/




















































































































































































































































































































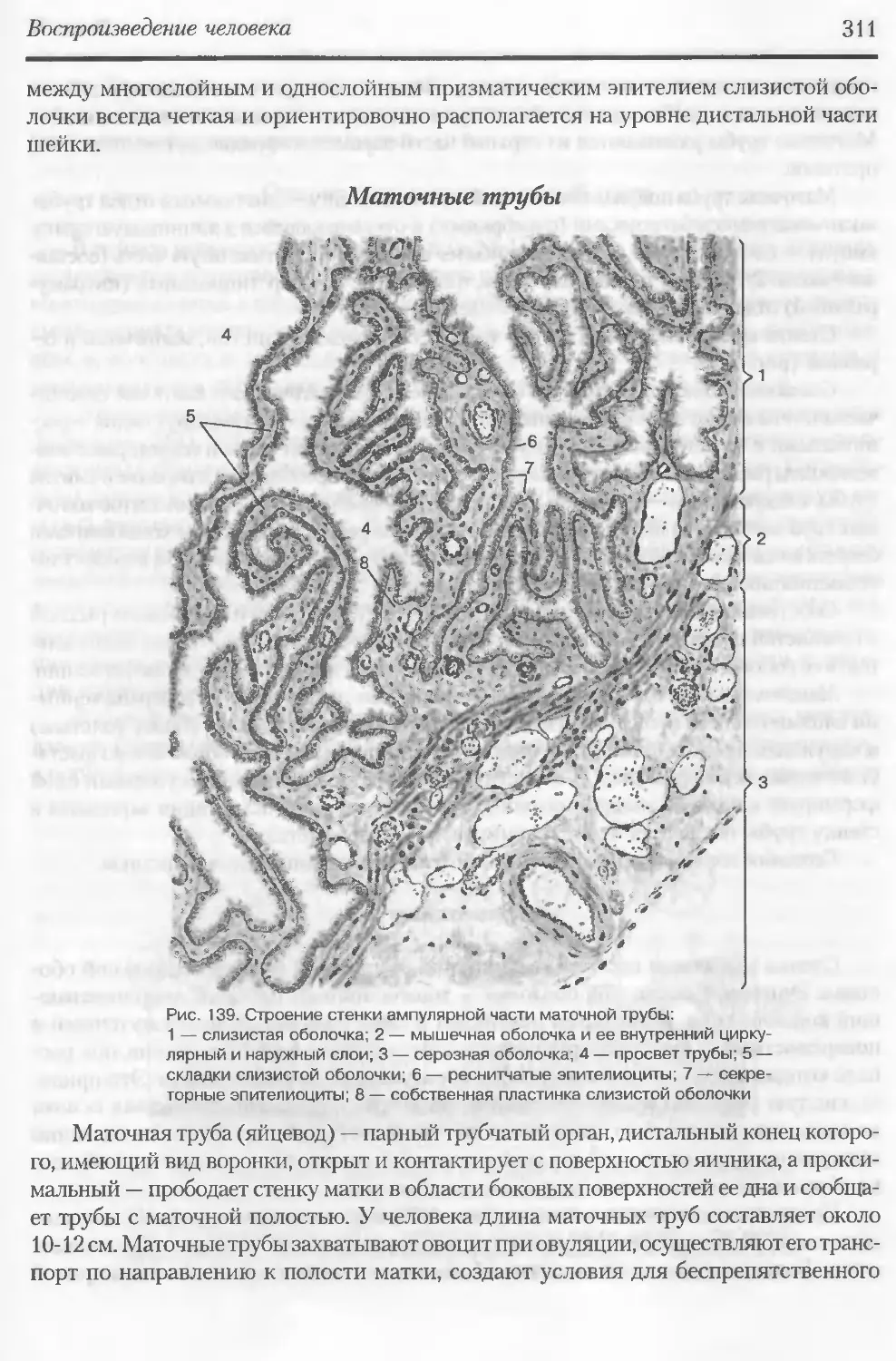








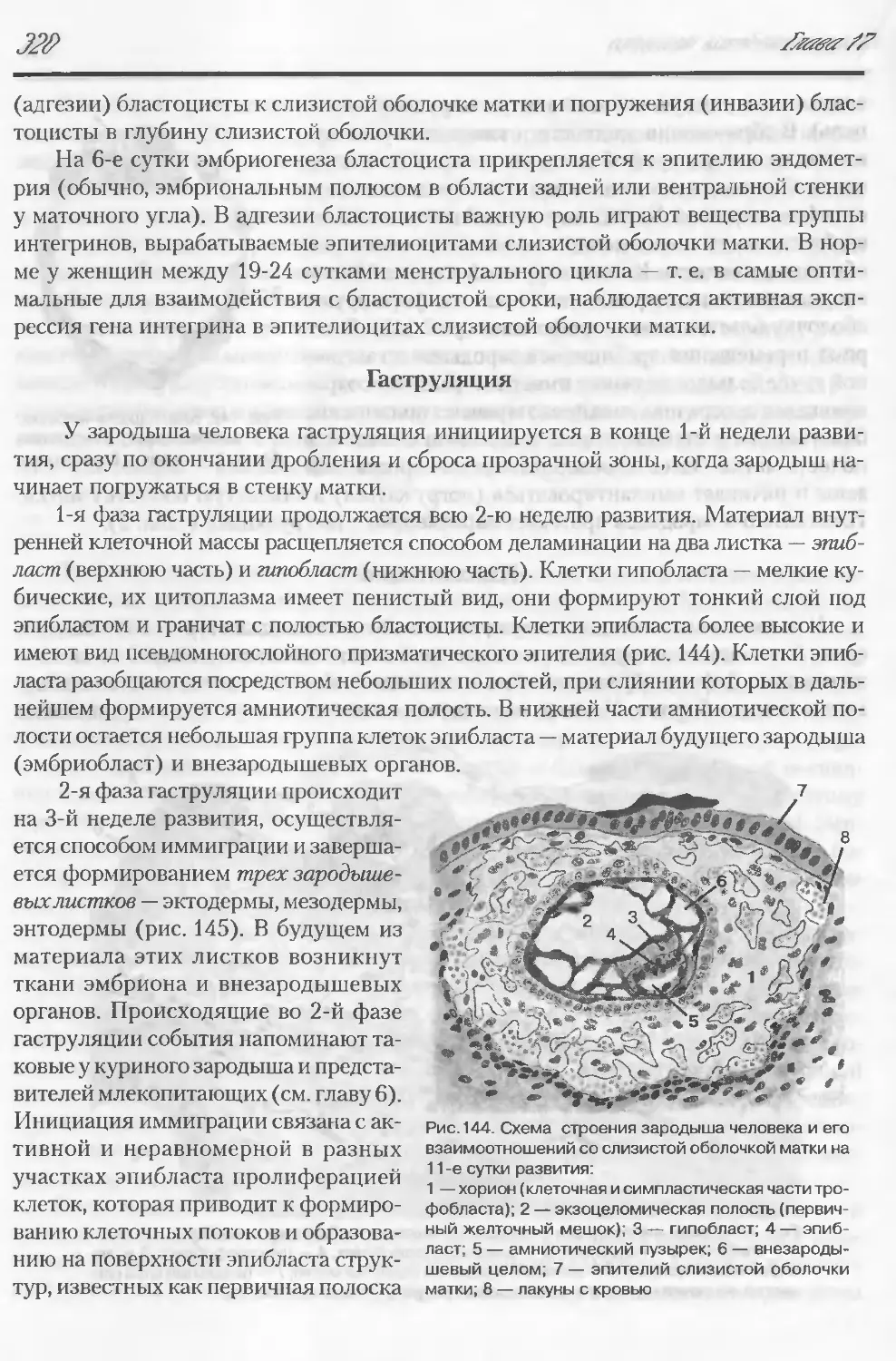















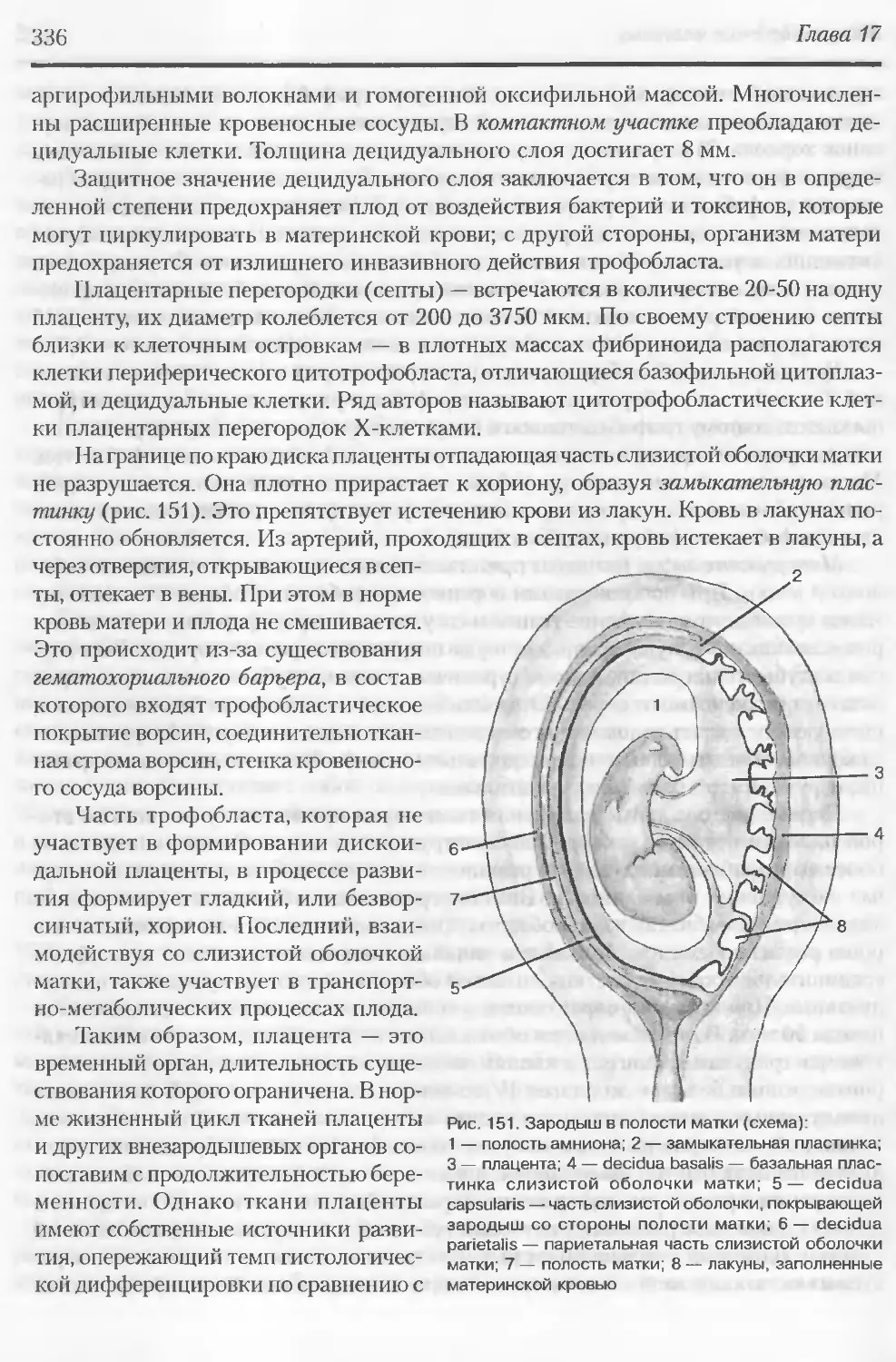




























Автор: Данилов Р.К. Клишов А.А. Боровая Т.Г.
Теги: анатомия анатомия человека сравнительная анатомия медицина гистология
ISBN: 5-93979-061-5
Год: 2003




Текст
Gi" 0('
A 8
r
T
r
Р.К. Данилов, А.А. Клишов, T.r. Боровая
-
'6
" ......
.... "9' О , . ,
lJ
. .
.. f1
, > . I
. ,
.. .
. .
.
. "
.р. ф ,
)
::--
...........
.
,
.....
.
Мультимедиа
ПJ}оrра мм а
k обучения
.. на СО ДИске
и ресурсы Интернет
А1шсросКDп и 'Fшpam pa. /J1ша;tuЮta J1. JI.
..
Учебная литература
для студентов медицинских вузов
K. Данилов,
А.А. Клишов,
T.f. Боровая
fистолоrия человека
в мультимедиа
Рекомендовано Управлением научных
и образовательных медицинских учреждений
Министерства здравоохранения
Российской Федерации в качестве учебника
для студентов медицинских вузов
51 {Q
.
БИБЛИОТЕКА
С.МI,.Петербурrско
rOCYA8pCfS8HHOi
медмцм 'H ко й
академии
.
CaHKT Петербурr
ЭЛБИ СПБ
2003
<О'\"\(О7)
,(l,J\'O:>
ББК
УДК 611/018
Д19
Данилов Р .К., КIIИШОВ А.А.. Боровая Т Т. rистолоrия человека в мультимедиа.
Учебиикдля студентов медицинских ВУЗОВ. СП6.: ЭЛБИ СПб_. 200З_ Зб2 с., ппл.
Это учебный комплекс, который включаеТ материалы, изложенные в виде
текста, а также на оптическом НQС(1теле (CD ROM) с ИСПОЛЬ30ВaJlИем: KOM
пьютерlIОЙ rрафики. анимации, аудио и ВИДСОТеХНИКИ, цифровой MaKpo и
микросъсмки анатомических и rиСТОЛOl"ических преПараТОВ. Наличие поис
копой системы ресурсов Интернет по аIlатомии, цитолоrии, эмбриолоrии и
rистолоrии 06еспечинает дистанционную поддсржку самостоятельнои рабо
ты путем обновления и дополнсния учебных электронных компонентов и
Дllалоrа с создателями учебника. Все это позволяет в paMк:l.'\: новой учебной
протраммы по ruстолorии (2002 Т.) дать сведения о важнейших биолоrn:чес
IШХ процессах, а также использовать мировые образоваТL'JIЫlые ресурсы для
достижения конечной цели получение знаний и умений.
Для курсантов, слушателей и иностранных воеННОСТIужащих, обучающихся
в ВоеННО "'1е,ДИЦIlНСКОЙ <l.кздсмии, и студентов медицинских институтов, пре
подавателей, аспирантов и врачей лаборантов-
Рецензенты: профессор, академик РАМН О.В.Волкова,
профессор, заслуженный деятель науки РФ в.д.Новиков,
профессор, ЧJlен корреспондент ВМедА и.В. rайворонский,
профессор, заслуженный работник высшей школы рф
Э.И.Вд.лькuвич
.
И'i..'iюстраЦl1U, вuдeoфраL.'lreJ)пы, текст!" u ajhpte ти уЧЕбною 1СО.ип
r "\.., _ . 1. "1r i:
.Ieкea '1Iе MOZym иыmь ри.эи<:. еНЫIJ .:'астu U Bocпp.o Qenы с UCllOлъзоваlluем
элп-тронных WlU U1lbI,X среdcпю,1 вк.I1РЧая Ф01fIО1\опlЧJоо uе u запись на .VlV
1luтный 1lOCuтCIb, без пllChveHH lO ра1рefflekия.. dвторов..;за UС1С.'iIOЧeНllе.V тех
суучаев, КOlда эmо необ.хООUJЮ обfjчающеvуся для реUlенШl тестов, сuту{щu
ОННЫ1: задач u J(01lтpa'ibHbL1. вопросов в pCLWlcaX учебных пршра,v...v_
ISBN 5 93979 061 5
@"ЭЛБИ СП6", 2003
@ Коллектив Авторов, 2003
ПРЕДИСЛОВИЕ
3
Развитие совремеШlOrо общества характеризуется повсеместным внедрением
во псе сферы деятельности человека ИllформаЦIIОlllrыл и коммуникационных Tex
нолоrиЙ, Естественно, что в сфере образования компьютерные технолоrии стали
неотъемлемым компонентом педаrоrическоrо процесса, предложив новые подхо
ды к изучению учебных матерналов, а также доступ к международным и нацио
на.1ьным uбразовательным ресурсам коллективною пользовання.
Учебный комплекс "rИСТОJlоrия человека в М'уЛЬТИ lепиа" это объединение
классичсскоrо учебника ПО rистолоrии, элеКТрОНllоrо учебника на оптическом
носителе (CD ROM), ресурсов Интернет по анатомии, цитолоrии, эмбриолоrии
11 rистолоrии и дистанционноЙ поддержки самостоятельноЙ работы путем обнов
леЮIЯ электронных компонентов и диалоrа с авторами труда с помощью вирту
3.%HOrO сервера для мuрфолоrов (http://wW\v\vin.wplus.net/pp/MediaMedic), Все
ЭТО позволяет B fCCTe с теоретичеСКIIМИ знаНИЯ),III, изложенными в привычном
для студента виде, с помощью компьютерной rрафики, видео, анимации, rиперс
сылок, поиска по ключевым словам и др., дать сведения о важнсliших биолоrи
ческих процессах в рамках изучаемоrо предмета, а также использовать мировые
оuразовательные ресурсы для ДОСТIIЖСНIIЯ конечной uеЛII получение знаний и
умений,
В учебнике наряду с фундаментальными проблемами большое внимание yдe
ляется прикладным военно меДИЦИIIСКИМ аспектам rистолоrии. Время показало,
что вопросы реактивиости и реrенерации тканеЙ при действии экстремальных
факторов среды. в том числе ш'нестрельноrо снаряда, не потеряли свою актуаль
ность Б связи С локальными конфликтами и распространением оружия среди Ha
селения. В текстовой части учебноrо комплекса наиболее сложно орrанизован
ные rистолоrические структуры и биолоrические процессы иллюстрированы
схемами. Однако- основной иллюстративный материал содержится на оптичес
ком носнтеле (CD ROM), Послсдний включает библиотеку анатомических и rи
столоrических прспаратов, схемы и электронные МlIкрофотоrрафllИ. словари Tep
"IIНОВ, предметныЙ указатель, план работы по конкретным темам, мультимедиа
тесты, учебные задачи, литературу, звуковые комментарии, анимацию важней
ШIIХ биолоrических процессов, видеофраrменты и краткую теоретическую ин
формацию о развитии, строении и функции все>. opraHoB и тканеЙ человека (Bce
ro более 1000 фаЙлов). ПОИСКОDая система ПОЗDоляет находить в сети Интернет
ресурсы по биЬлоrии человека, Быстрый поиск информации внутри электронно
ro учебника производится как по ключевым словам, так и по произвольно Ha
бранному в строке поиска тер IIIНУ.
4
Чтобы начать работу с оптическим носителем информации необходимо запус
тить проrрамму Setup в операционной среде Windows 3.1, 95, 98. Для этоrо кноп
кой Пуск открыть [лавное меню, переместить указатель мыши к строке ВЫПОk
нить и щелкнуть кнопкой мыши появится подменю Запуск проrраммы. В
текстовое поле Открыть необходимо внести строку: d:\mia\setup.exe и нажать кноп
ку ОК (при использовании CD ROM в дисководе с литерой "D"). Можно также
использовать кнопку Обзор для поиска необходимоrо файла. Далее ответить на
вопросы и следовать инструкциям проrраммы установки. Для запуска проrраммы
необходимо дважды нажать на иконку, появившуюся в результате инсталляции про
rpаммы, или строку "Cells and Tissues". Подробная инструкция по работе находит
ся в разделе "МедиаТур" оптическorо носителя.
Для реализации приложения «[истолоrия человека в мультимедиа в среде
Windows использован пакет проrрамм Multimedia Development Kit Version 1.0 фир
мы Мiсrosоft Corporation.
БЛАFОДАРНОСТИ
5
АВторы выражают rлубокую блаroдарность доцентам [-Я. [рафовой, Ю.К Хи
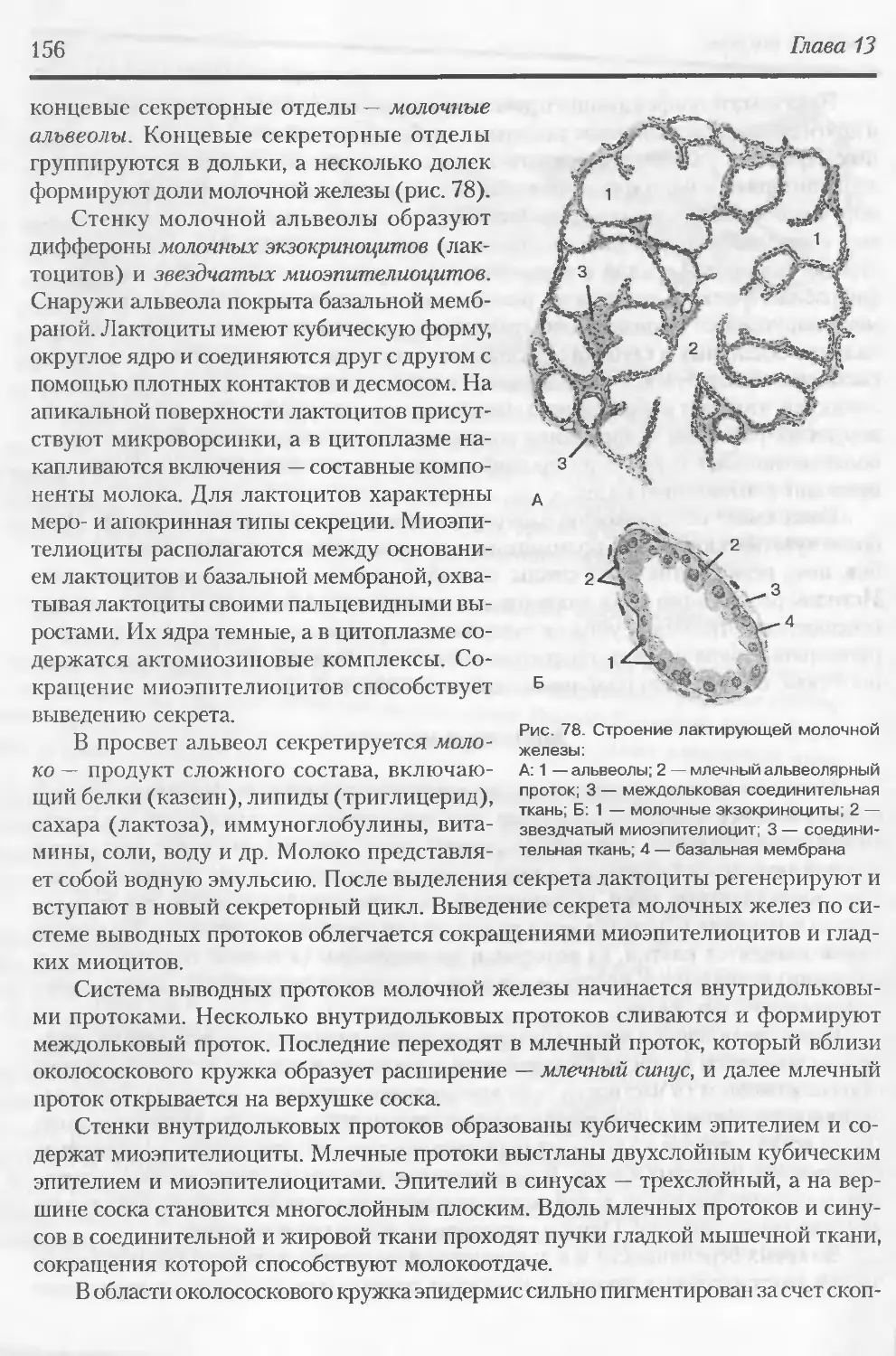
ловой, профессору B.r. [ололобову, старшему преподавателю Б.А [риrорян, всему
коллективу кафедры rистолоrии и эмбриолоrии Военно медицинской академии за
помощь в подrотовке препаратов к съемке, активное участие в обсуждении текстов
rлав учебника, за поиск материалов из киноархива кафедры и выбор фраrментов
фильмов, снятых доцентом З.Ф. Шавлаевым, машинописные работы и MHoroe дpy
roe, что было необходимо при подrотовке учебных материалов. Мультимедиа тесты
по пищеварительной системе составлены коллективом авторов под руководством
проф. Р.К Данилова. Некоторые схемы и рисунки с авторскими изменениями пере
несены из учебника профессора АА Клишова "[истолоrия человека" (из работ
В.Ф. Ивановой, В.В. [риroрьева, [ертвика иРокка, с.и. Щелкунова, Н.А Юриной,
л-с. Румянцевой, ю.и. Афанасьева, ЛЛ. Бобовой, B.r. Елисеева, АР. Далмане,
се Михайлова, B.r. Ясвоина, А Stevens, J. Lowe, J. Sobotta и др.),что, по мнению
авторов, необходимо для внедрения достижений науки в учебный процесс. Авторы
использовали изображения уникальных rистолоrических препаратов из личных KOk
лекций преподавателей и ученых (п.и. [алаховой, Л-В. Диденко, АЛ. Зашихина,
А.В. Павлова, Ас. Ростовщикова, В.Н. Семки на, [Н. Суворовой, Ю.А Хорошко
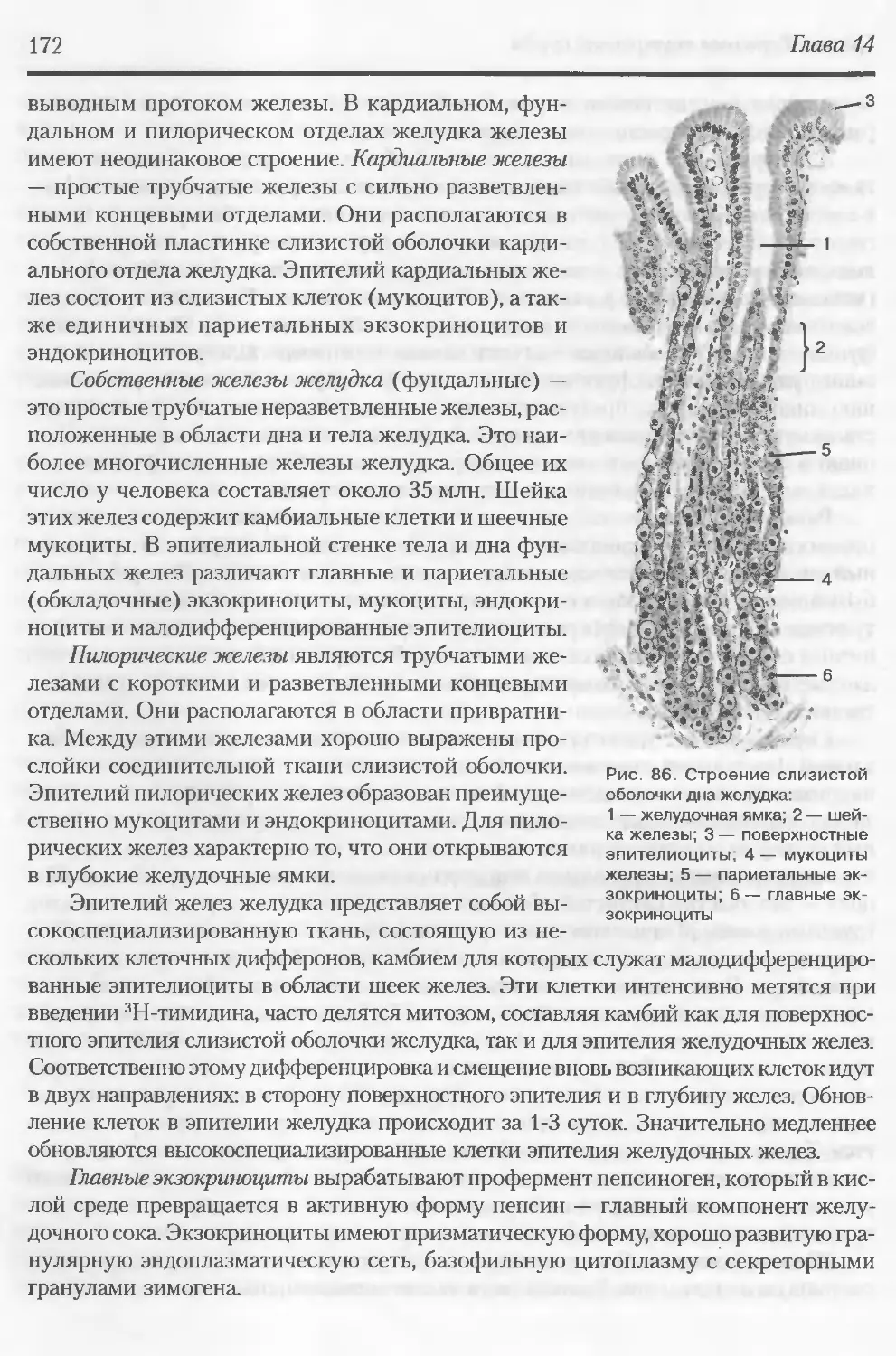
ва, М.и. Шашириной и др.), которые были заинтересованы во внедрении результа
тов научной работы в учебный процесс. Кафедры ВМедА любезно предоставили ви
деоматериалы по некоторым клиническим разделам. Часть иллюстраций выполнена
художником В.А Тюлюкиным.
Блаrодарим директора фирмы "Иста ВидеоТест" B.r. Пантелеева за техничес
кое сотрудничество и предоставление в аренду необходимой аппаратуры и про
rpaMM для цифровой обработки изображений микропрепаратов. Блаrодарим пре
подавателя ВМедА кандидата медицинских наук В.В. Ващенкова, оказавшеrо
помощь при освоении проrраммы и сотрудничество на первых этапах работы над
учебником. Блаrодарим профессоров О.В. Волкову, з.и. Вальковича, В.К Вери
на, В-Д. Новикова, Ю.В. Поrорелова, А.А Стадникова, АВ. Павлова, АН. [aHC
бурrскоrо, АЛ. Зашихина, СЛ. Кузнецова, Н.В. Ямщикова, преподавателей BMe
дА, студентов и курсантов за конструктивные советы при обсуждении концепции
данноrо учебноrо комплекса. Блаrодарим профессора И.В. [айворонскоro за пре
доставленные для съемки анатомические препараты.
Особую блаrодарность выражаем научным редакторам доцентам В.В. MOk
чановой и И.А: Одинцовой, труд которых во MHoroM способствовал ясности изло
жения учебноro материала.
Авторы уверены в том, что компьютерные технолоrии найдут свое место
6
в педаrоrическом процессе. Создание отечественных приложений позволит OCHac
тить персональные компьютеры и компьютерные классы, даст возможность бли
же познакомиться с принципами разработки мультимедийных при.iюжений и oцe
нить их эффективность в учебном процессе.
Возможно, что у читателя возникнет желание участвовать в развитии данноrо
проекта, обсудить и высказать критические замечания все будет принято с бла
rодарностью (E mail: rdanilov@mail.wplus.net).
ВВЕДЕНИЕ В rистолоrию
7
rЛАВА 1.
ПРЕДМЕТ И ЗАДАЧИ rnстолоrии. УРОВНИ
СТРУКТУРНО ФУНКЦИОНАЛЬНОЙ
орrлнизАЦИИ живоrо
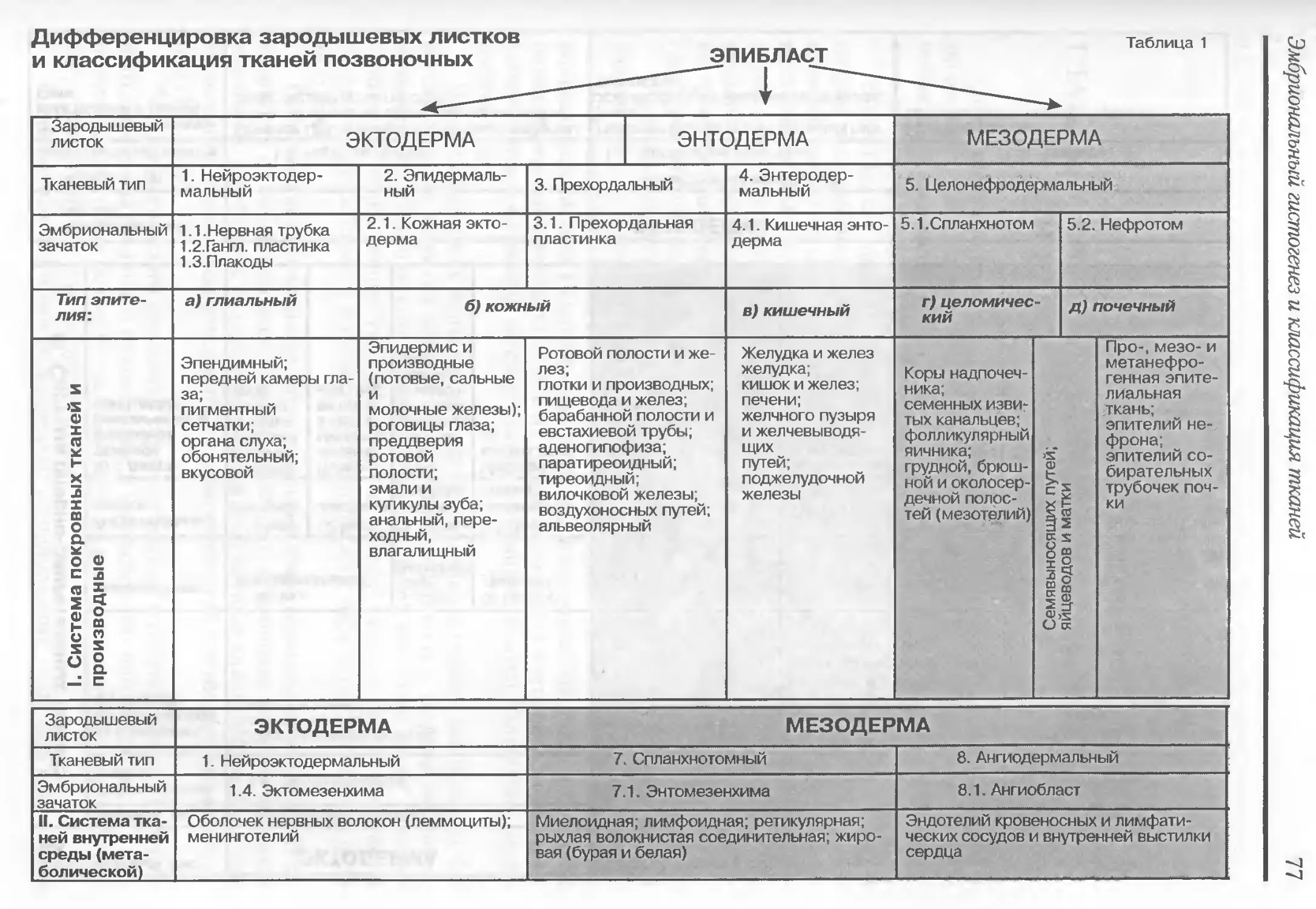
[истолоrия это наука, изучающая закономерности развития, строения и
функции тканей, а также межтканевые взаимодействия, в историческом и индиви
дуальном развитии человека и мноrоклеточных орrанизмов. Объект rистолоrии
ткани представляют собой филоrенетически сложившиеся, топоrрафически и
функционально связанные клеточные системы и их производные, из которых об
разованы opraHbI.
Как учебная дисциплина rистолоrия включает несколько разделов: 1) цито
лоrию учение о клетке; 2) эмбриолоrию науку о развитии зародыша, законо
мерностях закладки и образования тканей и opraHOB; 3) общую rистолоrию уче
ние о развитии, структуре и функциях тканей; 4) частную rистолоrию, изучающую
микроскопическое строение opraHOB и систем opraHoB.
Орrанизмы человека и животных являются целостными биолоrическими сис
темами, в которых условно можно выделить несколько взаимосвязанных, взаимо
действующих и соподчиненных уровней орrанизации молекулярный, субклеточ
ный, клеточный, тканевый и орrанный. Каждый из этих уровней обладает известной
автономностью и включает структурные единицы нижележащих уровней.
Орrанизменный уровень собственно орrанизм формируется как целост
ная биолоrическая система в процессе индивидуальнorо развития, именуемоrо
онтоrенезом.
Орrанный уровень включает комплекс взаимодействующих тканей в процессе
выполнения ими функций, свойственных данному конкретному opraHY или систе
ме opraHoB.
Тканевый уровень объединяет клетки и их производные. В состав тканей MOryT
входить клетки различной rенетической детерминации, однако основные свойства
тканей определяются ведущими клетками.
Клеточный уровень представлен основной структурно функциональной еди
ницей ткани клеткой и ее производными.
Субклеточный уровень включает структурно функциональные компоненты (KOM
партменты) клетки плазмолемму, ядро, цитозоль, орrанеллы, включения и др.
Наконец, молекулярный уровень характеризуется молекулярным составом клеточ
ных компонентов и механизмами их функционирования.
Представления об уровнях орrанизации и взаимосвязях различных уровней
позволяют рассматривать орrанизм как целостную и в то же время сложную
8
Fлава 1
иерархически соподчиненную систему. Структурные компоненты различных
уровней орrанизации живоrо являются объектом изучения разных медико био
лоrических дисциплин. В последние юды большое развитие получил комплекс
ный подход к изучению животных орrанизмов с использованием Bcero арсенала
методов и средств, которыми данные дисциплины располаrают. Это позволило
планировать и выполнять фундаментальные исследования и достиrнуть BЫCOKO
ro уровня знаний о структурно функциональной орrанизации живой материи, в
том числе орrанизма человека.
rлавное содержание rистолоrии как науки и учебной дисциплины составляют
закономерности rистоrенеза, морфофункциональной орrанизации, реактивности
и реrенерации тканей, выявленные на основе изучения большоrо фактическоrо
материала. Наиболее важное место среди теоретических достижений rистолоrии
занимают клеточная теория, теории зародышевых листков, эволюции тканей, rис
тоrенеза и реrенерации.
Актуальными задачами rистолоrии являются:
разработка общей теории rистолorии, отражающей эволюционную динами
ку тканей и закономерности эмбриональноrо и постнатальноrо rистоrенеза;
изучение rистоrенеза как комплекса координированных во времени и про
странстве процессов пролиферации, дифференциации, детерминации, интеrрации,
адаптивной изменчивости, проrраммированной rибели клеток и др.;
выяснение механизмов rомеостаза и тканевой реryляции (нервной, эндокрин
ной, иммунной), а также возрастной динамики тканей;
изучение закономерностей реактивности и адаптивной изменчивости кле
ток и тканей при действии неблаюприятных эколоrических факторов и в экстре
мальных условиях функционирования и развития, а также при трансплантации;
разработка проблемы реrенерации тканей после повреждающих воздействий
и методов тканевой заместительной терапии;
раскрытие механизмов молеКУЛЯРlю rенетической реryляции клеточной диф
ференцировки, наследования rенетическоrо дефекта развития систем человека, разра
6отка методов rенной терапии и трансплантации стволовых эмбриональных клеток;
выяснение процессов эмбриональноrо развития человека, критических пе
РИОДОВ развития, воспроизводства и причин бесплодия.
Изучение rистолоrии в медицинских вузах должно формировать у будущих Bpa
чей представление 06 уровнях структурно Функциональной орrанизации орrанизма
человека, их взаимосвязи и преемственности. rлубокие знания структуры и функции
орrанизма человека на всех уровнях ero орrанизации крайне необходимы COBpeMeHHO
му врачу, поскольку только на их основе возможно проведение квалифицированноrо
анализа этиопатоrенеза заболеваний и назначение патоrенетически обоснованной Te
рапии. Для медицины будущею, которая должна стать профилактической, знания о
структурных основах и закономерностях обеспечения устойчивости и надежности жи
вых систем (в том числе тканей) особенно важны, поскольку проrрессивное разви
тие цивилизации неизбежно влечет за собой появление новых факторов, неблаroпри
ятно воздействующих на животные орrанизмы, в том числе и человека.
Развитие микроскопии и открытие 'КЛетки
9
fЛАВА2.
РАЗВИТИЕ МИКРОСКОПИИ И ОТКРЫТИЕ
КЛЕТКИ. МЕТОДЫ ИССЛЕДОВАНИЯ
В rистолоrии
rистолоrия как самостоятельная наука выделилась в начале XIX века. Преды
сторию rистолоrии составили результаты мноrочисленных макроскопических (ви
зуальных) исследований составных частей различных животных и растительных
орrанизмов. Решающее значение для становления rистолоrии как науки о cтpoe
нии тканей имело изобретение микроскопа, первые образцы KOToporo были созда
ны в начале ХУН века (r. и З. Янсены, r. rалилей и др.). Одно из самых ранних
научных исследований с помощью микроскопа собственной конструкции провел
анrлийский ученый Роберт [ук (1635 1703). Он изучал микроскопическое CTpoe
ние мноrих предметов. Все изученные объекты Р. [ук описал в книrе "Микроrра
фия или некоторые физиолоrические описания мельчайших тел, выполненные при
посредстве увеличительных стекол...", изданной в 1665 [. Из своих наблюдений
р. [ук сделал вывод о широком распространении пузырьковидных клеток, или яче
ек, в растительных объектах и впервые предложил термин "клетка".
В 1671 [. анrлийский ученый Н. rрю (1641 1712) в своей книrе "Анатомия pa
стений" писал о клеточном строении как о всеобщем принципе орrанизации pac
тительных орrанизмов. Н. rрю впервые ввел в употребление термин "ткань" для
обозначения растительной массы, поскольку последняя напоминала по своей мик
роскопической конструкции ткани одежды. В том же [оду итальянец Дж. Мальпи
rn: (1628 1694) дал систематическое и детальное описание ячеистою (клеточноrо)
строения различных растений. В дальнейшем постепенно накапливались факты,
свидетельствующие о том, что не только растительные, но и животные орrанизмы
состоят из клеток Во второй половине ХУН века А. Левенrук (1632 1723) открыл
мир микроскопических животных и впервые описал красные кровяные тельца и
мужские половые клетки.
На протяжении Bcero ХУНI века происходило постепенное накопление фак
тов о клеточном строении растений и животных. Клетки животных тканей ПОk
робно исследовали и описали чешский ученый Ян Пуркиня (1787 1869) и ею уче
ники в начале XIX века.
Большое значение для развития знаний о микроскопическом строении opra
низмов имело дальнейшее усовершенствование микроскопов. В ХУI 11 веке мик
роскопы производились уже в большом количестве. В Россию они впервые были
привезены из rолландии Петром 1. Позднее при Академии наук в Петербурrе
была орrанизована мастерская по изrотовлению микроскопов. Для развития
10
Fлава 2
микроскопии в России MHoroe сделал М.В. Ломоносов, предложивший ряд Tex
нических усовершенствований конструкции микроскопа и ero оптической сис
темы. Вторая половина XIX века знаменательна бурным усовершенствованием
микроскопической техники. Были созданы новые конструкции микроскопов, и,
блаrодаря изобретению иммерсионных объективов (водная иммерсия стала при
меняться с 1850 r., масляная с 1878 r.), разрешающая способность оптических
приборов увеличилась в десятки раз. Параллельно с совершенствованием мик
роскопа развивалась и техника приrотовления микроскопических препаратов.
Если раньше объекты исследовали под микроскопом сразу после их выделения
из растений или животных без какой либо предварительной подrотовки, то Te
перь стали прибеrать к разнообразным методам их обработки, которые позволя
ли сохранять структуру биолоrических объектов. Были предложены разные спо
со бы фиксации материала. В качестве фиксирующих средств нашли применение
хромовая, пикриновая, осмиевая, уксусная и друrие кислоты, а также их смеси.
Простой и во мноrих случаях незаменимый фиксатор формалин впервые
был применен для фиксации биолоrических объектов в 1893 r.
Изrотовление препаратов, приrодных для исследования в проходящем свете,
стало возможным после разработки методов заливки кусочков в плотные среды, что
облеrчало получение тонких срезов. Изобретение специальных конструкций для
резки микротомов В лаборатории Я. Пуркиня значительно улучшило технику
изrотовления rистолоrических препаратов. В России первый микротом сконструи
ровал киевский rистолоr П.И. Перемежко. Для усиления контрастности структур
стали прибеrать к окрашиванию срезов различными красителями. Первым rистоло
rическим красителем, окрашивающим ядра клеток, нашедшим широкое примене
ние (начиная с 1858 r.), был кармин. Друrой ядерный краситель rематоксилин
стал применяться с 1865 r., однако долrое время ero свойства не были оценены в ПОk
ной мере. Ко второй половине XIX века уже употребляли' анилиновые красители,
были разработаны метод импреrнации тканей нитратом серебра (К rольджи, 1873)
и окраска нервной ткани метиленовым синим (А.с. Доrель, А.Е. Смирнов, 1887).
Блаrодаря фиксации биолоrическоrо материала и получению из Hero тончайших OK
рашенных срезов исследователи конца XIX века имели возможность значительно
rлубже проникнуть в тайны строения тканей и клеток, на основе чеrо был сделан ряд
величайших открытий. Так, в 1833 r. Р. Браун открыл постоянный компонент клет
ки ядро. В 1861 r. М. Шультце утвердил взrляд на клетку, как на "комочек прото
плазмы с лежащим внутри Hero ЯДрО I". rлавными составными частями клетки CTa
ли считать ядро и цитоплазму. В 70 x rодах XIX века rруппой исследователей
одновременно инезависимо друr от друrа был открыт непрямой способ деления кле
ток кариокинез, или мит03. В работах ид. Чистякова (1874), о. Бючли (1875),
Э. Страсбурrера (1875), В. Майзеля (1875), П.И. Перемежко (1878), В. Шлейхера
(1878), В. Флемминrа (1879) и др. были описаны и проиллюстрированы все стадии
непрямоrо клеточноrо деления. Это открытие имело большое значение для разви
тия знаний о клетке. Оно послужило также основой для более rлубокоrо изучения
TaKoro важнейшеrо биолоrическоrо процесса как оплодотворение. Изучение митоза
и оплодотворения при влекло особое внимание исследователей к ядру клетки и BЫ
Развитие микроскопии и открытие клетки
11
яснению ero значения в процесе передачи наследственных свойств. В 1884 [. о. [ep
твиr и Э. Страсбурrер независимо друr от друrа высказали rипотезу о том, что xpo
матин является материальным носителем наследственности. Объектом пристальноrо
внимания ученых стали хромосомы. Наряду с изучением ядра клетки, тщательному
анализу была подверrнута и цитоплазма.
Успехи микроскопической техники обусловили открьпие в цитоплазме орrанелл
постоянных и высокодифференцированных ее элементов, имеющих определенное
строение и выполняющих жизненно важные для клетки функции. В 1875 76 rr. He
мецким биолоrом о. rертвиrом и бельrиЙским ученым Ван Бенеденом был открыт
клеточный центр, или центросома; а в 1898 [. итальянским ученым К rольджи BHYT
риклеточный сетчатый аппарат (комплекс rольджи). в 1897 [. К. Бенда в животных
клетках, а в 1904 [. Ф. Мевес в растительных клетках описали хондриосомы, KOТO
рые позднее стали называться митохондриями.
Таким образом, к концу XIX века на основе успешноro развития микроскопи
ческой техники и анализа данных о микроскопическом строении клетки был Ha
коплен колоссальный фактический материал, позволивший выявить ряд важней
ших закономерностей в строении и развитии клеток и тканей. В это время учение о
клетке выделилось в самостоятельную биолоrическую науку цитолоrию.
Методы исследования в rистолоrии
Методы исследования в rистолоrии включают приrотовление rистолоrических
препаратов и их изучение с помощью световых или электронных микроскопов. rис
толоrические препараты представляют собой мазки, отпечатки opraHoB, пленочные
препараты, тонкие срезы кусочков opraHoB, окрашенные тем или иным красителем
(исследуются также нативные неокрашенные срезы), помещенные на предметное
стекло, заключенные в бальзам и покрытые тонким покровным стеклом.
Для изroтовления rистолоrическоro препарата необходимо после взятия Ma
териала произвести ero фиксацию в том или ином фиксаторе (формалине, спирте,
а для электронной микроскопии в rлутаровом альдеrиде и четырехокиси осмия).
Делается это для предотвращения процессов аутолиза и сохранения структуры opra
на, близкой к прижизненной. Далее следуют этапы обезвоживания кусочка opraHa
в спиртах возрастающей концентрации и в ксилоле с целью уплотнения тканей,
что необходимо для изrотовления тонких срезов. Для придания кусочку opraHa
еще большей плотности и rомоrенности, обеспечивающей высококачественную
резку, проводят ero заливку в орrаническую среду парафин, целлоидин (для CBe
товой микроскопии) и орrанические смолы (эпон, аралдит, дуркупан) для элек
тронно микроскопическоrо исследования.
Существуют также физические способы фиксации материала, наиболее pac
пространенным из которых является быстрое замораживание кусочка opraHa с по
мощью ЖИДКQro азота и друrих средств. Для резки замороженноrо материала ис
пользуют специальные приборы криостаты, или замораживающие микротомы.
Толщина срезов, предназначенных для световой микроскопии, не должна
превышать 4 5 мкм, для электронной 50 60 нм (такие ультратонкие срезы
12
Fлава 2
изrотавливают на специальном приборе ультратоме, используя стеклянные или
алмазные ножи и автоматический режим резки). .
После получения срезов их помещают на предметные стекла, далее следуют
этапы освобождения срезов от заливочной среды (при световой микроскопии) и
окраски для придания срезам контрастности. Среди rистолоrических красителей
наиболее часто употребляется сочетание rематоксилина, маркирующеrо ядро (кис
лотные молекулы), и эозина, избирательно окрашивающеrо белковые молекулы
(цитоплазматический краситель).
По окончании окрашивания срезы заключают в консервирующие среды (Ka
надский, кедровый бальзамы) и накрываются покровным стеклом.
Основным методом rистолоrnческоrо исследования клеток, тканей и opraнoB явля
ется световая микроскопия. В световом микроскопе для освещения объекта использу
ются лучи видимою спектра. Современные световые микроскопы позволяют получать
разрешение порядка 0,2 мкм (разрешающая способность микроскопа это то наимень
шее расстояние, при котором две рядом расположенные точки видны как отдельные).
Разновидности световой микроскопии фазово контрастная, интерференционная, по
ляризационная, темнопольная и др.
Фазово конmрасmная микроскопия метод изучения клеток в световом мик
роскопе, снабженном фазово контрастным устройством. Блаrодаря смещению фаз
световых волн в микроскопе такой конструкции повышается контрастность CTPYK
тур исследуемоrо объекта, что позволяет изучать живые клетки.
Инmерференционная микроскопия. В интерференционном микроскопе па
дающие на объект световые пучки раздваиваются один пучок проходит через
объект, друrой идет мимо. При последующем воссоединении пучков возни
кает интерференционное изображение объекта. По сдвиrу фаз одноrо пучка
относительно друrоrо можно судить о концентрациях различных веществ в ис
следуемом объекте.
Поляризационная микроскопия. В микроскопах этоrо типа световой пучок раз
лаrается на два луча, поляризованных во взаимно перпендикулярных плоскостях.
Проходя через структуры ткани со строrой ориентацией молекул, лучи запаздыва
ют друr относительно друrа вследствие неодинаковоrо их преломления. Возника
ющий при этом сдвиr фаз является показателем двойною лучепреломления кле
точных структур (таким способом были исследованы, например, миофибриллы).
Люминесценmная, Wlи флюоресценmная, микроскопия метод rистолоrическою
анализа с помощью люминесцентноrо микроскопа, в котором используется явление
люминесценции (свечения) веществ при действии на них коротковолновых лучей
(ультрафиолетовою света, рентrеновских лучей). Некоторые биолоrические соеди
нения, присутствующие в клетках, характеризуются спонтанной флюоресценцией
при попадании на клетку ультрафиолетовых лучей. Для выявления же большинства
друrих соединений клетки обрабатываются специальными флюорохромами (флюо
ресцеином, акридином оранжевым, корифосфином). С помощью флюорохромов
исследуют, например, содержание в клетках нуклеиновых кислот. При окраске aK
ридином ДНК дает красно зеленое свечение, а РНК оранжевое.
Люминесцентный микроскоп широко используется также для изучения им
Развитие микроскопии и открытие 1Ulетки
13
мунофлюоресценции. Иммуиофлюоресценция позволяет исследовать в клетке co
держание очень малых количеств белка. Препарат предварительно обрабатывают
антителами к исследуемому белку, меченными флюоресцирующим красителем.
Ультрафиолетовая микроскопия метод изучения клеток с помощью микро
скопов, В которых для освещения объекта используют ультрафиолетовые лучи (длина
волны которых равна 210 275 нм). Такие микроскопы имеют большую, чем обыч
ные световые микроскопы, разрешающую способность. Для наблюдения за объек
том требуется специальная аппаратура электронно оптический преобразователь,
который предохраняет opraH зрения от действия ультрафиолетовых лучей.
Электронная микроскопия. В электронных микроскопах используют пучок
электронов, длина электромаrнитной волны которых в 100 000 раз короче длины
волны видимоrо света. Разрешающая способность электронноrо микроскопа в сотни
и тысячи раз превышает обычные оптические приборы и равна 0,5 1 нм, а COBpe
менные меrавольтные электронные микроскопы дают увеличение до 1 000 000 раз.
е помощью электронных микроскопов получены мноrочисленные данные об уль
траструктуре клеток. Разновидностью электронной микроскопии является скани
рующая электронная микроскопия (СЭМ), при которой изучаются поверхност
ные структуры клеток.
Цитоспектрофотометрия метод изучения химическою состава клетки, oc
нованный на избирательном поrлощении теми или иными веществами лучей с оп
ределенной длиной волны. По интенсивности поrлощения света, которая зависит
от концентрации вещества, производится количественное определения ero coдep
жания в клетке.
Радиоавтоzрафия важный информативный метод, позволяющий изучать pac
пределение в клетках и тканях веществ, в состав которых искусственно введены
радиоактивные изотопы ен, I1С, 32р и др.). Введенный в орrанизм животноrо (или
в среду культивирования клеток) изотоп включается в соответствующие CTPYKTY
ры (например, меченый тимидин в ядра клеток, синтезирующих днк). Метод
основан на способности включенных в клетки изотопов восстанавливать бромис
тое серебро фотоэмульсии, которой покрывают срезы ткани или клетки. Образую
щиеся после проявления фотоэмульсии зерна серебра (треки) служат своею рода
автоrрафами, по локализации которых судят о включении в клетку примененных
веществ. Применение меченных тритием предшественников нуклеиновых кислот
(тимидина, аденина, цитидина, уридина) позволило выяснить мноrие важные ac
пекты синтеза днк, рнк и клеточных белков.
[исто и uммуноцитохuмические методы. В их основе лежит применение хи
мических реакций для выявления распределения химических веществ в CTPYK
турах клеток, тканей и opraHoB. Современные rистохимические методы позволя
ют обнаруживать аминокислоты, белки, нуклеиновые кислоты, различные виды
уrлеводов, липидов и др. Для выявления специфических белков используют им
муноцитохимические реакции. Для этоrо получают специфические сыворотки, co
держащие антитела (например, против белка микротрубочек тубулина). Далее хи
мическим путем соединяют эти антитела с флюорохромом (или друrим маркером).
Если меченые антитела нанести на rистолоrический срез, они вступают в
14
[Лава 2
соединение с соответствующими белками клетки и возникает специфическое CBe
чение, видимое в люминесцентном микроскопе. Современные иммуноцитохими
ческие методы, помимо флюорохромов, используют друrие самьте разнообразные
специфические маркеры, позволяющие качественно и количественно оценивать
содержание в клетке исследуемых соединений. Модификацией рассматриваемоrо
метода является введение меченых антител в цитоплазму живых клеток с помо
щью микроманипуляторов.
Метод культуры 'КЛеток, тка1lей заключается в выращивании клеток и тканей
вне орrанизма в искусственных питательных средах (в условиях in vitro). Для по
лучения изолированных клеток производят предварительную обработку материа
ла ферментами трипсином или коллаrеназой. Метод позволяет изучать реакции
клеток на различные воздействия, механизмы реryляции пролиферации, диффе
ренцировки и rибели. Особенно важное значение данный метод имеет для эмбрио
лоrических и цитофизиолоrических исследований, а также для трансплантации
эмбриональных клеток при лечении врожденных и приобретенных дефектов об
мена веществ.
Микроскопическая хируршя 'КЛетки совокупность методических приемов,
осуществляемых с помощью специальноrо прибора микроманипулятора. Этот
при бор позволяет производить различноrо рода тончайшие операции на клетке
(введение веществ, удаление или пересадка структурных компонентов клетки, Ha
несение уколов, разрезов и пр.) и нашел широкое применение в эмбриолоrии.
Цейтраффер1lая, или замедле1-l1lая, МИКрО1Ш1l0 или видеосъемка изучение жи
вых клеток Такой способ позволяет проследить за медленно протекающими измене
ниями клеток
Метод фраКЦИО1lирова1lИЯ (дифференциальноrо центрифуrирования) клеток
Ero суть заключается в получении из клеток изолированных структурных компо
нентов. Основан на разных скоростях осаждения этих компонентов при вращении
rOMoreHaToB клеток в ультрацентрифуrах. Данный метод сыrpал и иrрает очень важ
ную роль в изучении химическоrо состава и функциональных свойств субклеточ
ных элементов прежде Bcero, орrанелл.
КО1lфОКШlЬ1lая микроскопия современныЙ метод, использующий в качестве
осветителя лазерный луч, который последовательно сканирует всю толщину пре
парата. Информация о плотности объекта по каждой линии сканирования переда
ется в компьютер, [де специальная проrрамма осуществляет трехмерную peKOHCT
рукцию исследуемоrо объекта.
Следует отметить, что здесь приведены лишь самые основные, необходимые
для rистолоrическorо образования врачей всех профилей методы rистолоrическо
[о анализа. Существуют мноrие десятки друrих методов, с которыми можно озна
комиться в специальной rистолоrической и цитолоrической литературе.
Краткий исторический очерк
15
rЛАБАЗ.
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК
РАЗБИТИЯ УЧЕНИЯ О КЛЕТКЕ И ТКАНЯХ
Развитие методов микроскопии, возникновение rистолоrических школ послу
жило основой для перехода от периода накопления фактов к периоду формирова
ния научных теорий и концепций. Большое влияние на развитие учения о клетке и
тканях в XIX веке оказали работы М. Шлейдена (1804 1881), Ф. Лейдиrа (1821 1908),
И. Мюллера (1801 1858), Т. Шванна (181O 1882), Р. Вирхова (1821 1902), Р. Келли
кера (1817 1905) и др. Хотя мноrие исследователи высказывали положение о кле
точном строении орrанизмов, только Т. Шванн в своей моноrрафии "Микроскопи
ческое исследование о соответствии в структуре и росте животных и растений" (1839)
ясно сформулировал основные положения клеточной теории. Важнейший вывод дaH
ной теории состоял в том, что клетки представляют собой элементарные универ
сальные структурные единицы всех растений и животных.
Вскоре после опубликования книrи Т. Шванна австрийский rистолor А. Kek
ликер распространил положения клеточной теории на ранние стадии эмбриональ
Horo развития орrанизма. В 1841 1844 П. он показал, что сперматозоид и яйцо яв
ляются клетками. Из клеток состоит и орrанизм (зародыш), возникающий в ходе
дробления оплодотворенной женской половой клетки.
Параллельно с развитием клеточной теории складывались представления о том,
что клетки в составе орrанизма образуют системы более BbIcoKoro порядка TKa
ни. В 1801 r. французский анатом М.Ф.К Биша (1771 1802) на основе микроско
пических исследований предложил первую классификацию тканей. Ero ученик
КМайер ввел термин "rистолоrия" в изданном в 1819 r. труде "О rистолоrии и HO
вом подразделении тканей человеческоrо тела".
Во второй половине XIX века стало общепризнанным, что клетки в составе
мноrоклеточных животных существуют не как самостоятельные, изолированные
единицы, а как части тканей. Эмпирически было выделено 4 типа тканей: эпители
альные, соединительные, мышечные и нервная. Эта классификация тканей нашла
отражение в учебных руководствах Ф. Лейдиrа (1853) и А. Келликера (1855).
В России первые самостоятельные кафедры rистолоrии и эмбриолоrии были уч
реждены в 60 x roдах XIX века в Медико хирурrической (ныне Военно медицинс
кой) академии, Московском и Петербурrском университетах. Несколько позже они
были основаны в Казанском, Киевском и Харьковском университетах. Первыми про
фессорами руководителями кафедр rистолоrии были Н.М. Якубович, А.И. Бабу
хин, Ф.В. Овсянников, Ас. rолубев, П.И. Перемежко, Н.А Хржонщевский. С появле
нием кафедр начали формироваться и первые rистолоrические научные школы
Петербурrская, Московская, Харьковская и др.
16
Fлава 3
/1
Петербурrская (ленинrрадская) школа rистоло
rOB формировалась в стенах Медико хирурrической
академии, так как здесь в числе одной из первых в
России была основана кафедра rистолоrии. Впервые
в России систематическое преподавание rистолоrии
и эмбриолоrии было введено в академии КМ. Бэром,
руководившим кафедрой сравнительной анатомии и
физиолоrии в 1841 1852 п. Первым профессором,
специализировавшимся в области rистолоrии, был
Н.М. Якубович (1817 1879), он и возrлавил образо
вавшуюся самостоятельную кафедру rистолоrии Me
дико хирурrической академии (в 1868 r.).
Становление кафедры как учебноrо и научноrо
подразделения академии проходило под PYKOBOД
ством профессоров ф.н. 3аварыкина (1835 1905)
и м-д. Лавдовскоrо (1846 1903). МД. Лавдовский
вместе с московским профессором Ф.В. Овсянни
ковым (1826 1906) был редактором первоrо отече
cTBeHHoro руководства "Основания к изучению мик
роскопической анатомии человека и животных"
(1887 1888).
Почти 20 лет (с 1903 по 1922 п.) начальником
кафедры rистолorии был член корреспондент Poc
сийской академии наук профессор А.А. Максимов
(1874 1928), который является признанным OCHO
воположником учения о крови и соединительной
ткани. Он экспериментально обосновал унитарную ,:1, .....".-;::J;:..
теорию кроветворения, первым применил метод
прижизненных окрасок для цитолоrических иссле
дований, создал образцовый кабинет для культур
тканей и выполнил с помощью этих методов ряд BЫ
дающихся работ.
Особенно оживилась работа с приходом в aKaдe
мию в 1922 r. профессора А.А. 3аварзина (1886
1945) выдающеюся биолоrа и CTpaCTHoro патрио
та нашей Родины, ставшеrо впоследствии
академиком АН и АМН СССР, лауреатом Сталинс
кой премии СССР 1 й степени. Уже в 30 e rоды А.А. 3аварзин на основе rлубокоrо
сравнительно rистолоrическоrо изучения нервной системы сформулировал прин
цип параллелизма тканевых структур, переработанный позднее в теорию тканевой
эволюции. Он сделал вывод, что все животные имеют общий принцип тканевой opra
низации и состоят из 4 тканевых систем. Это связано с тем, что всякий орrанизм
находится в одинаковых условиях взаимодействия с окружающей средой и ВЫПОk
няет 4 наиболее общие функции защитную, внутреннею обмена и постоянства
''1\11
t ;. .« '.
"\
, ;';\
."""'
i: ,-
t.
, {.
h .f
Александр Александрович
Максимов
(1874-1928)
... . t....
,"i)lJ{... ...,,:,\- ..
. ; .
\
"
,
,;:
f..
--',
,'1.
,/
'.
'.
,
Алексей Алексеевич
Заварзин
(1886 1945)
Краткий исторический очерк
\<
17
внутрснней среды, движения, реактивности. А.А. 3a
варзин обосновал морфофункциональную классифи
кацию тканей. Теория А.А. 3аварзина называется Te
', .1 орией параллельных рядов тканевой эволюции.
lii1 Идеи А.А. 3аварзина оказали большое влияние
на развитие rистолоrии и мноrих друrих медико био
лоrическихдисциплин. Теория параллелизмов в раз
витии тканей нашла дальнейшее обоснование в pa
ботах сотрудников и учеников А.А. 3аварзина
(академик АН СССР Ю.А. Орлов, академик АМН
СССР H.f. Хлопин, члены корреспонденты АМН
СССР Ф.М. Лазаренко, СИ. Щелкунов, [С CTpe
лин, профессора Е.С Данини. [в. Ясвоин. А.А. Бра
ун, дс Сутулов И др.).
С 1936 r. по 1955 r. кафедру академии возrлав
лял академик АМН СССР, лауреат Сталинской пре
мии СССР 1 й степени профессор НТ. Хлопин
(1897 1961). Им и ero школой (член корреспондент
.....
>\( АМН СССР A.f. Кнорре, профессора Я.А. винни
.n ков, шд. fалустян, Н.И. fриroрьев, Н.Н. Кочетов, А.С. Лежава, вл. Михайлов,
...... В.Е. Цымбал, Н.А. Шевченко и др.) разработана теория диверrентноro развития
'I) тканей. Соrласно этой теории, эволюционное развитие тканей происходит прин
ципиально так же, как и орrанизмов, подчиняясь тем же основным закономерно
стям, что и целые орrанизмы. Как известно, орrанизмы развиваются диверrент
но, Т.е. расхождением признаков, блаrодаря чему и возникает мноrообразие форм.
Диверrентная эволюция тканей имеет, однако, свою специфику, в силу чеrо MHO
rообразие тканей оrраничено известными пределами признаков, свойственных 4
основным системам тканей. H.f. Хлопиным была предложена rенетическая клас
сификация тканей, обоснованная в монorрафии "Общебиолorические и экспе
риментальные основы rистолоrии" (1946).
Теории параллельноrо и диверrентноrо развития тканей дополняют друr дpy
ra, отражая разные стороны сложнorо процесса эволюции структур TKaHeBoro ypOB
ня орrанизации. Теория диверrентной эволюции раскрывает направление разви
тия тканей, связанное с rенетическим проrраммированием пути развития тканей.
Теория параллельных рядов развития тканей отражает результат и возможности
адаптивных изменений тканей при их функционировании в сходных условиях вза
имодействия орrанизмов с внешней средой.
В 1957 1977 rr. под руководством члена корреспондента АМН СССР профес
сора СИ. Щелкунова (1904 1977) кафедра разрабатывала различные вопросы rис
тоrенеза, реактивности и реrенерации тканей в условиях нормы и патолоrии. Были
изучены закономерности реактивных изменений тканей при опухолевом росте и
после действия различных экстремальных факторов. Соrласно взrлядам СИ. Щеk
кунова, развитие тканей определяются следующими основными принципами эво
люционной rист -'j1е fPffl1f<' U (наследственной обусловленностью),
C'Mkt'IIp.T p(\yprol(a"
rCH; уца ре T.eH O"
tlfJаИUIIНGКО"
,"'1 ·
..,
:;.. .
вi:
'-,4
".
.., Н.... J
fi:J>'........, " .
--'.- .
.. "d"
...
)
Николай rриrорьевич
Хлопин
(1897 19б1)
18
Fлава 3
rетерохронией (или разновременностью развития),
интеrрацией (или взаимозависимостью и взаимо
обусловленностью раЗБИТИЯ).
Дальнейшее развитие учения о rистоrенезе и
реrенерации получило в работах члена корреспон
дента АМН СССР профессора A.f. Кнорре (1914
1981) и профессора А.А. Клишова (1930 1991). По
А.С Кнорре, элементарными компонентами rисто
rенеза являются: клеточное размножение, клеточ
ный рост, клеточные перемещения (миrрации),
дифференциация клеток и их неклеточных произ
водных, межклеточные и межтканевые взаимодей
ствия (корреляции), отмирание клеток (клеточный
некробиоз и некроз). В ходе эмбриональноrо rис ...
тоrенеза образуются различные ткани, входящие в
состав opraHoB и систем opraHoB.
Профессор А.А. Клишов, развивая научные
традиции и достижения своих учителей, внес HO
Бое в разработку проблем rистоrенеза, реактивно
сти и реrенерации тканей, сделав акцент на изучении этих процессов в экспери
ментальных условиях. Под ero руководством были развернуты исследования
закономерных процессов rистоrенеза и реrенерации различных тканей с исполь
зованием современных методов исследования. Это позволило сформулировать
концепцию системно структурной орrанизации rистоrенеза, соrласно которой
развитие тканей продолжается до тех пор, пока живет орrанизм. [истоrенез вклю
чает взаимодействующие и взаимоперекрывающиеся базисные процессы дe
терминацию, пролиферацию, дифференциацию, интеrрацию и адаптацию кле
ток В восстановительном процессе ведущую роль иrрают закономерности
нормальноrо перманентноrо rистоrенеза, определяющие развитие и состояние
ткани до повреждения. Иными словами, после повреждения ткани продолжают
СБое развитие, но в иных, экстремальных условиях.
СущественныЙ вклад в разработку проблем rистоrенеза и реrенерации различ
НЫХ тканей внесли отечественные rистолоrи А.Н. Миславский, Б.И. Лаврентьев,
H.r. Колосов, Б.В. Алешин, АЛ. Поленов, В.И. Архипенко, П.В. Дунаев, H.f. Xpy
щов и др.
Проблема реактивности тканей, особенно соединительной ткани, явилась преk
метом исследований B.f. Елисеева и ero школы (Ю.И. Афанасьев, юя. Копаев,
НА Юрина и др.). Закономерности реактивности, компенсации и адаптации TKa
ней изучены КА. Зуфаровым и ею учениками.
Реrенерация тканей одна из актуальных медико биолоrических проблем, в
разработку которой внесли определенный вклад А.Н. Студитский, ВЛ. Михай
лов, Л.Н. Жинкин, [.с. Стрелин и др. Эмбриолоrические исследования А.Н. Ce
верцова, ПЛ. Иванова, Д.П. Филатова, п.r. Светлова, Б.П. Токина, A.f. Кнорре,
О.В. Волковой и др. являются достойным продолжением К.lJаСfических работ KO
. Щ ". н . .... _\
., н,,
...i'"
7<
, ,
i -,
L./" ' '
Алексей Андреевич
Клишов
( 1930 1991)
Краmкий исmорический очерк
19
рифеев эволюционной эмбриолоrии XIX столетия. Исследования Д.Н. HaCOHO
ва, вя. Александрова, А.А. Заварзина (мл.), п.п. Румянцева и др. расширили
представления о функциональном значении орrанелл, цитохимии, цитофизио
лоrии, клеточной адаптации и др_
20
цитолоrия
rЛАВА 4.
ОСНОВЫ ОБЩЕЙ цитолоrии
Цитолorия наука об общих закономерностях развития, строения и функций
клеток Клетка (лат. cellula) это микроскопической величины живая система,
оrраниченная биолоrической мембраной, состоящая из ядра и цитоплазмы, обла
дающая свойствами раздражимости и реактивности, реryляции состава BHYTpeH
ней среды и самовоспроизводства. Клетка является основой развития, строения и
функций всех животных и растительных орrанизмов (рис. 1). Как обособленная
единица живою она обладает признакам и индивидуальною целою. В то же время
в составе мноюклеточных орrанизмов клетка является структурной и функцио
нальной частью целоrо. Если в одноклеточных орrанизмах клетка выступает в роли
индивидуума, то в мноrоклеточных животных орrанизмах различают соматичес
кие клетки, составляющие тело орrанизма, и половые клетки, обеспечивающие
воспроизведение орrанизмов.
Современная цитолоrия представляет собой науку о природе и филоrенети
ческих связях клеток, основах их функций и специальных свойств. Следует OTMe
тить особое значение цитолоrии для медицины, так как в основе развития патоло
rических состояний лежит, как правило, патолоrия клетки.
Несмотря на крупные достижения в области современной биолоrии клетки,
непреходящее значение для развития идей о клетке имеет клеточная теория.
В 1838 r. немецкий зоолоr исследователь Т. Шванн впервые указал на rOMo
лоrичность, или сходство, клеток растительных и животных орrанизмов. Позже он
сформулировал 1Ulеточ1l.УЮ теорию строения орrанизмов. Поскольку при созда
нии этой теории Т. Шванн широко использовал результаты наблюдений HeMeЦKO
ro ботаника М. Шлейдена, последнеrо по праву считают соавтором клеточной Teo
рии. Стержнем теории Шванна Шлейдена является тезис о том, что клетки
представляют собой структурно функциональную основу всех живых существ.
В конце XIX столетия немецкий патолor Р. Вирхов пересмотрел и дополнил
клеточную теорию собственным важным выводом. В книrе "Целлюлярная патоло
rия, как учение, основанное на физиолоrической и патолоrической rистолоrии"
(1855 1859), он обосновал фундаментальное положение о преемственности кле
точноrо развития. Р. Вирхов в противоположность Т. Шванну, отстаивал взrляд на
образование новых клеток не из цитобластемы бесструктурной живой субстан
ЦИИ, а путем деления предсуществующих клеток (Omnis cellula е cellula). Лионс
кий патолоr Л. Барр подчеркнул специфичность тканей, дополнив: "Каждая клет
ка от клетки той же природы" .
Первое положение клеточной теории в ее современной трактовке rласит клет
ка является элементарной структурно функциональной единицей живой материи.
Основы общей цumОЛОlUU
21
Второе положение
указывает на то, что
клетки разных орrаниз
мов rомолоrичны по
своему строению. [OMO
лоrичность подразуме 5
вает сходство клеток по
основным свойствам и
признакам и отличие
по второстепенным. [o "
молоrичность строения
определяется общекле
точными функциями,
которые направлены на 2
поддержание жизни
клеток и их воспроиз
водство. В свою очередь, 1
разнообразие в строении
является результатом
Функциональнойспеци
ализации клеток, в OCHO 8
не которой лежат моле
кулярные механизмы
активации и репрессии 12
reHoB, составляющие по
нятие "клеточная дeTep
ми нация" .
Третье положение
клеточной теории заклю
чается в том, что различ
ные клетки происходят
путем деления исходной
материнской клетки.
Новейшие достижения биолоrии, связанные с научно техническим проrрес
сом, дали новые доказательства правильности клеточной теории как одной из
важнейших закономерностей развития живоrо.
11
Рис. 1. Схема строения животной клетки:
1 ядро; 2 плазмолемма; З микроворсинки; 4 рибосомы; 5
rранулярная зндоплазматическая сеть; 6 rладкая эндоплазматичес
кая сеть; 7 комплекс rольджи; 8 лизосома; 9 митохондрия; 10
клеточный центр; 11 микротрубочки; 12 десмосома
Морфофункциональные системы клетки
В орrанизме клетки объединяются в функциональные системы ткани. Извес
тно около 1SQ разновидностей клеток. Форма, размеры, внутренняя структура и фун
кции клеток весьма разнообразны. Размеры клеток человека колеблются от 5 до 100
мкм И более. По форме клетки MorYT быть окруrлыми, кубическими, призматичес
кими, веретеновидными, плоскими, звездчатыми и т. п. Различают одноядерные и
22
Fлава 4
мноroядерные клетки. Последние возникают в результате ацитокинетических мито
зов или путем слияния мноrих клеток (симпласты). При делении клеток MOryT Bpe
менно сохраняться цитоплазматические мостики (участки относительно широких ци
топлазматичеСЮIХ соединеНШ':'I между соседними клетками) так возникают синцитии.
Несмотря на большое разнообразие формы, размеров, способов взаимосвязи и
функций тканевых клеток, большинству из них присущи важнейшие общебиолоrи
чески и эволюционно обусловленные свойства: rенетическая индивидуальность и
способность передавать ее поколениям, реактивность и раздражимость, обмен вe
ществ, и, наконец, подвИЖ1l0сть. Эти четыре свойства живоrо обеспечиваются сле
дующиr. и комплексами взаимосвязанных и взаимодействующих структур, или сис
темами, клетки: покров ной, или поrpаничной, системой (плазмолемма), системой
восприятия, трансформации и передачи сиrналов (рецепторно трансдукторной),
внутренней метаболической средой (орrанеллы, ядро, включения и др.) и опорно
двиrательной системой.
Покровная система клетки (плазмолемма)
Плазмоле Iма (клеточная мембрана) обеспечивает дискретность живоrо веще
ства за счет разrраничения ero с внешней средой (микроокружением), rенетичес
кую индивидуальность, присущую клеткам данной особи, а также транспорт Be
ществ из клетки и в клетку. Данные ФУНКIlиональные свойства плазмолеммы свя
заны с ее молеку
лярной орrаниза
цией.
Плазмолемма
образована бимоле
кулярным слоем
полярных липидов
(преимущественно ,
фосфолипидов f 11 ' I
леЩ1Тина и цефали
на) и встроенными б < . \ , I / I I I I
в Hero молекулами .
rлобулярных беk . '.' , f
ков (рис. 2). rидро 5 I , , , I
фобные хвосты ли . .,.
пидных молекул...
спрятаны от BOД
ных сред rиалоп
лазмы и внешней
7
, .'/'
.
I 3
\ ., <" . ' .' .
-:. . .
4 . _ .
среды и направ
лены друr к друry
(6), а rидрофиль
ные rоловки (5)
Рис. 2. Мозаичная модель клеточной мембраны и поверхности мембраны по ли
нии замораживания скалывания (стрелка):
1 интеrральный белок; 2 полисахариды rликокаликса; З полуинтеrраль
ные белки; 4 периферический белок; 5 слой rидрофильных rоловок липид
ных молекул; б rидрофобные концы бислоя липидных молекул; 7 эпитоп
молекулы rистосовместимости I ro класса
Основы общей циmолоzии
23
обращены в сторону содержащих воду фаз. В отдельных участках бислоя фосфоли
пидов присутствуют молекулы холестерина, придающие мембране жесткость. В этих
участках мембрана, как правило, малоэластична, в связи с чем здесь не происходят
процессы эндо и экзоцитоза. Обращенные в межклеточную среду rоловки отдель
ных фосфолипидных молекул связаны с молекулами олиrосахаров (2) элемента
ми Vlи-ко-кали-кса. Липидные молекулы плазмолеммы обеспечивают ее основные фи
зико химические свойства, в первую очередь, текучесть мембраны, допускающую
свободное перемещение составляющих ее молекул.
Белковые компоненты плазмолеммы представлены собственно интеrральны
ми, или трансмембранными (1), полуинтеrральными (3), и периферическими (4)
белками. Интеrральные белки полностью (трансмембранно) располаrаются в би
липидном слое, их молекулы в своем составе имеют алифатические (липофиль
ные) аминокислоты, которые поrружены в липидный слой, И наружные rидрофиль
ные концы, с помощью которых белковые молекулы образуют связи с остатками
сахаров rликокаликса и периферическими белками. Полуинтеrральные белки по
rружены в бимолекулярный слой липидов частично. Весь набор белковых моле
кул распределен в мембране мозаично и леrко перемещается в ее плоскости с уча
стием элементов цитоскелета, которые образуют связи с интеrральными белками.
Периферические белки располаrаются вне липидноro бислоя в rиалоплазме и
непрочно связаны с внутренней поверхностью плазмолеммы.
Белковые молекулы выполняют ряд специальных функций рецепции, TpaHC
мембранных переносчиков, ферментативную.
Fли-ко-кали-кс образован уrлеводными участками rликолипидов и rликопротеи
нов плазмолеммы. Он придает мембране дополнительную механическую прочность,
обеспечивает адrезивные свойства (способность плазмолеммы взаимодействовать
с мембранами друrих клеток и межклеточным веществом), участвует в распозна
вании родственных клеток, рецепции специфических 2
сиrналов. В электронном микроскопе rликокаликс
имеет вид рыхлоrо слоя умеренной электронной плот
ности, покрывающеrо внешнюю поверхность клеточ
ной мембраны.
Со стороны внутренней поверхности плазмо
леммы располаrается тонкий кортикальный слой rи
алоплазмы (кортекс) с множеством микрофиламен з
тов (преимущественно актиновых). Этот слой свя
зан с периферическими белками плазмолеммы и ци
тоскелетом (рис. 3). Он также дает плазмолемме He 4
который запас прочности и участвует в поддержа
нии клеткой ее формы.
С молекулярной орrанизацией плазмолеммы
тесно связан!=> такое важнейшее свойство живоrо как
rенетическая индивидуальность. Последняя обеспе
чивается следующими факторами и механизмами:
1) непрерывностью плазмолеммы и ее реrенерацией,
Рис. З. Схема строения кортекса
клетки:
1 клеточная мембрана; 2 aK
тиновые филаменты; 3 актин
связывающие белки (линкерные);
4 внутренняя поверхность плаз
молеммы
24
Fлава 4
2) наличием в составе плазмолеммы молекулы rи
стосовместимости (rликопротеина), которая опре
деляет rенетические отличия клеток одноrо opra
низма от друrоrо (за исключением клеток у MOHO
зиroтных близнецов). Молекулу rистосовмести
мости 1 ro класса, содержат все клетки особи. Дан
ная молекула состоит из: 1) интеrральноrо TpaHC
мембранноrо белка, часть KOToporo находится в
цитоплазме, друrая пронизывает плазмолемму
и последняя наиболее длинная часть молекулы
находится в rликокаликсе; 2) периферическоrо
мембранноrо белка с малой молекулярной массой;
3) короткой молекулы белка, которая нековален
тно связывается с петлями внеклеточной части ин
теrральноrо трансмембранноrо белка. Именно
последняя часть молекулы (обычно это пептид из
9 аминокислот) является фраrментом нормально
ro белка клетки данной особи (рис. 4). Он и pac
познается как "свой" клетками иммунной систе
мы человека. В случае мутации на месте белка rистосовместимости появляется
белок с иной структурой молекулы (например, кодируемый вирусом,) и в ответ
на это возникает иммунная реакция со стороны орrанизма, направленная на унич
тожение данной клетки. Так сохраняется rенетическая индивидуальность клеток
и, следовательно, орrанизма.
Транспорт ионов и макромолекулярных соединений через плазмолемму про
исходит разными путями. Растворенные в жидкой среде вещества проникают че
рез клеточную мембрану либо сами без переносчиков (или носителей), либо с
помощью переносчиков, или носителей (рис. 5). Транспорт без носителей называ
ется пассивным (непосредственным транспортом) и осуществляется через каналы
мембран, т. е. в тех белоксодержащих участках, которые проницаемы для малых
молекул (воды, мочевины, ионов) и действуют подобно молекулярным ситам, а
также через липидную фазу мембраны (рис. 5, стрелки 1 и 2). В последнем случае
липидная фаза служит растворителем для ряда веществ (простые и сложные эфи
ры, жирные кислоты и др.).
Однако большинство веществ проникают через плазмолемму с помощью TpaHC
портных систем, или переносчиков (носителей). Это специфические мембранные
белки rруппы интеrральных, или функциональные комплексы липопротеидов,
которые связываются и трансмембранно переносят молекулы субстратов. Простей
шим примером транспорта с помощью носителя является облеrченная (опосредо
ванная) диффузия (рис. 5, стрелка 3). В этом процессе носитель облеrчает пере
нос какоrо либо вещества через мембрану в направлении rpадиента концентраций
без затраты энерrии.
Для осуществления процесса активноrо транспорта против rрадиента KOH
центрации (из области с низкой концентрацией в область с высокой концентраци
1
,
, I '
n -.;,
'" ' '" ",
I!>" " ' ,1!,
, /iii ( .,. .' >'J
.. ' , ' ''''..i:;, ....1;'
..., ....... " ,
,О ....... ,,, 1\
. ' ,:i.-. '-....,
; .
../
:,/ '
Рис. 4. Строение внеклеточноrо дo
мена молекулы rистосовместимости
I ro класса: два cerMeHTa альфа пе
тель формируют желобок (1), в KOTO
ром располаrается нековалентно свя
занная с молекулами альфа петель
короткая цепь пептида эпитоп
(2) животной клетки
ОС1l0вы общей цuтОЛОlUU
2S
ей), требуется не только
носитель, но и источник
энерrии, которым обыч
но является аденозинт
рифосфат (АТФ). Актив
ный транспорт может
служить для переноса ok
Horo вещества в одном
направлении (рис. S,
стрелка S), либо для пе
реноса двух веществ в
противоположных (или в
том же самом) направле
ниях. В последнем случае
перенос веществ называ
ется сопряженным aK
тивным транспортом
(рис. S, стрелки 6а и 6б).
В отличие от TpaHC
порта низкомолекуляр
ных соединений, макромолекулярные соединения транспортируются с помошью про
цессов эндоцитоза (в клетку) и экзоцитоза (из клетки).
Эllдоцuтоз это транспорт макромолекул через плазмолемму. COOTBeTCTBeH
но arperaTHoMY состоянию поrлощаемоro вещества выделяют пиноцитоз (захват и
транспорт клеткой жидкости или растворенных в жидкости соединений) и фаrо
цитоз (захват и транспорт твердых частиц). Эндоцитоз бывает неспецифический и
специфический.
Неспецифический эндоцитоз осуществляется без участия рецепторных белков
плазмолеммы (рис. 6). Первым этапом неспецифическоro эндоцитоза в случае Tpaнc
порта твердых частиц является адrезия (прилипание) частиц к внешней поверхности
плазмолеммы (важную роль в этом процессе иrpает rликокаликс). Второй этап по
rpужение частиц в клетку путем инваrинации плазмолеммы. Адrезия и поrpужение
происходят в тех участках плазмолеммы, которые свободны от холестерина, т. е. наи
менее жесткие, и к которым со
стороны цитоплазмы прилежит
слой белка клатрина (см. ниже).
После отшнуровки участка
плазмолеммы с твердыми час
тицами образуется внутрикле
точный пузырек эндосома.
Перемещение эндосомы в rиа
лоплазме осуществляется с по
мощью элементов цитоскелета.
Дальнейшая судьба эндосом
1 2
3
48
Рис. 5. Схема трансмембранноrо переноса веществ.
Стрелки (1 6) указывают пути и направления движения веществ через
nлазмолемму
а . CI О о.. .
.. .. .
:. " .е.
о е.
о
Цитоплазма
. .
. D 6).
Рис. 6. Схема эндоцитоза. Последовательные этапы aдre
зии nоrлощаемых частиц, инваrинации плазмолеммы, форми
рования пузырька и отделения ero от плазмолеммы. Фузоrен
ные белки (стрелки)
26
Fлава 4
может быть различна. Наиболее часто эндосомы подверrаются процессу внутрикле
точноrо переваривания: к эндосоме подходят и сливаются с ней перв чные лизосо
мы формируется фаrолизосома, в которой под действием rидролитических фермен
тов лизосом происходит химическое расщепление макромолекул до мономерных
соединениЙ. По мере расщепления (переваривания) макромолекул от мембраны фа
roлизосомы отшнуровываются фраrменты, которые встраиваются в плазмолемму и
восполняют ее дефицит, ранее образовавшийся при отшнуровке эндосомы.
Процесс пиноцитоза подразделяется на микро и макропиноцитоз. При мик
ропиноцитозе начальным этапом является образование инваrинации плазмолем
мы, в которой находится часть жидкой среды. Образующиеся затем по аналоrии с
эндосомами пиносомы представляют собой небольшие пузырьки (везикулы), KO
торые по мере продвижения по цитоплазме MorYT сливаться в более крупные
мультипиноцитозные образования. Поrружение капли жидкости при микропино
цитозе происходит не в случайных участках плазмолеммы, а в тех областях, KOTO
рые имеют со стороны rиалоплазмы тонкий слой особоrо белка клатрина. В этих
участках, как правило, отсутствует холестерин, что делает мембрану податливой к
инваrинации. Коrда от мембраны отшнуровывается пиносома (пиноцитозный пу
зырек, или везикула) по периферии она имеет слой клатрина, в связи с чем пузы
рек именуют окаймленным.
Макропиноцитоз отличается от микропиноцитоза тем, что с помощью довольно
длинных выростов плазмолеммы клетка активно захватывает фраrменты жидкой
среды. Макропиноцитоз в связи с этим именуется еще рофеоцитозом. После CMЫKa
ния конца выроста с соседним участком плазмолеммы образуется крупная пиноци-
тозная вакуоль. Таким образом, при макропиноцитозе процесс поrлощения клеткой
жидкости происходит более интенсивно.
е
.ее
Экзоцuтоз транспорт
ный процесс, имеющий проти
воположное эндоцитозу Ha
правление (рис. 7). Путем
экзоцитоза из клетки удаля
ются некоторые продукты Me
таболизма, непереваренные и
вредные для нее вещества, а из
железистых клеток продук
ты их секреции. Экзоцитозные
пузырьки, содержащие перечисленные соединения, приближаются к внутренней
поверхности плазмолеммы и сливаются с ней, формируя сообщение с внешней cpe
дой. Полаrают, что в процессе слияния мембран важная роль принадлежит oco
бым, так называемым фузоrенным, плазмолеммальным белкам (от латинскоrо
fusio слияние), которые концентрируются в участках контактов экзоцитозных
везикул с плазмолеммой.
ВажнеЙшее своЙство клетки и ее плазмолеммы формирование межклеточ-
ных соединений (контактов).
Простой lIесnециализироваllllЫЙ (адrезионный) контакт (рис. 8 А) образуется
.е
Цитоплазма
Рис. 7. Схема экзоцитоза. Последовательные этапы: прибли
жение пузырька к цитолемме, слияние мембраны пузырька с
цитолеммой, высвобождение содержимоrо пузырька во BHe
шнюю среду. Фузоrенные белки (стрелки)
Основы общей цитолоlUИ
27
1
д
2 2
щ
W З
, '"
w
na
з
5
з
4
4
Б
r
в
Рис. 8. Схема строения межклеточных контактов:
Д простой контакт (стрелки движение веществ по межклеточному пространству); Б ПЛОТНЫЙ KOH
такт (стрелки зоны сближения внешнених слоев плазмолеммы); В заякоривающий контакт (1
цитоплазматические мембраны, 2 связывающие трансмембранные белки кадrерины, 3 внутри
клеточные белки сцепления винкулин, 4 фибриллярные белки цитоскелета, 5 участки сцепления
связывающих белков с помошью катионов кальция); r щелевой контакт (1 цитоплазматические мем-
браны, 2 коннексоны, 3 каналы коннексонов)
за счет ЭЛементов rликокаликса трансмембранными rликопротеинами (Kaдre
ринами) взаимодействующих мембран. Слои rликокаликса удерживают мембра
ны клеток на расстоянии около 10-20 нм, оставляя свободной межклеточную щель
для транспортных процессов ионов и низкомолекулярных соединений. Обращен-
ные в сторону межклеточной щели молекулы кадrеринов связываются катионами
кальция. Простые контакты не обеспечивают высокой прочности межклеточных
взаимодействий. Иноrда плазмолеммы контактирующих клеток в области просто-
ro контакта образуют интердиrитации (взаимные пальцевидные внедрения участ
ков цитоплазмы), которые придают контакту большую прочность.
Плотный (запирающий) контакт характерен для клеток однослойных эпите-
лиев (см. рис. 8 Б). При формировании плотноrо контакта внешние слои мембран
в отдельных участках максимально сближаются. Б точках соприкосновения мемб
ран располаrаются интеrральные белки плазмолемм соседних клеток Б ряде слу-
чаев (в эпителии кишечноrо типа) плотные контакты формируют сплошные поло
сы, получившие название замыкающих пластинок. Эти контакты, помимо прочноrо
соединения клеток, изолируют межклеточные щели и делают их плохо проницае
мыми для ионов И молекул.
3ая-коривающий контакт. Б отличие от двух предыдущих в ero образовании
кроме клеточных мембран участвуют фибриллярные элементы цитоскелета (см.
рис. 8 Б). К этому виду соединений принадлежат десмосомы и полудесмосомы, в
формировании которых задействованы промежуточные филаменты цитоскелета.
Десмосома. В межклеточной щели в области десмосомы располаrается элект-
POHHO-ПЛОТНчIЙ слой, образованный взаимодействующими молекулами интеrраль-
ных rликопротеинов (десмоrлеинов) плазмолемм соседних клеток С помощью Ka
тионов кальция молекулы десмоrлеина сцеплены в межклеточном пространстве.
Со стороны rиалоплазмы в зоне десмосомы располаrается электронно плотный
28
rлава 4
слой белка десмоплакина, в который вплетаются промежуточные филаменты
цитоскелета. Десмосомы являются характерными контактами эпителиальных, эн
дотелиальных клеток, кардиомиоцитов и друrих, обеспечивая прочное сцепление
взаимодействующих структур.
Щелевой контакт. В отличие от всех рассмотренных выше он представляет
собой коммуникационное (обменное) соединение клеток (см. рис. 8 [). Через
щелевой контакт происходит прямой обмен химическими веществами между
клетками. Плазмолеммы соседних клеток в зоне щелевоrо контакта сближены
до 2 3 нм. Метод замораживания скалывания демонстрирует присутствие в
межклеточной щели контакта мелких частиц, имеющих внутри канальцы это
коннексоны (от анrл. connection соединение). В составе разных щелевых KOH
тактов насчитывается от нескольких единиц до нескольких тысяч коннексонов.
Коннексоны насквозь пронизывают плазмолемму и в мембранах соседних кле
ток соединяются "стык в стык". В результате образуются сквозные каналы, co
общающие между собой внутренние среды контактирующих клеток. KOHHeKCO
ны MorYT временно закрываться, оrраничивая активность обменных процессов
между клетками.
Таким образом, плазмолемма иrрает роль прочноrо механическоrо и биоло
rическоrо покрова, изолируя внутреннее содержимое клетки от внешней среды.
Эту функцию совместно с плазмолеммой выполняют rликокаликс и цитоплаз
матический кортекс актиновых филаментов. Кроме TOro, плазмолемма обеспе
чивает процессы переноса веществ из клетки во внеклеточную среду и наобо
рот из внеклеточной среды внутрь клетки, а также является носителем
информации о rенетической принадлежности особи.
Внутренняя метаболическая среда клетки и ее компартменты
Внутренняя среда клетки цитоплазма сложно орrанизованная система, вклю
чающая ядро, мембранные инемембранные орrанеллы, включения, которые находят
ся во взвешенном состоянии в rиалоплазме. Последняя представляет собой rель с из
меняющейся в зависимости от функциональноrо состояния клетки степенью вязкости.
В составе rиалоплазмы находятся структурные и ферментные белки клетки,
различные метаболиты, ионы. Здесь присутствуют ферменты, участвующие в син
тезе аминокислот, нуклеотидов, жирных кислот, биосинтезе сахаров. В rиалоплаз
ме происходят процессы rликолиза и синтез части АТФ, модификация ферментов
(например, фосфорилирование), приводящая к их активации, либо инактивации.
В rиалоплазме начинается ряд биосинтетических процессов, которые в дальней
шем продолжаются в той или иной внутриклеточной системе.
В электронном микроскопе rиалоплазма выrлядит rомоrенной и характеризу
ется низкой электронной плотностью. Меrавольтная электронная микроскопия об
наруживает в ней микротрабекулярную сеть, состоящую из тончайших фибрилл,
пересекающих rиалоплазму в различных направлениях. В ячейках этой сети pac
полаrаются орrанеллы, а в ее "узлах" фиксированы полисомы. Микротрабекуляр
ная сеть rиалоплазмы образует связи с микротрубочками и микрофиламентами
Основы общей цuтОЛОlUU
29
опорно двиrательной системы клетки и совместно с этими элементами участвует
в перемещении и функционировании внутриклеточных структур.
Орrанеллы это обязательно присутствующие в клетке ультрамикроскопичес
кие структуры, представляющие собой отrраниченные от окружающей rиалоплаз
мы биолоrической мембраной отсеки, или компартменты (мембранные орrанеллы),
и расположенные в rиалоплазме рибонуклеопротеиды, биополимеры и их комплек
сы (немембранные орrанеллы).
С точки зрения цитофизиолоrии целесообразно рассмотреть строение и функ
ции внутриклеточных структур в связи с их участием в метаболических процес
сах, таких как хранение и передача поколениям rенетической информации, синтез
и транспорт веществ, энерrообеспечение метаболических проuессов и др.
Ядро
Ядро важнейший компонент, с деятельностью KOToporo связаны хранение
rенетической информации, размножение клеток и передача rенетическоrо матери
ала поколениям, участие в синтезе белков.
В неделящихся клетках (интерфазе клеточноrо цикла) ядро хранит закодиро
ванную в ДНК хромосом информацию о белковом синтезе и обеспечивает синтез
тех белковых молекул, которые необходимы клетке в проuессе ее роста, диффе
ренцировки и физиолоrической реrенерации; в этот период в ядре синтезируются
участвующие в образовании белка рибосомальная, информационная и трансфер
ная РНК, формируются субъединицы рибосом. При подrотовке клетки к делению
ядро удваивает rенетическую информацию о белковом синтезе, создавая ее точ
ную копию для передачи дочерним клеткам.
В интерфазе в составе ядра обнаруживаются оболочка (нуклеолемма), xpOMa
тин, нуклеоплазма, ядрышко.
Ядерная оболочка представляет собой часть внутриклеточной мембранной сис
темы (совместно с rранулярной и аrpанулярной эндоплазматической сетью). Она
состоит из BHYTpeHHero и наружноrо листков, между которыми находится щелеВИk
ное пространство перинуклеарное, ширина KOToporo варьирует в зависимости от
функциональной активности клетки. Наружный листок нуклеолеммы со стороны
rиалоплазмы окружен сетью виментиновых промежуточных филаментов и имеет
на своей поверхности свободные рибосомы, прикрепленные к ней большими субъе
диницами. Внутренний листок rладкий, не содержит рибосом, образует связи с
пластинкой (ламиной) ядра и участвует в фиксации интерфазных хромосом. Лист
ки нуклеолеммы выполняют по отношению к ядру две важные функuии формо
образовательную и рецепторно барьерно транспортную.
Перинуклеарное пространство прерывается множеством пор. Они представляют
собой сквозные туннели, сообщающие содержимое ядра и rиалоплазмы. Стенка пор
образована ившимися листками нуклеолсммы (рис. 9). В области пор отсутствует
rетерохроматин. Диаметр ядерных пор около 90 нм, а численность варьирует в за
висимости от активности биосинтетических процессов в клетке. Поровые туннели за
полнены поровыми комплексами, состоящими из трех компонентов: поровых колец,
30
Тлава 4
", :! ,;.: ;,,"".''';:'4-... . «цент р альныхспицицен
. 7> <л.....,. ..... .> ,.,... .l >'" ..'" >.
....) ,.:;;' .. .. " ;' '<\ " .. ,. ".,,' "' ,, ', ; Т р альной rpaH . улы. Все
;., ., .. >f' \"" . ,;:" .". ' 1tIi", :tf...,,
,: ." t..,. ",,,.. t...... " ;...,.,'. " '...":<it\:,...}: ;., ; ',) компоненты поровоrо
.....""...., '«"'-' 1." .. ""' E "'" ,,.,...,..,.., i"" б
..' , : !I'. 'I".:t"-'«J .' .:>,! :K,;.;I:,'4 ';". комплекса елковые
'...,,' ,........;r':....f....". ,;..) 5:: \< f. ;.,, :<.:. ., j производные, обеспечи
. ' ,,:\:t' .\': '. :; . ,i....}.';,'?' -p. "'f: " ./ ,-: вающие строю избира
!.; " . 'h:' J . . .f' ", 2 ;';' «. j }" . l тельный транспорт Maк
.i. :: . ," .. c.'''; ' ", "....:. н <, ромолекул в ядро и из
, ,I,..,! "" , " .
,.. ,"'<, , ,'" liI ядра.
'\." / :-. .\.. . ... _H, '! \ :;.,! I ' 8 пр е р :::тъ :е : ь
...а: " '" "'....., , \
4 ;;. 4' , диафраrмы, состоящие
". : из восьми субъединиц
,,' II'";. . :1'...... (периферических rpa
Рис. 9. Ультраструктуры клеточноrо ядра: нул). В составе порово
1 ядрышко; 2 rетерохроматин; 3 ядерная оболочка; 4 ядерные ro комплекса выделя
поры ются два поровых коль
ца цитоплазматическое и ядерное, берущие начало соответственно от наруж
Horo и BHYTpeHHero листков ядерной оболочки (рис. 10). От белковых rранул
поровых колец к центру поры отходят множественные тонкие фибриллы, сходя
щиеся к центральной rрануле и формирующие диафраrму. Ядерные поры уча
ствуют в избирательном транспорте веществ между ядром и rиалоплазмой: все
ядерные белки поступают в ядро из rиалоплазмы, а все формы РНК транспорти
руются в rиалоплазму из ядра. В этом процессе комплекс поры выступает как
молекулярный механизм, исполняющий не только роль переносчика, но и сорти
ровщика молекулы, подлежащей транспорту.
:;+ "'"
i<, ': ":""" :' .
.....':-...:,.......""
....... -,t.a ...-._..-
: . :,; ':> ;S' : '
2
Рис. 10. Схема строения nopoBoro комплекса:
1 внешняя ядерная мембрана; 2 внутренняя ядерная мембрана; 3
центральная rранула; 4 периферические rранулы; 5 диафраrма
Через поры в ядро
свободно поступают
ионы, сахара, нуклеоти
дЫ, АТФ, некоторые rop
моны. Белки, транспор
тируемые в ядро, имеют
определенные последо
вательности аминокис
лот последовательнос..
ти ядерной локализации.
Рецепторы ядерных пор
узнают и пропускают эти
молекулы в ядро.
К внутреннему лист
ку нуклеолеммы на всем
протяжении за исключе
нием ядерных пор приле
жит тонкий пласт белко
ОС1Ю6Ы общей цитолоlUИ
2
31
вых филаментов (ядерная ламина). Блаrодаря пос
ледней ядро не утрачивает своей формы даже после
удаления оболочки ядра. От ядерной ламины в rлубь
ядра отходит сеть белковых филаментов, которая
служит основой для хромосом. В области ядрышка
формируется остов концентрически ориентирован
ных фибриллярных белков (рис. 11).
Хроматин это вещество, в состав KOToporo BXO
ДЯТ ДНК и белки. Различают rетерохроматин спи
рализованные участки хромосом и эухроматин дec
пирализованные участки хромосом. По соотношению
эу и rетерохроматина в ядре можно судить о степени
функциональной активности клетки. Например, в Ma
лодифференцированных и активно rотовящихся к про
Рис. 11. Схема строения ядерноrо лиферации клетках основная часть ядра занята эухро
Р:: ; ая оболочка (внутренняя матином, лишь у BHyтpeHHero листка кариолеммы и в
поверхность); 2 ядерные поры; отдельных областях центральной части ядра присут
3 элементыядернойламины;4 ствует rетерохроматин (хромоцентры). В высокодиф
связывающий лам ин; 5 собствен Ф
но ядерный матрикс (интерхрома еренцированных клетках хроматин конденсирован и
тиновый ламин); 6 фиброзный oc выrлядит в виде широкоЙ периферической зоны и Mac
тов ядрышка (ядрышковый ламин) сивных хромоцентров. Во всех клетках, независимо от
уровня их дифференцировки, в период интерфазы постоянно существуют конденси
рованные участки хроматина конститутивный хроматин соответствующие цeHT
po и теломерным участкам хромосом. В клетках женских особей к катеrории консти
ryтивноrо хроматина принадлежат тельца Барра (половой хроматин) диаметром около
0,5 мкм, являющиеся спирализованной Х хромосомой. В rpанулоцитах крови тельца
Барра представлены небольшими добавочными фраrментами ядер ("барабанные па
лочки").Тельца Барра обнаруживаются в 70 75 % интерфазных ядер соматических
клеток женскою орrанизма. Ero определение имеет большое практическое значение в
судебно медицинской практике.
Ядрышко в световом микроскопе выrлядит в виде мелкой темно окрашиваю
щеЙся частицы. Количество и размеры ядрышек в клетках варьируют в зависимости
от функциональной активности. В клетках, продуцирующих большое количество
белка, размер ядрышка может занимать до 25 % Bcero объема ядра.
Ядрышко образовано специализированными участками хромосом (13, 14, 15,
21 и 22), называемыми ядрышковыми орrанизаторами. Функции ядрышка COCTO
ят В синтезе рибосомальной РНК и образовании так называемых предшественни
КОВ большой и малой субъединиц рибосом. Дальнейшее созревание предшествен
ников в субъединицы происходит в rиалоплазме, куда их предшественники
попадают из ядра через поровые комплексы.
На электррнных микрофотоrpафиях в составе ядрышка различают три зоны: сла
боакрашенный компонент, содержащий ДНК из области ядрышковorо орrанизато
ра хромосом; центральный фибриллярный компонент, состоящий из множества TOH
КИХ (около 5 нм) рибонуклеопротеиновых фибрилл и представляющий собой
32
rлава 4
РНК транскрипты; периферический rpанулярный компонент, в состав KOToporo BXO
дЯТ предшественницы зрелых субъединиц рибосом. Варьирование размеров ядрышка
связано преимущественно с изменением доли rранулярноrо компонента.
НУЮlеоплазма жидкое внутреннее содержимое ядра, состоит из воды, ионов,
rликопротеинов, содержит РНК и ферментные белки.
Орlанеллы синтеза и транспорта биополu.меров
К орrанеллам данной rруппы относятся эндоплазматическая сеть, рибосомы,
комплекс fольджи, лизосомы, пероксисомы. Они осуществляют синтез орrаничес
ких соединений, их транспорт в процессе химической доработки из одноrо участка
канальцевой сети в друrой. накопление, перемещение, упаковку и экзоцитоз rOTo
вых продуктов синтеза.
Эидоплазматическая сеть и рибосомы. Эндоплазматическая сеть представле
на канальцами и цистернами, которые анастом03ИРУЮТ и формируют в rиалоплазме
трехмерную сеть. В состав сети входят rpанулярные (содержащие на внешних поверх
ностях мембран рибосомы) и arpанулярные (без рибосом) участки.
Рибосомы синтезируют все разнообразие клеточных белков. На светооптичес
ком уровне рибосомы неразличимы, об их количестве в клетке можно судить по
интенсивности окраски цитоплазмы общеrистолоrическими (базофилия) или спе
циальными rистохимическими реактивами и флюорохромами, маркирующими
РНК На субмикроскопическом уровне рибосомы выrлядят как осмиофильные
черные точки (диаметром около 20 25 нм), а их рабочие комплексы полисомы
как rруппы, или розетки, осмиофильных точек
Компоненты рибосом создаются в разных участках клетки: рибосомальные РНК
синтезируются в ядрышке; рибосомальные белки в цитоплазме. Последние посту
пают в ядро, rде комплсксируются с молекулами РНК и объединяются в рибосо
мальные субъединицы. Затем субъединицы РНК транспортируются из ядра через
поры и находятся в цитоплазме либо в диссоциированном (неактивном), либо acco
циированном друr с друrом (активном) состоянии.
Работающие орrанеллы состоят из двух ассоци
ированных (малой и большой) субъединиц, которые
удерживаются в обратимо связанном состоянии с
помощью катионов маrния (рис. 12). Большую
субъединицу рибосом образуют разные молекулы
РНК, имеющие сложную вторичную и третичную
структуру, в комплексе с рибосомальными протеи
нами. Большая субъединица значительно крупнее
малой и имеет форму полушара. Малая субъедини
ца выrлядит в виде маленькой шапочки. При acco
циации субъединиц в рибосому происходит законо
мерное взаимодействие их поверхностей.
Между субъединицами работающей рибосомы
имеет место cTporoe "разделение труда" малая
/
. )
" ",.
" ' } .....
........ ......-,
')
'" ...
у
'r'
.>(
2
,
-<'*..jj
Рис. 12. Схема строения рибосомы:
1 малая субьединица; 2 большая
субьединица
Основы общей цumолоlUU
зз
субъединица ответственна за связывание информационной РНК, большая ведает
образованием полипептидной цепи. В клетке нефункционирующие рибосомы Haxo
дятся в диссоциированном состоянии, в связи с чем получают возможность постоян
но обмениваться субъединицами и постоянно обновляться. В рабочем режиме рибо
сомы (от 3 дО 20 ЗО в rpуппе) образуют стабильный комплекс полисому, в котором
они связаны нитью информационной РНК
О степени развития в клетке rранулярной эндоплазматической сети можно cy
дить по базофилии цитоплазмы, обусловленной присутствием большоrо количе
ства рибосом; аrранулярные участки эндоплазматической сети на светооптичес
ком уровне не обнаруживаются. В большинстве клеток преобладает rранулярная
сеть, и оба вида сети имеют диффузную орrанизацию их элементы располаrают
ся в rиалоплазме свободно, без какой либо упорядоченности. Синтез белка в rpa
нулярной сети происходит на рибосомах и полисомах, а ее каналы и цистерны яв
ляются вместилищем и транспортными маrистралями для перемещения белка в
коплекс rольджи для доработки.
Ширина и количество канальцев и цистерн сети в клетках варьируют в зави
симости от их функциональною состояния при повышении функциональных
наrрузок на клетку канальцы и цистерны сети становятся множественными и зна
чительно расширяются. Канальцы эндоплазматической сети непосредственно свя
заны с перинуклеарным пространством клетки.
Значение rранулярной эндоплазматической сети состоит в синтезе мембран
ных белков и белков, предназначенных "на экспорт" и необходимых друrим клет
кам, либо используемых во внеклеточных физиолоrических реакциях. Этот вид
сети присутствует во всех клетках орrанизма человека (кроме зрелых спермиев),
однако наиболее развит в тех клетках, которые специализированы на синтезе боль
ших количеств белковых молекул. Таких видов клеток в орrанизме человека cpaB
нительно HeMHoro. Примером являются плазмоциты, синтезирующие антитела (или
иммуноrлобулины); клетки поджелудочной железы, вырабатывающие комплекс
белковых пищеварительных ферментов (панкреатический сок); rепатоциты, син
тезирующие широкий спектр белков плазмы крови, свертывающей и противосвер
тывающей систем, а также некоторые друrие клетки. В этих клетках канальцы сети
располаrаются упорядоченно (в некоторых случаях cTporo параллельно) в виде
так называемой эрrастоплазмы.
В малодифференцированных и неспециализированных клетках rранулярная
эндоплазматическая сеть, как правило, слабо развита, в структуре клеток преобла
дают свободные поли и рибосомы, обеспечивающие синтез белков, необходимых
клетке для роста и дифференцировки.
Аrранулярная эндоплазматическая сеть имеет вид коротких канальцев и пузырь
ков (везикул), которые диффузно располаrаются по всей rиалопла:зме.В большинстве
клеток элементы аrранулярной сети, как правило, немноrочисленны. В клетках, Bыpa
батывающих стероидные rормоны (клетки надпочечников, половых желез). arpaHY
лярная сеть хорошо развита и ее мноючисленные пузырьки занимают большие пло
щади, либо образуют муфты BOKpyr липидных включений предшественников
стероидных [ормонов. В мембранах сети находятся ферменты стероидоrенеза.
34
Fлава 4
ПОМИМО стероидоrенеза, она участвует в синтезе и метаболизме липидов, полисаха
ров, триrлицеридов, процессе детоксикации продуктов метаболизма лекарственных
препаратов и эндоrенных клеточных ядов. В канальцах аrpанулярноЙ сети депониру
ются большие запасы катионов кальция.
Комплекс rольджu. Представлен сплющенными цистернами (или мешками), co
бранны:\-ш в стопку. Каждая цистерна HeMHoro изоrнута и имеет выпуклую и воrнyтyю
поверхности. СредниЙ диаметр цистерн составляет около 1 мкм. В центре цистерны ее
мембраны сближены, а на периферии часто формируют расширения, или аМIrjЛЫ, от
которых отшнуровываются пузырьки. Пакеты плоских цистерн количеством в cpeд
нем около 5 10 фор:\-шруют диктиосому. Кроме цистерн, в комплексе rольджи присут
ствуют транспортные и секреторные пузырьки.
В диктиосоме в соответствии с направлением кривизны изоrнутых поверхнос
теЙ цистерн различают две поверхности. Выпуклая поверхность называется незре
лоЙ, или цис поверхностью. Она обращена к ядру или к канальцам rранулярноЙ
эндоплазматическоЙ сети и связана с последнеЙ пузырьками, отшнуровывающи
мися от rранулярноЙ сети и приносящими молекулы белка в диктиосому на дозре
вание и оформление в мембрану_
Противоположная TpaHC
поверхность диктиосомы Bor
нута. Она обращена к плазмо
лемме и именуется зрелоЙ
потому, что от ее мембран OT
шнуровываются секреторные
пузырьки, содержащие [OTO
вые к выведению из клетки
продукты секреции (рис. 13).
Комплекс rольджи уча
ствует в накоплении продук
тов, синтезированных в эндоп
лазматическоЙ сети, в их
химическоЙ перестроЙке и co
зревании. В цистернах комп
лекса rольджи происходит
синтез полисахаридов, их KOM
плексирование с белковы:\-ш
молекула:\-ш. Одна из rлавных
функциЙ комплекса rольд
жи фор:\-шрование [отовых
секреторных продуктов, KOTO
рые выводятся за пределы клетки путем экзоцитоза. ВажнеЙшими для клетки функ
циями комплекса rольджи также являются обновление клеточных мембран, в том числе
и участков плазмолеммы, а также замещение дефектов плаэмолеммы в процессе ceK
реторноЙ деятельности клетки. Комплекс rольлжи считается источником обраэова
ния первичных лизосом, хотя их ферменты синтезируются и в rpанулярноЙ сети.
/0
Встраивание НОВЫХ
фраrмеНТО8 клеточных
мембран
@
.
@ .
.-.. .
. ..
\
Лиэосомы
Секреция экзоцитозом
,-
.. протеолиз, СОрТИрО8ка белков Jf .::.. ::..... ..
.. пипидов, присоединение уrпеводныхостатков ."
( .:.
."
Мед G
... . )
.. . .. ..
Присоединение уrлеводных
остатков
Цис G
( .. -
:::." :
Формирование белков
:р
.. .. .. "," .. ..
u
о
Синтез лнпндов
Рис. 13. Схема метаболическоrо пути веществ в структурах KOM
плекса rольджи:
АЭС аrранулярная, rэс rранулярная эдоплазматическая
сеть; TpaHC , Meд и Цис G поверхности диктиосомы
Основы общей цuтОЛОlUU
35
Лuзосом.ы представляют собой внутриклеточно формирующиеся секреторные
вакуоли, заполненные rидролитическими ферментами, необходимыми для процес
сов фаrо и аутофаrоцитоза. На светооптическом уровне лизосомы можно инден
тифицировать и судить о степени их развития в клетке по активности rистохими
ческой реакции на кислую фосфатазу ключевой лизосомальный энзим.
При электронной микроскопии ли30сомы определяются как пузырьки, orpa
ниченные от rиалоплазмы мембраной. Условно выделяют 4 основных вида лизо
сом: первичные и вторичные лизосомы, аутофаrосомы и остаточные тельца.
Первичные лизосомы это мелкие мембранные пузырьки (средний диаметр
их составляет около 100 нм), заполненные rOMoreHHbIM мелкодисперсным coдep
жимым, представляющим собой набор rидролитических ферментов. В лизосомах
идентифицированы около 40 ферментов (протеазы, нуклеазы, rликозидазы, фос
форилазы, сульфатазы), оптимальный режим действия которых рассчитан на кис
лую среду (рН 5). Лизосомальные мембраны содержат специальные белки носи
тели для транспорта из лизосомы в rиалоплазму продуктов rидролитическоrо
расщепления аминокислот, сахаров и нуклеотидов. Мембрана лизосом устойчи
ва по отношению к rидролитическим ферментам.
Вторичные лизосомы образуются при слиянии первичных лизосом с эндо
цитозными либо с пиноцитозными вакуолями. Иными словами, вторичные ли
зосомы это внутриклеточные пищеварительные вакуоли, ферменты которых
поставляются первичными лизосомами, а материал для переваривания эндо
цитозной (пиноцитозной) вакуолью. Строение вторичных лизосом весьма раз
нообразно и изменяется в процессе rидролитическоrо расщепления содержи
Moro. Ферменты лизосом расщепляют попавшие в клетку биолоrические
вещества, в результате чеrо образуются мономеры, которые транспортируются
через мембрану лизосомы в rиалоплазму, rде утилизируются или включаются в
разнообразные синтетические и метаболические реакции.
Если взаимодействию с первичными лизосомами и rидролитическому расщеп
лению их ферментами подверrаются собственные структуры клетки (стареющие
орrанеллы, включения и пр.), формируется аутофаrосома. Аутофаroцитоз является
естественным процессом в жизнедеятельности клетки и иrpает большую роль в об
новлении ее структур при внутриклеточной реrенерации.
Остаточные тельца это одна из финальных стадий существования фаrо и
аутолизосом и обнаруживаются при незавершенном фаrо или аутофаrоцитозе и
впоследствии выделяются из клетки путем экзоцитоза. Они имеют уплотненное
содержимое, часто наблюдается вторичная структуризация непереваренных co
единений (например, липиды образуют сложные слоистые образования).
Перmcсuсом.ы (или микротельца) это вакуоли (0,3 1,5 мкм в диаметре), OKPy
женные мембраной. Внутреннее содержимое пероксисомы матрикс представ
лен мелкоrранулярным содержимым с нуклеоидом (сердцевиной) в центре. В HYK
леоиде часто J;!ИДНЫ кристаллоподобные структуры, которые состоят из упорядоченно
упакованных фибрилл и трубочек Пероксисомы обычно локализуются вблизи MeM
бран rранулярной эндоплазматической сети. Последние являются местом их обра
зования, хотя часть ферментов пероксисом синтезируется в rиалоплазме.
36
Fлава 4
В пероксисомах обнаружены ферменты, связанные с метаболизмом перекиси BOДO
рода. Это ферменты, ведущие окислительное дезаминирование аМИНОК!1СЛОТ (окси
дазы, уратоксидазы) с образованием вредной для клетки перекиси водорода, и фер
мент каталаза, разрушающая перекись.
Таким образом, эти орrанеллы, разрушающие орrанические соединения с об
разованием своеобразноrо клеточноrо яда в виде перекиси водорода, OДHOBpeMeH
но снабжены собственной защитой в виде нейтрализующих перекись ферментов.
Клеточные включения
Включения это непостоянные структуры клетки, которые появляются в ней
и исчезают в процессе метаболизма. Различают трофические, секреторные, экск
реторные и пиrментные включения.
[руппа трофических включений объединяет уrлеводные, липидные и беk
ковые включения. Наиболее распространенным представителем уrлеводных
включений является rликоrен полимер rлюкозы. На светооптическом ypOB
не наблюдать включения rликоrена можно при использовании rистохимичес
кой ШИК реакции. В электронном микроскопе rликоrен выявляется как осми
офильные rранулы, которые в клетках, rде rликоrена MHoro (rепатоцитах),
сливаются в крупные конrломераты rлыбки.
ЛипидllЪLМИ включениями наиболее боrаты клетки жировой ткани липоци
ты, резервирующие запасы жира для нужд Bcero орrанизма, а также стероидпроду
цирующие эндокринные клетки, использующие липид холестерин для синтеза CBO
их ropMOHOB. На ультрамикроскопическом уровне липидные включения имеют
правильную окруrлую форму и в зависимости от химическоrо состава характери
зуются высокой, средней или низкой электронной плотностью.
Белковые включения, например, вителлин в яйцеклетках, накапливается в ци
топлазме в виде rранул.
СекретОРllые включения представляют собой разнообразную rpуппу. CeKpeTOp
ные включения синтезируются в клетках и выделяются (секретируются ) в просветы
протоков (клетки экзокринных желез), в межклеточную среду (rормоны, нейроме
диаторы, факторы роста и др.), кровь, лимфу, межклеточные пространства (roрмо
ны). На ультрамикроскопическом уровне секреторные включения имеют вид мемб
ранных пузырьков, содержащих вещества разной плотности и интенсивности
окраски, что зависит от их химическоrо состава.
ЭкскреmОрllые включения это, как правило, продукты метаболизма клетки,
от которых она должна освободиться. К экскреторным включениям относятся TaK
же инородные включения случайно, либо преднамеренно (при Фаrоцитозе бак
терий, например,) попавшие в клетку субстраты. Такие включения клетка лизиру
ет с помощью своей лизосомальной системы, а оставшиеся частицы выводит
(экскретирует) во внешнюю среду. В более редких случаях попавшие в клетку areH
ты остаются неизменными и MorYT не подверrнуться экскреции такие включе
ния более правильно именовать чужеродными (хотя чужеродными для клетки яв
ляются и включения, которые она лизирует). '
Основы общей цuтОЛОlUU
37
Пuzм.е1l.т1l.ые включения хорошо выявляются как на светооптическом, так и
на ультрамикроскопическом уровнях. Очень характерный вид они имеют на элек
тронных микрофотоrрафиях в виде осмиофильных структур разных размеров
и формы. Данная rруппа включений характерна для пиrментоцитов. Пиrменто
циты, присутствуя в дерме кожи, защищают орrанизм от rлубокоrо проникнове
ния опасноrо для Hero ультрафиолетовоrо излучения, в радужке, сосудистой обо
лочке и сетчатке rлаза пиrментоциты реrулируют поток света на фоторецепторные
элементы rлаза и предохраняют их от перераздражения светом. В процессе CTa
рения очень мноrие соматические клетки накапливают пиrмент липофусцин, по
присутствию KOToporo можно судить о возрасте клетки. В эритроцитах и симп
ластах скелетных мышечных волокон присутствуют соответственно rемоrлобин
или миоrлобин пиrменты переносчики кислорода и уrлекислоты.
МитОХОllдрии
Это орrанеллы энерrообеспечения метаболических процесов в клетке. Раз
меры их варьируют от 0,5 до 5 7 мкм, количество в клетке составляет от 50 до 1000
и более. В rиалоплазме митохондрии распределены обычно диффузно, однако в
специализированных клетках сосредоточены в тех участках, rде имеется наиболь
шая потребность в энерrии. Например, в мышечных клетках и симпластах боль
шие количества митохондрий сосредоточены вдоль рабочих элементов сократи
тельных фибрилл. В клетках, функции которых сопряжены с особо высокими
энерrозатратами, митохондрии образуют множественные контакты, объединяясь
в сеть, или кластеры (кардиомиоциты и симпласты скелетной мышечной ткани).
В клетке митохондрии выполняют функцию дыхания. Клеточное дыхание
это последовательность реакций, с помощью которых клетка использует энерrию
связей орrанических молекул для синтеза макроэрrических соединений типа АТФ.
Образующиеся внутри митохондрии молекулы А ТФ переносятся наружу, обме
ниваясь на молекулы АДФ, ,...,....,."",6...........r-. :;.:-:" < ....... .. , '
-
находящиеся вне митохонд <>.. ,..." ,: .." ., . .....-;
u iirf!" ....... ," '0 п .,..
рии. В живои клетке мито ,:"::: ,. .":..- :.. }J , ' "
хондрии MorYT передвиrать : .. ' . >
ся с помощью элементов >"ч',
цитоскелета. . .
На ультрамикроскопи
ческом уровне стенка мито
хондрии состоит из двух
мембран наружной и BHYT
ренней. Наружная мембрана
имеет относительно ровную
поверхность,. внутренняя
образует направленные в
центр складки, или кристы
(рис. 14). Между наружной
38
Тлава 4
и внутренней мембранами возникает неширокое (около 15 нм) пространство, KO
торое называется наружной камерой митохондрии; внутренняя мембрана оrрани
чивает внутреннюю камеру. Содержимое наружной и внутренней камер митохон
дрии различно, и так же, как и сами мембраны, существенно отличается не только
по рельефу поверхности, но и по ряду биохимических и функциональных призна
ков. Наружная мембрана по химическому составу и свойствам близка к друrим
внутриклеточным мембранам и плазмолемме. Ее характеризует высокая проница
емость, блаrодаря наличию rидрофильных белковых каналов. Эта мембрана имеет
в своем составе рецепторные комплексы, распознающие и связывающие вещества,
поступающие в митохондрию. Ферментный спектр наружной мембраны небоrат:
это ферменты метаболизма жирных кислот, фосфолипидов, липидов И др. [лавной
функцией наружной мембраны митохондрии является отrраничение орrанеллы от
rиалоплазмы и транспорт необходимых для осуществления клеточноrо дыхания
субстратов.
Внутренняя мембрана митохондрий в большинстве клеток тканей различных op
raHoB формирует кристы в виде пластин (ламеллярные кристы), что значительно YBe
личивает площадь поверхности внутренней мембраны. В последней 20 25 % всех беk
ковых молекул составляют ферменты дыхательной цепи и окислительноrо фосфори
лирования. В эндокринных клетках надпочечников и половых желез митохондрии уча
ствуют в синтезе стероидных ropMOHOB. В этих клетках митохондрии имеют кристы в
виде трубочек (тубул), упорядоченно расположенных в определенном направлении.
Поэтому кристы митохондрий В стероидпродуцирующих клетках названных opraHoB
именуются тубулярными.
Матрикс митохондрии, или содержимое внутренней камеры, представляет
собой rелеобразную структуру, содержащую около 50 % белков. Осмиофильные
тельца, описанные при электронной микроскопии, это резервы кальция. MaT
рикс содержит ферменты цикла лимонной кислоты, катализирующие окисление
жирных кислот, синтез рибосом, ферменты, участвующие в синтезе РНК и ДНк.
Общее число ферментов превышает 40.
Помимо ферментов, матрикс митохондрии содержит митохондриальную ДНК
(митДНК) и митохондриальные рибосомы. Молекула митДНК имеет кольцевид
ную форму. Возможности внутримитохондриальноrо белковorо синтеза оrраниче
ны здесь синтезируются транспортные белки митохондриальных мембран и He
которые ферментные белки, участвующие в фосфорилировании АДФ. Все
остальные белки митохондрии кодируются ядерной ДНК, и их синтез осуществ
ляется в rиалоплазме, и в дальнейшем они транспортируются в митохондрию.
Жизненный цикл митохондрий в клетке короткий, поэтому природа наделила их
двойственной системой воспроизводства помимо деления материнской мито
хондрии, возможно образование нескольких дочерних орrанелл путем почкования.
Опорно двиrательная система клетки
Опорно двиrательная система и ее компоненты в качестве каркаса клетки обеспе
чивают противодействие внешним физическим факторам и вместе с тем леrко пере
Основы общей цитОЛОlИИ
39
страиваются и изменяют форму клетки, участвуют в реryляции потоков rиалоплазмы
и движении орrанелл.
К компонентам клетки с опорно двиrательной функцией относятся промежуточ
ные филаменты, микрофиламенты, микротрубочки и их специализированные произ
водные (микроворсинки, стереоцилии, реснички и жryтики). С деятельностью этих
структур связано выполнение практически всех клеточных функций.
Про.м.ежуточные фWlа.м.енты
Промежуточные филаменты построены из фибриллярных белковых MOHOMe
ров. Их пространственная конструкция напоминает плетение каната толщиной
около 8 1 О нм. В клетке они локализуются в виде трехмерной сети преимуществен
но в околоядерной области и собраны в пучки, которые направляются к периферии
клетки. Здесь они входят либо в состав десмосом и полудесмосом (в клетках эпите
лиальных тканей), либо направляются в отростки нервных клеток Эти части цитос
келета характерны для всех видов клеток, однако особенно хорошо развиты в клет
ках, испытывающих механические наrрузки, например, в клетках эпидермиса,
мышечных клетках, нейронах. В rруппу промежуточных филаментов входит несколь
ко родственных белков, однако в разных клетках это, как правило, разные белки. В
клетках мезенхимноrо происхождения (соединительнотканных, эндотелиальных,
клетках крови) промежуточные филаменты состоят из вUМe1lтИllа. В мышечных клет
ках белок промежуточных филаментов называется деСМИ1l0М (в поперечноисчерчен
ных мышечных волокнах десминовые филаменты входят в состав Z линий). В ней
ранах промежуточные филаменты поддерживают форму отростков нервных клеток
и фиксируют трансмембранные белки ионных каналов. В клетках эпидермиса про
межуточные филаменты, связываясь с друrими белками, формируют poroBoe веще
ство, представляющее собой мощный защитный слой кожи, непроницаемый для
мноrих водорастворимых опасных для орrанизма соединений. Наконец во всех клет
ках в составе ядра находятся белки ядерной пластинки (ламины). В отличие от yc
тойчивых промежуточных филаментов цитоплазмы слои филаментов ядерной пла
стинки леrко разбираются во время митотическоrо деления клетки.
Основными функциями промежуточных филаментов являются: опорная, под
держание формы клетки, участие в формировании межклеточных соединений типа
десмосом и полудесмосом, специальные функции в различных типах клеток
МUКрОфWlа.м.енты и их проuзводные
Это нитевидные сократимые образования толщиной около 5 нм, состоят из
белка актина и являются универсальными элементами цитоскелета. В цитоплазме
актиновые микрофиламенты располаrаются поодиночке, либо в виде сетки и пуч
ков, а с внутрен ей стороны плазмолеммы образуют сrущение кортикальный
слой клетки, или кортекс (см. рис. 3). В последнем актиновые филаменты обра
зуют сеть с помощью связывающих (линкерных) белков, одним из которых яв
ляется филамин. Филаменты актиновоrо кортекса фиксируются к плазмолемме
40
rлава 4
с помощью плазмолеммальных интеrральных белков интеrринов. В специали
зированных участках плазмолеммы и адrезионных контактов актин может CTa
новиться трансмембранным белком.
Во всех клетках актиновые филаменты взаимодействуют с модифицирован
ной формой миозина, представленной мономерной структурой МUНUМUОЗUНОМ
Минимиозин соединен с клеточными орrанеллами и облеrчает их транспорт, а также
перемещение везикул вдоль актиновых филаментов. При полимеризации актино
вых микрофиламентов возникают локальные перемещения цитоплазмы, необхо
димые для движения клетки.
Специализированными производными микро
филаментов являются mu-кровОРСU1l1Ш и их уплотнен
ные комплексы стереоцилии. Микроворсинки
это тонкие (0,1 мкм) и длинные (около 1 мкм) BЫ
росты апикальной (верхушечной) части клеток
(рис. 15). Внутри каждой микроворсинки располаrа
ется пучок актиновых микрофиламентов в количе
стве 20 30. Одними концами филаменты закрепле
ны на вершине микроворсинки, а нижняя часть
филаментов вплетается в актиновый кортекс. Фила
менты связываются в пучок поперечно расположен
ными белковыми молекулами (связками) фасцина и
фимбрина. В составе микроворсинок обнаружен и
сократительный белок минимиозин, который вызы
вает укорочение и удлинение микроворсинок.
Основными функциями микрофиламентов явля
ются: поддержание формы и придание жесткости клет
ке (осуществляется кортексом); участие в формиро
вании межклеточных соединений, участие в
транспортных процессах эндо , пино , экзоцитозе
(осуществляется кортексом); участие в процессах пе
ремещения клеточных орrанелл, транспортных и ceK
реторных пузырьков (осуществляется актиновыми
микрофиламентами с минимиозином, ассоциирован
ным с поверхностью указанных структур) и в образо
вании микроворсинок и стереоцилий, в формирова
нии специализированных для мышечных структур акто миозиновых сократительных
комплексов, а также в образовании клеточной перетяжки при цитотомии.
5
з
4
Рис. 15. Схема строения микро
ворсинки:
1 плазмолемма; 2 актиновые
филаменты; 3 актин связываю
щие белки (фасцин и фимбрин);
4 латеральный заякоривающий
белок (минимиозин); 5 аморф
ный материал верхушки микро
ворсинки; б спектрин; 7 KOp
текс субмембранный актин,
связанный спектрином; 8 цито
кератиновые филаменты
Клеточный центр и elO пРОUЗ60дные
Клеточный центр состоит из двух центриолей и центросферы. Основу цeHT
риоли составляют девять триплетов микротрубочек, расположенных по окружно
сти И формирующих полый цилиндр (рис. 16). Диаметр цилиндра центриоли co
ставляет около 0,15 0,2 мкм, длина от 0,3 до 0,5 мкм. Одна из микротрубочек
Основы общей цuтОЛОlUU
41
каждоrо триплета (микротрубочка А) состоит из
13 протофиламентов, две друrие (В и С) peдy
цированы и содержат по 11 протофиламентов.
Все микротрубочки триплета плотно прилежат
друr к друry. Каждый триплет по отношению к
радиусу формируемою ими цилиндра микротру
бочки располаrается под уrлом около 40 rраду
сов. В составе центриоли микротрубочки связа
ны поперечными белковыми мостиками, или
ручками. Последние отходят от А микротрубоч
ки и одним концом обращены в сторону центра
центриоли, друrим к С микротрубочке cocek
Hero триплета. Каждый триплет центриоли с
внешней стороны связан с белковыми тельцами
шаровидной формы сателлитами, от которых
в rиалоплазму расходятся микротрубочки, формирующие центросферу. BOKpyr
каждой центриоли обнаруживается тонковолокнистый матрикс, а сами триплеты
поrружены в аморфный материал умеренной электронной плотности, называемыЙ
муфтой центриоли.
В интерфазноЙ клетке присутствует пара (дочерняя и материнская) центриолеЙ,
или диплосома, которая чаще располаrается вблизи комплекса rольджи рядом с яk
ром. В диплосоме продольная ось дочернеЙ центриоли направлена перпендикулярно
продольной оси материнской. Дочерняя центриоль в отличие от материнской не име
ет перицентриолярных сателлитов и центросферы.
Центриоли выполняют в клетке функции орrанизации сети цитоплазматичес
ких микротрубочек (как в покоящихся, так и делящихся клетках), а также образу
ют микротрубочки для ресничек специализированных клеток
Мuкротрубочкu присутствуют во всех животных клетках за исключением эрит
роцитов. Они образованы полимеризованными
молекулами белка тубулина, который представ
ляет собой rетеродимер, состоящий из двух
субъединиц альфа и бета тубулина. При по
лимеризации альфа субъединица одноrо белка
соединяется с бета субъединицей следующеrо.
Так формируются отдельные протофиламенты,
которые, объединяясь по 13, формируют полую
микротрубочку, внешний диаметр которой co
ставляет около 25 нм, а внутренний 15 нм
(рис. 17).
Каждая микротрубочка имеет растущий
плюс конец и медленно растущий минус конец.
Микротрубочки один из наиболее динамичных
элементов цитоскелета. Во время наращивания
длины микротрубочки присоединение тубулинов
с
8 !
. '\
r 1 t;9)
[ А ,
c1i!
\O %
Рис. 1 б, Схема строения центриоли в жи
вотной клетке (соответствует формуле
(9хЗ)+О): поперечный и продольный cpe
зы микротрубочек А,В и С
Б
Рис, 17. Схема строения микротрубочки:
А продольное сечение, формирование
протофиламентов из молекул а (CBeT
лые) и 13 (темные) субъединиц тубулина;
Б поперечное сечение 1 3 протофила
ментов по окружности микротрубочки
42
Тлава 4
происходит на растущем плюс конце. Разборка микротрубочек наиболее часто проис
ходит с обоих концов. Белок тубулин, формирующий микротрубочки, не является co
кратительным белком, и микротрубочки не наделены способностью к сокращению и
передвижению. Однако микротрубочки цитоскелета принимают активное участие в
транспорте клеточных орrанелл, секреторных пузырьков и вакуолей. Из препаратов
микротрубочек отростков нейронов (аксонов) были выделены два белка кинезин и
динеин. Одним концом молекулы этих белков ассоциированы с микротрубочкой, дpy
rим способны связываться с мембранами орrанелл и внутриклеточных везикул. С
помощью кинезина осуществляется внутриклеточный транспорт к плюс концу мик
ротрубочки, а с помощью динеина в обратном направлении.
Реснички и Жlутики являются про из водными микротрубочек в клетках эпи
телия воздуховодных путей, женскоrо половоrо тракта, семявыносящих путей,
сперматозоидах.
Ресничка представляет собой тонкий ци
линдр С постоянным диаметром около 300 нм
(рис. 18). Это вырост плазмолеммы (аксолем
ма), внутреннее содержимое KOToporo aKCOHe
ма состоит из комплекса микротрубочек и He б
большоrо количества rиалоплазмы. Нижняя
часть реснички поrружена в rиалоплазму и об
разована базальным тельцем. Микротрубочки
располаrаются по окружности реснички парами
(дуплетами), повернутыми по отношению к ее
радиусу под небольшим уrлом около 10 rpa
дусов. В центре аксонемы расположена цeHT
ральная пара микротрубочек. Формула микро
трубочек в ресничке описывается как (9х2)+2.
В каждом дуплете одна микротрубочка (А) яв
ляется полной, т. е. состоит из 13 субъединиц,
вторая (В) неполной, т. е. содержит только 11
субъединиц. А микротрубочка имеет динеино
вые ручки, направленные к В l'vlИкротрубочке co
седнеrо дуплета. С помощью нектин связываю
щеro белка микротрубочка А соединяется с микротрубочкой В соседнеrо дуплета.
От А микротрубочки к центру аксоне мы отходит радиальная связка, или спица,
которая оканчивается rоловкой на так называемой центральной муфте. Последняя
окружает центральную пару микротрубочек. Центральные микротрубочки в отли
чие от периферических дуплетов микротрубочек располаrаются отдельно друr от
друrа на расстоянии около 25 нм.
Базальное тельце реснички состоит из 9 триплетов микротрубочек. A и В микро
трубочки триплетов базальноrо тельца, продолжаясь в A и В микротрубочки дупле
тов аксонемы, составляют вместе с ними единую структуру.
Реснички не содержат в своем составе сократительных белков, но при этом
совершают однонаправленные биения, не изменяя своей длины. Это происходит
А В
2
Рис. 18. Схема строения реснички на
поперечном срезе:
1 плазмолемма; 2 дуплет микро
трубочек; 3 центральный дуплет мик
ротрубочек; 4 центральная капсула
(муфта); 5 радиальная спица (связ
ка); б нектин связывающий белок;
7 динеиновые ручки
Основы общей цитОЛОlИИ
43
за счет смещения пар микротрубочек относительно друr друrа (продольноrо сколь
женил дуплетов) в присутствии АТФ.
Система реактивности клетки
(восприятия, трансформации и передачи сиrналов)
Комплекс структур, обеспечивающий клетке свойства раздражимости и peaK
тивности, тесно связан с плазмолеммой. Он представлен рецепторами, интеrpаль
ными белками переносчиками плазмолеммы, белковыми насосами и rликопротеи
нами rликокаликса. Эти структуры выполняют функции восприятия (рецепции) и
передачи (трансдукции) сиrналов.
Роль мноrих рецепторов заключается в передаче rормональных сиrналов
внутрь клетки на специальные белки ферменты, которые участвуют в формиро
вании общих и специфических ответов клеток на rормональные стимулы. Чаще
Bcero в роли TaKoro фермента выступает аде1lилилциклаза, инициирующая пре
вращение АТФ в циклический аденозинмонофосфат (цАМФ). Последний явля
ется активатором друrих ферментных систем, катализирующих специфические
внутриклеточные ответы соответственно характеру поступившеrо стимула. В про
цессе трансдукции сиrналов
принимают участие интеr rp' Рц
ральные белки плазмолем Ац
мы, так называемые G беЛ1(И
(рис. 19). При связывании
лиrанда (молекул ropMoHoB, ' G б
трансмиперов, ионов и др.) .
с рецепторной частью этorо rдф rдф'
белка возникает передача rp
активирующеrо или подав W'ц Ац
ляющеrо стимула на фер Специфический ..... ,
менты цитоплазмы. Так за функциональный ответ
внутриклеточных систем
пускается каскад внутри . " G . б
клеточных биолоrических ,
процессов, реализующийся rтф
визмененияхвнутриклеточ Рис. 19. Схема рецепторно трансдукторной системы клетки:
Horo метаболизма, делении, rp ropMoH; Рц рецептор; Ац аденилилциклаза; rдф rya
росте или rибели клеток. нидиндифосфат; rтф rуанидинтрифосфат; G б G белок
Таким образом, рецепторная функция плазмолеммы адаптирует клетку к BHe
шним условиям, способствует восприятию реrулирующих факторов и сохранению
постоянства внутриклеточноrо rомеостаза и жизнеспособности.
Кроме рецепторов, расположенных в плазмолемме, существует большая rруп
па внутрикле)"очных рецепторов, например, в цитоплазме к стероидным ropMo
нам, рецепторы на мембранах митохондрий, комплекса rольджи, ядра и др. Все они
участвуют в метаболических реакциях клетки, иrрая важную роль в трансмемб
ранном переносе веществ.
rp
Рц Ац
G б
./J.
44
Fлава 4
С помощью рецепторов обеспечивается специфический, или рецепторно опос
редованный, эндоцитоз. При специфическом эндоцитозе клетка избирательно по
rлощает те вещества (лиrанды), к которым имеет рецепторы в составе плазмолем
мы. Рецепторы, связывая лиrанд, способны активно смещаться в плазмолемме и
накапливаться в зонах эндоцитозных ямок. BOKpyr эндоцитозной ямки и В после
дующем BOKpyr эндосомы концентрируется слой белка клатрина, роль KOToporo
состоит в том, чтобы препятствовать слиянию эндосом. В процессе продвижения
эндосом по клетке клатриновая оболочка исчезает и отдельные эндосомы получа
ют возможность сливаться друr с друrом и формировать вакуоли. Обязательным
при рецепторно опосредованном эндоцитозе является возвращение рецептор со
держащеro фраrмента мембраны эндосомы в состав плазмолеммы.
Воспроизведение клеток
Период жизни большинства клеток тканей человека колеблется в широких пре
делах.
Следует различать клетки с короткой продолжительностью жизни. Последняя
равна времени от одною деления до друrorо и включает период подrотовки к мито
зу авmосuнmеmuческую интерфазу, и собственно митоз. Однако в rистоrенезе боль
шинство клеток после определенноrо числа делений переходит в lеmеросuнmеmuчес
-кую интерфазу, которая включает время роста, дифференцировки, функционирования,
старения и смерти (рис. 20). При этом продолжительность жизни клетки возрастает.
Например, клетки нервной ткани живут долrо, сравнимо с продолжительностью жиз
ни орrанизма.
СПEQИАЛН3АUИSI
СМЕРТЬ
КЛЕТКИ
МИТОТИЧЕСКИЙ uикп
ЖИЗНЕННЫЙ цикл КЛЕТКИ
Рис. 20. Схема жизненноrо цикла клетки:
G, пресинтетический период; S синтетический период; G 2 постсинтетический период
ОС1106Ы общей цитолоlUИ
4S
Основным способом деления животных клеток является митоз. Митозу предше
ствует автосинтетическая интерфаза. В последней выделяются три периода: 1) G, (от
анrл. gap промежуток ) постмитотический, пресинтетический, во время KOToporo
отсутствует синтез ДНК; 2) S синтетический, на протяжении KOToporo в XpOMOCO
мах клеточноrо ядра осуществляется синтез новой молекулы ДНК; 3) G2 премито
тический, постсинтетический, в течение KOToporo клетка rотовится к митозу. ПРОДОk
жительность указанных периодов в интерфазе различных клеток неодинакова.
S период следует считать одним из ключевых периодов, так как без синтеза ДНК He
возможны митотическое деление и последующее образование дочерних клеток, иден
тичных по объему rенетической информации исходной материнской клетке.
В процессе подrотовки клетки к митозу во время S периода интерфазы происхо
дит удвоение молекул ДНК ЭТО явление принято называть репликацией ДН К. После
окончания S периода количество ДНК в ядре становится равным 4 с, Torдa как coдep
жание ДНК в одном ядре в неделящихся диплоидных клетках составляет 2 с.
Следовательно, после завершения митоза в дочерние клетки попадают по одной po
дительской и комплементарной ей новой (дочерней) цепи ДНк. Так в каждой дочерней
клетке сохраняется исходная двухцепочечная структура молекулы ДНК rенетичес
кая копия родительской.
Цикл клеточной репродукции реryлируется мноrочисленными BHe и внутрикле
точными механизмами. К внеклеточным относятся влияния на клетку цитокинов,
факторов роста, rормональных и нейроrенных стимулов. Роль внутриклеточных
реryляторов иrpают специфические белки цитоплазмы. В течение каждоrо клеточно
ro цикла существуют несколько критических точек, соответствующих переходу клет
КII из одноrо периода цикла в друтой. Ключевое значение в прохождении каждою пе
риода цикла и в подrотовке клетки к вступлению в следующий период имеет сочетанное
влияние внутриклеточных белков (Gl ЦИКЛИНЫ, S циклины, М циклины И др.).
Таким образом, все критические точки цикла клеточной репродукции Haxo
дятся под контролем комплекса внутриклеточных специализированных белков.
Мутации reHoB, кодирующих некоторые из них, называются онкоrенными. Напри
мер, в норме белок PS3 ("блюститель reHoMa") воспринимает нарушение CTPYKTY
ры ДНК и останавливает клетку в G, или G2 периодах цикла. В случае, если He
возможна репарация ДНК, то белок PS3 инициирует rибель клетки путем апоптоза.
Существуют и друrие белки, участвующие в реrуляции синтеза и репарации ДНК,
блаroдаря которым прерывается клеточный цикл (например, за счет блока расхож
дения сестринских хроматид в анафазе митоза).
В митозе выделяют четыре фазы: профаза, метафаза, анафаза и телофаза. Каж
дая из этих фаз характеризуется определенными изменениями в структуре ядра и
цитоплазмы делящейся клетки (рис. 21).
Профаза. В профазе происходит конденсация хромосом, и они становятся ви
димыми при световой микроскопии. Хромосомы по мере компактизации ДНП при
обретают строение хорошо окрашивающихся нитей. Число хромосом равно 4п, что
соответствует количеству ДНК 4с. В связи с инактивацией reHoB в области ядрыш
KOBOro орrанизатора и уrнетением синтеза РНК в профазе отмечается исчезнове
ние ядрышек Ядерная оболочка постепенно распадается на фраrменты и мелкие
46
Fлава 4
, I ( '. I
. " . \
/v : ';\( " r:. l )
, . , ' i \j j
,,"" " J J
о\. f\,....,. ..."-....",......-..
', \ ..?"' J ,
\ / Б
мембранные пузырьки. При
этом к противоположным по
люсам клетки расходятся
центриоли. В сателлитном
участке материнской центри
оли начинается образование
микротрубочек, из которых
формируются нити веретена
деления.
Метафаза. Характерным
событием для метафазы явля
ется перемещение хромосом в
экваториальную плоскость Be
ретена. Здесь они располаrа
ются строю закономерно, об
разуя метафазную пластинку
(при взrляде на веретено деле
ния сбоку). Если рассматри
вать rруппу метафазных хромосом со стороны полюсов веретена, то отчетливо BЫ
ступает фиrура, напо:\шнающая звезду (так называемая материнская звезда). В этот
период можно определить число, форму и размеры хромосом (d XpOMOCOM, двой
ных хромосом), составляющих метафазную пластинку. К концу метафазы продоль
ные половинки хромосом (сестринские хроматиды) обособляются на всем протя
жении, кроме зоны первичной перетяжки.
Для каждоrо вида животных характерно cTporo постоянное число хромосом в
соматических клетках. Для человека оно равно 46. По длине хромосом различают
чередование окрашенных и неокрашенных участков. При этом каждая хромосома
отличается неповторимым рисунком дифференциальной окраски. Хромосомы че
ловека подразделяются на 7 rрупп по их размерам и особенностям строения (А, В,
С, D, Е. F. G) и каждая хромосома имеет свой номер. Совокупность признаков cтpo
ения хромосом, их размеров и числа составляет то, что называют кариотипом.
А1lафаза включает процесс расхождения хромосом к полюсам делящейся клет
ки. Механизм движения хромосом объясняется rипотезой скользящих нитей, соrлас
но которой состоящие из микротрубочек нити веретена, взаимодействуя друr с дpy
rOM и с сократительными белками, тянут хромосомы к полюсам. Скорость движения
хромосом достиrает O,2 O,S мкм/мин, а вся анафаза продолжается 2 3 мин. Анафаза
заканчивается перемещением двух идентичных наборов хромосом (S XpOMOCOM, или
одиночных хромосом) к полюсам, rде они сближаются, образуя фиryры, напомина
ющие по внешнему виду (если смотреть со стороны полюса) звезды. Эти фиryры
называют дочерними звездами. Так как хромосомные звезды образуются у каждоrо
из полюсов, данную стадию митоза иноrда называют стадией двойной звезды (диас
тер), или стадией дочерних звезд.
Телофаза конечная стадия митоза, в течение которой на полюсах веретена
реконструируются дочерние ядра. Перестройка телофазных хромосом напомина
.-i
(Q Y€i бf
f!i J (
\ , .' у
/ {::':" 1 ' " I
А }
/';'f f ) :/ " .
( /J:1/ ) .. , (с' ) }
(. '{' I (,
! 'v" l'
\ . f ).
'\......,' . ". /
,.r'" / ' '
r <L .
,.' J ....,J
'. .
в ,
'....,
> ... \/
"---'"'--\ ., " fio-
..S9 t /' () ."
() 'A. / ;" '
Рис. 21. Фазы митоза:
А профаза; Б метафаза; В анафаза; r телофаза
r
Основы общей циmолоlUИ
47
ет процессы их изменения в профазе, но происходящие в обратном направлении.
При взаимодействии хромосом с мембранными пузырьками цитоплазмы форми
руется ядерная оболочка. С переходом хромосом в интерфазное состояние образу
ются новые ядрышки. Телофаза завершается разделением тела клетки цитото
мией, или цитокинезом, что приводит к образованию двух дочерних клеток
Часть клеток может выходить из цикла репродукции и вступить на путь диф
ференцировки. Некоторые клетки MorYT выходить из клеточною цикла в Gl пери
оде или после S периода и находиться в покое (Gо период). Такие покоящиеся клет
ки сохраняют способность к делению и MOryT снова входить в цикл размножения.
Атипические миmозы. Наряду с типичными картинами митоза нередко наблю
даются атипические митозы, возникновение которых обусловлено теми или ины
ми отклонениями в деятельности указанных выше основных клеточных компонен
тов участников митоза. Так, если по каким либо причинам клеточный центр не
разделится надвое, то образуется однополюсное веретено деления. Такой митоз
называют однополюсным, униполярным (от лат. unus один) или моноцентри
ческим (от rpеч. monos один). При этом варианте митоза хромосомы не pacxo
дятся к полюсам веретена, и деление клетки не завершается плазмотомией. Hepek
ко встречаются мноrополюсные митозы, в процессе которых образуется несколько
клеточных центров (до S 6 и больше). Это связано с делением клеточноrо центра
(центросомы) не на две, как обычно, а на большее число центриолей. По числу
полюсов различают мноrополюсные, или мультиполярные (от лат. multum MHO
ro), или полицентрические (от rреч. poly MHoro), а митозы называют ТРИ J KBak
ри , reKca и т. д. полярными. Чаще Bcero мноrополюсный митоз не завершается
плазмотомией, вследствие чеrо образуются мноюядерные rиrантские клетки. Если
же наступает цитотомия, то дочерние клетки содержат неравноценный rенетичес
кий материал (анэуплоидия).
Ряд алкалоидов (колхицин) избирательно действует на веретено деления, в
результате чеrо исключается возможность расхождения хромосом в анафазе. В этом
случае течение митоза приостанавливается на стадии метафазы. Указанная разно
видность атипическоrо митоза в литературе известна под названием колхицино
Boro митоза, или К митоза. Колхицин используют в экспериментальной цитоло
rии при подсчете количества митозов в тканях. Фибриллярные структуры веретена
деления нарушаются также под действием бета меркаптоэтанола, эфира, HarpeBa
ния и повышенноrо давления.
Отклонения от нормальною течения митоза MOryT быть связаны с изменениями
структуры хромосом. Эти изменения называются хромосомными аберрациями (от
лат. aberratio уклонение). Различают несколько разновидностей хромосомных
аберраций: 1) слипание хромосом; 2) разрыв хромосомы на фраrменты; 3) выпаде
ние участка хромосомы при ее разрыве (нехватка, или делеция); 4) транслокация
обмен обломками между двумя хромосомами; 5) инверсия переворачивание на
1800 фраrмента хромосомы, расположенною между двумя переломами; 6) дуплика
ция удвоение какоrо либо участка хромосомы; 7) отставание хромосом во время
анафазноrо их перемещения с образованием хромосомных мостиков между pacxo
дящимися к полосам комплексами хромосом. Из обособившихся при этом хромосом
48
Тлава 4
MOryт возникать маленькие ядра кариомеры. Хромосомные аберрации возникают
при действии на клетки ионизирующеrо излучения. .
Эllдомитоз. Эндомитозом (от rpеч. endon внутри) называют вариант митоза,
коrда репликация хромосом не сопровождается исчезновением ядерной оболочки и
образованием веретена деления. При эндомитозе в одних случаях хромосомы выяв
ляются, В друrих же они не видны. Путем повторных эндомитозов количество
хромосомных наборов в клеточном ядре может значительно увеличиваться. Ядро
приобретает rиrантские размеры. Эндомитоз лежит в основе полиплоидии.
ПОЛИ11JlOидией (от rреч. poly MHoro и ploos складывать) называют такое
состояние клетки, коrда в ней в результате предшествующих эндомитозов оказы
вается более двух rаплоидных наборов хромосом. Полиплоидизация, в отличие от
митоза, осуществляется без снижения специфических клеточных функций и свой
ственна поли функциональным элементам (клеткам печени, сердца, слюнных же
лез и др.). В зависимости от числа хромосомных наборов в полиплоидных клетках
их называют три (при 3), тетра (при 4), пента (при 5) и т. д. плоидными. Поли
плоидные клетки отличаются rиrантскими размерами. Они довольно часто BCTpe
чаются в опухолевых тканях, а также в тканях, подверrнутых действию проникаю
щей радиации.
Среди модификаций митоза имеется еще один особый ero вариант, называемый
мейозом (от rреч. meiosis уменьшение). В результате мейоза происходит уменьше
ние числа хромосом вдвое (от диплоидноrо к rаплоидному). Этот способ клеточно
[о деления характерен для половых клеток.
Амитоз прямое деление клетки (ядра). При этом происходит перешнуровыва
ние или фраrментация ядра без выявления хромосом и образования веретена деле
ния. Одной из форм амитоза может быть сеrpеrация reHoMoB множественное пере
шнуровывание полиплоидноrо ядра с образованием мелких дочерних ядер. Как
правило, амитоз встречается в полиплоидных, отживающих или патолоrически изме
ненных клетках и ведет к образованию мноrоядерных клеток. В последние rоды факт
существования амитоза как способа нормальной репродукции клеток отрицается.
Рост и диффереНЦИРОБка клеток
Синтетическая деятельность клеток проявляется в изменении их величины.
При усилении синтеза наблюдается увеличение клеточных размеров и, наоборот,
при снижении синтетической активности может происходить их уменьшение. Все
клетки в той или иной степени способны к росту. Однако их рост оrраничен опре
деленными рамками. Некоторые клетки, например яйцевые, блаrодаря накопле
нию в них желтка, MOryT достиrать оrромной величины. Значительно превышают
обычные размеры rиrантские клетки, образующиеся особенно часто в опухолевых
тканях. Вместе с тем, для клеток некоторых типов (например, для клеток крови)
характерна стабильность средних размеров. Обычно клеточный рост сопровожда
ется преимущественным увеличением объема цитоплазмы, тоrда как величина ядра
изменяется в меньшей степени. На протяжении онтоrенеза строение клеток непре
рывно изменяется. Проrрессирующие изменения клеток, связанные с появлением
Основы общей цитолоlUИ
49
морфолоrических и функциональ
ных различий между ранее индиффе
рентными (однородными) клетщ ми
и обусловленные их специализацией
в процессе развития, называют диф
ференцировкой клеток Биохимичес
кой основой этоrо процесса является
синтез специфичес'/{их бел'/{ов и друrих
веществ. Молекулярные основы син
б \ Сплаисииr
теза елков складываются из TpaHC
САР
крипции первичной структуры MaT \
. Молекула мРНК
ричной РНК на основе информации ",САР
ДНК rена, кодирующей области re "
нов экзонные области; П р оцессин "-
Транспорт
ra мРНК, в результате KOToporo из HO в цитоплазму
вообразованной цепи удаляются He
смысловые последовательности HYK Рис. 22. Схема транскрипции rенетическоrо кода с MO
леотидов (интроны), перехода HOBO лекулы ДНК на молекулу матричной РНК. Объяснение
образованной мРНК в цитоплазму и в тексте
трансляции синтеза белка на аппарате синтеза белков клетки (рис. 22).
Дифференцировка сопровождается качественными, количественными и Bpe
менными параметрами, т. е. характеризуется изменениями клеточной структуры,
темпом развития (ускоренная или замедленная) и степенью (малодифференциро
ванные высокодифференцированные клетки). Усложнение структуры клетки co
провождается следующими изменениями: приобретением определенной формы и
размеров ядра и клетки; сдвиrом ядерно цитоплазменноrо отношения в связи с
более значительным ростом цитоплазмы по сравнению с ядром; развитием opra
нелл; образованием специализированных клеточных структур; синтезом специфи
ческих включений; образованием межклеточноrо вещества; появлением межкле
точных взаимодействий и установлением межклеточных и специализированных
контактов. Так, эпителиальные клетки приобретают кубическую, призматическую
или плоскую форму. Клетки тканей внутренней среды более разнообразны по фор
ме. Некоторые соединительнотканные клетки вырабатывают межклеточное веще
ство. Мышечные клетки содержат миофибриллы. Между нейронами формируют
СЯ синаптические контакты (подробно см. соответствующие rлавы).
Для разных тканевых клеток характерны определенные взаимоотношения
между процессами дифференцировки и деления. Однако в целом по мере повы
шения степени дифференцировки способность клеток к делению закономерно
уменьшается.
В rистоrенезе клетки определенноro цитотипа интеrpируются, частично теряя
автономность, JIРИСУЩУЮ ранней стадии (пролиферативной), вследствие формиро
вания реryляторных механизмов, оказывающих влияние на цитодифференцировку.
Клетки в составе тканей мноrоклеточных орrанизмов имеют различную ПрОДОk
жительность жизни. В некоторых тканях, например в эпителиях, смена клеточноrо
----- ............
-----
......
---
ЯАРО
""'-
/"
/
i
.;.
"' ,"
"-
з
\
\
\.
/
/5'
.
'.
!Первичная мРНК
{ Ш
I 5' CAP и поли А соеnинение
Транскрипция
САР
мм
n
()
мм
I
I
АААА/
/
/
./
ЦИТОПЛАЗМА
50
Fлава 4
состава происходит довольно быстро. По данным ряда исследователей, жизнь эпи
телиальных клеток (в частности, в кишечном эпителии) измеряет я нескольки
ми сутками. В друrих тканях (например, в мышечной и нервной) продолжитель
ность жизни клеток сравнима с продолжительностью жизни орrанизма. Так или
иначе, клеточный состав орrанизма не остается постоянным, он непрерывно из
меняется в результате rибели части клеток и замены их новыми клеточными по
колениями. В нормальных физиолоrических условиях rибели клеток предшеству
ют процессы старения. Они сопровождаются снижением функциональной
активности, а также появлением ряда дистрофических изменений. В общих чер
тах rибель клеток характеризуется сморщиванием ядра и клетки в целом, диф
фузной окрашиваемостью и исчезновением специализированных структур, pac
падом клетки на фраrменты, которые фаrоцитируются соседними клетками или
макрофаrами.
Реактивные изменения клеток
При действии разнообразных механических, химических, физических или био
[енных факторов имеют место реактивные изменения структуры и функций кле
ток Достаточно сильные раздражители вызывают состояние клетки, поrраничное
со смертью. Для обозначения такой предельной степени еще обратимоrо повреж
дения клетки Д.Н. Насонов и ВЯ. Александров (1934) предложили термин "па
ранекроз" (от rреч. para около и nekros мертвый). Это явление с физико хими
ческой точки зрения характеризуется подавлением rранулообразования,
диффузным окрашиванием цитоплазмы, уменьшением дисперсности коллоидов,
повышением вязкости, сдвиrом реакции цитоплазмы в кислую сторону и обрати
мостью этих изменеНИI':'I в начальных фазах действия areHToB.
Представляют интерес реактивные изменения клеток и клеточных орrанелл,
которые развиваются при действии различных факторов, сопутствующих боевому
поражен ию.
Повреждающее действие ионизирующей радиации на клетку связано в OCHOB
ном С ионизацией воды, входящей в ее состав, при которой образуются биолоrи
чески активные радикалы, вызывающие повреждение белков клеточных мембран.
Более чувствительны и быстрее подверrаются денатурации белки. входящие в co
став ферментов, особенно тех, которые содержат сульфrидрильные rруппы. Hapy
шение внутриклеточных окислительно восстановительных процессов приводит к
накоплению биолоrически активных метаболитов денатурированных белков, KO
торые вызывают дополнительные повреждения клеток
В клетках наиболее чувствительными к действию радиации являются мито
хондрии центры окислительно восстановительных реакций. Они набухают, их
матрикс просветляется, кристы укорачиваются, сrлаживаются и полностью исче
зают. Позднее митохондрии распадаются. Вследствие деструкции и rибели мито
хондрии в клетке уrнетается синтез АТФ.
Весьма чувствительна к повреждению как аrранулярная, так и rранулярная
части эндоплазматической сети. Цистерны и канальцы этих орrанелл вначале pac
ОСН0вы общей цитолоши
51
ширяются, затем фраrментируются. Количество рибосом при облучении снижает
ся, соответственно уменьшается содержание РНП в цитоплазме белоксинтезиру
ющих клеток. Число лизосом может быть увеличенным, они rруппируются BOKpyr
разрушающихся орrанелл, происходит аутофаrия продуктов распада. В ядрах кле
ток отмечаются слипание и перераспределение хроматина, возможна отслойка Ha
ружной ядерной мембраны с образованием перинуклеарных вакуолей. При этом
может наступить распад ядер и rибель клеток
Ионизирующая радиация оказывает повреждающее действие на клетку и во
время деления. Клетки в интерфазе, будучи облученными, внешне MorYT выrля
деть нормальными. Повреждение выявляется при последующем делении, коrда
появляются аномальные фиrуры митоза. Митотические хромосомы изменяют
форму, возникают их разрывы иноrда с последующим неправильным соединени
ем фраrментов. Наблюдаются аномалии веретена деления, оно может иметь три
и более полюсов. В связи с этим в анафазе расхождение хромосом к полюсам OKa
зывается неравномерным и некоторые хромосомы отстают, образуя XpOMOCOM
ные мостики. В друrих случаях хромосомы реплицируются, а деления ядра не
происходит, в результате чеrо образуются клетки с крупными полиплоидными
ядрами.
Излучение, воздействуя на rенетический аппарат клетки, может вызывать MY
тации. Измененные при этом rены в процессе митоза удваиваются и дочерние клет
ки будут обладать качествами клеток, испытавших лучевое воздействие. Возник
шая таким путем в клетках орrанизма мутация, может дать начало злокачественной
опухоли.
Достоверным признаком rибели клетки является нарушение структуры ядра.
Различают следующие формы изменения ядра: пикноз, лизис, рексис.
ПU'К1l0З (от rpеч. pyknos уплотнение) характеризуется интенсивным окраши
ванием rомоrенной массы ядерноrо вещества (rиперхроматоз), еro уплотнением и
сморщиванием вследствие потери воды. Лизис (от rреч. lysis растворение), наобо
рот, сопровождается набуханием ядер и их слабой окрашиваем остью с последую
щим полным растворением хроматина, что обозначается как хроматолиз. При этой
форме rибели клеточные ядра напоминают тени нормальных ядер. Кариорексис (от
rpеч. rhexis разрыв) отличается раздроблением хроматина на отдельные rлыбки,
после чеrо происходит обычно их растворение. Одновременно с этим в цитоплазме
rибнущих клеток отмечается появление вакуолей. При этом цитоплазма постепенно
теряет способность окрашиваться rистолоrическими красителями. Распадающиеся
клетки удаляются путем автолиза (самопереваривания под действием литических
ферментов) или путем фаrоцитоза, или в результате автолиза и фаroцитоза.
Кроме Toro, существует явление запроzраммирова1l.1ЮЙ шбели ето'К (апоптоз),
которое возникает в результате запуска собственной проrраммы самоуничтожения
при участии внутренних и внешних по отношению клетки факторов. В составе плаз
молеммы иденшфицированы рецепторы rибели (например Fas, ТNF и друrие), важ
нейшая функция которых связана с передачей цитотоксических сиrналов внутрь
клетки. Внутриклеточными ферментными белками, запускающими апоптоз (фраr
ментацию ДНК, нарушение структурных протеинов, активацию киназ и клеточноrо
52
rлава 4
цикла), являются ферменты семейства цистеин содержащих протеаз, именуемых Kac
пазами. При действии лиrанда на рецептор возникает реакция, в резул;ьтате которой
запускается каскад реакций, результирующим итоrом которою является rибель клет
ки.
Самым ранним морфолоrическим проявлением апоптоза является появление в
ядре резко очерченных уплотненных rOMoreHHbIx масс хроматина с внутренней CTO
роны ядерной оболочки. Наступает ядерная и цитоплазматическая фраrментация.
В дальнейшем фраrменты клетки поrлощаются соседними клетками. При этом при
знаки воспаления отсутствуют. Апоптоз наблюдается как в эмбриональном, так и в
постнатальном rистоrенезе.
Таковы в общих чертах основные проявления жизнедеятельности клеток При
веденные выше материалы свидетельствуют о неразрывном единстве структуры и
функции клеточных орrанелл. В живых клетках структурные компоненты ВЫПОk
няют определенные функции и эти функции имеют вполне соответствующие им
морфолоrические эквиваленты. Клетка блаrодаря тесному взаимодействию всех
структурных компонентов, представляет собой целостную биолоrическую систе
му на всех этапах жизненною цикла.
3акmoчение. Рассматривая клетку как элементарную единицу живой материи с
общебиолоrических позиций, важно определить ее положение и роль в составе иерар
хически наиболее высокоорrанизованной биолоrической системы орrанизма. Здесь
клетка выступает как ведущая структурно функциональная единица самостоятель
Horo уровня структурной орrанизации живоrо ткани. В ткани каждый тип клеток
запроrраммирован на выполнение ряда специальных функций. Чтобы выполнять
эти функции в соответствии с потребностями и адаптивными свойствами ткани,
клетка должна активно воспринимать тканевое окружение, реаrировать на Hero и
изменять свою функциональную активность в зависимости от общетканевоrо [OMe
остаза. Для этоrо в клетке значительная ее часть представлена мембранными CTPYK
турами, важнейшей из которых является плазмолемма. Будучи поrpаничным мемб
ранным комплексом клетки, плазмолемма с помощью рецепторной системы
связывает внутриклеточную среду с тканевой; обеспечивает межклеточные отноше
ния и взаимодействие реryляторных механизмов ткани с внутриклеточными CTPYK
турами и является важнейшей частью системообразующеrо механизма rистоrенеза.
Воспринимая изменения тканевой среды (микроокружения), плазмолемма переда
ет эту информацию по трансдукторной сети внутрь клетки на ключевые функцио
нальные комплексы (энзимные белки, депо кальция, ядро и пр.), которые обеспечи
вают перестройку физиолоrической активности клетки. Блаrодаря тесному
посредническому контакту плазмолеммы с тканевой средой, с одной стороны, и пря
мой связи с ключевыми внутриклеточными функциональными комплексами, с дpy
rой, клетка представляет собой целостную, устойчивую и, вместе с тем, необычайно
динамичную биолоrическую систему. Ей свойственны все черты живоrо, а именно
zенетическая индивидуальность и способность передавать ее будущим поколениям,
реактивность, обмен веществ и подвUЖ1l0сть.
ЭМБриолоrия
53
rЛАВА5.
ОСНОВЫ ОБЩЕЙ ЭМБриолоrии
Эмбриолоrия это наука о закономерностях эмбриональною развития заро
дыша. Термин "эмбриолоrия" возник от rреческою словосочетания ет bryo, что
означает "в оболочках". Эмбрион, или зародыш, это орrанизм, развивающийся
под покровом яйцевых оболочек или внутри материнскOl"О орrанизма в специали
зированном opraHe матке. У человека развивающийся орrанизм до 8 й недели
эмбриоrенеза называется зародышем, далее плодом. В задачи эмбриолоrии BXO
дИТ изучение развития зародыша от момента оплодотворения до рождения (BЫ
лупления из яйцевых оболочек или выхода из материнскою орrанизма), а также
изучение проrенеза процесса образования мужских и женских половых клеток
Медицинская (клиническая) эмбриолоrия изучает закономерности эмбриональ
Horo развития человека, причины нарушений эмбриоrенеза и механизмы возник
новения уродств, а также пути и способы влияния на эмбриоrенез.
Эмбриональное развитие, или эмбриоrенез, это сложный и длительный MOp
фоrенетический процесс, в ходе KOToporo из отцовской и материнской половых кле
ток формируется новый мноюклеточный орrанизм, способный к самостоятельной
жизнедеятельности в условиях внешней среды. Чтобы представить масштаб процес
сов, происходящих в развитии человека, достаточно вспомнить, что яйцеклетка ди
аметром 0,15 мм оплодотворяется спермием диаметром 0,005 мм, общая масса оп
лодотворенноrо яйца составляет Bcero лишь 5x10 9 r. Доношенный плод рождается
со средним размером 500 мм и массой 3400 r. От зиюты до рождения масса плода
возрастает примерно в миллиард раз.
Эмбриолоrические исследования домикроскопическоrо периода давали лишь
общую картину развития орrанизмов и не моrли раскрыть суть зачатия и развития
эмбриона и плода. С общебиолоrических позиций, однако, эти исследования OKa
зали существенное влияние на последующую трактовку мноrих научных фактов,
открытых с помощью микроскопических методов исследования.
Развитие эмбриолоrии как науки
История эмбриолоrии тесно связана с борьбой двух течений, зародившихся еще
в античные времена преформuзма и эпИlеиеза. Преформизм, означающий предоб
разование, утверждает, что развитие орrанизма является лишь ростом имеющеrося
зародыша. Теqретиком прсформизма является Ш. Бонне (1740 1793), утверждав
ШИЙ, что все opraHbI тела настолько тесно связаны между собой, что невозможно
допустить существование TaKoro момента, Korдa тот или друrой из них отсутствовал
бы. с позиций преформизма, вопрос заключался лишь в том, rде находится этот
54
rлава 5
зародыш. По мнению овистов (М. Мальпиrи),
зародыш находится в женской половой клет
ке, а по мнению анималькулистов в мужс
кой половой клетке. Сторонники эпиrенеза,
например, ж. Бюффон (1707 1788), отрица
ли предопределение, однако не смоrли ПОk
твердить свои убеждения фактами. Спор раз
решил русский академик К Вольф (17ЗЗ
1794), опубликовавший в 1759 [. диссертацию
"Теория зарождения", в которой доказал, что
для развития зародыша необходимы женские
и мужские половые клетки. К. Вольф экспе
риментально обосновал концепцию эпиrене
за учение о развитии, соrласно которому
новые разнородные части орrанизма появля
ются из исходноrо однородноrо материала
яйца под влиянием факторов, стоящих над за
родышем (иными словами, происходит HOBO
вобразование структур). Данная концепция
укрепилась блаrодаря работам х. Пандера
(1794 1865) и К Бэра (1792 1876).
Идеи преформизма вновь стали обсуж
даться в литературе, коrда развитие зародышей начали изучать методами молеку-
лярной биолоrии. Так, по мнению А. Спирито (1984), в яйцеклетке содержится не
анатомическая, а химическая миниатюра взрослоrо орrанизма (различия химичес
KOro состава разных участков яйца и в последующем цитоплазмы клеток зароды
ша, которые морфолоrически идентичны).
Становление эмбриолоrии как науки и систематизация фактическorо матери
ала связаны с именем ординарноro профессора Медико хирурrической академии
К Бэра. Он выявил, что в процессе эмбриональноrо развития раньше Bcero обна
руживаются общие типовые признаки, а затем появляются частные признаки клас-
са, отряда, семейства и, в последнюю очередь, признаки рода и вида. Данное зак
лючение было названо правилом Бэра. Соrласно этому правилу, развитие орrанизма
происходит от общеrо к частному. К Бэр указал на образование в эмбриоrенезе
двух зачатковых листков, описал хорду и др.
В развитии сравнительной эмбриолоrии ведущее место принадлежит русскому
эмбриолоry А.О. Ковалевскому (1840 1901). Он изучал мноrочисленных предста
вителей типов первично и вторичноротых и установил единый план развития
мноrоклеточных животных ланцетника, асцидий, червей, кишечнополостных.
А.О. КовалеВСКИII обосновал теорию зародышевых листков как образований, лежа
щих в основе развития всех мноrоклеточных орrанизмов. Опираясь на работы
А.О. Ковалевскоrо, немецкий биолоr э. fеккель (18З4 1919) сформулировал OCHOB
ной биоzеllетичес1СИЙ закон, который rласит, что онтоrенез есть краткое повторение
филоrенеза. Это означает, что в индивидуальном развитии можно наблюдать
'{".,;;.
" r!
A.
,r 1
,
'.
i
J
...
'
j;
{,
1
?'<
.. ..{::<
Карл Максимович
Бэр
(1792 1876)
Основы общей .мБРИОЛОlИИ
55
предковые признаки (или палинrенезы) например. образование у эмбрионов мле
копитающих зародышевых листков, хорды, жаберных щелей и др. Однако в ходе
эволюции появляются новые признаки ценоrенезы (образование провизорных,
или внезародышевых, opraHoB у рыб, птиц и млекопитающих). Явление повторения
в ходе эмбриональноrо развития высших орrанизмов тех или иных признаков более
низкоорrанизованных животных получило название рекапитуляция. Примерами pe
капитуляции в эмбриоrенезе человека являются смена трех форм скелета (хорда,
хрящевой скелет, костный скелет), образование и сохранение до трехмесячною воз
раста плода хвоста, развитие практически сплошною волосяноrо покрова (на 5 M
месяце внутриутробноrо развития), образование жаберных щелей и др.
Учение о рекапитуляции развил А.Н. Северцов (18бб 19Зб), который сформу
лировал положение о том, что онтоrенез не только повторяет филоrенез, но и творит
ero (теория филэмбриоrенезов). Так, если изменение индивидуальною развития идет
путем добавления новых стадий к предковым это надставка, или анаболия; изме
нения, начинающиеся со средних стадий, называются отклонением, или девиацией;
наконец, развитие может измениться с самых ранних стадий, Torдa это архаллаксис
(древний). В последнем случае определить предковые признаки в индивидуальном
развитии практически невозможно.
Большой вклад в развитие эмбриолоrии внесли ПЛ. Иванов (1878 1942) aB
тор теории о ларвальных и постларвальных cerMeHTax первичноротых, П.r. Светлов
(1892 1974) автор теории о критических периодах эмбриоrенеза и друrие исследо
ватели.
етодыисследования
Существует множество методов исследования, среди которых выделяются сле
дующие: наблюдение за жuвъlм.и зародышами с применением кино и видеосъемки
(используется в основном в эксперименте). Для этоrо применяется специальная
микрофотоустановка, соединяющаяся с термокамерой, в которой развивается за
родыш. При изучении развития куриною эмбриона, например, в скорлупе проде
лывается окошечко, которое закрывается прозрачной пластинкой. Данный метод
позволил проследить и уточнить динамику изменения формы и размеров зароды
шей в процессе развития.
Метод изучения фиксированных срезов зародышей с помощью световой и элект
ронной микроскопии, rисторадиоавтоrрафии, rисто и иммуноцитохимии. Эти Me
тоды позволяют анализировать тканевые и внутриклеточные изменения в динами
ке развития частей зародыша. С помощью rисто и иммуноцитохимических методов
исследуются биохимические процессы, происходящие в клетках зародышей, син
тез ДНК, РНК, белков, специфических рецепторных белков и др. С применением
этих методов была получена важная информация о клеточной и тканевой диффе
ренцировке в развитии эмбрионов и плодов.
Метод.маркировки, предложенный в 1925 r. В. Фоrтом (1888 1941), позволяет
изучать перемещения клеток в развивающемся зародыше. Для этою применяются
нетоксичные для зародышей маркеры (например, нейтральный красный, частицы
56
rлава 5
древесноrо уrля), а также антитела к определенным белкам. При применении aH
тител используется их свойство соединяться с флюоресцирующим красителем и
белками зародыша. С помощью флюоресцентной микроскопии прослеживается
распределение красителя и исследуется динамика белковоrо синтеза в развиваю
щихся тканях зародыша.
Методы МИКРОXUРУРZИИ разрабатывались в начале ХХ века представителями
школы [. Шпемана (1869 1941). Они включали: снятие оболочек яиц животных,
пересадку частей одноrо зародыша друюму и др. Данные методы используются
также для изучения последствий разрушения (например, с помощью лазернorо
луча) частей зародыша или ero отдельных клеток. Трансплантацию как раЗНОВИk
ность микрохирурrии используют для выявления путей миrрации клеток и источ
ников развития тканей. При этом пересаживают участок зародыша, например пе
репела, в тот же участок куриноrо зародыша на место удаленною участка. Ядра
клеток перепела имеют характерную структуру и поэтому отличимы от ядер кле
ток зародыша курицы.
Эксплантация иссечение небольшоrо участка зародыша и выращивание ero
на искусственной среде. С помощью этою метода можно получать информацию об
источниках развития тканей из данною участка зародыша и выявлять rистоrене
тические закономерности развития.
Трансплантация ядер метод, позволяющий клонировать зародышей. Ha
пример, пересадка ядер из клеток эпителия кишки rоловастика шпорцевой ля
ryшки в икринку ляrушки, ядро которой было инактивированно ультрафиолето
вым лучом, привела к появлению новых особей (опыты [ердон). Данные опыты
заложили основу клонирования высших позвоночных и способствовали появле
нию (в 1997 [.) знаменитой овцы Долли. Подобные эмбриолоrические экспери
менты убедительно показали, что ядра соматических клеток содержат полный
набор rенетической информации для развития HOBoro орrанизма.
Новейшим достижением экспериментальной эмбриолоrии явилась разработ
ка метода экстракорпорШlЪНОlO оплодотворения. Пересадка зародышей, зачатых в
пробирке, в матку составляет основу лечения бесплодия. В 1973 [. л. Шеттлз
(США) извлек предовуляторную яйцеклетку из яичника бесплодной женщины и
оплодотворил ее сперматозоидами мужа. Так было положено начало технике пере
садки зародышей человека с целью лечения бесплодия. Однако только в 1978 [. в
Великобритании в результате успешной пересадки в матку бесплодной женщины
зародыша человека на стадии 8 бластомеров после 2,5 суток культивирования по
явился первый в мире "пробирочный" ребенок массой 2700 r .
Половые клетки
В отличие от соматических, половые клетки (raMeTbI от слова "брачующие
ся") специализированы на воспроизводстве поколений орrанизмов 1:1 имеют поло
винный (rаплоидный) rенетический набор (1с1п, или 23 хромосомы у человека).
Различают мужские и женские половые клетки, которые несут rенетическую
информацию по отцовской и материнской линиям. В половых клетках у человека
Основы общей эмбриолоlUИ
57
присутствуют 22 аутосомы и 1 половая хромосома,
которая обозначается как Х или У у мужчин и Х
у женщин. При слиянии мужской и женской поло
вых клеток в процессе оплодотворения образуется
диплоидная клетка зиrота, дающая начало всем
клеткам HOBoro орrанизма. В эмбриоrенезе челове
ка половые клетки впервые обнаруживаются в жеk
точной энтодерме, т. е. внеrонадно, а затем миrри
руют в закладку половых желез.
При первом знакомстве со строением зрелых 111
мужских и женских половых клеток, обращает на
себя внимание их высокая функциональная целесо
образность. Сперматозоиды, которые должны быть
высокомобильны В женском половом тракте, преk
ставляют собой маленькие клетки, практически ли Рис. 23. Схема строения спермато
зоида:
шенные цитоплазмы и состоящие из ядросодержа 1 rоловка; 11 шейка; 111 хвост;
щей части, или rоловки, несущей rенетический 1 акросома; 2 ядро; 3 про
ксимальная центриоль; 4 мито
материал, и opraHa передвижения хвоста, или жry хондрии; 5 дистальная центриоль;
тика (рис. 23). Никаких иных субклеточных элемен 6 осевая нить
тов, кроме митохондрий, дающих энерrию для передвижения, акросомальной Ba
куоли с протеолитическими ферментами для растворения оболочек яйцеклетки, и
проксимальной центриоли, сперматозоиды не имеют. Общая длина спермия co
ставляет около 60 мкм, ИЗ которых на долю хвоста приходится 55 мкм.
На ультрамикроскопическом уровне в roловке сперматозоида различимы aK
росомальная и постакросомальная зоны, в хвосте промежуточный, rлавный и
концевой отделы. Большую часть rоловки занимает ядро, заполненное конденси
рованным хроматином. Пространство между ядром и передним участком плазмо
леммы спермия занято акросомальной вакуолью специализированной лизосо
мой, содержащей rруппу ферментов лизинов оболочек яйцеклетки: акрозин
(разрушает прозрачную зону овоцита), пенетраза (диссоциирует клетки лучисто
ro венца), rиалуронидаза (расщепляет rиалуроновую кислоту), кислая фосфатаза
(разрушает форсхолин при прохождении спермиев через плазмолемму овоцита).
Шейка спермия представляет собой короткий отдел, в котором находятся прокси
мальная центриоль и 9 сеrментированных колонн.
Промежуточный отдел содержит аксонему и 9 продольно ориентированных
элементов цитоскелета, состоящих из кератиноподобных белков и являющихся про
должением сеrментированных колонн. Кнаружи от этих волокон располаrаются
митохондрии. Аксонема построена по принципу реснички, биения которой ини
циируются катионами кальция, находящимися в окружающей среде, и МИТОХОНk
риальной АТФ спермия. В концевом отделе хвоста элементы цитоскелета исчеза
ют, и аксонема прикрыта лишь плазмолеммой. У человека дуплеты аксонемы в
концевом отделе распадаются на 18 одиночных микротрубочек.
По выходе из яичка спермии морфолоrически сформированы, но функциональ
но незрелы они неподвижны, не способны к оплодотворению и, помимо этоrо,
I {
11 {
2
3
4
5
6
58
rлава 5
являются носителями ряда специфических антиrенов, инактивация которых (как
и приобретение подвижности и оплодотворяющей способности капацитации)
происходит в системе семявыносящих путей мужскоrо половоrо тракта.
Яйце1Ulеmка в отличие от сперматозоидов
крупная и неподвижная (рис. 24). У боль
шинства млекопитающих и человека ее раз
меры достиrают 1 oo 200 мкм. Женская поло
вая клетка является хранительницей запаса
трофических соединений, необходимых для
paHHero развития зародыша, и "запаса" цитоп
лазматических структур для формирования
первых rенераций зародышевых клеток
бластомеров. У человека крупные размеры
яйцеклетки не связаны с большими запаса
ми желтка яйцеклетка человека бедна жел
тком (олиrолецитальная).
На ультрамикроскопическом уровне яй
цеклетка человека имеет крупное окруrлое
ядро, в котором преобладает эухроматин, и ци
топлазму с умеренным количеством митохон
дрий с немноrочисленными ламеллярными кристами. Хорошо развита система бел
KOBoro синтеза из за высокою содержания рибо и полисом, коротких канальцев
rранулярной эндоплазматической сети. В цитоплазме редко и диффузно располо
жены немноrочисленные скопления вителлиновых rранул коллекторов трофи
ческих включений.
Следует особенно подчеркнуть, что у высших плацентарных млекопитающих
и человека женские половые клетки не существуют изолированно. За исключени
ем самых ранних стадий развития (первичных половых клеток, овоrоний) они Ha
ходятся в тесном контакте с соматическими клетками яичника (фолликулярными
эпителиоцитами и соединительнотканными клетками), которые формируют BOK
pyr каждой половой клетки эпителиальную и соединительнотканную оболочки.
Комплекс "половая клетка соматические оболочки" именуется оварuалы-lым фол
лuкулом., или ово соматическим (фолликулярным) rистионом. Строение овариаль
ною фолликула усложняется в процессе овоrенеза (см. rлаву 17).
Рис. 24. Схема строения яйцеклетки:
1 ядро; 2 митохондрии; 3 вителлино
вые rранулы
Общая характеристи'Кll развития позвоначнъlX
59
rЛАВА6.
ОБЩАЯ ХАРАКТЕРИСТИКА
ИНДИВИДУАЛьноrо РАЗВИТИЯ
ПОЗВОНОЧНЫХ
Знакомство с эмбриоrенезом позвоночных позволяет оценить с позиции cpaB
нительной эволюционной rистолоrии постепенное изменение основных эмбрио
лоrических и rистоrенетических процессов и, в то же время, преемственность этих
изменений в их общебиолоrической основе, экстраполировать некоторые этапы
эмбриоrенезов на развитие человека. В эксперименте на животных можно модели
ровать различные условия развития, изучать действие тератоrенных веществ на
opraHo и rистоrенезы, а также критические периоды развития позвоночных.
Периодизация индивидуальноrо развития
Индивидуальное развитие, или онтоrенез, начинается с момента оплодотворе
ния и завершается смертью орrанизма. Принято различать эмбриональный (заро
дышевый, пренатальный) и постэмбриональный (постнатальный) периоды инди
видуальноrо развития.
Эмбриональный период развития (эмбриоrенез), в свою очередь, можно пред
ставить в виде ряда последовательно сменяющих друr друrа биолоrических про
цессов оплодотворения, дробления, возникновения бластулы и rаструлы, обо
собления комплекса зачатков opraHoB и тканей, rисто и орrаноrенеза.
Оплодотворение это слияние мужской и женской половых клеток и образо-
вание одноклеточноrо орrанизма зиrоты. При оплодотворении происходит вос-
становление диплоидноrо набора хромосом, и образующаяся зиrота приобретает
свойство тотипотентности способность дать начало всему разнообразию клеток
и тканей будущеrо орrанизма.
С помощью ферментов акросомы спермий локально разрушает оболочки яй
цеклетки и происходит слияние ero плазматической мембраны с плазматической
мембраной яйцеклетки. Затем в овоплазму проникает rоловка и промежуточная
часть спермия. Это происходит в результате взаимодействия между рецептора:\ш
клеточных оболочек сперматозоида и яйцеклетки. После проникновения rенети
ческоro материала сперматозоида в яйцеклетку возникает новая одноклеточная
живая система зиrота с диплоидным содержанием хромосом.
Дробление серия повторяющихся митотических делений зиroты и ее дочер
них клеток бластомеров, без последующеrо роста их размеров до размера MaTe
ринской клетки. Новые клетки не расходятся, а тесно прилежат друr к друrу. Ритм
60
Fлава 6
дробления зависит от вида
животноrо и колеблется от
десятков минут до десяти и
более часов. Темпы дробле
ния не сохраняются посто
янными и реrулируются
мноrими факторами. При
радиальном способе дробле
ния первая и вторая полосы
(борозды) дробления проходят в меридианальноЙ плоскости, но полосы дробле
ния находятся под прямым уrлом друr к друrу. Плоскость TpeTbero дробления
лежит под прямым уrлом к плоскостям первых двух дроблен иЙ и rлавноЙ оси
ЯI':'ща (широтно или экваториально). Чередование меридианальных и широтных
полос дробления вызывает увеличение числа бластомеров. У некоторых позво
ночных появляются танrенциальные полосы дробления, проходящие параллель
но поверхности скопления клеток (рис. 25). Характер дробления определяется
количеством желтка и разным распределением ero в цитоплазме яЙцеклетки (rи
потеза О. rертвиrа). Маложелтковые (алецитальные и олиrолецитальные), а TaK
же uзолецuталъ1lыe яЙцеклетки (клетки с небольшим количеством и paBHOMep
ным распределением желтка) дробятся полностью (rолобластически) и
равномерно. Однако следует понимать, что бластомеры при, казалось бы, внеш
не одинаковых размерах MorYT отличаться своими биохимическими своЙствами,
и понятие равномерности условно. В случае высокоrо содержания желтка в яЙ
цеклетке rОБОрЯТ о мноrожелтковых, или полuлецuталъ1lы,' яЙцеклетках, в KOTO
рых желток сосредоточен в веrетативноЙ, а орrанеллы в анимальноЙ частях.
Дробление в обоrащенноЙ включениями веrетативноЙ части цитоплазмы яЙцек
летки происходит более медленно. Дробление зародыша при этом, полное, но He
равномерное, и бластомеры на веrетативном полюсе отличаются большими раз
мерами, чем на анимальном (мезолецитальная яЙцеклетка). В случае очень
больших запасов белково липидных включениЙ в яЙцеклетке roворят о телоле
цuталЪ1l0Й или резко телолецитальноЙ яЙцеклетке. В этом случае дробится лишь
часть анимальноrо полюса яЙцеклетки, дробление частичное, или меробластu
чеС1Сое (дискоидальное, поверхностное).
у плацентарных млекопитающих яЙцеклетка маложелтковая вторично оли
rолецитальная и изолецитальная. Дробление полное, однако по характеру CTpoe
ния бластомеров и закономерностям появления новых бластомеров оно относится
к неравномерному и асинхронному.
В результате дробления возникает мнorоклеточныЙ зародыш, именуемыЙ бла
стулой. Бластула бывает в виде целобластулы с большим бластоцелем, если
дробление полное и равномерное; амфибластулы, коrда дробление полное, но He
равномерное, вследствие чеrо бластоцель располаrается эксцентрично. В бласту
ле различают стенку бластодерму и полость бластоцель, заполненную жиk
костью. В свою очередь в бластодерме выделяются крыша (анимальныЙ полюс
дробления), дно (веrетативныЙ полюс дробления), краевая зона, расположенная
CD& @
д
Б
в
Рис. 25.Способы дробления зиrоты:
Д полное равномерное; Б полное неравномерное; В He
полное, частичное,дискоидальное
Обшря характеристика развития пОЗ60UОЧll.blХ
61
между двумя вышепоименованными частями бластулы. Если дробление частич
ное, затраrивающее только часть (анимальную ) яйцеклетки (дискоидальное дроб
ление), то это приводит К возникновению мноrослойной структуры, напоминаю
щей диск (дискобластула). У млекопитающих в результате полноrо асинхронноrо
дробления возникает зародышевый пузырек, или бластоциста.
[лавным итоrом процесса дробления является увеличение числа клеток заро
дыша до такою критическоrо значения, при котором в клеточных пластах начина
ют возникать механические напряжения, инициирующие направленные переме
щения клеток в определенные участки зародыша. Продолжение активной
пролиферации клеток в развитии зародыша является одним из механизмов кле
точных транслокаций и, в частности, laструляции.
Ib.струл.яция результат активною деления клеток, роста и направленных пере
мещений (миrраций) клеточных потоков с формированием мноrослойноrо зароды
ша, или rаструлы, (возникновением послойно
расположенных, отделенных друr от друrа OT
четливой щелью, зародышевых листков: наруж
Horo эктодермы' среднеro мезодермы' BНYТ
peHHero эuтодермы). Перемещение клеток
происходит в cтporo определенной области за
родыша в области ceporo серпа. Последний
был описан В. Ру в 1888 r. В оплодотворенном
яйце амфибии серый серп выявляется как OKpa
шенная область на стороне, противоположной
проникновению спермия. В этом месте, как по
лаrают, локализуются факторы, необходимые
для rаструляции.
у разных представителей позвоночных ra
струляция совершается несколькими OCHOBHЫ
ми способами: путем ИU6аlUuации (впячива
ния), uмм.иzрации (перемещения части клеток
внутрь зародыша), эпиболии (обрастания), дe
лам.иuации (расщепления). Способы rаструля
ции зависят от типа яйцеклетки (рис. 26). При
любом способе rаструляции в качестве ведущих
сил выступают неравномерная пролиферация
клеток в разных частях зародыша, уровень об
менных процессов в клетках, расположенных
в разных частях зародыша. активность амебо
идных движений клеток, а также индуктивные
факторы (белки, нуклеопротеиды, стероиды и
др.). В результате rаструляции обособляются
основные зачатки opraHoB и тканей.
Следующим периодом эмбриоrенеза явля
ется rисто и орrаноrенез дифференцировка
А
r
Рис. 26. Способы rаструляции:
А инваrинация; Б эпиболия, или обра
стание; В иммиrрация; r деламина
ЦИЯ, или расщепление
62
Fлава 6
различных тканей и opraHOB орrанизма из материала зародышевых листков и эмб
риональных зачатков.
Образование зародышевых листков и эмбриональных
зачатков тканей в эмбриоrенезе у некоторых позвоночных
В результате rаструляции возникает мноrослойный зародыш. Несмотря на различ
ные способы rаструляции после выделения материала зародышевых листков по оси за
родыша находится материал хорды, который подстилает нервную пластинку, слева и
справа от хорды располаrается материал мезодермы. Все это характеризует осевой Koм.п
леке зачатков. В дальнейшем происходит формирование зачатков opraнoB, представля
ющих собой пространственно локализованные rpуппы стволовых клеток источников
развития тканей. Закономерности дифференцировки клеточноrо материала зачатков
можно проследить в эмбриоrенезе наиболее изученных животных.
Ланцетник
Классическим объектом эмбриолоrических исследований ланцентник, подроб
но изученный А.о. Ковалевским. Ланцетник является представителем класса xop
довых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне
в теплых морях. Свое название получил из за формы, напоминающей ланцет (хи
рурrический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиrо и изолецитальная, размером 110 мкм, ядро pac
полаrается ближе к анимальному полюсу. Оплодотворение наружное. Дробление
зиrоты полное, почти равномерное, синхронное и заканчивается образованием бла
стулы. В результате чередования меридианальных и широтных борозд дробления
формируется однослойная бластула с полостью, заполненной жидкостью блас
тоцелем. Бластула сохраняет полярность, ее дно представляет собой веrетативную,
а крыша анимальную части; между ними находится краевая зона.
При rаструляции происходит впячивание веrетативной части бластулы в ани
мальную. Впячивание постепенно уrлубляется и, наконец, формируется двустенная
чаша с широко зияющим отверстием, ведущим в новообразованную полость заро
дыша. Такой способ rаструляции называется uuваlUuацuей. Так бластула превраща
ется в rаструлу. В ней материал зародыша оказывается дифференцированным на
наружный листок эктодерму, и внутренний энтодерму. Полость чаши именует
ся zaстроцелем., или полостью первичной кишки, которая сообщается с внешней cpe
дой посредством бластопора, что соответствует заднепроходному отверстию. В бла
стопоре различают дорсальную, вентральную и две латеральных ryбы. В результате
инваrинации центр тяжести зародыша смещается, и зародыш поворачивается блас
топором кверху. Постепенно происходит смыкание краев бластопора и удлинение
зародыша. Топоrpафия клеток в составе ryб бластопора определяет развитие разных
частей зародыша. При rаструляции из BHYTpeHHero листка rаструлы обособляются
хорда и мезодерма, которые располаrаются между экто и энтодермой. Завершается
rаструляция образованием oceBoro комплекса зачатков и дальше обособлением
Обшря характеристика развития позвО1ЮЧllЫХ
63
зачатков opraHoB (рис. 27). Хорда ин
дуцирует развитие нервной трубки из
материала дорсальной эктодермы. Эта
часть эктодермы утолщается, образу
ется llервllQЯ пластИllка (нейроэкто
дep 1a), которая проrибается по cpek
ней линии и превращается в желобок
2 Края желобка постепенно смыкаются
в нервную трубку. Оставшаяся часть
эктодермы кожная, срастается над
llерв1l0Й трубкой. Однако на самом пе
реднем и заднем концах зародыша He
рвная трубка некоторое время сооб
щается с внешней средой с помощью
двух отверстий нейропоров. В пос
ледующем происходит разделение Me
8 зодермы на спинные сеrменты СОМИ
ты, количество которых нарастает от
1S пар дО 60 6S пар у взрослоrо лан
цетника. Часть латерально располо
женной мезодермы не сеrментирует
9
ся и расщепляется на наружный (па
риетальный) и внутренний (висце
ральный) листки спла1LXНотома. Эти
листки растут между экто и энтодер
мой и, достиrнув середины на BeHT
ральной стороне зародыша под ки
шечноЙ трубкой, срастаются, форми
руя единую вторичную полость цe
лом. На переднем конце зародыша воз
никает уrлубление (ротовая бухта)
растущее по направлению к передне
му отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепоrо KOH
ца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с
внешней средой. Аналоrичныl процесс протекает на заднем конце зародыша. По
бокам rоловноro отдела зародыша также возникает контакт кожноii эктодермы и ки
шечной энтодеРI\IЫ. В месте этоrо контакта происходит прорыв. Так полость пере
дней кишки сообщается с внешней средой (формируется жаберный аппарат). После
этоro зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миrрации бластомеров по
зволили выделить определенные области зародыша на ранних стадиях развития
(зиrоты бластулы), которые позднее развиваются в зародышевые листки и
эмбриональные зачатки opraHoB и тканей. Эти области были названы презумптив
lIЬ/.МИ (предполаrаемыми) участками, или зачатками.
А
Б
Рис, 27. Схема образования rаструлы ланцетника:
А продольное сечение; Б поперечное сечение; 1
эктодерма; 2 энтодерма; 3 rастроцель; 4 блас
топор; 5 нервная пластинка; 6 зачаток хорды; 7
мезодерма, 8 кожная эктодерма; 9 кишечная эн
тодерма
7
64
Тлава 6
Таким образом, на примере развития ланцетника можно наблюдать закономер
ное перемещение клеток и формирование зародышевых листков, oceBoro комплекса
зачатков и обособление зачатков opraHoB и тканей.
Птицы
Классическим объектом эмбриолоrических исследований является куриный эм
брион. Яйцеклетка у птиц боrата желтком пОЛWlеu,италънQЯ. Диаметр яйцеклетки
курицы достиrает 35 мм, а у страуса до 100 мм. В отложенном птичьем яйце Haxo
дится не яйцеклетка, а мноrоклеточный зародыш на стадии бластулы или ранней
laструлы. К яйцеклетке относится то, что мы называем желтком. Яйцеклетка, про
двиrаясь по яйцеводу, оплодотворяется. Диаметр овулировавшеrо яйца составляет
50 мкм И постепенно увеличивается до 6 мм, а затем и до 35 мм. Желточный MaTe
риал синтезируется в печени, переносится в фолликулярные клетки, а затем в ци
топлазму яйцеклетки. У рептилий и птиц наблюдается полиспермия и, хотя с женс
ким пронуклеусом сливается один мужской, избыточные спермии мероциты
(блуждающие ядра) способствуют переработке желтка.
Дробление зиюты птиц неполное, неравномерное, частичное, дискоидальное,
так как дробится всею 1/500 часть зиrоты на анимальном полюсе. Первые три бо
розды дробления радиальные, затем появляются широтные и танrенциальные
полосы. При дроблении возникают бластомеры различной величины, и в конеч
ном итоrе формируется мноrослойная дискобластула, распластанная на нераздро
бившемся желтке.
Как отмечал ось выше, к моменту откладки яйца зародыш находится в первой
фазе rаструляции. Это проявляется обособлением внезародышевой энтодермы
путем деламинаu,ии дискобластулы и возникновением двуслойноrо зародыша. Так
возникают две части: эпибласт наружная и zuпобласт внутренняя части дис
кобластулы (рис. 28). Периферическая часть rипобласта в дальнейшем образует
внезародышевую энтодерму, которая обрастает желток, формируя желточный Me
шок провизорный (временный) opraH, выполняющий трофическую и KpOBeTBOp
ную функции. В стенке желточною мешка обнаруживаются первичные половые
клетки (BHeroHaдHoe происхождение).
Наиболее важные морфоrенетические процессы развертываются в эпиблас
те источнике развития всех трех зародышевых листков. Здесь возникают кле
точные потоки вследствие асинхронноrо дробления клеток и их направленноrо
перемещения. Клетки перемещаются в течение 3 4 часов инкубации. Схема дви
жения клеточных потоков выrлядит следующим образом: клетки латеральных
частей эпибласта перемещаются в хвостовую часть зародыша и, встречаясь здесь,
движутся далее по средней линии эпибласта к rоловному концу. К 12 часам кле
точный поток принимает вид первичной полоски. Второй поток клеток (более
медленный) распространяется со стороны rоловноrо конца зародыша и движет
ся по средней части эпибласта навстречу первому потоку. Место встречи двух
потоков оnозначается как zензеновский, или первичный, узелок важнейшиЙ
rистолоrичсский маркер. На вершине rензеllОВСКОЮ узелка появляется уrлубле
Общая характеристика развития nозв01lачнЪLХ
65
ние nервичная ямка, а по средней линии первичной полоски первичная бо
роздка, как продолжение первичной ямки. Обе эти структуры рассматриваются
как rомолor бластопора. К 16 часам инкубации все описанные структуры хорошо
выявляюся на фиксированных препаратах.
Дальнейшие перемещения клеток uм.миzрация, характеризуют вторую фазу
rаструляции. В области переднеrо края rензеновскоrо узелка клетки, подворачи
ваясь, проходят через ямку и распространяются в направлении rоловноrо конца
зародыша, формируя rоловной отросток зачаток хорды (нотохорд). В свою оче
редь, зачаток хорды индуцирует раннюю детерминацию клеток нервной пластин
ки в составе эпибласта. Одновременно с материалом хорды из противоположноrо
", ""' ......................
// \ >llT' K :
/ f / , '. \
I I " " , '.
I I " \ I I
, \ :;:::,: / I 3
: \ \ \ ,,',' ' 4
I ' \ .. 5
\ '...
'\ \ б
\
\
\
\
\
\ 7
\ - .., I
. \ /
4
2
1
Б
8
9
1
/
/
б 7
/
.::
' , ?'
.
... ..... с..,. ... .... с_. .... ..... ......... .с:... .с.... с... c. с-;,. .i&!.)o с.. <11(....
1
/
/
s
9
Рис. 28. rаструлsщия у птиц:
А презумптивные зачатки в зародышевом диске; Б саrиттальный разрез; В поперечный разрез
кзади от rензеновскоrо узелка; 1 эктодерма; 2 нервная пластинка; 3 хорда; 4 rензеновский
узелок; 5 первичная ямка; б первичная полоска; 7 первичная бороздка; 8 rоловной отросток
(нотохорд); 9 энтодерма
66
[лава 6
края rензеновскоro узелка и передней части первичной полоски в иммиrрацию
вовлекаются новые клетки, однако последние включаются в состав rипобласта [o
ловной части зародыша. При этом происходит деформация rензеновскоrо узелка
и укорочение первичной полоски. Все эти процессы протекают в течение 8 10 ча
сов инкубации. Клетки оставшейся части первичной полоски представляют co
бой материал COMaTO и спланхномезодермы, Через 15 часов инкубации эти клетки
приходят в движение и миrрируют внутрь вдоль первичной полоски, распростра
няясь над ПlПобластом по бокам от удлиняющейся хорды.
Таким образом, в результате им
миrрации из эпибласта выселяются
материал хорды, прехордальной пла
стинки (а, по некоторым данным, Ma
териал зародышевой энтодермы),
мезодермы, что сопровождается дe
формацией, укорочением и исчезно
вением rензеновскоrо узелка и пер
вичной полоски. Оставшийся MaTe
риал эпибласта представляет собой эктодерму, которая в своем составе содержит
клетки будущей нервной трубки. Хорда индуцирует развитие HepBHoro желобка, CBO
рачивающеrося затем в трубку и поrружающеrося под эктодерму (рис. 29).
Клеточный материал мезодермы дифференцируется на сомиты, латерально
расположенные 1lефроmОJИЫ и наиболее латерально лежащие спла1lX1l0mомы. Co
миты и нефРОТО:\IЫ это сеrментированные части мезодермы, спланхнотом He
сеrментированная. Последний в дальнейшем расслаивается на висцеральный и па
риетальный листки со щелевидноЙ целомической полостью между ними. Под
хордой располаrается энтодерма, которая сворачивается в кишечную трубку, сооб
щающуюся с желтком. Так возникают эмбриональные зачатки opraHoB и тканей.
Наиболее сложна топorрафическая дифференцировка мезодермы. В составе соми
та различают следующие части: с'КЛероmом вентромедиальную часть, дерматом
дорсолатеральную часть и миотом дорсомедиальную часть, Клетки склеротома
интенсивно размножаются, выселяются из состава сомита, окружают хорду и диф
ференцируются в хрящевые клетки (возникают сложные формообразовательные
процессы закладки позвонков, ребер, лопаток и друrих костей скелета). Дерматом
является источником развития соединительных тканей кожи, а миотом источ
ником развития скелетной мышечной ткани.
Параллелыю образованию тела зародыша происходит образование внезаро
дышевых оболочек, или провизорных (временных) opraHoB. У зародышей высших
позвоночных имеются 4 вида оболочек aM1lUOll (водная), XOPUOll (серозная обо
лочка), aJUlallmouc и желmОЧllЫЙ мешок
, ,,1 3
. . '--:': < - ., , . ..
...,...; ' r ,'i.' "'c 0'0" .,
. 1:. . "11:" ..., - - - -. "' _....
. ,1!:" ' .' . ., , .,,/ · .., ,- - , "'-<'/:."
.... ..A"' r! 4 ...« ..... .-; :,I:r i..r l; . ;...
:'}...r:;,,- ...(.,... _"".T : ":::1.: 6.. '" .. ;,. <. >,.!..,
: tr;4 g '( . :.!!: ' .-" i {; :_:' «. . ..r ::
,... ..... .... <i I..... j;!. ""''tJ.3r.. :) "'".............. .......-... ............ w '-..- ;".J
.:.. ; : 'I.: : -,,_.J': . ..)....---- "'....-,.... .:.... ...1J' ...........
.'. -
о" .. . »-...........2
Рис. 29. Осевой комплекс: 1 нервная трубка; 2 xop
да; 3 мезодерма
Млекопитающие
Млекопитающие произошли от яйцекладущих предков рептилий и перешли
в процессе эволюции к живорождснию. Эмбрион вынашивается в специальном
Общая характеристика развития позв01lОЧНЪLX
67
opraHe материнскоrо орrанизма матке, [де он обеспечивается питательными Be
ществами и кислородом за счет материнскоrо орrанизма.
OpraHoM, осуществляющим связь зародыша с материнским орrанизмом, яв
ляется плацента. Вследствие Toro, что в эмбриоrенезе млеко питающих отпадает
необходимость накопления питательноrо материала в яйце, возникает неболь
шая, диаметром 100 200 мкм вторично изолецитальная и олиrолецитальная яй
цеклетка.
Оплодотворение у плацентар
ных млеко питающих внутреннее.
Дробление зиrоты полное, He
равномерное, асинхронное, в pe
зультате чеrо вначале возникает
скопление бластомеров в виде TY
товой яrоды МОРУЛЫ, или плот
ною шарика. Последующее дроб
ление приводит к возникновению
зародыша с полостью бластоци
сты. В бластоцисте выделяются
две части внутреннюю клеточ
ную массу, или эмбриобласт, и Ha
ружный слой клеток трофоб
ласт (рис. 30). Клетки трофобла
ста способны секретировать жиk
кость в полость бластоцисты и вза
имодействовать со слизистой обо
лочкой матки, участвуя в имплан
тации зародыша.
Имплантация, или внедрение,
зародыша в матку совершается в
течение 2 x суток и включает две
Рис. 30. Схема paHHero развития плацентарных млекопи фазы: прилипание (адrезия) и про
тающих:
1 первая полоса дробления; 2 асинхронное дробле никновение (инвазия). BOKpyr эм
ние бластомеров; 3 обрастание темных бластомеров бриобласта рано образуется внеза
эмбриобласта светлыми бластомера трофобласта; 4
морула плотное скопление бластомеров; 5 бластоци родышевая мезенхима, которая
ста, формирование полости; 6 трофобласт; 7 эмбри подстилает трофобласт изнутри.
облает Обе структуры ответственны за
развитие хориона ворсинчатой оболочки, которая с участком слизистой оболоч
ки матки формируют плаценту.
Только после имплантации зародыша становятся возможными развитие дpy
rих внезародышевых opraHoB и переход к rаструляции, rисто и орrаноrенезу. Раз
витие желточноrо мешка, хотя и происходит, функцию питания зародыша он не
выполняет. В стенке желточноrо мешка обнаруживаются первичные половые клет
ки, стволовые клетки крови, а также сосуды. Амнион, аллантоис и желточныЙ Me
шок развиваются из клеток эмбриобласта. Амнион появляется до возникновения
2
3
4
7
68
rлава 6
первичной полоски. С возникновением амниотическоrо пузырька оставшийся Ma
териал эмбриобласта расщепляется способом делами нации на наружную эпиб
ласт, и внутренюю lUпобласm клетnчные массы.
После выделения внезародышевых opraHoB, которые MOryт обеспечить ПрОДОk
жение развития зародыша, заметно активизируются пролиферативные и миrpацион
ные процессы в эпибласте. Они практически аналnrичны тем, которые происходят в
эпибласте куриноrо зародыша. Формируются первичная полоска и rензеновский узе
лак, клетки которых закономерно миrpируют в пространство между эпи и rип06лас
том. Так возникает I\fНОroслойный зародыш, в составе KOTopOro далее выделяются э-к
mодерма с нервной пластинкой, хордомезодерма и Эllmодерма (в rоловную часть
последней встраивается материал прехордальной пластинки). В последующем проис
ходят сеrментация и нейруляция.
Таким образом, в развитии позвоночных появлению тканей предшествуют про
цессы, связанные с дроблением, rаструляцией и обособлением эмбриональных за
чатков тканей. Данный временной отрезок можно определить как проrистоrенез.
После закладки комплекса эмбриональных зачатков тканей начинается период rис
тоrенеза и орrаноrенеза. Осуществляется дифференцировка и специализация кле
точноro материала эмбриональных зачатков, что составляет основу rистоrенеза
возникновения и развития тканеЙ. При этом происходит анатомо rистолоrическое
формирование практически всех opraHoB, характерных для взрослой особи.
у большинства позвоночных локализация зачатков тканей и opraHoB в составе
зародышевых листков и развитие из них конкретных производных совпадает. Так,
ОЖ1lая э-ктодерма является истnчником развития кожноrо эпителия. Нервная трубка
содержит стволовые клетки для развития тканевых элементов нервной системы. Из
-кишечной ЭllmодерМbl развивается эпителиальная выстилка кишечной трубки, КРУП
ные железы пищеварительноrо тракта (печень, поджелудочная железа). Мезодерма
содержит в себе стволовые клетки для большоrо числа производных скелетной
мышечной ткани (миотомы сомитов), соединительных тканей кожи (дерматомы co
митов), скелетных тканей (склеротомы сомитов) и др. Как правило, развитие боль
шинства тканей и opraHoB не завершается к рождению, а продолжается в постна
тальном периоде онтоrенеза. Классификация тканей и закономерные процессы
развития стволовых клеток зачатков тканей и opraHoB, составляющих суть эмбрио
нальноrо rисто и орrаноrенеза, рассматриваются в следующих rлавах.
[истorенез протекает и в провизорных opraHax (рис. 31). Он отличается рядом
специфических особенностей, в частности, опережающим развитием тканей BHe
зародышевых оболочек Несмотря на асинхронность развития тканей зародыше
вых и внезародышевых opraHoB, тем не менее на всех стадиях развития зародыш
является целостным орrанизмом. Взаимосвязь и взаимообусловленность ero час
тей усиливаются по мере повышения структурной орrанизации. Знания общих за
кономерностей индивидуальноrо развития, филоrенетической обусловленности
этих процессов у человека позволят будущим врачам при обрести навыки эволю
ционноrо подхода к оценке общебиолоrических явлений, в том числе и патолоrии,
умения применять их для профилактики, диаrностики и лечения различных Hapy
шений нормальноrо развития человека.
Обшр.я характеристи1((J. развития позвО1ЮЧ1iblХ
69
3
2
6
7
А
Б
в
Рис. 31. Схема развития провизорных opraHoB у млекопитающих:
А. Б, в три последовательные стадии:
1 тело эмбриона; 2 туловищная складка; 3 амниотические складки; 4 амниотическая оболочка;
5 желточный мешок; б аллантоис; 7 хорион.
Сплошная толстая линия трофобласт и эктодерма; сплошная тонкая линия энтодерма; пунктирная
мезодерма
"
70 ОБЩАЯ rистолоrия (УЧЕНИЕ о ТКАНЯХ)
fЛАВА 7.
ЭМБРИОНАЛЬНЫЙ fИСТОfЕНЕ3
И КЛАССИФИКАЦИЯ ТКАНЕЙ
В развитии низших и высших позвоночных отчетливо прослеживается единая
общебиолоrическая закономерность, выражающаяся в появлении зародышевых ли
стков и обособлении основных зачатков opraHoB и тканей. Процесс образования TKa
ней из материала эмбриональных зачатков составляет суть учения о rистоrенезах.
Эмбриональный rистоrенез. Определение понятия "ткань"
Эмбриональный rистоrенез, по определению А.А. Клишова (1984), это KOM
плекс координированных во времени и пространстве процессов пролиферации,
клеточноrо роста, миrрации, межклеточных взаи:-.юдействий, дифференциации,
детерминации, проrраммированной rибели клеток и некоторых друrих. Все назван
ные процессы в той или иной мере протекают в зародыше, начиная с самых ранних
стадий ero развития.
Пролиферация. Основной способ деления тканевых клеток это мumоз. По
мере увеличения числа клеток возникают клеточные rруппы, или популяции, объе
диненные общностью локализации в составе зародышевых листков (эмбриональ
ных зачатков) и обладающие сходными rистоrенетическими потенциями. Клеточ
ный цикл реryлируется мноrочисленными BHe и внутриклеточными механизмами.
К внеклеточным относятся влияния на клетку цитокинов, факторов роста, ropMo
нальных и нейроrенных стимулов. Роль внутриклеточных реrуляторов иrрают спе
цифические белки цитоплазмы. В течение каждоro клеточноrо цикла существуют
несколько критических точек, соответствующих переходу клетки из одноrо пери
ода цикла в друrой. При нарушении внутренней системы контроля клетка под вли
янием собственных факторов реrуляции элиминируется апоптозом, либо на HeKO
торое время задерживается в одном из периодов цикла.
Метод радиоrрафическоrо анализа клеточных циклов в различных тканях выя
вил особенности соотношения клеточной репродукции и дифференцировки. Напри
мер, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд про
лиферирующих клеток, за счет которых обеспечивается непрерывное возникновение
новых клеток взамен поrибающих, то эти ткани относятся к об1l0вляющu.м.ся. Друrие
ткани, например, некоторые соединительные, характеризуются тем, что в них YBe
личение количества клеток происходит параллельно с их дифференцировкой, клет
ки в этих тканях характеризуются низкой митотической активностью. Это pacmy
щuе ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы
репродукции заканчиваются в период эмбриональноro rистоrенеза (коrда форми
ЭмбрИО1ЮJlЬНЫЙ w.cmоzен.ез и 'ЮlO.сcuфика.цuя. тканей
71
руется основной запас стволовых клеток, достаточный для последующеrо развития
ткани). Поэтому она отнесена к стаБUJlЬНЬtМ (стационарным) тканям. Продолжи
тельность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно Ha
блюдается обновление внутриклеточных структур (внутриклеточная физиолоrичес
кая реrенерация).
Клеточный рост, миrрация и межклеточные взаимодействия. Рост клеток
проявляется в изменении их размеров и формы. При усилении функциональной
активности и внутриклеточных биосинтезов наблюдается увеличение объема клет
ки. Если объем клетки превышает некую норму, то rоворят о ее w.пертрофии, и
наоборот, при снижении Функщюнальной активности происходит уменьшение
объема клетки, а при переходе некоторых нормативных параметров возникает aт
рофuя. клетки. Рост клетки не беспределен и определяется оптимальным ядерно
цитоплазменным отношением.
Важное значение для rистоrенеза имеют процессы перемещения клеток. Миz
рация клеток наиболее характерна для периода rаструляции. Однако и в период
rисто и орrаноrенеза происходят перемещения клеточных масс (например, CMe
щения миобластов из миотомов в места закладки скелетных мышц; движение кле
ток из HepBHoro rребня с образованием спинномозrовых rанrлиев и нервных спле
тений, миrрация rоноцитов и т. д.). Миrрация осуществляется с помощью
нескольких механизмов. Так, различают хемотаксис движение клеток в направ
лении rрадиента концентрации какorо либо химическоrо areHTa (перемещения
спермиев к яйцеклетке, предшественников Т лимфоцитов из KOCTHoro мозrа в зак
ладку тимуса).
ю.птотаксис механизм перемещения клеток по rpадиенту концентрации aд
rезионной молекулы (движение клеток протока пронефроса у амфибий по rрадиен
ту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование
коrда в какой либо преrраде остается один канал для перемещения (описан у рыб
при образовании плавников).
Контактное Иlllибирование этот способ перемещения наблюдается у клеток
HepBHoro rребня. Суть способа заключается в том, что при образовании ламелло
подии одной клеткой и контакта ее с друrой клеткой, ламеллоподия прекращает
рост и постепенно исчезает, но в друrой части миrрирующей клетки при этом фор
мируется новая ламеллоподия.
В процессе миrрации клеток важную роль иrрают межклеточные взаимодей
ствия. Существует несколько механизмов TaKoro взаимодействия (контактною и дис
тантною). Выделяется большая rpуппа молекул 1Ulеточной адzезии (МКА). Так, Kaдre
рины это Са2+ зависимые МКА, отвечают за межклеточные контакты при
образовании тканей, за формообразование и др. В молекуле кадrерина различают BHe
клеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный
домен ответственен за адrезию клеток с одинаковыми кадrеринами, а внутриклеточ
ный за форму клетки. Друrой класс МКА это иммуноrлобулиновое суперсемей
ство Са2+ независимых МКА, обеспечивающих, например, адrезию аксонов к capKO
лемме мышечных волокон, или миrрацию нейробластов вдоль радиальных rлиоцитов
72
Fлава 7
в закладке коры большоrо мозrа и др. Следующий класс МКА это мембранные фер
менты rликозилтранферазы. Последние по типу "ключ замок" соединяются с уrле
водными субстратами rликозаминоrликанами надмембранноro комплекса клетки,
осуществляя таким образом прочное сцепление клеток
Кроме механизмов межклеточноrо взаимодействия, существуют механизмы
взаимодействия клеток с субстратом. Они включают формирование рецепторов
клетки к молекулам внеклеточноrо матрикса. К последним относят производные
клеток, среди которых наиболее изученными адrезионными молекулами являют
ся коллаrен, фибронектин, ламинин, тенасцин и некоторые друrие. Коллаzе1ibl, cpe
ди которых различают несколько десятков типов, входят в состав межклеточноrо
вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр.
ФибрО1iектU1i, секретируемый клетками, является связывающей молекулой меж
ду миrрирующей клеткой и межклеточным матриксом. ЛамU1iU1i компонент ба
зальной мембраны, также связывает миrрирующие клетки с межклеточным MaT
риксом (справедливо по отношению к эпителиоцитам инейробластам ).
Для осуществления связи миrрирующих клеток с межклеточным матриксом
клетки формируют специфические рецепторы. К ним относятся, например, сииде
каи, который обеспечивает контакт эпителиоцита с базальной мембраной за счет
сцепления с молекулами фибронектина и коллаrена. и1iтеzрu1iы клеточных поверх
ностей связывают с внеклеточной стороны молекулы внеклеточноrо матрикса, а BHYT
ри клетки белки цитоскелета (например, актиновые микрофиламенты). Так B03
никает связь внутри и внеклеточных структур, что позволяет клетке использовать
для перемещения собственный сократительный аппарат. Наконец, существует боль
шая rруппа молекул, формирующих клеточные контакты, осуществляющие KOMMY
никацию между клетками (щелевые контакты), механическую связь (десмосомы,
плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции
[ормонов и факторов роста (ФР). Последние это вещества, оказывающие стиму
лирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним
относятся, например, Ф Р, полученный из тромбоцитов и влияющий на переход кле
ток в фазу размножения (rладких миоцитов, фибробластов, rлиоцитов); эпидермаль
ный ФР стимулирует пролиферацию эпителиоцитов, производных эктодермы;
Ф Р фибробластов стимулирует пролиферацию фибробластов. Особо выделяется
большая rруппа пептидов (соматотропины, соматомедины, инсулин, лактоrен), вли
яющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также при
нимают кооперативное участие в процессах развития клеток и тканей. К ним OTHO
сятся, например, бета интерферон и трансформирующий ФР. Последний, однако,
в отношении разных клеточных типов действует по разному: блокирует размно
жение мноrих типов эпителиоцитов, но стимулирует размножение соединитель
нотканных клеток
Дифференциация это стойкое структурно функциональное преобразование
клеток в различные специализированные клетки. Дифференцировка клеток биохи
мически связана с синтезом специфических белков, а цитолоrически с образовани
Эм-бриО1ШЛЪНЫЙ m.стОle1lез и 'К.llO.сcuфикация тканей
73
ем специальных орrанелл и включений. IIри дифференцировке клеток происходит
избирательная активация reHOB. Важным показателем клеточной дифференцировки
является сдвиr ядерно цитоплазменноrо отношения в сторону преобладания разме
ров цитоплазмы над размером ядра. Дифференцировка происходит на всех этапах OH
тоrенеза. Особенно отчетливо выражены процессы дифференциации клеток на этапе
развития тканей из материала эмбриональных зачатков. Специализация клеток обус
ловлена их детерминацией.
Детерминация это процесс определения пути, направления, проrраммы раз
вития материала эмбриональных зачатков с образованием специализированных
тканей. Детерминация может быть оотипической (проrраммирующей развитие из
яйцеклетки и зиrоты орrанизма в целом), зачатковой (проrpаммирующей разви
тие opraHoB или систем, возникающих из эмбриональных зачатков), тканевой (про
rраммирующей развитие данной специализированной ткани) и клеточной (про
rраммирующей дифференцировку конкретных клеток). Различают детерминацию:
1) лабильную, неустойчивую, обратимую и 2) стабильную, устойчивую и необра
тимую. При детерминации тканевых клеток происходит стойкое закрепление их
свойств, вследствие чеrо ткани теряют способность к взаимному превращению
(метаплазии). Механизм детерминации связан со стойкими изменениями процес
сов репрессии (блокирования) и экспрессии (деблокирования) различных reHOB.
Клеточная rи6ель широко распространенное явление как в эмбриоrенезе,
так и в эмбриональном rистоrенезе. Как правило, в развитии зародыша и тканей
rибель клеток протекает по типу апоптоза. При мерами проrраммированной rибе
ли являются rибель эпителиоцитов в межпальцевых промежутках, rибель клеток
по краю срастающихся небных переrородок. Проrраммированная rибель клеток
хвоста происходит при метаморфозе личинки ляrушки. Это примеры морфоrене
тической rибели. В эмбриональном rистоrенезе также наблюдается rибель клеток,
например при развитии нервной ткани, скелетной мышечной ткани и др. Это при
меры rистоrенетической rибели. В дефинитивном орrанизме путем апоптоза по
rибают лимфоциты при их селекции в тимусе, клетки оболочек фолликулов яич
ников в процессе их отбора для овуляции и др.
Понятие о диффероне. По мере развития тканей из материала эмбриональ
ных зачатков возникает клеточное сообщество, в котором выделяются клетки раз
личной степени зрелости. Совокупность клеточных форм, составляющих линию
дифференцировки, называют диффероном-, или lистОlенетическим. рядом-. Диффе
рон составляют несколько rрупп клеток: 1) стволовые клетки, 2) клетки предше
ственники, 3) зрелые дифференцированные клетки, 4) стареющие и отмирающие
клетки. Стволовые клетки исходные клетки rистоrенетическоrо ряда это ca
моподдерживающаяся популяция клеток, способных дифференцироваться в раз
личных направлениях. Обладая высокими пролиферативными потенциями, сами
они (тем не менее) делятся очень редко.
Клетки предшественники (полустволовые, камбиальные) составляют следу
ющую часть rистоrенетическоrо ряда. Эти клетки претерпевают несколько цик
лов деления, пополняя клеточную совокупность новыми элементами, и часть из
них затем начинают специфическую дифференцировку (под влиянием факторов
74
rлава 7
микроокружения). Это популяция коммитированных клеток, способная диффе
ренuироваться в определенном направлении.
Зрелые функционирующие и стареющие клетки завершают rистоrенетичес
кий ряд, или дифферон (рис. 32). Соотношение клеток различной степени зрелос
ти в дифферонах зрелых тканей орrанизма неодинаково и зависит от основных
закономерных процессов физиолоrической реrенерации, присущих конкретному
виду ткани. Так, в обновляющихся тканях обнаруживаются все части клеточноrо
дифферона от стволовой до высокодифференцированной и rибнущей. В типе
растущих тканеЙ преобладают процессы роста. Одновременно в ткани присутству
ют клетки среднеЙ и конечной частей дифферона. В rистоrенезе митотическая aK
тивность клеток постепенно снижается до низкой или крайне низкой, наличие CTBO
ловых клеток подразумевается только в составе эмбриональных зачатков. Потомки
стволовых клеток некоторое время существуют как пролиферативный пул ткани,
но их популяция быстро расходуется в постнатальном онтоrенезе. В стабильном
типе тканей имеются лишь клетки высокодифференцированной и rибнущей час
теЙ дифферона, стволовые клетки обнаруживаются лишь в составе эмбриональ
ных зачатков и полностью расходуются в эмбриоrенезе.
Cf G[JOU<>t
Начальная
камбиальная
часть дифферона
Средняя
дифференцированная
часть дифферона
Конечная BblCOKO
дифференцированная
часть дифферона
Рис. 32. Схема орrанизации клеточноrо дифферона:
Столбики пролиферативная активность клеток
Изучение тканей с позиций их клеточно дифферонноrо состава позволяет
различать M01-l0дuффеРО1l1lые (например, хрящевая, плотная оформленная co
единительная и др.) и полuдuффеРО1-l1-lые (например, эпидермис, кровь, рыхлая
волокнистая соединительная, костная) ткани. Следовательно, несмотря на то,
что в эмбриональном rистоrенезе ткани закладываются как монодифферонные,
в дальнейшем большинство дефинитивных тканей формируются как системы
Эмбриональный zuстоl.ен.ез и классификация тканей
75
взаимодействующих клеток (клеточных дифферонов), источником развития KO
торых являются стволовые клетки разных эмбриональных зачатков.
Ткань это фWlо и оuтОlеuетuчески сложившаяся систе.ма клеточuых
диффероuов и их uеклетОЧUblХ производUblХ, Фуuкции и реlеuераторuая cпo
собuость которой определяется lистОlеuетическu.ми свойства.ми ведущеlО кле
тОЧUОlО диффероuа.
Ткань является структурным компонентом opraHa и в то же время частью одной
из четырех тканевых систем покровных, тканей внутренней среды, мышечных и
невральных.
Классификация тканей
с позиции филоrенеза предполаrается, что в процессе эволюции орrанизмов
как у беспозвоночных, так и позвоночных образуются 4 тканевых системы, обеспе
чивающие основные функции орrанизма: покровные, отrраничивающие от внеш
ней среды; внутренней среды поддерживающие rомеостаз; мышечные отвеча
ющие за движение, и нервные за реактивность и раздражимость. Объяснение
этому феномену дали А.А. Заварзин и H.r. Хлопин, которые заложили основы
учения об эволюционной и онтоrенетической детерминации тканей. Так, было
выдвинуто положение о том, что ткани образуются в связи с основными функция
ми, обеспечивающими существование орrанизма во внешней среде. Поэтому из
менения тканей в эволюции идут параллельными путями (теория параллелизмов
А.А. Заварзина). Однако диверrентный путь эволюции орrанизмов ведет к воз
никновению все большеrо разнообразия тканей (теория диверrентной эволюции
тканей H.r. Хлопина). Из этorо следует, что ткани в филоrенезе развиваются и
паршuzельныии рядами. и дивеРl.ентно.
Диверrентная дифференциация клеток в каждой из четырех тканевых систем в
конечном итоrе привела к большому разнообразию видов тканей, которые rистоло
rи в последующем стали объединять в системы или l.pyппъt тканей. Однако стало
ясно, что в ходе диверrентной эволюции ткань может развиваться не из одноrо, а из
нескольких источников. Выделение OCHOBHOro источника развития ткани, дающеrо
начало ведущему клеточному типу в ее составе, создает возможности для классифи
кации тканей по rенетическому признаку, а единство структуры и функции по
морфофизиолоrическому. Однако из этоrо не следует, что удалось построить COBep
шенную классификацию, которая была бы общепризнанной.
Большинство rистолоrов в своих работах опираются на морфофункциональ
ную классификацию А.А. Заварзина, сочетая ее с rенетической системой тканей
НТ Хлопина.
В основу известной классификации А.А. Клишова (1984) положена эволюци
онная детерминированность четырех систем тканей, развивающихся у животных
разных типов параллельными рядами, вместе с орrаноспецифической детерминаци
ей конкретных разновидностей тканей, образующихся диверrентно в онтоrенезе.
Автор в системе эпителиальных тканей выделяет 34 ткани, в системе крови, соеди
нительных и скелетных тканей 21 ткань, в системе мышечных тканей 4 ткани и
76
rлава 7
в системе нервных и нейроrлиальных тканей 4 ткани. Эта классификация включа
ет практически все конкретные ткани человека.
В качестве общей схемы (табло 1) приведен вариант классификации тканей по
морфофизиолоrическому принципу (rоризонтальное расположение) с учетом ис
точника развития ведущею 1<ЛетОЧ1lОlО дифферО1lа конкретной ткани (расположе
ние по вертикали). Здесь даны представления о зародышевом листке, эмбриональ
ном зачатке, тканевом типе большинства известных тканей позвоночных в
соответствии с представлениями о четырех тканевых системах. В приведенной
классификации не отражены ткани внезародышевых opraHoB, которые обладают
рядом особенностей.
Таким образом, иерархические отношения живых систем в орrанизме крайне
сложны. Клетки, как системы первоrо порядка, формируют дифферо1lы. Последние
образуют тка1lИ как мозаичные структуры или являются единственным дифферо
ном данной ткани. В случае полидифферонной структуры ткани необходимо BЫ
делить ведущий (основной) клеточный дифферон, который во MHoroM определяет
морфофизиолоrические и реактивные свойства ткани. Ткани формируют системы
следующеrо порядка орю1lы- В них также выделяется ведущая ткаиь, обеспечи
вающая rлавные функции данноrо opraHa. Архитектоника opraHa определяется ею
МОрфОфУ1lКЦИО1lалЬ1lЪLМИ едИ1lИЦами и lистИО1lами. Системы opraHoB являются об
разованиями, включающими все нижележащие уровни с их сqбственными закона
ми развития, взаимодействия и функционирования.
Все перечисленные структурные компоненты живою находятся в тесных вза
имоотношениях, rраницы условны, нижележащий уровень является частью выше
лежащеrо и так далее, составляя соответствующие целостные системы, высшей
формой орrанизации которой является орrанизм животных и человека.
Дифференцировка зародышевых листков
и классификация тканей позвоночных
Зародышевый
листок
Тканевый тип
1. Нейроэктодер
мальный
Эмбриональный 1.1.Нервная трубка
зачаток 1.2.rанrл. пластнка
1.З.плакоды
Тип эпите
лия:
:S:
':S:
Q)
ж
C'CI
10-
)(
jj
ж
111
О
а.
о Q)
с jj
C'CI Ж
::Е r::t
Q) о
ti
:S: :S:
CJ8.
..: с
Зародышевый
листок
Тканевый тип
Эмбриональный
зачаток
11. Система тка-
ней внутренней
средЬ! (MeTa
60лическойl
а) rлиальный
Эпендимный;
передней камеры rла
за;
пиrментный
сетчатки;
opraHa слуха;
обонятельный;
вкусовой
ЭКТОДЕРМА
.....
ЭКТОДЕРМА
ЭПИБЛАСТ
+
I
2. Эпидермаль
ный З. Прехордальный
2.1. Кожная экто З. 1. Прехордальная
дерма пластинка
б) кожный
Эпидермис и
производные
(потовые, сальные
и
молочные железы);
роrовицы rлаэа;
предцверия
ротовой
полости;
эмали и
кутикулы эуба;
анальный, пере
ходный,
влаrалищный
1. Нейроэктодермальный
1.4. Эктомезенхима
Оболочек нервных волокон (леммоциты);
менинrотелий
Ротовой полости и же
лез;
rлотки и производных;
пищевода и желез;
барабанной полости и
евстахиевой трубы;
аденоrипофиза;
паратиреоидный;
тиреоидный;
вилочковой железы;
воздухоносных путей;
альвеолярный
ЭНТОДЕРМА
4. Энтеродер
мальный
Таблица 1
w
о
о'
;ж:
о'
$::!,
N
i
-е..
i:;
J;::
;ж:
,
МЕ30ДЕРМА
6. Целонефродермальный
4.1. Кишечная энто 5.1.Спланхнотом
дерма
В) кишечный
Желудка и желез
желудка;
кишок и желез;
печени;
желчноrо пузыря
и желчевыводя
щих
путей;
поджелудочной
железы
МЕ30ДЕРМА
7. Спланхнотомный
7.1. Энтомезенхима
Миелоидная; лимфоидная; ретикулярная;
рыхлая волокнистая соединительная; жиро-
вая (бурая и белая)
5.2. Нефротом
r) целомичес- д) почечный
кий
Коры надпочеч
ника;
семенных изви
тых канальцев;
фолликулярный
яичника;
rрудной, брюш-
ной и околосер-
дечной полос
тей (меЗ0Т !1ИЙ)
)s...
о)
!>.:S:
1::",
i
o:s;
800
ха
А:З:
0000
0:0)
:::::!'
O»S
Uo:
Про-, мезо- и
метанефро-
rенная эпите-
лиальная
,ткань;
эпителий не-
фрона;
эпителий co
бирательных
трубочек поч
ки
8, Анrиодермальный
8.1. Анrиобласт
Эндотелий кровеносных и лимфати
ческих сосудов и внутренней выстилки
сердца
-...J
-...J
Зародышевый лис ЭКТОДЕРМА МЕ30ДЕРМА
ток
Тканевый тип 1. Нейроэктодермальный 6. Сомитный 9.Хордоидный
Эмбриональный 1.4. Эктомезенхима 6.1. Дерматом 6.2. Склеротом Хорда
.. зачаток
>S
Ф Соединительная склеры; Плотная неоформлен Плотная оформленная co
::1: а) с опорно- Хордальная
C'II механической костная черепа; ная соединительная единительная 1l<aHb.
функцией; дентин и цемент зуба хрящевые: rиалиновая. эла
)( стическая И ВОЛОКНИQТая;
:ii костные: пластинчатая и pe
::1:
JI тикулoqpиброзная
r:;
ф
... 2. Эпи
C'II Тканевый тип 1. Нейро 3. Прехор
L..
S эктодермальный дep дальный 6 Сомитный 7 СпланхнотомНЫЙ
111 мальный
ct
I
О Эмбрионалbl-lЫЙ 1. 1.HepB 1.2.rанrл. 2.1.Кож 3. 1.прехор 7.2. Миокардиальная пла
::1: ная труб 6.3. Миотом 7.1. Энтомезенхима
а. зачаток плаC1li1Нка ная эк дальная пла стинка
О ка тодерма стинка
1:
О б) с двиrа- Мионев Исчер Мио Миоидные Скелетная; Висцеральная; Сердечная:
C'II тельной ральная ченные эпители клетки тиму переднеro отдела пи. сосудистая сократительный,
::s функцией радужки миоциты альные са щеварительной труб проводящий и
ф (мышечные и и цили в COCTa слюнных, ки; креторный диффероны
... миоидные) apHoro ве OE[a потовых лимфатических cep
CJ
s тела нов НС и молоч дец;
() ных же электрических opra-
лез нов
-..J
00
Зародышевый ЭКТОДЕРМА
листок
Тканевый тип 1. Нейроэктодермальный
Эмбрионалbl-lЫЙ зачаток 1.1. Нервная трубка 1.2. rанrлиозная пластинка 1.3. Плакоды
IV. Система невраль OpraHoB ЦНС и нейросекреторные нейроны; Чувствительных и веrетативных raHr rанrлиев rоловы;
ных тканей и нейро- сетчатой оболочки rлаза; шишковиднои же лиев; спиральноrо и вестибулярноrо
rлия лезы; задней доли rипофиза хромаффинная 1l<aHb надпочечников;
rломусов
:?
о:,
'-J
Система nокровllЫХ ткаllей и их nроизводllыe
79
rЛАВА 8.
СИСТЕМА ПО КРОВНЫХ ТКАНЕЙ
И ИХ ПРОИ3ВОДНЫЕ
Эпителиальные ткани древнейшие rистолоrические структуры, которые в
фило и онтоrенезе возникают первыми. Основное свойство эпителиев nOlpa
llИЧ1l0стъ. Эпителиальные ткани (от rреч. epi над и thele кожица) располаrа
ются на rраницах двух сред, отделяя орrанизм или орrаны от окружающей среды.
Эпителии, как правило, имсют вид клеточных пластов и образуют наружный
покров тела, выстилку серозных оболочек, просветов opraHoB, сообщающихся с
внешней средой во взрослом состоянии или в эмбриorенезе. Через эпителии ocy
ществляется обмен веществ между орrанизмом и окружающей средой. Важной фун
кцией эпителиальных тканей является защита подлежащих тканей орrанизма от
механических, физических, химических и друrих повреждающих воздействий. He
которые эпителии специализированы на выработке специфических веществ pe
ryляторов деятельности друrих тканей орrанизма.
Производными покровных эпителиев являются железистые эпителии. Особый
вид эпителия эпителий opraHoB чувств.
Эпителии развиваются с 3 4 й недели эмбриоrенеза человека из материала всех
зародышевых листков. Некоторые эпителии, например эпидермис, формируются как
полидифферонные ткани, так как в их состав включаются клеточные диффероны,
развивающиеся из разных эмбриональных источников (клетки Ланrерrанса, мела
ноциты и др.).
В классификациях эпителия по происхождению за основу, как правило, бе
рется источник развития ведущеrо клеточнorо дифферона дифферОllа эnите
лиоu,итов. Цитохимическим маркером эпителиоцитов являются белки u,ито
кератиllы' образующие тонофиламенты. Цитокератины характеризуются
большим разнообразием и служат диаrностическим маркером KOHKpeTHoro вида
эпителия.
Различают эктодермальные, энтодермальные и мезодермальные эпителии. В за
висимости от эмбриональноrо зачатка, служащеrо источником развития ведущеrо
клеточноrо дифферона, эпителии подразделяются на типы: эпидермальный, энтеро
дермальный, целонефродермальныЙ, эпендимоrлиальный и анrиодермальный (Хло
лин H.r., 1946).
По rистолоrическим признакам строения ведущею (эпителиальноrо) клеточ
Horo дифферона различают однослоЙные и мноrослойные эпителии (рис. 33). Ok
нослойные эпителии по форме составляющих их клеток бывают плоские, кубичес
кие, призматические или цилиндрические.
Однослойные эпителии подразделяют на однорядные, если ядра всех клеток
80
Fлава 8
лежат на одном уровне, и мноrорядные, в
которых ядра расположены на разных ypOB
нях, т. е. в несколько рядов.
Мноrослойные эпителии подразделяют
ся на ороrовевающие и неороrовевающие.
Мноrослойные эпителии называют плоски
ми, учитывая форму клеток наружноrо слоя.
Клетки базальноrо и друrих слоев MorYT
иметь при этом цилиндрическую или непра
вильную форму. Кроме названных, выделя
ют еще переходный ЭПIJJ-елий, строение KO
Toporo меняется в зависимости от степени
ero растяжения.
На основе данных об орrаноспецифи
ческой детерминации эпителии подразде
ляются на следующие типы: кожный, ки
шечный, почечный, целомический и ней
роrлиальный. В составе каждоrо типа раз
личают несколько разновидностей эпите
лиев с учетом их строения и функций.
Эпителии перечисленных типов стойко
детерминированы. Однако при патолоrии
возможна трансформация одноrо вида
эпителия в друrой, но лишь в пределах Ok
Horo TKaHeBoro типа. Например, среди
эпителиев кожноrо типа мноrорядный
мерцательный эпителий воздухоносных
путей может переходить в мноrослойный плоский. Такое явление получило Ha
звание м-етаплазия.
Несмотря на разнообразие строения, выполняемых функций и происхожде
ния из разных источников, все эпителии имеют ряд общих признаков, на основе
которых их объединяют в систему или rруппу эпителиальных тканей. Эти общие
морфофункциональные признаки эпителиев следующие.
Большинство эпителиев по своей цитоархитектонике представляют собой ok
нослойные или мноrослойные пласты плотно сомкнутых клеток Клетки соедине
ны с помощью межклеточных контактов. Эпителий находится в тесных взаимо
действиях с подлежащей соединительной тканью. На rранице между этими тканями
имеется базШlЪ1laЯ м-ем-браllа (пластина). Эта структура участвует в формирова
нии эпителиально соединительнотканных взаимоотношений, выполняет функции
прикрепления с помощью полудесмосом эпителиоцитов, трофическую и барьер
ную. Толщина базальной мембраны обычно не превышает 1 мкм (рис. 34). Хотя в
некоторых opraHax ее толщина значительно возрастает. Электронно микроскопи
чески в составе мембраны выделяют светлую (расположенную ближе к эпителию)
и темную пластинки. Последняя содержит коллаrен IV ro типа, обеспечивающий
з
2
4
5
б
Рис. ЗЗ. Схема расположения эпителиоцитов в
различных видах эпителия:
1 однослойный плоский эпителий; 2 OДHO
слойный кубический эпителий; 3 однослой
ный цилиндрический эпителий; 4 мноroряд
ный мерцательный эпителий: 5 мноroслой
ный плоский ороrовевающий эпителий; б пе
реходный эпителий
Система покровllЫХ тканей и их проuзводllые
81
' + " ';"-,,t ....t;щ '"' , '"
;., ... .," Jl *;;''': .fi' . ..'! ;
:, < :. <.""',,, "F '"
' < '""- : - . '1, rf- .. . '. ' a::': :i
>': \. .,. , , ,"' " "'t: ,
'.. .......". " , .,. !' .".9i'''*!..
J ; ';, ...... ......... /" / " . ...... '.."
tf "-:...,.' ' ' , ' '-." ., ---:-
lf ; "\\ "." "1.: , '''''''. -:."
!..;' . ' ....> ..,: '.... t., ,. .. о
, , ?.. ,, . "' . 4." :
':>,1 ,1 :"'i . ',>.r ',' ,
t. ..., '"1"
н. '''1""'';1'"
;;';' '"7"
.. i. .r;
, 'Т' ----
е>
.,.(., ,, '
, ....
.....
J П
.,
' } '
< ,
. "
.IFO
)i;".. {
" . ,> .
,
.... .
Рис. 34. Ультраструктура базальной мембраны эпидермиса:
1 светлая пластинка; 2 темная пластинка; 3 полудесмосо,
мы эпителиоцитов; 4 сосочковый слой дермы
"
механические свойства мем
браны. С помощью адrезив
ных белков фибронектина
и ламинина осуществляется
при крепление эпителиоци
тов к мембране. Через баз(Щь
ную мембрану путем диффу
зии веществ происходит
питание эпителия. Базаль
ную мембрану рассматрива
ют в качестве барьера для po
ста эпителия вrлубь. При
опухолевых разрастаниях
эпителия она разрушается,
что позволяет измененным
раковым клеткам врастать в
подлежащую соединитель
ную ткань (fаршин B.r., 1939).
Эпителиоциты обладают zemeРOnОЛЯp1l0cmъю. Строение апикальной и базальной
частей клетки разное. В мноrослойных пластах клетки различных слоев отличаются
дрyr от друrа по структуре и функциям. Это называют вертикальной анизоморфией.
Эпителии обладают высокой способностью к реzеllерации за счет МИТ030В KaM
биальных клеток В зависимости от местоположения камбиальных клетЬк в эпите
лиальных тканях различают диффузный и локализованный камбий.
Эпителии кожноrо типа
Эпителии кожноrо типа развиваются из кожной эктодермы и прехордальной
пластинки. Из КОЖ1l0Й эктодермы возникают: мноroслойный плоский ороrовеваю
щий эпителий кожи (эпидермис), мноrослойный плоский неороrовевающий эпите
лий роrовицы, эпителий преддверия ротовой полости, эпителии слюнных, потовых,
сальных и молочных желез, переходный эпителий мочевыводящих путей и др.
Из прехордШZЪ1l0Й пластИllКИ развиваются мноrослойный плоский HeoporoBe
вающий эпителий пищевода, мнorорядный мерцательный эпителий воздухонос
ных путей, однослойный альвеолярный эпителий леrких, эпителий щитовидной,
околощитовидной, вилочковой желез и передней доли rипофиза (см. табл. 1).
По своему строению эпителии кожноrо типа MOryT быть мноrослойные, MHO
rорядные и однослойные. Мноrослойные эпителии состоят из нескольких клеточ
ных слоев, из которых лишь базальный слой прилежит к базальной мембране. Клет
ки базальноrо слоя эпителиоциты способны интенсивно делиться митозом.
Они служат источником пополнения клеточноrо состава вышележащих слоев. Ба
зальные эпителиоциты имеют призматическую форму. По мере смещения этих кле
ток в поверхностные слои они постепенно уплощаются. В мноrослойном плоском
ороrовевающем эпителии поверхностный слой образуют poroBbIe чешуйки.
82
rлава 8
Рис. 35. Слои и клеточные
диффероны эпидермиса:
А: 1 базальный слой; 2
шиповатый слой; 3 зерни
стый слой; 4 блестящий
слой; 5 роrовой слой; 6
базальные малодифферен
цированные эпителиоциты;
7 клетка Ланrерrанса; 8
роrовая чешуйка; Б: 9 Me
ланоцит; 1 О внyrриэпидер
мальный лимфоцит; 11
клетка Меркеля
Поrраничное положение
большинства эпителиев обус
ловливает определенную цито
архитектонику ткани, а также
специфические особенности
внутренней структуры клеток и
их объединения за счет форми
рования различных типов меж
клеточных контактов.
Эпидермис является наибо
лее типичной разновидностью
среди по кровных эпителиев
(рис. 35). Это полидифферонная
ткань. Эпителиальный дифферон
развивается из материала кожной
эктодермы, отличается стойкой
детерминированностью. Диффе
роны меланоцитов, клеток Лан
repraHca и клеток Меркеля разви
ваются из иных источников (см.
строение кожи). Эпителиальный
дифферон формирует MHoro
слойный пласт ороrовевающих
клеток (мноrослойный плоский
ороrовевающий эпителий). В нем
различают слои: базшlьны,' шиповаты,, зернистый и роювой. В базальном слое Ha
ходятся малодифференцированные клетки (базальные эпителиоциты) призматичес
кой формы, которые путем митотическоrо деления обеспечивают обновление кле
точноrо состава ткани. После митоза эти клетки перемещаются в вышележащий
шиповатый слой, образуя клетки мноrоуюльной формы. Клетки шиповатою слоя
(шиповатые, крылатые, или остистые, эпителиоциты) имеют в цитоплазме специа
лизированные структуры то1юфuламенты. При световой микроскопии arperaTbI
тонофиламентов описывают как тонофибршuzы. За счет опорных свойств последних
достиrается механическая прочность клеточною пласта. Между клетками образу
ются связующие комплексы, или межклеточные контакты дecMocoMы.
Следующую стадию дифференцировки составляют уплощенные эпителиоци
ты зернистоrо слоя. В цитоплазме этих клеток кроме тонофиламентов синтезиру
ются и накапливаются белки фWlaUРИ1/. и кератоли1/.И1/.. Ядра зернистых клеток
постепенно пикнотизируются, орrанеллы распадаются под влиянием внутрикле
точных ферментов.
Блестящий слой хорошо выявляется только в эпидермисе ладоней и подошв при
световой микроскопии. Ero образуют плоские постклеточные структуры кератино
цитыI в которых ядра и орrанеллы исчезают. Из последних образуются poroBbIe чешуй
ки поверхностноro слоя. Они имеют вид 14 rpанника. Между чешуйками находится цe
10
А
б
Система пОКРО6НЫХ тканей и их пРОИЗ60дные
83
ментирующее вещество, боrатое липидами (церамиды и др.). PoroBbIe чешуйки имеют
плотную оболочку (толщиной 1S нм), образованную кератолинином (инволюкрином),
ковалентно связанным с оболочкой чешуйки. Содержшюе чешуйки заполнено фиБРИk
лами зрелоrо кератина, который характеризуется водонерастворимостью и высокой стой
костью к химическим areHTaM. Созревание кератина это аrpеrация филаментов и обо
rащение серой за счет образования внутримолекулярных поперечных дисульфидных
связей. Этот процесс инициируется филarrpином и происходит при переходе эпителио
цитов из зернистоro слоя в роrовой. Самые поверхностные слои чешуек постепенно yт
рачивают связи дрyr с друrом и слущиваются.
Разновидностями мноrослойных эпителиев являются кубические и призмати
ческие эпителии, например, выводных протоков слюнных желез и некоторых дpy
rих opraHoB, а также мноrослоЙныЙ плоский неороrовеващиЙ эпителий роrовицы.
Последний состоит из базальноrо, шиповатоro и слоя плоских эпителиоцитов.
ОсобыЙ вид переходный эпителий мочевыводящих путеЙ. Он образован ба
зальным, промежуточным и поверхностным слоями. БазальныЙ (камбиальный) слой
образован мелкими эпителиоцитами. Полиrональные эпителиоциты располаrают
ся в промежуточном слое, а крупные 2 3 ядерные эпителиоциты в поверхност
ном слое. При растяжении мочевоrо пузыря ero стенка уплощается и эпителий pac
тяrивается, становится тонким, двуслойным и наоборот, при сокращении эпителий
утолщается. Эпителиоциты промежуточноrо слоя, не теряя связи с базальной мемб
раноЙ, становятся rрушевидными, а поверхностные куполообразные.
МНОlOрядный эпителий (ложномноrослоЙный) содержит клетки разной формы
Производными эпителиальноrо дифферона являются реснитчатые, вставочные эпи
телиоциты, бокаловидные экзокриноциты и эндокриноциты. Располаrаются все клет
ки на базальной мембране (см. рис. 33). Но вследствие разной высоты ядра эпители
оциов находятся на разных уровнях, что создает впечатление мноrослойности.
В кишечном типе эпителиев эпителиальный дифферон развивается из MaTe
риала кишечной энтодермы. Наиболее общий rистолоrический признак эпителиев
кишечноrо типа однослойность и высокопризматическая форма эпителиоциов
(см. рис. 33). Вместе с тем
каждая разновидность ки
шечных эпителиев имеет
свои орrаноспецифические
особенности строения, фун
кции И rистотопоrрафии.
Примером эпителия дaHHO
ro типа является всасываю
щий эпителий слизистой
оболочки тонкой кишки
(рис. 36). Это однослойный
цилиндрический эпителий
Эпителии кишечноrо типа
,/ .
I i ....... . " ",
':1 I .......' .
, ,,-' ;; G. li '.r ' I{\ "\..:.1 R
tTh' .,<j\ ' '. \.:.'/v, Qc I' !". Ой
i . ... " {'
..... '" -...........",-........ A... ........!
.... ,:;:... ...s;--:.,' (у <:IP 3
.4 .' €J <D
............2
Рис. Зб. Схема строения эпителия кишечноrо типа:
1 призматический эпителиоцит; 2 бокаловидный экзокрино
цит; 3 базальная мембрана
84
Fлава 8
с rетерополярностью различным строением базальной и апикальной частей кле
ток На апикальной поверхности клеток имеются m-икровОРСИll:ки, которые образу
ют щетОЧllУЮ каем-ку. При этом всасывающая поверхность увеличивается в 25 30
раз. В надмембранном комплексе rликокаликсе располаrаются ферменты при
стеночноrо пищеварения. Для эпителиев, образующих выстилку пищеваритель
Horo канала, характерно сильное развитие плотных межклеточных контактов за
пирающеrо типа, блаroдаря чему эпителиальный слой выполняет барьерную фун
кцию. Внутрь орrанизма вещества поступают не по межклеточным пространствам,
которые прочно переrорожены замыкательными поясками, а непосредственно че
рез сами клетки эпителия.
В составе эпителия существует друrая разновидность клеток бокаловuд1lые
ЭК30КрU1l0цuты это слизистые внутриэпителиальные одноклеточные железы. В
их цитоплазме содержится MHoro слизистоrо секрета, ядро оттеснено в базальную
часть.
Эпителий состоит из разных по степени созревания клеток: стволовых, камби
альных, малодифференцированных, дифференцированных (зрелых) и заканчива
ющих жизненный цикл. Стволовые клетки способны к диверrентной дифферен
цировке и формированию дифферонов каемчатых, апикально зернистых
эпителиоцитов, бокаловидных экзокриноцитов и эндокриноцитов.
В процессе физиолоrической реrенерации эпителиальный пласт обновляется
в течение 3 5 суток
К эпителиям кишечноrо типа относятся также эпителиальные ткани, COCTaB
ляющие основную массу печени и поджелудочной железы. Эпителии этих opraHoB
развиваются в эмбриоrенезе из общеrо с кишечным эпителием энтодермальноrо
зачатка и представляют особые разновидности эпителиев кишечноro типа. В их
структуре важный rистолоrический признак расположение клеток в виде плас
та наблюдается лишь на ранних этапах rисто и орrаноrенеза. В процессе после
дующеrо rистоrенеза их эпителии приобретают специфические для желез черты
строения, расположения и функции.
Эпителии почечноrо типа
Почечный тип эпителиев развивается из мезодермы, точнее 1lефротом-ов cer
ментных ножек Эпителии почечноrо типа образуют стенку мочевых канальцев поч
ки. По своему строению это однОСЛОЙ1lые эпителии. По длине мочевых канальцев
эпителий имеет разное строение. Эпителиоциты MOryT быть плоские, кубические,
призматические и различаются по функции. Одни из них выполняют функцию филь
трации, друrие всасывающую, третьи секреторную фУНКllИЮ. В зрелом орrаниз
ме митотическая активность эпителиоцитов крайне низкая.
Эпителии целомическоrо типа
Целомический тип эпителия m-езодерм-алЬ1l0lO происхождения, как правило, oд
НОСЛОЙIIЫЙ плоский или призматический, выполняет разrраничительную, барьер
Система пoKpoвHыx тканей и их производныe
8S
, "'
..............
/, '\
, \ .-:
У" / . \
/.\. \
i О . "-
., r--,
\ /
, , \ )
("r:: ' ,
/ '----. G '<
' } J?2\, "G:) . ' \
. \ / -,;
, '
ную, секреторную и друrие функ
ции. Целомические эпителии раз
виваются из материала BHYTpeH
ней выстилки сплахнотома, фор
мирующеrо целом (вторичную по
лость тела). Наиболее xapaKTep
ным эпителием среди тканей дaH
Horo типа является мезотелий
(рис. 37). Он покрывает серозные
оболочки листки брюшины и
плевры, эпикард и перикард. Me
зотелий однослойный плоский
эпителий, состоящий из плоских
эпителиоцитов (мезотелиоцитов).
При световой микроскопии плос
костных препаратов видны клеточ
ные rраницы. Последние хорошо
выявляются при импреrнации co
ля ми серебра. На первый взrляд Рис. 37. Мезотелий (пленочный препарат)
клетки мезотелиалыюro пласта мало чем отличаются друr от друrа. Однако MeTO
дами rисторадиоавтоrрафии показано, что в составе дифферона плоских эпители
оцитов имеются различно дифференцированные клетки, что определяется терми
ном lетероморфия. Есть эпителиоциты, делящиеся митозом (камбиальные), ДBY
ядерные и мноrоядерные клетки, есть и rибнущие клетки. Между мезотелиоцита
ми имеются контакты типа десмосом. За счет этоrо мезотелиоциты интеrрирова
ны в единую клеточную систему, имеющую вид пласта. Вместе с тем мезотелий в
связи с еro положением в своеобразной внутренней изофизиолоrической среде
утратил некоторые свойства поrраничных тканей.
rлавные функции мезотелия покровная, разrраничительная, секреторная.
Блаroдаря последней мезотелиальная выстилка создает необходимые условия для
скольжения соприкасающихся opraHoB (например, opraHoB брюшной полости) друr
относительно друrа. Наличие мезотелия на поверхности внутренних opraHoB пре
пятствует образованию спаек, которые оrраничивали бы движения opraHoB брюш
ной полости, леrких и сердца.
Физиолоrическая реrенерация мезотелия протекает довольно интенсивно за
счет диффузно расположенных в пласте камбиальных эпителиоциов. Для мезоте
лия характерно слущивание клеток На их место наползают новые, возникающие в
результате деления камбиальных клеток
К целомическим эпителиям, кроме мезотелия, относятся эпителии opraHoB
половой системы выстилающие извитые семенные канальцы, фолликулярный,
эпителий семявыносящих путей, эпителии матки и маточных труб, а также эпите
лий коры надпочечников. Каждая из этих тканевых разновидностей характеризу
ется специфическими особенностями детерминации, пролифераuии, дифференuи
ровки и взаимодействия клеток
86
Fлава 8
Эпителии нейроrлиальноrо типа
Нейроrлиальный тип эпителия
развивается из 1lейроэктодермы,
выстилает полости мозrа и HeKOTO
рых opraHoB чувств. По строению он
однослойный, плоский, кубический
или цилиндрический, выполняет
вспомоrательную для нервных TKa
ней функцию. Иноrда этот тип эпи
телиев называют эпендимоrлиаль
ным, так как одной из ero раЗНОВИk
ностей является эnе1lдима, или
эnе1lдUМ1lЫЙ эпителий, образующий
выстилку центральноrо канала
спинноrо мозrа и желудочков [o .. , "
ловноro мозrа (рис. 38). ЭпеНДИ 1 Рис. 38. Эпендимный эпителий
ный эпителий однослойный призматический эпителий. Некоторые авторы pac
сматривают эпендиму как вспомоrательную ткань нервной системы и не относят
ее к эпителиям. Кроме эпендимноro эпителия среди эпителиев нейроrлиальноrо
типа различают: эпителиЙ мозrовых оболочек, периневральный эпителий, эпите
лий передней камеры rлаза, хрусталиковый эпителий, пиrментныЙ эпителий ceT
чатки rлаза, эпителий opraHa слуха, обонятельный эпителий, вкусовой эпителий,
хромаффинный эпителий мозrовorо вещества надпочечников и адреналовых op
[анов (параrанrлиев).
Пиzме1lт1lЫЙ эпителий сетчатки rлаза представляет собой однослойный эпи
телий, состоящий из клеток полиrональной формы. В процессе дифференцировки
клетки этой ткани накапливают пиrментные включения, необходимые для ВЫПОk
нения эпителием ero функции поrлощения световых лучей и изоляции CBeTO
воспринимающих клеток друr от друrа.
ПерИ1lевралЪ1lЫЙ эпителий однослойный плоский эпителий, окружающий He
рвные стволы и выстилающий так называемые периневральные пространства
(щели).
Морфофункциональная и rистоrенетическая характеристика нейроrлиальных эпи
телиев затруднена недостаточными сведениями в отношении степени их детермина
ции, особенностей пролиферации и дифференцировки клеток, а также реактивности
этих тканей.
Продолжается дискуссия о природе и системной принадлежности сосудистою
Э1lдотелия. H.r. Хлопин относил эндотелий к эпителиям, выстилающим в виде OДHO
слойноrо пласта внутреннюю поверхность кровеносных, лимфатических сосудов и cep
дца (эпителий анrиодермальноrо типа). Н.А. Шевченко выделил эндотелий в особый
тип тканей. с.и. Щелкунов на основе поведения эндотелия в культуре рассматривает
сосудистый эндотелий как разновидность тканей внутренней среды. Участие эндотелия
rемокапилляров в трансмембранном переносе веществ функционально роднит ero с Tкa
. / ...
__О . .,
\\;., . .,: ,
".:t!l.""' " "
Iii., ',!;/W .-'
, . " ' ""'
7') Wt
]b' '
. ,;.A .", ' " , ' ", ' " "
. #t '
"& . <.. '
JJ..lio
. .....-.r
-i:); "
<;; \".>."
i !i.
7i r
.il '.'-"'.,
. '" .jjf .
, A-t
t .t' j
.:.:.&
I"t....-\Н'\ l.jh> ,. ,
. ; v ,.IV"''' \L} -w;.,
', .... . ;
#
1-'
iiI,..
,. , : .;
.., .
Система nокров1lъа ткаllей и их nроизводllые
87
нями внутренней среды с трофической функцией и поэтому строение эндотелия pac
сматривается в друroм разделе.
в
Рис. 39. Схема различных
типов секреции:
А мерокринный; Б апок
ринный; В rолокринный
Железистые эпителии
А
Эпителии, обладающие способностью вырабатывать
секреты или инкреты, называются железистыми. Они яв
ляются основным тканевым компонентом желез. Напри
мер, эпителии кожноrо типа дают производные в виде по
товых, сальных, слюнных (серозных, слизистых), I\ЮЛОЧ
ных, слезных желез.
Клетки эпителия желез lЛаllдулоциты весьма раз
нообразны по размерам, форме, ультраструктуре. ОНИ BЫ
рабатывают секреторные продукты различноrо химическо
ro состава. В rландулоцитах происходят следующие про
цессы: 1) поступление исходных для синтеза продуктов, 2)
синтез секрета, 3) созревание секрета и формирование ceK
реторных rранул, 4) накопление секрета, 5) выведение ceK
рета. Периодические структурно функциональные измене
ния секреторной клетки в процессе ее секреции называют
секретОРllЫМ ЦИКЛО},/.. В зависимости от способа выделения
секрета различают следующие типы секреций: мерОКрИll
llая секреция, при которой выход секрета из клетки проис
ходит без повреждения плазмолеммы; аnОКрИllllая ceKpe
ция выход секрета осуществляется путем отрыва BЫpOC
тов с секретом от апикальной поверхности клетки и lOЛОК
рИllllая секреuия, коrда образование секрета связано с апоп
тозом, разрушением клетки при явлениях пикноза ядра и
жировою пере рождения цитоплазмы (рис. 39).
Б
Физиолоrическая реrенерация эпителиев
Физиолоrическая реrенерация обновление клеток в составе эпителиальных
тканей в процессе их нормальною функuионирования. Это динамический процесс,
включающий как разрушение клеток, так и их репродукцию. Эпителиальные клет
ки сравнительно быстро изнашиваются, так как они испытывают значительное вли
яние внешних факторов в связи с тем, что большинство этих тканей занимает по
rраничное положение. Эпителии, как правило, обладают хорошо выраженной
реrенерационной способностью, выработанной в процессе эволюции, и относятся
к об1l0вляющuм.ся тканям. В эпителиях клеточное обновление происходит за счет
митотическою деления камбиальных клеток Однако следует подчеркнуть, что каж
дая разновидность эпителиев характеризуется специфическими особенностями
пролиферации, локализации камбиальных клеток и закономерностями дифферен
цировки и интеrрации клеток
88
rлава 9
rЛАВА 9.
СИСТЕМА ТКАНЕЙ ВНУТРЕННЕЙ
(МЕТАБОЛИЧЕСКОЙ) СРЕДЫ
Понятие "ткани внутренней среды" объединяет разнообразные по строению и
функциям ткани, которые не rраничат с внешней средой и полостями внутренних
opraHoB. Тер:\IИН этот нельзя считать вполне удачным, так как внутреннюю среду
орrаНИЗ:\1а образуют не только кровь, соединительные и скелетные ткани, но и раз
личные эпителии в составе внутренних opraHoB, мышечные ткани, а также ткани
нервной системы. Поэтому целесообразно включить в данную rруппу только те
ткани, которые по своим основным физиолоrическим свойствам создают e1l.ympe1l.
1I.ЮЮ среду для метаболических реакций, обеспечивают постоянство состава BHYT
ренней среды орrанизма, выполнение защитных функций орrанизма. В первую
очередь это относится к системе крови и некоторым разновидностям соединитель
ной ткани. Все указанные ткани являются производными мезе1lxu.мы. Друrие про
изводные мезенхимы соединительные ткани с выраженной опорно механичес
кой функцией и rладкая мышечная ткань вместе с мышечными тканями иноrо
происхождения будут рассмотрены в следующей rлаве.
Для большинства производных мезенхимы характерны следующие общие при
знаки: аполярность клеток, боrатство межклеточным веществом, выполнение за
щитных и трофических функций. Кроме Toro, структурные элементы ткани Haxo
дятся в тесном взаимодействии с кровеносными сосудами
Понятие "мезенхима". Ретикулярная ткань
Термин "мезенхима" (rpеч. Mesos средний, епсЬуmа заполняющая масса)
был предложен братьями [ертвиrами (1881). Это один из эмбриональных зачат
ков (по некоторым представлениям эмбриональная ткань), представляющий
собой разрыхленную часть среднею зародышевоrо листка мезодермы. Клеточ
ные элементы мезенхимы (точнее, энтомезенхимы) образуются в процессе диффе
ренцировки дерматома, с'КЛеротома, висцералЪ1l.0lO и париеталЪ1l.0lO листков спла1l.
Хllотома. Кроме тою, существует эктомезе1lXU.Ма (нейромезенхима), развивающаяся
из rанrлиозноЙ пластинки.
Мезенхима состоит из отростчатых клеток, сетевидно соединенных своими OT
ростками (рис. 40). Клетки MorYT высвобождаться от связей. амебоидно переме
щаться и фаrоцитировать инородные частицы. Вместе с межклеточной жидкостью
клетки мезенхимы составляют внутреннюю среду зародыша. По мере развития за
родыша в мезенхиму миrрируют клетки иноrо происхождения, нежели из пере
численных выше эмбриональных зачатков, например, клетки нейробластическоrо
Система тканей внутренней среды
89
мало чем отличаются друr от
друrа, и только очень чув
ствительные методы иссле
дования (иммуноцитохими
ческие, электронно микро
скопические ) выявляют в
составе мезенхимы клетки
различной природы.
Клетки мезенхимы об
наруживают способность к
ранней дифференцировке.
Например, в стенке желточною мешка 2 недельноrо эмбриона человека из состава
мезенхимы выделяются первичные клетки крови zемоцитъt, друrие формиру
ют стенку первuчнЪ/.х сосудов, третьи являются источником развития ретuкуляр
ной ткани остова кроветворных opraHoB. В составе провизорных opraHoB мезен
хима очень рано претерпевает тканевую специализацию, являясь источником
развития соединительных тканей.
Мезенхима существует только в эмбриональном периоде развития человека.
IIосле рождения в орrанизме человека сохраняются лишь малодифференцирован
ные (полипотентные) клетки в составе рыхлой волокнистой соединительной TKa
ни (адвентициальные клетки), которые MorYT диверrентно дифференцироваться в
различных направлениях, но в пределах определенной тканевой системы.
Ретикулярная ткань. Одним из производных мезенхимы является ретикуляр
ная ткань, которая в орrанизме человека сохраняет мезенхимоподобное строение.
Она входит в состав кроветворных opraHoB (KpacHoro KocTHoro МОЗI'а, селезенки,
лимфатических узлов) и состоит из звездчатых ретикулярных клеток, вырабатыва
ющих ретикулярные волокна (разновидность арrирофильных волокон). Ретикуляр
ные клетки неоднородны в функциональном отношении. Одни из них менее диффе
ренцированы и выполняют камбиальную роль. Друrие способны к фаrоцитозу и
перевариванию продуктов распада тканей. Ретикулярная ткань как остов KpOBeTBOp
ных opraHoB принимает участие в кроветворении и иммунолоrических реакциях,
выполняя роль микроокружения для дифференцирующихся клеток крови.
дифферона, миrрирующие
миобласты закладки скелет
ных мышц, пиrментоциты и
др. Следовательно, с опреде
ленной стадии развития за
родыша мезенхима пред
ставляет собой мозаику кле
ток, возникших из разных
зародышевых листков и эм
бриональных зачатков TKa
ней. Однако морфолоrичес
ки все клетки мезенхимы
с;- ;i9'&' \ 8
'? [., ' ' . r -'"
,...<J . :\ :), @ ;.
. .., 1" OJ
. i :- {
/" 2 t "e't ,
I , (f)" ' .
. , ' , ' &\ " '" " " , .
(f .'
.i', ' ):;.э ,,!
', е" \, :"" ТТ'I
( I , 1 @ '\!Y
) 1\"
.у
r
f.:/
("
s е",
4
,.:.;,
\
\
,
... \
"
Рис. 40. Мезенхима:
1 отростчатые клетки; 2 кровеносный сосуд и интраваскуляр
ный rемопоэз
90
Тлава 9
Кровь и лимфа как ткани
Кровь ткань внутренней среды защитно трофической функции, состоящая
из жидкorо межклеточноrо вещества (плазмы), постклеточных структур (эритро
цитов И тромбоцитов) и клеток как периферической крови и лимфы, так и клеток
на всех стадиях cBoero развития в кроветворных opraHax. Клеточные и посткле
точные структуры периферической крови называются форменнъt.М.и элементами.
Объем крови в орrанизме человека равен 5 5,5 л (или около 7% массы тела), при
этом форменные элементы составляют 40 45%, а плазма 55 60%.
Кровь выполняет следующие функции: 1) трофическую перенос питательных
веществ ко всем клеткам и тканям; 2) дыхательную rазообменную, или транспорт
кислорода к тканям и удаление из орrанизма уrлекислоты; 3) защитную (фаrоцитоз,
выработка антител); 4) реryляторную транспорт ropMoHoB и друrих ryморальных
факторов реryляции; 5) rомеостатическую поддержание физико химическоrо по
стоянства состава внутренней среды орrанизма.
ПлаЗ}fа крови это жидкое межклеточное вещество (рН 7,34 7,36), в котором
во взвешенном состоянии находятся форменные элементы крови. 93% плазмы co
ставляет вода, остальное белки (альбумины, rлобулины, фибриноrен и десятки
друrих), лишщы, уrлеводы, минеральные вещества. При свертывании крови фибри
HoreH переходит в HepaCТBO
римый белок фибрин. Oc
тавшаяся жидкая часть А.
пла3 fЫ после свертывания
фибриноrена называется
сывороткой. В сыворотке
содержатся антитела (им
муноrлобулины).
Форменные элементы
крови представляют собой
rетероморфную систему,
состоящую из различно
дифференцированных в
структурно функциональ
ном отношении элементов
(рис. 41). Объединяют их
общность rистоrенеза и co
вместное пребывание в пе
риферической крови.
Эритроциты челове
ка красные кровяные эле
менты, имеющие форму
двояковоrнутых дисков,
что на 20 30% увеличивает
площадь их поверхности.
о
' о , . ,, :' . . ; . ;. I : . } , .. ; . :' , : , . ' , ' . " , . : , . . t . . . ? з "".':"
. .. / ;' ё
о .9 8 """ '.. о
tP ........ 1.. . O
. ' о () Q О
i 10 0 . . О, О '
. 'iWg "
О .Р'" . '7 _1 4
О , О " ';";;' '
"о о ,,' о gci g
Рис. 41. Форменные элементы крови:
1 эритроцит: 2 сеrментоядерный нейтрофильный rранулоцит;
3 палочкоядерный нейтрофильный rранулоцит: 4 юный нейтро
фильный rранулоцит: 5 эозинофильный rранулоцит; 6 базо
фильный rранулоцит; 7 средний лимфоцит: 8 малый лимфоцит:
9 моноцит: 1 О кровяные пластинки
Система тка1lей 61lутре1l1lей среды
91
у друrих позвоночных (рыбы, амфибии, птицы и др.) это ядросодержащие клет
ки. В мазках крови эритроциты имеют окруrлую форму. Диаметр эритроцитов чело
века 7 8 мкм (в среднем 7,5 мкм), толщина в краевой зоне 2 2,5, а в центре
1 мкм. Наряду с эритроцитами 1l0рМОЦИmaм.и, которые составляют около 75 %,
бывают макроциты (диаметр 8 9 мкм), ZUla1lтоциты (12 мкм), микроциты (5 6 мкм).
При некоторых заболеваниях крови наблюдаются явления пОЙКUJlоцитоза изме
нение формы эритроцитов, а также а1luзоцитоза изменение размеров.
Количество эритроцитов в 1 л крови составляет 4 5,5x1012 у мужчин и 3,7
4,9х10 12 У женщин. Число эритроцитов может изменяться при разных физиолоrи
ческих состояниях орrанизма и реrиональных особенностях проживания. Стой
кое повышение их числа называется эритроцитозом, уменьшение эритропе1lией.
Диаrностическое значение имеет скорость оседания (аrrлютинация) эритроцитов
(СОЭ). в норме у мужчин СОЭ равна 4 8 мм в час, у женщин 7 10 мм в час.
Покровная и рецепторно трансдукторная системы эритроцита характеризуют
ся рядом особенностей. Плазмолемма имеет толщину 20 нм. В ней хорошо развиты
транспортные процессы за счет ионных насосов, каналов и белковых переносчиков.
Она обладает избирательной проницаемостью, обеспечивает перенос кислорода, ДBY
окиси уrлерода, ионов натрия и калия, но не препятствует соединению rемоrлобина
с окисью уrлерода (yrapHbIM rазом). Свойства плазмолеммы позволяют эритроциту
без повреждения проходить через капилляры, диаметр которых меньше диаметра
самою эритроцита. fликокаликс плазмолеммы, образованный rликолипидами и rли
копротеинами, содержит аrrлютиноrены А и В, определяющие rрупповую принад
лежность крови. Наличие в rликокаликсе аrлютиноrена резус фактора, определя
ет принадлежность человека к резус положительной (86 % людей имеют этот фактор)
или резус отрицательной популяциям.
Рецепторную функцию выполняют трансмембранные rликопротеины lJlИ
КОФОРИ1lЫ, обеспечивающие индивидуальные для каждоrо человека антиrенные
характеристики эритроцитов.
Двояковorнутая форма эритроцита поддерживается блаrодаря белкам опор
но двиrательной системы, в частности спектрИ1lа, формирующеrо в примембран
ном пространстве эритроцита сеть филаментов, и некоторых друrих белков.
Основную массу эритроцита составляют вода (66%) и белок rемоrлобин (33%).
Под электронным микроскопом содержимое эритроцитов выrлядит очень плотным.
В нем определяются мноrочисленные rранулы rемоrлобина диаметром 4 5 нм. [e
моrлобин дыхательный пиrмент. Белковая часть ею называется rлобин, железо
содержащая часть reM, который составляет 4 5% от массы rемоrлобина и придает
желтую окраску эритроциту. fемоrлобин леrко присоединяет кислород воздуха, пре
вращаясь в оксиrемоrлобин. Это происходит в капиллярах леrких. В онтоrенезе свой
ства rемоrлобина, меняются, в связи с чем различают rемоrлобин эмбриональный
(фетальный) и rемоrлобин взрослых. Блаюдаря накоплению rемоrлобина при эрит
ропоэзе эритроциты и выполняют дыхательную функцию. Наряду с транспортом
кислорода и друrих веществ (аминокислот, антител, токсинов) эритроциты перено
сят двуокись уrлерода из тканей в леrкие. Наличием rемоrлобина обусловлена окси
филия эритроцитов, т. е. сродство к кислым красителям.
92
Fлава 9
В rипотонической среде rемоrлобин выходит из эритроцитов в результате по
ступления в них воды и разрыва оболочки. Выход rемоrлобина называется [eMO
лизом. Некоторые вещества (например, фенилrидразин) вызывают rемолиз. Пос
ле удаления из эритроцита rемorлобина остается строма бесцветная масса (или
"тень" эритроцита).
Количество циркулирующих в орrанизме эритроцитов составляет около 2S
ЗОх10 12 . Появлению эритроцитов в крови предшествует длинный путь эритроци
топоэза. В кровь поступают наряду со зрелыми эритроцитами и молодые, бедные
rемоrлобином формы ретикулоциты, составляющие 1 2%.B них сохраняются
некоторые орrанеллы, которые при окраске мазков метиленовым синим выявля
ются В виде базофильных сетчатых структур. Возрастание числа ретикулоцитов
наблюдается при rипоксии, кровопотере и др.
Эритроциты живут в крови от 70 до 120 суток Продолжительность жизни эрит
роцитов может сокращаться до 24 суток (например, при занятии "моржеванием").
Ежесуточно распадается около 200 млрд. эритроцитов. Разрушение их происхо
дит В основном В селезенке и красном костном мозrе. Специальные клетки MaK
рофаrи захватывают эритроциты и обеспечивают реутилизацию (повторное ис
пользование) железа при развитии новых rенераций эритроцитов.
Лейкоциты. Лейкоцитами называют белые клетки крови. Число их в 1 л крови
во MHoro раз меньше, чем эритроцитов и составляет 4 9x109. Они подразделяются на
две rруппы: зернисmые лейкоциты (rранулоциты) и незернисmые лейкоциты (arpa
нулоuиты). Для зернистых лейкоцитов характерны следующие основные особенно
сти: наличие в цитоплазме специфической зернис'rОСТИ и сеrментация ядра. ПО OK
раске зернистости rранулоциты подразделяются на 3 вида: нейmрофилыlе,'
эо3uнофшlыlе,' базофшlыlle.. Основная функuия rранулоцитов участие в защит
ных реакциях орrанизма в соединитель
ных тканях. Схематично жизненный путь
rранулоцитов можно представить в виде
трех этапов: развитие в костном мозrе,
кратковременная циркуляция в KpOBeHOC
ном русле, пребывание в тканях. fлавное
назначение rpанулоuитов попасть в TKa
ни, rде они и выполняют свои функции.
нейmрофшlыlеe lраllулоциты (llейт
рофШlЫ). Нейтрофильные rранулоциты
составляют 6S 70% от общеrо числа лей
коцитов. Это окруrлые подвижные клет
ки диаметром 8 10 мкм (рис. 42). Рецеп
торно трансдукторная система клетки
воспринимает цитокины и передает сиr
налы опорно двиrательной системе, что
обусловливает направленное амебоидное
перемещение клетки в очаr воспаления.
Поэтому форма клетки может быть из
Рис. 42. Схема ультраструктуры сеrментоядерно
ro нейтрофильноrо rранулоцита:
1 азурофильные rранулы; 2 специфические
rранулы
Система тканей внутренней среды
93
менчивой. Цитоплазма клетки слабо оксифильна. Непосредственно под плазмо
леммой цитоплазма бедна орrанеллами, что способствует образованию псевдопо
дий. В остальной части цитоплазмы содержатся орrанеллы, включения rликоrена
и мноrочисленные rранулы числом до 200, которые воспринимают и кислые, и oc
новные красители. fранулы нейтрофилов, как правило, шаровидной формы. Элек
тронная плотность их различна. fранулы подразделяются на два типа: азурофUJlЪ
llble и специфические. Азурофильные (неспецифические, первичные) rранулы с
электронно плотной сердцевиной диаметром 0,4 0,8 мкм возникают первыми и co
ответствуют лизосомам. В них содержатся катионные белки, лизоцим, миелопе
роксидаза и др. Это система внутриклеточноrо переваривания инородных тел.
Специфическая (вторичная) зернистость появляется позднее, составляет 80 90%
от общеrо количества rранул. В специфических rранулах (с электронно прозрач
ным содержимым) диаметром 0,1 0,3 мкм определяются высокая активность ще
лочной фосфатазы, коллаrеназа, лизоцим, обладающий антибактериальным свой
ством и др. Эти вещества участвуют как во внутри , так и во внеклеточных реакциях.
Кроме TOro, описаны rранулы, которые участвуют в процессах миrрации rрануло
цита через стенку капилляров.
Наличие в нейтрофильных rранулоцитах rидролитических и окислительных
ферментов связано с фаrоцитарной активностью этих клеток (около 80% клеток
обладают этим свойством). и.и. Мечников называл нейтрофилы микрофаzQМИ.
Так, одна клетка может поrлотить в среднем до 9 кишечных палочек
В большей части нейтрофилов имеется сеrментированное ядро, состоящее из 3 5
cerMeHToB, соединенных узкими перемычками. 3 5 % клеток содержат палочковидное
ядро. В очень небольшом количестве (до 0,5%) в кровь попадают и юные нейтрофилы
с бобовидным ядром. В сеrментоядерных нейтрофилах у женщин определяется поло
вой хроматин, имеющий форму барабанной палочки (X xpOMOCOMa).
Продолжительность жизни нейтрофилов составляет около 8 суток, из них в
крови они циркулируют 8 12 часов.
Увеличение количества нейтрофилов
при мышечной работе, заболеваниях и эк
стремальных состояниях орrанизма назы
вают лейкоцитозом. При этом возрастает
доля малодифференцированных палоч
коядерных и юных форм, что называется
сдвиrом влево.
ЭОЗИНОфUJlъные zранулоциты (эозино
фUJlЫ) окруrлые клетки диаметром 1 o
12 мкм (рис. 43). Они встречаются в пери
ферической крови реже, чем нейтрофилы,
составляя 1 5% от общеro числа лейкоци
тов. Ядро эозинофилов имеет, как правило,
двухсеrментную конфиryрацию. XapaKTep
ным признаком эозинофильных лейкоци
тов является крупная эозинофильная
2
Рис. 43. Схема ультраструктуры эозино
фильноrо rранулоцита:
1 мелкие азурофильные rранулы; 2 спе
цифические rранулы
94
rлава 9
зернистость, равномерно расположенная в цитоплазме. rранулы имеют окруrлую
или овальную форму, окрашиваются эозином в ярко красный цвет. Мелкие азуро
фильные (неспецифические, первичные) rранулы (около 5%) диаметром до 0,4 мкм
содержат кислую фосфатазу, пероксидазу и друrие ферменты это лизосомы.
Крупные эозинофильные rранулы это специфические (вторичные), размером
до 1,5 мкм и числом около 200 содержат крuсmШUlOuд. В нем находится rлавный
щелочной (основной) белок и друrие ферменты, которые участвуют в антипарази
тарной функции клеток При контакте с паразитом эозинофилы секретируют co
держимое rранул, в частности rлавный щелочной белок, который разрушает обо
лочку паразита.
Количество эозинофилов значительно увеличивается (до 20 40%) при ak
лерrических состояниях (например, при бронхиальной астме). Так, при аллер
rических состояних и воспалении эозинофильные лейкоциты активно переме
щаются к источнику раздражения, например, к rистамину, выделяемому туч
ными клетками, поrлощают и разрушают ero, уменьшая степень выраженности
местных реакций в тканях. Кроме Toro, эозинофилы связывают комплексы aH
тиrен антитело.
Фаrоцитарная активность клеток ниже, чем у нейтрофилов. Длительность пре
бывания клеток в крови составляет з 1 О часов. Затем они миrрируют в ткани и
находятся там до 2 недель. Удаление стареющих эозинофилов происходит через
эпителий кишечной трубки. При проникновении паразитов в пищеварительную
трубку миrрация эозинофилов возрастает, как и увеличивается продукция этих
клеток в костном мозrе, что сопровождается эозинофилией.
Базофилы/.ъtе lра-н.улоцumъt (базофилъt). Эти клетки имеют 8 10 мкм в диамет
ре (рис. 44). Количество их в крови невелико от О до 0,5 1%. Ядро базофилов
слабодольчатое или S образное, плохо различимо из за базофильной зернистости
цитоплазмы, которая окрашивается MeTaxpo 1
матически, Т.е. не в тот цвет, каким обладает
краситель. Так, при окраске азуром (синяя
краска) зерна приобретают фиолетовый цвет.
Метахромазия характерна для структур, co
держащих сульфатированные rликозаминоr
ликаны. rранулы базофилов содержат rепа
рин, rистамин, серотонин, пероксидазу и др.
Размер rранул около 0,8 1 мкм. Они Heo
динаковы по степени зрелости. Некоторые
rранулы являются более светлыми, как бы
пустыми. При действии стресс факторов
происходит деrрануляция базофилов. Этот
процесс возникает при взаимuдействии pe
цепторов базофила с иммуноrлобулином
(IgE). Друrой вид rранул азурофильные,
немноrочисленны это лизосомы. Содержи
мое rранул вызывает ряд физиолоrических
Рис. 44. Схема ультраструктуры базофильно
ro rранулоцита:
1 азурофильные rранулы; 2 специфичес
кие rранулы
Система тканей внутренней среды
95
реакций тканеЙ. Так, rепарин препятствует свертыванию крови. Остальные веще
ства повышают проницаемость сосудов и влияют на сократимость rладких миоци
тов сосудистоЙ стенки.
Хемотаксические факторы обеспечивают направленную миrрацию базофилов
к очаrу воспаления. Базофилы циркулируют в крови до 1 суток, миrрируют в TKa
ни, [де продолжительность их жизни определяется несколькими сутками. Увели
чение числа базофилов в крови наблюдается при некоторых заболеваниях систе
мы крови и др.
Рис. 45. Схема ультраструктуры моноцита:
1 азурофильные rранулы
Моноциты самые крупные клетки
крови (аrранулоциты), имеющие слабоба
зофильную цитоплазму и бобовидное
ядро (рис. 45). Диаметр их 16 20 мкм. В
крови человека они составляют 6 8 % от
общеrо числа леЙкоцитов. В цитоплазме
моноцитов находятся мелкие азурофиль
ные зерна типа лизосом, MHoro вакуолей
и пузырьков, мелкие митохондрии и дpy
rие орrанеллы. Азурофильные rранулы
дают положительную реакцию на перок
сидазу и кислую фосфатазу. Моноциты
миrрируют из кровяноrо русла в ткани и
превращаются в свободные макрофаlU, rде
и выполняют свои мноrочисленные фун
кции. Для них характерно наличие большо
[о числа лизосом.
Функция моноцитов защитная, т. е. фаrоцитоз бактерий и продуктов pac
пада тканей. Моноциты выполняют также трофическую функцию, так как при
распаде они выделяют вещества, необходимые для друrих клеток. Моноциты OT
носятся К макрофаrической системе орrанизма, называемой также МОНОНУЮlеар
ной фаlOцитарной системой (МФС). Эта система включает несколько раЗНОВИk
ностей макрофаrов: rистиоциты в соединительной ткани, купферовские клетки
в печени, альвеолярные макрофаrи в леrких, фиксированные макрофаrи в селе
зенке и лимфатических узлах, остеокласты в костной ткани, rлиальные MaKpo
фаrи в центральной нервной системе и др. Моноциты осуществляют макрофа
rальную функцию в крови. Разновидность макрофаrов антиzенпредставляющие
'КЛетки находятся в качестве сопутствующею дифферона в покровных тканях
и участвуют в иммунных реакциях. Они захватывают чужеродные вещества, пе
рерабатывают их с помощью протеолитических ферментов, миrрируют в лимфа
тические узлы и вступают в кооперативные взаимодействия с лимфоидными клет
ками, формируя таким образом иммунный ответ на антиrены. При некоторых
патолоrических состояниях в opraHax обнаруживаются макрофаrи, содержащие
в своей цитоплазме инородные частицы, например, пылевые клетки в леrком, MHO
rоядерные клетки при длительно протекающем воспалении и др. Время пребы
вания моноцитов в крови от 1,5 до 5 суток.
96
Тлава 9
ЛИМфОЦИТЫ это аrранулоциты. Коли
чество их составляет 20 30% от общеrо чис
ла лейкоцитов. Большая часть лимфоцитов
(98%) находится в тканях. Различают малые
(диаметр около 6 мкм), средние (6 8 мкм) и
большие лимфоциты (8 12 мкм). В сосуди
стой крови преобладают малые лимфоциты.
Лимфоциты имеют базофильную цитоплаз
му, окружающую в виде узкоrо ободка плот
ное окруrлое ядро (рис. 46). Различают 4
вида лимфоцитов: малые светлые, малые
темные, средние и большие лимфоциты. Ta
кое подразделение недостаточно для точно
ro определения функциональных особенно Рис. 46. Схема ультраструктуры лимфоцита
стей разных видов лимфоцитов. При нечетких rистолоrических различиях они Becь
ма дифференцированы в отношении роли в защитных иммунных реакциях. Bыдe
лены два основных вида лимфоцитов: Т лимфоциты и В лимфоциты. Развитие
методов иммуноцитохимии позволило различать субпопуляции Т лимфоцитов,
отличающиеся набором тех или иных рецепторов (например, СО+4, СО+8 и др.).
Кроме Toro, отдельную rруппу составляют нулевые лимфоциты (не содержат Map
кируемые рецепторы), которые не относятся ни к B , ни к Т лимфоцитам. Среди
них основную популяцию составляют большие rранулярные лимфоциты, облада
ющие цитотоксическим свойством (натуральные киллеры, NК клетки).
Развиваются все лимфоциты из стволовой кроветворной клетки в красном KO
стном мозrе. Однако Т лимфоциты в последующем созревают в тимусе, тоrда как
В лимфоциты (от bursa Fabricius фабрициева сумка у птиц) после дифференци
ровки в красном костном мозrе оседают в тимуснезависимых зонах селезенки и
лимфатических узлов. Каждый из этих видов лимфоцитов подверrается еще более
узкой специализации, участвуя в иммунных реакциях как структурные элементы
клеточнorо и rуморальноrо иммунитета (см. ниже).
Общее количество лимфоцитов в орrанизме orpoMHo (1 4х109/л). Доволь
но интенсивно идет и репродукция лимфоцитов (на 1 Kr массы тела за 1 ч обра
зуется до 3 млн. лимфоцитов). 65 75% лимфоцитов крови относятся к долrо
живущим клеткам (продолжительность жизни от нескольких месяцев до 5 лет),
15 35% клеток к короткоживущим (продолжительность жизни от несколь
ких часов до 5 суток). Т лимфоциты долrоживущие. В лимфоциты KOpOT
коживущие. Для лимфоцитов характерны циркуляция и рециркуляция BЫ
ход из крови в ткани, переход из ткани и uиркуляция в составе лимфы,
возвращение в ткани.
Кровяные пластинки. Кровяные пластинки, или тромбоциты, представляют
собой свободно циркулирующие в крови безъядерные фраrменты uитоплазмы rи
rантских клеток KpacHoro KOCТHoro мозrа меlй1<арuоцuтов (рис. 47) . Размер пла
стинок 2 3 мкм. Количество их в 1 л составляет 200 300x109. Каждая кровяная
пластинка состоит из двух частей: центральной зернистой zра-н.уломера (xpOMO
Система тканей внутренней среды
97
мера) и периферической lИаломера.
В rрануломере имеются электронно
плотные rранулы (диаметр 0,2
0,5 мкм) нескольких видов, светлые
вакуоли, а также единичные митохон
дрии и rлыбки rликоrена. Самые круп
ные rранулы содержат фибриноrен,
тромбоrлобулин, фактор свертывания
У, переносчик фактора VHI CBepTЫBa
ния и др. Мелкие rранулы включают
в себя rистамин, серотонин, ионы
кальция и маrния, А ТФ и др. Суще
ствуют также немноrочисленные лизосомы. rиаломер в основном формируется эле
ментами опорно двиrательной системы микротрубочками, микрофиламентами
и промежуточными филаментами. Кроме тою, выявляются производные комплекса
rольджи в виде трубочек
у тромбоцитов функционально ведущей системой является рецепторно транс
дукторная. Рецепторы реryлируют уrнетение слипания пластинок или активацию
тромбоцитов и участие в процессе свертывания крови. При активации пластинок
на их поверхности образуются отростки, называемые "усиками", с помощью KOTO
рых кровяные пластинки формируют конrломераты. BOKpyr тромбоцитарных KOH
rломератов возникают нити фибрина. Выделяющийся из кровяных пластинок co
кратимый белок тромбостенин вызывает сжатие фибриновою cryCТKa.
Различают 5 видов кровяных пластинок: юные, зрелые, старые, деrенеративно
измененные и rиrантские. Продолжительность жизни кровяных пластинок COCTaB
ляет 5 8 сут.
Реrуляция количества кровяных пластинок в крови включает механизмы, YBe
личивающие объем и общую массу меrакариоцитов в красном костном мозrе.
Форменные элементы крови в норме находятся в определенных количествен
ных соотношениях, что называют lеМОlрам.м.ой. Лейкоцитарной формулой называют
соотношение доли различных клеточных форм лейкоцитов в крови взрослоrо чело
века в условиях нормы. Общее количество лейкоцитов в 1 мкл крови: 4 9 тыс., из
них: нейтрофилы составляют 65 70%, эозинофилы 1 5%, базофилы 0,5 1 %, лим
фоциты 20 30%, моноциты 6 8%.
Лимфа. Форменные элементы лимфы представлены в основном лимфоцитами,
а также моноцитами. Лимфа из капилляров поступает в лимфатические сосуды, про
текает через лимфатические узлы, rде обоrащается клетками крови (аrранулоцита
ми), и, поступая в крупные лимфатические сосуды, вливается в кровь. Таким обра
зом, между кровью и лимфой существуют определенные взаимодействия.
Рис. 47. Схема ультраструктуры тромбоцита
Кроветворные ткани и кроветворение в костном мозrе
Эмбриональный источник развития кроветворных тканей мезенхима. В
эмбриоrенезе можно различать 3 периода rемопоэза: 1) внезародышевый, или
98
Fлава 9
мезобластический (1 2 мес), 2) rепато тимо лиенальный (2 5 мес), 3) меДУk
ло тимо лимфоидный (5 10 мес).
Постэмбриональным rемопоэзом, или физиолоrической реrенерацией крови,
называют кроветворение во взрослом орrанизме.
Кроветворение в желточном мешке. В конце 2 й начале 3 й недели эмбриоrене
за в мезенхиме стенки желточноrо мешка образуются кровяные островки, в составе KO
торых клетки дифференцируются на плоские эндотелиальные и окрyrлые клетки. Пос
ледние преобразуются в стволовые кроветворные клетки. При внезародышевом
кроветворении из стволовых клеток формируются первичные эритробласты МRюлоб
ласты. Они делятся внутри сосудистоrо русла (u-н.траваскуляр-н.о). Небольшая часть
меrалобластов превращается в безъядерные первичные эритроциты МRlШlоциты. Об
разуется также незначительное количество вторичных эритроцитов меньшей величи
ны, чем меrалоциты. Экстраваскулярно дифференцируется часть первичных лейкоци
тов (rpанулоцитов нейтрофилов и эозинофилов). Из желточноrо мешка стволовые
кроветворные клетки по развивающимся сосудам расселяются по opraHaM зародыша.
Кроветворение в печени. На 2 M месяце эмбриоrенеза печень становится цeH
тром кроветворения. Источником rемопоэза здесь служат стволовые "poвeтвop
-н.ые клет"и. Кроветворение в печени происходит э"стравас"уляр-н.о. Из стволовых
кроветворных клеток образуются эритроциты, зернистые rранулоциты (нейтро
филы и эозинофилы) И меrакариоциты. Зернистые лейкоциты развиваются здесь
укороченным путем и не имеют четкой специфической зернистости. К концу эмб
риоrенеза человека кроветворение в печени постепенно прекращается.
Кроветворение в селезенке. На 4 5 M месяцах эмбриоrенеза человека селе
зенка становится универсальным opraHoM rемопоэза, в котором экстраваскулярно
образуются все клетки крови. Позднее процессы эритро и rранулоцитопоэза в ce
лезенке уrасают, но усиливается образование незернистых лейкоцитов.
Кроветворение в красном костном мозrе и тимусе. Постепенно центральным op
raHOM кроветворения становится красный костный мозr. Строму ero вначале образует
мезeю:uм.а, которая позднее преобразуется в рети'К!jЛЯР-н.ую т"ань. Ретикулярная ткань,
в трехмерной сети которой происходит развитие эритроцитов, rpанулоцитов, MOHO
цитов и меrакариоцитов, называют мueлоид-н.ой тканью. Миелоидная ткань специа
лизированная rемопоэтическая ткань KpacHoro костною мозrа. Она обеспечивает раз
витие стволовых клеток и всех форменных элементов крови. Наряду с миелоидной к
кроветворным тканям относится лимфouд-н.ая ткань, которая развивается в лимфати
ческих узлах, селезенке и друrих лимфоидных opraHax, составляющих лимфоидную
систему. Здесь в сети ретикулярной ткани происходит образование лимфоцитов, плаз
матических клеток, удаление клеток и продуктов их распада.
К центральным opraHaM кроветворения относится тимус, в котором на 2 M Me
сяце эмбриоrенеза начинают дифференцироваться лимфоциты тимуса. В дальней
тем они расселяются по периферическим opraHaM лимфоидной системы.
Кроветворение в лимфатических узлах начинается с 4 ro месяца эмбриоrенеза
после миrрации стволовых кроветворных клеток
В соответствии с унитарной теорией кроветворения А.А. Максимова, суще
ствует единый источник развития для всех клеток крови. Исходной клеткой для
Система тка'Н.ей в'Н.утре'Н.'Н.ей среды
99
всех ростков кроветворения является стволовая кроветвор'Н.ая клетка, сходная
по своему строению с малым лимфоцитом. А.А. Максимов (1911) писал, что ин
дифферентные блуждающие клетки, или лимфоциты в широком смысле, oдape
ны очень большой потенцией развития: "Это индифферентная мезенхимная блуж
дающая клетка, лимфоцит, является общей родоначальницей всех элементов
крови...Попадая в блаroприятные условия, она проявляет свою потенцию разви
тия, причем в зависимости от условий, направление развития и продукты ero по
лучаются очень разнообразными". Унитарная теория кроветворения была разви
та в трудах А.А. 3аварзина, H.r. Хлопина, А.Н. Крюкова, М.И. Аринкина и др.
Метод селезеночных колоний, разработанный канадскими учеными Тиллом и
МакКуллохом (1961), прозволил идентифицировать вид клеток, являющийся ис
точником развития клеток эритроидноrо, rранулоцитарноrо и меrакариоцитар
Horo рядов. Эту клетку, которая rистолоrически сходна с малым темным лимфо
цитом, авторы назвали коло'Н.иео6разующей единицей (КОЕ).
В развитии клеток крови условно выделяются классы клеток По мере перехо
да клеток из класса в класс, в каждом из них все более отчетливо обнаруживаются
rемопоэтические клеточные диффероны, которые характеризуются определенны
ми rистолоrическими признаками. Однако клетки первых трех классов по своему
строению идентичны. Только методы иммуноцитохимии позволяют различать клет
ки по наборам клеточных рецепторов, что является показателем диверrентной диф
ференцировки стволовой клетки.
В общем виде развитие клеток крови происходит в следующей последователь
ности (рис. 48).
I й класс плюрипотентные клетки это стволовые кроветворные клетки
(СКК). Стволовая клетка является общим самоподдерживающимся предшествен
ником всех клеток крови, включая все виды иммунокомпетентных клеток Пола
rают, что каждая из стволовых клеток способна проделать по меньшей мере 100
митозов, т. е. потомками одной стволовой клетки можно было бы обеспечить всю
кроветворную систему. Однако стволовые клетки после цикла пролиферации в эм
бриоrенезе переходят в состояние покоя. Она лишена каких либо специфических
признаков строения и локализуется в миелоидной ткани среди популяции лимфо
цито или моноцитоподобных элементов. Она может с током крови миrрировать
по тканям орrанизма. Объективным методом обнаружения и количественноrо учета
стволовых клеток является метод селезеночных колоний. Стволовые клетки co
ставляют около 0,1% популяции кроветворных элементов.
II й класс. Стволовые клетки под влиянием ряда факторов (тромбопоэтичес
кий, ИЛ 7 и др.) диверrентно дифференцируются в двух направлениях: полу
стволовые, или мультипотентные, клетки предшественники миелопоэза (KOE
rэмм) и мультипотентные клетки предшественники лимфопоэза (КОЕ Л).
В составе колоний эти клетки имеют оrраниченные возможности к самоподдер
жанию (около 3 4 недель). однако этоrо достаточно для поддержания физиоло
rической реrенерации крови.
III й класс. Из мультипотентных клеток предшественников миелопоэза
(КОЕ rэмм) в результате диверrентной дифференцировки, происходящей под
100
Fлава 9
влиянием ряда факторов микроокружения, возникают слсдующие клеточные
линии: а) родоначальные (проrениторные) клетки, или клетки предшественни
ки, эритропоэза (БОЭ Э, от анrл. burst взрыв) и развивающиеся из них KOE
Э; б) общие родоначальные клетки rранулоцито и моноцитопоэза (KOE rMo).
Последние в процессе дальнейшеI':" диверrентной дифференцировки под влияни
ем факторов микроокружения формируют родоначальные клетки для нейтро
фильных (rранулоцит стимулирующий фактор), эозинофильных (ИЛ S) и ба
зофильных (ИЛ 3) rранулоцитов (KOE rH, КОЕ Эо, КОЕ Б) и моноцитов
(KOE Mo, фактор моноцит колониестимулирующий).
Мультипотентные клетки лимфопоэза (КОЕ Л) под влиянием дифференци
ровочных факторов микроокружения (ИЛ 7, ИЛ 6) развиваются в родоначальные
клетки Т и B лимфоцитов.
Мультипотентные клетки КОЕ rэмм (при участии тромбопоэтина и ИЛ
11) являются источником развития родоначальной клетки для меrакариоцитов
(KOE Mer).
Таким образом, важнейшее свойство, которое приобретают в миелопоэзе и лим
фопоэзе кровеобразующие клетки это формирование рецеnmОрllO mрансду'Кmор
r:::\r:::\
\J \::/
11
IС:)
/
(:;;\
II
@ко:
ш i J7\
\V
IV V Дифференцировка rистолоrически идентифицируемых клеток
VI
Рис. 48. Схема постэмбриональноrо кроветворения (объяснение в тексте)
Система тканей внутренней среды
101
ной системы, реаrирующей на конкретные факторы дифференцировки (эритропо
этин, тромбопоэтин, колониестимулирующие факторы, интерлейкины ИЛ и др.),
вырабатываемые кроветворным микроокружением и клетками друrих opraHOB. Все
это при водит к тому, что в клетках появляются rистолоrические маркеры, на OCHO
ве которых можно с большой вероятностью отнести ту или иную клетку к KOHKpeT
ному rемопоэтическому ряду (дифферону).
IV й класс клеток rистолоrически распознаваемые клетки кроветворной TKa
ни это пролиферирующие клетки ("бласты"). Они способны к пролиферации и диф
ференцировке.
V й класс созревающие клетки ("про циты") и VI класс зрелые клетки
периферической крови.
Эрuтроцuтопоэз. Эритроцитопоэз начинается со стволовой кроветворной
клетки. Через стадию колониеобразующей мультипотентной клетки (KOE [ЭММ)
формируются бурстобразующая (БОЭ Э) и далее колониеобразующая единица
эритроцитов (КОЕ Э). Клетки этих колоний чувствительны к факторам реryля
ции пролиферации и дифференцировки. Например, эритропоэтин, вырабатывае
мый клетками почки, стимулирует пролиферацию и дифференцировку клеток в
эритробласты.
В IV й класс включаются базофильный. полихроматофильный и оксифильный
эритробласты. Проэритроциты, потом ретикулоциты составляют V й класс и, HaKO
нец, формируются эритроциты (VI й класс). В эритропоэзе на стадии оксифильно
ro эритробласта происходит выталкивание ядра. В целом цикл развития эритроцита
до выхода ретикулоцита в кровь продолжается до 12 суток Общее направление эрит
ропоэза характеризуется следующими основными структурно функциональными из
менениями: постепенным уменьшением размеров клетки, накоплением в цитоплаз
ме rемоrлобина, редукцией орrанелл, снижением базофилии и повышением
оксифилии цитоплазмы, уплотнением ядра с последующим ею выделением из co
става клетки. В эритробластических островках эритр06ласты поrлощают путем мик
ропиноцитоза железо, поставляемое макрофаrами, для синтеза rемоrлобина.
Развитие эритроцитов происходит в миелоидной ткани красною костною моз
ra. В периферическую кровь поступают только зрелые эритроциты и HeMHoro pe
тикулоцитов.
Состояние, при котором содержание rемоrлобина в крови значительно сниже
но, называется анемией. Оно бывает связано либо с уменьшением числа эритроци
тов, либо с понижением содержания rемоrлобина в них, и возникает в результате
ряда причин: rенетических (например, серповидноклеточная анемия, связанная с
нарушением синтеза rемоrлобина и распадом эритроцитов), кровопотери, воздей
ствия rемолитических ядов, вызывающих распад эритроцитов, дефицита железа
или витамина В 12 . В норме потребность в эритроцитах обеспечивается за счет раз
множения клеток IV V ю классов. Этот процесс называется lOмопластическим re
мопоэзом. При резком дефиците эритроцитов, вызванном кровопотерей или дpy
rими факторами, rомопластическоrо rемопоэза оказывается недостаточно
Эритроциты начинают развиваться путем деления клеток I III ro классов. Такой
процесс называется zетеропластическим rемопоэзом.
102
Fлава 9
rра"'улоцuтопозз. Образование rpанулоцитов происходит в миелоидной TKa
ни KpacHoro KocTHoro мозrа. Исходная стволовая клетка превращается в мульти
потентную клетку предшественник миелопоэза (КОЕ rэмм) и далее под воз
действием колониестимулирующих факторов дифференцируется в общую
родоначальную клетку для rранулоцитов и моноцитов (KOE rMH). в дальней
шем в результате диверrенции возникают родоначальные клетки для rранулоци
тов (KOE rH), которые дифференцируются в идентифицируемые миелобласты
(lV й класс клеток). В ряду дальнейшей клеточной дифференцировки (У й класс
клеток) различают стадии: промиелоцита, миелоцита, метамиелоцита. Начиная
со стадии промиелоцита, клетки подразделяются на 3 разновидности: нейтро
фильные, эозинофильные, базофильные. Более отчетливо это подразделение
можно провести на стадии миелоцитоВ, коrда в клетках накапливается достаточ
ное количество специфической зернистости. До стадии миелоцитов включительно
клетки rранулоцитопоэза делятся митозом. Метамиелоциты митозом уже не дe
лятся. В этих клетках ядро при обретает вначале палочковидную, а затем cerMeH
тированную форму.
Общее направление дифференцировки клеток rранулопоэза характеризует
ся: постепенным уменьшением размеров клетки, снижением базофилии цитоп
лазмы, появлением в цитоплазме специфических rранул, уменьшением разме
ров ядра, появлением сеrментированности ядра и ero уплотнением, сдвиrом
ядерно цитоплазменноrо отношения в сторону преобладания размеров цитоплаз
мы над размерами ядра.
В периферическую кровь поступают зрелые rранулоциты (УI й класс клеток)
нейтрофилы, эозинофилы и базофилы, а также небольшое количество малодиф
ференцированных (юных) rранулоцитов. Физиолоrическая реrенерация обеспе
чивается делением преимущественно клеток V ro класса миелоцитов.
Тро:м.боцuтопозз. Мультипотентные клетки (КОЕ rэмм) миелопоэза под
влиянием факторов микроокружения и тромбопоэтина дифференцируются в po
доначальные клетки меrакариоцитарноrо ряда (KOE Mer). Далее этот ряд вклю
чает следующие формы: меrакариобласт, промеrакариоцит, меrакариоцит, TpOM
боциты.
Меla1Сарuобласт крупная полиплоидная клетка. По мере увеличения степе
ни плоидности до 32 64 n она приобретает rиrантские размеры. Наиболее диффе
ренцированная клетка этоrо ряда m-еЮ1Сариоцит имеет базофильную цитоп
лазму с мноrочисленными азурофильными rранулами. Происходит значительное
увеличение размеров ядра вследствие ero полиплоидизации и сеrментации. Mera
кариоциты находятся в миелоидной ткани KpacHoro KocTHoro мозrа. От поверхно
сти цитоплазмы этих клеток по каналам аrранулярной эндоплазматической сети
отшнуровываются небольшие фраrменты, превращающиеся в 1Сровяные пластuн
1СИ. Последние попадают в кровяное русло. Основное проявление дифференциров
ки клеток при тромбоцитопоэзе сводится к увеличению размеров меrакариоблас
тов, полиплоидизации, появлению в цитоплазме азурофильной зернистости,
отшнуровыванию фраrментов цитоплазмы путем образования впячиваний плаз
молеммы, отделению от меrакариоцитов кровяных пластинок, попадающих в кровь.
Система тканей внутренней среды
103
МОll0цитопозз. Моноцитопоэз образование моноцитов происходит в Kpac
нам костном мозrе из стволовых клеток через стадии КОЕ rэмм, далее KOE
[Мо, затем KOE Мо, монобласта, промоноцита и моноцита. Конечной стадией диф
ференцировки клеток моноцитарноrо ряда является не моноцит, а макрофаz
(мононуклеарный фаrоцит), который находится вне сосудистоrо русла, Диффе
ренцировка клеток при моноцитопоэзе характеризуется: увеличением размеров
клетки, приобретением ядра бобовидной формы, снижением базофилии цитоплаз-
мы, превращением моноцита в макрофаr.
Лu.мфоцuтопозз и u.м.муноцuтопозз. Лимфоидная ткань у человека имеется в
составе лимфатических узлов, селезенки, МИНДалин, аппендикса и в друrих лимфо-
идных образованиях по ходу пищеварительною тракта. В лимфоидной ткани про
исходит лимфопоэз. Исходными клетками лимфопоэза являются стволовые клетки
KpacHoro костнorо мозrа. Через стадию мультипотентных клеток (КОЕ-Л) они
дифференцируются в родоначальные про- Т-и про В лимфобласты и далее в Т-и B
лимфобласты, T и В пролимфоциты И Т- и В лимфоциты.
В лимфоцитопоэзе в тимусе возникают субпопуляции Т клеток с различными
рецепторами (так называемая антиrеннезависимая пролиферация и дифференци
ровка). Т лимфоциты участвуют в формировании 'КЛеmочною uм.м.унитета. Дpy
rой ряд дифференцировки в лимфопоэзе приводит к образованию из В лимфоци
тов через стадии плазмобласта и проплазмоцита плазматических клеток
(плазмоцитов). Эти клетки вырабатывают антитела, обеспечивая lУМОРалъный им
мунитет. Подробнее образование иммунокомпетентных клеток и их участие в раз
витии воспаления рассматриваются ниже.
Из лимфобластов образуются большие, средние и малые лимфоциты. Этот
ряд дифференцировки сопровождается уменьшением размеров клеток, уплотне
нием ядер, снижением митотической активности. Малые лимфоциты способны
к "бласттрансформации" своеобразной дедифференцировке с последующей
повторной их дифференцировкой. Явление бласттрансформации открыто
А.А. Максимовым (1902).
Возрастные изменения и реактивность системы крови
Количество лейкоцитов достиrает уровня, свойственноrо взрослому человеку,
к 14 15 rодам. При этом оно снижается от 10 30 тыс в 1 мкл У новорожденноrо до
5 8 тыс в 1 мкл У взрослоrо. На 1 2 M rодах жизни лимфоциты (65%) преобладают
над нейтрофилами (25%). К 4 MY rоду эти показатели выравниваются. К периоду
половоrо созревания соотношение нейтрофилов (65%) и лимфоцитов (25%) CTa
новится обратным и достиrает нормы взрослоrо. Количество эритроцитов у HOBO
рожденноrо составляет 6 7 млн В 1 мкл. Нормы взрослоrо оно достиrает к периоду
половоrо созревания.
Все виды ионизирующею излучения, применяемые в клинике (лечебное тоталь
ное облучение орrанизма при множественных опухолях, трансплантации KocTHoro
мозrа и др.), MOryт привести к развитию радиационных повреждений кроветворных
тканей. Эти поражения объясняются высокой чувствительностью к лучевой энерrии
104
Fлава 9
клеток, проходящих митотический uикл в проuессе rистоrенеза крови. Под воздей
ствием радиаuии наступают подавление пролифераuии, дифференuировки, миrpаuии
развивающихся клеток крови, нарушение интеrpаuионных взаимоотношений элемен
тов микроокружения, rибель малодифференuированных клеток предшественников и
бластных форм, что приводит К значительному снижению интенсивности или прекра
щению rемопоэза. После облучения возможна реrенераuия миелоидной и лимфоид
ной тканей в opraHax кроветворения, что осуществляется за счет пролифераuии co
хранившихся стволовых кроветворных клеток Это постепенно приводит к
восстановлению rемопоэза. Показано, что для восстановления кроветворения ДOCTa
точно 0,1% стволовых кроветворных клеток Реrенераторные сдвиrи при несмертель
ном облучении в костном мозrе выявляются уже через 1 2 ч после поражения. rисто
rенез крови после лучевоro повреждения может пойти с отклонениями и привести к
развитию лейкоза.
Соединительные ткани
В rруппу соединительных тканей входят разновидности волокнистых тканей
(рыхлая соединительная, плотные фиброзные оформленная и неоформленная) и
соединительные ткани со спеuиальными свойствами (ретикулярная, жировая и др.).
Все они имеют единый эмбриональный источник развития мезею:uм.у. Среди TKa
ней этой rpуппы отчетливо выраженная метаболическая функция присуща рыхлой
волокнистой соединительной ткани.
Рыхлая волокнистая соединительная ткань
Это одна из наибо
лее распространенных в
орrанизме тканей (рис. 49).
Она сопровождает все KpO
веносные и лимфатические
сосуды, периферические
нервы, образует строму
внутренних opraHOB, за
полняет промежутки меж
ду орrанами, входит в co
став кожи и т. д. Рыхлая co
единительная ткань COCTO
ит из клеток и межклеточ
Horo вещества. В разных
opraHax клеточный состав
и межклеточные CTPYKTY
ры рыхлой соединитель
ной ткани имеют HeKOTO
рые особенности. Поэтому
2
-l
5
-/
}
,
'
:.J i : ,," J "
<J' -<i- p
' , (. '$.
, 3
" ' . \ f '\ 'с ..... ,......
.... \.'" . , ...., ," W'... ,......
, < <':."' I - . .
. }- '\ , ./>'
""., "' / j ,'l!':'$
'" V" ......,." /
!
'{
4
Рис. 49. Рыхлая волокнистая соединительная ткань:
1 фибробласты; 2 эндоплазма; 3 лимфоцит; 4 макрофаr;
5 коллаrеновое волокно; 6 эластическое волокно
Система тка1iей вЩjтре1i1iей среды
105
понятие "соединительная ткань" является очень обобщенным. По существу, в каж
дом opraHe имеется своя соединительная ткань, максимально приспособленная к
выполнению функции данноrо KOHKpeTHoro opraHa.
В составе рыхлой соединительной ткани находятся клетки различной rистоrе
нетической детерминации. Среди них различают фибробласты и фиброциты (Be
дущий клеточный дифферон), rистиоциты макрофаrи и антиrенпредставляющие
клетки, пиrментные клетки (меланоциты), тканевые базофилы (тучные клетки, лаб
роциты), перициты и адвентициальные клетки, жировые клетки (липоциты), плаз
матические (плазмоциты), клетки крови (rpанулоциты, моноциты, лимфоциты).
Адвентициальные клетки и перициты. Адвентициальные клетки наиме
нее дифференцированные, но rистолоrически распознаваемые клетки рыхлой co
единительной ткани. Располаrаются клетки nериваСКУЛЯР1iО, они подвижны, име
ют веретенообразную форму, их цитоплазма слабобазофильна, ядро овальное и
обычно rиперхромное. Адвентициальные клетки делятся митозом. По мере ди
верrентной дифференцировки они превращаются в фибробласты, миофиброб
ласты, миофиброкласты и липоциты. Следовательно, в рыхлой волокнистой co
единительной ткани существует совокупность клеток возрастающей степени
зрелости от камбиальной формы до фиброцита, что составляет фибробластичес
кий ряд, или фибробластический диффеРО1i.
Тесно связаны со стенкой KpoBeHocHoro сосуда микроциркуляторноrо русла
перициты. Эти клетки располаrаются между листками базальной мембраны эндо
телия кровеносных сосудов, что оrраничивает их подвижность. Клетки имеют OT
ростчатую форму, в цитоплазме хорошо развита опорно двиrательная система, что
придает клеткам способность к сокращению и реrуляции просвета rемокапилляра.
Фибробласты ведущие клетки рыхлой соединительной ткани, продуциру
ющие компоненты межклеточноrо вещества (рис. 50). Это отростчатые, BepeTeHO
образные или распластан
ные клетки размером OKO
ло 20 мкм. В них хорошо
развиты орrанеллы BHYT
ренней метаболической
среды. Ядро фибробласта
овальной формы, coдep
жит равномерно распы
ленный хроматин и 2 З
ядрышка. Цитоплазма OT
четливо подразделяется
на интенсивно окрашен
ную Э1iдоnлазму и слабо
окрашенную экmоnлQЗМУ.
Цитоплазма фиброблас
тов (особенно молодых)
базофильна. В ней выяв
ляется хорошо развитая
Рис. 50. Схема строения фибробласта и образование межклеточноrо
вещества:
1 молекулы тропоколлаrена; 2 rликозаминоrликаны; 3 прото
фибрилла; 4 коллаrеновая фибрилла; 5 молекулы эластина; б
эластическая фибрилла
106
rлава 9
эндоплазматическая сеть с большим количеством рибосом, прикрепленных к MeM
бранам в виде цепочек по 10 30 rранул. Такая ультраструктура rранулярной эн
доплазматической сети характерна для клеток, активно синтезирующих белок "на
экспорт". Имеются также мноrочисленные свободные рибосомы, хорошо разви
тый комплекс rольджи. Митохондрии крупные, количество их невелико. Цито
химическими методами показано наличие в цитоплазме фибробластов ферментов
rликолиза и rидролитических ферментов лизосом (особенно коллаrеназы). Me
нее активны окислительные ферменты митохондрии.
Опорно двиrательная система клетки обеспечивает их подвижность, изменение
формы, прикрепление к субстрату, механическое натяжение пленки, к которой клет
ка прикрепляется в культуре. На клеточной поверхности имеется MHoro микровор
синок и пузырчатых выростов. Фибробласты во взвешенном состоянии в жидкой
среде имеют шаровидную форму. Распластанным фибробласт становится после при
липания к твердой поверхности, по которой он передвиrается за счет псевдоподий.
Основная функция фибробластов синтез и секреция белков и rликозаминоrли
канов, идущих на формирование компонентов межклеточноrо вещества соединитель
ной ткани, а также выработка и секреция колониестимулирующих факторов (rpaHY
лоцитов, макрофаrов). Фибробласты долrое время сохраняют способность к
пролиферации. Фибробласты, закончившие цикл развития, называются фиброциmа
М,И. ЭТО долrоживущие клетки. Цитоплазма клеток обедняется орrанеллами, клетка
уплощается, пролиферативный потенциал падает. Однако клетка не теряет способ
ность участвовать в реryляции обменных процессов в ткани.
Межклеточное вещество. Состоит из фибриллярноrо и основною (аморфно
ro) компонентов. Методами rистоавторадиоrрафии с введением меченых аминокис
лот ен пРОЛИН, ЗН rлицин и др.) установлено, что в полисомах фибробластов про
исходит синтез молекул белка. Фибробласты одновременно MorYT синтезировать
несколько типов специфических белков и rликозаминоrликаны. Для синтеза белка
коллаrена имеет существенное значение наличие витамина С, при недостатке KOTO
poro коллаrеноrенез резко тормозится. Интенсивнее идет синтез межклеточноrо Be
щества в условиях пониженной концентрации кислорода. Одновременно с синте
зом коллаrена фибробласт разрушает примерно 2/3 этоrо белка с помощью фермента
'КОJUlаzеllазы, что препятствует преждевременному склерозированию ткани.
Синтезированные молекулы проколлаrена выводятся на поверхность фиброб
ластов путем экзоцитоза. При этом осуществляется переход белка из растворимой
формы в нерастворимую mpOnOKOJUlazell. Объединение молекул тропоколлаrена
в надмолекулярные структуры 'КОJUlаzе1l0вые фибриллы происходит в непос
редственной близости от клеточной поверхности блаrодаря действию особых Be
ществ, выделяемых клеткой. В частности, на поверхности фибробластов обнару
жен белок фибрОllе'КmИll, выполняющий адrезивную и друrие функции.
Последующие этапы фибриллorенеза происходят путем полимеризации и arpera
ции тропоколлаrена на ранее образованных фибриллах. При этом созревание KOk
лаrеновых волокон может протекать и без прямой связи с фибробластами.
[ликозаминоrликаны являются реryляторами коллаrенообразования и входят
в состав OCHoBHoro (аморфноrо) компонента межклеточноrо вещества.
Система тканей внутренней среды
107
Фибриллярный компонент межклеточноrо вещества рыхлой соединительной
ткани включает три типа волокон коллаrеновые, эластические и ретикулярные.
Они имеют сходный механизм образования, однако отличаются друr от друrа по
химическому составу, ультраструктуре и физическим свойствам. Белок коллаrен
идентифицируется по аминокислотному составу и последовательности располо
жения аминокислот в молекуле коллаrена. В зависимости от вариации аминокис
лот в полипептидной цепи, иммунных свойств, молекулярной массы и др. разли
чают 14 и более разновидностей коллаrеновых белков, которые входят в состав
соединительной ткани opraHoB. Все они составляют 4 основных типа, или класса,
коллаrена.
Коллаrен I ro типа встречается в соединительной и костной тканях, а также в
склере и роrовице rлаза; II ro типа в хрящевых тканях; III ro типа в стенке
кровеносных сосудов, в соединительной ткани кожи плода; IV ro типа в базаль
ных мембранах.
Коллаrеновые волокна. Образуются коллаrеновые волокна при полимериза
ции молекул тропоколлаrена, имеющих длину около 280 нм и толщину 1,4 нм.
Молекулу формируют три полипептидных цепочки, содержащих около тысячи
аминокислотных остатков. Из них 1/3 составляет rлицин, 1/3 приходится на
пролин или лизин И остальное надруrие аминокислоты. Молекулы тропоколла
reHa в коллаrеновом волокне продольно ориентированы. Они сдвинуты друr OTHO
сительно друrа на 1/4 своей длины. За счет этorо в протофибриллах коллаrеново
ro волокна возникает поперечная исчерченность с периодом повторяемости темных
и светлых участков в 64 70 нм. При участии rликозаминоrликанов и протеоrлика
нов из протофибрилл коллаrена формируются фибриллы толщиной 50 100 нм, а
затем и волокна с поперечником 1 3 мкм. Коллаrеновые волокна располаrаются в
различных направлениях, образуя подобие войлока. Они обладают малой растя
жим остью и большой прочностью на разрыв. Так, совокупность коллаrеновых BO
локон с поперечным сечением 1 см 2 может выдержать наrрузку до 500 Kr.
Эластические волокна. Это волокна диамером 0,2 10 мкм. В фиробластах синте
зируются молекулы белка эластина, содержащеrо аминокислоты: лизин, пролин,
rлицин, лейцин и в меньшей степени оксипролин и друrие. Внеклеточное формиро
вание эластических волокон происходит в два этапа: 1) расположение фибрилл в виде
пучка, 2) пропитывание этоrо пучка аморфным веществом. Молекулы эластина pac
полаrаются в фибриллах без определенной ориентировки (как молекулы в резине).
Эластические волокна обладают большой растяжимостью и сравнительно малой проч
ностью. Модуль упруrости их 4 6 Kr /см 2 . Они обычно анастомозируют друr с друтом,
образуя широкопетлистую сетьЛереходными формами развития эластическоrо BO
локна являются окситалановые и элауниновые волокна.
Ретикулярные, или ретикулиновые, волокна. Ретикулярные волокна имеют ди
аметр 0,1 2 мкм. Волокна эти называют также арrирофильными, так как они отли
чаются сродством к солям серебра и образованы коллаrеном 111 типа. В нем повы
шено содержание цистеина и rексозамина. Таких аминокислот, как пролин и
оксипролин меньше, чем в тропоколлаrене. Под электронным микроскопом в pe
тикулярных волокнах обнаружена периодическая исчерченность. Ретикулярные
108
rлава 9
волокна не перевариваются трипсином. Они образуют обычно сетчатые структуры
типа решетки (отсюда их друrое название решетчатые волокна). Ретикулярные
волокна входят в состав базальных мембран, располаrаются BOKpyr сосудов, в том
числе капилляров, нервных волокон, входят в состав сарколеммы мышечных воло
кон, вместе с ретикулярными клетками формируют остов кроветворных opraHoB.
ОСНОВНОЙ (аморфный) компонент межклеточноrо вещества это микроско
пически бесструктурная основа, в которой находятся клетки и волокна соединитель
ной ткани. Здесь осуществляются метаболические процессы. Биохимически это
полужидкий вязкий zель, состоящий из макромолекул, преимущественно полисаха
ридов, и большоrо количества тканевой жидкости. Полисахаридный компонент oc
HOBHoro вещества присутствует в виде rиалуроновой кислоты (rликозаминоrлика
на) длинной молекулы, которая в водных растворах образует изrибы, занимая
участок диаметром 400 нм И при этом связывает очень большой объем жидкости.
Соседние молекулы rиалуроновой кислоты формируют сеть, в петлях которой Ha
ходится тканевая жидкость. [ликозаминоrликаны связывают межклеточную воду,
реryлируют осмотическое давление и ионный состав OCHoBHoro вещества.
[ликозаминоrликаны бывают двух видов: сульфатuрова1-l1-lые (rепаринсульфат,
хондроитинсульфат, дерматансульфат); и 1-lесульФатuрова1-l1-lые rиалуроновая
кислота. Сульфатированные rликозаминоrликаны в норме соединены с белками и
образуют протеоrликаны. Этот процесс реryлируют тучные клетки. В составе oc
новноro вещества также обнаружены липиды, альбумин, rлобулины, минеральные
вещества и др. Таким образом, молекулы rликозаминоrликанов формируют сети,
ячейки, каналы, по которым циркулирует тканевая жидкость, и это является моле
кулярным барьером для бактерий и вирусов.
Выработка фибриллярноrо и OCHoBHoro компонентов межклеточноrо веще
ства rлавное проявление дифференцировки клеток фибробластическоrо ряда.
АдшIOЦИТЫ. Жировые клетки ади, поциты развиваются из адвентициаль
ных клеток Это крупные шаровидные клетки диаметром 30 50 мкм. В цитоплазме
адипоцитов накапливаются липидные включения в виде мелких капель, которые
позднее сливаются в одну большую каплю. Ядро при этом оттесняется на перифе
рию, и цитоплазма составляет лишь узкий ободок Обезжиренная клетка на rисто
лоrическом срезе напоминает по виду перстень. Под электронным микроскопом в
жировых клетках определяются слабо развитые цитоплазматическая сеть, комплекс
[ольджи и митохондрии. Адипоциты накапливают жир как трофическиЙ резервный
материал. Жировые клетки MOryT освобождаться от включений. При этом они CTa
новятся трудно отличимыми от клеток фибробластическоrо ряда.
Жировые клетки встречаются среди фибробластов рыхлой соединительной
ткани в незначительном количестве. В тех случаях, коrда они образуют большие
скопления, то roворят уже не об отдельных клетках, а о жировой ткани.
Пиrментоциты. В рыхлой волокнистой соединительной ткани обнаруживаются
клетки, цитоплазма которых содержит зерна пиrмента мела1-lU1-lа. Среди этих
клеток различают синтезирующие пиrмент меланоциты и фаrоцитирующие ro
товый пиrмент, например, фибробласты и макрофаrи. Ткань с большим количе
ством меланоцитов встречается у человека в радужке и сосудистоЙ оболочке rлаза,
Система тканей внутренней среды
109
в соединительноткаНIlЫХ слоях сильно пиrментированных участков кожи, а также
в родимых пятнах. Меланоциты являются производными HepBHoro rребня, имеют
отростчатую или веретеновидную форму, подвижны, функция и форма клеток MO
жет меняться в зависимости от ryморальных и нервных факторов. Клетки MorYT
втяrивать свои отростки или вытяrивать их, соответственно меняется окраска opra
на или, например, в opraHe зрения происходит защита фоточувствительноrо OTpO
стка нейрона от воздействия света.
Сказанным не исчерпывается все разнообразие клеточных форм, имеющихся
в составе рыхлой соединительной ткани.
В рыхлой соединительной ткани постоянно находятся клетки, являющиеся
потомками стволовой кроветворной клетки. Это rистиоциты макрофаrи, анти
rенпредставляющие клетки, тканевые базофилы (тучные клетки), плазмоциты,
клетки крови (rранулоциты, моноциты, лимфоциты).
rистиоциты макрофаrи. Они составляют
10 20% от Bcero клеточноrо состава рыхлой co
единительной ткани. Размер клеток 12
25 мкм. Макрофаrи, находящиеся в спокойном
состоянии, называют lистиоцитами, оседлыми
макрофаrами или блуждающими клетками в
покое (рис. 51). Подвижные макрофаrи, не
имеющие определенной локализации в ткани,
называют свободными макрофаrами. Ядро MaK
рофаrов темное, окруrлое, содержит крупные
rлыбки хроматина. Цитоплазма макрофаrов
четко контурирована. В ней содержатся боль
шое количество вакуолей фаroсом и лизосом,
комплекс [ольджи, мноrочисленные пиноци Рис. 51. Схема ультраструктуры макрофаrа:
тозные пузырьки. Остальные орrанеллы разви 1 комплекс rольджи; 2 лизосомы
ты умеренно. Хорошо развитая опорно двиrательная система способствует миrpа
ции клеток и фаrоцитозу инородних частиц. По характеру и количеству
ультраструктур выделяются макрофarи секреторноlO и фаlOцитарноlO видов. У пер
вых в цитоплазме преобладают секреторные вакуоли, у вторых лизосомальный
аппарат. Источником образования макрофаrов являются моноциты крови.
Особая разновидность макрофаroв принимает участие в качестве aHmUle1lпpeJcтaв
JlЯющей клетки и тем самым являются участниками кооперации Т и B лимфоцитов
при иммунном ответе на чужеродные вещества. Макрофarи: нейтрализуют токсины, MO
ryт накапливать витальные красители при введении их в кровь. Они проявляют анти
бактериальные свойства, выделяя лизоцим, кислые rnдролазы, лактоферрин и др., обла
дают антиопухолевой активностью, выделяя фактор некроза опухолей. Факторы роста
макрофаroв влияют на пролиферацию эпителиальных клеток, пролиферацию и диф
ференцировку фибробластов, новообразование кровеносных сосудов и др.
Способность к Фаrоцитозу является общебиолоrическим свойством мноrих TKa
невых клеток Однако только те клетки, которые способны захватывать и фермента
тивно перерабатывать в своей цитоплазме бактерии, инородние частицы, токсины
110
Fлава 9
и др., следует относить к макрофаzuческой системе орrанизма. Учение о макрофаrи
ческой системе заложил И.И. Мечников (1882), который в экспериментах на бес
позвоночных обнаружил подвижные клетки, накапливающиеся около инороднеrо
тела. Именно эти клетки были названы макрофаrами. Кроме макрофаrов rистиоци
тов в состав макрофаrической системы орrанизма входят макрофаrи печени (звезд
чатые макрофаrоциты, остеокласты, rлиальные макрофаrи, макрофаrи KpOBeTBOp
ных opraHoB, макрофаrи леrкоrо и др.). Реrуляция макрофаrической системы ocy
ществляется как местными так и центральными (нервная и эндокринная системы)
механизмами.
Тканевые базофилы (тучные клетки, лаб
роциты, rепариноциты) развиваются из CTBO
ловых кроветворных клеток. Клетки окруrлой
или овальной формы размером от 20 до зо
100 мкм, располаrаются преимущественно
вдоль мелких кровеносных сосудов. Они име
ют небольшое плотное ядро и зернистую цитоп
лазму (рис. 52). Наиболее характерный признак
тучных клеток это наличие в цитоплазме MHO
rочисленных rранул, диаметр которых 0,3
0,7 мкм, обладающих свойством метахромазии
Рис. 52. Схема улыраструктуры TKaHe
(окрашиваться не в цвет красителя). В rранулах Boro 6азофила:
содержатся rепарин, rистамин, хондроитинсуль 1 ядро; 2 rранулы различной плот
фаты, rиалуроновая кислота, серотонин, xeMO ности
таксические факторы для эозинофильных и нейтрофильных rpанулоцитов и др. При
деrрануляции тучных клеток выделяется rепарин, препятствующий свертыванию
крови. Выход биоrенных аминов сопровождается изменением проницаемости [eMa
TOTKaHeBoro барьера. Кроме Toro, тучные клетки вырабатывают цитокины, участву
ющие в иммунных процессах. Тучные клетки размножаются крайне редко.
Плазматические клетки. Это короткоживущие клетки, развивающиеся из B
лимфоцитов. Они имеют окруrлую форму, диаметр 10 12 мкм (рис. 53). OKpyr
лое ядро содержит плотный хроматин, распо
ложенный по радиусам, поэтому ядро имеет
вид "колеса со спицами". Цитоплазма плазмо
цитов сильно базофильна, содержит MHoro ри
бонуклеопротеида и цистерны rранулярной эн
доплазматической сети, расположенные цирку
лярно. Околоядерный участок клетки coдep
жит развитый комплекс rольджи, центриоли и
окрашивается менее базофильно ("дворик").
Иноrда определяются конrломераты иммуно
rлобулинов в виде оксифильных телец Pycce
ля. Клетки участвуют в формировании ryMO
ральноrо иммунитета, продуцируя иммуноrло
булины (антитела).
Рис. 5З. Схема улыраструктуры плазмати
ческой клетки:
1 светлый участок цитоплазмы ("дворик");
2 rранулярная эндоплазматическая сеть
Система ткан.ей 6н.утрен.н.ей среды
111
Рыхлая соединительная ткань является также местом постоянноrо пребывания
rpанулоцитов крови.
Жировая ткань
Значительные скопления жиро
вых клеток (адипоцитов), встречаю
щиеся среди друrих клеток в рыхлой
соединительной ткани, составляют
жировую ткань (рис. 54). Различают
два вида жировой ткани: белую и бу ".
рую. Белая жировая ткань состоит из
адипоцитов, содержащих обычно
одну крупную каплю жира. По фор
ме это перстневидные жировые
клетки. Адипоциты образуют комп
лексы, составляющие так называе
мые жировые дольки. Белая, или
обычная, жировая ткань встречает
ся у взрослых людей в подкожной
жировой клетчатке, в сальнике и Рис. 54. Белая жировая ткань:
строме внутренних opraHOB. 1 адипоциты; 2 кровеносный капилляр
Бурая жировая ткань xapaK
терна лишь для paHHero детскоrо возраста. Она состоит из адипоцитов, coдep
жащих мелко дисперrированные включения жира. Бурую окраску эти клетки
имеют блаrодаря присутствию желтых пиrментов ЦИТОХРОМОВ.
Функция жировой ткани трофическая, связанная с обеспечением энерrети
ческоrо запаса и резерва воды в орrанизме. Значительна роль жировой ткани TaK
же в процессах теплореryляции.
l
i..-"""
:
1 2
\ j
'" 4 } , .
\. , 4i
)
\L.
,
\! . ""
') t
. ,
....
"
\,
. {'
41,
,
.........."
, (
'-
Эндотелий
Эндотелий это пласт клеток, выстилающий кровеносные сосуды и эндо
кард, лимфатические сосуды. Источником развития эндотелия является мезен
хима спланхнотома (анrиобласт), в составе которой имеются стволовые клет
ки, дающие популяцию эн.дотелиобластО6. Дифференцировка последних
протекает в связи с ростом сосудов. Клетки митотически размножаются и пос
ле ряда митотических циклов постепенно уплощаются, вытяrиваются по оси
развивающеrося капилляра, формируют межклеточные контакты и создают He
прерывную выстилку сосудов. В ходе дальнейшей цитодифференцировки в co
ответствии с орrанной спецификой возникают специализированные формы эн
дотелиоцитов.
Эн.дотелиоциты полиrональные уплощенные клетки длиной 20 150 и ши
риной 10 20 мкм (рис. 55). Это, как правило, одноядерные диплоидные клетки
112
Тлава 9
толщиной 3 5 мкм в области
ядра и О, 1 0,4 мкм на перифе
рии. Электронно микроско
пически в каждом эндотели
оците выделяют три зоны:
а) ядерную и орrанелл; б) пе
риферическую; в) KOHTaKT
ную, и три поверхности лю
минальную, аблюминальную
и контактную. Форма ядер
овальная с мноrочисленными
инваrинациями ядерной обо
лачки в зависимости от растяжения сосудистой стенки. В околоядерной области CKOH
центрированы орrанеллы. В сосудах микроциркуляторноrо отдела KpoBeHocHoro pyc
ла в периферической зоне эндотелиоцитов находятся транспортные структуры, уча
ствующие в трофических и метаболических процессах: микропиноцитозные пузырь
ки, трансэндотелиальные каналы, фенестры, поры люки. Микропиноцитозные пу
зырьки обеспечивают транспорт веществ, рецепторно связывающихся с поверхнос
тью клетки. Фенестры это трансэндотелиоцитарные каналы диаметром от 30 до
80 нм, закрытые однослойными диафраrмами. Поры являются сквозными OTBep
стиями истонченных участков эндотелиоцита, непосредственно соединяющими про
свет сосуда с периэндотелиальным пространством.
Люминальная поверхность эндотелиоцита (обращенная к крови) образует мик
ровыросты, складки и микроворсинки. Латеральные поверхности имеют специа
лизированные межклеточные контакты простые, сложные (по типу интердиrи
тации), плотные и щелевые. Аблюминальная (базальная) поверхность клетки
формирует контакты с перицитами, rладкими миоцитами и фибриллярными CTPYK
турами соединительной ткани. Базальная мембрана является непрерывным слоем
электронно плотноro фибриллярноrо материала толщиной зо зоо нм, по CTpoe
нию близка к базальной мембране однослойных эпителиев. Однако в СИНУСОИk
ных и лимфатических капиллярах базальная мембрана отсутствует.
Реrенерация эндотелия осуществляется за счет митотическоrо деления и миr
рации эндотелиоцитов. Делящиеся клетки распределены в эндотелиальном плас
те диффузно.
Рис. 55. Схема ультраструктуры эндоте
лиоцита:
1 контактная поверхность; 2 люми
нальная поверхность; 3 аблюминаль
ная поверхность; 4 rликокаликс; 5
базальная мембрана; б клеточный
центр; 7 ядро
2
3
5
Взаимодействия клеток системы крови и соединительной
ткани в условиях воспаления и реrенерации тканей
Рыхлая соединительная ткань это система мноrих клеточных дифферонов,
или rистоrенетических рядов диверrентных линий клеточной дифференциров
ки. Несмотря на большое разнообразие клеточных форм, все они составляют еди
ную систему, выполняющую защитную и трофическую функции. Между кровью и
соединительной тканью существуют тесные взаимосвязи и постоянный обмен кле
точными элементами.
Система тканей внутренней среды
113
Структурно фУНКЦИОlIалыюй единицей соединительной ткани считается lИС
тион. Он включает участок микроциркуляторноrо русла с окружающими ero клет
ками и межклеточными структурами. Рыхлая соединительная ткань находится в
динамических взаимодействиях с друrими тканями, в частности, с эпителиальны
ми, ретикулярной, эндотелиальной, жировой. пиrментной, плотными волокнис
тыми соединительными тканями.
Воспаление и реrенерация. При действии повреждающих areHToB (механи
ческих, химических, бактериальных и друrих) в рыхлой соединительной ткани раз
вивается сложная сосудисто тканевая защитно приспособительная реакция вoc
паление. При воспалении наблюдаются как общие, так и местные изменения.
Местные проявления реакции орrанизма в очаrе воспаления включают несколько
взаимосвязанных фаз: 1) альтерация (повреждение) тканей; 2) высвобождение фи
зиолоrически активных веществ так называемых медиаторов воспаления; 3) co
судистая реакция с экссудацией. включающая изменение кровотока в микроцир
куляторном русле, повышение проницаемости сосудов; 4) резорбция продуктов
распада тканей; 5) пролиферация клеток с образованием "rрануляционной ткани"
и последующей реrенерацией тканей. Завершается воспаление образованием зре
лой волокнистой соединительной ткани.
При описании воспаления обычно выделяют три фазы: лейкоцитарную, с Пре
обладанием в очаrе воспаления нейтрофильных rpанулоцитов; макрофаrическую,
коrда продукты распада активно резорбируются макрофаrами; фибробластическую,
на протяжении которой на месте повреждения разрастается соединительная ткань.
Лейкоцитарная фаза воспаления характеризуется передвижением нейтрофиль
ных rранулоцитов в очаr распада ткани на месте ее повреждения. Пусковым Mexa
низмом воспаления является выброс медиаторов и цитокинов (rистамина, cepOTO
нина, лизосомных rидролаз и друrих биолоrически активных веществ). Источником
выделения медиаторов являются тканевые базофилы (тучные клетки), лейкоци
ты, кровяные пластинки, макрофаrи и лимфоциты. При этом развивается комп
лекс сосудистых изменений, включающий повышение проницаемости микроцир
куляторноrо русла, экссудацию жидких составных частей плазмы, эмиrрацию
клеток крови. Уже через 6 ч от начала воспаления образуется лейкоцитарный ин
фильтрат. Нейтрофильные rранулоциты проявляют высокую фаrоцитарную aK
тивность, поrлощая rлавным образом микроорrанизмы (отсюда их название мик
рофаrи). Часть нейтрофилов при этом распадается, выделяя большое количество
лизосомных rидролаз. Это способствует очищению очаrа воспаления от повреж
денных тканей.
Макрофаrическая фаза воспаления протекает при явлениях активизации MaK
рофаrов как reMaToreHHbIx (возникающих из моноцитов крови), так и rистиоrен
ных (оседлых макрофаrов rистиоцитов). Макрофаrи энерrично фаrоцитируют
продукты тканевorо распада. Вместе с тем они вырабатывают вещества стиму
ляторы восстановительных процессов в очаrе воспаления.
Фибробластическая фаза является завершающей фазой воспаления. Она xapaк
теризуется пролиферацией (размножением) клеток фибр06ластическоrо ряда и их пе
редвижением к воспалительному очаry. Поскольку к этому времени заканчивается
114
Тлава 9
в основном очищение места повреждения от продуктов TKaHeBoro распада, фибробла
сты заполняют бывший дефект ткани. Они интенсивно вырабатывают межклеточное
вещество. При этом образуются вначале тонкие арrирофильные, а позднее и коллаre
новые волокна. Вместе с клетками эти волокна отrpаничивают воспалительный очаr
от неповрежденной ткани. Развитие фибробластов постепенно приводит к замеще
нию воспалительноrо очаrа соединительной тканью. При значительном дефекте TKa
ни на месте очаrа воспаления формируется рубец. При наличии инородноrо тела BOK
pyr Hero образуется соединительнотканная капсула, отчетливо выраженная на 5 7 e
сутки от начала воспаления. Относительно источников развития фибробластов в оча
re повреждения существуют разные rипотезы. Так различают две субпопуляции фиб
робластов, имеющие разные источники и отличающиеся неодинаковой продолжитель
ностью жизни (KopOTKO И долrоживущие фибробласты).
Фибробласты, которые развиваются из стволовых кроветворных клеток это
короткоживущая популяция фибробластов защитно трофическоrо типа, участвую
щая в процессах воспаления, заживления ран и т. д. Друrие фибробласты происхо
дят от стволовых стромальных клеток (механоцитов) KocTHoro М:озrа. Это популя
ция долrоживущих фибробластов опорноrо типа с преимущественно механическими
функциями. Кроме Toro, существуют специализированные формы фибробластов
фиброкласты и миофибробласты, развивающиеся из адвентициальных клеток Фиб
рокласты обеспечивают перестройку соединительной ткани путем разрушения меж
клеточноrо вещества. В цитоплазме клеток обнаруживается хорошо развитый аппа
рат лизиса коллаrеновых фибрилл. Клетки выявляются преимущественно в области
формирования рубцовой ткани после повреждения opraHoB. Миофибробласты OT
личаются от фибробластов большим содержанием сократительных филаментов (aK
тина rладкомышечноrо типа). Они также участвуют в реrенерации путем KOHTpaK
ции краев раны.
Система оnорнъа и двиютеЛЪ1lЪLX тка1lей
115
rЛАВА 10. СИСТЕМА ОПОРНЫХ И двиrАТЕЛЬНЫХ
ТКАНЕЙ
Опорно двиrательная тканевая система выполняет опорно механическую и co
кратительную функции. Первая присуща разновидностям соединительных TKa
ней, в которых преобладают коллаrеновые или оссеиновые волокна. Источник раз
вития этих тканей мезе1lXИМа. Вторая сократительная функция присуща
большой rруппе специализированных на функции сокращения мышечным тканям.
Эмбриональными источниками развития данной rруппы тканей являются кожная
эктодерма, нейроэктодерма, тканевые зачатки в составе мезодермы, энтомезенхима.
Ткани с опорно механической функцией
Плотные волокнистые соединительные ткани
в этих тканях волокнистые структуры межклеточноrо вещества значительно
преобладают по своей массе над клетками. В зависимости от характера расположе
ния коллаrеновых волокон различают плотные волокнистые 1lеофор.мле1l1lыe и офор
мле1l1lыe соединительные ткани.
Различают также эластические co
единительные ткани.
Плотная волокнистая неофор
мленная соединительная ткань об .. .......,. з
..... ..... " .,.,......."..
разует сетчатый слой кожи, капсу >..... '!or",
лы opraHoB. Толстые пучки колла
rеновых волокон формируют здесь
трехмерную сеть (вязь).
Плотная волокнистая оформ
леннаясоединительнаятканьхарак
теризуется закономерным (парал
лельным) расположением коллаrе
новых пучков, между которыми pac
полаrаются фиброциты (рис. 56).
Примером такой ткани может быть
ткань, входящая в состав сухожилия.
Пучки коллаrеновых волокон при
дают opraHaM высокую механичес
кую прочность.
..........-.......................
,
...........
...........
......... .......
,.....
fIIIIP ,....
....
1 , ..........
, .
2 ___......... ......
..... ..
.........
................
........ ........
.....,
;., ф........ Q .. ........ , ..." р> . '"'
.' "
Рис. 56. Плотная волокнистая оформленная соединитель
ная ткань:
1 фиброциты; 2 пучки коллаrеновых волокон; 3 про
слойки рыхлой волокнистой соединительной ткани
116
Fлава 10
Скелетные ткани
Это хрящевые и костные ткани мезенхимноrо происхождения. Каждая из них в
свою очередь подразделяется на 3 разновидности. Хрящевые ткани это rиалино
вая, эластическая и волокнистая. Среди костных тканей различают костную денти
ноидную, костную ретикулофиброзную (rpубоволокнистую) и костную пластинча
тую ткани. Хрящевые и костные ткани выполняют опорную, механическую и
защитную функции, а также участвуют в водно солевом обмене орrанизма как ис
точники кальция и фосфора.
Хрящевые ткани
Хрящевые ткани состоят из крупных клеток хондробластов и хондроцитов,
а также плотноro межклеточноrо вещества сложноrо химическоrо состава. 70 80%
массы хрящевых тканей составляет вода, 1 o 15% орrанические вещества и 4 7%
минеральные соли. Межклеточное вещество содержит хондриновые фибриллы и
хондромукоид.
ruстozенез хрящевых тканей (хонi)РОluстОlенез). Хрящевые ткани разви
ваются из мезенхимы. Начинается хондроrенез с уплотнения мезенхимы на месте
будущеЙ хрящевой ткани и образования хондроrенноro участка. Клетки в составе
TaKoro участка интенсивно делятся митозом, сближаются друr с друrом, увеличи
ваются в размерах. Опорную функцию хондроrенные клетки выполняют за счет
собственноrо BHYTpeHHero напряжения, или Typropa.
На следующей стадии rистоrенеза хрящевые клетки начинают продуцировать меж
клеточное вещество. Формируется первичная хрящевая ткань. Происходит существен
ная перестройка внутреннеЙ орrанизации хондробластов, в которых развивается бе
локсинтезирующий аппарат (rранулярная эндоплазматическая сеть, комплекс
[ольджи). Хондробласты осуществляют синтез двух основных компонентов межкле
точноro вещества специфических коллаrеновых белков (II ro типа), которые фор
мируют фибриллы толщиной 10 20 нм, и
rликозаминоrликанов (рис. 57). Хондроб
ласты, начавшие синтез специфических
белков, сохраняют способность к репли
кации ДНК и MOryт делиться митозом. За
счет деления клеток масса первичной хря
щевой ткани увеличивается.
Следующая стадия rистоrенеза хря
щевых тканей характеризуется дальней
шей дифференцировкой хондробластов,
которые начинают секретир<?вать сульфа
тированные rликозаминоrликаны. В меж
клеточном веществе накапливаются про
теоrликаны соединение неколлаrено
вых белков с rликозаминоrликанами (XOH
. 2
Рис. 57. Схема улыраструктуры хондробласта:
1 rранулярная эндоплаэматическая сеть; 2
комплекс rольджи
Система оnорнъа и двиzaтеЛЪ1lЪLX тка1lей
117
дромукоид). Белки составляют 10 20%, а rликозаминоrликаны 80 90%. Большая
часть последних представлена хондроитинсульфатом (сульфатированным rликозами
ноrликаном), который окрашивается метахроматично. Поскольку хрящ не содержит
кровеносных сосудов, питание ткани происходит путем диффузии. Межклеточная
жидкость при этом иrpает ведущую метаболическую роль в проведении веществ к клет
кам (кислорода, ионов и др.). В центре хряща нередко создаются условия ухудшенной
трофики. В этих участках происходят rибель хрящевых клеток и межклеточноrо Be
щества и отложение солей кальция (асбестовая дистрофия хряща).
С увеличением массы межклеточною вещества синтетическая активность XOH
дробластов уменьшается. Блокируется и их способность к синтезу ДНК Хондроб
ласты превращаются в хондроциты зрелые хрящевые клетки. Последние распо
лаrаются обычно rруппами по 2, 4 или 8 клеток в общей полости. Это так
называемые изоrенные rруппы клеток, или "rнезда клеток". Как одиночные XOHk
роциты, так и их изоrенные rpуппы окружены слоем уплотненною оксифильноrо
межклеточною вещества, называемоrо "капсулой". Кнаружи от капсулы находит
ся слой базофильною вещества, содержащеrо rликозаминоrликаны, в том числе
свободную хондроитинсерную кислоту. Периферические слои этих клеточных Tep
риторий (или хондриновых шаров) снова оксифильны блаrодаря наличию здесь
rлобулярных белков.
Хрящевые клетки, располаrающиеся в rлубине развивающейся хрящевой TKa
ни, сохраняют некоторое время способность делиться митозом и синтезировать
межклеточное вещество, обеспечивая внутренний, И1lтерстициалЪ1lЫЙ, рост.
Наиболее распространена в орrанизме талИ1l0вая хрящевая ткань, определяе
мая на суставных поверхностях костей, на концах ребер, в стенке rортани и брон
хов. В нативном состоянии она выrлядит прозрачной, стекловидной. Хондрино
вые фибриллы имеют показатель преломления такой же, как и у OCHoBHoro
вещества, и потому они не видны. Выявляются эти фибриллы с помощью поляри
зационноrо микроскопа.
Эластическая хрящевая ткань встречается в ушной раковине, надrортаннике, в
составе стенки средних бронхов. В межклеточном веществе этой ткани преобладает
сеть эластических волокон. Последние
имеют толщину O,3 5 мкм и построены из
белка эластина. Эластическую хрящевую
ткань иноrда называют еще сетчатоЙ.
ВОЛОК1lистая хрящевая ткань входит
в состав межпозвоночных дисков, лонно
[о сочленения, встречается в местах при
крепления сухожилий и связок к rиали
новому хрящу и костям. Межклеточное
вещество содержит упорядоченно распо
ложенные коллаrеновые волокна, как и в
плотной оформленной соединительной
ткани, но клетки здесь хрящевые, а не
фиброциты (рис. 58). Коллаrеновые
ЧО)
",,)
. .; ,
Рис. 58. Волокнистая хрящевая ткань:
1 пучки коллаrеновых волокон; 2 хондроциты
.4';
"
Q
""
@
') ,
,'" .
,'"
,е.е
.
@
,,'"
'... '
.;...,
, " ",, ' ,j1, , . ,, ' 2" " ,
'
@ , ,
. - 1)';
z'
I
,,
,:.;)
@
118
Fлава 10
белки представлены преимущественно I bIM типом и незначительным количеством
11 ro типа.
Реzенерацuя хрящевых тканей. Хрящевые ткани способны к реrенерации. Важ
ную роль при этом иrрает llадхрЯЩlluца, rдe располаrаются камбиальные клетки. За
счет пролиферации этих клеток и их дифференцировки в хондробласты, образующие
межклеточное вещество, происходит заполнение дефекта.
Костные ткани
Костные ткани полuдuффеРОllllые ткани и состоят из клеток различной rис
тоrенетической детерминации (остеобластов, остеоцитов и остеоклаСТОБ) и очень
плотноrо межклеточноrо вещества, содержащеrо большое количество минераль
ных солей. Костные ткани выполняют опорную функцию. Они входят в качестве
rлавноrо CTPYKTYPHOro компонента Б состав скелета. Блаroдаря высокому coдep
жанию минеральных солей (до 65 70% сухой массы) костные ткани активно уча
ствуют в реryляции минеральноrо обмена. Между костными и кроветворными TKa
нями складываются особые взаимодействия, обеспечивающие блаrоприятное
микроокружение для пролиферации и дифференцировки клеток крови.
По степени упорядоченности расположения коллаrеновых волокон, которые в
костной ткани называются оссеиновыми, различают ретикулофиброзную (rрубо
волокнистую) и пластинчатую костные ткани. Кроме Toro, существует дентиноид
ная костная ткань (дентин зуба), а также цемент зуба.
ruстоzенезкостных тканей (остеоzuстоzенез). Источником развития KOC
тных тканей скелета человека служит m-езе1lXU.Ма склеротома. Костные ткани чере
па раЗБиваются из эктомезс1lXU.МЫ. Различают два способа развития костных TKa
.... .....;;", ;"" '1 ,.......,.., , ........-:--.. ней: остеоrистоrенез, протекающий
.. "'*' ..а il .. 1?JIO> """", ."..... непос р е д ственно в мезенхиме , и
;: """ .., .JIIIIi<
., .. .... """'.. остеоrистоrенез, источником KOTO
. , .' .....
, .. #,. ..,' ......,. _ poro является также мезенхима, но
.... о .., /' ' { : ' , . : Т : :уо:т:::е ; т::: а :Л=
........... '@." " '. .... . ,,1:'... #' принципиальны.
2 Остеоrистоrенез начинается с
· ,'...... А' . '... .... .. появления в мезенхиме скелетоrен
/ \ , .... ных участков с более плотным pac
.. "'.. ... ...
" ."" ........ .... положением клеток, среди которых
имеются стволовые клетки, диффе
ренцирующиеся в митотически деля
щиеся преостеобласты. Последние
., начинают вырабатывать межклеточ
ное вещество (рис. 59). Затем преос
.. rеобласты дифференцируются в oc
теобласты, которые постепенно Te
ряют способность делиться митозом.
,> , .,. _ .. ...... ...._ " . . 9.... ..,
, ,...,.. fiP fllJ ....... "-. ,{ _
.. 09 " ' .. /'"' ...........
. .. ......'.. -,А Ф,. (d ...... "
".' . '. .4 .. . "41" "C'
'* , · " / J' .......
, . .."'''' .....
Рис. 59. Развитие костной ткани:
1 остеобласты; 2 остеоциты; 3 костные балки; 4
остеокласт
.
Система оnорнъа и двиzaтеЛЪ1lЪLX тка1lей
119
Остеобласты это клетки, вырабатывающие межклеточное костное вещество.
Форма их зависит от функциональноrо состояния и бывает кубическая, цилиндри
ческая или отростчатая. Диаметр 15 20 мкм. Ядро имеет окруrлую или овальную
форму. В цитоплазме хорошо развита rpанулярная эндоплазматическая сеть, что Ha
ходится в связи С интенсивной продукцией этими клетками белков. Хорошо развит
и комплекс [ольджи, rде происходит синтез rликозаминоrликанов. В цитоплазме
остеобластов определяется высокое содержание щелочной фосфатазы. Все это сви
детельствует о высокой синтетической активности остеобластов и продукции opra
ническоrо матрикса остеоида.
Механизм внутриклеточноrо транспорта и выведения белковых макромоле
кул в остеобластах принципиально сходен с тем, что имеет место в фибробластах и
хондробластах. В общих чертах сходно протекают и первые фазы фибриллоrенеза.
Относительное количество оссеиновых (коллаrеновых) фибрилл в межклеточном
веществе костных тканей такое же, как и в хрящевых тканях, и составляет около
30% сухой массы. Оссеиновые фибриллы характеризуются высоким содержанием
орrаническоrо фосфата, что способствует процессам минерализации костной TKa
ни. Основное аморфное вещество костной ткани оссеомукоид содержит XOHk
роитинсульфаты, иrрающие роль активных накопителей и переносчиков ионов
кальция, а также белки неколлаrеновой природы (остеокальцин, остеопонтин, KO
стные морфоrенетические протеины, остеонектин и др.). Они обладают свойства
ми реryляторов минерализации, факторов роста, остеоиндуктивных веществ, ми
ToreHHbIx факторов, реrуляторов темпа образования коллаrеновых фибрилл. Это
также способствует минерализации костной ткани.
Непосредственно процесс минерализации костной ткани начинается после Ha
копления остеобластами большоrо количества щелочной фосфатазы. Под деЙстви
ем этоrо фермента rлицерофосфаты крови расщепляются на уrлеводы и фосфор
ную кислоту. Фосфорная кислота соединяется с ионами кальция, образуя
фосфорнокислый кальций, который вместе с уrлекислым кальцием формирует
кристаллы lUдроксиаnатита. Размер кристаллов: от 20 40 нм до 150 нм в длину И
от 1,5 до 75 нм в толщину. Иrольчатые и пластинчатые кристаллы апатита обнару
живаются как внутри оссеиновых фибрилл, повторяя их периодическую исчер
ченность, так и между оссеиновыми фибриллами.
Пропитанное минеральными солями межклеточное вещество костной ткани
имеет вид костных перекладин. Остеобласты располаrаются обычно на их поверх
ности. Некоторые остеобласты по мере роста и увеличения массы костной ткани
оказываются замурованными в толще костных перекладин. Здесь остеобласты пре
вращаются в зрелые высокодифференцированные клетки костной ткани ocтeo
циты. Последние имеют отростчатую форму, темное компактное ядро и слабоба
зофильную цитоплазму. Остеоциты представляют собой rетероморфную популяцию
клеток. Одни из остеоцитов имеют развитые мембранные структуры в цитоплазме
друrие находятся на различных стадиях деструкции. Остеоциты располаrаются
в костных полостях, или лакунах. Тонкие отростки остеоцитов проходят в KOCТ
ных канальцах, пронизывающих межклеточное вещество. При помощи этих Ka
нальцев происходит обмен веществ между остеоцитами и кровью.
120
Fлава 10
Остеоциты не делятся, но участвуют в процессах метаболизма, обновления меж
клеточных структур и поддержании ионноrо баланса орrанизма на определенном
уровне. Для ионноrо rомеостаза орrанизма немаловажен факт, что общая поверх
ность соприкосновения тканевой жидкости с пропитанным минеральными соля
ми межклеточным веществом костей у человека достиrает 5000 м 2 . Функция OCTe
оцитов. уже не способных вырабатывать межклеточное костное вещество, сводится
к участию в обменно транспортных процессах, реrуляции минеральноrо состава
костной ткани.
Развитие костной ткани на месте хряща протекает несколько сложнее, чем oc
теоrистоrенез, совершающийся непосредственно в мезенхиме. В этом случае раз
витию костной ткани предшествует образование хрящевой модели трубчатой KOC
ти, выполняющей опорную функцию на докостной стадии формирования скелета.
Исходными клетками являются камбиальные клетки надхрящницы адвеитuцu
алЪ1lые. При подрастании к надхрящнице кровеносных сосудов и улучшении усло
вий трофики И оксиrенации эти клетки дифференцируются не в хондробласты, а в
остеобласты, вырабатывающие межклеточное вещество ретикулофиброзной KOCT
ной ткани. Они образуют подобие костной манжетки, окружающей хрящевую MO
дель будущей трубчатой кости. Так возникает перихондральная костная ткань и
надкостница. Окруженные костной тканью хрящевые клетки, утратившие связь с
источником питания, подверrаются деrенерации. В возникшие полости деrенери
рующеrо хряща из надкостницы врастают кровеносные сосуды с расположенными
BOKpyr них камбиальными клетками. Некоторые из них превращаются в остеобла
сты, обусловливающие энхондральное развитие ретикулофиброзной костной TKa
нп. Клетки, которые замуровываются в межклеточное вещество, дифференциру
ются в остеоциты, а периферически расположенные клетки остеобласты
размножаются и продолжают синтез и секрецию компонентов межклеточноrо Be
щества. Все эти процессы первоначально протекают в середине хряшевой модели
трубчатой кости (диафизе) и распространяются в проксимальном идистальном
направлениях.
В зоне контакта хрящевой и костной
тканей можно выделить зоны неизменен
Horo хряща, размножающихся хондроци
тов, формирующих клеточные колонки,
зону деrенерации и замещения хряща KO
стной тканью. Зона размножающихся
хрящевых клеток определяет зону роста
будущей кости и важна для формирова
ния вектора роста кости.
Одновременно с формированием pe
тикулофиброзной костной ткани, coдep
жащеЙ остеобласты и остеоциты, возни
кает друrоЙ rистоrенетический тип кле
ток остеокласты (рис. 60). Это круп Рис. 60. Схема ультраструктуры остеокласта:
ные мноrоядерные (до 20 1 00 ядер) клет 1 ядра; 2 щеточная каемка
Система OпOP1l.bLX и двиютелъ1l.ЪLX тка1l.ей
121
ки размером до 100 мкм В диаметре являются производными стволовой KpOBeTBOp
ной клетки. Цитоплазма остеокластов оксифильна со слабо развитой эндоплазма
тической сетью. Хорошо развит комплекс rольджи. в цитоплазме MHoro лизосом,
содержащих кислую фосфатазу, коллаrеназу, карбоанrидразу и друrие ферменты.
Особенно MHoro лизосом в той части цитоплазмы остеокластов, которая обращена
к разрушаемой ткани. На этой поверхности имеются мноrочисленные выросты ци
топлазмы, образующие подобие "щеточной (rофрированной) каемки". Остеоклас
ты специализированы на "внеклеточной работе" лизосом: rидролитические фер
менты из них выходят и резорбируют межклеточное вещество. Методами микро
киносъемки показано, что остеокласты подверrают деминерализации и разруше
нию оссеиновые волокна и аморфное вещество, а затем макрофаrи фаroцитируют
остатки орrаническоrо субстрата. Остеокласты разрушают хрящевую ткань и pe
тикулофиброзную костную ткань, формируя каналы для врастающих сосудов и
проникновения остеобластов.
Последующие стадии rистоrенеза складываются из процессов новообразования
костной ткани, ее разрушения остеокластами и перестройки ремоделирования. Важ
ным фактором rистоrенеза пластинчатой костной ткани, входящей в состав трубчатой
кости, является вектор роста кости. Он определяет направление движения остеоклас
тов, следовательно, формирования каналов и врастание в них кровеносных сосудов
(по вектору роста). Кровеносный сосуд, в свою очередь, определяет упорядоченное
(концентрическое) расположение остеобластов BOKpyr себя. При этом остеобласты
синтезируют межклеточное вещество, оссеиновые волокна KOToporo упорядоченно (па
раллельно) располarаются возле остеобласта и при минерализации формируют KOcт
1I.ую ruюcmи1l.КУ, толщиной з 1 О мкм. Соседняя костная пластинка содержит оссеино
вые фибриллы, которые располаrаются под утлом по отношению к первым.
На протяжении rистоrенеза и всей возрастной динамики костной ткани в ней
происходит непрерывная перестройка блаroдаря соrласованной деятельности oc
теобластов и остеоцитов, образующих межклеточное вещество, а также остеоклас
тов, разрушающих костную ткань, что необходимо для процессов ее самообновле
ния. Так происходит смена rенераций костных пластинок и формирующихся
структурно функциональных единиц ocme01l.0B, достиrается упорядоченность
расположения последних, следовательно, высокая механическая прочность KOCT
ной ткани и кости как opraHa (см. кость).
Дентиноидная костная ткань отличается отсутствием тел костных клеток в TOk
ще межклеточноrо вещества. Дентин это вещество, состоящее из коллаrеновых
волокон и OCHoBHoro аморфноrо вещества, пропитанноrо минеральными солями.
Образующие дентин зуба клетки одО1l.тобласты (точнее их ядросодержащая
часть) расположены вне дентина в пульпе зуба. Дентин пронизан дентинными Ka
нальцами, в которых проходят отростки одонтобластов. Сходное строение имеет
цемент зуба.
Ретикулофиброзная (rpубоволокнистая) костная ткань характеризуется беспо
рядочным расположением оссеиновых фибрилл в виде толстых, плотных пучков BO
локон И OCHoBHoro аморфноrо вещества. Такая костная ткань образует кости в зароды
шевом и раннем постнатальном периодах. У ВЗрОCJIоrо человека она сохраняется лишь
122
[лава 10
на :\lecTe прикрепления сухожилий к
кости, в зарастающих швах черепа, а
также в составе TKaHeBoro pereHepa
та на месте переломов костей.
Пластинчатая костная ткань
отличается упорядоченным распо
ложением оссеиновых фибрилл в
составе костных пластинок Пос
ледние образуют расположенные
один за друrим слои пропитанно
ro соля:\1И кальция фибрилл, обра
зованных остеобластами. Слои
И:\fеют толщину от 3 7 до несколь t
ких сотен микрометров (рис. 61). .(
Каждая костная пластинка COCTO
ит из параллельно ориентирован
ных тонких оссеиновых (коллаrе
новых) волокон (коллаrен I ro
типа). Но коллаrеновые волокна
двух прилежащих друr к друrу KO
стных пластинок ориентированы
под разными уrлами. Костная пла Рис. 61. Пластинчатая костная ткань:
СТlIнка соединяется с соседней 1 остеоциты; 2 оссеиновые фибриллы; 3 отростки
пластинкой коллаrеновыми фиб остеоцитов
риллами. Так создается прочная волокнистая основа кости. Костные пластинки
располаrаются концентрически BOKpyr сосудов, то есть формируют ocmeoHы
структурно фvнкциональные единицы пластинчатой кости как opraHa. Кроме это
[о существуют наружные и внутренние окружающие и вставочные пластинки
трубчатой кости (см. ниже).
Реlе1lерацuя. В реrенерации костной ткани участвуют детерминированные oc
TeoreHHbIe элементы в составе надкостницы, механоциты KocTHoro мозrа, которые
размножаются и дифференцируются в остеобласты. Продуцируя межклеточное Be
щество, остеобласты дифференцируются в остеоциты и образуют ретикулофиб
розную костную ткань. Кроме Toro, адвентициальные клетки волокнистой соеди
нительноЙ ткани надкостницы также принимают участие в реrенерации костной
ткани. Однако дифференцировка их во MHoroM зависит от микроокружения, BHe
тканевых и BHeopraHHbIx факторов (например, от репозиции отломков, неподвиж
ности отломков, оксиrенации места перелома и др.).
Дифференцировка адвентициальных клеток возможна в трех направлениях:
oCTeoreHHoM, хондроrенном, фибробластическом. Этим определяется соотношение
различных видов тканей в pereHepaTe. При преимущественно остеобластическом
rистоrенезе формируется ретикулофиброзная костная ткань, которая постепенно
ремоделируется с образованием костной ткани, напоминающей по своему CTpoe
нию пластинчатую.
, .
\!
).. Ч..V '.
,. о" [,. r
JJ
r u/ З
,, /
)
1,
(
' 2
) .
(I"i. ....
,..
1
"
1'""fIitj
{ fj I ('
l
<
.'
.,......
l....
'1
";'
.
. . :
, :.
..'
.
'.
..\ 1
.
J
"'"
'-
i . 'л
'
..." 1''' <,-
r:.
,.
..:J.'
Система ОnОр1l.ъа и двиютелъ1l.ЪLX тканей
123
Ткани с двиrательной функцией
Мышечные ткани сократимые структуры, участвующие в выполнении фун
кции движения наиболее характерной функции животноrо орrанизма. Двиrа
тельные процессы мноrообразны и специфичны для различных уровней орrаниза
ции (субклеточноrо, клеточноrо, TKaHeBoro, opraHHoro и орrанизменноrо).
Источниками развития мышечных тканей и миоидных клеток позвоночных яв
ляются участки мезодермы, эктодермы и мезенхима. По классификации H.r. Хло
пина, все мышечные ткани делятся на 5 самостоятельных типов: 1) соматическоrо
типа; 2) целомическоrо типа: 3) rладкую внутренностей и сосудов; 4) нейральноrо
происхождения; 5) миоэпителиальные элементы.
Скелетная мышечная ткань
Тистozен.ез скелетной мышечной ткани позвоночных и человека это про
цесс преобразования структур от исходной клеточной формы орrанизации до сим
пластической, представляющей, по мнению А.А. Заварзина, высшую ступень диф
ференцировки определенной rистолоrической структуры. Вся скелетная мышеч
ная ткань развивается из единоrо источника миотомов мезодермы. Миотомы
мезодермы представляют собой дорсомедиальную часть сомита дорсальной мезо
дермы. Стволовые клетки миотомов (промиобласты) последовательно проходят
следующие стадии: миобластическую, миосимпластическую, мышечных трубочек
(миотуб), молодых и зрелых мышечных волокон.
Промиобласты в результате диверrентной дифференцировки дают начало сим
пластическому и клеточному (миосателлитоциты) дифферонам. Развитие этих
дифферонов протекает в единстве, в результате чеro формируется клеточно симп
ластическая система мышеч1l.0е волокно.
Симпластическая часть волокна развивается в результате слияния одноядер
ных миобластов на стадии закладки мышцы. Это клетки со слабо базофильной
цитоплазмой, немноrочисленными орrанеллами. Рецепторно трансдукторная
система миобластов позволяет узнавать друrие миобласты, выстраиваться в цe
почку, сливаться и формировать первые миосимпласты (рис. 62). В дальней
шем в периферической части миосимпласта синтезируются актиновые и мио
зиновые филаменты, которые объединяются в поперечноисчерченные
миофибриллы. Миосимпласт удлиняется, дифференцируется в мышечную TPy
бочку. В последней продолжается синтез сократительных белков, увеличивает
ся число миофибрилл, орrанелл. Кроме Toro, миосателлитоциты, которые при
лежат к покровной системе миосимпласта или мышечной трубочки, интенсивно
размножаются и после митоза одна из дочерних клеток сливается с симпласти
ческой частью формирующеrося мышечноrо волокна. Этим механизмом обес
печивает оптимальное ядерно саркоплазматическое отношение для контроля
синтеза белков.
Развивающаяся базальная мембрана покрывает снаружи как миосателлито
циты, так и симпластическую часть волокна. Так две структуры объединяются
124
Fлава 10
в единую структурно функциональную
единицу, состоящую из специализиро А
ванной на сокращении симпластичес
кой части и камбиальной клеточной.
В дальнейшем в связи с миофиБРИk
лоrенезом ядра симпласта постепенно
перемещаются на периферию, а миофиб Б
риллы занимают центральную часть.
Возникает молодое мышечное волокно.
При этом уменьшается пролифератив
ная активность миосателлитоцитов, из
меняется их структура ядро уплотня В
ется за счет появления rетерохроматина,
цитоплазма обедняется орrанеллами,
клетка уплощается и переходит в COCTO
яние пролиферативноrо покоя.
Стадия зрелоrо мышечною волокна
протекает в связи с иннервацией и xa
r
рактеризуется при обретением опреде
ленных rистохимических и физиолоrи
ческих свойств. Появляются медленные
оксидативные (красные), быстрые rли
колитические (белые, быстро утомляю
щиеся), оксидативно rликолитические
(красные, быстрые, более стойкие к
утомлению) мышечные волокна.
Строение. Структурно функциональной единицей поперечнополосатой CKe
летной мышечной ткани является мышечное волокно. Волокно может достиrать
12 см в длину, содержит большой объем саркоплазмы и сотни ядер. Каждое BO
локно покрыто сарколеммой, состоящей из двух слоев: BHYTpeHHero плазмо
леммы толщиной 8 1 О нм И внешнеrо базальной мембраны толщиной 30 40 нм.
Между плазмолеммой и базальной мембраной имеется пространство шириной
15 25 нм. Кроме тою, в базальную мембрану вплетаются ретикулярные волокна.
Значительный объем саркоплазмы занимают сократительные орrанеллы .м.и
офибрwulы. Каждая миофибрилла состоит из большоrо числа правильно чередую
щихся темных и светлых полос (дисков). В поляризованном свете темные диски
обнаруживают двойное лучепреломление, поэтому называются анизотропными (A
дисками). Светлые диски таким свойством не обладают и называются изотропны
ми (1 дисками). Каждая миофибрилла образована пучком параллельно идущих ми
офиламентов (рис. 63). А диски состоят из толстых и тонких миофиламентов, а
I диски только из тонких. Тонкие филаменты (5 8 нм) образованы белками aK
тином, тропомиозином, тропонином, а толстые (10 12 нм) миозином, белками
M и Н полос и друrими. Тонкие филаменты располаrаются между толстыми, об
разуя rексаrональное расположение.
' .
,:J .--:":":.: _ .. -.:.: . :::::.:....:.. Y/:i'
. ....,.. , ,С" ..'" ' ) .
." .__.:::... 'l. " () у.-.:..-...... .....
." "-'. ...
з
Рис. 62. rистоrенез скелетной мышечной ткани:
А миобласты; Б миосимпласт; В мышечная
трубочка; r зрелое мышечное волокно; 1 мио
сателлитоцит; 2 ядро сим пласта; 3 миофиб
риллы
Система опорных и двиzaтелъныx тканей
125
Структурно функциональ
ной единицей миофибриллы яв
ляется саркомер. Условная фор
мула саркомера 1/2 I диска +
А диск + 1/2 I диска. Линия
сшивки соседних саркомеров co 2
ответствует Z линии (телофраr
ме), которая состоит из белков
альфа актинина, десмина, вимен 3
тина. У позвоночных длина cap
комера равна 2 3 мкм. Средняя
часть миозиновоrо диска, куда не
доходят актиновые миофиламен
ты, более светлая и называется Н
полоской. Ее пересекает М линия
(мезофраrма), скрепляющая ми
озиновые нити посередине capKO
мера. В подмембранном слое сим
пласта обнаружены белки винку
лин и спектрин, входящие в co
став скелета симпласта.
Компоненты метаболической Рис. 63. Схема строения и взаиморасположения миофиб
рилл, сакоплазматической сети и Т системы в поперечно
среды симпласта хорошо выраже полосатом мышечном волокне:
ны. В rистоrенезе с возрастанием 1 миофибриллы; 2 саркоплазматическая сеть; 3
поперечная трубочка; 4 триада; 5 толстые миофила
степени зрелости симпластов Ha менты; 6 тонкие миофиламенты; 7 телофраrма; 8
блюдается увеличение числа мито мезофраrма
хондрии, которые ориентируются по бокам Z линии между миофибриллами и под
сарколеммой. rранулы rликоrена, липидные капли формируют скопления между ми
офибриллами и под сарколеммой. Содержание миоrлобина (связывающий кислород
пиrмент) варьирует в зависимости от образа жизни животноrо. Рибосомы представ
лены в виде полисом. Небольшое число лизосом принимают участие в процессах BHYТ
рисимпластической реrенерации. Клеточный центр в сим пласте отсутствует.
Саркоплазматическая сеть и Т трубочки развиваются параллельно. После
дние это инваrинации плазмолеммы, которые опоясывают каждый саркомер. В
продольном направлении BOKpyr каждой миофибриллы идут канальцы саркоплаз
матической сети. Так формируются продольная и поперечная системы, которые на
срезах видны как триады. 1jJиада это комплекс, состоящий из поперечной TPy
бочки и профилей двух цистерн саркоплазматической сети, расположенных сим
метрично по обе стороны от Т трубочки. В цистернах саркоплазматической сети
накапливаются ионы кальция, необходимые для сокращения миофибрилл.
В позднем онтоrенезе происходит ряд ультраструктурных изменений в клетках
и симпластах. Наиболее значимы утолщение базальной мембраны, дезорrаниза
ция миофибрилл и Z линии. возникновение скоплений митохондрий под сарколем
мой, отделение миосателлитоцитов от симпласта и переход их в интерстициальное
4
7
8
126
Fлава 10
пространство. И ннервация мышечных волокон осуществляется двиrательными ней
ронами передних poroB спинноrо мозrа, которые формируют нервно мышечные си
напсы примерно в центральной части волокна.
Реle'llерация. Для успешной реrенерации мышечной ткани необходимо coxpa
нение напряжения мышцы, восстановление кровоснабжения и нервной связи. Oc
новным источником реrенерации являются миосателлитоциты. После активации
последних происходит их митотическое деление, возникают миобласты, которые
претерпевают дифференцировку, сливаются друr с друrом и формируют симплас
ты. Развитие симпластов продолжается с участием размножающихся миосателли
тоциов, часть которых сливается с растущими симпластами. Так формируются HO
вые клеточно симпластические системы мышечные волокна.
Сердечная мышечная ткань
rистоzеllез. Источники развития сердечной мышечной ткани находятся в пре
кардиальной мезодерме. В rистоrенезе возникают парные складчатые утолщения
висцеральноrо листка спланхнотома миоэпикардиалъные пластинки, содержа
щие стволовые клетки сердечной мышечной ткани. Последние путем диверrент
ной дифференцировки дают начало следующим клеточным дифферонам: рабочим,
ритмзадающим (пейсмекерным), про водящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани кардиомиобласты характери
зvются рядом признаков: клетки уплощены, содержат крупное ядро, светлую ци
топлазму, бедную рибосомами и митохондриями. В дальнейшем происходит раз
витие комплекса rольджи, rранулярной эндоплазматической сети. В кардиомиоб
ластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки
обладают высоким пролиферативным потенциалом. После ряда митотических цик
лов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинает
ся саркомероrенез. В цитоплаЗ lе кардио IИОЦИТОВ увеличивается число полисом,
канальцев rранулярной эндоплазматической сети, накапливаются rранулы rлико
reHa, возрастает объем актомиозиновоrо комплекса. Кардиомиоциты сокращают
ся, но не теряют способность к дальнейшей пролиферации и дифференцировке.
Развитие сократительноro аппарата Б позднем эмбриональном и постнатальном
периодах происходит путем надставки новых саркомеров и наслоения вновь син
тезированных миофиламентов. Дифференцировка кардиомиоцитов сопровожда
ется увеличением числа митохондрий, распределением их у полюсов ядер и между
миофибриллами и протекает параллельно со специализацией контактирующих
поверхностей клеток Кардиомиоциты путем контактов "конец в конец", "конец в
бок" формируют клеточные комплексы сердечные мышечные волокна, и в цe
ЛО I ткань представляет собой сетевидную структуру.
Строение. Структурно функциональные единицы волокон кардиомиоциты
это клетки, имеющие вытянутую прямоуrольную форму (рис. 64). Длина рабочих Kap
диомиоцитов составляет 50 120 мкм, а ширина 15 20 мкм. OДHO ДBa ядра распола
l'аются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов зани
мают поперечноисчерченные миофибриллы, аналоrичные таковым в симпластах CKe
Система опорных и двиzaтелъных тканей
127
летномышечноrо волокна. Однако каналы
саркоплазматической сети и Т системы менее
отчетливо выражены. КардиомиоЦlПЫ отли
чаются большим количеством митохондрий,
расположенных тесными рядами меж.цу мио
фибриллами. Снаружи миоциты покрыты
сарколеммой, в составе которой выделяются
плазмолемма и базальная мембрана. XapaK
терной особенностью ткани является наличие
вставочных дисков на rpанице между KOHTaK
тирующими кардиомиоцитами. Вставочные
диски пересекают волокно в виде волнистой 12
или ступенчатой линии и включают межкле
точные контакты от простых, по типу дeCMO 7
сом и до щелевых (нексусов).
Часть кардиомиоцитов на ранних этапах
кардиомиоrенеза являются сократительно
секреторными. В дальнейшем в результате ди
верrентной дифференцировки возникают
"темные" (сократительные) и "светлые" (про
водящие) миоциты, в которых исчезают ceK
реторные rpанулы, тоrда как в предсердных
миоцитах они сохраняются. Так формирует
ся дифферон эндокринных кардиомиоцитов.
Эти клетки содержат центрально расположен
ное ядро с дисперrированным хроматином,
1 2 ядрышками. В цитоплазме хорошо развиты rpанулярная эндоплазматическая сеть,
диктиосомы комплекса fольджи, в тесной связи с элементами KOToporo находятся MHO
rочисленные секреторные rpанулы диаметром около 2 мкм, содержащие электронно
плотный материал. В дальнейшем секреторные rpанулы обнаруживаются под capKO
леммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный
пептидный ropMoH кардuодuлатин циркулирует в крови в виде кардионатрина, KOТO
рый вызывает сокращение rладких миоцитов артериол, увеличение почечноrо KpOBO
тока, ускоряет клубочковую фильтрацию и выделение натрия из орrанизма.
Кардиомиоциты проводящей системы rетероморфны. В них слабо развит мио
фибриллярный аппарат, расположение миофиламентов в составе миофибрилл pыx
лое, Z линии имеют неправильную конфиrypацию, эндоплазматическая сеть слабо
развита, находится на периферии миоцитов, число митохондрий незначительное. По
мере расположения этих кардиомиоцитов в проксимо дистальном направлении co
ответственно движению импульсов от синусно предсердноrо узла, через предсерд
но желудочковый узел, пучок fиса, ero ножки и клетки Пуркиня к рабочим миоци
там проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим
кардиомиоцитам.
Реzенерация. В rистоrенезе сердечной мышечной ткани специализированный
2
3
4
5
3
Рис. 64. Схема ультраструктуры сократитель
Horo кардиомиоцита:
1 сарколемма; 2 миофибрилла; 3 ми.
тохондрия; 4 поперечная трубочка; 5 ar
ранулярная эндоплазматическая сеть; 6
лизосома; 7 вставочный диск; 8 контакт
по типу щелевоrо вставочноrо диска; 9
зона прикрепления миофибрилл; 10 дec
мосома; 11 rранулы rликоrена; 12 Me
зофраrма; 13 телофраrма
128
rлава 10
камбш':', не возникает. Поэтому реrенерация ткани протекает на основе внутрикле
точных rиперпластических процессов. Вместе с тем для кардиомиоцитов млеко
питающих, приматов и человека характерен процесс полuплоuдuзацuu. Например,
у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов CTa
новятся TeTpa и октоплоидными. Полиплоидные кардиомиоциты возникают за
счет ацитокинетическоrо митоза, что приводит к мноrоядерности.
В условиях патолоrии сердечно сосудистой системы человека (ревматизм,
врожденные пороки сердца, инфаркт миокарда и друrие) важная роль в компенса
ции повреждений кардиомиоцитов принадлежит внутриклеточной реrенерации,
полиплоидизации как ядер, так и кардиомиоцитов.
fладкая мышечная ткань
rистоzеиез. Это ткань энтомезенхимноrо происхождения, которая делится на
два вида: висцеральную и сосудистую. В эмбриональном rистоrенезе даже элект
ронно микроскопически трудно отличить мезенхимные предшественники фибро
бластов от rладких миоцитов. В малодифференцированных rладких миоцитах раз
виты rpанулярная эндоплазматическая сеть, комплекс rольджи. Тонкие филаменты
ориентированы вдоль длинной оси клетки. По мере развития размеры клетки и чис
ло филаментов в цитоплазме возрастают. Постепенно объем цитоплазмы, занятый
сократительными филаментами, увеличивается, расположение их становится все
более упорядоченным. Пролиферативная активность rладких миоцитов в миоrене
зе постепенно снижается. Это происходит в результате увеличения продолжитель
ности клеточноrо цикла, выхода клеток из цикла репродукции и перехода в диффе
ренцированное состояние. Однако и в дефинитивном состоянии в rладкой мышечной
ткани клеточная реrенерация в виде размножения миоцитов полностью не прекра
щается. Существуют данные о том, что пролиферация и дифференцировка в боль
шей степени свойственна субпопуляции малых (по размерам) rладких миоцитов.
Строение. Структура дефинитивных rладких МИОЦИТОВ (леиuмиuцитов), входя
щих в состав внутренних opraHoB и стенки сосудов, имеет MHoro 06щеrо, но в то же Bpe
мя характеризуется rетероморфией. Так, в стенках вен и артерий обнаруживаются OBO
идные, веретеновидные, отростчатые миоциты длиной 1 0 40 MI<М, доходящие иноrда до
140 мкм. Наибольшей длины rладкие миоциты достиrают в стенке матки до 500 мкм.
Диаметр миоцитов колеблется от 2 до 20 мкм. В зависимости от характера внутрикле
точных биосинтетических процессов различают КОllтраюпUЛЪ1iые и секреториые мио
циты. Первые специализированы на функции сокращения, но вместе с тем сохраняют
секреторную активность. Плазмолемма расслабленной клетки имеет ровную поверх
ность, а при сокращении становится складчатой. В центре клетки имеется палочковид
ное ядро, которое при сокращении клетки спиралевидно изrибается. Практически все
ядра миоцитов содсржат диплоидное количество днк. rладкая ЭlЩоплазматическая сеть
занимает примерно 2 7% объема цитоплазмы, а rpанулярная сеть в контрактильных
миоцитах выражена плохо. МитохоlЩРИИ мелкие, сферические или овоидные, располо
жены у полюсов ядра. Характерной чертой rладких миоцитов является наличие множе
ства впячиваний (кавеол) плазмолеммы, содержащих ионы кальция.
Система опорных и двиzaтелъных тканей
129
Секреторные миоциты (синтетические) по своей ультраструктуре напомина
ют фибробласты, однако содержат в цитоплазме пучки тонких миофиламентов,
расположенные на периферии клетки. В цитоплазме хорошо развиты комплскс
rольджи, rранулярная эндоплазматическая сеть, MHoro митохондрий, rранул rли
KoreHa, свободных рибосом и полисом. По степени зрелости такие клетки относят
к малодифференцированным.
Сократительный аппарат миоцитов представлен тонкими актиновыми филамен
тами (rладкомышечным альфа актином), связанными с тропомиозином. Толстые
нити состоят из миозина, мономеры KOToporo располаrаются вблизи филаментов
актина. Соотношение актиновых и миозиновых филаментов в rладком миоците co
ставляет 12 к 1. Важным компонентом контрактильною аппарата МIIОЦИТОВ явля
ются электронно плотные структуры тельца прикрепления, расположенные CBO
бодно в цитоплазме (плотные тельца) или тесно связанные с плазмолеммой (рис. 65).
Основными белковыми компонентами плотных телец являются альфа актинин, aK
тин (немышечный) и кальпонин, что позволяет расссматривать их как функциональ
ный эквивалент Z линий миофибрилл скелетной мышцы. Актиновые филаменты
фиксируются на плотных тельцах. Промежуточные филаменты, включающие дec
мин и виментин, обеспечивают связи между плотными тельцами и плазмолеммой,
образуя прикрепительные пластины. Сократительные белки формируют решетча
тую структуру, закреплснную по окружности плазмолеМ:\IЫ, поэтому сокращение BЫ
ражается в укорочении клетки, которая приобретает складчатую форму, тorдa как в
состоянии покоя клетка вытянута. При возникновении нервною импульса, распро
страняющеrося по IIлазмолемме миоцита, происходит повышение уровня внутри
клеточноrо Са 2 +, который поступает в цитоплазму из кавеол, отшнуровывающихся в
цитоплазму в виде пузырьков. Высвобождение ионов калЬЦIlЯ приводит к каскаду
реакций, в результате KOTOpO
ro происходит полимериза
ция миозина и образование
перекрестных связей миозина
вдоль актиновых филаментов
по мере развития мышечноrо
сокрашения. Расслабление
мышцы возникает при BOC
становлении концентрации
исходноrо уровня Са 2 + BHYT
ри клетки путем ero псреме
щения внутрь саркоплазма
тической сети. При этом об
разоваnшиеся в присутствии
ионов кальция связи между
актином и миозином наруша
ются, акто миозиновый KOM
плекс распадается, rладкий
миоцит расслабляется.
А
Акто мнознновые
KOMlUleKCbl
Визнкулы
Рис. 65. Схема строения и сокращения rладкоrо миоцита:
А миоциты В состоянии релаксации; Б сокращенные rладкие
миоциты
130
rлава 10
rладкие МИОЦИТЫ синтезируют протеоrликаны, rликопротеиды, проколлаrен,
проэластин, из которых формируются коллаrеновые и эластические волокна и oc
новное вещество межклеточноrо матрикса.
Взаимодействие миоцитов осуществляется с помощью цитоплазматических MO
стиков, взаимных впячиваний, нексусов, десмосом или простых участков мемб
ранных контактов клеточных поверхностей.
Рezен.ерацuя. rладкая мышечная ткань висцеральноrо и сосудистоrо видов об
ладает значительной чувствительностью к воздействию экстремальных факторов.
В активированных миоцитах возрастает уровень биосинтетических процессов,
морфолоrическим выражением которых являются синтез сократительных белков,
укрупнение и rиперхроматоз ядра, rипертрофия ядрышка, возрастание показателей
ядерно цитоплазменноrо отношения, увеличение количества свободных рибосом и
полисом, активация ферментов, аэробноrо и анаэробноrо фосфорилирования, MeM
бранною транспорта. Клеточная реrенерация осуществляется как за счет дифферен
цированных клеток, обладающих способностью вступать в митотический цикл, так
и за счет активизации камбиальных элементов (миоцитов малоrо объема).
При действии ряда повреждающих факторов отмечается фенотипическая
трансформация контрактильных миоцитов в секреторные. Данная трансформация
часто наблюдается при повреждении интимы сосудов, формировании ее rиперп
лазии при развитии атеросклероза.
Мышечная ткань радужки и цилиарноrо тела относится к четвертому типу COKpa
тимых тканей, развивается из зачатка нервной системы. В ряду позвоночных мышеч
ные элементы радужки обнаруживают разнообразную диверrентную дифференциров
ку. Так, мионевральная ткань рептилий и птиц представлена исчерченными MHoro
ядерными волокнами, имеющими большое сходство с мускулатурой скелетноrо типа.
у млекопитающих и человека основной структурно функциональной единицей мышц
радужки является rладкий одноядерный миоцит, или м-иoпиl.Мeнmoциm (рис. 66). Пос
ледний имеет пиrментирован ." , ,
ное тело, содержащее ОДНО
ядро, вынесенное за пределы
веретеновидной сократимой
части. Цитоплазма клеток co ,'"
держит большое число мито
хондрий И пиrментные [paHY
ЛЫ, которые сходны по разме
рам и форме с rpанулами пиr
MeHTHoro эпителия.
Миофиламенты в миопиr
ментоцитах делятся на тонкие
(7 нм) и толстые (15 нм), по
размерам и расположению Ha
поминают миофиламенты
Мионевральная ткань
"'-:' " , :. ,
. t-'"
!\
""
"
""..'"
"tf!*ox, j у!..
. ,!I! ' ",,:
, '", r >
"""
.....
'
,
,
..
'""-
""
"
";::t;
-2
"1
1
"
,'"'" ""
'
t
r-r"
'''''' 1!
.., "
.
,.,.
;.....А '." .,...
Рис. 66. Ультраструктура миопиrментоцита:
1 ядро; 2 миофиламенты
Система OпOPllbLX и двиютелыlLx ткаllей
131
rладких миоцитов. Каждый миопиrментоцит окружен базальной мембраной. Возле
цитоплазматических отростков миоцитов обнаруживаются безмиелиновые нервные
волокна.
Реlенерация. В немноrочисленных работах показана низкая реrенерационная
активность после повреждения или ее отсутствие.
Миоидные клетки
Это большая rенетически разнообразная rруппа клеток Источниками разви
тия I\ПIОИДНЫХ клеток являются энтомезенхима мезодермы, эктодерма, нейроэкто
дерма и прехордальная пластинка.
Проuзводные энтомезенxuмы. Наиболее полно изучены миофибробласты. Клет
ки обнаруживаются в rpануляционной ткани заживающих ран. По ультраструктуре
характеризуются наличием в цитоплазме большоrо числа сократимых филаментов,
заНИ lающих 1/3 2/3 части объема цитоплаз:\IЫ, типичных для rладких :\ШОЦlпов плот
ных телец и палочковидноrо ядра. Миофибробласты взаимодеЙствуют путем дeCMO
сомоподобных и щелевых контактов. Цитоплазма связывает антитела к миозину rлад
ких мышц. Наряду с rладкими миофиламентами в цитоплазме миофибробластов об
наруживаются rранулярная эндоплазматическая сеть и комплекс rольджи, которые
участвуют в выработке коллаrена III ro типа.
Миоэндокринные. Это комплекс клеток, которые наряду с сократительными
филаментами в цитоплазме содержат эндокринные rранулы. Наиболее полно изу
чены миоэндокринные клетки юкстаrломерулярноrо комплекса почки, вырабаты
вающие рен ин и почечный эритропоэтический фактор. Имеются данные о нали
чии ренин секретирующих миоцитов в стенке матки.
Производные эктодермы. К этой rруппе относятся пюэпителиоциты слюн
ных, потовых, молочных желез и миоидные эпителиоциты реrенерирующеrо эпи
дер ми са. В составе желез миоэпителиоциты, как и rландулоциты, развиваются из
стволовых клеток путем диверrентной дифференцировки.
Миоидные эпителиоциты обнаруживаются в первые дни реrенерации эпидер
миса кожи в ero базальной части в области края раны. Они содержат в цитоплазме
плотно расположенные и ориентированные вдоль клетки миофиламенты, coxpa
няют связь с базальной мембраноЙ с помощью полудесмосом.
Производные нейроэктодермы. В эту rруппу включают миоидные клетки, Ha
поминающие по строению волокна скелетноЙ мускулатуры. Последние обнаруже
ны в составе шишковидной железы, мозжечка, rоловноrо мозrа и друrих. ЭТО MHO
rоядерные клетки, содержащие исчерченные миофиламенты.
К проuзводным прехордальной пластинки относятся миоидные клетки ти
муса. Последние расположены преимущественно в мозrовом веществе дольки,
миофибриллы формируют саркомеры; клетки содержат 1 2 ядра. Установлено Ha
личие большоrо числа I\IИОИДНЫХ клеток тимуса у больных миастениеЙ. Для мио
идных клеток независимо от источника развития (если он установлен) характерно
наличие в цитоплазме большою числа сократимых фила ментов rладких и попе
речноисчерченных.
132 Fлава 11
rЛАВА 11. СИСТЕМА НЕВРАЛЬНЫХ ТКАНЕЙ
И НЕйроrлия
Ткани нервной системы выполняют важнейшую функцию орrаНИЗ lа функцию
реактивности, основанную на способности нервных клеток воспринимать раздраже
ние, вырабатывать и передавать нервные импульсы. Они участвуют в получении, xpa
нении и переработке информации из внешней и внутренней среды орrанизма, обеспе
чивают реryляцию и интеrpацию деятельности всех opraHoB и систем человека.
В каждой части нервной системы клеточный состав нервной ткани и ее морфо
функциональные особенности неповторимы. Нервная ткань коры большоrо мозrа,
нервная ткань спинною мозrа, нейросекреторная ткань rипоталамуса, нервная ткань
ствола мозrа, нервная ткань веrетативных rанrлиев и друrих частей нервной систе
мы все это разновидности тканеЙ нервноЙ системы с достаточно четкими, специ
фическими (орrанотипическими) и стойко закрепленными признаками.
Особую rруппу вспомоrательных тканей в нервной системе образует нейро
rлия, или макроrлия, и ее разновидности (эпендима, астроrлия, олиrодендроrлия
и др.). Кроме Toro, в состав орrаноспецифическоЙ нервноЙ ткани входит микро
rлия, представленная диффероном макрофаrов. Мнorие авторы рассматривают
нейроrлию как составную часть нервной ткани, употребляя при этом термин "He
рвная ткань" в единственном числе.
rистоrенез
Источником развития нервной ткани и нейроrлии является нервная пластин
ка 18 суточноrо зародыша человека. После нейруляции из нее образуются нервная
трубка и rанrлиозные пластинки. Кроме Toro, в развитии черепных нервов ПрИНIf
мают участие плакоды утолщения эктодермы по бокам краниальной части бу
дущей нервной трубки. Эмбриональные зачатки состоят из малодифференциро
ванных клеток медуллобласmов (матричных клеток). На ранних этапах
rистоrенеза происходит дстерминация и диверrентная дифференцировка клеток,
в результате чею возникают два направления их развития: нейробластическое и
rлиобластическое. Из мезенхимы возникают клетки микроrлии ЦНС.
В неЙроrистorенезе различают стадии медуллобластов, неЙробласта, молодо
ro нейрона и зрелоrо неЙрона. Медуллобласты интенсивно делятся митозом. CTa
дия нейробласта характсризуется миrрацией клеток, при этом необратимо блоки
руется способность клеток к пролиферации. В цитоплазме нейробластов
определяются хорошо развитая rранулярная эндоплазматическая сеть, комплекс
rольджи и митохондрии. Начинается синтез специфических белков нсрвных кле
Система невралъных тканей и нейроlЛИЯ
133
ток, входящих в состав нейрофиламентов и микротрубочек. IIоявляется конус po
ста аксона.
Стадия молодоrо нейрона характеризуется ростом отростков, увеличением
объема клетки, образованием хроматофильной субстанции и появлением первых
синапсов. Дифференцировка нейробластов в нейроны происходит rруппами (rнез
дами), так что все их аксоны растут в виде пучка нервных волокон в одном направ
лении, образуя в дальнейшем проводящие пути и нервы.
Важной особенностью rистоrенеза является запроrpаммированная rибель нейро
нов по типу апоптоза. Например, в rистоrенезе спинноro мозrа позвоночных до 40
50% нервных клеток передних poroB rибнет после завершения пролиферативной фазы.
Самой продолжительной стадией является стадия зрелоrо нейрона, на протя
жении которой нейрон приобретает свою окончательную форму и специфическую
rистохимическую орrанизацию. Наряду с дифференцировкой нейронов происхо
дит все более rлубокая их интеrрация в составе рефлекторных дуr. Между нейро
нами устанавливаются мноrочисленные синаптические связи. Сложный характер
приобретают взаимодействия между нервными и rлиальными клетками.
Строение
Нейроны. Нервные клетки, нейроны, или нейроциты ведущий клеточный
дифферон нервной ткани. Клетки осуществляют рецепцию сиrнала, передачу ero
друrим нервным клеткам или клеткам эффекторам с помощью нейромедиаторов.
Нейроны отличаются большим разнообразием своих размеров, формы, строения,
функции и реактивности. Они занимают определенное место в составе рефлектор
ных дуr, представляющих материальный субстрат рефлексов. В связи с этим по
функциональным свойствам различают чувствительные (рецепторные), вставоч
ные (ассоциативные) и двиrательные (эффекторные) нейроны.
По rистолоrическим признакам нервные клетки подразделяются на звездчатые,
пирамидные, веретеновидные, паукообразные и др. (рис. 67). На форму клеток ВЛИЯ
ют число отростков и способы их отхождения от тела нейрона. Тело нервной клетки
содержит нейроплазму и обычно одно ядро. Размер тела варьирует в широких преде
лах от 5 до 130 мкм. Отростки имеют длину от нескольких микрометров до 1 1,5 м.
По количеству отростков выделяют нейроны униполярные (с одним OTpOCT
ком), псевдоуниполярные, биполярные (с двумя отростками) и мультиполярные
(с числом отростков более двух). Отростки нервных клеток специализированы на
выполнение определенных функций и потому подразделяются на два вида. Одни
из них называются дендритами (от dendron дерево), поскольку они сильно BeT
вятся. Эти отростки воспринимают раздражение и проводят импульсы по направ
лению к телу нейрона. Отростки друrоro вида называются аксонами. Они ВЫПОk
няют функцию отведения нервных импульсов от тела нейрона. Нервные клетки
имеют несколько дендритов, но один аксон.
Ядро нервной клетки крупное, крутлое, содержит деконденсированный хроматин.
В ядре определяется OДHO ДBa крупных ядрышка. Большинство ядер содержит дипло
идный набор хромосом. В некоторых типах нейронов (rрушевидные нейроны
134
Fлава 11
мозжечка) обнаруживаются по
липлоидные ядра со степенью
полиплOIIДИИ до 4 8 п. Ядро ней
рона осуществляет реrуляцию
синтеза белков в клетке. Для He
рвных клеток характерен BЫCO
кий уровень синтеза РНК и беk
ков. В нейроплазме имеются xo
рошо развитые элементы BHYT
рен ней метаболической среды
(rpанулярная эндоплазматичес
кая сеть с большим количеством
рибосом,МИТОХОНДРИИ, коплекс
[ольджи). При световой микро пирамидный
скопии В ней ро плазме выявляет нейрон rрушевидный
нейрон
ся хроматофильная субстанция,
или субстанция Ниссля, что свя
зано с наличием в нейроплазме
рнк. Субстанция Ниссля явля
ется основным белоксинтезиру
ющим компонентом нервной
клетки. Она располarается чаще
Bcero BOкpyr ядра, но встречает
ся и на периферии тела нейрона,
а также в дендритах. У места OT
хождения аксона (в аксонном
холмике) и по ходу аксона суб
станция НИСCJIя не определяет
ся. В зависимости от функцио
нальноrо состояния нейрона Be
личина и расположение rлыбок
субстанции Нисслямоryтзначи Рис. 67. Виды нейронов в тканях opraHoB нервной системы
тельно меняться. Исчезновение субстанции называют хроматолизом.
В цитоплазме нервных клеток выявляются компоненты опорно двиrательной
системы (микротрубочки, ПРОl\fежуточные филаменты нейрофиламенты и мик
рофиламенты). Нейрофиламенты это фибриллярные структуры диаметром 6
10 нм, состоящие из спиралевидно расположенных молекул кислых белков. Мик
ротрубочки цилиндрические структуры диаметром 24 нм. Под световым
микроскопом эти структуры не видны. Однако при импреrнации препаратов нервной
ткани солями серебра происходят аrреrация нейрофиламентов, осаждение на них
металлическоrо серебра, и Torдa нитчатые структуры становятся видимыми (рис. 68).
Такие искусственно аrpеrированные образования описаны под названием 1iейрофuб
рШUlы. Проходят они В теле нейрона в разных направлениях, а в отростках парал
лельно продольной оси, обеспечшзая ток аксоплазмы в двух направлениях. В нейро
L...J
2S.,.m
клетка зерно
мозжечка
интернейрон
бипопярный
нейрон
Система lI.евралъ1lых т-ка1lей и 1lейроlЛИЯ
135
плазме выявляются центриоли.
Основная часть белков нейроплаз
мы постоянно обновляется. Пока
зано непрерывное смещение аксоп
лазмы по направлению от тела
клетки к терминальным разветвле
ниям аксона (антероrрадный
транспорт). Токаксоплазмыпроис
ходит со скоростью около 2 5 мм в
сутки. Кроме медленноrо переме
щения аксоплазмы, существует Me
ханизм быстроrо перемещения беk
ков по отросткам нервных клеток Рис. 68. Мультиполярный нейрон:
Структурную основу быстроrо (от 1 ядро; 2 ядрышко; 3 нейрофибриллы
400 до 2000 мм в сутки) транспорта веществ от тела по отросткам составляют мик
рофиламенты и нейротрубочки. В аксонах и дендритах нейронов наблюдается TaK
же ретроrрадный транспорт, коrда макромолекулярный материал от периферичес
ких частей отростков доставляется в тело нейрона. Непрерывное обновление белков
в нервных клетках рассматривают как своеобразную модификацию физиолоrичес
кой реrенерации (внутриклеточную) в стабильной клеточной популяции нейронов.
Секреторные нейроны. В некоторых ядрах переднеrо rипоталамуса rоловно
[о мозrа (например, в супраоптических и паравентрикулярных) имеются клеточ
ные системы, состоящие из специализированных нейронов крупных ceKpeTOp
ных неЙронов. Последним присущи типичные для нейронов орrанеллы. Они
подверrаются воздеЙствию друrих нейронов через синаптические контакты. Ok
нако их ответы наряду с деполяризацией мембран и освобождением нейромедиа
тора включают также выделение в кровь или тканевые жидкости пептидных 1lей
рОlOрмои0в. По внешнему виду эти клетки сходны с мультиполярными нейронами.
Они имеют несколько коротких дендритов и один аксон. На дендритах и теле ceK
реторных нейронов выявляются мноrочисленные синапсы места переключения
импульсов от нейронов, расположенных в ядерных центрах roловноrо мозrа. В
цитоплазме и по ходу аксона секреторных нейронов определяются rpанулы нейро
секрета (например, окситоцин и вазопрессин). [ранулы неЙросекрета выводятся в
кровь или жидкость желудочков мозrа. Секреторные нейроны rипоталамуса уча
ствуют во взаимодействиях нервной и rуморальной систем реryляции.
НеЙРОlЛИЯ. В процессе развития тканей нервной системы из материала нервной
трубки, а также HepBHoro rребня происходит развитие rлиобластов. Результатом rли
областической диффсренцировки является образование неЙроrлиальных клеточных
дифферонов. Они выполняют опорную, разrраничительную, трофическую,
секреторную, защитную и друrие функции. Нейроrлия создает постоянную, стабиль
ную внутреннюю среду для нервной ткани, обеспечивая тканевый rомеостаз и HOp
мальное функционирование нервных клеток По строению и локализации клеток
различают эпендимную rлию, астроцитную rлию и олиrодендроrлию (рис. 69). He
редко эти разновидности rлии объединяют обобщенным понятием "макроrлия".
''\.
............,
1
2"" '_
"
: 1 .
! . i -....
"
............
/.
з
136
[лава 11
k".
Эпендимная rлия имеет эпители
оидное строение. Она выстилает цeH
тральный канал спинною мозrа и моз
[овые желудочки. В качестве эпендим
Horo эпителия эта разновидность
нейроrлии относится к нейроrлиаль
ному типу эпителиальных тканей. Bы
пячивания мяrкой оболочки мозrа в
просвет ero желудочков покрыты
эпендимоцитами кубической формы.
Они принимают участие в образова
нии спинномозrовой жидкости. В
стенке III ro желудочка мозrа Haxo В
дятся специализированные клетки Рис, 69. Виды rлиоцитов:
тттциты, обеспечивающие связь А волокнистый aCTpo
цит; Б протоплазма
между содержимым желудочка и KpO тический астроцит; В
вьюзасчетультрафильтрацииэлемен микроrлиоциты; r
тов спинномозrовой жидкости. олиrодендроrлиоциты;
Д Д эпендимоциты
Астроцитная rлия является
опорной структурой (каркасом) спинноrо и rоловноrо мозrа. В астроцитной rлии
различают два вида клеток: протоплазматические и волокнистые астроциты.
Первые из них располаrаются преимущественно в сером веществе мозrа. Они име
ют короткие и толстые, часто распластанные отростки. Вторые находятся в
белом веществе мозrа. Волокнистые астроциты имеют мноrочисленные OTpOCТ
ки, содержащпе арrирофильные фибриллы. За счет этих фибрилл формируются
rлиальные остов и разrраничительные мембраны в нервной системе, поrранич
ные мембраны BOKpyr кровеносных сосудов и так называемые "ножки" астроцит
ных отростков на кровеносных сосудах.
ОЛИlOдендрОlJlUЯ состоит из различно дифференцированных клеток олиrоден
дроцитов. Они плотно окружают тела нейронов и их отростки на всем протяжении
до концевых разветвлений. Есть несколько видов олиrодендроцитов. В opraHax цeH
тральной нервной системы олиrодендроrлия представлена мелкими отростчатыми
клетками, называемыми lJlиоцитами. BOKpyr тел чувствительных нейронов спинно
мозrовых rанrлиев находятся lJluоциты laНlJlUЯ (мантийные rлиоциты).
Отростки нервных клеток сопровождают нейролеммоциты, или шванновские
клетки. Источник их развития в периферических нервах, по данным некоторых
авторов, эктомезенхима нервною rребня.
Функции олиroдендроrлиоцитов мноrообразны и чрезвычайно важны для HOp
мальной деятельности нервных клеток Они обеспечивают трофику нейронов.
В единой метаболической системе "нейрон rлия" происходит взаимообмен HeKO
торыми ферментами, белками и рнк. Олиrодендроциты иrрают существенную
роль в процессах возбуждения и торможения нейронов и проведения по их OTpOC
ткам нервных импульсов. Так, нейролеммоциты совместно с отростками нейронов
образуют миелиновые и безмиелиновые нервные волокона периферической He
А
Б
11tt ..
' ..
fI
";,
........
Система 7lевралъ71ыхx т'/{аней и нейроvlUЯ
137
рвноЙ системы, выполняя при этом роль изоляторов, препятствующих рассеива
нию импульсов. Олиrодендроциты принимают участие в реrуляции водно соле
Boro баланса в нервной системе. Они MorYT набухать, перераспределять ионы и т. д.
Специализированные rлиоциты нервных окончаний участвуют в процессах рецеп
ЦИИ, а также в передаче HepBHoro импульса на рабочие структуры.
Помимо макроrлии в нервной системе есть еще МИ1<рОlЛИЯ. Источником ее разви
тия является мезeнxuма, а клетки микроrлии представляют собой rлиальные MaKpo
фаrи и относятся к нейроrлии лишь на основании rистотопоrpафии. Клетки микро
rлии MoryT размножаться, проявлять фаrоцитарную активность, синтезировать не
своЙственные орrанизму антиrены. что наблюдается при некоторых заболеваниях.
Нервные вОЛО1<:1lа и нервные о'/{ончания. Синапсы. Взаимодействия между
rлиальными и нервными клетками отчетливо проявляются в процессах развития
и структурной орrанизации нервных волокон. Нервным волокном называется OT
росток нервной клетки, окруженныЙ rлиальной оболочкой. Непосредственно сам
отросток называют еще осевым ЦШlИндром, а клетки rлиальной оболочки нейро
лем..моцuт{l;'.1.И. Различают i\Iиелиновые (мякотные) и безмиелиновые (безмякот
ные) нервные волокна.
В безмиелиновых нервных волокнах отростки нервных клеток поrружены в
уrлубления на поверхности нейролеммоцитов, имеющих вид желоба (рис. 70).
ПоrруженныЙ в тело rлиальной клетки нервный отросток оrраничен как собствен
ноЙ плазмолеммой, так и внешней мембраной нейролеммоцита. Он как бы ПОk
вешен на двухлистковой ее складке. Эти складки мембран (своеобразные ультра
структурные "брыжейки") называют меза'/{сонQМИ. БезмиеЛIIновые волокна MorYT
включать несколько осевых цилиндров.
Миелиновое нервное волокно состоит из HepBHoro отростка и неиролеммоцитов
(швашювских клеток). Осевой цилиндр не просто поrpужен в цитоплазму нейролем
моцита, а окружен спиральной слоистой оболочкой (миелином), образованной HaMa
тыванием мезаксонов неЙролем:\-юцитов при их вращении BOKpyr отростка нервной
клетки (рис. 71). В миелино "
"
вой оболочке обнаружены ли "
пиды, щелочной белок миели
на, маркерный белок S 100 и др.
Высокое содержание липидов
(почти 2/3 массы миелина) ]
выявляется при обработке пре
паратов четырехокисью осмия,
окрашивающей миелиновую
оболочку в темно коричневый
цвет. По ходу миелиновоrо BO
локна имеются сужения уз
ловъtе пepexвaтъt (перехваты
Ранвье). Они соответствуют
rранице смежных нейролем
моцитов. Каждый межузловой
2
.
4
.
..
138
Fлава 11
пульс проводится со CKOpOC
Рис, 71, Строение миелиновоrо HepBHoro волокна:
тью 1 2 м/с. Сложные взаи 1 цитоплазма нейролеммоцита; 2 осевой цилиндр; 3 узло
моотношения между HepBHЫ вой перехват (перехват РанвЬе)
ми И rлиальными клетками складываются при формировании чувствительных нервных
окончаний (реuепторов) и двиrательных нервных окончаниЙ (эффекторов).
Нервные окончания концевой аппарат нервных волокон, формирует меж
нейрональные контакты, или си'Напсы, рецептор'Ные (чувствительные) окончания
и двиютель'Ные (эффекторные) окончания.
Синапс (от synapsis соединение) специализированный для передачи He
рвных импульсов контакт между двумя нейронами или между нейроном и эффек
тором (рис. 72). Процессы возбуждения нейронов. возникновение импульсов и pac
пространение их по отросткам связаны с изменениями в плазмолемме. Она является
структурной основой возникновения и передачи потенциалов действия. Плазмо
лемма имеет существенные особенности строения и функции в участках, входя
щих в состав синапсов.
Межнейрональные синапсы
бывают нескольких видов: aKCOCO
матические (между аксоном OДHO
[о нейрона и телом друrоrо нейро
на); аксодендритические (между
аксоном одноrо нейрона и дендри
том друrоrо нейрона); aKcoaKCO
нальные (между аксонами двух
нейронов). Описаны также синап
сы соматосоматические, дeHДpo
дендритические и др.
Все синапсы по механизму пе
редачи импульсов между нервными
клетками подразделяются на 3 типа:
синапсы с химической передачей,
электротонические и смешанные си
напсы. ТипичныЙ синапс с химичес
кой передачей состоит из пресинап
тической и постсинаптической час
Teii, а также синаптической щели.
Пресинаптическая часть включает
cerMeHT оболочки волокна
представлен одним нейролем
моцптом. Мпелиновые волок
на толще безмиелиновых.
Скорость проведения HepBHO
[о и шульса по ним составля
ет s 120 м/с, Torдa как по без
миелиновым волокнам IIM
2 3
",, I
"
. ('.");-11
'."... , ..," ',," ' ..:i"
.......
А
Б
4
Рис. 72. Схема строения химическоrо синапса:
1 пресинаптическая часть; 2 синаптическая щель; 3
постсинаптическая часть; 4 синаптические пузырьки;
А постсинаптическая часть в виде однородноrо элект
ронно плотноrо уплотнения; Б то же в виде дискретных
образований
Система невральныx тканей и нейроVlИЯ
139
концевое расширение аксона, оrpаниченное пресинаптической мембраной. Специфи
ческими структурами этой части являются си-н.аптические пузырьки, содержащие ней
ромедиаторы. Пузырьки бывают со светлым и электронно плотным содержимым и
называются в связи с этим аrранулярными и rpанулярными. По форме они подразде
ляются на круrлые и уплощенные. На внутренней поверхности пресинаптической MeM
браны расположены конусовидные электронно плотные образования пресинапти
ческие уплотнения. В цитоплазме пресинаптической части имеются МИТОХOlщрии. Си
наптическая щель размером 20 30 нм содержит филаменты, связывающие наружные
слои плазмолеммы контактирующих нейронов.
Постсинаптическая часть в составе плазмолеммы BToporo нейрона имеет pe
цепторы к медиатору, который выделяется в синаптическую щель при деполяри
зации мембраны первоrо нейрона. Внутренняя поверхность постсинаптической
мембраны характеризуется наличием электронно плотноrо сл(jя цитоплазмы пo
стсинаптические уплотнения.
Механизм химической передачи нервных импульсов через синапс сводится к
тому, что содержимое синаптических пузырьков (медиатор) поступает в виде неболь
ших порций (квантов) в синаптическую щель и взаимодействует затем с рецептор
ными белками постсинаптической мембраны. Это вызывает деполяризацию мемб
раны и возбуждение следующеrо нейрона. Ультраструктурные особенности синапса
и механизм передачи импульсов определяют строryю однонаправленность переда
чи импульсов, что лежит в основе проведения импульсов по рефлекторным дуrам.
В зависимости от Toro, какое вещество выполняет роль нейромедиатора, синап
сы подразделяются на холинерrические (медиатор ацетилхолин), адренерrичес
кие (адреналин и норадреналин), дофаминерrические (дофамин), серотонинерrи
ческие (серотонин), пептидерrические (медиаторы пептиды и аминокислоты,
например, мет энкефалин, rамма аминомасляная кислота, rлицин и др.).
Нейрохимические синапсы подразделяются в функциональном отношении на
две противоположные по своему значению rруппы возбуждающие и тормозные.
Свойства этих синапсов зависят как от медиаторов, так и от ультраструктурных
особенностей синапсов. Так, некоторые медиаторы (например. rлутамат) xapaKTep
ны для возбуждающих синапсов, а в тормозных синапсах медиатором является [ам
ма аминомасляная кислота. Предполаrают, что в возбуждающих синапсах к пост
синаптической мембране прилежит электронно плотное вещество, в связи с чем
синапс приобретает асимметричное строение. В пресинаптических частях таких
соединений содержатся круrлые синаптические пузырьки. Тормозные синапсы име
ют симметричное строение. Синаптическая щель у них сужена и в пресинаптичес
кой части содержатся уплощенные синаптические пузырьки.
Кроме нейрохимических синапсов между нервными клетками (преимуществен
но между дендритами или телами нейронов) возникают электротонические синап
сы. Последние у млекопитающих встречаются редко и по строению соответствуют
щелевым контактам. Они проводят возбуждение блаrодаря формированию TpaHC
мембранных каналов коннексонов.
Каждый нейрон на своей поверхности имеет orpoMHoe количество (до 10000)
синапсов. Интеrрация импульсов в пределах отдельноrо нейрона происходит так:
140
Тлава 11
от синапсов, расположенных на дендритах и теле, по ero плазмолемме передается
импульс к аксонному холмику (reHepaTopHoMY пункту нейрона), rдe путем сумми
рования всех возбуждающих и тормозных импульсов возникает результирующий
nоmе1l.ЦИал дейсmвия.
Синаптические структуры обладают высокой чувствительностью к действию
токсических факторов, в том числе психотропных отравляющих веществ. Их из
менения иrрают важную роль в механизмах реактивности нервных тканей.
Рецепmор1l.ые нервные окончания подразделяются на две rруппы: экстерорецеп
торы, воспринимающие раздражения из внешней среды, и интерорецепторы, служа
щие для восприятия раздражен ий из внутренней среды орrанизма. В зависимости
от специфичности раздражения, воспринимаемоrо рецептором, различают следую
щие чувствительные нервные окончания: механорецепторы, барорецепторы, xeMO
рецепторы, терморецепторы, болевые рецептuры (ноцицепторы). Все рецепторы по
особенностям их строения подразделяют на свободные и несвободные нервные окон
чания. Первые из них состоят только из конечных разветвлений периферическоrо
отростка чувствительноro нейрона; вторые имеют в составе рецептора кроме He
pBHoro отростка rлиальный компонент, а нередко и соединительнотканную капсулу
(рис. 73). В последнем случае несвободные рецепторные окончания называют ин
капсулированными. К числу таких рецепторов относятся пластинчатые, осязатель
ные тельца, концевые колбы, мышечные веретена и др.
Эффекторные нервные окончания подразделяются на двиrательные и ceKpeTOp
ные. Двиrательный, или моторный, эффектор передает нервные импульсы на рабочие
opraHbI и ткани. В скелетных мышцах они называются нервно мышечными (aKCOMЫ
шечными) окончаниями (рис. 74). Терминальное разветвление аксона двиrательноrо
нейрона заканчивается булавовидным расширением. Оно соответствует пресинапти
ческой части HepBHO мышечноrо синапса, coдep
жит скопление синаптических пузырьков и мито
хондрий, оrpаничено пресинаптической мембра
ной xapaKTepHoro строения. Плазмолемма
мышечноrо волокна в этой области образует MHO
rочисленные складки и уrлубления. В постсинап
тической части волокна находится зернистая cap
коплазма с большим количеством митохондрий и
овальных ядер. Синаптическая щель имеет шири
ну 50 100 нм. У человека медиатором в HepBHO
мышечных синапсах является ацеmWlXОЛИ1l.. Mo
торные окончания в rладкой мышечной ткани, а
также секреторные эффекторы имеют вид тонких
пучков аксонов или их одиночных терминалей,
прилежащих к клеткам иннервируемых тканей.
возрасm1l.ые изменения нервных тканей
связаны с постепенным уменьшением запаса
нервных клеток, особенно чувствительных
нейронов, а также снижением уровня метабо
Рис. 73. Схема строения инкапсулиро
BaHHoro HepBHoro окончания:
1 отросток нервной клетки; 2 rли
альный компонент капсулы; 3 соеди
нительнотканный компонент капсулы
Система llеврШZЪ1iЪLX т'Каllей и 1I.ейроlЛИЯ
141
лических процессов, что выражается в законо
мерном накоплении включениЙ липофУСЦИllа
("пиrмент изнашивания") в нейроплазме.
Реlеllерацuя. Нервные ткани в составе op
rаНОБ относятся к стабильным тканям, так как
нейроны митозом не делятся. Физиолоrическая
и репаративная реrенерация происходит путем
частичной полиплоидизации ядер, BOCCTaHOB
ления синапсов после их повреждения, роста
поврежденных отростков, а rлавное путем об 4 5
новления химических и метаболических компо
Рис. 74. Схема ультраструктуры HepBHO
нентов нейронов при внутриклеточном обмене мышечноrо синапса:
веществ. На месте дефекта в нервной ткани раз 1 отросток MOTopHoro нейрона; 2 ней
ролеммоцит; 3 синаптические пузырьки;
растается lIейроrлия. Она является менее диф 4 пресинаптическая часть; 5 синапти
ференцированной тканью, клетки которой спо ческая щель; б постсинаптическая часть;
собны делиться митозом. Описаны rлиальные 7 саркоплазма мышечноrо волокна
клетки, обладающие высокими потенциями к размножению и развитию. Эти клет
ки принимают активное участие в восстановительных процессах нервноЙ ткани.
При повреждениях, приводящих к нарушению целостности нервных волокон
(оrнестрельные раны, разрывы), их периферические части распадаются на фраr
менты осевых цилиндров и миелиновых оболочек, поrибают и фаrоцитируются
макрофаrами (уоллеровская деrенерация осевых цилиндров). В сохранившейся ча
сти HepBHoro волокна начинается пролиферация нейролеммоцитов, формирующих
цепочку (бюнrнеровская лента), вдоль которой происходит постепенный рост oce
вых цилиндров. Таким образом, нейролеммоциты являются источником факторов,
стимулирующих рост oceBoro цилиндра. При отсутствии препятствий в виде оча
rОБ воспаления и соединительнотканных рубцов возможно восстановление иннер
вации тканей. Реrенерация нервных отростков идет со скоростью 2 4 мм в сутки.
В условиях лучевоrо воздействия происходит замедление процессов репаративно
ro rистоrенеза, что обусловлено в основном повреждением нейролеммоцитов и
клеток соединительной ткани в составе нерва. Способность нервных волокон к pe
rенерации после повреждения при сохранении целостности тела нейрона исполь
зуется в микрохирурrической практике при сшивании дистальноrо и проксималь
HOro отростков поврежденноrо нерва. Если это невозможно, то используют протезы
(например, участок подкожной вены), куда вставляют концы поврежденных He
рвов (футлериз). Реrенерацию нервных волокон ускоряет фактор роста нервной
ткани вещество белковой природы, выделенное из тканей слюнных желез и из
змеиноrо яда.
3
2
""tQj]) '.",
,: ': @;9::'
""',:: ',::
==- ='
142 Fлава 12
fЛАВА 12. РЕАКТИВНОСТЬ И РЕfЕНЕРАЦИЯ ТКАНЕЙ
Реактивность тканей
Важнейшее свойство живоrо отвечать комплексом изменений метаболизма,
двиrательной активности, размножения или rибели и др. на внешние и BHYTpeH
ние воздействия. Восприятие, трансформация и передача последних осуществля
ется рецепторно трансдукторной системой клетки, элементы которой связаны с
внутренней средой клетки. На клеточном уровне орrанизации живоrо первично
формируется каскад реакций, распространяющихся на тканевый, орrанный и opra
низменный уровни.
Под реактивностью тканей с rистоrенетических позиций следует понимать из
менения основных закономерных сторон развития ткани пролиферации, диффе
ренцировки, интеrрации клеток, межклеточных взаимодействий и друrих законо
мерных процессов rистоrенеза, происходящих под действием внешних для ткани
факторов. При самых разнообразных воздействиях (травмы, ожоrи, стрессовые си
туации и т. п.) ткани, для которых в норме характерно клеточное обновление путем
митоза, реаrируют прежде Bcero понижением или повышением степени пролифера
тивной активности клеток Уrнетение митозов или, наоборот, "вспышки" митоти
ческой активности непременные показатели реактивности таких тканей. В HeKO
торых тканях результатом реактивноrо изменения пролиферации клеток являются
эндомитоз и амитоз, образование двуядерных и мноrоядерных клеток
Наряду с повышением уровня дифференцировки клеток, реактивность тканей
может характеризоваться и явлениями дедифференцировки клеток Дедифферен
цировка клеток это упрощение их внутренней структуры, связанное с BpeMeH
ным снижением уровня дифференцировки (специализации) клеток Дедифферен
цированные клетки приобретают способность к пролиферации. В них активизи
руется синтез ДНК и общих неспецифических белков. Дедифференцировка как
реактивно приспособительное изменение клеток сопровождается увеличением
относительных объемов ядер и ядрышек, увеличением числа свободных рибосом,
исчезновением специальных орrанелл и включений в цитоплазме, редукцией MeM
бран комплекса rольджи, уменьшением числа митохондрий и друrими признака
ми. Под дедифференцировкой следует понимать структурно адаптационную pe
акцию клетки, а не обратное ее развитие.
При изменении функциональной наrрузки на ткани реактивно приспособитель
ные изменения клеток проявляются в метаболических сдвиrах трофических Ha
рушениях типа rипертрофии, дистрофии, атрофии, rипо или rиперсекреции и т. д.
Реактивность и реzенерация тканей
143
При rипертрофии клеток наблюдается rиперплазия внутриклеточных структур (ми
тохондрий, эндоплазматической сети, орrанелл) как морфолоrический эквивалент
повышенной функциональной активности.
Реактивность тканей характеризуется также изменениями межклеточных вза
имодействий. При действии повреждающих факторов в тканях может возникнуть
состояние дезинтеrрации клеток (например, нарушение межнейронных связей при
интоксикациях, дискомплексации эпителиев при нарушении контактов между эпи
телиоцитами и т. д.). Как проявление реактивности тканей следует рассматривать
прorраммированную rибель клеток за счет активации внутренней проrраМ:\1Ы ca
моуничтожения или ее запуска внешними стимулами, например факторами, Bыpa
батываемыми клетками микроокружения.
Реактивные изменения тканей зависят, конечно, во MHoroM от силы раздражите
ля повреждающеrо фактора. Существенное значение в реактивности тканей име
ет наследственная природа клеток самой ткани, так как каждая ткань отличается
rенетически детерминированным диапаЗОНО:\1 изменчивости или нормы адаптивной
реакции, обеспечивающей нормальное функционирование ткани. После воздействия
повреждающих факторов реактивные изменения клеток MOryT выходить за пределы
диапазона изменчивости, xapaKTepHoro для нормальноrо функционирования ткани.
Однако обычно резко выраженные вначале реактивные изменения клеток постепен
но сrлаживаются, и структурно функциональные свойства их приближаются к HOp
ме, обеспечивая адаптацию ткани к функционированию в новых условиях.
Физиолоrическая реrенерация тканей
На протяжении всей жизни орrанизма в тканях происходят процессы изнаши
вания и отмирания клеток (физиолоrическая деrенерация) и замены их новыми (фи
зиолоrическая реrенерация). Физиолоrическая реrенерация может быть внутрикле
точной (обновление орrанелл) и клеточной (обновление на уровне клеток за счет
пролиферации камбиальных или дифференцированных клеток). Для каждой ткани
характерны специфические особенности морфолоrических проявлениЙ физиолоrи
ческой реrенерации на клеточном и субклеточном уровнях. Если понимать физио
лоrическую реrенерацию тканей как процесс клеточноrо обновления, то к лабиль
ным (или обновляющимся) тканям следует отнести кроветворные ткани, кишечный
эпителий, эпидермис, рыхлую соединительную ткань и некоторые друrие. Для них
характерен высокий уровень пролиферативной активности клеток.
Ряд тканей отличаются сочетанием клеточной и внутриклеточноЙ форм физио
лоrической реrенерации (эпителий печени, почек, леrких, эпителии эндокринных
opraHoB, rладкая мышечная ткань и друrие).
Сердечная мышечная ткань и нервная ткань характеризуются внутриклеточной
формой физиолоrической реrенерации. В этих тканях, не имеющих камбиальных
клеток, происходит непрерывное обновление внутриклеточных ультраструктур.
Физиолоrическая реrенерация тканей это одно из проявлений сложноrо про
цесса постнатальноro rистоrенеза. Для физиолоrической реrенерации свойственна
rенетическая детерминированность составляющих ее процессов пролиферации
144
Тлава 12
клеток, их дифференцировки, роста, интеrpации и функциональной адаптации. За
кономерности постнатальноrо rистоrенеза обусловливают не только физиолorичес
кую реrенерацию тканей, но и все стороны их возрастной динамики.
Реrенерационный rистоrенез
В ответ на действие экстремальноrо фактора и нарушение тканевой орrаниза
ции opraHa возникает комплекс реакций с вовлечением всех структурных уровней
орrанизации живоrо. Можно лишь условно выделить те процессы, которые xapaK
терны для TKaHeBoro уровня а именно, процессы реrенерационноrо rистоrенеза.
Сразу же после повреждения в тканях развиваются реактивные изменения, co
провождающиеся нарушениями пролиферации, дифференцировки и интеrрации
клеток Если поврежденные клетки не адаптируются к новым условиям, наступает
их распад, rибель и элиминация. Формы проявления реrенерационноro rистоrене
за (например, клеточное размножение или rиперплазия внутриклеточных CTPYK
тур) обусловлены закономерными процессами эмбриональноrо rистоrенеза и спе
цифичны для каждой ткани. В обновляющихся тканях, для нормальноrо rистоrенеза
которых характерна пролиферация клеток путем митоза, и в процессах pereHepa
ции основная роль принадлежит митотическому делению клеток Реrенерацион
ный rистоrенез растущих тканей включает процессы как клеточной пролифера
ции, так и внутриклеточноrо увеличения структурных компонентов (орrанелл).
Реrенерационный rистоrенез стационарных тканей происходит за счет внутрикле
точных репаративных процессов (увеличение количества орrанелл, рост OTpOCT
ков И образование синаптических структур в нервных клетках).
Таким образом, изучение условий успешной реrенерации тканей возможно на
путях более rлубокоrо изучения rистоrенезов, ибо оптимизация посттравматичес
кой реrенерации должна проводиться с учетом особенностей физиолоrической
реrенерации конкретной ткани. Так, например, бесполезно стимулировать нейро
ны к митозу, если этот процесс им несвойственен. Напротив, стимуляция митозов
в обновляющихся тканях вполне оправданна.
В поврежденном opraHe процесс реrенерации включает всеrда комплекс меж
тканевых взаимодействий (корреляций). В ходе реrенерации складываются слож
ные взаимоотношения между эпителиями, соединительными и нервными тканя
ми. Воспалительные разрастания соединительной ткани в значительной степени
определяют исход восстановительноrо процесса. Взаимодействия различных TKa
ней с нервной, эндокринной, сосудистой, иммунной системами оказывают суще
ственное влияние на характер их реактивности и реrенерации.
Ткани, являясь составными частями opraHoB, в своих реrенеративных процес
сах подчинены не только собственно тканевым, но и opraHHbIM закономерностям.
Реализация способностей тканей к посттравматической реrенерации осуществля
ется в системе opraHa на основе межтканевых корреляций.
ЧАСТНАяrистолоrия
145
ВВЕДЕНИЕ В rистолоrию OprAHOB
И СИСТЕМ ЧЕЛОВЕКА
Частная rистолоrия это учение о тканях и особенностях их развития, CTpoe
ния, функций и межтканевых взаимодеЙствиЙ в составе opraHoB и систем opraHoB.
Орrаноrенез протекает на основе взаимодсЙствия rистоrенезов, в процессе KOToporo
формируются структурно функциональные единицы opraHa. Для обозначения пос
ледних предложен ряд понятиЙ (функциональные элементы, микрорайоны, реrио
ны, модули, ансамбли, rистионы и др.). Поскольку в состав структурно функцио
нальных единиц opraHoB входят клетки разной тканевой природы и разные ткани,
необходимы знания о тканевой специфике, источниках их развития, этапах rистоrе
неза и реrенерации.
Функции каждorо opraHa мноrообразны. Но всеrда существует ведущая, KOTO
рая осуществляется в рамках четырех основных свойств живоrо, рассмотренных
как в курсе цитолоrии, так и общей rистолоrии. Этот же принцип сохраняется в
курсе частноЙ rистолоrии. Все opraHbI человека можно объединить в 4 ведущие
функциональные системы, в состав которых входят комплексы opraHoB, связан
ных между собой общностью выполняемой ведущей функции: это система покро
ва орrанизма и производные, система opraHoB внутренней среды, связанная с BЫ
полнением метаболических функций, система opraHoB, обеспечивающих реактив
ные свойства, опорно двиrательная система человека.
В отдельную rруппу выделяются opraHbI репродуктивной системы человека,
которые рассматриваются в заключительном разделе "Эмбриоrенез человека".
146 Fлава 13
rЛАВА 13. СИСТЕМА ПОКРОВА
Кожа
OpraHoM системы покрова орrанизма является -кожа. Ее общая площадь у взрос
лоrо человека составляет около 1 ,5 2 м 2 . Ведущая функция кожи барьерная, пре
дохранение rлубжележащих тканей и opraHoB от механических, химических, фи
зических и друrих внешних воздействий. Эта функция осуществляется с помощью
различных механизмов. Кислая реакция эпидермиса и плотность ею poroBoro слоя
препятствуют проникновению в орrанизм микробов. Мощный роrовой слой зат
рудняет всасывание и потерю орrанизмом жидкости, что крайне важно для под
держания физиолorическоrо водно электролитноrо баланса (при поражении кожи
при обширных ожоrах наступает быстрое обезвоживание орrанизма). Вещества,
растворимые в липидах (хлороформ, эфир, фенол), а также отравляющие веще
ства кожною действия (иприт, люизит и др.), напротив, сравнительно леrко про
никают в кожу, т. К. хорошо смешиваются с кожным салом, покрывающим поверх
ность кожи, и липидной субстанцией, располаrающейся между роrовыми
чешуйками эпидермиса. Блаrодаря наличию в эпидермисе меланоцитов кожа за
щищает более rлубоко расположенные ткани от неблаrоприятноrо влияния ульт
рафиолетовых лучей. Защитная функция кожи проявляется также в аллерrичес
ких реакциях при попадании в орrанизм чужеродных белков антиrенов.
Кроме ведущей барьерной функции, кожа участвует в защите орrанизма от
rипо и rипертермии. Около 82% всей тепловой отдачи орrанизма происходит че
рез кожную поверхность путем конвекции, лучеиспускания, испарения. При Hapy
шении функции термореrуляции (например, при длительной работе в резиновом
комбинезоне) возникает переrpевание орrанизма (тепловой удар). Кожа участву
ет в выполнении выделительной функции вместе с потом через кожу в сутки
удаляется около 500 мл воды, а также ряд продуктов обмена веществ (белки, хло
риды, молочная кислота и др.). Экскреторная функция кожи усиливается при бо
лезнях opraHoB выделительной системы.
Наличие в коже сосудистых сетей позволяет ей выполнять роль депо крови
сосуды дермы в случае их расширения MOryT вместить до 1 л крови. Кожа прини
мает активное участие в обмене витаминов. Под действием ультрафиолетовых лу
чей в эпителиоцитах синтезируется витамин Д, препятствующий развитию рахи
та. Кожа представляет собой orpoMHoe рецепторное поле в ее отдельных участках
на 1 см 2 находится около 200 болевых, 25 тактильных, 17 терморецепторов.
С лечебной целью используется тесная взаимосвязь различных топоrрафичес
ких областей кожи с внутренними орrанами при физиотерапии, иrлоукалывании,
Система покрова
147
массаже. По этой же причине при патолоrии различных opraHOB возникает rипер
чувствительность в тех или иных участках кожи зонах Захарьина [еда. Мноrие
кожные и инфекционные заболевания сопровождаются появлением характерных
сыпей, имеющих важное диаrностическое значение.
Кожа и особенно ее производные (волосы, молочные железы) участвуют в раз
витии вторичных половых признаков.
Кожа образует производные, или придатки. в виде желез (потовых, сальных,
молочных), волос и ноrтей.
В различных областях тела кожа отличается по толщине, окраске, рельефу повер
хности, степени развития волос, содержанию потовых и са.1Jьных желез, микроскопи
ческой структуре тканей. Различают толстую и тонкую кожу. Толстая кожа покрывает
лишь ладони и подошвы, остальная поверхность тела покрыта тонкой кожей. Следует
отметить, что указанные понятия отражают лишь толщину эпидермиса, а не всей кожи
в целом. В среднем толщина кожи колеблется от 0,5 мм до 4 5 мм.
ruстоzеllез. В коже как в opraHe выделяют два слоя: наружный, которыЙ обра
зован мноrослойным ПЛОСКИ 1 ороroвеваЮЩИ 1 эпителием эпидер шсом, и BHYT
ренний, состоящий из волокнистоЙ соединительноЙ ткани. ПоследниЙ именуется
собственно дермой. В эпидермисе ведущим клеточным диффероном является эпи
телиальный, который развивается из кожной эктодермы. Источником развития
тканей дермы служит мезенхима дерматомов сомитов.
В первые недели Э lбриоrенеза эпителий кожи состоит из одноrо слоя плоских
клеток, к концу 2 ro месяца становится двухслойным, а на 3 I месяце мноrослоЙ
ным. На 3 M месяце эмбриоrенеза вследствие усложнения эпидермо дермальных вза
имодействий в коже появляются базальная мембрана, зачатки желез, волос, ноrтеЙ.
Параллельно с развитием эпидермиса осуществляются rистоrенетические процес
сы в соединительной ткани кожи, формируются слои дермы (сосочковый и сетча
тый), возникает подкожная жировая клетчатка (rиподерма).
В течение 3 ro и 4 ro месяцев на ладонях и подошвах, включая пальцы, появ
ляются поверхностные rребешки и бороздки. Характер возникающих при этом узо
ров имеет наследственную при роду и не меняется в течение жизни человека. Это
используется в методе дактилоскопии, применяемом в криминалистике. На 3 4 M
месяцах в эпидермис проникают пиrментные клетки меланобласты и меланоци
ты, происходящие из материала HepBHoro rребня, затем производные стволовой
кроветворной клетки клетки Ланrерrанса, а на 5 M месяце подрастают нервные
окончания чувствительных нейронов, позиционную информацию которьш пере
дают клетки Мсркеля нейроэндокринные клетки АРUD серии. .
[исто и орrаllоrенез кожи и ее производных характеризуется индуктивными
взаимодействиями эпидермиса и дермы. Вначале в качестве индуктора выступает
мезенхима, затем усиливается роль эпидермиса, приводящая к значительному yc
ложнению структуры и функции кожи как opraHa.
Строе1lие. Эпидер 1ИС полидифферонная ткань. Эпителиоциты формиру
ют мноrослойный ороrовевающий пласт клеток, в котором различают два OCHOB
ных функциональных слоя ростковый и рorовой. Эти слои подразделяются на
большее количество клеточных слоев в зависимости от толщины эпидермиса и
148
rлава 13
друrих rистотопоrрафических
особенностей кожи. В lIаибо
лее толстом эпидермисе ладо
ней и подошв, испытьшающем
сильные воздействия внешних
факторов, насчитывается 5
слоев клеток: базальный, ши
поватыЙ, зернистыЙ, блестя
щиЙ (выявляется только при
световой микроскопии) и po
rовой (рис. 75). В составе TOH
коЙ кожи блестящий слой OT
сутствует.
В эпидермисе присутству
ют 4 клеточных дифферона:
эпителиоциты, меланоциты,
клетки Ланrерrанса и клетки
Меркеля. Около 85% от обще
[о числа клеток составляют
эпителиоциты, или кератино
циты. Подразделение эпидер
миса на слои обусловлено Bep
тикально направленноЙ диф
ференцировкоЙ эпителиоци
тов в составе эпидеРМOJlЪ1IO пролифератИ61IOЙ едИ1lИЦЫ rистиона эпидермиса. Пос
ледниЙ представлен колонкой эпителиоцитов всех слоев эпидермиса, возникших
в результате пролиферации и дифференцировки одной стволовой клетки. Про
странственно rистион имеет вид шестиуrольной колонки и по площади равен ok
ноЙ роrовой чешуйке. В rистионе также присутствует клетка Ланrерrанса. Эпите
лиальные клетки rистиона имеют рецепторы к ряду факторов, реrулирующих их
развитие эпидермальному фактору роста и фактору роста кератиноцитов, сти
мулирующим пролиферацию, к кеЙлонам, тормозящим пролиферацию эпителио
цитов.
Мела1l0циты отростчатые пиrментные клетки нейроrлиальной природы,
расположенные в базальном слое эпидермиса. Их предшественниками являются
клетки меланобласты, миrрирующие в состав эпителия из HepBHoro rребня и моза
ично встраивающиеся среди эпителиоцитов. Количество меланоцитов может дoc
тиrать 10% и более от общеrо числа клеток базальноrо слоя. В их цитоплазме BЫ
являются rранулы темно коричневоrо пиrмента меланина, а цитохимическим
маркером меланоцитов служит фермент тирозиназа. Синтез меланина происхо
дит в специальных мембранных орrанеллах меланосомах. КожныЙ пиrмент из
меланоцитов выделяется в межклеточное пространство и эндоцитозом поступает
в эпителиоциты, концентрируясь в зоне ядер и защищая клетки от MYTareHHoro
влияния ультрафиолетовых лучей. Меланин может также передаваться по OTpOCT
.,,'0
,
8
Рис, 75. Строение эпидермиса и дермы толстой кожи:
1 базальный слой эпидермиса; 2 шиповатый слой; 3 зер
нистый слой; 4 блестящий слой; 5 роrовой слой; б co
сочковый слой дермы; 7 сетчатый слой; 8 потовая железа;
9 подкожная жировая клетчатка
Система покрова
149
кам в клетки подлежащей соединительной ткани дермы в меланодесмоциты, или
меланофоры. Меланин существует в двух формах эумеланина (черноrо) и фео
меланина (красною). Первый является фотопротектором. Феомсланин не облада
ет таким свойством и преобладает в коже рыжеволосых людей, которые чувстви
тельны к УФ облучению.
Количество пиrмента меняется в зависимости от мноrих внешних и внутренних
факторов. Сильная защитная пиrментация кожи (например при заrаре) развивается
при действии ультрафиолетовых лучей. Пиrментация наблюдается также при бере
менности. В условиях патолоrии из меланоцитов образуются злокачественные опухо
ли меланомы.
КлеткиЛанzерюнса (внутриэпидермальные макрофаrи, антиrенпредставляющие
клетки) это отростчатые клетки, расположенные в ростковом слое эпидермиса.
Они имеют костномозrовое происхождение, способны миrpировать из эпидермиса
в дерму и реrионарные лимфатические узлы, участвуя, таким образом, в формиро
вании иммунных реакциЙ. Признается также их влияние на численность эпителио
цитов в составе эпителиальноrо rистиона. С возрастом количество клеток Ланrер
raHca в эпидермисе уменьшается вплоть до полноrо их исчезновения.
Клетки Меркеля (осязательные) окруrлой или овальной формы, располаrа
ются в базальном слое эпидермиса, участвуют в осуществлении кожной чувстви
тельности. Имеют нейроrлиальную природу и проникают в эпидермис вместе с
врастающими чувствительными нервными волокнами. Встречаются в rлубоких
слоях эпидермиса человека (в основном в коже пальцев, кончика носа, эроrенных
зонах). По размерам превосходят эпителиоциты и пальцевидными выростами по
средством десмосом контактируют с ними. Цитоплазма клеток светлая, с YMepeH
ным количеством орrанелл, в базальной части содержит осмиофильные rранулы.
К клетке подходят чувствительные нервные окончания, и формируется комплекс
клетки Меркеля с нервной терминалью. Помимо рецепторной функции, клетки
Меркеля синтезируют нейропептиды (эндорфины, мет энкефалин, вазоактивный
кишечный полипептид и друrие rруппы интерлейкинов), стимулирующие иммун
ные процессы в орrанизме. Поэтому данные клетки относят к диффузной нейро
эндокринной системе орrанизма.
Покровные свойства кожи формируются эпидермальным диффероном за счет
присущих ему механизмов физиолоrической реrенерации. Друrие клеточные диф
фероны с их иммунными, иммуномодулирующими, рецепторными, защитными
свойствами, необходимы для функционирования эпителиоцитов и кожи в целом.
Дерма (собственно кожа). Эта часть кожи имеет толщину 1 2 мм (на подошвах
и ладонях 3 мм) и состоит из двух соединительнотканных слоев сосочковою и
сетчатоrо. Под дермой располаrается подкожная жировая клетчатка zиподерма.
Сосочковый слой образован рыхлой волокнистой соединительной тканью, Bдa
ющейся в эпидермис в виде сосочков. От эпидермиса соединительная ткань сосоч
KOBoro слоя отrраничена базальной мембраной. В сосочковом слое содержатся клет
ки ведущеrо фибробластическоrо ряда и друrие клеточные диффероны (макрофаrи,
тканевые базофилы, пиrментные клетки меланофоры), присущие рыхлой волок
нистой соединительной ткани. В межклеточном веществе рыхлоЙ соединительной
150
rлава 13
ткани сосочковоrо слоя беспорядочно располаrаются тонкие коллаrеновые, рети
кулярные и эластические волокна. В сосочковом слое MHoro кровеносных сосудов,
обеспечивающих трофику эпидермиса. Есть тут и пучки rладких мышечных кле
ток, сокращение которых обусловливает явления так называемой "ryсиной кожи".
При образовании "rусиной кожи" уменьшается приток крови к коже и понижается
отдача тепла. Сосочковый слой дермы определяет рисунок rребешков и бороздок
на поверхности кожи.
Сетчатый слой дермы образован плотной волокнистой неоформленной соеди
нительной тканью. Мноrочисленные пучки коллаrеновых волокон формируют
переплетения наподобие сети, строение которой зависит от функциональной Ha
rрузки на кожу. Сетчатый слой сильно развит в участках кожи, испытывающих
постоянное давление, и менее развит в тех участках, rде кожа подверrается значи
тельному растяжению. Сетчатый слой обусловливает прочность всей кожи. Кле
точный состав сетчатоrо слоя менее разнообразен, чем сосочковоrо слоя. Здесь в
основном встречаются клетки фибробластическоrо дифферона (фибробласты,
фиброциты). В сетчатом слое находятся концевые (секреторные) отделы потовых
и сальных желез, а также корни волос. Пучки коллаrеновых волокон из сетчатою
слоя продолжаются в подкожную жировую клетчатку. Толщина последней варьи
рует в различных участках тела и у разных людей, достиrая иноrда З 10 см и бо
лее. Функции подкожной жировой клетчатки депо жировой ткани, амортизация
кожи при механических повреждениях, участие в термореryляции.
Кровоснабжение кожи обеспечивается развитием нескольких сосудистых спле
тений, залеrающих на разных уровнях. Различают rлубокую (на rранице подкож
ной жировой клетчатки и сетчатоrо слоя) и поверхностную (подсосочковую) apTe
риальные сети. От подсосочковой сети отходят капилляры, снабжающие кровью
сосочковый слой дермы. Капиллярные сети окружают корни волос, потовые и саль
ные железы. Из капилляров кровь поступает в венозные поверхностные и rлубо
кие подсосочковые сплетения и далее в rлубокое дермальное венозное сплете
ние. Лимфатические сосуды образуют также два сплетения.
Иннервация кожи осуществляется разветвлениями цереброспинальных и Be
rетативных нервов, образующих субэпидермальное и дермальное нервные сплете
ния. В коже находится orpoMHoe число чувствительных нервных окончаний. CBO
бодные нервные окончания являются термореuепторами и ноuиuепторами (боле
выми рецепторами). В дерме располаrается большая rруппа инкапсулированных
нервных окончаний (пластинчатые тельца, концевые колбы, осязательные тельца
и др.), выполняющие функцию механореuепции.
Производные кожи
Потовые железы
Потовые железы производные эпителиальноrо дифферона, простые нераз
ветвленные трубчатые железы. Состоят из длинноrо конuевою ceKpeTopHoro OTдe
ла, закрученною в виде клубка, и прямоrо или слеrка извитоrо выводною протока.
Система покрова
151
Концевые отделы желез находятся в rлубоких частях сетчатоrо слоя дермы на
rранице этоrо слоя с подкожной жировоЙ клетчаткой. Выводные протоки OTKpЫ
ваются на поверхности эпидермиса потовой порой или впадают в волосяные BO
ронки вместе с выводными протоками сальных желез.
Потовые железы закладываются как впячивания эпидермиса в подлежащую
соединительную ткань. Вначале эти тяжи не имеют просветов, последние появля
ются лишь на 7 M месяце эмбриоrенеза. Потовые железы начинают функциониро
вать после рождения. В концевом отделе потовой железы, диаметр KOToporo около
30 мкм, эпителиальную стенку формируют дпффероны ЭК30КрИ1l0цитов (ceKpeTOp
ных клеток) и миоэпителоцитов (способных к сокращению). Первые имеют куби
ческую или коническую фОр:\1У. В их апикальной части накапливаются вакуоли с
жидким содержимым. Различают светлые и темные экзокриноциты. Первые Bыд
ляют преимущественно воду, вторые орrанические вещества. Для большинства
потовых желез характерен мерокринный тип секрецип.
Миоэпителиоциты в концевых отделах потовых желез располаrаются между
основаниями секреторных клеток и базальной мембраной. За счет сокращения этих
клеток облеrчается выведение секрета.
Стенка выводноrо протока состоит из двухслойноrо кубическоro эпителпя. Про
низывая эпидермис с плотно расположенными клетками, выводноЙ проток приоб
ретает штопорообразныЙ ход. В этой части ero стенка образована плоскими клетка
ми. В коже человека насчитывается 3 млн потовых желез, а в коже ладоней и подошв
на 1 см 2 поверхности открывается свыше 300 желез. Общая секреторная поверхность
эпителия всех потовых желез составляет примерно 5 м 2 . Пот содержит 98% воды и
2% орrанических и неорrанических веществ (хлориды, мочевина, аммиак, молочная
кислота и др.). За сутки выделяется 500 600 мл пота. Однако при тяжелой работе, в
сильную жару и при лихорадке количество выделяемоrо пота может возрастать до
2 4 и даже до 10 л. В условиях почечной недостаточности потовые железы MorYT в
известноЙ мере КОl\шенсировать нарушенную выделительную функцию почек
Сальные железы
СалЪ1lые железы также производные эпителиальноro дифферона, простые раз
ветвленные альвеолярные железы, почти всеrда связанные с волосами. Секрет саль
ных желез (кожное сало) служит жировой смазкой для эпидермиса и волос. За CYT
ки сальными железами человека выделяется около 20 r секрета. Жировая смазка
смяrчает кожу, делает ее непрошщаемой для воды, а также препятствует развитию
микробов на поверхности кожи, поскольку кожное сало имеет кислую реакцию. Саль
ные железы располаrаются в составе кожи более поверхностно, чем потовые их
концевые отделы лежат на rранице сосочковоrо и сетчатоrо слоев дермы. Выводной
проток сальной железы короткий, открывается, как правило, в волосяную воронку.
Стенка протока образована мноrослойным плоским эпителием. Больше Bcero саль
ных желез в коже rоловы. Они отсутствуют на ладонях и подошвах.
Железы состоят из различно дифференцированных эпителиальных клеток
салЪ1lЪ/Х ЭК30КрИ1l0цитов, или себоцитов (от sebum сало). На периферии железы
152
Fлава 13
на базальной мембране располаrаются камбиальные клетки. По мере дифферен
цировки себоциты накапливают в цитоплазме жировые включения и смещаются
по направлению к центру железы. Здесь происходит rибель себоцитов по механиз
му апоптоза. Это характеризует rолокринный тип секреции железы
Имеются данные о том, что физиолоrическая реrенерация сальной железы
ПрОIIСХОДИТ за счет формирования новых концевых отделов путем почкования.
Волосы
Волосы это ороroвевшие эпителиальные придатки кожи, покрывающие почти
всю кожную поверхность, за исключением ладонеЙ, подошв, боковых поверхностей
пальцев и др. Плотность расположения волос неодинакова. Различают три вида BO
лос: длинные, щетинистые, пушковые. Длинные волосы это волосы rоловы, боро
ды, усов, подмышечных впадин и лобка. Щетинистые волосы волосы бровей, pec
ниц, наружноrо слуховоro прохода, преддверия носовой полости. Пушковые волосы
покрывают остальную поверхность тела. Длина волос от нескольких миллимет
ров до 1,5 м, толщина от 0,005 до 0,6 мм. Волосы выполняют в основном защит
ную функцию ресницы защищают rлаза от попадания пыли и от механических
повреждениЙ. Волосы участвуют также в тактильной чувствительности.
Развитие волоса начинается на 3 M месяце эмбриоrенеза, коrда эпидермис в
виде тяжей врастает в подлежащую соединительную ткань. При этом образуются
волосяные фолликулы, из которых растет волос. Первоначальный волосяной по
кров (лануrо) исчезает еще до рождения, заменяясь пушковыми волосами (велюс).
После рождения новые волосяные фолликулы не образуются, а рост дефинитив
ных волос происходит за счет ранее возникших фолликулов.
В волосе различают стержень, выступающий над поверхностью кожи, и корень,
расположенный в толще кожи. Корень волоса оканчивается утолщением волося
ной луковицей и располаrается в особом футляре волосяном фолликуле (рис. 76).
Последний состоит из наружноrо и BHYTpeHHero эпителиальных корневых влаrалищ
и соединительнотканной волосяной сумки. На нижнем конце корня в волосяную
луковицу в виде сосочка вдается рыхлая соединительная ткань, выполняющая TpO
фическую роль. Волосяная луковица это матрица волоса. Она состоит из эпители
альных камбиальных клеток, обеспечивающих рост корня волоса и BHyтpeHHero эпи
телиальноrо KopHeBoro влаrалища. Кроме этих клеток, в матрице волоса имеются
меланоциты, синтезирующие пиrмент.
В корне волоса различают центральную часть мозrовое вещество, перифе
рические слои корковое вещество и кутикулу. Мозrовое вещество состоит из по
лиrональных эпителиоцитов, расположенных в виде монетных столбиков. В этих
клетках накапливаются rранулы трихоrиалина, а затем мяrкий кератин. KOpKO
вое вещество волоса на протяжении большей части корня и всею стержня состоит
из плоских роrовых чешуек, содержащих твердый кератин. Из них же (кутикуляр
ных эпителиоцитов), налеrающих друr на друrа в виде черепицы, состоит кутику
ла волоса. Разная толщина волос и прочность связаны с особенностями KopKOBoro
вещества волоса и ero кутикулы. Внутреннее корневое влаrалище окружает корень
Система покрова
153
волоса наподобие муфты. В нем
различают наружный эпителиаль
ный слой (бледный слой rенле),
средний эпителиальный слоЙ (rpa
нулосодержащий слой rексли) и
кутикулу, прилеrающую к кутику
ле корня волоса.
Наружное эпителиальное KOp
невое влаrалище это непосреk
ственное продолжение в rлубь дep
мы pOcTKoBoro слоя эпидермиса.
Волосяной фолликул у повер
хности кожи образует раСllIире
ние воронку, куда впадают про
токи сальных желез.
Волосяную сумку образуют Ha
ружный продольный и внутренний
циркулярный слои коллаrеновых
волокон. На rранице между воло
сяной сумкоЙ И волосяноЙ лукови
цей имеется базальная мембрана.
Корень волоса снабжен мыш
цей (состоит из rладкой мышечноЙ
ткани эктомезенхимноrо проис
хождения), поднимающей волос.
Одним концом она вплетена в BO
лосяную сумку, друrИ:\1 в сосоч
ковый слой дермы. При сокраще
нии мышцы волос принимает пря
мое положение, поднимаясь над поверхностью кожи ("волосы встают дыбом").
Существуют значительные половые различия в степени развития волос.
Волосы растут за счет пролиферации клеток матрицы и последовательной их Дlfф
ференцировки в кератинсодержащие клетки и далее в роroвые чешуйки. Последние
постепенно смещаются по направлению из области корня волоса в стержень. Скорость
роста волос у человека в среднем 0,4 мм в сутки. Продолжительность жизни волоса на
rолове 2 4 [ода. Прекращение пролиферации клеток матрицы означает остановку po
ста волоса. Затем пролиферация клеток матрицы возобновляется, начинается рост
HOBoro волоса, а старый волос при этом выпадает. Так пр()исходит смена волос.
4
з
{,
,:
J
, -$r t_
2
Рис. 76, Строение волоса и сальной железы:
1 волосяная луковица; 2 волосяной сосочек; 3 Ha
ружное эпителиальное влаrалище; 4 сальная железа
Ноrти
НОl.ти это poroBbIe эпителиальные придатки кожи. Составными частями Hor
тя служит плотная роrовая пластинка, лежащая на HorTeBoM ложе, и HorтeBoe ложе,
у основания и с боков оrpаничешюе кожными складками ноrтевыми валиками
154
rлава 1 3
(рис. 77). В ноrтевой пластинке раз
личают корень, тело, край. Корень
1l0lmя это задняя часть ноrтевой
пластинки, луночка ноrтя. Здесь
находится матрица ноrтя камби
альная часть HorтeBoro ложа. Эпи
телий HorтeBoro ложа имеет такое
же строение, как ростковый слой
эпидермиса. НOlmевая пласmUll:ка
образована роrовыми чешуйками, Рис. 77. Строение нопя:
содержащими твердый кератин. 1 нопевая пластинка; 2 нопевое ложе; 3 нопевой
Рост ноrтя происходит медленно со валик; 4 матрица нопя
скоростью около 1 мм в неделю за счет пролиферации клеток матрицы (онихоблас
тов) ноrтя. После размножения эти клетки дифференцируются и подверrаются opo
rовению. При ЭТО 1 они смещаются в виде роrовых чешуек в состав ноrтевой плас
тинки. Соединительнотканная часть HorтeBoro ложа образует продольные бороздки
и rребешки. Здесь проходят кровеносные сосуды. После повреждения ноrтя проис
ходит ero реrенерация, если клетки матрицы сохранили жизнеспособность.
:... : . ;
{ ''''''--:'' ,.....""i, '" ,
2 ,.' .. .' ':Qn
(",' О ".-.
, ' '"
1 '. . ...: . ....,:v.,1t...: ..
б ' Ac.fi., ;",
o., .. '.:
...
4
Реактивность и реrенерация кожи и ее производных
Реактивные свойства кожи отчетливо проявляются при действии факторов
внешней и внутреннеЙ среды. В эпидер шсе хорошо выражены процессы клеточ
Horo обновления (физиолоrическая реrенерация). ВысокиЙ уровень пролифера
ции эпителиоцитов отмечается в области матриц волос, а также в концевых OTдe
лах потовых и сальных желез.
Выделяются две разновидности стволовых клеток эпителиальноrо дифферо
на. Одни из них располаrаются в базалЬНОI\-1 слое эпидермиса и являются источни
ком развития дифферона ороroвеваlOЩИХ эпителиоцитов эпителиалыю пролифе
ративной единицы в физиолоrических условиях. Эти клетки обеспечивают также
реrенерацию при поверхностных повреждениях эпителиальноrо покрова кожи.
Вторая разновидность стволовых клеток эпителия локализуется в наружном
эпителиальном корневом влаrалище, непосредственно под устьями протоков саль
ных желез. Эти клетки являются источником развития поверхностных эпителиоци
тов, ЭПlпелиоцитов волоса и ero эпителиальных влаrалищ, эпителиоцитов желез, а
также участвуют в реrенерации кожи при ее rлубоких и обширных повреждениях.
При травматических и ожоrовых повреждениях кожи репаративная pereHepa
ЦlIЯ происходит путем стяпшания краев раны за счет развития rрануляционной
ткани и ЭПlпелизации раневой поверхности. Важную камбиальную роль в эпите
лизаЦlШ раневой поверхности иrраlOТ малодифференцированные клетки KOHцe
вых отдел()в потовых желез и наружных эпителиальных корневых влаrалищ воло
са. Это учитывается при взятии кожных лоскутов для трансплантации после
срезания TOHKoro лоскута кожи возникшая раневая поверхность достаточно быст
ро покрывается новым эпидсрмисом.
Система покрова
155
в результате повреждающеrо действия ядерною взрыва, отравляющих веществ
и друrих факторов возникают тяжелые и rлубокие поражения кожи и ее ПРОИЗВОk
ных. При этом удлиняется некротическая стадия paHeBoro процесса, тормозятся
лейкоцитарная и макрофаrальная реакции, снижается действие факторов клеточ
Horo и ryморальноrо иммунитета. В связи со значительным повреждением стволо
вых и камбиальных клеток в эпидермисе и дерме, подкожной жировой клетчатке,
уrнетением пролиферации и pereHepaTopHbIx процессов снижается число клеток
фибробластическorо дифферона, резко падает митотическая активность в эпидер
мисе, нарушаются формирование rрануляционной ткани и эпителизация ран. При
наличии обширных и rлубоких ожоrов изменяется соотношение реrенерационных
rистоrенезов эпидермиса и волокнистой соединительной ткани в процессе зажив
ления ран, что ведет к образованию обезображивающих рубцов. При действии иони
зирующей радиации повреждение кожною эпителия развивается быстрее в наи
более активных в отношении пролиферации участках волосяных влаrалищ, что
приводит к выпадению волос.
Кожа имеет неодинаковую радиочувствительность в разных ее отделах. Наи
более чувствительна кожа подмышечных впадин, паховых складок, локтевых crи
бов, шеи; резистентна кожа спины, разrибательных поверхностей конечностей.
Исходом реrенерации кожи является, как правило, образование pereHepaToB в виде
соединительнотканноrо рубца, не содержащеrо производных кожи (желез, волос)
pereHepaTa рубцовоrо типа, покрытою эпителием. Реже образуется pereHepaT кож
ною типа, по строению приближающийся к интактной коже.
Молочные железы
Молочные железы традиционно рассматриваются в разделе "Половая система",
однако, являясь производными кожи, они сохраняют в строении основные черты
cBoero происхождения. Источником развития молочных желез служат утолщения
кожной эктодермы (молочные точки), от которых в подлежащую мезенхиму Bpac
тают эпителиальные тяжи, ветвящиеся в своей терминальной части. Это начина
ется на 6 й неделе и продолжается вплоть до 6 ro месяца эмбриоrенеза. На концах
тяжей находятся клетки, из которых в последующем (в период беременности и
лактации) развиваются молочные альвеолы (концевые секреторные отделы альве
олярно трубчатых желез).
Строение молочной железы различно в детском, юношеском, детородном, CTap
ческом возрастах, во время беременности и лактации и зависит от rормональною
статуса женщины (в частности от концентрации половых ropMoHoB). Так, в He
активном состоянии в половозрелом возрасте в молочной железе различимы доли
(от 15 до 25), каждая из которых uкружена соединительной тканью с различным
содержанием жировых клеток. В соединительной ткани проходят млечные пpoтo
КИ числом, равным числу долей, которые в терминальной части разветвляются на
внутридолевые протоки. Однако секреторные отделы отсутствуют.
Во время беременности и в лактирующей молочной железе от терминальных
частей внутридолевых протоков начинают развиваться трубчато альвеолярные
156
Тлава 13
концевые секреторные отделы МОЛОЧ1lые
алъвеолы. Концевые секреторные отделы
rруппируются в дольки, а несколько долек
формируют доли молочной железы (рис. 78).
Стенку молочной альвеолы образуют
диффероны МОЛОЧ1lЪ/Х Э'КЗОКрИ1l0цитов (лак
тоцитов) и звездчатъ/Х миоэпителиоцитов.
Снаружи альвеола покрыта базальной мемб
раной. Лактоциты имеют кубическую форму,
окруrлое ядро и соединяются друr с друroм с
помощью плотных контактов и десмосом. На
апикальной поверхности лактоцитов присут
ствуют микроворсинки, а в цитоплазме Ha
капливаются включения составные компо
ненты молока. Для лактоцитов характерны
Mepo и апокринная типы секреции. Миоэпи
телиоциты располаrаются между основани
ем лактоцитов и базальной мембраной, OXBa
тывая лактоциты своими пальцевидными BЫ
ростами. Их ядра темные, а в цитоплазме co
держатся актомиозиновые комплексы. Co
кращение миоэпителиоuитов способствует
выведснию секрета.
Рис. 78. Строение лактирующей молочной
В просвет альвеол секретируется моло железы:
КО продукт сложноrо состава, включаю А: 1 альвеолы; 2 млечный альвеолярный
щиЙ белки (казеин), липиды (триrлицерид), проток; з междольковая соединительная
ткань; Б: 1 молочные экзокриноциты; 2
сахара (лактоза), иммуноrлобулины, вита звездчатый миоэпителиоцит; 3 соедини
мины, соли, воду и др. Молоко представля тельная ткань; 4 базальная мембрана
ет собой водную эмульсию. После выделения секрета лактоциты реrенерируют и
вступают в новый секреторный цикл. Выведение секрета молочных желез по си
стеме выводных протоков облеrчается сокращениями миоэпителиоцитов и rлад
ких миоцитов.
Система выводных протоков молочной железы начинается внутридольковы
ми протоками. Несколько внутридольковых протоков сливаются и формируют
междольковый проток. Последние переходят в млечный проток, который вблизи
околососковоrо кружка образует расширение млеч1lЫЙ СИ1lУС, и далее млечный
проток открывается на верхушке соска.
Стенки внутридольковых протоков образованы кубическим эпителием и co
держат миоэпителиоциты. Млечные протоки выстланы двухслойным кубическим
эпителием и миоэпителиоцитами. Эпителий в синусах трехслойный, а на Bep
шине соска становится мноrослойным плоским. Вдоль млечных протоков и сину
сов В соединительной и жировой ткани проходят пучки rладкой мышечной ткани,
сокращения которой способствуют молокоотдаче.
В области околососковоro кружка эпидермис сильно пиrментирован за счет скоп
"'I
J -r,") ,
..........t:'" "..: .. - ) "
?i. \: .; ';
! 1 . .
. ,-
. ._' 1
3
:! * '.
. h', .
. ,2_. "'..
"' " . J\
')" " 1
'" ,.t l ;;:. .
3/ ' . 1.
..'\
А
Б
r:. "'"
4 /2
2 . '" 't}'л
.' . Q"'. 3
\ e..:: , " ; " ' '''''' ',
;' (," . 4
" ' I
1 "\t? "
. " о'
,.
Система по'/(рова
157
ления меланоцитов. В сосочковом слое дермы находятся МНОl'Oчисленные инкапсу
лированные нервные окончания (механорецепторы). Секреторная активность лак
тоцитов реryлируется эстроrенами, проrестеРОНОI\1 и др. l'Oрмонами. С возрастом по
мере уменьшения синтеза половых ropMoHoB происходит постепенная инволюция
молочных желез.
158 [лава 14
fЛАВА 14. СИСТЕМА OPfAHOB ВНУТРЕННЕЙ СРЕДЫ,
СВЯЗАННАЯ С МЕТАБОЛИЗМОМ
Пищеварительный комплекс opraHoB
в орrаНИЗ1\lе человека пищеварительныЙ комплекс opraHoB иrрает исключи
тельную роль, поскольку обеспечивает поддержание трофики и жизнедеятельнос
ти всех клеток и тканеЙ. OpraHbI пищеварительноrо комплекса осуществляют Me
ханическую переработку и химическое расщепление компонентов пищи до более
простых соединениЙ, способных всасываться в кровь и лимфу и усваиваться Bce
ми клетка:\ш орrанизма для поддержания их жизнедеятельности и выполнения
специальных функций.
OpraHbI пищеварительноrо комплекса являются производными эмбриональноЙ
пищеварительной трубки, в котороЙ различают три отдела. Из переднеrо (rоловно
[о) отдела развиваются opraHbl ротовоЙ полости, rлотка и пищевод; из среднеrо (тy
ловищноrо) желудок, тонкая кишка, толстая кишка, печень и желчныЙ пузырь,
поджелудочная железа; из заднеrо каудальная часть пря:\юй кишки. Для каждоrо
из перечисленных opraHoB характерны специфические структурно функциональные
черты, детеР:\lИнированные эмбриональными зачатками тканеЙ и opraHoB.
Развитие и общий план строения пищеварительной трубки
Основные opraHbI пищеварительноrо комплекса образуются в процессе раз
вития эмбриональноЙ кишечноЙ трубки, которая вначале слепо заканчивается
на rоловном и хвостовом концах и соединяется с желточным мешком посредством
желточноrо стебелька. Позднее у зародыша образуются ротовая и анальная бух
ты. Дно этих бухт, соприкасаясь со стенкоЙ первичноЙ кишки, образует ротовую
и клоачную перепонки. На 3 4 й неделе эмбриоrенеза ротовая перепонка проры
вается. В начале 3 4 ro месяца происходит прорыв клоачноЙ перепонки. Кишеч
ная трубка становится открытоЙ с обоих концов. В краниальной части переднеЙ
КIIШКИ появляются пять пар жаберных карманов. Эктодерма ротовоЙ и анальной
бухт служит исходным материалом для развития мноrослоЙноrо плоскоrо эпи
телия преддверия POTOBOi,i полости и каудальноЙ части прямоЙ кишки. Кишеч
ная энтодерма является источником образования эпителия слизистоЙ оболочки
и желез rастроэнтеральноrо отдела пищеварительноЙ трубки. Соединительнот
канные и rладкомышечные тканевые элементы пищеваритсльных opraHoB фор
1\IИРУЮТСЯ IIЗ l\IсзеНХll1\lЫ, а однослойный плоскиЙ эпителиЙ серозной оболочки
Система Орlа1юв внутренней среды
159
из висцеральноrо листка спланхнотома. Поперечнополосатая мышечная ткань,
присутствующая в составе отдельных opraHoB пищеварительной трубки, разви
вается из митомов. Элементы нервной системы являются производными нервной
трубки и rанrлиозной пластинки.
Стенка пищеварительной трубки на всем протяжении имеет общий план CTpO
ения. Ее образуют следующие оболочки: слизистая с подслизистой основой, MЫ
шечная и наружная (серозная или адвентициальная). Слизистая оболочка состоит
из эпителия, собственной соединительнотканной пластинки и мышечной пластин
ки. Последняя присутствует не во всех opraHax. Называется эта оболочка слизис
той в связи с тем, что ее эпителиальная поверхность постоянно увлажнена слизью,
выделяемой слизистыми клетками и мноrоклеточными слизистыми железами. Под
слизистая основа представлена рыхлой волокнистой соединительной тканью. В ней
находятся кровеносные и лимфатические сосуды, нервные сплетения и скопления
лимфоидной ткани. Мышечная оболочка образована, как правило, двумя слоями
rладкой мышечной ткани (внутренним циркулярным и наружным продоль
ным). В межмышечной соединительной ткани проходят кровеносные и лимфати
ческие сосуды. Здесь же располаrается нервное сплетение. Наружная оболочка
бывает или серозной, или адвентициальной. Серозная оболочка состоит из мезоте
лия и соединительнотканной основы. Адвентuцuалъная оболочка образована толь
ко рыхлой соединительной тканью.
Производные переднеrо отдела пищеварительной трубки
OpraHbI ротовой полости (ryбы, щеки, десны, зубы, язык, слюнные железы,
твердое нёбо, мяrкое нёбо, миндалины) выполняют следующие основные функ
ции: механическая переработка пищи; химическая обработка пищи (смачивание
слюной, переваривание уrлеводов амилазой и мальтозой слюны); деrустация пищи
с помощью opraHa вкуса; rлотание и проталкивание пищи в пищевод. Кроме Toro,
некоторые opraHbI полости рта (например, миндалины) выполняют защитную фун
кцию, препятствуя проникновению в орrанизм микробов, участвуют в формиро
вании иммунноrо ответа орrанизма.
Ротовая полость
Слизистая оболочка ротовой полости состоит из мноrослойноrо плоскоrо эпи
телия кожною типа, развивающеrося из прехордальной пластинки, и собственной
соединительнотканной пластинки. Степень развития этих тканевых структур Heo
динакова в различных участках слизистой оболочки. Поверхность эпителия по
стоянно увлажняется за счет слюны, которая выделяется мноrочисленными мел
кими и крупными слюнными железами ротовой полости.
Эпителий слизистой оболочки включает rенетически различно детерминиро
ванные клетки. Это во MHoroM определяет сходство эпителия слизистой оболочки с
эпидермисом. Поверхностный слой эпителия образован плоскими клетками,
а в отдельных местах роювыми чешуйками. Эти эпителиальные клетки и poroBbIe
160
Fлава 14
чешуйки слущиваются в ротовую полость. Обновление клеточною состава эпите
лия происходит довольно интенсивно (за 4 6 суток). В целом за каждые пять минут
с поверхности эпителия ротовой полости слущивается до 0,5 млн клеток Столько
же клеток образуется вновь в результате деления базальных эпителиоцитов. Кроме
ведущею эпителиальноrо клеточноrо дифферона в эпителии слизистой оболочки
ротовой полости встречаются меланоциты, клетки Ланrерrанса и клетки Меркеля.
Эпителий в области rуб
постепенно переходит от эпи
дермиса (в кожной части) с ти
пичным строением к эпителию
слизистой оболочки ротовой
полости. В ryбе различают кож
ную, промежуточную (переход
ную, красную кайму) и слизис
тую части (рис. 79). В переХОk
ной части в эпителий высоко
вдаются соединительноткан
ные сосочки, содержатся MHO
rочисленные капилляры. Про
текающая в них кровь просве
чивает через тонкий слой
эпителия и придает этой части
ryб красноватый цвет. Здесь OT
сутствуют потовые железы, а по
краям ryб сохраняются сальные
железы.
В слизистой части эпителий ryб отличается значительной толщиной. В ПОk
слизистой основе располаrаются lуб1-/.ые железы смешанноrо (слизисто белково
[о) характера.
Основу щеки составляет поперечнополосатая мышечная ткань. Снаружи щека
имеет кожный покров, изнутри выстилается слизистой оболочкой, по строению
сходной с таковой у ryб. В подслизистой основе располаrаются концевые отделы
смешанных (белково слизистых) желез.
Слизистая оболочка десен с помощью плотных пучков коллаrеновых волокон
собственной пластинки неподвижно соединена с костями верхней и нижней челю
стей. Она содержит мноrочисленные нервные окончания.
Тубы, щекu, десны
Б
в
А
Рис.79.Строениеrубы:
А кожная часть; Б промежуточная часть; В слизис
тая часть; 1 мноrослойный плоский эпителий; 2 соб
ственная пластинка слизистой оболочки; 3 волосяной
мешок; 4 слюнные железы; 5 поперечнополосатая
мышца
Зубы
Зубы участвуют в механической обработке пищи: плоские резцы и клыки KO
нической формы откусывают пищу, малые и большие коренные зубы с коронками
Система орюнов внутренней среды
161
кубической формы и жевательными буrра
ми перетирают ее при еде. Зубы имеют важ
ное значение для артикуляции.
rucтo и ОрШ1l0lеllез зубов. У челове
ка различают две смены зубов выпадаю
щие, или молочные (20), и постоянные (32).
Развитие молочных зубов начинается в KOH
це 2 ro месяца эмбриоrенеза. В это время
эпителий ротовой полости врастает в виде
зубной пластинки в подлежащую мезенхиму
(рис. 80). На передней поверхности зубной
пластинки возникают эпителиальные зуб
ные почки по числу закладок зубов, BOKpyr
которых возникает уплотнение мезенхим
ных клеток зубные мешочки. Взаимодей
ствие двух эмбриональных зачатков приво
дит к изменению формы зубной пластин
ки она постепенно трансформируется в
структуру в виде бокала, внутри котороro
концентрируются мезенхимные клетки в
форме сосочка. Последний оказывает индук
тивное влияние надифференцировку клеток Рис. 80. Закладка зуба:
эпителиальноrо зубноrо бокала, в котором 1 мноrослойный плоский эпителий pOTO
вой полости; 2 эпителиальный зубной бо
топоrpафически выделяются внутренний и кал (эмалевый opraH); 3 энамелобласты;
наружный эмалевый эпителий и клетки про 4 мезенхимный эубной сосочек
межуточноro слоя. Внутренний эмалевый эпителий обращен к мезенхимному co
сочку, наружный образует "стенку" зубноrо бокала и тонкой клеточной ножкой oc
тается некоторое время связанным с эпителием ротовой полости; клетки промежу
точноrо слоя располаrаются между двумя первыми, при обретают звездчатую форму
и оттесняются друr от друrа накапливающейся здесь жидкостью.
Внутренний эмалевый эпителий отделяется от мезенхимноrо сосочка базаль
ной мембраной. Ero клетки дифференцируются в энамелобласты (амелобласты)
клетки образователи эмали. Образование базальной мембраны индуцирует диф
ференцировку расположенных рядом мезенхимных клеток в одонтобласты (дeH
тинобласты). Последние, в свою очередь, влияют на развитие энамелобластов.
Энамелобласты имеют вытянутую цилиндрическую форму, в них постепенно
происходит перемещение ядер из базальной части клеток в апикальную, посколь
ку В базальных частях клеток формируются эмалевые призмы, слой которых co
ставляет ЭМШlЪ зуба. Начинается кальцинация эмали. Каждый энамелобласт Bыpa
батывает одну эмалевую призму.
Клетки, прилежащие к энамелобластам, одонтобласты начинают секрети
ровать дентин во встречном с образованием эмали направлении. По мере развития
зуба массы эмали и дентина увеличиваются, и ряды клеток удаляются друr от дpy
ra. Энамелобласты при этом отодвиrаются наружу, а одонтобласты внутрь
2
3
4
t,
162
Fлава 14
развивающеrося зуба (рис. 81). К
моменту прорезывания молочных
зубов ядросодержащие части энаме
лобластов редуцируются, остаются
лишь тесно прилежащие друr KДPy
ry эмалевые призмы, покрытые KY
тикулоЙ, образованной остатками
промежуточных звездчатых клеток
и наружноrо эмалевою эпителия.
Последние постепенно уменьшают
ся в размерах и деrенерируют; кле
точная ножка, связывающая зача
ток зуба с эпителием полости рта,
фраrментируется и полностью ис
чезает. Зубной зачаток оказывается
поrpуженным в костную ткань че
люсти. Формирование эмали и дeH
тина распространяется от вершины
будущеrо зуба к боковым поверхно
стям. Клетки зубноrо мешочкадиф
ференцируются в цементобласты,
и незадолrо до прорезывания зуба
в области будуших корней форми
руют цемент. Клетки центральных
участков мезенхимных сосочков об
разуют пульпу зуба внутреннюю
рыхлую соединительную ткань
зуба, боrатую сосудами. Из клеток наружноrо слоя мезенхимноrо зубноrо мешочка
образуется зубная связка (периодонт), соединяющая зуб с альвеолой челюсти. Зубные
альвеолы формируются из окружающей зубные зачатки мезенхимы параллельно с фор
мированием зубов. Таким образом, в составе зуба эмаль имеет эпителиальную приро
ду. Все остальные части зуба (дентин, цемент, пульпа зуба), а также связочный аппа
рат это производные мезенхимы.
Закладка постоянных зубов происходит на 4 5 M месяцах эмбриоrенеза, коrда от
зубной пластинки начинают формироваться вторые эмалевые зачатки. Развитие их про
исходит принципиально так же, как и молочных зубов.
Строеll.ие зуба. В составе зуба различают коронковую, шеечную и корневую
части. Коронка выступает над десной, а шейка и корень поrружены в ткани зубной
альвеолы. Внутри зуба находится полость, заполненная пульпой. Коронку зуба
образуют эмаль, дентин и пульпа. Эмаль производное дифферона энамелоблас
тов. Структурными элементами эмали являются эмалевые прuзмы диаметром з
5 мкм. Они имеют S образно изоrнутый ход. В состав призмы входят орrаничес
кие вещества в виде субмикроскопической фибриллярной сети (филаментов
промежуточноrо типа), уrлеводы, кристаллы минеральных солей (фосфат каль
2
. . /. \,, . . '.'\.. О,' Н , . \,;'
:- : ,;.. . .' jI i , ;;..," ,'ii.. . & \, ,:
... .tf.l.:.. " J".f , - ..' ,'-..... t .'= \
.,;}..;... .;: : ..., . ....:. . ...,.---: _С ,"'"< .,, _ '
..j .'': _.,}- I ?:1; (.. . . . }. '4 ""'; < " ,_ ....\
'. .,,.; '.) ? . '.1 ", " ..., 4
.. . . .... .; .>;: .;...' ). : i :':: ..; :. ':\'"
у;," :' -.... ., " ,) ';"'" :о ...."...... .....
.1. f:: ,,,.:. < ' : .':l{: . '::- :;;:
/.. ';::J(: t ;. r.': .} ,_, '\, tf. .
1 -.:..1 Q # {а... 1.". .. .. ;:; : .. _А\
J'::' ..: ..!-; ",; ii :.l-;>i >&' , ,:': "". :;l ::
:'; :t. ,! ' 'li:; fi ij;; ; :. _ # з
,;; .:: t . :. ;/:'1:': ;".:: x ' )-:!,:::"5}.:
:..;. () '.../ -= )-1;' 4'j( ,. __.... - . -. ... : :
. ;:'Ч . /'.: /.' (\" . i' 4' ':; ); ,. > ' i..; :
.:'i :"":;.. '. ; P} .:;:f: i I JI.;'. t..
. ./' .:'1!: . : :' I,;",. . ,,-!, !y. ...
:: ., :::.. :,,' i, ',:1 1 "',-;:;,," '. '" , . '';r r, , ...
. .' .' . .1':> I . ; i 1 ;;' ,. ' -.. 't' .п,,- ..... *'"'
,; '" , . , " "..., ';"\i.:i.:':\ >; . ' ', ,'.. . jO- ., , . ,
1. < ..t; .:: . '" / ?::' ;:'f\. :..:'-.. .-(-"-:....\_ ,. ;J. ..... ;... :''#.-
.. t9 , .,; ,..' 1!:';",' , ''\1,. '>', , .. "..:} r.{ "
.;. . ,r"" '" . .. .:;,;:p. .:" ,t,...> "Ч А . }
:,.1. 1:' <Е. j<"\.;.. /r ;;: : \;.,' --. ";. __ . .
\/ .. ..ф. ;#. o; .-..::..:- f.:.'.....\.. ;:,l:..;,) . \ , .. t
/(H ? f t lrt : .f.
/. . :" ..: '. '\?-: '"i:::;:I:':: { ""t'; ); :::f" ; '"
< .. 4;', '. ," . x?,f:tl ; i;!.:' \ ; A[: ?J : t ,k
5
Рис, 81. rистоrенез зуба:
1 энамелобласты; 2 эмаль; 3 одонтобласты; 4
дентин; 5 пульпа зуба
Система 0PZa1IOR внутренней среды
163
ция в форме Пlдроксиапатита, фторид кальция). Доля последних равна 96 97%
массы эмали. Эмалевые призмы объединяются с помощью менее обызвествленно
ro межпризменноrо вещества и покрывают коронку зуба в виде эмали.
По твердости эмаль близка к кварцу. Снаружи эмаль покрыта тонкой кутику
лой, которая постепенно стирается при приеме пищи. Несмотря на то, что эмаль
это неклеточная структура, которая не содержит кровеносные сосуды, для нее xa
рактерен обмен веществ. Транспорт веществ в эмаль осуществляется эмалевой
жидкостью через межпризменные необызвествленные пространства. При HeДOCTaT
ке питательных веществ и витаминов эмаль разрушается.
Дентин ведущая ткань зуба, состоит из коллаrеновых фибрилл и склеи
вающеrо их вещества с большим количеством солей кальция. В дентине мине
ральные соли составляют 72%, а орrанические вещества 28%. Вещество дeH
тина пронизано дентинными канальцами, или трубочками. В них проходят
длинные отростки одонтобластов, расположенных в периферическом слое пуль
пы зуба. В дентинных канальцах проходят также безмякотные нервные волок
на. За счет этих канальцев осуществляются трофические процессы. В обмене
веществ дентина большое значение имеют так называемые интерrлобулярные
пространства необызвествленные участки в виде шарообразных полостей.
Блаrодаря таким участкам rраница между дентином и эмалью становится He
ровной, фестончатой, что обеспечивает прочное соединение двух тканей. Меж
ду одонтобластами, располаrающимися в периферических участках пульпы, и
дентином находится полоса необызвествленноrо матрикса, называемая пpeдeH
тином. За счет последующеrо отложения солей в предентине происходит аппо
зиционный рост дентина и рост зуба.
Цемент своеобразная костная ткань, покрывающая шейку и корень зуба.
В нем содержится 30% орrанических и 70% неорrанических веществ. Различают
две разновидности цемента: бесклеточный и клеточный. Бесклеточный цемент c!J
стоит из аморфною вещества и коллаrеновых волокон, которые переходят в пери
одонт и далее в костную ткань альвеол челюстей, прочно закрепляя зуб в ero ячей
ке. Клеточный цемент содержит цементоциты и по строению соответствует
rрубоволокнистой костной ткани. В составе цемента нет кровеносных сосудов,
поэтому трофические процессы в нем обеспечиваются за счет кровоснабжения пе
риодонта путем диффузии.
Пулъпа зуба (зубная мякоть) располаrается в полости зуба и в корневых KaHa
лах. Корневые каналы свободно открываются в зубную альвеолу. Пульпа зуба обра
зована рыхлой волокнистой соединительной тканью. Периферическое положение в
пульпе занимают одонтобласты. В промежуточном и l1ентральном слоях пульпы зуба
находятся адвентициальные клетки, фибробласты, макрофаrи, арrирофильные и
коллаrеновые волокна. В пульпе зуба разветвляются мноrочисленные кровеносные
сосуды, а также нервные волокна с чувствительными нервными окончаниями.
С возрастом уменьшается содержание орrанических веществ в эмали, дентине
и l1ементе зуба, а в связи с нарастающими склеротическими изменениями сосудов
пульпы ухудшаются кровоснабжение и трофика всех ero частей.
Репаративная реrенерация зуба возможна лишь в оrраниченных пределах.
164
Fлава 14
Эмаль после повреждения не восстанавливается. Дентин образуется медленно и в
очень небольшом количестве за счет дифференцировки одонтобластов. Цемент зуба
реrенерирует слабо.
Язык
Это мышечныЙ opraH, который участвует в механической обработке и проrла
тывании пищи, ее вкусовом восприятии и в артикуляции. Основу языка составля
ет поперечнополосатая мышечная ткань, волокна которой располаrаются в трех
взаимно перпендикулярных плоскостях. Подобно расrтоложение мышечных BO
локон и прикрепление языка только в области ero корня создают условия для ero
свободных движений, что важно для членораздельной речи (артикуляции). Ha
пример, врожденная короткая уздечка языка затрудняет формирование речевых
навыков у ребенка.
языК покрыт слизистой оболочкой. На верхней и боковоЙ поверхностях языка
она сращена с мышцами, на нижней переходит в подслизистую основу. Слизистая
оболочка состоит из мноrослоЙноrо плоскоrо частично ороrовевающеrо эпителия и
собственной пластинки, представленноЙ рыхлой соединительной тканью. На дop
сальной поверхности языка слизистая оболочка образует сосочки: нитевидны,' lpU
бовидныe и желобоваты.. У детей имеются еще листовидные сосочки. Все сосочки
покрыты мноrослоЙным плоским эпителием и имеют основу из рыхлой волокнис
той соединительноЙ ткани. Самые мноrочисленные нитевидные сосочки языка.
Они равномерно покрывают ею верхнюю поверхность, концентрируясь вблизи rpa
ницы с корнем языка, имеют коническую форму и не содержат вкусовых луковиц.
На поверхности нитевидных сосочков эпителий частично oporoBeBaeT. При HeKOTO
рых заболеваниях ороrовение эпителия сосочков слизистой оболочки языка усили
вается образуется так называемый налет.
В rрибовидных сосочках через эпителий просвечивают кровеносные каПИk
ляры, что придает им красный цвет. rрибовидные сосочки немноrочисленны,
располаrаются на спинке языка, сосредотачиваясь на кончике и по краям язы
ка. Они крупнее нитевидных, имеют форму rриба с широкой вершиной и узким
основанием. В эпителии содержат небольшое количество вкусовых луковиц.
Желобоватые сосочки (сосочки, окруженные валиком) в количестве 6 12 pac
положены между телом и корнем языка. Это самые крупные сосочки языка. По
форме они напоминают rрибовидные сосочки, но крупнее и как бы "утоплены" в
слизистую таким образом, что BOKpyr них образуются rлубокие желобки (рис. 82).
В этих желобках застаивается слюна с частицами пищи, что создает блаrоприят
ные условия для восприятия вкуса пищи клетками вкусовых луковиц, которых
очень MHoro в эпителии боковых поверхностей сосочков. В основания желобков
открываются выводные протоки белковых желез Эбнера. Секрет этих желез IIрО
мывает желобки и освобождает их от слущивающеrося эпителия, скаIIливающей
ся здесь пищи и микробов.
Листовидные сосочки располаrаются рядами по правому и левому краям языка.
В пространства между сосочками открываются выводные протоки белковых желез,
Система ОРlaнов внутренней среды
165
концевые отделы которых распо
лаrаются между мышечными BO
локнами языка. В эпителии боко
вых поверхностей листовидных
сосочков залеrают мноrочислен
ные вкусовые луковицы. У взрослых
листовидные сосочки редуцируют
ся, в местах расположения белко
вых желез развиваются жировая и
лимфоидная ткани.
В языке находится 3 вида же
лез белковые, слизистые, CMe
шанные. Белковые железы про
стые трубчатые разветвленные.
Заложены в основании желобова
тых и листовидных сосочков, в
толще языка. Слизистые железы
простые, трубчато альвеолярные
разветвленные железы, располо
жены в корне языка и вдоль ero боковых поверхностей. Протоки открываются в
крипты язычной миндалины. Смешанные железы залеrают в переднем отделе язы
ка. Их короткие протоки открываются вдоль складок слизистой оболочки под язы
ком, секреторные отделы располаrаются в мышечной ткани языка.
А
Рис. 82. Строение дорсальной поверхности языка:
А слизистая оболочка; Б мышечная оболочка; 1
желобоватый сосочек; 2 валик; 3 желобок; 4 MHO
rослойный плоский неороrовевающий эпителий; 5 соб
ственная пластинка слизистой оболочки; б вкусовые
луковицы; 7 концевые отделы слюнных белковых желез;
8 поперечнополосатая мышечная ткань
Большие слюнные железы
Помимо множества мелких слюнных желез, расположенных в слизистой обо
лочке щек, и желез языка, в полости рта находятся большие слюнные железы (OKO
лоушные, поднижнечелюстные и подъязычные), являющиеся производными эпи
телия слизистой оболочки ротовой полости. Они закладываются на 2 M месяце
эмбриоrенеза в виде парных плотных тяжей, врастающих в соединительную ткань.
В начале З rо месяца в закладках желез появляется просвет. От свободных концов
тяжей отпочковываются мноrочисленные выросты, из которых формируются аль
веолярные или трубчато альвеолярные концевые отделы. Их эпителиальная BЫC
тилка вначале образована малодифференцированными клетками. Позднее в ceK
реторном отделе в результате диверrентной дифференцировки исходной клетки
появляются мукоциты (слизистые клетки) и сероциты (белковые клетки), а TaK
же миоэпителиоциты. В зависимости от количественноrо соотношения этих кле
ток, характера выделяемою секрета и друrих структурно функциональных особен
ностей различают концевые (секреторные) отделы трех типов: белковые (серозные),
слизистые (мукоидные) и смешанные (белково слизистые).
В составе выводных путей слюнных желез различают вставочные и исчерчен
ные (или слюнные трубки) отделы внутридольковых протоков, междольковые про
токи, а также общий выводной проток По механизму секреции все большие
166
rлава 14
слюнные железы являются мерокринными. Слюнные железы вырабатывают ceK
реты, поступающие в ротовую полость. В различных железах секреторный цикл,
состоящий из фаз синтеза, накопления и выделения секрета, протекает reTepox
ронно. Это обусловливает непрерывную секрецию слюны.
Слюна представляет собой смесь секретов всех слюнных желез. Она содержит
99% воды, соли, белки, муцины, ферменты (амилаза, мальтаза, липаза, пептидаза,
протеиназа и др.), бактерицидное вещество лизоцим и друrие. В слюне содержат
ся слущенные клетки эпителия, лейкоциты и пр. Слюна увлажняет пищу, облеrчает
жевание и rлотание пищи, а также способствует артикуляции. Слюнные железы BЫ
полняют экскреторную функцию, выделяя из орrанизма мочевую кислоту, креати
нин, железо и др. Эндокринная функция слюнных желез связана с выработкой ин
сулиноподобноro вещества, фактора роста нервов, фактора роста эпителия и друrих
биолоrически активных соединений. У человека за сутки выделяется от 1 до 1,5 л
слюны. Слюноотделение повышается при стимуляции парасимпатических и YMeHЬ
шается при стимуляции симпатических нервных волокон.
Околоушные железы. Это белковые слюнные железы, состоящие из мноrочис
ленных долек В дольках железы различают концевые секреторные отделы (ацину
сы, или альвеолы), вставочные протоки, исчерченные слюнные трубки. В концевых
секреторных отделах эпителий представлен клетками двух типов: сероцитами и ми
оэпителиоцитами (рис. 83). Сероциты имеют форму конуса с четко выраженными
апикальной и базальной частями. Окруrлое ядро занимает почти срединное поло
жение. В базальной части располаrаются хорошо развитые rpанулярная эндоплаз
матическая сеть и комплекс fольджи. Это указывает на высокий уровень синтеза
белка в клетках. В апикальной части сероцитов концентрируются специфические
секреторные rранулы, содержа
щие амилазу и некоторые друrие
ферменты. Между сероцитами
выявляются межклеточные ceK
реторные канальцы. Миоэпители
оциты охватывают ацинусы напо
добие корзинок и лежат между oc б
нованиями сероцитов и базаль
ной мембраной. В их цитоплазме
имеются сократительные фила
менты, сокращение которых спо
собствует выделению секрета.
Вставочные отделы BЫBOД
ных протоков начинаются непос
редственно от концевых отделов.
Они имеют небольшой диаметр,
сильно разветвлены, выстланы
низким кубическим эпителием,
среди которых есть малодиффе
ренцированные камбиальные
1
, ' ,, ' , 'б , ! , ', ....; / . , " ' 4 : " ." , "',е ' ': , ' '''., ,,' ' " ,'
", О'' '..' 0)1 ","'
'. о ')(,t} &" ... '! с ' " ,О
:С?,С , i<Э ; ' ) ! 'C ..
., f"A' ' e/o '14" .S .,' .
О...," . ,: 1 1 ' ' /."".«! 'f .,. р
'( , > ," 9 ф\lЬil е I i" Q , "'""..'> "
'" . ' "\"_ \ ..(Q'Э 3 I) / . .
е .. .... ' ........,..:;:. ..
'е : .. ...... ........ '/'" ..I.!jJ->.: , ....
f):, . ' .. " ...o' ........ . I
4J ......;... . f.fI!.. ,:::,'e \ . -, . ф о ({) g .
,. "...... di# с:;> t 1" ...,. ф' If) .е
., .. ...E) J.. \ ti),.-at 1) 4
." ф' ........' "'$\..
i f) e 0<0" ( fЭ ' 7е::, . ; '. ,.
5 . f.ф; .h :Ф 4
е' \ , .... '. .. W,'\...
.. 0 ,.... .
2
Рис. 83. Строение околоушной слюнной железы:
1 концевой секреторный отдел; 2 вставочный проток;
3 исчерченный проток: 4 внутридольковый выводной
проток; 5 серозные rландулоциты; б миоэпителиоциты
Система Opla1IOB внутренней среды
167
клетки. Здесь, а также в исчерченных протоках встречаются миоэпителиоциты. Ис
черченные протоки имеют больший диаметр, широкий просвет и выстланы цилин
дрическим эпителием с выраженной оксифилией цитоплазмы. В базальной части
клеток выявляется исчерченность, обусловленная заКОНО:\1ерным расположением
митохондрий и rлубокими складками плазмолеммы. Эти клетки осуществляют
транспорт воды и ионов. В выводных протоках поодиночке или rруппами встреча
ются эндокринные клетки серотониноциты.
Поднижнечелюстные железы. По составу секрета эти железы относятся 1<
смешанным. Их концевые секреторные отделы бывают двух видов: белковые и бел
ково слuзистые. Преобладают белковые ацинусы, устроенные так же, как и в OKO
лоушной железе. Смешанные концевые отделы включают сероциты, составляю
щие так называемые серозные полулуния, и мукоциты. Имеются также и миоэпите
лиоциты. Мукоциты выrлядят более светлыми по сравнению с сероцитами. Ядро в
этих клетках лежит у основания, оно уплощено, а слизистый секрет занимает боль
шую часть цитоплазмы. Вставочные отделы короткие. Хорошо развиты исчерчен
ные протоки. Клетки исчерченных протоков синтезируют инсулиноподобный фак
тор и друrие биолоrически активные вещества. Эпителий междольковых прото
ков по мере увеличения калибра постепенно становится мноroслойным
Подъязычные железы. Это альвеолярно трубчатые железы, вырабатывающие
слизисто белковый секрет с преобладанием мукоида. В них имеются секреторные
отделы трех типов: белковые, слизистые и смешанные. Основную массу составляют
смешанные концевые отделы, образованные мукоцитами и полулуниями из сероци
тов. Вотавочные и исчерченные протоки в подъязычной железе развиты слабо.
Твердое и .мЯlкое нёбо
16ердое нёбо состоит из костной основы и плотно сращенной с ней слизистой
оболочки. Подслизистая основа здесь отсутствует. Слизистая оболочка включает
мноrослойный плоский неороrовевающий эпителий и собственную соединитель
нотканную пластинку, коллаrеновые волокна которой вплетаются в надкостницу.
МЯlкое нёбо и язычок имеют сухожильно мышечную основу, покрытую слизи
стой оболочкой. В них хорошо выражена подслизистая основа. На передней (pOTO
вой) поверхности эпителий мноrослойный плоский неороrовевающий. На зад
ней (носовой) поверхности мяrкоrо нёба и язычка эпителий мноrорядный
мерцательный. Основу язычка составляет поперечнополосатая мышечная ткань,
мышечные волокна в которой образуют ветвления и анастомозы.
Мяrкое нёбо и язычок участвуют в акте rлотания, закрывая собой носоrлотку
и препятствуя тем самым попаданию пищи в полость носа.
Миндалины
На rpанице ротовой полости и rлотки в составе слизистой оболочки расположены
скопления лимфоидной ткани в виде миндалин (нёбных, rлоточной, язычной, труб
ных, ropTaHHblx). Вместе они составляют лимфоэпителиальное rлоточное кольцо
168
Fлава 14
Пироrова. Миндалины выполня
ют защитную функцию, обезвре
живая микробы, попадающие с
воздухом и пищей. Миндалины
относятся к кроветворной и им
мунной системе и обеспечивают
развитие лимфоцитов.
Миндалины закладываются
[етерохронно в период с 3 ro по
S й месяц эмбриоrенеза. Mexa
низм их развития сводится к вза \.
имодействию эпителия, ретику .
лярной ткани и лимфоцитов. В Me ,
стах закладки нёбных и rлоточной
миндалин эпителий вначале MHO
roрядный мерцательный. Позднее ..
все миндалины оказываются по ..'.
крытыми мноroслойным плоским
неороrовевающим эпителием. В
подлежащую ретикулярную ткань
вселяются лимфоциты (преиму
щественно Т лимфоциты). Они
образуют мноroчисленные лuмфа
тические узелки, отделенные друr
от друrа тонкими прослойками co
единительной ткани. От поверхностноro эпителия образуются yrлубления, называ
емые криптами (рис. 84). Разветвляясь в rлубине миндалины, они образуют вторич
ные крипты. Эпителий в области крипт заселяется лимфоцитами и rpанулоцитами.
Инфильтрируя эпителий, лейкоциты обычно выходят на ero поверхность и фаroци
тируют попадающие сюда микробы. На дне крипт в миндалинах (например, в языч
ной) или BOKpyr миндалин (например, нёбных) открываются протоки небольших
слюнных желез, секрет которых способствует очищению крипт от продуктов фаrо
цитарной деятельности лейкоцитов, пищевых частиц и т. п.
"
.......\
I':;'
.,,
&. i";:;
"
';\:'
, 4
': '
. .'
:. ?
..С". ..
i'"
;.;.
ff'"
..i"" .....
"., ;
," '''''''''''''
"
,,: . ..
. \
-..1"' "!'''''' '''.
.
"
.... "'" 7',, -.
Рис. 84. Строение нёбной миндалины:
1 мноrослойный плоский неороrовевающий эпителий;
2 инфильтрация эпителия лимфоцитами; 3 крипта; 4
лимфатический узелок
rлот"а и пищевод
В rлотке перекрещиваются дыхательный и пищеварительный пути. Различа
ют носовую, ротовую и rортанную части rлотки. В носовой части слизистая обо
лочка покрыта мноroрядным мерцательным эпителием. В ротовой и rортанной ча
стях rлотки эпителий мноrослойный плоский неороroвевающий. Подслизистая
основа содержит концевые отделы слизистых желез. Мышечная оболочка преk
ставлена перечнополосатой мышечной тканью. Внутренний слой ее имеет продоль
ное направление волокон, а наружный циркулярное. Наружная адвентициаль
ная оболочка rлотки состоит из рыхлой соединительной ткани.
Система орюнов внутренней среды
169
Пищевод. Источником развития эпителия пищевода является материал пре
хордальной пластинки. Остальные ткани стенки пищевода за некоторым исклю
чением (см. ниже) развиваются из мезенхимы. Выстилка пищевода сначала преk
ставлена однослойным цилиндрическим эпителием. Затем этот эпителий
становится двухслойным. Сильно разрастаясь, он полностью закрывает просвет
пищевода. Последующий апоптоз клеток ведет вновь к образованию просвета, а
выстилка пищевода к З му месяцу эмбриоrенеза состоит из мноrорядноrо Mepцa
тельноrо эпителия. С 6 ro месяца эмбриоrенеза эпителий преобразуется в MHoro
слойный плоский с островками мерцательных или ороrовевших клеток Постепенно
усложняется строение и всех остальных тканей в составе стенки пищевода.
В стенке пищевода взрослою человека различают слизистую оболочку, ПОk
слизистую основу, мышечную и адвентициальную оболочки (рис. 85). Слизис
тая оболочка состоит из мноrослойноrо плоскоrо неороrовевающеrо эпителия,
собственной соединительнотканной и rладкомышечной пластинок В эпителии
ведущий клеточный дифферон
представлен эпителиоцитами,
находящимися на последова
тельных стадиях дифференци
ровки. Вместе с тем встречают
ся немноrочисленные клетки
Ланrерrанса и эндокринные
клетки. В собственной плас
тинке слизистой обuлочки пи
щевода в верхней и нижней ero
трети расположены 1(ардиалъ
ные железы пищевода, которые
по своему строению схожи с Ta
ковыми желудка. Это простые
разветвленные трубчатые же
лезы, концевые отделы KOTO
рых образованы однорядным
кубическим эпителием, в COCTa
ве KOToporo MHoro мукоцитов и
разновидностей эндокриноци
тов. Из этих желез возникают
кисты и опухоли.
Появление кардиальных
желез в пищеводе объясняется
меторизисом. Этим термином
В.М. Шимкевич (1908) обозна
чил смещение rраниц между
производными двух соседних
эмбриональных зачатков. По
лаrают, что в области перехода
1 ,""""
А
. . } 'б '.'..
. ,... ' .
,". - '... '
. :i :н J.....,. "::' ... з
!>'....'oj ,. .1 -... '" ...... $'""' , " A. ,""
, /:a&" ';' i."f :: .r': j;:---";'",
.; " ;; :'; ' 4 ,: ,'; .." ; . .> : 4
", . :' л 'l.piJ. ':," .
..., . t "'I:'f'; "'.....,.:. ':.',,' '. -
j - dP;, ';,,-,,;;,' . . ';.::
.' ' ;' " ' ' ..:'. '-: .:-; .
q.',ft;'l'i .. ,? .' .
'. , ,- У'.::':' 5
e:j-- , , i'#f'; ' t'.. ::. : .
>.f!I.' F . "... ....w...
... ..
.. ;: : '" . :..... ..
'"" .. ,
"".
............. '6 ...'
Б
в{
,
.-, '4"
I'J;'
;. ..'
Рис. 85. Строение стенки пищевода (нижняя треть):
А слизистая оболочка; Б подслизистая основа; В MЫ
шечная оболочка; 1 мноrослойный плоский HeoporoBeBa
ющий эпителий; 2 собственная пластинка слизистой обо
лочки; 3 мышечная пластинка слизистой оболочки; 4
собственные железы пищевода; 5 rладкие миоциты слоев
мышечной оболочки; б выводной проток собственной же-
лезы пищевода; 7 кардиальная железа пищевода
170
Тлава 14
пищевода в желудок материал кишечной энтодермы смещается в сторону пище
вода и дает начало железам желудочноrо типа в пределах слизистой оболочки
пищевода.
Мышечная пластинка слизистой оболочки состоит из продольно ориентиро
ванных пучков rладких мышечных клеток, изменение тонуса которых способству
ет прохождению пищи по пищеводу в желудок
В ПОДСЛИЗИСТOI':'f основе располаrаются собственные слизистые железы пище
вода производные эпителия пищевода. Это альвеолярно трубчатые железы. Их
концевые отделы образованы мукоцитами, постоянно выделяющими слизь. Pac
ширенные выводные протоки выстланы низким цилиндрическим эпителием, пе
реходящим в мноrослойный плоский эпителий. Среди ero клеток встречаются эн
докринные клетки серотониноциты.
Мышечная оболочка пищевода в верхней трети образована поперечнополоса
той мышечной тканью миотомноrо происхождения, в нижней трети rладкой Me
зенхимной природы, а в средней трети и той, и друrой мышечными тканями.
Эти ткани расположены в виде BHYTpeHHero циркулярноrо и наружноrо продоль
Horo слоев. Утолщения BHYTpeHHero циркулярною слоя мышечной оболочки об
разуют сфинктеры пищевода: верхний на уровне перстневидною хряща ropTa
ни и нижний в области перехода пищевода в желудок Мышечная ткань иннер
вируется преимущественно волокнами блуждающеrо нерва. Сокращение ее спо
собствует продвижению пищи из пищевода в желудок
Адвентициальная оболочка пищевода образована рыхлой волокнистой соеди
нительной тканью, содержащей кровеносные сосуды, нервы и нервные сплетения.
Реrенерация пищевода как физиолоrическая, так и репаративная выражена дoc
таточно хорошо. После хирурrических операций на пищеводе в связи с опух:олевы
ми процессами, ожоrами, ранениями и т. д. происходит обычно полное восстановле
ние эпителия и соединительнотканных структур в составе стенки пищевода.
Производные среднею и заднею отделов пищеварительной трубки
Средний, или rастроэнтеральный, отдел пищеварительной трубки включает
желудок, тонкую и толстую кишки, печень и желчный пузырь, поджелудочную
железу. В этом отделе происходят переваривание пищи под действием ферментов
желудочноrо и кишечноrо соков и всасывание необходимых для орrанизма пита
тельных веществ.
Желудок
Желудок выполняет ряд важных функций, связанных с химической перера
боткой пищи. Здесь под влиянием желудочною сока начинается активное хими
ческое расщепление пищи. Компонентами желудочноrо сока являются пепсин,
липаза, химозин, а также соляная кислота и слизь. Основной фермент желудочно
ro сока пепсин расщепляет сложные белки пищи на простые белки. Происходит
это только в кислой среде, что обеспечивается выработкой соляной кислоты Липа
Система OPZa11.06 6нутренней среды
171
за участвует в расщеплении жиров. Химозин вырабатывается в желудке только в
раннем детском возрасте он створаживает молоко.
Для нормальной деятельности слизистой оболочки желудка необходима защи
та ее от повреждающеrо действия соляной кислоты. Эту функцию выполняет слизь,
в состав которой входит нейтрализующее кислоту вещество (бикарбонат). Кроме
секреторной функции, желудок выполняет и экскреторную функцию, состоящую в
вьщелении через стенку в полость желудка ряда конечных продуктов обмена белков
(мочевины, аммиака и др.), а также солей тяжелых металлов. В желудке происходит
всасывание некоторых веществ (воды, спирта, солей, сахара и др.). Всасывательная
функция слизистой оболочки желудка, однако, оrраничена. Следует отметить еще
защитную (барьерную) функцию эпителия желудка, препятствующую проникнове
нию микробов в кровь, предупреждающую самопереваривание; моторную, осуще
ствляемую за счет сокращения мышечной оболочки, что важно для перемешивания
пищи и продвижения ее в двенадцатиперстную кишку. Эндокринная функция же
лудка имеет большее значение для реryляции пищеварения.
Развитие. Желудок закладывается на 4 й неделе эмбриоrенеза, но основные
процессы rистоrенеза происходят в течение 2 ю месяца. В это время энтодермаль
ный эпителий становится однослойным высокопризматическим. На протяжении
6 10 й недель формируются производные эпителия железы. Однако и к MOMeH
ту рождения процесс дифференцировки желудочных желез не завершается. Mы
шечная оболочка развивается из мезенхимы. Висцеральный листок спланхнотома
дает мезотелий. Окончательноrо развития желудок со всеми ero оболочками дoc
тиrает к 10 12 rодам.
Строение. В желудке взрослоrо человека различают следующие отделы: карди
альный, фундальный, тело желудка и пилорический отдел. Стенка желудка состоит
из слизистой оболочки, подслизистой основы, мышечной и серозной оболочек Сли
зистая оболочка желудка толщиной около 1 мм имеет неровную поверхность. Слож
ный рельеф ее обусловлен наличием складок, полей и ямок По малой кривизне же
лудка складки имеют продольное направление ( желудочная дорожка). Желудочные
поля это участки слизистой оболочки с отrpаниченными посредством бороздок
rpуппами желез. Желудочные ямки представляют собой мноrочисленные уrлубле
ния эпителия, в которые открываются по 2 3 железы. Общее число ямок достиrает
почти 3 млн. Внутренняя поверхность желудка покрыта однослойным высокоприз
матическим эпителием кишечною типа (рис. 86). Все клетки эпителия п06epXН0C
тные эпителиоциты, постоянно выделяют слизеподобный секрет. Слой слизи за
щищает слизистую оболочку от механических воздействий пищи и препятствует
самоперевариванию тканей желудочным соком. При действии раздражающих Be
ществ (спирта, кислот и др.) количество вьщеляемой слизи значительно увеличива
ется. Таким образом, поверхностный эпителий желудка представляет собой orpoM
ное железистое поле. Активная поверхность слизистой оболочки желудка во MHoro
раз увеличивается блаrодаря наличию мноrочисленных и разнообразных по CТPYK
туре желудочных желез собственных, пилорических и кардиальных.
Железы желудка. В желудочных железах различают шейку и rлавную часть,
состоящую из тела и дна. fлавная часть является секреторным отделом, а шейка
172
Fлава 14
ВЫВОДНЫМ протоком железы. В кардиальном, фун
дальном и пилорическом отделах желудка железы
имеют неодинаковое строение. КардuалЪ1lые железы
простые трубчатые железы с сильно разветвлен
ными концевыми отделами. Они располаrаются в
собственной пластинке слизистой оболочки карди
альноro отдела желудка. Эпителий кардиальных же
лез состоит из слизистых клеток (мукоцитов), а TaK
же единичных париетальных экзокриноцитов и
эндокриноuитов.
Со6сmве1l1lые железы желудка (фундальные)
это простые трубчатые неразветвленные железы, pac
положенные в области дна и тела желудка. Это наи
более мноrочисленные железы желудка. Общее их
число у человека составляет около ЗS млн. Шейка
этих желез содержит камбиальные клетки и шеечные
мукоциты. В эпителиальной стенке тела и дна фун
дальных желез различают rлавные и париетальные
(обкладочные)экзокриноциты,мукоциты,эндокри
ноuиты и малодифференцированные эпителиоuиты.
ПWlOрuческuе желеэы являются трубчатыми же
лезами с короткими и разветвленными концевыми
отделами. Они располаrаются в области привратни
ка. Между этими железами хорошо выражены про
слойки соединительной ткани слизистой оболочки.
Эпителий пилорических желез образован преимуще
ствепно мукоцитами и эндокриноuитами. Для пило
рических желез характерно то, что они открываются
в rлубокие желудочные ямки.
Эпителий желез желудка представляет собой BЫ
сокоспециализированную ткань, состоящую из He
скольких клеточных дифферонов, камбием для которых служат малодифференциро
ванные эпителиоциты в области шеек желез. Эти клетки интенсивно метятся при
введении зн тимидина, часто делятся митозом, составляя камбий как для поверхнос
THOro эпителия слизистой оболочки желудка, так и для эпителия желудочных желез.
Соответственно этому дифференцировка и смещение вновь возникающих клеток идут
в двух направлениях: в сторону поверхностноrо эпителия и в rлубину желез. Обнов
ление клеток в эпителии желудка происходит за 1 з суток Значительно медленнее
обновляются высокоспециализированные клетки эпителия желудочных. желез.
Fлав1lые Э'КЗОКрU1l0цumы вырабатывают профермент пепсиноrен, который в кис
лой среде превращается в активную форму пепсин rлавный компонент желу
дочноrо сока. Экзокриноциты имеют призматическую форму, хорошо развитую rpa
нулярную эндоплазматическую сеть, базофильную цитоплазму с' секреторными
rранулами зимоrена.
.......3
l ; ",)j!o#,r;
(''''t:-' _ '''I-Зi,''О:'''\
, ...",,' ,\' - " .
I .
j (\ :#,\,",,! .,:;,,1..
fI\ .- -"!.':w .С.... . :'s-".
..* '" ",' ' , . ;':1
'\ s.. т.. 81
,.,o ,\ .t' ) .......;,.L 1
'- :dt_i ; J;.: :
"t , 'C' , '\';''ii,,- .,:;; 8,.
:: , ? , f: ): } 2
1 !, 1 ,w/; .
.....t ..,.-".,'- .
tt:6 .'" !' *
I> l ' 'i :'.
'$У. .. i r-' :' ..
',.\." I''''->' 5
.. T;'-):j_ 't<:
' . rl ,,!':.' \:i
)': ,i!i\(it i; '
-(" .\- .",О{I' 1 'о
. .1:"1 ' \11 :''','
, .,f. "-" t }\- _\..
;'l ,, ' ' :"t.'(.. 4
6-- t.!"r. _;8 . ' .
.'(; t . ,..:
:.. ;: i\ ti ;It, :':
'. ..;.. 'f ;1. & \!<
.. >,\'.. ,1! , , ., ':"!: . J'i)
..,."''f: ,.. :;;;. 6
..; , :: ;;!.. !'i\.
, .. ... . . ,,,,,.} .
..,... . \ "
t \, .;.&''' ...r..
'\.. . ,.,.IW,
Рис. 86. Строение слизистой
оболочки дна желудка:
1 желудочная ямка; 2 шей.
ка железы; 3 поверхностные
эпителиоциты; 4 мукоциты
железы; 5 париетальные эк
зокриноциты; 6 rлавные эк
зокриноциты
Система орzтЮ6 61lутре1l1J.ей среды
173
ПарuеталЪ1lые ЭКЗ0крuuоцuты крупные, окруrлой или неправильно уrло
ватой формы клетки, расположенные в составе стенки железы кнаружи от rлав
ных экзокриноцитов и мукоцитов. Цитоплазма клеток резко оксифильна. В ней
содержатся мноrочисленные митохондрии. Ядро лежит в центральной части клет
ки. В цитоплазме есть система внутриклеточных секреторных канальцев, пере
ходящих в межклеточные канальцы. В просвет внутриклеточных канальцев BЫC
тупают мноrочисленные микроворсинки. По секреторным канальцам из клетки
на апикальную ее поверхность выводятся ионы Н. и CI , образующие соляную
кислоту. Париетальные клетки секретируют также внутренний фактор Кастла,
необходимый для всасывания витамина B t2 в тонкой кишке.
Мукоцuты слизистые клетки призматической формы со светлой цитоплаз
мой и уплотненным ядром, смещенным в базальную часть. При электронной микро
скопии в апикальной части слизистых клеток выявляется большое количество ceK
реторных rpанул. Располаrаются мукоциты в rлавной части желез, преимущественно
в теле собственных желез. Функция клеток выработка слизи.
ЭuдОКрU1l0циты желудка представлены несколькими клеточными дифферона
ми, для названия которых приняты буквенные сокращенные обозначения (ЕС, ECL,
С, Р, D, А и др.). Для всех этих клеток характерна более светлая цитоплазма, чем у
друrих эпителиальных клеток. Отличительным признаком эндокринных клеток
является наличие в цитоплазме секреторных rранул (рис. 87). Поскольку rpанулы
способны восстанавливать нитрат серебра, эти клетки называют арrирофильны
ми. Они интенсивно окрашиваются также бихроматом калия, с чем связано друrое
название эндокриноцитов энтерохромаффинные. На основании строения ceK
реторных rранул, а также с учетом их биохимических и функциональных свойств
эндокриноциты классифицируются на He
сколько видов.
ЕС КJlеткu самые мноrочисленные, распо
лаrаются в теле и дне железы, между rлавны
ми экзокриноцитами и секретируют cepOTO
нин имелатонин. Серотонин стимулирует
секреторную деятельность rлавных экзокри
ноцитов И мукоцитов. Мелатонин участвует в
реrуляции биолоrических ритмов функцио
нальной активности секреторных клеток в за
висимости от световых циклов.
ECL КJlemKU вырабатывают rистамин, KOTO
рый действует на париетальные экзокриноци
ты, реryлируя продукцию соляной кислоты.
С КJlеткu называют rастринпродуцирую
щими. В большом количестве они встречаются
в пилорических железах желудка. [астрин сти
мулирует деятельность rлавных и париеталь --:;'",Р:.-. ....;
ных экзокриноцитов, что сопровождается уси Рис. 87. Секторные rранулы (стрелки) в эн
ленной выработкой пепсиноrена и соляной докриноците железы желудка
-,.,
..;1'
. .
:
. J IJ-" I
1 . -, \
5'- "''''':.. '
f;"
I -
.....
"';,. i'
" .;' > r"f;
. "- ..
" ,.,t.'
").'t.-:.
S: 't
...,..-
...
...
.{
. ,
-, .
"'- f
? -:1
....
у,
.. .""...
..
\
:;, .
174
Fла6а 14
кислоты. У людей с повышенной кислотностью желудочноrо сока отмечается YBe
личение количества G клеток и их rиперфункция. Имеются данные о том, что G
клетки вырабатывают энкефалин морфиноподобное вещество, впервые обнару
женное в мозl}' и участвующее в реryляции чувства боли.
Р 'КЛеm1(U секретируют бомбезин, который усиливает сокращения rладкой MЫ
шечной ткани желчноrо пузыря, стимулирует выделение соляной кислоты парие
тальными экзокриноцитами.
D 'КЛеm1(U вырабатывают соматостатин инrибитор roрмона роста. Он yrHe
тает синтез белков.
ВИЛ 'КЛеm1(U продуцируют вазоинтестинальный пептид, расширяющий KpO
веносные сосуды и снижающий артериальное давление. Этот пептид стимулирует
также выделение ropMoHoB клетками островков поджелудочной железы.
А 'КЛеm1(U синтезируют энтероrлюкаrон, расщепляющий rликоrен до rлюкозы
подобно rлюкаrону А клеток островков поджелудочной железы.
В большинстве эндокриноцитов секреторные rранулы находятся в базальной
части. Содержимое rранул выделяется в собственную пластинку слизистой оболоч
ки и далее попадает в кровеносные капилляры.
Мышечная пластинка слизистой оболочки образована тремя слоями rладких
миоцитов.
Подслизистая основа стенки желудка представлена рыхлой волокнистой co
единительной тканью с сосудистыми и нервными сплетениями.
Мышечная оболочка желудка состоит из трех слоев rладкой мышечной ткани:
наружноro продольноrо, среднеrо циркулярноro и BHYTpCHHero с косым направлени
ем мышечных пучков. Средний слой в области привратника утолщен и образует пи
лорический сфинктер. Серозная оболочка желудка образована поверхностно лежа
щим мезотелием, а ее основу составляет рыхлая волокнистая соединительная ткань.
В стенке желудка расположены подслизистое, межмышечное и подсерозное
нервные сплетения. В rанrлиях межмышечноrо сплетения преобладают веrетатив
ные нейроны I ro типа, в пилорической области желудка больше нейронов II ro
типа. К сплетениям идут проводники от блуждающеrо нерва и из поrраничноrо
симпатическоrо ствола. Возбуждение блуждающеrо нерва стимулирует секрецию
желудочноrо сока, тоrда как возбуждение симпатических нервов, наоборот, yrHe
тает желудочную секрецию.
Тонкая кuшка
В тонкой кишке различают три переходящих друr в друrа отдела: двенадцати
перстную, тощую и подвздошную кишки. В тонкой кишке происходят дальнейшее
переваривание пищи, предварительно обработанной в ротовой полости и желудке,
и всасывание продуктов расшепления белков. жиров и уrлеводов. За счет COKpa
щений мышечной оболочки тонкая кишка выполняет механическую функцию, про
двиrая химус в каудальном направлении.
В переваривании пищи участвуют ферменты поджелудочной железы и экзок
РIIНОЦИТЫ эпителия тонкой кишки. Белки расщепляются под действием ферментов
Система ОрЮ1l0в в1lутре1l1lей среды
175
энтерокиназы, пептидазы, трипсина и др. Жиры переваривает липаза, а уrлеводы
амилаза, мальтоза, сахароза, лактоза, фосфатаза. Кроме химической обработки пищи
и всасывания продуктов расщепления, тонкой кишке свойственна эндокринная фун
кция. Эндокриноциты кишечноrо эпителия вырабатывают большую rpуппу биоло
rически активных веществ серотонин, мотилин, rистамин, секретин, энтероrлю
каroн, холецистокинин, rастрин и инrибитор rастрина. По образному выражению
А.М. Уrолева, двенадцатиперстная кишка является rипофизом пищеварительной си
стемы. Однако [ормоны и биолоrически активные амины, синтезируемые в кишеч
нике, имеют не только местное значение для пищеварительной функции орrанизма,
но мноrие из них иrрают важную общеметаболическую роль. Клетки, синтезирую
щие эти roрмоны, относятся к диффузной эндокринной системе орrанизма.
Развитие. Эпителий тонкой кишки начинает развиваться из кишечной энто
дермы на 5 й неделе эмбриоrенеза. К 8 й неделе он из однослойноrо кубическоrо
превращается в призматический эпителиЙ. В течение З rо месяца формируются
ворсинки и крипты. Дифференцировка
эпителиоцитов характеризуется после
довательным появлением столбчатых
(каемчатых и бескаемчатых) эпителио
цитов, бокаловидных экзокриноцитов и
эндокринных клеток. Среди последних
преобладают ЕС клетки. Параллельно с
развитием эпителия rистоrенетические
процессы происходят в остальных
структурах, составляющих стенку TOH
кой кишки, источниками развития KO
торых являются мезенхима и висце
ральный листок спланхнотома.
Строение. Стенка тонкой кишки
включает слизистую оболочку, подслизи
стую основу, мышечную и серозную обо
лочки. Слизистая оболочка состоит из
эпителия, соединительнотканной и rлад
комышечной пластинок. Рельеф слизис
той оболочки неровныЙ блаroдаря нали
чию складок, ворсинок и крипт (рис. 88).
КИШСЧ1lые вОРСИ1lКИ это выросты сли
зистой оболочки в просвет тонкоЙ киш
ки. КИШСЧ1lые -крипты трубчатые уrлуб
ления эпителия в собственной пластин
ке слизистоЙ оболочки. Количество ки
шечных ворсинок в тонкой кишке чело
века исчисляется несколькими милли
онами. Соотношение числа ворсинок и
крипт приблизительно равно 1:4. Высота
, :.... .. . ,А.
. .. / .
/':\ f. ':' ,
',' '",..... ..., .
А. .... ,;7" <r" . '; ,. -
)1" '. ":J .' \. !.."!.. ...., '" .."..
'.. .";1 ....\ 1':-.0'10" ...."'. .-:'-....:t-
Iр , \. \;, , :: , :t 1I.!:.t "" i :. , ..! ..
t:: 1 ":' .. .. JI '.' f. J.
' tI: '. ., .. , .."
t "; ' , " ; \ .'..\t.: \ :.i t'::; i :. \ ;1:
,.....! . .', t' ' " t' t ...
\ :,:. \<;':; . :, \ tj ' ,\'i :. $::
'. с:::. :.' r . .:. ,. J . . ' ' l '
'0' i;:',,:' :... 11:'- . ' J "; !.t ;.' ..
... '\ ." Jt.. . ".. . --"
'- ,\, ,,'. ' E ....;>i! .',. "
i- дt , Й :. Ч 1i,t , з:: --{.: ; ' .;. , : , '
f;f ,''.'.\ V,:, (i -,j 1\ . 1 :"!:
\ . , \ :O j' , ' , t:' l:: , ? \t :i.t \ y: l : @ " ,
о f. .. [\ '" . '. , " .... . .
... ..' "'... ... .. 1., . _ .__
-, .,..; i. '5." ...:[ .':" !';:" "':.
'J f,\ "i'i'-" " ' '.' 'i .:.... ;t , .#
tО:. ' Ч,: ;' \ : t .:;' \ ,',1 ' \ .., f :'1
, \ ,t' '\1' . :t '. O....' :
t ,: ',.l!'"I.'\; , .' n ..;..-' '. ! '. "::
1. ,; 1':' , .iH. ( "' , .', .' .'
,'J:.. .. , t 1t:} 13- f .:', .... .. . '.
\,' . i7 ;'-; iJ " :4 " ',: з: , " , . , < . {,\\ } '
'" . 1" ,. . · ..' ... ," " .
. \: ".;?'I:f . : ., . .). ':?:\ , '! l
! , .... "' . 2'; ). ",:,' <\ .
. ',е!'"",; ';А " .\.'
\ . '\ t ",, 'i :.. '4 \
i J 8. -м \ \. -I' \I...';
{I 'f "". .(' '\' . t..J-"!"'"
! . _ , i \ d.f;-; ; ;..;': I
l! ',,\-..\ ...;:..;;.... ,' ("'.f!:['
;-:;"- r X:J <'"( ..: :. :: 3
, 'lc --.;.. . , " ,.: i'i.'"" . itJ:.. 4,<. . if.j;,
. .":. .." . \ "",.' .' ,,' :,.,.-, 'р .
.,: :. ,''C:)'''' " /"'::' :;r:..;"',: ';;ic,.t!,' .f,
'".. . _ -- ....: : . ...:.,: ..;. ... 5 1' .. :. .. \:.l,"
I "": .""'''''' '''''I!I!'C:. ..:)._-
: { 1; -, 4
q",{....". ..... , ....., . ...... ........
.й; р : :."-..5
2
Рис. 88. Строение стенки тонкой кишки:
1 ворсинка; 2 крипта; 3 подслизистая осно-
ва; 4 мышечная оболочка; 5 серозная оболочка
176
Fлава 14
ворсинок около 1 мм. В двенадцатиперстной кишке они широкие и короткие. В TO
щей и подвздошной кишках ворсинки более высокие, но они несколько тоньше. В TOH
кой кишке общее число ворсинок превышает 1,4 млн.
Поверхность ворсинок образует одн.ОСЛОЙ1lЫЙ nрuзматuческий каемчатый эnu
телий, а основу составляет рыхлая волокнистая соединительная ткань, содержащая
кровеносные капилляры и один лимфатический капилляр. В строме ворсинок при
сутствуют rладкие мышечные клетки, которые обеспечивают до 4 6 сокращений
ворсинки в минуту, а также плазматические и тучные клетки, B и Т лимфоциты,
макрофаrи. Укорочение ворсинок, сжатие просвета их капилляров и ритмическое
проталкивание крови в капиллярах стромы ворсинок имеют существенное значение
в процессах всасывания и транспорта питательных веществ. В процессах BcaCЫBa
ния активная роль принадлежит, прежде Bcero, эпителию, покрывающему ворсин
ки, который составляет единую систему с эпителием крипты.
Всасывание питательных веществ происходит лишь после ферментативноrо
расщепления белков до аминокислот, жиров до моноrлицеридов, жирных кис
лот, rлицерина и уrлеводов до rлюкозы. Аминокислоты и rлюкоза, пройдя через
эпителиоциты ворсинки и подверrаясь при этом дальнейшим превращениям, по
ступают в кровеносные капилляры ворсинок. Всасывание жиров сопровождает
ся также сложными превращениями в эпителиоцитах с последующим поступле
нием липидов в лимфатические капилляры.
В кишечных криптах находятся камбиальные, а также стволовые клетки, за
счет которых происходит обновление клеточноrо состава эпителия крипт и BOp
синок. Фиrуры митоза встречаются преимущественно в средней части крипт. В
кишечном эпителии четко разrраничены места расположения малодифферен
цированных пролиферирующих и специализированных функционально актив
ных клеток. В процессе развития в результате диверrентной дифференцировки
из общеrо для всех клеток стволовой клетки образуется несколько различ
ных по структурно функциональным свойствам клеточных дифферонов: CTOk
бчатые эпителиоциты (каемчатые, всасывательные), бокаловидные экзокрино
циты (слизистые), клетки Панета и эндокриноциты. Миrрация этих клеток от
места пролиферации идет в двух направлениях. Первые два вида смещаются в
эпителий, покрывающий ворсинки. Клетки Панета и эндокриноциты миrриру
ют на дно крипт.
Столбчатые эnuтелuоцuты наиболее мноrочисленные клетки кишечноrо
эпителия, выполняющие основную всасывательную функцию кишечника. Эти
клетки составляют около 90% общеrо числа клеток кишечною эпителия. XapaK
терной чертой их дифференцировки является образование щетОЧ1l0Й каемки из
плотно расположенных микроворсинок на апикальной поверхности клеток. Дли
на микроворсинок около 1 мкм, диаметр, примерно, 0,1 мкм. Общее число микро
ворсинок на поверхности одной клетки колеблется в широких пределах от 500
до 3000. Микроворсинки покрыты снаружи rликокаликсом, который адсорбирует
ферменты, участвующие в пристеночном (контактном) пищеварении. За счет мик
роворсинок активная поверхность всасывания кишки возрастает в 30 40 раз.
Между эпителиоцитами в их апикальной части хорошо развиты контакты типа
Система орюнов внутренней среды
177
адrезивных поясков и плотных контактов. Базальные части клеток контактируют с
боковыми поверхностями соседних клеток посредством интердиrитаций и дeCMO
сом, а основание клеток прикрепляется к базальной мембране полудесмосомами.
Блаroдаря наличию этой системы межклеточных контактов кишечный эпителий
выполняет важную барьерную функцию, предохраняя орrанизм от проникновения
микробов и чужеродных веществ.
Бокаловидllые Э'К30КрИll0циты это по сути дела одноклеточные слизистые же
лезы, расположенные среди столбчатых эпителиоцитов. Они вырабатывают уrле
водно протеиновые комплексы муцины, выполняющие защитную функцию и спо
собствующие продвижению пищи в кишечнике. Количество клеток возрастает по
направлению к дистальному отделу кишечника. Форма клеток меняется в различ
ные фазы ceKpeTopHoro цикла от призматической до бокаловидной. В цитоплазме
клеток развиты комплекс rольджи и rранулярная эндоплазматическая сеть цeHT
ры синтеза rликозаминоrликанов и белков.
Клетки Панета, или экзокриноциты с ацидофильными rpанулами, постоянно
находятся в криптах (по 6 8 клеток) тощей и подвздошной кишок Общее число их
примерно 200 млн. В апикальной части этих клеток определяются ацидофильные
секреторные rpанулы. В цитоплазме выявляются также цинк, хорошо развитая rpa
нулярная эндоплазматическая сеть. Клетки выделяют секрет, боrатый ферментом
пептидазой, лизоцим и др. Полаrают, что секрет клеток нейтрализует соляную кис
лоту содержимоrо кишечника, участвует в расщеплении дипептидов до аминокис
лот, обладает антибактериальными свойствами.
Э1lдОКрИ1l0циты (энтерохромаффиноциты, арrентаффинные клетки, клетки
Кульчицкоrо) базальнозернистые клетки, расположенные на дне крипт. Они
хорошо импреrнируются солями серебра, имеют сродство к солям хрома. Среди
эндокринных клеток различают несколько видов, секретирующих различные [op
моны: ЕС клетки вырабатывают мелатонин, серотонин и вещество Р; S клетки
секретин; ЕСL клетки энтероrлюкаrон; I клетки холецистокинин; D клет
ки вырабатывают сома.тостатин, ВИП вазоактивные интестинальные пеп
тиды. Эндокриноциты составляют около 0,5% общеrо числа клеток кишечнorо
эпителия. Обновляются эти клетки значительно медленнее, чем эпителиоциты.
Методами rисторадиоавтоrрафии установлено очень быстрое обновление клеточ
Horo состава кишечноrо эпителия. Происходит это за 4 5 сут в двенадцатиперст
ной кишке и несколько медленнее (за 5 6 сут) в подвздошной кишке.
Собственная пластинка слизистой оболочки тонкой кишки состоит из рыхлой
волокнистой соединительной ткани, в которой определяются макрофаrи, плазма
тические клетки и лимфоциты. Имеются также как одиночные (солитарные ) лим
фатические узелки, так и более крупные скопления лимфоидной ткани arpera
ты, или rрупповые лимфатические узелки (пейеровы бляшки). Эпителий,
покрывающий последние, имеет ряд особенностей строения. В ero составе Haxo
дятся эпителиоциты с микроскладками на апикальной поверхности (М клетки).
Они формируют эндоцитозные везикулы с антиrеном и экзоцитозом переводят ero
в межклеточное пространство, [де располаrаются лимфоциты. Последующее раз
витие и образование плазматических клеток, выработка ими иммуноrлобулинов
178
Fлава 14
нейтрализует антиrены и микроорrанизмы кишечноrо содержимоrо. Мышечная
пластинка слизистой оболочки представлена rладкой мышечной тканью.
В подслuзucтой 0C1l0ве двенадцатиперстной кишки находятся дуоденальные (брун
неровы) железы. Это сложные разветвленные трубчатые слизистые железы. OCHOB
ной вид клеток в эпителии этих желез слизистые rландулоциты. Выводные протоки
этих желез выстланы каемчатыми клетками. Кроме Toro, в эпителии дуоденальных
желез встречаются клетки Панета, бокаловидные экзокриноциты и эндокриноциты.
Секрет этих желез участвует в расщеплении yrлеводов и нейтрализации соляной кис
лоты, поступающей из желудка, механической защите эпителия.
Мышечная оболочка тонкой кишки состоит из
BHYTpeHHero (циркулярною) и наружноrо (про
дольноrо) слоев rладкой мышечной ткани. В ДBe
надцатиперстной кишке мышечная оболочка TOH
кая и ввиду вертикальною расположения кишки
практически не участвует в перистальтике и про
движении химуса. Снаружи тонкая кишка покры
та серозной оболочкой.
Толстая кишка
В толстой кишке происходят интенсивное Bca
сывание воды, переваривание клетчатки с участи
ем бактериальной флоры, выработка витамина К
и комплекса витаминов В, выделение ряда Be
ществ, например, солей тяжелых металлов. В TOk
стой кишке всасывается около 7 л жидкости в CYT
ки (около 1 л слюны, 2 л желудочноrо сока, 0,5 л
поджелудочноrо сока, 1 л желчи, 1 л кишечноrо
сока и 1 2 л выпиваемой воды).
Развитие. Эпителий толстой кишки имеет
энтодермальное происхождение, кроме той части,
которая входит в состав выстилки промежуточной
и кожной зон прямой кишки. Здесь эпителий име
ет эктодермальный rенез. Остальные ткани име
ют мезенхимное (соединительные ткани и rлаk
кая мышечная ткань) и целомическое (мезотелий)
происхождения.
Строение. Стенка кишки образована слизистой
оболочкой, подслизистой основой, мышечной и ce
розной оболочками (рис. 89). В толстой кишке вop
синки отсутствуют, но крипты сильно развиты. Сли
зистая оболочка имеет мноrочисленные складки.
Эпителий слизистой оболочки толстой киш
ки однослойный призматический. Он состоит из
.
,-.:ч,. 1. и lit
« !.: t f! .j.t}.!} : \
" /1 l f ' { r,::, at
1 f,f {t . '\. <1 ц , j ,(
t ) ff. il' ' ::. , ! i 1
· 11 ' t · .
1: 1-.,. ' .
/ ., } I 1-!. .
, "i':fi Ф i,Al' :> i
\, . .. с.
:>- :' i'l i=i,": , . , У' , '
0,,_ .' 't ..... "
...... ......'t.' .......,
........ ,,,,..... ,(! " .4. .........
.., .,... .' .", '--,'"
: <!с., . t""1i Jf::::";;' .f:.;;.!
.. f"I' ><'",. '......
. ,. , fJl "'"
2 , ' "" v 10-- t_.J ".....
\.)'-""' ""'., :,,' ;
, "::'"-( );:, . ;.. ... .......
.. ....... "(,I. ' ,
V.. _. '\.... ...... .'1< \..,
." " 8. - .'_ ' ,, O# '.-c:
... \S ,." . ' ,; " .'$.,..... "" '"? ....'
_,,'#- .... _.4 _ ..A_ '-" .., '*" ... itlI<
... , " , ,' '.. \> р.; ,41-". "
. " -.. ' J!r.". '" l' '",0 -
) _ _ , -' I' _ _ .,*";",, _ _ ,"
> .. ','" .. \> ,,:''.А!.: *"."
.. 615 _ J ,_. ,V"t "W .:в:"I: .
ft-:'<. f :.,_, .,:.n: ,'!.. " ot . ·
з .f .,:. '''v tf',."...
. .,> .. ... >,,,;;, ; : /,'
-, . .:- e,.,., ... ..,. ; _ . .....
'..... ' :..: . ,.'.?:' . ' _ _',,,О
... -;",_. f' 't"'X.
/ ,.;:-. .. ;>'" .... ...., . , '
..' ":":.. .....: .
4 ...... .... " ....
, ...
,
.".
Рис. 89. Строение стенки толстой
кишки:
1 крипта; 2 подслизистая основа;
3 мышечная оболочка; 4 серозная
оболочка
Система орюнов внутренней среды
179
столбчатых эпителиоцитов (каемчатых и бескаемчатых), бокаловидных экзокрино
цитов, отдельных эндокриноцитов (преимущественно EC и ЕСL клеток) и мало
дифференцированных (камбиальных) клеток Наличие orpoMHoro количества бока
ловидных экзокриноцитов находится в корреляции с необходимостью выделения
слизистоrо секрета, который облеrчает проведение пищевых масс по кишечнику. В
собственной пластинке слизистой оболочки сильно развит защитный механизм от
микробов. Здесь имеются мноrочисленные лимфоидные скопления. Мышечная пла
стинка представлена циркулярным (внутренний) и продольным (наружный) слоя
ми rладких миоцитов.
В рыхлой волокнистой соединительной ткани подслизистой основы распола
rаются нервные и сосудистые элементы, скопления липоцитов.
Мышечная оболочка толстой кишки отличается от таковой у тонкой кишки:
сплошной циркулярный слой покрыт снаружи несплошным (в виде трех отдельных
лент) и более коротким продольным слоем. За счет разницы длины мышечных слоев
по ходу кишки формируются вздутия юустры, необходимые для медленноro пере
мещения содержимоro кишки, более полноrо всасывания из Hero воды и формирова
ния каловых масс. В серозной оболочке возникают скопления жировой ткани.
Физиолоrическая и репаративная реrенерация тканей стенки толстой кишки
происходит достаточно интенсивно. Клеточный состав эпителия обновляется ПОk
ностью за 4 S сут.
При травматических ранениях, химических ожоrах и, особенно, при лучевом по
вреждении в кишечнике резко замедляются или прекращаются проuессы пролифера
ции камбиальных клеток в кишечных криптах и железах, нарушаются дифференци
ровка эпителиоuитов и rлаIЩУЛОЦИТОВ, их интеrрация, резко замедляется миrpация
клеточных элементов из крипт на поверхность ворсинок, отмечаются уплощение, rи
бель и слущивание покровноrо секретирующеro эпителия. При этом обнажается co
единительнотканная строма складок, ворсинок, нарушается защитная функция эпи
телиальной выстилки. Репаративные процессы начинают развиваться в криптах и
железах, rде из сохранившихся камбиальных клеток за счет их пролиферации и пос
ледующей дифференuировки постепенно восстанавливается эпителиальный пласт.
Червеобразный отросток
Червеобразный отросток представляет собой дивертикул слепой кишки. Слизис
тая оболочка ero включает каемчатый призматический эпителий с хорошо развитыми
криптами (рис. 90). Кроме каемчатых и бескаемчатых столбчатых эпителиоцитов, в
эпителии встречаются бокаловидные экзокриноциты, клетки Панета и ЭIЩокриноци
ты (EC , s , D и друrие виды). Собственная пластинка слизистой оболочки образова
на рыхлой волокнистой соединительной тканью. В ней, а также в подслизистой OCHO
ве располаrаются крупные скопления лu.м.фоидной ткани. Иноrда лимфатические
узелки, сливаясь дрyr с друroм, полностью окружают просвет отростка и даже пере
крывают ero. При попадании в просвет и ткани червеобразноro отростка микробов
может произойти воспаление, что потребует хирурrическоrо удаления еro (аппеIЩЭК
томии). Лимфоидная ткань аппендикса участвует в защитной функции орrанизма
180
[лава 14
вместе с друrими лимфоидными обра
зованиями в составе стенки кишечной
трубки (см. выше).
1 2
,.:.:;' .,, '< ? /. j E't .
. : ;'. .s;.-:.;o,.;!;..... , ''''''', i 1.Ji'
;с. :... . . .....,.. .4>...... ... 'Ф: .... 0)"', -' .. у
:J; .,: rJ ; : .. .\.. J..:\ .. . '.'. -1'"'
\: . ' jil: 0 .. fj /;; ' l i.. п :: _: ..' ' '., t ';.; ;
..:;; ,, . ... : t}, .,; . "а, '
. ;.<>;>; !:! . ",.:t.,""?" ';;{ t . .. 3"':' :. '
t:.:;{ _ -.':'z .. ...F:., 1 . .' .'.- "Тс'
Каудальная часть прямой кишки ....:.;1;. :: '. -Ф ' t-I!.' : '11.,1' . " .: .,;.:t.
. .... .1,\0. '. .... '5,;f'.:". l,ft : -'V
составляет задний отдел пищевари- :..i....) ,: ".; ,_..:':-:< '" . tфi (t <'. ...'
тельно и U трубки. В анальнои U ее части ".:<'\8.r;: :!!'I' ;N. ;: 'flC. :- t;;J ",.
. ()... .....-;"у. f". Z?:J:':"'" " " ,""'-1""..'"''
различают столбчатую, промежуточ- f.... Ж fii. }' ; ; ;:: . , .::;. ...,: ;'\ -: t > , "' ' ;
...... ." r .'... .(.. ..':C'':;I< '" 'Ii'." .. ..... ;. : .. /'
С б ..' .. r .."'" t. .:!:;,........" . Uf" \- . ..... ......... ."",t..." .
ную И кожную зоны. лизистая о o ':., ,.::,.. r ',: if ,; ;., <' ;;, ". . .'. . .' : . t ;>' I
., #. .. '.:: ,J.;..\:} .j,./f!!!t>.I' :: ". " ' . "'f::'.
лочка в этих зонах состоит из эпите ;. :.... ;:"';' ;... .:м:"'.:.. 'i.. . ,:
лия, собственнои U И мыIечноиu u плас . ,'" . . . !: l' ..z 1"'; .. . \ 'ttf{! i :" -;'
"t": ;.' \." .. . y.. t }?t;g, t1:: .j:/!;.{.. -". '\[1 1
Э б "' \ . ., . .+,-40!" g. ; / r.... . f'". > ,.,.;"", . \ о.
тинок. пителий в стол чатой зоне ..' . .' . ,.: .. _"' I''''''.-J,' .. .. .,.:. -. . .,'. u'.'.
'.'- . .l'<';.? :",:а":\:."",,, ,,:,,!,,,, , ч,:,, (" '
;: E:;o = :i /' :-.;': (:.' : ':< ':k.;;\, \ \.
. . <. _.,.__.:':." \о. ';'Ф '\. ... :'. \:, :( .
мноrослойный плоский О р оrовеваю '. ",', '.' ..' '. . ....;. .:.
'-о ;o'\t f14'-' ..
щий. В собственной пластинке распо ..... . , :. "'. ;r.:.i , .-: '.. ::, , . ; . ,:..
ложены одиночные лимфатические .. ..."., .'.. -.....;;"'. _ .
узелки. В связи со слабым развитием .. ...... «' y -';;'.;: ': :: :.........->5
мышеной пластинки здесь нет четко ."-..: :.. ..
"-- ....' '"
ro разrраничсния собственной плас .
Рис. 90. Строение стенки червеобразноrо отростка:
тинки слизистоЙ оболочки и подсли 1 однослойный призматический эпителий; 2
зистой основы. Они содержат TOHKO крипта; 3 собственная пластинка слизистой обо
стенные lеморроuдалЪ1lые веиы. В ре- лочки; 4 подслизистая основа; 5 мышечная обо
лочка; 6 серозная оболочка; 7 лимфатический
зультате варикозноrо расширения узелок в подслизистой основе; 8 лимфатические
этих вен и их выпячивания в просвет узелки в собственной пластинке слизистой оболочки
анальноrо канала возникает заболевание rеморрой.
Мышечная оболочка содержит внутренний (циркулярный) и наружный (про
дольный) мышечные слои. За счет утолщения циркулярноrо слоя формируются
сфинктеры прямой кишки. Наружный сфинктер образован поперечнополосатой
мышечной тканью. Серозная оболочка покрывает прямую кишку лишь в верхнем
отделе. В нижнем отделе кишки вместо серозной оболочки имеется соединитель
нотканная оболочка.
Прямая кuшка
6
Поджелудочная железа
Поджелудочная железа состоит из экзокринной И эндокринной частей. Экзок-
ринная часть выполняет внешнесекреторную функцию, связанную с выработкой naи
креатuческою сока. Он содержит пищеварительные ферменты, трипсин, химотрип
син, карбоксилазу, липазу, амилазу и др. Панкреатический сок по выводным протокам
поступает в двенадцатиперстную кишку, rде ero ферменты участвуют в расщепле
нии белков, жиров и уrлеводов. Эндокринная часть вырабатывает юр.мо1lы (инсу
Система ОРШНОВ внутренней среды
181
лин, rлюкаroн, соматостатин, панкреатический полипептид и др.), реrулирующие
уrлеводный, жировой и белковый обмен веществ в орrанизме.
Развитие. Энтодермальный зачаток железы появляется в конце 3 й недели
эмбриorенеза. Вначале образуются дорсальное и вентральное выпячивания CTeH
ки туловищной кишки, которые вскоре срастаются в одну закладку opraHa. На 3 M
месяце эмбриоrенеза энтодермальный материал подразделяется на экзокринную
и эндокринную части железы. Из эпителиальных разрастаний зачатка железы CHa
чала образуется система выводных протоков, а затем концевые секреторные OT
делы пан'/(реатическuе ацинусы. Последними из камбиальных клеток терминаль
ных участков выводных протоков развиваются эндокринные элементы поджелу
дочной железы '/(лет'/(и остров'/(ов Ланzершнса. rистоrенез как экзокринноrо
(ацинарноrо), так и эндокринноrо (инсулярноrо) эпителиев происходит в услови
ях взаимодействия этих тканей с мезенхимой. В паренхиме железы развиваются
мноrочисленные кровеносные капилляры. К моменту рождения обе части подже
лудочной железы приобретают достаточно дифференцированное строение. OДHa
ко и в постнатальном периоде происходят процессы дальнейшеro структурно фун
кциональноro усложнения этоrо opraHa.
Строение. В железе различают rоловку, тело и хвост. Посредством протоков
поджелудочная железа сообщается с двенадцатиперстной кишкой. Поджелудоч
ная железа состоит из эпителиальной паренхимы и соединительнотканной cтpo
мы. Паренхима подразделена на дольки, между которыми находятся прослойки
рыхлой соединительной ткани, кровеносные сосуды, нервы.
Экзокринная часть. Это сложная альвеолярно трубчатая железа, имеющая доль
чатое строение. Основную ее массу образуют структурно функциональные едини
цы пан'/(реатuчес'/(uе ацинусы, отделенные друr от друrа прослойками рыхлой co
единительной ткани. Ацинус
состоит в основном из пaH'/(pe
атичес'/(uxэк.зо,/(риноцитов, или
ациноцитов, имеющих форму
усеченноrо конуса. Базальные
части этих клеток широкие,
апикальные суженные. Pac
полаrаясь в один ряд, 8 12 аци
ноцитов, а также несколько
клеток вставочноrо отдела фор 4
мируют ацинус, имеющий вид
окруrлоro или овальноrо обра
зования. В центре ацинуса Ha
ходится небольшой просвет
(рис. 91). На поверхности кле
ток, обращенной в просвет, име
ются короткие микроворсинки.
Боковые поверхности рядом
лежащих клеток в апикальной
б
Рис. 91. Строение экзо и эндокринноrо отделов поджелудо'l
ной железы:
1 ацинус; 2 внутридольковый выводной проток; 3
междольковый выводной проток; 4 островок Ланrерrанса;
5 кровеносный капилляр; б центроацинозные клетки
182
Тлава 14
их части соединены при помощи плотных контактов и десмосом. Базальная часть
ациноцитов отличается базОфWlUей. Здесь расположена развитая rpанулярная эн
доплазматическая сеть, в которой синтезируются ферменты панкреатическою сока.
Имеются также мноrочисленные митохондрии. Эту часть клетки называют rOMO
rенной зоной. Апикальная часть ациноцитов О'КСUфWlъ'Н.а здесь в форме зимоrена
накапливаются синтезированные в клетке пищеварительные ферменты трипси
HoreH, химотрипсиноrен, прокарбоксипептидазы и др. Окруrлое ядро располаrается
обычно в средней части ацинарных клеток
Структура ациноцитов подверrается изменениям в различные фазы ceKpeTOp
Horo цикла. При юлодании количество rpанул в цитоплазме увеличивается. Через
несколько минут после приема пищи отмечается интенсивный выход секреторных
rранул из клеток Выделение секрета происходит путем мерокринной секреции.
В состав ацинуса кроме ceKpeTopHoro входит и вставоч'Н.ый отдел в боль
шинстве случаев часть клеток вставочноrо протока как бы вдвинута внутрь ацину
са. При этом на срезе в центре ацинуса видны мелкие клетки, образующие стенку
вставочною отдела. Их называют це'Н.троацu'Н.оз'Н.ымu эпuтелuоцuтамu. Форма их
неправильная, уплощенная. Узкий слой цитоплазмы окружает овальное ядро. На
поверхности клеток, обращенной в просвет ацинуса, образуются немноючислен
ные микроворсинки. Вставочные отделы переходят в межацинозные протоки, BЫ
стланные однослойным кубическим эпителием. Соединяясь вместе, межациноз
ные протоки образуют внутридольковые протоки, переходящие в междольковые.
Последние впадают в общий выводной проток поджелудочной железы. Он OTKpЫ
вается в двенадцатиперстную кишку. Стенка ero выстлана однослойным высоким
призматическим эпителием. В устье протока имеется rладкомышечный сфинктер.
По ходу эпителиальной выстилки протоков поджелудочной железы встречают
ся слизистые бокаловидные экзокриноциты и эндокриноциты. Последние выраба
тывают ropMoHbI холецистокинин и панкреозимин, которые стимулируют про
дукцию пищеварительных ферментов экзокринной частью поджелудочной железы,
увеличивают секрецию желчи, сокращают мышечную ткань желчноrо пузыря, воз
буждают моторику кишки и т. д.
Эндокринная часть. Островки состоят из эпителиальных клеток пa'Н.'Кpea
тuчес'Кux э'Н.до'Крu'Н.оцuтов, или инсулоцитов. Величина островков, их форма и чис
ло входящих в состав клеток очень различны. Общее количество островков в ПОk
желудочной железе достиrает 1 2 млн. Средний размер островка О, 1 0,3 мм. Общий
объем эндокринной части составляет около 3% всею объема железы. Островки про
низаны кровеносными капиллярами, окруженными перикапиллярным простран
ством. Эндотелий капилляров имеет фенестры, облеrчающие поступление юрмо
нов от инсулоцитов В кровь через перикапиллярное пространство.
В островков ом эпителии различают 5 видов клеток: А клетки, В клетки,
D клетки, ВИП клетки, РР клетки.
А 'Юlет'Кu (альфа клетки, или ацидофильные инсулоциты) это крупные OK
руrлые клетки с бледным крупным ядром и цитоплазмой, содержащей ацидофиль
ные rранулы. rранулы обладают и арrирофилией. В состав этих rранул входит rop
мон lJlю'КаlO'Н., расщепляющий rликоrен и повышающий содержание сахара в крови.
Система ОрЮН0в в1lутре1l1lей cpeды
183
А клетки рассеяны по всему островку, образуя нередко небольшие скопления в цeH
тральной части. Они составляют около 20 25% от всех инсулоцитов.
В клетки (бета клетки, или базофильные инсулоциты) имеют кубическую или
призматическую форму, крупное темное, боrатое rетерохроматином ядро. Доля
В клеток достиrает 70 75% от общею числа инсулоцитов. В цитоплазме В клеток
накапливаются осмиофильные rранулы, содержащие ropMoH И1lСУЛИ1l. Инсулин pe
ryлирует синтез rликоrена из rлюкозы. При недостатке продукции инсулина rлюко
за не превращается в rликоrен, содержание ее в крови повышается и создаются усло
вия для развития заболевания, называемоrо сахарным диабетом.
D клетки (дельта клетки, или дендритические инсулоциты) составляют 5 10%
среди всех островковых клеток Форма их иноrда звездчатая с отростками. В цитоп
лазме определяются rранулы средних размеров и плотности. В rранулах накаплива
ется ropMOH соматостатИ1l. Он тормозит секрецию инсулина и rлюкаrона, снижает
продукцию ряда ropMoHoB желудочно кишечноrо тракта rастрина, секретина, эн
тероrлюкаrона, холецистокинина и др., подавляет секрецию соматотропноrо ropMo
на в rипофизе.
ВИП клетки (арrирофильные клетки) встречаются в островках внебольшом
количестве. В цитоплазме выявляются плотные арrирофильные rранулы, coдep
жащие вазоактив1lый и1lтести1lшlъ1lый полипептид. Он обладает выраженным co
судорасширяющим дейтвием, снижает кровяное давление, yrHeTaeT секрецию co
ляной кислоты В желудке, стимулирует выделение rлюкаrона и инсулина.
РР клетки полиrональной формы инсулоциты, расположенные преимуще
ственно по периферии островка. Количество их 2 5% от общеrо числа клеток OCT
ровка. В цитоплазме РР клеток выявляются мелкие rранулы, содержащие пaH'Кpea
тичес'Кий полипептид. Основная роль панкреатическоrо полипептида в орrанизме
реryляция скорости и количества экзокринной секреции поджелудочной железы и
желчи в печени. Таков клеточный состав OCTpoBKoBoro эпителия, представляющеrо
собой мозаику диверrентно развивающихся клеточных дифферонов.
В дольках поджелудочной железы встречаются еще аци1l0з1l0 и1lсуляр1lыe клет
КИ, в цитоплазме которых одновременно содержатся rранулы, характерные как для
ацинозных, так и для островковых клеток
Ткани поджелудочной железы иннервируют блуждающий и симпатический
нервы. В интрамуральных веrетативных rанrлиях находятся холинерrические и
пептидерrические нейроны, волокна которых заканчиваются на клетках ацинусов
и островков. Между нервными клетками rанrлиев и остров ковы ми клетками YCTa
навливается тесная связь с образованием нейроинсулярных комплексов.
С возрастом в поджелудочной железе постепенно уменьшается количество oc
тровков. В островках наблюдаются закономерные возрастные изменения клеточ
ных взаимоотношений, заключающиеся в быстрой смене после рождения преоб
ладан ия А клеток над В клетками на преобладание В клеток над А клетками у
взрослых. Затем происходит постепенное увеличение количества А клеток, KOTO
рое наряду с одновременным, хотя и незначительным, уменьшением числа В кле
ток приводит нередко в пожилом и особенно старческом возрасте вновь к преобла
данию А клеток над В клетками.
184
rлава 14
Реlенерация. В эмбриorенсзе островки растут блаrодаря пролиферации ис
ходных клеток преДlIIеСТRенниц и их диверrентной дифференцировке в COOTBeT
ствующие клеточные диффероны. У взрослых физиолоrическая реrенерация аци
нозных и островковых клеток происходит в основном путем внутриклеточноrо
обновления орrанелл. Митотическая активность клеток в связи с высокой специа
лизациеЙ низкая. После резекции части или повреждения opraHa наблюдается He
которое повышение уровня пролиферативной активности клеток ацинусов, про
токов и островков, последующее образование новых ацинусов. Однако ведущей
формой реrенсрации экзокринной части железы является реrенерационная rипер
трофия. Восстановительные процессы в эндокринной части железы происходят за
счет пролиферативной активности инсулоцитов и клеток эпителия протоков пу
тем ацино инсулярной трансформации.
Печень
Печень самая крупная железа человека ее масса составляет около 1,5 Kr.
Она выполняет мноrообразные функции и является жизненно важным opraHoM.
Чрезвычайно важными для поддержания жизнеспособности орrанизма являются
метаболические функции печени, в связи с чем ее называют биохимической лабо
раторией орrанизма. В печени образуется желчь, необходимая для всасывания
жиров и стимуляции перистальтики кишечника. В сутки выделяется около 1 л
желчи. Печень является opraHoM, выполняющим роль депо крови. В ней может
депонироваться до 20% всей массы крови. В эмбриоrенезе печень выполняет KpO
ветворную функцию.
Развитие. Зачаток печени возникает в конце 3 й недели эмбриоrенеза из эн
тодермальной выстилки вентральной стенки средней кишки. Выпячивание этой
стенки разрастается, образуя эпителиальные тяжи в мезенхиме брыжейки. Позднее
тяжи подразделяются на краниальный и каудальный отделы, из которых COOTBeT
ственно формируются печень и желчный пузырь с протоками. В rистоrенезе про
исходит rетерохронная диверrентная диффсренцировка печеночных эпителиоци
ТОЕ (rепатоцитов) и эпителиоцитов желчных проточков (холанrиоцитов). Начиная
со второй половины эмбриorенеза, в печени формируются структурно функцио
нальные единицы печеночные дольки. Образование долек это результат слож
ных взаимодеЙствий между эпителием и внутрипеченочной соединительной TKa
нью с развиваюшимися синусоидными кровеносными капиллярами.
Строение. В печени различают эпителиальную паренхиму и соединительнот
канную строму. Структурно функциональными единицами печени являются ne
че1l0Чllые долыcu числом около 500 тыс. Печеночные дольки имеют форму шести
rpaHHbIx пирамид с диаметром до 1,5 мм и несколько большей высотой, в центре
котороЙ находится центральная вена (рис. 92). В связи с особенностями rемомик
роциркуляции rепатоциты в разных частях дольки оказываются в различных yc
лов иях обеспечения кислородом, что отражается на их строении. Поэтому в доль
ке выделяются центральная, периферическая и находящаяся между ними проме
жуточная зоны. Особенностью кровоснабжения печеночной дольки является то,
Система OpZa1-Ю6 6нутренней среды
185
.,.:-.. ,i\: :L. ..,;,"{ ...,...,.;,.,..:.t!Y..(i .,";''' -J_. ' i*:;'l. __J- . ' -:.t...:.,, "" C ""
< ::;. : '::" ::.; : :. .: ?;.. : :.;. : :., ..;: z \'
. ..:; . .' -..." 11'._ #", "'Ф ""О .';...--
"'....:;. '- .......,.,"'".... . ...").;...(f',..t".......-...... ........ () . ", "
:- :<tc...*.k , '''' ''"..;'t/ > ... :.,..-::.. ........ f! J l' .. ;:--'l...
t'f'." ..е>" ... ,..........T...4. r.... <' ........,, 1..1" .. ....
'r : '" ;Jf',"1 ...': ;::;. :fi;:.., I ", :
'.. ...A :j"'.. :::...':'.....:.},;. : 5 \'''-:
1;"':; ....... 1':"'- . : ;....'\...""1-:*":'...f.....'": ..'" :... : '" ;, \::.z. / ., , ....:
,.,,'" -... .... ........;..-i'..."'....:',.'!,. ..;." -;".. .: ,," '\ 1 ,
. , .. .: .. ;...;......:.....::..; :.....-'1... ........,:... : '1 о.'....
. ............ ...;...."#'"::.1f.'... .. ...:.#'..:,;... " .... ..t::r '- " .... '!"
1.":;, >О,....,)........ ->< """" ...cJ'",I.",'i. ,.:""" .. 4
.. .. ">i,." ..,, , : '!:... .' '<'''-..'' ,..."1-"""....... .. ,( . .
iII#....""'. ,.,...... 1:.t""'...-:....",.з..... "".... н "," . Ii; .... .' f
.. j#< .: :«-'<}:.... ; .;;:;. . ...."'-;.:.;...11..'('&.,;" ...:'",1." ......., \.
'. c ...:t.l''': .,"'... 'f'" ,..."';,,;,:r.....ч..,.. . >t..r:.. ........ . '" .
."" .. ".:""j. ..: f::r..;' ...---..:::r:b...: .....,: :..,.. .....'.. ri ,'"
:t >iJ, . :!,;:::.;;.:(,: .....: .: : :' ,.:.. .-"'\.:" ,,' : .:. ,\\:). J., ............... 6
p.(!''' , l.. ..:;.; ..."' ' 'J::;j ,I-'. : "' .:: ,., ,., ':." .
,.......;/r: . , 1':"",., ",-" !.:\ ..: . ..- ." :.. .''f....'..:.. .. ........: ,
..", ..r:....:,t H \-:.... #!Io"'"". '.... -I". f. ... ... ".,. ......... ....." ..........
: ;'1:"':.,/ :\.. ......'..... Jf::' ;:"''':'j (,,, , . I.:: .. .., .. . ; :: :
'!Ii "',. "... .. ..Ф..... ' :,,4':" ..".- .. :... J{o ... ..." o.- '"
....._ :: :. J « :u">,: . \.. {! . r ""4tl : :'" " ;:-: \". ,.; 2
:t "';. ' ..i' -.IJ..?": -\.,,:: (::....;: ..\i ;, : I :,.:: .
_. .. ...::....) ; ..' ,:-<\!:; ; ..ti;tf'i:: . .. #, .. . Ji
1
что отходящие от вокруrдолько
вой артерии и вены внутридоль
ковые артерия и вена сливаются
и далее смешанная кровь по [e
мокапиллярам перемещается в
радиальном направлении по Ha
правлению к центральной вене.
Внутридольковые rемокапилля
ры идут между печеночными
балками (трабекулами). Они
имеют диаметр дО ЗО мкм И OT
носятся К синусоидному типу Ka
пилляров.
Таким образом, по внутри
дольковым капиллярам смешан
ная кровь (венозная из систе
мы воротной вены и артериаль
ная из печеночной артерии)
течет от периферии к центру
дольки. Поэтому rепатоциты периферической зоны дольки оказываются в более
блаrоприятных условиях снабжения кислородом, чем таковые в центре дольки.
По междольковой соединительной ткани, в норме слабо развитой, проходят
кровеносные и лимфатические сосуды, а также выводные желчные протоки. Как
правило, междольковая артерия, междольковая вена и междольковый выводной
проток идут вместе, образуя так называемые триады печени. Собирательные вены
и лимфатические сосуды проходят в некотором отдалении от триад.
Эпителий печени состоит из zепатоцитО6, составляющих 60% всех клеток пе
чени. С деятельностью rепатоцитов связано выполнение большей части функций,
свойственных печени. При этом нет строrой специализации между печеночными
клетками и потому одни и те же rепатоциты вырабатывают как экзокринный ceK
рет (желчь), так и по типу эндокринной секреции мноrочисленные вещества, по
ступающие в кровоток
rепатоцитbl имеют неправильную мноrоуrольную форму. Средний диаметр
клеток 20 25 мкм. Различают апикальную (билиарную) поверхность rепатоци
та, обращенную в просвет желчноrо капилляра, и базальную (васкулярную) по
верхность в сторону синусоидноro капилляра. Своими латеральными поверх
ностями rепатоцитоты формируют печеночные балки. В центральной части клетки
лежит OДHO ДBa окруrлых ядра. Часть из них представляет собой крупные, поли
плоидные ядра. Причем число таких ядер увеличивается с возрастом и может дoc
тиrать в старости 80%. В цитоплазме хорошо развита rранулярная эндоплазмати
ческая сеть, участвующая в синтезе белков крови. Метаболизм уrлеводов связан с
rладкой эндоплазматической сетью, которая рассеяна в цитоплазме в виде трубо
чек и пузырьков. Вблизи этих элементов rладкой эндоплазматической сети выяв
ляются rранулы rликоrена. Цитоплазма rепатоцитов изобилует митохондриями,
Рис. 92. Строение печени:
1 печеночные балки; 2 внутридольковый кровеносный
капилляр; З центральная вена; 4 междольковая артерия;
5 междольковая вена; б междольковый желчный проток
186
Тлава 14
число которых в одной клетке превышает 1000. Комплекс rольджи хорошо развит.
Встречаются пероксисомы, лизосомы, а также различные включения (жировые,
пиrментные и др.). Количество включений в rепатоцитах находится в связи с фа
зами пищеварения. После приема пищи резко возрастает количество rликоrена,
увеличивается число липидных включений. Для печени характерен выраженный
суточный ритм: синтез и выделение желчи интенсивнее происходят днем, а rлико
reH в большом количестве накапливается ночью. Больше rликоrена образуется в
клетках, расположенных около центральной вены, а желчи в rепатоцитах на пе
риферии дольки.
rепатоциты располаrаются обычно в виде двух тесно прилеrающих друr к дpy
ry рядов, образуя при этом печеночные балки. Между апикальными (билиарными)
поверхностями двух rепатоцитов образуется щелевидное пространство с диамет
ром 0,5 1 мкм. Эти межклеточные узкие щели называют желчными каnWlЛЯрами.
Последние начинаются слепо и в своей начальной части собственной стенки не
имеют. Однако ближе к периферии дольки формируются канальцы rеринrа жеk
чные проточки, стенка которых представлена как rепатоцитами, так и эпителио
цитами проточков (холанrиоцитами). По мере увеличения калибра стенка проточ
ка становится сплошной, выстланной однослойным эпителием, в составе KOToporo
располаrаются малодифференцированные камбиальные холанrиоциты. По проточ
кам желчь попадает в междольковые желчные протоки, выстланные однослойным
кубическим эпителием. При обычных методах окраски желчные капилляры не
выявляются, но обнаруживаются при импреrнации солями серебра, rистохимичес
кой реакцией на щелочную фосфатазу и друrими методами.
Таким образом, вырабатывая желчь, печень функционирует как ЭКЗ0кринная
железа. Вместе с тем она выделяет в кровь такие вещества, как rлюкоза, мочевина,
белковые фракции и др., что характеризует печень как эндокринный орюн. Из rепа
тоцитов эти вещества поступают через базальную (синусоидную) поверхность клет
ки. Между rепатоцитом и rемокапилляром здесь располаrается перисинусоидное npo
сmрансmво Диссе, в которое rепатоцит выделяет белки, rлюкозу, мочевину и друrие
вещества в процессе осуществления метаболических функций. В печеночной доль
ке существуют две системы, не связанные между собой и действующие по принципу
противотока: желчеотводящая, по которой желчь идет от центра на периферию доль
ки, и кровеносная, по которой кровь движется от периферии к центру дольки. Меж
ду желчными и кровеносными капиллярами нет непосредственноrо соединения, и в
условиях нормы желчь не поступает в кровоток Просвет желчноrо капилляра явля
ется замкнутым блаrодаря наличию между образующими ero соседними rепатоци
тами межклеточных контактов нескольких типов плотных, щелевых и десмосом.
В просвет желчноrо капилляра выступают микроворсинки, образованные на били
арной поверхности rепатоцитов.
Базальная поверхность rепатоцитов обращена в сторону перисинусоидноrо про
странства Диссе. В это пространство выступают также мноroчисленные микровор
синки, что увеличивает активную поверхность rепатоцитов. Само перисинусоидное
пространство, представляет собой узкую щель (шириной 0,2 1 мкм). Если одну CTeH
ку ею образует базальная поверхность rепатоцитов, то друrую стенка СИНУСОИk
Система орюнов внутренней среды
187
Horo rемокапилляра. В пространстве Диссе находятся жидкость, боrатая белками, а
также арrирофильные фибриллы, единичные фибробласты, отростки звездчатых кле
ток и др. В нем обнаружены особые мелкие клетки перuсuнусоидШlb1iые лuпоцuты,
или 1<.Леткu Ито. Они обладают способностью накапливать в цитоплазме липиды и
депонировать жирорастворимые витамины. Эти клетки называют также жиронакап
ливающими, или жирозапасающими, клетками. Их рассматривают как особый тип co
единительнотканных интерстициальных клеток С функциями клеток связывается
синтез и секреция белков коллаrена и участие в развитии цирроза печени. В периси
нусоидальном пространстве располarаются рit 1<.Леткu, относящиеся к большим rpa
нулярным лимфоцитам (натуральные киллеры), которые выделяют вещества, стиму
лирующие пролиферацию rепатоцитов, участвуют в защитной функции.
Стенка внутридольковых синусоидов выстлана эндотелием, в котором, кроме
плоских и тонких эндотелиоцитов, имеются мноrочисленные вкрапления более
крупных звездчатых клеток Последние известны под названием звездчатые MaK
рофаzoцuты, или 1<.Леткu Купфера. Это производные моноцитов крови и представ
ляют собой печеночные макрофаrи. В цитоплазме этих клеток MHoro пиноцитоз
ных и фаroцитозных пузырьков, плотных телец (вторичных лизосом). Печеночные
макрофаrи способны поrлощать из крови циркулирующие вещества, накапливать
их в цитоплазме, захватывать и переваривать бактерии, обломки эритроцитов. Они
способны к амебоидному движению и MorYT выходить в просвет синусоидов. Ha
бухая, эти клетки выполняют роль сфинктеров синусоидных капилляров.
Эндотелиоциты соединяются в пласт при помощи плотных межклеточных KOH
тактов. В выстилке синусоидных капилляров обнаружено наличие мелких OTBep
стий, посредством которых сообщаются между собой просвет синусоидов и про
странство Диссе. Поры имеют диаметр около 100 нм. Участки истонченной
цитоплазмы эндотелиоцитов, rде концентрируются эти отверстия, называют cи
тoвидHыии пластинкамu. Они иrpают роль фильтра. В стенке внутридольковых
синусоидных кровеносных капилляров на большом протяжении отсутствует ба
зальная мембрана, что облеrчает проникновение веществ из крови в перисинусо
идное пространство и в обратном направлении.
Эндотелий синусоидных rемокапилляров, печеночные макрофаrи, структуры
в перисинусоидном пространстве составляют вместе zепатоzематическuй барьер,
или rистион, через который происходит обмен веществ между эпителием печени и
кровью.
Наряду с классическими представлениями о строении печеночной дольки,
имеются и друrие трактовки ее rистоархитектуры. Так, соrласно одной из rипотез,
элементами дольки являются не печеночные балки, а пластины, состоящие из Ok
Horo слоя rепатоцитов. Печеночные пластины отroраживают, как стенками, цилин
дрические синусоидные пространства (лакуны), по которым протекает кровь.
Кроме классических печеночных долек, описаны так называемые портШlьные
дольки и печеночные aциHycы. Центром портальной дольки признается триада, а
периферическими ориентирами являются центральные вены трех смежных долек
В целом портальная долька имеет форму треуrольника. В ее пределах кровь течет
по направлению от центра на периферию. ПечеlIОЧНЫЙ ацинус образуют cerMeHTbI
188
Fлава 14
двух соседних классических долек, расположенных между близлежащими цeHT
ральными венами. Ацинус имеет ромбовидную форму. У острых уrлов ромба Haxo
дятся центральные вены, а у тупоrо триада.
Эти представления о структурно функциональных единицах печени помоrа
ют понять особенности поражений разных отделов печеночной дольки в условиях
патолоrии.
Возрастные изменения печени характеризуются понижением метаболической и
пролиферативной активности rепатоцитов, накоплением в их цитоплазме липофус
цина и дистрофическими явлениями. Между печеночными дольками разрастается
соединительная ткань. Иноrда это сопровождается явлениями цирроза печени.
Реактивность и реzенерацuя печени. Ткани печени отличаются высокой чув
ствительностью к действию повреждающих факторов. Действие ОВ, ионизирую
щей радиации, комбинированных повреждений приводит к резкому нарушению
кровообращения в печени, связанноrо с ею особенностями в этом opraHe. Наруша
ется интеrpация rепатоцитов в составе печеночных балок, в клетках снижается KO
личество rликоrена, изменяется активность окислительно восстановительных фер
ментов, подавляется фаrоцитарная активность печеночных макрофаrов. На месте
rибнущих rепатоцитов разрастается рыхлая волокнистая соединительная ткань.
Эпителий печени проявляет способность к физиолоrической и репаративной pere
нерации. При удалении в эксперименте на животных до 70% массы печени уже через
2 недели происходит полное восстановление. Этот феномен наблюдается каждый
раз при мнorократных резекциях, проводимых с интервалом около месяца. Однако
высокая реrенерационная способность печени не характерна для человека. В целом,
rепатоциты и холанrиоциты относятся к растущей клеточной популяции.
Желчевыводящие пути и желчный пузырь
Желчевыводящие пути пред ставлены внутрипеченочными и внепеченочны
ми желчными протоками. Первые из них это междольковые желчные протоки, в
которые желчь поступает из желчных капилляров. Стенка междольковых желч
ных протоков состоит из однослойноrо кубическою или цилиндрическою (в бо
лее крупных протоках) эпителия и TOHKoro слоя рыхлой соединительной ткани.
Вне печеночные желчевыводящие пути включают печеночные, пузырный и общий
желчный протоки. Стенка их состоит из слизистой, мышечной и наружной оболо
чек Просвет протоков выстилает высокий призматический эпителий, в котором,
наряду с каемчатыми призматическими, эпителиоцитами имеются бокаловидные
экзокриноциты и единичные эндокриноциты. В мышечной оболочке при впаде
нии протоков в желчный пузырь и двенадцатиперстную кишку находятся сфинк
теры, реryлирующие поступление желчи в эти opraHbI.
Желчный пузырь. Стенка состоит из слизистой, мышечной и аДвентициаль
ной оболочек (рис. 93). Слизистая оболочка образует мноrочисленные складки и
крипты. Высокопризматический поверхностный эпителий обладает способностью
всасывать воду и соли из желчи, что приводит к повышению концентрации желч
Horo пиrмента, холестерина и солей желчных кислот в пузырной желчи. В составе
Система орюнов внутренней среды
189
эпителия присутствуют поверхност
ные эпителиоциты, бокаловидные эк
зокриноциты, продуцирующие слизь,
и базальные клетки (камбиальные). В
собственной соединительнотканной
пластинке слизистой оболочки распо
лаrаются жировые, плазматические и
тучные клетки. Мышечная оболочка
желчноro пузыря состоит из преиму
щественно циркулярно расположен
ных rладких мышечных клеток Co
кращение мышечной ткани реryлиру
...... < ;'-й -.
ется ropMoHoM холецистокинином, '
который вырабатывается эндокрино . ., '. ..;; 4
цитами кишечноrо эпителия. Желчь Рис. 93. Строение стенки желчноrо пузыря:
поступает в кишечник порциями. Aд 1 поверхностный эпителий; 2 рыхлая волокнис
вентициальная оболочка желчноrо тая соединительная ткань собственной пластинки
слизистой оболочки; 3 мышечная оболочка; 4
пузыря представлена волокнистой co серозная оболочка
единительной тканью. Со стороны брюшной полости стенку желчноrо пузыря по
крывает серозная оболочка.
r ' _ ; 1
.\ <9
) "' " , ," , " ;. """" . ......
'- '- '\; с,...",. 8' ......-: ..., ] - ..;..
:;.-l' /..,:'!'" . ;...."\i3 !:.7 ( '\ ) ,tv:
-;: ""...." ,,"__ ""'.._"' .J .,.' .__' "". ."
'J, ........ ". 't", ( .'
.. ...... .,..., "", ., I ,f
,."_ fV. (.. -' .',. I" _ \' ", ...
\....J.(/(; tt r r""j ii '
, .. '<'o$t',,' fs '"--1' ../{.
... .... .1, :-r.... ".. _ _ _. . "......... : ·
..... . _..... ;
со .... ,,""V\ .......-r" ,.., _. ".
.. .. .{. ) : _:1
... .......--..,.;.. ...< " .,...
.. .....,..' ,7 'lt"" 2
.; .' o"',.... ..
': ':S ' ..;"
..
" ,., : '1)..
. .
'.
.
:.
3
-?: /
Сердечно сосудистый комплекс opraHoB
Сердечно сосудистый комплекс opraHoB включает сердце, артерии, сосуды
микроциркуляторноrо русла, вены, лимфатические сосуды. Сердце и замкнутая
сеть сосудов обеспечивают циркуляцию крови в орrанизме и транспорт лимфы 1<
сердцу. Деятельность сердечно сосудистоrо комплекса направлена на поддержа
ние метаболизма и постоянства внутренней среды орrанизма из крови к тканям
и клеткам поступают питательные вещества, кислород, биолоrически активные Be
щества, реrулирующие их развитие и функции; в кровь и лимфу удаляются He
нужные клеткам шлаки и продукты их специальной деятельности.
Развитие. Источником развития кровеносных сосудов является мезенхима.
Первые сосуды возникают вне орrанизма зародыша в стенке желточноrо мешка и
хориона в начале 3 й недели эмбриоrепеза. Первоначально образуются скопления
клеток мезенхимы, именуемые кровяными островками. Периферические клетки oc
тровков уплощаются И, соединяясь друr с друrом, формируют примитивные сосуды
в виде эндотелиальных трубок Uентрально расположенные мезенхимоциты диф
ференцируются в первичные клетки кропи (начальный интраваскулярный этап KpO
ветворения). В теле зародыша сосуды появляются позже, также из мезенхимы пу
тем разрастания ее клеток по стенкам щелевидных пространств зародыша. В конце
3 й недели устанавливается сообщение между первичными кровеносными cocyдa
ми внезародышевых opraHoB и тела зародыша. После начала циркуляции крови CTPYК
тура сосудов заметно усложняется в соответствии с реrиональными условиями re
:\fодинамики. В составе стенок сосудов, ПОМИI\Ю эндотелия, развиваются друrие ткани
190
Fлава 14
(происходящие также из мезенхимы), которые, объединяясь, формируют BHYTpeH
нюю, среднюю, и наружную оболочки сосудов.
Закладка сердца возникает в начале 3 й недели развития в виде парных мезен
химных трубок После их слияния начинается дифференцировка тканей BHYTpeH
ней оболочки сердца эндокарда. Средняя и наружная оболочки сердца форми
руются также IIЗ парных миоэпикардиальных пластинок фраrментов правоrо и
левоrо висцеральных листков спланхнотома. Миоэпикардиальные пластинки при
ближаются к закладке эндокарда, окружают ее снаружи, и далее, сливаясь, диффе
ренцируются в тканевые элементы мио и эпикарда.
Артерии
Артерии сосуды, обеспечивающие продвижение крови от сердца к микроцир
куляторному руслу. По величине диаметра они подразделяются на артерии малою,
среднеrо и крупною калибра. Стенка всех артерий состоит из трех оболочек: BHYT
ренней (tunica intima), средней (tunica media) и наружной (tunica externa). TKaHe
вый состав и степень развития этих оболочек в артериях разноrо калибра неодина
ковы, что связано с rемодинамическими условиями и особенностями функций,
выполняемых сосуда ш тех или иных отделов артериальноrо русла. По количествен
ному соотношению эластических и мышечных элементов в средней оболочке сосуда
различают артерии эластическоrо, смешанноrо (мышечно эластическоrо) и мышеч
ною типов.
Артерии эластическою типа (аорта и леrочная артерия) выполняют транспорт
ную функцию и функцию поддержания давления крови в артериальной системе во
время диастолы сердца. Стенка их испытывает ритмические изменения кровяноrо
давления. Кровь в эти сосуды поступает под высоким давлением (120 130 мм рт. ст.)
И со скоростью около 1 м/с. В этих условиях вполне оправдано сильное развитие эла
стическоrо каркаса стенки, который позволяет растяrиваться сосудам во время систо
лы и принимать исходное положение во время диастолы. Возвращаясь в исходное по
ложение, эластичная стенка таких сосудов способствует тому, что последовательно
выбрасываемые из желудочков сердца порции крови превращаются в непрерывный
кровоток Внутренняя оболочка сосудов эластическою типа (на примере аорты) co
стоит из эндотелия, подэндотелиальноrо слоя и сплетения эластических волокон.
В подэндотелиальном слое определяются малодифференцированные звездчатые клет
ки рыхлой соединительной ткани, отдельные rладкие мышечные клетки, большое KO
личество rликозаминоrликанов. С возрастом здесь отмечается накопление холестери
на. В средней оболочке аорты имеется до 50 эластических окончатых мембран (точнее
эластических окончатых цилиндров разных диаметров, вставленных друr в друrа), в
отверстиях которых располаrаются rладкие мышечные клетки и эластические волок
на. Наружная оболочка состоит из рыхлой волокнистой соединительной ткани, co
держащеЙ сосуды сосудов и нервные стволики.
Артерии смешанною (Мblшечно эластическою) типа характеризуются пример
но равным количеством мышечных и эластических элементов в составе средней обо
лочки. Между rладкими миоцитами лежат rycTbIe сети эластических фибрилл.
Система орюнов внутренней cpeды
191
На rpанице внутренней и средней
оболочек отчетливо выражена
внутренняя эластическая мембра
на. В наружной оболочке coдep
жатся пучки rладких мышечных
клеток, а также коллаrеновых и
эластических волокон. К артери
ям данноrо типа относятся COH
ная, подключичная и друrие.
Аpmериимыечноlo типа BЫ
полняют не только транспортную,
но и распределительную функ
ции, реrулируя приток крови к
opraHaM в условиях разных физи
олоrических наrpузок (это, так Ha
зываемые, opraHHbIe артерии).
Артерии мышечноrо типа coдep .....
жат в средней оболочке rладкие
миоциты (рис. 94). Это позволя
ет артериям реryлировать приток
Рис. 94. Строение стенки артерии мышечноrо типа:
крови К opraHaM и поддерживать 1 эндотелий; 2 подэндотелиальный слой; 3 BHyтpeH
наrнетание крови, что важно для няя эластическая мембрана; 4 мышечная оболочка: 5
кровоснабжения opraHoB, распо наружная эластическая мембрана; 6 наружная оболочка
ложенных на большом удалении от сердца. Артерии мышечноrо типа MOryT быть
крупноrо, среднеrо и малою калибров. Внутреннюю оболочку стенки этих артерий
образуют эндотелий, лежащий на базальной мембране, подэндотелиальный слой и
внутренняя эластическая мембрана, однако в мелких артериях внутренняя эласти
ческая мембрана выражена слабо. Средняя оболочка образована rладкой мышечной
тканью с небольшим количеством фибробластов, коллаrеновых и эластических BO
локон. rладкие миоциты располаrаются в средней оболочке по полоrой спирали.
Вместе с радиально и дуrообразно расположенными эластическими волокнами ми
оциты создают единый пружинящий каркас, который препятствует спадению apTe
рий, обеспечивая их зияние и непрерывность кровотока. На rранице между средней
и наружной оболочками имеется наружная эластическая мембрана. Последняя OT
носится к наружной оболочке, состоящей из рыхлой соединительной ткани. Колла
reHoBbIe волокна имеют косое и продольное направление. В наружной оболочке ap
терий мышечною типа проходят питающие их кровеносные сосуды и нервы.
С помощью растровой электронной микроскопии показано, что внутренняя
поверхность эндотелия артерий имеет мноrочисленные складки и уrлубления, раз
нообразные по форме микроскопические выросты. Это создает неровный и слож
ный микрорельеф внутренней (люминальной) поверхности сосудов. Такой мик
рорельеф увеличивает свободную поверхность соприкосновения эндотелия с
кровью, что имеет трофическое значение и создает блаrоприятные условия для re
модинамики.
4
5
.... ....
} ,
:.
."""",
---
-.1
"Ib
..
....
liiI
d)
192
Fлава 14
Сосуды микроциркуляторноrо русла
По мере уменьшения Ka
либра артерий все оболочки
их стенок становятся тоньше.
Артерии постепенно перехо
дят в артериолы, с которых Ha
чинается микроциркулятор
ное сосудистое русло (МЦР).
Через стенки ero сосудов ocy
ществляется обмен веществ
между кровью и тканями, по
этому микроциркуляторное
русло именуется обменным
звеном сосудистой системы.
Постоянно происходящий об
мен воды, ионов, микро И
макромолекул между кровью,
тканевой средой и лимфой,
представляет собой процесс
микроциркуляции, от состоя 2
ния которою зависит поддер
жание постоянства внутритка
HeBoro и внутриорrанноrо ro Рис. 95. Сосуды микроциркуляторноrо русла:
М 1 артериола; 2 rемокапилляр; З венула
меостаза. В составе ЦР раз
личают артериолы, прекапилляры (прекапиллярные артериолы), rемокапилляры,
посткапилляры (посткапиллярные венулы) и венулы (рис. 95).
Артерuолы мелкие сосуды диаметром 50 100 мкм, постепенно переходящие
в капилляры. Основная функция артериол реryлирование притока крови в OCHOB
ное обменное звено МЦР rемокапилляры. В их стенке еще сохраняются все три
оболочки, свойственные более крупным сосудам, хотя они и становятся очень TOH
кими. Внутренний просвет артериол выстлан эндотелием, под которым лежат еди
ничные клетки подэндотелиальноrо слоя и тонкая внутренняя эластическая мемб
рана. В средней оболочке спиралевидно располаrаются rладкие миоциты. Они
образуют всею 1 2 слоя. rладкие мышечные клетки имеют непосредственный KOH
такт с эндотелиоцитами, блаюдаря наличию перфораций во внутренней эластичес
кой мембране и в базальной мембране эндотелия. Эндотелио миоцитарные KOHTaK
ты обеспечивают передачу сиrналов от эндотелиоцитов, воспринимающих изменение
концентраций биолorически активных соединений, реryлирующих тонус артериол,
на rладкомышечные клетки. Характерным для артериол является также наличие мио
миоцитарных контактов, блаrодаря которым артериолы выполняют свою роль "Kpa
нов сосудистой системы" (Сеченов И.М.). Артериолы обладают выраженной COKpa
тительной активностью, называемой вазомоцией. Наружная оболочка артериол
чрезвычайно тонка и сливается с окружающей соединительной тканью.
, .. /
' \ , ('< ", ( -,
) \ Ф.-; ,(. \
... .:1;' , .. \
ft;) lj", '. t: ' . :
t ..... ''\' t "'.-' е
e) (.". .f:<' I с
."".,., ." 1
3 ...... } t;" ...:;: \ J ,"';''' ...
( .
. ' ,, ' "''''''''''' -) ' ,, { ,
..., '. . ...,... "
.. " ' "".: " .
:. ' .: "'<'
;;) ;" r '.''i'
; . :' , ' , '(,
...... 4" ",.,}... ' ....
;/ ... . .JO''? .J
. . <?.
" .
Система оршнов внутренней среды
193
ПрекаnWUlЯрЫ (прекапиллярные артериолы) тонкие микрососуды (диамет
ром около 15 мкм), отходящие от артериол и переходящие в rемокапилляры. Их
стенка состоит из эндотелия, лежащеrо на базальной мембране, rладкомышечных
клеток, расположенных поодиночке и наружных адвентициальных клеток В Mec
тах отхождения от прекапиллярных артериол кровеносных капилляров имеются
rладкомышечные сфинктеры. Последние реrулируют приток крови к отдельным
rруппам rемокапилляров и при отсутствии выраженной функциональной наrруз
ки на opraH большая часть прекапиллярных сфинктеров закрыта. В области сфин
ктеров rладкие миоциты формируют несколько циркулярных слоев. Эндотелио
циты имеют большое количество хеморецепторов и образуют множество контактов
с миоцитами. Эти особенности строения позволяют прекапиллярным сфинктерам
реаrировать на действие биолоrически активных соединений и изменять приток
крови в rемокапилляры.
rе.мокапWl.1lЯРЫ. Наиболее тонкостенные сосуды микроциркуляторною pyc
ла, по которым кровь транспортируется из артериальноrо звена в венозное. Из это
[о правила есть исключения: в клубочках почек rемокапилляры располаrаются
между приносящими и выносящими артериолами. Такие атипично расположен
ные кровеносные капилляры образуют сети, называемые чудесными. Функциональ
ное значение rемокапилляров чрезвычайно велико. Они обеспечивают направлен
ное движение крови и обменные процессы между кровью и тканями. По диаметру
rемокапилляры подразделяются на узкие (5 7 мкм), широкие (8 12 мкм). синусо
идные (20 30 мкм и более с меняющимся по ходу диаметром) и лакуны.
Стенка кровеносных капилляров состоит из клеток эндотелиоцитов и пери
цитов, а также неклеточноrо компонента базальной мембраны. Снаружи капилля
ры окружены сетью ретикулярных волокон. Внутренняя выстилка rемокапилляров
образована однослойным пластом плоских эндотелиоцитов. Стенку капилляра в
поперечнике образуют от одной до четырех клеток Эндотелиоциты имеют полиrо
нальную форму, содержат, как правило, одно ядро и все орrанеллы. Наиболее xapaK
терtlыми ультраструктурами их цитоплазмы являются пиноцитозные везикулы.
Последних особенно мною в тонких периферических (марrинальных) частях кле
ток Пиноцитозные везикулы связаны с плазмолеммой наружной (люминальной) и
внутренней (аблюминальной) поверхностей эндотелиоцитов. Их образование OTpa
жает процесс трансэндотелиальною переноса веществ. При слиянии пиноцитозных
пузырьков формируются сплошные трансэндотелиальные канальцы. Плазмолемма
люминальной поверхности эндотелиальных клеток покрыта rликокаликсом, ВЫПОk
няющим функцию адсорбции и активною поrлощения из крови продуктов обмена
веществ и метаболитов. Здесь эндотелиальные клетки образуют микровыросты, чис
ленность которых отражает степень функциональной транспортной активности re
мокапилляров. В эндотелии rемокапилляров ряда opraHoB наблюдаются "отверстия"
(фенестры) диаметром около 50 65 нм, закрытые диафраrмой толщиной 4 6 нм. Их
присутствие облеrчает течение обменных процессов.
Эндотелиальные клетки обладают динамическим сцеплением и непрерывно сколь
зят одна относительно друrой, образуя интердиrитации, щелевые и плотные KOHTaK
ты. Между эндотелиоцитами в rемокапиллярах некоторых opraHOB обнаруживаются
194
Тлава 14
щелевидные поры и прерывистая базальная мембрана. Эти межклеточные щели слу
жат еще одним из путей транспорта веществ между кровью и тканями.
Снаружи от эндотелия располаrается базальная мембрана толщиной 25 З5 нм.
Она состоит из тонких фибрилл, поrруженных в rомоrенный липопротеиновый
матрикс. Базальная мембрана в отдельных участках по длиннику rемокапилляра
расщепляется на два листка, между которыми лежат перициты. Они оказываются
как бы "замурованными" в базальной мембране. Полаrают, что деятельность и из
менение диаметра кровеносных капилляров реrулируется, блаrодаря способности
перицитов набухать и отбухать. Аналоrом наружной оболочки сосудов в reMoKa
пиллярах служат адвентициальные (периваскулярные) клетки вместе с преколла
rеновыми фибриллами и аморфным веществом.
Для rемокапилляров характерна opraH
ная специфичность строения. В этой связи
различают три типа капилляров: 1) непре
рывные, или капилляры соматическоrо
типа, располаrаются в мозrу, мышцах,
коже; 2) фенестрированные, или капилляры
висцеральноrо типа, располаrаются в эн
докринных opraHax, почках, желудочно ки
шечном тракте; 3) прерывистые, или каПИk
ляры синусоидноrо типа, располаrаются в
селезенке, печени (рис. 96).
В rемокапиллярах соматическою типа
эндотелиоциты соединены друr с друrом с по
мощью плотных контактов и образуют сплош
ную выстилку. Базальная мембрана их также
непрерывная. Присутствие подобных каПИk
ляров со сплошной эндотелиальной ВЫСТИk
кой в мозrу, например, необходимо для надеж
ности rемато энцефалическоrо барьера. [eMO
капилляры висцераЛЪ1lOl0 типа выстланы
эндотелиоцитами с фенестрами. Базальная
мембрана при этом непрерывная. Капилляры
этою типа характерны для opraHoB, в которых обменно метаболические отношения
с кровью более тесные эндокринные железы вьщеляют в кровь свои rормоны, в
почках из крови фильтруются шлаки, в желудочно кишечном тракте в кровь и лим
фу всасываются продукты расщепления пищи. В преръtвистых (си1lусоид1lы)) reMo
капиллярах между эндотелиоцитами имеются щели, или поры. Базальная мембрана
в этих участках отсутствует. Такие rемокапилляры присутствуют в opraHax KpOBe
творения (через поры в их стенке в кровь поступают созревшие форменные элемен
ты крови), печени, которая выполняет множество метаболических функций и клет
ки которой "нуждаются" в максимально тесном контакте с кровью.
Количество rемокапилляров в разных opraHax неодинаково: на поперечном
срезе в мышце, например, на 1 мм 2 площади насчитывается до 400 капилляров,
2
в
6
Рис. 96. Схема строения стенки rемокапил
ля ров разных типов:
А капилляр HenpepbIBHoro типа: Б Ka
пилляр фенестрированноrо типа; В Ka
пилляр прерывистоrо типа; 1 эндотели
оцит; 2 перицит; 3 базальная мембра
на; 4 плотный контакт; 5 фенестра; 6
пора
Система орюнов внутренней среды
195
Torдa как в коже Bcero 40. В обычных физиолorических условиях до 50 % reMo
Ф К ""
капилляров являются не ункционирующими. оличество открытых капилля
ров зависит от интенсивности работы opraHa. Кровь протекает через капилляры со
скоростью 0,5 мм/с под давлением 20 40 мм рт. ст.
Пост"апWlЛЯРЫ, или посткапиллярные венулы, это сосуды диаметром около
12 30 мкм, образующиеся при слиянии нескольких капилляров. Посткапилляры
по сравнению с капиллярами имеют больший диаметр и в составе стенки чаще
встречаются перициты. Эндотелий фенестрированноrо типа. На уровне посткапил
ляров происходят также активные обменные процессы и осуществляется миrра
ция лейкоцитов.
Веиулы образуются при слиянии посткапилляров. Начальным звеном BeHY
лярноrо отдела МЦР являются собирательные венулы. Они имеют диаметр OKO
ло 30 50 мкм и не содержат в структуре стенки rладких миоцитов. Собиратель
ные венулы продолжаются в мышечные, диаметр которых достиrает 50 1 00 мкм.
В этих венулах имеются rладкомышечные клетки (численность последних YBe
личивается по мере удаления от rемокапилляров), которые ориентированы чаще
вдоль сосуда. В мышечных венулах восстанавливается четкая трехслойная CTPYK
тура стенки. В отличие от артериол, в мышечных венулах нет эластической MeM
браны, а форма эндотелиоцитов более окруrлая. Венулы отводят кровь из каПИk
ляров, выполняя отточно дренажную функцию, выполняют вместе с венами
депонирующую (емкостную) функцию. Сокращение продольно ориентирован
ных rладких миоцитов венул создает некоторое отрицательное давление в их про
свете, способствующее "присасыванию" крови из посткапилляров. По венозной
системе вместе с кровью из opraHoB и тканей удаляются продукты обмена Be
ществ.
[емодинамические условия в венулах и венах существенно отличаются от Ta
ковых в артериях и ар тер иолах в связи с тем, что кровь в венозном отделе течет с
небольшой скоростью (1 2 мм/с) и при низком давлении (около 10 мм рт. ст.).
В составе микроциркуляторноro русла существуют также артеpuоло венулярные
анастомозы, или соустья, обеспечивающие прямой, в обход капилляров, переход KpO
ви из артериол в венулы. Путь кровотока через анастомозы короче транскапиллярно
ro, поэтому анастомозы называют шунтами. Различают артериоло венулярные aHac
томозы rломусноrо типа и типа замыкающих артерий. Анастомозы rломусноrо типа
реryлируют свой просвет посредством набухания и отбухания эпителиоидных rло
мусных Е клеток, расположенных в средней оболочке соединяющеrо сосуда, образу
ющеrо нередко клубочек (rломус). Анастомозы типа замыкающих артерий содержат
скопления rладких мышечных клеток во внутренней оболочке. Сокращение этих ми
оцитов И их выбухание в просвет в виде валика или подушечки MOryT уменьшить или
полностью закрыть просвет анастомоза. Артериоло венулярные анастомозы реryли
руют местный периферический кровоток, участвуют в перераспределении крови, Tep
мореryляции, реryляции давления крови. Различают еще атипические анастомозы (по
лушунты), В которых соединяющий артериолу и венулу сосуд представлен коротким
rемокапилляром. По шунтам протекает чистая артериальная кровь, а полушунты. бу
дучи rемокапиллярами, передают в венулу смешанную кровь.
196
Fлава 14
Вены
Вены в целом сходны по строению с артериями, однако особенности rемоди
намики (низкое давление и медленное движение крови в венах) придают CTPYK
туре их стенки ряд особенностей. По сравнению с артериями одноименные вены
имеют больший диаметр (в венозном звене сосудистоrо русла находится около
70% всей крови), тонкую, леrко спадающуюся стенку, слабо развитый эластичес
кий компонент, более слабо развитые rладкомышечные элементы в средней обо
лочке, хорошо выраженную наружную оболочку. Вены, расположенные ниже
уровня сердца, имеют полулунные клапаны. rраницы между оболочками в венах
менее отчетливы по сравнению с артериями. Внутренняя оболочка вен состоит
из эндотелия и подэндотелиальноrо слоя. Внутренняя эластическая мембрана
слабо выражена. Средняя оболочка вен представлена rладкими мышечными клет
ками, которые не образуют сплошноrо слоя, как в артериях, а располаrаются в
виде обособленных пучков, отделенных прослойками волокнистой соединитель
ной ткани. Эластических волокон мало. Наружная адвентициальная оболочка
представляет собой наиболее толстый слой стенки вены. Она содержит коллаrе
новые и эластические волокна, сосуды, питающие вену, и нервные элементы. Тол
стая адвентиция вен, как правило, непосредственно переходит в окружающую
рыхлую соединительную ткань и фиксирует вену в соседних тканях.
В зависимости от степени развития мышечных элементов вены подразделяют
ся на безмышечllые и мышечllые. Безмышечные вены располаrаются в участках op
[анов с плотными стенками (твердая мозrовая оболочка, кости, трабекулы селе
зенки), в сетчатке rлаза, плаценте. В костях и трабекулах селезенки, например,
стенки вен сращены своей наружной оболочкой с интерстиuиальной тканью opra
нов и, таким образом, не спадаются.
Строение стенки вен безмышечноro типа достаточно простое эндотелий, OKPy
женный слоем рыхлоЙ соединительной ткани. rладкомышечных клеток в стенке нет.
В венах мышечноro типа rладкомышечные клетки имеются во всех трех оболоч
ках. Во внутренней и наружной оболочках пучки rладких миоцитов имеют продоль
ное направление, в средней циркулярное. Мышечные вены подразделяются на He
сколько видов. Вены со слабым развитием мышечных элементов это мелкие вены
верхней части туловища, по которым кровь движется, rлавным образом, вследствие
собственной силы тяжести; вены со средним развитием мышечных элементов (Mek
кие вены, плечевая, верхняя полая вены). В составе внутренней и наружной оболочек
этих вен присутствуют единичные продольно ориентированные пучки rладкомышеч
ных клеток, а в средней оболочке циркулярные пучки rладких миоцитов, разделен
ные рыхлой соединительной тканью. Эластических мембран в структуре стенки нет, а
внутренняя оболочка по ходу вены образует немноroчисленные полулунные склад
ки 'КЛаnаllЫ, свободные края которых направлены к сердцу. В основании клапанов
находятся эластические волокна и rладкомышечные клетки. Предназначение клапа
нов препятствовать обратному току крови под влиянием ее собственной силы тя
жести. Клапаны открываются по ходу кровотока. Наполняясь кровью, они перекры
вают просвет вены и препятствуют обратному движению крови.
Система орюнов внутренней среды
197
Лимфатическая система про
водит лимфу от тканей в венозное
русло. Начинается она лuмфока
пWUlЯрами, которые представляют
собой слепо начинающиеся упло
щенные канальцы. Стенка их об
разована только эндотелием. Ба
зальной мембраны и перицитов
нет (в отличие от кровеносных Ka
пилляров). Эндотелий лимфати
ческих капилляров связан с OKPy
жающей соединительной тканью
пучками якорных, или стропных,
филаментов, препятствующих
спадению капилляров. Между эн
дотелиоцитами имеются щели.
Диаметр лимфатических капилляров (20 ЗО мкм) может изменяться в зависимости
от степени наполнения их лимфой. Лимфокапилляры в виде сети пронизывают все
opraHbI и ткани, за исключением мозrа, rлазноrо яблока, BHyтpeHHero уха, печеночных
долек, лимфоидной ткани селезенки, лимфатических узлов, миндалин, KOCTHoro моз
ra, аденоrипофиза, плаценты и некоторых друrих opraHoB.
Лимфатические капилляры выполняют дренажную функцию, участвуя в про
цессах всасывания фильтрата плазмы крови из соединительной ткани. На поверх
ности эндотелиоцитов, обращенной в сторону интерстициальной соединительной
ткани, присутствуют микровыросты, а транспортные пиноцитозные везикулы Ha
правлены в сторону просвета капилляра. Через так называемый посткапиллярный
отдел (который отличается от лимфокапилляров наличием клапанов) лимфати
ческие капилляры постепенно переходят в лимфатические сосуды малою, cpeДHe
ro и крупноrо калибров. В структуре стенки лимфатических сосудов мною общеrо
Вены с сильным развитием
мышечных элементов это круп
ные вены нижней части тулови
ща, например, нижняя полая вена
(рис. 97). Во внутренней оболоч
ке и адвентиции этих вен присут
ствуют множественные продоль
ные пучки rладких миоцитов, а в
средней оболочке циркулярно
расположенные пучки. Имеется
хорошо развитый клапанный ап
парат.
Лимфатические сосуды
1
.. ,'....
. .... --- .. ""'.;'" ".,' 2
,..
"-: '.,. . : . . "' >
.... ' ........................ . . ............. з
..... '1<-'"9, , '. ",.,."...
... ., .
...,.... __,.r- ff . -.
о. ....... ".,....,... .. ...
..... -:..
......
...
If
.... k... . 8*« ·
, еФ.
t .. '. .J/fI'I' ..... ,,"
""._< т '.; -<"'f'!j .,;t;
....> ' '>;; А ..' '" .ора"
"---,, - ..!";". -
j "'.. ....0'."."...
. ....r' ,,,.:., &1
.. t '.." '-
. : /i '< .'Д ",., ) ..:\.
. j . , .."... ."
... , . .. ",.. $о . ,. .\.., ..
- -f. J. : " ._ f '>. . /.
{ ,- К..
" ,, ;}:,,: j.!" .' $ j . ..;:; "'.
1Iijit''ti; '''' ...,-#<. . ......
..., ' f 't. ' .""" j!,'. .",
_ ';. ' ._..'" A'. .:, .
.' ..."... ' . /!;.
9.. .. . ",", """"
(,. . ,....."" '-----". .. 1 . \
't., ... '! .... ":"-" J .
'.. '. ,'" ..'
,) "., \ { .. "'
. . ) ..t) , :. ' " .., '..
1 , "'. ." .\ ' 8..
., '. \t . " "О "'_.
.. . ' ., ..'
,..,.' .'/
.,. . ..:.
. ,
5
.
,
;..
..
&" ,
'1 .
,
.
Рис, 97. Схема строения стенки нижней полой вены:
1 эндотелий; 2 подэндотелиальный слой; 3 средняя
оболочка; 4 наружная оболочка; 5 продольно располо
женные пучки rладких миоцитов
198
lлава 14
с венами, что объясняется сходными условиями лимфо И rемодинамики (низкое
давление, малая скорость протекания, направление тока от тканей к сердцу).
Различают лимфатические сосуды Jиышеч1l0lO и безмышеч1l0lO типов. Средние и
крупные лимфатические сосуды имеют в составе стенки три хорошо развитые обо
лочки (внутреннюю, среднюю и наружную). Внутренняя облочка лимфатических
сосудов образует мноrочисленные складки клапа1lЫ. Расширенные участки лим
фатических сосудов между соседними клапанами называются лимфа1llUО1lG.МU. Cpeд
няя оболочка более выражена в сосудах нижних конечностей. По ходу лимфатичес
ких сосудов расположены лимфатические узлы. Протекая через них, лимфа
обоrащается лимфоцитами. Особенностью строения стенки крупных лимфатичес
ких сосудов (rрудноrо протока и правоrо лимфатическоrо протока) является xopo
шо развитая наружная оболочка, которая в З 4 раза толще внутренней и средней
оболочек вместе взятых. В наружной оболочке проходят продольные пучки rладко
мышечных клеток По ходу rрудноrо протока имеется до 9 полулунных клапанов.
И1luервацuя сосудов. Веrетативные нервные волокна сопровождают сосуды,
заканчиваясь на их стенке рецепторами и эффекторами. Чувствительные нервные
окончания отличаются мнorообразием форм и большой протяженностью, залеrая
во всех трех оболочках сосудов. Эффекторные нервные волокна заканчиваются на
rладких мышечных клетках сосудов нервно мышечными синапсами.
Возрастuые измеuеuuя сосудов. В течение всей жизни происходит непрерыв
ная перестройка сосудистой системы в связи с изменением условий их функциони
рования. С возрастом стенка сосудов уплотняется вследствие разрастания соедини
тельнотканных структур, атрофии клеток средней оболочки и появления известковых
отложений. При нарушении структурной целостности тканей внутренней оболочки
сосудов (эндотелия, рыхлой волокнистой соединительной ткани) и изменении их
метаболизма возможно развитие атеросклероза. При этом во внутренней оболочке
сосудов происходит накопление холестерина и образование атеросклеротических
бляшек Подобные изменения в венечных (коронарных) артериях приводят к ише
мической болезни сердца. С возрастом нередко наблюдаются изменения стенки вен
и лимфатических сосудов, приводящие к их варикозным расширениям.
Реzен.ерацuя сосудов. Рост и реrенерация капилляров происходят за счет образо
вания эндотелиальных выпячиваний в виде почек по принцИIТУ "эндотелий от эндоте
лия" и формирования внутри этих выростов полости с протекающей в ней кровью. При
ранениях стенки кровеносных сосудов (оruестрельные раны, действие ударной волны,
сдавления, переrpузки и др. факторы) посттравматический mстоrенез приводит к He
полноценной реrенерации эндотелия, волокнистой и rладкомышечной тканей оболоч
КИ сосудов, нарушению межтканевых корреляций в области дефекта стенки, замеще
нию ero, в основном, соединительной тканью, что способствует образованию
посттравматических аневризм, а также к заращению просвета маmстральных сосудов.
Сердце
Сердце центральный opraH системы KpOBO и лимфообращения. Блаroдаря
способности к сокращениям, сердце приводит в движение кровь.
Система орюнов внутренней среды
199
Стенка сердца состоит из трех оболочек: .. .."....
эндокарда, миокарда и эпикарда (рис. 98).
Эндокард. Во внутренней оболочке cep
дца различают следующие слои: эндотелий,
выстилающий изнутри полости сердца, и ero
базальную мембрану; подэндотелиальный
слой, представленный рыхлой соединитель
ной тканью, в которой MHoro малодиффе .'
ренцированных клеток; мышечно эласти
ческий слой, состоящий из rладкой мышеч
ной ткани, между клетками которой в виде
rустой сети располаrаются эластические
волокна; наружный соединительнотканный
слой, состоящий из рыхлой соединительной
ткани. Эндотелий и подэндотелиальный . ,"'\:;,:: "
слои аналоrичны внутренней оболочке co
судов, мышечно эластический является "эк
вивалентом" С р едней оболочки, а на ру жный
Рис. 98. Строение части стенки сердца:
соединительнотканный слой аналоrичен Ha 1 эндокард; 2 проводящие волокна Пур
ружной (адвентициальной) оболочке cocy киня; 3 миокард; 4 эпикард
дов. Поверхность эндокарда идеально rладкая и не препятствует свободному дви
жению крови. В предсердно желудочковой области и у основания аорты эндо
кард образует дупликатуры (складки), именуемые клапанами. Различают
предсердно желудочковые и желудочково сосудистые клапаны. В местах при
крепления клапанов имеются фиброзные кольца. Клапаны сердца это плотные
пластинки волокнистой соединительной ткани, покрытые эндотелием. Питание
эндокарда происходит путем диффузии веществ из крови, находящейся в полос
тях предсердий и желудочков.
Миокард (средняя оболочка сердца) мноrотканевая оболочка, состоящая из
поперчнополосатой сердечной мышечной ткани, межмышечной рыхлой соедини
тельной ткани, мноrочисленных сосудов и капилляров, а также нервных элемен
тов. Основной структурой является сердечная мышечная ткань, в свою очередь
состоящая из клеток, формирующих и проводящих нервные импульсы, и клеток
рабочею миокарда, обеспечивающих сокращение сердца (кардиомиоцитов). Cpe
ди клеток, формирующих и проводяших импульсы, В проводящей системе сердца
различают три вида: Р клетки (клетки пейсмекеры), промежуточные клетки и клет
ки (волокна) Пуркиня.
Р клетки клетки водители ритма, располаrаются в центре синусноrо узла
проводящей системы сердца. Они имеют полиrональную форму и детерминиро
ваны на спонтанную деполяризацию плазмолеммы. Миофибриллы и орrанеллы
общеrо значения в клетках пейсмекерах выражены слабо. Промежуточные клетки
неоднородная по составу rруппа клеток, передают возбуждение от Р клеток к
клеткам Пуркиня. Клетки Пуркиня клетки с небольшим количеством миофиб
рилл и полным отсутствием Т системы, с большим по сравнению с рабочими
,. t'#:''<tlI ........... 1
.'... . ,,,,' ;.. ... ./
_, ,4 ....'*,\:1...-. .' .... /"
' ' 'v,""........ .: :;..,::>0-..:-
.... < ....
" . t.14, _\
. $-. ,7 .
", ,-
.". ..;. . fi- ..... '.. ''''' .. 't.."
... ,'-- '- ..; '" '\:.
., .. ' .....
2
.
...
. .."I> ,.."-
...........
..
.;.
."
W'.<" )1
....
...
.... .
.... ..."
,
'",.
з
.
..... ..-.c.. 'о
... -
, .' ....' \.o..1i... ' ....' ...
r ";:.- ............... ..........._.:.:... . 4
, __,,' 'Iii;
,... ' . .;:r:
:. -..
't' "':""...._:: ....:....... -А '.,".: <=i-:: ,,,..
..... -.:;; , ..
.......... ...
200
Тлава 14
сократительными миоцитами количеством циоплазмы. Клетки Пуркиня переда
ют возбуждение от промежуточных клеток к сократительным клеткам миокарда.
Они входят в состав пучка rиса проводящей системы сердца. Неблаrоприятное
влияние на клетки пейсмекеры и клетки Пуркиня оказывают ряд лекарственных
препаратов и друrие факторы, способные привести к возникновению аритмий и
блокады сердца. Наличие в сердце собственной проводящей системы чрезвычай
но важно, поскольку она обеспечивает ритмичную смену систолических COKpa
щений и диастол камер сердца (предсердий и желудочков) и работу ero клапан
Horo аппарата.
Основную массу миокарда составляют сократительные клетки сердечные
миоциты, или кардиомиоциты. Это клетки вытянутой формы с упорядоченной си
стемой поперечноисчерченных миофибрилл, расположенных на периферии. Между
миофибриллами находятся митохондрии с большим количеством крист. В миоци
тах предсердий Т система выражена слабо. Слабо развита в кардиомиоцитах rpa
нулярная эндоплазматическая сеть. В центральной части миоцитов располаrается
ядро овальной формы. Иноrда встречаются двуядерные кардиомиоциты. В мышеч
ной ткани предсердий присутствуют кардиомиоциты с осмиофильными ceKpeTOp
ными rранулами, содержащими натрийуретический пептид. В кардиомиоцитах оп
ределяются включения rликоrена, служащеrо энерrетическим материалом сердеч
ной мышцы. Содержание ero в миоцитах левоrо желудочка больше, чем в друrих
отделах сердца. Миоциты рабочеrо миокарда и про водящей системы соединяются
между собой посредством вставочных дисков специализированных межклеточ
ных контактов. В области вставочных дисков прикрепляются актиновые сократи
тельные миофиламенты, присутствуют десмосомы и щелевые контакты (нексусы)
Десмосомы способствуют прочному сцеплению сократительных миоцитов в фун
кциональные мышечные волокна, а нексусы обеспечивают быстрое распростране
ние волн деполяризации плазмолемм с одной мышечной клетки на друrую и суще
ствование сердечноrо мышечною волокна как единой метаболической единицы.
Характерным для миоцитов рабочеrо миокарда является присутствие aHaCTOMO
зирующих мостиков взаимосвязанных фраrментов цитоплазм мышечных кле
ток разных волокон с находящимися в них миофибриллами. Тысячи таких мости
ков превращают мышечную ткань сердца в сетчатую структуру, способную синх
ронно и эффективно сокращаться и выбрасывать из полостей желудочков
необходимые систолические объемы крови. После пере несенных обширных ин
фарктов миокарда (острых ишемических некрозов стенки сердца), коrда диффуз
но поражаются мышечная ткань сердца, система вставочных дисков, анастомози
рующих мостиков и проводящая система, возникают нарушения ритма работы cep
дца вплоть до фибрилляции. В этом случае сократительная деятельность сердца
превращается в отдельные несоrласованные подерrивания мышечных волокон и
сердце не в состоянии выбрасывать нужные систолические порции крови в пери
ферическую циркуляцию.
Миокард состоит в целом из высокоспециализированных клеток, утративших
способность делиться митозом. Лишь в определенных участках предсердий наблю
даются митозы кардиомиоцитов (Румянцев пл., 1982). Вместе с тем, для миокарда
Система орюнов внутренней среды
201
характерно наличие поли11Jloидныx миоцитов, что значительно усиливает ero рабо
чий потенциал. Явление полиплоидности наиболее часто наблюдается при компен
саторных реакциях миокарда, коrда повышается наrрузка на сердце, и при патоло
rии (недостаточности сердечных клапанов, заболеваниях леrких и др.). Сердечные
миоциты в этих случаях резко rипертрофируются, и стенка сердца в том или ином
отделе утолщается. В миокардиальной соединительной ткани заключена боrато раз
ветвленная сеть кровеносных и лимфатических капилляров, что обеспечивает по
стоян но работающую сердечную мышцу питанием и кислородом. В прослойках co
единительной ткани имеются плотные пучки коллаrеновых волокон, а также
эластические волокна. В целом, эти соединительнотканные структуры составляют
опорный скелет сердца, к которому прикрепляются сердечные мышечные клетки.
Сердце opraH, обладающий способностью к автоматизму сокращений. Оно
может функционировать в известных пределах автономно. Однако в орrанизме
деятельность сердца находится под контролем нервной системы. В интрамураль
ных нервных узлах сердца находятся чувствительные веrетативные нейроны (клет
ки Доrеля Il ro типа), малые интенсивно флюоресцирующие клетки МИФ клет
ки и эффекторные веrетативные нейроны (клетки Дorеля I ro типа). МИФ клетки
рассматриваются как вставочные нейроны.
Эпикард наружная оболочка сердца представляет собой висцеральный ли
сток околосердечноЙ сумки (перикарда). Свободная поверхность эпикарда выстла
на мезотелием так же, как и поверхность перикарда, обращенная в перикардиаль
ную полость. Под мезотелием в составе этих серозных оболочек находится
соединительнотканная основа из рыхлой волокнистой соединительной ткани.
Дыхательный комплекс opraHoB
Большинство метаболических процессов в клетках и тканях орrанизма ocy
ществляется с участием кислорода. Поступление кислорода из атмосферноrо воз
духа в кровь называется внешним дыханием и происходит в дыхательном КОМП
лексе opraHoB. Общая площадь дыхательной поверхности приблизительно равна
160 м 2 , что в 80 раз больше поверхности всей кожи, а общее число rазообменных
пузырьков (альвеол) в леrких составляет 300 млн.
Различают также внутреннее дыхание rазообмен (кислородом и уrлекисло
той) между кровью и клетками, в котором дыхательный комплекс непосредствен
Horo участия не принимает, однако внутреннее дыхание невозможно без внешнею.
Кроме ведущей функции rазообмена, дыхательный комплекс выполняет ряд
нереспираторных функций, связанных с кровообращением и обменом веществ: леr
кие участвуют в липидном обмене, активации ряда биолоrически активных веществ,
выработке факторов свертывания крови, в реryляции тепловоrо баланса; выполня
ют защитную функцию, участвуя в иммунных реакциях и очищая воздух от пыли и
микробов. Важную роль в физиолоrии дыхательною комплекса opraHoB и физиоло
rии всею орrанизма иrрает эндокринный аппарат эпителия воздухоносных путей.
Дыхательный комплекс opraHoB включает воздухоносные пути и респиратор
ный отдел.
202
rлава 14
Развитие. Источником развития трахеи, бронхов и респираторноrо отдела леr
ких служит материал вентральной стенки передней кишки, который является про
изводным nрехордШlЫЮЙ пластинки. На 3 й неделе эмбриоrенеза в ней появляется
мешковидное выпячивание, которое в нижней части делится на два зачатка пра
Boro и левоrо леrких.
В развитии леrкоrо различают три стадии. Первая стадия (железистая) OXBa
тывает период эмбриоrенеза с S й недели до 4 ro месяца. В это время зачаток леr
ких напоминает трубчатую железу. На этой стадии формируются воздухоносные
пути. Вторая каналикулярная стадия (4 6 й месяцы) характеризуется разви
тием респираторных бронхиол. Происходит это при явлениях интенсивной про
лиферации капилляров. Третья стадия (альвеолярная) протекает с 6 ro месяца и
до рождения. На протяжении этой стадии образуются альвеолярные ходы и альве
олы. При этом имеют место тесные взаимодействия между леrочными капилляра
ми и альвеолярным эпителием.
Развитие эпителия (прехордальноrо по происхождению) в воздухоносных пу
тях И В респираторном отделе сопровождается образованием нескольких клеточных
дифферонов (реснитчатые эпителиоциты, бокаловидные экзокриноциты, эндокри
ноциты, респираторные эпителиоциты и др.), взаимодействующих между собой в
процессе выполнения функций. Из мезенхимы, окружающей бронхиальное дерево,
дифференцируются волокнистая соединительная ткань, rладкая мышечная ткань,
rиалиновая и эластическая хрящевые ткани бронхов, а также капиллярная сеть, оп
летающая альвеолы. Нервные элементы являются производными нервной трубки.
В течение Bcero эмбриоrенеза альвеолы находятся в спавшемся состоянии.
После рождения ребенка при первом вдохе происходит их наполнение воздухом,
расширение и расправление.
Воздухоносные пути
Воздухопроводящую функцию выполняют следующие opraHbI: носовая по
лость, носоrлотка, [ортань, трахея, бронхи (внелеrочные и внутрилеrочные). В воз
духоносных путях вдыхаемый воздух очищается от пыли, увлажняется, темпера
тура ero приближается к температуре тела.
Полость носа
в ней различают преддверие, дыхательную и обонятельную области. Преддве
рие выстлано мноrослойным плоским ороrовевающим эпителием с короткими
щетинистыми волосками, очищающими воздух от пыли, который rлубже CTaHO
вится неороrовевающим, а волосы и железы исчезают. Дыхательная область BЫC
тлана слизистой оболочкой состоящей из мноrорядноrо столбчатоrо реснитчатоrо
эпителия и соединительнотканной собственной пластинки. В эпителии домини
руют реснитчатые эпителиоциты и бокаловидные экзокриноциты (рис. 99).
Реснитчатые эпителиоциты имеют на апикальной поверхности по lS 20 ресни
чек длиной до 7 мкм. Бокаловидные экзокриноциты вырабатывают слизь. Слизис
Система орюнов внутренней среды
203
тый секрет вьщеляют также желе
зы, которые открываются на повер
хности эпителия. На увлажненной
слизью поверхности эпителия oce
дают и прилипают частицы пыли и
микроорrанизмы. Блаrодаря бие з
ниям ресничек мерцательноrо эпи
телия, они удаляются из воздухо
носных путей (мукоцилиарный
механизм очищения воздуха). Соб
ственная пластинка слизистой обо
лочки носовой полости боrата спле
тениями кровеносных сосудов,
участвующими в соrревании Bдыxa
емоro воздуха.
В придаточных пазухах (rайморовых, лобных) эпителий имеет строение,
сходное с эпителием дыхательной части носовой полости. Мноrорядный pec
нитчатый эпителий в пазухах менее высокий и содержит меньшее количество
бокаловидных клеток.
' i:.v,
. ..:,<It
"'Ф'
, .
с;::::>
.'
е'
...
< . с7
' ,f
"'''4''<
Рис. 99. Мноrорядный мерцательный эпителий:
1 реснитчатые эпителиоциты; 2 бокаловидные экзо
криноциты; 3 вставочные эпителиоциты
rOpтaHb
Стенка rортани состоит из слизистой, фиброзно хрящевой и адвентициальной
оболочек. Слизистая оболочка включает мноrорядный столбчатый реснитчатый эпи
телий и собственную пластинку, которая состоит из рыхлой волокнистой неоформ
ленной соединительной ткани, содержащей MHoro эластических волокон. Фиброз
но хрящевая оболочка ВЫПолняет роль каркаса rортани. Блаrодаря наличию в ней
rиалиновой и эластической хрящевых тканей стенки rортани не спадаются при BДO
хе. К хрящам прикрепляются наружные и внутренние мышцы rортани. Адвентици
альная оболочка образована волокнистой соединительной тканью.
Слизистая оболочка rортани имеет некоторые особенности строения в облас
ти истинных и ложных roлосовых связок. rолосовые связки представляют собой
складки слизистой оболочки, выступающие в просвет rортани. Основу ложных
rолосовых связок составляет рыхлая волокнистая соединительная ткань с белко
во слизистыми железами и скоплениями лимфоидной ткани. В составе истинных
rолосовых связок, кроме соединительной ткани, имеются поперечнополосатые
мышцы, сокращение которых изменяет величину rолосовой щели и степень натя
жения или расслабления roлосовых связок. Истинные rолосовые связки покрыты
мноrослойным плоским неороrовевающим эпителием.
Надzoртанни'К, отделяющий [ортань от rлотки, имеет в качестве внутренней
опоры эластический хрящ. Слизистая оболочка надrортанника на передней ero по
верхности содержит мноrослойный плоский неороrовевающий эпителий. Эпите
лий, покрывающий нижнюю часть задней поверхности надrортанника, MHoro
рядный столбчатый реснитчатый. у основания надrортанноrо хряща в собственной
204
[лава 14
пластинке слизистой оболочки MHOrO белково слизистых желез и скоплений лим
фоидной ткани.
Стенка трахеи состоит из CJIизистой
оболочки, подслизистой основы, волок
нисто мышечно хрящевой и адвентици
альной оболочек (рис. 100). Эпителий
CJluзuстой оболочки мноrорядный CTOk
бчатый реснитчатый. В нем различают
реснитчатые эпителиоциты, бокаловид
ные ЭКЗ0КРИНОЦИТЫ, базальные (низкие)
и вставочные (высокие) эпителиоциты и
эндокриноциты. Наиболее мноrочислен
ные реснитчатые эпителиоциты на апи
кальной поверхности имеют 200 250 pec
ничек, которые участвуют в мукоцилиар
ном механизме очищения воздуха. Бока
ловидные ЭКЗ0КРИНОЦИТЫ эндоэпите
лиальные железы, форма которых зави 3
сит от секреторною цикла. Ядро распо
лarается в базальной части и уплощено.
Комплекс rольджи участвует в выработ
ке секрета (слизи). Эндокриноциты Tpa
Рис. 100. Строение стенки трахеи:
хеи относятся к диффузной эндокринной 1 слизистая оболочка; 2 подслизистая основа;
системе орrанизма и продуцируют пеп 3 волокнисто мышечно хрящевая оболочка
тидные rOpMOHbI и биоrенные амины (норадреналин, дофамин, серотонин, кальцито
нин и др.), имеющие важное значение для местных и общеметаболических процессов.
Они располаrаются поодиночке или небольшими rpуппами, в базальной части coдep
жат секреторные rранулы. Базальные клетки широким основанием с помощью полу
десмосом прикрепляются к базальной мембране, а с соседними клетками соединяют
ся посредством десмосом. Они не достиrают поверхности эпителия. Вставочные BЫ
сокие клетки напротив, располаrаются в толще эпителия от базальной мембраны до
просвета трахеи. На апикальной поверхности содержат микроворсинки, поrруженные
в rликокаликс. Цитоплазма электронно плотная из за большоrо содержания рибосом.
Источником диверrентной дифференцировки реснитчатых, бокаловидных и
эндокринных клеток являются базальные (низкие) и вставочные (высокие) клет
ки, выполняющие роль камбия для всею эпителия. Среди эпителиоцитов встреча
ются антиrенпредставляющие клетки Ланrерrанса, длинные отростки которых
проникают между эпителиоцитами. Эти клетки относятся к дифферону KOCTHO
мозrовоrо происхождения.
Собственная пластинка слизистой оболочки образована рыхлой волокнистой
соединительной тканью, в которой содержатся эластические волокна, пучки rлад
Трахея
,50 r :'i> .1 1'.'
k I 1 ', I j ( 1"
,1 ." .. I ),' ..''1', '. J ".
f.11t1:'f'I-,;,.I.,I, <r,.... io .!;.t- .,.. '4'J.....
-J " .. <>:'" Ь'..;tф
;.;;"'.... ... .-"."'.......!<J..... ...: ":' ."tJt/"1'.
. ,,' ". . . : . .... - .,'.. /"t'" ..
...... "'.......А.. ..,...r .....t.... .
:""0"..... . .......,-r: "'. 0').1'" " :..,
4t "" "" . .. ...... "... .. ;".;*'"'" .. ...." ..
.. -....... -;. . . .. "
.. ...,.,:.. .....
.
.....
[
..... ..................
...- ..... .., , ....""""",,,,,," ,,,,,,, J'
....... ..... .;;::w ...,,""""
.... ,-- ...... ...... .
2 .... .. .,............, ..:.... J ' .,..", ;::. ....l... ;....
-i': 7" ; r --,. '" ':.. : " #,"
.. \. . tui ,_ ..'I"....,,' ." " , .,'
....'. .';....., 'A'C- tf'; а:;... :i> f
.....-....,........,. ...о....." Ia.. .i' ,,1'-.,
-
(;'1:/':-.::::...
..,..
...
....
...'.
- *=-
. ....
r
:..
","
\-;
(."'
. .
......'
Система орюнов внутренней среды
205
ких мышечных клеток и лимфоидные узелки В подслизистой основе расположе
ны концевые отделы белково слизистых желез. Подслuзистая основа без резкой
rраницы переходит в волокнисто мышечно хрящевую оболочку, которая состоит
из 16 20 незамкнутых сзади колец rиалиновоrо хряща. На задней стенке свобод
ные концы этих образований соединены пучками rладких мышечных клеток. Это
способствует беспрепятственному прохождению пищевых комков по пищеводу, pac
положенному вплотную позади трахеи. В вертикальном направлении "кольца" Tpa
хеи связаны друr с друrом плотной волокнистой соединительной тканью. AдвeH
тициалъная оболочка трахеи образована рыхлой волокнистой соединительной
тканью, которая соединяет трахею с прилежащими орrанами и тканями cpeдocтe
ния. Трахея разделяется на две ветви правый и левый rлавные бронхи.
Леrкие
В состав леrких входят внутрилеroчные бронхи разноrо калибра и респира
торный отдел, представленный ацинусами. Система внутрилеrочных бронхов Ha
чинается с долевых бронхов, на которые деляется rлавные (внелеroчные) бронхи.
ВllутрuлеlОЧllые бро1LXU
Войдя в леrОЧIlУЮ паренхиму, долевые бронхи дихотомически делятся на брон
хи крупноrо, среднеrо и мелкоrо калибра. Конечным звеном внутрилеrочнorо брон
хиальноro дерева являются терминальные бронхиолы.
В rлавных бронхах стенка по строению сходна с трахеей. Волокнисто хряще
B<u:! оболочка, однако, в отличие от трахеи состоит не из полуколец, а из замкнутых
хрящевых колец.
Стенка внутрилеrочных бронхов также состоит из слизистой оболочки с ПОk
слизистой основой, волокнисто хрящевой и адвентициальной оболочек. Слизистая
оболочка включает эпителий, собственную и мышечную пластинки. Последняя об
разована косоциркулярными пучками rладких миоцитов.
По мере дихотомическоrо деления и уменьшения калибра внутрилеrочных
бронхов появляются существенные отличия в строении их оболочек. Так, MHoro
рядный мерцательный эпителий слизистой оболочки крупных и средних бронхов
переходит в однослойный кубический в мелких, количество бокаловидных кле
ток при этом уменьшается. В подслизистой основе бронхов постепенно исчезают
мноrоклеточные слизисто белковые железы. Изменения волокнисто хрящевой обо
лочки сводятся к тому, что в ее составе по мере дихотомическоrо деления бронхов
уменьшается содержание хрящевой ткани, поскольку стенка средних и мелких
бронхов, расположенных в rлубине паренхимы леrких, должна быть более подат
ливой при расправлении и спадении альвеол. Если в стенке крупных бронхов rиа
линовая хрящевая ткань образует протяженные пластинки, соединенные друr с дpy
[ом коллаrеновыми волокнами и пучками rладких миоцитов, то в бронхах среднеrо
калибра хрящ представлен мелкими островками и становится эластическим. HaKO
нец, в мелких бронхах, лежащих наиболее rлубоко, хрящевая ткань исчезает. Вместе
206
Fлава 14
с тем, по мере деления бронхов и уменьшения их калибра становится более выражен
ной циркулярная rладкомышечная пластинка, располаrающаяся на rpанице слизис
той оболочки и подслизистой основы. В стенках мелких бронхов она занимает значи
тельную часть. rладкомышечная ткань стенки мелких бронхов высокочувствительна
к концентрации СО ъ увеличение ero концентрации в воздухе вызывает расширение
мелких бронхов. В связи с сокращением мышечных клеток слизистая оболочка Mek
ких бронхов образует мноrочисленные складки. На всем протяжении бронхиальноrо
дерева в составе слизистой оболочки и подслизистой основы присутствуют скопле
ния лимфоидноЙ ткани, осуществляющей местные защитные реакции.
Бронхиальное дерево леп<их заканчивается термuишшнымu 6ро1lXUОЛамu. Их
эпителий становится низким кубическим и, кроме реснитчатых, бокаловидных и
базальных клеток, содержит секреторные клетки Клара, или бронхиолярные эк
зокриноциты. Последние в апикальной части накапливают специфические ceKpe
торные rранулы, являющиеся компонентом сурфактанта, а также содержат фер
менты, инактивирующие токсины. Под базальной мембраной эпителия в тонкой
собственной пластинке слизистоЙ оболочки продольно идут пучки эластических
волокон, между которыми располаrаются rладкие мышечные клетки.
Физиолоrическая реrенерация бронхиальноrо эпителия происходит за счет
митотическоrо деления базальных клеток Время обновления клеток эпителиаль
Horo пласта составляет около недели.
Респираторный отдел леllCUX
Структурно функциональноЙ единицей
респираторноrо отдела леrких является ацииус.
Этим термином обозначают систему, состоящую
из респираторных бронхиол 1 3 ro порядков,
альвеолярных ходов и альвеолярных мешочков.
Между воздухом внутри альвеол и кровью, Ha
ходящейся в капиллярах, оплетающих альвео
лярную стенку, происходит rазообмен.
Ацинус начинается респираторной брон
хиолой 1 ro порядка, которая в свою очередь
дихотомически делится на респираторные
бронхиолы 2 ro, а затем 3 ro порядков. Пос
ледние разветвляются на альвеолярные ходы,
заканчивающиеся двумя тремя сферическими
альвеолярными мешочками (рис. 101). Чис
ленность альвеол последовательно возраста
ет и, если в стенках репираторных бронхиол
еще имеются участки, состоящие из однослой
ною кубическою эпителия и тонкой прослой
ки коллаrеновых волокон и rладких миоцитов,
rде не происходит rазообмен, то альвеолярные
:--- ...."':'":' '''";!.. '''''&#'' ...."
, :О"'&> .... \:I };.'
: ''i..;.>t.i '',. ., . ' .:;
:::$;. > : : ', J J.J , ' , ..ё
.:.. .Э' .. 'l :; , , , .&;..-;
... t. . , . ' У6 '''''''''' '''
_.... ...... .\.: 1. '"
.; ,o' :'. . ' \Ъ.:. . J" Ч
... ,.,., .. t..J1. _. €.. -.
\, , ' . ' , . :;" f"5 ::*. " ) 11:"-
'{ (:: .... " ) f"" '''0''''''
> .,.. ..\'ti---;j -;< e.;' .
O ....{,:::. ...... tJ ,"" '.;.i-\ .)
,- "...', 2 !;.....'.& ..."
.. ", ,,. "'0" 1
O l( ' j, 1''( "А , 'j ......;;. , , ''\
, . 1 ..,.... , 4(" .
\.. f . ", t") J' /' '\
. з .\.. а' "r'- --" . ;. .
..., · ff
.t... <\Ч .. " ." .--', r..
' . .. 3 "'1.".......
.. ""-'''' ",. -<
..--: ...... (. """'"":.... tt: "';, .
Рис. 1 О 1. Строение ацинуса леrкоrо:
1 респираторная бронхиола; 2 альве
олярный ход 3 альвеолярные мешочки
Система ОрZа1юв внутренней среды
207
мешочки имеют стенку, сплошь состоящую из альвеол. По форме ацинус напоми
нает пирам иду или конус, в вершину, KOToporo входит респираторная бронхиола.
12 18 ацинусов образуют леlO'lJ.НУЮ дольку. Ацинусы отделены друr от друrа соеди
нительнотканными прослойками.
Важнейшим структурным элементом леrочноrо ацинуса является Шlьвеола. Cpeд
ний диаметр альвеол у взрослоrо человека 260 290 мкм. Альвеолы тесно прилежат
друr к друry. Между ними определяются тонкие межальвеолярные переrородки, по
которым проходят кровеносные капилляры. Имеются также эластические и ретику
лярные волокна, оплетающие альвеолы, и придающие им упруrость. В переroродках
между альвеолами обнаруживаются отверстия диаметром 10 15 мкм. Это так назы
ваемые альвеолярные поры Кона, создающие возможность проникновения воздуха
из одной альвеолы в друrую. Эластический каркас и rладкие мышечные клетки в
леrочных ацинусах участвуют в реryляции поступления воздуха в альвеолы.
Изнутри альвеолы выстланы однослойным плоским эпителием. Альвеолярная
выстилка включает несколько клеточных дифферонов. Респираторные плоские эпи
телиоциты (альвеолоциты 1 ro типа) это полиrональной формы клетки. В них
различают две части: более толстую ядросодержащую и тонкую безъядерную (плас
тинчатую). Околоядерная часть имеет толщину около 5 мкм. Толщина пластинча
той части не более 0,2 мкм. Орrанеллы располаrаются около ядра. Через пластинча
тую часть цитоплазмы происходит rазообмен, и в ней MHoro пиноцитозных
пузырьков. Респираторные эпителиоциты лежат на тонкой базальной мембране.
Своей пластинчатой частью они прилежат к базальным участкам эндотелиальных
клеток кровеносных капилляров. В этих участках базальные мембраны альвеоляр
ноro эпителия и эндотелия MOryT сливаться, блаrодаря чему аэроrематический барь
ер (барьер "воздух кровь") оказывается чрезвычайно тонким (около 0,5 мкм). Это
блаroприятствует rазообмену. Обмен rазов между альвеолярным воздухом и KpO
вью происходит путем диффузии в связи с разницей парциальноro давления 02 и
С0 2 В альвеолярном воздухе и в крови. Респираторные эпителиоциты являются BЫ
сокоспециализированными клетками, утратившими способность делиться митозом.
Аэроzематическим. барьером называется комплекс элементов стенки альвеолы и
KpoBeHocHoro капилляра, который преодолевают rазы (02 И С0 2 ) в процессе внеш
Hero дыхания (рис. 102). В ero состав входят слой сурфактанта, цитоплазматические
,-t
. , ':" .
' ,
. .::.'
т-
Рис. 102. Строение межальвеолярной переrородки: 1
1 респираторные плоские эпителиоциты; 2 большой rранулярный эпителиоцит; 3 rемокапилля
ры; 4 альвеолярный макрофаr
4
208
rлава 14
пластинчатые части респираторных эпителиоцитов, общая с эндотелиоцитами ба
зальная мембрана, аблюминальная и люминальная поверхности эндотелиоцитов re
мокапилляра, стенка эритроцита (если слияния базальных мембран нет, то CTPYKТY
ра барьера усложняется между двумя базальными мембранами располаrается
тонкая соединительнотканная прослойка).
Большие (rранулярные) эпителиоциты (альвеолоциты 2 ro типа) выполня
ют важную секреторную функцию. Они несколько крупнее респираторных эпи
телиоцитов. Их диаметр равен 8 12 мкм. Эти клетки имеют овальную или поли
rональную форму и короткие отростки. В цитоплазме MHoro различных орrанелл.
Характерной особенностью является наличие в их цитоплазме пластинчатых oc
миофильных телец включений сурфактанта. Пластинчатые тельца выделяют
ся из клетки путем экзоцитоза. Слой сурфакmанmа покрывает внутреннюю по
верхность стенки альвеол и состоит из двух фаз поверхностной мембранной
(апофаза), представленной молекулярной пленкой фосфолипидов, и жидкой (rи
пофаза), содержащей липиды, белки, полисахариды, воду и др., которая заполня
ет неровности и пространства между эпителиоцитами. Толщина сурфактантноrо
слоя 20 30 нм. Сурфактант имеет важное функциональное значение блаrодаря
тому, что участвует в поддержании поверхностноrо натяжения альвеол, предох
раняет их от спадания при выдохе, препятствует транссудации жидкости в про
свет альвеол, выполняет защитную функцию, обладая бактерицидностью. В HOp
ме синтез сурфактанта начинается еще в эмбриоrенезе, и, если к рождению
сурфактант в леrких плода отсутствует (так называемый, врожденный дистресс
синдром), ребенок не может сделать самостоятельный первый вдох, поскольку
альвеолы оказываются слипшимися из за отсутствия сурфактанта.
Большие эпителиоциты являются одновременно секретирующими и проли
ферирующими клетками. Между респираторными и большими эпителиоцитами
образуются межклеточные соединения типа плотных контактов. Кроме описанных
выше клеток, в стенке альвеол и в rипофазе обнаруживаются альвеолярные MaKpo
фаrоциты. Это производные моноцитов. Очищая вдыхаемый воздух, альвеоляр
ные макрофаrи выполняют функцию защиты.
Иннервация ле?ких. К бронхиальному дереву подходят симпатические и па
расимпатические нервы. Нервные импульсы, идущие по парасимпатическим He
рвным проводникам (ветви блуждающеrо нерва), вызывают сокращение rладких
мышц бронхов, а раздражение симпатических волокон, напротив, вызывает pac
слабление мышц.
Возрастные изменения. В постнатальном периоде проrpессирующе увеличи
вается дыхательная поверхность леrких. В пожилом возрасте происходит снижение
rазообменной функции в связи с постепенным разрастанием соединительнотканной
стромы леrких.
Ре?енерацuя леrких связана в основном с явлениями компенсаторной rиперт
рофии клеток альвеолярной выстилки. Показано, что большие эпителиоциты 2 ro
типа MOryT делиться митозом. Реrенерация леrких связана также с пролифераци
ей и миrрацией клеток бронхиальною эпителия, который врастает в зону повреж
дения и участвует в формировании альвеолоподобных структур.
Система орюнов внутренней среды
209
Плевра
Леrкие снаружи покрыты плеврой, называемой висцеральной, в отличие от па
риетальной плевры, которая выстилает изнутри стенку rpудной клетки. Висцераль
ная плевра состоит из мезотелия и подлежащей соединительной ткани. На rpанице
между ними имеется базальная мембрана. Мезотелий, кроме поrраничной, ВЫПОk
няет секреторную функцию, вырабатывая небольшой объем серозной жидкости в
плевральную полость. Среди соединительнотканных структур встречаются колла
reHoBbIe, эластические и ретикулиновые волокна. В леrочной плевре присутствуют
и rладкомышечные клетки. Висцеральная плевра плотно сращена с тканями леrко
ro. Плевра участвует в создании условий для облеrчения дыхательных движений
(экскурсии) леrких.
Выделительный комплекс opraHoB
Выделительный комплекс включает парный opraH почку и внепопечные MO
чевыводящие пути (мочеточники, мочевой пузырь, мочеиспускательный канал).
OpraHaM выделения принадлежит важнейшая роль в метаболических процессах
орrанизма и поддержании постоянства ero внутренней среды (rомеостаза). Почки,
фильтруя плазму крови, в которую постоянно выделяют продукты метаболизма
все клетки орrанизма, очищают орrанизм от конечных продуктов обмена и способ
ствуют сохранению физиолоrическоrо водно электролитноrо и кислотно щелоч
Horo баланса. Помимо функции мочеобразования почки синтезируют ряд биоло
rически активных соединений, которые имеют важное значение не только для фи
зиолоrии выделительной системы, но и общеметаболических процессов орrанизма.
В юкстаrломерулярном rистионе почки образуется белок ренин, который способ
стствует повышению артериальноrо давления, участвует в pery ляции секреции аль
достерона в надпочечниках, активации реабсорбции электролитов в почечных Ka
нальцах. Кроме Toro, здесь синтезируется ropMOH эритропоэтин, стимулирующий
дифференцировку клеток эритроидноrо ряда.
В интерстициальных эндокриноцитах почки синтезируются вещества rруппы
простаrландинов, вызывающие расширение артериол в почках и орrанизме в целом.
Развитие. Источником развития opraHoB выделения служит материал нефро
томов (cerMeHTHbIx ножек). У зародыша человека нефротомы сеrментированы лишь
в краниальном и туловищном отделах. В каудальном отделе тела находится не раз
деленный на cerMeHTbI нефроrенный зачаток. Материал нефротомов дифференци
руется reTepoxpOHHO. На 3 й неделе эмбриоrенеза из 8 10 передних (roловных) He
фротомов формируется предпочка, или пронефрос. Она состоит из трубочек
(протонефридий), один конец которых слепо замкнут и обращен к целому, а дpy
roй обращен в сторону сомитов, rде канальцы, соединяясь, образуют мезонефраль
ный (вольфов) проток Вскоре (через 40 ч) предпочка, не функционируя, редуциру
ется. Остается лишь мезонефральный проток, растущий в каудальном направлении.
На следующей стадии образуются канальцы первичной почки (мезонефроса)
из 25 туловищных нефротомов. Их дифференцировка происходит на протяжении
210
rлава 14
2 ro месяца эмбриоrенеза. Одним своим концом канальцы (метанефридии) соеди
няются с мезонефральным протоком, а противоположным (слепым концом) они
контактируют с клубочком капилляров, образуя BOKpyr Hero двухслойную капсу
лу. На 2 M месяце эмбриоrенеза мезонефрос достиrает максимальноrо развития.
Мезонефрос в эмбриоrенезе функционально активен и период ero экскреторной
деятельности прерывает начало функционирования окончательной почки. Процесс
мочеобразования в мезонефросе медленный. Это связано с низким артериальным
давлением у плода. Отсутствие петли fенле в нефроне мезонефроса обусловлива
ет rипотонию мочи. С 3 ro по 5 й месяц мезонефрос постепенно деrенерирует. Oc
тавшиеся канальцы используются для формирования семявыносящих протоков и
канальцев у плодов мужскоro пола и оболочек для половых клеток у плодов женс
Koro пола.
В период, коrда функционирует мезонефрос, начинается дифференцировка
несеrментированной части нефроrенноrо зачатка в каудальной части тела. Здесь
формируется вторичная, или окончательная, почка м.етанефрос.
Кроме нефроrенноrо зачатка, в развитии вторичной почки большая роль при
надлежит выросту стенки м.езонефральноlO протока. Этот вырост образуется в виде
слепоro выпячивания от стенки мезонефральноro протока в том месте, rде после
дний впадает в клоаку. Далее выпячивание стенки растет по направлению к нефро
reHHoMY зачатку, и из Hero в дальнейшем формируются мочеточник, почечная ло
ханка с почечными чашечками, а от последних возникают выросты, превращаю
щиеся в собирательные трубочки. Эти трубочки выполняют роль индуктора при
развитии канальцев в нефроrенном зачатке. Из последнеro образуются скопления
клеток, которые превращаются в замкнутые пузырьки. Разрастаясь в длину, пузырь
ки превращаются в слепые почечные канальцы, которые в процессе роста S образно
изrибаются. При взаимодействии стенки канальца, прилежащей к слепому выросту
собирательной трубочки, происходит объединение их просветов. Противоположный
слепой конец почечноro канальца приобретает вид двуслойной чаши, в уrлубление
которой врастает клубочек артериальных капилляров. Здесь формируется сосудис
тый клубочек почки, который вместе с капсулой образует почечное тельце.
Почки
Почки располаrаются в забрюшинном пространстве поясничной области. CHa
ружи почка покрыта соединительнотканной капсулой. Почка состоит из KOpKOBO
ro и мозrовоrо вещества. fраница между этими частями неровная, так как CTPYK
турные компоненты KopKoBoro вещества вдаются в мозrовое в виде колонок, а
мозrовое вещество проникает в корковое, образуя мозrовые лучи (рис. 103).
Основной структурно функциональной единицей почки является нефрон.
Нефрон представляет собой эпителиальную трубочку, которая начинается слепо в
виде капсулы почечноrо тельца, далее переходящей в канальцы разноro калибра,
впадающей в собирательную трубочку. В каждой почке имеется около 1 2 млн He
фронов. Длина канальцев нефрона составляет 2 5 см, а общая длина всех каналь
цев в обеих почках достиrает 100 км.
Система ОрЮ1l0в в1lутре1l1lей среды
211
в нефроне различают капсулу клу
бочка почечноrо тельца, проксималь
ный, тонкий и дистальный отделы.
Почечное тельце состоит из клубоч
ковой капиллярной сети и эпителиаль
ной капсулы. В капсуле различают Ha
ружную и внутреннюю стенки (листки).
Последняя вместе с эндотелиоцитами
клубочковой капиллярной сети форми
рует lематО1lефрuдuалЬ1lЫЙ lUCmU01l.
Клубочек капиллярной сети располо
жен между приносящей и выносящей
артериолами. Приносящая артериола
чаще дает четыре разветвления, которые
распадаются на 50 1 00 капилляров.
Между ними имеются мноючисленные
анастомозы. Эндотелий капилляров
клубочковой сети состоит из плоских
эндотелиоцитов с мноrочисленными
фенестрами в цитоплазме размером OKO
ло 0,1 мкм. Фенестрированные (оконча
тые) эндотелиоциты представляют co
бой своеобразное сито. Снаружи от эн
дотелиоцитов располаrается общая для
эндотелия и эпителия внутренней cтeH
ки капсулы базальная мембрана, толщи
ной около 300 нм. Для нее характерно
трехслойное строение. Эпителий BHYT
ренней стенки капсулы охватывает со всех сторон капилляры клубочковой сети.
Состоит он из одною слоя клеток, называемых подоцитами. Подоциты имеют слеr
ка вытянутую неправильную форму. Тело подоцита имеет 2 3 крупных длинных OT
ростка, называемых цитотрабекулами. От них в свою очередь отходит MHoro мелких
отростков цитоподий. Цитоподии представляют собой узкие цилиндрические об
разования (ножки) с утолщениями на конце, посредством которых они прикрепля
ются к базальной мембране. Между ними имеются щелевидные пространства разме
ром 30 50 нм. Эти щели имеют определенное значение в процессах фильтрации при
образовании первичной мочи. Между петлями капилляров клубочковой сети Haxo
дится разновидность соединительной ткани (мезанrий), содержащая волокнистые
структуры и мезанrиоциты.
Эпителий наружной стенки капсулы клубочка состоит из одною слоя плоских
эпителиоцитов. Между наружной и внутренней стенками капсулы имеется полость,
в которую поступает первичная моча, образующаяся в результате клубочковой
фильтрации (рис. 104).
Процесс фильтрацuu является перl3ЫМ этапом мочеобразования. Фильтруются
"i!.. 'tJ..\: \ 1 ':'\ А':" ;"iЦн f t\ ,'6, . ;
' ..''"' " #; !l ' f "" <V
' ,, '; , l , : ::, ": ..::\ , ti i; l; ': :""'
{i . w - - ; I f/; . ,,- <}; y. ;. !.:J1
:. '" "";" ,д ;[, '..' ::т. , ' j i': H ' :. ' , . , t".. " ""' ,, " , , ; J '
',.; .о',:" 'j' ':,-:Jiii! , :" ;" 0 <:.. t..,.... J
2' ,; "Н-, ) . f ;'1 " ' '.С.., .,'
'? ' )(:,: , '; "I:: ::Т J(I . ; \ , '" ';' 't.
_'\.'E, :i'.: : -: '3" ,-' ,"
} н . . \, ( ._ .'" ..' . "" <
o:\,!' '. ' ,.,..:;IJ.' . ." ',. , :"
" " , [{ ) '.'1-.. .....f!" ',,!
" "','" "::,', . , ,;}". /: ct ;" 1',
"o . ;?!i. " ;O'o;_, ;' ,,fr.o'
" Q . \\У'1>: p \. ' ',;", '14,',,#)..
f::'. i: ': ' :j . /,\ ifr i
,!,; p(' [ . <o ,'\! r;9;;.: t .'
"t ," ':,g i.;;Ч" ,\6/'.
" " ! ,,, ,," :\ '" , " " ,.j ,:: 1< , '
jO' .-,' К, Ч '1
{;(..l."', , :8f' . \ '!
,О ; iJi;i:! . i ' , .У=--- ,<
,':, '\ \ ,,, ' .,' /;
'\ " ", ' , "' , !' , k' '" '. # '
'V, "'q '1: , f'
t Ч .' ''t};. , .1
у.
А
Б
Рис. 103. Строение почки:
А корковое вещество; Б мозrовое вещество;
1 почечное тельце; 2 мозrовые лучи; 3 Ka
нальцы разноro калибра; 4 кровеносные сосуды
212
Fлава 14
практически все компоненты
плазмы крови, за исключением
Еысокомолекулярных белков и
форменных элементов крови.
Жидкость из просвета капилля
ра проходит через фенестриро
ванные эндотелиоциты, базаль
ную мембрану и между цитопо
диями подоцитов С их MHoro
численными фильтрационны ,.
ми щелями, при крытыми диаф
раrмами, в полость капсулы
клубочка. rематонефридиаль
ный rистион проницаем для
rлюкозы, мочевины, мочевой
КИСЛОТЫ,креатинина,ХЛОРИДОВ
и низкомолекулярных белков.
Эти вещества входят в состав
ультрафильтрата первичной
мочи. Большое значение для
эффективной фильтрации име
ет разность диаметров принося
щей и выносящей клубочковых
артериол, что создает высокое
фильтрационное давление (70
80 мм рт. ст.), а также большое
количество капилляров (около
SO 60) в составе клубочка. Во
взрослом орrанизме в течение
суток образуется около 1 SO
170 л первичноrо фильтрата
(мочи). Столь эффективная
фильтрация плазмы, осуществ
ляемая почками практически
беспрерывно, способствует MaK
симальному удалению из орrанизма вредных продуктов метаболизма шлаков. Сле
дующим этапом мочеобразования является обратное всасывание (реабсорбция ) He
обходимых орrанизму соединений (белков, rлюкозы, электролитов, воды) из пер
вичноro фильтрата с образованием окончательной мочи. Процесс реабсорбции про
исходит в канальцах нефрона.
В про-кcuмалъном. отделе нефрона различают извитую и прямую части канальца.
Это самый протяженный участок канальцев (около 14 мм). Диаметр проксимальноrо
извитоrо канальца составляет SO 60 мкм. Здесь происходит облиrатная реабсорбция
орrанических соединений по типу рецепторно опосредованноrо эндоцитоза с участи
б f 1
l
1 " >.:
'
,,'х '.
!; !"-.i
..,<F"';
i
5
\
"
'" ,
J
"
,
\}
\'
.
. .
.........7
.'".<1
з
,
со)
.
.'
. ...
t 11.
]
I
)
.........4
Рис. 104. Схема строения нефрона:
1 клубочковая капиллярная сеть; 2 эпителиальная кап
сула; 3 проксимальный отдел нефрона; 4 тонкий отдел
нефрона; 5 дистальный отдел нефрона; б юкстаrломе
рулярный комплекс; 7 собирательная трубочка
Система орюнов внутренней среды
213
ем энерrии митохондрий. Стенка проксимальноrо Ka
нальца состоит из однослойноrо кубическоrо микро
ворсинчатоrо эпителия (рис. 105). На апикальной по
верхности эпителиоцитов находятся мноrочисленные 1
микроворсинки длиной 1 3 мкм (щеточная каемка).
Число микроворсинок на поверхности одной клетки
2
достиrает 6500, что увеличивает активную всасываю
щую поверхность каждой клетки в 40 раз. В плазмо
лемме эпителиоцитов между микроворсинками име
ются уrлубления с адсорбированными макромолеку
лами белков, из которых формируются транспортные :5
пузырьки. Общая поверхность микроворсинок во всех
нефронах составляет 40 50 м 2 . Второй характерной
особенностью строения клеток эпителия проксималь
Horo канальца является базальная исчерченность эпи 4 ' M """"""'"
телиоцитов, образованная rлубокими складками плаз
молеммы и закономерным расположением между ними
мноrочисленных митохондрий (базальный лабиринт).
Плазмолемма эпителиоцитов базальноrо лабиринта
обладает свойством транспорта натрия из первичной
мочи в межклеточное пространство.
71J1lКUЙ отдел нефрона, или пerпля МЮlе, состоит из нисходящей тонкой части и
восходящей толстой части. Нисходящая часть имеет диаметр 1З 15 мкм. Стенка нис
ходящей части образована плоскими, а восходящей части кубическими эпителио
цитами, в которых выявляются немноrочисленные микроворсинки. В восходящей ча
сти происходит активная реабсорбция электролитов (натрия). Вслед за натрием
происходит пассивное перемещение воды из нисходящей части в интерстициальное
пространство и далее в rемокапилляры. Клетки нисходящей части плоские и xapaK
теризуются присутствием множественных пиноцитозных пузырьков.
Дистальный отдел нефрона подразделяется на две части прямую и извитую.
Диаметр канальца составляет 20 50 мкм. В этой части нефрона завершается обрат
ное всасывание электролитов и воды (факультативная реабсорбция), а также ceKpe
тируются катионы водорода, придающие окончательной моче кислую реакцию и за
щитные антимикробные свойства. Клетки, выстилающие стенку дистальноrо отдела,
имеют кубическую форму, в них выявляются складки базальной плазмолеммы, между
которыми залеrают удлиненные митохондрии. Эпителий дистальноrо отдела отли
чается высокой активностью rидролитических и rликолитических ферментов. Про
цесс реабсорбции реryлируется rормонами rипоталамуса (антидиуретическим rop
моном) и надпочечников (минералокортикоидами). Нарушения биосинтеза этих
roрмонов приводит к возникновению несахарноro мочеизнурения.
Дистальные отделы нефронов впадают в собирательные тру6ач:кu. Их стенка
образована кубическим и призматическим эпителием, в составе KOToporo различа
ют светлые и темные клетки. Светлые участвуют в реабсорбции электролитов и
воды с помощью водных каналов, которые формируются интеrральным белком
I ' I . 1 ,
{:. ' . .. ;..:
1 "ii ;' , : I "e
,,-:". ..:. .-..,. :
R:c; i'
./
( r'v \
\\ ....... .
\: 'О..... I
" - .
...
...M?:нo1It:
..............r.........
:t])
o ..... ...
!€J<>
!' е'.
\Ф /
"',.. . a.-"
Рис. 105. Схема строения эпители
оцитов различных отделов нефрона
и собирательной трубочки:
1 проксимальный отдел; 2 TOH
кий отдел; 3 дистальный отдел;
4 собирательная трубочка
214
rлава 14
плазмолеммы аквапорином. Темные секретируют катионы водорода, подкисля
ющие окончательную мочу.
Сосуды почки. Сосуды почки имеют своеобразную архитектонику в связи
с наличием двух типов нефронов: корковых и околомозrовых (юкстамедулляр
ных). Корковые нефроны почти полностью располаrаются в корковом веще
стве почки. Они имеют короткий тонкий отдел нефрона. Юкстамедуллярные
нефроны расположены в основном в мозrовом веществе, rде проходят их длин
ные тонкие отделы. Корковые нефроны составляют 80% от общеrо количества
всех нефронов. Юкстамедуллярные нефроны в норме не участвуют в процессе
мочеобразования.
Кровь поступает в почку через почечную артерию, переходящую в междоле
вые артерии и далее в дуювые артерии. От дуrовых артерий отходят радиальные
междольковые артерии, дающие приносящие клубочковые артериолы, капилляры
клубочковой сети, выносящие клубочковые артериолы. Выносящие артериолы
распадаются на перитубулярную (вторичную) капиллярную сеть, оплетающую Ka
нальцы нефрона, в которых осуществляется реабсорбция компонентов первичной
мочи. Далее капиллярная сеть переходит в венулярное звено, междольковые, дуrо
вые, междолевые вены и, наконец, в почечную вену. В комплексе эти сосуды co
ставляют кортикальный Kpyr кровообращения в почке.
Существует и второй юкстамедуллярный Kpyr. Он начинается принося
щими артериолами, отходящими от проксимальной части междольковых apTe
рий. Эти артериолы идут к юкстамедуллярным клубочкам, образуя здесь сеть
капилляров. Из клубочков выходят выносящие артериолы, из которых кровь
поступает в прямые артерии и далее непосредственно в прямые вены, не обра
зуя вторичной капиллярной сети. Прямые вены переходят далее в дуrовые, затем
в междолевые вены, несущие кровь в почечную вену. Юкстамедуллярное KpO
вообращение короче кортикальною и является своеобразным шунтом, по KOTO
рому кровь может частично сбрасываться, минуя корковое вещество. При этом в
корковых нефронах фильтрационное давление остается относительно стабиль
ным (70 80 мм рт. ст.). Данный путь имеет важное значение, т. ко при подъеме
артериальною давления (при физических наrрузках, например) резко возраста
ет приток крови к почкам.
Сосудистая система почки находится под реryлирующим влиянием как общих
нейроэндокринных механизмов, так и местных, среди которых наибольшее значе
ние имеют rормоны юкстаrломерулярноrо rистиона почки и интерстициальных
эндокриноцитов.
Клетки юкстаrломерулярноrо rистиона вырабатывают фермент ренин, необ
ходимый для образования анrиотензина, обладающеrо сильным сосудосуживаю
щим действием. Кроме Toro, ренин участвует в реrуляции водноrо и минеральноrо
обмена, оказывая влияние на продукцию альдостерона надпочечниками. Клетки
юкстаrломерулярноrо rистиона также вырабатывают эритропоэтин, следователь
но, это своеобразный эндокринный комплекс, называемый иноrда "ренинсекрети
рующим комплексом", или "ренинанrиотензинной системой". ЮкстаlЛомеруляр
'Н.ЫЙ lUCтUOH включает следующие клетки: миоидные эндокриноциты при носящих
Система орzшюв внутренней среды
215
клубочковыхартериол;эпителиоциты
плотноrо пятна дистальноrо отдела
нефрона; юкставаскулярные клетки
(клетки [урмаrтиrа) и мезанrиоциты
7
(рис. 106).
Миоидные эндокриноциты (юк
стаrломерулярньre, околоклубочковые
клетки) это видоизмененные rладко
мышечные клетки, средней оболочки
приносящей клубочковой артериолы в
месте контакта ее с плотным пятном ди
стальноrо канальца. В их цитоплазме З
накапливаются секреторные rранулы,
содержащиеренин.Этобарорецепторы.
Клетки плотною пятна находятся
в участке стенки дистальноrо канальца,
лежащем между приносящей и вынося
щей клубочковыми артериолами у co
Рис. 106. Схема строения почечноrо тельца и юкстаr
судистоrо полюса почечноrо тельца. ломерулярноrо rистиона:
Эти эпителиоциты числом 15 40 более 1 приносящая артериола; 2 выносящая артерио
высокие, прилежат к очень тонкой пре ла; З капилляры сосудистоrо клубочка; 4 полость
капсулы; 5 проксимальный отдел; 6 дистальный
рывистой базальной мембране, нередко отдел; 7 миоидные эндокриноциты; 8 эпителио
имеют отростки цитоплазмы, идущие циты плотноrо пятна; 9 юкставаскулярные клетки
между юкставаскулярными клетками. Полаrают, что эпителиоциты плотноrо пятна
воспринимают изменения концентрации натрия в крови и в жидкости, омывающей
плотное пятно, то есть являются осморецепторными клетками. Эту информацию они
передают миоидным эндокриноцитам, которые реryлируют выработку ренина.
Юкставаскулярные клетки ([урмаrтиrа) заполняют треуюльное пространство,
оrраниченное приносящей и выносящей клубочковыми артериолами и плотным
пятном. Мезанrиальные клетки располаrаются между капиллярами клубочковой
сети. Они способны к фаrоцитозу, синтезу межклеточноrо вещества, обладают co
кратительной активностью и поддерживают форму клубочка.
К эндокринному комплексу почки относятся также интерстициальные клет
ки, которые расположены в строме мозrовоrо вещества. Своими отростками они
оплетают канальцы петли нефрона и кровеносные капилляры перитубулярной сети.
Интерстициальные клетки вырабатывают одну из разновидностей простаrланди
нов с антиrипертензивным действием. Нарушение деятельности эндокринноrо ап
парата почки может привести к развитию артериальной rипертензии.
Юкстаrломерулярный rистион и интерстициальные эндокриноциты почек pe
rулируют внутрипочечное кровяное давление, что имеет важное значение для фун
кции почки фильтрация в почечных клубочках происходит лишь при давлении
крови в капиллярах клубочков корковых нефронов около 70 90 мм рт. СТ.; резкие
колебания внутрипочечноrо давления недопустимы и крайне неблаrоприятны для
функционирования выделительной системы.
2
9
8
4
216
Fлава 14
Мочевыводящие пути
к 1\I0чевыводящим путям относят почечные чашечки, лоханки, мочеточники,
мочевоЙ пузырь и мочеиспускательный канал. Строение этих opraHoB в общих чер
тах сходно, так как их стенка состоит из слизистой оболочки, подслизистой OCHO
вы, мышечной и наружной оболочек Эпителий мочевыводящих путей называется
переходнымо Он относится к эпителиям кожноrо типа. В эпителии различают ба
зальные клетки, выполняющие роль камбия, и более дифференцированные повер
хностные клетки. При этом около половины поверхностных клеток являются по
липлоидными.
В стенке почечных чашечек описаны особые rладкие мышечные клетки вo
дители ритма (пейсмекеры). За счет ритмическorо сокращения этих клеток моча
порциями поступает из собирательных трубочек и происходит опорожнение по
чечных чашечек
В мочеточниках rладкая мышеч
ная ткань образует в верхней полови
не два слоя: внутренний продольный '. '.\
и наружный циркулярный. В ниж
ней части мочеточников добавляется
еще один продольный слой, располо
женный снаружи от циркулярноrо.
В мочевом пузыре слизистая обо
лочка приспособлена к растяжению,
связанному с периодическим накопле
нием мочи (рис. 107). ЭпителиЙ при
этом меняет свою rистолоrическую
картину от растянутоrо двухслойноrо
до псевдомноrослойноrо в спавшемся
пузыре. В слизистой оболочке мuчевu
ro пузыря сильно развиты сосудистые
подэпителиальные сплетения. Mы
шечная оболочка мочевоrо пузыря co
стоит из трех слоев: BHYTpeHHero, Ha
ружноrо с продольным расположени
ем rладких миоцитов и среднеrо
циркулярноrо. В шейке мочевоrо пу
зыря имеется мышечный сфинктер.
Наружная оболочка образована соеди
нительной тканью, а в области дна
серозной оболочкой. Мочевой пузырь
иннервируется симпатическими и па
расимпатическими, а также спиналь
ными нервами. В нем имеется MHOro
веrетативных нервных rанrлиев.
o ''''; ? '
'. ,,,,,,,.,..- , 1
:... '" f.:;:" -:" . . /
f ;;. = ,,; _' _/, "\<. -
.-....- ''',' ''';' .' >o'i!l >о '"
: :' l ' ::.jfr't.i ", J. (:':","
'" ., о' ". .' ,. ,А > '.
.. :'. :... . .'. '. : : .,!1. ., '
.... , ", ,}\:. . 2 ';'
-:.... . : '<.ii! \ . \
,.,' . I ..... ," o .
.. ..., z.::."': , ",a:.: ;;' ... :' ,
, ....:.. , " 't?o4':';,;Jt!.:"",' .. ,.'" ,., ..,. о'.
..... . . ...,. ; «'!-.f f "'. - - "".' .
" # ..tQ-........ , - , .,..::...."> ._'J1 , '( .." , ' ......,., . , ' , }
,А .f'''' -111., "': ......; ./
. : .' ,._:;.:?:.......... i " :&.. . . Б
. ': ( :.....8 '.. '. _ .- 7'4t-!"'"'"
""", ....."1> t"Н'_....,/
., 0'0 .... : - d .:-...... ..' '-1_1
" , " .\' ,,0' J. '_ ,.
",' ....... :. ..:. ';;'ф ,...;...... ; '..
'], "", , '1I{r"..". '.
,. '"'..': и.. ' ," . _ ';. _, ! "1"-
: ., __- ;:' ;. t- $ _y'
; . . ,,- "" . '" .# ts;-"':
.ft}: А.'II ".... ...... 1:....:' ... ... ":;...," ,, ';rI ;J..-... f1....,
"..... !'......I. .....J.u.. " ... :3"
"" "' ....:,-, ...,:, '" ' ?Jf; I",.j.;y> v В
.. ....... r....,. " ,F< \.":,,;-_,, _ . .,"
'1"", ... """'.'r '!Jiт... з...
.'" f ' о .. . .; "
..... I "1 '...
....,,0 .. ii ... ...".
(;?
д
...
.
...
'.
t'O
"
}r
1!i:'Ir'.. .
Рис. 107. Строение стенки мочевоrо пузыря:
Д слизистая оболочка; Б подслизистая основа; В
мышечная оболочка; r адвентициальная оболочка;
1 переходный эпителий; 2 собственная пластинка
слизистой оболочки; 3 пучки rладких миоцитов
Система орюнов внутренней среды
217
Стенка мужскоro мочеиспускательною канала состоит из слизистой и мышеч
ной оболочек, а женскоrо слизистой, мышечной и адвентициальной оболочек
Эпителий слизистой оболочки из переходноrо постепенно трансформируется в
мноrослойный плоский неороrовевающий. В составе эпителия встречаются скоп
ления слизистых клеток Собственная пластинка слизистой оболочки содержит
уретральные слизистые железы. Мышечная оболочка включает внутренний про
дольный и наружный циркулярный слои rладких миоцитов. При прохождении MO
чеполовой диафраrмы мужская уретра окружается поперечнополосатой мышеч
ной тканью наружноro сфинктера мочевоrо пузыря. Женская уретра в средней своей
части окружается поперечнополосатой мышечной тканью наружноrо сфинктера.
Возрастные изменения. В постнатальном периоде продолжаются рост и раз
витие нефронов. При этом увеличиваются их длина и толщина. В связи с этим на
единицу массы почечной ткани у взрослоrо приходится в 1 О раз меньше почечных
телец по сравнению с новорожденным.
Реактивность и ре енерацuя. Реактивные изменения почек при действии эк
стремальных факторов (переохлаждение орrанизма, отравление ядовитыми веще
ствами, действие проникающей радиации, ожоrи, травмы и др.) весьма разнооб
разны с преимущественным поражением сосудистых клубочков или эпителия
различных отделов нефрона вплоть до rибели нефронов.
Реrенерация нефрона происходит более полно при внутриканальцевой rибели
эпителия. Наблюдаются клеточная и внутриклеточная формы реrенерации. Эпи
телий мочевыводящих путей обладает достаточно выраженной восстановительной
способностью.
Аномалии мочевыделительной системы, орrаноrенез которой достаточно сло
жен, являются одним из наиболее частых пороков развития. Причинами их обра
зования MorYT быть как наследственные факторы, так и действие различных по
вреждающих факторов ионизирующеrо излучения, алкоrолизма и наркомании
родителей и др. Вследствие ТOfО, что нефроны и собирательные трубочки имеют
разные источники развития, неправильность или отсутствие их контактов друr с
друroм приводит к патолоrии развития почек (поликистоз, rидронефроз, аrенезия
почек и др.).
218 Fлава 15
fЛАВА 15. СИСТЕМА РЕАКТИВНОСТИ
К системе реактивности орrанизма человека принадлежат орrаны, осуществ
ляющие восприятие всех внешних и внутренних сиrналов, их анализ и адекватную
конкретноЙ обстановке реryляцию жизнедеятельности, а также интеrрацию функ
ций opraHoB и систем орrанизма. Систему реактивности представляют орrаны им
мунноЙ защиты, эндокринные железы, нервная система с ее периферическим ceH
сорным аппаратом. Эти три части орrанизма объединяются в единую нейро эн
докринно иммунную систему, поскольку их деятельность взаимно соrласована и
зависима. Так. нейропептиды, синтезируемые эндокринными нейронами, влияют
на активность иммунокомпетентных клеток, а биолоrические активные вещества
иммунокомпетентных клеток оказывают влияние на клетки и ткани, сходные с Ta
ковыми для ropMOHoB эндокриноцитов и пептидов нейронов.
Иммунный комплекс opraHoB
Иммунный комплекс opraHoB включает вилочковую железу (тимус), лимфа
тические узлы, селезенку, лимфоидные образования в стенке пищеварительноrо
тракта и в друrих opraHax и красный костный мозr, rде развиваются все клетки
крови, в том числе осуществляющие иммунный надзор.
Несмотря на топоrрафическую разобщенность, эти орrаны вместе с кровью и
лимфой образуют единую в функциональном отношении систему, обеспечиваю
щую поддержание процессов кроветворения и иммунной защиты. Орrаны KpOBe
творения представляют собой открытую систему с постоянным перемещением
клеток крови.
Различают центральные и периферические орrаны кроветворения и иммуно
rенеза. К центральным opraHaM относят красный костный мозr и вилочковую же
лезу. К периферическим кроветворным и иммунным opraHaM принадлежат лим
фатические узлы, селезенка, миндалины и друrие лимфоидные образования в
составе слизистных оболочек opraHoB.
Красный костный МОЗf
Красный костныЙ мозr центральный rемопоэтический opraH. В нем Haxo
дится основная часть стволовых кроветворных клеток и происходит развитие кле
ток миелоидноro и лимфоидноrо рядов, осуществляется антиrеннезависимая диф
ференцировка В лимфоцитов (рис. 108).
В эмбриоrенезе человека костный мозr появляется впервые на 2 3 M месяцах в
Система реактивности
219
.".,.\,) ',;? I; ,' : 'f ;: . ",С:,:
. ; ' . Jk '"h.. ' " ,, 7.. 'f,t'J},,].P
. .: ._ O..,.,," . .. .... ........ .. . ;.
. ',,\t..'}" . .. '....' "У\.....' " .-.".
l ' ; { .. i' "E и 2
1 t-,...1"{' :'> (.1 ';fi4,r:, ';;,i. :i...
. , ..\. y.. l . . .. "' ..
,i ]/*.? ..,; : f '
С . . ...,t....,. (J- . t;....' . \. "f ..... ...... {
,,' . '< t" '," " .''itt;, 1f!I!!i" '>:'N '> ,..
.f: ";-; rл& . -1 : ' .; I r:...... ; ti'.. : "!. . ": . ......
,-":, ,'Ф' ' .,,"., ,..,.' .......... ,, . . .. ,.
":; <;, ':,(' ".. , ," .. r l" h:... """' 1J.'".c... чУ "if' Qi' ',' 1
"V 'f"!' '" '.. /;:.. i!r' / " ..., , :} .. l ) ': ,0'" f
.', ".-' ,,'...., .. ( . ., >WJ ' .'.,. ". . Т '
,,' а:;. iIi- ,'.. "'I \." ., y," 1iiIIii<" I!i> ..
' . .,. . @ '"10 '\ "'.. '"'= :. ,, ::. . . -.:.. .
· " ;' "..> ,,",,. . 1 i"" r ...'I/e...':"'" f""".. .. '.'::
. ..... ,.". , "'.. ../...,;; , .. tj . t
.. . ' ,,'" ",', "> ".;:-,,,: , ,'c. .... ... .. (..
.. . \. .., ....; 1< .... .; '" 't; .' : . . ". . " N
,'1. . ,. pr":':;;' :',lf/ '> " '"'>- ..": .. ", '<".':-'" ' ,
" : ' }'.<': ;:rt :( i':7 N j:,; , :; : \.
- ,. ,.,""",$, "r - " <", J -j".. ' , \ ..... .' >,
V ; ,_:';'С', ,...> /.<, ",' '\.. ;:..' .' ,,' 'f::. ' ___ . .., "'-
_ ,"' 'I"Y'.....,..",, ', ) ....., :"'"' .,..... . ": I ".."...c.,g..?;;;
- ' : "p" , ? , "\ ,ii d., "C{ J. ,,> \
/ ':, ,'! #t-.. '\" '" .,\":;')
/' ... .((--;::.:;":. ::«Lt ?' :::Jfil;(i> Kii' ".,, ;;...,,
5 ; - O' " ;... ,..../q < !>{"
\:".' ._ ".: ;t ;.:;r"'" :&'">; : ?: 'c (!'.; ' jf r_
..... "; .. "'""" ';""" ,'''"'''.v-
"... .. ;.,.>i?;::.;;_:., '!.;;':
.... ..;/ ...... . :t . ..")
. -.; ; i?: : .:
плоских костях и позвон
ках, на 4 M месяце в труб
чатых костях конечностей.
Различают красный Kocт
ный M03l и желтый кост
ный M03l. Красный костный
мозr находится в эпифизах
трубчатых костей, в ryбча
том веществе плоских KOC
тей, в лопатках, rpудине, по-
звонках, костях черепа. He
смотря на такое paccpeДOTO
чение, функционально он 4
тесно взаимосвязан блаro-
даря постоянной миrpации
клеток и наличию общих
механизмов реryляции про
цессов кроветворения.
Масса KOCTHoro мозrа
1,6-3,7 Kr, что составляет 3-
6% от массы тела. Красный
костный мозr имеет TeMHO
Рис. 108. Строение KOCTHoro мозrа:
красный цвет, Консистен- 1 эритробласт; 2 миелоцит; 3 меrакариоцит; 4 суносоид
ция ero полужидкая. Это ный капилляр; 5 эритроциты
позволяет делать из Hero тонкие мазки, изучение которых имеет большое диаrнос
тическое значение в клинике.
Строма KpacHoro KocTHoro мозrа образована костными перекладинами, иду--
щими от эндоста. Между ними располаrается ретикулярная ткань. Последняя co
стоит из трехмерной сети rетероморфных ретикулярных клеток фибробластичес--
Koro вида (фибробласты KocTHoro мозrа). Они вырабатывают межклеточное
вещество, включающее ретикулярные волокна и амфорный компонент с большим
содержанием rликозаминоrликанов, ростовые факторы (интерлейкины). Кроме ре-
тикулярных клеток к стромальным клеточным элементам относятся остеобласты,
входящие в состав эндоста и способные влиять на пролиферацию rемопоэтичес
ких клеток, адвентициальные малодифференцированные клетки, сопровожда
ющие кровеносные сосуды, жировые клетки. Все эти клетки развиваются в резуль
тате диверrентной дифференцировки стромальной стволовой клетки и иrрают роль
микроокружения для развивающихся клеток крови.
Строма KpacHoro KocTHoro мозrа пронизана кровеносными сосудами микро
циркуляторноro русла. В основном это капилляры СЩlусоидноrо типа с диамет
ром около 30 мкм.
В петлях ретикулярной ткани KpacHoro KocTHoro мозrа расположено множе
ство кроветворных клеток (в том числе стволовых кроветворных, клеток предше
ственников миело и лимфопоэза, клеток rранулоцитарноrо, эритроцитарноrо,
з
220
Fлава 15
лимфоцитарноrо, моноцитарноrо и тромбоцитарноrо рядов на различных стадиях
дифференцировки ).
Количество стволовых кроветворных клеток в красном костном мозrе наиболь
шее по сравнению с друrими кроветворными орrанами (50 на 105 клеток). KOHцeH
трация стволовых кроветворных клеток вблизи эндоста в 3 раза больше, чем в дpy
rих участках KOCТHoro мозrа. Именно здесь наиболее интенсивно идет KpOBeTBope
ние, что связывается с выработкой остеобластами интерлейкинов и повышенным
содержанием кальция.
Развивающиеся клетки крови располаrаются в красном костном мозrе rpуппа
ми (островками, "rнездами"), представляющими собой диффероны, или rистоrе
нетические ряды клеточной дифференцировки. Эритробласты находятся вблизи
макрофаrов, содержащих железо фаroцитированных эритроцитов, и получают от
них железо, необходимое для построения rемоrлобина. Созревающие rранулоци
ты образуют островки, подобно эритроидным клеткам, с тем, однако, отличием,
что они не имеют связи с макрофаrами.
Клетки тромбоцитарноrо ряда (меrакариобласты и меrакариоциты) локали
зуются преимущественно вблизи кровеносных синусоидов. Отростки цитоплаз
мы меrакариоцитов при этом проникают через поры в стенке синусоидов внутрь
сосудов, И от них отделяются фраrменты цитоплазмы в виде кровяных пластинок
(тромбоцитов). Последние тут же поступают в кровоток
В красном костном мозrе обычно BOKpyr кровеносных сосудов встречаются
небольшие rруппы лимфоцитов и моноцитов. Среди множества кровяных кле
ток в красном костном мозrе больше Bcero зрелых клеточных форм или близ
ких К состоянию зрелости (эритробластов, метамиелоцитов и др.). В случае He
обходим ости, например, при кровопотере, они MorYT быстро завершить
дифференцировку и перейти в кровоток. В нормальных условиях через стенку
синусоидных капилляров MorYT проникать лишь зрелые формы клеточных диф
феронов.
Желтый костный МОЗl расположен в диафизах трубчатых костей. Он представ
лен преимущественно жировой тканью. В жировых клетках содержится пиrмент
липохром, имеющий желтый цвет. Желтый костный мозr рассматривается как KpO
ветворный резерв, и в случае больших кровопотерь он начинает функционировать
как кроветворный opraH. Желтый и красный костный мозr это два Функцио
нальных состояния одноrо KpoBeTBopHoro opraHa.
Красный костный мозr очень чувствителен к действию радиации, интоксика
ций бензолом, толуолом и друrими ядами. Особенно уязвимы при этом "бласт
ные" клеточные формы. Происходит опустошение KocTHoro мозrа и в результате
остается лишь ретикулярная строма. Отмечаются выраженные изменения KOCTHO
ro мозrа, связанные с превращением миелоидной ткани в жировую. а в старческом
возрасте в слизистую, желатинозную ткани.
Реzеllерацuя. Костный мозr обладает высокой реrенерационной способностью.
После удаления части KocTHoro мозrа или после облучения ионизирующей радиа
цией происходит ero восстановление за счет заселения KocTHoro мозrа циркулиру
ющими в крови стволовыми клетками. Необходимым условием при этом является
Система реактивности
221
Тимус центральный opraH лим
фоидноrо кроветворения и иммунной
защиты орrанизма. В тимусе происхо
дит антиrеннезависимая дифферен
цировка костномозrовых предше
ственников Т лимфоцитов в иммуно
компетентные клетки Т лимфоци
ты. Последние осуществляют реакции
клеточноrо иммунитета и участвуют
в реryляции ryморальноrо иммуните
та, что происходит, однако, не в тиму
се, а в периферических opraHax KpOBe
Рис. 109. Строение тимуса:
творения и иммунной защиты. Кроме А корковое вещество; Б мозrовое вещество; 1
TOrO, в экстрактах тимуса обнаружено rемокапилляры; 2 тельце rассаля
более 20 биолоrически активных веществ, в том числе дистантноro действия, что
позволяет отнести тимус к железам эндокринной системы.
Развитие. Тимус закладывается на 2 M месяце эмбриоrенеза в виде небольших
выпячиваний стенок З й и 4 й пар жаберных карманов. На 6 й недсле зачаток желе
зы имеет отчетливо выраженный эпителиальный характер. На 7 й неделе он утрачи
вает связь со стенкой roловной кишки. Эпителий закладки железы, образуя BЫpOC
ты в мезенхиму, приобретает сетевидное строение. Вначале плотная эпителиальная
закладка железы разрыхляется блаrодаря заселению ее лимфоцитами. Число их бы
стро нарастает, и железа при обретает структуру лимфоэпителиалъноl.O орюна. Bpac
тающая мезенхима с кровеносными сосудами подразделяет тимус на дольки. В каж
дой дольке различают корковое и мозroвое вещество. В rистоrенезе тимуса в мозrовом
веществе долек образуются слоистые эпителиальные образования эпителиальные
жемчужины, или тельца [ассаля. В их составе определяются плотные эпителиаль
ные клетки, концентрически наслаивающиеся друr на друrа (рис. 109).
Строение. Снаружи вилочковая железа покрыта соединительнотканной капсу
лой. Отходящие от нее переroродки септы подразделяют тимус на дольки. Oc
нову дольки составляют отростчатые эпителиальные клетки эпителиоретикуло
циты, в сетевидном остове которых находятся тимические лимфоциты (тимоциты).
Источником развития Т лимфоцитов являются костномозroвые стволовые KpOBe
творные клетки. Далее предшественники Т лимфоцитов (претимоциты) поступают
с кровью в тимус И превращаются здесь в лимфобласты. В корковом веществе тиму
са одни из них под действием выделяемых эпителиальными клеткаМII пептидных
ropMoHoB тимозина, тимопоэтина и др., а также макрофаrов превращаются в
сохранение жизнеспособности CTpO
мальных клеток. В клинике широко
при меняют различные методы TpaHC
плантации костноro мозrа.
Тимус
.
. ...', ....t .....
.' .:е.-. '- '...", .-'
. '1->" '\1" 'с
, , .. " :..... - '"<J. .,.. .
...' '$ ." .J, .
tJ' ..:; I>t- .. f. tt' 1 :
Jij , ';tf:..... ".4$' '1f-
1 .!IJ ,.(j 'r ; {", ' ,
' ""Il 1.,- "', ;l., ,;! ". . f. 6,.
:i '" . . .. , . fo" ....
, ,,'';A f'- >' +."_: '" ; .,..{
,.. ' , ", ' "" . ..,1 '.У V . " ''' ;
.. ;'" м--," -'о, fP -:",...... .C; .. :'"
{. .e. ("'. ','Е', . '.j'..,.
!II ......'"i'" -'..." ",. ..{>-...,.. ".. - ' ,...,,+ -- -.
.,,:;"$. ._'t ..:-::. .-! #' /j . :;z 1ft '.,
t:;.,' " \ ?. ,\'"
" . . "'; r:э
2 i " / i tI' .
)
А
i!(
V'- Б
Э:.
'- ... 8,
.;" 9 ... ,.
;:' # .. .. ;p . . .
..... .,.,... '. , '.
'. ;;..,. 1 ..."\ ос . "",.Iti."....
. ,'\
.. -'.
." ';:
.. .: . .(
....."'"
222
Fлава 15
антиrен реактивные Т лимфоциты приобретают рецепторы к cTporo определен
ным антиrенам. Они выходят из тимуса, не попадая в мозrовое вещество, и заселяют
тимусзависимые зоны лимфатических узлов и селезенки. Здесь в периферических
opraHax иммуноrенеза происходит их дальнейшее созревание в Т киллеры (цито
токсические), Т хелперы, после чеro они способны к рециркуляции, клонированию
(пролиферации), образованию клеток памяти.
Друrие Т лимфобласты превращаются в аутоиммунокомпетентные клетки,
реактивные к аутоантиrенам. Они подверrаются разрушению путем апоптоза (при
мерно 95% клеток) и фаrоцитируются макрофаrами.
Процесс специализации Т лимфоцитов в корковом веществе долек тимуса про
исходит в условиях, предохраняющих от избыточноrо действия на них антиrенов.
Это достиrается за счет образования zем.атотuмическоlO lUстиона (барьера), COCTO
ящеrо из эндотелия rемокапилляров с отчетливо выраженной непрерывной базаль
ной мембраной, перикапиллярноrо пространства с межклеточным веществом и MaK
рофаrами, а также из эпителиоретикулоцитов с их базальной мембраной. Корковое
вещество тимуса имеет самостоятельное микроциркуляторное русло. Напротив, в
мозrовом веществе тимуса капиллярная сеть не выполняет барьерной роли, и через
эндотелий этих капилляров зрелые лимфоциты MorYT уходить и возвращаться в ти
мус, т. е. рециркулировать. В тимусе эти лимфоциты находятся в мозrовом веше
стве. Зрелые лимфоциты покидают тимус через стенку посткапиллярной венулы.
Мозrовое вещество тимуса имеет более светлую окраску, так как лимфоцитов
здесь меньше, чем в корковом веществе. Эпителиальный остов выступает более OT
четливо, а эпителиоретикулоциты здесь более крупные и мноroчисленные. В Cpek
ней части мозroвоrо вещества встречаются слоистые эпителиальные тельца юссаля.
С возрастом количество и размеры их увеличиваются. В строме тимуса кроме эпите
лиоретикулоцитов находятся макрофаrи, дендритные клетки костномозrовоrо про
исхождения, нейроэндокринные клетки про из водные HepBHoro rребня, а также
миоидные клетки.
Возрастные изменения и реактивность. После 20 лет происходит возрастная
инволюция тимуса. Это сопровождается уменьшением количества лимфоцитов и раз
витием жировой ткани. В тех случаях, коrда вилочковая железа не подверrается об
ратному развитию, возникает пониженная сопротивляемость орrанизма к инфекци
ям и интоксикациям.
Ткани тимуса являются высокореактивными структурами. При действии по
вреждающих факторов (интоксикации, травмы и др.) наблюдаются выброс Т лим
фоцитов в кровь И массовая их rибель преимущественно в корковом веществе.
Возникающая при таких стресс воздействиях инволюция тимуса называется Bpe
менной, быстропроходящей, или акцидентальной. Наблюдаемые при этом явле
ния лимфоцитолиза и фаrоцитоза макрофаrами rибнущих лимфоцитов paCCMaT
риваются как возможные способы высвобождения факторов роста и ДНК,
необходимых для восстановительных процессов в тканях. С rибелью лимфоцитов
в этих условиях связывают также селекцию Т лимфобластов. Реактивные измене
ния ВИЛОЧКОRОЙ железы находятся в тесной корреляции с функциональными из
менениями коры надпочечников и уровнем rлюкокортикоидов в крови. В тимусе
Система реактивности
223
выявлены адренерrические и холинерrические нервные волокна, входящие в opraH
вместе с сосудами.
Лимфатические узлы и лимфоидные образования в орrаиах
Лимфатические узлы это
opraHbI, расположенные по ходу
лимфатических сосудов. Размер их
O,5 1 см, форма чаще окруrлая,
овальная или бобовидная. Распола
rаются они обычно реrионарно,
rруппами. С выпуклой стороны
узла в Hero входят приносящие 2
лимфатические сосуды, а с проти
воположной, называемой BopOTa
ми, выходят выносящие лимфати
ческие сосуды. Кроме Toro, в BOpO
та входят артерии и нервы и BЫXO
ДЯТ вены. Общее число лимфати
ческих узлов достиrает 1 ТЫС., что
составляет около 1 % массы тела.
Лимфатические узлы выполняют 4
роль активноrо биолоrическоrо
фильтра, в котором задерживается
и фаrоцитируется до 99% всех ино
родных частиц и бактерий.
Различают неспецифическую
защитную функцию лимфатичес
ких узлов за счет элиминации микробов из лимфы и специфическую, выражаю
щуюся в иммунном ответе на антиrены. Эти opraHbI выполняют и кроветворную
функцию. Хотя стволовые клетки в них практически отсутствуют, но пролифера
ция лимфобластов, дифференцировка В лимфоцитов в плазмоциты происходит.
Лимфа, протекая через лимфатические узлы, обоrащается лимфоцитами.
Развитие. Зачатки лимфатических узлов появляются в конце 2 ro начале
3 ro месяца эмбриоrенеза в виде скоплений мезенхимы по ходу лимфатических
сосудов. Вскоре из мезенхимы образуется ретикулярная ткань, составляющая
строму opraHa. К концу 4 ro месяца в закладки узлов вселяются лимфоциты и
формируются скопления первичные узелки без центра размножения. OДHO
временно появляется подразделение opraHa на корковое и мозroвое вещество.
Строение. В лимфатическом узле имеются следующие структурные КОI\ШО
ненты: капсула, содержащая MHoro коллаrеновых волокон (в области ворот в кап
суле есть и rладкие миоциты), трабекулы перекладины из соединительной TKa
ни, которые, анастомозируя друr с друrом, образуют каркас узла, ретикулярная
ткань, заполняющая все пространство, оrраниченное капсулоЙ и трабекулами.
224
Fлава 15
В лимфатическом узле различают периферическое корковое вещество и централь
ное мозrовое вещество. Между этими частями выделяют еще паракортикальную
зону (рис. 110).
В корковом веществе расположены скопления лимфоидной ткани в виде BTO
ричных узелков. Это окруrлые образования диаметром до 1 мм. Центральную часть
узелка называют центром размножения, или реактивным центром. Здесь происхо
дит антиrензависимая пролиферация В лимфоцитов и дифференцировка их в преk
шественники плазматических клеток. Кроме Toro, в центре размножения находят
ся дендритные клетки костномозrовоrо происхождения, которые на своих отростках
удерживают антиrены, активирующие В лимфоциты, макрофаrи моноцитарноrо
rенеза, фаrоцитирующие поrибающие апоптозом аутоиммунные В лимфоциты,
антиrены и инородные частицы. По периферии вторичноrо узелка расположена
корона полулунной формы, состоящая из малых лимфоцитов (рециркулирующих
В лимфоцитов, В клеток памяти, незрелых плазматических клеток). На rранице
центра размножения короны обнаруживаются Т лимфоциты (хелперы), которые
способствуют развитию В лимфоцитов в иммунобласты. Последние миrрируют в
мозrовые тяжи, отходящие от паракортикальной зоны и узелков внутрь мозrовоrо
вещества.
Лимфатические узелки являются динамическими структурами. Они то обра
зуются, то исчезают. В процессе изменения структуры лимфатических узелков раз
личают несколько стадий (формирование светлоrо центра размножения, появле
ние BOKpyr центра короны из малых лимфоцитов и др.). Период их существования
2 З сут.
паракоpmикaJlыlя ЗОllа лимфатическоrо узла находится на rpанице между KOp
ковым и мозrовым веществом. Она называется тимусзависимой зоной, или Т зоной,
так как при удалении тимуса происходит ее исчезновение. В паракортикальной зоне
осушествляются бласттрансформация Т лимфоцитов, их пролиферация и превраще
ние в специализированные клетки системы иммунитета. Здесь MHoro дендритных кле
ток. Они появляются В результате миrpации из тканей системы покрова орrанизма
внутриэпидермальных макрофаroв. На своей поверхности они несут антиrены и преk
ставляют их Т лимфоцитам (хелперам). Кроме Toro, в этой зоне находятся особые Be
нулы, выстланные эндотелиоцитами кубической формы. Через стенку этих венул про
исходит переход T и В лимфоцитов из крови В строму лимфатическоrо узла.
Мозювое вещество лимфатических узлов является местом созревания плазма
тических клеток. Вместе со вторичными узелками KopKoBoro вещества мозroвые тяжи
составляют тимуснезависимую зону, или В зону, лимфатических узлов. Мозroвые
тяжи кроме B лимфоцитов и плазмоцитов содержат Т лимфоциты И макрофаrи.
Лимфа протекает через лимфатические узлы по синусам пространствам, co
держащим ретикулярную ткань, и orраниченным капсулой и трабекулами с одной
стороны и узелками и мозrовыми тяжами с друrой. Различают краевые, проме
жуточные и воротный синусы. Лимфа из последнеrо по лимфатическому сосуду
выходит в области ворот. В просвете синусов обнаруживаются ретикулярные клет
ки, макрофаrи, лимфоциты, плазматические клетки.
Возраст1lые uзмен.е1lUЯ. С возрастом лимфатические узелки и их центры раз
Система реактивности
225
множения постепенно исчезают, понижается фаrоцитарная активность макрофаrов,
разрастается соединительная ткань трабекул, развиваются явления атрофии узлов
и замещения их жировой тканью.
Реактивность и реlенерация. Лимфатические узлы весьма реактивные
структуры. Они чувствительны к действию различных повреждающих факторов
(радиации, инфекции, интоксикации и др.). Реrенерация их возможна, если coxpa
нены при носящие и выносящие лимфатические сосуды и, хотя бы частично, рети
кулярная ткань, пролиферация клеток которой сопровождается заселением CTBO
ловыми клетками и последующей их дифференцировкой.
Селезенка
Селезенка периферический opraH кроветворной и иммунной систем. Кроме
выполнения кроветворной и защитной функций, она участвует в процессах rибели
эритроцитов, вырабатывает вещества, уrнетающие эритропоэз, депонирует кровь.
Развитие. Закладка селезенки происходит на 5 й неделе эмбриоrенеза обра
зованием плотноrо скопления мезенхимы. Последняя дифференцируется в рети
кулярную ткань, прорастает кровеносными сосудами, заселяется стволовыми KpO
ветворными клетками. На 5 M месяце эмбриоrенеза в селезенке отмечаются
процессы миелопоэза, которые к моменту рождения сменяются лимфоцитопоэзом.
Строение. Селезенка снаружи 1
; ::т::: о : :: ;:л::з ,;:, ; .: " ': /. : . ". ?-., ,:;,<.
'\' ' . ..',' :.k,, , ,'и-:. .'_c.,- "'1I...
ткани и rл ад ких мио ц итов (р ис. 111 ) . &"=i".' :'\!J!':;< ". 'i. .: ,:.;' r.".", ,<' '\:,..: ..:; -С;;,;
кап: :: ;;,ь::ь.:::::; :;.::: ;i ;'; ',;71i i ' :
между собой. В них также есть волок
нистые структуры и rладкие миоциты.
Капсула и трабекулы образуют опор
но сократительный аппарат селезенки.
Он составляет 5 7% объема этоrо opra
з
на. Между трабекулами находится
пульпа (мякоть) селезенки, основу KO
торой составляет ретикулярная ткань. б
Стволовые кроветворные клетки опре
деляются в селезенке в количестве,
7
примерно, З,5 в 105 клеток Различают
белую и красную пульпы селезенки.
Белая пульпа селезенки это co
вокупность лимфоидной ткани, KOTO
рая образована лимфатическими
узелками (В зависимые зоны) и лим
фатичсскими периартериальными
влаrалищами (Т зависимые зоны).
4
5
. ..
. ",-,-
Рис. 111. Строение селезенки:
1 капсула; 2 красная пульпа; 3 лимфатический
узелок; 4 центральная артерия; 5 Т зависимая
зона; б трабекулярная артерия; 7 трабекулярная
вена
226
Fлава 15
Белая пульпа при макроскопическом изучении срезов селезенки выrлядит в виде
светло серых окруrлых образований, составляющих 1/5 часть opraHa и распреде
ленных диффузно по площади среза.
Лимфатическое периартериальное влаrалище окружает артерию после выхода ее
из трабекулы. В ero составе обнаруживаются антиrенпреДставляющие (дендритные)
клетки, ретикулярные клетки, лимфоциты (преимущественно Т хелперы), макрофа
rи, плазматические клетки. Лимфатические первичные узелки по своему строению
аналоrичны таковым в лимфатических узлах. Это окруrлое образование в виде скоп
ления малых В лимфоцитов, прошедших антиrеннезависимую дифференцировку в
костном мозrе, которые находятся во взаимодействии с ретикулярными и дендритны
ми клетками. Вторичный узелок с rерминативным центром и короной возникает при
антиrенной стимуляции и наличии Т хелперов. В короне присутствуют В лимфоци
ты, макрофаrи, ретикулярные клетки, а в rерминативном центре В лимфоциты на
разных стадиях пролиферации и дифференцировки в плазматические клетки, Т xek
перы, дендритные клетки и макрофаrи.
Краевая, или марrинальная, зона узелков окружена синусоидальными каПИk
лярами, стенка которых пронизана щелевидными порами. В эту зону Т лимфоци
ты миrрируют по rемокапиллярам из периартериальной зоны и поступают в сину
соидные капилляры.
Красная пульпа совокупность разнообразных тканевых и клеточных CTPYK
тур, составляющих всю оставшуюся массу селезенки, за исключением капсулы,
трабекул и белой пульпы. Основные структурные компоненты ее ретикулярная
ткань с клетками крови, а также кровеносные сосуды синусоидноro типа, образую
щие причудливые лабиринты за счет разветвлений и анастомозов. В ретикуляр
ной ткани красной пульпы различают два типа ретикулярных клеток малодиф
ференцированные и клетки фаrоцитирующие, в цитоплазме которых MHoro фаrосом
и лизосом. Между ретикулярными клетками располаrаются клетки крови эрит
роциты, зернистые и незернистые лейкоциты. Часть эритроцитов находится в co
стоянии деrенерации или полнorо распада. Такие эритроциты фаrоцитируются
макрофаrами, переносящими затем железосодержащую часть rемоrлобина в Kpac
ный костный мозr для эритроцитопоэза.
Синусы в красной пульпе селезенки представляют часть сосудистоrо русла,
начало которому дает селезеночная артерия. Далее следуют cerMeHTapHbIe, трабе
кулярные и пульпарные артерии. В пределах лимфоидных узелков пульпарные
артерии называются центральными. Затем идут кисточковые артериолы, артери
альные rемокапилляры, венозные синусы, пульпарные венулы и вены, трабекуляр
ные вены и т. д. В стенке кисточковых артериол есть утолщения, называемые rиль
зами, муфтами или эллипсоидами. Мышечные элементы здесь отсутствуют. В
эндотелиоцитах, выстилающих просвет rильз, обнаружены тонкие миофиламен
ты. Базальная мембрана очень пористая. Основную массу утолщенных rильз co
ставляют ретикулярные клетки, обладающие высокой фаrоцитарной активностью.
Полаrают, что артериальные rильзы участвуют в фильтрации и обезвреживании
артериальной крови, протекающей через селезенку.
Венозные синусы образуют значительную часть красной пульпы. Их диаметр
Система реактивности
227
12 40 мкм. Стенка синусов выстлана эндотелиоцитами, между которыми имеются меж
клеточные щели размером до 2 мкм. Они лежат на прерывистой базальноЙ мембране,
содержащей большое количество отверстий диаметром 2 6 мкм. В некоторых местах
поры в базальной мембране совпадают с межклеточными щелями эндотелия. Блаrо
даря этому устанавливается прямое сообщение между просветом синуса и ретикуляр
ной тканью красной пульпы, и кровь из синуса может выходить в окружающую их
ретикулярную строму. Важное значение для реryляции кровотока через венозные си
нусы имеют мышечные сфинктеры в стенке синусов в месте их перехода в вены. Име
ются также сфинктеры в артериальных капиллярах. Сокращения этих двух типов
мышечных сфинктеров реryлирует кровенаполнение синусов. Отток крови из микро
циркуляторноrо русла селезенки происходит по системе вен возрастающеrо калибра.
Особенностью трабекулярных вен являются отсутствие в их стенке мышечноrо слоя
и сращение наружной оболочки с соединительной тканью трабекул. Вследствие этоrо
трабекулярные вены постоянно зияют, что облеrчает отток крови.
Возрастные изменения. С возрастом в селезенке отмечаются явления aTpo
фии белой и красной пульпы, уменьшается количество лимфатических фоллику
лов, разрастается соединительнотканная строма opraHa.
Реактивность и реzенерация. fистолоrические особенности строения селе
зенки, ее кровоснабжения, наличие в ней большоrо количества крупных расши
ренных синусоидных капилляров, отсутствие мышечной оболочки в трабекуляр
ных венах следует учитывать при боевой травме. При повреждении селезенки
мноrие сосуды пребывают в зияющем состоянии, и кровотечение при этом caMO
произвольно не останавливается. Эти обстоятельства MOryT определить тактику
хирурrических вмешательств. Ткани селезенки очень чувствительны к действию
проникающей радиации, к интоксикациям и инфекциям. Вместе с тем они облада
ют высокой реrенерационной способностью. Восстановление селезенки после TpaB
мы происходит в течение 3 4 недель за счет пролиферации клеток ретикулярной
ткани и образования очаrов лимфоидноrо кроветворения.
Кроветворная и иммунная системы чрезвычайно чувствительны к различным
повреждающим воздействиям. При действии экстремальных факторов, тяжелых
травмах и интоксикациях в opraHax происходят значительные изменения. В KOCT
ном мозrе уменьшается число стволовых кроветворных клеток, опустошаются лим
фоидные орrаны (тимус, селезенка, лимфатические узлы), уrнетается кооперация
T и В лимфоцитов, изменяются хелперные и киллерные свойства Т лимфоцитов,
нарушается дифференцировка В лимфоцитов.
Иммунитет и ero виды
Иммунитет (от лат. immunitas осв бождение, избавление от чеrо либо)
невосприимчивость орrанизма к инфекцИQННЫМ инеинфекционным areHTaM и
веществам, обладающим антиrенными свойсТвами. Существует иммунитет врож
денный, нестерильный и стерильный, поствакцинальный, клеточный, ryмораль
ный и др. В реакциях иммунитета rлавным образом участвуют специализирован
ные лимфоциты, плазматические клетки и макрофаrи.
228
Тлава 15
Чужеродные для орrанизма человека белки, полисахари/l,Ы и их синтетичес
кие аналоrи, имеющие бактериальное, тканевое, вирусное или искусственное про
исхожденпе, называются антИlеuам.и. При их попадании в орrанизм возникает
иммунныЙ ответ, что выражается в продукции антител, появлении клеток эффек
торов и формировании пммунолоrическоЙ памяти на данный чужеродный объект.
Антитело это молекулы белка, присутствующие в сыворотке крови, KOTO
рые обладают способностью соединяться и прочно связываться с антиrеном. Для
каждоrо антиrена существует свое антитело. Природа создала несколько химичес
ки различных видов (классов) антител, или иммунorлобулинов: [gG, IgA, IgM, IgD,
IgE. Наиболее часто встречающаяся разновидность IgG активен в нейтрали
зации бактериальных токсинов и в защите от вирусов. Слюна и слизь боrаты IgM
(он леrко преодолевает клеточные барьеры). IgA располаrается на поверхности
слизистых оболочек, предупреждает проникновение инфекции во внутреннюю
среду орrанизма. Друroй белковый компонент крови опсоuины' обволакивая бак
терии, облеrчают их фаrоцитирование. rруппа белков сыворотки крови 'Компле
мент, фиксируется в комплексе антиrен антитело, что приводит к активации раз
рушающих стенку бактерии ферментов.
В зависимости от механизма уничтожения антиrена различают 'КЛеточный и
lуморшlьuый иммуuитет. Ведущие клетки, ответственные за специфическую им
мунореактивность, это T и В лимфоциты. Иммунокомпетентность T и В лим
фоциты приобретают в результате rенетически запроrраммированной антиrенне
зависимой пролиферации и дифференцировки, которая совершается в тимусе для
Т лимфоцитов и В красном костном мозrе для В лимфоцитов. Под влиянием фак
торов микроокружения п в результате rенетической изменчивости участков [eHO
ма, ответственных за молекулярную орrанизацию вариабельной части клеточноrо
рецептора, эти клетки на своей поверхности приобретают специальные рецепто
ры, реаrирующие с антиrенами. Соrласно клонально селекционной теории, в opra
низме существует множество T и В лимфоцитов (1012 лимфоцитов), запроrрам
мированных отвечать на один, определенный антиrен.
В периферических иммунных и кроветворных opraHax при встрече с антиrена
ми происходят антиrензависимая пролиферация и дифференцировка T и В лим
фоцитов. Пролиферации предшествует бласттрансформация лимфоцитов явле
ние, впервые описанное А.А. Максимовым (1902). После пролиферации
бласттрансформированных Т лимфоцитов возникают: 1) Т клетки киллеры (ци
тотоксические) основные эффекторные клетки иммунитета, они разрушают чу
жеродные клетки мишени и обеспечивают rенетическое постоянство внутренней
среды орrанизма, защищая ero от вторжения чужеродных клеток, новообразова
ний, аутоиммунных заболеваний; 2) Т клетки хелперы, участвующие в процессах
межклеточной кооперации с В лимфоцитами, без чеrо не возможна трансформа
ция В лимфоцитов в плазмоциты; 3) Т клетки супрессоры, блокирующие антите
лообразование B лимфоцитами при аутоиммунных реакциях.
Микроокружением для лимфоцитов Т зоны являются особые макрофаrи aH
тиrенпредставляющие (дендритные) клетки. Эти клетки распознают и фаrоцитиру
ют антиrены и в виде фраrментов (эпитопов) выделяют их на поверхности своих
Систем.а реактивности
229
отростков. Блаrодаря этому облеrчается распознавание антиrена лимфоцитами, что
и индуцирует пролиферацию Т лимфоцитов. Помимо иммунокомпетентных кле
ток и макрофаrов, в иммунных реакциях принимают участие тканевые базофилы и
эозинофилы. Тканевые базофилы выделяют rистамин, серотонин и rепарин, оказы
вающие стимулирующее действие на клетки, которые уча ствуют в иммунных peaK
циях. Эозинофилы разрушают и фаrоцитируют комплексы антиrен антитело, BЫ
полняя тем самым свойственную им дезинтоксикационную функцию.
Клеточные основы кооперации в иммунных реакциях
Реактивное свойство орrанизма, которое проявляется на клеточном уровне opra
низации живоrо и заключается в распозновании "своих и чужих" клеток, обеспечи
вается Т лимфоцитами. Молекулярные механизмы "узнавания" заключаются в том,
что практически все клетки орrанизма позвоночных экспрессируют молекулу rис
тосовместимости, которая находится на поверхности клетки и доступна для опозна
вания специализированными клетками иммунной системы. В связи с тем, что моле
кула в составе клетки одноrо орrанизма, будучи внесенная в друrой, вызывает
реакцию отторжения, она получила название антиrен rистосовместимости, или rлав
ная молекула rистосовместимости. Последняя бывает двух классов 1 и 11.
Молекула tUстосовм.естuмости I lO класса является rликопротеином. Она COCTO
ит из трех частей: а) трансмембранноro белка (тяжелой цепи), который пронизывает
плазмолемму клетки и имеет внутриклеточную, цитоплазматическую и внеклеточную
части (домены); б) белка малой молекулярной массы (эпитопа), расположенноrо между
петлями внеклеточной части трансмембранноro белка; в) периферическоrо белка. [ены,
кодирующие молекулярную орrанизацию трансмембранноrо белка, расположены на
6 й хромосоме в области, которая получила название rлавный комплекс rистосовмес
тимости (анrл. МНС Major Histocompatibility Complex).
Основная биолоrическая роль молекулы rистосовместимости I ro класса дать
возможность Т лимфоцитам с помощью своих Т клеточных рецепторов распознать
"свои и чужие" клетки. Информацию об этом несет разнообразный по составу и после
довательности расположения аминокислот низкомолекулярный белок эnиrrюп. Это
фраrмент, возникший из белкрв, присутствующих внутри клетки. Среди них MOryT
быть: а) фраrменты собственных белков, Torдa они не вызывают ответной реакции со
стороны специализированных Т лимфоцитов; б) молекулы белка, кодируемые виру
сом; в) молекулы белка, кодируемые мутантным [еномом раковой клетки орrанизма.
В случае появления двух последних происходит реакция клетки, несущей чужеРОk
ный эпитоп, с Т лимфоцитом, который называется цитотоксическим.
Цитотоксические лимфоциты имеют рецепторы, с помощью которых осуществ
ляется мониторинr практически всех клеток орrанизма (за исключеним тех, KOTO
рые отделены специальными rематотканевыми rистионами). Они имеют специфи
ческие рецепторы Т клеточные, с помощью которых взаимодействуют с эпитопом
тканевой клетки, и особые рецепторы, представленные поверхностными rликопро
теинами, которые выявляются моноклональными антителами CD8 (от анrл. cluster
of differentiation по номенклатуре, принятой в иммуноцитохимии рецепторов). Если
230
rлава 15
Т лимфоциты маркируются моноклональным антителом CD8, то они называются
СD8+Т лимфоцитами, цитотоксическими, или киллерами. Последние связывают
ся только с молекулой rистосовместимости I ro класса. В случае обнаружения чу
жеродноrо эпитопа происходит молекулярное взаимодействие в области контакта
рецепторов, что является сиrналом к запуску механизмов разрушения клетки, син
тезирующей чужеродный белок Это проявляется в секреции в межклеточное про
странство белков перфоринов, которые в виде трансмембранных белков встраива
ются в плазмолемму чужеродной клетки, формируют поры, при водящие к
нарушению rомеостаза, набуханию и распаду клетки. Кроме тою, цитотоксические
лимфоциты способны активизировать своими биолоrически активными вещества
ми ферменты апоптоза как при проникновении их в клетку через поры, так и через
рецепторы чужеродной клетки к индукторам апоптоза. Существуют и друrие Mexa
низмы цитотоксическоrо действия данной субпопуляции лимфоцитов на мишени.
В отличие от реакций клеточноrо типа существует друrой тип реакции иммун
ных клеток, выражающийся в выработке антител в жидкие среды орrанизма. Pe
цепторная система клеток (макрофаrов, B и Т лимфоцитов), участвующих в им
мунных реакциях данноrо типа, представлена молекулами lUстосовместимости
II lO класса. Последняя состоит из двух трансмембранных белков, между петлями
которых располаrается фраrмент антиrена эпитоп, или антиrенная детерминан
та. В отличие от описанною выше процесса возникновения эпитопа в составе MO
лекулы rистосовместимости 1 ю класса, во втором случае эпитоп появляется как
результат двух процессов: а) фаrоцитоза антиrена макрофаrом (антиrенпредстав
ляющей клеткой), внутриклеточноrо взаимодействия антиrена с лизосомами MaK
рофаrа и возврата фраrмента антиrена в виде эпитопа на клеточную поверхность;
б) рецепторно опосредованноrо эндоцитоза В лимфоцитами pacTBopeHHoro анти
[ена, ero модификации и возврата в виде эпитопа на клеточную поверхность.
Следовательно, принципиальные отличия в орrанизации молекулы rистосов
мести мости II ro класса заключаются в том, что не все клетки синтезируют дaH
ный класс молекул, а эпитоп имеет внеклеточное происхождение.
С молекулой 11 ro класса MorYT взаимодействовать только те лимфоциты, KOTO
рые имеют на своей поверхности друrой специальный рецептор класса CD4. К
ним относятся СD+4Т лимфоциты, или Т хелперы. Последние вместе с антиrенп
редставляющими клетками и В лимфоцитами отвечают за формирования иммун
ной реакции, которая проявляется в синтезе uм.м.УНОVlOбулинов, или антител, цир
кулирующих в крови И тканевой жидкости (ryморальный иммунитет).
Межклеточные взаимодействия протекают следующим образом. Т хелпер с по
мощью двух своих рецепторов Т клеточноrо (молекулы II ro класса) и CD4 свя
зывается с молекулой 11 ro класса rистосовместимости макрофаrа (антиrенпредстав
ляющей клетки). Если Т клеточный рецептор с одной стороны и эпитоп молекулы
11 ro класса макрофаrа с друrой стороны не комплементарны, то происходит актива
ция и пролиферации Т хелпера. Аналоrично происходит взаимодействие хелпера с
В лимфоцитом. Если эпитоп молекулы II ro класса В лимфоцита не соответствует
Т клеточному рецептору хелпера, то последний вырабатывает лимфо'кины' которые
вызывают пролиферацию и дифференцировку B лимфоцита в плазматические клет
Система реактивности
231
ки. Последние специализированы на синтезе в большом количестве К7Iеточных pe
цепторов к конкретному антиrену и секреции рецепторов в виде иммуноrлобулинов
в кровь, лимфу, тканевую жидкость. В свою очередь рецепторы связывают циркули
рующие в жидкостях орrанизма антиrены, предупреждают дальнейшее развитие за
болевания. Итак, uм.мУНОlJlOбулины это секретируемые плазматическими К7IeTKa
ми клеточные рецепторы к определенным антиrенам. СD4+Т лимфоциты
называются помощниками (хелперами), так как без их участия невозможна актива
ция, пролиферация и дифференцировка В лимфоцитов в плазматические К7Iетки.
Таким образом, иммунный комплекс opraHoB действует как целое. Блаrодаря вза
имодействию с эндокринной, нервной системами и местными факторами реryляции,
происходит пролиферация и дифференцировка лимфоидных К7IeToK, упорядоченная
их миrpация, осуществляемая через кровь и лимфу. Медленная МUlрация К7leТOK xa
рактерна для стволовых К7IeToK, T и В лимфоцитов, заселяющих периферические
opraHbI иммуноrенеза. Быстрая миzрация характерна для рециркулирующих долro
живущих лимфоцитов памяти. Количество рециркулирующих К7IeToK составляет 1010.
Они миrpируют из крови В периферические opraHbI иммуноrенеза и обратно. Рецир
куляция Т лимфоцитов обеспечивает поддержание иммунноrо rомеостаза.
Эндокринный комплекс opraHoB
в системе реактивности орrанизма реryляторные эффекты, вызываемые эндок
ринными механизмами, отличаются относительно медленным развитием и ПрОДОk
жительностью действия.
Эффектором в данном случае выступают особые секреторные продукты эндок
ринных клеток zoрмоны, которые ryморальным путем достиrают своих К7IeToK
мишеней и оказывают на них стимулирующие или тормозные влияния. OpraHbI,
которые образуют и секретируют в кровь, лимфу и меЖК7Iеточную среду [ормоны,
обладающие высокой биолоrической активностью, контактным и дистантным воз
действием на процессы жизнедеятельности друrих К7IeToK и тканей орrанизма назы
ваются эндокринными железами. Помимо эндокринных желез, в орrанизме челове
ка существует orpoMHoe количество одиночных эндокриноцитов, расположенных в
составе эпителиальных тканей кожи, дыхательной, пищеварительной и выделитель
ной систем, составляющие так называемую диффузную эндокринную систему.
Свойствами образовывать и выделять roрмоны обладают секреторные ядра мозrа
(rипоталамуса), rипофиз, эпифиз, щитовидная железа, околощитовидные железы,
надпочечники, островки поджелудочной железы, половые железы. Эндокринные же
лезы не имеют выводных протоков и выделяют roрмон во внутреннюю среду орrаниз
ма. Термин "ropMoH" (от rpеч. hormau возбуждаю) был предложен Э.r. Старлинrом
(1905). [ормоны обладают рядом уникальных свойств специфичностью действия,
высокой биолоrической активностью, дистантностью действия. Дистантное (через
кровь, лимфу, меЖК7Iеточное вещество) влияние [ормонов на К7Iетки мишени имену
ется эндокринным (или телекринным), воздействие на рядом расположенные К7Iетки
друrоro типа паракринным, а на собственную метаболическую и пролиферативную
активность аутокринным.
232
!Лава 15
По химическому строению roрмоны подразделяются на производные амИ1-l0
кислот (адреналин и норадреналин и др.), пептиды (наиболее мноrочисленный
класс, насчитывающий более 50 ropMOHOB, среди которых инсулин, rлюкаrон,
инrибин, rастрин и др.), стероид1-lblе (половые, коры надпочечников и др.), 1-le1-la
сыще1-l1-lые ЖUР1-lые кислоты (простаrландины). По физиолоrическому действию
различают пусковые rормоны и roрмоны исполнители. Пусковыми rормонами яв
ляются нейроroрмоны rипоталамуса и roрмоны rипофиза, они стимулируют или
тормозят синтез и секрецию roрмонов в друrих железах внутреннеЙ секреции. [op
моны исполнители действуют непосредственно на обменные процессы в клетках
и тканях мишенях.
Биолоrическое влияние ropMoHoB осуществляется, rлавным образом, посреk
ством реryляции синтеза белков в клетках мишенях. Последние бывают ЮрМО1-lЗа
висuмые, которые функционируют только в присутствии конкретноro ropMoHa, и
ЮРМО1-lчувствитеЛЪ1-lые MOryT функционировать без rормональноЙ стимуляции,
но их деятельность все же контролируется ropMOHOM.
КлеТКИ l\шшени активно захватывают и аккумулируют roрмон с помощью спе
цифическоrо для данноro ropMoHa белка рецептора (избирательность связывания
ropMoHa). Рецепторы MoryT быть внутриклеточные (для ropMoHoB, проникающих в
цитозоль), или располаrаться в виде интеrральных белков цитолеммы (для ropMo
нов, не проникающих в клетку). В последнем случае необходимы дополнительные
механизмы передачи rормональноrо сиrнала к внутриклеточным элементам. Пере
дача стимула внутрь клетки осуществляется вторичными медиаторами, или посред
никами (цАМФ, цrмФ, ионы кальция и др.). При соединении roрмона с рецепто
ром происходит активация rормонзависимоro фермента плазмолеммы адеНИЛИk
циклазы. Последняя активирует в цитоплазме образование из А ТФ
внутриклеточноro посредника циклическоrо аденозин монофосфата (цАМ Ф). Дa
лее следует взаимодействие посредника с внутриклеточным рецептором и переме
щение комплекса цАМ Ф рсцептор в ядро и возникновение новых синтезов. При этом
ускоряется течение обменных реакций в клетке.
Стероидные [ормоны способны проходить через плазмолемму и взаимодей
ствовать с внутриклеточными рецепторами. Они MoryT действовать и на rенети
ческиЙ аппарат клеток мишеней.
[ормоны обладают высокой биолоrической активностью, хотя и продуциру
ются В очень малых количествах. При введении извне эффективными являются
чрезвычайно малые концентрации roрмонов.
Необходимый уровень ropMoHoB в орrанизме поддерживается реrулирующи
ми системами на основе прямых и обратных связей. Эти системы реrуляции обес
печивают динамическое равновесие при действии ropMoHoB с различной или даже
противоположной направленностью (стимуляторов и инrибиторов функций).
Эндокринные железы образуют единую реryлирующую систему с rлубокими
и сложными функциональными взаимосвязями и взаимодействиями. Вместе с тем
в rистолоrическом и топоrрафическом отношениях эндокринные железы весьма
разнообразны. Они сосредоточены в разных отделах тела, имеют неодинаковое cтpo
ение и происхождение.
Система реактивности
233
Общим признаком всех эндокринных желез является высокая степень их Bac
куляризации. Обычно каждая железистая клетка в их составе контактирует с ce
тью кровеносных капилляров. rемокапилляры в эндокринных opraHax относятся
к синусоидному типу.
OpraHbI с эндокринной функцией подразделяются на две rpуппы. Одни из них
являются чисто эндокринными железами, выполняющими исключительно внутри
секреторную функцию, друrие относятся к смешанным opraHaM, осуществляю
ЩИМ наряду с внутренней, еще и внешнюю секрецию, либо друryю функцию (напри
мер, половые железы).
По происхождению, rистоrенезу и rистолоrическим признакам эндокринные
opraHbl классифицируют на три rруппы: бран.xuоzенная zpyппa (от rреч. branchia
жабры) железы, происшедшие из rлоточных карманов аналorов жаберных
щелей (щитовидная железа, околощитовидные железы); zpyппa надпочечников (KOp
ковое и мозroвое вещество надпочечников, параrанrлии); zруппа.мозloвыпридат
ков (rипоталамус, rипофиз и эпифиз).
Поскольку эндокринные железы составляют единую в функциональном отноше
нии реryлирующую систему, существует друrая классификация, в которой учитены
межорrанные связи и иерархическая зависимость эндокринных opraHoB (Б.В. Алешин):
1. Центральные звенья эндокринноro комплекса желез (реrулируют деятель
ность большинства периферических эндокринных желез):
1) rипоталамус (нейросекреторные ядра);
2) rипофиз (аденоrипофиз и нейроrипофиз);
3) эпифиз.
Па. Периферические аденоrипофиззависимые эндокринные железы и эн
докриноциты:
1) щитовидная железа (тироциты);
2) надпочечники (корковое вещество);
3) roнады (яички, яичники).
Пб. Периферические аденоrипофизнезависимые эндокринные железы и эн
докриноциты:
1) кальцитониноциты щитовидной железы;
2) околощитовидные железы;
3) мозrовое вещество надпочечников и параrанrлии;
4) эндокринные клетки островков Ланrерrанса поджелудочной железы;
5) нейроэндокриноциты в составе неэндокринных opraHoB, АРUD серия эн
докриноцитов.
Центральным звеном в системе эндокринноЙ реryляции является rипоталамо
rипофизарный нейросекреторный комплекс. НеЙросекреторные ядра rипоталамуса
и эпифиз находятся в тесных связях с друrими отделами rоловноrо мозrа и зани
мают в физиолоrическом отношении промежуточное положение между нервной и
эндокринной системами. Они выполняют роль нейроэндокринных трансмиттеров
(переключателеЙ), объединяя нервную и эндокринную системы в единую нейро
эндокринную систему. rипоталамус является высшим центром реrуляции BereTa
тивных функций орrанизма.
234
Fлава 15
fипоталамус
[ипоталамус, называемый "эндокринным мозrом", осуществляет реrуляцию
всех opraHOB эндокринноro комплекса. Он занимает подбуrровую область проме
жуточноrо мозrа и является одновременно нервным образованием и эндокринной
частью мозrа.
[ипоталамус связан с rипофизом посредством rипофизарной ножки (рис. 112).
Последняя на дистальном конце образует заднюю долю rипофиза (нейроrипофиз).
Кпереди от rипофизарной ножки утолшение дна III ro желудочка мозrа образует
1 2
/
'
А
Б
в
,(
j.
.
з
I
<
I'
/4
.'
r
.
....
'
Рис. 112. Схема rипоталамо rипофизарной нейросекреторной системы:
А rипоталамус; Б rипофизарная ножка; В аденоrипофиз; r нейроrипофиз; 1 супраоптичес
кое ядро; 2 паравентрикулярное ядро; 3 аркуатное ядро; 4 вентромедиальное ядро; 5 первич
ная капиллярная сеть; б вторичная капиллярная сеть; 7 средняя доля; 8 аксовазальный синапс
Система реактивности
235
срединное возвышение (медиальную эминенцию). Здесь эпендимоциты выстила
ют полость желудочка, а rлиоциты дифференцируются в танициты, которые свои
ми отростками контактируют с первичной капиллярной сетью, связанной, в свою
очередь, со вторичной капиллярной сетью передней доли rипофиза. Танициты, по
rлощая вещества из спинномозrовой жидкости, транспортируют их в просвет KpO
веносных сосудов.
Нервные и нейросекреторные клетки rипоталамуса располаrаются в виде ядер,
число которых превышает 30 пар. В rипоталамусе различают передний, средний (Me
диобазальный, или туберальный) и задний отделы. Передний отдел zuпоmaламуса
содержит парные супраоптические и паравентрикулярные ядра. Супраоптические
ядра располаrаются над перекрестом зрительных нервов, а паравентрикулярные в
стенке III ro желудочка. Они образованы крупными нейросекреторными клетками.
В телах и отростках этих клеток определяются секреторные rранулы, называемые
иноrда rомориположительными, так как они хорошо окрашиваются по методу [OMO
ри. Нейриты крупных нейросекреторных клеток идут через медиальную эминенцию
и rипофизарную ножку в заднюю долю rипофиза. Они образуют lипоталамО lипо
физарный тракm, состоящий из супраоптико rипофизарноro и паравентрикуло rи
пофизарноrо пучков.
Нейросекреторные клетки указанных ядер вырабатывают нейроrормоны
вазопрессин (антидиуретический [ормон) и окситоцин. Первый из них образуется
преимущественно в клетках супраоптических ядер, а второй (окситоuин) В па
равентрикулярных ядрах переднеro rипоталамуса.
Средний (медиобазальный) отдел lипоталамуса содержит ряд ядер, состоящих
из мелких нейросекреторных клеток и адренерrических нейронов различноrо раз
мера. Обычно мелкие нейросекреторные клетки лежат на периферии ядер. Среди
ядер медиобазальноro rипоталамуса наибольшее значение имеют аркуатные (инфун
дибулярные) и вентромедиальные ядра, расположенные в области ceporo буrра. Бо
лее крупные из них вентромедиальные ядра. Нейросекреторные клетки этих ядер
вырабатывают аденоrипофизарные [ормоны. Нейриты клеток в срединном возвы
шении формируют аксовазальные синапсы на петлях первичной капиллярной сети.
rипоталамические нейроrормоны являются низкомолекулярными белками
(олиrопептидами), которые или стимулируют, или уrнетают синтез и секрецию
соответствующих [ормонов клетками аденоrипофиза. rипоталамические нейроrор
моны называют также РUЛИЗИНl факторами, или РUЛИЗИ1/.l lOрмонами (от анrл.
release освобождать). Нейроrормоны, стимулирующие освобождение тропных
[ормонов rипофиза, называются либеринами. Для нейроrормонов инrибиторов
освобождения rипофизарных [ормонов предложено обозначение статины.
Среди либеринов различают: сомаТОТРОПИIl рилизинr фактор соматолиберин;
тиреотропин рилизинr фактор тиролиберин; Актr рилизинr фактор кортико
либерин; рилизинr фактор фолликулостимулирующеro [ормона фоллиберин; ри
лизинr фактор лютеинизирующеrо [ормона люлиберин; пролактин рилизинr фак
тор пролактолиберин; рилизинr фактор меланоцитостимулирующеrо [ормона
меланолиберин. Среди статинов выделяют: соматотропин инrибирующий фактор
соматостатин; пролактининrибирующий фактор пролактостатин; инrибирующий
236
Fлава 15
фактор меланоцитостимулирующеrо ropMoHa меланостатин. Либерины истатины
поступают в аденоrипофиз через кровоток от первичной капиллярной сети в медиаль
ной эминенции, далее в портальные вены и во вторичную капиллярную сеть rипофи
за. Синусоидные капилляры этой сети, окружая аденоuиты, обеспечивают воздействие
нейроrормонов на клетки аденоrипофиза.
Функции rипоталамуса находятся под контролем rоловнorо мозrа. В ею раз
ных частях находятся нейроэндокринные клетки, которые вырабатывают lI.ейро
пептиды (более 50). Среди них, например, энкефалин, является интернейрональ
ным медиатором, под воздействием которою по цепочке нейронов происходит
выработка нейроаминов серотонина и норадреналина нейронами лимбической
системы и норадреналина нейронами ретикулярной формации. Нейроамины вли
яют на секрецию rипоталамических нейроrормонов. Действие последних стиму
лирует или тормозит активность аденоцитов rипофиза. Так возникает тесная фун
кциональная связь нервной и эндокринной систем, обеспечивающая контроль,
интеrpацию и реактивность живоrо.
fипофиз
[ипофиз (питуитарная железа) вместе с rипоталамусом составляет rипотала
мо rипофизарную нейросекреторную систему. Он является мозювым придатком.
В rипофизе различают аде1l.0zuпофиз (переднюю долю, промежуточную и тубераль
ную части) и lI.ейроzuпофиз (заднюю долю, воронку).
Развитие. Аденоrипофиз развивается из эпителия крыши ротовой полости.
На 4 Й неделе эмбриоrенеза образуется эпителиальное выпячивание в виде rипо
физарноrо кармана (карман Ратке), из которою вначале формируется железа с BHe
шним типом секреuии. Затем проксимальный отдел кармана редуuируется, и aдe
номер становится обособленной эндокринной железоЙ. Нейроrипофиз образуется
из материала инфундибулярной части дна 111 ro желудочка мозrа и имеет нейраль
ное происхождение. Эти две разные по происхождению части вступают в контакт,
образуя rипофиз.
Строение. Аденоrипофиз состоит из эпителиальных тяжей трабекул. Меж
ду ними проходят синусоидные капилляры. Клетки представлены хромофильны
ми и хромофобными эндокриноцитами. Среди хромофильных эндокриноцитов
различают ацидофильные и базофильные эндокриноциты.
Ацидофuлыl.еe Эll.дОКрИ1l.0циты это клетки средних размеров, окруrлой или
овальноЙ формы, с хорошо развитой rранулярной эндоплазматической сетью. Ядра
находятся в центре клеток Они содержат крупные плотные rранулы, окрашива
ющиеся кислыми красителями. Лежат эти клетки по периферии трабекул и co
ставляют 30 35% от общеrо количества аденоцитов в передней доле rипофиза.
Имеется две разновидности ацидофильных эндокриноцитов: соматотропоциты,
вырабатывающие ropMoH роста (соматотропин), и лактотропоциты, или MaMMOT
ропоциты, продуцирующие лактотропный ropMoH (пролактин). Соматотропин сти
мулирует процессы роста всех тканеЙ и opraHoB. При rиперфункции соматотропо
цитов может развиться акромеrалия и rиrантизм, а в условиях rипофункции
Система реактИ8иости
237
замедление роста тела, что ПрИБОДИТ к rипофизарной карликовости. Лактотроп
ный [ормон стимулирует секрецию молока в молочных железах и проrестерона в
желтом теле яичника.
Базофuльuые Э1lдОКрИ1l0циты это крупные клетки, в цитоплазме которых есть
rранулы, окрашивающиеся основными красителями (анилиновым синим). Они
составляют 4 1 0% от общеrо числа клеток в передней доле rипофиза. В rранулах
содержатся rликопротеины. Базофильные эндокриноциты подразделяются на ти
ротропоциты и rонадотропоциты.
Тиротропоциты это клетки с большим числом плотных мелких rранул, OK
рашивающихся альдеrидфуксином. Они вырабатывают тиреотропный [ормон. При
недостатке в орrанизме тиреоидных [ормонов тиротропоциты трансформируются
в клетки тиреоидэктомии с большим числом вакуолей. При этом усиливается про
дукция тиротропина.
rонадотропоциты окруrлые клетки, в которых ядро смешено к периферии.
В цитоплазме есть макула светлое пятно, [де расположен комплекс rольджи.
Мелкие секреторные rранулы содержат rонадотропные [ормоны. При HeДOCTaT
ке в орrанизме половых [ормонов в аденоrипофизе появляются клетки KaCTpa
ции, для которых характерна кольцевидная форма из за наличия крупной BaKYO
ли в цитоплазме. Такая трансформация rонадотропной клетки связана с ее
rиперфункцией. Различаются две rруппы rонадотропоцитов, вырабатывающих
либо фолликулостимулирующий, либо лютеинизирующий ropMoHbI.
Кортикотропоциты это клетки неправильной, иноrда отростчатой формы.
Они разбросаны по всей передней доле rипофиза. В их цитоплазме определяются
секреторные rранулы в виде пузырька с плотной сердцевиной, окруженноrо мемб
раной. Между мембраной и сердцевиной имеется светлый ободок Кортикотропо
циты вырабатывают AKTr (адренокортикотропный [ормон), или кортикотропин,
активируюший клетки пучковой и сетчатой зон коры надпочечников.
Хромофоб1lые эuдОКрИ1l0циты составляют SO 60% от общеrо числа клеток aдe
ноrипофиза. Они располаrаются в середине трабекул, имеют небольшие размеры,
rранул не содержат, цитоплазма их слабо окрашивается. Это сборная rруппа кле
ток, среди которых находятся молодые хромофильные клетки, еще не накопившие
rранул секрета, зрелые хромофильные клетки, уже выделившие секреторные [pa
нулы, и резервные камбиальные клетки.
Таким образом, в аденоrипофизе обнаруживается система взаимодействующих
клеточных дифферонов, формирующих ведущую эпителиалыl.юю ткань этой час
ти железы.
Средll.ЯЯ (промежуточная) доля rипофиза у человека развита слабо, составляя
2% от общеrо объема rипофиза. Эпителий в этой доле однородный, клетки боrаты
мукоидом. Местами есть коллоид. В промежуточной доле эндокриноцитами Bыpa
батываются мелаllОЦИТОСТИМУЛИРУЮЩИЙ [ормон и липотропный ropMoH. Первый
адаптирует сетчатку rлаза к зрению в сумерках, а также активизирует кору надпо
чечников. Липотропный l'OpMOH стимулирует жировой обмен.
Влияние нейропептидов rипоталамуса на эндокриноциты осуществляется с по
мощью системы rипоталамо аденоrипофизарноro кровообращения (портальной).
238
Тлава 15
в первичную капиллярную сеть срединноrо возвышения секретируются rипотала
мические нейропептиды, которые далее по портальной вене попадают в аденоrипо
физ и в ero вторичную капиллярную сеть. Синусоидные капилляры последней pac
положены между эпителиальными тяжами эндокриноцитов. Так нейропептиды
rипоталамуса воздействуют на клетки мишени аденоrипофиза.
Нейроzunофиз имеет нейроrлиальную природу, не является rормонпродуциру
ющей железой, а выполняет роль нейроrемальноrо образования, в котором накапли
ваются ropMoHbI некоторых нейросекреторных ядер переднеrо rипоталамуса. В зад
ней доле rипофиза идут мноroчисленные нервные волокна rипоталамоrипофизарноro
тракта. Это нервные отростки нейросекреторных клеток супраоптическоrо и пара
вентрикулярноrо ядер rипоталамуса. Неройны этих ядер способны к нейросекре
ции. Нейросекрет (трансдуктор) транспортируется по нервным отросткам в заднюю
долю rипофиза, [де выявляется в виде телец Херринrа. Аксоны нейросекреторных
клеток заканчиваются в нейроrипофизе нейрососудистыми синапсами, посредством
которых нейросекрет поступает в кровь. Нейросекрет содержит два [ормона: анти
диуретический (AДr), или вазопрессин (он действует на нефроны, реrулируя об
ратное всасывание воды, а также суживает сосуды, повышая кровяное давление);
окситоцин, стимулирующий сокращение rладких мышц матки. Лекарственный пре
парат, полученный из задней доли rипофиза, называется питуитрином и использу
ется для лечения HecaxapHoro мочеизнурения. В нейроrипофизе имеются tIейроrли
альные клетки, называемые питуицитами.
Реактивность lипотала fO lипофuзарной системы. Боевые травмы и сопро
вождающие их стрессы при водят к сложным нарушениям нейроэндокринной pery
ляции rомеостаза. При этом неЙросекреторные клетки rипоталамуса усиливают BЫ
работку нейроroрмонов. В аденorипофизе уменьшается количество хромофобных
эндокриноцитов, что ослабляет репаративные процессы в этом opraHe. Число базо
фильных эндокриноцитов нарастает, а в ацидофильных эндокриноцитах появляют
ся крупные вакуоли, свидетельствующие об их напряженном функционировании.
При длительном лучевом поражении в железах внутренней секреции возникают дe
структивные изменения секреторных клеток и уrнетение их функции.
Эпифиз
Эпифиз (верхниЙ мозrовоЙ придаток, пинеальная, или шишковидная, железа)
расположен между передними буrрами четверохолмия. Это неЙроэндокринный
opraH, реrулирующий физиолоrические ритмы, так как секреция ero ведущеrо [op
мона изменяется от времени суток У некоторых позвоночных (рыб, амфибий и
др.) это фоторецепторный opraH.
Развитие. В эмбриоrенезе человека эпифиз закладывается на 5 6 й неделе как
выпячивание крыши промежуточноrо мозrа. В ero состав включается также субко
миссуральныЙ opraH производное эпендимы III ro желудочка мозrа.
Строение. Эпифиз покрыт соединительнотканной капсулой, от которой OT
ходят переroродки. Они делят паренхиму на дольки, состоящие из тяжей пинеаль
ных клеток (или пинеалоцитов) и клеток нейроrлии rлиоцитов.
Система реактивности
239
ПинеШLOцитъt полиrональные клетки с отростками, которые посредством була
вовидных расширений контактируют с капиллярами. В цитоплазме клеток содержат
ся крупные митохондрии, эндоплазматическая сеть, комплекс fольджи, везикулы и
др. Среди пинеалоцитов выделяют светлые и темные клетки. Предполаrают, что это
клетки одноrо типа, но находящиеся в различном функциональном состоянии.
тлиоциты имеют длинные отростки и выполняют опорную функцию.
Пинеалоциты выделяют следующие ropMoHbI: серотонин, действующий на
rладкую мускулатуру сосудов, повышая кровяное давление, служит медиатором в
центральной нервной системе, кроме Toro, он является предшественником мелато
нина; мелатонин действует как антаrонист меланоцитостимулирующеrо ropMo
на аденоrипофиза, и наряду с серотонином участвует в механизме "биолоrических
часов" (днем в эпифизе преобладает синтез серотонина, ночью мелатонина); aд
реноrломерулотропин ropMoH, реryлирующий секрецию альдостерона в клубоч
ковой зоне коры надпочечников.
Эпифиз вырабатывает антиrипоталамические факторы (антиroрмоны), оказы
вающие действие на rипофиззависимые эндокринные opraHbI. Действие это обрат
ное (тормозящее, инrибирующее) тропным ropMoHaM аденоrипофиза. Важное зна
чение имеет выработка пинеалоцитами антиrонадотропина, который тормозит
секрецию лютропина в аденоrипофизе, т. е. выполняет роль roнадостатина. Антиrо
надотропин эпифиза rонадолиберин rипоталамуса, действуя как roрмоны антаrо
нисты, совместно осуществляют реryляцию rонадотропной функции rипофиза.
Таким образом, имеется определенная связь эпифиза с rипоталамо rипофи
зарной системой. В целом эндокринная система находится под реrулирующим вли
янием нервной системы.
UЦитовиднаяжелеза
Щитовидная железа вырабатывает тиреоидные ropMoHbI тироксин и ТРИЙОk
тиронин, а также тирокальцитонин (кальцитонин).
Тироксин и трийодтиронин являются мощными стимуляторами окислитель
ных процессов в клетках, причем трийодтиронин в S 1 О раз активнее тироксина.
Эти rормоны усиливают обмен веществ, синтез белков, rазообмен, обмен' уrлево
дов и жиров. Тиреоидные rормоны оказывают значительное влияние на развитие,
рост и дифференцировку клеток и тканей. Они ускоряют развитие костной ткани.
Особенно большое влияние ropMoHbI щитовидной железы оказывают на rистоrе
нез нервной ткани. При недостаточности щитовидной железы тормозится диффе
ренцировка клеток и тканей rоловноrо мозrа, нарушается психическое развитие
человека. Тиреоидные rормоны оказывают стимулирующее действие на pereHepa
ционные процессы в тканях. В состав тироксина и трийодтиронина в качестве обя
зательноro инrредиента входит йод, поэтому поступление йода с питьевой водой и
пищей в орrанизм необходимо для нормальной деятельности щитовидной желе
зы. Не содержащий йода третий ropMoH щитовидной железы тирокальцитонин
участвует в реryляции кальциевоrо и фосфорноrо обмена.
Развитие. Зачаток щитовидной железы появляется на 4 й неделе эмбриоrенеза
240
Fлава 15
в виде выпячивания вентральной стенки rлоточной кишки между I й и II й пара
ми жаберных карманов. Выпячивание это превращается в эпителиальный тяж с
утолщением на конце. Щитовидная железа вначале закладывается как экзокрин
ная железа. Позднее проток, соединяющий железу с rлоткой, редуцируется, и от
этоrо протока остается лишь foramen cecum. Эпителиальный тяж раздваивается на
конце. В течение 3 ro месяца по ходу эпителиальных тяжей образуются перетяж
ки. В четкообразных cerMeHTax этих тяжей появляются просветы. По мере уrлуб
ления перетяжек эпителиальные тяжи распадаются на отдельные фолликулы и кле
точные островки. В конце 3 ro месяца эпителий фолликулов дифференцируется.
Клетки еro фолликулярные эндокриноциты (тироциты) начинают вырабаты
вать roрмоны, которые накапливаются в полости фолликула. Образование новых
фолликулов и их развитие происходят [етерохронно. К моменту рождения OTMe
чается наличие фолликулов с коллоидом ("коллоидный тип строения") и без KOk
лоида с десквамацией эпителия ("десквамативный тип строения"). Между фолли
кулами встречаются межфолликулярные островки клеток В процессе развития
железы наряду с дифференцировкой эпителия происходит разрастание мезенхи
мы, преобразующейся в соединительную ткань. Формируется строма железы, co
держащая rустую сеть капилляров. В строму проникают нервные волокна.
В зачаток железы врастают также производные 5 й пары жаберных карманов
клетки так называемых ультимобранхиальных телец. Это С клетки, вырабатыва
ющие кальцитонин. Это клетки нейроэктодермальной природы, и в закладку щи
товидной железы они внедряются через ультимобранхиальные зачатки.
Таким образом, в формировании структурно функциональных единиц щито
видной железы участвуют следу
ющие клеточные диффероны: это
ведущие клетки эпителия фолли
кулов фолликулярные эндок
риноциты, вырабатывающие ти
роксин и трийодтиронин; каль
цитониноциты, или С клетки,
продуцирующие кальцитонин и
друrие пептиды (соматостатин,
тиролиберин, серотонин и др.).
Cntpoeuue. )Келеза, состоя
щая из двух долей, снаружи по
крыта соединительнотканной
капсулой, от которой отходят пе
реrородки, подразделяющие па
ренхиму на дольки. CTPYKTYPHO
функциональной единицей щи
товидной железы является фол
ЛUКУЛ (рис. 113). Средний диа
метр фолликулов около 50 мкм.
Форма их преимущественно OK
фОё
....,:.. ем. ,.' #')
.,. ..,.,... . ., . e '1
., .' #, «"" .tJ .,_
. . v .... - -';'" .
; 1t i/f) :. 1 .
'ф ."", . е,е .. 3
· . ......, .".1',0 · ..
' (J' \ ., 'III.!O. 4
Q ..1..\. о. . ' 5
: ,,,",' ' 2
If .. . : , '0)1
- t& . t. .. ',\ ·
...., . .J;.б ' .' :)'J
". -: -О"" \ , if' 3
Рис. 113. Строение щитовидной железы:
1 фолликул; 2 интерфолликулярный островок; 3 ти
роциты; 4 коллоид: 5 rемокапилляры
Система реактивности
241
руrлая. Фолликулы представляют собой замкнутые пузырьки. Стенка их образова
на однослойным эпителием, состоящим из фоJulикулярныx эндокриноцитов (тиро
цитов). Среди этих клеток в виде небольших скоплений находятся С клеткu. Они
MOryT быть и около фолликулов, и между фолликулами.
Полость фолликула заполнена продуктом секреции тироцитов коллоидом,
содержащим белки тироrлобулины. Снаружи фолликулы оплетают сети KpOBe
носных и лимфатических капилляров. Между соседними фолликулами встреча
ются интерфолликулярные островки, состоящие из малодифференцированных эн
докриноцитов.
Фолликулярные эндокриноциты имеют кубическую форму и окруrлое ядро.
На апикальной поверхности их имеются микроворсинки. В цитоплазме хорошо
развиты орrанеллы, обеспечивающие синтез белков. MHoro свободных рибосом,
образующих полисомы. Соседние эндокриноциты в стенке фолликула соединя
ются при помощи плотных контактов, десмосом и интердиrитаций.
Особенностью rистофизиолоrии щитовидной железы является различно Ha
правленное движение секреторных продуктов: вначале в полость фолликула, а за
тем в противоположном направлении в кровь. Происходит это блаrодаря актив
ной деятельности фолликулярных эндокриноцитов. Секреторный цикл этих клеток
состоит из следующих фаз: поrлощение исходных веществ, синтез секрета, Bыдe
ление ero в полость фолликула в виде коллоида, иодирование коллоида, эндоци
тоз иодированноrо коллоида и ero модификация и выведение ropMoHa через ба
зальную часть клетки в окружающие ткани и кровеносные и лимфатические
капилляры. Выработка тиреоидных ropMoHoB начинается с синтеза тиреоrлобули
на в базальной части эндокриноцитов. Содержащие тиреоrлобулин продукты син
теза из эндоплазматической сети поступают в комплекс rольджи и далее в виде
секреторных rранул путем экзоцитоза выводятся в полость фолликула. К нейоди
рованному тиреоrлобулину присоединяется сначала один атом йода, а затем и BTO
рой, В результате чею образуются MOHO и дийодтиронины. Последующая комп
лексация дает тетрайодтиронин, или тироксин. При отщеплен ии одноrо атома йода
от тироксина образуется трийодтиронин.
В нормальных условиях процессы образования коллоида и ero резорбции ypaвHO
вешены. Это равновесие, однако, нарушается при rипо и rиперфункции щитовидной
железы. При rипофункции задерживается выведение ropMoHa. При этом фолликулы
обычно имеют большие размеры, в полости фолликула коллоида MHoro, он ryстой, не
имеет резорбционных вакуолей, эпителий представлен плоскими эндокриноцитами.
При rиперфункции, наоборот, усиливается выведение rормона коллоида в кровь. KOk
лоид разжижается, количество ею невелико, эпителий стенки фолликула становится
высоким призматическим.
Парафолликулярные эндокриноциты (С клетки, или кальцитониноциты) BЫ
рабатывают белковый ropMoH кальцитонин. Он понижает уровень кальция в крови
и является антаrонистом паратирина. Кальцитонин действует на остеокласты KO
стной ткани, уменьшая их резорбтивную активность. С клетки это более CBeT
лые и более крупные, чем фолликулярные эндокриноциты, и доля их составляет
0,1%. Обычно они лежат поодиночке или мелкими rруппами. В цитоплазме
242
Fлава 15
парафолликулярных эндокриноцитов MHoro арrирофильных или осмиофильных
секреторных rранул, содержащих кальцитонин и друrие пептиды.
Строма железы состоит из рыхлой волокнистой соединительной ткани, в KO
торой часто обнаруживаются тучные клетки, макрофаrи, лимфоциты. Фиброблас
ты щитовидной железы обладают рядом уникальных свойств, что определяется
особенностями рецепторно трансдукторной системы клеток В частности, они MO
ryT участвовать в развитии воспаления наряду с иммунокомпетентными клетка
ми. Хорошо развиты капилляры фенестрированноrо типа. Нервные волокна co
держат пептиды, характерные для С клеток
Реактивность. Физиолоrическая реrенерация щитовидной железы происхо
дит путем обновления клеток эпителия фолликулов за счет их митоза. Источни
ком развития новых фолликулов MOryT быть интерфолликулярные клеточные oc
тровки. Интоксикации, травмы, аутоиммунные процессы, наследственные факторы
и др. MorYT приводить к тиреотоксикозу или к rипотиреозу. Следует иметь в виду,
что для нормальнorо функционирования щитовидной железы необходимо ДOCTa
точное поступление в орrанизм йода. Щитовидная железа обладает высокой спо
собностью К реrенерации после травм. Примером аутоиммунноrо заболевания
щитовидной железы является болезнь Хашимото. Она возникает вследствие Toro,
что тирorлобулин проникает в строму щитовидной железы и, являясь антиrеном,
вызывает иммунную реакцию орrанизма. При этом сильно разрастается строма
железы, она инфильтрируется лимфоцитами и плазмоцитами, а фолликулы щито
видной железы содержат мало коллоида и постепенно атрофируются.
Околощитовидные железы
Функция околощитовидных желез состоит в выработке полипептидноro [op
мона паратирина (паратrормона), который участвует в реryляции обмена каль
ция и фосфора в орrанизме. Паратирин повышает содержание кальция в крови.
rиперкальциемический эффект паратирина обусловлен активацией остеокластов
и подавлением остеоцитов, что приводит к резорбции кости и выходу кальция в
кровь, усилению всасывания кальция в кишечнике и ускорению реабсорбции каль
ция в почках. Кроме паратирина на содержание кальция в орrанизме влияет каль
цитонин щитовидной железы. Взаимодействие этих [ормонов с противоположным
действием обеспечивает КaJIьциевый и фосфорный rомеостаз в орrанизме.
Развитие. Околощитовидные железы производные III й и IV й пар жабер
ных карманов. эпителиальная выстилка которых имеет прехордальный rенез. На
S 6 й неделе эмбриоrенеза образуются четыре зачатка желез в виде эпителиаль
ных почек На 7 8 й неделе эти почки отшнуровываются от стенок жаберных Kap
манов, присоединяясь к задней поверхности щитовидной железы. В процессе rис
тоrенеза эпителия околощитовидных желез составляющие ero клетки становятся
все более дифференцированными, их размеры увеличиваются, количество rлико
[ена в них уменьшается, цитоплазма приобретает светлую окраску. Они называют
ся rлавными паратироцитами. У S месячноrо плода rлавные паратироциты диф
ференцируются на светлые и темные паратироциты. На десятом [оду жизни
Система реактИШlOсти
243
появляется следующий вид эпителиальных клеток желез ацидофильные, или
оксифильные, паратироциты. В виде единичных включениЙ в паренхиме около
щитовидных желез MorYT находиться С клетки, вырабатывающие кальцитонин.
Строение. Паренхима железы образована эпитеЛИaJlЫ-lЪLМИ трабекулами, клеточ
ными тяжами и реже комплексами в виде фолликулов с оксифильным содержи
мым. Нежные прослоЙки соединительноЙ ткани, содержащие rycTbIe сети KpOBeHOC
ных капилляров, делят железу на мелкие дольки. Ведущим клеточным диффероном
среди железистых клеток составляют тaв7-1ыe паратироциты. Это клетки полиrональ
ноЙ формы, в светлоЙ цитоплазме которых определяются включения rликоrена и ли
пидов. Размеры клеток колеблются от 4 до 10 мкм. Среди rлавных паратироцитов раз
личают активные (темные) и неактивные (светлые) формы. В активных клетках
сильнее развиты орrанеллы, в неактивных больше липидных капель и rликоrена.
По соотношению двух видов паратироцитов можно судить о функциональноЙ актив
ности железы. Обычно на один темныЙ приходится 3 5 светлых паратироцитов.
Среди rлавных паратироцитов в паренхиме околощитовидноЙ железы BCTpe
чаются скопления оксифильных (ацидофильных) паратироцитов. Эти клетки круп
нее rлавных, в цитоплазме их содержится большое количество оксифильных зе
рен. Последние при электронноЙ микроскопии являются митохондриями, которые
занимают большую часть цитоплазмы. При этом секреторные rранулы не выявля
ются. Предполаrают, что ацидофильные паратироциты это стареющие, деrене
ративно измененные формы rлавных паратироцитов.
В железах у лиц пожилою возраста обнаруживаются фолликулы с коллоидопо
добным содержимым. [ормон в фолликуле не обнаружен.
Секреторные rранулы выводятся из клетки экзоцитозом. Уменьшение KOHцeHT
рации кальция и фосфора приводит к активации синтеза паратrормона. Рецептор
но трансдукторная система клетки воспринимает уровень внеклеточноrо кальция,
и происходит активация ceKpeTopHoro цикла клетки и секреция ropMoHa в кровь.
Реактивность. Разрастание эпителия околощитовидной железы, приводящее
к ее rиперфункции, вызывает нарушение процесса обызвествления костноЙ ткани
(остеопороз, остеомаляция) и выведение кальция и фосфора из костей в кровь.
При этом происходят резорбция костноЙ ткани, нарастание количества остеоклас
тов, разрастание фиброзноЙ ткани. Кости становятся хрупкими, что при водит К
повторным их переломам.
[ипофункция околощитовидноЙ железы (травмы, удаление во время операции,
инфекции) вызывает повышение нервно мышечноЙ возбудимости, ухудшение COKpa
тительноЙ способности миокарда, судороrи в связи с недостатком кальция в крови.
Надпочечники
Надпочечники представляют собоЙ парные железы, состоящие из корковою и
мозrовоrо вещества. Каждая из этих частей является самостоятельной железой
внутренней секреции, вырабатывающей свои ropMoHbI реrуляторы защитно при
способительных реакций орrанизма. Корковое вещество это аденоrипофиззави
симая, а мозrовое вещество аденоrипофизнезависимая эндокринные железы.
244
Fлава 15
Развитие. Корковая часть надпочечников развивается на S й неделе эмбриоrе
неза из участка целомическоro эпителия в области корня брыжейки у краниальноro
полюса правой и левой первичной почки. Отсюда происходит друrое название коры
надпочечников интерреналовое тело. Вначале образуется первичная (или феталь
ная) кора, состоящая из крупных ацидофильных эндокриноцитов. Начиная с 10 й
недели эмбриоrенеза из Toro же источника формируется дефинитивная кора надпо
чечников за счет мелких базофильных эндокриноцитов, окружающих снаружи пер
вичную кору. В эмбриоrенезе толщина фетальной коры значительной превосходит
таковую дефинтивной коры. Однако после рождения эндокриноциты фетальной
коры поrибают механизмом апоптоза, поэтому толщина коры уменьшается. Полное
развитие коры происходит после половоrо созревания.
Источник развития KopKoBoro вещества надпочечников находится вблизи от
зачатка [онад, с чем связана способность клеток сетчатой зоны коры вырабатывать
андроrеНIIЫЙ roрмон близкий по свойствам тестостерону.
Мозrовое вещество надпочечников образуется несколько позже (на 6 7 й He
деле эмбриоrенеза) из общеrо с симпатическими rанrлиями зачатка HepBHoro
rребня. Симпатобласты миrрируют в интерреналовое тело, размножаются, фор
мируя мозrовые эндокриноциты хром аффинной ткани.
Строение. Надпочечники покрыты соединительнотканной капсулой, под KO
торой находится слой малодифференцированных клеток Корковое вещество co
стоит из системы эпителиальных тяжей. Между ними по соединительнотканным
прослойкам проходят KpOBeHOC
ные капилляры.
В коре надпочечника разли
чают три зоны: клубочковую,
пучковую и сетчатую (рис. 114).
Клубочковая, или наружная,
зона располаrается под капсулой.
Корковыеэндокриноциты(адре
нокортикоциты) образуют здесь А
аркады, или клубки. В цитоплаз
ме этих клеток хорошо развита
аrранулярная эндоплазматичес
кая сеть, что вообще характерно
для клеток, синтезирующих CTe
роидные [ормоны. Эндокрино
циты клубочковой зоны выраба
тывают м.uнералокортuкоuдъt
(альдостерон и друrие). Альдос
терон реryлирует уровень натрия
в орrанизме, предотвращая ero
выведение. Эндокриноциты клу
бочковой зоны получают сиrна
лы об изменениях уровня натрия
. . У' . ......., .. - . . '. ' ''''':''
<..........,;,.". , ' ,:",;, 'r- ", "'...... '''1'..;"'"' ". F, "..L(" \..:.: .:.. 1
.: ;Q:t ::{.--:" ..": " '.. -:ij, t't-... 'J{J.;; A'i'''' "v . '
{ ..;.......;,,:..Ф ..&J"..;,#. "; >;" 4!t "'.o:_ ..
)..7;, '" . :. :: \ f) J 7
''';''''' , ' /..q:.W. .....'.. '. ' .... ,
11 ;':' f.."' '-,M..-- ---; <-1It r..J......"':"**. ;"9;" "'''' ...'; ';.,".'!-;-:;,
-- . '*. .. 111 '; ... yv4..\.... ",;o.l::;'''<f''' 2
'-1 . :{;" :;;,:: [ tJ :. _. f :; /: +1"1 " :
1 "'''-';:'' 1; ' ",",,!" .:,-}t, , ""..>,; " , 51..
..... ... "'"
......... ,.",.. 1:" ., ..,
: ; ; .:. ' J ''!''IF''' 1
,
\
...... .<-'it
\ ..
">,: .: 1!: "1 '"
.
1.'
З
......
ff ".-#
.; : :I j.-. ': -. :!r;, -; ; '«, .;: ,_ ,, .
{ ,,', , .,... . .: #-!,,,,,,,",",.. ,'.:....' {' , 4
Б f, ' :( ',,': . ;:; : : ' ;; : :: 1
,.' ,', '.,...... " .. :,jJ!.......::J
......... $- :"- ...'.. ,, !; :r"' 'a_,:
...... , ,'" i '\"" '. {. " j' ", ' , , "" ' ",.,<х ..;:.."" .'....:: "
_ '-i ,.f. A }} '-- --..,: "' !t' >. '\ -1
' " " '<#; , ';' ' ' , ' , ' ' ': " " ' # )i...>... ,. .
1t;J;;--;.;, 4- " ........,.. ..... .J_ ,
>t. ,. . \.:J<; ",.J
J! '" i : ": IF :i:. . "s
"''0.0 - . ('!I"".:!J"':
','.' А
t::: ;
Рис. 114. Строение надпочечника:
А корковое вещество; Б мозrовое вещество; 1
капсула; 2 клубочковая зона; 3 пучковая зона; 4
сетчатая зона
Система реактивН0сти
245
в крови через анrиотензиновую систему, и таким образом их деятельность оказыва
ется связанной с функцией юкстаrломерулярноrо rистиона почки. Минералокорти
коиды влияют на водно солевой обмен, усиливают воспаление и образование KOk
лаrена. В клубочковой зоне отмечается MHoro митозов эпителиальных клеток На
rpанице между клубочковой и пучковой зонами расположен суда1l0фоб1lЫЙ слой.
Здесь также находятся малодифференцированные эпителиоциты.
Пучковая ЗО1lа состоит из светлых кубических или призматических эндокри
ноцитов, образующих тяжи, или пучки, направленные перпендикулярно к поверх
ности надпочечника. В цитоплазме клеток определяются капли липидов, после pa
створения которых образуются вакуоли, а клетки приобретают вид ryбки (отсюда
происходит друrое название клеток пучковой зоны спонrиоциты). В цитоплаз
ме клеток хорошо развита rладкая эндоплазматическая сеть. Число рибосом опре
деляет темный или светлый вид клетки. Это зависит от фазы ceKpeTopHoro цикла.
По мере синтеза roрмона цитоплазма просветляется и [ормон выводится из клет
ки. Митохондрии эндокриноцитов пучковой зоны содержат кристы в виде пузырь
ков, извитых и ветвящихся трубочек В них определяются ферменты, которые обес
печивают превращение холестерина в lлюкокортикоид1lыe юрмо1lы кортикосте
рон, кортизол (rидрокортизон), кортизон. Эти [ормоны реrулируют обмен
уrлеводов, уменьшают проницаемость тканей, ослабляют воспаление, фаrоцитоз,
коллаrенообразование. rлюкокортикоиды увеличивают содержание rликоrена в
скелетных мышцах, печени и миокарде, а также способствуют образованию rлю
козы за счет тканевых белков. rлюкокортикоиды повышают сопротивляемость opra
низма к действию повреждающих факторов. Вместе с тем они ослабляют процес
сы иммуноrенеза и применяются, в частности, для подавления реакций HeCOBMec
тимости тканей при трансплантации opraHoB.
В отличие от клеток клубочковой зоны эндокриноциты пучковой зоны явля
ются аденоrипофиззависимыми клетками. Их деятельность стимулируется AKTr
аденоrипофиза и кортиколиберином rипоталамуса.
Сетчатая ЗО1lа коры надпочечников состоит из эндокриноцитов, образующих
рыхлую сеть. Эндокриноциты здесь имеют меньшие размеры, чем в пучковой зоне.
Форма их разнообразна. В цитоплазме определяется меньше липидных включе
ний, чем в эндокриноцитах пучковой зоны, и преобладают свободные рибосомы.
На rранице с мозrовым веществом встречаются крупные ацидофильные эндокри
ноциты, образующие Х зону (остатки фетальной коры).
В сетчатой зоне вырабатываются половые стероиды андроrенный [ормон (близ
кий по химической структуре и свойствам к тестостерону яичек), эстроrен и проrес
терон. Секреторная активность сетчатой зоны, так же как и пучковой, контролиру
ется rипоталамо rипофизарной системой.
Мозювое вещество надпочечников состоит из рыхлоrо скопления и тяжей круп
ных окруrлых клеток, называемых мозroвыми эндокриноциами, или хромаффи
ноцитами, BOKpyr которых располаrаются померживающие нейроrлиальные клет
ки. Такое название этим клеткам дано в связи с тем, что при обработке их растворами
бихромата калия образуется осадок восстановленных окислов хрома. Мозrовые
эндокриноциты вырабатывают катехолами1lы адреналин и норадреналин. В связи
246
Fлава 15
с этим различают два вида клеток: светлые эндокриноциты, или эпинефроциты,
вырабатывающие адреналин, и темные эндокриноциты, или норэпинефроциты,
продуцирующие норадреналин. В цитоплазме этих двух видов клеток содержатся
мноrочисленные секреторные rранулы.
Эпинефроциты содержат электронно плотные rpанулы, окруженные мембраной.
Эти клетки не флюоресцируют в ультрафиолетовых лучах, не дают реакции с йодом
и серебром. Норэпинефроциты, напротив, отличаются от первых тем, что содержат
в цитоплазме большие "окаймленные rранулы" с очень плотной сердцевиной, флю
оресцируют в ультрафиолетовых лучах, дают реакцию с йодом и серебром. В составе
секреторных rpанул разных типов кроме катехоламинов присутствуют белки, липи
ды, опиодные пептиды (энкефалины, эндорфин) и др.
Адреналин и норадреналин оказывают сходное физиолоrическое действие,
вызывая сужение сосудов и повышая артериальное давление. Вместе с тем в их
действии есть некоторые различия. Адреналин является ropMoHoM, а HopaдpeHa
лин медиатором при передаче нервных импульсов с постrанrлионарноrо симпа
тическоrо нейрона на иннервируемые эффекторные структуры. Адреналин повы
шает уровень rлюкозы в крови за счет мобилизации ее из печени, норадреналин
оказывает слабое действие на эти обменные реакции. Сосуды rоловноrо мозrа и
скелетных мышц под влиянием адреналина расширяются, тоrда как норадреналин
вызывает сосудосуживающий эффект. Адреналин усиливает работу сердца, уча
щает сердцебиение, а норадреналин замедляет сердцебиение. Адреналин не влия
ет на секрецию тиролиберина и roнадолиберина, а норадреналин усиливает ceKpe
цию этих ropMoHoB и т. д.
Эндокриноциты мозrовоrо вещества надпочечников являются видоизменен
ными симпатическими нейронами и их секреторная активность находится под
контролем симпатической нервноЙ системы. Вырабатываемые мозrовыми эндок
риноцитами катехоламины поступают в кровь. Между тяжами хромаффинных
клеток проходят кровеносные сосуды и капилляры синусоидноrо типа, выстлан
ные фенестрированными эндотелиоцитами. КаждыЙ эндокриноцит, с одной CTO
роны, контактирует с артериальным капилляром, а с друrоЙ с венозным сину
соидом. При этом синтезированные катехоламины поступают в венозные
синусоиды. В мозrовое вещество входят кровеносные сосуды, пронизывающие
кору надпочечников и приносящие секреторные продукты корковых эндокрино
цитов. Кроме Toro, в мозrовом веществе присутствуют мультиполярные неЙро
ны веrетативноЙ нервноЙ системы.
Парazа1llЛUU, подобно мозroвому веществу надпочечников, состоят из хромаф
финной ткани, развивающейся из симпатобластов HepBHoro rpебня. Различают брюш
ные, аортальные, каротидные, внутриорrанные (в сердце, коже, яичках, матке и т. д.)
параrанrлии. Снаружи они окружены соединительной тканью, прослойки которой
проникают между тяжами lра1lуляр1lыx Э1lдО-КРU1l0цumов. Последние диаметром 1 o
15 мкм имеют овальную или окруrлую формы и содержат специфические rранулы
разноro размера, в которых находятся катехоламины. Эндокриноциты окружены
поддерживающими клетками неЙроrлиальноrо происхождения. Капилляр синусо
идноrо типа с фенестрированными эндотелиоцитами прилежит к rруппе эндокри
Система реактивности
247
НОЦИТОВ В тоЙ части, rде отсутствуют поддерживающие клетки. Иннервация opraHa
осуществляется симпатическоЙ нервноЙ системоЙ.
Реактивность и реlенерацuя. При стрессах, сопровождающихся сильными
эмоциональными реакциями страха или ярости, преобладает активность симпати
ческоЙ нервноЙ системы над парасимпатическоЙ. При этом повышается не только
активность постrанrлионарных симпатических неЙронов, но и секреция клеток
мозrовоro вещества надпочечника. В кровь поступают большие количества HOpaд
реналина и адреналина. В результате учащаются и усиливаются сокращения cepk
ца, повышается артериальное давление, увеличивается объем циркулирующей KpO
ви в сосудах мышц и центральной нервноЙ системы, в кровь из печени выбрасыва
ются запасы rлюкозы. Усиленное выделение адреналина и норадреналина клетками
мозrовоrо вещества надпочечника происходит и рефлекторно при резком охлаж
дении, боли и друrих видах стресса.
Физиолоrическая реrенерация коры надпочечника осуществляется при учас
тии субкапсулярных клеток и клеток суданофобной зоны, которые находятся под
контролем AKTf аденоrипофиза. При удалении одноrо надпочечника наблюдает
ся компенсаторная rипертрофия и rиперплазия железистых клеток друrоrо надпо
чечника.
Эндокринные клетки в составе неэндокринных opraHoB. Диффузная
эндокринная система (АРUD серия)
Название rруппы эндокринных клеток, составляющих АРUD серию, проис
ходит от первых букв анrлийских слов amine precursor uptake and decarboxylation,
что означает поrлощение и декарбоксилирование аминов и их предшественни
ков. Это название отражает основной признак клеток данноЙ серии способность
накапливать предшественники биоrенных аминов, декарбоксилировать их и BЫ
рабатывать Биоlенныe амины и полипептидныe loрмоны. Последние иrрают важ
ную роль в процессах местной тканевой реrуляции и TKaHeBoro rомеостаза.
Для клеток АРUD серии характерно присутствие в них нейроаминов (напри
мер, серотонина), катехоламинов и друrих биоrенных аминов, которые выявляются
блаrодаря специфической флюоресценции после обработки в парах формальдеrида
и просмотра препаратов в ультрафиолетовых лучах; поrлощение предшественников
аминов при введении их в орrанизм (например, 5 окситриптофана, диоксифенила
ланина и друrих); декарбоксилирование аминов. Пептидные ropMoHbI выявляются
преимущественно иммуноцитохимическими методами.
В клетках АР U D серии определяется высокое содержание ряда ферментов (эс
тераз, холинэстераз, альфа rлицерофосфатдеrидроrеназ). Иными словами, клетки
данной серии сочетают в себе признаки как нервных, так и эндокринных 'КЛеток
Под электронным микроскопом в базальной части этих клеток выявляется
большое количество секреторных rранул, белковая сердцевина которых окружена
мембраной (рис. 115). Клетки АРUD серии дают специфическую реакцию при
обработке их антисыворотками к биоrенным аминам и полипептидным ropMoHaM.
Особенно MHoro клеток АРUD серии в эпителии желудочно кишечноrо тракта
248
Fлава 15
t
(эндокринные клетки разных типов),
rде они образуют так называемую Э1l.
терИ1l.0вую lОрJИО1l.аль1l.УЮ систему.
[истоrенетические и ПIстофизи
олоrические различия не дают OCHOBa
ний для объединения всех так назы
ваемых апудоцитов в единую (rенети
чески) клеточную систему. Несмотря
на сходство по некоторым, rлавным
образом ПIстохимическим. признакам,
эндокринные клетки неэндокринных
opraнoB являются составными элемен
тами соответствующих тканей, обра
зуя в этих тканях линии диверrентной
клеточной дифференцировки, или
клеточные диффероны. Поэтому раз
ли чают клетки АРUD серии: произ
водные 1I.е'Йроэктодермы (нейrоэндок
риноциты rипоталамуса, эпифиза,
пептидерrические нейроны ЦН С и
ПНС); производные кожн.ой эктодер
мы (клетки Меркеля, эндокриноциты
АРUD серии адеНОПIпофиза); произ
водные кишечuой Э1l.тодеРМbl (ЭНlI.ОК
риноциты rастроэнтеропанкреатичес
кой системы); проuзвод-н.ые .мезодермы
(клетки ЛеЙll.llrа, эндокриноциты теки
фолликула яичника) и др.
Таким образом, для эндокриноци
тов АРUD серии, несмотря на различные источники их происхождения, характерно
наличие в цитоплазме как неЙроамина (серотонина ), так и пеПТИll.ноrо ropMoHa. И тот,
и друrоЙ секреторныЙ продукт оказывает дистантное или местное (паракринное) воз
деЙствие на клетки мишени, расположенные в данном или друrом opraHe.
.;....; . . { '"E ' .,..tr ,
,< '"", t" . , ",I 1 ....
,, ,, ,:.:; # . It
· ". \..::.,"'" ,
t-.:.." '. tt:'I . ... i.,.' .;' >.. ·
j'''''r: !:'" w
. /'). . .':.. ..... Ir :i -..li'
J , : .. ri' ",_.\... ' . . ..l';' , \"
, .ii .'". i.\! J .... '. < 'k'l '/ , t
; "r. ." . ..t '., , ь '; ' !Iit
.' ' . . fl. f1 '
1..,"'.. ...... . ,. ... k. ./
... '\.' . - . .r'''.l.;.. ..."'( 1
* ОС... v" .....3 _ /'_. . , , , } ,4. .. . . ; ... '" \t
. .... , ..':.'!.'.fJ;r' .c....1/t. A r.. ;..",, :; :<:iI... :.'" {
-! ' ",.,, #.... , - С;;., .:: ,t' A'('!I "
А
,.
. . ,.
. :..
.. .:
-.. . 0/'
.. '!!I'""''4It . .
I ",
..& ......
. "'; ,о ·
...,,;4tr.
..
I
"
.. "
.. '1:, ....
Э. ''о . '- '
, .; 1, '
, ,"f,,
... .,.
<
1 ..
. tJi.
:,., . .....'
.'
Б
Рис. 115. Секреторные rранулы в эндокриноцитах же
лудка:
А ЕСL клетка; Б G клетка
Невральный комплекс opraHoB
Реактивные своЙства, присущие орrанизму человека, заключаются в способ
ности воспринимать раздражения и отвечать на них адекватными реакциями. Это
стало возможным блаrодаря развитию (наряду с вышеперечисленными иммунны
ми и эндокринными орrанаl\lИ) комплекса связанных между собоЙ невральных об
разованиЙ, претерпевших длинную эволюцию, в состав KOToporo входят как спе
циализироваНlIые орrаны, так и одиночные клетки и их rруппы.
[истолоrическими элементами, способными воспринимать раздражения
и возбуждаться в ответ на стимулы, являются нервные клетки, или неЙроны.
Система реактив-н.ости
249
Последние формируют нервные ткани, каждой из которых присущи особен
ности функциональной специализации в составе конкретных opraHoB.
Большинство авторов, однако, не разделяет подобную точку зрения. ДеЙстви
телыю, рассматривая нервную систему с позиции взаимосвязи ее ведущих rисто
лоrических элементов, можно сделать вывод о существовании в орrанизме единой
-н.ейро-н.1/.0Й сети. Это связано с особенностями нейроrистоrенеза, коrда часть He
рвной клетки перикарион находится в одном opraHe, а отростки простпраются
далеко за пределами этоrо opraHa. Кроме Toro, отростки нейронов, входя в друrие
opraHbI нервной системы, формируют синаптические связи с множеством друrих
нейронов, а с помощью эфферентных волокон взаимодействуют с исполнитель
ными орrанами. Такое объединение нейронов разной локализации в единую сис
тему является rистофизиолоrической основой для реrуляции всех жизненно важ
ных процессов в орrанизме человека и ero взаимодействия с внешней средой.
Нервная система функционирует на основе рефлексов ответных реакций opra
низма, осуществляемых при посреДCfве центральноЙ нервной системы. И.М. Сеченов
впервые в книrе "Рефлексы rоловноrо мозrа" (1863) обосновал положение о том, что все
акты сознательной и бессознательной жизни по способу происхождения представляют
собой рефлексы. Морфолоmческим субстратом рефлексов являются рефлекторныедуm,
состоящие из цепи афферентных, ассоциативных и эффекторных неЙронов.
Нервную систему в анатомическом отношении делят на центральную и пери
ферическую. Центральная нервная система состоит из rоловноrо и спинноrо моз
[а. К периферической нервной системе относятся нервы, отходящие от rоловноrо
и спинною мозrа (черепные и спинномозювые), нервные узлы и окончания пери
ферических нервов. Нервная система в физиолоrическом отношенпи подразделя
ется на соматическую (цереброспинальную) и веrетативную (автономную). Пер
вая иннервирует все тело, за исключением внутренних opraHoB, сосудов и желез,
вторая реryлирует деятельность висцеральных opraHoB.
Источником развития нервных тканеЙ и нейроrлии opraHoB нервной системы
является материал нейроэктодермы (нервной пластинки). Стромальные элемен
ты и оболочки имеют мезенхимное происхождение. После нейруляции образуют
ся нервная трубка и две rанrлиозные пластинки. Из туловищною отдела нервной
трубки развивается спинной мозr, из краниальноrо rоловной мозr. Из rанrлиоз
ных пластинок образуются спинномозrовые, симпатические и интрамуральные
узлы, а также хромаффинная ткань надпочечников, параrанrлиев и неЙроэндок
ринные клетки АРUD серии.
Параллельно с развитием rоловноrо и спинноrо мозrа происходит формиро
вание спинномозrовых и периферических веrетативных rанrлиев, [де также диф
ференцируются нейроны и клетки rлии.
Спинной МОЗf
Спинной мозr расположен в позвоночном канале, является частью централь
ной нервной системы.
РаЗбитие. Источником развития спинноrо мозrа является туловищный
250
Тлава 15
отдел нервной трубки. Кроме Toro, в развитии opraHa принимает участие Me
зенхима. Эмбриональная нервная трубка, вначале состоящая из одноrо слоя при
зматических клеток, становится вскоре мноrослойной блаrодаря интенсивно
му митотическому делению клеток (медуллобластов, или матричных клеток).
В результате митозов и дивсрrентной дифференцировки матричные клетки пре
вращаются в нейробласты, rлиобласты (спонrиобласты) и эпендимобласты.
Вскоре в стенке нервной трубки формируются три слоя: эпендимный, плаще
воЙ и краевая вуаль. Внутренний эпендимный слой, образует выстилку
центральноrо канала спинноrо мозrа. Плащевой слой образован нейробласта
ми и rлиобластами, которые миrрировали из эпендимноrо слоя. Из клеток пла
щевоrо слоя в дальнейшем образуется серое вещество спинноrо мозrа. Нейро
бласты в плащевом слое митозом уже не делятся. Они дифференцируются,
образуя отростки, в нейроны.
rлиобласты, выселяясь в толщу плащевоrо слоя, сохраняют способность дe
литься митозом. Из отростков нервных клеток и rлиобластов формируется наруж
ный слой нервной трубки краевая вуаль. Позднее этот слой образует белое Be
щество спинноrо мозrа.
Дифференцировка нейронов спинноrо мозrа происходит [етерохронно. Пер
выми развиваются двиrательные (моторные) нейроны передних poroB спинноrо
мозrа, затем ассоциативные (комиссуральные, канатиковые, пучковые) и после
дними нейроны студенистоrо (роландова) вещества, а также клетки Реншоу (TOp
мозные неЙроны в передних porax спинноrо мозrа).
Наряду с дифференuировкой неЙронов в rистоrенезе спинноrо мозrа YCTaHaB
ливаются связи между нервными клетками, а также между нейронами и исполни
тельными структурами в составе формирующихся рефлекторных дуr. Отсутствие
межклеточных контактов у части нейронов является одной из причин массовой их
rибели по механизму апоптоза.
Процесс межклеточной интеrрации сопровождается образованием клеточных
скоплениЙ, называемых ядрами. Образующие их нейроны имеют ветвящиеся дeH
дриты, тоrда как неЙриты (аксоны) объединяются в компактные пучки. В процес
се развития аксоны двиrательных нейронов выходят из спинноrо мозrа, образуя
передние корешки, и направляются в закладки скелетных мышц. Задние pora фор
мируются центральными отростками чувствительных нейронов спинномозrовых
rанrлиев, которые вступают во взаимодействия с нейронами ceporo вещества спин
Horo мозrа. Миелинизация отростков нейронов начинается с 4 ro месяца эмбрио
rенеза и продолжается после рождения. При этом rлиобласты (спонrиобласты) диф
ференцируются на астроциты и олиюдендроциты.
Развитие спинною мозrа тесно связано с процессом межнейронной интеrра
uии с неЙронами rоловноrо мозrа. МезеНХИ 1а формирует ткани мозrовых оболо
чек (см. ниже), кровеносные сосуды.
Строение. В спинном мозrе различают серое и белое вещество. На попереч
ном срезе спинною мозrа серое вещество имеет вид буквы Н. Выделяют передние
(вентральные), боковые, или латеральные (нижние шейные, [рудные, два пояснич
ных), и задние (дорсальные) pora ceporo вещества спинноrо мозrа.
Система реактивности
251
Серое вещество представлено телами нейронов и их отростками, нервными
окончаниями с синаптическим аппаратом, MaKpo и микроrлией и сосудами.
Белое вещество окружает снаружи серое вещество и образовано пучками мя
котных нервных волокон, которые формируют проводящие пути на протяжении
Bcero спинною мозrа. Эти пути направляются в сторону rоловноrо мозrа или нис
ходят из нею. Сюда же относятся волокна, направляющиеся в выше или нижеле
жащие сеrменты спинноrо мозrа. Кроме Toro, в белом веществе находятся aCTpo
циты, отдельные нейроны, rемокапилляры.
В белом веществе каждой половины спинноrо мозrа (на поперечном срезе)
различают три пары столбов (канатиков): задний (между задней срединной пере
rородкой и медиальной поверхностью заднеrо pora), боковой (между передним и
задним роrами) и передний (между медиальной поверхностью переднеrо pora и
передней срединной щелью).
В центре спинноrо мозrа проходит канал, выстланный эпендимоцитами, среди
которых различают малодифференцированные формы, способные, по данным HeKO
торых авторов, к миrpации и дифференцировке в нейроны. В нижних cerMeHTax спин
Horo мозrа (поясничном и сакральном) после половоrо созревания происходит про
лиферация rлиоцитов и зарастание канала, образование интраспиналъноlO орюна. В
составе последнеrо находятся
rлиоциты и секреторные клетки,
вырабатывающие вазоактивный
нейропептид. OpraH подверrает
ся инволюции после 36 лет.
Нейроны ceporo вещества
спинною мозrа являются мулъ
типолярныи.. Среди них разли
чают нейроны с немноrочислен
ными слабоветвящимися дeHД
ритами, нейроны с ветвящимися
дендритами, а также переходные
формы.
В зависимости от Toro, куда
идут отростки нейронов, Bыдe
ляют: внутренние нейроны, OT
ростки которых заканчиваются
синапсами в пределах спинною
мозrа; пучковые нейроны, ней
рит которых идет в составе пуч
ков (проводящих путей) в дpy
rие отделы спинноrо мозrа или
в rоловной мозr; корешковые
нейроны, аксоны которых поки
дают спинной мозr в составе пе
редних корешков (рис. 116).
А
Б
л
6
8
Рис. 116. Схема соматической и веrетативных рефлекторных
дуr:
А соматическая дуrа; Б веrетативные дуrи; 1 спинной
мозr; 2 спинномозrовой узел; 3 паравертебральный узел;
4 превертебральный узел; 5 интрамуральный узел; 6
чувствительный нейрон; 7 вставочные нейроны; 8 дви
rательные нейроны
252
Fлава 15
На поперечном срезе нейроны rруппируются в ядра, в составе которых находятся
сходные по строению и функции нейроны. На продольном срезе эти нейроны pac
полаrаются послойно в виде колонки, что отчетливо видно в области заднеrо pora.
Нейроны каждой колонки иннервируют cTporo определенные области тела. О за
кономерностях rруппировки нейронов и их функциях можно судить по пластинам
Рекседа (1 Х). В центре заднеrо рorа располаrается собственное ядро заднеrо pora,
у основания заднеrо pora трудное ядро (Кларка), латеральнее и несколько rлуб
же располаrаются базилярные ядра, в промежуточной зоне медиальное проме
жуточное ядро. В дорсальной части заднеrо pora из rлубины кнаружи последова
тельно располаrаются мелкие нейроны студневидноrо вещества (роландова), дa
лее мелкие нейроны ryбчатой зоны и, наконец, поrраничная зона, содержащая
мелкие нейроны.
Аксоны чувствительных нейронов из спинальных rанrлиев входят в спинной
мозr через задние корешки и далее в краевую зону, тде делятся на две ветви: KOpOT
кую нисходящую И длинную восходящую. По боковым ответвлениям от этих BeT
вей аксона импульсы передаются на ассоциативные нейроны ceporo вещества. Бо
левая, температурная и тактильная чувствительность проецируется на нейроны
студневидною вещества и собственною ядра заднеrо pora. В составе студневидно
ro вещества находятся интернейроны, продуцирующие опиоидные пептиды, KOTO
рые влияют на болевые ощущения (так называемые "ворота боли"). Импульсы от
внутренних opraHoB передаются на нейроны ядер промежуточной зоны. Сиrналы
от мышц, сухожилий, суставных капсул и др. (проприорецепция) направляются
на ядро Кларка и друrие ядра. Аксоны нейронов этих ядер формируют восходя
щие проводящие пути.
В задних porax спинноrо мозrа MHoro диффузно расположенных нейронов,
аксоны которых заканчиваются в пределах спинноrо мозrа той же или противопо
ложной стороны ceporo вещества. Аксоны этих нейронов выходят в белое веще
ство и тут же делятся на нисходящую и восходящую ветви. Распространяясь на
уровне 4 5 спинномозrовых cerMeHToB, эти ветви образуют в совокупности соб
ственные пучки белоrо вещества, непосредственно прилежащие к серому веществу.
При этом различают задний, боковой и передний собственные пучки. Все эти пуч
ки белоrо вещества относятся к собственному аппарату спинноrо мозrа. От aKCO
нов, входящих в состав собственных пучков, отходят коллатерали, заканчивающи
еся синапсами на двиrательных нейронах. Блаrодаря этому создаются условия для
лавинообразноrо нарастания числа нейронов, передающих импульсы по рефлек
торным дуrам собственноrо аппарата спинною мозrа.
В передних porax располаrаются двиrательные (моторные) нейроны. Их aKCO
ны выходят из спинноrо мозrа по передним корешкам и в составе периферических
нервов достиrают скелетных мышц, rдe заканчиваются на каждом мышечном BO
локне нервно мышечным синапсом (моторной бляшкой).
Моторные нейроны (мотонейроны) это крупные мультиполярные клетки (диа
метр 1 oo 140 мкм). Общее их количество составляет 2 З млн. Каждый моторный ней
рон иннервирует от единиц до тысяч мышечных волокон, образуя двиrательную еди
ницу. На поперечном срезе спинноrо мозrа моторные нейроны rpуппируются в ядра;
Система реактИ61юсти
253
на продольном в колонки. Моторные нейроны каждой колонки Иlшервируют толь
ко одну мышцу. Различают медиальную, центральную и латеральную rpуппы (ядра)
моторных нейронов. Медиальная rpуппа неЙронов иннервирует мышцы туловища,
центральная мышцы тазовоro и плечевоrо пояса, латеральная мышцы конечнос
тей. НеЙроны в функциональном отношении подразделяются на альфа мотонейроны
большие, альфа мотонейроны малые и rамма l\ютонеЙ:роны. Альфа мотонеЙроны боль
шие передают импульсы на .:жстрафузальные мышечные волокна, вызывая быстрые
фазические сокращения. Альфа мотонеЙ:роны малые поддерживают тонус скелетных
мышц. rамма мотонеЙроны направляют аксоны к интрафузальным мышечным волок
нам нервно мышечноrо веретена. Синапсы покрывают примерно 50% поверхности тела
и дендритов мотонеЙрона. Среднее число синапсов на моторном нейроне спинноrо
мозrа человека достиrает 25 35 тыс. Одновременно функционально активными I\Ю
ryT быть тысячи синапсов. КаждыЙ моторный нейрон испытывает множественное
влияние, идущее от нейронов спинальноro и супраспинальноrо уровней. При этом
возможно возвратное торможение MOTopHoro нейрона блаroдаря тому, что коллате
раль аксона MOTopHoro нейрона контактирует с вставочным нейроном (клеткоЙ PeH
шоу), а аксоны клетки Реншоу оканчиваются на теле MOTopHoro нейрона.
Наряду с соматическими рефлекторными дуrами через спинной мозr прохо
дят веrетативные рефлекторные дуrи, относящиеся к автономноЙ (веrетативной)
нервной системе.
СПИННОМОЗfовые узлы
Спинномозroвые узлы расположены в межпозвонковых отверстиях по ходу
задних корешков спинноrо мозrа. Они покрыты соединительнотканной капсулой.
В пределах узла тела псевдоуниполярных нейронов лежат на периферии rруппа
ми, разделенными прослойками соединительной ткани, тоrда как нервные волок
на проходят через центральную часть. В rистоrенезе из биполярных нейронов фор
мируется клетка, отростки которой (аксон и дендрит) тесно сближаются (клетка
приобретает вид псевдоуниполярной), а затем Т образно расходятся. Оба OTpOCТ
ка становятся миелинизированными и называются аксонами. Отросток, который
вступает в спинной мозr (возникшиЙ из аксона биполярноro неЙрона) принято
называть центральным, а направляющийся на периферию и заканчивающийся pe
цеп тором (возникший из дендрита) периферическим.
Тела неЙронов имеют rрушевидную или окруrлую форму, размеры тел нейро
нов вариабельны от 30 до 120 мкм в диаметре. В цитоплазме крупных нейронов
хорошо развит синтетический аппарат. BOKpyr неЙронов находятся мноroчислен
ные ПlИоциты rанrлия (мантийные клетки), отростки которых формируют капсу
лу. Между телом нейрона и rлиоцитами формируются мноroчисленные инваrина
ции плазмолеммы. Здесь возможны метаболические взаимообмены rлиоцита и
нейрона. BOKpyr отростков неЙронов располаrаются леммоциты, формирующие
миелиновую оболочку, и фибробласты соединительной ткани. цеllтршlыlеe oт
ростки чувствительных нейронов формируют задние корешки и передают нервные
импульсы на ассоциативные неЙроны спинноro мозrа. Перuферические отростки
254
Тлава 15
в cocTdBe смешанных спинномозrовых нервов направляются на периферию и в
opraHax формируют рецепторы.
[оловной мозr
rоловной мозr является высшим центром реrуляции всех жизненных функ
ций орrаНИЗl\lа. Состоит из ствола мозrа и плащевой части.
Развитие. Краниальный отдел нервной трубки в эмбриоrенезе человека ПОk
разделяется на три l\Iозrовых пузыря (передний, средний и ромбовидный). В даль
нейшем за счет складок и изrибов формируются пять отделов мозrа: конечный и
промежуточныЙ l\Iозr, средний сохраняется неделимым, ромбовидный подразделя
ется на продолrоватый и задний мозr. Конечный мозr вначале непарный в дальней
шем формирует два полушария большоrо мозrа. Производные среднеrо и ромбо
видноro мозrовых пузырей образуют ствол мозrа и являются древними образовани
ями, в которых сохраняется ПРИНllИП сеrментарности. Последний исчезает в
производных проыежуточноrо и конечноrо мозrа, как образованиях филоrенетичес
ки более молодых, в которых концентрируются IIнтеrративные функции.
Стволовыми клетками для развития нервной ткани и нейроrлии rоловноrо моз
[а служат матричные клетки, расположенные в эпендимном слое желудочков мозrа.
Матричные клетки интенсивно делятся митозоы, миrрируют за пределы эпендим
Horo слоя, дифференцируются на нейробласты и rлиобласты (спонrиобласты), даю
щие позднее соответственно нейроны и клетки rлии. Часть клеток остается на месте
и формирует эпендимную выстилку желудочков мозrа. Пролиферация и дифферен
цировка матричных клеток в разных отделах rоловноrо мозrа происходят [eTepox
ронно. При формировании коры важную роль в миrрации нейробластов иrpают pa
диальные rлиоциты (мюллеровы волокна). Тела клеток располаrаются в эпендимном
слое, а отроски простираются до наружной поверхности нейроэпителиальной BЫC
тилки нервной трубки. Именно по отросткам радиальных rлиоцитов происходит
направленная миrрация нервных клеток из rлубоких в поверхностные слои форми
рующихся корковых структур мозrа. Из мезенхимы развиваются сосуды, а из CTBO
ловых кроветворных клеток клетки микроrлии.
Ствол lолов1l0l0 .м.ОЗlа
К стволу rоловноrо мозrа относят продолrоватый мозr, мост, 1\юзжечок и обра
зования среднеrо и промежуточноrо мозrа. По мере перехода спинноrо мозrа в про
долrоватый теряется характерная форма ceporo вещества спинноrо мозrа, но прин
цип локализации ядер по функции сохраняется: чувствительные ядра занимают
дорсальную, а двиrательные вентральную части ствола. Между ними располаrа
ется ретикулярная формация.
В чувствительных ядрах находятся нейроны, которые аналоrичны не аффе
рентным псеВДОУIlИПОЛЯРНЫМ нейронам спинальноrо rанrлия, а нейронам ядер
задних poroB спинноrо мозrа. Эти пучковые нейроны получают импульсы от ней
рОIlОВ спинальных rанrлиев по отросткам, идущим в составе КЛИllовидноrо и TOH
Система реактивности
255
кою пучков, а также от чувствительных нейронов таких rанrлиев, как полулун
ный, коленный, каменистый, яремный, верхний пучковидный.
В двиzaтелъных ядрах ствола мозrа находятся мультиполярные нейроны, ocy
ществляющие моторную иннервацию скелетных мышц rоловы и шеи. Нейроны
веrетативных ядер продолrоватоrо и среднеrо мозrа направляют свои аксоны в Be
rетативные rанrлии.
В ассоциативных (пере1Ulючателъных) ядрах MHoro нейронов, которые обеспе
чивают переключение импульсов из спинноrо мозrа и ствола мозrа на нейроны
коры и в обратном направлении. Ствол rоловноrо мозrа связан с периферически
ми орrанами чувствительными и двиrательными волокнами черепных нервов. Ап
парат черепных нервов вполне аналоrичен собственному аппарату спинноrо моз
[а. Собственный внутренний аппарат ствола rоловною мозrа обеспечивает связи
между ею частями.
Рефлекторные дуrи построены не из простой цепочки нейронов, а включают
rpуппы нейронов, в которых возможно осуществление пре и постсинаптическоrо
торможения и модуляция с помощью нейропептидов интернейронов потока аффе
рентных и эфферентных сиrналов. fруппировки нейронов являются общим прин
ципом внутренней орrанизации корковых и ядерных структур мозrа. Объединять
нейроны в единую функциональную систему MOryT коллатерали афферентноrо OT
ростка и интернейроны (продуцирующие нейропептиды), модифицирующие про
ведение HepBHoro импульса.
Различные части ствола rоловноrо мозrа тесно взаимосвязаны блаrодаря Ha
личию BHYTpeHHero собственноrо рефлекторноrо аппарата. Важная роль в YCTaHOB
лении этих взаимосвязей принадлежит также аппарату двусторонних связей спин
ною мозrа и стволовой части rоловноrо мозrа, который включает восходящие и
нисходящие пути.
В состав ствола мозrа входит ретикулярная формация восходящая диффуз
но активирующая система rоловноrо мозrа. В ее сети располаrаются мультиполяр
ные нейроны (размером от 5 до 120 мкм) с маловетвящимися отростками. Ретику
лярная формация получает импульсы от афферентных путей, но сами импульсы
проходят через нее в 4 S раз медленнее, чем через прямые пути. Отростки нейро
нов ретикулярной формации направляются в кору большоrо мозrа, мозжечка, в
ядра ствола мозrа, rдe формируют синапсы (холин , aдpeH , дофаминэрrические и
др.) с нейронами. Так осуществляется интеrративная функция ретикулярной фор
мации. Нисходящие волокна нейронов ретикулярной формации взаимодейству
ют с моторными нейронами спинноrо мозrа. При этом они тормозят их активность.
Считается, что ретикулярная формация принимает участие в формировании эмо
ций, восприятии боли, осуществляет контроль стереотипных движений, тонуса
мышц.
Филorенетически новой надстройкой в ростральной части ствола является пpo
межуточный МОЗl (таламический мозr и rипоталамическая область). Составная часть
1:aJl<\мическоI'О МОЗУ<\ 1:QЛ<\МУС (ЗрИ"Iельный бууор) состоит практически из ceporo
вещества, разделенноrо прослойками белою вещества на ядра. К ним подходят BOC
ходящие (афферентные) проводящие пути и формируются синапсы с нейронами
256
Fлава 15
таламуса. Отростки последних образуют связи (лучистый венец) с нейронами rипо
таламуса, нейронами коры большоrо мозrа и нейронами различных центров проме
жуточноrо мозrа.
rипоталамическая область ствола мозrа высший веrетативный центр, coдep
жащиЙ: комплекс ядер (более 40), тесно связанных с ЦНС с помощью афферент
ных и эфферентных путей. rипоталамус участвует в реryляции температуры, KpO
вяною давления, водною, жировоrо обмена и др_ Среди ядер rипоталамуса особо
выделяются те, которые синтезируют нейроrормоны. В отличие от нейронов, Ha
пример, коры большою мозrа, нейроны rипоталамических ядер обладают двойной
чувствительностью: к действию нейромедиаторов, выделяемых в области межней
рональных синапсов; к действию физико химических факторов окружающей cpe
ды (температура, осмотическое давление, концентрация rлюкозы и др.).
Мозжечок. Мозжечок крупная часть rоловноrо мозrа, rде в процессе rистоrе
неза наблюдается перемещение ceporo вещества изнутри кнаружи. Так возникают
полушария с большим числом борозд и извилин. Белое вещество заключено в rлу
бине opraHa, rде располаrаются ядра мозжечка. Он связан со стволом мозrа 3 парами
ножек. Мозжечок выполняет роль системы, координирующей и контролирующей
сложные автоматически выполняемые движения человека. Это достиrается с помо
щью мноrочисленных афферентных и эфферентных связей нервных клеток мозжечка
с нейронами ядер ствола мозrа и спинноrо мозrа. Например, сиrналы от опорно дви
rательноrо аппарата приходят в мозжечок по спинно мозжечковым путям от HepBHO
мышечных и сухожильных веретен. Мозжечок интеrрирует информацию, поступа
ющую из спинноrо мозrа, opraHoB равновесия (вестибулярных ядер), ретикулярной
формации и коры большоrо мозrа (через нейроны ядер моста). Нейроны ядер моз
жечка, в свою очередь, связаны синаптическими связями с нейронами ядер таламуса
и нейронами ретикулярной формации ствола мозrа. Так мозжечок иrpает роль KOH
тролирующеrо устройства, прослеживающею каждое движение.
Развитие. Мозжечок образуется за счет разрастания дорсолатеральной CTeH
ки нервной трубки в области заднеrо мозrа. Кора в эмбриоrенезе формируется OT
носительно поздно. rладкая на ранних стадиях развития область закладки мозжечка
в последующем становится складчатой. Борозды и извилины на разрезе имеют xa
рактерную для мозжечка картину "древо жизни". От эпендимноrо слоя матрич
ных клеток нервной трубки путем миrpации по отросткам радиальных rлиоцитов
формируется наружный клеточный слой мозжечка. Отсюда клетки, дифференци
руясь в нейробласты, миrрируют в обратном направлении, встречаясь с нейробла
стами следующей волны миrрации из эпендимноrо слоя. Такая встречная миrра
ция двух потоков нейробластов способствует по мере их созревания формированию
послойной структуры коры мозжечка и образованию характерных для нее BHYT
ренних межнейрональных связей.
Строение. В мозжечке различают кору и белое вещество. В коре мозжечка раз
личают три слоя: молекулярный (наружный), rанrлионарный (средний) и зернис
тый (внутренний).
В 'ЮUlJlUОllар1l0М слое лежат rрушевидные нейроны (клетки Пуркиня). От Bep
шины тела этих клеток отходят 2 З дендрита, которые дают первичные, вторичные
Система реактивности
257
и третичные ветви (рис. 117).
На дендритах имеются MHoro
численные синапсы в виде ши
пиков (до 40 тыс. на одной клет
ке). Размер тела rpушевидных
нейронов 60х35 мкм. Распо
лarаются они в один ряд. От oc
нования этих клеток отходит aк
сон в сторону зернистоrо слоя,
образующий начальное звено
эфферентных тормозных пyrей
мозжечка. В зернистом слое от
них формируются возвратные
коллатерали, которые контакти
руют с соседними rpушевидны
ми нейронами. Два три дендри
та rpушевидноrо нейрона Ha
правляются в молекулярный
слой и располаrаются водпой
плоскости, перпендикулярной
длине извилины.
В молекулярном слое раз
личают корзинчатые и звеЗk
чатые клетки. Корзинчатые
клетки лежат в нижней трети
молекулярноrо слоя непосредственно над слоем rpушевидных нейронов. Аксоны
кuрзинчатых клеток длинные, и образуют мноrочисленные коллатерали. за счет
которых формируются "корзинки" BOKpyr тел rрушевидных нейронов. Одна KOp
зинчатая клетка может установить связь через коллатерали аксона с 240 rpуше
видными нейронами.
Звездчатые клетки лежат выше корзинчатых и бывают двух видов: небольших
размеров с короткими и крупные нейроны с длинными отростками. Первые YCTa
навливают контакты с дендритами rpушевидных нейронов. Вторые, наряду с этим,
MorYT направлять аксон в состав корзинки BOKpyr тела rрушевидных нейронов. Kop
зинчатые и звездчатые клетки представляют собой ассоциативные нейроны и OKa
зывают тормозное действие на rрушевидные нейроны.
В .зернистом слое rлавный вид клеток это клетки зерна (зерновидные нейро
вы) с небольшим ядром (5 8 мкм в диаметре) и узким ободком цитоплазмы. Ней
рон имеет 3 6 тонких дендритов с концевыми веточками наподобие птичьей лапки.
Здесь моховидные волокна, входящие в мозжечок, формируют синапсы, и вся CTPYK
тура называется "клубочком мозжечка". Аксон клетки зерна идет в молекулярный
слой, rде Т образно делится. Параллельно идущие волокна аксона образуют синап
тические связи с дендритами rрушевидных, корзинчатых и звездчатых нейронов.
Длин.! параллельных нервных волокон клеток зерен достиrает 3 мм. В зернистом
v J
Д Б
Рис. 117. Схема синаптических связей нейронов в коре мозжеч
ка:
Д поперечный срез извилины; Б продольный срез извили
ны; 1 rрушевидный нейрон; 2 корзинчатый нейрон; 3
зерновидный нейрон; 4 звездчатый нейрон (клетка rольджи);
5 моховидное волокно; б лиановидное волокно
258
Fлава 15
слое есть еще звездчатые нейроны (клетки [ольджи), тормозные. Различают три вида
этих нейронов: звездчатые с коротким нейритом, звездчатые с длинным нейритом и
веретенообразные rоризонтальные. Дендриты звездчатых нейронов распространя
ются в молекулярном слое, а нейрит вплетается в "клубочки мозжечка", тормозя пе
редачу HepBHoro импульса rрушевидному нейрону. Нейроны с длинными OTpOCTKa
ми обеспечивают связь между разными областями коры мозжечка (вертикальная
сочетательная система мозжечка). [оризонтальные нейроны лежат между зернис
тым и rанrлионарным слоями. Они имеют длинные rоризонтально идущие дендри
ты, а аксон их уходит в белое вещество, отдав коллатерали в зернистом слое.
Белое вещество находится под корой мозжечка и составляет мозrовое тело.
В rлубине белоrо вещества локализуются ядра мозжечка. Белое вещество coдep
жит афферентные и эфферентные волокна, которые направляются в ствол мозrа в
составе ножек мозжечка.
Афференmные волокна мозжечка. Различают два вида волокон, по которым
возбуждающие импульсы поступают в кору мозжечка. Это моховидные и лиано
видные волокна.
Моховидные (мшистые) волокна берут начало от спинно мозжечковых путей,
вестибулярных ядер, ретикулярной формации ствола и др. Они заканчиваются в
"клубочках мозжечка". Один аксон передает импульсы примерно на 10 клеток зе
рен, а те, в свою очередь, на мноrие rрушевидные нейроны. Большее число импуль
сов от нейронов коры большоrо мозrа поступает в мозжечок по аксонам нейронов
собственных ядер моста по мосто мозжечковому пути.
Лиановидные (лазящие) волокна проникают в мозжечок в составе оливо моз
жечковых путей. Нейроны оливы продолrоватоrо мозrа, в свою очередь, через цепь
нейронов среднеrо мозrа получают управляющие импульсы от базальных ядер KO
нечноrо мозrа. Послед\iие являются центрами, обеспечивающими выполнение He
произвольных движений, реryлирующие тонус скелетных мышц. Лазящие волок
на стелятся по дендритам rрушевидных нейронов. По ним импульсы проецируются
в кору мозжечка на определенные rрушевидные нейроны, т. е. "точка в точку". По
ступающая информация в коре мозжечка перерабатывается и на основе этоrо про
изводится коррекция двиrательных актов (оказывается тормозное действие).
Эфференmные волокна мозжечка. Эти волокна формируют пути, составленные
нейритами rрушевидных нейронов и нейронами ядер мозжечка. Один rрушеВИk
ный нейрон своим аксоном формирует синаптические связи с десятками нейронов
ядер мозжечка. В свою очередь с одним нейроном ядра мозжечка связываются бо
лее 800 rрушевидных нейронов. Через аксоны нейронов ядер мозжечка импульсы,
сформированные в коре мозжечка (тормозные), направляются в таламические ядра,
которые, в свою очередь, связаны моторными областями коры, а также на нейроны
ретикулярной формации ствола мозrа.
Мозжечок сам не сохраняет память о двиrательных актах, он только их pery
лирует. Команды на двиrательные реакции идут по пирамидным и экстрапираМИk
ным путям.
Клетки коры мозжечка очень чувствительны к действию интоксикации. BЫK
лючение мозжечка из ero контролирующей деятельности сопровождается расстрой
Система реактивности
259
ством координации движений. Ярким примером этоrо является нарушение KOOp
динации движений при алкоrольном опьянении.
Dzuя в м-озжечке представлена волокнистыми и протоплазматическими астроци
тами, а также олиrодендроцитами. В rанrлионарном слое есть особые rлиальные клет
ки, отростки которых идут в молекулярный слой. Это так называемые берrмановские
волокна. Они поддерживают дендриты rрушевидных нейронов. Микроrлия в виде rли
альных макрофаrов представлена в молекулярном и rанrлионарном слоях.
При осуществлении двиrательных актов соматическая нервная система вклю
чает ряд рефлекторных аппаратов: собственный аппарат спинноrо мозrа; собствен
ный аппарат ствола rоловноrо мозrа; аппарат двусторонних связей спинноro мозrа и
ствола мозrа, а также коры rоловноrо мозrа. Особую роль в реryляции двиrательной
активности выполняет в составе сложной системы этих аппаратов мозжечок
При осуществлении сложных двиrательных актов различные отделы ствола ro
ловноro мозrа связаны между собой непрерывными замкнутыми круroвыми нейрон
ными цепями, по которым импульсы длительное время циркулируют. Большую роль
в этих процессах иrpает лимбическая система (rиппокамп, лобно теменная кора, XBO
статое ядро и т. д.).
Кора БОЛЬШОlО M03la
Кора представлена слоем ceporo вещества толщиной 3 5 мм. В коре насчитывают
до 15 и более млрд. нейронов, а число rлиоцитов в rоловном мозrе более 100 млрд.
Развитие. Формирование коры больших полушарий происходит путем зако
номерной миrрации нейробластов эпендимноrо слоя вдоль вертикально ориенти
рованных радиальных rлиоцитов. Первыми возникают наиболее поверхностный и
rлубокий слои коры. Затем возникают очередные последовательные волны миrра
ции rрупп нейробластов, которые дифференцируются в нейроны V ro, затем IV ro
слоя и д. т. Таким образом, нейробласты очередной волны миrрации преодолевают
слой нейронов, возникших от более ранней волны миrрации. Это создает послой
ную (экранную) цитоархитектонику коры большоrо мозrа.
Между нейронами устанавливаются сложные взаимосвязи в соответствии с
их местом в составе рефлекторных дуr. Формируются ядерные и экранные нервные
центры. Тесные взаимоотношения в ходе rистоrенеза складываются между нейро
нами и клетками rлии.
Строение. Все нейроны коры мультиполярны.. Среди них по форме клеток раз
личают пирамидные и непирамидные (звездчатые, корзинчатые, веретенообразные,
паукообразные и rоризонтальные) неЙроны. Пирамидные нейроны, наиболее xapaK
терны для коры, имеют тело, по форме напоминающее пирамиду, вершина которой
обращена к поверхности коры. От основания пирамидноЙ клетки отходит аксон, име
ющий коллатерали (возвратные, rоризонтальные, косые). От вершины и боковых
поверхностей тела отходят длинные дендриты (апикальные и базальные). Апикаль
ные дендриты rруппы нейронов объединяются в дендритические пучки. На повер
хности дендритов одноrо пирамидноro нейрона может быть до 4 6 тыс. специаль
ных рецепторных аппаратов шипиков. Наличие в последних актомиозиновоrо
260
Тлава 15
комплекса позволяет изменять площадь синаптическоrо контакта, следовательно,
влиять на синаптическую связь.
Величина тела пирамидных клеток варьирует от 10 до 150 мкм. Различают
малые, средние, крупные и rиrантские пирам иды. I1ирамидные клетки это эф
ферентные неЙроны коры, их аксонные коллатерали образуют 3/4 всех синапсов в
коре.
Звездчатые неЙроны имеют тело в форме звезды. Дендриты отходят во все cтo
роны от тела звездчатоrо неЙрона. Они в большинстве случаев короткие и лишены
шипиков. Аксоны звездчатых клеток образуют сложные разветвления около клет
ки. Это так называемые околоклеточные паутинообразные аксонные сети. Эти клет
ки встречаются в нижних слоях коры.
Корзинчатые клетки (мелкие и крупные), расположены во II м и III M слоях
коры, своими мноrочисленными отростка 1И образуют синаптические связи с Te
лами пирамидных нейронов V ro слоя. Клетки содержат медиатор (rAMK), KO
торыЙ тормозит передачу возбуждения.
Нейроrлиоморфные клетки встречаются во всех слоях коры. Это мелкие муль
типолярные нейроны с короткими ветвяшимися дeHД
ритами и аксонами.
Биполярные нейроны малочисленная rруппа кле
ток, от тела которых отходят аксон и дендрит. В целом
соотношение между пирамидными и друrими форма:\1И
неЙронов равно 85:15, то есть в пользу пирамиднЫХ неЙ
ронов.
ЦlIтоархитектоника. В двиrательной зоне коры
различают шесть основных слоев: молекулярныЙ, Ha
ружныЙ зернистыЙ, пирамидныЙ, внутренниЙ зернис
тыЙ, rанrлионарныЙ, слоЙ полиморфных клеток
(рис. 118).
В первом (наружном) }ilолекуляр'юм слое почти нет
тел неЙронов. Обнаруживаются единичные rоризон
тально ориентированные неЙроны, танrенциальные
ветвления нервных волокон нижележащих неЙронов
и rлиальные клетки.
ВтороЙ, или наружный зернистый, слоЙ содержит
мелкие звездчатые и пирамидные неЙроны размером
около 1 О мкм. Аксоны этих нейронов оканчиваются в
III M, IV M И VI M слоях коры, а дендриты поднимают
ся в молекулярный слой.
ТретиЙ слоЙ это слой средних и крупных пира
мидных нейронов. Аксоны этих клеток образуют acco
циативные нервные волокна, идущие через белое Be
щество и соединяющие соседние участки коры.
ЧетвертыЙ, или внутренний зернистый, слой coдep
жит в основном мелкие звездчатые нейроны. Аксоны
T ; ": '..;' . .... :; ' ,, <,;'::"; }1
. ' ' . . l'
;. i:l}; l L llL ' 1 1;r;L \l:i }2
,1 1. i:, ", .. , I ' ," i /11 ; 11. J }
..' J: i; .! (\:1'.&.:. t, 1 : К::;
tl t;J fJ з
l l A', {: \:!/J }
" ,":, 1).!: :t'i,!.\( 4
X\JiJ.'J41 ii' . }
; :: :4i' 'j 5
, ,.'Zlj1 1j'
f; f 'y
k.. " , . , >:.... .:А.. .. .
}\ / J):: б
:-.. ,-:_._ .... , V: ."..:.:
Рис. 118.Послойноерасположе
ние нейронов в коре полушарий
большоrо мозrа:
1 молекулярный; 2 наруж
ный зернистый; 3 пирамид
ный; 4 внутренний зернистый;
5 rанrлионарный; 6 поли
морфных нейронов
Система реактивности
261
этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих
слоев коры. Этот слоЙ сильно развит в зрительной и слуховой зонах коры. Он co
стоит из сенсорных звездчатых нейронов, имеющих мноrочисленные ассоциатив
ные связи с нейронами друrих типов.
Пятый zатлионарный слой образован большими пирамидными неЙрона
ми (клетками Беца). Апикальные дендриты нейронов направляются в молекуляр
ный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и
проекционные нервные волокна, и прежде Bcero пирамидные пути.
Шестой слой слой полиморфных нейронов содержит также мною эффе
рентных ПИр<iМИДНЫХ нейронов. Кроме Toro, здесь есть веретенообразные нейро
ны. Дендриты нейронов шестоrо слоя пронизывают всю толщину коры, достиrая
молекулярною слоя.
Миелоархитектоника. Это закономерное расположение нервных волокон бе
лою вещества. Различают ассоциативные волоюш, которые связывают между co
боЙ корковые области одноrо и Toro же полушария или объединяют нейроны в
функциональный комплекс (например, зрительный анализатор). Источником ac
социативных волокон являются пирамидные нейроны I1 ro и III ю слоев. Аксоны
пирамидных нейронов направляются rоризонтально или уходят в белое вещество
и вновь возвращаются в кору.
Комиссуральные (КШ/JLOзалЪ1iые) вОЛО1СН.а связывают между собой полушария, объе
диняя их функциональную активность. Эти волокна проникают в кору через мозоли
стое тело и заканчиваются во всех слоях коры.
Проекционные волокна подразделяются на афферентные и эфферентные. Пер
вые (восходящие) связывают подкорковые центры полушарий и ствола мозrа с
нейронами коры. Войдя в кору, проекционные волокна взаимодействуют с деНk
ритами и телами нейронов практически всех слоев. Восходящие волокна имеют
большой диаметр и толстую миелиновую оболочку. Вторые эфферентные (нис
ходящие) волокна, образованы аксонами нейронов коры и направляются к ПОk
корковым центрам и в спинной мозr.
Закономерностями послойною (экранноrо) расположения нейронов и волокон
не исчерпывается нейронная орrанизация коры большою мозrа. Например, в ceHCO
моторной коре описаны вертикальные колонки, характеризующиеся rнездным
расположением эфферентных пирамидных нейронов и дендритными пучками. Сле
довательно, существует иная (вертикальная) орrанизация нейронов коры. Все это
вместе с физиолоrическими экспериментами по характеристике импульсной актив
ности нейронов, ин нервирующих вибриссы или сетчатку rлаза, создали преДПОСЫk
ки для обоснования принципа модульной орrанизации неокортекса.
Модуль динамичная, относительно автономная самоорrанизующаяся нейрон
ная структура, которая возникает под влиянием афферентных импульсов и TOpMO
зящеrо действия интернейронов. Он имет форму цилиндра (колонки) диаметром до
500 мкм С вертикальной ориентацией длинной оси. Примером модульной орrаниза
ции является локальная нейронная сеть. Это rруппа пирамидных нейронов III ro и
V ro слоев, апикальные дендриты которых собраны в пучок (до 20 дендритов). Между
дендритами возникают контакты щелевоrо типа. Такое rруппирование и взаимосвязь
262
Тлава 15
нейронов позволяет синхронизировать их работу, создавать единый вход для прихо
дящих импульсов. Результатом этою является формирование эфферентноrо импуль
cHoro "залпа".
Чрезвычайно большое значение для понимания основ деятельности мозrа
имеют межнейрональные взаимодействия, которые обеспечиваются большой
rруппой UllтерllеЙрО1l0в. С помощью иммуноцитохимических методов удалось
идентифицировать интернейроны с различными медиаторами норадренерrи
ческие, серотонинерrические, rАМКерrические, rлицинерrические и пептидер
rические. Последние в качестве медиатора используют более SO биолоrически
активных пептидов. Например, они продуцируют: тиреотропный ropMoH, COMa
тостатин, вещество Р, лютропин, ВИП, метионин энкефалин, лейцин энкефалин,
бета эндорфин, rастрин, холецистокинин, нейротензин, бомбезин, инсулин, бра
дикинин, AKTr, пролактин, вазопрессин, окситоцин. Нейропептид вещество Р
служит медиатором, специфически связанным с передачей информации о боли
от периферических болевых рецепторов в центральную нервную систему. Ней
роны, содержащие энкефалины, реrулируют поступление болевых сиrналов в ro
лов ной мозr. Эндорфины и энкефалины морфиноподобные вещества, выраба
тываемые нейроэндокринными клетками, рассеянными по разным отделам
rоловноrо мозrа.
rлутаминовая и аспараrиновая аминокислоты оказывают сильное возбужда
ющее действие на большинство нейронов. [АМК относится к обычным тормоз
ным медиаторам rоловноrо мозrа. Не менее трети синапсов мозrа используют в
качестве медиатора r АМк. rлицин служит тормозным медиатором в спинном мозI}'.
В тканях rоловноrо мозrа найдены простаrландины, которые оказывают разнооб
разное возбуждение и тормозное действие на нейроны. Интернейроны и их медиа
торы оказывают модифицирующее влияние на деятельность пирамидных нейро
нов внутри локальных связей.
МаКРОlJlUЯ представлена различными видами клеток Астроциты выполняют
опорную И трофическую функции в сером и белом веществе, формируют поrpа
ничные rлиальные мембраны. Эпендимная rлия выстилает желудочки мозrа. Оли
rодендроrлия формирует оболочки нервных волокон, участвует в метаболических
процессах.
МUКрОlJlUЯ в виде rлиальных макрофаrов костномозrовоrо происхождения уча
ствует в фаrоцитозе деrенерирующих нейронов.
Оболочки спинноrо и rоловноrо мозrа
rоловной и спинной мозr покрывают мяrкая (сосудистая), паутинная и твердая
оболочки (рис. 119). Они обеспечивают защитную, в том числе механическую (фик
сация мозrа в черепе и позвоночном канале) функции, участвуют в циркуляции цe
реброспинальной жидкости. Мяrкая и паутинная оболочки продолжаются вдоль He
рвов в виде периневрия.
Мяzкая мозzoвая оболочка непосредственно прилежит и сращена с тканями моз
ra, корешков нервов и повторяет в rоловном мозrе ход борозд и извилин. Строма
Система реактивности
263
нейроrлиальноrо проис
хождения менинrотели
ем. Сосуды стромы, прони
кающие в мозr, окружены
элементами rематоэнцефа
лическоro rистиона (барь 9
10
ера) астроцитами, ножки
которых BOKpyr сосудов
формируют непрерывную ;
муфту.
Таким образом, ножки
астроцитов и их базальная
мембрана являются rрани
цей между нервной тканью
и мозrовыми оболочками
(наружная rлиальная MeM
брана).
Паутинная оболочка
расположена между твердой и мяrкой оболочками. Она покрывает полностью по
верхность rоловноrо и спинноrо мозrа. В rоловном мозrе, однако, она не проника
ет в ero уrлубления. Над последними возникают подпаутинные цистерны, rде цир
кулирует цереброспинальная жидкость.
Снаружи паутинная оболочка выстлана однослойным плоским нейроrлиальным
эпителием, под которым располаrается 5 8 слоев уплощенных фибробластоподобных
клеток менинrоцитов. Цитоплазматические отростки последних и коллаrеновые
фибриллы образуют трехмерную сеть паутинных трабекул, которые прикрепляются к
наружной поверхности мяrкой мозrовой оболочки. В сети находятся макрофаrи, лим
фоциты, тучные клетки и крупные кровеносные сосуды, ветви которых проникают в
мяrкую мозrовую оболочку. Выросты паутинной оболочки в венозные синусы TBep
дой мозrовой оболочки, наиболее крупные из которых называются пахионавы rpaHY
ляции, служат для оттока цереброспинальной жидкости в венозный кровоток
16ердая оболочка образована плотной волокнистой соединительной тканью.
Между твердой и паутинной оболочками находится субдуральное пространство.
Оно содержит небольшое количество цереброспинальной жидкости и продолжа
ется в виде периневралЬН IХ пространств вдоль нервных стволов. Стенки этих про
странств выстланы однослойным плоским нейроrлиальным эпителием. Снаружи
оболочки представлена
рыхлой неоформленной
соединительной тканью с
большим количеством KpO
веносных сосудов и He
рвных волокон. Снаружи
строма покрыта однослой
ным плоским эпителием
.
;:: '
.' . x-f. :
- ". ...",........
.. f?.'
. .... I
7.' . _;- :. ..' .','; 1
:>, ",
8
264
Fлава 15
от твердой оболочки СПlшноrо мозrа находится эпидуральное пространство, за
полненное ЖИРОВОI':! тканью. Напротив, твердая оболочка rоловноrо мозrа плотно
сращена с надкостницей черепных костей, в связи с чем в черепе отсутствует эпи
дуральное пространство.
Кровеносные сосуды, проникающие в ткань rоловноrо мозrа, идут по каналам,
выстланным мяrкой мозrовой оболочкой. BOKpyr крупных сосудов имеется пери
васкулярное пространство. Оно сообщается с субарахноидальным пространством
и содержит цереброспинальную жидкость. BOKpyr кровеносных капилляров TaKO
ro пространства нет. Содержимое кровеносных капилляров отделено от ткани ro
ловноrо мозrа lемаmОЭ1lцефалuческuм lUcтU01l0M (барьером). Последний образу
ют: непрерывный слой эндотелия капилляров с базальной мембраной, при этом
эндотелиоциты соединены протяженными плотными межклеточными KOHTaKTa
ми; периваСКУЛЯРНdЯ поrраничная rлиальная мембрана, образованная ножками
астроцитов, которая в виде непрерывной муфты окружает капилляры мозrа.
Через rематоэнuефалический барьер из крови в мозr не проникают некоторые
лекарственные препараты, антитела и друrие крупномолекулярные вещества, Tor
да как rазы и мелкие молекулы, необходимые для питания нервной ткани, диф
фундируют через Hero.
Цереброспинальная жидкость, мяrкая и паутинная оболочки мозrа покрыва
ют roловноЙ и спинной мозr, выполняя роль rидравлическоrо амортизатора. С по
мощью отверстий в крыше четвертоrо желудочка пространства в оболочках мозrа
соединяются послсдовательно с полостями мозrовых желудочков. Исследование
цереброспинальной жидкости имеет большое диаrностическое значение в клини
ке. Местом образования ее в основном являются сосудистые сплетения, выступа
ющие в просвет всех четырех мозrовых желудочков.
Сосудистое сплетение снаружи покрыто однослоЙным кубическим эпителием
нейроrлиальноrо происхождения. Строма сплетения состоит из сuединительной
ткани, сосудов и нервов. На поверхности сплетения располаrаются макрофаrи
(клетки Кол мера).
В нервной системе постоянно происходит циркуляция цереброспинальной жидко
сти. Переход ее в кровь происходит в выростах паутинной оболочки (пахионовых rpa
нуляuиях), выступающих в венозные синусы твердой мозrовой оболочки. Следует OT
метить, что в центральной нервной системе нет лимфатических сосудов, которые моrли
бы отводить избыток жидкости, и потому роль арахноидальных ворсинок очень велика.
Транспортировка ликвора между полостью 111 ro желудочка и первичной капиллярной
сетью медиальной эминенции rипоталамуса осуществляется при активном участии Ta
ницитов клеток эпендимной выстилки. Для них характерно наличие длинных oтpoc
тков, обеспечивающих контакт с первичной капиллярной сетью. В цитоплазме таници
тов описана система мембранных полостей и пузырьков, с помощью которых
осуществляется внутриклеточный транспорт не только ликвора, но и мноrих roрмонов.
Периферические нервы
Периферические нервы состоят из пучков миелиновых и безмиелиновых He
Система реа'/{тивности
265
рвных волокон, одиночных неЙронов или их скоплений и оболочек Тела нейронов
находятся в сером веществе спинноrо и rоловноrо мозrа и спинномозrовых узлах
(rанrлиев). В составе нервов находятся ЧУВСТIЗительные (афферентные) и двиrатель
ные (эфферентные) нервные волокна, но чаще те и друrие. Между нервными волок
нами располаrается эндоневрий, представленный нежными прослойками рыхлой BO
локнистой соединительной ткани с сосудами.
Периневрий одевает отдельные пучки нервных волокон. Он содержит 5 6 пластов
однослойною эпителия эпендимоrлиальною типа, лежащеrо на базальной мембране,
разделенною прослойками рыхлой волокнистой соединительной ткани. Периневрий
является продолжением эпителия мозювых оболочек По жидкости периневральноrо
пространства MOryт распространяться вирусы (например, бешенства).
Наружная оболочка эпиневрий представляет собой поверхностную соеди
нительнотканную оболочку нерва, состоящую из плотной соединительной ткани с
кровеносными и лимфатическими сосудами, нервными окончаниями.
Одиночные нейроны и их скопления в составе нервов, как правило, встреча
ются в веrетативной нервной системе.
Веrетативная нервная система
Веrетативная нервная система является частью единой нервноЙ системы. Она
иннервирует внутренние opl'aHbI, кровеносные сосуды, железы, принимает учас
тие в иннервации скелетных мышц, реrулирует процессы кровообращения, дыxa
ния, об:\fена веществ, питания, выделения, термореrуляции и Ilр. Ее называют aB
тономноЙ, но автономность этой системы, хотя и функционирует она независимо
от сознания, относительна, так как все стороны ее деятельности находятся под KOH
тролем коры большоrо мозrа. И соматическая, и автономная системы построены
по одной схеме, но развиваются диверrентно: соматическая система вместе с opra
нами движения, а автоно:\шая вместе с внутренними орrанами.
Веrетативная нервная система подразделяется на симпатическую и парасимпа
тическую. Стимуляция симпатической нервной системы увеличивает частоту и силу
сердечных сокращений., вызывает сужение сосудов внутренних opraHoB, повышает
артериальное давление, расширяет бронхи, зрачки, снижает тонус желудочно кишеч
Horo тракта, оказывает адаптационно трофическое влияние на ткани. Стимуляция
парасимпатической нервной системы снижает силу и частоту сердечных сокраще
ний, снижает артериальное давление, при водит к усилению перистальтики кишеч
ника и др. Веrетативная нервная система подютавливает и обеспечивает соматичес
кие эффекты соответствующими метаболическими процессами.
В веrетативноЙ нервной системе различают центральные и периферические
отделы. ЦентРШlьные отделы симпатической нервной системы представлены яk
рам и боковых poroB тораколюмбальноrо отдела спинноrо мозrа. В парасимпати
ческой нервной системе центральные отделы включают ядра среднеrо и продолrо
BaToro мозrа, а также ядра боковых poroB сакральноrо отдела спинноrо мозrа.
Парасимпатические волокна краниобульбарпоrо отдела выходят в составе III й
VII й, IХ й и Х й пар черепных нервов.
266
Fлава 15
Перuферuчес'Кuе отделы веrетативной нервной системы образованы HepBHЫ
ми стволами, rанrлиями и сплетениями.
Веrетативные рефлекторные дуrи начинаются чувствительным нейроном, тело
KOToporo лежит в спинномозrовом узле (rанrлии), как и в соматической рефлек
торной дуrе (см. рис. 116). Ассоциативные нейроны находятся в боковых porax
спинноrо мозrа. Здесь нервные импульсы переключаются на промежуточные пре
rанrлионарные нейроны, отростки которых покидают центральные ядра и дости
rают веrетативных rанrлиев, rде передают импульсы на двиrательный нейрон. В
связи с этим различают нервные волокна преrанrлионарные и постrанrлионарные.
Первые из них покидают центральную нервную систему в составе вентральных
корешков спинномозrовых нервов и черепных нервов. Как в симпатической, так и
в парасимпатической системах преrанrлионарные нервные волокна принадлежат
холинерrическим нейронам. Аксоны нейронов, расположенных в веrетативных raH
rлиях, называются постrанrлионарными. Они не образуют прямых контактов с эф
фекторными клетками. Их терминальные отделы по своему ходу формируют pac
ширения варикозности, в составе которых находяся пузырьки медиатора. В
области варикозности нет rлиальной оболочки и нейромедиатор, выделяясь в OK
ружающую среду, влияет на эффекторные клетки (например, на клетки желез, rлаk
кие миоциты и др.).
В периферических rанrлиях симпатической нервной системы, как правило, Ha
ходятся адренерrические эфферентные нейроны (за исключением нейронов, имею
щих синаптические связи с потовыми железами, rде симпатические нейроны явля
ются холинерrическими). В парасимпатических rанrлиях эфферентные нейроны
всеrда холинерrические.
Fа1ilЛUU представляют co
бой скопления мультиполяр
ных нейронов (от нескольких
клеток до десятков тысяч). Эк
страорrанные (симпатические)
rанrлии имеют хорошо Bыpa
женную соединительноткан
ную капсулу, как продолжение
периневрия (рис. 120). Пара
симпатические rанrлии Haxo
дятся, как правило, в интраму
ральных нервных сплетениях.
rанrлии интрамуральных спле
тений, как и друrие веrетатив
ные узлы, содержат веrетатив
ные нейроны местных рефлек
торных дуr. Мультиполярные
нейроны диаметром 20 35 мкм
расположены диффузно, каж
дый нейрон окружен rлиоцита
",.... .,....... ....,., (
."..............,.-:"" =-...."<... ::.., -', - J. 1/
. >'''-'' . . ',. " ,,...,.....-'У '/'-I. '/' t!-M
/.....,. .'" '.. 'c,, "':::r '// 7',/
,.'..у .,,,.,...i..' '. ,;. ;r: 't'. :;! / /., j1
" '" :«"' . ,,,- , '.,( "('//'''iI / J
'" "'"', :'\ ;:, -;;';',, ..' :\ I(/ '1/, - t
j'. .. 1\ 4> .. ," ,., "I"{ ,
1/ , 1." , "" ' .' , .. tI ;, ,.,rto, h " ...t ': r Л,I , 1.;
I .: .., .'.. э, ,'I.t .'" '4i q ., \, !
,../...'If >,,;,..) ...:. "'", ......... _'-.:;.." ..\\.
,1 11 ,,;'. ".......,. ,/ ","',.:; ,,, ;... "<f\.<,. I
1(. .. . .....".." " .AIi!"'(J "'!' 1' .." '"',........,...... "'1 ''9- . \ .
1"1"", "" . "", .' ..;... ';'fIr ,.,.... ;:"'1' " '\'
11- ,. JlW" .:Jt' . (, :;\... .:_; "' ' ""'....-; :. : "" -""
/' ) ","ff',. .:t.... ;".,-,. .. t,:; 1;0, '.} 1
, _:- -. \ A -. " .. f/;" . ;:". ,. V
j l. '\"'. \i !I . J' ..,....' 4.. >,:.,.'
. "с' '* &tt "'\, , ,....'" '" 1 , ;"4.». f ., .JVi lIt"
' \ O&\ t.... f'i ,. .. " "А '.., ."."\ ",' '" "' . ::; .... , ";,,,
. .. , ..,1,">1. ..{ '" "' ' "" "Y .;> 'A:'!"Jo! "' .Y.:!"/
( , \ ". " , t. ' ,",,\, ."\':'" { ... '''' \ tIi " .' ..... "". 2
I .., 1'. ''0....\ '. ". <,... (- > "" '..' !
. \ '"" '.. + ...,.., ... 1I..""t \'\1' . ... 'fJ -,"-'-' ... ' А '
'. .. > ....... . .. .,. ... .. "'. '-. ,и
\i'4".,.... .,{! .. .; :( $ '" i-:: .,. -!t" \,.'
. '-<" .h.t.' ..' '." ;, "\ , " ".' ...,,"1. ,1
\:r"""",>"'JiJ. ,._. , '\}'''\ ''' ....1j:O ". '-"\. / '
,...;-. '\:i: "t y ';\1 .. ii ... >!oe ' ..."'! * .""_",' '\-t1....
\. ,о:' "... _. ,;. ....'4; о "':' \ .":. ..... I
\..:!J'''fa''«f;-<.t ..",-. ...,; '", 'I'" ,h> .;' "iI" /.1" .
,- .&'-'> It '" ...,:с; $< . . '.. ,.'
,, ..'"".'4) ь" .... "". ";.... .. J, __ ,...
,....\-'iA> f?:-e '(t:" "' ,i.. "'t':: * : ",, /./;I/
',,"':--;' .E. ....<:-. ' ,.,,".";'
......,iiI) -- ,, Ч;{"1r- "*' "",,""
........... ..... -=- .:.=;
Рис. 120. Строение симпатическоrо rанrлия (В окраске reMa
токсилин и эозин):
1 соединительнотканная капсула; 2 диффузное распо
ложение нейронов
Система реактивности
267
ми rанrлия. Кроме Toro, описаны нейроэндокринные, хеморецепторные, биполяр
ные, а у некоторых позвоночных и униполярные нейроны. Б симпатических raHr
лиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ клетки) с
короткими отростками и большим количеством rранулярных пузырьков в цитоп
лазме. Они выделяют катехоламины и оказывают тормозящее влияние на переда
чу импульсов с преrанrлионарных нервных волокон на эфферентный симпатичес
кий нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов веrетативных rанrлиев различа
ют: двиrательные (клетки Доrеля I ro типа), чувствительные (клетки Доrеля II ro
типа) и ассоциативные (клетки Доrеля III ro тип::!). Двиrательные нейроны имеют
короткие дендриты с пластинчатыми расширениями ("рецептивные площадки").
Аксон этих клеток очень длинный, уходит за пределы rанrлия в составе постrанr
лионарных тонких безмиелиновых нервных волокон и оканчивается на rладких
миоцитах внутренних opraHoB. Клетки I ro типа называют длинноаксонными ней
ронами. Нейроны II ro типа равноотростчатые нервные клетки. От их тела OTXO
дят 2 4 отростка, среди которых различить аксон трудно. Не разветвляясь, OTpOCT
ки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные
окончания, а аксон оканчивается на телах двиrательных нейронов в соседних raH
rлиях. Клетки II ro типа являются чувствительными нейронами местных BereTa
тивных рефлекторных дуr. Клетки Доrеля III ro типа по форме тела похожи на
веrетативные нейроны II ro типа, но их дендриты не выходят за пределы rанrлия, а
нейрит направляется в друrие rанrлии. Мноrие исследователи считают эти клетки
разновидностями чувствительных нейронов.
Таким образом, в периферических веrетативных rанrлиях имеются местные
рефлекторные дУlИ, состоящие из чувствительных, двиrательных и, возможно, ac
социативных веrетативных нейронов.
Интрамуральные веrетативные rанrлии в стенке пищеварительноrо тракта OT
личаются тем, что в их составе, кроме двиrательных холинерrических нейронов,
имеются тормозные нейроны. Они представлены адренерrическими и пуринерrи
ческими нервными клетками. Б последних медиатором является пуриновый HYK
леотид. Б интрамуральных веrетативных rанrлиях встречаются также пептидер
rические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд
друrих пептидов, с помощью которых осуществляются нейроэндокринная реryля
ция и модуляция деятельности тканей и opraHoB пищеварительной системы.
Реrенерация в нервной системе
Нервные ткани необыкновенно чувствительны к снабжению кислородом. Если
доставка кислорода задержится более, чем на одну минуту, нервные клетки будут
не в состоянии выполнять свои функции.
Различные виды боевой травмы (радиационные поражения, ударная волна,
термические воздействия, ОБ и др.) вызывают нарушения проницаемости стенок
кровеносных сосудов, структуры и функций нейронов в opraHax центральной и
периферической нервной системы. Б нервных клетках происходят разрушение
268
Fлава 15
хроматофильной субстанции, вакуолизация цитоплазмы, уменьшение количества
РНП, расширение цистерн эндоплазматической сети и комплекса rольджи, YMeHЬ
шение числа рибосом, набухание митохондрий с деформацией и разрушением
крист, кариопикноз и кариолизис. Наиболее чувствительными к повреждению яв
ляются нейроны пирамидноrо, rанrлионарноrо и полиморфноrо слоев коры полу
шариЙ большоrо мозrа, корзинчатые клетки и rрушевидные клетки мозжечка, неЙ
роны подбуrровоЙ области и ствола мозrа. При этом возможно нарушение
про ведения нервных импульсов в области синапсов, дезинтеrрации синапсов.
Посттравматический rистоrенез тканей opraHoB нервноЙ системы имеет свои
особенности в связи с отсутствием камбиальных элементов для нейронов. Проли
ферация соединительнотканных элементов, клеток нейроrлии, формирование KOk
лаrеновых волокон, межклеточные и межтканевые отношения в процессе rистоrе
неза приводят к образованию рубцов сложноrо TKaHeBoro состава.
В последние roды развивается идея постнатальноrо нейроrенеза, которая зак
лючается в том, что в opraHax нервноЙ системы продуцируются новые неЙроны в
течение всеЙ жизни особи. Местом, rде возн кают новые неЙроны, является субвен
трикулярная зона (лобная, височная, теменная области) rоловноrо мозrа, откуда
возникшие нейроны миrрируют в разные области rоловноro мозrа. Возможно появ
ление новых нейронов и в rиппокампе. Все это имеет перспективу для лечение забо
леваниЙ человека. связанных с rибелью неЙронов (например, болезни АльцrеЙмера,
сопровождающейся слабоумием).
Сенсорный комплекс opraHoB
Сенсорный комплекс opraHoB (сенсорная система) специализирован на BOC
приятие раздражениli внешнеЙ и внутренней сред и передачу информации в кору
rоловноrо мозrа. Ранее было отмечено, что все клетки орrанизма реаrируют на воз
действия, что относится к общей неспецифической реакции. Однако наряду с He
специфическоЙ реакцией в клетках наблюдаются специфические реакции на KOH
кретные внешние или внутренние стимулы с помощью системы восприятия,
трансформации и передачи сиrнала (рецептор ной системы клетки). Специализи
рованные клетки, воспринимающие и кодирующие энерrию, поступающую из
внешней среды (экстерорецепторы), входят в состав opraHoB чувств.
OpraHbI чувств представляют собоЙ анализаторы, под которыми, по и.п. Пав
лову, надо понимать сложный комплекс структур, включающих три тесно связан
ные в rистолоrическом и функциональном отношениях части: периферическую
часть, осуществляющую восприятие раздражения и трансформацию ero в нервныЙ
импульс; промежуточную часть, передающую импульс по проводящим путям И ПОk
корковым образованиям, и центральную часть кору roловноrо мозrа, rде проис
ходит высший анализ и синтез ощущений.
OpraHbI чувств подразделяются на экстерорецепторы и интерорецепторы
на основе Toro, какие раздражители внешней или внутренней среды они BOC
принимают. Термин "орrапы чувств" относится в большей степени к экстероре
цепторам.
Система реактивuости
269
В зависимости от особенностей развития, строения и функций HepBHoro и rли
альноrо компонентов opraHbI чувств подразделяются на три rруппы.
К первой rруппе относят opraH зрения и opraH обоняния. Эти opraHbl чувств
имеют особое происхождение в эмбриоrенезе, так как они являются производны
ми нервной трубки. В строении их нервною компонента (сетчатка, обонятельные
луковицы) отражается принцип экранною послойноrо расположения нейронов.
Рецепторные клетки, воспринимающие колебания световых волн или действие
молекул пахучих веществ, в этих opraHax чувств являются нервными клетками.
Они называются uейросеuсорuыми, или первичночувствующими.
Ко второй rруппе относят орrап слуха, opraH равновесия и opraH вкуса. В этих
opraHax чувств сильно развиты rлиальные сенсоэпителиальные клетки произ
водные плакод, которые воспринимают действие раздражителя и передают воз
буждение чувствительным нервным клеткам. Это сеuсоэпителиалъuые, или BTO
ричночувствующие, рецепторы. Все дифференцированные клетки opraHoB чувств
этой rруппы содержат на апикальной поверхности, кроме мноrочисленных МIIКрО
ворсинок (стереоцилий), реснички (киноцилии).
Третью rруппу opraHoB чувств составляют рецепторные инкапсулированные и
неинкапсулированные тельца периферические части анализаторов осязания
давления и др.
OplaH зрения
OpraH зрения rлаз представляет собой периферическую часть зрительно
ro анализатора. Пuсредством opraHa dреНИЯ человек получает 80 8S% информации
об окружающем мире. Зрение важнейший физиолоrический процесс, с помо
щью KOToporo создается представление о величине, форме и цвете предметов, о
взаимном их расположении и расстоянии. Эта информация пuзволяет человеку
ориентироваться в окружающем пространстве.
OpraH зрения состоит из rлазноrо яблока и вспомоrательною аппарата (веки, слез
ные железы, rлазодвиrательные мышцы). В rлазном яблоке различают три оболочки:
наружная склера и прозрачная ее часть роruвица; средняя сосудистая оболочка
с ее производными ресничным (цилиарным) телом и радужной оболочкой; BHYT
ренняя сетчатая оболочка (или сетчатка). Кроме Toro, в rлазном яблоке имеются
хрусталик, стекловидное тело, жидкость передней и задней камер rлаза. В функцио
нальном отношении выделяют несколько аппаратов: рецепторныЙ (сетчатая оболоч
ка), диоптрический, или светопреломляющиЙ (роювица, хрусталик, стекловидное тело,
жидкость передней и задней камер rлаза), аккомодационныЙ (радужная оболочка, pec
ничное тело) и вспомоrательный аппарат.
Развитие. [лаз развивается из нескольких источников. Сетчатка и зрительный
нерв формируются из HepBHoro и неЙроrлиальноrо материала выпячиваний стенки
переднеrо мозrа, которые имеют вид rлазных пузырей, позднее преобразующихся в
rлазные бокалы. Хрусталик развивается из эктодермалыюrо материала хрусталико
вой плакоды. Сосудистая оболочка и ее производные радужка и ресничное тело, а
также собственное вещество роrовицы и склера развиваются из мезенхимы.
270
Тлава 15
Мышцы, расширяющие и суживающие зрачок, образованы мионейральной тканью.
Большую роль в процессах развития rлаза иrрают индуктивные взаимодействия
материала различных эмбриональных зачатков.
Рецепторный аппарат rлаза. Сетчатка развивается из стенки rлазноro бокала.
Это внутренняя оболочка rлаза, состоящая из светочувствительноrо и пиrментно
ro листков, соответствующих внутреннему и наружному листкам стенки rлазноrо
бокала. По своему происхождению сетчатка является специализированной частью
мозrовой коры, вынесенной на периферию. На 4 й неделе эмбриоrенеза зачаток
сетчатки состоит из однородных малодифференцированных клеток На 5 й неделе
появляется разделение сетчатки на два слоя: наружный (от центра rлаза) ядер
ный, и внутренний слой, не содержащий ЯДер. Наружный ядерный слой выполня
ет роль матричной зоны, rде наблюдаются мноrочисленные митозы. В начале 6 й
недели из матричной зоны начинают выселяться нейробласты, образующие BHYT
ренний слой. В конце 3 ro месяца четко дифференцируется слой крупных rанrли
озных нейронов. Отростки последних проникают в краевую зону, образуя самый
внутренний слой нервных волокон, которые врастают в rлазной стебелек и фор
мируют зрительный нерв. В последнюю очередь в сетчатке дифференцируется Ha
ружный слой, состоящий из палочковидных и колбочковидных зрительных кле
ток Происходит это незадолrо до рождения. Помимо нейробластов в матричном
слое сетчатки образуются rлиобласты источники развития клеток rлии. Высоко
дифференцированными среди них становятся мюллеровы волокна, пронизываю
щие всю толщу сетчатки.
Строение. В сетчатке rлаза разли
чают зрительную (оптическую) и сле
пую части. ЗрительнаЯ'часть приобрета
ет сложный клеточный состав, тоrда как
слепая часть сетчатки, на которую не
попадают прямо световые лучи, OCTaeT
ся состоящей из двух пластов кубичес
Koro rлиальноro эпителия. rраница меж
ду зрительной и слепой частями сетчат
ки неровная. Она носит название зубча
Toro края.
Сетчатка сформированноrо rлаза в
оптической своей части имеет послойное
строение, характерное для экранных He
рвных центров. В ней различают три слоя
нейронов и два слоя межнейрональных
связей, а также слои, образованные rли
альными элементами (рис. 121). Bcero в
сетчатке выделяют 1 О слоев: пиrментный
эпителий сетчатки, слой палочек и KOk
бочек, наружная поrраничная мембрана,
наружный ядерный слой, наружный ceT
I
.
Рис. 121. Схема строения и синаптических связей
нейронов сетчатки rлаза:
1 палочковидная нейросенсорная клетка; 2
колбочковидная нейросенсорная клетка; 3 бипо
лярный нейрон; 4 rанrлиозный нейрон; 5 rори
зонтальный нейрон; 6 амакринный нейрон
Система реактивности
271
чатый слой, внутренний ядерный слой, внутренний сетчатый слой, rанrлионарный
слой, слой нервных волокон, внутренняя rлиальная поrраничная мембрана.
Наружный и внутренний ядерные слои, а также rанrлионарный слой COOTBeT
ствуют расположению тел нейронов, а сетчатые слои их синаптическим KOHTaK
там. Самый наружный слой сетчатки образован пиrментным эпителием. Пиrмен
тоциты имеют форму шестиrранных призм. Основание их прилежит к стеКЛОВИk
ной мембране сосудистой оболочки rлаза. От вершины клеток отходят отростки в
виде "бороды", содержащие пиrмент меланин. На свету количество пиrмента в OT
ростках увеличивается, в темноте он перемещается из отростков в тело клетки.
Пиrментосодержащие отростки этих клеток окружают палочки и колбочки нейро
сенсорных клеток и отделяют их друr от друrа, препятствуя рассеиванию света, а
также обеспечивают оптическую защиту от яркою света. Кроме Toro, пиrментный
эпителий обеспечивает транспорт метаболитов, кислорода из сосудистой оболоч
ки, фаrоцитирует деrенерирующие диски наружных cerMeHToB нейросенсорных
клеток и др.
Нейросенсорные клетки имеют периферический и центральный отростки. Тела
этих нейронов лежат в наружном ядерном слое сетчатки. Периферический OTpOC
ток нейрона имеет форму или палочки, или колбочки, что определяет их название.
Палочковые 'КЛетки состоят из ядросодержащей части и фоторецепторной час
ти палочки. В палочке различают наружный и внутренний cerMeHTbI. BHYTpeH
ний cerMeHT содержит мноrочисленные митохондрии и полирибосомы, комплекс
fольджи и элементы эндоплазматической сети. В наружном cerмeHTe, имеющем
цилиндрическую форму, находится множество сдвоенных поперечных мембран,
расположенных в виде стопки уплощенных мембранных пузырьков. Эти замкну
тые мембранные структуры называют дисками. Они образуются за счет rлубоких
складок плазмолеммы у основания наружноrо cerMeHTa. Диски составляют дина
мическую систему с высокой степенью обновляем ости. Через каждые 40 мин воз
никает новый диск, а ранее образовавшиеся смещаются к свободному концу cer
мента. Там диски фаrоцитируются клетками пиrментною эпителия. В мембранах
наружноrо cerMeHTa палочковых клеток содержится зрительный пиrмент родоп
син, который состоит из белка опсина в сочетании с ретинолом (альдеrидом вита
мина А). Между наружным и внутренним сеrментами определяются филаменты,
типичные для ресничек, с базальным тельцем во внутреннем cerMeHTe. Это OTpa
жает эволюционное происхождение cerMeHTOB как видоизмененных ресничек Дли
на палочковой клетки достиrает 60 мкм. Общее их количество в сетчатке около
130 млн. Палочки являются рецепторами черно белоrо (сумеречнorо) света, тоrда
как цветное зрение связано с функцией колбочек
Колбочковые 'КЛетки отличаются от палочковых некоторыми особенностями CTpO
ения наружноrо и BHyтpeHHero cerMeHToB, а также свойствами зрительноrо пиrмента.
Наружные cerMeHTbI колбочек состоят из полудисков. Во внутреннем cerMeHTe имеет
ся эллипсоид, состоящий из липидной капли, окруженной скоплением митохондрий.
В колбочках полудиски содержат зрительный пиrмент йодопсин. Мембраны полу
дисков здесь не обновляются подобно дискам в палочках. Имеет место лишь молеку
лярное обновление белков в составе мембран. Общее количество колбочковых клеток
272
Fлава 15
около 7 млн. Колбочковые клетки бывают трех типов, способных воспринимать три
основных цвета (красный, синий и зеленый). Трехкомпонентная теория цветною зре
ния впервые была выдвинута М.В. Ломоносовым в 1756 r.
Механизм фоторецепции связан с распадом молекул родопсина и йодопсина
при действии световой энерrии. Это запускает цепь биохимических реакций, KO
торые сопровождаются изменением проницаемости мембран в палочках и колбоч
ках и возникновением потенциала действия. После распада зрительноrо пиrмента
следует ero ресинтез, что происходит в темноте и при наличии витамина А Heдo
статок в пище витамина А может приводить к нарушению сумеречноrо зрения (KY
риная слепота). Цветовая слепота (дальтонизм) объясняется rенетически обуслов
ленным отсутствием в сетчатке одноrо или нескольких типов колбочек
Возбуждение нейросенсорной клетки передается посредством центральноrо OT
ростка на 2 й биполярный нейрон. Тела биполярных нейронов лежат во внутреннем
ядерном слое сетчатки. В этом слое, кроме биполярных неЙронов, находятся ассоци
ативные нейроны еще двух типов: rоризонтальные и амакринные. Биполярные ней
роны соединяют палочковидные и колбочковидные зрительные клетки с нейрона
ми rанrлионарноrо слоя. При этом колбочковидные клетки контактируют с
биполярными нейронами в соотношении 1:1, Torдa как с одной биполярной клеткой
образуют соединения несколько палочковидных клеток. rоризонтальные нервные
клетки имеют MHoro дендритов, с помощью которых контактируют с центральными
отростками фоторецепторных клеток Аксон rоризонтальных клеток также вступа
ет в контакт с синаптическими структурами между рецепторной и биполярной клет
ками. Здесь возникают множественные синапсы своеобразноrо типа. Передача им
пульсов через такой синапс и далее с помощью rоризонтальных клеток может
вызывать :эффект латеральноrо торможения, что увеличивает контрастность изоб
ражения объекта. Сходную роль выполняют амакринные нейроны, расположенные
на уровне BHYTpeHHero сетчатоrо слоя. У амакринных нейронов нет аксона, но есть
разветвленные дендриты. Тело нейрона иrpает роль синаптической поверхности.
ю:н.lJlИОllарllblе клетки образуют слой TaKoro же названия. Это наиболее круп
ные нервные клетки сетчатки. Они составляют З й компонент нейронной цепи.
Аксоны этих клеток дают слой нервных волокон, формирующих зрительный нерв.
Поддерживающие элементы в сетчатке представлены rлиальными клетками
(мюллеровыми волокнами) и астроцитами. Мюллеровы волокна это крупные ней
роrлиальные клетки с отростками, которые располаrаются вертикально по всей TOk
щине сетчатки, оплетают нейроны сетчатки, выполняя поддерживающую и трофи
ческую функции. Ядра клеток располаrаются на уровне BHyтpeHHero ядерною слоя.
Наружные отростки клеток заканчиваются мноrочисленными цитоплазматически
ми выростами (микроворсинками), которые формируют наружную поrраничную
мембрану, а внутренние завершаются на rpанице со стекловидным телом (форми
руя внутренюю поrpаничную мембрану).
В сетчатке есть желтое nят1l0 с центральной ямкой. Это место наилучшеrо ви
дения. Здесь мною колбочковых нейронов. Имеется также слепое пятно, которое co
ответствует месту выхода зрительноrо нерва.
Диоптрический аппарат rлаза. РOlовuца. Прозрачная часть наружной фиб
Система реактивности
273
розной оболочки rлаза называется роroвицей. Ее строма развивается из мезеНХII
мы. Передняя и задняя поверхности роrовицы покрыты эпителиальными ткаIlЯ
ми. Передний эпителий роrовицы мноrослойный плоский неороroвевающий эпи
телий, лежащий на базальной мембране, является продолжением эпителия
конъюнктивы. Эпителий пронизан мноrочисленными нервными окончаниями, YB
лажняется секретом слезных желез. Под эпителием располаrается передняя по
rраничная пластинка толщиной 6 9 мкм, представляющая собой наружную часть
стромы роrовицы. Она участвует в защите rлаза от травмы и проникновения бак
терий. Ее повреждение затрудняет последующую реrенерацию роrовицы.
Собственное вещество роroвицы состоит из параллельно лежащих коллаrено
вых волокон, образующих пластинки. Между ними определяется аморфное веще
ство, боrатое rликозаминоrликанами, и небольшое число отростчатых плоских
клеток фибробластическоrо дифферона. Компоненты основнorо вещества обеспе
чивают прозрачность стромы роroвицы. Собственное вещество роrовицы ПрОДОk
жается н склеру плотную непрозрачную соединительнотканную оболочку rлаз
Horo яблока, выполняющую опорную роль. Переход прозрачной роrовицы в
непрозрачную склеру происходит в области лимба. На rранице между стромой po
roвицы и задним эпителием роrовицы находится задняя поrраничная пластинка
(десцеметова мембрана) с упорядоченным расположением фибрилл. Заднюю по
верхность роroвицы выстилает эпителий, состоящий из одноrо слоя плоских по
лиrональных клеток нейроrлиальноrо происхождения. Эпителий участвует в пи
тании роrовицы (путем диффузии веществ из жидкости передней камеры rлаза).
При травме роrовицы проявляет слабую реrенераторную способность.
Воспалительные процессы в роroвице, сопровождающиеся врастанием в нее OT
сутствующих В норме кровеносных сосудов, MOryT приводить К помутнению ее (бель
мо) и полной слепоте. В настоящее время успешно проводится rомотрансплантация
роrовицы. Это объясняется тем, что роrовица лишена кровеносных сосудов и сюда
не поступают клетки крови, участвующие в отторжении трансплантата.
Хрусталик. Хрусталик развивается из материала эктодермальной хрустали
ков ой плакоды, преобразующейся под влиянием rлазноrо бокала в хрусталиковый
пузырек На 5 й неделе эмбриоrенеза хрусталиковыЙ,.пузырек отшнуровывается
от эктодермы.
Передняя стенка хрусталиковоrо пузырька состоит из однослойноro кубическою
эпителия, заднюю стенку образуют удлиненные клетки хрусталиковые волокна. По
мере их роста полость пузырька исчезает. В центре развивающеroся хрусталика из
первичных хрусталиковых волокон формируется ядро хрусталика. При этом xpYCTa
ликовые волокна теряют ядра, а процесс физиолоrической реrенерации белков ocy
ществляется на долrоживущих рнк. ДальнейшиЙ рост хрусталика связан с образова
нием вторичных хрусталиковых волокон за счет пролиферации клеток, находящихся
в экваториальной области. Цитоплазма хрусталиковых волокон содержит прозрачное
вещество белок кристаллин. Внутри хрусталика сосудов и нервов нет. Хрусталик
покрыт прозрачной капсулой. На передней поверхности хрусталика под капсулой co
храняется однослойный эпителий, называемый эпителием передней сумки. XpYCTa
лик поддерживается в rлазу в определенном положении посредством ресничноrо
274
Fлава 15
пояска, состоящеro из нитей цинновой связки. Нити прикрепляются с одной стороны к
ресничному телу, а с друroй к капсуле хрусталика в области ero экватора. Измене
ние степени натяжения цинновой связки сопровождается изменением кривизны по
верхностей хрусталика, имеющеrо форму двояковыпуклой линзы. При этом меняется
преломляющая сила хрусталика, блаroдаря чему возможна аккомодация rлаза, т. е.
способность четкоro видения различно удаленных предметов.
Стекловидное тело. В эмбриоrенезе различают три стадии формирования CTeK
ловидноrо тела: первичное стекловидное тело, состоящее из мезенхимных клеток,
проникающих в полость rлазноro бокала, которые дифференцируются в rиалоциты;
вторичное стекловидное тело, для KOToporo характерны редукция сосудов и продук
ция прозрачноro вещества нейроrлиальным эпителием внутренней оболочки rлаза;
и сформированное стекловидное тело. Оно представляет собой желеобразное про
зрачное вещество, содержащее около 99% воды и плотный остов из белка витреина и
rиалуроновой кислоты. Стекловидное тело представляет собой основную светопре
ломляющую среду rлаза. Вместе с тем это и CBoero рода амортизатор, способствую
щий созданию внутриrлазноrо давления. Стекловидное тело является метаболичес
ки активным веществом, участвующим в трофических процессах сетчатки. В CTeK
ловидном теле отсутствуют нервы и сосуды. При электронной микроскопии
установлено наличие в нем фибриллярных структур, образующих нитчатый остов,
rиалоцитов, макрофаrов и лимфоцитов.
Сосудистая оболочка. Питание rлаза ocy
ществляет средняя сосудистая оболочка.
В сосудистой оболочке различают надсосуди
стую, сосудистую, сосудисто капиллярную
пластинки, которые состоят из рыхлой волок
нистой соединительной ткани с большим KO
личеством меланоцитов. Сосудистая и cocy
дисто капиллярная пластинки содержат apTe
рии, вены и капиллярные сети. Здесь проис
ходит переход крови из артериальноrо в Be
нозное русло. На rpанице между сосудистой
оболочкой и пиrментным слоем сетчатки Ha
ходится мембрана Бруха, представляющая co
бой тонкий (1 4 мкм) слой коллаrеновых и
эластических волокон. Через нее происходит
диффузное питание сетчатки.
Аккомодационный аппарат rлаза. Pec
ничное тело и радужка выполняют функцию
аккомодации rлаза, блarодаря чему изменяют
ся кривизна хрусталика и величина зрачка и
создаются условия для четкоrо изображения
предмета на сетчатке (рис. 122).
Радужка. Это производное сосудистой
и сетчатой оболочек Радужка располаrается
6
Рис. 122. Схема строения переднеrо отдела
rлазноrо яблока:
1 роrовица; 2 передняя камера rлаза;
3 радужка; 4 задняя камера rлаза; 5
хрусталик; 6 цилиарное тело; 7 склера;
8 стекловидное тело
Система реактивности
275
перед хрусталиком, отделяет переднюю камеру rлаза от задней. Она имеет вид пла
стинки, в центре которой находится круrлое отверстие зрачок Величина ero
постоянно меняется. Радужка выполняет роль диафраrмы, реrулирующей CBeTO
вой поток. В ней различают 5 слоев. Спереди она покрыта однослойным плоским
нейроrлиальным эпителием, переходящим с задней поверхности роrовицы. Под
эпителием располаrается наружный поrраничный слой, состоящией из соедини
тельной ткани с варьирующим числом пиrментоцитов. Средний слой сосудис
тый. Последний прилежит к заднему поrраничному слою (по своему строению прак
тически не отличается от строения переднеrо слоя). Пятый слой это задний
эпителий. Он состоит из двух клеточных дифферонов. Непосредственно к задне
му поrраничному слою прилежат видоизмененные мюллеровы клетки (непиrмен
тированные), а снаружи находятся пиrментоциты продолжение пиrментноrо слоя
сетчатки.
Более rлубокий слой rлиальноrо эпителия в радужке подверrается сложной
перестройке с образованием здесь мионейральной ткани. Из этой ткани построе
ны мышцы, суживающая (циркулярная) и расширяющая зрачок
Реснuчное, Шlи ЦШlиарное, тело. В основе ресничноrо тела лежит ресничная
(или аккомодационная) мышца. Она состоит из rладких мышечных клеток (произ
водное эктомезенхимы HepBHoro rребня). Пучки rладких миоцитов располаrаются в
меридианальном, радиальном и циркулярном направлениях. Сокращение мышцы
вызывает расслабление цинновой связки. При этом хрусталик становится выпук
лым, и ero преломляющая сила увеличивается. От поверхности ресничноrо тела OT
ходят 70 80 отростков. Строма ресничных отростков состоит из рыхлоЙ волокнис
той соединительной ткани, большоrо числа капилляров с фенестрированными
эндотелиоцитами. Отростки покрыты rлиальным эпителием, состоящим из двух кле
точных дифферонов: внутренний образован непиrментированными цилиндричес
кими клетками аналоrами мюллеровых радиальных rлиоцитов (волокон) сетчат
ки (радиальной rлии); наружный слой является продолжением пиrментноrо слоя
сетчатки rлаза. В области ресничноro тела поисходит выработка водянистой влш-и.
Последняя содержит большую часть растворимых веществ плазмы крови, но очень
мало белка. Она rипертонична по отношению к плазме крови, в ней нет фибриноrе
на. В норме образуется и всасывается около 2 мм 3 жидкости В 1 мин. Важное клини
ческое значение имеет сохранение путеЙ оттока жидкости, так как вода несжимаема.
Нарушение оттока приводит к повышению внутриrлазноrо давления (в норме 20
25 мм рт.ст.) и нарушению функции сетчатки. Через трабекулярные пространства
уrла rлаза влаrа всасывается в веноЗ1lЫЙ синус склеры (шлеммов канал), выходит под
конъюнктиву rлазноrо яблока и попадает в водные вены.
Вспомоrательный аппарат rлаза. К нему относят веки, слезные железы, rлаз
ные мышцы. Веки развиваются из кожных складок, образующихся кверху и книзу от
rлазноrо бокала. Они растут по направлению друr к друry и спаиваются своим эпи
телиальным покровом. К 7 MY месяцу эмбриоrенеза спайка исчезает. Задняя повер
хность век покрыта слизистой оболочкой, называемой конъюнктивоЙ. Эпителий
конъюнктивы мноrослойный плоский. В ero составе есть бокаловидные экзокрино
циты, вырабатывающие слизь. В толще век имеются плотная соединительная ткань
276
Тлава 15
(тарзальная пластинка), кольцевая мышца, сальные железы. По краю век располаrа
ются ресницы и: ресничные железы, представляюшие собой видоизмененные пото
вые железы.
Слезный аппарат rлаза образован слезными железами, слезным мешком и слез
HO HOCOBЫM протоком. СлеЗll.ые железы это серозные сложные альвеолярно труб
чатые железы, секрет которых содержит 98% воды, 1,5% хлорида натрия, 0,5% аль
бумина и слизи. В слезной жидкости имеется бактерицидное вешество лизоцим.
Стенки слезноrо мешка и слезно носовоro протока выстланы ДBYX и мноrоряд
ным эпителием. У медиальноrо уrла rлазной шели расположено слезное мясцо.
Слезная жидкость постоянно увлажняет поверхность роrовицы, выполняя зашит
ную функцию.
ТлаЗll.ые мышцы участвуют в осушествлении rлазодвиrательной функции, что
особенно важно при бинокулярном зрении. По развитию, строению и функции они
мало чем отличаются от обычных поперечнополосатых скелетных мышц.
rистофизиолоrия зрения. Фоторецепторный (световоспринимаюший), диопт
рический (светопреломляюший), аккомодационный и вспомоrательный аппараты
тесно взаимосвязаны в функциональном отношении. Блаrодаря соrласованному их
взаимодействию световые лучи проходят через роrовицу, переднюю камеру rлаза,
зрачок, хрусталик, стекловидное тело, сетчатку и возбуждают фоторецепторные клет
ки. На сетчатке возникает изображение предметов. Импульсы с клеток фоторецеп
торов через цепь нейронов сетчатки передаются по аксонам rанrлиозных клеток в
наружное коленчатое тело основной подкорковый центр зрительноrо анализато
ра. Для Hero характерна послойная структура Часть волокон зрительноrо нерва ПОk
ходит к верхним буrоркам четверохолмия. Далее от подкорковых зрительных цeHT
ров афферентные импульсы поступают в зрительную область коры, [де и происходят
окончательный анализ и синтез информации.
РеактU811.0сть. Большую опасность для opraHa зрения представляет действие
лазерноrо излучения и радиоволн СВЧ поля. Наиболее чувствительными к дей
ствию СВЧ поля являются хрусталик и стекловидное тело ввиду недостаточной
термореrуляции этих структур. Лазерное облучение может вызвать сильное по
вреждение opraHa зрения, так как оптически прозрачные среды rлаза (роrовица,
жидкость камер rлаза, хрусталик, стекловидное тело) способны фокусировать CBe
товые лучи. В связи с этим плотность энерrии на сетчатой оболочке увеличивается
в несколько раз. Сетчатка превращается как бы в экран, на котором проецируется
изображение cBeToBoro пятна лазерноrо излучения. В зависимости от величины
падающей энерrии лазерное излучение видимой области может вызывать как Bpe
мен ное ослепление, так и необратимую потерю зрения из за сильноrо ожоrа ceT
чатки и повреждения зрительноrо нерва.
OpzaH обоняния
OpraH обоняния является хеморецептором. Он воспринимает действие моле
кул пахучих веществ. Это самый древний вид рецепции. В составе обонятельноrо
анализатора различают три части: обонятельную область носовой полости (пери
Система реактивности
277
4
ферическая часть), обонятельную луковицу
(промежуточная часть), а также обонятельные
центры в коре больших полушарий rоловною
мозrа.
Развитие. Источником образования всех
частей opraHa обоняния являются нервная
трубка, симметричные локальные утолщения
эктодермы оБО1lЯтелъные плакоды, располо
женные в области передней части rоловы за
родыша и мезенхuм.а. Материал плакоды впя
чивается в подлежащую мезенхиму, формируя
обонятельные мешки, связанные с внешней
средой посредством отверстий (будушие ноз
дри). В составе стенки обонятельноrо мешка
находятся стволовые клетки, которые на 4 M
месяце эмбриоrенеза путем диверrентной
дифференцировки развиваются в нейросен
сорные (обонятельные) клетки, поддержива
ющие и базальные эпителиоциты. Часть кле
f""
ток обонятельноrо мешка идет на построение
обонятельной (боуменовой) железы.
у основания носовой переrородки форми
руется вомероназальпый (якобсонов) opraH,
нейросенсорные клетки KOToporo реаrируют
на феромоны.
Строение. Обонятельная выстилка периферической части обонятельною aHa
лизатора находится на верхней и частично средней раковинах носовой полости
(рис. 123). Общая площадь ее около 10 см 2 . Обонятельная область имеет эпители
оподобиое строение. От подлежашей соединительной ткани рецепторная часть обо
нятельноrо анализатора отrраничена базальной мембраной. Обонятельные нейро
сенсорные клетки имеют веретенообразную форму с двумя отростками. По форме
они делятся на палочковидные и колбочковидные. Общее число обонятельных кле
ток у человека достиrает 400 млн при значительном преобладании количества па
лочковидных клеток
Периферический отросток обонятельной нейросенсорной клетки длиной 15
20 мкм имеет на конце утолшение, называемое обонятельной булавой. На oKpyr
лой вершине обонятельных булав имеются обонятельные волоски ahme1l1-Lbl в
количестве 10 12. Длина их достиrает 2 3 мкм. Антенны имеют ультраструктуру,
характерную для ресничек, т. е. содержат 9 периферических и 2 центральные спа
ренные протофибриллы, отходящих от типичных базальных телец. Антенны co
вершают непрерывные автоматические движения маятникообразноrо типа. Bep
шина антенн перемещается по сложной траектории, блаrодаря чему увеличивается
возможность их контакта с молекулами пахучих веществ. Антенны поrружены при
этом в жидкую среду, представляющую собой секрет трубчато альвеолярных
_.-:. .X'..-
'
Рис. 123. Схема строения обонятельной
выстилки:
1 нейросенсорная клетка с ресничками;
2 опорные клетки; 3 базальные клетки;
4 обонятельные реснички; 5 обоня
тельная слизь
278
Fлава 15
обонятельных желез (боуменовых). Для них характерен мерокринный тип ceKpe
ции. Секрет этих желез увлажняет поверхность обонятельной выстилки.
Центральный отросток обонятельной нейросенсорной клетки аксон, направ
ляется в промежуточную часть opraHa обоняния обонятельную луковицу и YCTa
навливает там синаптическую связь в виде клубочка с митральными нейронами. В
обонятельной луковице различают следующие слои: 1) слой обонятельных клу
бочков, 2) наружный зернистый слой, З) молекулярный слой, 4) слой митральных
клеток, 5) внутренний зернистый слой, 6) слой центробежных волокон.
Центральный отдел opraHa обоняния локализуется в тnnокампе и в rиппокампо
вой извилине коры большоrо мозrа, куда направляются аксоны митральных клеток и
формируют синаптические связи с нейронами.
Таким образом, opraH обоняния (обонятельная область носовой полости и обо
нятельная луковица), подобно opraHY зрения, имеет слоистое расположение ней
ронов, что характерно для экранных нервных центров.
Поддерживающие эпителиоциты обонятельной области высокопризмати
ческие клетки с микроворсинками, располаrаются в виде мноrорядноrо эпители
альноro пласта, обеспечивая пространственную орrанизацию нейросенсорных кле
ток. Некоторые из этих клеток являются секреторными, а также обладают
фаrоцитарноЙ способностью. Базальные эпителиоциты кубической формы явля
ются малодифференцированными (камбиальными) и служат источником образо
вания новых клеток обонятельной выстилки. Цикл физиолоrической реrенерации
нейросенсорных клеток оценивается в ЗО З5 суток
Боуменовы железы состоят из секреторных и миоэпителиальных клеток CeK
рет желез растворяет пахучие вещества, которые взаимодействуют с рецепторно
трансдукторной системой ресничек нейросенсорной клетки. Это вызывает изме
нение мембранноrо потенциала, который передается через цепь нейронов в
центральную часть opraHa обоняния.
OpzaH слуха u равновесия
OpraH слуха и равновесия осуществляет восприятие звуков, линейных, уrло
вых ускорений и земноrо притяжения. Состоит из трех частей: наружноrо уха (уш
ная раковина, наружный слуховой проход и барабанная перепонка), среднеrо уха
(полость, rде находятся слуховые косточки, слуховая труба) и BHYTpeHHero уха (KO
стный и перепончатый лабиринты).
Ушная раковина образована эластическим хрящом, переходящим в хрящ Ha
ружноrо слуховоrо прохода, покрыты м кожей. В коже с волосами, сальными желе
зами присутствуют особые видоизмененные потовые железы (церуминозные),
выделяющие ушную серу, которая обладает бактерицидными свойствами. Бара
банная перепонка толщиной 0,1 мм состоит из соединительнотканной пластинки,
покрытой снаружи мнorослойным плоским эпителием, а изнутри слизистой обо
лочкой, в составе котороЙ эпителий представлен одним пластом кубических или
плоских эпителиоцитов.
Среднее ухо выстлано слизистой оболочкой, полость ero соединяется с внеш
Система реактивности
279
ней средой при помощи слуховой (евстахиевой) трубы, имеющей однослойное эпи
телиальное покрытие. Кубический эпителий трубы при хронических воспалитель
ных проuессах может трансформироваться в мноrослойный плоскиЙ. В слизистой
оболочке и костных стенках среднеrо уха проходят ветви (лицевоrо, языкorлоточ
Horo, блуждающеro) нервов. С помощью слуховой трубы выравнивается давление
в среднем ухе, что улучшает звукопроведение.
Внутреннее ухо располаrается в каменистой части височной кости, включает
костный и перепончатый лабиринты, формы которых повторяют друr друrа. Пере
пончатыЙ лабиринт это замкнутая система трубочек, мешочков, заполненная
жидкостью эндолимфой. Между перепончатым и костным лабиринтами Haxo
дится перилимфатическое пространство, заполненное перилимфой. Перепончатый
лабиринт делится на слуховую (улитка) и вестибулярную (opraH равновесия) час
ти. Последняя состоит из трех полукружных каналов и двух отолитовых opraHoB
эллиптическоrо и сферическоrо мешочков.
Развитие. ИСТОЧНИКО 1 развития перепончатой части BHYTpeHHero уха являют
ся слуховые плакоды парные утолщения эктодермы на уровне развивающеrося зад
Hero мозrовоrо пузыря. На 3 й неделе эмбриоrенеза происходит впячивание матери
ала слуховых плакод в подлежащую мезенхиму с образованием слуховых ямок
В дальнейшем материал слуховых плакод полностью поrpужается во внутреннюю
среду и отшнуровывается от эктодермы. Возникают слуховые пузырьки. Каждый
слуховой пузырек имеет стенку из мноroрядноro эпителия и полость, заполненную
эндолимфой. В дальнейшем пузырек перетяrивается на две части: вестибулярную
(маточка с полукружными каналами) и мешочек с закладкой улитковоro канала. По
зднее улитка разрастается и отделяется от мешочка. Внутренняя выстилка всех этих
образований состоит из rлиальноrо эпителия. На характер дифференцировки кле
ток оказывает индуктивное влияние контакт эпителия с эмбриональным слуховым
нервным rанrлием, который подразделяется на rанrлий преддверия (вестибулярный)
и rанrлий улитки (слуховой rанrлий). В определенных участках маточки, мешочка,
ампул полукружных каналов, а также в улитке образуются рецепторные зоны, co
держащие чувствительные клетки, специализированные на восприятие звуковых,
rpавитационных и вибрационных раздражителей. Происходит это на 3 M месяце эм
бриоrенеза. Клеточный состав, структура и функция эпителия неодинаковы в раз
личных участках улитковоrо канала. [истоrенез эпителия вестибулярноro аппарата
характеризуется образованием желатинообразноrо тела купола rpебешков и oco
бых кристаллов отолитов. Параллельно с rистоrенезом эпителия перепончатоro
лабиринта изменяется окружающая лабиринт мезенхима, в результате редукции KO
торой образуются перилимфатические полости.
Строение opzaHa равновесия и zравитации. Вестибулярная часть перепон
чатоrо лабиринта состоит из мешочка, маточки и трех полукружных каналов. На
месте соединения их с маточкой имеются расширения, называемые ампулами. В
ампулах рецепторные участки имеют вид rpебешков (рис. 124). В мешочке и Ma
точке они представлены слуховыми пятнами. В этих участках эпителий поляризо
ван. Вся остальная часть вестибулярноrо перепончатоrо лабиринта выстлана плос
ким однослойным эпителием. Эпителий пятен состоит из сенсоэпителиальных
280
Fлава 15
клеток и расположенных меж
ду ними опорных клеток. Над
поверхностью эпителия имеет
ся отолитовая мембрана, coдep
жащая кристаллы уrлекислоrо
кальция (отолиты, или OTOKO
нии). Отолитовая мембранаяв
ляется продуктом секреции
опорных клеток. Мембрана как
бы плавает в эндолимфе. При 2
движении rоловы она может
смещаться. Соединенные с MeM
браной киноцилии сенсоэпите
лиальных клеток смещаются в Рис, 124. Схема строения рецепторных зон вестибулярноrо
сторону стереоцилий или от аппарата:
А rребешок; Б пятно; 1 волосковые сенсорные клет
них, что вызывает COOTBeT ки; 2 опорные клетки; 3 купол; 4 отолитовая мембра
ственно возбуждение или TOp на; 5 соединительная ткань
можение соответствующих нейронов, иннервирующих рецепторные клеки.
Различают два типа сенсоэпителиальных клеток. Клетки I ro типа имеют ши
рокое основание и кувшинообразную форму. На апикальной поверхности их име
ется кутикула и БО 80 неподвижных волосков стереоцилий, а также полярно к
пучку стереоцилий располаrается один подвижный волосок киноцилия. К OCHO
ванию волосковых клеток подходят чувствительные нервные окончания, образу
ющие футляр в виде чаши. Клетки II ro типа имеют цилиндрическую форму и Me
нее обширный и даже точечный контакт с нервными окончаниями.
rистолоrически пятна в мешочке и маточке мало чем отличаются друr от дpy
ra. Тем не менее они имеют различное функциональное значение. Пятно мешочка
воспринимает вибрационные колебания. Пятно маточки место восприятия раз
дражений, связанных с вертикальными изменениями положения тела, т. е. измене
ниями в положении тела по отношению к rравитационному полю Земли. Таким
образом, пятно маточки это рецептор rравитации.
Fребеш'Кu в ампулах полукружных каналов устроены принципиально так же,
как и пятна. В их составе имеются сенсоэпителиальные клетки и поддерживаю
щие эпителиоциты. Вместо отолитовой мембраны на поверхности rребешка обра
зуетсяжелатинообразное(студневидное) тело купол. Купол продуктrолок
ринной секреции поддерживающих эпителиоцитов. При движении rоловы и
ускоренном уrловом вращении тела купол с rребешком отклоняется, тоrда как эн
долимфа в полукружных каналах в силу инерции сохраняет некоторое время He
подвижность. Все это смещает купол относительно rребешка. Смещаясь, купол
тянет киноцилию или в сторону стереоцилий, или в противоположную от них CTO
рону. Это изменяет импульсную активность нейронов, ин нервирующих сенсоэпи
телиальные клетки. При этом происходит передача импульсов с последующей реф
лекторной реакцией соответствующих мышц. У человека утрата вестибулярной
функции вызывает неустойчивость позы.
2
Система реактивности
281
Таким образом, rребешки это рецепторы ускорения при поступательном ro
ризонталыюм движении и уrловоrо ускорения при вращении тела. РецепторныЙ
эпителий отделен от подлежащей соединительноЙ ткани базальной мембраной.
В соединительной ткани MHoro кровеносных капилляров и нервных волокон.
OpraH слуха. Периферическая часть слуховоrо анализатора находится в улит
ковом канале перепончатоrо лабиринта BHYTpeHHero уха. Спиральный (или корти
ев) opraH расположен по всей длине улитковorо канала. Он осуществляет воспри
яти е звуковых колебаний.
Строение. На поперечном срезе улитковый канал имеет форму треуrольника,
вершиной обращенноrо к центральному костному стержню улитки. Улитковый
канал, имеющий длину около 3,5 см, по спирали делает 2,5 завитка, слепо закан
чиваясь на верхушке улитки. Канал заполнен эндолимфой. Снаружи от улитково
ro канала находятся перилимфатические пространства, называемые лестниuами:
сверху вестибулярная (преддверная), снизу барабанная (тимпанальная) Bec
тибулярная лестница отделяется от полости среднеrо уха овальным окном, в KOTO
ром располаrается основание стремечка. Барабанная лестниuа отделяется от поло
сти среднеrо уха посредством мембраны круrлоrо окна.
Стенка улитковою канала, обращенная к вестибулярной лестнице, называет
ся вестибулярной (или реЙснеровой) мембраной. Она представлена соединитель
нотканноЙ тонкофибриллярноЙ пластинкой, покрытой со стороны улитковоrо Ka
нала однослойным плоским rлиальным эпителием, а со стороны вестибулярной
перилимфатической лестницы эндотелием.
Боковая стенка улитковоrо канала выстлана так называемой сосудистой полос
кой, эпителий которой участвует в продукции эндолимфы. Эпителий представляет
собой мноrорядный rлиальныЙ эпителиЙ, среди клеток KOToporo различают плос
кие светлые и высокие отростчатые призматические клетки. Последние в своеЙ uи
топлазме содержат мноrочисленные митохондрии и выrлядят темными клетками.
Среди клеточных дифферонов сосудистоЙ полоски обнаружены нейроэндокрино
циты АРUD серии, вырабатывающие серотонин, мелатонин, адреналин и др., KOTO
рые участвуют в реryляции объема эндолимфы. Здесь же идут мноrочисленные KpO
веносные капилляры. С сосудистой полоской связывают насыщение эндолимфы
кислородом, создание определенноrо ионноrо состава и объема эндолимфы и др.,
что необходимо для нормальноЙ функции opraHa.
Стенка улитковоrо канала, примыкающая к барабанной лестнице, имеет очень
сложное строение, так как на неЙ расположен спиральныЙ opraH рецептор звука
(рис. 125). Основу этоЙ стенки улитковою канала составляет базилярная мембра
на. Ее поверхность, обращенная в барабанную лестницу, покрыта тонким слоем
эндотелия. Базилярная мембрана это соединительнотканная пластинка. В виде
спирали она тянется вдоль всею канала улитки. Структурными элементами бази
лярноЙ мембраны являются тонкие коллаrеновые волокна слуховые струны.
Общее число их достиrает 24 000. Они располаrаются между спиральноЙ костной
пластинкой, отходящей от осевою стержня улитки, и спиральной связкой, распо
ложенноЙ на наружной стенке улитки. Длина слуховых струн неодинакова: у oc
нования улитки они короче (100 мкм), а на вершине улитки в 5 раз длиннее
282
rлава 15
(500 мкм). Диаметр струн
1 2 !\1КМ. Коллаrеновые BO
локна ( струны) окружены ro
моrенным основным проме
жуточным веществом. Бази
лярная мембрана со стороны
улитковоrо канала покрыта
поrраничноЙ базальноЙ MeM
браноЙ, на которой лежит
эпителиальноrлиальныЙ спи
ральныЙ opraH.
СпuрШlЪ1l.ЫЙ орю1l. образо
ван рецепторными волоско
вы ш сенсорны ш эпителио
цитами и опорными эпители
оцитами, имеющими высокую
степень структурно функцио
нальноЙ специализации. Pe
цепторные клетки подразде
ляются на внутренние и наружные волосковые сенсорные эпителиоциты. BHYT
ренние сенсорные эпителиоциты имеют кувшинообразную форму. Их ядра лежат
в расширенноЙ базальной части клетки. На поверхности суженной апикальной ча
сти клетки имеются кутикула и проходящие через нее 30 60 коротких микровыро
стов (слуховых волосков), расположенных в 3 4 ряда. Это стереоцилии. В отличие
от кпноцилий они неподвижны. Общее число внутренних сенсорных эпителиоци
тов около 3500. Располаrаются они в один ряд вдоль Bcero спиральноrо opraHa.
Их поддерживают внутренние фаланrовые эпителиоциты.
Наружные сенсорные эпителиоциты имеют цилиндрическую форму. На апикаль
ной поверхности этих клеток имеется кутикула, через которую проходят слуховые
волоски стереоцилии. Стереоцилии в количестве 60 70 образуют щеточку, распо
лаrаясь несколькими рядами. На поверхности волосковых сенсорных клеток среди
стереоцилий киноцилии отсутствуют, но от них сохраняются базальные тельца. CBO
ими вершинами стереоцилии прикрепляются к внутренней поверхности покровной
(желатинозноЙ) мембраны, нависающей над спиральным opraHoM. Окруrлым OCHO
ванием наружные волосковые эпителиоциты опираются на поддерживающие эпи
телиоциты, контактируя с чувствительными нервными волокнами нейронов спи
ральноrо rанrлия. Наружные волосковые клетки лежат в виде трех параллельных
рядов по всеЙ длине спиральноrо opraHa. Общее их число 12 20 тыс. Наружные ceH
сорные эпителиоциты воспринимают звуки большой интенсивности, внутренние
MorYT воспринимать и слабые звуки. Сенсорные эпителиоциты, находящиеся на Bep
шине улитки, воспринимают низкие звуки, клетки у основания высокие.
К базальноЙ поверхности сенсорных эпителиоцитов подходят афферентные
волокна преимущественно спиральноro rанrлия и эфферентные волокна оливо
кохлеарных путей, которые формируют здесь синаптические контакты. BHYTpeH
11
Рис. 125. Схема строения улитковоrо канала:
1 покровная мембрана; 2 туннель; З внутренняя волос
ковая клетка; 4 наружные волосковые клетки; 5 клетки стол
бы; б наружные фаланrовые клетки; 7 базилярная пластин
ка; 8 вестибулярная мембрана; 9 вестибулярная лестница;
1 О улитковый канал; 11 барабанная лестница; 12 костный
спиральный выступ; 13 спиральная связка; 14 лимб
Система реактивности
283
ние сенсорные эпителиоциты имеют преимущественно афферентную иннервацию,
а наружные преимущественно эфферентную иннервацию. Роль последней зак
лючается в торможении и модуляции HepBHoro импульса.
Опорные эпителиоциты спиральноrо opraHa отличаются выраженной дивер
rентной дифференцировкой. Различают несколько разновидностей этих клеток:
внутренние фаланrовые эпителиоциты, внутренние и наружные столбовые эпите
лиоциты (клетки столбы), наружные фаланrовые эпителиоциты (клетки Дейтер
са), наружные поrраничные эпителиоциты (клетки [ензена), наружные поддержи
вающие эпителиоциты (клетки Клаудиуса). Название "фаланrовые" для
внутренних и наружных поддерживающих эпителиоцитов дано в связи с тем, что
эти клетки имеют тонкие пальцевидные отростки. Посредством этих отростков
волосковые сенсорные эпителиоциты отделяются друr от друrа. В отличие от BO
лосковых эпителиоцитов все опорные эпителиоциты непосредственно прилежат к
базальной мембране, rде обнаруживаются мноrочисленные полудесмосомы. Через
всю цитоплазму фаланrовых эпителиоцитов проходит пучок фибрилл, начинаю
щихся от полудесмосом и образующих на апикальной поверхности расширенную
плоскую пластинку. Между этой пластинкой и апикальной частью волосковых
клеток имеются плотные контакты типа замыкающей полоски. Пучки фибрилляр
Horo вещества проходят и через клетки столбы.
fистофизиолоmя слуха. Звуки определенной частоты, воспринятые наружным
ухом и переданные через слуховые косточки и овальное окно перилимфе барабан
ной лестницы, вызывают колебания базилярной мембраны. В ответ на частоту звука
возникают колебания определенных участков спиральноro opraHa. Они восприни
маются волосковыми сенсорными эпителиоцитами блаroдаря тому, что их волоски
смещаются относительно покровной мембраны, в которую их кончики поrpужены.
Это приводит к возбуждению сенсорных эпителиоцитов и к изменению импульса
ции в афферентных нервных веточках, оплетающих основания волосковых ceHCOp
ных эпителиоцитов. От афферентных окончаний на волосковых клетках нервные
импульсы передаются нейронам спиральноro rанrлия. Один нейрон может получать
импульсы от мноrих волосковых сенсорных эпителиоцитов. Затем импульсы пере
даются по аксонам нейронов спиральноrо rанrлия, которые формируют улитковый
нерв, в слуховые ядра ствола мозrа и далее в слуховую область коры большоro мозrа
(верхнюю височную извилину).
Реактивность тканей opzaHa слуха и равновесия. Внутренние и наружные
сенсорные волосковые клетки отличаются неодинаковой чувствительностью к
повреждающим воздействиям. Так, при введении больших доз стрептомицина по
вреждаются внутренние, а при введении больших доз хинина наружные волос
ковы е клетки. Кратковременное и длительное воздействие звуков большой интен
сивности может приводить к профессиональной туrоухости. Сочетание шума и
вибрации, взрывная волна, сотрясение при водят к кровоизлияниям в барабанную
полость, полукружные каналы, в область пятна и rребешка. При этом происходят
деrенеративные изменения отолитовоrо аппарата и ампульных rребешков, дeCT
рукция сенсорных и поддерживающих эпителиоцитов спиральноrо opraHa, что
проявляется в нарушении слуха и вестибулярных расстройствах.
284
Тлава 15
Opzau вкуса
Периферическая часть BKycoBoro
анализатора выполняет функцию xeMO
рецептора. Она представлена совокуп С
ностью вкусовых почек (или луковиц),
расположенных в мноrослойном эпите
лии сосочков языка. Общее число BKY
совых почек варьирует от 2 до 9 тысяч.
Развитие. Существует два взrляда
на rистоrенез вкусовых почек Соrласно
первому из них, клетки вкусовых почек
развиваются за счет трансформации ба
зальною слоя эпителия языка под воз
действием подрастающих вкусовых He
рвных волокон. По второму взrляду,
вкусовые почки формируются из элемен
тов эмбриональной нервной rлии ней
ролеммоцитов терминальных окончаний
нервных волокон язычноrо, языкоrло
точноrо и блуждающеrо нервов.
Строение. Вкусовая почка имеет
эллипсоидную форму. Клетки в ней ле
жат наподобие ломтиков апельсина. Рис. 126. Схема строения вкусовой почки:
Различают сенсоэпителиальные клет 1 и 2 поддерживающие эпителиоциты (темные и
ки, темные поддерживающие и базаль светлые); з сенсоэпителиальная клетка; 4 ба
зальный эпителиоцит; 5 вкусовая пора; 6 He
ные (камбиальные) эпителиоциты рвные волокна; 7 синаптический контакт; 8 ceK
(рис. 126). Почка содержит ЗО 60 эпи рет
телиоцитов. Вершина почки сообщается с поверхностью эпителия языка при по
мощи отверстия вкусовой поры, которое ведет внебольшое уrлубление, называе
мое вкусовой ямкой. Сенсорные эпителиоциты имеют веретенообразную форму со
светлой или темной цитоплазмой. В цитоплазме в околоядерной и базальной час
тях находятся пузырьки с электронно плотным содержимым. На апикальном KOH
це клетки имеют до 40 50 тонких микроворсинок вкусовых штифтиков. Длина
микроворсинок З 5 мкм, а диаметр 0,2 мкм. Посредством этих микроворсинок
вкусовые клетки воспринимают раздражение веществ. Молекулы этих веществ aд
сорбируются на мембранах микроворсинок в активных центрах. Они избиратель
но реаrируют на действие веществ, обладающих rорьким, кислым, сладким и соле
ным вкусом. В передней части языка находятся сладкочувствительные клетки, а в
задней rорькочувствительные. В базальной части клеток формируются синапсы
с афферентными волокнами (1 6 на одну клетку).
Возбуждение вкусовых клеток передается на дендриты чувствительных ней
ронов, тела которых лежат в каменистом и коленчатом rанrлиях. Чувствительные
нервные волокна, заходя внутрь почки, контактируют с боковыми поверхностями
Система реактивности
285
сенсоэпителиальных клеток. В своеобразных синаптических контактах определя
ется ацетилхолинэстераза, принимающая участие в проведении нервных импуль
сов. Далее импульсы передаются на нейроны BKycoBoro ядра продолrоватоrо l'юз
[а, нейроны таламуса и, наконец, на нейроны вкусовой области коры мозrа.
Наряду с вкусовыми клетками, вкусовые почки содержат базальные и поддер
живающие эпителиоциты. Последние это, как правило, разнообразной формы
клетки со светлой или темной цитоплазмой. Клетки выполняют секреторную фун
кцию, выделяя секрет во вкусовую пору, и не имеют синаптических связей с He
рвными волокна:\1И. На апикальной поверхности поддерживающих клеток имеют
ся микроворсинки. Базальные эпителиоциты имеют небольшие размеры,
цитоплазма и орrанеллы развиты слабо. Они не достиrают вкусовой поры и не име
ют синаптических связей с нервными волокнами. Предполаrается их участие в фи
зиолоrической реrенерации сенсоэпителиальных и опорных клеток. КлеточныЙ
состав вкусовой почки обновляется каждые 10 12 суток. С возрастом количество
вкусовых клеток уменьшается.
Opla11 осязания
OpraH осязания воспринимает тактильные раздражения, возникающие в резуль
тате прикосновения или давления предметов на кожу. Афферентная иннервация кожи
осуществляется нервными волокнами, идущими от чувствительных неЙронов спип
номозrовых rанrлиев. Дендриты чувствительных неЙронов формируют осязатель
ные рец пторы, которые находятся в коже. Кожные рецепторы воспринимают не
только механические раздражения, но и температурные, химические, электричес
кие. В коже, помимо тактильных и температурных рецепторов, имеются также боле
вые рецепторы. Физиолоrи полаrают, что рецептор это не только и не просто BOC
принимающий аппарат, а творческая часть афферентною отдела рефл кторноЙ дуrи
Различают свободные и несвободные нервные окончания. Первые состоят
только из oceBoro цилиндра дендрита чувствительноrо нейрона. Вторые вклю
чают и клетки rлии нейролеммоциты (шванновские). Если у последних появля
ется соединительнотканная капсула, то такие нервные окончания называются ин
капсу лированными.
Болевые раздражения (ноцицепция) воспринимают свободные чувствительные
нервные окончания, пронизывающие толщу эпидермиса (рис. 127). Нервные вuлuк
на распространяются между эпителиоцитами, при этом утрачивают миелиновую
оболочку. Прикосновения, даже самые нежные, в первую очередь воспринимаются
чувствительными нервными окончаниями, оплетающими корни волос. В эпидер
мисе имеются клетки Меркеля. Они принимают участие в восприятии прикоснове
ний, поскольку тесно связаны с сетевидными концевыми разветвлениями чувстви
тельных нервов. Кроме Toro, клетки Меркеля синтезируют специфические для He
рвных клеток маркеры (нейрофиламенты, неЙрональные клеточные адrезионные MO
лекулы и др.). На основании присутствия в цитоплазме клеток нейропептидов они
относятся к диффузной эндокринной системе. Мет энкефалин, вырабатываемыЙ
клетками Меркеля, стимулирует иммунные реакции орrанизма.
286
Fлава 15
3
Типичными инкапсулиро
ванными орrанами осязания
(механорецепторами) являются
осязательные тельца МеЙС1iера,
расположенные в сосочковом
слое кожи пальцев, rуб, век, по
ловых opraHoB. Они имеют диа
метр около 100 мкм, снаружи OK
ружены соединительнотканной
капсулой. В составе этих телец
нейроrлиальные клетки образу
ют внутреннюю колбу BOKpyr
концевоrо утолщения чувстви
тельноrо HepBHoro волокна, KO
торое располаrается параллель
но поверхности кожи.
Специальным рецептором
Рис. 127. Свободные нервные окончания в эпителии poro
давления (барорецептором) яв вицы:
ляются пластинчатые тельца 1 эпителий; 2 соединительная ткань; 3 нервные
Фатер ПаЧU1iU. Они лежат rлу окончания
боко В подкожном слое в области пальцев, наружных половых opraHoB, стенки MO
чевоrо пузыря, капсуле внутренних opraHoB и др. Диаметр тельца достиrает 1 мм.
Для них характерно наличие мноrослойной пластинчатой соединительнотканной
капсулы (наружной колбы) и комплекса нейроrлиальных клеток (внутренней KOk
бы). В центре внутренней колбы проходит концевая веточка чувствительноrо He
pBHOro волокна.
1ельце РУФФU1iU лежит в rлубоких слоях кожи, например, подошвы стопы. Ди
аметр до 1 мм. Афферентное волокно образует наподобие кустика из немиели
низироваНIIЫХ веточек, которые оканчиваются уплощенными терминалями. Co
единитеЛЫIOтканная капсула хорошо выражена. Реаrируют на смещение кожи и
давление.
Концевые '/(олбы Краузе обнаруживаются в конъюнктиве, языке, наружных
половых opraHax. Диаметр до 150 мкм. Имеют тонкую капсулу, мноrочислен
ные разветвления афферентноrо окончания располаrаются в виде колбы. Это Me
ханорецептор.
При действии на opraHbI осязания и давления в механорецепторах кожи энер
шя раздражителя трансформируется в нервное возбуждение, которое по цепи ней
ронов передается от периферической части кожнorо анализатора в ero корковую
часть в заднюю центральную извилину. В верхней части ее проецируется чув
ствительность кожи Hor, в средней рук и туловища, а в нижней кожи rоловы.
.........
;
".,<;;;::;.;'
ОпОрlю двиzaтеЛЪ1lая система
287
rЛАВА 16. опоРно двиrАТЕЛЬНАЯ СИСТЕМА
Опорно двиrательную систему человека образуют костный скелет и скелет
ные мышцы. Блаrодаря способности к сокращению мышцы приводят в движение
кости скелета, в результате чеrо тело человека или ero части MoryT перемещаться в
пространстве и выполнять ту или иную работу. Сокращение мышц происходит под
влиянием нервных импульсов, поступающих из центральной нервной системы. CKe
летные мышцы являются одним из rлавных эффекторных аппаратов нервной сис
темы, что убедительно показано физиолоrами. И.М. Сеченов писал: "Все беско
нечное разнообразие внешних проявлений мозrовой деятельности сводится
окончательно к одному лишь явлению мышечному движению". Кроме KOCTHoro
скелета и мускулатуры, к системе opraHoB движения и опоры относятся суставы,
хрящи, сухожилия, связки, фасции.
Кости
rлавная функция костей обеспечение твердой опоры человеческоrо тела.
Наряду с этой механической функцией кости принимают также участие в мине
ральном обмене, поскольку в них содержится основной запас кальция, фосфора и
др. минеральных веществ. В костях находится красный костный мозr основной
opraH кроветворения. Кость opraH, построенный преимущественно из костной
ткани. В состав каждой кости входят также еше ряд тканей, находящихся в опреде
ленных соотношениях.
Для примера рассмотрим строение трубчатой кости, а именно бедренной кости
человека. Она состоит из пластинчатой костной ткани, надкостницы (периоста), эн
доста, суставных хрящей, синовиальноrо эндотелия, сосудов и нервов. Полость ди
афиза, а также пространства ryбчатоrо вещества эпифизов заполнены костным МОЗ
rOM. Компактное вещество кости представлено пластинчатой костной тканью.
Снаружи диафиза кости имеется надкостница (периост), далее идут наружные OK
ружающие (rенеральные) пластинки. Изнутри со стороны костномозrовой полости
располаrаются внутренние окружающие (rенеральные) пластинки, покрытые эндо
стом. Основную же часть трубчатой кости, расположенную между наружными и
внутренними окружающими пластинками, составляют остеоны и заполняющие про
межутки между ними вставочные пластинки (остаточные остеоны).
Остеоп это трехмерная цилиндрическая система концентрически располо
женных костных пластинок и остеоцитов, окружающих центральный канал OCTeo
на (рис. 128). В костных пластинках оссеиновые фибриллы плотно и параллельно
прилежат друr к друrу. Костно пластинчатые цилиндры как бы вставлены один
288
Fлава 16
в друrоЙ. В соседних
концентрических KOCT
ных пластинках оссеи
новые фибриллы идут
под друrим уrлом. Бла
rодаря этому достиrа
ется исключительная
прочность остеонов.
Сложная конструкция
остеонов образуется в
процессе rистоrенеза
костноЙ ткани и ее по
стоянной перестройки.
Часть остеонов разру
шается. Остатки их co
.... ' ," .,. . C " ... . " t .
,.... " "'" , , '. "" _......;,.r... , .;(: ".J "', ..... 1".. ' .
- ........ "'- .... .......,. . .. --- ...
..,. ,:"!'o . 1.;. , , tt< , .. , ..:",.,.." ....... , '.. . , ./'.'.
... ...,. '-.J.. "' ""'-"""." -_ ,...._ .с.. ,- А .. '" .. .. ...-* _ .,' '--"'
.....""-::..:. :...............,..... ....-: :"'.};. f , 'r. '... ..4..._ """' , . -
'o ," r ,:: ,'< ,' 'I'r....._,;. ,., о. " ,.,-#JI' \. .,
.. v ...... ."", ......""...". _ \ -\ ...,;...... .. 4 .."
#':" . ........-... ...... .. ". .-.: ..,.- . .j
7=f , ;&'.J/I' . "' .... '" ...,.,... '.:\..
' \j p..... &!.: ... .;; .]!E ..... '..-:.... ' ..::: ...,;: - 'i;... ' , i ,.
,. .,. r::, ....... .. ""'""'!"....." " '" у.." ". ...-:_ л......"
. ''''';' ",,,,,:,," :'(;.,. :,, ...,.;< _" ..... ,', . . A """, '". .
.. :-...... "'" . -'-.... . . '..,. ,,"' . ""'tI':.' , .' 1fН '
..... .;/"-'" "," ...... __,- . . .' 1W'..:.o! .. .., .
. --:.:.. '... "" \ ..... 'Н. "', .. "4< ... 2'" ..... ... ,'"
" og\ " ... M"""' , ... . " ' '< , '
"... .... , ... ,, .l: ... т." . .> /;;.. :. '
t- ..... , t:-.'..ф"f; ...: < 1 '.. " --; , q- "'P" -' "'" "'.
.....,..-...;.'"'40.-. .. ......... .... .... ..."....,... ..K ,.. :""O'- 1.4I
. ... . !Ii " ..........,........ ..?..... "- ,. ',J .... 'it '
-- 'w . .....'"... .1--..0« ... .. ..... ...
.!-...".... ! . "": """.., , W.J..f!f , ..-.:
........- .. ; :;:::., '* . ' ;.... 'i; 4 ;'
";<' "' , , ...;;... .;"'.. ..... ,оД ,:f:;<; , . .. . , ". ;; .rf
;;:'" , ''' j) : : ' .. ...{':'-:. . ..., .......'):a'l:# ...,....".
!,:-: :;.,, .. .:/:- .', < ..,.. -:-;<" r .I!
:?';': .. ' , .. ' , ;. "/, 'Н',,, ,,: , ..! "!t , r J ,
..... /..' i'7I. .-М;': I "" ., . . " ......; !f)l ..,.::,
.. , ';\f" ""' :- - ,!,'.. ":' , . :.'.. ," ...
::r;!! \' ; :;;v: ;J .,,""" :}.....,..:;:
"!.. ..;I'( .;.$.r,<,p ,. " <, " ',"t ._ .,<';1
. . \: ;:;' .'f":,' .. I .. .. ">.-. . # '... ...... ;
. ,', :.;...'\. , ,,,,'f:, ,J[ , ""' , . "AJt!' "":'. " .. ... ,. y """М_ , .;"Ч
';;""" '"2 :: . ,"'" "'J4'!j '{:').sM' . ' .' .' '. . ..A'
/.: 1i'fl!. . , . 1"o-. ,,: "i!f!1g;fi'i. 'f
.....""' .'j . ,..,...i.'. ....ч .......< .>.v::-'.w......, , ......
.j... ' 'J ..j.'....::8fi .......... ;;; + ---: 2 .:::9'. .'
ставляют вставочные
Рис, 128, Строение остеона по данным сканирующей электронной мик
пластинки. Наряду с роскопии:
этим возникают новые 1 центральный канал остеона; 2 лакуны остеоцитов; 3 канальцы
остеоны. Источником отростков остеоцитов
их служат камбиальные клетки, расположенные в рыхлой соединительной ткани BOK
pyr сосудов в каналах остеонов. Большую роль в процессе перестроЙки и особенно в
механизмах рецепции физических наrрузок отводят пьезоэлектрическим эффектам.
При crибании костных пластинок на их поверхности возникают + и заряды. Пола
rают, что положительный заряд вызывает дифференцировку остеокластов, а отри
цательный заряд остеобластов. Таким образом, в костной ткани rармонично про
текают процессы созидания и разрушения, блаroдаря этому достиrаются механи
ческая прочность и физиолоrическая реrенерация кости.
Рост трубчатых костей в длину заканчивается обычно к 20 rодам жизни. До
этоrо времени функционирует метазпuфuзар1l.ая пластu1l.1Ш роста, расположенная
между эпифизом и диафизом. В метаэпифизарной пластинке различают поrранич
ную зону, расположенную ближе к костной ткани эпифиза. Эту зону называют также
зоной покоящеrося хряща. Далее выделяют зону пролиферирующеrо молодоrо хря
ща, или зону столбчатых клеток Здесь образуются новые хондробласты для заме
ны тех хрящевых клеток. которые отмирают у диафизарной поверхности пластин
ки. Следующая зона в метаэпифизарной пластинке называется зоной созревающеrо
хряща, или зоной пузырчатых клеток Она характеризуется разрушением XOHДpO
цитов с последующим энхондральным окостенением. Выделяют еще зону оБызе
ствления хряща. Она непосредственно rраничит с костной тканью диафиза. В нее
проникают капилляры и oCTeoreHHbIe клетки. Последние превращаются в остеоб
ласты, образующие на диафизарной стороне метаэпифизарной пластинки костные
перекладины.
Таким образом. интерстициальный рост хряща на эпифизарной стороне MeTa
эпифизарной пластинки отодвиrает эпифиз от диафиза, но метаэпифизарная пла
стинка не увеличивается в толщине, так как со стороны диафиза она постоянно
ОпОр1-l0 двиzaтеЛЪ1-lая система
289
подверrается резорбции и замещается костной тканью. За счет этоrо и происходит
рост трубчатых костей в длину.
Реrенерация кости после
перелома. При переломах костей
наблюдаются прямые и непря
мые повреждения тканей, так как
разрываются связанные с костью
соединительные, мышечные TKa
ни, кровеносные сосуды и нервы.
В область перелома изливается
кровь, которая свертывается, об
разуя crYCТOK На некотором pac
стоянии от места перелома KOC
ти остеоциты в поврежденных
Рис. 129, Реrенерация кости после перелома:
остеонах rибнут и часть костной 1 наружная костная мозоль; 2 внутренняя костная
ткани отмирает. Реrенерация мозоль; 3 надкостница; 4 костный мозr; 5 отми
происходит путем образования рающие участки кости
новой костной ткани между отломками и BOKpyr них. В процессе формирования
pereHepaTa различают наружную (периостальную), межотломковую (интермеди
арную) и внутреннюю (эндостальную) ero части (рис. 129).
Источником развития периостальной части pereHepaTa служат остеоrенные
клетки надкостницы. Интермедиарная часть образуется за счет oCTeoreHHbIx эле
ментов, сохранившихся в каналах остеонов. Источником эндостальноrо остеоrис
тоrенеза являются остеоrенные клетки в составе эндоста и KocTHoro мозrа. Кроме
детерминированных остеоrенных клеток в формировании KocTHoro pereHepaTa при
ни мают участие камбиальные (адвентициальные) клетки, сопровождающие pac
тущие сосуды, которые MOryT дифференцироваться в остеобласты.
В наружных частях pereHepaTa за счет пролиферации эндотелиоцитов идет рост
сосудов, но медленный. Это отражается на дифференцировке ocтeoreHHbIx клеток: в
отсутствие капилляров (и кислорода) остеorенные клетки превращаются в XOHДpO
циты, В результате чеrо в наружных частях pereHepaTa образуется хрящ. Если каПИk
ляры прорастают в pereHepaT быстро, то в присутствии кислорода из остеоrенных
клеток сразу возникает ретикулофиброзная костная ткань. По мере перестройки pe
[енерата хрящ замещается костью rубчатоrо типа. Костные перекладины вблизи от
костных отломков прочно скрепляются с ними и между собой. В rубчатой костной
ткани между перекладинами образуются полости (туннели) за счет резорбции Mep
твых участков кости остеокластами. В образующиеся полости (туннели) врастают
капилляры и остеобласты. Выстилающие изнутри эти полости остеобласты путем
аппозиционнorо роста образуют слоистую структуру стенок За счет заполняющей
ся изнутри концентрическими слоями костной ткани стенки полостей становятся
толще, а просвет суживается. Таким образом возникают остеоны (rаверсовы систе
мы). Внутри остеонов имеется один или два кровеносных сосуда, доставляющих пи
тательные вещества для остеоцитов, окруженных пятью шестью концентрически
лежащими костными пластинками. Процесс внутренней перестройки pereHepaTa
290
Тлава 16
в области переЛО:\fа кости с превращением ретикулофиброзной костной ткани в пла
стинчатую продолжается долrо, шюrда несколько месяцев. Однако в конце концов в
результате этою сложноrо процесса в месте перелома кости, как правило, происхо
дит полное восстановление конфиryрации'и архитектоники кости. Тесное соприкос
новение и жесткая фиксация отломков кости способствуют более быстрому процес
су реrенерации.
Реrенерация при оrнестрельном повреждении кости имеет ряд особенностей.
Оrнестрельный снаряд при ударе о кость вызывает в последней возникновение
трещин и множества осколков, что замедляет процесс реrенерации. В эксперимен
тальных и клинических работах показано, что сохранившие жизнеспособность oc
TeoreHHbIe клетки костных осколков способны к пролиферации, дифференциации
и формированию ретикулофиброзной костной ткани, являясь источником пост
травматической реrенерации костной ткани.
Непременным участником реrенерации являются макрофаrи костномозrово
ro происхождения остеокласты. Последние участвуют в ремоделировании KOCТ
ной ткани.
Соединения костей
Различают непрерывные (синостозы, синхондрозы и синдесмозы) и прерывные
(диартрозы) соединения костей. Сино
стозы это неподвижные плотные co
единения, осуществляемые за счет KO
стной ткани (кости таза). Синхондро
зы соединения костей при помощи
хряща (например, межпозвонковые
диски). Синдесмозы фиброзные co
единения (рис. 130). К ним относятся
связки, межкостные перепонки и швы.
Прерывные соединения (диартрозы)
называют синовиальными соединсни
ями, или суставами.
В суставе различают суставные
поверхности сочленяющихся костей,
суставную сумку и суставную по
лость. Суставные поверхности костей покрыты слоем rиалиновоrо хряща, KOTO
рый не имст надхрящницы. Суставная капсула состоит из наружноrо фиброзноrо
слоя и внутренней синовиальной оболочки". Фиброзный слой капсулы представ
лен плотной волокнистой соединительной тканью. Соединение надкостницы Ok
ной из сочленяющихся костей с надкостницей друrоЙ осуществляется переплета
ющимися пучками коллаrеновых и эластических волокон, направленных вдоль
продольной оси сустава.
Синовиальная оболочка состоит из поверхностноrо и rлубокоrо волокнистых
коллаrеново эластических слоев, содержащих сосуды и нервные волокна, и покров
....... t ...... ,,' ..
.
>1
......
... .... '-
, . ' -......
. . 2
,. ;: , : 3
....
....
.... -.v- .....
...... ....
."'\.; >"
;,'.... ,
........
........,
..,.
Рис. 130. Строение связки:
1 пучки коллаrеновых волокон: 2 эластические
волокна; 3 прослойка рыхлой волокнистой соеди
нительной ткани с кровеносными сосудами
ОпОр1l0 двиzaтеЛЪ1lая система
291
Horo слоя, выстилающеrо суставную полость. Покровный слой представлен СИ1l0
виалъ1lым. Э1lдотелием. Он состоит из синовиоцитов специализированных кле
ток фибробластическоrо ряда. Различают макрофаrоподобные (фаrоцитарные)
синовиоциты и синовиальные фибробласты (или секреторные синовиоциты). MaK
рофаrоподобные синовиоциты (антиrенпредставляющие клетки) очищают coдep
жимое синовиальной полости от продуктов распада. Секреторные синовиоциты
вырабатывают синовию липкую прозрачную жидкость, содержащую rиалуро
новую кислоту. Синовия выполняет роль смазки, уменьшающей трение сочленяю
щихся в суставе костей.
Суставная полость представляет собой rерметически закрытое щелевидное
пространство, заполненное синовиальной жидкостью. Среди добавочных образо
ваний суставов различают складки синовиальной оболочки, внутрисуставные хря
щи (диски, мениски) и связки.
Хрящи MorYT быть представлены rиалиновой (суставные поверхности костей,
хрящи rортани и бронхов), эластической (ушная раковина, надrортанник) или BO
локнистой (межпозвонковые и суставные диски) тканями. На поверхности хряща
располаrается волокнистая соединительная ткань надхрящница, в составе KOTO
рой проходят сосуды и нервы, а также находятся малодифференцированные XOHk
роrенные клетки, составляющие камбий (рис. 131). За счет деления этих клеток
происходит рост хряща путем наложения (аппозиционный). Внутренний (интер
стициальный) рост осуществляется путем деления хондробластов. В центре хря
ща располаrаются изоrенные rруппы хондроцитов, которые утратили способность
делиться. В хряще нет сосудов, и пи
тание ero происходит диффузно. С воз
растом MorYT возникнуть rибель XOHД
роцитов и отложение солей кальция.
Хрящ применяется с целью трансплан
тации, в нем нет сосудов с кровью и он
не подверrается быстрому отторжению
из за несовместимости тканей.
Реrенерация хряща происходит за
счет деления камбиальных клеток Hak
хрящницы и синтеза межклеточноrо
вещества хондробластами.
Мышцы
Каждая скелетная мышца преk
ставляет собой активный opraH дви
жения, построенный из мноrих TKa
ней, rлавной из которых является
Хрящи
1
2
з
Рис. 1 3 1. Строение эластическоrо хряща:
1 надхрящница; 2 сеть эластических волокон;
3 хондроциты
292
Fлава 16
поперечнополосатая мышечная
ткань. Kpo:\fe Toro, в состав мыш
цы входят рыхлая и плотная co ... .............
единительные ткани, сосуды и
нервы (рис. 132). Функциональ
ное значение мышцы как opraHa
состоит в ее способности COKpa
щаться и изменять при этом свои
размеры. Это свойство мышцы
обусловлено сократительной спо
собностью скелетной мышечной
ткани, что в свою очередь связа
но с сокращением мышечных BO
локон в результате укорочения
миофибрилл в пределах каждоrо
саркомера. Мышечные волокна
являются основными рабочими
элементами мышцы.
Различают красные и белые мышечные волокна. Четкие rраницы между Kpac
ными и белыми мышечными волокнами отсутствуют, поэтому их подразделение
производится на основе сопоставления количественных показателей их CTPYKTYP
ных компонентов. Так, красные мышечные волокна (нередко обозначаемые миона
ми I ro типа) содержат больше саркоплазмы и, соответственно, меньшее количе
ство миофибрилл. В саркоплазме определяются мноrочисленные митохондрии,
которые отличаются высокой активностью окислительных ферментов (в частно
сти, сдr сукцинатдеrидроrеназы). СаркоплаЗl\fа характеризуется повышенным
содержанием миоrлобина. Диаметр красных мышечных волокон несколько MeHЬ
ше, чем средний диаметр белых мышечных волокон. BOKpyr красных мионов ин
тенсивно развита капиллярная сеть. По данным rисторадиоrрафии, для этих MЫ
шечных волокон характерен более высокиЙ уровень синтеза белков. На красных
мышечных волокнах некоторых мышц (например rлаза) определяются нетипич
ные множественные моторные бляшки.
Белые мышечные волокна (или мионы II ro типа) содержат большее число ми
офибрилл, расположенных в виде столбиков, или колонок, образующих на попе
речном срезе широкие поля. Саркоплазма образует узкие прослойки между мио
фибриллярными колонками. В неЙ мало митохондрий. Невысоко содержание и
миоrлобина. Диаметр белых мышечных волокон больше, чем красных. На каждом
белом мышечном волокне имеется лишь одна моторная бляшка типичноrо CTpoe
ния. Белые мионы сокращаются быстрее, чем красные.
Между красными и белыми мышечными волокнами имеются переходпые фор
мы промежуточные волокна. Мышцы человека, как правило, смешанные по co
ставу мионов разных типов, но каждая из них имеет свой рисунок, определяемый
процентным соотношением числа красных, белых и промежуточных мышечных
волокон.
..
8 "
.1#......."
'...... . .416"....
с
.......
...,
., ""
.......
. .' ,.'.\,"'"""' , ,< . "" . '"
? 1 .;.. ,.-ш.ш:.. ....О"" ' \ . е,,"
.................. ,,,,,, .., ""''''' .....'''!i, . 3 0\...
.. р'''''" .........-;о. . , ;.. (V, . "'i," ':
f , ,. 1 ",) 1,1 .'4
I 2 "с \,\.......". " ":";
':. .'в' ",\! I ,:-
.,..: ," {\ .. ,,.....
..Jf..../.J. ....... .....
...... .... 'l .. ..'--
'. .
...
,
___3., t<' " ;..:.
v"';-" ' \,",,",-3J U- с9 ..
,........ ":'.. . . .
....
.
"
" .j: .
;;;;;;""
JJI;
.
,.. ....... .:.....
....
"!-
Рис. 132. Прослойки рыхлой волокнистой соединительной
ткани между пучками мышечных волокон (перимизий):
1 артерия; 2 вена; 3 лимфатический сосуд
ОпОР1LO двиzaтелъная система
293
Важная роль в построении мышцы как opraHa принадлежит соединительной
ткани, которая объединяет мышечные волокна в пучки, проводит кровеносные
сосуды и нервы, а также обеспечивает прикрепление мышцы к костям. Рыхлая
соединительная ткань внутри мышечных пучков называется эндомизием. Пучки
мышечных волокон соединяются между собой также прослойками рыхлой co
единительной ткани, которую называют перu.мизием. Снаружи мышца покрыта
плотной соединительнотканной оболочкой эпu.мuзием, или фасциеЙ. Внутри
мышечная соединительная ткань обеспечивает развитие rустоЙ капиллярноЙ сети
BOKpyr каждою мышечноrо волокна. Блаrодаря эластическим своЙствам она уча
ствует в процессах, обусловливающих расслабление мышцы после ее сокраще
ния.
Иннервация мышц. Афферентные нервные волокна в скелетных мышцах име
ют разветвленные окончания в эндомизии. Они подходят также к сложноустроен
ным мышечным рецепторам типа мышечных и сухожильных веретен.
Мышечные веретена представляют собой инкапсулированные рецепторные
образования веретеновидноЙ формы длиноЙ 3 5 мм и толщиноЙ около 0,2 мм.
В пределах скелетных мышц они располаrаются продольно, и при удлинении всей
мышцы растяrиваются. Количество их в составе отдельных мышц измеряется дe
сятками или сотнями. Больше мышечных веретен содержится в мышцах кисти,
требующих тонких движений. Каждое веретено содержит rруппу (около 10) MЫ
шечных волокон, называемых интрафузальными. в отличие от экстрафузальных
волокон, находящихся вне веретен. Под соединительнотканноЙ капсулой Bepe
тена находится жидкость, омывающая интрафузальные мышечные волокна. Пос
ледние подразделяются на два типа: волокна с ядерной сумкой (ЯС волокна) и
волокна с ядерной цепочкой (ЯЦ волокна). Эти обозначения отражают характер
расположения в них ядер. ЯЦ волокна тоньше и короче и представлены в боль
шем количестве. Если ЯС волокон обычно от 1 до 4, то ЯЦ волокон может быть
до 1 О в одном мышечном веретене.
К мышечному веретену подходят афферентные и эфферентные нервные BO
локна. К ЯС волокнам подходят миелиновые афферентные нервные волокна, об
разующие окончания аннулоспиральноrо типа (первичные окончания). ЯЦ во
локна получают афферентную иннервацию как от Toro же первичноrо окончания,
так и от более тонких афферентных волокон (вторичные окончания). Первичное
окончание реаrирует на степень и скорость растяжения мышц, а вторичное толь
ко на степень растяжения. Импульсы по аффереНТНЫl\f нервным волокнам по
ступают в спинной мозr, [де афферентные нейроны передают их на [aMMa MOTO
нейроны.
Наряду с афферентной иннервацией интрафузальные мышечные волокна имеют
и эфферентную иннервацию, с помощью котороЙ они находятся в определенной CTe
псни натяжения. Эфферентные нервные окончания образуют нервные волокна [aM
ма мотонеЙронов. Такая сложная система иннервации мышечных веретен не06ходи
ма для автоматическоrо контроля ими за степенью и скоростью растяжения скелетных
мышц. Мышечные веретена сиrнализируют в центральную нервную систему о любых
рассоrласованиях между степенью растяжения интрафузальных и экстрафузальных
294
Тлава 16
мышечных волокон, на что следует ответная реакция, корриrирующая сокращение
мышцы в целом. Мышечные веретена обеспечивают "мышечное чувство" вместе с дpy
rими афферентными окончаниями из rpуппы проприоцепторов, которые участвуют в
реryляции движений и позы, а именно: сухожильные и суставные рецепторы.
СУХОЖШlъные рецепторы инкапсулированноrо типа находятся в местах соеди
нения мышц с их сухожилиями, а также в апоневрозах. Они имеют лишь афферен
тную иннервацию. Суставные рецепторы по строению очень похожи на сухожиль
ные веретена. В суставных капсулах MHoro свободных нервных окончаний,
пластинчатых телец и друrих механорецепторов.
Скелетные мышцы как эффекторные аппараты нервной системы содержат
большое количество эфферентных нервных волокон. На каждом мышечном BO
локне имеются двиrательные нервные окончания концевые моторные бляшки.
Посредством этих образований нервные импульсы передаются с MOTopHoro ней
рона на мышечное волокно. Моторная бляшка (нервно мышечное соединение, или
аксо мышечный синапс) состоит из HepBHoro полюса (концевоrо ветвления aKCO
на) и мышечноrо полюса части мышечноrо волокна, к которой прилежит терми
наль аксона. Последняя содержит митохондрии, синаптические пузырьки с aцe
тилхолином и пресинаптическую мембрану, а снаружи нейролеммоциты
(шванновские клетки). Синаптическая щель в аксомышечном синапсе имеет вид
уrлублений с первичными и вторичными складками. Постсинаптическая мембра
на представляет собой часть плазмолеммы мышечноrо волокна, через которую по
тенциал действия распространяется по мембранам трубочек Т системы. Это изме
няет содержание кальция в саркоплазматической сети и приводит к сокращению
миофибрилл.
Моторные бляшки бывают типичные и rроздевидные. На красных мионах оп
ределяются rроздевидные моторные бляшки. Для белых мышечных волокон xa
рактерны типичные моторные бляшки. Обычно двиrательное нервное волокно раз
ветвляется на несколько десятков веточек, каждая из которых заканчивается
моторной бляшкой. Совокупность мышечных волокон, иннервируемых одним
моторным нейроном, составляет вместе с этим нейроном нейромоторную единицу.
Рост скелетных мышц. В период постнатальноrо роста орrанизма вместе с
удлинением костей происходит рост в длину и толщину скелетных мышц. Масса
скелетных мышц увеличивается также при интенсивных физических упражнени
ях. При этом рост мышц обусловлен утолщением (rипертрофией) мышечных BO
локон. Количество же их существенно не меняется. В основе rипертрофии мышеч
ных волокон лежит увеличение числа миофибрилл и друrих орrанелл. Новые
миофибриллы образуются из миофиламентов, которые строятся в свою очередь из
синтезируемых сократительных белков. Рост мышечных волокон в длину осуще
ствляется в результате слияния с ними миосателлитоцитов. Удлинение миофиб
рилл достиrается при стройкой к их концам новых саркомеров. Это происходит в
местах прикрепления мышечных волокон к сухожилиям.
Реzеllерацuя скелетllЫХ мышц. После повреждения скелетных мышц часть MЫ
шечных волокон в области травмы подверrается разрушению. Продукты их распада
резорбируются путем фаrоцитоза и аутолиза (рис. 133). На месте травмы разраста
ОпОр1ю двuютеЛЪ1lая cucтeJ'da
295
· J t v>c.:* .:F:J. .
. . :.. ...,!:. ." j .
:} " : "Y; * ':;.. .'': й ' ' r 1 ?
'1" t.1. 1.;'t'" . ,__:. r 1' '!. -:' it-
.... J ..... . ..J;-:$t!, v .".." .'> .. \ .;::..
.. "''t > .h, ",; ' r"ц'" '" "":. r,,, ),."" ."".. .,
..( . "..;'P d;':i #" i."" \" "!\I. 'f, _ . ....... "" ......
.<'.....i!i: ....: ,..., f;'" ' " '" .'ti.. ft . ':
1''J1iI' .. о;7.", ... 4 -.:-.. ).- ...
Ji ..... . ... I:: € < "".... " '.&..-: .......;;. ' ;j :
,$,; . . . :.:;;,, ;; $. r .<.' > "\ _ . t '", __i!.. . '
. i\ ..... . ' -р. ... .... '" "<.- " .. :.; 'I:"
<;<>.,.... .,:, .rд; ........ . ... (' :\'.;, ';,;" . .\" : . (
:'tI .. . " '\iл!t . .' " ..'t"..........'. f!'!.....-:'
' ' r '. : ' I,;;.; м:l t: f{( r. . , & <.; .. . "
". .. "t: . :; . ";;..';--: . .:. " ,
",' " 7..' .,.,? '" . '..iJ 1
.: ., ... .:;1:-.t : ', " ",.. 't'
. ..,, ,,' . '''' '.}y. ; j" >; . .'- '" "!"
... ..;:: 't": ""; .4f"r., .
Рис. 133. Макрофаr. участвующий в резорбции поврежненноrо
мышечноrо волокна:
1 лизосомы макрофаrа; 2 отростки макрофаrа; 3 мышеч
ное волокно
ется rрануляционная ткань, в
которую врастают мышечные
элементы pereHepaTa. Источ
ником восстановления мышц
являются в основном миоса
теллитоциты, способные дe
литься митозом и превра
щаться в миобласты. После
дние сливаются друr с дpy
rOM, формируя миосимплас
ты. В ходе последующей их
дифференцировки образуют
ся новые мышечные трубоч
ки и зрелые мышечные BO
локна. Процесс восстановле
ния поврежденной мышцы
продолжается около месяца.
Для реrенерации скелетной
мышцы имеет важное значение восстановление иннервации (нервно трофическое
влияние), кровоснабжения и функциональноrо натяжения. После тяжелых травм
значительные дефекты скелетных мышц полностью не восстанавливаются. Обычно
в таких случаях место мышечной травмы заполняется фиброзной рубцовой тканью.
Этот процесс, как правило, сопровождает заживление Iышечной раны после orHe
. ".... ' стрельноrо повреждения.
" ., ...
ii 2
.. '! r
i/I
о, а:"
'"4;_
I "
...;-
..... .
..,,-... ...
....
d( ,О
"',,"
,'
. ..... ........ ......-.,: ,, ...,.-
..."
..
..
, .
. .
.
. . \
1\
. :
,
.
..
.
'..
...
.'
..., .....
".. r
,, "C.t ) . ·
, '.. .., {
...;.;.;. / J .. i
.... . ,
'. (..
'"
,
.
..
...
;
:
. .
:.
' ...
'.
--
'..
.
.
... ..;.........4
.... 4!!' ..,,)..
.
...
.
,
'
.
.j/it
..
Рис. 134. Строение сухожилия (поперечный срез):
1 пучки 1 ro порядка; 2 сухожильные клетки; 3 эн
дотеноний между пучками 2 ro порядка; 4 перитеноний
Сухожилия
11
с помощью сухожилий обеспе
чивается связь скелетных мышц с
костями. Сухожилия образованы
плотной оформленной соедини
тельной тканью (рис. 134). В cyxo
жил иях коллаrеновые пучки Haxo
дятся постоянно в состоянии натя
3 жения. В процессе rистоrенеза cy
хожилиЙ при дифференцировке cy
хожильных клеток образуются KOk
лаrеновые волокна, располаrающи
еся параллельно друr друrу. Каж
дыЙ пучок коллаrеновых волокон
отделяется от сосеДllеrо слоем cy
хожильных клеток (тендоцитов).
В uелом этот комплекс клеток и BO
локон называется сухожильным
296
[Лава 16
пучком 1 lO порядка. [руппа пучков 1 ro порядка, окруженная прослойками pыx
лой соединительной ткани, составляет сухожильный пучок 2 ro порядка. OTдe
ляющие эти пучки прослойки рыхлой соединительной ткани называют эндоте
нонием. [руппа сухожильных пучков 2 ro порядка образует сухожильный пучок
З rо порядка. Он окружен плотноЙ соединительнотканной оболочкой перите
нонием. Несмотря на сильное развитие коллаrеновых структур, оттесняющих на
второй план клетки, сухожилия способны к рсrенерации. Камбиальным источ
ником для llосстановительных процессов служат малодифференцированные фиб
робласты и адвептициальныс клетки в составе эндотенония.
Общие закономерности реrенерации орсанов. Боевые повреждения, как правило,
сопровождаются нарушеним целостности покрова орrанизма, opraHoB внутренней среды
и опорно двиrательной систе:\IЫ. При этом формируется дефект орсана, очаr некроза, яв
ления дезинтеrрации клеток, тканевых и орrанных структур. На это орrанизм отвечает пе
рестроЙкой систем защиты и восстановления, происходит заживление раны, в котором раз
личают несколько фаз.
Первая фаза paHeBoro процесса фаза воспаления характеризуется травматичес
юш отеком тканей, повышением сосудистой проницаемости, ацидозом, миrрацией лейко
цитов, тучных клеток и макрофаrов. Происходит очищение раны путем фаrоцитоза и ли
зиса некротизированных тканей.
Во второЙ фазе раневorо процесса фазе реrенерации происходит развитие rpaHY
ляционноЙ ткани, постепенно заполняющеЙ раневой дефект. Основными структурами этой
ткани являются фибробласты, межклеточное вещество и капилляры. Клетки фиброблас
тическоrо дифферона rрануляционноЙ ткани отличаются от фибробластов нормальной
соединительноЙ ткани высокой функциональной активностью. Они синтезируют белки и
rликозаминоrликаны, образуя коллаrеновые волокна. В развитии и созревании rрануля
ционноЙ ткани важную роль иrрают также макрофаrи, тучные и плазматические клетки.
[рануляционная ткань впоследствии преобразуется в рубцовую соединительную ткань.
Третья фаза раневосо процесса фаза реорrанизации рубца характеризуется про
rpессирующим уменьшением числа кровеносных сосудов и клеточных элементов (фиброб
ластов, макрофаrов, тучных клеток) при явлениях нарастания общей массы коллаrеновых
волокон. Параллельно с созреванием rрануляционной ткани и ее превращением в рубцовую
ткань происходит эпителизация раны. Эпителизация раны и созревание rрануляшюнной
ткани cTporo соответствуют во вре:\lени.
В зависш,юсти от характера и величины ранения, особенностей реактивности орrанизма
и друrих условий раневой процесс протекает по разному. При небольшом объеме поражения
имеет место заживление раны псрвичным натяжением. Воспаление и замещение дефекта в
тканях следуют непосредственно за их травматическим отеком и не сопровождаются HaCHoe
нием. К концу первой недели раневой процесс в основном завершается. Если же объем пора
жения велик и края раны оказываются на более или менее значительном расстоянии друс от
друса. то заживление раны осуществляется через напюение с образованием хорошо развитоЙ
rpануляционной ткани с последующим ее рубцеванием. Заживление раны происходит BTO
ричным натяжением со значительной продолжительностью фаз раневorо процесса.
Заживление раны первичным и вторичным натяжением имеет количественные, но не
качественные различия. Механизмы реrенерации при этом принципиально сходны и вклю
чают воспаление, разрастание соединительной ткани, эпителизацию. Знание ключевых зве
ньев ресенераЦIIОННОСО процесса позволяет целенаправленно вести поиск средств реryля
ции заживления ран 11 разработке способов тканевой терапии.
Воспроизведение человека
297
rЛАВА 17. ВОСПРОИЗВЕДЕНИЕ ЧЕЛОВЕКА.
ОСНОВЫ МЕДИЦИНСКОЙ ЭМБРиолоrии
Развитие половых клеток (проrенез)
Развитие половых клеток называ
ется проrенезом, или rаметоrенезом, и
происходит в половых железах. Суть
rаметоrенеза состоит в образовании из
диплоидных стволовых предшествен
ников половых клеток высокодиффе
ренцированных клеток (сперматозои
дов и яйцеклеток) с rаплоидным набо
ром хромосом.
Развитие мужских половых кле 2
ток сnермаmОlенез, включает четыре
фазы: размножения, роста, созревания,
формирования (рис. 135). В результа
те сперматоrенеза возникают клетки
сперматозоиды (спермии), содержа з
щие x или У половую хромосому.
Развитие женских половых кле
ток овozенез, состоит из трех фаз: раз
множения, роста, созревания. В резуль
тате овоrенеза образуются яйцеклетки
(овоциты), содержащие Х половую
хромосому.
РИс.135. Схема развития половых клеток:
В основном мужской и женский ra А сперматоrенез; Б овоrенез; фазы: 1 раз
метоrенезы протекают однотипно. В множения; 2 роста; 3 созревания; 4 форми
фазе раЗМ1l0жения исходные половые рования
клетки (сперматоrонии в сперматоrенезе и овоrонии в овоrенезе) в ходе клеточ
HOro цикла синтезируют ДНК и митотически делятся; их размножение протекает на
основе общих закономерностей, характерных и для соматических клеток В процес
се сперматоrенезаделится только часть сперматоrоний, так называемые светлые спер
матоrонии типа А, которые являются полустволовыми клетками, дающими начало
сперматоrониям типа В. Сперматоrонии типа В проходят несколько митотических
циклов, при этом не происходит полноЙ цитотомии сперматоroниЙ 11 они остаются
связанными цитоплазматичеСКИl\lИ мостиками. Фаза размножения сперматоrоний
!\
/\ 1\
/i i l 1\ i\
... .1..
;\
J\ /\
1 t t 1
4[
А
.
/\
. .
/\ 1\
....
JJl \
.
(л
Б
298
Тлава 17
контролируется естественньши митоrена ш [ормоном аденоrипофиза фоллитро
пином и интерлеiiкином 1, вырабатываеl\IЫМ клетками Сертоли и клетками Лейди
[а (интерстициальньши эндокриноцитами) яичек
В мужском орrаНlIзме фаза размножения сперматorониЙ, как и сперматоrенез
в целом, начинается в процессе половorо созревания (в репродуктивном возрас
те). В женскоы орrаНИЗ :lе фаза размножения овоrенеза инициируется и заверша
ется еще в эмбриоrенезе. К окончанию данноЙ фазы в каждом яичнике плода зак
ладывается около 6 7 млн овоrониЙ.
Фаза роста в сперматоrенезе выделяется в определенной степени условно, посколь
ку она не связана, как в женском rаметоrенезе, с накоплением питательных веществ
для будущеrо зародыша, и по этоЙ причине ее часто объединяют с третьей фазой спер
матоrенеза (фазой созревания) в одну, так называемую, мейотuческую фазу. В мейо
тической фазе половая клетка (именуемая первичным сперматоцитом, или спермато
цитом 1 ю порядка) проходит длинную профазу первоrо мейотическоrо деления,
которая у человека продолжается около 22 суток Рост характеризуется небольшим
увеличением объема сперматоцитов. Напротив, в фазе роста женская половая клетка
значительно увеличивает свои размеры в результате синтеза и накопления желтка (ви
теллина) в цитоплазме. В овоrенезе фаза роста более продолжительна, чем в сперма
тоrенезе, и включает малый и большой рост. Малый рост женские половые клетки
претерпевают в эмбриоrенезе, а в большой рост вступают после половоrо созревания
девочки (в репродуктивном возрасте).
Фаза созреваll.UЯ наиболее продолжительная фаза rаметоrенеза. В овоrенезе
она начинается в эыбриоrенезе (практически одновременно с началом малоrо poc
та половых клеток). К рождению девочки фаза созревания половых клеток (OBO
цитов) В ее яичниках приостанавливается и возобновляется лишь после наступле
ния половой зрелости. В фазе созревания как мужские, так и женские половые
клетки проходят мейоз особый вид деления, в ходе которою содержание XpOMO
сом в их ядрах сокращается наполовину и составляет 23.
Перед вступлением в мейоз диплоидные половые клетки с rенетическим набо
ром 2c2n (сперматоrонии типа В и овоrонии) в синтетическом периоде клеточноrо
цикла удваивают количество ДНК и, соответственно, количество субъединиц
хромосом. Их ядерная формула может быть представлена как 4c2n.
Собственно мейоз включает два последовательных деления созревания, про
текающих без интерфазы и количественноrо изменения rенетическоrо материала.
Первое деление именуется редукционным, второе эквационным.
В сперматоrенезе исходная половая клетка, вступающая в мейоз, носит назва
ние сперматоцита 1 ro порядка (первичноrо сперматоцита), в овоrенезе овоцита
1 ro порядка.
Ответственным этапом мейоза является профаза первоrо деления. В сперма
TO и овоrенезе она включает стадии лептотены, зиrотены. пахитены, диплотены и
диакинеза.
При этом в пахитене происходит обмен rенами и rруппами [енов между rомоло
rичными хромосомами (кроссинrовер). Значение последнеrо состоит в формирова
НIШ качественноrо разнообразия rенофонда половых клеток и в последующем
Воспроuзведе1luе челове",а
299
развивающихся из них орrанизмов. Следует отметить, что в профазе 1 ro деления мей
оза мноrие половые клетки поrибают из за сложности происходящих процессов.
В сперматоrенезе профаза непосредственно продолжается в последующие стадии
первоro деления меЙоза. В овоrенезе половые клетки останавливаются в стадии диаки
неза под влиянием мейоз инrибирующей субстанции и MOryT пребывать в ней разное
число лет. В этой связи стадия диакинеза в овоrенезе именуется сто.цuан.арн.ой сто.дueй
профазы первоro деления мейоза. Разные женские половые клетки выходят из стацио
нарноro состояния и продолжают свое развитие в разные периоды репродуктивноrо
возраста, мноrие поrибают, так и не реинициировав мейоз. Фактором, стимулирующим
продолжение мейоза, является мейоз стимулирующая субстанция, которая как и мей
оз инrибирующая, синтезируется соматическими (фолликулярными) клетками овари
альных фолликулов, в окружении которых развиваются женские половые клетки.
В процессе первоro мейотическоrо деления в каждую дочернюю клетку pacxo
дится по одной roмолоrичной двойной хромосоме от каждоrо бивалента. Иными
словами, каждая дочерняя клетка получает rаплоидный набор хромосом, в связи с
чем первое деление именуется редУ"'ЦUО1l1lЪLМ. Каждая из хромосом этих клеток,
однако, состоит из двух хроматид (ядерная формула клеток 2с1п). В сперматоrе
незе телофаза завершается неполной цитотомией и образующиеся клетки спер
матоциты 2 ro порядка также остаются связанными друr с друrом цитоплазма
тическими мостиками (формируется синцитий).
В овоrенезе в результате первоrо деления мейоза образуется один овоцит 1 ro
порядка и одна абортивная дочерняя клетка, именуемая редукционным тельцем,
для которой характерно практически полное отсутствие цитоплазмы. Это тельце,
как правило, остается расположенным рядом с овоцитом.
Далее следует второе деление созревания э",ваЦUО1mое, протекающее как
обычный митоз. Однако, в отличие от чередования митозов соматических клеток
здесь отсутствует отчетливая интерфаза и клетки переходят от первоrо деления
мейоза ко второму делению мейоза без деконденсации хроматина и удвоения co
держания ДНК В образующиеся дочерние клетки (в сперматиды при сперматоrе
незе и в овоциты 2 ro порядка при овоrенезе) расходятся хроматиды от каждой из
метафазных хромосом, таким образом, клетки получают истинно rаплоидный Ha
бор rенетическоrо материала (ядерная формула сперматид и овоцитов 2 ro поряд
ка 1с1п).
В результате мейоза в сперматоrенезе из одной исходной сперматоrонии обра
зуется 4 дифференцированные половые клетки сперматиды, которые теряют
синцитиальные связи. Половина образующихся сперматид содержит У половую
хромосому, друrая половина X XpOMOCOMY.
В овоrенезе, как указано выше, овоциты продолжают мейоз лишь при действии
мейоз стимулирующей субстанции. Первое деление мейоза и второе деление до
стадии метафазы женская половая клетка проходит, находясь в яичнике. На CTa
дии метафазы BToporo деления мейоза овоцит 2 ro порядка покидает яичник (OBY
лирует) и претерпевает aHa и телофазу в маточной трубе (превращаясь в резуль
тате активирующеrо влияния сперматозоидов в зрелую rаплоидную половую
клетку, или яйцеклетку). Если контакта со спермиями не происходит, овоцит 2 ro
300
Fлава 17
порядка так и не завершает мейоз и поrибает. В овоrенезе каждая материнская клет
ка при каждом делении мейоза дает лишь одну полноценную половую клетку; BTO
рая клетка оказывается абортивной и называется редукционным тельцем. PeДYK
ционное тельце, которое образуется в результате первоrо деления мейоза, при
втором делении также делится (но пеобязательно), давая два дочерних тельца. Ta
ким образом, в результате мейоза в овоrенезе образуется лишь одна дифференuи
рованная яйцеклетка и три редукционных тельца.
В фазе фор шрования сперматоrенеза сперматиды трансформируются в спер
матозоиды из части центриоли образуется жryтик, сбрасывается большая часть
цитоплазмы (что важно для обеспечения подвижности, а также для проникнове
ния спермия в яйцеклетку при оплодотворении), синтезируются фермеНТЫ ЛI1ЗИ
ны, оформляется акросомальная вакуоль, изменяется форма и объем ядра, KOHдeH
сируется и компактно упаковывается хроматин.
OpraHbI мужской половой системы
к opraHaM мужской половой системы относятся [онады (яички, или семенни
ки), выполняющие репродуктивную и эндокринную функции, добавочные opra
ны семявыносящие пути, железы. половой член.
Закладка яичек у человека появляется на 28 ЗО е сутки эмбриоrенеза в виде
парных утолщений целомическоrо эпителия на медиовентральной поверхности
первичных почек (мезонефросов). Здесь формируются половые валики, в которые
миrрируют первичные половые клетки lO7-lOциmы. С З3 З5 х суток начинается
фОР lIIрование и врастание в мезенхиму половых валиков клеточных тяжей из цe
ломическоrо эпителия и клеток канальцев первичной почки клеток сети. К KOH
цу 5 й началу 6 й недели формируется rонадная бластема, в состав которой BXO
дят rоноциты, клетки целомическоrо эпителия, канальцев первичной почки и
мезенхимы. Вплоть до начала 7 й недели rонада человека rистолоrически не диф
ференцирована по половому признаку (стадия индифферентной roнады).
Индифферентная [онада начинает развиваться по мужскому типу с 7 й недели,
коrда наблюдается формирование закладки семенных канальцев в виде половых тя
жей. Это происходит под влиянием экспрессии факторов У хромосомы. Первично
дифференцируются клетки Сертоли (поддерживающие эпителиоциты, сустентоци
ты), далее клетки Лейдиrа (интерстициальные эндокриноциты). [оноциты под вли
янием факторов, инrибирующих развитие мюллеровых протоков, секретируемых клет
ками Сертоли, дифференцируются в сперматоrонии, а мезенхимные клетки формируют
характерную для семенников соединительнотканную строму и сосуды. Между поло
выми тяжами и эпителием [онады возникает белочная оболочка, которая разделяет
яичко на отсеки, содержащие отдельные половые тяжи. На 7 M месяце эм6риоrенеза,
яички опускаются в мошонку и покрываются соответствующими оболочками.
Яички
Яичко парный opraH эллипсоидной формы, расположенный у человека в MO
Воспроизведение человека
301
шонке. От белочной оболочки в rлубь яичка отходят переrородки септы, которые
разделяют ero на 200 300 долек коническоЙ формы. В области верхнезаднеrо края
яичка септы сходятся, врастая в соединительнотканное средостение яичка. В cpeдo
стение проникают нервы и сосуды.
В каждой дольке яичка располаrаются 1 4 извитых семенных канальца длиной
от 30 до 80 см, в стенке которых происходит сперматоrенез. На верхушках долек
извитые канальцы продолжаются в короткие прямые канальцы, которые переходя т
в сеть семенника, а из сети берет начало система семявыносящих протоков.
Строение извитоrо ce
менноro канальца. Снаружи
извитой семенной каналец or
раничен базальной l\Iембра
ной. На ней располаrается ok
нослойный эпителий цело
мическоrо типа, состоящий из
клеmок Серmоли (рис. 136).
Последние имеют дреВОВИk
ную форму и широким OCHO
ванием прикреплены к базаль
ной мембране. Апикальные
части клеток Сертоли образу
ют множество анастомозиру
ющихдруrс друroм отросщов,
между которыми, как в KapMa
нах, располаrаются развиваю
щиеся половые клетки. Про
странства между базальными
частями клеток Сертоли в co
вокупности именуются ба
зальным отсеком канальца и
Рис. 136. Строение яичка:
заняты сперматоrониями раз 1 семенной извитой каналец; 2 клетки Лейдиrа; 3 клетки
ных типов здесь происходит Сертоли
процесс их размножения. Центральный (противоположный базальному) отдел каналь
ца назван адлюминальным отсеком здесь находятся более развитые формы поло
вых клеток, начиная со стадии сперматоцитов 1 ro порядка. Лдлюминальный отсек
изолирован от базальноro с помощью плотных контактов между отростками соседних
клеток Сертоли. Такая изоляция необходима потому, что в процессе развития (начи
ная со стадии сперматоцитов 1 ro порядка), коrда происходит кроссинroвер XpOMO
сом, половые клетки становятся носителями неизвестных орrанизму анmиzенов и ДОk
жны быть изолированы от системы крови и лимфы. Это достиrается путем их "поме
щения" в адлюминальный отсек Клетки Сертоли являются ведущими в создании le
маmоmесmикулярноzo барьера (ПБ), или rистиона, предотвращающеrо иммунную pe
акцию орrанизма на развивающиеся половые клетки. В состав ПБ входят: система
плотных контактов между отростками клеток Сертоли, базальная мембрана извитоro
. "." ..!!
1# .. .... .. . 9J..$!
",. .. . .. 4....."" .. 4:i.'- ..
. .. .,';. * .' \ 72.......
. . -.А. .." ...,.., . .... / t I ..
I.':t, IW' .- .с.. . ."... А.7..... . 11 ..",,'
"""\.. ;.$" . ... ; '..... e е. \- ":"..
" . . 'V\" ., .; . " '. .,.''' '' " . а #.
. ..., 'iUJ'В:. ' . ,Ж/, .
" ;1'6", . ..., . ' ' "t d. , ." ' . " .-
. V .J -.:JJI',.. .8' ';v .. ." .. *'л#.
.. .. .l , .. i ,}t?'8a.. .',0 . , . .,.. r ..
-"-- .' . .f!j; ''Ыf..ш, ,е ' . . .. '. .. " " . t;, . "
t' ,.; . . !f;" . .( .;.... " е-.tO , '"
. lko'fE".{jj.- " .. 1Р:. "\ . .. ,.
.l!i'-. / C "'...) ... .QIf"' ,Ji ""Ю " ,.
: {J WO:: ...,0' -: '1'. .. . '1.. t;: ..
" 1". ,. 1> .i..."a . ., ..\>« . ... <9,. . L .. 'f'io ;. 3
.- . . . . Ii> .... 'o 4'.. '
-.).."'. .. .... .... 'Д. :-!1/' _.. ...
'I" J. Ф. \""-'" . .. #: .. . ..
1 . 11 rt . I r ,, I&,-> . ,.
. ' ... > . ,. ". ''' i{]
i'l '''''''. 'i (,. . '"
#,,' k , .,..... ";:.. \, ... '.ti.
2 U . ::. ,. t' ". .....i:..".
- . 'i. . I ... 5'.
.1 · . t,1t . . е . ,#0. '"' . "." "" t
t!J';' .,., ..' "", . '. ',' '" "4 .'"
{/ , "'.. (jII@ ф ,j.. '." i/!" ,.,
"с . ,,; \,tg;,..9. ,," ," Ii! .. ". .....
6", o" . ..." ... ..... ;",.Дj'. 1;,
...,... '.f'i', ., f.O .',., . {\. /:.
... .'. "".. . .... ''' ''' .. ' ,f!P . .. . 1J ......">.,, . t> ,......' . ......., 4.'
... ." -.. .. "'l. '" '" '. Д .".r...........
., \0_" l' · """'; . ,, !;'j. . ,
,,1If iJ .. .. ..!i' r.J..".,. . о
"'".' "4!' .. f!!)r ""-V"" ., . >'.
.. !' ..' ./ .. .. '\. ' ..' .
i 1. , } .'.' ..... '-,1P.: l5i.. '\'\ tf,
· " k ф ' . ...!:/ S ,;;.' ,'. \
& b . ijC'f''' ...,:>..
->' . . . р _" " _о .,
. . ,,--:' "t". '"
,#'!' \\t"1
....
302
Тлава 17
семенною канальца, миоидные клетки, расположенные BOKpyr извитых канальцев, He
большое количество интерстициальной соединительной ткани, базальная мембрана
rемокапилляров, эндотелиальные клетки rемокапилляров.
Полаrают, что сперматоrонии типа В, инициирующие рост и fейоз и приобре
тающие антиrенные свойства, перемещаются из базальноrо в адлюминальный OT
сек по способу "шлюзования" в результате перестройки системы плотных KOH
тактов между отростками клеток Сертоли.
В извитом семенном канальце сперматоrенез протекает неравномерно BOk
нами, поэтому на rистолоrических препаратах opraHa на одном срезе канальца не
всеrда можно наблюдать все стадии развития половых клеток Цикл развития муж
ских половых клеток у человека продолжается около 74 суток
Пространства между петлями извитых канальцев заняты рыхлой волокнис
той соединительной тканью (интерстициальной тканью, стромой), содержащей
большое количество кровеносных и лимфатических сосудов, нервных окончаний,
а также клетки Лейдиrа. Клетки Лейдиza являются мезенхимными производны
ми. Это клетки окруrлой формы диаметром 10 15 мкм с оксифильной цитоплаз
мой, располаrающиеся между извитыми канальuами небольшими rруппами или
одиночно. Они синтезируют мужские половые ropMoHbI тестостерон, диrидро
тестостерон, андростендион, а также небольшие количества эстроrенов. Помимо
половых ropMoHoB, клетки Лейдиrа вырабатывают вещества, оказывающие пара
кринное воздействие на клетки Сертоли; небольшие количества окситоцина, сти
мулирующеrо сокращения перитубулярных миоидных клеток извитых семенных
канальцев, а также интерлейкин 1, действующий как фактор роста на спермато
rонии типа В.
Се,МявЫll0сящие пути и железы
Источником развития семявыносящих путей служат: часть канальцев первич
ной почки (вольфова тела), из которых развиваются выносящие канальцы придат
ка яичка; верхняя часть мезонефральноrо протока, материал которой идет на обра
зование протока придатка яичка; нижняя часть мезонефральноrо протока, дающая
семявыносящий проток; выросты мочеполовоrо синуса, из которых развиваются
семенные пузырьки и предстательная железа.
у эмбриона на ранней стадии развития закладываются две пары протоков: мезо
нефральные (или вольфовы) и парамезонефральные (или мюллеровы). При разви
тии мужскоi:i половой системы парамезонефральные протоки редуцируются. От них
остаются лишь маленький пузырек (rидатида) и рудиментарное образование "мужс
кая маточка". Формирование половых путей по мужскому типу определяет первич
ный мужской половой ropMoH белковой природы инrибин.
Развитие семя выносящих путей начинается с соединения BepxHero отдела Me
зонефральноrо протока с семенными извитыми канальцами яичка. Осуществляет
ся это посредством части канальцев первичной почки, образующих прямые каналь
цы и сеть канальцев в средостении яичка.
Семявыносящие пути выстилает эпителий. В различных отделах семявынося
Воспроизведение человека
303
щих путей строение и функция эпителия имеют свои особенности, обеспечивающие
транспорт и дозревание сперматозоидов. Поскольку спер lатозоилы, образующиеся
в яичке, подвижностью не обладают, то продвижение их по се:\IЯПЫНОСЯЩИМ путям
происходит за счет давления новообразующихся порций сперматозоидов, а также за
счет движений ресничек эпителиальных клеток и перистальтических сокращений
rладких мышечных элементов в составе стенки протоков.
Придаток яичка (эпидидuмис) состоит из rоловки, тела и хвоста. [оловка об
разована 12 15 выносящими канальцами. Просвет их имеет неровный контур, так
как выстилающий изнутри канальцы однослойный эпителий состоит из клеток раз
ной высоты. Более высокие клетки на апикальной поверхности имеют мерцатель
ные реснички и называются реснитчатыми эпителиоцитами. Они способствуют про
движению по канальцам спер:\1атозоидов. Низкие клетки, расположенные в "ямках"
эпителия, имеют микроворсинки. Для них характерны апокринный тип секреции и
реабсорбция жидкости из просвета.
Проток придатка яичка, образующий своими мноrочисленными извивами тело
придатка, выстлан двухрядным эпителием. В эпителии различают призматичес
кие и вставочные эпителиоциты. На свободной поверхности призматических кле
ток имеются стереоцилии немерцающие волоски. Вставочные эпителиоциты
являются камбиальными для призматических ЭПIпелиоцитов. Блаrодаря ceKpeTOp
ной деятельности эпителия протока придатка сперматозоиды дозревают и покры
ваются тонким слоем секрета (rликокаликсом). Значение rликокаликса состоит в
предотвращении иммунных реакций при попадании сперматозоидов в женские по
ловые пути. В стенку выносящих канальцев и протока придатка входят также MЫ
шечная оболочка, состоящая из rладких миоцитов, и наружная соединительнот
канная оболочка. В связи с более низкой температурой в яичках, чем общая
температура тела. кислой реакциеЙ и вязкостью среды сперматозоиды пребывают
здесь в состоянии относительною анабиоза.
Се-мявыносящий проток. Продвижение спермы от придатка яичка осуществ
ляется по семя выносящему протоку. Стенка ero состоит из слизистой, мышечной
и адвенпщиальной оболочек Слизистая оболочка образует 4 6 продольных скла
док, в связи с чем просвет протока на поперечном срезе имеет звездчатую форму.
Эпителий слизистой оболочки однослойныЙ мноrорядный. Высокие призматичес
кие клетки эпителия имеют стереоцилии. Мышечная оболочка представлена Tpe
мя слоями: наружным и внутренним с продольным расположением rладких ми
оцитов и средним циркулярным. Перистальтические сокращения мышечной
оболочки семявыносящеrо протока способствуют продвижению сперматозоидов
в семяизверrающий проток
Се-менные пузырьки это парные добавочные железы половоrо аппарата. Они
разиваются как выпячивания стенки семявыносящеrо протока, прилеrающие к oc
нованию предстательной железы. Слизистая оболочка семенных пузырьков обра
зует первичные, вторичные и третичные складки, местами срастающиеся между
собой. Выстилающий сетчато ячеистую структуру слизистой оболочки эпителий
однослойный высокий призматический, а иноrда и мноrорядныЙ. Функция ceMeH
ных пузырьков это выработка слабощелочноrо слизистоrо секрета, в котором
304
[лава 17
спер IaТОЗОИДЫ при обретают подвижность. Секрет, примешиваясь к сперме, нейт
рализует и разжижает ее, а также обеспечивает сперматозоиды энерrетическим Ma
териалом фруктозой. За счет жидкости из каждоrо пузырька объем эякулята
.
увеличивается на 1 3 мл.
предстателыlяжелезаa (пpocтaтa) это железисто мышечный opraH, ox
ватывающиЙ со всех сторон верхнюю часть мочеиспускательноrо канала (ypeT
ры). В состав железы входят 30 50 простатических желез трубчато альвеоляр
ной структуры мелкие слизистые, подслизистые и rлавные, или наружные,
обеспечивающие поступление основной части секрета. Концевые отделы и BЫ
водные протоки впадают в мочеиспускательный канал справа и слева от ceMeH
Horo буrорка. В эпителии концевых отделов различают высокие секретирующие
клетки и мелкие вставочные клетки. В выводных протоках эпителий мноrоряд
ный призматический. Секрет предстательной железы нейтрализует кислую pe
акцию спермы и стимулирует двиrательную активность сперматозоидов. CrycT
ки секрета иноrда пропитываются минеральными солями и в старческом возрасте
превращаются в простатические конкреции, или камни предстательной железы.
Предстательная железа содержит rладкую мышечную ткань, на долю которой при
ходится от 1/3 до 1/2 объема opraHa. Мощный мышечный аппарат, окружающий
железистые структуры предстательной железы, обеспечивает своим сокращени
ем выбрасывание секрета в момент эякуляции. В жидкости предстательной же
лезы и семенных пузырьков содержатся простаrландины, повышающие жизне
способность' и функциональную активность половых клеток Кроме Toro, они
вызывают сокращение rладкой мускулатуры, обладают антивоспалительным дей
ствием, влияют на синапсы в центральной нервной системе, модулируют функ
цию некоторых эндокринных opraHoB. В предстательной железе вырабатывается
также фактор, стимулирующий рост нервных волокон.
В месте впадения семявыносящих протоков в мочеиспускательный канал Ha
ходится семенной буюро-к. Это утолщение слизистой оболочки, покрытое переХОk
ным эпителием, содержит мноrочисленные нервные окончания. Раздражение их
вызывает эрекцию ceMeHHoro буrорка, что предотвращает затекание эякулята в
мочевой пузырь. В верхнюю часть мочеиспускательноrо канала открываются про
токи булъбоуретралъ1/.ЪLX желез, выделяющих слизистый секрет.
Половой член
Основу половоrо члена образуют два дорсальных пещеристых (кавернозных)
тела, окруженные фиброзной оболочкой. Третье пещеристое тело, называемое rуб
чатым, расположено вентральнее по средней линии. На дистальном конце оно pac
ширяется, образуя rоловку половоrо члена. В середине rубчатоrо тела проходит
мочеиспускательный канал. Кавернозные тела имеют вид сообщающихся лакун,
заполненных кровью. Внутренние стенки лакун выстланы эндотелием. В лакуны
кровь поступает из концевых артерий, а оттекает по тонкостенным венам в веноз
ные сплетения. Во время эрекции происходит сжатие вен и отток крови из лакун
пещеристых тел прекращается.
Воспроизведеuие человека
305
В мочеиспускателЪ1IOМ канале эпителий имеет неодинаковое строение. Он яв
ляется переходным в простатической части, мноrорядным призматическим в
перепончатой части, однослойным призматическим в пещеристоЙ части, затем
снова мноrорядным призматическим и далее Мllоrослойным плоским. В эпителии
MHoro бокаловидных клеток. Имеются также уретральные железы. вырабатываю
щие слизь.
Реryляция ФУНКЦИЙ яичек
Функциональная активность яичек реryлируется сложной системой нейроим
муноэндокринных факторов.
Нейросекреторные клетки мелкоклеточных ядер rипоталамуса вырабатывают
ropMoH люлиберин, который в пульсирующем режиме с интервалом в 2 ч выделя
ется в кровоток. Люлиберин активирует rонадотропоциты аденоrипофиза. KOTO
рые, в свою очередь, импу льсно секретируют в кровь rонадотропные [ормоны (фОk
ли И лютропин), воздействующие на клетки мишени яичек.
Ведущая роль в реrуляции сперматоrенной функции яичек принадлежит
Фоллuтропuну. Основными клетками мишенями, опосредующими стимулиру
ющие эффекты фОЛЛИТрОПИНd на сперматоrенез, являются клетки Сертоли. Под
воздействием фоллитропина клетки Сертоли синтезируют иrибин, активин, aH
дроrенсвязывающиЙ белок, трансферрин и цитокины. [ормоны инrибин и aK
тивин являются физиолоrическими антаrонистами. Инrибин по принципу OT
рицательной обратной связи подавляет секрецию фоллитропина в аденorипо
физе и, таким образом, тормозит сперматоrенез. Действие активина
противоположное, однако менее отчетливо выражено, чем у инrибина. AHДpO
rенсвязывающий белок поддерживает высокую (в сотни раз большую по cpaB
нению с перитубулярными, или околоканальцевыми, областями) концентрацию
тестостерона, которая необходима для физиолоrическоrо течения спеР!\lатоrе
неза. Тестостерон стимулирует выработку андроrенсвязывающеrо белка (таким
образом он участвует в поддержании постоянства собственной высокой KOH
центрации в семенных извитых канальцах). Тестостерон влияет на спермати
ды, которые он стимулирует к трансформации в сперматозоиды (инициирует
фазу формирования). Важнейший физиолоrический эффект тестостерона co
стоит также в репрессии reHoB апоптоза в половых клетках, что крайне важно
для полноценноrо сперматоrенеза и образования нужноrо для оплодотворения
количества зрелых спермиев.
Продукция тестостерона в клетках Лейдиrа в значительной степени подконт
рольналютропuну аденоrипофиза, хотя определенное паракринное влияние на эту
функцию оказывает также инсулиноподобный фактор роста, синтезируемый клет
ками Сертоли.
Трансферрин это естественный митorен, стимулирующиЙ пролиферацию
сперматоroний, мейоз и спермиоrенез.
В реrуляЦIШ сперматоrенеза важное место принадлежит нервным и иммун
ным факторам.
З06
Fлава 17
OpraHbI женской половой системы
Женская половая система состоит из половых желез (яичников) и opraHoB
половоrо тракта (маточные трубы, матка, влаrалище). Яичники выполняют reHe
ративную (созревание половых клеток) и эндокринную (синтез женских половых
ropMoHoB) функции. Маточные трубы обеспечивают транспорт половых клеток и,
если произошло оплодотворение, зародыша в матку. Матка является opraHoM,
rде зародыш развивается и пребывает до рождения.
Источником развития яичников являются парные утолщения целомическоrо
эпителия на медиальной поверхности первичной почки (мезонефроса), именуе
мые половыми валиками. На 5 6 й неделе половые валики заселяются первичны
ми половыми клетками (rоноцитами), а со стороны мезонефроса в половые вали
ки врастают клетки канальцев первичной почки. Пролиферируя, эти клетки
формируют клеточные тяжи и канальцы внутрияичниковой сети (rete ovarii).
Миrрировавшие в закладку rOHaд половые клетки активно размножаются и
после завершения циклов деления дифференцируются в овоrонии. Важнейшим
событием половой дифференцировки яичника является вступление овorоний в
мейоз. Основополаrающую роль в инициации мейоза иrрает сеть яичника ее
клетки вырабатывают мейоз индуцирующую субстанцию.
К 7 й неделе развития яичник отделяется уrлубляющимися бороздами от мезо
нефроса, начинают фОР 1Ироваться ворота opraHa. Через них в rOHaдy проникают
кровеносные и лимфатические сосуды, нервные волокна. К этому времени происхо
дит компартментализация яичника в ero составе обособляются овариальные фОk
ликулы с половыми клетками и интерстициальная соединительная ткань. Каждая
половая клетка окружается co б
матическими клетками произ
водными сети, образуются при
митивные овОфОЛЛИ-КУЛЯРllые
lUстИО1IЫ примордиальные
фолликулы. Оболочка таких
фолликулов состоит из одноrо
слоя плоских эпителиальных
клеток; внутри фолликула pac
полаrается овоцит 1 ro порядка
в диктиотене профазы 1 ro деле
ния мейоза.
Снаружи яичник покрыт
однослойным кубическим (за
чатковым) эпителием целоми
ческоrо происхождения. Под
эпителием располаrается соеди
Яичники
2
!-. .:.-::.
. . 9
.7
10
.. .. .......
Рис. 137. Схема развития овариальных фолликулов:
1 покровный эпителий яичника: 2 овоrонии; З выселе
ние клеток целомическоrо эпителия; 4 канальцы внутриова
риальной сети; 5 примордиальный фолликул; б первич
ные фолликулы; 7 вторичный фолликул; 8 третичный фол
ликул; 9 желтое тело; 10 атретический фолликул
Воспроизведение человека
307
нительнотканная белочная оболочка. В дефинитивном яичнике различают KOpKO
вое и мозroвое вещество. Корковое вещество занимает периферическую часть и co
держит фолликулы, в которых находятся половые клетки (рис. 137). Мозrовое Be
щество находится в центре. В нем проходят крупные орrанные сосуды и нервы.
ФОЛЛИ"УЛOlе1lез. Фолликулоrенезом называется поступательное развитие
фолликулов от стадии примордиальных до преовуляторных (третичных) фолли
кулов, или rраафовых пузырьков. Термином "овариальный фолликул" обознача
ется клеточно тканевый комплекс (ово соматический rистион), состоящий из жен
ской половой клетки и окружающих ее соматических тканей производных rete
ovarii. Фолликулоrенез возникает в орrанизме женщины лишь с наступлением пе
риода половоrо созревания. Этапность фолликулоrенеза заключается в развитии
примордиальноrо фолликула в первичный, первичноrо во вторичный и далее
вторичноrо в третичный фолликул, или rраафов пузырек (см. рис. 137). В про
цессе фолликулоrенеза половая клетка претерпевает стадию большоrо роста, и на
финальных стадиях фолликулоrенеза возобновляет мейоз.
Развитие примордиальных фолликулов в первичные характеризуется следую
щими признаками: а) возрастают размеры овоuита в связи с накоплением вителли
на и развитием орrанелл; б) фолликулярные клетки, окружающие овоцит, проли
ферируют, увеличиваются их число и размеры, они становятся кубическими или
призматическими, однослойный эпителий дифференцируется в мноrослойный. Так
образуется мноrослойная эпителиальная оболочка фолликулов (фолликулярный
эпителий, или зернистый слой). Как и в примордиальном фолликуле, овоцит пер
вичноrо фолликула пребывает в состоянии блока мейоза, несмотря на то, что ero
рост продолжается и даже активируется. В первичном фолликуле с двухслойным
фолликулярным эпителием происходит образование прозрачной зоны овоцита, в
состав которой входят кислые и нейтральные rликозаминоrликаны. Прозрачную зону
пронизывает множество каналов разной величины, в которых проходят uитоплаз
матические отростки фолликулярных клеток к овоциту, обеспечивающие полноцен
ную трофику половой клетки.
В процессе образования первичноrо фолликула BOKpyr зернистоrо слоя распо
лаrаются соединительнотканные клетки и создают внешнюю соединительноткан
ную оболочку фолликула теку. Формирование теки знаменует становление pe
rиональноrо кровоснабжения и иннервации фолликула.
В стадии вторичноrо фолликула в фолликулярном эпителии активизируются
пролиферативные процессы, тека дифференцируется на внутреннюю и наружную
части, совершенствуется нейро трофическое обеспечение, формируется фоллику
лярная полость, в мноrослойном фолликулярном эпителии обособляется область,
несущая половую клетку, яйценосный буlOрОК, или кумулюс.
Третичные фолликулы (rраафовы пузырьки, преовуляторные фолликулы) xa
рактеризуются высшей степенью развития и подrотовкоЙ фолликула к овуляции.
Фолликул имеет крупные размеры, большую полость, заполненную жидкостью, ис
тонченную стенку. Овоцит 1 ro порядка со стороны полости окружен одним тремя
слоями фолликулярных клеток лучистым венцом, и постепенно отходит от KYMY
люса. Сосуды теки характеризуются кровенаполнением. В овоците третичноrо
308
Тлава 17
фолликула репнициируется мейоз. Завершается первое и начинается второе эк
вационное деление меЙоза, возникает овоцит 2 ro порядка и первое редукционное
тельце. Общая продолжительность развития примордиалыюrо фолликула ПО CTa
дин преовуляторноro фолликула у человека составляет около 120 суток
В фолликулах, стенка которых состоит из фолликулярноrо эпителия и теки, ocy
ществляется синтез половых ropMoHoB. ОбразующиЙся эстрадиол частично TpaHC
ПОРТllруется в кровь, частично выделяется в полость фолликула. От внутрифолли
кулярной концентрации эстраДllола во MHoroM зависит судьба развивающеroся
фолликула чем выше эта концентрация, тем фолликул имеет больше возможнос
тей продолжить развитие и достиrнуть стадии зрелоrо rpаафова пузырька. Фолли
кулы с низким реrиональным уровнем эстрадиола (фолликулы с андроrенным про
филем), как правило, подверrаются атрезии (поrибают).
Оптимальные условия для развития женских половых клеток создает leMamo
фОЛJlИКУЛЯрllЫЙ барьер (шстUОll), в которыЙ входят: эндотелиоциты rемокапилля
ров теки, базальная мембрана эндотелия, интерстициальные элементы теки, базаль
ная мембрана фолликулярноrо эпителия, фолликулярные клетки купулы и
лучистоrо венца, прозрачная зона.
Овуляция представляет собоЙ rормонально зависимыЙ процесс разрыва стенки
третичноrо (преовуляторноrо) фолликула и выхода женскоЙ половоЙ клетки в Ma
точную трубу. В момент овуляции овоцит 2 ro порядка находится на стадии метафа
зы BToporo деления меЙоза. Если овоцит 2 ro порядка не оплодотворяется, он поrи
бает, так и не закончив меЙоз. Завершение меЙоза возможно лишь при условии
оплодотворения (под влиянием активирующеrо начала мужскоЙ половоЙ клетки).
Развитие желтОlО тела (лютеОlеllез). После овуляции стенка фолликула
частично спадается, фолликулярныЙ эпителий и внутренняя часть теки собирают
ся в складки, исчезают фраrменты разrраничивающеЙ их базальной мембраны. По
лость фолликула заполняется оставшеЙся фолликулярноЙ жидкостью и формен
ными элементами крови. Происходит размножение эпнтелиоцитов зернистоrо слоя
и клеток внутреннеЙ части теки. Сосуды и клетки внутреннеЙ части теки прорас
тают в толщу фолликулярноrо эпителия. Далее желтое тело вступает в стадию же
лезистоrо метаморфоза эпителиоциты зернистоrо слоя rипертрофируются, Ha
капливают орrанеллы стероидноrо синтеза и дифференцируются в лютеоциты.
Увеличение размеров и количества клеток, а также врастание стромальных компо
нентов теки в область бывшеrо фолликулярноrо эпителия приводят к закрытию
полости и формированию соединительнотканноrо центра желтоro тела.
Следующая стадия активноrо функционирования желтых тел продолжа
ется в зависимости от Toro, наступает оплодотворение или нет. В случае оплодо
творения желтое тело существует и секретирует проrестерон в течение нескольких
месяцев (до начала проrестеронообразования в плаценте) и именуется желтым Te
лом беременности. Если беременность не наступает, желтое тело функционирует
лишь в течение нескольких (4 7 и) суток и подверrается инволюции. У человека
соединительнотканные рубцы (белые тела) свидетельства лютеолиза coxpa
няются в течение мноrих месяцев и лет жизни, и по их количеству в яичнике мож
но косвенно судить о числе овуляциЙ.
Воспроизведение человека
309
. Атрезuя (zuбель) фоллuкулов. Различают атрезию малых (примордиаль
ных первичных, или однослойных) и крупных (имеющих зернистый слой) фол
ликулов. Атрезия фолликулов ранних стадий развития называется деrенерацион
ной, поскольку возникает деструкция и rи6ель овоцита и ею окружения. Атрезия
фолликулов с зернистым слоем протекает иначе здесь формируются атретичес
кие тела, клетки которых активно продуцируют андроrены и небольшие количе
ства эстроrенов.
Матка
,
Стенка матки состоит из трех оболочек: внутренней слизистой, или эндо
метрия, средней мышечной, или миометрия, и наружной серозной, или пери
метрия.
Эндомеmрий является наиболее динамичной оболочкой, так как циклически
перестраивается под влиянием разных концентраций половых ropMoHoB. Он co
стоит из однослойноrо призматическоrо эпителия целомическоrо типа и c06CTBeH
ной пластинки слизистой оболочки, содержащей маточные железы. Циклические
изменения эндометрия человека и высших приматов сопровождаются маточными
кровотечениями, в связи с чем половые циклы именуются менструальными
(рис. 138). В межменструальный период эн 1 1
дометрий имеет толщину около 1 2 мм. Bco
ставе эпителия присутствуют три вида кле ,\п); _::::' ':О.;./>:i
ток реснитчатые эпителиоциты, слизистые > :::аJ!:::' ,;: r{::::Я
экзокриноциты и эндокриноциты. О:'. T\ ",V':'IJ..
Маточные железы, являясь производны ...;;..02{rO,:" 11':} { .. . '-';:' , ",:,<;:
ми эпители эндометрия, залеrаю в rлубине '.". :.f;:.:':'.:,.q':'.' i : CI' li:;': So. i
собственнои пластинки слизистои оболочки. "\: :>" О ' '..,.: 4 ;;; ",--,>:,,:;:"'.::: .':; :'::". , .
Численность и плотность расположения желез f41 . .::-t::' 'i\;,':; :'i . fi "":::: r.;1 /)" ":: :; ;:i'
варьирует в зависимости от фазы половоrо :,. Ч:' J i '''' : ::Ji:: и:
цикла. Развитие и функционирование желези "c=;..,j li '; :,'C:,:': '1 ,...,; %.щ
cToro аппарата матки в большей степени ПОk ':;..};. .;.:-:( t Й
контрольно roрмону желтоrо тела проrесте -:" 3 . ; . ...> U \?:::?;i
. .<.' 0'0 Q. . .' .0." :. '.. .;);;,\..
рону, поэтому максимальное развитие желез ;;.;: . .:4 :. \\ . .:....;i:'>\ /:r
iI:- ". .о.'}).;.... . ... ..,
отмечается во 2 й половине менструальноrо :' 4f"P ::; :';) ' .;i"';' ; \;';:': !;
цикла, коrда концентрация этоrо ropMoHa oco ; ,:: ( 1 ::( )[j.' i' t;;.t;i
бенно высока. Эта часть цикла в связи с актив );::\. ::i:. 9 !.'. :щ
ной секреторной деятельностью маточных же :-: 1:; :' : : 1t;..
лез пол у чила название сек р ето р ной Ф азы. . \f:.1; ; ; ';;: . ':;':
. ...-.0 .......'r., . ....-...J.:;;r--_....
Маточны железы относятся к простым труб .Б...,',.... . . ,
чатым железам. В постменструальный период
они имеют прямолинейный ход, а в предмен
струальном периоде удлиняются и штопо
ро06разно закручиваются. Клеточный состав
желез аналоrичен составу эпителия слизистой
Рис. 138. Строение стенки матки:
А эндометрий в 1 й половине менструаль.
Horo цикла; Б эндометрий во 2 й полови
не менструальноrо цикла; 1 покровный
эпителий; 2 маточные железы; 3 сосу.
ды; 4 миометрий
310
Fлава 17
оболочки матки. Маточные железы с секреторными клетками покровноro эпителия
образуют маточную жидкость, которая представляет собой белково rликозаминоr
ликановый комплекс.
Собственная пластинка слизистой оболочки матки состоит из рыхлой волокни
стой соединительной ткани. В период секреторной фазы цикла вся толща собствен
ной пластинки пронизана мноrочисленными маточными железами их донные OT
делы достиrают миометрия. В течение половоro цикла значительные изменения
претерпевает клеточный состав соединительной ткани эндометрия. В предимплан
тационный период клетки собственной пластинки проявляют признаки децидуали
зации располаrаются rруппами, увеличиваются в размерах, накапливают rлико
reH, в них появляются рецепторы к ropMoHaM, они формируют множественные
контакты типа нексусов и десмосом. Дифференцировка децидуальных клеток rop
монозависимый процесс ее активируют ropMoHbI rруппы проrестерона и HeKOTO
рые биолоrически активные соединения (rистамин, простаrландины), синтезируе
мые в эндометрии и бластоцисте.
Слизистая оболочка матки боrато васкуляризирована. При менструации наруж
ный слой эндометрия, именуемый функциональным, некротизируется и отторrает
ся. Внутренний базальный слой, с сохраняющимися в нем донными отделами Ma
точных желез остается и участвует в процессе физиолоrической реrенерации
внутренней оболочки матки в постменструальном периоде.
Мuометрuй состоит из трех слоев rладкой мышечной ткани, между которыми
располаrаются прослойки рыхлой соединительной ткани. Из за отсутствия ПОk
слизистой основы миометрий неподвижно соединен с базальным слоем собствен
ной пластинки слизистой оболочки матки. Внутренний мышечный слой, располо
женныЙ под слизистой оболочкой, состоит из косо ориентированных пучков
rладких миоцитов, в среднем слое они имеют циркулярное направление, а в Ha
ружном субсерозном также косопродольное направление, противоположное
направлению во внутреннем слое. Между слоями мышечной ткани нет резких rpa
ниц. Здесь находятся крупные кровеносные сосуды. При сокращении матки cocy
ды пережимаются, что предотвращает кровотечение при менструации и родах. Эс
TporeHbI повышают электрическую возбудимость rладких миоцитов, а проrестерон,
напротив, увеличивает пороr возбудимости данных клеток
Перuметрий серозная оболочка матки, покрывает значительную часть opraHa,
за исключением передней и боковых поверхностей надвлаrалищной области. В фор
мировании периметрия участвуют мезотелиЙ и рыхлая волокнистая соединитель
ная ткань.
Шейка матки представляет собоЙ нижнюю суженную часть и имеет вид MЫ
шечноrо цилиндра. В центре шейки проходит шеечный, или цервикальный канал,
который начинается в полости тела матки внутренним зевом. Дистальная часть
шейки матки выступает во влаrалище и заканчивается наружным зевом. Шейка
матки состоит из тех же оболочек, что и тело. Канал шейки матки выстлан OДHO
слойным призматическим эпителием, который в области дистальной (влаrалищ
ной) части шейки соединяется с мноrослоЙным плоским неороrовевающим эпите
лием. Последний продолжается в эпителиЙ слизистой оболочки влаrалища. [раница
Воспроизведеllие человека
311
между мноrослойным и однослойным призматическим эпителием слизистой обо
лочки всеrда четкая и ориентировочно располаrается на уровне дистальной части
шейки.
Маточные трубы
3
.{
,
. .,
(,
#',
Рис. 139. Строение стенки ампулярной части маточной трубы:
1 слизистая оболочка; 2 мышечная оболочка и ее внутренний цирку
лярный и наружный слои; 3 серозная оболочка; 4 просвет трубы; 5
складки слизистой оболочки; 6 реснитчатые эпителиоциты; 7 ceKpe
торные эпителиоциты; 8 собственная пластинка слизистой оболочки
Маточная труба (яйцевод) парный трубчатый opraH, дистальный конец KOTOpO
ro, имеющий вид воронки, открыт и контактирует с поверхностью яичника, а прокси
мальный прободает стенку матки в области боковых поверхностей ее дна и сообща
ет трубы с маточной полостью. У человека длина маточных труб составляет около
1 o 12 см. Маточные трубы захватывают овоцит при овуляции, осуществляют ero тpaHC
порт по направлению к полости матки, создают условия для беспрепятственноrо
312
Fлава 17
продвижения спермиев навстречу овоциту, обеспечивают среду, необходимую для оп
лодотворения и дробления зародыша, транспортируют зародыш в полость матки.
Маточные трубы развиваются из верхней части парамезонефральных (мюллеровых)
протоков.
Маточная труба подразделяется на 4 отдела: воронку дистальный отдел трубы,
заканчивающийся бахромками (фимбриями) и открывающийся в яичниковую сумку,
ампулу следующую за воронкой наиболее широкую и протяженную часть (COCTaв
ляет около 2/3 длины трубы), перешеек, или истмус, и интерстициальный (интраму
ральный) отдел, прободающий стенку матки.
Стенка маточной трубы состоит из трех оболочек: слизистой, мышечной и ce
розной (рис. 139).
Слuзuсто.я оболочка состоит из однослойноrо призматическоrо эпителия целоми
ческоrо типа и собственной пластинки. Эпителий образован клетками двух видов pec
нитчатыми и секреторными. По ходу маточной трубы реснитчатые и секреторные эпи
телиоциты располаrаются неравномерно реснитчатые преобладают в воронке и ампуле
трубы, а секреторные в области перешейка. Для секреторных эпителиоцитов маточ
ных труб характерны апо и мерокринный типы секреции. Основными компонентами
секрета являются преаль6умины, трансферрин, rлобулин и липопротеиды, а также rли
козаминоrликаны, простаrландины, утероrлобин.
Собственная пластинка слизистой оболочки труб тонкая и образована рыхлой
волокнистой соединительной тканью. Помимо типичных для этой ткани видов кле
ток, в ее составе обнаруживаются клетки, способные к децидуальной трансформации.
Мышечная оболочка маточных труб образована двумя нерезко разrраниченны
I\Ш слоями rладкой мышечной ткани внутренним циркулярным (более толстым)
и наружным продольным (более тонким). Толщина мышечной оболочки возраста
ет от воронки к перешейку. В области перешейка внутренний циркулярный слой
фОР lИрует круrовую мыщцу маточной трубы. В случае имплантации зародыша в
стенку трубы последняя леrко травмируется и разрывается.
Серозная оболочка представлена мезотелием и соединительной тканью.
Влаzалuще
Стенка влаrалища состоит из слизистой, мышечной и адвентициальной обо
лочек. Эпителий слизистой оболочки мноrослойный плоский неороrовеваю
щий кожноrо типа. В эпителии различают 3 слоя: базальный, промежуточный и
поверхностный. Клетки поверхностноrо слоя содержат мною rликоrена, при pac
паде KOToporo под влиянием микробов образуется молочная кислота. Это прида
ет кислую реакцию и бактерицидные свойства слизи. Подслизистая основа
во влаrалище отсутствует. Мышечная оболочка состоит из продольно и частично
циркулярно идущих пучков rладкой мышечной ткани. Адвентициалъная оболоч
ка образована волокнистой соединительной тканью.
Преддверие влаrалища, а также малые ryбы выстланы мноrослойным плоским
эпителием. В преддверии открываются две бартолиниевы железы, выделяющие
слизь. Большие rубы представляют собоЙ складки кожи с прослойками жировой
Воспроизведение человека
313
ткани. Клитор состоит из двух пещеристых тел. IЪловка ero покрыта мнorослоЙ
ным плоским частично ороrовевающим эпителием. Содержит большое количество
чувствительных нервных окончаний.
Овариально менструальный цикл
в период репродуктивной зрелости (с 13 15 до 40 45 лет) орrанизм женщины
периодически подrотавливается к зачатию и вынашиванию плода. Физиолоrичес
ки это проявляется в циклических перестройках репродуктивной системы, имену
емых овариально менструальными циклами. Циклические процессы, происходя
щие в яичниках и матке, взаимосвязаны, синхронизированы во времени и
контролируются rипоталаl\ю rипофизарной нейросекреторной системой.
В овариально менструальном цикле выделяют фолликулярную и лютеиновую
фазы (рис. 140). В фолликулярной фазе цикла под влиянием люлиберина и rOHa
дотропных ropMoHoB аденоrипофиза в яичнике развиваются вторичные (антраль
ные) фолликулы, а также один третичный, или предовуляторный, фолликул, KO
торый подверrнется овуляции. В лютеиновой фазе цикла из оболочек проовули
ровавшеrо (разорвавшеrося) фолликула развивается желтое тело, клетки KOTOporo
вырабатывают проrестерон. Эстроrены, а затем проrестерон, последовательно
воздействуют на эндометрий и вызывают ero преrравидарную перестройку (от лат.
gravidarum беременность), которая заключается в rипертрофии и децидуализа
ции слизистой оболочки матки, усилении ее кровоснабжения, развитии и ceKpe
ции маточных желез.
В стенке матки в течение овариально менструальноro цикла различают MeHCTpy
альную (десквамативную), постменструальную (пролиферативную) и предменстру
альную (секреторную, преrравидарную) фазы. В менсmруGЛЫЮЙ фазе происходит
жт
ПФ
жт
о
5
ело
314
Fлава 17
отторжение (десквамация) поверхностноrо функциональноrо слоя эндометрия, KO
торое сопровождается маточным кровотечением. Продолжительность менструальной
фазы цикла у человека в среднем составляет около 4 x суток К концу менструальной
фазы в орrаНlIЗме возрастает секреция фоллитропина, что стимулирует в яичнике рост
вторичных фолликулов. Синтезируемые фолликулами эстроrены индуцируют pere
нерацию функциональноrо слоя эндометрия. В результате активной пролиферации
клеток донных отделов маточных желез не только полностью восстанавливается же
лезистый эпителий матки (достраиваются тела, шейки и устья желез), но и практичес
ки заново создается слизистая оболочка. Наиболее активно пролиферативные про
цессы происходят на 5 11 e сутки после окончания менструации, затем слизистая
оболочка матки в течение 3 4 x суток (11 14 e сутки цикла) пребывает в состоянии
относительноro покоя.
В постмеllструалыюй фазе цикла эндометрий не достиrает необходимоro ypOB
ня развития, так как маточные железы недостаточно дифференцированы, клетки
желез практически не вырабатывают слизь и содержат мало rликоrена. Эпителий
слизистой оболочки представлен призматическими клетками, среди которых мало
реснитчатых и доминируют малодифференцированные безреснитчатые. Большин
ство эпителиальных клеток обнаруживают признаки слабой секреторной актив
ности. Средняя продолжительность менструальной и постменструальной фаз цикла
у человека составляет около двух недель.
Следующая предмеllструалЪ1lая фаза продолжается также около 14 и суток
и знаменует собой завершение подroтовки репродуктивной системы к беременно
сти. Клетки желтоrо тела начинают вырабатывать проrестерон. Прorестерон сти
мулирует преrравидарную перестройку слизистой оболочки матки. Под ero влия
нием увеличиваются в размерах, приобретают характерную извитую форму и
ветвятся маточные железы. Их клетки накапливают rликоrен, набухают и начина
ют активно секретировать в полость матки ryстую слизь, которая необходима для
имплантации, трофики и активации развития зародыша. В составе эпителия YBe
личивается численность реснитчатых эпителиоцитов, большинство из них ceKpe
торно активны. Существенно возрастает кровоснабжение эндометрия. В соедини
тельной ткани собственной пластинки слизистой оболочки под влиянием
проrестерона дифференцируются децидуальные клетки. К концу предменструаль
ной фазы цикла толщина эндометрия увеличивается более, чем в два раза по cpaB
нению с толщиной в начале постменструальной фазы. Если происходит оплодот
ворение, то rипертрофированное состояние эндометрия поддерживается в течение
6 8 недель, что способствует развитию плаценты. Если оплодотворение не HaCTY
пает, функциональный слой слизистой разрушается и во время менструации OT
торrается.
Менструальная фаза и менструальное кровотечение индуцируются rормональ
ными перестройками и связанными с ними нарушениями кровоснабжения эндо
метрия. Отсутствие зачатия является основной причиной прекращения rормональ
ной деятельности желтоrо тела. Дефицит проrестерона приводит к резкому спазму
артериальных сосудов функциональноrо слоя эндометрия, развитию ишемии и
некроза этоrо слоя. Собственная пластинка слизистой оболочки матки инфильт
Воспроизведение человека
315
рируется лейкоцитами. Через некоторое время спиральные артерии функциональ
Horo слоя расслабляются, однако из за нарушения целостности их стенки, которое
возникло в результате спазма, в строму выходят первые порции крови, знаменую
щие начало менструации. В течение десквамативной фазы цикла весь Функцио
нальный слой эндометрия некротизируется, постепенно отторrается и вместе с KpO
вью удаляется из полости матки. Менструальную фазу цикла в rормональном
отношении рассматривают как стадию покоя, поскольку в это время желтое тело
не функционирует, а новая популяция вторичных фолликулов потенциальных
источников эстроrенов, еще не начала развитие. Как только падение KOHцeHTpa
ции проrестерона в периферической крови достиrает критическоrо уровня, в яич
никах под влиянием выброса фоллитропина активируется рост новой rруппы BTO
ричных фолликулов И начинает повышаться концентрация эстроrенов. В матке
вновь инициируется восстановительный процесс и подютовка слизистой оболоч
ки к имплантации зародыша.
Эмбриональное развитие человека
Онтоzенез, или индивидуальное развитие, включает пренатальный (внутри
утробный) период, который длится примерно 280 суток, или 1 О лунных месяцев, и
постнатальный (внеутробный) период, продолжительность KOToporo у разных
людей варьирует и во MHoroM определяется как внутренними, так и внешними по
отношению к человеку факторами.
Изучение пренатальноrо развития человека (эмбриоrенеза) встречается с ря
дом трудностей, связанных не только с получением необходимоrо для исследова
ний материала, но этическими и релиrиозными нормами, существующими в обще
ственном сознании. Ранние зародыши человека "редкая находка". Только в 1944 r.
впервые был исследован 7,5 суточный зародыш человека, а в 1946 r. 2 5 суточ
ные зародыши. Наиболее полная коллекция человеческих эмбрионов находится в
институте Карнеrи (Балтимор, США). Описания ранних эмбрионов человека даны
rертиrом, Рокком и Стритером. В отечественной эмбриолоrии ранние стадии раз
вития зародыша человека изучены и описаны А.[ Кнорре (зародыш "BMA l") и
БЛ. Хватовым (зародыш "Крым"). Развитие технолorии искусственною оплодот
ворения позволило в деталях исследовать механизмы оплодотворения и дробле
ния зиrоты у человека.
Оплодотворение (фертилизация)
у человека оплодотворение внутреннее. По клиническим наблюдениям, зача
тие чаще всею происходит у женщин вплоть до 2 й недели после менструации, хотя
друrие авторы указывают в качестве наиболее подходящих для зачатия сроков 11
17 e сутки менструальноrо цикла.
В результате rаметоrенеза у человека образуется rенетически однородная по
пуляция овоцитов (яйцеклеток), содержащих 22 соматические и одну половую
Х хромосомы; и два рода спермиев с различными rенетическими характеристиками
316
Fлава 17
(22+ Х и 22+ У). После
дние образуются в paB
ном количестве, поэтому
яйцеклетка имеет стати
стически равные возмож
ности на встречу как с
Х , так и с У спермием и,
соответственно, ожидает
ся рождение мальчиков и
девочек в равном COOTHO
шении. Однако физиоло
rические условия опло
дотворения вносят по
правку в эти результаты
(100: 106 в пользу рож
дения мальчиков).
Процесс направлен
Horo персмещения спер
миев по opraHaM женско
ro половоrо тракта из
влаrалища в маточную
трубу продолжается OKO
ло 1 О часов и является,
по сути, преодолением
клетками с оrраничен
ными метаболическими
потенциями orpoMHoro расстояния. Блаrодаря тому, что в эякуляте содержится в
среднем около 200 300 млн сперматозоидов, существует высокая вероятность тоro,
что небольшая часть спермиев (около 1 % от первоначальноrо числа) сохранит жиз
неспособность, достиrнет маточной трубы и будет участвовать в оплодотворении.
Скорость самостоятеЛЬНОl'О передвижения спермиев очень невелика около 2
4 мм/мин.
Женская половая клетка при овуляции попадает в маточную трубу блаrодаря
набуханию фимбрий и их тесному контакту с поверхностью яичника.
При взаимодействии сперматозоидов с орrанами женскоrо репродуктивноrо
тракта поисходит их 'Капацитация приобретение оплодотворяющей способнос
ти. При капацитации под влиянием секреторных продуктов женскоrо половоrо
тракта с поверхности сперматозоидов удаляются вещества, которые блокируют pe
цепторно трансдукторную систему сперматозоида, взаимодействующую с повер
хностью женской половой клетки. Собственно процесс оплодотворения условно
разделяют на фазы дистантноrо и KOHTaKTHoro взаимодействия и завершается
оплодотворение активацией метаболизма зиrоты.
В фазе дистантною взаимодействия происходит встреча половых клеток (ra
мет) в половых путях женщины. Важными механизмами дистантноro взаимодей
r
д
в
Е
Рис. 141. Оплодотворение яцеклетки (схема):
А первое деление мейоза, формирование первоrо полярноrо тельца;
Б овоцит после овуляции в блоке метафазы 2 ro деления мейоза, KOH
такт со спермием приводит к снятию блока мейоза и ero завершению;
В выделение BToporo полярноrо тельца, образование rаплоидноrо
женскоrо пронуклеуса, митотическое деление первоrо полярноrо тель'
ца; r сближение мужскоrо и женскоrо пронуклеусов, репликация xpo
мосом, подrотовка к митозу (первому делению дробления); Д митоз
зиrоты реплицированные хромосомы располаrаются в экваториаль,
ной плоскости, объединение мужскоrо и женскоrо rенетическоrо MaTe
риала (синкарион); Е анафаза митоза, при завершении деления зи
rOTbl возникнут две диплоидные клетки первые бластомеры
Воспроизведеиие человека
317
ствия являются положительные xeMO и реотаксис, а также электростатическое
взаимодействие [амет (на близком расстоянии).
В фазе контактною взаимодействия спермиЙ разрушает оболочки овоцита
лучистый венец, прозрачную зону и плазмолемму (рис. 141). В процессе оплодотво
рения не должно про изойти полиспермии проникновения в женскую половую
клетку нескольких сперматозоидов. Полаrают, что первый этап взаимодействия спер
миев с женской половой клеткой это механическое удаление части клеток лучис
TOro венца, которое осуществляют биения жryтиков спермиев. Дальнейшие собы
тия KOHTaKTHoro взаимодействия связаны со взаимодействием рецепторов двух
клеток, акросомной реакцией сперматозоида и кортикальной реакцией женской по
ловой клетки. При контакте с женской половой клеткоЙ, под действием активирую
щих субстанций (одна из которых фертилизин), инициируется активное поступ
ление катионов кальция в rоловку спермия. В результате происходят очаrовые
слияния плазмолеммы овоцита и акросомалыюй мембраны сперматозоида и их раз
рушение с появлением микроперфораций. Через образовавшиеся микроотверстия
выделяются фермеIlТЫ ЛИЗИНЫ сперм ия (rиалуронидаза, трипсиноподобный фер
мент и др.), которые разобщают контакты между клетками лучистоro венца, а также
между ними и плазмолеммой овоцита. Диссоциация лучистоrо венца проrpессирует
и наконец обнажается небольшой участок более rлубоко расположенной прозрач
ноЙ зоны. Выделяемый акросомой спермия акрозин разрушает в этом участке rли
козаминоrликаны прозрачной зоны и образует "окно", через которое сперм ий может
проникнуть к женской половой клетке. Пенетрация прозрачной зоны продолжается
около 20 минут. После разрушения участка прозрачной зоны сперм ий попадает в
заполненное жидкой средой перивителлиновое пространство между прозрачной зо
ной и плазмолеммой овоцита. В месте соприкосновения roловки спермия с плазмо
леммой овоцита цитоплазма женской половой клетки формирует выступ буюрок
оплодотвОРe1lИЯ (в этом участке овоцита активируется полимеризация актина) и здесь
же происходит слияние внешних мембран женской и мужской [амет. Слившиеся
участки мембран затем разрушаются и через образовавшееся отверстие спермий про
никает в женскую половую клетку. Ero плазмолемма при этом "сползает" и закрыва
ет дефект, образовавшийся в плазмолемме овоцита. Из цитоплазматических CTPYK
тур спермия KpUMe ядра в овоцит попадают проксимальная центриоль и шейка (хвост
остается снаружи и отпадает). Блаrодаря тому, что участок мембраны, привнесен
ный в плазмолемму овоцита спеРJl.шем, хорошо про ни цаем для катионов натрия,
последние начинают активно поступать внутрь женской половой клетки и изменя
ют ее мембранный потенциал. В течение очень KopoTKoro времени (около 1/10 сек)
мембранный потенциал овоцита резко падает, и женская половая клетка становится
невосприимчивоЙ к контактам с друrими спермиями. Затем наступает кортикалъ
иая реакция ОВОЦlпа. Это происходит в результате поступления катионов кальция 8
женскую половую клетку, что вызывает СЛlIяние мембран кортикальных rранул с
плазмолеммой овоцита и экзоцитоз их ферментов в перивителлиновое простран
ство. При этом прозрачная зона уплотняется, утолщается, теряет рецепторные беk
ки к сперматозоидам. Так возникает оболочка оплодотворения, препятствующая про
никновению друrих спермиев в овоцит.
318
rлава 17
в момент встречи со спермием овоцит находится в блоке метафазы BToporo мейо
тическоrо деления. После проникновения спермпя в овоплазму женская половая клетка
завершает второе деление созревания. При этом выделяется полярное тельце с лиш
ними хромосомами. Пока овоцит завершает мейоз, про нуклеус спермия окруrляется
и принимает вид интерфазною. В нем происходит синтез ДНК, и пронуклеус приоб
ретает набор двойных (реплицированных) хромосом. Ядро женской половой клетки
по завершении BToporo деления мейоза претерпевает точно такие же изменения. За
тем оба пронуклеуса переходят в профазу митоза. Центриоль, привнесенная сперми
ем, делится, формируя две центросомы. Последние прикрепляются к веретену деле
ния, образующемуся между пронуклеусами и, таким образом, хромосомы мужскоrо и
женскою пронуклеусов оказываются расположенными в экваториальной плоскости
происходит метафаза митоза. Далее следуют aHa и телофаза зиrота завершает пер
вое деление дробления, в результате KOToporo образуются первые две дочерние клет
ки бластомеры каждая с диплоидным набором хромосом.
Дробление
Дробление представляет собой серию митотических делений зиrоты с образова
нием мноrих дочерних клеток (бластомеров) меньшею размера. Митотические дe
ления зиrоты, а в последующем бластомеров происходят с увеличением числа
клеток, но без увеличения их массы, поэтому именуются дроблением. У человека
дробление не имеет принципиальных отличий от TaKOBoro у друrих представителей
позвоночных, однако протекает юраздо медленнее. Дробление полное, или rолобла
стическое (борозды дробления проходят через весь зародыш), неравномерное (в pe
зультате дробления образуются дочерние клетки бластомеры неравной величи
ны) и асинхронное (разные бластомеры дробятся с различной скоростью, поэтому
зародыш на отдельных стадиях дробления содержит нечетное число клеток). Пер
вое деление дробления продолжается в среднем около 30 часов, последующие бо
лее кратковременны (около 20 24 часов). В процессе дробления зародыш перемеща
ется по маточной трубе и на 6 e сутки развития попадает в полость матки.
Бластомеры первой rенерации у человека, как и зиrота, тотипотентны (каж
дый бластомер способен развиться в полноценный орrанизм). До стадии 8 бласто
меров клетки зародыша формируют рыхлую неоформленную rpуппу, и только после
третьею деления устанавливают между собой плотные контакты, образуя компак
тный клеточный шар из 16 бластомеров, именуемый м-орулой. Компактизация co
здает условия для развития наружной клеточной массы и внутренней клеточной
массы. Последняя это материал будущеrо тела зародыша (эмбриобласта) и BHe
зародышевых opraHoB. Бластомеры наружной клеточной массы мелкие и MHoro
численные (их примерно в 1 О раз больше, чем клеток внутренней клеточной Mac
сы), являются источником развития трофобласта.
Коrда морула попадает в проксимальный отдел маточной трубы и далее в
полость матки, через ее прозрачную зону начинает проникать содержащаяся в Ma
точной трубе и матке жидкость. Происходит кавитация морулы. Сначала жидкость
накапливается между клетками и образует небольшие промежутки, которые затем
Воспроизведение человека
319
сливаются в единую полость внутри морулы (бласто
цель). В образовании жидкости и кавитации участву
ют также клетки трофобласта, секретирующие жиk
кость. С момента появления полости зародыш имену
ется бласmоцисmой. Клетки внутренней клеточной Mac
сы бластоцисты локализованы на одном из полюсов и
обращены в полость. Клетки наружной клеточной Mac
сы уплощаются и, оrраничивая полость, формируют
оболочку бластоцисты трофобласт (рис, 142), В пе
риод перемещения дробящеrося зародыша по маточ
ной трубе большое значение имеет тот факт, что coxpa
няющаяся прозрачная зона предотвращает прилипание
бластоцисты к стенкам трубы и зародыш попадает в
полость матки. Здесь он освобождается от прозрачной
зоны и начинает имплантироваться (поrружаться) в слизистую оболочку матки.
Имплантация зародыша протекает параллельно с rаструляцией (табл. 3).
Имплантация
1
:.t .. ' .. .' / .. /
/ "" (T ' "
" :.,,,= .. . ,.
........ tQY'
1'<
"
3
&'
,
2
. O
-
Рис. 142. Бластоциста человека:
1 эмбриобласт; 2 трофо-
бласт; 3 полость бластоцисты
у человека имплантация поzружная, или интерстициальная (рис. 143), За одни
сутки бластоциста поrружается в эндометрий почти наполовину, а через 40 часов
полностью. Дефект, образовавшийся в слизистой оболочке матки, восстанавлива
ется в течение 5 суток. Условно имплантация состоит из двух фаз прилипания
,' -T""""--..
. .. _.J.f!'IД"" -'\, -'"' .,'. - )..
";\"!'iw.' . ,...........;,;:::::J. "1.. / ' " 4
".\_ ,< -:!. . .... ; \-=: \ ,.
;/.;, :'$" _____ 2 <F , <,...
$?, ',,.,.,,, , jo-;_".т-; 1 . ., и& '.' .J"'б
t1 -';;-,:Z ::'" "" <"'--фJ....... ./ы .--'" ,. < \ ..,r
.. /"......J< ,", '" '" jr. ' ... '.. ..... ) ,.
"" -: ."; ,, t$ V. ,"Ф -" (flJ---. - \ 8fI
"',,' ') .Q.i"\.....@ r ...... е ::, . Ii'H) ' ,cr,'.
$;!': (b ":. ,'- (1 у'1 з -: i J7.; d ' " \ ' I
,..rra' t;> \ - 4t,.'t / \$& (::' \ \.7! ! /,
1.." . 0...:' t?" .' , '0:;(" \ ' ""' ' '" '1..1
li:iI ,,-'\ , . '!i / ". ;-.-:\ \ .. /? ,f ".,"to l 7
, ..... '.. ;'0$'-..';,\ ) 6'\.}, *' " ,...... [ l' '..
...... .., & ;.а: 8' а .' '" "'...... ,..." ..
'\< а. \.. r .-.." ,: r.. ,,<tt-., ..;.:: "1,..... """..,.....
.'" .. '\ \,р ,........ ",,," -' :"'" "",""t:" ,,!", ....., .. ;............."........""...
'\.. . .,,, j -:. ...... JII.i':: .. "'..... .... ..... "" ." ",:" """.". '" >6' .... I
., & У ...... \. .\t ;:;"'" it '" ..,. у .. . "".... ........, J 6' """.,. .... .# .. .
.. '. ,,-,,, '.. ;;.9-" "'" ...., .... "" ...... . . ....
\( ",.':'"" ::! ."..,........."", " "'" <9> '" N.... . "-1
. , ... ш..... ... ..." ,,'"
f' h .. ". .". '"' : "'" ....., .-'01> ... ... ., <,щ., 8 <IP . .. '
(;...i. \;.. "" 4 .<39 б '. "4:/ . ... j _ 1
f .<OiI:>' "".,..,,' " I 9
.... "'., <" , .,. OfI>. ,
...."" е/ е"...,е JtJI ... tJ> '1 '"
.... е 6\""';;;"' ... I .. J
" fIJ;
Рис. 143. Схема имплантации бластоцисты человека на 7,5 сутки развития:
1 эпибласт; 2 rипобласт; 3 симпластотрофобласт; 4 цитотрофобласт; 5 aM
ниотический пузырек; б эпителий слизистой оболочки матки; 7 базальный слой сли
зистой оболочки матки; 8 кровеносный сосуд; 9 маточная железа
.J.?t!?
U«'t5'a' /?
(адrезии) бластоцисты к слизистой оболочке матки и поrружения (инвазии) блас
тоцисты в rлубину слизистой оболочки.
На 6 e сутки эмбриоrенеза бластоциста прикрепляется к эпителию эндомет
рия (обычно, эмбриональным полюсом в области задней или вентральной стенки
у маточноrо уrла). В адrезии бластоцисты важную роль иrрают вещества rруппы
интеrринов, вырабатываемые эпителиоцитами слизистой оболочки матки. В HOp
ме у женщин между 19 24 сутками менструальноrо цикла т. е. в самые опти
мальные для взаимодействия с бластоцистой сроки, наблюдается активная эксп
рессия [ена интеrрина в эпителиоцитах слизистой оболочки матки.
rаструляция
у зародыша человека rаструляция инициируется в конце 1 й недели разви
тия, сразу по окончании дробления и сброса прозрачной зоны, коrда зародыш Ha
чинает поrружаться в стенку матки.
1 я фаза rаструляции продолжается всю 2 ю неделю развития. Материал BHYT
ренней клеточной массы расщепляется способом деламинации на два листка эпиб
ласт (верхнюю часть) и тпобласт (нижнюю часть). Клетки rипобласта мелкие KY
бические, их цитоплазма имеет пенистый вид, они формируют тонкий слой под
эпибластом и rpаничат с полостью бластоцисты. Клетки эпибласта более высокие и
имеют вид псевдомноrослойноrо призматическоro эпителия (рис. 144). Клетки эпиб
ласта разобщаются посредством небольших полостей, при слиянии которых в даль
нейшем формируется амниотическая полость. В нижней части амниотической по
лости остается небольшая rруппа клеток эпибласта материал будущеrо зародыша
(эмбриобласт) и внезародышевых opraHoB.
2 я фаза rаструляции происходит
на З й неделе развития, осуществля
ется способом иммиrрации и заверша
ется формированием трех зародые
въ/Х листков эктодермы, мезодермы,
энтодермы (рис. 145). В будущем из
материала этих листков возникнут
ткани эмбриона и внезародышевых
opraHoB. Происходящие во 2 й фазе
rаструляции события напоминают Ta
ковые у куриноrо зародыша и предста
вителей млекопитающих (см. rлаву 6).
Инициация иммиrрации связана с aK
тивной и неравномерной в разных
участках эпибласта пролиферацией
клеток, которая приводит к формиро
ванию клеточных потоков и образова
нию на поверхности эпибласта CTPYK
тур, известных как пеРВИЧllая полоска
7
....i'''''" J :' 'ii 11'-<-
,.' ./ <!.- 11. '. 8
"_ . l' ...........J. .- .,
.11 ../'... "<C:.:.z . . . .... $\,} '" <::::C) . 1 .... " .' ";а
r ""> 77? IJ. ', . .' """"'.....
· ., . t'lI ''{i«'' . .. ":/' . ;' Q."
", 1 ( , ; ..." "" 2 3 '"" O '1\ . ? .
" r " t1I
..' /,. . .', 6
. -..С> . i "$ I ""-;"1'.
. '" ',. /.:,
,....)е. H:t.. .''1;'' 1. Jf
,;) 1'\ . ,\(oo. ('J" c . ."1 ,;"
'I",,_.. . ,... ... ,,<!!.: w... . " 'I!.
fI4. 't\'-.) '\i ? 'о. IoIIowrt.l" F1'";.. j.-'--:. {IW"' (':>;;:#.
J ," /.) ""'i. .... ,$:"; .1\. .. 5 -' -1 (1f"it. .
... ., . : .. C ..,. ..7:ff'l (. , .,.,
.. . .'.', l [;; -:" 1 r\ 1ft.\ € ...I: '
". -. . "'\... -: \ . пf:.:.)r · '" ,
О: .. ,, .., ..:-"'. A!. ., 4р I
.. . ......i ... -.. -. #!'
РИС.144. Схема строения зародыша человека и ero
взаимоотношений со слизистой оболочкой матки на
11 e сутки развития:
1 хорион (клеточная и симпластическая части TpO
фобласта); 2 экзоцеломическая полость (первич
ный желточный мешок); 3 rипобласт; 4 эпиб
ласт; 5 амниотический пузырек; б внезароды
шевый целом; 7 эпителий слизистой оболочки
матки; 8 лакуны с кровью
Воспроизведение человека
321
и rензеновский узелок. Окруrлый и плоский зародышевыЙ диск во 2 ю фазу racT
руляции превращается в вытянутый с расширенным краниальным и более узким
каудальным концами.
Особое морфоrенетическое значение в раннем эмбриоrенезе принадлежит roлов
ному отростку (нотохорду). fоловной отросток растет в краниальном направлении
между эпи и rипобластом, пока не достиrнет прехордальной пластинки, которая яв
ляется "индикатором" будущеro рта зародыша. fоловной отросток дает начало разви
тию хорды зародыша своеобразноrо клеточноrо стержня, который определяет пер
вичную ось эмбриона и придает ему "жесткость". Хорда формирует ось скелета
fjP
.'(}
(,()
.
6(' ')
fj'"
.. .<fo
()
'"
о'
@
8 р
, 1i 4' o. :.
09 a V Q
\ ф..,О'
'0 е(Р
If> ,
Рис. 145. Схема строения 15 суточноrо зародыша человека:
1 хорион; 2 соединительная ткань хориона; 3 амниотическая
полость; 4 полость желточноrо мешка; 5 энтодерма; 6 выселя
ющийся материал мезодермы; 7 эктодерма
322
[лава 17
зародыша человека и является основой развития костей oceBoro скелета (позвоночни
ка, ребер, rpудины, черепа). BOKpyr хорды в будущем сформируется позвоночный столб.
Нотохорд оказывает важное индуцирующее влияние на дифференцировку
прилежащеrо к нему сверху участка эктодермы в нервную пластинку и далее в
нервную трубку, из которой будут развиваться rоловной и спинной мозr. К концу
3 й недели эмбриоrенеза хорда почти полностью сформирована и простирается от
орофаринrеальной мембраны до каудальноrо конца зародыша.
Нейруляция
Развитие нервной трубки (нейруляция) включает образование нервной плас
тинки, HepBHoro желобка и замыкания последнеrо в нервную трубку.
Как только возникает нотохорд, эктодерма, расположенная над ним, начинает
утолщаться и формировать 1/.ерв1/.УЮ плаcmu1/.КУ. Первым признаком дифференци
ровки нейроэктодеР 1Ы является удлинение клеток этой области, и клетки припод
нимаются над остальной частью эктодермы. Нервная пластинка занимает около 50%
всей площади эктодермы. Материал нервной пластинки первоначально возникает
вблизи rензеновскоrо узелка, затем, коrда rоловной отросток удлиняется, формиро
вание нервной пластинки продолжается в краниальном направлении и, в конце KOH
цов, онадостиrает орофаринrеальной мембраны. Приблизительно, на 18 e сутки He
рвная пластинка проrибается вдоль своей продольной оси и образует нервный же
лобок с возвышениями по обе стороны желобка нервными валиками. В конце 3 й
недели в середине эмбриона нервный желобок превращается в нервную трубку и
далее этот процесс распространяется в каудальном и краниальном направлениях.
Однако в краниальной области смыкание желобка в трубку происходит более быст
рыми темпами. К концу 4 й недели нервная
трубка полностью сформирована (рис. 146). В ro
ловном конце, [де будет формироваться rолов
ной мозr, нервная трубка широкая и стенка ее TOk
стая, в ней контурируются вздутия и сужения, co
ответствующие будущим отделам roловноro моз
ra. В каудальном направлении трубка сохраняет
цилиндрическую форму и постепенно суживает
ся. Два открытых конца нервной трубки (спере
ди и сзади) именуются передним и задним ней
ропорами. В эмбриоrенезе млекопитающих, бла
rодаря наличию неЙропоров, через нервную труб
ку некоторое время "протекает" аМНlIотическая
жидкость. Позже нейропоры смыкаются. Как
только нервные валики сливаются и образуют He
рвную трубку, нейроэктодермальные клетки
(клетки нервноro rpе6ня), располаrающиеся меж
ду нервной трубкой и эктодермальным эпители
ем миrрируют в стороны от нервной трубки и
Рис. 146. Поперечный срез зародыша че
ловека на 21 28 e сутки развития:
1 эктодерма; 2 нервная трубка; 3
хорда; 4 энтодерма; 5 мезодерма;
б листки спланхнотома; 7 целом; 8
полость желточноrо мешка; 9 полость
амниона
Воспроизведение человека
323
Мезодерма, расположенная латерально от
хорды, образует широкие полосы вдоль каж
дой стороны хорды и нервной трубки зароды
ша, именуемые несеrментированной дорсаль
ной мезодермой. В результате индуктивноrо
влияния хорды и нервной трубки дорсальная
часть мезодермы подверrается сеrментации на
сомиты. Первая пара сом-итов (от rреч. soma
тело) развивается на 20 e сутки в шейном OT
деле зародыша; последующие пары формиру
ются в кранио каудальном направлении (приблизительно, по три пары сомитов в
сутки) вплоть до конца 5 й недели эмбриоrенеза (рис. 147). В конечном итоrе воз
никает от 42 до 44 пар со штов (4 пары затылочных, 7 шейных, 12 [рудных, 5 пояс
ничных, 5 крестцовых и 8 10 копчиковых сомитов). В последующем 1 я паразаты
лочных и 5 7 я пары копчиковых сомитов исчезают. В этот период развития число
сомитов часто используется в качестве критерия для определения возраста заро
дыша (табл. 2).
Под влиянием веществ, вырабатываемых клетками хорды и нервной трубки
(эмбриональная индукция), расположенные вентрально в составе СОi\lИТОВ клетки
начинают митотически делиться, становят
ся полиморфными и распределяются BOK
pyr хорды. В совокупности эти клетки Ha
зываются склеротом-ом-. В последующем
стволовые клетки в составе склеротома
дифференцируются в хондробласты и уча
ствуют в образовании осевою скелета (по
звонков, ребер, черепа и т. п.). После завер
шения миrрации клеток склеротома OCTaB
шиеся клетки сомита формируют ДBYX
слойную трубку с наружным слоем дep
м-атом-ом- и внутренним м-иотом-ом-. Из
дерматома в дальнейшем будут развивать
ся соединительные ткани кожи, из миото
ма поперечнополосатая скелетная MЫ
шечная ткань.
далее по всему зародышу. Позднее эти клетки
в разных участках эмбриона дадут начало He
скольким клеточным дифферонам, в том числе
пиrментным клеткам, клеткам l\Iозrовоrо веще
ства надпочечников, клеткам периферической
нервной системы.
Сеrментация мезодермы
Рис. 147. Схематическое изображение за
родыша человека на 25 e сутки развития:
1 rоловной конец зародыша; 2 первая и
вторая rлоточные дуrи; 3 сомиты; 4 XBO
стовой конец зародыша; 5 формируюший
ся пупочный канатик; 6 аллантоис; 7 жел
точный мешок; 8 полость амниона
Таблица 2. Корреляция числа СОМИТОВ
и возраста эмбриона (В сутках)
Приблизительный Количество
возраст зародыша пар сомитов
(сутки)
20 1 4
21 4 7
22 7 10
23 10 13
24 13 17
25 17 20
26 20 23
27 23 26
28 26 29
30 34 35
324
Тлава 17
ПЛОДНЫЙ период
ПЛОДНЫЙ период развития человека начинается с 8 й недели и характеризует
ся значительными преобразованиями в opraHax и системах плода, а также в ero
внезародышевых opraHax и в орrанизме матери (табл. 3).
Таблица з. Краткий календарь эмбриональноrо развитиs:l человека
N2 Возраст Морфоrенетические процессы
пл
1 2 3
1. 24 ч после овуляции, раз Происходит оплодотворение, возникает одноклеточный заро
мер 0,1 0,5 мм. дыш. После серии митотических делений зиrоты и ее дочерних
бластомеров образуется морула, которая на 3 4 e сутки покида
4 e сутки после оплодот ет маточную трубу и переходит в полость матки. В моруле появля
ворения, длина 0,1 O,2 ется полость, зародыш превращается в бластоцисту. Клетки бла
мм. стоцисты дифференцируются в эмбрио и трофобласт, что явля
ется первым проявлением дифференцировки в эмбриоrенезе.
2. 5 6 e сутки после опло Бластоциста начинает поrружаться в rлубину слизистой оболоч
дотворения, длина О, 1 ки. Эмбриобласт путем деламинации расщепляется на верхнюю
0,2 мм. и нижнюю клеточные массы (будущие эпи и rипобласт) т. е. ини
циируется 1 я фаза rаструляции. Формируются ворсины хорио
1 3 e сутки после оплодот на. Кровеносные сосуды появляются в области хориона, окружа
варения, длина 0,2 мм. ющеrо эмбрион, в стенке желточноrо мешка начинается rемопо
эз. Эмбрион связан соединительным стебельком с развивающей
16 e сутки после опло ся плацентой.
дотворения, длина 0,4 На 1 3 e сутки появляется первичная полоска, что характеризу
мм. ет начало 2 й фазы rаструляции.
В результате 2 й фазы rаструляции на 16 e сутки формируется
3 слойный зародыш.
3 17 19 e сутки после оп Зародыш rрушевидной формы с широким rоловным концом.
лодотворения, длина Утолщение эктодермы формирует нервную пластинку и далее
1 1,5 мм. нервный желобок. Возникают клетки крови эмбриона и параллель
но происходит образование сосудов. Появляются сомиты как уп
21 22 e сутки после оп лотнения мезодермы по внешней стороне HepBHoro желобка. На
лодотворения, длина данной стадии обнаруживаются три пары сомитов. Первичная по
1,5 3 мм, число соми лоска составляет от 1/4 до 1/3 длины зародыша. Появляются BTO
тов 4 12. ричные ворсинки хориона, содержащие сосуды. В стенке желточ
HOro мешка обнаруживаются rемопоэтические клетки OДHOBpeMeH
но с эндотелиальными клетками, которые формируют KpOBeHOC
ные сосуды новых первичных клеток крови. Начинается процесс
формирования сердца в виде двух пока раздельных миоэндокар
диальных трубок. Возникает и быстро редуцируется предпочка.
К 21 22 суткам происходит замыкание нервной трубки и слияние
миоэндокардиальных трубок. Эмбрион растет и удлиняется, YBe
личивается желточный мешок. Продолжается ростнервноroжелоб
ка, возникают участки замыкания желобка в нервную трубку. В Kpa
ниальной части зародыша обнаруживаются клетки HepBHoro rреб
ня. Заметно возникновение зачатков rлаз и opraHa слуха. Миоэн
докардиальные трубки сливаются, формируют единую S образную
трубку, инициируются сокращения сердечных мышечных клеток.
Воспроизведение человека
325
4. 4 я неделя после опло В начале 4 й недели эмбрион принимает С образную форму,
дотворения, длина от возникают 3 4 пары жаберных Ayr, почки верхних конечностей,
2,5 до 5 мм, число соми формируются закладки rлаз и ушей. Передняя часть нейропора
тов от 13 20 до 30АО. закрывается, передняя часть HepBHoro желобка замыкается на
стадии 20 сомитов. Сердечномышечная трубка имеет S образ
6 я неделя после опло ную форму, ритмично сокращается, становится заметным пере
дотворения, длина от мещение крови в разных участках тела зародыша.
7 9 до 1O 13 мм. Имеются закладки верхних конечностей, намечаются закладки
НО!'; формирование эпителиальноro покрова кожи эмбриона. Ha
чинается развитие костей и мышц из материала сомитов. Крани
Рис. 148 A B. альная часть нервной трубки дифференцируется на три отдела
передний, средний, задний. Обнаруживаются первичная ротовая
бухта и закладка языка. Происходит развитие закладки щитовид
ной железы и лимфатической системы. Продолжается формиро
вание леrких. В пищеварительной трубке возникает утолщение,
соответствующее поджелудочной железе, продолжается разви
тие тонкой кишки, желчноrо пузыря, а также закладывается пер
вичная почка. Печень крупная, заметна в rоловной части эмбрио
на. Развивающиеся сосуды печени соединяются с сосудами пла
центы через пупочный канатик. Хориальные ворсинки обеспечи
вают связь и обмен с кровью материнскоro орrанизма.
В начале 6 й недели у зародыша обозначен шейный изrиб, ro
лова располаrается под прямым уrлом к туловищу, вырисовыва
ется первичное лицо. Возникает ротовое уrлубление эктодермы
(стомодеум) по направлению к кишечной трубке. Развиваются
ушные раковины. Смыкаются веки, хорошо выявляются две сим
метричные носовые ямки. В верхних конечностях различаются cer
менты плеча, предплечья, кисти. В нижних конечностях начинает
ся формирование стоп и иннервации. Продолжается формиро
вание мышц спины. Проrрессирует развитие почек.
К концу 6 й недели появляются нижнечелюстные и лицевые
мышцы. Возникают зубные пластинки и зачатки зубов. Формиру
ется rипофиз. Начинают развиваться трахея и бронхи, в леrких
выделяются бронхо леrочные cerMeHTbI. Образуются 4 камеры
сердца. Формируется rрудобрюшная преrрада диафраrма. Pa
стущая кишка зародыша проникает в пупочный канатик (физио
лоrическая rрыжа зародыша). Происходит отделение rOHaд от Me
зонефроса и их заселение первичными половыми клетками. ОТ
ствола аорты отделяется леrочный ствол. Почки начинают Bыpa
батывать мочу; появляются половой буrорок, уроrенитальная и
анальная мембраны.
5. 6,5 7,5 недель после оп В rоловном мозrе хорошо различимы полушария. К началу 8 й
лодотворения длина от недели руки удлиняются и сrибаются в локтях и перекрещивают
17 22 до 26 31 мм. ся на животе. Завершается критический период развития ниж
них конечностей, различимы пальцы стоп, но между пальцами Hor
8 9 я недели, длина от сохраняются перепонки.
31 до 50 мм, масса около В полушариях rоловноrо мозrа появляются первые извилины,
1Or. и начинают формироваться структуры BHyтpeHHero уха. Эмбри
он совершает непроизвольные движения. Возникает межпред
РИс.148 r. сердная переrородка. Кишка начинает миrрировать из пупочно
ro канатика в брюшную полость, печень занимает вентральную
часть живота. Уроrенитальная мембрана дифференцируется по
326
Тлава 17
мужскому или женскому типу, становятся различимы яички и яич
ники. Происходит перфорация анальной мембраны. Внешний
вид rоловы становится характерным для человека, rолова oKpyr
лена и выпрямлена. Веки и ушные раковины более развиты; к
концу 8 й недели веки объединяются, и rлаза наполовину зак
рыты, сетчатка пиrментирована, полностью сформировано Ha
ружное ухо. Завершается критический период развития сердца.
Верхняя ryба зародыша сформирована, возникают вкусовые лу
ковицы языка, развиваются эпителиальные закладки зубов. Ko
сти твердоrо неба начинают сливаться. Кишка миrрирует в по
лость тела. В длинных костях появляются первичные очаrи OKOC
тенения, замещающие хрящ на кость. Это обычно начинается в
верхних конечностях.
У девочек формируется клитор, у мальчиков половой член.
Верхние и нижние конечности хорошо сформированы, пальцы yд
линяются и все разделены и расставлены. Присутствовавший pa
нее хвост деrенерирует.
6. 10 11 недель В результате выделения назолакримальной области и интермак
после оплодотворения, силлярноrо cerMeHToB лицо зародыша становится похожим на
длина 50 70 мм, масса лицо человека. Конечности хорошо развиты во всех своих частях.
20 40 r. Наружные rениталии дифференцированы по половому при знаку.
Начинают расти ноrти. Кожа плода тонкая и очень чувствитель
12 13 я недели после оп ная. Плод начинает издавать звуки, так как развиваются rолосо
лодотворения, длина 70 вые связки. Начинается закладка волосяных фолликулов.
90 мм, масса 45 60 r. Увеличивается масса мозrа, совершенствуется ero структура.
В ротовой полости присутствуют закладки 20 зубов. Кишка начи
нает перемещаться из пупочноrо канатика в брюшную полость.
Печень начинает секретироватьжелчь. Поджелудочная железа за
вершает развитие, клетки панкреатических островков дифферен
цированы и вырабатывают инсулин. Завершает развитие щито
видная железа.
Объем амниотической жидкости составляет около 50 мл.
На 12 13 й неделе плод начинает двиrаться. Хорошо прослу
шиваются сердцебиения плода. Руки приобретают конечные про
порции, однако ноrи еще короткие. Плод может совершать дви
жения rубами, челюстью. Наружные половые opraHbI развиты.
Сердце функционирует и перекачивает около 28 л крови в сутки.
В леrких дифференцируются бронхи, которые заполнены амнио
тической жидкостью. Продолжается развитие пищеварительной
системы. Завершается формирование селезенки, в которой про
исходит rемопоэз и образование антител.
7. 14 15 я недели после оп Веки сомкнуты, ноздри открыты, ryбы также приоткрыты, на rоло
лодотворения,длина100 ве и бровях имеется короткий пушок. Плод становится еще более
130 мм, масса 80 150 r. подвижным. Двиrает rоловой, ноrами, челюстью. rолова и шея ВЫП
рямляются, так развиваются и сокращаются мышцы спины. Haд
16 18 недель после опло бровные дуrи выдаются вперед, ушные раковины прижаты к черепу.
дотворения, длина от 140 Размер rоловы становится пропорциональным размеру туловища,
до 170 мм, масса от 200 так как тело плода интенсивно растет в длину. Хорошо развиты KO
дО 300 r. нечности. Продолжается развитие всех висцеральных систем.
Плод продолжает рост, но новые структуры тела не возникают.
Лицо выступает более вперед, чем в стороны. rлаза сформиро
ваны, возникает рефлекс морrания. Уши занимают обычное по
ложение. Продолжается развитие пальцев и костей конечностей.
Воспроизведение человека
327
утолщается пупочныи канатик, плацента достиrает размеров, co
ответствующих размеру плода.
Продолжается развитие всех opraHoB. Начинается процесс ми
елинизациинервов,функционируеткровообращениеплода.Вки
шечнике накапливается меконий продукт клеточной rибели,
деятельности пищеварительных желез и амниотическая жидкость.
К 18 й неделе плод хорошо сформирован. Веки сомкнуты, ноз
дри И ryбы приоткрыты. Это период наиболее бурноrо роста пло
да. Плод начинает фазно спать и двиrаться, но "предпочитает
спать". На rолове появляются временные волосы лануrо, фор
мируются ресницы. В области шеи и rруди развивается буровая
жировая ткань. Образуется плодовая смазка, покрывающая все
тело. Развиты матка и яичники, последние содержат половые клет
ки. Плацента полностью сформирована, увеличивается вдиамет
ре, но не в толщине.
8 20 24 недели после опло Плод совершает сосательные и rлотательные движения. Длина
дотворения, длина 190 ero Hor пропорциональна туловищу. К 24 й неделе opraHbI слуха и
230 мм, масса 450 820 r. зрения плода MOryт воспринимать соответствующие раздраже
ния, вкусовые рецепторы в области рта и ryб обладают чувстви
26 28 я недели после оп тельностью. OpraH слуха способен воспринимать звуки из внеш
лодотворения, длина от ней среды и возникают рефлекторные мышечные движения пло
240 до 270 мм, масса от да. Начинается развитие сосудистой системы леrких, в последних
900 до 1300 r. формируются альвеолы и начинается продукция сурфактанта. По
являются закладки постоянных зубов. Подкожная жировая клет
29 32 я недели после оп чатка слабо развита.
лодотворения, длина от На 26 28 й неделе у плода исчезает лануrо за исключением
280 до 300 мм, масса плеч. Формируется волосяной покров rОЛО9Ы. Возникает жиро
1400 2100 r. вая смазка кожи, выполняющая изолирующую функцию и являю
щаяся источником энерrии. rлаза частично открыты, есть ресни
цы. Реrистрируются сосательные и rлотательныедвижения. Боль
шие полушария мозrа растут, прикрывая остальные участки моз
ra, увеличивается количество извилин коры rоловноrо мозrа.
Леrкие rOToBbI к rазообмену. Кроветворение сосредотачивается
в красном костном мозrе. Яички спущены в мошонку.
На 29 32 й неделе замедляются темпы роста плода. Плод прак
тически не двиrается. rлаза некоторое время открыты и закрыва
ются во время сна. Цвет rлаз обычно rолубой, способность к пиr
ментации радужки еще не полностью развита, мышцы радужки
рефлекторно реаrируют на свет. Создается подкожная жировая
клетчатка, что делает окраску кожных покровов более светлой.
Продолжается быстрый рост мозrа, формируются множествен
ные извилины коры больших полушарий. Создаются мноrочислен
ные интернейрональные связи. Начинает развиваться иммунная
система. В конце периода волосы rоловы утолщаются, ноrти на
пальцах Hor полностью сформированы. Вследствие оrраниченно
ro маточноro пространства, ноrи сrибаются и принимают поло
жение, известное как "поза плода".
9 34 36 я недели после оп Плод поворачивается rоловой ко входу в малый таз. Maccaero под
лодотворения, длина от кожной жировой кле1'lатки увеличивается до 15% от массы тела, что
310 до 340 мм, масса от необходимо для подцержания постоянства температуры плода.
2200 до 2900 r. Руки поднимаются и прижимаются к rруди и шее. Кожа становится
328
Тлава 17
38 я неделя после опло светлой, блаrодаря поверхностному расположению кровеносных co
дотворения, длина 360 судов.
мм, масса 3400 r. На 36 й неделе увеличивается подкожный жировой слой плода.
В кишке аккумулируется некоторое количество мекония, который
обычно выделяется вскоре после рождения. Полость матки orpa
ничивает движения плода, и ero конечности плотно прижаты к тy
ловищу. Проrрессирует оссифицирование костей.
На 38 й неделе череп не тверд, так как 5 костных пластин раз
делены родничками и MOryт смещаться при родах, способствуя
изменению формы и удлинению rоловы, что необходимо для об
леrчения рождения без нарушения мозrа плода. После рождения
восстанавливается окруrлая форма rоловы. Леrкие увеличивают
продукцию сурфактанта, живот плода большой и круrлый, выпук
лый в области печени. Кожа становится толще и каждый день плод
прибавляет 14 r жира. Плод полностью соответствует морфоло
rическому статусу ребенка. 15% составляет жир, 80% KOToporo
находится в подкожной клетчатке и 20% BOKpyr opraHoB. Начи
наются схватки.
Примечание: эмбриолоrи отсчитывают возраст зрелоro плода, равный 266 суткам,
или 38 неделям, начиная от момента оплодотворения и образования зиrоты, т. е. от
дня первой стадии эмбриоrенеза. В акушерской практике дату рождения ребенка
принято определять по дню последней менструации у женщины, т. е., рождение pe
бенка ожидается через 280 суток (или 40 недель) после первоrо дня последней MeH
струации.
Воспроизведение человека
329
2
А
4
4
7
5
Б
в
8
r
Рис. 148. Изменение внешнеrо вида эмбриона человека на ранних стадиях развития:
А 28 суток развития (стадия 25 сомитов); Б 5 недель развития; В б недель развития; r 8 недель
развития; 1 зрительная плакода; 2 слуховая плакода; 3 жаберные дуrи; 4 сомиты; 5 пупоч
ный канатик; б сердечный выступ; 7 закладка верхней конечности; 8 закладка нижней конечности;
9 хвост; 1 О шейный изrиб; 11 формирующийся слуховой проход 12 формирование пальцев
рук; 1 3 формирование пальцев Hor
Воспроизведение человека
ЗЗ1
Имплантация зародыша активизирует пролиферативные и миrрационные про
цессы в эмбриобласте. Это приводит к развитию друrих внезародышевых opraHoB
амниона, желточноrо мешка, аллантоиса и хориона (в период с 7 x по 14 e сутки
эмбриоrенеза ).
Амнион (водная, амниотическая оболочка), представляет собой полый opraH
(мешок), заполненный жидкостью (околоплодными водами), в которой находит
ся и развивается зародыш. Основная функция амниона выработка ОКОЛОПЛОk
ных вод, которые обеспечивают оптимальную среду для развития зародыша и пре
дохраняют ero от высыхания и механических воздействий. Амнион возникает из
материала эпибласта путем образования в ero толще полости амниотическоrо
пузырька. В процессе развития эпителий амниона (сначала однослойный плоский)
на З м месяце эмбриоrенеза преобразуется в призматический. Располаrается эпи
телий на базальной мембране, под которой находится более плотный слой соеди
нительной ткани. Далее располаrается ryбчатый слой рыхлой волокнистой соеди
нительной ткани, простраI-:!2твенно связанный со стромой rладкоrо и ворсинчатоrо
хориона.
Эпителиоциты амниона обладают секреторной (в плацентарной части) и Bca
сывающей (во внеплацентарной части) активностью. Амниотическая жидкость
постоянно обменивается, имеет сложный химический состав, изменяющийся в ходе
развития плода. Помимо указанных выше функций, амниотическая жидкость имеет
важное значение для формообразовательных процессов развития ротовой и HO
совой полостей, opraHoB дыхания, пищеварения. Количество вод с течением бере
менности увеличивается и к родам достиrает O,5 1 ,5 л, коррелируя с длиной и Mac
сой плода и сроком беременности. В околоплодных водах MorYT определяться
клетки эпидермиса, эпителия ротовой полости и ваrинальноrо эпителия плода, ЭШI
телия пуповины и амниона, продукты секреции сальных желез, пушковые волосы.
Желточный мешок. Желточный мешок у человека (пупочный, или пуповин
ный пузырек) рудиментарное образование, утратившее функцию вместилища
питательных веществ. До 7 8 й недели эмбриоrенеза основная ero функция KpO
ветворная. Кроме Toro, в стенке желточноrо мешка появляются первичные поло
вые клетки rонобласты, которые миrрируют в Hero из области первичной полос
ки. Источниками развития тканей желточноrо мешка являются внезародышевая
энтодерма и внезародышевая мезенхима. Стенка желточноrо мешка выстлана жеk
точным эпителием особым подтипом эпителия кишечноrо типа. ЭпителиЙ co
стоит из одноrо слоя кубических или плоских клеток энтодермальноrо происхож
дения со светлой цитоплазмой и круrлыми интенсивно красящимися ядрами. После
формирования туловищной складки желточныЙ мешок связывается с полостью
средней кишки посредством желточноrо стебелька. Позднее желточный мешок
обнаруживается в составе пупочноro канатика в виде узкоЙ трубочки.
Аллантоис. В середине 1 ro месяца внутриутробноrо развития за счет проли
ферации эпителия каудальноrо участка желточноrо пузырька возникает алланто
ис. Аллантоис врастает в амниотическую ножку. Стенка аллантоиса состоит из oд
нослойноrо призматическоrо эпителия, клетки KOToporo имеют умеренно
выраженную оксифилию цитоплазмы. Не являясь мочевым мешком (как это было
ЗЗ2
Fлава 17
у яЙцекладущих), аллантоис со своими сосудами, которые связываются с ворсин
ками хори она, обеспечивает питание развивающеrося зародыша.
Пупочный канатик. Амниотическая ножка, посредством которой зародыш с 15
суток развития связан с хорионом, постепенно трансформируется в пупочный KaHa
тик Кроме мезенхимы амниотической ножки в формировании канатика принима
ют участие аллантоис со своими сосудами, желточный стебелек Снаружи амниоти
ческая ножка покрывается амниотической оболочкой, эпителий которой в области
пупочноrо отверстия срастается с эпителием кожи плода. Так формируется наруж
ное эпителиальное покрытие пупочною канати
ка. Строму пупочноrо канатика составляет особый
вид соединительной ткани, не встречающийся в
орrанизме человека после ero рождения. Это cтy
де'Нистая, или слизистая, тка'Нь (вартонов cтy
день). ИСТОЧНИКО 1 развития этой ткани является
внезародышевая мезодерма эмбриобласта. Ткань
отличается боrатством OCHoBHoro вещества и раз
личными клетками производными мезенхимы
(фибр06ласты, миофи6р06ласты, rладкие миоци
ты). Волокнистый компонент представлен слабо.
Вартонов студень предохраняет пупочные
сосуды (две пупочные артерии, по которым Te
чет венозная кровь от плода, и одну вену, по KO
тороЙ течет насыщенная кислородом кровь к пло
ду) от сжатия, обеспечивая упруюсть канатика.
ПупочныЙ канатик доношенноrо плода имеет
длину 40 50 см, диаметр около 1,5 см. При
крепляется он к вентральной стенке тела плода
к месту, имеющему кольцевидную форму и Ha
зываемому пупком.
Плацента
Плацента это основное связующее звено
матери и плода, относится к ворсинчатому reMo
хориальному типу. Развивающийся троф06ласт
разрушает ткани слизистой оболочки матки и
сосуды, формируются лакуны, куда изливается
артериальная кровь матери и далее кровь из ла
кун по венозной системе оттекает И3 матки.
Плацента человека дискоидальная, ее
структурно функциональной единицей является
котuледо'Н (котиледон rреч. щупальцы полипа).
Последний представлен стволовой, или якорной,
ворсинкой, которая срастается посредством пери
4
. .. .. ._ O
.. ..
..... .. -- --........ ..., ......." ... ..... -"
............ . .. .. .
''-о.. " __ T_
."" ${ ... -- .,. ", .,
'.., 5
"
"8
. "" '"
. . ; .....
.. . .:. ,,!'. ..,:. ".
. п : ." '-.6';' ',. .. .. ;.
1b' ; q. ".- ,
.. .::; < ... .. ..
. . ( ' ....'" .......,......,. .....'
" Qa o..jj "'.._t:- ........................
. : - --..y,; ,
.. е _ ,. ... 9 -''''' "',« "-.t:
. . ........ .ф.. - ..:
о. ........,... "
Рис. 150. Схема образования и CTpO
ения вторичных ворсин хори она:
1 соединительнотканная строма
ворсинки; 2 цитотрофобласт; 3
симпластотрофобласт; 4 амниоти-
ческий эпителий; 5 соединитель
нотканная пластинка хориона; б
кровяные лакуны; 7 свободная
(вторичная) ворсинка; 8 стволовая
(якорная) ворсинка; 9 базальная
пластинка эндометрия; 10 маточ
ная железа
Воспроизведение человека
333
ферическоrо цитотрофобласта с материнскими тканями, и свободными ворсинка
ми, колеблющимися в материнской крови лакун вторичными, третичными BOp
синками (рис. 150). Название opraHa происходит от лат. placenta пироr, лепешка,
оладья. В конце беременности плацента представляет собой мяrкий диск диаметром
15 18 см, толщиной в центральной части 2 4 см, массой около 500 600 [. Общая по
верхность хориальных ворсинок достиrает 16 м 2 , что значительно больше поверх
ности всех леrочных альвеол, а площадь их капилляров 12 м 2 . Обычно плацента
локализуется в матке на ее передней или задней поверхности, иноrда в области дна.
В плаценте различают две поверхности. Поверхность, которая обращена к плоду,
называется плодной. Она покрыта rладким амнионом, через который просвечива
ют крупные сосуды.
Материнская поверX1l.0стъ плаценты обращена к стенке матки. При ее внеш
нем осмотре обращает внимание серо красный цвет и шероховатость. Здесь пла
цента разделяется на котиледоны.
Плодная часть плаценты формируется в следующей последовательности. Tpo
фэктодерма бластоцисты при попадании зародыша в матку на 6 7 e сутки развития
дифференцируется в трофобласт, обладающий свойством прикрепляться к ВЫСТИk
ке матки. При этом клеточная часть трофобласта дифференцируется на две части
наряду с клеточной составляющей, снаружи возникает симпластическая часть TpO
фобласта. Именно последняя вследствие своею более дифференцированною cocтo
яния способна обеспечить имплантацию и подавить иммунную реакцию материнс
Koro орrанизма на внедрение rенетически чужеродною объекта (бластоцисты) в
ткани. За счет развития и ветвления симпластотрофобласта возникают первичныe
вОРСИ1lКИ, что увеличивает площадь соприкосновения трофобласта с тканями матки.
При имплантации в зародыше возрастают пролиферативные процессы, возникает
внезародышевая мезенхима, которая изнутри выстилает цитотрофобласт и являет
ся источником развития соединительной ткани в составе ворсинок Так формиру
ются вторичные ворсинки. На этой стадии трофобласт принято называть хорионом,
или ворсинчатой оболочкой.
Продолжающаяся плацентация и развитие аллантоиса и ero сосудов приводят к
тому, что кровеносные сосуды на 3 й неделе развития прорастают во вторичные BOp
синки. Дальнейшее ветвление ворсинок еще больше увеличивает площадь сопри
косновения плодной части плаценты с материнской кровью за счет формирования
третИЧНЪL"С, или терминшlъных' ворсинок, содержащих кровеносные сосуды плода.
При этом общая длина ворсинок достиrает почти 50 км. Эпителиальные клетки BOp
синок на апикальной поверхности имеют микроворсинки, образующие щеточную
кайму. Длина микроворсинок колеблется от 0,5 до 2 мкм. Щеточная кайма прини
мает участие в транспорте специфических веществ. В ней выявлены иммуноrлобу
лин, железо, трансферрин, ферритин, витамин В12, фолаты, кальций, аминокислоты,
rлюкоза, кортикостерон, липопротеины соединения, обеспечивающие работу
транспортных систем. В щеточной кайме присутствуют также рецепторы к юрмо
нам инсулину, соматомедину, эпидермальному фактору роста, хориальному [OHa
дотропину. rруппу прочих рецепторов составляют бета адренерrические, холинер
rические и опиатные. Также в области щеточной каймы выявляются высокая
ЗЗ4
Тлава 17
активность ферментов фосфатаз, пептидаз, rалактозилтрансферазы, rамма rлю
таминтранспептидазы, MHoro белков и антиrенов и таких небелковых компонентов,
как липиды, уrлеводы и сиаловые кислоты.
Симпластический покров ворсинок образует мноrочисленные выпячивания паль
цевидной формы. Симпластотрофобласт содержит MHoro орrанелл, секреторных и
осмиофильных rpанул. Ядра здесь, в основном, овальные, очень плотные, особенно по
периферии, содержат компактный хроматин, располаrаются неравномерно. В симп
ластотрофобласте хорошо развита ЭlЩоплазматическая сеть, как rpанулярная, так и
аrpанулярная, встречаются свободные полисомы. Митохондрии небольшие, а в еди
нице объема их больше, чем в клетках цитотрофобласта. MHoro мелких и крупных
осмиофильных rpанул. Число rpанул rликоrена незначительно. Выявлен хорошо раз
витый комплекс [ольджи, MHoro пиноцитозных пузырьков и др.
Кроме цитотрофобласта (слоя Ланrrанса) и симпластическоrо трофобласта, KO
торые составляют основу трофобластическоrо покрытия хориальных ворсинок, в
хорионе человека существует вневорсинчатый, или периферический, как ero иноrда
называют дополнительный цитотрофобласт. К нему относят клеточные колонны,
клетки базальной пластинки (базальной зоны) плаценты, клеточных островков и
плацентарных переюродок, цитотрофобласт, замурованный в фибриноидные Mac
сы, а также цитотрофобласт rладкоrо хориона (внеплацентарноrо).
В конце второй недели беременности при контакте трофобласта с тканями сли
зистоЙ оболочки матки в ее базальной части (место формирования базальноrо слоя,
содержащею децидуальные клетки см. ниже) происходит образование базаль
ной зоны (пластинки) хориона (от анrл. "basal plate"). В базальной зоне, определя
е},.юй как место тесною контакта плодных и материнских тканей, сосуществуют
клетки двух разных rенотипов, формируя, таким образом, базйлыl.йй lИсmиок Ба
зальная зона представляет собой область плацентарною при крепления к стенке
матки и вместе с плацентой отделяется при родах. В базальном слое эндометрия и
в миометрии выявляются клетки интерстициальноrо цитотрофобласта. Также об
наруживается и внутрисосудистый, или эндоваскулярный, цитотрофобласт, KOTO
рый проникает в стенки спиральных артерий матки.
Клеточные островки это окруrлые образования диаметром от 200 до
1000 мкм. Их основу составляет плотный rомоrенный фибриноид, в котором по
стоянно встречаются клетки цитотрофобласта.
Соединительная ткань стромы ворсинок хориона относится к особоЙ форме pыx
лой волокнистоЙ соединительноЙ ткани. Она характеризуется бедностью клеточны
ми элементами и обилием основною вещества. В неЙ MHoro rиалуроновой и ХОlЩрои
тинсернистой кислот, с которыми связана проницаемость rематоплацентарноrо
барьера. Здесь встречаются клетки фибробластическоrо дифферона, адвентициаль
ные, rбtaтоrенные эле},.lенты, макрофаrи (клетки Кащенко [офбауэра). Особо следу
ет подчеркнуть опережающее развитие соединительноЙ ткани внезародышевых opra
нов по сравнению с таковоЙ в зародышевых opraHax в эмбриоrенезе человека.
Следовательно, в развитии хориона условно можно вьщелить 3 стадии: 1) раз
растание трофобласта в ходе имплантации, дифференцировку ею на симпластичес
кую и клеточную части и образование первичных ворсинок; 2) возникновение BHe
Воспроизведение человека
335
зародышевой мезенхимы, подстилающей изнутри трофобласт и являющейся источ
ником развития соединительнотканной стромы стенки амниона и вторичных BOp
синок хориона; 3) разрастание и проникновение в соединительнотканную основу
сосудов, формирование терминальных ворсинок Все эти сложные этапы преобра
зования трофобласта и формирования сосудистой системы хориона позволили BЫ
делить этап плацентации как критический период в развитии плацентарных млеко
питающих и человека. У человека это 5 6 я недели развития. Формирование
плаценты заканчивается в конце 3 ro месяца развития. В ходе беременности проис
ходит быстрый рост плаценты. У 3 недельноrо зародыша плацента занимает 1/15
часть внутренней поверхности матки, у 8 недельноrо 1/3, У 5 недельноrо 1/2.
Ворсинка в процессе беременности претерпевает ряд rистолоrических измене
ний. Так, на 4 M месяце беременности цитотрофобласт распадается на отдельные rруп
пы клеток, поэтому трофобластическое покрытие ворсинок трансформируется He
которые участки ворсинок оказываются покрытыми только СlIмпластотрофобластом.
Местами изчезает и симпластотрофобласт, и тоrда на ero месте откладываются фиб
риноподобные массы продукт реакции сыворотки крови 11 распадающеrося симп
ластотрофобласта (фибриноид Ланrrанса).
Материнская частъ плаценты представлена видоизмененной отпадающей обо
лочкой матки. При проникновеНИII ворсинок трофобласта в слизистую оболочку
матки происходит разрушение тканей и сосудов ее rипертрофированноrо эндомет
рия возникают лакуны крови, в которые поrружены ворсинки хорпона. Рост BOp
сии В rлубь стенки матки, однако, оrраничен, и самые rлубокие слои эндометрия
остаются свободными от ворсин. Эта часть эндометрия называется базалЫ-lЪLМ слоем,
строму ее составляет волокнистая соединительная ткань, в которой обнаруживают
ся крупные, боrатые rликоrеном децидуальные клетки. Клетки имеют четкие rpани
цы, окруrлые ядра, оксифильную цитоплазму.
Первые децидуальные изменения эндометрия происходят у женщин во BTO
рой половине каждоrо овариально менструальноrо цикла. Однако значительно
более выраженная децидуальная реакция эндометрия наблюдается только в слу
чае наступления беременности. Вначале среди клеток базальноrо слоя стромы
эндометрия преобладают фибробласты. Они продолrоваты, в их цитоплазме xo
рошо развиты комплекс rольджи и канальцы эндоплазматической сети. Далее
соединительнотканные клетки слизистой оболочки матки превращаются в деци
дуальные. При этом они окруrляются, увеличиваются в несколько раз, достиrая
иноrда 50 мкм. В них изменяется обмен веществ, о чем можно судить по появля
ющимся rранулам rликоrена и каплям лиПlIДОВ, а также по способности синтези
ровать особые белки коллаrен IV ro типа, десмин, ВIII\lентин. В цитоплаз lе
децидуальных клеток содержится редкая эндоплазматическая сеть и MHoro ми
тохондрий, заметное развитие получает комплекс rольджи. Образование деци
дуальноrо слоя быстро завершается, и уже к концу 2 й недели с момента опло
дотворения происходит завершение перестройки эндометрия. В это время в
децидуальном слое различают rлубокий rубчатый и поверхностный компактныЙ
участки. Fубчатый участок имеет рыхлое строение, в нем lИorO полостеЙ, обра
зуемых остатками желез эндометрия. Редкие децидуальные клетки разделяются
ЗЗб
Fлава 17
арrирофильными волокнами и rомоrенной оксифильной массой. Мноrочислен
ны расширенные кровеносные сосуды. В компактном участке преобладают дe
цидуалыlеe клетки. Толщина децидуальноro слоя достиrает 8 мм.
Защитное значение децидуальноrо слоя заключается в том, что ОН в опреде
ленной степени предохраняет плод от воздействия бактерий и токсинов, которые
MorYT циркулировать в материнской крови; с друrой стороны, орrанизм матери
предохраняется от излишнеrо инвазивноrо действия трофобласта.
Плацентарные переrородки (септы) встречаются в количестве 20 50 на одну
плаценту, их диаметр колеблется от 200 до 3750 мкм. По своему строению септы
близки к клеточным островкам в плотных массах фибриноида располаrаются
клетки периферическorо цитотрофобласта, отличающиеся базофильной цитоплаз
мой, и децидуальные клетки. Ряд авторов называют цитотрофобластические клет
ки плацентарных переroродок Х клетками.
На rранице по краю диска плаценты отпадающая часть слизистоЙ оболочки матки
не разрушается. Она плотно прирастает к хориону, образуя замыкательную плас
тИ1l:КУ (рис. 151). Это препятствует истечению крови из лакун. Кровь в лакунах по
стоянно обновляется. Из артериЙ, проходящих в септах, кровь истекает в лакуны, а
через отверстия, открывающиеся в сеп
ты, оттекает в вены. При этом в норме
кровь матери и плода не смешивается.
Это происходит из за существования
lематохорuалЬНОlО барьера, в состав
KOToporo входят трофобластическое
покрытис ворсин, соединителыlткан
ная СТРШ1а ворсин, стенка KpOBeHOCHO
[о сосуда ворсины.
Часть троф06ласта. которая не
участвует в формировании дискои
дальной плаценты, в процессе разви
тия формирует rладкий, или безвор
синчатыЙ, хори он. Последпий, взаи
юдсЙствуя со слизистой оболочкой
матки, также участвует в транспорт 5
но метаболических процессах плода.
Таким образом, плацента это
временный opraH, длительность суще
ствования KOToporo оrраничена. В HOp
ме жизненныЙ цикл тканеЙ плаценты
и друrих внезародышевых opraHoB co
поставим с продолжительностью бере
менности. Однако ткани плаценты
имеют собственные источники ра."1ВИ
тия, опережающиЙ темп rистолоrичес
кой дифференцировки по сравнению с
Рис. 151. Зародыш в полости матки (схема):
1 полость амниона; 2 замыкательная пластинка;
3 плацента; 4 decidua basalis базальная плас
тинка слизистой оболочки матки; 5 decidua
capsularis часть слизистой оболочки, покрывающей
зародыш со стороны полости матки; б decidua
parietalis париетальная часть слизистой оболочки
матки; 7 полость матки; 8 лакуны, заполненные
материнской кровью
Воспроизведение человека
337
тканями плода. Сосудисто капиллярная система плацснты принадлежит плоду, ее
деятельность направлена на обеспечение потребностей плода, но и сами ворсинки
полностью снабжаются всеми необходимыми веществами для неоrраниченноrо 11
самостоятельноrо существования. Следствием Bcero перечисленноro является тот
факт, что плод может поrибнуть, а ворсинки хориона при этом сохраняют свою жиз
неспособность и MOryT перерождаться.
Важнейшие функции хориона можно разделить на 2 катеrории: по отношению
к орrанизму плода и по отношению к орrанизму матери. К первым относятся:
а) трофическая, транспортная (перенос аминокислот, уrлеводов, жиров, минера
лов); б) осуществление rазообмена; в) защитная, выражающаяся в формировании
биолоrическоrо барьера, фаrоцитозе, протеолитическоЙ активности клеток, анти
оксидантной способности, детоксикации; [) реryляция водно солевоrо обмена. Kpo
ме перечисленных функций, различают специальные влияние на развитие opra
нов плода (эндокринных желез, печени, почек, леrких и др.).
Вторую катеrорию функций по отношению к орrанизму матери, представ
ляют: а) функции, обеспечивающие имплантацию (протеолитическая и фаrоци
тарная); б) синтез веществ, способствующих перестройке opraHoB и систем MaTe
ри; в) иммунодепрессивная функция; [) анти и rемостатическая функция;
д) эндокринная функция.
Одной из наиважнейших функций хориона является трофическая. Показано,
что плацента избирательно проницаема для некоторых веществ. Так, rрибки и бак
терии не проходят через rематоплацентарный барьер в физиолоrических услови
ях, но при этом барьер проницаем для альбуминов, аминокислот, уrлеводов, вита
ми нов, иммуноrлобулинов G, антибиотиков, а также большинства вирусов.
Последние наиболее опасны для развивающеrося плода.
Эндокринная функция плаценты заключается в выработке хориальноrо rOHa
дотропина (XrT) и плацентарноrо лактоrена, или хориальноrо COMaTOMaMMOT
ропина (ХСМТ). [ормоны иммуноцитохимически идентифицированы в цитоп
лазме симпластотрофобласта. XrT стимулирует и пролонrирует деятельность
желтоrо тела и участвует в имплантации, действуя на иммунные процессы MaTe
ринскоrо орrанизма в качестве супрессора (уrнетателя). ХСМТ обладает COMa
тотропным, лактоrенным и лютеотропным действием. Кроме этих важнейших [op
монов, в плаценте обнаружены адренокортикотропный [ормон, меланотропный,
тиреотропный ropMoHbI, ренин, рилизинr факторы, аналоrичные таковым rипо
таламуса, анrиоrенные вещества, факторы роста, стероидные [ормоны (проrес
терон, эстроrены и андроrены). Поверхность ворсин хориона содержит вещества,
влияющие на свертываемость крови. Плацента имеет большие запасы тромбоп
ластических веществ. При нарушении баланса последних возникает послеродо
вое кровотечение.
Важнейшей функцией плаценты, направленной в сторону материнскоrо opra
низма, является им.мунодепрессивная. Трофобласт и в последующем плацента син
тезируют биолоrически активные вещества, влияющие на течение иммунных про
цессов. Так, наивысшая концентрация супрессоров иммунных реакций (XrT и
ХСМТ) обнаруживается на поверхности ворсинок хориона. Иммуномаскирующим
338
Fлава 17
действием обладают rликопротеины фибриноида Ланrrанса. При этом определен
ный вклад при вносят избирательный транспорт и связывание биолоrически aK
тивных веществ, фаrоцитоз иммунореактивных клеток и ryморальных факторов.
Амниотические воды содержат антитела, направленные против антиrенов крови
матери.
Орrанизм матери и плода тесно взаимосвязанные биолоrические системы.
Важнейший механизм взаимноrо влияния нейроrуморальный. В данном Mexa
низме выделяются рецепторные (воспринимающие информацию), реrуляторные
(перерабатывающие ее) и исполнительные составляющие. В матке расположены
чувствительные нервные окончания, которые воспринимают информацию о co
стоянии плода. В эндометрии обнаружены xeMO , MexaHO и терморецепторы, CBO
бодные нервные окончания, расположенные в стенке маточных вен базальной час
ти эндометрия. Раздражения рецепторов матки вызывают изменения интенсивно
сти дыхания, артериальноrо давления в орrанизме матери, направленные на
обеспечение оптимальных для плода условий развития. rипоталамическая и He
рвная системы матери это основные системы реrуляции взаимоотношений ее
орrанизма и плода. При беременности создается новое эндокринное равновесие,
соответствующее тому или иному периоду развития плода. Например, размеры
rипофиза беременной увеличиваются почти в два раза, в структуре субпопуляций
передней доли возрастают число и размеры базофильных эндокриноцитов, OTBeT
ственных за выработку лютропина, поддерживающеrо развитие желтоrо тела бе
ременности. После 3 ro месяца беременности возрастает число ацидофильных эн
докриноцитов, выделяющих пролактин, активизирующий биосинтез молока в MO
лочных железах. Активизация кортикотропоцитов усиливает секрецию клетками
коры надпочечника [ормона, обеспечивающеrо повышенный минеральный обмен.
Активизация клеток средней доли rипофиза вызывает характерную пиrментацию
беременных. Это только некоторые примеры эндокринной перестройки в орrаниз
ме беременной женщины.
В орrанизме плода также имеются рецепторы, воспринимающие сиrналы об
изменениях в орrанизме матери, а также звуки внешней среды. Нервные прибо
ры обнаружены в стенке пупочных сосудов, коже и opraHax пищеварения плода.
Двиrательные реакции плода наблюдаются на 2 3 M месяце развития, opraH слуха
к этому времени вполне сформирован и плод воспринимает звуки.
Роды заключительный акт развития плода. При наступлении родов плодные
оболочки вдвиrаются в шейку матки, при этом разрывается амниотическая оболоч
ка и амниотическая жидкость изливается наружу. К этому времени лонное сочлене
ние подrотовлено к тому, чтобы расшириться и пропустить rоловку плода (выработ
ка [ормона релаксина этому способствует). После выхода младенца из родовых путей
он остается связанным с плацентой пупочным канатиком, который пережимается и
перерезается акушером. Затем происходит отторжение плаценты (последа), что co
провождается кровотечением, быстро прекращающимся за счет сокращения матки
и пережатия сосудов, проходящих в миометрии. Слизистая оболочка матки путем
клеточной реrенерации восстанавливает свою целостность. Циклические изменения
эндометрия матки восстанавливаются после завершения лактации.
Воспроизведение человека
339
Критические периоды и аномалии развития
Эмбриональное развитие возможно лишь при оптимальном сочетании BHYT
ренних и внешних условий. Каждая последующая эмбриоrенеза вытекает из пре
дыдущей и из имеющихся в данный момент условий развития. Если KaKoro либо
внешнеrо или BHYTpeHHero условия недостает, или если появляется необычный BHe
шний фактор, способный кардинально повлиять на ход развития плода, эмбриоrе
нез может отклониться от нормальноro пути.
Результаты классических исследований эмбриолоrа П.f. Светлова (1960)
указали на два критических периода в развитии плацентарных млекопитающих
это имплантация и плацентация. В критические периоды зародыш или плод CTa
новится высоко чувствительным и лабильным по отношению к действию BHe
шних факторов. В эти периоды непосредственной причиной аномалии может по
служить либо остановка развития той или иной системы орrанизма, либо
нарушение координации в скорости компенсаторных ответных реакций систем
развивающеrося плода. Чем на более ранней стадии cBoero развития находится
эмбрион, тем сильнее отличается ero ответная реакция на действие патоrенноrо
фактора от реакции систем взрослоrо орrанизма.
В раннем эмбриоrенезе человека закладывается большая часть аномалий и
уродств развития (табл. 4).
Таблица 4. Сроки возникновения некоторых аномалий развития эмбрионов и плодов
человека
Форма аномалии Возраст
Анэнцефалия отсутствие rоловноrо мозrа,
свода костей черепа ......................................... ... ........................ ....... 26 суток
Менинrомиелоцеле выбухание оболочек
и тканей спинноro мозrа сквозь дефект в позвоночнике ...................... 28 суток
Экстрофия мочевоrо пузыря отсутствие нижней
части передней брюшной стенки и передней стенки
мочевоro пузыря. Дефект брюшной стенки замещен
задней стенкой мочевоrо пузыря с отверстиями мочеточников ........... 30 суток
rуба расщепленная (заячья) дефект развития верхней ryбы ............ 36 суток
Атрезия и свищи прямой кишки, возникновение
диафраrмальной rрыжи, дефект межжелудочковой
переrородки сердца ... ........................... ............ ............................. .... 6 недель
Расщелина верхней челюсти и неба .................................................... 9 1 О недель
rрыжа пуповинная (омфалоцеле) из за дефекта
брюшной стенки наружу выходят внутренние opraHbI,
покрытые истонченными тканями пуповины ........................................ 10 недель
rипоспадия отсутствие у детей мужскоrо пола
дистальной части мочеиспускательноrо канала,
а ero наружное отверстие находится в мошонке
или промежности; у детей женскоrо пола
отверстие мочеиспускательноrо канала открывается
во влаrалище ..... ...... ............................................. .......... ... .... ...... ....... 12 недель
Крипторхизм задержка опущения яичек в МОШQНКУ ......................... 7 10 месяцев
340
Fлава 17
В онтоrенезе человека к критическим периодам относят: 1) оплодотворение;
2) имплантацию (7 8 e сутки эмбриоrенеза); 3) развитие oceBoro комплекса зачат
ков opraHoB и плацентацию (3 8 я недели); 4) развитие roловноrо мозrа (15 20 я
недели); 5) формирование основных систем орrанизма (20 24 я недели); 6) рожде
ние; 7) период до 1 rода); 8) половое созревание (11 16 лет).
Около 10% причин смерти детей после рождения обусловлено врожденной
патолоrией. Статистика утверждает, что у 2,5% новорожденных реrистрируются
резко выраженные аномалии развития. Столь высокая частота пороков обуслов
лена сложностью эмбриональноrо развития, наличием нескольких критических
периодов в эмбриоrенезе, коrда зародыш наиболее подвержен влиянию TepaToreH
ных факторов, а также современным неблаroприятным состоянием эколоrической
среды. По данным медицинской статистики, около 20% врожденных пороков обус
ловлены нарушением структуры reHoB rенопатиями; 10% нарушением CTPYK
туры хромосом (хромосомными аберрация ми) и 10% пере несен ной внутриут
робной инфекцией. Остальную долю составляют пороки невыясненной этиолоrии
Методы iJuazHocmUKU и меры профuлактики аномалий развития челове
ка. С целью выявления аномалий развития человека современная медицина pac
полаrает рядом методов (неинвазивных и инвазивных). Так, всем беременным жен
щинам дважды (16 24 недели и 32 36 недель) про водят ультразвуковое
исследование, что позволяет обнаружить ряд аномалий развития плода и ero opra
нов. На 16 18 й неделе беременности с помощью метода определения содержания
альфа фетопротеина в сыворотке крови матери можно выявить пороки развития
ЦНС (в случае увеличения ero уровня более, чем в 2 раза) или хромосомные aHO
малии, например синдром Дауна трисомия хромосомы 21 или друrие трисомии
(об этом свидетельствует уменьшение более, чем в 2 раза, уровня исследуемоrо
вешества).
АМНlIоцентез инвазивный способ исследования, при котором через брюш
ную стенку матери производят взятие околоплодных вод (обычно на 16 й неделе
беременности). В дальнейшем производят хромосомный анализ клеток амниоти
ческой жидкости и друrие исследования.
Используется также визуальный контроль развития плода с помощью лапа
роскопа, введенноrо через брюшную стенку матери в полость матки (фетоскопия).
Существуют и друrие способы диаrностики аномалий развития плода. Однако
основной задачей медицинской эмбриолоrии является предупреждение их разви
тия. С этой целью разрабатываются методы rенетическоrо консультирования и
подбора супружеских пар. Методы искусственной инсеминации половыми клет
ками от заведомо здоровых доноров позволяют избежать наследования ряда He
блаroприятных признаков. Развитие rенной инженерии дает возможнось корри
rировать локальные повреждения rенетическоrо аппарата клетки. Так, существует
метод, сущность KOToporo заключается в получении биоптата яичка у мужчины с
rенетически обусловленным заболеванием. Внесение в сперматоrонии нормаль
ной ДНК, а затем трансплантация сперматоrоний в предварительно облученное
яичко (для уничтожения rенетически дефектных половых клеток). последующее
размножение трансплантированных сперматоroний при водит к тому, что вновь
Воспроизведение человека
341
образованные сперматозоиды освобождаются от rенетически обусловленноrо дe
фекта. Следовательно, такие клетки MorYT дать нормальное потомство при опло
дотворении женской половой клетки.
Метод криоконсервации спермы позволяет длительно сохранять оплодотво
ряющую способность сперматозоидов. Это применяется для сохранения половых
клеток мужчин, связанных с опасностью облучения, ранения и др.
Метод искусственноrо оплодотворения и переноса эмбрионов (экстракорпо
ральное оплодотворение) применяется для лечения как мужскоrо, так и женскоrо
бесплодия. Для получения женских половых клеток используют лапароскопию.
Специальной иrлой прокалывают оболочку яичника в области расположения пу
зырчатоrо фолликула, аспирируют овоцит, который в дальнейшем оплодотворя
ется спермиями. Последующее культивирование, как правило до стадии 2 4 8 бла
стомеров, и перенос зародыша в матку или маточную трубу обеспечивает ero
развитие в условиях материнскоrо орrанизма. При этом возможна транспланта
ция зародыша в матку "сурроrатной" матери.
Совершенствование методов лечения бесплодия и профилактики аномалий
развития человека тесно переплетаются с морально этическими, юридическими,
социальными проблемами, решение которых во MHoroM зависит от вековых тради
ций тою или иноrо народа. Это является предметом специальноrо исследования и
обсуждения в литературе. Успехи клинической эмбриолоrии и репродуктолоrии,
однако, не MOryT существенно повлиять на рост народонаселения в силу высокой
стоимости лечения и методических трудностей при работе с половыми клетками.
Поэтому профилактическая работа врача, основанная на знаниях процессов эмб
риоrенеза, составляет основу той деятельности, которая направлена на оздоровле
ние и численный рост населения. Для рождения здоровоrо потомства немаловаж
но соблюдать правила здоровоrо образа жизни и отказаться от вредных привычек,
а также проводить комплекс тех мероприятий, которые находятся в компетенции
медицинских, общественных и образовательных учреждений.
Таким образом, в результате изучения эмбриоrенеза человека и друrих позво
ночных установлены основные механизмы образования половых клеток и их сли
ян ия С возникновением одноклеточной стадии развития зиrоты. Последующее
развитие зародыша, имплантация, формирование зародышевых листков и эмбри
ональных зачатков тканей, внезародышевых opraHoB показывают тесную эволю
ционную связь И преемственность развития представителей различных классов жи
BOTHoro мира. Важно знать, что в развитии зародыша существуют критические пе
риоды, коrда резко возрастает риск внутриутробной rибели, либо развития по
патолоrическому пути. Знание основных закономерных процессов эмбриоrенеза
позволяет разрешить ряд проблем медицинской эмбриолоrии предупреждение
аномалий развития и уродств плода, лечение бесплодия, осуществлять комплекс
тех мероприятий, которые предупреждают rибель плодов и новорожденных.
342
ЗАКЛЮЧЕНИЕ
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ КУРСА rистолоrии
в учебнике материалы изложены с учетом проявлений основных свойств живоrо:
поzpа1lUЧ1locmи, обмена веществ (метаболизма), реактшлюсти и д6Иzameлыюй акmИ61l0
сти. Эти свойства прослеживаются на структурных уровнях орrанизации живоrо от кле
точноrо до орrанизменноrо. Такой подход к изложению учебных материалов, на наш
взrляд, позволяет нarлядно проследить преемственность развития и усложнение opra
низации клеток, тканей, орrанизма в целом в рамках четырех свойств живоrо.
Современная rистолоrия и ее уrлубление в область молекулярной биолоrии не
умаляют те orpoMHbIe достижения, которые создали фундамент для cOBpeMeHHoro
понимания строения и функционирования живых систем орrанизма человека. За
время развития rистолоrии накоплен и обобщен оrpомный фактический материал
по микроскопическому и субмикроскопическому строению клеток и тканей и выk
винут ряд научных теорий. Наиболее важная из них 'КЛеточная теория оказала
большое влияние на развитие всех биолоrических наук, в том числе и rистолоrии.
Это широкое фундаментальное обобщение в биолоrии и в наши дни служит проч
ной основой для разработки целоrо ряда актуальных медико биолоrических про
блем (рост, дифференцировка, реrенерация, воспаление, иммунитет и др.).
Теория зародышевых листков, основы которой были заложены К. Бэром,
АО. Ковалевским и И.И. Мечниковым, прочно вошла в эмбриолоrию и rистолоrию.
Она имеет большое значение для теоретической и практической медицины. Обособ
ление зародышевых листков, детерминация в них зачатков являются первой ступе
нью дифференциации и специализации клеточноrо материала зародыша и необхо
димой предпосылкой последующеrо возникновения из этоrо материала специали
зированных в различных направлениях тканей. Исторически возникшая тканевая
специфичность производных различных зародышевых листков накладывает отпе
чаток на свойства тканей в условиях нормы, реrенерации и опухолевоrо роста, что
необходимо учитывать при анализе мноrих медико биолоrических проблем.
Создание п.r. Светловым теории критических периодов развития имеет боль
шое значение для разработки вопросов профилактики патолоrии беременности,
пренатальной смертности, предупреждения эмбриопатий, аномалий развития и
наследственных болезней. Изучение в разделе эмбриолоrии общих закономернос
тей индивидуальноro развития, филоrенетической обусловленности их у человека
имеет целью при вить будущим врачам навыки эволюционноrо подхода к оценке
общебиолоrических явлений, в том числе и вопросов патолоrии.
Блаroдаря классическим работам АА Заварзина, H.r. Хлопина и их школ co
здано эволюционное направление в rистолоrии, сформулированы теория паралле
лизма развития тканевых систем и теория диверrентной эволюции тканей. Обосно
Медицинское значение "урса lистолоlUИ
343
вано положение о том, что с повышением орrанизации животных в ходе эволюции
все более оrраничиваются возможности взаимопревращения разнородных тканей
друr в друrа. Основные положения эволюционной rистолоrии явились преДПОСЫk
кой для построения теории lисmОlенеза. [истоrенез следует рассматривать как диа
лектическое единство взаимодействующих факторов: внутренних (наследственность
клеток и ткани, или rенетическая детерминация), проrраммирующих направление и
характер развития, и внешних (изменчивость, или фактор адаптации), обеспечива
ющих модуляцию клеточных фенотипов за счет различных механизмов реryляции
(нервной, rормональной, иммунной и др.). Структурно функциональным выраже
нием данноrо взаимодействия служат процессы пролиферации, дифференциации,
интеrрации, rибели клеток и др. в развивающихся тканях.
При изучении закономерностей rистоrенеза следует учитывать, что всякая ткань
развивается не изолированно, а в тесном окружении и в постоянном взаимодействии
со мноrими тканями соответствующих opraHoB и систем орrанизма. В связи с этим в
современной rистолоrии особое значение приобрела теория межтканевых корреля
ций, касающаяся Bcero мноюобразия связей, определяющих развитие тканей, opra
нов и орrанизма в целом.
Знание закономерностей rистоrенеза и особенностей ею проявления в разные MO
менты функционирования тканей необходимо для понимания проблем патолоrии, так
как закономерности нормальною rистоrенезаслужат фундаментом при выяснении сущ
ности патолоrических процессов.
Подход к проблеме реrенерации с позиций учения о rистоrенезе позволяет
приложить общие закономерности развития тканей не только к физиолоrической,
но и к репаративной реrенерации.
С позиций rистоrенетической концеПllИИ реrенерации суть дела состоит в He
обходимости выяснения диапазона детерминированной изменчивости тканей и
межтканевых корреляций, свойственных нормальному развитию, и условий про
явления этих закономерностей после повреждения opraHoB. Закономерности rис
TO и орrаноrенеза и межтканевых взаимодействий основа для понимания Mexa
низмов реrенерации opraHoB и тканей. Формы проявления реrенерации (клеточное
размножение или rиперплазия внутриклеточных структур) обусловлены rистоrе
незом и специфичны для каждой ткани. Знание этих закономерностей необходи
мо для изыскания эффективных способов реrуляции заживления ран.
Наряду с важным значением теоретических аспектов учения о тканях, COBpe
менная rистолоrия предоставляет для медицины оrромный фактический матери
ал прикладноrо плана. Значение этоrо материала трудно переоценить, ибо без зна
ния структурных основ клеточноrо и тканевою уровней не может быть полноrо
понимания механизмов патолоrических процессов.
Уже первые занятия по rистолоrической технике в курсе rистолоrии позво
ляют приобрести определенные навыки взятия материала для исследования, ero
фиксации и последующей обработки, так как в практической деятельности врача
может возникнуть необходимость самостоятельно делать биопсии opraHoB и TKa
ней для экспресс диаrностики и определения тактики лечения. Следует иметь в
виду, что rистолоrические, rистохимические и электронно микроскопические
344
Заключение
методы широко применяются в научно исследовательской работе практически
по всем медицинским и военно медицинским специальностям.
Материалы по цитолоrии и реактивности клеток и их орrанелл при действии
неблаrоприятных факторов различной природы, о сущности изменений клеточ
НOI':'! мембраны, орrанелл, rенетическоrо материала, а также метаболизма и трофи
ки клеток, состояния клеток после повреждения, изменения клеток во время про
хождения митотическоrо цикла, особенно при воздействии MYTareHHbIx факторов,
возможности внутриклеточной реrенерации и др. имеют прямое отношение к по
ниманию вопросов патолоrии.
Четкие представления о локализации камбиальных элементов, темпах проли
ферации клеток и ее реrуляции необходимы будущим врачам для Toro, чтобы про
думанно проводить хирурrические и терапевтические вмешательства на opraHax,
выстланных или покрытых эпителиями. Не нуждается в пояснении необходимость
четкою знания структурно функциональных особенностей различных клеток KpO
ви для прочтения лабораторных анализов и постановки диаrноза заболевания.
Современные представления о T и В лимфоцитах и их популяциях, мононукле
арной макрофаrальной системе, кооперациях и трансформациях клеток в процес
се ryморальною и клеточноrо иммунитета способствуют пониманию медицинс
ких проблем иммунолоrическоrо надзора в орrанизме, трансплантации opraHoB и
тканей, опухолевоrо роста и др. Важно понимать взаимоотношения местных и цeH
тральных факторов реryляции морфофункциональных единиц opraHoB особенно
в связи с учением о диффузной эндокринной системы.
Изучение патолоrии не входит в задачу нормальной rистолоrии. Однако зна
ние закономерностей развития структур клеточною, тканевою и opraHHoro ypOB
ней орrанизации тела человека в эмбриоrенезе и в постнатальном онтоrенезе, а TaK
же знание особенностей строения, функций, реактивности и реrенерации тканей с
учетом их медицинскоrо значения является необходимой основой для успешною
овладения клиническими дисциплинами, для последующеrо умения распознавать,
анализировать и правильно трактовать патолоrические изменения тканей и овла
девать дифференциальной диаrностикой важным компонентом клиническоrо
мышления врача.
Алфавитный указатель
345
Алфавитный указатель
Б
А
А клетки 174, 183
адвентициальная оболочка 159, 205, 312
адвентициальные клетки 120
аденилилциклаза 43
аденоrипофиз 236
актина белок 39
аллантоис 66
альвеола 207
аминокислоты 232
амитоз 48
амнион 66
амниотическая жидкость 331
анастомозирующие мостики 200
анафаза 46
анемия 101
анизоцитоз 91
антенны 277
антиrенпредставляющая клетка 109
антиrены 228, 301
антитела 230
антитело 228
апокриннаясекреция 87
апоптоз 73
артерии
мышечноro типа 191
смешанноrо (мышечно эластическоrо)
типа 190
эластическоrо типа 190
артериоло венулярные анастомозы 195
артериолы 192
ассоциативные (переключательные) ядра
255
ассоциативные волокна 261
астроциты
волокнистые 136
протоплазматические 136
атрезия (rибель) фолликулов. 309
атрофия 71
ацетилхолин 140
ацидофильныеэндокриноциты 236
ацинозно инсулярные клетки 183
ацинус 206
аэроrематический барьер 207
базальная мембрана 80
базалыIйй rистион 334
базальный слой 335
базофилия 182
базофильные rранулоциты (базофилы) 94
базофильные эндокриноциты 237
белая пульпа 225
белковые железы 165
белое вещество 251
белые мышечные волокна 292
биorенетический закон 54
биоrенные амины 247
бластопор 62
бластоциста 67, 319
бластула 60, 64
бокаловидные экзокриноциты 84, 177
большие слюнные железы 165
бранхиоrенная rруппа 233
6yropoK оплодотворения 317
6уль60уретральные железы 304
быстрая миrрация 231
в
Б клетки 183
вазоактивный IIнтестинальный
полипептид 183
ведущая ткань 76
ведущий клеточный дифферон 76
веки 275
венозный синус склеры 275
венулы 195
вены
безмышечные 196
мышечные 196
виментин 39
БИП клетки 174, 183
включения
белковые 36
липидными 36
пиrментные 37
экскреторные 36
вкусовая пора 284
вкусовая ямка 284
вкусовые луковицы 165
внутреннее ухо 279
внутренняя среда для метаболических
346
Алфавиmный указаmель
реакций 88
водители ритма 216
водянистая влаrа 275
возрастные изменения нервных тканей 140
возрастные изменения сосудов 198
волокна мозжечка
афферентные 258
эфферентные 258
волосы 152
корень 152
стержень 152
волосяная луковиuа 152
волосяная сумка 153
воспаление 11 3
вставочные диски 127
вставочные протоки 166
вставочный отдел 182
вторичная почка 210
вторичные ворсинки 333
r
rанrлии 266
rанrлионарные клетки 272
rанrлионарный слой 256, 261
rаптотаксис 71
rастроцель 62
rаструла ранняя 64
rаструляция 61
[аустры 179
rель 108
rематонефридиальный rистион 211
rематотестикулярный барьер (ПБ) 301
rематотимический rистион 222
rематофолликулярный барьер (rистион)
308
rематохориальный барьер 336
rематоэнцефалический rистион (барьер)
264
reMorpaMMa 97
rемокапилляры 193
висцеральноrо типа 195
прерывистые (синусоидные) 195
соматическоrо типа 194
rеморроидальные вены 180
rемоциты 89
rенетическая индивидуальность 52
rензеновский узелок 64
rепатоrематический барьер 187
rепатоциты 185
rетероморфия 85
rетеропластический rемопоэз 101
rетерополярность 81
rетеросинтетическая интерфаза 44
rиаломер 97
rиrантоциты 91
rидроксиапатита кристаллы 119
rипертрофия 71
rипобласт 64, 68, 320
rиподерма 149
rипоталамо rипофизарный тракт 235
rиппокамп 278
rистион 113
rистиоциты 109
rисто и иммуноцитохимические методы
13
rистоrенез 123
rистоrенез костных тканей
(остеоrистоrенез) 118
[истоrенез хрящевых тканей
(хондроrистоrенез) 116
rистоrенетическая концепция 343
rистоrенетический ряд 73
rистосовместимость I ro класса 229
rистосовместимость II ro класса 230
rлавные паратироциты 243
rлавные экзокриноциты 172
rлазные мышцы 276
rландулоциты 87
rликозаминоrликаны
несульфатированные 108
сульфатированные 108
rликокаликс 23
rликофорины 91
rлиоциты 136, 239
rлия 259
rлюкаrон 183
rлюкокортикоидные roрмоны 245
roлокринная секреция 87
rомопластический rемопоэз 101
rоноuиты 300
[ормоны 180, 231
rрануломер 96
rранулоцитопоэз 102
rранулоциты
базофильные 92
нейтрофильные 92
Алфавитный указатель
347
эозинофильные 92
rранулы
азурофильные 93
специфические 93
rpанулярныеэндокриноциты 246
rребешки 280
rруппа мозrовых придатков 233
rруппа надпочечников 233
rруппы тканей 75
ryбные железы 160
ryбчатый участок 335
ryморальный иммунитет 103
д
двиrательные ядра 255
деламинация 61
деламинация дискобластулы 64
дентин 161, 163
дерма 149
дерматом 66, 323
десмин 39
десмосома 27
детерминация
зачатковая 73
клеточная 73
оотипическая 73
тканевая 73
дискобластула 64
дифферон 73
эндокринных кардиомиоцитов 127
эпителиоцитов 79
дробление 59
дыхание 37
клеточное 37
Е
ЕС клетки 173
ЕСL клетки 173
ж
железистые эпителии 87
желтое пятно 272
желточный мешок 64, 66
желтый костный мозr 219, 220
желчные капилляры 186
3
замыкательная пластинка 336
запроrраммированная rибель клеток
(апоптоз) 51
зародышевые листки 320
звездчатые макрофаrоциты 187
звездчатые миоэпителиоциты 156
зернистые лейкоциты 92
зубная пластинка 161
зубная связка 162
зубные альвеолы 162
зубные мешочки 161
зубы 160
И
изолецитальные яйцеклетки 60
иммиrрация 61, 65
иммунитет
ryморальный 228
клеточный 228
ИМJ\lуноrлобулины 230, 231
иммунодепрессивная функция 337
имплантация 67
инваrинация 61
индивидуальность 22
иннервация сосудов 198
инсулин 183
интеrрины 72
интернейроны 262
интерстициальный рост 117
интерференционная микроскопия 12
интраваскулярное деление 98
интраспинальный opraH 251
исчерченные слюнные трубки 166
к
капацитация 316
кардиальные железы 169, 172
кардиодилатин 127
кардиомиобласты 126
кардиомиоциты 126, 200
кариорексис 51
катехоламины 245
кератолинин 82
кишечная энтодерма 68, 83, 248
кишечные ворсинки 175
кишечные крипты 175
клапаны 196, 198, 199
клетки Ито 187
клетки Купфера 187
348
Алфавuтll.ЫЙ указатель
клетки Ланrерrанса 149
клетки Лейдиrа 302
клетки Меркеля 149
клетки островков Ланrерrанса 181
клетки Панета 177
клетки Пуркиня 199
клетки Сертоли 301
клетки мишени
rормонзависимые 232
rормончувствительные 232
клеточная теория 20
клеточный иммунитет 103
клеточный центр 40
клубочковая (наружная) зона 244
кожа 146
кожная эктодерма 68, 81, 248
колбочковые клетки 271
колбы Краузе 286
коллаrеназа 106
коллаrеновые фибриллы 106
коллаrены 72
колониеобразующая единица 99
колонка 252
комиссуральные (каллозальные) волокна
261
компактный участок 336
коr.шлемент 228
контакт
заякоривающий 27
плотный 27
простой, неспециализированный 26
щелевой 28
контактное инrибирование 71
конфокальная микроскопия 14
концевые моторные бляшки 294
концевые секреторные отделы 166
корень нопя 154
корковое вещество 224
кортикальная реакция 317
кортикотропоцитыI 237
костная пластинка 121
котиледон 332
красная пульпа 226
красные мышечные волокна 292
красный костный мозr 219
крипты 168, 178
кристаллоиды 94
критические периоды 339
критические точки 45
кровяные пластинки 102
крупные скопления лимфоидной ткани
179
л
ламинин 72
леrочная долька 207
лейкоцитарная формула 97
лейкоцитоз 93
либерины 235
лизис 51
лизосомы 35
лимб 273
лимфа 224
лимфанrионы 198
лимфатические сосуды
безмышечноrо типа 198
мышечноrо типа 198
лимфатические узелки 168. 224
лимфоидная ткань 98
лимфокапилляры 197
лимфокины 230
лимфоцитопоэз и иммуноцитопоэз 103
лимфоэпителиальный opraH 221
листки спланхнотома 88
висцеральный 88
париетальный 88
люминесцентная. или флюоресцентная,
микроскопия 12
лютропин 305
м
макроrлия 262
макрофаr 103
макрофаrи
секреторные 109
фаrоцитарные 109
макрофаrическая система 110
макроциты 91
материнская поверхность плаценты 333
матка 67
меrакариобласт 102
меrакариоцит 102
меrалобласты 98
меrалоциты 98
медиаторы 232
медленная миrрация 231
Алфавит1lЫЙ указатель
349
медуллобласты 132
мезаксон 137
мезенхима 88, 98, 104, 115, 137, 277
мезенхима склеротома 118
мезодерма 61, 68. 88
мезонефральный проток 21 О
мезотелий 85
мейоз 48
мейотическая фаза 298
меланин 108, 149
меланоциты 148
мембрана Бруха 274
менструальная фаза 313
меробластическое дробление 60
мерокринная секреция 87
мероциты 64
местные рефлекторные дуrи 267
метанефрос 210
метаплазия 80
метафаза 46
метод изучения фиксированных срезов
зародышей 55
метод культуры клеток, тканей 14
метод маркировки 55
метод фракционирования 14
метод экстракорпоральноro
оплодотворения 56
методы микрохирурrии 56
меторизис 169
миrрация клеток 71
миелоидная ткань 98
микроворсинки 40, 84
микроrлия 137, 262
микроскопическая хирурrия клетки 14
микротельца 35
микрофаrи 93
микроциты 91
минералокортикоиды 244
минимиозин 40
миометрий 31 О
миопиnментоцит 130
миотом 66. 323
миофибриллы 124
миоциты
контрактильные 128
секреторные 128, 129
миоэпикардиальные пластинки 126
миоэпителиоциты 165
митоз 45. 70
млечные протоки 155
млечный синус 156
мноrорядный эпителий 83
модуль 261
мозrовое вещество 224, 245
мозжечок 256
молекулы клеточной адrезии 71
молоко 156
молочные альвеолы 155, 156
молочные железы 155
молочные экзокриноциты 156
мононуклеарная фаrоцитарная система 95
моноцитопоэз 103
морула 67, 318
морфофункциональные единицы 76
мочевой пузырь 216
мочеиспускательный канал 217. 305
мочеточник 216
мукоциты 165, 173
мультиполярные 251, 259
мышечная оболочка 159, 178, 312
мышечное волокно 123
мышечные веретена 293
мяrкая мозrовая оболочка 262
мяrкое нёбо 167
н
надrортанник 203
надхрящница 118
незернистые лейкоциты 92
нейроrипофиз 236, 238
нейроrормоны 135
нейролеммоциты 136, 137
нейромоторная единица 294
нейронная сеть 249
нейропептиды 236
нейропоры 63
нейросенсорные 269
нейрофибриллы 134
нейроэктодерма 86
нейроэктодермы 248
нейроэндокринная система 233
нейтрофильные rpанулоuиты
(нейтрофилы) 92
ненасыщенные жирные кислоты 232
нервная пластинка 63, 322
нервная трубка 63
350
Алфавитный указатель
нервные клетки 247
нервные окончания
свободные 285
нефрон 210
дистальный отдел 213
тонкий отдел 213
нефротомы 66, 84, 209
нопевая пластинка 154
нопи 153
нормоциты 91
нуклеоплазма 32
о
обмен веществ 22, 52, 342
обновляющиеся ткани 70, 87
оболочка оплодотворения 317
обонятельная луковица 278
обонятельные плакоды 277
овариальный фолликул 58
овоrенез 297
овофолликулярные rистионы 306
однослойные эпителии 84
однослойный призматический каемчатый
эпителий 176
одонтобласты 121, 161
окончания
двиrательные 138
рецепторные 138, 140
олиrодендроrлия 136
онтоrенез 315
оплодотворение 59
опсонины 228
осевой комплекс зачатков 62
осевой цилиндр 137
остеобласты 118
остеокласты 120
остеоны 121, 122
остеоциты 119
п
палочковые клетки 271
панкреатические ацинусы 181
панкреатические экзокриноциты 181
панкреатические эндокриноциты 182
панкреатический полипептид 183
панкреатический сок 180
параrанrлии 246
паракортикальнаязона 224
париетальные экзокриноциты 173
паутинная оболочка 263
пепсин 170
пептиды 232
первичная почка 209
передний отдел rипоталамуса 235
переходный эпителий 83
периметрий 31 О
перимизий 293
периневральный эпителий 86
периневрий 265
перисинусоидальные липоциты 187
периферические отделы 266
периферические отростки 256
перициты 105
пероксисомы 35
перфорины 230
петля [енле 213
печеночные ацинусы 187
печеночные балки 186
печеночные дольки 184
пиrментный эпителий 86
пикноз 51
пилорические железы 172
пинеалоциты 239
плацента 67, 330
поверхностные эпителиоциты 171
поrружная имплантация 319
подслизистая основа 159, 178, 205, 312
пойкилоцитоз 91
полилецитальные яйцеклетки 60
полипептидные [ормоны 247
полиплоидизация 128
полиплоидные миоциты 201
половые валики 300
половые стероиды 245
поляризационная микроскопия 12
поровые кольца 30
портальные дольки 187
поры Кона 207
посткапилляры 195
постменструальная фаза 314
постсинаптические уплотнения 139
потенциал действия 140
потовые железы 150
предентин 163
предменструальная фаза 314
предпочка 209
Алфавит'Ный указатель
ЗS1
предстательная железа (простата) 304
презумптивные участки 63
прекапилляры 193
преостеобласты 118
преформизм 53
прехордальная пластинка 81, 202
придаток яичка (эпидидимис) 303
проекционные волокна 261
прозрачная зона 307
производные мезодермы 248
промежуточный мозr 255
пронефрос 209
пространство Диссе 186
пульпа зуба 162, 163
пучковая зона 245
р
Р клетки 174, 199
радиоавтоrрафия 13
радужка 274
развитие желтоro тела (лютеоrенез) 308
растущие ткани 70
реrенерация 81
реrенерация костной ткани 122
редукционное деление 299
рекапитуляция 55
ресничное (цилиарное) тело 275
ретикулоциты 92
ретикулярная ткань 89, 98
ретикулярная формация 255
рефлексы 249
рецепторно трансдукторная система 100
речепторы
сенсоэпителиальные 269
рилизин rормоны 235
рилизинr факторы 235
роrовица 272
роды 338
РР клетки 183
с
С клетки 241
сальные железы 151
сальные экзокриноциты 151
свободные макрофаrи 95, 109
себоциты 151
секреторные отделы
белково слизистые 167
белковые 167
секреторный цикл 87, 241
семенной буrорок 304
семенные пузырьки 303
семявыносящий проток 303
серое вещество 251
серозная оболочка 159, 178, 312
серозные полулуния 167
сероциты 165
сетчатая зона 245
синапсы 138
синаптические пузырьки 139
синовиальный эндотелий 291
синусы 226
склеротом 323
слезные железы 276
слепое пятно 272
слизистая оболочка 159, 175, 204, 312
слизистые железы 165
слой полиморфных нейронов 261
слуховые плакоды 279
слуховые пятна 279
слюна 166
смешанные железы 165
собирательные трубочки 213
собственные железы желудка 172
соляная кислота 170
соматостатин 183
сомиты 63, 66, 323
сосочки 164
rрибовидные 164
желобоватые 164
нитевидные 164
сосудистый эндотелий 86
сперматоrенез 297
сперматозоиды 57
спиральный opraH 282
спланхнотом 63
среднее ухо 278
средний отдел rипоталамуса 235
средняя доля 237
стабильные ткани 71
статины 235
стационарная стадия 299
стволовая кроветворная клетка 99
стекловидное тело 274
стенка первичных сосудов 89
стероидные ropMoHbI 232
352
Алфавитный указатель
столбчатые эпителиоциты 176
строма 219
студенистая (слизистая) ткань 332
суданофобный слой 245
сурфактант 208
сухожильные рецепторы 294
сфинктеры пищевода 170
т
танициты 136
твердая оболочка 264
твердое нёбо 167
тека 307
телолецитальная яйцеклетка 60
телофаза 46
тельца [ассаля 222
тельца Мейснера 286
тельца Фатер Пачини 286
тельце Руффини 286
теория rистоrенеза 343
теория зародышевых листков 54, 342
терминальные бронхиолы 206
ткани 76
внутренней среды 75
монодифферонные 74
мышечные 75
нервные 75
покровные 75
полидифферонные 74
развитие
диверrентно 75
параллельными рядами 75
соединительные
неоформленные 115
оформленные 115
эластические 115
хрящевая
волокнистая 117
rиалиновая 117
эластическая 117
тонофибриллы 82
тонофиламенты 82
трансплантация ядер 56
трансферрин 305
третичные (терминальные) ворсинки 333
триада 125
триады печени 185
тромбоцитопоэз 102
тропоколлаrен 106
трофобласт 67
трофэктодерма 330
у
узловые перехваты 137
ультрафиолетовая микроскопия 13
уретральные железы 305
ушная раковина 278
ф
фаза
дистантноrо взаимодействия 316
KOHTaKTHoro взаимодействия 317
фаза размножения 297
фаза созревания 298
фазово контрастная микроскопия 12
фибробластический дифферон 105
фибронектин 72, 106
фиброциты 106, 115
филаrrрин 82
фолликулоrенез 307
фолликулы 240
фолликулярныеэндокриноциты 241
фоллитропин 305
форменные элементы 90
х
хемотаксис 71
химозин 171
хордомезодерма 68
хорион 66, 67
хроматин 31
хромофобные эндокриноциты 237
хрусталик 273
ц
цейтрафферная, или замедленная,
микрокиносъемка 14
целом 63
цемент 162, 163
цементобласты 162
цементоциты 163
центральные отделы 265
центральные отростки 253
центроацинозные эпителиоциты 182
цинновая связка 274
Алфавитный у-казателъ
353
цитокератины 79
цитоспектрофотометрия 13
ч
чувствительные ядра 254
ш
шейка матки 310
шунты 195
щ
щеки 160
щелочная фосфатаза 119
щеточная каемка 84
э
эквационное деление 299
экзокринная железа 186
экзокриноциты 151
экзоцитоз 26
эксплантация 56
экстраваскулярное кроветворение 98
эктодерма 61, 68
эктомезенхима 88, 118
эктоплазма 105
электронная микроскопия 13
эмалевые призмы 162
эмаль зуба 161
эмбриобласт 67
эмбрион 53
эмбриональное развитие 53
энамелобласты 161
эндокринные клетки 247
эндокринный opraH 186
эндокриноциты 177
эндокриноциты желудка 173
эндометрий 309
эндомизий 293
эндоневрий 265
эндоплазма 105
эндотелиобласты 111
эндотелиоциты 112
эндоцитоз 25
энтериновая rормональная система 248
энтодерма 61, 68
эпендима 86
эпибласт 64, 68, 320
эпиболия 61
эпиrенез 53
эпидермально пролиферативная единица
148
эпидермис 82
эпиневрий 265
эпителиальная ткань 237
эпителиальные трабекулы 243
эпителиоподобноестроение 277
эпитоп 229
эритропения 91
эритроцитоз 91
эритроцитопоэз 101
ю
юкстаrломерулярный rистион 214
я
ядерная оболочка 29
ядра 252
ядро 29
язык 164
яйцеклетка 58
яйценосный буrорок 307
354
Литература
ЛИТЕРАТУРА
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К, Уотсон Дж. Молекуляр
ная биолоrия клетки (т. 1 3): Пер. с анrл. М.: Мир, 1994.
Алмазов И.В., Сутулов л.с. Атлас по rистолоrии и эмбриолоrии. М.: Медици
на, 1978.
Брусиловский АИ., Роrажинская КК, Хачатурян Е.К, Килачицкая И.Р. PYCCKO
латинская терминолоrия по rистолоrии, цитолоrии и эмбриолоrииj Под ред.
проф. Ю.И. Афанасьева. Симферополь, 1987.
Волкова О.В., Пекарский М.И. Эмбриоrенез и возрастная rистолоrия внутренних
opraHoB человека. M.: Медицина,1976.
rистолоrия. А Хэм, Д. Кормак М.: Мир, т. 1 5, 1982 1983.
rистолоrия, цитолоrия и эмбриолоrия: Атлас j Под ред. О.В. Волковой, Ю.К Елец
Koro. М.: Медицина, 1996.
rистолоrия. Н.А Юрина, АИ. Радостина. М.: Медицина, 1996.
rистолоrия. Введение в патолоrию: Учебник jПод ред. э.r. Улумбекова и Ю.А Че
лышева. М.: [еотар, 1997.
rистолоrияjПод ред. Ю.И. Афанасьева, Н.А Юриной. М.: Медицина, 5 e изд.,
1999.
Данилов Р.К, Клишов А.А rистолоrия: Учебные материалы к лекциям и практи
ческим занятиям. СПб.: ВМедА, 1995.
Данилов Р.К, Клишов АА, Боровая Tr. rистолоrия человека в мультимедиа.
СПб.: Эскулап, 1997.
Данилов Р.К, Боровая Tr. Общая и медицинская эмбриолоrия. СПб.: СпецЛит,
2003.
Елисеев B.r., Афанасьев Ю.И., Котовский Е.Ф. Атлас микроскопическоrо и ульт
рамикроскопическоro строения клеток, тканей и opraHoB. М.: Медицина, 1970.
Карлсон Б. Основы эмбриолоrии по ПэттенуjПер. с анrл. М.: Мир, 1983.
Клишов АА rистоrенез и реrенерация тканей. Д: Медицина, 1984.
Клишов А.А rистолоrия человека: Учебник л.: ВМедА, 1989.
Кнорре Ar. Краткий очерк эмбриолоrии человека: 2 e изд. Л.: Медицина, 1967.
Кнорре Ar. Эмбриональный rистоrенез (морфолоrические очерки). л.: Меди
цина, 1971.
Кузнецов СЛ., Мушкамбаров Н.Н., rорячкина вл. Атлас по rистолоrии, цитоло
rии и эмбриолоrии. М.: МИА, 2001.
Лабораторные занятия по курсу rистолоrии, цитолоrии и эмбриолоrииjПод ред.
Ю.И. Афанасьева и АН. Яцковскоrо. М.: Медицина, 1999.
Международная rистолоrическая номенклатура (Под общей редакцией проф.
Ю.И. Афанасьева). М: Медицина, 1987.
Новиков ВД., Правоторов [В., Труфакин В.А Словарь по rистолоrии. Новоси
бирск: Новосибирский медицинский институт, 1998.
Общая и клиническая эмбриолоrияjПод ред. проф. Э.И. Вальковича. Д: ЛПМИ,
1989.
Литература
355
Пэттен Б.М. Эмбриолоrия человека/Пер. с aHr. М.: Медrиз, 1959.
Соловьев Б.А, Биноrрадов АФ. Аномалии развития человека. Учебное пособие.
Тверь: РИО TfMA, 1998.
Фалин Л.И. Атлас по эмбриолоrии человека. М.: Медицина, 1976.
Щелкунов с.и. Основные принципы клеточной дифференuировки. М.: Меди
цина, 1977.
Amenta P.S. Elias Pauly's Histology and Ниmап Microanatomy. N.Y Toronto:john
Wiley and Sons, 1987.
Clara М., Herschel К, Ferner Н. Atlas der normalen Microskopichen Anatomie des
Menshen. Leipzig, 1973.
Gartner L.P., Hiatt J.L. Color Atlas of Histology. Баltimorе: Williams and Wilkins,
1990.
Gartner L.P., Нiattj.L., Strumj.M. Histology. Williams &Wilkins, 1988.
Кimball's Бiоlоgу Pages. URL: http:j /www.ultrапеt.соm/ jkimЬаll/БiоlоgуРаgеs/
Krstic R.V. Ниmап Microscopic Anatomy. Ап Atlas for Students ofMedicine and Вiology.
Беrliп НеidеlЬеrg N. У.: Springer, 1991.
Moore R.L. Беfоrе We Are Боrn. Баsiс Embryology and Бirth Defects/ Third Edition.
Philadelphia: WБ. Saunders Соmрапу, 1989.
Sadler т.W Langman's Medical Embryology / Seventh Edition. Баltimоrе: Williams
and Wilkins, 1995.
Stevens А, LoweJ.S. Histology. Mosby, 1993.
356
Содержание
Содержание
ПРЕДИСЛОВИЕ .........................................................................................................3
БЛАrОДАРНОСТИ ...................................................................................................5
ВВЕДЕНИЕ В rистолоrию .................................................................................7
rЛАВА 1. ПРЕДМЕТ И ЗАДАЧИ rистолоrии.
УРОВНИ СТРУКТУРНО фУНКцИОНАЛьНОЙ
орrАНИЗАЦИИ живоrо ................................................................................... 7
rЛАВА 2. РАЗВИТИЕ МИКРОСКОПИИ И ОТКРЫТИЕ КЛЕТКИ.
МЕТОДЫ ИССЛЕДОВАНИЯ
В rистолоrии ........................................................................................................9
Методы исследования в rистолоrии ............................................................... 11
rЛАВА 3. КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК РАЗВИТИЯ
УЧЕНИЯ О КЛЕТКЕ И ТКАНЯХ ..................................................................... 15
ЦИТОЛ О rИЯ ................................................................................................................... 20
rЛАВА 4. ОСНОВЫ ОБЩЕЙ цитолоrии ............................................20
Морфофункциональные системы клетки ................................................. ...21
Покровная система клетки (плазмолемма) ............................................22
Внутренняя метаболическая среда клетки и ее компартменты ......28
Ядро ............................................................ ...................................................... 29
Орrанеллы синтеза и транспорта биополимеров ............................ 32
Клеточные включения ...............................................................................36
Митохондрии ................................................................................................37
Опорно двиrательная система клетки ..................................................... 38
Промежуточные филаменты ...................................................................39
Микрофиламенты и их производные ..................................................39
Клеточный центр и ero производные ..................................................40
Система реактивности клетки
(восприятия, трансформации и передачи сиrналов) ..........................43
Воспроизведение клеток ................................................................................44
Рост и дифференцировка клеток ................................................................48
Реактивные изменения клеток ....................................................................50
ЭМБРиолоrия ........................................................................................................... 53
rЛАВА 5. ОСНОВЫ ОБЩЕЙ ЭМБРиолоrии ....................................53
Развитие эмбриолоrии как науки .................................................................... 53
Методы исследования ..........................................................................................55
Половые клетки ......................................................................................................56
rЛАВА 6. ОБЩАЯ ХАРАКТЕРИСТИКА ИНДИВИДУАЛьноrо
РАЗВИТИЯ ПОЗВОНОЧНЫХ ..........................................................................59
Периодизация индивидуальноrо развития .................................................. 59
Содержание
357
Образование зародышевых листков и эмбриональных зачатков
тканей в эмбриоrенезе у некоторых позвоночных ....................................62
Ланцетник ........................................................................................................... 62
П тицы ................................................................................................................... 64
Млекопитающие ...............................................................................................66
ОБЩАЯ rистолоrия (УЧЕНИЕ О ТКАНЯХ) ..........................................70
rЛАВА 7. ЭМБРИОНАЛЬНЫЙ rистоrЕНЕ3
и КЛАССИФИКАЦИЯ ТКАНЕЙ .....................................................................70
Эмбриональный rистоrенез. Определение понятия "ткань" ................. 70
Классификация тканей ........................................................................................75
rЛАВА 8. СИСТЕМА ПОКРОВНЫХ ТКАНЕЙ
И ИХ ПРОИ3ВОДНЫЕ .........................................................................................79
Эпителии кожноrо типа ......................................................................................81
Эпителии кишечноrо типа ................................................................................. 83
Эпителии почечноrо типа ...................................................................................84
Эпителии целомическоrо типа .. ....................................................................... 84
Эпителии нейроrлиальноrо типа ..................................................................... 86
Железистые эпителии ..........................................................................................87
Физиолоrическая реrенерация эпителиев ................................................... 87
rЛАВА 9. СИСТЕМА ТКАНЕЙ ВНУТРЕННЕЙ
(МЕТАБОЛИЧЕСКОЙ) СРЕДЫ ......................................................................88
Понятие "мезенхима". Ретикулярная ткань ................................................88
Кровь и лимфа как ткани .................................................................................... 90
Кроветворные ткани и кроветворение в костном мозrе .......................... 97
Возрастные изменения и реактивность системы крови ....................... 103
Соединительные ткани ..................................................................................... 104
Рыхлая волокнистая соединительная ткань ....................................... 104
Жировая ткань ............................................................................................... 111
Эндотелий ............................................................................................................. 111
Взаимодействия клеток системы крови и соединительной ткани
в условиях воспаления и реrенерации тканей ......................................... 112
rЛАВА 10. СИСТЕМА ОПОРНЫХ
И двиrА ТЕЛЬНЫХ ТКАНЕЙ ........................................................................ 115
Ткани с опорно механической функцией .................................................. 115
Плотные волокнистые соединительные ткани .................................. 115
Скелетные ткани ............................................................................................ 116
Хрящевые ткани ....................................................................................... 116
Костные ткани ........................................................................................... 118
Ткани с двиrательной функцией ................................................................... 123
Скелетная мышечная ткань ....................................................................... 123
358
Содержание
Сердечная мышечная ткань ....................................................................... 126
rладкая мышечная ткань ............................................................................ 128
Мионевральная ткань .................................................................................. 130
Миоидные клетки .......................................................................................... 131
rЛАВА 11. СИСТЕМА НЕВРАЛЬНЫХ ТКАНЕЙ
И НЕйроrлия ...................................................................................................... 132
rистоrенез .............................................................................................................. 132
Строение ........................ ........................................................................................ 133
rЛАВА 12. РЕАКТИВНОСТЬ И РЕrЕНЕРАЦИЯ ТКАНЕЙ ............. 142
Реактивность тканей ................................................ ......................................... 142
Физиолоrическая реrенерация тканей ....................................................... 143
Реrенерационный rистоrенез ......................................................................... 144
ЧАСТНАЯ rИ СТОЛ О rИЯ ...................................................................................... 145
ВВЕДЕНИЕ В rистолоrию OprAHOB
И СИСТЕМ ЧЕЛ О ВЕКА .................................................................................... 145
r ЛАВА 13. СИСТЕМА ПО КРО ВА ................................................................. 146
Кожа ......................................................................................................................... 146
Производные кожи ............................................................................................. 150
Потовые железы ............................................................................................. 150
Сальные железы ............................................................................................. 151
Волосы ...................................................... . ........................................................ 152
Ноrти ........................................................ .. ........................................................ 153
Реактивность и реrенерация кожи и ее производных ..................... 154
Молочные железы ......................................................................................... 155
rЛАВА 14. СИСТЕМА OprAHOB ВНУТРЕННЕЙ СРЕДЫ,
СВЯЗАННАЯ С МЕТАБОЛИЗМОМ ........................................................... 158
Пищеварительный комплекс opraHoB ......................................................... 158
Развитие и общий план строения пищеварительной трубки .......158
Производные переднеrо отдела пищеварительной трубки ........... 159
Ротовая полость ........................................................................................ 159
rубы, щеки, десны ..................................................................................... 160
Зубы ......... ........................................... .......................................................... 160
Язык .............................................................................................................. 164
Большие слюнные железы .................................................................... 165
Твердое и мяrкое нёбо ............................................................................ 167
Миндалины ................................................................................................ 167
rлотка и пищевод ..................................................................................... 168
Производные среднеrо и заднеrо отделов
пищеварительной трубки ........................................................................... 170
Желудок ....................................................................................................... 170
Содержание
359
Тонкая кишка ............................................................................................. 174
Толстая кишка ........................................................................................... 178
Червеобразный отросток ....................................................................... 179
Прямая кишка ........................................................................................... 180
Поджелудочная железа .......................................................................... 180
Печень .......................................................................................................... 184
Желчевыводящие пути и желчный пузырь .................................... 188
Сердечно сосудистый комплекс opraHoB .................................................. 189
Артерии ................................. .............................. .............................................. 190
Сосуды микроциркуляторноrо русла .................................................... 192
Вены ................................................................................................................... 196
Лимфатические сосуды ............................................................................... 197
Сердце ........................................................... .............................................. ....... 198
Дыхательный комплекс opraHoB ................................................................... 201
Воздухоносные пути ............................................................................. ....... 202
Полость носа ..... ........................... .... ..... ................................ ..................... 202
[ортань ............................................... ........................................................... 203
Трахея ............................................. .............................................................. 204
Леrкие ............... ............................................................................ ..................... 205
Внутрилеrочные бронхи . ....................................................................... 205
Респираторный отдел леrких ............................................................... 206
Плевра ............ ....... ......................... .................................................. ............ 209
Выделительный комплекс opraHoB .............................................................. 209
Почки ........................................................................................... ................. 21 О
Мочевыводящие пути ............................................................................. 216
[ЛАВА 15. СИСТЕМА РЕАКТИВНОСТИ ................................................. 218
Иммунный комплекс opraHoB ........................................................................ 218
Красный костный мозr ........................................ ........................................ 218
Тимус.................................................................................................................. 221
Лимфатические узлы и лимфоидные образования в opraHax ...... 223
Селезенка .......................................................................................................... 225
Иммунитет и ero виды ................................................................................. 227
Клеточные основы кооперации в иммунных реакциях .................. 229
Эндокринный комплекс opraHoB .................................................................. 231
[ипоталамус ..................................................................................................... 234
[ипофиз ............................................................................................................. 236
Эпифиз ................. ...... .................... ........ ........... .... ............... .................. ........... 238
Щитовидная железа ..................................................................................... 239
Около щитовидные железы ........................................................................ 242
Надпочечники .................. ............................................................................... 243
360
Содержание
Эндокринные клетки в составе неэндокринных opraHoB.
Диффузная эндокринная система (АРUD серия) ........................... 247
Невральный комплекс opraHoB ..................................................................... 248
Спинной мозr .................................................................................................. 249
Спинномозrовые узлы ................................................................................. 253
rоловной мозr .................................................................................................. 254
Ствол rоловноrо мозrа ........................................................................... 254
Кора большоrо мозrа ............................................................................... 259
Оболочки спинноrо и rоловноrо мозrа ................................................. 262
Периферические нервы ............................................................................... 264
Веrетативная нервная система ................................................................. 265
Реrенерация в нервной системе ............................................................... 267
Сенсорный комплекс opraHoB ........................................................................ 268
OpraH зрения .............................................................................................. 269
OpraH обоняния ........................................................................................ 276
OpraH слуха и равновесия ..................................................................... 278
OpraH вкуса ................................................................................................ 284
OpraH осязания ......................................................................................... 285
rЛАВА 16. ОПОРНо ДВИrАТЕЛьНАя СИСТЕМА ............................287
Кости .................................................................................................................. 287
Соединения костей ....................................................................................... 290
Хрящи ...................................................... . ......................................................... 291
Мышцы .............................................................................................................. 291
Сухожилия ....................................................................................................... 295
rЛАВА 17. ВОСПРОИЗВЕДЕНИЕ ЧЕЛОВЕКА.
ОСНОВЫ МЕДИЦИНСКОЙ ЭМБРиолоrии ................................... 297
Развитие половых клеток (проrенез) . ......................................................... 297
Орrаны мужской половой системы . ....................................................... 300
Яички ............................................................................................................ 300
Семя выносящие пути и железы ......................................................... 302
Половой член ............................................................................................. 304
Pery ляция функций яичек ......................................................................... 305
Орrаны женской половой системы ......................................................... 306
Яичники ....................................................................................................... 306
Матка ............................................................................................. ............... 309
Маточные трубы ....................................................................................... 311
Влаrалище .................................................................................................. 312
Овариально менструальный цикл .......................................................... 313
Эмбриональное развитие человека .............................................................. 315
Оплодотворение (фертилизация) ........................................................... 315
Содержание
361
Дробление ........................................................................................................ 318
Имплантация .................................................................................................. 319
rаструляция ..................................................................................................... 320
Нейру ляция .........................................................._.......................................... 322
Сеrментация мезодермы ............................................................................. 323
Плодный период ................................................................................................. 324
Внезародышевые орrаны. Плацента .. .......................................................... 330
Плацента ...... ..................................................................................................... 332
Критические периоды и аномалии развития ........................................... 339
ЗАКЛЮЧЕНИЕ
МЕДИЦИНСКОЕ ЗНАЧЕНИЕ КУРСА rистолоrии ................... 342
АЛ ФАВИТНЫЙ УКАЗАТЕЛЬ ............................................................................. 345
ЛИТЕРАТУРА .......................................................................................................... 354
Учебная литература
для студентов медицинских вузов
Р.К Данилов,
А.А. Клишов,
T.r. Боровая
rистолоrиячеловека
в мультимедиа
Издательство 000 ЭЛБИ СПб..
ИД N 01520 от 14.04.00
Санкт Петербурr. Лабораторный пр., 23.
E mail: an@elbi.spb.su
(812) 322 92 57; 322 92 58
Подписано в печать 07.04.03. Формат 70х100'/;6'
Печать офсетная. Тираж 2000 экз. Заказ .N9 2742.
Отпечатано с диапозитивов в ФfУП Печатный двор.
Министерства РФ по делам печати, телерадиовешания
и средств массовых коммуникаций.
197110, Санкт Петербурr. Чкаловский пр., 15.
rИ( ТОtJ1().r Иfl
.
ч J :.J] ()IJ I It
-
J :J ..
"\
...... .. ..
......
4
.ol
1
'
&-4
," .
. 1 '
, I
,. ".
.
15'
\
I 1
.,
... I J 1
"..WI . . 1
.IUJJ' ' l'
", .. , r
r . I
" . J
,1..1 ..' . . .".. .,'"
11. . J 1. 1 А 1.
...1... .. R
I I
{.I
.1
v
f
'1 .1.
.
,
Данилов Ревхать Константинович, доктор медицинских наук,
профессор, заслуженный работник высшей школы РФ, Соросовский
профессор, лауреат конкурса 'Трант Санкт Петербурrа", npoвeдeHHoro
в 2002 r. Международной Проrраммой Образования в области точных
наук (ISSEP), ученик профессора д'А.Клиwoва, заведующий кафедрой
rистолоrии и эмбриолоrии Военно-медицинской академии (с 1991 r.),
автор учебных пособий, моноrрафий, ответственный редактор nepBoro
отечественноro двухтомноrо труда ВРуководство по rистолоrии"
и дрyrих уче6но--методических материалов.
Клишов Алексей Андреевич (19ЗО 1991), доктор медицинских наук,
профессор, заведовал кафедрой rистолоrии и эмбриолоrии
Военно медицинской академии с 1978 по 1991 roды, автор ряда
моноrрафий, учебника 'Тистолоrия человека", словарей терминов по
цитолоrии и эмбриолоrии.
Боровая Татьяна reHHaAbeBHa член корреспондент РАМН, доктор
медицинских наук, профессор кафедры rистолоrии и эмбриолоrии
педиатрическоrо факультета Российскоrо rосударственноrо
медицинскorо университета (Москва), лауреат конкурса 'Трант Москвы"
(2002 r.) в области наук и технолоrий в сфере образования, автор
уче6но методических пособий и моноrрафии.
;"11' f' """ir
i 11 j 111 1-
9 785939 790611