












































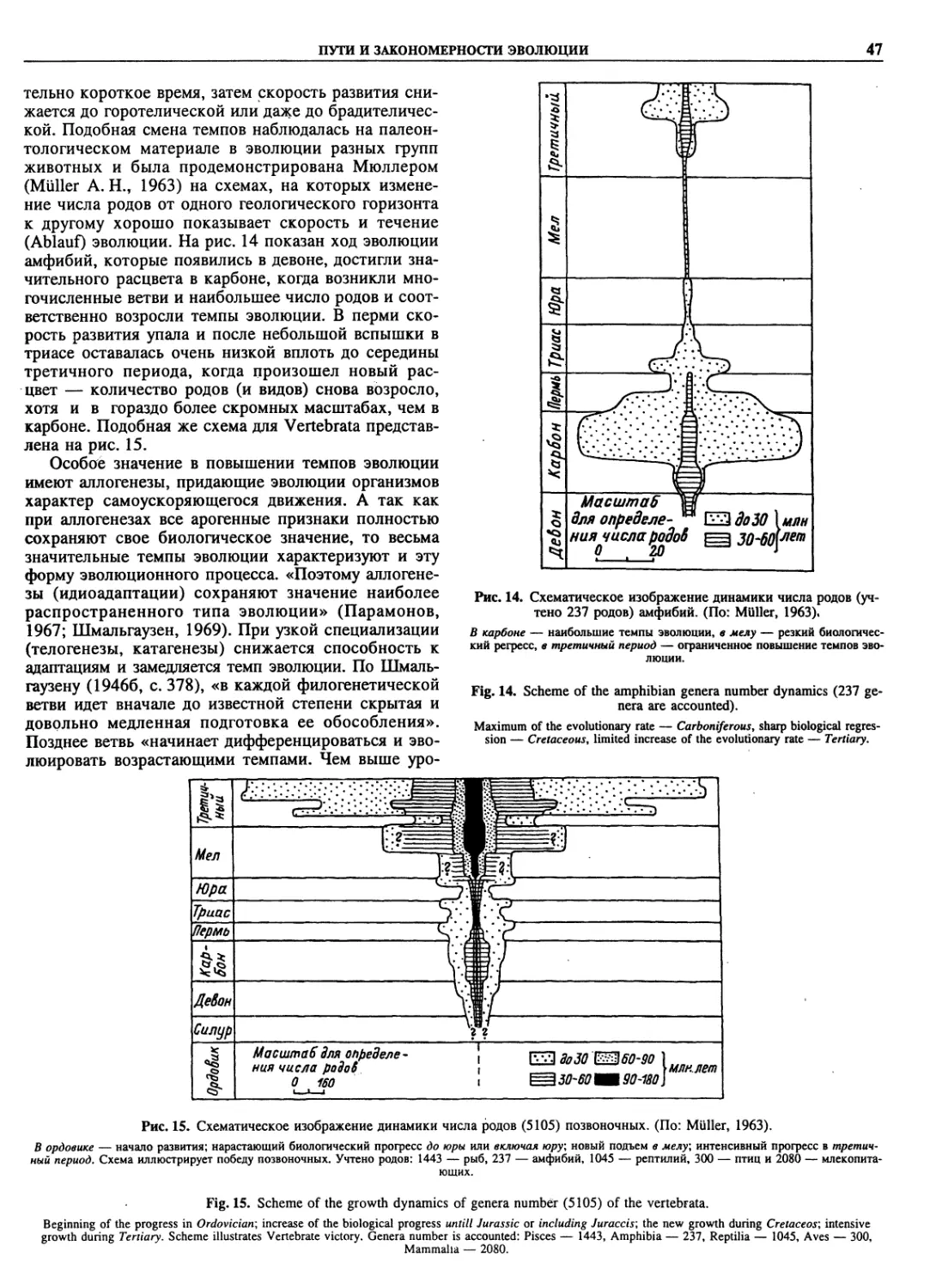



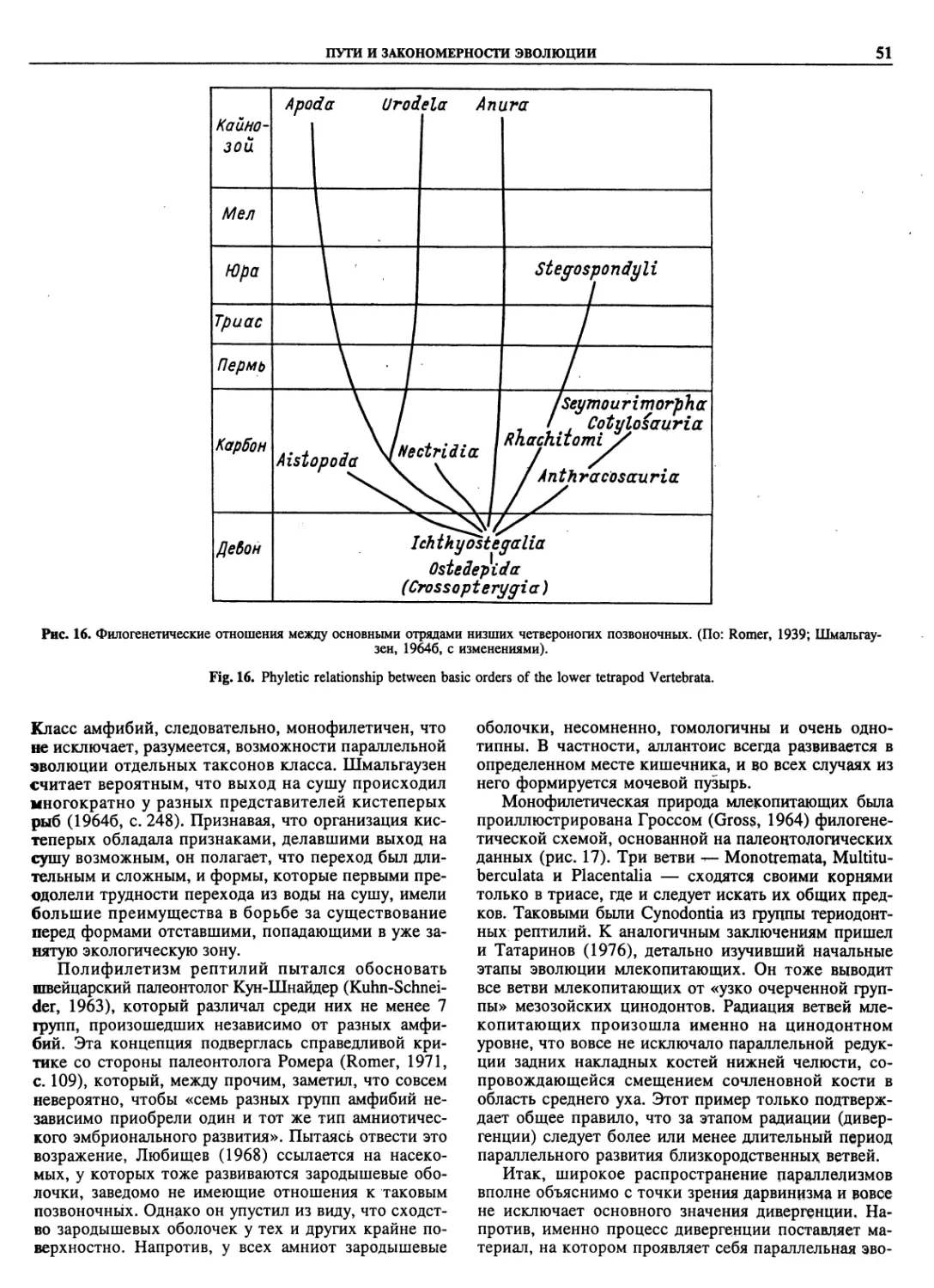
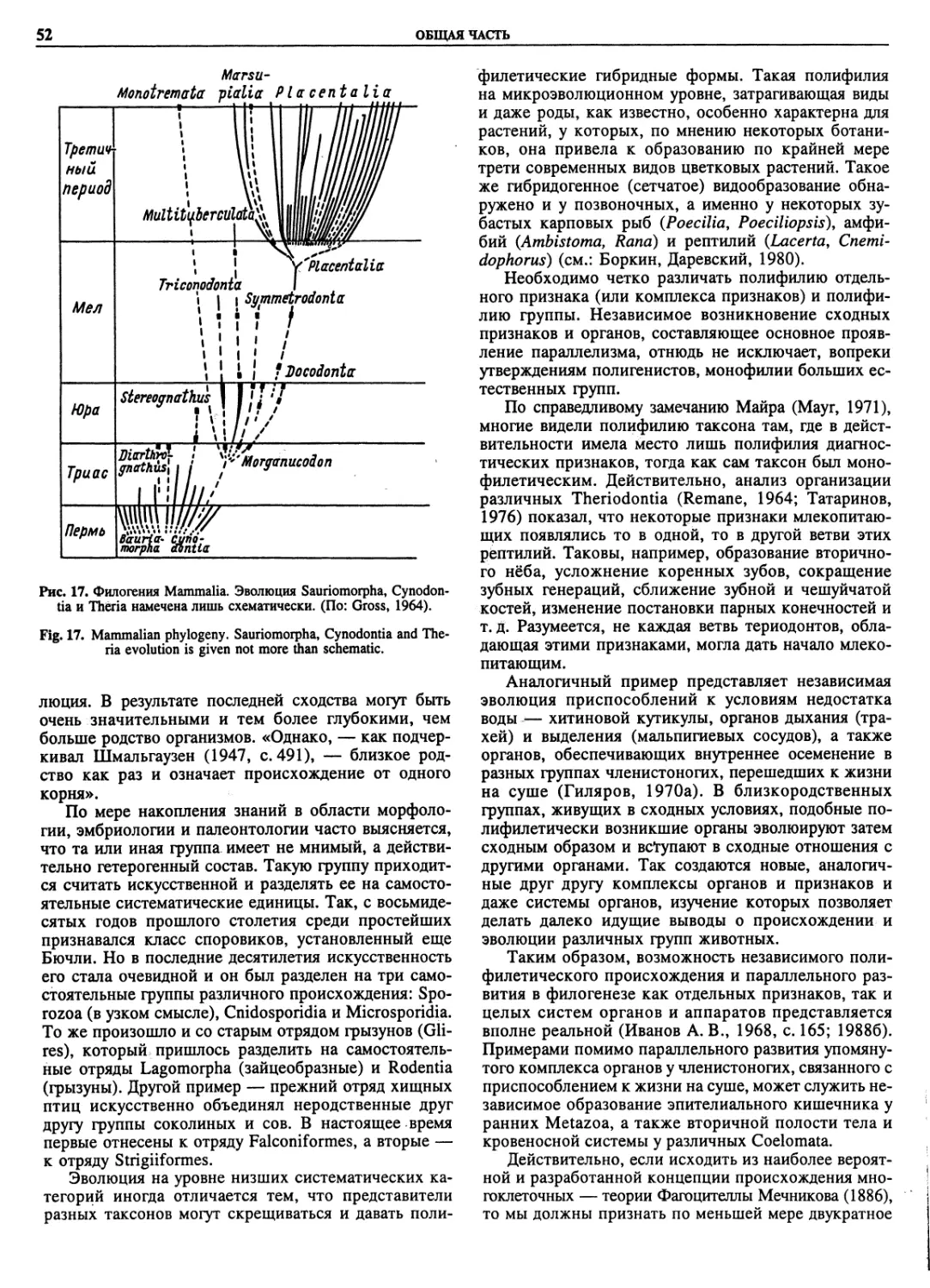





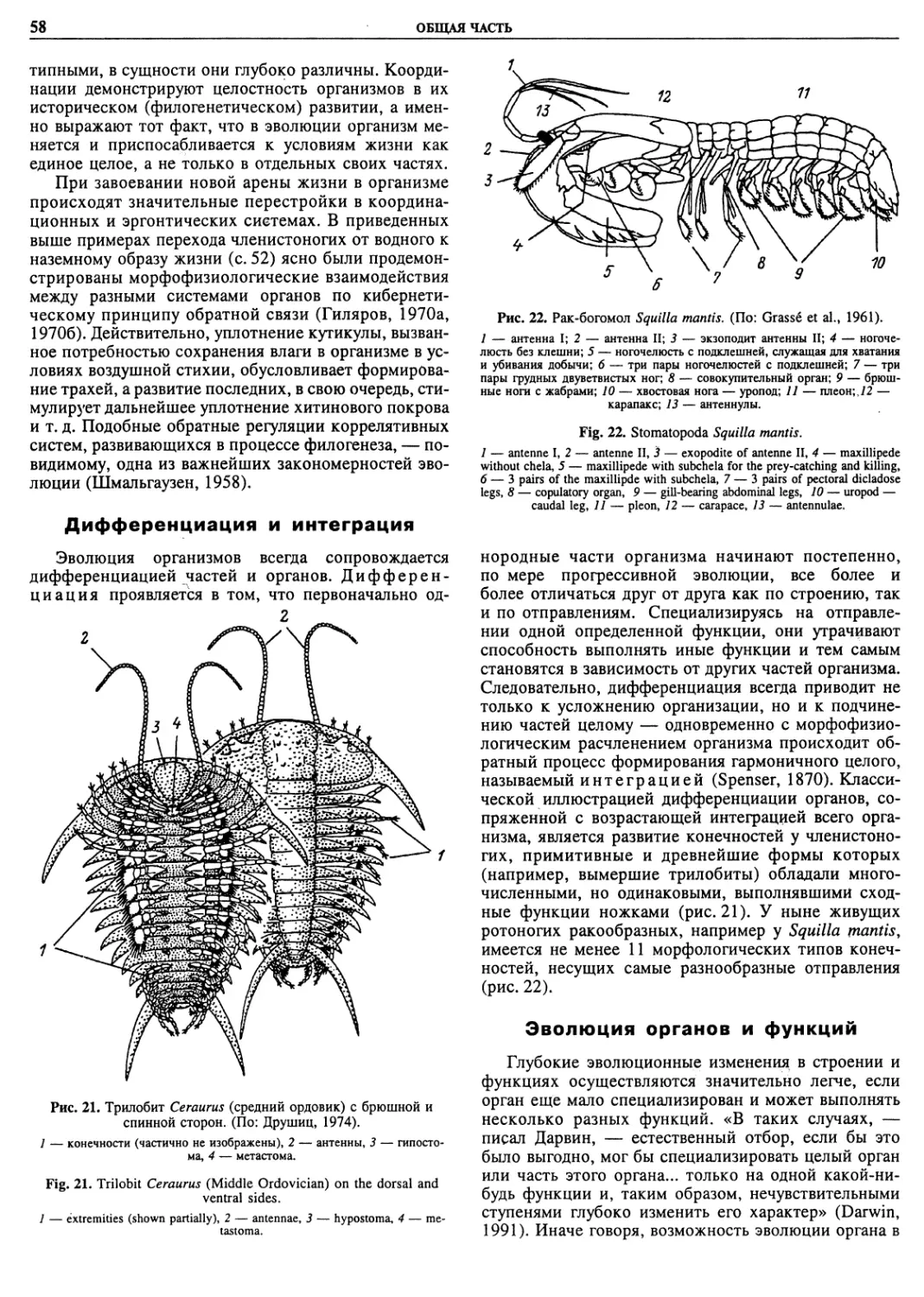





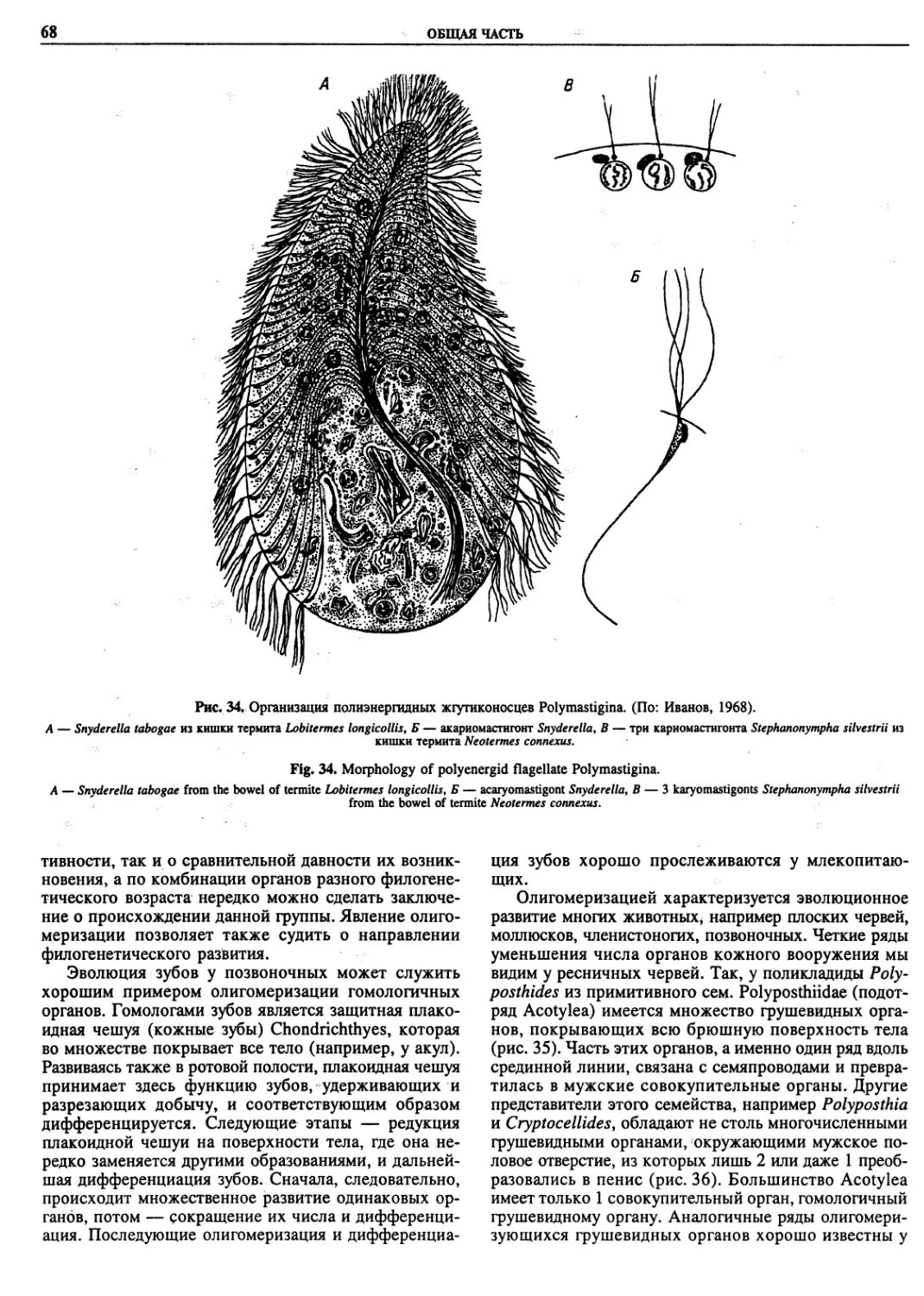
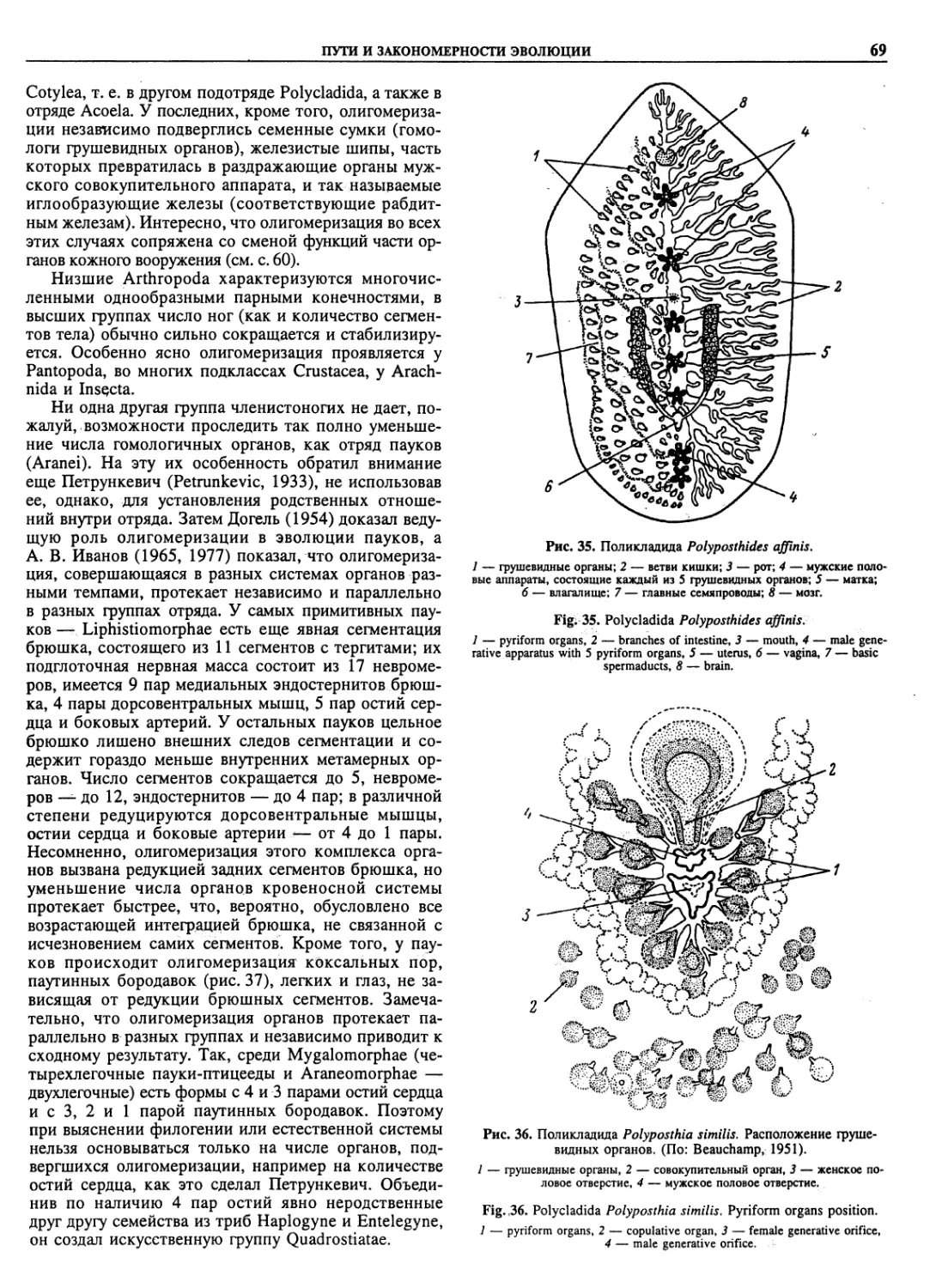





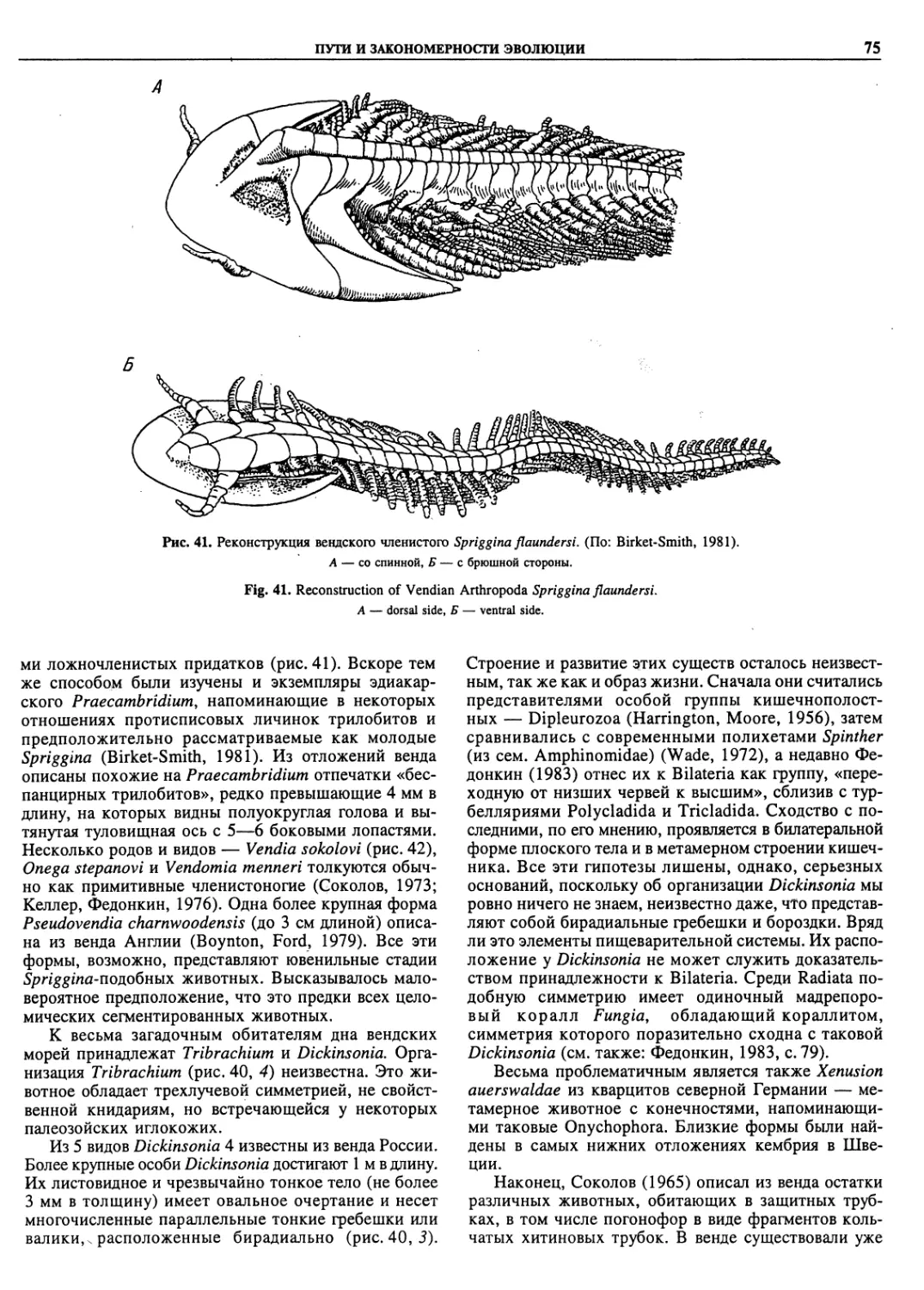

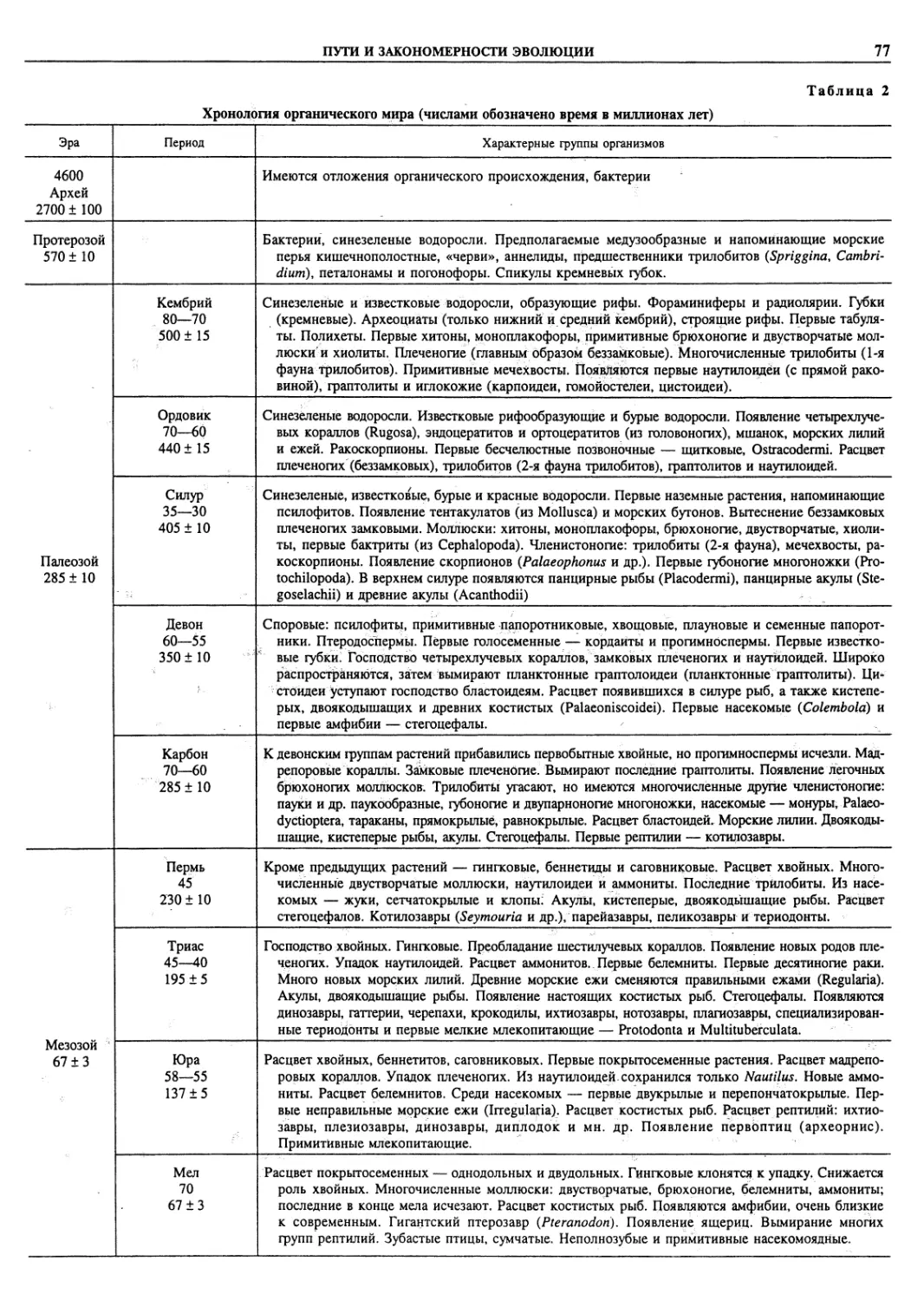












































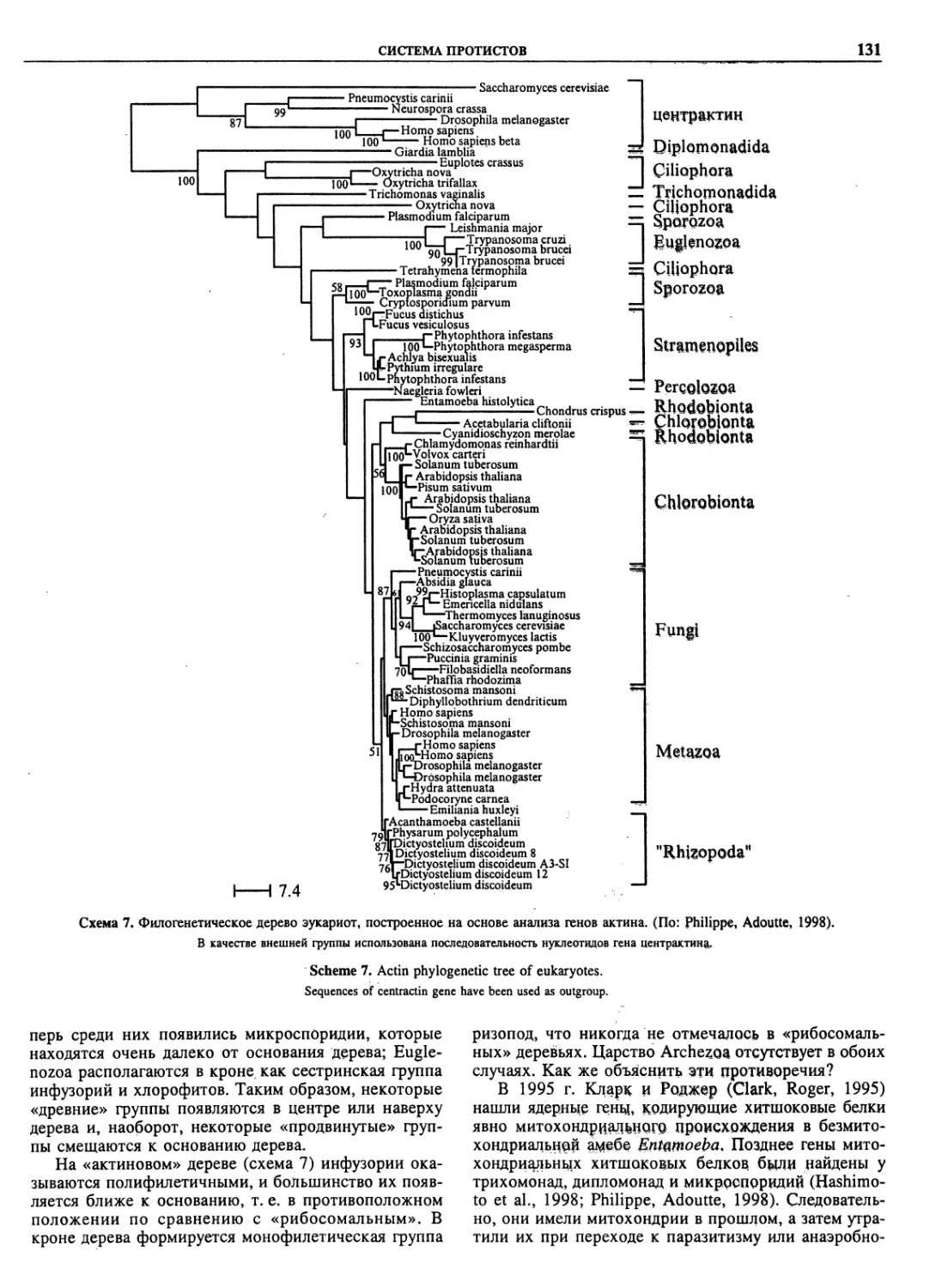
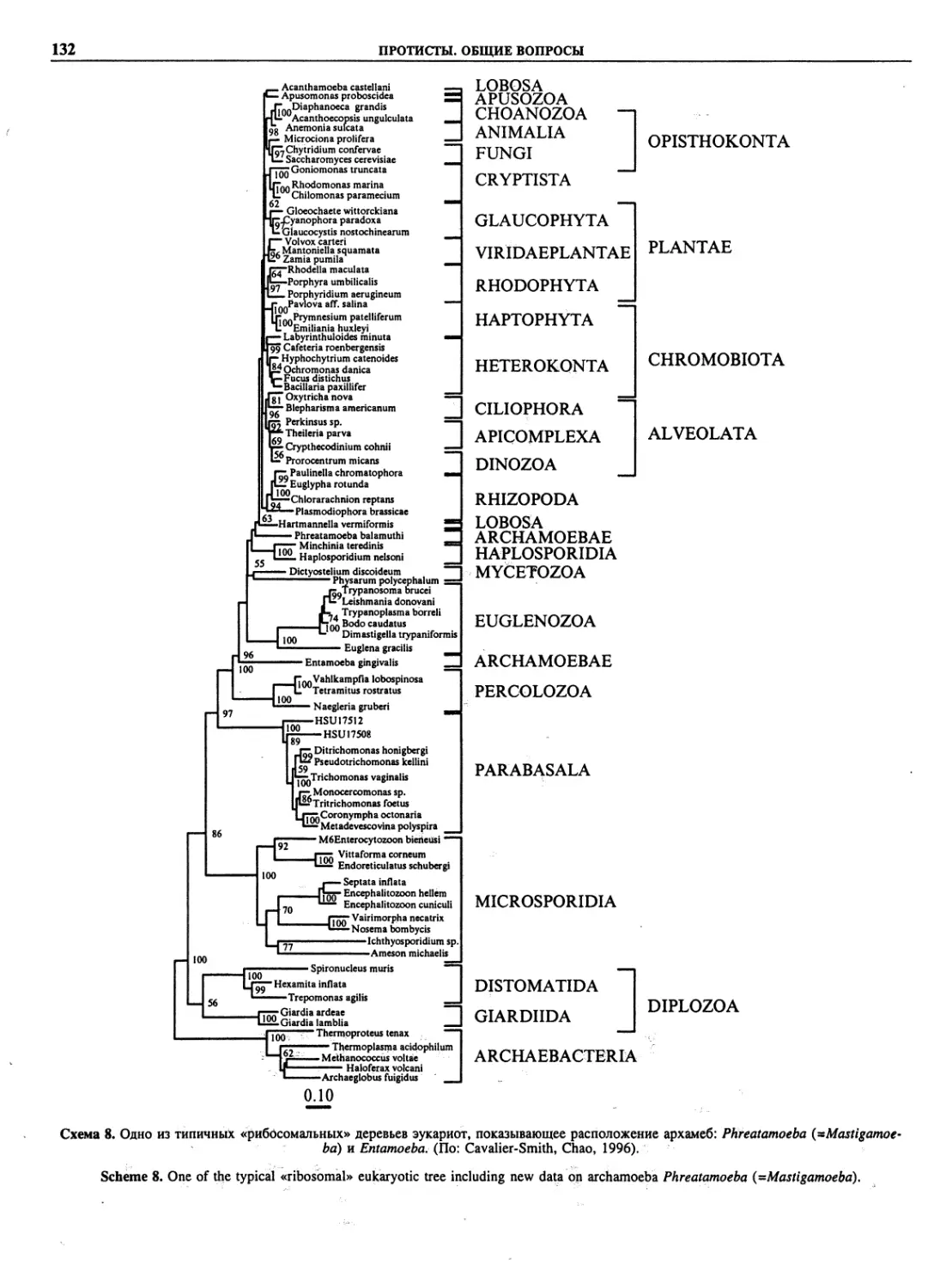














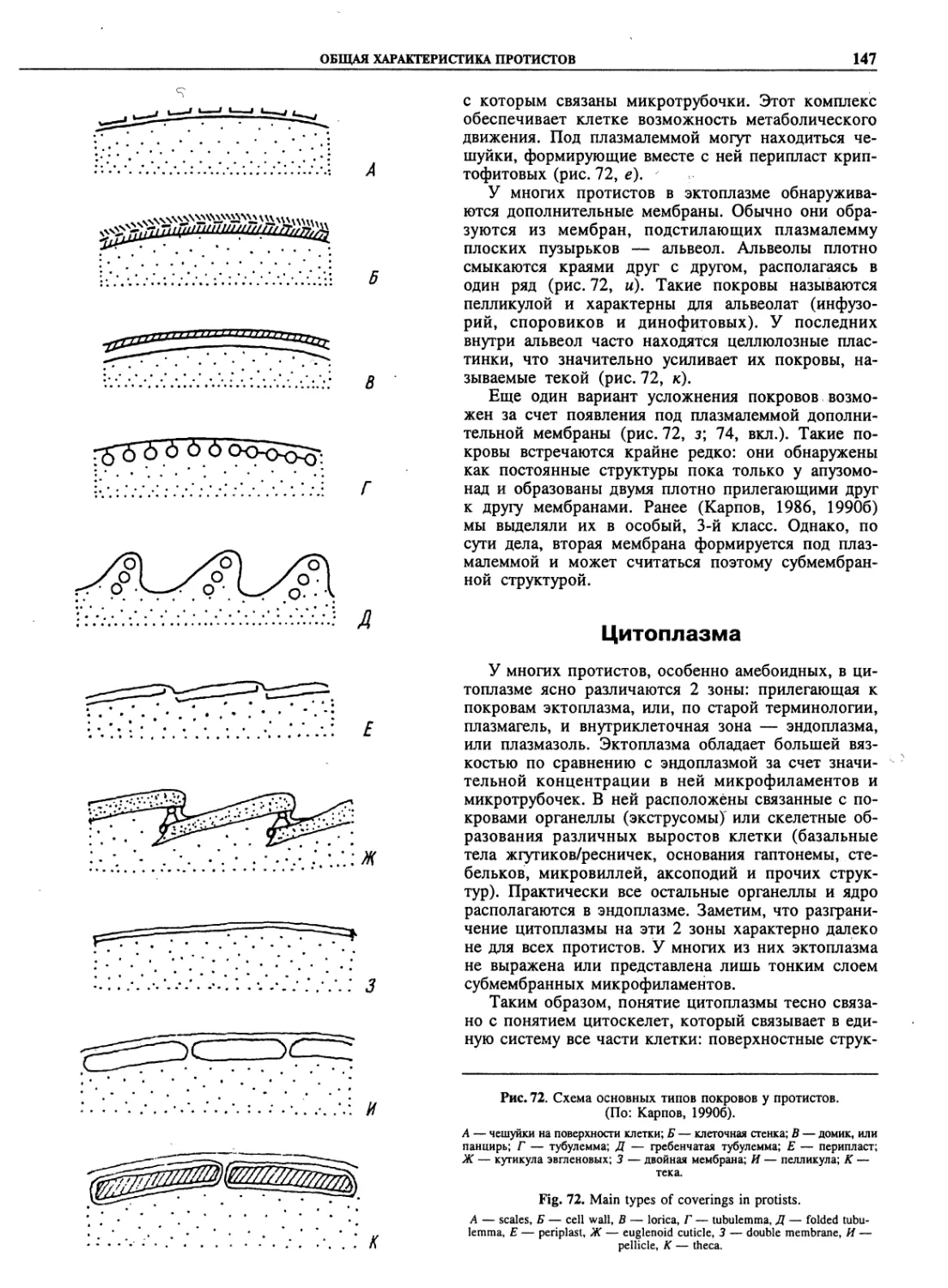

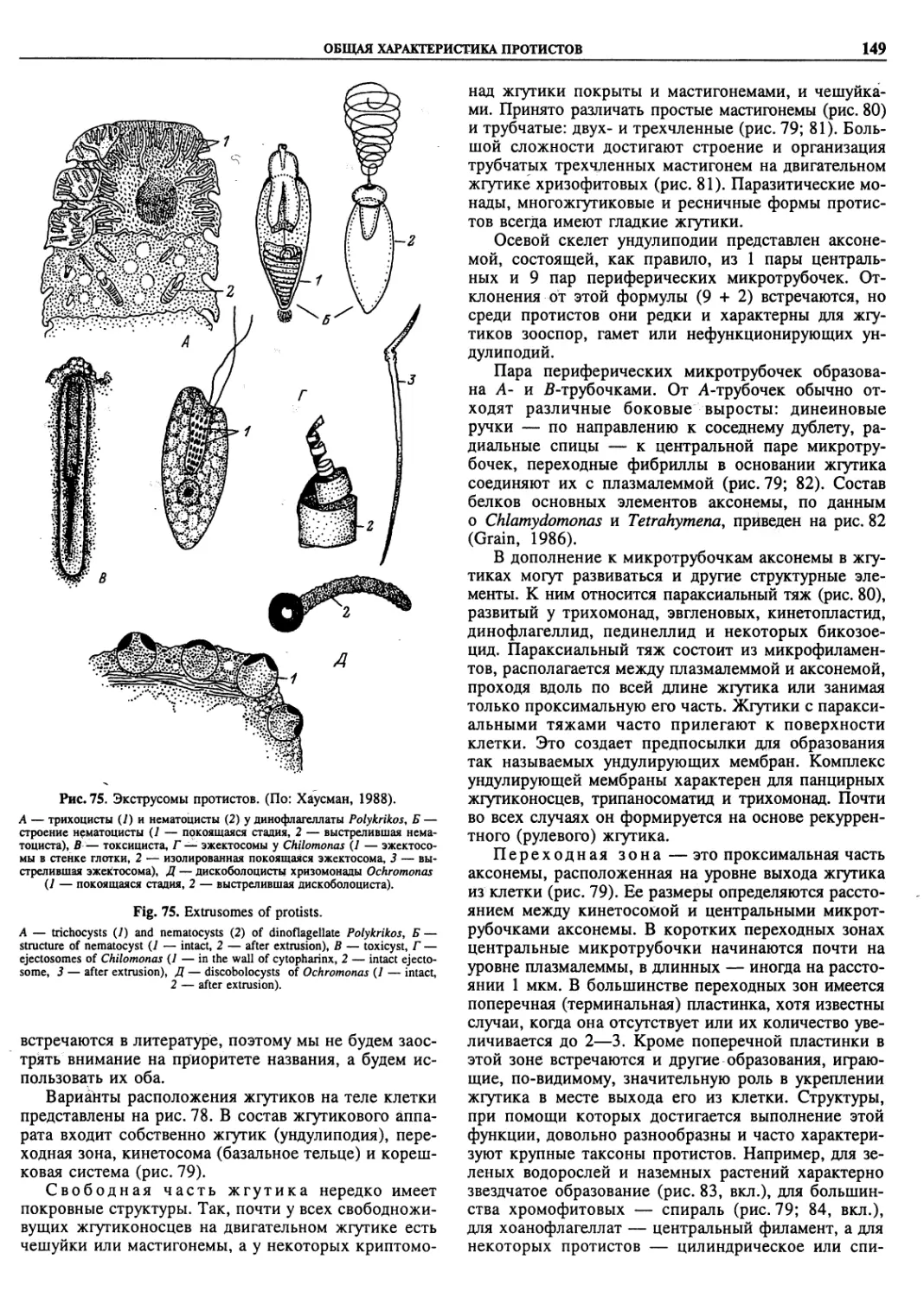



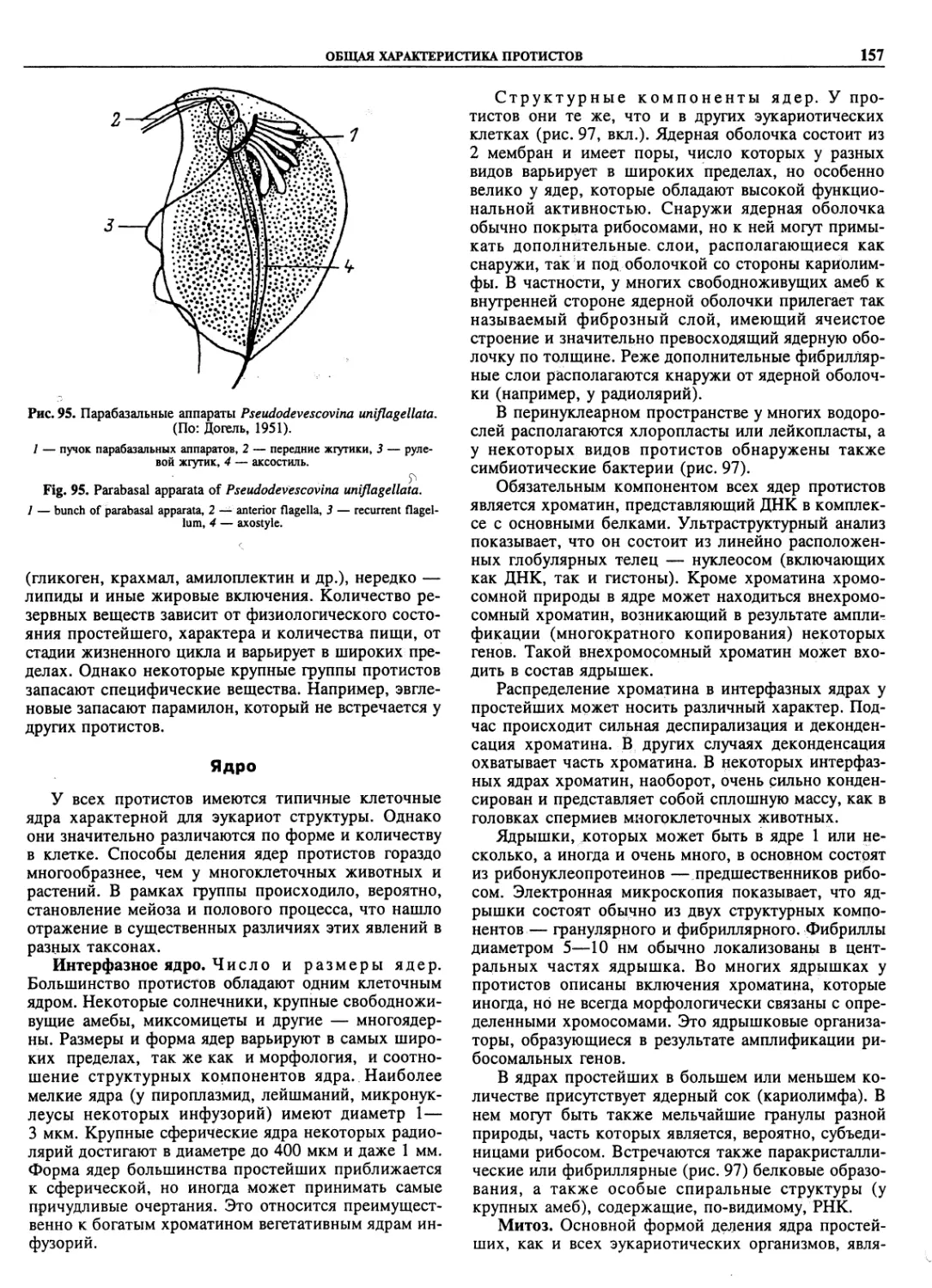

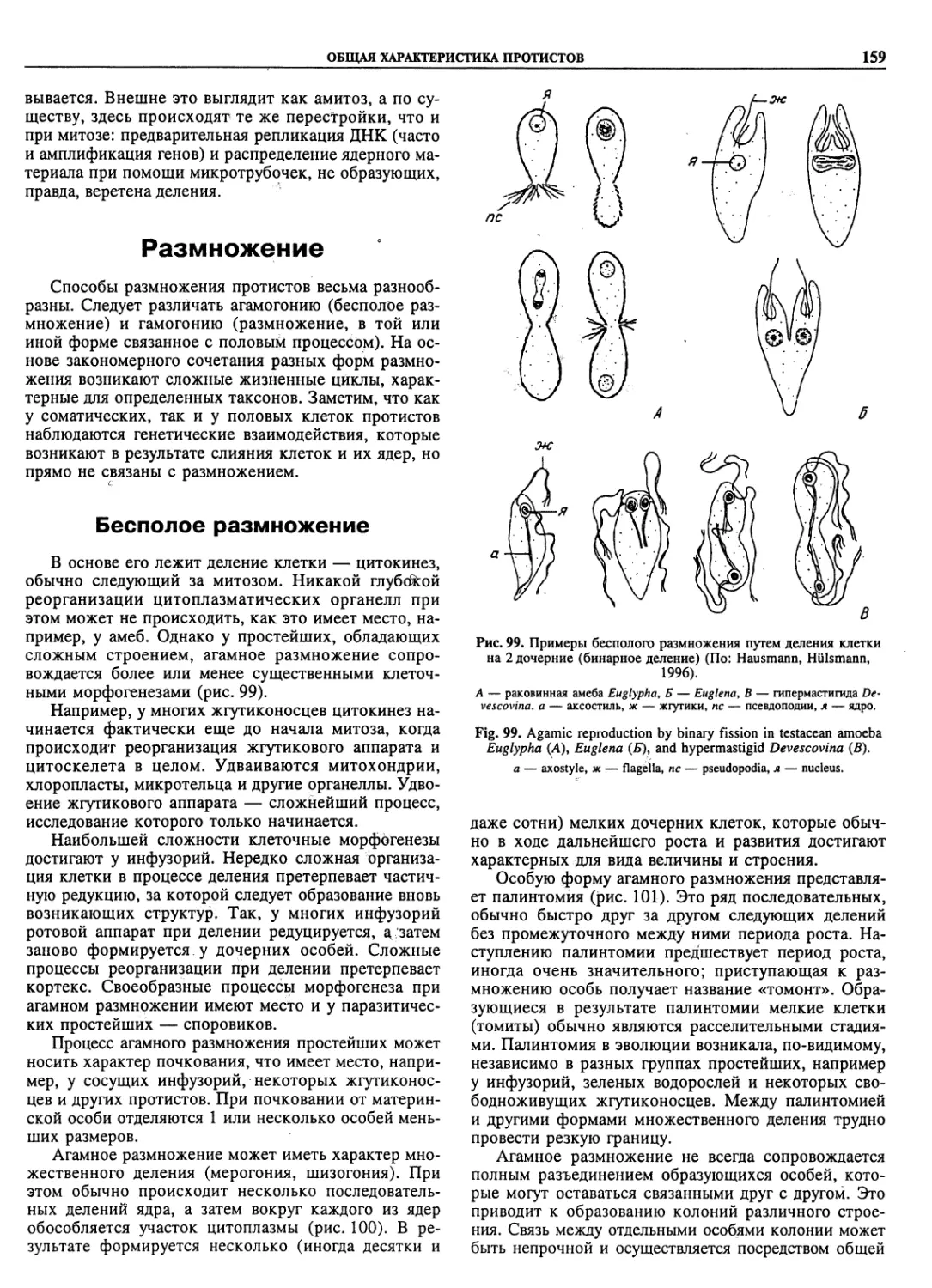
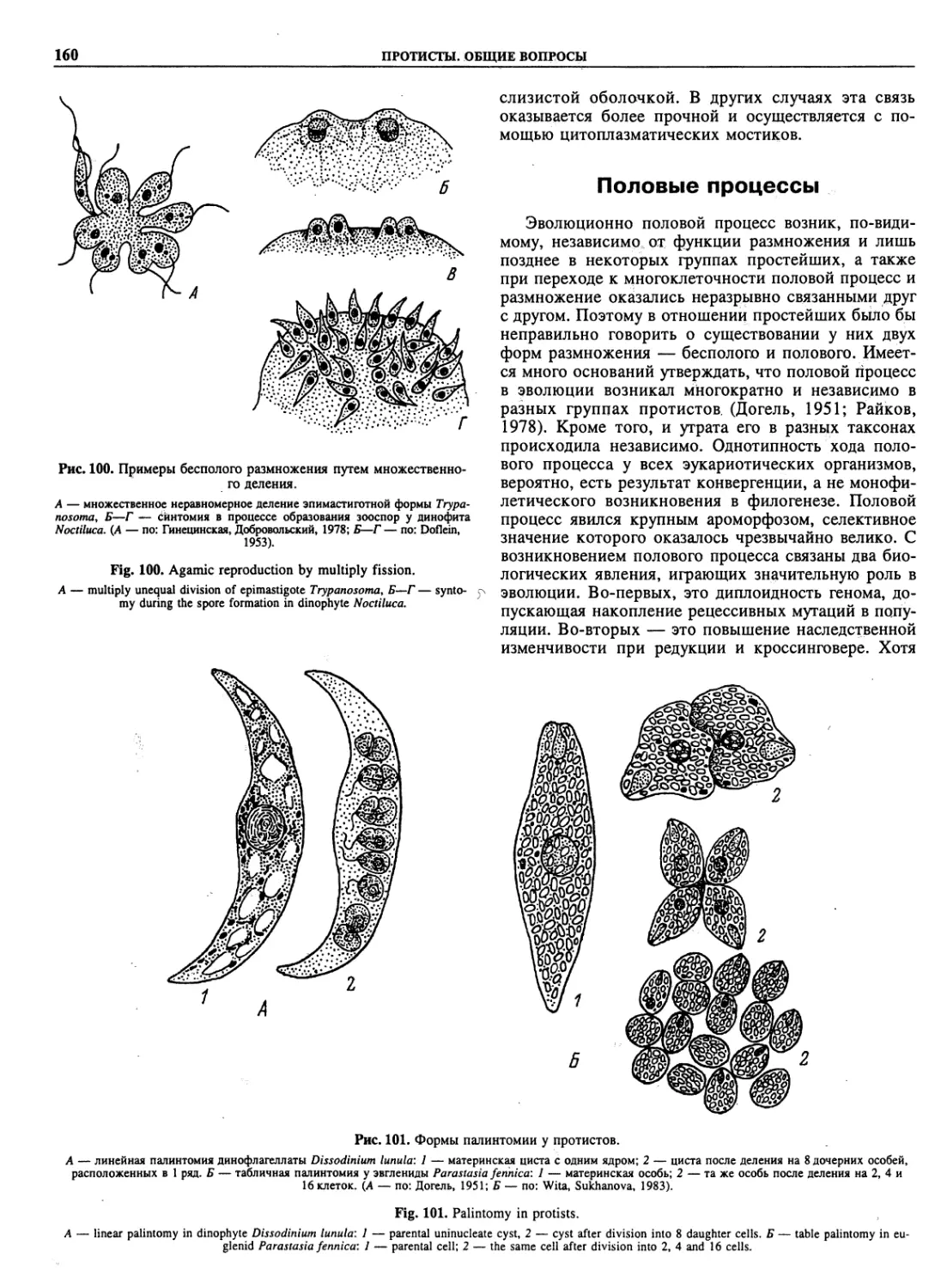
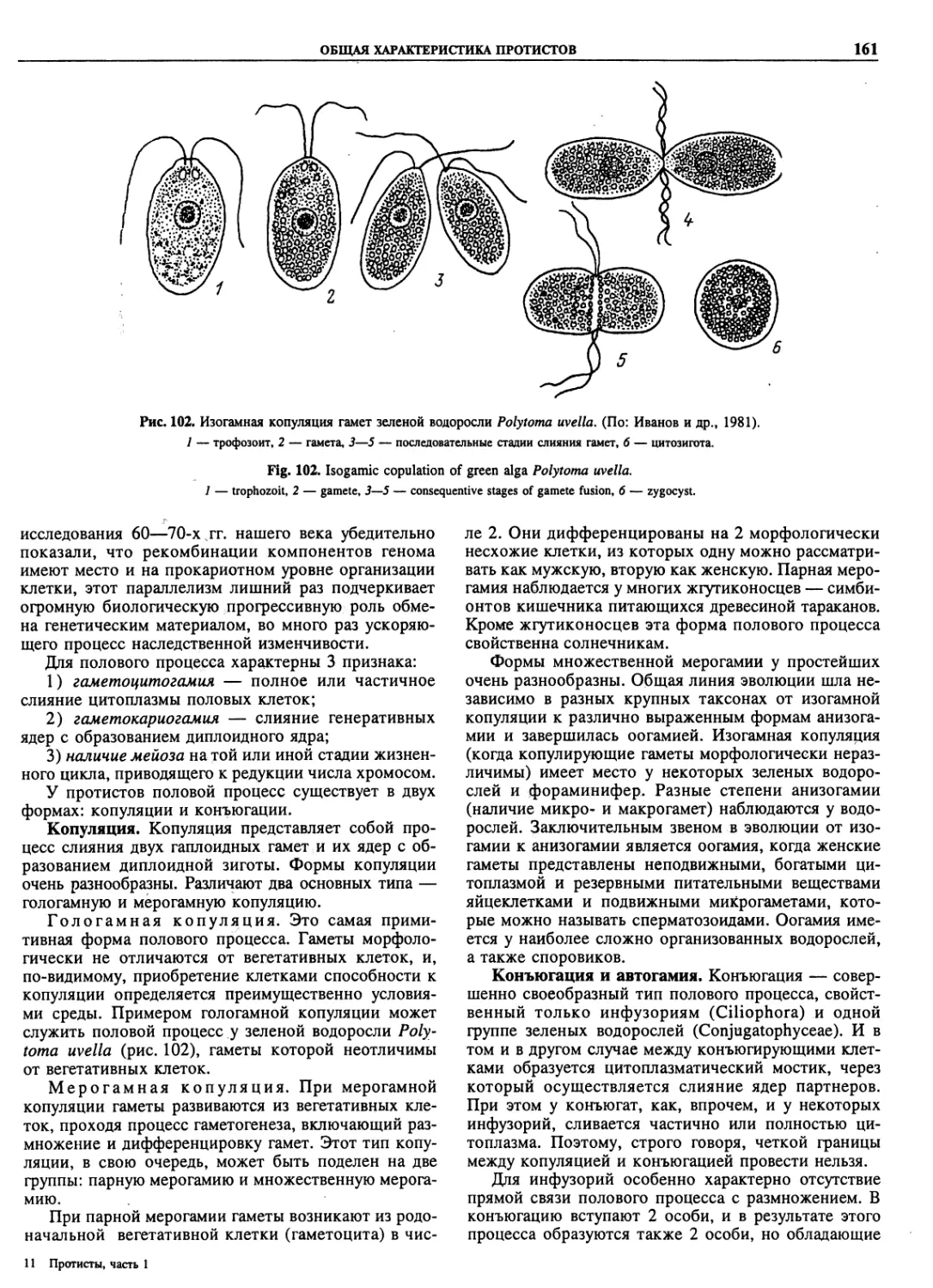



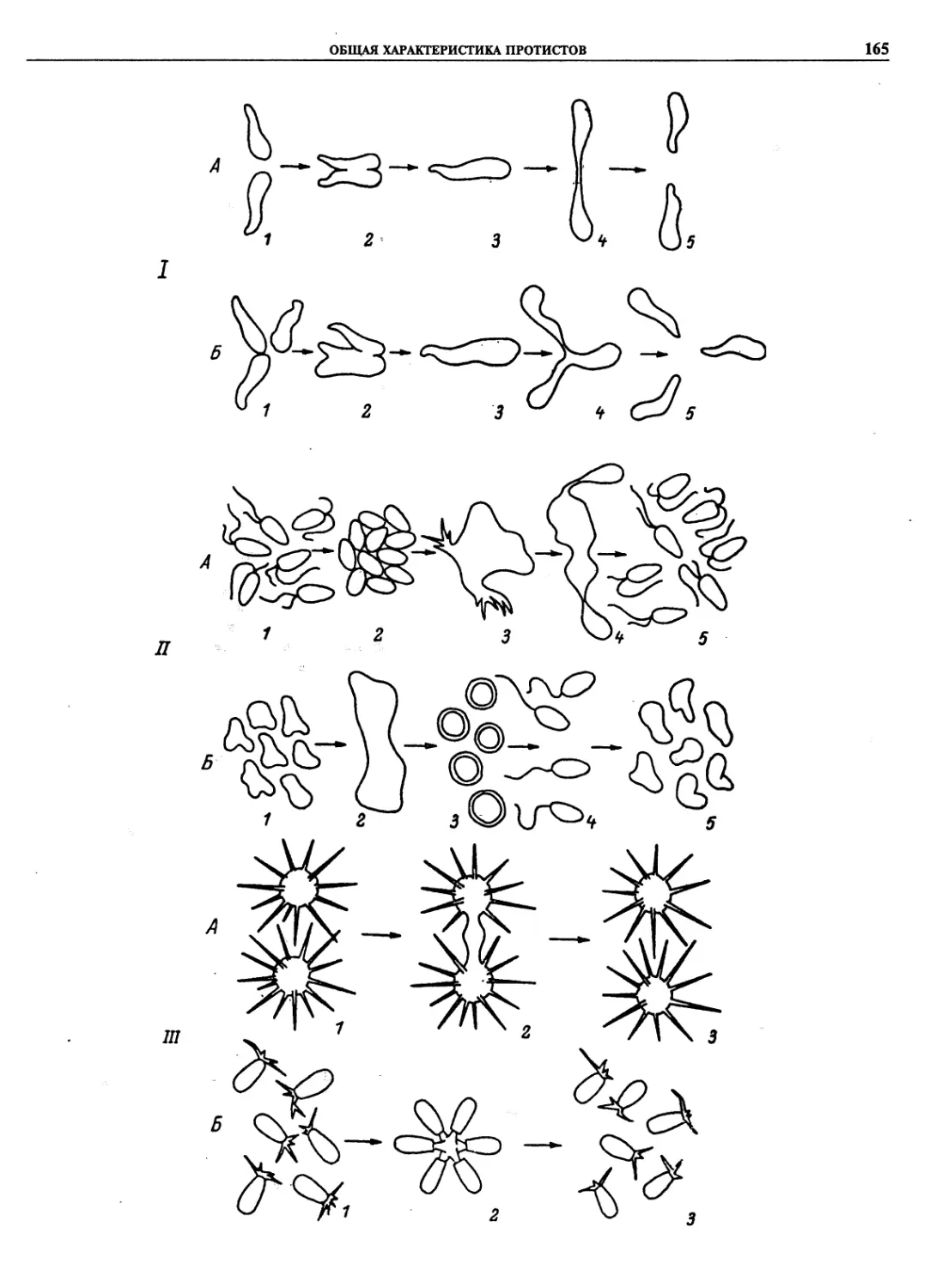

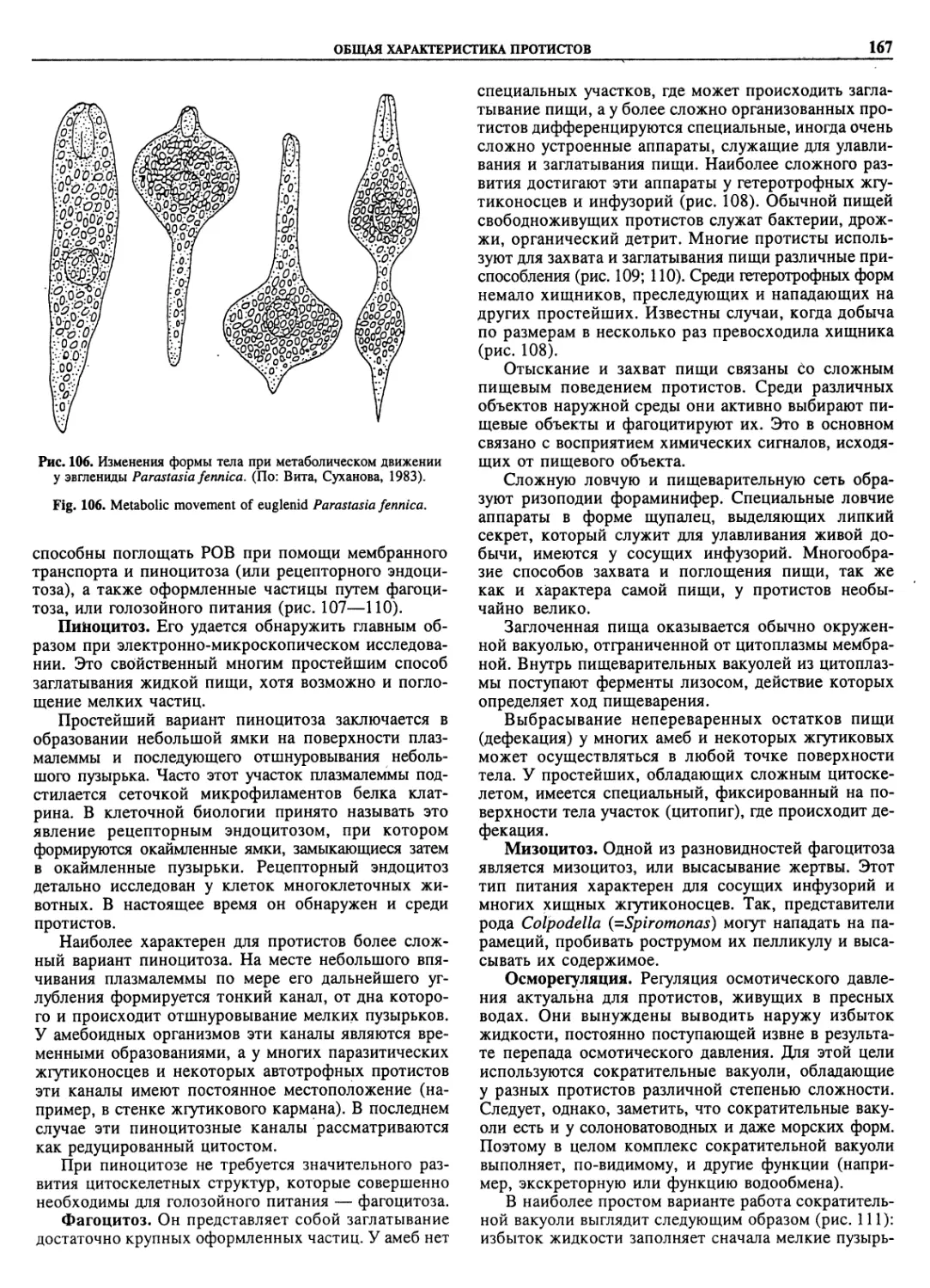
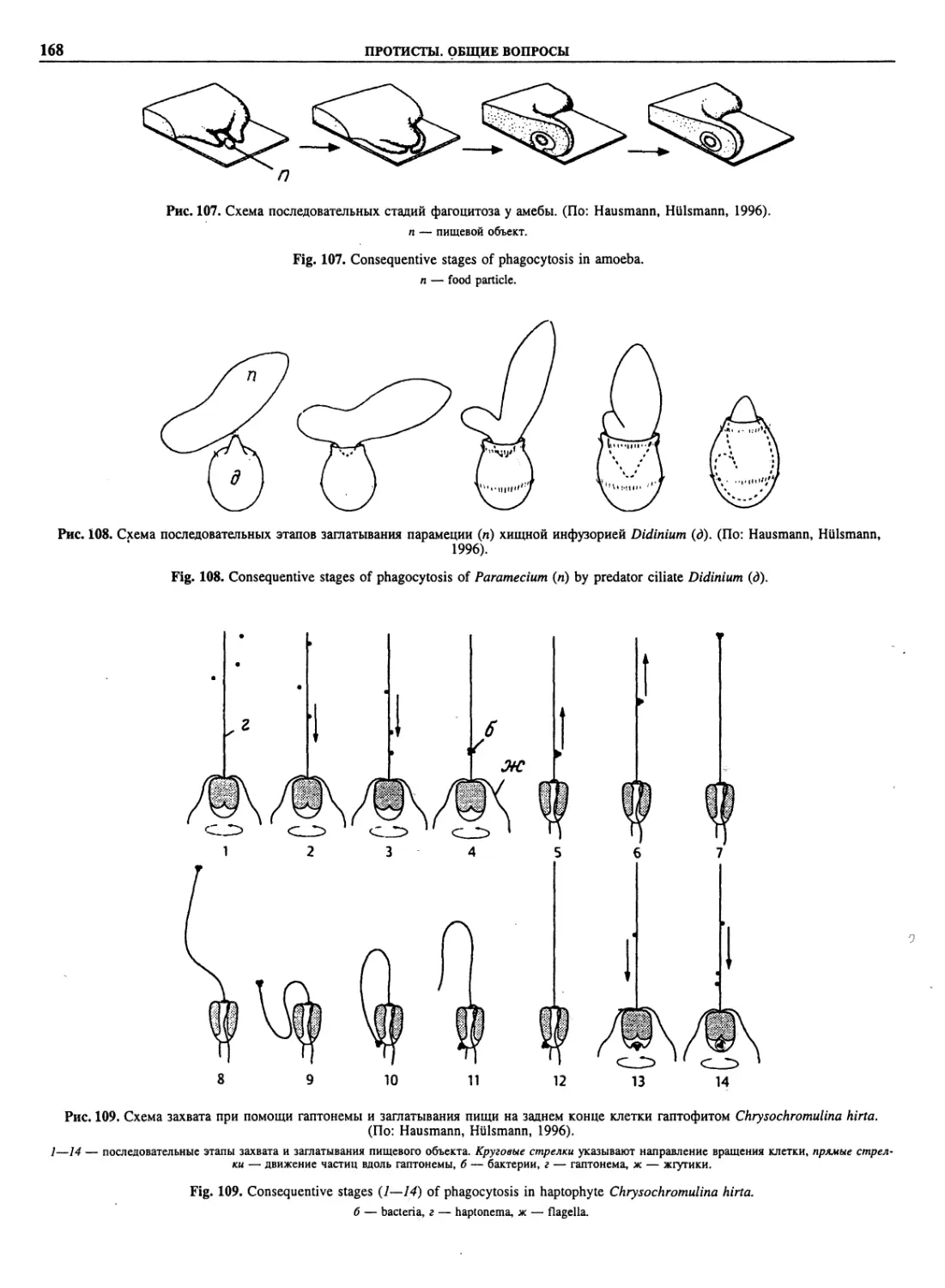
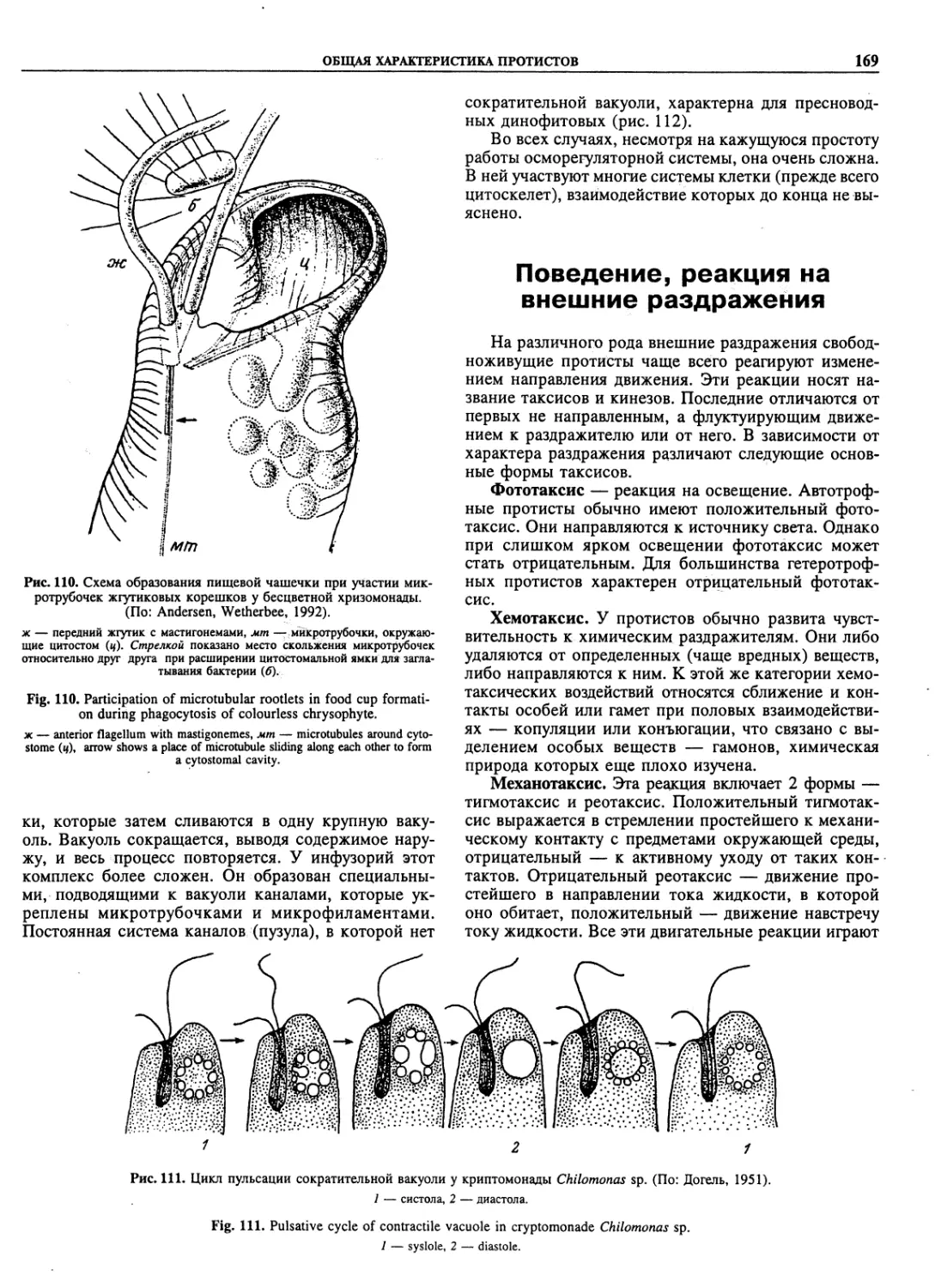

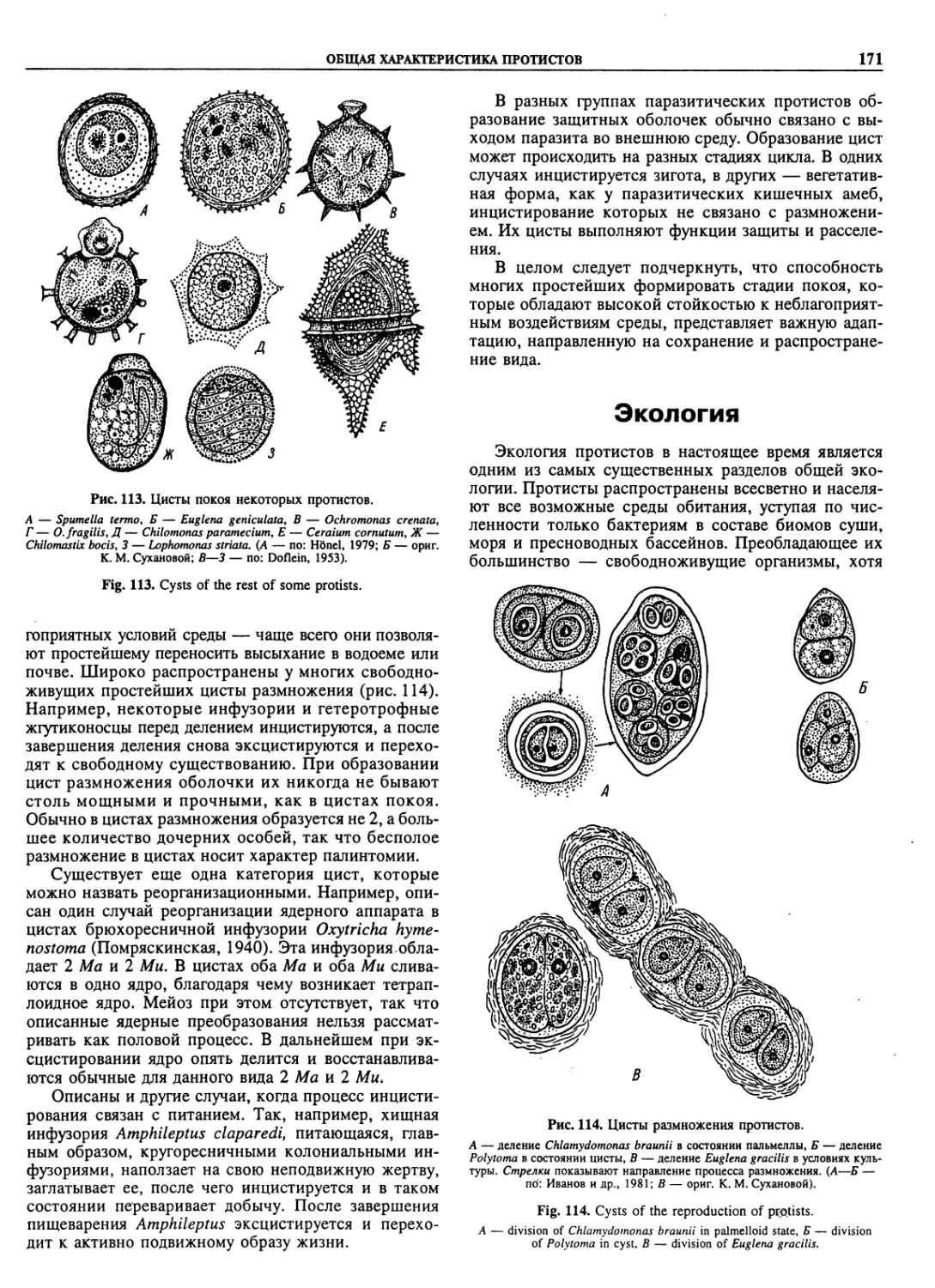





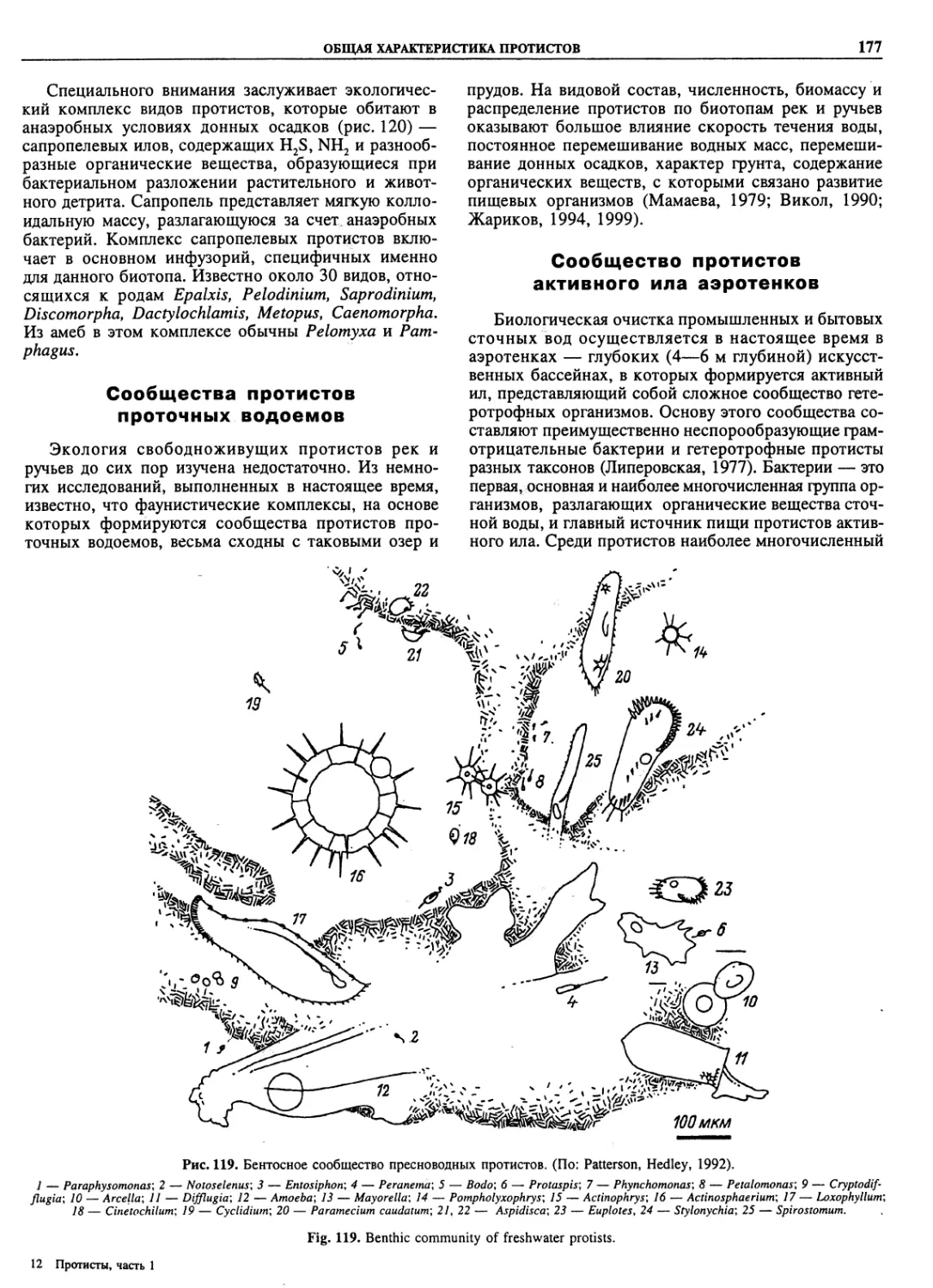















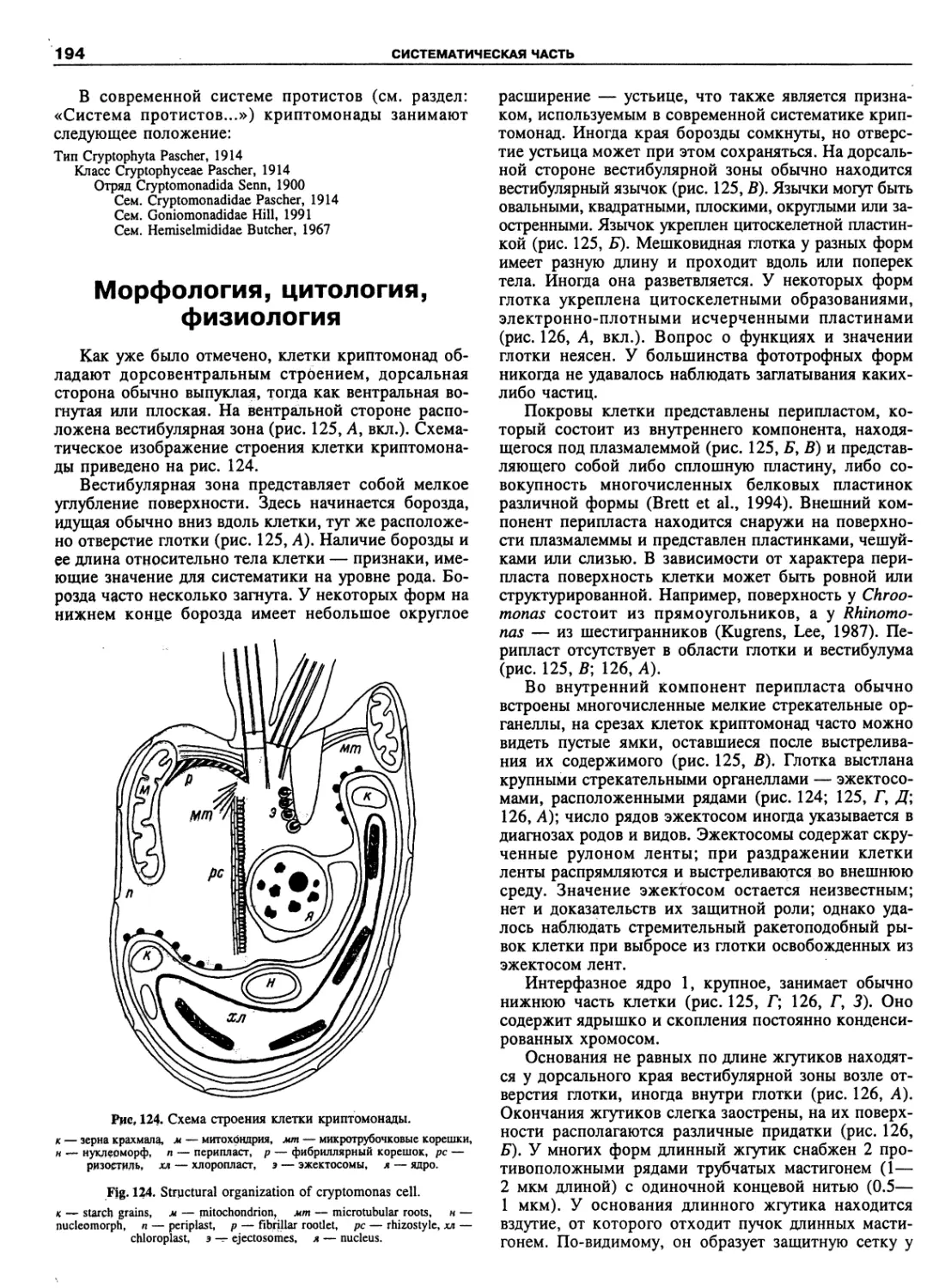


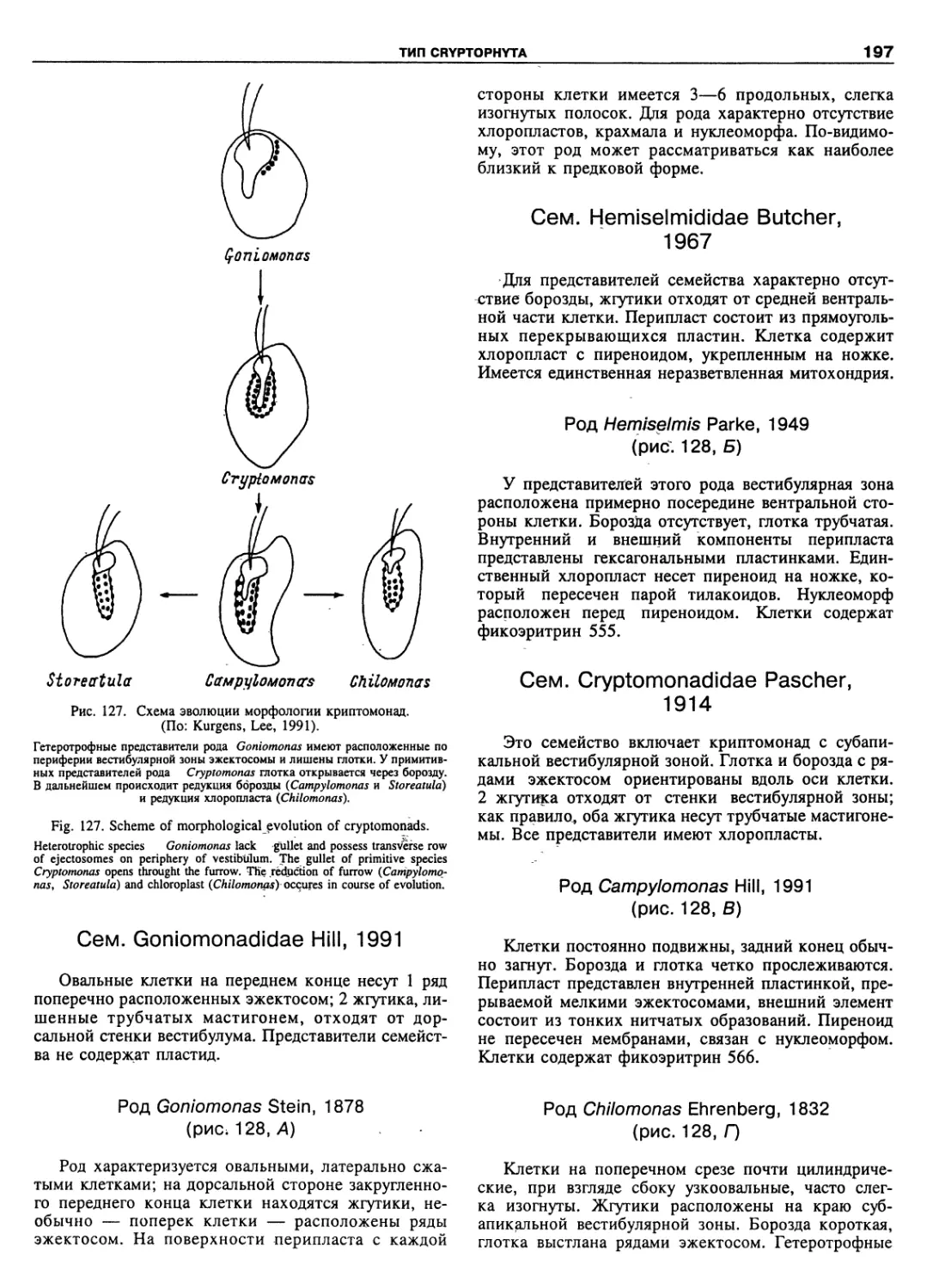





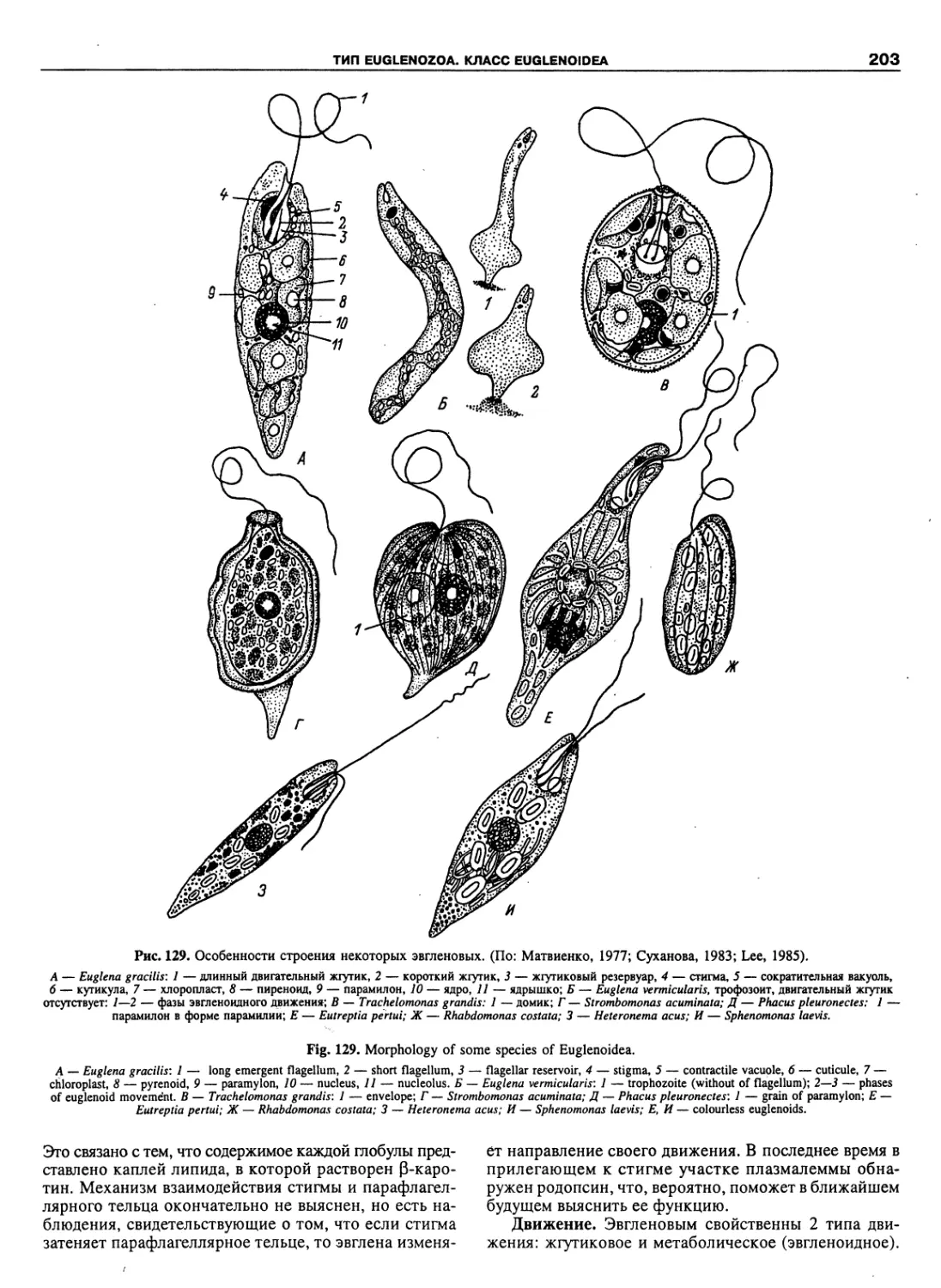
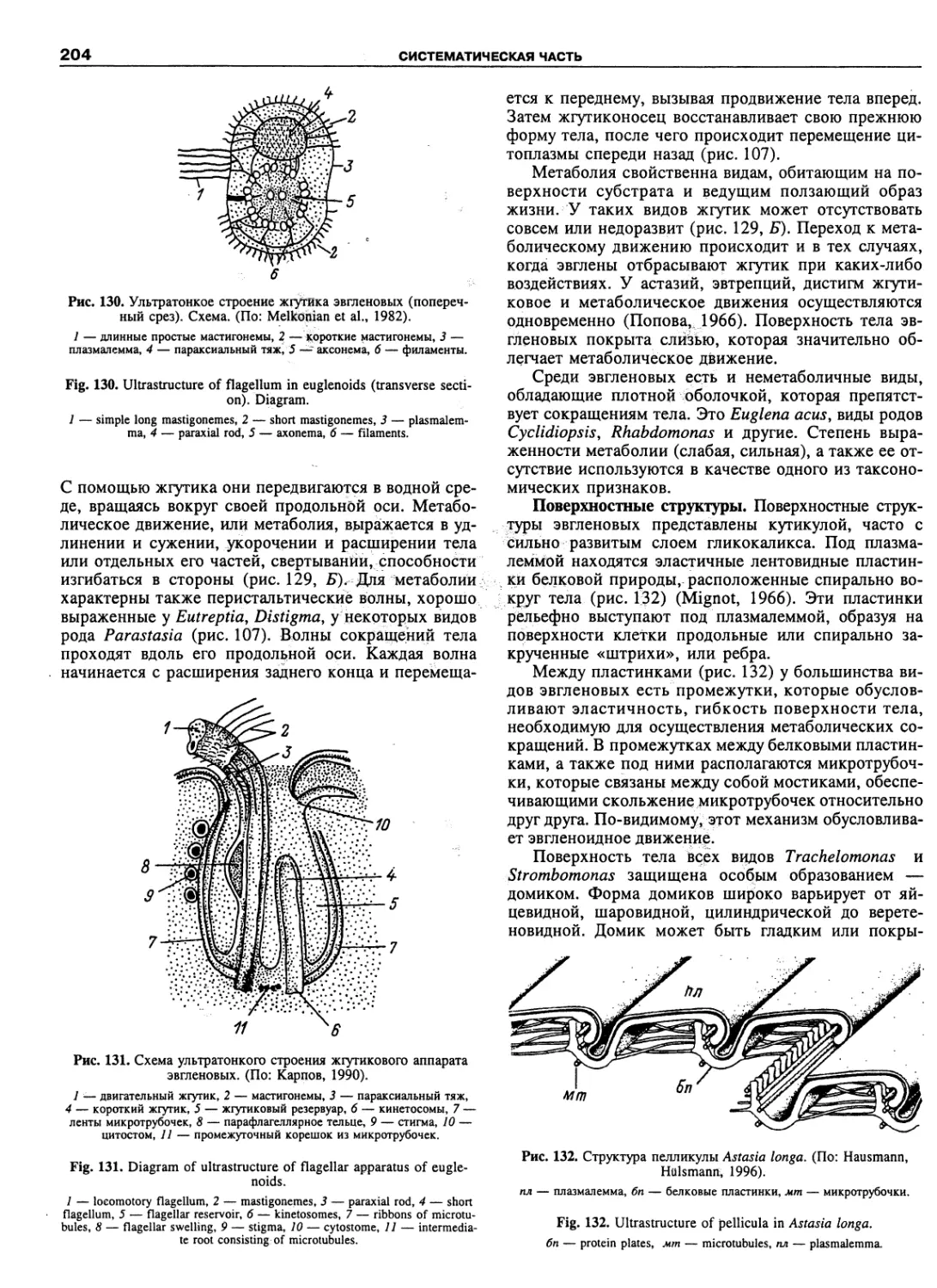



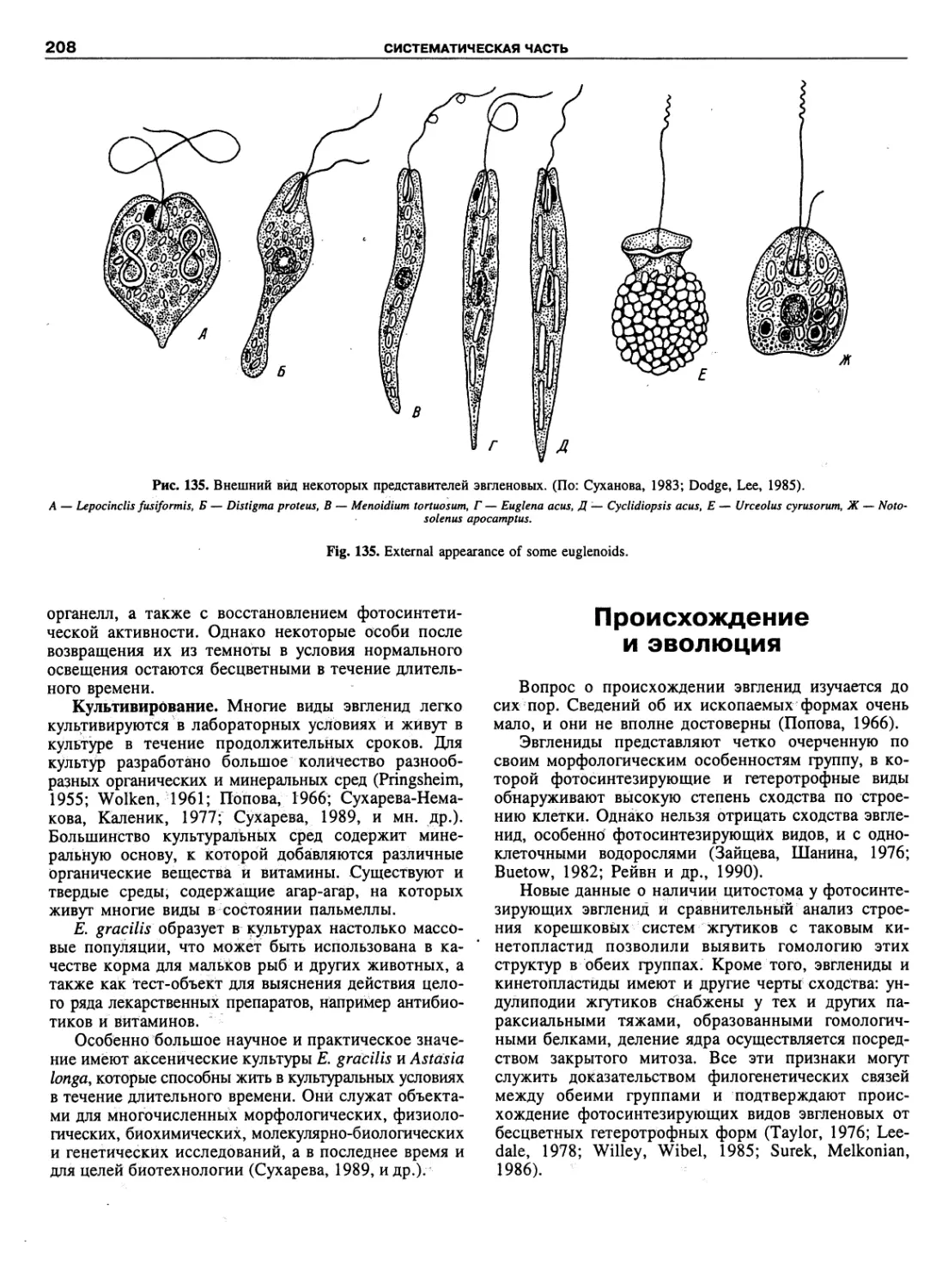





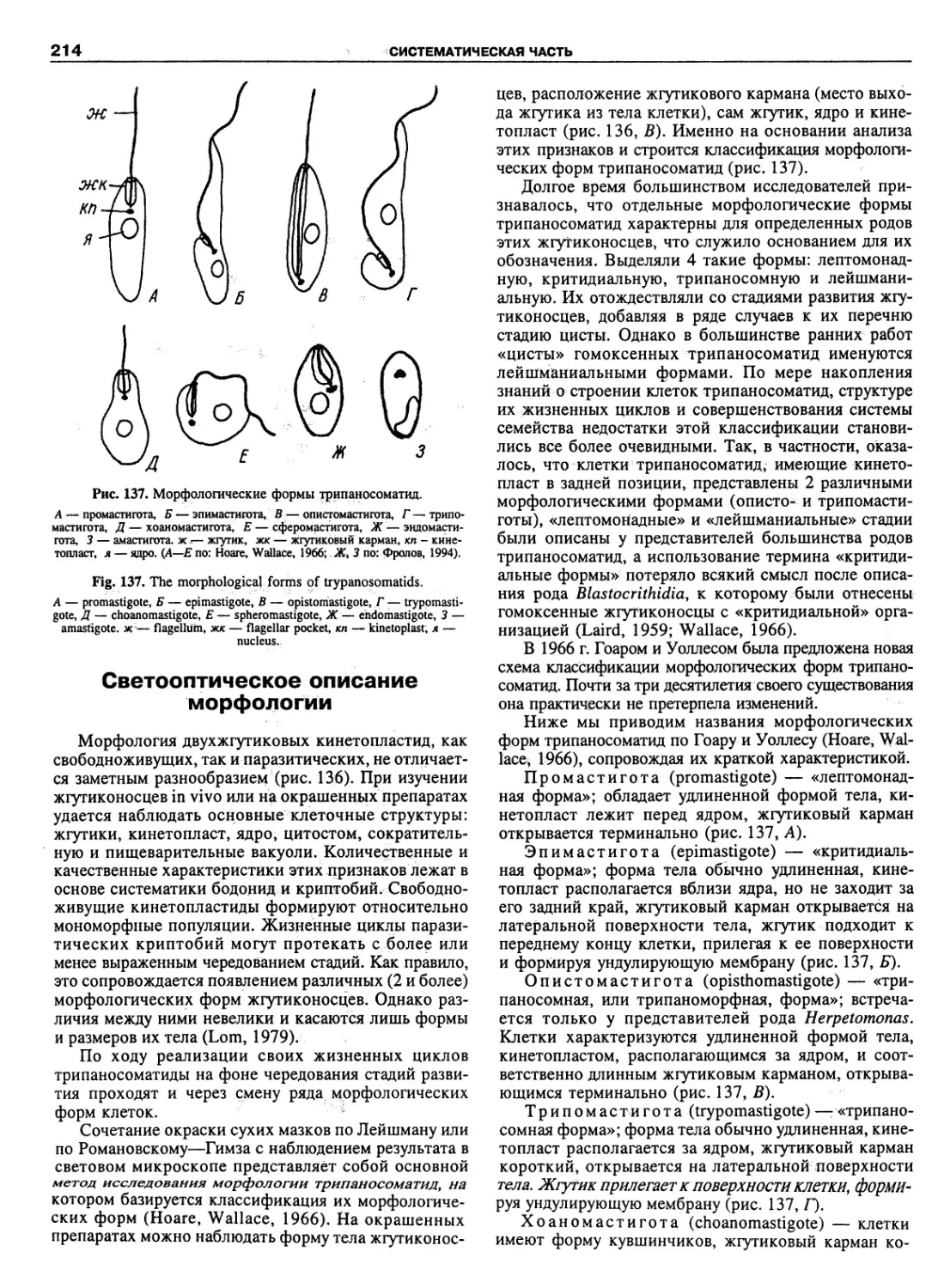
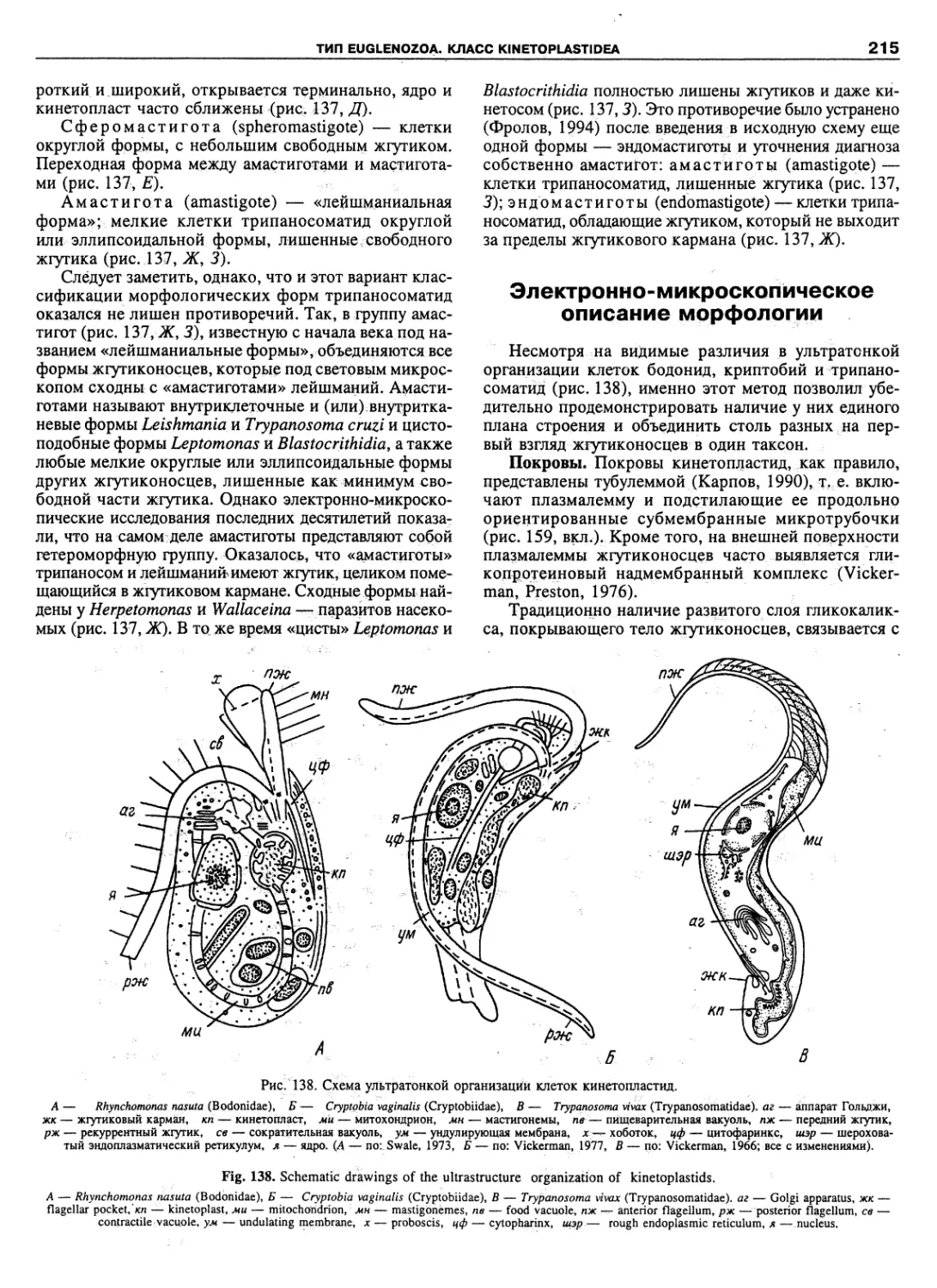





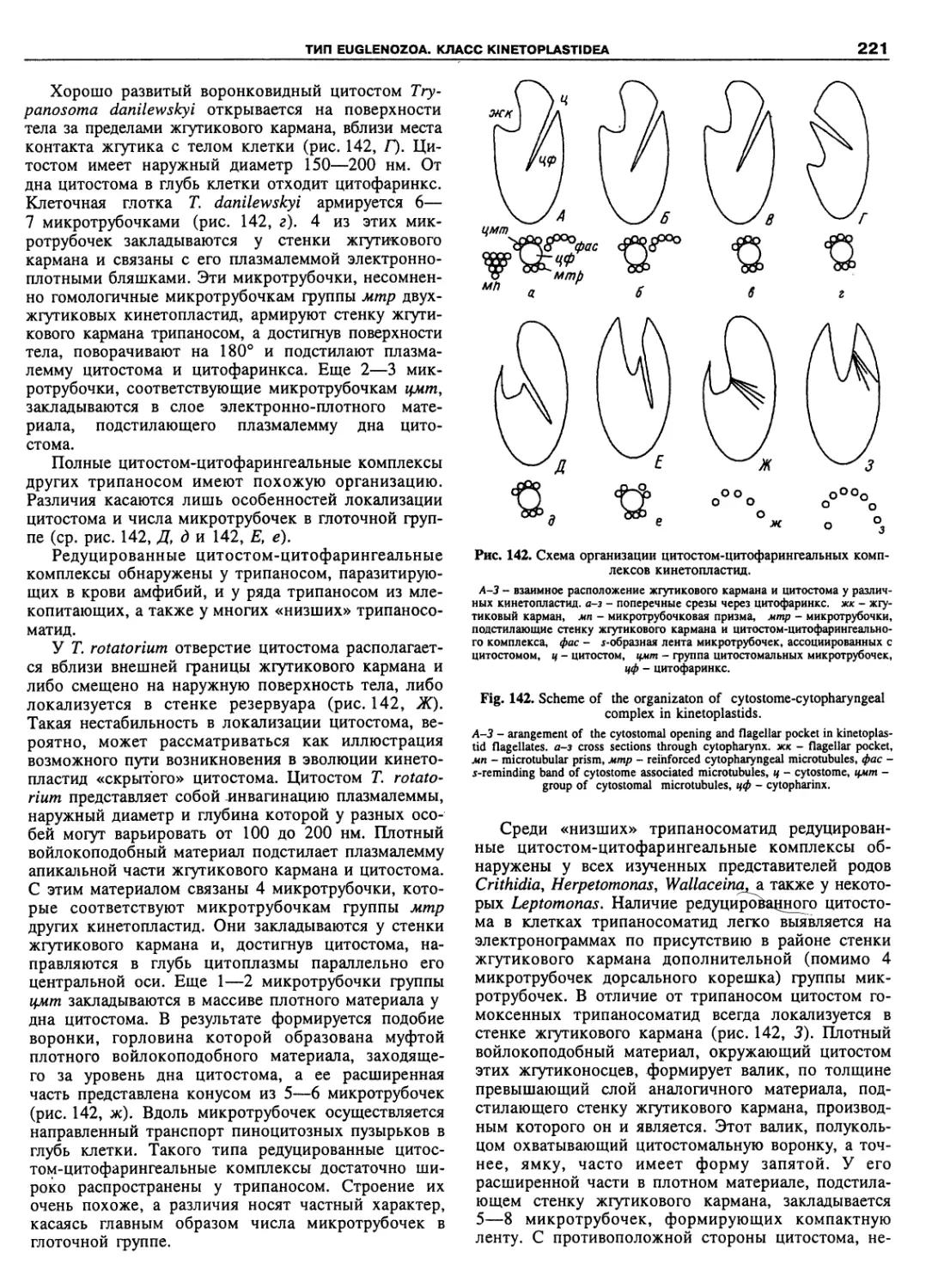





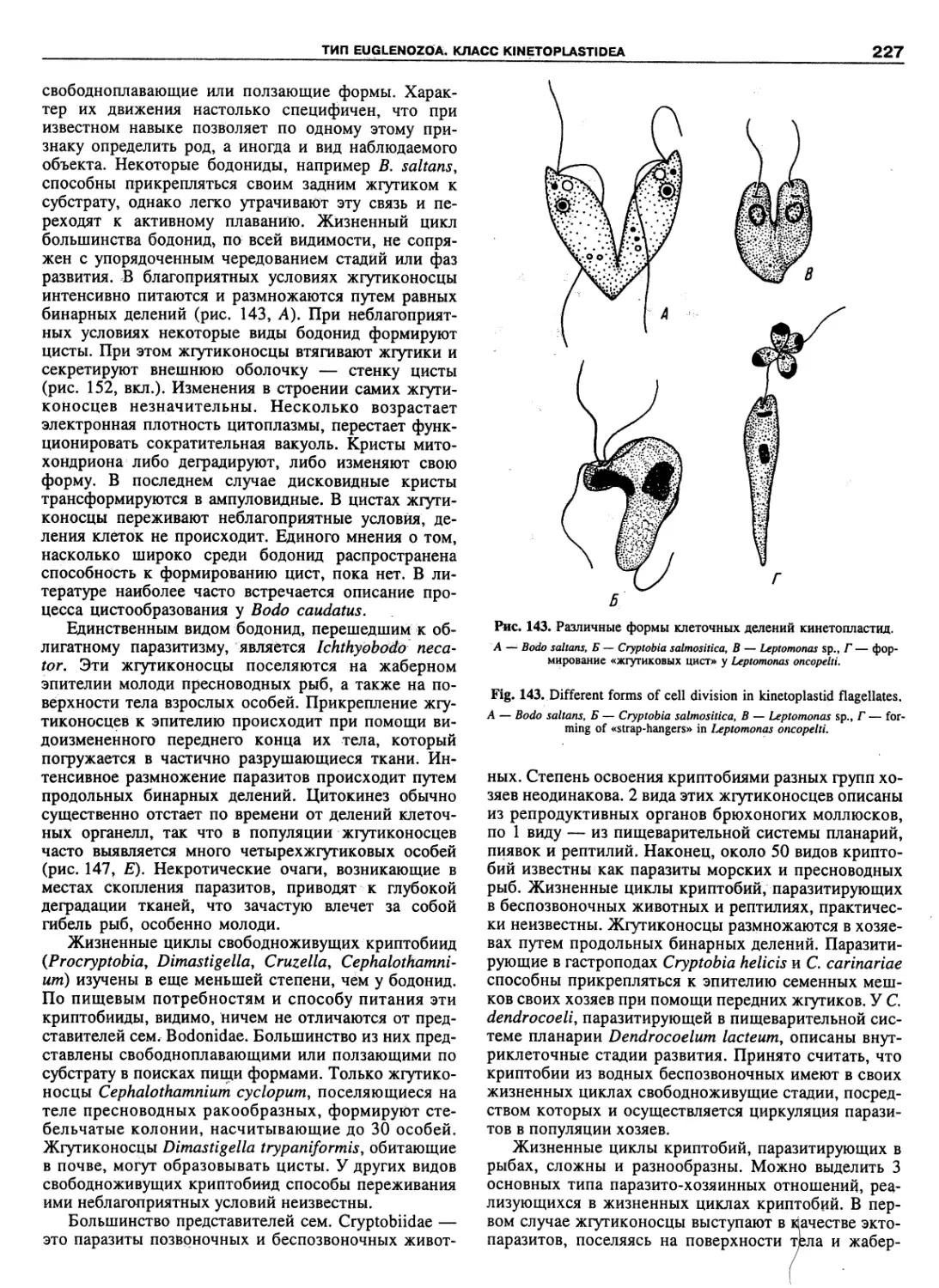









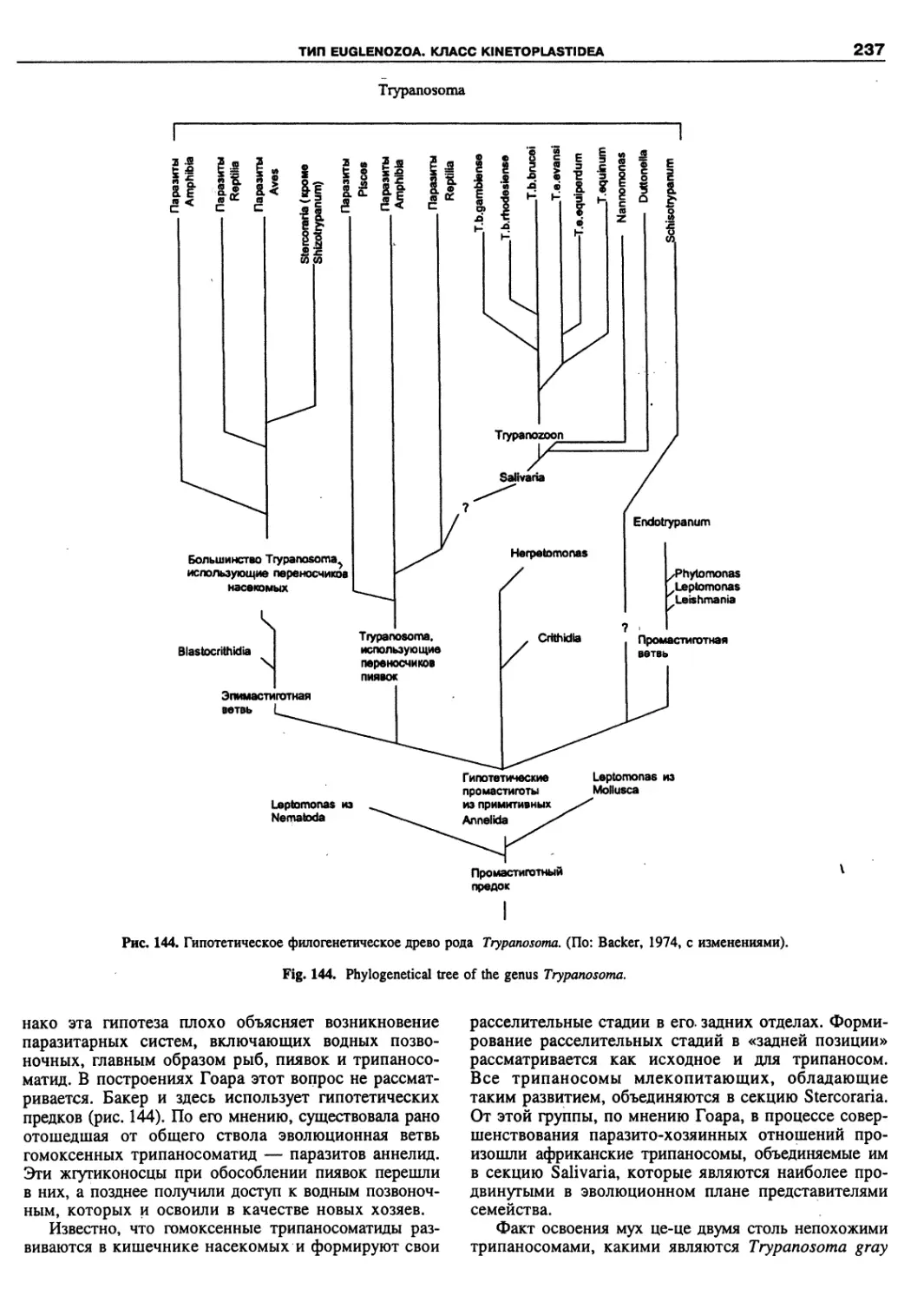



















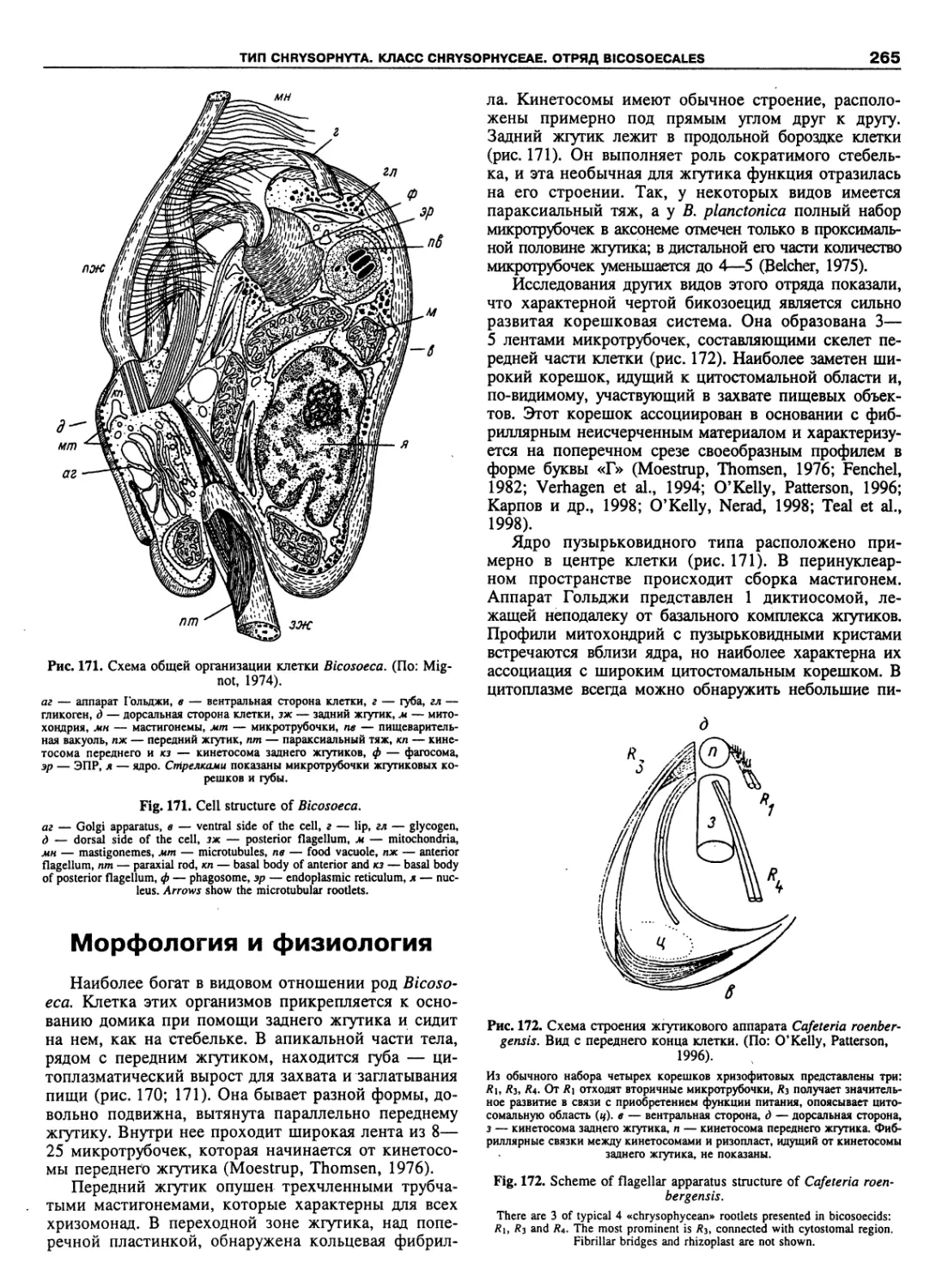

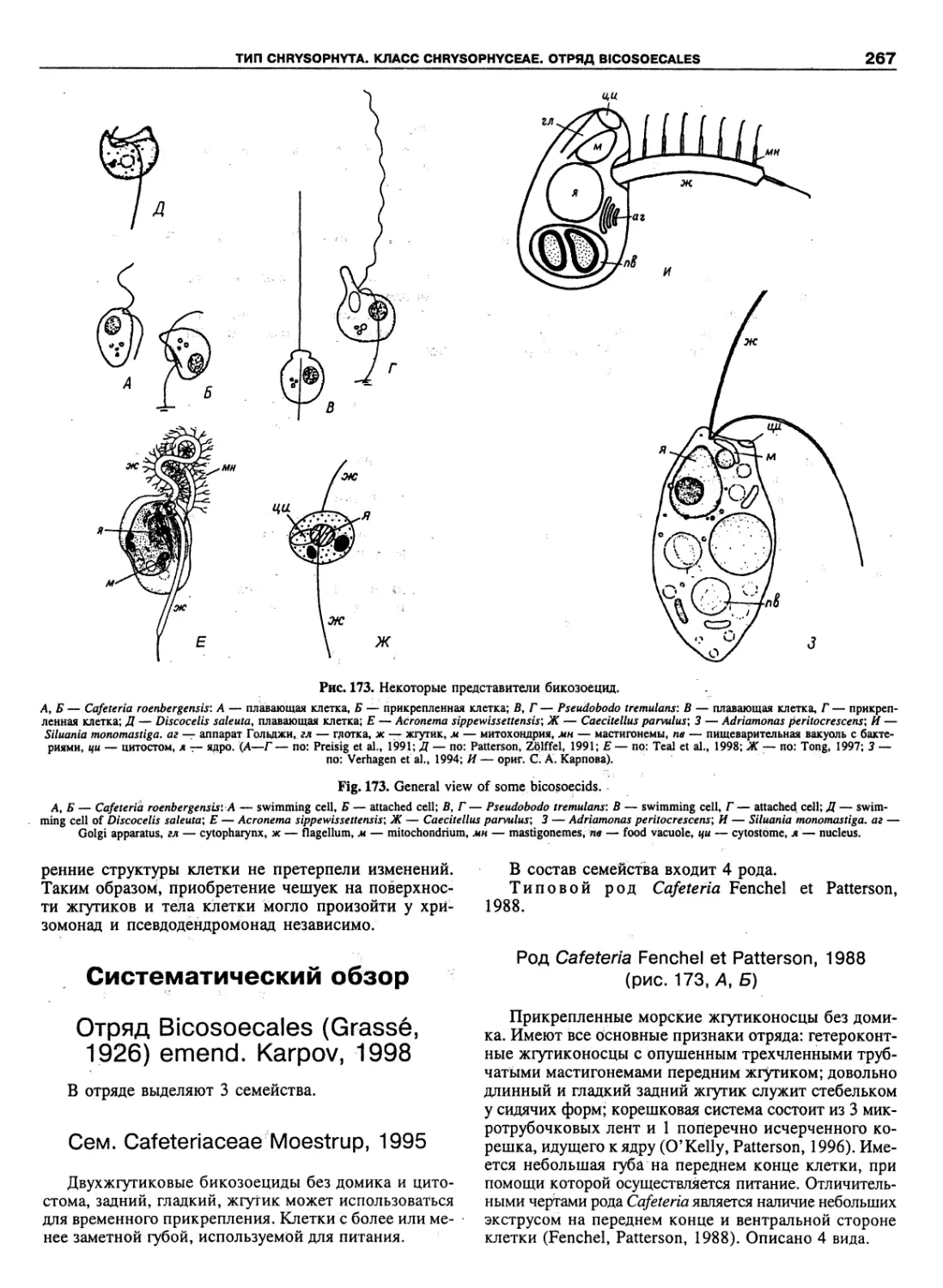



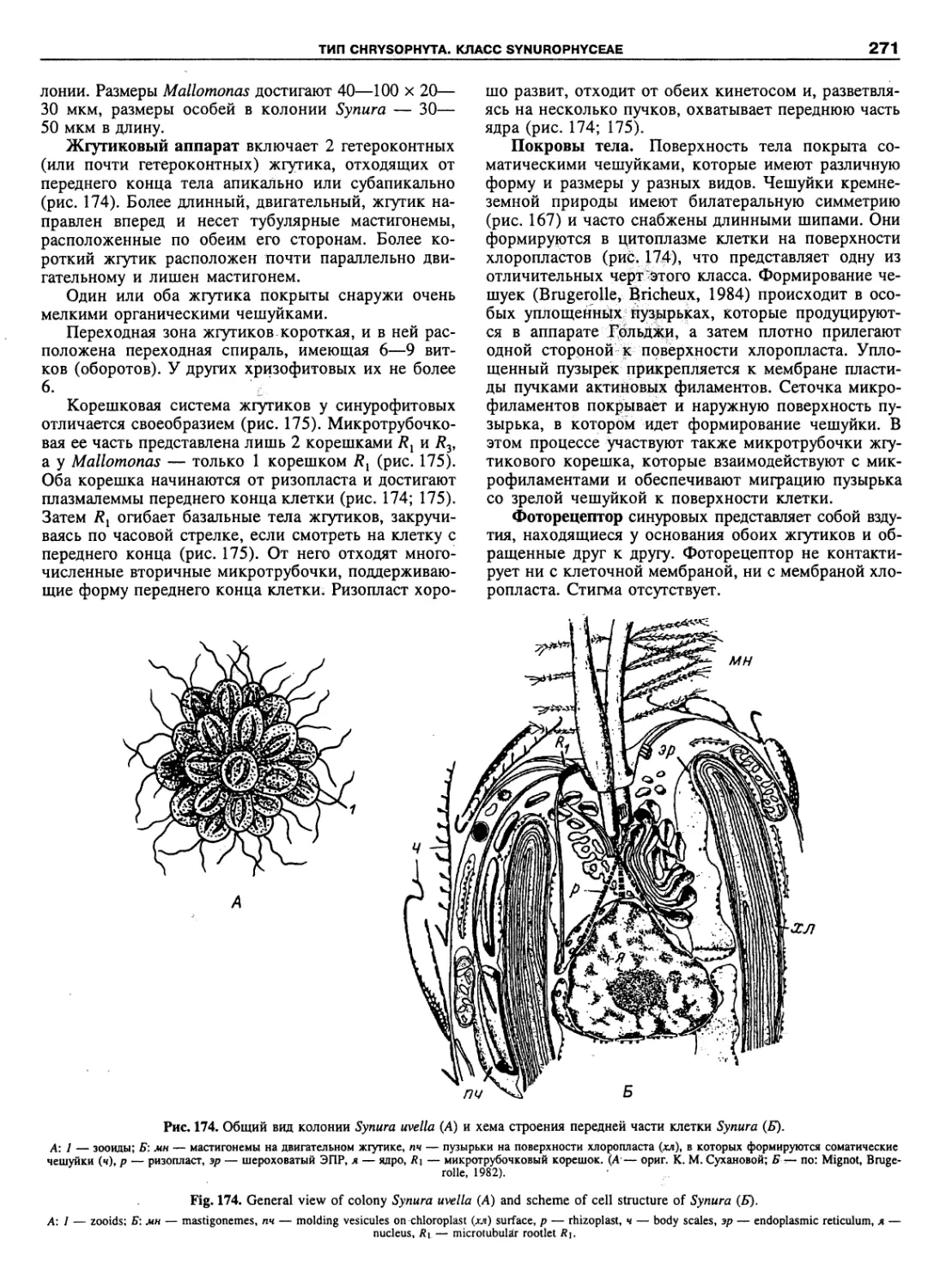














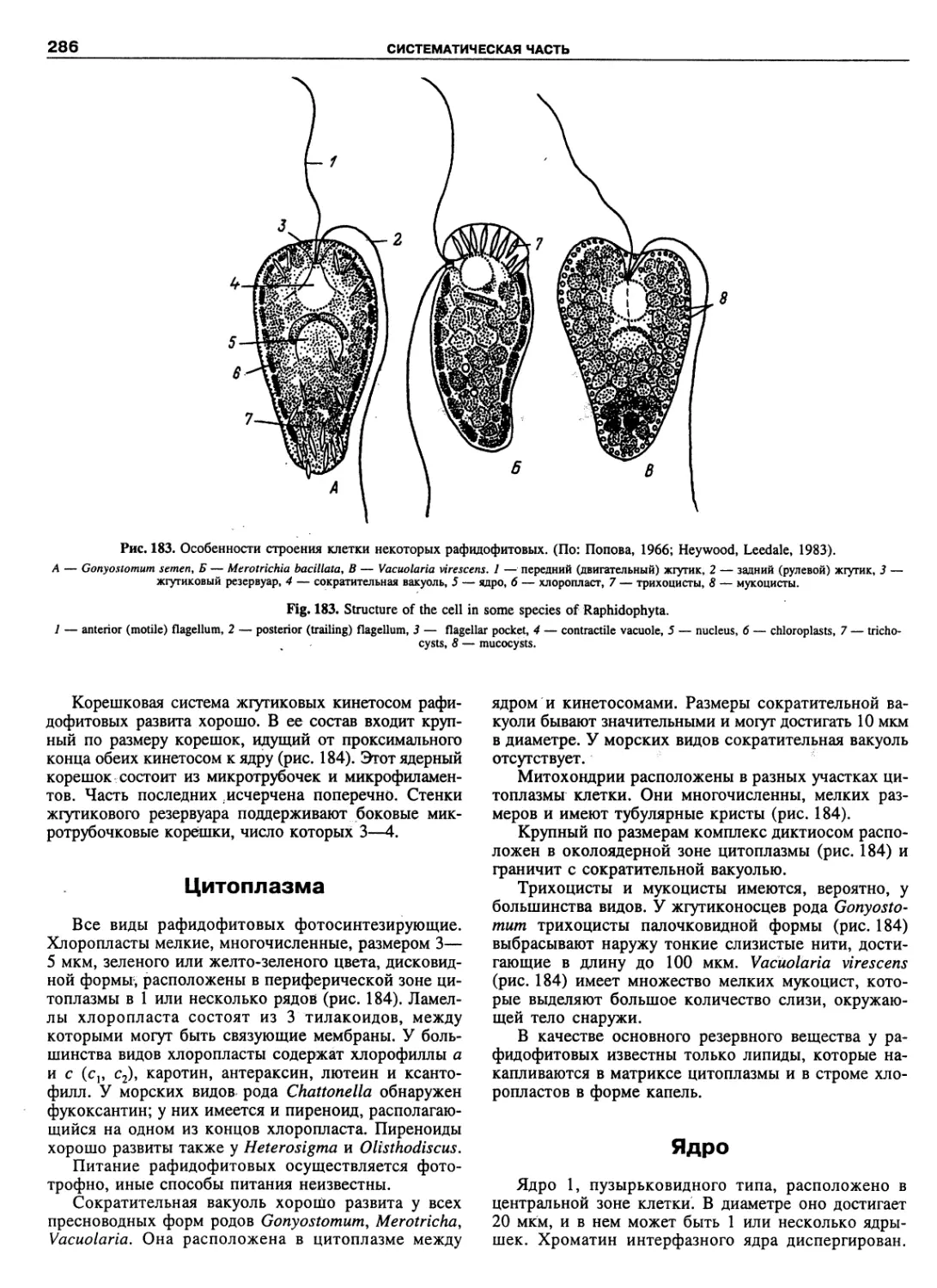










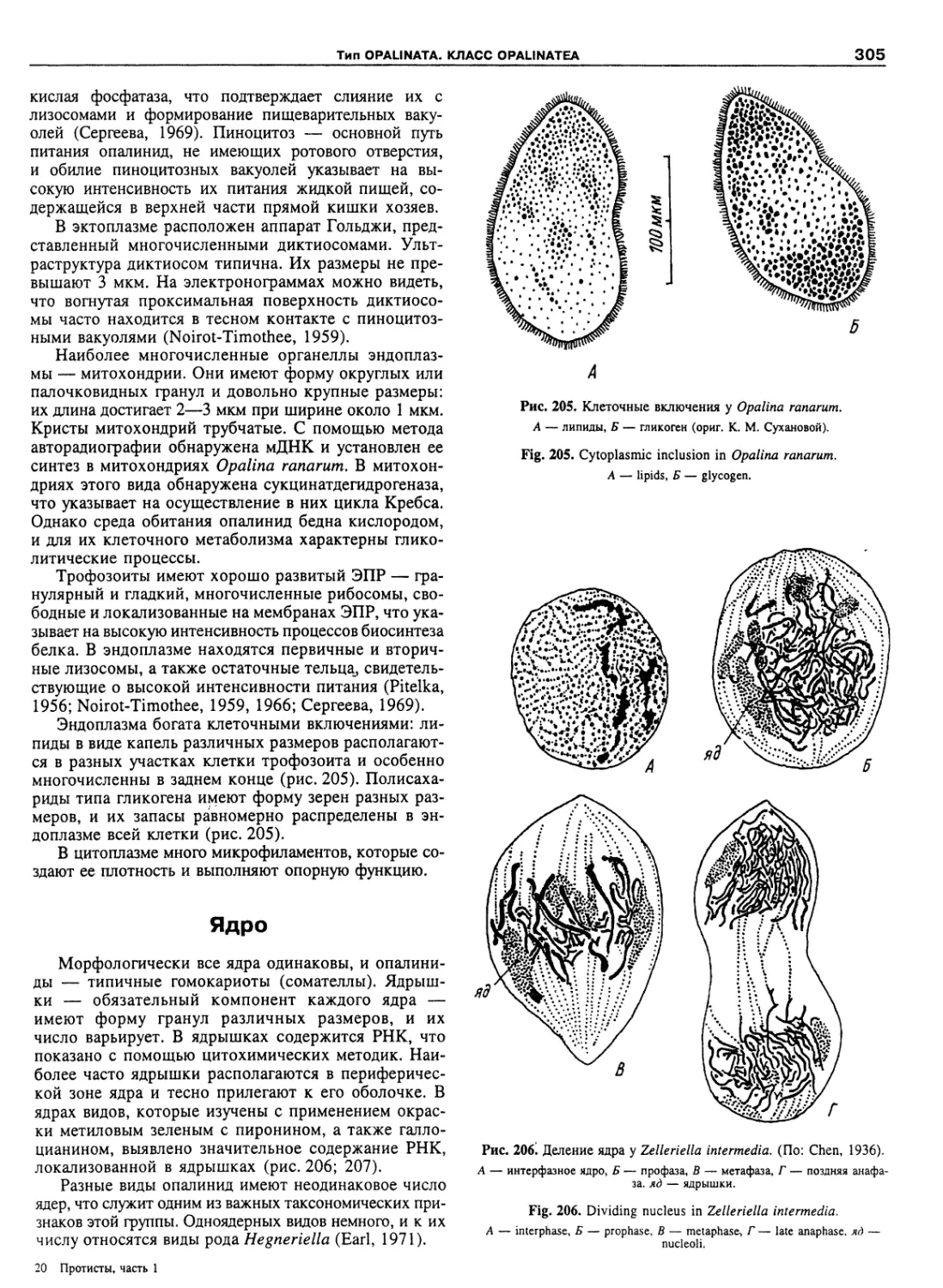
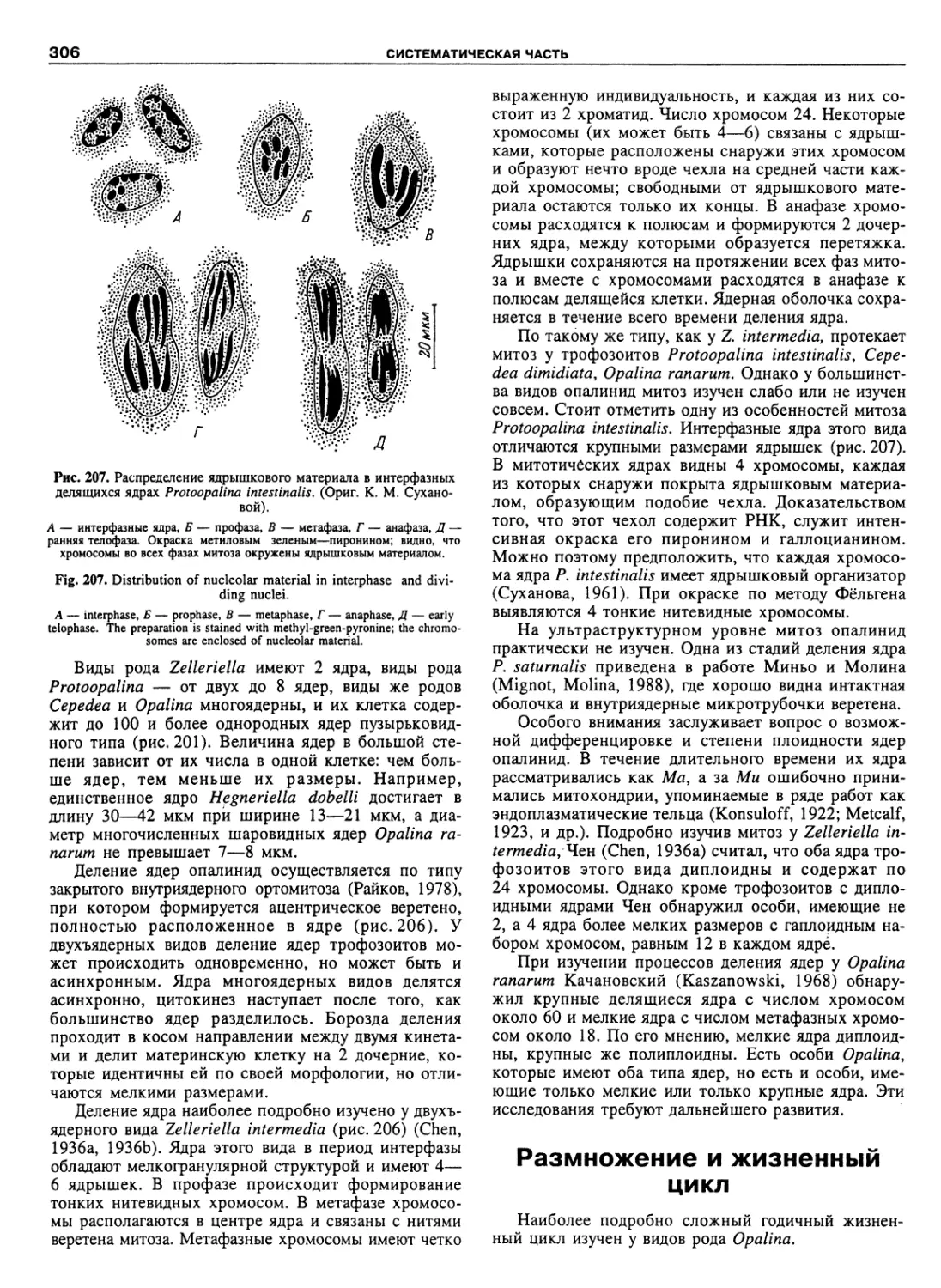
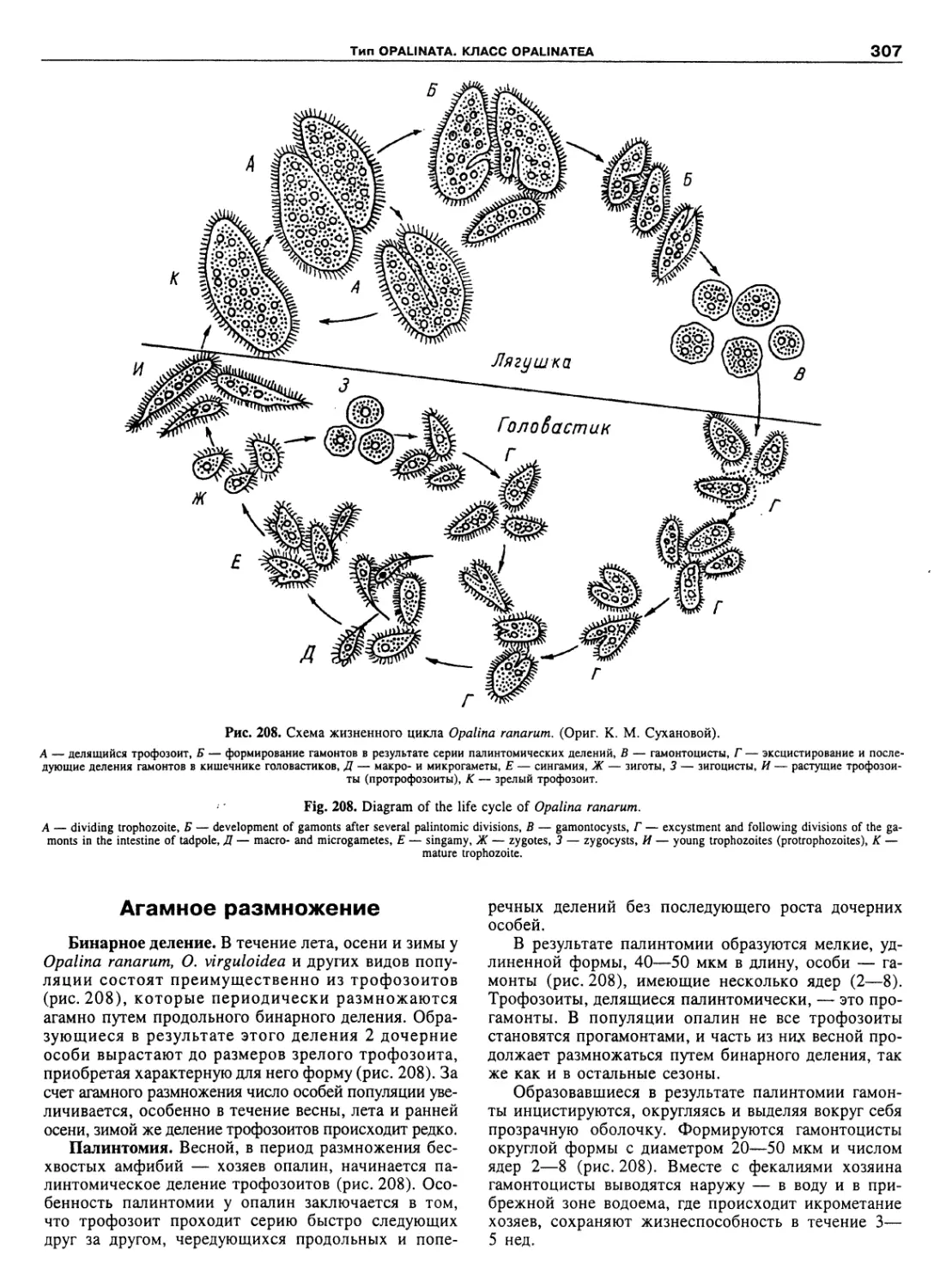

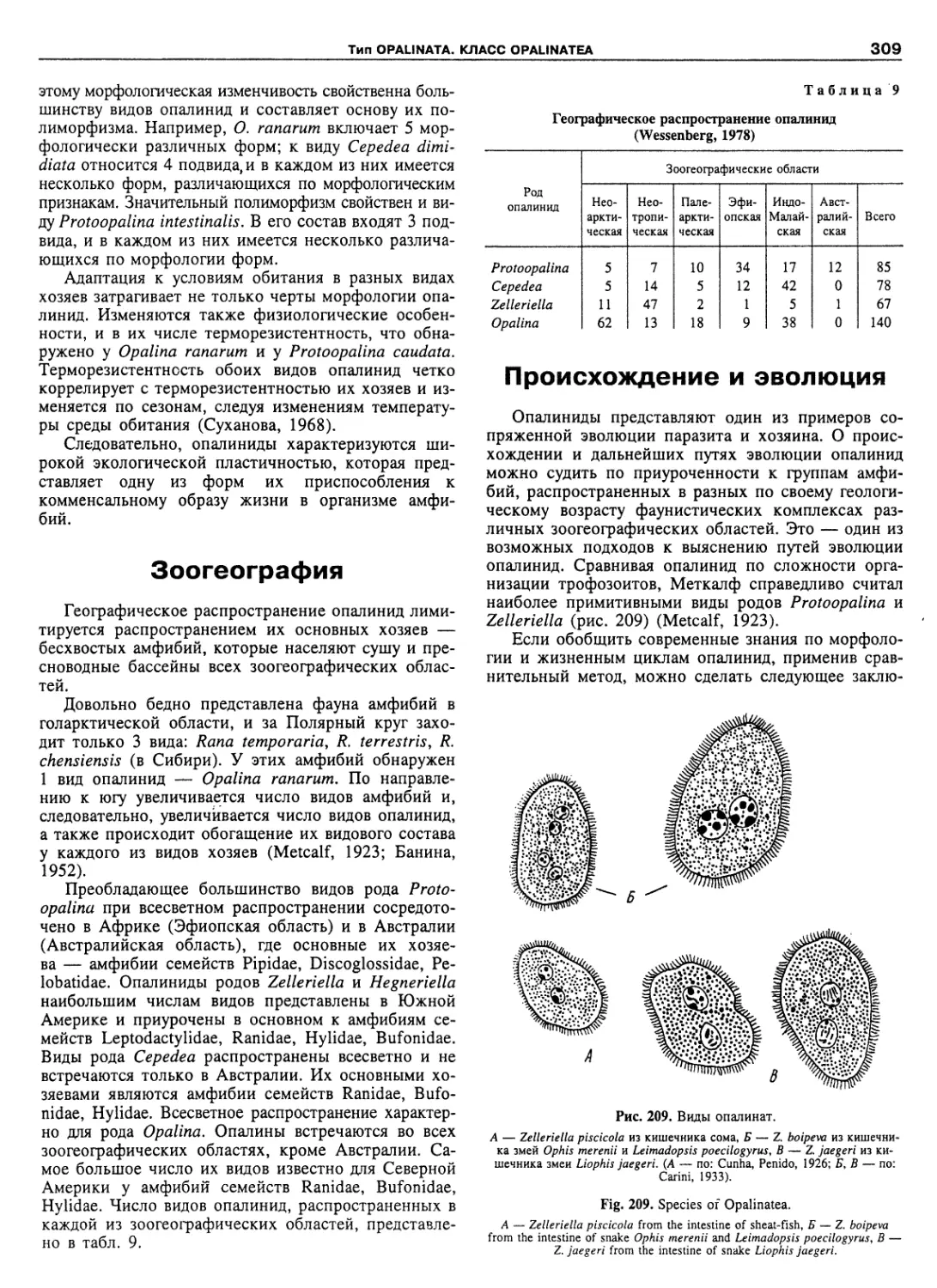









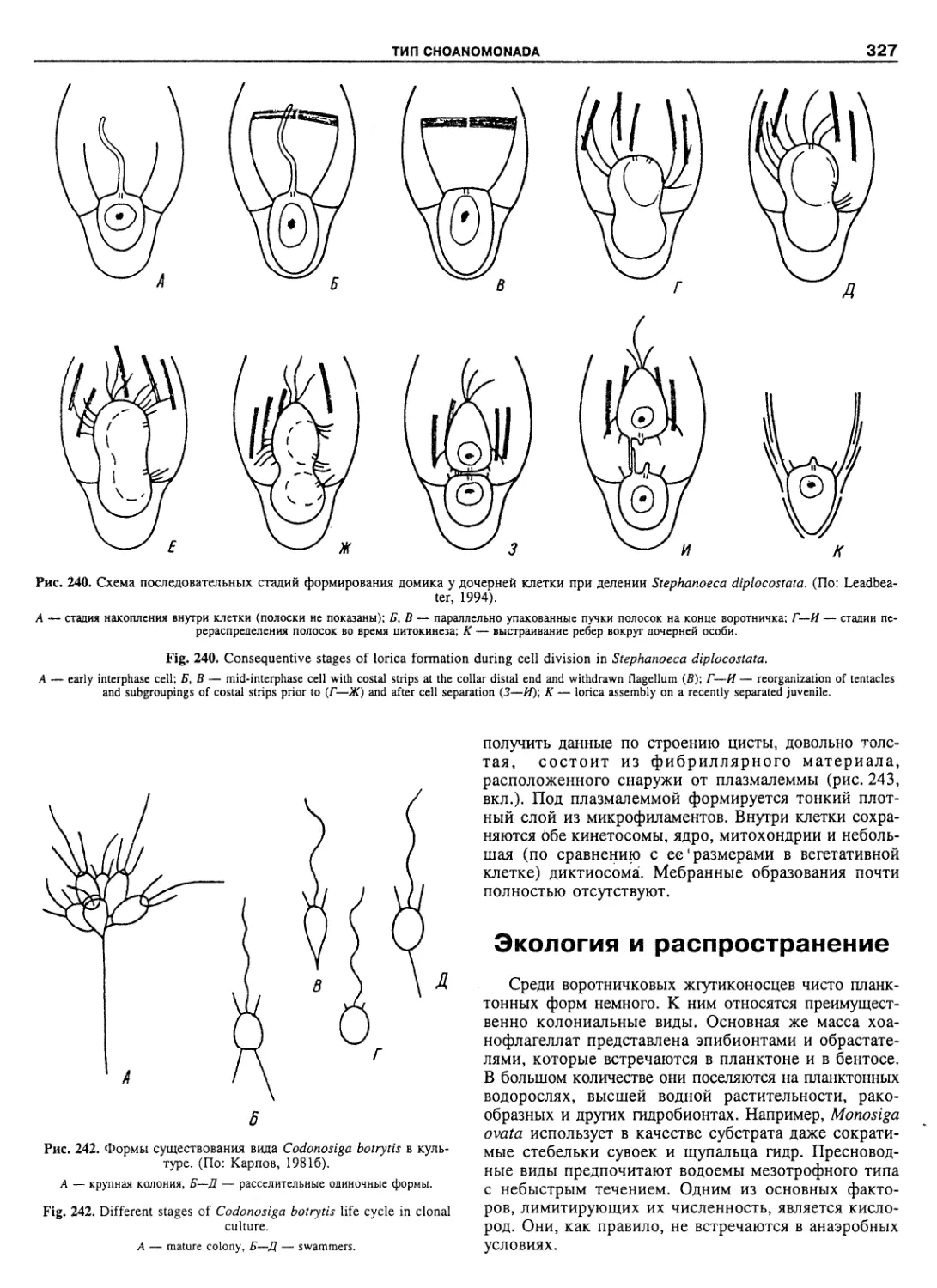
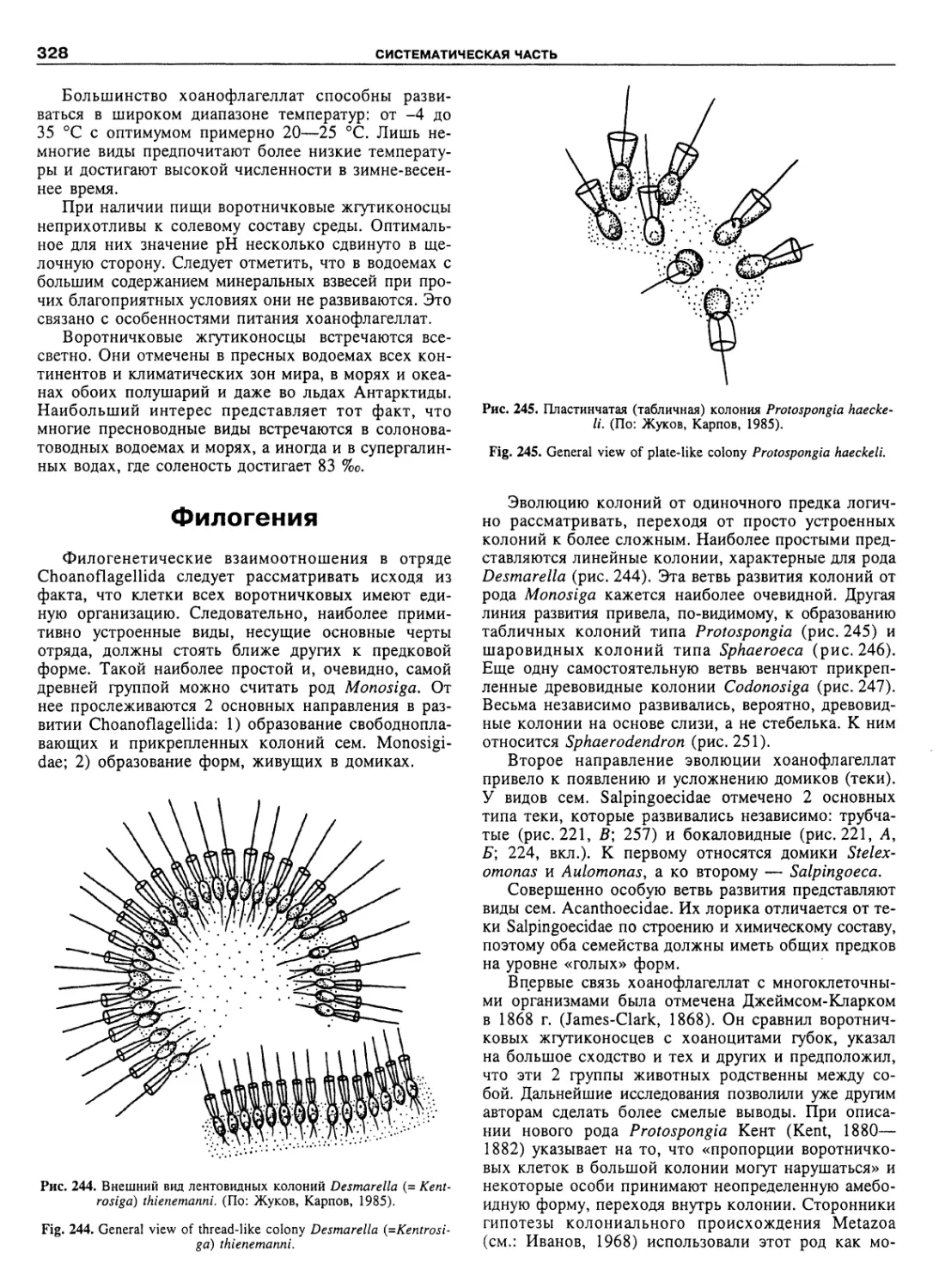









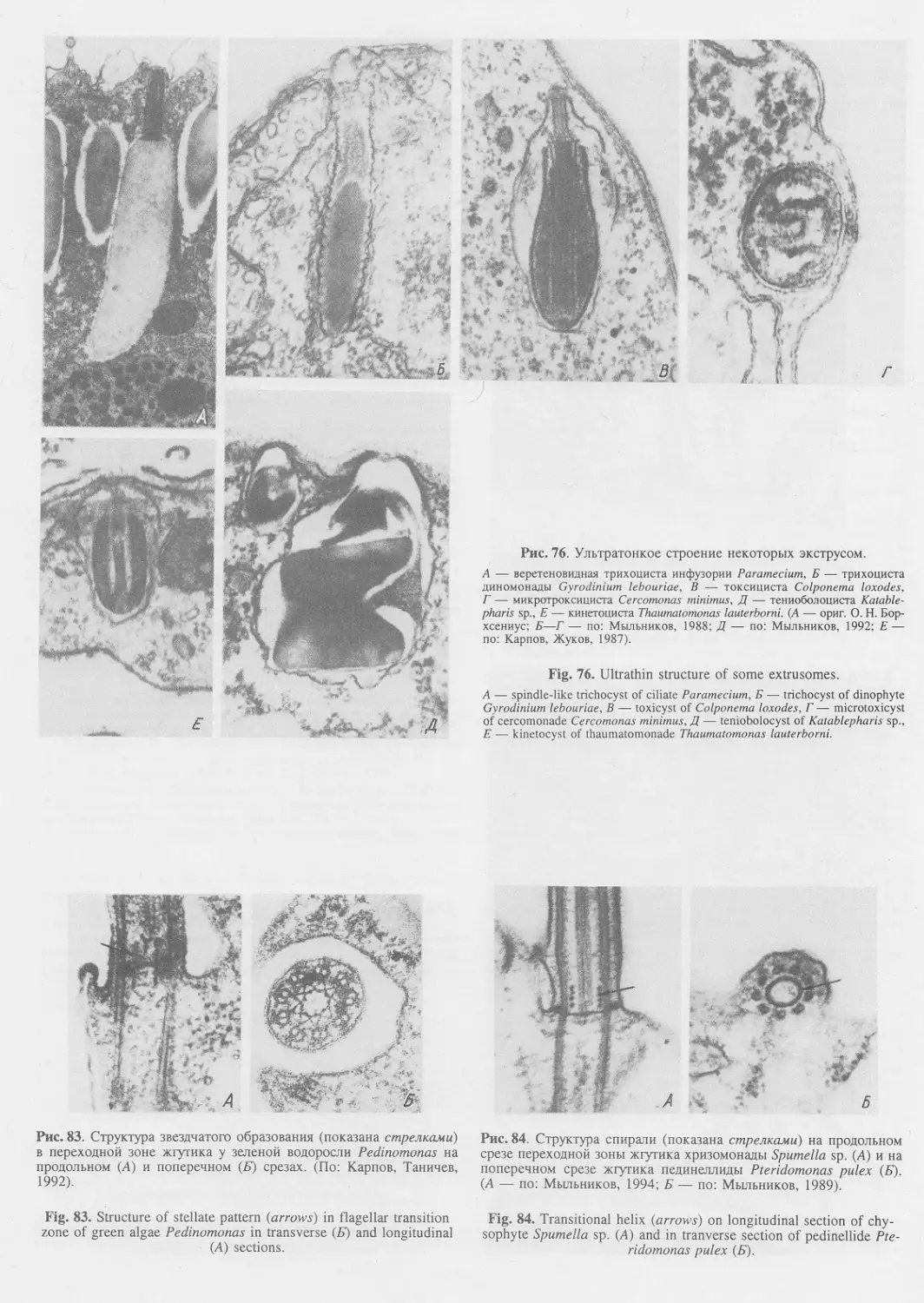
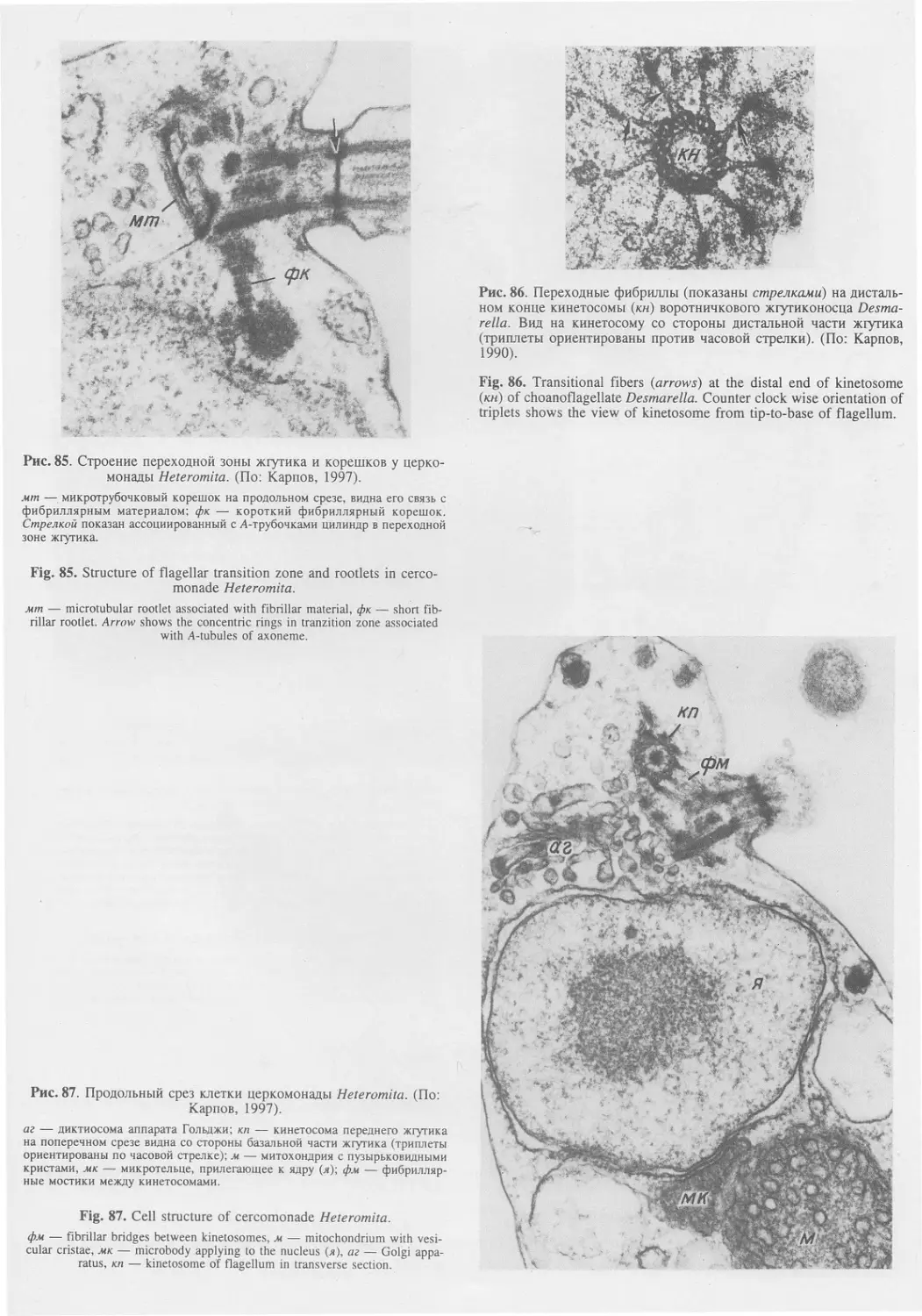
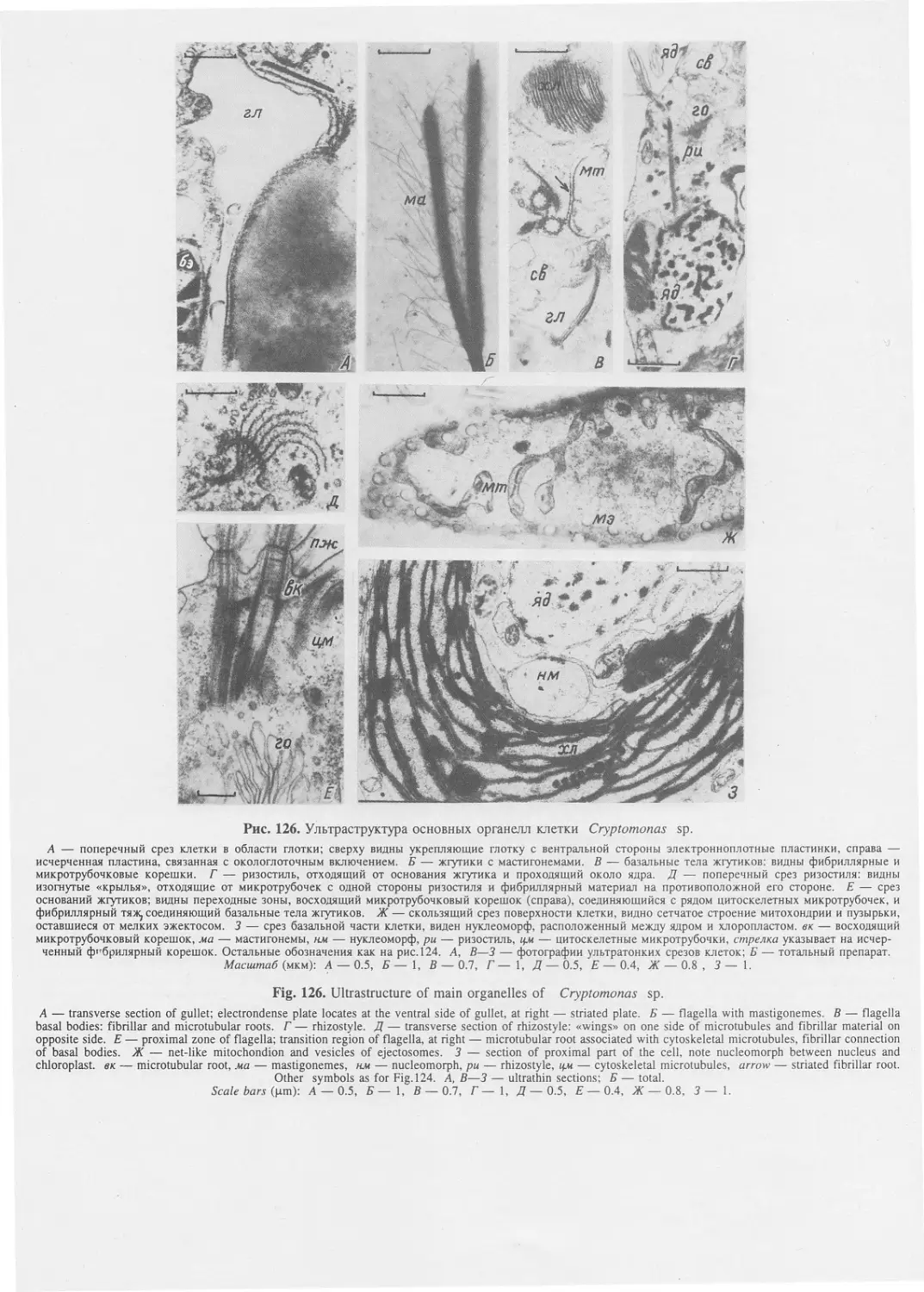
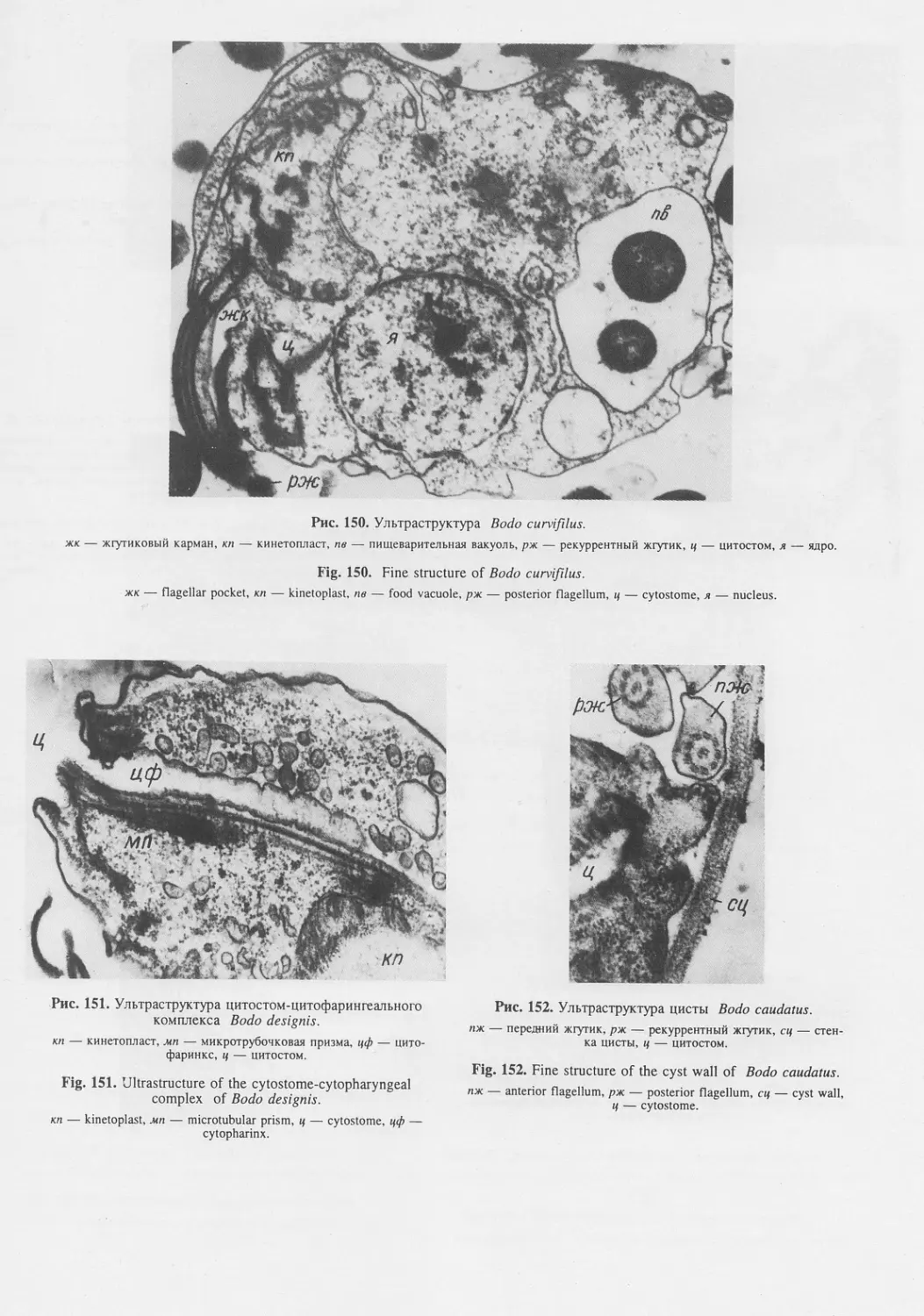
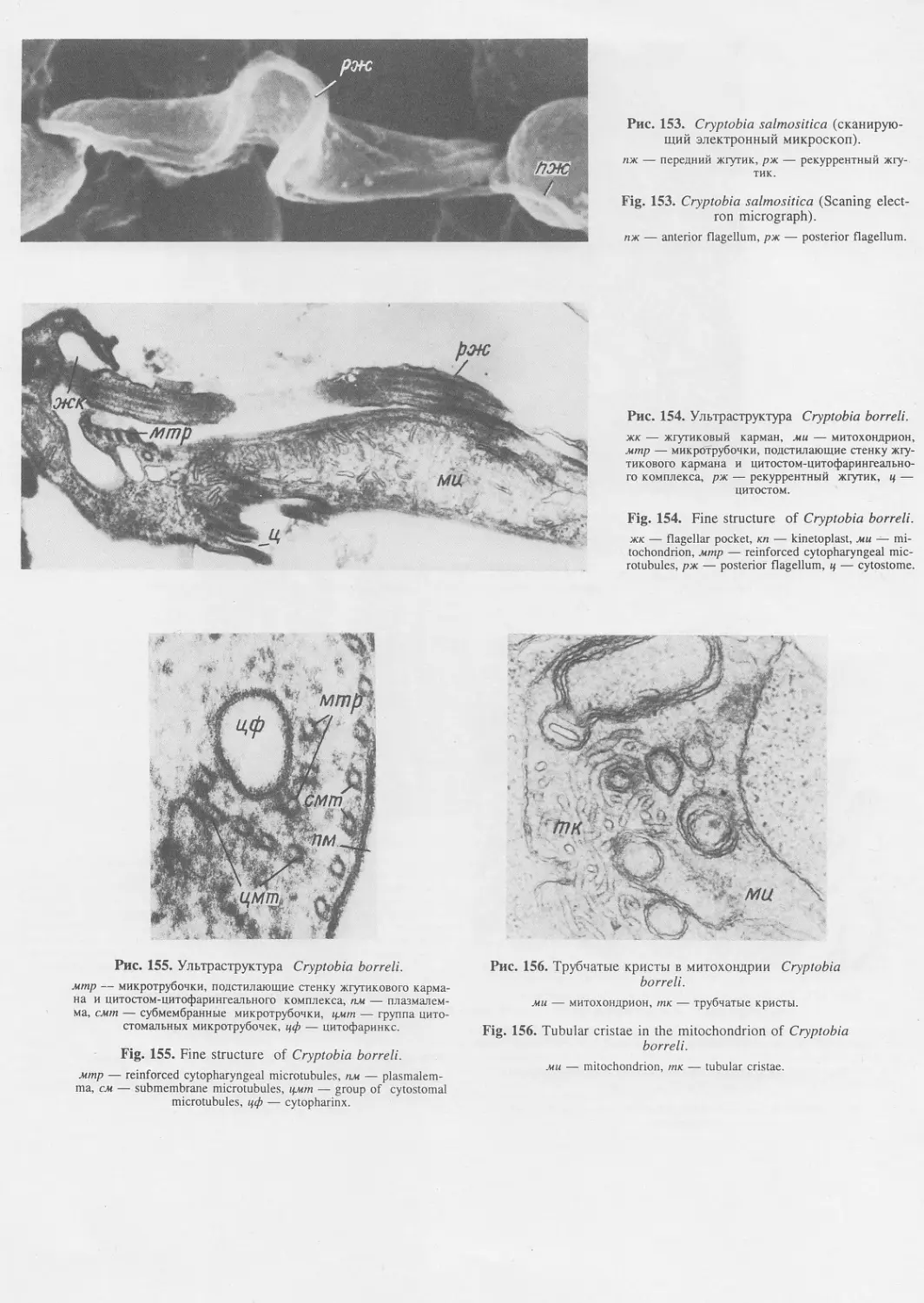
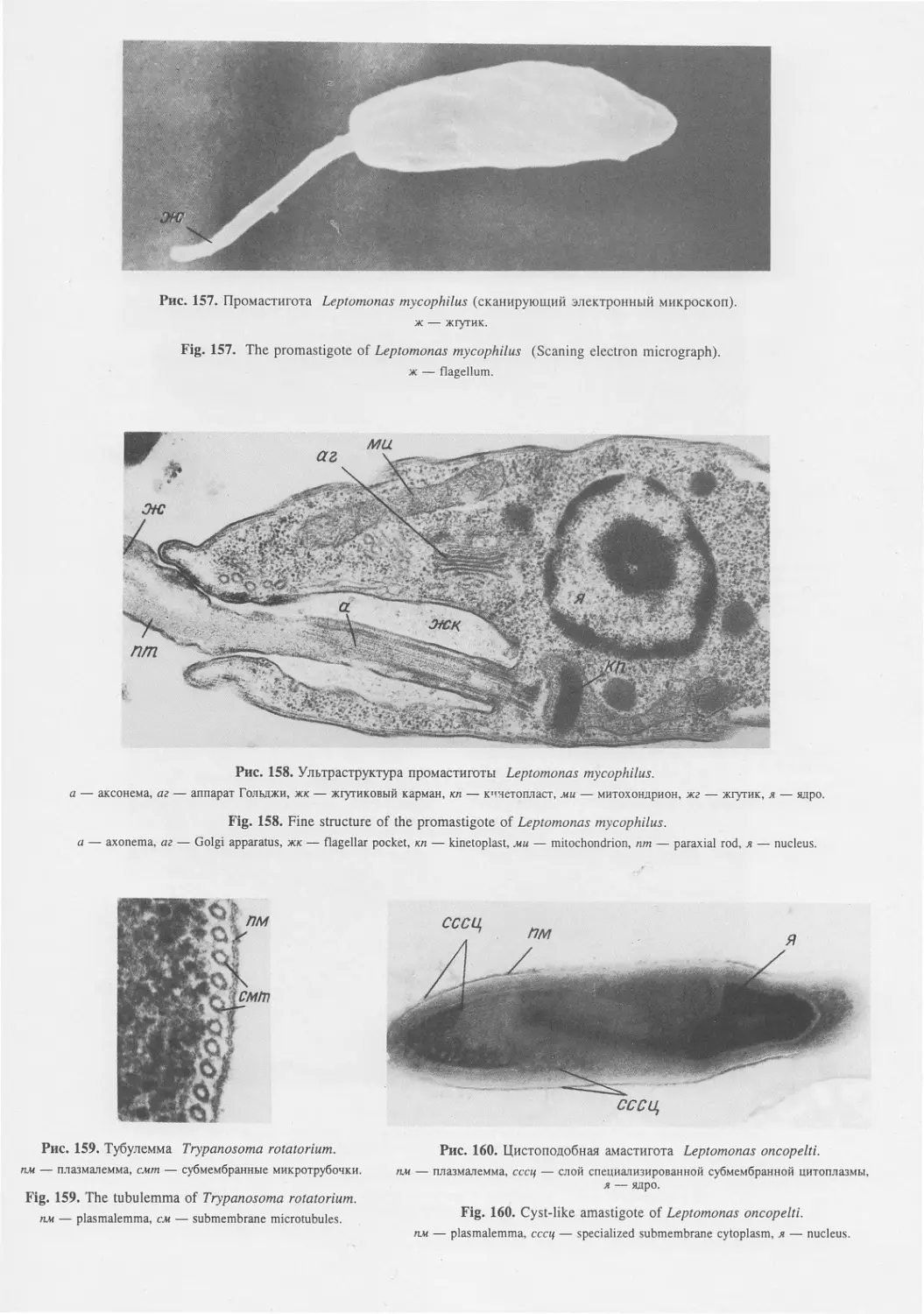

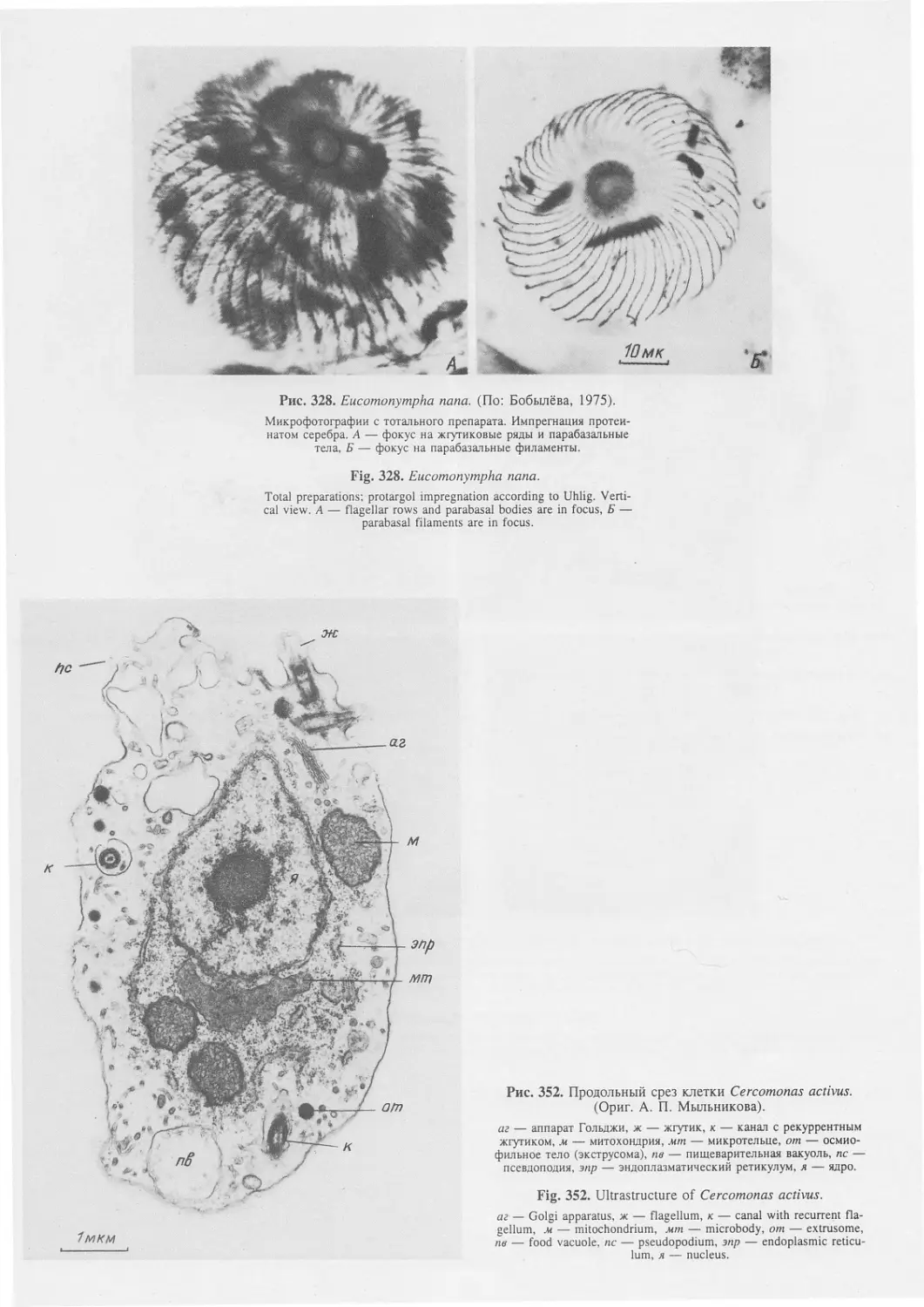
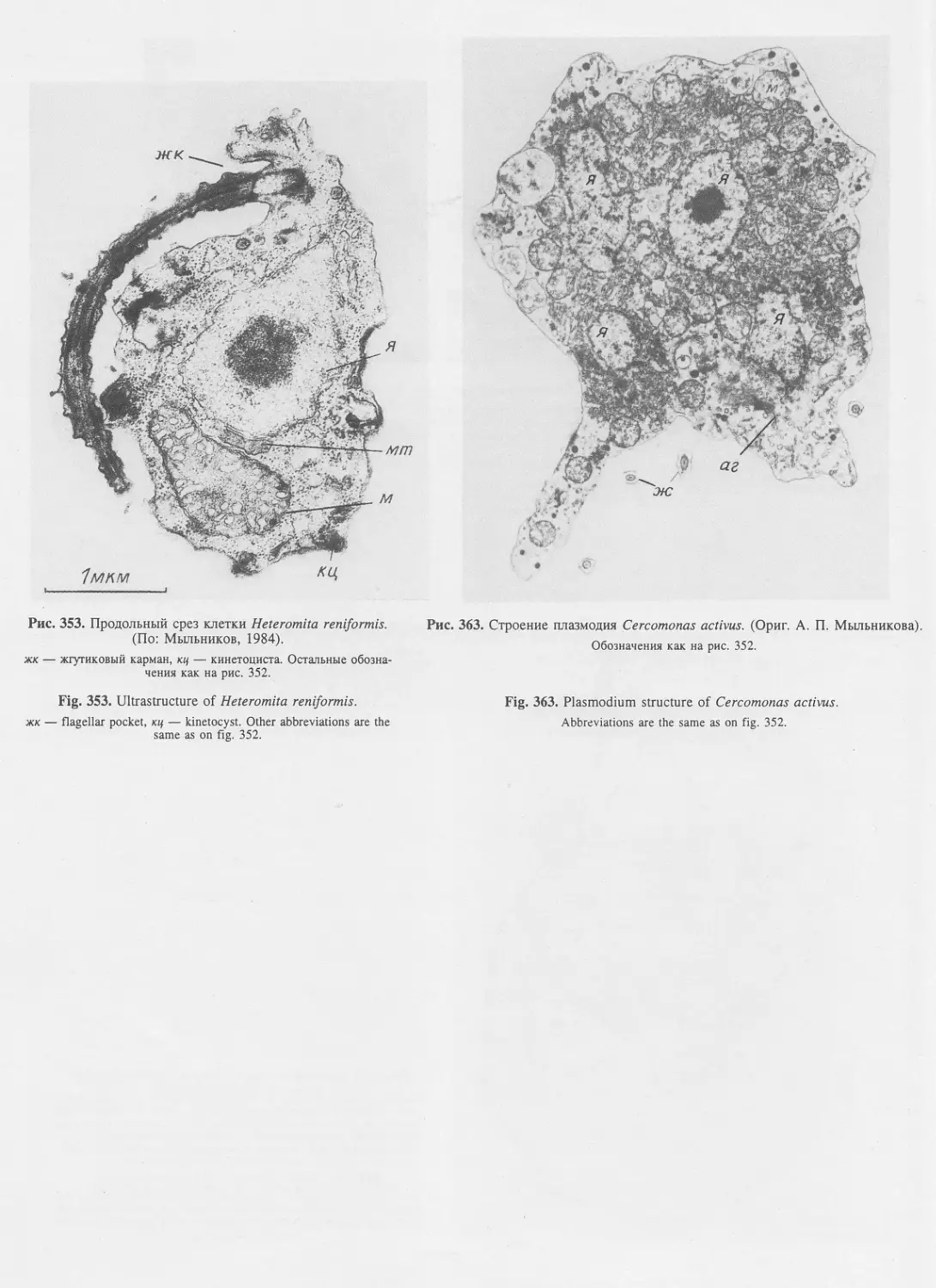
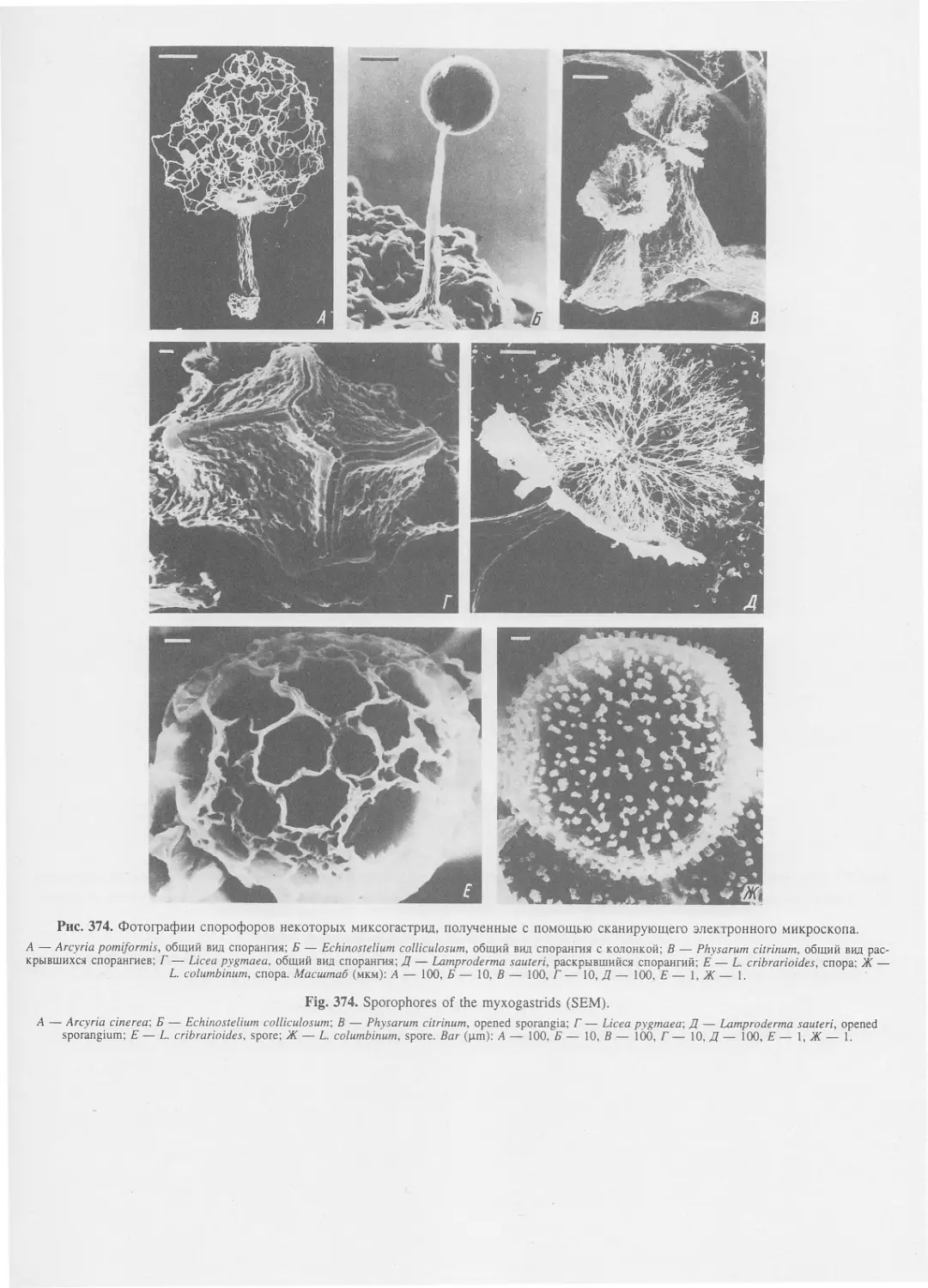
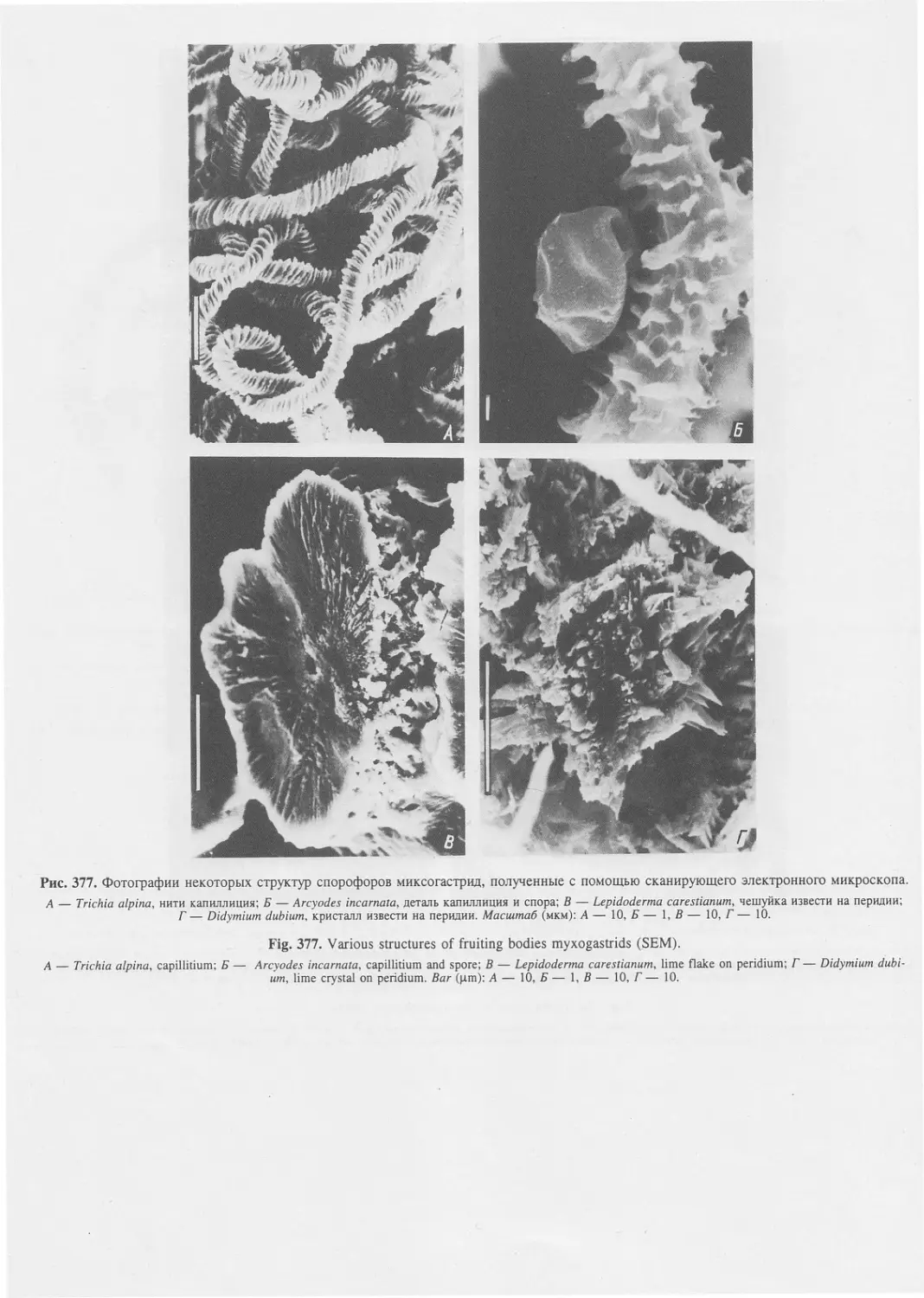






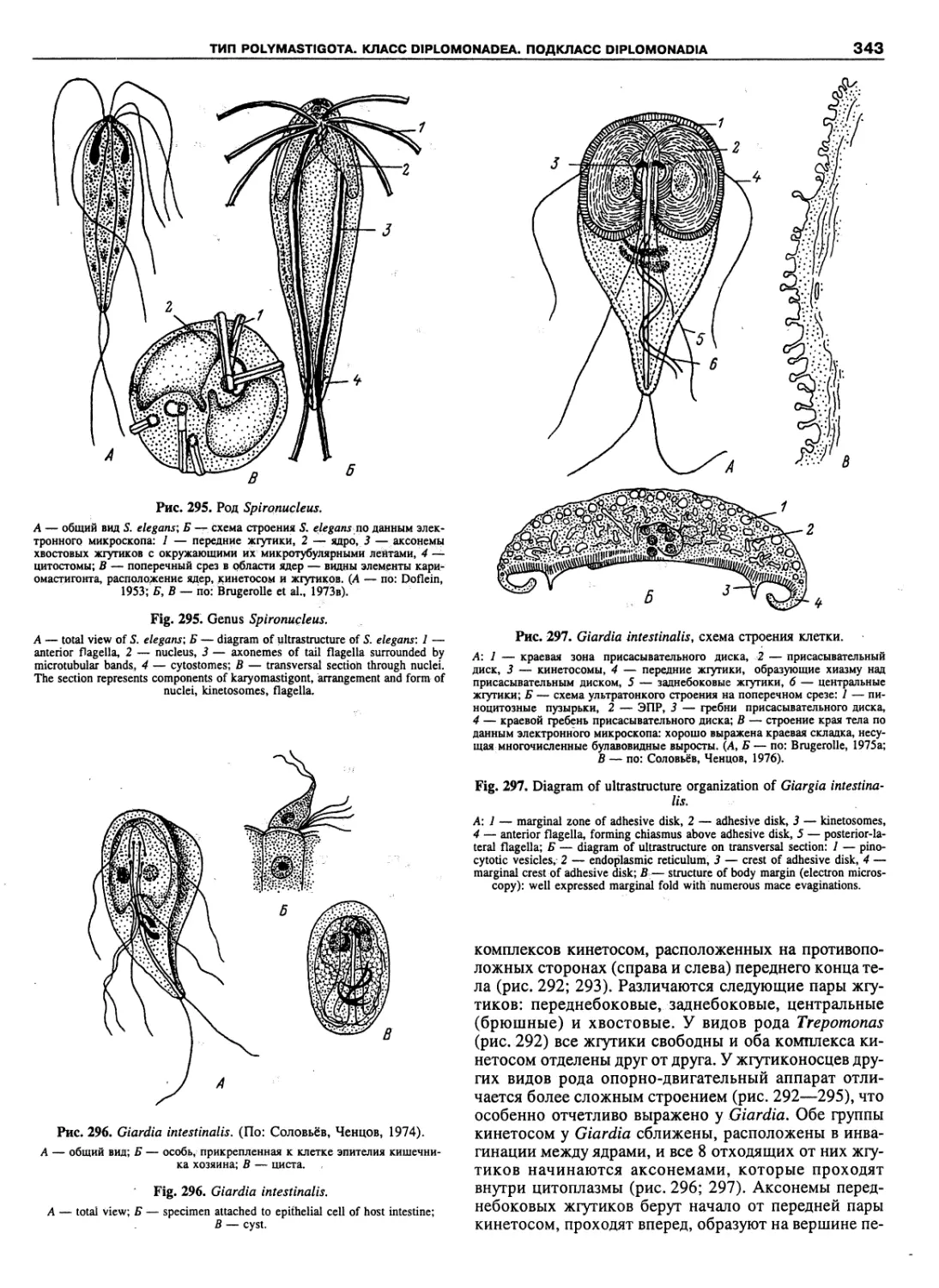






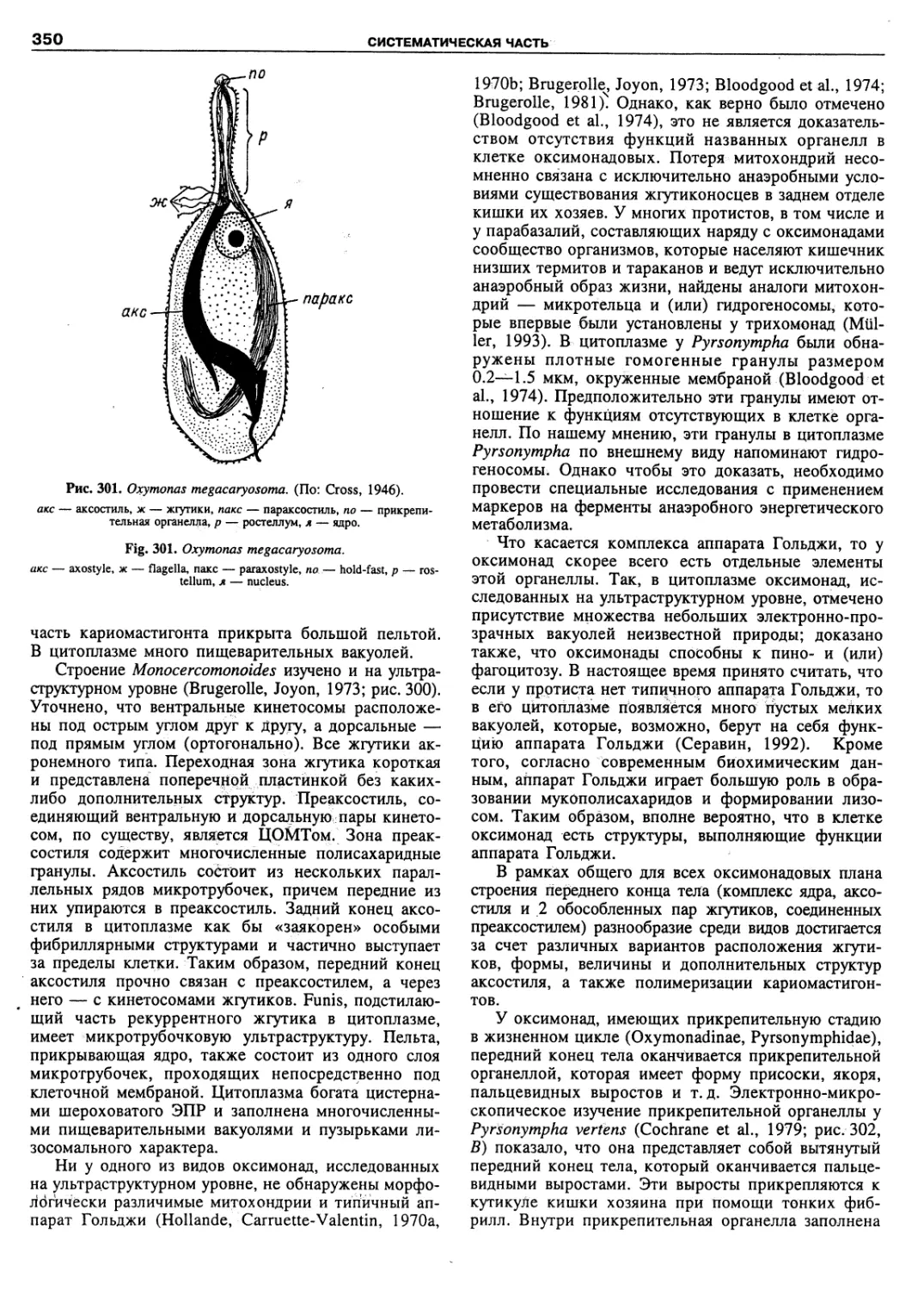
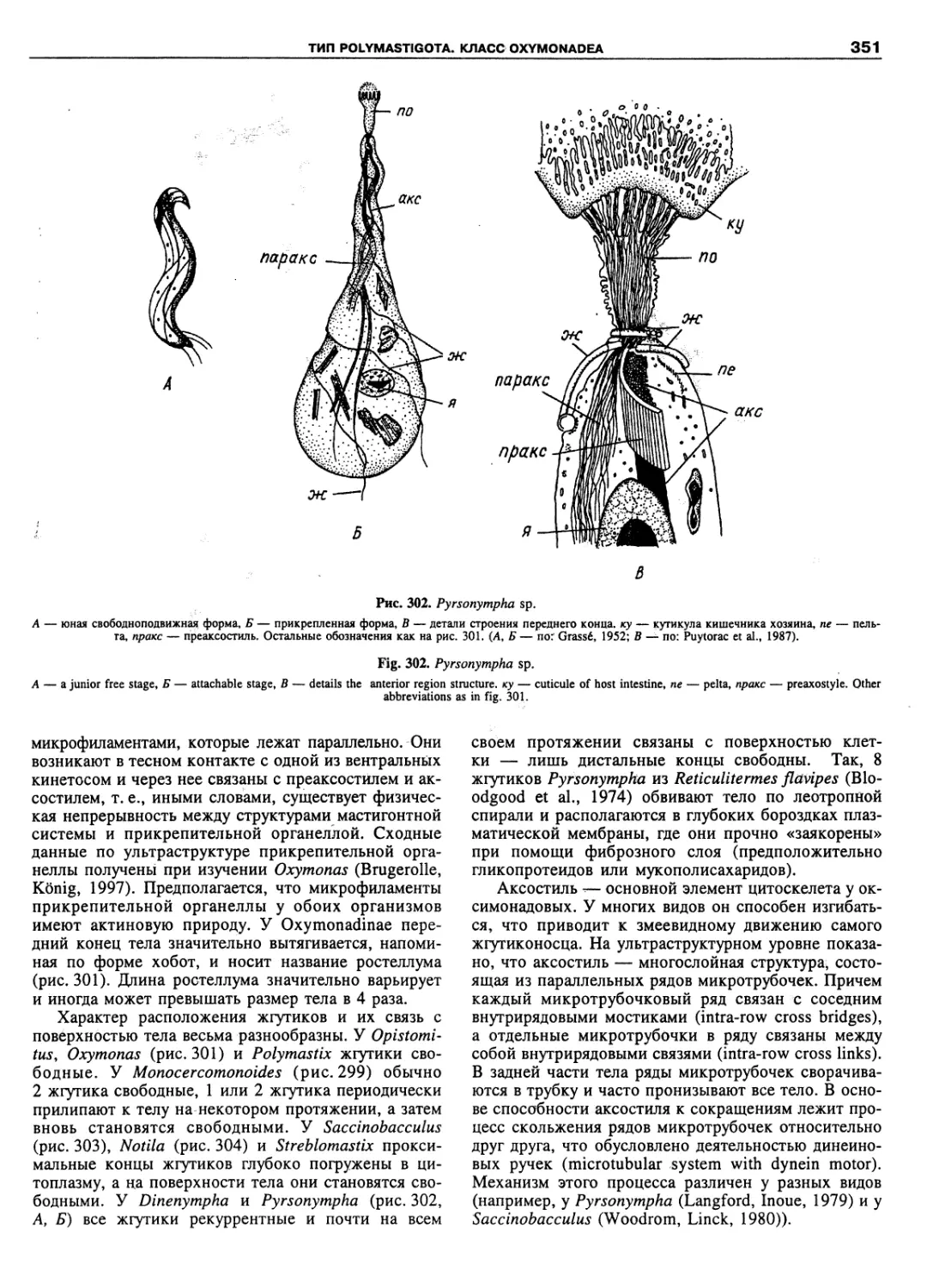
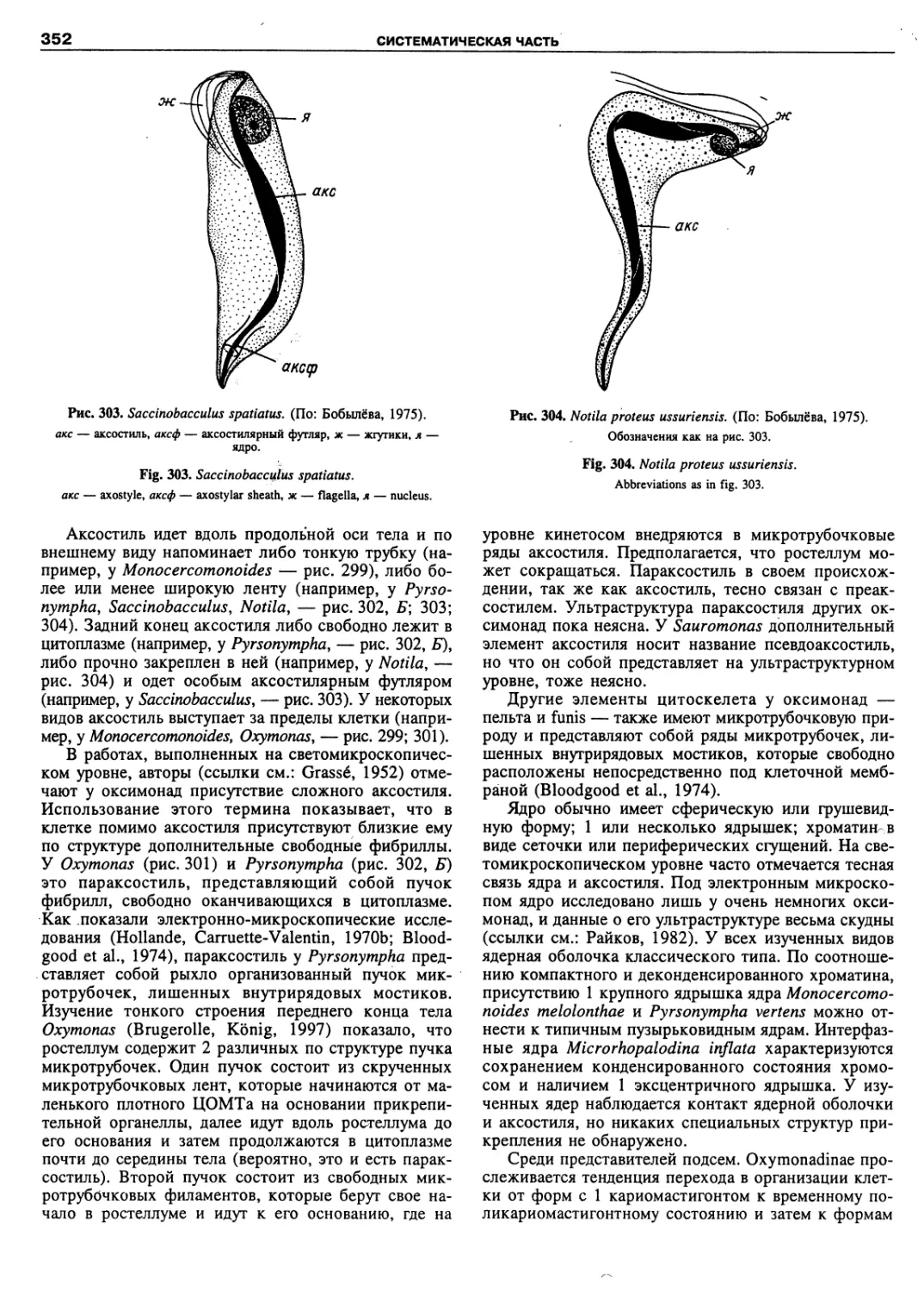









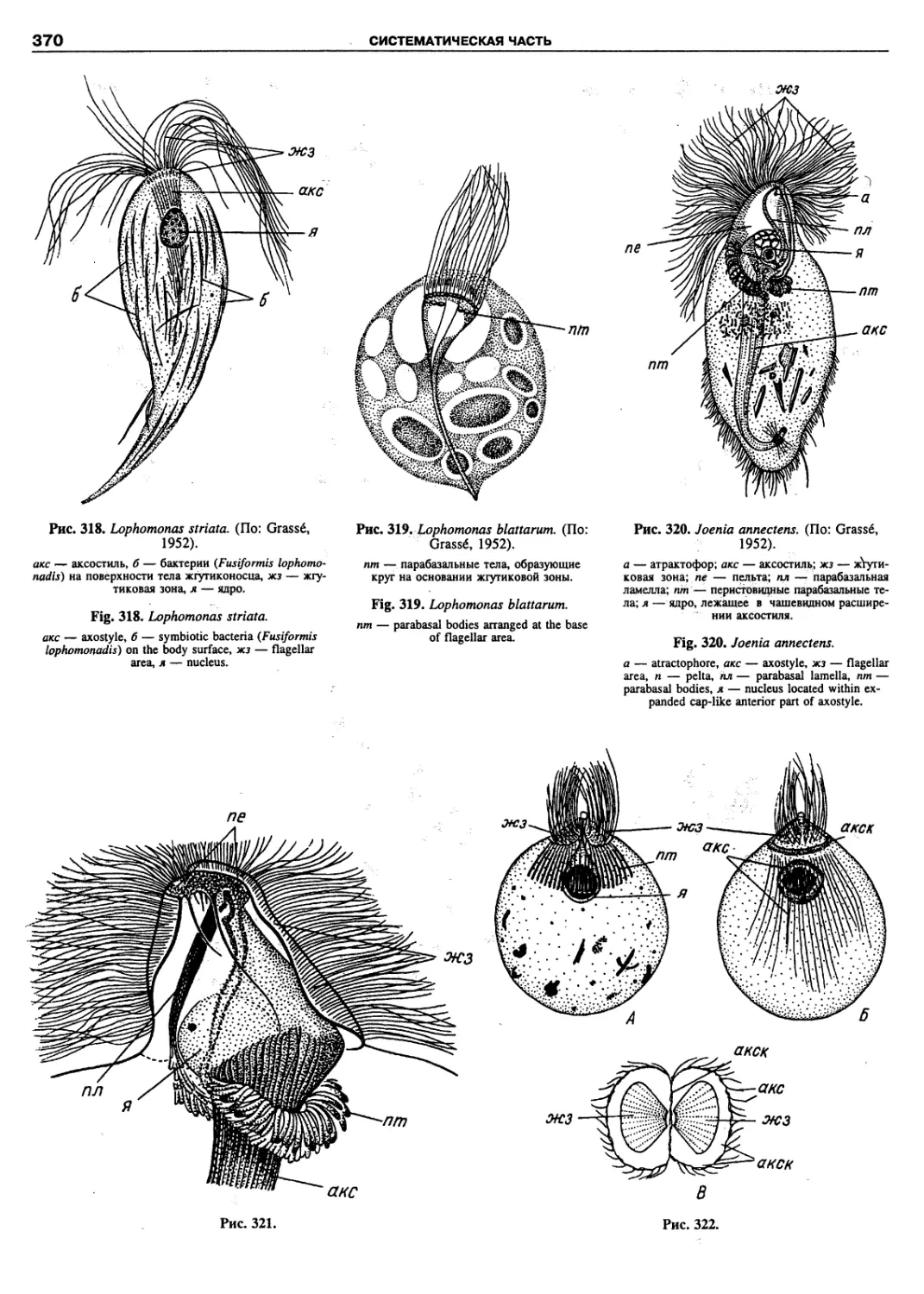
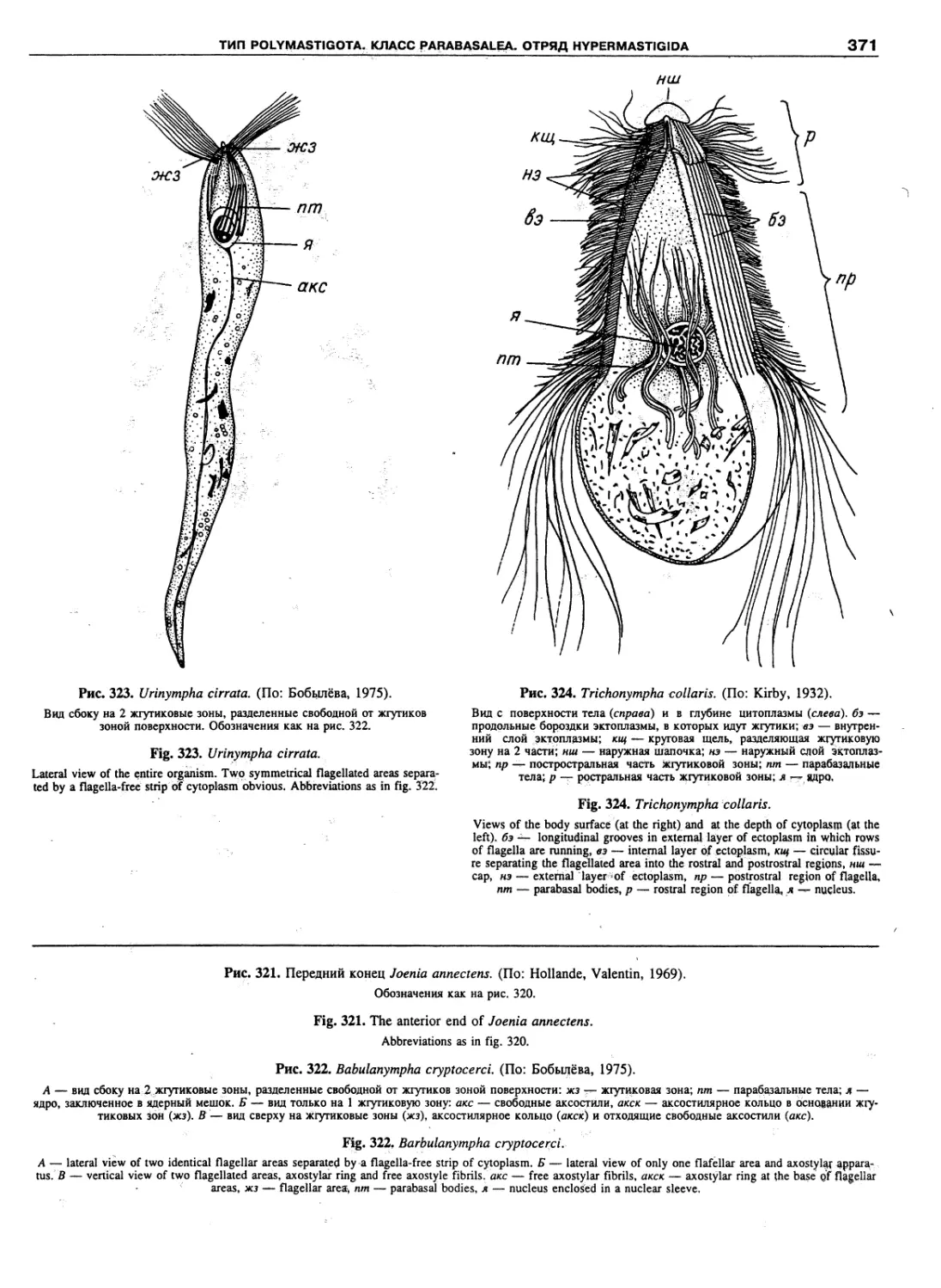

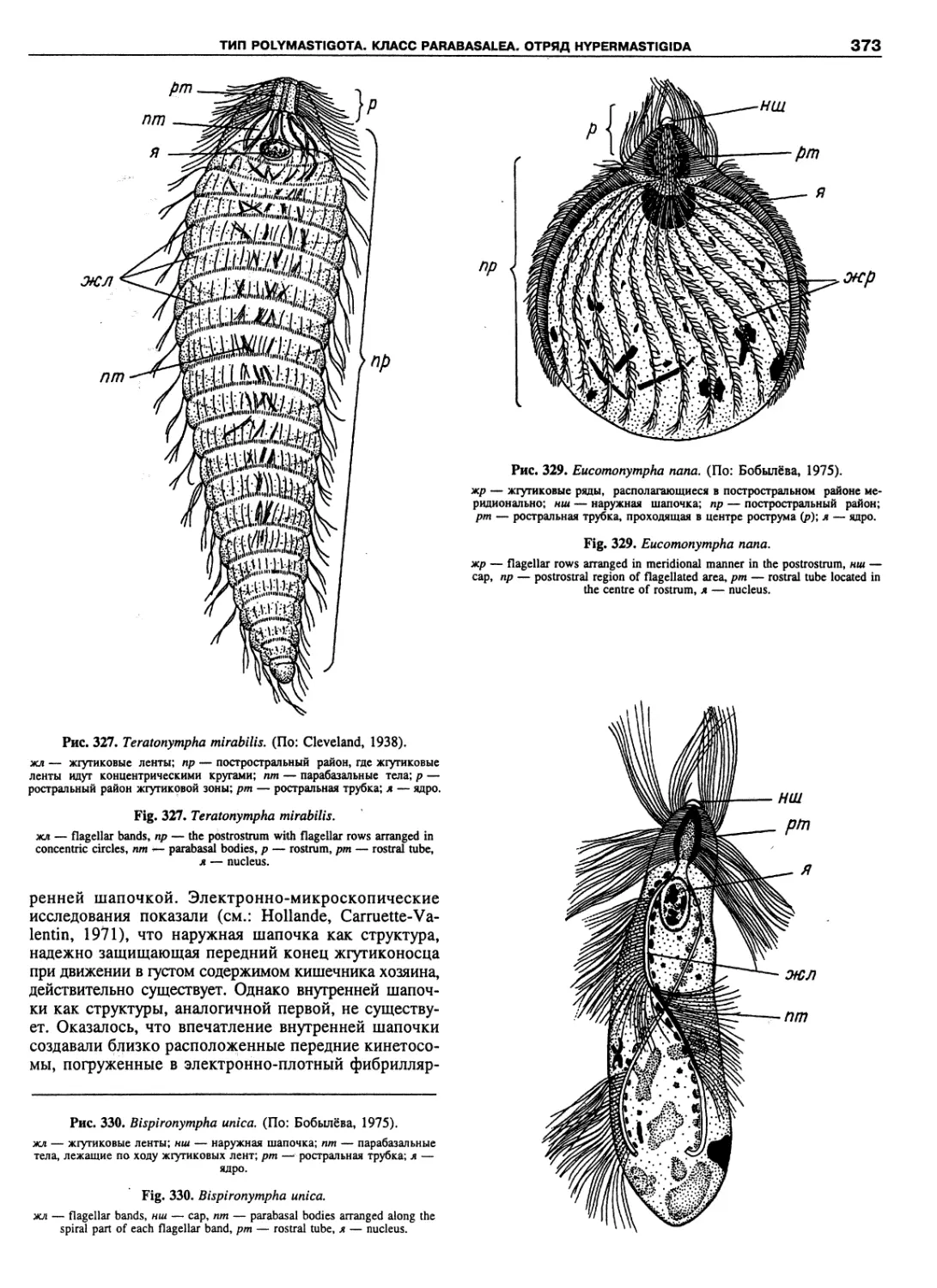

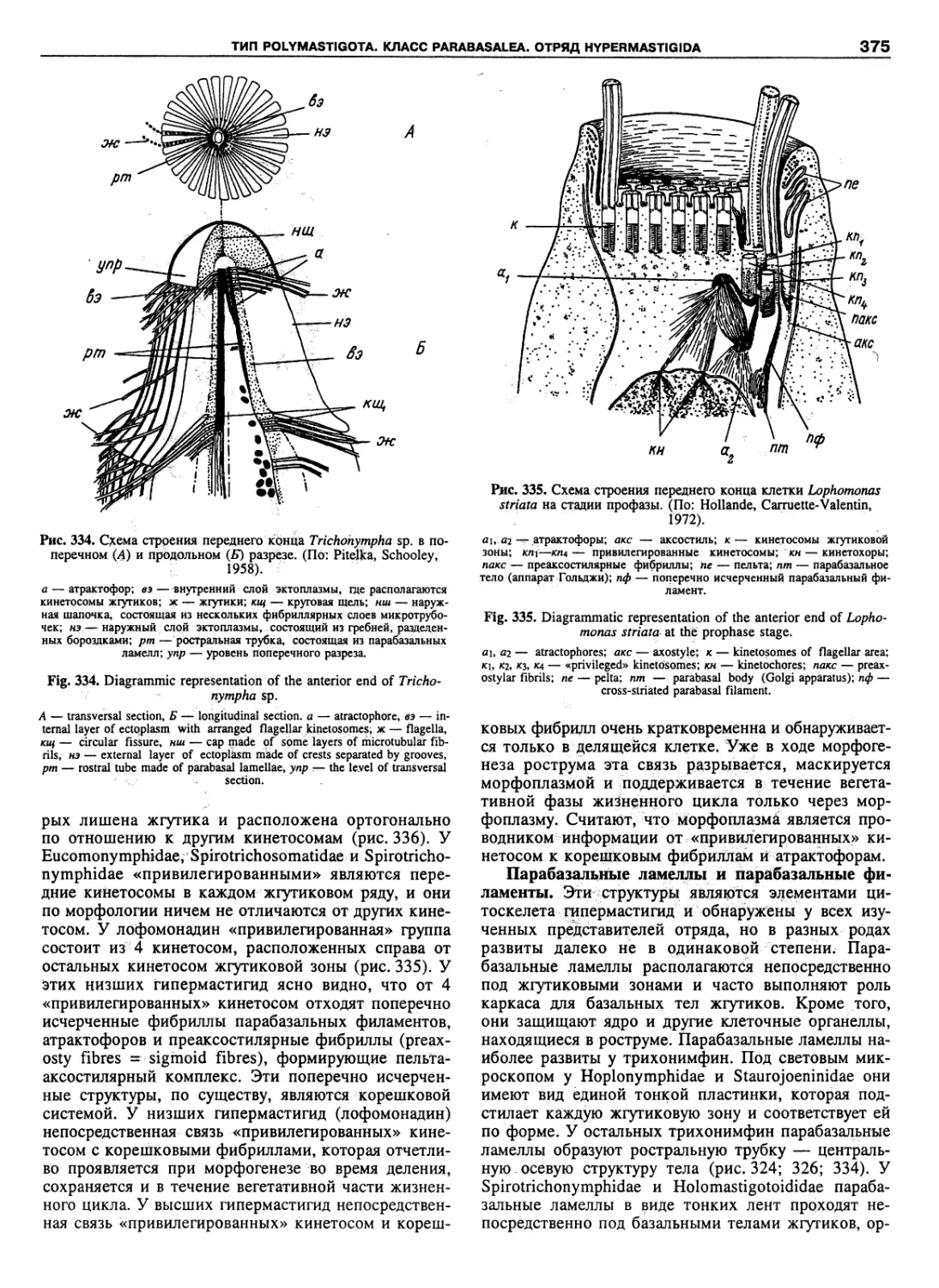
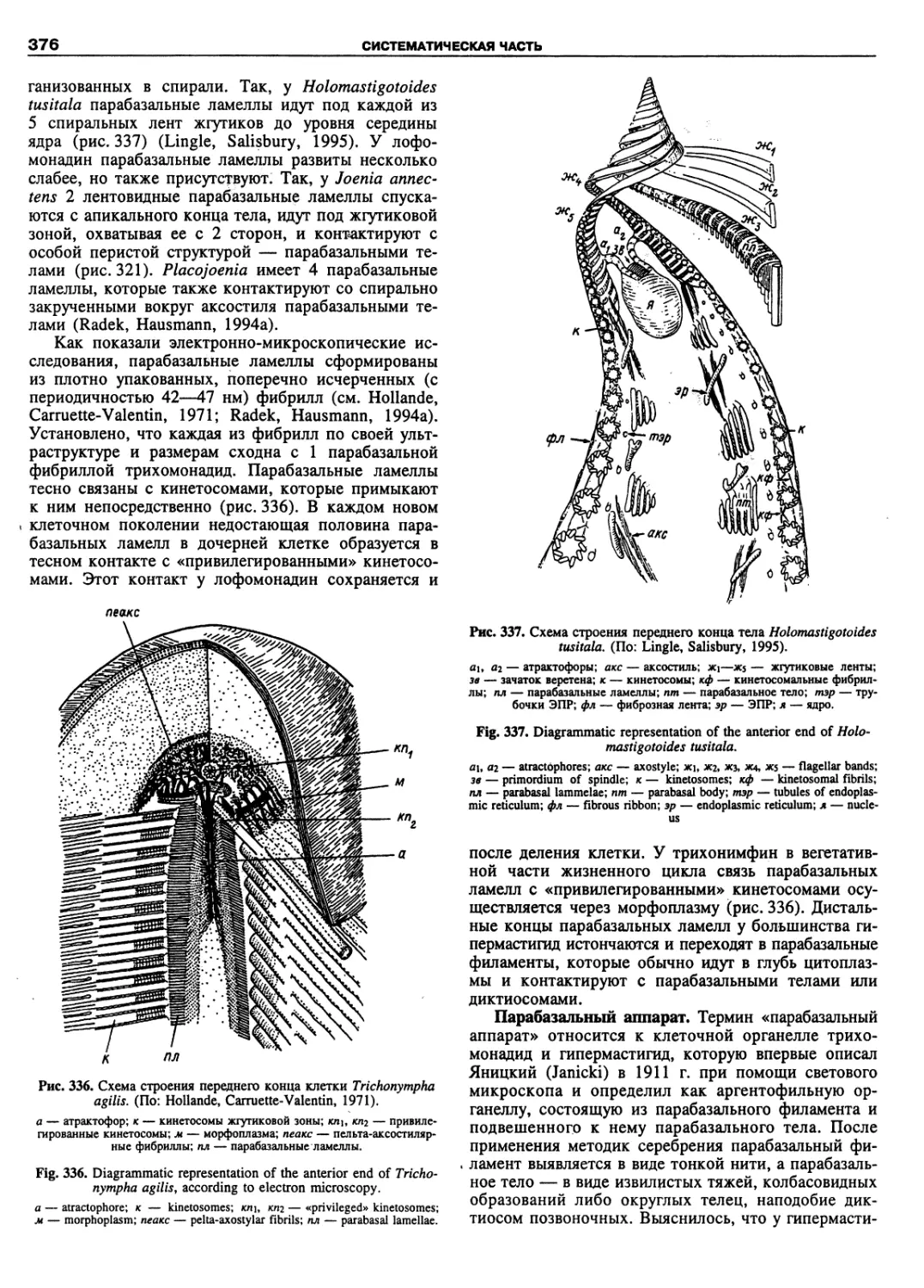





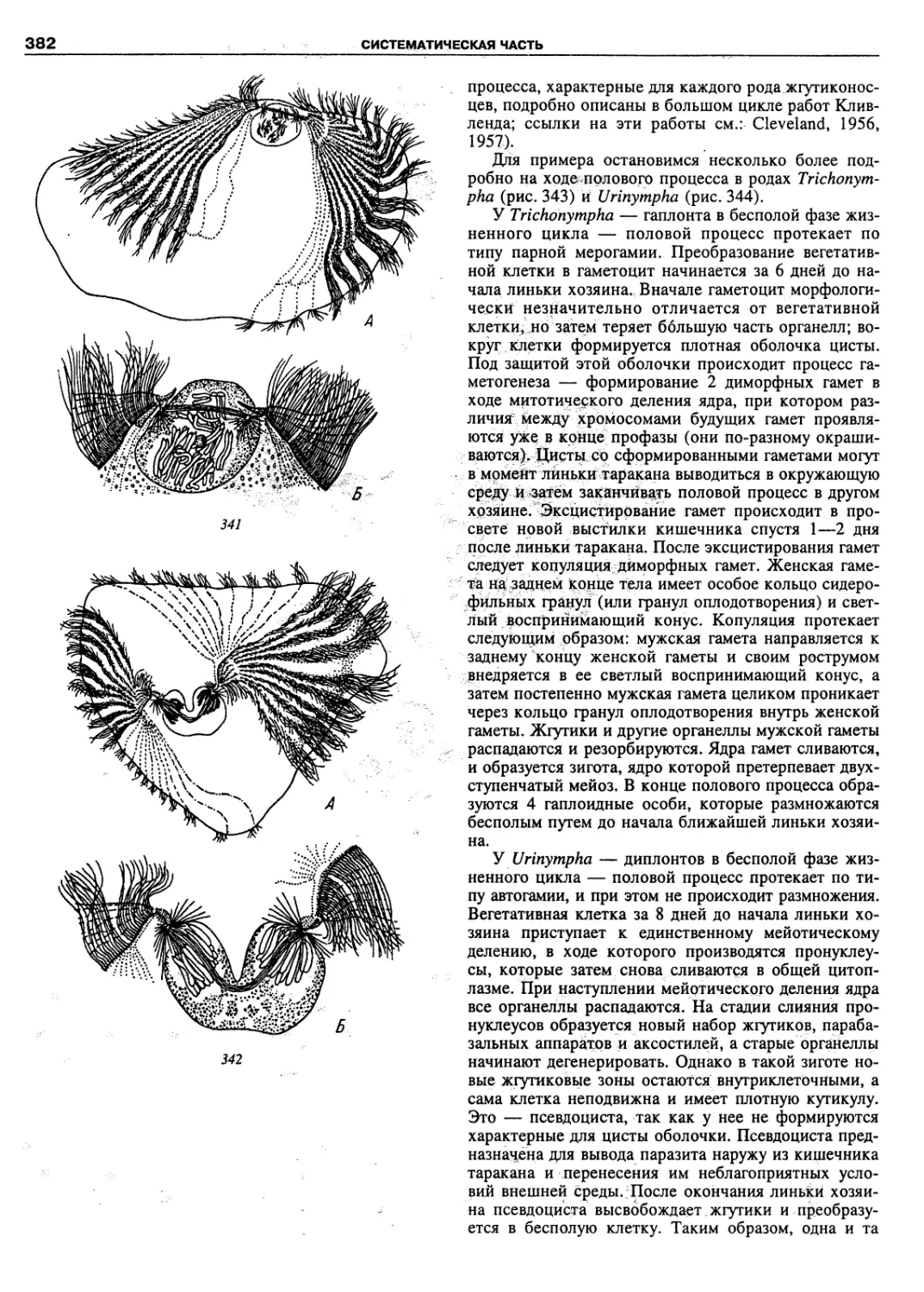
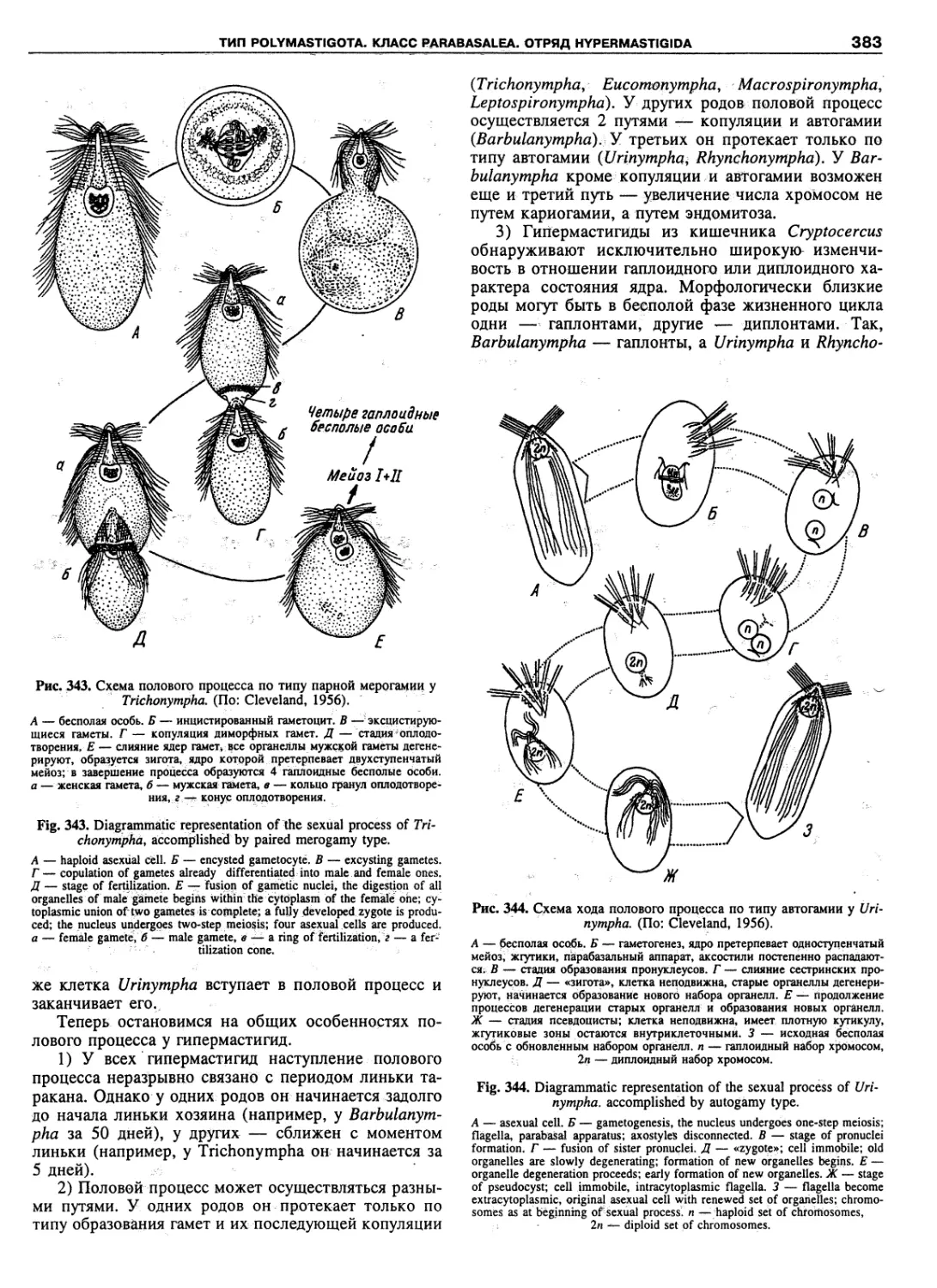

















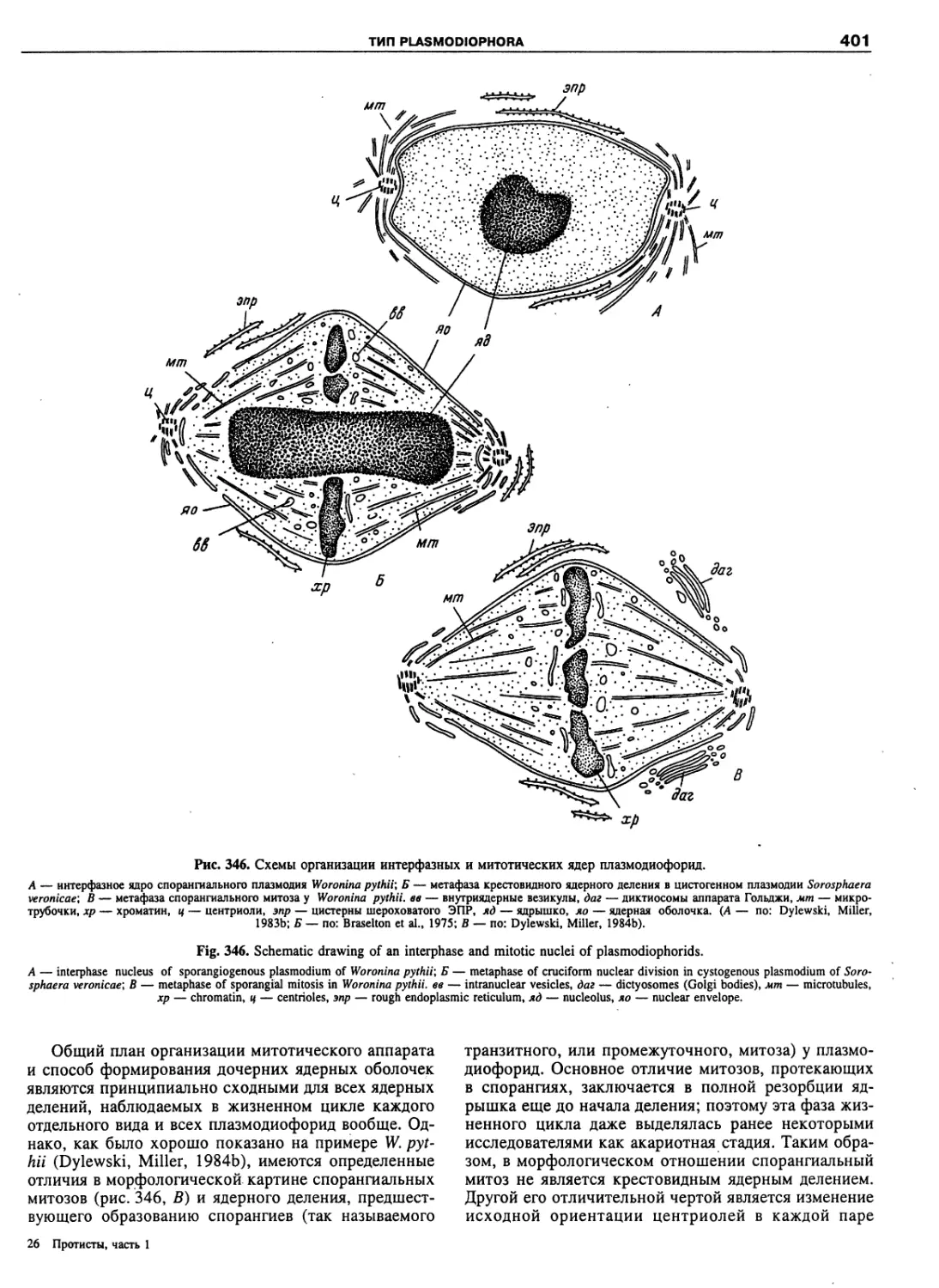






























































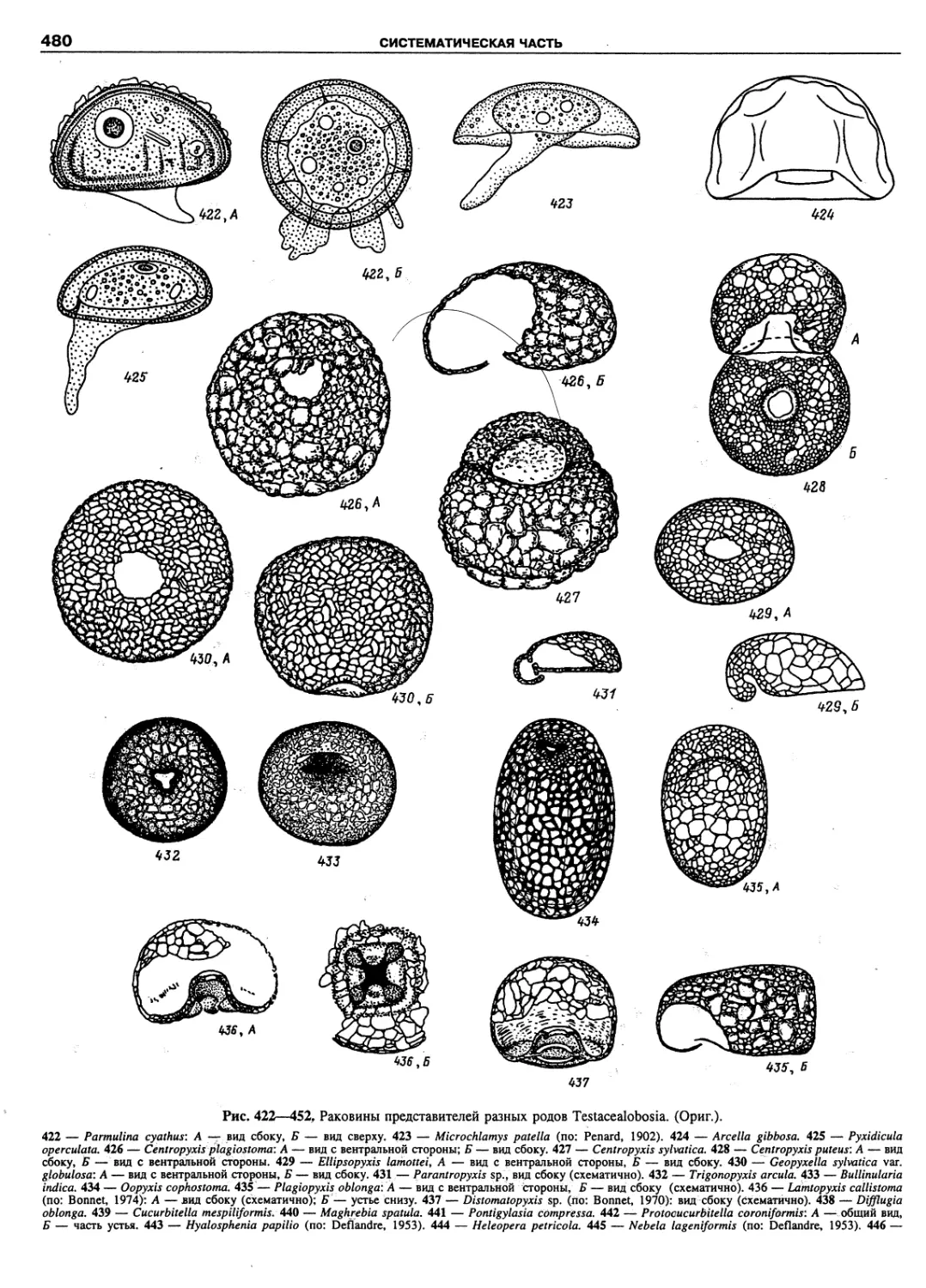
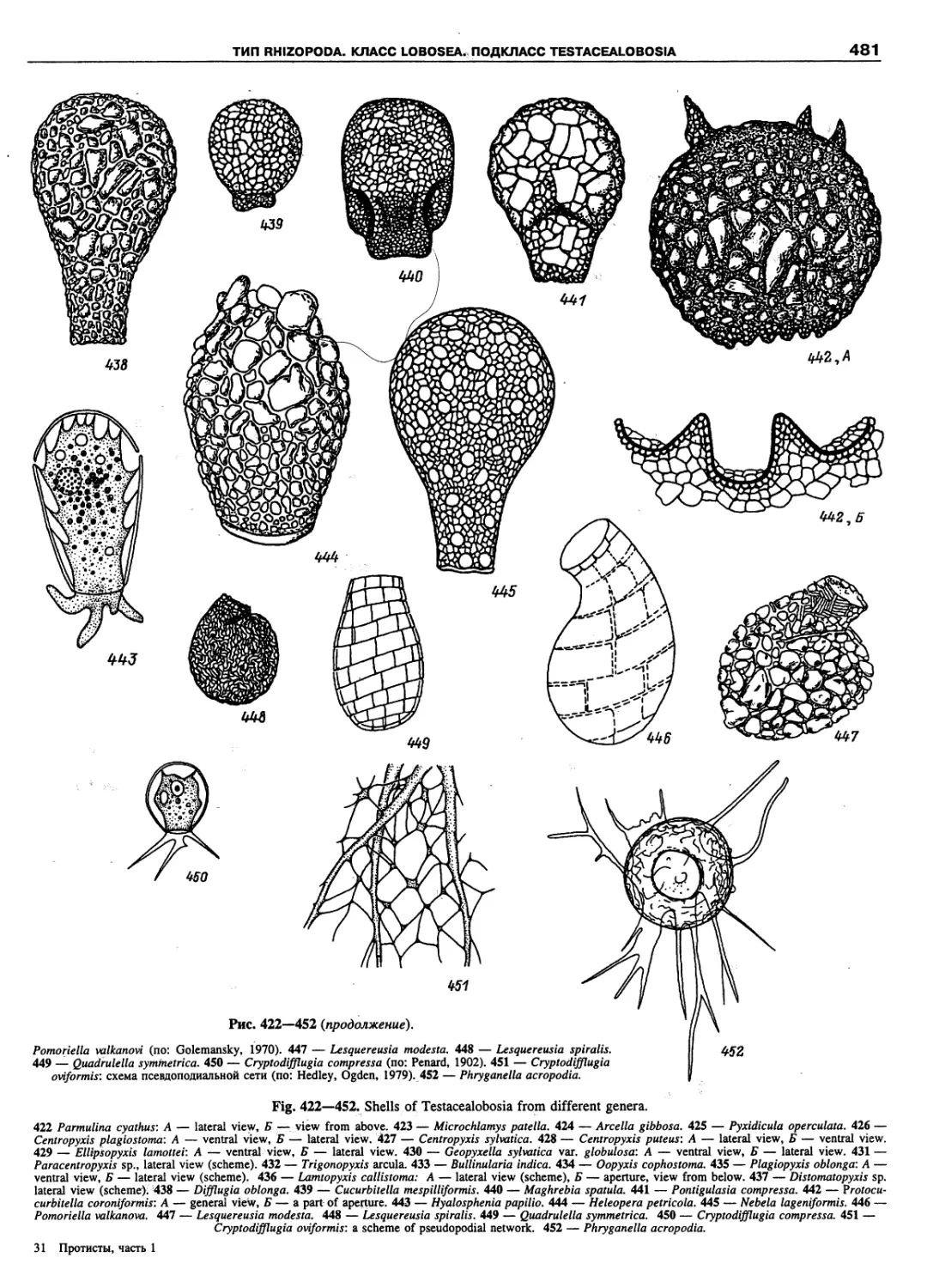





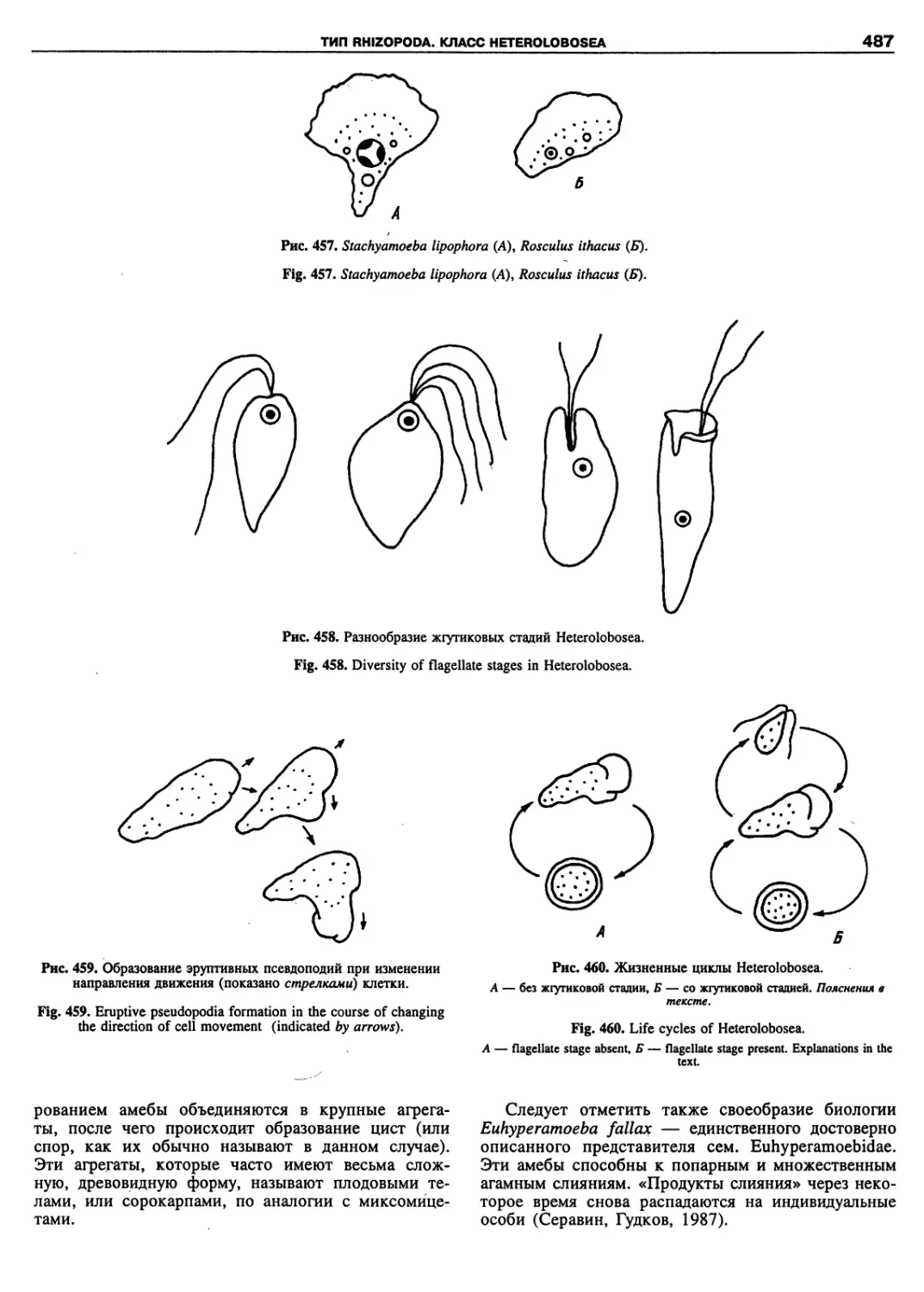








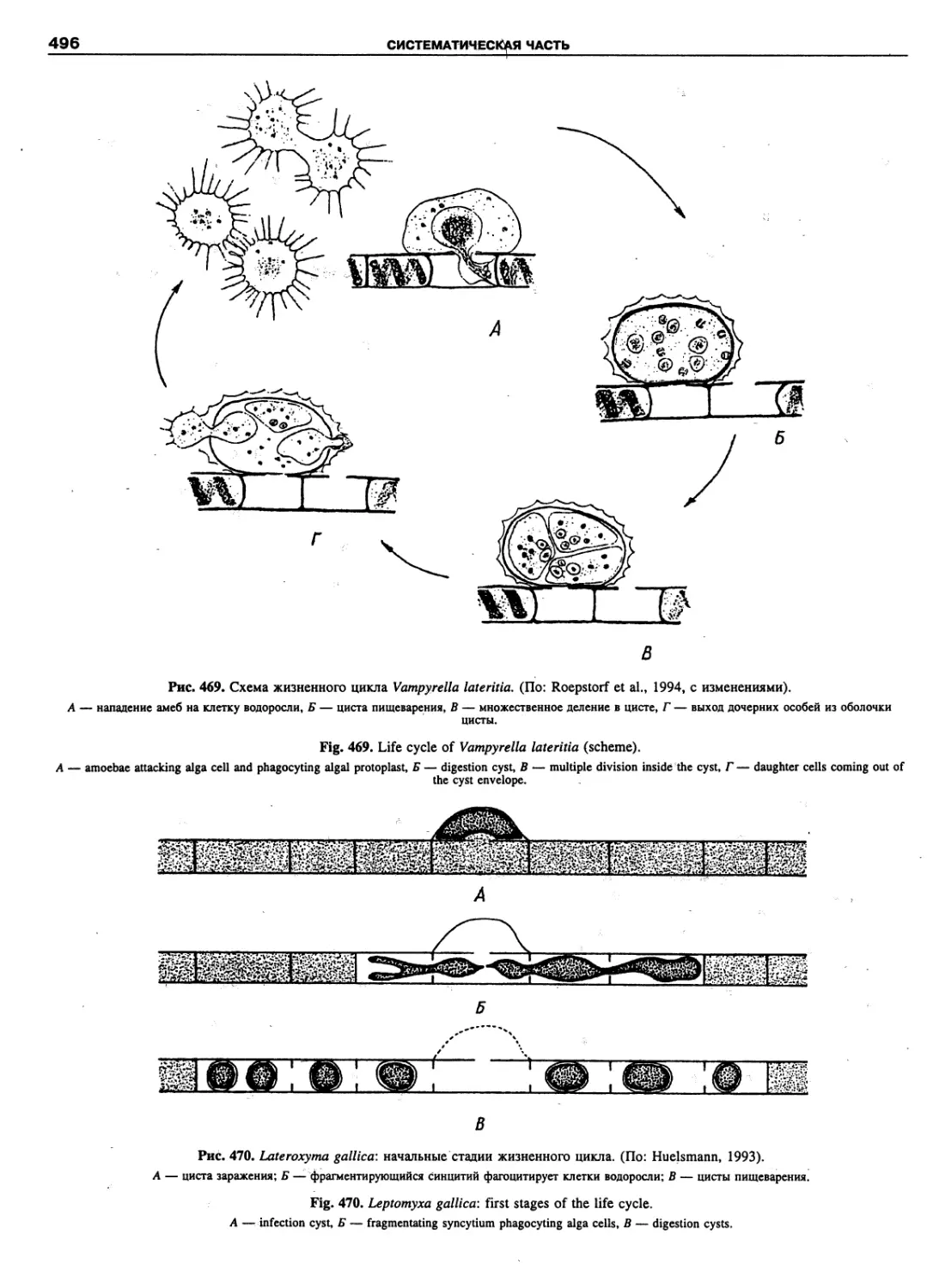




























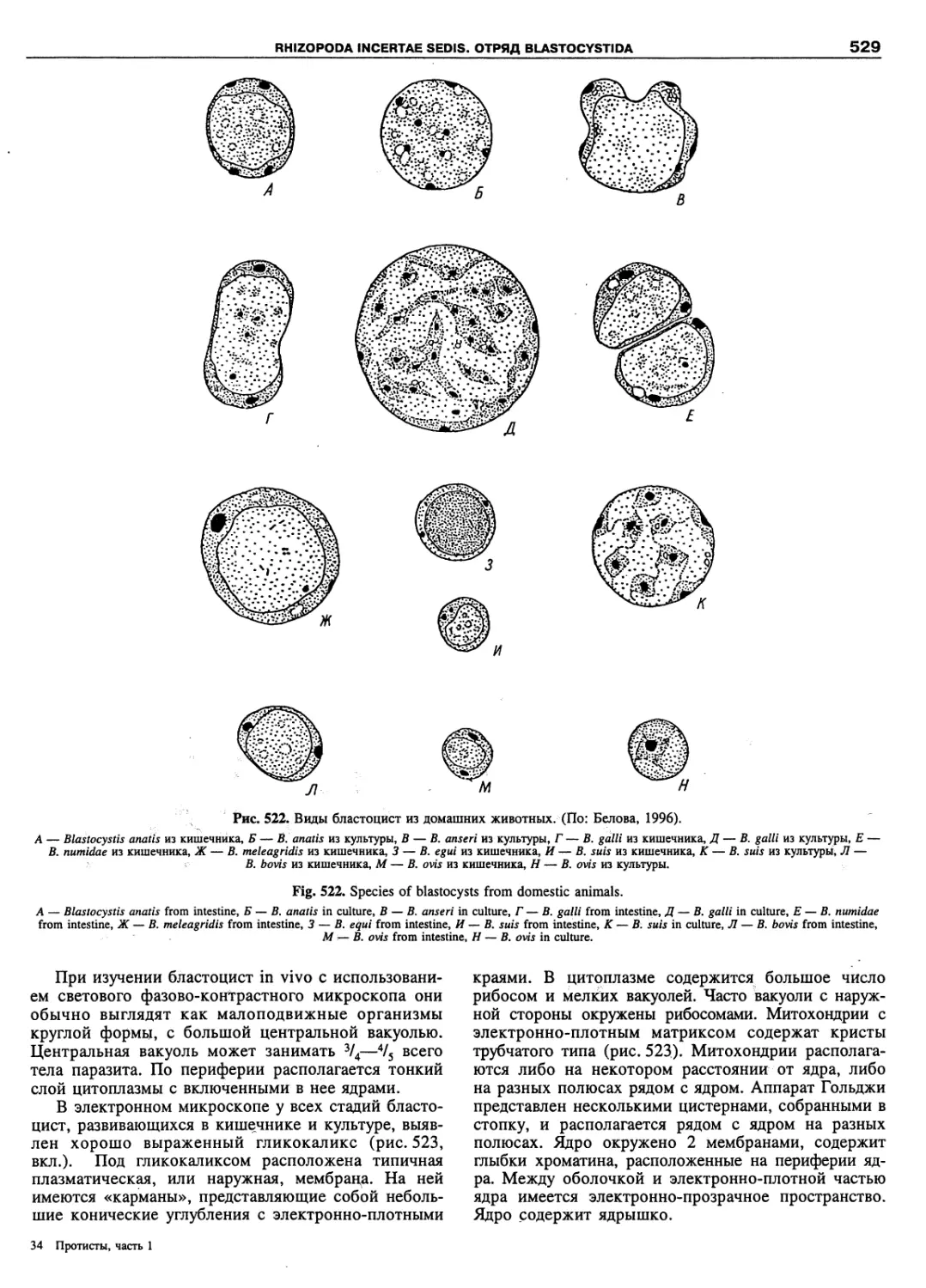
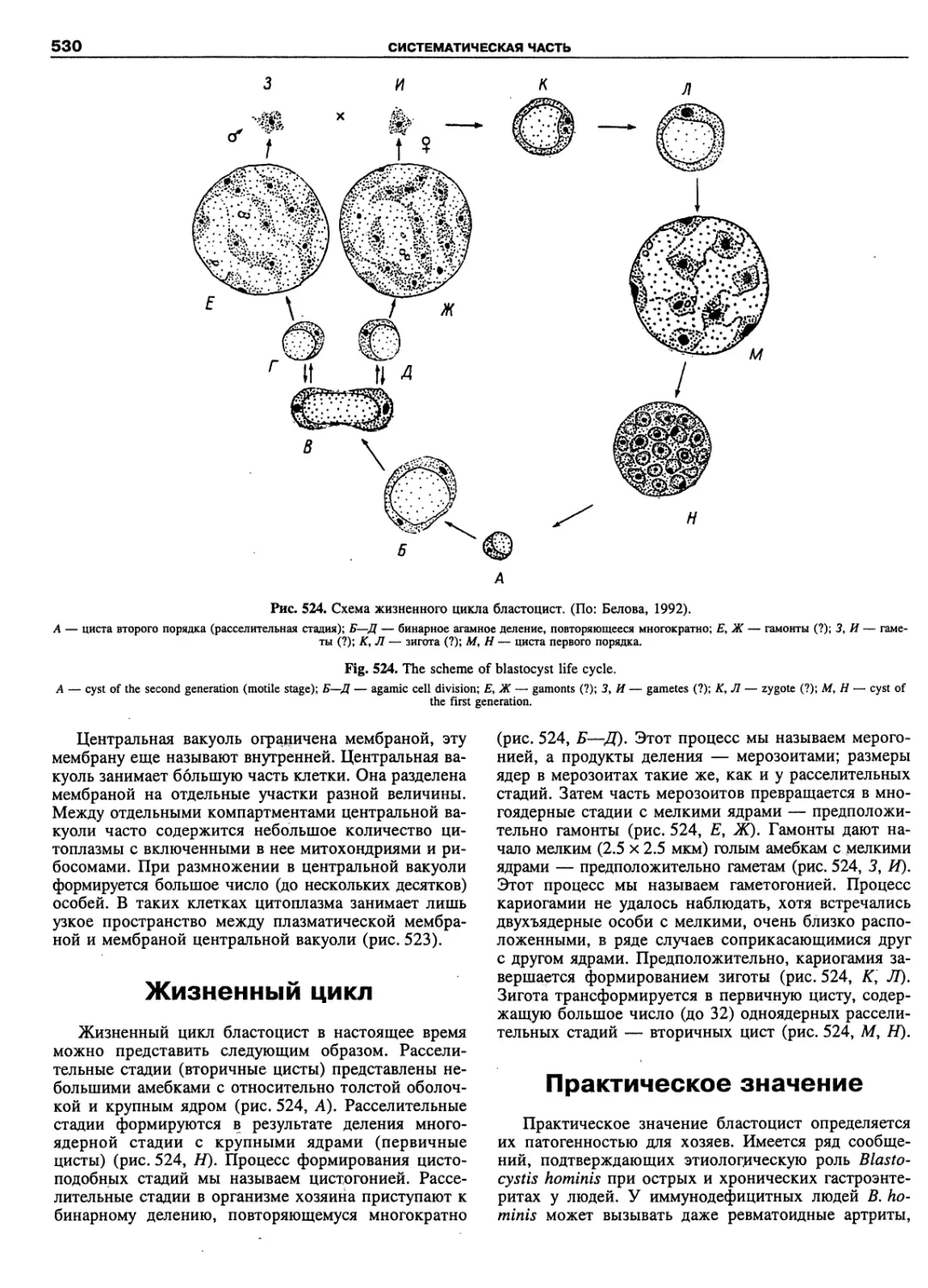










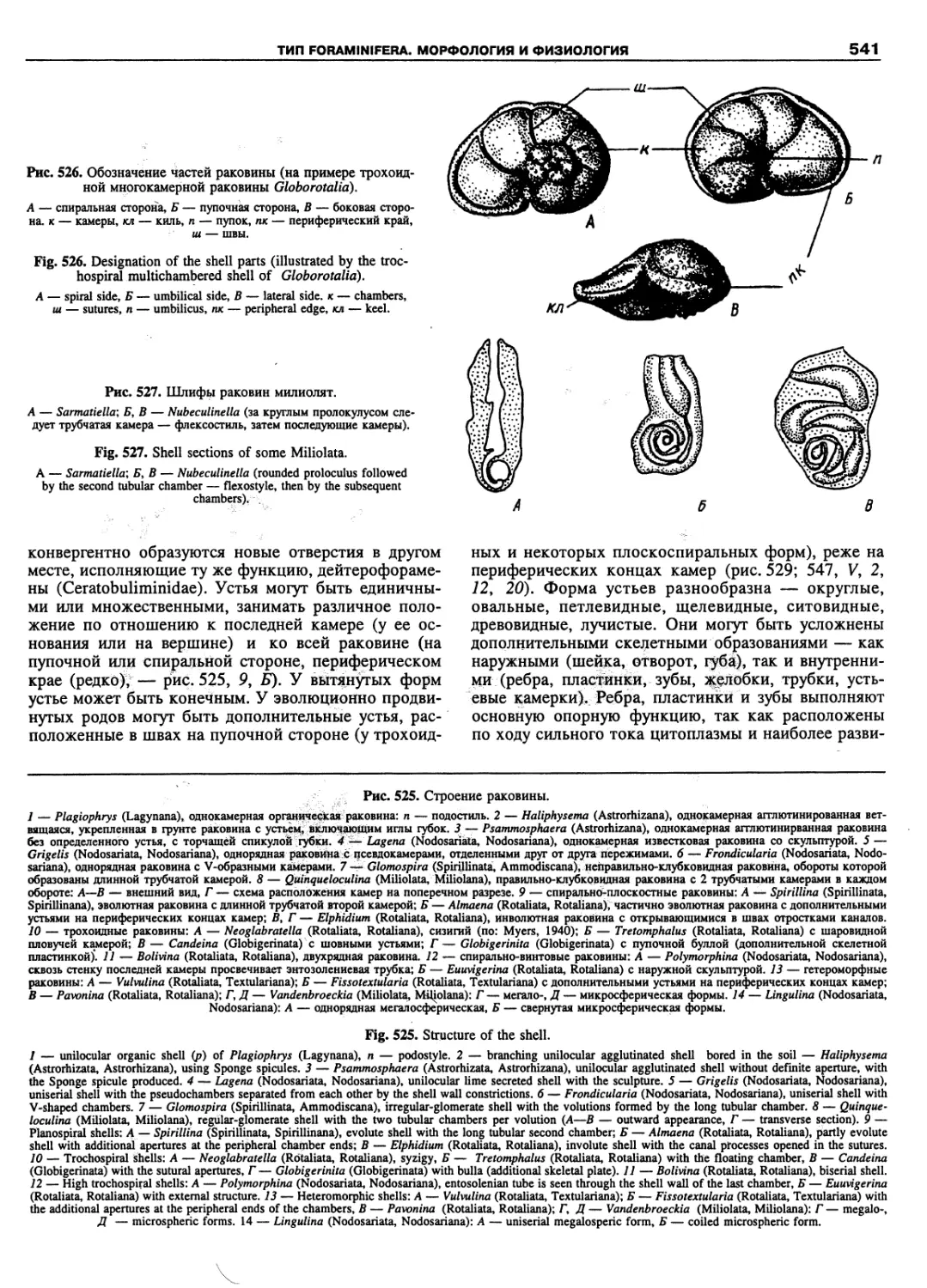
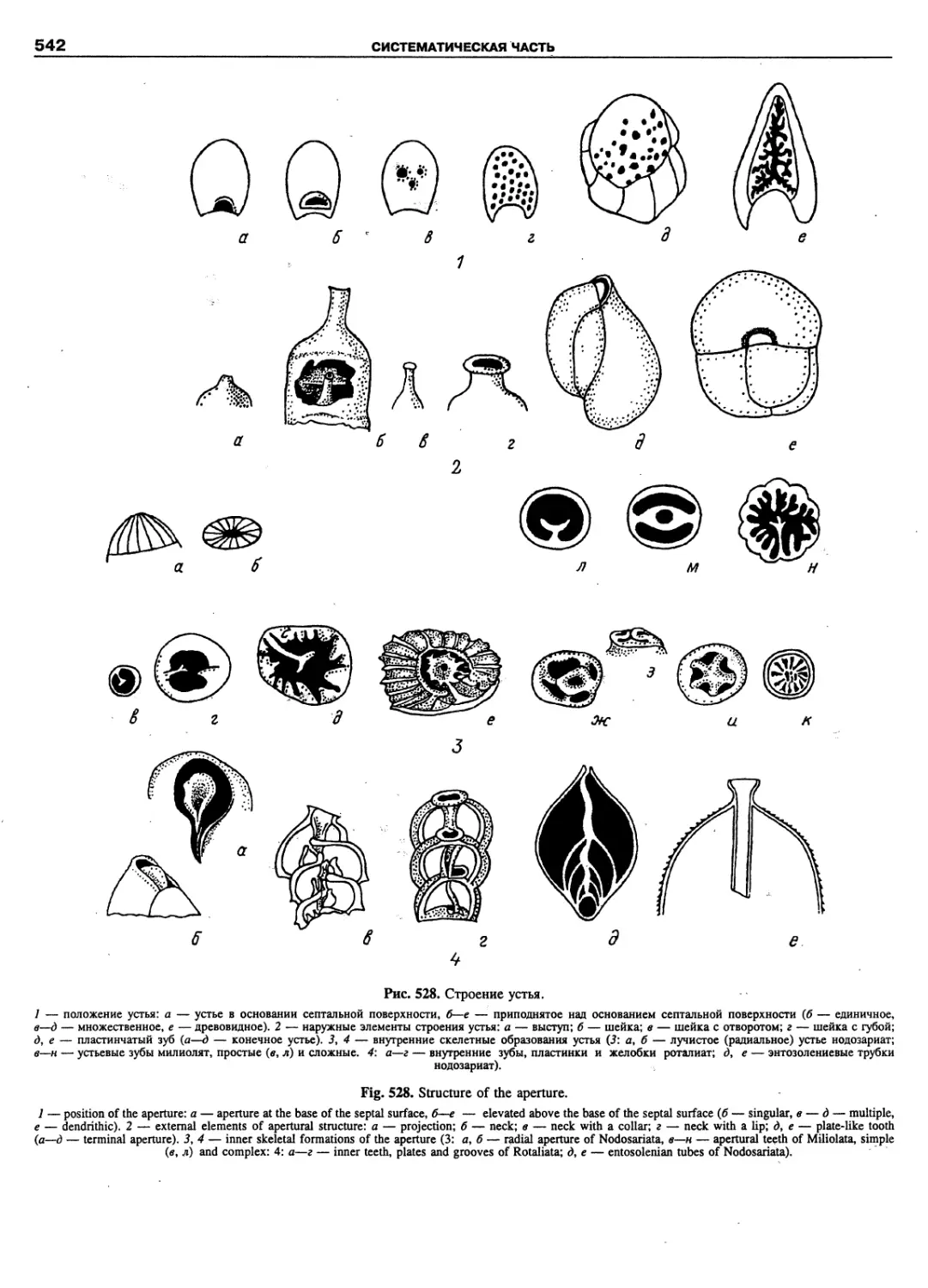















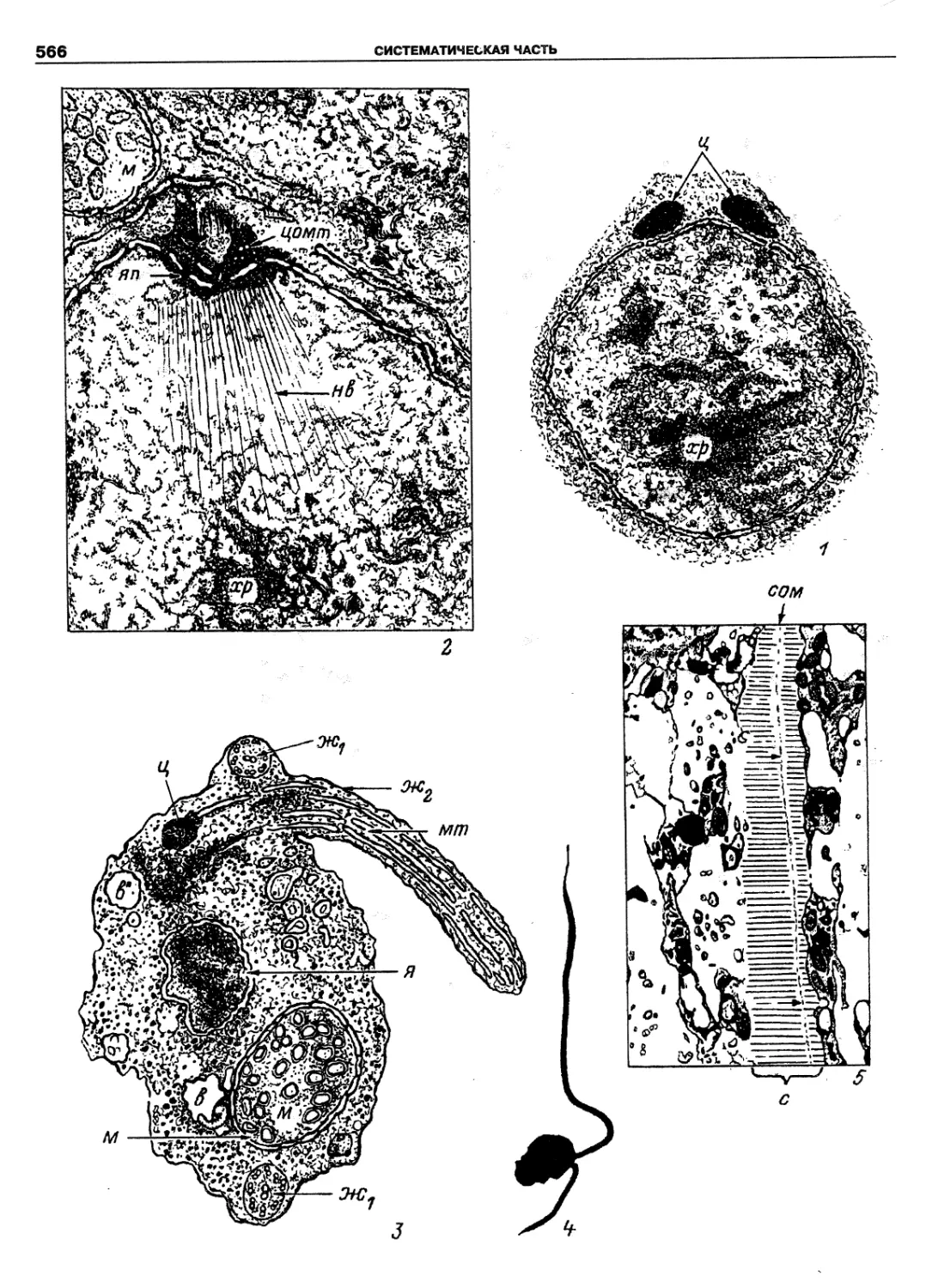

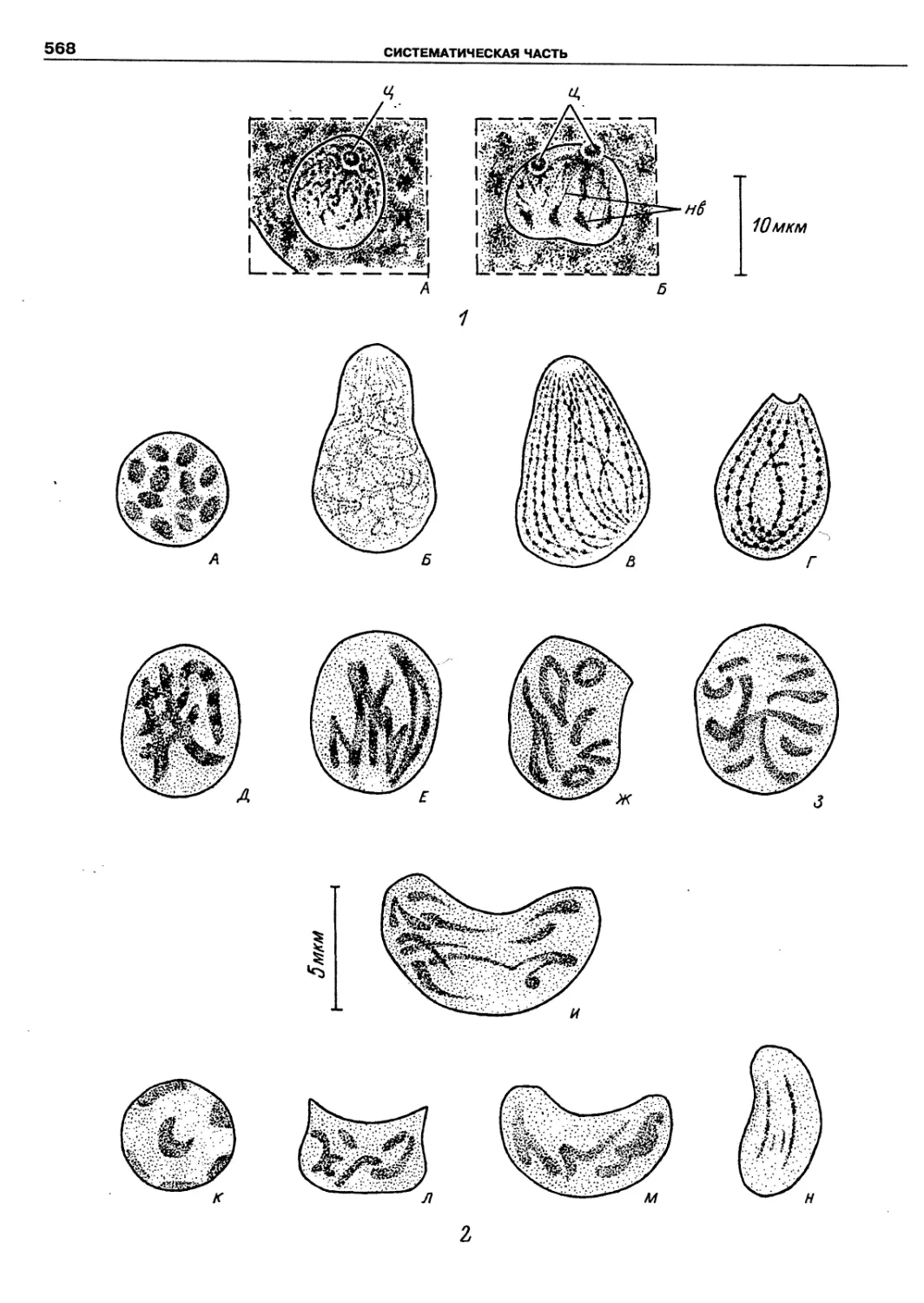

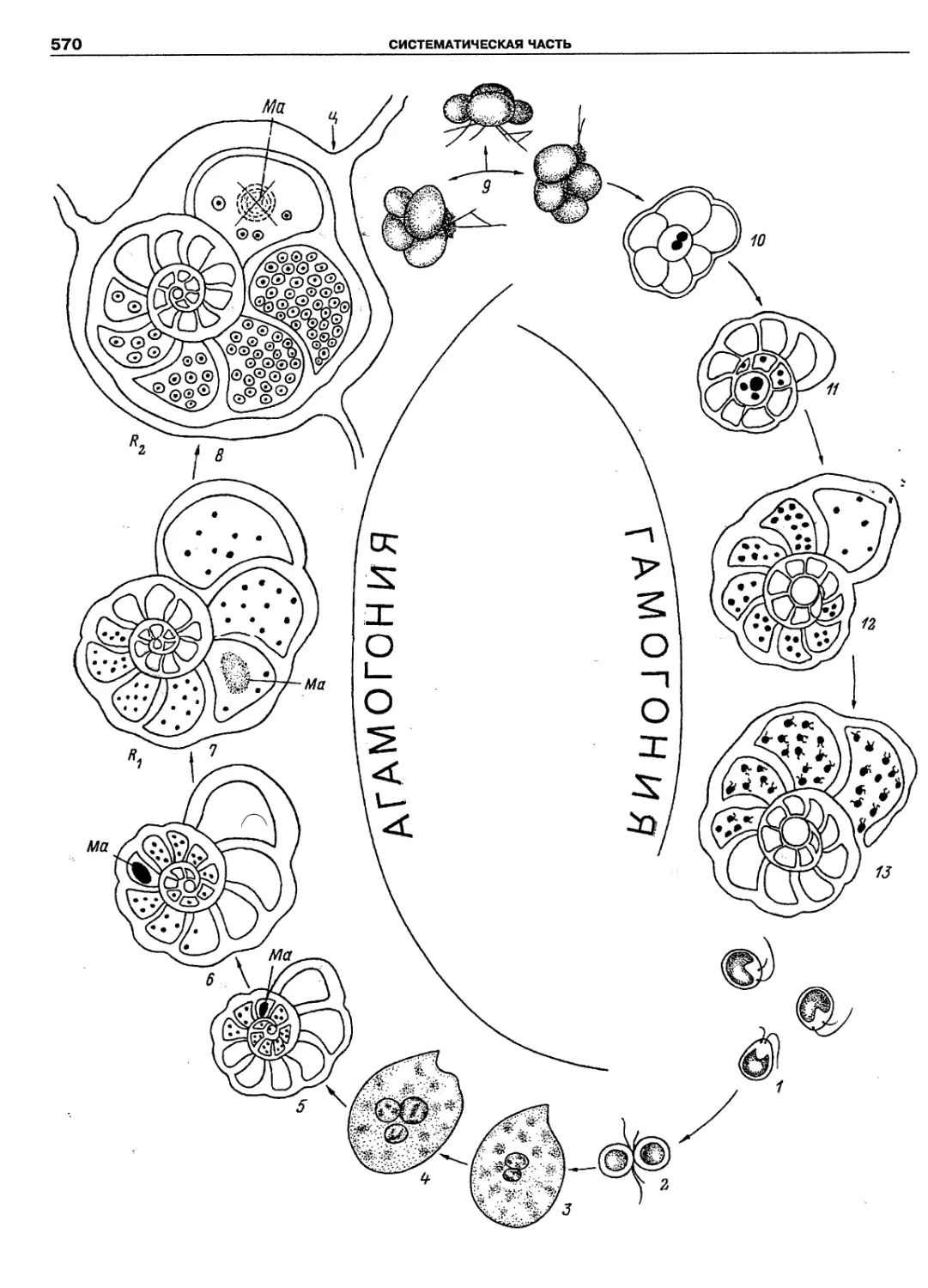


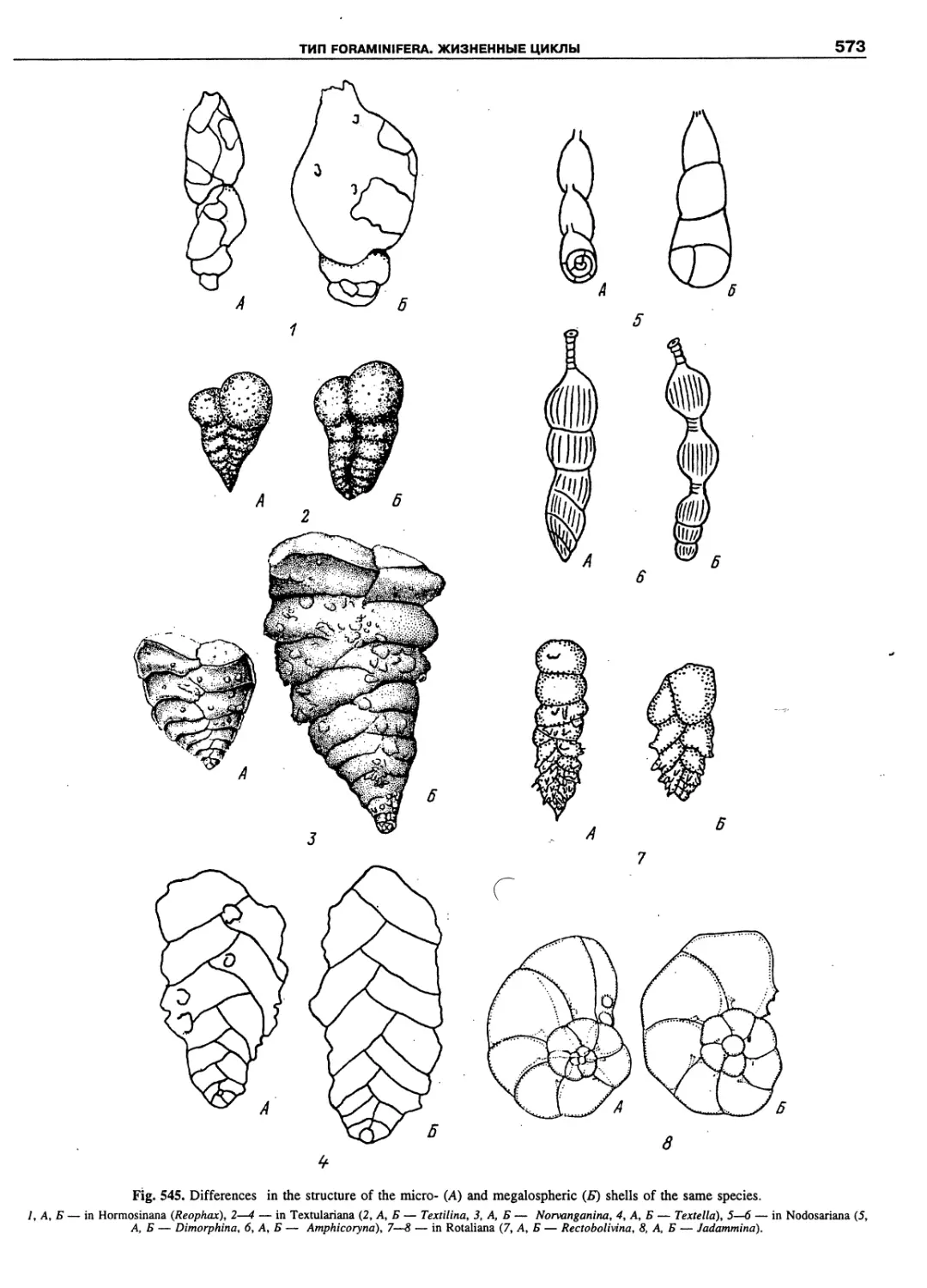









































































































Автор: Пугачев О.Н.
Теги: protozoa систематика животных специальные зоологические науки биология зоология российская академия наук руководство по зоологии протисты анатомия эукариотов
ISBN: 5-02-025864-4
Год: 2000




Thermus thermophilus
Anacystis nidulans
--------- Escherichia soli
Sulfolobus solfataricus
Methanococcus vannielii
Halobacterium halobium
Glugea atherinae
~ Ameson michaelis
Pleistophora sp.
— ----- Enterocytozoon salmonis
Enterocytozoon bienusi
— Encephalitozoon cuniculi
------- Vairimorpha necatrix
Nosema trichoplusiae
Coronympha octonaria
___Trichomonas vaginalis
___ Tritrichomonas foetus
I__ Monocercomonas sp.
___ Hexamita sp.
___ Glardia
Glardia
Glardia
_ Physarum polycephalum
Euglena gracilis
____Trypanosoma brucei
~___Bodo caudatus
Vahlkampfia lobospinosa -
Naegleria gruberi
histolytica -
gingivalis
discoideum
muris
lamblia
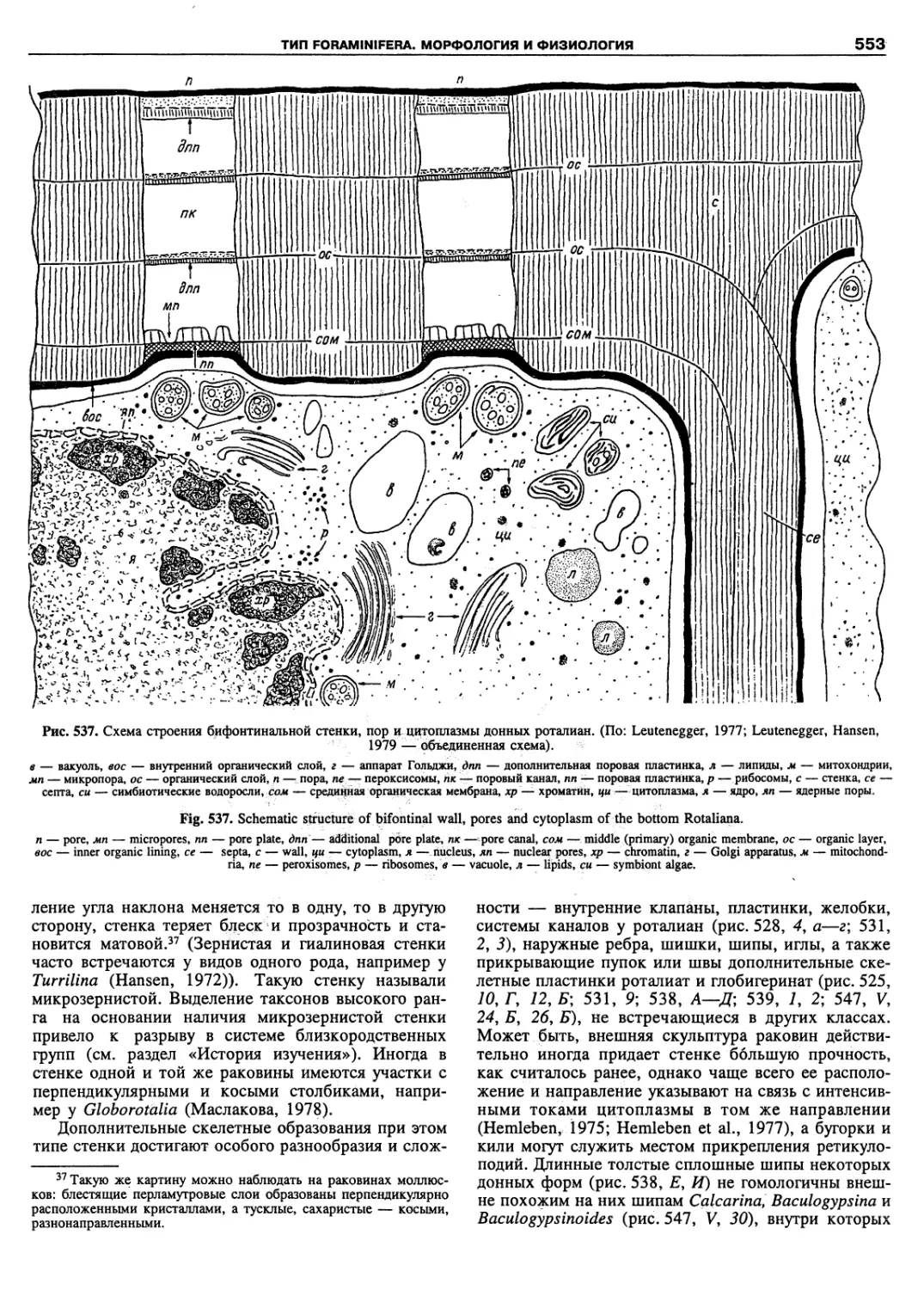
ardeae
___ Entamoeba
'__Entamoeba
. Dictyostelium
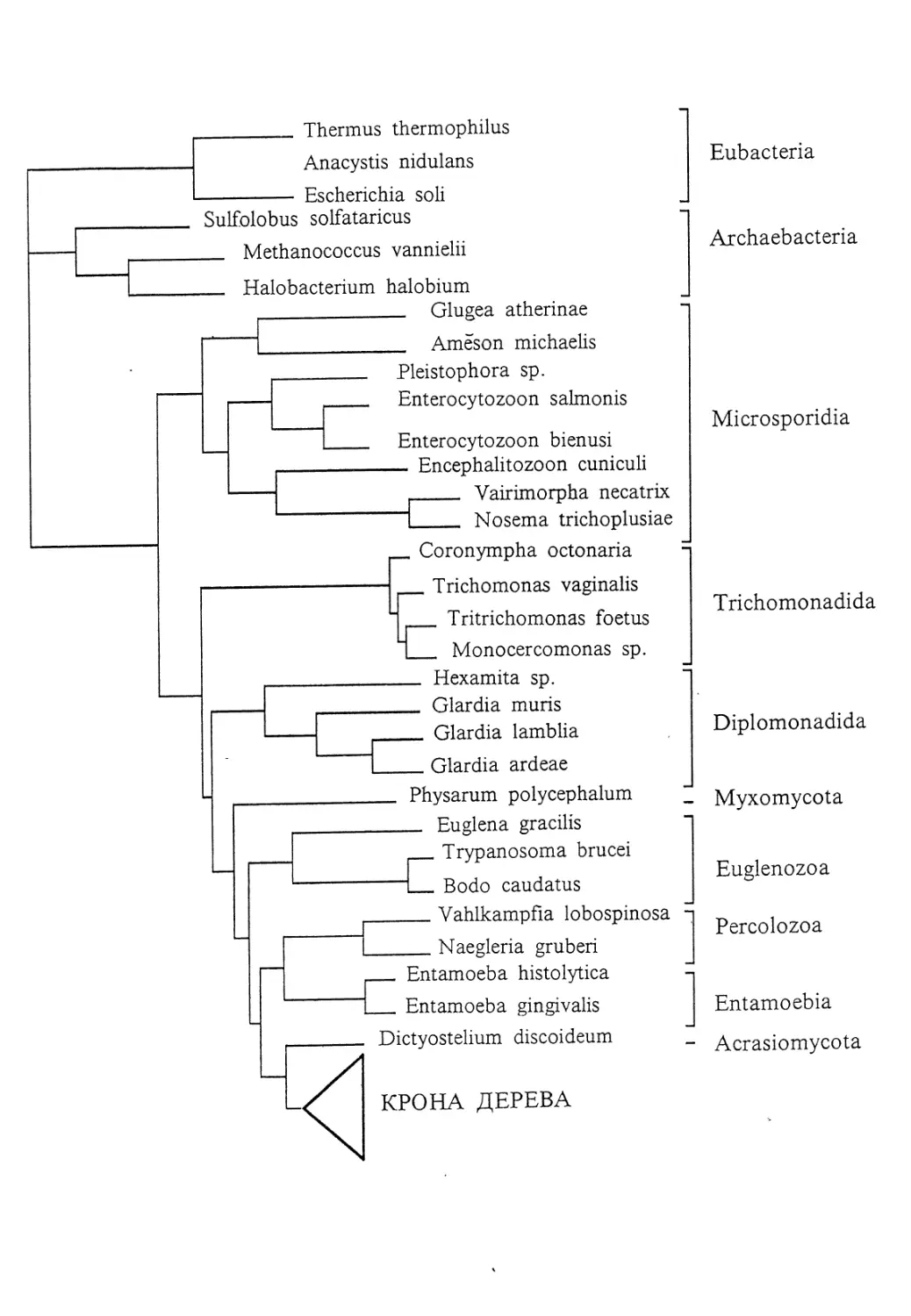
Eubacteria
Archaebacteria
Microsporidia
Trichomonadida
Diplomonadida
Myxomycota
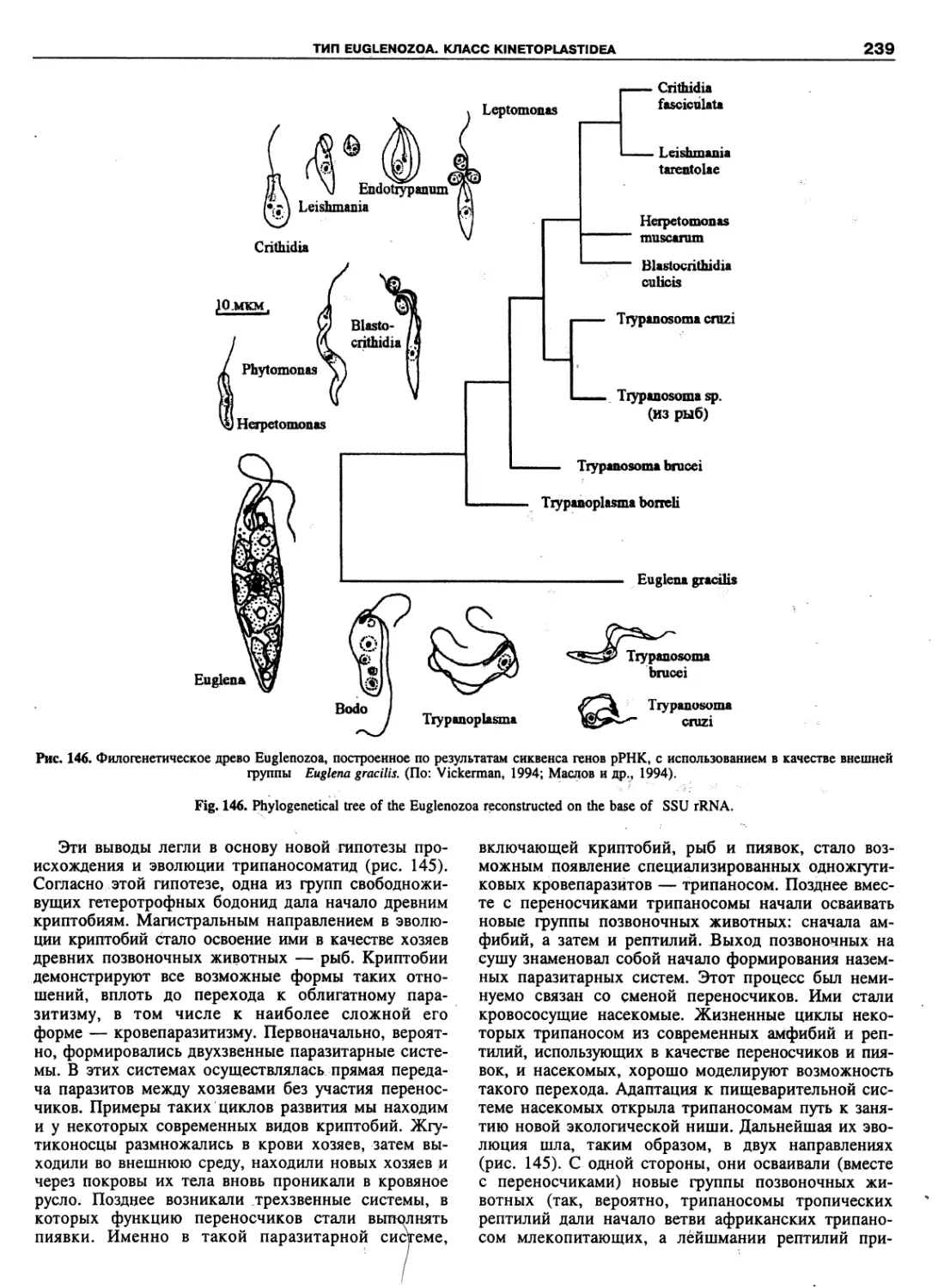
Euglenozoa
Percolozoa
Entamoebia
Acrasiomycota
КРОНА ДЕРЕВА
п
III chlorophyta (62) I
<3 chlorarachniophyta (HM)
эд charophyta (19)
chlorobionts
land plants (45)
==z& haptophyta (6)
--------«=^ chlorarachniophyta (5)
Euglyphina (2)
heterokont algae (32)
— Hyphochytrium (hyphochytriomycetes)
O oomycetes (3)
labyrinthulida (3)
--------Cafeteria (oicosoecida) _
ciliata (26)
—dinophyta (7)
-----Perkinsus
stramenopiles
alveolata
—glaucophyta (2)
-------=33 Acanthamoeba (3)
---32^ cryptophyta (HM)
rhodophyta (49)
choanoflagellida (2)
bilateria (43)
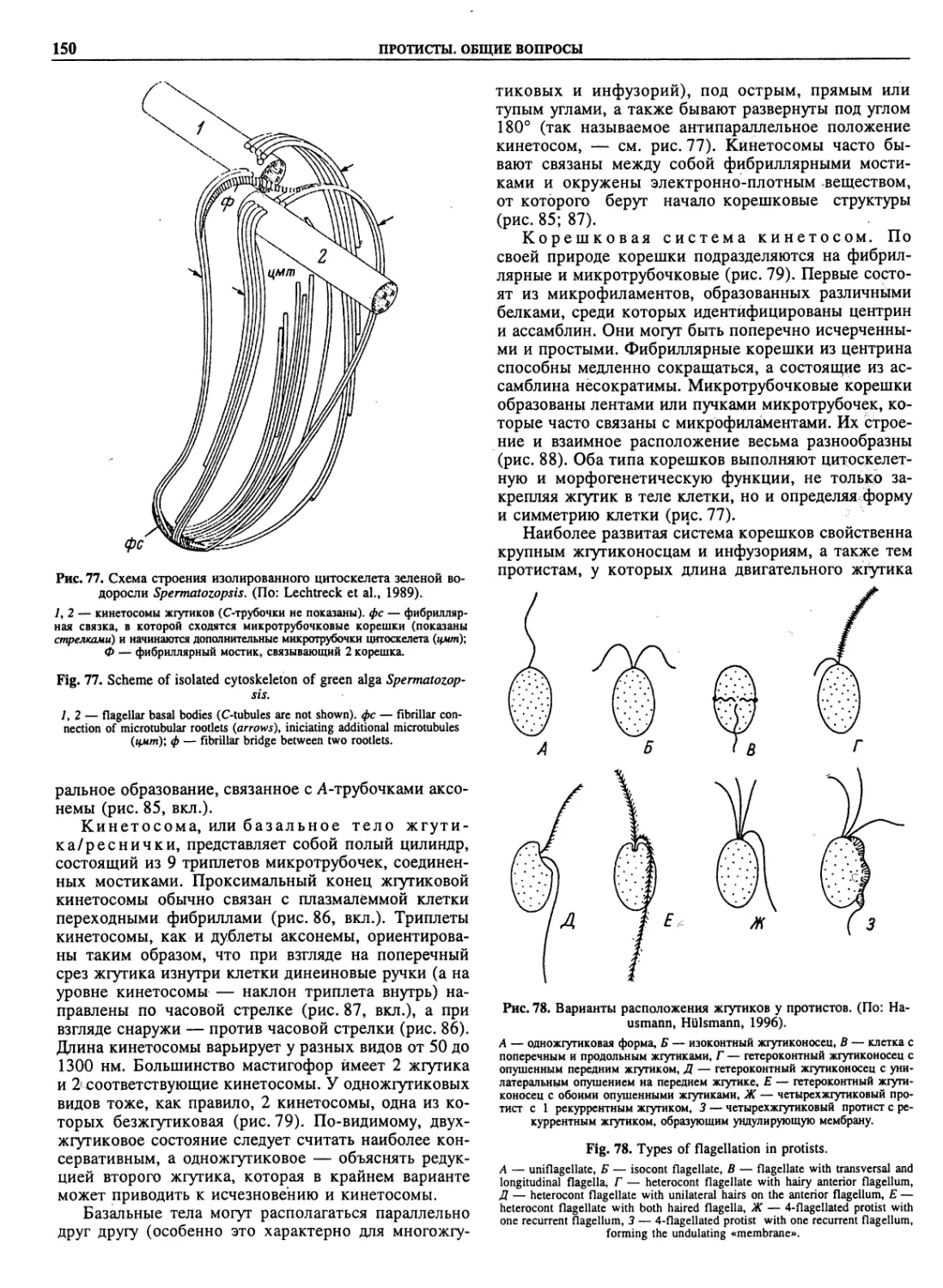
radiata (11)
animalia
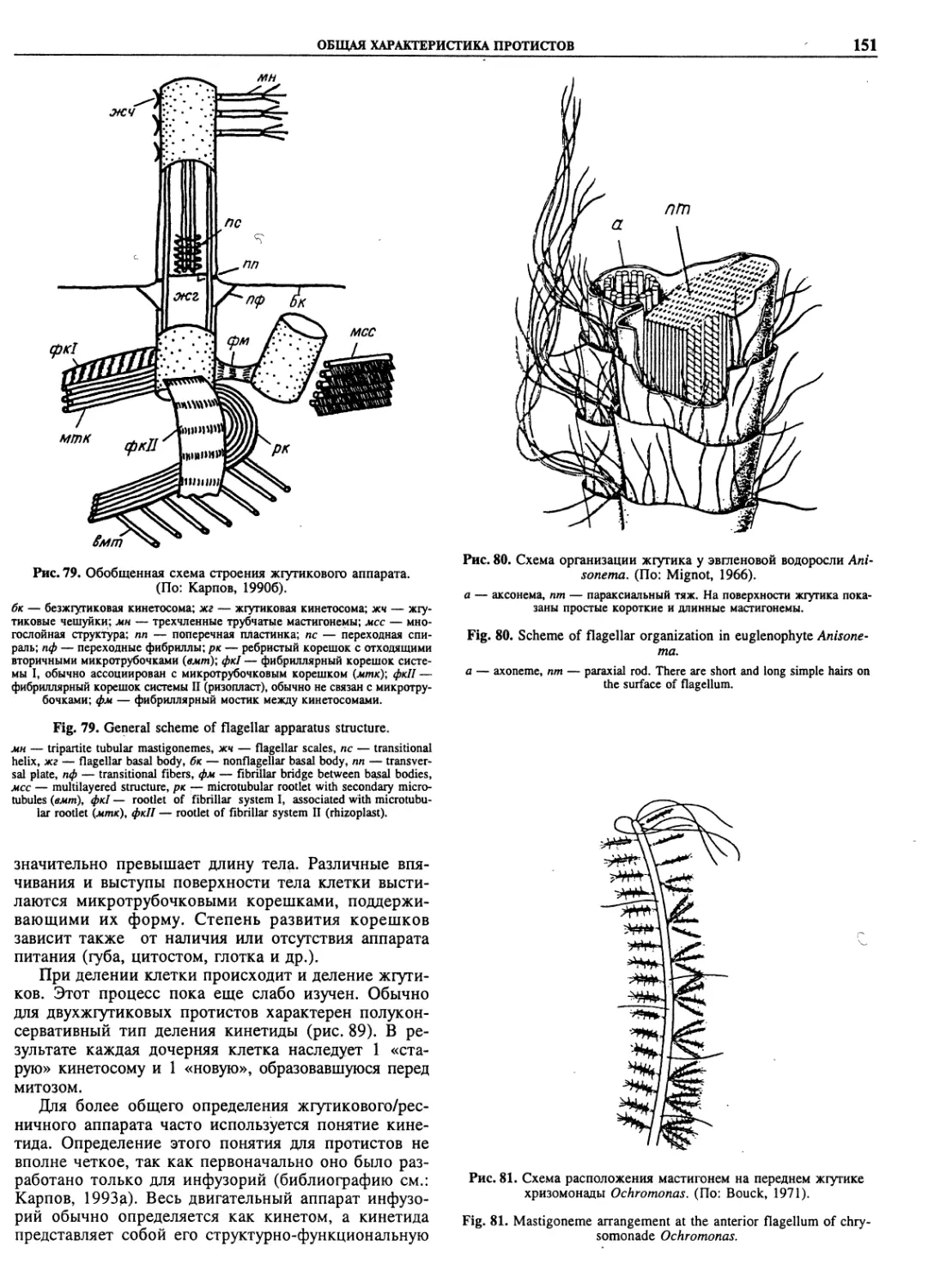
basidiomycetes (38)
ascomycetes (81)
chytridiomycetes (4)
zygomycetes (8)
fungi
СВЕТЛОЙ ПАМЯТИ
Б. Е. Быковского, А. В. Иванова, Д. В. Наумова,
Ю. И. Полянского, О. А. Скарлато,
А. А. Стрелкова,
СТОЯВШИХ У ИСТОКОВ ЭТОГО ИЗДАНИЯ
RUSSIAN ACADEMY OF SCIENCES
Zoological Institute
PROTISTA
PART 1
HANDBOOK ON ZOOLOGY
ST. PETERSBURG
«NAUKA»
2000
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Зоологический институт
ПРОТИСТЫ
ЧАСТЬ 1
РУКОВОДСТВО по зоологии
е
САНКТ-ПЕТЕРБУРГ
«НАУКА»
2000 ’
УДК 593.1
ББК 28.691
П82
Протисты: Руководство по зоологии. — СПб.:
Наука, 2000. — Ч. 1. — 679 с.
ISBN 5-02-025864-4
Монография представляет собой первую часть руковод-
ства по зоологии, посвященного протестам. Она содержит
вводный раздел, в котором рассмотрены вопросы развития
представлений о систематике и эволюции животного мира. В
основных главах книги содержатся общая характеристика и
система протестов, обсуждаются пути их эволюции. Наи-
большее внимание уделено истории изучения, таксономии,
морфологии и другим биологическим аспектам традиционно
протозоологических групп, входящих в состав типов: Crypto-
phyta, Euglenozoa, Chrysophyta, Haptophyta, Raphidophyta,
Saprolegnia, Opalinata, Choanomonada, Polymastigota, Plas-
modiophora, Mycetozoa, Rhizopoda и Foraminifera. Библиогр.
2654 назв. Ил. 551. Схем 10. Табл. 9.
Для зоологов всех специальностей, преподавателей
вузов и студентов.
Protista: Handbook on zoology. — St. Petersburg:
Nauka, 2000. — Pt 1. — 679 p.
ISBN 5-02-025864-4
The monograph written by the leading Russian protistolo-
gists is demoted to protists, with emphasis on the protozoologi-
cal groups. The first part comprises the history of zoology, the
pathways of evolution, taxonomy and classification of the ani-
mal world. The general part is devoted to the characterization
of protists, their classification and trends of evolution. The
main part of the volume includes the systematic part, compri-
sing the history, taxonomy, morphology and other biological
aspects of the phyla: Cryptophyta, Euglenozoa, Chrysophyta,
Haptophyta, Raphidophyta, Saprolegnia, Opalinata, Choano-
monada, Polymastigota, Plasmodiophora, Mycetozoa, Rhizo-
poda, and Foraminifera.
Главный редактор
директор Зоологического института РАН
академик РАН, проф. А. Ф. АЛИМОВ
Editor-in-Chief
Director of Zoological Institute (Russian Academy of Science)
Prof. A. F. ALIMOV, Member of Russian Academy of Science
Редакционная коллегия:
T. В. БЕЙЕР, М. В. КРЫЛОВ, Л. Н. СЕР АВИН,
Я. И. СТАРОБОГАТОВ
Научный редактор:
С. А. КАРПОВ
Ответственный редактор:
С. Д. СТЕПАНЬЯНЦ
Рецензенты:
Д. В. ОСИПОВ, И. Б. РАЙКОВ
Editorial Board:
Т. V. BEYER, М. V. KRILOV, L. N. SERAVIN,
Ya. I. STAROBOGATOV
Scientific Editor:
S. A. KARPOV
Demanding editor:
S. D. STEPANJANTS
Reviewers:
D. V. OSIPOV, I. B. RAYKOV
Издание осуществлено при поддержке Российского фонда фундаментальных исследований
по проекту № 97-04-62048
Без объявления
ISBN 5-02-025864-4
© Российская академия наук, 2000
© Зоологический институт РАН, 2000
© Коллектив авторов, 2000
© Е. И. Кудина, оформление, 2000
ПРЕДИСЛОВИЕ
Издание фундаментальных сводок по основным
современным научным проблемам — одно из важ-
нейших условий успешного развития науки. Без них
невозможны ни новые открытия и обобщения, ни
разработка практических научных задач, связанных с
развитием народного хозяйства и экономики государ-
ства.
Зоология имеет первостепенное значение как
одна из фундаментальных наук, составляющая основу
для развития общей биологии, экологии, медицины,
ветеринарии, паразитологии, сельского хозяйства,
рыбного и охотничьего хозяйств, охраны природы и
многих других дисциплин. Это обстоятельство было
главной причиной издания больших руководств-сво-
док по зоологии.
В Германии с 1859 г. издается многотомная серия
«Bronns Klassen und Ordnungen des Tierreichs», про-
должающая выходить и в настоящее время. Там же с
1923 г. и до настоящего времени публикуется «Hand-
buch der Zoologie» под редакцией Кюкенталя и Крум-
баха (Kiikenthal, Krumbach); с 1965 г. издается много-
томный учебник «Lehrbuch der speziellen Zoologie»,
написанный Кестнером (A. Kaestner). Во Франции на-
чиная с 1948 г. опубликовано 27 томов «Traite de Zoo-
logie» под редакцией Грассэ (Р. Grasse). Наконец, в
США издано 7 томов по зоологии беспозвоночных
животных («The Invertebrates»; 1-й том появился в
1940 г.), принадлежащих перу Хайман (L. Hyman).
В отечественной зоологии в этом отношении дело
обстоит явно неблагополучно. Несмотря на наличие
большой армии квалифицированных зоологов и вы-
дающиеся достижения в области зоологической науки,
у нас нет солидного руководства, отвечающего совре-
менному уровню знаний.
Первая попытка подготовить и издать «Руководст-
во по зоологии» в нашей стране не была осуществле-
на полностью. Вместо 7 запланированных томов
вышло только 4 (1937—1951), последний из них —
около 45 лет назад. Не были опубликованы тома по
таким важным в теоретическом и практическом от-
ношениях группам, как паукообразные, ракообраз-
ные, амфибии, рыбы, рептилии, млекопитающие.
Ныне содержание изданных томов устарело. Это
связано с тем, что в послевоенное время зоология зна-
чительно шагнула вперед. Были открыты и описаны
новые отряды, классы и типы животных, внесены су-
щественные изменения в классификацию животного
мира, проведены обширные сравнительно-физиологи-
ческие, гистологические, генетические и экологичес-
кие исследования наземной, пресноводной фауны и
фауны Мирового океана. Очень бурно стала разви-
ваться протозоология, изучающая не только однокле-
точных животных, но и водоросли, и низшие грибы.
Сейчас в протозоологии происходит настоящая рево-
люция, ведущая к синтезу знаний по всем эукариотам
и созданию единой системы органического мира. В па-
разитологии большие успехи достигнуты гельминто-
логами; возникла новая дисциплина — экологическая
паразитология. В области медицинской паразитоло-
гии успешно развивается важное учение о природных
очагах трансмиссивных заболеваний. Возникла также
отечественная почвенная зоология. Наконец, решают-
ся новые неотложные задачи, связанные с проблемой
сохранения среды и существующего разнообразия жи-
вотного населения.
Отечественными зоологами созданы и развиты
концепции, нередко противоречащие теориям и ги-
потезам, которые господствуют за рубежом. На запа-
де они остаются по существу неизвестными и потому
подчас не попадают в круг вопросов, обсуждаемых
мировой наукой.
Таким образом, потребность в издании руководст-
ва по зоологии давно назрела. Такое руководство,
получившее первоначально название «Основы зооло-
гии», было задумано еще в 1978 г. Инициаторами вы-
ступили два института Российской академии наук —
Зоологический институт (ЗИН, Санкт-Петербург) и
Институт эволюционной морфологии и экологии жи-
вотных (ныне — ИПЭЭ, Москва). По замыслу авто-
ров, руководство должно стать настольной книгой
для ученых-зоологов всех специальностей, главным
источником для получения справок и первоначаль-
ных основных сведений по разным группам живот-
ных. Аналогичное значение оно должно иметь и для
исследователей самых различных областей биологии,
а также для преподавателей университетов, медицин-
ских, ветеринарных, сельскохозяйственных и педаго-
гических институтов, средних школ и, конечно, для
аспирантов и студентов биологических специально-
стей.
В начале 90-х гг. были подготовлены первые
тома этого издания. В частности, были написаны
6
ПРЕДИСЛОВИЕ
первый из двух томов по протестам (ред4
| Ю. И. Полянский!), том по губкам и кишечнополост-
ным (ред. С. Д. Степаньянц), том по плоским чер-
вям (ред. |А. В. Иванов |). Однако все ухудшающееся
с годами экономическое состояние отечественной
науки отразилось и на издательском деле. Так, за-
вершенный и принятый в издательство «Наука» в
1992 г. первый том по протистам пролежал без
движения до 1998 г. В последние годы появилась
возможность публиковать некоторые наиболее зна-
чимые рукописи при финансовой поддержке Россий-
ского фонда фундаментальных исследований. Бла-
годаря этому были получены средства на издание
первого тома из числа готовых — по протистам
как самостоятельной коллективной монографии.
Монография «Протесты. Руководство по зооло-
гии» включает помимо описаний двух больших
групп Protozoa — жгутиконосцев и саркодовых и
ряда групп Chromista обширную общую часть, со-
стоящую из разделов «Краткая история зоологии»,
«Пути и закономерности эволюции», «Систематика
и ее задачи», «Система животного мира», «Система
протистов и проблемы их мегасистематики», «Пути
эволюции протестов», «Общая характеристика про-
тестов».
Авторы и редколлегия надеются, что предлагаемая
монография послужит толчком для продолжения под-
готовки к печати и публикации автономных выпус-
ков по отдельным группам животных.
Авторский коллектив выражает благодарность
А. А. Голикову за помощь при компьютерном наборе
и в составлении указателей.
Редколлегия
PREFACE
The publication of fundamental reviews on major
themes of present-day research is one of the essential
prerequisites of the successful development of science.
Zoology is of paramount importance as one Of the fun-
damental sciences contributing towards general biology;
ecology, medicine, veterinary science, parasitology, ag-
riculture, fisheries and game husbandry, nature conser-
vation and many other disciplines. This fact was one of
the reasons for the initiation of a series of large handbo-
oks on zoology.
In Germany, the multivolume series, «Bronns Klas-
sen und Ordnungen des Tierreichs», was first published
in 1859; the «Handbuch der Zoologie», edited by
Kiikenthal and Krumbach, was published in 1923; the
multivolume manual «Lehrbuch der speziellen Zoolo-
gie» by Kaestner has been published since 1965. In
France, 27 volumes of «Traite de Zoologie», edited by
P. Grasse, have been published since 1948. Finally, in
the USA, 7 volumes on invertebrate zoology entitled
«The Invertebrates» have been published by L. Hyman —
the 1st volume appeared in 1940. In Russia, however,
no fundamental manuals conforming to the modern
level of knowledge have been published in spite of the
large number of highly qualified zoologists and the out-
standing achievements in the field of zoology.
A first attempt to prepare and publish «The Manual
on Zoology» in Russia was not completed. Instead of
the 7 volumes originally planned only 4 were published
between 1937 and 1951, the most recent of which appe-
ared approximately 45 years ago. Volumes on taxa of
theoretical and applied importance, such as arachnids,
crustaceans, amphibians, fishes, reptiles and mammals
were never published.
The volumes in this series that were published have
now become outdated. Zoology has advanced dramati-
cally in the post-war years. New orders, classes, and
phyla have been defined and described. Essential amen-
dments have been made in classification of the animal
world. Extensive comparative studies have been conduc-
ted in physiology, histology, genetics and ecology of
terrestrial fauna, as well as of fauna of fresh waters and
the world’s oceans. Protistology, which incorporates not
only unicellular animals, but also algae and the lower
fungi, developed vigorously. Radical changes have now
taken place in protistology, which have lead to a synt-
hesis of knowledge on all eukaryotes and the estab-
lishment of a comprehensive classification of the orga-
nic world. In the field of parasitology great advances
have been achieved by helminthologists. A new discip-
line, ecological parasitology, has been established. In
the field of medical parasitology the important theory of
natural foci of transmissive diseases has been developed
successfully. Eventually new urgent tasks related to
conservation of the environment and existing diversity
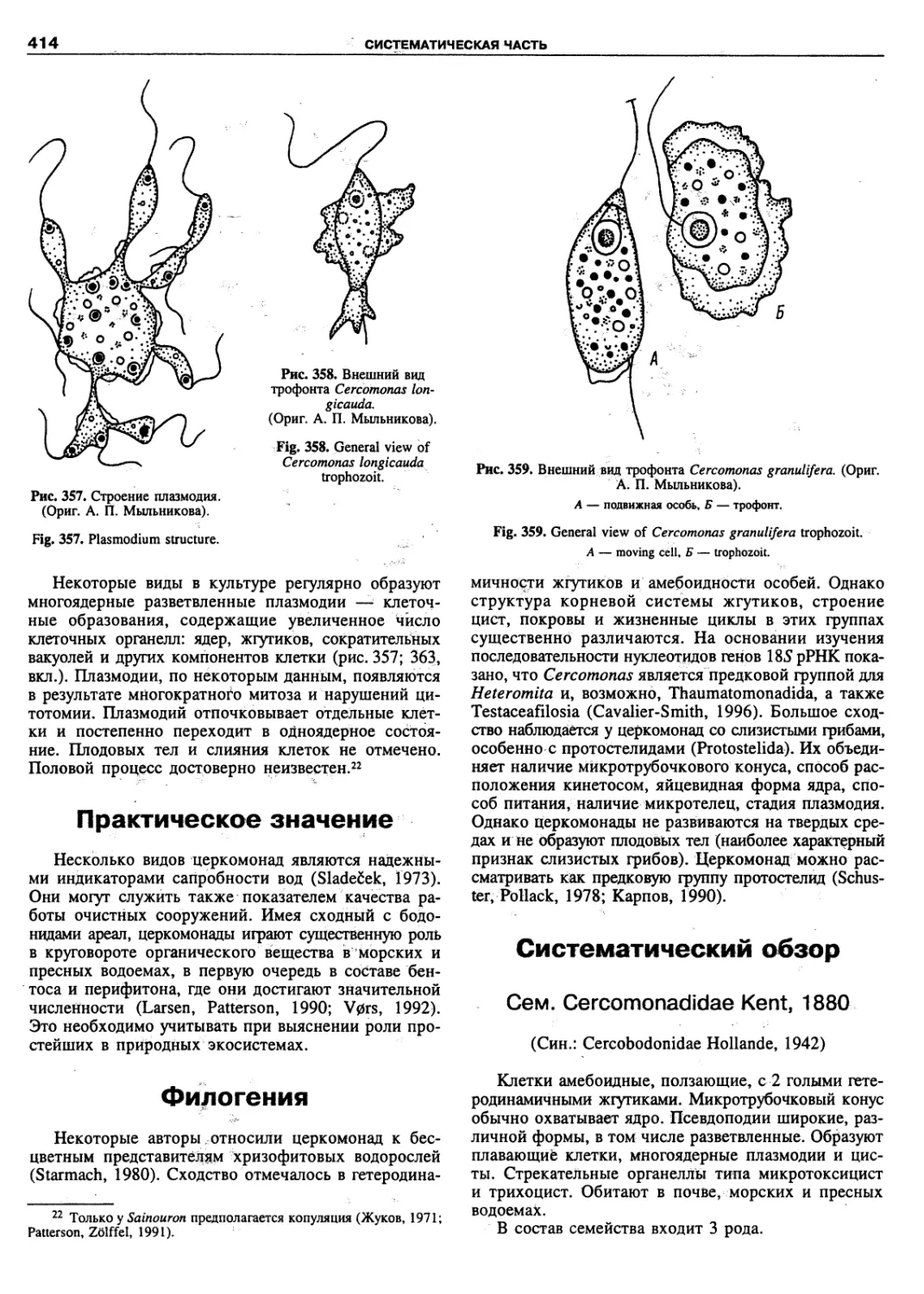
of the animal population are being resolved.
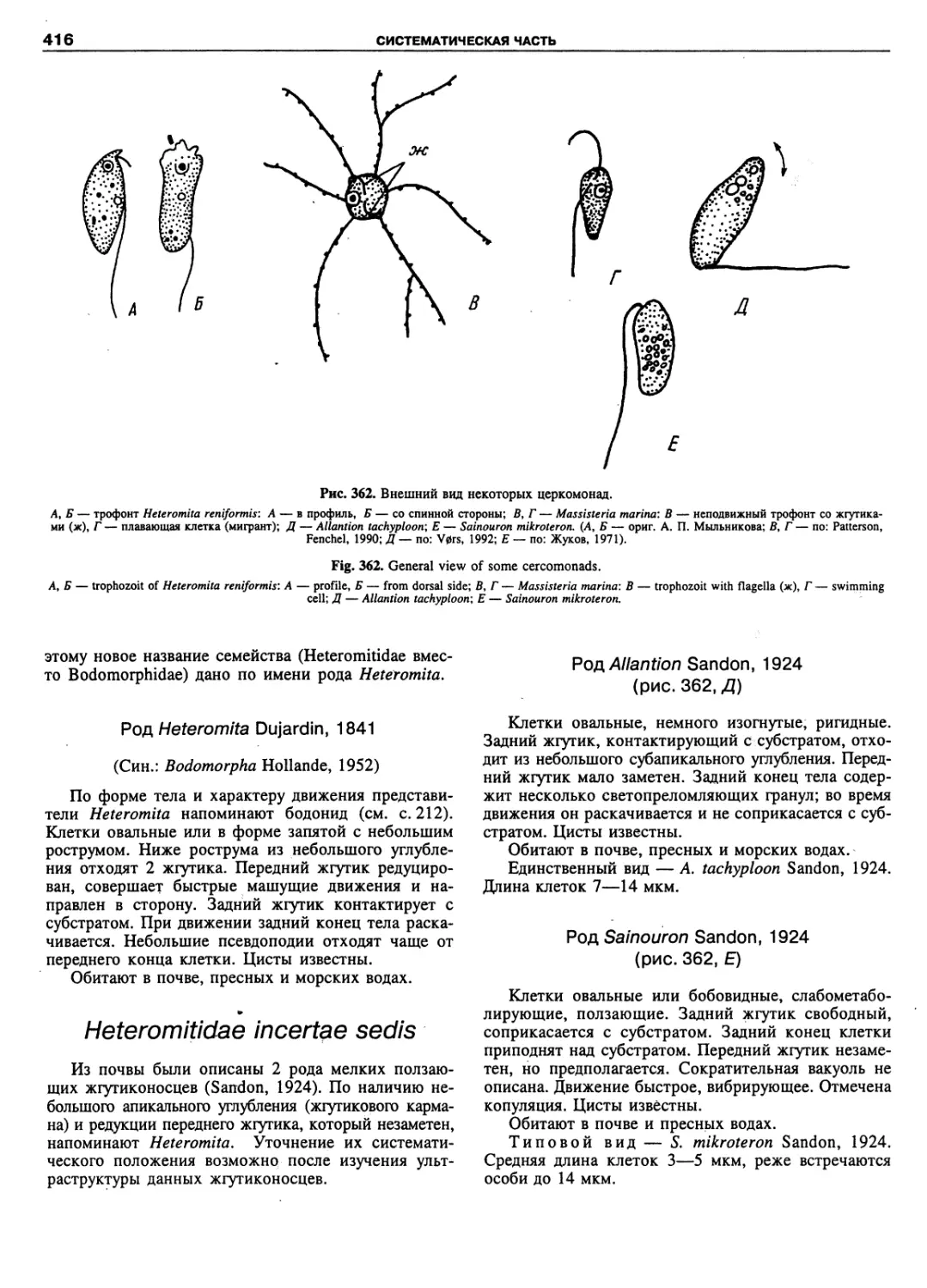
Russian zoologists have put forward and developed
scientific concepts, sometimes contradicting theories
and hypotheses accepted abroad. Many of these con-
cepts remain unknown in the West and elsewhere and,
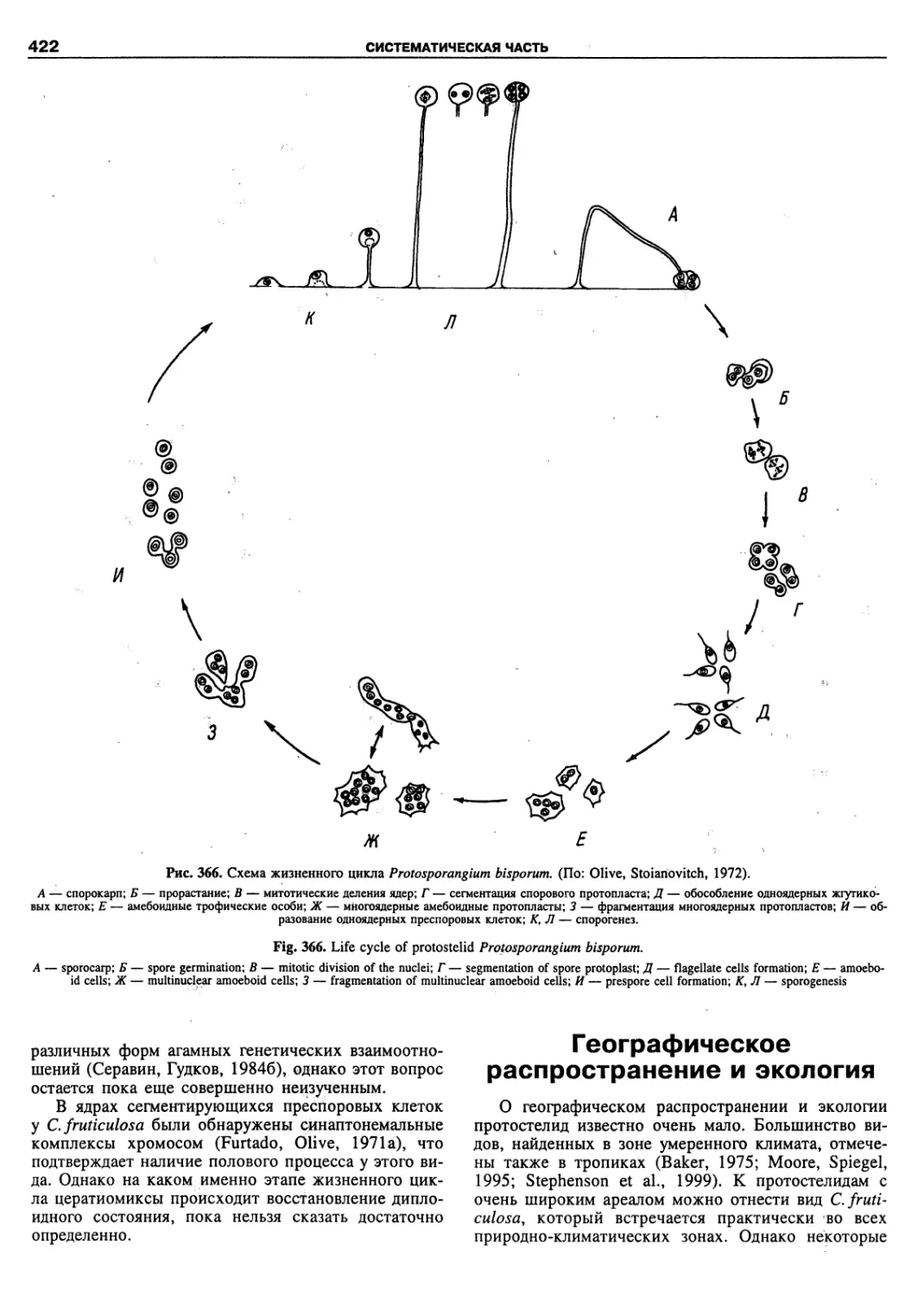
therefore, are often ignored by scientists outside Russia.
Thus the time is right for the publication of a guide
on zoology. Such a guide entitled «The Bases of Zoolo-
gy» was planned in 1978 by two institutes of the Russi-
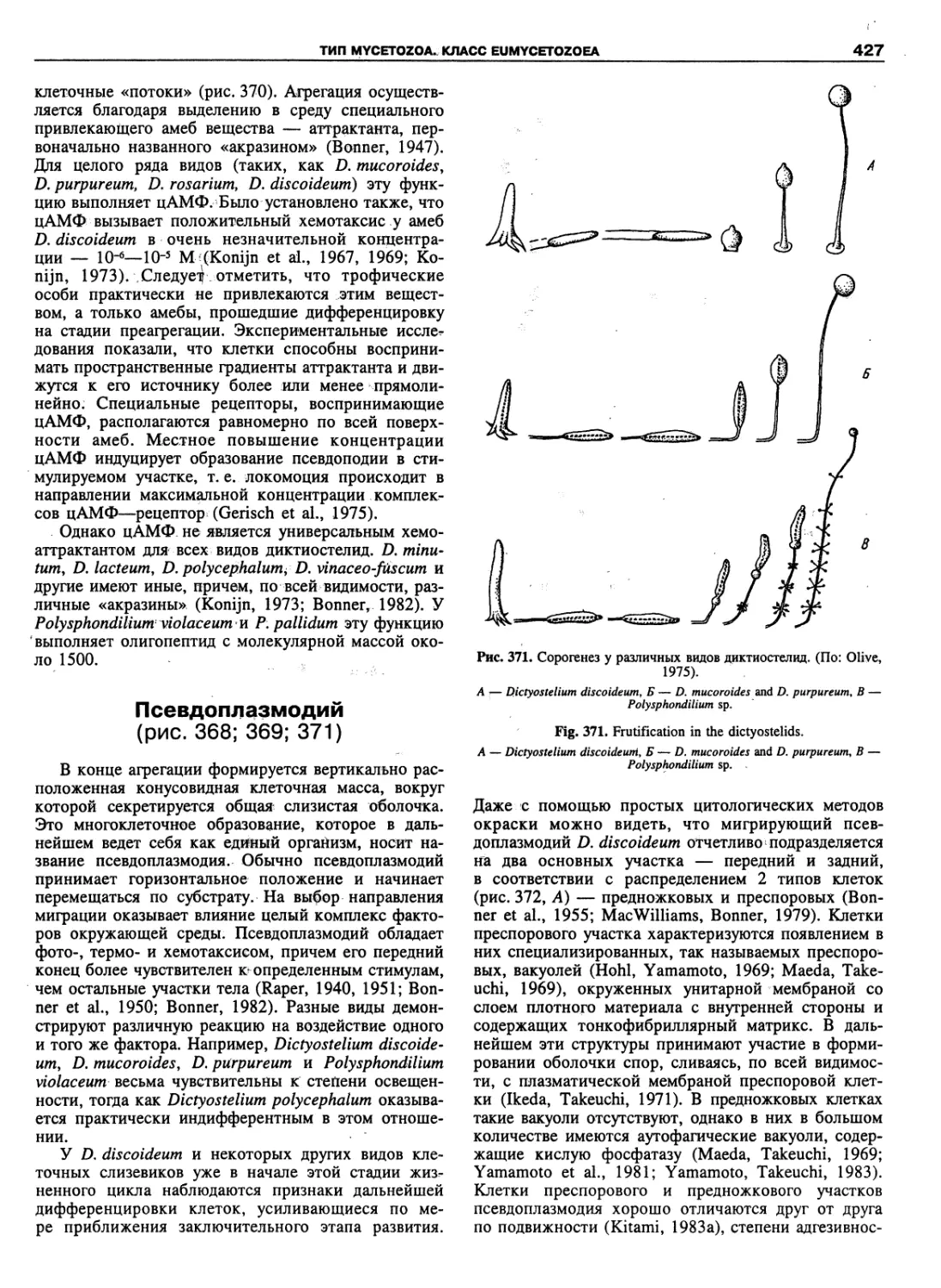
an Academy of Sciences: the Zoological Institute (St.
Petersburg) and the Institute of Animal Evolutionary
Morphology and Ecology (Moscow). It was the authors’
intention that «The Bases of Zoology» should become a
manual for zoologists of all specialities, and a major
source of basic reference information on different ani-
mal taxa. This manual is important also for researchers
in different fields of biology and for teachers in univer-
sities, medical, veterinary, agricultural, and pedagogical
institutions, secondary schools as well as for students of
biology, undergraduate and graduate.
In the early 1990s the first volumes of this edition
were prepared. The first of two volumes on protozoa
(Ed.| G. I. Poljansky]), a volume on sponges, and coelen-
terates (Ed. S. t). Stepanjants), and a volume on plathel-
minthes (Ed.|A. V. Ivanov|) were completed. However,
the gradually deteriorating economic situation of Russi-
an science also affected publishing activities. The first
volume on protozoa completed and adopted for publica-
tion in 1992 remained unpublished until 1998. Recently,
owing to support provided by the Russian Foundation
for Basic Research, an opportunity has arisen for the
publication of several of the most significant manusc-
ripts. As a result, funds were obtained for the publica-
tion of the first volume on Protozoa as a separate mo-
nograph by a number of authors.
The monograph «Protists: Handbook on Zoology»
comprises a large general part including sections on
the «Essay on the history of zoology», «Pathways
8
PREFACE
and principles of the evolution», «Systematics and its
purposes», «Classification of the animal world» and
the sections devoted to the general characterization of
protists, their classification and trends of evolution.
The main part of the volume is devoted to systema-
tics, comprising the history, taxonomy, morphology
and other biological aspects of the phyla: Cryptophy-
ta, Euglenozoa, Chrysophyta, Haptophyta, Raphidophyta,
Saprolegnia (=Oomycetes), Opalinata, Choanomonada,
Polymastigota, Plasmodiophora, Mycetozoa, Rhizopo-
da, and Foraminifera. All other groups of protists will
be included in a second volume.
The authors and the editorial board hope that this
monograph will serve as an impetus for the continued
preparation and publication of other volumes, each on a
separate animal group.
The Editorial Board
ОБЩАЯ ЧАСТЬ
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
Основные задачи зоологии
Зоология (от греческого «zoo» — животное и
«logos» — слово, наука) — наука о животных. Это —
часть биологии, изучающая многообразие животного
мира, строение и жизнедеятельность животных, их
распространение, связь со средой обитания, законо-
мерности индивидуального и исторического разви-
тия. Зоология связана с производственной деятель-
ностью человека; она ставит задачу освоения, рекон-
струкции и охраны животного мира Земли.
По задачам исследования зоология распадается на
ряд основных дисциплин. Систематика животных
имеет целью описание многообразия видов, система-
тизацию их по признакам сходства и различия, уста-
новление иерархии таксонов. Систематика стремится
к построению естественной системы, отображающей
пути исторического развития животного мира. Мор-
фология исследует строение животных — внешнее и
внутреннее. Последнее относится к области анато-
мии. .Гистология — также часть морфологии — ис-
следует микроскопическое строение животных. Срав-
нительная морфология изучает строение животных и
отдельные системы их органов в плане сравнения у
разных систематических групп, устанавливая при
этом гомологии и аналогии. Эмбриология животных
изучает индивидуальное развитие (онтогенез) от яйца
до взрослого организма. Филогенетика исследует
пути эволюции животного мира. Экология животных
ставит задачи изучения их взаймоотношений между
собой, с другими организмами и со средой обитания.
Зоогеография — раздел зоологии (пограничный с
географией), исследующий распределение животных
в разных регионах планеты, на суше и в воде, а также
факторы, определяющие это распространение. Палео-
зоология изучает вымерших животных прежних гео-
логических эпох. В этом она тесно связана с филоге-
нетикой и эволюционной морфологией. Палеозооло-
1 Этот раздел был написан[ю. И. Полянским] в 1990 г. В це-
лом мы оставили оригинальный текст автора без изменений. В
ряде случаев в него внесены небольшие дополнения в виде сносок.
Раздел «Учение Дарвина» написан Э. И. Колчинским. В разделе не
приводится библиография в традиционном понимании, но все ци-
тируемые автором работы можно найти в обзорных изданиях (см.:
«Литература», с. 114). Редколлегия.
гические исследования базируются на материалах
окаменелостей, отпечатках и реконструкциях по
ним. Разработка новых методов палеозоологических
исследований позволила в ряде случаев восстановить
не только внешний вид, но и внутреннюю организа-
цию ископаемых животных, как это было сделано,
например, шведским ученым Э. Стеншио для некото-
рых бесчелюстных.
Физиология животных, которая исторически воз-
никла как одна из ветвей зоологии, развилась в са-
мостоятельную биологическую науку, изучающую
функции животного организма.
Зоологию можно классифицировать и по объекту
исследования: на протозоологию — науку об однокле-
точных организмах, гельминтологию — о паразити-
ческих червях, малакологию — о моллюсках, карци-
нологию — о ракообразных, арахнологию — о пауко-
образных, акарологию — о клещах, энтомологию — о
насекомых, ихтиологию — о рыбах и рыбообразных,
батрахологию — о земноводных, герпетологию — о
пресмыкающихся, орнитологию — о птицах, териоло-
гию — о млекопитающих и т. п.
В зоологии используют разнообразные методы ис-
следования, общие для биологических дисциплин:
традиционные, морфологические, кариологические,
методы молекулярной биологии и другие.
Зоология тесно связана с другими биологически-
ми науками, а также с медициной и ветеринарией.
Некоторые разделы зоологии входят как составляю-
щая часть в такие комплексные дисциплины, как па-
разитология, гидробиология, эпизоотология, эпиде-
миология.
Зоология имеет широкое практическое примене-
ние в различных областях деятельности человека. В
области медицинской и ветеринарной паразитологии
зоология изучает животных — паразитов человека,
домашних и полезных животных. Зоологические ис-
следования лежат в основе организации мероприятий
по борьбе с животными-вредителями сельского и
лесного хозяйства. Многие беспозвоночные живот-
ные — моллюски, ракообразные, насекомые — ис-
пользуются человеком как пищевые объекты и для
технических целей. Эколого-зоологические исследо-
вания лежат в основе разработки мер по воспроиз-
водству рыбных запасов, регулированию численности
12
ОБЩАЯ ЧАСТЬ
объектов охотничьего хозяйства, акклиматизации и
интродукции полезных животных.
За последние десятилетия выделились некоторые
специальные области зоологии, представляющие
значительный теоретический и практический инте-
рес. Почвенная зоология изучает своеобразные поч-
венные биоценозы (работы академика М. С. Гиляро-
ва и др.). Исследуется фауна пещер. Изучается
псаммофильная фауна моря и пресных водоемов —
биоценоз, поверхностного слоя песка (шведский
ученый Б. Сведмарк, немецкий ученый А. Ремане и
др.). Значительный интерес представляют исследо-
вания фауны обрастаний подводных предметов в
пресной и морской воде, что имеет большое прак-
тическое значение при создании подводных соору-
жений и в кораблестроении.
Зоология, как и другие биологические науки, вы-
росла из запросов практики и на разных этапах исто-
рии человечества находила и находит широкое прак-
тическое применение.
Животные с самого начала появления человека на
Земле были объектом охоты, необходимым условием
существования и развития первобытного человеческо-
го общества. Со времени среднего палеолита осталось
довольно много изображений в виде наскальных ри-
сунков на стенах пещер, служивших людям жилищем.
Эти изображения были созданы свыше 50 тысяч лет до
н. э. По ним можно судить, на каких животных охо-
тился человек каменного века: среди них были мамон-
ты, шерстистые носороги и некоторые другие, ныне
вымершие, виды. Приручение животных произошло
значительно позднее. Оно относится к концу палео-
лита и приобрело массовый характер в неолитическую
эпоху (5—6 тыс. лет до н. э.), когда человек стал за-
ниматься наряду с рыбной ловлей и охотой земледе-
лием и скотоводством. Это произошло в первую оче-
редь в очагах древней культуры, которые, как показа-
но Н. И. Вавиловым, стали центрами происхождения
культурных растений. Первым животным, которое
приручил человек, была собака.
Знания человека о мире животных значительно
возросли в период формирования великих древних
культурных центров человечества — Древнего Егип-
та, Вавилона, Индии, Китая. В некоторых дошедших
до нас древних папирусах Египта (например, в знаме-
нитом «папирусе Эберса», написанном приблизи-
тельно за 1500 лет до н. э.) содержатся многочислен-
ные данные о животных. Информация о животных и
растениях тесно переплеталась со сведениями по ме-
дицине, анатомии человека. Последняя в Древнем
Египте достигла значительного совершенства, что
было связано с бальзамированием трупов. Однако все
эти довольно многочисленные знания о животном
мире носили разрозненный характер, не были объ-
единены в систему и поэтому не могут рассматри-
ваться как начало науки зоологии.
Античная древность
Зоология как наука возникла в Древней Элладе и
связана с именами знаменитых философов и ученых
Древней Греции: Гераклита (ок. 530—470 до н. э.),
Демокрита (ок. 460—370 до н. э.) и в особенности ге-
ниального ученого и философа Аристотеля (ок. 384—
322 до н. э.).
Гераклит — древнегреческий философ-материа-
лист, представитель античной диалектики, признав-
ший материальность мира и его непрерывное движе-
ние и развитие, рассматривал жизнь как этап этого
всеобщего процесса.
Демокрит — великий мыслитель древней Эллады,
учение которого об атомах как основных элементах
материального и духовного мира явилось одной из
первых материалистических всеобъемлющих концеп-
ций философии. Источником и основой жизни, в том
числе и животных организмов, Демокрит признавал
мельчайшие круглые гладкие атомы, по сравнению
с другими атомами — особенно подвижные. Живые
существа — лишь временные сочетания различной
формы атомов. Эта наивная материалистическая кон-
цепция представляет, однако, большую ценность, ибо
дает единую картину материальной природы мира. В
трудах Демокрита мы не находим ни описаний живот-
ных, ни попыток их классифицировать.
Аристотель был величайшим мыслителем антич-
ного мира, труды которого оказали огромное влияние
на развитие естествознания, в том числе и зоологии.
В течение более полутора тысяч лет (до XVII в.) они
являлись энциклопедией знаний о живой природе. В
сочинениях Аристотеля охвачены все отрасли совре-
менной ему науки — психология, логика, история,
политика, этика, эстетика. Много внимания уделено
естествознанию, в том числе ботанике и зоологии.
Сочинения Аристотеля, опубликованные им при
жизни, не сохранились. Известны лишь фрагменты,
приводимые древними авторами, а также рукописи
некоторых его сочинений, опубликованные спустя
300 лет после его смерти Андроником Родосским.
Биологические воззрения Аристотеля изложены в
трех его произведениях: «История животных», «О
частях животных» и «О возникновении животных».
Они распадаются на несколько самостоятельных раз-
делов, которые называются «книгами»; всего их на-
считывается 19. Аристотель — создатель первой в
истории науки системы животного мира. Он приво-
дит описание 454 видов животных и группирует их в
систематические категории более высокого порядка,
которые называет родами («эйдос»). Последние объ-
единяются в «Высшие роды», которые примерно со-
ответствуют современным классам. Наконец, «Выс-
шие роды» объединены в несколько групп еще боль-
шего таксономического значения. Нужно иметь в
виду, что описываемые Аристотелем отдельные фор-
мы животных не полностью соответствуют современ-
ному пониманию вида и в большинстве случаев близ-
ки, скорее, к понятию рода (например, оса — это не-
сколько видов ос, комнатная муха — несколько видов
мух, дрозд — несколько видов птиц и т. п.). Система
Аристотеля сыграла исключительную роль в разви-
тии зоологии. Она служила основой зоологических
знаний в течение полутора тысячелетий, поэтому
считаем необходимым привести ее в настоящей
главе. При этом мы воспроизводим (с сокращениями
и изменениями) таблицу из работы Л. А. Зенкевича
«История системы беспозвоночных» (1937), в кото-
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
13
Таблица 1
По Аристотелю Современные названия
А Животные, снабженные кровью. Позвоночные.
I Живородящие четвероног Млекопитающие (без кито-
гие. образных).
II Животные с двумя ногами и двумя крыльями. Птицы.
III Яйцекладущие четвероно- Земноводные и пресмыкаю-
гие и безногие с легоч- ным дыханием. щиеся.
IV Живородящие безногие, жи- вут в воде, дышат лег- кими. Китообразные.
V Безногие, покрытые чешуей и дышащие жабрами. Рыбы.
Б Животные без крови. Беспозвоночные.
I Мягкие животные Malakia. Головоногие моллюски.
Ноги окружают голову. Ли- шены твердого скелета. Имеют чернильный ме- шок.
II Мягкопанцирные с боль- шим количеством ног Malakostraca. Высшие ракообразные.
III Членистые с большим ко- Часть членистоногих (в т. ч.
личеством ног. насекомые и многонож- ки), кольчатые черви.
IV Безногие с твердым панци- Моллюски (кроме голово-
рем-раковиной Ostraco- ногих), иглокожие, кишеч-
derma. Мягкое тело внут- нополостные, губки, не-
ри твердой ломкой рако- которые асцидии и усо-
вины. ногие. Часть брюхоногих.
1 Раковинные.
2 Морские ежи.
3 Морские звезды.
4 Безраковинные свободно- Голотурии.
о живущие.
5 Безраковинные прирастаю- щие. Губки, актинии.
рой удачно сопоставлены подразделения аристоте-
левской системы с современными представлениями и
номенклатурой (табл. 1).
Знакомство с системой Аристотеля показывает,
насколько глубоко он понял основные черты архи-
тектоники животных и в ряде случаев сгруппировал
их в таксоны, приближающиеся к современным. При
этом Аристотель учитывал не какой-либо один или
немногие признаки, а всю совокупность основных
признаков.
Наряду с созданием системы Аристотелю принад-
лежат важные обобщения в области зоологии. На ос-
нове изучения морфологии животных им была выска-
зана идея о единстве их строения. Основание для
этого важнейшего заключения дали главным образом
позвоночные. Аристотелем был разработан ряд важ-
нейших сравнительно-анатомических обобщений, в
том числе учение о корреляциях (соотношении орга-
нов), которое получило дальнейшее развитие лишь в
XIX в. Аристотель различает аналогичные и.гомоло-
гичные органы. Очень важны наблюдения его над эм-
бриональным развитием животных, которые он про-
водил главным образом на курином зародыше. Эти
наблюдения дали Аристотелю основания для выводов
о том, что органы вначале имеют простое строение, а
позже претерпевают постепенное усложнение, и о
том, что зародыши разных животных похожи друг на
друга гораздо больше, чем взрослые формы.
Всеобъемлющий синтезирующий ум гениального
мыслителя глубоко проник в понимание ряда законо-
мерностей живой природы. Он признавал раздельное
существование материи и духовного начала. Мате-
рия — пассивная субстанция. Духовное начало, дей-
ствуя на материю, превращает ее в действительность.
Характерно для Аристотеля представление о непре-
рывной цепи, градации форм от более простых к
более сложным и совершенным. Эту идею он приме-
нял и к организмам. Таким образом, все его мировоз-
зрение носит телеологический характер. Иерархию
вещей в природе Аристотель не рассматривает как
нечто эволюционно развивающееся во времени. Это —
градация вечных форм, стремящихся к единой цели.
Римский период античной культуры в развитии
зоологии, как и биологии в целом, внес мало принци-
пиально нового. Среди римских ученых и философов
видное место занимает Тит Лукреций Кар (ок. 99—
55 до н. э.). Его философская поэма «О природе
вещей» представляет выдающееся поэтическое про-
изведение, а также важный философский трактат. В
ней получает дальнейшее развитие античный матери-
алистический атомизм. Живые организмы возникли в
результате сочетания первичных телец (атомов). По
Лукрецию, вначале возникали неприспособленные к
жизни организмы (безротые, уроды всякого рода и
т. п.). Они быстро погибали, пока в силу того же слу-
чайного сочетания атомов не возникли существа,
приспособленные к условиям среды. В этом можно
видеть предвестника учения о борьбе за существова-
ние и естественном отборе. Все эти представления
Лукреция облечены в увлекательную форму.
Широкой известностью пользовался Гай Плиний
Секунд Старший (ок. 23—79), знатный римлянин,
погибший при извержении Везувия. Он создал своего
рода энциклопедию естественных наук, опубликовав
«Естественную историю» в 37 томах. Это сочинение,
охватывающее все области естествознания, в том
числе и зоологию, которой отведено 4 тома, пред-
ставляет собой чисто компилятивный труд, в котором
суммирована вся античная литература. Плиний ука-
зывает, что использовал сочинения 146 римских и
327 иноземных авторов. В «Естественной истории»
нет оригинальных мыслей или обобщений. В ней до-
бросовестно переписано все, что было известно в I в.
н. э., и наряду с действительными фактами имеется
немало фантастики. Сочинения Плиния тем не менее
сыграли положительную роль, и к ним часто обраща-
лись ученые средневековья.
Важную роль в развитии зоологии сыграл также
знаменитый римский врач и естествоиспытатель
Клавдий Гален (ок. 130—200). Его труды, из которых
до нас дошла лишь небольшая часть (основная масса
их погибла во время пожара храма Мира в Риме),
посвящены вопросам анатомии человека и животных.
В них делается попытка связать форму органа с его
функцией. В его трудах много верных наблюдений.
14
ОБЩАЯ ЧАСТЬ
Он показал, что артерии наполнены кровью, доказал
проводящую возбуждение роль нервов (путем их пе-
ререзки). До XVII в. (до работ Гарвея) физиологичес-
кие представления Галена пользовались широким
признанием.
Средние века
Эпоха средневековья датируется со взятия и раз-
грабления Рима вестготами Алариха, что произошло в
4ДО г. Этот бтап истории человечества, продолжав-
шийся свыше 1000 лет (до XV—XVI вв.), в области
общественно-экономических отношений характери-
зуется феодальным строем, а в области идеологии —
господством1 католицизма в Западной Европе и пра-
вославия в России. В естествознании, и в том числе в
зоблогии, этот длительный период истории человечес-
тва знаменуется забвением античной философии и ес-
тествознания. За весь период можно указать лишь на
немногочисленные отдельные проблески в развитии
науки; большая часть которых безжалостно подавля-
лась. Хотя в этот период творения Аристотеля и от-
части Плиния широко использовались, но в большин-
стве случаев бралось только то, что не противоречило
церковным догматам. Одним из методов борьбы с не-
угодными идеями во вторую половину средневековья
на Западе были костры инквизиции.
Характерным представителем раннего средневе-
ковья был Блаженный Августин (354—430). Глубоко
философски образованный епископ, знаток античной
философии, в своих многочисленных сочинениях
развивал учение о всеобщей гармонии в природе как
проявлении божественной мудрости.
Важную роль в сохранении античной науки и в
развитии некоторых сторон естествознания сыграли
арабы. Проникнув в VII и VIII вв. на Южное побе-
режье Средиземного моря, достигнув Испании, поко-
рив Персию, Палестину, Египет, арабы принесли Ев-
ропе восточную культуру и ассимилировали вместе с
тем античную философию и науку. Арабы дали ряд
выдающихся мыслителей и ученых. Врач, философ и
натуралист Авиценна (Ибн Сина, 980—1037) был глу-
боким знатоком Аристотеля. В его «Каноне» кроме
изложения Аристотеля и Галена находим оригиналь-
ные сведения по медицине, зоологии, ботанике, а
также призыв обратиться к изучению природы. Еще
большее значение имели в XIII в. труды Авероса
(Ибн Рашида, 1120—1198). Будучи также натуралис-
том, врачом и философом, он высказывал некоторые
весьма прогрессивные мысли о вечности материи и
ее развитии. Особенно большую славу создал ему
«Большой комментарий» (1193), в котором наряду с
толкованием Аристотеля высказывается ряд смелых
и прогрессивных мыслей о развитии мира.
Большой популярностью в XIII в. пользовался
Альберт Великий (граф Альберт фон Болыптетт, род.
между 1193 и 1205, ум. 1280). Епископ, блестящий
лектор, плодовитый писатель (литературное наследие
Альберта Великого — 21 том) — он с позиций като-
лицизма освещает проблемы всех наук, широко ис-
пользуя при этом труды античных философов. В спе-
циальном трактате «De animalibus» немало места уде-
ляется им зоологии. Частично это пересказы Аристо-
теля и Плиния, частично — фантастические сведе-
ния, широко использовавшиеся в средние века (опи-
саны «единорог» с лошадиным туловищем, оленьей
головой и закрученным рогом, «пегас» с лошадиным
туловищем, бычьей головой и орлиными крыльями,
превращение лошадиного волоса в волосатика и
т. п.).
Весьма распространенными в средние века были
различного рода сборники (их авторство трудно уста-
новить), носившие название «Физиологус», «Бестиа-
рий». Такого рода сборники издавались начиная со II
и до XVI в. Переводы их проникали и в Россию. В
этих сборниках наряду с некоторыми правильными
сведениями о жизни и строении животных имелось
множество фантастических данных, например, опи-
сывались гуси, которые родятся на деревьях. На этом
основании диких гусей стали считать постными и
употреблять в пищу в постные дни. Как это ни пара-
доксально звучит, но понадобилась особая булла
папы Иннокентия III (1161—1216), чтобы заставить
считать гусей скоромной пищей. Этот исторический
анекдот показывает уровень развития естествознания
в средние века.
Таким же низким уровнем характеризуется и ряд
других, чисто компилятивных произведений средне-
вековья о живой природе. К их числу следует отнести
произведения Фомы из Кантимпрэ (1210?—1263),
написавшего обширную компилятивную энциклопе-
дию «О природе вещей» (20 книг), сочинение «Книга
природы» Конрада фон Магенбёрга (1309—1374).
Последняя книга (написанная не по-латыни, а по-не-
мецки) пользовалась особенно широким распростра-
нением и выдержала множество изданий. Имея в ос-
нове произведения Фомы из Кантимпрэ, она содер-
жит описание 450 видов животных. В значительной
мере и это произведение наряду с описаниями дейст-
вительно существующих животных полно фантасти-
ки и суеверий.
Особое место в философии и естествознании
средних веков занимает Роджер Бэкон (R. Bacon)
(1214/19—1290/94?). Он призывал к опыту и наблюде-
нию как критериям истины. Основное произведение
Бэкона — «Opus majus», состоящее из семи частей,
включает как философские проблемы (преимущест-
венно изложение основных идей античной филосо-
фии), так и основы наук о природе, главным образом
математики, астрономии, географии, зоологии. Вся
шестая часть этого опуса идет под девизом «без опы-
та ничто не может быть вполне познано» («Sine ехре-
rimentia nihil sufficienter sciri potest»). Идеи P. Бэкона
намного опередили его эпоху. Отцы церкви (он при-
надлежал к монашескому ордену францисканцев)
дважды сажали его в тюрьму, где он просидел в
общей сложности 24 года.
Во второй половине средневековья, и главным об-
разом в конце его, в связи с началом роста городов во
многих странах Европы возникают университеты.
Хотя наука во многих университетах носила типич-
ные черты средневековой схоластики, тем не менее
их возникновение имело важное положительное зна-
чение, ибо способствовало объединению ученых,
обучению молодежи и т. п. Наиболее древние универ-
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
15
ситеты Европы возникли в Болонье (1158), Монпелье
(1180), Париже (1200), Оксфорде (1200—1229), Не-
аполе (1224), Риме (1245), Лиссабоне (1290).
XV—XVIII века
Этот период в истории биологических наук начи-
нается эпдхой Возрождения (XV—XVI вв.), ознаме-
новавшейся ломкой феодальных общественно-эконо-
мических отношений и началом развития капитализ-
ма. Про этот период бурного роста науки и искусства
Энгельс писал: «Это был величайший прогрессивный
переворот, пережитый тогда человечеством, — эпоха,
которая нуждалась в титанах и которая породила ти-
танов по силе мысли, страстности и характеру, по
многосторонности и учености...» Великие географи-
ческие открытия обогатили естествознание огром-
ным количеством новых фактов. Развитие мореплава-
ния, торговли усилило связи между народами. Шел
процесс формирования национальных государств.
Резко возросли научные связи. В большинстве госу-
дарств Европы возникают национальные Академии
наук и Научные общества (среди последних особенно
большую роль сыграло Королевское Общество в
Лондоне). Академии и Общества начинают создавать
периодические издания (типа «Proceedings», «Trans-
actions», «Известия» и т. п.). Естествоиспытатели ак-
тивно участвуют в экспедициях во все страны мира,
по всем вновь открытым континентам, собирая и изу-
чая новых, ранее не известных представителей фауны
и флоры. При академиях и университетах возникают
музеи, ботанические сады и зоопарки.
Особо можно выделить период с XVI по XVII в.,
который характеризуется накоплением новых знаний
в области биологии. До создания системы животного
мира в зоологии дело доходит значительно позднее,
чем в ботанике, где признаки более наглядны и до-
ступны. Одним из ярких представителей этого пери-
ода в развитии зоологии явился швейцарский натура-
лист Конрад Геснер (Conrad Gesner, 1516—1565). Им
создана пятитомная энциклопедия зоологии (свыше
4000 страниц). В труде Геснера более 1000 рисунков.
При сравнении трудов Геснера с аналогичными рабо-
тами средневековых авторов виден большой прогресс
(это касается как объема материала, так и характера
его изложения). Все описания сделаны по единому
плану, включающему название животного, распрост-
ранение, описание строения и связей со средой, а
также указание на практическое значение для чело-
века. В произведении Геснера приводится много соб-
ственных наблюдений, но все же в значительной
части оно построено на компиляции преимуществен-
но античных литературных источников. Наряду с
описаниями существующих животных сохраняется
довольно значительное количество всякого рода
мифических существ (таковы были традиции почти
полуторатысячной давности!). Никакой системы и
даже попытки ее построения в трудах Геснера нет.
Животные располагаются в алфавитном порядке.
Сведения о внутреннем строении почти отсутствуют.
Среди ученых XVI в. наряду с Геснером следует на-
звать еще несколько имен видных зоологов, труды
которых менее энциклопедичны, чем таковые Гесне-
ра. Гийом Ронделэ (Guillame Rondelet, 1507—1666) —
профессор из Монпелье, известен как автор большой
работы «О морских рыбах». Однако кроме ихтиоло-
гического материала в его сочинении рассматривают-
ся моллюски, черви, ластоногие. Особенно подробно
на основании собственных наблюдений им описаны
средиземноморские рйбы.
Пьер Белон ^Pierre Belon, 1518—1564) известен
своими орнитологическими и ихтиологическими ра-
ботами. Им написаны «Естественная история птиц» и
«Ихтиология» (в последнюю входили не только
рыбы, но и другие водные животные). В его сочине-
ниях много довольно Точных данных по анатомйи, в
особенности птиц. Системы животных еще нет. ‘Зна-
чительная часть материала была собрана им лично во
время многочисленных путешествий по Европе и
Африке. (Можно было бы назвать и еще целый ряд
зоологов XVI в., занимавшихся сбором и накоплени-
ем материала, но не поднимавшихся до сколько-ни-
будь крупных обобщений и до построения системы
животного мира.) К этому же периоду относятся про-
изведения многих натуралистов, предпринявших пу-
тешествия во вновь открытые страны и описавших их
животный мир.
Для развития зоологии, как и других биологичес-
ких наук, большое значение имели труды английско-
го философа Фрэнсиса Бэкона (Francis Bacon, 1561—
1626) — сторонника экспериментального метода,
врага схоластики и дедукции. Бэкон считается твор-
цом индуктивного и экспериментального методов в
науке. Изложенные в его знаменитом произведении
«Великое восстановление» («Instauratio magna») взгля-
ды призывали к преобразованию наук на основании
строгого и точного опыта. Философия Бэкона оказа-
ла огромное влияние на развитие естественных наук
в конце XVI—начале XVII в.
Значительное влияние на развитие зоологии в
конце XV, XVI и начале XVII в. оказали работы ана-
томов, которые, изучая анатомию человека, иссле-
довали также анатомию животных. В ряд этих ана-
томических трудов следует прежде всего поставить
работы гениального мыслителя, художника и естест-
воиспытателя эпохи Возрождения Леонардо да Винчи
(Leonardo da Vinci, 1452—1519) — автора трудов по
анатомии человека и животных, сопровождавшихся
прекрасными рисунками. Леонардо да Винчи стре-
мился не только изобразить строение органа, но и
понять его функцию. По сравнению с античными ав-
торами анатомия человека и животных сделала боль-
шие успехи в XVI, а также начале XVII в. благодаря
трудам итальянцев Везалия (A. Vesal, 1514—1564)
(«Семь книг о строении человеческого тела»), Фал-
лопия (G. Fallopio, 1523—1562), Евстахия (В. Eustac-
hio, 1510—1574), Фабриция (I. Fabricius, 1537—
1619) и испанца Сервета (М. Serveto, 1509—1553).
Серия трудов этой блестящей плеяды анатомов созда-
ла предпосылки для дальнейшего проникновения не
только в строение, но и в функции животного орга-
низма.
В 1628 г. знаменитый английский врач и естество-
испытатель Вильям Гарвей (William Harvey, 1578—
1657) описал большой и малый круг кровообращения
16
ОБЩАЯ ЧАСТЬ
й изложил учение о кровообращении, что оказало ог-
ромное влияние на дальнейшее развитие физиологии,
медицины и зоологии. Ему же принадлежат эмбрио-
логические исследования развития цыпленка в яйце и
млекопитающих (изучалось развитие косуль по мате-
риалам королевской охоты). Гарвеем был сформули-
рован (в работе «Исследования о нарождении живот-
ных», 1666) знаменитый афоризм «всякое животное
из яйца» («omne animal ovo»). При этом нужно иметь
в виду, что «яйцо» Гарвея вовсе не было яйцевой
клеткой (которая была открыта позже), а представля-
ло собою зародыш млекопитающих на ранних стади-
ях развития.
В начале XVII в. произошло открытие микроскопа,
имевшее огромное значение для развития биологи-
ческих наук. Голландский естествоиспытатель-люби-
тель Левенгук (Antony van Leewenhoek, 1632—1723)
открывает целый новый неизвестный мир микроско-
пических существ, большинство которых являются
животными (простейшие, коловратки, мелкие нема-
тоды и т. п.). Ему же принадлежит описание сперма-
тозоидов человека. Эти открытия произвели большое
впечатление на современников. Итальянец Мальпиги
(Marcello Malpighi, 1628—1694) сделал исключитель-
но много в познании тонкого строения растений и
животных (как позвоночных, так и беспозвоночных).
Ему, в частности, принадлежит открытие капилляров,
которые не были еще известны Гарвею. Голландец
Сваммердам (Jan Swammerdam, 1637—1680) извес-
тен своими монографическими описаниями анатомии
беспозвоночных. Он широко пользовался микроско-
пической техникой.
В XVI—XVII вв. крупные успехи делает эмбрио-
логия животных, начиная с работ Фабрициуса (см.
выше). Исследования Гарвея, Сваммердама, Мальпиги,
Валлиснери (Antonio Vallisnieri, 1661—1730), Гарт-
секера (Nicolas Hartsoeker, 1656—1725) и ряда дру-
гих ученых приводят к формулировке двух, в значи-
тельной мере противоположных, концепций инди-
видуального развития: преформизма и эпигенеза.
Преформисты утверждали, что развитие зародыша
сводится к простому «разворачиванию» уже сущест-
вующей в яйце или сперматозоиде организации. Таким
образом, онтогенез представляет собой чисто коли-
чественные преобразования. Сторонниками префор-
мизма были Левенгук, Сваммердам, философ Лейб-
ниц и многие другие. Среди преформистов существо-
вало две группы. Одни (овулисты) полагали, что
зародыш заключен в яйце и сперматозоид дает лишь
толчок для его роста (Сваммердам, Валлиснери).
Другие (анималькулисты — Гартсекер, Левенгук и
др.) считали, что зародыш заключен в сперматозоиде
и что он растет и питается за счет яйца. В XVIII в.
преформизм нашел яркое выражение в трудах швей-
царца Шарля Боннэ (Charles Bonnet, 1720—1793). В
противоположность преформистам эпигенетики ут-
верждали, что зародыш развивается из неорганизо-
ванной массы, в которой по мере развития лишь по-
степенно возникают структуры. Из ранних эмбриоло-
гов к этой точке зрения примыкал Гарвей.
Теория эпигенеза была развита несколько позд-
нее, в XVIII в., в трудах выдающегося эмбриолога,
работавшего в России, Каспара Фридриха Вольфа
(Caspar Friedrich Wolff, 1733—1794). Теория пре-
формации, по существу, отрицала развитие как ка-
чественное новообразование. Эпигенез же приводил
многих исследователей к признанию некоего немате-
риального организующего начала, которое направля-
ет индивидуальное развитие по определенному пути в
сторону усложнения организации.
Накопление фактических знаний о многообразии
животного мира, его строении и индивидуальном раз-
витии приводит в конце XVII и первой половине
XVIII в. к первым попыткам создания системы, вы-
ходящей за границы аристотелевских представлений.
В этом плане существенное значение имели работы
двух англичан, выполненные в тесном сотрудничест-
ве, — Рея (John Ray, 1627—1705) и Виллоуби (Fran-
cis Willugby, 1635—1672). Рей известен как бота-
ническими, так и зоологическими трудами. В его
совместных с Виллоуби зоологических сочинениях,
основанных в значительной мере на собственных
экспедиционных исследованиях, дается классифика-
ция животных, во многом основанная (в отношении
позвоночных) на анатомических признаках (строение
органов дыхания, сердца) (1693, 1710). Однако глав-
ной заслугой Рея является попытка сформулировать
понятие вида как совокупности особей, похожих друг
на друга, как дети на родителей. В этом отношении
он явился предшественником знаменитого шведского
естествоиспытателя Карла Линнея (Carl von Linnae-
us, 1707—1778). Роль последнего в развитии бота-
ники и зоологии чрезвычайно велика. В вышедшей
первым изданием в 1735 г. «Системе природы» («Syste-
ma naturae») была четко установлена иерархия систе-
матических единиц (вид, род, порядок, класс) и со-
здана бинарная латинская номенклатура. Это сразу
внесло порядок в тот номенклатурный хаос, который
царил в ботанике и зоологии. Способ описания видов
в форме кратких диагнозов позволил наглядно пред-
ставить многообразие организмов и стимулировал
дальнейшее развитие систематики. Линней более
подробно разработал ботаническую систему. Что ка-
сается зоологии, то он разделил царство животных на
6 классов: млекопитающие, птицы, гады, рыбы, насе-
комые, черви. Таким образом, система беспозвоноч-
ных у Линнея была еще очень несовершенной. Лин-
ней непрерывно развивал и улучшал созданную им
«Систему природы», которая при его жизни вышла
12 изданиями (13-е издание было уже посмертным).
Общее число описываемых им видов животных в 10-м
издании было 4200, из них 1222 приходилось на поз-
воночных, 1936 —- на насекомых, 674 — на моллюс-
ков и плеченогих и лишь 376 — на всех остальных
беспозвоночных. Линней -— яркий представитель ме-
тафизической методологии в биологии, основными
положениями которой было представление о неиз-
менности природы и имманентно присущей организ-
мам целесообразности, отражающей мудрость твор-
ца. В своем знаменитом произведении «Философия
ботаники» («Philosophia botanica», §157) в 1751 г.
Линней писал: «Видов насчитывается столько, сколь-
ко разных форм создано в самом начале». Хотя в
конце жизни Линней допускал возникновение новых
видов в результате скрещивания, эти допущения не
меняли его общей метафизической концепции.
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
17
XVIII в. в истории зоологии характеризуется
дальнейшим быстрым накоплением фактов в систе-
матике, эмбриологии, сравнительной анатомии, воз-
никновением новых направлений исследования и за-
рождением идеи развития органического мира, кото-
рая станет господствующей лишь во второй половине
XIX в. На развитие прогрессивных направлений в
зоологии, как и биологии в целом, большое влияние
оказали французские философы-материалисты, из-
вестные под именем энциклопедистов, так как боль-
шинство из них было связано с изданием во Франции
знаменитой энциклопедии («Энциклопедия, или Тол-
ковый словарь наук, искусств и ремесел» — «Encyc-
lopedic, ou Dictionnaire de sciences, des arts et des me-
tiers», 35 томов, 1751—80 гг.). Французский матери-
ализм XVIII в. был в основном механистическим
материализмом. Тем не менее в борьбе с пережитка-
ми феодализма, католицизма и метафизикой он играл
огромную прогрессивную революционизирующую
роль. Наибольшее значение имели труды философов
Ламетри (J. La Metrie, 1709—1751), Дидро (D. Dide-
rot, 1713—1784), Гольбаха (Р. Н. Holbach, 1723—
1789), Гельвеция (С. Helvetius, 1715—1751), Робинэ
(J.-B. Robinet, 1735—1820) и ряда других. В произ-
ведениях французских материалистов XVIII в. неред-
ко высказывается мысль об изменяемости видов (осо-
бенно отчетливо — у Дидро). Среди французских
зоологов XVIII в. исключительно большой попу-
лярностью пользовался Жорж Бюффон (G. Buffon,
1707—1788). Его 36 томов «Естественной истории»
включали обширный материал по описанию преиму-
щественно млекопитающих и птиц. Его описания
животных включают огромный экологический мате-
риал и больше напоминают популярные очерки. В
вопросе об изменяемости видов Бюффон занимал не-
определенную, колеблющуюся позицию. К концу
жизни он, однако, признал существование эволюции
и рассматривал, например, животных Нового Света
как произошедших от животных Старого Света. При-
чину этих изменений он видел в непосредственном
влиянии климатических условий.
В XVIII в. в широких масштабах продолжается
изучение фауны на всех материках земного шара.
В качестве примера можно назвать экспедиции
русских ученых С. П, Крашенинникова (1711—1755),
Г В. Стеллера (1709—1746), И. Г, Гмелина (1709—
1755) в Восточную Сибирь и на Камчатку, во время
которых была описана ныне вымершая стеллерова
корова. К этому же периоду относятся и первые зоо-
логические исследования экспериментального харак-
тера. Трамблэ (A. Trembley, 1710—1784) публикует
свои наблюдения над размножением и регенерацией
гидр. Спалланцани (L. Spallanzani, 1729—1799) про-
водит ряд экспериментальных исследований по размно-
жению животных, в частности, он впервые осущест-
вляет искусственное оплодотворение у рыб (1787).
Ему же принадлежит и ряд экспериментов над про-
стейшими, у которых он наблюдает деление, инцис-
тирование и выход из цист. Спалланцани, опираясь
на опыты, выступает против широко распространен-
ного в XVIII в. представления о самозарождении.
Резко возрастает объем знаний о микроскопи-
ческих организмах. В этом плане особенно большое
2 Протесты, часть 1
значение имели работы Отто Фридриха Мюллера
(О. F. Muller, 1730—1784), описавшего около 400 ви-
дов микроскопических существ («Animalcula infuso-
ria», 1786), разных простейших, коловраток и дру-
гих. Шарль Боннэ, о преформистских взглядах кото-
рого уже было сказано, открывает партеногенез тлей.
В книге «Созерцание природы» (1764) он строит лест-
ницу существ, в основе которой находятся воздух,
вода и огонь, затем следуют неорганические субстан-
ции, далее — растения в восходящем порядке услож-
нения, далее — животные, человек и на вершине ее
бесплотные существа — ангелы различных степеней.
Эта лестница существ отнюдь не означала развития, а
давала представление о проявлении божественного
начала в природе. Во второй половине XVIII в. рабо-
тало много выдающихся зоологов, исследования ко-
торых были посвящены отдельным группам живот-
ных, их анатомии, систематике, биологии. Среди них
были Реомюр (R. A. Reaumur, 1683—1757), автор
семитомного сочинения «Мемуары по изучению ис-
тории насекомых» (1734—1742), Фабриций (J. Fabri-
cius, 1745—1898), опубликовавший 18 томов по сис-
тематике насекомых и значительно усовершенство-
вавший их систему.
XIX век
Конец XVIII и начало XIX в. в истории зоологии
характеризуются появлением ряда крупных исследо-
ваний, которые значительно продвинули познание
животного мира. Вместе с тем благодаря этим иссле-
дованиям ярко проявились две противоположные
идеологии: метафизическая, утверждавшая неизме-
няемость живой природы, и учение о развитии орга-
нического мира. К первой из названных концепций
относятся труды знаменитого французского ученого
Жоржа Кювье (Georges Cuvier, 1769—1832, и его
школа) — выдающегося сравнительного анатома, па-
леонтолога и вместе с тем яркого и воинствующего
представителя учения о неизменяемости форм в при-
роде. В пятитомном курсе сравнительной анатомии
(1799—1805) и четырехтомном «Царстве животных»
(1817—1829) Кювье было разработано учение о кор-
реляции органов, основанное на представлении о
целостности организма и взаимосвязанности его час-
тей. Очень важным явилось введение понятия типа,
которое основывалось на строении нервной системы.
Кювье установил 4 типа: позвоночные, моллюски,
членистоногие и лучистые. Кювье обосновал понятие
типа, которое он называл «ветвью» (embranchement),
само же слово «тип» было введено позже (1825) его
учеником Бленвиллем (A. Blainville, 1778—1850).
Изучая ископаемые фауны, Кювье видел их от-
личия от представителей современного животного
мира. Чтобы сочетать эти факты с учением о не-
изменности животного мира, Кювье вынужден был
создать теорию катастроф, согласно которой фауна
в прежние геологические периоды практически
уничтожалась грандиозными катаклизмами. Ученик
Кювье д’Орбинъи (A. D. d’Orbigny, 1802—1857) позже
дополнил эту теорию учением о повторных актах
творения.
18
ОБЩАЯ ЧАСТЬ
Современник Кювье, французский зоолог Жоф-
фруа Сент-Илер (Е. Geoffroy Sant-Hilair, 1772—
1844) в отличие от учения Кювье о четырех типах
строения животных развил идею о единстве плана
строения всех животных, причем сравнительно-ана-
томические представления он связывал с единством
происхождения животных, с их эволюцией, осущест-
влявшейся под прямым воздействием внешней среды.
Между Кювье и Жоффруа Сент-Илером произошло
серьезное столкновение, которое выразилось в дли-
тельном диспуте, проходившем в Париже в Музее ес-
тественной истории в 1830 г. Идее единства плана
строения (идея типа) сочувствовал и великий поэт и
естествоиспытатель Иоганн Вольфганг Гёте (J. W. von
Goethe, 1749—1832).
Большой вклад в развитие зоологии беспозвоноч-
ных внес современник Кювье Жан Батист Ламарк
(J. В. de Lamarck, 1744—1829). Ему принадлежит и
термин «беспозвоночные». В опубликованной се-
митомной (1815—1822) «Системе беспозвоночных
животных» Ламарк устанавливает 10 классов беспоз-
воночных (вместо двух имевшихся у Линнея) —
инфузории, полипы, лучистые, черви, насекомые,
паукообразные, ракообразные, кольчецы, усоногие,
моллюски, что представляло большой шаг вперед в
развитии зоологии. Ламарк также явился творцом пер-
вой эволюционной теории, изложенной в его «Фило-
софии зоологии» (1809).
В начале XIX в. рядом анатомов и эмбриологов
были получены данные о наличии в эмбриогенезе выс-
ших животных стадий, несущих признаки более
низко организованных групп (например, жаберные
щели у наземных позвоночных и т. п.), которые под-
тверждали идею единства плана строения всех жи-
вотных. Эти факты позже получили объяснение с по-
зиций эволюционной теории. Меккелем (J. Meckel,
1781—1833) и некоторыми другими исследователями
они трактовались как проявление предустановленно-
го общего плана строения. Противником постоянства
видов выступал ученый из Московского университета
К. Ф. Ру лье (1814—1858), призывавший изучать жи-
вотный организм в тесной связи с условиями среды.
Рулье явился одним из зачинателей экологии.
Важнейшим событием в истории зоологии и био-
логии в целом во второй четверти XIX столетия яви-
лось создание клеточной теории, которая была под-
готовлена в области ботаники Шлейденом (М. Schlei-
den, 1804—1881) и обоснована Шванном (Th. Schwann,
1810—1882) в его знаменитой работе «Микроскопи-
ческие исследования о соответствии в строении и
росте животных и растений» (1839). Развитие знаний
о строении клетки позволило Зибольду (С. Siebold,
1804—1885) сформулировать в 1845 г. представление
о простейших как об одноклеточных организмах и
выделить их в самостоятельный тип. Этим было окон-
чательно отвергнуто распространенное и развитое
Эренбергом (С. G. Ehrenberg, 1795—1876) учение о
простейших как о совершенных организмах, отли-
чающихся от других животных лишь мелкими разме-
рами. Эти взгляды Эренберг развил в монографии
«Животные инфузории как совершенные организмы»
(1838), пользовавшейся широкой популярностью.
Против них выступил французский протозоолог Дю-
жарден (F. Dujardin, 1801—1860), утверждавший, что
тело простейших состоит из полужидкого вещест-
ва — «саркоды» и не имеет сложной организации.
Дюжарден опроверг представления о фораминиферах
как о мелких головоногих моллюсках и отнес их к
простейшим.
В первой половине XIX в. было выполнено боль-
шое количество работ по отдельным группам живот-
ных и сделано много зоологических открытий.
В области эмбриологии русский академик К. М. Бэр
(1792—1876) опубликовал 2 тома «Истории развития
животных» (т. I, 1828; т. II, 1837). В этих классичес-
ких исследованиях заложены основы современной
эмбриологии. Бэр описал развитие основных органов
цыпленка, развил учение о зародышевых листках (у
позвоночных), дал эмбриологическое обоснование
теории типов. Рудольфи (К. Rudolphi, 1771—1832) в
серии работ положил начало науке о паразитических
червях (гельминтологии). Мильн-Эдвардс (Н. Milne-
Edwards, 1800—1885) опубликовал ряд крупных сво-
док по ракообразным (1828—1849) и коралловым
полипам (1858—1860) и значительно усовершенство-
вал систему беспозвоночных. Лейкарт (R. Leuckart,
1822—1898) выделил иглокожих из установленных
Кювье Radiata, отделив их от кишечнополостных.
Большой вклад в сравнительную анатомию внес Мюл-
лер (J.Muller, 1801—1858). Cape (М. Sars, 1808—
1869) ввел понятие «чередование поколений» у сци-
фоидных медуз (1835—1937). Стенструп (J. Steenst-
rup) в 1842 г. описал чередование поколений и
развитие сосальщиков. Все эти исследования прихо-
дили во все большее противоречие с метафизической
концепцией неизменяемости видов. Эта концепция
была разрушена гениальным творением Чарлза Дар-
вина «Происхождение видов» (1859), положившим
начало новой эпохе в развитии биологии.
Учение Ч. Дарвина
Эволюционные представления Дарвина стали
формироваться во время кругосветного путешествия
на корабле «Бигль» (1831—1836). Однако потребова-
лось более двух десятилетий кропотливого труда по
сбору литературных и музейных материалов, а также
оригинальных экспериментальных исследований для
всестороннего обоснования и развития гипотезы ес-
тественного отбора. В доэкологическую и догенети-
ческую эру биологии Дарвин смог показать наследст-
венный характер значительной части внутривидовой
изменчивости и избирательный характер выживае-
мости организма в результате сложных взаимодейст-
вий с абиотическими и биотическими факторами.
Особое значение имела работа Дарвина над учением
об искусственном отборе как главном методе выведе-
ния новых пород животных и сортов культурных рас-
тений. Используя в качестве модели механизм дейст-
вия искусственного отбора и опираясь на данные
самых различных отраслей биологии того времени,
Дарвин сформулировал гипотезу о причинах эволю-
ции животных и растений. Он предполагал, что она
осуществляется посредством естественного отбора,
главнейшими факторами которого выступают неоп-
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
19
ределенная изменчивость, геометрическая прогрессия
размножения и борьба за существование. И хотя в
распоряжении Дарвина еще не было прямых доказа-
тельств существования естественного отбора в при-
роде, он привел такое количество косвенных данных,
что выдвинутая им гипотеза убедила биологов в ре-
альности исторических изменений органических
форм.
Учение о естественном отборе объяснило причи-.
ны формирования видов со всем комплексом слож-
ных структур и функций, присущих каждому из них,
а следовательно, и причины современного многооб-
разия органического мира. В рамках этого учения
проблема органической целесообразности, и прежде
всего приспособленности организмов к условиям су-
ществования, получила естественнонаучное истолко-
вание. Вместе с тем был вскрыт и относительный
характер органической целесообразности, что в ко-
нечном счете вело к элиминации телеологии из био-
логии в целом и из зоологии в том числе. Дарвинизм
прокладывал пути и для изучения основных филоге-
нетических закономерностей эволюции, и к решению
вопросов о движущих силах прогрессивной эволю-
ции, неравномерности темпов эволюции, ее необра-
тимости и т. д. Особое значение имеет обоснованный
Дарвином принцип расхождения признаков, согласно
которому борьба за существование и естественный
отбор ведут ко все нарастающей дивергенции ранее
сходных форм, а следовательно, и к постепенному
увеличению разнообразия органического мира. Прин-
цип дивергенции объяснял и существующую иерар-
хию таксонов и позволял провести их естественную
классификацию, построенную на филогенетических
связях. Важное значение имела и дарвиновская кон-
цепция происхождения человека, в которой отмеча-
лись специфические особенности движущих сил ан-
тропогенеза, обусловивших становление характерных
человеку свойств.
Последарвиновский период
развития зоологии
(вторая половина XIX века)
Дарвинизм, утвердивший исторический метод в
биологии и раскрывший основной фактор эволю-
ции — естественный отбор, создал новую методоло-
гическую материалистическую основу биологической
науки и поставил перед всеми направлениями зооло-
гии новые задачи. Новая, прогрессивная методология
вызвала настоящий взрыв исследований. Старые, тра-
диционные разделы зоологии — систематика, фау-
нистика, зоогеография, сравнительная анатомия,
эмбриология животных — получили свое новое со-
держание. Перед ними встали и новые задачи. Возни-
кает целый ряд новых зоологических дисциплин, вы-
званных к жизни дарвинизмом. К их числу относятся
филогенетика, изучающая пути исторического разви-
тия животного мира, эволюционная эмбриология,
эволюционная палеозоология и другие. Эволюцион-
ное учение за короткий срок принимается всеми
биологами. Споры возникают не о факте эволюции
организмов, а о факторах эволюции. В этом плане
материалистическая дарвиновская концепция исто-
рического развития встретила со стороны некоторых
зоологов противодействие и выдвинула другие, анти-
дарвиновские концепции автогенетического толка.
На основе дарвинизма перестраивается система жи-
вотного мира, перед которой возникает новая зада-
ча — построение естественной системы, отражаю-
щей исторический путь развития животного мира.
Огромное значение в этом направлении имели иссле-
дования немецкого «апостола дарвинизма» Эрнста
Геккеля (Е. Haeckel, 1834—1896). В целом ряде спе-
циальных работ, посвященных радиоляриям, меду-
зам, губкам, сифонофорам, в книгах «Систематичес-
кая филогения» (т. I—III, 1894—1896), «Всеобщая
морфология организмов» (1866) и других трудах Гек-
кель последовательно развивает филогенетическую
зоологию, усовершенствует систему животных, создает
теорию происхождения организмов от колоний про-
стейших (теория гастреи, 1873 г.), формулирует уче-
ние филогенеза. Ему вместе с Ф. Мюллером (F. Mul-
ler, 1822—1897) принадлежит создание биогене-
тического закона, устанавливающего закономерную
связь между онтогенезом и филогенезом. Пропаганде
дарвинизма Геккель посвятил также несколько попу-
лярных книг («Мировые загадки», «Миротворение»),
которые пользовались широчайшей известностью и
были переведены на многие языки (в том числе и на
русский). Некоторые из построений Геккеля оказа-
лись недостаточно обоснованными, однако его роль в
пропаганде дарвинизма и внедрении эволюционной
теории в конкретные зоологические исследования ве-
лика. Усовершенствование зоологической системы в
свете идеи эволюции осуществляется и другими зоо-
логами, работающими во второй половине XIX в.
Среди них Томас Гексли (Т. Huxley, 1825—1895) —
пропагандист и защитник дарвинизма в Англии. На-
ряду с первоклассными исследованиями в области
зоологии, палеонтологии, сравнительной анатомии,
он предложил систему, включавшую 8 типов, которая
сыграла большую историческую роль. В ней иглоко-
жие рассматривались отдельно от кишечнополост-
ных, низшие черви — от кольчатых. Однако иглоко-
жих Гексли поставил в непосредственную близость с
червями на основании кажущегося сходства личинок.
Заметную роль в развитии зоологии и системы сыг-
рал выдающийся английский зоолог Рей-Ланкестер
(Е. Ray Lankester, 1847—1929). Он разбил червей на
3 типа, отнес кишечнодышащих и оболочников к
хордовым (1877). Ему принадлежит известное девя-
титомное руководство по зоологии (1900—1909).
Большое значение для развития зоологии имело со-
здание Бронном (Н. Вгопп, 1800—1862) многотомной
серии (ее издание началось в 1859 г.) «Классы и от-
ряды животного мира», в значительной своей части
посвященной разработке зоологической системы. Су-
щественное влияние на развитие зоологии в этот пе-
риод оказали работы Ланга (A. Lang, 1855—1914),
разработавшего теорию происхождения кольчатых
червей от турбелляриц, плоскцх червей от гребневи-
ков и создавшего учебник сравнительной анатомии
беспозвоночных (1888—1894). Большое значение
для создания системы одноклеточных организмов
20
ОБЩАЯ ЧАСТЬ
имели работы Бючли (О. Biitschli, 1848—1920), авто-
ра капитальной сводки по простейшим (1889). Среди
обобщающих руководств конца XIX и начала XX в.
следует также упомянуть многотомное французское
издание Делажа (J. Delage, 1854—1920) и Эруара
(Е. Herouard, 1858—1932) «Руководство конкретной
зоологии» («Traite de zoologie concrete», 1897—
1901), составленное на основе создания обобщенных
«морфологических типов», характерных для крупных
таксонов.
Разработка филогении и системы животных была
неразрывно связана со сравнительно-анатомическими
исследованиями, которые привели к установлению
некоторых существенных морфофизиологических за-
кономерностей эволюционного процесса. Сравни-
тельно-анатомические работы переплетаются с палео-
зоологическими. Палеонтология из науки, описываю-
щей ископаемые организмы, превращается в науку,
раскрывающую конкретные пути органической эво-
люции. На эволюционной основе был построен курс
сравнительной анатомии позвоночных Гегенбаура
(К. Gegenbaur, 1826—1903), изданный в 1867 г., на
котором воспитывалось не одно поколение зоологов.
Дорном (A. Dhom, 1840—1909) был установлен прин-
цип мультифункциональности органов и смены функ-
ций в процессе эволюции. Клейненберг (N. Kleinen-
berg, 1842—1897) разрабатывает принцип субститу-
ции органов (1886). Большое значение для анализа
эволюции животных имело разработанное основате-
лем эволюционной палеонтологии В. О. Ковалевским
(1842—1883) на материале ископаемых копытных
учение об адаптивных и инадаптивных путях эволю-
ции. Последователь Ковалевского — Долло (L. Dollo,
1857—1931) обосновал важный принцип необрати-
мости эволюции. Коп (Е. Соре, 1840—1897) устано-
вил принцип неспециализации, согласно которому
филогенетические ветви берут начало от неспециали-
зированных форм (1875).
Характерной чертой истории зоологии второй
половины XIX в. было создание сравнительной эм-
бриологии. Возникновение и развитие ее связано с
именами русских зоологов-эмбриологов А. О. Кова-
левского (1840—1901) и И. И. Мечникова (1845—
1916). Их исследования позволили распространить
учение о зародышевых листках на всех беспозвоноч-
ных. Наряду с этим Ковалевский решил ряд важных
вопросов положения в системе до того «загадочных»
по своему филогенетическому положению групп. Наи-
более ярким стало доказательство связи асцидий с
хордовыми, так как у личинок асцидий были обнару-
жены нервная трубка и хорда (1868). Вслед за Кова-
левским и Мечниковым и другие исследователи нача-
ли энергично разрабатывать эмбриологию разных
групп животных, причем особенно много в этом на-
правлении было сделано русскими зоологами (В. В. За-
ленский, 1847—1918; Н. В. Бобрецкий, 1843—1907;
С. М. Переяславцева, 1849—1903; А. А, Коротнев,
1854—1915; КН. Давыдов, 1877—1960; В.М.Шим-
кевич, 1858—1923 и др.). Эти исследования соста-
вили прочный фундамент для совершенствования
системы и филогении животного мира.
Быстрыми темпами развиваются систематика, фа-
унистика, зоогеография. Эти исследования в свете
дарвинизма также приобретают новое содержание,
ибо в них ставятся вопросы происхождения совре-
менных фаун в связи с их геологическим прошлым и
историей материков и морей. Во второй половине
XIX в. начинается систематическое изучение фауны
Мирового океана. Особенно большое значение в
этом плане имела кругосветная английская экспеди-
ция на корабле «Челленджер» (1872—1876). Зооло-
гические сборы этой экспедиции обширны, и они
были обработаны лучшими зоологами мира. Число
описанных видов животных быстро возрастает. Лин-
нею было известно 4200 видов, в первой половине
XIX в. их число возросло до 48 тысяч, а к концу века
превысило 412 тысяч.
В конце XIX в. начинает развиваться экология
животных. Термин этот был предложен Геккелем в
1866 г. для раздела биологии, изучающего взаимодей-
ствие животных и растений с окружающей их физи-
ко-химической и органической средой. Среди первых
крупных зоологов-экологов, основателей экологии,
следует назвать Мебиуса (К. Mobius, 1825—1908),
разработавшего на примере анализа устричных банок
понятие биоценоза и предложившего впервые этот
термин (1877), ученика Рулье Н. А. Северцова (1827—
1885), автора известного исследования «Периодичес-
кие явления в жизни зверей, птиц и гад Воронежской
губернии. Разсуждение, написанное для получения
степени магистра зоологии Николаем Северцовым по
наблюдениям, сделанным в 1844—53 годах» (М. : ти-
пография А. Евреинова, 1855). В эти же годы возни-
кает в качестве самостоятельной отрасли паразитоло-
гия, основателем которой следует считать Лейкарта
(R.Leuckart, 1822—1898); его известная книга «Об-
щая естественная история паразитов...» появилась в
1879 г.
Зоология в XX веке
и современные тенденции
ее развития
В XX в. все больший размах принимают фаунис-
тические исследования ранее не изучавшихся регио-
нов и сред обитания — абиссаль Мирового океана,
Антарктика, Арктика. Резко возрастает общее число
описанных видов, достигающее, по данным разных
авторов, свыше полутора миллионов. Но и в настоя-
щее время инвентаризация фауны Земли далеко не
завершена. Чрезвычайно много для зоологии дало
изучение фауны Мирового океана. В этих исследо-
ваниях участвовали десятки экспедиционных судов
всех стран. Особенно большое значение имели эк-
спедиции советского судна «Витязь» и датского «Га-
латея». С их работами связаны крупные зоологичес-
кие открытия. По материалам экспедиции на «Витя-
зе» А. В. Иванов (1906—1992) описал в 1960 г. новый
тип животных — погонофор, обладающих крайне
своеобразной организацией. Датские исследователи
Лемхе и Вингстранд во время экспедиции на «Гала-
тее» обнаружили в абиссали Восточной части Тихого
океана и описали примитивных моллюсков неопилин
(класс моноплакофоры, Monoplacophora) (Lemche,
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
21
Wingstrand, 1957, 1960). У юго-восточных берегов
Африки была поймана и в 1938 г. описана Смитом
(G. Smith) кистеперая рыба латимерия. До этой на-
ходки кистеперые рыбы были известны лишь из па-
леозойских и мезозойских отложений.
Особенно велик объем работ, выполненных в XX в.
по изучению насекомых (число описанных видов
близко к миллиону). Это связано как с огромной их
ролью в биосфере, так и с непосредственным практи-
ческим значением (вредители сельского хозяйства,
паразиты, переносчики заболеваний и т. п.). Исследо-
вание насекомых составляет самостоятельную обшир-
ную область зоологии — энтомологию.
В XX в. работа по изменению и усовершенствова-
нию системы животного мира продолжается, и она
еще далека от завершения (см. раздел «Система жи-
вотного мира»). На основе сравнительно-анатомичес-
ких, эмбриологических и палеонтологических иссле-
дований значительно возросло число выделенных выс-
ших таксонов — типов и классов. Кювье установил
4 типа, а, например, В. Н. Беклемишев (1890—1963)
в «Основах сравнительной анатомии беспозвоноч-
ных» (1964) выделяет 16 типов. Ряд современных
зоологов насчитывает их еще больше. Увеличению
числа крупных таксонов способствовало также ис-
пользование в зоологических исследованиях новых
методов, в результате чего были изучены ранее неиз-
вестные особенности организации животных. Осо-
бенно большое значение имела для развития зооло-
гии электронная микроскопия. Благодаря широкому
применению электронного микроскопа наши знания
об организации животных, в особенности низших
животных (в том числе одноклеточных), значительно
углубились и во многом изменились. В частности,
принимавшийся со времен Зибольда (С. Siebold, 1804—
1885) тип простейших (одноклеточных) в настоящее
время большинством исследователей делится на не-
сколько типов (о системе простейших см. раздел
«Система протестов и проблема их мегасистема-
тики»).
Во второй половине XX в. наметилась тенденция
объединять типы животных в еще более крупные
группировки, отражая этим основные этапы филоге-
нии. Если в отношении позвоночных животных ос-
новные пути филогенеза могут считаться установлен-
ными, то в отношении беспозвоночных имеет место
огромное разнообразие мнений и идут оживленные
дискуссии^ Опубликован ряд обобщающих трудов,
обсуждающих проблемы филогении. Основные рас-
хождения в построении системы базируются на раз-
ном понимании узловых точек филогенеза животного
мира. К их числу относятся проблемы происхожде-
ния многоклеточности, происхождения вторичной
полости тела и кровеносной системы (монофилети-
ческое или полифилетическое), соотношение вторич-
нополостных и паренхиматозных животных, первич-
ность или вторичность радиальной симметрии ки-
шечнополостных.
При изучении конкретных путей эволюции жи-
вотного мира сталкиваются две противоположные
тенденции. Значительная часть зоологов, исходя из
дарвиновских принципов естественного отбора, при-
нимает монофилетический характер филогенеза, счи-
тая, что дивергенция есть основной путь эволюции.
Другие в большей или меньшей степени стоят на по-
зициях полифилии — параллельного и конвергент-
ного развития (см. с. 48—53).
Большое влияние на развитие зоологии в XX в.
оказывает издание многотомных руководств, являю-
щихся сводом знаний по отдельным группам живот-
ных. С 1923 г. в Германии первоначально под редак-
цией Кюкенталя (W. Kiikenthal, 1861—1922) стало
издаваться «Руководство по зоологии» («Handbuch der
Zoologie»), которое не завершено и по сей день. Во
Франции под руководством Грассэ с 1948 г. выходит в
свет многотомный «Трактат по зоологии» (Р. Р. Gras-
se — «Traite de Zoologie») — издание также еще не за-
кончено. В США в 1940 г. Хайман (L. Hyman, 1888—
1969) начала публиковать многотомное руководство
«Беспозвоночные». Вышло 6 томов (последний — в
1959 г.), охватывающих примерно половину беспозво-
ночных. Издание прекратилось в связи с кончиной
автора. В СССР в 1937 г. начало издаваться много-
томное «Руководство по зоологии» под редакцией
Л. А. Зенкевича (1889—1970). Вышло всего 4 тома
(последний — в 1951 г.). Издание до конца не доведе-
но. Все эти публикации играют большую положитель-
ную роль, особенно в подготовке кадров ученых.
В зоологии XX в. энергично разрабатывается про-
блема вида. Важное место в этих исследованиях за-
нимает учение о популяциях как реальных формах
существования вида и элементарных единиц, в не-
драх которых начинается процесс микроэволюции —
видообразования. Вопросы структуры вида непосред-
ственно соприкасаются с популяционной генетикой и
эволюционной экологией животных, изучающей эко-
логические механизмы микроэволюции: многочис-
ленные работы Н. В. Тимофеева-Ресовского (1900—
1981), Майра (Е.Мауг, 1904—1970), С, С Шварца
(1919—1976) и других. Традиционные методы систе-
матики обогащаются новыми подходами и контакта-
ми с другими биологическими науками — биохи-
мией, молекулярной биологией, цитологией. Так,
А. И. Белозерский (1905—1972) изучал нуклеотидный
состав ДНК разных видов. Эти работы продолжают-
ся. Все шире в зоологии используется кариологичес-
кий анализ. Изучение числа и структуры хромосом
позволяет делать важные выводы в области система^
тики главным образом близких видов (Я. Н. Ворон-
цов). Широко используются серологические методы.
Наряду с микроэволюцией в зоологии, в особен-
ности в первой половине XX в., энергично разраба-
тываются проблемы макроэволюции, закономерности
эволюционных преобразований крупных таксонов. В
этом плане особенно большую роль сыграли работы
А. Н. Северцова (1866—1936) и его обширной школы
эволюционных морфологов, посвященные морфофи-
зиологическим закономерностям эволюции. Установ-
ление разных типов эволюционных изменений —
ароморфозов, идиоадаптаций, общей дегенерации —
позволило осуществить глубокий анализ путей эво-
люции позвоночных животных (объекты исследова-
ний Северцова), а затем распространить его на об-
ширный мир беспозвоночных. Не меньшее значение
для зоологии имела разработка Северцовым пробле-
мы соотношения онтогенеза и филогенеза.
22
ОБЩАЯ ЧАСТЬ
Среди учеников А. Н. Северцова особая роль при-
надлежит И, И, Шмальгаузену (1884—1963), который
связал основные закономерности эволюции с данны-
ми современной генетики и разработал учение о раз-
ных формах естественного отбора.
В познании морфофизиологических законов эво-
люции животных существенную роль сыграли работы
В. А. Догеля (1882—1955), в первую очередь его уче-
ние о роли полимеризации (увеличения) и олигоме-
ризации (уменьшения) числа гомологичных органов в
эволюции животных (1954). В морфологии животных
одним из важнейших обобщений в XX в. следует
считать учение 77. 77. Иванова (1878—1942) о первич-
ной и вторичной гетерономное™ метамерных живот-
ных.
Эмбриология животных в XX в. наряду с накопле-
нием и углублением материала 4ю морфологии онто-
генеза характеризуется казуальным, эксперименталь-
ным и функциональным анализом индивидуального
развития. Начало этому направлению, получившему
название механики и физиологии развития, положил
в Германии Вильгельм Ру (W.Roux, 1850—1924),
сформулировавший представления о самодифферен-
цировке и зависимой дифференцировке (1888). Про-
должение работ в этом направлении связано с имена-
ми многих исследователей: Шпемана (G. Spemann) и
Голыпфретера (J. Holtfreter) в Германии, Уоддингто-
на (С. Waddington) в Англии, Чайлда (Ch. Child) в
США, Вольфа (Е. Wolff) и Гальена (L. Gallien) во
Франции, Браше (G. Braschet) в Бельгии, Д. Л. Фила-
това и 77. Г. Светлова в бывшем СССР. 77. Г. Свет-
ловым (1892—1974) разработано важное обобще-
ние — учение о критических периодах в онтогенезе.
В текущем столетии широкое развитие получили
разделы зоологии (как в силу их теоретического ин-
тереса, так и практической значимости для медицины
и ветеринарии), предметом изучения которых явля-
ются паразитические животные. Число зоопаразити-
ческих исследований, равно как и число паразитоло-
гов, велико. В зависимости от объекта зоопаразито-
логия распадается на 3 раздела — протозоологию
(изучает паразитических простейших), гельминтоло-
гию (все группы паразитических червей) и арахноэн-
томологию (паразитические членистоногие и пере-
носчики различных заболеваний).
Параллельно с изучением отдельных групп пара-
зитов паразитология решает общие проблемы эколо-
гического и физиологического характера, что связано
с качественным своеобразием паразитов по сравне-
нию со свободноживущими (непаразитическими) жи-
вотными. Оно определяется тем, что средой обитания
паразита является другой живой организм. В XX в.
вопросы общей паразитологии разрабатывались во
многих странах. Очень широко эти исследования раз-
вернулись в бывшем СССР, где возникли крупные на-
учные паразитологические школы. Среди них школа
Е. Н. Павловского (1884—1965), разработавшего уче-
ние об организме как среде обитания, учение о при-
родной очаговости трансмиссивных заболеваний и
выдвинувшего понятие паразитоценоза; школа эколо-
гической паразитологии В. А. Догеля, изучавшего за-
висимость паразитофауны, взятой в целом, от усло-
вий окружающей хозяина среды и от физиологичес-
кого состояния хозяина; школа В. Н. Беклемишева,
изучавшего экологию переносчиков трансмиссивных
заболеваний; школа А. 77. Маркевича (Украина), ко-
торый развивал паразитоценологическое направле-
ние, ставящее задачей всестороннее изучение взаимо-
действия паразитов, населяющих хозяина.
Современная зоология изучает систематику, мбр-
фологию, экологию, этологию (поведение), геогра-
фию животных. Зоология неразрывно связана с эво-
люционной теорией, ибо изучает пути филогенеза
животных. Исследование популяций устанавливает
тесные связи зоологии с генетикой. Тесные связи су-
ществуют также между зоологией и цитологией,
молекулярной биологией. Физиология животных, ко-
торая возникла исторически как ветвь зоологии, вы-
делилась в самостоятельную науку, изучающую функ-
ции животного организма. Зоология входит как со-
ставная часть в такие комплексные дисциплины, как
гидробиология, паразитология, эпизоотология. Она
тесно связана с задачами сельского хозяйства (борьба
с вредителями), медицины, ветеринарии.
Развитие зоологии в России
В предыдущем разделе дан краткий очерк исто-
рии развития зоологии в мире. При этом упомина-
лись и некоторые этапы, проходившие в дореволюци-
онной России, а также бывшем СССР, и соответст-
венно были названы имена крупных отечественных
ученых. Очевидно, что развитие зоологии в нашей
стране неразрывно связано с мировой наукой. Одна-
ко представляется целесообразным дать более развер-
нутый очерк развития зоологии в России и республи-
ках бывшего СССР.
В XVII в. начинается планомерное изучение фауны
России, которое осуществляется благодаря экспеди-
циям. К числу важнейших работ этого периода отно-
сятся, например, исследования Камчатки, которые
проводились Г, В. Стеллером, С. 77. Крашениннико-
вым и И. Г. Гмелиным в составе знаменитой экспеди-
ции В, Беринга. Большое значение имели экспедиции
С. Г. Гмелина по изучению фауны юга России и Да-
гестана. И. И. Лепехин изучал животный мир севера
Европейской России и Сибири. В познание отечест-
венной фауны большой вклад внесли экспедиции и
работы П. С. Палласа, его учеников 77. И. Рычкова и
В. Ф. Зуева.
К первой половине XIX в. относятся экспедици-
онные исследования К. М. Бэра, охватившие Новую
Землю, Финский залив, Чудское озеро, Волгу^ К
этому периоду относятся и экспедиции А. Ф. Мид-
дендорфа на Кольский полуостров, в Сибирь и на
Белое море, в Ферганскую долину.
В Петербургском университете в это время рабо-
тали видные зоологи — С. С. Куторга, К. Ф. Кесслер
(особенно большое значение имели исследования
Кесслера в области ихтиологии).
Уже в начале XIX в. возникают первые русские
научные общества (Общество любителей естествоз-
нания, антропологии и этнографии при Московском
университете; Западное новороссийское общество
естествоиспытателей; Общество испытателей приро-
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
23
ды Харьковского университета; Московское общест-
во испытателей природы; Санкт-Петербургское об-
щество естествоиспытателей и др.). Географическое
общество организует экспедиции Н. М. Пржевальско-
го, А. П. Федченко и П. К. Козлова, которые знако-
мятся с ранее почти неизученной фауной Централь-
ной Азии.
Положительную роль в развитии зоологии, как и
других биологических наук, в России сыграли съезды
русских естествоиспытателей и врачей. С 1867 по
1913 г. состоялось 13 таких съездов.
Во второй половине XIX—XX в. начинается изу-
чение фауны морей, омывающих берега России. Осо-
бенно большой вклад в этом направлении внесли ра-
боты Н. М. Книповича по Баренцеву, Черному, Азовско-
му и Каспийскому морям, С. А. Зернова по Черному
морю, К. М, Дерюгина по Баренцеву, Белому и дальне-
восточным морям. Развитию зоологических исследо-
ваний способствовала организация биологических
станций, в особенности морских. Первыми были орга-
низованы Севастопольская (1871) и Соловецкая (1881)
станции.
Вторая половина XIX в. характеризуется распро-
странением в России учения Дарвина и его даль-
нейшей разработкой. Весьма значительны успехи в
области эмбриологии животных: кроме А. О. Кова-
левского и И. И. Мечникова, о которых уже было
сказано, на этом поприще трудятся такие крупные
ученые, как В. В. Заленский, В. Н. Ульянов, Н. В, Боб-
рецкий, В, М. Шимкевич и другие. Русская эмбрио-
логическая школа занимает ведущее место в мире.
Углубляются исследования в области фаунистики и
систематики различных групп животных. Н. В. На-
сонов, который в течение ряда лет был директором
Зоологического музея Академии наук в Петербурге,
в 1911 г. основывает издание серии монографий
под общим заглавием «Фауна России и сопредель-
ных стран» (издание продолжается и поныне).
Появляются теоретические исследования в области
систематики (А. П. Семенов-Тян-Шанский). М. А. Менз-
бир публикует ряд крупных работ в области зоогеогра-
фии. В конце XIX—XX в. появляется ряд руководств
и учебников, способствовавших распространению и
развитию зоологических знаний. Назовем среди них
«Биологические основы зоологии» (1900) и «Сравни-
тельную анатомию позвоночных» (1905) В. М. Шим-
кевича, «Учебник зоологии и сравнительной анато-
мии» (1905) и «Краткий курс энтомологии» (1890,
1912) Н. А. Холодковского. В. А. Вагнер начинает ис-
следования в области сравнительной зоопсихологии.
В Петербурге успешно работает в области протисто-
логии В. Т. Шевяков.
В царской России развитие зоологии (это справед-
ливо и для других естественных наук) было сосредото-
чено главным образом в немногих университетских
городах. Во всей огромной Сибири существовал лишь
один центр науки — Томский университет. Академия
наук в то время играла в развитии зоологии относи-
тельно небольшую роль, которая сводилась преиму-
щественно к работе в области систематики и фаунис-
тики в Зоологическом музее Санкт-Петербурга в так
называемой Особой зоологической лаборатории, воз-
главляемой в конце XIX в. А. О. Ковалевским.
Расцвет зоологии начинается в нашей стране в
двадцатые годы двадцатого столетия. Возникает мно-
жество новых научных центров на всей территории
тогдашнего Советского Союза, ставятся новые задачи
исследования, в науку вносится элемент плановости.
Разработка научных проблем связывается с народно-
хозяйственными задачами.
Быстрыми темпами в университетах, а также в
вузах другого профиля (педагогических, лесотехни-
ческих, сельскохозяйственных) происходит подго-
товка кадров зоологов. За это время количество оте-
чественных зоологов возросло примерно в 7—8 раз.
Число учреждений различного профиля, где осу-
ществляется научная работа в области зоологии, сей-
час велико. В системе Российской академии наук та-
ковыми являются Зоологический институт с музеем
(Санкт-Петербург), Институт эволюционной морфо-
логии и экологии животных им. А. Н. Северцова (ИПЭЭ,
Москва). Ряд институтов РАН в Москве решают раз-
личные проблемы зоологии: это Палеонтологический
институт, Лаборатория гельминтологии, Институт
биологии развития, Институт океанологии, Институт
биологии внутренних вод (Борок, Рыбинское водо-
хранилище), Институт экологии Волжского бассейна
(Тольятти), Мурманский морской биологический ин-
ститут (Восточный Мурман), Институт биологии
Южных морей (Севастополь, теперь Украина), Био-
логический институт Сибирского отделения РАН
(Новосибирск), Институт биологии моря (Дальневос-
точное отделение РАН, Владивосток), Институт эко-
логии растений и животных (Екатеринбург). Зооло-
гические учреждения имеются и в составе Академий
наук СНГ — это или специальные институты, или
комплексные институты зоологии и ботаники или
биологии. Отделы или лаборатории зоологии входят
также в состав филиалов РАН.
Зоологические исследования морских животных
проводятся Всероссийским научно-исследовательс-
ким институтом рыбного хозяйства и океанографии
(ВНИРО, Москва) совместно с филиалами, имеющи-
мися на всех отечественных морях. Государственный
научно-исследовательский институт озерного и реч-
ного рыбного хозяйства (ГОСНИОРХ, С.-Петербург)
аналогичную работу проводит на пресных водоемах.
Животные, вредящие в сельском хозяйстве (насеко-
мые, грызуны), изучаются Всероссийским институтом
защиты растений (ВИЗР, С.-Петербург) с его перифе-
рийными учреждениями. Разностороннюю работу в
области зоологии ведут биологические станции, мно-
гочисленные заповедники и заказники, находящиеся
под началом различных ведомств. Научная работа в
области зоологии ведется и на кафедрах университе-
тов, педагогических и сельскохозяйственных вузов.
Успехи зоологии в 20—50-е гг. характеризуются
созданием обобщающих сводок и руководств по раз-
личным разделам зоологии. В течение 1937—1951 гг.
вышли 4 тома коллективного труда «Руководство по
зоологии». Большую роль сыграли двухтомная «Срав-
нительная анатомия беспозвоночных» В. А. Догеля
(1938), «Основы сравнительной анатомии беспозво-
ночных» В. Н. Беклемишева (1944), «Курс общей эн-
томологии» Б.Н.Шванвича (1949). Обобщающими
трудами явились также «Курс сравнительной анато-
Z.4
ОБЩАЯ ЧАСТЬ
мии позвоночных животных» В. М. Шимкевича (ряд
изданий, первое из них вышло в 1905 г.) и «Основы
сравнительной анатомии позвоночных животных»
И, И. Шмальгаузена (ряд изданий, первое из них по-
явилось в 1923 г.).
Ряд крупных работ отечественных зоологов был
посвящен общим вопросам филогении. В их числе
назовем монографии Н. А. Ливанова «Пути эволюции
животного мира» (1955), Д. М. Федотова «Эволюция
и филогения беспозвоночных животных» (1966),
И. И, Шмальгаузена «Происхождение наземных поз-
воночных» (19646), А. В. Иванова «Происхождение
многоклеточных» (1968).
Углубленно изучаются морфофйзиологические за-
кономерности эволюции животного мира. К этой, пог-
раничной с эволюционной теорией, области знаний от-
носятся исследования А. Н. Северцова иИ. И. Шмаль-
гаузена. В разработку эволюционной морфологии
позвоночных большой вклад внесли также Б. С. Мат-
веев, С. Г. Крыжановский, С. В. Емельянов, Б. А. Дом-
бровский, М. М. Воскобойников, Д. К. Третьяков,
В. Г. Касьяненко, Н. С. Хранилов и многие другие.
Большой интерес представили работы П. П. Ива-
нова, предложившего теорию первичной гетероном-
ности метамерных животных. Особенно важными для
понимания закономерностей эволюции животных на
уровне тканей явились исследования А. А. Заварзина,
разработавшего учение о параллелизме развития гис-
тологических структур, а также работы Н. Г. Хлопина
по эволюционной гистологии.
В эмбриологии животных большое значение име-
ли исследования 77.77. Иванова и его монография «Об-
щая и сравнительная эмбриология» (1937), а также
труды А. А. Захваткина («Сравнительная эмбриоло-
гия низших беспозвоночных», 1949), Г. А. Шмидта
(«Эмбриология животных», т. 1 и 2, 1951—1953),
О, М. Ивановой-Казас (шеститомная «Сравнительная
эмбриология беспозвоночных животных», 1975—
1981) и работы московской школы эмбриологов под
руководством Л. В. Белоусова.
Большое теоретическое и практическое значение
имели исследования Б. Л. Астаурова и его школы по
экспериментальному партеногенезу и андрогенезу у
насекомых. Впервые у животных удалось получить
амфидиплоиды. Б. П. Токин развил учение о сомати-
ческом эмбриогенезе как особой форме развития при
вегетативном размножении.
Большие успехи были достигнуты в изучении мор-
фологии, систематики и фаунистики отдельных групп
животных. Зоологическим институтом (ЗИН РАН)
опубликовано около 300 томов в сериях «Фауна Рос-
сии», затем «Фауна СССР» и позже «Фауна России и
сопредельных стран»; «Определители по фауне СССР»
и позже «Определители по фауне России» и др.
Кроме того, ЗИН РАН и Академиями СНГ выпущено
много «региональных фаун», рассматривающих жи-
вотный мир отдельных районов суши и моря. К их
числу относится, например, серия «Фауна Украины»,
публикуемая Институтом зоологии АН Украины.
В области изучения простейших широко известны
работы В. А. Догеля и его школы. Книга В. А. Догеля
«Общая протистология» (1951) была удостоена Ле-
нинской премии. Большой вклад в познание парази-
тических простейших домашних животных внесен
В. Л. Якимовым и его школой, паразитических про-
стейших человека — Г. В. Эпштейном, П. Г. Серге-
евым, Ш. Д. Мошковским. В ЗИН РАН изучались и
изучаются как морские, так и паразитические про-
стейшие (А. А. Стрелков, М. Г. Петрушевская, В. В. Ре-
шетняк, А. В. Янковский, С. С. Шульман, В. В. Миха-
левич и др.). В Лаборатории протозоологии этого ин-
ститута под руководством М. В. Крылова развиваются
исследования низших трипаносоматид — паразитов
насекомых (А. О. Фролов, С. А. Подлипаев и др.).
В Лаборатории зоологии беспозвоночных
(БИНИИ СПбГУ, Старый Петергоф) под руководст-
вом профессора Л. Н. Серавина исследование про-
тестов ведется по трем основным группам: инфузории
(С. И. Фокин), амебы (А. В. Гудков) и жгутиконосцы
(С. А. Карпов). Кроме того, сукцессии морских про-
тестов перифитона изучаются А. И. Раилкиным и его
учениками. В Лаборатории кариологии одноклеточ-
ных животных того же института профессор Д. В. Оси-
пов руководит группой по изучению эндосимбионтов
инфузорий (М. С. Раутиан, О. В. Борхсениус, И, И. Скоб-
ло и др.). В ЦИН РАН под руководством!#. Б. Райкове^
иследуются различные аспекты жизнедеятельности
инфузорий (Г. А. Сергеева, Н. Н. Бобылёва, В. Г. Кова-
лёва), амеб (А. Л. Юдин, В. А. Сопина), споровиков
(Т. В. Бейер) и миксоспоридий (А. В. Успенская), их
морфология и жизненные циклы. Фауна бесцветных
жгутиконосцев, а также их систематика и ультраструк-
тура успешно изучаются Б. Ф. Жуковым, А. П. Мыльни-
ковым и другими в ИБВВ РАН (Борок). Еще один
центр по изучению фауны протистов формируется в
настоящее время в Лимнологическом институте Си-
бирского отделения РАН (Иркутск). Исследованиями
водорослей и бесцветных протистов Байкала занима-
ются Н.Н. Бондаренко, А. И. Таничев и другие. Изу-
чению жгутиковых водорослей и некоторых зоо-
споровых грибов посвящены работы Б. В. Громова и
О. В. Гавриловой (СПбГУ). Эвгленовые водоросли и
раковинные корненожки изучаются в СПбГПУ под ру-
ководством К. М. Сухановой, а амебы — на кафедре
зоологии беспозвоночных СПбГУ (А. В. Смирнов). Изу-
чению кинетопластид посвящены работы московских
авторов: В. М. Сафьяновой, В. Д. Каллиниковой и других.
В МГУ значительные успехи достигнуты в исследова-
нии инфузорий (И. В. Бурковский и его ученики), а
также в изучении почвенных простейших (ТО. Г. Гель-
цер и Г. А. Корганова). Микроспоридии изучаются
И. В. Исси и ее учениками (ВИЗР). Фауна и экология ин-
фузорий исследуется В. Н. Жариковым и его ученика-
ми в Институте экологии волжского бассейна РАН
(Тольятти). Общими вопросами систематики, таксо-
номии, филогении протистов, а также вопросами
эволюции эукариотной клетки занимаются Л. Н. Се-
равин, Я. И. Старобогатов, О. Г. Кусакин, А. Л. Дроз-
дов, С. А. Карпов.
Большой вклад внесен отечественными биологами
в изучение губок. Фаунистические, морфологические
и эмбриологические исследования проводились и про-
водятся 77. Д. Резвым, В. М. Колтуном (ЗИН РАН),
О. П. Коротковой, О. М. Ивановой-Казас, С. М. Еф-
ремовой (СПбГУ), М. А. Гуреевой, Л. И. Ивановой
(СПбГПУ), А. В. Ересковским (СПбГУ), Р. П. Анаки-
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
25
ной (ЗИН РАН) и другими. Общим вопросам биоло-
гии губок, их филогении и месте в системе эукариот
посвящены работы П. Д. Резвого, С. М. Колтуна и
Л. Н. Серавина (БИНИИ СПбГУ).
Ряд крупных монографий, касающихся системати-
ки, эволюции и фаунистики гидроидных, сифонофор
и сцифоидных принадлежат Д. В. Наумову, А. К. Лин-
ко, Н. В. Куделину и С. Д. Степаньянц (ЗИН РАН).
Аберрантных гидроидных рода Polypodium изучает
Е. В. Райкова (ЦИН РАН). Дальневосточной фауной
гидроидных, а также морфологией и классификацией
нематоцист занимается О. В. Шейко (Петропав-
ловск-Камчатский). Вопросами эволюции клеточ-
ных структур сцифоидных занимаются О. В. Чага
(СПбГУ) и Г. О. Напара (ЦИН РАН). Anthozoa ис-
следуются С. Д. Гребельным и Н. Н. Слепковой (ЗИН
РАН). Н. Н. Марфенин и его ученики (МГУ) решают
разные вопросы, касающиеся проблемы колониаль-
ное™ не только у гидроидных, но и у кораллов и
мшанок. Школы петербургских и московских эмбри-
ологов ведут многолетние исследования по эмбриоге-
незу и дифференциации клеточного состава тканей у
гидроидных (Д. Г. Полтева и В. В. Донаков, СПбГУ)
и изучению ростовых процессов и морфогенеза у
колониальных гидроидных (Л. В. Белоусов, МГУ).
Организация и регенерация гидр изучалась И. И. Ка-
наевым; исследования по таксономии и фаунистике
гидр с использованием кариосистематики начаты в
ЗИН РАН.
Л. Н. Серавиным возобновлены исследования мор-
фологии, систематики и эволюции Ctenophora.
По свободноживущим ресничным червям, их мор-
фологии и происхождению крупные исследования
выполнены Н. В. Насоновым, В. Н. Беклемишевым,
А. В. Ивановым, Ю. В. Мамкаевым и Б. И, Иоффе
(ЗИН РАН), Л. А. Евдониным, О. А. Тимошкиным
(Лимнологический ин-т, Байкал) и другими. Плас-
тичность нервной системы турбеллярий исследована
К. К Богутой (БИНИИ СПбГУ).
Подробно изучаются паразитические черви, имею-
щие большое практическое значение как паразиты
человека и животных. Под руководством К. И, Скря-
бина в течение многих лет проводилось планомерное
исследование фауны гельминтов в разных районах
России и сопредельных государств. Бурно развивает-
ся и эколого-фаунистическое направление в пара-
зитологии, созданное В. А. Догелем. Трудами этих
выдающихся исследователей и их многочисленных
учеников и коллег заложены основы современных
представлений и по отдельным группам гельминтов.
Школой К. И. Скрябина издается многотомная сводка
«Трематоды животных и человека» (вышло 28 то-
мов). Т. А. Гинецинская (СПбГУ) издала моногра-
фию, посвященную происхождению, эволюции и
жизненным циклам трематод. Специальные исследо-
вания по трематодам птиц опубликованы И. Е. Бы-
ховской-Павловской (ЗИН РАН) и М. М. Белополь-
ской (СПбГУ). Исследования в этом, направлении
активно развиваются и в настоящее время. В СПбГУ
различными аспектами биологии, развития и эво-
люции трематод занимаются А. А. Добровольский,
И. А. Тихомиров, Г. Л. Атаев, А. И. Гранович и дру-
гие. В Институте паразитологии РАН (Москва) ак-
тивно работают А. А. Шигин, Л. В. Филимонова,
С. А. Беэр. Морские трематоды изучаются К. В. Га-
лактионовым (ЗИН РАН). Ленточным червям посвя-
щены многочисленные работы А. А. Спасского,
М. Н. Дубининой, М. М. Белопольской и др. Эти ис-
следования продолжаются А. К. Галкиным (ЗИН РАН).
В Москве под редакцией К. И. Скрябина выходит мно-
готомная серия «Основы цестодологии». Многопла-
новые исследования ленточных червей проводятся
В. И. Фрезе и его сотрудниками (Институт паразито-
логии). Интересные работы по тонкой морфоло-
гии цестод и их развитию выполняются Б. И. Купер-
маном, В. Г. Давыдовым и рядом других исследовате-
лей в ИБВВ. В Новосибирске проблемами
систематики и эволюции цестод занимается В. Д. Гу-
ляев, а в Магадане — К.В. Регель. В изучение моно-
генетических сосальщиков большой вклад был вне-
сен Б. Е. Быховским (ЗИН РАН). Его работы продол-
жаю т| А. В. Zyceg|, Т. А. Тимофеева, П. И. Герасев.
Своеобразные паразиты беспозвоночных живот-
ных — ортонектиды исследуются Г. С. Слюсаревым.
Круглые черви, весьма разнообразные по своей эко-
логии, изучены неравномерно. Свободноживущим не-
матодам посвящено сравнительно немного работ,
среди которых большой интерес представляют иссле-
дования И. Н. Филипьева, Е. С. Кирьяновой, С. Я. Цало-
лихина, Т А. Платоновой, В. В. Гальцовой и А. Ю. Рыс-
са (ЗИН РАН), В В. Малахова, А. В. Чесунова (МГУ),
В. Г. Гагарина (ИБВВ), В. О. Мокиевского (ИОАН).
Нематодам, паразитирующим в животных, посвящена
огромная литература. Под редакцией К. И. Скрябина
издана серия монографий «Основы нематодологии».
В этой серии опубликованы работы А. А. Мозгового,
Т. И. Поповой, М. Д. Сонина, Р. С. Шульца и других.
В области нематод — паразитов растений обобщаю-
щие работы опубликованы А. А. Парамоновым и его
школой. Эволюционная морфология, система и фи-
логения нематод разрабатываются В. В. Малаховым.
В изучение типа Rotifera большой вклад внесли
Г. А. Маркевич и Л. А. Кутикова (ЗИН РАН).
Пелагические и донные немертины изучались
В. С. Короткевич (ЗИН РАН), а в настоящее время и
их исследованием занимается А. В. Чернышов (ИБМ).
Среди кольчатых червей полихеты подробно изу-
чены П. В. Ушаковым, В. В. Хлебовичем (ЗИН РАН),
В. А. Свешниковым (МГУ), Г. Н. Бужинской (ЗИН
РАН), В. Г. Аверинцевым (ММБИ), Ю. С. Миничевым,
О. В. Бубко, А. Б. Цетлиным и др. Крупные иссле-
дования по малощетинковым червям принадлежат
О. В. Чекановской, Т. С. Перель, И. И. Малевичу,
Н. П. Финогеновой (ЗИН РАН) и др. По пиявкам об-
ширные исследования выполнены В. Д. Зеленским,
Е. И. Лукиным, В. М. Эпштейном и другими. Мшанки
довольно полно изучены Г. А. Клюге, Е. И. Андросо-
вой, В. Н. Гонтарь (ЗИН РАН), А. Н. Островским
(СПбГУ) и др.
Огромный объем работ выполнен отечественными
исследователями членистоногих. Ряд крупных моно-
графических работ посвящен низшим ракам (С. С. Смир-
нов, В. М. Рылов, Е. В. Боруцкий, К. А. Бродский,
В. С. Шувалов, М. С. Кос, В. В. Гептнер (МГУ),
Е. Л. Мархасева и Н. В. Вышкварцева (ЗИН РАН) и
др.). Из высших раков подробно изучены бокоплавы и
26
ОБЩАЯ ЧАСТЬ
равноногие раки (Е. Ф, Гурьянова, Н. Л. Цветкова и
А. А. Голиков (ЗИН РАН), О, Г Кусакин (ИБМ, Вла-
дивосток) и др.). Эуфаузиевые, кумовые раки и кап-
реллиды изучены Н. Б, Ломакиной и С. В. Василенко
(ЗИН РАН). Глубоководные ракообразные исследова-
лись Я. А. Бирштейном и М. Е. Виноградовым (ИОАН).
Паразитическим ракообразным посвящены работы
А. П. Маркевича и |А. В. Гусе<?а|(ЗИН РАН).
Общие вопросы системы и филогении хелицеровых
освещены в трудах В. Б. Дубинина. Большое количест-
во исследований посвящено клещам, в особенности
иксодовым и аргасовым (Б. И. Померанцев, Г В. Сердю-
кова, В. И. Волгин, Н. А. Филиппова, Н. Г. Брегетова,
И. Г Галузо, А. А. Захваткин, В. Б. Дубинин, Е. Н. Пав-
ловский, Ю. С. Балашов — ЗИН РАН и др.). Галлооб-
разующие клещи растений изучаются В. Г. Шевченко
(СПбГУ). Менее подробно изучены пауки (С. А. Спас-
ский, П. И. Марковский, В. П. Тыщенко (СПбГУ) и др.),
скорпионы (В. А. Редикорцев, Е. Н. Павловский) и
другие непаразитирующие паукообразные.
Огромное количество работ посвящено насеко-
мым. Обобщающие исследования по систематике, пале-
онтологии, филогении и метаморфозу насекомых принад-
лежат А. П. Семенову-Тян-Шанскому, Н. А. Холодковско-
му, Б. Б. Родендорфу, Н. В. Кулагину, И. И. Ежикову,
Э. Г. Беккеру, В. В. Алпатову, Б. Н. Шванвичу, Д. Г. Ша-
рову, А. А. Захваткину, А. В. Мартынову, Е. С. Смир-
нову, Г. Я. Бей-Биенко и др. Важные проблемы диа-
паузы, холодоустойчивости и влияния фотопериода
на развитие насекомых освещены в исследованиях
И. В. Кожанчикова, Н. И. Калабухова, А. С. Данилев-
ского, Л. К. Лозина-Лозинского, Д. М. Штейнберга,
В. П. Тыщенко, Т. В. Кинд, Е. Б. Виноградовой, В. А. За-
славского, В. П. Семьянова, В. А. Масленниковой и
других. Многочисленны исследования, посвященные
отдельным группам насекомых. Упомянем ряд круп-
ных работ по прямокрылым и таракановым Г. Я. Бей-
Биенко, Л. Л. Мищенко. По двукрылым обобщающие
работы принадлежат А. А. ШтакелЪбергу, Н. Г. Ол-
суфьеву, А. С. Мднчадскому, А.В. Гуцевичу, И. А. Руб-
цову, П. П. Перфильеву, А. С. Лутта, В. Ф. Зайцеву,
В. Н. Танасийчуку, Э. П. Нарчук (ЗИН РАН) и другим.
Ручейники детально изучены С. Г. Лепневой, тли —
А. К. Мордвилко, Г. X. Шапошниковым, А. В. Стеколь-
щиковым, В. Г. Кузнецовой (ЗИН РАН). Крупные
работы по перепончатокрылым выполнены В. В. По-
повым, В. В. Гусаковым, С. И. Малышевым, В. И. То-
биасом, В. А. Тряпицыным, Ю. А. Песенко, Д. Р. Кас-
паряном (ЗИН РАН). По полужесткокрылым известны
работы И. М. Кержнера (ЗИН РАН), по цикадовым —
А. Ф. Емельянова (ЗИН РАН). Жесткокрылые изучены
Г. Г. Якобсоном, М. Е. Тер-Минасян, О. Л. Крыжанов-
ским, Г. С. Медведевым (ЗИН РАН) и другими; че-
шуекрылые — Н. Я. Кузнецовым, А. С. Данилевским,
В. И. Кузнецовым, А. А. Загуляевым и другими. Насе-
комыми, обитающими в почве, занимались М. С. Гиля-
ров, С. Я. Кузнецов. Изданы также десятки определите-
лей и региональных сводок, посвященных насекомым.
Из трудов по морским брюхоногим и пластинча-
тожаберным моллюскам упомянем работы К. М. Де-
рюгина, А. Н. Голикова, Ю. И. Галкина (ЗИН РАН),
Л. И. Москалева (ИОАН), Б. И. Сиренко, Э. Н. Егоро-
вой, 3. А. Филатовой, О. А. Скарлато (ЗИН РАН),
А. И. Кафанова (ИБМ) и других. В изучение пресно-
водных моллюсков большой вклад внесен В. И. Жа-
диным, А. Ф. Алимовым, Я. И. Старобогатовым
(ЗИН РАН) и др. Наземные моллюски исследованы
Е. С. Ромельмейром, И. М. Лихаревым, Я. И. Старо-
богатовым, А. А. Шилейко (ИОАН). Головоногие
моллюски изучаются К. Н. Несисом и Ч. М. Нигма-
тулиным. Эндемичная фауна моллюсков Байкала
изучалась М. М. Кожовым и Г Г Мартинсоном.
Иглокожие в сравнительно-анатомическом и палео-
зоологическом аспектах изучались Д. М. Федотовым.
По систематике этой группы работали А. М. Дьяконов,
3. И. Баранова, Г. М. Беляев и продолжают работать
А. Н. Миронов (ИОАН), Е. Н. Грузов, И. С. Смирнов,
А. В. Смирнов (ЗИН РАН) и другие.
Литература по позвоночным животным отечест-
венных регионов колоссальна. В области ихтиологии
следует прежде всего упомянуть исследования Л. С. Берга
по системе рыб и рыбообразных и его трехтомную,
ставшую классической, монографию «Рыбы пресных
вод СССР и сопредельных стран» (1948—1949).
Опубликованы монографии и определители рыб Се-
верных морей (Н. М. Книпович, А. П. Андрияшев), Даль-
невосточных морей (П. Ю. Шмидт, А. П. Андрияшев,
Г. У. Линдберг, А. Я. Линдберг, А. Я. Таранец, В. В. Бар-
суков, В. В. Федоров и др.), Черного и Аральского
морей (А. Н. Световидов, Г. В. Никольский). По рыбам
Антарктики опубликовано много работ А. П. Андрия-
шевымиА. В. Балушкиным (ЗИН РАН). Значительный
вклад в эмбриологию рыб сделан Ю. Н. Городиловым
(БИНИИ СПбГУ). Весьма обширная литература по-
священа вопросам экологии рыб, их миграции и дина-
мике численности. Большие монографические иссле-
дования в области ихтиологии созданы В. К. Солдато-
вым и Е. К. Суворовым. Интересные исследования
посвящены развитию рыб и вопросам этапности разви-
тия рыб (С. Г. Крыжановский, В. В. Васнецов). Разрабо-
таны методы регуляции полового цикла рыб и стиму-
лирования размножения (Я. Л. Гербильский, Б. Н. Ка-
занский, Г. М. Персов, И. А. Баранникова — СПбГУ, и
др.). Все эти исследования тесно связаны с вопросами
практики рыболовства и рыборазведения.
Фауна земноводных и пресмыкающихся пред-
ставлена довольно полно в трудах А. В. Никольского,
С. А. Чернова, П. В. Терентьева, Л. И. Хозацкого,
А. Г. Банникова, А. А. Емельянова, И. С. Даревского,
Н. Б. Ананьевой, Л. Я. Боркина (ЗИН РАН) и других.
В области орнитологии создан ряд крупных моно-
графий и определителей (например, «Полный опреде-
литель птиц СССР» С. А. Бутурлина и Г. П. Дементь-
ева, 1934—1941; «Птицы Советского Союза» — шес-
титомная монография под ред. Г. П. Дементьева и
Н. А. Гладкова и др.). Большое значение для развития
орнитологии имеют исследования П. П. Сушкина,
Б. К. Штегмана, Е. В. Козловой, Л. А. Портенко,
А. И. Иванова, К. А. Юдина, В. Д. Ильичева, В. М. Лос-
кота, Р. Л. Потапова (ЗИН РАН) и других по функ-
циональной морфологии, систематике, зоогеографии
птиц. Много работ посвящено изучению миграции
птиц (А. Я. Тугаринов, Э. В. Кумари, Е. В. Козлова,
А. Б. Кистяковский, В. Р. Дольник, К. В. Большаков —
ЗИН РАН, и др.) и другим сторонам экологии птиц
(А. С. Мальчевский — СПбГУ, В. Р. Дольник), а также
КРАТКАЯ ИСТОРИЯ ЗООЛОГИИ
27
вопросам социодемографических связей у некоторых
видов оседлых птиц.
Разносторонне изучена фауна млекопитающих.
Укажем на многотомную монографию С. И, Огнева
(1886—1951) «Звери СССР и прилежащих стран».
Много исследований посвящено отдельным группам
млекопитающих. Особенно подробно изучались гры-
зуны ввиду их практического значения. Этому вопросу
посвящены публикации Б. С. Виноградова, Б. К. Феню-
ка, Н. И. Калабухова, Н. П. Наумова, А. Н. Формозова,
И. М. Громова (ЗИН РАН), Ю. М. Ралль, Б. М. Касат-
кина, О. В. Петрова, П. К. Смирнова, Н. Н. Воронцова
(ИБР) и многих других. Хищные млекопитающие изу-
чались Г А. Новиковым, копытные — И. И. Соколо-
вым, ластоногие — Н. А. Смирновым, К, К. Чапским,
А. С. Соколовым, китообразные — А. Г. Томилиным,
С. Е. Клейненбергом, М. М. Слепцовым, А. В. Яблоковым
(ИБР), рукокрылые—А. П. Кузякиным и П. П. Стрелко-
вым (ЗИН РАН). Акклиматизации и промысловой тех-
нологии посвящены работы А. А. Насимович, Г Г До-
пелъмайра, Б. М. Житкова, Н. П, Лаврова, А. А. Слуд-
ского и других.
Изучение животных в отечественных регионах со-
провождается анализом распространения и проис-
хождения фаун. Именно такой характер в отношении
наземных животных носят работы 77. 77. Сушкина,
А. Н. Формозова, И. Г. Пидопличко, Н. К Верещаги-
на, Я. И. Старобогатова (ЗИН РАН) и других. Ши-
рокие зоогеографические обобщения в отношении
морской фауны имеются в работах К М. Дерюгина,
Л. А. Зенкевича, Е. Ф. Гурьяновой, О, Г. Кусакина,
К Н. Несиса, А. И. Кафанова и других.
Широкое и разностороннее развитие в нашей стра-
не получила экология животных. Она развивается и
как аутэкология, экология популяций и биоценология.
Большое значение имели обобщающие работы Д Н. Каш-
карова («Основы экологии животных», 1946), Н. П. Нау-
мова («Экология животных», 1955). Полевым методам
в экологии посвящен ряд работ Г А. Новикова. Экс-
периментальную направленность имеют работы
И. Д. Стрельникова, Н. И. Калабухова, В. С. Ивлева,
И. В. Кожанчикова. А. С. Данилевский, В. П. Тыщенко
и их школа на материале насекомых разработали уче-
ние о фотопериодизме. Популяционный экологичес-
кий анализ успешно развивали С. С. Шварц («Эво-
люционная экология животных», 1969) и его ученики.
Вопросам продукционной биологии водных орга-
низмов и экологической физиологией занимается
школа Г У. Винберга, идеи которого успешно развива-
ет А. Ф. Алимов.
Отечественная палеозоология также получила ши-
рокое развитие. Основные работы в области палеозоо-
логии беспозвоночных принадлежат Н. Н. Яковлеву,
Д. Д. Раузер-Черноусовой, Л. Ш. Давиташвили, А. Г. Эбер-
зину, В. Е. Руженцову, Н. Н. Субботиной, Б. К. Лиха-
реву, Б. С. Соколову, И. Т. Журавлевой, М. А. Федон-
кину, А. Б. Ивановскому, Е. С. Краснову и многим
другим. В области палеозоологии позвоночных работа-
ли Д. В. Обручев, А. А. Борисяк, Ю. А. Орлов, А. П. Быст-
ров, В. И. Громов, Л. И. Хозацкий, Л. П. Татаринов,
Э. И. Воробьева и другие. Широко известна 15-том-
ная монография «Основы палеонтологии» под редак-
цией ТО. А. Орлова.
Зоологические научные
общества.
Международные конгрессы
Во многих развитых странах в начале и середине
XIX в. возникают научные зоологические общества,
объединившие ученых зоологов и сыгравшие значи-
тельную положительную роль в развитии зоологии. В
ряде стран зоологические общества возникли как са-
мостоятельные организации, в других же они вошли
как подразделения (секции, отделения и т. п.) в био-
логические общества более широкого профиля. Зоо-
логические общества возникают в 1826 г. в Лондоне
(Zoological Society of London), в 1875 г. в Калькутте
(Zoological Society of Calcutta), в 1876 г. во Франции
(Societe Zoologique de France), в 1890 г. в Германии
(Deutsche Zoologische Gesellschaft), в 1895 г. в Нью-
Йорке (Zoological Society of New York). В России
объединения зоологов обычно в виде отделений вхо-
дили и входят в состав обществ более широкого
профиля. В 1805 г. в Москве возникло Общество ис-
пытателей природы, существующее и поныне. В 60—
70-е гг. прошлого столетия по рекомендации Всерос-
сийских съездов естествоиспытателей и врачей (они
проходили регулярно с 1867 по 1913 г.) во многих
университетских городах при университетах органи-
зуются Общества естествоиспытателей (в Петербурге
в 1868 г., Казани в 1869 г., Одессе в 1870 г., Томске
в 1889 г.), имевшие в своем составе зоологические
отделения. Большинство этих обществ функциониру-
ет и в настоящее время.
Наряду с зоологическими во многих странах были
созданы научные общества, охватывающие отдельные
направления зоологии. Например, Энтомологическое
общество Франции в 1832 г. (Societe Entomologique
de France), Энтомологическое общество в Лондоне в
1833 г. (Entomological Society of London), Британс-
кое объединение орнитологов (British Ornithological
Union) в 1859 г., Русское энтомологическое общест-
во в 1859 г. Большое число научных обществ по от-
дельным зоологическим дисциплинам возникло за
последние два десятилетия в разных странах, в том
числе и в России (паразитологические, ихтиологи-
ческие, герпетологические и др.). Большинство науч-
ных обществ издает журналы и разного рода непери-
одические издания (монографии, руководства по со-
биранию коллекций и т. п.). t
Международные зоологические конгрессы, необ-
ходимость в которых назрела во второй половине
XIX в., начали созываться с 1889 г. По настоящее
время состоялось 17 Конгрессов. Места и годы их ра-
боты следующие: 1-й — 1889 г., Париж; 2-й —
1892 г., Москва; 3-й — 1895 г., Лейден; 4-й — 1898 г.,
Кембридж; 5-й —1901 г., Берлин; 6-й — 1904 г.,
Берн; 7-й — 1907 г., Бостон; 8-й — 1910 г., Грац; 9-й —
1913 г., Монако; 10-й — 1927 г., Будапешт; 11-й —
1930 г., Падуя; 12-й — 1935 г., Лиссабон; 13-й —
1948 г., Париж; 14-й — 1953 г., Копенгаген; 15-й —
1958 г., Лондон; 16-й — 1963 г., Вашингтон; 17-й —
1972 г., Монако.
Первые зоологические конгрессы были относи-
тельно небольшими собраниями ученых. На первом
28
ОБЩАЯ ЧАСТЬ
из них, в Париже, присутствовало около 200 человек
и было сделано 38 докладов. На самом большом по
числу участников — 16-м Конгрессе в 1963 г. в Ва-
шингтоне присутствовало свыше 2 тысяч зоологов;
кроме ряда выступлений на пленарных заседаниях
доклады были сделаны на 29 специальных симпозиу-
мах и 16 секциях. Общее число прочитанных докла-
дов приближалось к 1000. Сопоставление этих цифр
указывает на огромный количественный рост зооло-
гических кадров, произошедший за последние 80 лет
развития зоологии. 17-й Конгресс, проходивший в
Монако, был относительно немноголюдным, и на нем'
были заслушаны только некоторые общие доклады, а
также обсуждался вопрос — имеет ли смысл созы-
вать зоологические конгрессы, или рациональнее пе-
рейти на более узко профилированные форумы, охва-
тывающие лишь конкретные разделы зоологии. Были
высказаны разные мнения, и вопрос остался нере-
шенным. Время и место очередного Конгресса так и
остались не назначенными. С тех пор Международ-
ные зоологические конгрессы не собирались. Между-
народное общение зоологов спонтанно пошло по
линии созыва более специализированных форумов.
Бесспорно, что Международные зоологические кон-
грессы сыграли в развитии зоологии большую поло-
жительную роль, обеспечив широкие контакты между
зоологами, без чего развитие науки на современном
этапе невозможно.
В бывшем Советском Союзе состоялись 4 Всесо-
юзных зоологических съезда. Первый происходил в
Ленинграде в 1922 г. (308 членов, 267 докладов),
второй — в 1925 г. в Москве (411 членов, 367 докла-
дов), третий — в 1927 г. в Ленинграде (758 членов,
401 доклад) и четвертый — в 1930 г. в Киеве
(882 члена, 417 докладов). Далее общезоологические
съезды перестали созываться, и в настоящее время
общение между зоологами России и стран СНГ осу-
ществляется на более узко специализированных фо-
румах (съезды и совещания энтомологов, гидробиоло-
гов, орнитологов, териологов, паразитологов, по
борьбе с вредителями и т. п.).
Зоологические музеи,
зоопарки, заповедники,
национальные парки
В развитии зоологии как науки, а также в пропа-
ганде зоологических знаний среди населения большую
роль играют зоологические музеи, зоопарки, заповед-
ники, национальные парки. Особенно большое значе-
ние имеют зоологические музеи, где сосредоточены
зоологические коллекции и ведутся научные работы,
главным образом по систематике и фаунистике. Во
многих странах зоологические музеи связаны с круп-
ными исследовательскими учреждениями — зоологи-
ческими институтами. Это принято в большинстве
развитых стран. Зоологические музеи и институты на-
чали возникать еще в XVII в. Наиболее крупные зоо-
логические музеи имеются в Париже, Вене, Праге,
Берлине. В Лондоне и Вашингтоне зоологические от-
делы входят в состав музеев естественной истории
(Natural History Museum) и включают кроме зоологии
геологию, минералогию и палеонтологию.
Старейший в России Зоологический музей, на ос-
нове которого образовался Зоологический институт
РАН, возник в Петербурге на базе Кунсткамеры Петра I
и в настоящее время представляет собой один из круп-
нейших зоологических музеев в мире. Официальной
датой его открытия считается 4 (16) июля 1832 г., когда
академик Ф. Ф. Брандт доложил Конференции Рос-
сийской академии наук о готовности Зоологического
музея принять посетителей. В России и в странах СНГ
при Академиях наук и университетах также имеются
аналогичные учреждения — преимущественно в сто-
лицах. Среди них важное место занимает основанный
в 1971 г. Зоологический музей при Московском госу-
дарственном университете. Крупный музей существует в
столице Украины Киеве при Институте зоологии Ук-
раинской академии наук.
Немалую роль для зоологии играют зоологические
сады и парки, имеющиеся во многих странах. В част-
ности, они важны для сохранения редких и вымира-
ющих видов, занесенных в «Красные книги».
В наше время, когда вопросы охраны природы
стоят особенно остро, для сохранения и изучения жи-
вотного мира особое значение приобретают заповед-
ники и заказники. В странах СНГ в настоящее время
имеется более 150 заповедников. Такое число запо-
ведников нельзя считать достаточным, однако они иг-
рали и продолжают играть большую положительную
роль, в том числе в спасении и последующем размно-
жении таких близких к исчезновению видов живот-
ных, как соболь, бобр, сайгак, зубр и другие.
Библиография
по истории зоологии
Подробная аннотированная библиография по зоо-
логии и истории зоологии имеется в книге В. И. Ле-
вина, В. Г. Левина и Д. В. Лебедева «Путеводитель
для биологов по библиографическим изданиям»:
Справочное пособие (М.: Наука, 1978), что позволяет
указать здесь только главнейшие библиографические
источники. Наиболее полным библиографическим
справочником по зоологии является «Zoological Re-
cord», издающийся с 1865 г. в Лондоне. Полная биб-
лиография имеется в зоологических разделах свод-
ных справочных изданий «Реферативный журнал.
Биология» (издается с 1954 г.; изд-во ВИНИТИ РАН,
Москва) и «Biological abstracts» (издается с 1926 г.;
Филадельфия). Биографические сведения о естество-
испытателях с середины XIX в. до 40-х гг. текущего
столетия имеются в словаре Поггендорфа (Poggen-
dorf J. С. Biographisch-literarisches Handworterbuch
zur Geschichte der exakten Wissenschaften. Bd 1—6.
Leipzig; Berlin, 1859—1940). Об истории Зоологичес-
кого института РАН и об исследовательской работе
его сотрудников см.: «Зоологический институт Ака-
демии наук СССР. 150 лет» (Л.: Наука, 1982);
А. Ф. Алимов, В. Н. Танасийчук, С. Д. Степаньянц.
«Коллекции Зоологического института РАН — осно-
ва для изучения биологического разнообразия» (Зоол.
журн. 1999. № 9).
ПУТИ Л ЗАКОНОМЕРНОСТИ эволюции
29
ESSAY ON THE HISTORY OF ZOOLOGY
SUMMARY
Zoology (Greek: «zoo» — animal; «logos» — scien-
ce) is a part of Biology. It involves some general bran-
ches of science, i. t. systematics, morphology, histology,
comparative morphology, embriology, phylogenetics,
zoogeography, paleozoology, physiology, etc.
Zoology might be classified on the base of the subject
of investigation: protozoology, helmintology, malacolo-
gy, carcenology, arachnology, entomology, ichtiology,
batrachology, herpetology, ornitology, teriology, etc.
The last decade some special lines of particular the-
oretical and practical interest were arisen in zoology,
i. e. soil zoology, cave-fauna investigations, interstitial
freshwater- and marine-fauna investigations; fouling
fauna investigations, etc.
The knowledge of animals grew through all the pe-
riod when ancient-human culture had risen — in Anci-
ent Egypt, Vavilon, India, China.
The origin of Zoology as a science during the Anti-
que Times is reviewed in this chapter.
The Zoology in the Middle Ages (Bacon), in XV—
XVIII centuries, in the first and the second halfs of
XIX century with a special attention paid to the
«Darvinist» and «post-Darvinist» times is observed.
The Zoology in XX century and its contemporary
trends are described. A special place is occupied with
the detailed description of Zoology in Russia.
Zoological Scientific Societies, International Zoo-
logical Congresses, Zoological Museums, Zoological
Parks and reservations of the world are described.
This chapter contains only general bibliographical
sources.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
О становлении современной
теории эволюции
Формирование теории эволюции — дарвинизма,
или синтетической теории эволюции (СТЭ), проис-
ходило во второй четверти XX в. Его основой стал
широкий синтез знаний о факторах, движущих силах
и закономерностях эволюции, добытых к тому време-
ни разными отраслями биологии. Синтез осуществ-
лялся на основе дарвиновской идеи о естественном
отборе как главной причине приспособительных пре-
образований в органическом мире.
Основные проблемы СТЭ были рассмотрены в
книгах Добжанского (Dobzhansky, 1937), Шмальгау-
зена (1938, 1939а, 19396, 1946а), Лукина (1940),
С. А. Северцова (1941), Хаксли (Huxley, 1942), Май-
ра (Мауг, 1942), Симпсона (Simpson, 1944), Парамо-
нова (1945), Стеббинса (Stebbins, 1950), Ренша (Rensch,
1954), а также в серии статей Г. Ф. Гаузе, С. М. Гер-
шензона, Н. П. Дубинина и Н. В. Тимофеева-Ресов-
ского, опубликованных в 1935—1948 гг.
Не касаясь подробностей (см.: Завадский, 1971;
Галл, Георгиевский, 1973; «The evolutionary...», 1980;
Мауг, 1982; Завадский и др., 1983; Воронцов, 1984;
Полянский, 1988), отметим главные особенности в
формировании СТЭ, необходимые для понимания
сущности современного эволюционизма.
Его фундаментом стали результаты эксперимен-
тальных исследований основных факторов эволюции:
изменчивости, наследственности, изоляции, дрейфа
генов, борьбы за существование и естественного от-
бора. Вначале это были‘исследования закономернос-
тей генетической изменчивости, приведшие к форми-
рованию генетики природных популяций (Baur, 1924;
Четвериков, 1926; Тимофеева-Ресовская, Тимофеев-
Ресовский, 1927; Fisher, 1930; Дубинин, 1931, 1932;
Дубинин, Ромашов, 1932; Sturtevant, Dobzhansky,
1936; Гершензон, 1941, и мн. др.). Изучались законо-
мерности мутаций, фонд рецессивных мутаций в по-
пуляциях, эволюционное значение гетерозиготности
и полиморфизм как способ поддержания гетероген-
ности популяции. Одновременно исследовались зако-
номерности модификационной изменчивости по мор-
фологическим, физиологическим, биохимическим и
поведенческим признакам и выяснялось эволюцион-
ное значение ненаследственной изменчивости (Кир-
пичников, 1935; Лукин, 1936; Камшилов, 1939; Гаузе,
1940, и др.).
В тесной связи с названными направлениями были
развернуты работы в области экспериментальной сис-
тематики (микросистематики); они показали, что вид
имеет сложную структуру и содержит генетически
разделяющиеся группы, приспособленные к опре-
деленным условиям существования: географические
и экологические расы, местные популяции, а также
внутрипопуляционные формы различной природы
(Clausen, 1921; Turreson, 1922; Вавилов, 1926, 1931;
Розанова, 1928, и др.). Изучались и различные формы
изоляции внутривидовых групп.
Для понимания механизмов адаптивных преобра-
зований популяций особое значение имели исследо-
вания динамики численности и плотности популя-
ций, внутривидовой и межвидовой конкуренции,
сложных взаимодействий между популяциями хищ-
ника и жертвы, паразита и хозяина и т. д. (Сукачев,
1927; Гаузе, 1934; Северцов С. А., 1941). Были про-
ведены разнообразные экспериментальные работы,
направленные на выяснение творческой роли естест-
венного отбора в изменении генетического состава
30
ОБЩАЯ ЧАСТЬ
популяций и в образовании новых видовых форм
(подробнее см.: Галл, 1983).
Таким образом, в центре названных исследований
было изучение причин, вызывающих преобразование
популяций, т. е. причин так называемой микроэволю-
ции. Однако неверно было бы сводить СТЭ лишь к
теории микроэволюции. На самом деле успехи в по-
знании генетических и экологических факторов мик-
роэволюции дали возможность объяснить причинную
обусловленность макроэволюции, т. е. эволюции на
надвидовом уровне, и ее закономерностей. В состав
современного эволюционизма как неотъемлемая его
часть вошло учение А. Н. Северцова—И. И. Шмаль-
гаузена о морфологических закономерностях макро-
эволюции: представления о главных направлениях
Эволюционного процесса, филэмбриогенезах, моду-
сах эволюции органов и т. д. Объединение генетико-
экологических данных о факторах микроэволюции с
данными филогенетической систематики, палеонто-
логии, морфологии, эмбриологии обеспечило сущес-
твенный прогресс в познании закономерностей мак-
роэволюции. Было принято, что нет оснований раз-
рывать процессы микро- и макроэволюции (Simpson,
1944). Существует единый процесс эволюции —
адаптивные преобразования популяции — и его ре-
зультаты: непосредственные и отдаленные, неболь-
шие по масштабу и огромные. Предположения же
о существовании каких-то особых причин макро-
эволюции, которые якобы сразу приводят к возни-
кновению новых видов и более крупных таксонов, до
сих пор не имеют надежных доказательств. Сторон-
ники СТЭ исходят из предположения, что вся эволю-
ция от доклеточных существ до высших многокле-
точных животных имеет приспособительный харак-
тер и вполне объясняется действием естественного
отбора.
Сущность СТЭ заключается в следующих положе-
ниях: 1) популяция является элементарной единицей
эволюции; 2) мутации (генные, хромосомные, геном-
ные) поставляют основной материал для эволюции;
3) элементарное эволюционное явление состоит в на-
правленном необратимом изменении генофонда по-
пуляций; 4) естественный отбор выступает как глав-
ный фактор эволюции; 5) видообразование завер-
шается формированием изоляционных механизмов
между новым и предковым видами; 6) вид представ-
ляет собой сложную политипическую систему, состо-
ящую из популяций и различных внутрипопуляцион-
ных групп; 7) существует единство механизмов микро-
и макроэволюции.
Факторы микроэволюции
и видообразование
Под микроэволюцией понимаются адаптивные
преобразования, протекающие еще в рамках вида.
Микроэволюция — статистический процесс, который
охватывает множество организмов, способное дать
материал для действия естественного отбора. Такое
множество получило название местной (локальной)
популяции.
В зависимости от сложности организации живот-
ных или растений, их места в цепях питания, степени
приспособленности к условиям среды и от ареала
средний размер популяции может быть очень раз-
ным: он может исчисляться триллионами особей у
микроорганизмов и сотнями и даже десятками у
высших позвоночных животных. Но эволюционный
процесс всегда происходит в недрах популяции. Из-
менение ее генетического состава оказывается эле-
ментарным эволюционным явлением (Тимофеев-Ре-
совский, 1958; Тимофеев-Ресовский и др., 1969). По-
этому популяция и признана в СТЭ элементарной
единицей эволюции. Исследования генетического со-
става природных популяций различных видов, осо-
бенно плодовой мушки — дрозофилы, существенно
углубили дарвиновские представления о наслед-
ственной изменчивости как источнике материала
для отбора. Всецело подтвердилось предположение
Четверикова (1926) о том, что природные популя-
ции буквально насыщены рецессивными мутациями,
представляющими обильный материал для формиро-
вания новых адаптаций. Тем самым было окончатель-
но устранено существенное возражение дарвинизму
(Берг, 1922), заключающееся в том что мутации на-
столько редки и так вредны, что не могут обеспечить
появление существующего разнообразия адаптивных
форм.
Действительно, большинство мутаций вредно. Од-
нако их отрицательное значение далеко не всегда
проявляется, так как в популяциях они чаще всего
находятся в гетерозиготном состоянии. При постоян-
ном же процессе перекомбинирования мутаций, про-
исходящем при скрещивании организмов с различны-
ми генотипами, вредные свойства мутаций нередко
погашаются и комбинации разных мутаций могут
стать основой для формирования новых, полезных
признаков. Материалом для эволюции панмиктичес-
ких организмов, как правило, служат не вновь возни-
кающие мутации, а мутации, уже прошедшие отбор
на обезвреживание в различных генотипических сре-
дах и оказавшиеся в таких системах гетерозигот, где
их фенотипическое проявление стало адаптивным
(Шмальгаузен, 19466).
Было доказано, что материалом для адаптивных
преобразований могут служить не только мелкие, но
и крупные мутации. Так, при исследовании при-
родных популяций дрозофилы Дубинин и Тиняков
(1946) показали, что концентрации тех или иных пе-
рестроек хромосом меняются в зависимости от сезо-
на. Следовательно, мутанты с разным типом хромо-
сомных перестроек отличаются по плодовитости и
выживаемости при разных температурах. При этом
большей жизнестойкостью отличаются гетерозиготы.
Влияние хромосомного полиморфизма на географи-
ческое распространение вида было вскрыто в работах
Добжанского и его сотрудников (см.: Dobzhansky,
1970). К настоящему времени накапливаются прямые
и косвенные доказательства того, что макромутанты
могут дать начало крупным морфологическим преоб-
разованиям не только у растений (Brucher, 1941;
Stubbe, Wettstein, 1941; Schwanitz, 1974; Тахтаджян,
1983), но, вероятно, и у насекомых и позвоночных
(Goldschmidt, 1940; Britten, Davidson, 1971; Eldredge,
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
31
Gould, 1972; Carson, 1980; Britten et al., 1982; Ворон-
цов, 1984; Татаринов, 1985, 1986).
Были получены также экспериментальные доказа-
тельства возможности образования новых жизнеспо-
собных форм на базе крупных хромосомных и геном-
ных мутаций. Классическими примерами эволюции в
результате геномных мутаций и отдаленной гибриди-
зации стали гибриды: ржано-пшеничный (Мейстер,
1927), капустно-редечный (Карпеченко, 1927), алычи
с терном (Рыбин, 1936), а также аллотетраплоидная
форма тутового шелкопряда (Астауров, 1940, 1968).
Имеется много данных о широком распространении
полиплоидии и гибридизации как факторов видооб-
разования у растений и животных (см.: Боркин, Да-
ревский, 1980; Bullini, 1994; Даревский, 1995).
Однако как бы ни был велик фенотипический эф-
фект мутаций, сами по себе они не могут предо-
пределить ни темп, ни направление эволюции. За-
крепление мутаций в популяции, формирование на ее
основе новой адаптивной нормы происходят под кон-
тролем отбора. Более того, без отбора популяция
быстро бы деградировала, так как ее адаптивная
норма была бы разрушена мутационным процессом,
идущим в разнообразных направлениях.
Признавая мутационный процесс главным постав-
щиком материала для естественного отбора, совре-
менные эволюционисты по-новому поставили вопрос
об эволюционном значении ненаследственной, моди-
фикационной изменчивости. Отбор действует не на
отдельные наследственные признаки организма, а на
весь фенотип в целом. Поэтому для выживаемости
организма отнюдь не безразлична его способность
приспособительно реагировать на изменения внеш-
ней среды. Сама способность образовывать адаптив-
ные (необратимые и обратимые, морфологические и
физиологические) модификации, — конечно, не какое-то
изначальное свойство живого, а результат предшест-
вующего отбора на приспособляемость. Эта способ-
ность наследственно закреплена и обусловлена нор-
мой реакции организма. Но при наличии адаптивных
реакций на изменение среды эффективность и ско-
рость отбора значительно возрастают (Камшилов,
1939; Гаузе, 1940; Waddington, 1942). Как показал
И. И. Шмальгаузен в теории стабилизирующего от-
бора, модификации, в сущности, предваряют эво-
люцию, прокладывая путь будущим, наследственно
закрепленным преобразованиям популяций. Способ-
ность к адаптивным модификациям нередко захваты-
вает целые комплексы органов и тем самым облегча-
ет становление новых, иногда очень сложных при-
способлений к изменившейся среде.
Среди факторов микроэволюции особенно спор-
ным остается эволюционное значение дрейфа генов
(Wright, 1931), или генетико-автоматических процес-
сов (Дубинин, Ромашов, 1932).2 Такие явления, как
основание новой популяции несколькими особями,
несущими лишь часть генофонда исходного вида
2 Дрейфом генов называют изменения частот генов в популя-
ции под действием случайных факторов. Так, при резком сокраще-
нии численности популяций под влиянием стихийных бедствий те-
ряется значительная часть аллелей, усиливается гомозиготность,
увеличивается вероятность проявления в фенотипе редких мута-
ций.
(«принцип основателя» Э. Майра), селективная экви-
валентность нескольких генотипов, наличие беспо-
лезных или даже вредных признаков, связанных кор-
реляциями с полезными признаками (коррелятивная
изменчивость Ч. Дарвина), бесспорно, вносят сущест-
венные элементы неопределенности в процессы диф-
ференциации популяций и видов. Возрастает вероят-
ность и случайного исчезновения отдельных мутаций
при резких изменениях численности в небольших
изолированных популяциях. Создатели концепции
прерывистого равновесия (Gould, Eldredge, 1977;
Stanley, 1979) утверждают даже, что быстрые морфо-
логические изменения на основе дрейфа генов в пе-
риферических изолятах являются главным механиз-
мом видообразования.
Однако дрейф генов не может действовать вопре-
ки естественному отбору. Это еще раз недавно было
доказано при изучении полиморфизма белков в при-
родных популяциях. Оказалось, что организмы, отли-
чающиеся по составу и структуре белков, обладают
различной устойчивостью к температурам, болезням
и т. д. Таким образом, стало ясно, что эволюция на
молекулярном уровне также носит адаптивный, а не
случайный характер, как это утверждали сторонники
нейтральной, так называемой недарвиновской эволю-
ции (King, Jukes, 1969; Fitch, Margolish, 1970; Kimu-
ra, 1983).
В работах Сукачева (1927), Гаузе (1934), С. А. Се-
верцова (1941), Завадского (1954) и некоторых дру-
гих эволюционистов дальнейшее развитие получило
дарвиновское представление о борьбе за существова-
ние как главной предпосылке для действия естествен-
ного отбора. Борьбой за существование со времен
Дарвина называют комплекс сложных внутрипопуля-
ционных и межвидовых взаимоотношений, в которые
вступает каждый организм, а также его отношения к
абиотическим факторам среды. Особенно велико
здесь значение внутривидовой конкуренции, развер-
тывающейся между близкородственными формами за
пищу, территорию, полового партнера и т. д. Борьба
за существование состоит из множества компонентов
(конкуренция, кооперация, хищничество, реакции
организма на абиотические факторы), которые нахо-
дятся в многообразных и противоречивых взаимодей-
ствиях. И хотя процесс борьбы за существование во
всей полноте разыгрывается в экосистемах, его эле-
ментарные следствия приурочены прежде всего к по-
пуляции. Избирательная элиминация — закономер-
ный итог борьбы за существование.
Центральное место в современном эволюциониз-
ме занимают исследования по естественному отбору.
Его эффективность, продемонстрированная на самых
разнообразных объектах, проявляется в возникнове-
нии меланистических форм и ядоустойчивых рас
насекомых, в формировании наследственно закреп-
ленных приспособлений у растений к повышенным
концентрациям вредных соединений в почвах и в
атмосфере, в усилении в сотни раз устойчивости
микроорганизмов к антибиотикам и т. д. Было дока-
зано и прямое участие отбора в процессах видообра-
зования. Так, при отборе дрозофил на максимальное
и минимальное число щетинок через тридцать поко-
лений были получены новые формы, фактически не
32
ОБЩАЯ ЧАСТЬ
скрещивающиеся друг с другом (Thoday, Gibson,
1962).
Было доказано, что под контролем естественного
отбора находятся все признаки и свойства организмов
и популяций, а также сами факторы эволюции (Завад-
ский, Колчинский, 1977). Оказалось, что естествен-
ный отбор контролирует темп и характер мутацион-
ной изменчивости. Рецессивность, доминантность —
это итоги деятельности отбора, направленного на ус-
транение вредного эффекта мутаций., образование мо-
билизационного резерва, делающего вид эволюционно
пластичным, на формирование адаптивного фенотипа,
его упрочение и защиту от дезорганизующего действия
новых мутаций.
Был выяснен селективный механизм поддержания
невысокой численности в популяциях бабочек, под-
ражающих несъедобным насекомым. Оказалось, что
их малочисленность является необходимым условием
выживания, так как при большей численности исчез
бы «эффект примеси» и птицы начали бы отличать
имитаторов от моделей. Численность подобных видов
регулируется летальными генами, концентрация ко-
торых может быть весьма высокой (гибнет до 60—
80 % зародышей в каждом поколении). Но имен-
но эта постоянная высокая смертность является важ-
ной адаптацией вида. У многих животных и растений
были обнаружены гены, летальные или семилеталь-
ные в гомозиготном состоянии, а в гетерозигот-
ном — повышающие жизнеспособность, иммунитет,
экологическую пластичность и другие важнейшие
свойства. Поэтому отбор часто сохраняет генетичес-
кие системы, способные поддерживать состояние
адаптивной гетерозиготности, и действует против го-
мозиготных особей.
При изучении генетического полиморфизма попу-
ляции удалось выяснить и роль отбора, поддерживаю-
щего постоянство численных отношений между раз-
личными формами в популяциях. Однако при извест-
ных обстоятельствах устойчивый (сбалансированный)
полиморфизм может смениться переходным (транзи-
тивным), при котором отбор будет способствовать пре-
обладанию одной (или нескольких) формы над други-
ми, вплоть до полной перестройки состава популяций
(Ford, 1964).
Создатели современного дарвинизма глубоко
обосновали понятия о движущих (накапливающая,
интегрирующая) и поддерживающих (нормализую-
щая, центростремительная, стабилизирующая) фор-
мах естественного отбора. Особенно важной стала
теория стабилизирующего отбора И. И. Шмальгаузе-
на, объясняющая механизм повышения автономиза-
ции онтогенеза и повышения надежности реализации
адаптивной нормы. Были полнее охарактеризованы
условия и факторы, вызывающие действия отбора в
одном направлении (ортоселекция). Изучение этой
формы отбора устранило возражения сторонников
ортогенеза.
Были выделены и такие формы естественного от-
бора, как дизруптивньщ, частотно-зависимый, сба-
лансированный и катастрофический. Мазер (Mather,
1955) и Тудей (Thoday, Gibson, 1962) проанализиро-
вали особенности действия отбора одновременно не-
скольких дискретных фенотипов в популяции (диз-
руптивный отбор), в результате чего в недрах одной
популяции происходит дивергенция форм. В исследо-
ваниях Бритье и Тесье (L’Heritier, Teissier, 1937) был
открыт частотно-зависимый отбор, при котором цен-
ность генотипа определяется частотой его нахожде-
ния в популяции. Оказалось, что преимущественное
сохранение гетерозигот отнюдь не является необхо-
димым условием поддержания устойчивого равнове-
сия генов в популяции. Напротив, оно может сохра-
няться и при снижении адаптивной ценности гетеро-
зигот по сравнению с гомозиготами в том случае,
если селективное преимущество имеет ген с низкой
частотой. В работе Льюиса (Lewis, 1962) было пока-
зано, что даже при катастрофическом отборе, когда в
результате стихийного бедствия погибает практичес-
ки весь вид, единственный выживший организм может
дать начало новому виду.
Однако сколь ни важны вышеназванные факторы
эволюции, процесс видообразования может считаться
завершенным и необратимым только в тех случаях,
когда формируются какие-то барьеры, препятствую-
щие обмену генетической информацией между ис-
ходной и новыми формами. Поэтому различные формы
изоляции (географическая, экологическая, морфоло-
гическая, физиологическая и генетическая) — необхо-
димые факторы эволюции (Мауг, 1974). Они действу-
ют не только у организмов с облигатным половым
размножением, но и у форм с различными формами
обмена генетической информацией внеполовым путем
(Серавин, Гудков, 1984).
Открытие горизонтального переноса генетической
информации между разными, подчас очень отдален-
ными организмами (подробнее см.: Хейсин, 1984;
Lewin, 1984) стимулирует поиск механизмов изоля-
ции, обеспечивающих стабильность видов в условиях
проникновения в их генофонд перемещающихся ге-
нетических элементов других видов (внутрихромо-
сомных транспозонов и им подобных сегментов
ДНК, внехромосомных плазмид), которые способны
интегрировать в геном хозяина вирусы (Дубинин,
1980). Во всех случаях обмена генетической инфор-
мацией внеполовым путем правильнее, видимо, гово-
рить не о механизмах изоляции, а о факторах, обес-
печивающих устойчивость генофонда (Полянский,
1986). Главным фактором, препятствующим фикса-
ции чужеродных генов в генофонде вида, является
отбор, который позволяет закрепление только генов,
дающих крупный выигрыш виду в борьбе за сущест-
вование. Это хорошо показывают процессы бурного
формирования среди бактерий штаммов, содержащих
плазмиды с детерминантами резистентности к анти-
биотикам.
Для более четкого понимания взаимоотношений
между основными факторами эволюции важное зна-
чение имеет кибернетическая теория эволюции, раз-
работанная в 50—60-х гг. Шмальгаузеном (1968). В
предложенной им схеме регулирующих механизмов
эволюции популяция рассматривалась как объект уп-
равления, регулятором которой выступает биогеоце-
ноз. Естественный отбор обеспечивает прямую связь
между ними, осуществляемую через процессы диффе-
ренциальной выживаемости и размножаемости осо-
бей. Здесь информация передается от зиготы одного
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
33
поколения к половым клеткам другого. Возникающие
при этом искажения информации обеспечивают при-
ток новых мутаций. Механизм обратной связи дейст-
вует через реализацию наследственной информации в
онтогенезе и проявляется в каждой особи, влияющей
через процессы борьбы за существование на состав
биогеоценоза.
В настоящее время различается несколько спо-
собов видообразования (Завадский, 1968; Тимофеев-
Ресовский и др., 1969; Мауг, 1974; White, 1978; Во-
ронцов, 1984). По пространственным отношениям
видообразование подразделяется на аллопатри-
ческое, связанное с пространственной изоляцией
отдельных популяций данного вида, и симпатри-
ческое, которое происходит на основе территори-
ально единой популяции при существовании в ней
нескольких четко различающихся форм особей (диз-
руптивный отбор). По размерам исходных преобразо-
ваний, по времени изоляции новой формы и темпам
самого процесса К. М. Завадский предложил различать
медленное видообразование путем накопления в
популяции многих мелких адаптивных изменений
(микроаккумулятивное) и быстрое, неоформоген-
ное видообразование — на основе размножения резко
измененных мутантных форм, с самого начала репро-
дуктивно изолированных от исходного вида. Иногда
этот способ видообразования называют также «мгно-
венным» или «внезапным». Предлагается также вы-
делять три типа видообразования по способу взаимо-
действия генетической информации: путем разделе-
ния исходного вида на несколько рас с последующей
их адаптивной радиацией и превращением в новые
виды (расщепляющееся видообразование,
сегрегациогенез); путем образования нового
вида на основе объединения генетической инфор-
мации двух или нескольких исходных видов в ре-
зультате отдельной гибридизации, трансдукции,
трансформации, симбиогенеза и пр. (синтезоге-
не з); путем преобразования исходного вида в один
дочерний (ортоселекция, филетическое ви-
дообразование).
Называют и другие способы видообразования с
помощью интрогрессии, т. е. путем приобретения
одним видом в результате гибридизации и отбора от-
дельных признаков других видов (Бобров, 1944; An-
derson, 1949), и видообразование, основанное на спо-
собности «генетически агрессивного вида» захваты-
вать при гибридизации целые комплексы геномов
других видов (Harlan, Wet, 1963). Относительно не-
давно была выделена специфическая форма видооб-
разования — кредитогенез (Боркин, Даревский,
1980), когда в ходе становления нового вида исполь-
зуется лишь часть генетической информации одного
из видов, ставших основой для образования новых
гибридных форм. Установлен и особый тип видооб-
разования на островах, начинающийся с залета или
заноса на отдаленные острова единичных особей
(«принцип основателя» Э. Майра).
Таким образом, в рамках современной СТЭ не
только были экспериментально изучены основные
факторы микроэволюции, но и описаны разнообраз-
ные способы видообразования, идущего под направ-
ляющим действием естественного отбора.
3 Протисты, часть 1
Основные закономерности
филогенеза
Еще Дарвин (Darwin, 1859) подчеркивал необхо-
димость изучения морфологических закономерностей
эволюции. Важно, конечно, построить генеалогичес-
кую схему животных, без этого любая теория эволю-
ции осталась бы абстрактным учением без конкрет-
ного содержания. Однако не менее существенно от-
крыть те общие принципы, по которым совершаются
филогенетические превращения органов и их функ-
ций и развитие целых организмов. Эта задача, су-
щественно интересующая филогенетику, представля-
ет собой и наиболее общий раздел эволюционной
морфологии. Начало такого рода исследованиям
было положено еще в XVIII в. в трудах Вик-д’Азира
(F. Vicq-d’Azyr), защищавшего идею о коррелятив-
ных зависимостях между органами одного организма
(см. с. 57). Позднее эта идея была разработана знаме-
нитым Кювье в его законе корреляций (Cuvier, 1801).
Приблизительно через 100 лет после Вик-д’Азира о
морфологических закономерностях, но уже в истори-
ческом, дарвиновском освещении, писал великий
русский палеонтолог В. О. Ковалевский (1874), обос-
новавший учение об адаптивных и инадаптивных из-
менениях органов. Но особенно широкое изучение
морфологических закономерностей эволюции было
осуществлено уже в начале нашего века А. Н. Север-
цевым и его учениками. Северцов создал свою школу
сравнительной анатомии. Основные проблемы, кото-
рые он поставил и разработал, — учение о биологи-
ческом и морфологическом прогрессе, типы, или мо-
дусы, филогенетического преобразования органов,
коррелятивное развитие органов, теория филэмбрио-
генеза. И. И. Шмальгаузен был блестящим продолжа-
телем исследований А. Н. Северцова в этой области.
В ряде последующих разделов в сжатом виде
будут изложены сведения о главных морфологичес-
ких закономерностях эволюционного развития, кото-
рыми руководствуется в своей работе каждый зоолог.
Приспособительный характер
эволюции
В борьбе за существование выживают и оставляют
потомство те особи, которые обладают особенностя-
ми, полезными для вида. Эволюционные изменения
поэтому всегда являются приспособлениями к усло-
виям среды. Соответственно одна из основных задач
эволюционной зоологии состоит в изучении конкрет-
ных путей и направлений филогенеза в зависимости
от образа жизни и условий существования изучаемых
животных.
Однако адаптивные признаки организмов весьма
разнородны и далеко не равноценны по эволюцион-
ному значению. Наиболее обычны узкие приспособ-
ления к строго определенным и ограниченным усло-
виям среды, носящие характер узкой специализации.
Они могут ограничить возможности дальнейшей про-
грессивной эволюции и называются идиоадапта-
ция ми (Северцов, 1939). Это частные приспосо-
34
ОБЩАЯ ЧАСТЬ
бительные изменения, при которых жизнедеятель-
ность животного остается на прежнем уровне, не по-
вышаясь и не понижаясь.
Другие эволюционные изменения имеют более
общее значение и не только позволяют животному
обитать в разнообразных условиях среды, но и обес-
печивают высокую пластичность организации. Про-
цесс такого приспособления был изучен Северцовым
(1939) и назван морфофизиологическим про-
грессом, или ароморфозом. Он-затрагивает жиз-
ненно важные органы (органы питания, дыхания,
кровообращения, нервную систему, органы чувств и
т. д.) и ведет к общему подъему организации и жизне-
деятельности организма. Ароморфоз дает возмож-
ность менять место обитания и образ жизни и, следо-
вательно, открывает широкую дорогу дальнейшим
эволюционным преобразованиям. Путем ароморфо-
зов, вероятно, совершался эволюционный переход к
видам, давшим затем начало новым большим систе-
матическим группам.
У млекопитающих, например, морфофизиологи-
ческий прогресс коснулся различных систем органов:
в частности, их конечности стали мультифункцио-
нальными, что обеспечило возможность самых раз-
личных функциональных изменений в филогенезе.
После достижения теплокровности и развития волося-
ного покрова млекопитающие оказались способными
к захвату новых сред обитания. В результате эти жи-
вотные расселились по всем континентам и океанам и
овладели разнообразными экологическими нишами.
Для этого типа эволюционных преобразований
Тахтаджян (1951) предложил этимологически более
правильное название — арогенез (аналогичное
термину адаптациогенез). Впрочем, этот термин
целесообразно употреблять для обозначения самого
эволюционного процесса, а для крупных морфологи-
ческих и физиологических адаптаций лучше приме-
нять северцовский термин ароморфоз. Сходным об-
разом не следует смешивать понятия идиогенез и
идиоадаптация.
Один из классических примеров арогенеза — эво-
люция сердца у позвоночных (рис. 1). Как известно,
для рыб характерно двухкамерное сердце — венозная
кровь попадает в цельное предсердие. Из него кровь
проталкивается последовательно в желудочек, аорту
и жаберные сосуды. В жабрах она окисляется, обога-
щается кислородом и затем разносится по всем орга-
нам тела. У земноводных предсердие уже разделено
неполной продольной перегородкой на две полови-
ны, причем правая по-прежнему принимает венозную
кровь, тогда как левая — артериальную. Однако оба
типа крови частично смешиваются в неразделенном
еще желудочке, хотя к голове из него направляется
преимущественно окисленная кровь, а к остальным
частям тела — главным образом венозная. У пресмы-
кающихся при совершенном разделении предсердий
желудочек разделен еще не полностью и артериаль-
ная кровь занимает почти целиком левую, а веноз-
ная — правую половину сердца. Наконец, у птиц и
млекопитающих сердце стало четырехкамерным и
артериальная кровь полностью изолирована от веноз-
ной.. У тех и других это состояние было достигнуто
независимо; у птиц сохранилась и функционирует
Рис. 1. Арогенная эволюция сердца у позвоночных. Схема строе-
ния сердца и артериальных дуг. (По: Kuhn, 1961).
А — рыбы, Б — хвостатые амфибии после метаморфоза (у бесхвостых ам-
фибий III дуга аорты редуцирована), В — рептилии, Г — млекопитающие.
1 — правая аорта, 2 — жаберные капилляры, 3 — головные артерии, 4 —
левая аорта, 5 — аорта, 6 — венозный синус, 7 — желудочек, 8 — пред-
сердие, 9 — легочная артерия, 10 — боталлов проток, 11 — легочные ка-
пилляры, 12 — правое предсердие, 13 — левое предсердие, 14 — левый
желудочек, 15 — правый желудочек. I—IV — дуги аорты.
Fig. 1. Arogenic evolution of the vertebrate heart. Scheme of the heart
and aortic arches morphology.*
A — Fishes, Б — Tailed amphibians after metamorphosis (III aortic arch is
reduced in the tailless amphibians), В — Reptiles, Г — Mammals. 1 — right
aorta, 2 — branchial capillaries, 3 — cephalic arteries, 4 — left aorta, 5 —
aorta, 6 — sinus venosus, 7 — ventricle, 8 — atrium, 9 — pulmonary artery,
10 — ductus Botalli, 11 — pulmonary capillaries, 12 — right atrium, 13 — left
atrium, 14 — left ventricle, 15 — right ventricle. I—IV — aortic arches.
правая, а у млекопитающих — левая дуга аорты. В
обоих классах прогрессивное развитие сердца соче-
талось с усовершенствованием органов дыхания. В
результате независимо развились более совершенные
процессы окисления крови и снабжения тканей и ор-
ганов кислородом. Произошло общее усиление ос-
новных функций организации, а физиологическая
координация органов стала многообразнее и совер-
шеннее. В конце концов в силу физиологических
корреляций и другие системы органов — пищевари-
тельный, нервный аппараты, органы чувств и прочие
получили стимул к прогрессивному развитию.
* Here and below reference is given in the language of the origi-
nal.
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
35
На широкое распространение ароморфозов в раз-
ных классах простейших указали Догель (1951) и
Полянский (1970). Ограничимся лишь одним приме-
ром. Переход к организации инфузорий сопровож-
дался морфофизиологическим прогрессом, который
выразился прежде всего в умножении (полимери-
зации) жгутиков, давших сплошной ресничный по-
кров. Это привело к значительному увеличению
подвижности инфузорий, повысило эффектив-
ность разыскивания пищи и ловли добычи и обес-
печило быструю ретировку животного из зоны с
неблагоприятными факторами среды. У Hypotricha
движения стали гораздо совершеннее после образова-
ния брюшных цирр. Часть ресничек затем дала нача-
ло специализированному ротовому аппарату, а у
равноресничных инфузорий появился особый па-
лочковый аппарат клеточной глотки. Последнее осо-
бенной сложности достигло у Entodiniomorpha, у
которых появились глоточные системы специальных
волокон, скелетных пластинок и некоторые другие
образования. Прогрессивным приобретением явилась
и выработка интенсивного циклоза эндоплазмы, спо-
собствующего пищеварительным процессам в пище-
вых вакуолях. Появлением клеточной порошицы и
развитием клеточной задней кишки с ее аппаратом
сократительных волоконец (у Entodiniomorpha)
заканчивается этот ряд прогрессивных изменений
пищеварительной системы. Сократительные вакуо-
ли, обеспечивающие осморегуляцию и экскрецию, у
инфузорий более совершенны, чем у жгутиконосцев.
То же можно сказать и в отношении их чувствитель-
ных органелл, и особенно сложных систем кинето-
сом, и проводящих, опорных и сократительных во-
локнистых структур поверхностной зоны цитоплаз-
мы — кортекса.
Ароморфозом Догель считает и наличие у инфузо-
рий двух типов ядер с разными функциями. Макро-
нуклеус представляет собой высокополиплоидное
ядро с большим количеством нуклеиновых кислот и
способен поддерживать высокий уровень метаболиз-
ма в теле животного. Все эти прогрессивные особен-
ности инфузорий значительно повысили их жизнеде-
ятельность, обеспечили их биологический прогресс
и, следовательно, не могут рассматриваться иначе,
как ряд ароморфозов (Догель и др., 1962).
Очень важным ароморфозом было возникновение
многоклеточной организации, характерной для Meta-
zoa. По наиболее вероятным представлениям предка-
ми многоклеточных животных были воротничковые
жгутиконосцы, Choanoflagellata. Усиление физио-
логической связи между клетками колонии (инте-
грация) постепенно привело к появлению индивиду-
альности целой колонии. Аналогичная постепенная
интеграция колоний хорошо известна у ряда одно-
клеточных организмов животной и растительной
природы и особенно хорошо прослеживается у коло-
ниальных и шаровидных Phytomonadida (в семейст-
вах Eudorinidae и Volvocidae, — см.: Захваткин, 1949).
Таким образом, индивидуальность метазоона не соот-
ветствует таковой моноэнергидного простейшего; ме-
тазоон — это индивид иного, более высокого уровня.
Существенным арогенным процессом, разыграв-
шимся при становлении многоклеточных, по мнению
Шмальгаузена (19466), было также разделение функ-
ций между клетками. Первый его этап проявился в
морфологической и физиологической дифференциа-
ции на соматические и половые клетки; последние
стали обеспечивать размножение. Второй этап за-
ключался в дифференциации соматических клеток на
периферические жгутиковые киноциты и внутренние
амебообразные фагоциты; соответственно у первых
Metazoa образовались два первичных органа: наруж-
ный слой локомоторных жгутиковых клеток — ки-
нобласт — и внутренняя пищеварительная клеточная
масса — фагоцитобласт, т. е. общий зачаток энто- и
мезодермы (Мечников, 1886; Беклемишев, 1964;
Иванов, 1968).
Весьма крупным ароморфозом у низших беспоз-
воночных было также образование кишечника за
счет фагоцитобласта у Radiata и у низших Bilateria.
Выработка билатеральной симметрии у Metazoa и
прогрессивное развитие мезодермы и ее дериватов —
опорной соединительной ткани, мускулатуры, крове-
носной системы и полости тела — были последую-
щими важными этапами арогенной эволюции.
Характерные арогенные приспособления проявля-
лись в эволюционном развитии всех больших таксо-
нов Metazoa. Так, в становлении членистоногих ос-
новными ароморфозами были развитие из кутикулы
наружного хитинового скелета, формирование чле-
нистых конечностей, отделов тела, в том числе слож-
ной головы, а также совершенных органов чувств,
нервной системы, сердца и т. д.
Ароморфозы, сопровождающиеся возникновени-
ем новых органов и их систем, открывающих пути
развития в новом направлении, было предложено на-
зывать кайнотропозами (Киршенблат, 1969). Мож-
но сослаться, например, на образование глотки у ки-
шечнополостных и у плоских червей, целома у Coe-
lomata или амбулакральной системы у иглокожих.
Появление этих органов открыло совершенно новые
возможности эволюции.
Идиоадаптации, подобно ароморфозам, обеспечи-
вают процветание данной группы животных. Однако
будучи приспособлениями к частным экологическим
условиям, они не имеют характера универсальных
адаптаций, способных значительно повысить общую
организацию, хотя строение и функции отдельных ор-
ганов могут достигать очень высокого совершенства.
К идиоадаптациям относятся, например, приспособ-
ления различных жизненных форм: ползающих, пла-
вающих или сидячих — прикрепленных к грунту или
живущих в защитных трубках или домиках животных,
симбионтов и паразитов, обитающих на теле или в
теле своих хозяев, и т. п. Покровительственная окрас-
ка и мимикрия также являются идиоадаптациями. Не-
сомненно, идиоадаптации представляют наиболее
обычные и характерные проявления приспособитель-
ной эволюции, посредством которых достигается
экологическая дивергенция, или адаптивная
радиация (термин принадлежит Осборну, — Os-
born, 1905).
Однако по степени специализации идиоадаптации
весьма различны. Это вызвало необходимость разли-
чать среди них два разных типа — алломорфозы и
теломорфозы (Шмальгаузен, 1939—1946).
36
ОБЩАЯ ЧАСТЬ
Рис. 2. Ленивец Bradypus tridactylus. Пример идиоадаптации типа
теломорфоза (специализация к узким условиям существования)
(По: Grasse, 1955).
Fig. 2. The Sloth Bradypus tridactylus. Idioadaptation of the telo-
morphosis type example (specialization to the narrow environment).
Алломорфозами были названы приспособле-
ния, лишенные особенностей узкой специализации и
не закрывающие возможностей для более широких
эволюционных преобразований (т. е. ароморфозов).
При алломорфозах органы животного сохраняют еще
значительную пластичность и мультифункциональ-
ность, т. е. основные условия для успешного завоева-
ния новой среды. Хорошей иллюстрацией могут слу-
жить становление и эволюция класса птиц. Их пред-
ками были мезозойские рептилии — псевдозухии,
которые лазали по ветвям деревьев или совершали
длинные прыжки и имели соответствующие алломорф-
ные приспособления. От прыжков, через планирова-
ние, они пришли к полету и приобрели организацию
летающих птиц.
Теломорфозы, напротив, представляют узкие
приспособления к условиям более ограниченной,
более специальной среды. Среди млекопитающих те-
ломорфозы можно проиллюстрировать на примере
форм, ведущих древесный (например, коала, или ле-
нивцы, — рис. 2) или роющий (кроты) образ жизни.
Муравьеды, способные питаться только мелкими об-
щественными насекомыми (муравьями и термитами),
представляют другой яркий пример. Множество при-
меров различных теломорфозов можно найти среди
беспозвоночных.
Иногда у Metazoa, ведущих неподвижный, при-
крепленный образ жизни, и особенно у паразитов,
т. е. у организмов, проникших в новую среду, част-
ные приспособления выражаются в регрессивной
эволюции, которая характеризуется исчезновением
(редукцией) ряда органов и нередко приводит к край-
нему упрощению всего организма. Впрочем, морфо-
логическая деградация отнюдь не означает биологи-
ческий регресс, но знаменует особый, нередко очень
эффективный путь к достижению биологического ус-
пеха. Общеизвестные примеры таких адаптаций, на-
зываемых катагенезами (Парамонов, 1967), ка-
таморфозами (Шмальгаузен, 1969), представлены
паразитическими корнеголовыми ракообразными
(Sacculina, — рис. 3; Peltogaster и др.), эндопарази-
тическими брюхоногими моллюсками (Parentero-
xenos, Entoconchidae, — рис. 4, и др.) и многими дру-
гими паразитами. Однако даже в этих крайних случа-
ях морфологический регресс одних органов сопро-
вождается прогрессивным развитием других (напри-
мер, органов прикрепления и половой системы).
Как частную форму катагенеза следует, по-види-
мому, рассматривать недоразвитие организма вслед-
ствие сохранения отношений со средой, свойствен-
ных его личинке или иной юной стадии онтогенеза.
Недоразвиваются те органы, которые нормально
(например, у близкородственных животных) форми-
руются в конце онтогенеза. Организм как бы останав-
ливается в развитии, но становится половозрелым.
Эти адаптации обозначаются как гипоморфозы
(Шмальгаузен, 1939а, 19396). Гипогенез (см. с. 37)
приводит к развитию неотении и прогенезу. Нео-
тенией называются те случаи, когда животное в
какой-то мере еще сохраняет личиночные признаки в
период половой зрелости (Kollmann, 1882). Примеры
неотении хорошо известны у хвостатых и бесхвостых
амфибий (Molge, Pelobates, Alytes), у которых она
может быть вызвана искусственно. При плохих усло-
виях жизни в результате задержки роста и превраще-
ния органов (кроме половых) из личинок иногда раз-
виваются мелкие неотенические особи с характерной
личиночной окраской и жаберным дыханием. Посто-
янная неотения, при которой половозрелое животное
сохраняет все типичные личиночные черты и лишено
полностью дефинитивных органов, называется про-
ге незом. Классический его пример представляет
общеизвестный аксолотль (Siredon pisciformis) — ос-
тановившаяся в своем развитии, водная, дышащая
жабрами, половозрелая личиночная форма хвостатой
амфибии (Ambistoma ligrinum), живущей на суше и
обладающей легкими.
Рис. 3. Корнеголовый рак Sacculina carcini на крабе Carcinus тае-
nas. Изображена часть всасывающих ветвящихся отростков тела
паразита. (По: Бирштейн, 1968).
1 — тело паразита
Fig. 3. Sacculina carcini on the crab Carcinus maenas. The part of
the sucker branching processes of the parasite body is shown.
1 — parasite body.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
37
Рис. 4. Брюхоногий моллюск Parenteroxenos dogieli, паразитиру-
ющий в полости тела голотурии Cucumaria japonica. Схема строе-
ния. (По: Иванов, 1949).
1 — стенка кишки хозяина, 2 — сифон, 3 — семенник, 4 — коконы с
яйцами, 5 — ложная мантия, 6 — перитонеум хозяина, 7 — внутренност-
ный мешок, 8 -— яичник, 9 — передний конец, 10 — яйцевод, 11 — женское
половое отверстие.
Fig. 4. Parasitic Gastropoda Parenteroxenos dogieli into Cucumaria
japorirca (Holothuroidea) body. Morphological scheme.
1 — host intestine wall, 2 — siphon, 3 -— testicle, 4 — coccons with eggs,
5 — pseudopallium, 6 — host peritoneum, 7 — entralis pouch, 8 — ovary,
9 — anterior end, 10 — oviduct, 11 — female sexual orifice.
Гипогенез и неотения приводят к утрате узкоспе-
циализированных признаков и таким образом созда-
ют возможности для прогрессивной эволюции в любом
новом направлении. Так, например, длительное неза-
растание лобного шва на черепе человека представ-
ляет необходимое условие для продолжения роста го-
ловного мозга. По мнению некоторых ученых (Balck,
1914; Иванов П. П., 1928а), аналогичные явления не-
доразвития органов (обозначаемые как случаи фета-
лизации) сыграли значительную роль в филогенети-
ческом становлении человека.
Таким образом, благодаря неотеническому раз-
витию и размножению в личиночном состоянии
(педогенезу) создаются возможности освобождения
организма от чрезмерного груза специализации.
Очевидно, в ряде случаев неотенические формы,
вступив на новый путь прогрессивной эволюции,
дали начало новым крупным таксонам животных.
На допущении этой возможности построены многие
гипотезы происхождения некоторых групп живот-
ных. Долгое время, например, было распространено
убеждение в неотенической природе коловраток,
предками которых считались трохофоры — личин-
ки аннелид (Hatschek, 1878). Ресничные черви
иногда производятся от планул — личинок книда-
рий (Беклемишев, 1964), а низшие насекомые —
аптериготы от шестиногих личинок двупарноногих
многоножек (Мечников, 1874а; Давыдов, 1942). Ап-
пендикулярии рассматриваются некоторыми зооло-
гами как измененные неотенические личинки асци-
дий (Willey, 1894; Brien, 1948) или бочёночников
(Garstang, 1928; Bone, 1960). Наконец, Гарстанг даже
выводил хордовых из организации диплеурулы —
личинки иглокожих.
Впрочем, в большинстве своем концепции неоте-
нического происхождения больших таксонов мало-
убедительны, а некоторые, как например гипотеза
Гарстанга о происхождении позвоночных, слишком
умозрительны. Их трудно обосновать, ибо ни сравни-
тельная анатомия, ни палеонтология не могут дать
никаких указаний о происхождении животных, под-
вергшихся постоянной неотении, поскольку они вне-
запно утратили взрослую фазу. Правда, в некоторых
случаях получение полезных данных можно ожидать
от сравнительно-эмбриологических исследований.
Действительно, о том, что аксолотль есть прогенети-
ческая личинка амбистомы, известно лишь благодаря
тому, что при некоторых условиях первая форма пре-
вращается во вторую. Не будь этого, было бы невоз-
можно установить их кровное родство.
Гарстанг и Де Бир (Garstang, 1928; De Beer, 1958)
придерживаются крайней точки зрения, полагая, что
значительные эволюционные изменения происходили
только на почве неотении и педоморфоза и что от-
сутствие переходных форм между типами и классами
животного мира объясняется исчезновением в онто-
генезе взрослых форм вследствие неотении.
«Вряд ли у нас есть основание расширять значение
процесса катаморфоза, и в частности гипоморфоза,
как это делает Гарстанг, — замечает И. И. Шмальгау-
зен (1969, с. 421). — Однако мы не будем полностью
отрицать возможность частичного обращения разви-
тия, или „омоложения”, специализированного орга-
низма... Если же говорить о некоторых признаках спе-
циализации, то возможность их утраты может иметь
очень большое значение именно для прогрессивной
эволюции организмов». В пользу истинности этих
слов свидетельствует следующее соображение. По-
скольку Земля населена не живыми примитивными
схемами, а специализированными (в том или ином от-
ношении) организмами, прогрессивная эволюция воз-
можна лишь благодаря их способности к некоторой
деспециализации. Эволюционное значение неотении
было продемонстрировано на примере Dinophilida.
Неотеническое происхождение этих олигомерных ан-
нелид, которых часть зоологов выделяли в особый
класс, предполагалось уже давно (Мечников, 1886;
Ливанов, 1940), но лишь в наше время выяснилось,
что между динофилидами и педогенетическими ли-
чинками полихет из сем. Eunicidae имеется целый ряд
переходных форм — Apodotrocha, Dorvilleidae и др.
(Westheide, 1985).
38
ОБЩАЯ ЧАСТЬ
Рис. 5. Скелет ирландского оленя Megaloceras giganteus. При-
мер идиоадаптации типа гипёргенеза (гипертрофия специализиро-
ванных органов). (По: Быстров, 1957).
Fig. 5. Megaloceras giganteus skeleton. Idioadaptation of hypergene-
sis type example (hypertrophy of the specialized organs).
Среди идиоадаптаций следует также различать
ги пер генез, проявляющийся в чрезмерном развитии
в каком-то одном направлении отельных частей тела
или органов. Этот путь эволюции рассматривается
как гипертрофия органов, возникающая в результате
их узкой и односторонней специализации. Таковы,
например, гигантские клыки вымерших саблезубых
тигров — Machairodus, служившие для умерщвления
крупных толстокожих растительноядных млекопита-
ющих (носорогов и мастодонтов), или большие рога
самцов ископаемого ирландского оленя — Megaloce-
ras giganteus, развившиеся в результате полового от-
бора (рис. 5). При незначительном изменении усло-
вий существования такая чрезмерная специализация
приводит к вымиранию данной группы животных.
Общее увеличение размеров тела в процессе эво-
люции также рассматривается как характерное про-
явление гиперморфоза. Этот несомненно выгодный
признак свойствен некоторым высшим процветаю-
щим группам. Среди водных беспозвоночных наи-
большими средними размерами тела отличаются го-
ловоногие моллюски (среди которых есть гигантские
формы), вымершие хелицеровые членистоногие —
эуриптериды, мечехвосты, некоторые высшие рако-
образные и, наконец, некоторые эндопаразитические
черви (из нематод и цестод).
Известные особенности организации членистоно-
гих исключили возможность появления среди обита-
ющих на суше паукообразных, многоножек и насеко-
мых очень крупных форм (хотя из отложений карбо-
на известны некоторые представители вымершего
отряда насекомых — Palaeodictyoptera до 60 см дли-
ной). Самые крупные формы наземных животных от-
носятся к высшим группам позвоночных. Мощность
животного возрастает с увеличением размеров тела, а
вероятность нападения хищников понижается. Дейст-
вительно, от нападения хищных птиц погибают пре-
имущественно мелкие млекопитающие. Крупные раз-
меры, таким образом, повышают биотический потен-
циал гигантских форм. И хотя почти все гигантские
наземные позвоночные вымерли (стегоцефалы палео-
зоя, гигантские рептилии мезозоя, гигантские птицы
и млекопитающие третичного времени), в каждом
классе Tetrapoda в период его расцвета гигантизм
проявлялся достаточно ярко. Среди современных
высших позвоночных гиганты сохранились в классе
млекопитающих (китообразные, слоны и др.).
Следует подчеркнуть, что А. Н. Северцов пра-
вильно противопоставил ароморфозы всем другим
формам адаптаций. Причина этому ясна. Всякая
вновь достигнутая арогенная организация, будучи в
отличие от идиогенной не только прогрессивной,
но и универсальной, открывает пути к проникнове-
нию в новые среды обитания. Поэтому ароморфоз
всегда представляет собой важный исторический
этап, после которого следует новая вспышка адап-
тивной радиации.
Универсальная арогенная организация рекапиту-
лируется в онтогенезе в виде палингенезов, связан-
ных морфогенетическими корреляциями (см. с. 54) с
новыми онтогенетическими изменениями — фил-
эмбриогенезами (Шмальгаузен, 1964а). Органи-
зация взрослого животного (фенотип), подпадая
под действие естественного отбора, выступает как бы
в роли «пробного камня», на котором проверяются
перестройки онтогенеза, а в конечном итоге этих
процессов создаются частные адаптации — алломор-
фозы. Последние, как можно видеть, достигаются
путем отбора и элиминации неудачных онтогенезов и
связанных с ними неприспособленных взрослых
форм.
Общие соотношения между арогенезами и аллоге-
незами, а также катагенезами хорошо иллюстрируют-
ся известной схемой А. Н. Северцова (рис. 6). Чаще
всего период арогенеза процветающих форм при их
непрерывной дифференциации и дивергенции сменя-
ется аллогенной эволюцией, а затем телогенезом. Эта
последовательность была подтверждена и А. С. Се-
верцовым (1973), сравнившим филогенезы членисто-
ногих, кистеперых рыб, птиц и млекопитающих. Он
же показал, что когда алломорфная специализация
достигается в адаптивной зоне, которая граничит с
другой зоной, не занятой высокоорганизованными
формами, то результатом конкуренции обычно быва-
ют захват этой соседней зоны и проявление нового
арогенеза. Предшествующая фаза аллогенеза при
этом выступает как необходимая предпосылка к но-
вому арогенезу, так как при ее медленном течении
она в достаточной степени обеспечивает осущест-
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
39
Рис. 6. Схема эволюционных преобразований. (По: Северцов, 1939).
1 — ароморфозы: подъем на более высокий уровень (плоскости // и ZZZ), 2 — идиоадаптации: отклонения в пределах данной плоскости, 3 — теломорфо-
зы: специализации, 4 — регресс катаморфоз: спуск на нижележащую плоскость (Z).
Fig. 6. Scheme of the evolutionary transformations.
I — aromophosis: ups on the higher level (planes ZZ and ZZZ), 2 — idioadaptations: departures within the given plane, 3 — telomorphosis: specialization, 4 — reg-
ressive katamorphosis: down on the low plane (Z).
вление необходимых координаций алломорфных
приспособлений и глубоких арогенных перестроек
организации.
Дивергенция (радиация),
конвергенция и параллелизм
Эволюция в основном является монофилетичес-
кой, т. е. развитие происходит от общего корня. Еще
Дарвин очень убедительно показал, что естественные
группы организмов (виды, роды и т. д.) развиваются
путем дивергенции из одного общего ствола в резуль-
тате вымирания менее приспособленных промежу-
точных форм (рис. 7). Этим английским словом (di-
vergence) Дарвин обозначал расхождение, или укло-
нение, признаков и их носителей — организмов. По-
этому было бы совершенно неверно рассматривать
термин Дарвина как адекватный «раздвоению», или
«дихотомии». При дивергенции не обязательно вы-
живают только две эволюционные ветви, очень часто
их больше. Вследствие этого термин Осборна (Os-
born, 1905) «адаптивная радиация», выражающий то
же явление, образнее и потому предпочтительнее
(см.: Парамонов, 1967, с. 419).
Однако ^говоря о монофилетическом развитии, мы
отнюдь не предполагаем развития от одной пары пра-
> Класс
2^й 3-й 4- а
777-----ч Отрлды
Семеистба
. Роды
Рис. 7. Схема дивергентного развития видов. (По: Парамонов, 1967).
Диергентная эволюция вида приводит к разнообразию видов, родов, семейств, отрядов, классов и типов. Пунктиром отмечено развитие видов, не оставив-
ших потомства.
Fig. 7. Scheme of the divergent species evolution.
Divergent species evolution gives rise to the species, genera, families, orders, classes and types diversity. Development of the species remained without offspring
is dotted.
40
ОБЩАЯ ЧАСТЬ
Рис. 8. Глаза типа глазного пузыря на разрезе.
А — Helix (Gastropoda), Б — Vanadis (Polychaeta), В — человек (Mamma-
lia). 1 — роговица, 2 — хрусталик, 3 — зрительные клетки, 4 — зритель-
ный нерв, 5 — пигментные клетки, 6 — сетчатка, 7 — добавочная сетчат-
ка, 8 — секреторные клетки, 9 — оптический ганглий, 10 — стекловидное
тело, 11 — центральная ямка, 12 — склера, 13 —цилиарная связка, 14 —
радужная оболочка, 15 — сосудистая оболочка.
Fig. 8. Eyes of the eye-bladder type in section.
A — Helix (Gastropoda), Б — Vanadis (Polychaeta), В — Homo sp. (Mamma-
lia). 1 — cornea, 2 — lens, 3 — optic cells, 4 — optic nerve, 5 — pigmental
cells, 6 — retina, 7 — adventitious retina, 8 — secretory cells, 9 — optical
nerve-knot, 10 — vitreous body, 11 — central pit, 12 — sclera, 13 — ciliary
retinaculum, 14 — iris, 15 — choroid.
родительских особей, но признаем происхождение от
группы родственных форм. Такими группами, как уже
говорилось (см. с. 30), являются популяции, т. е. сово-
купности особей, которыми реально представлены в
природе все группы живых организмов. Результатом
дивергентного развития всегда бывает возрастающее
расхождение эволюционных ветвей. Впрочем, сплошь
и рядом наблюдаются случаи вторичного сходства в
организации неродственных форм, уже давно разо-
шедшихся вследствие дивергенции. Это сходство, на-
зываемое схождением признаков, или конвергенцией,
развивается как приспособление к одинаковым усло-
виям среды и часто диктуется внешними по отноше-
нию к организму, чисто физическими или химически-
ми законами или условиями среды. Как известно,
орган зрения — глаза возникают совершенно незави-
симо в самых различных группах животных (у про-
стейших, книдарий, различных червей, моллюсков,
членистоногих, позвоночных). Несмотря на это, глаз
обычно устроен в общем по одному плану, а именно
по типу camera obscura, или по схеме фотографичес-
кого аппарата. В хорошо развитом глазу имеются в оп-
ределенном сочетании роговица, зрачок, хрусталик,
стекловидное тело, сетчатка, зрительный нерв и дру-
гие части, развитие и взаимное расположение которых
определено оптическими законами — распростране-
ния, преломления и отражения света (рис. 8). При пе-
реходе к водному образу жизни наземные позвоноч-
ные приобретают много общих признаков с рыбами и
становятся внешне очень похожими на них. Таковы из
пресмыкающихся мезозойские ихтиозавры (рис. 9), а
среди млекопитающих — китообразные. Другой при-
мер: при сидячем образе жизни у водных беспозвоноч-
ных вне зависимости от их родства и систематичес-
кого положения вырабатываются сходные черты
строения — тело приобретает признаки радиальной
симметрии, на полюсе, обращенном от субстрата, во-
круг рта развиваются щупальца, питание принимает
пассивный характер и часто осуществляется подгоном
ресничками взвешенных в воде пищевых частиц к ро-
товому отверстию, анальное отверстие смещается на
ротовой полюс тела, а кишечник становится петлеоб-
разным, часто развивается способность к бесполому
размножению. Эти признаки характерны для ряда не-
родственных, неподвижно сидящих животных, и не-
удивительно, что внешний облик их бывает очень
сходным. Сходство такого типа имеется между Вгуо-
zoa и Pterobranchia, между Poly chaeta S edentaria и
Pogonophora, а также между многими другими живот-
ными (рис. 10).
Очень часто одинаковые признаки развиваются не-
зависимо, но в более или менее родственных ветвях в
результате эволюции в сходной среде. Это явление
именуется параллельным развитием. Например, раз-
ные отряды паукообразных — Aranei, Solifugae, Pseudo-
scorpiones, Acari и др. — представляют собой самостоя-
тельные ветви эволюции, давно обособившиеся друг
от друга. Но они связаны общим происхождением, ко-
рне. 9. Ихтиозавр Stenopterygius quadriscissus. Скелет с отпечатками контуров тела на породе. Длина около 2 м. (Диас Виртемберга).
Fig. 9. The Ichthyosaur Stenopterygius quadriscissus. Skeleton with the body outline imprint on the rock. Length about 2 m.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
41
Рис. 10. Сходные жизненные формы Protozoa и Metazoa. (По: Remane, 1956).
А — панцирный жгутиконосец Leptodiscus, Б — гидроидная медуза Нотоеопета, В — инфузория Stentor, Г — коловратка Ptygura, Д — сосущая инфузо-
рия Dendrosoma, Е — гидроидный полип Syncoryne.
Fig. 10. The similar life forms of Protozoa and Metazoa.
A — loricate flagellata Leptodiscus, Б — hydromedusa Нотоеопета, В — infusorian Stentor, Г — Ptygura (Rotifera), Д — suctoria infusorian Dendrosoma,
E — hydroid Syncoryne.
торым объясняются многие общие черты их строения
и определяются их в общем одинаковые эволюцион-
ные возможности. Именно поэтому независимо при-
обретаемые ими органы дыхания — воздухоносные
трубочки, или трахеи, оказываются очень сходными.
Очень интересный случай параллельной эволю-
ции представляет брюхоногий моллюск Berthelinia
Птах (рис. И) с захлопывающейся двустворчатой ра-
ковиной, очень похожей на таковую моллюсков Bi-
valvia (Kawaguti, Baba, 1959). Хорошо прослежива-
ются параллелизмы на палеонтологическом материа-
ле. Очень демонстративен, например, параллелизм в
эволюции конечностей у вымерших южноамериканс-
ких копытных из сем. Protheroteridae и у предков ло-
шадей. В обоих случаях совершенно независимо, но
очень сходно в результате приспособления к бегу
происходит постепенное усиление третьего и редук-
ция второго и четвертого пальцев конечности.
Представление о дивергенции неизбежно связано
с понятием о монофилии (см. с. 48). Филогенетичес-
кое развитие прогрессивной группы, происходящей
от одного корня и ставшей на путь аллогенеза, идет
по линиям приспособления к разным жизненным
средам. Вследствие этого образуются разные эколо-
гические формы (например, гидро-, амфи-, хтоно-,
дендробионты и т. д.), возникает специализация (по-
являются теломорфозы, катаморфозы и т. д.). В раз-
ных классах животных параллельно происходит мно-
гогранная дивергенция («радиация»), приводящая к
появлению форм со сходным образом жизни, что, ес-
тественно, связано с возникновением конвергенций и
параллелизмов (см.: Парамонов, 1945). Разумеется,
во всех этих случаях пути эволюции определяются
естественным отбором. Так, если представители раз-
личных групп млекопитающих независимо перешли
к жизни в воде, то параллельное приобретение при-
способлений к плаванию стало для этих животных
биологической неизбежностью — без них успех в
борьбе за существование невозможен.
Приведем другой пример. Различные группы чле-
нистоногих (Onychophora, Myriapoda, Insecta, Arach-
nida), перешедшие от водного к наземному образу
жизни независимо, прежде всего нуждались в вы-
работке вполне определенных приспособлений для
защиты тела от высыхания, обеспечения тканей и ор-
ганов кислородом; появилась необходимость изме-
нений в азотном обмене в связи с ограниченным ба-
лансом воды, а также возникла потребность перехода
от наружного к внутреннему осеменению (Гиляров,
1970а). У всех перечисленных членистоногих пере-
ход к жизни на земле, по-видимому, совершался
через промежуточный этап обитания в почве и со-?
провождался уплотнением хитинового покрова и об-
разованием новых органов воздушного дыхания вза-
мен редуцировавшихся жабр.
Рис. 11. Брюхоногий моллюск Berthelinia Umax (Opisthobranchia
Saccoglossa) с двустворчатой раковиной. (По: Kawaguti, Baba,
1959).
А — слева, Б — со спинной стороны. 1 — просвечивающий мускул-замы-
катель раковины.
Fig. 11. Berthelinia Umax (Opistobranchia Saccoglossa) with the bi-
valved shell.
A — at the left, Б — on the back side. 1 — adductor muscle is transparent.
42
ОБЩАЯ ЧАСТЬ
Уплотнение хитинового покрова ограничивает воз-
можность кожного дыхания и потому стимулирует об-
разование впяченных внутрь тонкостенных органов в
виде легких или трахей. Формирование же последних,
в свою очередь, было непременным условием для даль-
нейшего уплотнения покровов, которые в конце кон-
цов стали непроницаемыми для воды и газов. Анало-
гичные приспособления к наземной жизни испытали в
процессе эволюции и высшие позвоночные — Amnio-
ta, у которых тоже имели место уплотнение покровов,
редукция жабр и развитие органов воздушного дыха-
ния— легких (Гиляров, 19706, 1975).
Таким образом, конвергентное развитие и парал-
лелизм являются естественным следствием конвер-
гентных или параллельных направлений отбора.
О направленности эволюции
В филогенезе самых различных групп животных
при арогенных и аллогенных изменениях легко заме-
тить последовательность развития определенных при-
знаков. Так, у позвоночных эволюция сердца идет от
двухкамерного сердца рыб к трехкамерному сердцу
амфибий. У рептилий начинает развиваться четырех-
камерное сердце, характерное для птиц и млекопи-
тающих. То же мы замечаем и в эволюции других
систем органов позвоночных. У лошадиных кроме
постепенного развития однопалости происходит уве-
личение размеров тела и моляризация ложнокорен-
ных зубов. У хоботных отмечается последовательное
умножение гребней на задних коренных зубах и раз-
витие бивней и хобота. В эволюции членистоногих в
разных их группах мы видим постепенную стабилиза-
цию сегментов тела, дифференциацию конечностей,
все возрастающую цефализацию и т. д.
Все эти проявления параллелизма создают впечат-
ление направленности эволюционного процесса и
используются как доказательство «Автономического
ортогенеза» (Берг, 1922) или «тенденции к совер-
шенствованию». В действительности же если у жи-
вотного возникают некоторые наследственные изме-
нения, то в силу преемственности генотипического
развития в последующих поколениях возникают из-
менения такого же рода — более слабые или, напро-
тив, выраженные сильнее. При скрещиваниях эти из-
менения комбинируются.
Анализ эволюции конечностей у лошадей ясно
показал, что чем короче боковые пальцы, тем лучше
приспособленность к бегу, например у Pliohippus по
сравнению с Miohippus. Отсюда следует, что отбор
пойдет в сторону редукции боковых пальцев. В фило-
генезе лошадей, следовательно, нет никакой «внут-
ренней автономной тенденции». Эволюция их конеч-
ностей определялась исключительно направлением
отбора. «Лошадиный ряд, — пишет А. А. Парамонов
(1945, с. 348), — от Эогиппуса (Hyracotherium пот.
validae) до Лошади вовсе не является ортогенезом в
смысле предопределенности развития. „Ряд” получа-
ется, если мы отвлечемся от всех боковых вымерших
ветвей. Это — линия победителей в борьбе за су-
ществование, которую мы можем проследить в гуще
„родословного древа Лошади”. Поэтому следует гово-
рить не об ортогенезе, а об ортоселекции. То, что мы
называем направленностью эволюции, следовательно,
есть результат направленного естественного отбора».
Из сказанного видно, что сравнительное изучение
параллелизмов и конвергенций представляет боль-
шой интерес для сравнительной морфологии, физио-
логии, филогенетики и систематики (см.: Догель,
1937). О степени родства животных групп, очевидно,
можно судить не только по их гомологичным орга-
нам, но и по многим аналогиям, возникшим в резуль-
тате параллельной эволюции. Так, щетинки аннелид-
ного типа (т. е. образуемые каждая одной хетогенной
клеткой на дне кожного впячивания) возникли неза-
висимо у Echiurida, Annelida, Chaetognatha, Pogono-
phora (а также у некоторых Mollusca и Tentaculata).
Однако они представляют собой проявление парал-
лельного течения эволюции перечисленных групп
Coelomata и, следовательно, говорят о монофилети-
ческом происхождении всех этих животных (Ива-
нов А. В., 1975а, 1976).
Вся совокупность наших знаний о животном мире
свидетельствует о том, что прогресс является глав-
ным направлением эволюции. В истории биологии не
было недостатка в попытках объяснить направленное
течение эволюционного процесса действием некото-
рых внутренних факторов, присущих живым сущест-
вам. Ламарк (Lamarque, 1809, — цит. по: Lamarque,
1959) приписывал всем организмам изначально свой-
ственное им непознаваемое стремление к совершен-
ствованию. Очень близких взглядов, в сущности, при-
держивался и Берг (1922), пытавшийся объяснить
филогенез внутренними имманентными закономер-
ностями (гипотеза номогенеза). Негели (Nageli, 1884)
полагал, что процесс эволюции покоится на направ-
ленных изменениях структуры наследственного ве-
щества — идиоплазмы.
Однако причины прогрессивной направленности
эволюции полнее всего могут быть поняты в свете
теории Дарвина. Как было показано выше, приспосо-
бительную эволюцию было бы неправильно сводить
только к арогенезу, или морфофизиологическому
прогрессу, ибо обычно она осуществляется также
путем аллогенеза, телогенеза, катагенеза и гипогене-
за. В связи с этим необходимо вслед за А. Н. Север-
цовым (1925) четко различать два разных, хотя и
тесно связанных явления: прогресс морфофизиологи-
ческий и прогресс биологический.
Морфофизиологический прогресс харак-
теризуется усложнением строения, повышением диф-
ференциации, усилением и совершенствованием
функций, целостности (Шмальгаузен, 1938) и гомео-
стаза (Huxley, 1942), приводящих к подъему общего
уровня жизнедеятельности. Биологический про-
гресс является результатом успеха данной группы
организмов в борьбе за существование. Он определя-
ется увеличением количества популяций данного
вида и входящих в них особей, расширением ареала
и распадением таксона на подчиненные системати-
ческие группы.
Морфологический прогресс животных, разумеет-
ся, сопровождается изменениями и усложнениями их
образа жизни и всей жизненной обстановки. Ароген-
ному и, следовательно, прогрессивному развитию
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
43
прежде всего подвержены органы и системы большо-
го физиологического значения, а именно органы дви-
жения, полость тела, органы, связанные с обменом
веществ (пищеварительные, дыхательные, кровенос-
ные, выделительные), а также органы чувств и нерв-
ный аппарат. Концентрация нервных и чувствитель-
ных элементов на переднем полюсе тела у примитив-
ных билатеральных беспозвоночных имела большие
биологические преимущества и в конце концов при-
вела к развитию органов чувств и головного мозга и
к образованию головы (процесс цефализации), в об-
ласть которой переместилось и ротовое отверстие.
В состав головы у высших сегментированных живот-
ных (Annelida, Arthropoda) вовлекается то или иное
количество передних сегментов туловища с их пар-
ными конечностями и нервными центрами. В ре-
зультате формируется голова очень сложной мор-
фологии. Аналогичное образование сложной по
сегментарному составу головы, сопровождающееся
возникновением головного мозга и головных органов
чувств, имело место и у предков позвоночных. По
Шмальгаузену (1964а, 1969), органы чувств Vertebra-
ta возникли еще у их предков, обитавших в воде.
Действительно, многочисленные глазки, расположен-
ные в нервной трубке, и ямка Гатчека имеются уже у
ланцетника (Branchiostoma). У бесчелюстных органы
чувств (глаза, органы обоняния, слуха, равновесия и
вкуса) развивались прогрессивно и достигли большо-
го совершенства у рыб. Замечательно, однако, что
при переходе позвоночных из водной стихии на
сушу все эти органы отнюдь не деградировали, а,
наоборот, приобрели особенно большое значение и
соответственно достигли высшего совершенства.
Таким образом, по Шмальгаузену, органы чувств,
приобретаемые в воде, не теряют своего значения и в
других средах обитания, становясь постоянными ат-
рибутами и при обновлении организации. То же
самое справедливо и для многих других систем орга-
нов, которые имеют большое физиологическое зна-
чение.
Естественно, что накопление арогенных приспо-
соблений неизбежно связано и с развитием новых фи-
зиологических и онтогенетических корреляций между
органами и их системами. В онтогенезе отбираются и
накапливаются различные регулирующие механизмы,
которые повышают общий уровень целостности орга-
низма и обеспечивают полное развитие и координа-
цию всех специфических частей развивающегося и
взрослого животного. Поэтому постепенно онтогенез,
как и собственно организм, становится все более неза-
висимым от условий среды. Таким образом, при мор-
фофизиологическом прогрессе приспособленность к
среде, численность вида и его распространение воз-
растают* т. е. имеет место биологическое процветание
вида.
К биологическому прогрессу (по А. Н. Северцову)
ведут следующие пути.
1. Морфофизиологический прогресс, или орогенез,
т. е. приспособления, повышающие энергию жизне-
деятельности взрослых потомков.
2. Аллогенез и другие идиоадаптации, т. е. при-
способительные изменения, при которых общая жиз-
недеятельность взрослых потомков не меняется.
Ъ.Ценогенез, или приспособления зародышей и
личинок, не изменяющие энергии жизнедеятельно-
сти взрослых потомков, но увеличивающие их число.
(Впрочем, ценогенезы, представляющие приспособ-
ления в онтогенезе, могут иметь и более широкий
характер, и в этом случае они приближаются к
арогенезу. Таковы, например, зародышевые оболоч-
ки насекомых и наземных позвоночных (Amniota),
обусловившие эволюционный выход на сушу их об-
ладателей. Плацента млекопитающих — другой при-
мер этого рода.)
4. Общая дегенерация, т. е. приспособления взрос-
лых потомков, сопровождаемые снижением энергии
жизнедеятельности.
Однако нередко биологический прогресс сменяет-
ся биологической стабильностью, когда приспособ-
ленность пребывает более или менее длительное
время на одном уровне, а численность особей остает-
ся постоянной. При биологическом регрессе, напро-
тив, происходит снижение приспособленности орга-
низма, так как по темпу его эволюция отстает от из-
менений среды. Элиминация возрастает, численность
особей падает, т. е. вид приближается к вымиранию.
Из сказанного ясно, что наибольшее значение в
эволюции принадлежит биологическому прогрессу,
связанному с морфофизиологическим прогрессом.
Однако в ряде случаев биологический прогресс соче-
тается с морфофизиологическим регрессом, как это
наблюдается в эволюции большинства паразитичес-
ких, а также сидячих животных. Среди более прими-
тивных Metazoa есть обширные группы — классы и
даже типы, все представители которых ведут парази-
тический образ жизни (многие классы плоских чер-
вей, скребни и др.); они сравнительно мало подверг-
лись морфологическим упрощениям. Но представи-
тели более высоко организованных групп, перейдя
к паразитизму (например, некоторые брюхоногие
моллюски или ракообразные), испытали крайнюю
степень упрощения организации. Интересно, что
строение паразитических простейших, напротив, зна-
чительно усложняется под влиянием паразитизма
(Полянский, 1970).
Важнейшими факторами арогенной эволюции ор-
ганизмов, несомненно, являются условия биотичес-
кой среды. Эта истина, ясно высказанная еще Дарви-
ном, была рассмотрена в свете учения В. И. Вернадс-
кого об эволюции биосферы (Шмальгаузен, 1939а,
19396; Завадский, 1958, 1968; Яблоков, 1968; Кол-
чинский, 1990). По этому поводу Камшилов (1974)
совершенно справедливо писал: «...в результате жиз-
недеятельности происходят медленные, но постоян-
ные перестройки биотических отношений, законо-
мерно изменяется среда жизни каждого вида. Таким
образом, изменения условий жизни, ведущие к пре-
образованию организмов, оказываются неизбежным
следствием самой жизни... Жизнедеятельность орга-
низмов оказывается весьма существенной, если не
самой главной причиной преобразования живых су-
ществ». Иначе говоря, эволюция жизни на уровне био-
сферы выступает как саморазвитие (Вяккерев, 1972;
Сутт, 1977).
Итак, главную форму, или направление, эволюции
представляет арогенез, приводящий к появлению
44
ОБЩАЯ ЧАСТЬ
новых крупных таксонов — отрядов, классов, типов.
Вопрос о движущих силах этой так называемой мак-
роэволюции решается современной СТЭ в том смыс-
ле, что они тождественны таковым микроэволюции
(см. с. 30). Опираясь на хорошо разработанную кон-
цепцию происхождения видов на основе естественно-
го отбора в популяциях, можно предполагать, что
иных: движущих сил эволюции не существует.
Особое внимание привлекла к себе проблема так
называемого неограниченного прогресса эво-
люции, или эпиморфоза, приведшего к появле-
нию на Земле человека (Шмальгаузен, 1939а, 19396;
Huxley, 1963). Все остальные линии эволюции рас-
сматривались при этом как ограниченные в своих
возможностях. Завадский (1958) считал неограничен-
ный прогресс высшим проявлением арогенеза. Ан-
тропологи Рогинский и Якимов (1968) правильно
подчеркнули, что это понятие уникально и примени-
мо только для заключительных этапов эволюции че-
ловека разумного, Homo sapiens.
Откуда эта уникальность? Почему переход к выс-
шей форме движения материи осуществился лишь
один раз? Шмальгаузен, Завадский, Яблоков и другие
ученые видят главную причину в том, что в осталь-
ных арогенных линиях эволюции проявили себя ог-
раничения специализации, исключившие возмож-
ность дальнейшего существенного прогресса. Только
отряд приматов избежал этого и на одной из своих
ветвей дал в конце концов гоминид.
Здесь уместно кратко коснуться еще вопроса, о
природе тех ограничений, которыми лимитируются
некоторые прогрессивные направления эволюции.
Нередко они связывались с идеей целеустремлен-
ности эволюции, или стремления ее к конечной цели
(Lamarque, 1809, — цит. по: Lamarque, 1959; Baer,
1876), с концепцией «органического роста» (Eimer,
1888, 1897) или с законом «автономического ортоге-
неза» (Берг, 1922). Некоторые палеонтологи (Соре,
1904; Deperet, 1907) объясняли ограниченность эво-
люции предустановленной тенденцией развития в
сторону узкой специализации, ведущей рано или позд-
но к вымиранию филетических ветвей. Подобные ре-
шения вопроса, по существу, есть признание предо-
пределенности эволюции и неизбежно приводят к те-
леологии.
Совсем другая трактовка прогрессивной эволю-
ции, связанная с признанием ограниченности воз-
можных направлений развития, дается в современ-
ном дарвинизме. По словам И. И. Шмальгаузена
(1939а, 19396, с. 13), «неизбежная ограниченность
эволюции многих филогенетических ветвей, выра-
жающаяся в ограниченном формообразовании, в ог-
раничении распространения в пространстве и во
времени, является вместе с тем условием для неог-
раниченной эволюции всего мира организмов^ иду-
щих ко все более высоким формам жизни... Нет
никаких данных, указывающих на то, что эволюция
в целом идет к какому-то концу. Наоборот, непре-
рывно возникающие более совершенные формы вы-
тесняют все устаревшее, уже отжившее. Именно в
этом устранении менее совершенных форм мы и
видим условие для возможности расцвета новых,
более высоко стоящих форм».
Эволюция всегда проявляется как возникновение
и накопление новых приспособлений, новых конст-
рукций и процессов, нового поведения и образа
жизни.
Итак, эволюция не есть развертывание или реали-
зация заранее предустановленных программ и воз-
можностей. Естественный отбор может не только со-
здавать новые структуры и функции и интегрировать
новые адаптации, но и направлять эволюционный
процесс по новым путям (Шмальгаузен, 1969).
Необратимость эволюции
Эволюция — процесс необратимый, как всякое
развитие. Первая формулировка этого закона принад-
лежит самому Дарвину, который в XI главе своей
книги «Происхождение видов...» писал: «Если вид
исчез с лица Земли, то нет оснований думать, что та
же самая, тождественная форма когда-либо появится
вновь». В более узком смысле, а именно в отношении
эволюции отдельных органов животного, принцип
необратимости позднее был обоснован палеонтоло-
гом Долло (Dollo, 1893).
Закон Дарвина легко понять, ибо при полном вы-
мирании данного вида исчезает та генетическая линия,
которую он представляет. Условия существования
также никогда не остаются неизменными, постоянно
меняются и отношения вида со средой, изменяется,
следовательно, и вид.
Согласно принципу Долло, орган, подвергшийся
редукции, никогда не появляется вновь, если даже по-
томки попали в прежние экологические условия и
вновь возникла потребность в утраченном органе. В этом
случае развивается новый орган, функционально заме-
щающий старый, но формирующийся из совсем дру-
гих зачатков и, следовательно, ему не гомологичный.
Так, в юре и мелу появились приспособившиеся к
водной жизни черепахи (Protiosphargis, Archelon и
некоторые другие) с весьма облегченным панцирем,
прорезанным широкими отверстиями. Это были по-
томки триасовых форм, отличавшихся мощным
сплошным панцирем. В олигоцене и миоцене водные
черепахи вернулись к жизни на суше, однако потреб-
ность в защитном панцире была удовлетворена у них
не восстановлением старого, а развитием совершенно
нового, вторичного щита (рис. 12). Впрочем, извест-
ны и случаи «возврата» утраченного органа или при-
митивного признака. Несколько примеров такого
рода были проанализированы Сушкиным (1915).
У человека, например, лобная кость (в норме непар-
ная) иногда оказывается парной, как у большинства
млекопитающих. У пингвинов цевка (кость задней
конечности) отличается особенностями, едва ли не
более примитивными, чем у первоптицы — Archaeor-
nis. Добавим еще один пример — самым примитив-
ным представителем класса ресничных червей (Tur-
bellaria) свойствен орган равновесия — статоцист,
расположенный в области головного мозга. Неожи-
данно этот орган обнаружен и у Lurus — одной из
форм, принадлежащих к «высшему» отряду — Neor-
habdocoela, и можно думать, что это результат ново-
образования уже утраченного предками органа (Ах,
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
45
Рис. 12. Эволюция щита у водных черепах. (По: Ziegler, 1972).
А — Proterochersis (триас); Б — Thalassemys (юра); В — Taxochelys (мел); Г — Protosphargis (мел); Д — Psephophorus (третичный период); Е — Dermo-
chelys (современный); Ж — то же, рудиментарный первичный панцирь. Для обитания в воде панцирь облегчается путем образования отверстий (ряд А—Г).
При возврате на сушу в прибойную зону (Д) развивается вторичный панцирь из мелких кожных окостенений, который при вторичном переходе к жизни в воде
(£, Ж) редуцируется.
Fig. 12. Shell evolution of the marine turtle.
A — Proterochersis (Trias), Б — Thalassemys (Jurassic), В — Taxochelys (Cretaceous), Г — Protosphargis (Cretaceous), Д — Psephophorus (Tertiary), E '•— Der-
mochelys (Recent), Ж — the same, rudimentary primary shell. Shell is lighten by the fontanelles formation in aquatic environment (А—Г row). Recur to the coast
intertidal zone of land (Д) brings to the secondary shell with the small dermal ossifications development It is reduced in the secondary going to the aquatic environment
(&Ж).
1961). Во всех подобных случаях, однако, речь может
идти не о настоящем возврате к уже пройденному
этапу филогенетического развития, а о гетерохронии
(см. с. 54) в эмбриональном развитии некоторых
органов. С этой точки зрения развитие цевки пингви-
нов просто задерживается на очень ранней и прими-
тивной еще ступени онтогенеза (т. е. имеет место не-
доразвитие органов типа гипогенеза). Легко пред-
ставить себе, что такого рода задержки в развитии
отдельных органов устраняют те черты их специали-
зации, которые оказываются излишними и даже вред-
ными для организма, попавшего в новые условия су-
ществования.
О темпах эволюции
Общая картина современного животного и расти-
тельного населения Земли свидетельствует о крайне
различной скорости эволюции в разных группах.
Рядом с высокоразвитыми формами — человеком,
позвоночными, насекомыми, головоногими моллюс-
ками — обитают такие организмы, как книдарии,
гребневики, губки, простейшие и прокариоты.
Разнообразие факторов, влияющих на темпы эво-
люции, очень велико. Среди них можно отметить и
процессы, протекающие в популяциях, в частности
изменения в их генетическом составе, и такие изме-
нения среды, как появление и исчезновение организ-
мов, служащих пищей, или хищников и паразитов, и,
наконец, изменения температуры, солености воды,
состава атмосферы, уровня моря, солнечной радиа-
ции и т. д. По данным генетики, частота мутаций за-
висит от естественных изменений среды. Повышение
мутирования ведет к генетическому многообразию
популяции, которое может увеличить темпы эволю-
ции в том случае, если, например, стабилизирующий
отбор сменится движущей формой отбора. Может,
кроме того, возрасти и модификационная изменчи-
вость.
Большое значение для темпов эволюции имеют
размеры популяции. В малых популяциях условия не
способствуют возникновению благоприятных комби-
наций путем скрещивания различающихся особей, а
в больших популяциях остается слишком мало шан-
сов для их сохранения. Благоприятные условия для
эволюции имеются в популяциях среднего размера.
Однако, согласно Райту (Wright, 1931), оптимальное
сочетание условий складывается в локальных средах
и даже в малых популяциях при отсутствии полной
изоляции. Между популяциями происходит постоян-
ный обмен эмигрантами, увеличивающий вероят-
ность комбинирования, а неполная изоляция способ-
ствует закреплению удачных комбинаций. Что каса-
ется результатов естественного отбора, то они более
ощутимы в больших популяциях (Шмальгаузен,
1969).
Накоплено много данных по эволюции растений и
животных, позволяющих различать две основные фор-
46
ОБЩАЯ ЧАСТЬ
мы видообразования: внезапную и постепенную. Вне-
запное видообразование сопряжено с быстрой пере-
стройкой всего генома, как это происходит, например,
при образовании полиплоидных форм. Таким путем
новый вид возникает за несколько поколений. Поли-
плоидные виды довольно обычны среди растений. Из-
вестны и другие случаи быстрого образования новых
видовых форм, связанные с частичной перестройкой
хромосом (фрагментацией и слиянием), как было по-
казано генетиками в многочисленных, экспериментах
с плодовыми мушками Drosophila.
Однако более широко распространено посте-
пенное видообразование. В настоящее время пале-
онтологические материалы по некоторым видам на-
столько полны, что позволяют установить время, не-
обходимое для их становления. Известно, например,
что формирование некоторых видов верхнеплиоцено-
вых двустворчатых моллюсков заняло несколько мил-
лионов лет, но наряду с этим в четвертичном периоде
один новый вид рода Paphia в Черном море возник
лишь за 6000 лет. Аналогичные данные получены для
нового подвида водяной полевки Arvicola terrestris,
для его образования понадобилось 300—350 тыс. лет,
а новый вид зубра Bison выделился за 10—16 тыс.
лет. Образование подвидов хохлатого жаворонка —
Galerida cristata и славки — Prinia gracilis продол-
жалось, по-видимому, 5—10 тыс. лет.
Имеются данные и о длительности существования
таксонов выше вида. Так, геологический возраст не-
которых родов двустворчатых моллюсков (Bivalvia)
измеряется сотнями миллионов лет. Впрочем, дли-
тельность существования многих родов Bivalvia со-
ставляет от 25 до 50 млн лет. Для родов отряда гры-
зунов (Rodentia) установлена продолжительность не
более 30, а для родов хищных млекопитающих (Car-
nivora) — 25 млн лет (см.: Simpson, 1948; Яблоков,
Юсуфов, 1976). Однако во многих классах наряду с
медленно развивающимися формами существуют и
быстро эволюирующие.
«Даже случайный наблюдатель истории развития
животного мира будет поражен тем, — пишет Симп-
сон (Simpson, 1948, с. 195), — как на общем фоне
стандартной для данной группы, средней скорости
эволюции то тут, то там появляются отдельные линии,
эволюирующие с необычайной быстротой, в то время
как некоторые другие линии изменяются так медлен-
но, как будто вовсе не эволюируют». Иначе говоря,
быстрый, средний и замедленный темпы эволюции
часто свойственны близкородственным филогенети-
ческим ветвям. Например, эволюция предков лоша-
дей протекала гораздо быстрее в период между сред-
ним и поздним эоценом, а позднее наступило замед-
ление эволюции, сменившееся новым ускорением к
раннему олигоцену и далее — от раннего к среднему
миоцену.
Внимательное изучение показывает, что средняя,
или стандартная, скорость эволюции (горотели-
ческая по Симпсону) обычно значительно варьиру-
ет от одной большой группы к другой. Так, у млеко-
питающих групповая скорость оказалась значительно
выше, чем у двустворчатых моллюсков. Выходящие
из стандарта низкие скорости эволюции были назва-
ны Симпсоном брадителическими, а более вы-
сокие— тахителическими (см. рис. 13).
Классические примеры брадителии известны в
классе двустворчатых моллюсков. Можно утверж-
дать, что ни один из уже вымерших родов не просу-
ществовал дольше 275 млн лет; эти невысокие темпы,
очевидно, соответствуют очень низкому уровню горо-
телической скорости в классе Bivalvia. Однако неко-
торые современные роды существуют уже более
400 млн лет (т. е. по крайней мере начиная с силура).
Это отдельные, наиболее медленно изменяющиеся,
т. е. брадителические, линии развития. Таковы ныне
живущие роды — Nucula, Leda. Modiolus, Pteria,
Lima и Ostraea.
У животных, перешедших к быстрой эволюции
(тахителии), эта особенность наблюдается сравни-
Адаптивная зона
перемещаетацпопу -
ляции горотелические
а
Гахителическая
Bemtb
Переход через порог
от перспективной
адаптации к пост-
адаптации
Адаптивная зона
стабильная^ популяции
брадителические
Время
Рис. 13. Схема, иллюстрирующая понятия горотелии, брадителии и тахителии. (По: Simpson, 1948).
Fig. 13. Scheme of horotely, bradytely, tachytely concepts.
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
47
тельно короткое время, затем скорость развития сни-
жается до горотелической или даже до брадителичес-
кой. Подобная смена темпов наблюдалась на палеон-
тологическом материале в эволюции разных групп
животных и была продемонстрирована Мюллером
(Muller А. Н., 1963) на схемах, на которых измене-
ние числа родов от одного геологического горизонта
к другому хорошо показывает скорость и течение
(Ablauf) эволюции. На рис. 14 показан ход эволюции
амфибий, которые появились в девоне, достигли зна-
чительного расцвета в карбоне, когда возникли мно-
гочисленные ветви и наибольшее число родов и соот-
ветственно возросли темпы эволюции. В перми ско-
рость развития упала и после небольшой вспышки в
триасе оставалась очень низкой вплоть до середины
третичного периода, когда произошел новый рас-
цвет — количество родов (и видов) снова возросло,
хотя и в гораздо более скромных масштабах, чем в
карбоне. Подобная же схема для Vertebrata представ-
лена на рис. 15.
Особое значение в повышении темпов эволюции
имеют аллогенезы, придающие эволюции организмов
характер самоускоряющегося движения. А так как
при аллогенезах все арогенные признаки полностью
сохраняют свое биологическое значение, то весьма
значительные темпы эволюции характеризуют и эту
форму эволюционного процесса. «Поэтому аллогене-
зы (идиоадаптации) сохраняют значение наиболее
распространенного типа эволюции» (Парамонов,
1967; Шмальгаузен, 1969). При узкой специализации
(телогенезы, катагенезы) снижается способность к
адаптациям и замедляется темп эволюции. По Шмаль-
гаузену (19466, с. 378), «в каждой филогенетической
ветви идет вначале до известной степени скрытая и
довольно медленная подготовка ее обособления».
Позднее ветвь «начинает дифференцироваться и эво-
люировать возрастающими темпами. Чем выше уро-
Рис. 14. Схематическое изображение динамики числа родов (уч-
тено 237 родов) амфибий. (По: Muller, 1963),
В карбоне — наибольшие темпы эволюции, в мелу — резкий биологичес-
кий регресс, в третичный период — ограниченное повышение темпов эво-
люции.
Fig. 14. Scheme of the amphibian genera number dynamics (237 ge-
nera are accounted).
Maximum of the evolutionary rate — Carboniferous, sharp biological regres-
sion — Cretaceous, limited increase of the evolutionary rate — Tertiary.
Рис. 15. Схематическое изображение динамики числа родов (5105) позвоночных. (По: Muller, 1963).
В ордовике — начало развития; нарастающий биологический прогресс до юры или включая юру; новый подъем в мелу; интенсивный прогресс в третич-
ный период. Схема иллюстрирует победу позвоночных. Учтено родов: 1443 — рыб, 237 — амфибий, 1045 — рептилий, 300 — птиц и 2080 — млекопита-
ющих.
Fig. 15. Scheme of the growth dynamics of genera number (5105) of the vertebrata.
Beginning of the progress in Ordovician; increase of the biological progress untill Jurassic or including Juraccis; the new growth during Cretaceos; intensive
growth during Tertiary. Scheme illustrates Vertebrate victory. Genera number is accounted: Pisces — 1443, Amphibia — 237, Reptilia — 1045, Aves — 300,
Mammalia — 2080.
48
ОБЩАЯ ЧАСТЬ
вень ветви, тем быстрее она специализируется». Симп-
сон (Simpson, 1948) указывал, что начальная эволю-
ция новой ветви протекает очень быстро и что тахи-
телия может быть одной из возможных причин появ-
ления пробелов в палеонтологической летописи у
основания этой ветви.
В связи с проблемой понижения темпа эволюции
уместно вспомнить о реликтовых животных (см.
также с. 76). Стабильность окружающей среды спо-
собствует во многих случаях чрезвычайно длительно-
му сохранению некоторых видов в почти неизменном
состоянии (в результате доминирования стабилизиру-
ющего отбора). Классический пример реликтовой
формы, остающейся неизменной со времен ордовика
до наших дней (т. е. в течение более 450 млн лет),
представляет делающее норку в песчаном морском
грунте плеченогре Lingula. Архаический моллюск
Neopilina galatheae. открытый в 1952 г. в восточной
части Тихого океана, оказался представителем ниж-
непалеозойского класса Monoplacophora (кембрий,
нижний девон — возраст 550—400 млн лет). Пять
видов палеозойского рода корабликов, Nautilus, дожи-
ли до наших дней. Реликтовым, несомненно, являет-
ся и класс мечехвостов (Xiphosura). Шесть современ-
ных видов этих морских членистоногих обитают в
прибрежных водах Японии, Индо-Вест-Пацифики и у
атлантических берегов Северной и Центральной
Америки. Остатки мечехвостов, близких к современ-
ным, находят начиная с триаса (200 млн лет). Из дру-
гих общеизвестных реликтов напомним о современ-
ной кистеперой рыбе Latimeria chalumnae, впервые
обнаруженной близ берегов Юго-Восточной Африки
в 1938 г. До того кистеперые (Crossopterygii) были
известны в ископаемом состоянии из девонских и ка-
менноугольных отложений (возраст 300—400 млн
лет).
Особую проблему составляет вопрос о скоростях
филогенетических изменений органов. Анализ его
сделан Воронцовым (1963, 1999) и Лукиным (1964).
Разумеется, степень эволюционных изменений строе-
ния и функций органов определяется действующими
формами отбора. Стабильность органа, естественно,
обеспечена исторически сложившимися корреляция-
ми и координациями, «охраняющими» адаптивность
данного органа. Когда стабильный орган становится
на путь филогенетических изменений, морфогенети-
ческие связи должны перестроиться. Очевидно, про-
блема неравномерности филогенетических измене-
ний органов и их систем может быть решена лишь
при внимательном изучении эволюции конкретных
групп животных и условий, в которых она протекает.
Результат неравномерной эволюции органов выража-
ется в явлениях гетеробатмии (см. с. 66).
Главную причину неравномерности эволюции раз-
ных систем органов Е. И. Лукин видит в том, что не-
которые из них играют ведущую роль в эволюции как
биологически и физиологически важные и поэтому
изменяются в первую очередь. Первой ведущей систе-
мой Лукин считает нервную, второй — двигательную,
затем следуют остальные системы. Половой аппарат
оказывается на последнем месте.
Таким образом, различия в темпах эволюции от-
дельных органов Лукин связывает с их экологичес-
ким значением. Можно лишь отметить, что порядок
«очередности» преуспевающих систем органов не во
всех группах животных одинаков. Нервная система и
соответственно поведение животного действительно
часто выходят на передний план. Но во многих груп-
пах ведущими системами становятся другие. Не вда-
ваясь в детали, можно сослаться, например, на веду-
щую эволюционную роль пищеварительной системы,
особенно передней кишки у нематод (Парамонов,
1967), коловращательного аппарата у коловраток,
амбулакральной системы и скелета у иглокожих, щу-
пальцевого аппарата у погонофор (Иванов А. В.,
1959) и половой системы у турбеллярий (Ливанов,
1955).
Монофилия и полифилия
Успехи зоологии и палеонтологии последних де-
сятилетий сопровождались попытками некоторых
выдающихся ученых обосновать полифилетическое
происхождение таких больших групп, как Metazoa,
Bilateria, Coelomata, Arthropoda, Amphibia, Reptilia,
Mammalia и других. Предлагались разные варианты
полифилетизма, уживающиеся с самыми различными
представлениями о факторах эволюции.
Если исключить концепции, использующие поня-
тия полифилии и параллельной эволюции как аргу-
менты против СТЭ, то можно утверждать, что мнение
большинства зоологов и ботаников склоняется к при-
знанию монофилетической природы естественных
групп.
Сторонники так называемой широкой монофилии
считают вслед за Симпсоном (Simpson, 1961) группу
монофилетической, даже если она объединяет не-
сколько параллельных ветвей, возникших независи-
мо от одной группы того же или более низкого ранга
(Негге, 1964; Тимофеев-Ресовский и др., 1969; Мауг,
1971). Они подчеркивают, что в этих случаях нет ос-
нований говорить о полифилии, поскольку в геноти-
пе параллельных стволов имеются общие комплексы
генов, способных сходно мутировать; отбор сохраня-
ет эти гомологические мутации.
Напротив, приверженцы строгой монофилии (среди
них Северцов А. Н., 1939; Шмальгаузен, 19466,19646;
Шаров, 1971; Лукин, 1975; Воробьёва, 1981; и мн.
др.), следуя за Дарвином, видят в естественном таксо-
не только потомков одного вида. Согласно этим авто-
рам, признание крупной полифилетической группы
всегда недостаточно обосновано и чаще всего прини-
мается вследствие неполноты палеонтологической ле-
тописи или ошибочных теоретических соображений.
Целый ряд серьезных аргументов против расши-
ренной трактовки монофилии по Симпсону высказали
И. И. Шмальгаузен, А. Ремане, Е. И. Лукин, А. Г. Ша-
ров и многие другие.
Особенно важное обстоятельство отметил еще в
1946 г. Шмальгаузен: параллелизмы, на которые опи-
раются полигенисты, проявляются лишь у близкород-
ственных организмов после периода начальной ди-
вергенции и потому свидетельствуют в пользу моно-
филии. Параллельные стволы начинаются от одного
родоначального вида.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
49
То же самое, по существу, писал и Ремане (Rema-
ne, 1964), утверждая, что палеонтологи говорят о по-
лифилии обычно в тех случаях, когда уходящие в
глубь веков линии ископаемых форм приводят к
одной общей исходной форме «не так скоро, как это
ожидалось». Наличие параллельных эволюционных
стволов не означает еще их независимого возникно-
вения, а свидетельствует лишь о том, что разветвле-
ние стволов надо искать в более древних слоях.
Впрочем, и длительное параллельное развитие сход-
ных стволов часто оказывается мнимым, поскольку
оно зависит от способа классификации ископаемых
форм. Можно, например, присоединить ископаемые
формы к современным группам, расширив их диаг-
ноз, или, наоборот, создать для них новые таксоны. В
зависимости от этого отдельные таксоны можно
трактовать то как монофилетические, то как поли-
филетические.
Отсутствие или крайнюю редкость ископаемых
«недостающих звеньев» между материнским и дочер-
ним таксонами часто также приводят как аргументы
в пользу полифилии. Однако в немногих случаях,
когда переходная форма сохраняется в ископаемом
состоянии, сомнений в монофилии дочернего таксо-
на не возникает. Пример этому можно найти в классе
птиц; между современными птицами и рептилиями
известна соединяющая их юрская первоптица — Аг-
chaeornis. Среди беспозвоночных подобные связую-
щие формы обнаруживаются гораздо реже, поскольку
в кембрии существовали уже почти все типы и боль-
шинство классов. Возможно, к переходным формам
относится докембрийская Spriggina — сегментиро-
ванное донное животное с некоторыми признаками
аннелид и трилобитов.
Редкость ископаемых переходных форм была
объяснена Симпсоном (Simpson, 1948). Возникнове-
ние нового большого таксона обычно связано с за-
хватом новой арены жизни, что всегда сопряжено с
переходом через неустойчивое состояние: в новой
обстановке старые приспособления утрачивают свое
значение, а новые лишь зарождаются. Поэтому усло-
вия жизни промежуточной зоны очень тяжелы для
эволюирующей группы и резко снижают ее числен-
ность, в то время как давление отбора значительно
ускоряет темп эволюции. Переход через эту зону
могут совершить немногие из параллельно эволюиру-
ющих ветвей — только обладающие достаточно вы-
сокими темпами эволюции. В результате сохранение
их в ископаемом состоянии представляется крайне
маловероятным. Симпсон, таким образом, вопреки
собственным аргументам в пользу «широкой моно-
филии» объяснил монофилетическую природу зарож-
дающегося большого таксона и подтвердил мнение
Шмальгаузена о том, что длительно существующие
родственные параллельные ветви возникают в резуль-
тате сравнительно недолгого периода дивергенции,
или адаптивной радиации.
Большинство зоологов принимают монофилию
Metazoa. Напротив, для многоклеточных растений —
Metaphyta, которые, впрочем, никогда не рассматри-
ваются как единая систематическая единица, совер-
шенно очевидно полифилетическое происхождение.
Разные Щ4пы водорослей возникли от различных жгу-
4 Протесты, часть 1
тиконосцев и имеют свою независимую эволюцион-
ную историю. При этом эволюция разных ветвей Me-
taphyta протекала в некоторых отношениях парал-
лельно, что привело к формированию аналогичных
ступеней морфологической дифференциации в раз-
ных филогенетических ветвях (Полянский В. И., 1956).
Однако ряд авторов поддерживали идею о совер-
шенно независимом происхождении разных типов
многоклеточных животных от различных протестов
(Pantin, 1951, 1960; Boyden, 1953; Greenberg, 1959;
Nursall, 1962). В частности, Гринберг полагает, что
представители типов Porifera, Cnidaria, Ctenophora и
Plathelminthes имеют лишь то общее, что построены
в виде двухслойного мешка; больше их ничто не объ-
единяет. Отсутствие общего предка у Porifera и Eume-
tazoa, по мнению Гринберга, — очевидный факт, по-
скольку у губок есть хоаноциты, которых нет у осталь-
ных многоклеточных животных, и налицо крайнее
своеобразие зародышевых листков. Не менее резко
обособлены и другие типы. Планула и гаструла в
цикле развития представляют только «технически не-
избежный результат» любого дробления яйца. Корен-
ные расхождения между типами проявляются и в осо-
бенностях симметрии стадий дробления, и в органи-
зации взрослых животных.
Аргументы Гринберга, на наш взгляд, неубеди-
тельны. Таким существенным стадиям развития, как
паренхимула, планула и гаструла, он отказывает в
каком-либо эволюционном значении, а особенности
проморфологии считает неизменными, сложившими-
ся еще у одноклеточных предков.
Старая точка зрения, что губки Porifera чужды по
происхождению остальным Metazoa (Sollas, 1884;
Minchin, 1900; Hadzi, 1944), позже опровергалась
(Беклемишев, 1964; Иванов А. В., 1968, 1971; Codre-
anu, 1970; Salvini-Plawen, 1978). Действительно, в
пользу монофилии губок, а также всех Metazoa сви-
детельствуют следующие общие их черты.
1. Однотипный жизненный цикл, состоящий из че-
тырех периодов: а) метагамного, б) эмбриогенеза и
роста, в) прогамного—мейотического и г) сингам-
ного.
2. Однотипность гаметогенеза и организации
гамет.
3. Гомология зародышевых листков.
4. Одинаковая организация жгутиковых клеток.
Последнее сходство выражается в наличии жгути-
ковых воротничковых клеток — хоаноцитов не толь-
ко у губок, но и у представителей самых различных
групп высших многоклеточных животных.
Замечательно также, что строение кинетиды (цен-
триолярно-жгутикового комплекса микротрубочек) в
жгутиковых клетках губок (особенно в локомотор-
ных жгутиковых клетках бластулообразных личинок)
и многих других групп Metazoa одно и то же.
Другим ярким примером полифилетического тол-
кования эволюции группы служит гипотеза Беклеми-
шева (1964) о независимом возникновении шести вы-
деляемых им ветвей Bilateria, а именно Scolecida,
Trochozoa, Actinotrochozoa, Brachiopoda, Chaetognat-
ha и Deuterostomia (cm. c. 53). При таком толковании,
во-первых, первичноротые (Protostomia) искусствен-
но разрываются на две независимые части (Scolecida
50
ОБЩАЯ ЧАСТЬ
и Trochozoa) и разрушается хорошо обоснованное
единство животных со спиральным дроблением (Spi-
ralia). Во-вторых, разъединяются родственные друг
другу мшанки (Bryozoa) и плеченогие (Brachiopoda),
близость которых хорошо обоснована сравнительно-
анатомически. Гораздо правдоподобнее выглядит объ-
яснение происхождения всех Bilateria их общностью
(Hyman, 1951; Ливанов, 1955; Федотов, 1966; Иванов
А. В., 1975а).
Идея о полифцлии членистоногих возникла в кон-
це прошлого столетия (Kennel, 1891, и др.), позднее ее
поддерживали многие авторы (Иванов П. П., 1933;
Tiegs, Manton, 1958; Федотов, 1966; Manton, 1977).
Критическая оценка их взглядов содержится в работах
Бирштейна (1948), Зивинга (Siewing, 1960), Шарова
(1966), Лукина (1975) и Ивановой-Казас (1979).
По мнению полигенистов, современные членисто-
ногие представлены несколькими группами, незави-
симо возникшими от разных аннелид. Такими само-
стоятельными группами Ментон, например, считает:
1) Uniramia — членистоногих с первично неветвисты-
ми конечностями (Onychophora, Myriapoda и Insecta),
2) Crustacea, 3) Chelicerata и 4) Trilobita. Многочис-
ленные различия в строении и положении органов
(фасеточных глаз, мандибул, конечностей, незамкну-
той кровеносной системы, сложной головы и мозга)
принимаются за изначальные особенности организа-
ции, унаследованной от разных предков. Однако в ре-
зультате анализа этих различий и собственных иссле-
дований Crustacea Зивинг (Siewing, 1960) очень убеди-
тельно показал, что все они гораздо правдоподобнее
объясняются как следствие параллельного развития,
имевшего место уже после «артроподизации», т. е.
после становления типа членистоногих. Зивинг разли-
чает две фазы артроподизации. Во время первой фазы
образовались открытая кровеносная система, спинное
сердце с остиями, мозговой комплекс, наружный хи-
тиновый скелет и расчлененные конечности. После
этого произошло отделение боковой ветви онихофор.
Вторая фаза заключалась в формировании фасеточ-
ных глаз, дальнейшей дифференциации конечностей,
завершении цефализации и в образовании головной
капсулы. Все эти арогенные процессы привели к появ-
лению трилобитообразных форм. В заключение своей
очень интересной статьи Зивинг приводит гипотети-
ческое, но вполне правдоподобное монофилетическое
древо всех членистоногих.
Как известно, у членистоногих количество лар-
вальных сегментов, закладывающихся в онтогенезе,
значительно варьирует. П. П. Иванов, автор теории
первичной гетерономности сегментированных живот-
ных, считал этот признак очень устойчивым и важ-
ным в филогенетическом отношении. Насчитывая у
ракообразных 3, у трилобитов и хелицеровых 4, а у
Antennata (вместе с Onychophora) 5 передних лар-
вальных сеплецтов, он высказал предположение о по-
лифилетическом происхождении Arthropoda тремя
независимыми стволами (Иванов П. П., 1933).
Однако нет оснований утверждать, что количест-
во ларвальных сегментов всегда соответствует числу
сегментов в теле предковых форм.
Естественнее рассматривать первичную гетеро-
номность как сосуществование в одном онтогенезе
двух разных способов развития сегментов (Ливанов,
1955; Иванова-Казас, 1978). Синхронная закладка
ларвальных сегментов, правда, отражает филогенети-
ческий процесс возникновения метамерии, связанной
с развитием параподий и упорядоченным расположе-
нием септ и мышечных пучков, и может, следова-
тельно, рассматриваться как первичный способ раз-
вития. Однако при значительном удлинений тела и
умножении сегментов он становится явно недоста-
точным и дополняется новым способом — последо-
вательным образованием постларвальных сегментов
из задней зоны роста (Беклемишев, 1964). Такого
рода изменения в ходе онтогенеза естественно рас-
сматривать как «рационализацию» развития (Шмаль-
гаузен, 19466). С другой стороны, постоянство числа
ларвальных сегментов в разных ветвях членистоногих
не абсолютно. У онихофор, вопреки мнению П. П. Ива-
нова, который опирался на старые данные, все сег-
менты закладываются последовательно, начиная с пе-
редних (Manton, 1949), и можно думать, что новый
и прогрессивный способ развития здесь полностью
вытеснил первичный тип развития (Иванова-Казас,
1979).
Родство трилобитов и хелицеровых очень правдо-
подобно. У первых развивалось 5 ларвальных сегмен-
тов, у мечехвостов вследствие редукции антеннально-
го сегмента их осталось лишь 4. С другой стороны, у
паукообразных, по новым данным, налицо 6 ларваль-
ных сегментов, что явилось следствием вторичного
уподобления 2 передних постларвальных сегментов
ларвальным. Эти примеры хорошо иллюстрируют
возможность вторичных изменений количества лар-
вальных сегментов в процессе филогенеза.
Гипотеза полифилетической природы класса ам-
фибий особенно защищается шведским палеонтоло-
гом Ярвиком (Jarvik, 1942, 1964, 1980), который по-
лагает, что отряд хвостатых амфибий, Urodela, про-
изошел от вымерших кистеперых рыб из отряда
Porolepiformes, тогда как предками бесхвостых ам-
фибий, Anura, якобы были представители другого от-
ряда кистеперых — Osteolepiformes. От них же про- j
изошли рептилии и млекопитающие. Ярвик объеди- j
няет даже Porolepiformes и Urodela в единую группу
Urodelomorpha. Такая полифилия вытекает из якобы
полной независимости эволюции кардинальных при-
знаков в строении лицевой части черепа, конечнос-
тей, позвонков, аппарата звукопередачи и других ор-
ганов в этих двух группах. Однако исследования
Шмальгаузена и его учеников, обобщенные им в мо-
нографии «Происхождение наземных позвоночных» ]
(19646), опровергают аргументы Ярвика и других за- |
щитников полифилетической природы земноводных. !
Было, в частности, доказано, что сходство в строении
упомянутых сложных органов и комплексов невоз-
можно объяснить независимым происхождением. Это
общие признаки всех амфибий, унаследованные ими
от предков, давших начало путем адаптивной радиа-
ции различным ветвям современных земноводных.
Предками амфибий были только Osteolepiformes,
но после периода дивергенции отдельные ветви земно-
водных эволюировали параллельно: бесхвостые от
батрахоморфных лабиринтодонтов, а хвостатые и без-
ногие — от лепоспондиальных стегоцефалов (рис. 16).
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
51
Рис. 16. Филогенетические отношения между основными отрядами низших четвероногих позвоночных. (По: Romer, 1939; Шмальгау-
зен, 19646, с изменениями).
Fig. 16. Phyletic relationship between basic orders of the lower tetrapod Vertebrata.
Класс амфибий, следовательно, монофилетичен, что
не исключает, разумеется, возможности параллельной
эволюции отдельных таксонов класса. Шмальгаузен
считает вероятным, что выход на сушу происходил
многократно у разных представителей кистеперых
рыб (19646, с. 248). Признавая, что организация кис-
теперых обладала признаками, делавшими выход на
сушу возможным, он полагает, что переход был дли-
тельным и сложным, и формы, которые первыми пре-
одолели трудности перехода из воды на сушу, имели
большие преимущества в борьбе за существование
перед формами отставшими, попадающими в уже за-
нятую экологическую зону.
Полифилетизм рептилий пытался обосновать
швейцарский палеонтолог Кун-Шнайдер (Kuhn-Schnei-
der, 1963), который различал среди них не менее 7
групп, произошедших независимо от разных амфи-
бий. Эта концепция подверглась справедливой кри-
тике со стороны палеонтолога Ромера (Romer, 1971,
с. 109), который, между прочим, заметил, что совсем
невероятно, чтобы «семь разных групп амфибий не-
зависимо приобрели один и тот же тип амниотичес-
кого эмбрионального развития». Пытаясь отвести это
возражение, Любищев (1968) ссылается на насеко-
мых, у которых тоже развиваются зародышевые обо-
лочки, заведомо не имеющие отношения к таковым
позвоночных. Однако он упустил из виду, что сходст-
во зародышевых оболочек у тех и других крайне по-
верхностно. Напротив, у всех амниот зародышевые
оболочки, несомненно, гомологичны и очень одно-
типны. В частности, аллантоис всегда развивается в
определенном месте кишечника, и во всех случаях из
него формируется мочевой пузырь.
Монофилетическая природа млекопитающих была
проиллюстрирована Гроссом (Gross, 1964) филогене-
тической схемой, основанной на палеонтологических
данных (рис. 17). Три ветви Monotremata, Multitu-
berculata и Placentalia — сходятся своими корнями
только в триасе, где и следует искать их общих пред-
ков. Таковыми были Cynodontia из группы териодонт-
ных рептилий. К аналогичным заключениям пришел
и Татаринов (1976), детально изучивший начальные
этапы эволюции млекопитающих. Он тоже выводит
все ветви млекопитающих от «узко очерченной груп-
пы» мезозойских цинодонтов. Радиация ветвей мле-
копитающих произошла именно на цинодонтном
уровне, что вовсе не исключало параллельной редук-
ции задних накладных костей нижней челюсти, со-
провождающейся смещением сочленовной кости в
область среднего уха. Этот пример только подтверж-
дает общее правило, что за этапом радиации (дивер-
генции) следует более или менее длительный период
параллельного развития близкородственных ветвей.
Итак, широкое распространение параллелизмов
вполне объяснимо с точки зрения дарвинизма и вовсе
не исключает основного значения дивергенции. На-
против, именно процесс дивергенции поставляет ма-
териал, на котором проявляет себя параллельная эво-
52
ОБЩАЯ ЧАСТЬ
Marsu-
Monotremata pialia Placentalia
Трети*
ный
период
Мел
Multiti/berculata
Triconoaonta
/
/
/
f Docodonta
Юра
Stereognathus | I V
Триас
/ ^Morjanucoion
/' t
! /
Пермь
Bauria- Curio-
morpha dontla
Рис. 17. Филогения Mammalia. Эволюция Sauriomorpha, Cynodon-
tia и Theria намечена лишь схематически. (По: Gross, 1964).
Fig. 17. Mammalian phylogeny. Sauriomorpha, Cynodontia and The-
ria evolution is given not more than schematic.
люция. В результате последней сходства могут быть
очень значительными и тем более глубокими, чем
больше родство организмов. «Однако, — как подчер-
кивал Шмальгаузен (1947, с. 491), — близкое род-
ство как раз и означает происхождение от одного
корня».
По мере накопления знаний в области морфоло-
гии, эмбриологии и палеонтологии часто выясняется,
что та или иная группа имеет не мнимый, а действи-
тельно гетерогенный состав. Такую группу приходит-
ся считать искусственной и разделять ее на самосто-
ятельные систематические единицы. Так, с восьмиде-
сятых годов прошлого столетия среди простейших
признавался класс споровиков, установленный еще
Бючли. Но в последние десятилетия искусственность
его стала очевидной и он был разделен на три само-
стоятельные группы различного происхождения: Spo-
rozoa (в узком смысле), Cnidosporidia и Microsporidia.
То же произошло и со старым отрядом грызунов (Gli-
res), который пришлось разделить на самостоятель-
ные отряды Lagomorpha (зайцеобразные) и Rodentia
(грызуны). Другой пример — прежний отряд хищных
птиц искусственно объединял неродственные друг
другу группы соколиных и сов. В настоящее время
первые отнесены к отряду Falconiformes, а вторые —
к отряду Strigiiformes.
Эволюция на уровне низших систематических ка-
тегорий иногда отличается тем, что представители
разных таксонов могут скрещиваться и давать поли-
филетические гибридные формы. Такая полифилия
на микроэволюционном уровне, затрагивающая виды
и даже роды, как известно, особенно характерна для
растений, у которых, по мнению некоторых ботани-
ков, она привела к образованию по крайней мере
трети современных видов цветковых растений. Такое
же гибридогенное (сетчатое) видообразование обна-
ружено и у позвоночных, а именно у некоторых зу-
бастых карповых рыб (Poecilia, Poeciliopsis), амфи-
бий (Ambistoma, Rana) и рептилий (Lacerta, Cnemi-
dophorus) (см.: Боркин, Даревский, 1980).
Необходимо четко различать полифилию отдель-
ного признака (или комплекса признаков) и полифи-
лию группы. Независимое возникновение сходных
признаков и органов, составляющее основное прояв-
ление параллелизма, отнюдь не исключает, вопреки
утверждениям полигенистов, монофилии больших ес-
тественных групп.
По справедливому замечанию Майра (Мауг, 1971),
многие видели полифилию таксона там, где в дейст-
вительности имела место лишь полифилия диагнос-
тических признаков, тогда как сам таксон был моно-
филетическим. Действительно, анализ организации
различных Theriodontia (Remane, 1964; Татаринов,
1976) показал, что некоторые признаки млекопитаю-
щих появлялись то в одной, то в другой ветви этих
рептилий. Таковы, например, образование вторично-
го нёба, усложнение коренных зубов, сокращение
зубных генераций, сближение зубной и чешуйчатой
костей, изменение постановки парных конечностей и
т. д. Разумеется, не каждая ветвь териодонтов, обла-
дающая этими признаками, могла дать начало млеко-
питающим.
Аналогичный пример представляет независимая
эволюция приспособлений к условиям недостатка
воды— хитиновой кутикулы, органов дыхания (тра-
хей) и выделения (мальпигиевых сосудов), а также
органов, обеспечивающих внутреннее осеменение в
разных группах членистоногих, перешедших к жизни
на суше (Гиляров, 1970а). В близкородственных
группах, живущих в сходных условиях, подобные по-
лифилетически возникшие органы эволюируют затем
сходным образом и вступают в сходные отношения с
другими органами. Так создаются новые, аналогич-
ные друг другу комплексы органов и признаков и
даже системы органов, изучение которых позволяет
делать далеко идущие выводы о происхождении и
эволюции различных групп животных.
Таким образом, возможность независимого поли-
филетического происхождения и параллельного раз-
вития в филогенезе как отдельных признаков, так и
целых систем органов и аппаратов представляется
вполне реальной (Иванов А. В., 1968, с. 165; 19886).
Примерами помимо параллельного развития упомяну-
того комплекса органов у членистоногих, связанного с
приспособлением к жизни на суше, может служить не-
зависимое образование эпителиального кишечника у
ранних Metazoa, а также вторичной полости тела и
кровеносной системы у различных Coelomata.
Действительно, если исходить из наиболее вероят-
ной и разработанной концепции происхождения мно-
гоклеточных — теории Фагоцителлы Мечникова (1886),
то мы должны признать по меньшей мере двукратное
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
53
происхождение эпителиального кишечника: во-пер-
вых, у общего предка кишечнополостных и, во-вто-
рых, у архаических Bilateria. На самом же деле можно
говорить даже о многократном появлении кишки.
Мы не имеем никаких фактических данных, указыва-
ющих на характер становления полостного кишечни-
ка у Coelenterata в их филогенезе, но низшие Turbel-
laria дают богатый материал, поскольку признаки не-
зависимого возникновения эпителизованной кишки
наблюдаются у них в пределах нескольких примитив-
ных отрядов (Nemertodermatida, Catenulida, Acoela,
Prolecithophora, — см.: Иванов, Мамкаев, 1973). Не-
которые авторы (Ах, 1961, с. 12, и ряд других) счита-
ют независимое образование кишечника «невероят-
ным и методически необоснованным» предположени-
ем, исходя из постулатов теории Гастреи. Трудно,
однако, усмотреть что-либо невероятное или методи-
чески необоснованное в эпителизации первоначально
аморфной пищеварительной паренхимы, содержащей
блуждающие фагоциты. Этот переход, в сущности, не
так уж сложен, фактически он совершается у некото-
рых турбеллярий в течение жизни особи многократно
как смена морфофизиологических состояний кишки.
Можно считать установленным, что обширная
полость тела у приапулид, несмотря на некоторые
гистологические отличия, имеет много особенностей,
общих с целомом кольчатых червей и вторичноротых
(Беклемишев, 1944). У немертин полость хоботного
влагалища тоже обладает рядом характерных призна-
ков вторичной полости тела, ее жидкость играет роль
гидростатического скелета, дающего опору сокраща-
ющимся мускульным волокнам в стенках хоботного
влагалища (Hyman, 1951; Clark, 1964). Перикард и
целом моллюсков, по-видимому, не гомологичны це-
лому аннелид и имеют независимое от него проис-
хождение (Salvini-Plawen, 1968, 1972). Теперь вряд
ли можно сомневаться, что целомы различных Coelo-
mata не всегда гомологичны. Можно ли, например,
рассматривать обширный нерасчлененный целом
эхиурид как наиболее примитивный и выводить из
него метамерный целом полихет? Старое представле-
ние об эхиуридах как о вторично упрощенных по-
томках полихет не подтверждается эмбриологически,
а эволюция в обратном направлении от эхиурид к по-
лихетам крайне сомнительна. Все это заставляет со-
гласиться с тем, что филогенетическое становление
нерасчлененного и метамерного целомов — незави-
симые процессы (Clark, 1979). По-видимому, то же
самое следует признать и в отношении целома вто-
ричноротых, щупальцевых и, возможно, погонофор.
Как показал Кларк (Clark, 1964), появление сплош-
ного целома было неизбежным приспособлением к
копанию норки у червеобразного животного, но ме-
тамерный целом необходим животным, длительно ро-
ющимся в грунте, тогда как димерный или тример-
ный целом свойствен формам, которые обитают в
трубке или домике и получают пищу и кислород по-
средством щупалец (см. также: Старобогатов, 1983).
К последним принадлежат Tentaculata, древние Deu-
terostomia и Pogonophora.
Наконец, уместно вспомнить многочисленные па-
раллелизмы в эволюции нервной системы. Примитив-
ные ее формы встречаются у низших представителей
различных беспозвоночных (например, у Coelentera-
ta, Turbellaria, Nemertini, Annelida, Phoronida, Hemi-
chordata, Pogonophora, Echinodermata) в виде сплете-
ния нервных клеток (плексуса), расположенного в
толще эпидермиса. Для начальных стадий эволюции
нервного аппарата характерны три процесса — кон-
центрация и централизация нервных элементов, по-
гружение их в глубь тела и формирование мозга
(Hanstrom, 1928; Ливанов, 1941; Беклемишев, 1964;
Reisinger, 1972). Различные этапы этих процессов
прослеживаются параллельно у турбеллярий Acoela,
Prolecithophora и Catenulida, что свидетельствует о
независимой эволюции нервной системы в этих отря-
дах, происходящей весьма сходными путями (Бекле-
мишев, 1964; Иванов, Мамкаев, 1973). Независимые,
но совершенно аналогичные этапы организации нерв-
ной системы (кожный плексус, погружение его и
концентрация элементов в ортогональную решетку,
образование брюшного и головного мозга) известны
также у немертин, моллюсков и кольчатых червей
(Бубко, Миничев, 1972).
Соотношение онтогенеза
и филогенеза
Уже в начале прошлого столетия зоологи, изучав-
шие эмбриональное развитие позвоночных животных
(Meckel, 1821, и др.), были убеждены в существова- -
нии закономерного соответствия между стадиями ин-
дивидуального развития и ступенями повышения ор-
ганизации взрослых форм. Однако К. Бэр пришел к
выводу, что зародыши высших животных сходны от-
нюдь не со взрослыми предками, а лишь с их эмбри-
онами. Дарвин, много размышлявший над этой проб-
лемой, в «Происхождении видов...» писал: «Интерес
к эмбриологии значительно повысится, если мы будем
видеть в зародыше более или менее затемненный
образ общего предка (во взрослом или личиночном
состоянии) всех членов одного и того же большого
класса» (Darwin, 1991, с. 388).
Вскоре эта идея была развита Фрицем Мюллером
в его известной книге «За Дарвина» (Muller F., 1864).
На примере развития некоторых ракообразных Мюл-
лер ясно продемонстрировал, что эволюция новых
форм совершается или путем уклонения от развития
предков на средних стадиях онтогенеза, или добав-
лением новых особенностей в его конце. Однако в
«биогенетическом законе» Геккеля (Haeckel, 1866)
трактовка соотношения фило- и онтогенеза приняла
весьма односторонний характер. В первоначальной
формулировке геккелевский закон означал, что он-
тогенез есть краткое, быстро протекающее воспроиз-
ведение (рекапитуляция) филогенеза, обусловленное
физиологическими функциями наследственности
(размножение) и адаптаций. По Геккелю, это воспро-
изведение тем полнее, чем в большей степени сохра-
няется первоначальное развитие предков (палинге-
нез). Но оно часто нарушается вторичными зароды-
шевыми приспособлениями (ценогенезами), которые
лишь затрудняют филогенетическое исследование.
Таким образом, Геккель представлял себе онтогенез
54
ОБЩАЯ ЧАСТЬ
механистически — как ряд «упрощенных» и «уко-
роченных» палингенезов. Новые изменения лишь
прибавляются к ранее существовавшим палингенезам
в качестве надставок. Развитие есть суммирование
надстраивающихся палингенезов, которые затем ото-
двигаются в глубь онтогенетического развития, В резуль-
тате Геккель утверждал, что «филогенез есть механи-
ческая причина онтогенеза», и, объясняя рекапитуля-
цию палингенезов, ссылался на «консервативную
наследственность» (Haeckel, 1875). Эти наивные
представления оказались очень живучими и отчасти
сохранились до настоящего времени.
Было показано, однако (Кремянский, 1941; Пара-
монов, 1967; Шмальгаузен, 1969), что палингенезы
проявляются вследствие сохранения отбором именно
тех морфофункциональных конструкций, которые
имеют жизненно необходимое значение. Так, у всех
зародышей позвоночных хорда и нервная трубка
формируются вовсе не в силу некоей «консерватив-
ной наследственности», а потому, что они жизненно
необходимы, так как коррелятивно связаны с заклад-
кой и развитием позвоночника, черепа, головного и
спинного мозга. Именно поэтому отбор шел на со-
хранение геномов, определяющих закладку и разви-
тие хорды и нервной трубки.
Дарвин и Мюллер в противоположность Геккелю
рассматривали онтогенез как активный фактор пере-
стройки направлений филогенеза. Эта концепция
оказалась весьма плодотворной и полностью подтвер-
дилась дальнейшим развитием биологии. Развивая и
дополняя ее, А. Н. Северцов (1910, 1939) разработал
современную теорию филэмбриогенеза, которая объ-
ясняет действительные отношения фило- и онтогене-
за и раскрывает творческую роль последнего в эволю-
ции организмов. Северцов предложил различать сле-
дующие характерные черты изменений онтогенеза.
1. Палингенезы — признаки относительной река-
питуляции организации предков (в их онтогенезе или
во взрослом состоянии).
Рис. 18. Филэмбриогенез по типу анаболии. (По: Северцов,
1939).
А — взрослая особь Belone acus; Б — голова взрослого Hemirhamphus;
В — малек В. acus; Г — то же, несколько более поздний.
Fig. 18. Phylembryogenesis after anaboly type.
A — matured Belone acus, Б — matured Hemirhamphus head, В — Belone
acus hutchling, Г — the same, more late larva.
2. Ценогенезы — провизорные приспособления к
зародышевому или личиночному существованию.
3. Гетерохронии — приспособительные измене-
ния во времени закладки и развития органа. Они
могут проявляться как в ускорении закладки (акселе-
рации), так и в запаздывании ее (ретардации).
4. Гетеротопии — изменения по месту закладки
и развития органа.
5. Филэмбриогенезы — изменения, которые воз-
никают у эмбрионов, но переходят во взрослое состо-
яние и служат источником для новых путей филоге-
неза.
В основе теории филэмбриогенеза лежит пред-
ставление о том, что все существенные новые направ-
ления эволюции животных первоначально проявля-
ются в онтогенезе предков.
Филэмбриогенезы возникают на разных стадиях
эмбрионального развития. Соответственно Северцов
различал анаболии (добавление новой стадии в
конце морфогенеза какого-либо органа с соответст-
вующим удлинением его онтогенетического разви-
тия), которые проявляются лишь в конце онтогенеза
как его надставки, девиации (уклонение в разви-
тии, изменение характера жизненного цикла без его
удлинения), возникающие на средних этапах эмбрио-
генеза, и архаллаксисы — онтогенетически
самые ранние филэмбриогенезы, представляющие
собой изменение самих зачатков.
Анаболии, или надставки, наиболее распростра-
нены. К уже законченному онтогенезу добавляется
новый этап развития и дифференциации органа. Это
связано, разумеется, со сложностью предшествующе-
го развития, не допускающей серьезных изменений
на его ранних и средних стадиях. Так, многие изме-
нения в строении скелета и мускулатуры и в распре-
делении периферических кровеносных сосудов у поз-
воночных происходят на самых поздних стадиях он-
тогенеза. Как пример типичной анаболии Северцов
рассматривал эволюцию очень длинных челюстей у
саргана, Belone acus (рис. 18, А). У этой морской
рыбы из сем. Scombresocidae на стадии раннего маль-
ка рекапитулируется состояние короткорылых скомб-
резоцид (рис. 18, В), а на следующей стадии нижняя
челюсть резко удлиняется и возникает состояние, ха-
рактерное для рода Hemirhamphus (рис. 18, Б). Таким
образом, ясно, что эволюция челюстей у саргана со-
вершалась путем добавления к онтогенезу вначале
стадии «удлиненной нижней челюсти» (тип Hemir-
hamphus), а затем и стадии «взрослого саргана».
Прекрасный пример эволюции путем анаболии
представляет краб-разбойник, или пальмовый вор, —
Birgus latro (Иванов А. В., 19556). Это хорошо при-
способленное к наземному образу жизни и к воздуш-
ному дыханию ракообразное (рис. 19, В) принад-
лежит к сем. Coenobitidae (отряд Decapoda), очень
близкому к ракам-отшельникам — Paguridae. Бли-
жайший сородич пальмового вора — Coenobita, в от-
личие от него будучи настоящим сухопутным отшель-
ником, прячет еще брюшко в пустую раковину брю-
хоногого моллюска. Birgus же использует раковину
только в молодости.
Самки Birgus, несущие яйца на брюшке, живут
очень скрытно в сырых норках, но покидают их для
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
55
Рис. 19. Пальмовый вор Birgus latro.
А — личинка зоэа, Б — стадия Coenobita, В — взрослый самец.
(По: Бирштейн, 1968).
Fig. 19. Robber crab Birgus latro.
A — zoea larva, Б — Coenobita stage, В — matured male.
миграции в море в период вылупления личинок.
В воде же из яйца выходит планктонная личинка
зоэа (рис. 19, А), весьма похожая на такую же личин-
ку Coenobita и раков-отшельников. Затем следует ряд
пелагических личиночных стадий, в том числе мизид-
ная личинка, и наконец подрастающее животное пе-
реходит к жизни на дне. Тут оно, по существу, пре-
вращается в рака-отшельника и прячет асимметрич-
ное брюшко в раковину моллюска. Короткая стадия
отшельника вначале протекает в море, затем живот-
ное выходит на сушу, превращаясь в стадию Coenobi-
ta (рис. 19, Б). После одной из ближайших линек мо-
лодой Birgus оставляет раковину, далее с каждой
линькой он утрачивает внешность рака-отшельника и
постепенно приобретает характерные морфологичес-
кие, физиологические и экологические особенности
взрослого пальмового вора.
Таким образом, у Birgus налицо очень полная ре-
капитуляция филогенеза. Повторяются не только
онтогенетические особенности предков (развитие в
море, серия планктонных личинок), но и ряд взрос-
лых анцестральных состояний, отвечающих орга-
низации и образу жизни современных Paguridae и
Coenobita. На значительном протяжении эволюция
Birgus несомненно заключалась в добавлении к пер-
воначальному онтогенезу последовательных надста-
вок: после стадии Paguridae — стадия Coenobita, после
нее — стадия Birgus. Характерно, что каждый из этих
этапов был связан с очень существенными изменени-
ями образа жизни, поведения, морфологии и физио-
логии, но не повлиял на ранние и средние стадии он-
тогенеза.
По Догелю (1951), эволюция простейших совер-
шалась на основе филэмбриогенезов в той же мере,
56
ОБЩАЯ ЧАСТЬ
что и у многоклеточных. Чаще всего наблюдается
анаболия, реже девиация, еще реже архаллаксис.
Хороший пример эволюции путем филэмбриоге-
незов представляют сосущие инфузории (Suctoria),
ведущие сидячий образ жизни. Большинство из них
прикрепляется стебельком, у многих тело одето
чашевидной раковиной, реснички отсутствуют. Мо-
нотомическое деление протекает без каких-либо
признаков рекапитуляции. Однако при почковании
(наружном или внутреннем) образуются бродяжки,
несущие реснички, которые рекапитулируют стадию
свободноплавающего инфузориеобразного предка.
По круговому расположению ресничек и по строе-
нию стебелька некоторые авторы сравнивают их с
кругоресничными инфузориями (Peritricha), другие,
ссылаясь на особенности инфрацилиатуры у бродя-
жек и некоторые другие черты, выводят Suctoria от
Рис. 20. Типы филэмбриогенезов в развитии дериватов кожи поз-
воночных. (По: Шмальгаузен, 1969).
А — развитие чешуи акулы — исходный тип развития (онтогенетические
стадии 1—6); Б — развитие чешуи рептилий (стадии онтогенеза 7—72), на
одной из средних стадий происходит девиация, способ развития чешуи
резко меняется (указано стрелкой)', В — развитие пера птиц (стадии 7—11,
13—15), вначале развитие идет как при развитии чешуи рептилий, но после
стадии 77 специфически изменяется путем анаболии; Г — развитие волоса
млекопитающих (стадии 16—21), на стадии 77 происходит изменение — ар-
халлаксис, ведущее к образованию волоса.
Fig. 20. Phylembriogenesis types presented in development of der-
mal derivatives in Vertebrata.
A — development of the shark dermal denticle as an initial type of development
(ontogenetic stages 7—6); Б — development of the Reptilian homy scale (on-
togenetic stages 7—12), deviation takes place at the one of the middle stages,
the mode of scale development changes abruptly (arrow)', В — bird feather
development (stages 7—77, 13—15), initial the same as in reptilian scale, after
77 stage it changes specifically by anaboly; Г— development of mammalian hairs
(stages 16—21), on stage 77 archallaxis bringing hair formation takes place.
примитивных равноресничных инфузорий. Как бы то
ни было, перед нами пример эволюции по типу ана-
болии (положительной и отрицательной). Очевидно,
предок сукторий, перейдя к сидячему образу жизни,
приобрел раковину и сосательные трубочки, а может
быть и стебелек (положительная анаболия), и парал-
лельно утратил рот, глотку и реснички (отрицатель-
ная анаболия).
Отмечены примеры филэмбриогенезов также у
Peritricha и некоторых других инфузорий, у целого
ряда Radiolaria и разных представителей Mastigopho-
га (Догель и др., 1962).
Подробный анализ с позиций морфофизиологи-
ческих закономерностей эволюции для паразитичес-
ких простейших со сложным жизненным циклом, от-
носящихся к классу споровиков (класс Sporozoa, тип
Apicomplexa), проведен в работах Каллиниковой (1991).
Последовательные стадии сложного жизненного цик-
ла споровиков с чередованием стадий бесполого (ме-
рогония) и полового (гамогония и спорогония) раз-
множения она рассматривает как целостный онто-
генез, сравнимый с онтогенезом многоклеточных
животных.
Сравнивая жизненные циклы разных групп споро-
виков, Каллиникова полагает, что большая часть их
прогрессивных эволюционных преобразований осу-
ществлялась путем надставок (анаболий). Таков, на-
пример, путь от относительно простого жизненного
цикла грегарин (Gregarinea) к сложному жизненному
циклу моноксонных и гетероксонных кокцидий.
В эволюции некоторых групп споровиков, а именно
гемогрегарин и гемоспоридий (Haemosporidia), имели
место девиации. Наконец, возникновение такой абер-
рантной группы кровяных паразитов позвоночных,
как пироплазмиды (Piroplasmida), связано, по-види-
мому, с изменениями типа архаллаксиса.
Примером девиации может служить образование
чешуи рептилий (Матвеев, 1932). Исследование эм-
бриогенеза чешуи ящерицы Lacerta muralis показало,
что вначале он протекает по типу развития плакоид-
ной чешуи далеких рыбообразных предков (рис. 20,
А). Потом, на одной из средних стадий, происходит
отклонение от этого пути. Прогрессивным преобра-
зованиям подвергается не соединительно-тканный со-
сочек, дающий у низших рыб дентиновый плакоид-
ный зуб, а эктодермальный слой, образующий рого-
вую чешую нового, «рептильного» типа (рис. 20, Б),
На хороший пример девиации указал сам А. Н. Се-
верцов (1939). Среднее ухо высших позвоночных го-
мологично каналу брызгальца (spiraculum) низших
рыб. Ранние стадии его развития очень сходны с раз-
витием спиракулярного мешка рыб, но дальнейшее
развитие этих двух образований протекает совершен-
но по-разному. У упомянутого выше саргана развитие
органа обоняния сначала совпадает с таковым других
костистых рыб, но на средних стадиях эмбриогенеза
разрастание одной из складок обонятельного мешка
препятствует образованию мостика, разделяющего
единую первичную ноздрю на две. Это отклонение от
обычного хода развития приводит к отсутствию у
саргана задней ноздри.
Развитие конечностей ракообразных, вероятно,
тоже можно толковать как пример девиации. На ран-
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
57
них стадиях развития (метаморфоза) они обычно
имеют еще характер примитивных двуветвистых пла-
вательных ножек, но позднее изменяют свое развитие
в сторону дефинитивного состояния. Таковы, напри-
мер, мандибулы и максиллы личинки-науплиуса.
Архаллаксис можно проиллюстрировать на при-
мере эмбрионального развития волоса у млекопитаю-
щих. Оно начинается с образования небольшого
скопления соединительно-тканных клеток под комп-
лексом эпидермальных клеток (рис. 20, Г). За счет
этого скопления формируется луковица волоса. Сле-
довательно, зачаток волоса состоит из эктодермаль-
ного и мезодермального сгущения клеток и в таком
виде вполне подобен и, очевидно, гомологичен зачат-
ку плакоидной чешуи хрящевых рыб. Однако далее
прогрессивно развивается не мезодермальный компо-
нент зачатка, как у рыб, а эктодермальный, который
быстро растет, удлиняется и в виде стержня волоса
врастает в кожу, одеваясь волосяной сумкой.
Хорошо изученное развитие легких у скорпионов
и пауков также протекает по типу архаллаксиса. Лег-
кие сначала закладываются как зачатки типичных ко-
нечностей, несущие зачаточные жаберные лепестки
на задней поверхности. Но на очень ранних стадиях
их эмбриогенеза развитие резко меняет свое направ-
ление в сторону формирования легкого.
Само собой разумеется, что истинные рекапитуля-
ции анцестральных состояний могут сохраняться при
эволюции по типу анаболии, поскольку в основном
онтогенез остается неизменным. При девиации река-
питуляцию можно ожидать лишь на ранних стадиях
развития органа до появления отклонения от первич-
ного хода онтогенеза. И очевидно, что при архаллак-
сисе последовательность онтогенетических событий
ничего не говорит о филогении, поскольку онтогенез
изменен с первых своих стадий.
Впрочем, в некоторых случаях предполагаемого
филэмбриогенеза типа девиации или архаллаксиса
трудно быть уверенным в том, что фактически на-
блюдаемое раннее изменение развития не есть ре-
зультат вторичного смещения в глубь онтогенеза ана-
болии, т. е. что оно не является гетерохронией.
Кроме того, говоря о филэмбриогенезах, нельзя
забывать, что причиной морфологической эволюции
являются отнюдь не перестройки в онтогенезе сами
по себе, а мобилизованные естественным отбором ге-
нотипические изменения. Понятия архаллаксис, де-
виация и анаболия характеризуют только те стадии
онтогенетического развития, на которых начинают
фенотипически проявляться наследственные измене-
ния.
Морфофизиологические
проблемы эволюции
Выяснение основных принципов филогенетичес-
ких преобразований (модусов) органов, общих для
всех групп животных, составляет особую проблему
морфофизиологических закономерностей эволюции.
Как уже было отмечено, первые серьезные наблюде-
ния и обобщения в этой области принадлежат самому
Дарвину и Ковалевскому (1874). В XX в. особенно
много в этом направлении было сделано А. Н. Север-
цовым и его школой.
Принцип корреляций
Животный организм представляет собой единое
целое, в котором все части и органы взаимосвязаны.
Когда в эволюционном процессе изменяется строе-
ние и функция одного органа, то это неизбежно вле-
чет коррелятивное изменение и в других органах, в
процессе онтогенеза связанных с первым физиологи-
чески и морфологически. Закон корреляции, или со-
относительного развития органов, в общей форме
был открыт Вик^д’Азиром и развит Кювье (Cuvier,
1801). Пользуясь им, нередко удается реконструиро-
вать организацию целого ископаемого животного по
его частям, например по скелету.
А. Н. Северцов (1939) называл координация-
ми наличие в одном организме всегда сосущест-
вующих и связанных друг с другом морфофизиоло-
гических особенностей строения; их сочетание, ха-
рактерное для данной монофилетической группы,
по-видимому, складывается и изменяется в процессе
эволюции. Таковы основные признаки многих типов,
например хордовых — Chordata, а именно хорда,
спинная нервная трубка, жаберные щели, брюшное
сердце. Эта координированная система признаков ха-
рактерна для всех животных, принадлежащих к типу
хордовых, разумеется, за исключением форм, подверг-
шихся резким вторичным изменениям. Координиро-
ванные признаки членистоногих (Arthropoda) — на-
личие сегментации тела, хитиновые покровы, чле-
нистые конечности, далеко зашедшая цефализация,
сложный головной мозг, брюшная нервная лестница,
или нервная цепочка, развитие мускульных пучков
вместо кожно-мускульного мешка, открытая крове-
носная система и т. д. По Северцову, главный крите-
рий координации •— сопряженное изменение призна-
ков в ходе эволюции. Координации представляют
собой результат естественного отбора, т. е. составля-
ют особую категорию исторически сложившихся со-
отношений.
Что касается корреляций, то, по Дарвину, они
проявляются в ходе онтогенеза. Их роль в филогенезе
исключительно велика и была особенно подробно
изучена Шмальгаузеном (1938, 1964а, 1969).
Главные типы корреляций определяются как фор-
мой связи между коррелированными органами и дру-
гими системами организма, так и характером осущест-
вления изменений в них. При геномных корреляциях
зависимость определяется наследственными фактора-
ми (генами) через биохимические процессы в клетках
и осуществляется в частях организма, не связанных
друг с другом непосредственно. Морфогенетические
корреляции характеризуются взаимодействием зачат-
ков развивающегося эмбриона при их контакте друг с
другом через переход веществ от одной части к дру-
гой. Наконец, эргонтические (или функциональные)
корреляции определяются взаимной зависимостью де-
финитивных функций взрослого животного.
Хотя факторы, лежащие в основе координаций и
корреляций, на первый взгляд представляются одно-
58
ОБЩАЯ ЧАСТЬ
типными, в сущности они глубоко различны. Коорди-
нации демонстрируют целостность организмов в их
историческом (филогенетическом) развитии, а имен-
но выражают тот факт, что в эволюции организм ме-
няется и приспосабливается к условиям жизни как
единое целое, а не только в отдельных своих частях.
При завоевании новой арены жизни в организме
происходят значительные перестройки в координа-
ционных и эргонтических системах. В приведенных
выше примерах перехода членистоногих от водного к
наземному образу жизни (с. 52) ясно были продемон-
стрированы морфофизиологические взаимодействия
между разными системами органов по кибернети-
ческому принципу обратной связи (Гиляров, 1970а,
19706). Действительно, уплотнение кутикулы, вызван-
ное потребностью сохранения влаги в организме в ус-
ловиях воздушной стихии, обусловливает формирова-
ние трахей, а развитие последних, в свою очередь, сти-
мулирует дальнейшее уплотнение хитинового покрова
и т. д. Подобные обратные регуляции коррелятивных
систем, развивающихся в процессе филогенеза, — по-
видимому, одна из важнейших закономерностей эво-
люции (Шмальгаузен, 1958).
Дифференциация и интеграция
Эволюция организмов всегда сопровождается
дифференциацией частей и органов. Дифферен-
циация проявляется в том, что первоначально од-
2
Рис. 21. Трилобит Ceraurus (средний ордовик) с брюшной и
спинной сторон. (По: Друшиц, 1974).
7 — конечности (частично не изображены), 2 — антенны, 3 — гипосто-
ма, 4 — метастома.
Fig. 21. Trilobit Ceraurus (Middle Ordovician) on the dorsal and
ventral sides.
7 — extremities (shown partially), 2 — antennae, 3 — hypostoma, 4 — me-
tastoma.
Рис. 22. Рак-богомол Squilla mantis. (По: Grasse et al., 1961).
7 — антенна I; 2 — антенна II; 3 — экзоподит антенны II; 4 — ногоче-
люсть без клешни; 5 — ногочелюсть с подклешней, служащая для хватания
и убивания добычи; б — три пары ногочелюстей с подклешней; 7 — три
пары грудных двуветвистых ног; 8 — совокупительный орган; 9 — брюш-
ные ноги с жабрами; 10 — хвостовая нога — уропод; 77 — плеон; 72 —
карапакс; 13 — антеннулы.
Fig. 22. Stomatopoda Squilla mantis.
1 — antenne I, 2 — antenne II, 3 — exopodite of antenne II, 4 — maxillipede
without chela, 5 — maxillipede with subchela for the prey-catching and killing,
6 — 3 pairs of the maxillipde with subchela, 7 — 3 pairs of pectoral dicladose
legs, 8 — copulatory organ, 9 — gill-bearing abdominal legs, 10 — uropod —
caudal leg, 77 — pleon, 72 — carapace, 73 — antennulae.
неродные части организма начинают постепенно,
по мере прогрессивной эволюции, все более и
более отличаться друг от друга как по строению, так
и по отправлениям. Специализируясь на отправле-
нии одной определенной функции, они утрачивают
способность выполнять иные функции и тем самым
становятся в зависимость от других частей организма.
Следовательно, дифференциация всегда приводит не
только к усложнению организации, но и к подчине-
нию частей целому — одновременно с морфофизио-
логическим расчленением организма происходит об-
ратный процесс формирования гармоничного целого,
называемый интеграцией (Spenser, 1870). Класси-
ческой иллюстрацией дифференциации органов, со-
пряженной с возрастающей интеграцией всего орга-
низма, является развитие конечностей у членистоно-
гих, примитивные и древнейшие формы которых
(например, вымершие трилобиты) обладали много-
численными, но одинаковыми, выполнявшими сход-
ные функции ножками (рис. 21). У ныне живущих
ротоногих ракообразных, например у Squilla mantis,
имеется не менее 11 морфологических типов конеч-
ностей, несущих самые разнообразные отправления
(рис. 22).
Эволюция органов и функций
Глубокие эволюционные изменения в строении и
функциях осуществляются значительно легче, если
орган еще мало специализирован и может выполнять
несколько разных функций. «В таких случаях, —
писал Дарвин, — естественный отбор, если бы это
было выгодно, мог бы специализировать целый орган
или часть этого органа... только на одной какой-ни-
будь функции и, таким образом, нечувствительными
ступенями глубоко изменить его характер» (Darwin,
1991). Иначе говоря, возможность эволюции органа в
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
59
различных направлениях прежде всего обеспечива-
ется его мультифункциональностью. Эта идея
была затем подхвачена Дорном (Dohrn, 1875), кото-
рый ввел понятие о смене функций, и широко
использована А. Н. Северцовым (1939) для опре-
деления общих принципов изменений органов в про-
цессе эволюции. (Например, даже такие специализи-
рованные органы, как ноги лошади, великолепно
приспособленные для бега, служат также для защиты
и выражения эмоций.)
Северцов различал следующие модусы (формы,
или способы) филогенетического органогенеза.
1. Усиление функции. Часто главная функция эво-
люирующего органа подвергается усилению или бла-
годаря изменениям в строении клеток и тканей, или
вследствие увеличения числа его составных частей.
Так, усиление функции мускула происходит в резуль-
тате превращения гладких мускульных волокон в по-
перечно-полосатые (что, между прочим, происходит
в самых разнообразных группах животного мира, на-
чиная с книдарий). Двигательная активность и мус-
кульная сила животного при этом резко возрастают,
и оно получает преимущества в борьбе за существо-
вание. Усиление деятельности различных желез, в
частности млечной железы млекопитающих, образу-
ющейся из маленьких трубчатых железок, — другой
пример того же рода. У китообразных плавательная
функция значительно усилилась вследствие вторич-
ного умножения фаланг пальцев. Наконец, у некото-
рых кишечнополостных функция ловли добычи уси-
лилась благодаря полимеризации разветвлений на
ловчих щупальцах (некоторые Anthomedusae, Cte-
nophora). Характерен этот модус эволюции и для Pro-
tozoa. Так, у многих инфузорий путем объединения
нескольких ресничек образуются их пучки, цирры —
более мощные двигательные органеллы (рис. 23). У та-
ких инфузорий постепенно усиливается работа со-
кратительных вакуолей, которые к тому же могут ум-
ножаться в числе.
Принцип усиления функций чрезвычайно широко
распространен и лежит в основе многих арогенных
процессов. В этих случаях усиление функций осу-
ществляется благодаря коренным изменениям в стро-
ении органа. Убедительным примером может служить
эволюция сердца у позвоночных, рассмотренная выше
(с. 34).
2. Ослабление функции. Представляет противопо-
ложный предыдущему принцип филогенетического
развития органа. Каждая функция может постепенно
ослабевать, а при регрессивной эволюции органа и
целиком исчезнуть. Примерами ослабления функции
могут служить широко распространенные рудимен-
тарные органы.
3. Расширение функции (Plate, 1912) или увеличе-
ние числа функций. Прогрессивное развитие органа
нередко сопровождается появлением новых физиоло-
гических отправлений. Парные плавники рыб воз-
никли как органы планирования в воде, но с развити-
ем специальной мускулатуры стали активными руля-
ми глубины, а также органами плавания или опоры и
движения по дну. Хорошими примерами этого модуса
являются случаи вторичного увеличения функций ан-
тенн у некоторых членистоногих. Так, у веслоногих
(Copepoda) длинные антенны I помимо основной чув-
ствительной функции служат усами для парения в
воде, а у самцов они изогнуты и приспособлены для
удержания самки при спаривании. У водяных блох
(Cladocera) антенны II — не только чувствительные
конечности, но также и органы парения и плавания.
Среди насекомых также известны примеры расшире-
ния функций антенн. Так, у некоторых кобылок (Ас-
ridiella, Acrida и др.) сильно уплощенные и расши-
ренные усики вытягиваются в покое вперед, как бы
удлиняя узкое тело, что делает его еще более похо-
жим на лист злака. В этом случае усики кроме своих
обычных функций приобрели новую функцию, дела-
ющую покровительственную форму насекомого более
совершенной. Наездник из сем. Braconidae — Hab-
robracon использует усики не только по «прямому
назначению», но и для механической опоры на ко-
леблющемся субстрате. Известно, что огромные уш-
ные раковины африканского слона (Loxodonta) слу-
жат не только резонаторами, но и как опахала, помо-
гающие животному отдавать избыточное тепло.
4. Уменьшение числа функций. Этот модус проти-
воположен предыдущему и чаще всего является след-
ствием усиления или смены функции. Так, у ластоно-
гих и в еще большей степени у китообразных исчезла
не только главная функция конечностей предков —
хождение, но и ряд побочных. Обычно сужение функ-
ций приводит к значительной специализации органа.
5. Активация функции. Иногда орган, представля-
ющий первоначально совершенно пассивное образо-
вание, в дальнейшем превращается в активно дейст-
вующий. Хорошей иллюстрацией этого модуса явля-
ется развитие подвижности челюстных костей у змей
(рис. 24). Как известно, змеи произошли от рептилий
с неподвижными верхними челюстями; возникшие
новые отношения оказались очень выгодными, по-
скольку подвижность maxillae (и других элементов)
позволяет проглатывать целиком крупную добычу, а
развивающаяся одновременно подвижность ядовитых
зубов усиливает функцию защиты и нападения.
Прекрасный пример активации функций отмечен
у кругоресничных инфузорий (Peritricha) — их при-
крепительный отдел тела, вначале неспособный к со-
кращению, в процессе эволюции приобрел сложное
строение и превратился в сократительный стебелек
(рис. 25).
Рис. 23. Брюхоресничная инфузория Stylonychia, ползущая по
субстрату при помощи цирр. Вид сбоку. (По: Biitschli; 1880—
1882).
1 — сократительная вакуоль, 2 — адоральные мембраны, 3 — цирры.
Fig. 23. Infusoria Stylonychia creeping up substratum with cilia,
viewed from the side.
7 — contractile vacuole, 2 — adoral membrane, 3 — cirri.
60
ОБЩАЯ ЧАСТЬ
Рис. 24. Череп змеи. Схема. (По: Северцов, 1939).
А — при закрытой пасти, Б — при открытой пасти. 1 — ядовитый зуб;
2—6 — кости: 2 — верхнечелюстная, 3 — поперечная, 4 — крыловид-
ная, 5 — чешуйчатая, 6 — квадратная.
Fig. 24. Scheme of a snake cranium.
A — with closed jaw, Б — with opened jaw. 1 — fang, 2 — maxillary bone,
3 — ectopterygoid bone, 4 — pterygoid bone, 5 — squamosal bone, 6 — quad-
rate bone.
Рис. 25. Сувойка Vorticella. (По: Remane et al., 1976).
1 — макронуклеус, 2 — микронуклеус, 3 — перистом, 4 — преддверие,
5 — сократительная вакуоль, 6 — клеточная глотка, 7 — мионемы, 8 —
пищеварительные вакуоли, 9 — ундулирующие мембраны адорального ап-
парата, 10 — мионема стебелька.
Fig. 25. Peritrichous infusorian Vorticella.
1 — macronucleus, 2 — micronucleus, 3 — peristome, 4 — funnel, 5 — con-
tractile vacuole, 6 — cellular farynx, 7 — myonemes, 8 — digestive vacuoles,
9 — undulating membrane of the adoral apparatus, 10 — mioneme of pedicle.
6. Иммобилизация функции, напротив, представ-
ляет собой модус перехода от активной к пассивной
функции. Многочисленные примеры вторичной утра-
ты подвижности элементов скелета у различных чле-
нистоногих и позвоночных хорошо его иллюстриру-
ют. Сюда относятся случаи слияния сегментов в
тагмы и члеников конечностей у первых и слияние
костей у вторых. В частности, у птиц в результате
приспособления к полету срастаются некоторые поз-
вонки, кости тела, задних конечностей и черепа (уси-
ление опорного значения позвоночника и облегчение
массы тела).
7. Смена функций (Dohrn, 1875). Перемена образа
жизни или менее резкие изменения условий сущест-
вования могут вызвать утрату главной функции орга-
на и превращение одной из второстепенных функций
в главную. Такая смена функций сопровождается со-
ответствующей перестройкой морфологии органа.
Этому модусу эволюции органов придается большое
значение, поскольку он хорошо объясняет возмож-
ность качественных функциональных изменений.
Примеры смены функций многочисленны, в частнос-
ти, у растений, у которых большинство органов от-
личается значительно меньшей специализацией.
Приведем несколько более ярких примеров, чтобы
отметить широкое распространение этого модуса в
разных группах животных.
У простейших известно несколько примеров, ка-
сающихся эволюции ресничек инфузорий (Догель,
1951). У Spathidium на переднем конце тела в неко-
торых рядах реснички утратили свою локомоторную
функцию и превратились в чисто осязательные орга-
неллы. Сосущие инфузории Ephelota обладают двумя
типами придатков — сосательными трубочками, слу-
жащими, как у остальных представителей Suctoria,
для высасывания добычи, и длинными хватательными
щупальцами, удерживающими жертву. Щупальца
представляют собой модификацию трубочек и разви-
лись путем смены функций. Эти и некоторые приве-
денные выше примеры показывают, что в принципе в
эволюции органелл Protozoa проявляются те же моду-
сы филогенетического развития, что и в эволюции
органов у Metazoa (см.: Gelei, 1950).
У низших ресничных червей из отрядов Acoela и
Polycladida многочисленные вначале, ядовитые орга-
ны «кожного вооружения» в процессе эволюции раз-
ных форм частично превращаются в половые органы,
а именно в предстательные железы, семенные сумки,
органы раздражения партнера по копуляции и, нако-
нец, в мужские совокупительные органы (см.: Ива-
нов, Мамкаев, 1973). У Xenoprorhynchus steinboecki
(из отряда Lecithoepitheliata), напротив, мужской копу-
лятивный орган отделяется от полового аппарата и
путем смены функций превращается в ядовитый сти-
лет, служащий для умерщвления добычи (рис. 26).
Среди брюхоногих моллюсков, перешедших к па-
разитированию в иглокожих, большим своеобразием
отличается Paedophoropus dicoelobius, обитающий в
водных легких голотурий Eupyrgus. У самок этого не-
большого моллюска, уже утратившего под влиянием
паразитического образа жизни раковину, мантию и
спиральное закручивание туловища (пример ослабле-
ния функций и редукции органов), первично локомо-
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
61
Рис. 26. Общий план строения турбеллярий Prorhynchus (А) и Xenoprorhynchus steinboecki (Б).
1 — половой атрий, 2 — семенник, 3 — семенной пузырек, 4 — простатические железы, 5 — простатический пузырек — ядовитый орган, 6 — мешок
пениса, 7 — стилет, 8 — ротовое отверстие, 9 — ротовая полость, 10 — мозг, 11 — глотка, 12 — яйцепровод, 13 — генито-интестинальный проток,
14 — кишка, 15 — яичник, 16 — ядовитая железа.
Fig. 26. Morphology of turbellaria Prorhynchus (A) and Xenoprorhynchus steinboecki (Б).
1 — sexual atrium, 2 — testis, 3 — testicular bladder, 4 — prostate, 5 — prostate bladder — poison organ, 6 — penis sac, 7 — stilet, 8 — mouth, 9 — buccal
cavity, 10 — encephalon, 11 — pharinx, 12 — oviduct, 13 — genital-intestinal duct, 14 — intestine, 75 — ovary, 16 — poison gland.
торный орган — нога — превращен в обширную вы-
водковую сумку, в которой развиваются эмбрионы
(рис. 27).
Примитивные двуветвистые конечности членисто-
ногих были гомономными и обладали несколькими
основными функциями — локомоторной, жеватель-
ной и дыхательной. В процессе эволюции различных
ветвей членистоногих эти конечности сильно измени-
лись. В зависимости от положения на том или ином
отделе тела и преимущественно путем смены функций
они резко дифференцировались как функционально,
так и морфологически. В результате, например, у
высших ракообразных (Malakostraca) мы видим чрез-
вычайное разнообразие конечностей (рис. 22).
У части перепончатокрылых насекомых (Нуте-
noptera aculeata) есть жало, связанное с ядовитыми
железами. Оно образовалось из элементов яйцеклада
самки. Последний, в свою очередь, сформировался у
предков перепончатокрылых из конечностей 8-го и
9-го сегментов брюшка. В обоих случаях эволюция
органа сопровождалась сменой функций.
Конечности млекопитающих служат в основном
для хождения. Помимо того, они могут выполнять по-
бочные функции — прыгания, лазания, бега, рытья,
ползания, плавания, хватания, чесания, чистки шерс-
ти, защиты. Они чрезвычайно мультифункциональны
и обычно очень пластичны, т. е. легко поддаются из-
менениям и перестройкам в зависимости от условий
среды и образа жизни. Любая их дополнительная функ-
ция может стать главной, что можно видеть в разных
отрядах млекопитающих.
Специализация органа на выполнение одной функ-
ции уменьшает, разумеется, вероятность развития по
принципу смены функции. Впрочем, у пингвинов даже
такая специализированная конечность, как крыло, из
органа полета превратилась в орган плавания.
Нельзя не отметить, что посредством смены функ-
ций во многих случаях возникали важнейшие ароген-
ные изменения организации. При выходе рыбообраз-
ных предков Tetrapoda на сушу брызгальце (spiracu-
lum) превратилось в среднее ухо. При становлении
организационного типа млекопитающих челюстные
сочленовные кости трансформировались в слуховые
косточки среднего уха — наковальню (incus) и моло-
точек (malleus).
Развивая концепцию смены функций, А. Н. Север-
цов обратил внимание на роль экологических условий
в осуществлении этого модуса и ясно показал связь между
исключительным разнообразием направлений эволю-
ции, к которым он приводит, и адаптивной радиацией.
8. Фиксация фаз. Иногда работа, выполняемая
органом, слагается из нескольких последовательных
фаз, и в процессе эволюции одна из них может стать
постоянной, т. е. фиксируется как главная. Стопохо-
дящие млекопитающие при беге сначала приподнима-
ются на пальцах (рис. 28). Через фиксацию этой про-
межуточной фазы движения ноги установились посто-
янное хождение и бег на пальцах, свойственные всем
копытным. Некоторые зоологи рассматривают этот
модус как частный принцип разделения функций.
9. Выпадение промежуточных функций — модус
(как и предыдущий), распространенный сравнитель-
но мало. Северцов иллюстрировал его эволюцией
квадратной кости (quadratum) при переходе от Therio-
morpha к Mammalia. У высших рыб с костным че-
репом и у низших наземных позвоночных нижняя
челюсть подвешивалась к черепу посредством quadra-
tum и articulare. Но у рептилий, предков млекопи-
тающих, эти две кости вышли из состава челюстной
дуги, так что нижняя челюсть — dentale — стала
вторично причленяться к височной области черепа
(рис. 29). Следовательно, на этой стадии эволюции
62
ОБЩАЯ ЧАСТЬ
Рис. 27. Паразитический брюхоногий моллюск Paedophoropus di-
coelobius. Самка. Схема. (По: Иванов, 1937).
1 — вершина внутренностного мешка, 2 — печень, 3 — почка, 4 — яич-
ник, 5 — рудиментарная мантийная полость, 6 — церебральный ганглий,
7 — рудиментарные щупальца, 8 — мускулы, 9 — хобот, 10 — пищевод,
11 — глотка, 12 — ротовое отверстие, 13 — половой вырост с половым
отверстием, 14 — педальная железа, 75 — нога, 16 — выводковая камера,
17 — коконы с зародышами.
Fig. 27. Parasitic Gastropoda Paedophoropus dicoelobius. Female.
Scheme.
7 — crop of entrails pouch, 2 — liver, 3 — kidney, 4 — ovary, 5 — rudimen-
tary pulmonary cavity, 6 — cerebral ganglion, 7 — rudimantary tentacles, 8 —
muscles, 9 — proboscis, 10 — esophagus, 77 — pharynx, 72 — mouth, 13 —
penis with the sexual orifice, 14 — pedal gland, 75 — leg, 16 — broot cham-
ber, 77 — cocoons with embrios.
articulate и quadratum утратили свою промежуточную
функцию поддерживания нижней челюсти. Однако
они не исчезли, подобно другим костям нижней че-
люсти (spleniale, coronoideum), но, изменив функцию,
превратились в слуховые косточки среднего уха.
10. Разделение функций и органов. Примером
этого модуса может служить расчленение первично
непарного спинного плавника рыб, который распада-
ется у некоторых форм на отделы, несущие разные
функции: спинной и анальный плавники являются
рулями, а хвостовой — главным двигательным орга-
ном (рис. 30).
Другой общеизвестный пример представляет пре-
образование брюшной ползательной ноги в венец
хватательных околоротовых рук и воронку у голово-
ногих моллюсков (Cephalopoda).
11. Симиляция функций, по определению А. Н. Се-
верцова, представляет собой уподобление друг другу
первоначально несходных органов. Таково сходство
по строению и функциям позвонков у угрей, безногих
ящериц и змей, ползающих на брюхе. Этот процесс
связан с вторичным увеличением числа гомодинам-
ных метамеров, в данном примере — позвонков. По
Махотину (1940), такова же природа умножения
шейных позвонков у некоторых птиц и рептилий, а
также сегментов тела у некоторых многоножек (Му-
riapoda).
12. Субституция органов (Kleinenberg, 1886) —
замещение в ходе эволюции одного органа другим.
Если орган исчезает или редуцируется, то он часто
заменяется другим, берущим на себя выполнение той
же функции. Классический пример этого модуса —
замещение у позвоночных хорды сначала хрящевыми,
а потом костными позвонками (рис. 31). Не менее ил-
люстративным случаем субституции органов является
замена редуцирующихся кровеносных сосудов у пия-
вок лакунарными каналами целомической полости,
несущими кровь. У щетинковой пиявки, Acanthobdel-
la, еще хорошо развиты кровеносные сосуды, лежа-
щие в сравнительно обширном целоме. У Rhynchob-
dellaea (Piscicola и др.) имеются лакунарные каналы,
но сохранились еще спинной и брюшной сосуды. На-
конец, у Gnathobdellaea последние полностью заме-
щены лакунами. У офиур Euryalae целом вытеснен
бурсами, которые образовали третичную полость
тела (рис. 32).
13. Физиологическая субституция. Под этим на-
званием Федотов (1936) описал замещение функции
одного органа другим, который морфологически упо-
добляется первому. Так, у живородящих офиур раз-
витие зародышей протекает в бурсах — кожных впя-
чиваниях, замкнутых со стороны целома (Stegophiura
nodosa и др.). Но у Ophionotus hexact is личинки вы-
нашиваются в полости тела, которая, следовательно,
берет на себя одну из функций бурс. Последние, од-
нако, не редуцируются и сохраняют остальные свои
функции (половую и дыхательную).
14. Субституция функций — утрата в ходе эво-
люции одной из функций и замещение ее другой,
биологически равноценной. Часто можно наблюдать
замещение функции одного органа аналогичной функ-
цией, которую выполняет другой орган, расположен-
ный в другом месте и развивающийся из другого ма-
териала. В качестве поясняющего примера А. Н. Север-
цов привел изменения, происходящие у рептилий при
переходе от ходьбы с помощью ног к ползанию по-
средством волнообразного изгибания тела, в котором
участвуют позвоночник, мускулатура туловища и
ребра. Этот процесс разыгрывался в классе рептилий
не один раз. Примеры подобного рода можно при-
вести для различных групп беспозвоночных. Так,
ленточные черви, гирокотилиды и амфилиниды под
влиянием паразитического образа жизни полностью
утратили кишечник. Однако функции принятия пита-
тельных веществ, их всасывания и химической пере-
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
63
в
Рис. 28. Движение задней конечности при ходьбе млекопитающих. (По: Северцов, 1939).
А — медведь: стопоходное движение, Б — тигр: пальцеходное движение, В — лошадь: унгулиградное движение. 1—4 — последовательные фазы движе-
ния.
Fig. 28. Movements of the mammalian hind limbs during walking.
A — bear: plantigrade locomotion, Б — tiger: digitigrade locomotion, В — horse: unguligrade locomotion. 1—4 — sequential phases of locomotion.
Рис. 29. Голова эмбриона млекопитающего. Схема. (По: Port-
mann, 1951).
1—7 — кости: 1 — межчелюстная; 2 — носовая; 3 — верхнечелюстная;
4 — лобная; 5 — скуловая; 6 — теменная; 7 — чешуйчатая, squamosum;
8 — наковальня, incusquadratum; 9 — стремечко, stapes-hyomandibulare;
10 — молоточек, malleus-articulare; 11 — processus stylloideus; 12 — гиоид;
73 — гортанные хрящи; 14 — барабанная кость, angulare; 75 — нижняя
челюсть, dentale; 16 — меккелев хрящ.
Fig. 29. Head of the mammalian embryo. Scheme.
7—7 — bones: 7 — premaxillary, 2 — nasal, 3 — maxillary, 4 — frontal,
5 — jugal, 6 — parietal, 7 — squamosal, <8 — incus, 9 — stapes, 10 — malle-
us, 77 — processus stylloideus, 72 — hyoid, 73 — laryngeal cartilages, 14 —
tympanic bone, 75 — low jaw, 16 — Meckel’s cartilage.
64
ОБЩАЯ ЧАСТЬ
Рис. 30. Разделение первично непарного спинного плавника рыб.
(По: Северцов, 1939).
А—В — последовательные стадии эволюции.
Fig. 30. Division of the fish dorsal fin, originally unpaired.
A—В — the successive stages of evolution.
работки выполняются физиологически специализи-
рованными покровами (погруженным эпидермисом,
несущим микротрихии). Аналогичный пример пред-
ставляют погонофоры, тоже утратившие кишечник.
Основным источником энергии для них являются ор-
ганические вещества, синтезируемые их симбионта-
ми — автотрофными серными бактериями. Послед-
ние обитают в трофосоме — особом органе хозяина,
который образуется за счет целомического эпителия
(Иванов А. В., 1960). Таким образом, как у ленточ-
ных червей, так и у погонофор функции исчезнувше-
го кишечника более или менее полно выполняются
органами совсем иной природы.
Примеры физиологической субституции известны
и у простейших. Так, у сосущих инфузорий (Suctoria)
исчезают ротовое отверстие и глотка, поскольку пи-
тательные вещества поглощаются сосательными тру-
бочками. У других инфузорий (Hypotricha, Ophryo-
scolecidae) защитная функция выполняется утолщен-
ной пелликулой, а трихоцисты исчезают.
Впрочем, учение А. Н. Северцова о модусах эво-
люции органов до сих пор представляет, по его же
словам (1939, с. 415), пока только «новое и широкое
поле исследований, изучение которого едва намечает-
ся». После Северцова эта проблема специально изу-
чалась лишь немногими зоологами, которые показали
некоторую искусственность северцовской классифи-
кации модусов и сложную связь, существующую меж-
ду ними.
Вспомним, что в основе всех модусов филогенети-
ческого органогенеза лежат совершенно реальные яв-
ления — мультифункциональность органов и изме-
нения их функций. Самые частые и элементарные
изменения функций — их усиление или ослабление.
Очевидно, каждая функция на каком-то этапе эво-
люции появляется как новое свойство органа, затем
усиливается, более или менее совершенствуется и ус-
ложняется. Ослабевающая функция в конце концов
исчезает. Эти элементарные явления составляют сущ-
ность всех модусов. Так, модус смены функций есть
усиление одной функции и одновременное ослабле-
ние другой. Модус уменьшения числа функций пред-
ставляет собой исчезновение некоторых из них, а
модус увеличения числа функций — зарождение
новых функций (Махотин, 1940).
Сходным образом проявляют себя элементарные
изменения функций и в тех модусах, в которых коор-
динированно эволюируют по крайней мере два раз-
ных органа. При субституции функций, например, за-
меняемый орган более или менее редуцируется, т. е.
испытывает ослабление функции, а замещающий, на-
оборот, — усиление функции или смену функций. Из
сказанного ясно, что модусы весьма неравноценны.
Некоторые из них, например усиление или ослабление
Рис. 31. Поперечные разрезы через осевой скелет различных поз-
воночных. (По: Северцов, 1939).
А — первая стадия; Б — вторая стадия (круглоротые, хрящевые ганоиды);
В — третья стадия (цельноголовые, селяхии, двоякодышащие); Г — четвер-
тая стадия (костистые рыбы, амфибии, амниоты); Д — пятая стадия, хря-
щевой туловищный позвонок; Е — схема строения грудного позвонка мле-
копитающего. 1 — мезодермальная ткань, 2 — хорда, 3 — первичная обо-
лочка хорды, 4 — спинная дуга, 5 — вторичная оболочка хорды, 6 —
брюшная дуга, 7 — остистый отросток, 8 — поперечный отросток, 9 —
сочленовный отросток, 10 — тело позвонка.
Fig. 31. Transversal sections through the axial skeleton of different
vertebrates.
A — the first stage; Б — the second stage (Cyclostomes, Chondrosteans); В —
the third stage (Selachians, Chimaeras, Dipneustomorpha); Г— the forth stage
(Teleosts, Amphibians, Amniotes); Д — the fifth stage, cartilaginous dorsal ver-
tebra; E — scheme of the mammalian dorsal vertebra morphology. 1 — meso-
dermal tissue, 2 — chorda, 3 — primary chordal membrane, 4 — neural arch,
5 — secondary chordal membrane, 6 — hemal arch, 7 — neural spine, 8 —
transverse processus, 9 — zygapophysis, 10 — centrum.
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
65
функций, достаточно просты и входят в состав более
сложных модусов.
Все известные ему модусы А. Н. Северцов разде-
лил на две группы в зависимости от количественного
или качественного характера функциональных изме-
нений.3 Однако, строго говоря, без четкого устано-
вления гомологии сравниваемых функций решить во-
прос, с какими их изменениями (количественными
или качественными) мы имеем дело, невозможно.
Между тем проблема гомологии функций до сих пор
остается неразработанной.
И тем не менее можно утверждать, что гомологич-
ные функции могут выполняться только гомологичны-
ми органами. Следовательно, даже «одинаковые» функ-
ции, выполняемые негомологичными органами, воз-
никли независимо, и называть их «сохраняющимися»
и качественно неизменными невозможно.
Рассмотрим с точки зрения гомологии функций
замещение хорды позвонками. Хорда не гомологична
позвоночнику, и, следовательно, опорная функция
последнего возникла независимо. Да и полного сход-
ства между опорной функцией того и другого образо-
вания нет. Хорда у низших хордовых, в частности у
низших рыб, представляет собой упругий, поддаю-
щийся сгибанию при сокращении мускулатуры тела,
но самостоятельно распрямляющийся стержень, тогда
как позвоночник всегда является цепью метамерных,
подвижно сочлененных твердых позвонков. Позвон-
ки двигаются один относительно другого посредством
мышц, которые прикрепляются к их отросткам, т. е.
их опорная функция в принципе отличается от тако-
вой хорды. В данном случае скорее можно говорить
о субституции функций, нежели о субституции орга-
нов. Костные позвонки развиваются путем окостене-
ния хрящевых, и считать это превращение субститу-
цией органов тоже не приходится.
Другой известный пример субституции органов —
возникновение дыхания посредством сети кровенос-
ных сосудов на пальцах безлегочных саламандр (Pletho-
dontidae). Предполагается, что в связи с редукцией
легких функцию дыхания взяли на себя совершенно
другие органы. В действительности же кожное дыха-
ние вообще широко распространено даже у хвостатых
амфибий и часто более или менее вытесняет легочное.
Поэтому более вероятно, что предками безлегочных
саламандр были формы, обладавшие как легочным,
так и кожным дыханием. Своеобразный дыхательный
орган Plethodontidae, следовательно, образовался как
локальное усиление побочной функции кожи. Таким
образом, если ограничиваться сравнением с ближай-
шими предками, этот пример следует рассматривать
как частный случай смены функций.
Аналогичное возражение можно привести и в от-
ношении обычно упоминаемого примера субституции
органов у растений, а именно образования листо-
подобных органов филлодиев и филлокладиев из
черешков и стеблей у кактусоподобных суккулентов.
Постулируется переход очень важной функции фота-
3 Выражение Северцова «качественные изменения» не следует
понимать буквально, т. е. в том смысле, что одна функция приоб-
ретает качественно новые признаки. Речь идет по большей части о
смене функций, возникших независимо.
5 Протисты, часть 1
Рис. 32. Полость телау офиур. (По: Федотов, 1939).
А — Разрез тела офиуры со вторичной полостью тела; Б — то же, Еигуа-
1ае с третичной полостью тела, образовавшейся путем вытеснения целома
полостями бурс. 1 — целом, 2 — бурса, 3 — кишечник, 4 — стенки
тела.
Fig. 32. Basketstar body cavity.
A — section of basketstar body with the secondary cavity; Б — the same,
Euryalae with the third cavity formed with coelom displacement by cavities
of bursa. 1 — coelom, 2 — bursa, 3 — intestine, 4 — body wall.
синтеза от листьев, которые исчезли, к стеблям и
субституция одного органа другим. Однако у многих
травянистых растений фотосинтезирующими являют-
ся не только листья, но в какой-то мере и зеленые
стебли, и не исключена возможность, что филлодии
и филлокладии образуются как результат смены
функций.
В случае физиологической субституции (по Д. М. Фе-
дотову) невозможно представить себе «переход» уже
резко выраженной у некоторых офиур функции вына-
шивания зародышей от эктодермальных бурс к цело-
му. Естественнее допустить, что живорождение раз-
вивалось у различных офиур независимо — у одних
в бурсах, у других в целоме. Следовательно, в этом
примере мы имеем дело не с количественными, а с ка-
чественными изменениями функций. Иными словами,
перед нами модус субституции функций, т. е. развитие
аналогичных функций в разных (не гомологичных)
органах.
Итак, можно сделать вывод, что, во-первых, особо-
го модуса физиологической субституции не сущест-
вует, во-вторых, принципы субституции органов и
субституции функций лучше объединить в один модус
замещения органов и функций.
Что касается принципа смены функций (по Дорну),
то, хотя к этому модусу иногда относят, по-видимо-
му, очень разные процессы, в вышеприведенных ти-
пичных примерах действительно имеет место усиле-
ние одной побочной функции и ослабление главной.
Впрочем, иногда, как в случае превращения ноги
брюхоногого моллюска Paedophoropus в инкубацион-
ный орган, приходится допускать появление совер-
шенно новой функции. В принципе можно говорить
о процессе возникновения новых функций так же,
66
ОБЩАЯ ЧАСТЬ
Рис. 33. Нефромиксий полихеты Alciope contrainii. Протонефри-
дий с приросшей к нему половой воррнкой. (По: Догель, 1938).
1 — половая воронка; 2 — соленоциты; 3, 4 нефридиальный канал.
Fig. 33. Alciope contrainii nephromixium. Protonephridium with the
accreted sexual funnel.
1 — generative funnel; 2 — solenocytes; 3, 4 — nephridial canal.
как говорится об образовании новых органов (см.:
Иванов А. В., 1937; Getei, 1950).
Наконец, принцип фиксации промежуточных фаз
некоторые авторы (Яблоков, Юсуфов, 1976) не без
основания рассматривают как частный вариант более
общего модуса разделения функций и органов.
Таким образом, общее число модусов должно
быть несколько сокращено. С другой стороны, эта
область эволюционной морфологии настолько мало
разработана, что можно ожидать открытия еще неиз-
вестных модусов. Так, судя по некоторым данным
сравнительной анатомии беспозвоночных, вероятно,
целесообразно установление нового модуса — слия-
ния (компликации) органов, противоположного мо-
дусу разделения органов и функций. Сущность его
состоит в слиянии двух или нескольких различных
органов предка с разными отправлениями в один
сложный орган потомка, который обладает своей,
более или менее специфической, функцией (Ива-
нов А. В., 1937). Таковы, например, нефромиксии
кольчатых червей, образовавшиеся путем слияния эк-
тодермального прото- или метанефридия с мезодер-
мальным целомодуктом (рис. 33). Такие «составные»
органы очень распространены; они усложняются в
процессе эволюции за счет присоединения к ним ме-
зодермальных клеток, которые формируют соедини-
тельную ткань, а также кровеносных сосудов (тра-
хей), мускульных и нервных элементов.
Эволюция координированных
органов
Морфофизиологические преобразования одного
органа не протекают изолированно и зависят от изме-
нения строения и функций ряда других органов (фи-
логенетическая корреляция, или координация, см.
с. 57). В частности, эволюционная перестройка одно-
го органа нередко делает излишней деятельность дру-
гого органа и становится условием его редукции (та-
ковы типичные случаи субституции функций, напри-
мер исчезновение хорды при развитии позвонков).
Часто, напротив, прогресс одного органа освобожда-
ет другой только от части его побочных функций,
создавая тем самым для него условия прогрессивного
развития в новом направлении (Рыдзюнский, 1939).
К числу более сложных и потому еще недостаточ-
но выясненных морфофизиологических преобразова-
ний в системах органов относятся принципы концен-
трации, компенсации органов и гетеробатмии. Пер-
вые два имеют отношение к координации отдельных
систем органов, последний представляет результат
координации разных систем.
1. Концентрация функций (Махотин, 1940) прояв-
ляется в эволюции серий сходных (обычно гомоди-
намных) органов, причем анцестральная функция
концентрируется на нескольких из них, в связи с чем
происходит ее усиление, остальные же органы меня-
ют ее на другую функцию или редуцируются. Хоро-
шими примерами концентрации функций Махотин
считает развитие задних прыгательных ног саранчр-
вых, кузнечиков и сверчков и передних хватательных
ног водяных клопов (Nepa, Naucoris) и богомолов.
Очевидно, под эту категорию явлений могут быть
подведены почти все случаи дифференциации конеч-
ностей у членистоногих, параподий у полихет, а также
любых других метамерных органов.
2. Компенсация органов и функций (Воронцов,
1961, 1967, 1999). Часто в процессе адаптации вида
прогрессивно развивается лишь одна часть системы
органов. Так, у ряда грызунов специализация к опре-
деленному сорту пищи больше касается строения же-
лудка, чем зубов. У других форм, напротив, большим
изменениям подвергается зубная система. Благодаря
такой «компенсации» органов, или дополнению одних
органов другими, всегда остается резерв неспециали-
зированных структур, которые могут быть использо-
ваны при изменении направления естественного от-
бора.
3. Гетеробатмией был назван результат неравно-
мерного темпа эволюции различных органов и систем
организма, выражающийся в неодинаковом уровне их
развития. Это явление, сначала отмеченное Тахтад-
жяном (1954, 1966) для растений, у которых коорди-
нация между частями тела и органами слабее, оказа-
лось характерным и для животных. У представителей
тропических растений — Trochodendracea и Tetra-
dendracea — высокоспециализированный цветок со-
четается с весьма примитивной проводящей сосудис-
той системой. У магнолий же отношения обратные.
Среди животных проявление гетеробатмии в яркой
форме может быть отмечено, например, у Onychop-
hora, у которых большая часть систем органов нахо-
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
67
дится еще на примитивном аннелидном уровне (гомо-
номность сегментации, эластичная кутикула, кожно-
мускульный мешок, сегментарные целомодукты и
др.), тогда как одновременно имеются высокоразви-
тое спинное сердце, ротовые органы и очень специа-
лизированные формы живорождения.
Полимеризация и олигомеризация
Важные морфологические закономерности эволю-
ции представляют также явления полимеризации
(умножения) и олигомеризации (уменьшения
числа) гомологичных органов. Они тесно связаны с
дифференциацией эволюирующего организма, пред-
ставляют явное проявление его интеграции и в фило-
генезе обычно сопряжены друг с другом. По сущест-
ву, поли- и олигомеризация являются модусами фи-
логенетического развития органов, но не единичных,
а координированных и относятся вместе с концент-
рацией органов, функциональной компенсацией и ге-
теробатмией к одному кругу закономерностей.
Полимеризация как характерный процесс эволю-
ционного усложнения организации одноклеточных
животных была открыта В. А. Догелем в 1929 г. Под
этим термином Догель (1951) понимал не что иное,
как развитие у простейших многих гомологичных и
равноценных органоидов. Он полагал, что полимери-
зация представляет важную закономерность в эволю-
ции одноклеточных организмов, но встречается и у
некоторых низших многоклеточных, проявляясь у
них в умножении числа гомологичных органов.
Умножение ядер представляет собой наиболее
обычное проявление полимеризации у Protozoa. Оно
начинается как простое деление моноэнергидной
клетки: делится ядро, но процесс размножения не
заканчивается — цитоплазма остается неразделен-
ной. При повторных ядерных делениях образуется
несколько моноэнергидных ядер в общей цитоплаз-
матической массе: так складывается тип организации
полиэнергидных простейших. Аналогичным образом
развиваются и весьма обычные у Metazoa многоядер-
ные симпластические образования.
У Mastigophora очень распространено умноже-
ние двигательных органелл или целых их комплек-
сов (кинетид), трофических структур (хроматофо-
ров, пиреноидов) и других образований. В этих
случаях общая организация весьма усложняется, но
животное остается моноэнергидной клеткой. Неред-
ко полимеризация затрагивает в равной степени
как органеллы, так и ядерный аппарат. Тогда воз-
никает организация, характерная для многих Poly-
mastigida, у которых к каждому ядру тяготеет его
собственная многожгутиковая кинетида и вместе с
ним составляет одну энергиду — так называемый
кариомастигонт (рис. 34).
Увеличение числа гомологичных органелл у Proto-
zoa может происходить или путем новообразования,
или посредством разделения заранее имевшихся за-
чатков. Первый способ имеет место, например, при
полимеризации пульсирующих вакуолей у инфузо-
рий, а второй — при умножении комплекса двига-
тельных органелл (кинетиды) у жгутиконосцев. В не-
которых случаях полимеризация есть результат не
доведенного до конца деления клетки, что особенно
ясно видно у таких форм, как колониальный панцир-
ный жгутиконосец Polykrikos или паразитический
жгутиконосец Lamblia (Догель, 1954).
О причинах преобладания у Protozoa процессов
полимеризации убедительные соображения выска-
заны Полянским и Райковым (1977). Авторы связыва-
ют этот факт с уровнем организации полимеризую-
щихся элементов — клеточных органоидов, который
приближается к молекулярному уровню. Стабиль-
ность и жесткость, характерные для молекулярной
конструкции, почти исключают возможность услож-
нения внутреннего строения органоидов клетки, при
котором организм мог бы обходиться немногими, но
высокоразвитыми органами. Остается лишь один
путь интенсификации функций органелл — их умно-
жение. Однако с появлением у простейших «органо-
идов второго порядка», т. е. структур, состоящих из
многих «элементарных органоидов», в эволюции на-
чинают занимать видное место процессы олигомери-
зации, т. е. уменьшения числа гомологичных струк-
тур. Обычно это происходит у наиболее развитых
форм, в частности у Infusoria, у которых очень отчет-
ливо прослеживаются олигомеризация и дифферен-
циация цирр, слагающихся из многих ресничек, и
олигомеризация кортикальных элементов, в частнос-
ти соединение соматических кинетосом в пары в
пределах каждой кинеты.
Идея о закономерном характере уменьшения ко-
личества однозначных органов в процессе эволюции
издавна как бы витала в воздухе. Однако высказыва-
ния авторов (например, Вгопп, 1858) по большей
части носили чисто умозрительный характер, были
далеки от эволюционной трактовки и оставались не-
достаточно обоснованными фактическими материа-
лами. Только Догелю (1936, 1954) удалось детально
проанализировать эту закономерность в эволюцион-
ном аспекте во многих группах Metazoa и выяснить
не только те биологические и физиологические пре-
имущества, к которым приводит процесс олигомери-
зации, но и изучить вопрос о природе соотношения
между возрастающей целостностью прогрессивно
развивающегося организма и дифференциацией его
частей и органов.
Как подчеркнул Догель, при значительных эволю-
ционных превращениях, как правило, происходят
большие перестройки организации, ведущие к возник-
новению новых органов. Особенно способствует но-
вообразованию органов перемена образа жизни, на-
пример переход от малоподвижного существования к
подвижному или от водного к наземному. Новые ор-
ганы формируются часто в большом числе, слабо раз-
виты, весьма однородны и нередко располагаются
беспорядочно (принцип множественной закладки
органов). По мере дифференциации они принимают
определенную локализацию, а число их все более
уменьшается и становится более или менее постоян-
ным для данной группы животных. Многие зоологи
вслед за Догелем видят в явлении олигомеризации су-
щественные моменты, проливающие свет на важные
вопросы филогенетики. По множественному или уже
олигомеризованному состоянию гомологичных орга-
нов нередко удается судить как о степени их прими-
68
ОБЩАЯ ЧАСТЬ
Рис. 34. Организация полиэнергидных жгутиконосцев Polymastigina. (По: Иванов, 1968).
А — Snyderella tabogae из кишки термита Lobitermes longicollis, Б — акариомастигонт Snyderella, В — три кариомастигонта Stephanonympha silvestrii из
кишки термита Neotermes connexus.
Fig. 34. Morphology of polyenergid flagellate Polymastigina.
A — Snyderella tabogae from the bowel of termite Lobitermes longicollis, Б — acaryomastigont Snyderella, В — 3 karyomastigonts Stephanonympha silvestrii
from the bowel of termite Neotermes connexus.
тивности, так и о сравнительной давности их возник-
новения, а по комбинации органов разного филогене-
тического возраста нередко можно сделать заключе-
ние о происхождении данной группы. Явление олиго-
меризации позволяет также судить о направлении
филогенетического развития.
Эволюция зубов у позвоночных может служить
хорошим примером олигомеризации гомологичных
органов. Гомологами зубов является защитная плако-
идная чешуя (кожные зубы) Chondrichthyes, которая
во множестве покрывает все тело (например, у акул).
Развиваясь также в ротовой полости, плакоидная чешуя
принимает здесь функцию зубов, удерживающих и
разрезающих добычу, и соответствующим образом
дифференцируется. Следующие этапы — редукция
плакоидной чешуи на поверхности тела, где она не-
редко заменяется другими образованиями, и дальней-
шая дифференциация зубов. Сначала, следовательно,
происходит множественное развитие одинаковых ор-
ганов, потом — сокращение их числа и дифференци-
ация. Последующие олигомеризация и дифференциа-
ция зубов хорошо прослеживаются у млекопитаю-
щих.
Олигомеризацией характеризуется эволюционное
развитие многих животных, например плоских червей,
моллюсков, членистоногих, позвоночных. Четкие ряды
уменьшения числа органов кожного вооружения мы
видим у ресничных червей. Так, у поликладиды Poly-
posthides из примитивного сем. Polyposthiidae (подот-
ряд Acotylea) имеется множество грушевидных орга-
нов, покрывающих всю брюшную поверхность тела
(рис. 35). Часть этих органов, а именно один ряд вдоль
срединной линии, связана с семяпроводами и превра-
тилась в мужские совокупительные органы. Другие
представители этого семейства, например Polyposthia
и Cryptocellides, обладают не столь многочисленными
грушевидными органами, окружающими мужское по-
ловое отверстие, из которых лишь 2 или даже 1 преоб-
разовались в пенис (рис. 36). Большинство Acotylea
имеет только 1 совокупительный орган, гомологичный
грушевидному органу. Аналогичные ряды олигомери-
зующихся грушевидных органов хорошо известны у
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
69
Cotylea, т. е. в другом подотряде Polycladida, а также в
отряде Acoela. У последних, кроме того, олигомериза-
ции независимо подверглись семенные сумки (гомо-
логи грушевидных органов), железистые шипы, часть
которых превратилась в раздражающие органы муж-
ского совокупительного аппарата, и так называемые
иглообразующие железы (соответствующие рабдит-
ным железам). Интересно, что олигомеризация во всех
этих случаях сопряжена со сменой функций части ор-
ганов кожного вооружения (см. с. 60).
Низшие Arthropoda характеризуются многочис-
ленными однообразными парными конечностями, в
высших группах число ног (как и количество сегмен-
тов тела) обычно сильно сокращается и стабилизиру-
ется. Особенно ясно олигомеризация проявляется у
Pantopoda, во многих подклассах Crustacea, у Arach-
nida и InsQcta.
Ни одна другая группа членистоногих не дает, по-
жалуй, возможности проследить так полно уменьше-
ние числа гомологичных органов, как отряд пауков
(Aranei). На эту их особенность обратил внимание
еще Петрункевич (Petrunkevic, 1933), не использовав
ее, однако, для установления родственных отноше-
ний внутри отряда. Затем Догель (1954) доказал веду-
щую роль олигомеризации в эволюции пауков, а
А. В. Иванов (1965, 1977) показал, что олигомериза-
ция, совершающаяся в разных системах органов раз-
ными темпами, протекает независимо и параллельно
в разных группах отряда. У самых примитивных пау-
ков — Liphistiomorphae есть еще явная сегментация
брюшка, состоящего из 11 сегментов с тергитами; их
подглоточная нервная масса состоит из 17 невроме-
ров, имеется 9 пар медиальных эндостернитов брюш-
ка, 4 пары дорсовентральных мышц, 5 пар остий сер-
дца и боковых артерий. У остальных пауков цельное
брюшко лишено внешних следов сегментации и со-
держит гораздо меньше внутренних метамерных ор-
ганов. Число сегментов сокращается до 5, невроме-
ров — до 12, эндостернитов — до 4 пар; в различной
степени редуцируются дорсовентральные мышцы,
остии сердца и боковые артерии — от 4 до 1 пары.
Несомненно, олигомеризация этого комплекса орга-
нов вызвана редукцией задних сегментов брюшка, но
уменьшение числа органов кровеносной системы
протекает быстрее, что, вероятно, обусловлено все
возрастающей интеграцией брюшка, не связанной с
исчезновением самих сегментов. Кроме того, у пау-
ков происходит олигомеризация коксальных пор,
паутинных бородавок (рис. 37), легких и глаз, не за-
висящая от редукции брюшных сегментов. Замеча-
тельно, что олигомеризация органов протекает па-
раллельно в разных группах и независимо приводит к
сходному результату. Так, среди Mygalomorphae (че-
тырехлегочные пауки-птицееды и Araneomorphae —
двухлегочные) есть формы с 4 и 3 парами остий сердца
и с 3, 2 и 1 парой паутинных бородавок. Поэтому
при выяснении филогении или естественной системы
нельзя основываться только на числе органов, под-
вергшихся олигомеризации, например на количестве
остий сердца, как это сделал Петрункевич. Объеди-
нив по наличию 4 пар остий явно неродственные
друг другу семейства из триб Haplogyne и Entelegyne,
он создал искусственную группу Quadrostiatae.
Рис. 35. Поликладида Polyposthides affinis.
1 — грушевидные органы; 2 — ветви кишки; 3 — рот; 4 — мужские поло-
вые аппараты, состоящие каждый из 5 грушевидных органов; 5 — матка;
6 — влагалище; 7 — главные семяпроводы; 8 — мозг.
Fig; 35. Polycladida Poly posthides affinis.
1 — pyriform organs, 2 — branches of intestine, 3 — mouth, 4 — male gene-
rative apparatus with 5 pyriform organs, 5 — uterus, 6 — vagina, 7 — basic
spermaducts, 8 — brain.
Рис. 36. Поликладида Polyposthia similis. Расположение груше-
видных органов. (По: Beauchamp, 1951).
7 — грушевидные органы, 2 — совокупительный орган, 3 — женское по-
ловое отверстие, 4 — мужское половое отверстие.
Fig. ,36. Polycladida Polyposthia similis. Pyriform organs position.
7 — pyriform organs, 2 — copulative organ, 3 — female generative orifice,
4 — male generative orifice.
70
ОБЩАЯ ЧАСТЬ
Рис. 37. Параллельная олигомеризация паутинных бородавок у
пауков. Схема. (По: Иванов, 1965).
А — Liphistius, Б — Heptathele, В — Atypus, Г — большинство Mygalomor-
phae, Д — Barychelidae, Е — Stegodyphus, Е' — то же, Ж — Araneus, 3 —
Uroctea, И — некоторые Entelegynae, К — Zodariidae, Л — Loxosceles,
М — многие Haplogynae, Н — Heardrotarsus ( А—В — Liphistiomorphae;
Е, Е' — Cribella; Ж—К — Entelegynae; Л—Н — Haplogynae).
Fig. 37. Parallel oligomerization of the spider warts (Araneida).
Scheme.
A — Liphistius, Б — Heptathele, В — Atypus, Г — the majority of Mygalomor-
phae, Д — Barychelidae, E — Stegodyphus, E — the same, Ж — Araneus,
3 — Uroctea, И — some Entelegynae, К — Zodariidae, Л — Loxosceles, M —
many of Haplogynae, H — Heardrotarsus (A—В — Liphistiomorphae; E, E —
Cribella', Ж—К — Entelegynae; Л—Н — Haplogynae).
Таким образом, лишь для небольших и цельных
групп пауков, в которых олигомеризация представля-
ет единый процесс, число олигомеризующихся орга-
нов приобретает основное значение для понимания
филогении.
Что касается простейших, то для них характерно
сложное сочетание и взаимодействие поли- и олиго-
меризации.
В эволюции инфузорий разыгрываются сложные
процессы олигомеризации в системе кортикального
слоя, происходящие как минимум на двух уровнях
(Полянский, Райков, 1977). На организменном уровне
имеет место дифференциация сложного ресничного
покрова с образованием ресничных и безресничных
зон, т. е. олигомеризация путем редукции целых кинет
или их отрезков. Так образуются, например, реснич-
ные венчики у Didinium, брюшное ресничное поле у
многих ползающих инфузорий (Loxodes, Trachelone-
та и др.), ротовые и околоротовые ресничные образо-
вания и чувствительные спинные щетинки Hypotricha.
На органоидном уровне элементарным процессом
олигомеризации является соединение соматических
кинетосом в пары в пределах каждой кинеты. Оно
происходит уже у самых примитивных инфузорий
(например, у Karyorelictida, обладающих еще макро-
нуклеусом, близким к диплоидному). У высших ин-
фузорий происходит дальнейшая олигомеризация —
из 2 кинетосом 1 теряет ресничку. Последующие
этапы развития сложных ресничных органоидов мем-
бранелл и цирр тоже включают как минимум два эле-
мента олигомеризации — процесс соединения ресни-
чек в органеллу высшего порядка й параллельный
процесс редукции фибриллярных отростков кинето-
сомы.
Догель (1954) установил следующие три типа оли-
гомеризации гомологичных органов.
1. Посредством полной редукции части гомоло-
гичных органов. Этот тип, в свою очередь, подразде-
ляется на несколько групп. Одну из них составляют
многочисленные мелкие образования, которые внача-
ле развиваются по всему телу, затем сохраняются в
более или менее измененном виде лишь на опреде-
ленных его участках. Таковы чешуя различной при-
роды у позвоночных, а также их перья и волосы. Во
вторую труппу входят все случаи постепенной редук-
ции множественных парных органов у метамерных
животных. Пример, приведенный Догелем, — 2 пары
нефридиев ракообразных, которые представляют
сохранившуюся часть изначально множественных ме-
тамерных органов. Особый характер носит редукци-
онная олигомеризация органов третьей группы, свя-
занная с развитием асимметрии у брюхоногих мол-
люсков. Это прежде всего — редукция первично левых
органов мантийного комплекса. Существуют и другие
формы редукционной олигомеризации.
2. Путем слияния зачатков гомологичных орга-
нов. Таково слияние зубов у двоякодышащих рыб
(Dipnoi) или слияние у низших рыб парных нёбно-
квадратных и меккелевых хрящей в непарные эле-
менты.
3. Посредством изменения функции части гомо-
логичных органов. Сюда Догелем были отнесены:
дифференциация в различных направлениях груше-
видных органов у турбеллярий (см. выше), превраще-
ние части яичников в желточники у плоских червей,
превращение части эстетов у хитонов (Polyplacopho-
га) в глаза и многие другие примеры.
О значении
палеонтологической летописи
Очень важным материалом для изучения эволюци-
онной истории животных являются их ископаемые
остатки: они позволяют наблюдать эволюционные из-
менения животных и прослеживать их смену во вре-
мени при условии, разумеется, что их родство и пре-
емственность истолкованы правильно. Однако непол-
нота палеонтологической летописи весьма велика.
Очень многие группы животных, лишенные скелета,
не сохраняются или оставляют лишь следы своей
жизнедеятельности. Чаще всего поэтому зоолог, за-
нимающийся систематикой и филогенезом, вынужден
ограничиваться данными биологических наук, изуча-
ющих ныне живущих животных, — в первую очередь
сравнительной анатомии, эмбриологии и экологии.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
71
К сожалению, имеется очень мало палеонтологи-
ческих данных о самом длительном первоначальном
периоде эволюции жизни, протекавшем на необозри-
мом протяжении архейской и протерозойской эр.
Первые остатки организмов хорошей сохранности
начинают попадаться лишь в нижнепалеозойских
слоях. Впрочем, существует ряд косвенных указаний^
свидетельствующих о наличии очень примитивных
форм жизни уже в архейскую эру, которая занимала
огромный промежуток времени в истории Земли (от-
ложения возрастом от 2.6 до 4.5 млрд лет).
1. Нахождение в породах архейского возраста в
Европе и Северной Америке отложений графита, со-
стоящего в основном из углерода. Возможно, они
произошли из остатков морских водорослей, так как
углерод выделяется из углекислоты лишь фотосинте-
зирующими растениями. После разложения отмер-
ших водорослей под влиянием бактерий мог остаться
углерод, который в процессе метаморфизма превра-
тился в графит. Поэтому многочисленные залежи
графита рассматриваются некоторыми палеонтолога-
ми как хорошее (хотя и косвенное) свидетельство су-
ществования растений в архее.
2. Частичное окисление археозойских железных
руд, приведшее к образованию гематитов и магнети-
тов, требует допущения присутствия свободного кис-
лорода, что, в свою очередь, указывает на наличие
уже в то время растений.
3. Наконец, в архейских толщах встречаются из-
вестняки, которые, как правило, образуются в резуль-
тате жизнедеятельности организмов.
Все это, вместе взятое, делает предположение о
присутствии растительных организмов в архее очень
вероятным.
О необъятности времени, протекшего с тех пор,
как появилась на Земле жизнь, отчасти дает пред-
ставление так называемая годичная календарная схема
Флюгеля (рис. 38), в которой все время существова-
ния нашей планеты представлено условно в виде
12 месяцев одного года. При этом на долю фанерозоя
приходится не более 2 последних месяцев (большая
часть ноября и декабрь), тогда как длительность до-
кембрийского периода оказывается в 6 раз больше.
Первые остатки организмов (если воспользоваться
схемой Флюгеля) появились в начале марта, a Euka-
ryota — только в начале сентября. Metazoa известны
только с конца октября.
Было предложено множество гипотез происхож-
дения жизни на Земле (см. обзоры: Опарин, 1957,
1980; Rutten, 1973; Folsom, 1982; Уголев, 1985).
Предполагается, что наша планета существует уже
около 4.7 млрд лет и что первые живые организмы
появились на Земле около 3.8 млрд лет назад. Весьма
распространены представления о происхождении
жизни в результате очень длительной (около 1 млрд
лет) «химической эволюции» (Опарин, 1924; Розано-
ва, 1928; Folsom, 1982, и др.), когда из неорганичес-
ких компонентов первобытной атмосферы под воз-
действием энергии ультрафиолетовых лучей, тепла и
электричества возникали и затем эволюировали раз-
нообразные органические соединения. Из комбина-
ций последних постепенно развивались сложные сис-
темы органических соединений, обладающие некото-
рыми основными физиологическими свойствами, ко-
торые способствовали их превращению в первых
протобионтов. Были осуществлены некоторые лабо-
раторные опыты, которые как будто подтверждали
эти предположения. Так, например, оказалось, что,
воздействуя световой и электрической энергией на
смеси соответствующих газов, находящихся над по-
верхностью воды, можно получить не только малые
органические молекулы, но и содержащие углерод
полимерные материалы в виде мельчайших сферул
(см.: Folsom, 1982). Предполагается, что множество
подобных сферических «предклеток» должно было
находиться в первичных водоемах.
Происхождение первых клеточных организмов
пытались объяснить две гипотезы. Опарин (1924)
приписывает основное значение в образовании клет-
ки белкам, а возникновение генетического аппарата
наследственной передачи признаков следующему по-
колению относит к более поздним этапам эволюции
(см. также: Fox, Dose, 1975). По другой гипотезе,
первоосновой жизни была нуклеиновая кислота — «го-
лый ген», обеспечивший построение и эволюцию
белков (Mliller G., 1969; Florkin, 1975, и др.). Все эти
проблемы недавно были критически проанализирова-
ны Уголевым (1985), который показал необходи-
мость рассматривать развитие жизни как закономер-
ный физиологический процесс.
Как бы то ни было, самые древние микроскопи-
ческие органические остатки, похожие на предпола-
гаемые «предклетки», были найдены в кремнистых
сланцах раннего докембрия Южной Африки (серии
Онвервахт и Фйг-Три из системы Свазиленд), датиру-
емые 3—3.7 млрд лет (см.: Rutten, 1973; Schopf,
1975; Ford Т. D., 1979). Они обнаружены на тонких
шлифах и в материале, полученном посредством ма-
церации, имеют сферическую или нитчатую форму и
размеры от 1 до 20 мкм. С известной долей вероят-
ности их можно сравнивать с прокариотными орга-
низмами, однако их биогенная природа все же оста-
лась недоказанной.
Несомненные ископаемые организмы были от-
крыты в породах среднего докембрия в сланцах фор-
мации Ганфлинг в Канаде (Tyler, Barghoom, 1954).
Им около 2000 млн лет. Разнообразные по форме —
сферические, удлиненные или нитчатые, простые или
разветвленные, они большей частью одноклеточные
или состоят из немногих клеток (Barghoorn, Tyler,
1965). Очень сходная микрофлора была обнаружена
также в Калифорнии (1075 млн лет), Центральной
Австралии и Северной Франции (700 млн лет). В Рос-
сии она была описана из венда (позднего докембрия)
Тимофеевым (1969, 1974). После изучения этих ис-
копаемых организмов с помощью электронного ми-
кроскопа их прокариотная природа стала более оче-
видной (Oehler, 1976). Многие формы оказались бак-
териями или синезелеными водорослями (Barghoorn,
1971; Крылов, 1972; Flugel, 1978; Келлер, 1979). От-
крытие этих древних существ относится к выдаю-
щимся достижениям современной палеонтологии.
Весьма обычными ископаемыми докембрия явля-
ются строматолиты, представляющие собой большие
слоистые куполообразные столбчатые разветвленные
отложения извести — продукта жизнедеятельности
72
ОБЩАЯ ЧАСТЬ
Декабрь
Ноябрь
r 570
О
S'
X
Октябрь
1000-
Сентябрь
’а
Абгуст
о
«о
Июль
---------2000
Июнь
Маи
Е
возраст
(млн JJtm)✓ТЗЗПТеТЖ;/nuv/wd
' 14 X1
Третичный
период
№
61-
137
195
230
Metazoa
285-
> 350
Eukaryota
905-
\ 990
500-
570
Мел
Юра
Триас
Пермь
Карбан
Дебон
Силур
Ордобик
Кембрии
Современность
,, 31.XI
-----Homo---------
**s
. 17X1
Aves-------
* т. 7J.Zff
=f=— Mammalia—-----
л 4-,- 33al
•*--Reptiha-------
'а Декабрь 1.ХД
* , 29Л
-•—Amphibia------
. 29Л
-•—Pisces------
<ъ
ь
Ноябрь ЮЛ
Апрель
--------2700
Март
Февраль
Рис. 38. Понятие геологического времени и деление истории Земли по формациям. Условный календарный год, облегчающий представ-
ление о длительности абсолютной протяженности геологических периодов. (По: Flugel, 1978).
Fig. 38. Geological time concept and the Earth history division in formations. The conventional calendar year promoted view of the persis-
tence of the absolute time of geological period.
сообществ синезеленых водорослей (рис. 39). Древ-
нейшие из них — известные системы Булавайо в
Юго-Восточной Африке — датируются периодом
времени от 3100 до 2700 млн лет. Подобные по-
стройки Cyanophyta образуют и поныне (в Западной
Австралии, Северной Америке и на Багамских остро-
вах) (см.: Вологдин, 1963). Докембрийские стромато-
литы обычно развивались в морских мелководьях и
иногда формировали рифы.
Таким образом, эволюция первобытных Ргокагуо-
ta продолжалась в течение огромного периода време-
ни (вероятно, начиная от 3750 до 1600 млн лет). За
это время у протобионтов возникли и эволюировали
генетический и белоксинтезирующий механизмы,
сложился передающийся наследственно анаэробный
тип обмена веществ. Протобионты, вероятно, были
гетеротрофными организмами. Питаясь органически-
ми веществами абиогенного происхождения, они
ПУГИ И ЗАКОНОМЕРНОСТИ эволюции
73
получали энергию путем анаэробной диссимиляции,
или брожения. Атмосферный кислород тогда еще не
существовал, атмосфера состояла из воды, аммиака,
углекислоты и, возможно, водорода и метана. Позд-
нее в результате жизнедеятельности широко распро-
страненных автотрофных фотосинтезирующих орга-
низмов, в первую очередь синезеленых водорослей
(Суanophyta), в атмосфере стал постепенно накапли-
ваться свободный кислород. Когда его количество до-
стигло 0.01 от современного его содержания (что, ве-
роятно, имело место около 1000 млн лет назад, —
см. рис. 38), появились организмы с энергетически
гораздо более совершенным обменом веществ, сопро-
вождавшимся аэробной диссимиляцией — процессом
дыхания. Несомненно, это был весьма важный этап в
истории жизни на Земле. Современное содержание
кислорода в воздухе было достигнуто лишь в конце
палеозоя. В раннем рифее (1600—1350 млн лет), по-
видимому, появились уже и Eukaryota (Розанова, 1928;
Тимофеев, 1969; Barghoorn, 1971; Соколов, 1981;
Видаль, 1984). На это указывает состав разнообраз-
ной микрофлоры формации Битер Спрингс в Австра-
лии, Лахадинской серии в Якутии и серии Балт в Се-
верной Америке. У некоторых одноклеточных орга-
низмов этого времени были отмечены признаки ядра,
находящегося в процессе деления (Nucellosphaeridi-
ит, Cariosphaeroides и др.). Ядро первобытной эука-
риотной клетки, скорее всего, возникло как результат
прогрессивной эволюции ее энергетических органелл
и генетического аппарата. Но в настоящее время ши-
роким признанием пользуется новая симбиогенети-
ческая гипотеза, рассматривающая митохондрии и
пластиды как видоизменения различных симбиоти-
ческих прокариотных клеток (Маргелис, 1983). Как
бы то ни было, возникновение первичных Eukaryota
предуготовило расцвет в рифее эукариотного планк-
тона и появление многоклеточных растений и живот-
ных.
Из верхнего докембрия — венда известны сфери-
ческие органические тела — мегасфероморфы, пред-
положительно, планктонные организмы, достигаю-
щие величины 5 мм и больше. Они обнаружены в
скалистых породах Канады, Швеции, Франции, Рос-
сии, Ирана, Индии и Австралии, датируемых в 600—
800 млн лет. Представителем этого типа микрофос-
силий, по-видимому, является и «маленький медузо-
ид» Chuaria, описанный Соколовым (1973) из венда
России. К какой группе — Prokaryota или Eukaryo-
ta — относятся мегасфероморфы, неизвестно, но их
появление совпадает с вендским расцветом Metazoa.
Что касается животного населения Земли, то оно
могло появиться лишь в среднем протерозое (рис. 38).
Хорошо известен поразительный факт, до сих пор не
получивший вполне удовлетворительного объясне-
ния, что в позднепротерозойских породах отсутству-
ют твердые скелетные части животных (обзор гипо-
тез см.: Шевырев, 1962). Все не очень разнообразные
остатки докембрийской фауны представлены только
следами их жизнедеятельности или отпечатками их
мягких частей. К древнейшим следам животных (в
возрасте около 1000 млн лет) относятся обнаружен-
ные в Центральной Австралии и на Южном Урале
червеобразные экскременты каких-то червей-ило-
Рис. 39. Строматолиты — постройка сообществ синезеленых во-
дорослей Linella avis из отложений венда. (По: Крылов, 1972).
А — часть строматолита, Б — шлиф строматолита.
Fig. 39. Stromatoliths — constructions of the blue-green algae Linel-
la avis from the Vendian.
A — part of construction, Б — stromatolith saw cut.
едов, так называемые Vermiculites. Вполне достовер-
ные остатки многоклеточных животных известны
только начиная с верхнего протерозоя — венда, да-
тируемые в пределах от 680 ± 20 до 570 ±10 млн лет.
Все более древние находки пока сомнительны и дол-
жны быть проверены (Соколов, 1965).
Первые несомненные остатки вендских многокле-
точных животных были обнаружены в Намибии (Юго-
Западная Африка) (Giirich, 1930). Затем очень богатая
вендская морская фауна была открыта австралийским
палеонтологом Сприггом (Sprigg, 1947) в Эдиакаре
(Южная Австралия).
Здесь было собрано около 1600 образцов —
большей частью отпечатки медузоподобных существ,
полипов и колоний полипов, похожих на морские
перья, а также форм, отдаленно напоминающих
кольчатых червей и даже трилобитообразных чле-
нистоногих.
Эдиакарские находки не единичны. Отпечатки
лишенных скелета животных вендского времени были
обнаружены также в Англии, Африке (Намибия) и
Восточной Польше. В семидесятых годах они были
найдены и на территории бывшего Советского Сою-
за — на Зимнем и Летнем берегах Белого моря, в
Якутии, западной Латвии и в других местах. Бело-
морская фауна венда оказалась богаче и представи-
тельнее эдиакарской и характеризует больший пери-
од времени (Соколов, 1976, 1981).
Среди беспозвоночных венда весьма обычными
оказались медузоподобные существа (Glaessner, 1958,
1963, 1971; Федонкин, 1983, 1987). Это округлые,
овальные, дисковидные отпечатки с концентрически-
ми, радиальными и лопастными образованиями.
Среди них, по-видимому, имеются формы, которые
довольно уверенно можно отнести к гидроидным или
сцифоидным медузам, есть также слепки, полу-
чившиеся в результате наполнения кишки медузоид-
74
ОБЩАЯ ЧАСТЬ
Рис. 40. Ландшафт дна вендского моря. (По: Биологический энциклопедический словарь, 1986, с изменениями).
1 — Vendia sokolovi, билатерально-симметричное животное неясного систематического положения; 2 — Nemiana simplex, радиально-симметричный орга-
низм; 3 — Dickinsonia costata; 4 — Tribrachium heraldicum; 5 — Spriggina flaundersi; 6 — Vendotaenia, слоевищная водоросль; 7 — Charniodiscus opposi-
te, морское перо (?); 8 — Chamia masoni, морское перо.
Fig. 40. Landscape of the Vendian Sea bottom.
1 — Vendia sokolovi — bilateral-symmetrical organism, incertae sedis; 2 — Nemiana simplex — radial-symmetrical organism; 3 — Dickinsonia costata; 4 — Trib-
rachium heraldicum; 5 — Spriggina flaundersi; 6 — Vendotaenia — frond algae; 7 — Charniodiscus oppositus — sea pen (?); 8 — Charnia masoni — sea pen.
ного существа песком (Wade, 1968, 1972). Однако
для большей части медузоподобных форм нет бес-
спорных доказательств принадлежности к книдариям
(Ford, 1979).
Вторую важную группу вендской фауны составля-
ют формы, охарактеризованные как Pennatulida —
морские перья (Glaessner, Wade, 1968). Полной уве-
ренности в этом, однако, нет. По мнению других ав-
торов (Jenkins, Gehling, 1978), относящиеся сюда
роды *— Arborea, Charnia (рис. 40, 8), Charniodiscus
(рис. 40, 7), Rangea и Pteridinium — с равным осно-
ванием могут рассматриваться как большие перооб-
разные водоросли или как совершенно неизвестные
вымершие кишечнополостные.
Внешне похожи на морские перья не менее таин-
ственные петалонамы (Petalonamae), найденные в по-
родах верхнего протерозоя Юго-Западной Африки, а
также в кварцитах Паунд в Южной Австралии. Пета-
лонамы послужили объектами крайне фантастичес-
ких филогенетических спекуляций Пфлюга (Pflug,
1970—1973). До сих пор их организация и система-
тическое положение остаются неизвестными.
Третья группа животных определена Глесснером
и Уэдом как Annelida. Одно из этих животных —
Spriggina flaundersi (рис. 40, 5) недавно изучено мето-
дом отливок из латекса, изготавливаемых по естест-
венным отпечаткам тела животного на горной поро-
де. Оказалось, что это животное не может быть при-
числено ни к аннелидам, ни к членистоногим, но по
некоторым признакам занимает промежуточное по-
ложение (Birket-Smith, 1981). Длинное сегментиро-
ванное тело Spriggina было защищено непарными
спинными и парными брюшными кутикулярными
пластинками. Большая подковообразная голова обла-
дала «глазами» и вентральными «антеннами». Каж-
дый сегмент был снабжен спинной и брюшной пара-
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
75
Б
Рис. 41. Реконструкция вендского членистого Spriggina flaundersi. (По: Birket-Smith, 1981).
А — со спинной, Б — с брюшной стороны.
Fig. 41. Reconstruction of Vendian Arthropoda Spriggina flaundersi,
A — dorsal side, Б — ventral side.
ми ложночленистых придатков (рис. 41). Вскоре тем
же способом были изучены и экземпляры эдиакар-
ского Praecambridium, напоминающие в некоторых
отношениях протисписовых личинок трилобитов и
предположительно рассматриваемые как молодые
Spriggina (Birket-Smith, 1981). Из отложений венда
описаны похожие на Praecambridium отпечатки «бес-
панцирных трилобитов», редко превышающие 4 мм в
длину, на которых видны полуокруглая голова и вы-
тянутая туловищная ось с 5—6 боковыми лопастями.
Несколько родов и видов — Vendia sokolovi (рис. 42),
Onega stepanovi и Vendomia menneri толкуются обыч-
но как примитивные членистоногие (Соколов, 1973;
Келлер, Федонкин, 1976). Одна более крупная форма
Pseudovendia charnwoodensis (до 3 см длиной) описа-
на из венда Англии (Boynton, Ford, 1979). Все эти
формы, возможно, представляют ювенильные стадии
5/?п££1па-подобных животных. Высказывалось мало-
вероятное предположение, что это предки всех цело-
мических сегментированных животных.
К весьма загадочным обитателям дна вендских
морей принадлежат Tribrachium и Dickinsonia. Орга-
низация Tribrachium (рис. 40, 4) неизвестна. Это жи-
вотное обладает трехлучевой симметрией, не свойст-
венной книдариям, но встречающейся у некоторых
палеозойских иглокожих.
Из 5 видов Dickinsonia 4 известны из венда России.
Более крупные особи Dickinsonia достигают 1 м в длину.
Их листовидное и чрезвычайно тонкое тело (не более
3 мм в толщину) имеет овальное очертание и несет
многочисленные параллельные тонкие гребешки или
валики,. расположенные бирадиально (рис. 40, 5).
Строение и развитие этих существ осталось неизвест-
ным, так же как и образ жизни. Сначала они считались
представителями особой группы кишечнополост-
ных — Dipleurozoa (Harrington, Moore, 1956), затем
сравнивались с современными полихетами Spinther
(из сем. Amphinomidae) (Wade, 1972), а недавно Фе-
донкин (1983) отнес их к Bilateria как группу, «пере-
ходную от низших червей к высшим», сблизив с тур-
белляриями Polycladida и Tricladida. Сходство с по-
следними, по его мнению, проявляется в билатеральной
форме плоского тела и в метамерном строении кишеч-
ника. Все эти гипотезы лишены, однако, серьезных
оснований, поскольку об организации Dickinsonia мы
ровно ничего не знаем, неизвестно даже, что представ-
ляют собой бирадиальные гребешки и бороздки. Вряд
ли это элементы пищеварительной системы. Их распо-
ложение у Dickinsonia не может служить доказатель-
ством принадлежности к Bilateria. Среди Radiata по-
добную симметрию имеет одиночный мадрепоро-
вый коралл Fungia, обладающий кораллитом,
симметрия которого поразительно сходна с таковой
Dickinsonia (см. также: Федонкин, 1983, с. 79).
Весьма проблематичным является также Xenusion
auerswaldae из кварцитов северной Германии — ме-
тамерное животное с конечностями, напоминающи-
ми таковые Onychophora. Близкие формы были най-
дены в самых нижних отложениях кембрия в Шве-
ции.
Наконец, Соколов (1965) описал из венда остатки
различных животных, обитающих в защитных труб-
ках, в том числе погонофор в виде фрагментов коль-
чатых хитиновых трубок. В венде существовали уже
76
ОБЩАЯ ЧАСТЬ
Рис. 42. Предполагаемое членистоногое Vendia "sokolovi (венд).
(По: Келлер, 1979).
Fig. 42. Assumed Arthropoda Vendia sokolovi (Vendian).
многочисленные и разнообразные группы многокле-
точных растений — Metaphyta (Соколов, 1979).
Предполагается, что все докембрийские Metazoa,
вероятно за исключением Spriggina и Praecambridi-
ит, были фильтраторами микропланктона. Однако,
вероятно, существовали и роющиеся в грунте детри-
тоядные животные, судя по оставленным ими следам
и норкам (Seilacher, 1956; Webby, 1970).
Таким образом, вендо-эдиакарская фауна была
разнообразна, богата и содержала уже некоторые вы-
сокоразвитые формы. Часть вендских животных, ве-
роятно, будет отнесена к известным типам и классам,
другая часть, возможно, принадлежит к неизвестным
еще, вымершим группам. Вендская радиация Metazoa
несомненно представляет важнейший этап в эволю-
ции животного мира.
Общую картину развития органического мира в
протерозое рисует Келлер (1979, с. 71): «В теплых
морях верхнего рифея до эпохи великого лапландс-
кого оледенения повсеместно были распространены
синезеленые водоросли, создавшие в карбонатных
толщах строматолитовые постройки. Животный мир
этого времени изучен слабо. В вендских отложениях
к послеледниковому времени произошло быстрое
расселение беспозвоночных животных, образующих
устойчивую, очень разнообразную по своему система-
тическому составу ассоциацию видов. Составляющие
ее формы относятся к организмам, лишенным твер-
дого скелета в виде панциря или раковины. В морях
геологического прошлого эта фауна появилась 600—
620 млн лет назад и существовала около 50 млн лет.
Многие типы и семейства, свойственные этой фауне,
мы находим и в более молодых кембрийских отложе-
ниях. Фитолиты в венде отходят на второй план, по-
являются и развиваются своеобразные растительные
сообщества, получившие название „вендотениевой
флоры”».
На границе протерозоя и кембрия картина резко
меняется — как бы внезапно появляются представи-
тели почти всех основных групп беспозвоночных;
причем многие из них обладают уже известковым,
кремневым и органическим скелетом. Создается впе-
чатление, что в протерозое существовали какие-то
условия, препятствовавшие образованию скелета,
или, быть может, способствовавшие его разрушению
после гибели животного. Одна из гипотез заключает-
ся в предположении, что свойства воды докембрийс-
ких океанов исключали возможность построения жи-
вотными минерального скелета; таким свойством мог
быть, например, недостаток насыщения морской
воды известью вследствие перенасыщения ее хлором
или углекислым газом. Этому предположению, одна-
ко, противоречит образование в протерозое упомяну-
тых выше мощных карбонатных отложений синезе-
леных водорослей (Виноградов, 1940). Впрочем, в
верхнем венде существовали уже организмы с крем-
невым и фосфатным скелетом.
Итак, в кембрийских слоях уже встречаются ос-
татки представителей разных типов беспозвоночных.
В кембрии существовали фораминиферы, радиоля-
рии и другие простейшие, а также кремневые хубки.
И те и другие несомненно появились гораздо раньше.
Особенно характерны археоциаты — своеобразные,
напоминающие губок, неподвижно прикрепленные
морские животные с известковым скелетом, обитав-
шие в мелководных морях, где они создавали мощ-
ные береговые и барьерные рифы (Вологдин, 1963).
Типичными кембрийскими формами были также та-
буляты и строматопораты, обладающие известковым
скелетом. Существовали уже разнообразные моллюс-
ки — двустворчатые, брюхоногие, хиолиты — и жи-
вотные, напоминающие онихофор. Членистоногие
были представлены трилобитами, мечехвостами, ра-
кообразными (остракодами) и некоторыми группами
неясного систематического положения. Характерно,
что в фауне нижнего кембрия по числу родов и более
высоких таксонов выделялись только археоциаты,
плеченогие и трилобиты и явно проявлялась последо-
вательность в появлении групп — сначала развива-
лись археоциаты, губки и плеченогие, затем первые
трилобиты, хиолитиды и брюхоногие моллюски (см.:
Шёвырев, 1962). В конце кембрия появились первые
головоногие моллюски (наутилоидеи), а также грап-
толиты и прикрепленные иглокожие. В кембрии, по-
видимому, не было еще позвоночных и наземных
членистоногих.
Паукообразные (настоящие наземные скорпионы)
и первые примитивные насекомые появились лишь в
девоне. Низшие позвоночные — бесчелюстные (Ag-
natha), а именно щитковые — Ostracodermi, впервые
встречаются в слоях нижнего ордовика, а настоящие
рыбы — в отложениях верхнего силура. Остатки пер-
вых млекопитающих — Protodonta и Multitubercula-
ta — известны из триаса, а человек (Homo habilis)
появился только в начале четвертичного периода
около 2.7—2.5 млн лет назад. Появление Homo sapi-
ens произошло примерно 200 тыс. лет назад. Общая
картина смены флоры и фауны на протяжении палео-
зойской, мезозойской и кайнозойской эр представле-
на в табл. 2.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
77
Таблица 2
Хронология органического мира (числами обозначено время в миллионах лет)
Эра Период Характерные группы организмов
4600 Архей 2700 ± 100 Имеются отложения органического происхождения, бактерии
Протерозой 570 ± 10 Бактерии, синезеленые водоросли. Предполагаемые медузообразные и напоминающие морские перья кишечнополостные, «черви», аннелиды, предшественники трилобитов (Spriggina, Cambri- dium), петалонамы и погонофоры. Спикулы кремневых губок.
Палеозой 285 ± 10 Кембрий 80—70 500 ± 15 Синезеленые и известковые водоросли, образующие рифы. Фораминиферы и радиолярии. Губки (кремневые). Археоциаты (только нижний и средний кембрий), строящие рифы. Первые табуля- ты. Полихеты. Первые хитоны, моноплакофоры, примитивные брюхоногие и двустворчатые мол- люски и хиолиты. Плеченогие (главным образом беззамковые). Многочисленные трилобиты (1-я фауна Трилобитов). Примитивные мечехвосты. Появляются первые наутилоидёи (с прямой рако- виной), граптолиты и иглокожие (карпоидеи, гомойостелеи, цистоидеи).
Ордовик 70—60 440 ±15 Синезеленые водоросли. Известковые рифообразующие и бурые водоросли. Появление четырехлуче- вых кораллов (Rugosa), эндоцератитов и ортоцератитов (из головоногих), мшанок, морских лилий и ежей. Ракоскорпионы. Первые бесчелюстные позвоночные — щитковые, Ostracodermi. Расцвет плеченогих (беззамковых), трилобитов (2-я фауна трилобитов), граптолитов и наутилоидей.
Силур 35—30 405 ± 10 Синезеленые, известковые, бурые и красные водоросли. Первые наземные растения, напоминающие псилофитов. Появление тентакулатов (из Mollusca) и морских бутонов. Вытеснение беззамковых плеченогих замковыми. Моллюски: хитоны, моноплакофоры, брюхоногие, двустворчатые, хиоли- ты, первые бактриты (из Cephalopoda). Членистоногие: трилобиты (2-я фауна), мечехвосты, ра- коскорпионы. Появление скорпионов (Palaeophonus и др.). Первые губоногие многоножки (Рго- tochilopoda). В верхнем силуре появляются панцирные рыбы (Placodermi), панцирные акулы (Ste- goselachii) и древние акулы (Acanthodii)
Девон 60—55 350 ±10 Споровые: псилофиты, примитивные папоротниковые, хвощовые, плауновые и семенные папорот- ники. Птеродоспермы. Первые голосеменные — кордайты и прогимноспермы. Первые известко- вые губки. Господство четырехлучевых кораллов, замковых плеченогих и наутилоидей. Широко распространяются, затем вымирают планктонные граптолоидеи (планктонные граптолиты). Ци- стоидеи уступают господство бластоидеям. Расцвет появившихся в силуре рыб, а также кистепе- рых, двоякодышащих и древних костистых (Palaeoniscoidei). Первые насекомые (Colembola) и первые амфибии — стегоцефалы.
Карбон 70—60 285 ±10 К девонским группам растений прибавились первобытные хвойные, но прогимноспермы исчезли. Мад- репоровые кораллы. Замковые плеченогие. Вымирают последние граптолиты. Появление легочных брюхоногих моллюсков. Трилобиты угасают, но имеются многочисленные другие членистоногие: пауки и др. паукообразные, губоногие и двупарноногие многоножки, насекомые — монуры, Palaeo- dyctioptera, тараканы, прямокрылые, равнокрылые. Расцвет бластоидей. Морские лилии. Двоякоды- шащие, кистеперые рыбы, акулы. Стегоцефалы. Первые рептилии — котилозавры.
Мезозой 67 ±3 Пермь 45 230 ± 10 Кроме предыдущих растений — гингковые, беннетиды и саговниковые. Расцвет хвойных. Много- численные двустворчатые моллюски, наутилоидеи и аммониты. Последние трилобиты. Из насе- комых — жуки, сетчатокрылые и клопы: Акулы, кистеперые, двоякодышащие рыбы. Расцвет стегоцефалов. Котилозавры (Seymouria и др.), парейазавры, пеликозавры и териодонты.
Триас 45—40 195 ±5 Господство хвойных. Гингковые. Преобладание шестилучевых кораллов. Появление новых родов пле- ченогих. Упадок наутилоидей. Расцвет аммонитов. Первые белемниты. Первые десятиногие раки. Много новых морских лилий. Древние морские ежи сменяются правильными ежами (Regularia). Акулы, двоякодышащие рыбы. Появление настоящих костистых рыб. Стегоцефалы. Появляются динозавры, гаттерии, черепахи, крокодилы, ихтиозавры, нотозавры, плагиозавры, специализирован- ные териодонты и первые мелкие млекопитающие — Protodonta и Multituberculata.
Юра 58—55 137 ±5 Расцвет хвойных, беннетитов, саговниковых. Первые покрытосеменные растения. Расцвет мадрепо- ровых кораллов. Упадок плеченогих. Из наутилоидей-сохранился только Nautilus. Новые аммо- ниты. Расцвет белемнитов. Среди насекомых — первые двукрылые и перепончатокрылые. Пер- вые неправильные морские ежи (Irregularia). Расцвет костистых рыб. Расцвет рептилий: ихтио- завры, плезиозавры, динозавры, диплодок и мн. др. Появление первоптиц (археорнис). Примитивные млекопитающие.
Мел 70 67 ±3 Расцвет покрытосеменных — однодольных и двудольных. Гингковые клонятся к упадку. Снижается роль хвойных. Многочисленные моллюски: двустворчатые, брюхоногие, белемниты, аммониты; последние в конце мела исчезают. Расцвет костистых рыб. Появляются амфибии, очень близкие к современным. Гигантский птерозавр (Pteranodon). Появление ящериц. Вымирание многих групп рептилий. Зубастые птицы, сумчатые. Неполнозубые и примитивные насекомоядные.
78
ОБЩАЯ ЧАСТЬ
Таблица 2 (продолжение)
Эра Период Характерные группы организмов
Кайнозой 70—67 Палеоге- новый 41 Палео- цен Широкое распространение тропической й субтропической флоры современного типа. Расцвет не- правильных морских ежей. Много двустворчатых и брюхоногих моллюсков. Кальмары и кара- катицы. Исчезновение последних белемнитов. Архаические млекопитающие: однопроходные, сумчатые, насекомоядные, лемуры, тупайи, долгопяты.
Палеоге- новый 41 25 ± 1 Эоцен Флора та же. Вымирание большинства мезозойских рептилий. Птицы современного типа. Развитие плацентарных млекопитающих. Расцвет копытных и хищных. Лемуры. Долгопяты. Приматы — амфипитеки.
Олиго- цен Флора та же. Вымирание древних млекопитающих. Низшие узконосые обезьяны. Приматы: амфипитеки, парапитеки, проплиопитеки.
Неогено- вый 25 Миоцен Флора та же. Расцвет третичных млекопитающих. Креодонты уступают место хищным. Развитие парнокопытных, носорогов, хоботных, лошадиных (мерихиппус, гиппарион). Среди приматов — дриопитеки.
Плиоцен Расцвет луговой растительности. Прогресс однодольных. Фауна беспозвоночных близка к совре- менной. Появление змей, развитие ящериц, крокодилов, черепах. Птицы близки к современным. Эволюция хоботных, носорогов, лошадей. Появление австралопитеков.
Четвертичный 1.7 Флора современная. Вымирание гигантских млекопитающих и развитие современных форм: лоша- дей (Equus), слонов, мамонтов, саблезубых тигров. Современные человекообразные обезьяны. Вымирание/австралопитеков. Появление Homo habilis и Н. erectus. Появление, широкое распро- странение и вымирание неандертальцев (Homo sapiens neanderthalensis). Появление и господство человека разумного (Homo sapiens sapiens).
В систематике и при филогенетических исследо-
ваниях (в частности, палеонтологии) особо важная
роль принадлежит сравнительному методу изучения,
широко используемому и за пределами этих наук, так
же как и вне естествознания.
Для отдельных частей организма или для органов
сравниваемых животных устанавливаются морфоло-
гические ряды, в которых крайние члены обычно
сильно различаются, но связаны друг с другом проме-
жуточными ступенями. Получается более или менее
постепенный переход от одного крайнего члена ряда
к другому, иллюстрирующий, как могла протекать
эволюция сравниваемых органов. Однако ряд может
быть искусственным и может не отражать действи-
тельный путь эволюции, если члены ряда подобраны
случайно. Составленный таким образом морфологи-
ческий ряд — в зависимости от природы исходного
материала — может быть сравнительно-анатомичес-
ким, онтогенетическим или палеонтологическим. Од-
нако ни один из них, взятый в отдельности, не может
претендовать на значение генеалогического ряда (не-
редко даже палеонтологический ряд, если он состав-
лен не из последовательных по времени захоронения
ископаемых остатков), до тех пор пока не будет под-
твержден другими рядами. Иными словами, истинный
филогенетический ряд устанавливается лишь путем
синтеза данных трех морфологических наук — срав-
нительной анатомии, эмбриологии и палеонтологии,
т. е. посредством так называемого метода тройного
параллелизма (Haeckel, 1894), который позволяет ус-
тановить филогенетические связи между современны-
ми и вымершими группами животных.
Бурное развитие биохимии и молекулярной био-
логии существенно расширило арсенал методов уста-
новления филогенетических отношений между раз-
личными таксонами. Например, успешно применяют-
ся способы выявления наиболее близких в биохи-
мическом отношении млекопитающих по степени их
реагирования на чужую сыворотку крови. Сравнение
последовательности аминокислот молекулы белка,
например гемоглобина, у разных групп позвоночных
стало важным методом для коррекции филогенети-
ческих древ. При этом активно используется пред-
ставление о так называемых молекулярных часах, ход
которых основан на случайных мутациях, которые
накапливаются с постоянной скоростью. Так, было
выяснено, что аминокислотные последовательности
двух альфа-цепей глобинов у человека и японской
макаки различаются по 4 сайтам, что составляет 2.8 %
от общего количества сайтов, у человека и собаки
соответственно по 23 сайтам (16.3 %), у человека и
акулы по 74 сайтам (55.2 %). Таблицы различий ами-
нокислотных последовательностей (в процентах) в
альфа-цепях глобина дают следующие величины: че-
ловека и карпа 48.6, тритона 44.0, курицы 24.8, ехид-
ны 26.2, кенгуру 19.1 (Кимура, 1985). Это в целом
соответствует принятым схемам филогенетических
отношений, отражающих время дивергенции таксо-
нов.
Наконец, геносистематика сделала попытку на
базе изучения изменений последовательности нуклео-
тидов установить «окончательную истину молекуляр-
ной эволюции», т. е. выразить различие и сходство
между всеми группами организмов от бактерий до че-
ловека в процентах гомологичных последовательно-
стей нуклеотидов. Были предложены следующие спо-
собы изучения эволюции генома: сравнительные ис-
следования нуклеотидного состава ДНК у различных
групп организмов; исследования степени метилирова-
ния нуклеотидов и наборов изоплит; изучение соот-
ношений фракций с разной степенью повторяемости
и чередования; молекулярная гибридизация ДНК
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
79
Рис. 43. Непрерывные ряды изменений раковины палюдин от Viviparus neymari до V. hoernesi (плиоценовые отложения Славонии).
(По: Неймар, цит. по: Шмальгаузен, 1969).
Fig. 43. Continuous rows of Viviparus shells changes (from V. neymari to V. hoernesi) (Pliocene, Slavonian).
(Медников, 1983). Однако при исследовании процент-
ного содержания гуанин + цитозин в ДНК оказалось,
что нуклеотидный состав ДНК варьирует у разных
групп по-разному. Наиболее лабильными по этому
признаку оказались микроорганизмы, а у высших ор-
ганизмов этот показатель теряет свое значение. Этот
метод эффективен на уровне выяснения отношений
между макротаксонами, например при сравнении
растений с грибами, многоклеточных и одноклеточ-
ных животных ит. д. Малоэффективными оказались
попытки установить филогенетические отношения по
степени метилирования нуклеотидов и наборов изо-
плит, соотношению фракций с разной степенью по-
вторяемости и чередования. Более перспективными
оказались методы молекулярной гибридизации, поз-
воляющие работать со столь близкими в системати-
ческом отношении объектами, как штаммы и биоти-
пы бактерий, экологические расы рыб и даже расы
человека и т. д. •
Несмотря на скепсис некоторых приверженцев
традиционных способов изучения филогенетических
закономерностей, новые методы оказались чрезвы-
чайно плодотворными. Путем измерения различий в
иммунологических реакциях или в молекулярных
структурах были существенно уточнены данные о
расхождении многих филогенетических линий позво-
ночных. В частности, оказалось, что дивергенция
эволюционных линий человека и человекообразных
обезьян произошла не 20 млн лет назад, как это было
общепринято, а значительно позже. Крупные чело-
векообразные обезьяны и гиббоны дивергировали
10 млн лет назад, а общий предок человека, шимпан-
зе и гориллы жил всего лишь 6—8 млн лет назад.
Более того, новейшие открытия Australopithecus ana-
mensis и A. afarensis показали, что становление ос-
новных черт организации Ното приходится как раз
на время дивергенции крупных человекообразных
обезьян и австралопитековых. Оказалось, что само
выделение Homo sapiens в самостоятельное семей-
ство неоправданно с точки зрения геносистематики,
так как по ДНК и белкам человек гораздо ближе к
шимпанзе и горилле, чем последние к орангутану.
Более того, различия между человеком и шимпанзе
на молекулярном уровне не превышают различий,
существующих обычно между видами-двойниками
(Goodman, 1974 et al.).
Если филогенетический ряд подтверждается мате-
риалами и других биологических наук (экологии,
этологии, физиологии, генетики, биохимии), то его
достоверность, как особенно подчеркивал Федотов
(1966), значительно возрастает. К сожалению, более
или менее полными (хотя еще и недостаточными)
данными располагает пока только сравнительная ана-
томия. Эмбриональное развитие многих форм остает-
ся еще неизученным, а палеонтологические остатки
по многим группам животных, как уже сказано выше,
вообще отсутствуют.
Настоящие филогенетические ряды ископаемых
предков удается получить лишь в редких случаях —
когда имеется особенно полный и богатый палеонто-
логический материал из последовательных слоев оса-
дочных пород. Таковы некоторые ряды третичных
моллюсков: Viviparus (рис. 43), Gyraulus, Didacna и
других. С очень высокой степенью достоверности ре-
конструирована генеалогия предков лошадей и сло-
нов. Впрочем, сравнительно хорошо изучена эволю-
ция многих млекопитающих: хищных, китообразных,
приматов и человека.
Остановимся вкратце на эволюции лошадей —
наиболее специализированной ветви непарнокопыт-
ных, которая прослежена почти от начала третичного
периода до четвертичного, т. е. на протяжении не-
скольких десятков миллионов лет (Ковалевский, 1874;
Osborn, 1918; Abel, 1929; Matthew, 1926; Simpson,
1951). Отдаленные предки лошади — Hyopsodus, оби-
тавшие во влажных лесах эоцена, были небольшими,
80
ОБЩАЯ ЧАСТЬ
Рис. 44. Гиракотерий Hyracotherium (эоцен). Реконструкция.
(Рис. В. А. Ватагина из: Мензбир, 1934).
Fig. 44. Hyracotherium (Eocene). Reconstruction.
Рис. 45. Филогения сем. лошади, Equidae. (По: Simpson, 1951).
7 — Hyracotherium (Eohippus), 2 — Orohippus, 3 — Epihippus, 4 — Meso-
hippus, 5 — Mio hippus, 6 — Parahippus, 7 — Meryhippus, 8 — Pliohippus,
9 Equus, 10 — Palaeotherium, 11 — Anchitherium, 12 — Hypqhippus,
13 — Megahippus, 14 — Archaeohippus, 15 — Hipparion, 16 — Neohipparion,
17 — Naiumhippus, 18 — Hippidion, 19 — Callippus, 20 — поедатели травы,
27 — поедатели ветвей.
Fig. 45. Horse phylogeny, Equidae.
20 — graminivorous, 27 — ramivorous.
Рис. 46. Параллельная эволюция передней конечности Condilart-
ra (A), Equidae (Б—Е) и Litbptema (Ж—3).
А — Phenacodus, Б — Eohippus, В — Мiohippus, Г — Parahippus, Д —
Pliohippus, Е — Equus, Ж — Diadiaphorus, 3 — Thoatherium. I—V —
пальцы пятипалой конечности.
Fig. 46. Parallel evolution of the Equidae (Б—E) and Litopterna
(Ж, 3} forelimbs.
I— V — fingers of pentadactylic forelimb.
величиной_с зайца, животными, приспособленными к
беганию по топкой почве лесных болот. Они облада-
ли 4 пальцами на передних и 3 пальцами на задних
ногах. Пища их состояла из мягкой растительности
подлеска, и зубы были бугорчатыми. Начиная от этих
примитивных копытных, к современной лошади
ведет ряд ископаемых промежуточных форм: от
Eohippus {Hyracotherium, — рис. 44) в Америке и Ев-
ропе — через Orohippus — Mesohippus — Meryhip-
pus — Pliohippus к Equus. Но кроме этих основных
рядов было открыто много боковых слепых ветвей,
например линия Miohippus, Anchitherium и другие.
Представители примитивных родов вели лесной образ
жизни, подобно Hyopsodus, но более поздние формы
приспособились к существованию на обширных, от-
крытых пространствах, к бегу по плотной почве и к
питанию жесткой степной растительностью (рис. 45).
В связи с этим у них происходили постепенное уве-
личение размеров тела (олигоценовый Mesohippus,
например, достиг уже 48 см в высоту) и существенные
изменения конечностей и зубов. Боковые пальцы ног
постепенно редуцировались при одновременном уси-
лении среднего, третьего пальца, произошли редук-
ция локтевой кости и прирастание ее к лучевой, ре-
дукция и прирастание малой берцовой и большой
берцовой костей (рис. 46). Коронки коренных зубов
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
81
Рис. 47. Эволюция хоботных Proboscidea. (По: Парамонов, 1945).
1 — Mammirthus primigenius, 2 — Elephas maximus, 3 — Loxodonta africana pumilio, 4 — L. africana, 5 — Parelephas jeffersonii, 6 — Stegodon ganesa, 7 —
Stegomastodon arizonae, 8 — Archidiskodon imperator, 9 — Anancus arvernensis, 10 — Tetralophodon campester, 11 — Cuvieronius humboldtii, 12 — Mammut
americanutn, 13 — M. borsoni, 14 — Trilophodon giganteus, 15 — Dibelodon andium, 16 — Deinotherium gigantissimum, 17 — Serridentinus productus", 18 —
Rhynchotherium tlascalae, 19 — Palaeomastodon berdnelli, 20 — Phiomia osborni, 21 — Moeritherium lyonsi.
Fig. 47. Evolution of proboscideans.
(моляров) увеличились и усложнились. Верхнеэоце-
новый Epihippus имел уже гребешки на коренных
зубах, образовавшиеся слиянием бугров. Затем этот
процесс моляризации распространился и на ложно-
коренные зубы. Основной ряд предков лошади, обна-
руженный в Америке, по-видимому, несколько раз
становился источником появления лошадиных в Ев-
ропе. Так, от американского трехпалого рода Meso-
hippus возник европейский Anchitherium, а трехпа-
лый Meryhippus дал европейского Hipparion. Однако
европейские ветви лошадиных быстро вымирали.
Менее детально известна эволюция хоботных,
прослеженная, начиная с эоценового меритерия —
Moeritherium (рис. 47). Она характеризуется посте-
пенным увеличением размеров тела, прогрессивным
развитием хобота и своеобразной специализацией
зубной системы. Из резцов у настоящих слонов раз-г
вивается только одна верхняя пара, которая образует
огромные бивни. Четырехбугорчатые коренные зубы
резко уменьшаются в числе, превращаясь в шестибу-
6 Протисты, часть 1
горчатые, а позднее — в многобугорчатые у масто-
донтов и в складчатые зубы у слонов (рис. 48). У ма-
монта и современного индийского слона число скла-
док достигает 27—29.
Изучение вымерших животных позволяет во мно-
гих случаях установить ясную родственную связь между
типами, классами и отрядами в виде промежуточных
форм. Особенно демонстративны в этом отношении
ископаемые «связующие звенья» между классами
позвоночных (см.: Быстров, 1942, 1957; Kuhn-Schny-
der, 1963; Шмальгаузен, 19646). Среди них прежде
всего заслуживают упоминания пресноводные девон-
ские кистеперые рыбы из отряда Osteolepiformes,
очень близкие к древнейшим наземным позвоноч-
ным — стегоцефалам и обладавшие уже внутренними
ноздрями — хоанами, легкими и лопастевидными пар-
ными плавниками (Шмальгаузен, 19646). Их потом-
ки — Ichthyostegalia (верхний девон) представляли
следующее соединительное звено, которое, однако, на-
ходилось уже в пределах класса амфибий (см. рис. 16).
82
ОБЩАЯ ЧАСТЬ
II
Рис. 48. Палеонтологический ряд хоботных, представленный че-
репами с очертаниями головы (/) и коренными зубами (II). (По:
Шмальгаузен, 1969).
А — Moeritherium (верхний эоцен), Б — Palaeomastodon (нижний олиго-
цен), В — Trilophodon (миоцен), Г — Mastodon (димовий), Д — Stegodon
(плиоцен), Е — Elephas (современный).
Fig. 48. Paleontological row of the Proboscidea represented with cra-
nium with the head outlines {I) and molar teeth (II).
Это были первые стегоцефалы (рис. 49) с настоящи-
ми пятипалыми конечностями, но сохранившие еще
много чисто рыбьих черт — каналы боковой линии,
пронизывающие кости крыши черепа, отсутствие
овального окошечка, внутреннего уха, кости жабер-
ной крышки (praeoperculum и suboperculum), характер-
ный хвостовой плавник и другие (Save-Soderbergh,
1932).
В нижней перми появились сеймурии — Seymou-
riamorpha — первые и наиболее примитивные репти-
лии (Ефремов, 1946; Быстров, 1957), настолько близ-
кие еще к стегоцефалам, что некоторые палеонтологи
относят их к амфибиям. Действительно, они облада-
ли еще сошниковыми и нёбными зубами, тяжелым
стегальным черепом с теменным отверстием (рис. 50),
следами боковой линии и одним тазовым позвонком;
их личинка развивалась в воде, претерпевая метамор-
фоз, подобно земноводным. Однако сеймурии имели
уже один затылочный мыщелок, подвижное сочлене-
ние в области передних шейных позвонков и более
совершенные пояса конечностей, т. е. признаки,
свойственные пресмыкающимся.
Хорошо известны также переходные формы, свя-
зывающие рептилий и млекопитающих (см. рис. 17). В
триасу в различных эволюционных линиях териодонт-
ных пресмыкающихся — Cynodontia (рис. 51) — стали
параллельно развиваться признаки, сближающие их с
Mammalia; началось преобразование некоторых
костей нижней челюсти в слуховые косточки
среднего уха; прогрессировала дифференциация
зубов (см. с. 51). Мозаика признаков этих форм поз-
воляет различать границы между двумя классами толь-
ко условно, как статистическое состояние (см. с. 92).
В настоящее время сделано уже 6 находок остат-
ков ископаемых первоптиц, или ящерохвостых птиц, —
Archaeornithes в юрских Золенгофенских литограф-
ских сланцах (Франконская Юра в Германии) (см.:
Flugel, 1978). Наиболее полно сохранившиеся экзем-
пляры позволили установить 2 рода — Archaeopteryx
(«лондонский экземпляр») и Archaeornis («берлин-
ский экземпляр», — рис. 52). После открытия «лон-
донского экземпляра» (в 1861 г.) одни авторы отнес-
Рис. 49. Ichthyostega (верхний девон). (По: Шмальгаузен, 19646).
А — скелет, Б — реконструкция внешнего вида.
Fig. 49. Ichthyostega (upper Devonian).
A — skeleton, Б — reconstruction of the exterior habitus.
ПУТИ И ЗАКОНОМЕРНОСТИ эволюции
83
л
Б
Рис. 50. Череп Seymouria (нижняя пермь). (По: Шмальгаузен, 19646).
А — сверху, Б — снизу. 1 — квадратная кость, 2 — чешуйчатая кость, 3 — ушная вырезка, 4 — скуловая кость, 5 — слезно-носовой проток, 6 — межче-
люстная кость, 7 — сошник, 8 — нёбная кость, 9 — птеригоид; 10 — парасфеноид, 11 — окно для челюстной мышцы.
Fig. 50. Cranium of Seymouria (lower Permian).
A — dorsal view, Б — ventral view. 1 — quadrate, 2 — squamosal, 3 — otic notch, 4 — jugal bone, 5 — nasolacrimal duct, 6 — premaxillary, 7 — vomer,
8 — palatine, 9 — pterygoid, 10 — parasphenoid, 11 — subtemporal fossa for adductor.
ли его к рептилиям по длинному хвосту (Wagner,
1861), другие по перьям и крыльям — к настоящим
птицам (Owen, 1863). Последнее мнение было под-
тверждено при изучении других находок (Dames,
1883). Промежуточный характер Archaeomithes лучше
всего выражен Шмальгаузеном (1947, с. 455—456) в
следующих словах: «...сходство со скелетом некото-
рых Coelurosauria так велико, что их, наверное, от-
несли бы к одной группе, если бы не самые настоя-
щие птичьи перья, которые покрывали все тело...
Именно присутствие покрова из перьев, имевших на
крыльях характерное расположение маховых перьев,
и заставляет смотреть на это животное как на птицу.
В строении скелета имеется, конечно, большое сход-
ство с современными птицами, но все же в некото-
рых отношениях он ближе к скелету псевдозухий и
динозавров. Таз птичий... В отличие от птиц грудные
позвонки... не срастались, имелся длинный хвост из
большого числа позвонков (18—20), грудина была
слаба... и не имела киля. Все три пальца передней
конечности имели полное число фаланг (2, 3, 4) и
были снабжены когтями; кости пястья между собой
не срастались. Имелись брюшные ребра (как у псев-
дозухий и динозавров). Из этого сопоставления уже
ясно, что эта птица была, по сути, ближе к рептили-
ям, у нее отсутствовало многое из того, что характер-
но для птицы как летающего организма».
В отношении происхождения полета первоптиц
были высказаны две гипотезы: или планирующий
полет с дерева, на которое животное сначала залезав
ло, или вначале короткий разбег на земле, а потом
затяжной прыжок, переходящий в планирующий
полет с заключительным приземлением на задние ко-
нечности (Ostrom, 1974).
Данный раздел был подготовлен около десяти лет
назад. За прошедший период появилось немало работ
по проблемам эволюции, многие из которых до сих
пор продолжают вызывать дискуссии среди биологов.
Основная трудность эволюционной теории — отыс-
кание причин, позволяющих виду переходить из
одной экологической зоны в другую, поскольку при
этом вид должен выйти на некоторое время из-под
контроля естественного отбора. Часть эволюционис-
тов считает, что в действительности такие переходы
Рис. 51. Реконструкция Cynognathus (верхняя пермь), совмещающего черты рептилий и млекопитающих. (По: Шмальгаузен, 1969).
Fig. 51. Reconstruction of Cynognathus (?Permian) combined features of Reptilia and Mammalia.
84
ОБЩАЯ ЧАСТЬ
Рис. 52. Сравнение скелетов первоптицы Archaeomis siemensi (Archaeopteryx lithographica nom. valide) (А, юра) и голубя (Б). (По: Re-
mane et al., 1976).
Fig. 52. Comparison of the sceletons of Archaeornis siemensi (Archaeopteryx lithographica nom. valide) (Jurassic) (A) and Columba (Б).
возможны в тех случаях, когда обнаруживается
достаточно широкая пластичность онтогенетических
реакций; другие полагают, что такой переход может
быть осуществлен только с помощью преадаптаций;
третьи склоняются к тому, что условия переходной
экологической зоны способствуют вскрытию мобили-
зационного резерва изменчивости и отбор имеет дело
с чрезвычайно широким спектром форм.
Не достигнуто еще согласие в понимании отноше-
ния элементарных факторов эволюции к естественно-
му отбору. Большинство современных биологов ука-
зывают, что их нельзя сводить лишь к мутационному
процессу, популяционным волнам и изоляции. Необ-
ходимо учитывать эволюционное значение поведения
животных, роль фенотипа в эволюции, структуру по-
пуляций, различные способы размножения и т. д. Про-
должаются дискуссии вокруг концепции нейтральной
эволюции Дж. Кинга, М. Кимуры и Т. Ота, согласно
которой большинство нуклеотидных замен в ДНК и
соответствующих им изменений первичной структуры
белков не имеет селективного значения и происходит
в результате случайного закрепления «нейтральных
мутаций». Дискуссионными остаются проблемы раз-
личных форм видообразования (симпатрического,
аллопатрического, парапатрического и т. д.), о меха-
низмах внутривидовой конкуренции, о групповом от-
боре и его роли в эволюции. К нерешенным еще до
конца относятся вопросы, возникающие в связи с не-
полнотой палеонтологической летописи^ природой
крупных филетических хиатусов и происхождени-
ем высших таксонов, возможностями и причинами
внезапных видообразований, сальтационного преоб-
разования органов, огромных ускорений эволюцион-
ного процесса на относительно малых отрезках геоло-
гического времени.
В эволюционной теории только начинается разра-
ботка проблемы особенностей эволюционного про-
цесса в различных таксономических группах (эволю-
ция эволюции), общих тенденций и закономерностей
в эволюции биосферы, эволюции других экосистем,
что также вызывает новые дискуссии.
Однако столь значительное количество нерешен-
ных проблем отнюдь не является свидетельством сла-
бости СТЭ (как пытаются утверждать некоторые ее
оппоненты). В действительности же они подчеркива-
ют ее творческий, динамический характер. Их реше-
ние успешно осуществляется в рамках той научной па-
радигмы, ядром которой по-прежнему остается дарви-
новское учение о естественном отборе как главном
механизме эволюции живого.
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
85
PATHWAYS AND PRINCIPLES OF THE EVOLUTION
SUMMARY
The present evolutionary theory is based upon synthesis of know-
ledge of factors, moving forces and regularities of the evolution. The
synthesis, realised in the second quarter of the 20th century, was star-
ted from the following principles: 1) populations are elementary units
of the evolution; 2) mutations (of gene, chromosome, and genome)
provide the major material for the evolution; 3) elementary evolution
phenomenon involves directional and irreversible change of gene pool
of populations; 4) natural selection acts as the major factor of the
evolution; 5) speciation completes with formation of isolation mecha-
nisms between a new and ancestral species; 6) a species exists as a
complex polytypic system consisting of populations and intrapopulati-
on groups; 7) there is no principle difference between the mechanisms
of micro- and macroevolution. These principles were verified by nu-
merous experimental studies. Evidence of emergence of new viable
forms, isolated from the initial ones, was obtained. Different forms of
natural selection (stabilising, leading, disruptive, frequency-dependent,
catastrophic) and different modes of speciation (allopatric, sympatric,
neoformogenic, abrupt, etc.) were defined and analyzed.
The study of phylogenetic regularities of the evolution has a par-
ticular role in the modern evolutionary theory. The study of these is
conducted by means of correction of traditional research methods
(comparative morphology, embryology, and palaeontology) and by
means of elaboration of new approaches based upon achievements of
genetics, phenogenetics, development mechanics, biochemistry, palaeo-
biochemistry, etc. This allows not only studying phylogenetic regula-
rities, but also discussing causes of macroevolution, which is of adap-
tive character. Biological progress, as was shown by A. N. Severtzov,
may proceed by means of different morphophysiological adaptations
(aromorphosis, idioadaptation, degeneration, cenogenesis), sharply
differing as regards their evolution perspective. Subsequently, mor-
phophysiological and ecological criteria were verified for defining the
major pathways of macroevolution: arogenesis, allogenesis, catagene-
sis, hypogenesis, hypergenesis, etc. Evolution is mostly monophyletic
in the broad sense, which suggests a possibility of origin of a taxon
from a group of related forms. At the same time we cannot exclude a
possibility of polyphyly in the origin of some taxa. Adaptive radiation
is of primary importance, but various cases of convergence and paral-
lelisms are widely spread, and the cases of symbiogenesis and reticu-
late formation of new taxa are also possible.
Evolution has directional character. Its major features are irregular
rate and irreversibility. Examples of bradytelic, horotelic and tachytelic
rates can be found in every large taxonomic group. The theory, pro-
posed by A. N. Severtzov for phyloembryogeneses, offers a new inter-
pretation of relationship between phylogenesis and ontogenesis. It re-
veals diverse ways of rearrangement of ontogenesis (archalaxes, devi-
ations and anabolies), which constitute basis of phylogenetic changes.
It is necessary to take into account, along with phyloembryogeneses,
the changes in ontogenesis, such as palingeneses, cenogeneses, hete-
rochronies and heterotopies. Equally important for understanding the
regularities of macroevolution are ideas of modi of formation of or-
gans, differentiation and integration in the evolution of ontogenesis,
the role of coordinations and correlations in the transformation of
organs and functions (polymerisation and oligomerization, extension
and decrease of the number of functions, activation and immobi-
lisation of functions, substitution of functions and organs, fixation of
phases, etc.). The present knowledge of palaeontological records, data
of comparative embryology and morphology are the major sources for
the reconstruction of phylogenetic relations. But the modern develop-
ment of genosystematics and molecular biology made it possible to
correct the notions of the pathways ahd regularities of the evolution.
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ4
Систематика (от греч. «systema»), или таксономия
(от греч. «taxis» — расположение и «nomos» — за-
кон), — наука о классификации организмов. Впро-
чем, эти термины рассматривались как синонимы
лишь до недавнего времени. Ныне многие биологи
предпочитают ограничивать таксономию как науку о
теоретических основах классификации, а системати-
ку определяют более широко — как науку о разнооб-
разии живого мира. По Симпсону (Simpson, 1961),
систематика есть научное исследование различных
организмов, их разнообразия, а также всех взаимоот-
ношений между ними.
Систематика — наука синтетическая и строит свои
выводы на данных морфологии, эмбриологии, пале-
онтологии, физиологии, биохимии, экологии, этоло-
гии и генетики. Возможности и достижения система-
тики в значительной степени определяются успехами
этих наук.
Первая задача, неизбежно возникающая перед ис-
следователем любой биологической проблемы, за-
ключается в описании и классификации относящих-
ся к ней явлений. В области ботаники и зоологии
этот первый шаг осуществляет систематика, описыва-
4 Этот и следующий раздел («Система животного мира») были
написаны А. В. Ивановым в 1990 г. незадолго до смерти, поэтому
мы сочли необходимым оставить текст автора без изменений. При-
мечания и комментарии редактора даны в сносках к соответствую-
щим местам текста.
ющая и классифицирующая все растения и всех жи-
вотных. Общее представление о масштабах этой ра-
боты дает грубый подсчет растительных и животных
форм, обитающих на Земле. Ныне описано около
500 тыс. видов растений. С 1753 по 1942 г. вместе с
синонимами и подвидами в науку было введено более
1 млн названий для явнобрачных и тайнобрачных
растений (Mayr et al., 1956). Число описанных жи-
вотных значительно больше 1 млн (табл. 3). «Число
именованных животных, — пишет Майр (Mayr et al.,
1956, с. 141), — включая подвиды, составляет более
2 млн, и ежегодно описывается примерно 10 тыс.
новых форм. Для одних лишь насекомых уже исполь-
зовано 1.5 млн названий (Metcalf, 1940). Если при-
нять, что число всех существующих видов насекомых
равно, вероятно, 3 млн (Silvestri, 1929) и что каждый
вид имеет в среднем 5 морфологически различных
фаз развития, то для характеристики всех фаз всех
видов насекомых в конечном итоге потребуется 15 млн
описаний! Когда мы представим себе, что эти 3 млн
видов необходимо распределить в рамках более вы-
соких категорий, выражая при этом их естественные
взаимоотношения и анализируя структуру популяции
каждого данного вида, мы сможем до некоторой сте-
пени понять грандиозность задач, стоящих перед сис-
тематикой лишь одной группы».
Задачи эти настолько сложны, а связанная с ними
литература настолько обширна, что ныне даже очень
86
ОБЩАЯ ЧАСТЬ
квалифицированный систематик не способен це-
ликом охватить какую-либо крупную группу живот-
ных. Неизбежным следствием этого является все воз-
растающая специализация среди зоологов-система-
тиков.
Основным понятием в систематике является вид
(species), представляющий конкретную форму сущес-
твования организмов. Жизнь существует только в
виде индивидов или особей (и их объединений — ко-
лоний и кормусов), принадлежащих к тем или иным
отдельным видам. О содержании понятия вид много
писалось (см., напр.: Kein, 1958; Завадский, 1968;
Мауг, 1968, 1974); здесь мы приведем его краткое оп-
ределение по Гептнеру (1976).
Таблица 3
Число современных описанных видов животных
Название группы Число видов Автор Название группы Число видов Автор
Protozoa 27100 Grell, 1980a, 1980b * Myzostomida 150 Hartmann-Schroder, 1982
* Placozoa 2 Иванов A. B., 1973 Oligochaeta 3 500 ’
Porifera 5 000 Kilian, 1980 Hirudinea 400 Лукин, 1976
2000 Колтун, 1987 * Onychophora 90
Cnidaria -7 700 Tardigrada 400 Moritz, 1982
* Scyphozoa 200 Werner, 1984 Pentastomida 80
* Cubozoa 16 Степаньянц (рукопись) Arthropoda -863 000
Hydrozoa 1800 * Xiphosura 5 Fage, 1949
Anthozoa 4 850 Arachnida 57 000
* Mesozoa 80 Pantopoda 500 Mayr, 1971
* Orthonectida 20 Иванов А. В. (рукопись) Crustacea 20000
* Rhombozoa 60 Chilopoda 2 800
* Ctenophora ~80 Werner, 1984 Diplopoda 7 200 Kaestner, 1959
Plathelminthes -16 800 Pauropoda 362
* Xenoturbellida 1 Westblad, 1949 Symphyla 120 /
Turbellaria 3 400, Insecta 775 000
* Gnathostomulida 80 Odening, 1984 Tentaculata -4 800
* Aspidogastraea 40 Тимофеева* * Phoronidea 10 Emig, 1985
Trematoda 7 200 Odening, 1984 Bryozoa 4 500 Андросова*
Monogenea 2500 Гусев* * Brachiopoda 280
* Gyrocotylida 10 Odening, 1984 * Chaetognatha 50 Kaestner, 1963
* Amphilinida 10 Дубинина, 1982 * Pogonophora 150 Иванов A. B., 1988a
Cestoda 3 500 Гинецинская, Доброволь- * Hemichordata 81
ский, 1978 * Enteropneusta 60
Nemathelminthes -22700 * Pterobranchia 2 Kaestner, 1963
* Gastrotricha 350 * Planctosphaerea 1
Nematoda -20000 Hartwich, 1984 Echinodermata 5 980
Rotifera -2000 Crinoidea 620
* Gordiacea 250 Holothuroidea 1 100
Kinorhyncha 100 Echinoidea 860 Kaestner, 1963
* Priapulida 15 Asteroidea 1500
* Loricifera 10 Иоффе* Ophiuroidea 1900
Acanthocephala 750 Chordata ~43 770
* Kamptozoa 100 Hartwich, 1984 * Acrania 30 Deckert, 1967
Nemertini 850 Odening, 1984 Tunicata 2040
Mollusca 114499 Ascidiacea 1900
* Caudofoveata 83 * Pyrosomida 10
* Solenogastres 176 Иванов Д. Л., 1986 * Salpae 25 Gunther, 1967
Polyplacophora 850 Сиренко* * Doliolida 15
* Monoplacophora 15 * Appendiculariae 100
Gastropoda 110000 Vertebrata 41700 Mayr, 1971
* Scaphopoda 350 Kilian, 1982 I * Cyclostomata 46 Sterba, 1967
Bivalvia 20000 Pisces -20600
Cephalopoda 750 Amphibia -2500 Наумов, Карташев, 1979
* Echiurida 140 Gruner, 1982 Reptilia -8 600 Mayr et al., 1956
* Sipunculida 250 Мурина, 1977 Aves 8 590
Annelida 17 050 [ Mammalia 3700—4 000
Polychaeta 13 000
Примечание. Плюсом («+») отмечены личные сообщения специалистов; звездочкой (*) — группы, в состав которых входит небольшое число видов
(от 1 до 400).
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
87
Вид — это генетически ограниченная группа по-
пуляций, особи которых характеризуются только им
присущими, общими признаками и способны свобод-
но скрещиваться и производить нормальное плодови-
тое потомство. По так называемой биологической
концепции вида (Тимофеев-Ресовский, 1965; Мауг,
1982), основной особенностью вида считается нали-
чие репродуктивной изоляции особей, обеспечиваю-
щее «защиту генофонда». Поэтому возникают особые
затруднения при установлении вида у бесполых,
партеногенетических или самооплодотворяющихся
организмов. У этих форм (например, у облигатно
агамных протестов) видом приходится считать сово-
купность клонов и линий, сходных морфологически,
физиологически и экологически (см., напр.: Полян-
ский Ю. И., 1986, 1992). Каждый вид занимает свой
ареал — определенное географическое пространст-
во. Каждый вид отличается от других видов того же
рода рядом морфологических и физиологических
особенностей; различия по этим признакам создают
разрывы (хиатусы) между родственными видами.
Межвидовые скрещивания в природе относительно
редки и не нарушают самостоятельности видов. Каж-
дый вид, таким образом, вполне реален. Границы
между двумя видами неразличимы лишь в том случае,
если один из них является дочерним, находящимся
еще в процессе отделения от материнского вида.
Встречаются, правда, практически едва различимые
«виды-двойники», обладающие такими отличиями в
кариотипе или в половом поведении, которые исклю-
чают появление плодовитого гибридного потомства.
Эти виды находятся в начальной стадии обособления
и являются генетически изолированными при отсут-
ствии каких-либо других различий.
Новый вид обычно происходит в результате дли-
тельной эволюции от другого вида путем его превра-
щения или распадения — дивергенции — на два или
несколько самостоятельных видов. Новые виды могут
возникать также путем гибридизации или полиплои-
дизации, особенно у растений. Сложившийся вид
обычно подвергается действию стабилизирующего
отбора и обладает относительной неизменностью, во
всяком случае в течение времени, равного продолжи-
тельности человеческой истории.
Будучи определенным качественным этапом эво-
люционного процесса, вид может рассматриваться в
известном смысле как основная единица живой при-
роды (Remane, 1956; Гептнер, 1976). Вместе с тем
вид большей частью политипичен, т. е. распадается
на внутривидовые систематические категории, среди
которых основным является подвид, или геогра-
фическая раса (Семенов-Тян-Шанский, 1910).
Характерно, что особи одного подвида обладают
признаками, постепенно переходящими в признаки
другого подвида, и хиатус между подвидами отсутст-
вует. В пограничных зонах их ареалов, где особи
подвидов встречаются, обычна гибридизация между
ними. Образование подвидов, в сущности, есть нача-
ло дивергенции, а сам подвид, чаще всего, — заро-
ждающийся вид.
Систематика — самая древняя из биологических
дисциплин. Первая классификация животного мира
была создана еще в IV столетии до н. э. великим
Таблица 4
Приблизительное количество видов в группах высших рангов
Группа Число видов
Protozoa 27 100
Metazoa 1 104271
Phagocytellozoa 2
Enantiozoa 5 000
Eumetazoa 1099269
Mesozoa 80
Radialia 7780
Bilateria 1091 360
Scolecida 41 200
Coelomata 1 050 160
Trochozoa 995 509
Tentaculata 4 800
Chaetognatha 50
Pogonophora 150
Deuterostomia 49750
Animalia -1131000
Аристотелем. Начало систематики как науки, одна-
ко, датируется серединой XVIII в., когда вышел зна-
менитый труд Карла Линнея — «Systema naturae»
(первое издание в 1735 г.). Линнеевское и послелин-
неевское время было первым периодом расцвета этой
науки.
Во второй половине XIX столетия, после выхо-
да в свет «Происхождения видов» Дарвина (Darwin,
1859), в зоологии возникло множество новых про-
блем, оказавшихся чрезвычайно важными. Это об-
стоятельство отодвинуло систематику на задний
план, хотя сам Дарвин широко использовал выводы
систематики в своих обобщениях и был всемирно
известен как систематик усоногих ракообразных. В
конце XIX и в начале XX в. систематика претер-
пела период особенных сомнений и колебаний и
даже некоторого упадка. Ясного представления о
том, что такое вид, еще не существовало. Нередко
делались попытки дискредитировать систематику
как науку; указывалось, в частности^ что задачи
систематики сводятся лишь к регистрации фактов,
что систематика должна только описывать и назы-
вать животных, чтобы уметь их различать. Утверж-
далось даже, что систематика, в сущности, не наука;
ее сравнивали иногда с простым коллекциониро-
ванием, например с филателией. Скепсис к систе-
матике отчасти был вызван явным разочарованием
в многочисленных филогенетических гипотезах и
спекуляциях, которые расцвели пышным цветом к
концу XIX и в начале XX столетия, но не были
аргументированы ни экологически, ни физиологи-
чески и потому по большей части носили крайне
умозрительный и даже фантастический характер.
Особенно скептически были настроены физиологи,
биохимики и «механики развития», видевшие в
систематике, как и в других «описательных нау-
ках» — морфологии, экологии и зоогеографии, «от-
жившую» область знания в противоположность «точ-
ным», экспериментальным наукам. Эти утвержде-
ния, охотно повторяющиеся иногда даже в наши дни,
основаны на наивном представлении, что наука оп-
88
ОБЩАЯЧАСТЬ
ределяется приемами исследования, а не теми зада-
чами, которые она перед собой ставит.
«К счастью, — пишет В.Г. Гептнер (1956), —
систематики оказались дальновиднее своих хулите-
лей и продолжали упорно работать, настойчиво соби-
рая фактический материал и разрабатывая теорети-
ческие основы своей науки, и прежде всего — про-
блему вида». К началу XX столетия в хаосе мнений и
предположений начали обозначаться основные кон-
туры современной систематики. В первой четверти
XX в. определилось уже ее основное понятие — вид,
а сама систематика превратилась в хорошо разрабо-
танную биологическую дисциплину. Было достигнуто
и ясное разграничение разных проявлений внутриви-
довой изменчивости, правильно понята географичес-
кая изменчивость и оценена роль географического
фактора в процессе видообразования (Семенов-Тян-
Шанский, 1910). Возникла и укрепилась концепция
«широкого», или политипического, вида как системы
подчиненных подвидовых единиц — разновидностей.
При этом пришлось отказаться от чисто морфологи-
ческого толкования сущности вида. Именно в этот
период сложилась новая, так называемая биологичес-
кая трактовка вида, представляющая собой одно из
важных обобщений современной биологии. Достиже-
ния современной систематики и новые проблемы,
вставшие перед ней, тесно связали ее с дарвинизмом,
экологией и популяционной генетикой и превратили
из вспомогательной отрасли знания, какой она пред-
ставлялась раньше, в важнейшую общебиологичес-
кую науку.
В наше время изучение проблем эволюции требу-
ет широкого привлечения систематического материа-
ла, а анализ проблем видообразования основывается
на данных систематики; Центральный ее вопрос —
определение природы вида — представляет собой в
то же время основную биологическую проблему.
Вместе с тем по-прежнему одной из главных
задач систематики остается изучение и описание
различных представителей и групп животного мира
и целых фаун. Другая ее задача, как и раньше, —
создание классификации, т. е. построение иерархи-
ческой системы соподчиненных таксонов. Класси-
фикация должна быть естественной, т. е. должна
строиться на генеалогически важных признаках и,
следовательно, отражать родственные отношения
животных групп. За последнее время заметно воз-
росло и практическое значение систематики. Мно-
гие группы животных, еще недавно считавшиеся
безразличными для человека или по крайней мере
малозначительными, теперь оказались важными в
практическом отношении. Число таких групп не-
прерывно растет. Здравоохранение, защита живот-
ных и растений от паразитов и вредителей, борьба
с паразитами — переносчиками болезней человека
и животных, карантинная служба, рыбное, охот-
ничье, промысловое хозяйства, сельское и лесное
хозяйства, рыборазведение, разведение сельскохо-
зяйственных и пушных животных и многие другие
отрасли народного хозяйства нуждаются в деталь-
ном изучении фауны в целом и в разработке сис-
тематики самых различных групп организмов. Ряд
неотложных практических задач встает перед со-
временной систематикой и в связи с новыми про-
блемами защиты и охраны среды и животного на-
селения на территории бывшего СССР и за его
пределами.
Мы очень далеки еще от точной оценки объема и
состава мировой фауны. Почти полвека тому назад
Гептнер (1956) с полным основанием мог утверждать,
что более или менее точное число существующих
видов установлено только для класса птиц и что в
отношении остальных групп, в том числе млекопита-
ющих, можно говорить лишь о весьма приблизитель-
ных цифрах. С тех пор мало что изменилось. Были
открыты некоторые новые группы животных, но не-
многие виды, к ним принадлежащие, не изменили за-
метно наши представления о суммарном числе видов
(табл. 3, 4). Все еще имеются большие группы бес-
позвоночных, в которых описано менее половины
действительно существующих видов. Таковы, по-ви-
димому, Monogenea, Nematoda, Acari, а также Pogo-
nophora и другие. Особенно слабо изученным счита-
ется видовой состав свободноживущих круглых чер-
вей (Nematoda). Одни авторы насчитывают около
14 000 видов (Цалолихин, 1980), тогда как другие
(см., напр.: Парамонов, 1962), опираясь на непрекра-
щающийся поток новоописаний, увеличивают это
число в 50 раз (до 700 000). Характерно, что систе-
матика подобных групп до сих пор остается недоста-
точно разработанной и требует серьезной ревизии с
позиций учения о политипическом виде. Как показы-
вает практика, в результате такой ревизии всегда
имеет место значительное сокращение числа видов.
Так, в 20-х гг. среди млекопитающих насчитывали до
6000 видов, а к 1953 г. — уже только 3200—3400. В
начале нашего столетия некоторые орнитологи раз-
личали около 28 000 видов птиц, но после разработ-
ки видовой структуры этого класса число видов со-
кратилось до 8590 (Mayr et al., 1956). Что касается
насекомых, то, по одной, крайней точке зрения, из-
вестно лишь 10—15 % существующих видов — ут-
верждение явно абсурдное. Так, Ирвин и Скотт
(Erwin, Scott, 1980) считают даже, что общее число
видов насекомых составляет около 1млн. По-видимо-
му, гораздо ближе к истине предположение Крыжа-
новского (1958) о том, что в большинстве стран уже
описано около 50 % видов. Таким образом, вероятно,
на Земле обитает 2—2.5 млн видов насекомых. Это
самый обширный класс животного мира; следующим
по объему, очевидно, должен считаться класс нема-
тод (табл. 3).
С другой стороны, на всех «этажах» главной сис-
темы мы встречаемся с совсем небольшими группами
рангом не ниже подкласса, которые пережили когда-
то пору своего расцвета и ныне представлены всего
1—20 видами (Placozoa, Planctosphaerea, Pogonopho-
ra, Afrenulata, Monoplacophora, Priapulida, Xiphosura,
Phoronida и др.) или несколькими десятками видов
(Mesozoa, Ctenophora, Onychophora, Enteropneusta и
др.), а все вместе — не более чем .400 видами (в
табл. 3 эти группы отмечены звездочкой). Характер-
ные особенности этих филогенетических реликтов были
проанализированы Родендорфом (1959). В наше вре-
мя количество таких групп возросло до 46 за счет
добавления типов Placozoa и Pogonophora (с 2 груп-
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
89
пами Frenulata и Afrenulata), классов Xenoturbellida,
Gnathostomulida, Caudofoveata, Solenogastres, Mono-
placophora, Myzostomida, Planctosphaerea и подклас-
сов ракообразных — Cephalocarida, Mystacocarida,
Ascothoracida и других.
По справедливому мнению Родендорфа, общей
чертой всех реликтовых групп является их крайняя
систематико-филогенетическая обособленность и вы-
сокий систематический ранг. Все они, кроме того,
отличаются малым количеством подчиненных таксо-
нов — классов в типах, отрядов в классах, семейств
в отрядах и т. д. В ряде случаев филогенетические
реликты отличаются прогрессивными чертами, позво-
лившими им дожить до наших дней. Таковы, например,
приспособления к живорождению у онихофор, скорпи-
онов и акул или скрытный образ жизни — в грунте
(Gnathostomulida, Gastrotricha, Priapulida, Cephalocari-
da, Mystacocarida), в норках, пустых раковинах и труб-
ках (Echiurida, Sipunculida, Enteropneusta), в собствен-
ных защитных домиках и трубках (Phoronida, Ptero-
branchia, Pogonophora).
Все реликтовые группы очень примитивны, ис-
ключительная древность этих форм очевидна в тех
случаях, когда имеются твердые скелетные части,
сохраняющиеся в ископаемом состоянии (Monopla-
cophora, Xiphosura, Brachiopoda, Pogonophora). Так,
современных плеченогих насчитывается всего 280
видов, а ископаемых (начиная с нижнего кембрия)
описано около 10 000.
Две группы — примитивные погонофоры и совер-
шенные головоногие моллюски выделяются как будто
бы несоответствием между высотой организации и
количеством видов.
Действительно, Pogonophora представляют одну
из самых древних групп беспозвоночных, но до сих
пор сохраняют довольно значительное систематичес-
кое разнообразие (в их составе имеется 2 подкласса,
5 отрядов, И семейств и более 25 родов). Остатки
трубок погонофор известны из отложений верхнего
протерозоя (Соколов, 1965). Однако вопреки такой
древности, непрекращающиеся описания новых видов
погонофор заставляют предполагать, что 150 видов,
описанных до сих пор, составляют не более 10 % су-
ществующих форм. Причина этого разнообразия и
изобилия видов, возможно, состоит в том, что пого-
нофоры, во-первых, защищены хитиновой трубкой,
отчасти погруженной в ил, и, во-вторых, не имеют
серьезных пищевых конкурентов, так как питаются
преимущественно за счет органических веществ, син-
тезируемых их симбионтами — хемотрофными сер-
ными бактериями.
Головоногие проделали длительную прогрессив-
ную эволюцию, начавшуюся еще в кембрии с очень
примитивных Nautiloidea, и дали в течение фанерозоя
громадное разнообразие форм и не менее 10 000 видов.
Современные же головоногие, несмотря на совер-
шенную организацию, тем не менее сравнительно ма-
лочисленны (750 видов). Относительное количество
известных современных и вымерших животных пока-
зано на рис. 53.
Рассмотрим некоторые особенности сравнитель-
ной морфологии, связанные с теми общими задачами,
которые стоят перед систематикой и сравнительной
морфологией, а именно с построением естественной
системы животного мира.
Широко распространено ложное мнение, что сис-
тематика представляет собой давно пройденную сту-
пень науки; некоторые склонны считать систематику
и морфологию бесполезными для дальнейшего разви-
тия биологии; указывают даже, что имеет смысл за-
ниматься только экспериментальными науками —
физиологией, биохимией, молекулярной биологией,
генетикой и т. д. Это мнение обычно основано на
убеждении, что путь к познанию сложного состоит
только в расчленении его на элементарные части и
процессы, объяснимые с точки зрения физики и
химии, и что сложное познается по свойствам его
частей (см.: Энгельгардт, 1970).
Несостоятельность этого принципа исследования,
обычно называемого редукционизмом, в отношении
биологических явлений отмечалась неоднократно
(см., напр.: Любищев, 1977). Не выдерживает крити-
ки и наивная классификация наук по технике и при-
емам исследования. Значительная роль перечислен-
ных выше «экспериментальных» биологических наук
в развитии научного, технического и экономического
прогресса несомненна, но без систематики и, следо-
вательно, без сравнительной морфологии все они, в
сущности, бессильны, однако это, к сожалению, редко
осознается.
Интересные суждения по этому вопросу мы нахо-
дим у Ремане (Remane, 1956), который совершенно
справедливо подчеркнул, что все наши наблюдения и
эксперименты мы производим на отдельных особях,
поскольку жизнь существует только в виде отдельных
живых существ. Следовательно, индивиды, или особи,
являются нашими первичными объектами исследова-
ния. Наблюдение и эксперимент тем менее надежны,
чем меньше особей было изучено. Если отвергнуть
систематику с ее выводами, то, строго говоря, полу-
ченные при изучении индивидов результаты прило-
жимы только к ним одним, и всякое познавательное
научное значение этих результатов прекращается
сразу со смертью изученных индивидов. Действитель-
но, если, например, физиолог открыл какое-то явле-
ние в нерве лягушки, то из его открытия самого по
себе еще никак не вытекает, что это явление свойст-
венно также и другим лягушкам, и тем более всем
амфибиям. Если и можно сделать такой вывод, то
лишь после того, как будет определена видовая при-
надлежность исследованной лягушки, изучены срав-
нительно-анатомически и систематически другие ам-
фибии и показана степень их родства. Другими сло-
вами, наш физиолог может обобщать результаты своего
труда, только опираясь на громадную работу, уже
проделанную систематиками и сравнительными ана-
томами. Между тем мало кто из биологов-экспери-
ментаторов останавливается перед широкими обоб-
щениями результатов своего исследования; обычно
они, не задумываясь, считают их действительными
для многих миллионов особей, большого числа про-
шедших, настоящих и будущих поколений, всего
вида, даже семейства, отряда и т. д.
Биохимик, публикующий, например, работу «О
пищеварительных ферментах виноградной улитки —
Helix pomatia», пользуясь видовым названием — Helix
90
ОБЩАЯ ЧАСТЬ
Protozoa
Рис. 53. Относительное количество известных современных и вымерших видов животных. (По: Schnyder, 1967, с небольшими измене-
ниями).
10 равен 3500 видам. Зачерченная часть сектора пропорциональна числу ископаемых видов.
Fig. 53. Relative number of the known present and fossil animal species.
1 is equal to 3500 species; the shaded part of the sector is proportional to the fossil species.
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
91
pomatia, распространяет результаты своей работы на
все особи, которые систематик отнес бы к этому
виду. Между тем если бы не было систематики, загла-
вие его статьи должно было бы выглядеть примерно
так: «О пищеварительных ферментах 10 животных
со спиральной раковиной, собранных 2 августа
1969 г. в саду в окрестностях Киева». Автор должен
был бы подчеркнуть еще, что «ныне эти животные
уже не существуют», так как с их смертью значение
исследования сильно падает.
Дело в том, что понятие «вид» (species), вырабо-
танное систематикой, играет роль до некоторой сте-
пени такого же средства обобщения, какое в физике
и химии имеет понятие «элемент», или «чистое со-
единение» (Remane, 1956).
Однако в систематике существуют и другие при-
емы обобщения. Обратимся к примеру, приводимому
Ремане. Во всех учебниках по генетике утверждается,
что у бабочек (отряд Lepidoptera) женские особи ге-
терогаметны, т. е. откладывают яйца двух сортов — с
Х-хромосомой и Y-хромосомой. Было исследовано
около 50 видов, каждый в единичных экземплярах.
Отряд бабочек содержит более 100 000 видов, следо-
вательно, соотношение исследованного материала с
тем, на который распространено это обобщение,
равно 1 :2000. Имеют ли право генетики делать такое
обобщение?
Практически, однако, к подобному приему при-
бегают очень часто, и он оправдывает себя в тех
случаях, когда выполняются условия, эмпирически
установленные систематиками. Оказывается, дей-
ствительно нет нужды изучать всех представителей
данной систематической группы, можно ограни-
читься небольшим их числом, если наблюдаемое
явление или свойство присуще наиболее удаленным
друг от друга ее членам. Следовательно, выбор объ-
ектов для любого биологического исследования дол-
жен быть согласован с данными естественной сис-
темы.5
Систематика определяет также границы, до кото-
рых может быть распространено обобщение. Одну из
обычных ошибок биологов, недостаточно знакомых с
эволюционной системой, представляет необоснован-
ное перенесение свойств низших организмов на выс-
шие или неродственные им формы. Нельзя, напри-
мер, приписывать некоторые особенности организа-
ции гидры всем кишечнополостным или низших
позвоночных — млекопитающим и человеку.
Таким образом, естественная система играет фун-
даментальную роль в вопросах общей биологии.
Только естественная система и понятие о виде дают
возможность биологам всех направлений уверенно
ориентироваться в бесчисленном множестве явлений
в мире организмов и делать из их сопоставления вы-
воды общего характера. Без их помощи стремление
биологов понять общие закономерности было бы
5 Я. И. Старобогатов считает, что главная задача системати-
ки — построение системы с прогностическими свойствами. В
этом смысле естественная система — это система с максимально
возможной прогностической силой. Более подробно об этих про-
блемах, а также о необходимости наряду с естественной и искус-
ственных систем см.: Старобогатов, 1989, — примеч. редактора.
столь же безнадежно, как исследование процессов на
берегу моря путем изучения каждой отдельной пес-
чинки, которая исследовалась бы структурно, хими-
чески, физически, геологически, минералогически и
т. д. Великий немецкий поэт и натуралист-мыслитель
Гёте, прекрасно понимавший обобщающую силу сис-
тематики и сравнительной морфологии, писал по
этому поводу: «Можно выпить все море, пока будут
изучены все индивидуальные явления — наблюдены,
изменены, взвешены и описаны».
Именно эта обобщающая сила систематики и срав-
нительной морфологии дает замечательные возмож-
ности предсказывать еще неизученные структуры,
процессы и явления. Предположим, говорит Ремане,
что в наши руки попал экземпляр неизвестного еще
науке животного, например жука-плавунца из рода
Dityscus. Даже один сухой экземпляр достаточен,
чтобы сделать длинный ряд сбывающихся предсказа-
ний о новом виде, предсказаний, относящихся к
строению (анатомии, гистологии) всех его систем ор-
ганов, к множеству деталей его эмбрионального раз-
вития, к его физиологии, строению, экологии и физио-
логии его личинки и куколки и т. д., даже к природе
его паразитов. Дело в том, что о строении, развитии
и жизни всех стадий Dityscus, в частности D. margi-
natus, написаны уже толстые книги и почти все, со-
держащееся в них, приложимо и к нашему виду.
Подобного рода предсказания и их последующее
фактическое подтверждение — самое обычное, по-
вседневное явление в систематике и сравнительной
морфологии; оно воспринимается как нечто само
собой разумеющееся и никого не удивляет. Между
тем если вдуматься, — это поразительное дости-
жение в биологии, поскольку точность возможных
предсказаний в систематике и сравнительной морфо-
логии сопоставима только с той, которая достигнута
в генетике. Никакие другие биологические науки не
могут делать подобных предсказаний. Это замеча-
тельное свойство естественной системы обусловлено
прежде всего тем, что она отражает эволюционное
родство.
Несмотря на большие успехи, современная систе-
матика все еще представляет собой «недостроенное
здание». По ряду принципиальных проблем в ней
уживаются одновременно самые противоречивые мне-
ния. Мы остановимся здесь лишь на некоторых общих
вопросах, главным образом на отношениях филоге-
нии и системы, определении понятия признак и оп-
ределении уровней развития систематики.
Теория происхождения видов Ч. Дарвина, пред-
ставляющая синтез всех биологических знаний сере-
дины прошлого века, объяснила многие, непонятные
до того особенности живых существ. Прежде всего,
она установила кровное родство всего органического
мира, основываясь на глубоком сходстве строения и
отправлений самых разнообразных организмов. Тео-
рия Дарвина объяснила и характерную черту мира
живых существ — прерывистость его многообразия,
т. е. расчленение его на обособленные систематичес-
кие группы. Нашли свое объяснение также и причи-
ны давно известной иерархической субординации,
т. е. соподчиненности систематических подразделе-
ний. Дарвин убедительно доказал, что естественные
92
ОБЩАЯ ЧАСТЬ
группы организмов развивались путем расхождения
признаков (дивергенции) из одного общего ствола
вследствие вымирания менее приспособленных про-
межуточных форм. Отдельные группы животных воз-
никали монофилетически от ограниченной совокуп-
ности близкородственных форм (в современном по-
нимании — популяций). Все это очень хорошо
объясняло, почему естественная система организмов,
отражающая реально существующие в природе отно-
шения, обладает иерархическим строем и системати-
чески может быть представлена в виде ветвящегося
древа. Однако это важнейшее положение дарвинизма
решительно отвергается его противниками, для взгля-
дов которых характерен полный отрыв филогении от
системы, что, впрочем, не препятствует им упорно
именовать систему, предлагаемую ими самими, ес-
тественной. При этом предполагается, что система
должна создаваться на основании «объективных»
сходств и различий между животными независимо от
каких бы то ни было гипотез об их происхождении.
Так, например, Смирнов (1969) утверждает, что
^...систематические группировки, установленные при
классификации по сходству, вовсе не должны иметь
значение филогенетических, так как сходство еще не
указывает на кровное родство». Однако особенно
последовательно отстаивал идею «независимости»
системы от филогении Любищев (1923). Свою статью
он заключает следующими словами: «Мы вправе счи-
тать эволюцию доказанной... но в вопросах систе-
матики мы не можем пользоваться ее языком... Пост-
роение системы на философии Дарвина оказалось
иллюзией: надо строить систему, отрешившись от
эволюционного подхода». Любищев, таким образом,
не скрывает своего отрицательного отношения к дар-
винизму.
Невозможно согласиться с этим антиэволюцион-
ным подходом к построению системы, так же как
нельзя принять и иные системы, предлагаемые други-
ми противниками дарвинизма, например сетчатую
схему эволюции Лотси или схему Л. С. Берга в виде
«пшеничного поля».
В действительности филогения и система тесно
связаны и первая определяет построение второй, как
правильно подчеркивал это в специальной статье
Зенкевич (1939). Это не означает, разумеется, что
систематика и филогенетика — одно и то же. Тесная
связь между ними гораздо сложнее, чем это может
показаться на первый взгляд. Основные задачи фило-
генетики и систематики различны. В идеале филоге-
нетика стремится установить непрерывную линию
эволюции организмов — от предков к потомкам, в
которой по самой природе эволюционного процесса
нельзя установить четких границ между материнским
и дочерним таксонами. Это хорошо показывает при-
веденный выше (с. 82) пример Seymouriamorpha —
промежуточной группы, которую трудно отнести как
к амфибиям, так и к рептилиям. Именно такие слу-
чаи и подтверждают правильность филогенетических
выводов. Систематика, напротив, направляет свои
усилия на определение границ между таксонами, и
это ей удается лучше всего именно в тех случаях,
когда филогенетическая линия оказывается с разры-
вами, ограничивающими таксоны (см.: Скарлато,
Старобогатов, 1974). Филогенез есть процесс, проте-
кающий во времени, система — отражение статичес-
кого состояния (Hennig, 1950).
Из сказанного вытекает, что границы таксонов,
устанавливаемые систематикой, до известной степени
условны, что, однако, нисколько не умаляет ни их
естественности, ни их познавательного и практичес-
кого значения.
Дарвин писал, что «всякая истинная классифика-
ция есть генеалогическая». Поэтому в естественной
системе особенное значение должны иметь признаки,
которые указывают на кровное родство. Биологи, го-
ворящие об «объективности» и независимости систе-
мы от филогении, естественно придают всем призна-
кам одинаково важное значение независимо от их от-
ношения к филогенезу. Признание равноценности
всех признаков естественно вытекает из пренебреже-
ния к различиям между гомологиями, аналогиями,
конвергенциями и параллелизмами. Особенно рекла-
мировали принцип равноценности всех признаков
сторонники фенетической, или так называемой нуме-
рической, систематики (Sokal, Sneath, 1963, и др.).
Систематические группы — таксоны предлагается
«создавать» не по генетическим признакам, а по
числу общих признаков. Для подсчета совпадающих
признаков применяется современная вычислительная
техника, при этом считается, что чем больше призна-
ков учтено, тем лучше, — характер этих признаков
остается в тени.
Нумерическое направление возникло как реакция
на распространившиеся в систематике субъективность
и произвол и проявилось в использовании матема-
тических и статистических приемов для исследования
биологических объектов. К сожалению, теоретичес-
кий уровень фенетической систематики оказался не-
достаточным для создания филогенетической систе-
мы, хотя она и претендовала на ведущую роль. «Все
нумерические системы, — пишет К. А. Юдин (1974), —
содержат в себе некий неучитываемый и неустрани-
мый математическим методом элемент искусственно-
сти». Вина падает не на математический метод, а на
лежащие в основе нумерического направления устаре-
лые принципы диагностической систематики.
На количественных показателях и признаках, ото-
рванных от филогении, основан и метод математи-
ческого таксономического анализа Смирнова (1969).
Значение признака оценивается им не путем установ-
ления гомологии, а по степени его распространения
среди сравниваемых видов данной группы — чем у
большего числа видов он выражен, тем значительнее
его систематический вес. Можно, однако, возразить,
что сходство проявляется не только благодаря насле-
дованию какого-либо признака от общих предков, но
и в результате параллельной эволюции. Метод Смир-
нова, таким образом, позволяет лишь уточнять сход-
ства, но ничего не говорит об их природе.
Имеются и другие современные направления (на-
пример, кариологические и биохимические), осно-
ванные, подобно фенетической систематике, на при-
менении какого-либо одного метода. Их кажущаяся
связь с современной СТЭ и с генетикой, в сущности,
формальна. Системы, которые ими создаются, пост-
роены по одиночным признакам или их комплексам
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
93
и потому лишены истинного филогенетического со-
держания. Несмотря на это, они представляют значи-
тельное подспорье в работе систематика.
Выводы эволюционной морфологии и системати-
ки получают большую убедительность, если подтвер-
ждаются данными физиологии и биохимии. Уверен-
ность в правильности разделения билатеральных жи-
вотных — Bilateria — на Protostomia и Deuterostomia
возросла, когда стало известно, что главным источни-
ком энергии мышечного сокращения у первых явля-
ется артининфосфорная, а у вторых — креатинфос-
форная кислота. Такого рода данные очень полезны
для систематики, но не могут быть решающими.
Между тем среди физиологов и биохимиков встреча-
ется мнение, что биохимические методы, и в частно-
сти, серологические и иммунологические, могут пол-
ностью раскрыть родственные связи между группами
животных и тем самым сделать работу сравнитель-
ного анатома и систематика излишней. Американ-
ский систематик Сокэл — лидер фенетической сис-
тематики — дошел до крайности, утверждая, что в
будущем «музейная морфология» отомрет совсем и
ее заменит одна химия. Такое мнение явно абсурдно.
Однако поскольку нам известны случаи, когда биохи-
мия действительно подтверждает выводы системати-
ки, то очень важно выяснить, всегда ли мы можем на
нее положиться и при каких обстоятельствах. Обсуж-
дению этого вопроса посвящена очень интересная
статья Ремане (Remane, 1967), к которой мы и обра-
тимся.
Молиш (Molisch, 1933) рассмотрел распростране-
ние химических веществ в растительном мире и по-
казал, что многие из них, органические и неоргани-
ческие, характерны для определенных семейств рас-
тений. Таким образом, существует корреляция между
химизмом и естественной системой. Однако это
справедливо лишь для одной части веществ. Другие
вещества рассеяны столь беспорядочно в разных се-
мействах, что приходится признать их независимое
многократное появление у разных растительных ор-
ганизмов. То же справедливо и для многих веществ,
встречающихся у животных. Так, хитин и гликоген
свойственны различным неродственным группам жи-
вотных и, более того, встречаются и у грибов. Дыха-
тельными пигментами крови у близкородственных
животных могут быть различные вещества. Кроме
позвоночных гемоглобин встречается независимо в
самых различных группах — у личинок насекомых
(Chironomus и др.), некоторых моллюсков (Planorbis
и др.), аннелид (олигохеты, пиявки, Capitellidae, Аге-
nicola и др.), погонофор. У полихет помимо гемогло-
бина в крови встречается и другое железосодержащее
вещество — зеленый хлорокруорин. Интересно, что
в пределах одного рода Spirorbis в крови разных
видов мы находим то гемоглобин (5. corrugatus), то
хлорокруорин (S. borealis), то вообще бесцветную
кровь (5. milliaris), а у Serpula в крови одной особи
содержится как гемоглобин, так и хлорокруорин. У
неродственных друг другу членистоногих и моллюс-
ков имеется гемоцианин, содержащий медь и облада-
ющий в окисленном состоянии синеватым цветом,
впрочем, у них же нередок и гемоглобин. Среди ра-
кообразных жаброногие, ракушковые, усоногие обла-
дают гемоглобином, а десятиногие и ротоногие — ге-
моцианином. Возникает вопрос: при таком мозаич-
ном распространении дыхательных пигментов в жи-
вотном царстве можно ли говорить о каком-либо их
филогенетическом значении?
Большую роль в подтверждении филогенетическо-
го родства играет серодиагностика. В ряде случаев
этот метод показал высокую корреляцию между хи-
мическим и морфологическим родством. Так, филоге-
нетическое древо приматов, полученное на основе се-
рологических данных, совпадает в общем с построе-
ниями систематики. Корреляция в этом случае очень
высока, но не абсолютна.
Изучение эстераз печени многих млекопитающих
показало, что виды одного рода обладают близкими
ферментами. Однако обнаружились и несоответст-
вия. Южноамериканская обезьяна Lagottrix имеет эс-
теразы, идентичные с человеческими, тогда как между
человеком, орангутаном и гиббоном в этом отноше-
нии есть существенные различия. Таким образом, за-
ключает Ремане, изучение эстераз обнаружило как
далеко идущие совпадения с естественной системой,
так и большие различия.
Пэнтин (Pantin, 1966) выделяет три типа морфо-
логического сходства между организмами: наряду с
гомологией и аналогией — еще и сходство в ультра-
структурах, из которых построены клеточные орга-
неллы, которые обнаруживаются только с помощьк)
электронного микроскопа и представляют прояв-
ление «химической идентичности». Это сходство
различных, неродственных организмов выражается в
совершенно одинаковом расположении в их идентич-
ных структурах элементов (или их комплексов) в виде
мембран, полосок исчерченности фибрилл, трубочек
или в виде решеток, весьма напоминающих простран-
ственное размещение молекул в кристаллах (рис. 54,
вкл.). Такого рода структурное сходство целиком за-
висит от чисто физических свойств и не обусловлено
действием отбора. В его основе лежит абсолютная
идентичность молекул химических веществ, свойства
которых остаются одними и теми же во всех организ-
мах.6 Отсюда вывод — присутствие одинаковых ве-
ществ в разных организмах не обязательно свиде-
тельствует об их родстве.
Однако как только мы обращаемся не к отдель-
ным молекулам, а к целому классу веществ с очень
большими и сложными молекулами, а именно к бел-
кам или нуклеиновым кислотам, мы встречаемся с
бесконечными вариациями внутри него, разнообразие
которых не уступает разнообразию видов среди орга-
низмов. Очевидно, молекула ДНК каждого вида дол-
жна быть специфична для него, а различия между
ДНК близких видов должны быть меньше, чем у
форм неродственных (Белозерский, 1964). На этом
постулате основаны интересные и многообещающие
попытки современных биохимиков определить сте-
6 Этот вывод, разумеется, не относится к более сложным
структурам, наблюдаемым с помощью электронного микроскопа
органеллам и состоящим из них внутриклеточным аппаратам. Их
особенности в ряде случаев имеют значение очень существенных
систематических признаков и важны в филогенетическом отноше-
нии.
94
ОБЩАЯ ЧАСТЬ
пень родства различных групп животных по свойст-
вам их нуклеиновых кислот. Такая возможность
стала реальной благодаря методу молекулярной гиб-
ридизации ДНК, разработанному Хойером, Байтовом
и Маккарти (Hoyer et al., 1964) и основанному на
том, что в известных условиях денатурированная
ДНК одного вида может образовывать гибридные
комплексы с таковой другого вида. При этом процент
гибридизации пропорционален проценту гомологич-
ных нуклеотидных последовательностей по которо-
му, по-видимому, можно судить о степени генетичес-
кой близости рассматриваемых видов. Таким спосо-
бом удалось установить степень родства некоторых
вирусов, бактерий, растений и животных (Медников
и др., 1973). Когда гибридизация ДНК горбуши (On-
corhynchus gorbuscha) с ДНК другой особи этого же
вида была принята за 100 %, оказалось, что для пары
горбуша—кета (Oncorhynchus keta) число «сходных
генов» составило 98 %, между тем как для пары гор-
буша—форель (Salmo trutta facia) — только 79.2 %.
Различия между сигами и лососевыми, выраженные в
числе гомологичных нуклеотидных последователь-
ностей или в числе гибридных комплексов, указали
на родство межсемейственного ранга, тогда как зоо-
логи обычно считают эти группы подсемействами.
Хрящевых ганоидов — Chondrostei, по данным этого
метода, следует рассматривать как самостоятельный
класс — доля гомологий в ДНК осетра и акулы ока-
залась приблизительно того же уровня, что и у кури-
цы и черепахи, а также у человека и лосося. Оказы-
вается, кроме того, что степень вариабельности со-
става ДНК у разных представителей данного таксона
может служить одним из критериев его эволюцион-
ного возраста (Антонов, 1968).
В последнее время делаются попытки определе-
ния филогенетического родства путем сравнения пер-
вичной структуры ДНК отдельных генов и рРНК.
Возможно, что при дальнейшей разработке этого ме-
тода могут быть получены достоверные результаты
благодаря практически неограниченному набору нук-
леотидных позиций в гомологичных молекулах.
Таким образом, в тех случаях, когда исследуется
ДНК, биохимия способна дать важные указания на
филогенетическое родство. Во многих случаях эти
данные самостоятельного значения не имеют, и, оче-
видно, каждый раз необходимо выяснять, имеет ли
изучаемый биохимический признак филогенетичес-
кое значение, или он лишен его, будучи лишь проявле-
нием независимо возникшей химической аналогии.
Ничего исключительного в этом нет. Морфологичес-
кие признаки тоже требуют дифференциальной оцен-
ки — одни представляют собой гомологии, свидетель-
ствующие о родстве, другие развились независимо.
Следует всегда помнить, что гомологичными могут
быть только те функции или биохимические процес-
сы, которые протекают в гомологичных органах.
В истории систематики ясно различаются по край-
ней мере три периода, которым приблизительно соот-
ветствуют разные уровни в современной систематике
(Mayr et al., 1956). Глубина систематической разра-
ботки любой группы животных в большей мере зави-
сит от степени ее изученности. Наряду с хорошо ис-
следованными группами, такими как классы и отряды
позвоночных, некоторые отряды насекомых и мол-
люсков, до сих пор еще имеется значительное число
недостаточно изученных групп. Для многих из них нет
пока удовлетворительных сведений ни по физиологии,
ни по экологии, нередко отсутствуют данные об эм-
бриональном развитии. Поэтому и систематическое
изучение разных групп находится на весьма различ-
ных уровнях и современная систематика представляет
смесь различных уровней и состояний.
Юдин (1974) различает три главных уровня совре-
менной систематики, соответствующих в общем трем
историческим периодам развития этой науки.
1. Диагностический уровень (a-систематика по
Майру) проявляет себя, когда группа недостаточно
изучена или когда для чисто практических целей
приходится сознательно упрощать систему, выбирая
удобные для определения диагностические признаки.
Этот уровень таксономической обработки представ-
ляет прямое наследие додарвиновской систематики,
для которой характерно использование легко наблю-
даемых морфологических признаков. Диагностичес-
кие системы иногда как будто не противоречат прин-
ципу выживания наиболее приспособленных особей
в ходе эволюции, но признать их филогенетическими
нельзя.
2. Классический филогенетический уровень до-
стигается в тех случаях, когда группа изучена на-
столько, что есть возможность опираться на дарви-
новскую теорию естественного отбора и применять
метод тройного параллелизма, т. е. использовать дан-
ные сравнительной анатомии, эмбриологии и палео-
зоологии. В историческом аспекте он способствовал
появлению систематики последарвиновского перио-
да, начиная с филогенетических древ и классификации
Геккеля (h-систематика по Майру). Фактическим ма-
териалом для построения системы служат преобразо-
вания органов в морфологических рядах. Экологичес-
кие и физиологические данные почти не учитываются.
Поэтому классические системы все же остаются не-
обоснованными в аспекте учения о естественном от-
боре. Увы, до сих пор систематическое изучение гро-
мадного большинства животных групп находится на
этом уровне.
3. Морфобиологический уровень (е-систематика
по Майру) представляет собой закономерный резуль-
тат развития эволюционной и сравнительной морфо-
логии, и возник он на почве исследований В. А. Ко-
валевского, Луи Долло, А. Н. Северцова, И. И. Шмаль-
гаузена. Этот уровень характеризуется объединением
классической филогенетики с учением об эволюции
адаптаций (Ковалевский, Долло), а следовательно, с
физиологией и экологией и с морфобиологической
теорией инициации Северцова (отсюда предложен-
ное К. А. Юдиным название уровня). Системы, по-
строенные на морфобиологическом принципе, тесно
связаны с современным дарвинизмом. Их задача —
представить филогенез как приспособительный про-
цесс, направляемый естественным отбором (Шмаль-
гаузен, 1964а, 19646, 1969). Для этого синтезируются
данные сравнительной морфологии (анатомии, гисто-
логии, цитологии), функциональной морфологии, а
также эмбриологии, палеозоологии, экологии и это-
логии. Синтез должен обнаружить причинные связи
СИСТЕМАТИКА И ЕЕ ЗАДАЧИ
95
между образом жизни, строением и функцией орга-
нов и их систем и историческим становлением изуча-
емых групп животных.
Исследования в этом духе, касающиеся позвоноч-
ных, несомненно ушли вперед, но и в изучении
беспозвоночных наметились существенные сдвиги и
возможности, поскольку появились новые принци-
пиально важные труды об эволюции отдельных сис-
тематических групп, основанные на обширдых срав-
нительно-физиологических и экологических данных.
Таковы многие систематические работы, появившие-
ся в отечественных и зарубежных исследованиях, ка-
сающиеся самых различных животных. Перечислять
их здесь нет смысла. Большие успехи сравнительной
эмбриологии беспозвоночных, а также электронно-
микроскопические исследования значительно им
способствовали. Глубокий морфофизиологический
и экологический анализ проблемы выхода на сушу
позвоночных (Шмальгаузен, 1964а, 19646) и чле-
нистоногих (Гиляров, 1970а), исследование путей
эволюции типов животного мира (Ливанов, 1955;
Беклемишев, 1964; Федотов, 1966, и др.) с позиций
морфобиологической теории Северцова также весьма
способствовали развитию современной морфобио-
логической систематики, в которой наиболее полно
сливаются система и филогения, представляя собой
залог ее будущих успехов.
Итак, в наше время вся система животных перест-
раивается по генеалогическому принципу, хотя во
многих своих частях она остается еще искусствен-
ной. Многие группы животных все еще недостаточно
изучены для создания эволюционной классификации.
Однако современная система стремится к тому, чтобы
во всех частях стать естественной. Если обнаружива-
ется, что генетически разные группы объединяются в
системе в одну группу, то это значит, что классифи-
кация в этом месте в дальнейшем будет исправлена.
В самом историческом развитии систематических
построений существует своя неизбежная последова-
тельность. Лишь самый первый, приближенный эскиз
классификации создается эмпирически, т. е. незави-
симо от филогенетических соображений, и только в
этом случае можно говорить о «независимости систе-
мы и филогении». Однако эта независимость, вынуж-
денная и временная, существует только до появления
эволюционных выводов, опирающихся на изучение
гомологий.
Теоретические соображения, на которые всегда
опиралась практическая деятельность систематиков
(т. е. основы таксономии), значительно менялись с
течением времени. По-видимому, можно назвать
только пять основных концепций классификации (Мауг,
1971), а именно: эссенциализм, номинализм, фене-
тизм, кладизм и эволюционную классификацию. Пер-
вые три теории относятся преимущественно к додар-
виновскому периоду биологии, две последние — к
последарвиновскому.
Эссенциализм' (сюда прежде всего относятся
постулаты системы Аристотеля) пытается свести раз-
нообразие организмов к некоторым основным типам,
стоящим на разных уровнях. Предполагается, что все
члены таксона отражают одну и ту же естественную
сущность (essentiale nature) или, иначе говоря, соот-
ветствуют одному и тому же типовому плану строе-
ния. Нередко поэтому эссенциализм именуется типо-
логией. Изменчивость считается чем-то незначитель-
ным, а постоянство таксонов, а также разрывы между
ними явно преувеличиваются. Главный порок эссен-
циализма состоит в отсутствии каких-либо критериев
существенных (типовых) свойств организма, устанав-
ливаемых поэтому совершенно произвольно. Эссен-
циализм господствовал в течение многих веков, был
принят Линнеем и до сих пор защищается некото-
рыми таксономистами (Borgmeier, 1957; Thompson,
1962, и др.).
Номинализм постулирует, что в действитель-
ности существуют только особи. Все другие группи-
ровки — роды, семейства, отряды и т. д. — являются
мнимыми понятиями, созданными умом человека.
Основная ошибка этой концепции заключается в
ложном толковании причинной связи между сходст-
вом и родством. Игнорируется тот факт, что каждая
группа организмов, родственных по происхождению,
обладает единством наследственной ДНК, определя-
ющей общность их признаков. Так, птицы вовсе не
есть искусственный продукт, созданный человечес-
ким разумом, но представляют собой реальную, су-
ществующую в природе группу родственных живот-
ных. Члены этого таксона принадлежат к нему вовсе
не в силу своего сходства друг с другом, как это при-
нимается номиналистами, а поскольку обладают
общей наследственностью (Simpson, 1961).
Представители фенетической, или нумеро-
логической, систематики (Sokal, Sneath, 1963)
приписывают всем признакам одинаковую ценность,
не учитывая ни мозаичную эволюцию, ни гомоло-
гии, ни узкие адаптации, ни параллельное и кон-
вергентное развитие. Иногда систематикам-фенети-
кам удается создать классификацию, сходную с
системой, основанной на эволюционном подходе.
Но это объясняется только тем, что два организма
всегда тем более сходны, чем ближе они по про-
исхождению. Критику фенетической систематики
можно найти у Симпсона (Simpson, 1961), Майра
(Мауг, 1971) и Юдина (1974).
Представители этого направления считают, что
вообще нет необходимости в теоретическом толкова-
нии систематики. Предполагается, что при «разумной
оценке достаточного числа признаков» естественная
система возникает автоматически. Действительно,
систематик-практик часто действует по этому эмпи-
рическому принципу, не размышляя, по крайней
мере при создании первого приближенного эскиза
классификации. Однако он не может не осознавать,
что полученная таким путем классификация остается
бессмысленной без эволюционного толкования, пред-
ложенного Дарвином.
В концепции кладизма ранг таксонов и клас-
сификации целиком определяется «давностью про-
исхождения от общего предка». Ранг таксона якобы
зависит только от положения точек ветвления на
филогенетическом древе. Кладисты не учитывают,
что истинное родство определяется двумя различ-
ными явлениями — генетической близостью и ге-
неалогическим родством. Эти два понятия совпада-
ют по своему значению только у очень близких
96
ОБЩАЯ ЧАСТЬ
tri punctata
car dint
calloptera
canalinea
replete dreyfusi
funebris
annulirnana
rubrifro ns
testacea
' quinoria
MICODROSOPHILA
robusta
melanica
virilis
Phloridosa
pinicola
im-migrans
/2APRI0NUS
Dorsilopha
MICRODROSOPHILA
LEUCOPHEN !}A
DEITOPSOMYIA
IDIOM VIA и некоторые
другие габайские роды
/ у гадайские
Drosophila
'sCAPTOMYZA
bryani
cancellata \
victoria—2^ v
Pholodoris
wtllistoni
saltans t melanogaster
\! / .obscura
chymomyza4<
Sophophora
Рис. 55. Филогения высших позвоночных. (По: Мауг, 1971).
1 — точка расхождения крокодилов и других рецентных рептилий; 2 —
точка расхождения крокодилов и птиц, которая приходится на гораздо
более позднее время.
Fig. 55. Phylogeny of the higher vertebrates.
1 — the divergence point of crocodiles and other recent Reptilia; 2 — the
divergence point of crocodiles and birds which falls on the much more late time.
родственников. Но в филогенезе, который разыгры-
вается в потоке тысяч и миллионов генераций и
сопровождается бесчисленными мутациями, реком-
бинациями и отбором, невозможно выразить истинное
родство только генеалогически. Основным критери-
ем в этих случаях следует считать не расположение
на генеалогическом древе, а степень генетического
сходства. Если эволюционная линия подвергается
сильному давлению отбора и вследствие этого резко
дивергирует от генеалогически ближайших родичей,
то «сестринские линии» могут стать настолько раз-
личными, что считать их близкими будет бессмыс-
ленно. Так, крокодилы, с точки зрения кладизма,
ближе всего к птицам (ибо и те и другие возникли
от псевдозухий, — рис. 55). Однако по «кровному
родству», т. е. по генному составу, крокодилы тем
не менее ближе к другим рептилиям, нежели к
птицам (Мауг, 1971).
Определяя ранг таксонов исключительно по месту
ветвления, кладисты полностью забывают о различи-
ях в темпах эволюции. Дистанция генеалогическая
при этом ошибочно отождествляется с дистанцией
генетической. Наиболее последовательный защитник
кладизма — Хенниг (Hennig, 1966, с. 236) писал, что
таксоны в их иерархии «подчинены друг другу в со-
ответствии с временной дистанцией между момента-
ми их возникновения и настоящим временем... сопод-
чинение групп устанавливается по их относительно-
му возрасту...». Следуя Хеннигу, таксоны, которые
возникли посредством ответвления в докембрийское
время, следует возводить в ранг типа, образовавшиеся
DROSOPHILA
Рис. 56. Филогения сем. плодовых мушек, Drosophilidae. (По:
Мауг, 1971).
Многие роды представляют собой специализированные концевые точки
некоторых групп видов Drosophilidae.
Род — прописные буквы; подрод — первая буква прописная, осталь-
ные — строчные; вид — все буквы строчные.
Fig. 56. Fam. Drosophilidae phylogeny.
Many genera represent specialized terminal points of some Drosophilidae spe-
cies groups. Genus — the capital letters; subgenus:—the capital letter for the
first one, small leters for the others; species — the all small letters.
Рис. 57. Филогения Tetrapoda. (По: Мауг, 1971).
Новые классы возникли путем ответвления дочерних линий, в то время как
родительская линия продолжается без существенных изменений. 1 — Cros-
sopterygii; 2 — Amphibia; 3 — Reptilia; 4 — Mammalia; 5 — Aves (2 —
дочерняя линия 1; 3 — дочерняя линия 2; 4 и 5 — дочерние линии 3).
Fig. 57. Phylogeny of tetrapods.
New classes developed by ramification of the daughter lines, while parental line
extends without significant changes. 1 — Crossopterygii, 2 — Amphibia, 3 —
Reptilia, 4 — Mammalia, 5 — Aves (2 — daughter line 1; 3 — daughter line 2;
4 and 5 — daughter lines 3).
СИСТЕМА ЖИВОТНОГО МИРА
97
между кембрием и девоном — в ранг класса, между
началом карбона и пермью, — в ранг отряда и т. д.
«В таком случае млекопитающих следовало бы назы-
вать отрядом... Marsupialia и Placentalia низводились
бы до семейства, а отряды плацентарных стали бы
трибами». Абсурдность этих заключений очевидна
(см.: Мауг, 1971).
Необоснованность постулатов кладизма хорошо была
продемонстрирована на примере детально изучен-
ной эволюции сем. плодовых мушек, Drosophilidae
(рис. 56). Большая часть их представителей произош-
ла путем дивергенции от единой филогенетической
линии, медленно изменявшейся во времени. Линии, ди-
вергирующие из одной точки ствола, расходятся и из-
меняются в разной степени, вплоть до образования
особых родов. Виды рода Drosophila представляют
собой сложную мозаику признаков своих близких ро-
дичей.
Шаров (1965) справедливо заметил, что, по пред-
ставлениям кладистов, родительский таксон исчезает,
дав начало путем расщепления двум дочерним таксо-
нам (сестринским группам по Хеннигу). В действи-
тельности же это бывает редко. Напротив, обычно
родительский таксон также продолжает существовать
еще очень долго, иногда более чем 100 млн лет
(рис. 57). Новая группа в этом случае становится как
бы «сестринской группой» своего родительского так-
сона.
Эволюционная классификация, подобно
эмпирическим схемам, опирается на факт существова-
ния в природе реальных групп, таких как птицы, насе-
комые, летучие мыши, жуки и т. п. Но эволюционная
систематика отличается от эмпирических схем тем,
что объясняет природу этих групп и использует эти
объяснения для усовершенствования классификации.
Такое объяснение эволюционной систематики было
дано Дарвином (Darwin, 1859) в «Происхождении ви-
дов...»: «Я полагаю, что (в нашей классификации) за-
ключено нечто большее, чем простое сходство, и что
близость происхождения — единственная известная
причина сходства живых существ».
Таким образом, можно сказать, что эволюционная
классификация — закономерный продукт системати-
ки современного морфобиологического уровня.
SYSTEMATICS AND ITS PURPOSES
SUMMARY
Systematics (Greek: «systema»), or taxonomy (Greek: «taxis» —
arrangement, «nomos» — law) is a science dealing with classification
of organisms. Some years ago these terms were supposed to be syno-
nims. Now major biologists suppose taxonomy to be a science dealing
with the theoretical base of classification.
Systematics involves scientific investigations of different orga-
nisms, their diversity and all the kinds of interrelations between them.
The general concept of systematics is based upon a species as a
concrete form of existence of living bodies. According to V. Geptner
«species is a group of populations, limited genetically, which individu-
als might be characterized with the peculiarities inherent in them
only». If the «biological conception of species» is being concerned
(Timofeev-Ressovsky, 1965; Mayr, 1982) the general species’ peculiari-
ty is believed to lie in the reproductive isolation of the individuals, that
ensures «the genofund safety».
Systematics is the eldest among the biological sciences. The first
classification of fauna was developed by Great Aristotel (IV cent.
before n. e). The starting point of systematics as a serious science
dates back to mid-XVII century, when the «Systema nature» by Carl
Linnaeus (1735) was published.
One of the general tasks of systematics appears to be describing
different representatives and groups of fauna. The other task is the
elaboration of classification, i. e. the hierarchic system of subordinate
taxa. The classification must be natural that means it should be based
on genealogically important characters.
The natural system plays an important role in Fundamental Bio-
logy. «The Theory of the Species’ Origin» by Ch. Darvin explains
many different questions within systematics and fundamental biolo-
gy. Discussion dealing into different anti-Darvin’s opinions, such
as the theories of independence of system from phylogeny elaborated
by E. Smirnov and by A. Ljubischev; the theory of L. Bergh; the nu-
merical systems; cladistical conception, etc. is included into this chap-
ter.
СИСТЕМА ЖИВОТНОГО МИРА7
К истории большой системы
животного мира
В отечественной зоологической литературе есть
подробные очерки истории классификации живот-
ных Зенкевича (1937) и Плавилыцикова (1941), кото-
рые отражают все основные концепции, существовав-
шие до конца тридцатых годов нашего столетия. По-
этому здесь можно ограничиться главным образом
рассмотрением систем, которые появились в последу-
ющее время.
7 См. сноску 4 (с. 85).
Современная зоология все еще не может похва-
литься существованием общепринятой «большой
системы», т. е. классификации таксонов, стоящих по
рангу выше класса. Если в вопросе о числе, объеме и
составе классов достигнуто относительное единство
взглядов, то в отношении более высоких таксонов —
типов, надтипов, разделов — мнения все еще резко
расходятся.
Тем не менее в первой четверти XX в. намети-
лись два основных направления в разработке зоо-
логической системы, расходящихся, во-первых, в
оценке систематического значения таких эмбриоло-
гических признаков, как способы образования де-
финитивного рта и вторичной полости тела, и, во-
7 Протисты, часть 1
98
ОБЩАЯ ЧАСТЬ
вторых, в различном толковании природы тех групп
Bilateria, которые лишены целома.
Сторонники старой школы, продолжавшей тради-
ции Каруса, Гексли, Рея Ланкестера, Геккеля, Ланга,
Бючли, Гертвига, Деляжа и Эруара, полагали, что
одинаковые способы развития органов могли сло-
житься независимо в разных группах Bilateria и что,
следовательно, эти признаки лишены большого сис-
тематического значения. Такое допущение позволяло
сближать до некоторой степени группы со схизоцель-
ным и энтероцельным целомом, или с первичным или
вторичным ртом, и не препятствовало последователь-
ному соблюдению монофилетического принципа при
построении системы.
В частности, принималось как нечто бесспорное,
что более просто организованные черви, а именно
Plathelminthes, Nemathelminthes и Nemertini, часто
объединяемые под названием Scolecida, представляют
собой наиболее примитивные группы Bilateria. Счи-
талось, что от них произошли все Coelomata, т. е.
принимался постулат о монофилетической природе
всех Bilateria. К последним детищам этой школы от-
носятся классификации, принятые в многотомных за-
рубежных руководствах — Handbuch der Zoologie
(издатели W. Kiikenthal и Th, Krumbach) и Traite de
Zoologie (издатель P.-P. Grasse).
К ним тесно примыкают также системы Кэно и
Бриана. Однако последние резко отличаются призна-
нием дифилетического происхождения Bilateria и от-
рицанием примитивности Scolecida (или Acoelomata),
которые трактуются как деградировавшие Coelomata.
В результате все Bilateria считаются целомическими
животными. В основе этих представлений лежит ста-
рая идея Седжвика (Sedgwick, 1884) о дифилетизме
Coelomata, произошедших от кишечнополостных
якобы двумя независимыми стволами (см. с. 100).
Названия этих стволов — Hyponeura и Epineu-
га — принадлежат Ламеру (Lameere, 1915). Hyponeu-
ra отличаются расположением центральной нервной
системы вдоль брюшной стороны тела и соответству-
ют всем билатеральным беспозвоночным (Ipvertebrata
по Ламарку). Epineura имеют спинную нервную сис-
тему и объединяют только хордовых (Prochordata и
Vertebrata). Hyponeura и Epineura вовсе не соответст-
вуют Protostomia и Deuterostomia, как может пока-
заться на первый взгляд; в состав Hyponeura входят
такие типичные вторичноротые, как иглокожие и
полухордовые. Таким образом, системы Кэно и Бри-
ана лишь внешне похожи на классификации Гатчек-
Гроббеновского типа.
Классификации второго типа берут свое начало
от старой системы Гатчека (Hatschek, 1888), развитой
в начале XX столетия Гроббеном (Grobben, 1908) и
самим Гатчеком (Hatschek, 1911). Характерная их
черта — резкое разделение всех Bilateria на две боль-
шие ветви — Protostomia и Deuterostomia по способу
образования рта взрослого животного и по типу фор-
мирования целома (рис. 58). У первичноротых рот
развивается за счет бластопора, а у вторичноро-
тых — на переднем конце тела независимо от блас-
топора. Кроме того, у первичноротых целом форми-
руется телобластическим способом — пара мезоблас-
тов сначала дает мезодермальные полоски, в которых
А
ZYQONEURA Ambulacralia
сЪ°гй°п™'
METERAXONIA
(~ Bilateria)
PROTAXONIA
(-Coelenterata)
Б
PROTOSTOMIA DEUTEROSTOMIA
COELOMATA
(-Bilateria)
COELENTERATA
(-Diploblastica)
Рис. 58. Взаимоотношения основных групп Eumetazoa.
А — по: Hatschek, 1888; Б — по: Grobben, 1908.
Fig. 58. Relations of the main Eumetazoa groups.
затем схизоцельно появляются целомические полос-
ти. У Вторичноротых целом возникает из выпячива-
ний первичной кишки, т. е. энтероцельно. Позднее
были найдены и другие, как казалось, принципиаль-
ные различия между Proto- и Deuterostomia, в том
числе в первичном типе дробления яйца, плане стро-
ения нервной системы, биохимии мышечного сокра-
щения, и разделение всех Bilateria на две ветви стало
едва ли не общепринятым. Однако, как будет видно
далее, оно привело к весьма двусмысленной ситуа-
ции.
Объединение всех высших животных, обладаю-
щих по крайней мере в зародышевом состоянии вто-
ричной полостью тела — целомом, в одну обширную
группу Coelomata было осуществлено Геккелем. В
знаменитой работе о теории Гастреи он писал: «Если
усматривать главное значение в целоме и связанной с
ним сосудистой, или кровеносной, системе, то основ-
ная группа многоклеточных распадается на две раз-
личные группы: с одной стороны, — на низших
животных с зародышевыми листками, но лишенных
целома и гемолимфы: зоофиты и животные, не име-
ющие целома (Acoelomata, плоские черви); с дру-
гой — на высших многоклеточных, имеющих целом
и гемолимфу (Coelomata), и происшедших из них че-
тыре высших типа животных: иглокожие, членисто-
ногие, моллюски и позвоночные» (цит. по рус. изда-
нию: Геккель, Мюллер, 1940, с. 221; см. также: Haec-
kel, 1872, с. 465, 468).
Это совершенно правильное толкование понятия
Coelomata быстро вошло в зоологию. Однако основ-
ные постулаты появившейся вскоре цикломерной ги-
потезы Седжвика (Sedgwick, 1884) заставили многих
зоологов отойти от первоначальных представлений о
систематическом составе Coelomata. Седжвик сделал
первую попытку решить проблему происхождения
Coelomata с помощью энтероцельной теории, кото-
рая к тому времени казалась уже хорошо обоснован-
ной (Мечников, 18746; Hertwig, Hertwig, 1882). Сег-
ментацию аннелид и хордовых Седжвик выводил не-
зависимо из радиальной симметрии (цикломерии)
кишечнополостных. Такое решение оставляло, одна-
ко, происхождение Scolecida без объяснения, и по-
следователи Седжвика были вынуждены признать
этих, в действительности примитивных червей за вто-
рично упрощенных Coelomata.
Уже в конце XIX в. идеи Седжвика получили ши-
рокое распространение: появились варианты цикло-
мерной гипотезы (Beneden, 1891; Masterman, 1898), а
СИСТЕМА ЖИВОТНОГО МИРА
99
HYPONEURA EPINEURA
Acoelomata
COELOMATA
ctenophora:
Chidaricc
Spotigia —
Рис. 59. Филогенетические отношения между основными группа-
ми животных. (Составлено по: Cuenot, 1952).
Fig. 59. Phylogenetic relations between the main animal groups.
в большом английском руководстве «Treatise on Zoo-
logy» (Lankester, 1900) Eumetazoa были разделены
только на две большие группы: Enterocoela ^Coelen-
terata) и Coelomacoela (=Coelomata). Таким образом,
понятия Bilateria и Coelomata полностью совпали.
В XX в., несмотря на полную очевидность прими-
тивности паренхиматозных и первичнополостных форм,
концепция Седжвика упорно держалась в системах и
филогенетических схемах различных школ (не только
в системе Claus—Grobben—Kuhn, 1932 и Remane et
al., 1976, но также Cuenot, 1952, — рис. 59; Brien,
1961, 1963, 1972). В частности, ее принял Гроббен,
обосновавший деление Bilateria на две ветви — Proto-
stomia и Deuterostomia.
Системы Старой школы
Рассмотрим вначале классификацию Кюкенталя
(Kiikenthal, 1923), которая была положена в основу
многотомного немецкого руководства «Handbuch der
Zoologie», основанного этим автором в 20-х гг. и вы-
ходящего до сих пор.
Система Кюкенталя—Крумбаха (Kiikenthal, 1923)
Подцарство Protozoa
Тип Protozoa
Подцарство Metazoa
Надраздел Parazoa
Тип Porifera
Надраздел Eumetazoa
Раздел Radiata
Тип Coelenterata
Тип Ctenophora
Тип Mesozoa, или Planuloidea
Раздел Bilateria
Тип Vermes
Подтип Amera
Ветвь Platodes, включая Nemertini
Ветвь Nemathelminthes (классы: Rotatoria, Gastro-
tricha, Kinorhyncha, Nematodes, Nematomorpha,
Acanthocephala)
Ветвь Kamptozoa
Подтип Polymera-Annelida
Подтип Oligomera
. Ветвь Tentaculata (Phoronida, Bryozoa)
Ветвь Brachiopoda
Ветвь Chaetognatha
Ветвь Branchiotremata (Pterobranchia, Enteropneusta)
Тип Echinodermata
Тип Mollusca
Тип Arthropoda
Тип Chordata
Эта система почти полностью совпадает со старой
классификацией Бючди, принятой им в «Лекциях по
сравнительной анатомии» (Biitschli, 1910). Однако
4 типа Protozoa, принятые Бючли, низведены до ранга
класса. Сборный тип Vermes, фигурировавший во всех
старых системах, расчленен на 3 подтипа — Amera,
Oligomera и Polymera. В особенности искусственным
представляется подтип Oligomera, в котором объеди-
нены столь разнородные животные, как Tentaculata,
Chaetognatha и Branchiotremata (=Hemichordata). На-
конец, занижен ранг таких групп, как Platodes, Ne-
mertini, Nemathelminthes, Chaetognatha, Tentaculata,
Branchiotremata, ибо большинство зоологов уже давно
считают их по крайней мере типами.
Принципы этой школы еще явно чувствуются и в
другом большом современном руководстве по зооло-
гии, публикуемом во Франции с 1948 г., но еще не за-
конченном. Речь вдет о «Traite de Zoologie», издавае-
мом Грассе (Р.-Р. Grasse). В вышедших уже 40 книгах
мы находим такую последовательность материала.
Система П- Грассе (Р.-Р. Grass^, 1948)
Тип Protpzoa
Подтип Rhizoflagellata
Надкласс Flagellata (Phytomonadina, Xanthomonadina, Dino-
flagellata, Ebriadina, Silicoflagellata, Chrysomonadina, Zoo-
flagelli4a)
Тип Spongiaria
Тип Plathelminthes (классы: Turbellaria, Temnocephala, Monogenea,
Cestodaria, Cestoda, Trematoda)
Тип Mesozoa (классы: Orthonectida, Dycyemida)
Тип Acanthocephala
Тип Nemertini
Тип Nemathelminthes (классы: Nematoda, Rotifera, Gastrotricha, Ki-
norhyncha)
Тип Annelida (классы: Polychaeta, Oligochaeta, Hirudinea, Myzosto-
mida, Sipunculida, Priapulida, Endoprocta, Phoronida, Bryozoa,
Brachiopoda)
Тип Pogonophora
Тип Mollusca (классы: Aplacophora, Polyplacophora, Monoplaco-
phora, Bivalvia, Gastropoda, Scaphopoda, Cephalopoda)
* Onychophora8
* Tardigrada
* Pentastomida
Тип Arthropoda
Подтип Trilobitomorpha (Trilobita и др.)
Подтип Chelicerata (Merostomata, Arachnida, Pantopoda)
Подтип Mandibulata (Crustacea, Myriapoda, Insecta)
Тип Echinodermata
Тип Stomochordata (Enteropneusta, Pterobranchia, Graptolitoidea)
Тип Tunicata (Appendicularia, Thaliacea, Ascidiacea)
Тип Cephalochordata
Тип Vertebrata
Как видим, в «Traite de Zoologie», несмотря на
фундаментальность и несомненные высокие достоин-
ства этого руководства, нет вполне упорядоченной
системы. Изолированно, по существу, рассматрива-
ются 17 типов (Embranchements), начиная с Protozoa
и кончая Vertebrata. Деление Bilateria на Proto- и Deu-
terostomia отсутствует. Особенно удивляет то обстоя-
тельство, что классы Phoronida, Bryozoa и Brachiopo-
8 Звездочкой (*) обозначены группы неясного происхождения.
100
ОБЩАЯ ЧАСТЬ
da попали в состав типа Annelida вместе с Priapulida
и Sipunculida. В этом характерном сочетании сказы-
вается влияние старой классификации с ее концеп-
цией типа червеобразных — Vermidea. Достаточно
вспомнить, например, систему Деляжа и Эруара (De-
lage, Herouard, 1897), в которой Vermidea искусст-
венно объединяют с Gephyrea (т. е. Priapulida, Sipun-
culida и Echiurida), считавшихся родственными An-
nelida, вместе с Bryozoa, Axobranchia (Phoronida +
Pterobranchia), Trochelminthes (Rotifera + Gastrotri-
cha), Kinorhyncha, Chaetognatha и Brachiopoda. Ранг
Spongiaria, Onychophora, Tardigrada и Pentastomida
не указан. Необычно выделение Tunic-ata, Cephalo-
chordata и Vertebrata в качестве самостоятельных типов,
причем Chordata считаются надтипом. Неясно, в какой
части этой системы будут помещены Chaetognatha,
когда издание будет закончено.
В системе выдающегося бельгийского зоолога П. Бри-
ана, лежащей в основе его двухтомного учебника
«Elements de Zoologie, notions d’anatomia comparee»
(Brien, 1961, 1963), сохранен еще старинный тип
Vermes, но наряду с этим фигурируют Hypo- и Epi-
neura.
Вместе с тем признается целоматная природа ско-
лецид, которые якобы претерпели вторичную дегра-
дацию организации (тип Parannelida). В системе Бри-
ана, таким образом, уживаются рядом как традиции
систематической зоологии конца XIX столетия, так и
«новая» гипотеза о деградации Scolecida, несмотря на
то что последняя имеет логическую связь с учением о
Proto—Deuterostomia, которое самим Брианом отвер-
гается (см. ниже, а также систему Ремане — с. 104).
Следуя взглядам своих учителей Бенедена и Ламера,
Бриан поддерживает цикломерную теорию последне-
го (Lameere, 1915) и его учение о дифилетическом
происхождении Coelomata от архаических коралло-
вых полипов (см. с. 98).
Система Бриана (Brien, 1961, 1963)
Protozoa
Тип Flagellata
Тип Rhizopoda
Тип Mycetozoa
Тип Sporozoa
Тип Infusoria
Metazoa
Diploblastica, s. Acoelomata
Тип Spongiaria
Тип Coelenterata (Hydrozoa, включяя Acalephae, Antho-
zoa, Ctenophora)
Triploblastica, s. Coelomata
Hyponeura
Тип Vermes
Подтип Annelida (включая Echiurida, Sipunculida, Pri-
apulida).
Подтип Parannelida (Nemertini, Gastrotricha, Rotifera)
Подтип Nemathelminthes (Nematoda, Gordiacea, Acan-
thocephala, Echinoderida)
Подтип Plathelminthes
Подтип Mesozoa
Тип Vermidea
Подтип Kamptozoa
Подтип Molluscoidea (Phoronida, Bryozoa, Brachiopo-
. da, Chaetognatha) .
Подтип Enteropneusta (Pterobranchia, Balanoglossi,
Graptolitha, Pogonophora)
Тип Echinodermata
Тип Mollusca
Тип Arthropoda s. 1.
Подтип Onychophora
Подтип Tardigrada
Подтип Pentastomida
Подтип Arthropoda s. str.
Epineura
Тип Prochordata (Tunicata, Cephalochordata)
Тип Vertebrata
Итак, система Бриана весьма своеобразна. Кроме
указанных выше особенностей необходимо отметить
еще следующие ее черты. Все классы Protozoa возведены
в ранг типа. Spongiaria тесно примыкают к Coelenterata,
одним из классов которых считаются Ctenophora. Извест-
ный исследователь губок, глава европейской школы
спонгиологов Бриан упорно придерживался так назы-
ваемой целентератной гипотезы происхождения гу-
бок, берущей начало от Геккеля (Haeckel, 1872,1874),
который рассматривал первичную губку Olynthus
как полипообразную гастреаду, перешедшую к сидя-
чему образу жизни, и гомологизировал ее централь-
ную полость с гастроцелем, а устье — с первичным
ртом. Данные современной сравнительной эмбриоло-
гии, однако, резко противоречат этим представлениям
(Codreanu, 1970; Иванов А. В., 1971; Salvini-Plawen,
1978).
Тип Vermes разделен у Бриана на 5 подтипов. Из
них, очевидно, только Annelida представлены пер-
вичными формами, остальные подтипы несут якобы
все более и более резкие следы упрощения и деграда-
ции в последовательности: Parannelida—Nemathel-
minthes—Plathelminthes—Mesozoa. Впрочем, даже
среди Annelida три класса, составлявшие некогда
группу Gephyrea, по мнению Бриана, уже отличаются
вторичной утратой сегментации. Очень своеобразен
подтип Parannelida, содержащий весьма разнородные
классы — Nemertini и часть первичнополостных чер-
вей. В тип Vermidea включены Chaetognatha и вто-
ричноротые животные — Hemichordata и Pogonophora.
Наконец, подчеркнем сближение Tunicata и Cepha-
lochorda под общим названием Prochordata в проти-
вовес типу Vertebrata.
Гатчек-Гроббеновская школа
Дифилетические системы
Начнем с классификаций, допускающих дифиле-
тическое происхождение Bilateria, так как именно
эти системы появились раньше других и составляют
основное ядро Гатчек-Гроббеновской школы. Это преж-
де всего системы Гроббена и Гатчека, сравнительно
мало изменившиеся в учебнике Клауса, Гроббена и
Кюна (Claus et al., 1932). Большинство последовате-
лей Гроббена настолько резко обособляло первично-
ротых от вторичноротых, что в поисках их предков
спускалось по филогенетическому древу до уровня
кишечнополостных и даже простейших. Среди со-
временных систем сюда относятся классификации,
предложенные авторами «Руководства по зоологии»
(Зенкевич, 1937), Д. М. Федотовым (1966 и более
ранний вариант) и А. В. Ивановым (1957). Сюда же,
по-видимому, примыкает и система Кестнера (Kaest-
ner, 1965).
СИСТЕМА ЖИВОТНОГО МИРА
101
Scolecida рассматриваются этими системами как
низшие и более примитивные Protostomia, a Deute-
rostomia выводятся совершенно независимо от при-
митивных Metazoa.
В незаконченном советском «Руководстве по зоо-
логии» (Зенкевич, 1937) большая система животного
мира выглядела следующим образом (табл. 5).
В этой системе 13 типов. Термин Protostomia
употреблен в классическом его понимании, а вклю-
чение в состав Protostomia Tentaculata, что делается
далеко не всегда, правильно подчеркивает принци-
пиальные различия между олигомерными мшанками
и крыложаберными. Есть Parazoa, но отсутствуют
Eumetazoa. Недостатком следует считать также от-
сутствие понятий Radiata и Bilateria (или соответст-
венно Diplo- и Triploblastica) и неоправданно боль-
шое число «добавлений». Так, самые характерные,
основные классы старого типа Nemathelminthes —
Rotatoria, Gastrotricha, Kinorhyncha — оказались до-
бавлением к нему. Что же осталось в самом типе?
Только Nematoda, Nematomorpha, Acanthocephala.
Федотов (1966) в большой и интересной моногра-
фии «Эволюция и филогения беспозвоночных живот-
ных» разбирает проблемы филогении внутри отдель-
ных групп, но редко упоминает о большой системе.
Он твердо держится широко распространенного раз-
деления Bilateria на протостомий и дейтеростомий,
относя низших червей к первой ветви в соответствии
с классическим ее определением. Однако Федотов не
считает Scolecida видоизмененными Coelomata. Пого-
нофор он не находит возможным отнести ни к Pro-
tostomia, ни к Deuterostomia и помещает их между
теми и другими, что равносильно признанию третьей
независимой ветви Bilateria (рис. 60).
Таблица 5
Система, принятая в «Руководстве по зоологии»
(по: Зенкевич, 1937)
Protozoa I Protozoa
Metazoa Parazoa II Porifera
Eumetazoa III Coelenterata
Protostomia IV Plathelminthes V Nemathelminthes VI Nemertina Добавление к низшим чер- вям: классы Rota- toria, Gastro- tricha, Kino- rhyncha, Kamptozoa
VII Annelida Добавление к высшим чер- вям: классы: Si- punculidea, Priapulidea
VIII Arthropoda IX Mollusca X Tentaculata
Deuterostomia XI Enteropneusta XII Echinodermata XIII Chordata Добавление к Deuterostomia: класс Chae- tognatha
РROTOS ТО MIA PO$ONO - BE UTE ROS TOM IA
Rhizopod a
Рис. 60. Филогенетические отношения между основными группа-
ми животных. (Составлено по: Федотов, 1966).
Fig. 60. Phylogenetic relations between the main animal groups.
Современным вариантом дифилетической концеп-
ции Гатчека явилась предложенная А. В. Ивановым
(1957) филогенетическая схема беспозвоночных, в ко-
торой Protostomia выводились от мечниковской фаго-
цителлы независимо от кишечнополостных, a Deute-
rostomia (вслед за В. Н. Беклемишевым) от первичных
Coelenterata. Сколециды, в согласии с Гатчеком, рас-
сматривались как низшие протостомии. Несколько
позднее эта схема фигурировала в учебнике «Зоология
беспозвоночных» Догеля (1959).
Наконец, классификация Кестнера (Kaestner, 1959),
приведенная в прекрасном немецком руководстве
«Lehrbuch der speziellen Zoologie» (1959—1984) также
построена по образцу дифилетической системы, хотя
автор и полагает, что из трех возможных вариантов
толкования природы Bilateria (моно-, ди- и полифи-
летического) ни один пока не может привести в свою
пользу решающие аргументы (Kaestner, 1965). Во
Entoprocta
Memertini
Plathelminthes
Pogonophora
Chaetoynatha^J?
' Tentaculata
SPIRALIA
X ____ Диплеирула
рохофора Радиальное
Спиральное ' Bilaterogastraea дробление
d дробление s.Phagocytella
__Радиальное
дробление
Рис. 61. Филогенетическое дерево Metazoa.
(По: Gruner, 1980, с изменениями).
Fig. 61. Metazoa phylogenetic tree.
102
ОБЩАЯ ЧАСТЬ
введении к последнему изданию «Руководства» Кест-
нера Грунер (Gruner, 1980) приводит схему, помещен-
ную на рис. 61.
В классификации Кестнера интересна система Pro-
tozoa, очерк по которым написан Ветцелем (Kaestner,
1959). Подцарство Одноклеточных разделено на два
раздела — Cytomorpha и Cytoidea, а группы, обычно
принимаемые за классы, переведены в ранг типа. Кест-
нер приводит 27 типов и принимает деление Bilateria
на Proto- и Deuterostomia в обычном смысле.
Монофилетические системы
Вторую группу Гатчек-Гроббеновского типа со-
ставляют классификации, приписывающие всем Bila-
teria монофилетическое происхождение. Согласно
этим классификациям, самые примитивные Bilate-
ria — предшественники целомических Protostomia, а
Deuterostomia произошли или от очень примитивных
первичноротых, или от общих с ними предков. В эту
группу следует отнести системы Хайман, Ливанова,
Грассе, Ремане и А. В. Иванова (см. соответствующие
системы).
Классификация Хайман (Hyman, 1940) в ее все-
мирно известном руководстве по зоологии беспозво-
ночных «The Invertebrates» (6 томов которого выходи-
ли с 1940 по 1959 г.) отличается большой четкостью и
хорошо отражает состояние современной зоологии.
Хайман счастливо избегла неудобства употребления
понятия Protostomia как таксономической единицы,
разделив Coelomata на Schizocoela и Enterocoela (см.
рис. 62). Она приводит 23 типа и выделяет среди Me-
tazoa 3 надраздела. Coelomata (Eucoelomata у Хайман)
разделены на Schizocoela и Enterocoela по типу обра-
зования в онтогенезе целома группы, соответствую-
щие Coelomata, Trochozoa и Deuterostomia.
Система Хайман (Hyman, 1940)
I. Подцарство Protozoa
Тип Protozoa
П. Подцарство Metazoa
Ветвь A. Mesozoa
Тип Mesozoa
Ветвь В. Parazoa
Тип Porifera s. Spongiaria
Ветвь С. Eumetazoa
Ступень I Radiata
Тип Cnidaria s. Coelenterata
Тип Ctenophora
Ступень II Bilateria
A. Acoelomata
Тип Plathelminthes
Тип Rhynchocoela s. Nemertinea
B. Pseudocoelomata
Тип Aschelminthes
Тип Entoprocta
C. Eucoelomata
1. Schizocoela
Тип Bryozoa s. Polyzoa
Тип Phoronida
Тип Brachiopoda
Тип Mollusca
Тип Sipunculoidea
Тип Priapuloidea
Тип Echiuroidea
Тип Annelida
Тип Arthropoda
2. Enterocoela
Тип Chaetognatha
Тип Echihodermata
Тип Hemichordata
Тип Pogonophora
Тип Chordata
С появлением классификации Хайман закончи-
лась разработка характерного для середины XX сто-
летия типа системы.
В филогенетической схеме Ливанова (1955), при-
веденной в его книге «Пути эволюции животного
мира», к сожалению, мы снова встречаемся со ста-
рым типом Vermes, который автор делит на 4 подти-
па: Plathelminthes, Nemathelminthes, Nemertini и An-
nelides (рис. 63). Названия Protostomia и Deuterosto-
mia заменены соответственно новыми терминами —
Protencephala и Deutencephala, так как судьба бласто-
пора, по мнению автора, не так показательна, как
план строения нервной системы. Однако полного со-
ответствия тут нет, ибо у Ливанова деление на две
ветви ограничено лишь целомическими животными,
а низшие черви под понятие Protencephala не подпа-
дают. Оказывается, что нижняя граница Protencephala
должна проходить между подтипами аннелид и не-
мертин внутри единого типа червей.
Таким образом, Protencephala и Deutencephala Ли-
ванова отвечают хаймановским Schizocoela и Entero-
coela. Тип Pogonophora Ливанов не признает, считая
этих животных аберрантными аннелидами.
Классификация Грассе лежит в основе известного
двухтомного французского учебника трех авторов
(Grasse, Poisson, Tuzet, 1961). В самой системе раз-
дел Protostomia отсутствует (несмотря на наличие
раздела Deuterostomia), но на филогенетической схе-
ме, составленной Грассе (Grasse et al., 1961, с. 14), две
ветви Bilateria обозначены как Proto- и Deuterosto-
mia. Авторы различают 20 типов (Embranchements) и
5 групп incertae sedis приблизительно в этом же
ранге.
SCHIZOCOELA ENTEROCOELA
\ / Chaeto-
\ /,/9natha
Примитивные
Acoela
I
BILATERIA
^RADIATA
Planula
(Phaffocytella)
Рис. 62. Родственные отношения основных групп Metazoa. (По:
Hyman, 1940, с изменениями).
Fig. 62. Relationship between the main groups of Metazoa.
СИСТЕМА ЖИВОТНОГО МИРА
103
PROUHCEPHALA UEUTEHCEPHALK
Nemertini
Turbellaria
I RADIATA
PAR AZO A QASTRA'eADA
I I
Рис. 63. Родство основных групп животных. (По: Ливанов, 1955).
Fig. 63. Relationship between the main animal groups.
Система Грассе (Grasse et al., 1961)9
Unicellulaires
Тип Protozoa
Pluricellulaires или Metazoa
Diploblastica
Тип Spongiaria
Тип Cnidaria
Тип Ctenaria
Triploblastica
Acoelomata
Тип Plathelminthes (классы: Turbellaria, Cestodaria, Mo-
nogenea, Cestoda, Trematoda)
Incertae sedis (классы: Acanthocephala, Priapulida)
Тип Mesozoa (классы: Orthonectida, Dycyemida)
Тип Nemertini
Тип Nemathelminthes (классы: Nematoda, Nematomorpha)
Тип Nematorhyncha (классы: Gastrotricha, Kinorhyncha)
Тип Rotifera
Coelomata
Тип Annelida (Polychaeta, Oligochaeta, Hirudinea)
Родственные группы (Sipunculida, Echiurida)
Тип Lophophorata (Endoprocta, Phoronida, Ectoprocta,
Brachiopoda)
Тип Mollusca
Тип Arthropoda
Trilobitomorpha
Chelicerata
Mandibulata s. Antennata
Pararthropoda (разнородные архаические членистоно-
гие)
(Onychophora, Tardigrada, Pentastomida)
Тип Chaetognatha
Deuterostomia Epithelioneuria
Тип Echinodermata
Тип Stomochordata (Enteropneusta, Pterobranchia, Grapto-
litha)
Тип Pogonophora
Deuterostomia Epineura, s. Chordata
Тип Tunicata (Thaliacea, Appendicularia, Ascidia)
Тип.Cephalochordata
Тип Vertebrata
Одноклеточные в этой системе составляют один
тип, разделенный на 5 подтипов, причем каждый из
них соответствует классу в обычном понимании. Губки
тесно сближены с книдариями и гребневиками в со-
ответствии со взглядами одного из авторов учебника
(О. Tuzet), которые не без серьезных оснований оспа-
риваются многими современными зоологами (Лива-
нов, 1955; Беклемишев, 1964; Codreanu, 1970; Ива-
нов А. В., 1971; Salvini-Plawen, 1978). В связи с этим
нет и разделения Metazoa на надразделы (superdivisi-
ones) Parazoa и Eumetazoa. Вторичноротые понима-
ются в узком смысле (без включения в них Chaeto-
gnatha) и разделены на Epithelioneura, отличающиеся
эпителиальным расположением нервного аппарата, и
Epineura (они же Chordata) с погруженной спинной
центральной нервной системой. Epineura содержат
три типа: Tunicata, Cephalochordata и Vertebrata, ко-
торые обычно считаются только подтипами хордо-
вых. Наконец, следует отметить разделение явно ис-
кусственного типа Nemathelminthes, или (по другому
наименованию) Aschelminthes, на несколько самосто-
ятельных типов: Nemathelminthes в узком понимании,
Nematorhyncha, Rotifera и группу incertae sedis —
Acanthocephala. Последних, разумеется, тоже следует
рассматривать в ранге типа, как уже предлагали Скря-
бин и Шульц (1931) и Петроченко (1956).
Судя по филогенетическому древу Грассе (Grasse
et al., 1961, с. 14) и по расположению материала в
учебнике, низшие черви Acoelomata рассматриваются
как самые примитивные Bilateria — предки Protosto-
mia Coelomata. Этим система Грассе сближается с
классификацией Хайман и Ливанова.
Явно выраженным монофилетическим толковани-
ем эволюции Bilateria характеризуется также система
Ремане, вариант которой мы находим в учебнике
«Systematische Zoologie, Stamme des Tierreiches» (Re-
mane et al., 1976) (рис. 64). Главная ее особенность
(общая с системой Кэно и Бриана) заключается в
причислении низших червей — Scolecida (т. е. Plat-
helminthes, Nemathelminthes, Nemertini) к Coelomata
на том основании, что это якобы деградировавшие
потомки целомических форм. Поэтому в «линии»
(Linie) Protostomia наиболее примитивными считают-
ся Tentaculata, затем следуют Sipunculida, Scolecida,
Mollusca и Articulata (=Annelida + Arthropoda). К
ARCHICOELO-
MATA
I
RADI A TA
(Anthozotz)
Рис. 64. Филогения основных групп Metazoa. (По: Remane et al.,
1976, с изменениями).
9 Классы в ряде типов опущены.
Fig. 64. Phylogeny of the main Metazoa groups.
104
ОБЩАЯ ЧАСТЬ
альтернативному «ряду» (Reihe) Deuterostomia отно-
сятся Hemichordata, Pogonophora, Chaetognatha, Echino-
dermata и Chordata. Филогенетической основой сис-
темы Ремане является архицеломатная теория (Archi-
coelomatentheorie), по которой исходной и основной
группой всех Bilateria считаются гипотетические це-
ломические и тримерные Archicoelomata, давшие на-
чало как протостомиям, так и дейтеростомиям. Именно
поэтому олигомерные формы как среди первых, так
и среди вторых трактуются как наиболее примитив-
ные. В этом пункте взгляды Ремане и Бриана совпа-
дают.
Система Ремане (Remane et aL, 1976)
1. Подцарство Protozoa (классы: Flagellata, Rhizopoda, Sporozoa,
Cnidosporidia, Ciliata)
2. Подцарство Metazoa
A. Porifera
B. Cnidaria
C. Ctenophora
D. Coelomata
Линия Protostomia
I. Tentaculata
II. Sipunculata
III. Scolecida
1. Plathelminthes
2. Gnathostomulida
3. Nemertini
4. Aschelminthes
5. Kamptozoa
IV. Mollusca
V. Articulata
1. Annelida
2. Pentastomida
3. Tardigrada
4. Arthropoda
Protarthropoda (Onychophora)
Eu arthropoda
A. Trilobitomorpha
B. Chelicerata
C. Mandibulata
1. Crustacea
2. Antennata
a. Chilopoda
b. Progoneata
c. Insecta
Ряд Deuterostomia
I. Chaetognatha
II. Pogonophora
III. Hemichordata
IV. Echinodermata
V. Chordata
1. Tunicata
2. Copelata
3. Acrania
4. Vertebrata
К сожалению, в системе Ремане не указывается
таксономический ранг большинства групп, и о нем
можно только догадываться. Однако ясно, что Porife-
ra, Cnidaria и Ctenophora находятся в одном очень
высоком ранге (выше типа), наравне с Coelomata.
Последние, очевидно, по объему совпадают с Bilate-
ria. Итак, губки не противопоставляются остальным
Metazoa, а сближаются в общем с книдариями (Рема-
не является защитником старой Геккелевской целен-
тератной теории происхождения Porifera). Coelomata
разделены на 10 типов, причем Tentaculata рассмат-
риваются как примитивный тип Protostomia, a Chae-
tognatha и Pogonophora отнесены к Deuterostomia.
Одним из современных вариантов монофилети-
ческой системы является классификация Нильсена
(Nielsen, 1985) (рис. 65), построенная на постулатах
гипотезы Трохеи (Trochaea) (Nielsen, Norrevang, 1985).
По поводу этой концепции, однако, был сделан ряд
серьезных критических замечаний (Иванова-Казас,
Иванов, 1987). Чрезмерно большое значение в гипо-
тезе Трохеи придается личиночным признакам, и не-
которые личиночные формы (трохофора, торнария)
рассматриваются в ней как рекапитуляция гипоте-
тических стадий филогенеза. По Нильсену и Норре-
вангу, самые ранние Bilateria уже обладали пелаги-
ческой личинкой с эпителиальной кишкой, апикаль-
ным органом чувств и задним ресничным пояском —
археотрохом, который окружал бластопор и играл
роль локомоторного и подгоняющего пищевые части-
цы органа.
Гипотеза Трохеи является развитием теории Гаст-
реи и разделяет все ее слабые стороны, но если поня-
тие о Гастрее опирается на широкое распростране-
ние инвагинационной гаструлы во многих группах
Metazoa, то Трохея — существо, созданное йсключи-
тельно умозрительным путем, ни одно животное ни
на какой стадии развития не имеет сходства с ней.
По гипотезе Нильсена и Норреванга, ресничные
пояски трохофоры (прото-, мета-, тело- и гастро-
трох) являются характерными чертами личинки про-
тостомных Bilateria, дериватами гипотетического ар-
хеотроха. Однако подобное толкование неприемлемо
хотя бы потому, что прототрох, например, не гомоло-
гичен передней части археотроха, поскольку он раз-
вивается из бластомеров, относящихся ко всем четы-
рем квадрантам зародыша, а не только к квадранту В,
примыкающему спереди к бластопору. В действи-
тельности же ресничные пояски трохофорных личи-
нок возникали в процессе эволюции de novo, не имея
предшественника в форме гипотетического археотро-
ха. Предполагается, что, согласно этой гипотезе, на
филогенетической стадии Властей отделились ветви
Placozoa и Porifera, а на стадии Гастреи — Cnidaria.
Далее Гастрея приобрела археотрох и превратилась в
Трохею, которая затем дала начало Гастронейрону
(Gastroneuron) — общему предку всех протостомных
SPIRALIA
Articulata
Parenchymia
Bryozoa
Gastroneuron
CYRTOTRETA
BRACHIATA
Phoronida
Brachiopoda Enteropneusta
Echinodermata
Chordata
Notoneuron
ASCHELMIHTHES
Rotifera
Acanthocephala
Chaetognatha
Nematoda
Protornaea
Tornaea
Trochaea
________Cnidaria
Qastraea
Рис. 65. Филогения Metazoa согласно теории Трохеи. (По: Niel-
sen, 1985, с изменениями).
Fig. 65. Metazoa phylogeny according Trochaea theory.
СИСТЕМА ЖИВОТНОГО МИРА
105
Bilateria, или, как их предпочитает именовать Ниль-
сен, Gastroneuralia. Гастронейрон стал первым бенти-
ческим билатеральным животным, сохранившим пе-
лагическую личинку трохофорного типа.
От Трохеи же произошли и дейтеростомные Bila-
teria — Notoneuralia, но ранняя их эволюция была го-
раздо сложнее, и для ее объяснения понадобились ги-
потетические промежуточные формы: Проторнея
(Protornaea), наделенная 4 каналами акрогастера (от
нее якобы произошла боковая ветвь Ctenophora),
Торнея (Тогпаеа), созданная по образу торнарии —
личинки Enteropneusta, и, наконец, Нотонейрон (No-
toneuron) — предполагаемый общий предок Notoneu-
ralia. Нотонейрон, подобно Гастронейрону, приобрел
пелагобентический жизненный цикл, т. е. во взрос-
лом состоянии стал донным существом, развиваю-
щимся из пелагической торнариеобразной личинки.
Система Trochaeozoa (=Bilateria)
по Нильсену (Nielsen, 1985)
Trochaeozoa
Gastroneuralia (=Protostomia)
Spiralia
Articulata
Annelida s. 1. (вместе c Pogonophora)
Echiura
Gnathostomulida
Onychophora
Arthropoda s. 1.
Mollusca
Sipuncula
Parenchymia
Nemertini
Plathelminthes
Bryozoa
Entoprocta
Ectoprocta
Aschelminthes
Rotifera
Acanthocephala
Nematoda
Kinorhyncha
Loricifera
Priapulida
Gastrotricha
Notoneuralia (=Deuterostomia)
Brachiata
Pterobranchia
Phoronida
Echinodermata
Cyrtotreta
Enteropneusta
Chordata
Urochordata
Cephalochordata
Vertebrata
Несмотря на привлечение новых интересных дан-
ных по тонкой структуре и функциям личиночных
органов в разных группах Bilateria, гипотеза Трохеи
носит слишком умозрительный характер и не может
служить основой для построения системы. Класси-
фикация Trochaeozoa, предложенная Нильсеном,
вызывает ряд решительных возражений, полностью
проанализировать которые в коротком очерке невоз-
можно. Прежде всего вызывает сомнения общая ха-
рактеристика двух основных ветвей Trochaeozoa. «Луч-
шим и наиболее постоянным признаком Gastroneura-
lia» Нильсен считает строение и положение их нервной
системы. Гипотетическому предку — Гастронейро-
ну — он приписывает «типичную нервную систему»
с фронтальным мозгом и парой продольных брюш-
ных стволов, лучше всего представленную у совре-
менных аннелид — якобы самой примитивной груп-
пы Гастронейралий. Однако, как теперь известно,
очень примитивные формьх полихет — Dinophilidae
и Oweniidae — обладают интраэпидермальной нерв-
ной системой с многими продольными стволами (Буб-
ко, Миничев, 1972; Котикова, 1973; Лагутенко, 1985).
Очевидно, исходным прототипом для аннелид следу-
ет признать интраэпителиальный нервный плексус,
или ортогон, возникший независимо от погруженного
в паренхиму ортогона плоских червей.
Среди Notoneuralia исходный тип нервной систе-
мы якобы сохранился у Enteropneusta, обладающих
интраэпителиальным спинным нервным стволом, ко-
торый образует трубчатый мозг в области воротничка
(мезосомы). При этом, однако, забывается, что ки-
шечнодышащие имеют еще и брюшной ствол, связан-
ный кольцевой комиссурой между мезо- и метасомой
со спинным стволом. Такие же особенности нервной
системы обнаруживают и Pogonophora (Иванов А. В.,
1959; Salvini-Plawen, 1972).
Невозможно согласиться с тем, что у первобыт-
ных Gastroneuralia бластопор вытянулся в форме щели,
замкнулся в средней части и разделился на ротовое и
анальное отверстия. У большинства сравнительно
примитивных Bilateria (Polycladida, Nemertini, Sipun-
culida, Echiurida, Mollusca, многие Polychaeta) блас-
топор целиком превращается в рот, который затем
смещается на брюшную сторону и вперед из-за силь-
ного разрастания эктодермы в задней области спин-
ного меридиана. Именно таким путем совершается
переход от радиальной симметрии зародыша к дефи-
нитивной билатеральной симметрии (Иванова-Казас,
1983).
Все соображения Нильсена об эволюции Spiralia
проникнуты идеей вторичного упрощения организа-
ции нецеломических несегментированных червей.
Parenchymia (т. е. Nemertini и Plathelminthes), а также
Aschelminthes помещены в системе после Articulata,
среди которых наиболее примитивными считаются
Annelida.
Нильсен сталкивается с теми же трудностями, что
и другие филогенетики, выводящие Bilateria от Radiata
двумя отдельными стволами, поскольку такие груп-
пы, как Tentaculata, Chaetognatha и Pogonophora, не
могут быть безоговорочно причислены ни к Gastro-
neuralia, ни к Notoneuralia. Пренебрегая такими су-
щественными «нотонейральными» признаками Chae-
tognatha, как вторичный рот, энтероцельный целом и
олигомерное расчленение тела, Нильсен счел воз-
можным обосновать их «родство» с Rotifera и Acan-
thocephala наличием «внутриклеточной кутикулы»,
развитие которой в разных группах правдоподобнее
объяснить как результат независимой эволюции.
Также нельзя согласиться ни с соединением мша-
нок Ectoprocta с паренхиматозной группой Kampto-
zoa (=Entoprocta), ни со сближением Gnathostomulida
и Pogonophora с Annelida.
Однако настоящим «камнем преткновения» как
для гипотезы Трохеи, так и для системы Trochaeozoa
106
ОБЩАЯ ЧАСТЬ
PROCOELOMA1A
SCOLECIDA
AR С HI Bl LATER IА
Рис. 66. Родственные отношения основных групп Metazoa.
(По: Иванов, 1975а).
Fig. 66. Phylogenetic reflations between the main Metazoa groups.
является причисление к Notoneuralia Phoronida и
Brachiopoda, особенно если вспомнить, что у форо-
нид дефинитивный рот образуется из бластопора, а
анус возникает независимо от него, как у большинст-
ва низших Gastroneuralia.
Таким образом, собирающий пиЩу послеротовой
ресничный аппарат актинотрохи (личинки форонид)
нельзя рассматривать как дериват неотроха, который,
согласно гипотезе Трохеи, возник у Notoneuralia в
связи с появлением вторичного рта.
Особое место среди монофилетических схем за-
нимает классификация А. В. Иванова (1975а, 1976),
подробнее рассмотренная ниже (см. с. 109). Она от-
личается, во-первых, разделением всех Coelomata не
на 2, а на 5 параллельных ветвей (надтипов) и, во-
вторых, отсутствием термина Protostomia (рис. 66).
Общее понятие Protostomia тем не менее остается,
более примитивные первичноротые (Protostomia Aco-
elomata) представлены в системе как синоним под-
раздела Scolecida, а высшие (Protostomia Coeloma-
ta) — как синоним надтипа Trochozoa.
Полифилетическая система
В. Н. Беклемишева
До сих пор мы имели дело только с монофилети-
ческими и дифилетическими системами. Однако, по
справедливому замечанию многих зоологов, помимо
Protostomia и Deuterostomia существуют еще большие
группы Coelomata, не укладывающиеся в рамки ни
тех ни других. Такими, как бы «промежуточными»,
группами обычно называют Tentaculata (Phoronida,
Bryozoa, Brachiopoda) и Chaetognatha. Это обстоя-
тельство, так же как значительное организационное
их своеобразие, обнаруживающееся при детальном
проморфологическом анализе, заставило Беклемише-
ва предложить полифилетическую гипотезу проис-
хождения Bilateria. «Очевидно, — писал Беклемишев
(1964, с. 364), — животное царство широким фрон-
том, многими стволами поднялось с уровня организа-
ции Bilateria, и одни из этих стволов ближе между
собой, другие менее близки. В этом смысле проис-
хождение Bilateria является полифилетическим».
Система В. Н. Беклемишева (1944, 1964)
Подцарство I. Protozoa
Тип I. Protozoa
Подцарство II. Metazoa
Надраздел I. Enantiozoa
Тип II. Porifera
Надраздел II. Archaeocyatha
Тип III. Archaeocyatha
Надраздел III. Enterozoa
Раздел Radiata
Тип IV. Coelenterata
Подтип Cnidaria
Подтип Acnidaria
Раздел Bilateria
Подраздел Protostomia
Надтип Protrochozoa
Тип V. Scolecida
Подтип Plathelminthes
Подтип Acanthocephala
Подтип Nemathelminthes
Подтип Nemertini
Надтип Trochozoa
Тип VI. Mollusca
Тип VII. Articulata
Подтип Annelides
Подтип Tardigrada
Подтип Pentastomida
Подтип Onychophora
Подтип Arthropoda
НаДкласс Trilobitoidea (Trilobita, Pantopoda,
Chelicerata)
Надкласс Crustacea
Надкласс Atelocerata
Тип VIII. Prosopyga (Sipunculoidea)
Тип IX. Kamptozoa
Тип X. Podaxonia (Phoronoidea, Bryozoa)
Надтип Actinotrochozoa
Подраздел Brachiopoda
Тип XL Brachiopoda
Подраздел Deuterostomia
Тип XII. Pogonophora
Тип XIII. Hemichordata
Тип XIV. Chordata
Тип XV. Echinodermata
Подраздел Chaetognatha
Тип XVI. Chaetognatha
Таким образом, Беклемишев различает 6 незави-
симых стволов Bilateria (рис. 67):
1. Protrochozoa (т. е. нецеломические Protosto-
mia), предками которых предположительно были
неотенические планулы книдарий.
2. Trochozoa (целомические Protostomia), кото-
рые, вероятно, произошли от первичных плавающих
Coelenterata.
3. Actinotrochozoa (форониды и мшанки).
4. Deuterostomia, берущие начало от примитив-
ных гребневиков.
СИСТЕМА ЖИВОТНОГО МИРА
107
и
ча
q
<0
Ь
К
Cnida-
phora
ria
RAUTATA
Рис. 68. Полифилетическое происхождение многоклеточных орга-
низмов от протистов. Схема. (По: Nursall, 1959).
Fig. 68. Scheme of the Metazoa polyphyletic origin from Protista.
Рис. 67. Происхождение основных ветвей Bilateria. (По: Беклеми-
шев, 1944).
Fig. 67. Origin of the Bilateria main branches.
5. Brachiopoda, которые, вопреки широко рас-
пространенному мнению, вовсе не связаны с мшанка-
ми и форонидами.
6. Chaetognatha.
В системе Беклемишева, однако, вместо 6 подраз-
делов Bilateria имеется только 4, так как, вопреки
филогенетической концепции автора, Protrochozoa,
Trochozoa и Actinotrochozoa все же соединены в одну
группу Protostomia, очевидно, имеющую трифилети-
ческое происхождение. Наиболее вероятных предков
Brachiopoda и Chaetognatha, по Беклемишеву, также
следует искать среди примитивных кишечнополост-
ных. Происхождение же Trochozoa и Deuterostomia
трактуется в духе классической цикломерной теории,
основной постулат которой составляет признание го-
мологии периферических карманов (каналов) гаст-
ральной полости предков и метамерных целомичес-
ких мешков потомков. Однако энтероцельную приро-
ду целома автор признает только для Deuterostomia.
У Trochozoa, напротив, имеется схизоцельный целом,
учение о возникновении которого из первичной па-
ренхимы путем ее эпителизации настолько убеди-
тельно разработано автором, что с ним трудно не со-
гласиться. Таким образом, представление о схизо-
цельном целоме Trochozoa находится в противоречии
с основным постулатом цикломерной теории. Кроме
того, полностью отрывая Scolecida от Trochozoa, Бе-
клемишев разрушает понятие о монофилетическом
единстве Protostomia (=Spiralia).
Невозможно также согласиться с отрывом Brachio-
poda от Bryozoa и Phoronida. Родство всех этих групп,
входящих, по мнению большинства зоологов, в один
тип Tentaculata, хорошо обосновано сравнительно-ана-
томически (Kaestner, 1965; Emig, 1977) и не вызывает
сомнений даже при наличии значительных эмбрио-
логических различий между ними. Наконец, выделе-
ние палеозойских Archaeocyatha в самостоятельный
надраздел (superdivisio) представляется недостаточно
обоснованным. Мы слишком мало знаем об организа-
ции этих животных.
Идея полифилетического происхождения основ-
ных групп животного царства нашла свое крайнее
выражение в схеме, представленной на рис. 68 (см.:
Nursall, 1959).
О происхождении Coelomata
и толковании альтернатив
Protostomia — Deuterostomia
Из обзора современных зоологических систем хо-
рошо видно, что они расходятся главным образом в
определении систематического значения Protostomia
и Deuterostomia, зависящего в сильной степени от
того, как объясняется происхождение Scolecida и
Coelomata. Среди теорий, претендующих на решение
этого вопроса, следует отметить следующие.
1. Учение о полифилетической природе Bilateria,
разработанное Беклемишевым (1944) и поддержан-
ное его последователями (Шаров, 1965; Мельников,
1971; Миничев, Старобогатов, 1972). Bilateria рас-
сматриваются как серия самостоятельных ветвей,
развившихся от кишечнополостных (см. с. 106): Sco-
lecida, Trochozoa, Deuterostomia, Brachiopoda и Chae-
tognatha. Однако концепция Беклемишева о первич-
ной плагиоаксонии аннелид, лежащая в основе его
представлений о независимом происхождении Tro-
chozoa и Scolecida, была проанализирована в свете
данных сравнительной эмбриологии (Светлов, 1967;
Иванова-Казас, 1974) и не выдержала проверки.
Учение Беклемишева, как уже отмечалось, пред-
ставляет собой вариант цикломерной гипотезы. Из
него следует, что билатеральная симметрия основных
108
ОБЩАЯ ЧАСТЬ
ветвей Coelomata возникала несколько раз одновре-
менно с целомом. Понятие Protostomia Беклемише-
вым, в сущности, уничтожается (рис. 67), хотя он и
обозначает один из подразделов своей системы этим
названием. Сходство в спиральном дроблении Scole-
cida и Trochozoa считается возникшим конвергентно.
2. Дифилетическая гипотеза, или гипотеза неза-
висимого происхождения Proto- и Deuterostomia,
была развита Гатчеком (Hatschek, 1911), который по-
лагал, что первичноротые (=Ecterocoelida) произошли
от ктенофорообразных, а вторичноротые (=Enterocoe-
lida) — от актиниеподобных предков. При этом эво-
люция первичноротых объяснялась с позиций трохо-
форной теории (Hatschek, 1878; 1888), a Scolecida
считались примитивными предшественниками цело-
мических протостомий (рис. 58, А).
Независимое происхождение Hypo- и Epineura (не
совпадающих по составу с Proto- и Deuterostomia) от
близких друг другу кишечнополостных на основе
цикломерно-энтероцельной гипотезы признавалось
также Ламером (Lameere, 1915, 1930) и Кэно (Cue-
not, 1952). Ламер, развивший представления своего
учителя Бенедена (Beneden, 1891), полагал, что Нуро-
neura и Epineura возникли независимо от различных
актиниеподобных организмов, причем оральная по-
верхность предков стала якобы брюшной стороной у
первых и спинной — у вторых. Сходные представле-
ния защищал и Бриан (Brien, 1969). По мнению всех
этих бельгийских авторов, Scolecida — настоящие
гипомерные Coelomata, испытавшие сильное вторич-
ное упрощение организации.
3. Архицеломатная теория (Archicoelomatentheo-
rie), представляющая современную интерпретацию
старой цикломерной концепции Седжвика и системы
Гроббена, была высказана еще в конце прошлого
столетия (Masterman, 1898) и возрождена в середине
нашего века (Ulrich, 1949, 1973). Особенно пропаган-
дировалась она в Германии (Ах, 1961; Siewing, 1967,
1969, 1972, 1976; Remarie, 1973). По этой гипотезе,
Coelomata произошли от примитивных четырехлуче-
вых полипов, начавших ползать на ротовой стороне
тела (по Siewing, 1976 — от ползающей Benthogast-
гаеа). Предполагается далее, что гипотетические пер-
вобытные Archicoelomata вели полусидячую жизнь;
их тело состояло из трех отделов, каждый из которых
имел пару целомических мешков; второй отдел (ме-
зосома) был снабжен щупальцами. По Ульриху и Зи-
вингу, в современной фауне существуют реликтовые
группы, еще близкие к архицеломатам, — Tentaculata,
Pterobranchia и Pogonophora. От вымерших архицело-
мат произошли 2 главные ветви целомических жи-
вотных — Spiralia или Protostomia (с их деградиро-
вавшими потомками сколецидами) и Deuterostomia.
Эта гипотеза не согласуется со многими данными
сравнительной анатомии и эмбриологии (см. кри-
тические статьи: Hyman, 1959; Hartman, 1963; Ива-
нов А. В., 1968, 1975а, 19756, 1976; Codreanu, 1970;
Иванов, Мамкаев, 1973). В частности, невозможно
без нагяжек свести организацию всех современных
Coelomata к исходному трехмерному плану строения.
Вслед за Седжвиком и Гроббеном и их последовате-
лями архицеломатная теория считает всех Bilateria
настоящими Coelomata.
4. Теория Н. А.Ливанова. По мнению Ливанова
(1955), первыми целомическими животными были
аннелиды, произошедшие от немертинообразных
предков. От них получили свое начало как первично-
ротые (=Protencephala), так и вторичноротые (=Deuten-
cephala). Таким образом, Ливанов не относит низших
червей к Protostomia (рис. 63), что противоречит пер-
воначальному определению этого понятия (см.: Grob-
ben, 1908). В теории Ливанова сколециды занимают
приблизительно подобающее им место. Однако пред-
полагаемое родство немертин и аннелид очень спор-
но. Несмотря на это, в основе построений Ливанова
лежит правильная идея о монофилетической природе
Coelomata и о схизоцельном происхождении целома
(см.: Иванов А. В., 1976).
5. Теория Хайман (Hyman). Особого внимания за-
служивают филогенетические взгляды Хайман (Hyman,
1940), которая выводит Schizocoela (=Protostomia и
Scolecida) и Enterdcoela (=Deuterostomia) от «прими-
тивных ацелоидных Plathelminthes». Это монофиле-
тическая теория происхождения Bilateria и в то же
время — признание дифилетической природы цело-
ма у Coelomata.
Современные Scolecida (=Acoelomata и Pseudoco-
elomata) у Хайман представляют ранние и слепые
ветви протостомий (рис. 62). Против концепции Хай-
ман трудно было бы что-либо возразить, если бы она
не причисляла всех Целомат только к двум основным
группам — Schizocoela и Enterocoela (см. с. 102).
6. Монофилетическая схема эволюции Bilateria,
вытекающая из теории Фагоцителлы И. И. Мечнико-
ва (Иванов А. В., 1968, 1975а, 1975в, 1976), находит-
ся в гораздо большем соответствии с данными срав-
нительной морфологии и с результатами изучения
физиологии движения беспозвоночных и их экологии
(см.: Clark, 1964).
По этой гипотезе, первобытные билатерии — Аг-
chibilateria, вероятнее всего, находились на организа-
ционном уровне низших турбеллярий, но были лише-
ны свойственных им специализаций. От Archibilateria
развились далее все Scolecida и независимо от них —
первобытные целомические животные Procoelomata.
AIOTOMCURALIA
(Chordata)
I
HETERONEURAL1A
( Oligomera)
qASTRONEURALIA
(целомические)
qASTRONEURALlA
(Ирецеломи ческие)
TRTPLOBLASTICA
I
diploblastic a
Рис. 69. Филогения основных групп Metazoa. (По: Salvini-Pla-
wen, 1972).
Fig. 69. Phylogeny of the main Metazoa groups.
СИСТЕМА ЖИВОТНОГО МИРА
109
(Protostomia) (Deuterostomia)
ТЕ N ТА CULA ТА СОЕ LO МОРОНА
другие
SPIRALIA CHORDATA
POLY MERA
PLATHELMINTHES
(А тега)
DIPLOBLASTICA
Рис. 70. Филогения основных групп Metazoa. (По: Gutmann,
1979).
Fig. 70. Phylogeny of the main Metazoa groups.
Естественно считать, что все эти филогенетически
связанные друг с другом группы, а именно Archibila-
teria, Scolecida и Procoelomata, принадлежат к Proto-
stomia, так как должны обладать всеми характерны-
ми для первичноротых признаками (рис. 66). Что ка-
сается потомков Procoelomata, то среди них имеются
как истинные первичноротые, так и вторичноротые и
группы, по существу, не принадлежащие ни к тем ни
к другим (Иванов А. В., 1975а, 19756, 1976). Недавно
к сходным взглядам пришли также Кодреану (Codre-
anu, 1970) и Сальвини-Плавен (Salvini-Plawen, 1972)
(рис. 69).
Перейдем теперь к оценке современных представ-
лений о составе Proto- и Deuterostomia и о их месте
в системе. На рис. 70 в схематическом виде показа-
но, как в только что рассмотренных нами теориях
понимается отношение первичноротых и вторично-
ротых друг к другу и к их предполагаемым предкам.
Отметим, прежде всего, что все построения, основан-
ные целиком и отчасти на цикломерно-энтероцель-
ной теории, должны быть отброшены. Таковы все по-
лифилетические учения, рискующие выводить цело-
мат непосредственно от кишечнополостных, а также
варианты гипотезы Седжвика среди монофилетичес-
ких концепций. Нельзя согласиться ни с включением
Scolecida в состав Coelomata, ни с их полным отры-
вом от Trochozoa, ни, наконец, с исключением их из
состава первичноротых.
Таким образом, идея Гроббена о существовании
двух равноценных филетических ветвей Bilateria ^Coe-
lomata) находится ныне в двусмысленном положении,
чтобы не сказать — в состоянии глубокого кризиса.
В самом деле, с одной стороны, понятие Protostomia
(=Spiralia) вполне правомочно как удобное объедине-
ние очень больших, вполне реальных систематичес-
ких групп — Scolecida и Trochozoa. С другой сторо-
ны, Protostomia отнюдь нельзя сопоставить с Deute-
rostomia, в состав которых входят только некоторые
целомические животные. Очевидно, при построении
естественной системы понятие Protostomia не может
быть употреблено как название таксономической
группы. Действительно, в предлагаемой системе Pro-
tostomia распадаются на две группы различного так-
сономического ранга: Protostomia Acoelomata пред-
ставлены подразделом Scolecida, a Protostomia Coelo-
mata — надтипом Trochozoa. Напротив, Deutero-
stomia (в узком понимании этого названия) остаются
конкретной систематической группой, вполне сопоста-
вимой с другими большими группами целомических
животных, а между ними и с равными им по рангу —
с Trochozoa. Говоря образно, Protostomia — это
ствол дерева, тогда как Deuterostomia — лишь одна
из его ветвей (рис. 66).
Монофилетическое единство
Coelomata
Изучение проблемы Proto- и Deuterostomia все
еще затруднено недостаточностью наших знаний о
строении и развитии многих групп животных. В част-
ности, были бы весьма желательны данные, позволя-
ющие с большой уверенностью сделать выбор между
полифилетическим и монофилетическим толковани-
ем природы Coelomata. Впрочем, в наше время поя-
вилась возможность рассмотреть в новом свете этот
вопрос благодаря успехам экспериментальной зооло-
гии (физиологии движения и экологии Coelomata),
развитию электронной микроскопии и открытию среди
целомических животных совершенно нового органи-
зационного типа, представленного современными Po-
gonophora.
В организации этих животных есть многие сущест-
венные черты, общие с Trochozoa и, что особенно
важно, присущие также Tentaculata и Chaetognatha.
Таковы, например, щетинки аннелидного типа, спо-
собность образовывать хитин, общая связь гонад с
целомом и другие. Еще больше характерных сходств
обнаруживают погонофоры с Deuterostomia и, опять-
таки, — с Tentaculata и Chaetognatha (энтероцельный
целом; разделение тела на 2—4 отдела, причем туло-
вищный отдел — метасома — содержит одну пару
гонад; не ортогональный тип нервной системы и
т. д.). Очень важное сходство с Deuterostomia прояв-
ляется в асимметрическом развитии первой пары це-
ломических мешочков (протоцелей) погонофор —
особенность, общая с Echinodermata, Hemichordata и
Acrania (Иванов А. В., 1975а, 19756, 1975в). Все эти
сходства нельзя объяснить конвергентным развитием.
Одни носят, скорее, характер параллелизмов, часто
проявляющихся в родственных группах (например,
щетинки аннелидного типа или слоистая структура
кутикулы), другие несомненно представляют гомоло-
гии (например, упомянутая асимметрия в развитии
протоцелей).*
Результатом независимого конвергентного разви-
тия можно считать только возникновение полимер-
ного (многосегментного) состояния в туловище анне-
лид и в заднем отделе тела у погонофор. В обоих
случаях причиной возникновения сегментации, веро-
ятно, был активный роющий образ жизни, приводя-
щий к сегментарному расчленению первично нерас-
* Вопрос о положении погонофор в системе животного царст-
ва разбирается также в монографии Малахова и Галкина (1998).
по
ОБЩАЯ ЧАСТЬ
члененного целома (Clark, 1963, 1964). Развитие по-
лимерного состояния у Chordata, напротив, обычно
связывают с активным плаванием (Clark, 1964; Gut-
mann, 1972). Таким образом, полимерный план стро-
ения трижды возникал у Coelomata.
Вспомним, что уже давно отмечали явно «проме-
жуточный» характер организации Tentaculata, кото-
рые обнаруживают ряд важных сходств как с Proto-
stomia, так и с Deuterostomia (см., напр.: Hyman, 1959;
Беклемишев, 1964; Siewing, 1967, 1972).
Итак, ясно, что «промежуточные» группы Pogo-
nophora, Tentaculata и Chaetognatha по одним, очень
характерным признакам сходны с Trochozoa, по дру-
гим — с Deuterostomia. Сходства носят характер па-
раллелизмов и гомологий, что свидетельствует о фи-
логенетическом родстве всех сравниваемых групп.
Можно сказать, что все современные Coelomata про-
изошли от общих предков, вероятно, уже приблизив-
шихся к целомическому уровню организации (Ива-
нов А. В., 1975а). К такому же выводу о монофилети-
ческом единстве Coelomata пришел и Зивинг после
очень детального анализа сходств и различий между
разными группами целомических животных (Siewing,
1967, 1972, 1975).
Итак, в большой системе помимо новых достиже-
ний современной науки должны быть учтены следую-
щие моменты.
1. Особая судьба зародышевых листков у Porifera
и раннее обособление этой группы от общего ствола
Metazoa.
2. Примитивность Scolecida.
3. Единство Protostomia (=Spiralia).
4. Несопоставимость альтернатив Protostomia—
Deuterostomia.
5. Монофилетическое происхождение Coelomata.
6. Известный параллелизм в эволюции главных
стволов Coelomata.
Отвечающий этим требованиям вариант системы
(Иванов А. В., 1975а, 1976) был в свое время реко-
мендован для «Основ зоологии». С небольшими из-
менениями эта система имеет следующий вид.
Animalia
Подцарство A. Protozoa10
Тип 1. Sarcomastigophora
Тип 2. Apicomplexa
Тип 3. Cnidosporidia
Тип 4. Microsporidia
Тип 5. Ciliophora
Подцарство В. Metazoa
Надотдел I. Phagocytellozoa
Тип 6. Placozoa
Надотдел II. Enantiozoa (=Parazoa)
Тип 7. Porifera
Надотдел III. Eumetazoa (=Enterozoa)
Отдел I. Radiata (=Diploblastica)
Тип 8. Cnidaria
Тип 9. Ctenophora
Тип 10. Mesozoa
Отдел 2. Bilateria (=Triploblastica)
Подотдел a. Scolecida (=Protostomia Acoelomata)
Тип 11. Plathelminthes
Тип 12. Nemathelminthes
Тип 13. Nemertini
10 В настоящем издании принята иная, более современная сис-
тема простейших (см. с. 135).
Подотдел b. Coelomata
Надтип Trochozoa (Protostomia Coelomata)
Тип 14. Mollusca
Тип 15. Sipunculida
Тип 16. Echiurida
Тип 17. Annelida
Тип 18. Tardigrada
Тип 19. Pentastomida
Тип 20. Onychophora
Тип 21. Arthropoda
Надтип Tentaculata
Тип 22. Tentaculata
Надтип Chaetognatha
Тип 23. Chaetognatha
Надтип Pogonophora
Тип 24. Brachiata
Надтип Deuterostomia
Тип 25. Hemichordata
Тип 26. Echinodermata
Тип 27. Chordata
Животное царство (Regnum) — Animalia, или
Zoa, делится прежде всего на 2 подцарства (Subreg-
na): Protozoa — одноклеточные и Metazoa — много-
клеточные. Это разделение, предложенное Геккелем
(Haeckel, 1874), считается общепризнанным. К под-
царству Protozoa принадлежат 5 типов: Sarcomasti-
gophora, Apicomplexa, Cnidosporidia, Microsporidia и
Ciliophora. Следует, однако, иметь в виду, что эти
таксоны не соответствуют по своему рангу типам Me-
tazoa, так как установлены не по признакам промор-
фологии и общего плана строения, а преимуществен-
но по сложности организации клеточных органелл,
их производных и комплексов (Grell, 1980а, 1980b).
Тем не менее их естественность и правомочность
этим не нарушается, поскольку в новых макросисте-
мах (см. напр.: Серавин, 1980; Старобогатов, 1986)
понятия Protozoa вообще не существует, и все тради-
ционно относимые к ним группы распределены по
разным царствам, в которых принципы установления
высших таксонов могут быть иными, чем для Anima-
lia. Metazoa разделены на 3 надотдела (superdivisio-
nes).-Первый надотдел — Phagocytellozoa, Фагоци-
теллообразные — установлен А. В. Ивановым (1973)
на основании изучения Trichoplax adhaerens — наи-
более примитивного представителя современных Me-
tazoa. По организации Trichoplax очень близок к
фагоцителле, которая является, по И. И. Мечникову,
общим предком многоклеточных. Грелль (Grell, 1971)
установил для Trichoplax и его родичей новый тип —
Placozoa, который по-русски мы будем называть плас-
тинчатые. Второй надотдел — Enantiozoa, энантио-
зои (Delage, 1892), или Parazoa (Sollas, 1884). К нему
относится единственный современный тип Archaeo-
cyatha. Анализ современных исследований эмбрио-
нального развития губок (см.: Codreanu, 1970; Ива-
нов А. В., 1971; Иванова-Казас, 1975; Гуреева, 1976;
Salvini-Plawen, 1978, и др.) дает все основания под-
держивать учение Деляжа об инверсии зародышевых
листков у этих животных. Третий надотдел составля-
ют Eumetazoa — эуметазои (название принадлежит
Бючли, — см.: Butschli, 1910). Это животные в гро-
мадном большинстве с кишечником и с ротовым от-
верстием (Enterozoa, по терминологии Ланкесте-
ра, — см.: Lankester, 1877), с органами, построенны-
ми из тканей. Eumetazoa, в свою очередь, делятся на
2 отдела (divisiones): Radiata и Bilateria.
СИСТЕМА ЖИВОТНОГО МИРА
111
Radiata — радиальные животные. Этот отдел вы-
делен Шимкевичем (Schimkewitsch, 1891). Тело Ra-
diata во взрослом состоянии слагается из двух кле-
точных слоев — эпидермы и гастродермы, отвечаю-
щих зародышевым листкам — экто- и энтодерме
(отсюда их синоним Diploblastica, — Lankester, 1873).
Сюда относятся два типа: Cnidaria — стрекающие,
или книдарии, и Ctenophora — гребневики. Иногда
оба типа объединяются под старым названием Coe-
lenterata — кишечнополостные.
К Radiata мы причисляем провизорно (как допол-
нение) тип Mesozoa, содержащий два класса пара-
зитических животных — Orthonectida и Rhombozoa
(=Dicyemida). Первые обитают в различных морских
беспозвоночных, вторые — в головоногих моллюс-
ках. И те и другие отличаются сложным жизненным
циклом. Название Mesozoa было предложено Бенеде-
ном (Beneden, 1876), который видел в этих животных
формы, переходные от Protozoa к Metazoa. До сих
пор, однако, систематическое положение Mesozoa
неясно. Многие авторы считают их сильно упрощен-
ными плоскими червями, другие оценивают простоту
строения Mesozoa как первичную. Действительно, по
организации они стоят на уровне личинки книда-
рий — паренхимулы, а отсутствие у них мускульных
и нервных элементов и наличие бесполых генератив-
ных клеток свидетельствует против родства с Plathel-
minthes.
Bilateria (Hatschek, 1888), или Triploblastica (Lan-
kester, 1873), — билатеральные животные; обладают
двусторонней симметрией тела и тремя зародышевы-
ми листками.
Что касается систематического расчленения Bila-
teria, то, как мы видели, у разных авторов оно сильно
варьирует (см. выше). У многих зоологов это понятие
целиком совпадает с понятием Coelomata — взгляд,
основанный, как мы могли убедиться, на очень шат-
ких гипотезах. Однако еще Гексли (Huxley Th., 1864)
ясно показал, что среди червей следует различать
наиболее примитивную группу — Scolecida, которая
объединяет формы, еще лишенные вторичной полос-
ти тела и сегментации. Эта концепция должна быть
признана единственным возможным решением.
Итак, в состав Bilateria входят 2 подотдела (subdi-
visiones): Scolecida и Coelomata.
Низшие черви, или Scolecida (=Acoelomata, —
Schimkewitsch, 1891), с шестью типами, которые по
большей части совпадают с типами подраздела того
же названия в системе Грассе (см. с. 99). Однако мы
не включаем сюда Mesozoa, а гнатостомулид (Gna-
thostomulida) не считаем особым типом, как это дела-
ют некоторые авторы (Riedl, 1969; Sterrer, 1972; Rie-
ger, Mainitz, 1977), но причисляем к плоским червям
в качестве класса.
Обширный и, возможно, гетерогенный тип первич-
нополостных червей — Nemathelminthes (=Aschel-
minthes), фигурирующий в большинстве учебников и
руководств, разделен нами на несколько подтипов.
Попытки разбить его на несколько самостоятельных
типов предпринимались давно. Прежде всего, был вы-
делен тип скребней — Acanthocephala (Скрябин, Шульц,
1931). Серьезная реформа в классификации Nemathel-
minthes была произведена Грассе (Grasse et al., 1961),
который предложил различать 4 типа: Rotifera, Acan-
thocephala, Nemathelminthes (s. str.) (с классами Nema-
toda и Nematomorpha) и Nematorhyncha (с классами
Gastrotricha и Kinorhyncha). Однако два последних
таксона вряд ли можно признать достаточно обосно-
ванными — слишком разнородными являются объеди-
няемые в них классы. В классификации, принятой
нами, эти группы заменены двумя другими, по сущест-
ву, новыми группами, установленными и обоснован-
ными Малаховым (1980). Один из них, Nemathelmin-
thes (s. str.) (с тем же названием, что и у Грассе), объ-
единяет два несомненно близкородственных класса —
Nematoda и Gastrotricha. Другой таксон, которому Ма-
лахов придал ранг типа — Cephalorhyncha, содержит
классы Kinorhyncha, Gordiacea и Priapulida, обладаю-
щие рядом общих, весьма характерных признаков. К
ним, по-видимому, должен быть присоединен откры-
тый несколько лет назад (Kristensen, 1983) класс
Loricifera.
Внимательное рассмотрение диагнозов этих так-
сонов, однако, не позволяет признать в них настоя-
щие типы, сравнимые по рангу с таковыми других
Eumetazoa (табл. 6). Из таблицы ясно видно, сколь
немногочисленны серьезные различия между срав-
ниваемыми группами. Истинная природа мозга недо-
статочно ясна; нервные стволы во всех случаях могут
рассматриваться как сильно олигомеризованные ство-
лы ортогона. Иными словами, ни для одной из четы-
рех групп первичнополостных червей нельзя дать
полноценный диагноз типового ранга. Поэтому, по-
видимому, будет более правильным пока считать все
эти группы подтипами одного большого типа — Ne-
mathelminthes, все члены которого находятся на одном,
а именно первичнополостном, уровне эволюции (см.:
Беклемишев, 1964).
Ко вторичнополостным — Coelomata (Haeckel,
1874) принадлежат все остальные типы.
Из сказанного выше следует, что современные Co-
elomata представлены не двумя, а несколькими рав-
ноценными по рангу группами.
Одну из них составляет надтип (Superphylum)
Trochozoa — трохофорные, установленный В. Н. Бе-
клемишевым (1944). Трохофорные животные обладают
характерной личинкой — трохофорой. Надо отме-
тить, что Сальвини-Плавен резко отделяет моллюс-
ков от всех Trochozoa и сближает их со сколецидами,
полагая вслед за рядом авторов, что они произошли
от турбелляриеобразных предков, а целом приобрели
независимо. Эти положения обоснованы в блестящей
работе Сальвини-Плавена (Salvini-Plawen, 1972). Од-
нако мы не решаемся принять эту точку зрения, не-
смотря на большую ее соблазнительность, из-за нали-
чия у моллюсков типичной трохофоры.
Что касается эхиурид, то они часто рассматрива-
лись как вторично упрощенные потомки полимерных
кольчатых червей и присоединялись к типу Annelida
в качестве отдельного класса. Ныне эта точка зрения
признана ошибочной (Hyman, 1959; Кот, 1960; Clark,
1964, 1969; Kaestner, 1965). Первично нерасчленен-
ный целом эхиурид как нельзя лучше свидетельствует
о том, что сегментация отнюдь не представляет собой
первичную черту организации первобытных Coelo-
mata.
112
ОБЩАЯ ЧАСТЬ
Таблица 6
Сопоставление признаков типов, составленное по диагнозам В. В. Малахова (1980)
Rotifera (класс Rotifera) Acanthocephala (класс Acanthocephala) Nemathelminthes s. str. (классы: Nematoda, Gastrotricha) Cephalorhyncha (классы: Gordiacea, Kinorhyncha, Priapulida)
Интраэпительная пластинка Кутикула Продольная и кольцевая мускула- тура Эндон Пара продольных нервных ство- лов Схизоцель .. Сквозной кишечник Мастакс Протонефридии Клоака Раздельнополость Видоизмененное спиральное дроб- ление Бластопор — рот Интраэпительная пластинка Кутикула Хобот с влагалищем1 Продольная и кольцевая мускула- тура Эндон3 Пара продольных нервных ство- лов Схизоцель Протонефридии Клоака Раздельнополость Видоизмененное спиральное дроб- ление Кутикула Продольная мускулатура2 Нервное кольцо Ортогон или пара стволов Схизоцель Сквозной кишечник pharynx bulbosus Протонефридии Раздельнополость Гермафродитизм1 Билатеральное дробление5 Бластопор — рот—анус Кутикула Хобот (типа интроверт)1 Продольная и кольцевая мускула- тура Нервное кольцо4 Вентральный нервный ствол Схизоцель Сквозной кишечник pharynx bulbosus Протонефридии Раздельнополость Бластопор—анус
1 Признак, свойственный лишь данной группе.
2 У Gastrotricha есть слабо развитая кольцевая мускулатура.
3 Мозг расположен в задней части хоботного влагалища.
4 Нервное кольцо в передней части хобота.
5 Сильно видоизмененное спиральное дробление (Иванова-Казас, 1975).
Сипункулиды, вероятно, представляют реликто-
вую группу, близкую к вымершим первичным Trocho-
zoa с несегментированным целомом (Clark, 1964,
1969; Salvini-Plawen, 1972). Старые данные о том,
что у некоторых сипункулид мезодермальные полос-
ки временно сегментируются, не подтвердились (см.:
Hyman, 1959). Впрочем, у взрослых сипункулид от
туловищного целома обособлен небольшой передний
участок, охватывающий в виде узкого кольца перед-
ний конец кишечника и посылающий целомические
каналы в околоротовые щупальца. Таким образом,
можно говорить о наличии у сипункулид двух отде-
лов целома.
В состав типа Annelida входят классы Polychae-
ta, Dinophilida, Myzostomida, Oligochaeta и Hirudi-
nea. Олигомерные динофилиды и мизостомиды обла-
дают только ларвальными сегментами (Иванов П. П.,
1928а, 19286; Федотов, 1929) и характеризуются
несегментированным половым целомом, лишенным
опорной функции. Приняв Dinophylus за первичную
примитивную форму, мы не смогли бы понять эко-
логические и физиологические причины появления
целома у его предков. Передвижение посредством
мерцательных поясков не требует развития целома
с гидроскелетной функцией. Наличие у Dinophylus
и Myzostomida метамерии в некоторых системах
органов (нервный аппарат, нефридии), таким обра-
зом, естественно рассматривать как наследие от ан-
нелидных предков, т. е. от животных, прошедших
уже филогенетическую фазу роющих организмов,
когда появление расчлененного целома имело явные
селективные преимущества. Несовершенный несег-
ментированный целом характерен для ларвальных
сегментов некоторых примитивных полихет (Ива-
нов П. П., 1928; Беклемишев, 1964). Очень вероят-
но поэтому, что Dinophylus и Myzostomida про-
изошли от их личиночных форм (метатрохофор)
посредством неотении.
Tardigrada, Pentastomida и даже Onychophora
иногда включаются в состав Arthropoda, но многие
авторы рассматривают их как самостоятельные
типы, что представляется нам оправданным. Вряд
ли можно сомневаться, что артроподные черты в
этих группах развились параллельно и независимо
от членистоногих. Энтероцельное развитие целоми-
ческих мешочков у Tardigrada несомненно носит
вторичный характер и тем не менее указывает на
самостоятельный путь эволюции этой группы. Что
касается Pentastomida, то старое предположение об
их родстве с клещами (Leuckart, 1860, и др.) ка-
жется очень маловероятным (Cuenot, 1949; Osche,
1963; Kaestner, 1965).
Неправдоподобно и сближение пентастомид с
ракообразными (карповыми вшами — Branchiura),
основанное на сходстве ультраструктуры спермиев
у тех и у других. Поэтому мы присоединяемся к
мнению Беклемишева, Кестнера и Кено, защищаю-
щих типовую самостоятельность пятиусток. Panto-
poda условно помещены рядом с Chelicerata.
Труднейшую проблему, далекую от разрешения,
представляет вопрос о систематическом положении
Kamptozoa (это название принадлежит Кори, —
см.: Cori, 1929). Как показывает метаморфоз ли-
чинки, линия, соединяющая рот и анус взрослого
животного, является медиовентральной, тогда как у
Phoronida и Bryozoa это спинная сторона тела.
CLASSIFICATION OF THE ANIMAL WORLD. SUMMARY
113
Сближение Kamptozoa и Tentaculata поэтому ис-
ключается. Разделение тела на чашечку и стебелек
(а не на полипид и цистид), отсутствие целома,
развитие протонефридиев подтверждают это заклю-
чение. Неоднократно отмеченное сходство в строе-
нии личинки Kamptozoa и некоторых Bryozoa (Ива-
нов П. П., 1937; Nielsen, 1971) не может его поко-
лебать. Отнюдь не лучшим выходом из положения
было бы причисление Kamptozoa в ранге ^типа к
Scolecida, основанное на наличии спирального дроб-
ления, развития паренхимы за счет бластомера 4d
и присутствия протонефридиев (Hatschek, 1888; Mac-
Bride, 1914). Однако в спиральном гомоквадрант-
ном дроблении Kamptozoa есть много специальных
гомологий с таковым Polychaeta, а личинка Kamp-
tozoa очень близка к трохофоре. План строения
Kamptozoa, следовательно, можно рассматривать как
видоизменение организации трохофоры, приспособ-
ленной к сидячему образу жизни (Беклемишев,
1964). Присоединяясь к этому мнению, мы включа-
ем Kamptozoa провизорно в надтип Trochozoa. От-
сутствие целома вряд ли является препятствием, если
предположить, что у Kamptozoa он вторично исчез
в связи с сидячей жизнью.
Самостоятельный надтип, по нашему мнению,
должны составлять Tentaculata — щупальцевые.
Под этим названием Гатчек (Hatschek, 1888) объ-
единял Phoronida, Bryozoa и Brachiopoda. Между
всеми классами тентакулят существует несомненное
родство, что подтверждается сравнительно-анатоми-
чески и эмбриологически (Hyman, 1959; Ливанов,
1963; Kaestner, 1965). Вопреки этому, Беклемишев
(1964) резко обособляет Brachiopoda как самосто-
ятельный подраздел (subdivisio), т. е. как группу,
соответствующую по рангу Protostomia и Deutero-
stomia. Для обозначения щупальцевых было пред-
ложено несколько различных наименований: Lo-
phophorata (Hyman, 1959), Podaxonia (Беклемишев,
1944), Prosopygia (Ливанов, 1963), но мы не видим
ни в одной из них особых преимуществ перед ста-
рым гатчековским названием.
Как особую группу в ранге надтипа, по-видимо-
му, следует рассматривать Chaetognatha — щетин-
кочелюстных (Leuckart, 1854). Их тело состоит из
двух первичных отделов. Причисление Chaetognatha
к вторичноротым вряд ли оправдано (Беклемишев,
1964).
Четвертый надтип — Pogonophora (Johansson, 1937),
погонофоры. По основному плану строения и осо-
бенностям развития целома погонофоры ближе к
Deuterostomia, чем к другим Coelomata, но по спи-
ральному дроблению, образованию щетинок и связи
половой системы с целомом они сходны с Trochozoa
и отчасти с Tentaculata. Единственный тип погоно-
фор можно именовать Brachiata — брахиаты (Ива-
нов А. В., 1955а, 19556).
Наконец, еще один надтип составляют Deuterosto-
mia — вторичноротые, понимаемые в узком смысле
слова как объединение только явно родственных
типов: Hemichordata, Echinodermata и Chordata, в ос-
нове организации которых лежит тримерный план
строения.
CLASSIFICATION OF THE ANIMAL WORLD
SUMMARY
The present-day zoology can not afford boasting of the universal
«general system» of a taxon ranging higher than class.
In the early XX century two general directions of elaboration —
into zoological system were evident. These directions disagree with 2
points:
1. Over the evaluation of systematic significance of embriological
peculiarities, for instance the character of definitive mouth and the
coelom development.
2. Over the interpretation of nature of the groups devoid of coe-
lom such as Bilateria.
The Old School supported the opinion that the similar ways of
organs’ development might succeed into different Bilateria groups in-
dependently and as it takes place the characters appear to be of no
systematic importance. It allowed to pull together the groups with
schisocoelic and enterocoelic coelom or the ones with primary or se-
condary mouth. This position can not prevent from the monophyletic
principle being observed consistently during the system’s constructi-
on. Within the framework of this school some systems were elabora-
ted, such as the ones that Kukenthal—Krumbach used in «Handbuch
der Zoology», or P.-P. Grasse — in «Traite de Zoologie», or
P. Brien — in «Elements de Zoologie, notions d’anatomia comparee».
The classification of the Hatchek—Grobbens’s school permits di-
vision into: A — diphyletic systems, В — monophyletic systems,
C — polyphyletic systems. Within the first systems’ framework the
classifications elaborated by Zenkevich (1937), Fedotov (1966),
A. Ivanov (1957) and Kaestner (1965) are discussed. To illustrate the
second systems’ classifications those of Hyman (1940); Livanov
(1955); Grasse et al. (1961);. Remane et al. (1976); Nielsen (1985) are
adduced. As an example of the polyphyletic systems’ classification the
one by V. Beklemishev (1944, 1964) is discussed.
If Protostomia and Deuterostomia systematic importance is con-
cerned the presenuday zoological systems seem to be quite different.
What different ways these conceptions would explain Scolecida and
Coelomata origin.
6 concepts are named in this chapter:
1. Polyphyletic nature of Bilateria (V. Beklemishev).
2. Diphyletic hypothesis concerning the independent origin of
Protostomia and Deuterostomia (Hatschek, 1911).
3. Archicoelomate theory (Masterman, 1898; Ulrich, 1949, 1973).
4. Theory after Livanov (1955) based on monophyletic nature of
Coelomata and on schizocoelic origin of the coelom.
5. Conception of Hyman (1940) concerning the monophyletic ori-
gin of Bilateria and diphyletic nature of coelom of Coelomata.
6. Monophyletic scheme of the Bilateria evolution based on the
Phagocityella theory of I. Mechnikov.
Monophyletic unity of Coelomata is discussed.
The system of Animalia by A. V. Ivanov recommended for zoo-
logists is presented.
8 Протисты, часть 1
114
ОБЩАЯ ЧАСТЬ
ЛИТЕРАТУРА11
Антонов А. С. Степень вариабельности состава ДНК живот-
ных как критерий эволюционного возраста таксона // Проблемы
эволюции. Новосибирск, 1968. Т. I, С. 37—46.
Астауров Б. Л. Искусственный партеногенез у тутового шел-
копряда. М.; Л., 1940.
Астауров Б. Л. Цитогенетика развития тутового шелкопряда и
ее экспериментальный контроль. М.: Наука, 1968.
Беклемишев В. Н. Основы сравнительной анатомии беспозво-
ночных. М., 1944.
Беклемишев В. Н. Основы сравнительной анатомии беспозво-
ночных. 3-е изд. М„ 1964.
Белозерский А. Н. Состав нуклеиновых кислот и эволюци-
онная систематика И Молекулярная биология. М., 1964. С. 5—
13.
Берг Л. С. Номогенез или эволюция на основе закономерно-
стей. СПб., 1922.
Биологический энциклопедический словарь. М., 1986.
Бирштейн Я. А. Новые взгляды на филогению членистоно-
гих И Успехи соврем, биол. 1948. Т. 25, № 1. С. 143—149.
Бирштейн Я. А. Класс Ракообразные. Crustacea // Атлас бес-
позвоночных Каспийского моря. М., 1968. С. 118—120.
Бобров Е. Г. Об особенностях флоры эрратической области
(один из путей формообразования) И Советская ботаника. М.; Л.,
1944. Т. 2. С. 3—20.
Боркин Л. Я., Даревский И. С. Сетчатое (гибридогенное) ви-
дообразование у позвоночных // Журн. общ. биол. 1980. Т. 40,
№ 4. С. 465—506.
Бубко О. В., Миничев Ю. С. Нервная система Oweniidae (Po-
lychaeta) И Зоол. журн. 1972. Т. 1, № 9. С. 1288—1298.
(Быстров А. П.) Bystrow А. Р. Dekknochen und Zahne der Oste-
olepsis und Dipterus // Acta Zool. 1942. N 25, P. 263—289.
Быстров А. П. Прошлое, настоящее и будущее человека. М.,
1957.
Вавилов Н. И. Центры происхождения культурных растений.
Л., 1926.
Вавилов Н. И. Линнеевский вид как система // Тр. по прикл.
ботан., генет., селекц. 1931. Т. 26, № 3. С. 109—134.
Видаль Г. Древнейшие эукариотические клетки И В мире науки.
1984. № 13. С. 13—24.
Виноградов А. П. О причине отсутствия известковых скелетов
у докембрийских беспозвоночных И Докл. АН СССР. 1940. Т. 27,
№ 3. С. 230—233.
Вологдин А. Г. Земля и жизнь. М., 1963.
Воробьёва Э. И. Структурная перестройка позвоночных в
связи с выходом на сушу И Современные проблемы эволюционной
морфологии животных. М., 1981. С. 17—18.
Воронцов Н. Н. Неравномерность темпов преобразования
органов пищеварительной системы грызунов и принцип компен-
сации функций И Докл. АН СССР. 1961. Т. 136, № 6. С. 1494—
1497.
Воронцов Н. Н. Неравномерность темпов преобразования ор-
ганов и принцип компенсации функций К Зоол. журн. 1963. Т. 42,
№ 9. С. 1289—1305.
Воронцов Н.Н. Эволюция пищеварительной системы грызу-
нов. Новосибирск, 1967.
Воронцов Н.Н.Теория эволюции: истоки, постулаты и про-
блемы. М., 1984.
Воронцов Н. Н. Развитие эволюционных идей в биологии. М.:
Прогресс—Традиция, 1999.
Вяккерев Ф. Ф. Проблемы самодвижения в материалистичес-
кой диалектике. Л., 1972.
Галл Я. М. Исследования по естественному отбору # Развитие
эволюционной теории в СССР. Л., 1983. С. 221—252.
Галл Я. М., Георгиевский А. Б. Синтетическая теория эволю-
ции как современный этап развития дарвинизма И Биология в
щколе. 1973. № 3. С. 7—15; № 4. С. 5—14.
(Гаузе Г. Ф.) Gause G. F. The struggle for existence. Baltimore,
1934.
Гаузе Г. Ф. Роль приспособляемости в естественном отборе И
Журн. общ. биол. 1940. Т.1,№ 1.С. 105—120.
Геккель Е., Мюллер Ф. Основной биологический закон // Из-
бранные работы. М., 1940.
11 Литература к разделам «Пути и закономерности эволюции»,
«Систематика и ее задачи», «Система животного мира».
Гептнер В. Г. О числе видов в фауне СССР и об отношении
ее к мировой фауне И Зоол. журн. 1956. Т. 35, № 12. С. 1780—
1790.
Гептнер В. Г. Систематика биологическая И БСЭ. 3-е изд. М.,
1976. Т. 23. С. 1349—1405.
Гершензон С. М. Мобилизационный резерв внутривидовой из-
менчивости И Журн. общ. биол. 1941. Т. 2, № 1. С. 85—107.
Гиляров М. С. Закономерность приспособления членистоно-
гих к жизни на суше. М., 1970а.
Гиляров М. С. Закономерности и направления филогенеза //
Журн. общ. биол. 19706. № 2. С. 179—188.
Гиляров М. С. Общие направления эволюции насекомых и выс-
ших позвоночных // Зоол. журн. 1975. Т. 56, № 6. С. 822—831.
Гинецинская Т. А., Добровольский А. А. Частная паразитоло-
гия. М., 1978.
Гуреева М. А. Эмбриональное развитие и зародышевые листки
у губок// Тр. Ленинигр. о-ва естествоиспыт. 1976. Т. 84, № 1.
С. 10—25.
(Давыдов К. К.) Dawydoff С. С. Observation sur le developpe-
ment des Scalopendrelles (Symphyla) indohinoises // Bull. Biol. Fran-
ce Belgique. 1942. N 77. P. 1—28.
Даревский И. С. Эпистандартная эволюция и гибридогенное
видообразование у пресмыкающихся И Журн. общ. биол. 1995.
Т. 56, №3. С. 310—316.
(Догель В. A.) Dogiel V. A. Polymerization als ein Prinzip der
progressiven Entwicklung bei Protozoen // Biol. Zbl. 1929. N 49.
S. 451—469.
Догель В. А. Олигомеризация гомологичных органов как один
из процессов эволюции животных И Архив анат., гистол. и эмбри-
ол. 1936. Т. 15, № 3. С. 101—114.
Догель В.А. Некоторые перспективы развития советской срав-
нительной анатомии в связи с ее достижениями за последнее
время И Природа. 1937. № 11. С. 46—53.
Догель В. А. Сравнительная анатомия беспозвоночных. Ч. 1.
Л., 1938.
Догель В. А. Общая протистология. М., 1951.
Догель В. А. Зоология беспозвоночных. М., 1952.
Догель В. А. Олигомеризация гомологичных органов как один
из главных путей эволюции животных. Л., 1954.
Догель В. А. Зоология беспозвоночных. 5-е изд. М., 1959.
Догель В. А., Полянский Ю. И., Хейсин Е. М. Общая прото-
зоология. М.; Л., 1962.
Друшиц В. В. Палеонтология беспозвоночных. М., 1974.
Дубинин Н. П. Генетико-автоматические процессы и их значе-
ние для механизма органической эволюции И Журн. эксперим.
биол. 1931. Т. 7, вып. 5—6. С. 463^79.
Дубинин Н. П. О некоторых основных проблемах генетики И
Ботан. журн. 1932. Т. 1, вып. 2. С. 112—146.
Дубинин Н. П. Генетика и ее значение для человечества И
Вести. АН СССР. 1980. № 6. С. 73—81.
Дубинин Н. П., Ромашов Д. Д. Генетическое строение вида и
его эволюция И Биол. журн. 1932. Т. 1, вып. 5—6. С. 52—95.
Дубинин Н. П., Тиняков Г. Г. Сезонный цикл и концентра-
ция инверсий в популяциях И Докл. АН СССР. 1946. Т. 52, вып. 1.
С. 77—79.
Дубинина М. Н. Паразитические черви класса Amphilinida
(Plathelminthes). Л., 1982. (Тр. ЗИН АН СССР; Т. 100).
Ефремов И. А. О подклассе Batrachosauria — группе форм,
промежуточных между земноводными и пресмыкающимися # Изв.
АН СССР. 1946. № 6. С. 615—638.
Завадский К. М. О причинах выпада растений в гнездовых по-
севах различной плотности в зависимости от размеров гнезд и ус-
ловий минерального питания И Ботан. журн. 1954. Т. 39, № 4.
С. 515—544.
Завадский К. М. К пониманию прогресса в органической при-
роде И Проблемы развития природы и общества. Л.; М., 1958.
С. 79—120.
Завадский К. М. Вид и видообразование. Л., 1968.
Завадский К. М. Синтетическая теория эволюции и диалекти-
ческий материализм И Философские проблемы эволюционной тео-
рии. Ч. 2. М., 1971. С. 4—30.
Завадский К.М., Колчинский Э. И. Эволюция эволюции. Л.,
1977.
Завадский К. М., Колчинский Э. И., Ермоленко М. Т. Главные
этапы развития эволюционной теории в СССР И Развитие эволюци-
онной теории в СССР. Л., 1983. С. 8—42.
ЛИТЕРАТУРА
115
Захваткин А. А. Сравнительная эмбриология низших беспоз-
воночных. М., 1949.
Зенкевич Л. А. История системы беспозвоночных: Руководст-
во по зоологии. Т. 1. Беспозвоночные. М.; Л„ 1937.
Зенкевич Л. А. Система и филогения И Зоол. журн. 1939.
Т. 18, №4. С. 600—611.
Иванов А. В. Морфологические адаптации к паразитическо-
му образу жизни И Учен. зап. ЛГУ. 1937. № 13, Т. 3, вып. 4.
С. 53—94.
Иванов А. В. Строение и развитие эндопаразитического брю-
хоногого моллюска Parenteroxenos dogieli A. Ivanow (ceM.Entocen-
chidae). Сообщ. 2, 3 И Изв. АН СССР. 1949. № 2. С. 109—139.
Иванов А. В. О принадлежности класса Pogonophora к особо-
му типу вторичноротых — Brachiata A. Ivanov, phyl. nov. И Докл.
АН СССР. 1955а. Т. 100, № 3. С. 595—596.
Иванов А. В. Промысловые водные беспозвоночные. М., 19556.
Иванов А. В. Филогенез беспозвоночных животных // БСЭ.
2-е изд. М., 1957. Т. 45. С. 109—112.
Иванов А. В. Пуги эволюционного развития в классе Pogono-
phora // Тр. ИЭМЭЖ. 1959. Вып. 27. С. 274—278.
Иванов А. В. Погонофоры. М.; Л., 1960.
Иванов А. В. Пауки, их строение, образ жизни и значение для
человека. Л., 1965.
Иванов А. В. Происхождение многоклеточных животных. Л.,
1968.
Иванов А. В. Эмбриональное развитие губок (Porifera) и поло-
жение их в системе животного мира И Журн. общ. биол. 1971.
Т. 32, № 3. С. 557—572.
Иванов А. В. Trichoplax adhaerens — фагоцителлообразное
животное И Зоол. журн. 1973. Т. 52, № 8. С. 117—131.
Иванов А. В. О происхождении Coelomata // Журн. общ. биол.
1975а. Т. 36, № 5. С. 643—653.
Иванов А. В. Наблюдения над эмбриональным развитием Po-
gonophora. Сообщ. 2. О систематическом положении Pogonopho-
ra И Зоол. журн. 19756. Т. 54, № 8. С. 1125—1140.
(Иванов А. В.) Ivanov А. V. Embryonalentwicklung der Pogo-
nophora und ihre systematische Stellung // Ztschr. Zool. Syst.-Evol.
Forsch. 1975b. S. 10—44.
Иванов А. В. Соотношение между Protostomia и Deuterostomia
и система животного мира // Зоол. журн. 1976. Т. 55, № 6.
С. 1125—1137.
Иванов А. В. Олигомеризация и филогения в отряде пауков И
Значение процессов полимеризации и олигомеризации в эволю-
ции. Л., 1977. С. 50—52.
Иванов А. В. Тип Погонофоры (Pogonophora) И Жизнь живот-
ных. Т. 2. 2-е изд. М., 1988а.
(Иванов А. В.) Ivanov А. V. Analysis of the embryonic develop-
ment of Pogonophora in connection with problems of phylogenetics //
Ztschr. Zool. Syst.-Evol. Forsch. 19886. Bd 26, N 3. S. 161—185.
Иванов А. В., Мамкаев Ю. В. Ресничные черви, их происхож-
дение и эволюция. Л., 1973.
Иванов Д. Л. Филогения подкласса Caudofoveata // Тр. ЗИН
АН СССР. 1986. Т. 148. С. 12—32.
Иванов П. П. Эмбриональные черты в строении человека И
Человек. 1928а. № 2—4. С. 117—133.
(Иванов П. П.) Ivanoff Р. Р. Die Entwicklung der Larvalsegmen-
te bei den Anneliden // Ztschr. Morphol. Oekol. Tiere. 1928b. N 10.
S. 62—161.
(Иванов П. П.) Ivanoff P. P. Die Entwicklung von Limulus mol-
lucana // Zool. Jb. Anat. 1933. N 56. S. 163—348.
Иванов И. П. Общая и сравнительная эмбриология. М., 1937.
Иванова-Казас О. М. К вопросу о соотношении морфологи-
ческих осей у Spiralia И Зоол. журн. 1974. Т. 53, № 1. С. 5—18.
Иванова-Казас О. М. Сравнительная эмбриология беспозво-
ночных животных. Новосибирск, 1975.
Иванова-Казас О. М. Современное состояние теории первич-
ной гетерономности сегментов И Зоол. журн. 1978. Т. 57, № 11.
С. 1605—1617.
Иванова-Казас О. М. Сравнительная эмбриология беспозво-
ночных животных. Членистоногие. М., 1979.
Иванова-Казас О. М. Филогенетическое значение спирально-
го дробления И Биология моря, 1981. № 5. С. 3—14.
Иванова-Казас О. М. Становление билатеральной симметрии
в онтогенезе // Тр. ЗИН АН СССР. 1983. Т. 109. С. 90—110.
Иванова-Казас О. М., Иванов А. В. О теории Трохеи и о фи-
логенетическом -значении ресничных личинок И Биология моря.
1987. № 2. С. 5—21.
Иорданский И. Н. Развитие жизни на Земле. М., 1981.
Каллиникова В. Д. Эволюционные преобразования жизнен-
ных циклов Sporozoa и филэмбриогенезы И Успехи соврем, биол.
1991. Т. 111, вып. 1. С. 108—121.
Камшилов М. М. Отбор как фактор, меняющий зависимость
признака от изменения внешних условий И Докл. АН СССР. 1939.
Т. 23, № 4. С. 361—364.
Камшилов М. М. Эволюция биосферы. М., 1974.
Карпеченко Г. Д. Полиплоидные гибриды Raphanus sativus L.
и Brassica oleracea L. // Тр. по прикл. ботан., генет., селекц. 1927.
Т. 17, вып. 3. С. 305—410.
Келлер Б. М. Загадки верхнего докембрия // Природа. 1979.
№ 1. С. 66—75.
Келлер Б. М., Федонкин М. А. Новые находки окаменелостей
в Валдайской серии докембрия по р. Сюзьме // Изв. АН СССР.
Сер. геол. 1976. N 3. С. 38—44.
Кирпичников В. С. Роль ненаследственных изменений в про-
цессе естественного отбора И Биол. журн. 1935. Т. 4, вып. 5.
С. 775—801.
Киршенблат Я. Д. Кайнотропозы — одно из главных направ-
лении эволюционного процесса // Вести, зоологии. 1969. № 3.
С. 24—28.
(Ковалевский В.) Kowalewsky W. Monographic der Gattung
Antracotherrium Cuv. und Versiich einer natiirlichen Klassifikation
der fassilen Huftthiere // Paleograph. 1874. N. F. N 11. P. 133—285.
Колтун В. M. Надраздел Parazoa // Жизнь животных. T. 1. М.,
1987. С. 128—152.
Колчинский Э. И. Эволюция биосферы: историко-критичес-
кие очерки исследований в СССР. Л., 1990.
Котикова Е. А. Новые данные о нервной системе Archianneli-
da // Зоол. журн. 1973. Т. 52, вып. 11. С. 1611—1615.
Кремянский В. И. Переход от ведущей роли естественного от-
бора к ведущей роли труда И Успехи соврем, биол. 1941. № 14.
С. 356—371.
Крыжановский О. Л. К вопросу о числе видов насекомых в
фауне СССР и в мировой фауне (по поводу статьи проф. В. Г. Гепт-
нера «О числе видов в фауне СССР и об отношении ее к мировой
фауне» в «Зоологическом журнале») И Энтомол. обозр. 1958.
Т. 37, № 3. С. 661—669.
Крылов И. Н. На заре жизни. Органический мир докембрия.
Л., 1972.
Лагутенко Ю. П. О структурной организации нервного плексу-
са Овении (Polychaeta, Oweniidae) // Зоол. журн. 1985. Т. 64,
вып. 12. С. 1802—1807.
Ливанов Н. А. Класс Полихет (Polychaeta) # Руководство по
зоологии. Т. II. М.; Л., 1940. С. 10—136.
Ливанов Н. А. Основные этапы эволюции нервной системы И
Учен. зап. Казан, гос. ун-та. 1941. Т. 101, № 1. С. 79—142.
Ливанов Н. А. Пути эволюции животного мира. М., 1955.
Ливанов Н. А. Форониды, мшанки, брахиоподы И Тр. о-ва ес-
тествоиспыт. при Казан, гос. ун-те. Т. 66. 1963. С. 55—81.
Лукин Е. И. Про пичини залини в процесс! органично! сво-
люцц неспадковых зм!з спадковими з погляду теорп природного
добору U Учен. зап. Харьщв. госп. ун-та. 1936. Т. 6—7. С. 199—
209.
Лукин Е. И. Дарвинизм и географические закономерности в
изменении организмов. М.; Л., 1940.
Лукин Е. И. Различия в скорости эволюции разных систем ор-
ганов и приспособления к размножению и развитию животных И
Зоол. журн. 1964. Т. 45, № 8. С. 1106—1119.
Лукин Е. И. Дарвиновский принцип монофилии и проблема
уменьшения количества параллельных систематических групп в за-
висимости от повышения их ранга И Зоол. журн. 1975. Т. 54, № 1.
С. 5—19.
Лукин Е. И. Пиявки. Т. 1. Л., 1976. (Фауна СССР; Т 109).
Любищев А. А. О форме естественной системы организмов И
Изв. биол. НИИ Перм. ун-та. 1923. Вып. 2. С. 99—110.
Любищев А. А. Проблемы систематики И Проблемы эволю-
ции. Новосибирск, 1968. Т. 1. С. 9—29.
Любищев А. А. Редукционизм и развитие морфологии и систе-
матики И Журн. общ. биол. 1977. Т. 38, № 2. С. 245—263.
Малахов В. В. Cephalorhyncha — новый тип животного царст-
ва, объединяющий Priapulida, Kinorhyncha, Gordiacea, и система
первичнополостных червей И Зоол. журн. 1980. Т. 59, № 4. С. 485—
499.
Малахов В. В., Галкин С. В. Вестиментиферы — бескишеч-
ные беспозвоночные морских глубин. М.: КМК «Scientific Press
Ltd.», 1998.
Маргелис Л. Роль симбиоза в эволюции клеток. М., 1983.
116
ОБЩАЯ ЧАСТЬ
Матвеев В. С. Об эволюции кожных покровов позвоночных
путем эмбриональных изменений // Зоол. журн. 1932. № 11.
С. 38—59.
Махотин А. А. О взаимоотношениях типов филогенетических
изменений органов И Памяти акад. А. Н. Северцова. Т. 2, ч. 1. М.;
Л„ 1940. С. 209—250.
Медников Б. М. Исследования эволюции генома И Развитие
эволюционной теории в СССР. Л., 1983. С. 449—463.
Медников Б. М., Попов Л. С., Антонов А. С. Характеристика
первичной структуры ДНК как критерий для построения естест-
венной системы рыб И Журн. общ. биол. 1973. Т. 34, № 4. С. 516—
529.
Мейстер Г. К. Проблема межвидовой гибридизации в освеще-
нии современного экспериментального метода И Журн. по агроно-
мии Юго-Востока. 1927. Т. 4, № 1. С. 3—88.
Мельников О. Д. О первичной гетерономности сегментов тела
Articulata И Журн. общ. биол. 1971. Т. 32, № 5. С. 597—612.
Мензбир М. А. Очерк истории фауны европейской части
СССР (от начала третичной эры). М.; Л., 1934.
(Мечников И. И.) Metschnikoff Е. Apsilis lentiformis, ein Ra-
dertier // Ztschr. wiss. Zool. 1866. Bd 16. S. 346—356.
(Мечников И. И.) Metschnikoff E. Embryologie der doppeltfus-
sigen Myriapoden U Ztschr. wiss. Zool. 1874a. Bd 24. S. 253—283.
(Мечников И. И.) Metschnikoff E. Studien uber die Entwick-
lung der Medusen und Siphonophoren // Ztschr. wiss. Zool. 1874b.
Bd 21. S. 15—83.
(Мечников И. И.) Metschnikoff E. Embryologische Studien an
Medusen. Ein Beitrag zur Genealogie der Primitiv-Organe. Wien,
1886.
Миничев Ю. С., Старобогатов Я. И. Проблема торсионного
процесса и проморфологические перестройки у личинок трохо-
форных животных # Зоол. журн. 1972. Т. 51, № 10. С. 1437—1449.
Мурина В. В. Морские черви Сипункулиды арктических и бо-
реальных вод Европы. Л., 1977.
Наумов Н. П., Карташев Н. Н. Зоология позвоночных. Ч. 1,2.
М., 1979.
Опарин А. И. Происхождение жизни. М., 1924.
Опарин А. И. Возникновение и начальное развитие жизни. М.,
1957.
Опарин А. И. Современное представление о происхождении
жизни на Земле К Журн. всесоюз. хим. о-ва. 1980. Т. 25. С. 246—
252.
Парамонов А. А. Курс дарвинизма. М„ 1945.
Парамонов А. А. Основы фитогельминтологии. Т. 1. М., 1962.
Парамонов А. А. Пути и закономерности эволюционного про-
цесса (основные принципы филогенетики) // Современные пробле-
мы эволюционных теорий. Л., 1967. С. 342—441.
Петроченко В. И. Акантоцефалы (скребни) домашних и диких
животных. Т. 1. М., 1956.
Плавильщиков Н. Н. Очерки по истории зоологии. М., 1941.
Полянский В. И. Монадная форма строения водорослей и не-
которые вопросы их филогении И Ботан. журн. 1956. Т. 41, № 8.
С. 1095—1106.
Полянский Ю. И. О некоторых морфологических закономер-
ностях в эволюции паразитических животных И Паразитол. сб.
ЗИН АН СССР. 1969. Т. 24. С. 208—219.
Полянский Ю. И. О морфологических закономерностях эво-
люции простейших И Зоол. журн. 1970. Т. 49, вып. 4. С. 560—568.
Полянский Ю. И. Общее и частное в проблеме вида И Систе-
ма интеграции вида. Вильнюс, 1986. С. 7—22.
Полянский Ю. И. Дарвинизм: история и современность И Дар-
винизм: история и современность. Л., 1988. С. 5—15.
(Полянский Ю. И.) Polyansky G. I. Protozoology and the prob-
lem of species // J. Protozool. 1992. Vol. 39 (1). P. 177—180.
Полянский Ю. И., Райков И. Б. Полимеризация и олигомери-
зация в эволюции простейших И Журн. общ. биол. 1977. Т. 38,
№ 3. С. 326—335.
Рогинский Я. Я., Якимов В. П. Теоретическое наследие А. Н. Се-
верцова и некоторые проблемы антропогенеза И Журн. общ. биол.
1968, № 1. С. 68—72.
Родендорф Б. Б. Филогенетические реликты И Тр. ИЭМЭЖ.
1959. Вып. 27. С. 41—51.
Розанова М. А. О низших систематических единицах И Журн.
рус. ботан. о-ва. 1928. Т. 13, № 3—4. С. 329—342.
Рыбин В. А. Гибриды терна и алычи и проблема происхожде-
ния культурной сливы // Тр. по прикл. ботан. 1936. Т. 2, вып. 100.
С. 1—44.
Рыдзюнский А. Типы связей между преобразованиями орга-
нов в филогенезе животных (филогенетические корреляции орга-
нов) // Памяти акад. А. Н. Северцова. Т. 1. М.; Л., 1939. С. 231—
256.
Светлов П. Г. Отношения между морфологическими осями в
онтогенезе и филогенезе различных групп животных И Журн. общ.
биол. 1967. Т. 28, № 5. С. 567—578.
Северцов А. Н. Эволюция и эмбриология И Речи, произнесен-
ные в соединенных заседаниях обществ... М., 1910. С. 18—31.
Северцов А. Н. Главные направления эволюционного процес-
са И Прогресс, регресс и адаптация. М., 1925.
Северцов А. Н. Морфологические закономерности эволюции.
М.; Л., 1939.
Северцов А. С. Становление ароморфоза И Журн. общ. биол.
1973. Т. 34, № 1. С. 21—36.
Северцов С. А. Динамика населения и приспособительная эво-
люция животных. М.; Л., 1941.
Семёнов-Тян-Шанский А. С. Таксономические границы вида
и его подразделений // Зап. Рос. акад. наук. 1910. Т. 25, № 1. С. 1—
29.
Серавин Л. Н. Новые представления о системе одноклеточных
животных И Тр. ЗИН АН СССР. 1980. Т. 94.
Серавин Л. Е., Гудков А. В. Возможные формы агамных гене-
тических взаимоотношений у протистов и пути становления поло-
вого процесса И Цитология. 1984. № 1. С. 12—24.
Скарлато О. А., Старобогатов Я. И. Филогенетика и принци-
пы построения естественной системы И Тр. ЗИН АН СССР. 1974.
Т. 53. С. 30—46.
Скрябин К. И., Шульц Р. С. Гельминтозы человека. Ч. 2. М.;
Л., 1931.
Смирнов Е. С. Таксономический анализ. М., 1969.
Соколов Б. С. Древнейшие отложения раннего кембрия и Са-
беллитиды// Всесоюз. симпоз. по палеонтологии докембрия: Тез.
докл. Новосибирск, 1965. С. 78—91.
(Соколов Б. С.) Sokolov В. S. Vendian of Northern Eurasia И
Mem. Amer. Ass. Petrol. Geol. 1973. N 19. P. 204—218.
Соколов Б. C. Metazoa докембрия и вендо-кембрийский рубеж И
Палеонтол. журн. 1976. № 1. С. 3—18.
Соколов Б. С. Жизнь на древних континентах, ее становление
и развитие И Тр. Всесоюз. Палеонтол. о-ва. Л., 1981.
Старобогатов Я. И. Брахиоцельная (гидроцельная) гипотеза
происхождения целома// Тр. ЗИН АН СССР. 1983. Т. 109.
С. 111—122.
Старобогатов Я. И. К вопросу о числе царств эукариотных ор-
ганизмов // Тр. ЗИН АН СССР. 1986. Т. 144. С. 4—24.
Сукачев В. Н. К вопросу о борьбе за существованье между био-
типами одного и того же вида // Сб., поев. И. П. Бородину. Л.,
1927. С. 195—219.
Сутт Т. Проблема направленности органической эволюции.
Таллинн, 1977.
Сушкин П. П. Обратим ли процесс эволюции? // Новые идеи в
биологии. 1915. Вып. 8. С. 1—39.
Татаринов Л. П. Морфологическая эволюция териодонтов и
общие вопросы филогении. М., 1976.
Татаринов Л. П. Палеонтология и эволюционное учение. М.,
1985.
Татаринов Л. П. Сальтационизм и неокатастрофизм в совре-
менных теориях эволюции К Вести. АН СССР. 1986. № 6. С. 10—
22.
Тахтаджян А. Л. О границах, методах и символах филогенети-
ческих настроений в ботанике // Бюл. Моск, о-ва испытат. приро-
ды. 1947. Т. 52, № 3. С. 95—120.
Тахтаджян А. Л. Пути приспособительной эволюции расте-
ний // Ботан. журн. 1951. № 3. С. 231—286.
Тахтаджян А. Л. Вопросы эволюционной морфологии расте-
ний. М.; Л., 1954.
Тахтаджян А. Л. Система и филогения цветковых растений.
М ; Л., 1966.
Тахтаджян А. Л. Макроэволюционные процессы в истории
растительного мира И Ботан. журн. 1983. № 12. С. 1593—1603.
Тимофеев Б. В. Сфероморфиды протерозоя. Л., 1969.
Тимофеев Б. В. Микрофитофоссилии протерозоя — раннего
палеозоя СССР. М., 1974.
Тимофеев-Ресовский Н. В. Микроэволюция// Ботан. журн.
1958. № 3. С. 317—336.
Тимофеев-Ресовский Н. В. К теории вида И Тр. Ин-та биол.
(Урал. фил. АН СССР). 1965. Вып. 44. С. 11—25.
Тимофеев-Ресовский Н. В., Воронцов Н.Н., Яблоков А. В.
Краткий очерк теории эволюции. М., 1969.
(Тимофеева-Ресовская Е. А., Тимофеев-Ресовский Н. В.) ТЬ
mofeeff-Ressovsky Е. A., Timofeeff-Ressovsky N. W. Genetische
ЛИТЕРАТУРА
117
Analyse einer freilebenden Drosophila melanogaster Population //
Roux' Arch. Entomol. Org. 1927. Bd 109. S. 70—109.
Уголев A. M. Эволюция пищеварения и принципы эволюции
функций. Л., 1985.
Федонкин М. А. Органический мир венда И Итоги науки и
техники (Стратиграфия и палеонтология). 1983. Т. 12. С. 3—
127.
Федонкин М. А. Бесскелетная фауна венда и ее место в эво-
люции Метазоа. М.: Наука, 1987. (Тр. Палеонтол. ин-та; Т. 226).
(Федотов Д. М.) Fedotov D. M.Beitrage zur Kenntnis der Mor-
phologic der Myzostomidae // Ztschr. Morphol. Oecol. Tiere. 1929.
Bd 15, N 1/2. S 156—191.
Федотов Д. M. Морфологические закономерности эволюции в
применении к беспозвоночным И Изв. АН СССР. 1936. № 5.
С. 1015—1031.
Федотов Д. М. О субституциях вторичной полости тела у
офиур // Зоол. журн. 1939. Т. 18, № 6. С. 943—959.
Федотов Д. М. Эволюция и филогения беспозвоночных живот-
ных. М., 1966.
Хейсин Р. В. Непостоянство генома. М., 1984.
Цалолихин С. Я. Свободноживущие нематоды Байкала. Ново-
сибирск, 1980.
Четвериков С. С. О некоторых моментах эволюционного про-
цесса с точки зрения современной генетики И Журн. эксперим.
биол. 1926. Т. 2, вып. 1. С. 3—54.
Шаров А. Г. Происхождение и основные этапы эволюции чле-
нистоногих // Зоол. журн. 1965а. Т. 44, № 6. С. 803—817.
(Шаров А. Г.) Sharov A. G. Evolution and taxonomy И Ztschr.
Zool. Syst.-Evol. Forsch. 1965b. S. 349—558.
(Шаров А. Г.) Sharov A. G. Basic arthropodanstoox. Pergamon
Press, 1966. c
Шаров А. Г. Монофилия и полифилия в эволюции И Журн.
общ. биол. 1971. Т. 32, № 6. С. 667—678.
Шевырев А. А. Проблемы происхождения раннекембрийской
фауны И Палеонтол. журн. 1962. № 4. С. 43—57.
(Шимкевич В.) Schimkewitsch W. Versuch einer Klassification
des Thierreiches // Biol. Zbl. 1891. Bd 11, N 9—10. S. 291—294.
Шмальгаузен И. И. Организм как целое в индивидуальном и
историческом развитии. М.; Л., 1938.
Шмальгаузен И. И. Пути и закономерности эволюционного
процесса. М.; Л., 1939а.
Шмальгаузен И. И. Дарвинизм и теория направленной эволю-
ции // Зоол. журн. 19396. Т. 18, вып. 4. С. 544—556.
Шмальгаузен И. И. Проблемы дарвинизма. М., 1946а.
Шмальгаузен И. И. Факторы эволюции. М., 19466.
Шмальгаузен И. И. Основы сравнительной анатомии позво-
ночных животных. М., 1947.
Шмальгаузен И. И. Регулирующие механизмы эволюции И
Зоол. журн. 1958. № 37. С. 1291—1306.
Шмальгаузен И. И. Регуляция формообразования в индивиду-
альном развитии. М., 1964а.
Шмальгаузен И. И. Происхождение наземных позвоночных.
М., 19646.
Шмальгаузен И. И. Кибернетические вопросы биологии. Но-
восибирск, 1968.
Шмальгаузен И. И. Проблемы дарвинизма. Л., 1969.
Энгельгардт В. А. Интегратизм — путь от простого к сложно-
му в понимании явлений жизни // Изв. АН СССР. Сер. биол. 1970.
№ 6. С. 799—822.
Юдин К. А. О понятии «признак» и уровнях развития систе-
матики животных И Теоретические вопросы систематики и фило-
гении животных. Л., 1974. С. 5—29.
Яблоков А. В. О разных формах прогрессивного развития в
органическом мире И Проблемы эволюции. Новосибирск, 1968.
Т. 1. С. 98—115.
Яблоков А. В., Юсуфов А. Г. Эволюционное учение. М., 1976.
Abel О. Palaeobiologie und Stammesgeschichte. Jena, 1929. _
Anderson E. Introgressive hybridization. New York; Wiley,
1949.
Ax P. Verwandtschaftsbeziehungen und Phylogenie der Turbella-
rien // Ergebn. Biol. 1961. N 24. S. 1—68.
Baer К. E. V. Uber den Zweck in der Vorhangen der Natur. 1.
Halfte II Studien aus dem Gebiete der Naturwissenschaften. 1876.
S. 49—105.
Baick L. Uber die Korperlange der Niederlander und derem Zu-
nahme in den letzten Dezennien # Ztschr. Morphol., Anthropol. 1914.
Bd 18. S. 15—48.
Barghoorn E. S. The oldest fossily // Sci. Amer. 1971. N 224.
P. 30—42.
Barghoorn E.S., Tyler S. A. Microorganisms from the gunflint
chort H Science. 1965. N 147. P. 563—577.
Baur E. Untersuchungen uber das Wesen der Entstellung und der
Vererbung von Rassenunterschieden bei Antimhinum majus // Biblio-
theca genetica. 1924. Bd 4. S. 1—170.
Beauchamp P., de. A propose d’une Planaire du Congo // Beige
Rev. Zool. et Botan. Africaines. 1951. Vol. 45, f. 1—2, P. 90—98.
De Beer G. Embryos and Amertors. Oxford, 1958.
Beneden E., van. Recherches sur les Dicyemides // Bull. Belgique
Cl. Sci. ser. 2. 1876. Vol. 41, N 2. P. 1150—1205.
Beneden E., van. Recherches sur le developpement des Arachnac-
tis. Contribution a la morphologic des Cerianthides // Arch. Biol.
1891. N 11. P. 115—147.
Birket-Smith S.J. R. A reconstruction of the Pre-Cembrian
Spriggina II Zool. Jb. Anat. 1931. N 105. S. 237—258.
Birket-Smith S. J. R. Is Praecambridium a juvenile Spriggina?
1981.
Bone G. The origin of the Chordata // J. Linn. Soc. Lond. Zool.
1960. Vol. 44, N 29. P. 252—269. .
Borgmeier T. Basic question of systematics // Syst. Zool. 1957.
N 6. P. 53—69.
Boyden A. A. Comparative evolution with special reference to pri-
mitive mechanisms // Evolution. 1953. N 7. P. 21—30.
Boynton H. E., Ford T. D. Pseudovendia charnwoodensis, a new
Praecambrian for Fossil Arthropod from Charnwood Forest, Leicester-
shire II Mercian Geol. 1979. N 7. P. 175—177.
Brien P. Embranchement der Tuniciers H Traite de Zoologie.
1948. T. 2. P. 553—930.
Brien P. Elements de Zoologie et notions d’anatomie comparee.
Vol. 1. Desoer; Liege, 1961.
Brien P. Elements de Zoologie et notions d’anatomie comparee.
Vol. 2. Desoer; Liege, 1963.
Brien P. Principes de Phylogenese animale appliques a la interp-
retation des Metazoarres Triploblastiques // Memoires Acad. roy.
Belg. Cl. Sci. 1969. T. 8, f. 7. P. 3—110.
Brien P. Le fenillets embryonnaires des Sponges (Reponse an
prof. A. V. Ivanov) // Acad. Roy. Bull. Cl. Sci. 1972. Ser. 5, N 57.
P. 715—732.
Britten R., Davidson E. Repetitive and nonrepetitive DNA sequen-
ces and speculation of evolutionary novelty // Quart, rev. biol. 1971.
Vol. 46, N 1. P. 111—133.
Britten R., Davidson E., Dower G. et al. Genomic change and
morphological evolution // Life Sci. Rept. 1982. Vol. 22. P. 19—39.
Bronn H. Morphologische Studien fiber die Gestaltungs-Gesetze
der Naturkorper fiberhaupt und der organischen Insbesondere. Leipzig,
1858.
Brucher H. Vitalitumsteigerung bei Mutanten in kiinstlichen
Klima II Naturwiss. 1941. Bd 29. S. 422—423.
Bullini L. Origin and Evolution of animal hybrid species //
Trends in Ecology and Evolution. 1994. Vol. 9. N 11. P. 421—426.
Biitschli O. Vorlesungen fiber vergleichenden Anatomie. Berlin,
1910.
Carson H. L.A provocative view of the evolutionary process Ю
Richard Goldchmidt-Controversial Geneticistfand Creative Biologist
Boston. Stuttgart, 1980. P. 24—26.
Clark R. B. The evolution of the coelom and metameria segmen-
tation // The lower Metazoa. Univ. Calif, Press. 1963. P. 91—107.
Clark R. B. Dynamics in metazoan evolution. The origin of the
coelom and segments. Oxford, 1964.
Clark R. B. Systematics and phylogeny: Annelida, Echiura, Si-
puncula II Chem. Zool. 1969. N 4. P. 1—68.
Clark R. B. Radiation of the Metazoa U The origin of major inver-
tebrate groups. Acad. Press, 197?. P. 59—102.
Claus C., Grobben K., Kuhn A. Lehrbuch der Zoologie. 10.
Aufl. Berlin, 1932.
Clausen J. Studies on the collective species Viola tricolor L. //
Botan. Tidsskr. 1921. Vol. 37. P. 205—221.
Codreanu R. Grands problemes controverses de 1’evolution phyloge-
netique des Metazoaires // Ann. Biol. 1970. Vol. 9, N 11/12. P. 671—709.
Cope E. The primary factors of organic evolution. Chicago, 1904.
Cori C. J. Kamptozoa// Handbuch der Zoologie. Bd 2. 1929.
S. 1—64.
Cuenot L. Les Tardigrades. Les Pentastomides // Traite de Zoolo-
gie. 1949. N 6. P. 40—75.
Cuenot L. Phylogenese du regne animal // Traite de Zoologie.
1952. T. 1. P. 1—33.
Cuvier G. Lecons d’anatomie comparee. Vol. 1. Paris, 1801.
118
ОБЩАЯ ЧАСТЬ
Dames W. Uber eine tertiare Wirbelthier-fauna von der westlic-
hen Insel des Birket-el-Qurun in Fajum (Aegypten). Berlin, 1883.
Darwin Ch. The origin of species by means of natural selection.
London, 1859.
(Darwin Ch.) Дарвин Ч. Происхождение видов путем естест-
венного отбора. СПб., 1991.
Deckert К. Unterstamm Aerania И Urania Tierreich. 1967. N 3.
P. 39—40.
Delage Y. Embryogenie des Sponges. Development post larvaire
des Sponges siticenses et fibrenses marines et d’eau douce // Arch.
Zool. exp. gen. 1892. N 10. P. 345—498.
Delage Y., Herouard E. Traite de Zoologie concrete. Lecons pro-
fessees ala Sorbonn. T. 1—5. Paris, 1896—1899.
DepSret Ch. Les transformations du mond animal. Paris, 1907.
Dobzhansky Th. Genetics and origin of species. New York,
1937.
Dobzhansky Th. Genetics of the evolutionary process. New York;
London, 1970.
Dohrn A. Ursprung der Wirbelthiere und das Prinzip des Funkti-
onswechsels: Genealogische Skizzen. Leipzig, 1875.
Dollo L. Les lois de 1’evolution // Bull. Soc. Beige Geol. 1893.
N 7. P. 164—166.
Eimer T. Die Entstehung der Arten. 1. Teil. Jena, 1888.
Eimer T. Die Entstehung der Arten. 2. Teil. Leipzig, 1897.
Eldredge N., Gould S. Punctuated equilibria: an alternative to
phyletic gradualism // Models in Paleobiology. 1972. P. 82—115.
Emig С.C. Embryology of Phoronida // Amer. Zool. 1977.
Vol. 17, N 1. P. 184—193.
Erwin T. L., Scott J. C. Seasonal and size patterns, trophic struc-
ture and richness of Coleoptera in the tropical arboreal ecosystem //
Coleopterists Bull. 1980. Vol. 34, N 3. P. 305—322.
The evolutionary synthesis: perspectives on the unification of
biology. Cambridge; London, 1980.
Fage L. Classe des Merostomaces // Traite de Zoologie. 1949. T. 4.
P. 219—262.
Fisher R. The genetical theory of natural selection. Oxford, 1930.
Fitch W., Margolish E. The usefulness of amino acid and nucle-
otide sequences in evolutionary studies U Evolutionary biology. 1970.
Vol. 4. P. 67—109.
Florkin M. Ideas and experiments in the field of prebiological
chemical evolution H Comprehensive biochemistry. 1975. Vol. 29, pt
В. P. 231—260.
Flugel E. Palaeontologische Beitrage zur Evolution der Organis-
men II Evolution. G. Fischer, 1978. Taschenbiich., 748. S. 221—262.
(Folsom С.) Фолсом К. Происхождение жизни. М., 1982.
Ford Е. Ecological genetics. London etc., 1964.
Ford T. D. Precambrian fossils and the origin of the Phanerozoic
phyla U The origin of major Invertebrate groups. 1979. P. 21.
(Fox S., Dose К.) Фокс С., Дозе К. Молекулярная эволюция и
возникновение жизни. М., 1975.
Garstang W. The morphology of Tunicata and its bearing on the
phylogeny of the Chordata U Quart. J. Microsc. Sci. 1928. Vol. 72,
pt 1. P. 51—187.
Gelei J. Die Morphogenese der Einzeller mit Rusksicht auf die
morphogenetischen Prinzipien von Sewertzoff // Acta Biol. Acad. Sci.
1950. T. 1, f. 1—4. P. 69—134.
Ginsburg L. Les reptiles fossiles // Traite de Zoologie. 1970.
T. 14, f. 3. P. 1161—1332.
Glaessner M. F. The oldest fossil faunas of South Australia //
Geol. Rundschau. 1958. Vol. 47, N 2. P. 522—531.
(Glaessner M. F.) Глесснер M. Ф. Древнейшие бесскелетные
организмы И Природа. 1963. № 11. С. 73—78.
Glaessner М. F. Die Entwicklung des Lebens im Prakambrium
und seine geologische Bedeutung // Geol. Rundschau. 1971. Vol. 60.
P. 1323—1339.
Glaessner M. F., Wade M. The late Precambrian fossils from
Ediacana, South Australia // Paleontology. 1968. N 9. P. 599—628.
Goldschmidt R. The material basis of evolution. New Hakn,
1940.
Goodman M., Moore G., Farris W. Primate phylogeny from the
perspective of molecular systematics // Transplant. Proc. 1974. N 6.
P. 217.
Gould S., Eldredge N. Punctuated equilibria: the tempo and mode
of evolution reconsidered II Paleobiology. 1977. Vol. 3, N 2. P. 115—
151.
Grasse P.-P. Ordre des edentes. Formes actuelles // Traite de Zoo-
logie. 1955. T. 17, f. 11. P. 1183—1246.
Grasse P.-Р., Poisson R. A., Tuzet O. Zoologie. 1. Invertebras.
Masson; Paris, 1961.
Greenberg M. J. Ancestors, embryos and symmetry // Sy st. Zool.
1959. Vol. 8, N 4. P. 312—321.
Grell K. G. Trichoplax adhaerens F. E. Schulze und die Entstehung
der Metazoen U Naturw. Rundschau. 1971. Bd 22, N 4. S. 160—161.
Grell K. G. Unterreich Protozoa, Einzeller oder Urtiere. Jena,
1980a.
Grell K. G. I. Stamm Protozoa. II. Stamm Placozoa И Lehrbuch
der speziellen Zoologie. Jena, 1980b. Bd 1, T. 1. S. 157—250.
Grobben K. Die Systematische Einteilung des Tierreiches // Verh.
Zool.-Botan. Ges. Wien. 1908. Bd 58, N 10. S. 491—511.
Gross W. Polyphyletische Stamme in System der Wirbelthiere //
Zool. Anz. 1964. Bd 173, H. 1. S. 1—22.
Gruner H. E. Einfiihrung // Lehrbuch der Speziellen Zoologie.
Jena, 1980. Bd 1, T. 1. S. 15—156.
Gruner H. E. 14. Stamm Echiwida // Lehrbuch der Speziellen
Zoologie. Jena, 1982. Bd 1, T. 2. S. 261—275.
Gunther K. Unterstamm Tunicata // Urania Tierreich. 1967. N 3.
S. 15—38.
Giirich G. Der bislang altesten Spuren von Organismen in Sudaf-
rica U G. R. Int. Geol. Congr. 1930. P. 670—680.
Gutmann W. F. Die Hydroskelett-theorie. Aufsatze // Reden Sen-
ckenb. naturwiss. Ges. 1972.
Gutmann W. F. Das Analogie-Denken Darwins und die physika-
listische Evolutionstheorie H Biol. Rundschau. 1979. Bd 17, H. 2.
S. 84—99.
Hadzi J. Turbelarijska teorija knidarijev. Ljubljana, 1944.
Haeckel E. Generelle Morphologic der Organismen. Bd II. Berlin,
1866.
Haeckel E. Die Kalkschwamme. 1, 4 // Phylosophie der Kalk-
schwamme. 1872. S. 451—484.
Haeckel E. Die Gastraea-theorie, die phylogenetische Klassifizie-
rung des Tierreichs und die Homologie der Keimblatter // Jen. Ztschr.
Naturwiss. 1874. N 8. S. 1—55.
Haeckel E. Die Gastrula und die Entwicklung der Thiere // Jen.
Ztschr. Naturwiss. 1875. N 9. S. 402—408.
Haeckel E. Systematische Byogenie. T. 1. Berlin, 1894.
Hanstrom B. Vergleichende Anatomie des Nervensystems der
wirbellosen Tiere, unter Benicksichtigung seiner Funktion. Berlin,
1928.
Harlan J., Wet J. The compilospecies concept // Evolution. 1963.
Vol. 17, N 1. P. 17—22.
Harrington H. J., Moore R. C. Dipleurozoa H Treatise on Inver-
tebrate Paleontology. 1956. F24—F27.
Hartman W. D. A vritique of enterocoele theory // The lower Me-
tazoa. Univ. Calif. Press, 1963. P. 55—77.
Hartwich G. 9. Stamm Entoprocta; 10. Stamm Nemathelmint-
hes II Lehrbuch der Speziellen Zoologie. Jena, 1984. Bd 1. T. 2.
S. 455—542.
Hatschek B. Studien uber die Entwicklungsgeschichte der Anne-
liden II Arb. Zool. Inst. 1878. P. 1—128.
Hatschek B. Lehrbuch der Zoologie. I. Jena, 1888.
Hatschek B. Das neue zoologische System. Leipzig, 1911.
Hennig W. Grundziige einer Theorie der phylogenetischen Syste-
matic Berlin, 1950.
Hennig W. Phylogenetic systematics. Urbona, 1966.
L’HSritier Ph., Teissier G. Elimination des formes mutantes dans
les populations de Drosophiles. I. Cas des Drosophiles «bar» // C. r.
Soc. bid. 1937. Vol. 124. P. 880—882.
Herre W. Zum Abstammungsproblem von Amphibien und Tylo-
poden so wie uber Parallelbildungen und zur Polyphylierangen // Zool.
Anz. 1964. Vol. 173, N 1. P. 66—91.
Hertwig O., Hertwig K. Die Coelom-Theorie. Ein Versuch einer
Erklarung des mittleren Keimblates // Jen. Ztschr. Naturwiss. 1882.
N 15. S. 1—150.
Hoyer В. H., McCarthy B. J., Bolton E. T. A molecular approach
in the systematics of higher organisms // Science. 1964. Vol. 144, N 3621.
P. 459.
Huxley J. Evolution: the modern synthesis. London, 1942.
Huxley J. Evolution: the modern synthesis. London, 1963.
Huxley Th. H. Lectures on the elements of comparative anatomy.
London, 1864.
Hyman L. H. The Invertebrates: Protozoa through Ctenophora.
New York; London, 1940.
Hyman L. H. The Invertebrates: Platyhelmintes and Rhynchocala.
The acoelomata Bilateria. New York, 1951.
Hyman L.H. The Invertebrates. New York; London, 1959.
Jarvik E. On the structure of the snout of Grossopterygans and
lower Gnathostomes in general // Zool. Beitrag. 1942. N 21. P. 235—
675.
ЛИТЕРАТУРА
119
Jarvik Е. Specialisations in early vertebrates // Ann. Soc. Roy.
Zool. Belg. 1964. Vol. 94, N 1. P. 11—95.
Jarvik E. Basic structure and evolution of vertebrates. New York,
1980.
Jenkins R. J., Gehling J. G. A review of the frond-like fossils of
the Ediacara assemblage // Rec. South. Aust. Mus. 1978. Vol. 17,
N 23. P. 347—359.
Johansson К. E. Uber Lamellisabella zachsi und ihre systematis-
che Stellung // Zool. Anz. 1937. N 117. S. 23—26.
Kaestner A. Lehrbuch der speziellen Zoologie. Jena, 1959.
Kaestner A. Lehrbuch der speziellen Zoologie. Jena, 1963.
Kaestner A. Lehrbuch der speziellen Zoologie. Jena, 1965.
Kawaguti S., Baba K. A preliminary note on a two-valved sac-
coglosson Gastropod Tamanovalva limax n. g., n. sp. from Tamano-
Japan II J. Okayama Univ. 1959. N 5. P. 177—184.
(Kein А.) Кейн А. Вид и его эволюция. М., 1958.
Kennel J. Die Verwandschaftsverhaltnisse der Arthropoden. Dor-
pat, 1891.
Kielmeyer C. F. Uber die Verhaltnisse der organischen Krafte
untereinander in der Reihe der verschiedenen Organisationen // Tubin-
gen. 1814. 18 M. S. 38—41.
Kilian E. F. 3. Stamm Porifera // Lehrbuch der Speziellen Zoolo-
gie. Jena, 1980. S. 251—288.
Kilian R. 12. Stamm Mollusca // Lehrbuch der Speziellen Zoolo-
gie. Jena, 1982. S. 9—245.
Kimura M. The neutral theory of molecular evolution. London, 1983.
(Kimura M.) Кимура M. Молекулярная эволюция: теория ней-
тральности. М.: Мир, 1985. С. 88.
King J., Jusces Т. Non-Darwinian evolution // Science. 1969.
Vol. 164. P. 788—798.
Kleinenberg N. Die Entstehung der Annelids aus der Larve von
Lopadorhynchus // Ztschr. wiss. Zool. 1886. Bd 44. S. 1—227.
Kollmann J. Uberwintern von Frosch- und Triton-Larven und die
Umwandlung des mexikanischen Axolotl // Verh. naturforsch. Ges.
Basel. 1882. N 7. S. 387.
Korn H. Erganzende Beobachtungen zur Struktur der Larve von
Echiurus abyssalis Skor. # Ztschr. wiss. Zool. 1960. Bd 164, N 3/4.
S. 199—237.
Kristensen R. M. Loricifera. A new phylum with Aschelminthes
characters from the meiobesethons // Ztschr. Zool. Syst.-Evol. Forsch.
1983. Bd 21. S. 163—180.
Ktihn-Schneider E. Wege der Reptiliensystematik // Paleontol.
Ztschr. 1963. N 37. S. 61—87.
Kiikenthal W. Handbuch der Zoologie. Berlin, 1923.
(Lamarque G. В.) Ламарк Ж. Б. Философия зоологии И Из-
бранные произведения в двух томах. 1959. Т. 1. С. 171—843.
Lameere A. Une Theorie zoologique // Bull. Sci. France. Belgi-
que. 1915. N 49. P. 378—431.
Lameere A. Origin du coelome // Arch. Zool. Ital. 1930. Pt I.
P. 197—206.
Lankester E. R. On the primitive Af-layers of the embryo as the
basis of genealogical classification of animals etc. // Ann. Mag. Nat.
Hist. 1873. N 11. P. 321—338.
Lankester E. R. Notes on the embryology and classification on
the animal kingdom, comprising a revision in speculations relative to
origin and significance of the germ layers // Quart. J. micr. Sci. 1877.
N 17. P. 399—454.
Lankester E. R. The Enterocoela and the Coelomocoela // A trea-
tise on Zoology. London, 1900. P. 1—37.
Leuckart R. Zoologische Untersuchungen. 1854.
Leuckart R. Bau und Entwicklungsgeschichte der Pentastomen.
Leipzig, 1860.
Lewin B. Science as a way of knowing-molecular evolutional //
Amer. Zool. 1984. Vol. 24, N 3. P. 451—464.
Lewis H. Catastrophic selection as a factor in speciation // Evolu-
tion. 1962. Vol. 16, N 3. P. 257—271.
Linnaeus C. Systema naturae, sive regna tria naturae systematice pro-
posita per classes, ordines, genera et species. Lugduni Batavorum, 1735.
MacBride E. W. Text-book of embryology. Vol. I. Invertebrata.
London, 1914.
Manton S.M. Studien on the Onychophora. VII // Phil. Trans.
Roy. Soc. London. B. 1949. N 233. P. 483—580.
Manton S. M. The Arthropoda habits functional morphology and
evolution. Oxford, 1977.
(Margelis L.) Маргелис Л. Роль симбиоза в эволюции клетки.
М., 1983.
Masterman А. Т. On the theory of archimeric segmentation and
its bearing upon the phyletic classification of the Coelomata // Proc.
Roy. Soc. Edinb. 1898. N 22. P. 270—310.
Mather K. Polymorphism as an outcome of disruptive selection //
Evolution. 1955. Vol. 9, N 1. P. 52—61.
Matthew W. D. The evolution of the Horse; a record and its in-
terpretation II Quart. Rev. Biol. 1926. Vol. 1. P. 130—185.
Mayr E. Notes on the Polynesian species of Aplonis // Amer,
mus. nov. New York, 1942. N 1166.
(Mayr E.) Майр Э. Зоологический вид и эволюция. М., 1968.
(Mayr Е.) Майр Э. Принципы зоологической систематики.
М., 1971.
(Mayr Е.) Майр Э. Популяции, виды и эволюция. М., 1974.
Mayr Е. The growth of biological thought. Cambridge; London,
1982.
(Mayr E., Linsly E., Usiter R.) Майр Э., Линсли Э., Юзитер Р.
Методы и принципы зоологической систематики. М., 1956.
Meckel J. F. System der vergleichenden Anatomie. Bd I. Allge-
meine Anatomie. Halle, 1821.
Metcalf Z. P. How many insects are there in the world? U Ento-
mol. News. 1940. N 51. P. 219—222.
Mmchin E. A. Sponges // A Treatise on Zoology. 1900. P. 1—178.
Molisch H. Pflanzenchemie und Pflanzenverwandtschaft. Jena,
1933.
Moritz M. 16. Stamm Onychophora; 17. Stamm Tardigrada; 18.
Stamm Pentastomida // Lehrbuch der speziellen Zoologie. Jena, 1982.
S. 470—545.
Muller A. H. Lehrbuch der Palaozoologie. Bd I. Allgemeine Grund-
lagen. Jena, 1963.
Muller F. Fur Darwin. Leipzig, 1864.
(Muller G.) Мюллер Г. Углистые метеориты и возникновение
жизни И Возникновение жизни. 1969. С. 306—335.
Nageli С. К. U. Mechanisch-physiologische Theorie der Abstam-
mungslehre. Munchen, 1884.
Nielsen C. Entoproct life-cycles and the entoproct-ektoproct rela-
tionship. Univ. Copenhagen, Mar. biol. lab. // Ophelia. 1971. Vol. 9,
N 2. P. 209—341.
Nielsen C. Animal phylogeny in the light of the Trochaea theo-
ry # Biol. J. Linn. Soc. 1985. Vol. 25. P. 243—299.
Nielsen C., Norrevang A. The Trochaea theory: an example of
life cycle phylogeny // The systematics association. Spec. vol. 28. Ox-
ford, 1985. P. 28—41.
Nursall J. R. The origin of the Metazoa // Trans. Roy. Soc. Cana-
da. 1959. Vol. 53, N 3/5. P. 1—5.
Nursall J. R. On the origins of the major groups of animals //
Evolution. 1962. Vol. 16, N 1. P. 118—123.
Odening K. 7. Stamm Plathelminthes: 8. Stamm Nemertini //
Lehrbuch der speziellen Zoologie. Jena, 1984. S. 347—454.
Oehler D. Z. Transmission electron microscopy of organic micro-
fossils from late Precambrian Biter Springs Formation of Australia //
J. Paleontol. 1976. N 50. P. 90—106.
Osborn H. F. The ideas and terms of modern phylosophical ana-
tomy // Science. 1905. Vol. 5, N 21. P. 59—61.
Osborn H. F. Squidae of Oligocene, Miocene, and Pliocene of
North America, iconographic type revision # Mem. Amer. Nat. Hist.
1918. Vol. 2. P. 1—217.
Osche G. Die systematische Stellung und Phylogenie der Penta-
stomida II Ztschr. Morphol. Okol. Tiere. 1963. Vol. 52. S. 487—596.
Ostrom J. H. Archaeopteryx and the origin of flight // Quart. Rev.
Biol. 1974. N 49. P. 27—47.
Owen R. On the Archaeopteryx of von Meyer, with a description
of the fossil remains of the longtailedspecies from lithographic stone
of solen hafen // Phil. Transact. 1863. P. 33—47.
Pantin C.F. A. Organic design // Advanc. Sci. 1951. N 8. P. 137—
150.
Pantin C.F.A. Diploblastic animals // Proc. Lin. Soc. 1960.
N 171. P. 1—14.
Pantin C. F. A. Homology, analogy and chemical identity in Cni-
daria II Symp. Zool. Soc. London, 1966. N 16. (The Cnidaria and their
Evolution).
Petrunkevic A. An inquiry into the natural classification of spi-
ders, based on a study of their internal anatomy. New Hawen, 1933.
Pflug H. D. Zur fauna der Nama-Schichten in Siidwest-Africa //
Palaeontographica. A. 1970. Bd 1, N 134. S. 226—262; 1971. Bd 2,
N 135. S. 698—731; 1972. Bd 3, N 139. S. 134—170; 1973. Bd 4,
N 144. S. 166—202.
Plate L. Prinzipen der Systematik mit besonderem Beriicksichti-
gung des Systems der Tiere // Die Kultur der Gegenwart. 1912. Bd 4,
N 4. S. 92—164.
Portmann A. Ontogenesetypes und Cerebralisation in der Evolu-
tion der Vogel und Sauger // Rev. Suis. Zool. 1951. T. 58, fasc. 3.
P. 427—434.
120
ОБЩАЯ ЧАСТЬ
Reisinger Е. Die Evolution des Orthogons der Spiralier und das
Archicoelomatenproblem // Ztschr. Zool. Syst. Evolutionsforsch.
1972. Bd 10, H. I. S. 1—43.
Remane A. Die Grundlagen des natiirlichen Systems der verglei-
chenden Anatomie und Systematic. Leipzig, 1956.
Remane A. Das Problem Monophylie-Polyphylie mit besonderer
Beriicksichtigung der Phylogenie der Tetrapoden // Zool. Anz. 1964.
Bd 173. S. 23—49.
Remane A. Phylogenetische Methoden ausserhalb der mofpholo-
gischen Verwandtschaftsforschung // Zool. Anz. 1967. Bd 179, H. 1/2.
Remane A. Stellungshahm // Senckenb. nat. Ges. 1973. Bd 22.
S. 105—108.
Remane A., Storch V., Welsch U. Systematische Zoologie. Stam-
me des Tierreichs. Jena, 1976.
Remane A., Storch V., Welsch U. Systematische Zoologie. Stutt-
gart, 1986.
Rensch B. Neuere Probleme der Abstammungslehre. Die trans-
spezifische Evolution. Stuttgart, 1954. Bd 12.
Riedl R. Gnathostomulida from America // Science., 1969. N 163.
P. 445—452.
Rieger R., Mainitz M. Comparative fine structure study of the
body wall in Gnathostomulida and their phylogenetic position between
Platyhelminthes and Aschelminthes // Ztschr. Zool. Syst.-Evol.
Forsch. 1977. Bd 15, H. 1. S. 9—35.
(Romer A. S.) Ромер А. Ш. Палеонтология позвоночных. M.;
Л., 1939.
Romer A. S. Unorthodoxies in reptilian phylogeny // Evolution.
1971. Vol. 25, N 1.
(Rutten M.) Рутген M. Происхождение жизни. M., 1973.
Salvini-Plawen L. Die Funktion der Coelomtheorie in der Evolu-
tion der Mollusken // Syst. Zool. 1968. Bd 17, N 3. S. 192—208.
Salvini-Plawen L. Zur Morphologie und Phylogenie der Mollus-
ken // Ztschr. wiss. Zool. 1972. Bd 184, N 3/4. S. 205—391.
Salvini-Plawen L. On the origin and evolution of the lower Me-
tazoa II Ztschr. Zool. Syst-Evol. Forsch. 1978. Bd 16. S. 40—87.
Save-Soderbergh G. Notes on the Devonian Stratigraphy of East
Greenland // Meddelelser am Gronland. 1932. Bd 94, N 4.
Schopf J. W. Precambrian palaeobiology: problems and perspecti-
ves // Ann. Rev. Earth and Planetary Sci. 1975. N 4.
Schwanitz F. Genetik und Evolutionsforschung bei Pflanzen //
Die Evolution der Organismen. Bd 2. Stuttgart. 1974. S 1—189.
Sedgwick A. On the origin of metameric segmentation and some
other morphological questions // Quart. J. Mier. Sci. 1884. N 24.
P. 43—82.
Seilacher A. Der Beginn des Kambrian als biologische Wende //
Neues Jb. Geol. Palaeontol. Abhand. 1956. N 103. S. 155—180.
Siewing R. Zum Problem der Polyphylie der Arthropoden //
Ztschr. wiss. Zool. 1960. Bd 164, N 3/4. S. 238—270.
Siewing R. Diskussionbeitrag zur Phylogenie der Coelomata //
Zool. Anz. 1967. N 179. S. 132—170.
Siewing R. Lehrbuch der vergleichender Entwicklungsgeschichte
der Tiere. Hamburg; Berlin, 1969.
Siewing R. Zur Deszendenz der Chordaten. Erwiderung und Ver-
such einer Geschichte der Archecoelomaten // Ztschr. Zool. Syst.-
Evol. Forsch. 1972. Bd 10. S. 267—291.
Siewing R. Thoughts about the phylogenetic-systematic position
of Pogonophora // Ztschr. Zool. Syst.-Evol. Forsch. 1975. Sonderheft’.
S. 127—138.
Siewing R. Probleme und neuere Erkenntnisse in der Grossyste-
matik der Wirbellosen // Verh. Dtsch. Zool. Ges. 1976. S. 59—83.
Silvestri F. The relation of taxonomy to other branches of ento-
mology // The 4th Intern. Congr. of entomol. 1929. Vol. 2. P. 52—54.
Simpson G. G. Tempo and mode in evolution. New York, 1944.
(Simpson G. G.) Симпсон Д. Г. Темпы и формы эволюции. М.;
Л., 1948.
Simpson G. G. Horses. The story of the Horse Family. New York,
1951.
Simpson G. G. Principles of animal taxonomy. New York, 1961.
Sokal R. R., Sneath P. H. A. Principles on numerical taxonomy.
London, 1963.
Sollas W. £ On the development of Halisarca lobularis // Quart.
J. micr. Sci. 1884. N 24. P. 603—621.
Southward A. J., Southward E. C., Dando P. R., Rau G. H., Pel-
beck H., Flugel H. Bacterial symbionts and low 13 C/12 rations in
tissues of Pogonophora indicate unusual nutrition and metabolism //
Nature. 1981. Vol. 293, N 5834. P. 616—620.
(Spenser G.) Спенсер Г. Основания биологии. СПб., 1870.
Sprigg R. С. Early Cambrian jellyfishes from the Flinders Ranges,
South Australia // Trans. Roy. Soc. S. Austr. 1947. Vol. 71. P. 212—224.
Stanley S. Macroevolution: pattern and process. San Francisco,
1979.
Stebbins G. Variation and evolution in plants. New York, 1950.
Sterba G. Unterstamm Agnata // Urania Tierreich. Leipzig, 1967.
Bd 3. S.41—46.
Sterrer W. Systematics and evolution within the Gnathostomuli-
da // System. Zool. 1972. Vol. 21, N 2. P. 151—173.
Stubbe H., Wettstein F. Uber die Bedeutung von Klein- und
Grossmutation fur die Evolution // Biol. Zbl. 1941. Bd 61. S. 264—
297.
Sturtevant A., Dobzhansky Th. Inversion of the third chromoso-
me of wild races of Drosophila pseudoobscura and their use in the
study of the history of the species // Proc. Nat. Acad. Sci. U. S. 1936.
Vol. 22. P. 448—450.
Thoday J., Gibson J. Isolation by disruptive selection # Nature.
1962. Vol. 193. P. 1164—1166.
Thompson W. R. Evolution and Taxonomy // Studia Entomol.
1962. Vol. 5. P. 549—570.
Tiegs O. W., Manton S. M. The evolution of the Arthropoda //
Biol. Rev. Cambr. Philos. Soc. 1958. Vol. 33, N 3. P. 225—337.
Turresson G. The genotipical response of the plant species to the
habitat // Hereditas. 1922. Bd 3. S. 221—350.
Tyler S. A., Barghoorn E. S. Occurrence of structurally preserved
plants in precambrian rocks of the Canadian Schield // Science. 1954.
N 119. P. 606—608.
Ulrich W. Uber die systematische Stellung einer neuen Tierklasse
(Pogonophora) den Begriff der Archicoelomaten und die Einteilung
der Bilaterien // Strz. Dtsch. Akad. Wiss. Berlin Math. Naturwiss. KI.
Jg. 1949. N 2. S. 1—25.
Ulrich W. Archicoelomaten // Aufsatze Red. Senckenb. Nab. Ges.
1973. N 22. S. 7—50.
Waddington С. H. Canalisation of development and the inheritan-
ce of acquired characters # Nature. 1942. Vol. 150, N 3811. P. 563—
565.
Wade M. Preservation of soft bodied animals: Precambrian sands-
tones at Edialara, South Australia // Lethaia. 1968. N 1. P. 238—267.
Wade M. Hydrozoa and Scyphozoa and other medusoids from the
Precambrian Ediacara fauna South Australia // Paleontology. 1972.
N 15. P. 197—225.
Wagner J. A. Ein eneus, angeblich mit Vogelfedern versehenes
Reptil // Sitzungsler. Acad. Munchen. 1861. S. 146—154.
Webby B. D. Late Precambrian Trace fossils from New South
Wales U Lethaia. 1970. N 3. P. 79—109.
Werner B. 4. Stamm Cnidaria; 5. Stamm Ctenophora // Lehrbuch
der speziellen Zoologie. Jena, 1984. S. 11—335.
Westblad E. Xenoturbella bocki n. g., n. sp., a peculiar, primitive
Turbellarian type // Arc. Zool. (s. 2). 1949. N 1. P. 11—29.
Westheide W. The systematic position of the Diniphilidae and the
archiannelid problem // The origin and relationships of lower Inverteb-
rates. 1985. Vol. 28. P. 310—326.
Wetzel A. Technische Hydrobiologie. Trink-, Branch-, Abwasser.
Leipzig, 1969.
White M. Modes of speciation. San Francisco, 1978.
Willey A. Amphioxy and the ancestry of Vertebrates. New York,
1894.
Wright S. Evolution of mendelian populations // Genetica. 1931.
Vol. 16, N 1. P. 97—159.
ПРОТИСТЫ
ОБЩИЕ ВОПРОСЫ
СИСТЕМА ПРОТИСТОВ
И ПРОБЛЕМЫ ИХ МЕГАСИСТЕМАТИКИ
Эволюция представлений о системе протистов, с
которой следовало бы начать эту главу, весьма обсто-
ятельно изложена в книге Кусакина и Дроздова «Фи-
лема органического мира» (1994). Нет нужды повто-
рять описание истории развития многоцарственных
систем, поэтому здесь мы ограничимся лишь самыми
главными, на наш взгляд, вехами.
Понятие «протесты» было введено Геккелем в
1866 г. (Haeckel, 1866) для обозначения третьего цар-
ства (наряду с животными и растениями) живых ор-
ганизмов. Оно включало простейших, водоросли,
низшие грибы, а также прокариот и некоторых ни-
зших многоклеточных животных. Позднее, в процес-
се уточнения объема этого царства и обсуждения его
с другими биологами того времени Геккель опять
распределил протистов между царствами животных и
растений и вернулся фактически к аристотелевской
двухцарственной системе организмов.
Идею протистов (протоктистов — термин Хогга
(Hogg, I860)) возродил Коупланд (Copeland, 1956),
который вновь ввел это царство эукариот, но уже в
современном понимании, включив в него и настоя-
щие грибы. Протесты в современном объеме как
одно из 4 царств эукариот были введены Уиттекером
(Whittaker, 1969). Его идеи были развиты в серии
работ Корлисом (см.: Corliss, 1984), который вклю-
чил в это царство простейших, водоросли и зооспо-
ровые грибы.
Уже поверхностный взгляд на характеристику про-
тистов (см. раздел «Общая характеристика протис-
тов») дает представление о них как о гетерогенной и,
очевидно, сборной группе эукариот. И все же почти
через 100 лет после работ Хогга и Геккеля биологи
вернулись, хотя и на короткое время, к протистам как
к единому таксону. Почему это произошло? Основные
причины, по-видимому, заключались в следующем.
1. Расширение ультраструктурных исследований
выявило большое многообразие в организации клет-
ки простейших и водорослей. Уже в 60-е гг. разделе-
ние простейших на 4 группы — амебы, жгутиконос-
цы, инфузории и споровики — казалось весьма ус-
ловным. Так, солнечники и фораминиферы могли
быть отнесены к амебам с большой натяжкой. Доволь-
но рано было показано, что споровики разделяются
на три четко очерченных типа (Microsporidia, Мухо-
sporidia и Sporozoa), а жгутиконосцы, как показали
работы 70-х гг., настолько разнообразны, что иссле-
дователи вынуждены были пересмотреть не только
систему протистов, но и эукариот в целом. Только
инфузории претерпели наименьшие изменения и до
сих пор представляют собой целостный, хорошо
очерченный таксон с яркими отличительными при-
знаками.
2/Была обнаружена таксономическая и филогене-
тическая близость у некоторых групп простейших и
водорослей. Еще в 1973 г. Сли (Sleigh, 1973) в своей
книге «The biology of Protozoa» указывал на большое
сходство эвгленовых водорослей и кинетопластид,
что в настоящее время общепризнано. Однако эти от-
ряды помещались в разные классы вплоть до 1980 г.
Еще более неестественная ситуация складывалась с
зелеными водорослями, ксантофитовыми и хризофи-
товыми. Они имеют в своем составе как бесцветные,
так и окрашенные формы. По старой системе, бес-
цветные формы относились к простейшим (царство
животных), а организмы с хлоропластами — к водо-
рослям (царство растений). Поэтому все чаще авторы
вынуждены были рассматривать простейшие и водо-
росли в одном таксоне за пределами растений и жи-
вотных.
3. Возрождение симбиогенетических идей на кле-
точном уровне привело к представлениям о первич-
ности гетеротрофных форм в эволюции эукариот. В
самом деле, если хлоропласты могли быть приобре-
тены путем симбиоза, скажем, с цианобактериями, то
разумно предположить, что первыми эукариотами
были гетеротрофы. Дальнейшую же эволюцию эука-
риот уже невозможно было рассматривать отдельно в
рамках только простейших или только водорослей.
4. Вокруг вопроса о происхождении эукариотной
клетки были объединены усилия протозоологов, фи-
кологов, микологов, клеточных биологов. Благодаря
124
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Схема 1. Филогенетические взаимоотношения между основными группами живых организмов. (По: Whittaker, 1969).
Протисты являются переходной в эволюционном плане группой от прокариот к эукариотам с многоклеточной и мицелиальной организацией (животные,
растения и грибы).
Scheme 1. Phylogenetic relationships among the main groups of living organisms.
The protists are a transitional evolutionary group from prokaryotes towards multicellular and mycelial eukaryotes: animals, plants and fungi.
симбиогенезу — мощному направлению в биологии
последних лет — активизировалось изучение проис-
хождения клеточных органелл биологами разных
специальностей, что, в свою очередь, сделало насущ-
ным пересмотр системы простейших, водорослей,
низших грибов и всей системы эукариот в целом.
По этим причинам простейшие, водоросли и зоо-
споровые грибы рассматриваются в одной системе
координат, а до последнего времени многие авторы
считали их единым царством Protista (Corliss, 1984;
Карпов, 19906; Margulis, 1992).
По Уиттекеру (Whittaker, 1969), протисты пред-
ставляют собой переходную в эволюционном плане
группу эукариот от прокариотических организмов к
другим эукариотам с многоклеточной и мицелиаль-
ной организацией (схема 1). Три царства эукариот —
животные, растения и грибы — хорошо очерчены и
не имеют четкой границы только с протистами. Эти
три ветви представляют собой основные линии разви-
тия (специализации) живого в соответствии с типом
питания: из протистов, имеющих все типы питания,
вычленяются животные — гетеротрофы, растения —
автотрофы, грибы — сапрофиты. Для каждой из этих
групп характерны определенные признаки.
Животные — преимущественно многоклеточные
гетеротрофы с голозойным питанием; кристы в мито-
хондриях пластинчатые; обычно со сложным эмбри-
ональным развитием; мейоз гаметический.
Растения — преимущественно многоклеточные
автотрофы, содержат хлорофилл а и Ь; преимущест-
венно наземные; целлюлоза в клеточной стенке, крах-
мал запасается внутри хлоропластов; кристы в мито-
хондриях пластинчатые; доминирует спорический
тип мейоза.
Грибы — мицелиальные гетеротрофы с сапро-
фитным питанием; клеточная стенка из хитина; крис-
ты в митохондриях пластинчатые; обычен зиготичес-
кий мейоз.
Протисты — преимущественно одноклеточные,
микроскопические, но есть и крупные синцитиаль-
ные, ценобиальные и многоклеточные формы; под-
вижность за счет жгутиков, ресничек, псевдоподий,
поверхностных структур клетки; питание автотроф-
ное, гетеротрофное, миксотрофное; формы мей-
оза — все варианты; кристы в митохондриях самой
различной формы.
Развитие представления о протистах как о пере-
ходной группе между прокариотами и остальными
эукариотами привело к пониманию того, что в преде-
лах протистов происходило, вероятно, становление
не только типов питания, но и всех клеточных сис-
тем, которыми в дальнейшем стали «пользоваться»
растения, животные и грибы.
На уровне генома наблюдается становление ди-,
поли- и амфиплоидности, многоядерности и гетеро-
морфизма, всех типов распределения генетического
СИСТЕМА ПРОТИСТОВ
125
материала ядра при бесполом размножении (все типы
митозов).
На уровне белок синтезирующей систе-
мы как проявления генома происходит фактически
ее эукариотизация (Серавин, 1992). Например, в
пределах протистов встречаются разные константы
седиментации рибосом — от характерной для прока-
риот 70S до типично эукариотической 80S.
На уровне клетки наблюдается приобретение
митохондрий со всеми возможными типами крист,
всех вариантов хлоропластов, из которых только
один реализуется в наземных растениях; всех типов
клеточных покровов, органелл и структур цитоскеле-
та, которые только можно обнаружить в клетках
многоклеточных и грибов.
На уровне организма встречаются все возмож-
ные типы жизненных циклов и форм существования
клетки, все формы редукции генетического материа-
ла (гаметический, спорический, зиготический), а
также обе формы полового процесса (с одно- и двух-
ступенчатым мейозом); все типы клеточного движе-
ния и клеточного питания, различные варианты мно-
гоклеточное™.
Следовательно, с позиций филогенетической
систематики мы не можем принять концепцию так-
сона протистов (Patterson, 1994). Они не эквива-
лентны растениям, животным или грибам, а вклю-
чают их в себя. Поэтому лучше говорить о фило-
гении эукариот в целом, и о филогенетической
системе, охватывающей всех эукариот. С этой точки
зрения приводить систему только протистов было
бы бессмысленно.
Лидером в совершенствовании системы эукариот
является Кавалье-Смит, опубликовавший множество
компилятивных, иногда противоречивых работ по
проблемам их систематики и филогении, происхож-
дения эукариотной клетки, автогенеза и симбиогенеза
(см.: Cavalier-Smith, 1987—1998). Под влиянием Пат-
терсона и Кавалье-Смита Корлисс опубликовал сис-
тему, включающую 34 типа протистов, распределен-
ных по 5 царствам эукариот (Corliss, 1994). В ней
уже нет царства Protista как таксономической катего-
рии, а само имя «протесты» превратилось в имя на-
рицательное в том смысле, который мы применяем
теперь по отношению к водорослям.
Прежде чем перейти к последним достижениям в
построении системы эукариот, рассмотрим главные
критерии в таксономии протистов, на основе кото-
рых выделяются наиболее весомые признаки.
Морфологические признаки
в систематике протистов
Оценивая роль морфологических признаков в
систематике живых организмов, мы должны при-
знать, что как у животных, так и у растений они
являются ведущими в мега-, макро- и микросисте-
матике. Основные признаки протистов определяют-
ся их клеточным уровнем организации и сочетают
как морфологические, так и биохимические и мо-
лекулярно-биологические. В определении «веса»
признака, соответствующего тому или иному уров-
ню таксона, и состоит главная трудность таксоно-
мии протистов.
Применение электронного микроскопа позволило
изучить строение клетки протистов и определить
наиболее существенные морфологические признаки.
70-е гг. можно считать расцветом ультраструктурных
исследований протистов, после чего наступил период
обобщений и постепенного угасания интереса к этой
области в результате переключения на молекулярно-
биологические методы. Можно констатировать, что
ультраструктурная революция завершилась к концу
80-х гг., в результате чего сформировался основной
набор ультраструктурных, а также биохимических
признаков у протистов. Наиболее весомыми призна-
ками считают особенности цитоскелета, преимущест-
венно жгутикового аппарата, структуру псевдоподи-
альных выростов, покровов, строение хлоропластов,
форму крист митохондрий, наличие/отсутствие дик-
тиосом, наличие уникальных органелл. Из биохими-
ческих особенностей наиболее важны наборы основ-
ных и дополнительных пигментов у фототрофных
эукариот.
Итоги ультраструктурной революции можно сум-
мировать в виде нескольких положений.
1) Установлена консервативность формы крист
митохондрий (трубчатые и пластинчатые).
2) В строении жгутикового аппарата обнаружены
гомологичные корешки у инфузорий, зеленых водо-
рослей и высших растений, хризофитовых водоро-
слейи миксомицетов.
3) Обнаружены уникальные структуры и органел-
лы, свидетельствующие о многообразии протистов и
позволяющие более четко определить границы таксо-
нов.
4) Выдвинута и частично подтверждена гипотеза
симбиогенного происхождения клеточных органелл.
При построении системы эукариот были выделе-
ны следующие монофилетические группы:
1) зеленые водоросли и высшие растения — обра-
зуют одну филу эукариот по многим признакам и
объединяются в царство Viridiplantae;
2) эвгленовые и кинетопластиды — объединяются
в тип Euglenozoa;
3) показано, что две группы организмов, содер-
жащих хлорофилл b (хлорофитовые и эвгленовые),
представляют собой независимые линии эукариот;
4) гетероконты, или страминопилы, являются мо-
нофилетической группой (царство Chromista), куда
вошли все протесты, содержащие хлорофилл с, и
многие бесцветные организмы, относимые ранее к
зооспоровым грибам или простейшим, которые имеют
трубчатые мастигонемы на переднем жгутике и труб-
чатые кристы в митохондриях;
5) сформировано царство Archezoa (Archamoebae,
Microsporidia, Metamonada), в которое вошли орга-
низмы, лишенные всех основных клеточных органелл
и структур (хлоропластов, митохондрий, гидрогено-
сом, пероксисом и диктиосом).
Вместе с тем на фоне этих очевидных успехов
ультраструктурных исследований имели место и
явные неудачи. В качестве одного из негативных
последствий для таксономии отметим распростра-
126
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
нившееся заблуждение, что отсутствие типичных
для эукариотической клетки признаков означает ее
примитивность, а присутствие их — продвинутость.
Многие протисты (амебы) оказались настолько бед-
ны морфологическими признаками, что их положе-
ние в системе не удается установить. Гетероген-
ность протистов не уменьшилась, а, скорее, даже
увеличилась, так как выросло количество «ультра-
структурных индивидуальностей», которые невоз-
можно отнести к каким-либо известным таксонам.
Другими словами, росло число протистов неопреде-
ленного таксономического положения, несмотря на
то что их ультратонкое строение было известно.
Помимо этого, новые системы протистов оказыва-
лись в большей степени «таксономическими», чем
филогенетическими. Из-за отсутствия (за редкими
исключениями) ископаемых форм и эмбриогенеза у
протистов невозможно было установить последова-
тельность появления групп в эволюции. Невозмож-
ность использовать фактор времени не позволяла
определить направленность эволюции даже в одно-
родных таксонах, так как морфологические ряды
могли читаться в обоих направлениях.
Чтобы перейти к следующему этапу — созданию
действительно филогенетической системы, необходи-
мо ввести фактор времени, т. е. определить последо-
вательность возникновения этих таксонов.
Молекулярно-биологические
признаки в систематике
и филогении протистов
В основе использования молекулярно-биологичес-
ких признаков в систематике лежит идея о молеку-
лярных часах, согласно которой изменения генома в
процессе эволюции идут с одинаковой скоростью.
Обобщенно можно сказать, что филогенетическая
удаленность одной группы организмов от другой оп-
ределяется процентом несходства последователь-
ностей нуклеотидов одного и того же участка ДНК.
Вероятность накопления мутаций (в соответствии с
гипотезой нейтральной эволюции) должна быть оди-
накова в разных группах организмов. Однако в дейст-
вительности она, конечно, могла различаться как у
разных организмов, так и в разных участках молеку-
лы ДНК. Более того, накопление мутаций не может
идти с одинаковой скоростью в разные периоды вре-
мени даже в одной группе организмов. Например, в
эволюции класса Diplomonadea (дипломонады), кото-
рый признается монофилетичным (Brugerolle; 1976),
происходили резкие изменения (удвоение органелл),
что не могло не сказаться на скорости эволюции белок
синтезирующих систем (Siddal et al., 1992). Тем не
менее в течение последних 15 лет многие биологи
с энтузиазмом занимались секвенированием (опре-
делением последовательности нуклеотидов) ДНК
эукариот и конструировали филогенетические де-
ревья, отражающие эволюцию организмов как внутри
родов и семейств, так и в пределах всех про- и эука-
риот.
«Рибосомальные» деревья
Построение филогенетических деревьев на основе
молекулярных данных началось со сравнительного
анализа последовательностей ДНК, кодирующих бел-
ки гемоглобин (Zuckerkandl, Pauling, 1965) и цито-
хром с (Fitch, Margoliash, 1967). Это направление не
получило тогда дальнейшего развития. В 1977 г. был
предложен более универсальный маркер — последо-
вательность нуклеотидов генов 5S рРНК — для пост-
роения филогении организмов и параллельно откры-
ты архебактерии (Woese, Fox, 1977). Полученный
результат вселил уверенность в правильности вы-
бранного пути. Позднее было признано, что гены 5S
рРНК значительно беднее информативно, чем у РНК
малой субъединицы рибосом (16S у прокариот или ее
гомолог 18S у эукариот), длина которой больше, а
усовершенствованные методы секвенирования и об-
работки материала позволяли получать данные в
сравнительно короткие сроки. Еще более полную ин-
формацию несет участок ДНК, кодирующий 23(28)S
рРНК, но он был слишком велик для полного секве-
нирования. РНК малой субъединицы рибосом обла-
дает рядом преимуществ, которые предопределили ее
использование в качестве филогенетического мар-
кера в течение последних 15 лет. Она универсальна,
поскольку есть у всех организмов (прокариот и эука-
риот), а также в митохондриях и хлоропластах. Ее
гены функционально консервативны и почти наверня-
ка гомологичны у всех организмов. Предполагается
также, что возможность «горизонтального» переноса
ее генов в эволюции крайне маловероятна.
В 1986 г. Согин с соавт. (Sogin et al., 1986) по-
строили дерево жизни на основании последователь-
ностей нуклеотидов генов 18S рРНК, куда вошли и
9 протистов (схема 2). В результате было показано
молекулярное разнообразие среди протистов, кото-
рое оказалось даже больше различий между растени-
ями и животными. В то же время протисты, как и
положено первым эукариотам, оказались в основании
дерева, а многоклеточные — на его вершине, или
кроне. Это вполне укладывалось в прежние представ-
ления о протистах как о переходной группе между
прокариотами и многоклеточными, поэтому статья
предопределила структуру молекулярного филогене-
тического дерева на весь дальнейший период.
То что произошло в последующие 10 лет, можно
назвать молекулярным взрывом. Накопление данных
по сиквенсам (последовательностям нуклеотидов)
генов рРНК как прокариот, так и эукариот и их оценка
шли лавинообразно. Не было построено двух одина-
ковых деревьев даже в одной лаборатории. Методы
подсчета различий между сиквенсами все время со-
вершенствуются и, естественно, различаются у раз-
ных авторов. Кроме того, включение новых видов в
результате расширения исследований вносит откло-
нения в принятые схемы.
К настоящему времени получено примерно 2000
последовательностей нуклеотидов генов рРНК эука-
риот, которые хранятся в специально созданном ге-
нетическом банке. Параллельно с накоплением дан-
ных по последовательностям усложняются и методы
их обработки. Например, в настоящее время сущест-
система протестов
127
Схема 2. Одно из первых «рибосомальных» деревьев, построен-,
ное на основе дистанционного анализа последовательностей нук-
леотидов генов 16(18)S рРНК. (По: Sogin et al., 1986).
Нижняя ветвь — прокариоты, верхняя — эукариоты.
Scheme 2. One of the first ribosomal tree derived from a distance-
matrix analysis of alinged «core regions» of small-subunit rRNA.
Rattus norvegicus
Xenopus laevis
Artemia salina
Zea mays
Oryza sativa
Acanthamoeba castellanii
Saccharomyces cerevisiae
Paramecium tetraurelia
Tetrahymena thermophilia
Oxytricha nova
Stylonychia pustulata
Dictyostelium discoideum
Trypanosoma brucei
Euglena gracilis
Sulfolobus solfataricus
Halobacterium volcanii
•Anacystis nidulans
Escherichia coli
100
100
100
75____
99
86
57
Thermus thermophilus
TXT? Anacystis nidulans
1001---------- Escherichia coli
-— Sulfolobus solfataricus
------- Methanococcus vannielii
------7- Halobacterium halobium
— Glugea atherinae
— Ameson michaelis
Pleistophora sp.
qq] I-------Enterocytozoon salmonis
10()l---Enterocytozoon bienusi
---------!--Encephalitozoon cuniculi
---------|------- Vairimorpha necatri
1001----- Nosema irichoplush
|—Coronympha octonaria
100ч— Trichomonas vaginalis
76Ц—Tritrichomonas foetus
78*— Monocercomonas sp.
Hexamita sp.
Glardia muris
Glardia lamblia
Glardia afdeae
88
99
| Trypanosoma brucei
100— Bodo caudatus
67
98
100
Vahlkampfia lobospinosa
Naegleria gruberi -
-----1 Entamoeba histolytica
100 Entamoeba gingivalis
Dictyostelium discoideum
КРОНА ДЕРЕВА
Eubacteria
Archaebacteria
Microsporidia
Trichomonadida
Diplomonadida
Myxomycota
Euglenozoa
Percolozoa
Entamoebia
Acrasiomycota
I
III
I----1 1.8
Схема 3. Три зоны, обычно выделяемые в типичном «рибосомальном» дереве эукариот. (По: Philippe, Adoutte, 1995).
В основании дерева {зона Z) длинные ветви микроспоридий, дипломонад и парабазалий (царство Archezoa), порядок ветвления которых может меняться.
В средней части (зона II), также меняясь местами друг с другом, находятся эвгленовые и кинетопластиды, клеточные и неклеточные слизевики, амебофла-
геллаты. Все остальные эукариоты (зона III) формируют крону дерева.
Scheme 3. Three zones which are usually distinguished in typical «ribosomal» eukaryotic tree.
Zone I at the basis contains long branches of microsporidia and metamonada (kingdom Archezoa) in variable order. In the middle part (zone II), also being interc-
hanged the position with each other there are euglenozoa, cellular and non-cellular slime molds, and amoeboflagellates. All other eukaryotes (zone III) form a crown.
128
ПРОТЕСТЫ. ОБЩИЕ ВОПРОСЫ
81
"1 56
56
70188
chlorophyta (62)1
------£ 7 chlorarachniophyta (HM) (5)
charophyta(19)
land plants (45)
chlorobionts
93 J
Ю0 llir™ haptophyta (6)
8 ।—--------- in "S3 chlorarachniophyta (5)
Euglyphina (2)
89
78
8-
heterokont algae (32)
57
94
94
75 l-
96
821
Hyphochytrium (hyphochytriomycetes)
oomycetes (3)
ишмММШД labyrinthulida (3)
-------Cafeter*a (oicosoecida)
J ciliata (26)
dinophyta (7)
Perkinsus
apicomplexa (17)
stramenopiles
alveolata
cryptophyta (6)
glaucophyta (2)
<z3 Acanthamoeba (3)
—cryptophyta (HM)
87
«цщяд Plasmodium (9)
rhodophyta (49)
таа choanoflagellida (2)
Схема 4. Крона эволюционного дерева эукариот, построенного на
основе анализа 500 последовательностей нуклеотидов генов рРНК
малой субъединицы. (По: Van de Peer, De Wachter, 1997).
Количество последовательностей показано в скобках после каждого назва-
ния таксона. Нуклеоморф (НМ) криптомонад занимает указанное положе-
ние только в том случае, если не приведены данные по нуклеоморфу хлор-
арахниофитов.
Scheme 4. Evolutionary tree reconstructed from 500 SSU rRNA se-
quences from eukaryotes of «crown».
The number of sequences comprised in each claster is mentioned in parantheses
after the taxon name. The cryptomonads occupy the evolutionary position indi-
cated by a dotted line only when the chlorarachniophyte nucleomorph sequen-
ces are omitted from the set.
вует несколько достаточно сложных программ, кото-
рые стали доступны многим исследователям благода-
ря сети Internet. Несмотря на несоответствия получа-
емых деревьев, общее расположение ветвей было
примерно одинаково, что указывало на правильность
выбранного маркера. Обычно в каждом дереве выде-
лялось 3 зоны (Philippe, Adoutte, 1995): основание,
средняя часть и крона (схема 3).
В основании дерева эукариот, как правило, распо-
лагались длинные ветви микроспоридий, дипломонад
и парабазалий. Их порядок мог меняться, но они
всегда первыми ответвлялись от эукариотического
ствола и считались наиболее древними эукариотами,
составляющими царство Archezoa. В средней части
дерева, также с известной степенью ротации, обычно
находились эвгленовые и кинетопластиды, клеточные
и неклеточные слизевики и некоторые амебофлагел-
латные группы. Их ветви несколько короче, чем у
групп, расположенных в основании, что указывало на
более медленную эволюцию этих организмов. Нако-
нец, все остальные эукариоты формировали крону
дерева, в которой невозможно было определить поря-
док ветвления, а сами ветви были много короче преды-
дущих, что свидетельствовало о сравнительно недав-
нем происхождении этих групп.
Один из примеров расположения эукариот в кроне
показан на схеме 4 (Van de Peer et al., 1997). В целом
эта схема отражает основные результаты, получен-
ные к настоящему времени разными группами иссле-
дователей. Зеленые водоросли, харовые и высшие
растения образуют филу хлоробионтов; водоросли,
содержащие хлорофилл с (за исключением Crypto-
phyta), образуют вместе с бесцветными страминопи-
лами линию развития, относящуюся к хромистам; ин-
фузории, динофлагелляты и споровики формируют
устойчивую монофилетическую группу Alveolata; в
один большой кластер попадают воротничковые жгу-
тиконосцы, Metazoa и настоящие грибы. В других
схемах существенные отличия касались положения
красных водорослей, которые часто объединяются с
хлоробионтами, а также гаптофитовых и криптомо-
над.
Молекулярная догма
Выводы, сделанные на основании полученных ре-
зультатов, стали перерастать в догму, которая начала
определять обсуждение таких проблем, как проис-
хождение эукариотной клетки, ее органелл, полового
размножения и т. д. Это ясно видно на обобщенных
схемах развития эукариот (см.: Patterson, 1994; Sleigh,
1995; Margulis, 1996), совмещающих морфологичес-
кие и молекулярные данные (схема 5).
К самым древним эукариотам (Archezoa) относи-
ли Archamoebae, Microsporidia и Metamonada. Пер-
выми эукариотами считались представители царства
Archezoa, которые приобрели только ядро. Среди
них наиболее просто устроены Microsporidia, для ко-
торых характерны рибосомы прокариотного типа
(5S, 16S и 23S рРНК), закрытый внутриядерный
плевромитоз, отсутствие подвижных стадий в жиз-
ненном цикле, наличие хитина в клеточной стенке
(что свойственно также и архебактериям), отсутствие
митохондрий, пероксисом, хлоропластов, гидрогено-
сом, диктиосом и жгутиков. Тип Microsporidia пред-
ставлен только внутриклеточными паразитами.
На следующем этапе эукариотическая клетка при-
обретает жгутики и появляются архамебы и мета-
монады. Archamoebae являются свободноживущими
анаэробными амебофлагеллатами, содержащими 1 ка-
риомастигонт. Их жгутики малоподвижны или реду-
цированы, от 1 кинетосомы отходит к ядру конус из
микротрубочек. Тип митоза неизвестен. Archamoebae
включает отряды Pelobiontida (Pelomyxa palustris) и
Mastigamoebida (Mastigina, Mastigella).
Metamonada представлены паразитами или сво-
бодноживущими анаэробами. Они имеют 4 кинетосо-
СИСТЕМА ПРОТИСТОВ
129
ACANTHAREA
ACTINOPHYIDA
APUSOMONADIDA
CERCOMONADIDA
COPROMYXIDA
DESMOTHORACIDA
DIMORPHIDA
EBRIIDA
RAMICRISTATES
GRANULORETICULOSA
альвеолы
CILIATA
APICOMPLEXA
DINOFLAGELLATA
GYMNOSPHAERIDA -И трубчатые l\_ EUMYCETOZOA
HAPLOSPORIDA ----------
TRIPLOBLASTICA
кристы
VAMPYRELLIDA
THAUMATOMONADA
SPONGOMONADIDA
PSEUDODENDROMONADID
POLYCYSTINEA
PHAEODAREA
PARAMYXEA
KATHABLEPHARIDA
HAPTOPHYTA
CHRYSOPHYTA
AXODINA
SYNUROPHYTA
XANTHOPHYTA
RAPHIDOPHYTA
EUSTIGMATOPHYTA
BROWN ALGAE
DIATOMEA
PARMALEA
wHкр —
SPONGES
CHOANOFLAGELLATA
CHYTRIDA
EUFUNGI
RED ALGAE
VIRIDAEPLANTAE
PLASMODIOPHORIDA
CRYPTOMONADA
CYANOPHOREA
CENTROHELIDA
(митохондрии |
плоские
7
трехчленные
мастигонемы
BICOSOECIDA
LABYR1NTHULIDA
OOMYCETES
OPALINEA
дисковидные
кристы
EUGLENIDA
KIN ETO PL ASTI DA
NUCLEARIIDA
SCHIZOPYRENIDA
ACRASIDA
ELLOBIOPSIDA
SPIRONEMIDAE
KOMOKIIDA
XENOPHYOPHOREA
HYPERMASTIGIDA
[ ДИКТИОСОМЫ |
TRICHOMONADIDA
OXYMONADIDA
фибриллярный
цитоскелет
| жгутик I
? -
I ядро
DIPLOMONADIDA
RETORTAMONADIDA
PELOBIONTIDA
ENTAMOEBIDA
MICROSPORIDIA
Схема 5. Обобщенная схема формирования таксонов эукариот в процессе эволюции. (По: Patterson, 1994).
Scheme 5. The general scheme of eukaryotic evolution.
мы и 4 жгутика, которые организованы попарно, ре-
куррентная кинетосома ассоциирована по меньшей
мере с 1 микротрубочковым корешком; для них ха-
рактерен закрытый или полузакрытый митоз. Meta-
monada включает отряды Retortamonadida, Diplomo-
nadida (Giardia, Hexamita и Trichomonas имеют 16S
рРНК) и Oxymonadida.
С точки зрения морфолога, в царстве Archezoa
объединяются настолько разные организмы, что
вполне можно говорить об их независимом проис-
хождении. Бружероль (Brugerolle, 1991) считает, что
жгутиконосцы из архамеб и метамонад не имеют го-
мологичных структур. Цитоскелет архамеб может
быть как примитивным, так и регрессивным. Метамо-
нады могут иметь гомологичные жгутиковые структу-
ры, а могут и не иметь их. Основные признаки, по
которым выделяется царство — отсутствие митохон-
дрий и прочих органелл, могут быть вторичными.
Этого не исключал и Кавалье-Смит (Cavalier-Smith,
1993). Все представители царства — или паразиты,
или анаэробы, которым не нужны ни митохондрии,
ни хлоропласты, потому что они адаптированы к
очень специфическим условиям обитания. Кроме
того, почти во всех группах эукариот встречаются
безмитохондриальные организмы (Muller, 1998), од-
нако они появляются, на молекулярном дереве далеко
от основания и поэтому считаются утратившими ми-
тохондрии вторично. Архезои же претендовали на
9 Протисты, часть 1
исходное отсутствие митохондрий. И хотя молеку-
лярных данных по архамебам тогда не было, их все-
таки помещали в основание дерева (схема 5).
На следующем этапе эволюции происходит при-
обретение диктиосом: аппарат Гольджи формирует
стопки цистерн, что увеличивает эффективность его
работы. Таким образом появляются парабазалии
(Trichomonadida и Hypermastigida). Только после этого
эукариоты приобретают, наконец, митохондрии, и
вся дальнейшая их эволюция идет по типам крист
митохондрий, приобретению хлоропластов, трубча-
тых мастигонем и других органелл и структур (схе-
ма 5). Подобные схемы становления эукариотной
клетки стали включаться в учебники.
«Белковые» деревья
Параллельно с «рибосомальными» строились и «бел-
ковые» деревья, которые были основаны на последо-
вательностях ДНК, кодирующих различные широкорас-
пространенные белки: а- и (3-тубулины, актин, глицер-
альдегид-3-фосфатдегидрогеназу (GAPDH), фактор
элонгации a (EF-1 а) и хитшоковые белки (HSP 70). На
фоне «рибосомальных» деревьев они выглядели менее
представительно, появились позже и поэтому, естест-
венно, испытывали сильное влияние со стороны «ри-
босомальных» построений. В них подчеркивались
130
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
961_____
72
93
90
94
68
тт----------Entamoeba histolytica
—--^Dictyostelium discoideum
-Physarum polycephalum
-----Arabidopsis thaliana
Reticulomyxa filosa
Euglena gracilis
Trypanosoma cruzi
hlamydomonas reinhardtii
Naegleria gruberi
Tetrahymena pyriformis
Plasmodium falciparum
100.— Homo sapiens
Fl-Homo sapiens
67j । ....— Schizophyllum commune
L-51Lr—— Schizosaccharomyces pombe
’---------------Neurospora crassa
Entamoeba histolytica
—— Reticulomyxa filosa
---Chondrus crispus
Porphyra purpurea
.......Reticulomyxa filosa
— Physarum polycephalum
— Dictyostelium discoideum
y - Trichomonas vaginalis
"|i— Giardia lamblia
80*.... Hexamita sp.
p Caenorhabditis elegans
__r- Caenorhabditis elegans
95 99*-Caenorhabditis elegans
Г Caenorhabditis briggsae
UpDrosophila melanogaster
|g— Drosophila melanogaster
Homo sapiens
4 r— Drosophila erecta
*tp“ Gallus gallus
77 Gallus gallus
p—?“ Schizophyllum commune
“..........EncephalitoZoon hellem
—Pneumocystis carinii
r—— Schizosaccharomyces pombe
jr-.....— Geotrichum candiaum
oTl r Saccaromyces cerevisiae
100 *--Candida albicans
i---Collectotrichum graminicola
100“— Collectotrichum gloeosporioides
richoderma viride
Trichoderma viride
Penicillium digitatum
Neurospora crassa
Emericella nidulans
81 *- Acremonium chrysogenum
□
77
74
70
81
93
-Physarum polycephalum
r-Chlamyaomonas reinhardtii
76
98 75r Oryza sativa
Zea mays
Arabidopsis thaliana
I лл|"г Arabidopsis thaliana
Arabidopsis thaliana
ryza sativa
Zea mays
Arabidopsis thaliana
96**Arabidopsis thaliana
95p Porphyra purpurea
-------Porphyra purpurea
ц—Achlya kiebsiana
oj Ectocarpus variabilis
д Euplotes crassus
59 pEuplotes octocarinatus
Ip Stylonychia lemnae
ni Paramecium tetraurelia
1ПTetrahymena thermophila
69rr-Babesia bovis
|p-Plasmodium falciparum
97lp Eimeria tenella
SO^Toxoplasma gondii
• Naegleria gruberi
i—Euglena gracilis
I 94 rTrypanosoma brucei
I П-Trypanosoma cruzi
lOoLrLeishmania mexicana
1001- Leishmania major
а-тубулин
Rhodobionta
Mycetozoa
Trichomonadida
Diplomonadida
Metazoa
Fungi
Microsporidia
Fungi
Mycetozoa
Chlorobionta
Rhodobionta
Stramenopiles
Ciliophora
Sporozoa
Percolozoa
Euglenozoa
Схема 6. Филогенетическое дерево эукариот, построенное на основе анализа гена [3-тубулина. (По: Philippe, Adoutte, 1998).
В качестве внешней группы использована последовательность нуклеотидов гена а-тубулина.
Scheme 6. 0-tubuline phylogenetic tree of eukaryotes.
Sequences of a-tubuline gene have been used as outgroup.
только те аспекты, которые были в согласии с «рибо-
сомальными», а несоответствия объяснялись разными
причинами. Приведем 2 наиболее полных, недавно
опубликованных филогенетических дерева* (Philippe,
Adoutte, 1998): «тубулиновое» (схема 6) и «актино-
вое» (схема 7).
Очевидно несоответствие «тубулинового» дерева
«рибосомальному» (ср. схемы 3 и 4 со схемой 6). На
схеме 6 две амебы появляются в основании дерева;
дипломонады и трихомонады ответвляются намного
позже (как монофилетическая группа); грибы все
еще формируют сестринскую группу Metazoa, но те-
СИСТЕМА ПРОТИСТОВ
131
87
99
100
7.4
---------...... —........Saccharomyces cerevisiae
— Pneumocystis carinii
""" -.... Neurospora crassa
I-------j-----Drosophila melanogaster
i nn 1 r—Homo sapiens
1001-----Homo sapiens beta
----------Giardia lamblia
----------Euplotes crassus
|---Oxytricha nova
100*—— Oxytricha trifallax
...... Trichomonas vaginalis
..........--- Oxytricna nova
.... — Plasmodium falciparum
f—“ Leishmania major
....Г77П |-------------Trypanosoma cruzi
TnT-pTrypanosoma brucei
99| Trypanosoma brucei
—-------— Tetrahymena termophila
co j—Plasmodium falciparum
j 001"’Toxoplasma gondii
Cryptosporidium parvum
lOOp-Fucus distichus
i-Fucus vesiculosus
. Г7Т pPhytophthora infestans
93 J 100 PPhytophthora megasperma
4 j-Achlya bisexuaiis
if-Pythium irregulare
lOOLphytophthora infestans
......."Naegleria fowleri
.......---- Entamoeba histolytica
j—.......................... Chondrus crispus
*......- Acetabularia cliftonii “
-----Cyanidioschyzon merolae Z
Chlamydomonas reinhardtii
Volvox carteri
Solanum tuberosum
Arabidopsis thaliana
Pisum sativum
Arabidopsis thaliana
— Solanum tuberosum
Oryza sativa
Arabidopsis thaliana
Solanum tuberosum
“Arabidopsis thaliana
Solanum tuberosum -
—Pneumocystis carinii
r—Absidia giauca
87 6j 99pHistoplasma capsulatum
I Emencella nidulans
LJ 1--------Thermomyces lanuginosus
941 iSaccharomvces cerevisiae
1001—Kluyveromyces lactis
।---Schizosaccharomyces pombe
*]!—Puccinia graminis
7(jlf—Filobasidiella neoformans
‘—PhafTia rhodozima
гёо Schistosoma mansoni *
Diphyllobothrium dendriticum
’ Homo sapiens
^-Schistosoma mansoni
-Drosophila melanogaster
a’lomo sapiens
[ото sapiens
osophila melanogaster
osophila melanogaster
ira attenuata
iocoryne carnea
Emihania huxleyi
'Acanthamoeba castellanii
^nirPhysarum polycephalum
87|TDictyostehum discoideum
771Dictyostelium discoideum 8
76f“Dictyostelium discoideum A3-SI
IrDictyostelium discoideum 12
951,Dictyostelium discoideum
r-Phytophthora infestans
T-PhvtoDhthora megasperma
00
5
100
51
центрактин
=f Diplomonadida
I Ciliophora
_ Trichomonadida
— Ciliophora
=; Sporozoa
Euglenozoa
— Ciliophora
Sporozoa
Stramenopiles
— Percolozoa
Rhodobionta
Chlorobionta
Rhodobionta
Chlorobionta
Fungi
Metazoa
Rhizopoda"
Схема 7. Филогенетическое дерево эукариот, построенное на основе анализа генов актина. (По: Philippe, Adoutte, 1998).
В качестве внешней группы использована последовательность нуклеотидов гена центрактина.
Scheme 7. Actin phylogenetic tree of eukaryotes.
Sequences of centractin gene have been used as outgroup.
перь среди них появились микроспоридии, которые
находятся очень далеко от основания дерева; Eugle-
nozoa располагаются в кроне как сестринская группа
инфузорий и хлорофитов. Таким образом, некоторые
«древние» группы появляются в центре или наверху
дерева и, наоборот, некоторые «продвинутые» груп-
пы смещаются к основанию дерева.
На «актиновом» дереве (схема 7) инфузории ока-
зываются полифилетичными, и большинство их появ-
ляется ближе к основанию, т. е. в противоположном
положении по сравнению с «рибосомальным». В
кроне дерева формируется монофилетическая группа
ризопод, что никогда не отмечалось в «рибосомаль-
ных» деревьях. Царство Archez_oa отсутствует в обоих
случаях. Как же объяснить эти противоречия?
В 1995 г. Кларк и Роджер (Clark, Roger, 1995)
нашли ядерные гены, кодирующие хитшоковые белки
явно митохондриального происхождения в безмито-
хондриальной амебе Entamoeba. Позднее гены мито-
хондриальных хитшоковых белков, были найдены у
трихомонад, дипломонад и микроспоридий (Hashimo-
to et al., 1998; Philippe, Adoutte, 1998). Следователь-
но, они имели митохондрии в прошлом, а затем утра-
тили их при переходе к паразитизму или анаэробно-
132
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
97
--- 86
100
I— 56
Acanthamoeba castellani —
C— Apusomonas proboscides
J7 Diaphanoeca grandis
r{*- uAcanthoecopsis ungulculata ш„
98 Anemonia sulcata
f— Microciona prolifera ---
•hZ} Chy tridium confervae
I— Saccharomyces cerevisiae __
rj"OQ Goniomonas truncata
[Год Rhodomonas marina
Chilomonas paramecium
62 v
r— Gloeochaete wittorckiana
jr^2yanophora paradoxa
L- Glaucocystis nostochinearum
Г— Volvox carteri
k, Mantoniella squamata
Zamia pumila
ERhodella maculata
Porphyra umbilicalis
Porphyridium aerugineum
r00Paviova aff. salina
[r^Prymnesium patelliferum
u^Emiliania huxleyi
Labyrinthuloides minuta мм
Г99 Cafeteria roenbergensis
|p Hyphochytrium catenoides
WOchromonas danica
V-Fucus distichus
*—Bacillaria paxillifer
Oxytricha nova —
~• Blepharisma americanum
96 _ . , МММ
ktx Perkmsus sp. —
Theileria parva
Ir^Crypthecodinium cohnii =
° Prorocentrum micans
r~ Paulinella chromatophora мм
[L— E ugly ph a rotunda
П——Chiorarachnion reptans
piasmodiophora brassicae
H——Hartmannella vermiformis JJJjJ
.... Phreatamoeba balamuthi
I I "L. Minchinia teredinis яв
1222. Haplosporidium nelsoni
I ' Dictyosteiium discoideum """
—— Physarum polycephalum я
JaoTrypanosoma brucei
H-^Leishmania donovani
Л- Trypanoplasma borreli
Pqq Bodo caudatus
I jQQ l- Dimastigeila trypaniformis
. L»,"" Euglena gracilis иии
1 — Entamoeba gingivalis «=
lYooVahlkampfia lobospinosa
I LwTetramitus rostratus
—— Naegleria gruberi
r—— HSU17512 ""
--------HSU 17508
I 89
Ditrichomonas honigbergi
Pseudotrichomonas kellini
i|59
J —^Trichomonas vaginalis
I r-LMonocercomonas sp.
[p^Tritrichomonas foetus
Ir^Coronympha octonaria
Metadevescovinapolyspira ___
M6Enterocytozoon bieheusi
* -Тюб ^‘tta^orma corneum
- Endoreticulatus schubergi
r—— Septata inflata
Encephalitozoon hellem
Encephalitozoon cuniculi
Vairimorpha necatrix
"”ь—— Nosema bbmbycis
,, Ichthyosporidium sp.
Ameson michaelis
Spironucleus muris
Hexamita inflata
I - ...Trepomonas agilis ___
rrTT Giardia ardeae
1122.Giardia lamblia ___
Grj.. ''1 Thermoproteus tenax
— Thermoplasma acidophilum
Methanococcus voltae
f.....—... Haloferax volcani
I—-Archaeglobus fuigidus __
0.10
LOBOSA
APUSOZOA
CHOANOZOA
ANIMALIA
FUNGI
CRYPTISTA
GLAUCOPHYTA
VIRIDAEPLANTAE
RHODOPHYTA
HAPTOPHYTA
HETEROKQNTA
CILIOPHORA
APICOMPLEXA
DINOZOA
RHIZOPODA
LOBOSA
ARCHAMOEBAE
HAPLOSPORIDIA
MYCETOZOA
EUGLENOZOA
OPISTHOKONTA
PLANTAE
CHROMOBIOTA
ALVEOLATA
96
100
100
70
77
ARCHAMOEBAE
PERCOLOZOA
PARABASALA
MICROSPORIDIA
DISTOMATIDA
GIARDIIDA
DIPLOZOA
ARCHAEBACTERIA
Схема 8. Одно из типичных «рибосомальных» деревьев эукариот, показывающее расположение архамеб: Phreatamoeba (=Mastigamoe‘
ba) и Entamoeba. (По: Cavalier-Smith, Chao, 1996).
Scheme 8. One of the typical «ribosomal» eukaryotic tree including new data on archamoeba Phreatamoeba (=Mastigamoeba).
система протестов
133
му образу жизни, т. е. можно считать что приобрете-
ние митохондрий произошло до появления всех со-
временных эукариот.
Примерно в то же время (Cavalier-Smith, Chao, 1996)
были получены данные по последовательности нукле-
отидов рРНК у Phreatamoeba {-Mastigamoeba) — ти-
пичного представителя архамеб (схема 8). Эти данные
показывают, что Phreatamoeba располагается далеко
от основания дерева и, скорее, является сестринской
группой лобозных амеб, что вполне соответствует
морфологическим данным. В результате царство Ar-
chezoa было упразднено (Cavalier-Smith, 1998).
Таким образом, Archamoebae, как и в прежних
системах, могут быть включены в тип Rhizipoda под
названием Peloflagellata (Caryoblastea), а представи-
тели Metamonada (ретортамонады, дипломонады и
оксимонады) занимают свое обычное место среди по-
лимастигин (тип Polymastigota).
На пути к построению
истинного
филогенетического дерева
эукариот
Как же все-таки объяснить возникшие несоответ-
ствия между «рибосомальными» и «белковыми» де-
ревьями? Эти проблемы, а также многочисленные
«ловушки» при построении молекулярных деревьев
были подробно обсуждены в недавней статье фран-
цузских исследователей (Philippe, Adoutte, 1998). Не
углубляясь в детали анализа всех подводных камней
на пути построения «правильных» молекулярных де-
ревьев, остановимся лишь на тех из них, которые наи-
более существенны для нашей цели — определения
филогенетических и таксономических взаимоотно-
шений между крупными группировками протистов.
Оказалось, что наиболее сомнительны ветви, на-
ходящиеся в основании дерева. Они всегда длиннее
других, что свидетельствует о высокой скорости эво-
люции соответствующих групп. На языке молекуляр-
ной генетики это означает, что последовательности
генов рРНК у этих организмов мутационно насыще-
ны. При анализе групп с неравными скоростями эво-
люции проявляется феномен, который известен как
«long branch attraction» артефакт. Суть его состоит в
том, что виды с более высокими скоростями эволю-
ции имеют тенденцию объединяться друг с другом,
вместо того чтобы примыкать к их истинной сест-
ринской группе, и появляются на дереве ниже их ис-
тинного положения. По-видимому, это и было причи-
ной раннего ответвления архезоев на дереве.
Таким образом, базальная часть дерева оказывает-
ся асимметричной и искаженной из-за явления «long
branch attraction», в то время как апикальная его
часть обычно симметрична и правильно отражает ог-
ромную радиацию эукариот. Поэтому можно полу-
чить более совершенную картину филогении эукари-
от при сопоставлении только симметричных частей
деревьев. Сравнение крон «рибосомальных» и «бел-
ковых» деревьев магло бы дать хорошие результаты,
однако здесь встречается еще одна трудность молеку-
лярной филогении. Если разделение между предками
определенных видов произошло в короткое время, то
может оказаться, что молекулярные синапоморфные
признаки, которые позволили бы реконструировать
тип ветвления, еще не успели накопиться. Таким об-
разом, вместо последовательных ветвлений дерева мы
получим огромный «неразрешенный» кустарник, ги-
гантскую мультифуркацию. В принципе эта проблема
может быть преодолена, но «разрешение» ветвлений,
или минимальный временной промежуток, который
можно зарегистрировать, зависит от количества нук-
леотидов анализируемой последовательности. Напри-
мер, полная последовательность РНК малой субъе-
диницы рибосом дает надежное «разрешение» только
для видообразования, разделенного более чем 40 млн
лет. Отсутствие «разрешения» у Metazoa, таким обра-
зом, совместимо с палеонтологическими данными, в
соответствии с которыми главные типы многоклеточ-
ных появились в промежутке менее чем 20 млн лет, в
раннем кембрии. Подсчеты показывают, что при оцен-
ке филогении Metazoa 500 нуклеотидов дают «разре-
шение» в 140 млн лет, но требуется проанализировать
80 000 нуклеотидов для получения разрешения в
1 млн лет. При этом мы должны полагаться на со-
мнительное предположение о точной работе молеку-
лярных часов.
Показательны в этом отношении результаты рабо-
ты Филипа и Адута (Philippe, Adoutte, 1998) по ана-
лизу топологии молекулярных деревьев всех эукари-
от. Авторы исходили в своих рассуждениях из двух
положений: 1) при сравнении деревьев группы из ос-
нования и середины «рибосомального» дерева оказы-
вались в кроне «белкового», 2) в кроне невозможно
определить последовательность ветвления. Поэтому
очень трудно или даже невозможно найти наборы
молекулярных синапоморфных признаков, которые
позволили бы изобразить полностью «разрешенное»
дерево эукариот в форме последовательных ветвле-
ний. В связи с этим авторы задались вопросом: явля-
ется ли эволюционная история эукариот постепен-
ной, или имела место эволюционная радиация?
Если предположить возможность дивергенции эука-
риот в течение относительно короткого времени, то
станет ясно, почему до сих пор не были найдены
морфологические и молекулярные синапоморфии.
Авторы даже подсчитали, что эта радиация произош-
ла 700—1000 млн лет назад, и высказали предполо-
жение, что ей предшествовал длительный период ста-
новления эукариотной клетки, однако большинство
потомков (если не все) этого раннего периода вымер-
ли (схема 9). В течение этого времени было накопле-
но достаточное количество молекулярных и морфо-
логических синапоморфий в пределах некоторых
линий, что позволяет идентифицировать их как мо-
нофилетические группы. Если связать эти рассужде-
ния с представлениями о митохондриальном симбио-
зе, который предшествовал радиации, то такое объяс-
нение вполне возможно.
Таким образом, можно предположить, что все
ныне живущие эукариоты произошли от одного пред-
ка, получившего в виде прообраза митохондрий ка-
кие-то протеобактерии. Эта радиация произошла, по-
видимому, сравнительно недавно, когда насыщен-
134
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
ность кислорода в атмосфере достигла современного
уровня. Можно даже предположить, что полученное
преимущество было столь велико, что привело к ис-
чезновению всех первично амитохондриальных пред-
ков. Авторы называют эту гипотезу «большим мито-
хондриальным взрывом» и полагают, что все имею-
щиеся в настоящее время организмы возникли как
его следствие (Philippe, Adoutte, 1998).
Так ли это происходило в действительности, ска-
зать трудно. Для нас важно другое. Вслед за авторами
этой и других последних работ по филогении эукари-
от мы можем констатировать, что достигнуто согла-
шение по монофилии многих групп эукариот и это в
значительной степени облегчает задачу построения
истинной системы эукариот. Правда, как и до моле-
кулярных исследований, она лишь с большой натяж-
кой может называться филогенетической, так как
последовательность появления таксонов остается не-
Схема 9. Гипотеза «большого взрыва». (По: Philippe, Adoutte,
1998).
Современные эукариоты предположительно возникли в результате быстрой
радиации, связанной, вероятно, с приобретением митохондриальных симби-
онтов. Ранние (возможно, анаэробные) группы вымерли полностью. Ряд мо-
нофилетических групп высокого таксономического уровня можно иденти-
фицировать в кроне, но их взаимоотношения трудно установить. Среди них
выделяются только 2 группы очень высокого ранга. Одна объединяет грибы,
микроспоридии и животные, другая — зеленые растения и красные водоросли.
Scheme 9. The «big bang» hypotesis.
Extant eukaryotes are assumed to have arisen during a period of rapid diversi-
fication, perhaps correlated with the establishment of mitochondrial symbiosis.
Earliest (possibly anaerobic) groups have not left living representatives. A num-
ber of high level monophyletic groups can still be identified in the «crown»,
such as Alveolates or Stramenopiles, but their relationships are difficult to es-
tablish. Only two even higher level groups are suggested, that uniting Fungi,
Microsporidia and Animalia, and that uniting Rhodobionta and Chlorobionta.
известной. Радует, однако, тот факт, что морфологи-
ческая однородность нескольких групп протистов,
так же как и многие догадки морфологов, подтверж-
дены молекулярно-биологическими данными.
В целом можно констатировать, что большие на-
дежды, связанные с использованием нуклеиновых
кислот как надежных филогенетических маркеров,
пока не оправдались, но синтез двух разных подходов
дал результаты, которые не кажутся теперь противо-
речивыми.
В то же время признание монофилетичности мно-
гих (но далеко не всех) групп эукариот еще не озна-
чает фактического построения системы. Самое слож-
ное заключается в определении ранга этих группиро-
вок и их взаимоотношений друг с другом. Здесь
мнения могут быть самыми разными. В соответствии
с взглядами Филиппа и Адутта (Philippe, Adoutte,
1998) каждый из монофилетических таксонов (схема 9)
мог бы быть отдельным царством эукариот, хотя сами
авторы считают, что крупных группировок эукариот
не должно быть более 4—5. Паттерсон (1999) вы-
деляет около 70 групп эукариот (не считая сотни
других, имеющих неопределенное систематическое
положение). Их ранг варьирует от рода (например,
Trimastix) как минимум до царства (Opistocontha),
объединяющего всех многоклеточных животных
(включая губки и миксоспоридии), воротничковых
жгутиконосцев и грибы (включая микроспоридии).
Кавалье-Смит (Cavalier-Smith, 1998) предпочитает
выделять 5 царств эукариот: Plantae, Metazoa, Fungi,
Chromista и Protozoa. Маргулис (Margulis, 1996) до
сих пор считает приемлемым сохранение царства
Protista.
Среди современных отечественных систем выде-
ляется оригинальностью система Кусакина и Дроздо-
ва (1994, 1998), в которой насчитывается 11 царств
эукариот.
Таким образом, принятие определенной системы
эукариот, по-видимому, преждевременно.
По этой причине, вероятно, не принята единая ме-
тасистема протистов во втором издании американ-
ского «Атласа простейших», выход которого из печа-
ти был намечен на 1999 год.
Первоначально (5 лет назад) для нашего издания
была принята единая система протистов, в соответ-
ствии с которой были написаны все главы. По сути,
она представляла собой модифицированную систему
Корлисса (Corliss, 1994), в которой, правда, отсутст-
вовало царство Archezoa, а протесты распределялись
по 5 царствам эукариот: Animalia, Fungi, Plantae,
Chromista и Protozoa. Принятие этой системы не про-
тиворечит в целом современным представлениям о
филогенетической системе эукариот. Однако уже
сейчас просматриваются тенденции к объединению
грибов и животных, а царство простейших, без со-
мнения, является полифилетичным и будет пересмот-
рено. По-видимому, альвеолаты будут объединены с
эвгленозоями, а криптофитовых и гаптофитовых все
более сближают с растениями. Думается, что мета-
система эукариот будет принята в общем виде в бли-
жайшие 3—5 лет. Из приведенных рассуждений можно
сделать вывод, что любая современная мегасистема
будет временной. Поэтому здесь мы не приводим
СИСТЕМА ПРОТИСТОВ
135
общей системы эукариот, полагая даже, что это не
является необходимым при монографическом описа-
нии отдельных групп протистов. Отсутствие общей
системы не может повлиять ни на способ подачи ма-
териала, ни на рассуждения о филогении того или
иного таксона. В то же время большое внимание уде-
лено системе каждого типа протистов.
Классификация протистов,
рассматриваемых
в этой книге
В настоящем издании излагаются материалы не по
всем группам протистов. Ограниченный объем не
позволяет включить в него пединелломорфид, сол-
нечников, радиолярий, алвеолат, микроспоридий,
миксоспоридий и протистов неясного таксономичес-
кого положения. Предполагается, что все эти группы
будут рассмотрены во второй части «Протесты: Руко-
водство по зоологии». Не рассматриваются здесь и
такие традиционно непротозоологические группы,
как желтозеленые и диатомовые водоросли, гифохит-
ридиевые грибы и некоторые другие.
По этой же причине весьма кратко изложены ма-
териалы по так называемым фитофлагеллатам (гапто-
фиты, рафидофиты, эвгленофиты) и традиционно ми-
кологическим группам (оомицеты). В то же время по
возможности полно даны сведения по миксомицетам,
которые близки к гетеротрофным протистам.
Список таксонов протистов,
рассматриваемых в этой книге
Тип Cryptophyta Pascher, 1914
Класс Cryptophyceae Pascher, 1914
Тип Euglenozoa Cavalier-Smith, 1981
Класс Euglenoidea Biitschli, 1884
Класс Kinetoplastidea Honigberg, 1963
Тип Chrysophyta Pascher, 1914
Класс Chrysophyceae Pascher, 1914
Отряд Bicosoecales (Grasse) Karpov, 1998
Класс Synurophyceae Andersen, 1987
Класс Pelagophyceae Andersen et Saunders, 1993
Тип Haptophyta Christensen, 1962
Класс Haptophyceae Christensen, 1962
Тип Raphidophyta Chadefaud, 1950
Класс Raphidophyceae Chadefaud, 1950
Тип Saprolegnia Zerov, 1972
Класс Saprolegnea (=Oomycetes) Zerov, 1972
Класс Labyrinthomorphea Page, 1979
Отряд Labyrinthulida Cienkowski, 1867
Отряд Thraustochytrida Sparrow, 1943
Тип Opalinata Wenyon, 1926
Класс Proteromonadea Grasse, 1952
Класс Opalinatea Wenyon, 1926
«Chromista» incertae sedis: Spongomonada (Hibberd) Karpov,
1990; Pseudodendromonada Hibberd, 1985.
Тип Choanomonada Kent, 1880
Класс Choanomonadea Kent, 1880
Тип Polymastigota Biitschli, 1884
Класс Diplomonadea Wenyon, 1926
Класс Oxymonadea Grasse, 1952
Класс Parabasalea Honigberg, 1973
Отряд Trichomonadida Kirby, 1947
Отряд Hypermastigida Grassi et Foa, 1911
Тип Plasmodiophora Zopf,1884
Тип Mycetozoa de В ary, 1859
Класс Cercomonadea Mylnikov, 1986
Класс Eumycetozoea Zopf, 1884
Подкласс Pfotostelia Olive, 1970
Подкласс Dictyostelia Olive, 1970
Подкласс Myxogastria Olive, 1970
Mycetozoa incertae sedis: Hyperamoeba flagellata Alexeieff,
1926.
Тип Rhizopoda Siebold, 1845
Класс Lobosea Carpenter, 1861
Подкласс Gymnamoebia Haeckel, 1862
Подкласс Testacealobosia de Saedeleer, 1934
Класс Heterolobosea Page et Blanton, 1985
Отряд Acrasida Schrbter, 1886
Отряд Schizopyrenida Singh, 1952
Класс Peloflagellatea (=Caryoblastea) Goodkov et Seravin, 1988
Класс Filosea Leidy, 1879
Подкласс Aconchulinea de Saedeleer, 1934
Подкласс Testaceafilosia de Saedeleer, 1934
Класс Xenophyophorea Schulze, 1904
Rhizopoda incertae sedis: Komokiida Kamenskaya, 1992;
Athalamia Haeckel, 1862, Monothalamia Haeckel, 1862.
Тип Foraminifera D’Orbigni, 1826
Выделение хромобионтов, или гетероконтов, в са-
мостоятельное царство Chromista обсуждается уже
давно. В него обычно включаются фототрофы, содер-
жащие хлорофиллы а и с. В последние годы к ним
причисляются и некоторые гетеротрофные группы
протистов, имеющие характерное строение зооспор,
а именно гетероконтные жгутиконосцы с передним
опушенным жгутиком. Паттерсон (Patterson, 1989)
назвал их страминопилами (stramenopiles). Такое
строение имеют зооспоры, гаметы и вегетативные
клетки почти всех водорослей, содержащих хлоро-
филл с, и многих зооспоровых грибов. Кроме того,
эти признаки сочетаются с наличием трубчатых крист
в митохондриях и спирали в переходной зоне жгути-
ка. По этим признакам к хромистам, или хромофито-
вым в широком смысле, помимо водорослей относят-
ся зооспоровые грибы (оомицеты, траустохитридие-
вые и гифохитридиевые), опалины и протеромонады.
Некоторые авторы включают в эту группу и крип-
тофитовых, также имеющих хлорофилл с (Cavalier-
Smith, 1993). Однако их клетки устроены совершенно
по-другому: митохондрии с пластинчатыми кристами;
жгутики изоконтные, и обычно оба несут мастигонемы
иного строения, чем хромофиты; своеобразен и сам жгу-
тиковый аппарат; покровы уникальны для протистов;
особенности строения хлоропластов и наличие фико-
билинов также резко выделяют криптофитовых из про-
чих водорослей с хлорофиллом с, да и из всех эукари-
от. В таксономическом смысле криптофитовые так же
уникальны, как, например, воротничковые жгутико-
носцы.
В системе типа Chrysophyta в последнее время
произошли некоторые изменения (Preisig, 1995; Мое-
strup, 1995). Так, в класс Chrysophyceae включен отряд
бикозоецид, как это и было предложено ранее (Кар-
пов, 19906). Помимо класса Synurophyceae Прейсиг
(Preisig, 1995) предлагает включать в подтип хризофи-
товых класс Dictyochophyce^e Silva, 1980 с тремя отря-
дами, в которые входят пединелловые водоросли,
роды Rhizochromulina и Dictyocha. У всех этих орга-
низмов обнаружены наряду со жгутиками и аксопо-
136
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
дии, микротрубочковый скелет которых отходит от
ядерной оболочки (Moestrup, Thomsen, 1990; O’Kelly,
Wujek, 1995). Поэтому само по себе объединение их в
один таксон не вызывает сомнений. Однако по этому
признаку они сходны с актинофриидными солнечни-
ками и вместе с ними помещены нами в тип Pedinello-
morpha Karpov, 1990. Тип Pedinellomorpha будет рас-
смотрен во второй части «Протесты: Руководство по
зоологии» наряду с солнечниками и радиоляриями.
Таким образом, в нашей классификации приведена
лишь часть хромистов: Chrysophyta, Raphidophyta, Sap-
rolegnia (=Oomycetes), Traustochytridiomycetes, Laby-
rinthulida, Opalinata и Haptophyta, поэтому и сам термин
«хромисты» становится неформальным и является сино-
нимом хромофитов, или страминопилов. Отряды спон-
гомонад и псевдодендромонад не могут быть отнесены
к какому-либо таксону хромистов, поскольку имеют
признаки, присущие сразу нескольким типам. Поэто-
му обе группы оставлены в статусе отрядов неопреде-
ленного таксономического положения и помещены в
конце списка таксонов, относящихся к хромистам.
В настоящем издании имя Opalinata Wenyon, 1926
принято для типа, совпадающего по объему со Slopa-
linata Patterson, 1985, который объединяет протеро-
монад и опалин.
В тип миксомицетов введен новый для них класс —
Cercomohadea Mylnikov, 1986. Цитоскелет церкомо-
над близок по строению к таковому протостелид
(Мыльников, 1989; Карпов, 19906). Недавно исследо-
ванная Hyperamoeba flagellata (Карпов, Мыльников,
1997) представляет собой переходное звено между
церкомонадами и протостелидами (Карпов, 1997) и
помещена в группу миксомицетов неясного таксоно-
мического положения.
Rhizopoda до сих пор остаются наименее изучен-
ным типом, система которого так же несовершенна,
как и 10 лет назад. Некоторые авторы даже рассмат-
ривают всех ризопод как эукариот неопределенного
таксономического положения (Hausmann, Hiilsmann,
1996). Распределение этих протистов по типу псевдо-
подий, конечно, весьма условно и приводит к форми-
рованию явно сборных таксонов. Однако более со-
вершенной системы пока не существует, поэтому мы
взяли за основу систему Пэйджа (Page, 1987), в кото-
рую внесли некоторые изменения, кажущиеся нам
очевидными.
Так, класс Granuloreticulosea упразднен как явно
сборный. Недавние исследования представителей
рода Penardia из аталамид (Микрюков, Мыльников,
1996) показали, что они имеют совершенно необыч-
ное для гранулоретикулозных амеб строение: в част-
ности, кристы в митохондриях пластинчатые, а не
трубчатые. В результате был описан новый род Вог-
kovia (Микрюков, Мыльников, 1996), который будет
рассмотрен в протестах неясного таксономического
положения. Фораминиферы как наиболее четко
очерченный таксон выделены в самостоятельный тип.
Кардинальный пересмотр ризопод предпринят не-
давно Паттерсоном (Patterson, 1999). Он предлагает в
качестве основного признака использовать форму
крист митохондрий и выделяет довольно большую
группу амеб (Ramicristata), имеющих разветвленные
кристы. Однако эта точка зрения во многом спорна,
поэтому мы оставляем старую систему ризопод.
В типе Rhizopoda имеется довольно большая груп-
па неопределенного таксономического положения,
что неудивительно, так как весь тип является, по-ви-
димому, полифилетичным.
THE CLASSIFICATION OF PROTISTS
SUMMARY
The term «protista» was introduced by E. Haeckel in 1866 as
a name for the third kingdom (along with Animalia and Plantae) of
the organisms. It included firstly protozoa, algae, lower fungi, bac-
teria and spongi. Later Haeckel being influenced by other biologists
of that time came back to the Aristotel’s two-kingdom classification
of organisms. The term «protista» in the modern sense has been
reborn by Whittaker (1969). His ideas have been developed by
J. O. Corliss in a series of papers (1979—1987), where the inclusion
of the protozoa, algae and zoosporic fungi in this kingdom has been
corroborated.
The heterogeneity of protists was always clear, but,, nevertheless,
the above authors came back to the idea of protista as a single taxon
almost 100 years after Haeckel and Hogg. Why did this occur? The
main reasons are:
1. The evidence on the taxonomic and phylogenetic relationships
between some taxa of algae and protozoa, and heterogeneity of both
groups.
2. The revival of the symbiogenic ideas at the cellular level by
L. Margulis led to the estimation of heterotrophic protists as the most
ancient eukaryotes, from which the phototrophic ones evolved.
3. The collaboration of protozoologists, phycologists, mycologists
and cell biologists in the study of the origin of eukaryotic cell.
As a result, the protozoa, algae and zoosporic fungi have been
recognized as a single kingdom of eukaryotes up to the recent time
(Corliss, 1984; Karpov, 1990; Margulis, 1992, 1996).
The protists were considered as predominantly unicellular, mic-
roscopic, with some macroscopic syncitial, cenobial and multicellular
forms. They move using flagella, cilia and pseudopodia, more rarely
by gliding or metabolic, movement. The feeding is autotrophic, hete-
rotrophic and mixotrophic. They have all possible types of mitosis,
meiosis, and mitochondrial cristae of different shape.
If we accept the protists as a transitional group in evolution from
prokaryotes to other eukaryotes, we should assume that within the
limits of protists the development of all cellular eukaryotic classifica-
tion and modes of nutrition occurred, which the animals, fungi and
plants «use» in their life.
Indeed, it can be easily shown that it took place at the genome
level: the appearance of di-, poly- and amphiploid forms, the multi-
nucleiness, nuclear heteromorphism, and all types of separation of the
genomic material.
At the level of protein syntesis system as a genome reali-
zation: the development of rRNA from the 70S to 80S.
At the cellular level: the appearance of mitochondrion with all
crista shapes, all types of chloroplasts, cell coverings, organelles, and
cytoskeleton structures.
At the organism level: all possible types of life-cycle, forms of
cell existence, all types of genomic reduction (gametic, sporic and zygo-
tic), both types of meiosis (one- and two-step), all possible types of cell
movement and cell nutrition, and also different types of multicellula-
rity.
Thus, from the phylogenetic point of view we cannot accept the
protists as a taxon, because the protists are not equivalent to Plantae,
Animals, or Fungi, but they include all eukaryotes (Patterson, 1994).
From this point of view, there is no sense to give at present the clas-
sification of just protists.
Following this way J. O. Corliss, being influenced by T. Cavalier-
Smith and D. J. Patterson, published the classification including 34 phyla
of protists spread between 5 kingdoms of eukaryotes (Corliss, 1994).
THE CLASSIFICATION OF PROTISTS
137
THE MORPHOLOGICAL CHARACTERS
IN SYSTEMATICS OF PROTISTS
The morphological characters are essential for large eukaryotes
like plants, animals and fungi. For small prokaryotes the most impor-
tant characters are biochemical and physiological ones. In regard of
size the protists are located between small prokaryotes and large euka-
ryotes. Their main characters are determined by the cellular level of
organization and combine the morphological, biochemical and mole-,
cular characters. This is the great difficulty in protists taxonomy.
At present, the most important morphological characters are: fea-
tures of cytoskeleton (predominantly cilia/flagellar apparatus), structu-
re of pseudopodia, coverings, chloroplasts, cristae of mitochondria,
and the unique organelles and structures.
THE MOLECULAR CHARACTERS IN PROTISTS TAXONOMY
For the majority of protists no fossils have been recorded. For the
phylogeny reconstruction of organisms the study of their genome evo-
lution is widely accepted now. A recent understanding of molecular
phylogeny of protists based on the analysis of SSU rRNA gene sequ-
ences, as well as on gene sequences of some proteins, is presented at
the in time published book «Evolutionary Relationships Among Pro-
tozoa» (cds G. H. Coombs, K. Vickerman, M. A. Sleigh and A. War-
ren; 1998). Following the critical reviews by H. Philippe and A. Ado-
utte (1998) and Van de Peer and De Wachter (1997) we can conclude,
that molecular method does not show an order of branching in phylo-
genetic tree, so, we cannot say which eukaryotes are more ancient. At
the same time it confirms the existence of monophyletic taxa among
protists. This agrees in general with morphological data accumulated
during the last 30 years of ultrastructural investigations of eukaryotes
and there are no reasons do hot accept it.
GENERAL CLASSIFICATION OF EUKARYOTES
At present, there is almost no doubt in monophyly of chlorobi-
onts (vascular plants, chiorophytes, charophytes); biliphyta (red algae
and glaucophytes); fungi (true fungi, chytridiomycetes and microspo-
ridia); euglenozoa (kinetoplasids and euglenids); alveolates (apicom-
plexans, dinophytes and ciliates); polymastigines (=metamonads);
metazoa (animals and myxozoa); chromists, or heteroconts, including
besides of usual set of chlorophyll c containing algae the zoosporic
fungi (oomycetes, traustochytridiae and hyphochytridiae), opalines
and proteromonads (phylum Opalinata). Cryptophyta should not be
included in chromists as having different pigments, flagellar apparatus
structure, mitochondrial cristae, coverings, and extrusomes. The cryp-
tophytes are as unique as choanoflagellates.
Unfortunately, there are many other small groups of protists still re-
main, which may compose very heterogenous taxon Protozoa. Together
with hundreds of protists of uncertain affinities this does not permit us
to have a comprehensive general classification of eukaryotes at the mo-
ment. Modern general classifications deal with different number of
highest level taxa (from one kingdom Protista, — Margulis, 1996; to
5 kingdoms of eukaryotes, — Cavalier-Smith, 1998; to 11 kingdoms of
eukaryotes, — Kusakiri, Drosdov, 1994, 1998; and to appr. 70 ultra-
structural identities of eukaryotes, — Patterson, 1999).
TAXA CONSIDERED IN THIS VOLUME
For those reasons we do not accept here a general classification
of protists. At the same time, the classification is accepted for each
phylum of following list of phyla which are considered in this volume:
Phylum Cryptophyta Pascher, 1914
Class Cryptophyceae Senn, 1900
Phylum Euglenozoa Cavalier-Smith, Д 978
Class Euglenoidea Biitschli, 1884
Class Kinetoplastidea Honigberg, 1963
Phylum Chrysophyta Pascher, 1914
Class Chrysophyceae Pascher, 1914
Order Bicosoecales (Grasse) Karpov, 1998
Class Synurophyceae Andersen, 1987
Class Pelagophyceae Andersen et Saunders, 1993
Phylum Haptophyta Christensen, 1962
Phylum Raphidophyta Chadefaud, 1950
Phylum Saprolegnia Zerov, 1972
Class Saprolegnea (=Oomycetes) Zerov, 1972
Class Labyrinthomorphea Page, 1979
Order Labyrinthulida Cienkowski, 1867
Order Thraustochytrida Sparrow, 1943
Phylum Opalinata Wenyon, 1926
Class Proteromonadea Grasse, 1952
Class Opalinatea Wenyon, 1926
«Chromista» incertae sedis: Spongomonada (Hibberd) Karpov,
1990; Pseudodendromonada Hibberd, 1985.
Phylum Choanomonada Kent, 1880
Class Choanomonadea Kent, 1880
Order Monosigida Kent, 1880
Order Salpingoecida Kent, 1880
Order Acanthoecida Norris, 1965
Phylum Polymastigota Biitschli, 1884
Class Diplomonadea Wenyon, 1926
Order Retortamonadida Grass£, 1952
Order Diplomonadida Wenyon, 1926
Class Oxymonadea Grasse, 1952
Order Oxymonadida Grasse, 1952
Class Parabasalea Honigberg, 1973
Order Trichomonadida Kirby, 1947
Order Hypermastigida Grassi et Foa, 1911
Phylum Plasmodiophora Zopf, 1884
Phylum Mycetozoa de Bary, 1859
Class Cercomonadea Mylnikov, 1986
Class Eumycetozoea Zopf, 1884
Subclass Protostelia Olive, 1970
Subclass Dictyostelia Olive, 1970
Subclass Myxogastria Olive, 1970
Mycetozoa incertae sedis: Hyperamoeba flagellata Alexeieff,
1926.
Phylum Rhizopoda Siebold, 1845
Class Lobosea Carpenter, 1861
Class Heterolobosea Page et Blanton, 1985
Class Peloflagellata (=Caryoblastea) Goodkov et Seravin, 1988
Class Filosea Leidy, 1879
Class Xenophyophorea Schulze, 1904
Rhizopoda incertae sedis: Komokiida Kamenskaya, 1992;
Athalamia Haeckel, 1862; Monothalamia Haeckel, 1862.
Phylum Foraminifera D’Orbigni, 1826
The bicosoecids are included in the class Chrysophyceae (Karpov,
19906; Moestrup, 1995). Dictyochophyceae Silva, 1980 is now consi-
dered to include the pedinellids, Rhizochromulina and Dictyocha (Mo-
estrup, Thomsen, 1990; O’Kelly, Wujek, 1995). All of them have the
axopodia originating from the nuclear envelope. According to this
character they have been put in the phylum Pedinellomorpha Karpov,
1990, which also unites the actinophriid heliozoans, as they have the
same axopodial MTOC and the tubular, cristae in mitochondria (other
heliosoans have flagellated cells of different structure, and the flat
cristae).
The concept of kingdom (or subkingdom) Archezoa is not accepted
within these limits. At present-day classifications the microsporidia are
placed into the Fungi (Canning, 1998), Mastigamoeba (=Phreatamoe-
baj is a sister group of lobose amoebae (Cavalier-Smith, Chao, 1996).
In our classification the archamoebae are included in the phylum
Rhizopoda as Peloflagellata (Caryoblastea), and the so-called metamo-
nads are in their usual place in the phylum Polymastigota.
The phylum Mycetozoa includes also the class Cercomonadea
Mylnikov, 1986, as the cytoskeleton of theif flagellated cells has a
number of homologous elements, and they have similar biological
characters including the life-cycle features (Karpov, 1997).
The classification of rhizopods is based on the classification of
F. Page (1987). The new taxon Ramicristata (Patterson, 1999) is rather
discutive at the moment, therefore it is not accepted here. The taxon
Granuloreticulosea does not exist any more, as Penardia and Gymnoph-
rys are removed from athalamids (Mikrjukov, Mylnikov, 1996), and
the rank of Foraminifera is raised to a phylum.
The phylum Helipzoa contains only the organisms which have the
axopodia produced by either axoplast or centroplast, and with predo-
minantly flat cristae in mitochondria.
The Alveolata, Mixosporidia, Microsporidia, Heliozoa, Radiola-
ria, Pedinellomorpha and the protists of uncertain position will be
placed in the second volume of Protista. Some taxa, which are not
traditional for protozoologists (Xanthophyceae, Bacillariophyceae,
Hyphochytridia and some other minor groups), are also not included
in the present volume.
138
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
ПУТИ ЭВОЛЮЦИИ ПРОТИСТОВ
Основные направления и закономерности эволюции
Metazoa разработаны довольно хорошо: для подтверж-
дения этого стоит лишь вспомнить имена А. Н. Се-
верцова (1939,1945,1949), И. И. Шмальгаузена (1969,
1982) и В. А. Догеля (1954). Подобную же работу в от-
ношении Plantae проделал А. Л. Тахтаджян (1948, 1954,
1983, 1991). Начало изучению путей эволюции про-
тистов было положено Догелем (1929, 1951), который
показал, что прогрессивное развитие этих, еще прими-
тивных эукариот основано на полимеризации (умно-
жении) числа органелл и других клеточных компонен-
тов. Эта идея нашла развитие в книге А. А. Захваткина
(1949) и в ряде работ Ю. И. Полянского (Полянский,
1969, 1981а, 19816; Полянский, Райков, 1977).
За последние два десятилетия накоплены новые
интересные данные по электронной микроскопии и
молекулярной биологии различных протистов, кото-
рые обогатили наши знания о путях эволюции этих
организмов. В предлагаемом издании излагаются как
сделанные ранее, так и последние обобщения, касаю-
щиеся основных направлений эволюционного про-
цесса у низших эукариот.
В настоящее время можно выделить 6 основных
путей эволюции протистов: эукариотизацию, цитои-
дизацию, дезорганеллизацию, амебоидизацию, дипло-
идизацию и полицеллюляризацию. Рассмотрим вкрат-
це каждый из этих путей.
Эукариотизация
Этот процесс представляет собой постепенную ут-
рату протистами ряда оставшихся у них от прокари-
отных предков бактериальных признаков; наиболее
четко он прослеживается по изменению некоторых
свойств рибосом и особенностей строения их рРНК.
В связи с этим отметим, что в современных учебни-
ках и сводках по-прежнему подчеркивается большой
разрыв между признаками рибосом и их РНК у Рго-
karyota и Eukaryota (Бохински, 1987; Льюин, 1988;
Инге-Вечтомов, 1989, и др.). При этом указывается,
что константа седиментации (в единицах Сведберга)
у рибосом прокариотных организмов всегда равняет-
ся 70S, а у эукариотных — всегда 80S. Считается,
что рибосомы прокариот имеют три рРНК с констан-
тами седиментации 23S, 16S и 5S, тогда как у эука-
риот их 4: 28S, 18S, 5S и 5,8S рРНК. Последняя счи-
тается обязательной для всех эукариотных организ-
мов без исключения.
Константа седиментации молекул зависит в пер-
вую очередь от их массы. Следовательно, приведен-
ные данные свидетельствуют о том, что гомологич-
ные по НП 23S и 28S рРНК содержат разное коли-
чество нуклеотидов. Действительно, первая из них
имеет в среднем в молекуле около 2900 оснований, в
то время как 28S — не менее 4700; 16S рРНК содер-
жит приблизительно 1500 нуклеотидов, а гомологичная
ей 18S рРНК— до 1900. Кроме того, известно, что
прокариотные рРНК имеют участки (домены) с НП,
отсутствующие у эукариот; напротив, у эукариотных
рРНК появляются свои специфические домены.
В 80-е годы появились работы научной группы
Воссбринка (Vossbrinck, Woese, 1986; Vossbrinck et
al., 1987), в которых было показано, что у мельчай-
ших внутриклеточных паразитических протистов, от-
носящихся к типу Microsporidia, а именно у Vairimor-
pha necatrix, рибосомы, как и у типичных прокариот,
имеют константу седиментации 70S и содержат всего
три рРНК — 23S, 16S и 5S, тогда как 5,8S рРНК еще
отсутствует. 16S рРНК V. necatrix состоит всего из
1244 нуклеотидов, т. е. она даже меньше, чем у боль-
шинства бактерий. Более того, у нее отсутствует ряд
доменов, которые считаются типичными для других
эукариот, например расположенные в позиции нукле-
отидов 180—225 и 590—650. Учитывая вышеска-
занное, а также тот факт, что у микроспоридий нет
митохондрий, жгутиков и центриолей, Воссбринк с
соавторами пришли к выводу, что эти протисты воз-
никли задолго до всех остальных эукариот, а именно
тогда, «когда земная атмосфера была лишена свобод-
ного кислорода... до эволюции митохондрий и пре-
жде возникновения 5,8S рРНК у эукариот» (Vossb-
rinck et al., 1987, с. 413).
Однако вскоре в работе других авторов (Sogin et
al., 1989) было показано, что у паразитического ана-
эробного жгутиконосца Giardia lamblia (тип Polymas-
tigota) рибосомы также содержат 16S рРНК, причем
с характерным для бактерий количеством нуклеоти-
дов — 1453; ей присуще высокое содержание нукле-
отидов Г + Ц (75 %) — признак также прокариот-
ный. Кроме того, выяснилось, что эта рРНК у Giar-
dia имеет ряд сайтов бактериального типа, например
для связывания прокариотной рРНК. Правда, эука-
риотные признаки у 16S рРНК G. lamblia все же пре-
обладают. Поэтому, начиная с работы Согина с соавт.,
специалисты стали часто употреблять выражения
«188-подобная рРНК» и «288-подобная рРНК», когда
речь идет о рРНК эукариот с пониженной констан-
той седиментации и при наличии у них пока еще до-
статочно выраженных прокариотных свойств.
Опираясь на данные по НП генов 188-подобных
рРНК у разных эукариот, Согин с соавт. (Sogin et al.,
1989) построили молекулярно-биологическое филоге-
нетическое дерево. Мы модифицировали его в более
привычное для систематиков обычное филогенетичес-
кое дерево (схема 10). Под названием каждого кон-
кретного вида, у которого была исследована НП генов
188-подобной рРНК, дано название более крупного
таксона (типа или группы типов), к которому принад-
лежит данный организм. Ветви дерева, обозначенные
пунктирными линиями, взяты из молекулярно-биоло-
гических филогенетических деревьев, приведенных в
работе Шлегеля (Schlegel, 1991). Для краткости мы
будем называть филогенетическое дерево, приведен-
ное на схеме 10, деревом Согина—Шлегеля.
Перемещаясь вдоль ствола этого дерева снизу
вверх, можно обнаружить некоторые особенности
эукариотизации рибосом. Во-первых, у протистов
ПУТИ ЭВОЛЮЦИИ ПРОТИСТОВ
139
уже на довольно ранних этапах эволюции появляется
обособленная 5,8S рРНК, которая отсутствует у микро-
споридий. Во-вторых, наблюдается четкая картина
последовательного увеличения числа нуклеотидов, а
значит, и увеличения константы седиментации как у
288-пддобной, так и у 188-подобной рРНК. Если у
микроспоридий самая большая рРНК имеет констан-
ту седиментации 23S, как и у прокариот, то у пред-
ставителей подтипа Apicomplexa она равняется 23S—
24S, у Rhodophyta 25S, у Ciliophora 25S или 26S, и
лишь у некоторых голых амеб (например, у Acanth-
amoeba castellani), а также у высших животных до-
стигает величины 28S (см.: Серавин, 1992).
Как уже отмечалось, у дипломонад и микроспори-
дий в рибосомах присутствует 16S рРНК. У Apicom-
plexa, Rhodophyta и Ciliophora константы седимента-
ции гомологичных ей рРНК не превышают 17S—17,5S.
И только у высших Rhizopoda, Plantae и Metazoa она
равняется 18S (см.: Серавин, 1992).
Xenopus laevis
(Metazoa)
Ochromonas danica
(Chromophyta)
Saccharomyces ellipsoides
flFungi)
Zea mays
(Plantae)
Chlamydomonas reinhardtii
(Chlorophyta)
Acanthamoeba castellani
(Rhizopoda, Lobosea)
Plasmodium berghei
(Apicomplexa)
Prorocentrum micans
(Dinophyta)
Oxytricha nova
(Ciliophora)
Entamoeba histolitica
(Rhizopoda, Lobosea)
Rhodophyta
Euglena gracilis
(Euglenophyta)
Dictyostelium discoideum
z (Mycetozoa)
Physarum polycephalum
(Mycetozoa)
Naegleria gruberi
(Heterolobosea)
Giardia lamblia
(Diplomonada)
Vairimorpha necatrix
(Microsporidia)
Prokaryota
Схема 10. Филогенетическое дерево. (По: Sogin et al., 1989 и
Schlegel, 1991, модифицировано).
Scheme 10. Phylogenetic tree.
Необходимо подчеркнуть еще одно весьма важное
обстоятельство, упущенное из виду молекулярными
биологами. Имеющиеся данные достаточно четко по-
казывают, что в рибосомах одного и того же таксона
протистов эукариотизация по НП в разных рРНК
может идти с неодинаковой скоростью. Поэтому на
молекулярно-биологическом филогенетическом дереве,
которое построено Перассо с соавт. (Perasso et al.,
1990) на основании сравнения 288-подобных рРНК,
Rhodophyta расположены в верхней трети ствола. На
подобном же дереве Согина—Шлегеля (см. схему 10)
красные водоросли ответвляются от середины этого
ствола, однако на основании данных по НП генов 5S
рРНК Rhodophyta оказываются в самой нижней части
филогенетического дерева эукариот (Hori et al., 1985).
Этот факт нельзя признать простой случайностью
или объяснить методическими причинами. Эйден с
соавт. (Eyden et al., 1988) показали, что и в первич-
ной и во вторичной структурах 5S рРНК красной во-
доросли Porphyra umbilicalis имеется много прокари-
отных признаков. Исследователи подчеркивают, что
5S рРНК красных водорослей, пожалуй, стоит ближе
к прокариотным, чем эукариотным молекулам. В то
же время 18S- и 288-подобные рРНК этих организ-
мов обладают значительно более выраженными эука-
риотными свойствами.
Однако в этом отношении красные водоросли вовсе
не являются каким-то особым исключением. На моле-
кулярно-биологических филогенетических деревьях,
построенных по НП генов 23S—28S рРНК, 16S—18S
рРНК и 5S рРНК, весьма неодинаковое положение за-
нимают и другие крупные таксоны, например Ciliop-
hora, Euglenophyta и Fungi (Hori et al., 1985; Perasso et
al., 1989; Sogin et al., 1989; Schlegel, 1991). Важно от-
метить тот факт, что в крупных таксонах протистов (в
крупных ветвях филогенетического дерева) эукариоти-
зация рибосом протекает независимо и параллельно
(Серавин, 1992), что имеет принципиальное значение
для оценки всех нынешних теорий эволюции. Во вся-
ком случае, подобные факты свидетельствуют в пользу
того, что любые молекулярно-биологические данные,
как и любые морфологические данные, в том числе и
электронно-микроскопические, следует использовать
в систематических и филогенетических построениях с
осторожностью.
Имеющиеся факты позволяют прийти к заключе-
нию, что нет какой-то принципиальной разницы в
организации рибосом прокариот и эукариот. У про-
тистов можно найти весь ряд рРНК с константами
седиментации от 23S до 28S и от 16S до 18S, а в
рРНК низших эукариот могут сохраняться типично
прокариотные сайты.
Таким образом, совершенно ясно, что эукариот-
ные протесты произошли от прокариотных в процес-
се длительной эволюции, следы которой легко улав-
ливаются у современных низших форм.
Цитоидизация
Так называют эволюционный процесс формирова-
ния среди протистов сложных клеткоподобных орга-
низмов. В современных учебниках по зоологии бес-
140
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
позвоночных (см., напр.: Догель, 1981) простейших
называют одноклеточными, а в общебиологической
литературе специально подчеркивают, что они по
своему строению прямо сравнимы с отдельной клет-
кой Metazoa, В последней инструктивной статье
Международного комитета по систематике Protozoa
подчеркивается, что простейшие — принципиально
одноклеточные существа (Levine et al., 1980).
Против утверждения, что любых протистов можно
отождествлять с клеткой Metazoa, выступил еще До-
белл (Dobell, 1911). Он провел сравнение строения
тела инфузорий из отряда Entodiniomorpha со строе-
нием тела коловраток, имеющих подобные же разме-
ры, и нашел у них много сходных черт. На переднем
конце этих протистов есть такой же коловращатель-
ный аппарат, который, как и у коловраток, может
втягиваться внутрь тела. Ротовое отверстие инфузо-
рий ведет в глотку, которая у них также может иметь
несколько отделов. Затем следует аналог средней киш-
ки, которая обособлена от остальной цитоплазмы
специальными скелетными элементами; далее распо-
лагается задняя «кишка», заканчивающаяся анальным
отверстием. Имеются аналогии и в строении других
систем. Конечно же, невозможно даже представить
клетку с ротовым отверстием, обособленным пищева-
рительным трактом и анусом. Добелл пишет: «Ясно,
что протист не более гомологичен одной клетке ме-
тазоа, чем одному его органу, например мозгу или пе-
чени» (Dobell, 1911, с. 272).
Догель провел подобное сравнение инфузорий с
ресничными червями турбелляриями и пришел к по-
хожим выводам, которые он опубликовал в статье
«Простейшие как совершенные организмы» (Догель,
1925).
Немилов, участвуя в дискуссии о природе про-
стейших, писал: «В них (инфузориях) есть нечто от
клетки, но они, если можно так выразиться, «переро-
сли» клетку. «Ведущим» для них является не то, что
их можно приравнять к клетке, а высокая степень
дифференцировки при отсутствии расчлененности.
Недаром именно со стороны протистологов, которым
специфика этих организмов известна лучше, чем
кому-либо другому, было предложено называть их
«бесклеточными». Но в самом слове «бесклеточные»,
«неклеточные» есть некоторый элемент пройденной
стадии одноклеточности. Поэтому более удобно, как
представляется, название «надклеточные» организ-
мы» (Немилов, 1934, — цит. по: Вермель, 1970, с.
182—183).
Вообще на все возникающие здесь вопросы в прин-
ципе можно было бы отвечать относительно просто,
если бы дело касалось только инфузорий и их услож-
нений за счет возникновения новых, необычных для
клеток метазоа органелл. Однако усложнение самых
разных протистов часто происходит в результате ум-
ножения (полимеризации) практически всех структур
их тела (Догель, 1929).
Giardia lamblia, о которой уже шла речь, относит-
ся к классу Diplomonadea, что в переводе означает
«удвоенная монада». Действительно, тело этого про-
тиста состоит как бы из двух одинаковых, параллель-
но расположенных особей, но не разделенных плаз-
малеммой. Дипломонады представляют собой как бы
удвоенных, не разошедшихся в результате продоль-
ного деления ретортамонад (Захваткин, 1949). У ряда
Hypermastigida (класс Parabasalea) при индивидуаль-
ном развитии особи нередко наблюдается последова-
тельная полимеризация — удвоение^ утроение, вооб-
ще значительное умножение всех частей тела (обыч-
но кроме ядра). Однако внутри тела простейшего
такого разделения не возникает, хотя оно иногда
внешне выглядит как десяток жгутиконосцев, образу-
ющих единую цепочку (Захваткин, 1949).
Подобных сложных протистов, если они обладают
постоянной формой тела (жгутиконосцы, инфузории
и др.), предложено называть не клетками, а сомател-
лами (Захваткин, 1949), ибо это уже не клетки, а
клеткообразные существа (Серавин, 1991).
У протистов, для которых характерна амебоидная
активность (собственно амебы, некоторые жгутико-
носцы), в результате не доведенного до конца деле-
ния возникают многоядерные плазмодии. Впрочем,
они могут формироваться и иным способом. Так, де-
сятки особей амебы Euhyperamoeba fallax входят в
контакт друг с другом и сливаются в единый плазмо-
дий. Позднее он распадается на множество отдель-
ных амеб (Серавин, Гудков, 1983, 1999). Крупные
плазмодии миксомицетов могут образовываться бла-
годаря сочетанию двух процессов одновременно —
путем слияния отдельных особей и в результате деле-
ния ядер, не сопровождаемого плазмотомией. Плаз-
модий Fuligo septica в конечном итоге может дости-
гать длины 1.5 м и состоять из сотен тысяч амеб, не
отделенных друг от друга поверхностными мембрана-
ми (см. раздел «Класс Eumycetozoea»), и его уже
нельзя назвать одноклеточным организмом. Морские
многоядерные представители класса Xenophyophorea
могут иметь длину тела до 25 см; диаметр некоторых
вымерших многоядерных фораминифер равнялся
18 см (см. раздел «Тип Foraminifera).
Образование плазмоидальных форм довольно ши-
роко распространено в таких таксонах, как Chryso-
phyceae и Xanthophyceae (Вассер, 1989). Однако осо-
бой сложности с выраженным внешним расчленени-
ем тела достигают водоросли, имеющие сифональный
тип таллома. Они возникают независимо в типе Chlo-
rophyta (класс Siphonophyceae) и классе Xanthophy-
ceae (порядок Botrydiales).
Слоевище сифоновой водоросли Caulerpa prolife-
га достигает в длину полметра и более. Оно сильно
расчленено и по внешнему облику подобно высшему
растению. Стелющиеся по грунту ризомы — аналоги
стеблей — многократно ветвятся. Местами вниз от
них отходят корнеподобные ризоиды, а вверх — по-
беги, которые часто имеют вид листоподобных плас-
тинок; при этом тело каулерпы не разделено на клет-
ки. Ботаники называют сифоновые водоросли некле-
точными организмами (Виноградова, 1977; Догадина,
Матвиенко, 1977; Масюк, 1989). Действительно, не-
возможно этот организм, имеющий многие тысячи
ядер и столь сложно устроенный таллом, считать од-
ноклеточным.
Электронно-микроскопические исследования по-
казали, что какие бы усложнения и перестройки не
претерпевало тело протистов в процессе эволюции,
организмы типа плазмодиев, сомателл, сифоновых
ПУТИ ЭВОЛЮЦИИ ПРОТИСТОВ
141
слоевищ и т. п. остаются на клеточном уровне орга-
низации. Их тело одето единой эукариотной поверх-
ностной мембраной, они содержат эукариотное ядро
(ядра) и цитоплазму эукариотного типа, однако это
уже не одноклеточные организмы. Поэтому было
предложено (Серавин, 1984, 1991) называть таких
протистов сложными цитоидами, т. е. клеткообраз-
ными, ибо сложный цитоид обычно соответствует не
одной клетке Metazoa (эуциту), а полностью или час-
тично полимеризованной совокупности клеток, прав-
да, не отделенных друг от друга мембранами. Строго
говоря, саму клетку следует рассматривать как про-
стую, исходную форму цитоида, тогда как плазмодии,
сомателлы и т. п. — производными, сложными цито-
идами. Конечно, термин «клетка» настолько прочно
вошел в обиход науки и сыграл в развитии биологии
столь важную роль, что его необходимо сохранить,
тем более что среди протистов имеется довольно
много одноклеточных видов.
Резюмируя сказанное, следует подчеркнуть, что в
процессе эволюции в разных таксонах Protista неза-
висимо происходит сильное морфологическое услож-
нение (цитоидизация) организмов, что приводит к
возникновению сомателл, плазмодиев, сифоновых
слоевищ и т. п. Образно говоря, протесты стремятся
достигнуть качественно нового уровня организации,
не выходя за пределы клетки.
Дезорганеллизация
Процесс редукции и бесследного исчезновения
важнейших клеточных органелл (митохондрий, аппа-
рата Гольджи, жгутиков, хлоропластов) носит назва-
ние дезорганелл изации.
Согласно молекулярно-филогенетическому дере-
ву Согина—Шлегеля (схема 10), самыми древними
из современных протистов являются жгутиконос-
цы — дипломонады. Следовательно, все позднее
возникшие безжгутиковые виды лишены кинетид
вторично, т. е. такие организмы потеряли их в про-
цессе эволюции. Так, среди жгутиконосцев отряда
Trichomonadida (класс Parabasalea) жгутики отсутст-
вуют лишь у одного вида — Dientamoeba fragilis
(Came et al., 1974), который получил такое родовое
название потому, что перемещается с помощью
псевдоподий. Однако простейшее сохранило при
этом кинетосомы и связанный с ними опорный
микротрубочковый аппарат трихомонадной органи-
зации, тогда как представители типов Microsporidia
и Rhodophyta лишены не только собственно жгути-
ков, но также кинетосом и центриолей. Кинетиды
у них исчезли как морфологические элементы пол-
ностью и бесследно. Вот эти два способа редукции
жгутикового аппарата весьма широко встречаются у
протистов, причем потеря кинетид происходит неза-
висимо в самых разных таксонах. Нами были про-
анализированы данные многих авторов, касающиеся
последовательных рядов исчезновения жгутикового
аппарата в конкретных группах протистов (Сера-
вин, 1992). .
В типе Euglenozoa в разных группах, прямо не
связанных родством, появляются виды, лишенные ки-
нетид; такие простейшие перемещаются по субстрату
с помощью метаболического изменения формы их
тела (Серавин, Фролов, 1983). В классе Heterolobosea
протист Percolomonas — типичный жгутиконосец, а
вот Naegrelia может в определенные моменты жиз-
ненного цикла терять жгутики (вместе с кинетосома-
ми) и ползать, как амеба; наконец, представители от-
ряда Acrasida обычно нацело лишены каких-либо
следов кинетид. У Protostelida (тип Mycetozoa) есть и
жгутиковые и безжгутиковые виды, у последних ки-
нетиды исчезают бесследно. Свободноживущие пред-
ставители подтипа Apicomplexa из рода Colpodella —
типичные хищные жгутиконосцы, паразитические
Gregarinida сохраняют жгутики лишь у мужских гамет,
а у кровяных споровиков кинетиды имеют тенденцию
к исчезновению. Количество подобных примеров
можно было бы значительно увеличить, однако в
этом нет необходимости.
Самое важное из всего только что сказанного за-
ключается в том, что у целого ряда протистов кине-
тиды как морфологические образования исчезают в
процессе эволюции бесследно. Поэтому вряд ли
можно признать правильной точку зрения, что Mic-
rosporidia, Rhodophyta и Fungi являются первично-
безжгутиковыми (Старобогатов, 1986), тем более что
основные молекулярно-биологические данные свиде-
тельствуют в пользу того, что предки современных
протистов уже были жгутиконосцами (Adoutte et al.,
1989; Sogin et al., 1989).
Пластиды, как сейчас хорошо известно, могут в
процессе эволюции исчезать у тех или иных предста-
вителей практически во всех типах водорослей (Се-
равин, 1984; Вассер, 1989). Впрочем, это в полной
мере относится и к паразитическим высшим расте-
ниям (Тахтаджян, 1954). Сначала редуцируются хло-
рофиллы, так что пластиды превращаются в лейко-
пласты, а позднее исчезают и они. Таким образом,
хлоропласты как морфологические структуры в про-
цессе эволюции могут, исчезнув, не оставить после
себя никакого выраженного следа. Этот факт очень
важен, ибо, зная его, мы не имеем права утверждать,
что расположенные в нижней части филогенетичес-
кого дерева Согина—Шлегеля Metamonada и Micro-
sporidia действительно первично лишены пластид, а
не потеряли их в ходе эволюции. Данные микропале-
онтологии, между тем, свидетельствуют в пользу того,
что первичные эукариоты — акритархи (Acritarcha),
обитавшие на Земле свыше 1 млрд лет назад, были
планктонными и, скорее всего, фототрофными орга-
низмами (Тимофеев и др., 1976; Тимофеев, 1982; Ви-
даль, 1984; Янаускас, 1989). Самые древние из извес-
тных нам эукариотных организмов — дипломонады
являются, все без исключения, анаэробными протес-
тами, а поэтому неизбежно должны были потерять
пластиды, поскольку анаэробных фототрофов среди
эукариотных организмов не существует (Hook, Craw-
ford, 1976). Молекулярно-биологические исследо-
вания показывают, что частичная, весьма редуциро-
ванная информация о предковых пластидах может
длительное время сохраняться в ДНК типичных гете-
ротрофных простейших, например у Apicomplexa
(Wilson, 1993). Будем надеяться, что когда-нибудь ее
обнаружат и в геноме метамонад.
142
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
До самого последнего времени считалось, что у
всех Metamonada первично отсутствует и аппарат
Гольджи (Cavalier-Smith, 1987, 1989). Однако недав-
ние электронно-микроскопические исследования по-
казали, что у метамонады Giardia lamblia в период
формирования стенки цисты в цитоплазме появляют-
ся типичные диктиосомы (Reiner et al., 1990). По-ви-
димому, ближайшие предки современных протистов
имели не только жгутики, но также пластиды и дик-
тиосомы.
Хотя митохондрии принято считать обязательны-
ми компонентами клеток эукариот (Ченцов, 1984;
Биологический энциклопедический словарь, 1986),
благодаря электронно-микроскопическим исследова-
ниям было установлено, что эти органеллы отсутству-
ют более чем у 2000 видов протистов, к тому же от-
носящихся к разным, часто неродственным таксонам.
Кавалье-Смит (Cavalier-Smith, 1987, 1989) выде-
лил таксоны низших протистов, не имеющих мито-
хондрий, сначала в подцарство, а затем в особое цар-
ство Archezoa, в которое включил 4 типа (табл. 7),
причем он полагает, что представители двух из
них — Archamoeba и Metamonada, кроме того, пер-
вично лишены и аппарата Гольджи. Как уже было
указано, в отношении метамонад такая точка зрения
оказалась ошибочной.
Охарактеризуем вкратце основные таксоны архе-
зоев. Оказывается, что все они либо паразиты, либо
анаэробы, или же одновременно являются и парази-
тическими и анаэробными видами.
Между тем среди инфузорий (подтип Ciliophora) и
хитридиевых грибов (тип Chytridiomycota) также
имеется большое число анаэробных видов, часть ко-
торых одновременно является и паразитами.
Электронно-микроскопические исследования мно-
гих авторов (Yarlett et al., 1981, 1983, 1984; Van Brug-
gen et al., 1983, 1984, 1988; Finley, Fenchel, 1989; Fen-
chel, Finley, 1991; Серавин, 1992) показали, что и у
инфузорий, которые эволюционно не являются при-
митивными протистами, в анаэробных условиях, вооб-
ще при дефиците кислорода в среде, исчезают мито-
хондрии. Это — самое обычное явление (Серавин,
1992). Однако у большинства из них появляются но-
вые органеллы — гидрогеносомы, которые способны
утилизировать пируват с образованием АТФ, причем
конечным продуктом реакции является Н2. Поэтому в
цитоплазме таких безмитохондриальных протистов
Таблица 7
Подцарство (царство) Archezoa (по: Cavalier-Smith, 1987)
Тип Класс
1. Archamoebae Mastigamoeba Pelobiontea
(Pelomyxa, Entamoebidae)
2. Metamonada Anaxostylea (Diplomonadida, Retortomonadida) Axostylaria (Oxymonadida)
3. Microsporidia 4. Parabasalia (Trichomonadida, Hypermastigida)
нередко поселяются в качестве эндосимбионтов метано-
генные архебактерии (очень часто Methanobacterium
formicium), которые утилизируют образующийся во-
дород, продуцируя метан.
Фенчел и Финлей (Fenchel, Finley, 1991) полага-
ют, что гидрогеносомы инфузорий — преобразован-
ные митохондрии, поскольку они могут иметь двой-
ную наружную мембрану. Однако у Entodiniomorpha
доказать присутствие такой двойной мембраны не
удалось (Yarlett et al., 1984), а у паразитических хит-
ридиевых вокруг этой органеллы имеется всегда одна
мембрана (см.: Серавин, 1992). К тому же Фенчел и
Финлей (Fenchel, Finley, 1991) установили, что среди
свободноживущих анаэробных инфузорий есть виды,
у которых бесследно исчезли митохондрии; у них нет
ни гидрогеносом, ни метаногенных бактерий. Неко-
торые безмитохондриальные цилиаты, хитридиевые,
а также некоторые другие протисты (например, спо-
ровики пироплазмиды) одновременно полностью те-
ряют и аппарат Гольджи (см.: Серавин, 1992).
Таким образом, у разных протистов в процессе
эволюции могут бесследно исчезать не только жгути-
ки и хлоропласты, но и митохондрии.
Кавалье-Смит (Cavalier-Smith, 1987, 1989) счита-
ет наиболее примитивными эукариотами представи-
телей созданного им типа Archamoeba, потому что,
по его мнению, они первично лишены митохондрий,
гидрогеносом и аппаратаТольджи. Однако все пред-
ставители этого типа — анаэробы, а некоторые к
тому же и паразиты. Как показывает проведенный
органеллологический анализ (Серавин, 1992), арха-
мебы могли потерять все эти органеллы в результате
их специфической среды обитания. В пользу спра-
ведливости нашей точки зрения свидетельствуют сле-
дующие факты. Кавалье-Смит (Cavalier-Smith, 1987)
отнес к архамебам Pelomyxa palustris и Entamoebidae
(см. табл. 7). Между тем электронно-микроскопичес-
кие исследования показали, что у пеломиксы есть
диктиосомы, гидрогеносомы (Серавин, Гудков, 1987а,
19876), а также метаногенные эндосимбиотические
прокариоты, включая Methanobacterium formicium
(Van Bruggen et al., 1988). Таким образом, у пело-
миксы имеются все внутриклеточные структуры, ха-
рактерные для тех инфузорий, которые относительно
недавно перешли к анаэробному образу жизни, но
уже стали безмитохондриальными. Столь глубокие
аналогии свидетельствуют, что Pelomyxa palustris —
вовсе не архамеба, а протист, который, перейдя к
анаэробному образу жизни, потерял свои митохонд-
рии, но приобрел гидрогеносомы и метаногенных эн-
досимбионтов.
Молекулярно-биологические данные (Sogin et al.,
1989) свидетельствуют о том, что Entamoebidae от-
ветвились от общего ствола филогенетического дере-
ва где-то чуть ниже его середины (схема 10), т. е.
они — тоже не архамебы и не архезои, а протисты, в
процессе эволюции потерявшие митохондрии, аппа-
рат Гольджи и кинетиды. Поэтому Кавалье-Смит (Ca-
valier-Smith, 1989) вынужден был убрать их из со-
зданного им царства. Проведенный подробный орга-
нелл ологический анализ (Серавин, 1992) позволяет
думать, что царство Archezoa — искусственный так-
сон, в который произвольно включена часть протис-
ПУТИ ЭВОЛЮЦИИ ПРОТИСТОВ
143
тов, потерявших из-за своего образа жизни некото-
рые важнейшие органеллы.
Для того чтобы избежать тех филогенетических и
систематических ошибок, которые делают Кавалье-
Смит и некоторые другие исследователи, имеющие
дело с низшими эукариотами, было предложено вы-
делить в пределах цитологии (клеточной биологии)
частную науку — органеллологию, задача которой
заключается в изучении закономерностей возникно-
вения, развития, редукции и полного исчезновения
клеточных органелл (Серавин, 1992, 1993). Только
сравнительная и эволюционная органеллология поз-
волит исследователям достаточно верно установить,
первично или вторично лишены те или иные протес-
ты своих важнейших органелл.
Накопленные факты свидетельствуют в пользу
того, что все ныне существующие протесты про-
изошли от тех предковых эукариот, которые уже
имели все основные органеллы, т. е. митохондрии,
пластиды, жгутики и диктиосомы. Приспосабливаясь
к различным, неодинаковым экологическим нишам и
меняя в связи с этим свой образ жизни, отдельные
виды и даже более крупные таксоны протистов стали
терять имеющиеся или, напротив, приобретать новые
органеллы.
Дезорганеллизация — одно из важнейших направ-
лений эволюционного процесса у протистов, проявля-
ющееся независимо в разных ветвях филогенетичес-
кого дерева эукариот. Это весьма затрудняет построе-
ние общей системы Eukaryota и выяснение точных
филогенетических взаимоотношений между разными
(особенно крупными) таксонами протистов.
Поскольку существуют протесты, вторично ли-
шенные ряда или практически всех важнейших орга-
нелл, становится ясно, что происхождение Eukaryota
и происхождение их органелл — это две очень важ-
ные, но непосредственно прямо не связанные пробле-
мы (см.: Серавин, 1986).
Амебоидизация
Этот процесс характеризуется тем, что у отдель-
ных видов или более крупных таксонов и даже мак-
ротаксонов протистов возникает амебоидная актив-
ность. Она может проявиться в форме образования
псевдоподий (лобоподий, филоподий, аксоподий, ре-
тикулоподий) для парения в воде, захвата добычи, пе-
ремещения по субстрату или в форме плазмодизации,
т. е. формирования плазмодиев. Имеются виды, кото-
рые проявляют все основные способы амебоидной
активности.
Как уже говорилось выше, по современным пред-
ставлениям, наиболее примитивными протестами яв-
ляются жгутиконосцы — дипломонады. Для них не-
известна амебоидная активность. Отсутствует она и у
представителей таких типов, как Euglenophyta и Ki-
netoplastida (Sleigh, 1989). В пределах класса Hetero-
lobosea наблюдается плавный эволюционный переход
от типичных жгутиконосцев рода Percolomonas до
представителей отряда Acrasida, которые вообще ли-
шены жгутиковой фазы и представляют собой амебо-
идные организмы.
Эволюционные переходы от видов жгутиконос-
цев, не обладающих никакой амебоидной актив-
ностью, к жгутиконосцам, способным питаться фа-
готрофно с помощью одной или нескольких псевдо-
подий, а от них — к видам, перемещающимся по
субстрату амебоидным образом, наблюдаются в типе
Haptophyta, подтипе Dinophyta, в классах Chrysophy-
ceae, Xanthophyceae и др. (Вассер, 1989). Rhizopoda,
Myxozoa, Foraminifera, Radiolaria и другие типы про-
тистов представляют собой макротаксоны, состоя-
щие из чисто амебоидных организмов, происхожде-
ние которых от близких или дальних жгутиконосных
предков не вызывает сомнений (Sleigh, 1989; Карпов,
1990а).
Некогда весьма распространенная точка зрения,
что «амебы» — самые примитивные и наиболее
просто устроенные простейшие, оказалась совер-
шенно неверной. Амебоидность всегда возникает у
протистов в результате длительной эволюции, при-
чем независимо в разных макротаксонах. Достаточ-
но вспомнить, что самых крупных представителей
в группе протистов можно найти среди амебоидов,
а именно у Myzetozoa. Амебоидные организмы не-
которых групп способны образовывать невероятно
сложные, необыкновенно красивые скелеты; это в
первую очередь относится к радиоляриям. Форами-
ниферы образуют раковины такой формы и слож-
ности, что крупные формы ^тих протистов некогда
относили к моллюскам. Амебоиды чрезвычайно
разнообразны по своему строению, весьма эволюци-
онно стойки, размножаются быстро и в массовых
количествах. Неудивительно, поэтому, что многие
горные породы — известняки, радиоляриты и дру-
гие состоят преимущественно из раковинок и ске-
летов вымерших амебоидных протистов, т. е. во
многом определяют состав земной коры и даже ее
внешний облик (Серавин, 1984).
Амебоидизация протистов, осуществляется парал-
лельно и независимо во многих ветвях филогенети-
ческого дерева эукариот, однако не следует забывать,
что в некоторых крупных макротаксонах протистов
она вообще отсутствует (классы Euglenoidea, Kine-
toplastidea, подтип Ciliophora) или крайне редка
(Chlorophyta).
Диплоидизация
Диплоидизация — процесс независимого возник-
новения отдельных видов и групп протистов с поло-
вым процессом (мейозом) и наличием диплоидной
фазы в их жизненном цикле. В своей книге «Ядро
простейших» Райков (1978, с. 154) пишет: «По-види-
мому, половой процесс и сопряженный с ним мейоз
возникали в эволюции простейших многократно и
независимо. Другими словами, агамные жизненные
циклы, наблюдающиеся во всех низших группах про-
стейших (амебы, многие отряды Ж1угиконосцев), яв-
ляются первичными и им соответствует гаплоидное
состояние ядра... Подобные взгляды, утверждающие
полифилетизм мейоза и полового процесса у Proto-
zoa, разделяются в настоящее время большинством
специалистов (Wenrich, 1954; Догель и др., 1962;
144
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Иванов, 1968, 1970; Полянский, 1971, и др.)». Более
поздние исследования Сли (Sleigh, 1989) подтверди-
ли эти взгляды. Другими авторами также было пока-
зано, что у представителей древнейших протистов из
типа Metamonada половой процесс отсутствует (Sleigh,
1989; Серавин, Гудков, 1999).
Опираясь на сравнение аминокислотных последо-
вательностей белка фактора элонгации — 1L, группа
японских исследователей (Hasegawa et al., 1993) по-
строила своеобразное филогенетическое дерево эука-
риот. По данным этих авторов, самыми древними со-
временными протестами являются энтамебы, кото-
рые, как известно, не обладают половым процессом.
Как можно видеть, разные авторы, исходя из разных
посылок, помещают в основание филогенетического
дерева самые разные группы. Однако во всех этих
случаях здесь оказываются протесты, не обладающие
половым процессом или приобретающие его в ходе
длительной эволюции.
Райков (1978), Сли (Sleigh, 1989) и Серавин и
Гудков (1999) приводят много данных, свидетельству-
ющих о том, что половой процесс, т. е. и диплоиди-
зация протистов, происходили в разных таксонах не-
зависимо и неоднократно. Одно несомненно, что все
высшие эукариоты Phaeophyta, Plantae, Fungi, Meta-
zoa, а также достаточно высокоорганизованные про-
тесты подтипа Ciliophora, типов Foraminifera, Radio-
laria и другие обладают им (за редкими случаями его
вторичной утраты у немногих форм).
Полицеллюляризация
Для этого процесса характерно образование в раз-
ных таксонах протистов колониальных и многокле-
точных форм в результате механического объедине-
ния особей (например, слизью) вследствие не до
конца доведенного деления исходных индивидуумов
и на основе агрегации, т. е. сползания и объединения
в единый многоклеточный организм особей одного
вида, как это происходит у представителей подкласса
Dictyostelia.
Формирование колоний — процесс чрезвычайно
широко распространенный, можно сказать, весьма
обычный среди простейших и водорослей. Разные
пути их образования неплохо изучены (Захваткин,
1949; Иванов, 1968; Вассер, 1989; Sleigh, 1989; Масюк,
1989).
Истинные многоклеточные виды появляются неза-
висимо у Chrysophyceae, Xanthophyceae, Haptophyta,
Rhodophyta, несколько раз у Chlorophyta, а также у
Ascetospora, Labyrinthomorpha, Chytridiomycota,
Oomycetes и в некоторых других макротаксонах про-
тистов. Одноклеточные амебки подкласса Dictyoste-
lia и отряда Acrasida (класс Heterolobosea) группиру-
ются вместе и образуют многоклеточные псевдоплаз-
модии.
Возникает законный вопрос, почему же перечис-
ленные таксоны, несмотря на их многоклеточность,
большей частью относятся к царству Protista? Да потому,
что тело у их представителей состоит из одинаковых
вегетативных клеток. Эти организмы остаются на кле-
точном уровне организации потому, что их клетки не
дифференцированы и не формируют тканей.
В научной литературе принято дебатировать проб-
лему возникновения многоклеточное™ у животных
и растений как достижение нового уровня их сис-
темной организации. Однако, на самом деле, такой
проблемы вовсе не существует: многоклеточность,
как таковая, возникает легко, независимо и много-
кратно в самых различных ветвях филогенетическо-
го дерева эукариот, оставляя, тем не менее, такие
организмы большей частью на уровне организации
протистов. На самом деле есть другая чрезвычайно
важная проблема — возникновение у эукариот раз-
ноклеточности (дифференцированности) t. вегетатив-
ных клеток в пределах одного и того же организ-
ма. Такая гетероклеточность появляется в четкой
форме у Phaeophyta, Spongia, Metazoa и Plantae.
Как, по каким причинам, под влиянием каких
внешних и внутренних факторов возникает это яв-
ление? Ответ на этот вопрос невольно подменяется,
а потому и затушевывается, когда исследователи
вместо него в центр обсуждения ставят проблему
происхождения многоклеточное™.
Мы показали 6 важных путей, 6 направлений эво-
люционного процесса у протистов. Образно говоря,
они «стремятся» преобразоваться так, чтобы по своей
организации как можно дальше уйти от прокариот.
При этом они «пробуют» найти такие формы строе-
ния, которые позволили бььПм в результате перейти
на принципиально новый системный уровень органи-
зации. Отсюда появление сложно устроенных цитои-
дов путем умножения уже существующих внутрикле-
точных структур, или создание новых органелл, или
увеличение в составе организма числа одинаковых
вегетативных клеток (формирование колоний и ис-
тинно многоклеточных особей). Но все это так и не
привело к переходу этих организмов на новый уро-
вень системной организации. Выход на такой уровень
был осуществлен протестами в докембрии (венде),
когда возникли первые гетероклеточные организмы
(Федонкин, 1987). Однако при этом они перестали
быть протистами. Они превратились в метазои и мета-
фиты — организмы, которые пошли новыми путями
морфофункциональной эволюции, хотя на клеточном
уровне ими использовались частично и некоторые из
прежних направлений.
Полянский (1969, 1971, 1981а, 19816; Полянский,
Райков, 1977) многократно обращал внимание на
своеобразие путей и закономерностей морфофункци-
ональной эволюции простейших. Опираясь на имев-
шиеся данные, он писал: «Настоятельно необходи-
мым становится сейчас создание «частных теорий
эволюции», последующий синтез которых будет, ве-
роятно, призван поднять эволюционную теорию на
качественно новую ступень» (Полянский, 19816,
с. 156). Завадский и Колчинский (1977) четко пока-
зали необходимость таких теорий для понимания
эволюции самой эволюции живых организмов.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
145
THE PATHWAYS OF PROTIST EVOLUTION
SU M M
The six major mechanisms of the evolutionary process can be
distinguished in the present-day protists.
1. Eukaryotization — a gradual loss of some remains of prokaryotic
features. This is most obvious at the rRNA level: sets of 16S, 23S and
5S rRNA occur in lower forms (such as microsporidians) and sets of
18S, 28S, 5S and 5,8S rRNA are found in the higher protists (e. g.
Acanthamoeba castellani).
2. Cytoidization — formation of complex cellular (but not unicel-
lular) forms, such as somatella, plasmodia and siphonous talli, i.e. cyto-
ids resulting from the polymerization of organelles and other intracellu-
lar structures and also from agamic fusion of unicellular organisms be-
longing to the same species, or from incomplete division of protists.
3. Desorganellization — a partial or complete reduction of cell
organelles (mitochondria, dictyosomes, flagella, chloroplasts). In the
parasitic and anaerobic protists almost all these organelles can be re-
duced to challenge the investigator’s capacity to guess whether the
obvious absence of certain important organelles is primary or secon-
ARY
dary. We suppose that all the present-day heterotrophic protists, regard-
less of their systematic position, are of secondary origin, i.e. they must
have phototrophic ancestors.
4. Amoeboidization — origin of species and higher taxa with
amoeboid activity in different groups of protists (catching of prey with
pseudopodia, amoeboid locomotion, or forming plasmodia).
5. Polycellularization — producing colonial and multicellular or-
ganisms in different groups of protists by means of mechanical integ-
ration of organisms (e. g. with slime) due to incomplete division or
aggregation, i. e. by means of uniting independent organisms belon-
ging to the same species into a common single multicellular organism.
6. Diploidization — an independent origin of species and groups
of protists with the sexual process (meiosis) and the diploid stage
available in their life cycle.
It should be stressed that all six ways of evolution can run parallel
and independently in different branches of the phylogenetic tree of
protists.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
Протисты представляют собой обширную по ко-
личеству и многообразию видов группу эукариот. В
настоящее время насчитывается, по разным оценкам,
от 120 до 200 тыс. видов этих организмов. Причем с
полной уверенностью можно утверждать, что это
лишь меньшая часть действительно существующего в
биосфере многообразия протистов. Изучение видово-
го состава некоторых крупных таксонов только еще
началось, и описана лишь относительно небольшая
часть реально существующих видов. В качестве при-
мера можно указать тип микроспоридий. До настоя-
щего времени изучено несколько сот видов этих па-
разитических простейших. Но имеются основания
предполагать, что почти каждый вид насекомых и
других членистоногих имеют свой специфический
вид микроспоридий. Если это так, то число видов
только микроспоридий превысит миллион. Столь же
неполно описание видового многообразия и других
крупных таксонов протистов. Например, по некото-
рым оценкам, общее число видов только диатомовых
водорослей может насчитывать до 10 000 000! (An-
dersen, 1992). Одноклеточные организмы, имеющие
нерастворимый в морской воде скелет, представлены
преимущественно ископаемыми формами. Так, при-
мерно из 40 000 видов фораминифер 80 % являются
ископаемыми.
Протисты распространены в природе во всех воз-
можных для жизни средах обитания — океанах и
морях, пресных и солоноватых водах, почве. Среди
них много паразитов, причем их хозяевами являются
буквально все группы эукариот.
Велико практическое значение протистов в про-
дукции и деструкции органических веществ, в цепях
питания животных в морских и пресных водоемах, в
индикации степени органического загрязнения воды,
в биологической очистке сточных вод и самоочище-
нии водоемов.
10 Протисты, часть 1
К сожалению, как в отечественной, так и в зару-
бежной литературе очень мало уделяется внимания
характеристике этой группы эукариот. Пожалуй, наи-
более яркое и в то же время достаточно полное пред-
ставление о протистах, их роли в биосфере и жизни
человека дает книга Л. Н. Серавина «Простейшие...
что это такое?» (Серавин, 1984).
Морфология
Форма тела, размеры
Дать какое-либо общее описание формы тела про-
тистов невозможно, ибо она в высшей степени разно-
образна. Многие из них (например, амебы) постоян-
но меняют форму тела, образуя цитоплазматические
выросты — псевдоподии, служащие для захвата пищи
и движения (рис. 71) Встречаются формы, приближаю-
щиеся к шаровидной, причем от шаровидного тела
обычно отходят тонкие радиальные псевдоподии типа
аксоподий (рис. 71). Форма тела активно подвижных
свободноживущих простейших необычайно разно-
образна, что обусловлено характером движений, пи-
тания, особенностями среды обитания. У протистов,
ведущих сидячий образ жизни (кругоресничные ин-
фузории, солнечники, воротничковые жгутиконосцы,
пединеллиды), проявляются элементы радиальной
симметрии.
Многие представители протистов образуют коло-
нии, имеющие все возможные варианты форм: ли-
нейные, пластинчатые, шаровидные, древовидные.
Некоторые организмы (лабиринтулы) образуют сет-
чатые-колонии, внутри которых свободно переме-
щаются отдельные клетки. Встречаются и сетчатые
плазмодии — огромные многоядерные особи. Разме-
ры тела протистов варьируют в очень широких пре-
146
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 71. Многообразие форм протистов с различными типами
псевдоподий. (По: Hausmann, Hulsmann, 1996).
1 — конические псевдоподии Mayorella, 2 — радиальные аксоподии у Ас-
tinophrys, 3 — моноподиальная Saccamoeba, 4 — ламеллиподии Vannella,
5 — полиподиальная Amoeba, 6 — лобоподии у Nebela, 7 — Cyphoderia и
8 — Nuclearia с филоподиями, 9 — Stereomyxa с ветвящимися псевдоподия-
ми, 10 — ретикулоподии Allogromia.
Fig. 71. Diversity of protists with different pseudopodia.
1 — Mayorella with conical pseudopodia, 2 — Actinophrys with radial axopo-
dia, 3 — monopodial Saccamoeba, 4 — Vannella with lamellipodia, 5 — poly-
podial Amoeba, 6 — Nebela with lobopodia, 7 — Cyphoderia and 8 — Nucle-
aria with filopodia, 9 — Stereomyxa with branched pseudopodia, 10 — Allog-
romia with net of reticulopodia.
делах — от 1 мкм (споровики) до 10 и более метров
(бурые водоросли), т. е. масштаб вариаций составля-
ет 107.
Представления о протистах как об исключительно
одноклеточных организмах, мягко говоря, неточны.
Организация их тела очень многообразна. Это легко
видеть на примере водорослей, у которых разные ав-
торы выделяют от 7 до 14 типов организации талло-
ма, или тела водоросли (Масюк, 1993). Многие про-
стейшие существуют в виде больших многоядерных
плазмодиев, или симпластов, размеры которых дости-
гают десятков сантиметров и даже в редких случаях
могут занимать площадь около 5 м2. По своим раз-
мерам, сложности членения таллома и высокой спе-
циализации органов размножения некоторые водо-
росли являются фактически многоклеточными орга-
низмами. Однако большинство протистов все-таки
представлено одноклеточными формами. Поэтому
в общем морфологическом обзоре мы будем рас-
сматривать основные клеточные системы и органел-
лы на примере одноклеточного организма. В клеточ-
ной биологии формально выделяют 3 основные сис-
темы клетки: поверхностные структуры, цитоплазму
и ядро.
Поверхностные структуры
Как у всякой эукариотной клетки, в основе поверх-
ностных структур протистов лежит плазмалемма, об-
разованная трехслойной поверхностной мембраной и
прилегающим к ней снаружи слоем гликокаликса.
Очень многие протисты покрыты только плазмалем-
мой, но существует и немало организмов, обладаю-
щих дополнительными усложнениями покровов, ко-
торые призваны усиливать их защитную функцию
или выполнять функции движения, размножения,
поддержания формы клетки (цитоскелетная функ-
ция), питания, дыхания и даже энергетического обме-
на. Многообразие покровов протистов очень велико,
а их терминология сложна и часто запутана, так как
названия пришли из разных наук: фикологии, прото-
зоологии и микологии, а светооптические термины
переплетаются с ультраструктурными. Ревизия тер-
минологии и номенклатуры поверхностных структур
протистов проведена недавно коллективом авторов
(Preisig et al., 1994), которые, однако, не выработали
определенной концепции, а попытались описать все
многообразие покровов и привести синонимы их на-
званий.
Несмотря на большое разнообразие покровов про-
тистов, морфологические усложнения этих структур
можно разделить на 2 группы: 1) за счет образования
структурированных или аморфных надмембранных
слоев, 2) за счет изменения прилегающей к плазма-
лемме цитоплазмы (Карпов, 1986, 19906). Иногда эти
типы покровов могут сочетаться друг с другом.
1) Изменение покровов за счет надмембранных
слоев (рис. 72, а—в). Самое простое усиление покро-
вов этого типа связано с развитием гликокаликса. Он
может достигать значительной толщины у амеб, и,
по-видимому, на его основе формируются дискрет-
ные структуры — гликостили, а также тегумент и так
называемая кутикула, характерные для амебоидных
организмов. Довольно часто на поверхности плаз-
малеммы встречаются органические чешуйки различ-
ных форм и размеров (рис. 72, а\ 73, вкл.); изредка
обнаруживаются волоски, или соматонемы. Иногда
поверхность плазмалеммы покрывает дополнительная
мембрана — перилемма, встречающаяся у некоторых
инфузорий. К более мощным структурам этого типа
относятся клеточные стенки, характерные для мно-
гих водорослей и зооспоровых грибов (рис. 72, б), а
также домики, в которых живут клетки многих ризо-
под, хризомонад и воротничковых жгутиконосцев
(рис. 72, в). Клеточные стенки водорослей состоят
преимущественно из целлюлозы, а у некоторых зоос-
поровых грибов и микроспоридий — из хитина. До-
мики обычно имеют органическое происхождение, а
усиливаются за счет инкрустации солями железа,
кремния, стронция, кальция.
2) Изменение покровов за счет субмембранных
слоев цитоплазмы (рис. 72, г—к). Почти у всех кле-
ток протистов имеется субмембранный слой микро-
филаментов. Особенно хорошо он развит у амебоид-
ных организмов. В него входят актиновые и миозино-
вые филаменты, позволяющие клетке не только
менять форму, но и активно передвигаться.
Под плазмалеммой некоторых жгутиконосцев
можно обнаружить идущие вдоль клетки микротру-
бочки, которые обычно связаны с поверхностной
мембраной и образуют с ней единый комплекс — ту-
булемму (рис. 72, г). У опалин и протеромонад этот
тип покровов образует гребни и называется гребенча-
той тубулеммой (рис. 72, д). Эвгленовые водоросли
имеют более сложные покровы, называемые обычно
кутикулой (рис. 72, ж). Под плазмалеммой находится
плотный белковый слой в виде продольных полосок,
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
147
с которым связаны микротрубочки. Этот комплекс
обеспечивает клетке возможность метаболического
движения. Под плазмалеммой могут находиться че-
шуйки, формирующие вместе с ней перипласт крип-
тофитовых (рис. 72, е).
У многих протистов в эктоплазме обнаружива-
ются дополнительные мембраны. Обычно они обра-
зуются из мембран, подстилающих плазмалемму
плоских пузырьков — альвеол. Альвеолы плотно
смыкаются краями друг с другом, располагаясь в
один ряд (рис. 72, и). Такие покровы называются
пелликулой и характерны для альвеолат (инфузо-
рий, споровиков и динофитовых). У последних
внутри альвеол часто находятся целлюлозные плас-
тинки, что значительно усиливает их покровы, на-
зываемые текой (рис. 72, к).
Еще один вариант усложнения покровов возмо-
жен за счет появления под плазмалеммой дополни-
тельной мембраны (рис. 72, з; 74, вкл.). Такие по-
кровы встречаются крайне редко: они обнаружены
как постоянные структуры пока только у апузомо-
над и образованы двумя плотно прилегающими друг
к другу мембранами. Ранее (Карпов, 1986, 19906)
мы выделяли их в особый, 3-й класс. Однако, по
сути дела, вторая мембрана формируется под плаз-
малеммой и может считаться поэтому субмембран-
ной структурой.
Цитоплазма
У многих протистов, особенно амебоидных, в ци-
топлазме ясно различаются 2 зоны: прилегающая к
покровам эктоплазма, или, по старой терминологии,
плазмагель, и внутриклеточная зона — эндоплазма,
или плазмазоль. Эктоплазма обладает большей вяз-
костью по сравнению с эндоплазмой за счет значи-
тельной концентрации в ней микрофиламентов и
микротрубочек. В ней расположёны связанные с по-
кровами органеллы (экструсомы)' или скелетные об-
разования различных выростов клетки (базальные
тела жгутиков/ресничек, основания гаптонемы, сте-
бельков, микровиллей, аксоподий и прочих струк-
тур). Практически все остальные органеллы и ядро
располагаются в эндоплазме. Заметим, что разграни-
чение цитоплазмы на эти 2 зоны характерно далеко
не для всех протистов. У многих из них эктоплазма
не выражена или представлена лишь тонким слоем
субмембранных микрофиламентов.
Таким образом, понятие цитоплазмы тесно связа-
но с понятием цитоскелет, который связывает в еди-
ную систему все части клетки: поверхностные струк-
Рис. 72. Схема основных типов покровов у протистов.
(По: Карпов, 19906).
А — чешуйки на поверхности клетки; Б — клеточная стенка; В — домик, или
панцирь; Г — тубулемма; Д — гребенчатая тубулемма; Е — перипласт;
Ж — кутикула эвгленовых; 3 — двойная мембрана; И — пелликула; К —
тека.
Fig. 72. Main types of coverings in protists.
A — scales, Б — cell wall, В — lorica, Г — tubulemma, Д — folded tubu-
lemma, E — periplast, Ж — euglenoid cuticle, 3 — double membrane, И —
pellicle, К — theca.
148
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
туры, органеллы цитоплазмы и ядро. У протистов
можно найти все переходы в ^строении цитоскелета
от сеточки тонких актиновых филаментов амеб до
мощных тяжей микротрубочек и микрофиламентов у
трихомонад и эктофибриллярной системы инфузо-
рий.
Экструсомы. Эти органеллы представляют собой
тельца, окруженные мембраной, которая у зрелых эк-
струсом обычно контактирует с шгазмалеммой. В
ответ на различные внешние раздражения (механи-
ческие, химические, электрические и др.) они выбра-
сывают наружу свое содержимое. Различают несколь-
ко типов экструсом, распространенных среди про-
тистов (Hausmann, Hiilsmann, 1996).
Веретеновидные трихоцисты обнаружены
у инфузорий, динофитовых (рис. 75; 76, вкл.), эвгле-
нид и некоторых других жгутиконосцев. Эти органел-
лы имеют белковую природу, им свойственна перио-
дическая исчерченность. При выстреливании три-
хоциста увеличивается в длину примерно в 8 раз.
Трихоцисты выполняют, по-видимому, защитную
функцию.
Токсицисты представляют собой цилиндри-
ческие трубки. Трубка имеет плотную стенку, про-
свет которой занят внутренней, меньшей по диамет-
ру, трубкой. Последняя при выстреливании либо вы-
двигается телескопически из внешней трубки, либо
выворачивается, подобно пальцу перчатки (рис. 75;
76), и вонзается в тело жертвы. Токсицисты свойст-
венны хищным жгутиконосцам. Они содержат ядови-
тые вещества, способные обездвиживать и убивать
жертву (простейших и другие мелкие организмы).
Этот тип экструсом обнаружен у хищных эвгленид,
инфузорий и у хищного жгутиконосца Colponema
loxodes (рис. 76).
Микротоксицисты (гаптоцисты) обнаружены
у церкомонадид (рис. 76) и сукторий. Внутри пузырь-
ка находятся 2 вложенные друг в друга трубки или 2
конуса. Диаметр микротоксицист около 150 нм. Гап-
тоцисты у сосущих инфузорий крупнее, но принцип
действия их тот же: при выстреливании внутренняя
трубка выворачивается, как палец перчатки, и содер-
жимое микротоксицисты впрыскивается в тело жерт-
вы. Формирование этих органелл происходит в аппа-
рате Гольджи.
Мукоцисты (слизеносные тельца) типичны
для эвгленид, церкомонадид и некоторых амеб.
Они имеют шаровидную, веретеновидную или па-
лочковидную форму и открываются наружу порами.
Их слизистое содержимое при выходе наружу (вы-
стреливании) образует слизистый слой на поверх-
ности тела, который облегчает движения и имеет
защитное значение (особенно при формировании
оболочки цист).
Эжектосомы (тениоболоцисты) встречаются у
Cryptophyta, Prasinophyceae и хищного жгутиконосца
Katablepharis (рис. 75; 76). В спокойном состоянии
они напоминают рулоны свернутых лент, а при выст-
реливании образуют длинные трубки. Функция до
конца не выяснена.
Дискоболоцисты — органеллы некоторых ви-
дов хризомонадид (рис. 75). Форма их сферическая,
и на стороне, обращенной к плазмалемме, располо-
жено дисковидное уплотнение. Функция пока не вы-
яснена.
Нематоцисты обнаружены у ряда видов пан-
цирных жгутиконосцев и представляют собой стрека-
тельные капсулы, напоминающие книдоцисты книда-
рий. Верхняя часть капсулы — головка — имеет от-
верстие, закрытое слизистой пробочкой, а внутри
капсулы находится спирально свернутая стрекатель-
ная нить. Нематоцистам присуща, по-видимому, за-
щитная функция.
Кинетоцисты характерны для солнечников и
тауматомонад (рис. 76). Они имеют вид пузырьков с
достаточно сложным содержимым. Способ экструзии
и назначение этих органелл неизвестны, поскольку
выстрелившие кинетоцисты не обнаружены.
Рабдоцисты — палочковидные экструсомы,
обнаружены пока только у кариореликтид (Ciliata).
Механизм выстреливания соответствует таковому те-
лескопически выдвигающихся токсицист.
Цитоскелет. Цитоскелетные структуры протистов
весьма разнообразны как по строению и биохимичес-
кой природе, так и по выполняемым функциям.
Строго говоря, к цитоскелетным образованиям следу-
ет относить и все покровы (например, клеточную
стенку, или теку), которые часто определяют и под-
держивают форму клетки протиста. В основе скелет-
ных образований цитоплазмы лежат два типа струк-
тур — микрофиламенты и микротрубочки. Микро-
филаменты имеют диаметр 4—10 нм. Они могут быть
образованы разными белками — актином, миозином,
центрином, ассамблином, динеином, нексином, спаз-
мином и некоторыми другими, в том числе и еще не-
идентифицированными белками. Микрофиламенты
часто формируют как сократимые, так и несократи-
мые фибриллярные тяжи, связанные с ядром, плазма-
леммой, кинетосомами. Довольно часто они ассоции-
рованы с микротрубочками, обеспечивая движение
частиц цитоплазмы. Микротрубочки обычно имеют
диаметр 24—26 нм, хотя встречаются и более толст
тые — до 40 нм. Они состоят, как правило, из глобул
белков а- и Р-тубулина и формируются из этих гло-
бул путем самосборки. Микротрубочки выполняют в
клетке как опорную, так и сократительную функцию.
Они располагаются под покровами многих инфузо-
рий, жгутиконосцев, часто определяя форму клетки
(рис. 77), входят в состав ресничного и жгутикового
аппарата, аксоподий и прочих структур. За счет мик-
ротрубочек в процессе деления ядра формируется
митотическое веретено, посредством которого осу-
ществляется расхождение хромосом. Роль микротру-
бочек и микрофиламентов в жизни клетки протиста
чрезвычайно существенна и многообразна.
Жгутиковый/ресничный аппарат. Подвижность
очень многих протистов обеспечивается наличием
жгутиков или ресничек. И те и другие устроены оди-
наково, но исторически термин «реснички» появился
раньше для обозначения многочисленных подвижных
выростов (ундулиподий) на поверхности клеток ин-
фузорий. У большинства протистов количество жгу-
тиков невелико (от Г до 8 на клетку), хотя есть нема-
ло и многожгутиковых форм. Некоторые авторы счи-
тают, что правильнее называть все эти структуры
ресничками. Однако и тот и другой термины широко
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
149
Рис. 75. Экструсомы протистов. (По: Хаусман, 1988).
А — трихоцисты (7) и нематоцисты (2) у динофлагелляты Polykrikos, Б —
строение нематоцисты (7 — покоящаяся стадия, 2 — выстрелившая нема-
тоциста), В - —- токсициста, Г эжектосомы у Chilomonas (1 — эжектосо-
мы в стенке глотки, 2 — изолированная покоящаяся эжектосома, 3 — вы-
стрелившая эжектосома), Д — дискоболоцисты хризомонады Ochromonas
(7 — покоящаяся стадия, 2 — выстрелившая дискоболоциста).
Fig. 75. Extrusomes of protists.
A — trichocysts (7) and nematocysts (2) of dinoflagellate Polykrikos, Б —
structure of nematocyst (7 — intact, 2 — after extrusion), В — toxicyst, Г —
ejectosomes of Chilomonas (7 — in the wall of cytopharinx, 2 — intact ejecto-
some, 3 — after extrusion), Д — discobolocysts of Ochromonas (7 — intact,
2 — after extrusion).
встречаются в литературе, поэтому мы не будем заос-
трять внимание на приоритете названия, а будем ис-
пользовать их оба.
Варианты расположения жгутиков на теле клетки
представлены на рис. 78. В состав жгутикового аппа-
рата входит собственно жгутик (ундулиподия), пере-
ходная зона, кинетосома (базальное тельце) и кореш-
ковая система (рис. 79).
Свободная часть жгутика нередко имеет
покровные структуры. Так, почти у всех свободножи-
вущих жгутиконосцев на двигательном жгутике есть
чешуйки или мастигонемы, а у некоторых криптомо-
над жгутики покрыты и мастигонемами, и чешуйка-
ми. Принято различать простые мастигонемы (рис. 80)
и трубчатые: двух- и трехчленные (рис. 79; 81). Боль-
шой сложности достигают строение и организация
трубчатых трехчленных мастигонем на двигательном
жгутике хризофитовых (рис. 81). Паразитические мо-
нады, многожгутиковые и ресничные формы протис-
тов всегда имеют гладкие жгутики.
Осевой скелет ундулиподии представлен аксоне-
мой, состоящей, как правило, из 1 пары централь-
ных и 9 пар периферических микротрубочек. От-
клонения от этой формулы (9 + 2) встречаются, но
среди протистов они редки и характерны для жгу-
тиков зооспор, гамет или нефункционирующих ун-
дулиподий.
Пара периферических микротрубочек образова-
на А- и В-трубочками. От А-трубочек обычно от-
ходят различные боковые выросты: динеиновые
ручки — по направлению к соседнему дублету, ра-
диальные спицы — к центральной паре микротру-
бочек, переходные фибриллы в основании жгутика
соединяют их с плазмалеммой (рис. 79; 82). Состав
белков основных элементов аксонемы, по данным
о Chlamydomonas и Tetrahymena, приведен на рис. 82
(Grain, 1986).
В дополнение к микротрубочкам аксонемы в жгу-
тиках могут развиваться и другие структурные эле-
менты. К ним относится параксиальный тяж (рис. 80),
развитый у трихомонад, эвгленовых, кинетопластид,
динофлагеллид, пединеллид и некоторых бикозое-
цид. Параксиальный тяж состоит из микрофиламен-
тов, располагается между плазмалеммой и аксонемой,
проходя вдоль по всей длине жгутика или занимая
только проксимальную его часть. Жгутики с паракси-
альными тяжами часто прилегают к поверхности
клетки. Это создает предпосылки для образования
так называемых ундулирующих мембран. Комплекс
ундулирующей мембраны характерен для панцирных
жгутиконосцев, трипаносоматид и трихомонад. Почти
во всех случаях он формируется на основе рекуррен-
тного (рулевого) жгутика.
Переходная зона — это проксимальная часть
аксонемы, расположенная на уровне выхода жгутика
из клетки (рис. 79). Ее размеры определяются рассто-
янием между кинетосомой и центральными микрот-
рубочками аксонемы. В коротких переходных зонах
центральные микротрубочки начинаются почти на
уровне плазмалеммы, в длинных — иногда на рассто-
янии 1 мкм. В большинстве переходных зон имеется
поперечная (терминальная) пластинка, хотя известны
случаи, когда она отсутствует или их количество уве-
личивается до 2—3. Кроме поперечной пластинки в
этой зоне встречаются и другие образования, играю-
щие, по-видимому, значительную роль в укреплении
жгутика в месте выхода его из клетки. Структуры,
при помощи которых достигается выполнение этой
функции, довольно разнообразны и часто характери-
зуют крупные таксоны протистов. Например, для зе-
леных водорослей и наземных растений характерно
звездчатое образование (рис. 83, вкл.), для большин-
ства хромофитовых — спираль (рис. 79; 84, вкл.),
для хоанофлагеллат — центральный филамент, а для
некоторых протистов — цилиндрическое или спи-
150
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 77. Схема строения изолированного цитоскелета зеленой во-
доросли Spermatozopsis. (По: Lechtreck et al., 1989).
1,2 — кинетосомы жгутиков (С-трубочки не показаны), фс — фибрилляр-
ная связка, в которой сходятся микротрубочковые корешки (показаны
стрелками) и начинаются дополнительные микротрубочки цитоскелета (цмт)-,
Ф — фибриллярный мостик, связывающий 2 корешка.
Fig. 77. Scheme of isolated cytoskeleton of green alga Spermatozop-
sis.
1, 2 — flagellar basal bodies (C-tubules are not shown), фс — fibrillar con-
nection of microtubular rootlets (arrows), iniciating additional microtubules
(цмтУ, ф — fibrillar bridge between two rootlets.
ральное образование, связанное с А-трубочками аксо-
немы (рис. 85, вкл.).
Кинетосома, или базальное тело жгути-
ка/реснички, представляет собой полый цилиндр,
состоящий из 9 триплетов микротрубочек, соединен-
ных мостиками. Проксимальный конец жгутиковой
кинетосомы обычно связан с плазмалеммой клетки
переходными фибриллами (рис. 86, вкл.). Триплеты
кинетосомы, как и дублеты аксонемы, ориентирова-
ны таким образом, что при взгляде на поперечный
срез жгутика изнутри клетки динеиновые ручки (а на
уровне кинетосомы — наклон триплета внутрь) на-
правлены по часовой стрелке (рис. 87, вкл.), а при
взгляде снаружи — против часовой стрелки (рис. 86).
Длина кинетосомы варьирует у разных видов от 50 до
1300 нм. Большинство мастигофор имеет 2 жгутика
и Ъ соответствующие кинетосомы. У одножгутиковых
видов тоже, как правило, 2 кинетосомы, одна из ко-
торых безжгутиковая (рис. 79). По-видимому, двух-
жгутиковое состояние следует считать наиболее кон-
сервативным, а одножгутиковое — объяснять редук-
цией второго жгутика, которая в крайнем варианте
может приводить к исчезновению и кинетосомы.
Базальные тела могут располагаться параллельно
друг другу (особенно это характерно для многожгу-
тиковых и инфузорий), под острым, прямым или
тупым углами, а также бывают развернуты под углом
180° (так называемое антипараллельное положение
кинетосом, — см. рис. 77). Кинетосомы часто бы-
вают связаны между собой фибриллярными мости-
ками и окружены электронно-плотным веществом,
от которого берут начало корешковые структуры
(рис. 85; 87).
Корешковая система кинетосом. По
своей природе корешки подразделяются на фибрил-
лярные и микротрубочковые (рис. 79). Первые состо-
ят из микрофиламентов, образованных различными
белками, среди которых идентифицированы центрин
и ассамблин. Они могут быть поперечно исчерченны-
ми и простыми. Фибриллярные корешки из центрина
способны медленно сокращаться, а состоящие из ас-
самблина несократимы. Микротрубочковые корешки
образованы лентами или пучками микротрубочек, ко-
торые часто связаны с микрофиламентами. Их строе-
ние и взаимное расположение весьма разнообразны
(рис. 88). Оба типа корешков выполняют цитоскелет-
ную и морфогенетическую функции, не только за-
крепляя жгутик в теле клетки, но и определяя форму
и симметрию клетки (рис. 77).
Наиболее развитая система корешков свойственна
крупным жгутиконосцам и инфузориям, а также тем
протестам, у которых длина двигательного жгутика
Рис. 78. Варианты расположения жгутиков у протистов. (По: Ha-
usmann, Hiilsmann, 1996).
А — одножгутиковая форма, Б — изоконтный жгутиконосец, В — клетка с
поперечным и продольным жгутиками, Г — гетероконтный жгутиконосец с
опушенным передним жгутиком, Д — гетероконтный жгутиконосец с уни-
латеральным опушением на переднем жгутике, Е — гетероконтный жгути-
коносец с обоими опушенными жгутиками, Ж — четырехжгутиковый про-
тает с 1 рекуррентным жгутиком, 3 — четырехжгутиковый протает с ре-
куррентным жгутиком, образующим ундулирующую мембрану.
Fig. 78. Types of flagellation in protists.
A — uniflagellate, Б — isocont flagellate, В — flagellate with transversal and
longitudinal flagella, Г — heterocont flagellate with hairy anterior flagellum,
Д — heterocont flagellate with unilateral hairs on the anterior flagellum, E —
heterocont flagellate with both haired flagella, Ж — 4-flagellated protist with
one recurrent flagellum, 3 — 4-flagellated protist with one recurrent flagellum,
forming the undulating «membrane».
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
151
Рис. 79. Обобщенная схема строения жгутикового аппарата.
(По: Карпов, 19906).
бк — безжгугиковая кинетосома; жг — жгутиковая кинетосома; жч — жгу-
тиковые чешуйки; мн — трехчленные трубчатые мастигонемы; мсс — мно-
гослойная структура; пп — поперечная пластинка; пс — переходная спи-
раль; пф — переходные фибриллы; рк — ребристый корешок с отходящими
вторичными микротрубочками (вмт)\ фк! — фибриллярный корешок систе-
мы I, обычно ассоциирован с микротрубочковым корешком (мтк)\ фк11—
фибриллярный корешок системы II (ризопласт), обычно не связан с микротру-
бочками; фм — фибриллярный мостик между кинетосомами.
Рис. 80. Схема организации жгутика у эвгленовой водоросли Ani-
sonema. (По: Mignot, 1966).
а — аксонема, пт — параксиальный тяж. На поверхности жгутика пока-
заны простые короткие и длинные мастигонемы.
Fig. 80. Scheme of flagellar organization in euglenophyte Anisone-
ma.
a — axoneme, nm — paraxial rod. There are short and long simple hairs on
the surface of flagellum.
Fig. 79. General scheme of flagellar apparatus structure.
мн — tripartite tubular mastigonemes, жч — flagellar scales, nc — transitional
helix, жг — flagellar basal body, бк — nonflagellar basal body, nn — transver-
sal plate, пф — transitional fibers, фм — fibrillar bridge between basal bodies,
мсс — multilayered structure, рк — microtubular rootlet with secondary micro-
tubules (влт), фк! — rootlet of fibrillar system I, associated with microtubu-
lar rootlet (jwmx), фкП — rootlet of fibrillar system II (rhizoplast).
значительно превышает длину тела. Различные впя-
чивания и выступы поверхности тела клетки высти-
лаются микротрубочковыми корешками, поддержи-
вающими их форму. Степень развития корешков
зависит также от наличия или отсутствия аппарата
питания (губа, цитостом, глотка и др.).
При делении клетки происходит и деление жгути-
ков. Этот процесс пока еще слабо изучен. Обычно
для двухжгутиковых протистов характерен полукон-
сервативный тип деления кинетиды (рис. 89). В ре-
зультате каждая дочерняя клетка наследует 1 «ста-
рую» кинетосому и 1 «новую», образовавшуюся перед
митозом.
Для более общего определения жгутикового/рес-
ничного аппарата часто используется понятие кине-
тида. Определение этого понятия для протистов не
вполне четкое, так как первоначально оно было раз-
работано только для инфузорий (библиографию см.:
Карпов, 1993а). Весь двигательный аппарат инфузо-
рий обычно определяется как кинетом, а кинетида
представляет собой его структурно-функциональную
Рис. 81. Схема расположения мастигонем на переднем жгутике
хризомонады Ochromonas. (По: Bouck, 1971).
Fig. 81. Mastigoneme arrangement at the anterior flagellum of chry-
somonade Ochromonas.
152
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 82. Белки жгутиков и ресничек. (По: Grain, 1986).
вр — внутренние ручки образованы динеином и актиноподобным белком;
грс — головка радиальной спицы состоит из 5 белков различного м. в. (от
24 до 123 кд); мт — микротрубочки из тубулина; нр — наружные ручки
образованы динеином (м. в. 300 000 кд); нс — нексиновая связка; оцм —
оболочка центральных микротрубочек состоит из 23 белков различного
м. в. (от 14 до 360 кд); пм—- плазматическая мембрана, содержит основные
белки: 65 000 кд, 125 000 кд и тубулин; пф — 3 протофиламента образова-
ны белками с м. в. меньшим, чем у тубулинов; срс — стебелек радиальной
спицы состоит из 12 белков различного м. в. (от 20 до 124 кд); cml и cm2 —
центральные микротрубочки аксонемы.
Fig. 82. Proteins of flagella and cilia.
cml and cm2 — central tubules of axoneme; ep — inner dinein arms, нр —
outer arms, composed with dinein of 300000 kd; нс — nexin connection;
им — plasma membrane composed with main proteins of 65000 and
125000 kd; пф — three protofilaments contain proteins with molecular weight
less than tubulin; мт — tubulin microtubules; cpc — radial spoke shaft conta-
ins 5 different proteins with m. w. from 20 to 124 kd; грс — radial spoke head
contains 5 different proteins with m. w. from 24 to 123 kd; оцм — central
sheat contains 23 different proteins from 14 to 360 kd.
единицу. Для жгутиконосцев, имеющих 1—2 жгути-
ка, понятия кинетома и кинетиды совпадают. Наибо-
лее консервативная часть кинетиды — кинетосомы,
количество которых в кинетиде подавляющего боль-
шинства протистов равно 2. Двухкинетосомальное
состояние кинетиды распространено наиболее широ-
ко, хотя есть немало видов, имеющих 1 кинетосому.
Это так называемые истинно одножгутиковые формы
(в клетке 1 жгутик и 1 кинетосома) и некоторые мно-
гожгутиковые или ресничные (опалиниды), у кото-
рых имеются непарные кинетосомы.
Для многих полимастигин характерно объедине-
ние жгутиков с ядром в единый морфофункциональ-
ный комплекс. Его принято называть кариомастигон-
том. Обычно кариомастигонт содержит 4 кинетосо-
мы и 4 жгутика, расположенные попарно. Поскольку
пары кинетосом часто не связаны друг с другом мор-
фологически, можно считать, что кариомастигонт со-
держит две кинетиды и одно ядро.
Функции жгутика/реснички. Основная функция
жгутика — движение. В активной работе жгутика
движущим началом являются периферические мик-
ротрубочки и их ручки, обладающие АТФ-азной актив-
ностью. Центральные микротрубочки имеют опорное
значение. Формы движения жгутика различны, но
обычно это винтообразное движение, позволяющее
жгутиконосцу «ввинчиваться» в воду, совершая до
40 оборотов в секунду. У инфузорий и многожгути-
ковых протистов движение ресничек организовано
по типу метахрональных волн. Жгутики и реснички
часто используются также и для питания. При этом
поведение жгутика может меняться, а может оста-
ваться неизменным. Взаимодействие биения жгути-
ков, вызываемых ими токов жидкости и направления
движения клетки у различных протистов показано на
рис. 90. Например, у вольвоцид 2 одинаковых жгути-
ка проталкивают клетку вперед гребными движения-
ми, у гетероконтных жгутиконосцев задний жгутик
обычно пассивен, выполняя, по-видимому, рулевую
функцию. Многие хризомонады имеют редуцирован-
ный рулевой жгутик. Обычно в его основании распо-
ложено вздутие, представляющее часть фоторецеп-
торного аппарата, т. е. жгутики могут выполнять и
рецепторную функцию (см. также рис. 88).
Среди жгутиконосцев имеются виды, проводящие
большую часть жизненного цикла в прикрепленном
состоянии. В этот период жгутик теряет обычную для
него функцию движения и превращается в органеллу
прикрепления, стебелек или ножку (рис. 91). Еще
одна функция жгутика заключается в том, что он сво-
ими движениями очищает поверхность тела от нали-
пающих на него мелких посторонних частиц.
Эндоплазматические органеллы
В эндоплазме протистов находятся одно или не-
сколько ядер, а также все органеллы и структуры,
свойственные эукариотической клетке: ЭПР, рибосо-
мы, аппарат Гольджи, митохондрии, пероксисомы,
гидрогеносомы, пластиды (у автотрофных протис-
тов), лизосомы, пищеварительные вакуоли. Некото-
рые протисты обладают также специфичными только
для них органеллами, к которым относятся кинето-
пласт, пузула и ряд других.
Митохондрии. Они присутствуют в клетках боль-
шинства протистов. Их строение значительно разно-
образнее, чем у клеток многоклеточных животных и
растений (рис. 92; 93, вкл.). На мембранах митохонд-
рий осуществляются процессы окислительного фос-
форилирования, завершающиеся образованием АТФ.
Для многих одноклеточных протистов показано, что
в клетке находится лишь 1, иногда сильно ветвящая-
ся митохондрия. Внутренняя мембрана ее образует
впячивания внутрь митохондрии, называемые криста-
Рис. 88. Схема организации жгутиковых аппаратов различных хромистов. (По: Andersen, 1991).
А — хризофитовые, Б — бурые водоросли, В — пединелловые, Г — синурофитовые, Д — ксантофитовые, Е — оомицеты, Ж — рафидофитовые, 3 —
гаптофитовые. Пронумерованы гомологичные корешки. Показаны также мастигонемы (мн), вздутие в основании заднего жгутика (вз), фибриллярный ко-
решок (ризопласт — р), вторичные микротрубочки (вмт) и переходная спираль (пс).
Fig. 88. Flagellar apparatus organization in chromists.
A — chrysophytes, Б — phaeophytes, В — pedinellids, Г — synurophytes, Д — xanthophytes, E — oomycetes, Ж — raphidophytes, 3 — haptophytes. Homolo-
gous rootlets have the same number, мн — mastigonemes, вз — flagellar swelling, p — rhizoplast, вмт — secondary microtubules, nc — transitional helix.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
153
154
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 89. Схема трансформации жгутиков зеленой водоросли Neph-
roselmis в процессе деления клетки. Полуконсервативный тип ре-
пликации жгутиков. (По: Melkonian et al., 1987).
А — в интерфазе, Б — перед началом деления, В — во время митоза. 1 —
наиболее старая кинетосома материнской клетки; 2 — более молодая кине-
тосома материнской клетки; 21 — дочерняя кинетосома кинетосомы 7;
22 — дочерняя кинетосома кинетосомы 2; 12 — кинетосома дочерней клет-
ки, в которую превращается кинетосома 2 материнской клетки.
Fig. 89. Semiconservatiye type of flagellar replication during cell di-
vision of Nephroselmis.
A — interphase, Б — predivision cell with nascent kinetosomes, В — during
mitosis. 1 — the eldest kinetosome of parental cell; 2 — the younger kinetoso-
me of parental cell; 21, 22 — nascent kinetosomes; 12 — the parental kinetoso-
me no. 2 has been transformed to no. 1 kinetosome.
ми. Кристы различаются по форме, вариации кото-
рой можно описать тремя основными морфотипами:
трубчатые, пузырьковидные и пластинчатые (Сера-
вин, 1993). Форма крист митохондрий весьма кон-
сервативна в пределах крупных таксонов протистов и
используется в настоящее время в их таксономии.
У большинства протистов кристы трубчатой формы.
В матриксе митохондрий иногда встречаются крис-
таллические включения или фибриллярные образова-
ния. Внутри трубчатых крист некоторых хромофито-
вых обнаруживаются тонкие осевые нити. Природа
этих включений и филаментов неизвестна.
Гидрогеносомы. Многие виды паразитических и
свободноживущих протистов, обитающих в условиях
дефицита кислорода, обладают анаэробным обменом
и поэтому лишены митохондрий. Это — параба-
залии, дипломонады, оксимонады, микроспоридии,
некоторые инфузории и амебы. У трихомонад, гипер-
мастигин, анаэробных цилиат, некоторых жгути-
коносцев и хитридиевых грибов обнаружены гидро-
геносомы, впервые описанные около 20 лет назад
Мюллером и в дальнейшем детально изученные (см.:
Muller, 1993). Это особые органеллы в виде пузырь-
ков с 1 или 2 мембранами и гомогенным содержи-
мым. Они также содержат ферменты, окисляющие
пируваты и малаты с освобождением молекулярного
водорода. Представляя акцептор электронов в цепи
реакции окисления пировиноградной кислоты, водо-
род выполняет важную функцию в анаэробном мета-
болизме. Однако трихомонады могут существовать и
Рис. 90. Формы движения жгутиков, вызываемые при этом токи
жидкости и направление движения клетки у разных протистов.
(По: Vickerman et al., 1991).
А — динофлагеллята Ceratium; Б — критидия; В — трипаносома; Г, Д —
хризомонады Ochromonas и Bicosoeca; Е — дипломонада Trepomonas; Ж —
хламидомонада (движение вперед); 3 — хламидомонада (движение назад);
И — траектория плавающий эвглениды Rhadbomonas; К — трихомонада;
Л скользящее движение церкомонады; М — метахрональные волны опа-
лины. Стрелка внутри клетки указывает направление ее движения, пунк-
тирные стрелки — направление движения жидкости, сплошные и жирные
стрелки — направление биения жгутиков.
Fig. 90. Flagellar motion and cell locomotion.
Closed arrows, direction of motion of the waves; open arrows, direction of cell
movement; dashed arrows, water flow. A — dinophyte Ceratium; Б, В — kine-
toplastids Crithidia and Trypanosoma; Г, Д — chrysophytes Ochromonas and
Bicosoeca; E, Ж — breast-stroke flagellar motion in Trepomonas and Chlamy-
domonas; 3 — reversed swimming in Chlamydomonas; И — swimming path-
way of Rhabdomonas; К — trichomonade Trichomitus; Л — gliding motion of
Cercomonas; M — flagellar metachronal waves in Opalina.
в аэробных условиях. В этом случае гидрогеносомы
прекращают продуцировать водород и клетка пере-
ключается на кислородно-зависимый метаболизм
примитивного типа без митохондрий. Гидрогеносо-
мы, как правило, встречаются в клетках, не имеющих
митохондрий, однако бесцветный жгутиконосец Psal-
teriomonas lanterna имеет и митохондрии, и гидроге-
носомы (Broers et al., 1993). У этого жгутиконосца и
у многих анаэробных инфузорий гидрогеносомам со-
путствуют симбиотические метаногенные бактерии,
лежащие рядом с ними в цитоплазме.
Можно предположить, что гидрогеносомы него-
мологичны друг другу. Показано, что у цилиат они
появляются из митохондрий, у хитридиевых грибов —
из пероксисом, у парабазалий их происхождение
также связывают с митохондриями (Muller, 1998).
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
155
Рис. 91. Схема различных типов прикрепления жгутика трипано-
соматид к эпителию пищеварительного тракта насекомых. (Ориг.
А; О. Фролова и С. О. Скарлато).
1 — вплетение жгутика (ж) обычного строения в щеточную каемку (мв — микровилли), 2 — вплетение расширенного кончика жгутика (жр) в щеточную
каемку, 3 — вплетение отростков (жо) сильно измененного жгутика в щеточную каемку, 4 — прикрепление расширенного жгутика к эпикутикуле (эп) с
помощью полудесмосомы (nd).
Fig. 91. Different types of attachement of trypanosomatid flagella to the epithelium of insect intestine.
1 — penertation of normal flagellum (ж) among microvilli (ля), 2 — penertation of modified flagellum (жр) among microvilli, 3 — penertation of projections
(жо) of extremely modified flagellum (жр) among microvilli, 4 — attachment of modified flagellum to epicuticle (эп) with semidesmosome (nd).
В соответствии с другой точкой зрения, гидрогеносо-
мы могут иметь независимое и несимбиотическое
происхождение.
Пероксисомы. У всех протистов, имеющих мито-
хондрии, встречаются в цитоплазме и пероксисомы.
Это тельца округлой формы, ограниченные мембра-
ной, в которой заключен компактный гомогенный
матрикс. Их размеры обычно не превышают 1 мкм.
Пероксисомы отличаются содержанием оксидаз, ко-
торые продуцируют перекись водорода, каталазу и
ферменты глиоксилатного цикла. У них, как и у гид-
рогеносом, отсутствует ДНК и собственный синтез
белка.
Пластиды. Пластиды имеются у фототрофных и
близких к ним протистов и представлены хлороплас-
тами и лейкопластами. Последние обычно трактуют-
ся как редуцированные хлоропласты и характерны
для бесцветных (гетеротрофных) хризомонад. Они
располагаются обычно в перинуклеарном простран-
стве ядра, ограничены 1—2 мембранами и содержат
остатки тилакоидов и пигментные гранулы.
Хлоропласты на светооптическом уровне различа-
ются по форме, размерам и количеству в клетке.
Электронно-микроскопические исследования выяви-
ли существенные особенности в их строении, кото-
рые используются для характеристики крупных так-
сонов (рис. 94). Так, оболочка хлоропластов красных
водорослей, хлорофитов и высших растений состоит
из 2 мембран, у эвгленид и диномонад — из 3, а у
остальных протистов — из 4 мембран. У криптомо-
над между наружной и внутренней парами мембран
находится нуклеоморф с рибосомами и гранулами
полисахаридов.
Организация тилакоидов в хлоропласте также ока-
залась существенным признаком для фототрофных
протистов. Одиночные тилакоиды имеются у крас-
ных водорослей, у криптомонад они собраны попар-
но, а в хлоропластах зеленых водорослей тилакоиды
образуют стопки разного размера, подобные гранам
Рис. 92. Схема строения митохондрий с различными типами
крист. (По: Кусакин, Дроздов, 1994).
А — пластинчатые, Б — трубчатые, В — пузырьковидные, Г — ампуло-
видные кристы.
Fig. 92. Different shape of cristae in mitochondria.
A — plate-like, or lamellar; Б — tubular; В — vesicular; Г — ampula-like.
156
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
высших растений. У остальных водорослей тилакои-
ды собраны в стопки по 3.
В хлоропластах имеются центры синтеза полиса-
харидов — пиреноиды, которые не всегда морфоло-
гически выражены. Их строение довольно разнооб-
разно и может быть использовано для характеристи-
ки водорослей на уровне рода. Полисахариды обычно
откладываются в цитоплазме. Только у зеленых водо-
рослей они запасаются внутри хлоропласта, как у на-
земных растений.
Основными пигментами хлоропластов являются
хлорофиллы а, b и с. Хлорофилл с встречается в трех
формах: с7, с2 и сЗ, Для разных групп фототрофных
протистов характерны определенные наборы хлорофил-
лов. У зеленых водорослей и эвгленовых содержатся
хлорофиллы а и Ь, у красных водорослей — только
хлорофилл а, остальные имеют хлорофиллы а и с.
Из вторичных пигментов у водорослей встречают-
ся каротины и ксантофиллы, которые в больших кон-
центрациях могут маскировать зеленый хлорофилл и
придавать хлоропластам разнообразную окраску от
желто-зеленой до красновато-коричневой. У крипто-
монад имеются фикобилины. Эти пигменты, правда,
несколько иного состава, характерны также для крас-
ных водорослей и цианобактерий, где встречаются в
виде фикобилисом на поверхности тилакоидов.
Аппарат Гольджи обнаружен практически у всех
исследованных видов протистов. До изучения с по-
мощью электронного микроскопа эта органелла была
известна как парабазальное тельце. Функция его не
была определена, хотя некоторые исследователи
сравнивали его именно с аппаратом Гольджи (Догель,
1951). Парабазальное тельце наиболее сильно разви-
то у трихомонадид и гипермастигид — парабазалий
(рис. 95).
Наиболее часто аппарат Гольджи расположен по
соседству с ядром и представлен одной или несколь-
кими стопками плоских цистерн (диктиосом), окру-
женных мелкими пузырьками (рис. 96, вкл.). Однако
аппарат Гольджи не всегда встречается в виде диктио-
сом. У некоторых протистов он образован одиночны-
ми цистернами. Отсутствие диктиосом обычно тракту-
ется как примитивный признак. Вероятно, это вполне
справедливо, так как стопки цистерн аппарата Гольд-
жи работают эффективнее, чем одиночные цистерны.
Однако отсутствие диктиосом у современных протис-
тов не может однозначно свидетельствовать об их при-
митивности, поскольку диктиосомы — весьма лабиль-
ные образования, а их формирование и разборка в
большой степени зависят от внешних воздействий на
клетку (например, снижение концентрации кислорода
в среде) или от физиологических перестроек самого
протиста (переход к инцистированию).
Лизосомы и другие органеллы и включения. В
клетках протистов, как и в клетках многоклеточных
животных, присутствуют лизосомы. Эти цитоплазма-
тические тельца в форме мелких пузырьков (первич-
ные лизосомы) образуются в аппарате Гольджи. В
них локализованы пищеварительные гидролитичес-
кие ферменты. На электронограммах первичные ли-
зосомы выглядят как тельца округлой формы с диа-
метром меньше 1 мкм и с гомогенным внутренним
содержимым. Они расположены во всех участках
клетки. Вторичные лизосомы, или пищеварительные
вакуоли, хорошо выражены только у гетеротрофных
протистов, питающихся путем фагоцитоза.
В эндоплазме разных протистов в большем или
меньшем количестве присутствуют резервные пита-
тельные вещества, используемые в процессах мета-
болизма. Чаще всего это различные полисахариды
Рис. 94. Схемы расположения основных органелл, связанных с хлоропластами у красных водорослей и глаукофитовых (А), хлорофито-
вых (В), криптомонад (В), хризомонад (Г) и эвгленовых (Д). (По: Sleigh, 1989).
к — крахмал, л — лейкозин, нм — нуклеоморф, п — парамилон, с — стигма, хл — хлоропласт, я — ядро.
Fig. 94. Arrangement of membranes associated with chloroplast in Rhodophyta (A), Chlorophyta (B), Cryptophyta (B), Chrysophyta (Г), Eugle-
nophyta (Д).
нм — nucleomorph, n — paramylon, л — leucosin, хл — chloroplast, cm — stigma, к — starch, я — nucleus.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
157
Рис. 95. Парабазальные аппараты Pseudodevescovina uniflagellata.
(По: Догель, 1951).
1 — пучок парабазальных аппаратов, 2 — передние жгутики, 3 — руле-
вой жгутик, 4 — аксостиль.
Fig. 95. Parabasal apparata of Pseudodevescovina uniflagellata.
1 — bunch of parabasal apparata, 2 — anterior flagella, 3 — recurrent flagel-
lum, 4 — axostyle.
(гликоген, крахмал, амилоплектин и др.), нередко —
липиды и иные жировые включения. Количество ре-
зервных веществ зависит от физиологического состо-
яния простейшего, характера и количества пищи, от
стадии жизненного цикла и варьирует в широких пре-
делах. Однако некоторые крупные группы протистов
запасают специфические вещества. Например, эвгле-
новые запасают парамилон, который не встречается у
других протистов.
Ядро
У всех протистов имеются типичные клеточные
ядра характерной для эукариот структуры. Однако
они значительно различаются по форме и количеству
в клетке. Способы деления ядер протистов гораздо
многообразнее, чем у многоклеточных животных и
растений. В рамках группы происходило, вероятно,
становление мейоза и полового процесса, что нашло
отражение в существенных различиях этих явлений в
разных таксонах.
Интерфазное ядро. Число и размеры ядер.
Большинство протистов обладают одним клеточным
ядром. Некоторые солнечники, крупные свободножи-
вущие амебы, миксомицеты и другие — многоядер-
ны. Размеры и форма ядер варьируют в самых широ-
ких пределах, так же как и морфология, и соотно-
шение структурных компонентов ядра. Наиболее
мелкие ядра (у пироплазмид, лейшманий, микронук-
леусы некоторых инфузорий) имеют диаметр 1—
3 мкм. Крупные сферические ядра некоторых радио-
лярий достигают в диаметре до 400 мкм и даже 1 мм.
Форма ядер большинства простейших приближается
к сферической, но иногда может принимать самые
причудливые очертания. Это относится преимущест-
венно к богатым хроматином вегетативным ядрам ин-
фузорий.
Структурные компоненты ядер. У про-
тистов они те же, что и в других эукариотических
клетках (рис. 97, вкл.). Ядерная оболочка состоит из
2 мембран и имеет поры, число которых у разных
видов варьирует в широких пределах, но особенно
велико у ядер, которые обладают высокой функцио-
нальной активностью. Снаружи ядерная оболочка
обычно покрыта рибосомами, но к ней могут примы-
кать дополнительные, слои, располагающиеся как
снаружи, так и под оболочкой со стороны кариолим-
фы. В частности, у многих свободноживущих амеб к
внутренней стороне ядерной оболочки прилегает так
называемый фиброзный слой, имеющий ячеистое
строение и значительно превосходящий ядерную обо-
лочку по толщине. Реже дополнительные фибрилляр-
ные слои располагаются кнаружи от ядерной оболоч-
ки (например, у радиолярий).
В перинуклеарном пространстве у многих водоро-
слей располагаются хлоропласты или лейкопласты, а
у некоторых видов протистов обнаружены также
симбиотические бактерии (рис. 97).
Обязательным компонентом всех ядер протистов
является хроматин, представляющий ДНК в комплек-
се с основными белками. Ультраструктурный анализ
показывает, что он состоит из линейно расположен-
ных глобулярных телец — нуклеосом (включающих
как ДНК, так и гистоны). Кроме хроматина хромо-
сомной природы в ядре может находиться внехромо-
сомный хроматин, возникающий в результате ампли-
фикации (многократного копирования) некоторых
генов. Такой внехромосомный хроматин может вхо-
дить в состав ядрышек.
Распределение хроматина в интерфазных ядрах у
простейших может носить различный характер. Под-
час происходит сильная деспирализация и деконден-
сация хроматина. В других случаях деконденсация
охватывает часть хроматина. В некоторых интерфаз-
ных ядрах хроматин, наоборот, очень сильно конден-
сирован и представляет собой сплошную массу, как в
головках спермиев многоклеточных животных.
Ядрышки, которых может быть в ядре 1 или не-
сколько, а иногда и очень много, в основном сострят
из рибонуклеопротеинов — предшественников рибо-
сом. Электронная микроскопия показывает, что яд-
рышки состоят обычно из двух структурных компо-
нентов — гранулярного и фибриллярного. Фибриллы
диаметром 5—10 нм обычно локализованы в цент-
ральных частях ядрышка. Во многих ядрышках у
протистов описаны включения хроматина, которые
иногда, но не всегда морфологически связаны с опре-
деленными хромосомами. Это ядрышковые организа-
торы, образующиеся в результате амплификации ри-
босомальных генов.
В ядрах простейших в большем или меньшем ко-
личестве присутствует ядерный сок (кариолимфа). В
нем могут быть также мельчайшие гранулы разной
природы, часть которых является, вероятно, субъеди-
ницами рибосом. Встречаются также паракристалли-
ческие или фибриллярные (рис. 97) белковые образо-
вания, а также особые спиральные структуры (у
крупных амеб), содержащие, по-видимому, РНК.
Митоз. Основной формой деления ядра простей-
ших, как и всех эукариотических организмов, явля-
158
ПРОТЕСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 98. Диаграмма классификации митозов у протистов.
(По: Райков, 1994).
Fig. 98. Classification of metaphase figures in mitotic nucleus of
protists.
ется митоз. Главная черта митоза — закономерное
расхождение в дочерние ядра двух копий каждой
реплицированной хромосомы (хроматиды) — хоро-
шо известна. Этот важнейший биологический про-
цесс обеспечивает преемственность и непрерывность
наследственной информации. Хотя общая схема й
сущность митоза едины для всех эукариот, но в де-
талях процесс митоза может протекать по-разному.
Поведение и судьба участвующих в митозе компонен-
тов — ядерной оболочки, центриолей, ахроматино-
вого аппарата клетки — варьируют у протистов в
широких пределах. Здесь как бы осуществлялись раз-
ные попытки реализации митоза, из которых лишь
немногие были заимствованы наземными растениями,
настоящими грибами и животными.
Классификация типов митоза у протистов пред-
принималась неоднократно. По мере накопления зна-
ний она становилась все более дифференцированной.
Особое значение для углубления представлений о
многообразии форм митоза имела электронная мик-
роскопия, которая позволила раскрыть ряд ранее не-
известных фактов.
В настоящей книге мы принимаем классифика-
цию Райкова (1982, 1994), которая, в свою очередь,
представляет собой дальнейшее усовершенствование
классификации, предложенной в 1972 г. Холлан-
дом (Hollande, 1972). Мы ограничимся здесь только
общей схемой, детализация которой будет иметь
место при рассмотрении отдельных групп простей-
ших. Райков различает следующие 6 форм митоза
(рис. 98).
Открытый митоз
1. Открытый ортомитоз (эумитоз)
Полузакрытый митоз
2. Полузакрытый ортомитоз
3. Полузакрытый плевромитоз (парамитоз)
Закрытый митоз с внутриядерным веретеном
4. Внутриядерный плевромитоз
5. Внутриядерный ортомитоз
Закрытый митоз с внеядерным веретеном
6. Внеядерный плевромитоз
При открытом митозе в отличие от закрытого раз-
рушается ядерная оболочка, а ядрышки и хромосомы
оказываются свободно лежащими в цитоплазме. За-
крытым митозом именуют такие его формы, при ко-
торых ядерная оболочка сохраняется в течение всего
процесса. При ортомитозе в метафазе хромосомы
располагаются по экватору веретена, образуя мета-
фазную экваториальную пластинку. При плевромито-
зе экваториальная пластинка, как правило, не образу-
ется и ахроматиновое веретено представлено 2 полу-
веретенами, лежащими под углом друг к другу вне
или внутри ядра, в результате чего вся фигура деле-
ния приобретает асимметричный характер. Различ-
ные сочетания указанных черт и образуют те 6 кате-
горий митоза, которые названы выше.
Проблема амитоза. Амитоз, или деление ядра
простой перешнуровкой надвое без участия митоти-
ческого аппарата и видимой реорганизации хромати-
на, известен со времен светооптических исследова-
ний. Исследование ультраструктуры таких «амитоти-
чески» делящихся ядер показало, что многие формы
«амитоза» на самом деле являются закрытым мито-
зом. В результате к настоящему времени амитотичес-
ки делящиеся ядра известны лишь для очень немно-
гих протистов.
В культурах клеток воротничковых жгутиконос-
цев, некоторых эвгленид и Acanthamoeba castellani
всегда обнаруживается небольшой процент особей,
ядра которых делятся амитотически. В результате
клетка может содержать 2 или 3 часто неодинаковых
по размеру ядра. Цитокинез в таких случаях приво-
дит к образованию нежизнеспособных особей. Есть
тем не менее предположения, что это особая форма
размножения путем так называемых хромидий (Mar-
gulis, 1992), или генеративных тел (Карпов, Жгарев,
1987).
К другой группе относятся амитотически делящи-
еся ядра симбионтов. Известно, что таким образом
делятся ядра симбионтов некоторых динофитовых, а
также нуклеоморф криптофитовых, который по про-
исхождению считается остатком ядра симбионта. По-
видимому, сильная зависимость от клетки-хозяина
освобождает некоторых симбионтов от необходимос-
ти равного распределения генетического материала
между дочерними особями. В этих случаях следует
считать амитоз вторичным упрощением аппарата де-
ления ядра.
Вегетативные ядра инфузорий — Ма — богаты
хроматином. Форма их может быть разнообразной,
подчас причудливой (четковидной, лентовидной и др).
При делении инфузорий, обладающих Ма сложной
конфигурации, обычно происходит их конденсация и
они становятся яйцевидными. Видимого образования
хромосом нет. Однако ДНК проходит характерные
для митотического цикла стадии Gl, S (репликация
ДНК) и G2. В результате количество ДНК удваивает-
ся, после чего Ма обычно вытягивается и перешнуро-
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
159
вывается. Внешне это выглядит как амитоз, а по су-
ществу, здесь происходят те же перестройки, что и
при митозе: предварительная репликация ДНК (часто
и амплификация генов) и распределение ядерного ма-
териала при помощи микротрубочек, не образующих,
правда, веретена деления.
Размножение
Способы размножения протистов весьма разнооб-
разны. Следует различать агамогонию (бесполое раз-
множение) и гамогонию (размножение, в той или
иной форме связанное с половым процессом). На ос-
нове закономерного сочетания разных форм размно-
жения возникают сложные жизненные циклы, харак-
терные для определенных таксонов. Заметим, что как
у соматических, так и у половых клеток протистов
наблюдаются генетические взаимодействия, которые
возникают в результате слияния клеток и их ядер, но
прямо не связаны с размножением.
Бесполое размножение
В основе его лежит деление клетки — цитокинез,
обычно следующий за митозом. Никакой глубокой
реорганизации цитоплазматических органелл при
этом может не происходить, как это имеет место, на-
пример, у амеб. Однако у простейших, обладающих
сложным строением, агамное размножение сопро-
вождается более или менее существенными клеточ-
ными морфогенезами (рис. 99).
Например, у многих жгутиконосцев цитокинез на-
чинается фактически еще до начала митоза, когда
происходит реорганизация жгутикового аппарата и
цитоскелета в целом. Удваиваются митохондрии,
хлоропласты, микротельца и другие органеллы. Удво-
ение жгутикового аппарата — сложнейший процесс,
исследование которого только начинается.
Наибольшей сложности клеточные морфогенезы
достигают у инфузорий. Нередко сложная организа-
ция клетки в цроцессе деления претерпевает частич-
ную редукцию, за которой следует образование вновь
возникающих структур. Так, у многих инфузорий
ротовой аппарат при делении редуцируется, а затем
заново формируется у дочерних особей. Сложные
процессы реорганизации при делении претерпевает
кортекс. Своеобразные процессы морфогенеза при
агамном размножении имеют место и у паразитичес-
ких простейших — споровиков.
Процесс агамного размножения простейших может
носить характер почкования, что имеет место, напри-
мер, у сосущих инфузорий, некоторых жгутиконос-
цев и других протистов. При почковании от материн-
ской особи отделяются 1 или несколько особей мень-
ших размеров.
Агамное размножение может иметь характер мно-
жественного деления (мерогония, шизогония). При
этом обычно происходит несколько последователь-
ных делений ядра, а затем вокруг каждого из ядер
обособляется участок цитоплазмы (рис. 100). В ре-
зультате формируется несколько (иногда десятки и
Рис. 99. Примеры бесполого размножения путем деления клетки
на 2 дочерние (бинарное деление) (По: Hausmann, Hiilsmann,
1996).
А — раковинная амеба Euglypha, Б — Euglena, В — гипермастигида De-
vescovina. а — аксостиль, ж — жгутики, пс — псевдоподии, я — ядро.
Fig. 99. Agamic reproduction by binary fission in testacean amoeba
Euglypha (A), Euglena (Б), and hypermastigid Devescovina (B).
a — axostyle, ж — flagella, nc — pseudopodia, я — nucleus.
даже сотни) мелких дочерних клеток, которые обыч-
но в ходе дальнейшего роста и развития достигают
характерных для вида величины и строения.
Особую форму агамного размножения представля-
ет палинтомия (рис. 101). Это ряд последовательных,
обычно быстро друг за другом следующих делений
без промежуточного между ними периода роста. На-
ступлению палинтомии предшествует период роста,
иногда очень значительного; приступающая к раз-
множению особь получает название «томонт». Обра-
зующиеся в результате палинтомии мелкие клетки
(томиты) обычно являются расселительными стадия-
ми. Палинтомия в эволюции возникала, по-видимому,
независимо в разных группах простейших, например
у инфузорий, зеленых водорослей и некоторых сво-
бодноживущих жгутиконосцев. Между палинтомией
и другими формами множественного деления трудно
провести резкую границу.
Агамное размножение не всегда сопровождается
полным разъединением образующихся особей, кото-
рые могут оставаться связанными друг с другом. Это
приводит к образованию колоний различного строе-
ния. Связь между отдельными особями колонии может
быть непрочной и осуществляется посредством общей
160
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 100. Примеры бесполого размножения путем множественно-
го деления.
А — множественное неравномерное деление эпимастиготной формы Trypa-
nosoma, Б—Г — сйнтомия в процессе образования зооспор у динофита
Noctiluca. (А — по: Гинецинская, Добровольский, 1978; Б—Г— по: Doflein,
1953).
Fig. 100. Agamic reproduction by multiply fission.
A — multiply unequal division of epimastigote Trypanosoma, Б—Г — synto- у
my during the spore formation in dinophyte Noctiluca.
слизистой оболочкой. В других случаях эта связь
оказывается более прочной и осуществляется с по-
мощью цитоплазматических мостиков.
Половые процессы
Эволюционно половой процесс возник, по-види-
мому, независимо от функции размножения и лишь
позднее в некоторых группах простейших, а также
при переходе к многоклеточное™ половой процесс и
размножение оказались неразрывно связанными друг
с другом. Поэтому в отношении простейших было бы
неправильно говорить о существовании у них двух
форм размножения — бесполого и полового. Имеет-
ся много оснований утверждать, что половой процесс
в эволюции возникал многократно и независимо в
разных группах протистов. (Догель, 1951; Райков,
1978). Кроме того, и утрата его в разных таксонах
происходила независимо. Однотипность хода поло-
вого процесса у всех эукариотических организмов,
вероятно, есть результат конвергенции, а не монофи-
летического возникновения в филогенезе. Половой
процесс явился крупным ароморфозом, селективное
значение которого оказалось чрезвычайно велико. С
возникновением полового процесса связаны два био-
логических явления, играющих значительную роль в
эволюции. Во-первых, это диплоидность генома, до-
пускающая накопление рецессивных мутаций в попу-
ляции. Во-вторых — это повышение наследственной
изменчивости при редукции и кроссинговере. Хотя
Рис. 101. Формы палинтомии у протистов.
А — линейная палинтомия динофлагелляты Dissodinium lunula: 1 — материнская циста с одним ядром; 2 — циста после деления на 8 дочерних особей,
расположенных в 1 ряд. Б — табличная палинтомия у эвглениды Parastasia fennica: 1 — материнская особь; 2 — та же особь после деления на 2, 4 и
16 клеток. (А — по: Догель, 1951; Б — по: Wita, Sukhanova, 1983).
Fig. 101. Palintomy in protists.
A — linear palintomy in dinophyte Dissodinium lunula: ] — parental uninucleate cyst, 2 — cyst after division into 8 daughter cells. Б — table palintomy in eu-
glenid Parastasia fennica: 1 — parental cell; 2 — the same cell after division into 2, 4 and 16 cells.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
161
Рис. 102. Изогамная копуляция гамет зеленой водоросли Polytoma uvella. (По: Иванов и др., 1981).
1 — трофозоит, 2 — гамета, 3—5 — последовательные стадии слияния гамет, 6 — цитозигота.
Fig. 102. Isogamic copulation of green alga Polytoma uvella.
1 — trophozoit, 2 — gamete, 3—5 — consequentive stages of gamete fusion, 6 — zygocyst.
исследования 60—70-х гг. нашего века убедительно
показали, что рекомбинации компонентов генома
имеют место и на прокариотном уровне организации
клетки, этот параллелизм лишний раз подчеркивает
огромную биологическую прогрессивную роль обме-
на генетическим материалом, во много раз ускоряю-
щего процесс наследственной изменчивости.
Для полового процесса характерны 3 признака:
1) гаметоцитогамия — полное или частичное
слияние цитоплазмы половых клеток;
2) гаметокариогамия — слияние генеративных
ядер с образованием диплоидного ядра;
3) наличие мейоза на той или иной стадии жизнен-
ного цикла, приводящего к редукции числа хромосом.
У протистов половой процесс существует в двух
формах: копуляции и конъюгации.
Копуляция. Копуляция представляет собой про-
цесс слияния двух гаплоидных гамет и их ядер с об-
разованием диплоидной зиготы. Формы копуляции
очень разнообразны. Различают два основных типа —
гологамную и мерогамную копуляцию.
Гологамная копуляция. Это самая прими-
тивная форма полового процесса. Гаметы морфоло-
гически не отличаются от вегетативных клеток, и,
по-видимому, приобретение клетками способности к
копуляции определяется преимущественно условия-
ми среды. Примером гологамной копуляции может
служить половой процесс у зеленой водоросли Poly-
toma uvella (рис. 102), гаметы которой неотличимы
от вегетативных клеток.
Мерогамная копуляция. При мерогамной
копуляции гаметы развиваются из вегетативных кле-
ток, проходя процесс гаметогенеза, включающий раз-
множение и дифференцировку гамет. Этот тип копу-
ляции, в свою очередь, может быть поделен на две
группы: парную мерогамию и множественную мерога-
мию.
При парной мерогамии гаметы возникают из родо-
начальной вегетативной клетки (гаметоцита) в чис-
11 Протисты, часть 1
ле 2. Они дифференцированы на 2 морфологически
несхожие клетки, из которых одну можно рассматри-
вать как мужскую, вторую как женскую. Парная меро-
гамия наблюдается у многих жгутиконосцев — симби-
онтов кишечника питающихся древесиной тараканов.
Кроме жгутиконосцев эта форма полового процесса
свойственна солнечникам.
Формы множественной мерогамии у простейших
очень разнообразны. Общая линия эволюции шла не-
зависимо в разных крупных таксонах от изогамной
копуляции к различно выраженным формам анизога-
мии и завершилась оогамией. Изогамная копуляция
(когда копулирующие гаметы морфологически нераз-
личимы) имеет место у некоторых зеленых водоро-
слей и фораминифер. Разные степени анизогамии
(наличие микро- и макрогамет) наблюдаются у водо-
рослей. Заключительным звеном в эволюции от изо-
гамии к анизогамии является оогамия, когда женские
гаметы представлены неподвижными, богатыми ци-
топлазмой и резервными питательными веществами
яйцеклетками и подвижными микрогаметами, кото-
рые можно называть сперматозоидами. Оогамия име-
ется у наиболее сложно организованных водорослей,
а также споровиков.
Конъюгация и автогамия. Конъюгация — совер-
шенно своеобразный тип полового процесса, свойст-
венный только инфузориям (Ciliophora) и одной
группе зеленых водорослей (Conjugatophyceae). И в
том и в другом случае между конъюгирующими клет-
ками образуется цитоплазматический мостик, через
который осуществляется слияние ядер партнеров.
При этом у конъюгат, как, впрочем, и у некоторых
инфузорий, сливается частично или полностью ци-
топлазма. Поэтому, строго говоря, четкой границы
между копуляцией и конъюгацией провести нельзя.
Для инфузорий особенно характерно отсутствие
прямой связи полового процесса с размножением. В
конъюгацию вступают 2 особи, и в результате этого
процесса образуются также 2 особи, но обладающие
162
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
уже другим (гибридным) геномом. Число особей при
этом не увеличивается, но физиологическое состоя-
ние их меняется, а также изменяется генотип. Таким
образом, завершившая конъюгацию особь представ-
ляет собой в генетическом смысле новое поколение.
После соединения конъюгирующих особей диплоид-
ное генеративное ядро каждой из них — Ми — про7
делывает 2 мейотических деления, ведущих к редук-
ции числа хромосом. Из 4 гаплоидных -ядер обычно
3 ядра резорбируются в цитоплазме, а 1 претерпевает
третье деление (митоз), дающее начало 2 пронуклеу-
сам. Один из них мигрирует в клетку партнера и сли-
вается с его стационарным ядром. В результате этой
кариогамии в каждом из партнеров образуется дипло-
идное ядро — синкарион. Вскоре после его образо-
вания конъюганты (ставшие теперь эксконъюганта-
ми) расходятся. В дальнейшем синкарион делится
(число делений у разных видов различно), и часть
продуктов его деления преобразуется в Ма, другие
дают начало Ми. Что касается судьбы старого Ма, то
по ходу конъюгации он распадается на фрагменты и
постепенно резорбируется в цитоплазме.
У некоторых инфузорий наряду с конъюгацией
может происходить и самооплодотворение — автога-
мия. В отличие от конъюгации при этом не происхо-
дит временного соединения двух особей и все про-
цессы протекают в теле одной особи. Ход ядерных
процессов при автогамии такой же, как и при конъ-
югации. Важно отметить, что поскольку оба пронук-
леуса — суть продукты митотического деления одно-
го гаплоидного ядра, они генетически идентичны.
Следовательно, образующееся в результате их слия-
ния ядро по всем локусам будет гомозиготным.
Мейоз. Мейоз связан с двумя важнейшими биоло-
гическими функциями: во-первых, с уменьшением
вдвое числа хромосом (редукция), во-вторых, с ре-
комбинацией генного материала. Последнее происхо-
дит или на уровне целых хромосом в результате их
независимого распределения при мейотическом деле-
нии ядра, или в ходе кроссинговера, т. е. обмена
участками гомологичных (парных) хромосом.
Основные формы мейоза. Не рассматривая
детально цитологические особенности хода мейоза в
разных группах простейших, остановимся лишь на
главных его чертах.
У протистов встречаются 2 основные формы мей-
оза (рис. 103): с 2 делениями ядра (двухступенчатый)
и с одним делением (одноступенчатый).
Двухступенчатый мейоз наиболее схож с тем, что
имеет место у многоклеточных животных и растений.
Эта форма мейоза с различными модификациями
имеет место у многих простейших, у которых наблю-
дается половой процесс, — солнечников, форамини-
фер, инфузорий, некоторых зеленых водорослей и
опалин.
Мейоз с одним делением ядра (одноступенчатый)
протекает проще. В профазе гомологичные хромосо-
мы соединяются попарно, а затем, не расщепляясь,
отходят к полюсам ахроматинового веретена (рис. 103).
При одноступенчатом мейозе (в отличие от двухсту-
пенчатого) отсутствует обмен частями гомологичных
хромосом. Эта форма мейоза наблюдается у некото-
рых оксимонадид и гипермастигид, причем только у
тех, которые живут в кишечнике тараканов. Данные
о наличии одноступенчатого мейоза у споровиков и
динофитовых не подтвердились. И у тех и у других
обнаружены синаптонемальные комплексы, что сви-
детельствует о нормальной конъюгации хромосом
(Райков, 1995).
Обсуждая вопрос о происхождении и эволюции
мейоза, можно было бы считать одноступенчатый
мейоз примитивной исходной формой, на базе кото-
рой возник двухступенчатый мейоз. Это могло про-
изойти путем вовлечения в мейотический процесс од-
ного митотического деления. Однако в настоящее
время большинство исследователей придерживаются
мнения о вторичности одноступенчатого мейоза (Рай-
ков, 1995), встречающегося только у симбионтов дре-
воядных тараканов. У других оксимонадид и гипер-
мастигид имеет место двухступенчатый мейоз, отли-
чающийся, правда, некоторым своеобразием.
Предпосылкой к возникновению мейоза и редук-
ции могла явиться часто наблюдаемая при митозе тен-
денция гомологичных хромосом сближаться друг с
другом, образуя пары (биваленты). Этот процесс ши-
роко известен под названием конъюгации хромосом.
Он наблюдается не только у простейших, но и у мно-
гоклеточных животных и растений. Как уже упомина-
лось выше, мейоз и связанный с ним половой процесс
возникли независимо в разных стволах филогенети-
ческого дерева. Это находит свое подтверждение и в
многообразии форм мейоза^ простейших, хотя био-
логический смысл процесса остается неизменным.
Мейоз связан с соотношением гапло- и диплофа-
зы в жизненном цикле. По-видимому, у большинства
агамно размножающихся простейших имеется гапло-
идный (одинарный) набор хромосом в ядре и каждая
хромосома представлена в единственном числе. Весь-
ма вероятно, что среди агамных одноклеточных орга-
низмов имеются также полиплоидные формы с боль-
шим количеством гомологичных хромосом.
Место мейоза в жизненном цикле про-
стейших. Различают 3 основные формы соотноше-
ния гаплоидной и диплоидной фазы в жизненном
цикле простейших (рис. 104).
При зиготической редукции диплоидна лишь зиго-
та — продукт оплодотворения, слияния гамет. Уже
первое деление зиготы является мейозом, приводя-
щим к редукции числа хромосом. Все остальные
этапы жизненного цикла протекают в гаплоидной
фазе. Описанная форма соотношения гапло- и дипло-
фазы свойственна многим водорослям и гетеротроф-
ным жгутиконосцам, а также всем споровикам.
Гаметическая редукция является прямой противо-
положностью зиготической. Здесь все фазы жизнен-
ного цикла диплоидны, мейоз наблюдается при обра-
зовании гамет, которые являются единственной гап-
лоидной фазой в жизненном цикле. Гаметическая
редукция имеет место в жизненном цикле солнечни-
ков, инфузорий, некоторых жгутиконосцев.
Гетерофазное чередование поколений (споричес-
кая редукция) характеризуется закономерной сменой
диплоидной фазы гаплоидной. Типичный пример этого
явления демонстрируют фораминиферы. Из зиготы
развивается диплоидное поколение, размножающееся
агамно путем образования бесполых одноядерных
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
163
Рис. 103. Схема поведения хромосом в митозе (А), мейозе с двумя делениями ядра (К), мейозе с одним делением ядра (В), (По: Sleigh,
1989).
п — гаплоидное ядро, 2п — диплоидное ядро.
Fig. 103. Comparison of the behaviour of chromosomes in mitosis (A), two-division meiosis (£), one-division meiqsis (B),
n — haploid nucleus, 2n — diploid nucleus.
агамет. Их формирование происходит посредством
мейоза. Из гаплоидных агамет развивается следующее
поколение — гамонты. В конце жизни этого гапло-
идного поколения происходит формирование гамет
(без мейоза). Последние копулируют, и из зиготы раз-
вивается диплоидное поколение.
В ботанической литературе эту форму редукции
генетического материала принято называть споричес-
кой. Она предшествует появлению гаплоидного поко-
ления, развивающегося в виде гаплоидных спор, ко-
торые дают начало гаметофиту. Последний продуци-
рует гаметы, дающие в результате слияния зиготу, из
которой, в свою очередь, вырастает спорофит.
Одним из вариантов гетерофазного чередования
поколений можно считать соматическую редукцию,
характерную для зеленой водоросли Prasiola. Мейоз
происходит в вегетативных клетках диплоидного рас-
тения, где возникают участки гаплоидных клеток, да-
ющих впоследствии гаплоидные гаметы.
Другие формы генетических
взаимоотношений
Помимо различных форм полового процесса у
протистов встречаются и другие генетические взаи-
модействия клеток, при которых происходит обмен
целыми геномами, частями геномов или какими-либо
другими факторами, несущими генетическую инфор-
мацию (Серавин, Гудков, 1984). Такие явления следу-
ет называть агамными, так как они лишены одного
или двух признаков полового процесса, Они проис-
ходят в результате полного иди частичного, времен-
ного или постоянного слияния вегетативных клеток
(соматоцитогамия), при котором в одной цитоплазме
оказываются разнородные геномы (ядра) (рис. 105),
или даже в результате последующего слияния ядер
(соматокариогамия). Эти генетические взаимодейст-
вия обычно не связаны с размножением.
Соматоцитогамия. Естественные полные слияния
двух или нескольких вегетативных особей без после-
дующего слияния ядер описаны в разных таксонах
протистов (солнечники, лобозные и филозные амебы,
тауматомонады) и называются псевдокопуляцией. При
этом возникают временные двух-, трех- и многоядер-
ные клетки, которые затем фрагментируются на од-
ноядерные особи, генетически отличающиеся от ис-
ходных, так как каждая клетка содержит уже другое
ядро и цитоплазму. Какие именно генетические взаи-
модействия происходят при временном объединении
ядер в одной цитоплазме, в настоящее время неизвест-
но. Очевидно, однако, что псевдокопуляция приводит
к важным физиологическим перестройкам в резуль-
тате генетических взаимовлияний.
Различают 2 формы таких слияний: псевдокопу-
ляция, при которой полностью сливаются 2 (бинар-
164
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 104. Схема жизненных циклов протистов. (По: Sleigh, 1989).
а — агамный, б — с зиготической редукцией, в — со спорической редук-
цией (чередование поколений), г — с гаметической редукцией, к — конъ-
югация гамет, р — редукция числа хромосом. Одной линией обозначена
гаплофаза, двумя — диплофаза жизненного цикла.
Fig. 104. Four different patterns of life cycle in protists.
a — agamic and haploid, 6 — haploid with zygotic meiosis, в — haplodiplo-
id with intermedial meiosis and г — diploid with gametic meiosis, к — fertili-
sation, p — meiosis.
ная) или 3 (множественная) клетки, и плазмодиза-
ция — объединение множества клеток во временный
или постоянный плазмодий.
При неполном слиянии клеток (псевдоконъюга-
ции) также происходит временное объединение ци-
топлазмы двух или нескольких клеток с сохранением
их индивидуальности и последующим расхождением
тех же особей. Псевдоконъюгация известна у солнеч-
ников, филозных и лобозных амеб, апузомонад. Часто
она наблюдается при питании, когда 2 или несколько
особей захватывают одну крупную добычу и образуют
вокруг нее единую пищеварительную вакуоль (группо-
вое питание).
Значение генетических взаимовлияний при этих
процессах еще не изучено, однако ясно, что они зани-
мают важное место в жизни протистов. Опыты по под-
саживанию и удалению ядер у Amoeba proteus (Юдин,
1982) показывают, что у потомков этих амеб наблюда-
ется наследственная изменчивость по некоторым мар-
керным признакам. Можно предположить, что в пери-
од временного объединения ядер в одной цитоплазме
происходит обмен генетической информацией.
Соматокариогамия. Соматокариогамия встреча-
ется у протистов гораздо реже и еще менее изучена,
чем соматоцитогамия. Наиболее широко она рас-
пространена среди зигнемовых водорослей (класс
Conjugatophyceae), у которых процесс слияния гапло-
идных соматических клеток с последующим слия-
нием их ядер — весьма обычное явление. Подобное
явление также отмечается, хотя и редко, у несовер-
шенных грибов. Можно предположить, что соматока-
риогамия — довольно обычное для эукариот явление,
хотя наблюдается оно не часто.
Проблема происхождения
полового процесса
Существование спонтанного слияния клеток и их
ядер позволяет обсуждать и вопрос о возможном пути
появления полового процесса в эволюции. Для полной
картины становления полового процесса у протистов
не хватает фактически только одного признака типич-
ного полового процесса — мейоза. В этой связи инте-
ресны явления спонтанной гаплоидизации и сома-
тического (митотического) кроссинговера среди про-
тестов и некоторых эукариот. Так, диплоидные ядра
некоторых грибов (дейтеромицетов), образовавшиеся
в результате соматокариогамии, мшуг возвращаться в
исходное гаплоидное состояние в ряду митотических
делений. При этом с небольшой частотой встречается
и соматический кроссинговер, так как гомологичные
хромосомы могут конъюгировать попарно перед рас-
хождением в дочерние клетки. Так называемый ре-
комбинантный митоз, при котором происходит крос-
синговер между сестринскими хромосомами, встреча-
ется и в клетках многоклеточных животных. Таким
образом, в природе можно найти попытки перехода к
конъюгации сестринских хромосом при возврате^ к
гаплофазе, т. е. редукции числа хромосом.
Можно представить возникновение полового про-
цесса у протистов в виде эволюционной смены гапло-
фазных жизненных циклов (Серавин, Гудков, 1984).
1. Возникновение гетерокариотического жизнен-
ного цикла, при котором одна из стадий с 2 или не-
сколькими ядрами формируется в результате необра-
тимой соматоцитогамии.
2. Возникновение монокариотического жизненно-
го цикла, при котором появляются особи с диплоид-
Рис. 105. Схема основных типов и форм агамных слияний у простейших. (По: Серавин, Гудков, 1984).
I — псевдокопуляция: А — бинарная, Б — множественная; II — плазмодизация: А — временная, Б — постоянная; III — псевдоконъюгация: А — бинарная,
Б — множественная. I, А — бинарная псевдокопуляция у Euhyperamoeba fallax: 1 — сближение двух клеток, 2 — момент слияния, 3 — «объединенная»
амеба, 4 — разделение, 5 — образовавшиеся амебы; I, Б — множественная псевдокопуляция у Euhyperamoeba fallax: I — сближение и контакт трех клеток,
2 — момент слияния трех особей, 3 — «объединенная» амеба, 4 — разделение, 5 — образовавшиеся амебы. II, А — временная плазмодизация у ThaumatO’
monas lauterbomi: 1 — одиночные жгутиконосцы, 2 — агрегат из одноядерных особей, 3 — плазмодий, 4 — фрагментация плазмодия (деплазмодизация),
5 — образовавшиеся одиночные жгутиконосцы; II, Б — постоянная плазмодизация у Myxochrysis paradoxa: 1 — отдельные амебы, 2 — плазмодии, 3 —
цисты, 4 — зооспоры, 5 — образовавшиеся одиночные амебы. Ill, А — бинарная псевдоконъюгация у Actinophrys sp.: 1 — 2 отдельные особи, 2 — 2 особи,
связанные цитоплазматическим мостиком, 3 — те же особи, вновь обособленные; III, Б — множественная псевдоконъюгация у Euglypha rotunda: 1 — оди-
ночные особи, 2 — «розетка», 3 — вновь обособившиеся те же самые простейшие.
Fig. 105. Main types and forms of agamic fusion in protists.
I — pseudocopulation: A — binary, Б — multiply; Il — plasmodisation: A — temporary, Б — permanent; III — pseudoconjugation: A — binary, Б — multiply.
I, A — binary pseudocopulation in Euhyperamoeba fallax: 1 — cell attachment, 2 — fusion, 3 — «united» amoeba, 4 — fission, 5 — amoebae after fission; I, Б —
multiply pseudocopulation in Euhyperamoeba fallax: I — cell attachment, 2 — fusion of three cells, 3 — «united» amoeba, 4 — fission, 5 — amoebae after fission.
II, A — temporary plasmodisation in Thaumatomonas lauterborni: 1 — single flagellates, 2 — aggregation of inunucleate cells, 3 — plasmodium, 4 — plasmodium
fragmentation (deplasmodisation), 5 — single flagellates; II, Б — permanent plasmodisation in Myxochrysis paradoxa: 1 — single amoebae, 2 — plasmodia, 3 —
cysts, 4 — zoospores, 5 — single amoebae. Ill, A — binary pseudoconiugation in Actinophrys sp.: I — two separate cells, 2 — the same cells being connected with
cytoplasmic bridge, 3 — the same cells after separation; III, Б — multiply pseudoconjugation in Euglypha rotunda: 1 — single cells, 2 — «socket», 3 — separated cells.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
165
1
2
3
166
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
ными ядрами, образовавшимися при соматокариога-
мии.
3. Формирование гаплофазного цикла с зиготи-
ческой редукцией на основе спонтанной гаплоидиза-
ции и митотической рекомбинации.
Конечно, эта схема весьма гипотетична, однако
она указывает вполне вероятный путь становления
полового процесса у протистов. Если соматоцито-
гамия — широко распространенное явление среди
протистов, то соматокариогамия отмечается крайне
редко, что объясняется также и очень слабой изучен-
ностью этого явления. Наиболее слабо аргументи-
рован третий этап этого процесса, который прак-
тически не изучен у протистов. По-видимому, весьма
вероятно (и даже более вероятно) возникновение не
гаплофазного, а гетерофазного чередования поколе-
ний. Спонтанная гаплоидизация происходит (там, где
она отмечена) не сразу после соматокариогамии, а в
ряду поколений. Таким образом, какое-то время дол-
жен существовать ряд организмов с диплоидным ядром,
прежде чем популяция сможет вернуться в гаплоид-
ную фазу.
Так или иначе, большинство современных иссле-
дователей считают, что становление типичного поло-
вого процесса имело место в пределах протистов и
происходило, по-видимому, неоднократно и независи-
мо в разных группах, а возможно, и разными путями.
Органоиды
и формы движения
Формы движения простейших разнообразны, чему
соответствует и многообразие органоидов движения.
Как правило, различают 5 основных форм движе-
ния — амебоидное, жгутиковое, ресничное, метабо-
лирующее и скользящее. Кроме того, существуют и
некоторые более специализированные формы двига-
тельной активности. Жгутиковое и ресничное движе-
ния были рассмотрены ранее в связи со строением и
функцией жгутика/реснички, поэтому здесь их описа-
ние не приводится.
Амебоидное движение. Оно связано с формиро-
ванием псевдоподий, хотя само по себе образование
псевдоподий не всегда обусловливает движение, а
может быть связано с фагоцитарной активностью.
Различают несколько форм псевдоподий (рис. 71):
широкие с тупым концом лобоподии, тонкие невет-
вящиеся филоподии, тончайшие анастомозирующие
цитоплазматические выросты — ретикулоподии, или
ризоподии, и, наконец, тонкие нитевидные неанасто-
мозирующие аксоподии. Скелет лобоподий и фило-
подий представлен, как правило, актиновыми микро-
филаментами, внутри ретикулоподий обнаруживают-
ся одиночные неупорядоченные микротрубочки, а
осевой скелет аксоподий состоит из хорошо органи-
зованных микротрубочек. Эти существенные разли-
чия в строении псевдоподий связаны с различными
механизмами движения клетки. Например, солнечни-
ки используют для передвижения механизм сбор-
ки/разборки микротрубочек, в результате чего одни
аксоподии удлиняются, другие — укорачиваются, а
сама клетка перекатывается по субстрату. Некоторые
протесты эффективно используют аксоподии в ка-
честве гребных пластинок, активно передвигаясь в
толще воды.
Ретикулоподии практически не используются для
передвижения. Чаще всего они образуют ловчую сеть
и захватывают мелкие организмы для питания.
Филоподии наряду с лобоподиями чаще других
псевдоподий используются для движения. Но и здесь,
несмотря на их большое сходство в строении, меха-
низмы движения могут быть совершенно различны-
ми. Механизм собственно амебоидного движения пы-
тались выяснять в многочисленных исследованиях,
которые и в настоящее время еще далеки от заверше-
ния.
При амебоидном движении клетки могут образо-
вывать 1 псевдоподию или несколько, активно меняя
форму тела. Другие амебы могут передвигаться до-
вольно быстро, практически не изменяя формы тела.
Считается, что контакт с субстратом — необходимое
условие амебоидного движения. Наиболее популярна
в настоящее время гипотеза генерализованного сокра-
щения субмембранного корсета из микрофиламентов,
которая детально рассматривается в главе, посвящен-
ной лобозным амебам. Заметим только, что эта гипо-
теза объясняет только часть данных по амебоидному
движению. В настоящее время уже многие исследова-
тели приходят к мысли, высказанной Л. Н. Серавиным
более 30 лет назад (Серавин, 1967), что не существует
единого механизма амебоидного движения.
Метаболирующее движение. Метаболирующий
тип движения был выделен сравнительно недавно
(Серавин, Фролов, 1983). Он характерен для эвглено-
вых водорослей и осуществляется путем активного
скольжения белковых полос кутикулы относительно
друг друга. Внешне такое движение выглядит как пе-
ристальтические или волнообразные изгибания тела
клетки (рис. 106), за счет которых безжгутиковые
формы могут активно передвигаться.
Скользящее движение. Строго говоря, скользя-
щее движение характерно лишь для немногих специ-
ализированных групп протистов. Прежде всего это
диатомовые водоросли, лабиринтулы и грегарины.
Механизмы скользящего движения у них, по-видимо-
му, различны. Если скольжение грегарин по субстра-
ту осуществляется за счет волнообразных движений
их продольных складок, то внешне похожее движе-
ние клеток из двух других групп объяснить труднее.
У диатомовых водорослей большую роль в этом про-
цессе играет выделяемая ими слизь, а у веретеновид-
ных клеток лабиринтул существует, по-видимому,
иной механизм скольжения.
Захват пищи, питание
и осморегуляция
Дня протистов характерны 3 способа питания: ав-
тотрофный, гетеротрофный и миксотрофный. Чис-
тых автотрофов, по-видимому, нет, так как все про-
тесты, имеющие хлоропласты, весьма требовательны
к РОВ, т. е. используют их для питания наряду с
энергией солнечных лучей. Гетеротрофные протесты
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
167
Рис. 106. Изменения формы тела при метаболическом движении
у эвглениды Parastasia fennica. (По: Вита, Суханова, 1983).
Fig. 106. Metabolic movement of euglenid Parastasia fennica.
способны поглощать РОВ при помощи мембранного
транспорта и пиноцитоза (или рецепторного эндоци-
тоза), а также оформленные частицы путем фагоци-
тоза, или голозойного питания (рис. 107—110).
Пийоцитоз. Его удается обнаружить главным об-
разом при электронно-микроскопическом исследова-
нии. Это свойственный многим простейшим способ
заглатывания жидкой пищи, хотя возможно и погло-
щение мелких частиц.
Простейший вариант пиноцитоза заключается в
образовании небольшой ямки на поверхности плаз-
малеммы и последующего отшнуровывания неболь-
шого пузырька. Часто этот участок плазмалеммы под-
стилается сеточкой микрофиламентов белка клат-
рина. В клеточной биологии принято называть это
явление рецепторным эндоцитозом, при котором
формируются окаймленные ямки, замыкающиеся затем
в окаймленные пузырьки. Рецепторный эндоцитоз
детально исследован у клеток многоклеточных жи-
вотных. В настоящее время он обнаружен и среди
протистов.
Наиболее характерен для протистов более слож-
ный вариант пиноцитоза. На месте небольшого впя-
чивания плазмалеммы по мере его дальнейшего уг-
лубления формируется тонкий канал, от дна которо-
го и происходит отшнуровывание мелких пузырьков.
У амебоидных организмов эти каналы являются вре-
менными образованиями, а у многих паразитических
жгутиконосцев и некоторых автотрофных протистов
эти каналы имеют постоянное местоположение (на-
пример, в стенке жгутикового кармана). В последнем
случае эти пиноцитозные каналы рассматриваются
как редуцированный цитостом.
При пиноцитозе не требуется значительного раз-
вития цитоскелетных структур, которые совершенно
необходимы для голозойного питания — фагоцитоза.
Фагоцитоз. Он представляет собой заглатывание
достаточно крупных оформленных частиц. У амеб нет
специальных участков, где может происходить загла-
тывание пищи, а у более сложно организованных про-
тистов дифференцируются специальные, иногда очень
сложно устроенные аппараты, служащие для улавли-
вания и заглатывания пищи. Наиболее сложного раз-
вития достигают эти аппараты у гетеротрофных жгу-
тиконосцев и инфузорий (рис. 108). Обычной пищей
свободноживущих протистов служат бактерии, дрож-
жи, органический детрит. Многие протисты исполь-
зуют для захвата и заглатывания пищи различные при-
способления (рис. 109; 110). Среди гетеротрофных форм
немало хищников, преследующих и нападающих на
других простейших. Известны случаи, когда добыча
по размерам в несколько раз превосходила хищника
(рис. 108).
Отыскание и захват пищи связаны со сложным
пищевым поведением протистов. Среди различных
объектов наружной среды они активно выбирают пи-
щевые объекты и фагоцитируют их. Это в основном
связано с восприятием химических сигналов, исходя-
щих от пищевого объекта.
Сложную ловчую и пищеварительную сеть обра-
зуют ризоподии фораминифер. Специальные ловчие
аппараты в форме щупалец, выделяющих липкий
секрет, который служит для улавливания живой до-
бычи, имеются у сосущих инфузорий. Многообра-
зие способов захвата и поглощения пищи, так же
как и характера самой пищи, у протистов необы-
чайно велико.
Заглоченная пища оказывается обычно окружен-
ной вакуолью, отграниченной от цитоплазмы мембра-
ной. Внутрь пищеварительных вакуолей из цитоплаз-
мы поступают ферменты лизосом, действие которых
определяет ход пищеварения.
Выбрасывание непереваренных остатков пищи
(дефекация) у многих амеб и некоторых жгутиковых
может осуществляться в любой точке поверхности
тела. У простейших, обладающих сложным цитоске-
летом, имеется специальный, фиксированный на по-
верхности тела участок (цитопиг), где происходит де-
фекация.
Мизоцитоз. Одной из разновидностей фагоцитоза
является мизоцитоз, или высасывание жертвы. Этот
тип питания характерен для сосущих инфузорий и
многих хищных жгутиконосцев. Так, представители
рода Colpodella (=Spiromonas) могут нападать на па-
рамеций, пробивать рострумом их пелликулу и выса-
сывать их содержимое.
Осморегуляция. Регуляция осмотического давле-
ния актуальна для протистов, живущих в пресных
водах. Они вынуждены выводить наружу избыток
жидкости, постоянно поступающей извне в результа-
те перепада осмотического давления. Для этой цели
используются сократительные вакуоли, обладающие
у разных протистов различной степенью сложности.
Следует, однако, заметить, что сократительные ваку-
оли есть и у солоноватоводных и даже морских форм.
Поэтому в целом комплекс сократительной вакуоли
выполняет, по-видимому, и другие функции (напри-
мер, экскреторную или функцию водообмена).
В наиболее простом варианте работа сократитель-
ной вакуоли выглядит следующим образом (рис. 111):
избыток жидкости заполняет сначала мелкие пузырь-
168
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 107. Схема последовательных стадий фагоцитоза у амебы. (По: Hausmann, Hulsmann, 1996).
п — пищевой объект.
Fig. 107. Consequentive stages of phagocytosis in amoeba.
n — food particle.
Рис. 108. Схема последовательных этапов заглатывания парамеции (и) хищной инфузорией Didinium (д). (По: Hausmann, Hulsmann,
1996).
Fig. 108. Consequentive stages of phagocytosis of Paramecium (n) by predator ciliate Didinium (d).
Рис. 109. Схема захвата при помощи гаптонемы и заглатывания пищи на заднем конце клетки гаптофитом Chrysochromulina hirta.
(По: Hausmann, Hulsmann, 1996).
1—14 — последовательные этапы захвата и заглатывания пищевого объекта. Круговые стрелки указывают направление вращения клетки, прямые стрел-
ки — движение частиц вдоль гаптонемы, б — бактерии, г — гаптонема, ж — жгутики.
Fig. 109. Consequentive stages {1—14} of phagocytosis in haptophyte Chrysochromulina hirta.
6 — bacteria, г — haptonema, ж — flagella.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
169
Рис. 110. Схема образования пищевой чашечки при участии мик-
ротрубочек жгутиковых корешков у бесцветной хризомонады.
(По: Andersen, Wetherbee, 1992).
ж — передний жгутик с мастигонемами, мт — микротрубочки, окружаю-
щие цитостом (ц). Стрелкой показано место скольжения микротрубочек
относительно друг друга при расширении цитостомальной ямки для загла-
тывания бактерии (б).
Fig. 110. Participation of microtubular rootlets in food cup formati-
on during phagocytosis of colourless chrysophyte.
ж — anterior flagellum with mastigonemes, мт — microtubules around cyto-
stome (ц), arrow shows a place of microtubule sliding along each other to form
a cytostomal cavity.
ки, которые затем сливаются в одну крупную ваку-
оль. Вакуоль сокращается, выводя содержимое нару-
жу, и весь процесс повторяется. У инфузорий этот
комплекс более сложен. Он образован специальны-
ми, подводящими к вакуоли каналами, которые ук-
реплены микротрубочками и микрофиламентами.
Постоянная система каналов (пузула), в которой нет
сократительной вакуоли, характерна для пресновод-
ных динофитовых (рис. 112).
Во всех случаях, несмотря на кажущуюся простоту
работы осморегуляторной системы, она очень сложна.
В ней участвуют многие системы клетки (прежде всего
цитоскелет), взаимодействие которых до конца не вы-
яснено.
Поведение, реакция на
внешние раздражения
На различного рода внешние раздражения свобод-
ноживущие протесты чаще всего реагируют измене-
нием направления движения. Эти реакции носят на-
звание таксисов и кинезов. Последние отличаются от
первых не направленным, а флуктуирующим движе-
нием к раздражителю или от него. В зависимости от
характера раздражения различают следующие основ-
ные формы таксисов.
Фототаксис — реакция на освещение. Автотроф-
ные протесты обычно имеют положительный фото-
таксис. Они направляются к источнику света. Однако
при слишком ярком освещении фототаксис может
стать отрицательным. Для большинства гетеротроф-
ных протистов характерен отрицательный фототак-
сис.
Хемотаксис. У протистов обычно развита чувст-
вительность к химическим раздражителям. Они либо
удаляются от определенных (чаще вредных) веществ,
либо направляются к ним. К этой же категории хемо-
таксических воздействий относятся сближение и кон-
такты особей или гамет при половых взаимодействи-
ях — копуляции или конъюгации, что связано с вы-
делением особых веществ — гамонов, химическая
природа которых еще плохо изучена.
Механотаксис. Эта реакция включает 2 формы —
тигмотаксис и реотаксис. Положительный тигмотак-
сис выражается в стремлении простейшего к механи-
ческому контакту с предметами окружающей среды,
отрицательный — к активному уходу от таких кон-
тактов. Отрицательный реотаксис — движение про-
стейшего в направлении тока жидкости, в которой
оно обитает, положительный — движение навстречу
току жидкости. Все эти двигательные реакции играют
Рис. 111. Цикл пульсации сократительной вакуоли у криптомонады Chilomonas sp. (По: Догель, 1951).
1 — систола, 2 — диастола.
Fig. 111. Pulsative cycle of contractile vacuole in cryptomonade Chilomonas sp.
1 — syslole, 2 — diastole.
170
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 112. Схема каналов пузулы у динофлагеллаты Oodinium.
(По: Hausmann, Hulsmann, 1996).
ок — общая камера, ск — собирающие каналы, тп — текальные пластин-
ки.
Fig. 112. Scheme of pusula structure in dinoflagellate Oodinium.
ok — common chamber, ск — collecting channels, mn — thecal plates.
существенную роль в поведении простейших в естест-
венной среде обитания.
Геотаксис (гравитаксис). Для некоторых протис-
тов доказана их двигательная реакция в гравитацион-
ном поле в отношении направления силы тяжести.
Чаще геотаксис оказывается отрицательным — про-
тисты движутся в направлении, противоположном
вектору силы тяжести. Однако эта реакция очень из-
менчива и во многом зависит от физиологического
состояния клетки.
Термотаксис. Движение в направлении оптималь-
ной температуры при наличии температурного гради-
ента среды свойственно многим протистам. Нужно
иметь в виду, что понятие «оптимальная температу-
ра» в значительной степени условно, ибо оптимум
легко смещается в зависимости от предшествующего
температурного режима.
Гальванотаксис. В электрическом поле большин-
ство протистов обнаруживают определенную прост-
ранственную ориентировку. Например, особи Para-
mecium при наличии постоянного электрического
тока направляются прямо к катоду, где и образуют
скопление. Некоторые другие организмы (например,
Opalina, Chilomonas), напротив, движутся к аноду.
Причина этих различий неясна и требует дальнейших
исследований.
Жизненные циклы
Характерной чертой многих таксонов простей-
ших является наличие у них жизненных циклов, со-
провождающихся сменой морфологических форм, а
нередко и среды обитания, характера метаболизма.
Под термином «жизненный цикл» понимают сово-
купность следующих друг за другом этапов развития,
заключенных между произвольно взятыми стадиями.
Одна из них принимается за начальную и повторяет-
ся в каждом цикле. У многоклеточных считается, что
цикл начинается с оплодотворения яйцеклетки и за-
вершается достижением половой зрелости. Дня про-
тистов такое общее определение принять нельзя, ибо
у них имеются таксоны, в пределах которых полово-
го процесса нет. Однако у многих видов, обладающих
половым процессом, начальным этапом жизненного
цикла (аналогично многоклеточным) можно считать
образование зиготы. Жизненный цикл протистов сла-
гается из ряда клеточных поколений. Следовательно,
в пределах жизненного цикла имеется несколько кле-
точных циклов.
Жизненные циклы осуществляются в конкретных
условиях среды. Для паразитических протистов, об-
ладающих особенно сложными жизненными цикла-
ми, такой средой является организм хозяина. Измене-
ния условий внешней среды оказывают существенное
влияние на ход жизненного цикла. Вероятно, для
более точной характеристики последующих стадий
развития простейшего следует принять понятие цикл
развития, а реализацию его в конкретных условиях
среды именовать жизненным циклом. Необходимо
отметить, что в указанном смысле жизненные циклы
протистов (как свободноживущих, так и паразитичес-
ких) еще плохо изучены. Лучше других исследованы
в этом плане паразитические формы, в отношении же
свободноживущих (радиолярии, инфузории) эти све-
дения достаточно фрагментарны.
В ходе жизненных циклов протистов при наступле-
нии определенных условий формируются покоящиеся
стадии — цисты, что имеет место как у свободноживу-
щих, так и у паразитических форм. У последних ин-
цистирование обычно бывает связано с выходом из хо-
зяина в окружающую среду и с заражением новых осо-
бей хозяина.
У протистов, обладающих сложной организацией
(акантарии, солнечники, инфузории, некоторые жгу-
тиконосцы) при образовании цист имеет место дедиф-
ференцировка. Ресничные образования, скелет, орга-
ноиды захвата и приема пищи и другие, связанные с
активной жизнью, резорбируются, а иногда отбрасы-
ваются. Обычно происходит обезвоживание и уплотне-
ние цитоплазмы, исчезают вакуоли, в том числе пище-
варительные. Вокруг тела простейшего выделяются за-
щитные оболочки. Их, как правило, бывает не менее 2:
наружная — экзоциста и внутренняя — эндоциста. Чис-
ло защитных оболочек может быть и более 2. Обладая
слабой проницаемостью, оболочки служат не только ме-
ханической, но и химической защитой особи. В инцис-
тированном состоянии простейшие могут сохранять
жизнеспособность месяцами и даже годами, цисты
могут иметь различное физиологическое назначение.
Цисты покоя (рис. 113) служат для перенесения небла-
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
171
Рис. 113. Цисты покоя некоторых протистов.
А — Spumella termo, Б — Euglena geniculata, В — Ochromonas crenata,
Г— O.fragilis, Д — Chilomonas paramecium, E — Ceraium cornutum, Ж —
Chilomastix bocis, 3 — Lophomonas striata. (A — no: Hdnel, 1979; Б — ориг.
К. M. Сухановой; В—3 — по: Doflein, 1953).
Fig. 113. Cysts of the rest of some protists.
В разных группах паразитических протистов об-
разование защитных оболочек обычно связано с вы-
ходом паразита во внешнюю среду. Образование цист
может происходить на разных стадиях цикла. В одних
случаях инцистируется зигота, в других — вегетатив-
ная форма, как у паразитических кишечных амеб,
инцистирование которых не связано с размножени-
ем. Их цисты выполняют функции защиты и расселе-
ния.
В целом следует подчеркнуть, что способность
многих простейших формировать стадии покоя, ко-
торые обладают высокой стойкостью к неблагоприят-
ным воздействиям среды, представляет важную адап-
тацию, направленную на сохранение и распростране-
ние вида.
Экология
Экология протистов в настоящее время является
одним из самых существенных разделов общей эко-
логии. Протисты распространены всесветно и населя-
ют все возможные среды обитания, уступая по чис-
ленности только бактериям в составе биомов суши,
моря и пресноводных бассейнов. Преобладающее их
большинство — свободноживущие организмы, хотя
гоприятных условий среды — чаще всего они позволя-
ют простейшему переносить высыхание в водоеме или
почве. Широко распространены у многих свободно-
живущих простейших цисты размножения (рис. 114).
Например, некоторые инфузории и гетеротрофные
жгутиконосцы перед делением инцистируются, а после
завершения деления снова эксцистируются и перехо-
дят к свободному существованию. При образовании
цист размножения оболочки их никогда не бывают
столь мощными и прочными, как в цистах покоя.
Обычно в цистах размножения образуется не 2, а боль-
шее количество дочерних особей, так что бесполое
размножение в цистах носит характер палинтомии.
Существует еще одна категория цист, которые
можно назвать реорганизационными. Например, опи-
сан один случай реорганизации ядерного аппарата в
цистах брюхоресничной инфузории Oxytricha hyme-
nostoma (Помряскинская, 1940). Эта инфузория обла-
дает 2 Ма и 2 Ми. В цистах оба Ма и оба Ми слива-
ются в одно ядро, благодаря чему возникает тетрап-
лоидное ядро. Мейоз при этом отсутствует, так что
описанные ядерные преобразования нельзя рассмат-
ривать как половой процесс. В дальнейшем при эк-
сцистировании ядро опять делится и восстанавлива-
ются обычные для данного вида 2 Ма и 2 Ми<
Описаны и другие случаи, когда процесс инцисти-
рования связан с питанием. Так, например, хищная
инфузория Amphileptus claparedi, питающаяся, глав-
ным образом, кругоресничными колониальными ин-
фузориями, наползает на свою неподвижную жертву,
заглатывает ее, после чего инцистируется и в таком
состоянии переваривает добычу. После завершения
пищеварения Amphileptus эксцистируется и перехо-
дит к активно подвижному образу жизни.
Рис. 114. Цисты размножения протистов.
А — деление Chlamydomonas braunii в состоянии пальмеллы, Б — деление
Polytoma в состоянии цисты, В — деление Euglena gracilis в условиях куль-
туры. Стрелки показывают направление процесса размножения. (А—Б —
по: Иванов и др., 1981; В — ориг. К. M. Сухановой).
Fig. 114. Cysts of the reproduction of prQtists.
A — division of Chlamydomonas braunii in palmelloid state, Б — division
of Polytoma in cyst, В — division of Euglena gracilis.
172
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
многие виды протистов являются симбионтами чело-
века, животных и растений. Во всех природных эко-
системах протесты входят в состав сообществ (био-
ценозов) наряду с животными, растениями, грибами и
другими организмами, популяции которых сосущест-
вуют и взаимодействуют в пространстве и времени.
Первые исследования протистов, имеющие четко
выраженную экологическую направленность, отно-
сятся к концу XIX—началу XX столетия (Мережков-
ский, 1878; Метальников, 1911; Woodruff, 1912; Ефи-
мов, 1922; Гаузе, 1933, 1934, 1940, и др.). Особенно
существенный вклад в изучение экологии протистов
был внесен в последние 50 лет (Faure-Fremiet, 1950;
Догель, 1951; Stout, 1955; Полянский, 1957; Чорик,
1968; Суханова, 1968; Finley, 1969; Sladedek, 1963;
Мамаева, 1979; Бурковский, 1984; Fenchel, 1987;
Толмашева, 1990; Жуков, 1993; Атлас и определи-
тель пелагобионтов Байкала, 1995; Никитина, 1997;
Лихачёв, 1997; Раилкин, 1998; Жариков, 1999).
Изучение взаимодействий протистов с окружаю-
щей средой включает 3 основных уровня: экологию
особей, популяций и сообществ, в состав которых
протесты входят в качестве одной из многочислен-
ных облигатных групп. Населяя любой природный
биотоп, протесты ведут активный образ жизни толь-
ко в водной среде: это одна из главных особенностей
их экологии. К числу других специфических свойств
протистов относятся их микроскопические размеры
(что представляет известные трудности при изучении
экологии), высокая скорость размножения, сложные
жизненные циклы у многих видов, широкая моди-
фикационная изменчивость, часто встречающееся
расселение по микробиотопам. Необходимо отметить
также, что экология протистов находится в настоя-
щее время на самом раннем этапе ее изучения и до
сих пор в некоторых комплексных экологических ис-
следованиях мнргие группы протистов не учитыва-
ются.
Сообщества протистов
морей и океанов
В сравнении с другими средами обитания фауна
морских свободноживущих протистов — самая бога-
тая по видовому составу. Некоторые крупные таксо-
ны представлены только морскими организмами (фо-
раминиферы, радиолярии). Особое значение для мор-
ских организмов имеет соленость морской воды,
которая в большой мере определяет высокую степень
эндемизма морской фауны и отличает ее от фауны
пресных вод (Хлебович, 1974). По соотношению
ионов морская вода довольно близка к жидкостям
внутренней среды живых организмов, поэтому про-
тесты, как и множество других обитателей морей,
изоосмотичны по отношению к этой среде. Одной из
форм адаптаций к соленостному фактору может слу-
жить отсутствие у морских протистов активно функ-
ционирующих сократительных вакуолей — органелл
осморегуляции.
Протесты представляют многочисленную облигат-
ную группу организмов, населяющих две основные
экологические зоны морей и океанов: толщу воды —
пелагиаль и дно — бенталь (рис. 115). Вертикальную
экологическую зональность определяют изменения
абиотических и биотических условий существования
организмов, направленные от поверхности в глубину
и от прибрежных частей водного бассейна к откры-
тым. Выражена также горизонтальная зональность,
которая определяет различия в распределении сооб-
ществ организмов в прибрежных, эвтрофных, и в от-
крытых, удаленных от берега, олиготрофных зонах.
Протесты обладают разнообразными формами мор-
фологических и физиологических адаптаций к обита-
нию в каждой из этих зон морских бассейнов.
Протесты пелагиали. Для пелагических протис-
тов характерны приспособления, которые увеличива-
ют их флотационные способности, а также активные
движения в толще морской воды. К таким морфофи-
зиологическим адаптациям относятся разнообразные
выросты поверхности раковин в форме полых шипов
и радиально расположенных игол у фораминифер
(глобигеринаты), радиолярий, динофитовых (Cerati-
ит) и кокколитофорид. Эти приспособления увели-
чивают площадь поверхности тела, одновременно
уменьшая его удельную массу (рис. 116).
Свободному парению в морской воде способствуют
также отверстия во внутреннем скелете радиолярий и
их аксоподии, расположенные радиально. Такой же
адаптивный признак представляет собой медузовидная
форма тела ряда пелагических протистов, например
жгутиковых Leptodiscus и Medusochloris (рис. 116).
Ряд видов тинтиннид, активно передвигающихся в
воде, обладает тонкой прозрачной раковиной, имею-
щей длинную и узкую торпедообразную форму (До-
гель, 1951; Fenchel, 1987).
Флотационная способность радиолярий, динофи-
товых и ряда других протистов пелагиали значитель-
но увеличивается за счет вакуолизации цитоплазмы, а
также благодаря липидам, накапливающимся в ци-
топлазме в форме капель. Плавучесть планктонных
фораминифер (роталиаты) обеспечивается тончай-
шими ретикулоподиями диаметром менее 1 мкм, ко-
торые анастомозируют друг с другом и образуют во-
круг раковины густую сеть, снижая удельную массу
тела. Механизмом, регулирующим плавучесть фора-
минифер в планктоне на уровне клетки, служит уни-
кальная система криптосом и микровиллей цитоплаз-
мы (везикулярный ретикулум), которая способствует
изменению удельной массы тела путем замещения в
их структуре легких ионов хлоридов тяжелыми иона-
ми сульфидов. Бифонтинальная стенка раковин пе-
лагических роталиат обеспечивает легкость и проч-
ность самой раковины при малой ее толщине (Миха-
левич, 1992).
По длине тела пелагические протесты разделяются
на 3 группы: нанопланктон 2.0—20.0 мкм, микро-
планктон 20.0—200.0, мезопланктон 200.0 мкм и более.
К нанопланктону относится преобладающее число
видов гетеротрофных жгутиковых. Нанопланктон
морей разных широт представлен большим разнооб-
разием видов: например, в Черном море их известно
37, в Беринговом 19 (Моисеев, 1983), в Баренцевом 24
(Мыльников, Жгарев, 1984), в Японском 15 (Жуков,
1983). Особенно широким разнообразием видов жгу-
тиковых отличается нанопланктон шельфовой зоны:
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
173
Рис. 115. Основные экологические группы протистов. (По: Haus-
mann, Hulsmann, 1996).
1 — бентосные, 2 — обрастатели, 3 — эпибионты, 4 — интерстициаль-
ные, 5 — планктонные, 6 — нейстонные, 7 — водные, 8 — почвенные,
9 — эпифиты, 10 — эндофитобионты, 11 — эндозообионты.
Fig. 115. Main ecological groups of protists.
1 — benthos, 2 — periphyton, 3 — epibionts, 4 — interstitial, 5 — plank-
ton, 6 — neuston, 7 — water, 8 — soil, 9 — epiphytes, 10 — endophytobi-
ons, 11 — endozoobionts.
много видов хризофитовых из родов Paraphysomonas,
Ochromonas, Spumella (Monas), Pseudobodo, кинето-
пластид рода Bodo и мелких эвгленид. Численность
нанопланктона в шельфовой зоне достигает в среднем
1000 клеток/мл, но уменьшается с нарастанием глуби-
ны.
Жгутиковые протисты представляют важную группу
первичных консументов, с которых начинаются пи-
щевые цепи и сети многих протистов микро- и мезо-
планктона, а также мелких многоклеточных живот-
ных.
Микропланктон состоит из многих видов инфузо-
рий и динофитовых, эвгленид и диатомовых водоро-
слей. Для морского мезопланктона обычны также
крупные по размерам динофитовые. Это прежде
всего ночесветка, Noctiluca miliaris, достигающая в
диаметре 2 мм и распространенная преимущественно
в теплых морях (Догель, 1951). Ночесветка — гете-
ротрофный вид, пищу ее составляют протисты мик-
ропланктона, личинки морских беспозвоночных самых
мелких размеров, яйца ракообразных и рыб. Ноче-
светка обладает свойством биолюминесценции и, об-
разуя плотные скопления в поверхностных слоях
воды, вызывает наряду с другими динофитовыми и
бактериями свечение моря, особенно яркое в вечер-
ние и ночные часы. Из самых крупных известных
морских видов динофитовых можно назвать Protope-
ridinium claudicans и Р. divergens, размеры которых
составляют около 5 см. Оба вида обнаружены в от-
крытой части Черного моря, где популяции их осо-
бенно многочисленны. Инфузории в составе морско-
го мезопланктона встречаются достаточно редко.
Бентосные протисты. Сообщество бентосных
протистов Мирового океана не менее богато видами,
чем сообщество пелагиали. В состав его входят фора-
миниферы, инфузории, эвгленйды, кинетопластиды,
голые и раковинные амебы.
В состав морского бентосного сообщества входит
многочисленная группа протистов, состоящая из ин-
фузорий, эвгленид, кинетопластид, динофитовых,
хризофитовых, бикозоецид и амеб (рис. 117), но их
экология почти не изучена. В большинстве своем
жгутиковые протисты — неспецифические обитате-
ли бентоса, они входят в качестве облигатного звена
в пищевые цепи радиолярий, фораминифер, инфузо-
рий и мелких беспозвоночных, обитающих на поверх-
ности и в толще морских донных осадков.
Сообщества пресноводных
протистов
Пресноводные бассейны характеризуются широ-
ким разнообразием условий обитания: варьирует их
глубина, скорость течения воды, ее pH, содержание
газов (О2, СО2, H2S), органических веществ автохтон-
ной и аллохтонной природы.
Число видов протистов в пресных водоемах зна-
чительно меньше, чем в морских. Особенностью прес-
ных водоемов является отсутствие четких различий
между вертикальными зонами; соответственно нет
особой разницы между планктонными и бентосными
видами, отсутствует специфичная для морей интер-
стициальная фауна инфузорий. Тем не менее в круп-
ных озерах (Байкал, Ладожское, Онежское и др.)
можно достаточно отчетливо различить вертикаль-
ную зональность и выделить планктонное и бентос-
ное сообщества протистов (Гаевская, 1933).
Планктонное сообщество. Протозойный нано-
планктон представлен преимущественно гетеротроф-
ными жгутиковыми, среди которых много мелких эв-
гленид, хризофитовых, воротничковых, криптомона-
довых, бикозоецид, бодонид. Часто они образуют
скопления у поверхностной пленки воды, где концен-
трируются бактерии — их пищевые объекты. Следу-
ет отметить, что мелкие планктонные жгутиковые
протисты до сих пор изучены слабо, главным обра-
зом из-за трудностей методов сбора и изучения. От-
носительно недавно был опубликован довольно пол-
ный список видов жгутиконосцев и инфузорий Бай-
кала (Атлас и определитель пелагобионтов Байкала,
1995). Микропланктон озер и прудов включает гете-
ро- и фототрофных эвгленовых, хризофитовых, ди-
нофитовых (Ceratium), инфузорий, которые пред-
ставлены многочисленными видами (Чорик, 1968;
Мажейкайте, 1973; Мамаева 1979, и др.).
Кроме жгутиковых протистов и инфузорий в план-
ктоне пресных водоемов, особенно у поверхностной
пленки воды, развиваются популяции мелких амеб. К
условиям обитания в толще воды, где есть растения,
приспособлен один из видов диффлюгий — Difflugia
hydrostatica, популяции которой могут достигать вы-
сокой численности (Расе, 1982).
174
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Рис. 116. Некоторые виды морских планктонных жгутиконосцев.
А — Gymnodinium fuscum, Б — Cladopyxis brachiolatum, В — Acanthodinium caryophyllum, Г — Medusochloris phiale, Д — Syracosphaera apsteini — с ра-
ковинкой из бокаловидных кокколитов. (А—Г— по: Догель, 1951; Д— по: Голлербах, 1977).
Fig. 116. Planktonic marine flagellates.
A — Gymnodinium fuscum, Б — Cladopyxis brachiolatum, В — Acanthodinium caryophyllum, Г — Medusochloris phiale, Д — Syracosphaera apsteini with lori-
ca composed with coccolithes.
Сообщество обрастателей и эпибионтов. Обрас-
татели (перифитон) представлены большой группой
морских и пресноводных протистов, ведущих при-
крепленный образ жизни на твердых субстратах в
микробиотопах на камнях и скалах, на поверхности
детрита, гравия и искусственных субстратах. Эпиби-
онты — протисты, поселяющиеся на поверхности
тела разнообразных водных животных. Обрастатели
и эпибионты представляют обширное по количеству
видов и численности сообщество, относящееся в рав-
ной степени к планктону и бентосу.
Основу сообщества составляют прикрепленные
хоаномонады, кинетопластиды, церкомонады, бико-
зоециды, эвглениды, хризофитовые, а также инфузо-
рии (рис. 118). Экологические группы гетеротроф-
ных жгутиковых представлены прикрепленными (си-
дячими) и ползающими формами (Раилкин и др.,
1990). Эти же экологические группы могут быть при-
няты и для инфузорий.
Высокоспециализированную группу обрастателей
и эпибионтов самых разнообразных водных животных
представляют кругоресничные инфузории (перитри-
хи) (Банина, 1990). Известно около 100 видов одиноч-
ных й колониальных прикрепленных перитрих, кото-
рые широко распространены в пресных водоемах.
Прикрепленные протисты, относящиеся к разным
таксономическим группам, образуют основной комп-
лекс сообщества обрастателей и эпибионтов, обладая
многими сходными приспособлениями именно к при-
крепленному образу жизни. Второй комплекс сооб-
щества образуют неспецифические эврибионтные
виды протистов, ведущих подвижный образ жизни и
относящихся к разным таксономическим группам.
Биотопы обрастателей в водоемах многочислен-
ны, они представляют собой разнообразные природ-
ные и искусственные субстраты, отделенные друг от
друга пространствами, заполненными водой, что со-
ставляет специфику пространственной структуры по-
пуляций каждого вида данного сообщества. Подвиж-
ные животные — носители эпибионтов перемещают-
ся на значительные расстояния в вертикальном и
горизонтальном направлениях в любом водоеме, что
также составляет особенность структуры популяций
прикрепленных протистов.
Бентосное сообщество. Дно озер, прудов, рек,
канав и других водоемов покрыто слоем детрита, ко-
торый состоит из разлагающихся остатков растений
и животных. Литоральная зона озер часто имеет пес-
чаный грунт, который нередко свойствен и берегам
рек. В отличие от песчаного грунта морей в пресных
водоемах в нем развивается комплекс неспецифичес-
ких видов протистов (рис. 119). Донные органичес-
кие осадки (сапропель) разлагаются как в аэробных,
так и в анаэробных условиях. В них в зависимости от
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
175
Рис. 117. Некоторые виды морских бентосных жгутиконосцев и раковинных амеб.
А — Mastigamoeba psammobia; Б — М. simplex; В — Notoselenus tamanduensis: 1 — общий вид, 2 — поперечный срез; Г — Petalomonas baodicea; Д —
Р. raiula; Е — Ploeotia decipiens: 1 — общий вид, 2 — поперечный срез; Ж — Amphidinium herdmanii; 3 — Chardesia caudata; И — Aleptiella tricoronata;
К — Pseudocorythion acutum var. nipponicum; Л — P. wailesi; M — Corythionelloides fumiana. (А—Ж — no: Larsen, Patterson, 1990; 3, И — no: Goleman-
sky, 1970; K—M — no: Sbdzuki, 1979).
Fig. 117. Some marine benthic flagellates and testacean amoebae.
В, E; 1 — general view, 2 — transverse section.
176
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
преобладания тех или иных процессов развиваются
анаэробные или аэробные сапробионтные комплексы
протистов, бактерий, синезеленых водорослей и гри-
бов.
На поверхности детрита и в верхнем его слое до
глубины около 6 см существует множество микробио-
топов, в которых развивается целый комплекс видов
эвгленид, бодонид, хризофитовых, дипломонад, раз-
нообразных видов голых и раковинных амеб, солнеч-
ников и инфузорий. Раковинные амебы класса Lobo-
sea представляют облигатную группу пресноводного
бентоса, и наиболее широко в мезосапробных водо-
емах распространены виды родов Difflugia, Arcella,
Centropyxis, Cyclopyxis, а из класса Filosea — Cypho-
deria и Euglypha (Викол, 1990). Для пресноводного
микробентоса достаточно специфичны солнечники.
Значительную по численности группу микробентоса
представляют голые амебы разных систематических
групп (Mayorella, Vahlkamphia, Hartmanella, Pelo-
myxa palustris).
Широким обилием видов и многочисленностью
популяций отличается комплекс бентосных жгутико-
вых протистов. Среди них особенно много эвгленид
родов Euglena, Trachelomonas, Phacus, Peranema, Ur-
ceolus и других, преобладают миксотрофные виды,
гетеротрофные, питающиеся бактериями, частицами
детрита, и хищники, питающиеся мелкими жгутико-
выми и другими протистами, а также их цистами.
Много бодонид и церкомонадид. Очень разнообразны
хризофитовые, криптофитовые (особенно из рода
Chilomonas), динофитовые (Peridinium, Gymnodinium,
Ceratium).
Комплекс инфузорий аэробного сапропелевого
сообщества отличается большим разнообразием ви-
дов и высокой численностью. Эта зона пресных водо-
емов богата колониальными и одиночными, прикреп-
ленными и свободноплавающими кругоресничными
инфузориями (Peritricha), а также сосущими инфузо-
риями (Suctoria), которые ведут преимущественно
прикрепленный образ жизни. По сравнению с други-
ми пресноводными сообществами аэробный сапропе-
левый комплекс включает наибольшее количество
инфузорий, число видов которых достигает 100 и
более. В 1 м2 численность инфузорий может состав-
лять десятки миллионов экземпляров, а их био-
масса — сотни миллиграммов (Бурковский, 1984).
Рис. 118. Сообщество пресноводных протистов-эпибионтов. (По: Patterson, Hedley, 1992).
1 — Monosiga, 2 — Codonosiga, 3 — Anthophysa, 4 — Bicosoeca, 5 — Actinomonas, 6 — Paraphysomonas, 7 — Bodo, 8 — Rhynchomonas, 9 — Urceolus,
10 — Entosiphon, 11 — Clathrulina, 12 — Acanthocystis, 13 — Aspidisca,! 4 — Euplotes, 15 — Trithigmostoma, 16 — Acineta, 17 — Vorticella, 18 — Vagi-
nicola, 19 — Stichotricha, 20 — Stentor, 21 — Chilodonella.
Fig. 118. Epibiotic protist community.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
177
Специального внимания заслуживает экологичес-
кий комплекс видов протистов, которые обитают в
анаэробных условиях донных осадков (рис. 120) —
сапропелевых илов, содержащих H2S, NH2 и разнооб-
разные органические вещества, образующиеся при
бактериальном разложении растительного и живот-
ного детрита. Сапропель представляет мягкую колло-
идальную массу, разлагающуюся за счет анаэробных
бактерий. Комплекс сапропелевых протистов вклю-
чает в основном инфузорий, специфичных именно
для данного биотопа. Известно около 30 видов, отно-
сящихся к родам Epalxis, Pelodinium, Saprodinium,
Discomorpha, Dactylochlamis, Metopus, Caenomorpha.
Из амеб в этом комплексе обычны Pelomyxa и Рат-
phagus.
Сообщества протистов
проточных водоемов
Экология свободноживущих протистов рек и
ручьев до сих пор изучена недостаточно. Из немно-
гих исследований, выполненных в настоящее время,
известно, что фаунистические комплексы, на основе
которых формируются сообщества протистов про-
точных водоемов, весьма сходны с таковыми озер и
прудов. На видовой состав, численность, биомассу и
распределение протистов по биотопам рек и ручьев
оказывают большое влияние скорость течения воды,
постоянное перемешивание водных масс, перемеши-
вание донных осадков, характер грунта, содержание
органических веществ, с которыми связано развитие
пищевых организмов (Мамаева, 1979; Викол, 1990;
Жариков, 1994, 1999).
Сообщество протистов
активного ила аэротенков
Биологическая очистка промышленных и бытовых
сточных вод осуществляется в настоящее время в
аэротенках — глубоких (4—6 м глубиной) искусст-
венных бассейнах, в которых формируется активный
ил, представляющий собой сложное сообщество гете-
ротрофных организмов. Основу этого сообщества со-
ставляют преимущественно неспорообразующие грам-
отрицательные бактерии и гетеротрофные протисты
разных таксонов (Липеровская, 1977). Бактерии — это
первая, основная и наиболее многочисленная группа ор-
ганизмов, разлагающих органические вещества сточ-
ной воды, и главный источник пищи протистов актив-
ного ила. Среди протистов наиболее многочисленный
Рис. 119. Бентосное сообщество пресноводных протистов. (По: Patterson, Hedley, 1992).
1 — Paraphysomonas; 2 — Notoselenus; 3 — Entosiphon; 4 — Peranema; 5 — Bodo; 6 — Protaspis; 7 — Phynchomonas; 8 — Petalomonas; 9 — Cryptodif-
flugia; 10 — Arcella; 11 — Difflugia; 12 — Amoeba; 13 — Mayorella; 14 — Pompholyxophrys; 15 — Actinophrys; 16 — Actinosphaerium; 17 — Loxophyllum;
18 — Cinetochilum; 19 — Cyclidium; 20 — Paramecium caudatum; 21, 22 — Aspidisca; 23 — Euplotes, 24 — Stylonychia; 25 — Spirostomum.
Fig. 119. Benthic community of freshwater protists.
12 Протисты, часть 1
178
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
50 мкм
Рис. 120. Бентосное сообщество анаэробных протистов. (По: Patterson, Hedley, 1992).
1 — Mastigamoeba, 2 — Mastigella, 3 — Pelomyxa, 4 — Hexamita, 5 — Trepomonas, 6 — Trigonomonas, 7 — Brachonella, 8 — Metopus, 9 — Spirosto-
mum, 10 — Plagiopyla, 11 — Loxodes, 12 — Sarpodinium, 13 — Myelostoma, 14 — Caenomorpha.
Fig. 120. Benthic community of anaerobic protists.
и специфичный комплекс видов составляют инфузо-
рии класса Peritricha (Sessilida). Обычно в активном
илу можно обнаружить 30—40 видов прикрепленных
перитрих, а также ряд видов свободноплавающих ин-
фузорий (Банина, 1983, 1990). В состав сообщества
активного ила входит много видов саркодовых, сол-
нечников и гетеротрофных жгутиковых протистов
(Фауна аэротенков.., 1984).
Протисты грунтовых вод
Данные о протистах, населяющих грунтовые воды,
еще немногочисленны (Николюк, Гельцер, 1972; Ага-
малиев, 1990). Однако разнообразные виды их обнару-
жены в грунтовых водах, залегающих на небольшой
глубине. Это гетеротрофные жгутиконосцы (Bodo, Spu-
mella и др.), виды рода Amoeba, инфузории (Colpoda)
и другие. В грунтовые воды протисты попадают из
почвы, и в этой среде обитания развиваются преиму-
щественно влаголюбивые виды. Грунтовая вода — очень
непостоянная среда обитания: изменяется уровень ее
залегания, соленость, pH, газовый режим и температу-
ра в зависимости от типа й состояния близлежащего
водоема, а также от окружающей почвы. Поэтому фауна
протистов и их сообщества в грунтовых водах носят
временный характер. Специфичных для грунтовых
вод видов протистов пока не обнаружено.
Сообщество почвенных протистов
Протисты представляют наиболее многочислен-
ную, всесветно распространенную группу организмов
почвенной биоты, уступая по количеству видов толь-
ко бактериям. В почвах разных географических зон
протисты образуют сообщества, включающие в свой
состав жгутиконосцев, инфузорий и саркодовых, при-
способленных к своеобразным условиям, свойствен-
ным почве как среде обитания.
К числу облигатных видов протистов, адаптиро-
ванных к условиям обитания в почве, относятся ин-
фузории рода Colpoda, некоторые виды Hypotrichida
и Prostomatida. Специфичный многочисленный и
постоянный комплекс видов почвенной биоты со-
ставляют раковинные амебы родов Centropyxis, Plagio-
pyxis, Nebela, Difflugia, Arce Ila, Corythion, Euglypha,
Trinema и другие. Эти организмы распространены
всесветно, населяют разнообразные типы почв, но наи-
большее богатство видов и высокая их численность
характерны для почв лесов (Bonnet, Thomas, 1960;
Николюк, Гельцер, 1972; Лепинис и др., 1973; Гель-
цер, 1993; Корганова, 1997, и др.). Почвенным про-
тистам, и особенно раковинным амебам, принадле-
жит существенная роль в биологической индикации
типов почв, деструкции и минерализации органичес-
ких веществ, природном круговороте кальция и
кремния.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
179
Протисты — симбионты
В настоящее время известно свыше 10 000 ви-
дов симбионтов из разных групп протистов, что со-
ставляет около 10 % от их общего количества. В
современной экологии принято рассматривать не-
сколько форм симбиотических взаимоотношений
между совместно живущими организмами: ком-
менсализм, мутуализм, паразитизм и многие пере-
ходные между ними формы.
Симбиоз (греч. symbiosis) в самом общем виде
представляет собой любые формы совместного су-
ществования разноименных организмов, которые делят
между собой функции регуляции своих взаимоотно-
шений с внешней средой. Основой для формирова-
ния симбиотической системы могут служить пище-
вые, пространственные и другие взаимоотношения. В
природе известен факультативный симбиоз, когда
каждый из организмов при отсутствии партнера может
жить самостоятельно, и облигатный — когда партне-
ры не могут существовать друг без друга. Протистам
свойственны все типы симбиотических взаимоотно-
шений.
При комменсализме (лат. сот — вместе; mensa —
стол, трапеза) один из партнеров системы (коммен-
сал) возлагает регуляцию своих отношений с внеш-
ней средой на партнера (хозяина), но не вступает с
ним в тесную взаимосвязь. Комменсал обычно полу-
чает пользу для себя, а для хозяина он, по-видимому,
нейтрален. Комменсалами могут быть протисты, оби-
тающие на поверхности тела, жабр, в ротовой и дру-
гих полостях тела, а также во внутренних органах
животных. Примером могут служить кругоресничные
инфузории из рода Orbopercularia, обитающие в ро-
товой полости и пищеводе водных жуков. Они ис-
пользуют организм хозяина в качестве места обита-
ния, питаясь частью его пищи, но какой-либо пользы
или вреда жуку не приносят (Банина, 1990). Много-
численные комменсалы населяют рубец жвачных
(парнокопытных) млекопитающих. Они представ-
лены инфузориями и гетеротрофными жгутиконосца-
ми. И те и другие питаются бактериями и остатками
пищи хозяина, однако для хозяина, скорее всего, без-
различны, так как, с одной стороны, не наносят ему
вреда, а с другой — не помогают переваривать расти-
тельную пищу, не имея расщепляющих целлюлозу
ферментов. Такие ферменты вырабатывают много-
численные бактерии, населяющие рубец жвачных и
служащие пищей как хозяину, так и комменсалам.
Другие примеры комменсализма среди протистов
можно найти в систематической части этого издания.
Мутуализм также широко представлен среди про-
тистов. При этой форме симбиоза взаимоотношения
выгодны для обоих партнеров (лат. mutuus — взаим-
ный), и они уже не могут существовать друг без
друга. Классическим примером таких отношений
служит симбиоз термитов с гетеротрофными жгути-
коносцами из отрядов Trichomonadida, Hypermastigi-
da и Oxymonadida. Жгутиконосцы живут в задней
кишке термитов и расщепляют целлюлозу, которой
питается хозяин, до жирных кислот, всасываемых в
кишечнике термита (подробнее об этом см. соответ-
ствующие разделы настоящего издания). Другая груп-
па протистов — динофитовые — сожительствует с
кораллами. Динофлагеллаты, способные одновремен-
но к фототрофному и гетеротрофному питанию,
обеспечивают своих хозяев продуктами фотосинте-
за — углеводами. Кроме того, они способствуют
осаждению карбоната кальция, необходимого корал-
лам для формирования скелета (Бигон и др., 1989).
Паразитизм (греч. parasitos — нахлебник) пред-
ставляет собой антагонистическую форму взаимоот-
ношений. Почти все крупные группы протистов, за
исключением радиолярий и фораминифер, обладаю-
щих минеральным скелетом, имеют в своем составе
паразитические формы. Многие таксоны протистов
включают только паразитические виды.
Для паразита организм хозяина представляет среду
обитания; природная система микропаразит—хозяин
возникла в процессе длительной сопряженной эволю-
ции (коэволюция). Эта система подвергается естест-
венному отбору, становится устойчивой, и в природе
сохраняются оба вида — вид хозяина и вид паразита.
Паразитические протисты оказывают существенное
воздействие на отдельные особи, популяции хозяина,
часто снижая численность животных, а иногда и рас-
тений.
По локализации в организме хозяина различают
эктопаразитов, поселяющихся на поверхности тела, и
эндопаразитов, обитающих во внутренних органах,
тканях и клетках хозяина. Многие виды паразитов
специфичны к виду хозяина, его органам, тканям и
клеткам. Наибольшее число эктопаразитов известно
среди инфузорий. К ним относится целый ряд видов
таких родов, как Trichodina, Rhabdostyla, Capriniana,
Apiosoma, и других. Они прочно прикрепляются к
плавникам, жабрам и поверхности тела рыб, вызывая
патогенный эффект при интенсивном заражении. Эн-
допаразиты животных и человека населяют кишеч-
ный тракт, печень, органы мочеполовой системы,
кровь, головной и спинной мозг, глаза, а также другие
органы и ткани. Например, Entamoeba histolytica —
возбудитель амебной дизентерии — и инфузория Ba-
lantidium coli — возбудитель балантидиаза — лока-
лизуются в толстом кишечнике человека, изъязвляя
его стенку; Giardia (=Lamblia) intestinalis — возбуди-
тель лямблиоза — обитает в тонком кишечнике и
желчных протоках человека.
Особую группу эндопаразитов составляют виды,
обитающие внутри клеток и тканей. Это кокцидии,
токсоплазма, криптоспоридии и другие, микробиото-
пами которых служат клетки кишечного эпителия
животных и человека. Среди микропаразитов крови
выделяются 2 группы: паразиты плазмы крови (три-
паносомы, криптобии и др.) и внутриклеточные па-
разиты эритроцитов (малярийные плазмодии, пиро-
плазмиды, гемогрегарины).
Внутриклеточные паразиты — это одна из наибо-
лее высоко специализированных форм паразитизма
среди простейших. Для них характерны сложные
жизненные циклы, активное проникновение в клетку
хозяина, активная реакция клетки хозяина на внедре-
ние и последующее развитие в ней паразита.
Особую экологическую значимость у эндопарази-
тических простейших, и прежде всего у внутрикле-
точных паразитов со сложными жизненными цикла-
180
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
ми, имеют персистирующие стадии. Например, у кок-
цидий, криптоспоридий (Eimeria, Cryptosporidium и
др.) персистирование сопряжено с формированием
ооцист, которые выводятся из организма хозяина во
внешнюю среду и в течение длительного срока (око-
ло года) остаются жизнеспособными (Бейер, 1989;
Бигон и др., 1989).
Одну из важных особенностей экологии эндопара-
зитических простейших представляют способы рас-
пространения их в природе. Для ряда видов этих ор-
ганизмов известны способы передачи контактным
путем (например, таким путем передаются Trichomo-
nas foetus, Т. vaginalis и др.). Широко распространен
в природе способ передачи паразитических простей-
ших с помощью цист (лямблии, патогенные виды
амеб, инфузорий и др.). Наиболее сложный способ
передачи паразитов от одного хозяина к другому свя-
зан с переносчиком. Этот способ известен для маля-
рийных плазмодиев (переносчики — малярийные ко-
мары), пироплазмид и ряда других паразитических
простейших животных и человека.
Протисты — хозяева прокариот
и вирусов
Благодаря ультраструктурным, физиологическим
и биохимическим исследованиям протистов выявле-
ны многочисленные случаи формирования устойчи-
вых симбиотических систем. Материал по симбион-
там у протистов огромен. Среди них могут встречать-
ся бактерии, другие протисты и даже вирусы.
В настоящее время изучение симбиотических сис-
тем протистов и прокариот идет по двум направлени-
ям: 1) углубленные исследования ограниченного
круга широко используемых модельных объектов из
жгутиконосцев, инфузорий, солнечников и амеб с
применением всего арсенала современных методик;
2) сравнительно-эволюционный анализ симбионтов в
различных группах протистов.
В рамках этого раздела мы постараемся дать толь-
ко общее представление о встречаемости прокариот
и вирусов в клетках протистов. Более подробные све-
дения, касающиеся этой проблемы, можно найти в
недавно опубликованой сйодке (см.: Осипов и др.,
1997).
Встречаемость бактерий. Анализ, проведенный
по всем основным группам протистов, показал, что
среди прокариотических симбионтов встречаются
как эктобионты, так и эндобионты.
Эктобионты. Эктобиотические бактерии рас-
положены на поверхности клетки и ассоциированы с
ней на протяжении большей части жизненного цикла
протеста. Они обнаружены у многих жгутиконосцев
(особенно характерны для парабазалий и оксимонад),
а также некоторых инфузорий. Большинство эктоби-
онтов не определено до вида, и лишь в нескольких
случаях известны их точные видовые названия.
Характер взаимодействия эктобионтов с поверх-
ностными структурами весьма существенно различа-
ется у разных видов жгутиконосцев. Некоторые экто-
биотические бактерии несомненно реально участвуют
в движении клетки простейшего, хотя у большинства
оксимонад и гипермастигин основная нагрузка при
движении приходится, конечно, на жгутики. Среди
бактерий наиболее эффективными движителями явля-
ются спирохеты.
Эндобионты. Прокариотические эндобионты
обнаружены у более чем 100 видов амеб, жгутико-
носцев и инфузорий. Причем чаще всего они заселя-
ют цитоплазму (Осипов и др., 1997). Видовая принад-
лежность бактерий, за редкими исключениями, не
определена. Все они являются грамотрицательными,
т. е. не имеют муреина в клеточной стенке. Здесь они
могут встречаться как в симбионтофорной вакуоли,
так и свободно лежать в цитоплазме. Метаногенные
бактерии Methanobacterium formicicum ассоциирова-
ны с гидрогеносомами в цитоплазме Psalteriomonas и
некоторых анаэробных инфузорий.
Перинуклеарное пространство заселяется симби-
онтами значительно реже (Фокин, Карпов, 1995). Не-
смотря на то что перинуклеарное пространство и ше-
роховатый ЭПР представляют собой единый компарт-
мент клетки, у одних жгутиконосцев эндобионты
отмечены только в ЭПР, а у других — только в пери-
нуклеарном пространстве. Лишь у бесцветной хризо-
монады Paraphysomonas vestita бактерии найдены в
обоих отделах этого компартмента (рис. 121, вкл.).
Еще реже отмечаются прокариоты в нуклеоплазме,
причем они никогда не окружаются мембранными
структурами клетки-хозяина. Такого рода симбиоз ог-
раничивается, по-видимому, инфузориями (рис. 122,
вкл.), динофитовыми и некоторыми эвгленовыми.
У динофлагеллаты Woloszynskia pascheri обнару-
жены симбионты в хлоропластах и митохондриях
(Wilcox, 1985). Это уникальное среди эукариот явле-
ние, к сожалению, не было в дальнейшем исследовано.
Встречаемость вирусов. В большинстве как об-
зорных, так и оригинальных работ, посвященных вза-
имоотношениям вирусов и простейших, отмечен
только сам факт обнаружения вирионов в клетке
(Lemke, 1976; Терас, 1981; Wang, Wang, 1991; Lips-
comb, Riordan, 1995). Вирионы или вирионоподоб-
ные частицы обнаружены у хризофитовых (рис. 123,
вкл.), криптомонад, динофитовых и кинетопластид.
Протисты могут заражаться вирусами, поглощая зара-
женные бактерии, а также непосредственно получая
их из окружающей среды в процессе эндоцитоза. Ви-
русы, по-видимому, могут накапливаться в клетках
протестов, а также оказывать влияние на ход их мик-
роэволюционного процесса.
Адаптации протистов
к факторам внешней среды
Распространение протистов в природе, распреде-
ление по биотопам, устойчивость их природных со-
обществ и численность популяций в водных, почвен-
ных и наземных экосистемах зависят от многих аби-
отических и биотических факторов внешней среды.
К числу ведущих абиотических факторов, оказываю-
щих на протистов непосредственное воздействие
в течение всех фаз их жизненного цикла, относятся:
температура, соленость, влажность, газовый состав,
pH среды и другие. Биотические факторы включают
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
181
прежде всего пищу, конкурентные взаимоотношения
за пищевые ресурсы, хищников.
Абиотические факторы. Температура. Про-
тисты представляют одну из групп эктотермных орга-
низмов, которые могут существовать в широком диа-
пазоне толерантных температур от 0—4 до 30—35 °C.
Однако оптимальные температуры, при которых все
процессы жизнедеятельности у большинства протис-
тов протекают наиболее активно, соответствуют 20—
30 °C, а каждый вид имеет свой температурный опти-
мум. У протистов, обитающих в холодных северных
зонах, температурный оптимум размножения и других
процессов жизнедеятельности значительно ниже, чем
у протистов южных климатических зон (Смарагдова,
1941; Dragesco, 1973; Бурковский, 1984; Fenchel, 1987;
Хаусман, 1988, и др.).
Многие виды протистов в природе и в экспери-
ментальных условиях выдерживают действие экстре-
мальных низких и высоких температур и способны
переносить их длительное время. В термальных ис-
точниках Камчатки, Кавказа, Забайкалья, Италии,
Японии, США, в которых температура воды достига-
ет 30—56 °C, обнаружены мелкие гетеротрофные
жгутиконосцы, мелкие голые амебы, инфузории родов
Paramecium, Litonotus, Frontonia, Tetrahymena (Pres-
cott, 1957; Kudo, 1966; Лозина-Лозинский, 1961;
Kahan, Sharon, 1976, и др.).
Более высокие температуры (до 70—80 °C) вы-
держивают трофозоиты некоторых жгутиконосцев.
Однако особенно устойчивы к действию экстремаль-
ных высоких температур цисты многих протистов, в
частности Colpoda cucullus. При 70 °C они сохраня-
ют жизнеспособность в течение 26 ч, а сухие цисты,
находящиеся при 106 °C, — в течение 3 ч (Taylor,
Stricland, 1936).
Изучая С. maupasii, обитающих в крайнеаридной
почве пустыни Заалтайской Гоби, где дневная темпе-
ратура поверхности песка достигает 74 °C, Гельцер
(1990) обнаружил явление псевдокристаллизации
трофозоитов. Это явление представляет собой одну
из форм ксероанабиоза (ангидроанабиоз) и служит
защитной реакцией организма в ответ на действие ле-
тальной для трофозоитов температуры. Факты устой-
чивости протистов к действию столь высоких темпе-
ратур представляют одно из доказательств широких
пределов их адаптивных возможностей.
В стадии трофозоитов и цист протисты способны
переносить влияние отрицательных температур в
природе в пределах от -1 до -60, а в условиях экспе-
римента — от -1 до -269 °C. В природе температур-
ный минимум активной жизнедеятельности протис-
тов находится в пределах 0 °C, и зимой в водоемах,
под покровом льда, где температура воды не превы-
шает 2 °C, живет и развивается богатое видами сооб-
щество инфузорий и жгутиконосцев (Жинкин, 1930;
Золотарев, 1989, и др.). Трофозоиты и цисты многих
видов свободноживущих протистов и симбионтов
выдерживают замораживание при температурах -79,
-196, -269 °C в течение года и более (Лозина-Лозин-
ский, 1964; Sakai, 1964, и др.).
Адаптивные реакции протистов, возникающие в
ответ на изменения температуры окружающей среды,
разнообразны и затрагивают все функции жизнедея-
тельности. Смена температуры служит причиной из-
менений морфологии, функциональной активности
клетки-организма, течения жизненных и митотичес-
ких циклов, агамного и полового размножения. По-
казано, что при температуре ниже оптимальной зна-
чительно снижается скорость размножения Amoeba
proteus, Chilomonas paramecium, Euglena gracilis,
Paramecium caudatum, Colpoda maupasii и других.
Температура выше оптимальной также способствует
снижению скорости размножения протистов (Daniel,
Chalkley, 1932; Smith, 1940; Полянский Ю. И., 1957;
Franceschi, 1959; Ковалева, 1962; Соок, 1966, и др.).
Изменение скорости агамного размножения простей-
ших, подвергающихся постоянным колебаниям тем-
пературы, носит адаптивный характер и является по
своей природе адаптивной модификацией.
Способность простейших адаптироваться к сезон-
ным и суточным колебаниям температуры окружаю-
щей среды в природе, к меняющемуся температурно-
му режиму в условиях эксперимента зависит от их
тепло- и холодоустойчивости (терморезистентности).
Уровень терморезистентности находится в непосред-
ственной зависимости от тех температурных усло-
вий, к которым приспособлен в природе каждый вид
саркодовых, жгутиконосцев, инфузорий.
В качестве критерия для оценки уровня терморе-
зистентности в экспериментальных исследованиях
многих авторов принято время выживаемости про-
стейших (в мин) при действии сублетальных и ле-
тальных температур (Александров, 1948; Полянс-
кий Ю. И., 1957; Суханова, 1963, и др.). Результаты
многочисленных исследований показали, что одна из
важных закономерностей, которой подчинены изме-
нения терморезистентности простейших, выражается
в снижении их теплоустойчивости и повышении хо-
лодоустойчивости при адаптации к низким темпе-
ратурам. Соответственно повышение температуры
условий обитания сопряжено с повышением теплоус-
тойчивости и снижением холодоустойчивости про-
стейших.
Эта закономерность справедлива для эври- и сте-
нотермных видов свободноживущих, а также симбио-
тических протистов, обитающих в организме экто- и
гомотермных хозяев (Полянский Ю. И., 1959, 1963;
Лозина-Лозинский, 1961; Vogel, 1966; Суханова, 1968,
1973, и др.). В табл. 8 показаны изменения терморе-
зистентности парамеций при культивировании в усло-
виях разных температур в течение 3—8 нед.
Достаточно быстрая реакция простейших на смену
температурного режима, выражающаяся в изменении
их терморезистентности, представляет адаптивную
модификацию. При длительном содержании культур в
условиях повышенной или низкой температуры адап-
тивные модификации могут приобретать форму дли-
тельных модификаций, которым принадлежит особая
роль в приспособительных реакциях простейших к
среде обитания (Орлова, 1947; Полянский Ю. И., 1957;
Crippa-Franceschi, 1970).
Популяции, входящие в состав вида, различаются
по устойчивости к низким и высоким температурам.
Но и каждая популяция включает в свой состав груп-
пы особей — клоны, также неодинаковые по своей
терморезистентности (табл. 8). Гетерогенность попу-
182
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Таблица 8
Продолжительность выживания (в мин)
Paramecium caudatum при 40 °C после предварительного
культивирования в разных температурных условиях
(по: Полянский Ю. И., 1957 с изменениями)
Клон Продолжительность выживания (в мин) в зависимости от предварительного культивирования при температуре (в °C)
12—13 18—20 24—28
А 6.7 ±0.16 13.9 ±0.5 35.4 ±0.86
В 9.6 ±0.20 17.6 ±0.42 34.7 ± 0.30
10.6 ±0.30 19.1 ±0.30 42.1 ±0.77
6.1 ±0.05 19.3 ±0.70 31.6 ±0.69
Д 10.9 ±0.10 Нет данных 35.7 ±0.30
12.5 ±0.10 23.3 ±0.13 36.9 ± 0.20
ляции по отношению к температурному фактору спо-
собствует сохранению вида в природе: в случае вне-
запного, резкого изменения температуры среды оби-
тания может погибнуть часть особей, а оставшиеся,
более устойчивые особи (клоны) быстро адаптируют-
ся к изменившимся температурным условиям, начи-
нают размножаться, и численность популяции восста-
навливается.
Начиная с первых лет исследований температур-
ных адаптаций возник вопрос о природе терморе-
зистентности простейших и клеток многоклеточных
организмов. Были высказаны различные гипотезы. В од-
ной из них на основе полученных экспериментальных
данных сделано заключение об эпигеномной природе
терморезистентности простейших (Осипов, 1966).
Была также сформулирована гипотеза о зависи-
мости терморезистентности от особенностей функци-
онирования клеточных белков в условиях низких и
высоких температур (Ушаков, Кусакина, 1960; Уша-
ков, 1964, 1965; Александров, 1965; Суханова, 1968,
и др.). Однако белкщ связанные с терморезистент-
ностью клеток, в то время еще не были идентифици-
рованы.
В течение последних лет в клетках многих ор-
ганизмов (из простейших у инфузорий) обнаружены
гены теплового шока, отвечающие на температур-
ные воздействия. Изучена локализация этих генов в
хромосомах, их структурная организация и функци-
ональная активность в процессе синтеза РНК и
белков теплового шока. Эти белки обнаружены у
всех исследованных организмов. Синтез белков теп-
лового шока, наступающий в клетках в ответ на
температурное воздействие, имеет адаптивное зна-
чение. Белки теплового шока имеют широкий
спектр действия и синтезируются не только в мо-
мент температурного влияния, но и в нормальных
условиях жизнедеятельности клеток и организма в
целом (Заварзин и др., 1992).
Соленость. Фаунистические комплексы про-
тистов морей и пресных водоемов существуют в при-
роде раздельно, они не смешиваются, обладая высо-
кой степенью эндемизма, который обусловлен соле-
ностью воды. Граница раздела между пресноводной и
морской фауной определяется соленостью 5—8 %о.
Именно эта граница солености оказывается крити-
ческой: верхней для пресноводных и нижней для
морских организмов (Хлебович, 1962, 1974).
При солености воды в пределах 3—8 %о в приро-
де развивается и постоянно существует небольшой по
численности комплекс солоноватоводных видов ин-
фузорий и других протистов. Типичные морские
виды инфузорий обитают в воде с соленостью не ниже
10 %о, но для многих морских протистов соленость
ниже 21 %о неблагоприятна (Бурковский, 1984). Ряд
видов инфузорий обитает в природных гипергалин-
ных водоемах с соленостью воды до 150—250 %о
(Агамалиев, 1990).
Вместе с тем среди протистов есть немногочис-
ленный комплекс эвригалинных видов, способных к
обитанию в широком диапазоне солености водной
среды: в пресных, морских, солоноватоводных и ги-
пергалинных водоемах с меняющимся соленостным
режимом, что свидетельствует о широких адаптив-
ных возможностях протистов.
Способность амеб, жгутиконосцев и инфузорий
адаптироваться к изменению содержания солей в вод-
ной среде показана во многих экспериментальных
исследованиях. Одним из первых объектов этих ис-
следований стала инфузория Paramecium caudatum.
При культивировании в среде с повышенным содер-
жанием Са++ резистентность парамеций к этому хи-
мическому элементу значительно повышается по
сравнению с нормой, и уровень ее повышения зави-
сит от концентрации Са++ в среде (Орлова, 1947; Гри-
горян, 1964).
Повышение резистентности парамеций к Са+\ К+ и
другим ионам, применявшимся в экспериментах,
представляет собой адаптивные модификации и при
продолжительных сроках культивирования приобре-
тает характер длительных модификаций, имеющих
четко выраженный адаптивный эффект. Повышение
устойчивости к солям кальция, натрия, калия и другим
соединениям было установлено для многих видов ин-
фузорий, амеб и жгутиконосцев (Ирлина, 1964; Vogel,
1966; Полянский и др., 1967; Толмашова, 1990).
Наиболее высокая устойчивость к воздействию
солей свойственна простейшим, живущим при опти-
мальных или близких к ним условиях солености, к
которым каждый вид адаптирован в природе. Резуль-
тат опытов с перемещением Uronema marinum из
природной среды, где соленость равна 34 %о, в мор-
скую воду с соленостью 17 %о показал, что при сни-
женной вдвое солености инфузории прекращают раз-
множаться, при 8.8 %о погибает 50 % особей, а при
солености 3.5 %о через 4 сут погибает вся популяция
(Parker, 1976; Бурковский, 1984).
Итоги экспериментальных исследований показа-
ли, что у простейших повышение устойчивости к
солям сочетается с повышением их устойчивости к
действию других агентов, например к этанолу, темпе-
ратуре. В данном случае происходит неспецифичес-
кое повышение устойчивости к разным факторам,
которая, вероятно, обусловлена общими клеточными
механизмами и имеет адаптивное значение (Орлова,
1947; Полянский Ю. И., 1957; Сопина, 1963; Vogel,
1966; Суханова, 1968).
Изменение устойчивости протистов к содержанию
солей в водной среде представляет физиологическую
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОТИСТОВ
183
адаптацию. Это — одна из форм модификационной
изменчивости, которая определяется нормой реакции
генотипа. Одним из важных внутриклеточных меха-
низмов соленостной адаптации инфузорий и многих
видов морских беспозвоночных служит изменение
концентрации свободных аминокислот в клетках при
изменении осмомолярности внешней среды. Это —
феномен внутриклеточной изоосмотической амино-
кислотной регуляции (Заварзин и др., 1992).- Повы-
шение концентрации солей в водной среде приводит
к активации биосинтеза аминокислот, таких как ала-
нин, глицин, пролин, с которыми связано повышение
осмотического давления в клетке. Со свободными
аминокислотами сопряжена регуляция внутриклеточ-
ного обмена, изменяющегося в ответ на смену осмо-
молярности среды обитания.
Кроме свободных аминокислот у водных орга-
низмов существуют и другие механизмы внутрикле-
точной изоосмотической регуляции. К ним относят-
ся сахара и многоатомные спирты в клетках расте-
ний, неорганические ионы (Na+, К+, С1~) в клетках
морских моллюсков, изменение содержания кото-
рых компенсирует нарушения внутриклеточного ос-
мотического давления при смене осмомолярности
внешней среды.
Газовый состав среды обитания. Содер-
жание кислорода в среде обитания — экологический
фактор, который в значительной степени определяет
видовой состав, его разнообразие и численность лю-
бого природного сообщества протистов. Преобладаю-
щее большинство свободноживущих протистов —
аэробы, имеющие митохондрии и распространенные
в широком диапазоне вариаций содержания кислоро-
да в морях, пресноводных и почвенных биотопах.
По результатам исследований Бурковского (1984),
зона оптимальных значений содержания кислорода в
среде обитания у большинства изученных видов сво-
бодноживущих инфузорий находится в интервале 0—
10 мг/л. Для планктонных инфузорий этот интервал
составляет 0—5.0, для эпибионтных видов 6—12 мг/л;
для бентосных инфузорий, например сапропелевых и
интерстициальных видов, оптимум содержания кис-
лорода ниже и равен 0.1—1.0 мг/л.
Содержание кислорода в грунте дна водоемов
уменьшается от поверхности в глубину. Проникнове-
ние кислорода в грунт происходит путем диффузии и
движения воды. На поверхности грунта накапливает-
ся слой осадков, состоящий из органического мате-
риала — детрита, который подвергается бактериаль-
ному разложению.
По мере увеличения глубины залегания грунта со-
держание кислорода в нем резко уменьшается и увели-
чивается содержание СО2 и H2S. Поэтому оксифиль-
ные виды протистов, особенно инфузории, достигают
максимальной численности в верхней, окислительной
зоне, где окислительно-восстановительный потенциал
выше 200 м/В. В средней, восстановительной зоне
грунта, где окислительно-восстановительный потен-
циал составляет 0—200 м/В, число оксифильных видов
уменьшается. В более глубокой, сульфидной зоне, где
грунт черного цвета и окислительно-восстановитель-
ный потенциал ниже 0 м/В, по численности преобла-
дают анаэробные виды протистов.
Самые нижние слои грунта, лишенные О2 и с высо-
кими концентрациями H2S и СО2, населены облигат-
ными и факультативными видами анаэробных инфузо-
рий, жгутиконосцев и амеб, которые в этих условиях
достигают высокой численности популяций (Бурков-
ский, 1984; Fenchel, 1987). В отличие от оксифиль-
ных протистов большинство обитателей сульфидной
зоны не имеет митохондрий. К их числу относится
крупная по размерам амеба Pelomyxa palustris, инфузо-
рии родов Metopus, Caenomorpha, Parablepharisma,
Saprodinium, Myelostoma, Frontonia и другие. Сюда же
относятся свободноживущие жгутиконосцы родов
Trepomonas, Hexamita (дипломонадиды), а также много
других.
У свободноживущих протистов, особенно у инфу-
зорий, обитающих в анаэробных условиях, широко
распространены экто- и эндобиотические бактерии,
относящиеся к разным морфологическим типам. Каж-
дый вид инфузорий обладает одним видом бактерий.
Количество бактерий очень велико: например, у
одной особи инфузорий рода Sonderia на поверхно-
сти тела располагается до 105 клеток бактерий и
примерно такое же их количество находится в цито-
плазме. Показано, что бактериальные эндобионты
инфузорий родов Metopus и Caenomorpha относятся
к метанобразующим видам, подобно бактериям ци-
топлазмы инфузорий из рубца жвачных животных
(Van Bruggen et al., 1983). Реакции образования ме-
тана в бактериальной клетке сопровождаются выде-
лением кислорода (Хаусман, 1988).
У многих протистов, обитающих в анаэробных ус-
ловиях, в течение последних лет обнаружены особые
органеллы энергетического метаболизма — гидроге-
носомы. Впервые гидрогеносомы были обнаружены у
трихомонад, эндопаразитических жгутиконосцев жи-
вотных и человека (МйПег, 1975). Гидрогеносомы со-
держат ферменты, окисляющие пировиноградную
кислоту с образованием АТФ. В этих реакциях, где
конечным акцептором электронов служат протоны,
образуется молекулярный водород (МйПег, 1980; За-
варзин и др., 1992).
pH среды обитания. Пресноводные протисты
выдерживают широкий диапазон вариаций активной
реакции среды обитания от 3.5 до 8.5—9.0. Колеба-
ния значений pH морской воды происходят в более
узких пределах, и морские протисты развиваются
нормально при значениях pH 7.0—8.5. Однако влия-
ние pH на процессы жизнедеятельности, жизненные
циклы, размножение, инцистирование и эксцистиро-
вание протистов изучено еще недостаточно; мало
работ по изучению этого важного экологического
фактора в экспериментальных и полевых условиях.
Показано, например, что Euglena gracilis живет и
размножается в лабораторных культурах при вариа-
циях pH от 3.1 до 6.5 (Полянский В. И., 1948). При
низких значениях pH (3.5—5.0) в заболоченных поч-
вах обитают многие виды голых и раковинных амеб,
фотосинтезирующих и гетеротрофных жгутиконос-
цев, ряд видов инфузорий. В экспериментальных ус-
ловиях показано влияние pH на скорость эксцисти-
рования почвенных инфузорий Colpoda cucullus:
среда, имеющая pH 4.0—5.0, не влияет на время вы-
хода инфузорий из цист по сравнению с контролем;
184
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
в среде с pH 1.3—3.5 этот процесс значительно за-
медляется (Meier, 1979).
Большинство видов пресноводных инфузорий оби-
тает в биотопах, где pH воды составляет 6.0—8.0, и
сравнительно немногие виды обитают в условиях
меняющегося pH ниже или выше этого значения
(примерно в пределах 5 единиц, — Bick, Kunze,
1971). Бурковский (1984) отмечает, что свободно-
плавающие инфузории планктона и эпибионты оби-
тают в нейтральной и слабощелочной средах, инфу-
зории сапропелевых илов — в нейтральной и слабо-
кислой средах. При значениях pH ниже 5 обитают
инфузории в заболоченных биотопах, например в
сфагновых и торфяных болотах. Оптимальные значе-
ния pH для инфузорий большинства видов — около
7.0, и отклонения от оптимума в сторону снижения
или повышения изменяют скорость размножения, что
показано в работах разных авторов (см.: Бурковский,
1984).
Биотические факторы. В составе любого природ-
ного сообщества протисты живут в окружении и в
тесном взаимодействии с множеством других орга-
низмов. Бактерии и синезеленые водоросли, растения
и животные, образующие сообщество, создают ту
благоприятную биотическую среду, в которой про-
тисты могут осуществлять все функции своей жизне-
деятельности.
Главную форму связей между организмами природ-
ного сообщества составляют пищевые взаимоотноше-
ния, которые лежат в основе сложных циклов и
цепей питания. Пищевые и пространственные взаи-
моотношения представляют основу формирования
биотических комплексов, объединяющих виды сооб-
щества в единую биотическую макросистему, обли-
гатным звеном которой являются протисты.
Пищевой фактор. Способы питания простей-
ших весьма разнообразны. К ним относятся: авто-
трофный (голофитный), сапротрофный (сапрозой-
ный), миксотрофный и гетеротрофный (голозойный)
способы питания. Многим простейшим свойственна
широкая трофическая пластичность: каждый вид ис-
пользует в процессе питания не один, а несколько и
даже много пищевых объектов, спектр которых спе-
цифичен только для данного вида.
Сапрофитное питание осуществляется путем по-
глощения органических веществ, растворенных в
воде, с помощью активного или пассивного транс-
порта через поверхностную мембрану клетки —
плазмалемму. В состав веществ, растворенных в воде
океанов, морей, озер, рек и других водоемов, входят
белки, пептиды, аминокислоты, липиды, витамины, а
также продукты метаболизма водных организмов.
Сапрофитный способ питания свойствен в основном
многим мелким бесцветным жгутиконосцам, не име-
ющим цитостома, но среди инфузорий этот тип пита-
ния встречается редко.
Одно из главных направлений эволюционной эко-
логии простейших сопряжено с их адаптацией к пи-
танию твердой пищей, которое обеспечивает необхо-
димыми веществами все виды их жизнедеятельности
и осуществляется путем фагоцитоза. Процесс фаго-
цитоза широко распространен среди саркодовых, ин-
фузорий и жгутиконосцев.
У саркодовых фагоцитоз обеспечивается псевдо-
подиями самых разнообразных типов. С помощью
псевдоподий голые и раковинные амебы, форами-
ниферы, радиолярии и солнечники захватывают пи-
щевые частицы, направляют их в цитоплазму, где
формируется фагосома. После соединения фагосомы
с лизосомами образуется пищеварительная вакуоль
(вторичная лизосома), в которой происходит перева-
ривание пищи. У амеб (Amoeba proteus и других) на-
ряду с активным фагоцитозом происходит питание
путем пиноцитоза, особенно в среде, богатой раство-
ренными органическими веществами. Спектр пище-
вых объектов у саркодовых весьма разнообразен и
включает бактерии, разнообразные водоросли, грибы,
простейших, мелкий детрит и другие объекты.
Многие виды гетеротрофных жгутиконосцев и ин-
фузорий активно захватывают живые организмы и
питаются ими. К этой группе простейших относятся
бактериофаги и хищники. Среди бактериофагов много
седиментаторов, одну из особенностей которых со-
ставляет выделение большого количества слизи, спо-
собствующей слипанию бактерий и других пищевых
частиц. Образующиеся пищевые комки, которые у
инфузорий подгоняются к ротовому отверстию с по-
мощью движения ресничек перистома, оседают на
дне глотки, откуда затем поступают в цитоплазму. К
седиментаторам относятся свободноплавающие ин-
фузории родов Paramecium, Tetrahymena, Stentor и
ряд других, а также перитрихи — прикрепленные ин-
фузории родов Opercularia, Vorticella, Carchesium и
другие.
Все инфузории-седиментаторы имеют сложный
ротовой аппарат, снабженный ресничными комплек-
сами, часто типа мембранелл. Ресничные ряды созда-
ют постоянный ток воды вокруг цитостома, сортируют
и осаждают пищевые организмы и частицы, поступаю-
щие вместе с водой. Рот их нерастяжим и постоянно
открыт.
В качестве одного из типов питания у простейших
широко распространено хищничество, и хищники
известны во всех группах саркодовых, жгутиконосцев
и инфузорий. По отношению к жертвам различают
2 группы: неспециализированных и специализиро-
ванных хищников. Первые активно преследуют жер-
тву, ловят ее и заглатывают в живом состоянии. К
числу неспециализированных хищников относятся,
например, бесцветные эвглениды рода Регапета, ин-
фузории родов Didinium, Euplotes и другие.
Специализированные хищники известны среди ин-
фузорий и жгутиконосцев (Colpodella edax). Реснич-
ные инфузории родов Hemiophrys, Litonotus, Amphi-
leptus сначала убивают свою жертву или парализуют ее
с помощью экструсом (трихоцисты, токсоцисты) и
только после этого заглатывают. Сосущие инфузории
(Suctorida) также относятся к группе специализиро-
ванных хищников. Эти инфузории не имеют цитосто-
ма, но обладают щупальцами, с помощью которых они
ловят, убивают и высасывают добычу.
Фототрофное питание свойственно простейшим,
имеющим хлоропласты. К ним относятся многие
виды хризофитовых, эвгленофитовых, динофитовых,
криптофитовых жгутиковых протистов, которые по
типу питания близки к зеленым водорослям. Однако
ЛИТЕРАТУРА
185
среди эвгленовых существуют виды, обладающие
миксотрофным питанием, формы которого у зеленых
видов эвглен — фотосинтез и гетеротрофное пита-
ние — имеют важное значение в адаптации к меняю-
щимся условиям среды обитания (световой режим,
пища и др.).
В питании простейших могут принимать участие
эндобионты — бактерии и одноклеточные зеленые
водоросли. Они нередко встречаются в цитоплазме
инфузорий (Paramecium bursaria, Stentor polymorphus
и др.). Эндобионты выделяют в клетку хозяина цен-
ные питательные вещества — аминокислоты, сахара,
витамины и другие продукты фотосинтеза. Некото-
рые из них могут защищать своих хозяев от хищни-
ков, выделяя вещества, отпугивающие этих хищников
(Бурковский, 1984).
Наличие пищи, хищники, пищевые конкуренты
являются важнейшими биотическими факторами. От
наличия пищевых ресурсов зависят плотность попу-
ляций простейших в природных условиях, их относи-
тельная стабильность, а также сукцессии в сообщес-
твах. Экспериментальные исследования показали,
что сообщества простейших непостоянны и числен-
ность особей в них может меняться даже в течение
короткого времени. Причинами нестабильности могут
быть истощение пищевых ресурсов и появление хищ-
ников не только из числа простейших, но и много-
клеточных. Популяции простейших с особенно вы-
сокой численностью особей обитают в природных
биотопах, где отсутствуют многоклеточные, которые
питаются простейшими или вытесняют их потому,
что оказываются сильными конкурентами за пищу
(Хаусман, 1988).
К дефициту пищи в среде обитания простейшие
имеют ряд адаптаций, приобретенных в процессе
эволюции: переход на несвойственные им пищевые
объекты, включая каннибализм, образование цист, в
состоянии которых они могут находиться в течение
длительного времени, переживая неблагоприятные
для существования условия.
THE GENERAL CHARACTERIZATION OF PROTISTS
SUMMARY
The chapter includes a general description of protists in accor-
dance with the following scheme.
Introduction. Diversity of protists, their actuality and impor-
tance.
Morphology. Shape and dimensions, coverings, cytoplasmic
structures and organelles (extrusomes, cytoskeleton, flagellar/ciliary
apparatus and its functions, mitochondria, hydrogenosomes, peroxiso-
mes, plastids, Golgi apparatus, lysosomes and other organelles and
inclusions), nucleus (number and dimensions, structural compounds,
mitosis, amitosis).
Reproduction. Asexual reproduction: binary and multiple fis-
sion, colony division. Sexual reproduction: copulation, conjugation
and autogamy, main types of meiosis and its place in life cycles, other
genetic relationships (somatocytogamy, somatokaryogamy), sexual
process origin.
Movement. The moving cell structures and types of movement:
flagellar, amoeboid, metabolic, slime.
Feeding. Pinocytosis, phagocytosis, mysocytosis. Osmoregulation.
Behaviour. Phototaxis, chemotaxis, mechanotaxis, gravitaxis,
termotaxis, galvanotaxis. Kineses.
Life cycles.
Ecology. Marine communities: pelagic and benthic protists.
Freshwater communities: planctonic, benthic, river, fouling and epibi-
onts, of active sludge and air tanks, subsoil waters. Soil communities.
Symbionts. Protists are the hosts of prokaryotes and viruses.
Occurrence of bacteria (ectobionts, endobionts) and viruses.
ЛИТЕРАТУРА12
(Агамалиев Ф. Г.) Agamaliev F. G. Ciliates of the low-salinity
lagoons of the Caspian Sea // Arch. Protistenk. 1986. Bd 131, H. 162.
S. 201—214.
Агамалиев Ф. Г. Фауна и распределение инфузорий супрали-
торальной зоны Апшеронского побережья Каспия // Экология сво-
бодноживущих морских и пресноводных простейших. Л.: Наука,
1990. С. 58—63. (Протозоология; Вып. 13).
Александров В. Я. Специфическое и неспецифическое в реак-
ции клетки на повреждающие воздействия И Тр. Ин-та цитол., гис-
тол., эмбриол. 1948. Т. 3, № 1. С. 3—82.
Александров В. Я. О биологическом смысле соответствия
уровня теплоустойчивости белков температурным условиям сущес-
твования вида // Успехи соврем, биол. 1965. Т. 60, № 1 (4). С. 28—
44.
Атлас и определитель пелагобионтов Байкала. Новосибирск,
1995.
Банина Н. Н. Peritricha sessilida в биоценозе активного ила.
Л.: Наука, 1983. С. 87—116. (Протозоология; Вып. 8).
Банина Н. Н. Тип Инфузории — Ciliophora Doflein, 1941 //
Фауна аэротенков. Атлас. Л.: Наука, 1984. С. 136—186.
Банина Н. Н. Эволюция прикрепленных перитрих с адаптив-
но — экологической точки зрения # Экология свободноживущих
морских и пресноводных простейших. Л.: Наука, 1990. С. 99—
117. (Протозоология; Вып. 13).
12 Цитируемая и рекомендуемая литература к разделам «Систе-
ма протистов и проблемы их мегасистематики», «Пути эволюции
протистов», «Общая характеристика протистов».
Бейер Т. В. Клеточная биология споровиков — возбудителей
протозойных болезней животных и человека. Л., 1989.
Биологический энциклопедический словарь. М., 1986.
Бохински Р. Современные воззрения в биохимии. М., 1987.
Бурковский И. В. Экология свободноживущих инфузорий. М.:
Изд-во МГУ, 1984.
Вассер С. П. (ред.). Водоросли. Киев, 1989
Вермеяь Е. М. История учения о клетке. М., 1970.
Видаль Г. Древнейшие эукариотные клетки // В мире науки.
1984. Т. 4, № 1.
Викол М. М. Раковинные корненожки (Rhizopoda, Testacea) и
их роль в продукционно-деструкционных процессах водоемов //
Экология свободноживущих морских и пресноводных простей-
ших. Л.: Наука, 1990. С. 121—132. (Протозоология; Вып. 13).
Виноградова К. Л. Класс сифоновых (Siphonophyceae) //
Жизнь растений. М., 1977. Т. 3.
Виноградова К. Л. Красные водоросли И Определитель пре-
сноводных водорослей СССР. Л., 1980. Вып. 13.
(Гаевская Н. С.) Gajewskaja N. S. Zur Oekologie, Morphologie
und Systematik der Infusorien des Baikalsees // Zoologica. 1933.
Vol. 32.
Гаузе Г. Ф. Математический подход к проблемам борьбы за
существование // Зоол. журн. 1933. Т. 12, № 3. С. 3—16.
Гаузе Г. Ф. Экспериментальное исследование борьбы за су-
ществование между Paramecium caudatum, Р. aurelia, Stylonychia
mytilis И Зоол. журн. 1934. Т. 3,№ 1. С. 1—17.
Гаузе Г. Ф. Наблюдения над стабилизирующим отбором в
культурах солоноватоводных инфузорий рода Euplotes И Зоол.
журн. 1940. Т. 19, № 3. С. 363—378.
186
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Гельцер Ю. Г. «Псевдокристаллизация» — особый способ пе-
ренесения неблагоприятных условий существования у Colpoda ma-
upasii из крайнеаридных почв Заалтайской Гоби (МНР) И Цитоло-
гия. 1988. Т. 30, № 11. С. 1386—1389.
Гельцер Ю. Г. Простейшие (Protozoa) как компонент почвен-
ной биоты (систематика, экология). М.: Изд-во МГУ, 1993.
Григорян Д. А. Зависимость теплоустойчивости Paramecium
caudatum от концентрации ионов Са в среде И Цитология. 1964.
Т. 6, № 1. С. 105—109.
Громов Б. В. Строение бактерий. Л.: ЛГУ, 1985. 189 с.
Догадина Т.В., Матвиенко А. М. Желто-зеленые водоросли
(Xanthophyta) // Жизнь растений. М., 1977. Т. 3.
Догель В. А. Простейшие, как совершенные организмы // При-
рода. 1925. № 4—6.
(Догель В. A.) Dogiel V. A. Polymerization als Prinzip der prog-
ressiven Entwicklung bei Protozoen // Biol. Zbl. 1929. Bd 49, S. 451—
469.
Догель В. А. Зоология беспозвоночных. M., 1931.
Догель В. А. Общая протистология. М.: Сов. наука, 1951.
Догель В. А. Олигомеризация гомологичных органов как один
из главных путей эволюции животных. Л., 1954.
Догель В. А. Зоология беспозвоночных. М.: Высш, шк.,
1981.
Догель В. А., Полянский Ю. И., Хейсин Е. М. Общая прото-
зоология, М., Л.: Изд-во АН СССР, 1962.
Ефимов В. В. Вымерзание и переохлаждение простейших й
Архив Рус. протистол. о-ва. 1922. Т. 1. С. 153—168.
Жариков В. В. Инфузории — индикаторы сапробного состоя-
ния среды волжских водохранилищ й Биоиндикация, методы, при-
ложения. Тольятти : 1994. С. 88—99.
Жариков В. В. Кадастр свободноживущих инфузорий водо-
хранилищ Волги (состав, распределение по водохранилищам,
обзор методов исследований). Тольятти: ИЭВБ РАН, 1996.
Жариков В. В. Свободноживущие инфузории Волги: Авто-
реф. дис. ... докт. биол. наук. СПб., 1999.
Жариков В. В., Ротарь Ю. М., Быкова С. В. Биоиндикация
качества воды по инфузориям — индикаторам й Волжский бас-
сейн: экологическая ситуация и пути рационального природополь-
зования. Тольятти, 1996. С. 45—49.
Жинкин Л. Н. Жир и причина его образования у инфузории
Stentor polymorphus Shrbg. й Тр. Петергоф, естеств.-науч. ин-та.
1929. Т. 6. С. 199—215.
(Жинкин Л. Н.) Zhinkin L. N. Zur Frage der Reservestoffe bei
Infusorien // Ztschr. Morphol. (Okol. Tiere. 1930. Bd 18, N 1—2.
S. 199—207.
Жуков Б. Ф. К фауне зоофлагеллат Японского моря й Биоло-
гия моря. 1983. Вып. 1. С. 57—58.
Жуков Б. Ф. Атлас пресноводных гетеротрофных жгутиконос-
цев. Рыбинск, 1993.
Жуков Б. Ф., Карпов С. А. Пресноводные воротничковые жгу-
тиконосцы. Л.: Наука, 1985.
Жуков Б. Ф., Мыльников А.П. Фауна зоофлагеллат очист-
ных сооружений И Простейшие активного ила. Л.: Наука, 1983.
С. 27—42.
Завадский К. М., Колчинский Э. И. Эволюция эволюции. Л.,
1977.
Заварзин А. А., Харазова А. Д., Молитвин М. Н. Биология
клетки. СПб.: Изд-во СПбГУ, 1992.
Заика В. Е. Распределение морского микропланктона. Киев,
1976.
Захваткин А. А. Сравнительная эмбриология низших беспоз-
воночных. М., 1949.
Иванов А. В. Происхождение многоклеточных животных. Л.,
1968.
(Иванов А. В.) Ivanov А. V. On the independent and conver-
gent origin of sexual process // Monit. Zool. Ital., n. ser. 1970. Vol. 4,
P. 189—193.
Инге-Вечтомов С. Г. Генетика с основами селекции. M., 1989.
Ирлина И. С. О стойкости Paramecium caudatum, адаптиро-
ванных к различным температурам, к повреждающему действию
солей // Морфология и физиология простейших. М.; Л., 1963.
С. 92—101.
Карпов С. А. Строение покровов жгутиконосцев // Цитология.
1986. Т. 28, № 2. С. 139—150.
Карпов С. А. Анализ отрядов Phalansteriida, Spongomonadida и
Thaumatomonadida // Зоол. журн. 1990а. Т. 69. № 3. С. 5—12.
Карпов С. А. Система протистов. Омск, 19906.
Карпов С. А. Пути эволюции кинетиды у протистов // Вести.
СПбГУ. 1993а. Вып. 3, № 17. С. 28—37.
Карпов С. А. Ультратонкое строение бесцветного жгутиконосца
Thaumatomonas seravini й Цитология, 19936. Т. 35, № 9. С. 8—11.
(Карпов С. A.) Karpov S. A. (ed.). The biology of free-living
heterotrophic flagellates. St. Petersburg: Nauka, 1995. (Cytology;
Vol. 37, N 11).
(Карпов C. A.) Karpov S. A. Cercomonads and their relationship
to the Myxomycetes // Arch. Protistenk. 1997. Bd 148. P. 297—307.
Карпов С. А., Жгарев H. А. О способах размножения ворот-
ничковых жгутиконосцев й Зоол. журн. 1987. Т. 66, № 1. С. 130—
133.
Карпов С. А., Жуков Б. Ф. Ультратонкое строение бесцветно-
го жгутиконосца Pleuromonas jaculans Petty (Kinetoplastida, Zoo-
mastigophorea) // Простейшие активного ила. Л.: Наука, 1983.
С. 153—156.
(Карпов С. А., Жуков Б. Ф.) Karpov S. A., Zhukov B.F. Ultra-
structure and taxonomic position of Apusomonas proboscidea Alexei-
eff // Arch. Protistenk. 1986. Bd 131. P. 13—26.
Карпов С. А., Жуков Б. Ф. Цитологические особенности бес-
цветного жгутиконосца Thaumatomonas lauterborni / Цитология.
1987. Т. 29, № 10. С. 1168—1171.
Карпов С. А., Мыльников А. П. Биология и ультраструктура
бесцветных жгутиконосцев Apusomonadida ord. п. И Зоол. журн.
1989. Т. 68, № 8. С. 5—17.
(Карпов С. А., Мыльников А. П.) Karpov S. A., Mylnikov А. Р.
The ultrathin structure of colourless flagellate Hyperamoeba flagellata
with special reference to the flagellar apparatus // Eur. J. Protistol.
1997. Vol. 33. P. 349—355.
(Карпов С. А., Таничев А. И.) Karpov S. A., Tanichev A. I.
The ultrastructural study of green alga Pedinomonas tenuis Massjuk,
1970 with special reference to the flagellar apparatus // Arch. Protis-
tenk. 1992. Vol. 141, N 4. P. 315—326.
(Карпов С. А., Фокин С. И.) Karpov S. A., Fokin S. I. The
structural diversity of flagellar transitional zone in heterotrophic fla-
gellates and other protists // The biology of free-living heterotrophic
flagellates / Ed. S. A. Karpov. 1995. P. 1038—1052 (Cytology;
Vol. 37, N 11).
Ковалёва H. E. Влияние температуры культивирования на чув-
ствительность инфузорий к повреждающему действию рентгено-
вых лучей й Цитология. 1962. Т. 4, № 3. С. 306—317.
Корганова Г. А. Почвенные раковинные амебы (Protozoa, Tes-
tacea): фауна, экология, принципы организации сообществ. Авто-
реф. дис. ... докт. биол. наук. М., 1997.
Кусакин О. Г., Дроздов А. Л. Филема органического мира.
Часть 1: Пролегомены к построению филемы. СПб.: Наука, 1994.
Кусакин О. Г., Дроздов А. Л. Филема органического мира.
Часть II: Прокариоты и низшие евкариоты. СПб.: Наука, 1998.
Лепинис А. К., Гельцер Ю. Г., Чибисова О. И., Гептнер В. А.
Определитель Protozoa почв Европейской части СССР. Вильнюс,
1973.
Липеровская Е. М. Простейшие как биоиндикаторы степени
очистки й Итоги науки и техники. М.: 1977. Т. 4. С. 105—140.
Лозина-Лозинский Л. К. Устойчивость к различным агентам
парамеций, адаптированных к жизни в горячем радиоактивном ис-
точнике й Цитология. Т. 3, № 2. 1961. С. 154—166.
Лозина-Лозинский Л. К. Устойчивость насекомых к глубоко-
му охлаждению и внутриклеточному замерзанию И Клетка и темпе-
ратура среды. М.; Л.: Наука, 1964. С. 66—72.
Лозина-Лозинский Л. К. Очерки по криобиологии. Л.: Наука,
1972.
Львова Т. Г. Жизненный цикл и соленостные адаптации Mic-
ronephthys minuta Thell (Nephtuidae, Polychaeta) в Белом море: Ав-
тореф. дис. ... канд. биол. наук. Л., 1981.
Льюин Б. Гены. М., 1988.
Мажейкайте С. И. Класс Ресничные инфузории — Ciliata //
Определитель пресноводных беспозвоночных Европейской части
СССР. Л.: Гидрометизд., 1977. С. 46—97.
Макрушин А. В. Биологический анализ качества вод. Л., 1974.
Мамаева Н. В. Инфузории бассейна Волги. Л.: Наука, 1979.
Масюк Н. П. Строение тела особи й Водоросли. Справочник.
Киев : Наукова думка, 1989. С. 63—74.
Масюк Н. П. Эволюционные аспекты морфологии эукариоти-
ческих водорослей. Киев : Наукова думка, 1993.
Мережковский К. Этюды над простейшими животными Севе-
ра России // Тр. СПб. о-ва естествоисп. 1878. № 1.
Метальников С. И. К физиологии внутриклеточного пищева-
рения у простейших. СПб., 1911.
Микрюков К. А., Мыльников А. П. Новые данные о строении
и жизненном цикле аталамидных амеб (Protista, Athalamida) //
Зоол. журн. 1996. Т. 75, вып. 9. С. 1283—1293.
ЛИТЕРАТУРА
187
Михалевич В. И. Ультраструктура фораминифер / Проблемы
современной микропалеонтологии. Л.: Наука, 1990. С. 36—46.
Михалевич В. И. Макросистема фораминифер: Автореф.
дис. ... докт. биол. наук. СПб., 1992.
Моисеев Е. В. К изучению зоофлагеллат Черного моря // Се-
зонные изменения Черноморского планктона. М., 1983. С. 100—
112.
Мыльников А. П. Строение экструсом некоторых жгутико-
носцев // Цитология. 1988. Т. 30, № 12. С. 1402—1408.
Мыльников А. П. Биология и ультраструктура амебоидных
жгутиконосцев Cercomonadida ord. nov. И Зоол.журн. 1986. Т. 65.
€. 683—692.
Мыльников А. П. Корешковая система жгутиков церкомона-
дид // Информ, бюл. Л. : Наука, 1989а. № 81. С. 52—54.
Мыльников А. П. Ультраструктура и систематическое поло-
жение Pteridomonas pulex (Pedinellales). Борок : ИБВВ АН СССР,
19896. Деп. в ВИНИТИ 11.01.89. № 272-В 89.
Мыльников А. П. Строение стрекательных органелл хищного
жгутиконосца Katablepharis sp. И БВВ. Информ, бюл. 1992. № 95.
С. 28—33.
Мыльников А. П. Тонкое строение бесцветной хризомонады
Monas sp. // БВВ. Информ, бюл. 1994. № 96. С. 16—21.
Никитина Л. И. Почвенные инфузории Среднего Приамурья.
Хабаровск: Хабар, гос. ун-т, 1997.
Николюк В. Ф., Гельцер Ю. Г. Почвенные простейшие СССР.
Ташкент, 1972.
Определитель паразитов пресноводных рыб фауны СССР.
Т. 1—2 / Под ред. О. Н. Бауера. Л.: Наука. 1984.
Орлова А. Ф. Длительные модификации у Paramecium cauda-
tum и Paramecium multimicronucleatum И Зоол. журн. 1941. Т. 20,
№ 3. С. 341—370.
Орлова А. Ф. Об адаптациях и длительных модификациях у
инфузорий И Зоол. журн. 1947. Т. 26, № 6. С. 521—530.
Осипов Д. В. Анализ наследственных механизмов, определяю-
щих теплоустойчивость Paramecium caudatum И Генетика. 1966.
№1.0 119—131.
(Осипов Д. В., Карпов С. А., Смирнов А. В., Раутиан М. С.)
Ossipov D. V., Karpov S. A., Smirnov А. V., Rautian М. S. Prokari-
otic endo- and ectocytobionts in protists // Acta Protistol. 1997.
Vol. 37. P. 1—18.
Полевой В. В. Физиология растений. М., 1989.
Полянский В. И, Экспериментальное исследование изменчи-
вости Euglena gracilis Klebs в условиях культуры // Учен. зап.
ЛГПИ. 1948. Т. 70. С. 153—170.
Полянский Ю. И. Температурные адаптации у инфузорий. I.
Зависимость теплоустойчивости Paramecium caudatum от темпера-
турных условий существования // Зоол. журн. 1957. Т. 36, № 11.
С. 1630—1646.
Полянский Ю. И. Температурные адаптации у инфузорий. II.
Изменение теплоустойчивости и холодоустойчивости Paramecium
caudatum при культивировании при низких температурах И Цито-
логия. 1959. Т. 1, № 6. С. 714—727.
Полянский Ю. И. О способности инфузории туфельки (Para-
mecium caudatum) переносить отрицательные температуры // Acta
protozool. 1963. Vol. 1, № 1. Р. 165—175.
Полянский Ю. И. О некоторых морфологических закономер-
ностях эволюции паразитических животных // Паразитол. сб. Л.,
1969. Т. 25.
Полянский Ю. И. О своеобразных чертах прогрессивной эво-
люции на клеточном уровне организации // Журн. общ. биол. 1971.
Т. 32, № 5.
Полянский Ю. И. Качественные особенности микро- и макро-
эволюционных процессов на клеточном уровне организации И
Вести. ЛГУ. 1981а. № 21.
Полянский Ю. И. О специфике эволюционного процесса в
крупных таксонах (на примере эволюции простейших) // Пробле-
мы новейшей истории эволюционного учения. Л., 19816.
Полянский Ю. И., Райков И. Б. Полимеризация и олигомери-
зация в эволюции простейших // Журн. общ. биол. 1977. Т. 38,
№ 3. С. 325—335.
Полянский Ю. И., Суханова К. М., Сопина В. А., Юдин А. Л.
Устойчивость Amoeba proteus к действию летальной температуры
и этилового спирта И Изменчивость теплоустойчивости клеток жи-
вотных в онто- и филогенезе. М.; Л.: Наука, 1967. С. 43—62.
Помряскинская Н. Наблюдения над цистами брюхоресничной
инфузории Oxytricha hymenostoma // Учен. зап. ЛГПИ. 1940. Т. 30.
С. 93—132.
Раилкин А. И. Процессы колонизации и защита от обраста-
ний. СПб., 1998.
Раилкин А. И., Жуков Б. Ф., Золотарёв В. А. Фауна, эколо-
гия и распределение бесцветных гетеротрофных флагеллат на ис-
кусственных субстратах в Белом море // Экология свободноживу-
щих морских и пресноводных простейших. Л.: Наука, 1990.
С. 47—57. (Протозоология; Вып. 13).
Райков И. Б. Ядро простейших, морфология и эволюция. Л.,
1978.
(Райков И. Б.) Raikov I. В. The protozoon nucleus. Morphology
and evolution. Wien; New York: Springer. 1982. Vol. 9.
(Райков И. Б.) Raikov I. В. The diversity of forms of mitosis in
Protozoa: a comparative review. Eur. J. Protistol. 1994. Vol. 30.
P. 253—269.
(Райков И. Б.) Raikov I. В. Meiosis in protists: recent advances
and persisting problems. Eur. J. Protistol. 1995. Vol. 31. P. 1—7.
Северцов A. H. Морфологические закономерности эволюции.
M.; Л. : Изд-во АН СССР, 1939.
Северцов А. Н. Общие вопросы эволюции И Собр. соч. М.,
1945. Т. 3.
Серавин Л. Н. Двигательные системы простейших. Строение,
механохимия и физиология. Л.: Наука, 1967.
Серавин Л. Н. Простейшие... Что это такое? Л.: Наука, 1984.
Серавин Л. Н. Происхождение эукариотной клетки. IV. Общая
гипотеза аутогенного происхождения эукариот // Цитология. 1986.
Т. 28, № 9. С. 899—910.
Серавин Л. Н. Существует ли в действительности такой так-
сон — подцарство Protozoa? И Биол. науки. 1989. № 7. С. 4—13.
Серавин Л. Н. Основные этапы развития клеточной теории и
место клетки среди живых систем И Цитология. 1991. Т. 33,
№ 12. С. 3—27.
Серавин Л. Н. Эукариоты, лишенные важнейших клеточных
органелл (жгутиков, аппарата Гольджи, митохондрий) и главная
задача органеллологии // Цитология. 1992. Т. 34, № 5. С. 3—33.
Серавин Л. Н. Основные типы и формы тонкого строения
крист митохондрий; степень их эволюционной консервативности
(способности к морфологической трансформации) И Цитология.
1993. Т. 35, № 4. С. 3—12.
Серавин Л. Н., Гудков А. В. Спонтанные слияния клеток в
культуре морской амебы Hyperamoeba fallax U Цитология. 1983.
Т. 25, № 2. С. 194—199.
Серавин Л. Н., Гудков А. В. Основные типы и формы агамных
слияний клеток у простейших И Цитология. 1984. Т. 26, № 2.
С. 123—131.
Серавин Л. Н., Гудков А. В. Цитоплазматические гранулы, по-
добные микротельцам, у амебы Pelomyxa palustris # Цитология.
1987а. Т. 29, № 5. С. 600—603.
Серавин Л. Н., Гудков А. В. Аппарат Гольджи у амебы Pelo-
myxa palustris // ДАН СССР. 19876. Т. 296, № 1. С. 249—250.
Серавин Л. Н., Гудков А. В. Агамные слияния протистов и
происхождение полового процесса. СПб.; Омск, 1999.
Серавин Л. Н., Фролов А. О. Метаболирующее движение как
одна из основных форм клеточного движения // Цитология. 1983.
Т. 25, № 12. С. 1343—1352.
Смарагдова Н. П. Географическая изменчивость парамеций и
роль стабилизирующего отбора в происхождении географических
различий И Журн. общ. биол. М., 1941. Т. 2, № 1. С. 71—84.
Сопина В. А. Устойчивость эндопаразитических простейших
некоторых видов амфибий к этиловому спирту // Цитология. 1963.
Т. 5, № 3. С. 343—347.
Сорокин Ю. И. Зоофлагеллаты как компонент сообщества эв-
трофных и олиготрофных вод Тихого океана № Океанология. 1979.
Т. 19, № 3. С. 476—480.
Старобогатов Я. И. К вопросу о числе царств эукариотных
организмов // Систематика простейших и их физиологические
связи с низшими эукариотами. Л.: ЗИН АН СССР, 1986. С.
4—25.
Суханова К. М. Исследование температурных адаптаций и
особенностей морфологии некоторых видов инфузорий Astomata
из олигохет Восточного Мурмана // Морфология и физиология
простейших. М.; Л.: Наука, 1963. С. 75—90.
Суханова К. М. Температурные адаптации у простейших. Л.:
Наука, 1968.
(Суханова К. М.) Sukhanova К. М. Some mechanisms of adap-
tive cell reactions in agamic Protozoa // Progress in Protozoology.
Clermont, 1973. T. 1. P. 408—409.
Суханова И. H., Чебан Е. А. Гетеротрофный фитопланктон
Черного моря в ранневесенний период развития фитоцена // Океа-
нология. 1990. Т. 30, вып. 6.
Тахтаджян А. Л. Морфологическая эволюция покрытосемян-
ных. М., 1948.
188
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Тахтаджян А. Л. Вопросы эволюционной морфологии расте-
ний. Л., 1954.
Тахтаджян А. Л. Макроэволюционные процессы в истории
растительного мира И Бот. журн. 1983. Т. 68, № 12. С. 1593—1603.
(Тахтаджян A.) Thakhtajan A. Evolutionary trends in flowering
plants. New York, 1991.
Tepac Ю. X. Формы взаимоотношений простейших с вируса-
ми // Взаимоотношения простейших с вирусами. Л. : Наука, 1981.
С. 31—55.
Тимофеев Б. В. Микрофитофоссилии раннего докембрия. Л.,
1982.
Тимофеев Б. В., Герман Т. Н., Михайлова Н. С. Микрофито-
фоссилии докембрия, кембрия и ордовика. Л., 1976.
Толмашова Г. Т. Адаптивная изменчивость эвглен и астазий в
среде с повышенным содержанием кальция И Экология свободно-
живущих морских и пресноводных простейших. Л. : Наука, 1990.
С. 163—186. (Протозоология; Вып. 13).
Ушаков Б. П. Исследование теплоустойчивости клеток и про-
топлазматических белков пойкилотермных животных в связи с
проблемой вида // Клетка и температура среды. М.; Л.: Наука,
1964. С. 214—222.
Ушаков Б. П. Проблема сопряженного изменения белков в
процессе видообразования // Цитология. 1965. Т. 7, № 4. С. 467—
480.
Ушаков Б. П., Кусакина А. А. О лабильности и консерватив-
ности адаптации клеток животных, обнаруживаемой на белковом
уровне // Цитология. 1960. Т. 2, № 4. С. 428—441.
Фауна аэротенков: Атлас. Л., Наука, 1984.
Федонкин М. А. Бесскелетная фауна венда и ее место в эво-
люции метазоа. М.: Наука, 1987.
(Фокин С. И., Карпов С. A.) Fokin S. I., Karpov S. A. Bacte-
rial endocytobionts inhabiting the perinuclear space of Protista // En-
docytob. Cell Res. 1995. Vol. 11. P. 81—94.
Хаусман К. Протозоология. M.: Мир, 1988.
Хлебович В. В. Особенности состава водной фауны в зависи-
мости от солености среды И Журн. общ. биол. 1962. Т. 23, № 2
С. 90—97.
Хлебович В. В. Критическая соленость биологических про-
цессов. Л.: Наука, 1974.
Ченцов Ю. С. Общая цитология. М., 1984.
Чорик Ф. П. Свободноживущие инфузории водоемов Молда-
вии. Кишинев, 1968.
Шмальгаузен И. И. Проблемы дарвинизма. Л., 1969.
Шмальгаузен И. И. Пути и закономерности эволюционного
процесса. М., 1982.
Шульман С. С., Донец 3. С., Ковалёва А. А. Класс миксоспо-
ридий мировой фауны. Т. 1. Общая часть. СПб.: Наука, 1997.
Юдин А. Л. Ядерно-цитоплазматические взаимоотношения:
клеточная наследственность у амеб И Л.: Наука, 1982.
Янаускас Т. В. (ред.) Микрофоссилии докембрия СССР. Л.,
1989.
Adoutte A., Baroin A., Perasso R. Molecular phylogeny of uni-
cellular eukaryotes based on partial sequencies of ribosomal RNA //
Moll. Zool. 1989. Vol. 56. P. 225.
Andersen R. A. Diversity of eukaryotic algae // Biodiversity and
Conservation. 1992. Vol. 1. P. 267—292.
Andersen R. A., Wetherbee R. Microtubules of the flagellar ap-
paratus are active during prey capture in the chrysophycean alga Epi-
pyxis pulchra // Protoplasma. 1992. Vol. 166. P. 8—120.
Anderson C. L. Phylogenetic relationshis of the Myxozoa // Evo-
lutionary relationships among Protozoa / Eds G. H. Coombs et al.
London : Kluwer, 1998. P. 341—350.
Barr D. J. S. Evolution and kingdoms of organisms from the per-
spective of a mycologist // Mycologia, 1992. Vol. 84. P. 1—11.
Beakes G. W. Relationships between lower fungi and protozoa //
Evolutionary relationships among Protozoa / Eds G. H. Coombs et al.
London : Kluwer, 1998. P. 351—374.
(Begon M., Harper J. L., Townsend C. R.) Битон M., Хар-
пер Дж., Таунсенд К. Экология. Особи, популяции и сообщества.
М.: Мир, 1989. Т. 1.
Bick Н., Kunze S. Eine Zusammenstellung von autbkologischen
und saprobiologischen Befunden an Siipwasserciliaten # Hydrobiolo-
gia. 1971. Bd 58, N 3. S. 337—384.
Bonnet L., Thomas R. Thecamoebines du sol. Paris, 1960.
Bouck G. B. The structure, origin, isolation and composition of
the tubular mastigonemes of the Ochromonas flagellum // J. Cell Biol.
1971. Vol. 50. P. 362—384.
Broers С. A. M., Meijers H. H. M., Symens J. C., Stumm С. K.,
Vogels G. D., Brugerolle G. Symbiotic association of Psalteromonas
vulgaris n. spec, with Methanobacterium formicicum // Eur. J. Protis-
tol. 1993. Vol. 29. P. 98—105.
Brugerolle G. Contribution a 1’etude cytologique et phyletique
des diplozoaires (Zoomastigophorea, Diplozoa, Dangeard 1910). VI.
Caracteres generaux des diplozoaires Й Protistologica. 1975. T. 11.
P. 111—118.
Brugerolle G. Etude ultrastructurale du flagelle parasite Polymas-
tix melolonthae (Oxymonadida) // Protistologica. 1981. T. 17. P. 139—
145.
Brugerolle G. Flagellar and cytoskeletal systems in amitochond-
rial flagellates: Archamoeba, Metamonada and Parabasala // Protoplas-
ma, 1991. Vol. 164. P. 70—90.
Camp R. R., Mattern C. F. T., Honigberg В. M. Studies of Dien-
tamoeba fragilis Jepp and Dobell // J. Protozool. 1974. Vol. 21.
P. 69—82.
Canning E. U. Evolutionary relationships of Microsporidia //
Evolutionary relationships among Protozoa I Eds G. H. Coombs et al.
London: Kluwer, 1998. P. 77—90.
Cavalier-Smith T. Eukaryotes with no mitochondria // Nature.
1987. Vol. 326. P. 332—333.
Cavalier-Smith T. Archezoa and origin of Protozoa // Abstr. pap.
8th. Int. congr. protozool. Tsukuba, Japan, 1989.
Cavalier-Smith T. Kingdom Protozoa and its 18 phyla Й Micro-
biol. Rev. 1993a. Vol. 57. P. 953—994.
Cavalier-Smith T. The protozoan phylum Opalozoa // J. Euk.
Microb. 1993b. Vol. 40. P. 609—615.
Cavalier-Smith T. Neomonada and the origin of animals and fun-
gi // Evolutionary Relationships Among Protozoa / Eds G. H. Coombs
et al. London: Kluwer, 1998. P. 375—408.
Cook J. R. Adaptations to temperature in two closely related stra-
ins of Euglena gracilis // Biol. Bull. 1966. Vol. 131, N 1. P. 83—93.
Cook J. R. Influence of culture pH and phosphate on synchrony
of Euglena gracilis // Exper. Cell Res. 1971. Vol. 69. P. 207—211.
Copeland H. F. The classification of lower organisms. Palo Alto,
1956.
Corliss J. O. The kingdom Protista and its 45 phyla // BioSys-
tems. 1984. Vol. 17. P. 87—126.
Corliss J. O. An interim utilitarian («User-friendly») hierarchical
classification and characterization of the protists // Acta Protozoologi-
es 1994. Vol. 33. P. 1—51.
Corliss J. O. Classification of protozoa and protists: the current
status II Evolutionary Relationships Among Protozoa / Eds G. H. Co-
ombs et al. London: Kluwer, 1998. P. 409—448.
(Crippa-Franceschi J.) Криппа-Франчески T. Длительные мо-
дификации И Журн. общ. биол. 1970. Т. 31, № 5. С. 572—577.
Daniel G. Е., Chalkley Н. W. The influence of temperature upon
the process of division in Amoebae proteus // J. Coll. Comp. Physiol.
1932. Vol. 2, N3. P. 311—327.
Dobell С. C. The principles of protistology // Arch. Protistenk.
1911. Bd 23, S. 269—310.
Doflein F. Lehrbuch der Protozoenkunde. Jena, 1953.
Dragesco J. Quelques Donnee ecologiques sur les cilies libres de
1’Afrique // Amer. zool. 1973. Vol. 13. P. 231—232.
Evolutionary relationships among Protozoa / Eds G. H. Coombs
et al. London: Kluwer, 1998.
Eyden H., van den, Baere, de Wachter R. Sequence and secon-
dary structure of Porphyra umbilicali 5S rRNA. Relevance for the
evolutionary origin of red algae // Nucl. Acids Res. 1988. Vol. 16.
P.10919.
Faure-Fremiet E. Ecologie des psammophiles littoraux // Bull.
Biol. France Belg. 1950. Vol. 84, N 1. P. 35—75.
Felsenstein J. Numerical methods for inferring evolutionary trees.
Quart. Rev. Biol. 1982. Vol. 57. P. 379—404.
Fenchel T. Ecology of Protozoa. The Biology of Free-living pha-
gotrophic Protists. Berlin etc., 1987.
Fenchel T., Finley В. I. The biology of free-living anaerobic cili-
ates II Eur. J. Protistol. 1991. Vol. 26. P. 201—215.
(Finley H. E.) Финлей Г. E. Экология кругоресничных инфузо-
рий И Успехи протозоологии. Л., 1969. Т. 3. С. 191—193.
Finley В. I., Fenchel Т. Hydrogenosomes in some anaerobic pro-
tozoa resemble mitochondria // FEMS Microbiol. Lett. 1989. Vol. 65.
P. 311—314.
Franceschi J. Relacione tra e ritmo di scissione in colpoda
(C. maupasii e cucullus) // Boll. Zool. 1959. Vol. 26, N 2. P. 637—651.
(Genermont J.) Женермон Ж. Проблема длительных модифи-
каций у простейших // Журн. общ. биол. 1970. Т. 31,142 6. С. 661—
671.
ЛИТЕРАТУРА
189
Golemansky V. Adaptations morphologicues des Thecamoebiens
psammobiontes du psommal supralittoral des mer // Acta protozool.
1978. Vol. 17. P. 141—152.
Golemansky V. Thecamoebiens intersticiales du supralittoral sab-
leux des mers. Sofia, 1980.
Grain J. The cytoskeleton in protists: nature, structure and func-
tions II Int. Rev. Cytol. 1986. Vol. 104. P. 153—249.
Haeckel E. Generelle Morphologie der Organismen. Bd 2. Berlin,
1866. CLX. 462 S.
Hartwig E. Die Ciliaten des gezeiten — sandtrandes der Nordsee-
insel, sylt. // Microfauna des Meersbodens. 1973. Bd 18. S.,1—69.
Hasegawa M., Hashimoto T., Adachi J., Iwabe N., Miyata T.
Early branchings in the evolution of eukaryotes: ancient divergence of
Entamoeba that lack mitochondria reveales by protein sequence data //
J. Mol. Evol. 1993.Vol. 36, P. 380—388.
Hashimoto T., Sdnchez L. B., Shirakura T., Miiller M., Hasega-
wa M. Secondary absence of mitochondria in Giardia lamlia and Tri-
chomonas vaginalis revealed by valyl-tRNA synthetase phylogeny //
Proc. Nat. Acad. Sci. USA. 1998. Vol. 95. P. 6860—6865.
Hausmann K, Hiilsmann N. Protozoology. Stuttgart, New York:
Thieme, 1996.
Hogg J. On the distributions of a plant and an animal, and on a
fourth Kingdom of nature. Edinburg New Philos. J. (N. S.). 1860.
Vol. 12. P. 216—225.
Hollande A. Le deroulement de la cryptomitose et les modalites
de la segregation des chromatides dans quelques groupes de Protozoa-
ires II Annee biol. 1972. Vol. 11. P. 427—466.
Hook D.D., Crawford R.M.M. (eds.) Plant life in anaerobic
environments. Collingwood, 1978.
Hori H., Lim B.-L., Osawa S. Evolution of green plants as dedu-
ced from 5S RNA sequence II Proc.Nat. Acad. Sci. USA. 1985.
Vol. 82, P. 820—823.
Hungate R. E. The rumen microbial ecosystem // Ann. Rev. Ecol.
a Systematics. 1975. Vol. 6, N 1. P. 39—66.
Hiilsmann N., Hausmann K. Towards a new perspective in Pro-
tozoan Evolution // Europ. J. Protistol. 1994. Vol. 30. P. 365—374.
Jellery G. M. Extended low — temperature preservation of
human malaria parasites II J. Parasitol. 1957. Vol. 43, N 4. P. 488—
509.
Kahan D., Scharon R. Effect of temperature on growth, cell size,
and free amino acid pool of the thermophilic ciliate cyclidium citrul-
lus II J. Protozoology. 1976. Vol. 23, N 3. P. 478—481.
Kahl A. Ciliata librea et ectocommensalia // Die Tiere der Nord —
und Ostsee. Leipzig, 1933. Bd 11, S. 29—146.
Kudo R. R. Protozoology. IV ed. Illinois (USA): Springfield:
Thomas. 1966.
Lechtreck K.-F., McFadden G. I., Melkonian M. The cytoske-
leton of naked green flagellate Spermatozopsis similis: isolation,
whole mount electron microscopy, and immunological charac-
terization. Cell Motil. Cytosk. 1989. Vol. 14. P. 552—561.
Lemke P. A. Viruses in eukaryotic microorganisms И Ann. Rev.
Microbiol., 1976. Vol. 30. P. 105—145.
Levine N. D., Corliss J. €>., Cox F. E. G. et al. A new revised
classification of the Protozoa U J. Protozool. 1980. Vol. 27. P. 37—
58.
Lipscomb D.L., Riordan G. P. Ultrastructural examination of
virus-like particles in a marine rhizopods (Sarcodina, Protista) # Acta
Protozool. 1995. Vol. 34. P. 35-44.
Margulis L. Biodiversity: molecular biological domain, symbiosis
and kingdom origin // BioSystems. 1992. Vol. 27. P. 39—51.
Meier D. Untersuchungen uber der Dauercysten von Colpoda cu-
cullus O. F. Muller (Holotricha, Ciliata) gegeniiber der Einwirkung
einiger chemisches und physicalisches abiotischer Faktoren // Diss.
Univ. Hamburg. 1979.
Melkonian M., Reize I. B., Preisig H. R. Maturation of a flagel-
lum/basal body requires more than one cycle in algal flagellates: stu-
dies of Nephroselmis olivacea (Prasinophyceae) И Algal develop-
ment, molecular and celular aspects / Eds W.Wiessner et al. Berlin
etc.: Springer, 1987. P. 102—113.
Mignot J.-P. Structure et ultrastructure de quelques Euglenomo-
nadines U Protistologica, 1966. T. 2. P. 51—117.
Moestrup 0. Current status of chrysophyte ’splinter groups’: sy-
nurophytes, pedinellids, silicoflagellates // Chrysophyte algae: ecolo-
gy, phylogeny and development / Eds C. D. Sandgren et al. Cambrid-
ge : Univ. Press, 1995. P. 75—91.
Moestrup 0., Thomsen H. A. Dictyocha speculum (Silicoflagel-
late, Dictyochophyceae), studies of armored and unarmored stages.
Det Kongel Danske Vidensk Selskab. Biol Skri. 1990. Vol. 37.
P. 1—57.
Miiller M. Biochemistry of protozoan microbodies: peroxisomes,
a-glycerophosphate oxidase bodies, hydrogenosomes // Ann. Rev.
Microbiol. 1975. Vol. 20. P. 467-483.
Miiller M. The hydrogenosome // Soc. Gen. Microbiol. Symposi-
um. 1980. Vol. 30. P. 127—142.
Miiller M. The hydrogenosome. // J. Gen. Microbiol. 1993.
Vol. 139. P. 2879—2889.
O’Kelly C. J., Patterson D. J. The flagellar apparatus of Cafeteria
roenbergensis Fenchel et Patterson, 1988 (Bicosoecales-Bicosoecida).
Europ. J. Protistol. 1996. Vol. 32. P. 216—226.
O’Kelly C. J., Wujek D. E. Status of the Chrysamoebales (Chry-
sophyceae): observations on Chrysamoeba pyrenoidifera, Rhizochro-
mulina marina and Laginion delicatulum. // Chrysophyte algae: ecolo-
gy, phylogeny and development / Eds C. D. Sandgren et al. Cambrid-
ge: Univ. Press. 1995. P. 361—372.
Pace M. L. Planctonic ciliates: their distribution, abundance, and
relationship to microbial resources in a monomictic lake // Can. J.
Fish. Aquat. Sci. 1982. Vol. 39. P. 1106—1116.
Page F. C. The classification of «naked» amoebae (Phylum Rhi-
zopoda). # Arch. Protistenk. 1987. Bd 133. S. 199—217.
Parker J. G. Cultural characteristics of the marine ciliated proto-
zoan, Uronema marinum // J. exp. mar. Biol. Ecol. 1976. Vol. 24.
P. 213—226.
Patterson D. J. Stramenopiles: chromophytes from a protistan
perspective // The chromophyte algae: problems and perspectives /
Eds J. C. Green et al. Oxford : Clarendon Press, 1989. P. 357—379.
Patterson D. J. Protozoa: evolution and systematics. И Progress in
Protozoology, Proc. IX Intern. Congr. of Protozool., Berlin, 1993 I
Eds K. Hausmann, N. Hiilsmann). Stuttgart: Fischer, 1994. P. 1—
14.
Patterson D. J. The diversity of eukaryotes // Amer. Naturalist.
1999. (In press).
Perasso R., Baroin F., Qu L. H., Bechellerie J. P., Adoutte A.
Origin of algae U Nature. 1989. Vol. 339. P. 142—144.
Philippe H., Adoutte A. How far can we trust the molecular phy-
logeny of protists? // Verh. Dtsch. Zool. Ges. 1996. Vol. 89, N 2.
P. 49—62.
Philippe H., Adoutte A. The molecular phylogeny of Eukaryota:
solid facts and uncertainties // Evolutionary relationships among Pro-
tozoa / Eds G. H. Coombs et al. London : Kluwer, 1998. P. 25—56.
Preisig H. R. A modern concept of chrysophyte classification //
Chrysophyte algae: ecology, phylogeny and development / Eds
C. D. Sandgren et al. Cambridge : Univ. Press, 1995. P. 46—74.
Preisig H. R., Anderson O. R., Corliss J. O., Moestrup 0., Po-
well M. J., Roberson R. W., Wetherbee R. Terminology and nomen-
clature of protist cell surface structures. Protoplasma, 1994. Vol. 181.
P. 1—28.
Prescott D. M. Relation between multiplication rate and tempera-
ture in Tetrahymena pyriformis, strains HS and GE П J. Protozool.
1957. Vol. 4, N 4. P. 252—256.
Reiner D. S., McCaffery M., Gillin F. D. Sorting of cyst wall pro-
teins to a regulated seecretory pathway during differentiation of the
primitive eukaryote, Giardia lamblia // Eur. J. Cell Biol. 1990. Vol. 53.
P. 142—153.
Schlegel M. Protist evolution and phylogeny as discrened from
small subunit ribosomal RNA sequence comparisions // Eur. J. Protis-
tol. 1991. Vol. 27. P. 207—219.
Siddal M. A., Hong H., Desser S. S. Phylogenetic analysis of the
Diplomonadida (Wenyon, 1926) Brugerolle, 1975: evedence of hete-
rochrony in Protozoa and against Giardia lamblia as a «Missing link».
J. Protozool. 1992. Vol. 39, N 3. P. 361—367.
Sl&de&k V. Quantitative characteristic of Protozoan communities
in waste waters // Progress in Protozoology. Prague, 1963. P. 338—
340.
Slddetek V. System of water quality from biological point of
view // Ergebnisse der Limnologie. Stuttgart, 1973.
Sleigh M. A. The biology of Protozoa. London. 1973.
Sleigh M. A. Protozoa and other protists. London : Arnold, 1989.
Sleigh M. A. Progress in understanding the phylogeny of flagella-
tes II Cytology, 1995. Vol. 37, N 11. P. 985—1009.
Smith J. A. Some effects of temperature on the frequency of di-
vision and on the volume of starch and fat in Chilomonas parameci-
um H Biol. Bull. 1940. Vol. 79, N 3. P. 379—396.
Sogin M. L., Gunderson J. H., Elwood H. J., Alonso R. A., Plat-
tic D. A. Phylogenetic meaning of kingdom concept. A unusual riboso-
mal RNA from Giardia lamblia // Science. 1989. Vol. 243. P. 75—77.
Stout J. D. Environmental factors affecting the life history of
three soil species of Colpoda (Ciliata) // Transact. Roy. Soc. N.Z.
1955. Vol. 82, N 5. P. 1165—1188.
190
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ
Taylor С. V., Stricland A. G. Effect of high vacua and extreme
temperatures on cysts of Colpoda cucullus // Physiol. Zool. 1936.
Vol. 9, N 1. P. 15.
Taylor F. J. R. Symbiosos in marine Protozoa // Ecology of ma-
rine Protozoa. N. Y., Oxford : Oxford Univ. Press, 1990. P. 323—
340.
Tourasse N. J., Gouy M. Evolutionary relationships among pro-
tist phyla constructed from LSUrRNAs accounting for unequal rates
of substitution among sites // Evolutionary relationships among Pro-
tozoa I Eds G. H. Coombs et al. London : Kluwer, 1998. P. 57—
76.
Van Bruggen J. J. A., Stumm С. K., Vogels G. D. Symbiosis of
methanogenic bacteria and sapropelic protozoa // Arch. Microbiol.
1983. Vol. 136. P. 89—96.
Van Bruggen J. J. A., Zwart К. B., Van Assema R. M., Stumm
С. K., Vogels G. D. Methanobacterium formicicum, an endosymbiont
of the anaerobic ciliate Metopus striatus McMurrion // Arch. Microbi-
ol. 1984. Vol. 139. P. 1—7.
Van Bruggen J. J. A., Van Rens G. L. M., Geerman E. G. M.,
Zwart К. B., Stumm С. K., Vogels G. D. Isolation of methanogenic
endosymbyont of the sapropelic amoeba Pelomyxa palustris Greef II J.
Protozool. 1988. Vol. 35. P. 20—23.
Van de Peer Y., De Wachter R. Evolutionary relationships
among the eukaryotic crown taxa taking into account site-to-site rate
variation in 18S rRNA // J. Molec. Evol. 1997. Vol. 45. P. 619—
630.
Vickerman K. Revolution among Protozoa // Evolutionary Rela-
tionships Among Protozoa / Eds G. H. Coombs et al. London : Klu-
wer, 1998. P. 1—24.
Vickerman K., Brugerolle G., Mignot J.-P. Mastigophora. In:
Microscopic Anatomy of Invertebrates. Vol. 1. Protozoa / Eds
F. W. Harrison, J. O. Corliss. New York etc.: Wiley-Liss, 1991. P. 13—
160.
Vogel W. Uber die Hilze- und Kalteresistenz von Zoothamnium
hiketes Precht (Ciliata, Peritricha) // Zeitschr. wissensch. Zool. 1966.
Bd 173, N 3—4. S. 345—378.
Vossbrinck C. R., Woese R. Eukaryotic ribosomes that lack a
5,8S RNA H Nature. 1986. Vol. 320. P. 287—288.
Vossbrinck C. R., Maddox J. V., Friedman S., Debrunner-Vos-
sbrinck B.A., Woese C. R. Ribosomal RNA sequence suggest that
microsporidia are extremely ancient eukaryotes // Nature. 1987. Vol. 326.
P. 411—414.
Wang A. L., Wang С. C. Viruses of Protozoa. Ann. Rev. Micro-
biol. 1991. Vol. 45. P. 251—263.
Wenrich D. H. Comments on the origin and evolution of «sez» //
Sex in microorganisms. Waschington, 1954.
Whittaker R. H. New concept of kingdoms of organisms. Scien-
ce. 1969. Vol. 163. P. 150—160.
Wilcox L. W. Prokaryotic endosymbionts in the chloroplast stro-
ma of the dinoflagellate Woloszynskia pascheri U Protoplasma. 1985.
Vol. 135. P. 71—79.
Wilson I. Plastids better red than dead II Nature. 1993. Vol. 366.
P. 638.
Woodruff L.L. Observation on the origin and sequence of the
Protozoan fauna of hay infusions // J. Exper. Zool. 1912. Vol. 12, N 2.
P. 205—265.
Yarlett N., Hann A. C., Lloyd D., Williams A. G. Hydrogeneso-
mes in the rumen protozoan Dasytricha ruminantum Schuberg # Bio-
chem J. 1981. Vol. 200. P. 365—372.
Yarlett N., Hann A. C., Lloyd D., Williams A. G. Hydrogenoso-
mes in a mixed isolate of Isotricha prostoma and Isotricha intestinalis
from ovine rumen contents # Comp. Biochem. Physiol. 1983.
Vol. 74b. P. 357—364.
Yarlett N., Coleman G. S., Williams A. G., Lloyd D. Hydrogeno-
somes in known species of rumen entodinomorphid protozoa II FEMS
Microbiol. Lett. 1984. Vol. 21. P. 15—19.
СИСТЕМАТИЧЕСКАЯ
ЧАСТЬ
Тип CRYPTOPHYTA Pascher, 1914 — КРИПТОФИТОВЫЕ
ВОДОРОСЛИ, КРИПТОМОНАДЫ
Одноклеточные подвижные организмы с типичным дорсовент-
ральным строением. Имеют овальную, яйцевидную, почковидную
или бобовидную форму. Тело клетки сжато с боков и имеет выпук-
лую дорсальную и плоскую (или вогнутую) вентральную сторону,
передний край обычно косо срезан. Здесь расположена вестибу-
лярная зона, от которой вдоль тела клетки отходит косая борозда;
у начала борозды расположено отверстие глотки, уходящей внутрь
тела на различную глубину. В вестибулярной зоне находятся осно-
вания 2 жгутиков, из которых один несколько длиннее другого.
Глотка выстлана рядами крупных стрекательных органелл —
эжектосом. Преимущественно строгие фототрофы, но есть и гете-
ротрофные формы, способные заглатывать и переваривать клетки
бактерий. Клетки фототрофных крйптомонад могут иметь различ-
ную окраску — коричневую, сине-зеленую или красную за счет
различного содержания фотосинтетических пигментов, таких как
хлорофиллы а и с2, водорастворимые фикобилиновые пигменты
фикоцианин и фикоэритрин. Хлоропласты криптомонад имеют
оболочку из 4 мембран, из которых наружная является продол-
жением ядерной мембраны. Между 2 наружными и 2 внутренними
мембранами находится перипластидное пространство, в котором
располагаются рудиментарное ядро — нуклеоморф, цитоплазма
с рибосомами и запасные питательные вещества. Половой про-
цесс неизвестен. Одни формы всегда подвижны, даже при делении,
другие образуют скопления заключенных в слизь неподвижных
клеток. В редких случаях формируются круглые неподвижные
цисты.
История изучения
Первым ученым, исследовавшим криптомонад,
был Эренберг (Ehrenberg, 1830, 1831), описавший
6 фототрофных видов, отнесенных к предложенному
им роду Cryptomonas, и 1 гетеротрофный вид, Chilo-
manas paramaecium. Описания Эренберга, как обыч-
но в те времена, были очень краткими и носили
самый общий характер. В дальнейшем новые роды и
виды описали Пашер (Pascher, 1913, 1914), Скуйя
(Skuja, 1948, 1956) и другие. Подробный обзор морс-
ких форм сделал Батчер (Butcher, 1967). Разными ав-
торами было описано более 200 видов криптомонад.
Однако детали строения сложно организованных
клеток крйптомонад плохо различимы в световом
микроскопе, а описания многих таксонов, приводи-
мые в ботанической литературе, являются недоста-
точно четкими. Многие признаки весьма изменчивы,
без изучения культур установить природу организма
затруднительно. Поэтому неудивительно, что в тече-
13 Протисты, часть 1
ние последнего десятилетия в результате использова-
ния электронного микроскопа систематика крипто-
монад претерпела резкие изменения. Большое значе-
ние для развития современной систематики крипто-
монад имели работы Санторе (Santore, 1984, 1985,
1987), хотя не всегда его взгляды получали призна-
ние. Переописания ряда родов и описания новых так-
сонов крйптомонад осуществили Хилл и Ветерби
(Hill, 1991; Hill, Wetherbee, 1988, 1989, 1990). Эти
описания основаны преимущественно на данных
электронной микроскопии, но они иногда недоста-
точно конкретны и неоднозначны, поэтому и в наши
дни определение родовой, не говоря уже о видовой,
принадлежности этих организмов представляет зна-
чительные трудности. Поставив под сомнение право-
мерность использования ультраструктурных критери-
ев в таксономии, Новарино и Лукас (Novarino, Lukas,
1993) пересмотрели систему крйптомонад, значи-
тельно сократив число родов. Создание коллекций
типовых культур позволило в последнее время провес-
ти сравнение последовательностей генов 18S рРНК и
обосновать выдвинутую ранее гипотезу о происхож-
дении криптофит в результате симбиоза гетеротроф-
ных двухжгутиковых монад и коккоидных красных
водорослей (Marin et al., 1998).
Классификация
Криптомонады традиционно являются объектами
исследования в различных отделах биологии. В соот-
ветствии с зоологической системой (Хаусман, 1988)
они могут быть классифицированы следущим обра-
зом:
Тип Sarcomastigophora Honigberg et Balamuth, 1963
Подтип Mastigophora Diesing, 1866
Класс Phytomastigdphorea Calkins, 1909
Отряд Cryptomonadida Senn, 1900
Согласно ботанической классификации (Горлен-
ко, 1981; Саут, Уиттик, 1990), их включают в
Отдел Pyrrophyta Pascher, 1914
Класс Cryptophyceae Pascher, 1914
Порядок Cryptomonadales Pascher, 1914
194
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
В современной системе протистов (см. раздел:
«Система протистов...») криптомонады занимают
следующее положение:
Тип Cryptophyta Pascher, 1914
Класс Cryptophyceae Pascher, 1914
Отряд Cryptomonadida Senn, 1900
Сем. Cryptomonadidae Pascher, 1914
Сем. Goniomonadidae Hill, 1991
Сем. Hemiselmididae Butcher, 1967
Морфология, цитология,
физиология
Как уже было отмечено, клетки криптомонад об-
ладают дорсовентральным строением, дорсальная
сторона обычно выпуклая, тогда как вентральная во-
гнутая или плоская. На вентральной стороне распо-
ложена вестибулярная зона (рис. 125, А, вкл.). Схема-
тическое изображение строения клетки криптомона-
ды приведено на рис. 124.
Вестибулярная зона представляет собой мелкое
углубление поверхности. Здесь начинается борозда,
идущая обычно вниз вдоль клетки, тут же расположе-
но отверстие глотки (рис. 125, А). Наличие борозды и
ее длина относительно тела клетки — признаки, име-
ющие значение для систематики на уровне рода. Бо-
розда часто несколько загнута. У некоторых форм на
нижнем конце борозда имеет небольшое округлое
Рис, 124. Схема строения клетки криптомонады.
к — зерна крахмала, м — митохондрия, мт — микротрубочковые корешки,
н — нуклеоморф, п — перипласт, р — фибриллярный корешок, рс —
ризостиль, хл — хлоропласт, э — эжектосомы, я — ядро.
Fig. 124. Structural organization of cryptomonas cell.
к — starch grains, м — mitochondrion, мт — microtubular roots, н —
nucleomorph, n — periplast, p — fibrillar rootlet, pc — rhizostyle, хл —
chloroplast, э ejectosomes, я — nucleus.
расширение — устьице, что также является призна-
ком, используемым в современной систематике крип-
томонад. Иногда края борозды сомкнуты, но отверс-
тие устьица может при этом сохраняться. На дорсаль-
ной стороне вестибулярной зоны обычно находится
вестибулярный язычок (рис. 125, В). Язычки могут быть
овальными, квадратными, плоскими, округлыми или за-
остренными. Язычок укреплен цитоскелетной пластин-
кой (рис. 125, Б). Мешковидная глотка у разных форм
имеет разную длину и проходит вдоль или поперек
тела. Иногда она разветвляется. У некоторых форм
глотка укреплена цитоскелетными образованиями,
электронно-плотными исчерченными пластинами
(рис. 126, А, вкл.). Вопрос о функциях и значении
глотки неясен. У большинства фототрофных форм
никогда не удавалось наблюдать заглатывания каких-
либо частиц.
Покровы клетки представлены перипластом, ко-
торый состоит из внутреннего компонента, находя-
щегося под плазмалеммой (рис. 125, Б, В) и представ-
ляющего собой либо сплошную пластину, либо со-
вокупность многочисленных белковых пластинок
различной формы (Brett et al., 1994). Внешний ком-
понент перипласта находится снаружи на поверхно-
сти плазмалеммы и представлен пластинками, чешуй-
ками или слизью. В зависимости от характера пери-
пласта поверхность клетки может быть ровной или
структурированной. Например, поверхность у Chroo-
monas состоит из прямоугольников, а у Rhinomo-
nas — из шестигранников (Kugrens, Lee, 1987). Пе-
рипласт отсутствует в области глотки и вестибулума
(рис. 125, В; 126, А).
Во внутренний компонент перипласта обычно
встроены многочисленные мелкие стрекательные ор-
ганеллы, на срезах клеток криптомонад часто можно
видеть пустые ямки, оставшиеся после выстрелива-
ния их содержимого (рис. 125, В). Глотка выстлана
крупными стрекательными органеллами — эжектосо-
мами, расположенными рядами (рис. 124; 125, Г, Д\
126, А); число рядов эжектосом иногда указывается в
диагнозах родов и видов. Эжектосомы содержат скру-
ченные рулоном ленты; при раздражении клетки
ленты распрямляются и выстреливаются во внешнюю
среду. Значение эжектосом остается неизвестным;
нет и доказательств их защитной роли; однако уда-
лось наблюдать стремительный ракетоподобный ры-
вок клетки при выбросе из глотки освобожденных из
эжектосом лент.
Интерфазное ядро 1, крупное, занимает обычно
нижнюю часть клетки (рис. 125, Г; 126, Г, 3). Оно
содержит ядрышко и скопления постоянно конденси-
рованных хромосом.
Основания не равных по длине жгутиков находят-
ся у дорсального края вестибулярной зоны возле от-
верстия глотки, иногда внутри глотки (рис. 126, А).
Окончания жгутиков слегка заострены, на их поверх-
ности располагаются различные придатки (рис. 126,
Б). У многих форм длинный жгутик снабжен 2 про-
тивоположными рядами трубчатых мастигонем (1—
2 мкм длиной) с одиночной концевой нитью (0.5—
1 мкм). У основания длинного жгутика находится
вздутие, от которого отходит пучок длинных масти-
гонем. По-видимому, он образует защитную сетку у
ТИП CRYPTOPHYTA
195
отверстия глотки. Короткий жгутик несет короткие
мастигонемы (около 1 мкм длиной) с коротким и
длинным волосками на концах. На поверхности мем-
браны жгутиков иногда обнаруживаются чешуйки —
семиугольные розетки 140—170 нм в диаметре. Они
могут покрывать также й тело криптомонады. В от-
личие от других групп протистов у криптомонад наблю-
дается сильное варьирование характера жгутиковых
придатков. Приведенное выше описание справедливо
лишь в отношении некоторых форм. Варьирует
структура мастигонем, их расположение, кроме че-
шуек могут присутствовать иглоподобные крючко-
видные выросты или волоски. Систематическое зна-
чение подобных различий пока не установлено; во
всяком случае, в современных диагнозах родов крип-
томонад эти признаки не упоминаются.
Особенностью переходной зоны жгутиков крипто-
монад является наличие здесь 2 септ, из которых
верхняя находится на уровне плазмалеммы (125, В;
126, Е). Кинетосомы (базальные тела жгутиков) ассо-
циированы с фибриллярными и микротрубочковыми
жгутиковыми корешками (рис. 124; 126, В—Е). У раз-
ных форм эти структуры различаются. Характерен
для криптомонад ризостиль — лента микротрубочек,
соединенных сократимыми фибриллами. Ризостиль
может проходить в непосредственной близости от
ядра (126, Г) и достигать заднего конца клетки. Воз-
можно, что при этом он как бы стягивает клетку
вдоль ее длинной оси, что обусловливает искривлен-
ную форму тела у видов Campylomonas. На попереч-
ном срезе ризостиль образует изогнутую пластинку
(рис. 126, Г, Д). Микротрубочки ризостиля иногда
снабжены изогнутыми пластинками — крыльями и
соединены с дополнительным фибриллярным тяжем
(рис. 126, Д). У многих форм отмечен исчерченный
фибриллярный корешок, идущий вдоль поверхности
митохондрии (рис. 126, В). Восходящий микротру-
бочковый корешок соединяется с пластиной, образо-
ванной цитоскелетными микротрубочками (рис. 126,
Е).
Сократительная вакуоль находится у апикального
конца клетки, она открывается в область глотки око-
ло ее начала (рис. 125, В; 126, В). В передней части
клетки расположен аппарат Гольджи, образованный
1—2 диктиосомами, состоящими из 12—15 цистерн
(рис. 125, Г, Д; 126, Е).
Митохондрия, по-види^ому, 1, сильно разветвлен-
ная, с пластинчатыми, ламеллярными кристами, об-
разует периферическую и центральную сети; перифе-
рическая сеть расположена под плазмалеммой, цент-
ральная —соответственно в центре клетки (рис. 125,
В, Д; 126, В, X 3).
Большинство криптомонад осуществляют фото-
синтез и являются автотрофами. Редкие формы
(например, Chroomonas pochmannii) миксотрофны,
осуществляют фотосинтез и могут захватывать и пе-
реваривать клетки бактерий. Известно 2 рода гете-
ротрофных криптомонад — Goniomonas и Chilomo-
nas. Бактерии обнаруживаются в пищеварительных
вакуолях, но механизм их попадания в тело крипто-
монады до сих пор не описан.
Фототрофные формы содержат 1 или 2 хлоро-
пласта. Иногда один из хлоропластов образует 2 боль-
шие лопасти, соединенные тонким мостиком, часто
трудно установить, сколько именно этих органелл со-
держит клетка. В хлоропластах находятся специали-
зированные плоские мембранные цистерны — тила-
коиды, содержащие основные светособирающие пиг-
менты и компоненты электрон-транспортных цепей.
Тилакоиды обычно расположены попарно, иногда
в группах по 3 (рис. 125, Д). Клетки фототрофных
криптомонад могут иметь различную окраску — ко-
ричневую, сине-зеленую или красную за счет различ-
ного содержания фотосинтетических пигментов, та-
ких как хлорофиллы а и с2, водорастворимые фико-
билиновые пигменты фикоцианин и фикоэритрин.
Фикобилины состоят из а- и Р-субъединиц фико-
билинпротеинов и линейных тетрапирролов — хро-
мофоров. Эти пигменты расположены во внутритила-
коидном пространстве и не образуют каких-либо ви-
димых структур, подобных фикобилисомам красных
и синезеленых водорослей. Каждый вид содержит
только один из фикобилиновых пигментов, причем
существует несколько типов фикоцианина и фико-
эритрина, характеризующихся различными максиму-
мами поглощения света (в описаниях родов макси-
мум указан в нанометрах после названия пигмента).
Интересно отметить, что генетические исследования
свидетельствуют об отсутствии у криптомонад белков
фикоцианинов. Предполагается, что замена хромо-
фора у фикоэритрина мимикрирует спектроскопи-
ческие характеристики истинных фикоцианинов
(Marin et al., 1998). Представители одного рода обыч-
но содержат один определенный пигмент, хотя вопрос
о таксономическом значении пигментации у крипто-
монад является пока дискуссионным (Hill, Rowan,
1989). Нередко в хлоропласте находятся небольшие
липидные глобулы (рис. 126, 3),
В хлоропласте, в его выросте, или на стебельке
над ним может располагаться пиреноид. Это белко-
вое тело, содержащее основные ферменты фиксации
углекислоты. Пиреноиды иногда рассечены мембра-
нами тилакоидов. Крахмальная обкладка пиреноида
не всегда хорошо выражена, что затрудняет его обна-
ружение в световом микроскопе. У немногих форм
криптомонад в хлоропласте располагается глазок —
стигма, которая состоит из группы липидных глобул
на периферии хлоропласта, иногда в его выросте.
Фототрофные формы способны к фототаксису. Их
светочувствительность связывают с присутствием фла-
вопротеинов на жгутиках.
Хлоропласт окружен 4 мембранами. 2 внешние
мембраны принято обозначать как хлоропластный
ЭПР. Внешняя мембрана хлоропластного ЭПР пере-
ходит во внешнююю мембрану ядра (рис. 124; 126,
3). Перипластидный компартмент, ограниченный
хлоропластным ЭПР и 2 внутренними спаянными
мембранами, включает зерна крахмала, нуклеоморф
и цитоплазму с эукариотическими рибосомами.
Нуклеоморф — рудиментарное ядро эукариотиче-
ского фототрофного симбионта — может быть ассо-
циирован с пиреноидом, ядром, иногда располагается
в выросте хлоропласта, иногда — внутри пиреноида
(рис. 126, 3). Нуклеоморф обычно имеет округлую
или грушевидную форму, окружен двойной мембраной,
в которой обнаруживаются поры; он делится амито-
196
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тически, простой перетяжкой. В нуклеоморфе вы-
делены зоны скопления РНК, соответствующие зоне
ядрышка, и зоны скопления ДНК. Геном нуклеомор-
фа у исследованных в этом отношении форм пред-
ставлен 3 хромосомами и не является пассивным об-
разованием; некоторые его гены работают, в частнос-
ти гены 18S рРНК.
Клетка криптомонады состоит в отличие от боль-
шинства других протистов из 4 различных клеточных
компартментов, содержащих собственный геном: ци-
топлазматического, перипластидного, пластидного и
митохондриального, каждый из которых ограничен
двойной мембраной.
Размножение,
жизненный цикл
Размножение крйптомонад происходит путем про-
дольного деления клетки, делящиеся клетки часто со-
храняют подвижность. Митоз начинается с реплика-
ции кинетосом, ядро перемещается в передний конец
клетки. Ядерная оболочка в профазе фрагментирует-
ся и частично разрушается, в случае открытого мито-
за из хлоропластного ретикулума формируется до-
полнительная перинуклеарная оболочка. ЦОМТами
служат базальные тела, но на полюсах веретена рас-
полагаются исчерченные фибриллярные корешки и
нуклеоморфы. Цитокинез идет параллельно кариоки-
незу, борозда деления формируется от переднего к
заднему концу клетки. Каждая дочерняя клетка полу-
чает 1 материнскую кинетосому и 1, построенную de
novo.
Криптомонады — это гаплоидные организмы.
Половой процесс неизвестен. Однако описан вид
Proteomonas sulcata, имеющий популяции гаплоид-
ных и диплоидных клеток, которые различаются по
размерам, морфологий и организации жгутикового
аппарата (Hill, Wetherbee, 1986). В редких случаях в
жизненном цикле образуются круглые неподвижные
цисты, при прорастании которых происходит деле-
ние их содержимого и из цисты выходят 2 вегетатив-
ные подвижные клетки.
Экология
Криптомонады относятся к организмам-убиквис-
там. Они характеризуются значительной устойчи-
востью к загрязнению, встречаются даже в сточных
водах и навозной жиже. Способны адаптироваться к
резким изменениям условий среды: например, Chroo-
monas africana не теряет жизнеспособности при пе-
реносе из морской воды в пресную. Многие виды яв-
ляются морскими организмами, их разнообразие
больше такового пресноводных. Последние могут
быть найдены в планктоне. Особенно часто они насе-
ляют прибрежные участки водоемов, старицы, пруды
и болота. Обычно число клеток в сообществах неве-
лико, но иногда происходит массовое развитие крип-
томонад, они вызывают «цветение» воды, окрашивая
ее в тот или иной цвет. Известны виды Cryptomonas,
способные развиваться на поверхности снежного
покрова и подо льдами Арктики. Хороший рост
культур наблюдался при нейтральной или слабокис-
лой реакции среды; сильный свет тормозит их рост.
Филогения
В соответствии со схемой, предложенной Кюг-
ренсом и Ли (Kugrens, Lee, 1991), исходными в
эволюции морфологических типов крйптомонад счи-
таются формы, близкие к представителям рода Goni-
omonas (рис. 127). У них вестибулярно-глоточное
пространство не было разграничено и открывалось
непосредственно в окружающую среду. К настояще-
му времени общепризнано, что пластиды криптомо-
над появились в результате вторичного эндосимбио-
за, в котором эндобионт был фототрофным эукари-
отом. Этот этап эволюции связан с образованием
устойчивого симбиоза с эукариотами (предположи-
тельно из группы Rhodophyta) и уменьшением раз-
меров вестибулярного пространства. При этом про-
странство глотки, направленной внутрь клетки, со-
общалось с окружающей средой через борозду. В
дальнейшем происходили смыкание вентральных
стенок вестибулярной зоны и редукция борозды.
Глотка стала открываться в окружающую среду толь-
ко через вестибулярную зону, как у представителей
рода Chroomonas. Эжектосомы переместились с пе-
риферии вестибулярной зоны на мембраны, выстила-
ющие глотку, и приобрели вертикальное расположе-
ние относительно длинной оси клетки. В процессе
эволюции, по-видимому, произошли редукция хло-
ропласта и повторный переход к гетеротрофии, что
привело к возникновению рода Chilomonas, предста-
вители которого обладают сильно редуцированной
бороздой, развитой глоткой и лейкопластами. При-
веденная гипотетическая схема подтвержается ана-
лизом нуклеотидных последовательностей генов 18S
рРНК (Marin et al., 1998).
Таким образом, криптомонады являются организ-
мами-химерами. Только представители одного рода —
Goniomonas, не обладающие ни хлоропластами, ни
нуклеоморфом, возможно, являются потомками ис-
ходного организма-хозяина.
Систематический обзор
В настоящее время в качестве основных призна-
ков, которые могут быть использованы для классифи-
кации крйптомонад, рассматриваются особенности
организации вестибулярной зоны, борозды и глотки,
строение перипласта, характер организации пластид-
ного комплекса — число и форма хлоропластов, рас-
положение нуклеоморфа, наличие тех или иных пиг-
ментов. Эти признаки рассматриваются в качестве ро-
довых (Santore, 1984, 1985^ 1987; Hill, Wetherbee,
1988,1989; Hill, 1991). Ниже мы приводим характерные
признаки некоторых родов, изученных с использова-
нием современных методов, исходя при этом преиму-
щественно из представлений Хилла и Ветерби* Гипо-
тетические филогенетические связи различных родов
приведены на рис. 127.
ТИП CRYPTOPHYTA
197
QonioMonas
Storecrtula Самру1омопсга ChilOMonas
Рис. 127. Схема эволюции морфологии криптомонад.
(По: Kurgens, Lee, 1991).
Гетеротрофные представители рода Goniomonas имеют расположенные по
периферии вестибулярной зоны эжектосомы и лишены глотки. У примитив-
ных представителей рода Cryptomonas глотка открывается через борозду.
В дальнейшем происходит редукция борозды (Campylomonas и Storeatula)
и редукция хлоропласта (Chilomonas).
Fig. 127. Scheme of morphological evolution of cryptomonads.
Heterotrophic species Goniomonas lack gullet and possess transverse row
of ejectosomes on periphery of vestibulum. The gullet of primitive species
Cryptomonas opens throught the furrow. The reduction of furrow {Campylomo-
nas, Storeatula) and chloroplast (Chilomonas) occures in course of evolution.
Сем. Goniomonadidae Hill, 1991
Овальные клетки на переднем конце несут 1 ряд
поперечно расположенных эжектосом; 2 жгутика, ли-
шенные трубчатых мастигонем, отходят от дор-
сальной стенки вестибулума. Представители семейст-
ва не содержат пластид.
стороны клетки имеется 3—6 продольных, слегка
изогнутых полосок. Для рода характерно отсутствие
хлоропластов, крахмала и нуклеоморфа. По-видимо-
му, этот род может рассматриваться как наиболее
близкий к предковой форме.
Сем. Hemiselmididae Butcher,
1967
Для представителей семейства характерно отсут-
ствие борозды, жгутики отходят от средней вентраль-
ной части клетки. Перипласт состоит из прямоуголь-
ных перекрывающихся пластин. Клетка содержит
хлоропласт с пиреноидом, укрепленным на ножке.
Имеется единственная неразветвленная митохондрия.
Род Hemiselmis Parke, 1949
(рис. 128, Б)
У представителей этого рода вестибулярная зона
расположена примерно посередине вентральной сто-
роны клетки. Борозйа отсутствует, глотка трубчатая.
Внутренний и внешний компоненты перипласта
представлены гексагональными пластинками. Един-
ственный хлоропласт несет пиреноид на ножке, ко-
торый пересечен парой тилакоидов. Нуклеоморф
расположен перед пиреноидом. Клетки содержат
фикоэритрин 555.
Сем. Cryptomonadidae Pascher,
1914
Это семейство включает криптомонад с субапи-
кальной вестибулярной зоной. Глотка и борозда с ря-
дами эжектосом ориентированы вдоль оси клетки.
2 жгутика отходят от стенки вестибулярной зоны;
как правило, оба жгутика несут трубчатые мастигоне-
мы. Все представители имеют хлоропласты.
Род Campylomonas Hill, 1991
(рис. 128, В)
Клетки постоянно подвижны, задний конец обыч-
но загнут. Борозда и глотка четко прослеживаются.
Перипласт представлен внутренней пластинкой, пре-
рываемой мелкими эжектосомами, внешний элемент
состоит из тонких нитчатых образований. Пиреноид
не пересечен мембранами, связан с нуклеоморфом.
Клетки содержат фикоэритрин 566.
Род Goniomonas Stein, 1878
(рис. 128, А)
Род характеризуется овальными, латерально сжа-
тыми клетками; на дорсальной стороне закругленно-
го переднего конца клетки находятся жгутики, не-
обычно — поперек клетки — расположены ряды
эжектосом. На поверхности перипласта с каждой
Род Chilomonas Ehrenberg, 1832
(рис. 128, Г)
Клетки на поперечном срезе почти цилиндриче-
ские, при взгляде сбоку узкоовальные, часто слег-
ка изогнуты. Жгутики расположены на краю суб-
апикальной вестибулярной зоны. Борозда короткая,
глотка выстлана рядами эжектосом. Гетеротрофные
198
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 128. Схема строения представителей основных родов криптомонад.
А — Goniomonas, Б — Hemiselmis, В — Campylomonas, Г — Chilomonas, Д — Chroomonas, Е — Cryptomonas, Ж — Falcomonas, 3 — Geminigera,
И — Guillardia, К — Котта, JI — Pyrenomonas, М — Rhinomonas, Н — Rhodomonas, О — Storeatula, П — Teleaulax.
Fig. 128. Scheme of morphology of main genera.
организмы. Настоящие хлоропласты отсутствуют, вмес-
то них имеются лишенные пигментов пластиды — лей-
копласты. В остальном этот род схож с родом пресно-
водных фототрофных криптомонад Campylomonas.
Род Chroomonas Hansgirg, 1885 emend. Hill,
1991 (рис. 128, Д)
Клетки обычно бочонковидные, иногда слегка
сжатые, с уплощенной вентральной стороной. Часто
переходят в пальмеллоидное состояние. Глотка труб-
чатая, ветвящаяся, борозда отсутствует. Перипласт
образует прямоугольные пластинки. Клетки сине-зе-
леного цвета за счет присутствия фикоцианина 630.
Род Cryptomonas Ehrenberg, 1831 emend. Hill,
1990 (рис. 128, E)
Представители этого рода, обладая обычным для
криптомонад строением, характеризуются прежде всего
тем, что они дорсовентрально уплощены, обычно об-
разуют пальмеллоидные слизистые колонии, имеют
ТИП CRYPTOPHYTA
199
сложно устроенную систему борозда—глотка. Пери-
пласт состоит из более или менее овальных пластинок
внутреннего компонента и тонких фибрилл внеш-
него. Клетки обладают 2 хлоропластами; содержат
фикоэритрин 566. Пиреноид не пересечен тила-
коидами. Представители рода — обитатели пресных
вод.
Род Falcomonas Hill, 1991 (рис. 128, Ж)
Клетки имеют форму капли или запятой. Борозда
отсутствует. Перипласт образован пластинками. Пи-
реноид погружен в хлоропласт, но содержит инваги-
нацию перипластидной цитоплазмы с нуклеоморфом.
Характерной особенностью рода является наличие
необычного фикоцианина 569.
Род Geminigera Hill, 1991 (рис. 128, 3)
Клетки заметно сжаты с боков, борозда хорошо
выражена и ограничена многочисленными эжекто-
сомами. Особенностью рода является единственный
лопастной хлоропласт с 2 почковидными, располо-
женными на стебельках пиреноидами. Пиреноиды не
пересечены тилакоидами и окружены полукругом
крахмальных зерен. Нуклеоморф помещается в ос-
тровке перипластидной цитоплазмы в ее инвагинации
в ядре. Хлоропласт содержит фикоэритрин 545.
Род Guillardia Hill et Wetherbee, 1990
(рис. 128, И)
Клетки постоянно подвижны; имеется глотка, но
борозда отсутствует. Внутренний компонент пери-
пласта представляет собой пластину, находящуюся
под плазмалеммой, внешний компонент — относи-
тельно крупные, неправильной формы пластинки, об-
разованные более мелкими субъединицами. Всего 1 хло-
ропласт. Пиреноид не пересечен тилакоидами и погру-
жен в строму хлоропласта. Имеется фикоэритрин 545.
Род Котта Hill, 1991 (рис. 128, К}
Передний конец клетки закруглен, задний сужен и
иногда заострен. Борозда отсутствует. Перипласт обра-
зован внутренними и внешними гексагональными плас-
тинками. Пиреноид глубоко погружен в вентральную
область хлоропласта, не пересечен тилакоидами, внут-
ри находится нуклеоморф. Ядро лежит ниже пиренои-
да. Клетки содержат фикоцианин 645.
Род Pyrenomonas Santore,1984
(рис. 128, Л)
Клетки с короткой бороздой, глотка продольная.
Внутренний компонент перипласта представлен прямо-
угольными пластинками, внешний — фибриллярный.
Хлоропласт с 2 лопастями. Пиреноид расположен в
области соединения лопастей, он не пересечен тила-
коидами. Нуклеоморф находится в инвагинации ци-
топлазмы внутри пиреноида. Клетки содержат фико-
эритрин 545.
Род Rhinomonas Hill et Wetherbee, 1988
(рис. 128, M)
Клетки постоянно подвижные, апикальная часть
закруглена, вестибулярная область расположена при-
мерно на расстоянии V3 длины клетки от ее переднего
конца. Глотка трубчатая, с продольными рядами три-
хоцист, борозда отсутствует. Перипласт представлен
внутренним и внешним компонентами — это более
или менее гексагональные пластинки. Хлоропласт 1.
Пиреноид, в котором заключен нуклеоморф, не пере-
сечен тилакоидами. Представители рода — морские
организмы. Клетки содержат фикоэритрин 545.
Род Rhodomonas Karsten, 1898 emend.
Hill et Wetherbee, 1989 (рис. 128, H)
Клетки подвижные, но иногда оседают в культуре.
Борозда короткая, глотка слегка изогнута в сторону
вентральной стороны тела. Перипласт образован про-
дольными рядами прямоугольных пластинок. Пире-
ноид без тилакоидов, но пронизан инвагинацией пе-
рипластидной цитоплазмы с нуклеоморфом. Клетки
содержат фикоэритрин 545.
Род Storeatula Hill, 1991
(рис. 128, О)
Клетки лишены борозды. Глотка открывается на
дорсальной стороне вестибулярной зоны; она тянется
примерно до половины клетки и окружена по крайней
мере 4 рядами больших эжектосом. Внутренний эле-
мент перипласта — пластинка, внешний — тонкий,
фибриллярный. Хлоропласт 1, состоит из ряда ло-
пастей, с пиреноидом; нуклеоморф расположен в
углублении пиреноида. Клетки содержат фикоэритрин
545.
Род Teleaulax Hill, 1991
(рис. 128, /7)
Клетки подвижны и не образуют слизи, борозда
начинается от вестибулярной зоны, которая располо-
жена на расстоянии 1/4 от переднего конца клетки, и
тянется почти до ее конца. Борозда ограничена 4—
6 рядами эжектосом. Внутренний компонент пери-
пласта представлен пластинкой, прерываемой вези-
кулами мелких эжектосом. С поверхности клетка по-
крыта гептагональными чешуйками. Пиреноид в
центре хлоропласта, он не пересечен тилакоидами.
Ядро расположено центрально. Содержат фикоэрит-
рин 545.
200
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
THE PHYLUM CRYPTOPHYTA, PASCHER, 1914
SUMMARY
The cryptomonads are motile unicellular organisms with a convex
dorsal side and a concave or flat ventral side. The vestibulum is loca-
ted at the anterior part of the cell, the furrow starts from the vestibu-
lum. Gullet aperture is located at the base of the furrow. The gullet is
lined by the rows of the ejectosomes. The majority of cryptomohads
are phototrophic organisms; the heterotrophic forms Ccapabale to en-
gulf bacteria also exist. Main pigments are chlorophylls a, c2 and
water-soluble phycobiliprotein phycoherythrin. The latter is dispersed
homogenously within the intrathilacoidal space. Every species posses-
ses a sole type of phycobyliprotein, with a definite maximum of light
absorbance. Four membranes encircle the chloroplast, the outer mem-
brane covered with ribosomes is a continuation of outer nuclear mem-
brane.
Cryptomonads, besides the nuclear, mitochondrial, and chloro-
plast genomes contain the genome of a nucleomorph. The nucleo-
morph is situated within the periplastid space, sunounded by two
membranes, and possesses typical eukaryotic chromosomes with acti-
ve genes including the one of 18S rRNA. Sometimes the nucleomorph
demonstrates structural connections with the pyrenoid.
The cell is surrounded by periplast, its inner part consists of pro-
tein plates. Two flagella of different length and functions are attached
nearly at the right side of the vestibulum. Both flagella are covered by
tubular mastigonemes, their structure and position are very variable.
Sometimes the scales and needlelike protrusions are present. The po-
sition of two septae in the transition zone is typical of cryptomonads.
The rhizostyle, a microtubular band associated with contractile fibrils,
which stretches from the flagellar base towards the nucleus, is typical
of the flagellar root system. Golgi is composed of 1—2 dictyosomes;
it lies at the anterior part of the cell. The mitochondrion possesses flat
cristae. The nucleus contains permanently condensed chromosomes
and the nucleolus. Nuclear division occurs by means of open mitotic
spindle. Flagellar basal bodies serve as MTOC of the division spindle.
Cysts can be formed sporadically in the life cycle; two vegetative cells
appear after cyst germination. Sexual reproduction is unknown. Cryp-
tomonads are ubiquitous organisms, although marine species domina-
te. They can cause water blooms.
In accordance with modern system of protists, cryptomonads be-
long to the phylum Cryptomonada, including the class Cryptomonadea
with one order Cryptomonadida. The order contains three families.
The family Goniomonadidae with the unique genus Goniomonas is
characterized by the absence of chloroplast, nucleomorph, and starch-
like storage product. A row of ejectosomes lies across the anterior
end. Goniomonas species are considered as ancestral forms without a
phototrophic symbiont. The Family Hemiselmididae (genus Hemisel-
mis) is characterized by the absence of the furrow; the nucleomorph is
situated near the pyrenoid, the periplast consists of rectangular plates.
The majority of the genera (Campylomonas, Chilomonas, Chroomo-
nas, Cryptomonas, Falcomonas, Geminigera, Guillardia, Котта, Py-
renomonas, Rhinomonas, Storeatula, Teleaulax) belong to the family
Cryptomonadidae characterized by the subapical vestibulum, gullet
and furrow associated with the rows of ejectosomes, with chloroplasts,
and the nucleomorph. The characteristics of the vestibulum, the fur-
row, the gullet, the periplast structure, the number and shape of chlo-
roplasts, the position of nucleomorph, the presence of specific pig-
ments, the number of ejectosomes are valuable in the description of
genera and species. It is considered that ancestral forms of cryptomo-
nads were heterotrophic, similarly to Goniomonas. The relatedness of
ancestral forms with other groups of protists is unclear. Phototrophic
cryptomonads are most probably the result of symbiosis with rho-
dophyte algae. Common trend of the evolution of cryptomonads is
the reduction of the furrow together with a secondary loss of chloro-
plasts.
Л ИТЕРАТУPA
Горленко M. В. (ред.) Курс низших растений. М., 1981.
Саут Р., Уиттик А. Основы альгологии. М., 1990.
Хаусман К. Протозоология. М., 1988.
Brett S. J., Perasso L., Wetherbee R. Structure and development
of the cryptomonad periplast: a review // Protoplasma. 1994. Vol. 181.
P. 106—122.
Butcher R. W. Cryptophyceae // An introductory account of the
smaller algae of British coastal waters. Fishery Investigations /
HMSO. London, 1967. Ser. IV, pt IV. P. 1—54.
Ehrenberg E. G. Beitrage zur Kenntnis der Organisation der In-
fusorien und ihrer geographischen Verbreitung, besonders in Sibiri-
en // Abhandlungen der Koniglichen Akademie der Wissenschaften.
Physical. KI. Berlin, 1830. P. 1—88.
Ehrenberg E. G. Uber die Entwicklung und Lebensdauer der In-
fusionsthiere; nebst ferneren Beitragen zu einer Vergleichung der or-
ganischen Systeme // Abhandlungen der Koniglichen Akademie der
Wissenschaften. Physical. KI. Berlin, 1831. P. 1—154.
Hill D. R. A. A revised circumscription of Cryptomonas (Cryp-
tophyceae) based on examination of Australian strains // Phycologia.
1991. Vol. 30. P. 170—188.
Hill D. R. A., Rowan K. S. The biliproteins of the Cryptophy-
ceae // Phycologia. 1989. Vol. 28. P. 455—463.
Hill D. R. A., Wetherbee R. Proteomonas sulcata gen. et sp. nov.
(Crypthophyceae), a cryptomonad with two morphologically distinct
and alternating forms // Phycologia. 1986. Vol. 25. P. 521—543.
Hill D. R. A., Wetherbee R. The structure and taxonomy of Rhi-
nomonas pauca gen. et sp. nov. (Cryptophyceae) // Phycologia. 1988.
Vol. 27. P. 355—365.
Hill D. R. A., Wetherbee R. A reappraisal of the genus Rhodomo-
nas (Cryptophyceae) // Phycologia. 1989. Vol. 28. P. 143—158.
Hill D. R. A., Wetherbee R. Guillardia theta gen. et sp. nov.
(Cryptophyceae) // Can. J. Bot. 1990. Vol. 68. P. 1873—1876.
Kugrens P., Lee R. E. An ultrastructural survey of cryptomonad
periplast using quick-freezing freeze fracture techniques // J. Phycol.
1987. Vol. 23. P. 365—376.
Kugrens P., Lee R. E. Organization of cryptomonads // The bio-
logy of free-living heterotrophic flagellates // The Systematics Associ-
ation Special / Eds Patterson D. J. et al. Oxford, 1991. Vol. 45.
P. 219—233.
Marin B., Klinberg M., Melkonian M. Phylogenetic relationships
among the Cryptophyta: analyses of nuclear-encoded SSU rRNA se-
quences support the monophyly of extant plastid-containing lineages //
Protist. 1998. Vol. 149. P. 265—276.
Novarino G.F.L.S., Lucas I. A.N. Some proposals for a new
classification system of the Cryptophyceae // Bot. J. Linn. Soc. 1993.
Vol. 3. P. 3—21.
Pascher A. Cryptomonadinae // Die Susswasser-Flora Deutsch-
lands, Osterreichs und der Schweiz. Jena, 1913. H. 2. S. 97—114.
Pasher A. Flagellate I. (Farblose Flagellaten) // Die Susswasser-
Flora Deutschlands, Osterreichs und der Schweiz. Jena, 1914. H. 1.
S. 1— 27.
Santore U. J. Some aspects of the taxonomy in the Cryptophy-
ceae // New Phytologist. 1984. Vol. 98. P. 627—646.
Santore U.J. A cytological survey of the genus Cryptomonas
(Cryptophyceae) with comments on its taxonomy // Arch. Protistenk.
1985. Vol. 130. P. 1—52.
Santore U. j. A cytological survey of the genus Chroomonas with
comments on the taxonomy of this natural group of the Cryptophy-
ceae II Arch. Protistenk. 1987. Vol. 134. P. 83—114.
Skuja H. Taxonomie des Phytoplanctons einiger Seen in Uppland,
Schweden // Symb. Bot., Uppsal. 1948. Vol. 9. P. 183—238.
Skuja H. Taxonomische und biologische Studien fiber das Phy-
toplankton Schwedischen Binnengenwasser // Nova Acta Regiae Soc.
Sci., Uppsal. 1956. Vol. 16, N 3.
Тип EUGLENOZOA Cavalier-Smith, 1981 —
ЭВГЛЕНОЗОИ
Фототрофные и гетеротрофные формы с 1—2, реже с не-
сколькими жгутиками, имеющими простые мастигонемы и пара-
ксиальный тяж. Жгутики, как правило, выходят из глубокого
впячивания на переднем конце клетки и имеют 2—3 микро-
трубочковых корешка. Кристы в митохондриях преимуществен-
но пластинчатые (дисковидные), но могут принимать и трубчатую
форму. Деление ядра — закрытый внутриядерный митоз. Ци-
тостом имеется как у гетеротрофных, так и у фототрофных
форм. Свободноживущие и паразитические.
Класс Euglenoidea Biitschli, 1884 — Эвгленоидеи
В составе класса одиночные и колониальные, фототрофные
(зеленые) и гетеротрофные (бесцветные) протисты. Форма тела
широко варьирует от овальной, веретеновидной до листовидной и
игловидной. Длина тела 5—500 мкм и более. У большинства видов
2 гетероконтных гетеродинамичных жгутика, выходящих из дна
жгутикового резервуара на переднем конце тела. Жгутики с пара-
ксиальным тяжем, простыми мастигонемами, длинной переходной
зоной и 3 микротрубочковыми корешками. Поверхность тела по-
крыта кутикулой, некоторые виды имеют домик. Движение жгути-
ковое и метаболическое (эвгленоидное).
Фототрофные виды имеют фоторецептор, включающий стигму
и парафлагеллярное тельце. Хлоропласты в числе от 1 до несколь-
ких сотен, с оболочкой из 3 мембран. Тилакоиды в ламелле собра-
ны по 3. Основные пигменты хлоропластов: хлорофилл а, b и fl-
каротин. Имеются пиреноиды. Митохондрии с пластинчатыми
кристами. Способы питания: голофитное, сапрозойное, голозой-
ное, миксотрофное. Гетеротрофные и некоторые фотосинтезирую-
щие виды имеют цитостом. Основные резервные вещества — пара-
милон и липиды.
Эвглениды одноядерны. Хромосомы конденсированы на про-
тяжении всего жизненного цикла, при делении ядра ядрышко не
разрушается. Размножение агамное путем продольного бинарного
деления. В жизненном цикле обычны пальмеллевидные формы и
цисты. Класс объединяет около 1000 свободноживущих, паразити-
ческих и эпибионтных видов, которые распространены всесветно.
История изучения
Первое описание ряда основных видов и родов эв-
гленовых принадлежит Эренбергу (Ehrenberg, 1830).
К их числу относятся: представители родов Euglena,
Astasia, Distigma, Colacium, Trachelomonas. Фауна их
интенсивно изучалась и в последующие годы (Dujar-
din, 1841; Perty, 1852; Stein, 1878; Klebs, 1883, и др.),
что позволило выделить эту группу в отдельный
отряд в типе Protozoa. Накопление знаний по морфо-
логии и физиологии способствовало обоснованию
растительной природы эвгленовых.
Идея о выделении их в качестве одной из групп од-
ноклеточных жгутиковых водорослей принадлежит
Ценковскому (1870). Эвгленовые были включены в
систему споровых растений в качестве отдельного
типа (отдела) Euglenophyta Biitschli, 1884 с 1, обосно-
ванным позже, порядком (классом) Euglenales Popova,
1951. В настоящее время доминирует идея о первич-
ности гетеротрофных форм в эволюции эвгленовых.
Подавляющее большинство бесцветных эвгленид ис-
ходно не имеют хлоропластов, которые, по-видимому,
были приобретены позднее в результате симбиоза.
В современной системе протистов эвгленовые
вместе с кинетопластидами выделены в особый тип
Euglenozoa Cavalier-Smith, 1981 и представляют в
нем отдельный класс Euglenoidea Biitschli, 1884 (Ca-
valier-Smith, 1978; Corliss, 1984).
В настоящее время эвгленовые известны в качестве
одной из важнейших групп протистов, имеющих боль-
шое значение в природе, а также как объект для целого
ряда экспериментальных работ. Поэтому в течение
последних десятилетий много внимания уделено изу-
чению их фауны, описанию новых видов, родов, се-
мейств, издан ряд хорошо иллюстрированных посо-
бий, атласов, определителей (Св1ренко, 1938; Goidics,
1953; Попова, 1955, 1966;Mignot, 1966; Leedale, 1967,
1978, 1985; Асаул-Ветрова, 1975; Попова, Сафонова,
1976; Bourrelly, 1985; Willey et al., 1988; Triemer, Ott,
1990; Dawson, Walne, 1994; Лихачёв, 1997; Simpson,
1997). Было сделано подробное описание группы эн-
допаразитических эвгленовых из кишечника, полости
тела и яйцевых мешков пресноводных веслоногих рач-
ков (Michajlow, 1972, 1978).
Большие успехи достигнуты в культивировании
ряда видов эвгленовых, получены их аксенические
202
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
(безбактериальные) культуры (Pringsheim, 1946; Гол-
лербах, Полянский В. И., 1951; Wolken, 1961, и др.).
Массовые клональные культуры (особенно эвглен и
астазий) широко применяются в самых разнообраз-
ных исследованиях — морфологических, электрон-
но-микроскопических, биохимических, генетичес-
ких, молекулярно-биологических, а также в биотех-
нологии (Epstein, Allaway, 1967, 1968; Leedale, 1967;
Buetow, 1968, 1982; Зайцева, Шанина, 1976; Сухано-
ва, 1983, 1985). В течение всех последних лет под-
робно изучается значение эвгленовых в природе и их
экология (Винберг, 1966; Sladecek, 1973; Anderson,
1987; Larsen, Patterson, 1990; Simpson et al., 1996/97).
Классификация
Класс Euglenoidea Biitschli, 1884 объединяет 5 от-
рядов, к которым относится 18 родов и около
1000 видов.
Отряд Eutreptiida Leedale, 1967 — Эвтрептины
Отряд Euglenida Biitschli, 1884 — Эвглениды
Отряд Rhabdomonadida Leedale, 1967 — Рабдомонадины
Отряд Sphenomonadida Leedale, 1967 — Сфеномонадины
Отряд Heteronematida Leedale, 1967 — Гетеронематины
Морфология и физиология
Внешнее строение
Форма и размеры тела. Форма тела трофозоитов
широко варьирует: она может быть шаровидной, эл-
липсоидальной, листовидной, яйцевидной, игловид-
ной, веретеновидной (рис. 129). Последняя наиболее
типична и широко распространена в разных таксонах
класса. Передний конец тела более или менее закруг-
лен, задний же часто вытянут и заканчивается заост-
ренным отростком. Одну из особенностей эвглено-
вых составляет спиральная скрученность тела, выра-
женная неодинаково у разных видов, влияющая на
форму тела и обусловливающая спиральную симмет-
рию у ряда видов.
Жгутиковый аппарат. Большинство видов имеет
2 неравных по длине, гетеродинамичных жгутика, и
только у немногих, например у эвтрепций, оба Ж1уги-
ка практически одинаковы (рис. 129, Е). Есть также
виды с 1 или несколькими Ж1угиками.
Жгутики выходят наружу через отверстие, распо-
ложенное на переднем конце тела. Оно ведет в узкий
канал, который продолжается в углубление грушевид-
ной формы — жгутиковый резервуар. Выходящие на-
ружу жгутики несут длинные мастигонемы, располо-
женные в 1 ряд с одной стороны (рис. 130), а также ко-
роткие мастигонемы, расположенные спирально по
всей длине ундулиподии. Обязательная внутренняя
структура жгутиков — параксиальный тяж. В длинной
переходной зоне жгутиков часто отсутствует даже по-
перечная пластинка и только у Entosiphon обнаружена
мощная псевдозвездчатая структура.
2 кинетосомы расположены параллельно друг дру-
гу; от них отходят корешки, состоящие из микротру-
бочек. Все 3 корешка направлены в переднюю часть
клетки и проходят вдоль стенок Ж1угикового резерву-
ара (Farmer, Triemer, 1988), поддерживая его форму
(рис. 131). У некоторых видов в системе корешков
имеются фибриллярные элементы.
Жгутиковый резервуар эвгленовых отличается
крупными размерами и достаточно сложным упоря-
доченным расположением связанных с ним орга-
нелл. Он выстлан плазмалеммой, которая представ-
ляет собой продолжение плазмалеммы поверхности
тела. Жгутики отходят от дна резервуара, и длинный
двигательный жгутик проходит вдоль дорсальной
стенки резервуара. Второй, у большинства видов ко-
роткий жгутик, часто не выходящий за пределы ре-
зервуара, расположен вблизи вентральной его стенки
(рис. 131).
Сократительная вакуоль расположена в брюшной
стенке жгутикового резервуара, в который изливается
ее содержимое при сокращении. У фотосинтезирую-
щих эвглен (род Euglena) недавно обнаружен руди-
ментарный цитостом, который, вероятно, частично
сохраняет функциональную активность (как пиноци-
тозный канал), так как в этом месте обнаруживаются
лизосомы. Он открывается в дистальную часть резер-
вуара (рис. 131). Скрытый цитостом эвглен гомо-
логичен цитостому бесцветных видов эвгленовых
(рис. 110) и кинетопластид (Willey, Wibel, 1985;
Surek, Melkonian, 1986).
Среди эвгленовых есть ряд хищных гетеротроф-
ных видов, у которых развиты весьма сходные при-
способления для захвата пищевых организмов. Это
представители родов Peranema, Heteronema, Entosi-
phon, обладающие глоточным палочковым аппара-
том. У Р. trichophorum глоточный аппарат лежит в
вентральной части переднего конца клетки. Его стен-
ки укреплены 2 трихитами — глоточными палочками.
Трихиты состоят из многочисленных микротрубочек,
ассоциированных с фибриллярными структурами.
Весь этот ротоглоточный комплекс обеспечивает за-
глатывание как мелких, так и крупных пищевых объ-
ектов.
Глоточный палочковый аппарат сходного строе-
ния имеют Heteronema acus и другие виды этого рода,
а также виды рода Entosiphon. У Е. sulcatus глоточ-
ные структуры проходят почти по всей длине тела.
Они образованы тремя мощными пучками микротру-
бочек, с проходящими вдоль них фибриллярными
«парусами», обращенными во внутреннее простран-
ство глотки (Triemer, Fritz, 1986). Пищей этих хищ-
ных эвгленовых жгутиконосцев служат не только
мелкие, но и крупные простейшие, их цисты, а также
мелкие многоклеточные.
С жгутиковым аппаратом фототрофных эвгленид
тесно связан фоторецептор, состоящий из парафлагел-
лярного тельца и стигмы. Парафлагеллярное тельце
представляет собой расширенный участок длинного
(дорсального) Ж1угика (рис. 131) и расположено у его
основания в жгутиковом резервуаре. Напротив параф-
лагеллярного тельца, вблизи дорсальной стенки жгу-
тикового резервуара, расположена стигма (глазок).
Эта органелла состоит из отдельных глобул, каждая из
которых покрыта мембраной (рис. 131). Стигма окра-
шена либо в красный, либо в оранжево-красный цвет.
ТИП EUGLENOZOA. КЛАСС EUGLENOIDEA
203
Рис. 129. Особенности строения некоторых эвгленовых. (По: Матвиенко, 1977; Суханова, 1983; Lee, 1985).
А — Euglena gracilis: 1 — длинный двигательный жгутик, 2 — короткий жгутик, 3 — жгутиковый резервуар, 4 — стигма, 5 — сократительная вакуоль,
6 — кутикула, 7 — хлоропласт, 8 — пиреноид, 9 — парамилон, 10 — ядро, 11 — ядрышко; Б — Euglena vermicularis, трофозоит, двигательный жгутик
отсутствует: 1—2 — фазы эвгленоидного движения; В — Trachelomonas grandis: 1 — домик; Г — Strombomonas acuminata; Д — Phacus pleuronectes: 1 —
парамилон в форме парамилии; Е — Eutreptia pertui; Ж — Rhabdomonas costata; 3 — Heteronema acus; И — Sphenomonas laevis.
Fig. 129. Morphology of some species of Euglenoidea.
A — Euglena gracilis: 1 — long emergent flagellum, 2 — short flagellum, 3 — flagellar reservoir, 4 — stigma, 5 — contractile vacuole, 6 — cuticule, 7 —
chloroplast, 8 — pyrenoid, 9 — paramylon, 10 — nucleus, 11 — nucleolus. Б — Euglena vermicularis: 1 — trophozoite (without of flagellum); 2—3 — phases
of euglenoid movement. В — Trachelomonas grandis: 1 — envelope; Г — Strombomonas acuminata; Д — Phacus pleuronectes: 1 — grain of paramylon; E —
Eutreptia pertui; Ж — Rhabdomonas costata; 3 — Heteronema acus; И — Sphenomonas laevis; E, И — colourless euglenoids.
Это связано с тем, что содержимое каждой глобулы пред-
ставлено каплей липида, в которой растворен (3-каро-
тин. Механизм взаимодействия стигмы и парафлагел-
лярного тельца окончательно не выяснен, но есть на-
блюдения, свидетельствующие о том, что если стигма
затеняет парафлагеллярное тельце, то эвглена изменя-
ет направление своего движения. В последнее время в
прилегающем к стигме участке плазмалеммы обна-
ружен родопсин, что, вероятно, поможет в ближайшем
будущем выяснить ее функцию.
Движение. Эвгленовым свойственны 2 типа дви-
жения: жгутиковое и метаболическое (эвгленоидное).
204
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 130. Ультратонкое строение жгутика эвгленовых (попереч-
ный срез). Схема. (По: Melkonian et al., 1982).
1 — длинные простые мастигонемы, 2 — короткие мастигонемы, 3 —
плазмалемма, 4 — параксиальный тяж, 5 ~ аксонема, 6 — филаменты.
Fig. 130. Ultrastructure of flagellum in euglenoids (transverse secti-
on). Diagram.
1 — simple long mastigonemes, 2 — short mastigonemes, 3 — plasmalem-
ma, 4 — paraxial rod, 5 — axonema, 6 — filaments.
С помощью жгутика они передвигаются в водной сре-
де, вращаясь вокруг своей продольной оси. Метабо-
лическое движение, или метаболия, выражается в уд-
линении и сужении, укорочении и расширении тела
или отдельных его частей, свертывании, способности
изгибаться в стороны (рис. 129, Б). Для метаболии
характерны также перистальтические волны, хорошо
выраженные у Eutreptia, Distigma, у некоторых видов
рода Parastasia (рис. 107). Волны сокращений тела
проходят вдоль его продольной оси. Каждая волна
начинается с расширения заднего конца и перемеща-
Рис. 131. Схема ультратонкого строения жгутикового аппарата
эвгленовых. (По: Карпов, 1990).
1 — двигательный жгутик, 2 — мастигонемы, 3 — параксиальный тяж,
4 — короткий жгутик, 5 — жгутиковый резервуар, 6 — кинетосомы, 7 —
ленты микротрубочек, 8 — парафлагеллярное тельце, 9 — стигма, 10 —
цитостом, 11 — промежуточный корешок из микротрубочек.
Fig. 131. Diagram of ultrastructure of flagellar apparatus of eugle-
noids.
1 — loeomotory flagellum, 2 — mastigonemes, 3 — paraxial rod, 4 — short
flagellum, 5 — flagellar reservoir, 6 — kinetosomes, 7 — ribbons of microtu-
bules, 8 — flagellar swelling, 9 — stigma, 10 — cytostome, 11 — intermedia-
te root consisting of microtubules.
ется к переднему, вызывая продвижение тела вперед.
Затем жгутиконосец восстанавливает свою прежнюю
форму тела, после чего происходит перемещение ци-
топлазмы спереди назад (рис. 107).
Метаболия свойственна видам, обитающим на по-
верхности субстрата и ведущим ползающий образ
жизни. У таких видов жгутик может отсутствовать
совсем или недоразвит (рис. 129, Б). Переход к мета-
болическому движению происходит и в тех случаях,
когда эвглены отбрасывают жгутик при каких-либо
воздействиях. У астазий, эвтрепций, дистигм жгути-
ковое и метаболическое движения осуществляются
одновременно (Попова, 1966). Поверхность тела эв-
гленовых покрыта слизью, которая значительно об-
легчает метаболическое движение.
Среди эвгленовых есть и неметаболичные виды,
обладающие плотной оболочкой, которая препятст-
вует сокращениям тела. Это Euglena acus, виды родов
Cyclidiopsis, Rhabdomonas и другие. Степень выра-
женности метаболии (слабая, сильная), а также ее от-
сутствие используются в качестве одного из таксоно-
мических признаков.
Поверхностные структуры. Поверхностные струк-
туры эвгленовых представлены кутикулой, часто с
сильно развитым слоем гликокаликса. Под плазма-
леммой находятся эластичные лентовидные пластин-
ки белковой природы, расположенные спирально во-
круг тела (рис. 132) (Mignot, 1966). Эти пластинки
рельефно выступают под плазмалеммой, образуя на
поверхности клетки продольные или спирально за-
крученные «штрихи», или ребра.
Между пластинками (рис. 132) у большинства ви-
дов эвгленовых есть промежутки, которые обуслов-
ливают эластичность, гибкость поверхности тела,
необходимую для осуществления метаболических со-
кращений. В промежутках между белковыми пластин-
ками, а также под ними располагаются микротрубоч-
ки, которые связаны между собой мостиками, обеспе-
чивающими скольжение микротрубочек относительно
друг друга. По-видимому, этот механизм обусловлива-
ет эвгленоидное движение.
Поверхность тела всех видов Trachelomonas и
Strombomonas защищена особым образованием —
домиком. Форма домиков широко варьирует от яй-
цевидной, шаровидной, цилиндрической до верете-
новидной. Домик может быть гладким или покры-
Рис. 132. Структура пелликулы Astasia longa. (По: Hausmann,
Hulsmann, 1996).
пл — плазмалемма, бп — белковые пластинки, мт — микротрубочки.
Fig. 132. Ultrastructure of pellicula in Astasia longa.
6n — protein plates, мт — microtubules, пл — plasmalemma.
ТИП EUGLENOZOA. КЛАСС EUGLENOIDEA
205
тым шипами, бугорками, различными выростами, по-
рами, сетевидными утолщениями (рис. 129, Б), На
переднем конце домик имеет горлышко с отверстием
для выхода двигательного жгутика, а на заднем конце
у некоторых видов имеется хвостовидный отросток.
Форма и размеры домиков, их орнаментация специ-
фичны для вида и являются важными таксономически-
ми признаками. Стенки домиков представляют собой
утолщенный поверхностный слой, состоящий из уплот-
ненных слизистых веществ полисахаридной и белковой
природы. Целлюлозы в стенках домиков нет, они ин-
крустированы солями железа, которые придают им ха-
рактерный бурый или коричневый цвет разных оттен-
ков, делают их более жесткими и прочными.
Цитоплазма
Хлоропласты. Фотосинтезирующие виды эвглено-
вых имеют хлоропласты, которых нет у бесцветных
гетеротрофных видов. Однако у некоторых из них,
например у Khawkinea quartana, обнаружены руди-
ментарные пластиды. Число хлоропластов у разных
видов варьирует от 1—2 до нескольких сотен. Если
хлоропластов немного, то число их постоянно для
вида и может служить таксономическим признаком.
Форма хлоропластов разнообразна (рис. 94). Они
могут быть дисковидными, звездчатыми или лентовид-
ными. Оболочка хлоропластов состоит из 3 мембран.
Крупные хлоропласты несут пиреноиды, у мелких они
отмечены не всегда. На поверхности пиреноидов, обра-
щенной к цитоплазме, откладывается парамилон, син-
тез которого происходит вне хлоропластов.
Хлоропласты травянисто-зеленого цвета, содер-
жат хлорофиллы а и Ь, 0-каротин, диадиноксантин и
другие пигменты в малом количестве. Тилакоиды в
ламелле группируются по 3.
Митохондрии. Эти многочисленные органеллы
расположены в цитоплазме всех участков клетки. Им
свойственны пластинчатые кристы. Эвглениды, оби-
тающие в анаэробных условиях, лишены митохонд-
рий. Это продемонстрировано недавно на примере
Postgaardi mariagerensis (Simpson et al., 1996/1997).
Сократительная вакуоль. Чаще всего сократи-
тельная вакуоль 1, и расположена она в передней части
тела возле дорсальной стенки жгутикового резервуара.
Содержимое вакуоли изливается в полость жгутикового
резервуара, а из него — наружу.
Аппарат Гольджи. Диктиосомы имеют типичное
строение, и число их неодинаково у разных видов.
Часто они представлены стопками из 10—15 цистерн.
Резервные вещества. Основное резервное вещество,
специфичное для всех видов эвгленид, — полисахарид
парамилон. Центрами его синтеза в клетке служат пи-
реноиды, однако он образуется и у бесцветных форм.
По своей химической природе парамилон представляет
0-1,3-связанный глюкан, который откладывается в ци-
топлазме в виде зерен различной величины и формы,
специфичных для отдельных родов и видов. У одних
видов зерна парамилона мелкие и заполняют всю ци-
топлазму, например у Euglena gracilis, Astasia longa,
Parastasia fennica и других; у ряда видов кроме мелких
зерен формируются еще 1—4 крупных зерна, называемых
Рис. 133. Резервные полисахариды у Astasia longa. (Ориг. Сухано-
вой).
1 — парамилон, 2 — гликоген.
Fig. 133. Reserved polysaccharide in Astasia longa.
1 -г- paramylon, 2 glycogen.
парамилиями. Место расположения парамилий в ци-
топлазме строго постоянно и специфично для вида.
Форма парамилей может быть палочковидной, как на-
пример у Cyclidiopsis acus, Euglena acus или кольцевидной,
что характерно для родов Phacus, Lepocinclis. Парами-
лии свойственны слабометаболичным видам и неметабо-
личным видам с постоянной формой тела (рис. 129, Д).
У бесцветных эвгленовых кроме парамилона син-
тезируется гликоген и небольшие его запасы откла-
дываются в цитоплазме в виде мелких зерен (рис. 133).
Резервным веществом служит, также нейтральный жир,
который откладывается в цитоплазме в виде капель.
У некоторых эвглен (Euglena rubra) в качестве за-
пасного вещества отмечен гематохром, который на-
капливается в цитоплазме при температуре 27—30 °C
и дает клетке красную окраску.
Ядро
Эвгленовые —- одноядерные организмы. Ядро ча-
ще всего шаровидное, расположено либо в централь-
ной части клетки, либо несколько смещено в ниж-
нюю ее половину. В центре ядра находится крупное
ядрышко (рис. 129). Хромосомы концентрируются в
периферической зоне ядра. В интерфазном ядре хро-
мосомы имеют высокую степень конденсации, и по
своей ультраструктуре они сходны с митотическими
хромосомами (Райков, 1978). Деление ядра осуществ-
ляется по типу закрытого плевро- или ортомитоза.
Размножение
Основной формой размножения служит бинарное
деление, но известна и палинтомия, свойственная
всем известным видам Parastasia и некоторым пред-
206
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 134. Жизненный цикл Parastasia fennica (=Astasia norvegica). (По: Michajlow, 1964).
1 — трофозоит в кишечнике Eucyclops serrulatus, 2 — делящаяся особь в воде, 3 — стадии палинтомии, 4 — жгутиковые зооспоры, 5 — погибшая зооспора.
Fig. 134. The life cycle of Parastasia fennica (=Astasia norvegica).
1 — trophozoite in the intestine of Eucyclops serrulatus, 2— dividing specimen in water, 3 — palintomic division (some stages), 4 — flagellates, 5 — perish zo-
ospore.
ставителям Khawkinea. Перед делением ядро обычно
мигрирует к переднему концу клетки. Цитокинез у всех
эвгленид происходит в продольном направлении, и бо-
розда, разделяющая 2 дочерние особи, проходит от пе-
реднего конца тела к заднему. Фототрофным видам
свойствен циркадный ритм делений. Бесцветные виды
делятся асинхронно в течение всего времени суток. Де-
ление может происходить в подвижном состоянии или
в состоянии пальмеллы. При этом делящаяся особь вы-
деляет большое количество слизи, которая образует
прозрачную и довольно толстую оболочку.
Жизненные циклы
В жизненных циклах свободноживущих и при-
крепленных эвгленовых имеется несколько фаз: ак-
тивные трофозоиты, делящиеся особи, пальмеллевид-
ные формы и цисты покоя. В природных условиях в
течение теплых сезонов эти фазы периодически сме-
няют друг друга, но зимой большинство эвгленовых
находятся в состоянии цист покоя.
У метаболичных видов цисты покоя имеют шаро-
видную форму и покрыты плотной оболочкой муко-
полисахаридной природы (рис. ИЗ, Б). У видов, не
способных к метаболическим движениям (представи-
тели родов Cyclidiopsis, Phacus и др.), переход в по-
коящееся.состояние осуществляется просто путем от-
брасывания жгутика. Изменений формы тела при
этом не происходит.
Из немногочисленной группы эндопаразитических
эвгленовых жизненные циклы достаточно подробно
изучены у нескольких видов рода Parastasia (Michaj-
low, 1972, 1978; Вита, Суханова, 1983; Лихачёв,
1989). Все известные виды парастазий имеют слож-
ные и довольно однотипные жизненные циклы, сходные
с жизненным циклом Parastasia fennica (рис. 134).
Он включает в себя 2 фазы: трофическую, проходя-
щую в организме хозяина (рачка), и репродуктивную,
протекающую в воде. На репродуктивной стадии об-
разуются жгутиковые инвазионные клетки, похожие
на свободноживущих астазий. Регулярное чередование
трофической и репродуктивной фаз жизненного цик-
ла происходит только в течение весны, лета и ранней
осени; зимуют парастазии в организме хозяина или (в
редких случаях) в цистах покоя.
Экология и практическое
значение
Распространение и места
обитания
Эвгленовые распространены всесветно. Огромное
большинство их — обитатели пресных континен-
тальных водоемов, и, вероятно, нет такого водоема, в
котором нельзя было бы найти те или иные виды
этих протистов. В шельфовых зонах морей и океанов
ТИП EUGLENOZOA. КЛАСС EUGLENOIDEA
207
преобладают гетеротрофные эвглениды (Patterson,
Simpson, 1996).
Популяции эвгленовых наиболее многочисленны
в мелководных стоячих пресных водоемах лесной и
лесостепной зон. Это водоемы, в которых много
растительности, хорошо прогревается вода и содер-
жится много органических веществ. Многочисленны
эвгленовые и в удобряемых рыбоводных прудах, на
рисовых полях, а также в биологических прудах, где
происходит очистка сточных вод. В таких водоемах
они часто представляют доминирующую группу орга-
низмов микропланктона и микробентоса. Развиваясь
в массовом количестве, могут служить причиной зе-
леного и красного «цветения» воды.
Среди фотосинтезирующих эвгленид преобла-
дают одиночные свободноживущие виды, которые
ведут плавающий образ жизни: эвглены, факусы,
трахеломонасы и многие другие. Напротив, гетеро-
трофные эвглениды большую часть времени ведут
ползающий образ жизни, лишь ненадолго перехо-
дя к плаванию в поисках скоплений микроорганиз-
мов, которые служат им пищей. Таковы, в частно-
сти, Petalomonas, Anisonema, Peranema, Ploeotia и
Другие.
Небольшой группе эвгленовых свойствен непод-
вижный (на большей части жизненного цикла), при-
крепленный образ жизни. Сюда относится несколько
видов рода Colacium — эпибионтов ракообразных и
других планктонных животных. К внешним покро-
вам животных они прикрепляются передним концом
тела с помощью слизистого стебелька или подушеч-
ки, образуя ветвистые колонии. Cyclidiopsis acus спо-
собны временно прикрепляться к какому-либо субст-
рату с помощью слизи, выделяемой ими на заднем,
заостренном конце тела. В прикрепленном состоянии
они вегетируют, размножаются и могут образовывать
временные колонии.
Еще одну группу эвгленовых представляют эндо-
паразитические виды родов Astasia и Parastasia, оби-
тающие в период трофической фазы жизненного
цикла во внутренних органах пресноводных Copepo-
da (Michajlow, 1972; Лихачёв, 1989).
Влияние факторов
внешней среды
Пищевой фактор. Несмотря на то что эвглено-
вым присущи все способы питания, свойственные
фототрофным и гетеротрофным организмам, пище-
вой фактор, и прежде всего содержание в воде орга-
нических веществ, лимитирует их распределение по
биотопам и распространение. Способность многих
видов к миксотрофному или сапрофитному питанию
обусловливает массовое развитие их в водоемах или
в тех зонах водоемов, которые богаты органически-
ми веществами. Поэтому большинство эвгленид
может быть использовано в качестве индикаторных
организмов водоемов с повышенной степенью сап-
робности (Sladecek, 1973). Пищевыми объектами
фаготрофных эвгленовых (роды Peranema, Urceolus
и др.) служат бактерии, мелкие простейшие, водоро-
сли.
Температура. Популяции эвгленовых начинают
развиваться ранней весной, и их численность увели-
чивается с повышением температуры в июне. В сере-
дине лета численность популяций снижается, но в
конце лета и начале осени вновь значительно увели-
чивается. При этом время весеннего и осеннего мак-
симумов развития различается в разных климатичес-
ких зонах (Попова, 1966, и др.).
Снижение температуры воды осенью стимулирует
формирование цист покоя, которые переживают в
грунте водоемов зиму. Однако ряд холодолюбивых
видов остается в активном состоянии поздней осенью
и зимой подо льдом при температуре воды 4—6 °C.
Один из холодолюбивых видов — Euglena viridis —
образует многочисленные популяции поздней осенью
в сильно эвтрофированных мелких водоемах, а также
в биологических прудах. Популяции этого вида эв-
глен достигают численности до 1—3 и более милли-
онов особей в 1 л воды. Они локализуются в поверх-
ностном слое, где активно осуществляют фотосинтез,
улучшая кислородный режим биологических прудов
и природных водоемов, что особенно важно для про-
цессов минерализации органических веществ (Вин-
берг, 1966; Попова, 1966; Суханова, 1983).
Холодолюбивые Euglena viridis, Е. proxima, трахе-
ломонасы, перанемы и другие зимой могут вмерзать в
лед, не теряя при этом своей жизнеспособности.
Интересные данные о полиморфизме вида по
температурному оптимуму размножения получены
для 26 штаммов Е. gracilis из разных мест обитания
(Pringsheim, 1958). В пределах этих штаммов выде-
лены 3 группы, различающиеся по температурному
оптимуму размножения.
1. Эвглены прекращают размножение при 27 °C,
остаются зелеными, и у них сохраняется стигма.
2. Эвглены интенсивно размножаются при 34 °C,
но многие из них становятся бесцветными, теряя при
этом стигму.
3. Эвглены интенсивно размножаются при 34 °C,
остаются зелеными, и у них сохраняется стигма.
Данные этих экспериментов показывают, что в
природе существуют разные физиологические груп-
пировки (расы) в пределах вида Е. gracilis, адаптиро-
ванные к разным температурным условиям.
Свет. Продолжительность фаз клеточного и жиз-
ненного циклов, время размножения и другие жиз-
ненные функции фототрофных эвгленовых лимити-
руются наряду с температурой световым днем. Раз-
множение этих организмов происходит в темноте.
При отсутствии света многие зеленые виды стано-
вятся бесцветными, гетеротрофными и нуждаются в
органическом источнике углерода. Их морфология и
клеточный метаболизм претерпевают глубокие изме-
нения. У обесцвеченных в темноте эвглен (Ё. gracilis)
уменьшается содержание каротиноидов, резорбиру-
ются пиреноиды, в цитоплазме откладываются боль-
шие запасы парамилона, у многих особей исчезает
стигма. Обесцвеченные эвглены морфологически
сходны с Astasia longa, что послужило причиной
многих дискуссий о систематическом положении
этого вида (Pringsheim, 1958; Попова, 1966). Переме-
щение обесцвеченных эвглен из темноты на свет со-
пряжено с восстановлением хлоропластов и других
208
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 135. Внешний вид некоторых представителей эвгленовых. (По: Суханова, 1983; Dodge, Lee, 1985).
А — Lepocinclis fusiformis, Б — Distigma proteus, В — Menoidium tortuosum, Г — Euglena acus, Д — Cyclidiopsis acus, E — Urceolus cyrusorum, Ж — Noto-
solenus apocamptus.
Fig. 135. External appearance of some euglenoids.
органелл, а также с восстановлением фотосинтети-
ческой активности. Однако некоторые особи после
возвращения их из темноты в условия нормального
освещения остаются бесцветными в течение длитель-
ного времени.
Культивирование. Многие виды эвгленид легко
культивируются в лабораторных условиях и живут в
культуре в течение продолжительных сроков. Для
культур разработано большое количество разнооб-
разных органических и минеральных сред (Pringsheim,
1955; Wolken, 1961; Попова, 1966; Сухарева-Нема-
кова, Каленик, 1977; Сухарева, 1989, и мн. др.).
Большинство культуральных сред содержит мине-
ральную основу, к которой добавляются различные
органические вещества и витамины. Существуют и
твердые среды, содержащие агар-агар, на которых
живут многие виды в состоянии пальмеллы.
Е. gracilis образует в культурах настолько массо-
вые популяции, что может быть использована в ка-
честве корма для мальков рыб и других животных, а
также как тест-объект для выяснения действия цело-
го ряда лекарственных препаратов, например антибио-
тиков и витаминов.
Особенно большое научное и практическое значе-
ние имеют аксенические культуры Е. gracilis и A stasia
longa, которые способны жить в культуральных условиях
в течение длительного времени. Они служат объекта-
ми для многочисленных морфологических, физиоло-
гических, биохимических, молекулярно-биологических
и генетических исследований, а в последнее время и
для целей биотехнологии (Сухарева, 1989, и др.).
Происхождение
и эволюция
Вопрос о происхождении эвгленид изучается до
сих пор. Сведений об их ископаемых формах очень
мало, и они не вполне достоверны (Попова, 1966).
Эвглениды представляют четко очерченную по
своим морфологическим особенностям группу, в ко-
торой фотосинтезирующие и гетеротрофные виды
обнаруживают высокую степень сходства по строе-
нию клетки. Однако нельзя отрицать сходства эвгле-
нид, особенно фотосинтезирующих видов, и с одно-
клеточными водорослями (Зайцева, Шанина, 1976;
Buetow, 1982; Рейвн и др., 1990).
Новые данные о наличии цитостома у фотосинте-
зирующих эвгленид и сравнительный анализ строе-
ния корешковых систем жгутиков с таковым ки-
нетопластид позволили выявить гомологию этих
структур в обеих группах. Кроме того, эвглениды и
кинетопластиды имеют и другие черты сходства: ун-
дулиподии жгутиков снабжены у тех и других па-
раксиальными тяжами, образованными гомологич-
ными белками, деление ядра осуществляется посред-
ством закрытого митоза. Все эти признаки могут
служить доказательством филогенетических связей
между обеими группами и подтверждают проис-
хождение фотосинтезирующих видов эвгленовых от
бесцветных гетеротрофных форм (Taylor, 1976; Lee-
dale, 1978; Willey, Wibel, 1985; Surek, Melkonian,
1986).
ТИП EUGLENOZOA. КЛАСС EUGLENOIDEA
209
Систематический обзор
Отряд Eutreptiida Leedale, 1967
Включает 2 рода: Eutreptia Perty, 1852 и Distigma
Ehrenberg, 1830.
Представители рода Eutreptia — одиночные, силь-
но метаболичные организмы с веретеновидной фор-
мой тела (рис. 129, Е). Передний конец тела закруглен
или косо срезан, задний несет заостренный отрос-
ток, который у разных видов различается по длине.
2 почти равных по длине и толщине жгутика выхо-
дят из жгутикового резервуара; у основания каждого
из них расположен фоторецептор на уровне стигмы.
Хлоропласты имеют форму мелких пластинок, без
пиреноидов.
К роду Distigma относятся бесцветные гетеро-
трофные виды с 2 неравными по длине гетеродина-
мичными жгутиками. Стигмы нет.
У всех видов обоих родов парамилоновые зер-
на мелкие, многочисленные. Обитают в шельфовых
зонах морей, реже в пресноводных бассейнах
(рис. 135, Б).
Отряд Euglenida Biitschli, 1884
К этому таксону относится около 500 фотосин-
тезирующих, гетеротрофных, в разной степени мета-
бол ичных видов. Тело вытянуто в длину, несколько
асимметричное, спирально скрученное. Большинство
представителей этого отряда — одиночные организ-
мы, и только род Colacium включает колониальные
формы. Жгутиков 2: один длинный, двигательный,
выходит наружу из жгутикового резервуара, второй
короткий и наружу не выходит. У зеленых видов в
основании двигательного жгутика расположен фото-
рецептор (рис. 131; 135). Стигма оранжево-красного
цвета. Хлоропласты различной формы и размеров, у
большинства видов с двухскорлупчатыми пиреноида-
ми. Парамилон образуется вне хлоропластов, имеет
форму мелких и крупных кольцевидных или палоч-
ковидных зерен. Обитатели пресноводных бассей-
нов.
К отряду относится свыше 10 родов. Основные
из них: Euglena Ehrenberg, 1830; Colacium Ehren-
berg, 1833; Trachelomonas Ehrenberg, 1833; Astasia
Dujardin, 1841; Phacus Dujardin, 1841; Strombomonas
Deflandre, 1930; Parastasia Michajlow, 1972.
Большинство эвгленид — свободноживущие жгу-
тиконосцы, но род Colacium включает эпибионтов, а
к роду Parastasia относится около 20 эндопаразити-
ческих видов, обитающих во внутренних органах вес-
лоногих рачков.
Отряд Rhabdomonadida Leedale,
1967
Представлен небольшой группой мелких, до
30 мкм в длину, бесцветных эвгленовых с ригидным,
неметаболичным телом (рис. 129; 135). Жгутики ге-
теродинамичные и неравные по длине. 1 длинный,
выходит из жгутикового резервуара наружу и выпол-
няет двигательную функцию, второй очень короткий
и наружу не выходит. Жгутиковое отверстие распо-
ложено субапикально. Тело цилиндрическое или уд-
линенное, изогнутое, поверхность его ребристая.
Стигмы нет. Питание сапрофитное. Парамилона
много, иногда зерна его крупные. Резервных липи-
дов также много. Пальмеллевидные формы и цисты
не обнаружены. Обитают в эвтрофных пресных во-
доемах. Все виды свободноживущие.
К отряду относится свыше 30 видов. Основные
роды: Menoidium Perty, 1852; Rhabdomonas Fresenius,
1901; Gyropaigne Skuja, 1926; Rhabdospira Pringshe-
im, 1963.
Отряд Sphenomonadida Leedale,
1967
Бесцветные фаготрофные одиночные эвгленовые
с 1 или 2 гетеродинамичными жгутиками, неравны-
ми по длине (рис. 129; 135). Длинный жгутик двига-
тельный, направлен вперед, второй (если имеется)
короткий и неподвижный, проходит по боковой сто-
роне тела. Сплющенное, ригидное тело с нескольки-
ми килями или с продольными бороздками достигает
40 мкм в длину. Обычны в шельфовых зонах морей,
а также в эвтрофных пресных водоемах; свободно-
живущие, бентосные и планктонные виды.
В отряд входит свыше 20 видов. Основные ро-
ды: Anisonema Dujardin, 1841; Sphenomonas Stein,
1878; Petalomonas Stein, 1878; Notosolenus Stokes,
1890.
Отряд Heteronematida Leedale,
1967
Бесцветные фаготрофные эвглениды с удлинен-
ным пластичным телом до 300 мкм в длину, округ-
лым в поперечном сечении. Жгутиков 2, оба выходят
наружу из жгутикового отверстия, расположенного
субапикально (рис. 129). Жгутики неодинаковые по
длине, толщине и двигательной активности. Более
длинный и толстый — двигательный жгутик во
время движения направлен вперед, ригидный, дис-
тальный конец его совершает круговые или осцил-
лирующие движения. Второй жгутик короткий, тон-
кий, направлен назад, тесно прижат к поверхности
тела и несет рулевую функцию. Поверхность тела
ребристая. Имеется глоточный аппарат из 2 палочек,
связанных с цитостомом (Регапета), или же в фор-
ме трубки (Entosiphon). Экструсомы многочисленны,
крупных размеров. Стигма отсутствует. Распростра-
нены в морских, солоноватоводных и пресноводных
бассейнах.
Известно около 50 видов, относящихся к этому
отряду. Основные роды: Peranema Dujardin, 1841;
Heteronema Dujardin, 1841; Urceolus Mereschkowsky,
1870; Entosiphon (Dujardin) Stein, 1878.
14 Протисты, часть 1
210
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
THE PHYLUM EUGLENOZOA CAVALIER-SMITH, 1978
THE CLASS EUGLENOIDEA BUTSCHLI, 1884
SUM
The class includes not less than 1000 green and colourless speci-
es. All euglenoids are monads except Colacium, which form dendroid
colonies of cells. Most of the species are elongated, spindle shaped,
others are ovoid, leaf-shaped or spherical. The body of euglenoids
possesses bilateral or helical symmetry, with or without flattening.
They have 2 flagella per cell arising within reservoir — an invagina-
tion of anterior part of the cell.
In most species only one flagellum emerges from the reservoir;
the second flagellum is short, being entirely within the reservoir.
Some species have 2 heterodynamic flagella of equal length. The
emergent flagella of all species bear paraflagellar rods, and unilateral
arrays of mastigonems. The transition zone of flagella is long,
transversal plate is normally present. All green euglenoid and some
colourless species have orange-red stigma (eyespot). It is located
near the base of the reservoir. It is independent of the chloroplast
and contains derivatives of ^-carotene and some other carotenoid
pigments. Opposite the stigma a flagellar swelling is situated near
the base of the longer flagella. The stigma and flagellar swelling
are two structural and functional components of euglenoid photore-
ceptor.
All species of green euglenoids have chloroplasts varying in num-
ber from one to several hundreds, in the shape from plates and discs
to ribbons, in dimensions, which are characteristic of each species.
Some chloroplasts have pyrenoids, which vary in shape and can be
sheeted, naked, projected, etc. The lamellae of a chloroplast normally
consist of 3 or more thylakoids. The basic pigments of chloroplasts
are: chlorophyll a, b, p-carotene, diadinoxantin. Colourless euglenoids
are free of chloroplasts and leucoplasts.
The euglenoid cell is covered with cuticle which forms strips
consisting of protein. The pellicular strips are situated under the cell
membrane. They can be flexible or rigid in some species. In addition,
the surface of euglenoid body is covered with a thin layer of gly-
cocalyx. Some euglenids (Trachelomonas and Strombomonas) have
dense lorica consisting of mucilage, impregnated with ferric hydroxide.
Many submembrane mucilaginous bodies can be seen in all euglenoid
species.
MARY
All freshwater species of euglenoids have a contractile vacuole
situated near the flagellar reservoir, mitochondria with lamellar cris-
tae, well developed dictyosomes and numerous lysosomes. The cytop-
lasm of euglenoids is rich in paramylon (P-l,3-linked glucan), which
is stored as solid grains.
The interphase euglenoid cell has one nucleus of vesicular type. It
divides by a specific closed euglenoid orthomitosis. The nuclear enve-
lope and the nucleolus remain intact during the process of nuclear
division. Reproduction is only agamic, and occurs by longitudinal fis-
sion of the cell.
Euglenoid flagellates are primary free living protozoa. Marine and
parasitic species are quite rare. Fresh water euglenids inhabit all types
of water basins: lakes, ponds, small temporary water bodies, swamps,
and even wet soil. The main type of locomotion is swimming with
highly mobile flagella. They also often glide with anteriorly directed
flagella, creeping on the surface. The euglenids have a specific kind
of movement — metabolic shortening of their body or rapid change
of body shape, the «euglenoid movement».
Phagotrophic species have well developed cytostome-pharyngeal
complex for food ingestion. Some of them are predators. The majority
of euglenids are photosynthetic. However, in spite of their mode of
feeding, the rudimentary cytostome has been found in all investigated
species of phototrophic flagellates.
The euglenoids usually form very dense populations in natural
water basins and play a very important role in food chains and nets of
hydrobionts. They can be important organisms in diagnosis of water
pollution. Many species of euglenoids are cultivated in laboratory con-
ditions. They form mass populations in the laboratory cultures and are
used as food organisms for young fishes. Cultivation of Euglena gra-
cilis allows using the species as a test object for the elucidation of the
activity of many medicine preparations, antibiotics and vitamins inclu-
ded. The phylum Euglenophyta Biitschli, 1884 includes the class Eugle-
nophyceae Biitschli, 1884, comprising 5 orders: Eutreptiida Leedale,
1967; Euglenida Biitschli, 1884; Rhabdomonadida Leedale, 1967; Sphe-
nomonadida Leedale, 1967; Heteronematida Leedale, 1967. The descrip-
tion of the main genera is presented in the chapter.
ЛИТЕРАТУРА13
Асаул-Ветрова 3.1. Визначник евгленових водоростей Укра-
шении РСР. Кшв, 1975.
Вннберг Г. Г. Биологические пруды в практике очистки сточ-
ных вод, Минск, 1966.
(Вита И., Суханова К. М.) Wita I., Sukhanova К. М. Studies
on the biology and cytology of Parastasia fennica Michailow (Flagel-
lata, Euglenida), a parasite of Cyclopida (Copepoda) // Acta Protozool.
1983. Vol. 22, N 1. P.55—70.
Гинецинская T. А., Добровольский А. А. Частная паразитоло-
гия. M.: Высш, шк., 1978. Т. 1.
Голлербах М. М., Полянский В. И. Пресноводные водоросли
и их изучение. Определитель пресноводных водорослей СССР. М.,
1951.
Жуков Б. Ф. Атлас пресноводных гетеротрофных жгутико-
носцев (биология, экология, систематика). Рыбинск, 1993.
Зайцева Г.Н., Шанина Н. А. Биохимия генетической системы
ядер простейших. Кариология и генетика простейших. Л.: Наука,
1976. С. 33—63.
Карпов С. А. Система протистов. Омск, 1990.
Лихачёв С. Ф. Эндопаразитические эвгленовые жгутиконосцы.
Омск, 1989.
Лихачёв С. Ф. Эвгленовые жгутиконосцы рода Trachelomo-
nas Ehrenberg, 1883 из водоемов Омской области // Учен. зап.
биол. ф-та Омск. гос. пед. ун-та. Биол. сер. 1996а. Вып. 5.
С. 93—123.
Лихачёв С. Ф. Эвгленовые жгутиконосцы рода Phacus Du-
jardin, 1884 из водоемов Омской области // Учен. зап. биол.
13 Цитируемая и рекомендуемая литература к разделу «Класс
Euglenoidea».
ф-та Омск. гос. пед. ун-та. Биол. сер. 19966. Вып. 5. С. 124—
142.
Лихачёв С. Ф. Эвгленовые водоемов Омской области. Омск,
1997.
Полянский В. И. Экспериментальное исследование изменчи-
вости Euglena gracilis Klebs, в условиях культуры И Учен. зап.
ЛГПИ. 1948. Т. 70. С. 153—170.
Попова Т. Г. Эвгленовые водоросли: Определитель пресновод-
ных водорослей СССР. М.: Сов. наука, 1955. Вып. 7.
Попова Т. Г. Эвгленовые водоросли. М.; Л.: Наука, 1966.
(Флора споровых растений СССР; Т. 8, вып. 1).
Попова Т.Г., Сафонова Т. к. Эвгленовые водоросли. Л.:
Наука, 1976. (Флора споровых растений СССР; Т. 9, вып. 2).
Райков И. Б. Ядро простейших. Морфология и эволюция. М.:
Наука, 1978.
(Райков И. Б.) Raikov I. В. The protozoan nucleus. Morphology
and evolution. Wien; New York, 1982. (Cell Biology Monographs;
Vol. 9).
Рейвн П., Эверт P., Айкхорн С. Современная ботаника. М.:
Мир, 1990. Т. 1.
Сафонова Т. А. Отдел Эвгленовые водоросли (Euglenophyta) И
Жизнь растений. М.: Просвещение, 1977. Т. 3.
CeipeHKO Д. О. Eugleninae. Кшв, 1938.
Суханова К. М. Фауна и экология эвгленовых жгутиконосцев
сооружений биологической очистки сточных вод И Простейшие
активного ила. Л.: Наука, 1983. С. 43—54.
Суханова К. М. Морфофизиологический полиморфизм и кле-
точные мехайизмы его поддержания у эвгленовых жгутиконосцев //
Микроэволюция. М., 1985. С. 29—31.
Сухарева Н. Н. Биотехнология получения липидных препа-
ратов из культур жгутиконосцев // Простейшие — новые объекты
биотехнологии. Л.: Наука, 1989. С. 5—78.
ТИП EUGLENOZOA. КЛАСС KIN ETOPLASTIDEA
211
Сухарева-Немакова H. Н., Каленик Н. М. Пути синтеза и со-
став жирных кислот у эвгленид // Науч. докл. высш. шк. Биол.
науки. 1977. № 5. С. 5—18.
(Ценовский Л.) Cienkowsky L. Uber Palmallaceen und emige
Flagellaten // Arch, microsk. Anat. 1870. Bd 6, N 8. S. 421—438.
Bourrelly P. Les algues d’eau douce. Initiation a la systematique.
T. 3: Les alques Meues et rouges, les Euglenines, Peridiniens et Cryp-
tomona dines. 2nd ed. Paris, 1985.
Buetow D. E. Morphology and ultrastructure of Euglena // The
Biology of Euglena. London, 1968. Vol. 1. P. 109—184.
Buetow D. E. Phylogenetic origin of the chloroplast //J: Protozo-
ol. 1976. Vol. 23, N 1. P. 41—47.
Buetow D. E. The Biology of Euglena. Physiology. London, 1982.
Cavalier-Smith T. The evolutionary origin and phylogeny of
microtubules, mitotic spindles and eukaryote flagella // BioSystems,
1978. Vol. 10. P. 93—114.
Cavalier-Smith T. Eukaryote kingdoms: seven or nine? // Bio-
Systems. 1981. Vol. 14. P. 461—484.
Corliss J. O. The kingdom Protista and its 45 phyla // BioSys-
tems. 1984. Vol. 17. P. 87—126.
Dawson N. S., Walne P. L. Evolutionary trends in euglenoids //
Arch. Protistenk. 1994. Bd 144. S. 221—225.
Doflein F. Lehrbuch der Protozoenkunde. Jena, 1953.
Dujardin F. Histoire naturelle des Zoophytes-Infusoires. Paris, 1841.
Epstein H.T., Allaway E. Properties of selectivity starved Eugle-
na // Biochim., Biophys. Acta. 1967. Vol. 142, N 1. P. 142—207.
Farmer M. A., Triemer R. E. Flagellar systems in the euglenoid
flagellates // BioSystems. 1988. Vol. 21. P. 283—291.
Goidics M. The genus Euglena. Wisconsin, 1953.
Hoare C. The trypanosomes of mammals. Oxford: Blackwell Sci.
Publ., 1972.
Klebs G. Uber die Organization einiger Flagellatengruppen und
ihre Beziehungen zu anderen Infusorien // Untersuch. Bot. Inst. Tubin-
gen. 1883. Bd 1, H. 2.
Larsen J., Patterson D. J. Some flagellates (Protozoa) from tro-
pical marine sediments // J. Nat. Hist. 1990. Vol. 24, N 6. P. 801—937.
Leedale G. F. Euglenoid flagellates. New Jersey : Euglewood
Cliffs, 1967.
Leedale G. F. The nucleus in Euglena // The biology of Euglena.
New York; London, 1968. P. 185—243.
Leedale G. F. Phylogenetic criteria in euglenoid flagellates // Bio-
Systems. 1978. Vol. 10, N 1. P. 183—187.
Leedale G. F. Order 3. Euglenida Butschli, 1884. Illustrated guide
to the Protozoa. Lawrence : Allen Press, 1985. P. 41—53.
Michajlow W. Euglenoidina parasitie in Copepoda. S. 1.: Polish.
Acad. Sci. polon., 1972.
Michajlow W. Biologia parozytniczych Euglenoidina. Warszawa,
1978.
Mignot J.-P. Structure et ultrastructure de quelques euglenomona-
dines // Protistologica. 1966. Vol. 2. P. 51—117.
Perty M. Zur Kentniss kleinster Lebensformen. Berlin, 1852.
Pringsheim E. G. Mixotrophy in Flagellates // Planta. 1958.
Vol. 52, N 4.
Pringsheim E. G. Physiologische Varietaten bei Flagellaten //
Berl. Dtsch. Bot. Ges. 1955. Bd 8.
Pringsheim E. G. The biphasic or soil-water culture for growing
Algae and Flagellatae // J. Ecol. 1946. Vol. 3, N 2.
Simpson A. G. B. The identity and composition of Euglenozoa H
Arch. Protistenk. 1997. Bd 148. S. 318—328.
Simpson G. B., Hoff J., Bernard C., Burton H. R., Patterson
D. J. The ultrastructure and systematic position of the Euglenozoon
Postgaardi mariagerensis, Fenchel et al. // Arch. Protistenk. 1996/1997.
Bd 147. S. 213—225.
Slade&k V. System of water quality from the biological point of
view. Ergebnisse limnologie // Arch. Hydrobiol. 1973. Bd 7, N 1.
S. 1—218.
(Slade€ek V.). Сладенек В. Атлас сапробных организмов. Уни-
фицированные методы исследования качества вод. М., 1977.
Stein F. R. Der Organismus der Infusionsthiere. Leipzig, 1878. H. 3.
Surek B., Melkonian M. A cryptic cytostome is present in Eug-
lena II Protoplasma. 1986. Vol. 133. P. 39—49.
Suzaki T., Williamson R. Ё. Pellicular ultrastructure and eugleno-
id movement in Euglena ehrenbergii Klebs and Euglena oxyuris
Schmarda // J. Protozool. 1986. Vol. 33. P. 165—171.
Triemer R. E., Farmer M. A. An ultrastructural comparison of
the flagellar, feeding and mitotic apparatus in euglenoids and kinetop-
lastids // Protoplasma. 1991. Vol. 164. P. 91—104.
Triemer R. £., Fritz L. Structure and operation of the feeding
apparatus in a colorless euglenoid Entosiphon sulcatum // J. Protozool,
1986. Vol. 34. P. 154—162.
Triemer R.E., Ott D. W. Ultrastructure of Diplonema ambulator
Larsen et Patterson (Euglenozoa) and its relationship to Isonema U
Eur. J. Protistol. 1990. Vol. 25. P. 316—320.
Willey R. L., Wibel R. G. A cytostome cytopharynx in green eug-
lenoid flagellates (Euglenales) and its phylogenetic implications К
BioSystems. 1985. Vol. 18. P. 369—376.
Willey R. L., Walne P. L., Kivic P. Phagotrophy and the origins
of the euglenoid flagellates # CRC Critical Rev. in Plant Sci. 1988.
Vol. 7. P. 303—340.
Wolken J. J. Euglena. An Experimental organism for biochemical
and biophysical studies. Rutgers: State univ.; Inst.of microbiol.,
1961. Vol. XII.
Класс Kinetoplastidea Honigberg, 1963
Бесцветные Ж1угиконосцы, обладающие 2 гетеродинамными
(или единственным, направленным вперед) жгутиками, отходящими
от дна жгутикового кармана. Жгутики несут параксиальный тяж,
который проходит параллельно аксонеме. Покровы представлены
тубулёммой, степень целостности которой различна у отдельных
представителей кинетопластид. Единственная митохондрия, часто
имеющая сложную пространственную организацию, формирует
специализированный участок — кинетопласт, который содержит
всю или почти всю митохондриальную ДНК, или несколько диск-
ретных ДНК-содержащих участков. Кинетопласт, как правило,
тесно связан с базальной частью кинетосом жгутиков. Кинетоплас-
тиды имеют 1 пузырьковидное ядро, деление которого осуществля-
ется по типу закрытого внутриядерного плевромитоза, протекающе-
го без конденсации хромосом. Наличие полового процесса доказа-
но, однако частота и формы его реализации пока неизвестны.
Ротовой аппарат, включающий цитостом и цитофаринкс, стенки
которых подстилаются микротрубочками, редуцирован у ряда кро-
вепаразитов. Сократительная вакуоль, если имеется, открывается в
жгутиковый карман. Одиночные, в исключительных случаях коло-
ниальные организмы; в жизненных циклах ряда видов отмечено за-
кономерное формирование многоядерных особей. Свободноживущие
гетеротрофы, комменсалы позвоночных и беспозвоночных живот-
ных, паразиты животных и растений. Описано около 1000 видов.
В настоящее время к кинетопластидам относят 3
хорошо дифференцируемые группы жгутиконосцев:
бодонид, криптобий и трипаносоматид (рис. 136).
Бодониды, имеющие 2 свободных гетеродинамных
жгутика (рис. 136, А), в подавляющем большинстве
представлены свободноживущими формами, У крип-
тобий, также имеющих 2 жгутика, направленный
назад рекуррентный, или рулевой, жгутик прикреп-
лен к телу клетки на значительном его протяжении
(рис. 136, К; 153, вкл.). Большинство криптобий —
комменсалы или паразиты беспозвоночных и позво-
ночных животных. Трипаносоматиды имеют только
1 направленный вперед жгутик (рис. 136, В; 157, вкл.).
Все представители трипаносоматид — облигатные па-
разиты животных или растений.
История изучения
Поскольку многие представители трипаносоматид
паразитируют в человеке, домашних животных и куль-
турных растениях, зачастую вызывая их серьезные забо-
левания, эта группа кинетопластид всегда была в центре
212
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 136. Морфология кинетопластид в световом микроскопе.
А — Bodo saltans, Б — Cryptobia salmositica, В — Trypanosoma mega,
кп — кинетопласт, пж — передний жгутик, рж — рекуррентный жгутик,
св — сократительная вакуоль, ум — ундулирующая мембрана, я — ядро.
Fig. 136. Morphology of kinetoplastids in the light microscope.
A — Bodo saltans, Б — Cryptobia salmositica, В — Trypanosoma mega,
кп — kinetoplast, пж — anterior flagellum, рж — posterior flagellum, ce —
contractile vacuole, ум — undulating membrane, я — nucleus.
внимания ученых, являясь предметом многочислен-
ных и разносторонних исследований. Именно этим
объясняется богатая и динамичная история их изуче-
ния. Напротив, история изучения двухжгутиковых
кинетопластид представляет собой монотонную хро-
нологию описания их отдельных таксонов, начиная с
работы Эренберга (Ehrenberg, 1830), описавшего род
Bodo, и до наших дней. Эта группа кинетопластид долгое
время оставалась в тени, и сегодня наши знания об эко-
логии, жизненных циклах и даже видовом разнообра-
зии бодонид и криптобий очень бедны и фрагментар-
ны.
История изучения кинетопластид, вероятно, ведет
свой отсчет от работ Антони ван Левенгука. Однако
начало целенаправленного и интенсивного изучения
группы можно датировать лишь серединой XIX сто-
летия. Именно в это время в научной литературе на-
чинают появляться отдельные описания кинетоплас-
тид, а уже в скором времени этот процесс принимает
лавинообразный характер. Сразу вслед за свободно-
живущими Bodo (Ehrenberg, 1830) в поле зрения уче-
ных попали трипаносомы из крови бесхвостых амфи-
бий (Gluge, 1842), для которых и было предложено
(Gruby, 1843) родовое название Trypanosoma. В это
же время в литературе появляется описание крипто-
бий из репродуктивных органов моллюсков (Leidy,
1846). К началу XX в. наука уже располагала многи-
ми фактами нахождения трипаносом в крови позво-
ночных животных, в том числе и млекопитающих.
Так, в крови лошадей и верблюдов, болевших суррой,
были обнаружены жгутиконосцы, названные впослед-
ствии в честь их первооткрывателя Т. evansi (Evans,
1880), а в крови крыс из Индии была описана Т. 1е-
wisi (Lewis, 1878, 1879). Кроме того, оказалось, что
трипаносомовые могут паразитировать и в кишечни-
ке беспозвоночных животных, в частности насеко-
мых (Leidy, 1853, 1856; Kent, 1880). К этому периоду
относятся и пионерские работы русских ученых Мит-
рофанова (1883), Данилевского (1888) и Боровского
(Боровский, 1898, — цит. по: Сафьянова, 1982). Пос-
леднему принадлежит приоритетное описание клини-
ческой картины кожного лейшманиоза (болезни Бо-
ровского) и доказательство протозойной природы его
возбудителя Leishmania tropica. Несомненно, глав-
ным событием конца XIX в. стало многоплановое ис-
следование Trypanosoma brucei — возбудителя нага-
ны — распространенного в экваториальной Африке
тяжелого заболевания домашнего скота. В серии ис-
следований Давиду Брюсу (Bruce, 1895, 1897, 1902)
удалось доказать, что это заболевание вызывается
трипаносомами, которые передаются от больных жи-
вотных здоровым через укус кровососущих насеко-
мых — мух рода Glossina, и что резервуаром парази-
тов в природе служат дикие животные. Совокупность
этих открытий, по сути, определила всю дальнейшую
стратегию исследования трипаносоматид как возбуди-
телей опасных трансмиссивных заболеваний человека
и домашних животных, характеризующихся природ-
ной очаговостью. Работы Брюса явились своего рода
детонатором, предопределившим исследовательский
взрыв, пришедшийся на первое десятилетие XX в. В
период с 1900 по 1911 г. были выявлены возбудители
всех основных трипаносомозов и лейшманиозов на
континентах Старого и Нового Света, описаны новые
роды гетеро- и гомоксенных трипаносоматид, сведен-
ные наконец в одно сем. Trypanosomatidae (Grobben,
1905, — цит. по: Ноаге, 1972). Среди многочисленных
исследований жизненных циклов трипаносоматид сле-
дует выделить работу Клейна (Kleine, 1909), который
показал, что, проходя через переносчика, трипаносомы
претерпевают определенное закономерное развитие,
предшествующее формированию стадий, инфективных
для позвоночных животных. Важным достижением
этого периода явилась разработка метода лабораторно-
го культивирования трипаносоматид (Novy et al., 1907)
на искусственных кровьсодержащих средах (NN и
NNN среды).
Накопление в литературе большого фактического
материала не могло не повлечь за собой попыток к оп-
ределенным теоретическим обобщениям. На данном
этапе они вылились в полемику по вопросу о проис-
хождении трипаносоматид (Leger, 1904а, 1904b; Wood-
cock, 1906; Minchin, 1908; Leger, Dubosq, 1910). В этом
научном споре победу одержала гипотеза Леже, соглас-
но которой предками трипаносом и лейшманий были
гомоксенные жгутиконосцы из кишечника насекомых,
произошедшие, в свою очередь, от гипотетических сво-
бодноживущих лептомонасподобных предков (Leger,
Dubosq, 1910). Несколько позже такой свободноживу-
щий Ж1угиконосец был найден Вудкокком (Woodcook,
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
213
1914) и назван Proleptomonas faecicola. Лишь спустя
шестьдесят с лишним лет удалось показать, что Р. fae-
cicola имеет отличную от кинетопластид организацию
хондриома и двигательного аппарата и не может быть
отнесен к этой группе жгутиконосцев.
Следующий период в истории изучения кинето-
пластид охватывает первую половину XX столетия.
В это время появляется ряд крупных, обобщающих
сводок, среди которых, бесспорно, выделяются со-
ответствующие разделы в монографиях Веньона и
Грассе (Wenyon, 1926; Grasse, 1952). На этом этапе
начинается постепенное обособление научных школ,
специализирующихся на изучении отдельных групп
и даже отдельных видов трипаносоматид, которых
разделяли на основании таксономических, клиничес-
ких или географических критериев. Таким образом
возникли группы ученых, изучающих различные лей-
шманиозы, африканские и южноамериканский три-
паносомозы, трипаносомозы домашних животных и
т. д. Часть трипаносоматид, не представляющая прак-
тического интереса, оказалась при этом фактически
за чертой внимания исследователей.
Начало 60-х гг. открывает новейший период в
истории изучения кинетопластид. Бурное развитие
методов электронной микроскопии, биохимии и мо-
лекулярной биологии наряду с совершенствованием
традиционных методов ставит исследование этой
группы на качественно новый уровень. Так, выясне-
ние природы и организации кинетопласта уже на
ранних этапах этих исследований способствовало
определению групп жгутиконосцев, в том числе и
свободноживущих, родственных паразитическим
трипаносоматидам. На основании этих данных Хо-
нигберг в 1963 г. объединил бодонид, криптобий и
трипаносоматид в 1 таксон Kinetoplastida (Honig-
berg, 1963). Результатом дальнейшей интеграции на-
учных знаний стало появление фундаментального
многотомного издания «Биология кинетопластид»
(Biology of.., 1976, 1979), в написании которого
приняли участие практически все ведущие специа-
листы в данной области. Этот всеобъемлющий труд
служит отправной точкой для большинства совре-
менных исследований. Вероятно, уже на этом этапе
можно было ожидать и нового осмысления теорети-
ческих вопросов эволюции и филогении кинетоплас-
тид, поскольку была признана филогенетическая це-
лостность кинетопластид, как свободноживущих, так
и паразитических, и получены достаточно многочис-
ленные данные по сравнительной цитологии различных
представителей группы. Однако этого не произошло,
и в дальнейшем авторы подтвердили привержен-
ность традиционным представлениям о происхожде-
нии и эволюции группы.
80-е и начало 90-х гг. проходят под знаком интен-
сивного применения методов электронной микроскопии
и молекулярной биологии в исследовании кинетопластид.
Одним из важных событий последнего десятилетия
стало доказательство наличия генетических обменов у
трипаносоматид (Крылов и др., 1985; Tait, 1983, и др.).
Наконец, комплексное сравнительно-морфологическое
исследование кинетопластид позволило по-новому
взглянуть на филогению этой группы (Фролов, 1993).
Оказалось, что по целому ряду признаков трипаносо-
мы гораздо ближе к двухжгутиковым кинетопласти-
дам, чем «низшие» гомоксенные паразиты насекомых.
В течение следующих пяти лет эта гипотеза нашла свое
подтверждение в серии работ по сиквенсу генов рРНК
у различных представителей кинетопластид и эвглено-
вых жгутиконосцев.
В настоящее время кинетопластиды являются од-
ной из наиболее интенсивно изучаемых групп низ-
ших эукариот. Ежегодно в различных изданиях пуб-
ликуются сотни работ, посвященных исследованию
трипаносоматид и их ближайших родственников —
бодонид и криптобий.
Классификация
Принципы классификации кинетопластид основа-
ны главным образом на анализе морфологических
признаков, которые можно наблюдать в световом
микроскопе, — это форма и размеры тела жгутико-
носцев, местонахождение жгутикового кармана, ко-
личество и размеры жгутиков, расположение в теле
основных органоидов и т. п. Для паразитических ки-
нетопластид учитывается, кроме того, приурочен-
ность к определенным хозяевам или их группам. В
последнее время в классификации кинетопластид все
чаще используются данные об их ультраструктуре и
некоторые молекулярно-биологические характерис-
тики их изолятов.
В настоящее время в классе Kinetoplastidea выде-
ляют следующие таксоны.
Отряд Bodonida Hollande, 1952
Сем. Bodonidae Dophlein, 1901
Род Bodo (Ehrenberg, 1830) Stein, 1875
Род Parabodo Skuja, 1939
Род Ichthyobodo Pinto, 1928
Род Rhynchomonas Klebs, 1893
Род Rhynchobodo Lackey, 1940
Род Hemistasia Griessmann, 1913
Сем. Cryptobiidae Poche, 1911
Род Cryptobia Leidy, 1846
Род Cephalothamnium Stein, 1878
Род Procryptobia Vickerman, 1978
Род Dimastigella (Sandon, 1928) Vickerman, 1978
Род Cruzella Faria et Pinto, 1922
Отряд Trypanosomatida Kent, 1880
Сем. Trypanosomatidae Grobben, 1905
Род Blastocrithidia Laird, 1959
Род Crithidia Leger, 1902
Род Herpetomonas Kent, 1880
Род Leptomonas Kent, 1880
Род Wallaceina Podlipaev, Frolov et Kolesnikov, 1990
Род Rhinchoidomonas Patton, 1910
Род Phytomonas Donovan, 1909
Род Leishmania Ross, 1903
Род Endotrypanum Mesnil et Brimont, 1908
Род Trypanosoma Gruby, 1943
Морфология
Кинетопластиды представляют собой целостную
группу организмов, характеризующуюся своеобраз-
ным планом строения, главным признаком которого
является наличие в их клетках кинетопласта (Honig-
berg, 1963; Vickerman, Preston, 1976; Фролов, 1993;
Frolov, Karpov, 1995).
214
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 137. Морфологические формы трипаносоматид.
А — промастигота, Б — эпимастигота, В — опистомастигота, Г— трипо-
мастигота, Д — хоаномастигота, Е — сферомастигота, Ж — эндомасти-
гота, 3 — амастигота. ж — жгутик, жк — жгутиковый карман, кп - кине-
топласт, я — ядро. (А—Е по: Ноаге, Wallace, 1966; Ж, 3 по: Фролов, 1994).
Fig. 137. The morphological forms of trypanosomatids.
A — promastigote, Б — epimastigote, В — opistomastigote, Г — trypomasti-
gote, Д — choanomastigote, E — spheromastigote, Ж— ehdomastigote, 3 —
amastigote. ж — flagellum, жк — flagellar pocket, кп — kinetoplast, я —
nucleus.
Светооптическое описание
морфологии
Морфология двухжгутиковых кинетопластид, как
свободноживущих, так и паразитических, не отличает-
ся заметным разнообразием (рис. 136). При изучении
жгутиконосцев in vivo или на окрашенных препаратах
удается наблюдать основные клеточные структуры:
жгутики, кинетопласт, ядро, цитостом, сократитель-
ную и пищеварительные вакуоли. Количественные и
качественные характеристики этих признаков лежат в
основе систематики бодонид и криптобий. Свободно-
живущие кинетопластиды формируют относительно
мономорфные популяции. Жизненные циклы парази-
тических криптобий могут протекать с более или
менее выраженным чередованием стадий. Как правило,
это сопровождается появлением различных (2 и более)
морфологических форм жгутиконосцев. Однако раз-
личия между ними невелики и касаются лишь формы
и размеров их тела (Lorn, 1979).
По ходу реализации своих жизненных циклов
трипаносоматиды на фоне чередования стадий разви-
тия проходят и через смену ряда морфологических
форм клеток. '
Сочетание окраски сухих мазков по Лейшману или
по Романовскому—Гимза с наблюдением результата в
световом микроскопе представляет собой основной
метод исследования морфологии трипаносоматид, на
котором базируется классификация их морфологиче-
ских форм (Ноаге, Wallace, 1966). На окрашенных
препаратах можно наблюдать форму тела жгутиконос-
цев, расположение жгутикового кармана (место выхо-
да жгутика из тела клетки), сам жгутик, ядро и кине-
топласт (рис. 136, В). Именно на основании анализа
этих признаков и строится классификация морфологи-
ческих форм трипаносоматид (рис. 137).
Долгое время большинством исследователей при-
знавалось, что отдельные морфологические формы
трипаносоматид характерны для определенных родов
этих жгутиконосцев, что служило основанием для их
обозначения. Выделяли 4 такие формы: лептомонад-
ную, критидиальную, трипаносомную и лейшмани-
альную. Их отождествляли со стадиями развития жгу-
тиконосцев, добавляя в ряде случаев к их перечню
стадию цисты. Однако в большинстве ранних работ
«цисты» гомоксенных трипаносоматид именуются
лейшманиальными формами. По мере накопления
знаний о строении клеток трипаносоматид, структуре
их жизненных циклов и совершенствования системы
семейства недостатки этой классификации станови-
лись все более очевидными. Так, в частности, оказа-
лось, что клетки трипаносоматид, имеющие кинето-
пласт в задней позиции, представлены 2 различными
морфологическими формами (описто- и трипомасти-
готы), «лептомонадные» и «лейшманиальные» стадии
были описаны у представителей большинства родов
трипаносоматид, а использование термина «критиди-
альные формы» потеряло всякий смысл после описа-
ния рода Blastocrithidia, к которому были отнесены
гомоксенные жгутиконосцы с «критидиальной» орга-
низацией (Laird, 1959; Wallace, 1966).
В 1966 г. Гоаром и Уоллесом была предложена новая
схема классификации морфологических форм трипано-
соматид. Почти за три десятилетия своего существования
она практически не претерпела изменений.
Ниже мы приводим названия морфологических
форм трипаносоматид по Гоару и Уоллесу (Ноаге, Wal-
lace, 1966), сопровождая их краткой характеристикой.
Промастигота (promastigote) — «лептомонад-
ная форма»; обладает удлиненной формой тела, ки-
нетопласт лежит перед ядром, жгутиковый карман
открывается терминально (рис. 137, А).
Эпимастигота (epimastigote) — «критидиаль-
ная форма»; форма тела обычно удлиненная, кине-
топласт располагается вблизи ядра, но не заходит за
его задний край, жгутиковый карман открывается на
латеральной поверхности тела, жгутик подходит к
переднему концу клетки, прилегая к ее поверхности
и формируя ундулирующую мембрану (рис. 137, Б).
Опистомастигота (opisthomastigote) — «три-
паносомная, или трипаноморфная, форма»; встреча-
ется только у представителей рода Herpetomonas.
Клетки характеризуются удлиненной формой тела,
кинетопластом, располагающимся за ядром, и соот-
ветственно длинным жгутиковым карманом, открыва-
ющимся терминально (рис. 137, В).
Трипомастигота (trypomastigote) — «трипано-
сомная форма»; форма тела обычно удлиненная, кине-
топласт располагается за ядром, жгутиковый карман
короткий, открывается на латеральной поверхности
тела. Жгутик прилегает к поверхности клетки, форми-
руя. ундулирующую мембрану (рис. 137, Г).
Хоаномастигота (choanomastigote) — клетки
имеют форму кувшинчиков, жгутиковый карман ко-
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
215
роткий и широкий, открывается терминально, ядро и
кинетопласт часто сближены (рис. 137, Д).
Сферомастигота (spheromastigote) — клетки
округлой формы, с небольшим свободным жгутиком.
Переходная форма между амастиготами и мастигота-
ми (рис. 137, Е),
Амастигота (amastigote) — «лейшманиальная
форма»; мелкие клетки трипаносоматид округлой
или эллипсоидальной формы, лишенные свободного
жгутика (рис. 137, Ж, 3).
Следует заметить, однако, что и этот вариант клас-
сификации морфологических форм трипаносоматид
оказался не лишен противоречий. Так, в группу амас-
тигот (рис. 137, Ж, 3), известную с начала века под на-
званием «лейшманиальные формы», объединяются все
формы жгутиконосцев, которые под световым микрос-
копом сходны с «амастиготами» лейшманий. Амасти-
готами называют внутриклеточные и (или) внутритка-
невые формы Leishmaniaи Trypanosoma cruzi и цисто-
подобные формы Leptomonas и Blastocrithidia, а также
любые мелкие округлые или эллипсоидальные формы
других жгутиконосцев, лишенные как минимум сво-
бодной части жгутика. Однако электронно-микроско-
пические исследования последних десятилетий показа-
ли, что на самом деле амастиготы представляют собой
гетероморфную группу. Оказалось, что «амастиготы»
трипаносом и лейшманий имеют жгутик, целиком поме-
щающийся в жгутиковом кармане. Сходные формы най-
дены у Herpetomonas и Wallaceina — паразитов насеко-
мых (рис. 137, Ж). В то же время «цисты» Leptomonas и
Blastocrithidia полностью лишены жгутиков и даже ки-
нетосом (рис. 137,3). Это противоречие было устранено
(Фролов, 1994) после введения в исходную схему еще
одной формы — эндомастиготы и уточнения диагноза
собственно амастигот: амастиготы (amastigote) —
клетки трипаносоматид, лишенные жгутика (рис. 137,
3); эндомастиготы (endomastigote) — клетки трипа-
носоматид, обладающие жгутиком, который не выходит
за пределы жгутикового кармана (рис. 137, Ж).
Электронно-микроскопическое
описание морфологии
Несмотря на видимые различия в ультратонкой
организации клеток бодонид, криптобий и трипано-
соматид (рис. 138), именно этот метод позволил убе-
дительно продемонстрировать наличие у них единого
плана строения и объединить столь разных на пер-
вый взгляд жгутиконосцев в один таксон.
Покровы. Покровы кинетопластид, как правило,
представлены тубулеммой (Карпов, 1990), т. е. вклю-
чают плазмалемму и подстилающие ее продольно
ориентированные субмембранные микротрубочки
(рис. 159, вкл.). Кроме того, на внешней поверхности
плазмалеммы жгутиконосцев часто выявляется гли-
копротеиновый надмембранный комплекс (Vicker-
man, Preston, 1976).
Традиционно наличие развитого слоя гликокалик-
са, покрывающего тело жгутиконосцев, связывается с
Рис. 138. Схема ультратонкой организации клеток кинетопластид.
А — Rhynchomonas nasuta (Bodonidae), Б — Cryptobia vaginalis (Cryptobiidae), В — Trypanosoma vivax (Trypanosomatidae). аг — аппарат Гольджи,
жк — жгутиковый карман, кп — кинетопласт, ми — митохондрион, мн — мастигонемы, пв— пищеварительная вакуоль, пж — передний жгутик,
рж — рекуррентный жгутик, св — сократительная вакуоль, ум — ундулирующая мембрана, х,--^ хоботок, цф — цитофаринкс, шэр — шерохова-
тый эндоплазматический ретикулум, я — ядро. (4 — по: Swale, 1973, Б — по: Vickerman, 1977, В — по: Vickerman, 1966; все с изменениями).
Fig. 138. Schematic drawings of the ultrastructure organization of kinetoplastids.
A — Rhynchomonas nasuta (Bodonidae), Б — Cryptobia vaginalis (Cryptobiidae), В — Trypanosoma vivax (Trypanosomatidae). аг — Golgi apparatus, жк —
flagellar pocket, кп — kinetoplast, mu — mitochondrion, мн — mastigOnemes, ne — food vacuole, пж anterior flagellum, рж — posterior flagellum, ce —
contractile vacuole, ум — undulating membrane, x — proboscis, цф — cytopharinx, шэр— rough endoplasmic reticulum, л — nucleus.
216
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 139. Схема организации покровов кинетопластид.
А — неармированная плазмалемма бодонид, Б—Г — различные формы сег-
ментированной тубулеммы, Д — целостная тубулемма трипаносоматид.
вмт — вентральные субмембранные микротрубочки, дмт — дорсальные суб-
мембранные микротрубочки, им — плазмалемма, фас — j-образная лента
микротрубочек, ассоциированных с цитостомом.
Fig. 139. Scheme of the organizaton of kinetoplastid coverings.
A — simple plasmalemma of the some bodonid flagellates, Б-Г — various exam-
ples of the segmented tubulemma, Д — complete tubulemma of trypanosomatids.
вмт — ventrale microtubules, дмт — dorsal microtubules, им — plasmalemma,
фас — ^-reminding band of cytostome associated microtubules.
представителями кинетопластид, которые ведут пара-
зитический образ жизни (Vickerman, Preston, 1976).
Более того, до недавнего времени эту особенность
организации покровов кинетопластид рассматривали
исключительно с точки зрения адаптации их предста-
вителей к кровепаразитизму. Однако недавно глико-
каликс был описан Бружеролем (Brugerolle, 1985) и
Мыльниковым (1986) у бодонид из рода Rhynchobo-
do. У этих жгутиконосцев мощный слой гликокалик-
са, достигающий 30 нм толщины, покрывает всю по-
верхность их тела, выстилает жгутиковый карман и
переходит на плазмалемму свободной части жгутиков.
В другой группе двухжгутиковых кинетопластид —
у криптобий — в различной степени развитый глико-
каликс оказался обычным атрибутом наружной по-
верхности плазмалеммы. Причем отмечен он и у кро-
вепаразитов, и у жгутиконосцев, которые паразитиру-
ют в других органах хозяев. У ряда видов, например
у Cryptobia sabnositica, надмембранный комплекс до-
стигает толщины 30 нм и покрывает всю поверхность
клетки, жгутиковый карман и жгутики. У эндопарази-
тов, подобных С. vaginalis, напротив, гликокаликс
располагается на поверхности клеток жгутиконосцев
нерегулярно и не достигает высокой степени развития.
У трипаносоматид наличие выраженного слоя по-
верхностных гликопротеинов характерно в первую
очередь для кровяных стадий трипаносом. Особенно
мощного развития достигает гликокаликс, формиру-
ющийся на поверхности клеток кровяных стадий аф-
риканских трипаносом млекопитающих. Эти трипа-
носомы имеют мощный надмембранный комплекс,
достигающий толщины 15 нм и более. Однако при-
менение методов электронно-микроскопической ци-
тохимии позволило продемонстрировать наличие
гликопротеиновой «шубы» как у большинства других
трипаносом, так и у лейшманий и даже у гомоксен-
ных трипаносоматид. Известно, что, покрывая поверх-
ность клеток трипаносоматид, паразитирующих в
крови позвоночных животных, гликокаликс обеспе-
чивает защиту транспортных систем плазмалеммы и
поверхностных рецепторов от атак антител хозяина.
В этой связи интересны факты обнаружения развито-
го гликокаликса на поверхности клеток так называе-
мых низших трипаносоматид — паразитов кишечно-
го тракта насекомых. Так, у эндомастигот жгутико-
носцев рода Wallaceina плазмалемму покрывает мощный
слой структурированного гликокаликса, более напо-
минающий надмембранные комплексы ряда ризопод.
Жгутиконосцы Phytomonas serpens обладают разви-
тым гликокаликсом и при этом, подобно кровяным
трипаносомам, при чередовании стадий по ходу жиз-
ненного цикла сначала полностью освобождаются от
гликопротеиновой шубы, а затем вновь ее приобретают.
У свободноживущих бодонид (Bodo, Rhynchobodo,
Rhynehomonas) покровы представлены чередующими-
ся участками плазмалеммы и тубулеммы (рис. 139, А—
В). Замкнутое кольцо субмембранных микротрубочек
отмечено в отдельных участках тела только у Rhyncho-
bodo. Обычно под плазмалеммой у бодонид выявляют-
ся 2—3 группы микротрубочек. Ленты дорсальных и
вентральных микротрубочек закладываются у кинето-
сом жгутиков в виде одноименных кинетосомальных
корешков, третья — s-образная группа микротрубочек
(ФАС — рис. 139; 140) в своем происхождении связа-
на с элементами цитостом-цитофарингеального комп-
лекса. Наиболее развитые участки тубулеммы обнару-
живаются на переднем конце клеток жгутиконосцев.
У большинства видов свободноживущих бодонид
здесь представлены микротрубочки всех трех групп
(рис. 139, Б; 140). Наиболее многочисленны микро-
трубочки, армирующие дорсальную поверхность клет-
ки, — это микротрубочки дорсальной группы и ФАС.
Количество вентральных субмембранных микротру-
бочек обычно не превышает 4—5. Микротрубочки ци-
тостомальной группы ФАС подстилают плазмалемму
на незначительном протяжении и в посторальной об-
ласти тела погружаются в глубь цитоплазмы. У ряда
видов бодонид основные ленты субмембранных мик-
ротрубочек — дорсальная и вентральная могут быть
очень короткими, и тогда покровы жгутиконосцев на
большем своем протяжении формируются за счет про-
стой, неармированной плазмалеммы (рис. 139, А).
Покровы представителей другой группы двухжгутико-
вых кинетопластид — криптобий (рис. 139, Б—Г), с
одной стороны, очень близки к покровам свободножи-
вущих бодонид. Так, у Cryptobia intestinalis в форми-
ровании тубулеммы принимают участие все 3 перечис-
ленные выше группы микротрубочек. С другой сторо-
ны, большинство криптобий, например С. branchialis,
С. helicis, C.jubilans, С. vaginalis, и все кровепаразиты
утрачивают s-образную группу микротрубочек, при
этом у них резко возрастает число вентральных суб-
мембранных микротрубочек. Тубулемма этих жгути-
коносцев формируется за счет двух лент микротрубо-
чек — дорсальной и вентральной, разделенных участ-
ками неармированной плазмалеммы (рис. 139, В). У С.
borreli, С. helicis, С. sabnositica число дорсальных суб-
мембранных микротрубочек прогрессивно возрастает
по направлению к заднему концу клетки. При этом в
посторальной части тела этих жгутиконосцев обе лен-
ты смыкаются, формируя тубулемму с единственной
брешью, где в субмембранной зоне часто располагаются
гребни гладкого ЭПР (рис. 139, Г).
Ни на одной из стадий жизненного цикла покро-
вы трипаносоматид не бывают представлены одной
лишь плазмалеммой (рис. 139, Г—ДБ Для многих
видов трипаносоматид (все «низшие трипаносомати-
ды» и Leishmania) характерен цельный, замкнутый
ТИП EUGLENOZOA. КЛАСС KIN ETOP LASTIDEA
217
ряд субмембранных микротрубочек (рис. 139, Д).
Только у трипаносом в этом ряду имеется брешь
(рис. 139, Г). Подобно криптобиям, у трипаносом в
эту брешь заходят гребни ЭПР. Субмембранные мик-
ротрубочки трипаносоматид закладываются как еди-
ная группа в электронно-плотном материале, подсти-
лающем плазмалемму апикальной части жгутикового
кармана.
Все разнообразие в организации покровов кине-
топластид может быть представлено в виде эволюци-
онного ряда, отражающего степень развития тубулем-
мы (рис. 139). Полярные позиции в этом ряду занима-
ют покровы некоторых свободноживущих бодонид,
представленные на значительном своем протяжении
неармированной плазмалеммой, и целостная тубулем-
ма большинства трипаносоматид. Промежуточное
положение в этом ряду занимают покровы крипто-
бий. Для большинства из них характерно наличие
сегментированной тубулеммы, которая формируется
за счет 2 (иногда 3) групп субмембранных микротру-
бочек: дорсальной и вентральной. Вместе с тем неко-
торые криптобии демонстрируют тенденцию к фор-
мированию хотя и неполного, но единого ряда суб-
мембранных микротрубочек. Подобная тубулемма,
характеризующаяся наличием единственной бреши в
ряду субмембранных микротрубочек, встречается
только у трипаносом, которые по этому признаку за-
нимают промежуточное положение между криптоби-
ями и остальными трипаносоматидами.
Особо следует остановиться на покровах рассели-
тельных стадий кинетопластид, приспособленных для
переживания неблагоприятных условий окружающей
среды. Такие стадии описаны у свободноживущих бо-
донид и у паразитических гомоксенных трипаносома-
тид из насекомых. Известно, что некоторые виды сво-
бодноживущих бодонид могут образовывать настоящие
цисты. У свободноживущих гетеротрофных бодонид про-
цесс инцистирования связан с формированием внекле-
точной оболочки — стенки цисты (рис. 152, вкл.). За-
щитная оболочка формируется над плазматической
мембраной. Жгутиконосцы втягивают жгутики внутрь
клетки и округляются. Никаких существенных изме-
нений в морфологии субмембранной зоны у инцисти-
рованных особей жгутиконосцев выявить не удается.
Основным элементом поверхностных структур
цистоподобных расселительных стадий низших три-
паносоматид, который непосредственно граничит с
окружающей средой, является плазмалемма (рис. 160,
вкл.). У некоторых видов трипаносоматид на поверх-
ности плазмалеммы цистоподобных стадий может
присутствовать в той или иной степени выраженный
слой гликокаликса. Специфической чертой организации
этих стадий, кроме того, является наличие слоя специали-
зированной субмембранной цитоплазмы (рис. 160, вкл.),
в которую погружаются субмембранные микротрубочки.
Жгутики. Жгутики кинетопластид представляют
собой покрытые плазмалеммой цитоплазматические
выросты (рис. 140), отходящие от дна перманентной
инвагинации, формирующейся на теле жгутиконос-
цев, которая получила название жгутикового карма-
на. Все свободноживущие бодониды и все криптобии
имеют по 2 гетеродинамных жгутика, а все трипано-
соматиды — 1 направленный вперед двигательный
Рис. 140. Организация цитоскелета бодонид (схема).
вмт — вентральные субмембранные микротрубочки; вк — вентральный
кинетосомальный корешок; дмт — дорсальные субмембранные микротру-
бочки; дк — дорсальный кинетосомальный корешок; ж — жгутики; жк - жгу-
тиковый карман; мтр — микротрубочки, подстилающие стенку жгутикового
кармана и цитостом-цитофарингеального комплекса; фас — 5-образная лента
микротрубочек, ассоциированных с цитостомом; ц — цитостом; цмт — груп-
па цитостомальных микротрубочек; цф - цитофаринкс.
Fig. 140. Scheme of the organizaton of kinetoplastid cytoskeleton.
вмт — ventrale microtubules; вк — ventrale kinetosomale rootlet; дмт — dorsal
microtubules; дк — dorsal kinetosomale rootlet; ж — flagellum; жк — flagellar
pocket; мтр — reinforced cytopharyngeal microtubules; фас — 5-reminding
band of cytostome associated microtubules; ц — cytostome; цмт — group of cy-
tostomal microtubules; цф — cytopharinx.
жгутик (рис. 138). Передние жгутики бодонид и крип-
тобий свободные. Направленные назад рекуррентные
жгутики бодонид свободные, в то время как у крип-
тобий рекуррентные жгутики плотно прилегают к по-
верхности тела жгутиконосцев. В процессе движения
жгутиконосцев участок тела, прилегающий к рекур-
рентному жгутику, в ответ на его изгибания перио-
дически вытягивается в уплощенную складку — ун-
дулирующую мембрану, которая хорошо видна даже
под световым микроскопом. Единственный жгутик
трипаносоматид может либо быть свободным (это ха-
рактерно для видов, жгутиковый карман которых от-
крывается на апикальном конце клетки), либо, подобно
рекуррентному жгутику криптобий, прилегать к телу
клетки (если жгутиковый карман открывается на ла-
теральной ее поверхности). В последнем случае
также формируется ундулирующая мембрана, причем
в зоне контакта плазмалеммы жгутика и тела про-
стейшего у трипаносоматид обнаруживаются множес-
твенные точечные десмосомы.
Тонкое строение жгутика кинетопластид в целом
типично. В нем можно выделить 3 отдела: кинетосо-
му, переходную зону и собственно свободную часть
жгутика.
В неделящихся клетках кинетопластид обнаружи-
ваются 2 кинетосомы. Обычно они располагаются под
произвольным углом друг к другу, но чаще их про-
дольные оси параллельны друг другу. Обе кинетосо-
мы представляют собой цилиндры, стенки которых
сформированы за счет 9 триплетов микротрубочек.
У свободноживущих бодонид и у криптобий обе
кинетосомы являются «рабочими», т. е. связаны с
функционирующими жгутиками: одна — с направ-
ленным вперед, двигательным, другая — с рекуррент-
ным. У трипаносоматид одна из кинетосом является
«рабочей» — она формирует единственный, направ-
ленный вперед двигательный жгутик. Другая в неде-
лящихся клетках не связана со жгутиком. Эту кине-
тосому называют покоящейся, или сателлитной. Суб-
218
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 141. Схема организации переходных зон
в жгутиках кинетопластид.:
А, Г - Bodonidae; Б, Д - Cryptobiidae; В,Е- Тгура-
nosomatidae. А-В - организация переходных зон на
продольных срезах жгутиков, Г-Е - поперечные сре-
зы через терминальные структуры переходных зон.
ак - аксосома; бп - базальная пластинка, лф - луче-
видные филаменты, пм - плазмалемма, пп - парак-
сиальная пластинка, пс - периферическая спираль,
пт - параксиальный тяж, пц - периферический ци-
линдр, тп - терминальная пластинка.
Fig. 141. Scheme of the organizaton of flagel-
lar tranzition zone in kinetoplastid flagellates.
ак - axosome, 6n - basal plate, Лф - beam-like radial
filaments, пм plasmalemma, nn - paraxial plate, nc -
periferal spiral, пф - paraxial rod, пц - periferal cylin-
der, mn - terminal plate.
структура рабочей и сателлитной кинетосом одно-
типна. Известно, что при бинарном делении трипано-
соматид новый жгутик дочерней особи «отрастает»
от сателлитной кинетосомы (Sherwin, Gull, 1989),
после чего около рабочих кинетосом закладываются
новые сателлиты. У двухжгутиковых кинетопластид в
процессе деления обе кинетосомы удваиваются, фор-
мируя новые жгутики дочерних особей. Таким обра-
зом, в делящихся клетках кинетопластид могут выяв-
ляться одновременно 4 кинетосомы.
У свободноживущих бодонид и у криптобий
дифференцируются 2 микротрубрчковых корешка:
один связан с кинетосомой двигательного жгутика,
другой — с кинетосомой рекуррентного жгутика
(рис. 140). Первый, состоящий из 3—4 микротрубо-
чек, часто связанный в основании с плотной фиб-
риллярной пластинкой, проходит под дорсальной
стенкой жгутикового кармана и принимает участие в
формировании группы дорсальных субмембранных
микротрубочек. Около кинетосомы рекуррентного
жгутика закладывается микротрубочковый корешок,
связанный с вентральными субмембранными микро-
трубочками. Обычно он насчитывает всего 2 мик-
ротрубочки. Кроме основных микротрубочковых
корешков обе кинетосомы обычно формируют ко-
роткие фибриллярные шпоры. В районе локализа-
ции кинетосом также могут присутствовать отдель-
ные микротрубочки или их небольшие ассоциаты,
но их связь с кинетосомами не прослеживается.
Единственный кинетосомальный корешок, состо-
ящий из 3—4 сближенных микротрубочек, найден у
всех исследованных видов трипаносоматид. Отходя
от основания кинетосомы, он спирально огибает жгу-
тиковый карман, и его микротрубочки проникают в
муфту плотного, войлокоподобного материала, окру-
жающего апикальную часть жгутикового кармана. На
начальных этапах деления трипаносоматид, еще до
закладки новых сателлитных кинетосом, кинетосрма
отрастающего жгутика (бывшая сателлитная) форми-
рует собственный корешок.
Жгутики кинетопластид характеризуются наличи-
ем длинной переходной зоны, существенно выступа-
ющей за уровень дна жгутикового кармана. Эта часть
жгутикового аппарата имеет типичное для протистов
строение и представляет собой цилиндр, стенки ко-
торого построены из 9 периферических дублетов
микротрубочек (рис. 141). В переходной зоне жгути-
ков кинетопластид выявляются 2 поперечные струк-
туры, которые обычно называют базальной и терми-
нальной пластинками.
Базальная пластинка чаще всего имеет вид фиб-
риллярной шайбы, наружный диаметр которой со-
ответствует внутреннему диаметру кольца перифе-
рических дублетов, а диаметр ее центрального от-
верстия обычно составляет не менее 50—60 нм
(рис. 141, А—В).
Терминальная поперечная структура переход-
ной зоны жгутиков свободноживущих бодонид бо-
лее всего соответствует «поперечной пластинке»
(рис. 141, А, Г). Как правило, это диск, диаметр
которого близок к внутреннему диаметру кольца пе-
риферических дублетов переходной зоны (рис. 141,
Г). На верхней поверхности этого диска расположен
еще один диск (или валик) меньшего диаметра, ко-
торый можно рассматривать как аксосому (рис. 141,
А). Иногда такой валик располагается под терми-
нальной пластинкой. В обоих случаях с ним связаны
основания одной или обеих микротрубочек цент-
ральной пары аксонемы. Однако у некоторых бодо-
нид дополнительные уплотнения на поверхности
терминальной пластинки могут отсутствовать. На
уровне терминальной пластинки, между перифери-
ческими дублетами и плазмалеммой жгутика, у бодо-
нид находится так называемая параксиальная плас-
тинка, от которой берет начало параксиальный тяж.
По мере приближения к плазмалемме параксиальная
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
219
пластинка переходит в массивную фибриллярную
структуру. Оба жгутика обращены друг к другу па-
раксиальными пластинками, и их плазмалеммы в
этом месте часто плотно прилегают друг к другу.
Терминальная структура переходной зоны жгути-
ков многих видов криптобий, таких как С. intestina-
lis, С. jubilans, С. vaginalis, сходна с аналогичной
структурой свободноживущих бодонид. Здесь также
имеется параксиальная пластинка, и жгутики взаи-
модействуют друг с другом аналогичным образом.
Однако у целого ряда представителей криптобиид
терминальная пластинка в переходной зоне жгу-
тиков отсутствует (рис. 141, Б, Д). Терминальная
структура переходной зоны С. helicis, С. borreli, С.
salmositica представлена толстостенным фибрилляр-
ным «стаканом», дно которого обращено к устью
жгутикового кармана. От стенок этого «стакана» к
динеиновым ручкам периферических дублетов пе-
реходной зоны отходят лучевидные филаменты
(рис. 141, Д). От дна «стакана» берут начало цент-
ральные микротрубочки аксонемы, следовательно,
есть все основания рассматривать этот элемент как
аксосому.
Терминальная структура переходной зоны жгути-
ка трипаносоматид состоит из 2 элементов (рис. 141,
В, Е). Как правило, это хорошо оформленная аксо-
сома глобулярного типа и периферический цилиндр
или спираль, связанные с А-трубочкамиаксонемы.
Параксиальная пластинка у трипаносоматид отсутст-
вует, а параксиальный тяж начинается выше уровня
терминальной структуры переходной зоны.
Ундулиподия. На уровне терминальной структуры
переходной зоны в жгутиках кинетопластид заклады-
вается центральная пара микротрубочек аксонемы.
Аксонема на всем своем протяжении имеет класси-
ческое строение 9 + 2. Дистальная часть аксонемы
формируется при последовательной утрате централь-
ной пары микротрубочек и замене периферических >
дублетов на синглеты.
Среди распространенных аберраций аксонемаль-
ного комплекса жгутиков кинетопластид можно от-
метить обнаружение «голой» аксонемы в цитоплазме
клеток кинетопластид. Часто удается наблюдать раз-
мещение таких «внутриклеточных» аксонем вблизи
субмембранных микротрубочек и даже их встраива-
ние в тубулемму. Это явление, вероятно, отчасти объ-
ясняется тем, что микротрубочки аксонемы и суб-
мембранные микротрубочки имеют идентичную изо-
форму а-тубулина, что было показано на примере
Trypanosoma brucei (Schneider et al., 1987).
Характерной чертой в организации жгутиков ки-
нетопластид является наличие в них параксиального
тяжа — микрофиламентозной структуры со сложной
пространственной организацией, ассоциированной с
аксонемой. Среди протистов сходные структуры най-
дены в жгутиках эвгленовых жгутиконосцев, дино-
флагеллят и у некоторых других протистов (Cachon
et al., 1988; Карпов, 1990). Функция параксиальных
структур пока неясна. У двухжгутиковых кинетоплас-
тид параксиальный тяж является обязательным атри-
бутом обоих жгутиков. У трипаносоматид паракси-
альный тяж формируется в жгутиках, выполняющих
двигательную функцию, и отсутствует лишь в двига-
тельных жгутиках ряда симбионтсодержащих видов
(Freymuller, Camargo, 1981). В жгутиках, выполняю-
щих прикрепительную функцию, эта структура обыч-
но подвергается разборке.
Параксиальный тяж сопровождает аксонему на
большем ее протяжении, заканчиваясь несколько
ниже того уровня, с которого начинается реорганиза-
ция ее дистальной части. Таким образом, кончик
жгутика, часто слегка вздутый^ лишен основных
структурных элементов и, вероятно, представляет
собой наиболее пластичную его часть. Основными
элементами параксиального тяжа трипаносоматид яв-
ляются микрофиламенты диаметром 7 и 25 нм.
Параксиальный тяж в жгутиках кинетопластид за-
нимает константное положение по отношению к
плоскости, проходящей через пару центральных мик-
ротрубочек аксонемы, нумерация дублетов которой
подчиняется правилу Афцелиуса. В двигательных
(передних) жгутиках свободноживущих бодонид и
криптобий параксиальный тяж ассоциирован с 4-м и
7-м дублетами аксонемы, а в их рекуррентных жгути-
ках он морфологически связан либо с 3-м и 6-м, либо
с 5-м и 8-м дублетами. В единственном жгутике три-
паносоматид параксиальный тяж морфологически
связан с 4-м и 7-м дублетами.
Жгутики кинетопластид являются полифункцио-
нальными органеллами. Наиболее известным приме-
ром этого служит способность паразитических кине-
топластид преобразовывать свой жгутик из двигатель-
ной органеллы в специализированную органеллу
прикрепления, В зависимости от типа субстрата жгу-
тиконосцы используют либо принцип заякрривания
(это характерно для жгутиконосцев, которые закреп-
ляются на эпителиях, формирующих микроворсин-
ки), либо принцип присоски — при закреплении на
кутикулярных выстилках эпителия. Оба способа при-
крепления широко распространены у трипаносома-
тид, причем у разных стадий одного и того же вида
могут использоваться оба способа. У криптобий по
типу заякоривания происходит закрепление видоиз-
мененного жгутика Cryptobia sp. на эпителии сперма-
теки улитки Triadopsis multilineata, a Cryptobia bran-
chialis использует принцип присоски, закрепляясь на
эпителии жабр рыб.
Завершая рассмотрение организации двигательного
аппарата кинетопластид, можно сделать следующие
выводы. По своему строению жгутики кинетоплас-
тид образуют очевидный эволюционный морфологи-
ческий ряд, в котором крайние позиции занимают
свободноживущие Bodonidae и Trypanosomatidae, а
промежуточную — Cryptobiidae. Со свободноживу-
щими бодонидами криптобий сближает наличие 2
гетеродинамных жгутиков, однотипная организация
кинетосомальных корешков, наличие параксиальной
пластинки, расположение параксиальных тяжей в
обоих жгутиках и формирование межжгутиковых
контактов на уровне дистальной части переходных
зон. Криптобий и трипаносоматид сближает отсутст-
вие в переходных зонах их жгутиков терминальной
пластинки, наличие обособленной аксосомы, спо-
собность к трансформации двигательных жгутиков в
органы прикрепления. Наличие единственного 3 или
4 микротрубочкового кинетосомального корешка,
220
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ассоциированного с субмембранными микротрубоч-
ками, и жесткая связь параксиального тяжа с 4-м и
7-м дублетами аксонемы позволяют рассматривать
единственный жгутик трипаносоматид и двигатель-
ные (передние) жгутики бодонид и криптобий как
гомологичные структуры.
Ротовой аппарат. Потребление питательных ве-
ществ клетками кинетопластид может происходить
различными способами (Vickerman, Preston, 1976). У
многих кинетопластид этим целям служат, в част-
ности, специализированные клеточные структуры,
формирующие сложные цитостом-цитофарингеаль-
ные комплексы (рис. 151, вкл.; 154, вкл.; 155, вкл.).
В настоящее время такие комплексы описаны у всех
изученных представителей свободноживущих бодо-
нид, криптобий и у многих трипаносоматид. Только
среди трипаносоматид имеются жгутиконосцы, у ко-
торых ротовой аппарат полностью редуцирован.
Среди них — все представители африканских три-
паносом группы Salivaria, Leishmania и Endotrypanum,
Phytomonas, гомоксенные Blastocrithidia и часть Lep-
tomonas.
Среди всего многообразия цитостом-цитофарин-
геальных комплексов кинетопластид мы выделяем
2 основные их формы: полные (рис. 142, А—Е) и ре-
дуцированные (рис. 142, Ж, 3) комплексы. Последние
отличаются частичной утратой ряда элементов комп-
лекса, и в первую очередь отсутствием выраженной
клеточной глотки.
Наиболее развиты и сложно организованы пол-
ные цитостом-цитофарингеальные комплексы сво-
бодноживущих бодонид (рис. 151, вкл.). У этих жгу-
тиконосцев они приспособлены для заглатывания
крупных пищевых частиц, например бактерий. На-
ружный диаметр цитостома бодонид часто равен ди-
аметру наружного отверстия жгутикового кармана
или превышает его. Плазмалемма цитостома подсти-
лается плотным войлокоподобным материалом, ко-
торый обычно имеет сложную пространственную ор-
ганизацию и с которым связаны основания 4—5
групп микротрубочек (рис. 142, А, а). Общими для
всех изученных видов бодонид являются 2 основные
ленты микротрубочек, одна из которых (мтр) закла-
дывается в плотном материале у стенки жгутикового
кармана, другая (фас) — в аналогичном материале у
стенки цитостомальной воронки. 5—6 микротрубо-
чек группы мтр по восходящей спирали армируют
стенку жгутикового кармана, проникают в цитоплаз-
му переднего конца клетки, где делают поворот на
180° и затем армируют последовательно стенку ци-
тостомальной воронки и цитофаринкс. Отличительной
чертой этой группы является связь ее микротрубочек
с плазмалеммой жгутикового кармана, цитостома и
частично цитофаринкса посредством электронно-
плотных «бляшек». Группа микротрубочек фас за-
кладывается в плотном материале под плазмалеммой
цитостома. Эта лента по широкой спирали окружает
воронку цитостома и проксимальную часть цитофа-
ринкса и уходит в глубь клетки. Часто, на значительном
протяжении эта лента микротрубочек располагается
в субмембранной зоне жгутиконосцев. В массивах
плотного материала, подстилающего плазмалемму
цитостома, берут начало 4—5 цмт, которые закла-
дываются по отдельности и не формируют компакт-
ную ленту, цмт армируют стенки цитостома и цито-
фаринкса вместе с микротрубочками группы мтр
(рис. 142, а). Кроме перечисленных элементов в фор-
мировании цитоскелета цитостомального комплекса
ряда свободноживущих бодонид принимают участие
так называемые микротрубочковые призмы. Это
компактные группы, насчитывающие по 10—20 и
более микротрубочек, которые берут начало в плот-
ном материале у дна цитостома. Взаимное располо-
жение микротрубочек таково, что на поперечном
срезе призмы их профили образуют правильный тре-
угольник или трапецию (рис. 142, а). Микротрубоч-
ки в призме связаны друг с другом мостиками. Мик-
ротрубочковая призма сопровождает цитофаринкс
на всем его протяжении. Однако непосредственной
связи между этими структурами нет, и функции
таких призм пока неясны.
У криптобий цитостом-цитофарингеальные комп-
лексы организованы так же, как у свободноживущих
бодонид. Наиболее близка к бодонидному типу орга-
низация цитостом-цитофарингеального комплекса
Cryptobia intestinalis. Цитоскелет хорошо развитого
орального аппарата этого жгутиконосца включает все
3 основные группы микротрубочек: мтр, фас и цмт
(рис. 142, Б, б). Кроме того, цитостом и цитофаринкс
С. intestinalis окружены мощной фибриллярной муф-
той, имеющей периодическую исчерченность. У
большинства других криптобий цитостом-цитофа-
рингеальные комплексы по сравнению с бодонида-
ми как бы миниатюризованы. Цитостом открывается
на вентральной поверхности тела жгутиконосцев
(рис. 142, В, в). Его наружный диаметр несколько
меньше диаметра жгутикового кармана. От дна ци-
тостомальной воронки отходит трубчатый цитофа-
ринкс. Его диаметр существенно меньше, чем у бодо-
нид. Армирующие клеточную глотку микротрубочки
имеют двоякое происхождение. 5—6 микротрубочек
группы мтр прослеживаются от стенки жгутикового
кармана, с которой каждая из них связана бляшками
плотного войлокоподобного материала. Достигая
вентральной поверхности тела жгутиконосца, они по-
ворачивают на 180°, локализуясь в гребневидном вы-
пячивании тела жгутиконосца (рис. 154, вкл.). Так
же, как и у стенки жгутикового кармана, плотные
бляшки соединяют их здесь с плазмалеммой гребня.
Внутри гребня эти микротрубочки сопровождают 2
пластинки, расположенные по обе стороны от ленты.
Они состоят из плотного, войлокоподобного матери-
ала и ассоциированы с 2—3 дополнительными мик-
ротрубочками, представляющими группу с мт. Дости-
гая отверстия цитостома, оральный гребень встраива-
ется в его стенку. Теперь микротрубочки группы мтр
и цмт армируют цитостомальную воронку и далее —
канал цитофаринкса (рис. 155, вкл.). Ближе к центру
клетки часть этих микротрубочек исчезает, и здесь
цитофаринкс армируется всего 3—5 микротрубоч-
ками.
Среди трипаносоматид полный цитостом-цитофа-
рингеальный комплекс известен у трипаносом, пара-
зитирующих в крови рыб, у Shisotrypanum и одного
представителя «низших» гомоксенных трипаносома-
тид — Leptomonas nabicula (рис. 142, Г—Е).
ТИП EUGLENOZOA. КЛАСС KIN ETOPLASTIDEA
221
Хорошо развитый воронковидный цитостом Try-
panosoma danilewskyi открывается на поверхности
тела за пределами жгутикового кармана, вблизи места
контакта жгутика с телом клетки (рис. 142, Г). Ци-
тостом имеет наружный диаметр 150—200 нм. От
дна цитостома в глубь клетки отходит цитофаринкс.
Клеточная глотка Т. danilewskyi армируется 6—
7 микротрубочками (рис. 142, г). 4 из этих мик-
ротрубочек закладываются у стенки жгутикового
кармана и связаны с его плазмалеммой электронно-
плотными бляшками. Эти микротрубочки, несомнен-
но гомологичные микротрубочкам группы мтр двух-
жгутиковых кинетопластид, армируют стенку жгути-
кового кармана трипаносом, а достигнув поверхности
тела, поворачивают на 180° и подстилают плазма-
лемму цитостома и цитофаринкса. Еще 2—3 мик-
ротрубочки, соответствующие микротрубочкам цмт,
закладываются в слое электронно-плотного мате-
риала, подстилающего плазмалемму дна цито-
стома.
Полные цитостом-цитофарингеальные комплексы
других трипаносом имеют похожую организацию.
Различия касаются лишь особенностей локализации
цитостома и числа микротрубочек в глоточной груп-
пе (ср. рис. 142, Д, д и 142, Е, е).
Редуцированные цитостом-цитофарингеальные
комплексы обнаружены у трипаносом, паразитирую-
щих в крови амфибий, и у ряда трипаносом из мле-
копитающих, а также у многих «низших» трипаносо-
матид.
У Т. rotatorium отверстие цитостома располагает-
ся вблизи внешней границы жгутикового кармана и
либо смещено на наружную поверхность тела, либо
локализуется в стенке резервуара (рис. 142, Ж).
Такая нестабильность в локализации цитостома, ве-
роятно, может рассматриваться как иллюстрация
возможного пути возникновения в эволюции кинето-
пластид «скрытого» цитостома. Цитостом Т. rotato-
rium представляет собой .инвагинацию плазмалеммы,
наружный диаметр и глубина которой у разных осо-
бей могут варьировать от 100 до 200 нм. Плотный
войлокоподобный материал подстилает плазмалемму
апикальной части жгутикового кармана и цитостома.
С этим материалом связаны 4 микротрубочки, кото-
рые соответствуют микротрубочкам группы мтр
других кинетопластид. Они закладываются у стенки
жгутикового кармана и, достигнув цитостома, на-
правляются в глубь цитоплазмы параллельно его
центральной оси. Еще 1—2 микротрубочки группы
цмт закладываются в массиве плотного материала у
дна цитостома. В результате формируется подобие
воронки, горловина которой образована муфтой
плотного войлокоподобного материала, заходяще-
го за уровень дна цитостома, а ее расширенная
часть представлена конусом из 5—6 микротрубочек
(рис. 142, ж). Вдоль микротрубочек осуществляется
направленный транспорт пиноцитозных пузырьков в
глубь клетки. Такого типа редуцированные цитос-
том-цитофарингеальные комплексы достаточно ши-
роко распространены у трипаносом. Строение их
очень похоже, а различия носят частный характер,
касаясь главным образом числа микротрубочек в
глоточной группе.
Рис. 142. Схема организации цитостом-цитофарингеальных комп-
лексов кинетопластид.
А-З - взаимное расположение жгутикового кармана и цитостома у различ-
ных кинетопластид. а-з - поперечные срезы через цитофаринкс. жк - жгу-
тиковый карман, мп - микротрубочковая призма, мтр - микротрубочки,
подстилающие стенку жгутикового кармана и цитостом-цитофарингеально-
го комплекса, фас - j-образная лента микротрубочек, ассоциированных с
цитостомом, ц - цитостом, цмт - группа цитостомальных микротрубочек,
цф - цитофаринкс.
Fig. 142. Scheme of the organizaton of cytostome-cytopharyngeal
complex in kinetoplastids.
A-3 - arangement of the cytostomal opening and flagellar pocket in kinetoplas-
tid flagellates, а-з cross sections through cytopharynx, жк - flagellar pocket,
мп - microtubular prism, мтр - reinforced cytopharyngeal microtubules, фас -
5-reminding band of cytostome associated microtubules, ц - cytostome, цмт -
group of cytostomal microtubules, цф - cytopharinx.
Среди «низших» трипаносоматид редуцирован-
ные цитостом-цитофарингеальные комплексы об-
наружены у всех изученных представителей родов
Crithidia, Herpetomonas, Wallaceina, а также у некото-
рых Leptomonas. Наличие редуцированного цитосто-
ма в клетках трипаносоматид легко выявляется на
электронограммах по присутствию в районе стенки
жгутикового кармана дополнительной (помимо 4
микротрубочек дорсального корешка) группы мик-
ротрубочек. В отличие от трипаносом цитостом го-
моксенных трипаносоматид всегда локализуется в
стенке жгутикового кармана (рис. 142, 3). Плотный
войлокоподобный материал, окружающий цитостом
этих жгутиконосцев, формирует валик, по толщине
превышающий слой аналогичного материала, под-
стилающего стенку жгутикового кармана, производ-
ным которого он и является. Этот валик, полуколь-
цом охватывающий цитостомальную воронку, а точ-
нее, ямку, часто имеет форму запятой. У его
расширенной части в плотном материале, подстила-
ющем стенку жгутикового кармана, закладывается
5—8 микротрубочек, формирующих компактную
ленту. С противоположной стороны цитостома, не-
222
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
сколько ближе к заднему концу клетки, подобным
образом закладывается 1—2 микротрубочки. Объ-
единяясь, обе группы формируют «глоточный
желоб» (рис. 142, з), с которым связаны начальные
этапы транспортировки пиноцитозных пузырьков,
формирующихся у дна цитостомальной ямки. Мик-
ротрубочковый «желоб», вероятно, является функци-
ональным аналогом клеточной глотки, а точнее, ее
рудиментом, поскольку в его формировании прини-
мает участие часть элементов, которые имеются в
полном цитостом-цитофарингеальном комплексе.
Организация редуцированных цитостомальных комп-
лексов различается у отдельных представителей го-
моксенных трипаносоматид лишь по наборам мик-
ротрубочек в глоточной группе. Так, у Herpetomonas
muscarum количество микротрубочек в глоточной
группе достигает ГО, а у Wallaceina brevicula их 8.
Поскольку цитостом-цитофарингеальные комп-
лексы кинетопластид, как свободноживущих, так и
паразитических видов, являются гомологичными
структурами, все их многообразие может быть пред-
ставлено в виде эволюционного ряда (рис. 142). Оче-
видно, что первую позицию в этом ряду следует
отвести цитостом-цитофарингеальным комплексам
свободноживущих гетеротрофных бодонид, посколь-
ку организация трипаносоматид и криптобий так
или иначе связана с вторичной адаптацией их пред-
ставителей к паразитическому образу жизни. Следу-
ющую позицию в этом ряду занимают цитостомаль-
ные аппараты криптобий. У Cryptobia intestinalis со-
храняется типичный «бодонидный» тип строения
этого комплекса, однако большинство криптобий ут-
рачивают одну из основных групп микротрубочек,
армирующих цитофаринкс бодонид, — фас микро-
трубочки. Собственно, именно такой тип цитостом-
цитофарингеального комплекса формируется и у
трипаносом, паразитирующих в крови рыб. У крип-
тобий и трипаносом наблюдается пространственная
разобщенность цитостома и жгутикового кармана и
наличие только 2 групп микротрубочек, армирую-
щих стенки комплекса. В организации ротовых ап-
паратов остальных трипаносоматид ясно прослежи-
ваются 2 основные эволюционные тенденции: сме-
щение отверстия цитостома с наружной поверхности
тела на стенку жгутикового кармана и последова-
тельная редукция всего комплекса вплоть до полной
его утраты рядом трипаносом, лейшманий и гомок-
сенных «низших» трипаносоматид.
Кинетопласт — митохондрион. Уникальным при-
знаком в организации кинетопластид является нали-
чие в их клетке единственной гигантской митохондрии
со специализированным ДНК-содержащим участ-
ком — кинетопластом (рис. 161, вкл.). У некоторых
кинетопластид митохондрия может иметь простую
форму удлиненного эллипсоида, однако чаще мито-
хондрион представлен сетью кольцевых и продоль-
ных рукавов, которые могут ветвиться и анастомози-
ровать друг с другом. Митохондрйон кинетопластид
можно наблюдать в световом микроскопе, используя
методы специфического окрашивания митохонд-
риальных ферментов (например, тетранитроголубым
тетразолием) или применяя флюоресцентные краси-
тели типа родамина 123.
Как и у большинства других эукариот, внутренняя
мембрана митохондрий кинетопластид формирует
особые складки — кристы. Как правила, морфология
митохондриальных крист протистов высоко консер-
вативна и в ряде случаев характеризует рамки таксо-
нов высокого ранга (Taylor, 1978; Серавин, 1980;
Старобогатов, 1986; Карпов, 1990). Считается, что
для кинетопластид характерно наличие пластинча-
тых тарелковидных крист в митохондриях. Однако
некоторые виды кинетопластид демонстрируют уни-
кальные примеры диморфизма митохондриальных
крист. Так, наряду с типичными тарелковидными
кристами в митохондрии С. borreli часто обнаружи-
ваются ворсинчатые (длинные трубчатые) кристы
(рис. 156, вкл.). У С. jubilans в митохондрии обнару-
жены ампуловидные кристы, которые можно отнести
также к типу трубчатых крист. У трипаносоматиды
Crithidia fasciculata в митохондрии обнаружены
2 типа крист: тарелковидные и ворсинчатые; анало-
гичные кристы обнаружены и у Wallaceina brevicula.
У африканских трипаносом в ходе клеточной диффе-
ренцировки происходит регулярное чередование та-
релковидных и ворсинчатых крист, сопровождающе-
еся изменением типа метаболизма клеток. 2 типа
крист обнаруживаются в митохондриях Trypanosoma
danilewskyi. Изменение морфологии митохондриаль-
ных крист можно наблюдать при инцистировании не-
которых свободноживущих бодонид.
Трансформация митохондрий кинетопластид в
ходе цитокинеза изучена пока очень слабо. Нераз-
ветвленная; палочковидная митохондрия кровяных
стадий африканских трипаносом удваивается путем
простого продольного деления. Вероятно, подобным
образом происходит деление слабо ветвящихся ми-
тохондрий большинства двухжгутиковых кинето-
пластид. Что же касается ретикулярного митохонд-
риона трипаносоматид, то его преобразование в ходе
цитокинеза обычно связывают с появлением так на-
зываемых лабиринтовидных митохондрий. Их осо-
бенность заключается в том, что митохондриальный
матрикс на этой стадии оказывается пронизан мно-
жеством каналов, содержащих цистерны ЭПР. Одна-
ко сам механизм удвоения ретикулярного митохонд-
риона пока не прослежен.
Кинетопласт представляет собой расширенный
участок митохондриона, располагающийся у основа-
ния кинетосом жгутиков (рис. 138), в котором сосре-
доточена вся или почти вся митохондреальдая ДНК
(рис. 150; 158, вкл.). Кинетопласт стал известенгши-
рокому кругу исследователей вслед за введением в
лабораторную практику окраски по Романовскому—
Гимза. Первая догадка о природе кинетопласта и
введение этого термина в научную литературу при-
надлежат Алексееву (1917). Интересно, что его
предположения были подтверждены лишь через пол-
века, когда удалось увидеть кинетопласт в электрон-
ном микроскопе (Clark, Wallace, 1960; Steinert,
1960).
Митохондриальную ДНК, сосредоточенную в ки-
нетопласте, называют кинетопластной ДНК (кДЙК).
В клетках трипаносоматид на ее долю приходится до
20 % общего количества ДНК. Переплетающиеся
кольца кДНК формируют нуклеоид, который распо-
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
223
лагается в мешковидном вздутии митохондриона—
капсуле кинетопласта. У бодонид и криптобий она
ориентирована параллельно продольной оси клетки и
представляет собой дистальную часть колбасовидного
митохондриона (рис. 161, вкл.), а у трипаносоматид
капсула кинетопласта чаще располагается поперек
клетки (рис. 158, вкл.) и в ретикулярных митохонд-
рионах обычно представляет собой мешковидный
анастомоз, соединяющий латеральные рукава мито-
хондрии.
Викерман (Vickerman, 1977) выделяет 3 основ-
ные морфологические формы кинетопластов: панки-
нетопласты, поликинетопласты и эукинетопласты. В
панкинетопластах фибриллы кДНК располагаются
более или менее диффузно в митохондриальном мат-
риксе, капсула кинетопласта практически не выра-
жена. Вероятно, это наиболее примитивная форма
кинетопластии, встречающаяся у свободноживущих
бодонид и ряда криптобий. При поликинетопластии
кДНК образует несколько хорошо дифференцируе-
мых скоплений (рис. 138, F), часто приуроченных к
локальным вздутиям митохондрии. Поликинетоплас-
тия может сочетаться с панкинетопластией при че-
редовании стадий развития ряда бодонид и крипто-
бий. Эукинетопласты известны у представителей
всех 3 групп кинетопластид (рис. 158, вкл.; 161,
вкл.). В данном случае кДНК собрана в одном, спе-
циализированном участке митохондриона, располо-
женном у основания кинетосом жгутиков. У трипа-
носоматид известна только эта форма организации
кинетопластов (Каллиникова, 1977). Однако если
эукинетопласты свободноживущих бодонид и крип-
тобий в большинстве своем морфологически сход-
ны, то морфология кинетопластов трипаносоматид
практически не имеет аналогов среди других ки-
нетопластид. Она характеризуется компактной упа-
ковкой колец кДНК, при которой нуклеоид в форме
слегка вогнутого диска располагается обычно в цен-
тре капсулы, имеющей сходную форму. У трипаносо-
матид известны случаи полной утраты оформленного
нуклеоида (при сохранении капсулы кинетопласта).
Это явление получило название акинетопластии,
или, что более корректно, дискинетопластии. Спон-
танная дискинетопластия встречается у многих три-
паносоматид. Однако жизнеспособные дискинетоп-
ластные штаммы могут формироваться только в
популяциях африканских трипаносом. Наиболее ха-
рактерно это явление для трипаносом из подрода
Trypanozoon. У Т. evansi и Т. equiperdum существуют
как природные дискинетопластные штаммы, так
и штаммы, полученные в лабораторных условиях.
Структура нуклеоида изучена пока только у три-
паносоматид, у которых он формируется за счет
компактной упаковки мини- и максикольцевых мо-
лекул кДНК. Первых насчитывается десятки тысяч,
вторых — сотни на кинетопласт. Размеры мини-
и максикольцевых молекул достаточно специфичны
на уровне групп видов, иногда видов трипаносома-
тид.
Ядро. Интерфазные ядра. Интерфазные ядра
бодонид, криптобий и трипаносоматид (рис. 138;
150, вкл.; 158, вкл.) имеют во многом сходную орга-
низацию. В центре ядра или несколько в стороне от
него расположено ядрышко. Наиболее характерной
чертой интерфазных ядер кинетопластид является
конденсированный гетерохроматин, основная масса
которого располагается по периферии ядра, при-
легая к внутренней поверхности ядерной оболочки
(рис. 150; вкл.; 158, вкл.). Помимо пристеночных
массивов гетерохроматин встречается в кариоплазме
в виде отдельных глыбок. У бодонид гетерохроматин
иногда представлен несколькими плотными тяжами.
Подобная организация ядра отмечена и у ряда видов
криптобий.
Ядра цист и цистоподобных стадий. В
клетках инцистированных особей свободноживущих
бодонид морфология ядер не претерпевает каких-ли-
бо заметных изменений по сравнению с интерфазны-
ми ядрами активных жгутиконосцев.
Совершенно особой организацией отличаются
ядра цистоподобных амастигот, выполняющих функ-
ции расселительных стадий в жизненных циклах го-
моксенных трипаносоматид из родов Leptomonas и
Blastocrithidia. Их ядра имеют диаметр менее 2 мкм
и часто неправильную форму (рис. 160, вкл.). Харак-
терная черта в организации ядер цистоподобных
амастигот — это гиперконденсация хроматина. У
всех изученных Blastocrithidia, у Leptomonas oncopel-
ti и L. rigidus ядерный хроматин формирует так назы-
ваемую лабиринтовидную структуру. Основными эле-
ментами этой структуры являются тонкие (3—5 нм)
фибриллы, которые собираются в тяжи диаметром
20—25 нм. Последние формируют единую извитую
ленту, тесно прилегающую к ядерной оболочке и за-
нимающую большую часть внутриядерного простран-
ства.
Хотя ядра предшественников цистоподобных
амастигот, уже обладающие лабиринтовидными
структурами, продолжают делиться, в них не обнару-
жены ни характерная для митоза трипаносоматид де-
конденсация хроматина, ни оформленное микротру-
бочковое веретено, так что вопрос о способе деления
таких ядер пока остается открытым. В ядрах зрелых,
т. е. завершивших все деления, цистоподобных ста-
дий структура конденсированного хроматина не диф-
ференцируется (рис. 160, вкл.). Различия в организа-
ции ядер цист свободноживущих бодонид и цистопо-
добных стадий трипаносоматид подтверждают, что
происхождение этих стадий в обеих группах кине-
топластид, вероятнее всего, было независимым.
Митотические ядра. Деление ядер у кинето-
пластид протекает по типу закрытых внутриядерных
митозов и характеризуется полным отсутствиемспи-
рализованных хромосом на всех этапах митоза.
Деление ядра свободноживущего жгутиконосца
Boda curvifilus начинается с полной деконденсации
массивов пристеночного гетерохроматина. Карио-
плазма этих ядер более гомогенная, чем у интерфаз-
ных ядер. Ядрышковый материал разрыхляется, су-
щественно увеличиваясь в объеме. На более поздних
сроках в ядре формируется микротрубочковое вере-
тено (рис. 162, вкл.). Микротрубочковое веретено
В. curvifilus имеет очевидную биполярную организа-
цию. Центры организации микротрубочек не выраже-
ны. В формировании веретена принимают участие
полуверетена, закладывающиеся под углом друг к
224
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
другу. В поздней профазе и ранней метафазе в центре
ядра выявляются 2 группы микротрубочек по 13—
15 штук в каждой, располагающиеся под углом друг
к другу. В экваториальной плоскости делящегося
ядра формируется 10 пар кинетохоров. В начале ана-
фазы ядро принимает эллипсоидальную форму. Вере-
тено вытягивается, большинство микротрубочек про-
стираются между полюсами, которые занимают диа-
метрально противоположное положение (рис. 162,
вкл.). Кинетохоры начинают двигаться к полюсам де-
лящегося ядра, причем среди них можно выделить
6—7 «быстрых» кинетохоров и 3—4 «медленных». К
тому моменту, когда «быстрые» кинетохоры уже
практически достигают полюсов, «медленные» про-
ходят всего Ч3 дистанции. Сильно фрагментирован-
ный материал ядрышка сосредоточен вокруг цент-
ральной части веретена. Однако отдельные его фраг-
менты обнаруживаются у полюсов ядра вне связи с
центральным массивом. На стадии телофазы ядро
принимает гантелевидную форму. Кинетохоры исче-
зают. Некоторое время сохраняется центральная
часть веретена, микротрубочки которого располага-
ются в канале, соединяющем дочерние ядра. В обоих
ядрах выявляются компактные ядрышки и присте-
ночные сгустки конденсированного гетерохроматина.
Наконец ядра окончательно обособляются.
Среди криптобий морфология делящихся ядер
исследована у Cryptobia borreli (Скарлато, 1987;
Скарлато, Лом, 1997). Ранняя профаза характеризу-
ется полной деконденсацией гетерохроматина. Фор-
мирующееся затем веретено внутриядерных микро-
трубочек имеет «рыхлую» структуру. Митотическое
веретено представлено 3—4 пучками, содержащими
до 40 микротрубочек каждый. С микротрубочками
веретена связаны кинетохоры. Пока их число не
установлено, однако если судить по косвенным дан-
ным, в частности по числу микротрубочек веретена,
то. число кинетохоров в ядрах С. borreli должно
быть не менее десяти. В телофазе веретено микро-
трубочек сильно вытягивается и дочерние ядра рас-
ходятся, оставаясь соединенными длинным узким ка-
налом, в котором проходят микротрубочки веретена.
На этом этапе кинетохоры еще сохраняют связь с
веретеном, однако у внутренней мембраны дочерних
ядер уже выявляются массивы конденсированного
гетерохроматина, сегрегация которого происходит,
вероятно, без участия кинетохоров и микротрубочек
веретена. В культурах криптобий обнаружены мно-
гоядерные клетки, формирование которых предшес-
твует множественному делению жгутиконосцев.
Среди трипаносоматид митоз изучен у отно-
сительно небольшого числа видов, представляю-
щих, однако, все основные их группы: гомоксенных
«низших» трипаносоматид, трипаносом и лейшма-
ний.
Еще до формирования митотического веретена в
ранней профазе в ядрах жгутиконосцев происходит
деконденсация гетерохроматина и фрагментация яд-
рышка. У большинства трипаносоматид ядрышковый
материал в дальнейшем тесно прилегает к микротру-
бочкам веретена и при вытягивании последнего рас-
пределяется между дочерними ядрами. Однако у Try-
panosoma cruzi описано полное исчезновение ядрыш-
ка и формирование ядрышек de novo в каждом из
дочерних ядер (Solari, 1980).
Сформированное митотическое веретено в ядрах
трипаносоматид имеет хорошо выраженную биполяр-
ную организацию, причем у трипаносом бба полюса
и центральная часть «экваториальной пластинки»
лежат приблизительно в одной плоскости. Однако у
лейшманий и «низших» трипаносоматид полуверете-
на находятся под заметным углом друг к другу вплоть
до вытягивания веретена в анафазе—телофазе мито-
за. Количество микротрубочек, формирующих вере-
тено деления, различается у отдельных представите-
лей семейства. Менее 20 микротрубочек формирует-
ся в митотическом веретене некоторых Crithidia. От
20 до 30 микротрубочек имеют в своих митотичес-
ких веретенах большинство изученных видов гомо-
ксенных трипаносоматид из родов Blastocrithidia и
Wallaceina, а также гетероксенные Trypanosoma rho-
desiense, Т. danilewskyi, Т. rotatorium и Endotrypa-
num sp. Около 60 микротрубочек имеют веретена
лейшманий. И наконец, самое развитое митотическое
веретено, включающее около 120 микротрубочек, ха-
рактерно для делящегося ядра Trypanosoma cruzi»
Пространственная организация микротрубочек,
формирующих веретено, также различается у отдель-
ных видов трипаносоматид. Условно можно выделить
2 основные формы организации митотического вере-
тена у трипаносоматид — компактную и рыхлую. В
первом случае микротрубочки собраны в 1 компакт-
ный пучок, во втором формируют 2—3 пространст-
венно разобщенных пучка. У жгутиконосцев с про-
мастиготной организацией (у Leishmania, Wallaceina,
Crithidia) и у Endotrypanum (в культуре промастигот)
митотическое веретено имеет компактный вид, а у
жгутиконосцев с эпимастиготной и трипомастигот-
ной организацией обнаружены только «рыхлые» ве-
ретена. Важная черта организации митотических
ядер трипаносоматид — наличие «danse plaques»
(кинетохорподобных структур), которые ассоцииро-
ваны с микротрубочками митотического веретена.
Тонкое строение и молекулярная организация этих
структур, выясненные в основном благодаря работам
Соляри, свидетельствуют о том, что эти структуры
гомологичны кинетохорам других эукариот. Число
кинетохоров варьирует у различных представителей
трипаносоматид: у Trypanosoma cruzi их 10, у Leish-
mania 6, у Endotrypanum также 6, у Wallaceina 4, у
Crithidia fasciculata и Blastocrithidia triatomae 3. Из-
вестно, что при митозе эукариот кинетохорам, ассо-
циированным с отдельными хромосомами и микро-
трубочками веретена или с ядерной оболочкой, отво-
дится важнейшая роль в сегрегации генома (Райков,
1982). Иными словами, у «нормальных» эукариот
число кинетохоров и хромосом одинаково. Однако у
трипаносоматид это правило, видимо, нб^Зействует.
Морфологически установить взаимосвязь кинетохо-
ров и хромосом у этих жгутиконосцев невозможно,
так как последние в митозе полностью деспирализо-
ваны, т. е. во время деления ядра трипаносоматид к
его полюсам расходятся «пустые» кинетохоры. Толь-
ко связь тонких фибрилл (диаметром около 5 нм) с
этими структурами свидетельствует, что по крайней
мере часть генома распределяется между дочерними
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
225
ядрами классическим способом. Различными молеку-
лярно-биологическими методами показано, что ре-
альное количество интактных молекул ДНК или хро-
мосом у трипаносоматид исчисляется десятками и
даже сотнями на ядро (Van der Ploeg et al., 1984a,
1984b, 1989; Скарлато и др., 1990). Таким образом,
скорее всего, значительная часть генома трипаносо-
матид распределяется между дочерними ядрами без
участия кинетохоров. Возможно, эта роль отводится
ядерной оболочке или микротрубочкам веретена. Во
всяком случае, число кинетохоров в ядрах трипано-
соматид не тождественно числу хромосом и не позво-
ляет судить ни о количестве последних, ни о плоид-
ности самих ядер.
Несмотря на то что в представлениях об органи-
зации ядерного аппарата кинетопластид еще имеется
множество пробелов, некоторые выводы все же можно
сделать.
Так, с большой долей уверенности можно гово-
рить о том, что описанная выше форма митоза ха-
рактерна для кинетопластид в целом. В качестве
основных признаков, отличающих митоз кинетоплас-
тид, можно перечислить следующие.
1) Ядерная оболочка остается интактной в про-
цессе деления ядра.
2) На ранних этапах митоза происходит полная
деконденсация хроматина.
3) ЦОМТы митотического веретена, имеющего
биполярную организацию, морфологическими мето-
дами не выявляются.
4) Спирализация хромосом не происходит.
5) К полюсам делящегося ядра расходятся кине-
тохоры, ассоциированные с микротрубочками вере-
тена. Число кинетохоров прогрессивно убывает в
ряду Bodo curvifilus—Cryptobia borreli—Trypanosoma
cruzi—Leishmania и Endotrypanum—остальные трипа-
носоматиды.
Внутриклеточные мембранные системы. К этой
группе внутриклеточных структур у протистов обыч-
но причисляют всю совокупность секреторных, экск-
реторных и транспортных элементов, представлен-
ных окруженными цитоплазматическими мембрана-
ми каналами, цистернами, вакуолями и пузырьками.
Эндоплазматический ретикулум. Цистер-
ны и каналы ЭПР обоих типов пронизывают всю ци-
топлазму клеток кинетопластид. Обычно клетки ки-
нетопластид не богаты шероховатым ЭПР. Отдель-
ные фрагменты его каналов чаще сосредоточены
возле ядра, где отчетливо выявляется их связь с ядер-
ной оболочкой. Однако у ряда трипаносоматид этот
тип ретикулума развит очень хорошо. Так, у некото-
рых Phytomonas многочисленные каналы шерохова-
того ЭПР формируют сложный трехмерный многолу-
чевой комплекс. Хорошо развитая система каналов
этого типа обнаружена у кровяных стадий Trypanoso-
ma fallisi. В цитоплазме этой трипаносомы шерохо-
ватый ЭПР образует значительные по объему скопле-
ния параллельно ориентированных каналов, иногда
формирующих многорядные концентрические ок-
ружности. У этой же трипаносомы отмечена еще
одна интересная особенность. Отдельные цистерны
шероховатого ЭПР выстраиваются параллельно плаз-
малемме, подстилая субмембранные микротрубочки.
15 Протисты, часть 1
Сеть гладкого ЭПР в клетках кинетопластид, как
правило, хорошо развита. Его каналы встречаются
во всех регионах клетки, однако в двух местах —
вблизи комплекса Гольджи и в субмембранной
зоне — они особенно многочисленны. У многих
трипаносом отмечена приуроченность гребней глад-
кого ЭПР к субмембранной зоне в месте формиро-
вания ундулирующей мембраны. Впервые этот фено-
мен был обнаружен у трипаносом из секции Saliva-
ria (Taylor, Godfrey, 1969). Каналы гладкого ЭПР,
берущие свое начало от ядерной оболочки, подходят
к месту контакта жгутика с телом клетки и форми-
руют комплекс с 3—4 субмембранными микротру-
бочками по обе стороны от зоны контакта, лишен-
ной микротрубочек. Эти комплексы были названы
субпелликулярными органеллами. Позднее аналогич-
ные структуры были описаны под названием гребни
ретикулума у большинства трипаносом. Интересно,
что у гомоксенных трипаносоматид, обладающих ун-
дулирующей мембраной, эта структура отсутствует,
но зато она широко распространена у криптобий.
Функции субпелликулярных гребней гладкого ЭПР
пока неясны.
Комплекс Гольджи. Аппарат Гольджи у ки-
нетопластид имеет типичное строение. В зоне цито-
плазмы, лишенной рибосом, между ядром и кинето-
пластом располагаются 3—5, реже до 10 его цистерн,
полюса которых окружены многочисленными пу-
зырьками.
Типичным для свободноживущих бодонид следует
считать наличие в клетке единственной диктиосомы
комплекса Гольджи. Однако у Bodo caudatus в пери-
од их инцистирования в клетке обнаруживаются мно-
жественные диктиосомы. Сходную полимеризацию
диктиосом удается наблюдать у другого представите-
ля двухжгутиковых кинетопластид — Cryptobia sal-
mositica.
У трипаносоматид обычно имеется 1 диктиосома.
В этом смысле исключением являются Trypanosoma
theileri и Т. fallisi, у которых в клетке бывает 2 и
более диктиосомы. У последнего вида несколько дик-
тиосом располагаются вблизи друг от друга. Отмече-
но формирование ими пузырьков с электронно-плот-
ным содержимым, которые обнаруживаются также в
цитоплазме вокруг жгутикового кармана и в его про-
свете.
С деятельностью аппарата Гольджи традиционно
связывают наличие в цитоплазме трипаносоматид
мультивезикулярных тел — мембранных мешков, за-
полненных мелкими, окруженными мембранами пу-
зырьками. Однако их функции и природа пока не
изучены. То же самое можно сказать и об акантосо-
мах, или «окаймленных пузырьках». Это везикулы
диаметром около 50 нм, которые обычно располага-
ются в пространстве между дном жгутикового карма-
на и ядром. Окружающая их мембрана несет на
своей наружной поверхности сеточку из белка клат-
рина. Функции акантосом связывают с транспортам
протеинов.
Гликосомы. Одной из характерных черт в орга-
низации кинетопластид считается наличие в их ци-
топлазме электронно-плотных микротелец — глико-
сом, или пероксисомоподобных органелл (Vicker-
226
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
man, Preston, 1976; Brugerolle et al., 1979). Профили
гликосом на электронограммах клеток кинетоплас-
тид либо рассредоточены по всей цитоплазме, либо
группируются в каком-либо одном месте. Существует
точка зрения, что, подобно митохондриону, гликосо-
мы могут формировать в клетке единую ретикуляр-
ную структуру. Однако данные по ультратонкой орга-
низации трипаносоматид не подтверждают этого.
Гликосомы являются органеллами, играющими важ-
ную роль в утилизаций глюкозы. В них обнаружены
ферменты, участвующие в гликолизе, а также фер-
менты пиримидинового синтеза.
Сократительная вакуоль. Пульсирующая,
или сократительная, вакуоль является еще одним эле-
ментом эндомембранной системы. Сократительные
вакуоли найдены как у свободноживущих, так и у па-
разитических кинетопластид. Обычно роль сократи-
тельной вакуоли рассматривают в связи с регуляцией
осмотического градиента в клетках протистов, одна-
ко возможно, что у кинетопластид они могут выпол-
нять и другие функции, в частности экскреторную.
Морфология пульсирующих вакуолей и их деятель-
ность изучены пока очень слабо. Интересной осо-
бенностью сократительных вакуолей кинетопластид
является выведение их содержимого в полость жгути-
кового кармана, Морфологически система пульсиру-
ющей вакуоли включает в себя собственно резервуар
вакуоли и всю совокупность ветвящихся приводящих
каналов. Последние интересны тем, что их попереч-
ные профили по морфологии и размерам сходны с
акантосомами.
В н у т р и к л е т о ч н ы е с и м б и о н т ы. В нутрикле-
точные симбионты встречаются у протистов доволь-
но часто. Кинетопластиды в этом смысле не являются
исключением — прокариотные эндосимбионты опи-
саны у многих видов свободноживущих бодонид и у
криптобий. У трипаносоматид эндосимбионты встре-
чаются гораздо реже, чем у двухжгутиковых кинето-
пластид, однако именно в этой группе эндосимбион-
ты стали объектом специальных исследований, К
настоящему времени эндосимбионты описаны у
4 гомоксенных видов трипаносоматид из насеко-
мых — Blastocrithidia culicis, Crithidia deanei, C. de-
souzai и C. oncopelti, а также у Trypanosoma cobitis
из пресноводных рыб и Г. fallisi из амфибий. Взаи-
моотношение эндосимбионтов с клетками трипано-
сом пока не изучены. Что же касается трипаносома-
тид насекомых, то в отношении их эндосимбионтов
имеется довольно обширная литература. Исследова-
ние их морфологии, а именно организации клеточ-
ной стенки и протопласта, показало, что эндосимби-
онты трипаносоматид являются грамположительными
бактериями (Gutteridge, Macadam, 1971). Используя
различные антибиотики, удалось получить симбионт-
свободные штаммы жгутиконосцев из их исходных,
симбионтсодержащих культур (Chang, 1974). Это
позволило сравнить метаболизм симбионтсодержа-
щих и «чистых» штаммов жгутиконосцев (Chang,
Trager, 1974). В частности, оказалось, что лишенные
эндосимбионтов штаммы Blastocrithidia culicis и
Crithidia oncopelti являются ауксотрофами по гемину
в отличие от исходных геминнезависимых их штам-
мов. Предполагается, что кроме гемина эндосимби-
онты могут поставлять в клетки трипаносоматид ряд
витаминов и белковых кофакторов. Наличие симби-
онтов в клетках трипаносоматид существенно сказы-
вается на их морфологии. Так, жгутики симбионтсо-
держащих особей лишены параксиального тяжа
(хотя его специфические белки сохраняются), а
ветви митохондриона занимают субмембранную по-
зицию, вклиниваясь между микротрубочками. Одна-
ко эти изменения оказываются обратимыми. При
удалении симбионтов морфология трипаносоматид
приобретает характерные черты: появляется парак-
сиальный тяж, ветви митохондриона рассредоточива-
ются в цитоплазме (Freymuller, Camargo, 1981). Не-
давно методом сравнительного сиквенса генов рРНК
было показано, что симбионты всех 4 видов гомок-
сенных трипаносоматид чрезвычайно близки друг
другу (более 97 %) и имеют монофилетическое про-
исхождение (Du et al., 1994).
Кроме прокариотных эндосимбионтов в клетках
трипаносоматид описаны также вирусоподобные час-
тицы. Однако об их взаимоотношениях с клетками
жгутиконосцев практически ничего не известно. Мо-
линьо описал вирусоподобные частицы у промасти-
гот Leishmania hertigi — кровепаразитов крыс из Па-
намы. Вирусоподобные частицы размером 50—60 нм
находились в цитоплазме промастигот. При транс-
формации промастигот в амастиготы (при развитии в
позвоночном животном) эти частицы переставали вы-
являться. Через 3—5 дней после того как амастиготы
скармливались москитам, в кишечнике последних
промастиготы вновь имели в своей цитоплазме вирус-
ные частицы (Molyneux, Ashford, 1983).
Особенности биологии
и жизненные циклы
кинетопластид
Жизненные циклы двухжгутиковых кинетопластид
изучены пока еще очень плохо. В первую очередь это
относится к свободноживущим представителям груп-
пы.
Большинство бодонид обитает в постоянных и
временных водоемах, как пресных, так и с различ-
ным уровнем солености. По-видимому, многие виды
достаточно легко переносят переход из соленой
воды в пресную и наоборот. Бодониды в изобилии
встречаются в почве и охотно поселяются на различ-
ных органических субстратах (фекальные массы,
компосты и т. п.). Большинство бодонид питаются
бактериями, захватывая их мощно развитым цито-
стомом (Bodo designis, В, curvifilus). В пищевари-
тельных вакуолях жгутиконосцев можно встретить и
мелкие водоросли, и оформленные органические
частички. Есть среди бодонид и облигатные хищни-
ки (Rhynchobodo), добычей которых служат мелкие
жгутиконосцы. Вместе с тем ряд жгутиконосцев (на^
пример, В. caudatus) может сочетать бактериофагию
и хищничество. Экспериментально показано, что бо-
дониды могут длительное время питаться растворен)-
ными органическими веществами, сохраняя способ-
ность к размножению. Большинство бодонид — это
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
227
свободноплавающие или ползающие формы. Харак-
тер их движения настолько специфичен, что при
известном навыке позволяет по одному этому при-
знаку определить род, а иногда и вид наблюдаемого
объекта. Некоторые бодониды, например В. saltans,
способны прикрепляться своим задним жгутиком к
субстрату, однако легко утрачивают эту связь и пе-
реходят к активному плаванию. Жизненный цикл
большинства бодонид» по всей видимости, не сопря-
жен с упорядоченным чередованием стадий или фаз
развития. В благоприятных условиях жгутиконосцы
интенсивно питаются и размножаются путем равных
бинарных делений (рис. 143, А). При неблагоприят-
ных условиях некоторые виды бодонид формируют
цисты. При этом жгутиконосцы втягивают жгутики и
секретируют внешнюю оболочку — стенку цисты
(рис. 152, вкл.). Изменения в строении самих жгути-
коносцев незначительны. Несколько возрастает
электронная плотность цитоплазмы, перестает функ-
ционировать сократительная вакуоль. Кристы мито-
хондриона либо деградируют, либо изменяют свою
форму. В последнем случае дисковидные кристы
трансформируются в ампуловидные. В цистах жгути-
коносцы переживают неблагоприятные условия, де-
ления клеток не происходит. Единого мнения о том,
насколько широко среди бодонид распространена
способность к формированию цист, пока нет. В ли-
тературе наиболее часто встречается описание про-
цесса цистообразования у Bodo caudatus.
Единственным видом бодонид, перешедшим к об-
лигатному паразитизму, является Ichthyobodo песа-
tor. Эти жгутиконосцы поселяются на жаберном
эпителии молоди пресноводных рыб, а также на по-
верхности тела взрослых особей. Прикрепление жгу-
тиконосцев к эпителию происходит при помощи ви-
доизмененного переднего конца их тела, который
погружается в частично разрушающиеся ткани. Ин-
тенсивное размножение паразитов происходит путем
продольных бинарных делений. Цитокинез обычно
существенно отстает по времени от делений клеточ-
ных органелл, так что в популяции жгутиконосцев
часто выявляется много четырехжгутиковых особей
(рис. 147, Е). Некротические очаги, возникающие в
местах скопления паразитов, приводят к глубокой
деградации тканей, что зачастую влечет за собой
гибель рыб, особенно молоди.
Жизненные циклы свободноживущих криптобиид
(Procryptobia, Dimastigella, Cruzella, Cephalothamni-
um) изучены в еще меньшей степени, чём у бодонид.
По пищевым потребностям и способу питания эти
криптобииды, видимо, ничем не отличаются от пред-
ставителей сем. Bodonidae. Большинство из них пред-
ставлены свободноплавающими или ползающими по
субстрату в поисках пищи формами. Только жгутико-
носцы Cephalothamnium cyclopum, поселяющиеся на
теле пресноводных ракообразных, формируют сте-
бельчатые колонии, насчитывающие до 30 особей.
Жгутиконосцы Dimastigella trypaniformis, обитающие
в почве, могут образовывать цисты. У других видов
свободноживущих криптобиид способы переживания
ими неблагоприятных условий неизвестны.
Большинство представителей сем. Cryptobiidae —
это паразиты позвоночных и беспозвоночных живот-
Рис. 143. Различные формы клеточных делений кинетопластид.
А Bodo saltans, Б — Cryptobia salmositica, В — Leptomonas sp., Г— фор-
мирование «жгутиковых цист» у Leptomonas oncopelti.
Fig. 143. Different forms of cell division in kinetoplastid flagellates.
A — Bodo saltans, Б — Cryptobia salmositica, В — Leptomonas sp., Г — for-
ming of «strap-hangers» in Leptomonas oncopelti.
ных. Степень освоения криптобиями разных групп хо-
зяев неодинакова. 2 вида этих жгутиконосцев описаны
из репродуктивных органов брюхоногих моллюсков,
по 1 виду — из пищеварительной системы планарий,
пиявок и рептилий. Наконец, около 50 видов крипто-
бий известны как паразиты морских и пресноводных
рыб. Жизненные циклы криптобий, паразитирующих
в беспозвоночных животных и рептилиях, практичес-
ки неизвестны. Жгутиконосцы размножаются в хозяе-
вах путем продольных бинарных делений. Паразити-
рующие в гастроподах Cryptobia helicis и С. carinariae
способны прикрепляться к эпителию семенных меш-
ков своих хозяев при помощи передних жгутиков. У С.
dendrocoeli, паразитирующей в пищеварительной сис-
теме планарии Dendrocoelum lacteum, описаны внут-
риклеточные стадии развития. Принято считать, что
криптобии из водных беспозвоночных имеют в своих
жизненных циклах свободноживущие стадии, посред-
ством которых и осуществляется циркуляция парази-
тов в популяции хозяев.
Жизненные циклы криптобий, паразитирующих в
рыбах, сложны и разнообразны. Можно выделить 3
основных типа паразито-хозяинных отношений, реа-
лизующихся в жизненных циклах криптобий. В пер-
вом случае жгутиконосцы выступают в качестве экто-
паразитов, поселяясь на поверхности тела и жабер-
228
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ном эпителии главным образом пресноводных рыб.
Биология эктопаразитических видов криптобий сла-
бо изучена. Часть жгутиконосцев, например Cryp-
tobia carassi, поселяющиеся в слизи на поверхности
тела хозяев, являются, по сути, комменсалами. Одна-
ко жгутиконосцы, которые заселяют жаберный эпи-
телий рыб, например С. branchialis, могут быть при-
чиной заболевания и массовой гибели рыб, особенно
их молоди. Эти жгутиконосцы прикрепляются к по-
верхности ткани при помощи видоизмененного жгу-
тика и интенсивно размножаются путем продольных
бинарных делений. Дочерние особи прикрепляются к
поверхности эпителия и также делятся. Интенсивное
размножение паразитов, вероятно, приводит к бло-
каде поверхности жабр и соответственно нарушению
дыхания. Следует, однако, отметить, что, возможно,
ряд эктопаразитических криптобий, а с точки зрения
By (Woo, 1987) — большинство, на самом деле не
являются самостоятельными видами, а представляют
собой стадии развития жгутиконосцев, паразитирую-
щих в кровеносной системе рыб. Как мы увидим ни-
же, для такого предположения есть серьезные осно-
вания.
Вторая группа (около 7 видов криптобий) пред-
ставлена паразитами кишечника или (в редких случа-
ях) других внутренних органов рыб. Почти все они,
за исключением С. jubilans, описаны из морских рыб.
Жгутиконосцы размножаются путем продольных би-
нарных делений в содержимом кишечника, не при-
крепляясь к поверхности энтероцитов. Немногочис-
ленные экспериментальные данные свидетельствуют
о том, что передача криптобий от хозяина к хозяину
может происходить тремя основными путями: 1) не-
посредственно через воду, свободными жгутиконос-
цами; 2) через временных накопителей (беспозвоноч-
ных, служащих пищей для хозяев); 3) при хищни-
честве или каннибализме. Вероятно, большинство из
этих видов (С. dahlii, С, stilbia, С, intestinalis и т. п.)
непатогенны для своих хозяев, и часто их рассматри-
вают как эндокомменсалов рыб. Однако С. jubilans
высоко патогенна для своих хозяев — пресноводных
рыбок Herichthys cyanoguttatum. Кроме пищевари-
тельной системы эти жгутиконосцы инвазируют мно-
гие внутренние органы: печень, селезенку, яичники и
могут проникать в кровь, где размножаются в макро-
фагах. Однако в крови вне макрофагов паразиты от-
сутствуют.
Третья и самая большая группа криптобий, на-
считывающая, по разным данным, от 30 до 40 ви-
дов, включает облигатных кровепаразитов рыб. Эти
криптобии описаны и в морских и в пресноводных
хозяевах. Главной особенностью этих видов является
их гетероксенность. Переносчиками жгутиконосцев
служат пиявки. Таким образом, жизненный цикл
жгутиконосцев включает минимум две фазы. Одна из
них протекает в позвоночном, а другая — в беспоз-
воночном животном-хозяине. В крови рыб развитие
криптобий протекает по стандартной схеме: резко
нарастающая инвазия переходит в хроническую
форму, сопровождающуюся постепенным снижением
численности паразитов. Единичные особи обнаружи-
ваются в крови хозяев через год и более с момента
заражения. Размножение криптобий в крови рыб
обычно протекает по типу бинарных делений
(рис. 143, Б). Данные о возможности неоднократных
делений нуждаются в проверке. Бинарные деления
могут протекать по-разному у разных жгутиконос-
цев. By (Woo, 1987) выделяет 2 основные их формы.
В первом случае, как это описано у Cryptobia catas-
tomi, жгутиконосцы перед делением округляются и в
результате деления формируют 2 одинаковые клет-
ки. Другой тип деления описан у С. salmositica. В
этом случае жгутиконосцы перед цитокинезом как
бы вытягиваются в длину, деление начинается с за-
днего конца, а формирующиеся особи различаются
по форме тела (широкие и узкие). Кроме того, при
первом типе деления дочерние особи формируют по
1 новому жгутику, а другой получают от материнс-
кой особи. Второй тип деления характеризуется тем,
что одна из дочерних особей (узкая форма) форми-
рует оба жгутика заново. Многие кровепаразитичес-
кие криптобии, например С. borreli или С. salmosi-
tica, не обладают выраженной специфичностью по
отношению к позвоночным хозяевам. Однако есть
виды, которые используют, по-видимому, довольно
узкий круг хозяев. Примером такого рода может
служить С. catostomi.
О патогенности кровепаразитических криптобий
имеется много сведений, особенно в старой литера-
туре. Однако Лом (Lorn, 1979) и By (Woo, 1987)
полагают, что часто приписываемая криптобиазам
гибель рыб на самом деле происходит по другим
причинам. Тем не менее для некоторых видов уда-
лось проследить патогенное воздействие на хозяина,
в том числе и экспериментальными методами. При-
мер тяжелого течения криптобиаза демонстрирует С.
salmositica. Главными признаками острой формы за-
болевания служат экзофтальмия, спленомегалия, на-
растающая анемия и растяжение брюшины асцитной
жидкостью. Криптобиаз служит причиной гибели
молоди лососевых и особенно опасен в рыбоводчес-
ких питомниках, где может принимать характер эпи-
зоотии.
Развитие криптобий в переносчиках связано с
интенсивным размножением паразитов, но в отличие
от трипаносом практически не сопровождается чере-
дованием морфологических форм жгутиконосцев.
Так, С. beckeri, паразит морской рыбы Scorpaenicht-
hys marmoratus, попадая с кровью хозяина в пище-
варительную систему пиявки Malmiana diminuta,
претерпевает серию бинарных делений, и уже через
72 ч его инфективные стадии локализуются во вла-
галище хоботка. Эти стадии отличаются от жгутико-
носцев из кишечника пиявки лишь более мелкими
размерами. Темпы развития паразитов соответствуют
времени переваривания пищевого комка и в значи-
тельной степени зависят от окружающей температу-
ры. Так, жгутиконосцы Cryptobia bullocki формиро-
вали инфективные стадии при температуре воды
5 °C через 10 дней после попадания в морскую пи-
явку Calliobdella vivida, а при температуре 20 °C —
уже через 24 ч. Заражение новых особей рыб про-
исходит при нападении на них зараженных крипто-
биями пиявок. Со слюной пиявки в кровеносную
систему рыбы вводятся жгутиконосцы, которые, по-
видимому, тут же приступают к размножению.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
229
Очень важной чертой в биологии ряда кровепа-
разитических криптобий является способность к
прямой (без участия переносчика) передаче пара-
зитов от хозяина к хозяину. Типичный пример
такой передачи можно наблюдать у Cryptobia sal-
mositica. При совместном содержании зараженных
и незараженных особей радужной форели пример-
но у 80% последних можно обнаружить присут-
ствие криптобий в крови спустя четыре месяца
после начала эксперимента. Эта величина оказыва-
ется несколько ниже (50 %), если садок разделить
проволочной сеткой, препятствующей прямым кон-
тактам между зараженными и незараженными ры-
бами. Примерно через 6 нед после заражения хо-
зяина криптобии начинают появляться в слизи на
поверхности тела рыб. Эти «эктопаразитические»
формы, если их инокулировать в кровь хозяина, за-
ражают его, и дальнейшее развитие криптобий про-
текает по обычному сценарию. Однако каков естес-
твенный путь криптобий с поверхности тела ры-
бы реципиента в ее кровеносную систему, пока
неизвестно. Хотя биология трипаносоматид изу-
чена гораздо глубже и полнее, чем у остальных
представителей кинетопластид, остается немало от-
крытых вопросов, касающихся хода их жизненного
цикла. Так, применение электронного микроскопа,
особенно при изучении паразито-хозяинных отно-
шений трипаносоматид и беспозвоночных живот-
ных, позволило выявить ряд форм или стадий жгу-
тиконосцев, роль которых в жизненных циклах три-
паносоматид неясна. В первую очередь это касается
внутритканевых стадий развития паразитов и много-
ядерных, или «гигантских», форм. Кроме того, до
сих пор ничего не известно о формах полового про-
цесса и его месте в жизненном цикле этих жгутико-
носцев.
Размножение
трипаносоматид
Жизненные циклы трипаносоматид, как правило,
включают несколько стадий, принимающих участие
в эндогенной агломерации паразитов. Исключение
составляет лишь Trypanosoma equiperdum, вторично
утратившая переносчика и обладающая, по сути,
одним, растянутым во времени циклом размножения.
Стадии, на которых размножаются трипаносомати-
ды, представлены разными морфологическими фор-
мами. Часто клеточные деления предшествуют про-
цессам клеточной дифференцировки, связанным со
сменой стадий или морфологических форм жгутико-
носцев, или сопровождают их. Связь размножения
трипаносоматид с половым процессом пока не уста-
новлена. Здесь можно вспомнить, что до недавнего
времени трипаносоматид вообще относили к агам-
ным простейшим.
Формы делений у трипаносоматид весьма разно-
образны. В этой группе протистов встречаются и
бинарные и множественные деления, причем и те и
другие могут быть и эквивалентными и неравными.
Наиболее обычной формой деления у трипаносома-
тид являются эквивалентные бинарные деления, т. е.
деления, приводящие к образованию 2 приблизи-
тельно равных по размеру и сходных по морфологии
дочерних клеток (рис. 143, В). Чаще всего такое
деление начинается с изменений в организации дви-
гательного аппарата, затем происходит деление ки-
нетопласта и вслед за ним — митоз. По времени с
этим процессом совпадает начало цитокинеза, кото-
рый обычно связан с расхождением передних концов
клеток трипаносоматид, однако иногда наблюдается и
обратная картина. Деление клетки, начинающееся с
ее заднего конца, в ряде случаев, по-видимому, со-
пряжено с формированием гигантских многоаксо-
немных жгутов, объединяющих «грозди» жгутико-
носцев.
К неравным бинарным делениям относится «поч-
кование», описанное у гомоксенных трипаносоматид
в связи с формированием «strap-hangers» — цисто-
подобных стадий, прикрепляющихся к жгутику мате-
ринской особи (рис. 143, Г)- В данном случае оче-
редность дупликации органелл та же, что и в преды-
дущем случае, однако в процессе почкования сначала
всегда обособляется задний конец «почки», а затем
борозда деления смещается в сторону передних кон-
цов родительской и дочерней клеток. Неравные би-
нарные деления известны и у трипаносом. Например,
некоторые трипаносомы из крови рыб и амфибий,
попадая в пищеварительную систему пиявок, первое
время размножаются именно таким способом. В этом
случае делящаяся трипомастигота дает начало 2 мор-
фологически отличным формам: удлиненной и округ-
лой.
В литературе встречается описание бинарных де-
лений, характеризующихся поперечным цитокинезом.
Эта форма делений еще мало исследована. Известно,
что цитокинезу в данном случае предшествует миг-
рация органоидов к противоположным полюсам
клетки, вслед за чем она перешнуровывается при-
мерно посередине.
Большой интерес представляют множественные
деления трипаносоматид. Эти деления могут рас-
сматриваться как результат отставания процессов
цитокинеза от темпов удвоения клеточных органои-
дов. В жизненных циклах трипаносоматид регулярно
происходящие множественные деления описаны
только у трипаносом. Эндогенная агломерация пара-
зитов, осуществляемая путем множественных деле-
ний, характерна для представителей подрода Негре-
tosoma. У большинства этих трипаносом множест-
венные деления жгутиконосцев протекают в крови
или в других тканях позвоночных животных, однако
у Trypanosoma lewisi, кроме того, они происходят и
внутри клеток кишечного эпителия блох. Множест-
венные деления трипаносом в клетках кишечного
эпителия насекомых-переносчиков описаны также у
Т brucei и Т cruzi. Сходным образом в клетках
слюнных желез клопов размножаются Т. rangeli и
Phytomonas serpens. В лабораторных культурах мно-
жественное деление описано у Trypanosoma сопо-
rhini. В процессе множественного деления у три-
паносоматид формируются так называемые гигант-
ские клетки. Интерес к этим стадиям объясняется
их возможной причастностью к половому процес-
су.
230
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Развитие трипаносоматид
в позвоночных животных
В позвоночных животных развитие трипаносома-
тид приурочено главным образом к кровеносной
системе. Считается, что паразитирование жгутико-
носцев в тканях внутренних органов, не связанных
непосредственно с кровеносной системой, носит у
трипаносоматид характер вторичных адаптаций. Од-
нако в последнее время это положение было поко-
леблено. Выяснилось, что в южноамериканских сум-
чатых и грызунах некоторые трипаносомы, относя-
щиеся к подродам Schisotrypanum, Megatrypanum и
Herpetosoma, могут развиваться в пищеварительной
системе хозяев, а именно в их ректальных железах.
Если к тому же учесть многочисленные описания
трипаносоматид из кишечника различных рептилий,
то, как нам кажется, не следует исключать возмож-
ность реализации алиментарной фазы жизненного
цикла в позвоночном-хозяине, по крайней мере у
части трипаносоматид.
В кровеносной системе позвоночных животных
возможны 3 различных пути развития трипаносома-
тид. Первый из них представлен в жизненных циклах
Leishmania и Endotrypanum. Этот путь связан с пора-
жением паразитами форменных элементов крови хо-
зяев. 2 вида жгутиконосцев из рода Endotrypanum
развиваются в эритроцитах южноамериканских ле-
нивцев (Edentata). Эти трипаносоматиды изучены
очень плохо. Известно лишь, что в эритроцитах на
ранних стадиях инвазии у одного из видов обнаружи-
ваются эпи-, а у другого — трипомастиготы. Клиника
заболевания неизвестна.
Мишенью для лейшманий служат мононуклеар-
ные фагоциты (макрофаги) хозяев, в которых лейш-
мании размножаются в форме эндомастигот. Разру-
шение москитом при помощи хоботка эпидермаль-
ных клеток и введение в ранку слюны стимулируют
приток макрофагов к месту укуса. Судьба паразитов,
которых зараженный москит инокулирует в ткани
хозяина, различна. Жгутиконосцы, попавшие непос-
редственно в кровь, гибнут, как и многие особи,
захваченные фагоцитами. Только в нестимулирован-
ных макрофагах промастиготы выживают, трансфор-
мируются в эндомастиготы и приступают к размно-
жению, только внутри этих клеток паразиты могут
разноситься по организму хозяина. Сыворотка крови
одинаково губительна и для промастигот и для эндо-
мастигот лейшманий. Передача эндомастигот между
макрофагами происходит при разрушении заражен-
ных клеток и захвате эндомастигот новыми макро-
фагами. Лейшманиозы проявляются у позвоночных
животных в виде более или менее значительных не-
кротических очагов, образующихся в местах лока-
лизации паразитов. Существуют 2 основные формы
лейшманиозов — кожная, в результате которой по-
ражается ретикуло-эндотелиальная система кожи и
подкожная клетчатка, и висцеральная, характеризу-
ющаяся широким поражением организма хозяина,
включая практически все ^системы внутренних орга-
нов. Второй путь связан с развитием жгутиконосцев
в крови хозяина без поражения ими форменных эле-
ментов крови и клеток тканей внутренних органов.
По такому типу происходит развитие трипаносом в
низших позвоночных животных, а также трипаносом
подрода Megatrypanum в млекопитающих. Эта груп-
па циклов в настоящее время выглядит достаточно
целостной, однако нельзя забывать, что трипаносо-
мы из низших позвоночных животных изучены
очень поверхностно. Данный тип развития трипано-
сом в позвоночных-хозяевах сопряжен со слабо вы-
раженным плеоморфизмом кровяных стадий и их
низкой пролиферативной активностью. Так, у боль-
шинства трипаносом, паразитирующих в крови рыб
(Lorn, 1979), размножение в позвоночных-хозяевах
не происходит. Слабо выраженный плеоморфизм
трипаносом в данном случае, вероятно, связан с кле-
точным ростом паразитов. Если же размножение
происходит, то оно приурочено к кровеносному
плексусу какого-либо органа (сердце, почки, кост-
ный мозг) и осуществляется в форме бинарных де-
лений. Трипаносомы в крови амфибий и рептилий
могут размножаться и путем бинарных, и путем мно-
жественных делений, часто протекающих на фоне
чередования морфологических форм жгутиконосцев.
Однако и здесь размножение жгутиконосцев чаще
происходит не в периферической крови, а в капил-
лярах внутренних органов. У трипаносом амфибий
обнаружено наличие циркадных ритмов, связанных с
фотопериодичностью.
Трипаносомы из птиц демонстрируют чрезвычай-
но низкий уровень инвазии^ так что выявить их часто
удается лишь методом слепого посева культур на
NNN среду. Исключение составляют Trypanosoma
avium backeri и Т bouffardi; последний вид, правда,
выпадает из данной группы, так как может развивать-
ся за пределами кровеносной системы хозяев. Из
всех представителей подрода Megatrypanum только у
Trypanosoma theileri описаны стадии, размножающи-
еся в крови млекопитающих. Другие представители
этого подрода, по морфологии очень близкие к три-
паносомам из амфибий, обладают слабым «возраст-
ным» плеоморфизмом при хроническом течении ин-
вазии. В отношении Т. theileri в литературе имеется
очень много противоречивых данных. Часто зараже-
ние этими жгутиконосцами сопряжено со смешанной
инвазией протозойной (Theileria) или иной (Anoplas-
ma) природы. Так что размножение в форме амасти-
гот, приписываемое этой трипаносоме, требует про-
верки, так же как и наличие у данного вида экстра-
васкулярных стадий.
Третий путь развития трипаносоматид в позво-
ночных животных представлен в жизненных циклах
большинства трипаносом, паразитирующих в млеко-
питающих. Его характерной чертой является выход
трипаносом за пределы кровеносной системы и их раз-
витие в тканях различных внутренних органов своих
хозяев. Среди всего многообразия такого рода циклов
можно выделить 3 группы, различающиеся по особен-
ностям экстраваскулярного развития паразитов.
В первую группу мы объединяем жизненные цик-
лы жгутиконосцев из подрода Herpetosoma, за исклю-
чением Trypanosoma (Я.) lewisi и ряда близких к ней
видов. В данном случае инвазия позвоночных живот-
ных протекает в 3 этапа. На первом этапе (инкуба-
ционный период) трипаносомы размножаются путем
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
231
множественных делений в тканях различных внут-
ренних органов хозяина, локализуясь обычно в их
межклетниках. Второй этап, на котором жгутико-
носцы выходят в кровь, соответствует пику паразите-
мии. Этот этап ограничен во времени и постепенно
переходит в следующий, на котором происходит уга-
сание инвазии, вплоть до полного освобождения хо-
зяина от паразитов. Обычно при этом хозяева приоб-
ретают устойчивый иммунитет к данному виду трипа*,
носом.
Ко второй группе могут быть отнесены жизнен-
ные циклы трипаносоматид подрода Schisotrypanum.
Эти жгутиконосцы используют кровеносную систему
хозяев в двух целях: для расселения по его организму
и для аккумуляции расселительных стадий. Попада-
ющие в кровяное русло млекопитающего метацикли-
ческие трипаносомы проникают через стенки сосу-
дов в ткани различных органов, поражая отдельные
клетки. Здесь трипаносомы трансформируются в ок-
руглые эпимастиготы (сферомастиготы), которые де-
лятся, формируя микропопуляцию эндомастигот. По-
следние размножаются, насколько позволяют габа-
риты пораженной клетки, которая в конце концов
превращается в мембранный мешок, заполненный
паразитами, — «псевдоцисту». После разрушения
пораженной клетки эндомастиготы трансформиру-
ются в трипомастигот, которые возвращаются в кро-
вяное русло. Их популяция плеоморфна. Узкие три-
паносомы продолжают цикл развития в хозяине,
повторяя путь метациклических стадий. Широкие
формы трипаносом служат для заражения перенос-
чиков. Эти формы преобладают в крови хозяев при
хроническом течении заболевания. Интересно, что
ряд животных выступает в качестве биологических
фильтров именно этих стадий. В мышах, например,
формируются штаммы Trypanosoma cruzi, характери-
зующиеся присутствием в крови хозяина только ши-
роких форм трипаносом. Различные штаммы Т. cruzi
демонстрируют предрасположенность к развитию в
тканях определенных органов. Наиболее тяжелые
последствия у млекопитающих вызывает поражение
трипаносомами печени, сердечной мышцы и цент-
ральной нервной системы. Будучи высокопатогенны-
ми для человека, жгутиконосцы Т, cruzi, по-види-
мому, находятся в достаточно лояльных отношениях
с дикими млекопитающими, среди которых много
представителей реликтовых групп: сумчатые, броне-
носцы и муравьеды (Ноаге, 1972).
Трипаносом африканских млекопитающих обыч-
но причисляют к «чистым» кровепаразитам, так как
их размножение происходит в периферической кро-
ви хозяев. Однако эти трипаносомы встречаются в
тканях практически всех внутренних органов, а в
лимфоидной ткани и в спинномозговой жидкости
они могут и размножаться. Эти трипаносомы облада-
ют выраженным плеоморфизмом кровяных стадий.
Длинные узкие трипаносомы ответственны за размно-
жение паразитов. Широкие формы, процент которых
возрастает в процессе ремиссии паразитемии, явля-
ются предшественниками проциклических стадий и
служат для заражения переносчиков. Т. equiper-
dum — типично тканевой паразит. В кровяное русло
хозяина эти трипаносомы проникают только для рас-
селения по его организму. Развитие жгутиконосцев
происходит главным образом в слизистой гениталиев
и в межклетниках тканей этих органов. На поздних
стадиях заболевания трипаносомы проникают в сли-
зистые рта, носа, поражают конъюктиву глаз и лока-
лизуются в подкожной клетчатке.
Вопрос о специфичности трипаносом к позво-
ночным животным изучен очень слабо. Считается,
что большинство трипаносом млекопитающих обла-
дают более или менее широкой специфичностью.
Это демонстрируется, например, наличием лабора-
торных модельных животных (мыши, крысы, хомя-
ки), на которых удается вести практически любых
трипаносоматид млекопитающих. Более того, Т bru-
cei, например, может длительное время развиваться
в крови рептилий, сохраняя при этом способность к
заражению своих естественных хозяев — млекопи-
тающих.
Представления о специфичности трипаносом низ-
ших позвоночных животных крайне противоречивы.
Собственно говоря, здесь сталкиваются две крайнос-
ти. Одна — это ошибочные представления об узкой
специфичности этих трипаносом, формирующиеся в
силу использования традиционного подхода при их
описании по принципу «новый хозяин — новый вид
паразита», что особенно хорошо видно на примере
трипаносом рыб (Lorn, 1979). Другая крайность —
это виды-«гиганты», такие как Г. rotatorium или
Т. inopinatum, к которым автоматически относят
большинство трипаносом из Anura (Bardsley, Harmsen,
1973), описанных на всех континентах, где встреча-
ются эти позвоночные. В данном случае, очевидно,
сборные виды трипаносом демонстрируют ошибоч-
ную картину широкой специфичности. Достоверная
узкая специфичность обнаруживается у ряда трипа-
носоматид, паразитирующих в реликтовых группах
хозяев. Таковы, в частности, Endotrypanum из амери-
канских ленивцев или Trypanosoma gray из афри-
канских крокодилов.
Анализ литературы, посвященной вопросу специ-
фичности трипаносоматид к позвоночным животным,
позволяет сделать вывод о том, что чем ниже эволю-
ционный уровень позвоночных животных, тем более
узкую специфичность по отношению к ним проявля-
ют трипаносоматиды. Однако, учитывая, что как раз
в этих группах животных трипаносоматиды изучены
очень поверхностно, мы оставляем вопрос о специ-
фичности трипаносоматид к позвоночным животным
открытым.
Развитие трипаносоматид
в беспозвоночных животных
Среди беспозвоночных животных хозяевами три-
паносоматид являются в основном клещи, пиявки и
насекомые. Известны отдельные находки трипаносо-
матид в нематодах, моллюсках, коловратках и инфу-
зориях, однако в большинстве случаев это единичные
описания, требующие дополнительных подтвержде-
ний. Исключением в данном случае является Lepto-
monas, паразитирующий в макронуклеусах инфузо-
рий. Однако эти жгутиконосцы изучены очень плохо,
232
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
и характеризовать данную систему паразит—хозяин
пока не представляется возможным.
Из литературы известно, что клещи могут быть
хозяевами как гомоксенных трипаносоматид, так и
переносчиками трипаносом. Гомоксенные жгутико-
носцы представлены в этой группе хозяев единствен-
ным видом Blastocrithidia hyalommae, который опи-
сан из гемолимфы иксодовых клещей. В последнее
время, правда, многие исследователи все чаще схо-
дятся во мнении, что за самостоятельный вид в дан-
ном случае были ошибочно приняты стадии Trypa-
nosoma theileri, развивающиеся в переносчике. Раз-
витие жгутиконосцев в гемолимфе и предполагаемая
трансовариальная передача Blastocrithidia hyalommae
между клещами в целом нетипичны для гомоксен-
ных трипаносоматид. Поэтому при отсутствии досто-
верных экспериментальных исследований обсуждать
особенности развития В. hyalommae в клещах не
имеет смысла. В качестве переносчиков клещей ис-
пользуют некоторые трипаносомы, паразитирующие
в летучих мышах, птицах и млекопитающих. Пара-
зито-хозяинные отношения в этих системах пока не
изучены. Однако тот факт, что в жизненных циклах
трипаносом наряду с клещами в качестве основных
переносчиков почти всегда фигурируют насекомые,
с большой вероятностью свидетельствует о том, что
освоение трипаносомами клещей носило вторичный
характер.
Таким образом, пиявки и насекомые являются ос-
новными группами беспозвоночных животных, с ко-
торыми связано развитие трипаносоматид. В пиявках
паразитируют только гетероксенные трипаносомы из
водных позвоночных животных: рыб, амфибий и реп-
тилий. Независимо от вида позвоночного хозяина в
переносчиках-пиявках эти трипаносомы претерпева-
ют похожее развитие. Жгутиконосцы концентриру-
ются в желудке пиявок, где трипаносомы интенсивно
делятся, часто со сменой морфологических форм.
Проциклические стадии многих видов в начале эндо-
генной агломерации проходят через фазу множест-
венных делений с последующей дифференцировкой,
сопровождающейся бинарными делениями. В зависи-
мости от вида трипаносом эти деления могут проте-
кать у про- и эпимастигот или (редко) у трипомасти-
гот. В желудке и других частях средней кишки пия-
вок трипаносомы могут прикрепляться к щеточной
каемке кишечного эпителия, заякориваясь расширен-
ным кончиком своего жгутика среди микроворсинок
энтероцитов. Ряд трипаносом (вне связи с их приуро-
ченностью к какой-либо определенной группе позво-
ночных-хозяев) формируют метациклические стадии
непосредственно в желудке пиявок. Другие мигриру-
ют в передние отделы кишечника, и их метацикли-
ческие стадии локализуются во влагалище хоботка.
Здесь они могут прикрепляться к его кутикулярной
выстилке за счет внедрения в нее кончика жгутика, в
котором формируются полудесмосомы. Такой тип
контактов отмечен только для жгутиков трипаносом,
развивающихся в пиявках. Роль метациклических
стадий у трипаносом, развитие которых связано с пи-
явками, могут выполнять либо эпи-, либо трипомас-
тиготы. Метациклические стадии трипаносом форми-
руют в пиявках плеоморфные популяции. Этот фено-
мен, проявляющийся в наличии узких и широких
форм жгутиконосцев, пока не имеет убедительного
объяснения. Высказывается точка зрения, что в пере-
носчике таким образом закладывается плеоморфизм
будущих кровяных стадий. Однако экспериментально
эта идея пока не подтверждена. Единственным при-
мером формирования метациклических стадий в
слюнных железах пиявок до сих пор остается Trypa-
nosoma barbari, паразитирующая в тритонах Triturus
torosus. В пиявке Actinobdella sp. метациклические
стадии этих жгутиконосцев найдены в слюнных же-
лезах и во влагалище хоботка хозяина. Однако тон-
кие механизмы развития этих трипаносом в слюнных
железах неизвестны.
Подавляющее большинство трипаносоматид ис-
пользуют в качестве беспозвоночных-хозяев насеко-
мых. В настоящее время известно более 200 видов
гомоксенных и более 300 видов гетероксенных три-
паносоматид, которые в своем развитии связаны с
насекомыми. Среди насекомых хозяева трипаносо-
матид распределены по 10 отрядам. Однако возмож-
ность паразитирования трипаносоматид в Anoplura,
Homoptera, Orthoptera и Trichoptera требует подтвер-
ждения из-за единичности находок и неполноты
описаний. В насекомых из отрядов Hymenoptera, Le-
pidoptera, Mecoptera и Siphonaptera найдено около
40 видов трипаносоматид, что составляет менее
10% от общего числа паразитов, описанных в насе-
комых. Все эти жгутиконосцы изучены очень плохо,
кроме паразитов блох. Более 90 % трипаносоматид,
развитие которых протекает в насекомых, приуроче-
но к 2 отрядам Insecta — Diptera и Hemiptera, между
которыми эти жгутиконосцы распределяются при-
мерно поровну. Несмотря на экологические, пове-
денческие, анатомические и физиологические раз-
личия, существующие между насекомыми-хозяевами,
развитие в них трипаносоматид протекает очень по-
хоже. Исключением являются случаи, связанные с
механическим переносом трипаносом между позво-
ночными животными двукрылыми кровососущими
насекомыми. При этом развития жгутиконосцев в
переносчике не происходит, они лишь переживают
короткое время в капле крови< сохраняющейся в
хоботке насекомого (Ноаге, 1972). Во всех осталь-
ных случаях трипаносоматиды, попав в пищевари-
тельную систему насекомых, претерпевают опреде-
ленное развитие, связанное с закономерной сменой
стадий, которая проходит на фоне нескольких цик-
лов эндогенной агломерации, часто пространственно
разобщенных, и завершают свое развитие в насеко-
мых, формируя неразмножающиеся расселительные
(метациклические) стадии. В наиболее общем случае
все развитие трипаносоматид в насекомых ограниче-
но рамками их пищеварительной системы.
Целый ряд общих черт в строении пищеваритель-
ной системы у насекомых имеет первостепенное зна-
чение в освоении этой стации трипаносоматидами. К
таким чертам следует отнести наличие кутикулярных
выстилок в передних и задних отделах кишечника
насекомых, имеющих эктодермальную природу, на-
личие развитой щеточной каемки у энтероцитов эн-
тодермальной средней кишки и присутствие в этом
отделе перитрофической оболочки или ее аналогов.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
233
Заражение насекомых происходит в процессе их пи-
тания. Расселительные стадии трипаносоматид, за-
хваченные вместе с пищей, транзитом проходят кути-
кулизированные отделы передней кишки и попадают
в среднюю кишку. В двукрылых насекомых пищевой
комок, содержащий проциклические стадии трипано-
соматид, сразу изолируется здесь от стенки кишечни-
ка перитрофической оболочкой, имеющей форму
мешка или трубки. Эта оболочка секретируется на
границе передней и средней кишки специальными
клетками. Она представляет собой динамичное обра-
зование, в котором постоянно изменяются вязкость
самого секрета, длина «мешка», толщина его стенок
и т. д. Ультраструктура перитрофической оболочки
двукрылых насекомых обычно характеризуется нали-
чием 2 слоев: гомогенного основного вещества и слоя
по-разному ориентированных микрофибрилл. В дан-
ном отряде Insecta динамика формирования перитро-
фической оболочки и ее барьерная функция лучше
всего изучены у мух це-це и москитов. Развитие аф-
риканских трипаносом группы Trypanosoma brucei
связано с инвазией эктоперитрофического простран-
ства мух це-це и, следовательно, с преодолением пе-
ритрофической оболочки. Это может достигаться 2
способами: или непосредственной пенетрацией обо-
лочки, или через открытый дистальный конец этой
структуры, имеющей у глоссин форму трубки. Вто-
рой путь реализуется у Т. gray и Т. congolense. Одна-
ко для Г. brucei низкие значения pH среды в задних
отделах кишечника мух губительны. Проникновение
этих трипаносом в эктоперитрофическое пространст-
во происходит непосредственно через стенку перит-
рофической трубки. Причем единственным местом,
где это может происходить, является передний слабо
полимеризованный ее участок, с которым трипаносо-
мы контактируют очень короткий промежуток време-
ни. Именно этим объясняется очень низкая заражае-
мость мух це-це данным видом трипаносом. Дело в
том, что только у молодых, еще не питавшихся мух
этот участок достаточно обширный. У взрослых мух
перитрофическая оболочка практически непреодоли-
ма для трипаносом.
При заражении москитов лейшманиями наблюда-
ется похожее явление. На границе передней и сред-
ней кишки этих насекомых пищевой комок быстро
обволакивается вязким секретом. Эта перитрофичес-
кая оболочка непреодолима для лейшманий, и они
оказываются изолированными от просвета кишечни-
ка. Однако у различных видов москитов перитрофи-
ческая оболочка в процессе пищеварения ведет се-
бя неодинаково. У некоторых видов перитрофичес-
кий мешочек сохраняет свою целостность вплоть до
момента дефекации насекомых. Хотя лейшманий в
этом случае и могут претерпевать определенное раз-
витие в перевариваемом пищевом комке, тем не ме-
нее они лишены возможности завершить его долж-
ным образом и полностью выводятся из организма
хозяина. У других видов москитов еще до попадания
пищевого комка в задние отделы кишечника пери-
трофическая оболочка фрагментируется и лейшма-
нии оказываются в просвете кишки.
У клопов классическая перитрофическая оболоч-
ка отсутствует. Однако и в их кишечнике пищевые
массы изолируются от плазмалеммы микроворсинок
энтероцитов средней кишки. Прежде всего, для мор-
фологии этого отдела пищеварительной системы по-
лужесткокрылых характерно наличие дополнитель-
ной, внеклеточной мембраны, покрывающей плаз-
малемму апикальных концов энтероцитов, включая и
микроворсинки. Снаружи от дополнительной мемб-
раны в кишке клопов располагаются внеклеточные
мембранные образования, часто формирующие мощ-
ные упорядоченные слои. Такие экстраклеточные,
мембранные структуры топографически и, очевидно,
функционально соответствуют настоящей перитро-
фической оболочке насекомых. Трипаносоматиды
различным образом реагируют на присутствие экст-
раклеточных мембранных образований в кишке кло-
пов. Жгутиконосцы Blastocrithidia triatomae, напри-
мер, способны вызывать редукцию этих образований,
после чего атакуют незащищенные энтероциты хозя-
ев — триатомовых клопов (Schaub, 1992). Однако в
тех же клопах жгутиконосцы Trypanosoma cruzi не
могут преодолеть барьер из экстраклеточных мемб-
ран и проходят среднюю кишку хозяев транзитом.
В других группах насекомых влияние перитрофи-
ческих оболочек на ход развития трипаносоматид
пока не изучено.
Дня средней кишки насекомых характерна актив-
ная перистальтика, способствующая продвижению
пищевых масс, которые в этом отделе подвергаются
основной ферментативной обработке. Тот факт, что
в средней кишке насекомых большинство видов три-
паносоматид интенсивно размножаются, свидетель-
ствует о том, что здесь создаются оптимальные усло-
вия для их развития. Неудивительно поэтому, что
большинство трипаносоматид стремятся закрепиться
в данном отделе кишечника насекомых. Известно
(Догель, 1947), что такого рода проблемы у многих
кишечных паразитов решаются путем формирования
особых прикрепительных органов (органелл). В этом
смысле трипаносоматиды не являются исключением.
Однако природа и строение их прикрепительных
органелл весьма оригинальны. На определенном
этапе жизненного цикла трипаносоматид эту функ-
цию берет на себя их единственный жгутик, являю-
щийся специализированной органеллой движения.
Основной способ закрепления жгутиконосцев на по-
верхности кишечного эпителия средней кишки насе-
комых основан на заякоривании их в той или иной
степени видоизмененных жгутиков в зоне щеточной
каемки. Другим способом, позволяющим жгутико-
носцам задерживаться в средней кишке хозяев, явля-
ется их проникновение в толщу кишечного эпите-
лия. В ряде случаев проникновение в стенку кишки
связано с миграцией паразитов в гемолимфу и далее
к слюнным железам хозяев. Примеры такого рода
демонстрируют Т. rangeli, Т brucei и различные Phy-
tomonas. Однако у многих гомоксенных трипаносо-
матид проникновение в толщу кишечного эпителия
может служить для удержания паразитов в данном
отделе кишечника.
В задней части средней кишки определяются ос-
новные пути дальнейшего развития жгутиконосцев.
Одни виды приступают здесь к завершающей фазе
своего развития в насекомых, формированию рассе-
234
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
лительных стадий. Это происходит у трипаносом из
секции Stercoraria, ряда лейшманий и большинства
гомоксенных трипаносоматид (Wallace, 1966). Для
Trypanosoma cruzi и лейшманий показано, что запуск
процессов клеточной дифференцировки их метацик-
лических стадии индуцируется циклическим аде-
нозинмонофосфатом; В насекомых-переносчиках это
вещество присутствует в секрете мальпигиевых сосу-
дов, протоки которых открываются на границе сред-
ней и задней кишки.
Некоторые трипаносоматиды все свое развитие
осуществляют в задней кишке насекомых. Таковы, в
частности, представители рода Crithidia. Для закреп-
ления в этом отделе кишечника жгутиконосцы ис-
пользуют видоизмененные «присосковидные» жгути-
ки, прикрепляясь ими к кутикулярной выстилке эпи-
телия.
Трипаносоматиды, формирующие расселительные
стадии в передних отделах кишечника, мигрируют
из средней кишки к ротовому аппарату своих хозя-
ев. Здесь они локализуются на кутикулярной вы-
стилке эпителия, причем способ прикрепления иден-
тичен способу, который трипаносоматиды использу-
ют в ректуме хозяев.
Жгутиконосцы, формирующие расселительные
стадии в слюнных железах хозяев, проникают в эти
органы из гемолимфы. В процессе продвижения к
слюнным железам жгутиконосцам приходится актив-
но проникать в клетки и ткани различных органов
насекомых. Механизм проникновения до конца не-
ясен и, вероятно, индивидуален у отдельных жгутико-
носцев. Т. brucei проникает в энтероциты мух це-це
следующим образом. Сначала жгутиконосцы инду-
цируют формирование инвагинационного кармана на
апикальной поверхности энтероцита и вводят в него
свое тело. Затем они покидают этот карман и оказы-
ваются непосредственно в цитоплазме, откуда прони-
кают под базальную мембрану кишечника и выходят
в гемолимфу. Жгутиконосцы Т.rangeli и Phytomonas
serpens проникают сквозь стенку слюнных желез
клопов-хозяев одинаковым способом. Базальная
мембрана просто протыкается жгутиком, а у плазма-
леммы клеток слюнных желез жгутик индуцирует
формирование инвагинационного кармана, который
замыкается в паразитофорную вакуоль. Дифференци-
ровка расселительных стадий перечисленных видов
происходит в просвете слюнных желез хозяев. У Тгу-
panosoma brucei и Phytomonas nordicus этому пред-
шествует закрепление жгутиконосцев на щеточной
каемке эпителиальных клеток желез, которое проис-
ходит посредством заякоривания жгутика среди мик-
роворсинок. Процесс проникновения Trypanosoma
brucei в слюнные железы мух це-це пока неясен. Из-
вестно, что трипаносомы внедряются в стенку сред-
ней кишки, где размножаются путем множественных
делений, формируя так называемые цистоподобные
тела. После этого жгутиконосцы выходят в гемолим-
фу мух. Однако внутри клеток слюнных желез трипа-
носомы не обнаружены. Это заставляет предполо-
жить, что выход жгутиконосцев в гемоцель — яв-
ление в значительной степени спонтанное, а их
проникновение в слюнные железы осуществляется в
ходе миграции трипаносом через гипофаринкс и про-
токи желез. Возможно, что у разных штаммов Т. bru-
cei могут реализовываться оба пути. Жгутиконосцы
Т. rangeli, Phytomonas serpensи Ph. nordicus прони-
кают в слюнные железы из гемолимфы.
Воздействие трипаносоматид на насекомых-хозя-
ев изучено пока явно недостаточно. Основные факты,
имеющиеся на этот счет, суммированы в ряде обзо-
ров (Molyneux, 1977; Schaub, 1992). С нашей точки
зрения, можно выделить 3 основных типа воздейст-
вий трипаносоматид на насекомых.
Первый тип — это влияние жгутиконосцев на по-
ведение их хозяев. Так, развитие Trypanosoma rangeli
в триатомовых клопах и Т. congolense в мухах це-це
приводит к проявлению у их хозяев синдрома преры-
вистого питания. Прерывистое питание характерно и
для москитов, зараженных лейшманиями. Данный
эффект обычно связывают с двумя возможными при-
чинами. Обнаружено, что трипаносомы Т. congolense
в гипофаринксе мух це-це наряду с кутикулярной вы-
стилкой этого отдела пищеварительной системы при-
крепляются и к поверхности механорецепторов. Воз-
действие на механорецепторы, естественно, может
сказываться на поведении хозяев. Однако, возможно,
по крайней мере в случае с лейшманиями и москита-
ми, что прерывистое питание связано с закупоркой
паразитами канала пищеварительной системы. В дан-
ном случае атакующие позвоночных животных мос-
киты проходят все этапы акта кровососания, однако
в их пищеварительную систему кровь не поступает,
или сюда попадает лишь ничтожное ее количество. У
таких насекомых возникает необходимость в повтор-
ном питании и т. д.
Второй тип — это патогенное воздействие трипа-
носоматид на насекомых. Различные механизмы та-
кого воздействия паразитов на их хозяев хорошо из-
вестны из курса общей паразитологии (Догель, 1947).
Однако среди трипаносоматид факты существования
прямого патогенного воздействия на хозяина отмеча-
ются крайне редко (Schaub, 1992). В частности, из-
вестно, что Blastocrithidia triatomae вызывает высо-
кую смертность среди личинок Triatoma infestans,
связанную с нарушением их нормального пищеваре-
ния. Показано, что отчасти это связано с разрушени-
ем дистального отдела мальпигиевых сосудов и нару-
шением их секреторной деятельности.
Наконец, к третьему типу можно отнести опосре-
дованное патогенное воздействие трипаносоматид на
их хозяев. Например, активное проникновение жгу-
тиконосцев через стенку кишки в гемоцель открыва-
ет в ряде случаев доступ к гемолимфе хозяев предста-
вителям бактериальной кишечной флоры. Инвазия
гемолимфы бактериями обычно приводит к гибели
насекомых.
Способы передачи инвазии
Способы заражения животных и растений трипа-
носоматидами сводятся в основном к двум формам.
Заражение хозяев контаминативным путем свойст-
венно всем гомоксенным трипаносоматидам. Этим
же путем происходит заражение позвоночных живот-
ных трипаносомами, относящимися к секции Stereo-
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
235
raria. У гомоксенных трипаносоматид заражение но-
вых особей хозяев обычно происходит при их пита-
нии на контаминированном субстрате. У ряда видов в
жизненном цикле имеются цистоподобные стадии,
способные переживать во внешней среде не менее
года. Контаминативное заражение позвоночных жи-
вотных трипаносомами основано на активном про-
никновении их метациклических стадий через по-
врежденный эпидермис или слизистые хозяина в его
кровяное русло. Это может происходить при фекаль-
ной контаминации переносчиком покровов или сли-
зистых животного-реципиента, при поедании позво-
ночными животными переносчиков и при попадании
в пищеварительную систему или на слизистые других
органов позвоночного животного крови зараженных
особей.
Первый способ реализуется у Trypanosoma cruzi.
Эффективность трансмиссии паразитов обеспечива-
ется спецификой поведения триатомовых клопов в
процессе питания (Ноаге, 1972). Дело в том, что у
триатомин каждый акт кровососания сопровождается
или завершается дефекацией насекомых. Выходящие
с экскрементами клопа метациклические трипаносо-
мы оказываются, таким образом, вблизи ранки, в ко-
торую могут попадать в дальнейшем, например при
расчесывании места укуса. Таким же образом трипа-
носомы могут попадать на слизистые рта, носа и т. д.
Этому способствует еще и тот факт, что триатомовые
$лопы обычно питаются на лицевой поверхности го-
ловы позвоночных животных. Существует такж:е вер-
сия, что метациклические трипаносомы Т. cruzi
могут проникать в организм хозяина и через непо-
врежденный эпителий, но пока она не доказана.
Поедание переносчиков лежит в основе трансмис-
сии жгутиконосцев из подрода Herpetosoma и, веро-
ятно, трипаносом и лейшманий, паразитирующих в
рептилиях. Ранее считалось, что заражение трипано-
соматидами в данном случае идет через слизистые
ротовой полости хозяев. Однако пероральное зараже-
ние крыс культурой трипаносом, помеченных изото-
пом трития, показало, что основной мишенью для
метациклических стадий Trypanosoma lew is i служат
ткани желудка и кишечника, а не слизистые ротовой
полости.
Третий способ может реализовываться двумя раз-
личными путями: при половых контактах животных
и при циркуляции паразитов в системе хищник-
жертва. Венерическая трансмиссия паразитов досто-
верно установлена только у Т. equiperdum. Заражение
через кровь жертв в процессе питания, вероятно, рас-
пространено довольно широко, но изучено в отноше-
нии трипаносоматид пока очень плохо. Классичес-
кий пример, имеющийся во всех учебниках, — это
заражение летучих мышей-кровососов Desmodes го-
tundus жгутиконосцами Trypanosoma evansi в процес-
се питания вампиров кровью лошадей. Хищничество
и каннибализм также могут способствовать циркуля-
ции паразитов в популяциях хозяев. Примеры такого
рода известны для ряда трипаносом, однако стабиль-
ность такого типа передачи пока не доказана.
Инокулятивное заражение хозяев основано на ак-
кумуляции паразитов в районе ротового аппарата
переносчиков или в органах, непосредственно с ним
связанных (слюнные железы, зоб Diptera). Этим спо-
собом осуществляется заражение низших позвоноч-
ных животных трипаносомами через переносчиков-
пиявок. Таким же способом попадают в позвоноч-
ных животных трипаносомы, относящиеся к секции
Salivaria, Leishmdriia, Trypanosoma rangeli и жгути-
коносцы из рода Phytomonas, паразитирующие в тка-
нях растений. Перенос паразитов может осуществ-
ляться механически, или же формированию инфек-
тивных стадий предшествует определенное развитие
жгутиконосцев в переносчиках. Возникновение
механической .передачи паразитов, вероятно, сопря-
жено с выходом трипаносом за границы ареала ис-
ходного переносчика и вторичным включением в их
жизненный цикл неспецифических переносчиков.
Пример такого рода демонстрирует Trypanosoma
evansi. Выход этого вида на север Африки за грани-
цу обитания мух це-це и последующее распространение
трипаносом на широких пространствах Евроазиат-
ского и Американского континентов происходили,
вероятно, благодаря включению в их жизненный
цикл в качестве переносчиков слепней (Tabanidae).
Среда кишечника этих двукрылых губительна для
трипаносом, однако в ротовом аппарате насекомых
жгутиконосцы короткий период времени сохраня-
ют жизнеспособность. Прерывистый характер пи-
тания слепней, сопровождающийся частой сменой
жертв, оказался достаточным условием для устойчи-
вой циркуляции паразитов в популяции хозяев
(Ноаге, 1972).
За рамками рассмотренных примеров остался еще
1 способ передачи паразитов между их хозяевами.
Речь идет о трансовариальной трансмиссии трипано-
соматид у беспозвоночных животных и трансплацен-
тарной инвазии плода у позвоночных. До недавнего
времени постулировалось наличие трансовариальной
передачи в жизненных циклах 4 видов гомоксенных
трипаносоматид: Leptomonas pyrrhocorisв клопах
Pyrrhocoris apterus, L. rhinoestri в Rhinoestrus nivar-
letti (Oestridae), L. jaculum в клопах Nepa cinerea и
Blastocrithidia hyalommae в клещах из рода Hyalom-
ma. Однако сейчас уже очевидно, что визуальное об-
наружение жгутиконосцев в органах репродуктивной
системы насекомых-хозяев не является достаточным
основанием для установления факта трансовариаль-
ной передачи паразитов. Экспериментально транс-
овариальная передача пока ни для одного из перечис-
ленных видов не подтверждена.
Трансплацентарная инвазия плода позвоночных
животных описана у Trypanosoma brucei, Т. cruzi и
Leishmania sp. (висцеральная форма). Распространен-
ность этого явления среди диких животных пока
практически не изучена. Известно, что Trypanosoma
theileri осуществляет нормальное развитие в культуре
клеток плода коровы. Т. musculi в период репро-
дуктивной фазы цикла концентрируется на плаценте
в материнском организме, однако потомству мышей
не передается. Вероятно, следует заключить, что ни
трансовариальная передача трипаносоматид у беспоз-
воночных животных, ни трансплацентарная передача
у позвоночных животных не получили широкого рас-
пространения в данной группе паразитов. В тех слу-
чаях, когда такого рода передача все же имеет место,
236
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
она принадлежит к разряду сопутствующих явлений и
всегда подкрепляется одним из перечисленных выше
способов трансмиссии паразитов.
Филогения
В обширной и очень неоднородной по составу
группе низших эукариот, которых объединяют под
общим названием жгутиконосцы, кинетопластиды
занимают особое положение. Характерный план
строения их клетки, центральным элементом кото-
рого является кинетопласт, определяет четкие грани-
цы этого таксона и не оставляет сомнений в мрно-
филетическом происхождении его представителей.
Среди других жгутиконосцев наиболее близки к
кинетопластидам эвгленовые. В пользу этого свиде-
тельствует заметное сходство в организации цито-
скелета и двигательного аппарата у представителей
обеих групп и наличие в их жгутиках похожих па-
раксиальных структур, формирующихся к тому же
за счет гомологичных белков (Kivic, Walne, 1984).
Факт филогенетического родства кинетопластид и
эвгленовых жгутиконосцев подтверждается и моле-
кулярно-биологическими исследованиями по сравни-
тельному сиквенсу генов их рРНК. Считается, что
обе группы имеют общее и очень древнее происхож-
дение, а их обособление и последующая дивергенция
произошли еще до возникновения большинства ныне
существующих таксонов эукариот (Cavalier-Smith, 1993).
Гораздо меньше ясности в вопросе о филогенети-
ческих отношениях между отдельными группами
самих кинетопластид. Надо заметить, что со времени
сведения в один таксон бодонид, криптобий и трипа-
носоматид (Honigberg, 1963) и до начала 90-х гг. этот
вопрос не поднимался. А до работы Хонигберга он
просто не возникал, поскольку все 3 группы обычно
помещались в системе либо независимо друг от друга,
либо в ранге родов или семейств среди многих других
«зоомастигофор». Так что единственным вопросом,
который привлекал к себе внимание исследователей
все это время, был вопрос о происхождении и эволю-
ции трипаносоматид.
Интересно, что уже в начале XX столетия были
предложены, а затем отвергнуты практически все
возможные варианты гипотез происхождения Тгу-
panosomatidae и эволюции их отдельных групп. В
1904 г. в работе, посвященной изучению морфоло-
гии трипаноплазмы (=Cryptobia), французский зоо-
лог Леже (Leger, 1904а) высказал предположение,
что эти двухжгутиковые простейшие являются пред-
ками трипаносом. Такое предположение целиком
базировалось на обнаружении у трипаноплазмы «ун-
дулирующей мембраны». Однако в том же 1904 г. в
другой своей работе Леже (Leger, 1904b) частично
отказывается от этой гипотезы. Он считает, что, ве-
роятно, только небольшая часть видов трипаносом
связана в своем происхождении с трипаноплазмами,
тогда как большинство представителей семейства
{Trypanosoma, Leishmania) произошли от жгутико-
носцев из кишечника насекомых — таких как Lep-
tomonas и Crithidia (^Blastocrithidia). Идея дифи-
летического происхождения трипаносоматид неожи-
данно нашла признание у ряда исследователей того
времени. Альтернативная гипотеза была предложена
в 1908 г. немецким зоологом Минхиным (Minchin,
1908). Годом раньше этот исследователь описал на-
личие цист в жизненном цикле Trypanosoma gray из
крокодилов (как стало известно позднее — ошибоч-
но). Основываясь на этом факте, а также на том,
что, по его мнению, трипаносоматиды в целом более
специфичны по отношению к позвоночным живот-
ным, чем к беспозвоночным, Минхин заключает, что
именно позвоночные животные были первичными
хозяевами трипаносоматид. По его мнению, предки
трипаносоматид могли обитать в кишечнике древних
Vertebrata, откуда периодически попадали в крове-
носную систему хозяев. Когда они уже вполне адап-
тировались к кровепаразитизму, в циклы их разви-
тия смогли включиться в качестве переносчиков
кровососущие беспозвоночные. Интересно, что хотя
эта гипотеза не раз воскрешалась позднее многими
исследователями, самим автором она была категори-
чески отвергнута 6 лет спустя. Причем произошло
это под влиянием новой гипотезы Леже (Leger, Du-
bosque, 1910). К этому времени Леже окончательно
разочаровался в идее «трипаноплазменной» природы
трипаносоматид и утвердился в мнении, что предка-
ми кровяных жгутиконосцев могли быть только мо-
ногенетические паразиты кишечного тракта насеко-
мых. Именно эта гипотеза, причем с минимальными
изменениями, и составляла до недавнего времени ост
нову представлений о происхождении и эволюции
трипаносоматид. Гипотеза Леже, однако, не могла
ответить на вопрос о происхождении трипаносо-
матид, паразитирующих у насекомых, или, как их
стали называть, «низших» трипаносоматид. Эта роль
явно не подходила бодонидам и криптобиям, по-
скольку ни те ни другие не освоили в качестве
хозяев насекомых. Долгое время этот вопрос, правда,
не стоял остро, так как в литературе имелось описа-
ние «свободноживущей трипаносоматиды» Prolepto-
monas faecicola, найденной в почве, контаминиро-
ванной фекалиями овец (Woodcock, 1914). Однако
позднее выяснилось, что описанный Вудкокком жгу-
тиконосец не относится к кинетопластидам (Le Ray,
Vickerman, 1976). Именно этим обусловлено введе-
ние в более поздние схемы гипотетического предка
трипаносоматид (рис. 144). Предполагалось, что это
были свободноживущие лептомонасподобные орга-
низмы. Сначала они освоили в качестве хозяев бес-
позвоночных животных. Гоар (Ноаге, 1972) считает,
что это были специфичные паразиты насекомых.
Бакер (Baker, 1974) предполагает, что кроме насеко-
мых, а вероятнее, и до них, такие жгутиконосцы
освоили кольчатых червей, причем ту ветвь их эво-
люции, которая позднее дала начало насекомым.
Лептомонасподобные жгутиконосцы в процессе эво-
люции дивергировали, дав начало 2 основным лини-
ям трипаносоматид — промастиготной и эпимасти-
готной. Первая объединила гомоксенных Leptomonas,
Crithidia, Herpetomonas и дала начало гетероксенным
Leishmania и Phytomonas. Эпимастиготная линия че-
рез гомоксенных Blastocrithidia и, возможно, Rhyn-
choidomonas (эти жгутиконосцы пока не изучены)
привела к обособлению различных Trypanosoma, Од-
ТИП EUGLENOZOA. КЛАСС KIN ETOPLASTIDEA
237
Trypanosoma
Рис. 144. Гипотетическое филогенетическое древо рода Trypanosoma. (По: Backer, 1974, с изменениями).
Fig. 144. Phylogenetical tree of the genus Trypanosoma.
нако эта гипотеза плохо объясняет возникновение
паразитарных систем, включающих водных позво-
ночных, главным образом рыб, пиявок и трипаносо-
матид. В построениях Гоара этот вопрос не рассмат-
ривается. Бакер и здесь использует гипотетических
предков (рис. 144). По его мнению, существовала рано
отошедшая от общего ствола эволюционная ветвь
гомоксенных трипаносоматид — паразитов аннелид.
Эти жгутиконосцы при обособлении пиявок перешли
в них, а позднее получили доступ к водным позвоноч-
ным, которых и освоили в качестве новых хозяев.
Известно, что гомоксенные трипаносоматиды раз-
виваются в кишечнике насекомых и формируют свои
расселительные стадии в его. задних отделах. Форми-
рование расселительных стадий в «задней позиции»
рассматривается как исходное и для трипаносом.
Все трипаносомы млекопитающих, обладающие
таким развитием, объединяются в секцию Stercoraria.
От этой группы, по мнению Гоара, в процессе совер-
шенствования паразито-хозяинных отношений про-
изошли африканские трипаносомы, объединяемые им
в секцию Salivaria, которые являются наиболее про-
двинутыми в эволюционном плане представителями
семейства.
Факт освоения мух це-це двумя столь непохожими
трипаносомами, какими являются Trypanosoma gray
238
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Свободноживущие бодониды
Рис. 145. Гипотетическое филогенетическое древо кинетоплас-
тид, построенное по данным сравнительно-морфологического ис-
следования группы. (По: Фролов, 1993, с изменениями).
Fig. 145. Phylogenetical tree of the Kinetoplastida reconstructed on
the base of comparative morphological data.
и T. brucei, стал исходной точкой в альтернатив-
ной гипотезе происхождения африканских трипано-
сом, предложенной By (Woo, 1970)? Этот исследователь
в серии экспериментов доказал, что Т, brucei может
длительное время сохранять жизнеспособность в кро-
ви рептилий и в дальнейшем вновь заражать млеко-
питающих. Он предположил, что предками африкан-
ских трипаносом млекопитающих могли быть не ги-
потетические Stercoraria, а жгутиконосцы из крови
тропических рептилий, подобные Trypanosoma gray.
По его мнению, в паразитарную систему Reptilia—
Trypanosoma—Hirudinea сначала в качестве неоснов-
ного переносчика могли включиться Glossina. Расши-
ренйе трофических связей этих «кровососов позднее
привело к переносу паразитов рептилий на млекопи-
тающих, к обитанию в которых они в конце концов
приспособились. Недавнее происхождение этих пара-
зитарных систем объясняет высокую патогенность
трипаносом для целого ряда млекопитающих.
Существовали, однако, две точки зрения, отража-
ющие иное-понимание возможной эволюции трипа-
носоматид. Обе они не аргументированы и, строго
говоря, не являются даже гипотезами, но тем не
менее представляют большой интерес. Грассе, ана-
лизируя взгляды Леже, высказал предположение, что
эволюция трипаносоматид могла идти по пути по-
следовательного освоения жгутиконосцами позвоноч-
ных животных в строгом соответствии с возрастани-
ем их эволюционного ранга: от рыб к млекопитаю-
щим. Формирование наземных паразитарных систем
при этом сопровождалось бы сменой переносчиков:
пиявок заменяли кровососущие насекомые (Grasse,
1952). В то же время Уоллес не исключал, что пред-
ки трипаносоматид могли быть кровепаразитами. С
одной стороны, переход паразитов из крови отно-
сительно крупных позвоночных животных в более
мелких переносчиков легче поддается объяснению,
когда речь идет о неустоявшихся паразитарных сис-
темах. .С другой стороны, и лейшмании, и многие
трипаносомы хорошо растут в лабораторных культу-
рах и способны выживать и даже размножаться в
кишечнике и гемолимфе неспецифичных насекомых.
Напротив, гомоксенные трипаносоматиды в крови
позвоночных животных очень быстро гибнут (Walla-
ce, 1966).
К началу 90-х гг. был накоплен уже значительный
по объему материал, касающийся ультратонкой ор-
ганизации различных кинетопластид. Это позволило
по-новому подойти к решению вопроса о филогене-
тических отношениях между бодонидами, криптоби-
ями и трипаносоматидами.
В группах протистов, лишенных палеонтологи-
ческой летописи, филогенетические построения и
определение эволюционного ранга их отдельных
представителей могут основываться на результатах
сравнительно-морфологических исследований ныне
живущих организмов. В основе этого метода лежит
выявление исходного (анцестрального) плана строения
и анализ эволюционных рядов гомологичных призна-
ков.
Поскольку возникновение паразитизма в эволю-
ции эукариот носит вторичный характер, то естест-
венно рассматривать организацию свободноживущих
гетеротрофных бодонид как исходную в эволюции
кинетопластид. Тогда особенности организации крип-
тобий и трипаносоматид следует оценивать с точки
зрения эволюционных преобразований исходного
(бодонидного) плана строения их клеток.
Оказалось (Фролов, 1993), что по всем основным
признакам (организация покровов, двигательного ап-
парата, ротового аппарата и т. п.) кинетопластиды
образуют следующий эволюционный ряд: свободно-
живущие бодониды—криптобии-—трипаносомы (Try-
panosoma)—остальные трипаносоматиды (лейшмании,
эндотрипанум и все гомоксенные паразиты насеко-
мых). Причем криптобии и трипаносомы (особенно
трипаносомы рыб и Т. cruzi) демонстрируют удиви-
тельно высокий уровень гомологий.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
239
г----Crithidia
Crithidia
Leptomonas
fasciculata
----Leishmania
tarentolae
Herpetomonas
muscarum
Phytomonas
Blasto-
crithidia
Blastocrithidia
culicis
---- Trypanosoma cruzi
Trypanosoma sp.
(из рыб)
У Herpetomonas
Euglena
Trypanosoma brucei
> Trypanoplasma borreli
Euglena gracilis
Bodo
Tiypanoplasma
? Trypanosoma
brucei
Trypanosoma
cruzi
Рис. 146. Филогенетическое древо Euglenozoa, построенное по результатам сиквенса генов рРНК, с использованием в качестве внешней
группы Euglena gracilis. (По: Vickerman, 1994; Маслов и др., 1994).
Fig. 146. Phylogenetical tree of the Euglenozoa reconstructed on the base of SSU rRNA.
Эти выводы легли в основу новой гипотезы про-
исхождения и эволюции трипаносоматид (рис. 145).
Согласно этой гипотезе, одна из групп свободножи-
вущих гетеротрофных бодонид дала начало древним
криптобиям. Магистральным направлением в эволю-
ции криптобий стало освоение ими в качестве хозяев
древних позвоночных животных — рыб. Криптобии
демонстрируют все возможные формы таких отно-
шений, вплоть до перехода к облигатному пара-
зитизму, в том числе к наиболее сложной его
форме — кровепаразитизму. Первоначально, вероят-
но, формировались двухзвенные паразитарные систе-
мы. В этих системах осуществлялась прямая переда-
ча паразитов между хозяевами без участия перенос-
чиков. Примеры таких циклов развития мы находим
и у некоторых современных видов криптобий. Жгу-
тиконосцы размножались в крови хозяев, затем вы-
ходили во внешнюю среду, находили новых хозяев и
через покровы их тела вновь проникали в кровяное
русло. Позднее возникали трехзвенные системы, в
которых функцию переносчиков стали выполнять
пиявки. Именно в такой паразитарной системе,
включающей криптобий, рыб и пиявок, стало воз-
можным появление специализированных одножгути-
ковых кровепаразитов — трипаносом. Позднее вмес-
те с переносчиками трипаносомы начали осваивать
новые группы позвоночных животных: сначала ам-
фибий, а затем и рептилий. Выход позвоночных на
сушу знаменовал собой начало формирования назем-
ных паразитарных систем. Этот процесс был неми-
нуемо связан со сменой переносчиков. Ими стали
кровососущие насекомые. Жизненные циклы неко-
торых трипаносом из современных амфибий и реп-
тилий, использующих в качестве переносчиков и пия-
вок, и насекомых, хорошо моделируют возможность
такого перехода. Адаптация к пищеварительной сис-
теме насекомых открыла трипаносомам путь к заня-
тию новой экологической ниши. Дальнейшая их эво-
люция шла, таким образом, в двух направлениях
(рис. 145). С одной стороны, они осваивали (вместе
с переносчиками) новые группы позвоночных жи-
вотных (так, вероятно, трипаносомы тропических
рептилий дали начало ветви африканских трипано-
сом млекопитающих, а лёйшмании рептилий при-
240
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
способились к паразитированию в млекопитающих),
а с другой стороны — насекомых. Возникновение
гомоксенных трипаносоматид могло быть связано с
группами хищных насекомых, в частности клопов,
жертвами которых становились переносчики трипа-
носом. Возможно, что сами переносчики сочетали
оба типа питания и это приводило к периодической
изоляции паразитов от их хозяев — позвоночных
животных. Утрата позвоночного хозяина не всегда
приводит к гибели паразитов. Известно, например,
что Т. cruzi может передаваться от одной особи
клопов редувиид к другой при каннибализме. То что
эволюция гомоксенных трипаносоматид из насеко-
мых теснейшим образом связана с насекомыми —
переносчиками трипаносом, подтверждается и тем
фактом, что 90 % видов «низших» трипаносоматид
описано из двукрылых, клопов и блох, т. е. из тех
отрядов насекомых, в которых гематофагия получи-
ла наибольшее развитие и к которым приурочены
основные переносчики гетероксенных трипаносома-
тид.
Нетрудно заметить, что предложенная гипотеза
полностью противоречит прежним взглядам на эво-
люцию кинетопластид (ср. рис. 144 и 145). Филоге-
нетическое древо, построенное на основе гипотезы
Леже, оказывается попросту перевернутым. Новая ги-
потеза нашла подтверждение в недавней серии работ
разных исследователей (Fernandes et al., 1993; Mas-
lov et al., 1995, 1996), использовавших методы моле-
кулярной филогении (рис. 146). Полученные в этих
работах данные однозначно указывают на то, что
гетероксенные трипаносомы появились значительно
раньше, чем гомоксенные паразиты насекомых, ко-
торые вместе с Leishmania и Endotrypanum форми-
руют крону филогенетического древа Kinetoplastida.
Рис. 147. Представители сем. Bodonidae. (По разным источникам).
А — Bodo caudatus, Б — В. spora, В — В. angustatus, Г — Rhynchomo-
nas nasuta, Д — Bodo saltans, E — Ichthyobodo necator.
Fig. 147. The representatives of Bodonidae,
Систематический обзор
Отряд Bodonida Hollande, 1952
Сем. Bodonidae Dophlein, 1901
Кинетопластиды, обладающие 2 гетеродинам-
ными жгутиками; направленный назад рекуррент-
ный жгутик не прикреплен к поверхности тела
(рис. 147).
Род Bodo (Ehrenberg, 1830) Stein, 1875
(= Prowazekia Hartmann et Chagas)
Бодониды с единственным крупным кинетоплас-
том; рострум короткий; цитостом открывается лате-
ровентрально по отношению к жгутиковому карману;
форма тела эллипсоидальная, яйцевидная, тело часто
сплюснуто с боков; ряд видов способны формировать
цисты.
Типовой вид В. saltans (-Pleuromonas jacu-
lans) (рис. 136, A; 147, Д) характеризуется наличием
простых мастигонем на переднем (двигательном) жгу-
тике и способностью прикрепляться к субстрату кон-
чиком заднего жгутика.
Представители рода Bodo широко распростране-
ны во всех типах водоемов с различным уровнем
соленности, а также в почве; обычные представи-
тели копрозойной фауны. Длина тела у различных
видов варьирует от 4 до 20 мкм. По числу видов род
Bodo самый многочисленный в семействе, однако
валидность большинства описаний сомнительна. Наибо-
лее распространены в природе В. saltans (рис. 147,
Д), В. caudatus (рис. 147 А), В. angustatus (рис. 147,
В), В. des ignis, В, curvifilus, В. spora (рис. 147, В).
Род Parabodo Skuja, 1939
В основном характеристика совпадает с таковой
рода Bodo, но отверстие цитостома смещено к перед-
нему концу клетки, который при этом выглядит раз-
двоенным.
Выделение рода Parabodo носит в значительной
степени традиционный характер. Исследование ульт-
раструктуры Р. nitrophilus (Мыльников, 1986) не вы-
явилр/каких-либо отличий а организации этих жгути-
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
241
коносцев от типичных Bodo. 2 вида наиболее обыч-
ны в природе: Р. nitrophilus и Р. attenuatus.
Род Ichthyobodo Pinto 1928
(= Costia Leclercq, 1890)
Бодониды, ретикулярный митохондрион которых
содержит несколько кинетопластов. Эктопаразиты
рыб, прикрепляющиеся к эпителию хозяина видоиз-
мененным передним концом тела (рострумом). Един-
ственный вид L necator (= Costia necatrix) широко
распространен в пресных водоемах (рис. 147, Е). В
рыбоводческих хозяйствах костиаз может быть при-
чиной массовой гибели молоди рыб.
Род Rhynchomonas Klebs, 1893
Мелкие бодониды, обладающие хорошо развитым
цитоплазматическим хоботком, к которому прикреп-
лен короткий передний жгутик; цитостом открывает-
ся у основания хоботка.
Типовой вид Rh. nasuta (рис. 147, Г), обитает
как в пресной, так и в морской воде, копрофил, в
стоячих мезосапробных водоемах иногда обильно
развивается в поверхностной пленке. Оба жгутика
Rh. nasuta несут простые мастигонемы. Близкий
вид — Rh. metabolica — отличается активной мета-
болией тела. Ряд исследователей склонны рассматри-
вать оба названия как синонимы одного вида.
Род Rhynchobodo Lackey, 1940
(Phyllomitus Steiri, 1878; Cryptaulax
sensu Novarino et al., 1994)
Бодониды, цитостом которых открывается терми-
нально на вершине хорошо развитого рострума, ро-
струм содержит многочисленные экструсомы; плаз-
малемма покрыта толстым слоем гликокаликса. Пок-
ровы формируют спиральную бороздку, идущую от
переднего конца тела к заднему. Поли- и эукинето-
пластные виды.
До последнего времени жгутиконосцы Rhynchobo-
do, Cryptaulax и Phyllomitus часто рассматривались
как представители самостоятельных родов. Однако
особенности их организации, надежно выделяющие
этих жгутиконосцев среди остальных бодонид, не
позволяют разграничить их между собой. Обитают
как в пресных, так и в морских водах. Наиболее
достоверны описания 4 видов: Rh. agilis, Rh. armata,
Rh. taeniata, Rh. simius (Patterson, Simpson, 1996).
Род Hemistasia Griessmann, 1913
Свободноплавающие поликинетопластные жгути-
коносцы, с 2 почти равными жгутиками, выходящи-
ми из большого субапикального жгутикового карма-
на. Жгутики часто обвиваются вокруг тела клетки.
Детально описан 1 вид Н. phaeocysticola — актив-
но металлирующие, морские жгутиконосцы (Elbras-
chter et aL, 1996). Клетки с апикальным рострумом,
пронизанным цитофаринксом. Выстреленные экстру-
сомы имеют вид сетчатой трубки. Отличается от
представителей рода Rhynchobodo отсутствием спи-
ральной бороздки тела и выраженного гликокаликса
на поверхности тела.
Сем. Cryptobiidae Poche,
1911
Кинетопластиды, обладающие 2 гетеродинамны-
ми жгутиками, направленный назад рекуррентный
жгутик прикреплен к поверхности тела (рис. 148).
Род Cryptobia Leidy, 1846
(= Trypanoplasma Laveran et Mesnil, 1901)
Одиночные криптобииды с различной организа-
цией кинетопластов (пан-, поли-, эукинетопласты);
комменсалы и паразиты беспозвоночных и позвоноч-
ных животных.
Типовой вид С. helicis (рис. 148, Е); паразити-
рует в семенных мешках ряда видов гастропод (Pul-
monata). Так же в репродуктивных органах морских
пелагических моллюсков Carinaria паразитирует дру-
гой вид — Cryptobia carinariae. С. vaginalis паразити-
рует во влагалище пиявок Haemopis sanguisuga и Ш-
rudo medicinalis, a Cryptobia dendrocoeli — в кишеч-
нике планарии Dendrocoelium lacteum.
Более 50 видов криптобий — паразиты рыб
(Lorn, 1979). Около 7 видов паразитируют в задней
кишке морских костных рыб, среди них наиболее
изучены Cryptobia dahlii (рис. 148, А) и С. intestina-
lis (рис. 148, Ж). Более 40 видов криптобий обитают
в крови морских и пресноводных рыб. Среди наи-
более обычных видов можно отметить С. cyprini
(рис. 148, Г) и С. borreli (рис. 148, Д). Вероятно, все
кровепаразиты являются гетероксенными паразита-
ми, использующими в качестве переносчиков пия-
вок, у некоторых видов, например у С. salmositica,
возможна прямая трансмиссия паразитов через во-
ду. Несколько видов — С. carassii, С. branchialis
(рис. 148, Е, В), С. indica и другие — считаются
эктопаразитами, возможно, однако, что в ряде слу-
чаев в качестве самостоятельных видов среди экто-
комменсалов ошибочно описаны стадии развития
кровепаразитов.
Род Cephalothamnium Stein, 1878
(= Cephalothamnion Lemmerman, 1913)
Криптобииды, обладающие крупным эукинето-
пластом (Hitchen, 1974); формируют сидячие коло-
нии, в которых отдельные особи прикрепляются за-
днимихконцами тела и рекуррентными жгутиками к
секретируемому стебельку. Единственный представи-
16 Протисты, часть 1
242
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 148. Представители сем. Cryptobiidae. (По разным источникам).
А — Cryptobia dahlii, Б, В — С. branchialis, Г — С. ciprini, Д — С. borreli, Е — С. helicis, Ж — С. intestinalis, 3 — Dimastigella trypaniformis.
Fig. 148. The representatives of Cryptobiidae.
тель рода C. cyclopum — эктокомменсал некоторых
пресноводных ракообразных. Колонии этих жгутико-
носцев насчитывают до 30 особей. Питаются бакте-
риями.
Род Procryptobia Vickerman,
1978
Свободноживущие криптобииды, обладающие
крупным эукинетопластом; рекуррентный жгутик
прикрепляется к поверхности лишь в передней
части тела жгутиконосцев. 3 вида свободноживущих
криптобиид отнесены Викерманом (Vickerman, 1978)
к этому роду. Питаются бактериями, заглатывая их
через хорошо развитый цитостом.
Типовой вид Р. vorax. Могут образовывать
цисты.
Род Dimastigella (Sandon, 1928) Vickerman,
1978
Свободноживущие криптобииды с поликинето-
пластной организацией митохондриона; на перед-
нем конце тела расположен хорошо развитый рост-
рум.
Род включает 2 вида: D. trypaniformis (рис. 148, 3)
и D. mimosa. D. trypaniformis обитает в богатых орга-
никой почвах, a D. mimosa встречается в пресных во-
доемах. Оба вида питаются бактериями. Различаются
расположением отверстия цитостома: у D. trypanifor-
mis оно открывается у основания рострума, а у D. mi-
mosa — на его вершине. При неблагоприятных усло-
виях жгутиконосцы формируют цисты.
Род Cruzella Faria et Pinto, 1922
Морские криптобииды; форма тела яйцевидная;
рострум хорошо развит; характерна сильная метабо-
лия тела.
Единственный известный вид С. marina описан из
морской воды в окр. Рио-де-Жанейро. Размеры авто-
рами не приводятся. Для этого вида, вероятно, харак-
терна поликинетопластия.
Cryptobiidae incertae sedis
Род Phanerobia Skuja, 1948
Жгутиконосцы немного похожи на Dimastigella, од-
нако гораздо крупнее. Описан один вид — Ph. pelo-
phila. Для определения места этих жгутиконосцев
в системе необходимо исследование их ультраструкту-
ры.
ТИП EUGLENOZOA. КЛАСС KtNETOPLASTIDEA
243
Отряд Trypanosomatida Kent, 1880
Кинетопластиды с единственным, направленным
вперед жгутиком; тубулемма полная, только у трипа-
носом кольцо субмембранных микротрубочек разом-
кнуто; кинетопласты относительно небольшие и ком-
пактные; цитостом-цитофарингеальные комплексы у
большинства представителей в различной степени ре-
дуцированы.
Сем. Trypanosomatidae Grobben,
1905
Характеристика семейства соответствует таковой
отряда (рис. 149).
Род Blastocrithidia Laird, 1959
Гомоксенные трипаносоматиды, в жизненных
циклах которых имеются эпимастиготные формы.
Многие виды формируют цистоподобные амастиго-
ты. Паразиты насекомых. Жгутиконосцы рода Blas-
tocrithidia паразитируют в пищеварительной системе,
иногда в гемолимфе насекомых-хозяев. Среди хозяев
преобладают полужесткокрылые и двукрылые насе-
комые.
Многие представители этого рода, например В.
familiar is, В. miridarum (рис. 149, 3), В. triatbmae,
формируют «жгутиковые цисты» — мелкие цисто-
подобные стадии, отпочковывающиеся от материн-
ской эпимастиготы и прикрепляющиеся к ее жгути-
ку. В. triatomae оказывает патогенное воздействие
на своих хозяев — клопов редувиид, вызывая их
высокую смертность, особенно среди личинок кло-
пов.
Типовой вид В. gerridis, всесветно распро-
страненный паразит кишечного тракта клопов водо-
мерок.
Род Crithidia Leger, 1902
Гомоксенные трипаносоматиды, имеющие харак-
терную «кувшиновидную» форму тела и очень широ-
кий жгутиковый карман. Паразиты насекомых, все
развитие которых протекает в ректуме хозяев. Пара-
зитируют и в личинках и в имаго насекомых. В
жизненном цикле имеются только стадии, обладаю-
щие свободным жгутиком. Большинство крити-
дий — паразиты двукрылых и в меньшей степени —
полужесткокрылых насекомых. В качестве модель-
ных объектов в лабораториях часто культивируют С.
oncopelti (рис. 149, А) и С. lucilia (рис. 149, Е).
Типовой вид С. fasciculata (рис. 149, Д), все-
светно распространенный паразит комаров Culici-
dae. Заражение происходит через воду, где рассели-
тельные стадии критидий выживают до 10 дней, со-
храняя способность к размножению по крайней
мере первые 5 сут.
Род Herpetomonas Kent, 1880
Гомоксенные трипаносоматиды, в жизненных
циклах которых представлены про-, эндо- и описто-
мастиготные формы. Паразиты насекомых.
Практически всё валидные описания представи-
телей рода относятся к паразитам Diptera. Развитие
жгутиконосцев приурочено к задним отделам пище-
варительной системы хозяев, к ректуму и мальпиги-
евым сосудам. Цистоподобные стадии в жизненных
циклах Herpetomonas отсутствуют. Некоторые виды
могут сохранять свою жизнеспособность при повы-
шенной (до 37 °С1 температуре. Более того, повы-
шение температуры стимулирует появление в попу-
ляции жгутиконосцев опистомастигот.
Типовой вид Н. muscarum (рис. 149, Б), широ-
ко распространенный паразит кишечного тракта раз-
личных мух.
Род Leptomonas Kent, 1880
Гомоксенные трипаносоматиды, в жизненных
циклах которых чередуются промастиготные и амас-
тиготные формы. Паразиты беспозвоночных живот-
ных. Первым Leptomonas, описанным из насекомых,
был L. jaculum (рис. 149, И). Жизненный цикл этих
жгутиконосцев протекает в кишечнике водяных
скорпионов Nepa cinerea. Лептомонады заселяют
среднюю кишку, где, прикрепляясь жгутиками к
плазмалемме энтероцитов, интенсивно размножают-
ся. В задних отделах кишечника жгутиконосцы фор-
мируют розетки (ассоциаты лептомонад, соединен-
ных жгутиками). Деления в розетках завершаются
образованием мелких цистоподобных амастигот, вы-
полняющих роль расселительных стадий. Представи-
тели этого рода паразитируют в кишечнике различ-
ных насекомых, однако подавляющее большинство
Leptomonas приурочено к 2 их отрядам — Hemiptera
и Diptera. К этому роду относятся единственные
представители трипаносоматид, паразитирующие в
простейших, — L. karyophilus и L. ciliatorum из мак-
ронуклеусов инфузорий.
Типовой вид L. butschlii. Название вида дано
Кентом (Kent, 1880) для жгутиконосцев, описанных
из морской нематоды Trilobius gracilis. Существова-
ние этого вида, как и Leptomonas, описанных из мол-
люсков, требует подтверждения.
Род Wallaceina Podlipaev,
Frolov et Kolesnikov, 1990
{-Proteomonas Podlipaev,
Frolov et Kolesnikov, 1990)
Гомоксенные трипаносоматиды, в жизненных
циклах которых чередуются промастиготы и эндо-
мастиготы. Эндомастиготы 2 типов: мелкие, с длин-
ным петлеобразным жгутиковым карманом, огибаю-
щим ядро, и крупные, с коротким прямым жгутико-
вым карманом. Паразиты насекомых. Жгутиконосцы
244
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 149. Представители сем. Trypanosomatidae. (По разным источникам).
А — Crithidia oncopelti; Б — опистомастигота Herpetomonas muscarum; В — Phytomonas nordicus; Г— про- и эвдомастиготы Leishmania donovani; Д — Crithidia
fasciculata; Е — С. lucilia; Ж — про- и эндомастиготы Wallaceina brevicula; 3 — Blastocrithidia miridarum, эпимастигота с прикрепленными к жгутику
цистоподобными амастиготами; И — Leptomonas jaculum со жгутиком преобразованным в прикрепительную органеллу; К — Trypanosoma gallinarum; Л—
Т. varani; М —Т (Н.) lewisi; Н — Т. (5.) cruzi; О—Т. (D.) vivax; П — Т. (N.) congolense; Р — Т. (Т.) brucei; С — Т. (Т.) gambiense; Т — Т. (Г.) evansi.
Н—Т — для каждого из этих видов приведены широкие и узкие формы трипомастигот.
Fig. 149. The representatives of Trypanosomatidae.
A — Crithidia oncopelti; Б — Herpetomonas muscarum (opistomastigote); В — Phytomonas nordicus; Г — Leishmania donovani (promastigote and endomastigote);
д — Crithidia fasciculata; E — C. lucilia; Ж — Wallaceina brevicula (promastigote and endomastigote); 3 — Blastocrithidia miridarum (epimastigote with
«strap-hangers»); И— Leptomonas jaculum (flagellum transformed in the special attachment organella); К — Trypanosomagallinarum; Л — T. varani; M~ T. (H.) lewisi;
H— T. (S.) cruzi; О — T. (D.) vivax; П — T. (N.) congolense, P — T. (T) brucei; C — T. (T.) gambiense; T — T. (T.) evansi (H-T both slender and stumpy forms of
trypanosoms is shown).
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
245
паразитируют в ректуме хозяев, здесь же формируются
мелкие эндомастиготы, выполняющие функции рас-
селительных стадий. Другой представитель рода
W. brevicula (рис. 149, Ж) — паразит хищных клопов
из сем. Nabidae.
Типовой вид Ж inconstans из кишечника кло-
пов слепняков Calocoris sexguttatus.
Род Rhinchoidomonas Patton, 1910
Гомоксенные трипаносоматиды, имеющие в жиз-
ненном цикле трипомастиготы (?); паразитируют в
мальпигиевых сосудах двукрылых насекомых. Мор-
фология и жизненный цикл не изучены.
Род Phytomonas Donovan, 1909
Гетероксенные трипаносоматиды с промастигота-
ми и амастиготами в жизненном цикле; тело промас-
тигот часто винтообразно закручено. Паразиты расте-
ний, переносчики — клопы-фитофаги.
Наиболее важны жгутиконосцы, вызывающие за-
болевания культурных растений. Ph. francai парази-
тирует в млечниках маниота и кассавы (сем. Euphor-
biaceae). В Бразилии на плантациях могут вызывать
массовую гибель растений. Ph. leptovasorum парази-
тирует во флоэме, наводит серьезный ущерб планта-
циям кофейных деревьев (сем. Rubiaceae). Перенос-
чик — Lincus spatuliger (Hemiptera, Pentatomidae).
Phytomonas staheli описан из флоэмы масличных и
кокосовых пальм (сем. Агесасеае). Могут вызывать
массовую гибель деревьев. Переносчики — клопы
лигеиды (Lygaeidae). Необычный жизненный цикл
характерен для самого северного представителя ро-
да — Р. nordicus (рис. 149, В). Эти жгутиконосцы
паразитируют в кишечнике, слюнных железах и ге-
молимфе хищных клопов Troilus luridus (Pentatomi-
dae). По своей морфологии, особенностям развития
в насекомых — это типичные фитомоносы. Однако
в их жизненном цикле отсутствует второй хозяин —
растение. Стадии жгутиконосцев, которые развива-
ются в слюнных железах хозяев, со слюной вводятся
в гемолимфу насекомых-жертв Т. luridus. Таким об-
разом, каждый акт питания клопов сопровождается
их автоинвазией. Передача Phytomonas nordicus от
одного хозяина к другому происходит контаминатив-
ным путем.
Типовой вид Рй. davidi, паразит растений се-
мейств Аросуапасеае и Asclepiadaceae.
Род Leishmania Ross, 1903
Гетероксенные паразиты позвоночных животных
(рептилий и млекопитающих); переносчики — мос-
киты; в позвоночных хозяевах паразитируют в фор-
ме эндомастигот, в москитах развиваются промасти-
готы.
Типовой вид L. major Yakimoff, 1915.
Биология представителей рода Leishmania весьма
разнообразна. В рамках этого таксона обычно выде-
ляют группы видов, характеризующиеся общностью
циклов развития и различающиеся между собой по
тем или иным критериям. Одно из наиболее удач-
ных, с нашей точки зрения, решений этого вопро-
са — выделение 2 подродов: Leishmania (паразиты
млекопитающих) и Sauroleishmania (паразиты репти-
лий) (Сафьянова, 1982). Однако в последнее время в
литературе преобладает другой подход. По аналог
гии с трипаносомами представителей рода Leishma-
nia разделяют на 3 группы нетаксономического ран-
га (секции) в соответствии с местом развития их
инфективных стадий в переносчиках. Характеризуя
жизненные циклы лейшманий, мы уже обсуждали
недостатки такого подхода. Тем не менее, поскольку
данная схема в настоящее время принимается боль-
шинством специалистов, мы приводим ее в нашей
работе.
1. Промастиготы развиваются в задней и пря-
мой кишке москитов. Сюда относятся паразиты из
секции Hypopylaria (паразиты рептилий) — Leishma-
nia agamae, L. ceramodactyli, L. gymnodactyli, L. guli-
kae, L. senegalensis (описано еще несколько видов,
однако их валидность требует подтверждения).
2. Промастиготы развиваются в средней кишке
и пилорусе, после чего могут мигрировать в перед-
ние отделы пищеварительной системы москитов. К
этой группе относятся паразиты из секции Peripylaria
(паразиты рептилий и млекопитающих) — Leishma-
nia adleri, L. braziliensis, L. tarentolae.
3. Промастиготы из средней кишки мигрируют
в передние отделы пищеварительной системы мос-
китов. В эту группу включены паразиты из секции
Suprapylaria (паразиты млекопитающих) — Leishma-
nia aethiopica, L chagasi, L. donovani, L. enriettii,
L. infantum, L. hertigi, L. major, L. mexicana, L. tro-
pica.
Кроме того, в характеристике представителей ро-
да Leishmania традиционно используется географи-
ческий критерий. Известно, что по целому ряду при-
знаков лейшманий Старого и Нового Света сущест-
венно различаются между собой.
Лейшманий Старого Света
Возбудители кожных
лейшманиозов
Leishmania tropica Wright, 1903.
(= L. t. minor Yakimoff, 1915).
Распространение: Средиземноморье, Ближний
и Средний Восток, Индостан, Закавказье и Средняя
Азия.
Хозяева: кроме человека заражает собак и раз-
личных грызунов.
Переносчики: Phlebotomus intermedius, Ph. mi-
gonei, Ph. pessoai, Ph. sergenti, Ph. whitmani.
L. tropica — возбудитель «сухой» язвы антропо-
нозного кожного поздноизъязвляющегося лейшмани-
оза, в том числе и туберкулоидной формы. Инкуба-
ционный период длится от нескольких месяцев до
нескольких лет.
246
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Leishmania major Yakimoff, 1915.
(=L. tropica major Yakimoff, 1915).
Распространение: Монголия, Передняя и
Средняя Азия, Индия, Северная и Восточная Аф-
рика.
Хозяева: кроме человека заражает грызунов и
различные виды хищных млекопитающих.
Переносчики: Phlebotomus andrejvi, Ph. саиса-
sicus, Ph. duboscqi, Ph. mongolensis, Ph. papatasi.
L. major — возбудитель зоонозного кожного oc-
тронекротизирующегося лейшманиоза, «мокнущей»
язвы. Инкубационный период длится от нескольких
дней до нескольких недель.
Leishmania aethiopica Bray, Ashford et Bray,
1973.
Распространение: влажные горные районы
Кении и Эфиопии.
Хозяева: кроме человека описан у доманов.
Переносчики: Phlebotomus longipes, Ph. pedifer.
L. aethiopica — возбудитель кожного лейшманио-
за.
Возбудители висцеральных
лейшманиозов
Leishmania donovani (Laveran et Mesnil, 1903).
(=Piroplasma donovani Laveran et Mesnil, 1903).
Распространение: Индия.
Хозяева: кроме человека заражает овец, собак,
лошадей.
Переносчик: Phlebotomus argentipes.
L. donovani (рис. 149, Г) — возбудитель кала-аза-
ра — висцерального индийского лейшманиоза. Ин-
кубационный период длится от 10 дней до года. На-
иболее интенсивно поражаются селезенка, печень,
костный мозг и лимфатические узлы. При отсутст-
вии специфического лечения, как правило, заканчи-
вается летальным исходом.
Leishmania infantum Nicolle, 1908.
(= L. donovani infantum, Nicolle, 1908).
Распространение: Средняя Азия, Средизем-
номорье, Китай.
Хозяева: кроме человека заражает различные
виды хищных млекопитающих.
Переносчики: Phlebotomus ariasi, Ph. chinensis,
Ph. celiae, Ph. major, Ph. perniciosus.
L. infantum — возбудитель средиземноморско-
среднеазиатского висцерального лейшманиоза. Тече-
ние заболевания как при кала-азаре.
Leishmania archibaldi Castellani et Chalmers,
1919.
(=L. donovani archibaldi Castellani et Chalmers, 1919).
Распространение: Восточная Африка.
Хозяева: кроме человека заражает различные
виды хищных млекопитающих.
Переносчики: Phlebotomus martini, Ph. orien-
talis.
L. archibaldi — возбудитель восточно-африканс-
кого висцерального лейшманиоза.
Лейшмании Нового Света
Возбудители кожных
лейшманиозов
Leishmania braziliensis braziliensis Vianna, 1911.
(= L. americana var. flagellata Escomel, 1913; L. braziliensis
braziliensis Lainson et Shaw, 1972; L. tropica nbi. americana Laveran
et Nattan-Larrier, 1912; L. t. var. braziliensis Floch, 1954; L. wrightii
Pedroso et da Silva, 1910).
Распространение: Боливия, Бразилия, Вене-
суэла, Колумбия, Парагвай, Перу, Эквадор.
Хозяева: кроме человека заражает грызунов и
многих других млекопитающих.
Переносчики: Lutzomyia intermedia, L. paraen-
sis, L. pessoai, Psychodopygys wellcomei.
L. b. braziliensis — возбудитель кожно-слизйстого
лейшманиоза; поражаются кожа и слизистые носа с
последующим разрушением хрящевых тканей.
Leishmania braziliensis peruviana Velez, 1913.
Распространение: горные районы Перу.
Хозяева: кроме человека, возможно, заражает
собак.
Переносчики: Lutzomyia peruensis, L. verruca-
rum.
L. b. peruviana вызывает кожные язвы.
Leishmania braziliensis guyanensis Floch, 1954.
Распространение: Бразилия, Гвиана, Венесуэ-
ла, Суринам.
Хозяева: кроме человека заражает ленивцев и
муравьедов.
Переносчики: Lutzomyia anduzei, L. umbratilis.
L. b. guyanensis вызывает кожные язвы, может по-
ражать слизистую носоглотки.
Leishmania braziliensis panamensis Lainson et
Shaw, 1972.
Распространение: Гондурас, Коста-Рика, Па-
нама.
Хозяева: кроме человека заражает различные
виды грызунов и неполнозубых.
Переносчики: Lutzomyia gomezi, L. panamensis,
L. trapidoi, L. ylephiletrix.
L. b. panamensis — возбудитель типичного кожно-
го лейшманиоза.
Leishmania mexicana mexicana Biagi, 1953.
(=L. braziliensis mexicana Pessoa, 1961; L. mexicana Gamham,
1962; L. tropica mexicana Biagi, 1953).
Распространение: Бразилия, Гватемала, Мек-
сика.
Хозяева: кроме человека заражает грызунов.
ТИП EUGLENOZOA^ КЛАСС KINETOPLASTIDEA
247
Переносчик: Lutzomyia olmeca,
L. m. mexicana — поражает преимущественно
кожу ушных раковин; заболевание сопровождается
разрушением хрящевых тканей ушей, носа и гортани.
Leishmania mexicana pifanoi Medina et Romero,
1962.
(=L. braziliensis pifanoi Medina et Romero, 1962; L. diffusa
Adler, 1962; L. pifanoi Medina et Romero, 1962).
Распространение: Венесуэла.
Хозяева: кроме человека заражает грызунов.
Переносчик: неизвестен.
L. т. pifanoi — возбудитель «диффузной» формы
кожного лейшманиоза.
Leishmania mexicana amazoensis Lainson et Shaw,
1972.
(=L. braziliensis Nery-Gumaraes et da Costa, 1966).
Распространение: Боливия, Бразилия, Трини-
дад.
Хозяева: кроме человека заражает различные
виды грызунов и опоссумов.
Переносчик: Lutzomyia flaviscutellata.
L. т. amazoensis — вызывает неглубокие кожные
поражения.
Возбудители висцеральных
лейшманиозов
Leishmania chagasi Cunha et Chagas, 1937.
Распространение: Боливия, Бразилия, Гвате-
мала, Колумбия, Мексика, Парагвай, Сальвадор.
Хозяева: кроме человека поражает ряд видов
хищных млекопитающих.
Переносчик: Lutzomyia longipalpis.
Течение заболевания как при кала-азаре.
Род Endotrypanum Mesnil
et Brimont, 1908
Трипаносоматиды, паразитирующие в эритроци-
тах ленивцев (Edentata) в форме эпи- и трипомас-
тигот; в переносчиках-москитах развиваются в фор-
ме промастигот. Описаны 2 вида: Е. monterogei и
£. schaudinni. Биология изучена плохо, в культуре
существуют в форме промастигот.
Распространение: Центральная и Южная Аме-
рика.
Переносчики: москиты рода Lutzomyia.
Род Trypanosoma Gru by,
1843
Гетероксенные кровепаразиты позвоночных жи-
вотных; переносчики — пиявки и артроподы; в
жизненном цикле обязательно представлены эпи- и
трипомастиготы.
Потенциальные переносчики — комары {Си-
lex territans и др.) и пиявки (различные виды из
семейств Glossiphonidae, Hirudinidae, Ichthyobdelli-
dae). При развитии в культуре и в переносчиках
формирует различные морфологические формы:
про-, трипо-, эндо- и эпимастиготы.
Типовой вид: Т. rotatorium Gruby, 1843, рас-
пространенный всесветно паразит крови бесхвостых
амфибий.
Род Trypanosoma представляет собой весьма ге-
терогенную группу трипаносоматид. В 1964 г. Гоар
(Ноаге, 1964) предложил рассматривать в пределах
этого рода несколько подродов, объединяющих па-
разитов млекопитающих. Большая группа трипа-
носом из низших позвоночных и птиц (сама по
себе также крайне неоднородная) остается в подро-
де Trypanosoma. Ниже перечислены подроды трипа-
носом млекопитающих и приведены их краткие ди-
агнозы.
Секция Stercoraria
Развитие метациклических стадий трипаносом
происходит в задних отделах кишечника насекомых-
переносчиков.
Подрод Megatrypanum Ноаге,
1964
Крупные трипаносомы с расположенными вблизи
друг от друга ядром и кинетопластом, смещенными к
середине тела жгутиконосцев. В млекопитающих раз-
множаются эпимастиготы.
Подрод Herpetosoma Doflein,
1901
Трипаносомы средних размеров с субтерминаль-
ным кинетопластом, однако четко отстоящим от
конца клетки. В млекопитающих размножаются
амастиготы и (или) эпимастиготы.
Подрод Schisotrypanum Chagas,
1909
Мелкие трипаносомы, на сухих мазках имеющие
характерную С-образную форму тела. Кинетопласт
примыкает к заднему концу клетки. В млекопитаю-
щих размножаются как внутриклеточные эндомасти-
готы.
Секция Salivaria
Развитие трипаносом в переносчиках происходит
в «передней позиции», т. е. в слюнных железах или в
хоботке кровососущих насекомых. Основные пере-
носчики — двукрылые насекомые Glossina и различ-
ные Tabanidae.
248
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подрод Duttonella Chalmers, 1918
Трипаносомы с большим, терминально распо-
ложенным кинетопластом, свободный жгутик всегда
имеется. В переносчиках развиваются только в про-
босцисе.
Подрод Nannomonas Ноаге, 1964
Мелкие трипаносомы с небольшим, маргинально
расположенным кинетопластом. Развиваются в сред-
ней кишке и пробосцисе Glossina.
Подрод Trypanozoon Luhe, 1906
Трипаносомы с мелким субтерминальным кинето-
пластом. Развиваются в средней кишке и слюнных
железах Glossina.
Подрод Pycnomonas Ноаге, 1964
Широкие мономорфные трипаносомы с маленьким
субтерминальным кинетопластом и коротким сво-
бодным жгутиком. Развиваются в средней кишке, и в
слюнных железах Glossina.
Подрод Trypanosoma Gruby, 1843
Несмотря на практически всесветное распростра-
нение (только трипаносомы Reptilia в основном при-
урочены к зоне тропиков и субтропиков) и довольно
высокую частоту встречаемости в хозяевах, предста-
вители подрода Trypanosoma относятся к наименее
изученным трипаносомам. В рыбах, амфибиях и пре-
смыкающихся обычно паразитируют крупные трипа-
носомы с компактным кинетопластом. У многих опи-
сана сократительная вакуоль. Вероятно, эти трипано-
сомы непатогенны для своих хозяев. Как правило,
заболевание протекает в хронической форме. Чис-
ленность паразитов в периферической крови низкая,
и размножение трипаносом здесь наблюдается край-
не редко. Обычно если жгутиконосцы и размножают-
ся в позвоночном хозяине, то это происходит в ка-
пиллярах различных внутренних органов (почки, пе-
чень, сердце). Переносчики — пиявки.
В рыбах описано около 170 видов трипаносом. В
качестве примеров приведем Trypanosoma rajae из
крови скатов и Т. murmanense — паразита различных
видов костных морских рыб. В пресноводных рыбах
обычны Т danilewskyi, Т. percae, Т. carassii и другие.
Около 60 видов трипаносом описаны из амфибий.
Среди них — классические лабораторные объекты
Т. mega (рис. 136, В), Т rotatorium и Т. inopinatum.
Переносчики — пиявки, москиты, комары.
Из всей группы наименее изучены трипаносомы
рептилий (около 70 видов). Лишь Т. grayi (паразит
нильского крокодила), переносчиком которой явля-
ются мухи рода Glossina? хорошо знакома зоологам
(Ноаге, 1931). Несколько видов (Г. chelodina, Т. dia-
monia и др.) описаны из черепах. Наиболее вероят-
ные их переносчики — пиявки и москиты. Валидность
большинства видов трипаносом, описанных из крови
ящериц (рис. 149, Л) и змей, нуждается в подтверж-
дении.
В птицах описано около 80 видов трипаносом.
Переносчиками служат мошки, мокрецы, комары,
мухи-кровососки и клещи Dermanyssus gallinae. За-
ражение птиц трипаносомами происходит контами-
нативно — трипаносомы попадают в организм птиц
при поедании ими переносчиков, либо активным пу-
тем через кожу или слизистые, загрязненные фекали-
ями. Объектом многих исследований стала Trypano-
soma avium <— распространенный паразит различных
видов диких птиц. Ряд трипаносом, например Т. gal-
linarum (рис. 149, К), Т. numidae, Т. calmettei, извест-
ны как паразиты домашних птиц.
Секция Stercoraria
Подрод Megatrypanum Ноаге, 1964
Trypanosoma (Megatrypanum) theileri Laveran,
1902.
(= Crithidia haematopotae Jegen, 1924; C. pangonia Rodhain,
van den Branden et Bequaert, 1913; C. subulata (Leeger, 1904) Pat-
ton, 1907; Herpetomonas subulata Leeger, 1904; Leptomonas pango-
niae Rodhain, Rons, van den Branden et Bequaert, 1912; Trypanoso-
ma americanum Crawley, 1909; T. cephalophi Bruce, Harvey, Hamer-
ton, Davey et Bruce, 1915; T. falshawi Knuth, 1909; T.franki Frosch,
1909; T. himalayanum Lingard, 1906; T. indicum Lingard, 1907; T.
lingardi Blanchard, 1904; T. scheini Knuth, 1909; T theileri Bruce,
1902; T. tragelaphi Kinghorn, Yorke et Lloyd, 1913; T transvaaliense
Laveran, 1902; T. wrublewskii Wladimiroff et Yakimoff, 1908; Trypa-
nozoon theileri (Laveran, 1902) Luhe, 1906).
Распространение: всесветное.
Хозяева: паразитирует в крови большинства по-
род крупного рогатого скота; обычный кровепаразит
африканских антилоп.
Переносчики: слепни (Tabanidae) и, вероятно,
иксодовые клещи.
Обычно Т. (М.) theileri считается слабопатогенной
или непатогенной трипаносомой.
Trypanosoma (Megatrypanum) melophagium (Flu,
1908).
(-Crithidia melophagi Swingle, 1909; C. melophagia Hu, 1908;
Herpetomonas melophagia Doflein, 1916; Leptomonas melophagi
Mesnil, 1909; L. melophagia Dunkerley, 1913; Trypanosoma (Cysto-
trypanosoma) melophagi Brumpt, 1913; T. (C.) melophagia Brumpt,
1922; T. woodcocki Brumpt, 1913).
Распространение: всесветное.
Хозяева: паразитирует в крови овец.
Переносчик: Melophagus ovinus.
Проникновение трипаносом в хозяина происходит
при расчесывании овцами мест питания кровососок
или через слизистую полости рта. Т. (М.) melophagium
относится К непатогенным видам трипаносом.
Trypanosoma (Megatrypanum) cervi Kingston et
Morton, 1975.
Распространение: Северная Америка.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA
249
Хозяева: паразитирует в крови лосей и различ-
ных видов оленей.
Переносчики: слепни (Tabanida).
Развитие в хозяевах не изучено.
Подрод Herpetosoma Doflein,
1901
Trypanosoma (Herpetosoma) lewisi (Kent, 1880)
Laveran et Mesnil, 1901.
{-Herpetomonas lewisi Kent, 1880; Trypanosoma rattorum Bor-
ner, 1881; Trichomonas lewisi Crookshank, 1886; «Trypanomonas
murium» Danilewsky, 1888; T. lewisi Labbe, 1891; Trypanosoma le-
wisi Doflein, 1901; Trypanozoon lewisi Luhe, 1906; Trypanosoma lo-
gocaudense Lingard, 1906; T. eburneense Delanoe, 1915; Lewisonella
lewisi Chalmers, 1918).
Распространение: всесветное.
Хозяева: Паразитирует в крови крыс (Rattus rat-
tus и R. norvegicus).
Переносчики: блохи Nosopsyllus fasciatus и Xe-
nopsylla cheopis.
Обычно заражение крыс происходит при поеда-
нии ими зараженных блох. Короткий инкубацион-
ный период (около 6 дней) сменяется вспышкой .ин-
тенсивного размножения трипаносом в перифери-
ческой крови хозяина (около 10 дней). После этого
трипаносомы перестают размножаться, и по прошес-
твии нескольких недель или месяцев кровь хозяев
полностью очищается от трипаносом. Г. (Н.) lewisi
(рис. 149, Af), вероятно, непатогенна для своих хозя-
ев. Однако, в Индии описано несколько случаев тя-
желого трипаносомоза у людей, возбудителем кото-
рого считают данный вид трипаносом.
Trypanosoma (Herpetosoma) rangeli Tejera, 1920.
(=T. ariarii Groot, Renjifo et Uribe, 1951; T. cebus Floch et
Abonnenc, 1949; T. encomeli Yorke, 1920; T. guatemalense Leon,
1946).
Распространение: Центральная и Южная Аме-
рика.
Хозяева: кроме человека заражает домашних
кошек и собак, опоссумов, многие виды приматов и
других млекопитающих.
Переносчики: клопы сем. Reduviidae.
Инкубационный период длится обычно около
2 нед. В кровяном русле хозяев трипаносомы сохра-
няются до года и более. Т. (Н.) rangeli непатогенна
для своих хозяев.
Подрод Schizotrypanum Chagas,
1909
Trypanosoma (Schizotrypanum) cruzi Chagas, 1909.
(=T. deanei Coutinho et Pattolli, 1964; T. rhesii Terry, 1911;
T. triatpmae Kofoid et MacCulloch, 1916; T. vickersae Brumpt, 1909;
Schizotrypanum mexicana Perrin, 1950).
Возбудитель болезни Чагаса.
Распространение: Центральная и Южная Аме-
рика.
Хозяева: кроме человека заражает многие виды
млекопитающих — как диких, так и домашних.
Переносчики; клопы сем. Reduviidae, возмож-
но Cimex lectularis, и некоторые кровососущие Ly-
gaeidae.
Инкубационный период длится до 2 нед. Сначала
в месте проникновения трипаносом появляется вос-
палительный инфильтрат — характерная чагома.
Иногда возникают вторичные чагомы в подкожной
клетчатке. Болезнь Чагаса нередко осложняется ме-
нингоэнцефалитом. Поражаются многие внутренние
органы, особенно опасны поражения миокарда.
Больные умирают при явлениях сердечно-сосудистой
недостаточности. Даже при хроническом течении
болезни Чагаса без специального лечения благопо-
лучный исход крайне редок. По данным ВОЗ, в
Центральной и Южной Америке в 1982 г. Т. (S.)
cruzi (рис. 149, Н) было заражено не менее 20 млн
человек. Кроме Т, (S.) cruzi к этому подроду отно-
сится еще несколько видов трипаносом, главным об-
разом паразитов летучих мышей (Г. vespertilionis, Т.
phyllostomae, Т. hipposideri). Они характеризуются
практически всесветным распространением, но их
биология пока изучена слабо.
Секция Salivaria
Подрод Duttonella Chalmers, 1918
Trypanosoma (Duttonella) vivax Ziemann, 1905.
{=Duttonella vivax Chalmers, 1918; Trypanosoma angolens Bro-
den et Rodhain, 1910; T. bovis Kleine, 1910; T. caprae Kleine, 1910;
T. cazalboui Laveran, 1905; T. guyanense Leger et Vienne, 1919;
T. vivax Luhe, 1906).
Распространение: большая часть Африки и
Америки, Индия.
Хозяева: домашние и дикие парнокопытные,
верблюды, лошади.
Переносчики: Glossina fuscipes, G. longipalpis,
G. morsitans, G. pallidipes, G. palpalis, G. swynnertoni,
G. tachinoides, G. vanhofi; за пределами ареала Glossi-
na — слепни (Tabanidae).
Инкубационный период варьирует в зависимости
от вида хозяина и вирулентности штамма трипано-
сом. Т. (D.) vivax (рис. 149, О) патогенны для круп-
ного рогатого скота и овец. Заболевание часто со-
провождается летальным исходом. Дикие животные,
а также козы и верблюды переносят заболевание
относительно легко.
Trypanosoma (Duttonella) uniforme Bruce et al.,
1911.
{^Duttonella uniformis (Chalmers, 1918)).
Распространение: Африка.
Хозяева: те же, что и у Г. (D.) vivax.
Переносчики: Glossina fuscipes, G. morsitans,
G. palpalis, механические переносчики Stomoxys.
Течение болезни как при заражении Г. (D.)
vivax.
250
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подрод Nannomonas Ноаге, 1964
Trypanosoma (Nannomonas) congolense Broden,
1904.
{-Duttonella pecorum Chalmers, 1918; Trypanosoma berhei
Chardome et Peel, 1967; T. cellii Martoglio, 1911 pro parte; T. confu-
sum Montgomery et Kinghorh, 1909; T. dimorphon Laveran et Mesnil,
1904; T, dimorphon Luhe, 1906; T. frobeniusi Weissenborh, 1911; T.
mossoense Chardome et Peel, 1967; T. multiforme Kinghom et al.,
1913 pro parte; T. nanum Luhe, 1906; T. pecorum Bruce et al., 1910;
T ruande van Saceghem, 1921; T. somaliense Martoglio, 1911 pro
parte; T. urundiense Chardome et Peel, 1967; Trypanozoon congolense
(Luhe, 1906); T. dimorphon Luhe, 1906; T. nanum Luhe, 1906).
Распространение: тропическая Африка.
Хозяева: дикие и домашние парнокопытные,
африканский слон, собаки, лошади, верблюды.
Переносчики: Glossina austeni, G. brevipalpis,
G. fuscipes, G. longipalpis, G. morsitans, G. pallidipes,
G. palpalis, G. swynnertoni, G. tachiniodes, G. vanho-
ofi.
Инкубационный период при заражений Т. (N.)
congolense (рис. 149, Z7) длится от нескольких дней
до 2—3 нед. У домашних животных болезнь обычно
протекает остро, с летальным исходом. Свиньи от-
носительно резистентны к заболеванию.
Trypanosoma (Nannomonas) simiae Bruce et al.,
1912.
(=Duttonella simiae Chalmers, 1918; Trypanosoma ignotum Kin-
ghom et Yorke, 1912; T. porci Schwetz, 1932; T. rodhaini Walravens,
1924).
Распространение: в основном Восточная и
Центральная Африка.
Хозяева: бородавочники, домашние свиньи, од-
ногорбые верблюды, обезьяны.
Переносчики: Glossina austeni, G. brevipalpis,
G. fusca, G. fuscipleuris, G. longipalpis, G. morsitans,
G. pallidipes, G. palpalis, G. tabaniformis, G. tachinoi-
des, G. vanhoofi.
Инкубационный период длится от 5 до 15 дней.
Г. (N.) simiae патогенны для свиней и верблюдов.
Животные гибнут через несколько дней после зара-
жения.
Подрод Trypanozoon Luhe, 1906
Trypanosoma (Trypanozoon) brucei Plimmer et
Bradford, 1899.
(=T. pecaudi Laveran, 1907).
Возбудитель заболевания, известного под названи-
ем нагана.
Распространение: тропическая Африка.
Хозяева: дикие и домашние парнокопытные, ло-
шади, верблюды, а также кошки, собаки и грызуны.
Переносчики: Glossina morsitans, G. pallidipes,
G. swynnertoni.
T. (T.) brucei (рис. 149, P) — один из наиболее
важных в практическом отношении видов трипано-
сом. Очень тяжело переносят заболевание наганой
лошади, овцы, козы, верблюды. У крупного рогатого
скота болезнь обычно протекает в хронической
форме.
Trypanosoma (Trypanozoon) gambiense Dutton,
1902.
Возбудитель сонной болезни.
Распространение: Западная и Центральная Аф-
рика.
Хозяева: кроме человека может заражать до-
машних свиней.
Переносчики: Glossina fuscipes, G. palpalis, G.
tachinoides.
T. (T.) gambiense (рис. 149, С) вызывает относи-
тельно легкую форму сонной болезни людей. Инку-
бационный период длится около 3 нед. Течение бо-
лезни обычно хроническое. Зараженные люди могут
годами являться носителями возбудителя без прояв-
ления острых признаков болезни.
Trypanosoma (Trypanozoon) rhodesiense Stephens
et Fantham, 1910.
Возбудитель сонной болезни.
Распространение: Восточная и Южная Аф-
рика.
Хозяева: кроме человека заражает крупный и
мелкий рогатый скот, в природе резервуаром служат
антилопы.
Переносчики: Glossina morsitans, G. pallidipes,
G. swynnertoni.
Инкубационный период длится до 2 недель. Эта
форма сонной болезни протекает остро на фоне глу-
боких психических расстройств. Без интенсивной те-
рапии смерть наступает в течение первого года.
Trypanosoma (Trypanozoon) evansi (Steel, 1885)
Balbiani, 1888.
(= Haematomonas evansi Crookshank, 1886; H. evansis Blanc-
hard, 1888; Herpetomonas lewisi Danilewskyi, 1888; Spirochaeta
evansi Steel, 1885; Trypanosoma aegyptum Nattan-Larrier et Noyer,
1932; T. annamense Laveran, 1911; T. cameli Pricolo et Ferraro,
1914; T. elmassiani Lignieres, 1902; T. equinum Voges, 1901; T. hip-
picum Darling, 1910; T. marocanum Sergent, L’heritier et Belleval,
1915; T. ninae kohl-yakimovi Yakimoff, 1921; T. soudanense Laveran,
1907; T. venezuelense Mesnil, 1910).
Возбудитель заболевания, известного под названи-
ем сурра.
Распространение: Африка, Малая и Средняя
Азия, Индокитай, Центральная и Южная Америка,
Филиппины, Европа.
Хозяева: заражает многие виды диких и домаш-
них млекопитающих; возможно неспецифическое за-
ражение человека.
Переносчики: как правило, происходит меха-
нический перенос трипаносом кровососущими Dip-
tera. Широко известен пример передачи паразитов
кровососущими летучими мышами (Desmodes rotun-
das).
Инкубационный период при заражении Т. (Т.)
evansi (рис. 149, Г) длится от нескольких дней до 3
нед. Заболевание может протекать в острой и хрони-
ческой формах. В первом случае животные, как пра-
вило, гибнут. Особенно тяжело болеют лошади и вер-
блюды. Крупный рогатый скот переносит заболева-
ние относительно легко.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA. ЛИТЕРАТУРА
251
Trypanosoma (Trypanozoon) equiperdum Doflein, 1901.
(-Castellanella equiperdum Chalmers, 1918; Trypanosoma equi
Blacklock et Yorke, 1913; T. rougeti Laveran et Mesnil, 1901; Trypa-
nozoon equiperdum Luhe, 1906).
Возбудитель заболевания, известного под названи-
ем дурина.
Распространение: всесветное.
Хозяева: лошади, ослы и их гибриды.
Переносчики: трипаносомы передаются поло-
вым путем.
Болезнь протекает вяло, в хронической форме,
через 1—2 года в 50 % случаев может оканчиваться
гибелью животных.
Подрод Pycnomonas Ноаге,
1964
Trypanosoma (Pycnomonas) suis Ochmann,
1905.
Распространение: Африка.
Хозяева: дикие и домашние свиньи.
Перенбсчики: Glossina brevipalpis, G. van-
hoffi.
Инкубационный период длится около недели.
Заболевание у взрослых животных протекает вяло, у
поросят — в острой форме и может заканчиваться их
гибелью.
THE PHYLUM EUGLENOZOA CAVALIER-SMITH, 1991
THE CLASS KINETOPLASTIDEA HONIGBERG, 1963
SUM
Cinetoplastids represent an integrated group of organisms with a
characteristic basic structure, the central feature of which is the pre-
sence of a kinetoplast in the cell. In different versions of contempora-
ry classifications of protists the status of kinetoplastids varies from the
rank of an order to that of a phylum. In this chapter the classification
of the class Kinetoplastidea comprising three orders Bodonida, Cryp-
tobiida, Trypanosomatida, has been accepted. Bodonids are. mainly
free-living heterotrophic organisms. They have two heterodynamic
flagella, the recurrently directed one being free from the body. Most
of cfyptobiids are parasites. These flagellates also have two heterody-
namic flagella, but their recurrent flagellum is adherent alongside the
cell body. All trypanosomatids are obligatory parasites. They have
only one flagellum directed anteriorly.
A complete list of genera kinetoplastids is given and a detailed
description of the most important representatives of each genus is
cited.
It is well known that the origin and evolution of kinetoplastids
(especially Trypanosomatidae) has received much attention since the
beginning of this century. In this chapter I again assert the idea that
in the groups of protists lacking palaeontological records, phylogene-
tic conceptions and definitions of their evolutionary rank may be pro-
posed on the basis of comparative morphology studies of the orga-
nisms living nowadays. The application of this method to Protista is
based on the conception of relative evolutionary conservatism of cell
structures in protistan natural groups. Revealing of original general
structure and the analysis of evolutionary series of homologous featu-
res within such natural groups in some cases permits us to draw con-
clusions about the evolutionary position of their representative genera.
M
ARY
As the appearance of parasitism in eukaryotic evolution seems to
be secondary, it is natural to consider the free-living bodonid organi-
zation to be the original one in kinetoplastid evolution. Then the fea-
tures of cryptobiid and trypanosomatid organization may be conside-
red from the point of view of evolutionary reorganization of the initial
(bodonid) general cell structure. A significant part of this chapter is
devoted to the analysis of this reorganization.
The unique general cell structure with a unique organelle — a
kinetoplast as its central feature in free-living heterotrophic bodonids,
parasitic cryptobiids and trypanosomatids indicates that these flagella-
tes represent a natural group of organisms with direct phylogenetic
relationships. This means that from the point of view of facts available
at present there are no grounds for including hypothetical free-living
ancestors with the structure principally differing from that of modern
free-living biflagellate kinetoplastids into the evolution scheme of the
trypanosomatids.
The analysis of the evolutionary reorganizations of the original
«bodonid» plan of organization in different kinetoplastid groups illus-
trated by the evolutionary series of homologous features (coverings,
kinetosomal rootlets, flagellar transitional zones, cytostome-cytopha-
ryngeal complexes) shows that according to all these features the
cryptobiids occupy an intermediate position in the kinetoplastid evo-
lution between the free-living bodonids and the trypanosomes and can
be considered to be potential ancestors of the latter.
The recent molecular data based on ribosomal RNA sequences of
the euglenids (as outgroup), bodonids, cryptobiids and trypanosoma-
tids also support new ideas on kinetoplastid evolution (see for exam-
ple: Vickerman, 1994).
ЛИТЕРАТУРА14
(Алексеев A.) Alexeieff A. Sur la fonction glycoplastique du ki-
netoplaste (=kinetonucleus) chez les flagelles // C. R. Soc. Biol. 1917.
Vol. 80. P. 512.
Балашов Ю. С. Паразито-хозяинные отношения членистоно-
гих с наземными позвоночными. Л.: Наука, 1982. (Тр. ЗИН АН
СССР; Т. 97).
Белова Л. М. Множественные формы малатдегидрогеназы
(К. Ф. 1.1.1.37) у лептомонад (Mastigophora, Trypanosomatidae) //
Жизненные циклы простейших. Л.: Изд. ЗИН АН СССР, 1985.
С. 26—34. (Тр. ЗИН АН СССР; Т. 129).
Белова Л.М. Полиморфизм малатдегидрогеназы у Blastocrithi-
dia gerricola и его значение для таксономии // Систематика про-
стейших и их филогенетические связи с низшими эукариотами.
Л.: Изд. ЗИН АН СССР, 1986. С. 57—60. (Тр. ЗИН АН СССР;
Т. 144).
14 Цитируемая и рекомендуемая литература к разделу «Класс
Kinetoplastidea».
Гинецинская Т. А., Добровольский А. А. Частная паразитоло-
гия. М.: Высш, шк., 1978. Т. 1.
Горбунов П. С. Эндопаразитические жгутиконосцы рода Cri-
thidia (Trypanosomatidae, Zoomastigophorea) из пищеварительного
тракта шмелей // Зоол. журн. 1987. Т. 46. С. 1775—1780.
Данилевский Б. Сравнительно паразитологические исследова-
ния крови. I. Зоопаразиты крови птиц // Тр. Харьк. ун-та. 1888.
Вып. 1. С. 1—146.
Догель В. А. Курс общей паразитологии. 2-е изд. Л.: Учпедгиз,
1947.
Жуков Б. Г. Атлас пресноводных гетеротрофных жгутиконос-
цев (биология, экология, систематика). Рыбинск, 1993.
Каллшшкова В. Д. Клеточная органелла кинетопласт. Л.:
Наука, 1977.
Карпов С. А. Система протистов. Омск, 1990.
Карпов С., Жуков Б. Ультратонкое строение Pleuromonas ja-
culans Perty (Kinetoplastida, Zoomastigophorea) // Простейшие ак-
тивного ила. Л.: Наука, 1983. С. 53—156.
252
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Колесников А. А. Молекулярные параметры кинетопластной
ДНК трипаносоматид как таксономический признак И Цитология.
1993, С. 24—38.
Крылов М. В. Возбудители протозойных болезней домашних
животных и человека. В 2-х тт. СПб., 1994.
Крылов М. В. Определитель паразитических простейших.
СПб., 1996.
Крылов М. В., Самовар А. Г., Подлипаев С. А., Хаецкий
А. С. Исследование наличия генетического обмена в. жизненном
цикле Crithidia oncopelti (Protozoa, Kinetoplastmonada). Л.: Изд.
ЗИН АН СССР, 1985. С. 4—25. (Тр. ЗИН АН СССР; Т. 129).
Малышева М. Н., Скарлато С. О. Исследование ультратонкой
организации жгутиконосцев Leptomonas peterhoffi, культивируе-
мых на жидкой и твердой питательных средах // Цитология. 1989.
Т. 31. С. 267—272.
Малышева М. Н., Фролов А. О. Свободноживущие раздели-
тельные стадии трипаносоматид из водных насекомых // Паразито-
логия. 1993. Т. 27. С. 337—340.
Малышева М. Н., Фролов А. О. Цикл развития жгутиконосца
Proteomonas brevicula (Trypanosomatidae) в хищных клопах семей-
ства Nabidae (Hemiptera) // Паразитология. 1995. Т. 29. С. 289—
297.
Манукян И. А., Сафьянова В. М. Сравнительное изучение
ультраструктуры лептомонадных форм Leishmania tropica Wright,
L. donovani Laveran et Mesnil, а также лептомонад, выделенных от
рептилий и москитов И Мед. паразитол. и паразит, бол. 1968.
С. 319—323.
(Митрофанов П.) Mitrophanov Р. Beitrage zur Kenntnis der
Hamatozoen. Neue monadenformige Parasiten des Fischblutes, ihnen
ahnliche Organismen und ihre Beziehung zu den Blutelementen //
Biol. Cbl. 1883. Bd 3. S. 35—39.
Мыльников А. Ультратонкое строение жгутиконосцев Parabo-
do nitrophilus // Цитология. 1986. T. 28. C. 1056—1060.
(Мыльников A.) Mylnikov A. Ultrastructure of a colourless fla-
gellate, Phylomitus apiculatus Skuja 1948 (Kinetoplastida) // Arch.
Protistenk. 1986. Vol. 132. P. 1—10.
Подлипаев С. А. Новые виды низших трипаносоматид из полу-
жесткокрылых (Heteroptera) семейств Gerridae и Nabidae: стадии
их жизненных циклов в природе и при культивировании в лабора-
тории. Л.: Изд. ЗИН АН СССР, 1985. С. 35—47. (Тр. ЗИН АН
СССР; Т. 129).
Подлипаев С* А. Каталог мировой фауны простейших семей-
ства Trypanosomatidae // Тр. ЗИН АН СССР. 1990. Т. 217.
Подлипаев С. А., Фролов А. О. Описание и лабораторное
культивирование Blastocrithidia miridarum sp. n. (Mastigophora,
Trypanosomatidae) // Паразитология. 1987. T. 21. C. 545—552.
Подлипаев С. А., Фролов А. О., Колесников A. A. Proteomo-
nas inconstans n. gen., n.sp. (Kinetoplastida: Trypanosomatidae) пара-
зит клопа Calocoris sexgutatus (Hemiptera: Miridae) l Паразитоло-
гия. 1990. T. 24. C. 339—346.
(Подлипаев С. А., Малышева M. H., Колесников A.A) Podli-
paev S. A., Malysheva M.N, Kolesnikov A. A. Leptomonas rigidus
sp.n. (Trypanosomatidae) — a parasite of Saida littoralis L. (Hemip-
tera: Heteroptera) // Acta Protozool. 1991. Vol. 30. P. 121—127.
Сафьянова В. M. Проблема таксономии лейшманий // Лейш-
мании. Л.: Наука, 1982. С. 5—109. (Сер. «Протозоология»; Вып. 7).
Серавин Л. Н. Макросистема жгутиконосцев // Принципы по-
строения макросистемы одноклеточных животных. Л., 1980. С. 4—
22. (Тр. ЗИН АН СССР; Т. 94).
Серавин Л. Н. Основные типы и формы тонкого строения
крист митохондрий; степень их эволюционной стабильности (спо-
собность к морфологическим трансформациям) // Цитология.
1993. Т. 35. С. 3—34.
Скарлато С. О. Особенности тонкого строения ядра паразити-
ческого жгутиконосца Trypanoplasma borreli (Kinetoplastida) в ин-
терфазе и митозе // Докл. АН СССР. 1987. Т. 293. С. 220—221.
(Скарлато С. О., Лом И.) Skarlato S. О., Lorn J. Mitosis in the
flagellate Trypanoplasma borreli (Kinetoplastidea: Bodonida) // Eur. J.
Protistol. 1997. Vol. 33. P. 77—86.
Скарлато С. О., Малышева M. H. Ультраструктура гигантс-
ких многоядерных клеток паразитических жгутиконосцев Crithidia
oncopelti If Цитология. 1987. Т. 29. С. 1337—1342.
Скарлато С. О., Фролов А. О. Необычный способ деления у
трипаносоматид // Докл. АН СССР. 1988. Т. 298. С. 729—731.
(Скарлато С. О., Лом И., Нохинкова Е.) Skarlato S. О., Lom
J., Nohynkbva Е. Fine structural morphology of the nucleus of Try-
panosoma danilewskyi (Kinetoplastida, Trypanosomatina) during mi-
tosis // Arch. Protistenk. 1987. Bd 133. S. 3—14.
Скарлато С. О., Паршкова T. А., Фролов А. О. Электронно-
микроскопическое и молекулярно-биологическое исследование
ядра у низших трипаносоматид Blastocrithidia miridarum и Crithidia
brevicula И Цитология. 1990. Т. 32. С. 317—324.
Скарлато С. О., Сомова Н. В., Розанов Ю. М., Кудрявцев
Б. Н., Бобылёва Н. Н., Фролов А. О. Особенности организации
хромосомного аппарата жгутиконосцев Crithidia sp. // Цитология.
1998. Т. 40. С. 991—1005.
Сомова Н. В., Скарлато С. О., Фролов А. О. Хромосомы сво-
бодноживущих жгутиконосцев-кинетопластид // Докл. РАН. 1997.
Т. 357. С. 414—416.
Старобогатов Я. И. К вопросу о числе царств эукариотных ор-
ганизмов // Систематика простейших и их филогенетические связи
с низшими эукариотами. Л.: Изд. ЗИН АН СССР, 1986. С. 4—25.
(Тр. ЗИН АН СССР; Т. 144).
Фролов А. О. Жизненный цикл Leptomonas pyrrhocoris Z. (Ki-
netoplastida, Trypanosomatidae) // Зоол. журн. 1987a. T. 66. С. 5—
11.
Фролов А. О. Жизненный цикл Blastocrithidia miridarum (Ki-
netoplastida, Trypanosomatidae) И Зоол. журн. 19876. Т. 66. С. 655—
661.
Фролов А. О. Происхождение трипаносоматид // Паразитоло-
гия. 1993. Т. 27. С. 97—107.
Фролов А. О. Классификация морфологических форм жгути-
коносцев семейства Trypanosomatidae // Паразитология. 1994. Т. 28.
С. 261—269.
(Фролов А. О., Карпов С. A.) Frolov А. О., Karpov S. A. Com-
parative morphology of kinetoplastids H Cytology. 1995. T. 37. P. 1072—
1096.
Фролов А. О., Малышева М. Н. Crithidia allae sp. п. и Crithidia
brevicula sp.n. (Protozoa, Trypanosomatidae) из клопа Nabis brevis #
Зоол. журн. 1989a. T. 68. C. 5—10.
Фролов А. О., Малышева M. H. Цикл развития, паразитичес-
кого жгутиконосца Crithidia brevicula (Trypanosomatidae) в лабора-
торной культуре И Цитология. 19896. Т. 31. С. 971—975.
Фролов А. О., Малышева М. Н. Эндомастиготы — особый
тип разделительных стадий трипаносоматид рода Proteomonas //
Паразитология. 1992. Т. 26. С. 351—354.
Фролов А. О., Малышева М. Н. Описание Phytomonas nordi-
cus n. sp. (Trypanosomatidae) из хищного клопа Troilus luridus
(Hemiptera: Pentatomidae) // Паразитология. 1993. T. 21. C. 227—
232.
Фролов А. О., Подлипаев С. А. Необычный способ формиро-
вания инвазионных стадий у Leptomonas rigidus (Kinetoplastida:
Trypanosomatidae) // Паразитология. 1996. T. 30. С. 473—477.
Фролов А. О., Скарлато С. О. Свето- й электронно-микроско-
пические исследования Leptomonas pyrrhocoris Z. (Trypanosomati-
dae) // Паразитология. 1987. T. 21. С.’З—
Фролов А. О., Скарлато С. О. Локализация и способы закреп-
ления жгутиконосцев Blastocrithidia miridarum в пищеварительном
тракте клопов Adelphocoris quadripunctatus 7/ Паразитология. 1988.
Т. 22. С. 481—487.
Фролов А. О., Скарлато С. О. Электронно-микроскопическое
исследование жгутиконосцев Leptomonas jaculum из средней
кишки клопа Nepa cinerea И Паразитология. 1989. Т. 23. С. 383—
389.
Фролов А. О., Скарлато С. О. Структура розетковидных кле-
точных ассоциатов у низших трипаносоматид // Цитология. 1990.
Т. 32. С. 445—461.
Фролов А. О., Скарлато С. О. Описание Leptomonas mycophi-
lus sp.n. (Trypanosomatidae) — паразита клопа Phytocoris sp. (Mi-
ridae) // Паразитология. 1991. T. 25. C. 99—103.
Фролов А. О., Скарлато С. О. Тонкое строение и механизмы
адаптации низших трипаносоматид в полужесткокрылых насеко-
мых // Цитология. 1995. Т. 37. С. 539—560.
(Фролов А. О., Скарлато С. О.) Frolov A., Skarlato S. Unusual
pattern of mitosis in free-living flagellate Dimastigella mimosa (Kine-
toplastida) // Protoplasma. 1998. Vol. 201. P. 110—114.
Фролов А. О., Скарлато С. О., Шаглина Е. Г. Морфология
цистоподобных клеток жгутиконосцев Leptomonas jaculum // Ци-
тология. 1991. Т. 33. С. 960—969.
(Фролов А. О., Карпов С. А., Малышева М. Н.) Frolov А. О.,
Karpov S. A., Malysheva М. N. The ultrastructure of mitosis in the
free-living kinetoplastid Bodo curvifilus // Eur. J. Protistol. 1996.
Vol. 32. P. 498—505.
Фролов А. О., Малышева M. H., Подлипаев С. А. Необычный
способ формирования цистоподобных стадий у бластокритидий
(Kinetoplastida, Trypanosomatidae), паразитирующих в кишечном
ТИП EUGLENOZOA. КЛАСС KINETOPLASTIDEA. ЛИТЕРАТУРА
253
тракте клопов водомерок (Hemiptera, Gerridae) // Паразитология.
1997а. Т. 31. С. 356—363.
Фролов А. О., Малышева М. Н., Подлипаев С. А., Скарла-
то €. О. Электронно-микроскопическое исследование ядер про-
мастигот трипаносоматид Endotrypanum monterogei, Leishmania ta-
rentolae и Phytomonas sp. (Kinetoplastida, Trypanosomatidae) в ин-
терфазе и Митозе // Цитология. 19976. Т. 39. С. 278—284.
Фролов А. О., Мыльников А. П., Малышева М/Электронно-
микроскопическое исследование нового вида свободноживущего
жгутиконосца Dimastigella mimosa sp.n. // Цитология. 1997в. Т. 39.
С. 442—448.
Хаецкий А. С. Клонирование жгутиконосца Crithidia oncopelti
на плотной питательной среде неопределенного состава, не содер-
жащей гемин // Цитология. 1982. Т. 24. С. 211—214.
Adler S. Leishmania. Advances in Parasitology. London; New
York, 1964. Vol. 2. P. 35—96.
Alvarez P., Cortinas M. N., Musto H. The analysis of protein
coding genes suggest monophyly of Trypanosoma // Mol. Phylogen.
Evol. 1996. Vol. 5. P. 333—343.
Anderson B., Ellis R. Ultrastructure of Trypanosoma lewisi: fla-
gellum, microtubules and kinetoplast // J, Protozool. 1965. Vol. 12.
p. 483—499.
Angelopoulos E. Pellicular microtubules in the family Trypanoso-
matidae II J. Protozool. 1970. Vol. 17. P. 39—51.
Anez N., East J. Studies on Trypanosoma rangeli Tejera, 1920 II.
Its effect on feeding behaviour of triatomine bugs # Acta Tropica.
1984. Vol. 41. P. 93—95.
Attias M., Roitman I., Camargo E. P., Dollet M., De Sousa W.
Comparative analyse of the fine structure of four isolates of trypano-
somdtids of the genus Phytomonas // J. Protozool. 1988. Vol. 35.
P. 365—370.
Ayala S. C. Trypanosomes in wild California sandflies and extrin-
sic stages of Trypanosoma bufophlebotomi // J. Protozool. 1971.
Vol. 18. P. 433—436.
Baker J. The evolutionary origin and speciation of the genus Try-
panosoma# Soc. General Microbiol. Symp. 1974. Vol. 25. P. 343—
366.
Bardsley J., Harmsen R. The trypanosomes of Anura // Adv. Pa-
rasitol. 1973. Vol. 11. P. 1—72.
Becker C. D., Katz M. Transmission of the hemoflagellate Cryp-
tobia salmositica Katz 1951, by a rhynchobdellid vector. // J. Parasitol.
1965. Vol. 51. P. 95—99.
Bianchi L., Rondanelly E. G., Garosi G., Gerna G. Endonuclear
mitotic spindle in the leptomonad of Leishmania tropica # J. Parasitol.
1969. Vol. 55. P. 1091—1092.
Brack C. Elektronmikroskopische Untersuchungen zum Lebens-
zyklus von Trypanosoma cruzi. Unter besonderer Berucksichtigung
der Entwicklungsformen im Uebertrager Rhodnius prolixiis // Acta
trop. 1968. Vol. 25. P. 289—294.
Brazil R. P., Fiorihi J. E.> Faria de Silva P.M. Phytomonas sp.
a trypanosomatid parasite of tomato, jolated from salivary glands of
Phthia picta (Hemiptera: Coreidae) in Southeast Brazil // Mem. Inst.
Oswald© Cruz. 1990. Vol. 85. P. 239—240.
Breunig A., Konig H., Brugerolle G., Vickerman K., Hertel H.
Isolation and ultrastructural features of a new strain of Dimastigella
trypaniformis Sandon 1928 (Bodonina, Kinetoplastida) and compari-
son with a previously isolated strain // Eur. J. Protistol. 1993. Vol. 29.
P. 416—424. -
Brooker B. E. The fine structure of Crithidia fasciculata with spe-
cial reference to the organelles involved in the ingestion and digestion
of protein // Ztschr. Zellenforsch. Mikrosk. Anat. 1971a. Bd 116.
S. 532—541.
Brooker В. E. Fine structure of Bodo saltans and Bodo cauda-
tus (Zoomastigophora, Protozoa) and their affinities with the Try-
panosomatidae II Bull. Brit. Mus. Nat. Hist. 1971b. Vol. 20. P. 89—
102. ~
Brooker В. E. The cell coat of Crithidia fasciculata // Parasitolo-
gy. 1976. Vol. 72. P. 259—267.
Brooker В. E., Ogden C. G. Encystment of Bodo caudatus // Pro-
toplasma. 1972. Vol. 74. P. 397—409.
Brugerolle G. Des trichocystes chez les bodonides, un charactere
phylogenetique supplementaire entre Kinetoplastida et Euglenida //
Protistologica. 1985. T. 22. S. 339—348.
Brugerolle G., Lom J., Nohynkova E., Joyon L. Comparaison et
evolution des structures cellulaires chez plusieurs especes de bodoni-
des et cryptobiides appartenant aux genres Bodo, Cryptobia et Trypa-
noplasma (Kinetoplastida, Mastigophora) // Protistologica. 1979. T. 15.
S. 197—221.
Bruce D. Preliminary report on the tsetse fly disease or nagana in
Zululand. Durban: Bennett and David, 1895.
Bruce D. Further report on the tsetse fly disease or nagana in
Zululand. London: Harrison and Sons, 1897.
Bruce D. Note on the discovery of a new Trypanosoma // Proc.
Roy. Soc. 1902. Vol. 69. P. 496.
Burzel L. A. Observations on the proboscis-cytopharynx complex
and flagella of Rhynchomonas metabolica Pshenin 1964 (Zoomasti-
gophora: Bodonidae) // J. Protozool. 1973. Vol. 20. P. 385—393.
Biitschli С. M. Beitrage zur Kenntniss der Flagellaten und einiger
verwandten Organismen // Ztschr. Wiss. Zool. 1878. Bd 30. S. 205—
281.
Cachon J., Cachon M., Cosson M.-P., Cosson J. The paraflagel-
lar rod: a structure in search of a function // Biol. Cell. 1988. Vol. 63.
P. 169—181.
Cavalier-Smith T. Kingdom Protozoa and its 18 phyla # Micro-
biol. Rev. 1993. Vol. 57. P. 953—994.
Cavalier-Smith T., Chao E. Molecular evidence that free-living
zooflagellates span 3 kingdoms // Eur. J. Protistol. 1995. Vol. 31. A 2.
P. 110.
Cerisola J. A.4 Del Prado С. E., Rohwedder R., Bozzi-
ni J. P. Blastocrithidia triatomae n.sp. faund in Triatoma infestans
from Argentina // J. Protozool. 1971. Vol. 18. P. 503—506.
Chang К. P. Ultrastructure of symbiotic bacteria in normal and
antibiotic-treated Blastocrithidia culicis and Crithidia oncopelti // J.
Protozool. 1974. Vol. 21. P. 699—707.
Chang К. P. Red used growth of Blastocrithidia culicis and Cri-
thidia oncopelti freed of intracellular symbionts by chloramphenicol //
J. Protozool. 1975. Vol. 22. P. 271—276.
Chang К. P. Nutritional significance of symbiont bacteria in two
species of hemoflagellates // Science. 1974. Vol. 183. P. 531—532.
Chatterjee S. N., Sen Gupta P. C. Ultrastructure of the promasti-
gote of Leishmania donovani // Ind. J. Med. Res. 1970. Vol. 58.
P.70—76. .
Christophers S. R., Shortt H. E., Bairaud P. J. The morphology
and life cycle of the parasite of Indian kala-azar in culture // Ind. Med.
Res. Mem. 1926. Vol. 4. P. 19—53.
Clark T., Wallace F. A comparative study of kinetoplast ultra-
structure in the Trypanosomatidae // J. Protozool. 1960. Vol. 7.
P. 115—121.
Croft S. L. Ultrastructural study of the nucleus of Leishmania her-
tigi // Protistologica. 1979. T. 15. S. 103—110.
Current W. L. Cryotobia sp. In the snail Triadopsis multilineata
(Say): fine structure of attaced flagellates and their mode of attach-
ment to the spermatheca // J. Protozool. 1980. Vol. 27. P. 278—287.
Desser S. S. The ultrastructure of the epimastigote stages of Try-
panosoma rotatorium in the leech Batracobdella picta # Canad. J.
Zool. 1976. Vol. 54. P. 1712—1723.
De Souza W., Souto-Padron T. The paraxial structure of the
flagellum of trypanosomatidae // J. Parasitol. 1980. Vol. 66. P. 229—
235.
Du Y., Maslov D. A., Chang K.-P. Monophyletic origin of p-divi-
sion proteobacterial ehdosymbionts and their coevolution with insect
trypanosomatid protozoa Blastocrithidia culicis and Crithidia spp. //
Proc. Nat. Acad. Sci. USA. 1994. Vol. 91. P. 8437.
Dyrce L., Jacenir R. The development of extraintestinal cycle of
Trypanosoma cruzi in Triatoma infestans and Panstrongylus megis-
tus // An. Acad. Bras. Cienc. 1984. Vol. 56. P. 221—230;
Ehrenberg G.Infbzionstiere als vollkommene Organismen. Leip-
zig, 1830.
Elbraschter M., Schnepf E., Balzer I. Hemistasia phaeocysticola
(Scherffel) comb.nov., redescription of a free-living, marine, phago-
trophic kinetoplastid flagellate // Arch. Protistenk. 1996. Vol. 147.
P. 125—136.
Ellis D., Evans D. A., Stamford S. The penetration of the salivary
glands of Rhodnius prolixus by Trypanosoma rangeli // Ztschr. Para-
sitenk. 1980. Bd 62. S. 63—74.
Eperon S., McMahon-Pratt D.I. Extracellular cultivation and
morphological characterization of amastigote-like forms of Leishma-
nia panamensis and L. braziliensis // J. Protozool. 1989. Vol. 36.
P. 502—510.
Evans D. A., Ellis D. S. Recent observations on the behavior of
certain trypanosomes within their insect hosts // Adv. Parasitol. 1983.
Vol. 22. P. 1—42.
Evans G. Report on ’surra* disease in the Dera Ismail Khan dist-
rict. Punjab. Govt. Milit. Dept. 1880. N 493. P. 446.
Eyden В. P. Morphology and ultrastructure of Bodo designis
Skuja 1948 // Protistologica. 1977. T. 8. S. 169—179.
254
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Fantham Н. В., Herpetomonas pediculi nov. spec., parasitic in the
alimentary tract of Pediculus vestimenti, the human body laouse //
Proc. Roy. Soc. Ser. B. 1912. Vol. 84. P.505—517.
Farina M., Attias M., Souto-Padron T., De Souza W. Further
studies on the organization of the paraxial rod of trypanosomatids // J.
Protozool. 1986. Vol. 33. P. 552—557.
Feng L. C. The role of the peritrophic membrane in Leishmania
and trypanosome infections of sandflies. // Peking Nat. Hist. Bull.
1951. Vol. 19. P. 327—314.
Fernandes A. P., Nelson K., Beverley S. M. Evolution of nuclear
ribosomal RNAs in kinetoplastid protozoa: perspectives on the age
and origin of parasitism // Proc. Nat. Acad. Sci. USA. 1993. Vol. 90.
P. 11608—11612.
Frevert U., Reinwald E. Formation of filopodia in Trypanosoma
congolense by crosslinking the variant surface antigen // J. Ultr.-
Molec. Struct. Res. 1988, Vol. 99. P. 124—136.
Freymuller E., Camargo E. P. Ultrastructural differences be-
tween species of trypanosomatid with and without endosymbionts // J.
Protozool. 1981. Vol. 21. P. 175—182.
Freymuller E., Milder R., Jankevicius J. V., Jankevicius S. I.,
Camargo E. Ultrastructural studies on the the trypanosomatid Phyto-
monas serpens in the salivary glands of a phytophagous hemipteran //
J. Protozool. 1990. Vol. 37. P. 225—229.
Fuge H. Electron microscopic studies on the intra-flagellar struc-
tures of trypanosomes // J. Protozool. 1969. Vol. 16. P. 460—466.
Gardner P. J. Pellicle-associated structures in the amastigote
stage of Trypanosoma cruzi and Leishmania species // Ann. Trop.
Med. Parasitol. 1974. Vol. 68. P. 167—176.
Gibbs A. J. Leptomonas serpens n.sp. parasitic in the digestive
tract and salivary glands of Nezara viridula (Pentatomidae) and in the
sap of Solanum lycopersicum (tomato) and other plants // Parasitolo-
gy. 1957. Vol. 47. P. 297—303.
Gluge G. Ueber ein eigenthiimliches Entozoon im Blute des Fros-
ches II Arch. Anat. Physiol. Wiss. Med. Berlin, 1842. S. 148—164.
Grassg P. Trait£ de Zoologie. Paris, 1952. T. 1. Fasc. 1.
Griessmann K. Ueber marine Flagellaten // Arch. Protistenk.
1913. Bd 32. S. 1—78.
Gruby D. Recherches et observations sur une nouvelle espece
d’hematozoaire, Trypanosoma sanguinis // C. R. Acad. Sci. 1843. T. 17.
S. 1134—1148.
Gutteridge W. E., Macadam R. F. An electron microscopic study
of the bipolar bodies in Crithidia oncopelti // J. Protozool. 1971.
Vol. 18. P. 637—640.
Hecker H., Schwarzenbach M., Rudin W. Development and in-
teractions of Trypanosoma rangeli in and with the reduviid bug Rhod-
nius prolixus // Parasitol. Res. 1990. Vol. 76. P. 311—318.
Heywood P., Weinman D. Mitosis in the hemoflagellate Trypano-
soma cyclops // J. Protozool. 1978; Vol. 25. P. 287—292.
Heywood P., Weinman D., Lipman M. Fine structure of Trypa-
nosoma cyclops in non cellular culture U J. Protozool. 1974. Vol. 21.
P. 232—238.
Hitchen E. T. The fine structure of the colonial kinetoplastid fla-
gellate Cephalothamnium cyclopum Stein // J. Protozool. 1974. Vol. 21.
P. 221—231.
Ноаге C. Studies on Trypanosoma grai. 3. Life-cycle in the tse-
tse-fly and in the crocodile // Parasitology. 1931. Vol. 23. P. 449—
470.
Ноаге C. Morphological and taxonomic studies on mammalian
trypanosomes. Revision of the systematics // J. Protozool. 1964. Vol. 11.
P. 200—223.
Ноаге C. The classification of mammalian trypanosomes // Erg.
Microbiol. 1966. Vol. 39. P. 43—121.
Ноаге C. The trypanosomes of mammals. Oxford: Blackwell,
1972.
Ноаге C. A., Wallace F. G. Developmental stages of trypanoso-
matid flagellates, a new terminology // Nature. 1966. Vol. 212. P. 1385—
1386.
Honigberg B.M. A contribution to the systematics of the non
pigmented flagellates // Progress in Protozoology / Eds J. Ludvig, J.
Vavra. New York: Acad. Press, 1963. P. 68—69.
Ibrahim E., Molyneux D. H. Pathogenicity of Crithidia fascicula-
ta in the haemocoele of Glossina // Acta Tropica. 1987. Vol. 44.
P. 13—22.
Jadin J. Cytologie et cytophysiologie des Trypanosomatidae //
Acta Zool. Pathol. Antverp. 1971. Vol. 53. P. 5—168.
Janovy J., Lee K., Brumbaugh J. The differentiation of Herpeto-
monas megaseliae: ultrastructural observations // J. Protozool. 1974.
Vol. 21. P. 53—59.
Jensen C., Schaub G., Molyneux D. H. The effect of Blastocrit-
hidia triatomae (Trypanosomatidae) on the midgut of the reduviid bug
Triatoma infestans // Parasitology. 1990. Vol. 100. P. 1—9.
Kent W. S. A Manual of the Infusoria. London: Bogue, 1880;
Killik-Kendrick R. Biology of Leishmania in phlebotomine sand-
flies // Biology of the Kinetoplastida. Vol. 2 / Eds W. H. R, Lums-
den, D. A, Evans. London: Acad. Press, 1979. P. 395—460.
Killick-Kendrick R., Molyneux D. H., Ashford R.W. Leis-
hmania in phlebotomid sandflies. I. Modifications of the flagellum
associated with attachment to the mid-gut and oesophageal valve of
the sandfly // Proc. Roy. Soc. London. (B). 1974. Vol. 187. P. 409—
419.
Kivic P. A., Walne P. L. An evaluation of a possible phylogenetic
relationship between the Euglenophyta and Kinetoplastida // Origins
of Life. 1984,. Vol. 13. P. 269—288.
Kleine F. K.Weitere wissenschaftliche Beobachtungen uber die
Entwicklung von Trypanosomen in Glossinen // Dtschr. Med. Wschr.
1909. Bd 35. S. 924—978.
Laird M. Blastocrithidia n.g. (Mastigophora: Protomonadina) for
Crithidia (in part), with a subarctic record for B. gerridis (Patton) //
Canad. J. Zool. 1959. Vol. 37. P. 749—752.
Lake J. A., De la Cruz V. F., Ferreira P. C. G., Morel C., Sim-
pson L. Evolution of parasitism. Kinetoplastid protozoan history con-
structed from mitochondrial rRNA gene sequences // Proc. Nat. Acad.
Sci. USA. 1988. Vol. 89. P. 4779—4783.
Lambrecht F. L. Palaeoecology of tsetse flies and sleeping sick-
ness in Africa // Proc. Amer. Philosoph. Soc. 1980. Vol. 124. P. 367—
385.
Lauge G., Nishioka R. S. Ultrastructural study of the relations be-
tween Leptomonas oncopelti (Noguchi and Tilden), Protozoa, Trypa-
nosomatidae, and the rectal wall of adults of Oncopeltus fasciatus,
Dallas, Hemiptera, Lygaeidae // J. Morphol. 1977. Vol. 154. P. 291—
306.
Lavier G. L*evolution de la morphologie dans le genre Trypano-
soma // Ann. Parasitol. 1943. Vol. 19. P. 168—179.
Leger L. Sur la forme gregarienne de Herpetomonas // C. R.
Seanc. Soc. Biol. 1902. T. 54. P. 400-401.
Leger L. Sur la morphologie du Trypanoplasma du vari-
ous // C. R. Hebd. Seanc. Acad. Soc. Paris, 1904a. Vol. 138. P. 824—
825.
Leger L. Sur les affinites de 1’Herpetomonas sabulata et la phylo-
gene des trypanosomes // C. R. Seanc. Soc. Biol. 1904b. T. 56. S. 615—
617.
Leger L., Dubosq O. Selenococcidium intermedium Leg. et Dub.
et la sistematique des Sporozoaires // Arch. Zool. Exp. Gener. 1910.
T. 5. S. 615—617.
Lehmann D. L. Notes on the life cycle and infectivity of Trypa-
nosoma barbari // J. Parasitol. 1952. Vol. 38. P. 550—553.
Leidy J. Description of anew genus and species of Entozoa //
Proc. Acad. Nat. Sci. Phyladelphia. 1846. Vpl. 3. P. 100—101.
Leidy J. A flora et fauna within living animals // Smiths. Con.
Know. 1853. Vol. 5. P. 1—67.
Leidy J. A synopsis of Entozoa and some of their ectocongeners
observed by the author // Proc. Acad. Nat. Sci. Philadelphia. 1856.
Vol. 8. P. 42—58.
Le Ray D., Vickerman K. Are there any free-living trypanosoma-
tids? // J. Protozool. 1977. Vol. 24. P. 8A.
Lewis J. W., Ball S. J. Attachment of the epimastigotes of Trypa-
nosoma cobitis (Mitrophanow, 1883) to the crop wall of the leech
vector Hemiclepsis marginata // Ztschr. Parasitenk. 1979. Bd 60.
S.29—36.
Lewis J., Ball S. Observations on the division of the epimastigotes
of Trypanosoma cobitis // Arch. Protistenk. 1981. Bd 124. S. 337—
344.
Lewis T. R. The microscopic organisms found in the blood of
man and animals and their relation to disease // Ann. Rep. San. Com-
mis. G. I. C. 1878. N 14.
Lewis T. R. Flagellated organisms in the blood of healthy rats //
Quart. J. Microsc. Sci. 1879. Vol. 19. P. 109—113.
Lipa J. Blastocrithidia raabei sp.n., a flagellate parasite of Meso-
cerus marginalis L. (Hemiptera: Coreidae) // Acta Protozool. 1966.
Vol. 4. P. 19—23.
Lom J. Biology of the trypanosomes and trypanoplasms of fish //
Biology of the Kinetoplastida. Vol. 2 I Eds W. H. R. Lumsden,
D. A. Evans. London: Acad. Press, 1979. P. 269—337.
Lom J. Cryptobia branchialis Nic. from fish gills: ultrastructural
evidence of ectocomensal function // J. Fish Diseas. 1980. Vol. 3.
P. 427H436.
ТИП EUGLENOZOA. КЛАСС KINETOPLASTID£A. ЛИТЕРАТУРА
255
Lom J., Paulin J. J., Nohynkova Б. The fine structure of the fish
trypanosome, Trypanosoma danilewskyi: I. Presence of a cytopharyn-
geal complex in bloodstream trypomastigotes // Protistologica. 1980.
Vol. 16. P. 365—373.
Martin D. S., Desser S. S. A light and electron microscopic study
of Trypanosoma fallisi n.sp. in toads (Bufo americanus) from Algon-
quin Park, Ontario // J. Protozool. 1990. Vol. 37. P. 199—206.
Maslov D., Simpson L. Evolution of parasitism in kinetoplastid
Protozoa II Parasitology Today. 1995. Vol. 11. P. 30—32.
Maslov D. A., Lukes J., Jirku M., Simpson L. Phylogeny of try-
panosomes as inferred from the small and large subunit rRNAs impli-
cations for the evolution of parasitism in the trypanosomatid proto-
zoa // Mol. Biochem. Parasitol. 1996. Vol.75. P.197—205.
McGhee B., Cosgrove W. Biology and physiology of the lower
Trypanosomatidae // Microbiol. Rev. 1980. Vol. 44. P. 140—173.
Mehlhorn H., Schaub G., Peters W., Haberkom A. Electron
microscopic studies on Blastocrithidia triatomae Cerisola et al., 1971
(Trypanosomatidae) // Tropenmed. Parasitol. 1979. Bd 30. S. 289—
300.
Meyer H., De Souza W. On the fine structure of Trypanosoma
cruzi in tissue cultures of pigment epithelium from the chick embryo.
Uptake of melanin granules by the parasite // J. Protozool. 1973.
Vol. 20. P. 590—593.
Milder R., Deane M. P. The cytostome of Trypanosoma cruzi and
T. conorhini // J. Protozool. 1969. Vol. 16. P. 730—737.
Minchin E. A. Investigations on the development of trypanoso-
mes in tsetse flies and other Diptera // Quart. J. Microsc. Sci. 1908.
Vol. 52. P. 159—260.
Minchin E. A. The development of trypanosomes in the inverteb-
rate host // Nature (London). 1914. Vol. 94. P. 405—412.
Molyneux D. H. The fine-structure of the epimastigote forms of
Trypanosoma lewisi in the rectum of the flea Nosopsyllus fasciatus H
Parasitology. 1969. Vol. 59. P. 55—66.
Molyneux D. H. Vector-parasite relationships in the Trypanoso-
matidae // Adv. Parasitol. 1977. Vol. 15. P. 1—82.
Molyneux D. H. Host-parasite relationships of Trypanosomatidae
in vectors // Current topics in vector research. Vol. 1. New York: Prae-
ger, 1983. P. 117—148.
Molyneux D. H., Ashford R. The biology of Trypanosoma and
Leishmania, parasites of man and domestic animals. London: Taylor
and Francis, 1983.
Molyneux D. H., Killick-Kendrick R. Morphology, ultrastructure
and life cycles // The Leishmaniases in Biology and Medicine. Vol. 1.
London: Acad. Press, 1987. P. 121—176.
Nohynkova E. A new pathogenic Cryptobia from freshwater fis-
hes: light and electron microscopic study // Protistologica. 1984.
Vol. 20. P. 181—195.
Novarino G., Warren A., Kinner N. E., Harvey R. W. Protists
from a sewage contaminated aquifer on Cape Cod, Massachusetts //
Geomicrobiology. 1994. Vol. 12. P. 23—36.
Novy F. G., Mac Neal W. J., Torrey H. N. The trypanosomes the
mosquitoes and other insects // J. infect. Dis. 1907. Vol. 4. P. 223—
235.
O’Farell W. R. Preliminary note on a new flagellate, Crithidia
hyalommae sp. n., found in the tick Hyalomma aegyptium (Linnaeus,
1758) // J. Trop. Med. Hyg. |gl3. Vol. 16. P. 245—246.
Paterson D. J., Simpson A. G. B. Heterotrophic flagellates from
costal marine and hypersaline sediments in Western Australia // Eur.
J. Protistol. 1996. Vol. 32. P. 423—448.
Paterson W., Woo P. T. K. Electron microscopic observations of
the blood stream form of Cryptobia salmositica Katz 1951 (Kineto-
plastida: Bodonina) // J. Protozool. 1983. Vol. 30. P. 431—437.
Paterson W. B., Woo P. T. K. Ultrastructural studies on mitosis
in Trypanosoma danilewskyi (Mastigophora: Zoomastigophorea) //
Canad. J. Zool. 1984. Vol. 62. P. 1167—1171.
Peng P. L., Wallace F. G. The cysts of Blastocrithidia triatomae
Cerisola et al., 1971 II J. Protozool. 1982. Vol. 29. P. 464—4(>7.
Porter A. The life-cycle of Herpetomonas jaculum (Leger), para-
sitic in the alimentary tract of Nepa cinerea // Parasitology. 1910.
Vol. 2. P. 367—391.
Preston M. The form and function of the cytostome-cytopha-
rynx of the culture forms of the elasmobranch haemoflagellate Try-
panosoma raiae Laveran et Mesnil // J. Protozool. 1969. Vol. 16.
P. 320—333.
Rangel-Aldao R., Triana F., Fernandez V., Comach G., Abate
T., Montoreano R. Cyclic AMP as an inducer of the cell differentia-
tion of Trypanosoma cruzi // Biochem. Intern. 1988. Vol. 17. P. 337—
344.
Rowton E. D., Lushbaugb W. B., McGhee R. B. Ultrastructure of
the flagellar apparatus and attachment of Herpetomonas ampelophilae
in the gut and malpighian tubules of Drosophila melanogaster // J.
Protozool. 1981. Vol. 28. P. 297—301.
Shaub G. The effects of trypanosomatids on insects // Adv. Para-
sitol. 1992. Vol. 31. P. 255—304.
Schaub G., Boker C. Scanning electron microscopic studies of
Blastocrithidia triatomae (Trypanosomatidae) in the rectum of Tri-
atoma infestans (Reduviidae) // J. Protozool. 1986. Vol. 33. P. 266—
.270.
Schaub G., Losch P. Parasite/host — interrelationships of the try-
panosomatids Trypanosoma cruzi and Blastocrithidia triatomae and
the reduviid bug Triatoma infestans: influence of starvation of the
bug // Ann. Trop. Med. Parasitol. 1989. Vol. 83. P. 215—223.
Schaub G., Pretsch T. Ultrastructural studies on the excystment
of Blastocrithidia triatomae (Trypanosomatidae) // Trans. Roy. Soc.
Trop. Med. Hyg, 1981. Vol. 75. P. 168—171.
Schaub G., Schnitker A. Influence of Blastocrithidia triatomae
(Trypanosomatidae) on the reduviid bug Triatoma infestans: alterati-
ons in the Malpighian tubules // Parasitol. Res. 1988. Vol. 75. P. 88—
97.
Schaub G. A., Boker C. A., Jensen C., Reduth D. Cannibalism
and coprophagy are modes of transmission of Blastocrithidia triatomae
(Trypanosomatidae) between triatomines // J. Protozool. 1989.
Vol. 36. P. 171—175.
Schaub G., Neukirchen K., Golecki J. Attachment of Blastocrit-
hidia triatomae (Trypanosomatidae) by flagellum and cell body in the
midgut of the reduviid bug Triatoma infestans // Eur. J. Protistol.
1992. Vol. 28. P. 322—328.
Schlein Y., Jacobson R., Schlomai J. Chitinase secreted by Lei-
shmania functions in the sandfly vector // Proc. Roy. Soc. London.
(B). 1991. Vol. 245. P. 121—126.
Schneider A., Shervin T., Sasse R., Russel D., Gull K., See-
beck T. Subpellicular and flagellar microtubules of Trypanosoma bru-
cei brucei contain the same a-tubulin isoforms // J. Cell Biol. 1987.
Vol. 104. P. 431—438.
Schultz H., MacClure E. Elektronmikroskopische Untersuchung
des Trypanosoma cruzi mit besonderer Beriicksichtigung des Periplas-
ten und des Blepharoplasten // Ztschr. Zellenforsch. Mikroskop. Anat.
1961. Bd 55. S. 389-412.
Sherwin T., Gull K. The cell division cycle of Trypanosoma bru-
cei brucei: Timing of event markers and cytoskeletal modulations //
Phil. Transact. Roy. Soc. London, B. 1989. Vol. 323. P. 573—588.
Solari A. J. The 3-dimensional fine structure of the mitotic spin-
dle in Trypanosoma cruzi // Chromosoma. 1980a. Vol. 78. P. 23.9—
255.
Solari A. J. Function of the dense plaques during mitosis in Try-
panosoma cruzi II Exp. Cell. Res. 1980b. Vol. 127. P. 457—460.
Solari A. J. Nuclear ultrastructure during mitosis in Crithidia fas-
ciculata and Trypanosoma brucei // J. Protozool. 1982. Vol. 29. P. 330—
331.
Solari A. J. The ultrastructure of mitotic nuclei of Blastocrithidia
triatomae // Ztschr. Parasitenk. 1983. Vol. 69. P. 3—15.
Solari A. J. De Souza W. Presence and comparative behavior of
mitotic plaques in five species of Trypanosomatidae // Microsc. Elec-
tron. Biol. Cell. 1983. Vol. 7. P. 28—43.
Solari A. J. Tandler C. J., Duschak V. Fine structure and cyto-
chemistry of the mitotic plaques of Trypanosoma cruzi and Crithidia
fasciculata // J. Submicrosc. Cytol. 1985. Vol. 17. P. 583—591.
Steiger R. On the ultrastructure of Trypanosoma (Trypanozoon)
brucei in the course of its life cycle and some related aspects // Acta
Tropica. 1973. Vol. 30. P. 64—168.
Steinert M., Novikoff A. The existence of a cytostome and the
occurrence of pinocytosis in the trypanosome, Trypanosoma mega II J.
Biophis., Biochem. Cytol. 1960. Vol. 8. P. 563—569.
Tait A. Sexual processes in the Kinetoplastida // Parasitology.
1983. Vol. 86. P. 29—57.
Taylor A. E. F., Godfrey D. G. A new organelle of bloodstream
salivarian trypanosomes // J. Protozool. 1969. Vol. 16. P. 466—470.
Taylor F. J. R. Problems in the development of an explicit hypo-
thetical phylogeny of the lower eukaryotes // Biosystems. 1978.
Vol. 10. P. 67—89.
Tetley L., Vickerman K. Differentiation in Trypanosoma bru-
cei: host-parasite cell junctions and their persistence during acqui-
sition of the variable antigen coat // J. Cell Sci. 1985. Vol. 74. P. 1—
19.
Tieszen K., Molyneux D. Morphology and hostparasite relation-
ships of Crithidia flexonema (Trypanosomatidae) in the hindgut and
256
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Malpighian tubules of Gerris odontogaster (Hemiptera: Gerridae) // J.
Parasitol. 1989. Vol. 75. P. 441-448.
Tieszen K., Heywood P., Molyneux D. Ultrastructure and hostpa-
rasite association of Blastocrithidia gerridis in the ventriculus of Ger-
ris odontogaster (Gerridae: Hemiptera) // Canad. J. Zool. 1983. Vol. 61.
P. 1900—1909.
Tieszen K., Molyneux D., Abdel-Hafez S. Ultrastructure of cyst
formation in Blastocrithidia familiaris in Lygaeus pandurus (Hemiptera:
Lygaeidae) // Ztschr. Parasitenk. 1985. Vol. 71. P. 179—188.
Tieszen K., Molyneux D., Abdel-Hafez S. Host-parasite relation-
ships of Blastocrithidia familiaris in Lygaeus pandurus Scop. (Hemip-
tera: Lygaeidae) // Parasitology. 1986i Vol. 92. P. 1—12.
Tieszen K., Kenneth L., Molyneux D. H. Transmission and eco-
logy of trypanosomatid flagellates of water striders (Hemiptera, Ger-
ridae) // J. Protozool: 1989a. Vol. 36. P. 519—523.
Tieszen K., Molyneux D., Abdel-Hafez S. Host-parasite relation-
ships and cysts of Leptomonas lygaei (Trypanosomatidae) in Lygaeus
pandurus (Hemiptera: Lygaeidae) // Parasitology. 1989b. Vol. 98.
P. 395—400;
Urena F. Three-dimentional reconstructions of the mitotic spindle
and dense plaques in three species of Leishmania // Ztschr. Parasitenk.
1986. Bd 72. S. 299—306.
Valentin G. Ueber ein Entozoon im Blute von Salmo fario //
Arch. Anat., Physiol, wiss. Med. Berlin, 1841. S. 435—444.
Van der Ploeg L.H.T., Schwartz D. C., Cantor C. R.,
Borst P. Antigenic variation in Trypanosoma brucei analyzed by elec-
trophoretic separation of chromosome-sized DNA molecules II Cell.
1984a. Vol. 37. P. 77—84.
Van der Ploeg L. H. T., Cornelissen A. W. C. A., Barry J. D.,
Borst P. Chromosomes of Kinetoplastida // EMBO J. 1984b. Vol. 3.
P. 3109—3115.
Van der Ploeg L. H. T., Smith C. L., Polvere R. I., Gottesdie-
ner К. M. Improv^ separation of chromosome-sized DNA from Try-
panosoma brucei, stock 427-60 // Nucl. Acids Res. 1989. Vol. 17.
P. 3217—3227.
Vickerman K. Electron microscope studies on akinetoplastic try-
panosomes II J. Protozool. 1963. Vol. 10. (suppl. 15).
Vickerman Ki Polymorphism and mitochondrial activity in slee-
ping sicknes trypanosomes // Nature. 1965. Vol. 208. P. 762—774.
Vickerman K. The fine structure of Trypanosoma congolense in
its blood phase// J. Protozool. 1969. Vol. 16. P. 54—69.
Vickerman K. The diversity of the kinetoplastid flagellates // Biology
of the Kinetoplastida. Vol. 17 Eds W. H. R. Lumsden, D. A. Evans.
London etc.: Acad. Press, 1976. P. 1—34.
Vickerman K. DNA throughout the single mitochondrion of a ki-
netoplastid flagellate: observations on the ultrastructure of Cryptobia va-
ginalis (Hesse, 1910) // J. Protozool. 1977. Vol. 24. P. 221—233.
Vickerman K. The free-living trypanoplasms. Descriptions of
three species of the genus Procryptobia N. G. and redescription of
Dimastigella trypaniformis Sandon, with notes on their relevance to
the microscopical diagnosis of disease in man and animals // Trans.
Amer. Microsc. Soc. 1978. Vol. 97. P. 485—502.
Vickerman K. Phylum Zoomastigina. Class Kinetoplastida //
Handbook of Protoctista / Eds L. Margulis et al. Boston: Jones and
Bartlett, 1989. P. 215—238.
Vickerman K. Organization of the bodonid flagellates // The bio-
logy of free-living heterotrophic flagellates. The systematics associa-
tion special volume / Eds D. J. Paterson, J. Larsen. 1991. Vol. 45.
P. 159—184.
Vickerman K. The evolutionary expansion of the trypanosomatid
flagellates // Int. J. Parasitol. 1994. Vol. 24. P. 1317—1331.
Vickerman K., Preston T. M. Spindle microtubules in the divi-
ding nuclei of trypanosomes // J. Cell Sci. 1970. Vol. 6. P. 365—383.
Vickerman K., Preston T. M. Comparative cell biology of the
kinetoplastid flagellates // Biology of the Kinetoplastida. Vol. 1 / Eds
W. H. R. Lumsden, D. A. Evans. London etc.: Acad. Press, 1976.
P. 35—130.
Wail B., Oschman J. Structure and function of the rectum in
insects // Fortschritte der Zoologie. 1975. Vol. 23. P. 193—222.
Wallace F. G. The trypanosomatid parasites of Insects and Arach-
nids // Exp. Parasitol. 1966. Vol. 18. P. 124—193.
Walters L., Modi G., Tesh R., Burrage T. Host-parasite relation-
ship of Leishmania mexicana mexicana and Lutzomyia abonnenci
(Diptera: Psychodidae) // Amer. J. Trop. Med. Hyg. 1987. Vol. 39.
P. 294—314.
Walters L., Modi G., Chaplin G., Tesh R. Ultrastructural deve-
lopment of Leishmania chagasi in its vector, Lutzomyia longipalpis
(Diptera: Psychodidae) // Amer. J. Trop. Med. Hyg. 1989. Vol. 41.
P. 295—317.
Weinman D., Wite E., Antipa G. Trypanosoma lucnowi a new
species of trypanosome from Macaca mulata with observations on its
fine structure//J. Protozool. 1984. Vol. 31. P. 429—433.
Wenyon C. Protozoology. London, 1926.
Woo P.T.K. Origin of mammalian trypanosomes which develop
in the anterior-station of blood-sucking artropods // Nature. London,
1970. Vol. 228. P. 1059—1062.
Woo P. T. K. Cryptobia and Cryptobiasis in fishes // Adv. Parasi-
tol. 1987. Vol. 26. P. 199—237.
Woo P. T. K., Wehnert S. D. Direct transmission of a hemofla-
gellate, Cryptobia salmositica (Kinetoplastida: Bodonina) between ra-
inbow trout under laboratory conditions // J. Protozool. 1983. Vol. 30.
P. 334—337.
Woodcock H.M. The haemoflagellates: a review of present
knowledge relating to the trypanosomes and allied forms // Quart. J.
Microsc. Sci. (N. S.) 1906. Vol. 50. P. 151—168.
Woodcock H. M. Observations on coprozoic flagellates // Phylos.
Trans. Roy. Soc. Ser. B. London, 1914. Vol. 207. P. 375—412.
Zeledon R., Monge E. Natural immunity of the bug Tryatoma
infestans to the protozoan Trypanosoma rangeli // J. Invertebr. Pathol.
1966. Vol. 8. P. 420—424.
Тип CHRYSOPHYTA Pascher, 1914 - ХРИЗОФИТЫ
Преимущественно фототрофы (но есть и бесцветные формы),
с различной организацией таллома, а также одиночные и колони-
альные протисты. Большинство из них подвижные, но есть и при-
крепленные особи разнообразного вида и размеров. Жгутиковые
клетки обычно имеют 2 гетероконтных жгутика: двигательный пе-
редний несет трубчатые мастигонемы, задний жгутик гладкий,
часто прилегает к поверхности клетки, формируя фоторецептор-
ный аппарат. В переходной зоне жгутика расположена спираль.
Хлоропласты окрашены в желто-зеленый или золотисто-бурый
цвет, обычно располагаются в перинуклеарном пространстве, име-
ют оболочку из 3 мембран и содержат хлорофиллы а и с, а также
каротиноиды. Митохондрии с трубчатыми или пузырьковидными
кристами. Резервные вещества — хризоламинарин и липиды. Раз-
множение агамное, половой процесс изучен у немногих видов.
В систему типа включены 3 класса, которые объединяют
около 1000 видов, распространенных всесветно.
История изучения
Интенсивное изучение Chrysophyta началось во
второй половине XIX столетия, когда было сделано
описание многих одиночных и колониальных видов,
изучены особенности морфологии и биологии целого
ряда этих жгутиконосцев. Накопленные знания поз-
волили Штейну (Stein, 1878) выделить жгутиконос-
цев, окрашенных в золотистый цвет, в самостоятель-
ный таксон Chrysomonadina Stein, 1878 и включить
его в класс Mastigophora Diesing, 1856. Впоследствии
по мере описания новых видов и изучения морфологии
вегетативных форм и цист система этого таксона много
раз пересматривалась, что послужило причиной вы-
деления его в отряд Chrysomonadida Engler, 1898.
Система типа Chrysophyta подвергалась многим
изменениям и продолжает разрабатываться в настоя-
щее время, В соответствии с зоологической систе-
мой, принятой в 1980 г. (Levine et al., 1980), хризо-
фитам был придан ранг отряда Chrysomonadida En-
gler, 1898 в классе Phytomastigophorea Calkins, 1909.
В ботанической системе хризофиты выделены в са-
мостоятельный отдел Chrysophyta — золотистые во-
доросли, в состав которого было включено 5 классов
в соответствии с типом организации таллома (Матви-
енко, 1977). Класс Chrysopodophyceae объединяет
организмы, имеющие преимущественно амебоидный
таллом, класс Chrysomonadophyceae — хризофиты с
монадной организацией тела. Представители класса
Chrysocapsophyceae характеризуются пальмеллоидной
17 Протисты, часть 1
(=гемимонадной), класса Chrysosphaerophyceae —
коккоидной структурой тела; класс Chrysotrichophy-
сеае объединяет организмы с нитчатым, разнонитча-
тым и пластинчатым строением тела водоросли.
Однако начиная с середины 70-х гг. в результате
накопления данных по ультраструктуре Chrysophyta
стал развиваться другой подход к классификации
этих водорослей. Благодаря работам Д. Хибберда
(D. J. Hibberd), Г. Прейсига (Н. R. Preisig), О. Меструпа
(0. Moestrup) и Р. Андерсена (R. A. Andersen) глав-
ными признаками в макро- и мегасистематике Chry-
sophyta становятся особенности организации жгути-
ковой клетки. Это направление продолжает успешно
развиваться и в настоящее время, что привело к пере-
смотру существовавшей системы типа Chrysophyta и
принятию новой системы, опубликованной Прейси-
гом (Preisig, 1995).
«Class Chrysophyceae Pasher (1914, nom. descript.), Hibberd (1976,
nom. tipificatum)
Order 1. Bicosoecales Grassd, 1926 («Bicoecinae», nom. nud.)
Order 2. Chromulinales Pascher, 1910 (incl. Ochromonadales, Pas-
cher, 1910)
Order 3. Hibberdiales Andersen, 1989
Order 4. Hydrurales Pascher, 1931
Order 5. Sarcinochrysidales Gayral et Billard, 1977
Order 6. Chrysomeridiales O’Kelly et Billard, 1995
Class Dictyochophyceae Silva, 1980
Order 1. Pedinellales Zimmermann et al., 1984
Order 2. Rhizochromulinales O’Kelly et Wujek, 1994
Order 3. Dictyochales Haeckel, 1894 («Dictyocheae»)
Class Synurophyceae Andersen, 1987 (Cavalier-Smith, 1986: «Synurea»)
Order 1. Synurales Andersen, 1987»
В списке отсутствует класс Pelagophyceae Ander-
sen et Saunders, 1993 по причине задержки выхода в
свет всей книги «Chrysophyte algae», в которой опуб-
ликована эта система.
Класс Dictyochophyceae объединяет организмы с
аксоподиями, аксонемы которых берут начало от
ядерной оболочки. Эти протисты были включены ранее
в класс Pedinellophyceae Karpov, 1990 типа Pedinello-
morpha Karpov, 1990 (Карпов, 1990), который в данном
издании не рассматривается. Здесь приводится под-
робная характеристика класса Chrysophyceae и отряда
Bicosoecales, представители которого являются класси-
ческим объектом протозоологов. Для традиционно «аль-
гологических» таксонов даны только общие сведения.
258
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 163. Типы организации одиночных видов хризофитов. (По: Doflein, 1953; Матвиенко, 1977; Hibberd, Leedale, 1985).
Жгутиковые (монадные) формы: А — Chromulina psammobia (7 — передний, двигательный жгутик; 2 — хлоропласты; 3 — ядро; 4 — хризоламинарин),
Б — Spumella (Monas) guttula; амебоидные (ризоподиальные) формы: В — Brechmiella chrysphydra, Г — Ochromonas elegans (1 — псевдоподия с фагоци-
тозной вакуолью); коккоидная форма: Д — Chrysococcus ornatus; виды, имеющие домик: Е — Pseudokephyrion acutum, Ж — Stokesiella dissimilis (1 —
t домик).
Fig. 163. Some types of structure in individual species of Chrysophyta.
А, Б — Flagellate (monad) species: A — Chromulina psammobia (1 — anterior locomotory flagellum, 2 — chloroplasts, 3 — nucleus, 4 — chrysolaminarin),
Б — Spumella (Monas) guttula; В—Г — Amoeboid species: В — Brechmiella chrysohydra, Г — Ochromonas elegans (1 — pseudopodium with phagocytic va-
cuole); Д — coccoid species: Chrysococcus ornatus; E, Ж — species with shell: E — Pseudokephyrion acutum; Ж — Stokesiella dissimilis (1 — shell).
Классификация
В соответствии с принятой в настоящем издании
системой тип Chrysophyta разделяется на 3 класса:
Класс Chrysophyceae Pascher, 1914 — Хризофитовые
Класс Synurophyceae Andersen, 1987 — Синурофитовые
Класс Pelagophyceae Andersen, Saunders, 1993 — Пелагофитовые
В протозоологической литературе приняты окон-
чания отрядов «-ida», хотя это не требуется Между-
народным кодексом зоологической номенклатуры
(МКЗН), который определяет окончания до надсе-
мейства включительно. Тем не менее здесь мы будем
придерживаться традиционных «ботанических» окон-
чаний, так как только бикозоециды выделяются из
общей массы хризофитовых.
Ниже рассмотрены основные черты, присущие
каждому из трех классов, входящих в тип Chryso-
phyta.
Класс Chrysophyceae Pascher, 1914 — Хризофитовые
Представители класса несут все основные черты типа. Среди
них преобладают одноклеточные жгутиковые формы, однако
встречаются коккоидный, ризоподиальный, нитчатый и паренхи-
матозный типы организации тела. Клетки чаще всего покрыты
лишь плазмалеммой, хотя многие живут в домиках, или панцирях.
Монады обычно гетероконтные. Задний жгутик может редуциро-
ваться частично или полностью (остается лишь кинетосома). Клет-
ки содержат от одного до нескольких хлоропластов. Вспомога-
тельный пигмент — фукоксантин. Наиболее распространенный за-
пасной продукт — хризоламинарин. Размножение осуществляется
бесполым И подовым путями. Хризофитовые обитают обычно в
пресных водах, формируя большую часть планктона прохладных
водоемов.
Морфология и физиология
Форма и размеры тела
Одну из особенностей внешнего строения хризо-
фитовых составляет широкая вариабельность размеров
и формы тела (рис. 163). Постоянная форма тела при-
суща большинству видов, но некоторые из них имеют
амебоидную форму и способны к образованию псев-
доподий различного типа, имеются также плазмодии.
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE
259
Рис. 164. Некоторые типы колоний хризофитовых. (По: Doflein, 1953; Матвиенко, 1977; Hibberd, Leedale, 1985).
А — Uroglenopsis europea, Б — Hyalobryon ramosum, В — Heliapsis mutabilis, Г — Chrysidiastrum catenatum, Д — Chrysosphaerella brevispina (7 — ШИПИ-
ки соматических чешуек зооидов).
Fig. 164. Some types of colony in Chrysophyta.
A — Uroglenopsis europea, Б — Hyalobryon ramosum, В — Heliapsis mutabilis, Г — Chrysidiastrum catenatum, Д — Chrysosphaerella brevispina: 1 — thorns
of somatic scales in zooids.
Большинство хризофитовых — подвижные од-
ноклеточные и колониальные организмы (рис. 163;
164), но нередки и неподвижные, находящиеся на
большей части жизненного цикла в пальмеллоидном
состоянии, и им свойственна шаровидная или эллип-
соидальная форма тела.
Жгутиковый аппарат
Подвижные хризофитовые имеют 2 жгутика, отхо-
дящих от вершины переднего конца тела. Один из них
длинный, второй — очень короткий. Первый, двигатель-
ный жгутик, направлен вперед и несет трубчатые масти-
гонемы, расположенные с обеих сторон (рис. 165) или
только с одной стороны. Каждая мастигонема состоит
из короткого основания, или базальной части, длиной
0.2—0.3 мкм, длинного трубчатого стержня, достигаю-
щего в длину 0.7—0.8 мкм, и дистальной части, пред-
ставленной одним или несколькими филаментами.
Длинные и короткие филаменты расположены также
по обе стороны трубчатого стержня (рис. 166).
Второй жгутик короткий, без мастигонем. Он на-
правлен вдоль тела, и у некоторых видов поверхность
его покрыта мелкими чешуйками.
Переходная зона жгутиков короткая, в ней кроме
одной поперечной пластинки имеется еще и одинар-
ная спираль.
260
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 165. Схема строения переднего конца клетки хризофитовых.
(По: Hibberd, 1976; Карпов, 1990).
1 — передний жгутик с мастигонемами, 2 — короткий жгутик, 3 — па-
рафлагеллярное тельце, 4 — хлоропласт, 5 — стигма, 6 — ламелла хло-
ропласта, 7 — ядро, 8 — аппарат Гольджи (диктиосома), 9 — поперечно
исчерченный корешок (ризопласт), 10 — микротрубочковый корешок,
11 — сократительная вакуоль, 12 — митохондрия.
Fig. 165. Diagram of structure of anterior part in chrysophyte cell.
1 — anterior flagellum with mastigonemes, 2 — short flagellum, 3 — parafla-
gellar body, 4 — chloroplast, 5 — stigma, 6 — lamella of chloroplast, 7 —
nucleus, 8 — Golgi apparatus (dictyosome), 9 — rhizoplast, 10 — microtubu-
lar rootlet, 11 — contractile vacuole, 12 — mitochondrion.
Рис. 166. Строение трехчленной трубчатой мастигонемы хризофи-
товых. (По: Bouck, 1971).
1 — терминальные филаменты, 2 — трубчатый стержень с длинными и
короткими филаментами (волосками) с обеих сторон, 3 — базальная
часть.
Fig. 166. Structure of tripartite tubular mastigoneme of Chrysophyta.
1 — terminal filament; 2 — tubular shaft with long and short filaments, on
the right and left sides; 3 — basal part.
Корешковая система, укрепляющая жгутики, име-
ет черты, специфичные для хризофитовых. Она изу-
чена у ряда видов, в числе которых виды рода Ос-
hromonas (Hibberd, 1976). Особенностью корешковой
системы жгутиконосцев этого рода служит сильно
развитый поперечно исчерченный фибриллярный
корешок — ризопласт, идущий от кинетосом к ядру.
Он достигает ядерной оболочки и соединяет кинето-
сомы с ядром (рис. 165). Развиты также корешки,
состоящие из микротрубочек. Они расположены ла-
терально и проходят непосредственно под плазма-
леммой переднего конца клетки. Расположение мик-
ротрубочковых корешков весьма консервативно в
пределах класса (рис. 88). В наиболее типичном ва-
рианте от кинетосомы переднего жгутика отходят
2 корешка: 1-й опоясывает передний конец клетки
по периферии, давая начало вторичным микротру-
бочкам, 2-й, короткий корешок, проходит между ки-
нетосомами. От кинетосомы рекуррентного жгутика
также отходят 2 корешка: 3-й и 4-й. Они идут с
противоположных сторон кинетосомы навстречу друг
другу и, огибая углубление поверхности клетки, в
котором обычно находится стигма, образуют петлю.
Различия в строении корешковой системы характе-
ризуют хризофитовых как на уровне отрядов, так и
на уровне родов.
Амебоидные формы, образующиеся в жизненном
цикле многих видов, иногда обладают жгутиками на-
ряду с ризоподиями.
Поверхностные структуры
Основу структур поверхности тела хризофитовых
составляет плазмалемма с хорошо развитым слоем
гликокаликса. У ряда видов на поверхности нет
каких-либо дополнительных структур и тело очень
пластично. Поверхность тела других видов покрыта
тонкими кремнеземными, известковыми или органи-
ческими соматическими чешуйками. Немало также
форм, которым свойственны домики, или поверх-
ность тела их армирована какими-либо другими, спе-
цифичными для вида, структурами (рис. 167).
Цитоплазма
Хлоропласты. В цитоплазме фотосинтезирующих
хризофитовых имеется 1—2 удлиненных хлороплас-
та с хорошо выраженными пиреноидами (рис. 165;
168). Хлоропласта находятся в перинуклеарном про-
странстве и имеют собственную оболочку, состоя-
щую из 3 мембран. Ламеллы обычно образованы
3 тилакоидами, по периферии хлоропласта лежит
опоясывающая ламелла. Матрикс пиреноида свобо-
ден от тилакоидов. Комплекс пигментов хлоропластов
включает хлорофиллы а, с1 и с2, p-каротин и не-
сколько форм ксантофиллов, среди которых фуко-
ксантин, диадиноксантин, диатоксантин и одна форма
эхинеона. Окраска хлоропластов чаще всего желто-
коричневая, золотистая или желто-зеленая. Бесцвет-
ные представители хризофитовых (Spumella, Рага-
physomonas) содержат рудиментарный хлоропласт —
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE
261
Рис. 167. Соматические чешуйки хризофитовых. (По: Hibberd, Leedale, 1985).
А — Paraphysomonas vestita (7 — трофозоит, фагоцитирующий пищевой объект; 2 — чешуйки, х 2000), Б — Mallomonas caudata (7 — трофозоит; 2 —
чешуйка, х 5000), В — чешуйки Synura uvella (х 7500), Г — чешуйки Chrysosphaerella brevispina (1 — овальная чешуйка, х 10000; 2 — игла вместе с
овальной чешуйкой, х 5000).
Fig. 167. Somatic scales of Chrysophyta.
A — Paraphysomonas vestita (7 — trophozoite; 2 — scales, x 2000), Б — Mallomonas caudata (7 — trophozoite; 2 — body scale, x 5000), В — body scale of
Synura uvella, x 7500; Г — body scales of Chrysosphaerella brevispina, x 10000.
Рис. 168. Строение жгутиковой клетки хризофитовых. (По: Hibberd, Leedale, 1985).
А — Chrysococcus rufescens (схематизировано по электронограмме), Б — Microglena butcheri, В — Ochromonas tuberculatus, х 2000. (Б, В рисунки сде-
ланы с помощью светового микроскопа). 1 — передний жгутик с мастигонемами, 2 — короткий жгутик, 3 — стигма, 4 — хлоропласт, 5 — сократитель-
ная вакуоль, б — капли жира, 7 — аппарат Гольджи, 8 — ядро, 9 — митохондрии, 10 — хризоламинарин, 77 — жгутиковый резервуар, 72 — дискоболо-
цисты.
Fig. 168. Structure of some flagellate cells of Chrysophyta.
A — Chrysococcus rufescens (diagrammatic section based upon electron microscopy), Б — Microglena butcheri, В — Ochromonas tuberculata, x 2000. 7 — ante-
rior flagellum with mastigonemes, 2 — short flagellum, 3 — stigma, 4 — chloroplast, 5 — contractile vacuole, 6 — drops of lipid, 7 — Golgi apparatus, 8 — nucle-
us, 9 — mitochondria, 10 — chrysolaminarin, 77 — flagellar reservoir, 72 — discobolocysts.
262
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
лейкопласт, который также располагается в перинук-
леарном пространстве. Бикозоециды, однако, пол-
ностью лишены каких-либо следов хлоропластов.
Стигма. С одним из хлоропластов тесно связана
стигма (рис. 165; 168), имеющаяся у большинства ви-
дов. Она расположена в матриксе верхнего, более уз-
кого конца хлоропласта. Стигма состоит из одного
ряда липидных глобул, содержащих пигмент, близкий к
астаксантину — производному Р-каротина. Часто стиг-
ма находится в особом отростке хлоропласта, кото-
рый плотно прилегает к поверхности клетки. Именно
в этом месте находится парафлагеллярное тельце,
или вздутие в основании короткого жгутика. В том
месте, где расположено парафлагеллярное тельце, по-
верхность клетки обычно слегка вогнута (рис. 165).
Митохондрии. У хризофитовых митохондрии мел-
кие, многочисленные, расположенные во всем объ-
еме цитоплазмы. Кристы митохондрий пузырьковид-
ные (рис. 165).
Аппарат Гольджи. Эта органелла (рис. 165) пред-
ставлена 1 или, реже, несколькими диктиосомами,
расположенными в участке цитоплазмы между кине-
тосомами и ядром (Hibberd, 1976). Диктиосомы
крупные, имеют изогнутую форму, и их цистерны
окружены многочисленными пузырьками. Один из
типов пузырьков содержит материал, идущий на фор-
мирование мастигонем двигательного жгутика. Дик-
тиосомы также являются местом формирования со-
матических и жгутиковых чешуек у ряда видов.
Сократительная вакуоль. Все пресноводные виды
имеют 1 или несколько сократительных вакуолей,
локализованных в передней части тела. У морских
видов сократительная вакуоль отчетливо не выра-
жена.
Экструсомы. Эти органеллы представлены слизе-
носными тельцами и дискоболоцистами (рис. 75).
Первые особенно сильно развиты у видов, имеющих
слизистую оболочку.
Резервные вещества. Основное резервное вещес-
тво хризофитовых — хризоламинарин, или лейко-
зин, накапливается в цитоплазме в виде 1 или не-
скольких капель, приуроченных к нижней части тела
(рис. 168). В цитоплазме накапливаются также липи-
ды в виде мелких капель.
Ядро
Хризомонады — одноядерные жгутиконосцы.
Ядро расположено в переднем конце тела. Форма его
шаровидная или грушевидная, и более узкий конец
направлен к кинетосомам, что связано с особеннос-
тями его деления. Ядро пузырьковидного типа, с 1
ядрышком, занимающим центральную зону нуклео-
плазмы. В период интерфазы хроматин диспергиро-
ван. От наружной мембраны отпочковываются много-
численные пузырьки, участвующие в формировании
ближайшей к ядру цистерны диктиосомы. Наружная
мембрана ядра участвует и в формировании мастиго-
нем двигательного жгутика. Мастигонемы формиру-
ются в расширенных участках перинуклеарного про-
странства, а затем мигрируют к поверхности клетки
(Leedale, 1970; Farmer, 1980).
Процесс митоза изучен лишь у немногих видов
хризофитовых. Ультраструктурные исследования по-
казали, что деление ядра относится к типу открытого
ортомитоза (Райков, 1978). Наиболее детально он ис-
следован у Ochromonas danica (Slankis, Gibbs, 1972;
Bouck, Brown, 1973).
Перед митозом происходит удвоение кинетосом,
ризопласт разделяется на 2 части и каждая пара до-
черних кинетосом получает свой ризопласт. В про-
фазе митоза ядерная оболочка распадается и остают-
ся только части ее, контактирующие с хлоропласта-
ми. Веретено митоза образуется между ризопластами,
которые представляют центры формирования микро-
трубочек веретена. В метафазе образуется экватори-
альная пластинка, представленная мелкими компакт-
ными хромосомами без морфологически выраженных
кинетохоров. В составе веретена есть непрерывные и
хромосомные микротрубочки. В анафазе и телофазе
дочерние группы хромосом располагаются у полюсов
делящегося ядра. Формирование новых ядерных обо-
лочек в дочерних клетках происходит в телофазе.
Размножение и жизненные
циклы
Агамное размножение
Эта основная форма размножения присуща всем
хризофитовым и осуществляется путем деления мате-
ринской особи на 2 дочерние, а также путем распада
колоний и плазмодиев на части. Деление происходит
в подвижном или пальмеллоидном состоянии. В ре-
зультате бинарного деления, не доведенного до кон-
ца, образуются монотомические колонии преимущес-
твенно шаровидной и древовидной формы. Агамное
размножение происходит также с помощью жгутико-
вых и амебоидных зооспор.
Половой процесс
Половое размножение известно пока только для
немногих видов и изучено еще недостаточно. Проис-
ходит оно в форме изогамии.
Жизненные циклы
В жизненном цикле ряда видов происходит отчет-
ливо выраженная смена генераций, морфологически
и функционально отличающихся друг от друга. На-
пример, у Chrysamoeba radians в жизненном цикле
чередуются амебоидная и жгутиковая формы (рис. 169).
В сложном жизненном цикле Myxochrysis paradoxa
регулярно чередуются стадии многоядерного плазмо-
дия, жгутиковых и амебоидных зооспор (рис. 169).
Для хризофитовых характерны цисты покоя, ко-
торые очень разнообразны, специфичны для каждого
вида и образуются в результате как агамного размно-
жения, так и полового процесса (рис. 169). Типичная
циста хризофитовых состоит из кремниевых соедине-
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE
263
Рис. 169. Стадии жизненных циклов некоторых хрузофитовых. (По: Doflein, 1953; Матвиенко, 1977; Hibberd, Leedale, 1985).
А — Chrysamoeba radians (7 — ризоподиапьная форма; 2 — жгутиковая, плавающая форма), Б — Myxochrysis paradoxa (7 — растущий плазмодий. 2 —
зрелый плазмодий, 3 — образование амебоидных форм), В — Chromulina pascheri (7 — жгутиковая форма, 2 — амебоидная форма, 3 — циста).
Fig. 169. Stages of the life cycles of some chrysophytes.
A — Chrysamoeba radians (7 — rhizopodial form; 2 — flagellate, swimming form), Б — Myxochrysis paradoxa (7 — young plasmodium, 2 — mature plasmodi-
um, 3 — formation of amoeboid specimens), В — Chromulina pascheri (7 — flagellate, 2 — amoeboid form, 3 — cyst).
ний и обычно имеет отверстие, закрытое пробочкой.
В состоянии цист многие виды могут находиться в
течение длительного времени, особенно при небла-
гоприятных условиях внешней среды.
Экология, распространение
и практическое значение
До недавнего времени хризофитовые считались
пресноводными жгутиконосцами. Но в последние
годы появились данные о широком распространении
этих жгутиконосцев в морских бассейнах, где им
принадлежит значительная роль в планктонных со-
обществах, особенно в шельфовых зонах. Хризофи-
товые распространены всесветно, но наиболее мно-
гочисленны в водных бассейнах умеренного клима-
та. В почве встречаются редко.
Основная масса хризофитовых — обитатели
планктона водоемов, но среди них есть также нейс-
тонные и бентосные формы. Особенно много этих
жгутиконосцев в воде сфагновых болот, которые
имеют повышенную кислотность.
В морских и пресных водоемах хризофитовые встре-
чаются в течение всех сезонов, но максимум их развития
относится к холодному времени. В планктоне они доми-
нируют ранней весной, поздней осенью и зимой.
Размножаясь в огромном количестве, хризофиты
составляют в водоемах значительную долю биопро-
дукции, являясь пищевыми объектами для многих
морских и пресноводных животных. Иногда нейстон-
ные виды вызывают «цветение» воды. Например, при
массовом развитии пресноводного жгутиконосца
Chromulina rosanoffii вода окрашивается в золотистый
цвет — в 1 мм2 поверхностной пленки воды содержит-
ся около 23 000 особей хромулины (Сафонова, 1977).
264
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отряд Bicosoecales (Grassd,
1926) emend. Karpov, 1998 —
бикозоециды
Бесцветные гетероконтные хризофитовые с 1 или 2 жгутика-
ми, живущие в домиках или без домиков. Передний жгутик с 1 или
2 рядами трубчатых мастигонем свободен, задний гладкий, часто
прикрепляется дистальным концом к основанию домика или суб-
страту. Аппарат питания представлен специальным подвижным
выростом (губой) или цитостомом, ведущим в глотку. Жгутиковый
аппарат характеризуется развитой корешковой системой из мик-
ротрубочек «охромонадного» типа. Самый широкий корешок свя-
зан с цитостомальной областью, в основании ассоциирован с фиб-
риллярным материалом и в поперечном сечении имеет характер-
ный профиль в форме буквы «Г». К широкому корешку обычно
прилегает митохондрия, имеющая пузырьковидные или трубчатые
кристы. Среди бикозоецид есть одиночные и колониальные
формы, свободноплавающие и прикрепленные, морские и пресно-
водные.
Классификация
До недавнего времени в отряде выделялся 1 род
Bicosoeca (Bourrelly, 1951; Жуков, 1978). Все ранее
описываемые роды (Codomonas, Poteriodendron, Ste-
phanocodon) оказались различными стадиями в жиз-
ненном цикле того или иного вида Bicosoeca (Жуков,
1978). Сравнительно недавно были описаны и изуче-
ны новые роды и виды бесцветных жгутиконосцев из
морских и пресных вод. Многие из них оказались
близки Bicosoeca по строению клетки и справедливо
отнесены к бикозоецидам. Прежде всего это род Ca-
feteria (Fenchel, Patterson, 1988), насчитывающий
4 вида, представители которых лишены домиков, но
по ультратонкому строению клетки сходны с бико-
зоецидами. По ряду признаков к этому отряду следу-
ет отнести вид Discocelis saleuta, описанный Вере в
1988 г. (V0rs, 1988), 2 вида Pseudobodo (Preisig et al.,
1991), а также представителей родов Adriamonas
(Verhagen et al., 1994), Acronema (Teal et al., 1998) и
Caecitellus (O’Kelly, Nerad, 1998). Последняя ревизия
этого отряда была проведена после описания нового
рода и вида Siluania monomastiga Karpov, 1998, вклю-
ченного в новое сем. Siluaniaceae Karpov, 1998 (Кар-
пов и др., 1998).
Рис. 170. Внешний вид некоторых бикозоецид (А—С) и стадии де-
ления и развития Bicosoeca conica (Т—Щ). (По: Жуков, 1978).
А, Б — В. annulata; В—Ж — В. mitra (Г —- цисты внутри домика); 3, И —
В. depucquesiana; К — В. lacustris (особи, сидящие на Melosira); Л —
В. ovata; М — В. griessmanni; Н — В. synoica, сидящая на В. planctonica;
О — В. mediterranea (особи сидящие на нити Skeletonema); П—С — В. ех-
ilis; Т — взрослая питающаяся особь В. conica; У — начало деления; Ф —
завершение деления; X — выход дочерней клетки из домика; Ц — плаваю-
щая особь; Ч — клетка, прикрепившаяся к субстрату через 30 мин после
выхода из домика; Ш, Щ— фазы формирования домика через 1—1.5 ч пос-
ле прикрепления к субстрату.
Fig. 170. General view of some bicosoecids (A—C) and stages of
cell division and development of Bicosoeca conica (Т—Щ).
А, Б — B. annulata; В—Ж — В. mitra (Г — cysts inside the lorica); 3, И —
В. depucquesiana; К — В. lacustris (attached to Melosira); Л — B. ovata; M —
B. griessmanni, H — B. synoica (attached to B. planctonica); О — В. mediterra-
nea (attached to Skeletonema); П—C — B. exilis; T — adult trophozoit of B. co-
nica; У — first stage of cell division; Ф — last stage of cell division; X —
separation of daughter cell; Ц — swimming cell; 4 — cell attached to the sub-
stratum in 30 min after leaving the lorica; Ш, Щ — stages of lorica formation
in 60—90 min after attachment.
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE. ОТРЯД BICOSOECALES
265
Рис. 171. Схема общей организации клетки Bicosoeca. (По: Mig-
not, 1974).
аг — аппарат Гольджи, в — вентральная сторона клетки, г — губа, гл —
гликоген, д — дорсальная сторона клетки, зж — задний жгутик, м — мито-
хондрия, мн — мастигонемы, мт — микротрубочки, пв — пищеваритель-
ная вакуоль, пж — передний жгутик, пт — параксиальный тяж, кп — кине-
тосома переднего и кз — кинетосома заднего жгутиков, ф — фагосома,
эр — ЭПР, я — ядро. Стрелками показаны микротрубочки жгутиковых ко-
решков и губы.
Fig. 171. Cell structure of Bicosoeca.
аг — Golgi apparatus, в — ventral side of the cell, г — lip, гл — glycogen,
d — dorsal side of the cell, зж — posterior flagellum, м — mitochondria,
мн — mastigonemes, мт — microtubules, ne — food vacuole, пж — anterior
flagellum, nm — paraxial rod, кп — basal body of anterior and кз — basal body
of posterior flagellum, ф — phagosome, эр — endoplasmic reticulum, я — nuc-
leus. Arrows show the microtubular rootlets.
Морфология и физиология
Наиболее богат в видовом отношении род Bicoso-
eca. Клетка этих организмов прикрепляется к осно-
ванию домика при помощи заднего жгутика и сидит
на нем, как на стебельке. В апикальной части тела,
рядом с передним жгутиком, находится губа — ци-
топлазматический вырост для захвата и заглатывания
пищи (рис. 170; 171). Она бывает разной формы, до-
вольно подвижна, вытянута параллельно переднему
жгутику. Внутри нее проходит широкая лента из 8—
25 микротрубочек, которая начинается от кинетосо-
мы переднего жгутика (Moestrup, Thomsen, 1976).
Передний жгутик опушен трехчленными трубча-
тыми мастигонемами, которые характерны для всех
хризомонад. В переходной зоне жгутика, над попе-
речной пластинкой, обнаружена кольцевая фибрил-
ла. Кинетосомы имеют обычное строение, располо-
жены примерно под прямым углом друг к другу.
Задний жгутик лежит в продольной бороздке клетки
(рис. 171). Он выполняет роль сократимого стебель-
ка, и эта необычная для жгутика функция отразилась
на его строении. Так, у некоторых видов имеется
параксиальный тяж, а у Я planctonica полный набор
микротрубочек в аксонеме отмечен только в проксималь-
ной половине жгутика; в дистальной его части количество
микротрубочек уменьшается до 4—5 (Belcher, 1975).
Исследования других видов этого отряда показали,
что характерной чертой бикозоецид является сильно
развитая корешковая система. Она образована 3—
5 лентами микротрубочек, составляющими скелет пе-
редней части клетки (рис. 172). Наиболее заметен ши-
рокий корешок, идущий к цитостомальной области и,
по-видимому, участвующий в захвате пищевых объек-
тов. Этот корешок ассоциирован в основании с фиб-
риллярным неисчерченным материалом и характеризу-
ется на поперечном срезе своеобразным профилем в
форме буквы «Г» (Moestrup, Thomsen, 1976; Fenchel,
1982; Verhagen et al., 1994; O’Kelly, Patterson, 1996;
Карпов и др., 1998; O’Kelly, Nerad, 1998; Teal et al.,
1998).
Ядро пузырьковидного типа расположено при-
мерно в центре клетки (рис. 171). В перинуклеар-
ном пространстве происходит сборка мастигонем.
Аппарат Гольджи представлен 1 диктиосомой, ле-
жащей неподалеку от базального комплекса жгутиков.
Профили митохондрий с пузырьковидными кристами
встречаются вблизи ядра, но наиболее характерна их
ассоциация с широким цитостомальным корешком. В
цитоплазме всегда можно обнаружить небольшие пи-
Рис. 172. Схема строения жгутикового аппарата Cafeteria roenber-
gensis. Вид с переднего конца клетки. (По: O’Kelly, Patterson,
1996).
Из обычного набора четырех корешков хризофитовых представлены три:
R\, Ri, R4. От /?1 отходят вторичные микротрубочки, Яз получает значитель-
ное развитие в связи с приобретением функции питания, опоясывает цито-
сомальную область (ц). в — вентральная сторона, д — дорсальная сторона,
з — кинетосома заднего жгутика, п — кинетосома переднего жгутика. Фиб-
риллярные связки между кинетосомами и ризопласт, идущий от кинетосомы
заднего жгутика, не показаны.
Fig. 172. Scheme of flagellar apparatus structure of Cafeteria roen-
bergensis.
There are 3 of typical 4 «chrysophycean» rootlets presented in bicosoecids:
/?i, R3 and R4. The most prominent is Я3, connected with cytostomal region.
Fibrillar bridges and rhizoplast are not shown.
266
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
щеварительные вакуоли. В качестве запасного пита-
тельного вещества откладывается гликоген (рис. 171).
Домики бикозоек бывают различной формы: яй-
цевидные, бокаловидные, как правило, с широким
устьем (рис. 170). Они могут быть прозрачными или
желтовато-коричневыми в результате импрегнации
солями железа. Иногда в световой микроскоп видна
кольцевая исчерченность. Исследования ультраструк-
туры домиков показали, что у всех изученных видов
они образованы кольцевыми ребрами, которые связа-
ны между собой многочисленными тонкими нитями
(Kristiansen, 1972). На срезах хорошо видно, что у
одних видов стенка домика двухслойная, у других
трехслойная (Mignon 1974).
Питаются бикозоециды в основном бактериями и
мелкими водорослями, которые приносятся токами
воды на губу. Возможно, эти жгутиконосцы обладают
не только фагоцитозом, но и пиноцитозом, и мемб-
ранным транспортом, однако пока это не установлено.
Процесс фагоцитоза у прикрепленных жгутиконос-
цев выглядит следующим образом. Передний жгутик
создает направленные токи воды, которые приносят
мелкие частички к апикальному концу клетки. Пи-
щевой объект попадает на губу, захватывается ею и
транспортируется в цитоплазму клетки внутри пище-
вой вакуоли.
Размножение и стадии
жизненного цикла
У бикозоецид известно только бесполое размно-
жение. Клетки размножаются путем продольного де-
ления на 2 дочерние особи. Этот процесс можно
проиллюстрировать на примере В. conica (Жуков,
1978). В начале деления передний конец клетки
раздвигается и вырастает еще 1 пара жгутиков
(рис. 170). Когда бороздка деления достигает заднего
конца материнской клетки, особи разделяются, и в
этот момент в домике находятся 2 дочерние двух-
жгутиковые клетки: одна — прикрепленная, другая —
свободноплавающая. Последняя вскоре уплывает в
поисках подходящего для дальнейшего развития мес-
та. Весь процесс деления занимает около часа. Под-
вижная вегетативная клетка прикрепляется к субстрату
задним жгутиком, постепенно строит домик (рис. 170),
начинает активно питаться и при достижении раз-
меров взрослой особи делится, давая прикрепленную
и расселительную формы. У колониальных видов до-
черняя клетка не уплывает после деления, а цеп-
ляется задним жгутиком за край домика материн-
ской особи и, используя его как субстрат, формиру-
ет домик и стебелек. Электронно-микроскопически
процесс деления не изучен, поэтому неизвестны по-
ведение ядра во время деления, тип митоза и т. д.
Мы рассмотрели типичный для бикозоек жизнен-
ный цикл, в котором участвуют прикрепленная и рас-
селительная стадии. Прикрепленные формы могут
быть одиночными и колониальными. Среди колони-
альных видов есть и свободноплавающие, например
В, socialis. Другие жизненные формы практически
не встречаются. Образование спор отмечено только у
В. mitra (Жуков, 1978).
Экология
Бикозоециды распространены очень широко. Они
могут жить и в морях, и в пресных водах. Пресновод-
ные виды чаще встречаются в (3-мезосапробных водо-
емах, где в осенний период они достигают очень
большой численности (до 500 000 особей на литр),
т. е. составляют примерно половину всех планктон-
ных зоофлагеллат (Жуков, 1978). Таким образом,
они являются существенным звеном в цепи трофи-
ческих связей планктонных организмов. Подавляю-
щее большинство бикозоецид — прикрепленные
формы, поселяющиеся на водорослях, высшей вод-
ной растительности, детрите, планктонных ракооб-
разных и другом субстрате.
Филогения
Бикозоециды существенно отличаются от других
хризомонад полным отсутствием каких-либо следов
пластид. Вероятно, их можно считать исходно гете-
ротрофными и, следовательно, наиболее примитив-
ными хризофитами. Другой близкой к ним группой
являются псевдодендромонады (см. с. 317). Послед-
ние характеризуются такой же симметрией тела клет-
ки, как и бикозоециды, представлены преимущест-
венно прикрепленными формами и имеют большое
сходство в строении корешковой системы жгутиков.
Однако у псевдодендромонад нет домиков — поверх-
ность клетки покрыта чешуйками. Кроме того, они
лишены трубчатых мастигонем. Вместо губы у них
имеется постоянное отверстие — цитостом, ведущий
в хорошо выраженную глотку. Ранее эти признаки не
позволяли объединять обе группы в один таксон. В
настоящее время становится все более очевидным,
что они должны быть объединены в одну таксономи-
ческую группу.
Филогенетически бикозоецид легко связать и с
хризофитовыми, и с псевдодендромонадами, пред-
ставив первых в качестве предковых форм. Наибо-
лее просто устроены, по-видимому, Cafeteria, кото-
рые лишены домика и несут, как гетероконтные жгу-
тиконосцы, все основные черты хризофитовых. В
результате симбиоза с эукариотными водорослями эти
формы могли дать начало фотосинтезирующим хри-
зомонадам. Перейдя к свободноплавающему образу
жизни и автотрофному способу питания, они могли
утратить губу, усилить корешковую систему за счет
развития ризопласта, связывающего кинетосомы с
ядром, и начать синтезировать чешуйки по способу
формирования субъединиц домика. С другой сторо-
ны, приобретение домика, по-видимому, привело к
появлению рода Bicosoeca и утрате у части его ви-
дов, живущих в домиках с узким устьем, трубчатых
мастигонем. От предковых форм бикозоецид без до-
миков могли произойти и псевдодендромонады. При
этом, вероятно, появились соматические чешуйки и
были утрачены мастигонемы, а губа превратилась в
постоянный цитостом, укрепленный гомологичной
лентой микротрубочек. Переходной в этом отноше-
нии группой можно считать род Adriamonas. В нут-
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE. ОТРЯД BICOSOECALES
267
Рис. 173. Некоторые представители бикозоецид.
А, Б — Cafeteria roenbergensis: А — плавающая клетка, Б — прикрепленная клетка; В, Г — Pseudobodo tremulans: В — плавающая клетка, Г — прикреп-
ленная клетка; Д — Discocelis saleuta, плавающая клетка; Е — Acronema sippewissettensis: Ж — Caecitellus parvulus: 3 — Adriamonas peritocrescens: И —
Siluania monomastiga. аг — аппарат Гольджи, гл — глотка, ж — жгутик, м — митохондрия, мн — мастигонемы, пв — пищеварительная вакуоль с бакте-
риями, ци — цитостом, я г~~ ядро. (А—Г— по: Preisig et al., 1991; Д — по: Patterson, Zolffel, 1991; Е — по: Teal et al., 1998; Ж :— no: Tong, 1997; 3 —
no: Verhagen et al., 1994; И — ориг. С. А. Карпова).
Fig. 173. General view of some bicosoecids.
А, Б — Cafeteria roenbergensis: A — swimming cell, Б — attached cell; В, Г — Pseudobodo tremulans: В — swimming cell, Г — attached, cell; Д — swim-
ming cell of Discocelis saleuta: E — Acronema sippewissettensis: Ж — Caecitellus parvulus: 3 — Adriamonas peritocrescens: И — Siluania monomastiga. аг —
Golgi apparatus, гл — cytopharynx, ж — flagellum, м — mitochondrium, мн — mastigonemes, ne — food vacuole, ци — cytostome, я — nucleus.
ренние структуры клетки не претерпели изменений.
Таким образом, приобретение чешуек на поверхнос-
ти жгутиков и тела клетки могло произойти у хрй-
зомонад и псевдодендромонад независимо.
Систематический обзор
Отряд Bicosoecales (Grassd,
1926) emend. Karpov, 1998
В отряде выделяют 3 семейства.
Сем. Cafeteriaceae Moestrup, 1995
Двухжгутиковые бикозоециды без домика и цито-
стома, задний, гладкий, жгутик может использоваться
для временного прикрепления. Клетки с более или ме-
нее заметной губой, используемой для питания.
В состав семейства входит 4 рода.
Типовой род Cafeteria Fenchel et Patterson,
1988.
Род Cafeteria Fenchel et Patterson, 1988
(рис. 173, Д, Б)
Прикрепленные морские жгутиконосцы без доми-
ка. Имеют все основные признаки отряда: гетероконт-
ные жгутиконосцы с опушенным трехчленными труб-
чатыми мастигонемами передним жгутиком; довольно
длинный и гладкий задний жгутик служит стебельком
у сидячих форм; корешковая система состоит из 3 мик-
ротрубочковых лент и 1 поперечно исчерченного ко-
решка, идущего к ядру (O’Kelly, Patterson, 1996). Име-
ется небольшая губа на переднем конце клетки, при
помощи которой осуществляется питание. Отличитель-
ными чертами рода Cafeteria является наличие небольших
экструсом на переднем конце и вентральной стороне
клетки (Fenchel, Patterson, 1988). Описано 4 вида.
268
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Discocelis Vers, 1988
(рис. 173, Д)
Морской бесцветный жгутиконосец, лишенный
домика, но по многим признакам сходный с бикозо-
еками (Vers, 1988). Клетка дисковидной формы, с
велумом на переднем конце тела, который вполне
можно принять за «губу» бикозоек. Корешковая сис-
тема похожа на таковую бикозоек, но менее развита:
3 латеральных микротрубочковых корешка укрепля-
ют велум и переднюю часть клетки. К ядру прилега-
ет микротельце, характерное и для бикозоек. Пере-
дний жгутик короткий, задний — длинный, прилега-
ет к боковой поверхности клетки. Митохондрии с
пузырьковидными кристами. Отличие от других би-
козоецид состоит в отсутствии мастигонем на переднем
жгутике. Под плазмалеммой обнаружен необычный
паракристаллический слой. По периферии клетки
расположен ряд экструсом. Обитание Discocelis
между частицами донного песка, по-видимому, при-
вело к утрате передним жгутиком двигательной ак-
тивности, что вызвало его редукцию и, как следст-
вие, исчезновение мастигонем. В результате и ко-
решковая система Discocelis не такая мощная, как у
остальных бикозоецид. Описан 1 вид — D. saleuta
Vers, 1988.
Род Pseudobodo Griessmann, 1913
(рис. 173, В, Г)
Клетка почти сферической формы, с длинным (в
2—4 раза длиннее клетки), направленным вперед
жгутиком, который слегка изогнут у прикрепленной
особи и почти прямой у плавающей. Более короткий,
гладкий, жгутик направлен назад и располагается в
бороздке на теле клетки. Клетка прикрепляется к
субстрату кончиком этого жгутика, который может
сильно вытягиваться в особую прикрепительную ор-
ганеллу. Цитостомальная зона в виде губы находится
по соседству с основанием двигательного жгутика,
как у других бикозоецид. Ультраструктура изучена
недостаточно (Fenchel, 1982).
Род Асгопета Teal et al., 1998
(рис. 173, Е)
Бесцветный морской гетероконтный жгутиконо-
сец с опушенным трубчатыми мастигонемами пере-
дним и гладким задним жгутиками. Обычно плавает,
но может и прикрепляться задним жгутиком к суб-
страту. Питается бактериями, как и другие бикозоеци-
ды, но цитостом отсутствует. Оба жгутика имеют
хорошо выраженные акронемы. К ядру прилегает
небольшое парануклеарное тельце, экструсомы от-
сутствуют. От кинетосом отходят 3 типичных для
бикозоецид микротрубочковых корешка. Самый ши-
рокий из них направлен в сторону вентральной
бороздки. Кинетосома заднего жгутика направлена
влево. Внутри кинетосом обнаружены электронно-
плотные аморфные структуры. В жизненном цикле
имеется стадия покоя — округлившаяся клетка без
жгутиков, но и без клеточной стенки.
Единственный описанный вид A. sippewissettensis
характеризуется овальной или шаровидной формой и
небольшими размерами (5x4 мкм).
Сем. Bicosoecaceae Stein, 1878
Бикозоециды, живущие в домиках и не имеющие
цитостома.
В состав семейства входит 1 род.
Типовой род Bicosoeca (=Bicoeca) James-Clark,
1866.
Род Bicosoeca James-Clark, 1866
(рис. 170)
Одиночные или колониальные гетероконтные бес-
цветные жгутиконосцы, планктонные или прикреп-
ленные. Клетки находятся в домиках и прикреплены
задним жгутиком к их основанию. Корешковая система
жгутиков хорошо развита, состоит из 4—5 микротру-
бочковых корешков. В переходной зоне жгутика имеется
кольцевая фибрилла, расположенная над поперечной
пластинкой. Кристы в митохондриях пузырьковидные.
Морские и пресноводные. Всего описано 38 видов.
Сем. Siluaniaceae Karpov, 1998
Бикозоециды без домика с хорошо развитым ци-
тостомальным аппаратом.
В состав семейства входит 3 рода.
Типовой род Siluania Karpov, 1998.
Род Siluania Karpov, 1998
(рис. 173, И)
Одножгутиковые бикозоециды, питающиеся голо-
зойно. Жгутик расположен субапикально, цитостом
находится на апикальном конце клетки. В переход-
ной зоне жгутика имеется кольцевая фибрилла, рас-
положенная под поперечной пластинкой. К этому
роду относится единственный вид S. monomastiga
Karpov, 1998. Это планктонная, очень мелкая (1.5—
2.0 мкм) бикозоецида. Жгутик равен по длине телу
клетки, которое имеет яйцевидную форму; выходит
из небольшого углубления по соседству с цитосто-
мальным отверстием и несет на одной стороне труб-
чатые мастигонемы. Плавает довольно быстро, как
бы кувыркаясь. Обнаружен в холодных пресных водах.
Род Adriamonas Verhagen et al., 1994
(рис. 173, 3)
Свободноживущий почвенный жгутиконосец.
Клетка удлиненная, с 2 субапикальными и почти рав-
ными жгутиками с длинными акронемальными отрос-
ТИП CHRYSOPHYTA. КЛАСС CHRYSOPHYCEAE. ОТРЯД CH RO MU LIN ALES
269
тками. Мастигонемы на жгутиках отсутствуют. Ци-
тостом расположен в переднебрюшной части клетки.
По ультратонкой организации он имеет общие при-
знаки и с псевдодендромонадами, и с бикозоецидами.
От кинетосом отходят 3 микротрубочковых корешка
(1 дорсальный и 2 вентральных, или цитостомаль-
ных) и 1 фибриллярный, направленный к ядру. Дор-
сальный корешок формирует вторичные микротру-
бочки, укрепляющие дорсальную сторону клетки. В
проксимальной части широкого корешка имеется
фибриллярное уплотнение, а его дистальная часть ох-
ватывает воронку цитостома, укрепляя это постоян-
ное образование. Митохондрия с трубчатыми криста-
ми также ассоциирована с широким корешком. В пе-
редней части клетки обнаружены сократительная
вакуоль и 1—2 экструсомы. Описание дано по одно-
му виду A. peritocrescens (Verhagen et al., 1994). Ав-
торы относят этот вид к псевдодендромонадам, одна-
ко по ультраструктурным признакам он обнаруживает
большое сходство с Pseudobodo tremulans (С. А. Кар-
пов, неопубл, данные) и поэтому помещен в отряд
Bicosoecales.
Род Caecitellus Patterson et al., 1993
(рис. 173, Ж)
Мелкий морской гетероконтный жгутиконосец с
2 гладкими жгутиками и хорошо выраженным цито-
стомом, ведущим в глотку. Как правило, ползает по
субстрату, хотя может и недолго плавать. Питается
бактериями. Обычно не приклепляется к субстрату
задним жгутиком, что не характерно для других би-
козоецид. От кинетосом отходят 3 типичных для би-
козоецид микротрубочковых корешка. Кинетосома
заднего жгутика направлена влево. У этого организ-
ма отсутствуют фибриллярные корешки, экструсомы
и микротельце. Внутри кинетосом обнаружены элек-
тронно-плотные аморфные структуры.
Единственный изученный вид С. parvulus (Griess-
mann, 1913) Patterson et al., 1993 имеет небольшие
размеры (3—7 мкм), передний жгутик в 1.5—2.5 раза
длиннее тела клетки, оканчивается длинной акроне-
мой, задний жгутик в 2.5—3.5 раза длиннее тела
клетки, без акронемы (O’Kelly, Nerad, 1998).
Отряд Chromulinales Pascher,
1910
По классификации Прейсига {Preisig, 1995), этот
отряд объединяет и охромонад (Ochromonadales). Ранее
представители обеих групп разделялись исследовате-
лями лишь на основании разного количества жгути-
ков, видимых в световой микроскоп. Сейчас ясно,
что их типовые роды Chromulina и Ochromonas обла-
дают 2 жгутиками. Представители обоих родов сход-
ны во всех отношениях, различаясь фактически толь-
ко длиной рекуррентного жгутика (Preisig, 1995).
Представители отряда Chromulinales — преиму-
щественно монадные организмы, но встречаются
пальмеллоидные и ризоподиальные формы. Клетки
обычно голые, но есть несколько видов с простыми
органическими чешуйками. Прейсиг (Preisig, 1995)
выделяет в отряде 10 семейств: Chromulinaceae, Di-
nobriaceae, Paraphysomonadaceae, Chrysolepidomo-
nadaceae, Chrysamoebaceae, Myxochrysidaceae, Chry-
socapsaceae, Stichogloeaceae (=Chrysosphaeraceae),
Phaeothamniaceae и Chrysothallaceae (=Phaeoplaca-
ceae).
Ниже приводятся описания некоторых родов этого
отряда.
Род Chromulina Cienkowski, 1870
В составе рода около 120 видов. Это одиночные
свободноплавающие организмы, в жизненном цикле
которых есть амебоидные или пальмеллевидные стадии.
Двигательный жгутик 1, второй очень короткий, нахо-
дится постоянно в небольшом углублении на переднем
конце тела. Большая часть видов обитает в поверхнос-
тных зонах водоёмов и относится к нейстонным фор-
мам. Один из широко распространенных видов — Ch.
psammobia Faure-Fremiet, 1958 (рис. 163, А).
Род Chrysococcus Klebs,
1883
Подвижные одиночные жгутиконосцы, сходные с
представителями предыдущего рода (рис. 163; 168).
Имеют домик, гладкий или скульптурированный, жел-
тый или коричневый. Жгутиков 2, но только 1 выходит
наружу, второй — короткий и не выходит за пределы
углубления на переднем конце тела. Широко распрост-
ранены в пресноводном планктоне. Один из типичных
видов — Ch. rufescens Klebs, 1893.
Род Chrysamoeba Klebs,
1893
В жизненном цикле преобладает амебоидная стадия
с тонкими гранулированными ризоподиями (рис. 169).
Жгутиковая стадия сходна с таковой Chromulina. Жгу-
тиков 2, короткие, из них 1 выходит наружу, второй
виден только с помощью электронного микроскопа.
Размножение происходит на амебоидной стадии. Пре-
сноводные и морские виды. Типичный вид — Ch. ra-
dians Klebs, 1893.
Род Myxochrysis Pascher,
1916
Включает 1 вид — М. paradoxa Pascher, 1916. Это-
му виду свойственны плазмодиальные формы. Зрелые
плазмодии многоядерны, имеют хлоропласты, не-
сколько сократительных вакуолей, запасы хризолами-
нарина. Плазмодий покрыт плотной оболочкой, которая
содержит кальций и инкрустирована солями железа.
Ему свойственны короткие лопастные псевдоподии
(рис. 169). Жизненный цикл сложный, и в нем череду-
ются стадии одноядерных амеб, одножгутиковых зоо-
спор и плазмодиев (Hibberd, Leedale, 1985).
270
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отряд Hibberdiales Andersen,
1989
Включает 1 сем. Hibberdiaceae Andersen, 1989 с
1 монотипическим родом Hibberdia (типовой вид
Я. magna (Belcher) Andersen). Н, magna {-Chryso-
sphaera magna Belcher) имеет 2 стадии в жизненном
цикле: капсоидную колониальную неподвижную и
жгутиковую одиночную подвижную. Жгутик сохраня-
ется и у капсоидной формы, но плохо заметен и
почти не функционирует из-за обилия слизи. Жгути-
ковый аппарат Н. magna отличается от такового хро-
мулинид: кинетосомы расположены под тупым углом
друг к другу, а микротрубочковые корешки не фор-
мируют петлю под вторым жгутиком. Наряду с фу-
коксантином у этой водоросли имеется другой свето-
собирающий каротиноид — антераксантин. Это —
уникальный случай среди водорослей.
Отряд Hydrurales Pascher, 1931
Единственное сем. Hydruraceae включает 3 рода:
Hydrurus и Celloniella, имеющие капсоидную органи-
зацию, и Phaeodermatium с паренхиматозным строе-
нием тела. Их зооспоры отличаются от других хри-
зомонад тетраэдральной формой, с длинным пере-
дним жгутиком, для которого не выявлено наличие
трубчатых мастигонем, и коротким задним жгути-
ком, лишенным центральных микротрубочек. Жгути-
ковые корешки не образуют петлю под 2-м жгути-
ком. Уникальная форма зооспор поддерживается хо-
рошо развитым скелетом из микротрубочек —
Класс Synurophyce
Синуроф
Синурофитовые — свободноживущие фототрофные, одиноч-
ные и колониальные протисты пресных водоемов. Имеют 2 гетеро-
контных (или почти гетероконтных) жгутика, отходящих от перед-
него конца тела. Более длинный жгутик несет трубчатые мастиго-
немы, расположенные по обеим его сторонам. В переходной зоне
жгутиков есть спираль, в которой насчитывается 6—9 витков. Ко-
решковая система представлена фактически одним ризопластом и
отходящим от него микротрубочковым корешком R\. Поверхность
одного или обоих жгутиков покрыта органическими чешуйками;
поверхность тела покрыта кремнеземными билатерально-симмет-
ричными чешуйками, формирующимися на внешней поверхности
хлоропластов. Хлоропластов 2, они содержат хлорофилл а и ci, а
также фукоксантин. В ламелле хлоропласта 3 тилакоида; есть опо-
ясывающая ламелла. Фоторецептор представляет лишь вздутия,
расположенные на каждом жгутике по 1; он не связан с плазмалем-
мой и с мембраной хлоропласта; стигмы нет. Система сократи-
тельной вакуоли расположена в нижней части клетки. Митохонд-
рии с пузырьковидными кристами.
История изучения
Первое описание колониального вида Synura uvel-
la принадлежит Эренбергу (Ehrenberg, 1838). Пашер
(Pascher, 1913) выделил класс Chrysophyceae, куда
были включены синуровые водоросли. В последнее
время на основании данных электронной микроско-
производных 1-го корешка. Фоторецептор отсутству-
ет. Деление ядра идет по типу полузакрытого орто-
митоза.
Отряд Sarcinochrysidales Gayral et
Billard, 1977
В этот отряд включены хризофитовые, тяготею-
щие к бурым водорослям. Они исключительно морские
или солоноватоводные, не Имеют стоматоцист. По-
движные клетки не имеют стигмы; пиреноид стебель-
чатый, как у бурых водорослей; имеются пигменты:
диатоксантин й диадиноксантин; отсутствует пере-
ходная спираль, а поперечная пластинка находится
выше поверхности клетки. Несколько родов распре-
делены по 3 семействам: Ankylochrysidaceae O’Kelly
et Billard, 1995; Sarcinochrysidaceae Gayral et Billard,
1977 и Nematochrysopsidaceae Gayral et Billard, 1977.
Отряд Chrysomeridiales O’Kelly et
Billard, 1995
В этом отряде объединена довольно большая груп-
па родов, относимых ранее к отряду Sarcinochrysidales.
В нем выделено 2 семейства: Chrysomeridiaceae Bour-
relly, 1957 й Phaeosacciaceae Feldmann, 1949. Представи-
тели отряда в отличие от видов отряда Sarcinochrysida-
les лишены стебельчатого пиреноида, имеют стигму у
подвижных клеток, спираль в переходной зоне жгутика,
а поперечная пластинка находится на уровне плазмалем-
мы; различаются они и строением корешковой системы.
ае Andersen, 1987 —
итовые
пии и биохимии из этого класса был выделен и обо-
снован новый класс Synurophyceae (Andersen, 1987) с
2 новыми семействами, объединенными в 1 отряд.
Классификация
Класс Synurophyceae Andersen, 1987 включает
1 отряд — Synurales Andersen, 1987. В составе отря-
да — 2 новых семейства: Synuraceae Andersen, 1987
и Mallomonadaceae Andersen, 1987, которые насчи-
тывают более 30 видов, распространенных в пресных
водоемах.
Морфология и физиология
По особенностям общей морфологии синурофи-
товые сходны с хризофитовыми. Одиночные организ-
мы (род Mallomonas) и особи колониальных видов
(род Synura) имеют удлиненную форму тела, которое
суживается на заднем конце (рис. 167). Особи, обра-
зующие колонию, на заднем конце имеют стебелек, с
помощью которого они прикрепляются в центре ко-
ТИП CHRYSOPHYTA. КЛАСС SYNUROPHYCEAE
271
лонии. Размеры Mallomonas достигают 40—100 х 20—
30 мкм, размеры особей в колонии Synura — 30—
50 мкм в длину.
Жгутиковый аппарат включает 2 гетероконтных
(или почти гетероконтных) жгутика, отходящих от
переднего конца тела апикально или субапикально
(рис. 174). Более длинный, двигательный, жгутик на-
правлен вперед и несет тубулярные мастигонемы,
расположенные по обеим его сторонам. Более ко-
роткий жгутик расположен почти параллельно дви-
гательному и лишен мастигонем.
Один или оба жгутика покрыты снаружи очень
мелкими органическими чешуйками.
Переходная зона жгутиков короткая, и в ней рас-
положена переходная спираль, имеющая 6—9 вит-
ков (оборотов). У других хризофитовых их не более
6.
Корешковая система жгутиков у синурофитовых
отличается своеобразием (рис. 175). Микротрубочко-
вая ее часть представлена лишь 2 корешками Rx и R3,
а у Mallomonas — только 1 корешком R{ (рис. 175).
Оба корешка начинаются от ризопласта и достигают
плазмалеммы переднего конца клетки (рис. 174; 175).
Затем /?! огибает базальные тела жгутиков, закручи-
ваясь по часовой стрелке, если смотреть на клетку с
переднего конца (рис. 175). От него отходят много-
численные вторичные микротрубочки, поддерживаю-
щие форму переднего конца клетки. Ризопласт хоро-
шо развит, отходит от обеих кинетосом и, разветвля-
ясь на несколько пучков, охватывает переднюю часть
ядра (рис. 174; 175).
Покровы тела. Поверхность тела покрыта со-
матическими чешуйками, которые имеют различную
форму и размеры у разных видов. Чешуйки кремне-
земной природы имеют билатеральную симметрию
(рис. 167) и часто снабжены длинными шипами. Они
формируются в цитоплазме клетки на поверхности
хлоропластов (рис. 174), что представляет одну из
отличительных черт этого класса. Формирование че-
шуек (Brugerolle, Bricheux, 1984) происходит в осо-
бых уплощенных пузырьках, которые продуцируют-
ся в аппарате Гольджи, а затем плотно прилегают
одной стороной к поверхности хлоропласта. Упло-
щенный пузырек прикрепляется к мембране пласти-
ды пучками актиновых филаментов. Сеточка микро-
филаментов покрывает и наружную поверхность пу-
зырька, в котором идет формирование чешуйки. В
этом процессе участвуют также микротрубочки жгу-
тикового корешка, которые взаимодействуют с мик-
рофиламентами и обеспечивают миграцию пузырька
со зрелой чешуйкой к поверхности клетки.
Фоторецептор синуровых представляет собой взду-
тия, находящиеся у основания обоих жгутиков и об-
ращенные друг к другу. Фоторецептор не контакти-
рует ни с клеточной мембраной, ни с мембраной хло-
ропласта. Стигма отсутствует.
Рис. 174. Общий вид колонии Synura uvella (А) и хема строения передней части клетки Synura (Б).
А: / — зооиды; Б: мн — мастигонемы на двигательном жгутике, пч — пузырьки на поверхности хлоропласта (хл), в которых формируются соматические
чешуйки (ч), р — ризопласт, эр — шероховатый ЭПР, я — ядро, Ri — микротрубочковый корешок. (А — ориг. К. M. Сухановой; Б— по: Mignot, Bruge-
rolle, 1982).
Fig. 174. General view of colony Synura uvella (A) and scheme of cell structure of Synura (Б).
A: I — zooids; Б-. мн — mastigonemes, пч — molding vesicules on chloroplast (хл) surface, p — rhizoplast, ч — body scales, эр — endoplasmic reticulum, я —
nucleus, R\ — microtubular rootlet R}.
272
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 175. Общий вид Mallomonas caudata (А) и схема строения жгутикового аппарата у Mallomonas (Б),
А: 1 — кремнеземные щетинки, 2 — чешуйки, 3 — хлоропласт, 4 — вакуоль; Б: вз — вздутие на опушенном жгутике (часть фоторецептора), пс — пере-
ходная спираль, р — ризопласт, R\ — микротрубочковый корешок. (А — по: Doflein, 1953; Б — по: Beech, Wetherbee, 1990).
Fig. 175. General view of Mallomonas caudata (A) and scheme of cell structure of Mallomonas (Б).
A: 1, 2 — silica scales; 3 — chloroplast; 4 — vacuole. Б: вз — flagellar swelling, nc — transitional helix, p — rhizoplast, R\ — microtubular rootlet Яь
Хлоропласты. В клетке синурофитовых обычно
2 вытянутых в длину хлоропласта желто-зеленого
цвета. Ламеллы хлоропласта включают по 3 тила-
коида. Есть опоясывающая ламелла. Хлоропласты
содержат хлорофилл а и с/, в отличие от хризо-
фитовых у синурофитовых отсутствует хлорофилл
с2. Как правило, для синурофитовых свойствен каро-
тиноид фукоксантин; он обычен и для хризофито-
вых.
Ядро 1, по форме близкое к шаровидному, нахо-
дится в центре клетки. Митоз синурофитовых по-
лузакрытого типа, причем, как и у хризофитовых,
ЦОМТами митотического веретена служат ризоплас-
ты (Andersen, 1989).
Сократительная вакуоль. Система сократительной
вакуоли имеется у всех видов синурофитовых. Рас-
положена она в задней части клетки и включает цент-
ральный резервуар и прилегающие к нему вакуоли.
Резервные вещества представлены, вероятно, хри-
золаминарином, расположенным в вакуолях цито-
плазмы.
Размножение и жизненный
цикл
Для синурофитовых обычно агамное размножение
путем продольного бинарного деления клетки. Поло-
вое размножение изучено недостаточно, но у не-
скольких видов Mallomonas обнаружены изогаметы,
образующие при слиянии зиготу, которая инцистиру-
ется.
В жизненном цикле преобладает стадия активного
трофозоита, но у всех исследованных видов обнару-
жены типичные для хризомонадовых цисты. Для цист
характерны шаровидная форма и плотный покров из
перекрывающихся краями кремнеземных чешуек, ко-
торые образуют своего рода панцирь. У синуры и
малломонас в жизненном цикле известны также паль-
меллоидные стадии.
Экология
Синурофитовые — пресноводные протисты, вхо-
дящие в состав планктонных сообществ озер, прудов
и других водоемов. Они служат пищевыми объектами
для многих видов мелких животных: коловраток, вет-
вистоусых и веслоногих рачков, мелких личинок вод-
ных насекомых.
У колоний синура (Synura uvella) отчетливо выра-
жены весенний и осенний максимумы численности
популяций. «Цветение» воды прудов и прибрежных
участков озер вызвано массовым размножением этих
организмов: вода при этом приобретает золотистый
оттенок.
ТИП CHRYSOPHYTA. КЛАСС PELAGOPHYCEAE
273
Происхождение и эволюция
Вопросы происхождения синурофитовых еще не
могут быть решены прежде всего потому, что этот
класс выделен из состава хризофитовых сравнитель-
но недавно (Andersen, 1987). Сходство этих 2 групп,
несмотря на обнаруженные различия, касающиеся
фотосинтезирующих пигментов, структуры жгутико-
вого аппарата, фоторецептора, внешнего покрова те-
ла и ряда других особенностей, позволяет высказать
предположение о наличии общего предка у синуро-
фитовых и хризофитовых.
Систематический обзор
Класс Synurophyceae Andersen, 1987 включает 1
отряд — Synurales Andersen, 1987, который разделен
на 2 семейства: Synuraceae и Mallomonadaceae.
Сем. Synuraceae Andersen,
1987
В состав семейства входят колониальные жгути-
коносцы, относящиеся к родам Synura, Chlorodesmos,
Chrysodidymus, которые имеют шаровидные колонии,
образующиеся путем монотомии. Колонии желто-зе-
леного цвета.
Типовой род Synura Ehrenberg, 1838 (рис. 174).
Сем. Mallomonadaceae Andersen,
1987
Семейство включает одиночные виды, широко
распространенные в планктоне малых водоемов.
Это — мелкие жгутиконосцы родов Mallomonas, Cond-
radiella.
Типовой род Mallomonas Perty, 1929 (рис. 167).
Класс Pelagophyceae Andersen et Saunders, 1993 —
Пел агофитовые
В результате морфологических, ультраструктурных и молеку-
лярно-биологических исследований единственного вида Pelagomo-
nas calceolata выделены новое семейство, новый отряд и класс
(Andersen et al., 1993). Основные признаки класса: 1 жгутик; 1
кинетосома; отсутствие жгутиковых корешков; трубчатые двух-
членные мастигонемы на жгутиках; зубчатый параксиальный тяж;
необычные митохондрии и экструсома; тонкая органическая тека
на поверхности тела. Анализ последовательности нуклеотидов
генов 18S рРНК показал, что Р. calceolata относится к хризофито-
вым. В составе класса, по-видимому, имеется несколько видов, но
изучен только 1 морской вид.
Классификация
Класс Pelagophyceae Andersen et Saunders, 1993
является монотипическим.
Отряд Pelagomonadales Andersen et Saunders, 1993 — Пелагомона-
диды
Сем. Pelagomonadaceae Andersen et Saunders, 1993 — Пелаго-
монадацеи
Род Pelagomonas Andersen et Saunders, 1993 — Пелагомо-
нас
Вид Pelagomonas calceolata Andersen et Saunders, 1993
Морфология и физиология
Жгутиконосец имеет овальную форму тела, не-
сколько расширенную на переднем конце (рис. 176).
Размеры тела: 2—3 мкм в длину, 1.5 мкм в ширину.
Большая часть тела покрыта с наружной сторо-
ны тонкой плотной текой из органического матери-
ала.
Жгутиковый аппарат. Пелагомонас имеет 1 жгу-
тик, длина которого в 2 или 3 раза больше длины
тела. Жгутик, закреплен на заднем конце тела и рас-
положен в продольной жгутиковой бороздке. Он несет
тонкие волоски, расположенные по обеим его сторо-
18 Протисты, часть 1
нам. Волоски состоят из 2 частей: тубулярного ос-
нования и концевого филамента (рис. 177).
Проксимальный конец единственного базального
тельца жгутика опирается на клеточную мембрану
заднего конца тела, а правая сторона его тесно свя-
зана с ядром. Жгутиковые корешки отсутствуют, так
же как отсутствует и вторая кинетосома, которая
развивается только непосредственно перед делением
клетки.
В переходной зоне жгутика находится поперечная
пластинка, которая расположена немного выше уровня
клеточной мембраны и состоит из толстого плотного
Рис. 176. Внешний вид Pelagomonas calceolata (схема). (По: An-
dersen et al., 1993).
1 — жгутик, 2 — передний конец тела, 3 — задний конец тела, 4 —
брюшная сторона, 5 — спинная сторона.
Fig. 176. Diagram of external appearance of Pelagomonas calceolata.
1 —‘ flagellum, 2 — anterior part of the body, 3 — posterior part of the
body, 4 — ventral side, 5 — dorsal side.
274
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 177. Схема строения мастигонем у Pelagomonas calceolata.
(По: Andersen et al., 1993).
I — терминальный филамент, 2 — трубчатая часть мастигонемы, 3 — по-
верхность жгутика.
Fig. 177. Diagram of ultrastructure of mastigonemes in Pelagomo-
nas calceolata.
1 — terminal filament, 2 — tubular part of mastigoneme, 3 — surface of fla-
gellum.
кольца между аксонемой жгутика и его плазмалем-
мой и тонкого диска, лежащего внутри аксонемы.
Ниже этой типичной поперечной пластинки располо-
жена вторая пластинка, которая представляет собой
тонкий диск, лежащий во внутреннем пространстве
между микротрубочками аксонемы. 2 витка переход-
ной спирали расположены между поперечными плас-
тинками.
Жгутик имеет параксиальный тяж, поверхность
которого снабжена зубцами. Органеллы в клетке ори-
ентированы таким образом, что ядро находится с
правой стороны жгутиковой кинетосомы, а митохон-
дрия — с левой.
Органеллы цитоплазмы и ядро
В клетке имеется 1 митохондрия с электронно-
плотным матриксом и пузырьковидными кристами.
Диктиосома (аппарат Гольджи) 1 и занимает место
вблизи проксимальной части кинетосомы. Цистерны
диктиосомы часто расширены. Хлоропласт 1, золо-
тистого цвета, занимает большую часть спинной сто-
роны тела. Он окружен внешней мембраной ядерной
оболочки, что является одним из характерных признаков
хризофитовых водорослей. Ламеллы хлоропласта со-
стоят из 3 тилакоидов. Имеется опоясывающая ламелла.
Хлоропласт содержит целый ряд каротиноидов, к кото-
рым относятся фукоксантин (55 %) 19’- бутаноилок-
сифукоксантин (32), диадиноксантин (8), диатоксантин
(2 %). Стигма отсутствует. Для хлоропласта специфич-
ны 2 типа хлорофилла: а и с. Пиреноид отсутствует.
Резервные вещества типа хризоламинарина и ли-
пидов локализованы в 1 или нескольких вакуолях ци-
топлазмы. В кортикальной зоне цитоплазмы брюш-
ной стороны находится специфичная для данного
вида экструсома цилиндрической формы с грануляр-
ным материалом внутри (Andersen et al., 1993).
В клетке 1 сферическое ядро. Деление ядра и
формы размножения пелагомонаса пока не изучены.
Филогения
В результате анализа последовательности нуклеидов
генов 18S рРНК и исследования ультраструктурной ор-
ганизации клетки было показано, что класс Pelagop-
hyceae относится к хризофитовым водорослям и его
место — в типе Chrysophyta. Имеющиеся данные еще
не позволяют решить вопрос о филогенетических вза-
имоотношениях между Pelagophyceae и другими хро-
мофитовыми. Однако они дают основание для сравне-
ния ультраструктуры клеток сходных групп жгутико-
носцев, которые указывают на принадлежность этого
нового класса к филогенетической линии именно хри-
зофитовых водорослей (Andersen et al., 1993).
THE PHYLUM CHRYSOPHYTA PASCHER, 1914
SUMMARY
The chrysophytes are phototrophic and heterotrophic forms with
different types of body organisation. The heterocont flagellates nor-
mally have anterior moving flagellum with tubular mastigonemes, and
the recurrent smooth flagellum, which often applies to the body surfa-
ce and forms the photoreceptor. There is a transition helix inside the
flagella. Yellow-green or golden-brown chloroplasts are usually loca-
ted in the perinuclear space and have the envelope of three membra-
nes. The pigments are presented with chlorophylls a + c, and with
carotionoids. Mitochondria with vesicular cristae. The storage pro-
ducts are chrysolaminarine and lipids. The reproduction is predomi-
nantly agamic, the sexual process has been studied in few species.
There are 3 classes (Chrysophyceae Pascher, 1914; Synurophyceae
Andersen, 1987; Pelagophyceae Andersen et Saunders, 1993) in die phy-
lum, which unites approximately 1000 of species spread worldwide.
The description of morphology and physiology of chryso-
phytes includes: shape and dimensions of cells, flagellar apparatus
structure, coverings, chloroplasts, stigma, mitochondria, Golgi appara-
tus, contractile vacuole, extrusomes, storage products, nucleus and its
division.
Reproduction. Agamic reproduction, sexual process.
Ecology and practical significance.
In the systematic part descriptions of the following taxa are
presented. The class Chrysophyceae Includes: the order Bicosoecales
Grassd, Karpov, 1998; the order Chromulinales Pascher, 1910; the
order Hibberdiales Andersen, 1989; the order Hydrurales Pascher,
1931; the order Sarcinochrysidales Gayral et Billard, 1977; the order
Chrysomeridiales O’Kelly et Billard, 1995.
Primary attention has been given to the order Bicosoecales as the
traditional zoological taxon.
THE ORDER BICOSOECALES (GRASSE, 1926) KARPOV, 1998
’Chrysophycean’ heterotrophic flagellates without plastids, having
one or two flagella, with or without a lorica; feeding apparatus repre-
ТИП CHRYSOPHYTA. ЛИТЕРАТУРА
275
sented by lip or by permanent cytostome with cytopharynx; transi-
tional concentric rings, if present, is located above or under transverse
plate; the broadest microtubular rootlet are connected with the cy-
tostome region, often «L-shaped» in cross section and associated with
fibrillar material. Sedentary and planktonic, freshwater and marine.
According to the International Botanical Code of Nomenclature the
order Bicosoecales (Grassd, 1926) Karpov, 1998 includes 3 families:
Siluaniaceae Karpov, 1998 — bicosoecids without a lorica; a per-
manent cytostome with developed cytopharynx present.
3 genera: Siluania Karpov, 1998; Adriamonas Verhagen et al.,
1994; Caecitellus Patterson et al., 1993.
Cafeteriaceae Moestrup, 1995 — bicosoecids without a lorica
and cytopharynx.
4 genera: Cafeteria Fenchel et Patterson, 1988; Pseudobodo
Griessmann, 1913; Acronema Teal et al., 1998; Discocelis V0rs,
1988.
Bicosoecaceae Stein, 1878 — bicosoecids with a lorica and with-
out cytopharynx.
1 genus: Bicosoeca Stein, 1878.
The general characteristics and description of the classes Synu-
rophyceae Andersen, 1987 and Pelagophyceae Andersen et Saunders,
1993 have been done after Andersen and co-authors (Andersen et al.,
1987, 1993) and in accordance with the following scheme: a general
description, a historical review, classification, morphology and phy-
siology, reproduction and life-cycle, origin and evolution, ecology, a
taxonomic review with a description of families and genera.
ЛИТЕРАТУРА15
Жуков Б. Ф. Определитель бесцветных жгутиконосцев отряда
Bicosoecida Grassd et Deflandre (Zoomastigophorea, Protozoa) // Био-
логия и систематика низших организмов. Л. : Наука, 1978. С. 3—
28. '
Карпов С. А. Система протистов. Омск, 1990.
Karpov S. A. Flagellate phylogeny: ultrastructural approach //
Flagellates / Eds J. Green, B. S. C. Leadbeater. 1999. (In press).
(Карпов C. A.) Karpov S. A. Flagellate phylogeny: ultrastructural
approach // The Flagellates I Ed. by B. S. C. Leadbeater, J. C. Green. Lon-
don: Taylor and Francis Syst. Assoc. Spec. Publ., 2000. P. 336—360.
(Карпов С. А., Керсанах P., Вильямс Д. M.) Karpov S. A.,
Kersanach R., Williams D. M. Ultrastructure and 18S rRNA gene se-
quence of a small heterotrophic flagellate Siluania monomastiga gen.
et sp. nov. (Bicosoecida) // Eur. J. Protistol. 1998. Vol. 34. P. 415—
425.
Матвиенко A. M. Отдел Золотистые водоросли (Chrysophy-
ta) И Жизнь растений. М. : Просвещение, 1977. Т. 3. С. 93—103.
Мыльников А. П. Ультратонкое строение и систематическое
положение Histiona aroides (Bicoecales) // Ботан. журн. 1989. № 74.
С. 184—189.
Сафонова Т. А. Отдел эвгленовые водоросли (Euglenophyta) И
Жизнь растений. М.: Просвещение, 1977. Т. 3. С. 259—265.
Райков И. Б. Ядро простейших. Морфология и эволюция. Л. :
Наука, 1978.
Andersen R. A. Synurophyceae classis nov., a new class of
algae // Amer. J. Bot. 1987. Vol. 74, N 3. P. 337—353.
Andersen R.A.The Synurophyceae and their relationship to
other Golden algae // Nowa Hedwigia. 1989. Vol. 95, NLP. 1—
26.
Andersen R.A., Saunders G.W., Paskind M.P., Sexton J.P.
Ultrastructure and 18S rRNA gene sequence for Pelagomonas calceo-
lata gen. et sp. nov. and description of a new algae class, the Pelago-
phyceae classis nov. // J. Phycol. 1993. Vol. 29, N 6. P.701—715.
Belcher J. H. The fine structure of the loricate colourless flagel-
late Bicoeca planktonica Kisselew // Arch. Protistenk. 1975. Vol. 117.
P. 78—84.
Bouck G. B. The structure, origin, isolation and composition of
the tubular mastigonemes of the Ochromonas flagellum // J. Cell Biol.
1971. Vol. 50. P. 362—384.
Bouck G. B., Brown D. L. Microtubule biogenesis and cell shape
in Ochromonas. 1. The distribution of cytoplasmic and mitotic micro-
tubules // J. Cell. Biol. 1973. Vol. 56, N 2. P. 340—359.
Bourrelly P. Notes sur les Flagelles incolores # Arch. Zool. exp.
gen. 1951. Vol. 88. P. 73—84.
Brugerolle G., Bricheux G. Actin microfilaments are involved in
scale formation of the chrysomonad cell Synura // Protoplasma. 1984.
Vql. 123. P. 203—212.
Ehrenberg D. Ch. G. Infusionsthierreichen als vollkommene Or-
ganismen. Leipzig, 1838.
Farmer J. M. The Protozoa. Introduction to Protozoology. New
York, 1980.
Fenchel T. Ecology of heterotrophic microflagellates. I. Some
important forms and their functional morphology // Mar. Ecol. Prog.
Ser. 1982. Vol. 8. P. 211—223.
Fenchel T., Patterson D. J. Cafeteria roenbergensis nov. gen.,
nov. sp., a heterotrophic microflagellate from marine plankton // Mar.
Microb. Food Webs. 1988. Vol. 3. P. 9—19.
15 Литература к разделу «Тип Chrysophyta».
Flavin M., Nerad T. A. Reclinomonas americana n.g., n.sp., a
new freshwater heterotrophic flagellate // J. Euk. Microb. 1993.
Vol. 40. P. 172—179. • '
Hibberd D. J. The ultrastructure and taxonomy of the Chrysophy-
ceae and Prymnesiophyceae (Haptophyceae) a survey with some new
observations on the ultrastructure of the Chrysophyceae // Bot. J. Linn.
Soc. 1976. Vol. 72, N 1. P. 55—80.
Hibberd D. J., Leedale G. F. Order Chrysomonadida Hibberd,
Leedale, 1985 // Illustrated. Guide to the Protozoa. 1985. P. 54—70.
Kristiansen J. Structure and occurrence of Bicoeca cristallina,
with remarks on the taxonomic position on the Bicosoecales // Brit.
Phycol. J. 1972. Vol. 7. P. 1—12.
Leedale G. F. Phylogenetic aspects of nuclear cytology in the
algae // Ann. N. Y. Acad. Sci. 1970. Vol. 175, N 3. P. 420—453.
Levine N. D., Corliss J. О., Cox P. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R., Lom J„ Lynn D.,
Merinfeld E. G., Page F. C., Poljansky G., Sprague V., Vavra J.,
Wallace F. G. A newly revised classification of the Protozoa II j. Pro-
tozool. 1980. Vol. 27, N 1. P. 37—58.
Mignot J.-P. Etude ultrastructurale des Bicoeca, protistes fla-
gellds II Protistologica. 1974. Vol. 10. P. 543—565.
Moestrup 0., Thomsen H. A. Fine structural studies on the fla-
gellate genus Bicoeca. I. Bicoeca maris with particular emphasis to
±e flagellar apparatus // Protistologica. 1976. Vol. 12. P. 101—120.
O’Kelly C. J., Nerad T. Kinetid architecture and bicosoecid affi-
nities of the marine heterotrophic nanoflagellate Caecitellus parvulus
(Griessmann, 1913) Patterson et al., 1993 // Eur. J. Protistol, 1998.
Vol. 34. P. 369—375.
O’Kelly C. J., Patterson D. J. The flagellar apparatus of Cafeteria
roenbergensis Fenchel et Patterson, 1988 (Bicosoecales=Bicosoeci-
da) U Eur. J. Protistol. 1996. Vol. 32. P. 216—226.
Pascher A. Chrysomonadinae // Die Siisswasser-Flora Deutschlands,
Osterreichs und der Schweiz. Bd 2. Jena : Fischer, 1913. P. 7—95.
Preisig H. R. A modern concept of chrysophyte classification //
Chrysophyte algae: ecology, phylogeny and development / Eds
C. D. Sandgren et al. Cambridge : Univ. Press, 1995. P. 46—74.
Preisig H. R., Vors N., Hallfors G. Diversity of heterotrophic
heterocont flagellates // The biology of free-living heterotrophic fla-
gellates I Eds D. J. Patterson, J. Larsen. Oxford : Clarendon Press,
1991. P. 361—399. (Syst. Assoc. Special Vol.; N 45).
Slankis T., Gibbs S. P. The fine structure of mitosis and cell
division in the chrysophycean alga Ochromonas danica // J. Phycol.
1972. Vol. 8, N 2. P. 243—256.
Stein F. R. Der Organismus der Infusionsthiere. Bd 3. Leipzig,
1878.
Teal Т.Н., Guillemete T., Chapman M., Margulis L. Acronema
sippewissettensis gen. nov. sp. nov., microbial mat bicosoecid (Bico-
soecales = Bicosoecida) // Europ. J. Protistol. 1998. Vol. 34. P. 402—
414.
The Haptophyte Algae / Eds J. C. Green, B. S. C. Leadbeater.
Oxford : Clarendon Press, 1994. (Syst. Assoc. Special; Vol. 51).
Tong S.M. Heterotrophic flagellates and other protists from So-
uthampton water, U.K. // Ophelia. 1997. Vol. 47, N 2. P. 71—131.
Verhagen F.J.M., Zolffel M., Brugerolle G., Patterson D.J.
Adriamonas peritocrescens gen. nov., sp. nov., a new free-living soil
flagellate (Protista, Pseudodendromonadida incertae sedis) // Eur. J. Pro-
tistol. 1994. Vol. 30. P. 295—308.
V0rs N. Discocelis saleuta gen. nov. et sp. nov. (Protista incertae
sedis) — a new heterotrophic marine flagellate // Eur. J. Protistol.
1988. Vol. 23. P. 297—308.
Тип HAPTOPHYTA Christensen, 1962 — ГАПТОФИТЫ
Гаптофиты — мелкие, не более 30 мкм в длину, преимущест-
венно морские свободноживущие жгутиконосцы. Среди них пре-
обладают одиночные свободноплавающие организмы, но есть и не-
подвижные коккоидные, пальмеллоидные и колониальные формы.
Гаптофиты обладают уникальной нитевидной органеллой — гапто-
немой, которая расположена на переднем конце тела между 2 изо-
контными (или иногда гетероконтными) жгутиками и функцио-
нально тесно с ними связана. Поверхность тела, а часто и поверх-
ность жгутиков, а также гаптонемы покрыты органическими
чешуйками, состоящими из двухслойных пластинок, каждый слой
которых несет радиально расположенные фибриллы. Поверхность
тела кокколитофорид покрыта кальцифицированными пластинка-
ми — кокколитами.
Гаптофиты — фототрофные жгутиконосцы, имеют 1—2 хло-
ропласта золотисто-бурого цвета, содержат хлорофиллы а и с и
каротиноиды. Большинство видов гаптофитовых лишено стигмы.
Ядро 1, способ деления ядра — открытый ортомитоз. Размножают-
ся путем продольного бинарного деления. Половой процесс в
форме изогамии известен у нескольких видов. Распространены в
морском планктоне, солоноватоводных и пресноводных видов не-
много. К типу относится свыше 200 видов, принадлежащих к 1
классу, который включает 4 отряда.
История изучения
Начало изучения гаптофитовых относится к 1836 г.,
и первое сообщение об этих жгутиконосцах принад-
лежит Эренбергу, который открыл у них мельчайшие
поверхностные структуры — пластинки, состоящие
из извести, кокколиты (Ehrenberg, 1838). Несколько
позже, в 1858 г.,г появилась работа Хаксли, который
обнаружил кокколиты в морских глубинных осадках
(Huxley, 1858). Первое описание гаптонемы известно
из работы Шерффеля (Scherffel, 1901), который эту
органеллу назвал «третий жгутик», обнаружив ее у
Phaeocystis.
Первая опубликованная работа по системе кокко-
литофорид относится к началу XX столетия (Loch-
mann, 1902). Несколько позже начинается интенсив-
ное изучение гаптофитовых, появляются описания
новых видов, совершенствуется система. В 1910 г.
Пашер (Pascher, 1910) выделил отряд Isochrysidales в
системе хризофитовых с 2 изоконтными жгутиками,
а в 1930 г. вышла в свет первая монография (Schiller,
1930), посвященная этой особой группе жгутиконос-
цев. Дальнейшее изучение строения клетки, жгутико-
вого аппарата показало, что гаптонема — это не жгу-
тик, а новая органелла, присущая только гаптофито-
вым. Изучение ультраструктуры клетки гаптофито-
вых начинается с 1972 г.
Накопление знаний об особенностях организации
клетки гаптофитовых, их биологий, распространении
в морских и других водных бассейнах, их значении в
морских экосистемах, классификации, а также опи-
сания новых видов продолжаются и в настоящее вре-
мя.
Название класса гаптофитовые было введено в
1962 г. (Christensen, 1962). Позже этот класс был на-
зван примнезиофитовые — Prymnesiophyceae Hib-
berd, 1976. В системе класса Phytomastigophorea Cal-
kins, 1909 (Levine et al., 1980) гаптофитовым придан
ранг отряда Prymnesiida Hibberd, 1976; Дальнейшие
исследования привели к созданию новой системы,
которая была построена на основе главных призна-
ков, отличающих эту группу от других жгутиконос-
цев, и в частности от хризофитовых. К числу таких
признаков относятся: гаптонема, кокколиты и двух-
слойные фибриллярные чешуйки, покрывающие тело
жгутиконосцев снаружи (The Haptophyte Algae, 1994).
Классификация
В последнее время была предложена новая систе-
ма типа Haptophyta (Green, Jordan, 1994). В нем име-
ется 1 класс Prymnesiophyceae, в котором выделяют
2 подкласса: Prymnesiophycidae с порядком Prymne-
siales и Pavlovophycidae с порядком Pavlovales. Пред-
ставители бывшего порядка Isochrysidales образуют
сем. Isochrysidaceae в порядке Prymnesiales, а кокко-
литофориды распределены по 12 семействам порядка
Prymnesiales. Однако в настоящем издании мы при-
держиваемся более устоявшейся и привычной систе-
мы гаптофитовых.
Тип Haptophyta Christensen, 1962
Класс Haptophyceae Christensen, 1962
Отряд Isochrysidales Pascher, 1910
Отряд Coccosphaerales Lemmermann, 1908
Отряд Prymnesiales Papenfuss, 1955
Отряд Pavlovales Green, 1976
ТИП HAPTOPHYTA
277
Морфология и физиология
Внешняя морфология
Размеры и форма тела. Гаптофитовые — мел-
кие одиночные и реже колониальные жгутиконосцы
(рис. 178; 179). Большинство из них свободноплава-
ющие, но есть также коккоидные и пальмеллоидные,
неподвижные формы. Размеры тела не превышают
30 мкм в длину у одиночных особей, колониальные
формы могут достигать до 8.0 мм в диаметре (Phae-
ocystis pouchetii). Большинство видов имеет постоян-
ную форму тела, которая варьирует от шаровидной,
овальной до уплощенной. Однако в жизненных цик-
лах некоторых видов есть амебоидные стадии, отли-
чающиеся непостоянной формой тела (Pavlova).
Жгутиковый аппарат. 2 жгутика равной или иног-
да неравной длины отходят от вершины переднего
конца тела у шаровидных и овальных по форме тела
особей, а у уплощенных — сбоку (субапикально).
Жгутики не несут трубчатых мастигонем, но у не-
которых видов покрыты тонкими волосками. Между
жгутиками расположена гаптонема — специфическая
для данного типа органелла (рис. 178). Длина жгути-
ков и характер жгутикового движения широко варь-
ируют у разных видов.
Переходная зона жгутиков гаптофитовых, имею-
щая важное таксономическое значение, достаточно
разнообразна (Карпов, 1990). Для видов Pavlova и
Chrysochromulina характерна длинная переходная зо-
на, и в ней имеется 1 поперечная пластинка. Некото-
рые гаптофиты обладают 2 поперечными пластинка-
ми, которые соединяются центральным филаментом,
а в дистальной части переходной зоны находятся тон-
кие мостики между периферическими группами мик-
ротрубочек (Hibberd, Norris, 1984; Green, Hori,. 1994).
Переходная зона жгутиков Pleurochrysis имеет не-
сколько необычное строение: к периферическим
группам микротрубочек тесно прилегает цилиндр с
тонкими стенками, а в него вложен другой цилиндр,
обладающий толстыми стенками; поперечная плас-
тинка отсутствует (Inouye, Pienaar, 1985).
Корешковая система жгутиков отличается доволь-
но сложным строением и изучена у нескольких ви-
дов, относящихся к отрядам Pavlovales, Prymnesiales
и Coccosphaerales. У всех изученных видов рода Pav-
lova от кинетосом отходит 2 корешка (рис. 180).
Один из них расположен сбоку, проходит под плаз-
малеммой и состоит из 7 микротрубочек. 2-й коре-
шок направлен в глубь клетки и состоит из 2 микрот-
рубочек.
Виды, относящиеся к отряду Prymnesiales имеют
более сложную и разнообразную корешковую сис-
тему (рис. 181). У Chrysochromulina apheles и Ch.
parva имеется 4 корешка, состоящих из микротрубо-
чек. Один из них широкий, состоит из 5 микротру-
бочек, и в основании его находится тонкая пластин-
ка, а 2 других корешка, состоящих из 1 и 2 микрот-
рубочек, направлены к хлоропласту. Последний, 4-й
корешок представлен микротрубочками, проходящи-
ми вдоль одной из кинетосом (Moestrup, Thomsen,
1986).
Система жгутиковых корешков организована наи-
более сложно у видов отряда Coccosphaerales. Кокко-
литофориды имеют 3 корешка, которые образованы
микротрубочками. Один из корешков состоит из
20 микротрубочек, и от него под углом 90° отходит
100—200 вторичных микротрубочек. 2-й корешок
включает 4 микротрубочки, и от него отходят много-
численные вторичные микротрубочки. 3-й корешок
обладает более простым строением и состоит из 6—
7 микротрубочек (Inouye, Pienaar, 1988; Карпов,
1990).
Гаптонема. Эта особая органелла гаптофитовых
имеет форму тонкой нити и способна закручиваться
спирально у многих видов. Длина гаптонемы неоди-
накова у разных видов: гаптонема Chrysochromulina
достигает в длину 100 мкм при диаметре тела жгути-
коносца 8 мкм, у Isochrysis и Chrysotila длина ее со-
ставляет 1—2 мкм (рис. 182). У Dicrateria и Imanto-
nia гаптонема настолько укорочена, что может быть
видна только с помощью электронного микроскопа;
она имеет форму хоботка, расположенного между
2 жгутиками. По своей ультраструктуре гаптонема
значительно отличается от жгутика (рис. 182). В цен-
тре ее проходит лента из 6—8 микротрубочек, кото-
рые окружены каналом гладкой эндоплазматической
сети (Moestrup, Thomsen, 1986). Микротрубочки гап-
тонемы отходят от одной из кинетосом и потому ее
иногда считают одним из корешков жгутика, образо-
ванных микротрубочками, хотя и у самой гаптонемы
(рис. 180) обнаружены фибриллярные корешки (Hib-
berd, 1976; Карпов, 1990).
Гаптонема тесно соединена с жгутиками много-
численными фибриллами, и движения жгутиков взаи-
мосвязаны с функционированием гаптонемы (Green,
Jordan, 1994). Существует также мнение, что гапто-
нема выполняет роль прикрепительной органеллы
(Хаусман, 1988).
Поверхностные структуры. Поверхность тела боль-
шинства видов гаптофитовых покрыта органически-
ми чешуйками (пластинками), расположенными в 1
или несколько рядов (рис. 179). Дистальная поверх-
ность чешуйки несет концентрические фибриллы, а
также радиально расположенные фибриллы на про-
ксимальной поверхности. У некоторых видов слой
более мелких чешуек, чем на поверхности тела, пок-
рывает снаружи гаптонему.
Поверхностными структурами у кокколитофорид
служат довольно крупные, размером 0.25—3.0 мкм,
кокколиты (коккосферы). Основу кокколитов со-
ставляют органические чешуйки, на поверхности ко-
торых карбонат кальция откладывается в виде каль-
цита или арагонита, образуя ромбические кристаллы
или гексагональные призмы. Кокколиты очень разно-
образны по форме (рис. 178). Одной из их особен-
ностей является отсутствие или наличие внутреннего
канала. По этому признаку кокколиты делятся на 2
группы: сплошные — дисколиты и с осевым каналом
в центре — тремалиты.
Все типы чешуек, а также некоторые кокколиты
формируются внутри клетки, в аппарате Гольджи, а
затем транспортируются к поверхности клетки. Плот-
ный покров («стенка») коккоидных клеток и ните-
видных форм состоит из нескольких слоев тесно упа-
278
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 178. Особенности строения некоторых гаптофитовых. (По: Hibberd, Leedale, 1985).
А — Prymnesium parvum: 1 — жгутики, 2 — гаптонема, 3 — хлоропласты, 4 — ядро, 5 — аппарат Гольджи, 6 — капли жира, 7 -
Chrysochromulina ericina: В — Coccolithus pelagicus: Г — Ochrosphaera neopolitana: 1 — кокколиты; Д — Isochrysis galbana: E
1 — кокколиты; Ж — Pavlova gyrans: 1 — стигма.
Fig. 178. Cell morphology of some species of Haptophyta.
A — Prymnesium parvum: I — flagella, 2 — haptonema, 3 — chloroplasts, 4 — nucleus, 5 — Golgi bodies, 6 — drops of lipid, 7 —
Chrysochromulina ericina: В — Coccolithus pelagicus: Г — Ochrosphaera neopolitana: 1 — coccoliths; Д — Isochrysis galbana; E -
1 — coccoliths; Ж — Pavlova gyrans: 1 — stigma.
хризоламинарин; Б—
- Ophiaster hydroideus:
chrysolaminarin; Б —
Ophiaster hydroideus:
ТИП HAPTOPHYTA
279
J
Рис. 179. Колониальный гаптофит Phaeocystis pouchetii. (По: Hib-
berd, Leedale, 1985).
1 — колонии в слизистом футляре, 2 — изолированные особи колонии
(зооиды), 3 — подвижные формы с двумя жгутиками и гаптонемой (зоо-
споры), 4 — чешуйки (схематизировано по данным электронного микро-
скопа, х 100 000).
Fig. 179. Colonial species of Haptophyta — Phaeocystis pouchetii.
J — colonies enclosed in mucous envelope, 2 — cells of colony (zooids),
3 — moving cells with two flagella and haptonema (zoospores), 4 — scales
(electron micrograph, x 100000).
Рис. 180. Схема строения жгутикового аппарата рода Pavlova.
(По: Карпов, 1990).
1 — жгутики, 2 — гаптонема, 3 — микротрубочки гаптонемы, 4 — цис-
терна эндоплазматической сети, 5 — фибриллярные корешки, 6 — мик-
ротрубочковые корешки.
Fig. 180. Diagram of structure of flagellar apparatus in Pavlova
genus.
1 — flagella, 2 — haptonema, 3 — microtubules of haptonema, 4 — cistern
of endoplasmatic reticulum, 5 — fibrillar rootlets, 6 — microtubular rootlets.
Рис. 181. Обобщенная схема строения жгутикового аппарата у
гаптофитовых. (По: Andersen, 1991).
1—4 — микротрубочковые корешки, пронумерованные по аналогии с
хризофитовыми. га — гаптонема.
Fig. 181. General diagram of structure of flagella apparatus in Hap-
tophyta.
1—4 — microtubular rootlets number by analogy with Chrysophyta; га —
haptonema.
Рис. 182. Схема строения типич-
ной гаптонемы. (По: Inouye, Ка-
wachi, 1994).
А — поперечные срезы гаптонемы на
уровнях (а—и), показанных на про-
дольном срезе (Б). 1—9 — микротру-
бочки гаптонемы, которые реорганизу-
ются по мере приближения к дисталь-
ной части, л — кинетосома левого
жгутика, п — кинетосома правого жгу-
тика.
Fig. 182. Diagram of structure of
typical haptonema.
A — Transversal sections of haptonema
on the levels (a—w), shows on the longi-
tudinal section (F). 1—9 — microtubules
of haptonema which undergo reorganiza-
tion by measure approach to the distal
part, л — kinetosome of left flagellum,
n — kinetosome of right flagellum.
280
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
кованных чешуек (Leadbeater, 1994). Такие чешуйки
представляют собой сеть концентрических и ради-
ально расположенных целлюлозных фибрилл, кото-
рые у коккоидных особей, имеющих «стенки», пог-
ружены в желатинизированный слой полисахаридов
типа пектина (Hibberd, Leedale, 1985). Гаптофитовые
рода Pavlova не имеют типичных чешуек на поверх-
ности клетки.
Форма, размеры и ультраструктура чешуек гапто-
фитовых, включая и особенности строения кокколи-
тов у разных видов, имеют большое значение в пост-
роении системы типа.
Цитоплазма
Фототрофные виды имеют 1—2 хлоропласта
(рис. 178), окрашенных в золотисто-бурый цвет бла-
годаря содержанию хлорофиллов а и с (с19 с2), а
также каротиноидов с преобладанием фукоксантина.
Оболочка хлоропластов состоит из 4 мембран
(включая мембрану ЭПР, в расширении которого на-
ходится хлоропласт). Каждая ламелла хлоропласта
включает 3 тилакоида, опоясывающая ламелла от-
сутствует. Хлоропласт несет 1 пиреноид, наполови-
ну погруженный в строму самого хлоропласта. Мат-
рикс пиреноида пронизывают 2 ламеллы тилакоида.
Преобладающее большинство видов фототрофы, но
есть и небольшое число фаготрофных видов, например
в составе рода Chrysochromulina (Hibberd, Leedale,
1985). Стигма свойственна лишь немногим видам и
наиболее хорошо развита у жгутиконосцев рода Pav-
lova (рис. 178). Она находится внутри хлоропласта, в
одной из его долей, и состоит из 1 ряда окрашенных
в красный цвет глобул. Парафлагеллярное тельце
имеется только у 1 вида — Diacronema vlkianum.
Комплекс диктиосом локализован вблизи ядра и
часто имеет веерообразную форму. В цистернах дик-
тиосом синтезируются все виды чешуек, включая и
кокколиты. Формирование поверхностных чешуек —
одна из важных и специфических функций аппарата
Гольджи в клетке гаптофитовых.
Сократительная вакуоль свойственна пресновод-
ным видам и расположена в передней части тела.
Многочисленные экструсомы в периферическом
слое цитоплазмы представлены мукоцистами, которые
выбрасывают наружу слизь в форме тонких нитей.
В цитоплазме постоянно имеются включения хри-
золаминарина и липидов. Они накапливаются в
форме капель различных размеров, расположенных
во всех участках клетки.
Ядро и его деление
Гаптофитовые — одноядерные организмы. Ядро
пузырьковидного типа, часто расположено в средней
части клетки. Ядрышко 1 или несколько. Деление
ядра осуществляется путем открытого ортомитоза,
который подробно изучен у Prymnesium parvum, а
также у Chrysochromulina chiton и других. Перед де-
лением ядра к полюсам расходятся удвоившиеся к
этому времени кинетосомы жгутиков. Ядерная обо-
лочка фрагментируется в поздней профазе, и одно-
временно формируется веретено митоза. Оно сим-
метричное, с хорошо выраженными пучками мик-
ротрубочек. Метафазная пластинка плотная, и в ней
не удается различить отдельные хромосомы. Ядрыш-
ко в процессе митоза резорбируется и образуется
вновь в каждом дочернем ядре в телофазе после
образования новых оболочек дочерних ядер. Верете-
но митоза остается в цитоплазме и там резорбирует-
ся (Райков, 1978; Hori, Green, 1985; Green et al.,
1989)
Размножение и жизненные
циклы
Все гаптофитовые размножаются агамно путем
бинарного деления в подвижном и пальмеллоидном
состояниях. Например, колонии Phaeocystis pouchetii
размножаются путем фрагментации на части. Поло-
вой процесс с формированием изогамет известен для
немногих видов, но изучен пока недостаточно.
Достоверно формирование изогамет известно у
Hymenomonas carterae, в сложном жизненном цикле
которого чередуются неподвижная (пальмеллоидная)
и подвижная свободноплавающая стадии (Gayral,
Frensel, 1983; Hibberd, Leedale, 1985). У одного из
морских видов рода Ochrosphaera (рис. 178) в жиз-
ненном цикле обнаружены гаплоидные изогаметы,
которые сливаются попарно и образуют зиготу, пре-
терпевающую мейоз. В результате мейоза формиру-
ются 4 гаплоидные зооспоры. Они представляют рас-
селительную стадию и, развиваясь, превращаются в
обычные для вида вегетативные особи.
Экология и практическое
значение
Большинство видов гаптофитовых обитает в мор-
ских бассейнах. В солоноватоводных и пресных во-
доемах их немного. Морские виды населяют толщу
воды до глубины 150 м. Наибольшей численности
они достигают в планктоне морей южных и умерен-
ных широт, в северных морях их значительно мень-
ше.
Кокколитофориды — обитатели открытых зон
морей и океанов. Приуроченность их именно к этим
зонам обусловлена высокой концентрацией солей
кальция в морской воде, достаточной освещен-
ностью толщи воды, ее прозрачностью, что создает
оптимальные условия для фотосинтеза. Ряд кокколи-
тофорид и других видов обитают также в шельфо-
вых зонах морей.
С массовым развитием гаптофитовых, в частности
кокколитофорид, может быть связано явление «цве-
тения» морской воды. Причины цветения часто
обусловлены массовым развитием Phaeocystis pou-
chetii, которое приурочено к осени и весне. Доста-
точно крупные колонии этого вида, покрытые снару-
жи слизистой оболочкой (рис. 179) и окрашенные в
золотисто-бурый цвет, образуют в воде плотную сли-
зистую массу, которая представляет серьезное пре-
ТИП HAPTOPHYTA
281
пятствие для миграций морских животных. «Цветение»
воды, связанное с развитием токсичных видов из
родов Chrysochromulina и Prymnesium, приводит к
массовой гибели морских животных, особенно рыбы,
что может отрицательно сказываться на экономике.
Плотные популяции образует также Isochrysis gab
bana — один из видов морского планктона, который
вместе с другими видами служит пищей различным
морским животным, и в первую очередь личинкам
моллюсков (рис. 178).
Известковые панцири современных и ископаемых
кокколитофорид образуют мощные пласты известня-
ков. Некоторые известковые породы, в частности мел,
на 50—75 % состоят из кокколитофорид. В ископае-
мом состоянии кокколитофориды известны со вре-
мен кембрия. Наиболее подробно они изучены из юр-
ских и меловых отложений. Их ископаемые панцири
используются с целью определения геологического
возраста известковых пород, поскольку каждому гео-
логическому периоду развития Земли соответствует
определенная группа этих жгутиконосцев.
Из немногих пресноводных гаптофитовых извест-
ны Diacronema vlkianum, несколько видов Deropyxis
и Pavlova (рис. 178). Эти фототрофные виды обычны
в планктоне пресноводных и солоноватоводных бас-
сейнов.
Происхождение и эволюция
Среди окрашенных жгутиконосцев (хромистов)
гаптофиты представляют обособленную группу, так
как им свойственна специфическая для отряда орга-
нелла — гаптонема, которая, вероятно, представляет
собой дериват жгутика. Однако гаптофиты обнару-
живают существенное сходство с хризомонадами по
многим морфологическим признакам, в частности по
структуре жгутиков, ультраструктуре хлоропластов,
форме крист в митохондриях, а также по типам хло-
рофиллов. Хризомонады и гаптофиты — филогене-
тически родственные группы, к которым близки ди-
атомовые и бурые водоросли (Taylor, 1976; Серавин,
1980). Молекулярно-биологические данные, однако,
постоянно демонстрируют большое расстояние меж-
ду хризофитами и гаптофитами на филогенетическом
древе (Van de Peer, De Wachter, 1997). Морфологи
также находят все больше существенных различий
между этими группами.
Систематический обзор
К типу принадлежит 4 различающихся по морфо-
логическим признакам отряда, и ниже приведена их
краткая характеристика с составе класса гаптофито-
вых — Haptophyceae.
Отряд Isochrysidales Pascher, 1910
Отряд включает несколько мелких видов гаптофи-
товых, длина тела которых не превышает 10 мкм, с
гладкими, без мастигонем, изоконтными жгутиками,
отходящими от переднего конца тела и во время дви-
жения направленными вперед. Форма тела эллипсоид-
ная, слегка уплощенная в дорсовентральном направ-
лении. Тело метаболичное, покрыто мелкими овальными
чешуйками, и очень мелкие чешуйки покрывают ко-
роткую, сильно редуцированную гаптонему.
Хлоропластов 1—2, они расположены по бокам те-
ла и окрашены в золотисто-бурый цвет. Ядро сфери-
ческое, находится в центре тела. Фототрофы. Резерв-
ные вещества — жир и лейкозин. Размножение агамное
путем продольного бинарного деления, которое проис-
ходит как в подвижном, так и в неподвижном состоя-
нии. Изучение в культуре показало, что в период разм-
ножения образуется толстостенный спорангий, внутри
которого особь делится на 16 подвижных клеток. Все
известные виды отряда — морские.
Типовой вид — Isochrysis galbana Parke, 1949.
Жгутиконосец очень мелкий, длина тела его не превы-
шает 5—6 мкм, толщина 2.5—3.5, ширина 2—4 мкм.
Жгутики примерно в 2 раза длиннее тела или равны его
длине. Тело покрыто мелкими чешуйками 2 типов: че-
шуйки, имеющие размеры 0.3—04-х 0.2—0.3 мкм, и че-
шуйки размером около 0.06 мкм с утолщением в центре.
Такие же мельчайшие чешуйки покрывают гаптонему.
Отряд Coccosphaerales
Lemmermann, 1908
Отряд объединяет свыше 100 известных видов,
морфология клетки которых типична для гаптофито-
вых. Основной признак коккосферид (кокколитофо-
рид) — внешний покров, в состав которого входят
кокколиты, образованные на основе органических
чешуек. На поверхности этих чешуек карбонат каль-
ция кристаллизуется в виде кальцита или арагонита,
формируя ромбовидные кристаллы и гексагональные
призмы (Black, 1963; The Haptophyte Algae, 1994).
Кокколиты имеют большое значение для системати-
ки отряда, так как обладают широким разнообразием
форм, размеров и характера расположения у разных
видов. Многим кокколитофоридам свойственны жиз-
ненные циклы со сменой стадий, отличающихся по
признакам морфологии, формам размножения, под-
вижности, способности к инцистированию и т. д.
Ниже приведены краткие характеристики наиболее
широко распространенных родов, включающих зна-
чительное число видов и типичных для отряда.
Род Coccolithus Schwarz, 1932
К роду относятся морские планктонные кокколи-
тофориды, у которых в жизненном цикле чередуются
подвижная и неподвижная стадии. Внешний покров
неподвижной стадии представлен плаколитами, со-
стоящими из 2 дисков. Каждый диск образован пере-
крывающимися, радиально расположенными сегмен-
тами. В центре плаколита находится округлое отвер-
стие — центральная пора, края которой образованы
соединенными между собой дисками. Подвижные
особи несут на поверхности тела кокколиты (крис-
талл олиты), которые чередуются с обычного вида ор-
282
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тоническими чешуйками. В составе рода 1 вид С. ре-
lagicus (Wallich) Schiller, 1929.
В жизненном цикле этого вида 2 фазы — подвиж-
ная, называемая «Crystallolithus», и неподвижная —
«Coccolithus». Подвижная стадия имеет сферическое
или близкое к сферической форме тело, диаметр ко-
торого 8—18 мкм. Жгутики одинаковые или различа-
ющиеся по длине, гомодинамичные. Гаптонема спи-
рально закручена, ее длина 2—10 мкм, у ее основа-
ния расположено вздутие. Внутри гаптонемы обычно
находится 6 микротрубочек. Тело покрыто кокколита-
ми типа кристаллолитов, которые состоят из эллип-
соидальных пластинок, образованных ромбовидными
кристаллами кальцита. Кроме кокколитов в поверх-
ностных покровах имеется 2 типа органических че-
шуек: овальной формы, размером 2x1.5 мкм, и мел-
кие удлиненные пластинки размером 0.8 х 0.45 мкм.
Одна и та же особь может иметь покров из несколь-
ких слоев чешуек и кокколитов.
Неподвижная фаза жизненного цикла — «Cocco-
lithus» — сферической или близкой к сферической
формы. Тело неподвижных особей покрыто кокколи-
тами (плаколитом), состоящими из 2 дисков, которые
соединяются по краю центральной поры (см. выше).
Между кокколитами расположены органические че-
шуйки овальной формы, средний размер которых со-
ставляет 7.5 х 5.0 мкм и очень мелкие чешуйки около
2 мкм в диаметре (Hibberd, Leedale, 1985).
Подвижные и неподвижные формы — фототрофы,
и морфология их клеток сходна. В клетке имеется
2—4 хлоропласта удлиненной формы, расположенных
на ее боковых сторонах. Хлоропласты окрашены в
золотисто-бурый цвет, и каждый из них содержит пи-
реноид, наполовину выступающий в цитоплазму.
Ядро находится в центре клетки. На нижнем конце
клетки имеется глубокая щель. Размножение атомное.
С. pelagicus обычен в морском планктоне и пред-
ставляет единственный вид этого отряда в холодных
водах Северной Атлантики.
Род Ochrosphaera Schussing, 1930
В жизненном цикле кокколитофорид этого рода
доминирует неподвижная фаза, которая может быть
представлена либо одноклеточными особями, либо
мелкими сферическими колониями, поверхность те-
ла которых покрыта кокколитами. Одноклеточные
неподвижные особи лишены жгутиков. Они имеют
2 крупных хлоропласта золотисто-бурого цвета; каж-
дый хлоропласт несет крупный пиреноид. Ядро рас-
положено в центре клетки. Резервные вещества —
липиды и хризоламинарин. Размножение происходит
путем деления клетки внутри оболочки, покрытой
кокколитами, в результате чего формируются 2 или
4 подвижные дочерние особи.
У одного вида известно половое размножение, в
процессе которого формируются изогаметы. Дипло-
идная зигота, возникающая при слиянии изогамет,
претерпевает мейоз и делится на 4 подвижные вегета-
тивные клетки. К роду Ochrosphaera относится не-
сколько морских планктонных видов. Ниже приведе-
но краткое описание типового вида этого рода.
Ochrosphaera neopolitana Schussing, 1930.
Жизненный цикл вида включает неподвижные од-
ноклеточные особи сферической формы 4.7—6.0 мкм
в диаметре, и подвижные, такие же мелкие, сфери-
ческие особи. У подвижных особей есть 3 изоконт-
ных жгутика, длина которых 12—15 мкм. Гаптонема
сильно редуцирована. Внешний покров жгутиковых
особей состоит из чешуек почти сферической формы
(диаметр 0.5—0.8 мкм) и кокколитов 2 типов, кото-
рые различаются по форме и размерам. У жгутико-
вых особей кокколиты развиваются либо на протяже-
нии всего времени их жизни, либо непосредственно
перед превращением их в неподвижную безжгутико-
вую стадию (Hibberd, Leedale, 1985).
Отряд Prymnesiales Papenfuss, 1955
Очень типичный для гаптофитовых отряд, вклю-
чающий немногочисленных, преимущественно морс-
ких и солоноватоводных жгутиконосцев. Для них ха-
рактерна яйцевидная форма тела. Жгутики изоконт-
ные, длина их либо равна длине тела, либо в 1.5 раза
превышает ее. 1-й жгутик расположен под прямым
углом по отношению к переднему концу тела, 2-й
проходит по одной из боковых сторон. Гаптонема не
закручивается спирально, длина ее соответствует
V3 длины тела. Внешний покров тела представлен
органическими чешуйками. Хлоропластов 2, они на-
ходятся под плазмалеммой в периферическом слое
цитоплазмы. Ядро расположено в центре клетки.
Имеется сократительная вакуоль, но она почти не
функционирует. В периферической цитоплазме
много мукоцист (Green et al., 1982).
Типовой вид — Prymnesiumparvum Carter, 1937.
Подобно другим видам отряда, — это мелкий жгути-
коносец с размерами тела 8—15x4—10 мкм. Оба
жгутика одинаковы по длине, которая равна 12—
20 мкм. Гаптонема короткая, прямая, ее длина 3—
5 мкм. Тело покрыто плоскими органическими че-
шуйками овальной формы, расположенными в
2 слоя. Чешуйки 2 типов: во внешнем слое их разме-
ры равны 0.3—0.4 х 0.23—0.3 мкм, они имеют узкий
гибкий ободок; чешуйки внутреннего слоя с размера-
ми 0.3—0.36 х 0.26—0.32 мкм имеют широкий обо-
док, проходящий по их дистальной поверхности.
В жизненном цикле Prymnesium образуются оваль-
ные цисты, оболочка которых состоит из нескольких
слоев чешуек, покрытых снаружи кремнеземом. Это
единственный случай содержания кремнезема в че-
шуйках гаптофитовых.
Р, parvum обитает в планктоне солоноватоводных
бассейнов. Известен как токсический жгутиконосец,
который при массовом размножении вызывает гибель
рыбы в солоноватых водоемах севера Европы, Сред-
него Востока, и особенно Израиля.
Отряд Pavlovales Green, 1976
Гаптофитовые этого отряда имеют ряд специфи-
ческих, присущих только им признаков, и значитель-
но отличаются от других видов. Форма тела этих жгу-
ТИП HAPTOPHYTA. SUMMARY
283
тиконосцев варьирует от удлиненной до амебоидной,
органические чешуйки на поверхности тела отсутст-
вуют. Жгутиков 2, они гетероконтные и сильно раз-
личаются по длине. Оба жгутика отходят не от самой
вершины переднего конца тела, а несколько ниже, на
брюшной стороне сбоку.
Длинный жгутик направлен вперед, короткий —
назад. Гаптонема прямая, не закручена спирально,
длина ее варьирует у разных видов. В /клетке 1 хло-
ропласт желто-зеленого цвета. Стигма расположена
на переднем конце тела, интенсивно окрашена в
оранжевый цвет, не связана с хлоропластом. Ядро 1,
сферическое, находится в передней трети тела. Раз-
множение агамное, половой процесс неизвестен.
К отряду относится 2 рода, включающих свыше
10 видов планктонных морских, солоноватоводных и
пресноводных жгутиконосцев.
Род Pavlova Butcher, 1952
Включает несколько видов, которым свойственны
все основные признаки отряда. 2 жгутика, выходящие
субапикально сбоку, сильно различаются по длине, и
длинный, передний жгутик более толстый, чем ко-
роткий, задний. Короткая прямая гаптонема может
быть снабжена тонкими волосковидными отростками
на конце. Внешний покров тела тонкий, гладкий, об-
разован плазмалеммой, под которой расположены до-
вольно крупные полости. Хлоропласт 1, желто-зеле-
ного цвета, находится в центре клетки. Он удлинен и
состоит из 2 боковых долей, между которыми располо-
жен пиреноид. Стигма крупная, располагается вблизи
жгутиковых кинетосом. У пресноводных видов имеется
сократительная вакуоль. Размножаются путем про-
дольного бинарного деления (Green, 1980).
В состав рода входит 2 вида. Один из них —
Р. gyrans Butcher, 1952. Это очень мелкий жгути-
коносец: размеры его тела составляют 4.0—10.0 х
2.0—2.5 мкм. Форма тела варьирует от удлиненной
до амебоидной. Передний, длинный жгутик покрыт
слоем «войлока» и несет продольные ряды плотных
телец, имеющих размеры 0.03 х 0.025 мкм. Во время
движения жгутик принимает характерную s-образ-
ную форму. Короткий, задний жгутик достигает
V3 длины переднего жгутика и покрыт очень тонки-
ми волосками. Гаптонема тонкая, внутри снабжена
только 2 микротрубочками и хорошо развитым ка-
налом ЭПР.
Хлоропласт желто-зеленый, двухлопастной, с
центральным пиреноидом. Стигма ярко-красная, рас-
положена на одной из долей хлоропласта. Иногда
на заднем конце тела образуются тонкие филаменты,
представляющие, вероятно, слизь, выделяемую муко-
цистами.
Р. gyrans — планктонный вид, распространенный
в солоноватоводных бассейнах.
Род Diacronema Prauser, 1958
К роду относятся мелкие гаптофитовые с 2 гете-
роконтными жгутиками с гладкой поверхностью, и
каждый из них несет тонкий волосковидный отрос-
ток. Длинный, передний, двигательный жгутик обра-
зует характерную петлю, когда жгутиконосец не дви-
жется; задний жгутик короткий. Оба жгутика выхо-
дят на брюшной стороне тела субапикально сбоку.
Гаптонема прямая, короткая. Стигма крупная, крас-
ная, приурочена к передней трети тела.
В клетке 1 двухлопастной хлоропласт светло-жел-
того или оливкового цвета. Его узкие лопасти, распо-
лагающиеся по бокам клетки, соединяются на заднем
конце тела. Пиреноид отсутствует. Ядро сферичес-
кое, обычно находится в передней части тела. План-
ктонные морские и пресноводные жгутиконосцы.
Известен 1 вид рода — D. vlkianum Prauser, 1958.
Жгутиконосцы этого вида обладают широким оваль-
ным телом с размерами 3.5—7.5 х 4.0—5.0 х 1.5—
3.0 мкм. Длинный, передний жгутик достигает в
длину 7.0—10.0 мкм, и его свободный конец несет
тонкий волосковидный отросток длиной 0.5—1.5 мкм.
Задний жгутик более короткий и его длина состав-
ляет 6.0—9.0 мкм, а длина его волосковидного от-
ростка 2.0—3.5 мкм. Оба жгутика по всей длине
несут очень тонкие волоски. У основания заднего
жгутика есть небольшое расширение — парафлагел-
лярное тельце, расположенное над стигмой.
Гаптонема короткая, ее длина около 1 мкм, и на
свободном конце имеется небольшое расширение.
Вид пресноводный, обладает сократительной ва-
куолью. Резервное вещество — липиды в виде мелких
капель. На заднем конце клетки расположены 2 круп-
ные сферические вакуоли (Hibberd, Leedale, 1985).
THE PHYLUM HAPTOPHYTA CHRISTENSEN, 1962
SUMMARY
Most species of the phylum are marine planctonic flagellates, and
only a few species inhabit freshwater reservoirs. The majority of the
flagellates are monads, though some species have amoeboid, palmelloid
stages or coccoid and colonies of various dimensions and form. The
unicellular body of flagellates is spherical, oval, curved or flattened.
The very specific organelle of Haptophyta is haptonema, situated
between two flagella originating on the apex of the cell. The haptone-
ma is completely different from flagella in its structure and functions.
It consists of 6—8 single microtubules, which are surrounded by a
cistern of endoplasmic reticulum. In some species it is very long,
coiled or uncoiled, in others it is reduced. Some species may swim
with the flagella and haptonema.
The body of haptophyta is covered with one or several layers of
unmineralised scales (organic) with fibrills and radiating lines.
Coccolithophorids are covered with large unmineralised scales,
coccoliths, on die surface of which calcium carbonate is crystallized as
calcite and aragonite in the form of rhombohedral crystals of hexagonal
prisms. All types of scales are formed inside the cell, either in the dicty-
osomes of Golgi apparatus or endoplasmic vesicles and transported to
the body surface. Some genera and species (Pavlova) have no scales.
Freshwater species have a contractile vacuole.
Coloured species have 2 saucer-shaped chloroplasts per cell.
Each chloroplast possesses a single pyrenoid. The chloroplast lamellae
consist of 3 thylacoids, a girdle lamella is absent. Chloroplasts
284
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
аге golden-brown, contain chlorophyll а, с, and various carotenoids:
fucoxantin, diadinoxantin, diatoxantin, characteristic of Chrysophyta.
Most species are photoautotrophic, and others are phagotrophic.
All species reproduce by longitudinal fission. The sexual process also
takes place, however it has been studied insufficiently.
Some haptophytes, especially coccolithophorids, are the main
constituent of marine plankton, being very important in primary pro-
duction. Some marine benthic species form supralittoral bands consis-
ting mostly of chalk.
A description of the main genera is presented in this chapter.
ЛИТЕРАТУРА16
Карпов С. А. Система протистов. Омск, 1990. в
Райков И. Б. Ядро простейших. Морфология и эволюция. Л. :
Наука, 1978.
Серавин Л.Н. Макросистема жгутиконосцев // Принципы пост-
роения макросистемы одноклеточных животных. Л., 1980. С. 4—22.
Хаусман К. Протозоология. М.: Мир, 1988.
Black М. The fine structure of the mineral parts of Coccolithop-
horidae // Proc. Linn. Soc. Lond. 1963. Vol. 174. P. 41—46.
Christensen T. Alger. Botanik. Bd 2. Systematic Botanik. N 2.
Copenhagen: Munksgaard, 1962.
Ehrenberg D. Ch. G. Infusionsthierreichen als vollkommene Or-
ganismen. Leipzig, 1838.
Gayral R., Frensel J. Description, sexualite et cycle de develop-
ment d’une nouvelle coccolithophoracee (Prymnesiophyceae): Pleuro-
cystis pseudoroscoffensis sp. nov. // Protistologica. 1983. T. 19, N 2.
P. 245—261.
Green J. C. The fine structure of Pavlova hingius Green and pre-
liminary survey of the order Pavlovales (Prymnesiophyceae) // Brit.
Phycol. J. 1980. Vol. 15, N 1. P. 151—191.
Green J. C., Hibberd H. J., Pienaar R. N. The taxonomy of
Prymnesium (Prymnesiophyceae) including a description of a new cos-
mopolitan species P. patellifera sp. nov. and further observations on
P. parvum N. Carter // Brit. Phycol. J. 1982. Vol. 17, N 2. P. 363—382.
Green J. C., Hori T., Course P. A. An structural study of mitosis
in Chrysochromulina chiton (Prymnesiophyceae) // Phycologia. 1989.
T. 28, N 3. P. 318—330.
Green J. C., Jordan R. W. Systematic history and taxonomy //
The Haptophyte Algae I Eds J. C. Green, B. S. C. Leadbeater. Oxford,
1994. P. 1—21.
Green J. C., Leadbeater B. S. C. Chrysochromulina parkeae sp.
nov. (Haptophyceae) a new species recorded from S. W. England and
Norway // J. Mar. Biol. Ass. U. K. 1972. Vol. 52, N 3. P. 469—474.
Hibberd D. J. The ultrastructure and taxonomy of the Chrysophy-
ceae and Prymnesiophyceae (Haptophyceae) a survey with some new
observations on the ultrastructure.of the Chrysophyceae // Bot. J. Linn.
Soc. 1976. Vol. 72, N 1. P. 55—80.
Hibberd D. J., Leedale G. F. Order Prymnesiida Hibberd, Leeda-
le, 1985 // Illustrated Guide to the Protozoa. 1985. P. 74—88.
Hibberd D. J., Norris R. Ё. Cytology and ultrastructure of Chlo-
rarachnion reptans (Chlorarachniophyta divisio nova, Chlorarachniop-
hyceae classis nova) // J. Phycol. 1984. Vol. 20, N 3. P. 310—330.
Hori T., Green J. C. An ultrastructural study of mitosis in non-
motile coccolith-bearing cells of Emiliania huxleyi (Lohm) Hay et Moh-
ler (Prymnesiophyceae) // Protistologica. 1985. T. 21, N 1. P. 107—120.
Inouye j., Pienaar R. N. Ultrastructure of the flagellar apparatus
in Pleurochrysis (Class Prymnesiophyceae) // Protoplasma. 1985. Vol. 125,
Nl. P. 24—35.
Inouye J., Pienaar R. N. Light and electron microscope observa-
tions of the type species of Syracosphaera, S. pulchra (Prymnesiophy-
ceae) // Brit. Phycol. J. 1988. Vol. 23, N 2. P. 205—217.
Leadbeater B. S. C. Cell coverings // The Haptophyte Algae I
Eds J. C. Green, B. S. C. Leadbeater. Oxford, 1994. P. 23—46.
Levine N. D., Corliss J. O., Cox P. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R., Lom J., Lynn D.,
Merinfeld E. G., Page F. C., Poljansky G., Sprague V., Vavra J.,
Wallace F. G. A newly revised classification of the Protozoa // J. Pro-
tozool. 1980. Vol. 27, N 1. P. 37—58.
Lochmann H. Die Coccolithophoridae eine Monographic der coc-
colithobildenen Flagellaten // Arch. Protistenk. 1902. Bd 1. S. 89—
165.
Manton I. Further observations on the microanatomy of the hap-
tonema in Chrysochromulina chiton and Prymnesium parvum // Pro-
toplasma. 1968. Vol. 66. P. 35—58.
Manton L, Leedale G. F. Observations on the fine structure of
Prymnesium parvum Carter // Arch. Microbiol. 1963. Vol. 45, N 2.
P. 285—303.
Manton I., Peterfi L. S. Observation on the fine structure of coc-
coliths, scales and the protoplast of a freshwater coccolithophorid Hy-
menomonas roseola Stein, with supplementary observations on the
protoplast of Cricosphaera carteriae // Proc. Roy. Soc. London. Ser. B.
1969. Vol. 172, N 1. P. 1—15.
Moestrup 0., Thomsen H. A. Ultrastructure and reconstruction of
the flagellar apparatus in Chrysochromulina apheles sp. nov. (Prymne-
siophyceae) // Canad. J. Bot. 1986. Vol. 64, N 5. P. 593—610.
Parke M., Green J. C. Haptophyta // J. Mar. Biol. Assoc. U. K.
1976. Vol. 56, N 5. P. 527—594.
Pascher A. Chrysomonaden aus dem Hirschberger Grossteiche //
Monogr. Abh. Int. Rev. Ges. Hydrobiol., Hydrogr. 1910. Bd 1. S. 5—
66.
Pienaar R. N. Chrysophytes // Developments in marine biology.
Vol. 2. Phytoflagellates. New York, 1980. P. 213—242.
Scherffel A. Kleiner Beitrag zur Phylogenie einiger Gruppen nie-
derer Organismen // Bot. Zeitg. 1901. Bd 59. S. 143—158.
Scherffel A. Beitrag zur Kenntnis der Chrysomonadinen // Arh.
Protistenk. 1911. Bd 22, H. 2. S. 299—344.
Schiller J. Coccolithineae // L. Rabenhorst’s und Kryptogamenf-
lora von Deutschland, Ousterreich und der Schweiz. 2. Aufl. Leipzig:
Akad. Verlagsges., 1930. Bd 10, Abt. 2. S, 89—273.
Taylor F. T. R. Flagellate phylogeny: a study in conflicts // J. Pro-
tozool. 1976. Vol. 23, N 1. P. 28—40.
The Haptophyte Algae I Eds J. C. Green, B. S. C. Leadbeater.
Oxford, 1994.
Van de Peer Y., De Wachter R. Evolutionary relationships
among the eukaryotic crown taxa taking into account site-to-site rate
variation in 18S r RNA // J. Molec. Evol. 1997. Vol. 45. P. 619—630.
16 Цитируемая и рекомендуемая литература к разделу «Тип
Haptophyta».
Тип RAPHIDOPHYTA Chadefaud, 1950 -
РАФИДОФИТОВЫЕ
(Син.: Chloromonadophyta)
Тип объединяет небольшую группу морфологически сходных
жгутиконосцев с постоянной формой тела, которое достигает у
разных видов 30—100 мкм в длину. Тело уплощенное в дорсовен-
тральном направлении. Под плазмалеммой обычно располагаются
многочисленные вакуоли. 2 гетероконтных жгутика выходят из уг-
лубления (резервуара) на переднем конце тела. Двигательный жгу-
тик несет трубчатые мастигонемы. Фотосинтезирующие жгутико-
носцы, имеют многочисленные хлоропласты зеленого цвета. Стиг-
мы нет. Резервное вещество — липиды. Экструсомы 2 типов:
трихоцисты и мукоцисты. Ядро 1, делится путем полузакрытого
ортомитоза. Размножаются агамно с помощью бинарного деления.
Половой процесс обнаружен у 1 вида. Большинство рафидофито-
вых — пресноводные виды, но есть солоноватоводные и морские.
История изучения
Начало изучения рафидофитовых относится ко
второй половине XIX столетия. В 1892 г. Клебс вы-
делил отряд Chloromonadida, в который объединил
3 известных к этому времени рода — Gonyostomum,
Merotricha и Vacuolaria. Позже к этому отряду Бих-
лер добавил новый, выделенный им в 1936 г. род
Chattonella. Остальные 2 рода — Olisthodiscus и He-
terosigma получили обоснование в качестве отдель-
ных родов соответственно в 1937 и 1968 гг. после
детального изучения ультраструктуры их клеток
(Нага, Chihara, 1982, 1985; Hara et al., 1985; Vesk,
Moestrup, 1987).
В течение последних 10 лет эти жгутиконосцы
были объединены в отряд Raphidomonadida Heywood
et Leedale, 1983 в классе Phytomastigophorea Cal-
kins, 1909 (Levine et al., 1980). В системе однокле-
точных водорослей эта небольшая группа мелких
фототрофных жгутиковых известна под названием
хлоромонадовые — Chloromonadophyceae (Pascher,
1913; Матвиенко, 1977). Рафидофитовые в настоящее
время достаточно подробно изучены с помощью све-
тового и электронного микроскопа, что позволило
изменить их. таксономическое положение и выде-
лить в самостоятельный тип Raphidophyta Chadefa-
ud, 1950.
Классификация
Тип Raphidophyta включает 1 класс Raphidophy-
сеае с 1 отрядом Raphidomonadales, в котором известно
7 родов:
Род Merotricha Mereschkowsky, 1877 — Меротрихи
Род Vacuolaria Cienkowsky, 1880 — Вакуолярии
Род Gonyostomum Diesing, 1866 — Гониостомум
Род Chattonella Biechler, 1936 — Шаттонелла
Род Olisthodiscus Carter, 1937 — Олистодискус
Род Heterosigma Hada, 1968 — Гетеросигма
Род Fibrocapsa Toriumi et Takano, 1975 — Фиброкапса
Морфология и физиология
Внешнее строение
Форма и размеры тела. Все рафидофитовые —
одиночные одноклеточные протисты. Тело их упло-
щено в дорсовентральном направлении (рис. 183).
Посередине брюшной стороны проходит продольная
бороздка. На переднем конце тела она доходит до
жгутикового резервуара, ведущего в глубь цитоплаз-
мы и имеющего треугольную форму. Размеры тела
варьируют от 20 до 100 мкм, варьирует и форма те-
ла — от грушевидной до овальной и шаровидной.
Имеются пальмеллевидные формы.
Жгутиковый аппарат. 2 гетероконтных жгутика
выходят из резервуара, расположенного на вершине
переднего конца тела (Vacuolaria), либо сбоку тела,
под его передним краем (Merotricha). Двигательный
жгутик направлен вперед и несет трубчатые масти-
гонемы. Длина его либо равна длине тела, либо не-
сколько больше нее. Второй жгутик направлен назад
и выполняет рулевую функцию. Поверхность его
гладкая, без мастигонем. Переходная зона жгутиков
простая, короткая, в ней отсутствует переходная
спираль.
286
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 183. Особенности строения клетки некоторых рафидофитовых. (По: Попова, 1966; Heywood, Leedale, 1983).
А — Gonyoslomum semen, Б — Merotrichia bacillata, В — Vacuolaria virescens. 1 — передний (двигательный) жгутик, 2 — задний (рулевой) жгутик, 3 —
жгутиковый резервуар, 4 — сократительная вакуоль, 5 — ядро, 6 — хлоропласт, 7 — трихоцисты, 8 — мукоцисты.
Fig. 183. Structure of the cell in some species of Raphidophyta.
1 — anterior (motile) flagellum, 2 — posterior (trailing) flagellum, 3 — flagellar pocket, 4 — contractile vacuole, 5 — nucleus, 6 — chloroplasts, 7 — tricho-
cysts, 8 — mucocysts.
Корешковая система жгутиковых кинетосом рафи-
дофитовых развита хорошо. В ее состав входит круп-
ный по размеру корешок, идущий от проксимального
конца обеих кинетосом к ядру (рис. 184). Этот ядерный
корешок состоит из микротрубочек и микрофиламен-
тов. Часть последних исчерчена поперечно. Стенки
жгутикового резервуара поддерживают боковые мик-
ротрубочковые корешки, число которых 3—4.
Цитоплазма
Все виды рафидофитовых фотосинтезирующие.
Хлоропласты мелкие, многочисленные, размером 3—
5 мкм, зеленого или желто-зеленого цвета, дисковид-
ной формы; расположены в периферической зоне ци-
топлазмы в 1 или несколько рядов (рис. 184). Ламел-
лы хлоропласта состоят из 3 тилакоидов, между
которыми могут быть связующие мембраны. У боль-
шинства видов хлоропласты содержат хлорофиллы а
и с (ср с2), каротин, антераксин, лютеин и ксанто-
филл. У морских видов рода Chattonella обнаружен
фукоксантин; у них имеется и пиреноид, располагаю-
щийся на одном из концов хлоропласта. Пиреноиды
хорошо развиты также у Heterosigma и Olisthodiscus.
Питание рафидофитовых осуществляется фото-
трофно, иные способы питания неизвестны.
Сократительная вакуоль хорошо развита у всех
пресноводных форм родов Gonyostomum, Merotricha,
Vacuolaria. Она расположена в цитоплазме между
ядром и кинетосомами. Размеры сократительной ва-
куоли бывают значительными и могут достигать 10 мкм
в диаметре. У морских видов сократительная вакуоль
отсутствует.
Митохондрии расположены в разных участках ци-
топлазмы клетки. Они многочисленны, мелких раз-
меров и имеют тубулярные кристы (рис. 184).
Крупный по размерам комплекс диктиосом распо-
ложен в околоядерной зоне цитоплазмы (рис. 184) и
граничит с сократительной вакуолью.
Трихоцисты и мукоцисты имеются, вероятно, у
большинства видов. У жгутиконосцев рода Gonyosto-
mum трихоцисты палочковидной формы (рис. 184)
выбрасывают наружу тонкие слизистые нити, дости-
гающие в длину до 100 мкм. Vacuolaria virescens
(рис. 184) имеет множество мелких мукоцист, кото-
рые выделяют большое количество слизи, окружаю-
щей тело снаружи.
В качестве основного резервного вещества у ра-
фидофитовых известны только липиды, которые на-
капливаются в матриксе цитоплазмы и в строме хло-
ропластов в форме капель.
Ядро
Ядро 1, пузырьковидного типа, расположено в
центральной зоне клетки. В диаметре оно достигает
20 мкм, и в нем может быть 1 или несколько ядры-
шек. Хроматин интерфазного ядра диспергирован.
ТИП RAPHIDOPHYTA
287
Рис. 184. Схема ультратонкого строения переднего конца клетки
рафидофитовых. (По: Карпов, 1990).
1 — передний жгутик с трубчатыми мастигонемами, 2 — задний жгутик,
3 — кинетосомы, 4 — микротрубочковые корешки, 5 — фибриллярные
корешки, 6 — ядро, 7 — митохондрия с трубчатыми кристами, 8 — аппа-
рат Гольджи, 9 — хлоропласт, 10 — ламелла из трех тилакоидов, 11 —
рибосомы, 12 — экструсома.
Fig. 184. Diagram of ultrastructure of anterior part of Raphydophyta
cell.
1 — anterior flagellum with tubular mastigonemes, 2 — posterior flagellum,
3 — kinetosome, 4 — microtubular rootlets, 5 — fibrillar rootlets, 6 -r- nucle-
us, 7 — mitochondria with tubular cristae, 8 — Golgi apparatus, 9 — chlo-
roplast, 10 — lamella consists of three thylakoids, 11 — ribosomes, 12 —
trichocysts.
По данным биохимического анализа, в молекуле ядер-
ной ДНК Gonyostomum semen содержится 35 % гуа-
нина и цитозина, а у Vacuolaria virescens 34 % (Hey-
wood, Leedale, 1985).
Тип деления ядра — полузакрытый ортомитоз
(Райков, 1978). Он изучен у 2 видов: Vacuolaria vi-
rescens и Gonyostomum semen. При этом типе митоза
ядерная оболочка • сохраняется в течение всего вре-
мени от профазы до телофазы, но на обоих полю-
сах делящегося ядра в оболочке имеются отверстия.
Центры формирования микротрубочек находятся в
цитоплазме. Микротрубочки проходят в ядро через
полярные отверстия. Внутри ядра они образуют
двухполюсное симметричное веретено митоза, в ко-
тором имеются как хромосомные, так и непрерыв-
ные нити.
Метафазная пластинка выражена отчетливо, и за-
метна индивидуальность хромосом. У V. virescens
число метафазных хромосом равно 97—99, а у G. se-
men 65—75 при длине 1—12 мкм. Хромосомы ните-
видные, и у первого вида обнаружены кинетохоры,
при помощи которых хромосомы прикрепляются к
микротрубочкам веретена. Новая оболочка у дочер-
них ядер формируется в период телофазы (Heywood,
1974, 1978; Heywood, Leedale, 1985).
Размножение
Рафидофитовые размножаются агамным путем.
Бинарное деление происходит в подвижном состоя-
нии с сохранением жгутиков материнской особи и в
пальмеллевидном состоянии. Половой процесс, обна-
руженный у Chattonella subsalsa, пока не изучен.
Экология
Распространенные всесветно, рафидофитовые
обитают преимущественно в малых эвтрофирован-
ных водоемах, в сфагновых болотах и реже в круп-
ных озерах, морях и солоноватоводных бассейнах.
Они принадлежат к планктонным организмам, но
встречаются и в придонных зонах водоемов. Часто
обитают в воде с повышенной кислотностью. У неко-
торых видов, например Gonyostomum semen (рис. 183,
А), отчетливо выражены суточные вертикальные миг-
рации: утром жгутиконосцы локализуются в поверх-
ностных слоях, днем — в придонной зоне водоемов
(Попова, 1955; Heywood, Leedale, 1985).
Филогения
О филогении рафидофитовых сведений пока не-
много. Их наиболее близкими родственными группа-
ми, вероятно, можно считать хризофитовых и ксан-
тофитовых. На филогенетическую близость этих
групп указывает сходство ультраструктуры их клеток:
гетероконтные жгутики, тип мастигонем переднего
жгутика, ламеллы хлоропластов, каждая из которых
включает 3 тилакоида, хлорофиллы а и с, липиды в
качестве резервного вещества и ряд других особен-
ностей строения клетки (Нага et al., 1985).
Систематический обзор
Тип Raphidophyta представляет небольшую, вклю-
чающую около 20 видов, четко ограниченную по
морфологическим признакам группу фототрофных
протистов. В состав типа входит 7 родов, объеди-
ненных в 1 класс Raphidophyceae, в котором имеет-
ся только 1 отряд — Raphidomonadales.
Род Gonyostomum Diesing,
1886 (рис. 183, А)
Видам этого рода свойственно овальное или почти
округлое, уплощенное в дорсовентральном направле-
нии тело. На брюшной стороне четко выражена про-
дольная бороздка. 2 жгутика выходят в центре пере-
днего конца тела из резервуара треугольной формы.
Передний, двигательный жгутик по длине равен телу,
задний, рулевой жгутик превышает длину тела почти
в 2 раза. В периферической зоне цитоплазмы рас-
положены палочковидные трихоцисты, собранные в
группы на переднем и заднем конце тела. Хлороплас-
ты многочисленны и расположены в 1 ряд в перифе-
рической зоне цитоплазмы. Сократительная вакуоль
находится в передней части тела между ядром и жгу-
тиковым резервуаром. Аппарат Гольджи крупный и
занимает переднюю часть околоядерной зоны, тесно
прилегая к оболочке ядра.
288
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
В составе рода известно 4 вида: G. semen, G. la-
tum, G. intermedium, G. ovatum. Наиболее хорошо
изучен G. semen (рис. 183), достигающий 40—90 мкм
в длину. Этот вид обитает в мелких водоемах, в кото-
рых pH воды составляет 5.5—5.8.
Род Merotricha Mereschkowsky, 1877
(рис. 183, Б)
Меротрихам свойственно тело удлиненно-оваль-
ной формы с закругленным- цитоплазматическим вы-
ростом на переднем конце, в котором, локализуются
палочковидные трихоцисты. 2 жгутика, одинаковые
по длине, выходят сбоку на переднем конце тела из
небольшого углубления. Хлоропласты дискрвидные,
ярко-зеленые, занимают периферическую зону ци-
топлазмы. Их нет только в выросте переднего конца
тела, где находятся трихоцисты. Аппарат Гольджи в
форме палочковидной структуры находится вблизи
переднего конца ядра. К роду относится 1 вид — М.
bacillata (рис. 183). Размеры его особей составляют
40—50 х 18—20 мкм. Обитает в мелких пресных во-
доемах с pH воды ниже 6.0.
Род Vacuolaria Cienkowsky, 1880
(рис. 183, В)
Форма тела вакуолярий варьирует у разных видов
и в пределах одного вида от удлиненной цилиндри-
ческой до сферической. Значительно варьируют и
размеры тела. Передний конец скошен, и посереди-
не выходят 2 жгутика из небольшого резервуара тре-
угольной формы. Длина жгутиков одинакова и равна
длине тела. Хлоропласты многочисленны, располо-
жены во всем объеме цитоплазмы в несколько рядов.
Мукоцисты мелкие, сферические, сосредоточены в
периферической зоне .цитоплазмы; они образуют
слизистую оболочку пальмеллоидных форм. Сокра-
тительная вакуоль крупная, расположена в передней
трети тела. Аппарат Гольджи тесно прилегает к обо-
лочке передней части ядра.
В состав рода входит несколько видов, среди кото-
рых наиболее часто встречаются V. virescens (рис. 183), V.
viridis, V. penardii. Вакуолярии — мелкие жгутико-
вые протисты: V. virescens, например, имеет размеры
30—85 х 18—25 мкм. Места обитания — неглубокие
пресные водоемы.
Род Chattonella Biechler, 1936
Форма тела шаттонелл овальная, часто с хвосто-
вым отростком. 2 жгутика одинаковой длины, кото-
рая равна длине тела. Хлоропластов много, они рас-
положены в 1 ряд в периферической зоне цитоплаз-
мы. Палочковидные трихоцисты также локализуются
в периферической цитоплазме. Сократительная ваку-
оль отсутствует. К роду относится 1 вид — Ch. sub-
salsa. Это мелкие организмы, достигающие размеров
30—50 х 15—25 мкм и обитающие в солоноватых и
морских водоемах (Hollande, Enjument, 1956).
Род Heterosigma Hada, 1968
Жгутиконосцы овальной формы, их размеры
варьируют в пределах 11—20x9—12 мкм. Тело по-
крыто снаружи слоем слизи, уплощено в дорсовент-
ральном направлении, в центре брюшной стороны
проходит продольная бороздка. 2 почти одинаковых
по длине жгутика выходят субапикально из бокового
углубления на переднем конце тела. Двигательный
жгутик направлен вперед, второй, ригидный, —
назад. Жгутиконосец плавает, совершая вращатель-
ные движения вокруг продольной оси тела. Хлороп-
ластов 10—25, они имеют дисковидную форму и ок-
рашены в желто-коричневый цвет. Стигма и сокра-
тительная вакуоль отсутствуют. Ядро расположено в
центре клетки. Размножение агамное. Перед деле-
нием клетки теряют жгутики и округляются. Иног-
да округлившиеся клетки слипаются, образуют не-
подвижный агрегат, в состоянии которого делятся
(Нага, Chihara, 1987). К роду относится 1 вид —
Н. akashiwo Hada, 1968 (син.: Entomosigma akashiwo
Hada, 1968).
Род Olisthodiscus Carter, 1937
Мелкие, желто-зеленого цвета жгутиконосцы
15—25 мкм в длину, 10—16 мкм в ширину при тол-
щине 5—7 мкм. Тело уплощено в дорсовентральном
направлении, с выпуклой спинной и вогнутой брюш-
ной сторонами, внешний покров представляет собой
утолщенную стенку. По брюшной стороне проходит
продольная бороздка. 2 гетероконтных жгутика вы-
ходят из углубления, расположенного посередине пе-
реднего конца тела. 1 жгутик двигательный, в 1.0—
1.5 раза превышает длину тела, направлен вперед и
несет 2 ряда мастигонем. Второй жгутик погружен в
бороздку на брюшной стороне, имеет длину, состав-
ляющую 0.5—1.2 длины тела, и направлен назад.
Жгутиконосец движется плавно, причем брюшная
сторона направлена кверху. Хлоропласты дисковид-
ные; расположены в передней части клетки число их
6—13. Ядро каплевидное, шаровидное, находится в
центре клетки. Размножение агамное. Обитает в со-
лоноватых и морских водоемах. В состав рода входит
1 вид — О. luteus Carter, 1937 (Hara et al., 1985).
Род Fibrocapsa Toriumi et Takano, 1975
Одноклеточные мелкие, овальной формы морские
жгутиконосцы, размеры которых варьируют в преде-
лах 15—20x10—12 мкм. Тело покрыто тонкой
плазмалеммой, уплощено в дорсовентральном на-
правлении. На переднем конце — небольшое округ-
лое отверстие, из которого выходят 2 жгутика почти
одинаковой длины. Двигательный жгутик направлен
вперед и несет мастигонемы на двух его сторонах,
второй, ригидный, направлен назад и не несет масти-
гонем. Кинетосом 2, жгутиковые корешки фибрил-
лярного типа. Стигма, ризостиль и сократительная
вакуоль отсутствуют. В эндоплазме расположены:
ТИП RAPHIDOPHYTA. ЛИТЕРАТУРА
289
крупное ядро, большое число митохондрий, мукоцисты,
диктиосомы, ЭПР и другие включения. В эктоплазме
расположены дискоидальные хлоропласты, отделен-
ные друг от друга тонкими прослойками цитоплазмы.
Хлоропласты с многими тилакоидами, которые фор-
мируют 2 или 3 ламеллы. Внутри каждого хлороплас-
та 1 пиреноид, имеющий форму полукруглого выпук-
лого отростка, направленного к центру клетки.
В нижней части клетки много мукоцист, подо-
бных таковым Gonyostomum semen. Они локализова-
ны вблизи ядра, имеют плотный матрикс и окружены
одной мембраной. По периферии клетки, в эктоплаз-
ме, расположен фибриллярный материал. Размноже-
ние клеток путем деления происходит в подвижном
состоянии. Однако перед делением они могут терять
жгутики и объединяться в агрегаты, подобные плаз-
модию.
Fibrocapsa japonica Toriumi et Tokano, 1975 —
единственный вид рода в отряде Raphidomodales
(Hara, Chihara, 1985).
THE PHYLUM RAPHIDOPHYTA
SUMMARY
The raphidophytes are a small group of unicellular flagellates,
fresh-water and marine. They have two heterocont flagella arising
from invagination near the cell apex. The anterior flagellum bears
tubular hairs (mastigonemes). The recurrent flagellum is smooth. The
transition zone of flagella lacks transition helix. Among the flagellar
rootlets the nuclear one is most developed.
The body is flattened dorsoventrally, the furrow is on the ventral
surface. Body shape is pyriform, spherical or ovoid. All species con-
tain discoid chloroplasts. Chloroplast lamellae include 3 thylakoids.
The girdle lamella is present. Chlorophylls a and с, В — carotene,
antheraxanthin, lutein, xanthophyll are the pigments of chloroplasts,
which are usually bright, yellowish-green. The reserve material is oil.
The extrusomes are mucocysts and trichocysts. The flagellates have no
photoreceptor. A contractile vacuole occurs in fresh-water species.
The nucleus is large (about 20 pm). Nuclear division is studied by
means of electron microscope. Reproduction of cells is by binary fis-
sion.
Only 7 genera are known in the phylum Raphidophyta.
ЛИТЕРАТУРА
Матвиенко A. M. Отдел Золотистые водоросли (Chrysophy-
ta) // Жизнь растений. М., 1977. Т. 3. С. 93—103.
Попова Т. Г. Определитель пресноводных водорослей СССР.
Эвгленовые водоросли. М., 1955.
Райков И. Б. Ядро простейших. Морфология и эволюция. Л.:
Наука, 1978.
Нага Y., Chihara М. Ultrastructure and taxonomy of Chattonel-
la (Class Raphidophyceae) in Japan // Jap. J. Phycol. 1982. Vol. 30.
P. 47—56.
Hara Y., Chihara M. Ultrastructure and taxonomy of Fibrocapsa
japonica (Class Raphidophyceae) // Arch. Protistenk. 1985. Vol. 130.
P. 133—141.
Hara Y., Chihara M. Morphology, ultrastructure and taxonomy
of the Raphidophycean alga; Heterosigma akashiwo // Bot. Mag.
Tokyo. 1987. Vol. 100. P. 151—163.
Hara Y., Inouye I., Chihara M. Morphology and ultrastructure of
Olisthodiscus luteus (Raphidophyceae) with special reference to the
taxonomy // Bot. Mag. Tokyo. 1985. Vol. 98. P. 251—262.
Heywood P. Mitosis and cytokinesis in the chloromonadophycean
Goniostomum semen U J. Phycol. 1974. Vol. 10, N 2. P. 355—358.
Heywood P. Ultrastructure of mitosis in the chloromonadophycean
alga Vacuolaria virescens // J. Cell Sci. 1978. Vol. 31, N 1. P. 37—51.
Heywood P., Leedale G.F. Order Raphidomonadida Leedale,
1983 // Illustrated Guide to the Protozoa. 1985. P. 70—74.
Hollande A., Enjument M. Sur une invasion des eaux du port
d’ Alger par Chatonella subsalsa (Homeilia marina Sub.) Biechler. Re-
marques sur la taxicite Chloromonadine // Bull. Stn. Agr. Peche Cos-
tiglione (N. S.). 1956., Vol. 8, N 2. P. 271—280.
Levine N. D., Corliss J. О., Cox P. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R., Lom J., Lynn D.,
Merinfeld E. G., Page F. C., Poljansky G., Sprague V., Vavra J.,
Wallace F. G. A newly revised classification of the Protozoa // J. Pro-
tozool. 1980. Vol. 27, N 1. P. 37—58.
Pascher A. Chloromonadinae II Siisswasserflora Deutschlands.
1913. H. 2. S. 175—181.
Vesk M., Moestrup 0. The flagellar root system in Heterosigma
akashiwo (Raphidophyceae) // Protoplasma. 1987. Vol. 137. P. 15—28.
19 Протисты, часть 1
Тип SAPROLEGNIA Zerov, 1972 (=OOMYCETES) —
САПРОЛЕГНИИ
Зооспоры представлены бесцветными гетероконтными жгути-
коносцами с характерными чертами хромистов: передний жгутик
опушен трубчатыми мастигонемами, кристы в митохондриях пу-
зырьковидные, у некоторых видов имеются рудиментарные пластиды
и стигма. В переходной зоне жгутиков имеется двойная спираль.
Корешковая система весьма консервативна, состоит из 4 микрот-
рубочковых корешков, которые, по-видимому, гомологичны таковым
хризофитовых. Клеточная стенка содержит целлюлозу. Характерен
неклеточный мицелий, или внеклеточная цитоплазматическая сеть.
Паразитические или сапрофитные, морские, солоноватовод-
ные, пресноводные или почвенные организмы.
К типу относится 2 класса: Saprolegniea Zerov, 1972
и Labyrinthomorphea Page, 1979.
Подробно класс Saprolegniea здесь не рассматри-
вается. Приводятся только его общая характеристика
и обоснования, по которым этот таксон зооспоровых
грибов включен в царство Chromista.
Сапролегнии имеют зооспоры с характерными
чертами типа. В жизненном цикле доминирует дип-
лоидная стадия. Мицелий многоядерный, не септиро-
ванный. Половой процесс в форме оогамии. Парази-
тические или сапрофитные, пресноводные и почвен-
ные организмы.
Класс Labyrinthom
Лабирин
Лабиринтоморфы — колониальные морские протисты со
сложным жизненным циклом, включающим следующие фазы: ко-
лония — плазмодий, состоящий из внеклеточной эктоплазмати-
ческой сети трубок с веретеновидными и амебоидными клетками;
цисты; спороцисты; спорангии; зооспоры с 2 гетероконтными жгу-
тиками. Двигательный передний жгутик несет мастигонемы. Пере-
ходная зона жгутика короткая, с 1 переходной пластинкой и двой-
ной переходной спиралью. Зооспоры — единственная свободно-
плавающая фаза в жизненном цикле лабиринтоморф. Внешний
покров представлен чешуйками. Митохондрии с тубулярными
кристами, внутри каждой кристы проходит осевая нить.
Уникальная для лабиринтоморф органелла — сагеногенетосо-
ма (ботросома) выполняет функцию формирования внеклеточной
эктоплазматической сети. Размножение агамное путем бинарного
деления клеток. Деление ядра осуществляется посредством закрыто-
го внутриядерного плевромитоза. При формировании зооспор на-
блюдается двухступенчатый мейоз. Половое размножение не изучено.
Лабиринтоморфы — паразиты морских растений, а также ус-
триц.
Кроме сходства с хромистами по строению зоо-
спор сапролегнии имеют и другие черты, сближающие
их со многими водорослями из этого царства. Так, в
качестве запасного вещества у них откладывается
крахмал, а клеточная стенка состоит из целлюлозы.
Кроме того, как и для бурых водорослей, для них ха-
рактерен гаметангиальный мейоз, а у всех изученных
оомицетов обнаружен лейкопласт (К-тело). По после-
довательности нуклеотидов генов малой субъединицы
рРНК оомицеты, бурые водоросли и хризофитовые со-
ставляют одну филогенетическую линию эукариот. Есть
достаточно оснований полагать, что оомицеты близки
к содержащим хлорофилл с водорослям таксономи-
чески и филогенетически (Barr, 1992) и, по-видимому,
представляют потомков тех из них, которые утратили
хлоропласты при переходе к наземному или паразити-
ческому образу жизни.
Название класса дано по имени порядка Saproleg-
niales, который несет все основные признаки ормице-
тов, тогда как собственно оомицеты (порядок Оошу-
cetales) представляют собой довольно специализиро-
ванную группу.
orphea Раде, 1979 —
о м о р ф ы
История изучения
Лабиринтоморфы известны с 1867 г. по исследо-
ванию Ценковского (Cienkowski, 1867). Он устано-
вил название рода Labyrinthula. изучил и дал под-
робное описание 2 первых видов — L. vitellina и
L, macrocystis. С этого времени начинается изучение
лабиринтоморф, которое было направлено прежде
всего на поиски новых видов и изучение их морфо-
логии. В конце прошлого столетия был обнаружен
еще 1 вид рода — L. cienkowski Zopf, 1894, а затем
и L. zopfi Dangeard, 1910. В состав сем. Labyrinthu-
lidae Haeckel, 1834 был включен новый род Labyrin-
thomyxa Dubosq, 1921 с 1 видом — L. sauvageaui
(Dubosq, 1921).
ТИП SAPROLEGNIA. КЛАСС LABYRINTHOMORPHEA
291
В 1936 г. появилась работа с описанием нового
сем. Thraustochytriaceae Sparrow, 1936, в состав ко-
торого был включен род Thraustochytrium Sparrow,
1936 с 1 видом — Th. proliferum Sparrow, 1936. Позже
в это семейство был включен род Schizochytrium
Goldstein, Belsky, 1964 с видом Sch. aggregation Gold-
stein, Belsky, 1964. Оба рода вначале были отнесены
к сапролегниевым грибам (Phycomycetes) по особен-
ностям строения плазмодия и зооспор. Дальнейшие
исследования показали, что ультраструктура «ри-
зоидов» Thraustochytriaceae идентична таковой сети
трубок лабиринтулид. Оба эти образования полу-
чили название внеклеточной эктоплазматической
сети, с которой связаны мембранные органеллы —
сагеногенетосомы. Идентичной оказалась и хими-
ческая природа мембран. Поэтому сем. Thrausto-
chytriaceae было включено в состав лабиринтоморф
(Darely, Fuller, 1970; Perkins, 1972; Levine et al.,
1980).
Наибольшее число работ, посвященных изучению
лабиринтоморф, относится к 1975—1995 гг. В этот
период были обнаружены новые виды, изучены жиз-
ненные циклы ряда видов, строение колоний, мор-
фология и ультраструктурная организация клеток,
формы размножения, питание и особенности экологии.
Большинство исследований проведено на лабораторных
культурах разных видов, для которых были разрабо-
таны специальные методы культивирования (Hollan-
de, Enjumet, 1955; Goldstein, 1963; Perkins, 1968,
1972; Perkins, Amon, 1969; Grell, 1971; Schmoller,
1971; Schwab-Stey, Schwab, 1974; Puytorac et al.,
1987; Хаусман, 1988). Однако лабиринтоморфы изу-
чены еще недостаточно.
Классификация
В прежней системе простейших (Levine et al.,
1980) лабиринтоморфы. были выделены в самостоя-
тельный тип Labyrinthomorpha Page, 1980 и в его со-
став входил 1 класс Labyrinthulea Levine et Corliss,
1963 с 1 отрядом Labyrinthulida Cienkowski, 1867,
который включал 2 семейства: Labyrinthulidae Haec-
kel, 1834 и Thraustochytriidae Sparrow, 1936.
В соответствии с классификацией, принятой в на-
стоящем изданий, лабиринтоморфы отнесены к типу
Saprolegnia в ранге класса Labyrinthomorphea, в кото-
рый входит 2 отряда — Labyrinthulida и Thraustochyt-
rida, 2 семейства — Labyrinthulidae и Thraustochytrii-
dae, включающие 4 рода и около 30 видов.
Морфология и физиология
Строение колонии
Внеклеточная эктоплазматическая сеть. Вегета-
тивная фаза жизненного цикла — колония, которая
представляет собой сетчатый плазмодий (рис. 185;
186) и состоит из тонких, нитевидных трубок с на-
ходящимися, внутри них веретеновидными клетками.
Трубки ветвятся, анастомозируют друг с другом, об-
разуя сеть; на концах трубки также разветвляются.
Рис. 185. Общий вид колонии Labyrinthula algeriensis. (По: Hol-
lande, Enjumet, 1955).
Fig. 185. Total view of a colony of Labyrinthula algeriensis.
Диаметр трубок варьирует от 0.1 до нескольких мик-
рометров; длина их также различна (рис. 187).
Трубки представляют собой эктоплазматические
выросты находящихся в них клеток. Внешний пок-
ров трубок образован плазмалеммой, несущей на по-
верхности чешуйки. Поэтому весь комплекс ветвя-
щихся трубок получил название внеклеточной экто-
плазматической сети (Perkins, 1972). В трубках нет
органелл цитоплазмы, а внутри них находится волок-
нисто-зернистый матрикс, в котором расположены
цистерны, ограниченные мембранами (рис. 188).
Веретеновидные клетки. Как уже было упомяну-
то, эти клетки находятся внутри эктоплазматических
трубок. Их размеры небольшие: длина варьирует у
разных видов от 10 до 30 мкм. Внешний покров кле-
ток образован плазмалеммой. Оба компонента коло-
нии — эктоплазматическая сеть и находящиеся в ней
веретеновидные клетки, будучи непосредственно свя-
занными друг с другом, образуют одно функциональ-
ное целое. Клетки движутся внутри трубок в разных
направлениях, но они не имеют оформленных орга-
нелл движения — псевдоподий или жгутиков — и
передвигаются путем скольжения.
У Thraustochytrium motiyum Goldstein, 1963 и Schi-
zochytrium aggregatum скользящего движения верете-
новидных клеток не обнаружено в отличие от видов
рода Labyrinthula.
Форма веретеновидных клеток относительно пос-
тоянна. Это типичные эукариотические клетки с четко
оформленным ядром и сложной структурой цитоп-
лазмы (рис. 187). У их поверхности, непосредствен-
но под плазмалеммой, расположены особые, специ-
фические для лабиринтоморф органеллы, называемые
сагеногенетосомами (рис. 188). Они имеют довольно
сложную ультраструктуру. Каждая сагеногенетосома
(ботросома) состоит из электронно-плотного зернис-
того компонента (гранулярный агрегат) дисковидной
формы, внутри которого находится цистерна ЭПР.
В цистерне на границе с гранулярным компонентом
292
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 186. Развивающаяся колония Labyrinthula algeriensis. (По: Hollande, Enjumet, 1955).
Fig. 186. The younger colony of Labyrinthula algeriensis.
расположены мелкие пузырьки, которые сливаются
и образуют внутренние мембраны эктоплазматичес-
кой сети. Диаметр каждого пузырька около 17 нм.
Эктоплазматическая сеть колонии связана с 1 или
более (до 20) сагеногенетосомой (Perkins, 1972). Фун-
кция этих органелл — формирование внеклеточной
эктоплазматической сети.
Ядро 1, расположено в центре интерфазной клет-
ки. В нем имеется 1 крупное фельген-отрицательное
ядрышко, лежащее эксцентрично. В ядрышке преоб-
ладает грайулярный компонент. Ядерная оболочка
пронизана многочисленными порами со всеми эле-
ментами порового комплекса, свойственными ядру
эукариотических клеток. Перинуклеарные цистерны
сообщаются с каналами и цистернами ЭПР.
Гранулярный ЭПР хорошо развит. Некоторые цис-
терны его увеличены и имеют вид крупных пузырь-
ков неправильной формы с гомогенным содержимым.
Они образуют анастомозирующую сеть в цитоплазме.
Митохондрии многочисленны, в них довольно отчет-
ливо выражены тубулярные кристы. Внутри каждой
кристы проходит осевая нить. Аппарат Гольджи име-
ется в каждой клетке, причем диктиосомы распо-
ложены в разных участках цитоплазмы. Резервные
вещества представлены липидами в форме крупных
округлых капель, не окруженных мембранами и до-
Рис. 187. Схема строения колонии Labyrinthula caenocystis. (По:
Schwab-Stey, Schwab, 1973).
7 — клеточная, 2 — внутренняя мембраны, 3 — внешняя мембрана труб-
ки, 4 — сагеногенетосома, 5 — веретеновидная клетка.
Fig. 187. Diagrammatic representation of a colony of Labyrinthula
caenocystis.
1 — cell membrane, 2 — internal membrane, 3 — outer membrane of tubule,
4 — sagenogenetosoma, 5 — spindle cell.
Рис. 188. Схема ультратонкого строения сагеногенетосомы Laby-
rinthula minuta. (По: Perkins, 1972).
1 — клеточная мембрана, 2 — внутренняя мембрана, 3 — ряд мелких пу-
зырьков, 4 — ЭПР, 5 — внутренняя цистерна, б — мембрана трубки, 7 —
митохондрии, 8 — гранулярный компонент.
Fig. 188. Diagrammatic expression of ultrastructure of sagenogeneto-
soma of Labyrinthula minuta.
1 — cell membrane, 2 — internal membrane, 3 — a line of small vesicules,
4 — endoplasmic reticulum, 5 — internal cistema, 6 — membrane of tubula,
7 — mitochondria, 8 — granular component.
ТИП SAPROLEGNIA. КЛАСС LABYRINTHOMORPHEA
293
Рис. 189. Амебоидная клетка Labyrinthomyxa pholia. (По: Schmol-
ler, 1966).
Fig. 189. Amoeboid cell of Labyrinthomyxa pholia.
стирающими в диаметре 0.5—1.0 мкм (Perkins, Amon,
1969).
По всей вероятности, веретеновидные клетки пи-
таются сапрофитно: фагоцитоз и пиноцитоз у них не
обнаружены.
Амебоидные клетки. Кроме веретеновидных кле-
ток в колониях у многих видов, особенно в составе
родов Labyrinthomyxa и Thraustochytrium, имеются
амебоидные клетки (рис. 189) с тонкими или лопаст-
ными псевдоподиями, с небольшими размерами тела,
которое достигает у Labyrinthomyxa pholia 10—20 мкм
в длину и около 5 мкм в ширину. Цитоплазма их
четко разделена на экто- и эндоплазму. Ядро 1, ок-
руглое и занимает центральную часть клетки. Аме-
боидные клетки представляют активную стадию в
жизненном цикле Labyrinthomyxa. В отличие от ве-
ретеновидных клеток они фагоцитируют бактерии и
другие мелкие организмы.
Амебоидные клетки — это стадия жизненного цикла,
которая дает начало развитию колонии. Они инцис-
тируются, и в состоянии цисты происходит их диф-
ференцировка (рис. 190), приводящая к превраще-
нию амебоидной клетки в клетку веретеновидную,
что связано и с образованием внеклеточной экто-
плазматической сети (Schmoller, 1971).
Зооспоры. Размеры зооспор значительно меньше
размеров веретеновидных и амебоидных клеток: 3—
5 мкм в длину и 2—3 мкм в ширину. Они могут быть
удлиненной или грушевидной формы с несколько за-
остренным задним концом. Зооспора отличается посто-
янством формы, которая при движении не изменяется.
Одна из характерных особенностей зооспор (пла-
нонтов) — наличие 2 жгутиков (рис. 191). Жгутики
отходят на боковой стороне тела в передней его
трети, и у их основания находится небольшая инва-
гинация. Жгутики неравной длины: передний имеет
длину 13—15 мкм, задний 6—10 мкм. Передний
жгутик несет мастигонемы, расположенные либо
только с одной стороны, как у Labyrinthula sp., либо
с двух сторон. Задний жгутик лишен мастигонем и
имеет длинный тонкий отросток на дистальном конце.
Ультраструктура жгутиков типична и не имеет отли-
чий от таковой жгутиков других протистов.
Переходная зона жгутиков короткая, в ней распо-
ложена 1 переходная пластинка и двойная переход-
Рис. 190. Дифференцировка амебоидной клетки в веретеновид-
ную. (По: Schmoller, 1971).
А — вегетативная особь, Б — инцистированная амеба, В — веретеновид-
ная клетка в трубке эктоплазматической сети.
Fig. 190. Differentiation of amoeboid cell into spindle cell.
A — vegetative specimen, Б — encysted amoeba, В — spindle cell inside of
tubule of ectoplasmatic net.
ная спираль. Корешковая система кинетосом включа-
ет группы корешков, состоящих из микротрубочек и
1 ребристого корешка.
Рядом с кинетосомами, непосредственно под плаз-
малеммой, расположена стигма желто-оранжевого цве-
та. Стигма состоит из нескольких электронно-плот-
ных гранул, которые лежат рядом друг с другом и
могут, сливаясь, образовать 1 удлиненное тельце. Возле
стигмы находится 1 ряд из 17—21 параллельных
Рис. 191. Схема ультратонкого строения зооспоры. (По: Perkins,
1968).
1 — передний жгутик, 2 — мастигонемы, 3 — кинетосомы, 4 — митохон-
дрия, 5 — ядро, 6 — аппарат Гольджи, 7 — вакуоль, 8 — капля липида,
9 — задний жгутик.
Fig. 191. Diagrammatic representation of ultrastructure of zoospore.
1 — anterior flagellum, 2 — mastigonemes, 3 — kinetosomes, 4 — mitochon-
drion, 5 — nucleus, 6 — Golgi apparatus, 7 — vacuole, 8 — drop of lipid,
9 — posterior flagellum.
294
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
микротрубочек, идущих непосредственно от микрот-
рубочек кинетосомы переднего жгутика. Этим мик-
ротрубочкам принадлежит, вероятно, опорная роль.
Зооспора — одноядерная клетка. В ее цитоплаз-
ме имеется 1 митохондрия удлиненной формы с
трубчатыми кристами и расположенная между ядром
и стигмой (рис. 191). Аппарат Гольджи находится
рядом с ядром. Цитоплазма содержит вакуоли, и ре-
зервные вещества в ней представлены нейтральными
липидами.
Зооспоры выходят в воду путем разрыва оболоч-
ки спорангия и активно плавают в воде около 24 ч
с помощью переднего жгутика. Затем они оседают
на растения или животных (моллюсков), которые
служат им хозяевами, округляются, теряют жгутики
и дифференцируются в веретеновидные или амебо-
идные клетки, с которых начинается рост колонии.
Инцистирование зооспор не обнаружено, так же как
не обнаружена их копуляция. Следовательно, зоо-
споры нельзя принять за изогаметы — это истинные
споры, агамные клетки.
Рис. 192. Labyrinthula algeriensis. Формирование клеточных агре-
гатов в расширенных участках эктоплазматической сети. (По:
Hollande; Enjumet, 1955).
Fig. 192. Labyrinthula algeriensis. Forming of cell agregates inside
of delations parts of ectoplasmatic net.
Рис. 193. Labyrinthula algeriensis. Формирование зооспор. (По:
Hollande, Enjumet, 1955).
1 — спороциста, 2 — спорангий, 3 — зооспоры.
Fig. 193. Labyrinthula algeriensis.
Development of zoospores: 7 — sporocyste, 2 — sporangium, 3 — zoospores.
Размножение
и жизненный цикл
Веретеновидные и амебоидные клетки размножа-
ются путем бинарного деления, в результате которого
возрастает их число и размеры колонии увеличива-
ются. Деление ядра осуществляется посредством за-
крытого плевромитоза. Размножение клеток в коло-
ниях происходит постоянно. Оно подробно изучено у
нескольких видов, в том числе у Labyrinthomorpha
pholia. В культурах этого вида на агаровых пластин-
ках четко различаются 3 зоны — роста, размножения
и инцистирования. При развитии колонии в зоне
роста размножение осуществляется не только путем
деления амебоидных и веретеновидных клеток, но и
путем формирования значительных по длине трубок,
которые содержат эти же клетки и распадаются на
ряд фрагментов, дающих начало новым колониям.
В зоне инцистирования амебоидные клетки обра-
зуют цисты, в которых затем происходит дифферен-
циация, завершающаяся, как уже отмечалось выше,
формированием веретеновидных клеток (рис. 190).
Размеры колонии увеличиваются, а цисты, кроме
того, дают начало новым колониям. Цисты состоят из
1 или многих клеток, скапливающихся внутри трубок
эктоплазматической сети (Schmoller, 1966).
Скопления (агрегации) клеток внутри трубок се-
тевидного плазмодия происходят и при зооспоруля-
ции. В местах агрегаций трубки плазмодия расширя-
ются (рис. 192), и в них формируются спороцисты.
Число спороцист в одной агрегации варьирует — их
может быть 5 и более. В каждой спороцисте развивает-
ся несколько спорангиев. В результате деления клеток
спорангиев образуются зооспоры (рис. 193). Форми-
рование зооспор свойственно большинству изученных
видов рода Labyrinthula.
Наиболее подробно этот сложный процесс иссле-
дован на уровне электронного микроскопа у несколь-
ких видов (Hollande, Enjumet, 1955; Amon, Perkins,
1968; Perkins, Amon, 1969; Perkins, 1970). При фор-
мировании спороцист у Labyrinthula algeriensis и La-
byrinthula sp. веретеновидные клетки округляются и
размеры их несколько увеличиваются. В некоторых
колониях этот процесс протекает довольно синхрон-
но. Зооспоруляция сопровождается не только мито-
тическим делением клеток, но и мейозом. Первое де-
Рис. 194. Структура синаптонемальных комплексов в ядре Laby-
rinthula sp. Профаза мейоза I. (По: Moens, Perkins, 1969).
Fig. 194. Structure of synaptonemal complexes in the nucleus of La-
byrinthula sp. Prophase of meiosis I.
ТИП SAPROLEGNIA. КЛАСС LABYRINTHOMORPHEA
295
ление клеток при формировании зооспор — это мей-
оз, который протекает в 2 этапа. В профазе 1-го мейо-
тического деления, на стадии зигонемы и пахинемы,
возникают синаптонемальные комплексы (рис. 194).
Формирование синаптонемальных комплексов подроб-
но изучено у Labyrinthula sp. (Moens, Perkins, 1969).
У этого вида образуется 9 синаптонемальных комп-
лексов. Поскольку каждый из них объединяет пару
гомологичных хромосом (биваленты), то число гап-
лоидных хромосом у этого вида равно 9. Синаптоне-
мальные комплексы прикрепляются к ядерной мембра-
не (рис. 194). Они указывают на наличие полового
цикла у Labyrinthula sp. и позволяют различать мейо-
тическую профазу.
В профазе мейоза происходит формирование
центриолей на противоположных полюсах ядра.
Центриоли образуются только при зооспоруляции;
веретеновидные клетки в интерфазе центриолей не
имеют. Метафаза мейоза характеризуется частичной
резорбцией ядерной оболочки и внедрением микрот-
рубочек формирующегося веретена деления в нуклео-
плазму. Полной резорбции ядерной оболочки не
происходит. Возникает плотная метафазная пластин-
ка, и в ней с трудом можно различить отдельные
хромосомы, потому что они имеют форму очень
тонких нитей. Кинетохоры у хромосом не обнаруже-
ны. В анафазе хромосомы мигрируют к противопо-
ложным полюсам ядра. В телофазе происходит ре-
организация ядерной оболочки, оба дочерних ядра
полностью отделяются друг от друга и резорбируют-
ся микротрубочки веретена деления. Наступает ци-
токинез, и материнская клетка делится в попереч-
ном направлении с образованием перетяжки.
В итоге мейоза I спорангий содержит 2 одноядер-
ные клетки, которые претерпевают мейоз II. Второе
мейотическое деление протекает так же, как и пер-
вое, но без образования синаптонемальных комплек-
сов. После мейоза II спорангий содержит 4 гаплоид-
ные клетки. Третье деление клеток осуществляется
путем митоза, в результате которого образуется 8
гаплоидных зооспор, что служит отличительным при-
знаком изученного вида Labyrinthula sp. (Perkins,
1970). У L. algeriensis в 1 спорангии образуются 4 зоо-
споры (Hollande, Enjumet, 1955).
Экология
Все известные в настоящее время виды лабирин-
томорф — сапрозойные или паразитические орга-
низмы, обитающие на морских растениях, а также в
морских моллюсках (устрицах). Их колонии разру-
шают ткани хозяина, вызывая его гибель. Из мол-
люсков особенно интенсивно бывают заражены уст-
рицы; особую опасность для них представляет Labyrin-
thomyxa marina — колониальная форма с обилием
амебоидных клеток. Разрастаясь, колония разрушает
ткани моллюска, и он погибает. Этот вид может
быть причиной массовой гибели устриц в природных
местах обитания и в устричных хозяйствах. Отрица-
тельная роль лабиринтоморф возрастает еще и пото-
му, что они уничтожают водоросли, которые служат
пищей разнообразным морским животным. Массо-
вое поражение водорослей обнаружено по берегам
Атлантического океана в США. Однако географи-
ческое распространение лабиринтоморф пока еще
не изучено, так же как не изучены особенности
их экологии, включающие разнообразие адаптаций к
условиям окружающей среды. В литературе пока
имеются немногочисленные сведения об отношении
лабиринтоморф к температуре, освещению, солевому
составу среды и к пище, основанные на наблюдени-
ях в условиях лабораторной культуры (Goldstein,
1963).
Систематический обзор
К отряду Labyrinthulida Lankester, 1877 относится
1 семейство — Labyrinthulidae Haeckel, 1834, кото-
рое включает 2 рода. Отряд Thraustochytrida Sparrow,
1936 также содержит 1 семейство — Thraustochytrii-
dae Sparrow, 1936 с 2 родами.
Сем. Labyrinthulidae Haeckel, 1834
Двухжгутиковые зооспоры имеют глазок и вздутие
в основании заднего жгутика. Вегетативные клетки
без центриолей, внеклеточная сеть обычно хорошо
развита.
Род Labyrinthula Cienkowski, 1867
К этому роду относится около 15 видов, которые
являются паразитами или сапрофитами морских во-
дорослей.
Род Labyrinthomyxa Dubosq, 1921
В составе рода 5 видов — паразитов морских во-
дорослей и моллюсков, особенно устриц.
Сем. Thraustochytriidae Sparrow, 1936
Фоторецепторный аппарат у зооспор отсутствует;
в вегетативных клетках имеются кинетосомы. Вне-
клеточная сеть менее развита, чем у Labyrinthulidae.
В состав семейства входит 2 рода: Thraustochytri-
ит Sparrow, 1936 и Schizochytrium Goldstein et Bels-
ky, 1964, которые здесь не рассматриваются.
Представители обоих родов паразитируют на мор-
ских водорослях.
296
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
THE PHYLUM SAPROLEGNIA ZEROV, 1972
THE CLASS LABYRINTHOMORPHEA PAGE, 1979
SUM
The labyrinthomorphs are a small group of colonial protists with
the complex life cycle, including several phases, differing sufficiently
in their morphology and functional activity. Trophic phase of the life
cycle is plasmodium, which consists of extra-cellular net of tubes.
Inside the tubes there are spindle-like cells that move by sliding. The
mechanism of sliding movement is yet unknown. The spindle cells are
related to the internal walls of the tubules with the specific organel-
les — sagenogenetosomes (bothrosomes).
There are individual amoeboid cells, the specific stage in the life
cycle of some species of Labyrinthomorpha. The spindle-like and amo-
eboid cells multiply by binary fission. Double-stage meiosis is charac-
teristic of the zoosporogenesis in many species. As a result of meiosis
the formation of the motile biflagellated zoospores takes place.
Zoospores are often covered with scales and have two heterocont
flagella. The anterior one bears tubular mastigonems, which are typi-
cal of chromists. The transition zone of flagella has the double helix.
There are 2 microtubular rootlets passing from each basal body.
All species of labyrinthomorphids are parasitic or saprophytic,
marine and fresh-water organisms, inhabiting water plants and shell-
fish. Many species of Labyrinthomorpha are pathogenous: they dest-
M ARY
roy tissues of their hosts and can cause mass death of oysters, zostera,
laminaria, and other organisms.
The class Labyrinthomorphea Page, 1979 is divided into 2 orders:
Labyrinthulida Cienkowski, 1867 and Thraustochytrida Sparrow,
1943.
The order Labyrinthulida Cienkowski, 1867 includes one family
Labyrintholidae Haeckel, 1878. Zoospores have two flagella; they also
have the photoreceptor, consisting of stigma and swelling on the hin-
der flagellum. The extracellular net of plasmodium is well developed.
Vegetative amoeboid cells are usually without centrioles. There are
two genera in the family: Labyrinthula Cienkowski, 1867 (15 species)
and Labyrinthomyxa Dubosq, 1921 (5 species). All species are parasi-
tic or saprophytic organisms of marine algae. Among them there are
some pathogenic parasites of shell-fish, especially oysters. The order
Thraustochytrida Sparrow, 1943 also includes one family Thrausto-
chytridae Sparrow, 1936. Zoospores have no photoreceptor. Extracel-
lular net is weakly developed. The family includes 2 genera: Thraus-
tochytrium Sparrow, 1936 (3 species) and Schizochytrium Goldstein et
Belsky, 1964 (1 species). Species of both genera are parasites of ma-
rine algae.
ЛИТЕРАТУРА17
Хаусман К. Протозоология. М., 1988.
Amon J. Р., Perkins F. О. Structure of Labyrinthula sp. zoospo-
res // J. Protozool. 1968. Vol. 15, N 3. P. 543—547.
Barr D. J. S. Evolution and kingdoms of organisms from the per-
spective of a mycologist. Mycologia. 1992. Vol. 84. P. 1—11.
Cienkowski L. Ueber den Bau und Entwicklung des Labyrinthu-
len // Arch. mier. Anat. 1867. Bd 3. S. 274—308.
Darely W. M., Fuller M.S.Cell wall chemistry and taxono-
mic position of Schizochytrium // Amer. J. Bot. 1970. Vol. 57, N 6.
P. 761.
Doflein F. Lehrbuch der Protozoenkunde. Jena, 1953.
Dubocq O. Labyrinthomyxa sauvageau n. sp., Proteomyxee para-
site // Soc. biol. 1921. T. 84. P. 27—32.
Farmer J. N. The Protozoa. Introduction to Protozoology. St.
Louis etc., 1980.
Goldstein S. Development and nutrition of new species of Thra-
ustochytrium // Amer. J. Bot. 1963. Vol. 50, N 3. P. 271—279.
Goldstein S., Belsky M. Axenic culture studies of a new marine
phycomycete possessing an unusual type of asexual reproduction //
Amer. J. Bot. 1964. Vol. 51, N 1. P. 72—78.
Grell K. G. Protistologie. Berlin etc., 1968.
Grell K. G. Labyrinthula coenocystis (Proteomyxidea). Bewegung
und Fortpflanzung // Encyclopedia cinematographica. 1971. E 1172.
S. 2—8.
Hohl H. R. The fine structure of the slimeways in Labyrinthula //
J. Protozool. 1966. Vol. 13, N 1. P. 41—43.
Hollande A., Enjumet M. Sur L’evolution et la systematique des
Labyrinthulidae des Labyrinthula algeriensis nov. sp. // Ann. Sci. nat.
Zool. 1955. Vol. 17, N 2. P. 357—368.
Levine N. D., Corliss J. O. Two new subclassis of sarcodines, of
Labyrinthula subcl. nov. and Proteomyxida subcl. nov. // J. Protozool.
1963. Vol. 10, suppl. P. 27.
Levine N. D., Corliss J. О., Cox P. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R., Lom J., Lynn Z.,
Merinfeld E. G., Page F. C., Poljansky G., Sprague V., Vavra J.,
Wallace F. G. A newly revised classification of the Protozoa // J.
Protozool. 1980. Vol. 27, N 1. P. 37—58.
Moens P. B., Perkins F. O. Chromosome number of a small Protist:
Accurate Determination // Science. 1969. Vol. 166, N 5. P. 1289—1291.
Perkins F. O. Fine structure of zoospores from Labyrinthomyxa
sp. parasitizing the clam Macoma baltica // Science. 1968. Vol. 9, N 3.
P. 198—208.
Perkins F. O. Formation of centriole and centriole-like structures
during meiosis in Labyrinthula sp. (Rhizopodea, Labyrinthulida) // J.
Cell Sci. 1970. Vol. 6, N 4. P. 629—655.
Perkins F. O. The ultrastructure of Holdfasts, «rhizoids» and
«slime tracts» in Traustochytriaceous Fungi and Labyrinthula spp. //
Arch. Microbiol. 1972. Vol. 84, N 1. P. 95—118.
Perkins F. O., Amon J. P. Zoosporulation in Labyrinthula sp.; an
electron microscope study // J. Protozool. 1969. Vol. 16, N 2. P. 235—
257.
Perkins F. O., Menzel R. W. Ultrastructure of sporulation in the
Ouster pathogen Dermocystidium marinum // J. Invertebr. Path. 1967.
Vol. 9, N 2. P. 205—229.
Pokorny K. L. Labyrinthula// J. Protozool. 1967. Vol. 14, N 4.
P. 697—708.
Puytorac P., Grain J. P., Mignot J. Precis de Protistologie. Paris,
1987.
Schmoller H. Beitrag zur Kenntnis der Labyrinthula Entwick-
lung // Arch. Protistenk. 1966. Bd 109, N 2. S. 226—244.
Schmoller H. Die Labyrinthulen und ihre Beziehung zu den
Amoeben // Naturwiss. 1971. Bd 58, N 1. S. 142—146.
Schwab-Stey H., Schwab D. Uber die Feinstruktur von Labyrint-
hula coenocystis Schmoller nach Gefrieratzung // Protoplasma. 1973.
Bd 76, N 3. S. 455—464.
Schwab-Stey H., Schwab D. Scanning electron microscopic in-
vestigation on the motile stage of Labyrinthula coenocystis Schmol-
ler // Cytobiol. 1974. Vol. 8, N 3. P. 383—394.
Sparrow F. K. Biological observations on the marine fungi of
Woods Hole waters // Biol. Bull. 1936. Vol. 70, N 2. P. 236—263.
Valkanov A. Untersuchungen uber die Structur und den Entwick-
lungszyklus von Labyrinthodyction magnifilum Valk. // Arch. Protis-
tenk. 1972. Bd 114, N 4. S. 426—442.
Zopf W. Einige neue Beobachtungen an Labyrinthula Cienk. //
Beitr. Physiol. Morphol. niederer Organismen. 1894. Bd 4. S. 36—48.
17 Цитируемая и рекомендуемая литература к разделу «Тип
Saprolegnia».
Тип OPALINATA Wenyon, 1926 — ОПАЛ ИН АТЫ
Опалинаты — комменсалы кишечника преимущественно эк-
тотермных позвоночных. Гетеротрофные одноклеточные протис-
ты с уплощенным листовидным или цилиндрическим телом, несу-
щим 2 или много жгутиков (ресничек). В переходной зоне жгу-
тиков расположена двойная спираль. Покровы образованы
гребенчатой тубулеммой. Митохондрии с трубчатыми кристами.
Цитостом отсутствует, питание происходит путем пиноцитоза.
Ядра мономорфные, пузырьковидного типа, от 1 до многих. Деле-
ние ядра осуществляется путем закрытого внутриядерного орто-
митоза. Размножение агамное и половое. Мейоз гаметический,
двухступенчатый. Жгутиковые анизогаметы 1-ядерные. Жизнен-
ный цикл у большинства видов сложный, тесно связанный с жиз-
ненным циклом хозяина.
К типу относится 2 класса, в которых насчитыва-
ется около 400 видов.
Класс Proteromonadea
Протеромонады — мелкие одноклеточные орга-
низмы, длина тела которых не превышает 30 мкм.
Трофозоиты имеют 2—4 жгутика, отходящих от пе-
реднего конца тела; жгутиковые корешки состоят из
микротрубочек и соединены с аппаратом Гольджи.
Митохондрия 1. В гребнях тубулеммы проходит от
1 до 10 микротрубочек. Комменсалы кишечника рыб,
амфибий, рептилий и мелких млекопитающих.
История изучения
Протеромонады относятся к числу пока еще слабо
изученных жгутиковых протистов. Система класса и
его таксономическое положение были определены
только после изучения ультраструктуры клетки этих
организмов (Brugerolle, Joyon, 1975; Patterson, 1985;
Карпов, 1990). До исследования ультраструктурных
особенностей клетки протеромонады были отнесены
к кинетопластидам (Honigberg, 1963) на основании
внешнего сходства их единственной крупной мито-
хондрии с кинетопластом. Наиболее подробно изуче-
ны 2 вида протеромонад — Proteromonas lacertae vi-
ridis Grassi, 1879 и Karotomorpha bufonis Dobell,
1908. Класс требует дальнейшего изучения.
Классификация
Класс Proteromonadea Grasse, 1952
Отряд Proterpmonadida Grasse, 1952
Сем. Proteromonadidae Grasse, 1952
Род Proteromonas Kunstler, 1883
Род Karotomorpha Travis, 1954
Grasse, 1952 —
Протеромонады
В настоящее время известно около 10 видов про-
теромонад.
Морфология и физиология
Трофозоит
Форма и размеры тела. Для трофозоитов харак-
терна удлиненная форма тела с расширенным перед-
ним и заостренным задним концом (рис. 195). Разме-
ры трофозоитов варьируют у разных видов от 10 до
30 мкм в длину и от 1.5 до 6 мкм в ширину.
Жгутиковый аппарат. Одну из особенностей про-
теромонад составляет отсутствие жгутикового резер-
вуара — жгутики отходят непосредственно от перед-
него конца тела. У видов рода Proteromonas имеется
2 жгутика: передний и задний, или хвостовой (рис. 195).
Передний жгутик, длина которого превышает длину
тела более чем в 2 раза, утолщен за счет разрастания
цитоплазмы, окружающей аксонему (рис. 196). Зад-
ний жгутик тонкий и несколько короче переднего.
От кинетосомы заднего жгутика отходит 2 микротру-
бочковых корешка, и 1 корешок связан с кинетосо-
мой переднего жгутика (рис. 196). Корешки пересе-
кают аппарат Гольджи, проходят в бороздку ядра и
заканчиваются на поверхности митохондрии (Bruge-
rolle, Joyon, 1975).
У видов Karotomorpha все 4 жгутика направлены
назад (рис. 195), так же как и обе пары их кинето-
сом, расположенных по сторонам центральной оси
тела. От каждой пары кинетосом отходит по 2 мик-
ротрубочковых корешка, которые не связаны с аппа-
298
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 195. Особенности строения некоторых протеромонад.
А — Proteromonas lacertae viridis: 1 — передний жгутик, 2 — аппарат Гольджи, 3 — ядро, 4 — ядрышко, 5 — митохондрия, 6 — пищеварительные вакуоли,
7 — ризостиль, 8 — кинетосомы; Б — Karotomorpha fusiformis: В — циста Proteromonas lacertae viridis: 1 — ядро, 2 — митохондрия, 3 — оболочка
цисты. (А — по: Grassi, 1897; Б — по: Тимофеев, Шульман, 1962; В — по: Kulda, Nohynkova, 1978).
Fig. 195. Morphology of Proteromonadea.
A — Proteromonas lacertae viridis: 1 — anterior flagellum, 2 — Golgi apparatus, 3 — nucleus, 4 — nucleolus, 5 — mitochondrion, 6 — digestive vacuoles, 7 —
rhizostyl, 8 — kinetosomes; Б — Karotomorpha fusiformis: В — cyst of Proteromonas lacertae viridis: 1 — nucleus, 2 ~ mitochondrion, 3 — cyst wall.
ратом Гольджи; они идут вдоль боковой стороны ядра
и заканчиваются у поверхности клетки (рис. 197). В
переходной зоне жгутиков обоих видов находится двой-
ная спираль, расположенная над поперечной пластин-
кой. Эти структурные элементы переходной зоны
имеют большое значение при выяснении филогенети-
А
ческих взаимоотношений протеромонад с другими
таксонами жгутиконосцев.
Покровы. Для протеромонад характерны плот-
ные покровы, образованные гребенчатой тубулеммой
(Карпов, 1990). Продольные ребра тубулеммы слабо
развиты у Proteromonas, и в каждом из них проходит
только 1 микротрубочка (рис. 196). Особенность Karo-
tomorpha составляют сильно развитые ребра, идущие
вдоль тела и слегка изогнутые спирально (рис. 195;
197). Внутри каждого ребра находится лента из 10 со-
единенных между собой микротрубочек. Всего по по-
верхности клетки проходит 18—28 продольных ребер.
Среди жгутиконосцев гребенчатая тубулемма обнару-
жена еще только у опалинид.
Нижняя часть тела Proteromonas покрыта трубча-
тыми трехчленными волосками— соматонемами, ко-
торые подобны мастигонемам хризомонад (рис. 196;
198). Соматонемы прикрепляются к апикальной по-
Рис. 196. Особенности строения трофозоита Proteromonas lacer-
tae viridis из кишечника ящерицы. (По: Brugerolle, Joyon, 1975).
А — схема строения клетки, Б — поперечный срез переднего жгутика,
В — поперечный срез через тубулемму. аг — аппарат Гольджи, ак — ак-
сонема, жк — жгутиковые корешки, зж — задний жгутик, м — митохонд-
рия, мт — микротрубочки, проходящие в складках тубулеммы, пж — пе-
редний жгутик, пл — плазматическая мембрана, сн — соматонемы, я —
ядро.
Fig. 196. Morphology of trophozoite of Proteromonas lacertae viri-
dis from the intestine of a lizard.
A — diagram of the cell structure, Б — cross section through the anterior fla-
gellum, В — cross section through tubulemma. аг — Golgi apparatus, ак —
axonema, жк — flagellar rootlets, зж — posterior flagellum, м — mitochond-
rion, мт — microtubules inside the folds of tubulemma, пж — anterior fla-
gellum, пл — cytoplasmic membrane, ch — somatonemes, я — nucleus.
ТИП OPALINATA. КЛАСС PROTEROMONADEA
299
Рис. 197. Схемы сборки, миграции и крепления соматонем на по-
верхности клетки Proteromonas lacertae viridis. (По: Brugerolle,
Bardele, 1988).
аг — аппарат Гольджи, мт — микротрубочки, мф — микрофиламенты,
пл — плазматическая мембрана, сн — соматонемы, эр — ЭПР, я — ядро.
Fig. 197. Diagram of synthesis, migration, and attachement of soma-
tonemes to the surface of cell of Proteromonas lacertae viridis.
аг — Golgi apparatus, мт — microtubules, microfilaments, мф — microfila-
ments, пл — cytoplasmatic membrane, ch — somatonemes, эр — endoplas-
mic reticulum, я — nucleus.
верхности гребней тубулеммы (Brugerolle, Bardele, 1988).
Они синтезируются в цистернах аппарата Гольджи и
выявляются в его секретирующей зоне. Функция со-
матонем не изучена.
Митохондрии и другие органеллы цитоплазмы.
Протеромонады имеют 1 митохондрию крупных раз-
меров, напоминающую по форме кинетопласт жгу-
тиконосцев отряда Kinetoplastida. Митохондрия рас-
положена непосредственно под ядром и имеет фор-
му полукольца. Кристы митохондрии тубулярные
(рис. 196; 197), матрикс ее осмйофильный, плотный,
не окрашивающийся по Фёльгену и на электро-
Рис. 198. Схема ультратонкого строения трофозоита Karotomorp-
ha bufonis. (По: Brugerolle, Joyon, 1975).
А — расположение органелл в клетке: 1 — жгутики, 2 — кинетосомы,
3 — складки тубулеммы, 4 — аппарат Гольджи, 5 — ядро, б — митохонд-
рии, 7 — ЭПР, 8 — ризостйль; Б — поперечный срез через складки тубу-
леммы.
Fig. 198. Diagram of ultrastructure of trophozoite of Karotomorpha
bufonis.
A — arrangement of cell organelles: 1 — flagella, 2 — kinetosomes, 3 —
folds tubulemma, 4 — Golgi apparatus, 5 — nucleus, 6 — mitochondrion,
7 — endoplasmic reticulum, 8 — rhizostyl; Б — cross section through the
folds of tubulemma.
нограммах в нём не выявляются хромосомные фиб-
риллы.
Цитоплазма трофозоитов насыщена цистернами
гранулярной и гладкой ЭПР, большинство которых
локализуется в передней половине тела. В цитоплаз-
ме содержится много свободных рибосом. Аппарат
Гольджи находится в переднем конце тела над ядром
и обращен к ядру своей секретирующей поверхно-
стью (рис. 196; 197). Лизосомы, мелкие округлые тель-
ца, содержащие кислую фосфатазу, особенно- мно-
гочисленны в передней половине тела трофозоита.
В этой же зоне сосредоточены пищеварительные ва-
куоли, образующиеся путем слияния пиноцитозных
пузырьков. Пцноцитоз осуществляется путем по-
глощения жидкой пищи всей поверхностью тела и
представляет основной тип питания. Цитостом отсутст-
вует (Brugerolle, Joyon, 1975; Kulda, Nohynkova, 1978).
Цисты
В жизненном цикле протеромонад имеются цисты
покоя. Они мелкие, округлые, около 10 мкм в диа-
метре (рис. 195). Оболочка цист толстая и плотная. В
300
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 199. Жизненный цикл Proteromonas lacertae viridis. (По: Puy-
torac et al., 1987).
AP — агамное размножение, ВС — внешняя среда, ПР — половое размно-
жение. 1 — трофозоит, 2 — делящаяся особь, 3 — трофозоит, 4 — копуля-
ция, 5 — слияние особей и образование’зигоцисты, 6 — зигоциста, 7 — де-
ление ядер в зигоцисте, 8 — формирование зооспор и выход их во внеш-
нюю среду.
Fig. 199. The life cycle of Proteromonas lacertae viridis.
AP — agamic reproduction, BC — outside of host, ПР — sexual reproduction.
1 — trophozoite, 2 — dividing 'specimen, 3 — trophozoite, 4 — copulation,
5 — confluence of 2 specimens and zygocyst formation, 6 — zygocyst, 7 —
dividing nuclei in zygocyst, 8 — formation of zoospores, and their excystment.
цитоплазме цисты находится 1 ядро и митохондрия,
но отсутствуют аксонемы жгутиков и субмембранные
микротрубочки. У Proteromonas lacertae viridis обна-
ружены также зигоцисты, в которых происходит раз-
множение (рис. 199).
Размножение
и жизненный цикл
Протеромонады размножаются агамным и поло-
вым путем. При агамном размножении трофозоиты
претерпевают продольное бинарное деление, в ре-
зультате которого образуются 2 одинаковые дочер-
ние клетки. Процесс деления изучен пока только на
уровне светового микроскопа и только у Р. I. viridis
(Kulda, Nohynkova, 1978).
Митоз закрытый, протекающий полностью под
ядерной оболочкой (рис. 200). Он начинается с деле-
ния кинетосом, вслед за этим у делящейся клетки по-
является двойной набор жгутиков. Кинетосомы удва-
иваются, и дочерние кинетосомы мигрируют к проти-
воположным полюсам ядра. Они находятся в тесном
контакте с ядерной оболочкой и служат центрами де-
ления. Жгутиковые корешки резорбируются.
В профазе митоза видны тонкие нитевидные хро-
мосомы, начинается резорбция ядрышка. Образуется
внутриядерное веретено митоза с кинетосомами на
полюсах. В метафазе хорошо видны хромосомы,
расположенные в одной плоскости и образующие
экваториальную пластинку. Ядрышко резорбируется,
и хромосомы начинают расходиться к противопо-
ложным полюсам ядра; анафаза хорошо выражена.
Затем ядро растягивается, приобретает гантелеобраз-
ную форму и делится на 2 равные части, что свой-
ственно телофазе. Образуются 2 дочерних ядра,
после чего наступает цитокинез.
Во время телофазы митохондрия также делится
на 2 равные части, разделяется пополам и аппарат
Гольджи (Kulda, Nohynkova, 1978).
Половой процесс обнаружен пока только у вида
Р. I. viridis (Puytorac et al., 1987), популяции которо-
го обитают в просвете кишечника зеленой ящерицы,
Lacerta viridis. Основную массу особей в популяции
составляют активные трофозоиты, размножающиеся
агамно путем продольного бинарного деления (рис. 199).
Агамное размножение чередуется с половым процес-
сом, который протекает также в просвете кишечника
в форме гологамной копуляции 2 зрелых трофозои-
тов, морфологически не отличающихся друг от друга
(рис. 199). Трофозоиты соединяются попарно, инцис-
тируются, и в цисте происходит полное слияние ядер
и цитоплазмы (рис. 199).
После копуляции здесь же, в просвете кишечника
хозяина, образуется зигоциста. В ней происходит де-
ление синкариона и начинается формирование до-
черних клеток — будущих зооспор (рис. 199). В
таком состоянии зигоциста выводится во внешнюю
среду вместе с фекалиями хозяина, где в ней разви-
ваются 32 жгутиковые зооспоры (рис. 199).
Жизненный цикл Р. /. viridis относится к числу
дигенетических циклов — в нем имеется 2 поколе-
ния: агамное и половое. Большая часть жизненного
цикла протекает в просвете кишечника ящерицы и
представляет эндогенную часть цикла. Только одна
его фаза — циста размножения, или зигоциста, раз-
вивается во внешней среде. Она представляет экзо-
генную часть цикла и выполняет функцию распрост-
ранения протеромонад среди их хозяев — ящериц
(Puytorac et al., 1987). Мейоз и место его в жизнен-
ном цикле пока не изучены.
Культивирование
Для Proteromonas lacertae viridis разработаны 2
типа культур in vitro. Один из них представляет аксе-
нические культуры на жидкой питательной среде, в
которых плотность популяций достигает 106 организ-
мов/мл (Kulda, 1973). При втором способе культиви-
рования к. стерильной жидкой среде добавляют дрож-
жи Candida sp.
Тип OPALINATA. КЛАСС OPALINATEA
301
Рис. 200. Бинарное деление трофозоита Proteromonas Lacertae viridis. (По: Doflein, 1953).
А — трофозоит в период интерфазы, Б—Д — фазы митоза. 1 — жгутики, 2 — кинетосомы, 3 — ризопласт, 4 — аппарат Гольджи, 5 — ядро, 6 — яд-
рышко, 7 — митохондрия.
Fig. 200. Binary fission of trophozoite of Proteromonas Lacertae viridis.
A — trophozoite (interphase), Б—Д — phases of mitosis. 1 — flagella, 2 — kinetosomes, 3 — rhizoplast, 4 — Golgi apparatus, 5 — nucleus, 6 — nucleolus,
7 — mitochondrion.
Происхождение и эволюция
Ультраструктурная организация трофозоитов Pro-
teromonas и Karotomorpha позволяет судить о сход-
стве их с опалинатами и хризомонадами. Признака-
ми сходства с обеими группами могут служить пе-
реходная зона жгутика, в которой имеется двойная
переходная спираль; один тип покровов — гребен-
чатая тубулемма; трубчатые кристы в митохондриях
и один тип митоза (Patterson, 1985). Наличие со-
матонем наряду с другими признаками указывает
на возможное происхождение их от одной предко-
вой группы свободноживущих жгутиконосцев, кото-
рые могли быть подобны хризомонадам (Patterson,
1985).
Систематический обзор
2 рода сем. Proteromonadidae отличаются друг от
друга по числу жгутиков, ультратонкой организации
тубулеммы, форме и локализации аппарата Гольджи,
строению жгутиковых корешков (рис. 196; 197).
Род Proteromonas Kunstler, 1883
(Син.: Heteromita Grassi, 1881; Bodo Prowazek,
1904; Prowazekella Alexeieff, 1912; Prowazekia
Swezy, 1916; Schizobodo Chatton, 1917)
В составе рода имеется 4 вида, которые обитают в
кишечнике моллюсков, хвостатых амфибий, ящериц
и черепах, а также морских свинок. Все виды не па-
тогенны для своих хозяев. Типовой вид — Р. 1а-
certae viridis Grassi, 1879.
Род Karotomorpha Travis, 1934
(Син.: Tetramitus Belar, 1916;
Tetramastix Alexeieff, 1916)
Включает 3 достоверно известных вида: К. bufonis
Dobell, 1908, K.fusiformis Timofeeff et Schulman, 1962
и К. ovalis Timofeeff et Schulman, 1962. Первый вид оби-
тает в кишечнике амфибий, 2 последних — в кишеч-
нике пресноводных рыб (Хайбулаев, Шульман, 1984).
Класс Opalinatea Wenyon, 1926 — Опалинаты
Опалинаты — комменсалы прямой кишки эктотермных позво-
ночных; по морфологическим и физиологическим признакам пред-
ставляют достаточно однородную группу гетеротрофных протис-
тов. Хозяева — бесхвостые амфибии и значительно реже рыбы и
рептилии. Плоское листовидное или цилиндрическое веретеновид-
ное тело опалинат покрыто многочисленными, одинаковыми по
длине жгутиками (ресничками). Переходная зона жгутиков содер-
жит двухнитчатую спираль.
Покровы представлены гребенчатой тубулеммой. Цитостом от-
сутствует; основной способ питания — пиноцитоз. Митохондрии
302
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
многочисленные, с трубчатыми кристами. Ядра пузырьковидного
типа, однородные по структуре, их число варьирует у разных видов
от 1 до нескольких сотен. Размножение агамное и половое. Мейоз
гаметический, двухступенчатый. Формируются жгутиковые Ма и
Ми. Процесс их слияния — сингамия. Обычны гамонтоцисты и
зигоцисты. Жизненный цикл сложный и тесно связан с сезонными
циклическими изменениями хозяев.
История изучения
Первое описание опалинат принадлежит Левенгу-
ку и относится к 1683 г. В первой половине про-
шлого столетия был выделен род Opalina Purkinje et
Valentin, 1835. В этот род были включены гетеро-
трофные протисты, большинство из которых обита-
ет в кишечнике амфибий. С момента описания рода
началось интенсивное изучение фауны опалинат, их
морфологии и жизненных циклов.
Подробные описания большинства известных в
настоящее время родов, видов и жизненных циклов
опалинат принадлежат Меткалфу (Metcalf, 1909, 1923,
1940). Им был создан также первый атлас-определи-
тёль семейств, родов и видов опалинат, который не
потерял своей ценности и в наши дни.
Описание новых видов и внутривидовых форм опа-
линат продолжается до сих пор. К настоящему вре-
мени детально изучены жизненные циклы ряда видов,
особенно Opalina ranarum, О. obtrigonoidea, О. virgu-
loidea (Суханова^ 1953; Wessenberg, 1961, 1978). Ис-
следован процесс митоза и обнаружен мейоз в жиз-
ненном цикле О: ranarum (Kaszanowski, 1971; Patter-
son, 1985). Данные по морфологии клетки О. ranarum,
О. virguloidea, О. japonica, О. obtrigonoidea, Cepedea
dimidiata(Pitelka, 1956; Noirot-Timothee, 1959; Wessen-
berg, 1961; 1966, 1978; Patterson, 1985), Protoopalina
saturnalis (Mignot, Molina, 1988) получены благодаря
применению многих цитологических методик, вклю-
чая и электронную микроскопию.
Ряд работ посвящен вопросам экологии опалинат.
Наиболее подробно исследованы адаптации О. гапа-
Рис. 201. Трофозоиты опалинат разных видов.
А — Hegneriella dobelli, Б— Zelleriella macronucleata, В — Protoopalina caudata lata, Г — Cepedea dimidiata vulgaris, Д — Opalina triangulata subsp. viri-
dis. (A — no: Earl, 1971; Б — no: Metcalf, 1923; В—Д — ориг. K.:M. Сухановой).
Fig. 201. Trophozoites of different species.
Тип OPALINATA. КЛАСС OPALINATEA
303
гит к температуре среды обитания, внутривидовая
адаптивная изменчивость морфологических признаков,
взаимоприспособленность жизненных циклов опали-
нат к циклическим изменениям хозяина (Metcalf, 1940;
Суханова, 1968; Wessenberg, 1978).
Классификация
Класс Opalinatea Wenyon, 1926
Отряд Opalinida Poche, 1913
Сем. Opalinidae Claus, 1874
В состав семейства входит 5 родов, насчитываю-
щих порядка 400 видов.
Морфология и физиология
Внешнее строение
Форма и размеры тела. Большинство опалинид
обладает крупными размерами: длина тела трофозои-
тов некоторых видов достигает 1 мм. Форма тела ши-
роко варьирует у разных видов — от узкой веретено-
видной и округлой в поперечном сечёнии до оваль-
ной и широкой листовидной с плоским поперечным
сечением (рис. 201). У «плоских» видов тело сплю-
щено с боков, дорсальная сторона несколько выпук-
лая, а вентральная вогнута. Передний конец тела за-
круглен, утолщен, и край его образует специфический
для опалинид отдел — фалькс, разделяющий правую
и левую стороны тела (рис. 202). Задний конец тела
может быть округлым или заостренным.
Поверхностные структуры. Наиболее подробно
поверхностные структуры изучены у О. гапагит, О.
virguloidea, Protoopalina satufnalis (Pitelka, 1956;
Wessenberg, 1961; Noirot-Timothee, 1966; Patterson,
1985; Mignot, Molina, 1988). Снаружи тело покрыто
тубулеммой, которая образует складки, идущие вдоль
тела и занимающие промежутки между двумя сосед-
ними кинетами (рис. 203). Высота тубулярных греб-
ней 800—900 нм при ширине 50 нм. Внутри них
проходят микротрубочки — в 1 ряд по высоте складки.
Каждая микротрубочка соединена с соседней двумя
краевыми связками. В одном ряду может быть 13—
24 микротрубочки. Число гребней варьирует от 1 до
25, и они придают значительную прочность покровам.
При наблюдении с помощью светового микроскопа
тесно лежащие складки создают впечатление опалес-
ценции.
Жгутики. Тело опалинид покрыто многочислен-
ными жгутиками, длина которых одинакова и дости-
гает 10—12 мкм. Расположение жгутиков строго упо-
рядочено, и они образуют ряды — соматические кине-
ты, идущие вдоль тела и слегка изогнутые S-образно.
Все кинеты берут свое начало от фалькса (рис. 202).
Число кинет варьирует у разных видов, и, например,
у Opalina virguloidea от фалькса берет начало 210—
220 кинет, различающихся по своей длине. До задне-
го конца доходит 50—60 длинных кинет, а осталь-
ные — короткие и заканчиваются в передней полови-
не тела. В отличие от продольных соматических
Рис. 202. Схема расположения кинет у Opalina virguloidea (А, Б)
и у Protoopalina satumalis (В).
А — у растущего трофозоита, Б —в области фалькса, В — схема органи-
зации фалькса и прилегающих сЬматических кинет. Кинетосомы фалькса
(кф) окружены зоной фибриллярного материала (зф), от которого начина-
ются микротрубочки цитоскелета (мт) и с которой связаны продольные
ряды кинет (пк). к — кинеты, ф — фалькс, я — ядро. (А, Б — по: Wes-
senberg, 1961; В — по: Mignot, Molina, 1988).
Fig. 202. Diagram of arrangement of kineties in Opalina virguloidea
(А, Б), and in Protoopalina saturnalis (B).
A — protrophozoite, Б — falx zone, В — diagram of falx structure and adjo-
ing somatic kineties. The kinetosomes of falx (кф) are surrounded by zone of
fibrillar material (зф), from which microtubules of skeleton (мт), and longitu-
dinal rows of kineties (пк) originate, к — kineties, ф — falx, я — nucleus.
кинет ряды кинет фалькса идут в поперечном направ-
лении и занимают передний край тела (рис. 202).
Жгутики — основные органеллы движения всех опа-
линид. Биение жгутиков начинается от фалькса —
возникают метахрональные волны, которые распро-
страняются вдоль тела от переднего конца к заднему.
304
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 203. Процесс пиноцитоза у опалин.
А — пиноцитозные вакуоли в эктоплазме Opalina ranarum, Б — вакуолизация цитоплазмы у Zelleriella telmatobii. 1 — складки тубулеммы с проходящи-
ми в них микротрубочками, 2 — пиноцитозные каналы, 3 — пиноцитозные вакуоли. (А — по: Munch, 1970; Б — по: Metcalf, 1923).
Fig. 203. Pinocytosis in Opalina.
A — pinocytotic vacuoles in Opalina ranarum ectoplasm, Б — vacuolisation of cytoplasm of Zelleriella telmatobii. 1 — folds of tubulemma with microtubules in-
side, 2 — pinocytotic canals, 3 — pinocytotic vacuoles.
При движении тело трофозоита вращается вокруг
своей продольной оси и он медленно плавает в ки-
шечном содержимом. Эта группа кинетосом обособ-
лена от остального кинетома электронно-плотным
фибриллярным материалом, из которого начинаются
как соматические кинеты, так и микротрубочки тубу-
Рис. 204. Схема строения основания жгутика и кинетосомы Opali-
na ranarum. Продольный срез. (По: Wessenberg, 1966).
1 — центральные микротрубочки, 2 — складки тубулеммы, 3 — микротру-
бочки кинетосомы, 4 — фибриллярная зона, 5 — переходная пластинка,
6 — поперечно исчерченная фибрилла, 7 — плотные гранулы в центре ки-
нетосомы, 8 — переходные фибриллы дистальной части кинетосомы.
Fig. 204. Diagram of structure of flagella basis and kinetosome in
Opalina ranarum. Longitudinal section.
1 — central microtubules, 2 — folds of tubulemma, 3 — microtubules of kine-
tosoma, 4 — fibrillar zone, 5 — transitional plate, 6 — transversal striated
fibrille, 7 — dense granules in the center of kinetosome, 8 — transition fibres of
the distal part of the kinetosome.
леммы (Mignot, Molina, 1988). Жгутики оиалинид
имеют типичное ультратонкое строение, свойствен-
ное ресничкам и жгутикам других протистов.
В переходной зоне имеется двойная спираль, рас-
положенная над поперечной пластинкой (рис. 204).
Диаметр кинетосомы около 200 нм, длина около
1 мкм. В центральной части кинетосомы находятся
электронно-плотные гранулы (рис. 204). В одной ки-
нетосоме может находиться 72—96 таких гранул при
диаметре одной гранулы 25—30 нм. Функция этих
гранул пока неясна (Wessenberg, 1961; Patterson, 1985;
Mignot, Molina, 1988).
От каждой кинетосомы отходит 2 фибриллярных
корешка: длинный и короткий. Они направлены ла-
терально в сторону соседней кинетосомы. Длинные
корешки соединяют кинетосомы друг с другом, об-
разуя кинету. Корешки в кинете ориентированы в
сторону фалькса. От дистального конца кинетосомы
отходят и прикрепляются к плазмалемме 9 переход-
ных фибрилл, которые выполняют опорную функ-
цию (рис. 204). Проксимальный участок кинетосомы
имеет колесовидную структуру (Patterson, 1985).
Цитоплазма
Эктоплазма представляет поверхностный слой ци-
топлазмы, который у опалинид отчетливо выражен, и
в нем расположены кинетосомы жгутиков. Одну из
особенностей эктоплазмы составляет сильная вакуолиза-
ция. Вакуоли мелкие, уплощенной или округлой фор-
мы. С помощью светового микроскопа вакуолизация
эктоплазмы особенно хорошо видна у уплощенных
форм (рис. 203). По своей природе — это пиноцитоз-
ные вакуоли (Noirot-Timothee, 1959, 1966; Patterson,
1985). Они периодически образуются у поверхности
клетки, а затем проходят в более глубокие слои цито-
плазмы и, сливаясь вместе, образуют вакуоли более
крупных размеров (рис. 203). В вакуолях обнаружена
Тип OPALINATA. КЛАСС OPALINATEA
305
кислая фосфатаза, что подтверждает слияние их с
лизосомами и формирование пищеварительных ваку-
олей (Сергеева, 1969). Пиноцитоз — основной путь
питания опалинид, не имеющих ротового отверстия,
и обилие пиноцитозных вакуолей указывает на вы-
сокую интенсивность их питания жидкой пищей, со-
держащейся в верхней части прямой кишки хозяев.
В эктоплазме расположен аппарат Гольджи, пред-
ставленный многочисленными диктиосомами. Ульт-
раструктура диктиосом типична. Их размеры не пре-
вышают 3 мкм. На электронограммах можно видеть,
что вогнутая проксимальная поверхность диктиосо-
мы часто находится в тесном контакте с пиноцитоз-
ными вакуолями (Noirot-Timothee, 1959).
Наиболее многочисленные органеллы эндоплаз-
мы — митохондрии. Они имеют форму округлых или
палочковидных гранул и довольно крупные размеры:
их длина достигает 2—3 мкм при ширине около 1 мкм.
Кристы митохондрий трубчатые. С помощью метода
авторадиографии обнаружена мДНК и установлен ее
синтез в митохондриях Opalina ranarum, В митохон-
дриях этого вида обнаружена сукцинатдегидрогеназа,
что указывает на осуществление в них цикла Кребса.
Однако среда обитания опалинид бедна кислородом,
и для их клеточного метаболизма характерны глико-
литические процессы.
Трофозоиты имеют хорошо развитый ЭПР — гра-
нулярный и гладкий, многочисленные рибосомы, сво-
бодные и локализованные на мембранах ЭПР, что ука-
зывает на высокую интенсивность процессов биосинтеза
белка. В эндоплазме находятся первичные и вторич-
ные лизосомы, а также остаточные тельца, свидетель-
ствующие о высокой интенсивности питания (Pitelka,
1956; Noirot-Timothee, 1959, 1966; Сергеева, 1969).
Эндоплазма богата клеточными включениями: ли-
пиды в виде капель различных размеров располагают-
ся в разных участках клетки трофозоита и особенно
многочисленны в заднем конце (рис. 205). Полисаха-
риды типа гликогена имеют форму зерен разных раз-
меров, и их запасы равномерно распределены в эн-
доплазме всей клетки (рис. 205).
В цитоплазме много микрофиламентов, которые со-
здают ее плотность и выполняют опорную функцию.
Ядро
Морфологически все ядра одинаковы, и опалини-
ды — типичные гомокариоты (сомателлы). Ядрыш-
ки — обязательный компонент каждого ядра —
имеют форму гранул различных размеров, и их
число варьирует. В ядрышках содержится РНК, что
показано с помощью цитохимических методик. Наи-
более часто ядрышки располагаются в периферичес-
кой зоне ядра и тесно прилегают к его оболочке. В
ядрах видов, которые изучены с применением окрас-
ки метиловым зеленым с пиронином, а также галло-
цианином, выявлено значительное содержание РНК,
локализованной в ядрышках (рис. 206; 207).
Разные виды опалинид имеют неодинаковое число
ядер, что служит одним из важных таксономических при-
знаков этой группы. Одноядерных видов немного, и к их
числу относятся виды рода Hegneriella (Earl, 1971).
20 Протисты, часть 1
Рис. 205. Клеточные включения у Opalina ranarum,
А — липиды, Б — гликоген (ориг. К. М. Сухановой).
Fig. 205. Cytoplasmic inclusion in Opalina ranarum.
A — lipids, Б — glycogen.
Рис. 206. Деление ядра у Zelleriella intermedia. (По: Chen, 1936).
A — интерфазное ядро, Б — профаза, В — метафаза, Г — поздняя анафа-
за. яд — ядрышки.
Fig. 206. Dividing nucleus in Zelleriella intermedia.
A — interphase, Б — prophase, В — metaphase, Г — late anaphase, яд —
nucleoli.
306
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 207. Распределение ядрышкового материала в интерфазных
делящихся ядрах Protoopalina intestinalis. (Ориг. К. М. Сухано-
вой).
А — интерфазные ядра, Б — профаза, В — метафаза, Г — анафаза, Д —
ранняя телофаза. Окраска метиловым зеленым—пиронином; видно, что
хромосомы во всех фазах митоза окружены ядрышковым материалом.
Fig. 207. Distribution of nucleolar material in interphase and divi-
ding nuclei.
A — interphase, Б — prophase, В — metaphase, Г — anaphase, Д — early
telophase. The preparation is stained with methyl-green-pyronine; the chromo-
somes are enclosed of nucleolar material.
Виды рода Zelleriella имеют 2 ядра, виды рода
Protoopalina — от двух до 8 ядер, виды же родов
Cepedea и Opalina многоядерны, и их клетка содер-
жит до 100 и более однородных ядер пузырьковид-
ного типа (рис. 201). Величина ядер в большой сте-
пени зависит от их числа в одной клетке: чем боль-
ше ядер, тем меньше их размеры. Например,
единственное ядро Hegneriella dobelli достигает в
длину 30—42 мкм при ширине 13—21 мкм, а диа-
метр многочисленных шаровидных ядер Opalina ra-
narum не превышает 7—8 мкм.
Деление ядер опалинид осуществляется по типу
закрытого внутриядерного ортомитоза (Райков, 1978),
при котором формируется ацентрическое веретено,
полностью расположенное в ядре (рис. 206). У
двухъядерных видов деление ядер трофозоитов мо-
жет происходить одновременно, но может быть и
асинхронным. Ядра многоядерных видов делятся
асинхронно, цитокинез наступает после того, как
большинство ядер разделилось. Борозда деления
проходит в косом направлении между двумя кинета-
ми и делит материнскую клетку на 2 дочерние, ко-
торые идентичны ей по своей морфологии, но отли-
чаются мелкими размерами.
Деление ядра наиболее подробно изучено у двухъ-
ядерного вида Zelleriella intermedia (рис. 206) (Chen,
1936а, 1936b). Ядра этого вида в период интерфазы
обладают мелкогранулярной структурой и имеют 4—
6 ядрышек. В профазе происходит формирование
тонких нитевидных хромосом. В метафазе хромосо-
мы располагаются в центре ядра и связаны с нитями
веретена митоза. Метафазные хромосомы имеют четко
выраженную индивидуальность, и каждая из них со-
стоит из 2 хроматид. Число хромосом 24. Некоторые
хромосомы (их может быть 4—6) связаны с ядрыш-
ками, которые расположены снаружи этих хромосом
и образуют нечто вроде чехла на средней части каж-
дой хромосомы; свободными от ядрышкового мате-
риала остаются только их концы. В анафазе хромо-
сомы расходятся к полюсам и формируются 2 дочер-
них ядра, между которыми образуется перетяжка.
Ядрышки сохраняются на протяжении всех фаз мито-
за и вместе с хромосомами расходятся в анафазе к
полюсам делящейся клетки. Ядерная оболочка сохра-
няется в течение всего времени деления ядра.
По такому же типу, как у Z. intermedia, протекает
митоз у трофозоитов Protoopalina intestinalis, Cepe-
dea dimidiata, Opalina ranarum. Однако у большинст-
ва видов опалинид митоз изучен слабо или не изучен
совсем. Стоит отметить одну из особенностей митоза
Protoopalina intestinalis. Интерфазные ядра этого вида
отличаются крупными размерами ядрышек (рис. 207).
В митотических ядрах видны 4 хромосомы, каждая
из которых снаружи покрыта ядрышковым материа-
лом, образующим подобие чехла. Доказательством
того, что этот чехол содержит РНК, служит интен-
сивная окраска его пиронином и галлоцианином.
Можно поэтому предположить, что каждая хромосо-
ма ядра Р. intestinalis имеет ядрышковый организатор
(Суханова, 1961). При окраске по методу Фёльгена
выявляются 4 тонкие нитевидные хромосомы.
На ультраструктурном уровне митоз опалинид
практически не изучен. Одна из стадий деления ядра
Р. saturnalis приведена в работе Миньо и Молина
(Mignot, Molina, 1988), где хорошо видна интактная
оболочка и внутриядерные микротрубочки веретена.
Особого внимания заслуживает вопрос о возмож-
ной дифференцировке и степени плоидности ядер
опалинид. В течение длительного времени их ядра
рассматривались как Ма, а за Ми ошибочно прини-
мались митохондрии, упоминаемые в ряде работ как
эндоплазматические тельца (Konsuloff, 1922; Metcalf,
1923, и др.). Подробно изучив митоз у Zelleriella in-
termedia,Чен (Chen, 1936а) считал, что оба ядра тро-
фозоитов этого вида диплоидны и содержат по
24 хромосомы. Однако кроме трофозоитов с дипло-
идными ядрами Чен обнаружил особи, имеющие не
2, а 4 ядра более мелких размеров с гаплоидным на-
бором хромосом, равным 12 в каждом ядре.
При изучении процессов деления ядер у Opalina
ranarum Качановский (Kaszanowski, 1968) обнару-
жил крупные делящиеся ядра с числом хромосом
около 60 и мелкие ядра с числом метафазных хромо-
сом около 18. По его мнению, мелкие ядра диплоид-
ны, крупные же полиплоидны. Есть особи Opalina,
которые имеют оба типа ядер, но есть и особи, име-
ющие только мелкие или только крупные ядра. Эти
исследования требуют дальнейшего развития.
Размножение и жизненный
цикл
Наиболее подробно сложный годичный жизнен-
ный цикл изучен у видов рода Opalina.
Тип OPALINATA. КЛАСС OPALINATEA
307
Рис. 208. Схема жизненного цикла Opalina ranarum. (Ориг. К. М. Сухановой).
А — делящийся трофозоит, Б — формирование гамонтов в результате серии палинтомических делений, В — гамонтоцисты, Г — эксцистирование и после-
дующие деления гамонтов в кишечнике головастиков, Д — макро- и микрогаметы, Е — сингамия, Ж — зиготы, 3 — зигоцисты, И — растущие трофозои-
ты (протрофозоиты), К — зрелый трофозоит.
Fig. 208. Diagram of the life cycle of Opalina ranarum.
A — dividing trophozoite, Б — development of gamonts after several palintomic divisions, В — gamontocysts, Г — excystment and following divisions of the ga-
monts in the intestine of tadpole, Д — macro- and microgametes, E — singamy, Ж — zygotes, 3 — zygocysts, И — young trophozoites (protrophozoites), К —
mature trophozoite.
Агамное размножение
Бинарное деление. В течение лета, осени и зимы у
Opalina ranarum, О. virguloidea и других видов попу-
ляции состоят преимущественно из трофозоитов
(рис. 208), которые периодически размножаются
агамно путем продольного бинарного деления. Обра-
зующиеся в результате этого деления 2 дочерние
особи вырастают до размеров зрелого трофозоита,
приобретая характерную для него форму (рис. 208). За
счет агамного размножения число особей популяции уве-
личивается, особенно в течение весны, лета и ранней
осени, зимой же деление трофозоитов происходит редко.
Палинтомия. Весной, в период размножения бес-
хвостых амфибий — хозяев опалин, начинается па-
линтомическое деление трофозоитов (рис. 208). Осо-
бенность палинтомии у опалин заключается в том,
что трофозоит проходит серию быстро следующих
друг за другом, чередующихся продольных и попе-
речных делений без последующего роста дочерних
особей.
В результате палинтомии образуются мелкие, уд-
линенной формы, 40—50 мкм в длину, особи — га-
монты (рис. 208), имеющие несколько ядер (2—8).
Трофозоиты, делящиеся палинтомически, — это про-
гамонты. В популяции опалин не все трофозоиты
становятся прогамонтами, и часть из них весной про-
должает размножаться путем бинарного деления, так
же как и в остальные сезоны.
Образовавшиеся в результате палинтомии гамон-
ты инцистируются, округляясь и выделяя вокруг себя
прозрачную оболочку. Формируются гамонтоцисты
округлой формы с диаметром 20—50 мкм и числом
ядер 2—8 (рис. 208). Вместе с фекалиями хозяина
гамонтоцисты выводятся наружу — в воду и в при-
брежной зоне водоема, где происходит икрометание
хозяев, сохраняют жизнеспособность в течение 3—
5 нед.
308
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Как только вылупившиеся из икринок головасти-
ки начинают питаться (основная их пища — разно-
образные мелкие организмы), в кишечник вместе с
пищей попадают гамонтоцисты опалин. В кишечнике
головастика происходит эксцистирование и из цист
выходят гамонты, которые локализуются в прямой
кишке.
Половое размножение
Гаметогенез. При делении гамонтов (гаметогенез)
образуются гаметы 2 типов: Ма и Ми (рис. 208).
Мейоз гаметический, двухступенчатый. Особенность
его у Opalina ranarum заключается в том, что мейоти-
ческое деление претерпевает только 1 ядро гамонта,
остальные ядра в это время находятся в интерфазном
состоянии. Число бивалентов в мейотическом ядре
равно 8—9, число же хромосом в 1 диплоидном ядре
трофозоита порядка 18 (Kaszanowski, 1971).
Сингамия. Ма морфологически сходны с гамон-
тами, но каждая из них содержит 1 ядро. В отличие
от Ма Ми узкие, тонкие, снабжены длинным ните-
видным хвостовым отростком; жгутики их немного-
численны, движения медленные. Встречаясь с Ма,
Ми прикрепляется к ее поверхности хвостовым от-
ростком, который внедряется в Ма, и обе гаметы пол-
ностью сливаются друг с другом (рис. 208); сливают-
ся и их ядра. Так протекает у опалин анизогамная
копуляция (сингамия), в результате которой форми-
руется зигота округлой формы (рис. 208).
Зигота. Вскоре после образования зигота инцис-
тируется, превращаясь в зигоцисту, которая вместе с
фекалиями выходит в воду, но затем может вновь
попасть в кишечник головастика вместе с пищей
(рис. 208). Вероятно, не все зиготы выходят в воду,
часть их остается в кишечнике головастика и дает
начало новой популяции опалин. Развитие же зиго-
ты, снова попавшей из воды в кишечник, зависит от
возраста головастика. У молодых головастиков, не
приступивших еще к метаморфозу, зигота в своем
развитии повторяет стадии гамонта, а также гамет,
которые копулируют и вновь образуют зиготу. У го-
ловастиков, претерпевающих метаморфоз, зигота (в
кишечник головастика обычно попадает не 1, а не-
сколько зигоцист) дает начало новой популяции опа-
лин.
Развитие трофозоита
Из зигот в кишечнике головастика развиваются
протрофозоиты. Ядро каждого протрофозоита делит-
ся несколько раз без деления цитоплазмы и развива-
ется удлиненная клетка с несколькими ядрами, обра-
зующими 1 ряд вдоль средней оси тела (рис. 208).
Такое расположение ядер в 1 ряд сохраняется до
8—9-ядерной стадии, а затем ширина клетки увели-
чивается, число ядер возрастает, и они распределя-
ются по всему объему цитоплазмы. У молодых тро-
фозоитов формируется фалькс (рис. 202), увеличива-
ется число кинет и клетка приобретает характерную
для вида форму.
Когда метаморфоз головастика заканчивается, то
перед выходом его из воды часть сформированных
уже трофозоитов вновь превращается в прогамонтов,
которые делятся палинтомически. В результате деле-
ния формируются гамонты с несколькими ядрами.
Они инцистируются, превращаясь в гамонтоцисты,
которые вместе с фекалиями выходят из кишечника
в воду и заражают новые особи хозяев (Wessenberg,
1961, 1978).
Экология
Обитание в столь специфической среде, как про-
свет прямой кишки хозяина, связано с возникнове-
нием целого ряда адаптаций. Популяции опалинид
локализуются в передней, расширенной части пря-
мой кишки хозяина, что препятствует свободному
выносу их из кишечника вместе с фекалиями. Попу-
ляция опалинид представляет собой компактное
скопление особей.
Основная масса особей популяции — это зрелые,
периодически делящиеся трофозоиты. Уплощенная
форма тела, отсутствие цитостома, представляют со-
бой признаки адаптации к среде, содержащей раст-
воренные и взвешенные в жидком кишечном содер-
жимом органические вещества. Поэтому питание
опалинид осуществляется путем пиноцитоза и мик-
рофагоцитоза.
Жизненный цикл опалинид, точно приуроченный
к годичному жизненному циклу амфибий, также ука-
зывает на высокую степень их адаптации к условиям
обитания в организме хозяина. Весной, в период раз-
множения амфибий, наступает половое размножение
опалинид, что связано с особенностями их распрост-
ранения и с заражением нового поколения хозяев.
Широкому распространению способствует массовое
образование гамонтоцист и зигоцист, которые могут
длительное время сохранять жизнеспособность в при-
донном иле прибрежных зон водоемов, где обитают
головастики.
Сопряженность периода полового размножения
опалинид с размножением амфибий зависит в значи-
тельной степени от гормональной активности хозяев:
половое размножение опалинид происходит только в
половозрелых животных. Экспериментальные иссле-
дования показали, что палинтомическое деление и
инцистирование Opalina ranarum можно индуциро-
вать в разные сезоны путем инъекции амфибиям го-
надотропина, эстерона, тестостерона, суспензии ги-
пофиза (Суханова, 1953; El Mofty, Smith, 1964; Wes-
senberg, 1961, 1978).
Одна из важных особенностей экологии опали-
нид — их четко выраженная специфичность по отно-
шению к определенной группе хозяев, ограниченной
для большинства видов только Anura. Однако один и
тот же вид О. ranarum может обитать в разных видах
хозяев. Например, О. ranarum может быть комменса-
лом кишечника травяной, земляной и озерной лягу-
шек и обыкновенной и зеленой жаб.
Приспособление опалинид к условиям обитания в
каждом из видов хозяев приводит к межпопуляцион-
ным различиям по морфологическим признакам. По-
Тип OPALINATA. КЛАСС OPALINATEA
309
этому морфологическая изменчивость свойственна боль-
шинству видов опалинид и составляет основу их по-
лиморфизма. Например, О. ranarum включает 5 мор-
фологически различных форм; к виду Cepedea dimi-
diata относится 4 подвида, и в каждом из них имеется
несколько форм, различающихся по морфологическим
признакам. Значительный полиморфизм свойствен и ви-
ду Protoopalina intestinalis. В его состав входят 3 под-
вида, и в каждом из них имеется несколько различа-
ющихся по морфологии форм.
Адаптация к условиям обитания в разных видах
хозяев затрагивает не только черты морфологии опа-
линид. Изменяются также физиологические особен-
ности, и в их числе терморезистентность, что обна-
ружено у Opalina ranarum и у Protoopalina caudata.
Терморезистентность обоих видов опалинид четко
коррелирует с терморезистентностью их хозяев и из-
меняется по сезонам, следуя изменениям температу-
ры среды обитания (Суханова, 1968).
Следовательно, опалиниды характеризуются ши-
рокой экологической пластичностью, которая пред-
ставляет одну из форм их приспособления к
комменсальному образу жизни в организме амфи-
бий.
Зоогеография
Географическое распространение опалинид лими-
тируется распространением их основных хозяев —
бесхвостых амфибий, которые населяют сушу и пре-
сноводные бассейны всех зоогеографических облас-
тей.
Довольно бедно представлена фауна амфибий в
голарктической области, и за Полярный круг захо-
дит только 3 вида: Rana temporaria, R. terrestris, R.
chensiensis (в Сибири). У этих амфибий обнаружен
1 вид опалинид — Opalina ranarum. По направле-
нию к югу увеличивается число видов амфибий и,
следовательно, увеличивается число видов опалинид,
а также происходит обогащение их видового состава
у каждого из видов хозяев (Metcalf, 1923; Банина,
1952).
Преобладающее большинство видов рода Proto-
opalina при всесветном распространении сосредото-
чено в Африке (Эфиопская область) и в Австралии
(Австралийская область), где основные их хозяе-
ва — амфибии семейств Pipidae, Discoglossidae, Ре-
lobatidae. Опалиниды родов Zelleriella и Hegneriella
наибольшим числам видов представлены в Южной
Америке и приурочены в основном к амфибиям се-
мейств Leptodactylidae, Ranidae, Hylidae, Bufonidae.
Виды рода Cepedea распространены всесветно и не
встречаются только в Австралии. Их основными хо-
зяевами являются амфибии семейств Ranidae, Bufo-
nidae, Hylidae. Всесветное распространение характер-
но для рода Opalina. Опалины встречаются во всех
зоогеографических областях, кроме Австралии. Са-
мое большое число их видов известно для Северной
Америки у амфибий семейств Ranidae, Bufonidae,
Hylidae. Число видов опалинид, распространенных в
каждой из зоогеографических областей, представле-
но в табл. 9.
Таблица 9
Географическое распространение опалинид
(Wessenberg, 1978)
Род опалинид Зоогеографические области
Нео- аркти- ческая Нео- тропи- ческая Пале- аркти- ческая Эфи- опская Индо- Малай- ская Авст- ралий- ская Всего
Protoopalina 5 7 10 34 17 12 85
Cepedea 5 14 5 12 42 0 78
Zelleriella 11 47 2 1 5 1 67
Opalina 62 13 18 9 38 0 140
Происхождение и эволюция
Опалиниды представляют один из примеров со-
пряженной эволюции паразита и хозяина. О проис-
хождении и дальнейших путях эволюции опалинид
можно судить по приуроченности к группам амфи-
бий, распространенных в разных по своему геологи-
ческому возрасту фаунистических комплексах раз-
личных зоогеографических областей. Это — один из
возможных подходов к выяснению путей эволюции
опалинид. Сравнивая опалинид по сложности орга-
низации трофозоитов, Меткалф справедливо считал
наиболее примитивными виды родов Protoopalina и
Zelleriella (рис. 209) (Metcalf, 1923).
Если обобщить современные знания по морфоло-
гии и жизненным циклам опалинид, применив срав-
нительный метод, можно сделать следующее заклю-
Рис. 209. Виды опалинат.
А — Zelleriella piscicola из кишечника сома, Б — Z. boipeva из кишечни-
ка змей Ophis merenii и Leimadopsis poecilogyrus, В — Z. jaegeri из ки-
шечника змеи Liophis jaegeri. (А —• по: Cunha, Penido, 1926; Б, В — по:
Carini, 1933).
Fig. 209. Species of Opalinatea.
A — Zelleriella piscicola from the intestine of sheat-fish, Б — Z. boipeva
from the intestine of snake Ophis merenii and Leimadopsis poecilogyrus, В —
Z. jaegeri from the intestine of snake Liophis jaegeri.
310
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 210. Виды опалинат.
1 — Protoopalina intestinalis, 2'— Opalina virguloidea, 3 — О. ranarum. a — микрогаметы, б — сингамия, в — зиготы, г — зигоцисты. (7, 2 — по: Wes-
senberg, 1961; 3 — ориг. К. М. Сухановой).
Fig. 210. Species of Opalinatea.
a — microgametes, 6 — singamy, в — zygotes, г — zygocysts.
чение. У всех изученных видов половой процесс про-
текает однотипно в форме анизогамной копуляции
(рис. 210). Ма и Ми (рис. 210) сходны по своей мор-
фологии у разных видов, снабжены небольшим чис-
лом жгутиков и представляют гаметы флагеллоидного
типа с выраженными анцестральными признаками.
Зигота опалинид (рис. 210), образующаяся путем
сингамии, также снабжена жгутиками и вскоре после
формирования инцистируется (рис. 210). Метагамный
период развития зиготы характеризуется не только
инцистированием, но и тем, что все ее содержимое
переходит в особь первого поколения. Это показа-
тель наиболее простого и короткого пути перехода
от полового поколения к агамному.
Молодые трофозоиты, развивающиеся после экс-
цистирования зигоцист в кишечнике головастика, у
всех видов имеют мелкие размеры и одноядерны
(рис. 208). Следовательно, есть основание для пред-
положения о том, что одноядерная стадия в онтогене-
зе многоядерных опалинид рекапитулирует анцест-
ральную форму, по-видимому, довольно примитивно-
го одноядерного жгутиконосца. Среди современных
опалинид одноядерны только виды рода Hegneriella
(рис. 201).
Дальнейшее развитие одноядерного трофозоита в
онтогенезе происходит по пути увеличения размеров
клетки и умножения числа ядер. Примечательно, что
двухъядерные протрофозоиты многоядерных видов
сходны с протоопалинами (рис. 211), а протрофозо-
иты с 5—10 ядрами — с Protoopalina axonucleata, но
меньше по размерам (рис. 212). Митотическое деле-
ние ядер у опалин и цепедей не сопровождается
цитокинезом, и их число увеличивается до опреде-
ленного предела, после чего многоядерная клетка
делится пополам в продольном (косом) направлении.
Поэтому можно предположить, что в филогенезе
опалинат происходил процесс полимеризации ядер и
жгутиков при прогрессивном увеличении размеров
Тип OPALINATA. КЛАСС OPALINATEA
311
Рис. 211. Виды опалинат.
А — Protoopalina intestinalis subsp. minor, Б — двухъядерные протрофозо-
иты Opalina ranarum. (А — по: Банина, 1952; Б — ориг. К. М. Сухано-
вой).
Fig. 211. Species of Opalinatea.
A — Protoopalina intestinalis subsp. minor, Б — binucleate protrophozoites
of Opalina ranarum.
клетки-организма без увеличения размеров самих
ядер и органелл цитоплазмы и что опалинаты могут
иметь общего предка с одноядерными гетеротроф-
ными жгутиконосцами, у которых в процессе эволю-
ции отмечается переход от свободноживущего состо-
яния к облигатному комменсализму.
Из 5 родов опалинид Protoopalina считается наи-
более древним. Хозяева Protoopalina — амфибии
низших семейств группы Salientia, происхождение
которых относится к триасу. Достаточно древним
считается и род Cepedea, возникновение которого
относится к меловому периоду мезозойской эры
(Metcalf, 1923, 1940; Банина, 1952).
Анцестральные формы опалинат, по-видимому,
подобно современным протоопалинам, имели тело с
округлым поперечным сечением. Многоядерные Opa-
lina с уплощенным телом появились в процессе эво-
люции значительно позже, о чем можно судить и по
геологическому возрасту их хозяев.
Изучение ультраструктуры клетки Opalina rana-
rum, как уже было упомянуто выше, показало сход-
ство с протеромонадными жгутиконосцами родов
Proteromonas и Karotomorpha, причем сходные орга-
неллы обеих групп гомологичны. Гомология многих
ультраструктурных признаков может служить и до-
казательством единства происхождения протеромо-
надных жгутиконосцев и опалинат. В самом деле, в
пределах типа Opalinata происходит полимеризация
кинетиды: род Proteromonas имеет 2 жгутика, род
Karotomorpha 4, класс Opalinatea — много жгутиков.
На фоне полимеризации жгутиков увеличивается и
высота складок тубулеммы, что определяется коли-
чеством проходящих в них микротрубочек (от 1 у
Proteromonas до 10 у Karotomorpha и 25—30 у Opa-
lina). Происходит изменение структуры корешковой
системы, но двойная переходная спираль сохраняет-
ся у представителей обоих классов. Поэтому в целом
тип Opalinata представляется весьма однородным, а
по признакам строения крист в митохондриях и на-
Рис. 212. Виды опалинат.
А — Protoopalina axonucleata, Б — протрофозоиты Opalina ranarum.
(Ориг. К. М. Сухановой).
Fig. 212. Species of Opalinatea.
Б — protrophozoites of Opalina ranarum.
личию переходной спирали в жгутиках он, очевидно,
связан с другими таксонами царства «хромистов».
Систематический обзор
Сем. Opalinidae Claus, 1874
Род Hegneriella Earl, 1971
К этому роду относится 2 одноядерных вида опа-
линид с уплощенным телом листовидной формы
(рис. 201).
Род Protoopalina Metcalf, 1918
Известно около 70 видов этого рода, для кото-
рых характерна удлиненная веретеновидная форма
тела. Большинство Protoopalina имеет 2 ядра, они
пузырьковидные, одинаковые по размерам и струк-
туре (рис. 201). Однако в составе рода есть виды,
например Р. axonucleata, у которых число ядер
может достигать 8—10. Жизненный цикл протоопа-
лин изучен недостаточно, особенно гаметогенез и
последующее развитие зиготы в кишечнике головас-
тика.
Род, Zelleriella Metcalf, 1923
Опалиниды с уплощенным листовидным телом в
отличие от видов рода Protoopalina. Трофозоиты с
2 ядрами, довольно однотипны по морфологическим
признакам (рис. 201). К роду относится около 60 ви-
312
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
дов. Среди них несколько видов обитает в прямой
кишке рыб (Z. piseicola Cunha et Penido, 1926) и змей
(Z boipevae Carini, 1933 и Z. jaegeri Carini, 1933)
(рис. 209). Эти виды почти не изучены.
Род Cepedea Metcalf, 1923
В состав рода входит около 70 многоядерных
видов. Тело вытянуто в длину, округлое в поперечном
сечении, спирально закручено. Кинеты, расположен-
ные вдоль тела, также спирально изогнуты (рис. 201).
Большинство видов рода полиморфны по ряду мор-
фологических признаков.
Род Opalina Purkinje et Valentin, 1835
(рис. 201)
Известно около 80 видов опалин, обитающих
в прямой кишке амфибий, в основном рода Rana. В
отличие от рода Cepedea опалины обладают листо-
видным телом, уплощенным в поперечном сечении.
Все виды опалин многоядерны, и число ядер у зре-
лых трофозоитов может достигать 200 и более. Фор-
ма тела, размеры, число ядер у каждого из видов
опалин значительно варьируют, что указывает на
внутривидовой морфологический полиморфизм. Ря-
ды кинет спирально изогнуты.
THE PHYLUM OPALINATA WENYON, 1926
SUMMARY
Opalinates are endobiotic heterotrophic protists inhabiting the in-
testine (rectum) of fish, amphibians, and reptiles. The body of tropho-
zoites is flat or spindle-shaped. The body size varies from 20 to
400 pm. 2—4 flagella of proteromonads are located apically, many
flagella of opalines are organised in rows on the body surface. Flagel-
lar pocket and cytostomal structure are absent. The main mode of
nutrition is pinocytosis. Body surface is covered with folded tubulem-
ma. Transition zone of flagella has double stranded helix, and trans-
versal plate. Mitochondria with tubular cristae. Golgi apparatus is pre-
sent. Reproduction occurs by means of asexual and sexual processes.
Nuclear apparatus consists of one or many homomorphic nuclei. Mi-
totic nuclear spindle is located within the nucleus (closed mitosis).
The phylum Opalinata includes 2 classes: Proteromonadea Grass€,
1952 and Opalinatea Wenyon, 1926.
THE CLASS PROTEROMONADEA GRASSE, 1952
Proteromonadida Grassd, 1952, the single order of the class Protero-
monadea, comprises the single family Proteromonadidae, Grasse, 1952.
The family includes 2 genera: Proteromonas Kiinstler, 1883 and Karoto-
morpha Travis, 1934. Approximately 10 species of the family Proteromo-
nadidae are known today. All the species are uninucleate parasitic flagella-
tes. The trophozoites have 2 (Proteromonas) or 4 (Karotomorpha) hetero-
dynamic flagella without paraxial rods. Trophozoites of both genera have
spindle-shaped body narrowing posteriorly. The posterior part of the body
surface of Proteromonas is covered with somatonemes. The presence of
large single mitochondrion is characteristic of both genera.
The spherical or oval resistant cysts form both genera — Prote-
romonas and Karotomorpha. The trophozoites multiply by binary lon-
gitudinal fission. The sexual process is studied in Proteromonas lacer-
tae viridis. This process proceeds in a form of hologamic copulation
of two mature trophozoites, which are identical in their morphology.
The trophozoites join in pairs, and encyst. Both of the nuclei and
cytoplasm join inside the cyst. As soon as the zygocyste is formed, it
leaves the intestine of the host. Inside zygocyst 92 flagellate spores are
developed. Zygocyst is the only exogenic stage of P. lacertae viridis.
THE CLASS OPALINATEA WENYON, 1926
All species of Opalinatea are endobionts of the rectum of ecto-
thermic vertebrates: amphibians, fishes, reptiles. According to mor-
phological, physiological indications and unique life cycles, the class
Opalinatea is a sufficiently homogenous group of protists. They have
a flat, leaf-like or spindle-shaped body, which is covered with nume-
rous flagella (cilia). The folded tubulemma is well developed and con-
tains up to 30 microtubules inside each ridge. The mitochondria with
tubular cristae are numerous. The number of nuclei varies notably in
different species from 1 to 100 and more. Agamic and sexual repro-
duction present.
The life cycles of Opalina, Cepedea, Zelleriella, Protoopalina are
adapted to the breeding season of the amphibian (Anura) hosts. The
opalinids multiply during the year by binary longitudinal division of
the trophozoits cell. In spring, during the breeding season of amphibians,
many trophozoits (tomonts) undergo a series of divisions without continued
growth. The result of this palintomy is formation of small individual
cells — gamonts, which transform into gamontocysts. The gamontocysts
are released from the intestine of a host into the water. Newly hatched
tadpoles feed on algae, detritus and ingest opaline cysts with the food.
Excystment of the gamontocysts occurs in the gut of tadpoles and
gamonts go out of the cysts. The gamonts concentrate in the hindgut
of tadpoles. Gamont divides and two types of gametes are formed: Mi-
and Afa-gametes. The meiosis is prezygotic. Ma- and Л/Z-gametes fuse
completely. The zygote develops and incysts. Namely the zygote gives
a start for the new population of opalinids.
The class Opalinatea includes one order Opalinida Poche, 1913
with a family Opalinidae Claus, 1874. There are 5 genera in the family
uniting about 400 species: Hegneriella Earl, 1971; Protoopalina Met-
calf, 1918; Zelleriella Metcalf, 1923; Cepedea Purkinje et Valentin,
1835; Opalina Purkinje et Valentin, 1835.
Geographical distribution of opalines is limited by the distribution
of their hosts — Anura, inhabiting land, and fresh water basins, of all
zoogeographical zones. However, their fauna is more poor in the
North. The predominant number of species of Protoopalina are con-
centrated in Africa and Australia. The greatest number of species of
Zelleriella and Hegneriella inhabit in South America. The species of
Opalina inhabit all geographical zones of the world, with prevalence
in North America.
Opalinatea can be regarded as an example of co-evolution of an
endobiont and a host. Ontogenesis of Opalinatea includes the uninuc-
lear stage, which is characteristic of each species of the phylum. The
phenomenon permits suggesting that the mononuclear specimen reca-
pitulates an ancestral stage.
ЛИТЕРАТУРА18
Банина H. H. Протоцилиата амфибий СССР // Уч. зап. ЛГУ.
1952. Т. 28, № 141. С. 139—150.
18 Цитируемая и рекомендуемая литература к разделу «Тип
Opalinata».
Карпов С. А. Строение покровов жгутиконосцев // Цитология.
1986. Т. 28, №2. С. 139—150.
Карпов С. А. Система протистов. Омск, 1990.
Райков И. Б. Ядро простейших. Морфология и эволюция. Л.,
1978.
ТИП OPALINATA. ЛИТЕРАТУРА
313
Сергеева Г. И. Электронно-микроскопическое исследование
лизосом у Opalina ranarum Ehrbg. (Mastigophora) // Цитология.
1969. T. 11, №6. С. 714—720.
Суханова К. М. Влияние факторов внешней среды на жизнен-
ный цикл Opalina ranarum // Уч. зап. ЛГПИ. 1953. Т. 91. С. 31—69.
Суханова К. М. Морфологическое и цитохимическое исследо-
вание Protoopalina intestinalis // Цитология. 1961. Т. 3, № 5. С.
578—584.
Суханова К. М. Температурные адаптации у простейших. Л.,
1968.
Хайбулаев К. X., Шульман С. С. Тип Жгутиконосцы // Опре-
делитель паразитов пресноводных рыб. Л., 1984. Т. 1. С. 13—42.
Хаусман К. Протозоология. М., 1988.
Amaro A. «Cepedea marginata» sp. n., enterozoario de «Hyla
similis» Cochran (Sarcomastigophora, Opalinata) // Rev. Brasil. Biol.
1964. Vol. 24, N. 1. P. 5—10.
Amaro A. Sobre um novo Opalinideo, «Zelleriella freitasi» sp. n.
enterozoario de «Hyla fuscomarginata» Lutz do Brasil (Sarcomasti-
gophora, Opalinata) II Atas Soc. Biol. Rio de Janeiro. 1965. Vol. 9,
N 1. P. 1—3.
Amaro A. Sobre a sistematica da familia «Opalinidae» Claus, 1874
(Sarcomastigophora, Opalinata). Ill nota: Genro «Cepedea» Metcalf,
1920 // Atas Soc. Biol. Rio de Janeiro. 1966a. Vol. 10, N 2. P. 29—32.
Amaro A. Sobre a sistematica da familia «Opalinidae» Claus,
1874 (Sarcomastigophora, Opalinata). IV nota: Genero Opalina Pur-
kinje Valentin, 1835 // Atas Soc. Biol. Rio de Janeiro. 1966b. Vol. 10,
N 2. P. 33—34.
Belar K. Protozoenstudien // Arch. Protistenk. 1921. Bd 43, N 3.
S. 431—462.
Blanckart S. Die Oberflachenstrukturen von Paramecium sp. und
Opalina ranarum // Ztschr. wiss. Mikrosc. 1957. Bd 63, H. 2. S. 276—
287.
Brugerolle C., Joyon L. Etude cytologique ultrastructurale des
genres Proteromonas et Karotomorpha (Zoomastigophorea, Protero-
monmadida Grassd, 1952) // Protistologica. 1975. T. 11, N 4. P. 561—
546.
Carini A. Sobre uma nova Zelleriella (Z. jaegeri) de intestino de
uma cobra // Arch. Biol. 1933a. T. 16, N 174. S. 50—59.
Carini A. Zelleriella boipevae n. sp. de intestino de uma cobra //
Arch. Biol. 1933b. T. 16, N 174. S.60—65.
Chen T. T. Observations on mitosis in opalinids. 1. The behavior
and individuality of chromosomes and their significance // Proc. Nat.
Acad. Sci. USA. 1936a. Vol. 22, N 5. P. 594—601.
Chen T. T. Observations on mitosis in opalinids. 11. The associ-
ation of chromosomes and nucleoli II Proc. Nat. Acad. Sci. USA.
1936b. Vol. 22, N 5. P. 602—607.
Corliss J. O. The opalinid infusorians: Flagellates or Ciliates? // J.
Protozool. 1955. Vol. 2, N L.P. 107—114.
Corliss J. O. The kingdom protista and its 49 phyla // BioSystems.
1984. Vol. 17, N 1. P. 87—126.
Cunha A. M., Penido J. C. N. Nouveau protozoaire parasite des
poissons: Zeleriella piscicola n. sp. // Compt. Rend. Soc. Biol. 1926.
T.95. S. 1003—1005; 1010—1011.
Dobell C. Antony van Leuvenhoek and his «Little Animals». New
York, 1958.
Doflein F. Lehrbuch der Protozoenkunde. Jena, 1953.
Earl P. R. Hegneriella dobelli gen. n. (Opalinidae) from Bufo
valliceps and some remarks on the systematic position of the Opalini-
dae // Acta Protozool. 1971. Vol. 9, N 1. P. 41—48.
Earl P. R. Notes on the taxonomy of the Opalinids (Protozoa),
including remarks on continental drift II Trans. Amer. Mier. Soc.
1979. Vol. 98, N 4. P. 549—557.
El Mofti M. M., Smith J. D. Endocrine control of Encyststion in
Opalina ranarum parasitic in Rana temporaria // Exp. Parasitol. 1964.
Vol. 15, N 3. P. 185—199.
Gatenby J. B., King S. D. Opalina ranarum — a flagellate // Na-
ture. 1925. Vol. 116. P. 712.
Grasse P.-P. Contribution а Г etude des flagelles parasites // Arch.
Zool. Exp. Gen. 1926. Vol. 65, N 3. P. 345—602.
Grasse P.-P. Sur la cytologie du Flagelle parasite Proteromonas
lacertae-viridis Grassi // C. R. Assoc. Anat. Berdeaux. 1929. T. 18,
N 2. P. 267—275.
Honigberg В. M. Evolutionary and sistematic relationships in the
flagellate order Trichomonadida Kirby // J. Protozool. 1963. Vol. 10,
N 1. P. 20—63.
Kaszanowski A. Mitosis and Polyploidy in nuclei of Opalina ra-
narum II Experientia. 1968. Vol. 24, N 7. P. 846—847.
Kaszanowski A. Opalina ranarum Purkinje, Valentin: Meiosis and
dimorphysm of nuclear behavior meiosis II Acta Protozool. 1971.
Vol. 9, N 1. P. 105—106.
Khan M. C. Cepedea hylae sp. n. — as new Opalinid ciliate from
India // Trans. Amer. Microsc. Soc. 1962. Vol. 81, N 2. P. 205—207.
Konsuloff S. Untersuchungen liber Opalina // Arch. Protistenk.
1922. Bd 44, N 2. S. 285—345.
Kulda J. Axenic cultivation of Proteromonas lacertae viridis
(Grassi, 1879) // J. Protozool. 1973. Vol. 20, N 4. P. 536—537.
Kulda J., Nohynkova E. Proteromonadidae // Parasitic Protozoa /
Ed. J. P. Kreier. 1978. Vol. 2. P. 118—127.
Lynn D. N., Small E. B. Subphyllum Opalinata // Illustr. Guide to
the Protozoa. USA, 1985. P. 156—157.
Metcalf M. M. Opalina. Its anatomy and reproduction a descripti-
on of infection experiments and a chronological review of the litera-
ture // Arch. Protistenk. 1909. Bd 13, N 2. S. 195—375.
Metcalf M. M. The Opalinid ciliate infusorians // Bull. USA Nat.
Mus. 1923. Vol. 120, N 1. P. 1—484.
Metcalf M. M. Further studies on the opalinid ciliate infusorians
and their hosts // Proc. USA Nat. Mus. 1940. Vol. 87, N 3077.
P. 465—634.
Mignot J.-P., Molina A. Etude ultrastructurale de Protoopalina
saturnalis (Leger et Duboscq, 1904) Metcalf 1908, protiste parasite du
poisson marin Box boops L. 11 Arch. Protistenk. 1988. Bd 135.
S. 255—270.
Moskowitz N. Observations on some intestinal flagellates from
reptilian host (Squamata) // J. Morphol. 1951. Vol. 89, N 2. P. 257—
321.
Munch R. Endocytose Nachweis bei Opalina ranarum // Cytobio-
logie. 1970. Bd 2, H. 1. S. 108—122.
Neresheimer E. Die Fortpflanzung der Opalinen II Arch. Protis-
tenk. 1907. Bd 1. S. 1—42.
Nie D. Morphology and taxonomy of the intestinal Protozoa of the
guinea-pig, Cavia porcella // J. Morphol. 1950. Vol. 86, N 3. P. 381—
494.
Noirot-Timothee C. Rechearches sur Г ultrastructure d* Opalina
ranarum // An. Sci. nat. Zool., Biol. An. 1959. T. 1, ser. 12. P. 265—
281.
Noirot-Timothee C. Presence simultanee de deux types de vesi-
cules de micropinocytose chez Cepedea dimidiata (Protozoa Opalina-
ta) // C. R. Acad. Sci. Paris. 1966. T. 263, ser. D. P. 1230—1233.
Patterson D. J. The fine structure of Opalina ranarum (family
Opalinidae): Opalinid phylogeny and classification // Protistologica.
1985. T.21, N4. P. 413—428.
Pitelka D. R. An electron microscope study of cortical structure
of Opalina obtrigonoidea // J. Biophys.-Biochem. Cytol. 1956. Vol. 2,
N 3. P. 423—432.
Puytorac P., Grain J., Mignot J. P. Precis de protistologie. Paris,
1987.
Uttangi J. C. Some more binucleated Opalinid Ciliates (Protozoa)
from Indian frogs // J. Univ. Bombay. 1960. Vol. 29, N 3—5. P. 110—
133.
Wessenberg H. Studies on the cycle and morphogenesis of Opa-
lina // Univ. Calif. Berkley Publ. Zool. 1961. Vol. 61. P. 315—370.
Wessenberg H. Observations on cortical ultrastructure in Opali-
na // J. Microsc. 1966. Vol. 4, N 5. P. 471—492.
Wessenberg H. Opalinata // Parasitic Protozoa I Ed. J. P. Kreier.
1978. Vol. 2. P. 551—581.
314
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
«CHROMISTA» INCERTAE SEDIS
Отряд Spongomonadida (Hibberd)
Karpov, 1990 — Спонгомонады
Бесцветные, обычно колониальные жгутиконосцы; клетки с
1 или 2 апикально расположенными жгутиками, которые пол-
ностью или частично окружены цитоплазматическим выростом
апикальной части клетки. Колонии прикрепленные, матрикс коло-
нии образован слизью и железосодержащими гранулами эндоген-
ного происхождения. Микротрубочки жгутиковых корешков вее-
ром расходятся от развитых фибриллярных ЦОМТов, переходная
зона жгутика длинная, равна высоте цитоплазматического выроста.
Митохондрии с пузырьковидными кристами. Свободноживущие. В
отряде имеется 3 рода.
История изучения
Роды Rhipidodendron и Spongomonas описаны Штей-
ном (Stein, 1878) как колониальные бесцветные жгу-
тиконосцы с 2 равными жгутиками. Их таксономическое
положение и филогенетические связи всегда были не-
ясными: то они считались бесцветными хризомонада-
ми, то сближались с бодонидами и хоанофлагеллата-
ми. До последнего времени практически не изучались.
Первое серьезное исследование было предпринято
Хиббердом (Hibberd, 1976b). Изучив ультраструктуру
этих родов на примере S.uvella и Rhipidodendron splendi-
dum, автор показал, что они представляют хорошо очер-
ченную и своеобразную группу, которую невозможно
отнести ни к кинетопластидам, ни к хоанофлагеллатам,
ни к полимастигинам. Позднее (Hibberd, 1983) для них
был установлен особый отряд Spongomonadida Hib-
berd, 1983.
Еще 1 род этого отряда — Phalansterium был опи-
сан более ста лет назад (Cienkowsky, 1870). Сначала
Штейн (Stein, 1878) объединял его с другими колони-
альными бесцветными жгутиконосцами, затем Кент
(Kent, 1880—1882) отнес их к воротничковым жгути-
коносцам, так как на' переднем конце тела клетки
есть небольшой воротничок, как у хоанофлагеллат,
расположенный вокруг единственного жгутика. Это
толкование выглядело вполне обоснованным, потому
что такой воротничок можно было считать рудимен-
том нормального воротничка хоанофлагеллат, редук-
ция которого произошла из-за полного погружения
его в слизь колонии и смены функции.
Электронно-микроскопическое исследование
Ph. digitatum (Hibberd, 1983) показало, что клетка
этого вида устроена совсем иначе, чем у хоанофла-
геллат. Оказалось, что она обладает целым рядом
необычных черт, в частности пузырьковидными
кристами в митохондриях, своеобразным жгутико-
вым аппаратом, цитоплазматической воронкой вмес-
то воротничка. Это позволило Хибберду (Hibberd,
1983) выделить данный вид в самостоятельный отряд
Phalansteriida Hibberd, 1983.
Классификация
В результате детального сравнительно-морфоло-
гического анализа всех 3 родов (Карпов, 1990) было
показано, что Ph. digitatum принципиально не отли-
чается от спонгомонад и может быть отнесен к от-
ряду Spongomonadida в качестве семейства с единст-
венным родом Phalansterium. Таким образом, отряд
содержит 2 семейства: Spongomonadidae и Phalanste-
riidae.
Морфология
Особи колонии Rhipidodendron splendidum распо-
ложены на концах дихотомически ветвящихся сли-
зистых трубок (рис. 213). Максимальные размеры ко-
лонии достигают в высоту 440 мкм с кроной около
310 мкм в диаметре. Размеры клетки 20 х 4.5 мкм.
Колонии Spongomonas uvella обычно дисковидной
или шаровидной формы (рис. 214). Они также могут
достигать значительных размеров — до 370 мкм в
диаметре. Особи почти шаровидные, диаметром
около 10 мкм. Наиболее часто встречаются 4—16-
клеточные колонии.
У Phalansterium digitatum колонии древовидной
формы, достигают иногда 100 мкм в высоту (рис. 215).
Клетки размером 20 х 8 мкм, расположены на кон-
цах тонких, дихотомически ветвящихся стебельков.
Жгутик 1, в базальной части окружен небольшим ци-
топлазматическим воротничком. Особи с воротнич-
ком погружены в слизь, которая вместе с содержащи-
мися в ней коричневыми гранулами образует матрикс
колонии. Такой матрикс колонии характерен для
всех спонгомонад. Гранулы размером от 1 до 1.5 мкм
образованы солями железа. Они вырабатываются
внутри клетки, возможно при участии симбиотичес-
ких бактерий, и выделяются наружу, заполняя вместе
со слизью пространство между клетками.
Строение клеток родов Rhipidodendron и Spongo-
monas, несмотря на различия в их форме и разме-
рах, одинаково (рис. 216). 2 апикальных жгутика вы-
ходят из жгутикового кармана, один край которого
образует вокруг жгутиков вырост в виде совка около
5 мкм высотой. Оба жгутика гладкие, примерно оди-
наковой длины, без параксиальных структур, содер-
жат аксонемы с обычным набором микротрубочек
(9 + 2). Переходная зона длинная (около 1 мкм).
Кинетосомы расположены параллельно друг другу и
связаны между собой 2 фибриллярными мостиками
(рис. 216). От кинетосом отходят корешки в виде
одиночных микротрубочек и фибриллярного тяжа,
идущего к ядру. Микротрубочки расходятся веером,
не образуя при этом симметричной картины, что
связано, очевидно, с асимметрией всего переднего
конца клетки (рис. 216).
Радиально-симметричные клетки Phalansterium
имеют 1 жгутик, окруженный цитоплазматической
воронкой (рис. 217). Ее высота также не превышает
4—5 мкм, а в целом эту воронку легко представить
как сомкнувшийся краями совковообразный вырост
Spongomonas в результате редукции одного из жгути-
ков. Оставшийся единственный жгутик тоже гладкий,
имеет длинную переходную зону и обычный набор
микротрубочек в аксонеме. Второй жгутик редуциро-
«CHROMISTA» INCERTAE SEDIS. ОТРЯД SPONGOMONADIDA
315
Рис. 213. Общий вид клеток (А) и распределение зооидов в коло-
нии (5) Rhipidodendron. (По: Patterson, Headley, 1992).
з — зооиды, ж — жгутики.
Fig. 213. General view of the cell (A) and zooid arrangement in the
colony (Б) of Rhipidodendron.
з — zooids, ж — flagella.
Б
Рис. 214. Общий вид клеток (А) и распределение зооидов в коло-
нии (Б) Spongomonas. (По: Patterson, Headley, 1992).
з — зооиды, ж — жгутики.
Fig. 214. General view of the cell (A) and zooid arrangement in the
colony (F) of Spongomonas.
з — zooids, ж — flagella.
Рис. 216. Схема строения переднего конца клетки спонгомонад.
(По: Hibberd, 1976).
аг — аппарат Гольджи, аф — аморфный фибриллярный материал, ж —
жгутики, жк — жгутиковый карман, ик — исчерченный фибриллярный ко-
решок, иф — исчерченный фибриллярный материал, м — митохондрия с
пузырьковидными кристами, мт — микротрубочки, мтк — микротрубоч-
ковые корешки, ст — слизеносные тельца, у — углубление между жгути-
ком и цитоплазматическим выростом, фк — фибриллярный корешок,
фм — фибриллярный мостик между кинетосомами, цв — асимметричный
цитоплазматический вырост, я — ядро.
Fig. 216. Scheme of spongomonade cell structure.
аг — Golgi apparatus, аф — amorphous fibrillar material, ж — flagella, жк —
flagellar pocket, ик — striated fibrillar rootlet, иф — striated fibrillar material,
м — mitochondrion, мт — microtubules, мтк — microtubular rootlets, cm —
mucilage vesicles, у — pit, фк — fibrillar rootlet, фм — fibrillar bridge, цв —
anterior prolongation, я — nucleus.
вался, по-видимому, полностью, так как отсутствует
даже обычная в таких случаях безжгутиковая кинето-
сома. Корешковая система также представлена одним
фибриллярным корешком и радиально расходящими-
ся одиночными микротрубочками.
Строение других органелл у представителей всех
3 родов очень сходно. Покровы представлены только
плазмалеммой. Ядро пузырьковидного типа, с хоро-
шо выраженным ядрышком, лежит примерно в сере-
дине клетки. Митохондрии с пузырьковидными крис-
тами. Единственная диктиосома аппарата Гольджи
расположена между ядром и базальным комплексом
Рис. 215. Общий вид клеток (А) и распределение зооидов в коло-
нии (К) Phalansteriwn. (По: Patterson, Headley, 1992).
в — воронка, г — железосодержащие гранулы в матриксе колонии, ж —
жгутик, з — зооиды.
Fig. 215. General view of the cell (A) and zooid arrangement in the
colony (Б) of Phalansteriwn.
в — collar, г — iron containing spherules in the colony matrix, ж — flagel-
lum, з — zooids.
316
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 217. Схема строения апикального конца клетки Phalansteri-
um digitatum. (По: Hibberd, 1983).
аг — аппарат Гольджи, в — цитоплазматическая воронка вокруг жгутика,
м — митохондрия, мт — микротрубочки корешковой системы, расходя-
щиеся от электронно-плотных зон (х, у, z), фк — фибриллярный корешок.
Fig. 217. Scheme of apical end cell structure of Phalansterium digi-
tatum.
аг — Golgi apparatus, в — collar, м — mitochondrion, мт — microtubular
rootlets associated with circular'bands (x, y, z) concentric with the flagellar
base, фк — fibrillar rootlet.
жгутиков (рис. 216; 217). В крупных вакуолях базальной
части клетки содержатся электронно-плотные желе-
зосодержащие сферулы. Иногда рядом с ними, внут-
ри вакуолей, обнаруживаются бактерии. Под по-
кровами отмечаются также небольшие тельца, похо-
жие на мукоцисты. По-видимому, они продуцируют
слизь, объединяющую все компоненты колонии. В
базальной части клетки находится 1—2 сократитель-
ные вакуоли. Пищеварительных вакуолей не обнару-
жено.
Способы питания и размножения этих жгутико-
носцев не изучены.
Из других стадий жизненного цикла известны
только цисты.
Филогения
Филогенетические отношения между семейства-
ми спонгомонад можно объяснить, исходя из редук-
ции одного из жгутиков. Примеры полной редукции
жгутика (вместе с кинетосомой), хотя и не часто, но
встречаются среди протистов. Поэтому можно пред-
положить, что при утрате жгутика асимметричный
вырост замкнулся в цитоплазматическую воронку во-
круг единственного жгутика, а корешковая система
приобрела радиальную симметрию, как и вся клетка.
Среди прикрепленных клеток протистов двухжгути-
ковые формы всегда асимметричны. В качестве при-
меров можно привести представителей спонгомонад,
псевдодендромонад, бикозоецид, хризомонад. Од-
ножгутиковые же прикрепленные протисты всегда
радиально-симметричны: хоанофлагеллаты, пединел-
ловые водоросли, гелиофлагеллаты. По-видимому, это
общая закономерность для прикрепленных форм
протистов, которая может объяснить связь Phalans-
terium с другими спонгомонадами.
Спонгомонады, как и другие отряды бесцветных
жгутиконосцев, обладают целым комплексом своеоб-
разных черт, поэтому их трудно связать с какими-
либо зоофлагеллатами. Кинетопластиды и хоанофла-
геллаты являются четко очерченными таксонами, да
к тому же имеют пластинчатые кристы в митохонд-
риях. Отряд Pseudodendromanadida также очень свое-
образен. Хотя его представители имеют пузырьковид-
ные кристы в митохондриях, по строению цитоскеле-
та и общей симметрии клетки они резко отличаются
от спонгомонад.
Все отряды зоофлагеллат с трубчатыми или пу-
зырьковидными кристами в митохондриях принято
выводить из хризомонад. Среди последних есть и
бесцветные виды. Кроме того, эта группа водорослей
довольно многочисленна и сравнительно гетерогенна
по ультраструктурным признакам. Поэтому среди
них всегда можно обнаружить формы, сходные по
некоторым признакам с зоофлагеллатами. Однако и
в этом отношении спонгомонады являются исключе-
нием. Отсутствие фаготрофии наряду с продуцирова-
нием железосодержащих гранул свидетельствует о
примитивном способе обмена веществ. Возможно,
они являются очень древними эукариотами, хотя для
более обоснованного обсуждения этого предположе-
ния необходимы сведения о питании и размножении
спонгомонад.
Систематический обзор
Сем. Phalansteriidae Kent, 1880
Клетки радиально-симметричные, с 1 жгутиком.
На апикальном конце клетки имеется цитоплазмати-
ческая воронка, или воротничок. Единственный род
Phalansterium представлен 2 видами: Ph. digitatum и
Ph. consociatum.
Сем. Spongomonadidae Karpov, 1990
Клетки асимметричные, с 2 жгутиками. На апи-
кальном конце клетки совковообразный цитоплазма-
тический вырост.
Роды: Rhipidodendron и Spongomonas.
CHROMISTA» INCERTAE SEDIS. ОТРЯД PSEUDODENDROMONADIDA
317
Отряд Pseudodendromonadida
Hibberd, 1985 — Псевдодендромонады
Свободноживущие бесцветные жгутиковые с 2 почти равными
жгутиками. Свободноплавающие или прикрепленные формы на
стебельках. Прикрепленные организмы часто образуют дихотоми-
чески ветвящиеся колонии. Тело клетки характерной треугольной
формы: апикальный конец плоский, с одной его стороны на не-
большом возвышении находятся жгутики, на противоположной
стороне лежит хорошо развитый цитостом, переходящий в глотку.
Базальный конец тела закруглен. Клетка покрыта неминеральными
чешуйками, которые синтезируются в аппарате Гольджи. Жгутики
покрыты плотно прилегающими к их поверхности мелкими че-
шуйками. Один из них имеет хорошо выраженную акронему. Под
апикальной поверхностью клетки проходит широкий корешок из
микротрубочек, переходящий в скелет глотки. Митохондрии с пу-
зырьковидными кристами.
История изучения
Представители этого отряда стали известны недав-
но: Петерсеном и Хансеном был описан Cyathobodo
stipitatus (Petersen, Hansen, 1961). Затем Свэйл и Бел-
чер (Swale, Belcher, 1975) изучили ультраструктуру
еще 3 видов этого рода, считая их близкородственны-
ми бодонидам. Исследования были продолжены Хиб-
бердом (Hibberd, 1976а, 1985), изучившим ультраст-
руктуру Cyathobodo.
Род Pseudodendromonas сначала был ошибочно
описан как Dendromonas (Vlk, 1938), затем Бурели
(Bourrelly, 1953) исправил ошибку и описал его как
новый род и вид Pseudodendromonas vilkii. Миньо
(Mignot, 1974) исследовал его ультратонкое строение
и отнес к хризофитовым водорослям. Позднее изуче-
ние этого вида и 2 видов рода Cyathobodo продолжил
Хибберд (Hibberd, 1976а, 1985). Он обобщил данные
по обоим родам и пришел к выводу, что они образу-
ют особую группу жгутиконосцев ранга отряда.
Классификация
В настоящее время в отряд Pseudodendromonadida
входит 1 семейство Pseudodendromonadidae, включа-
ющее 2 рода: Cyathobodo с 9 видами и Pseudodendro-
monas с 3 видами.
Морфология
Строение клеток всех представителей отряда
идентично. Форма прикрепленных особей весьма
своеобразна: они сплющены латерально и при взгля-
де с боку похожи на треугольник, обращенный вер-
шиной вниз (рис. 218; 219). Апикальная поверхность
клетки плоская. На одном ее крае расположена не-
большая папилла, из которой выходят жгутики. На
противоположном крае находится цитостом, перехо-
дящий в глотку.
Снаружи тело клеток покрыто чешуйками. Разли-
чия в их форме являются важным признаком при
описании видов. Например, для Cyathobodo salpinx
характерно наличие 1 слоя чешуек, имеющих плос-
кое основание, из центра которого торчит трубчатая
Рис. 218. Внешний вид колонии Pseudodendromonas vlkii (А) и
отдельного зооида (Б). (По: Patterson, Headley, 1992).
ж — жгутики, ст — стебелек, я — ядро.
Fig. 218. General view of the colony (A) and separate zooid (Б) of
Pseudodendromonas vlkii.
ж — flagella, cm — stalk, я — nucleus.
стойка. Для Pseudodendromonas vilkii характерны
плоские овальные чешуйки с перекладинами, в не-
сколько слоев покрывающие клетку.
Оба жгутика примерно равной длины, один из них
имеет хорошо выраженную акронему. На поверхнос-
ти обоих жгутиков Cyathobodo обнаружены мелкие
(примерно 50 нм в диаметре) блюдцевидные чешуй-
ки, очень плотно прилегающие к плазмалемме. Жгу-
тики Pseudodendromonas не имеют чешуек.
Кинетосомы расположены почти параллельно
друг другу. От них отходит короткий поперечно ис-
черченный фибриллярный корешок и 2 корешка из
микротрубочек. Наиболее широкий микротрубочко-
вый корешок проходит под плазмалеммой апикаль-
ного конца клетки, прилегая своей внутренней сторо-
ной к митохондрии (рис. 219). Дистальный конец
этого корешка закручивается вокруг цитостома и на-
правляется в глубь клетки, укрепляя глоточный канал.
Другой микротрубочковый корешок проходит вдоль
боковой поверхности клетки.
Ядро пузырьковидного типа, немного вытянуто в
сторону кинетосом. С одной стороны к нему при-
легает диктиосома аппарата Гольджи, в которой син-
тезируются соматические чешуйки. Митохондрии с
пузырьковидными кристами. По периферии клетки
расположены запасные вещества в виде гранул глико-
гена.
Филогения
Миньо (Mignot, 1974) относит их к одному из по-
рядков гаптофитовых водорослей Isochrysidales,
представители которых не имеют гаптонемы, на жгу-
тиках нет трубчатых мастигонем, а тело клетки пок-
рыто чешуйками. Однако Хибберд (Hibberd, 1976,
1985) отметил их принципиальные отличия от гапто-
318
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 219. Схема ультратонкого строения клетки Pseudodendromonas vlkii. (По: Mignot, 1974).
аг — аппарат Гольджи; бп — базальная пластинка; гл — гранулы гликогена; ж — жгутики; м — митохондрия с пузырьковидными кристами; мтк — мик-
ротрубочковые корешки; ог — осмиофильные глобулы; пв — пищеварительная вакуоль; ст — стебелек; фк — фибриллярный корешок; ц — цитостом, ве-
дущий в глотку; я — ядро, в — вентральная, д — дорсальная сторона клетки.
Fig. 219. Scheme of cell structure of Pseudodendromonas vlkii. .
аг — Golgi apparatus, 6n — basal plate, гл — glycogen, ж — flagella, м — mitochondrium, мтк — microtubular rootlets, ог — osmiophilic globules, ne — food va-
cuole, cm — stalk, фк — fibrillar rootlet, ц — cytostome, я — nucleus, в — ventral, d — dorsal side of the cell. Flagellar and body scales are not shown.
монад в расположении клеточных органелл, строе-
нии аппарата Гольджи, форме чешуек и способе их
формирования.
Для понимания филогенетических и таксономи-
ческих связей псевдодендромонад необходимо срав-
нить их строение с морфологией некоторых бикозое-
цид, особенно представителей родов Caecitellus и Ad-
riamonas (см. разд. «Отряд Bicosoecales»). Для них
характерны наличие постоянного цитостома с раз-
витым цитофаринксом, 2 гладких жгутика, пузырько-
видные кристы в митохондриях, а также сходство
общей симметрии клетки, которая определяется на-
личием цитостомального корешка. До последнего
времени наличие цитофаринкса у псевдодендромонад
служило препятствием для объединения отрядов Bi-
cosoecales и Pseudodenromonadida в один таксон. Од-
нако описание типичной бикозоециды с развитыми
цитостомом и глоткой (Siluania) позволяет по-новому
взглянуть на эту проблему (Карпов и др., 1998). S. то-
nomastiga отличается от псевдодендромонад факти-
чески только наличием мастигонем на жгутике и от-
сутствием чешуек. Строение корешковой системы, ас-
социация широкого корешка с митохондрией, общая
симметрия клетки у представителей Siluania, Adriamo-
nas, Pseudodenromonas и Cyathobodo настолько похо-
жи, что предлагается объединить все эти роды в один
отряд Bicosoecida Karpov, 2000 (см.: Карпов, 2000).
Филогенетически оценить ситуацию можно следу-
щим образом. Если оставлять отряд бикозоецид
среди хризофитовых по признаку «наличие масти-
гонем», то наиболее примитивными следует считать
представителей родов Cafeteria и Pseudobodo, у кото-
«CHROMISTA» INCERTAE SEDIS. SUMMARY
319
рых отсутствует домик и еще нет цитостома. На сле-
дующем этапе эволюции был, по-видимому, приобре-
тен домик (Bicosoeca'). Другая линия эволюции связа-
на с приобретением постоянного ротового отверстия
и глотки (Siluania). Утрата мастигонем происходила,
вероятно, в разных таксонах независимо. Если же
считать первичными чешуйчатые формы псевдоденд-
ромонад, то придется предположить возникновение в
этой группе трубчатых трехчленных мастигонем и ут-
рату постоянного цитостома. Последнее для голозой-
но питающихся форм весьма сомнительно. Независи-
мое от хризомонад приобретение трубчатых мастиго-
нем также маловероятно.
Таким образом, вероятнее всего происхождение
псевдодендромонад связано с бикозоецидами или
общим предком этих двух групп. Предковая форма
могла обладать чешуйками и мастигонемами. Приоб-
ретение цитофаринкса и утрата мастигонем привели в
процессе эволюции, по-видимому, к формированию
группы псевдодендромонад.
Систематический обзор
В отряде имеется только 2 рода.
Род Cyathobodo Petersen et Hansen, 1961
(рис. 218, A)
Свободноплавающие или прикрепленные неветвя-
щимся стебельком длиной 4—50 мкм одиночные клет-
ки. Стебелек тонкий, 0.3—0.4 мкм в диаметре, состо-
ит из переплетающихся фибрилл, прикрепленных к
едва заметной пластинке в основании клетки. Разме-
ры клеток от 4.5 до 7 мкм в длину. У прикрепленных
особей задний конец заострен, у плавающих — за-
круглен. 2 почти равных жгутика 6—10 мкм длиной
примерно с 20 рядами очень мелких блюдцевидных
чешуек.
Род Pseudodendromonas Bourrelly, 1953
(рис. 218, Б)
Свободноплавающие клетки или прикрепленные,
дихотомически ветвящиеся зонтиковидные колонии
до 200 мкм высотой, с кроной около 100 мкм в диа-
метре. Клетки в колонии грушевидные или коничес-
кие с заостренным задним концом, размером от 6 до
10 мкм в длину, плавающие особи размером 6x6 мкм,
имеют закругленный задний конец. Стебелек полый,
с толстыми стенками. 2 почти равных жгутика дли-
ной 7.5—11 мкм лишены чешуек. Попытки сблизить
эти организмы с кинетопластидами (Swale, Belcher,
1975) несостоятельны, так как их клетки лишены ки-
нетопласта и в митохондриях имеются не пластинча-
тые, а пузырьковидные кристы.
Следует добавить, что в состав отряда некоторые
авторы (Verhagen et al., 1994; Patterson, 1999) вклю-
чают род Adriamonas Verhagen et al., 1994. Он опи-
сан по одному виду A. peritocrescens, который обна-
ружен в обызвествленных почвах пастбищ. Клетка у
этого жгутиконосца удлиненной формы, с 2 субапи-
кальными и почти равными жгутиками. У него име-
ется цитостом, который расположен в переднебрюш-
ной части клетки. Однако у представителей этого вида
отсутствуют характерные для псевдодендромонад че-
шуйки на жгутиках и теле клетки, а также стебелек.
По ультратонкой организации он имеет общие при-
знаки и с псевдодендромонадами, и с бикозоецида-
ми. Наличие цитостома и глотки не является теперь
препятствием для включения организма в отряд Bico-
soecales при сходстве других особенностей строения
(Карпов и др., 1998). Поэтому описание рода Adria-
monas Verhagen et al., 1994 приводится в разделе
«Отряд Bicosoecales».
«CHROMISTA» INCERTAE SEDIS
SUMMARY
THE ORDER SPONGOMONADIDA (HIBBERD) KARPOV, 1990
The spongomonads are colourless colonial flagellates, which
form dichotomously-branched digitate colonies. The single cells are
normally embedded within the distal end of each of the ultimate
branches. The cells produce endogenously small spherules composed
of iron compounds. These spherules together with mucilage
form matrix of a colony. There are one or two flagella at the apical
end of the cell, surrounded partly or totally with a cytoplasmic
collar.
The arrangement of organelles within the cell conforms more or
less to the radial symmetry. Microtubular flagellar rootlets pass radi-
ally from the special MTOC around the basal bodies. The fibrillar
rootlet associated with the Golgi apparatus is also present. The flagel-
lar transition zone is as long as the cytoplasmic collar. Mitochondria
with vesicular cristae. Free-living.
There are two families in the order: Spongomonadidae Karpov,
1990 with two genera — Rhipidodendron Stein, 1878, Spongomonas
Stein, 1878; and Phalansteriidae Kent, 1880 with one genus — Pha-
lansterium Cienkowsky, 1870.
The representatives of the order are quite poorly investigated.
We know only the cell structure and very little about their biology.
After the investigations of cell structure in Rhipidodendron splen-
didum, Spongomonas uvella, and Phalansterium digitatum D. J. Hib-
berd established two separate orders: Phalansteriida Hibberd, 1983,
and Spongomonadida Hibberd, 1983 (Hibberd, 1983). The detailed
comparison of ultrastructure in all tree genera has shown that they
have the same main characters (Karpov, 1990). Cell covered only with
the plasmalemma, the vesicular nucleus is located in the anterior part
of the cell, mitochondria with vesicular cristae, a dictyosome lies be-
tween nucleus and flagellar basal complex, there are spherules of iron
compounds in large vacoules, where the symbiotic bacteria also can be
found. One or two contractile vacoules are located in the posterior part
of the cell.
The flagellar apparatus structures also reveal many common cha-
racters: rootlets of single microtubules pass from striated MTOC late-
rally having a tendency to radial arrangement, one fibrillar rootlet is
directed to the nucleus and associated with the dictyosome. All repre-
sentatives have unusually long flagellar transition zone, which is con-
nected with the protection of basal part of undulipodia by cytoplasmic
collar (Phalansterium) or semicollar (spongomonads). The differences
between spongomonads and phalansteriids are connected with the
number of flagella. I supposed that the reduction of the second flagel-
lum in Phalansterium is connected with its more ancient sedentary
320
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
state. All these differences are regarded as those of the family level.
Spongomonads form a very peculiar group of protists, which can be
related with Chromista only on the basis of cristae shape.
THE ORDER PSEUDODENDROMONADIDA HIBBERD, 1985
The pseudodendromonads are solitary and colonial freshwater he-
terotrophic flagellates with 2 almost equal long flagella. The cell body
has the characteristic triangular profile. The apical end of the cell is
flat and both flagella emerge from one side of it. The well developed
cytostome leading into the pharynx is located opposite the flagella.
The basal end of the cell is rounded and attached to the stalk. Cells
are covered with unmineralized scales, synthesised in the Golgi appa-
ratus. Flagella also have one layer of scales, which are smaller than
the somatic ones.
The colonies are often attached to the substratum and are branc-
hed dichotomically. The stalks are rather wide and often with mucila-
ge and accumulated debris around them. This order uncludes two ge-
nera: Cyathobodo Petersen et Hansen, 1961 and Pseudodendromonas
Bourrelly, 1953.
The main characters of the internal structure are: both basal bodies
lie almost parallel to each other, they produce one short fibrillar cross-
striated rootlet, and two microtubular rootlets. One of the latter is
broader and passes beneath the plasmalemma to support the pharynx.
Another one underlies the lateral side of the cell. The nucleus of ve-
sicular type is slightly elongated towards the basal bodies. The Golgi
apparatus applies closely to its surface and takes part in the scale
production. The mitochondria have vesicular cristae.
Very little is known about their feeding and reproduction.
Their taxonomic and phylogenetic position is not clear at the mo-
ment. J.-P.Mignot (1974) put these flagellates into the haptophytes (Jso-
chrysidales), which have no haptonema, tubular mastigonemes, and the
body is covered with scales. Hibberd (1976, 1985), however, noticed
important differences between them in the organelle position, Golgi
apparatus structure, and the mode of scale formation. The relationship
between the pseudodendromonads and the bicosoecids is also discussed.
ЛИТЕРАТУРА19
Карпов С. А. Анализ отрядов Phalansteriida, Spongomonadida и
Thaumatomonadida И Зоол. журн. 1990. Т. 69, вып. 3. С. 5—12.
(Карпов С. A.) KarpovS. A. Flagellate phylogeny: ultrastructural
approach // The Flagellates / Ed. by B. S. C. Leadbeater, J. C. Green.
London: Taylor and Francis Syst. Assoc. Spec. Publ., 2000. P. 336—
360.
Bourrelly P. Flagell6s incolores rares ou nouveaux // Ost. bot.
Ztschr. 1953. Bd 100. S. 533—539.
Cienkowsky L. Ober Palmellaceen und einige Flagellaten //
Arch, mikrosk. Anat. 1870. Bd 7. S. 421—438.
Hibberd D. J. Observations on the ultrastructure of three new
species of Cyathobodo Petersen et Hansen (C. salpinx, C. intricatus
and C. simplex) and on the external morphology of Pseudodendromo-
nas vlkii Bourrelly // Protistologica. 1976a. Vol. 12, N 2. P. 249—
261.
Hibberd D. J. The fine structure of the colonial colourless flagel-
lates Rhipidodendron splendidum Stein and Spongomonas uvella Stein
with special reference to the flagellar apparatus // J. Protozool. 1976b.
Vol. 23. P. 374—385.
Hibberd D. J. Ultrastructure of the colonial colourless zooflagel-
lates Phalansterium digitatum Stein (Phalansteriida ord. nov.) and
Spongomonas uvella Stein (Spongomonadida ord. nov.) // Protistolo-
gica. 1983. Vol. 19. P. 523—535.
Hibberd D. J. Observations on the ultrastructure of new species
of Pseudodendromonas Bourrelly (P. operculifera and P. insignis) and
Cyathobodo Petersen et Hansen (C. peltatus and C. gemmatus), Pseudo-
dendromonadida ord. nov. / Arch. Protistenk. 1985. Vol. 129. P. 3—11.
Kent W. S. A manual of the infusoria. Vol. 1—3. London,
1880-1882.
Mignot J.-P. Etude ultrastructurale d’un protiste flagelle incolore:
Pseudodendromonas vlkii Bourrelly // Protistologica. 1974. Vol. 10.
P. 543—565.
Petersen J. B., Hansen J. B. On some neuston organisms. Ill //
Bot. Tidsskr. 1961. Vol. 57. P. 293—305.
Stein F. R. Der Organismus der Infusionstiere. Liepzig: Engel-
mann, 1878. Bd 3.
Swale E. M. F., Belcher J. H. A study of three new species of the
colourless scaly flagellate Cyathobodo Petersen et Hansen: C. reticu-
latus, C. crucifer and C. umbraculum // Arch. Protistenk. 1975.
Vol. 117. P. 69—275.
Vlk W. Uber den Bau der Geissel // Arch. Protistenk. 1938.
Vol. 90. P. 448—488.
19 Литература к разделу «„Chromista” incertae sedis».
Тип CHOANOMONADA Kent, 1880 — ВОРОТНИЧКОВЫЕ
ЖГУТИКОНОСЦЫ, или ХОАНОФЛАГЕЛЛАТЫ
(Син.: Craspedomonadines James-Clark, 1868)
Одиночные или колониальные гетеротрофные жгутиконосцы,
планктонные или прикрепленные. Клетки овальной или грушевид-
ной формы, размером от 3 до 25 мкм. На апикальном конце клет-
ки находится 1 жгутик, окруженный тентакулами (микровиллями),
образующими воротничок. В переходной зоне жгутика имеется
центральный филамент, микротрубочковые корешки расходятся
радиально от жгутиковой кинетосомы, кристы в митохондриях
пластинчатого типа. Особи могут жить в домиках или без них,
часто обладают хорошо развитым гликокаликсом. Ядро 1, распо-
ложено в центре или в передней половине клетки.
Размножаются, по-видимому, только бесполым путем преиму-
щественно за счет продольного деления клетки на 2 дочерние. В
жизненном цикле обычно есть 2 основные формы: прикрепленная
или свободноплавающая и стадия покоя — циста. Распространены
во всех районах земного шара; встречаются в пресных, солонова-
тых и морских водах. Свободноживущие.
Эта группа — одна из ведущих среди зоофлагеллат в планкто-
не, бентосе и обрастаниях. Их численность и биомасса в опреде-
ленные сезоны соизмеримы с таковыми бактерий и зоопланктона.
Обладая большой скоростью размножения и являясь активными
бактериофагами, воротничкЪвые жгутиконосцы играют значитель-
ную роль в жизни водоемов.
История изучения
Впервые воротничковые жгутиконосцы (или хоа-
нофлагеллаты) были описаны Эренбергом (Ehrenberg,
1838), ошибочно принявшим их за инфузорий из
рода Epistylis. Этот вид впоследствии был назван Со-
donosiga botrytis Stein, 1878. В 1868 г. Джеймс-Кларк
(James-Clark, 1868) описал еще несколько видов хоа-
нофлагеллат и впервые выделил их в особую группу
Craspedomonadines. Однако статус таксономической
категории этим простейшим — «Отряд Choanoflagel-
lata» — впервые придал Кент (Kent, 1880—1882). С
тех пор воротничковые жгутиконосцы изучаются как
единая, четко очерченная группа и альгологами, и зо-
ологами. Дальнейшие исследования хоанофлагеллат
можно разделить на 2 периода: светооптический (до
1954 г.) и электронно-микроскопический.
Использование только световой оптики не позво-
ляло выяснить многие морфологические особенности
этих мелких жгутиконосцев, в частности строение
21 Протисты, часть 1
воротничка. В первый период было описано много
новых видов и родов и начаты исследования питания
и размножения.
Применение электронного микроскопа дало воз-
можность установить строение воротничка, уточнить
способы захвата и заглатывания пищи, углубить
представления о способах размножения хоанофла-
геллат. В результате изучения органелл клетки были
сделаны важные обобщения, касающиеся не только
этой группы, но и жгутиконосцев в целом.
Первая работа на современном уровне была сдела-
на Хиббердом на Codonosiga botrytis (Hibberd, 1975).
Серию исследований по ультраструктуре и жизненно-
му циклу провели Ледбите с соавт. (Leadbeater, Mor-
ton, 1974; Leadbeater, 1977, 1983, 1991, 1994) и Кар-
пов с соавт. (Карпов, 1981, 1982а, 19826, 1985; Жу-
ков, Карпов, 1985; Карпов, Мыльников, 1993; Карпов,
Ледбите, 1997, 1998). Видовое многообразие хоаноф-
лагеллат, особенно, морских видов, исследовали
Трондзен (Throndsen, Г974), Томсен с соавт. (Thom-
sen, Buck, 1991, Thomsen et al., 1995, 1997) и Мар-
шан (Marchant, 1990).
Эти и другие работы подтвердили морфологичес-
кое единообразие в строении клетки хоанофлагеллат,
а также своеобразие и четкую очерченность этого так-
сона. В настоящее время совершенно неясны связи
этой группы с другими жгутиконосцами. Поэтому в
дальнейшем необходимо расширить ультраструктур-
ные исследования морских видов (акантоецид).
Спорным остается вопрос о родстве хоанофлагел-
лат и многоклеточных животных, особенно губок.
Однако здесь не хватает данных по строению жгути-
ковых клеток метазоев.
Классификация
Четкая очерченность таксона и своеобразие в
строении хоанофлагеллат побуждают исследователей
повышать ранг этого таксона до типа (Corliss, 1984),
322
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
надтипа (Карпов, 1990) и даже подцарства (Mohn,
1984).
В настоящем руководстве принимается ранг типа
Choanomonada, который представлен 1 классом Cho-
anomonadea Kent, 1880, содержащим единственный
отряд Choanoflagellida Kent, 1880. Отряд делится на
3 семейства, представители которых различаются по
наличию или отсутствию домиков и их строению.
Сем. Monosigidae Kent, 1880 — клетки без доми-
ков, часто погружены в слизь.
Сем. Salpingoecidae Kent, 1880 — клетки в доми-
ках из целлюлозы.
Сем. Acanthoecidae Norris, 1965 — клетки в до-
миках из кремниевых волокон.
Морфология
Клетки воротничковых жгутиконосцев независи-
мо от принадлежности к семейству, а также у оди-
ночных и колониальных видов устроены сходно
(рис. 220; 223, вкл.; 224, вкл.). Тело особи имеет
грушевидную или овальную форму. Сужение его пе-
редней части называется шейкой. Из апикальной
части шейки выходит жгутик, окруженный воро-
тничком. Многие хоанофлагеллаты имеют домики,
которые бывают 2 типов: прозрачные из целлюлозы
(тека, — рис. 221) и в форме корзинки из жестких
кремниевых полосок, или волокон (лорика, —
рис. 222). Особи без домиков обычно окружены
Рис. 220. Схема строения клетки хоанофлагеллат. (По: Жуков,
Карпов, 1985).
в — воротничок, ж — жгутик, зв — задние выросты, о — оболочка, пв —
пищеварительная вакуоль, св — сократительная вакуоль, т — тело клет-
ки, ш. — шейка клетки, я — ядро с ядрышком.
Fig. 220. Choanoflagellate cell structure.
в — collar, ж — flagellum, зв — posterior tentacles, о — envelope, пв — fo-
od vacuole, ce — contractile vacuole, m — body, iu — neck, я — nucleus.
Рис. 221. Клетки сальпингоецид в домиках разной формы. (По:
Жуков, Карпов, 1985).
А — Silpingoeca urnula, Б — S. megachelia, В — Aulomonas purdyi.
Fig. 221. Some salpingoecids.
слизью или тонкой оболочкой, которую можно об-
наружить только под электронным микроскопом.
Оболочка сохраняется у текатных и лорикатных хо-
анофлагеллат для фиксации клетки в домике.
Воротничковые жгутиконосцы снабжены наруж-
ными выростами, к которым относятся жгутик, ок-
ружающие его тентакулы воротничка, тентакулооб-
разные выросты в других местах клетки и цитоплаз-
матические мостики между соседними особями в
колониях некоторых видов.
Тентакулы воротничка представляют собой тон-
кие цитоплазматические выросты толщиной около
200 нм. Они примерно одинаковой длины и разделе-
ны равными промежутками, располагаясь вокруг жгу-
тика по окружности. Внутри каждой тентакулы про-
ходит пучок актиновых микрофиламентов, поддер-
живающий ее в расправленном состоянии (рис. 225,
вкл.; 226, вкл.).
Тентакулообразные выросты встречаются и в дру-
гих частях клетки. Чаще они располагаются на зад-
нем конце особи и служат для прикрепления к суб-
страту или образования стебелька. Иногда есть и бо-
ковые выросты. У текатных и лорикатных форм они
принимают участие в образовании домика. Тентаку-
лообразные выросты могут быть временными или
постоянными, а число их значительно меняется в
пределах онтогенеза одной клетки. Внутри них также
есть актиновые микрофиламенты, что свидетельству-
ет о гомологии всех тентакулообразных выростов Хо-
анофлагеллат. Тентакулы воротничковых сходны с
микрофиллями некоторых специализированных кле-
ток многоклеточных животных. И те и другие имеют
постоянную толщину по всей длине, расположены
примерно на одинаковом расстоянии друг от друга,
равны по длине и содержат микрофиламенты, состо-
ящие из актина.
Латеральные цитоплазматические мостики между
соседними клетками обнаружены в колониях Codono-
siga botrytis, Desmarella moniliformis и D. thienemanni.
Это временные образования, остающиеся, вероятно,
ТИП CHOANOMONADA
323
после не прошедшего до конца деления клеток. Внут-
ри мостика обычно имеется плотная перегородка, ко-
торая пронизана 1 или несколькими порами. Вероят-
но, цитоплазмы соседних клеток колонии еще неко-
торое время сообщаются между собой через поры.
Строение свободной части жгутика типично для
большинства подобных образований. Это длинный
цитоплазматический вырост апикального конца клет-
ки, внутри которого проходит аксонема, состоящая
из 2 центральных микротрубочек и 9 периферичес-
ких дублетов. У некоторых видов на жгутике обна-
ружены очень тонкие боковые волоски — простые
мастигонемы. Однако у большинства изученных во-
ротничковых жгутиконосцев мастигонемы отсутству-
ют. Строение переходной зоны хоанофлагеллат уни-
кально. Центральные микротрубочки расположены
значительно выше поперечной пластинки и связаны
с ней центральным филаментом (рис. 227, вкл.; 228,
вкл.).
Базальное тело жгутика, или кинетосома, состоит
из 9 триплетов периферических микротрубочек. Дис-
тальная часть кинетосомы соединяется с поверхност-
ной мембраной в основании жгутика при помощи хо-
рошо выраженных 9 переходных фибрилл (рис. 86).
Так устроена жгутиковая кинетосома. Ниже и пер-
пендикулярно к ней расположена безжгутиковая ки-
нетосома, не отличающаяся по строению от обычной
центриоли. Обе кинетосомы связаны друг с другом
фибриллярным мостиком. Базальный комплекс жгу-
тика, по-видимому, у всех хоанофлагеллат имеет дип-
лосомное строение, т. е. состоит из 2 ортогонально
расположенных кинетосом.
Корешковая система жгутиков воротничковых
жгутиконосцев представлена лентами микротрубо-
чек, которые радиально .расходятся от жгутиковой
кинетосомы и лежат в одной плоскости с ее про-
дольной осью (рис. 229—232, вкл.). Ленты микро-
трубочек начинаются от сателлитов — электронно-
плотных образований, расположенных вокруг кине-
тосомы. Число и форма сателлитов у разных видов
могут быть различны (рис. 229—231). Различается и
количество микротрубочек. Например, почти у всех
одиночных видов общее число микротрубочек в ко-
решковой системе не превышает 100, а у колониаль-
ных оно может достигать 200—250. Микротрубочки
расходятся под поверхностной мембраной, образуя
скелет передней половины клетки и одновременно
защищая снаружи ядро (рис. 225; 233). Проходя так-
же между основаниями тентакул воротничка, они
обычно контактируют с пучками их микрофиламен-
тов (рис. 233) и формируют, таким образом, единый
цитоскелетный комплекс. От безжгутиковой кинето-
сомы отходит тонкий поперечно исчерченный фиб-
риллярный корешок, с которым связана мембрана
сильно уплощенного мешочка. Весь этот фибрил-
лярно-мембранный комплекс направлен к диктиосо-
ме и обычно с ней связан (рис. 233).
Ядро воротничковых жгутиконосцев относится
к пузырьковидному типу (рис. 225). Оно лежит в
передней половине клетки. Ядрышко довольно круп-
ное, занимает центральное положение. Глыбки ге-
терохроматина немногочисленны, распределены по
всему объему ядра.
Рис. 222. Клетки акантоециды «Acanthoecopsis» (= Acanthocor-
bisl) unquiculata в домике из кремниевых полосок. (По: Thrond-
sen, 1974).
Fig. 222. Acanthoecid choanoflagellate «Acanthoecopsis» (=Acantho-
corbisT) unguiculata.
Между ядром и базальным комплексом жгутика
всегда расположена единственная диктиосома аппа-
рата Гольджи (рис. 225). Митохондрии с пластинча-
тыми кристами чаще находятся вблизи ядра. 1 или 2
сократительные вакуоли лежат в задней части клетки
недалеко от поверхности. У морских видов они могут
отсутствовать.
В задней же половине скапливаются пищевари-
тельные вакуоли и включения запасных питательных
веществ (липидов). У одиночных видов пищевари-
тельные вакуоли обычно мелкие. В них содержится
по 1—3 бактерии. Клетки колониальных видов часто
имеют крупные пищеварительные вакуоли (могут со-
держать до 20 бактерий). Эти различия связаны с
различиями в способах питания хоанофлагеллат (см.
ниже). Цитоплазма довольно плотная, с большим ко-
личеством рибосом^ У морского вида Choanoeca рег-
plexa были обнаружены немногочисленные симбио-
тические бактерии (Leadbeater, 1977).
Подводя итог морфологическому описанию воро-
тничковых жгутиконосцев, следует отметить, что все
они устроены весьма однообразно (рис. 225; 233).
Сначала это заключение было основано на светооп-
тических наблюдениях, а затем полностью подтвер-
дилось электронно-микроскопическими исследова-
ниями. Это особенно важно потому, что выделяемая
ранее только по наличию воротничка группа про-
стейших оказалась однородной и по многим другим
признакам: у всех хоанофлагеллат между ядром и
базальным комплексом жгутика лежит аппарат Голь-
джи; митохондрии с пластинчатыми кристами; жгу-
тик 1, кнутовидного типа; базальный аппарат имеет
диплосомное строение; в переходной зоне жгутика
расположен центральный филамент; корешковая
система жгутика образована радиально расходящи-
мися лентами микротрубочек и 1 фибриллярным ко-
решком. Практически все воротничковые жгутико-
324
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 233. Схема строения цитоскелета воротничковых жгутико-
носцев. (По: Карпов, Ледбите, 1998).
аг — диктиосома аппарата Гольджи; аф — актиновые, филаменты в тента-
кулах воротничка; бк — безжгутиковая кинетосома; жк — жгутиковая ки-
нетосома; мт — микротрубочки корешковой системы, которые связаны с
основаниями пучков микрофиламентов в тентакулах воротничка; пф — пе-
реходные фибриллы; пц — плоская цистерна, прилегающая к фибриллярно-
му корешку (фк) безжгутиковой кинетосомы; фм — фибриллярный мостик
между кинетосомами; ЦОМТ — центр организации микротрубочек кореш-
ковой системы (показан кольцевой ЦОМТ, который характерен для Mono-
siga ovata)', цф — центральный филамент в переходной зоне.
* Fig. 233. Scheme of choanoflagellate cytoskeleton structure.
аг — Golgi apparatus; аф — actin filaments in the tentacles; бк — nonflagellar
basal body; жк — flagellar basal body; мт — microtubular rootlets associated
with actin filaments; пф — transitional fibers; пц — flat cystem associated with
fibrillar rootlet (фк); фм — fibrillar bridge between basal bodies; ЦОМТ —
MTOC (circular ring is characteristic for Monosiga ovata); цф — central fila-
ment.
носцы способны образовывать временные или посто-
янные выросты, морфологически сходные с тентакулами
воротничка. Поразительное однообразие ультраструк-
турной организации хоанофлагеллат, вероятно, сви-
детельствует об их филогенетическом единстве.
Питание
Воротничок хоанофлагеллат всегда и справедли-
во считался органоидом питания. Однако объясняли
этот процесс по-разному. Одним из примеров может
служить работа Кента (Kent, 1880—1882). Автор по-
лагал, что пищевые частицы попадают внутрь во-
ротничка и заглатываются у основания жгутика. В
дальнейшем эти предположения не подтвердились.
Верным оказалось лишь то, что жгутик создает на-
правленные токи жидкости вокруг неподвижной
клетки (рис. 234). Движение жидкости осуществля-
ется таким образом, что увлекаемые им пищевые
частицы попадают на наружную поверхность воро-
тничка и прилипают к ней. В районе шейки про-
стейшего появляется язычковидная псевдоподия, ко-
торая растет по направлению к воротничку. Она захва-
тывает пищевые частицы с наружной поверхности
воротничка и, замыкаясь, образует пищевую вакуоль,
которая быстро втягивается в клетку и попадает в
ее заднюю часть, где и происходит переваривание
пищи.
Рис. 234. Схема токов жидкости, возникающих при пищевом бие-
нии жгутика хоанофлагеллат. (По: Жуков, Карпов, 1985).
Стрелками показано направление движения жидкости.
Fig. 234. Scheme of water currents around the cell coursed by feed
flagellar beating.
Сопоставление светооптических наблюдений с
электронно-микроскопическими исследованиями
этого процесса позволило обнаружить существенные
различия в питании хоанофлагеллат и выделить
3 основных способа захвата и заглатывания пищи
(Карпов, 19826).
Рис. 235. Три основных способа захвата и заглатывания пищи во-
ротничковыми жгутиконосцами. (По: Жуков, Карпов, 1985).
А — одноклеточные виды, Б — безжгутиковые формы, В — колониаль-
ные виды.
Fig. 235. Three main types of food capture in choanoflagellates.
A — single cell species, Б — aflagellate forms, В — colonial species.
ТИП CHOANOMONADA
325
Первый характерен для одиночных видов со жгу-
тиком (рис. 235, А; 236, вкл.). Пищевая псевдоподия
образуется в основании воротничка и движется вверх
по его наружной поверхности. Происходит слия-
ние цитоплазмы псевдоподии и тентакул воротничка.
При контакте с бактерией псевдоподия немного отги-
бается наружу, и между ее дистальной частью и воро-
тничком образуется пищевая чашечка, в которую по-
падают 1—2 бактерии. Затем чашечка замыкается и
образует пищеварительную вакуоль, которая быстро
втягивается в цитоплазму особи.
Принципиально иным способом питаются безжгу-
тиковые формы Choanoeca perplexa (рис. 235, Б). Их
воротничок значительно больше, чем у плавающих
жгутиковых клеток, и обычно широко раскрыт. Бак-
терии и пищевые частицы оседают на его внутренней
поверхности и постепенно концентрируются в осно-
вании воротничка. Периодически из апикальной части
клетки внутри воротничка появляются пищевые псев-
доподии, которые движутся вдоль его внутренней по-
верхности по направлению к бактериям и захватыва-
ют их в пищевую чашечку, впоследствии замыкающу-
юся и образующую пищевую вакуоль.
Третий способ питания характерен в основном
для колониальных хоанофлагеллат (рис. 235, В). Пи-
щевая псевдоподия образуется на уровне середины
тела и по мере движения вперед и в сторону растя-
гивает тонкую оболочку на переднем конце. В ре-
зультате в образовавшееся между оболочкой и по-
верхностью шейки пространство втягиваются бактерии
с нижней части воротничка. Пищевая псевдоподия
охватывает пищу и, замыкаясь, образует у основания
воротничка пищевую вакуоль. Последняя втягивается
в шейку клетки и транспортируется в заднюю поло-
вину особи. При таком способе питания может за-
хватываться по нескольку бактерий одновременно,
поэтому образуются крупные пищевые вакуоли.
Размножение и жизненные
формы
Бесполое размножение хоанофлагеллат осуществ-
ляется путем продольного деления клетки пополам.
Описанные в прошлом веке образование спор, конъ-
югация и поперечное деление клеток в дальнейшем
не были обнаружены.
Процесс деления изучен наиболее детально у Мо-
nosiga ovata (Карпов, Мыльников, 1993; Карпов,
Ледбите, 1997). Базальный аппарат жгутика удваива-
ется, а сам жгутик быстро втягивается внутрь клетки,
где происходит разборка аксонемы (рис. 237). Шейка
особи набухает, увеличиваясь в диаметре. Раздви-
гаются в стороны основания тентакул воротничка, и
последний, если смотреть на клетку сверху, превра-
щается из окружности в эллипс. Обе пары кинетосом
мигрируют в противоположные стороны к будущим
полюсам делящегося ядра, сохраняя при этом кореш-
ки из микротрубочек. Ядро вытягивается в направ-
лении, перпендикулярном продольной оси клетки.
Внутри него образуется микротрубочковое веретено,
которое направляет хромосомный материал к середи-
не ядра. На стадии поздней профазы кинетосомы на-
ходятся на полюсах ядра и уже сформированы новые
короткие жгутики, ядерная оболочка еще сохраняет-
ся, а митохондрия вплотную прилегает к поверхности
ядра. На стадии метафазы (рис. 238, вкл.) ядерная
оболочка прорывается на полюсах ядра и микротру-
бочки веретена соединяются с кинетосомами. После
этого начинается процесс расхождения хромосом и
дальнейшее разрушение ядерной оболочки. В анафа-
зе и ранней телофазе ядерная оболочка отсутствует
(рис. 239, вкл.) и восстанавливается только в поздней
телофазе. Хромосомный материал мигрирует вместе
с остатками микротрубочек вслед за кинетосомами,
которые постепенно перемещаются к середине апи-
кальной части клетки. Ветви митохондрии окружают
ядерный материал на всем протяжении митоза, а в
анафазе и телофазе частично выполняют функции
ядерной оболочки. Цитокинез завершается почти
сразу после формирования дочерних ядер, а жгутик
вырастает до нормальных размеров только после
окончательного отделения клеток друг от друга и пе-
ремещения кинетиды в центр воротничка.
Таким образом, метафазная фигура ядра у М. ova-
ta относится к типу ортомитоза, который за период
метафазы трансформируется из закрытого в полузак-
рытый (по классификации И. Б. Райкова, — см.:
Райков, 1982). Похожим образом протекает митоз у
морского воротничкового жгутиконосца Stephanoeca
diplocostata (Leadbeater, 1994).
Деление клетки S. diplocostata связано с форми-
рованием домика у дочерней особи (Leadbeater, 1994).
Образование домика происходит в 2 этапа: сначала
внутри клетки синтезируются и накапливаются крем-
ниевые полоски, а затем они группами выделяются
наружу и осуществляется сборка домика. Полоски син-
тезируются в так называемых пузырьках накопления,
которые ассоциированы с микротрубочками кореш-
ковой системы. По мере «роста» полосок пузырьки
вытягиваются вдоль микротрубочек, служащих опорным
и направляющим образованием. На заключительной
стадии формирования полоски собираются группами
по 5—8 штук на периферии клетки (рис. 240).
На втором этапе полоски выделяются наружу и
скапливаются в районе воротничка (рис. 240). При-
чем пакеты полосок будущих продольных и попереч-
ных ребер расположены взаимно перпендикулярно. В
дальнейшем, по мере процесса деления, полоски пре-
терпевают реорганизацию при помощи тентакул во-
ротничка. Концы полосок каким-то образом прикле-
иваются друг к другу, и корзинка приобретает окон-
чательную форму взрослой особи.
Среди представителей лорикатных морских хоа-
нофлагеллат (сем. Acanthoecidae) принято различать
2 типа деления клетки: нудиформное (nudiform) и
тектиформное (tectiform). В результате нудиформно-
го деления образуется дочерняя клетка без домика,
которая уплывает в поисках подходящего субстрата
для прикрепления. Такие клетки формируют домик
только после прикрепления к субстрату. У тектифор-
мных видов дочерняя клетка строит свой домик сразу
после деления (рис. 240) и только после этого уплы-
вает на поиски нового места для прикрепления.
Одним из эффективных способов увеличения чис-
ленности колониальных видов является деление
326
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 237. Стадии деления клетки Monosiga ovata. (По: Карпов, Ледбите, 1997).
А — интерфазная клетка, Б — втягивание жгутика и удвоение кинетосом, В — расхождение кинетосом, Г — стадия ранней метафазы, Д — стадия мета-
фазы, Е — телофаза с полным разрушением ядерной оболочки, Ж — формирование оболочки вокруг дочерних ядер, 3 — перемещение жгутиков внутрь
воротничков, И — полное расхождение дочерних клеток и развитие жгутиков нормальной длины.
Fig. 237. Stages of cell division in Monosiga ovata.
A — interphase, Б — flagellar involvement and replication of basal bodies, В — migration of basal bodies, Г — early metaphase, Д — mid-metaphase, E — te-
lophase with nuclear envelope disintegration, Ж — nuclear envelope formation around the daughter nuclei, 3 — flagellar movement inside the collar, И — comple-
te separation of daughter cells and formation of flagella of normal length.
целых колоний. Лентовидные колонии распадаются
на группы из нескольких особей. При этом нет како-
го-либо критического количества клеток, с которого
начинается фрагментация. Одной из возможных причин
деления колонии может быть механическое воздейст-
вие. Однако таким образом трудно объяснить де-
ление шаровидных колоний (рис. 241/вкл.). Сначала
колония принимает форму эллипсоида. Затем по его
малому экватору возникает перетяжка, которая пос-
тепенно увеличивается и приводит к формированию
2 дочерних колоний. Деление происходит независи-
мо от размеров, и при этом часто образуются 2 не-
равные колонии.
Интересно сопоставить между собой размножение
и расселение хоанофлагеллат. У одиночных жгутико-
носцев эти процессы протекают одновременно, т. е.
деление прикрепленной особи на 2 вызывает обра-
зование расселительной формы. Для колониальных
хоанофлагеллат характерно то, что расселительная
форма появляется не в результате деления, как у оди-
ночных, а более или менее независимо от этого про-
цесса. Из колоний выходят (отрываются), как прави-
ло, взрослые зооиды, морфологически неотличимые
от других особей. Расселительные формы очень
важны для прикрепленных колоний — таких как Со-
donosiga. У них имеется несколько одноклеточных
стадий с расселительной функцией (рис. 242). Для
плавающих колоний расселение не столь актуально,
и разнообразие одноклеточных форм у них невелико.
Все изученные воротничковые жгутиконосцы об-
разуют в своем жизненном цикле свободноплавающие
особи, лишенные домиков и служащие для расселе-
ния. Для них характерны1 тентакулообразные выро-
сты на заднем конце и активное движение в толще
воды, чаще всего задним концом вперед. Только у
одиночного вида Monosiga ovata клетки обычно пла-
вают передним концом вперед, что значительно уве-
личивает их скорость передвижения.
Наибольшее многообразие форм обнаружено при
исследовании культур прикрепленных колониальных
хоанофлагеллат из рода Codonosiga (рис. 242). У них
встречается до 3 расселительных форм и ряд стадий
в развитии колоний. Эти колониальные стадии зако-
номерно сменяют одна другую, чередуясь с одиноч-
ными, и образуют цикл развития. Вероятно, все отме-
ченные в культуре формы имеют место в природе и
сменяются в зависимости от изменений условий ок-
ружающей среды.
Пресноводные хоанофлагеллаты могут формиро-
вать при неблагоприятных условиях округлые цисты
с толстой оболочкой. По-видимому, это характерная
черта всех воротничковых, хотя данных по морским
видам пока нет. Оболочка цисты Desmarella moni-
liformis — единственного вида, у которого удалось
ТИП CHOANOMONADA
327
Рис. 240. Схема последовательных стадий формирования домика у дочерней клетки при делении Stephanoeca diplocostata. (По: Leadbea-
ter, 1994).
А — стадия накопления внутри клетки (полоски не показаны); Б, В — параллельно упакованные пучки полосок на конце воротничка; Г—И — стадии пе-
рераспределения полосок во время цитокинеза; К — выстраивание ребер вокруг дочерней особи.
Fig. 240. Consequentive stages of lorica formation during cell division in Stephanoeca diplocostata.
A — early interphase cell; Б, В — mid-interphase cell with costal strips at the collar distal end and withdrawn flagellum (В); Г—И — reorganization of tentacles
and subgroupings of costal strips prior to (Г—Ж) and after cell separation (3—И); К — lorica assembly on a recently separated juvenile.
Рис. 242. Формы существования вида Codonosiga botrytis в куль-
туре. (По: Карпов, 19816).
А — крупная колония, Б—Д — расселительные одиночные формы.
Fig. 242. Different stages of Codonosiga botrytis life cycle in clonal
culture.
A — mature colony, Б—Д — swammers.
получить данные по строению цисты, довольно толс-
тая, состоит из фибриллярного материала,
расположенного снаружи от плазмалеммы (рис. 243,
вкл.). Под плазмалеммой формируется тонкий плот-
ный слой из микрофиламентов. Внутри клетки сохра-
няются обе кинетосомы, ядро, митохондрии и неболь-
шая (по сравнению с ее'размерами в вегетативной
клетке) диктиосома. Мебранные образования почти
полностью отсутствуют.
Экология и распространение
Среди воротничковых жгутиконосцев чисто планк-
тонных форм немного. К ним относятся преимущест-
венно колониальные виды. Основная же масса хоа-
нофлагеллат представлена эпибионтами и обрастате-
лями, которые встречаются в планктоне и в бентосе.
В большом количестве они поселяются на планктонных
водорослях, высшей водной растительности, рако-
образных и других гидробионтах. Например, Monosiga
ovata использует в качестве субстрата даже сократи-
мые стебельки сувоек и щупальца гидр. Пресновод-
ные виды предпочитают водоемы мезотрофного типа
с небыстрым течением. Одним из основных факто-
ров, лимитирующих их численность, является кисло-
род. Они, как правило, не встречаются в анаэробных
условиях.
328
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Большинство хоанофлагеллат способны разви-
ваться в широком диапазоне температур: от -4 до
35 °C с оптимумом примерно 20—25 °C. Лишь не-
многие виды предпочитают более низкие температу-
ры и достигают высокой численности в зимне-весен-
нее время.
При наличии пищи воротничковые жгутиконосцы
неприхотливы к солевому составу среды. Оптималь-
ное для них значение pH несколько сдвинуто в ще-
лочную сторону. Следует отметить, что в водоемах с
большим содержанием минеральных взвесей при про-
чих благоприятных условиях они не развиваются. Это
связано с особенностями питания хоанофлагеллат.
Воротничковые жгутиконосцы встречаются все-
светно. Они отмечены в пресных водоемах всех кон-
тинентов и климатических зон мира, в морях и океа-
нах обоих полушарий и даже во льдах Антарктиды.
Наибольший интерес представляет тот факт, что
многие пресноводные виды встречаются в солонова-
товодных водоемах и морях, а иногда и в супергалин-
ных водах, где соленость достигает 83 %о.
Рис. 245. Пластинчатая (табличная) колония Protospongia haecke-
li. (По: Жуков, Карпов, 1985).
Fig. 245. General view of plate-like colony Protospongia haeckeli.
Филогения
Филогенетические взаимоотношения в отряде
Choanoflagellida следует рассматривать исходя из
факта, что клетки всех воротничковых имеют еди-
ную организацию. Следовательно, наиболее прими-
тивно устроенные виды, несущие основные черты
отряда, должны стоять ближе других к предковой
форме. Такой наиболее простой и, очевидно, самой
древней группой можно считать род Monosiga, От
нее прослеживаются 2 основных направления в раз-
витии Choanoflagellida: 1) образование свободнопла-
вающих и прикрепленных колоний сем. Monosigi-
dae; 2) образование форм, живущих в домиках.
Рис. 244. Внешний вид лентовидных колоний Desmarella (= Kent-
rosiga) thienemanni. (По: Жуков, Карпов, 1985).
Fig. 244. General view of thread-like colony Desmarella (=Kentrosi-
ga) thienemanni.
Эволюцию колоний от одиночного предка логич-
но рассматривать, переходя от просто устроенных
колоний к более сложным. Наиболее простыми пред-
ставляются линейные колонии, характерные для рода
Desmarella (рис. 244). Эта ветвь развития колоний от
рода Monosiga кажется наиболее очевидной. Другая
линия развития привела, по-видимому, к образованию
табличных колоний типа Protospongia (рис. 245) и
шаровидных колоний типа Sphaeroeca (рис. 246).
Еще одну самостоятельную ветвь венчают прикреп-
ленные древовидные колонии Codonosiga (рис. 247).
Весьма независимо развивались, вероятно, древовид-
ные колонии на основе слизи, а не стебелька. К ним
относится Sphaerodendron (рис. 251).
Второе направление эволюции хоанофлагеллат
привело к появлению и усложнению домиков (теки).
У видов сем. Salpingoecidae отмечено 2 основных
типа теки, которые развивались независимо: трубча-
тые (рис. 221, В; 257) и бокаловидные (рис. 221, А,
В; 224, вкл.). К первому относятся домики Stelex-
omonas и Aulomonas, а ко второму — Salpingoeca.
Совершенно особую ветвь развития представляют
виды сем. Acanthoecidae. Их лорика отличается от те-
ки Salpingoecidae по строению и химическому составу,
поэтому оба семейства должны иметь общих предков
на уровне «голых» форм.
Впервые связь хоанофлагеллат с многоклеточны-
ми организмами была отмечена Джеймсом-Кларком
в 1868 г. (James-Clark, 1868). Он сравнил воротнич-
ковых жгутиконосцев с хоаноцитами губок, указал
на большое сходство и тех и других и предположил,
что эти 2 группы животных родственны между со-
бой. Дальнейшие исследования позволили уже другим
авторам сделать более смелые выводы. При описа-
нии нового рода Protospongia Кент (Kent, 1880—
1882) указывает на то, что «пропорции воротничко-
вых клеток в большой колонии могут нарушаться» и
некоторые особи принимают неопределенную амебо-
идную форму, переходя внутрь колонии. Сторонники
гипотезы колониального происхождения Metazoa
(см.: Иванов, 1968) использовали этот род как мо-
ТИП CHOANOMONADA
329
дель перехода от колониальное™ к дифференциров-
ке клеток в соответствии с различными функциями,
а затем и к образованию двухслойного организма. В
общих чертах этот процесс мог выглядеть следующим
образом. Воротничковые клетки, получив достаточное
количество пищи, отбрасывали жгутик и воротничок,
переходили в амебоидное состояние и переползали
внутрь колонии для переваривания пищи. С течением
времени дифференцировка на наружные воротничко-
вые (эктодермальные) и внутренние амебоидные (энто-
дермальные) клетки завершилась и образовался пер-
вый двухслойный организм, от которого произошли
все современные многоклеточные животные.
Одним из главных аргументов в этой гипотезе
было приведенное выше описание Кента. При более
детальных наблюдениях за представителями рода
Protospongia (-Proterospongia) оказалось, что ни
амебоидных, ни других клеток, кроме воротничко-
вых, в колониях всех 5 видов этого рода не сущест-
вует. Нет амебоидных клеток и у других колониаль-
ных хоанофлагеллат. Таким образом, подобные мо-
дели не подтверждаются фактическим материалом.
Применение электронно-микроскопических мето-
дов показало, что воротничковые жгутиконосцы и
хоаноциты губок обладают рядом общих черт. Об-
щая морфология и симметрия клетки у них одинако-
вы. Сходно устроены тентакулы воротничка. Кристы
в митохондриях имеют пластинчатую форму. /Есть
определенное сходство и в структуре жгутикового
аппарата, хотя для более детального сравнения не
хватает данных по строению жгутиков у хоаноцитов.
Близость по многим признакам не может быть
случайной, поэтому многие авторы связывают проис-
хождение губок с воротничковыми жгутиконосцами.
Однако недавнее исследование хоаноцитов пресно-
водной губки Ephydatia fluviatilis (Карпов, Ефремо-
ва, 1994) показало, что имеются значительные отли-
чия в строении их жгутикового аппарата. Так, в пе-
реходной зоне жгутика хоаноцитов расположена
спиральная фибрилла, а не центральный филамент,
как у хоанофлагеллат. Кроме того, в основании жгу-
тика находится ямка, что характерно для инфузорий
и не встречается у хоанофлагеллат. Конечно, для
того чтобы сделать окончательный вывод о строении
жгутика у хоаноцитов губок, необходимы исследова-
ния и других видов, особенно морских представите-
лей. Тем не менее уже первые такие данные вызыва-
ют сомнения в близости хоанофлагеллат и губок.
Систематический обзор
Сем. Monosigidae Kent, 1880
Особи лишены домиков, окружены тонкой обо-
лочкой или слизью. Наряду с одиночными часто встре-
чаются колониальные виды. Колонии прикреплен-
ные и свободноплавающие. Среди них можно найти
все основные типы колоний: линейные (рис. 244),
плоскостные, или табличные (рис. 245), шаровидные
(рис. 246) и древовидные (рис. 247).
Встречаются в пресных и морских водах.
Рис. 246. Шаровидная колония Sphaeroeca volvox. (По: Жуков,
Карпов, 1985).
А — колония, Б — одиночная особь.
Fig. 246. Spherical colony of Sphaeroeca volvox.
A — colony, Б — single cell.
Рис. 247. Древовидная колония Codonosiga botrytis. (По: Жуков,
Карпов, 1985).
Fig. 247. General view of tree-like colony Codonosiga botrytis.
330
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 248. Колония Astrosiga. (По: Жуков, Карпов, 1985).
Fig. 248. General view of Astrosiga.
Род, Astrosiga Kent, 1880 (рис. 248)
Колонии звездчатого типа; клетки расположены
на концах довольно длинных стебельков, отходящих
от общего центра.
Род Cladospongia Iyengar et Ramanathan,
1940 (рис. 249)
Прикрепленные слизистые колонии, дихотоми-
чески ветвящиеся на концах. Клетки расположены
по периферии колонии, погружены в слизь и связа-
ны между собой цитоплазматическими выростами.
Прикрепленные организмы.
Монотипический род.
Род Codonocladium Stein,
1878
Колониальные прикрепленные жгутиконосцы;
клетки собраны в группы на конце общего сте-
белька. В отличие от стебелька Codonosiga стебелек
Codonocladium обильно ветвится на конце, образуя
вторичные стебельки разной длины. Особи обратно-
яйцевидной формы, с оттянутым задним концом.
Род Codonosiga James-Clark, 1866 (рис. 224,
вкл.; 247)
Колониальные прикрепленные жгутиконосцы;
клетки собраны в группы на конце общего стебелька
по 2—20.
Род Codonosigopsis Senn,
1900 (рис. 252)
Прикрепленные колонии, похожие на таковые ро-
да Codonosiga, но клетки имеют по 2 воротничка.
Род Desmarella Kent, 1880 (рис. 244)
Планктонные лентовидные, иногда разветвленные
колонии, клетки в которых соединяются боковыми
сторонами, часто при помощи цитоплазматических
мостиков. На заднем конце клетки могут присутство-
вать тентакулообразные выросты.
Род Diplosiga Frenzel, 1892 (рис. 253)
Одиночные прикрепленные жгутиконосцы без до-
мика, с 2 воротничками, из которых наружный коро-
че внутреннего.
Род Kentrosiga Schiller, 1953
(син.: Desmarella (Karpov et Coupe, 1998)).
Род Monosiga Kent, 1880 (рис. 250)
Одиночные формы без домика. Прикрепленные
формы могут иметь стебелек. Плавающие клетки спо-
собны передвигаться передним концом вперед. Посе-
ляются на различных субстратах, часто на планктон-
ных водорослях.
Род Protospongia Kent, 1880 (рис. 245)
Планктонные пластинчатые колонии; клетки со-
браны в группы по 4 и погружены в слизь.
Род Sphaeroeca Lauterborn, 1894 (рис. 246)
Планктонные шаровидные колонии; клетки рас-
положены в поверхностном слое слизистого шара с
выступающими наружу воротничками и жгутиками.
От базальной части клетки отходит тонкий отрос-
ток, соединяющий ее с центром колонии.
Род Sphaerodendron Zhukov,
1978 (рис. 251)
Прикрепленные колониальные жгутиконосцы.
Основание колонии составляет слизистый стебель;
на его расширенной вершине с помощью базальных
Рис. 249. Общий вид колонии Cla-
dospongia (А) и расположение кле-
ток в одной из ее ветвей (Б). (По:
Жуков, Карпов, 1985).
Fig. 249. General view of colony (A)
and cell arrangement in the branch
(5) of Cladospongia.
Рис. 250. Различная форма кле-
ток у Monosiga ovata. (По:
Жуков, Карпов, 1985).
Fig. 250. Cells of different shape
in culture of Monosiga ovata.
ТИП CHOANOMONADA
331
Рис. 251. Sphaerodendron. (По: Жу-
ков, Карпов, 1985).
Fig. 251. General view of Sphaero-
dendron.
Рис. 252. Codonosigopsis. (По:
Thomsen, Buck, 1991).
Fig. 252. General view of Codono-
sigopsis.
Рис. 253. Diplosiga. (По: Thomsen, Buck, 1991).
Fig. 253. General view of Diplosiga.
выростов фиксируются воротничковые клетки, кото-
рые образуют неправильной формы шар.
Некоторые авторы (Bourrelly, 1968) считают ро-
ды с 2 воротничками дискуссионными, полагая, что
наружный воротничок может быть на самом деле
расширенным устьем плотно прилегающего к клетке
тонкостенного домика или представлять собой псев-
доподиальные выросты. Для решения вопроса не-
обходимо изучение ультраструктуры представителей
родов Codonosigopsis и Diplosiga, а также рода Dip-
losigopsis из сем. Salpingoecidae.
Сем. Salpingoecidae Kent,
1880
В основном одноклеточные формы с домиком —
текой. Домики состоят из целлюлозы и имеют глад-
кую поверхность. Их форма различна и считается
видовым признаком. Тека может быть трубчатой,
колбовидной, похожей на удлиненную вазу и т. д.
(рис. 221). В верхней части есть устье, из которого
выступают воротничок и жгутик особи. Края устья
слегка расширены. Нижняя часть домика заострена
и постепенно переходит в стебелек, который бывает
разной длины. В месте прикрепления к субстрату он
образует небольшое расширение. Особи закрепляют-
ся внутри домика при помощи тонкой оболочки в
районе шейки.
Встречаются в пресных и морских водах.
Наиболее характерный и многочисленный по ви-
довому составу род Salpingoeca.
Род Aulomonas Lackey, 1942 (рис. 221, В)
Клетка расположена в верхней части прозрачного
трубчатого домика, прямого или слегка изогнутого.
Домик может быть инкрустирован снаружи прилип-
шими частицами. Планктонные формы.
Монотипический род.
Род Choanoeca Ellis, 1930 (рис. 254)
Клетки одиночные, находятся в колбовидном до-
мике на коротком стебельке. Воротничок длинный,
обычно широко раскрыт. Взрослые особи не имеют
жгутика, который появляется только в результате де-
ления у ювенильной клетки.
Род Diploeca Ellis, 1929 (рис. 255)
Домики с двойной стенкой, разделены на 2 отдела:
нижний и верхний. Наружный слой толстый, часто
окрашенный, формирует нижнюю часть домика;
внутренний — прозрачный — заключает особь и
формирует горлышко, края которого напоминают
второй воротничок.
Род Diplosigopsis France, 1897 (рис. 256)
Клетки с 2 воротничками находятся в домиках
различной формы с 1 устьем. Прикрепленные орга-
низмы.
Род Lagenoeca Kent, 1880
Свободноплавающие одиночные формы в прозрач-
ных или коричневатых домиках. Отсутствуют прикре-
пительные выросты в задней части домика. Толстые
постоянные выросты в задней части домика у некото-
рых видов не служат для прикрепления к субстрату.
Род Pashysoeca Ellis, 1930
Одиночные клетки находятся в колбовидных тол-
стостенных однослойных домиках. Последние могут
быть прозрачные или коричневые. Род требует пере-
смотра из-за большого сходства с Lagenoeca,
Рис. 254—257. Сальпингоециды. (По: Thomsen, Buck, 1991).
254 — Choanoeca, 255 — Diploeca, 256 — Diplosogopsis, 257 — Stelexomo-
nas.
Fig. 254—257. General view of some salpingoecids.
332
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Salpingoeca James-Clark, 1868 (рис. 221,
Д, Б] 224, вкл.)
Прикрепленные жгутиконосцы в однослойных
прозрачных бокаловидных домиках со стебельком
или без него.
Род Stelexomonas Lackey,
1942 (рис. 257)
Колониальные планктонные формы, клетки рас-
положены в верхних частях дихотомически ветвяще-
гося, прозрачного трубчатого домика. Домик может
быть инкрустирован снаружи прилипшими частицами.
Монотипический род.
Сем. Acanthoecidae Norris, 1965
Подавляющее большинство — одноклеточные ви-
ды с домиком — лорикой. Лорика в виде ажурной
корзинки вокруг особи построена из кремниевых по-
лосок, составляющих продольные и поперечные реб-
ра. Взаимное расположение ребер и форма лорики
различны и считаются видовыми признаками. Ульт-
раструктура клетки почти не изучена.
Характерные роды: Stephanoeca, Diaphanoeca. Жи-
вут только в морской воде.
Чтобы перейти к систематическому описанию ро-
дов семейства, необходимо ввести некоторые терми-
ны. Лорика часто имеет для прикрепления к субст-
рату ножку — педицел. Педицел может быть простой
(полоски ребер соединяются концами в основании
лорики), агрегированный (состоит из пучка поло-
сок), составной (полоски дивергируют от заднего
конца лорики).
Соединение полосок ребер друг с другом также
различно и имеет важное значение для классифи-
кации родов акантоецид. Принято различать Т-сое-
динение (продольные полоски упираются в поперечные
в виде буквы «Т»), соединение типа «конец-в-ко-
нец» (продольные полоски упираются в соединение
поперечных), У-соединение (продольная полоска от-
ходит примерно от середины другой, образуя букву
«У») и еще один тип соединения, когда несколько
продольных полосок упираются в одну поперечную.
Продольные ребра могут выступать за пределы
верхнего поперечного ребра. Если эти выступы мень-
ше длины продольной полоски, то их называют ши-
пами (spine), если больше — выступами (protrusion).
Род Acanthocorbis Hara et Takahashi, 1984
(рис. 258)
Лорика образована палочковидными полосками с
передними выступами и максимальным диаметром в
переднем поперечном ребре. Имеются многочислен-
ные продольные ребра (6—17) и 2 или более попе-
речных.
Род Acanthoeca Ellis, 1930 (рис. 259)
Лорика образована палочковидными полосками с
8—17 выступами; полоски расположены спирально.
Лорика, как правило, с агрегированным педицелом.
Размножение нудиформное.
Род Amoenoscopa Hara et Takahashi, 1987
(рис. 260)
Лорика образована толстыми палочковидными
полосками с 15—18 выступами, имеющими особые
трезубцы на концах. Имеется 2—3 поперечных ребра.
Педицел агрегированный.
Монотипический род.
Род Apheloecion Thomsen, 1973 (рис. 261)
Лорика образована палочковидными полосками с
шипами или выступами. Имеется 1 поперечное реб-
ро и 4—5 продольных. Педицел простой (от 1 до
нескольких полосок в длину).
Род Bicosta Leadbeater, 1978 (рис. 262)
Лорика состоит из 7 полосок, образующих 2 про-
дольных ребра (по 3 полоски в каждом). Педицел
простой, из 1 полоски. Поперечных ребер нет.
Род Calliacantha Leadbeater, 1978 (рис. 263)
Лорика состоит из узких полосок с 3—7 выступами
или шипами. 2 верхних поперечных ребра сближены.
Имеется 4—7 продольных ребер. Педицел простой.
Род Calotheca Thomsen et Moestrup, 1983
(рис. 264)
Лорика состоит из 6 продольных ребер, образо-
ванных широкими полосками с Т-соединением. Зад-
нее поперечное ребро редуцировано (У-соединения
лишены каждого второго «зигзага»). Клетка полно-
стью окружена перекрывающимися продольными по-
лосками. Поперечные полоски соединяются друг с
другом своеобразными сочленениями.
Монотипический тепловодный род, близкий к
Stephanacantha.
Род Campyloacantha Hara et Takahashi, 1987
(рис. 265)
Лорика образована палочковидными полосками. Пе-
реднее поперечное ребро состоит из характерных струк-
тур в виде буквы «Ь» (каждая представляет собой па-
ру слившихся полосок — поперечной и продольной).
Имеется 6—8 продольных ребер. Педицел простой.
Род Conion Thomsen, 1982 (рис. 266)
Лорика образована палочковидными полосками с
сильно перекрывающимися соединениями типа «ко-
нец-в-конец» и шипами. Имеется примерно 10 про-
дольных и 3 поперечных ребра. Педицел простой.
Монотипический род. Обнаружен только в водах
Северной Гренландии. Требуется его переописание и
уточнение связей с Cosmoeca, Diaphanoeca pedicella-
te и D. undulata.
Род Cosmoeca Thomsen, 1984 (рис. 267)
Лорика бочковидная, состоит из палочковидных
полосок с соединением типа «конец-в-конец». Могут
присутствовать шипы. Имеется 9—12 продольных и
2—4 поперечных ребра. Может присутствовать про-
стой педицел.
Род близок к Platypleura, от которого отличается
более широкими полосками.
ТИП CHOANOMONADA
333
Рис. 258—286. Представители основных родов акантоецид. (По: Thomsen, Buck, 1991).
258 — Acanthocorbis; 259 — Acanthoeca; 260 — Amoenoscopa; 261 — Apheloecion; 262 — Bicosta', 263 — Calliacantha; 264 — Calotheca; 265 — Campyloa-
cantha’, 266 — Conion; 267 — Cosmoeca; 268 — Crinolina; 269 — Crucispina', 270 — Diaphanoeca; 271 — Diplotheca', 272 — Kakoeca; 273 — Monocosta;
274 — Nannoeca; 275 — Parvicorbicula; 276 — Platypleura; 277 — Pleurasiga: P. minima (a), P. orculaeformis (6); 278 — Polyfibula; 279 — Polyoeca; 280 —
Saepicula, лорики с разными типами соединений полосок: а — «Т-соединение», б — соединение «конец-в-конец»; 281 — Saroeca; 282 — Savillea; 283 —
Spiraloecion; 284 — Stephanacantha; 285 — Stephanoeca; 286 — Syndetophyllum.
Fig. 258—286. General view of acanthoecids.
258 — Acanthocorbis; 259 — Acanthoeca; 260 — Amoenoscopa; 261 — Apheloecion; 262 — Bicosta; 263 — Calliacantha; 264 — Calotheca; 265 — Campyloa-
cantha; 266 — Conion; 267 — Cosmoeca; 268 — Crinolina; 269 — Crucispina; 270 — Diaphanoeca; 271 — Diplotheca; 272 — Kakoeca; 273 — Monocosta;
274 — Nannoeca; 275 — Parvicorbicula; 276 — Platypleura; 277 — Pleurasiga: P. minima (a), P. orculaeformis (6); 278 — Polyfibula; 279 — Polyoeca; 280 —
Saepicula, loricae with different types of strip junctions: a — «Т-junction», 6 — «end-to-end junction»; 281 — Saroeca; 282 — Savillea; 283 — Spiraloecion;
284 — Stephanacantha; 285 — Stephanoeca; 286 — Syndetophyllum.
Род Crinolina Thomsen, 1976 (рис. 268)
Лорика в форме юбочки, образована палочковид-
ными полосками с выступами. 12—16 продольных и
2 поперечных ребра. Переднее ребро меньшего ди-
аметра, чем заднее. Лорика не закрыта сзади, и про-
дольные ребра могут выступать за заднее поперечное
ребро в виде шипов или выступов. Клетка занимает
срединное положение в лорике (концы тентакул во-
ротничка достигают переднего поперечного ребра).
Род близок к Diaphanoeca.
Род Crucispina Espeland, 1986 (рис. 269)
Лорика состоит из палочковидных полосок, обра-
зующих Х-образную структуру (2 передних и 2 зад-
них расходящихся выступа), прикрепленную к одной
стороне маленькой камеры лорики.
Монотипический род теплых вод.
Род Diaphanoeca Ellis, 1930 (рис. 270)
Лорика колбовидная, образована палочковидными
полосками с выступами. Имеется 10—16 продольных
и 2—6 поперечных ребер. Максимальный диаметр
лорики на уровне второго спереди поперечного ребра.
Клетка находится в передней трети лорики. Иногда
имеется простой педицел.
Род включает большое разнообразие форм и нуж-
дается в пересмотре. Наиболее типичные виды рода
имеют 10 и более продольных ребер, а передние
334
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
выступы образованы 2 полосками. У D. pedicellata и
D. undulata только 5 продольных ребер, 1 выступ и
клетка находится в задней части лорики.
Род Diplotheca Valkanov, 1970 (рис. 271)
Лорика бочковидная, состоит из 2 камер. Перед-
няя камера образована толстыми полосками, у кото-
рых концы часто имеют вид лопаточки. К каждой пе-
редней поперечной полоске прикрепляется более чем
2 продольных ребра. Задняя камера образована мно-
гочисленными широкими перекрывающимися полос-
ками. Ряд поперечных ребер расположен в месте су-
жения лорики.
Монотипический род.
Род Какоеса Buck et Marchant, 1991
(рис. 272)
Лорика бочковидная, образована множеством па-
лочковидных полосок. Задний конец лорики состоит
из продольных и поперечных ребер, ориентирован-
ных параллельно и перпендикулярно к продольной
оси клетки, передняя часть — из спирально располо-
женных, поперечных ребер и продолжений продоль-
ных ребер. Клетка укреплена в лорике хорошо замет-
ной мембраной.
Монотипический род.
Какоеса похожа на Savillea. Они различаются по
расположению ребер в передней части лорики.
Род Monocosta Thomsen, 1979 (рис. 273)
Лорика состоит из единственного поперечного реб-
ра, образованного 5 палочковидными полосками.
Род монотипический. Найден только в прибреж-
ных водах Финляндии.
Род Nannoeca Thomsen, 1988 (рис. 274)
Лорика состоит из палочковидных полосок с ко-
роткими шипами. Имеется 6—10 продольных ребер,
каждое из которых состоит из 2 полосок, а также
2 поперечных ребра. Соединения полосок на перед-
нем конце лорики разного типа.
Монотипический род.
Род Parvicorbicula (Meunier) Deflandre, 1960
(рис. 275)
Лорика воронковидная, состоит из палочковидных
полосок с Т-соединениями. Передних шипов или вы-
ступов нет. Имеется 4—10 продольных и 2 попереч-
ных ребра. Клетка находится в задней части лорики.
Все виды этого рода можно разделить на 2 группы
по количеству продольных ребер.
Род Platypleura Thomsen, 1983 (рис. 276)
Лорика бочковидная, состоит из широких поло-
сок с соединением типа «конец-в-конец». Могут при-
сутствовать небольшие шипы. Имеется 10—12 про-
дольных и 2—3 поперечных ребра. Сложный педицел
(если имеется) обычно состоит из 4 расходящихся
полосок.
Род Pleurasiga Schiller, 1925 (рис. 277)
Лорика состоит из 8 продольных и 3 поперечных
ребер. Клетка расположена в задней части лорики.
Длинный жгутик выступает за пределы переднего по-
перечного ребра.
Большинство видов этого рода отнесено в насто-
ящее время к родам Parvicorbicula, Stephanoeca, Po-
lyfibula, Nannoeca. Еще 4 вида, изученных электронно-
микроскопически, вероятно, будут отнесены к Par-
vicorbicula. Если это произойдет, то в роде останется
один малоизвестный и не изученный электронно-мик-
роскопически типовой вид Р. orculaeformis.
Род Polyfibula Manton et Bremer, 1981
(рис. 278)
Лорика частично построена из модифицирован-
ных палочковидных полосок. Полоски 2 передних
поперечных ребер имеют центральное «гранение».
Имеется 6—8 продольных и 3 поперечных ребра.
Есть соединения Т-типа, иногда встречаются пе-
редние шипы. У некоторых видов есть простой пе-
дицел.
Род Polyoeca Kent, 1880
(рис. 279)
Клетки одиночные или объединены в линейные
или древовидные колонии. Лорика состоит из мно-
гочисленных палочковидных полосок с передними
11—17 выступами. 2 или более ленты поперечных
ребер с равными промежутками между ними охваты-
вают камеру лорики. 30—40 продольных ребер про-
должаются в агрегированный педицел. Размножение
нудиформное.
Монотипический род.
Род Saepicula Leadbeater,
1980 (рис. 280)
Лорика состоит из палочковидных полосок. Задняя
камера лорики образована многочисленными полос-
ками, которые объединены в 2 группы: одна почти
параллельна продольной оси клетки, другая поч-
ти перпендикулярна ей. Передняя камера состоит из
10 продольных ребер с коротко заостренными кон-
чиками; все продольные формы объединены одним
поперечным ребром. Наиболее часты соединения Т-
типа, но у некоторых экземпляров встречается со-
единение типа «конец-в-конец».
Род Saroeca Thomsen,
1979 (рис. 281)
Лорика воронковидная, состоит из палочковид-
ных полосок. Имеется 3—7 продольных ребер. Их
передние полоски заострены на обоих концах. Име-
ется 1—2 поперечных ребра. Переднее поперечное
ребро прикрепляется к продольным в точках мак-
симальной ширины передних полосок продольных
ребер. Педицел простой. Продольные полоски зна-
чительно выступают над передним поперечным реб-
ром.
Род Savillea (Ellis) Loeblich III, 1967
(рис. 282)
Лорика грушевидная или яйцевидная, состоит из
палочковидных полосок, образующих 2 слоя спиральных
ребер. Размножение нудиформное. В условиях «куль-
тур» 5. micropora может терять жгутики.
ТИП CHOANOMONADA. SUMMARY
335
Род Spinoeca Thomsen, Ostergaard
et Hansen, 1995
Лорика состоит из 4 продольных и 2 систем по-
перечных ребер. Каждое продольное ребро состоит
из 2 гетерополярных, дистально заостренных по-
лосок. Передние продольные полоски выступают в
виде шипов над передним поперечным ребром, кото-
рое состоит из 5 полосок длиной 4—5 мкм каждая.
Это поперечное ребро расположено выше соедине-
ния полосок продольного ребра примерно на V5 всей
длины передней продольной полоски. Пары попе-
речных полосок образуют крестовидные связи между
каждой соседней парой задних продольных ребер.
Один конец каждой поперечной полоски связан с
областью соединения продольных полосок, а другой
конец прикрепляется к соседней задней продольной
полоске на V5 ниже соединения продольных полосок.
Продольные полоски расходятся на обоих концах ло-
рики. Лорика открыта сзади. Деление тектиформное.
Род Spiraloecion Marchant et Perrin, 1986
(рис. 283)
Лорика состоит из палочковидных полосок, и
только задняя часть и педицел образованы более
толстыми и плоскими полосками. Лорика конусо-
видная, с выступами.
Монотипический род.
Род Stephanacantha Thomsen, 1983
(рис. 284)
Лорика состоит из широких полосок с передни-
ми Т- и Y-соединениями в средней части лорики.
Имеются передние поперечные полоски с шипом на
одном конце. Для одних видов характерен сложный
педицел, состоящий из 4 дивергирующих полосок,
у других видов лорика оканчивается некремниевым
уплощенным педицелом.
Род Stephanoeca Ellis, 1930
(рис. 285)
Лорика состоит из палочковидных полосок и раз-
делена поперечной перетяжкой на 2 камеры. Апи-
кальная часть передней камеры сужена, без шипов и
выступов. Несколько продольных и поперечных
ребер часто (по меньшей мере, на заднем конце)
располагаются не параллельно основным осям лори-
ки. Может присутствовать простой или агрегирован-
ный педицел.
Некоторые из описанных видов нуждаются в пе-
реисследовании.
Род Syndetophyllum Thomsen et Moestrup,
1983 (рис. 286)
Лорика состоит из широких полосок. Переднее
поперечное ребро образовано налегающими друг на
друга косыми и асимметричными полосками. Моди-
фицированные соединения типа «конец-в-конец» с
дополнительными поперечными полосками располо-
жены между теми из них, которые крепят продоль-
ные полоски. Педицел сложный, состоит из диверги-
рующих полосок.
Монотипический род.
Приведем также и названия других родов и видов,
которые встречаются в литературе, но являются си-
нонимами перечисленных выше.
Acanthoecopsis apoda (=Acanthocorbis apoda)\
Acanthoecopsis spiculifera (^Polyoeca dichotoma).
Campanoeca — скорее всего, идентичен Polyfibula
sphyrelata.
Microsportella (^Diaphanoeca).
Pseudomicrosportella — скорее всего, пустая ло-
рика Acanthocorbis unguiculata, временно занятая re-
тероконтным жгутиконосцем.
Sportelloeca (-Stephanoeca norrisii).
THE PHYLUM CHOANOMONADA KENT, 1880
SUMMARY
Choanoflagellates are unicellular or colonial flagellated protists
with cell size of 5—25 pm. They have rounded body and a collar,
which circles the single flagellum. They use the collar for feeding on
bacteria. Some of them have cellulose theca or siliceous lorica around
the cell body, but inner organization of cell is the same. They are
predominantly attached organisms living in fresh, marine and brackish
habitats.
This small group contains not more than 200 species, but is con-
sidered here as a separate phylum, because there are no other protists
with collar.
Choanoflagellates are one of the most important groups of proto-
zoans in both marine and freshwater ecosystems. Their abundance,
particularly in winter freshwater ponds, may compose a half of proto-
zoan nanoplankton as a whole. They are represented by colonial or
single cell species, and occur in all parts of the world. Freshwater
choanoflagellates often have cysts in their life-cycle.
All choanoflagellates are very conservative in their cell organiza-
tion: there is one vesicular nucleus, one mitochondrion with flat cris-
tae, one collar, one dictyosome, one flagellum, and two basal bodies.
Cytoskeleton is presented by flagellar rootlet microtubules, which pass
radially from the flagellar basal body and are connected with the actin
microfilaments of tentacles. There is a fibrillar rootlet, associated with
special cistern and originating from the non-flagellar basal body. Cho-
anoflagellates differ from each other by the development of microtu-
bular rootlet system, which depends on relative length of the flagel-
lum, and by the number of tentacles in the collar.
The unique characters for choanoflagellates are: presence of col-
lar, central filament in flagellar transition zone, and arrangement of
main organelles (basal bodies, dictyosome, nucleus) along the longitu-
dinal cell axis according to radial cell symmetry.
Choanoflagellates feed by the ingestion of bacteria. The flagellum
beats with the base to tip planar wave. This causes water flows around
the cell in the way that water is drawn through the collar between the
tentacles. Bacteria and other particles adhere to the outer surface of
the collar and are ingested by pseudopodia, which arise from the base
of the collar.
No sexual reproduction has been found.
The vegetative reproduction is represented by longitudinal divisi-
on of the cell and by colony division.
The cell division includes: basal bodies duplication, flagellar in-
volvement and disintegration, basal bodies separation and migration to
the future nuclear poles of the elongated nucleus. The metaphase fig-
ure shows semiopen orthomitosis. Two new short flagella arise in
early metaphase. In anaphase the nuclear envelope disintegrates and
nuclear material is separating. Flagellar bases reorient their position
from antiparallel during metaphase to approximately orthogonal in late
336
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
telophase, and nuclear envelopes appear de novo around the daughter
nuclei. After the separation of cell bodies the flagella grow up to the
normal length.
Reproduction by colony division usually resembles fragmentation
of thread-like or plate-shaped large colonies into smaller ones. The
spherical colonies of Sphaeroeca volvox divide by constriction of their
equator region into two smaller spheres.
TAXONOMY
The Phylum Choanomonada includes the class Choanomonadea
Kent, 1880 with the single order Choanoflagellida Kent, 1880. The
order consists of 3 families.
Monosigidae (=Codonosigidae) Kent, 1880 — naked cells often
immersed in mucilage;
Salpingoecidae Kent, 1880 — cells have thick coverings (theca)
composed of cellulose;
Acathoecidae Norris, 1965 — cells have basket-like lorica com-
posed of siliceous costae.
A description of the main genera has been prepared.
Choanoflagellates seem to be not related with any group of pro-
tists, but we can find choanocytes in some metazoans (Spongi, Cnida-
ria, Echinodermata), and propose therefore the relationship between
choanoflagellates and metazoans. This point of view is supported now
by molecular data (SSU rRNA gene sequence). On the other hand, the
details of cell structure are different in choanoflagellates and choano-
cytes. We can assume that «choano-cells» could appear in evolution
of different eukaryotic taxa independently. For the phylogeny it is
more essential to compare the choanoflagellates with larva epithelial
cell of sponges and other «lower» metazoans.
ЛИТЕРАТУРА20
Жуков Б. Ф., Карпов С. А. Пресноводные воротничковые жгу-
тиконосцы. Л.: Наука, 1985.
Иванов А. В. Происхождение многоклеточных животных. Л.:
Наука, 1968.
Карпов С. А. Ультратонкое строение воротничкового жгути-
коносца Sphaeroeca volvox // Цитология. 1981. Т. 23, № 9. С. 991—
996.
Карпов С. А. Ультраструктура воротничкового жгутиконосца
Monosiga ovata // Цитология. 1982а. Т. 24, № 4. С. 400—404.
Карпов С. А. Способы питания воротничковых жгутиконос-
цев // Вести. ЛГУ. 19826. № 21. С. 91—94.
Карпов С. А. Ультратонкое строение воротничкового жгути-
коносца Kentrosiga thienemanni // Цитология. 1985. Т. 27, № 8.
С. 947—949.
Карпов С. А. Система протистов. Омск, 1990.
Карпов С. А., Ефремова С. М. Ультратонкое строение жгути-
кового аппарата хоаноцита губки Ephydatia fluviatilis // Цитология.
1994. Т. 36, № 5. С. 403—408.
(Карпове. А., Коуп С. Дж.) Karpov S. A., Coupe S.J. A re-
vision of choanoflagellate genera Kentrosiga Schiller, 1953 and
Desmarella Kent, 1880 // Acta Protozool. 1998, Vol. 37 . P. 23—
27.
(Карпов С. А., Ледбите Б. C.) Karpov S. A., Leadbeater B. S. C.
Cell and nuclear division in freshwater choanoflagellate Monosiga
ovata // Eur. J. Protistol. 1997. Vol. 33. P. 323—334.
(Карпове. А., Ледбите Б.С.) Karpov S. A., Leadbeater
B. S. C. The cytoskeleton structure and composition in choanoflagella-
tes // J. Euk. Microbiol. 1998. Vol. 45, N 3. P. 361—367.
(Карпов С. А., Мыльников А. П.) Karpov, S. A., Mylnikov
A. P. Preliminary observations on the ultrastructure of mitosis in cho-
anoflagellates II Eur. J. Protistol. 1993. Vol. 29. P. 19—23.
(Райков И. Б.) Raikov I. В. The protozoan nucleus. Wien; New
York: Springer, 1982.
Corliss J. O. The kingdom Protista and its 45 phyla // Biosys-
tems.1984. Vol. 17. P. 87—126.
Ehrenberg D. Ch. G. Infusionsthierreichen als vollkommene Or-
ganismen. Leipzig, 1838.
Hibberd D. J. Observations on the ultrastructure of the choa-
noflagellate Codosiga botrytis (Ehr.) Saville-Kent with special reference
to the flagellar apparatus // J. Cell Sci. 1975. Vol. 17. P. 191—219.
James-Clark H. Conclusive proofs of the animality of the ciliate
sponges, and of their affinities with the infusoria flagellata // Ann.
mag. nat. Hist. Ser. 3. 1868. Vol. 19. P. 13—18.
Kent W. S. A manual of the infusoria. Vol. 1—3. London, 1880—1882.
Leadbeater B. S. C. Observations on the life-history and ultra-
structure of the marine choanoflagellate Choanoeca perplexa Ellis II J.
mar. biol. Ass. U. K. 1977. Vol. 57. P. 285—301.
Leadbeater B. S. C. Life history and ultrastructure of a new ma-
rine species of Proterospongia (Choanoflagellida) // J. mar. biol. Ass.
U. K. 1983. Vol. 63. P. 135—160.
Leadbeater B. S. C. Choanoflagellate organization with special re-
ference to loricate taxa // Free-living heterotrophic flagellates I Eds
D. J. Patterson, J. Larsen. Oxford: Oxford Univ. Press, 1991. P. 241—258.
Leadbeater B. S. C. Developmental studies on the loricate choa-
noflagellate Stephanoeca diplocostata Ellis. VIII. Nuclear division and
cytokinesis // Eur. J. Protistol. 1994. Vol. 30. P. 171—183.
Leadbeater B. S. C., Morton C. A light and electron microsope
study of the choanoflagellates Acanthoeca spectabilis Ellis and A. bre-
vipoda Ellis // Arch. Microbiol. 1974. Vol. 95. P. 279—292.
Marchant H. J. Grazing rate and particle size selection by the
choanoflagellate Diaphanoeca grandis from the sea-ice of Lake Saro-
ma, Hokkaido // Proc. NIPR Symp. Polar Biol. 1990. Vol. 3. P. 1—7.
Mohn E. System und Phylogenie der Lebewesen. Bd 1. Physika-
lische, chemische und biologische Evolution, Prokaryota, Eukaryota
(bis Ctenophora). Stuttgart, 1984.
Thomsen H. A., Buck, K. R. Choanoflagellate diversity with par-
ticular emphasis on the Acanthoecidae II Free-living heterotrophic fla-
gellates I Eds D. J.Patterson, J. Larsen. Oxford: Clarendon Press,
1991. P. 259—284.
Thomsen H. A., Ostergaard J. B., Hansen L. E. Loricate choa-
noflagellates from West Greenland (August 1988) including the desc-
ription of Spinoeca buckii gen. et sp. nov. // Eur. J. Protistol. 1995.
Vol. 31. P. 38—44.
Thomsen H. A., Garrison D. L., Kosman C. Choanoflagellates
(Acanthoecidae, Choanoflagellida) from the Weddell Sea, Antarctica,
taxonomy and community structure with particular emphasis on the
ice biota; with preliminary remarks on choanoflagellates from Arctic
sea ice (Northeast Water Polynya, Greenland) II Arch. Protistenk.
1997. Vol. 148. P. 77—114.
Throndsen J. Planktonic choanoflagellates from North Atlantic
waters // Sarsia. 1974. Vol. 56. P. 95—122.
20 Литература к разделу «Тип Choanomonada».
Рис. 54. Часть поперечного разреза стенки клеточной глотки
хищной инфузории Phascolodon vorticella. (По: Pantin, 1966).
1 — пластинки микротрубочек, 2 — наружные опорные палочки.
Fig. 54. Part of the transverse section of the cellular pharynx wall
of the predatory infusoria Phascolodon vorticella.
Рис. 73. Чешуйки на поверхности клетки Thaumatomonas. (По:
Карпов, Жуков, 1987).
сч — соматические чешуйки.
Fig. 73. Body scales on the cell surface of Thaumatomonas.
сч — body scales.
Рис. 74. Двойная мембрана, образующая покровы Apusomonas.
(По: Карпов, Мыльников, 1989).
дм — двойная мембрана, пл — плазмалемма.
Fig. 74. Double membrane of Apusomonas.
дм — double membrane, пл — plasmalemma.
Рис. 76. Ультратонкое строение некоторых экструсом.
А — веретеновидная трихоциста инфузории Paramecium, Б — трихоциста
диномонады Gyrodinium lebouriae, В — токсициста Colponema loxodes,
Г — микротроксициста Cercomonas minimus, Д — тениоболоциста Katable-
pharis sp., Е — кинетоциста Thaumalomonas lauterborni. (А — ориг. О. Н. Бор-
хсениус; Б—Г — по: Мыльников, 1988; Д — по: Мыльников, 1992; Е —
по: Карпов, Жуков, 1987).
Fig. 76. Ultrathin structure of some extrusomes.
A — spindle-like trichocyst of ciliate Paramecium, Б — trichocyst of dinophyte
Gyrodinium lebouriae, В — toxicyst of Colponema loxodes, Г — microtoxicyst
of cercomonade Cercomonas minimus, Д — teniobolocyst of Katablepharis sp.,
E — kinetocyst of thaumatomonade Thaumalomonas lauterborni.
Рис. 83. Структура звездчатого образования (показана стрелками)
в переходной зоне жгутика у зеленой водоросли Pedinomonas на
продольном (А) и поперечном (5) срезах. (По: Карпов, Таничев,
1992).
Рис. 84. Структура спирали (показана стрелками) на продольном
срезе переходной зоны жгутика хризомонады Spumella sp. (А) и на
поперечном срезе жгутика пединеллиды Pteridomonas pulex (Б).
(А — по: Мыльников, 1994; Б — по: Мыльников, 1989).
Fig. 83. Structure of stellate pattern (arrows) in flagellar transition
zone of green algae Pedinomonas in transverse (Б) and longitudinal
(A) sections.
Fig. 84. Transitional helix (arrows) on longitudinal section of chy-
sophyte Spumella sp. (A) and in tranverse section of pedinellide Pte-
ridomonas pulex (Б).
Рис. 86. Переходные фибриллы (показаны стрелками) на дисталь-
ном конце кинетосомы (кн) воротничкового жгутиконосца Desma-
rella. Вид на кинетосому со стороны дистальной части жгутика
(триплеты ориентированы против часовой стрелки). (По: Карпов,
1990).
Fig. 86. Transitional fibers (arrows) at the distal end of kinetosome
(kh) of choanoflagellate Desmarella. Counter clock wise orientation of
triplets shows the view of kinetosome from tip-to-base of flagellum.
Рис. 85. Строение переходной зоны жгутика и корешков у церко-
монады Heteromita. (По: Карпов, 1997).
мт — микротрубочковый корешок на продольном срезе, видна его связь с
фибриллярным материалом; фк —- короткий фибриллярный корешок.
Стрелкой показан ассоциированный с Л-трубочками цилиндр в переходной
зоне жгутика.
Fig. 85. Structure of flagellar transition zone and rootlets in cerco-
monade Heteromita.
мт — microtubular rootlet associated with fibrillar material, фк — short fib-
rillar rootlet. Arrow shows the concentric rings in tranzition zone associated
with A-tubules of axoneme.
Рис. 87. Продольный срез клетки церкомонады Heteromita. (По:
Карпов, 1997).
аг — диктиосома аппарата Гольджи; кп — кинетосома переднего жгутика
на поперечном срезе видна со стороны базальной части жгутика (триплеты
ориентированы по часовой стрелке); м — митохондрия с пузырьковидными
кристами, мк — микротельце, прилегающее к ядру (я); фм — фибрилляр-
ные мостики между кинетосомами.
Fig. 87. Cell structure of cercomonade Heteromita.
фм — fibrillar bridges between kinetosomes, м — mitochondrium with vesi-
cular cristae, мк — microbody applying to the nucleus (л), аг — Golgi appa-
ratus, кп — kinetosome of flagellum in transverse section.
Рис. 96. Строение диктиосомы эвгленовой водоросли.
(Ориг. С. А. Карпова).
Рис. 93. Ультратонкое строение крист митохондрий у раз-
ных протистов.
А — пластинчатые кристы в митохондриях Desmarella thienemanni,
Б — трубчатые кристы в митохондрии инфузории Paramecium du-
boscqui, В — пузырьковидные кристы в митохондрии Thaumatomo-
nas seravini (на поверхности митохондрии видно также формирова-
ние соматической чешуйки), Г — ампуловидные кристы в митохон-
дрии Opalina ranarum. (А — ориг. С. А. Карпова; Б — ориг.
С. И. Фокина: В — по: Карпов, 1993; Г— по: Карпов, 1990).
Fig. 93. Mitochondrium ultrastructure of some protists.
A — lamellar cristae of choanoflagellate Desmarella thienemanni, Б —
tubular cristae in ciliate Paramecium duboscqui, В — vesicular cristae
in Thaumatomonas seravini (formation of body scale is also visible on
the mitochondrium surface), Г — ampula-like cristae in Opalina rana-
rum.
Fig. 96. Dictyosome structure of euglenid.
Рис. 97. Строение ядра кинетопластиды Bodo saltans с симбиотической бактерией (с) в перинуклеарном пространстве и фибриллами
(стрелка) в кариоплазме. (Ориг. С. А. Карпова).
гх — гетерохроматин, яд ядрышко.
Fig. 97. Nuclear structure of Bodo saltans with symbiotic bacterium (c) in perinuclear space and unknown fibrills (arrow) in karyoplasm.
kx — heterochromatin, яд — nucleolus.
Рис. 121. Эндосимбиотические бактерии (б) в перинуклеарном пространстве и ЭПР гетеротрофного хризофита Paraphysomonas vestita.
(Ориг. С.А.Карпова и А.И.Таничева).
я — ядро.
Fig. 121. Endobiotic bacteria (б) in perinuclear space and endoplasmic reticulum of heterotrophic chrysophyte Paraphysomonas vestita.
я — nucleus.
Рис. 122. Симбиотические бактерии в макронуклеусе инфузории
Paramecium. (Ориг. С.И.Фокина).
Fig. 122. Endobiotic bacteria in macronucleus of Paramecium.
Рис. 123. Вирусоподобные частицы в цитоплазме гетеротрофного
хризофита Siluania. (Ориг. С.А.Карпова).
Fig. 123. Virus-like particles in heterotrophic chrysophyte Siluania.
Рис. 125. План строения и структура поверхности клетки криптомонады.
А — передний конец клетки Campylomonas sp.: видны вестибулярная зона и отверстие глотки; от правого края борозды отходят жгутики. Б — покровы
клетки Cryptomonas sp.: видны плазмалемма, внешний и внутренний элементы перипласта. В — срез в области вестибулярной зоны Cryptomonas sp. Г —
продольный срез клетки Cryptomonas sp. Д — поперечный срез клетки Cryptomonas sp. бэ — большая эжектосома, гл — глотка, го — аппарат Гольджи,
с« — сократительная вакуоль, мт — митохондрия, мэ — пузырьки мелких эжектосом, пе — перипласт, пж — переходная зона жгутика, пм —
плазмалемма, хл — хлоропласт, яд — ядро, яз — вестибулярный язычок. А — сканирующий электронный микроскоп, Б—Д — фотографии ультратонких
срезов клеток.
Масштаб (мкм): А — 3, Б — ОД, В — 0.4, Г— 1, Д — 0.6.
Fig. 125. Basic structures and architecture of cell surface.
A — anterior end of Campylomonas sp.: note vestibulum, and gullet apertura; flagella extend from right side of furrow. Б — cell surface of Cryptomonas sp.:
note plasmalemma, inner and outer periplast elements. В — ultrathin section of vestibulum zone of Cryptomonas sp. Г — longitudinal section of cell Cryptomonas
sp. Д — transverse section of Cryptomonas sp. бэ — big ejectosome, гл — gullet, го — Goldgi apparatus, ce — contractile vacoule, мт — mitochondrion,
мэ — small ejectosomes, ne — periplast, пж — transition region of flagella, пм — plasmalemma, хл — chloroplasts, яд — nucleus, яз — vestibular ligule. A —
SEM, Б—Д — ТЕМ.
Scale bars (ptm): A — 3, 5 — 0.7, В — 0.4, Г—1, Д — 0.6.
Рис. 126. Ультраструктура основных органелл клетки Cryptomonas sp.
А — поперечный срез клетки в области глотки; сверху видны укрепляющие глотку с вентральной стороны электронноплотные пластинки, справа —
исчерченная пластина, связанная с окологлоточным включением. Б — жгутики с мастигонемами. В — базальные тела жгутиков: видны фибриллярные и
микротрубочковые корешки. Г — ризостиль, отходящий от основания жгутика и проходящий около ядра. Д — поперечный срез ризостиля: видны
изогнутые «крылья», отходящие от микротрубочек с одной стороны ризостиля и фибриллярный материал на противоположной его стороне. Е — срез
оснований жгутиков; видны переходные зоны, восходящий микротрубочковый корешок (справа), соединяющийся с рядом цитоскелетных микротрубочек, и
фибриллярный тяж^ соединяющий базальные тела жгутиков. Ж — скользящий срез поверхности клетки, видно сетчатое строение митохондрии и пузырьки,
оставшиеся от мелких эжектосом. 3 — срез базальной части клетки, виден нуклеоморф, расположенный между ядром и хлоропластом, вк — восходящий
микротрубочковый корешок, ма — мастигонемы, нм — нуклеоморф, ри — ризостиль, цм — цитоскелетные микротрубочки, стрелка указывает на исчер-
ченный ф«гбрилярный корешок. Остальные обозначения как на рис. 124. А, В—3 — фотографии ультратонких срезов клеток; Б — тотальный препарат.
Масштаб (мкм): А — 0.5, Б — 1, В — 0.7, Г—1, Д — 0.5, £ — 0.4, Ж — 0.8 , 3—1.
Fig. 126. Ultrastructure of main organelles of Cryptomonas sp.
A — transverse section of gullet; electrondense plate locates at the ventral side of gullet, at right — striated plate. Б — flagella with mastigonemes. В — flagella
basal bodies: fibrillar and microtubular roots. Г — rhizostyle. Д — transverse section of rhizostyle: «wings» on one side of microtubules and fibrillar material on
opposite side. E — proximal zone of flagella; transition region of flagella, at right — microtubular root associated with cytoskeletal microtubules, fibrillar connection
of basal bodies. Ж — net-like mitochondion and vesicles of ejectosomes. 3 — section of proximal part of the cell, note nucleomorph between nucleus and
chloroplast, ex — microtubular root, ма — mastigonemes, нм — nucleomorph, pu — rhizostyle, — cytoskeletal microtubules, arrow — striated fibrillar root.
Other symbols as for Fig. 124. A, В—3 — ultrathin sections; Б — total.
Scale bars (pm): A — 0.5, 5—1, В — 0.7, 5—1, Д — 0.5, E—0.4, Ж — 0.8, 3—1.
Рис. 150. Ультраструктура Bodo curvifilus.
жк — жгутиковый карман, кп — кинетопласт, пв — пищеварительная вакуоль, рж — рекуррентный жгутик, ц — цитостом, я — ядро.
Fig. 150. Fine structure of Bodo curvifilus.
жк — flagellar pocket, кп — kinetoplast, ne — food vacuole, рж — posterior flagellum, ц — cytostome, я — nucleus.
Рис. 151. Ультраструктура цитостом-цитофарингеального
комплекса Bodo designis.
кп — кинетопласт, мп — микротрубочковая призма, цф — цито-
фаринкс, ц — цитостом.
Fig. 151. Ultrastructure of the cytostome-cytopharyngeal
complex of Bodo designis.
кп — kinetoplast, мп — microtubular prism, ц — cytostome, цф —
cytopharinx.
Рис. 152. Ультраструктура цисты Bodo caudatus.
пж — передний жгутик, рж — рекуррентный жгутик, сц — стен-
ка цисты, ц — цитостом.
Fig. 152. Fine structure of the cyst wall of Bodo caudatus.
пж — anterior flagellum, рж — posterior flagellum, сц — cyst wall,
ц — cytostome.
Рис. 153. Cryptobia salmositica (сканирую-
щий электронный микроскоп).
пж — передний жгутик, рж — рекуррентный жгу-
тик.
Fig. 153. Cryptobia salmositica (Scaning elect-
ron micrograph).
пж — anterior flagellum, рж — posterior flagellum.
Рис. 154. Ультраструктура Cryptobia borreli.
жк — жгутиковый карман, ми — митохондрион,
мтр — микротрубочки, подстилающие стенку жгу-
тикового кармана и цитостом-цитофарингеально-
го комплекса, рж — рекуррентный жгутик, ц —
цитостом.
Fig. 154. Fine structure of Cryptobia borreli.
жк — flagellar pocket, кп — kinetoplast, mu — mi-
tochondrion, мтр — reinforced cytopharyngeal mic-
rotubules, рж — posterior flagellum, ц — cytostome.
Рис. 155. Ультраструктура Cryptobia borreli.
мтр — микротрубочки, подстилающие стенку жгутикового карма-
на и цитостом-цитофарингеального комплекса, пм — плазмалем-
ма, смт — субмембранные микротрубочки, цмт — группа цито-
стомальных микротрубочек, цф — цитофаринкс.
Fig. 155. Fine structure of Cryptobia borreli.
мтр — reinforced cytopharyngeal microtubules, пм — plasmalem-
ma, cm — submembrane microtubules, цмт — group of cytostomal
microtubules, цф — cytopharinx.
Рис. 156. Трубчатые кристы в митохондрии Cryptobia
borreli.
ми — митохондрион, тк — трубчатые кристы.
Fig. 156. Tubular cristae in the mitochondrion of Cryptobia
borreli.
mu — mitochondrion, тк — tubular cristae.
Рис. 157. Промастигота Leptomonas mycophilus (сканирующий электронный микроскоп),
ж — жгутик.
Fig. 157. The promastigote of Leptomonas mycophilus (Scaning electron micrograph),
ж — flagellum.
Рис. 158. Ультраструктура промастиготы Leptomonas mycophilus.
a — аксонема, аг — аппарат Гольджи, жк — жгутиковый карман, кп — кинетопласт, ми — митохондрион, жг — жгутик, я — ядро.
Fig. 158. Fine structure of the promastigote of Leptomonas mycophilus.
a — axonema, аг — Golgi apparatus, жк — flagellar pocket, кп — kinetoplast, mu — mitochondrion, nm — paraxial rod, я — nucleus.
Рис. 159. Тубулемма Trypanosoma rotatorium.
пм — плазмалемма, смт — субмембранные микротрубочки.
Fig. 159. The tubulemma of Trypanosoma rotatorium.
пм — plasmalemma, cm — submembrane microtubules.
сссц
Рис. 160. Цистоподобная амастигота Leptomonas oncopelti.
пм — плазмалемма, сссц — слой специализированной субмембранной цитоплазмы,
я — ядро.
Fig. 160. Cyst-like amastigote of Leptomonas oncopelti.
пм — plasmalemma, сссц — specialized submembrane cytoplasm, я — nucleus.
Рис. 161. Кинетопласт Bodo designis.
кр — дисковидные пластинчатые кристы, ну — нуклеоид.
Fig. 161. Kinetoplast of Bodo designis.
кр — discoidal cristae, ну — nucleoid.
Рис. 162. Митоз Bodo curvifilus.
kx — кинетохоры, мтв — микротрубочки митотического веретена, я — ядро, яд — ядрышковый материал,
яо — интактная ядерная оболочка. (По: Фролов и др., 1996).
Fig. 162. Mitosis in Bodo curvifilus.
kx — kinetochore, мтв — microtubules of the mitotic spindle, я — nucleus, яд — nucleolus,
яо — intact nuclear envelope.
Рис. 223. Колония Codonosiga botrytis.
Прижизненный снимок. Интерференци-
онный контраст. (Ориг. S. J. Coupe).
в — воротничок, ж — жгутик, ст — общий
стебелек колонии.
Fig. 223. Alive colony of Codonosiga
botrytis.
в — collar, ж — flagellum, cm — com-
mon stalk of the colony.
Рис. 224. Две особи Salpingoeca spp.
Прижизненный снимок. Интерференци-
онный контраст. (Ориг. S. J. Coupe).
т — домик (тека), л — ядро.
Fig. 224. Two alive organisms of Salpin-
goeca spp.
m — theca, я — nucleus.
Рис. 225. Продольный срез клетки Monosiga ovata. (По:
Карпов, Ледбите, 1997).
аг — диктиосома аппарата Гольджи, бк — безжгутиковая кинето-
ма, ж — жгутик, м — митохондрии, мт — микротрубочки ко-
решковой системы, мф — пучки микрофиламентов в тектакулах
воротничка (те), пв — пищеварительная вакуоль, я — ядро с яд-
рышком.
Fig. 225. Ultrathin structure of Monosiga ovata.
аг — Golgi apparatus, бк — nonflagellar basal body, ж — flagel-
lum, м — mitochondria, мт — rootlet microtubules, мф — microfi-
laments in the tentacles (me), ne — food vacuole, я — nucleus.
Рис. 226. Поперечный срез тентакулы воротничка Monosi-
ga ovata при большом увеличении. (Ориг. С. А. Карпова).
Актиновые микрофиламенты внутри тентакулы (показаны стрел-
ками) хорошо видны при фиксации с применением таниновой
кислоты.
Fig. 226. Transverse section of collar tentacule of Monosiga
ovata at high magnification.
Arrows show actin microfilaments.
Рис. 227. Продольный срез переходной зоны жгутика Des-
marella thienemanni. (Ориг. С. А. Карпова).
Стрелкой показан центральный филамент.
Fig. 227. Longitudinal section of flagellar transition zone of
Desmarella thienemanni.
Arrow shows a central filament.
Рис. 229. Поперечный срез жгутиковой кинетосомы Mono-
siga ovata. (Ориг. С. А. Карпова).
Стрелкой показан кольцевой ЦОМТ корешковой системы.
Fig. 229. Transverse section of flagellar basal body of Mono-
siga ovata.
Arrow shows a circular MTOC of flagellar rootlet system.
Рис. 228. Поперечный срез переходной зоны жгутика
Sphaeroeca volvox. (Ориг. С. А. Карпова).
Стрелкой показан центральный филамент.
Fig. 228. Transverse section of flagellar transition zone of
Sphaeroeca volvox.
Arrow shows a central filament.
Рис. 230. Поперечный срез жгутиковой кинетосомы Des-
marella thienemanni. (Ориг. С. А. Карпова).
Стрелками показаны отдельные ЦОМТы корешковой системы.
Fig. 230. Transverse section of flagellar basal body of Des-
marella thienemanni.
Arrows show the separate MTOCs of flagellar rootlet system.
Рис. 231. Поперечный срез жгутиковой кинетосомы Spha-
eroeca volvox. (Ориг. С. А. Карпова).
Стрелками показаны 2 ЦОМТа корешковой системы.
Fig. 231. Transverse section of flagellar basal body of Spha-
eroeca volvox.
Arrows show 2 separate MTOCs of flagellar rootlet system.
Рис. 236. Образование пищевой чашечки у Monosiga
ovata. (Ориг. С. А. Карпова).
аг — аппарат Гольджи, б — бактерия, ж — жгутик, пс — пище-
вая псевдоподия, те — тентакулы воротничка, я — ядро.
Fig. 236. Food cap formation in Monosiga ovata.
аг — Golgi apparatus, 6 — bacterium, ж — flagellum, nc — food
pseudopodium, me — collar tentacles, я — nucleus.
Рис. 232. Тангентальный срез ЦОМТа (показан стрел-
кой) корешковой системы Sphaeroeca volvox. (Ориг.
С. А. Карпова).
Микротрубочки организованы в несколько вертикальных слоев.
Fig. 232. Oblique section of one of the MTOC (.arrow) of
flagellar rootlet system in Sphaeroeca volvox.
Microtubules are arranged in several layers.
Рис. 238. Стадия поздней метафазы Monosiga ovata с об-
разованием фенестр (ф) на полюсах веретена. (Ориг.
С. А. Карпова).
к — кинетосомы, оя — оболочка ядра, хр — хромосомы.
Fig. 238. Late metaphase in Monosiga ovata with fenestrae
(ф) formation at the nuclear poles.
x — basal bodies, оя — nuclear envelope, xp — chromosomes.
Рис. 239. Стадия телофазы Monosiga ovata. (Ориг.
С. А. Карпова).
Ядерный материал (ям) сгруппирован у кинетосом (к) и не окру-
жен оболочкой, ж — жгутик, м — митохондрия.
Fig. 239. Telophase in Monosiga ovata.
Nuclear material (ям) without envelope joins to basal bodies (к),
ж — flagellum, м — mitochondrium.
Рис. 241. Деление шаровидной колонии Sphaeroeca vol-
vox. (По: Жуков, Карпов, 1985).
Fig. 241. Division of spherical colony of Sphaeroeca volvox.
Рис. 243. Строение цисты Desmarella moniliformis. (Ориг.
С. А. Карпова).
к — кинетосомы, л — липидное включение, м — митохондрия,
оц — оболочка цисты, пл — плазмалемма, ф — фибриллярный
слой микрофиламентов под плазмалеммой, я — ядро.
Fig. 243. Cyst structure of Desmarella moniliformis.
к —. basal bodies, л — lipids, м — mitochondrium, оц — cyst wall,
пл — plasmalemma, ф — submembrane microfilaments, я — nucle-
us.
Рис. 328. Eucomonympha папа. (По: Бобылёва, 1975).
Микрофотографии с тотального препарата. Импрегнация протеи-
натом серебра. А — фокус на жгутиковые ряды и парабазальные
тела, Б — фокус на парабазальные филаменты.
Fig. 328. Eucomonympha папа.
Total preparations; protargol impregnation according to Uhlig. Verti-
cal view. A — flagellar rows and parabasal bodies are in focus, Б —
parabasal filaments are in focus.
1 мкм
Рис. 352. Продольный срез клетки Cercomonas activus.
(Ориг. А. П. Мыльникова).
аг — аппарат Гольджи, ж — жгутик, к — канал с рекуррентным
жгутиком, м — митохондрия, мт — микротельце, от — осмио-
фильное тело (экструсома), пв — пищеварительная вакуоль, пс —
псевдоподия, эпр — эндоплазматический ретикулум, я — ядро.
Fig. 352. Ultrastructure of Cercomonas activus.
аг — Golgi apparatus, ж — flagellum, к — canal with recurrent fla-
gellum, м — mitochondrium, мт — microbody, от — extrusome,
tie — food vacuole, nc — pseudopodium, эпр — endoplasmic reticu-
lum, я — nucleus.
Рис. 363. Строение плазмодия Cercomonas activus. (Ориг. А. П. Мыльникова).
Обозначения как на рис. 352.
Fig. 363. Plasmodium structure of Cercomonas activus.
Abbreviations are the same as on fig. 352.
Рис. 353. Продольный срез клетки Heteromita reniformis.
(По: Мыльников, 1984).
жк — жгутиковый карман, кц — кинетоциста. Остальные обозна-
чения как на рис. 352.
Fig. 353. Ultrastructure of Heteromita reniformis.
жк — flagellar pocket, кц — kinetocyst. Other abbreviations are the
same as on fig. 352.
Рис. 374. Фотографии спорофоров некоторых миксогастрид, полученные с помощью сканирующего электронного микроскопа.
А — Arcyria pomiformis, общий вид спорангия; Б — Echinostelium colliculosum, общий вил спорангия с колонкой; В — Physarum citrinum, общий вид рас-
крывшихся спорангиев; Г — Licea pygmaea, общий вид спорангия; Д — Lamproderma sauteri, раскрывшийся спорангий; Е — L. cribrarioides, спора; Ж —
L. columbinum, спора. Масштаб (мкм): А — 100, Б — 10, В — 100, Г — 10, Д — 100, Е — 1, Ж — 1.
Fig. 374. Sporophores of the myxogastrids (SEM).
A — Arcyria cinerea; Б — Echinostelium colliculosum; В — Physarum citrinum, opened sporangia; Г — Licea pygmaea; Д — Lamproderma sauteri, opened
sporangium; E — L. cribrarioides, spore; Ж — L. columbinum, spore. Bar (pm): A — 100, Б — 10, В — 100, Г— 10, Д — 100, E — 1, Ж — 1.
Рис. 377. Фотографии некоторых структур спорофоров миксогастрид, полученные с помощью сканирующего электронного микроскопа.
А — Trichia alpina, нити капиллиция; Б — Arcyodes incarnata, деталь капиллиция и спора; В — Lepidoderma carestianum, чешуйка извести на перидии;
Г — Didymium dubium, кристалл извести на перидии. Масштаб (мкм): А — 10, Б — 1, В — 10, Г— 10.
Fig. 377. Various structures of fruiting bodies myxogastrids (SEM).
A — Trichia alpina, capillitium; Б — Arcyodes incarnata, capillitium and spore; В — Lepidoderma carestianum, lime flake on peridium; Г — Didymium dubi-
um, lime crystal on peridium. Bar (pm): A — 10, Б — I, В — 10, Г — 10.
Рис. 394. Организация гликокаликса голых амеб.
А — аморфный (Thecamoeba striata)', Б — филаментозный (Amoeba proteus)', В — толстый, многослойный («кутикула») (Mayorella vespertilioides); Г —
пентагональные гликостили (Pessonella sp.); Д — спиральные гликостили (Paradermamoeba valamo); Е — микрочешуйки (Korotnevella bulla).
Fig. 394. Organization of glycocalix in naked amoebae.
A — amorphous (Thecamoeba striata)', Б — filamentous (Amoeba proteus)', В — thick multilayered («cuticle») (Mayorella vespertilioides); Г — pentagonal gly-
costyles (Pessonella sp.); Д — spiral glycostyles (Paradermamoeba valamo); E — microscales (Korotnevella bulla).
Рис. 396. Скопление микрофиламентов вокруг формирующейся пищевой чашечки у Pessonella sp.
Fig. 396. Association of microfilaments around forming food cup in Pessonella sp.
Рис. 399. Представители сем. Amoebidae.
A — полиподиальная форма Amoeba proteus, Б — та же клетка при быстрой локомоции принимает практически моноподиальную форму, В — Polychaos
annulatum (полиподиальная форма), Г — моноподиальная форма той же клетки при быстрой локомоции, Д — сотовый слой в ядре Amoeba proteus.
Fig. 399. Representatives of Amoebidae.
A — polypodial form of Amoeba proteus, Б — the same cell during fast locomotion aquires nearly monopodial form, В — Polychaos annulatum (monopodial
form), Г — monopodial form of the same cell during fast locomotion, Д — honeycomb-like layer in the nucleus of Amoeba proteus.
Рис. 400. Представители сем. Thecamoebidae.
А — Thecamoeba striata, Б — Paradermamoeba valamo, В — Pseudothecamoeba sp.
Fig. 400. Representatives of Thecamoebidae.
Рис. 401. Представители сем. Hartmannellidae.
A — Hartmannella sp., Б — Saccamoeba Umax.
Fig. 401. Representatives of Hartmannellidae.
Рис. 402. Представители сем. Paramoebidae.
A — Mayorella cantabrigiensis, Б — Korotnevella Stella.
Fig. 402. Representatives of Paramoebidae.
Рис. 404. Представители сем. Vannellidae.
A — Vannella simplex, Б — Platyamoeba stenopodia.
Рис. 403. Vexillifera bacillipedes (сем. Vexilliferidae).
Fig. 403. Vexillifera bacillipedes (Vexilliferidae).
Fig. 404. Representatives of Vannellidae.
Рис. 410. Flamella sp. (сем. Flabellulidae).
Рис. 411. Rhizamoeba sp. (сем. Leptomyxidae).
Fig. 410. Flamella sp. (Flabellulidae).
Fig. 411. Rhizamoeba sp. (Leptomyxidae).
Рис. 412. Leptomyxa reticulata (сем. Leptomyxidae).
Fig. 412. Leptomyxa reticulata (Leptomyxidae).
Рис. 415. Представители Testacealobosia. (Ориг.).
А — Difflugia crassa, Б — D. acuminata, В — D. urceolata, Г — D. claviformis; Д — Nebela sp., Е — Nebela sp., Ж — Centropyxis aculeata, 3 — Cochlio-
podium bilipibosum. A—Д, Ж — раковина; E — материал раковины.
Fig. 415. Representatives of Testacealobosia.
A — Difflugia crassa, Б — D. acuminata, В — D. urceolata, Г — D. claviformis, Д — Nebela sp., E — Nebela sp„ Ж — Centropyxis aculeata, 3 — Cochlio-
podium bilimbosum. А—Д, Ж — shell; E — shell material.
Рис. 416. Представители Testacealobosia. (Ориг.).
А — Lesquereusia spiralis, раковина; Б — L. spiralis, материал раковины; В — Netzelia luberculata, раковина; Г — Netzelia luberculata, материал ракови-
ны; Д — Arcella sp.; Е — Arcella sp.; Ж — Arcella sp., тангентальный срез стенки раковинки; 3 — Arcella discoides; И — Arcella sp., альвеолы, э — эпи-
подии, я — ядра.
Fig. 416. Representatives of Testacealobosia.
A — Lesquereusia spiralis, shell; Б — L. spiralis, shell material; В — Netzelia luberculata, shell; Г — N. luberculata, shell material; Д — Arcella sp.; E — Ar-
cella sp.; Ж — Arcella sp., transverse section of shell wall; 3 — Arcella discoides', И — Arcella sp., alveoli, э — epipodia, я — nuclei.
Рис. 508. Уроид Pelomyxa palustris, на поверхности кото-
рого располагается «щетка» гиалиновых выростов (вилли).
Масштаб: 12 мкм.
Fig. 508. Uroid of Pelomyxa palustris with hyaline villi.
Bar =12 pm.
Рис. 507. Общий вид движущейся Pelomyxa palustris.
Стрелкой показано направление движения, у — уроид.
Масштаб: 200 мкм.
Fig. 507. Pelomyxa palustris, general view of locomoting
specimen.
Arrow — direction of locomotion, у — uroid. Bar = 200 pm.
Рис. 509. Облигатные бактериальные эндоцитобионты Pelomyxa palustris.
A — скопление «крупных» бактерий по периферии гликогенового тела; Б — деление «мелких» форм в цитоплазме пеломиксы (гт — участок гликогено-
вого тела). Масштаб: 0.5 мкм.
Fig. 509. Bacterial endocytobionts of Pelomyxa palustris.
A — «large» endocytobionts, Б — division of «small» endocytobionts. (гт — glycogen body). Bar = 0.5 pm.
Рис. 510. Микротрубочки в эктоплазме Pelomyxa
palustris.
Масштаб: 0.5 мкм.
Рис. 511. Основание жгутика Pelomyxa palustris с кореш-
ковой системой микротрубочек.
Масштаб: 0.25 мкм.
Fig. 510. Microtubules in the ectoplasm of Pelomyxa
palustris.
Bar = 0.5 pm.
Fig. 511. Pelomyxa palustris, the base of flagellum with
microtubular rootlet system.
Bar = 0.25 pm.
Рис. 523. Ультратонкая организация бластоцисты Blastocystis galli. (По: Белова, 1996).
А — общий вид клетки, Б — фрагмент клетки при большем увеличении, аг — аппарат Гольджи, г — гликокаликс, ми — митохондрия, цв — централь-
ная вакуоль, я — ядро.
Fig. 523. The scheme of ultrastructure of Blastocystis galli.
A — general view of the cell, Б — fragment of the cell, аг — Golgi apparatus, г — surface coat, mu — mitochondria, цв — central vacuole, я — nucleus.
Рис. 536. Детали строения монофонтинальной и бифонтинальной стенки.
I — Deniostomina (Schlumbergerinana) (монофонтинальная стенка): А — внешний вид раковины; Б — игольчатая ультраструктура внутренней поверхности
стенки и зуба, х 10000; В — поперечный срез раковины, х 200; Г— микрогранулярная ультраструктура на поперечном срезе, х 3000. 2 — Borelis (Miliola-
na), х 220 (монофонтинальная стенка), снаружи видны отверстия псевдопор. 3 — отверстия псевдопор на поверхности стенки некоторых Nodosariata (моно-
фонтинальная стенка): А — Laevidentalina, х 7000; Б — Percultazonaria, х 3000; 4 — отверстия пор на поверхности стенки некоторых Globigerinata (бифон-
тинальная стенка): А — Globorotalia, х 1500; Б — Globigerinoides, х 4000 (краст). 5 — отверстия пор на поверхности Eponides (Rotaliana), х 1500 (бифон-
тинальная стенка). (7 — по: Михалевич и др., 1986; 2 — по: Михалевич, 1983; 3 — по: Loeblich, Tappan, 1988).
Fig. 536. Structural details of the monofontinal and bifontinal wall.
/ — Deniostomina (Schlumbergerinana) (monofontinal wall): A — outer view of the shell; Б — needle ultrastructure of the inner shell surface and tooth, x 10 000;
В — transverse section of the shell, x 200; Г — microgranular ultrastructure (transverse section), x 3000. 2 — Borelis (Miliolana), x 220 (monofontinal wall);
pseudopore openings seen from the outside. 3 — pseudopore openings at the shell surface of some Nodosariata (monofontinal wall): A — Laevidentalina, x 7000;
Б — Percultazonaria, x 3000. 4 — Pore openings at the shell surface of some Globigerinata (bifontinal wall): A — Globorotalia, x 1500; Б — Globigerinoides
x 4000 (crust). 5 — Pore openings at the shell surface of Eponides (Rotaliana), x 1500 (bifontinal wall).
Lcrgynana || Arcrnoscalccricrricz
Рис. 547.
V Rotate ata
IV. Nodosariata
Рис. 547 (продолжение).
Рис. 547. Характерные представители различных классов и подклассов фораминифер (сплошная линия — границы между классами,
пунктирная — между подклассами). (По: Михалевич, 1999, с изменениями).
/.Класс Astrorhizata. Подкласс Lagynana: 1 — Diplogromia; 2 — Lagynis; 3 — Phainogulmia; 4 — Xenothekella. Подкласс Ammoscalariana: 5, A, 5, Б —
Periptigma; 6 — Ammoscalaria. Подкласс Astrorhizana: 1 — Sorostomasphaera, 2—Thuramminopsis, 3—Halyphysema, 4—Lagenammina, 5 — Astrammina, 6, A,
6, Б — Schizammina (A — раковина, Б — внутреннее строение лабиринтовой стенки); 7 — Masonella (с канальцами в толстой стенке); 8 — Rhabdammina,
9 — Notodendrodes; 10—Saccorhiza. ZZ. Класс Spirillinata. Подкласс Ammodiscana: 1 — Ammodiscus, 2 — Repmanic. Подкласс Spirillinana: 3 — Archa-
ediscus (сечение), 4 — Howchinia (аксиальное сечение), 5 — Lasiotrochus (аксиальное сечение), 6 — Babelispirillic (аксиальное сечение), 7 — Coronipora,
8 — Trocholina, 9 — Spirillina, 10 — Spirotrocholina (10, A, 10, Б — вид раковины co спиральной стороны и сбоку, 10, В — схема каналов на аксиальном
разрезе); 11 —Annulopate Ilina', 12 — Patellina (вид с устьевой стороны, с Т-образной пластинкой). ZZZ. Класс Mi И о lata. Подкласс Schlumbergerinana: 1 —
Dentostomina’, 2 — Sigmoilopsis; 3—Ammomassilina. Подкласс Miliolana: 4 — Squamulic; 5 — Cornuspira; 6 — Cornuspiroides; 7 — Gheorgianina; 8 — Discos-
pirina; 9 — Spirophtalmidium; 10 — Ophtalmidium; 11—13 — Quinqueloculina (13 — схема расположения камер на поперечном сечении); 14 — Idalina
(поперечное сечение с начальными клубкообразными оборотами и поздними оборотами, объемлющими в одной плоскости); 15 — Articulina, 16 — Alveolic
(схема строения); 17, А, 17, Б — Hauerina (17, А — микросферическая форма, почти целиком плоскоспиральная с едва заметным начальным квинквелоку-
линовым отделом; 17, Б — мегалосферическая форма, почти целиком квинквелокулиновая); 18, А, 18, Б, 19 — Fisherinella; 20 — Laevipeneroplis; 21 —
Parasorites. IV. Класс Nodosariata. Подкласс Hormosinana: Z — Hormosinella; 2 — Saccamminis; 3, A, 3, Б, 3, В — Nodosinum (3, A — раковина; 3, Б —
вид радиального устья с продольными ребрами изнутри; 3, В — вид устья снаружи, сверху); 4 — Adelungia; 5 — Pseudopalmula. Подкласс Nodosariana: 6 —
Lagena; 7 — Parqfissurina; 8 — Syzrania; 9, 10 — Grigelis; 11 — Nodosaria; 12 — Multiseptida (схема продольного сечения); 13, A, 13, Б — Lingulina (13,
A — микросферическая, 13, Б — мегалосферическая формы); 14 — Marginulic; 15, А, 15, Б — Tristix; 16 — Kyphopixa; 17 — Dyofrondicularia; 18 —
Flabellina, 19 — Polymorphic; 20 — Laryngosigma; 21 — Lenticulic. V. Класс Rotaliata. Подкласс Textulariana: Z — Haplophragmoides; 2, A, 2, Б —
Fissotextularia (2, A — мегалоферическая, 2, Б — микросферическая формы); 3 — Pseudobolivina; 4 — Cuneolinella; 5, A, 5, Б — Clavulina (5, A — внешний
вид раковины; 5, Б — сечение начального отдела); 6 — Minouxia; 7 — Spiroplectinata; 8, А, 8, Б — Gaudryina; 9, А, 9, Б — Dobrogelina (9, А — вид со
спиральной; 9, Б — с пупочной стороны); 10, А, 10, Б — Asterotrochammina (10, А — вид со спиральной, 10, Б — с пупочной стороны); 11, А, 11, Б —
Arenagula (11, А — вид со спиральной, 11, Б — с пупочной стороны); 12 — Tiphotrocha (вид с пупочной стороны); 13 — Montsechiana. Подкласс Rotaliana:
14 — Bermudezinella; 15 — Elphidium; 16 — Bolivinellina; 17 — Brizalina; 18 — Sporobulimina; 19 — Pavonina; 20 — Pseudobuliminella; 21 — Reussella; 22 —
Eouvigerina; 23 — Euuvigerina; 24, A, 24, Б — Sestronophora (24, A — вид co спиральной, 24, Б — с пупочной стороны); 25 — Rectoeponides (вид с пупочной
стороны); 26, А, 26, Б — Discorbis (26, А — вид со спиральной, 26, Б — с пупочной стороны); 27 — фрагмент системы каналов Rotalia; 28, А, 28, Б —
Neocribrella (28, А — со спиральной, 28, Б — с пупочной стороны); 29 — Neoconorbina (вид с пупочной стороны); 30 — Baculogypsinoides (горизонтальное
сечение мегалосферической особи с каналами); 31 — Eulinderina (срединное сечение мегалосферической формы).
Fig. 547. Characteristic representatives of the different Foraminiferal classes and subclasses (unbroken line — boundary between the classes,
dotted line — between the subclasses).
/.Class Astrorhizata. Subclass Lagynana: 1 — Diplogromia; 2 — Lagynis; 3 — Phainogulmia; 4 — Xenothekella. Subclass Ammoscalariana: 5, А, Б —
Periptigma; 6 — Ammoscalaria. Subclass Astrorhizana: 1 — Sorostomasphaera; 2 — Thuramminopsis; 3 — Halyphysema; 4 — Lagenammina; 5 — Astrammina;
6, А, Б — Schizammina (A — the shell, Б — inner structure of the labyrinthic wall); 7 — Masonella (with canaliculi in the thick wall); 8 — Rhabdammina; 9 —
Notodendrodes; 10 — Saccorhiza. ZZ. Class Spirillinata. Subclass Ammodiscana: 1 — Ammodiscus; 2 — Repmanina. Subclass Spirillinana: 3 — Archaediscus
(section); 4 — Howchinia (axial section); 5 — Lasiotrochus (axial section); 6 — Babelispirillina (axial section); 7 — Coronipora; 8 — Trocholina; 9 — Spirillina;
10 — Spirotrocholina (А, Б — view of the shell from the spiral and lateral sides; В — scheme of the canals, axial section); 11 — AnnulopateIlina; 12 — Patellina
(view from the apertural side, with the T-shaped plate). ZZZ. Class Miliolata. Subclass Schlumbergerinana: 1 — Dentostomina; 2 — Sigmoilopsis; 3 — Ammo-
massilina. Subclass Miliolana: 4 — Squamulina; 5 — Cornuspira; 6 — Cornuspiroides; 7 — Gheorgianina; 8 — Discospirina; 9 — Spirophtalmidium; 10 —
Ophtalmidium; 11—13 — Quinqueloculina (13 — including the transverse section showing chamber arrangement); 14 — Idalina (transverse section showing initial
glomerate whorls and later ones, embracing and pianospiral); 15 — Articulina; 16 — Alveolina (schematic structure); 17, A, 17, Б — Hauerina (A — microspheric
form, nearly entirely pianospiral with the initial quinqueloculine part hardly seen, Б — megalospheric form, nearly entirely quinqueloculine); 18 A, 18 Б, 19 —
Fisherinella; 20 — Laevipeneroplis; 21 — Parasorites. IV. Class Nodosariata. Subclass Hormosinana: 1 — Hormosinella; 2 — Saccamminis; 3, A, 3, Б, 3,
В — Nodosinum (A — view of the shell, Б — view of the radial aperture with the inner longitudinal ribs, В — outer apertural view from the apertural side); 4 —
Adelungia; 5 — Pseudopalmula. Subclass Nodosariana: 6 — Lagena; 7 — Parafissurina; 8 — Syzrania; 9, 10 — Grigelis; 11 — Nodosaria; 12 — Multiseptida;
(scheme of the longitudinal section); 13, A, 13, Б — Lingulina (A — microspheric, Б — megalospheric forms); 14 — Marginulina; 15, A, 15, Б — Tristix; 16 —
Kyphopixa; 17 — Dyofrondicularia; 18 — Flabellina; 19 — Polymorphic; 20 — Laryngosigma; 21 — Lenticulina. V. Class Rotaliata. Subclass Textulariana:
1 — Haplophragmoides; 2, A, 2, Б — Fissotextularia (A — megalospheric, Б — microspheric forms); 3 — Pseudobolivina; 4 — Cuneolinella; 5 A, 5, Б —
Clavulic (A — outer view of the shell; Б — section of the initial part); 6 — Minouxia; 7 — Spiroplectinata; 8 A, 8, Б — Gaudryic; 9 A, 9, Б — Dobrogelina
(A — view from the spiral side, Б — view from the umbilical side); 10 A, 10, Б Asterotrochammina (A — view from the spiral side, Б — view from the umbilical
side); 11, A, 11, Б — Arenagula (A — view from the spiral side, Б — view from the umbilical side); 12 — Tiphotrocha (view from the umbilical side); 13 —
Montsechiac. Subclass Rotaliana: 14 — Bermudezinella; 15 — Elphidium; 16 — Bolivinellic; 17 — Brizalina; 18 — Sporobulimina; 19 — Pavonina; 20 —
Pseudobuliminella; 21 — Reussella; 22 — Eouvigerina; 23 — Euuvigerina; 24, A, 24, Б — Sestronophora (A — view from the spiral side, Б — view from the
umbilical side); 25 — Rectoeponides (view from the umbilical side); 26, A, 26, Б — Discorbis (A — view from the spiral side, Б — view from the umbilical side);
27 — fragment of Rotalia canal system; 28, A, 28, Б — Neocribrella (A — view from the spiral side, Б — view from the umbilical side); 29 — Neoconorbic
(view from the umbilical side); 30 — Baculogypsinoides, horizontal section of the megalospheric individual with the canals); 31 — Eulinderina (median section of
the megalospheric form).
Тип POLYMASTIGOTA Biitschli, 1884-
ПОЛ И МАСТИ ГОТЫ
К типу относятся гетеротрофные жгутиконосцы, имеющие 2 и
более жгутика и обладающие уникальным для типа комплексом
органелл — кариомастигонтом, который включает кинетосомы,
сложную систему жгутиковых корешков и ядро. В каждом классе
этого типа наблюдается полимеризация жгутиков или всего карио*
мастигонта. Полимастиготы — преимущественно комменсалы и
эндопаразиты органов пищеварения беспозвоночных и позвоноч-
ных животных, а также человека. Некоторые виды — патогенные
паразиты урогенитального тракта млекопитающих и человека.
Обитая во внутренних органах своих хозяев, полимастиготы обла-
дают многими адаптивными признаками, к которым относятся ка-
риомастигонт, ундулирующая мембрана у ряда видов, гидрогеносо-
мы, отсутствие митохондрий, а у многих видов и отсутствие дик-
тиосом аппарата Гольджи.
Свободноживущих видов немного, они — обитатели водоемов
с высоким содержанием органических веществ, преимущественно
анаэробы.
В тип включены 3 класса:
Класс Diplomonadea Wenyon, 1926 — Дипломонадеи
Класс Oxymonadea Grasse, 1952 — Оксимонадовые, или Оксимо-
нады
Класс Parabasalea Honigberg, 1973 — Парабазалии
Общее число видов полимастигот около 150.
Распространены всесветно.
Класс Diplomonadea Wenyon, 1926— Дипломонадеи
Класс объединяет мелких, не более 20 мкм в длину, гетеро-
трофных свободноживущих и паразитических видов с осевой (ак-
сиальной) симметрией. Специфическую морфофункциональную
особенность дипломонадей составляет кариомастигонт, включаю-
щий 1—4 жгутика с их кинетосомами и системой жгутиковых ко-
решков, а также ядро. Кариомастигонт имеет большое таксономи-
ческое значение, определяя, в частности, деление на отряды: в
отряд Enteromonadida выделены виды с 1 кариомастигонтом, отряд
Diplomonadida включает виды с 2 кариомастигонтами и соответст-
венно с двойным комплексом органелл цитоплазмы. В клетках
дипломонадей отсутствует аксостиль.
Питание осуществляется посредством фаго- и пиноцитоза. Раз-
множение агамное путем бинарного деления активного трофозоита
или в цисте. Деление ядра — закрытый плевромитоз (подкласс
Retortamonadia) или полузакрытый ортомитоз (подкласс Diplomo-
nadia). В классе около 70 видов, среди которых преобладают эндо-
паразиты животных и человека. Распространены всесветно.
История изучения
Несмотря на то что отряд Diplomonadida был вы-
делен в качестве самостоятельного таксона этого
ранга давно (Wenyon, 1926), жгутиконосцев, вклю-
ченных в него, в течение длительного периода отно-
сили к отряду Protomonadida Blochmann, 1895 (Dofle-
in, 1953). В этом отряде для дипломонадид было вы-
делено отдельное сем. Distomatidae Klebs, 1892 (син.:
Diplozoa Dangeard, 1910). Затем эта группа жгутико-
носцев была помещена в отряд Polymastigida Bloch-
22 Протисты, часть 1
mann, 1895, в котором оставалась до настоящего вре-
мени.
Значительные трудности в изучении морфологии,
функциональной активности, жизненных циклов и
таксономии связаны с очень мелкими размерами этих
жгутиконосцев. Поэтому прогресс во всех исследова-
ниях дипломонадид основан на применении методов
электронной микроскопии. В настоящее время ульт-
ратонкое строение трофозоитов известно для основ-
ных видов отряда, что отразилось и на его системати-
ке (Соловьёв, Ченцов, 1966, 1970; Brugerolle et al.,
1973а, 1973b, 1974; Brugerolle, 1974; Kulda, Nohyn-
kova, 1978).
Объединение родов Retortamonas и Chilomastix в
одно семейство, а затем и в один отряд основано на
их глубоком морфологическом сходстве, установлен-
ном в настоящее время на уровне как световой, так и
электронной микроскопии (Brugerolle, 1973, 1977).
Первоначально семейство было названо Embadomo-
nadidae Alexeieff, 1912, но затем получило современ-
ное название Retortamonadidae Wenrich, 1932 по на-
званию типового рода Retortamonas Grassi, 1879.
Большинство видов подкласса Retortamonadia ис-
следовано только под световым микроскопом, и
лишь у нескольких видов изучено ультратонкое
строение (Brugerolle, 1973, 1977). Разработаны мето-
ды культивирования этих простейших на жидких пи-
338
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тательных средах, но аксеничные культуры получить
не удается (Kulda, Nohynkova, 1978).
В 70-х гг. проведена ревизия отряда Diplomonadi-
da на основании данных электронной микроскопии
и установлена таксономическая позиция семейств,
подсемейств и родов (Brugerolle, 1975а, 1975b;
Kulda, Nohynkova, 1978). Основанием для включе-
ния в отряд сем. Enteromonadidae, к которому отно-
сятся виды с 1 кариомастигонтом, послужили дан-
ные по ультраструктуре жгутиконосцев рода Entero-
monas da Fonseca, 1915 (син.: Tricercomonas Wenyon
et O’Konnor, 1917). Эти данные показали высокую
степень их сходства с представителями одного из
типичных родов дипломонадид — Hexamita Dujar-
din, 1838 (Brugerolle, 1975а). Однако впоследствии
это семейство с относящимися к нему родами было
выделено в самостоятельный отряд в подклассе Dip-
lomonadia Wenyon, 1926.
На основе данных электронной микроскопии раз-
работана система сем. Hexamitidae Kent, 1880. Осо-
бенно подробно изучены эндопаразитические виды,
патогенные для животных и человека. Это, прежде
всего представители рода Giardia Kiinstler, 1882, для
которых известны также названия: Cercomonas Lambl,
1859; Hexamita Davaine, 1875; Megastoma Grassi, 1881;
Lamblia Blanchard, 1888.
Из видов рода Giardia наибольшее практическое
значение имеет G. intestinalis Alexeieff, 1914 (син.:
Cercomonas intestinalis Lambl, 1859; Megastoma ente-
rica Grassi, 1881; Lamblia intestinalis Blanchard,
1888; Giardia lamblia Stiller, 1915; G. enterica Kofo-
id, 1920). В современных литературных источниках
наиболее часто используются 2 названия вида: си-
нонимы Giardia intestinalis и Lamblia intestinalis, а
заболевание человека, вызванное этими жгутиконос-
цами, получило название жиардиазис, или лямблиоз.
Название «лямблия» дано по фамилии профессора
Харьковского университета В. Лямбля, который
впервые описал этого жгутиконосца (1859 г.). Пер-
вое наблюдение над жгутиконосцами из кишечника
человека принадлежит А. Левенгуку и относится к
1681 г.
Для исследования отряда Diplomonadida (особен-
но лямблий — род Giardia) большое значение имеют
культуры in vitro. В культурах детально изучены мор-
фология трофозоитов и цист, механизм прикрепле-
ния лямблий к субстрату, способ деления клетки (Ка-
рапетян, 1960; Соловьёв, 1963; Соловьёв, Ченцов,
1966; Шарапов, Соловьёв, 1976). Однако большинст-
во видов отряда изучено еще недостаточно.
Классификация
Класс Diplomonadea Wenyon, 1926
Подкласс Retortamonadia Grass6, 1952
Отряд Retortamonadida Grasse, 1952
Сем. Retortamonadidae Wenrich, 1932 (син.: Embadomo-
nadidae Alexeieff, 1917; 2 рода, около 40 видов).
Подкласс Diplomonadia Wenyon, 1926
Отряд Enteromonadida Brugerolle, 1975
Сем. Enteromonadidae Kulda, Nohynkova, 1978 (3 рода)
Отряд Diplomonadida Wenyon, 1926
Сем. Hexamitidae Kent, 1880 (3 рода)
Сем. Giardiidae Kulda, Nohynkova, 1978 (2 рода)
Retortomonadia отличаются от Diplomonadia нали-
чием цитостома, строением покровов, образованных
тубулеммой, и особенностями деления ядра.
Морфология и жизненный
цикл
Жизненный цикл Diplomonadea включает 2 фазы:
1) трофозоит, 2) цисту. Размножение путем бинарно-
го деления происходит как в фазе трофозоита, так и
на стадии цисты. Особенности морфологии, размно-
жения, жизненных циклов дипломонадей, их культи-
вирования in vitro, а также происхождение и эволю-
ция рассматриваются по каждому подклассу раздель-
но. Систематический обзор сделан также по каждому
подклассу.
Подкласс Retortamonadia Grassd, 1952
Морфология, размножение,
жизненный цикл
Ретортамонады — мелкие одноядерные жгутико-
носцы, не более 20—25 мкм в длину. Форма тела гру-
шевидная или овальная. Покровы представлены тубу-
леммой. На переднем конце тела находится 2—
4 жгутика, один из которых направлен назад и
проходит в углублении цитостома. Пузырьковидное
ядро прилегает к кинетосомам. Митохондрии и дик-
тиосомы аппарата Гольджи не обнаружены. Размно-
жение агамное, путем бинарного деления клетки. Де-
ление ядра осуществляется посредством закрытого
или полузакрытого плевромитоза. Цисты образуются
у всех видов. Почти все виды (за исключением одно-
го) являются непатогенными паразитами пищевари-
тельного тракта многих беспозвоночных, всех групп
позвоночных животных и человека.
Для трофозоитов характерна грушевидная или
овальная форма тела с закругленным передним и вы-
тянутым задним концом (рис. 287). Тело обычно уп-
лощено в дорсовентральном направлении. Цитостом
расположен на вентральной стороне тела. Он пред-
ставляет собой трубковидное углубление, или желоб
(рис. 287), и укрепляется микротрубочками и микро-
филаментами. Цитостом переходит в цитофаринкс,
на дне которого происходит эндоцитоз. Форма и раз-
меры цитостома и поддерживающих его структур раз-
личны у разных видов.
У жгутиконосцев обоих родов имеется 4 кинето-
сомы, расположенные попарно. От них отходят фиб-
риллярные и микротрубочковые корешки, часть ко-
торых направлена к ядру, а часть вдоль цитостома.
Цитостомальные корешки усиливаются дополнитель-
ными микротрубочками и микрофиламентами, фор-
мируя опорные структуры вдоль краев цитостома.
ТИП POLYMASTIGOTA, КЛАСС DIPLOMONADEA. ПОДКЛАСС RETORTAMONADIA
339
Рис. 287. Retortomonas intestinalis. (По: Kulda, Nohynkova, 1978).
A — трофозоит, Б — циста. 1 — ядро, 2 — передний жгутик, 3 — зад-
ний жгутик, 4 — цитостом, 5 — цитостомальные фибриллы, 6 — оболоч-
ка цисты.
Fig. 287. Retortomonas intestinalis.
А — trophozoite; Б — cyst. 1 — nucleus, 2 — anterior flagellum, 3 — recur-
rent flagellum, 4 — cytostome, 5 — fibrilles of cytostome, 6 — cyst wall.
У видов рода Chilomastix все 4 кинетосомы несут
жгутики, из которых 3 двигательных направлены впе-
ред, а 1 рекуррентный — назад и идет вдоль цитосто-
ма. У видов Retortamonas только 2 жгутика: перед-
ний, двигательный, и задний, рулевой (рис. 287; 288).
Задний жгутик часто имеет складки, увеличивающие
его поверхность. Основная пища ретортамонадид —
бактерий, скопления которых можно видеть на дне
цитостома и в пищеварительных вакуолях (Brugerol-
le, 1973, 1977).
Цитоплазма богата каналами ЭПР, пищеваритель-
ными вакуолями и мелкими гранулами запасных ве-
ществ (рис. 289). Митохондрии, диктиосомы и какие-
либо аксиальные элементы цитоскелета у ретортамо-
надид отсутствуют.
Формирование цист в жизненном цикле — об-
щая особенность видов ретортамонадид. Цисты мел-
кие, грушевидной или овальной формы (рис. 287; 288).
Размеры цист несколько различаются у разных видов
и достигают не более 7—10 мкм (рис. 287; 288).
Цисты бесцветные. Снаружи они покрыты довольно
толстой оболочкой, содержащей филаментозный ма-
териал. Оболочка плотная, сплошная, без отверстий
и мелких пор.
В цитоплазме цисты находятся все органеллы,
свойственные трофозоиту, и их расположение сходно
с таковым у трофозоита (рис. 288). Ядро также со-
храняет свое положение на переднем полюсе цисты.
Субмембранные микротрубочки в цисте отсутствуют.
Они резорбируются при инцистировании (Brugerolle,
1973). По своей природе — это цисты покоя. Они
выводятся из кишечника животных и человека вместе
с фекальными массами и представляют инфекцион-
ную стадию в жизненном цикле ретортамонадид.
Трофозоиты ретортамонадид (Retortamonas и Chi-
lomastix} размножаются в кишечнике хозяина агамно
путем бинарного деления клетки в продольном на-
правлении. Механизмы митозами цитокинеза изучены
недостаточно. Известно, что у Retortamonas митоз за-
крытого типа и все его фазы протекают под ядерной
оболочкой (рис. 290). Изучение ультраструктуры де-
лящейся клетки показало, что центрами веретена де-
ления служат кинетосомы, которые образуют 2 груп-
пы (по паре кинетосом в каждой) и находятся в тес-
ном контакте с ядерной оболочкой.
Митотическое веретено у Retortamonas внутри-
ядерное, и его микротрубочки прикрепляются к внут-
ренней ядерной мембране. У Chilomastix микротру-
Рис. 288. Виды рода Chilomastix.
А — трофозоит Chilomastix mesnili, Б — циста Ch. mesnili, В — Ch. gallinarum, Г — Ch. undulata. 1 — ядро, 2 — 3 свободных передних жгутика, 3 —
задний жгутик, 4 — цитостом, 5 — цитостомальные фибриллы, 6 — пищеварительные вакуоли, 7 — оболочка цисты. (А, Б — по: Doflein, 1953; В, Г —
по: Мыльников, Жуков, 1983).
Fig. 288. Some species of Chilomastix.
A — trophozoite of Chilomastix mesnili, Б — cyst of Ch. mesnili, В — Ch. gallinarum, Г — Ch. undulata. 1 — nucleus, 2 — three anterior flagella, 3 — recur-
rent flagellum, 4 — cytostome, 5 — fibrilles of cytostome, 6 — digestive vacuoles, 7 — cyst wall.
340
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 289. Схема ультраструктурной организации Retortamonas agi-
lis. (По: Brugerolle, 1977).
1 — передний жгутик; 2 — ядро; 3 — кинетосомы; 4 — задний жгутик;
5 — цитостом; 6 — цитофаринкс; 7 — ленты микротрубочек и исчерчен-
ные фибриллы, поддерживаюШие цитостом и окружающие цитофаринкс;
8 — микротрубочки тубулеммы; 9 — микрофибриллы, окружающие ядро;
10 — цистерны ЭПР; 11 — пищеварительные вакуоли.
Fig. 289. Diagrammatic expression of ultrastructure organization of
Retortamonas agilis.
1 — anterior flagellum, 2 — nucleus, 3 — kinetosomes, 4 — recurrent flagel-
lum, 5 — cytostome, 6 — cytopharynx, 7 — band of microtubules and strai-
ted fibrilles, supporting cytostome and surrounding cytopharynx, 8 — microtu-
bules of tubulemma, 9 — microfibrilles surrounding nucleus, 10 — endoplas-
mic reticulum, 11 — digestive vacuoles.
бочки веретена выходят наружу и прикрепляются
к кинетосомам. Это — полузакрытый митоз с цито-
плазматическим центром веретена деления (Bruge-
rolle, 1977). Хромосомы очень мелкие, их число не
всегда удается сосчитать, но известно, что у Ch. inte-
stinalis их 6, а у Ch. aulastomi 20—24 (рис. 290). Из-
менения клеточных органелл в период митоза изуче-
ны у Ch. intestinalis. В поздней профазе происходит
резорбция цитостома и жгутиков материнской клет-
ки. Эти органеллы формируются заново в ранней те-
лофазе (Brugerolle, 1973).
Интересные наблюдения за процессом деления
трофозоитов у представителей рода Retortamonas
были проведены с помощью светового микроскопа.
У них обнаружено нарушение обычного процесса
деления: в клетке делятся ядра и формируются пол-
ные наборы клеточных органелл у дочерних клеток,
но не происходит цитокинез. В результате образуют-
ся организмы с 2 ядрами и двойным набором клеточ-
ных органелл, напоминающие дипломонадид (Bis-
hop, 1934; Doflein, 1953).
Деление ретортамонадид осуществляется только в
фазе трофозоита; в цисте деления не происходит (Kul-
da, Nohynkova, 1978). Половой процесс не обнаружен.
Культивирование
Ряд видов Retortamonas и Chilomastix выделены в
культуры in vitro и живут в них при регулярной смене
культуральной среды в течение длительного времени.
Для культивирования чаще всего используются двух-
фазные среды, пригодные для роста кишечных амеб
и трихомонад. Наиболее часто используются моди-
фицированные среды на растворе Рингера и Ринге-
ра—Локка с добавлением сыворотки крови и содер-
жимого куриного яйца (Boeck, Drobohlav, 1925; Do-
bell, Leidlaw, 1926; Wenrich, 1947; Hilber et al., 1960).
Среды подвергаются стерилизации в автоклаве.
Для содержания культур необходимы следующие
условия. Культуральная среда должна иметь pH в
пределах 7.2—7.4. Жгутиконосцы, обитающие в ки-
шечнике млекопитающих, растут наиболее быстро
Рис. 290. Chilomastix aulastomi.
(По: Doflein, 1953).
А — трофозоит, Б — фазы деления.
Fig. 290. Chilomastix aulastomi.
А — trophozoite, Б — phases of cell di-
vision.
ТИП POLYMASTIGOTA. КЛАСС DIPLOMONADEA. ПОДКЛАСС DIPLOMONADIA
341
при 35—37 °C, виды из кишечника птиц — при 38—
40 °C, виды же из кишечника амфибий, рептилий и
беспозвоночных — при комнатной температуре в
анаэробных условиях. При комнатной температуре
способны размножаться также виды из кишечника
млекопитающих и птиц, но развитие культуры идет
значительно медленнее. В культурах ретортамонади-
ды регулярно образуют цисты.
Происхождение и эволюция
Представители отряда Retortamonadida имеют мор-
фологическое сходство с жгутиконосцами отрядов Dip-
lomonadida и Oxymonadida (класс Oxymonadea). К
числу сходных черт относятся: кинетосомы, располо-
женные двумя парами; цитостомы, которые поддер-
живаются фибриллярными структурами у ретортамона-
дид и дипломонадид. Митоз у ряда видов ретортамо-
надид, протекающий с образованием внутриядерного
веретена, сходен с митозом оксимонадид, а полузак-
рытый митоз сходен с митозом такого же типа у дип-
ломонадид. Кроме того, у жгутиконосцев этих 3 от-
рядов отсутствуют митохондрии и диктиосомы аппа-
рата Гольджи.
Сходство морфологических признаков может слу-
жить одним из доказательств существования единой
предковой группы, от которой берут начало жгутико-
носцы всех 3 отрядов. Виды предковой группы были,
вероятно, свободноживущими, и их дальнейшая эво-
люция проходила по линии адаптации к паразитичес-
кому образу жизни в кишечнике самых разнообраз-
ных животных. Филогенетические взаимоотношения
ретортамонадид требуют дальнейшего изучения.
Систематический обзор
Род Retortamonas Grassi, 1879
(Син.: Embadomonas Mackinnon, 1911; Waskia Wenyon et
O’Connor, 1917) (рис. 287)
Род включает жгутиконосцев с 2 жгутиками не-
равной длины. Цитостомальная зона овальной фор-
мы и поддерживается 2 фибриллами. Виды этого
рода обнаружены у человека, обезьян, крупного ро-
гатого скота, овец, морских свинок и кроликов.
Все эти жгутиконосцы не патогенны для своих хо-
зяев.
Род Chilomastix Alexeieff,
1912
(Син.: Macrostoma Alexeieff, 1909)
Для рода характерно наличие 4 жгутиков, из кото-
рых 3 передних, а 1 направлен назад. Цитостомаль-
ная зона в форме продольной щели (рис. 287) и под-
держивается 2 фибриллами. Виды Chilomastix обна-
ружены в кишечнике представителей всех классов
позвоночных, а также у насекомых (термиты) и коль-
чатых червей (пиявки). В кишечнике человека обна-
ружен 1 вид — Ch. mesnili Alexeieff, 1912. Так же
как и другие виды ретортамонадид, этот вид для че-
ловека не патогенен. К этому роду относится единст-
венный свободноживущий вид отряда — Ch. undulata
Skuja (рис. 287), который обитает в анаэробных усло-
виях лимносапробных водоемов.
Подкласс Diplomonadia Wenyon, 1926
Морфология и физиология
Трофозоит
Внешнее строение. Дипломонады — мелкие бес-
цветные жгутиконосцы, размеры тела которых варь-
ируют от 5 до 20 мкм в длину у разных видов. Форма
тела трофозоитов также варьирует и может быть гру-
шевидной, яйцевидной, овальной. Передний конец
тела у многих видов закруглен, а задний, или хвосто-
вой, конец вытянут и заострен (рис. 291). Тело вы-
пуклое с дорсальной стороны и плоское с вентраль-
ной. Обе стороны тела различаются по своему строению.
Вентральная сторона имеет более сложное строение
по сравнению с дорсальной. По ней вдоль централь-
ной оси тела проходит углубление — желоб, в кото-
ром размещается пара центральных жгутиков. У ряда
видов в задней части тела находятся цитостомы (2), а
у рода Giardia (Lamblia) большую часть поверхности
вентральной стороны занимает присасывательный диск
(см. с. 343).
Одним из основных признаков дипломонад являет-
ся осевая (аксиальная) симметрия большинства видов,
которая обусловлена локализацией в одной клетке 2
ядер и 2 наборов органелл опорно-двигательного ап-
парата (кариомастигонты). Исключение составляют
виды отряда Enteromonadida, трофозоиты которых
имеют только 1 кариомастигонт (рис. 291, В), кото-
рый по своей структуре сходен с кариомастигонтами
других дипломонадей.
Кариомастигонт представляет собой опорно-дви-
гательный аппарат. В состав одного кариомастигонта
входят жгутики, их кинетосомы, сложная система
жгутиковых корешков и 1 ядро, расположенное на
переднем конце тела и функционально связанное с
другими элементами комплекса (рис. 291; 292).
Число жгутиков у трофозоитов разных видов варь-
ирует от 1 (род Caviomonas) до 8 (сем. Hexamitidae).
У видов с 1 жгутиком, например у С. mobilis
(рис. 291; 292), жгутик, берущий начало от кинето-
сомы, которая расположена на переднем конце тела
над ядром, направлен назад и длина его превышает
длину тела. На переднем конце жгутик прилегает к
поверхности тела.
Виды сем. Hexamitidae с осевой симметрией тела
имеют 4 пары жгутиков, которые берут начало от 2
342
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 291. Морфология клетки Caviomonas и Enteromonas. (По:
Kulda, Nohynkova, 1978).
A — Caviomonas mobilis из кишечника морской свинки: 1 — ядро, 2 —
микротубулярная лента, 3 — жгутик; Б — Enteromonas hominis из кишеч-
ника человека: 1 — ядро, 2 — передние жгутики, 3 — хвостовой жгутик,
4 — микротубулярная лента; В — схема ультратонкой организации Entero-
monas sp.: 7 — передние жгутики, 2 — задний жгутик, 3 — лента микро-
трубочек, 4 — цистерны ЭПР.
Fig. 291. Cell morphology of Caviomonas and Enteromonas.
A — Caviomonas mobilis from intestinae of guinea pig: 1 — nucleus, 2 —
microtubular band, 3 — flagellum; Б — Enteromonas hominis from intestine
of a men: 7 — niicleus, 2 — anterior flagella, 3 — tail flagellum, 4 — micro-
tubular band; В — diagram of ultrastructure of Enteromonas sp.: 7 — anterior
flagella, 2 — recurrent flagellum, 3 — band of microtubules, 4 — endoplas-
mic reticulum.
Рис. 292. Особенности морфологии дипломонадей рода Trepomo-
nas. (По: Brugerolle et al., 1973).
А — морфология Т. rotans, Б — схема ультратонкого строения клетки
Trepomonas sp. 1 — передний жгутик, 2 — свободные задние жгутики,
3 — ядра, 4 — цитостомальные фибриллы, 5 — исчерченные ламеллы,
6 — микротубулярные фибриллы.
Fig. 292. Morphology of Diplomonadea of genus Trepomonas.
A — morphology of T. rotans; Б — diagram of ultrastructure of Trepomonas
sp. 7 — anterior flagella, 2 — free recurrent flagella, 3 — nuclei, 4 — cytos-
tomal fibrills, 5 — striated lamellae, 6 — microtubular fibrills.
Рис. 294. Род Octomitus.
Рис. 293. Морфология клетки Hexamita.
А — общий вид Н. inflata; Б — схема ультратонкого строения Hexamita
sp.; В — поперечный срез в области ядер (схема), видны все элементы ка-
риомастигонта, расположение кинетосом и жгутиков. (А — по: Doflein,
1953; Б, В по: Brugerolle, 1974). 1 — передние жгутики, 2 — ядра, 3 — за-
дние жгутики, 4 — цитостомы, 5 — цистерны гранулярного ЭПР.
Fig. 293. Morphology of Hexamita cell.
A — face view of H. inflata; Б — diagram of ultrastructure of Hexamita sp.;
В — transversal section through nuclei (diagram), illustrates all elements of
karyomastigont, arrangement of kinetosomes and flagella. 1 — anterior flagel-
la, 2 — nuclei, 3 — posterior flagella, 4 — cytostomes, 5 — cisterns of gra-
nular endoplasmic reticulum.
A — общий вид трофозоита О. muris; Б — циста; В — схема ультратонко-
го строения Octomitus sp.: 1 — передние жгутики, 2 — ядра, 3 — аксоне-
мы хвостовых жгутиков, 4 — цистерны гранулярного ЭПР, 5 — исчерчен-
ные ламеллы; Г — поперечный срез в области ядер — видны все элементы
кариомастигонта, расположение и форма ядер, жгутики и их кинетосо-
мы, межъядерные фибриллы. (А, Б — по: Doflein, 1953; В, Г — по: Bruge-
rolle et al., 1974).
Fig. 294. Genus Octomitus.
A — total view of trophozoite of O. muris; Б — cyst; В — diagram of ultrast-
ructure of Octomitus sp.: 7 — anterior flagella, 2 — nuclei, 3 — axonemes of
tail flagella, 4 — cisterns of granular endoplasmic reticulum, 5 — striated la-
mellae; Г — transversal section through nuclei: the section illustrates all ele-
ments of karyomastigont, arrangement and form of nuclei, flagella, kinetoso-
mes, intranuclear fibrills.
ТИП POLYMASTIGOTA. КЛАСС DIPLOMONADEA. ПОДКЛАСС DIPLOMONADIA
343
Рис. 295. Род Spironucleus.
А — общий вид S. elegans; Б -г схема строения 5. elegans по данным элек-
тронного микроскопа: 1 — передние жгутики, 2 — ядро, 3 — аксонемы
хвостовых жгутиков с окружающими их микротубулярными лентами, 4 —-
цитостомы; В — поперечный срез в области ядер — видны элементы кари-
омастигонта, расположение ядер, кинетосом и жгутиков. (А — по: Doflein,
1953; Я, В — по: Brugerolle et al., 1973в).
Fig. 295. Genus Spironucleus.
A — total view of S. elegans; Б — diagram of ultrastructure of S. elegans: 1 —
anterior flagella, 2 — nucleus, 3 — axonemes of tail flagella surrounded by
microtubular bands, 4 — cytostomes; В — transversal section through nuclei.
The section represents components of karyomastigont, arrangement and form of
nuclei, kinetosomes, flagella.
Рис. 296. Giardia intestinalis. (По: Соловьёв, Ченцов, 1974).
A — общий вид; Б — особь, прикрепленная к клетке эпителия кишечни-
ка хозяина; В — циста.
Fig. 296. Giardia intestinalis.
А — total view; Б — specimen attached to epithelial cell of host intestine;
В — cyst.
Рис. 297. Giardia intestinalis, схема строения клетки.
A: 1 — краевая зона присасывательного диска, 2 — присасывательный
диск, 3 — кинетосомы, 4 — передние жгутики, образующие хиазму над
присасывательным диском, 5 — заднебоковые жгутики, 6 — центральные
жгутики; Б — схема ультратонкого строения на поперечном срезе: 1 — пи-
ноцитозные пузырьки, 2 — ЭПР, 3 — гребни присасывательного диска,
4 — краевой гребень присасывательного диска; В — строение края тела по
данным электронного микроскопа: хорошо выражена краевая складка, несу-
щая многочисленные булавовидные выросты. (А, Б — по: Brugerolle, 1975а;
В — по: Соловьёв, Ченцов, 1976).
Fig. 297. Diagram of ultrastructure organization of Giargia intestina-
lis.
A: I — marginal zone of adhesive disk, 2 — adhesive disk, 3 — kinetosomes,
4 — anterior flagella, forming chiasmus above adhesive disk, 5 — posterior-la-
teral flagella; Б — diagram of ultrastructure on transversal section: 1 — pino-
cytotic vesicles, 2 — endoplasmic reticulum, 3 — crest of adhesive disk, 4 —
marginal crest of adhesive disk; В — structure of body margin (electron micros-
copy): well expressed marginal fold with numerous mace evaginations.
комплексов кинетосом, расположенных на противопо-
ложных сторонах (справа и слева) переднего конца те-
ла (рис. 292; 293). Различаются следующие пары жгу-
тиков: переднебоковые, заднебоковые, центральные
(брюшные) и хвостовые. У видов рода Trepomonas
(рис. 292) все жгутики свободны и оба комплекса ки-
нетосом отделены друг от друга. У жгутиконосцев дру-
гих видов рода опорно-двигательный аппарат отли-
чается более сложным строением (рис. 292—295), что
особенно отчетливо выражено у Giardia. Обе группы
кинетосом у Giardia сближены, расположены в инва-
гинации между ядрами, и все 8 отходящих от них Ж1у-
тиков начинаются аксонемами, которые проходят
внутри цитоплазмы (рис. 296; 297). Аксонемы перед-
небоковых жгутиков берут начало от передней пары
кинетосом, проходят вперед, образуют на вершине пе-
344
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
реднего конца тела перекрест (хиазма), огибают при-
сасывательный диск и выходят наружу в форме сво-
бодных жгутиков на уровне верхней границы ядер.
Аксонемы заднебоковых жгутиков направлены
назад, проходят в цитоплазме между ядрами и выхо-
дят наружу, образуя свободные жгутики несколько
ниже середины тела. Аксонемы центральных жгути-
ков проходят внутри цитоплазмы только на расстоя-
нии, равном длине ядер, и, выходя наружу, образуют
свободные жгутики, лежащие в продольном брюш-
ном желобе. При движении они изгибаются спираль-
но. Свободноподвижные хвостовые жгутики у Giar-
dia выходят наружу на заднем конце, а аксонемы их
проходят внутри цитоплазмы на протяжении почти
всей длины тела.
Жгутики и их кинетосомы тесно связаны с целым
рядом других органелл, выполняющих опорную фун-
кцию. К ним относятся и лентовидные структуры.
Переходная зона жгутиков короткая и содержит
1 переходную пластинку, которая не имеет дополни-
тельных структур. Корешковая система жгутиков
включает 2 корешка, состоящих из микротрубочек и
связывающих кинетосомы с ядром, и пластинку, ко-
торая проходит вдоль жгутикового канала и состоит
из комплекса микрофибрилл и микротрубочек (Кар-
пов, 1990). Поэтому аксонемы хвостовых жгутиков
хорошо видны с помощью светового микроскопа и в
ряде исследований были ошибочно приняты за аксо-
стили. В действительности же они по своей ультраст-
руктуре принципиально отличаются от микротубу-
лярных аксостилей Trichomonadida и Oxymonadida,
представляя оригинальные органеллы кариомастигон-
тов Diplomonadida.
Группы опорных микротрубочек расположены
также в околоядерной зоне. Это, как уже было от-
мечено, — корешковые структуры кинетосом. Из
них хорошо развиты микротрубочки, поддерживаю-
щие ядра сверху, а также межъядерные и поперечные
группы у основания ядер (рис. 293; 294). Имеются и
многочисленные субмембранные микротрубочки,
входящие в состав цитоскелета. Микротубулярная
природа присуща особым, специфичным для рода
Giardia медианным структурам, которые находятся в
центральной части тела под присасывательным дис-
ком (рис. 297). Функция этих структур пока еще не
вполне ясна.
К опорным органеллам относятся также исчер-
ченные фибриллы, проходящие вдоль тела у Trepo-
monas, и пучки микрофибрилл у Giardia, связанные
с нижним краем присасывательного диска (рис. 297).
Опорную роль играют и ламеллярные структуры. Ис-
черченные ламеллы хорошо развиты у Trepomonas
(рис. 292, Б). У Giardia хорошо развиты продольные
краевые гребни, идущие на протяжении всей длины
центральных жгутиков; они имеют уплощенную
форму.
Цитостомы. У каждой особи имеется 2 цитосто-
ма, расположенных по бокам заднего конца тела, ко-
торые образованы отверстиями микротубулярных лент
(рис. 292, Б*; 293; 295, А), выполняющих опорную
функцию.
Края и стенки цитостомов поддерживаются мик-
ротубулярными лентами, а также V-образными ис-
черченными ламеллами (модифицированные фиб-
риллы жгутиковых корешков) и межъядерными мик-
ротрубочками. Каждый цитостом имеет форму труб-
ки и от заднего конца тела проходит вперед до об-
ласти ядра.
Через цитостомы осуществляется питание жгути-
коносцев путем фагоцитоза. Они фагоцитируют бак-
терии и другие плотные пищевые частицы. Осуществ-
ляется также пиноцитоз. Жгутиконосцы родов Octo-
mitus и Giardia питаются только путем пиноцитоза,
поскольку они лишенцы цитостомов. У Giardia пиноцитоз
происходит на выпуклой спинной стороне (рис. 293, А;
294), а также в центральной части присасывательного
диска, которая свободна от фибриллярных структур.
Виды рода Octomitus поглощают жидкую пищу всей
поверхностью тела (Brugerolle et al., 1974; Соловьёв,
Ченцов, 1976; Kulda, Nohynkova, 1978).
Присасывательный диск Giardia. Большую часть
центральной поверхности тела жгутиконосцев рода
Giardia занимает присасывательный диск, выполня-
ющий функцию прикрепления их к микроворсинкам
щеточной каймы тонкого кишечника хозяина. Эта
органелла имеет округлую форму и представляет со-
бой впадину, ограниченную опорными микротубу-
лярными фибриллами (рис. 296, А; 297, А). Поверх-
ность диска покрыта плотной пелликулой, состоя-
щей из микротрубочек, которые упакованы
спирально и несут гребни, расположенные перпен-
дикулярно поверхности диска (рис. 297, Б). Наибо-
лее сильно развит краевой гребень, который имеет в по-
перечном сечении серповидную форму и образует
заостренный краевой выступ (борт) высотой около
0.3 мкм. С помощью этого краевого выступа, «крюч-
ка», жгутиконосцы закрепляются между микровор-
синками клеток щеточной каймы эпителия тонкого
кишечника хозяина (рис. 297, Б). В своей задней
части присасывательный диск образует выемку полу-
круглой (куполообразной) формы, которая переходит
в расширяющийся вентральный желоб. Этот желоб
представляет собой общий резервуар центральных
жгутиков (Brugerolle, 1975а; Соловьёв, Ченцов, 1976;
Гинецинская, Добровольский, 1978).
Органеллы цитоплазмы (цитозоля). В цитоплаз-
ме дипломонад сосредоточены каналы и цистерны
ЭПР. Особенно сильно развит гладкий ЭПР. Каналы
и цистерны этой органеллы либо расположены равно-
мерно в цитоплазме всей клетки (Giardia), либо лока-
лизованы в определенных ее участках. У рода Hexami-
ta основная масса каналов и цистерн концентриру-
ется вокруг цитостомов, а у Octomitus они окружают
аксиальную зону опорных органелл (рис. 294, В).
Рибосомы многочисленны, и большинство их сво-
бодно располагается в цитоплазме. Пищеварительные
вакуоли многочисленны, что указывает на высокую
интенсивность питания. Эндопаразитические виды
питаются жидкой, полностью переваренной пищей
хозяина, что можно видеть у Octomitus и Giardia. В
их цитоплазме обычно локализовано множество пи-
ноцитозных вакуолей, наполненных гомогенным со-
держимым. У Giardia больше всего пищеварительных
вакуолей находится в цитоплазме дорсальной сторо-
ны тела, где наиболее интенсивно происходит пино-
цитоз.
ТИП POLYMASTIGOTA. КЛАСС DIPLOMONADEA. ПОДКЛАСС DIPLOMONADIA
345
Отличительной чертой дипломонад служит отсут-
ствие митохондрий, аппарата Гольджи, аксостиля и
микротелец.
Ядро. Трофозоиты жгутиконосцев сем. Enteromo-
nadidae имеют 1 ядро овальной формы, расположен-
ное на переднем конце тела. Ядро пузырьковидного
типа (рис. 291, Б). Продольная ось ядра проходит в
дорсовентральном направлении, и впереди ядра раз-
мещаются кинетосомы.
У жгутиконосцев сем. Hexamitidae в период ин-
терфазы трофозоиты имеют 2 одинаковых ядра, рас-
положенных в цитоплазме передней трети тела сим-
метрично по отношению к его продольной оси. Оба
ядра однородны по своей структуре, и в каждом из
них видно крупное округлое ядрышко. Оболочка
ядер типична для эукариотических клеток и состоит
из 2 мембран с поровыми комплексами, разделенных
перинуклеарной цистерной.
Форма и размеры ядер специфичны для трофо-
зоитов каждого рода. У видов рода Trepomonas оба
ядра вытянуты в длину и изогнуты в соответствии
с закругленной формой переднего конца тела
(рис. 292, Б). Жгутиконосцам рода Octomitus свойст-
венны овальные по форме ядра, у которых внутрен-
няя поверхность изогнута так> что образуется не-
большая впадина с расположенными в ней кинетосо-
мами. Эта особенность хорошо видна на поперечном
срезе в области ядер (рис. 294, В). Ядрам жгутико-
носцев рода Spironucleus присуща S-образная форма.
Оба ядра соприкасаются своей внутренней поверх-
ностью, и края их налегают друг на друга (рис. 295).
Для видов рода Giardia характерны овальные по фо-
рме ядра, расположенные на расстоянии друг от дру-
га (рис. 294, А). В верхней части межъядерного про-
странства располагается кинетосомальный комплекс.
Циста
Формирование цист — общая особенность жиз-
ненного цикла преобладающего большинства видов
отряда Diplomonadida, но структура цист и процессы,
протекающие в них, изучены пока только у рода Gi-
ardia (Соловьёв, Ченцов, 1970; Scheffeld, Bjorvath,
1977). Цисты у жгутиконосцев рода Giardia мелкие,
овальные, длина их не более 10—12 мкм. Снаружи
циста покрыта плотной оболочкой, состоящей из фи-
ламентозного материала, который имеет невысокую
электронную плотность. Толщина оболочки 0.3—
0.4 мкм.
По своему строению циста значительно отличает-
ся от трофозоита (рис. 296, В). В цисте происходит
деление ядер и удвоение всех органелл клетки. Поэ-
тому в цисте содержится 4 ядра, а также удвоенный
комплекс жгутиков и кинетосом, которые погружа-
ются в цитоплазму при инцистировании. Аксонемы
центральных, заднебоковых и хвостовых жгутиков
образуют единый пучок, проходящий параллельно
продольной оси тела (рис. 296, В), а аксонемы
передних жгутиков расположены так же, как и у
трофозоитов, т. е. в передней части цисты.
Цитоплазма цисты вакуолизирована, содержит
многочисленные рибосомы, в ней находятся удвоен-
Рис. 298. Деление ядра.
А — схема деления Giardia duodenalis; Б — деление ядра Hexamita infla-
ta по типу полузакрытого ортомитоза: 1 — экстрануклеарные полуверете-
на, прикрепленные к кинетосомам, и микротрубочки, образующие их, вы-
ходят из полярных отверстий ядра. (А —по: Соловьёв, Ченцов, 1963; Б —
по: Brugerolle, 1974).
Fig. 298. Division of nucleus.
A — diagram of division of Giardia duodenalis; Б — division of nucleus in
Hexamita inflata; semi-open orthomitosis. 1 — extranuclear half-spindles at-
tached to kinetosomes and their microtubules go out of nuclear phenestrae.
ные медианные структуры. В процессе инцистирова-
ния присасывательный диск распадается на отдель-
ные фрагменты. Митохондрии и аппарат Гольджи в
цистах не обнаружены.
Цисты представляют инвазионную стадию в жиз-
ненном цикле эндопаразитических дипломонад и вы-
водятся из кишечника хозяина во внешнюю среду
вместе с фекальными массами.
Размножение
Все виды дипломонад размножаются агамным
путем. Половой процесс у них не обнаружен. Размно-
жение происходит в фазе трофозоита и в цисте. Спо-
соб размножения — продольное бинарное деление,
которое протекает по типу полузакрытого ортомито-
за с образованием 2 полуверетен у полюсов каждого
ядра (рис. 298).
Размножение трофозоита
Бинарное деление трофозоита изучено с помощью
светового микроскопа наиболее подробно у Giardia
(Lamblia) duodenalis (Соловьёв, 1963). Наблюдения в
культуре in vitro показали, что деление трофозоитов
этого вида продолжается 15—20 мин. При делении
346
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
тело трофозоита сильно расширяется, присасыватель-
ный диск резорбируется. Ядра делятся без разруше-
ний ядерной оболочки. В ядрах формируются хромо-
сомы, отчетливо видимые при светооптическом ис-
следовании; хромосом в каждом ядре 6. Затем ядра
смещаются к центру и располагаются одно за другим
вблизи центральной оси тела (рис. 298, А), после че-
го растягиваются, отодвигаясь в стороны. В каждой
половине клетки оказывается по 2 ядра,-из которых
одно произошло от одного, а другое — от другого ма-
теринского ядра. Дочерние ядра перемещаются в са-
гиттальную плоскость, и последующее деление ци-
топлазмы совершается не во фронтальной, а в сагит-
тальной плоскости (Соловьёв, 1963).
Обе дочерние клетки получают по 1 из каждой
пары жгутиков материнской особи, и дальше идет
восстановление недостающих жхугиков у дочерних
трофозоитов. Происходит также формирование но-
вых присасывательных дисков.
Исследование процесса деления дипломонад
было значительно дополнено данными электронной
микроскопии (Brugerolle, 1974, 1975а; Kulda, No-
hynkova, 1978). Деление трофозоита начинается с
деления кинетосом, которые группируются у проти-
воположных полюсов ядра и функционируют в ка-
честве центров деления (рис. 298, Б1), а ядерная мем-
брана сохраняется. Микротубулярные фибриллы ве-
ретена митоза формируются в ядре, выходят через
полярные отверстия наружу в цитоплазму и образу-
ют экстрануклеарные полуверетена, прикрепленные
к кинетосомам. В метафазе оба полюса ядра соеди-
няются внутриядерным фибриллярным интердесмо-
сом (= парадесмос, центродесмос). Затем наступает
анафаза, после которой начинается формирование
дочерних ядер с последующим цитокинезом.
Одной из особенностей деления Enteromonas слу-
жит довольно часто встречающееся отсутствие цито-
кинеза при удвоившихся ядрах и двойном наборе
других органелл клетки. Эта особенность рода Ente-
romonas, трофозоиты которого имеют 1 кариомасти-
гонт, важна для решения вопроса о происхождении
дипломонад.
Размножение в цисте
У всех изученных видов подкласса Diplomonadia
в цисте происходит бинарное деление, (т. е. это
цисты размножения). Детально оно изучено у рода
Giardia. Деление ядра происходит сразу после ин-
цистирования, и процесс деления сходен с делением
ядра трофозоита. Присасывательный диск во время
деления ядра резорбируется. В последующий период
развития цисты происходит удвоение мастигонтов, и
в зрелой цисте Giardia содержится 4 ядра и удвоен-
ный набор органелл кариомастигонта, но цитокинез
и формирование дочерних особей завершаются пос-
ле эксцистирования (Соловьёв, Ченцов, 1970; Bruge-
rolle, 1975а; Kulda, Nohynkova, 1978). В цистах Hex-
amita обнаружены 2 полностью сформированные до-
черние клетки. В каждой из них содержится 2 ядра
и 2 мастигонта, что указывает на возможность цито-
кинеза в цисте.
Культивирование
Культуры in vitro получены пока лишь для немно-
гих видов эндопаразитических дипломонад. Культуру
Giardia (Lamblia) intestinalis впервые получил Кара-
петян (1960), выделив трофозоиты из дуоденального
содержимого человека. В состав питательных сред,
выбранных для этой культуры, были включены соле-
вые растворы Ханкса и Эрла, куриный эмбриональ-
ный экстракт, сыворотка крови человека, перевар
Хоттингера и антибиотики: пенициллин, стрептоми-
цин. Лямблии росли в культуре вместе с дрожжевыми
грибками Candida quilliermondi, которые обитают в
двенадцатиперстной кишке человека. Культуры Giar-
dia intestinalis росли при 37 °C в анаэробных услови-
ях в течение 7 мес.
Этот же вид лямблий, а также G. duodenalis из
тонкого кишечника кролика, шиншиллы и кошки
культивировались в течение нескольких лет на моди-
фицированной среде Карапетяна (1962) с добавлени-
ем Saccharomyces cerevisiae и Candida sp. (Шарапов,
Соловьёв, 1976). Получены также аксенические куль-
туры Giardia intestinalis.
Большинство попыток культивировать эндопара-
зитические виды Hexamita пока не увенчались успехом.
Получены только временные культуры Н. salmonis и
Н. nelsoni. Такие же временные культуры получены
при попытках выращивания на курином эмбрионе
Spironucleus meleagridis и видов рода Enteromonas
(Hughes, Zander, 1954; Kulda, Nohynkova, 1978).
Разработана впервые методика культивирования
свободноживущих дипломонадей — обитателей во-
доемов с высоким содержанием органических ве-
ществ, обитателей сооружений биологической очист-
ки сточных вод (Жуков, Мыльников, 19836). К ним
относятся анаэробные виды: Trigonomonas tortuosa,
Т. inflata, Trepomonas agilis, T. rotans, Hexamita in-
flata, H. fissa, H. caudata, H. mutabilis. В качестве
оптимальной питательной среды для этих жгути-
коносцев предложена среда Пратта (KNO3 — 0.1,
MgSO4 — 0.01, КНРО3 — 0.01, FeCl3 — 0.001 г/л)
с добавлением 1 г пептона или мясо-пептонного
бульона на 1 л среды. Среда не должна содержать
кислорода, так как любая его концентрация губи-
тельна для анаэробов, pH должен быть равен 6.5—
7.8, а водородный показатель рН2 — 17—18 единиц.
Пищей для свободноживущих дипломонад в культуре
служат бактерии Klebsiella (=Aerobacter) aerogenes
или бактерии из очистных сооружений. Для культур
благоприятна температура 7—12 °C (Жуков, Мыль-
ников, 1983).
Экология
Многие виды рода Giardia патогенны для млеко-
питающих и человека. Из них наиболее широко из-
вестен один — G. intestinalis, паразитирующий в две-
надцатиперстной кишке и в других частях переднего
отдела тонкого кишечника человека. Этот жгутико-
носец известен в качестве возбудителя заболевания
человека, называемого жиардиазис, или лямблиоз,
которое при интенсивной инвазии выражается в рас-
ТИП POLYMASTIGOTA. КЛАСС DIPLOMONADEA. ПОДКЛАСС DIPLOMONADIA
347
стройстве деятельности кишечника. Есть данные о
том, что при лямблиозе на поверхности эпителия ки-
шечника, равной 1 см2, может находиться до 1 млн
жгутиконосцев. При таком скоплении G. intestinalis
в зоне их прикрепления цитоплазма клеток эпите-
лия вакуолизирована, клетки сдавлены и поврежде-
ны, микроворсинки укорочены и утолщены. В зонах
прикрепления лямблий клетки эпителия делятся быс-
трее, чем обычно. Во многих случаях заражение че-
ловека лямблиями протекает бессимптомно (Гине-
цинская, Добровольский, 1978; Kulda, Nohynkova,
1978).
Заражение человека происходит цистами лямб-
лий, которые попадают в кишечник вместе с пищей
или с водой. В кишечнике из цист, под воздействи-
ем ферментов хозяина, выходят трофозоиты, кото-
рые прикрепляются к щеточной кайме эпителия пе-
реднего отдела тонкого кишечника. Прикрепление
происходит с помощью присасывательного диска и в
этом процессе участвует также краевая складка тела
лямблий (рис. 296, F; 297, В), снабженная микро-
ворсинками (Соловьёв, Ченцов, 1976).
Питание лямблий осуществляется путем пиноци-
тоза. Насасыванию жидкой, переваренной пищи хо-
зяина из зоны щеточной каймы эпителия кишечника
способствует энергичное движение центральных жгу-
тиков (Соловьёв, Ченцов, 1976).
Лямблии отличаются исключительно высокой про-
дукцией цист: до 300 млн цист может выводиться в
течение суток во внешнюю среду вместе с фекалия-
ми зараженного человека. Эта особенность представ-
ляет одну из причин широкого распространения за-
раженности лямблиями среди людей всех стран
мира, и особенно среди детей: 50—80 % детей зара-
жены лямблиями (Kulda, Nohynkova, 1978).
О патогенности жгутиконосцев рода Giardia для
животных известно пока еще мало/ В имеющейся
литературе есть несколько работ с сообщениями
о заболевании лямблиозом кошек (G. cati), а также
крупного рогатого скота (G. bovis) с четко выра-
женной диарреей (Brightman, Slonka, 1976; Kulda,
Nohynkova, 1978). Достоверных сведений о заболе-
ваниях лямблиозом других животных в литературе
нет.
Происхождение
и эволюция
При построении системы отряда Diplomonadida,
существующей в настоящее время, был использован
принцип, основанный на гомологии клеточных
структур. Ключевая роль в эволюции класса принад-
лежит жгутиконосцам родов Enteromonas и Hexamita.
По структуре своей дипломонадной клетки Hexamita
представляет удвоенную клетку Enteromonas (рис. 291;
293). Высокая степень гомологии структур у этих
2 родов подтверждена на ультраструктурном уровне
(Brugerolle, 1975b). Возможно, что дипломонадиды,
которые известны в настоящее время в природе,
представляют результат незавершенного деления
клетки анцестральной формы, обладавшей сходст-
вом с Enteromonas. Вероятность такого предположе-
ния может быть подтверждена тем, что у ряда видов
энтеромонадид довольно часто встречаются удво-
енные организмы, образовавшиеся в результате не-
завершенного цитокинеза (Kulda, Nohynkova, 1978).
Эволюция жгутиконосцев сем. Hexamitidae шла
по пути удвоения кариомастигонтов и Других орга-
нелл клетки. Это — путь умножения ядер и орга-
нелл цитоплазмы, который свойствен многим груп-
пам простейших и обобщен в теории полимеризации
органелл Догеля (1951). Внутри семейства эволюция
шла, по всей вероятности, в направлении от прими-
тивных свободноживущих форм, сходных с видами
Trepomonas, к высокоспециализированным эндопа-
разитическим видам Giardia.
В эволюции паразитизма, развивающегося внутри
сем. Hexamitidae, отчетливо прослеживается ре-
дукция цитостома, приводящая к полному его исчез-
новению (Octomitus, Giardia). Прослеживается также
тенденция к уменьшению подвижности от быстро
плавающих свободноживущих Trepomonas до при-
крепленных к эпителию кишечника хозяина жиар-
дий (лямблий), обладающих мощным присасыватель-
ным диском. Примерный путь эволюции жгутико-
носцев сем. Hexamitidae отражает следующий ряд:
свободноживущие *—» эндопаразиты: Trepomonas —»
Hexamita —> Spironucleus —> Octomitus —» Giardia.
Diplomonadida присущи морфологические при-
знаки, которые сближают их с Retortamonadida и, в
меньшей степени, с Oxymonadida. Поэтому можно
предполагать, что жгутиконосцы этих 3 отрядов про-
исходят от общего предка (Brugerolle, 1977).
Систематический обзор
Отряд EnteromonadidaBrugerolle,
1975
В составе отряда выделено 1 семейство.
Сем. Enteromonadidae Kulda,
Nohynkova, 1978
Энтеромонадиды — мелкие жгутиконосцы, длина
тела которых около 5 мкм. Имеют 1 кариомастигонт
с числом жгутиков от 1 до 4 (рис. 291). Если число
жгутиков 3—4, то один из них хвостовой, укреплен-
ный лентой из микротрубочек и связанный с цито-
стомом.
В составе сем. Enteromonadidae известно 3 рода:
Enteromonas da Fonseca, 1915 (син. Tricercomonas
Wenyon et O’Connor, 1917); Trimitus Alexeieff, 1910
и Caviomonas Nie, 1950. В жизненном цикле энте-
ромонадид изучены трофозоиты, и только у рода
Enteromonas обнаружены цисты. Все виды семейст-
ва — непатогенные комменсалы самых разнооб-
разных животных и человека. Распространены все-
светно;
348
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отряд Diplomonadida Wenyon, 1926
Отряд включает 2 семейства.
Сем. Hexamitidae Kent, 1880
Это дипломонады с аксиальной симметрией, име-
ющие 8 жгутиков и 2 кариомастигонта. В жизнен-
ном цикле у всех видов известны трофозоиты и
цисты (рис. 292, А; 293, А; 295, А).
В состав семейства входит 3 основных рода: Hexa-
mita Dujardin, 1838; Spironucleus Lavier, 1936; Trepo-
monas Dujardin, 1841. Одним из характерных при-
знаков этих жгутиконосцев является наличие 2 ци-
тостомов, которые имеют форму трубок, проходят
вдоль тела, достигают заднего его конца, где распо-
ложены их отверстия и где происходит заглатывание
пищи. В составе семейства имеются свободноживу-
щие и эндопаразитические виды, распространенные
всесветно. Типовой род — Hexamita, к которому
относятся как свободноживущие, так и паразитичес-
Класс Oxymonadea Grasse,
или Окси
Оксимонадовые — жгутиконосцы, в основе организации кото-
рых лежит кариомастигонт с 2 парами жгутиков, соединенных
между собой паракристаллической структурой — преаксостилем и
аксостилем, передний конец которого входит в преаксостиль. В
основном это одноядерные организмы, но среди них отмечены и
роды, обладающие постоянной поликариомастигонтной организа-
цией; многие виды имеют тенденцию к временному поликариомас-
тигонтному состоянию. Аксостиль в некоторых родах является ак-
тивной локомоторной органеллой. У оксимонад нет митохондрий
и типичного аппарата Гольджи. На поверхности тела и в цитоплаз-
ме ряда оксимонад присутствуют симбиотические прокариотные
организмы — спирохеты и бактерии. Все оксимонады — парази-
тические организмы, обитающие в заднем отделе кишечника ни-
зших термитов и древесиноядных тараканов рода Cryptocercus (ис-
ключение составляет сем. Polymastigidae, часть видов которого
обитает и в других группах насекомых, а также в позвоночных).
Они либо находятся в просвете кишки в свободноподвижном со-
стоянии, либо прикрепляются к ее хитиновой выстилке при помо-
щи особой прикрепительной органеллы. Жизненный цикл у одних
оксимонад включает только свободноподвижную стадию, у дру-
гих — также и прикрепительную. Митоз оксимонад по своему
типу относится к закрытому внутриядерному плевромитозу. Окси-
монады, населяющие кишечник низших термитов, размножаются в
течение всего жизненного цикла только бесполым путем. Жизнен-
ный цикл оксимонад, паразитирующих в Cryptocercus, включает
обязательный половой процесс.
Определение видов оксимонад затрудняется маскировкой не-
зрелых форм одного рода под зрелые формы другого рода, измене-
нием внешнего вида особей, связанным с чередованием подвижной
и прикрепительной стадий, а также миграцией ядер, дегенерацией
и смещением органелл во время митоза в некоторых родах.
История изучения
История изучения оксимонадовых началась со
времени первого ‘описания кишечной фауны терми-
та, сделанного Лейди (Leidy) в 1877 г. Так, первыми
были описаны Pyrsonympha и Dinenympha из Reticuli-
termes ftavipes. Круг исследованных оксимонад рас-
ширялся по мере увеличения числа изученных видов
кие виды, патогенные для рыб (Я. salmonis), ящериц
(Я. parva) и устриц (Я. nelsoni). Род Spironucleus
включает только паразитические виды, патогенные
для индеек (S. meleagridis), голубей, мышей и рыб.
Свободноживущие виды родов Hexamita и Trepomo-
nas — обитатели пресных водоемов, анаэробы.
Сем. Giardiidae Kulda, Nohynkova, 1978
В состав семейства входят 2 рода: Giardia Kiinst-
ler, 1882 и Octomitus Prowazek, 1904.
Для жгутиконосцев обоих родов характерны
четко выраженная бинарная аксиальная симметрия
(рис. 294, А; 296, А) и отсутствие цитостомов. Аксо-
немы хвостовых жгутиков проходят внутри цито-
плазмы вдоль тела и только на заднем конце выходят
наружу, образуя свободные жгутики; питаются путем
пиноцитоза. Все виды семейства либо комменсалы
(род Octomitus), либо эндопаразиты кишечника жи-
вотных и человека. Распространены всесветно. Ти-
повой род — Giardia Kiinster, 1882.
1952 — Оксимонадовые,
монады
термитов. Оксимонады в составе кишечной фауны
таракана Cryptocercus были открыты позже (Cleve-
land et al., 1934). В настоящее время известно около
80 видов этого отряда (см. сводный каталог: Yamin,
1979).
Среди работ, имеющих первостепенное значение
для понимания плана строения и систематики окси-
монад, можно отметить работы, выполненные с при-
менением светового микроскопа: Дюбоска и Грассе
(Duboscq, Grassi) по Pyrsonymphidae; Кофоида и
Свези (Kofoid, Swezy) по Streblomastix и Microrho-
palodbur, Кросса (Cross) по Oxymonas\ Целифа (Ze-
liff) no Barroelkr, серию работ Кирби и Кливленда
(Kirby, Cleveland) по Saccinobacculinae (ссылки см.:
Grasse, 1952). Электронно-микроскопические иссле-
дования тонкого строения различных оксимонадо-
вых относятся к 70-м гг. нашего столетия. Они были
проведены в основном французскими исследователя-
ми (Hollande, Carruette-Valentin, 1970а, 1970b; Bru-
gerolle, Joyon, 1973; Bloodgood et al., 1974; Brugerol-
le, 1981).
Классификация
Современные оксимонадовые ранее наряду с раз-
личными трихомонадами, в том числе и населяющи-
ми кишечник низших термитов, входили в состав
отряда Polymastigina (Blochmann) Reichenow, 1928. За-
тем Кирби (Kirby, 1947) выделил трихомонад в са-
мостоятельный отряд. В 1952 г. Грассе (Grasse,
1952) выделил из оставшихся полимастигин два са-
мостоятельных отряда Oxymonadina и Pyrsonymphi-
па. Сем. Polymastigidae, включающее 2 рода — Мо-
nocercomonoides Travis, 1932 и Polymastix Biitschli,
ТИП POLYMASTIGOTA. КЛАСС OXYMONADEA
349
1884, заняло неопределенное систематическое по-
ложение. С появлением новых данных о строении
жгутиконосцев, паразитирующих в термитах и тара-
канах, Хонигберг (Honigberg, 1963а) предложил со-
здать отряд Oxymonadida Grasse, 1952, в который
включил 3 семейства: Oxymonadidae Kirby, 1928,
Pyrsonymphidae Grassi, 1892 и Monocercomonoididae
Honigberg, 1963; в последнем он оставил только род
Monocercomonoides.
Дальнейшие ультраструктурные исследования
представителей этих семейств (Hollande, Carruette-
Valentin, 1970а, 1970b; Brugerolle, Yoyon, 1973; Blo-
odgood et al., 1974; Brugerolle, 1981) подтвердили
принципиальное сходство в структурной организа-
ции клетки различных оксимонадовых и правиль-
ность выделения их в единую таксономическую
группу — отряд Oxymonadida (Levine et al., 1980).
Кроме того, сравнительное изучение тонкого строе-
ния Monocercomonoides и Polymastix показало, что
эти роды близки по своей организации и несомнен-
но составляют одно семейство, поэтому правомерно
вернуться к первоначальному названию семейства —
Polymastigidae.
В современной системе протистов оксимонады
имеют ранг класса и входят наряду с дипломонадами,
ретортамонадами и парабазалиями в тип Polymastigo-
ta Biitschli, 1884.
Класс Oxymonadea Grass6, 1952
Отряд Oxymonadida Grassd, 1952
Сем. Polymastigidae Biitschli, 1884 emend. Grassd, 1952
Сем. Oxymonadidae Kirby, 1928
Сем. Pyrsonymphidae Grassi, 1892
Морфология и физиология
Однотипная организация кариомастигонта у раз-
личных оксимонад была установлена еще на све-
томикроскопическом уровне (см.: Grasse, 1952). С
помощью электронного микроскопа была выяснена
ультраструктура всего комплекса органоидов карио-
мастигонта и подтверждена их тесная связь между
собой. Общий план строения переднего конца окси-
монад можно продемонстрировать на примере наи-
более примитивного представителя отряда из рода
Monocercomonoides (рис. 299). Трофозоит М. cavia
имеет почти круглое тело, снабженное 4 жгутиками,
которые разделены на 2 обособленные группы, со-
единенные между собой. 2 вентральных жгутика —
свободные, из 2 дорсальных один свободен, а другой
направлен назад и прилегает к телу на некотором
протяжении. Прилегающая к телу часть рекуррент-
ного жгутика подстилается в цитоплазме особой лен-
той (funis). Базальные тела вентральной и дорсаль-
ной групп жгутиков связаны между собой изогнутой
фибриллой — преаксостилем, от которого отходит
аксостиль, идущий вдоль продольной оси тела и име-
ющий однообразную толщину на всем протяжении.
Задний конец аксостиля несколько выступает за
пределы клетки. Ядро расположено в непосредствен-
ной близости от дорсальной пары жгутиков и аксо-
стиля — под преаксостилем. Оно округлой формы и
содержит крупную сферическую эндосому. Передняя
Рис. 299. Monocercomonoides cavia. Схематический рисунок по
данным световой микроскопии. (По: Kulda, Nohynkova, 1978).
акс — аксостиль; пдж — переднедорсальный жгутик; пе — пельта; пж —
2 передневентральных жгутика; пл — лента, подстилающая возвратный
жгутик (funis); пракс — преаксостиль; рж — рекуррентный жгутик; я —
ядро с эндосомой.
Fig. 299. Monocercomonoides cavia. Diagrammatic representation of
the organism, according to light microscopy.
акс — axostyle, пдж — anterior dorsal flagellum, ne — pelta, пж — two an-
terior ventral flagella, пл — funis, пракс — preaxostyle, рж — recurrent fla-
gellum, я — nucleus with endosome.
Рис. 300. Monocercomonoides sp. Схематический рисунок по дан-
ным электронной микроскопии. (По: Kulda, Nohynkova, 1978).
акс — аксостиль; к\,кг — кинетосомы передневентральной пары жгутиков,
кз, кд — кинетосомы дорсальной пары жгутиков, пв — пищеварительные
вакуоли; пг — полисахаридные гранулы; пе — пельта; пл — лента, подсти-
лающая возвратный жгутик (funis); пракс — преаксостиль; шэр — шерохо-
ватый эндоплазматический ретикулум.
Fig. 300. Monocercomonoides sp. Diagramatic reconstruction of ul-
trastructure of the organism, according to electron microscopy.
акс — axostyle, k\, K2 — kinetosomes of the anteroventral pair of flagella, ку,
ka — kinetosomes of the dorsal pair of flagella, ne — digestive vacuoles, m —
polysaccharide granules, ne — pelta, пл — funis, пракс — preaxostyle, шэр —
rough endoplasmic reticulum.
350
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 301. Oxymonas megacaryosoma. (По: Cross, 1946).
акс — аксостиль, ж — жгутики, паке — параксостиль, по — прикрепи-
тельная органелла, р — ростеллум, я — ядро.
Fig. 301. Oxymonas megacaryosoma.
акс — axostyle, ж — flagella, паке — paraxostyle, по — hold-fast, р — ros-
tellum, я — nucleus.
часть кариомастигонта прикрыта большой пельтой.
В цитоплазме много пищеварительных вакуолей.
Строение Monocercomonoides изучено и на ультра-
структурном уровне (Brugerolle, Joyon, 1973; рис. 300).
Уточнено, что вентральные кинетосомы расположе-
ны под острым углом друг к другу, а дорсальные —
под прямым углом (ортогонально). Все жгутики ак-
ронемного типа. Переходная зона жгутика короткая
и представлена поперечной пластинкой без каких-
либо дополнительных структур. Преаксостиль, со-
единяющий вентральную и дорсальную пары кинето-
сом, по существу, является ЦОМТом. Зона преак-
состиля содержит многочисленные полисахаридные
гранулы. Аксостиль состоит из нескольких парал-
лельных рядов микротрубочек, причем передние из
них упираются в преаксостиль. Задний конец аксо-
стиля в цитоплазме как бы «заякорен» особыми
фибриллярными структурами и частично выступает
за пределы клетки. Таким образом, передний конец
аксостиля прочно связан с преаксостилем, а через
него — с кинетосомами жгутиков. Funis, подстилаю-
щий часть рекуррентного жгутика в цитоплазме,
имеет микротрубочковую ультраструктуру. Пельта,
прикрывающая ядро, также состоит из одного слоя
микротрубочек, проходящих непосредственно под
клеточной мембраной. Цитоплазма богата цистерна-
ми шероховатого ЭПР и заполнена многочисленны-
ми пищеварительными вакуолями и пузырьками ли-
зосомального характера.
Ни у одного из видов оксимонад, исследованных
на ультраструктурном уровне, не обнаружены морфо-
логически различимые митохондрии и типичный ап-
парат Гольджи (Hollande, Carruette-Valentin, 1970а,
1970b; Brugerolle, Joyon, 1973; Bloodgood et al., 1974;
Brugerolle, 1981)' Однако, как верно было отмечено
(Bloodgood et al., 1974), это не является доказатель-
ством отсутствия функций названных органелл в
клетке оксимонадовых. Потеря митохондрий несо-
мненно связана с исключительно анаэробными усло-
виями существования жгутиконосцев в заднем отделе
кишки их хозяев. У многих протистов, в том числе и
у парабазалий, составляющих наряду с оксимонадами
сообщество организмов, которые населяют кишечник
низших термитов и тараканов и ведут исключительно
анаэробный образ жизни, найдены аналоги митохон-
дрий — микротельца и (или) гидрогеносомы, кото-
рые впервые были установлены у трихомонад (Mul-
ler, 1993). В цитоплазме у Pyrsonympha были обна-
ружены плотные гомогенные гранулы размером
0.2—1.5 мкм, окруженные мембраной (Bloodgood et
al., 1974). Предположительно эти гранулы имеют от-
ношение к функциям отсутствующих в клетке орга-
нелл. По нашему мнению, эти гранулы в цитоплазме
Pyrsonympha по внешнему виду напоминают гидро-
геносомы. Однако чтобы это доказать, необходимо
провести специальные исследования с применением
маркеров на ферменты анаэробного энергетического
метаболизма.
Что касается комплекса аппарата Гольджи, то у
оксимонад скорее всего есть отдельные элементы
этой органеллы. Так, в цитоплазме оксимонад, ис-
следованных на ультраструктурном уровне, отмечено
присутствие множества небольших электронно-про-
зрачных вакуолей неизвестной природы; доказано
также, что оксимонады способны к пино- и (или)
фагоцитозу. В настоящее время принято считать, что
если у протиста нет типичного аппарата Гольджи, то
в его цитоплазме появляется много пустых мелких
вакуолей, которые, возможно, берут на себя функ-
цию аппарата Гольджи (Серавин, 1992). Кроме
того, согласно современным биохимическим дан-
ным, аппарат Гольджи играет большую роль в обра-
зовании мукополисахаридов и формировании лизо-
сом. Таким образом, вполне вероятно, что в клетке
оксимонад есть структуры, выполняющие функции
аппарата Гольджи.
В рамках общего для всех оксимонадовых плана
строения переднего конца тела (комплекс ядра, аксо-
стиля и 2 обособленных пар жгутиков, соединенных
преаксостилем) разнообразие среди видов достигается
за счет различных вариантов расположения жгути-
ков, формы, величины и дополнительных структур
аксостиля, а также полимеризации кариомастигон-
тов.
У оксимонад, имеющих прикрепительную стадию
в жизненном цикле (Oxymonadinae, Pyrsonymphidae),
передний конец тела оканчивается прикрепительной
органеллой, которая имеет форму присоски, якоря,
пальцевидных выростов и т. д. Электронно-микро-
скопическое изучение прикрепительной органеллы у
Pyrsonympha vertens (Cochrane et al., 1979; рис. 302,
В) показало, что она представляет собой вытянутый
передний конец тела, который оканчивается пальце-
видными выростами. Эти выросты прикрепляются к
кутикуле кишки хозяина при помощи тонких фиб-
рилл. Внутри прикрепительная органелла заполнена
ТИП POLYMASTIGOTA. КЛАСС OXYMONADEA
351
В
Рис. 302. Pyrsonympha sp.
А — юная свободноподвижная форма, Б — прикрепленная форма, В — детали строения переднего конца, ку — кутикула кишечника хозяина, пе — пель-
та, пракс — преаксостиль. Остальные обозначения как на рис. 301. (А, Б — nor Grassd, 1952; В — по: Puytorac et al., 1987).
Fig. 302. Pyrsonympha sp.
A — a junior free stage, Б — attachable stage, В — details the anterior region structure, ку — cuticule of host intestine, ne — pelta, пракс — preaxostyle. Other
abbreviations as in fig. 301.
микрофиламентами, которые лежат параллельно. Они
возникают в тесном контакте с одной из вентральных
кинетосом и через нее связаны с преаксостилем и ак-
состилем, т. е., иными словами, существует физичес-
кая непрерывность между структурами мастигонтной
системы и прикрепительной органеллой. Сходные
данные по ультраструктуре прикрепительной орга-
неллы получены при изучении Oxymonas (Brugerolle,
Konig, 1997). Предполагается, что микрофиламенты
прикрепительной органеллы у обоих организмов
имеют актиновую природу. У Oxymonadinae пере-
дний конец тела значительно вытягивается, напоми-
ная по форме хобот, и носит название ростеллума
(рис. 301). Длина ростеллума значительно варьирует
и иногда может превышать размер тела в 4 раза.
Характер расположения жгутиков и их связь с
поверхностью тела весьма разнообразны. У Opistomi-
tus, Oxymonas (рис. 301) и Polymastix жгутики сво-
бодные. У Monocercomonoides (рис. 299) обычно
2 жгутика свободные, 1 или 2 жгутика периодически
прилипают к телу на некотором протяжении, а затем
вновь становятся свободными. У Saccinobacculus
(рис. 303), Notila (рис. 304) и Streblomastix прокси-
мальные концы жгутиков глубоко погружены в ци-
топлазму, а на поверхности тела они становятся сво-
бодными. У Dinenympha и Pyrsonympha (рис. 302,
А, Б) все жгутики рекуррентные и почти на всем
своем протяжении связаны с поверхностью клет-
ки — лишь дистальные концы свободны. Так, 8
жгутиков Pyrsonympha из Reticulitermes flavipes (Blo-
odgood et al., 1974) обвивают тело по леотропной
спирали и располагаются в глубоких бороздках плаз-
матической мембраны, где они прочно «заякорены»
при помощи фиброзного слоя (предположительно
гликопротеидов или мукополисахаридов).
Аксостиль основной элемент цитоскелета у ок-
симонадовых. У многих видов он способен изгибать-
ся, что приводит к змеевидному движению самого
жгутиконосца. На ультраструктурном уровне показа-
но, что аксостиль — многослойная структура, состо-
ящая из параллельных рядов микротрубочек. Причем
каждый микротрубочковый ряд связан с соседним
внутрирядовыми мостиками (intra-row cross bridges),
а отдельные микротрубочки в ряду связаны между
собой внутрирядовыми связями (intra-row cross links).
В задней части тела ряды микротрубочек сворачива-
ются в трубку и часто пронизывают все тело. В осно-
ве способности аксостиля к сокращениям лежит про-
цесс скольжения рядов микротрубочек относительно
друг друга, что обусловлено деятельностью динеино-
вых ручек (microtubular system with dynein motor).
Механизм этого процесса различен у разных видов
(например, у Pyrsonympha (Langford, Inoue, 1979) и у
Saccinobacculus (Woodrom, Linck, 1980)).
352
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 303. Saccinobacculus spatiatus. (По: Бобылёва, 1975).
акс — аксостиль, аксф — аксостилярный футляр, ж — жгутики, я —
ядро.
Fig. 303. Saccinobacculus spatiatus.
акс — axostyle, аксф — axostylar sheath, ж — flagella, я — nucleus.
Рис. 304. Notila proteus ussuriensis. (По: Бобылёва, 1975).
Обозначения как на рис. 303.
Fig. 304. Notila proteus ussuriensis.
Abbreviations as in fig. 303.
Аксостиль идет вдоль продольной оси тела и по
внешнему виду напоминает либо тонкую трубку (на-
пример, у Monocercomonoides — рис. 299), либо бо-
лее или менее широкую ленту (например, у Pyrso-
nympha, Saccinobacculus, Notila, — рис. 302, 5*; 303;
304). Задний конец аксостиля либо свободно лежит в
цитоплазме (например, у Pyrsonympha, — рис. 302, Б),
либо прочно закреплен в ней (например, у Notila, —
рис. 304) и одет особым аксостилярным футляром
(например, у Saccinobacculus, — рис. 303). У некоторых
видов аксостиль выступает за пределы клетки (напри-
мер, у Monocercomonoides, Oxymonas, — рис. 299; 301).
В работах, выполненных на светомикроскопичес-
ком уровне, авторы (ссылки см.: Grasse, 1952) отме-
чают у оксимонад присутствие сложного аксостиля.
Использование этого термина показывает, что в
клетке помимо аксостиля присутствуют близкие ему
по структуре дополнительные свободные фибриллы.
У Oxymonas (рис. 301) и Pyrsonympha (рис. 302, Б)
это параксостиль, представляющий собой пучок
фибрилл, свободно оканчивающихся в цитоплазме.
Как показали электронно-микроскопические иссле-
дования (Hollande, Carruette-Valentin, 1970b; Blood-
good et al., 1974), параксостиль у Pyrsonympha пред-
ставляет собой рыхло организованный пучок мик-
ротрубочек, лишенных внутрирядовых мостиков.
Изучение тонкого строения переднего конца тела
Oxymonas (Brugerolle, Konig, 1997) показало, что
ростеллум содержит 2 различных по структуре пучка
микротрубочек. Один пучок состоит из скрученных
микротрубочковых лент, которые начинаются от ма-
ленького плотного ЦОМТа на основании прикрепи-
тельной органеллы, далее идут вдоль ростеллума до
его основания и затем продолжаются в цитоплазме
почти до середины тела (вероятно, это и есть парак-
состиль). Второй пучок состоит из свободных мик-
ротрубочковых филаментов, которые берут свое на-
чало в ростеллуме и идут к его основанию, где на
уровне кинетосом внедряются в микротрубочковые
ряды аксостиля. Предполагается, что ростеллум мо-
жет сокращаться. Параксостиль в своем происхож-
дении, так же как аксостиль, тесно связан с преак-
состилем. Ультраструктура параксостиля других ок-
симонад пока неясна. У Sauromonas дополнительный
элемент аксостиля носит название псевдоаксостиль,
но что он собой представляет на ультраструктурном
уровне, тоже неясно.
Другие элементы цитоскелета у оксимонад —
пельта и funis — также имеют микротрубочковую при-
роду и представляют собой ряды микротрубочек, ли-
шенных внутрирядовых мостиков, которые свободно
расположены непосредственно под клеточной мемб-
раной (Bloodgood et al., 1974).
Ядро обычно имеет сферическую или грушевид-
ную форму; 1 или несколько ядрышек; хроматин в
виде сеточки или периферических сгущений. На све-
томикроскопическом уровне часто отмечается тесная
связь ядра и аксостиля. Под электронным микроско-
пом ядро исследовано лишь у очень немногих окси-
монад, и данные о его ультраструктуре весьма скудны
(ссылки см.: Райков, 1982). У всех изученных видов
ядерная оболочка классического типа. По соотноше-
нию компактного и деконденсированного хроматина,
присутствию 1 крупного ядрышка ядра Monocercomo-
noides melolonthae и Pyrsonympha vertens можно от-
нести к типичным пузырьковидным ядрам. Интерфаз-
ные ядра Microrhopalodina inflata характеризуются
сохранением конденсированного состояния хромо-
сом и наличием 1 эксцентричного ядрышка. У изу-
ченных ядер наблюдается контакт ядерной оболочки
и аксостиля, но никаких специальных структур при-
крепления не обнаружено.
Среди представителей подсем. Oxymonadinae про-
слеживается тенденция перехода в организации клет-
ки от форм с 1 кариомастигонтом к временному по-
ликариомастигонтному состоянию и затем к формам
ТИП POLYMASTIGOTA. КЛАСС OXYMONADEA
353
Рис. 305. Proboscidiella kofoidi. (По: Kirby, 1928).
акс — аксостиль, р — ростеллум, я — ядро.
Fig. 305. Proboscidiella kofoidi.
акс — axostyle, р — rostellum, я — nucleus.
с постоянной поликариомастигонтной организацией.
Так, Opisthomitus, Oxymonas (рис. 301) и Sauromo-
nas — в основном одноядерные формы, но в их жиз-
ненном цикле существуют особи с 2, 4 и 6 кариомас-
тигонтами. Виды родов Microrhopalodina (-Probossi-
diella) (рис. 305) и Barroella обладают постоянной
поликариомастигонтной организацией. При полиме-
ризации кариомастигонтов преаксостиль сильно
уменьшается, пельта и параксостиль редуцируются,
но аксостиль сохраняется и обычно соответствует
числу кариомастигонтов в клетке.
На поверхности тела большинства оксимонад най-
дены бактерии и спирохеты. Особенности прикрепле-
ния спирохет и палочковидных бактерий к клеточ-
ной мембране Pyrsonympha и их взаимоотношения с
хозяином подробно исследованы на ультраструктурном
уровне (Bloodgood et al., 1974; Bloodgood, Fitzharris,
1976). Оказалось, что образуются специализирован-
ные структуры в местах прикрепления прокариотов
к поверхности жгутиконосца, причем возникшая
мембранная специализация обеспечивается обоими
партнерами. Эти специализированные мембранные
структуры обеспечивают прочное механическое «за-
якоривание» спирохет и бактерий на поверхности
клетки жгутиконосца. Таким образом, возникает ас-
социация организмов, в основе которой лежат сим-
биотические отношения между партнерами.
У Polymastix на поверхности тела располагаются
многочисленные длинные грамположительные бакте-
рии рода Fusiformis (Brugerolle, 1981). Они прикреп-
ляются к плазматической мембране хозяина глико-
каликсом, причем в местах прикрепления обнару-
живается и слой электронно-плотного материала,
находящийся на внутренней стороне клеточной мем-
браны жгутиконосца. Особенности расположения и
прикрепления эктосимбионтов часто используются в
качестве таксономического признака. Разнообразные
симбиотические бактерии содержатся и в цитоплазме
оксимонад. Более подробно о симбиотических отно-
шениях между прокариотными организмами и жгути-
коносцами из кишечника термитов и тараканов см.
раздел «Отряд Hypermastigida».
Питание
Мелкие формы оксимонад, например Opisthomi-
tus, питаются осмотрофно, т. е. поглощая необходи-
мые для жизни вещества из ректальной жидкости не-
посредственно всей поверхностью тела. Большинство
оксимонад (в частности Pyrsonympha vertens, Micro-
rhopalodina inflata) питаются частицами древесины и
бактериями, содержащимися в заднем отделе кишеч-
ника хозяина-ксилофага. Пища попадает в организм
жгутиконосца посредством образования впячивания
во внутрь клеточной поверхности на заднем конце
его тела. Образуется пищевая вакуоль, содержащая
частицы древесины и кишечную жидкость с бактери-
ями. Питание идет в основном по типу фагоцитоза.
Однако возможен и пиноцитоз, поскольку в цито-
плазме найдены пузырьки и трубочки, указывающие
на поглощение ректальной жидкости. В пищевари-
тельной вакуоли Pyrsonympha vertens отмечено при-
сутствие кислой фосфатазы. (Ссылки на работы, по-
священные питанию оксимонад, см. в обзоре: Radek,
Hausmann, 1994b).
Размножение
Бесполое размножение
На светомикроскопическом уровне митоз описан
довольно давно у Oxymonas, Saccinobacculus и Pyr-
sonympha (ссылки см.: Grasse, 1952); отмечены наи-
более характерные его черты: полное сохранение
ядерной оболочки на протяжении всего митоза и
внутриядерное положение веретена и хромосом. На
ультраструктурном уровне митоз изучен пока только
у Pyrsonympha flage llata (Hollande, Carruette-Valentin,
1970b). Показано, что на внутренней стороне интак-
тной ядерной оболочки располагается 2 центриоляр-
ных пластинки, от которых тянутся микротрубочки,
образующие 2 полуверетена. Центриолярные плас-
тинки имеют тонко гранулярную структуру, т. е. они
23 Протисты, часть 1
354
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
являются, по существу, ЦОМТами. В ранней профазе
центриолярная пластинка всего 1 и хромосомы связа-
ны с ней посредством хромосомных пучков микро-
трубочек. Затем центриолярная пластинка разделяется
на 2 и образуются 2 полуверетена; одновременно
происходит постепенное разъединение сестринских
хроматид, которые на протяжении всего митоза силь-
но спирализованы. 2 полуверетена постепенно откло-
няются друг от друга; часть микротрубочек образует
нити центрального веретена, остальные прикрепляются
к кинетохорам хроматид. В конце митоза полуве-
ретена достигают диаметрально противоположной
позиции на ядерной оболочке и разделение хроматид
завершается. Механизм отклонения полуверетен обес-
печивается, по-видимому, как скольжением ЦОМТов
по внутренней оболочке ядра, так и процессом растя-
жения самой внутренней оболочки между 2 центрио-
лярными пластинками. Митоз оксимонадовых по
классификации типов митозов относится к закрыто-
му внутриядерному плевромитозу (см.: Райков, 1994).
У представителей подсем. Oxymonadinae в про-
фазе наблюдается миграция ядра в задний конец
клетки и его возвращение в телофазе в свое интер-
кинетическое положение, в котором оно находилось
перед началом цитокинеза. У этих жгутиконосцев в
профазе дегенерирует старый аксостиль, а у некото-
рых представителей также и жгутики. Плазмотомия
у всех оксимонад осуществляется продольно. Митоз
может происходить как в свободноподвижном, так и
в прикрепленном состоянии. Плазмотомия может
приводить к возникновению в жизненном цикле либо
свободноподвижной, либо прикрепленной стадии.
Оксимонады, паразитирующие в кишечнике низших
термитов, размножаются только бесполым путем.
Оксимонады, населяющие задний отдел кишечника
таракана Cryptocercus, размножаются бесполым пу-
тем большую часть своего жизненного цикла, кото-
рая проходит в нелиняющем хозяине. В период линь-
ки таракана у оксимонад наблюдается обязательный
половой процесс.
Половой процесс
У оксимонад, обитающих в кишечнике таракана
Cryptocercus, в период линьки хозяина происходит
половой процесс. У каждого из 3 родов — Oxymo-
nas, Notila и Saccinobacculus — половой процесс
имеет свои отличительные черты, присущие только
этому роду (ссылки см.: Cleveland, 1956, 1957), однако
можно выделить ряд черт, общих для всех оксимонад.
Во-первых, половой процесс может осуществлять-
ся двумя путями: 1) в результате образования гамет и
их последующей копуляции, 2) в результате автога-
мии (слияния сестринских ядер, образовавшихся в
ходе митоза гаметоцита, при котором не происходит
клеточного деления).
Во-вторых, отличительной чертой полового про-
цесса оксимонад является слияние аксостилей, кото-
рое наблюдается в ходе слияния гамет или автргамии.
В-третьих, у всех родов оксимонад происходит од-
ноступенчатый мейоз, сопровождающийся полной или
частичной реорганизацией всех клеточных органелл.
Рис. 306. Половой процесс по типу копуляции гамет или автога-
мии у Saccinobacculus. (По: Cleveland, 1956).
А — бесполая особь. Б — гаметоцит. В — гаметы, различающиеся между
собой только физиологически. Г — копуляция гамет. Д — образование
клетки с 8 жгутиками; произошло слияние цитоплазмы гамет, а также ак-
состилей; у пронуклеусов происходит лишь контакт ядерных оболочек. Е —
зигота — клетка округляется, кутикула утолщается, жгутики остаются внут-
ри клетки. Ж — зигота, которая претерпевает одноступенчатый мейоз; при
этом происходит дегенерация старых аксостилей и жгутиков и образование
нового набора органелл. 3 — образование в ходе цитокинеза 2 новых гап-
лоидных особей, п — гаплоидный набор хромосом, 2п — диплоидный на-
бор хромосом.
Fig. 306. Sexual process in Saccinobacculus, accomplished by a
complete gametogenesis or autogamy.
A — asexual cell. Б — gametocyte not differing morphologically from an
asexual cell. В — gametes differing from each other only physiologically. Г —
cytoplasmic fusion of gametes. Д — cell with 8 flagella; axostyles are fused
completely, but pronuclei contact only with their nuclear envelopes. E — zygo-
te: pronuclei fused; 8 flagella and 2 axostyles are seen retained inside the cell;
the cell rounds up. Ж — a single-step meiotic division occurs in the nucleus
following host moulting; axostyles are discarded and renewed; one set of flagel-
la is lost and new ones are produced. 3 — two haploid asexual cells with new
set of organelles, n — haploid chromosome set, 2n — diploid chromosome set.
В-четвертых, у всех названных родов половой
процесс растянут во времени (начинается примерно
за 40 дней до линьки таракана) и включает покоя-
щиеся стадии, когда никаких изменений не происхо-
дит в течение довольно долгого периода, а заверша-
ется спустя 1—2 дня после линьки хозяина.
В качестве примера остановимся более подробно на
ходе полового процесса у Saccinobacculus (рис. 306).
Бесполая особь (А) — гаплонт — в период подготов-
ки таракана к линьке (за 42 дня) преобразуется в га-
метоцит (В), который морфологически почти не от-
личается от нее; затем процесс может идти либо по
типу автогамии (Б, Д), либо по типу копуляции гамет
{Б—Г). В ходе ядерного (митоз) и цитоплазматичес-
кого деления формируются 2 гаметы (В), различаю-
щиеся между собой только физиологически. Во вре-
мя копуляции гамет (Г) их цитоплазма сливается; ак-
ТИП POLYMASTIGOTA. КЛАСС OXYMONADEA
355
состили также сливаются, а у пронуклеусов наблюда-
ется лишь соприкосновение ядерных оболочек (Д),
Стадия слияния (Д) наблюдается и в ходе автогамии.
На этой стадии половой процесс как бы замирает —
клетка, обладающая 8 жгутиками, аксостилем и 2 яд-
рами, которые соприкасаются своими оболочками
(Д), существует в кишечнике готовящегося к линьке
таракана около 35 дней. Затем за 4—5 дней до линь-
ки хозяина ядра сливаются и образуется зигота (Е),
которая имеет 8 жгутиков. На этой стадии Saccino-
bacculus переживает линьку хозяина; при этом клет-
ка округляется, покровы утолщаются. На следующий
день после линьки таракана ядро зиготы жгутиконос-
ца претерпевает одноступенчатый мейоз (X), кото-
рый сопровождается дегенерацией обоих аксостилей
и одного набора жгутиков и образованием заново жгути-
ков и аксостилей. Затем происходит цитокинез и образу-
ются 2 гаплоидные особи (3), размножающиеся бесполым
путем до наступления ближайшей линьки хозяина.
Половой процесс Oxymonas, которые являются
гаплонтами на бесполой стадии жизненного цикла,
подобен таковому Saccinobacculus, но отличается тем,
что при слиянии гамет наблюдается и одновременное
слияние пронуклеусов и аксостилей.
Половой процесс у Notila (рис. 304) — диплонтов
на бесполой стадии жизненного цикла — чрезвычай-
но своеобразен, хотя и здесь четко прослеживаются
вышеупомянутые общие черты этого процесса у ок-
симонад. Особенности полового процесса у Notila за-
ключаются в следующем.
1) Гаметоцит производит диплоидные «гаметы»,
которые копулируют, и при этом происходит слияние
цитоплазмы и аксостилей, а ядра не сливаются в те-
чение долгого периода.
2) После линьки таракана Ядра гамет приступают
к одноступенчатому мейозу, который сопровождается
реорганизацией всех клеточных органелл; при этом
цитокинез не наблюдается.
3) Сформировавшиеся в результате мейоза про-
нуклеусы попарно сливаются, и в одной клетке обра-
зуются 2 диплоидных ядра, каждое из которых имеет
полный набор органелл. Впоследствии такая клетка
испытывает цитокинез и дает начало 2 диплоидным
бесполым особям.
Эволюция
Эволюционные связи оксимонадовых с другими
жгутиковыми до настоящего времени остаются весь-
ма неясными. Наиболее близки Oxymonadida по
морфологическому плану организации клетки Retor-
tamonadida и Diplomonadida. Среди оксимонадовых
сем. Polymastigidae несомненно наиболее примитив-
ное. Можно предположить, что именно это семей-
ство находилось в начале филогенетического древа
отряда и от него эволюировали 2 другие ветви —
семейства Oxymonadidae и Pyrsonymphidae. В эво-
люции Oxymonadinae можно выделить морфологи-
ческий ряд форм, усложнение организации которых
связано с процессом полимеризации кариомастигон-
тов. В начале этого ряда находятся Opisthomitus, Ox-
ymonas и Sauromonas, имеющие в жизненном цикле
стадии с временной поликариомастигонтной органи-
зацией; замыкают ряд Proboscidiella и Barroella, ддя
которых характерна постоянная йоликариомастигон-
тная организация. Эволюция Saccinobacculinae, веро-
ятно, связана с развитием аксостиля как активной
локомоторной органеллы. Эволюция оксимонад ока-
залась тесно связанной исключительно с 2 родствен-
ными группами насекомых — низшими термитами
(семейства Calotermitidae, Termopsidae, Rhinotermiti-
dae) и тараканами рода Cryptocercus, Однако в про-
шлом оксимонадовые, вероятно, имели более широ-
кое распространение, чем теперь. На это указывает
современный круг хозяев наиболее примитивных ро-
дов — Monocercomonoides и Polymastix, в который
входят не только низшие термиты и тараканы Cryp-
tocercus, но и другие группы насекомых, а также
многоножки, амфибии, рептилии и млекопитающие.
Подавляющее большинство Oxymonadida наряду
с Hypermastigida входят в состав кишечной фауны
Cryptocercus и низших термитов и имеют симбиоти-
ческие отношения мутуалистического характера со
своими хозяевами. Подробнее об этом и о путях
заражения хозяев этими жгутиконосцами см. раздел
«Отряд Hypermastigida».
Систематический обзор
Отряд Oxymonadida Grasse, 1952
Сем. Polymastigidae Butschli, 1884
emend. Grasse, 1952
Мелкие оксимонады (до 10 мкм) с 1 кариомасти-
гонтом; свободноподвижные, прикрепленных форм
нет; 2 пары жгутиков, из которых 1 часто загибается
назад. Аксостиль тонкий, несократимый. Ядро сфе-
рической формы, с 1 крупным ядрышком. Часто при-
сутствует пельта. Способны образовывать цисты.
Паразиты многих групп насекомых, многоножек, ам-
фибий, рептилий и млекопитающих. Включает 2 рода.
Род Monocercomonoides Travis, 1932
Мелкие оксимонады, у которых рекуррентный
жгутик связан на значительном расстоянии с поверх-
ностью тела и от него отходит микротрубочковый ко-
решок — funis, подстилающий в цитоплазме этот
жгутик.
Всего около 20 видов, из них М. termitis найден в
термите Kalotermes sinaicus, а М. globus населяет ки-
шечник таракана Cryptocercus punctulatus, остальные
обитают в других группах насекомых, а также извес-
тны из рептилий, амфибий и млекопитающих.
Род Polymastix Butschli, 1884
Мелкие оксимонады. Все 4 жгутика свободные и
направлены вперед. Сохраняется рекуррентная мик-
ротрубочковая фибрилла — funis. Поверхность тела
356
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
покрыта многочисленными симбиотическими бакте-
риями рода Fusiformis.
Известно 5 видов, из них лишь 1 обитает в кишеч-
нике термита рода Neotermes, остальные — в других
группах насекомых и многоножках.
Сем. Oxymonadidae Kirby, 1928
Оксимонады с 1 или многими кариомастигонтами;
свободноподвижные и прикрепленные формы; жгути-
ки свободные либо на некотором расстоянии закреп-
лены в цитоплазме. Аксостиль у многих родов сокра-
тимый, часто с дополнительными структурами; зад-
ний конец аксостиля лежит свободно в цитоплазме,
либо закреплен в ней, либо выступает из клетки. Для
многих родов известны цисты; для видов, населяю-
щих Cryptocercus, известен половой процесс.
Паразиты низших термитов сем. Calotermitidae и
тараканов рода Cryptocercus. Включает два подсе-
мейства — Oxymonadinae и Saccinobacculinae.
Подсем. Oxymonadinae Kirby, 1928
emend. Cross, 1946
1 или много кариомастигонтов. Жгутики свобод-
ные. Свободноподвижные и прикрепленные формы;
ростеллум с прикрепительной органеллой. Ядро сфе-
рическое или грушевидное, с 1 и более ядрышками.
Аксостиль сократимый у большинства родов; лежит
свободно в цитоплазме либо выступает из клетки.
Часто присутствует параксостиль. Ядро в течение
профазы мигрирует в задний конец клетки и возвра-
щается к своему интеркинетическому положению, в
котором оно находилось перед началом цитокинеза.
Для некоторых родов известны цисты. Паразиты низ-
ших термитов сем. Calotermitidae. Включает 5 родов.
Род Opisthomitus Duboscq et Grasse,
1934
Мелкие формы (до 10 мкм); свободноподвижные.
Аксостиль тонкий, пронизывает задний конец тела,
несократимый. В основном одноядерные формы, но
не редки временные формы, содержащие 2 или 4 ка-
риомастигонта.
Известны О. avicularis из Calotermes flavicollis и
Opisthomitus sp. из Glyptotermes irridipennis.
Род Oxymonas Janicki, 1915
В основном одноядерные формы, но часто воз-
никают формы, содержащие 2 кариомастигонта; ре-
же встречаются особи с 4—6 кариомастигонтами.
Форма тела сферическая, грушевидная или удлинен-
ная. Имеются как мелкие, так и крупные формы (от
5 до 240 мкм). Свободноподвижные и прикрепленные
формы. Ростеллум в виде длинного тонкого трубча-
того отростка переднего конца тела, который закан-
чивается прикрепительной органеллой. Аксостиль
сложный, состоит из основной части различной
формы (тонкой ленты, стилета, кривой сабли и т. д.)
и передней части, продолжающейся в ростеллум и
возвращающейся в виде более или менее длинного
пучка фибрилл — параксостиля. Основная часть ак-
состиля лежит свободно в цитоплазме либо выступа-
ет из клетки. Аксостиль слабо сократимый. Для не-
которых видов известны цисты. У многих видов при-
сутствуют прокариотные экто- и эндосимбионты.
Наиболее многочисленный род по числу видов
(более 30). Паразиты тараканов рода Cryptocercus и
термитов сем. Calotermitidae.
Род Sauromonas Grasse et Hollande,
1952
Внешне сходен с Oxymonas. Аксостиль с допол-
нительным пучком фибрилл — псевдоаксостилем.
В жизненном цикле появляются формы с 2 симмет-
ричными зонами жгутиков. Способны образовывать
цисты.
Известно 2 вида — S. m’baikiensis и S. termitidis
из термитов рода Glyptotermes.
Род Proboscidiella Kofoid et Swezy,
1926
(Син.: Microrhopalodina Grassi etFoa, 1911)
Многоядерные формы; содержат в среднем от
5 до 13 кариомастигонтов, но в некоторых особях
их число достигает 50. Кариомастигонты располага-
ются в передней части* клетки в виде короны. Форма
тела грушевидная либо удлиненная. У самых мелких
форм длина тела до 23, у самых крупных — до
165 мкм. Свободноподвижные и прикрепленные
формы. Ростеллум в виде длинного тонкого отрост-
ка. Аксостиль имеет вид тонкой ленты, параксостиль
развит слабо.
Прокариотические эктосимбионты. Известно 4 вида
из термитов сем. Calotermitidae.
Род Barroella Zeliff, 1944
Многоядерные формы. Молодые особи по внеш-
нему виду напоминают Microrhopalodina; содержат
от 2 до 12 кариомастигонтов. Зрелые особи имеют
неправильную мешковидную форму; содержат мно-
жественные короны кариомастигонтов, либо карио-
мастигонты беспорядочно рассеяны в цитоплазме; в
последнем случае число ядер и аксостилей редко со-
впадает. Молодые особи возникают в результате от-
почковывания от крупного зрелого организма. Сво-
бодноподвижные и прикрепленные формы. Самые
мелкие экземпляры 27, самые крупные 224 мкм. Ак-
состиль с параксостилем.
На поверхности тела располагаются прокариоти-
ческие симбионты. Известны В. zeteki из термита
Calcaritermes brevicollis и В. coronaris из термита
Postelectrotermes howa.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA
357
Подсем. Saccinobacculinae Kirby, 1934
Одноядерные оксимонады; только свободнопод-
вижные формы. Проксимальные концы жгутиков по-
гружены в цитоплазму. Аксостиль широкий, лентовид-
ный, проходит через все тело, но не выступает за его
пределы; закреплен в цитоплазме. Аксостиль является
активной локомоторной органеллой. Форма тела варь-
ирует от сферической до удлиненной, что зависит от
положения аксостиля в момент прохождения через не-
го волны сокращения. Ядро сферической формы, с 1 яд-
рышком; хроматин сетчатый. Известен половой процесс.
Паразиты тараканов рода Cryptocercus. Включает 3 рода.
Род Saccinobacculus Cleveland, Hall, Sanders
et Collier, 1934
Самые мелкие формы 30, наиболее крупные —
около 100 мкм. Задний конец аксостиля окружен аксо-
стилярным футляром и не выступает за пределы клетки.
Известно 5 видов из тараканов Cryptocercus punc-
tulatus и С. relictus.
Род Notila Cleveland, 1950
Длина тела около 100 мкм. Задний конец широкого
лентовидного аксостиля лишен аксостилярного футляра.
Известен 1 вид — N. prbteus — из тараканов
Cryptocercus punctulatus и его подвид N. р. ussurien-
sis из С. relictus.
Род Paranotila Cleveland, 1966
Мелкие оксимонады (до 10 мкм). Задний конец
аксостиля не пронизывает клетку и лишен аксости-
лярного футляра.
Известен 1 вид — Р. lata из таракана Cryptocercus
punctulatus.
Сем. Pyrsonymphidae Grassi, 1892
Одноядерные, свободноподвижные и прикреплен-
ные формы. Жгутики свободные либо на значитель-
ном расстоянии прикреплены к поверхности тела. У
некоторых видов есть прикрепительная органелла.
Ядро грушевидное либо веретенообразное. Аксо-
стиль тонкий, длинный, сократимый; у некоторых
видов есть параксостиль.
Имеются экто- и эндосимбионты. Паразиты низ-
ших термитов семейств Rhinotermitidae и Termopsi-
dae. Включает 2 рода.
Род Pyrsonympha Leidy, 1877
Форма тела грушевидная или удлиненная, причем са-
мо тело может быть изогнуто или спирально закручено.
Длина тела самых мелких форм 40, самых крупных
150 мкм. Жгутики загнуты назад, закручены по лео-
тропной спирали и располагаются в углублениях или
желобках клеточной мембраны. Жгутики на всем своем
протяжений прикреплены к поверхности тела, сво-
бодны лишь их дистальные концы. Часто наблюдают-
ся особи с 8 жгутиками. В разных видах хозяев встреча-
ются формы с прикрепительной органеллой и без нее.
Ядрр грушевидной формы. Аксостиль тонкий, длинный,
сократимый, часто есть и параксостиль. На поверхно-
сти тела располагаются спирохеты, в цитоплазме —
симбиотические бактерии. Паразиты низших термитов
сем. Rhinotermitidae. Род Pyrsonympha насчитывает бо-
лее 20 видов и состоит из 2 подродов — Pyrsonympha
(аксостиль свободно лежит в цитоплазме) и Dinenympha
(аксостиль фиксирован сбоку на заднем конце тела).
Крупные Pyrsonympha, описанные впервые Лейди
(Leidy) в 1877 г. из Reticuliternies flavipes и позднее
Грассе (Grassd) из R. lucifugus под видовым названи-
ем Pyrsonympha vertens, вероятно, являются 2 разны-
ми видами, что вытекает из электронно-микроскопи-
ческих исследований этих пирсонимф (Hollande, Саг-
ruette-Valentin, 1970b; Bloodgood et al., 1974).
Род Streblomastix Kofoid et Swezy,
1919
Тонкие и длинные оксимонады (длина тела от 15
до 530 мкм). Тело спирально закручено; 4—8 глубо-
ких желобков разделяют ленты мионем, закрученные
по леотропной спирали. Жгутики свободные. Ядро
веретенообразное. На переднем конце тела прикре-
пительная органелла в виде присоски. Аксостиль
тонкий, есть параксостиль.
Паразиты низших термитов сем. Termopsidae. Из-
вестен 1 вид — S. strix из Zootermopsis angusticollis и
Z. nevadensis.
Класс Parabasalea Honigberg, 1973 — Парабазалии
Парабазалий — трихомонады и гипермастигиды (многожгути-
ковые жгутиконосцы) — комменсалы и эндопаразиты животных и
человека. Специфичность класса определяет особый, присущий
этим жгутиконосцам парабазальный комплекс органелл, включаю-
щий аппарат Гольджи (от 1 до многих диктиосом) и парабазальные
филаменты (от 1 до многих у разных видов) Число жгутиков у
разных видов от нескольких до многочисленных, отходящих от
переднего конца тела. У трихомонад развйта увдулирующая мемб-
рана. Корешковая система жгутиков имеет сходный тип строения
у трихомонад и гипермастигид и включает фибриллярные кореш-
ки, к числу которых относятся и парабазальные филаменты.
Для парабазалий специфичен пельта-аксостилярный комплекс,
органеллы которого — пельта и аксостиль состоят из многочис-
ленных микротрубочек. Ядро 1, расположено спереди в тесном
контакте с аксостилем. Кинетосомы, корешковая система жгути-
ков, пельта, аксостиль и ядро образуют сложный кариомастигонт
парабазалий. В их клетке отсутствуют митохондрии; имеются гид-
рогеносомы. Размножение агамное путем бинарного деления клет-
ки. Тип деления ядра — закрытый внеядерный плевромитоз.
Среди парабазалий есть целый ряд видов, патогенных для жи-
вотных и человека, вследствие чего исследование этой группы эн-
допаразитических жгутиконосцев имеет большое значение для ме-
дицины и ветеринарии. Общее число видов парабазалий — свыше
300. Распространены всесветно.
Класс Parabasalea Honigberg, 1973 подразделяется
на 2 отряда: отряд Trichomonadida Kirby, 1947 —
358
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Трихомонадиды, в состав которого входит 4 семейст-
ва и 32 рода, и отряд Hypermastigida Grassi et Foa,
1911 — Гипермастигиды, который включает 2 под-
отряда, 12 семейств и более 30 родов.
Отряд Trichomonadida Kirby, 1947 —
Трихомонадиды
Многочисленный по своему составу отряд включает комменса-
лов и эндопаразитических жгутиконосцев пищеварительной систе-
мы и других внутренних органов животных и человека. Трихомо-
надиды — одноклеточные простейшие, адаптированные к анаэ-
робным условиям обитания и обладающие сложным строением
клетки. Они имеют 4—6 жгутиков, лишенных мастигонем; один из
жгутиков направлен назад и образует ундулирующую мембрану. В
клетке у преобладающего большинства видов 1 кариомастигонт и
1 парабазальный аппарат. Система жгутиковых корешков включа-
ет 3 коротких фибриллярных и 2—3 поперечно исчерченных ко-
решка и 1 косту. Пельта-аксостилярный комплекс имеет микроту-
булярную природу. Аксостиль не относится* к системе жгутиковых
корешков. Среди органелл цитоплазмы имеются многочисленные
гидрогеносомы. Ядро 1, в своем расположении связано с аксости-
лем. Способ деления ядра — типичный внутриядерный плевроми-
тоз с внеядерным веретеном и атрактофорами. Размножаются бес-
полым путем.
К отряду относится около 200 видов, среди которых много па-
тогенных, в частности возбудителей трихомонозов животных и че-
ловека. Распространены всесветно.
История изучения
Начало изучения трихомонадид относится ко вто-
рой половине XVIII в. В 1773 г. появилась работа с
описанием Trichomonas tenax (О. F. Muller, 1773)
(Kazubski, 1990; Benchimol et al., 1993; Yemilohun,
1998)/бесцветного жгутиконосца, который был обна-
ружен в ротовой полости человека. Первое описание
рода Trichomonas Donne, 1836 и именно выделение
этих жгутиконосцев в самостоятельный род связано с
тем, что А. Донне обнаружил трихрмонад в урогени-
тальном тракте человека. Он установил их патоген-
ное значение, изучил морфологию и дал первое опи-
сание вида Т. vaginalis Donne, 1837.
В последующие годы появилось описание не-
скольких видов трихомонадид из кишечника грызу-
нов (Tritrichomonas caviae Davaine, 1875; Т. muris
Grassi, 1879) и бесхвостых амфибий (Trichomitus bat-
rachorum Petry, 1892; Tritrichomonas augusta Alexei-
eff, 1911), а также ряда других видов. С конца про-
шлого столетия начинается интенсивное изучение
трихомонадид, что обусловлено большим практичес-
ким значением тех их ввдов, которые представляют
возбудителей заболеваний (трихомонозов) позвоноч-
ных животных и человека. Изучение трихомонадид,
описание их новых родов и видов продолжается и в
настоящее время.
В 1926 г. эта группа паразитических жгутиконос-
цев была объединена в сем. Trichomonadidae Weny-
on, 1926 в составе отряда многожгутиковые — Poly-
mastigina (Blochmann) Reichenow, 1928. Накопление
знаний о трихомонадидах послужило основанием
для пересмотра системы семейства, и в результате
они были выделены в самостоятельный отряд Tricho-
monadida Kirby, 1947 с включением в него 4 се-
мейств.
Последующая ревизия системы отряда Trichomo-
nadida была осуществлена Хонигбергом (Honigberg,
1963). Он тщательно пересмотрел структуру всех
4 семейств, используя данные, полученные с по-
мощью светового и электронного микроскопов, внес
* новые предложения о комплексе признаков, на осно-
вании которых следует выделять таксоны трихомона-
дид с учетом эволюционного принципа.
В настоящее время изучению трихомонадид пос-
вящена огромная литература, разработаны методы
культивирования in vitro, что в первую очередь отно-
сится к патогенным видам, особенно таким как Tri-
chomonas vaginalis, Tritrichomonas foetus, T. suis, ко-
торые исследованы с применением многих современ-
ных цитологических методик. Наряду с подробным
анализом ультраструктурной организации клетки и ее
органелл подробно изучены особенности метаболи-
ческих процессов, антигенные свойства, патоген-
ность, симптомы, клиническая картина и лечение
трихомонозов животных и человека. Однако целый
ряд видов трихомонадид беспозвоночных, а также
рыб, амфибий, рептилий и диких птиц изучены еще
очень мало.
Классификация
В состав отряда Trichomonadida входит 4 семейст-
ва, каждое с 1 или. несколькими подсемействами, и
32 рода.
Сем. Monocercomonadidae Honigberg et Kuldova, 1969
Подсем. Monocercomonadinae Honigberg, 1963
Подсем. Protrichomonadinae Honigberg et Kuldova, 1969
Подсем. Dientamoebinae Honigberg, 1984
Подсем. Chilomitinae Honigberg, 1963
Подсем. Hypotrichomonadinae Honigberg, 1963
Сем. Devescovinidae Kirby, 1931
Подсем. Devescovinihae Kirby, 1931
Подсем. Gigantomonadinae Kirby, 1944
Сем. Calonymphidae Grassi, 1911
Сем. Trichomonadidae Chalmers et Pekkola, 1918
Подсем. Trichomonadinae Honigberg, 1963
Подсем. Tritrichomonadinae Honigberg, 1963
Подсем. Pentatrichomonadinae Honigberg, 1963
Морфология и физиология
Трихомонадиды — эндопаразитические бесцвет-
ные жгутиконосцы, обладающие, несмотря на боль-
шое разнообразие таксонов, многими сходными чер-
тами морфологии, клеточного метаболизма и жизнен-
ных циклов. Все они — одноклеточные животные,
размеры тела которых широко варьируют от 10 до
100 и более микрометров в длину. Столь же широко
варьирует и форма их тела — от округлой или гру-
шевидной до сильно удлиненной с заостренными в
разной степени передним и задним концами (рис. 307).
Жгутиковый аппарат. От переднего конца тела
отходит 4—6 жгутиков; один из них направлен назад.
Задний жгутик у некоторых видов (рис. 308) имеет
свободный, выступающий за пределы тела конец, у
других же он по всей длине прикреплен к поверхно-
сти тела. На протяжении заднего жгутика, той его
части, которая прикреплена к телу, у многих видов
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД TRICHOMONADIDA
359
Рис. 307. Основные типы организации трихомонадид.
А — Dientamoeba fragilis — амебоидная особь, Б — Parahistomonas wenrichi, В — Chilomitus caviae, Г — Tritrichomonas foetus. 1— передние жгутики,
2 — Ядро, 3 — парабазальное тело, 4 — задний жгутик, 5 — аксостиль, 6 —. гидрогеносомы, 7 — коста, 8 — ундулирующая мембрана. (А — по: Brugerol-
le, 1980; Б — по: Honigberg, Kuldova, 1969; В — по: Nie, 1950; Г— по: Doflein, 1953).
Fig. 307. Some basic morphological types of Trichompnadida.
A — Dientamoeba fragilis — amoeboid specimen, Б . Parahistomonas wenrichi, В — Chilomitus caviae, Г — Tritrichomonas foetus. 1 — anterior flagella, 2 —
nucleus, 3 — parabasal body, 4 — posterior flagellum, 5 — axostyle, 6 — hydrbgenosomes, 7 — costa, 8 — undulating membrane.
проходит ундулирующая мембрана, но у некоторых
видов она отсутствует.
Жгутики и их кинетосомы, группа корешковых
структур, отходящих от кинетосом, парабазальный
аппарат, а также пельта-аксостилярный комплекс
образуют мастигонтную систему, которая представ-
ляет основу структурной организации трихрмойад-
ной клетки. Каждая клетка-организм, за небольшим
исключением, имеет 1 кариомастигонтную систему.
Ультратонкое строение свободных передних жгу-
тиков типичное (рис. 308), так же как и строение ак-
Рис. 308. Схема ультратонкого строения Tritrichomonas muris.
(По: Селюкайте, 1977).
1 — жгутиковые кинетосомы, 2 — пельта, 3 — аксостиль, 4 — коста,
5 — рудиментарная коста, 6 — парабазальный филамент, 7 — ундулирую-
щая мембрана, 8 — свободный конец заднего жгутика, 9 — десмосомопо-
добные контакты, 10 — парабазальное тело (аппарат Гольджи), 11 — гид-
рогеносомы, 12 — параксостилярное кольцо, 13 — гребень, 14— ядро.
Fig. 308. Diagram of ultrastructure organization of Tritrichomonas
muris.
1 — kinetosomes of flagella, 2 — pelta, 3 — axostyle, 4 — costa, 5 — rudi-
mentary costa, 6 — parabasal filament, 7 — undulating membrane, 8 — free
end of posterior flagellum, 9 — desmosome-like contacts, 10 — parabasal
body (Golgi apparatus), 11 — hydrogenosomes, 12 — paraxostylar ring, 13 —
cresta, 14 — nucleus.
сонемы заднего жгутика, который у ряда видов распо-
лагается по краю ундулирующей мембраны. У жгути-
коносцев родов Trichomonas, Trichomitus и других ун-
360
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 309. Схемы строения кариомастигонта.
А — изолированный кариомастигонт Histomonas meleagridis: 1 — передний жгутик, 2 — кинетосомы, 3 — пельта, 4 — парабазальное тело, 5 — параба-
зальный филамент, 6 — аксостиль, 7 — ядро; Б — Devescovina lemniscata: 1 — передние жгутики, 2 — кинетосомы, 3 — ядро, 4 — парабазальное тело,
5 — гребень, 6 — аксостиль, 7 — задний жгутик; В — Tritrichomonas foetus: 1 — кинетосомы передних жгутиков, 2 — пельта, 3 — аксостиль, 4 — ядро,
5 — парабазальные филаменты, б — коста, 7 — гребень, 8 — сигмоидный филамент, 9 — левый филамент первой кинетосомы, 10 — межкинетосомаль-
ное тельце, 11 — гидрогеносомы, 12 — задний жгутик, 13 — ундулирующая мембрана, 14 — аксонема заднего жгутика. (А — по: Honigberg, Bennet,
1971; Б— по: Kirby, 1941; В — по: Honigberg, 1971).
Fig. 309. Diagram of karyomastigont structure.
A — isolated karyomastigont of Histomonas meleagridis: 1 — anterior flagellum, 2 — kinetosomes, 3 — pelta, 4 — parabasal body, 5 — parabasal filament, 6 —
axostyle, 7 — nucleus; Б — Devescovina lemniscata: 1 —. anterior flagella, 2 — kinetosomes, 3 — nucleus, 4 — parabasal body, 5 — cresta, 6 — axostyle, 7 —
posterior flagellum; В — Tritrichomonas foetus: 1 — kinetosomes of anterior flagella, 2 — pelta, 3.— axostyle, 4 — nucleus, 5 — parabasal filaments, 6 — cos-
ta, 7 — cresta, 8 — sigmoid filament, 9 — left filament of the first kinetosome, 10i— interkinetosomal body, 11 — hydrogenosomes, 12 — posterior flagel-
lum, 13 — undulating membrane, 14 — axoneme of posterior flagellum.
Аудирующая мембрана развита хорошо, идет от пере-
днего конца к заднему, занимая значительную часть
длины тела (рис. 309).
Плазмалемма этого органоида, идущего вдоль дор-
сальной стороны тела жгутиконосца, не сливается с
плазмалеммой клетки, а имеет с ней специальные
контакты, по-видимому, подвижные, которые осу-
ществляются с помощью плотного гомогенного ве-
щества (рис. 310). Нижняя, направленная к краю
клетки часть ундулирующей мембраны, образует
Рис. 310. Ультратонкое строение ундулиру-
ющей мембраны (схематизировано).
А — Tritrichomonas minuta, Б — Т. lacertae, В —
Т. augusta, Г — Т. foetus, Д — Т. muris, Е — Tri-
chomonas gallinae, Ж — Т. vaginalis. 1 — ундули-
рующая мембрана, 2 — аксонема заднего краевого
жгутика, 3 — продольные тяжи, 4 — параксиаль-
ный тяж, 5 — связующий компонент. (А—В — по:
Joyon et al., 1969; Г — по: Honigberg, 1971: Д —
по: Селюкайте, 1977, Е — по: Mattern et al., 1967;
Ж — по: Nielsen et al., 1966).
Fig. 310. Ultrastructural organization of undu-
lating membranes (diagram).
A — Tritrichomonas minuta, A — T. lacertae, Б —
T. augusta, В — T. foetus, Г — T. muris, Д — Tri-
chomonas gallinae, E — T. vaginalis: 1 — undula-
ting membrane, 2 axonema of posterior flagellum,
3 — longitudinal fibrills, 4 — paraxial body, 5 —
bounding component.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД TRICHOMONADIDA
361
Рис. 311. Цистоподобные стадии трихомонадид.
А — Tritrichomonas muris var. meriones; Б — T. muris var. hystrici; В — T. caviae; Г—Т. hominis; Д — T. muris: 1 — оболочка цистоподобной особи, 2 —
аксостиль, 3 — ядро 4 — коста, 5 — ундулирующая мембрана; Е — ультратонкое строение цистоподобной формы Т. muris (схематизировано по электро-
нограмме): 1 — внешняя оболочка; 2 — ундулирующая мембрана; 3 — жгутики, погруженные внутрь клетки; 4 — ЭПР; 5 — гидрогеносомы; 6 — аксо-
стиль (видны микротрубочки, образующие его стенку); 7 — гликоген; 8 — ядро; 9 —- коста; 10 — аппарат Гольджи. (А — по: Пак, 1960; Б — по: Dubinin,
1938; В — по: Wenyon, 1965; Г — по: Gwelessiany, 1929; Д, Е — по: Селюкайте, 1977).
Fig. 311. Stages of Trichomonadida similar to cysts.
A — Tritrichpmonas muris var. meriones; Б — T. muris var. hystrici; В — T. caviae; Г — T. hominis; Д —. T. muris: 1 — envelope, 2 — axostyle, 3 — nucleus,
4 — costa, 5 — undulating membrane; E — ultrastructure of cyst-like specimen of T. muris (diagram): 1 — envelope, 2 — undulating membrane, 3 — flagella in-
side the cell, 4 -^- endoplasmic reticulum, 5—hydrogenosomes, 6—axostyle composed by microtubules, 7—glycogen, 8 — nucleus, 9 — costa, 10 —
Golgi apparatus.
складку. Строение этой складки неодинаково у раз-
ных видов (рис. 310, Д); наибольшей сложности она
достигает у Tritrichomonas muris — обычной трихо-
монады кишечника грызунов. У большинства же
видов трихомонад ундулирующая мембрана представ-
ляет тонкий лентовидный органоид без дополнитель-
ных опорных<структур.
У Т. muris ундулирующая мембрана утолщена и
непосредственно под аксонемой жгутика проходит
2 продольных плотных тяжа. Вся остальная часть
ундулирующей мембраны занята кристаллическим
телом. Это — наиболее хорошо развитая часть орга-
ноида, которая вместе с продольными тяжами пред-
ставляет опорную структуру. Кристаллическое тело
Рис. 312. Histomonas meleagridis. (По: Honigberg, Bennet, 1971).
А — жгутиковая особь, Б — амебоидные особи. 1 — передний жгутик,
2 — ядро, 3 — парабазальное тело, 4 — аксостиль.
Fig. 312. Histomonas meleagridis.
А — flagellate specimen, Б — amoeboid specimens. 1 — anterior flagellum,
2 — nucleus, 3 — parabasal body, 4 — axostyle.
состоит из множества электронно-плотных, очень
тонких филаментов, расположенных друг по отно-
шению к другу под углом 90°. Филаменты находятся
на одинаковом расстоянии друг от друга и образуют
упорядоченную структуру, которая состоит из чере-
дующихся светлых и темных полос. В том месте, где
заканчивается ундулирующая мембрана, жгутик, иду-
362
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
щий по ее краю, остается свободным и выходит за
пределы клетки (рис. 309).
Кинетосомы всех жгутиков образуют кинетосо-
мальный комплекс, который включает 5 кинетосом.
4 из них расположены параллельно продольной оси
клетки и друг другу, давая начало свободным передним
жгутикам. 5-я кинетосома, соответствующая заднему
жгутику, расположена под косым углом по отноше-
нию к продольной оси клетки и к остальным кине-
тосомам. Форма кинетосом цилиндрическая, каждая
из них состоит из 9 триплетов микротрубочек. Пе-
реходная зона жгутиков трихомонадид короткая, с 1
переходной пластинкой, без других дополнительных
структур (Карпов,. 1990).
Корешковая система жгутиков образована тремя
короткими фибриллярными корешками. Из них наи-
более крупный — сигмоидный корешок, который
представляет ЦОМТ пельта-аксостилярного комплек-
са. Корешковую систему дополняют 2 парабазальные
фибриллы и коста, которые представляют собой по-
перечно исчерченные жгутиковые корешки (рис. 308;
309). Аксостиль не относится к корешковой системе
жгутиков (Карпов, 1990).
Коста отходит от базальной части кинетосомы
заднего жгутика. Проксимальная часть косты, приле-
гающая к кинетосоме, расширена. Коста слегка изо-
гнута, проходит вдоль всего тела по его дорсальной
стороне параллельно ундулирующёй мембране и под-
держивает ее. В состав косты входят белки с молеку-
лярной массой от 26 до 122 кДа; углеводов в ее со-
ставе нет (Monteiro-Leal et al., 1993). У некоторых
видов, например у Tritrichomonas muris, есть еще и
рудиментарная коста — тонкий филамент, идущий
параллельно косте в передней ее части. У этого вида
верхний конец косты контактирует с другой попереч-
но-полосатой структурой — гребнем.
Пельта-аксостилярный комплекс. Комплекс
включает 2 органоида, состоящих из микротрубочек:
пельту и аксостиль. Пельта — органелла серповид-
ной формы, поддерживающая кинетосомальный ком-
плекс (рис. 309). Микротрубочки расположены по
длине пельты, и ее расширенную часть, контактиру-
ющую с аксостилем, образуют 35—40 микротрубо-
чек, каждая по 20 нм в диаметре. Микротрубочки
расширенной части пельты входят внутрь головки
аксостиля и вместе с его микротрубочками образуют
зону тесного контакта обеих органелл.
Головка аксостиля — расширенная его часть —
имеет ложковидную форму. Аксостиль проходит по
всей длине тела, состоит из микротрубочек (каждая
по 20 нм в диаметре), которые располагаются в
1 ряд, плотно прилегая друг к другу и образуя плас-
тинку. Расширена только передняя часть аксости-
ля — головка, на большей же части его длины края
микротубулярной пластинки смыкаются и аксостиль
имеет форму трубки. Задний конец аксостиля выда-
ется наружу в виде заостренного стержня, покрытого
плазмалеммой; на границе его выхода из тела распо-
ложено параксостилярное кольцо. Аксостиль три-
хомонадид может совершать небольшие движения.
Внутренняя часть аксостиля наполнена гликогеном,
который представляет основное резервное вещество
клетки.
Органеллы цитоплазмы. У трихомонадид отсут-
ствуют митохондрии, что представляет одну из их
специфических особенностей. Постоянными орга-
неллами цитоплазмы клетки трихомонад являются
гидрогеносомы — плотные округлые микротельца
около 1 мкм в диаметре (рис. 308; 309; 315). Одна
группа гидрогеносом находится возле вогнутой по-
верхности косты, вторая расположена возле аксости-
ля. Гидрогеносомы покрыты плотной мембраной; их
внутреннее содержимое представляет собой мел-
козернистый матрикс с уплотненной центральной
частью. Морфогенез гидрогеносом, изученный у не-
скольких видов трихомонад, подобен таковому ми-
тохондрий и осуществляется путем деления с обра-
зованием 2 дочерних органелл. Гидрогеносомы тесно
связаны с ЭПР и другими органеллами клетки при
помощи пузырьков, осуществляющих транспорт ве-
ществ. На поверхностной мембране клетки трихомо-
над (Т. foetus и др.) обнаружен белок адгезии, ко-
торый концентрируется в пузырьках цитоплазмы, а
затем выделяется наружу. Предполагается, что он
способствует адгезии трихомонад к эритроцитам хо-
зяина. После установления контакта эти пузырьки
осуществляют обмен веществ между клетками хозяина
и паразита (Monteiro-Leal et al., 1993; Benchimol et
al., 1993, 1996; da Silva et al., 1999). Гидрогеносомы
продуцируют молекулярный водород в качестве ко-
нечного продукта ферментативного расщепления уг-
леводов (Muller, 1973).
Ядро. Ядро располагается в передней, расширен-
ной части аксостиля. По своей организации оно ти-
пично для эукариотической клетки и может быть от-
несено к хромосомному типу строения ядер простей-
ших (Райков, 1978).
Митоз. Деление ядра осуществляется по типу за-
крытого плевромитоза (рис. 316). Этот тип митоза
имеет ряд особенностей: 1) ядерная оболочка сохра-
няется на всех фазах деления; 2) формируется вне-
ядерное веретено, образующееся в цитоплазме между
2 атрактофорами («носители веретена»). Атрактофо-
ры имеют пластинчатую или палочковидную форму и
отходят от основания кинетосомального комплекса.
Веретено митоза имеет форму нити, которая соеди-
няет оба атрактофора, и известно под названием па-
радесмос и центродесмос.
С помощью электронного микроскопа наиболее
полно изучен митоз у Trichomonas vaginalis (Bruge-
rolle, 1975с; Puytorac et al., 1987; рис. 316, Б). У этого
вида атрактофоры пластинчатые и находятся в тес-
ном контакте с основаниями кинетосом. Нитевидное
центральное веретено представляет собой пучок мик-
ротрубочек. От нижних концов атрактофоров отхо-
дят хромосомные и астральные микротрубочки. Хро-
мосомы имеют кинетохоры мелких размеров, кото-
рые «встроены» в ядерную оболочку. Хромосомные
микротрубочки прикрепляются к кинетохорам, что
хорошо видно в метафазе. Расхождение хромосом в
анафазе происходит за счет микротрубочек веретена.
В телофазе ядро принимает гантелевидную форму и
перешнуровывается, образуя ядра 2 дочерних клеток.
Митотическое деление клетки трихомонадид со-
провождается сложной перестройкой цитоплазмати-
ческих органелл, и прежде всего системы мастигонта.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД TRICHOMONADIDA
363
Изучение деления клетки Tritrichomonas foetus в
культуре in vitro показало, что дупликация кинетосом
начинается в ранней профазе. В это же время форми-
руется новый парабазальный аппарат при сохранении
старого. После этого органеллы мастигонта делятся
на 2 группы. Одна группа содержит 2 передних сво-
бодных жгутика, старую косту, старый парабазальный
аппарат и ундулирующую мембрану. Во-вторую группу
входит 3-й свободный жгутик, новая коста и новый
парабазальный аппарат. Недостающие органеллы каж-
дого мастигонта восстанавливаются в процессе по-
следующего деления. В конце профазы формируется
внеядерное веретено митоза, исчезает ядрышко и ста-
новятся хорошо видимыми 6 хромосом. Аксостиль и
пельта во время деления материнской клетки подвер-
гаются резорбции, и у обеих дочерних клеток форми-
руются зачатки 2 дочерних аксостилей, имеющих в
этот момент форму запятой.
Цитокинез осуществляется в продольном направ-
лении (Kuldova, Kulda, 1974). По такому же типу
протекает митоз и у Histomonas meleagridis (Honig-
berg, Bennett, 1971).
Кроме монотомии известен и способ множествен-
ного деления у Tritrichomonas foetus и ряда других
видов. При таком способе размножения ядра делятся
без цитокинеза, в результате чего образуются много-
ядерные особи, которые имеют & и более ядер и до-
стигают в диаметре 30—35 мкм (рис. 315). В много-
ядерной клетке числу ядер соответствует и число
мастигонтов. Такие клетки сохраняют способность к
делению и потому получили название временных со-
мателл. Появлению многоядерных клеток трихомо-
над способствует воздействие разнообразных небла-
гоприятных внешних факторов. Например, у Т. foetus
они появляются в особенно большом количестве при
лечении больных животных противотрихоманозными
препаратами и в культурах на жидких питательных
средах (Тимофеев, Петровский, 1967). У людей,
больных трихомонозом, многоядерные клетки Tricho-
monas vaginalis образуются в большом количестве в
период лечения, а в. культурах in vitro — при добав-
лении в питательную среду различных солей, щело-
чей и других веществ, изменяющих ее оптимальный
солевой состав (Падченко, 1975).
Жизненные циклы
У большинства видов трихомонадид жизненные
циклы изучены слабо, и наиболее подробные сведе-
ния имеются только о трофозоитах, обитающих в ор-
ганизме хозяина. О стадиях жизненного цикла, про-
ходящих во внешней среде, известно мало, и до сих
пор остается дискуссионным вопрос о существовании
цист у, большинства видов. Решение этого вопроса
особенно важно в случае патогенных видов, так как с
ним связано выяснение путей распространения три-
хомонадид в природе и путей заражения животных и
человека.
Наиболее полно жизненный цикл изучен у пато-
генного для человека вида Т. vaginalis (Падченко,
1975; Honigberg, 1978b). Трофозоиты этого вида об-
ладают довольно широким морфологическим поли-
морфизмом как в урогенитальном тракте человека,
так и в культурах in vitro. В составе популяций этих
трихомонад имеются неделящиеся и делящиеся особи,
но ни в одной популяции не были обнаружены цисты.
Заражение человека происходит с помощью трофо-
зоитов, передающихся от больных трихомонозом
людей при половом контакте. Таким же путем про-
исходит заражение трихомонадами крупного рогато-
го скота (Tritrichomonas foetus) и свиней (Т suis).
У трихомонад, обитающих в пищеварительном
тракте различных животных и человека (Trichomonas
tenax, Т. gallinae, Pentatrichomonas hominis), а также
у девесковинид из кишечника термитов цисты неизвестны.
Заражение животных происходит трофозоитами. Боль-
шая литература существует о цистоподобных стади-
ях, или псевдоцистах, у трихомонадид, паразитирую-
щих в кишечнике грызунов (Tritrichomonas muris,
Т. caviae). Псевдоцисты (рис. 311) лишены плотной
оболочки, которая свойственна цистам других про-
стейших. Свободные передние жгутики, ундулирую-
щая мембрана и свободный конец аксостиля втянуты
внутрь клетки и находятся в цитоплазме псевдоцис-
ты. Роль псевдоцист в заражении грызунов не выяс-
нена (Дубинин, 1938; Пак, 1960; Селюкайте, 1977).
Псевдоцисты обнаружены и изучены у Trichomitus la-
certae. Их структура сходна с псевдоцистами трихо-
монад из кишечника грызунов (Brugerolle, 1973b).
Цисты, подобные цистам покоя других простей-
ших, известны у Т. batrachorum из кишечника амфибий
и у Monocercomonas tipulae, обитающего в кишечни-
ке личинок ручейников и у типулид (Brugerolle, 1973b).
Цисты этих 2 видов трихомонадид изучены с помощью
светового и электронного микроскопов. Форма цист
округлая, диаметр их у Т. batrachorum равен 5—
6 мкм. Оболочка цист плотная, довольно толстая
(0.15 мкм), состоит из микрофиламентов, а также
включает плотные осмиофильные гранулы. Поверх-
ностная мембрана клетки образует многочисленные
складки. Внутри оболочки цитоплазма клетки совер-
шает ротационные движения. Свободные жгутики и
ундулирующая мембрана втянуты внутрь клетки, ак-
состиль и пельта изогнуты, сохраняют свое положение
в клетке парабазальный филамент и аппарат Гольд-
жи. Цитоплазма инцистированной клетки сильно ва-
куолизирована, и особенно много вакуолей располо-
жено вокруг парабазального аппарата. Цисты устой-
чивы к факторам внешней среды.
Более сложный жизненный цикл свойствен Histo-
monas meleagridis (Honigberg, Bennett, 1971; Honig-
berg, 1978a). Этот вид трихомонадид паразитирует в
слепой кишке и печени куриных и гусиных птиц.
В его жизненном цикле не обнаружены цисты, но
существует 2 фазы: амебоидная и жгутиковая (рис. 312).
Первая инвазирует ткани слепой кишки и печени
птиц. Амебоидные гистомонады активно питаются,
поглощая клеточный материал, и размножаются пу-
тем бинарного деления. Размеры активных амебоид-
ных форм 10—15 мкм.
Жгутиковые формы Н. meleagridis обитают в
просвете слепой кишки птиц, и их движение среди
пищевых масс осуществляется с помощью псевдопо-
дий (лобоподий) и жгутика; в отличие от других
трихомонадид этот вид гистомонад имеет только
364
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
1 жгутик. Форма жгутиковых особей почти округ-
лая, 10—15 мкм в диаметре, они активно делятся
(рис. 312).
Амебоидные и жгутиковые особи повреждают тка-
ни печени и слепой кишки, и заболевание птиц гисто-
монозом протекает часто в острой, тяжелой форме.
Жизненный цикл Н. meleagridis протекает со сме-
ной хозяев, и вторым их хозяином служит нематода
Heterakis sp., которая паразитирует в слепой кишке
птиц. Эта нематода заглатывает гистомонад в качест-
ве пищевых объектов. В кишечнике нематод гисто-
монады не погибают, а проникают в клетки кишечно-
го эпителия, активно в них размножаются и разруша-
ют их. Трофозоиты гистомонад попадают в полость
тела нематоды и из нее — в клетки репродуктивных
органов.
У женских особей нематод трофозоиты мигриру-
ют в яичники и локализуются между оогониями. Раз-
вивающиеся ооциты оказываются зараженными гис-
томонадами, которые продолжают размножаться. У
зараженных мужских особей нематод гистомонады
также локализуются в гонадах и располагаются
между сперматозоидами. Поэтому возможно зараже-
ние женских особей нематод при копуляции с зара-
женными мужскими особями.
Заражение птиц происходит инвазированными яй-
цами нематод, в которых трофозоиты Histomonas me-
leagridis могут сохранять жизнеспособность в тече-
ние нескольких лет. В фекалиях птиц трофозоиты
остаются жизнеспособными в течение 2—3 ч, и путь
инвазии через фекалии значительна менее эффекти-
вен, чем через промежуточного хозяина. Заражение
птиц инвазированными яйцами нематод представляет
собой пример двойной инвазии: птицы одновременно
заражаются нематодами и гистомонадами.
Культивирование
При изучении трихомонадид культуры in vitro
имеют особое значение и широко используются с
целью диагностики трихомонозов животных и чело-
века, для электронно-микроскопических и биохими-
ческих работ, изучения антигенных свойств, форм
изменчивости* а также воздействия факторов внеш-
ней среды. Разработаны методы клонирования трихо-
монадид в культурах, изучения вирулентности й пато-
генности клонов, специфичности к хозяину разных
видов трихомонадид, действия лекарственных препа-
ратов.
Наиболее хорошо разработаны методы культиви-
рования и питательные среды для патогенных видов
трихомонадид: Trichomonas vaginalis, Т gallinae; Tri-
trichomonas foetus, T. suis, Histomonas meleagridis и
других. Эти культуры требуют совершенно опреде-
ленных условий, при которых возможно их длитель-
ное содержание: анаэробные условия, для некото-
рых — наличие СО2, температура 37 °C и другие.
Для культивирования Trichomonas vaginalis предло-
жено много разнообразных сред. Культуры этого вида
на минеральных средах известны с начала 20-х гг. на-
шего столетия. Известны также агнотобиотические
культуры, среда которых включает минеральные ком-
поненты и сыворотку крови млекопитающих. Однако
эти среды непригодны для длительного культивирова-
ния трихомонад, так как в них достаточно быстро
развиваются бактерии.
В настоящее время разработан целый ряд сред для
аксенических культур трихомонадид, а также полу-
- жидкие и плотные среды, содержащие разные коли-
чества агар-агара. Состав питательных сред различен.
Обычна печеночная среда с добавлением цистеина,
пептона и мальтозы, печеночная среда с добавлением
цистеина, триптозы и мальтозы, а также упрощенная
среда — сыворотка крови с триптиказой. В культу-
ральные среды добавляют антибиотики: пенициллин,
стрептомицин, неомицин и другие с целью уничтоже-
ния бактерий, что необходимо для длительного куль-
тивирования трихомонадид (Diamond, 1957; Терас, 1963;
Honigberg, 1978а, 1978b).
Для ряда видов трихомонадид аксенические куль-
туры еще не разработаны. Таких культур нет, напри-
мер; для Tritrichomonas muris, Histomonas meleagri-
dis, Dientamoeba fragilis и других. 2 последних вида
растут только на средах, содержащих бактерии, ко-
торые необходимы для питания этих жгутиконосцев
(Camp et al., 1974; Honigberg, 1978a).
В изучении морфологии, клеточного метаболизма,
патогенности и других особенностей биологии три-
хомонадид большое значение имеет замораживание
их культур при сверхнизких температурах от -79 до
-190 °C. Разработаны специальные режимы замора-
живания и оттаивания. Например, аксенические
культуры Trichomonas vaginalis замораживались до
температуры -190 °C с добавлением в них 5%
ДМСО в качестве криопротектора. При заморажива-
нии культур, находившихся в поздней логарифмичес-
кой или ранней стационарной фазах, выживало 80—
100% трихомонад (определение выживаемости про-
водилось с использованием теста движения в разные
сроки после пребывания их в условиях температур
жидкого азота, — Honigberg, 1978b). Размораживание
культур проводилось также ступенчато. В опытах ря-
да исследователей получены данные о том, что после
замораживания в жидком азоте и пребывания в таком
состоянии в течение 2 лет Т. vaginalis не только сохра-
няет жизнеспособность, но и не теряет патогенность.
Т. gallinae хорошо переносит замораживание в
жидком азоте с добавлением ДМСО или глицерина.
В культуральных средах* содержащих 5 % ДМСО,
эти трихомонады находились в замороженном со-
стоянии 12 лет, после чего оставалось живыми свыше
90 % особей. Они были подвижны и без каких-либо
морфологических изменений (Honigberg, 1978а).
Tritrichomonas foetus после замораживания в жид-
ком азоте также сохраняет свои патогенные свойства
после разных сроков пребывания в условиях сверх-
низких температур, если к культуральной среде до-
бавляют 5 % ДМСО. Результаты опытов с Т foetus
имеют особую практическую значимость для живот-
новодства: сперма быков, замороженная при темпе-
ратуре -190 °C и предназначенная для искусственно-
го осеменения, может служить источником инвазии
коров, если она получена от зараженных трихомона-
дами животных (трихомоноз — одна из причин сеп-
тических абортов у коров).
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД TRICHOMONADIDA
365
Замораживание при -79, -95 и -190 °C хорошо
переносят также Trichomonas tenax и Pentatrichomo-
nas hominis, если при охлаждении культур в них до-
бавляют ДМСО или глицерин (Honigberg, 1978b).
Происхождение и эволюция
Среди трихомонадид у основания филогенетиче-
ского древа отряда стоит сем. Monocercomonadidae,
ряд видов которого обладает многими примитивны-
ми чертами организации. К числу таких видов отно-
сятся: Monocercomonas colubrorum Hammerschmidt,
1844, Paratrichomonas wentrichi и Histomonas mele-
agridis (рис. 312). Эти виды — амебофлагеллаты с
хорошо выраженными псевдоподиями лобозного типа.
Они отличаются простым строением кариома-
стигонта, в котором структуры его не так тесно свя-
заны друг с другом, как у трихомонадид, имеющих
более высокую степень дифференциации клеточных
органелл (рис. 307; 315). В клетке этих видов имеется
1 кариомастигонт; структурные элементы его сход-
ны морфологически и, вероятно, гомологичны тако-
вым других трихомонадид (Honigberg, 1963).
Метод сравнительной морфологии позволяет про-
следить развитие элементов кариомастигонта в про-
цессе эволюции трихомонадид, которая проходила
по пути их приспособления к обитанию в кишечни-
ке и в других органах беспозвоночных и позвоноч-
ных хозяев. Одним из важнейших путей эволюции
трихомонадид служит полимеризация опорно-двига-
тельных органелл и ядер их клетки (Догель, 1951).
Процесс полимеризации проходил прежде всего по
линии умножения числа свободных жгутиков от 1 у
Histomonas meleagridis до 5 у Pentatrichomonas homi-
nis. При умножении числа жгутиков умножается и
число их кинетосом, но другие органеллы полиме-
ризации не подвергаются. Более сложный путь поли-
меризации у трихомонадид представляет умножение
2 и большего числа органелл на уровне моноэнер-
гидной клетки. Примером тому может служить Pse-
udodevescovina uniflagellata с увеличением числа па-
рабазальных аппаратов до 5—6 при 3 свободных
жгутиках. В этом процессе ядро участия не при-
нимает, и умножение органелл происходит путем их
деления или образования заново в одноядерной клетке.
Еще один путь полимеризации органелл — их де-
ление в пределах одной клетки, причем умножение
претерпевают несколько органелл и ядер. Такой путь
полимеризации свойствен, например, Coronympha
clevelandi, у которой в одной клетке образуется
16 кариомастигонтов, расположенных в виде венчика
на переднем конце тела (рис. 314). Следовательно,
Coronympha в процессе эволюции становится много-
ядерным организмом, стоящим на более высокой сту-
пени организации, которая получила название сома-
теллы (Догель, 1951).
Полимеризация органелл — путь прогрессивной
эволюции в отряде Trichomonadida. Она направлена
на усиление функциональной активности организма,
что связано и с увеличением размеров тела при пере-
ходе организма на более высокий уровень энергети-
ческого обмена.
Ниже в виде схемы представлены возможные пути
эволюционных взаимоотношений в отряде Trichomo-
nadida (Honigberg, 1963b).
Pentatrichomonadinae
?
Tritrichomonadinae Trichomonadinae
?
Trichomonadidae
?
Calonymphidae
?
Hypotrichomonadinae
?
Devescovinidae Monocercomonadinae
T
Chilomitinae
?
Monocercomonas
?
MONOCERCOMONADIDAE
Происхождение Trichomonadida от каких-либо сво-
бодноживущих форм установить довольно трудно.
Систематический обзор
Сем. Monocercomonadidae Honigberg
et Kuldova, 1969
В состав семейства входит 5 подсемейств и 10 ро-
дов. Моноцеркомонадиды характеризуются наиболее
примитивной организацией, разнообразием морфоло-
гии и жизненных циклов (Honigberg, Kuldova, 1969;
Camp et al., 1974). Среди них известны амебоидные
формы (Dientamoeba fragilis Jepps et Dobell, 1918, —
рис. 307), виды с одинаково хорошо развитыми аме-
боидными и жгутиковыми формами (Paratricho-
monas, Histomonas), а также виды с типичными для
трихомонадид жгутиковыми клетками (Chilomitus,
Hexamastix, Pseudotrichomonas, Tritrichomonas, —
рис. 307). Соответственно имеются и 2 формы движе-
ния: амебоидное с образованием псевдоподий и жгу-
тиковое с помощью передних и заднего жгутиков.
В клетке всех видов моноцеркомонадид содержится
1 кариомастигонт (рис. 307; 315) и число свободных
передних жгутиков у трофозоитов разных видов варьи-
рует от 1 до 5. Задний жгутик свободен по всей дли-
не, но у некоторых видов проксимальная часть его
прикреплена к дорсальной поверхности переднего
конца тела и проходит вдоль дополнительного фила-
мента (род Monocercomonas), Ундулирующая мембра-
на не у всех видов, но если она есть, то слабо развита
(род Hypotrichomonas). Коста отсутствует.
Аксостиль у большинства видов тонкий, палочко-
видный, головка его соединяется с пельтой. Параба-
зальный аппарат включает хорошо развитый фила-
мент и парабазальное тельце (аппарат Гольджи) па-
лочковидной, дисковидной или V-образной формы
(рис. 307; 315).
Моноцеркомонадиды распространены всесветно.
Они населяют пищеварительный тракт насекомых
(Pentatrichomastix parisii MacKinnen, 1913), рыб
(Protrichomonas legeri Alexeieff, 1911), амфибий
366
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
(Hexamastix batrachorum Alexeieff, 1912), рептилий
(Hypotrichomonas acosta Lee, I960), птиц (Parahisto-
monas wenrichi Honigberg, 1969), морских свинок
(Chilomitus caviae Da Fonseca, 1915), человека (Dien-
tamoeba fragilis).
Большинство видов семейства — непатогенные
обитатели пищеварительного тракта, за исключени-
ем Histomonas meleagridis (рис. 312). Этот вид, как
уже было отмечено, поражает слепую кишку и пе-
чень (чаще всего у индеек), и болезнь особенно ост-
ро протекает у молодых птиц, нередко вызывая их
гибель.
Сем. Devescovinidae Kirby, 1931
Включает 2 подсемейства и 8 родов. Девесковини-
ды — непатогенные комменсалы кишечника терми-
тов и тараканов рода Cryptocercus.
Большинству видов присуще удлиненное тело,
размеры которого широко варьируют от 6 до 140 мкм
в длину. Кариомастигонт 1, типичный по своему
строению, но есть ряд отличий, характеризующих
особенности семейства (Kirby, 1941). Передних жгу-
тиков 3, они одинаковой длины, и 1 задний жгутик,
имеющий значительную толщину и прикрепленный
к дорсальной стороне тела на небольшом участке
переднего конца (рис. 313). Остальная часть жгутика
свободна и идет вдоль тела по его дорсальной сто-
роне. Ундулирующей мембраны нет. В составе кари-
омастигонта есть особая пластинчатая структура —
гребень (рис. 309, £>), проходящий вдоль переднего
края тела по дорсальной стороне там, где прикреп-
ляется задний жгутик. Аксостиль и парабазальный
аппарат варьируют по форме и размерам; у многих
видов парабазальный филамент обвивается вокруг
аксостиля (рис. 309). Коста отсутствует.
Рис. 313. Gigantomonas herkulea. (По: Догель, 1951).
А — жгутиковая особь, Б — амебоидная особь. 7 — передние жгутики,
2 — ядро, 3 — аксостиль, 4 — креста, 5 — псевдоподии, 6 — пищевые
частицы.
Fig. 313. Gigantomonas herkulea.
А — flagellate specimen, Б — amoeboid specimen, 7 — anterior flagella, 2 —
nucleus, 3 — axostyle, 4 — cresta, 5 — pseudopodia, 6 — food particles.
Девесковиниды — ксилофаги. Они питаются мел-
кими частичками древесины, содержащимися в зад-
ней кишке хозяев. Если термитов лишить этих жгу-
тиконосцев, то они погибнут, поскольку ферменты
их кишечника не действуют непосредственно на^
клетчатку. Девесковиниды синтезируют ферменты,
расщепляющие целлюлозу, и только с помощью этих
жгутиконосцев она подготавливается к переварива-
нию ферментами хозяина (Догель, 1951). Питание
частицами древесины связано у некоторых видов с
переходом в амебоидную стадию, что наблюдал До-
гель (1916) у Gigantomonas herkulea (рис. 313).
У девесковинид нет цист, и заражение насекомых
ими происходит при питании фекалиями: капли фе-
калий, в которых содержатся трофозоиты жгутико-
носцев, выделяются из ануса и слизываются другими
особями термитов. Поэтому постоянно обновляются
популяции жгутиконосцев у имаго термитов и зара-
жаются молодые особи этих насекомых.
Среди трофозоитов, находящихся в задней кишке
хозяев, Догель (1951) обнаружил небольшое число
«мумифицированных» особей округлой формы с уп-
лотненной цитоплазмой и неподвижных. Такие особи,
как считает Догель, свободно проходят через желу-
док и тонкий кишечник, избегая каких-либо повреж-
дений за счет действия пищеварительных ферментов
хозяина. При линьке термитов удаляется вся выстил-
ка эпителия задней кишки, и вместе с ней удаляются
все обитающие там жгутиконосцы. После линьки хо-
зяина происходит новое его заражение жгутиконос-
цами.
Типовой род семейства — Devescovina (рис. 309,
Б).
Сем. Calonymphidae Grassi, 1911
В составе семейства 5 родов, к которым относят-
ся облигатные комменсалы заднего отдела кишечни-
ка термитов. В отличие от девесковинид они имеют
постоянную полимонадную организацию и у разных
Рис. 314. Coronympha octonaria. (По: Kirby, 1941).
7 — жгутики, 2 — комплекс кариомастигонтов, 3 — аксостили.
Fig. 314. Coronympha octonaria.
1 — flagella, 2 — complex of karyomastigonts, 3 — axostyles.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД TRICHOMONADIDA
367
видов в клетке содержится от 8 до 345 индивиду-
альных кариомастигонтов, расположенных спереди
(рис. 314). Для рода Calonympha характерны также
акариомастигонты, так как часть ядер у них не со-
единена с опорно-двигательными органеллами. У
этого рода аксостили всех мастигонтных систем со-
единяются в 1 плотный пучок, проходящий вдоль
тела.
По способу питания и другим особенностям био-
логии калонимфиды сходны с девесковинидами.
Типовой род семейства — Calonympha.
Сем. Trichomonadidae Chalmers
et Pekkola, 1918
Семейство объединяет 3 подсемейства и 8 родов.
Для этих жгутиконосцев характерно наличие 4—
6 Ж1угиков (рис. 307; 315), из которых 1 направлен
назад и проходит по краю ундулирующей мембраны,
варьирующей по степени развития у разных видов.
Ундулирующая мембрана поддерживается костой, ко-
торая располагается в краевой зоне цитоплазмы вдоль
линии прикрепления ундулирующей мембраны к
телу. Аксостиль, расширенная головка которого про-
должается в пельту, и парабазальный аппарат хорошо
развиты.
В состав подсемейства Trichomonadinae входит
4 рода. Типовой род — Trichomonas, к которому
относится Т. vaginalis (рис. 315) — патогенный пара-
зит урогенитального тракта человека. Этот вид три-
хомонадид уже около 150 лет привлекает к себе вни-
мание протозоологов и врачей и в настоящее время
изучен достаточно подробно. Кроме морфологии на
уровне светового и электронного микроскопов, кле-
точного метаболизма, жизненного цикла и разработ-
ки условий культивирования in vitro подробно изуче-
ны антигенные свойства Т. vaginalis, разработаны ме-
тоды иммунодиагностики (Терас, 1963, и ряд др.
работ).
Г. vaginalis имеет всесветное распространение, и
трихомоноз урогенитального тракта встречается до-
статочно часто. Например, в 1972 г. свыше 180 млн
женщин и мужчин во всех странах мира были больны
этой формой трихомоноза (Honigberg, 1978b). Отсю-
да — огромная практическая значимость изучения
Т. vaginalis, клинической картины заболевания,
средств лечения и профилактики.
Из других видов трихомонад, обитающих в орга-
низме человека, часто встречается Т. tenax. Этот вид,
патогенное значение которого пока неясно, обитает в
ротовой полости, и, по-видимому, относится к числу
комменсалов. Трихомонады ротовой полости, так же
как и большинство других видов рода, распростране-
ны всесветно, и в разных странах мира заражение
ими человека варьирует от 0 до 78 % (Honigberg,
1978b). Обычным комменсалом кишечника человека
считается Pentatrichomonas hominis Dayaine, 1860.
Еще один типичный и очень многочисленный род
семейства — Tritrichomonas. К нему относится 2 па-
тогенных вида сельскохозяйственных животных —
Т foetus и Т. suis. К этому же роду принадлежит и
Рис. 315. Морфология клетки типичных видов трихомонадид.
А — Trichomonas vaginalis. Б — Pentatrichomonas hominis. В — Tritrichomonas augusta. 1 — передние жгутики, 2 — ундулирующая мембрана, 3 — зад-
ний жгутик, 4 — коста, 5 — гидрогеносомы, б — ядро, 7 — аксостиль, 8 — парабазальное тело, 9 — парабазальнай филамент, 10 — пельта, 11 — пара-
ксиальное кольцо.
Fig. 315. Cell morphology in typical species of Trichomonadida.
A — Trichomonas vaginalis, Б — Pentatrichomonas hominis, В — Tritrichomonas augusta. 1 — anterior flagella, 2 — undulating membrane, 3 — posterior fla-
gellum, 4 — costa, 5 — hydrogenosomes, 6 — nucleus, 7 — axostyle, 8 — parabasal body, 9 — parabasal filament, 10 — pelta, 11 — paraxial ring.
368
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 316. Закрытый внутриядерный плевромитоз у трихомонадид.
А — анафаза митоза у Tritrichomonas muris, Б — ядро в стадии анафазы у
Т. caviae, В — схема митоза у Trichomonas vaginalis. 1 — внеадерное вере-
тено митоза (парадесмос); 2 — микротрубочки парадесмоса, 3 — хромо-
сомные микротрубочки; 4 — кинетохоры; 5 — атрактофоры; 6 — микро-
фибриллы, соединяющие атрактофоры с основаниями кинетосом; 7 — яд-
ро; 8 — хромосомы; 9 — ядерная оболочка. (А, Б — по: Doflein, 1953;
В — по: Puytorac et al., 1987).
Fig. 316. Close intranuclear pleuromitosis in Trichomonadida.
A — anaphase in Tritrichomonas muris, Б — nucleus in anaphase (T. caviae),
В — diagram of mitosis in Trichomonas vaginalis. 1 — extranuclear mitotic
spindle (paradesmos), 2 — microtubules of paradesmos, 3 — chromosome
microtubules, 4 — kinetochores, 5 — atractophores, 6 — microfibrilles con-
necting atractophores with kinetosomes, 7 — nucleus, 8 — chromosomes, 9 —
nuclear envelope.
еще одна группа непатогенных видов трихомонад,
обитающих в кишечнике грызунов (Г. muris и ряд
других), амфибий (Г. augusta) и насекомых (Г. aula-
codi). Все эти виды имеют всесветное распростране-
ние.
Отряд Hypermastigida Grass!
et Foa, 1911 — Гипермастигиды
Наиболее высокоорганизованная группа среди всех жгутико-
вых, для которой характерны следующие черты.
1) Наличие многочисленных жгутиков (от 10 до нескольких
тысяч), расположенных в определенном порядке в 1 или несколь-
ких зонах: по полному или неполному кругу, в виде 1 или несколь-
ких пластинок или, наконец, продольными или спиральными ряда-
ми. Каждая жгутиковая зона тесно связана с парабазальными ла-
меллами, парабазальными аппаратами (Janicki-type) и структурами
пельта-аксостилярного комплекса. Жгутиковая зона в комплексе с
вышеперечисленными органеллами образует мастигонтную систе-
му. Таких систем в клетке может быть 1 либо 2, изредка 4. В
организации клетки с 2 или 4 мастигонтными системами наблюда-
ется билатеральная симметрия, сочетающаяся с радиальной.
2) Наличие группы «привилегированных» кинетосом.
3) 1 ядро в клетке. Митоз протекает с сохранением ядерной
оболочки и внеядерным веретеном деления.
4) Присутствие атрактофоров — структур, формирующих ве-
ретено деления.
5) Существование множественного парабазального аппарата.
6) Наличие гидрогеносом. )
7) Наличие у некоторых гипермастигид цист и полового про-
цесса.
8) Все представители отряда — паразиты, населяющие ки-
шечник низших термитов и тараканов рода Cryptocercus (исклю-
чение составляет Lophomonas, паразитирующий в тараканах Blat-
ta, Blattella и Periplaneta). Большинство гипермастигид связаны
симбиотическими отношениями мутуалистического характера со
своими хозяевами, но некоторые являются настоящими паразита-
ми.
История изучения
Первое описание гипермастигид из кишечника
термита было сделано Леспесом (Lespes) в 1856 г. С
этого времени начались интенсивные исследования
кишечных фаун различных видов термитов. В та-
раканах гипермастигиды впервые были обнаружены
Кливлендом с соавт. (Cleveland et al., 1934), которые
описали кишечную фауну древесиноядного таракана
Cryptocercus punctulatus, обитающего в Северной
Америке. Жгутиконосцы из кишечника этого тара-
кана оказались морфологически и физиологичейси
близки жгутиконосцам из термитов. В конце 70-х гг.
стала известна кишечная фауна еще одного вида
этих реликтовых тараканов — Cryptocercus relictus,
обитающего на Дальнем Востоке (Бобылёва, 1975).
К 80-м гг. нашего столетия было исследовано
205 видов низших термитов (из 521 вида известных
науке) и 2 вида Cryptocercus (из 3 известных); опи-
сано более 200 видов гипермастигид (см. сводный
каталог: Yamin, 1979).
Среди работ, посвященных всестороннему изуче-
нию гипермастигид, первостепенное значение имеет
большой цикл работ американского ученого Клив-
ленда (Cleveland), выполненных в период с 1923 по
1966 г., и монография Грассе (Grasse, 1952). Эти
работы заложили прочный фундамент для последую-
щего электронно-микроскопического изучения ги-
пермастигид, которые являются весьма сложными и
трудными объектами для такого рода исследований.
Тонкое строение жгутика впервые было детально
изучено на гипермастигидах (Pitelka, Schooley, 1958;
Gibbons, Grimstone, 1960). Большое значение для
понимания общего плана строения клетки этой
группы организмов имели ультраструктурные иссле-
дования лофомонадин (Hollande, Valentin, 1969; Hol-
lande, Carruette-Valentin, 1972; Radek, Hausmann,
1994a) и трихонимфин (Hollande, Carruette-Valentin,
1971; Kubai, 1973; Lingle, Salisbury, 1995). Благода-
ря этим работам удалось не только показать природу
и ультраструктуру всех клеточных органелл гипер-
мастигид, но и выявить универсальный характер вза-
имосвязи между «привилегированными» кинетосома-
ми и корешковыми фибриллами, а также органи-
зующую роль этих кинетосом во время онтогенеза
клетки. Вопросы эволюции гипермастигид, их взаи-
моотношений с хозяевами и трансмиссии были под-
робно исследованы Догелем (1922, 1951, 1956) и
Кирби (Kirby, 1937, 1941, 1947, 1949).
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
369
Классификация
Морфология и физиология
С первых шагов изучения гипермастигид исследо-
ватели были единодушны в том, что эти своеобраз-
ные жгутиконосцы, населяющие кишечник термитов
и тараканов, представляют единую таксономическую
группу. Большинство авторов разделяло ее на
2 крупные эволюционные ветви — Lophomonadina и
Trichonymphina (Догель, 1922; Kirby, 1949; Honig-
berg, 1963b; Levine et al., 1980). Однако, по мнению
французских исследователей (Grasse, Hollande, 1942;
Hollande, Carruette-Valentin, 1971), гипермастигиды
распадаются на 3 эволюционные линии. Эти авторы
выделяют семейства Spirotrichonymphidae и Holoma-
stigotoididae в отдельную ветвь. Приводимая ниже
классификация отражает взгляды большинства про-
тозоологов на систематику этой группы.
Отряд Hypermastigida Grassi et Foa, 1911 — Гипермастигиды
Подотряд Lophomonadina Light, 1927 — Лофомонадины
Сем. Joeniidae Janicki, 1915
Сем. Lophomonadidae Kent, 1880
Сем. Kofoidiidae Light, 1927
Сем. Rhyzonymphidae Grasse et Hollande, 1951
Подотряд Trichonymphina Poche, 1913 — Трихонимфины
Сем. Hoplonymphidae Light, 1926
Сем. Staurojoeninidae Grassi, 1917
Сем. Trichonymphidae Kent, 1880
Сем. Eucomonymphidae Cleveland et al., 1934
Сем. Teratonymphidae Koidzumi, 1921
Сем. Spirotrichosomatidae Hollande et Carruette-Valentin, 1971
Сем Spirotrichonymphidae Grassi, 1917
Сем. Holomastigotoididae Janicki, 1915
Жгутиковый аппарат. У наиболее примитивных
гипермастигид подотряда Lophomonadina жгутики
собраны в одну зону, занимающую апикальное поло-
жение и имеющую весьма разнообразную форму.
Так, у Lophomonas жгутиковая зона имеет вид коро-
ны, у Joenia — форму парика, у Joenina — в виде
полумесяца, у Placojoenia — подковообразная и т. д.
(рис. 317; 318; 320; 321).
У гипермастигид из подотряда Trichonymphina
многочисленные жгутики расположены в основном в
2 билатерально-симметричных зонах, изредка в
4 идентичных симметричных зонах (Staurojoenini-
dae). У представителей Hoplonymphidae и Staurojoe-
ninidae жгутиковые зоны разделяются свободной от
жгутиков поверхностью; внутри каждой зоны жгути-
ки располагаются продольными рядами (рис. 322;
323). У представителей Trichonymphidae, Eucomo-
nymphidae, Teratonymphidae, Spirotrichosomatidae,
имеющих ростральную трубку, принято различать в
пределах жгутиковой зоны ростральную и построст-
ральную части. В ростральной части края 2 билате-
рально-симметричных зон тесно сближены и жгути-
ки расположены радиальными продольными рядами
(рис. 324). В постростральной части жгутики могут
быть расположены продольными (Trichonymphidae)
(рис. 324), меридиональными (Eucomonymphidae)
(рис. 328, вкл.; 329); концентрическими (Teratonym-
Рис. 317. Lophomonas blattarum.
А — структурная организация на светомикроскопическом уровне: акс — аксостиль; жз — жгутиковая зона; пв — пищеварительные вакуоли; я — ядро,
локализованное внутри чашевидного расширения аксостиля; ят — «ядерные трубочки». Б — диаграмма ультраструктурной организации околоядерного
района: аг — аппарат Гольджи, акс — аксостиль, г — гликоген, дят — дистальная часть «ядерной трубочки», к — кинетосомы, мт — микротрубочки
аксостиля, п — поры, пят — проксимальная часть «ядерной трубочки», р — рибосомы, я — ядро, яо — ядерная оболочка. (А — по: Beams et al., 1961; Б —
по: Beams, Sekhon, 1969).
Fig. 317. Lophomonas blattarum.
A — diagrammic representation of the organism according to light microscopy: акс — axostyle, жз — flagellar zone, ne — digestive vacuoles, я — nucleus located
within an expanded cap-like anterior part of axostyle, ят — «nuclear tubules». Б — diagram of the perinuclear region, according to electron microscopy: аг — Golgi
apparatus, акс — axostyle, г — glycogen, дят — distal parts of «nuclear tubules», к — kinetosomes, мт — axostylar microtubules, n — pores, пят — proximal
parts of «nuclear tubules», p — ribosomes, я — nucleus, яо — nuclear envelope.
24 Протисты, часть 1
370
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 318. Lophomonas striata. (По: Grassi,
1952).
акс — аксостиль, б — бактерии (Fusiformis lophomo-
nadis) на поверхности тела жгутиконосца, жз — жгу-
тиковая зона, я — ядро.
Fig. 318. Lophomonas striata.
акс — axostyle, б — symbiotic bacteria (Fusiformis
lophomonadis) on the body surface, жз — flagellar
area, я — nucleus.
Рис. 319. Lophomonas blattarum. (По:
Grassd, 1952).
nm — парабазальные тела, образующие
круг на основании жгутиковой зоны.
Fig. 319. Lophomonas blattarum.
пт — parabasal bodies arranged at the base
of flagellar area.
Рис. 320. Joenia annectens. (По: Grasse,
1952).
a — атрактофор; акс — аксостиль; жз — жгути-
ковая зона; пе — пельта; пл — парабазальная
ламелла; пт— перисГовидные парабазальные те-
ла; я — ядро, лежащее в чашевидном расшире-
нии аксостиля.
Fig. 320. Joenia annectens.
а — atractophore, акс — axostyle, жз — flagellar
area, n — pelta, пл — parabasal lamella, nm —
parabasal bodies, я — nucleus located within ex-
panded cap-like anterior part of axostyle.
Рис. 321.
Рис. 322.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
371
Рис. 323. Urinympha cirrata. (По: Бобылёва, 1975).
Вид сбоку на 2 жгутиковые зоны, разделенные свободной от жгутиков
зоной поверхности. Обозначения как на рис. 322.
Fig. 323. Urinympha cirrata.
Lateral view of the entire organism. Two symmetrical flagellated areas separa-
ted by a flagella-free strip of cytoplasm obvious. Abbreviations as in fig. 322.
Рис. 324. Trichonympha collaris. (По: Kirby, 1932).
Вид с поверхности тела {справа) и в глубине цитоплазмы (слева), бэ —
продольные бороздки эктоплазмы, в которых идут жгутики; вэ — внутрен-
ний слой эктоплазмы; кщ — круговая щель, разделяющая жгутиковую
зону на 2 части; нш — наружная шапочка; нэ — наружный слой эктоплаз-
мы; пр — постростральная часть жгутиковой зоны; пт — парабазальные
тела; р —- ростральная часть жгутиковой зоны; я — ядро,
Fig. 324. Trichpnympha collaris.
Views of the body surface (at the right) and at the depth of cytoplasm (at the
left), бэ — longitudinal grooves in external layer of ectoplasm in which rows
of flagella are running, вэ — internal layer of ectoplasm, кщ — circular fissu-
re separating the flagellated area into the rostral and postrostral regions, нш —
cap, нэ — external layerof ectoplasm, np — postrostral region of flagella,
nm — parabasal bodies, p — rostral region of flagella, я — nucleus.
Рис. 321. Передний конец Joenia annectens. (По: Hollande, Valentin, 1969).
Обозначения как на рис. 320.
Fig. 321. The anterior end of Joenia annectens.
Abbreviations as in fig. 320.
Рис. 322. Babulanympha cryptocerci. (По: Бобылёва, 1975).
A — вид сбоку на 2жгутиковые зоны, разделенные свободной от жгутиков зоной поверхности: жз жгутиковая зона; пт — парабазальные тела; я —
ядро, заключенное в ядерный мешок. Б — вид только на 1 жгутиковую зону: акс — свободные аксостили, акск — аксостилярное кольцо в основании жгу-
тиковых зон (жз). В — вид сверху на Жгутиковые зоны (жз), аксостилярное кольцо (акск) и отходящие свободные аксостили (акс).
Fig. 322, Barbulanympha cryptocerci.
А — lateral view of two identical flagellar areas separated by a flagella-free strip of cytoplasm. Б — lateral view of only one flafellar area and axostylar appara-
tus. В — vertical view of two flagellated areas, axostylar ring and free axostyle fibrils, акс — free axostylar fibrils, акск — axostylar ring at the base of flagellar
areas, жз — flagellar area, nm — parabasal bodies, я — nucleus enclosed in a nuclear sleeve.
372
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 325. Trichonympha ussuriensis. (По: Бобылёва, 1975).
пт — парабазальные тела, пф — парабазальные филаменты, я — ядро.
Fig. 325. Trichonympha ussuriensis.
пт — parabasal bodies, пф — parabasal filaments, я — nucleus.
phidae) (рис. 327) рядами, а также спиралями (Spiro-
trichosomatidae) (рис. 330—332). У гипермастигид
Spirotrichonymphidae и Holomastigotoididae спираль-
ные ленты жгутиков начинаются непосредственно на
переднем конце тела и охватывают тело жгутиконос-
ца несколькими леотропно либо дексиотропно за-
крученными спиралями (рис. 333).
У лофомонадин жгутик полностью свободен. У
трихонимфин для разных семейств характерно раз-
личное заглубление жгутика в цитоплазме: у Hoplo-
nymphidae и Staurojoeninidae проксимальные части
жгутиков заглублены в цитоплазме; у Trichonymphi-
dae и Eucomonymphidae он связан на значительном
протяжении с поверхностью тела. Эктоплазма у этих
гипермастигид имеет дольчатое или листовидное
строение, т. е. состоит из 2 слоев — внутреннего
и весьма значительного наружного. Во внутреннем
слое располагаются базальные тела жгутиков, кото-
рые проходят радиально. Наружный слой состоит
из параллельных складок или гребней, разделенных
глубокими бороздками. Жгутик появляется из цито-
плазмы на дне бороздки между эктоплазматическими
гребнями и проходит в ней значительное расстояние,
прежде чем стать свободным на поверхности тела
(рис. 334). Как показали электронно-микроскопичес-
кие исследования, в бороздке жгутик закреплен при
помощи особой системы филаментов (Hollande, Саг-
ruette-Valentin, 1971). У Spirotrichonymphidae и Ho-
lomastigotoididae жгутик одной своей стороной так-
же часто объединяется со стенкой тела.
Рис. 326. Передний конец Trichonympha major. (По: Бобылёва,
1975).
вэ — внутренний слой эктоплазмы, кщ — круговая щель, нш — наружная
шапочка, нэ — наружный слой эктоплазмы, рт — ростральная трубка,
я — ядро, ям — ядерный мешок.
Fig. 326. Anterior part of Trichonympha major.
вэ — internal layer of ectoplasm, кщ — circular fissure, нш — cap, нэ — ex-
ternal layer of ectoplasm, pm — rostral tube, я — nucleus, ям — nuclealar
sleeve.
Различные виды Trichonympha, Pseudotrichonym-
pha стали одними из первых объектов, у которых
было изучено тонкое строение жгутика (Pitelka,
Schooley, 1958; Gibbons, Grimstone, 1960). Эти дан-
ные по ультраструктуре жгутика стали классически-
ми, и их можно найти в любом руководстве по цито-
логии. Из особенностей ультраструктуры жгутика у
гипермастигид можно отметить следующие.
1) Атипично длинные кинетосомы (до 5 мкм) у
многих видов Trichonympha, Eucomonympha, Pseudo-
trichonympha, Spirotrichonympha (Hollande, Carruette-
Valentin, 1971), Placojoenia (Radek, Hausmann, 1994a).
2) Переходная зона без дополнительных струк-
тур.
3) Сложная система кинетосомальных фибрилл
(Gibbons, Grimstone, 1960; Lingle, Salisbury, 1995).
У всех гипермастигид каждая из кинетосом свя-
зана с соседними кинетосомами в жгутиковом ряду
и с 2 соседними кинетосомами ближайших жгутико-
вых рядов посредством особой системы фибрилл.
Таким образом, все кинетосомы жгутиковой зоны
связаны между собой.
«Привилегированные кинетосомы» и морфо-
плазма. На светомикроскопическом уровне все ис-
следователи отмечали, что передний конец тела три-
хонимфин прикрыт не только наружной, но и внут-
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
373
Рис. 327. Teratonympha mirabilis. (По: Cleveland, 1938).
жл — жгутиковые ленты; пр — постростральный район, где жгутиковые
ленты идут концентрическими кругами; пт — парабазальные тела; р —
ростральный район жгутиковой зоны; рт — ростральная трубка; я — ядро.
Fig. 327. Teratonympha mirabilis.
жл — flagellar bands, пр — the postrostrum with flagellar rows arranged in
concentric circles, nm >— parabasal bodies, p — rostrum, pm — rostral tube,
я — nucleus.
ренней шапочкой. Электронно-микроскопические
исследования показали (см.: Hollande, Carruette-Va-
lentin, 1971), что наружная шапочка как структура,
надежно защищающая передний конец жгутиконосца
при движении в густом содержимом кишечника хозяина,
действительно существует. Однако внутренней шапоч-
ки как структуры, аналогичной первой, не существу-
ет. Оказалось, что впечатление внутренней шапочки
создавали близко расположенные передние кинетосо-
мы, погруженные в электронно-плотный фибрилляр-
Рис. 330. Bispironympha unica. (По: Бобылёва, 1975).
жл — жгутиковые ленты; нш — наружная шапочка; пт — парабазальные
тела, лежащие по ходу жгутиковых лент; рт — ростральная трубка; я —
ядро.
Fig. 330. Bispironympha unica.
жл — flagellar bands, нш — cap, пт — parabasal bodies arranged along the
spiral part of each flagellar band, pm — rostral tube, я — nucleus.
Рис. 329. Eucomonympha папа. (По: Бобылёва, 1975).
жр — жгутиковые ряды, располагающиеся в постростральном районе ме-
ридионально; нш — наружная шапочка; пр — постростральный район;
рт — ростральная трубка, проходящая в центре рострума (р); я — ядро.
Fig. 329. Eucomonympha папа.
жр — flagellar rows arranged in meridional manner in the postrostrum, нш —
cap, np — postrostral region of flagellated area, pm — rostral tube located in
the centre of rostrum, я — nucleus.
374
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 331. Leptospironympha gigantea. (По: Cleveland, Day, 1958).
жл — жгутиковые ленты, располагающиеся в построструме леотропно за-
крученными спиралями; пр — постростральный район; р — ростральный
район жгутиковой зоны.
Fig. 331. Leptospironympha gigantea.
жл — flagellar band located in the rostrum leotropic spirals, np — postrosral
region of flagellar area, p — rostral region of flagellar area.
Рис. 332. Spirotrichosoma magnum. (По: Cleveland, Day, 1958).
акс — аксостили; вжл — вторичные жгутиковые ленты, отходящие от пер-
вичных и идущие до конца тела; пжл — первичные жгутиковые ленты, ох-
ватывающие тело по леотропной спирали; рт — ростральная трубка; я —
ядро.
Fig. 332. Spirotrichosoma magnum.
акс — axostyles, вжл — secondary flagellar bands arising from the primary
bands and stretching towards the body end, пжл — primary flagellar bands en-
veloping the body as leotropic spirals, pm — rostral tube, я — nucleus.
ный матрикс, который часто распадается на отдель-
ные тяжи, идущие в глубь цитоплазмы. Сопоставле-
ние свето- и электронно-микроскопических данных
привело к заключению, что эти структуры играют ис-
ключительно важную организующую роль в морфоге-
незе клетки во время деления. В контакте с ними в
каждом клеточном поколении формируются недоста-
ющая половина жтугиков, а также поперечно-исчер-
ченные фибриллы. Эта группа кинетосом была названа
«привилегированной», а окружающая их электронно-
плотная масса цитоплазмы — морфоплазмой (Hollan-
de, Carruette-Valentin, 1971). У таких трихонимфин,
как Hoplonymphidae и Staurojoeniriidae, эта группа
состоит из 4 кинетосом, которые лишены жгутика и
расположены между жгутиковыми зонами. У Tricho-
nymphidae в нее входит 2 кинетосомы, одна из кото-
Рис. 333. Spirotrichonmpha flagellatа. (По: Cleveland, 1958).
акс — аксостили; жл — жгутиковые ленты, начинающиеся на вершине
тела, идут леотропно закрученными спиралями до конца тела; пт — пара-
базальные тела; я — ядро.
Fig. 333. Spirotrichonympha flagellata.
акс — axostyles, жл — flagellar bands starting on the body top and follo-
wing leotropic spirals to the body end, nm — parabasal bodies, я — nucleus.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
375
Рис. 334. Схема строения переднего конца Trichonympha sp. в по-
перечном (4) и продольном (Б) разрезе. (По: Pitelka, Schooley,
1958).
а — атрактофор; вэ — внутренний слой эктоплазмы, где располагаются
кинетосомы жгутиков; ж — жгутики; кщ — круговая щель; нш — наруж-
ная шапочка, состоящая из нескольких фибриллярных слоев микротрубо-
чек; из — наружный слой эктоплазмы, состоящий из гребней, разделен-
ных бороздками; рт — ростральная трубка, состоящая из парабазальных
ламелл; упр — уровень поперечного разреза.
Fig. 334. Diagrammic representation of the anterior end of Tricho-
nympha sp.
A — transversal section, Б — longitudinal section, a — atractophore, вэ — in-
ternal layer of ectoplasm with arranged flagellar kinetosomes; ж — flagella,
кщ — circular fissure, нш — cap made of some layers of microtubular fib-
rils, нэ — external layer of ectoplasm made of crests separated by grooves,
pm — rostral tube made of parabasal lamellae, ynp — the level of transversal
section.
рых лишена жгутика и расположена ортогонально
по отношению к другим кинетосомам (рис. 336). У
Eucomonymphidae, Spirotrichosomatidae и Spirotricho-
nymphidae «привилегированными» являются пере-
дние кинетосомы в каждом жгутиковом ряду, и они
по морфологии ничем не отличаются от других кине-
тосом. У лофомонадин «привилегированная» группа
состоит из 4 кинетосом, расположенных справа от
остальных кинетосом жгутиковой зоны (рис. 335). У
этих низших гипермастигид ясно видно, что от 4
«привилегированных» кинетосом отходят поперечно
исчерченные фибриллы парабазальных филаментов,
атрактофоров и преаксостилярные фибриллы (ргеах-
osty fibres = sigmoid fibres), формирующие пельта-
аксостилярный комплекс. Эти поперечно исчерчен-
ные структуры, по существу, являются корешковой
системой. У низших гипермастигид (лофомонадин)
непосредственная связь «привилегированных» кине-
тосом с корешковыми фибриллами, которая отчетли-
во проявляется при морфогенезе во время деления,
сохраняется и в течение вегетативной части жизнен-
ного цикла. У высших гипермастигид непосредствен-
ная связь «привилегированных» кинетосом и кореш-
Рмс. 335. Схема строения переднего конца клетки Lophomonas
striata на стадии профазы. (По: Hollande, Carruette-Valentin,
1972).
аь 02 — атрактофоры; акс — аксостиль; к— кинетосомы жгутиковой
зоны; кт—кп4— привилегированные кинетосомы; кн— кинетохоры;
паке — преаксостилярные фибриллы; пе — пельта; пт — парабазальное
тело (аппарат Гольджи); пф — поперечно исчерченный парабазальный фи-
ламент.
Fig. 335. Diagrammatic representation of the anterior end of Lopho-
monas striata at the prophase stage.
ai, 02 — atractophores; акс — axostyle; к — kinetosomes of flagellar area;
Ki, К2, КЗ, K4 — «privileged» kinetosomes; кн — kinetochores; паке — preax-
ostylar fibrils; ne — pelta; nm — parabasal body (Golgi apparatus); пф —
cross-striated parabasal filament.
ковых фибрилл очень кратковременна и обнаруживает-
ся только в делящейся клетке. Уже в ходе морфоге-
неза рострума эта связь разрывается, маскируется
морфоплазмой и поддерживается в течение вегета-
тивной фазы жизненного цикла только через мор-
фоплазму. Считают, что морфоплазма является про-
водником информации от «привилегированных» ки-
нетосом к корешковым фибриллам й атрактофорам.
Парабазальные ламеллы и парабазальные фи-
ламенты. Эти структуры являются элементами ци-
тоскелета гипермастигид и обнаружены у всех изу-
ченных представителей отряда, но в разных родах
развиты далеко не в одинаковой степени. Пара-
базальные ламеллы располагаются непосредственно
под жгутиковыми зонами и часто выполняют роль
каркаса для базальных тел жгутиков. Кроме того,
они защищают ядро и другие клеточные органеллы,
находящиеся в роструме. Парабазальные ламеллы на-
иболее развиты у трихонимфин. Под световым мик-
роскопом у Hoplonymphidae и Staurojoeninidae они
имеют вид единой тонкой пластинки, которая под-
стилает каждую жгутиковую зону и соответствует ей
по форме. У остальных трихонимфин парабазальные
ламеллы образуют ростральную трубку — централь-
ную осевую структуру тела (рис. 324; 326; 334). У
Spirotrichonymphidae и Holomastigotoididae параба-
зальные ламеллы в виде тонких лент проходят не-
посредственно под базальными телами жгутиков, ор-
376
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ганизованных в спирали. Так, у Holomastigotoides
tusitala парабазальные ламеллы идут под каждой из
5 спиральных лент жгутиков до уровня середины
ядра (рис. 337) (Lingle, Salisbury, 1995). У лофо-
монадин парабазальные ламеллы развиты несколько
слабее, но также присутствуют. Так, у Joenia annec-
tens 2 лентовидные парабазальные ламеллы спуска-
ются с апикального конца тела, идут под жгутиковой
зоной, охватывая ее с 2 сторон, и контактируют с
особой перистой структурой — парабазальными те-
лами (рис. 321). Placojoenia имеет 4 парабазальные
ламеллы, которые также контактируют со спирально
закрученными вокруг аксостиля парабазальными те-
лами (Radek, Hausmann, 1994а).
Как показали электронно-микроскопические ис-
следования, парабазальные ламеллы сформированы
из плотно упакованных, поперечно исчерченных (с
периодичностью 42—47 нм) фибрилл (см. Hollande,
Carruette-Valentin, 1971; Radek, Hausmann, 1994a).
Установлено, что каждая из фибрилл по своей ульт-
раструктуре и размерам сходна с 1 парабазальной
фибриллой трихомонадид. Парабазальные ламеллы
тесно связаны с кинетосомами, которые примыкают
к ним непосредственно (рис. 336). В каждом новом
i клеточном поколении недостающая половина пара-
базальных ламелл в дочерней клетке образуется в
тесном контакте с «привилегированными» кинетосо-
мами. Этот контакт у лофомонадин сохраняется и
Рис. 336. Схема строения переднего конца клетки Trichonympha
agilis. (По: Hollande, Carruette-Valentin, 1971).
a — атрактофор; к — кинетосомы жгутиковой зоны; кп\, кпг — привиле-
гированные кинетосомы; м — морфоплазма; пеакс — пельта-аксостиляр-
ные фибриллы; пл — парабазальные ламеллы.
Fig. 336. Diagrammatic representation of the anterior end of Tricho-
nympha agilis, according to electron microscopy.
a — atractophore; к — kinetosomes; кп\, кпг — «privileged» kinetosomes;
м — morphoplasm; пеакс — pelta-axostylar fibrils; пл — parabasal lamellae.
Рис. 337. Схема строения переднего конца тела Holomastigotoides
tusitala, (По: Lingle, Salisbury, 1995).
аь 02 — атрактофоры; акс — аксостиль; Ж1—жз — жгутиковые ленты;
зв — зачаток веретена; к — кинетосомы; кф — кинетосомальные фибрил-
лы; пл — парабазальные ламеллы; пт — парабазальное тело; тэр — тру-
бочки ЭПР; фл — фиброзная лента; эр — ЭПР; я — ядро.
Fig. 337. Diagrammatic representation of the anterior end of Holo-
mastigotoides tusitala.
a\, аг — atractophores; акс — axostyle; жь ж?, жз, Ж4, ms — flagellar bands;
зв — primordium of spindle; к — kinetosomes; кф — kinetosomal fibrils;
пл — parabasal lammelae; nm — parabasal body; тэр — tubules of endoplas-
mic reticulum; фл — fibrous ribbon; эр — endoplasmic reticulum; л — nucle-
us
после деления клетки. У трихонимфин в вегетатив-
ной части жизненного цикла связь парабазальных
ламелл с «привилегированными» кинетосомами осу-
ществляется через морфоплазму (рис. 336). Дисталь-
ные концы парабазальных ламелл у большинства ги-
пермастигид истончаются и переходят в парабазальные
филаменты, которые обычно идут в глубь цитоплаз-
мы и контактируют с парабазальными телами или
диктиосомами.
Парабазальный аппарат. Термин «парабазальный
аппарат» относится к клеточной органелле трихо-
монадид и гипермастигид, которую впервые описал
Яницкий (Janicki) в 1911 г. при помощи светового
микроскопа и определил как аргентофильную ор-
ганеллу, состоящую из парабазального филамента и
подвешенного к нему парабазального тела. После
применения методик серебрения парабазальный фи-
< ламент выявляется в виде тонкой нити, а парабазаль-
ное тело — в виде извилистых тяжей, колбасовидных
образований либо округлых телец, наподобие дик-
тиосом позвоночных. Выяснилось, что у гипермасти-
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA 377
гид имеется большое число таких парабазальных ап-
паратов. Типичный парабазальный аппарат имеют
некоторые лофомонадины (например, Lophomonas) и
многие из трихонимфин (семейства Hoplonymphidae,
Staurojoeninidae, Trichonymphidae, Teratonymphidae),
причем парабазальные филаменты и парабазальные
тела более или менее свободно располагаются в глу-
бине цитоплазмы вокруг ядра (рис. 319; 322—325;
337). У других гипермастигид (семейства Eucomo-
nymphidae, Spirotrichosomatidae, Spirotrichonymphi-
dae, Holomastigotoididae) парабазальный аппарат
после применения методик серебрения выявляется в
виде тонких лент, подстилающих каждый жгутико-
вый ряд (они идентифицируются с парабазальными
филаментами), и округлых телец, следующих по ходу
этих лент (рис. 328, вкл.; 330; 333). У некоторых
гипермастигид парабазальный аппарат имеет более
сложную структуру. Так, у Joenia annectens параба-
зальный аппарат образует перистую структуру вокруг
аксостиля (рис. 320; 321). У Placojoenia парабазаль-
ный аппарат в 5—7 рядов обвивает акостиль по спи-
рали. Помимо парабазальных тел, связанных с пара-
базальным филаментом, в цитоплазме у Placojoenia
найдены свободные парабазальные тела.
Электронно-микроскопические исследования по-
казали, что 2 элемента парабазального аппарата имеют
разную ультраструктурную организацию. Парабазаль-
ный филамент, как уже было сказано, отходит от па-
рабазальных ламелл и идентичен им по ультраструк-
туре. Он может состоять из 1, 2 или нескольких по-
перечно исчерченных парабазальных фибрилл.
Парабазальное тело по своей ультраструктурной и
цитохимической организации является аппаратом
Гольджи (рис. 317, Б\ 335; 337). Для гипермастигид
характерно, что каждая клетка имеет обычно много-
численные, хорошо развитые и типичные по структу-
ре аппараты Гольджи. Обычно на одну диктиосому
приходится 25—28 плотно прилегающих друг к другу
цистерн, образующих замкнутые трубочки, от кото-
рых отделяются пузырьки и гранулы с электронно-
плотным содержимым. Парабазальный филамент
всегда располагается параллельно стопке цистерн.
Однако недавно появились данные о том, что дикти-
осомы не всегда связаны с парабазальными филамен-
тами. Так, у Holomastigotoides tusitala (Lingle, Salis-
bury, 1995) многочисленные диктиосомы идут по хо-
ду жгутиковых лент; они обнаруживают тесную связь
с микротрубочками аксостилей и ЭПР (рис. 337).
Пельта-аксостилярный комплекс. Под этим на-
званием объединены структуры, в основе организа-
ции которых лежат микротрубочки: аксостиль, пель-
та и пельта-аксостилярные фибриллы. Они наряду с
парабазальными ламеллами являются элементами ци-
тоскелета клетки. В делящейся клетке микротрубоч-
ковые структуры деполимеризуются и вновь образу-
ются в ходе морфогенеза в контакте с преаксостиляр-
ными фибриллами у Lophomonas (рис. 335), либо в
контакте с парабазальными ламеллами у Holomastigo-
toides (рис. 337), либо вблизи «привилегированных»
кинетосом.
У лофомонадин многослойные микротрубочковые
структуры (аксостиль) играют ведущую роль в фор-
мировании цитоскелета клетки. На ультраструктур-
ном уровне показано, что у Lophomonas. Joenia, Pla-
cojoenia аксостиль состоит из спирально закрученных
рядов микротрубочек. Отмечается, что внутри аксо-
стиля разбросаны гранулы гликогена. Аксостиль, про-
ходящий через все тело, является основным опорным
органоидом у лофомонадин. Так, у Lophomonas ак-
состиль имеет вид тонкой трубки, пронизывающей
тело, и весьма напоминает аксостиль трихомонадид
(рис. 317). У других родов — Joenia и Placojoenia —
аксостиль представляет собой массивную трубку,
имеющую на переднем конце расширение в виде ча-
ши (capitulum), где помещается ядро (рис. 320; 321).
Трихонимфины имеют многочисленные нитевид-
ные аксостили, которые свободно лежат в эндоплазме
вокруг ядра (рис. 322, Я; 323; 331; 333) либо объеди-
няются в пучок. У гипермастигид, населяющих ки-
шечник таракана Cryptocercus, аксостилярные фиб-
риллы образуют ядерный мешок — структуру, плотно
охватывающую ядро и поддерживающую его в опре-
деленном положении в клетке (рис. 326) (Cleveland et
al., 1934; Hollande, Carruette-Valentin, 1971). У Holo-
mastigotoides. обитающих в низших термитах сем. Но-
dotermitidae, аксостили также принимают участие в
поддержании ядра в фиксированном положении (Lin-
gle, Salisbury, 1995).
Слой микротрубочек, проходящих непосредствен-
но под клеточной мембраной апикального конца
клетки, образует пельту. У лофомонадин пельта разной
величины прикрывает передний конец тела (рис. 320).
У всех трихонимфин пельта и пельта-аксостилярные
фибриллы принимают участие в образовании наруж-
ной шапочки — структуры, прикрывающей передний
конец простейшего и защищающей его при движении
в густом содержимом кишечника хозяина (рис. 322, А;
324; 326; 329; 334; 336).
Атрактофоры. Термин «атрактофоры» («носите-
ли веретена») относится к своеобразным органеллам,
которые у гипермастигид при митозе формируют и
выносят веретено деления к ядру. Морфология этих
органелл и их роль при митозе была подробно иссле-
дована под световым микроскопом в большой серии
работ американского исследователя Кливленда (Cle-
veland et al., 1934; Cleveland, 1960), который называл
их центриолями по аналогии с их функциональной
значимостью в клетке. Однако последующие элект-
ронно-микроскопические исследования четко пока-
зали, что эти органеллы не имеют морфологии, ха-
рактерной для центриолей, но выполняют их функ-
цию в клетке (Hollande, Valentin, 1969; Hollande,
Carruette-Valentin, 1971, 1972; Radek, Hausmann,
1994a; Lingle, Salisbury, 1995). Атрактофоры харак-
терны для всех гипермастигид, но у одних они появ-
ляются в клетке лишь в период митоза, у других же
хорошо видны и в интерфазе. Так, у Hoplonymphidae
они имеют вид 2 лентовидных тяжей, идущих под па-
рабазальными ламеллами и оканчивающихся близ
ядра. У Trichonymphidae в интерфазе виден лишь
1 атрактофор в виде палочки в просвете ростральной
трубки (рис. 336). У Eucomonymphidae атрактофоры
появляются лишь перед началом митоза. Дистальные
концы атрактофоров у некоторых гипермастигид (на-
пример, у Barbulanympha) окружены атрактосфера-
ми. У большинства гипермастигид атрактосфер нет и
378
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
веретено деления образуется непосредственно между
дистальными концами атрактофоров. У Lophomonas,
Holomastigotoides дистальные концы атрактофоров и
в интерфазе соединены зачатком центрального вере-
тена (рис. 337).
На ультраструктурном уровне, как выяснилось, у
разных гипермастигид атрактофоры не однотипны по
своей организации. У Hoplonymphidae, Trichonymphi-
dae и Eucomonymphidae атрактофоры имеют микро-
фибриллярную структуру, являясь как бы частью
морфоплазмы (Hollande, Carruette-Valentin, 1971). У
Joenia, Placojoenia, Holomastigotoides и Spirotrichoso-
matidae атрактофоры имеют поперечно-периодичес-
кую структуру, как и парабазальные ламеллы, при-
датками которых они являются (Hollande, Valentin,
1969; Hollande, Carruette-Valentin, 1971; Radek, Haus-
mann, 1994a; Lingle, Salisbury, 1995). У Lophomonas
атрактофоры имеют также структуру поперечно ис-
черченных фибрилл, которые и в интерфазе находят-
ся в непосредственном контакте с «привилегирован-
ными» кинетосомами и постоянно соединены между
собой зачатком центрального веретена (рис. 335).
Атрактосферы в интерфазе имеют мелкозернис-
тую структуру и представляют собой внеядерный
ЦОМТ. Пучки микротрубочек центрального веретена
и хромосомальные фибриллы возникают из атракто-
сфер. У таких гипермастигид, как Lophomonas, Joe-
nia, Staurojoenia, Holomastigotoides и других, атрак-
тосфер нет и центральное веретено образуется прямо
между дистальными концами атрактофоров. В ходе
деления клетки четко прослеживается, что атракто-
форы вновь образуются и функционируют в контакте
с «привилегированными» кинетосомами; они являют-
ся элементами корешковой системы. У большинства
гипермастигид этот контакт разрывается уже в ходе
митоза и поддерживается в интерфазе через морфо-
плазму. Предполагается, что атрактофоры являются
связующим звеном между «привилегированными».ки-
нетосомами и формирующимся веретеном деления. По
этой гипотезе, атрактофоры; которые являются про-
изводными «привилегированных» кинетосом, переда-
ют информацию от этих кинетосом и находятся под
их контролем.
Ядро. Гипермастигиды —- одноядерные организ-
мы. Основные особенности ядра этих жгутиконосцев
были отмечены еще на светомикроскопическом уровне
(см.: Cleveland et al., 1934; Grasse, 1952; Райков,
1982). Ядро обычно шаровидной формы, содержит 1,
реже несколько ядрышек. У многих гипермастигид
(Barbulanympha, Тrichonympha, Spirotrichonympha,
Holomastigotoides) в интерфазе хромосомы находят-
ся в конденсированном состояний. Но у лофомона-
дин и некоторых трихонимфин (Urinympha, Stauro-
joenina) ядра относятся к пузырьковидному типу, при
котором хроматин находится в более или менее дис-
пергированном состоянии. Под электронным мик-
роскопом ядра гипермастигид изучены недостаточно,
но у исследованных объектов структурные компо-
ненты ядра — ядерная оболочка, ядрышко и хрома-
тин — в большинстве случаев имеют классическое
строение (см.: Райков, 1982). Однако у ядер некото-
рых лофомонадин обнаружено усложнение ядерной
оболочки. Так, у Joeninia и Holomastigotoides наруж-
ная сторона ядерной оболочки имеет слой микро-
фибрилл (Hollande, Valentin, 1969; Lingle, Salisbury,
1995). Особый случай разрастания наружной мемб-
раны ядерной оболочки отмечен у Lophomonas blat-
tarum (Beams, Sekhon, 1969; рис. 317, К). Здесь она
образует многочисленные, радиально расположенные
выросты (ядерные трубочки). Эти выросты сначала
тонкие и извилистые (проксимальная часть ядерной
трубочки), но постепенно расширяются и заканчива-
ются слепо замкнутым концом (дистальная часть).
Проксимальные концы ядерных трубочек несут на
наружной стороне своей мембраны многочисленные
рибосомы. Полость ядерных трубочек сообщается с
перинуклеарным пространством и заполнена элект-
ронно-плотным веществом. Вероятно, в «ядерных
трубочках» синтезируются при участии рибосом
какие-то вещества, которые накапливаются в их по-
лости. Ядерная оболочка у другой лофомонадины —
Placojoenia sinaica (Radek, Hausmann, 1994a) — об-
разует большую разветвленную сеть, причем этот
процесс затрагивает сразу обе мембраны. В перинук-
леарном пространстве ядерной оболочки найдены
гидрогеносомы; У трихонимфин ядерная оболочка
классического типа. Однако у Trichonympha ca^to яд-
ро окружено 1 или несколькими концентрическими
цистернами гранулярной ЭПР. Электронно-микро-
скопические исследования показали (см.: Райков,
1982), что у Trichonympha^ Barbulanympha и Spirot-
richonympha хромосомы в неделящемся ядре остают-
ся ясно очерченными, хотя и разрыхляются по срав-
нению с митотическими. Интерфазные хромосомы
этих жгутиконосцев постоянно закреплены своими
кинетохорами на внутренней стороне ядерной обо-
лочки. Кинетохоры в интерфазе имеют вид пласти-
нок из тонкофибриллярного материала, «встроенных»
между собственно телом хромосомы и внутренней
мембраной ядерной оболочки. Особенно компактны
интерфазные хромосомы Holomastigotoides. Показа-
но, что они состоят из 2 взаймозакрученных хро-
матид и окружены общим хромосомным матриксом
невысокой плотности, явственно отличающимся от
кариолимфы. Электронно-микроскопические иссле-
дования Holomastigotoides tusitala (Lingle, Salisbury,
1995) показали, что в интерфазной клетке между
дистальными концами атрактофоров присутствует
веретено. Оно, наподобие подушки, плотно прилега-
ет к ядерной оболочке, в которой видны «встроен-
ные» кинетохоры.
Цитоплазма. В цитоплазме гипермастигид при-
сутствуют многочисленные гидрогеносомы, которые
являются функциональными аналогами митохондрий
и типичны для протистов, ведущих анаэробный образ
жизни (см.: Серавин, 1992; Muller, 1993). Гидрогено-
сомы морфологически отличаются от митохондрий и
впервые были описаны у трихомонадид как микро-
тельца с относительно однообразным матриксом, ок-
руженные одинарной или, как показано для некото-
рых протистов, двойной мембраной. Гидрогеносомы
в основном разбросаны в эндоплазме. Однако у Holo-
mastigotoides tusitala они располагаются одним слоем
непосредственно под клеточной мембраной — между
ней и базальными телами жгутиков (Lingle, Salisbury,
1995). Отмечается, что гидрогеносомы у Joenia и Р1а-
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
379
cojoenia имеют кристаллоподобные включения (Hol-
lande, Valentin, 1969; Radek, Hausmann, 1994a). Гид-
рогеносомы играют важную роль в энергии анаэробного
гликолиза. Пируваты, образующиеся в ходе гликоли-
за, окисляются с образованием АТФ. В гидрогеносомах
найдены все энзимы, характерные для анаэробного
метаболического пути. В результате окислительно-
восстановительныхт процессов пируватов и малатов,
происходящих в гидрогеносомах, в эквимолярных ко-
личествах образуются углекислый газ, молекулярный
водород и ацетаты. Предполагается, что первые 2 из
конечных продуктов утилизируются симбиотическими
бактериями, содержащимися в цитоплазме многих ги-
пермастигйд, при этом в ходе метаболизма этих бак-
терий образуется метан (Ohkuma et aL, 1995).
Симбионты гипермастигид. Свободная от жгу-
тиков поверхность тела у многих гипермастигид по-
крыта эктобионтами. У жгутиконосцев Barbulanymp-
ha, Urinympha й Staurojoenina палочковидные бактерии,
прикрепленные к клеточной мембране, создают впе-
чатление исчерченности на поверхности^ тела (Cleve-
land et aL, 1934). Кроме палочковидных бактерий на
поверхности тела Barbulanympha обнаружены спиро-
хеты. У Joenia поверхность тела покрывают бактерии
(Schizophyta).
Исследования на электронно-микроскопическом
уровне показали, что в местах прикрепления эктоби-
онтов к клеточной мембране-возникают специализиро-
ванные структуры, прочно удерживающие эктобион-
тов на поверхности тела. Так, палочковидные бакте-
рии у Joenia покрыты толстым слоем гликокаликса,
который образует чехол вокруг бактерии, а в месте
прикрепления бактерии кроме этого слоя под кле-
точной мембраной жгутиконосца существует слой элек-
тронно-плотного материала (Hollande, Valentin, 1969).
Специализированные структуры прикрепления к
клеточной мембране развиваются и у палочковидных
бактерий — эктобионтов Barbulanympha и Urinym-
pha (Hollande, Carruette-Valentin, 1971; Bloodgood,
Fitzharris, 1976). Бактерии, покрытые снаружи слоем
гликокаликса, лежат в небольших нишах на клеточной
мембране жгутиконосца. Каждая такая ниша по дли-
не соответствует длине тела бактерии и подстилается
с внутренней стороны клеточной мембраны хозяина
слоем электронно-плотного материала. Кроме того,
бактерия, находящаяся в такой нише, с обеих сторон
закреплена в ней скобочками — расположенным
перпендикулярно к клеточной мембране электронно-
плотным слоем эктоплазмы хозяина. Спирохеты за-
крепляются на поверхности тела Barbulanympha при
помощи узкого носовидного выроста, который кон-
тактирует непосредственно с клеточной мембраной
жгутиконосца (Bloodgood, Fitzharris, 1976).
По-видимому, часть эктобионтов служит источ-
ником пищи для жгутиконосцев, так как они обна-
руживаются в фагоцитарных вакуолях. Но все же
роль эктобионтов в жизненном цикле гипермастигид
до конца неясна. Неизвестна и биохимическая сто-
рона симбиоза между гипермастигидами и различны-
ми прокариотическими организмами, прикрепленны-
ми к клеточной мембране.
В цитоплазме гипермастигид также найдены эн-
добиотические бактерии. Эти бактерии не связаны с
пищевыми вакуолями и обычно лежат свободно в
цитоплазме. Вероятно, некоторые из них относятся
к метаногенным бактериям, которые утилизируют
молекуляный водород и углекислый газ, образуя
метан (Ohkuma et al., 1995). Другие бактерии, веро-
ятно, вовлечены в метаболизм гликогена у гипермас-
тигид. Так, в цитоплазме Placojoenia найдены много-
численные цитоплазматические пластинки, которые и
послужили поводом дать такое название этому роду
жгутиконосцев (Radek, Hausmann, 1994а). В центре
каждой пластинки находится 1 или 2 палочковидные
бактерии, остальное пространство заполнено грану-
лами гликогена. Стенка бактерии снаружи окружена
маленькими частицами гликогена. Цитоплазматичес-
кие пластинки граничат с цистернами ЭЦР, ассоци-
ированного с рибосомами. В цитоплазме Placojoenia
между цитоплазматическими пластинками лежат еще
и спиралевидные бактерии. Эндобиотические бакте-
рии и гликоген в цитоплазме найдены и у Joenia
(Hollande, Carruette-Valentin, 1969). Однако не во
всех случаях ясно, связаны ли эндобиотические бак-
терии с метаболизмом клетки хозяина.
Питание
Гипермастигиды питаются главным образом ку-
сочками древесины, которые в изобилии имеются в
заднем отделе кишечника хозяина-ксилофага. Пище-
вые частицы захватываются на заднем, неструктури-
рованном, конце тела посредством инвагинаций, об-
разованных поверхностной мембраной клетки, либо
путем формирования псевдоподий (Cleveland et al.,
1934). В последние годы на Trichonympha было по-
казано, что тип псевдоподий при этом амебоидном
процессе зависит от размера пищевой частицы. До-
казано также, что для захвата и селективного про-
глатывания целлюлозы очень важны липкие свойства
поверхностной мембраны жгутиконосца. Описание
процесса эндоцитоза варьирует у разных авторов,
поскольку он, вероятно, специфичен для каждого
вида гипермастигид. Исследования процесса питания
у Joenia annectens показали, что пищевые частицы
прилипают к заднему концу тела, поверхность кото-
рого покрыта бактериями, и втягиваются во внутрь
без образования псевдоподий. Таким образом, пище-
вая частица оказывается покрытой бактериями, ко-
торые служат дополнительным источником питания.
Как показали электронно-микроскопические иссле-
дования, питание гипермастигид осуществляется по
смешанному типу, т. е. кроме фагоцитоза доказана
интенсивная пиноцитозная активность. Так, для 2 ви-
дов Joenia показано, что у них на уровне располо-
жения парабазальных тел в эктоплазме существуют
тысячи трубочек длиной 1 мкм и диаметром 60 нм,
которые лежат перпендикулярно клеточной мембра-
не (Hollande, Valentin, 1969). Активный пиноцитоз
обнаружен при помощи электронного микроскопа
также у Placojoenia (Radek, Hausmann, 1994а). Ко-
нечные продукты переваривания кусочков древесины
(углекислый газ, водород, различные ацетаты) хоро-
шо известны, но сам процесс энзиматической дегра-
дации целлюлозы, происходящий у жгутиконосцев,
380
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
еще недостаточно ясен. Возможно, эндобиотические
бактерии вносят свой вклад в процесс образования
различных целлюлаз.
Дефекационные процессы у гипермастигид еще
мало изучены. Показано, что у Joenia во время деле-
ния клетки идут активные дефекационные процессы
(Hollande, Valentin, 1969) (ссылки на работы по пи-
танию гипермастигид см.: Radek, Hausmann, 1994b).
Размножение и жизненные
циклы
Бесполое размножение
Деление клетки у гипермастигид изучено доста-
точно хорошо на светомикроскопическом уровне
(Cleveland et al., 1934; Grasse, 1952). На электронно-
микроскопическом уровне подробно изучен митоз
(Hollande, Valentin, 1969; Hollande, Carruette-Valen-
tin, 1971, 1972; Kubai, 1973), но морфогенез при де-
лении исследован современными методами еще недо-
статочно.
Митоз. Митоз гипермастигид на основании ори-
гинальных работ довольно подробно описан в отечест-
венной литературе (Райков, 1982, 1994), поэтому
приведены лишь основные особенности этого типа
митоза, который классифицируется как закрытый
внеядерный плевромитоз.
1) Полное сохранение ядерной оболочки на про-
тяжении митоза.
2) Внеядерное положение всего веретена деления,
включая нити центрального веретена и хромосомаль-
ные фибриллы, а также ЦОМТы. Формирующееся
веретено располагается латерально по отношению к
ядру. Хромосомы взаимодействуют с микротрубочка-
ми веретена через интактную ядерную оболочку. Для
этого типа митоза сформировалась удивительная сис-
тема связи кинетохоров с микротрубочками хромо-
сомальных фибрилл. Кинетохоры прорастают сквозь
ядерную оболочку, и к ним со стороны цитоплазмы
прикрепляется пучок хромосомальных микротрубочек.
Первоначальное расхождение кинетохоров после их
удвоения обеспечивается мембранным механизмом.
Дальнейшее расхождение прикрепленных к ним сес-
тринских хроматид происходит за счет хромосомных
пучков микротрубочек, а окончательное разделение
2 хромосомных наборов — за счет сильного роста в
длину центрального веретена.
3) Стадия экваториальной пластинки хромосом не
выражена, и анафаза следует непосредственно за про-
фазой.
Морфогенез. В качестве примера последователь-
ности событий при делении гипермастигид кратко
рассмотрим процесс деления Trichonympha magna
(Cleveland, 1960) (рис. 338—342); при этом восполь-
зуемся современной терминологией для органелл.
Деление начинается с продольного расщепления
рострума на 2 половины, которые постепенно откло-
няются друг от друга, оставляя между собой ясную
полосу безжгугиковой зоны (рис. 338). Во время это-
го процесса идет расщепление строго на 2 половины
всех органелл — наружной шапочки, рядов жгутиков,
парабазальных ламелл, составляющих стенку рост-
ральной трубки, парабазальных аппаратов (рис. 339;
340). Параллельно идет процесс удвоения «привиле-
гированных» кинетосом. В это время ядро (уже на
стадии профазы) мигрирует в передний конец клетки
(рис. 340).
Одновременно с процессом расщепления орга-
нелл идет рост атрактофоров. Как только длина ат-
рактофоров становится одинаковой, на их дистальных
концах появляется лучистость, состоящая из микро-
трубочек, которые растут, увеличиваются в размерах
и формируют центральное веретено и хромосомаль-
ные фибриллы (рис. 340). В это время ядро занимает
положение под веретеном (рис. 341). На этой стадии
начинается процесс образования новой, недостающей
половины органелл. В контакте с «привилегирован-
ными» кинетосомами в каждой дочерней половине
формируются кинетосомы жгутиков, парабазальные
ламеллы, атрактофоры. Далее центральное веретено
растет в длину и вплотную прилегает к ядру снаружи,
при этом целостность ядерной оболочки не наруша-
ется (рис. 341). Хромосомальные фибриллы приходят
в контакт с кинетохорами хромосом (в это время ки-
нетохоры уже разделились и начали отдаляться друг
от друга; каждая хромосома состоит уже из 2 сест-
ринских хроматид, — рис. 341, Я).
Митотические хромосомы трихонимф остаются
удлиненными на протяжении всех стадий митоза.
Стадия экваториальной пластинки у трихонимф не
выражена. Далее идет бурный рост недостающей по-
ловины органелл (рис. 342, А). Центральное верете-
но продолжает расти, и хромосомы «растаскиваются»
к полюсам и образуют 2 дочерние группы (рис. 342, Б).
Наконец, цитокинез завершает деление.
Вероятно, при морфогенезе «привилегированные»
кинетосомы являются координирующим центром,
обеспечивающим точное и упорядоченное распреде-
ление всех структур каждой мастигонтной системы.
Половой процесс
Гипермастигиды, населяющие кишечник низших
термитов и Cryptocercus, близки между собой в мор-
фофизиологическом отношении, но в характере раз-
множения жгутиконосцев существует глубокое разли-
чие. В термитах гипермастигиды размножаются толь-
ко агамным путем. При каждой линьке термиты
теряют и вновь приобретают всю кишечную фауну.
Гипермастигиды, паразитирующие в Cryptocercus, в
течение большей части жизненного цикла также раз-
множаются бесполым путем, но в период линьки хо-
зяина они претерпевают обязательный половой про-
цесс, в ходе которого у жгутиконосцев образуются
цисты и псевдоцисты. Часть из них вместе со старой
выстилкой кишечника таракана выводится наружу в
окружающую среду и служит для заражения молоди
таракана, а оставшаяся часть цист и псевдоцист в ки-
шечнике служит для увеличения массы симбионтов.
Половой процесс у гипермастигид носит столь
разнообразный характер, что дать его общую картину
весьма затруднительно (различные аспекты полового
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
381
Рис. 33&—340.
Рис. 338—342. Схематические рисунки различных стадий деления Trichonympha sp. (По: Cleveland, 1960).
338 — начальная стадия расщепления рострума на 2 половины. А — жгутиковые зоны постепенно отклоняются друг от друга. Б — в клетке еще 1 атракто-
фор; ядро уже на стадии профазы и начинает двигаться вперед. 339 — детали строения рострума на начальной стадии расщепления, а — атрактофор. 340 —
продолжение расщепления рострума на 2 половины. А — расщепление наружной шапочки; половины жгутиковых зон все дальше удаляются друг от друга.
Б — атрактофоры сравнялись по длине, и между их дистальными концами образуется центральное веретено; ядро на стадии профазы продвигается вперед.
341 — начало процесса образования новой недостающей половины органелл в дочерних особях. А — расхождение жгутиковых зон достигло максимума и
начинается образование новых органелл в контакте с «привилегированными» кинетосомами; ядро достигло центрального веретена, которое вдавливается
снаружи ядерной оболочки; начинается образование хромосомальных фибрилл. Б — то же, но более крупным планом. 342 — завершающая стадия деления
клетки. А — новая половина органелл завершает свой рост; ядро на стадии телофазы. Б — то же, но более крупным планом.
Fig. 338—342. Diagrammatic representation of cell division stages in Trichonympha sp.
338 — the early stage of the rostrum separation in two halves. A — the flagellar rows begin to separate into two groups leaving a clear aflagellate area between
them. Б — still one atractophore has remained in cell; the early prophase nucleus begins to move forth. 339 — details of the rostrum structure.at the early stage of
separation. Rostrum is opened up, atractophores (a) are not active. 340 — continued rostrum separation in two halves. A — formation of new halves of the cap;
flagellar rows move apart and produce a broad aflagellate region. Б — atractophores have become equal in length and formed a small central spindle between their
distal ends; the prophase nucleus continues to move forth. 341 — the early formation of a new half of organelles set during cell duplication. A — separation of
flagellar, areas is at its peak and new organelles are produced in contact with the «priveleged» kinetosomes; the nucleus reached the central spindle, which is forced
down outside the nuclear envelope; chromosomal fibrils arise. Б — the same, a close-up view. 342 — the completion of cell division. A — a new half of organelles
set completes growing; the nucleus is at telophase. Б — the same, a close-up view.
382
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
341
342
процесса, характерные для каждого рода жгутиконос-
цев, подробно описаны в большом цикле работ Клив-
ленда* ссылки на эти работы см.: Cleveland, 1956,
1957).
Для примера остановимся несколько более под-
робно на ходе полового процесса в родах Trichonym-
pha (рис. 343) й Urinympha (рис. 344).
У Trichonympha — гаплонта в бесполой фазе жиз-
ненного цикла — половой процесс протекает по
типу парной мерогамии. Преобразование вегетатив-
ной клетки в гаметоцит начинается за 6 дней до на-
чала линьки хозяина. Вначале гаметоцит морфологи-
чески незначительно отличается от вегетативной
клетки, но затем теряет большую часть органелл; во-
круг клетки формируется плотная оболочка цисты.
Под защитой этой оболочки происходит процесс га-
метогенеза — формирование 2 диморфных гамет в
ходе митотического деления ядра, при котором раз-
личия между хромосомами будущих гамет проявля-
ются уже в конце профазы (они по-разному окраши-
ваются). Цисты со сформированными гаметами могут
в момент линьки таракана выводиться в окружающую
среду и <затем заканчивать половой процесс в другом
хозяине. Эксцистирование гамет происходит в про-
свете новой выстилки кишечника спустя 1—2 дня
после линьки таракана. После эксцистирования гамет
следует копуляция диморфных гамет. Женская гаме-
та на заднем конце тела имеет особое кольцо сидеро-
фильных гранул (или гранул оплодотворения) и свет-
лый воспринимающий конус. Копуляция протекает
следующим /образом: мужская гамета направляется к
заднему концу женской гаметы и своим рострумом
внедряется в ее светлый воспринимающий конус, а
затем постепенно мужская гамета целиком проникает
через кольцо гранул оплодотворения внутрь женской
гаметы. Жгутики и другие органеллы мужской гаметы
распадаются и резорбируются. Ядра гамет сливаются,
и образуется зигота, ядро которой претерпевает двух-
ступенчатый мейоз. В конце полового процесса обра-
зуются 4 гаплоидные особи, которые размножаются
бесполым путем до начала ближайшей линьки хозяи-
на.
У Urinympha — диплонтов в бесполой фазе жиз-
ненного цикла — половой процесс протекает по ти-
пу автогамии, и при этом не происходит размножения.
Вегетативная клетка за 8 дней до начала линьки хо-
зяина приступает к единственному мейотическому
делению, в ходе которого производятся пронуклеу-
сы, которые затем снова сливаются в общей цитоп-
лазме. При наступлении мейотического деления ядра
все органеллы распадаются. На стадии слияния про-
нуклеусов образуется новый набор жгутиков, параба-
зальных аппаратов и аксостилей, а старые органеллы
начинают дегенерировать. Однако в такой зиготе но-
вые жгутиковые зоны остаются внутриклеточными, а
сама клетка неподвижна и имеет плотную кутикулу.
Это — псевдоциста, так как у нее не формируются
характерные для цисты оболочки. Псевдоциста пред-
назначена для вывода паразита наружу из кишечника
таракана и перенесения им неблагоприятных усло-
вий внешней среды. После окончания линьки хозяи-
на псевдоциста высвобождает жгутики и преобразу-
ется в бесполую клетку. Таким образом, одна и та
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
383
Рис. 343. Схема полового процесса по типу парной мерогамии у
Trichonympha. (По: Cleveland, 1956).
А — бесполая особь. Б — инцистированный гаметоцит. В —• эксцистирую-
щиеся гаметы. Г — копуляция диморфных гамет. Д — стадия оплодо-
творения, Е — слияние ядер гамет, все органеллы мужской гаметы дегене-
рируют, образуется зигота, ядро которой претерпевает двухступенчатый
мейоз; в завершение процесса образуются 4 гаплоидные бесполые особи.
а — женская гамета, б — мужская гамета, в — кольцо гранул оплодотворе-
ния, г — конус оплодотворения.
Fig. 343. Diagrammatic representation of the sexual process of Tri-
chonympha, accomplished by paired merogamy type.
A — haploid asexual cell. Б — encysted gametocyte. В — excysting gametes.
Г— copulation of gametes already differentiated into male and female ones.
Д — stage of fertilization. E — fusion of gametic nuclei, the digestion of all
organelles of male gamete begins within the cytoplasm of the female one; cy-
toplasmic union of two gametes is complete; a fully developed zygote is produ-
ced; the nucleus undergoes two-step meiosis; four asexual cells are produced,
a — female gamete, 6 — male gamete, в — a ring of fertilization, г — a fer-
tilization cone.
же клетка Urinympha вступает в половой процесс и
заканчивает его.
Теперь остановимся на общих особенностях по-
лового процесса у гипермастигид.
1) У всех гипермастигид наступление полового
процесса неразрывно связано с периодом линьки та-
ракана. Однако у одних родов он начинается задолго
до начала линьки хозяина (например, у Barbulanym-
pha за 50 дней), у других — сближен с моментом
линьки (например, у Trichonympha он начинается за
5 дней).
2) Половой процесс может осуществляться разны-
ми путями. У одних родов он протекает только по
типу образования гамет и их последующей копуляции
(Trichonympha, Eucomonympha, Macrospironympha,
Leptospironympha). У других родов половой процесс
осуществляется 2 путями — копуляции и автогамии
(Barbulanympha). У третьих он протекает только по
типу автогамии (Urinympha, Rhynchonympha). У Bar-
bulanympha кроме копуляции и автогамии возможен
еще и третий путь — увеличение числа хромосом не
путем кариогамии, а путем эндомитоза.
3) Гипермастигиды из кишечника Cryptocercus
обнаруживают исключительно широкуку изменчи-
вость в отношении гаплоидного или диплоидного ха-
рактера состояния ядра. Морфологически близкие
роды могут быть в бесполой фазе жизненного цикла
одни — гаплонтами, другие — диплонтами. Так,
Barbulanympha — гаплонты, a Urinympha и Rhyncho-
Рис. 344. Схема хода полового процесса по типу автогамии у Uri-
nympha. (По: Cleveland, 1956).
А — бесполая особь. Б — гаметогенез, ядро претерпевает одноступенчатый
мейоз, жгутики, парабазальный аппарат, аксостили постепенно распадают-
ся. В — стадия образования пронуклеусов. Г — слияние сестринских про-
нуклеусов. Д — «зигота», клетка неподвижна, старые органеллы дегенери-
руют, начинается образование нового набора органелл. Е — продолжение
процессов дегенерации старых органелл и образования новых органелл.
Ж — стадия псевдоцисты; клетка неподвижна, имеет плотную кутикулу,
жгутиковые зоны остаются внутриклеточными. 3 — исходная бесполая
особь с обновленным набором органелл, п — гаплоидный набор хромосом,
2п — диплоидный набор хромосом.
Fig. 344. Diagrammatic representation of the sexual process of Uri-
nympha. accomplished by autogamy type.
A — asexual cell. Б — gametogenesis, the nucleus undergoes one-step meiosis;
flagella, parabasal apparatus; axostyles disconnected. В — stage of pronuclei
formation. Г — fusion of sister pronuclei. Д — «zygote»; cell immobile; old
organelles are slowly degenerating; formation of new organelles begins. E —
organelle degeneration proceeds; early formation of new organelles. Ж — stage
of pseudocyst; cell immobile, intracytoplasmic flagella. 3 — flagella become
extracytoplasmic, original asexual cell with renewed set of organelles; chromo-
somes as at beginning of sexual process, n — haploid set of chromosomes,
2n — diploid set of chromosomes.
384
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
nympha — диплонты; Leptospironympha — гаплонты,
a Macrospironympha — диплонты.
4) Редукция числа хромосом может осуществ-
ляться путем двух- или одноступенчатого мейоза,
причем у морфологически близких родов тип мейоза
также варьирует. Так, у Barbulanympha двухступенча-
тый мейоз происходит на стадии зиготы, а у Urinym-
pha наблюдается одноступенчатый мейоз при обра-
зовании пронуклеусов в ходе автогамищ Leptospiro-
nympha — гаплонт с зиготическим одноступенчатым
мейозом, a Macrospironympha — диплонт с гамети-
ческим двуступенчатым мейозом.
5) Довольно пестрая морфологическая картина
наблюдается и при сравнении отдельных периодов
полового процесса у различных родов. В прогамном
периоде у одних родов гаметоцит, в который преоб-
разуется вегетативная клетка под влиянием гормона
линьки хозяина, морфологически почти не отличается
от бесполой клетки (например, у Barbulanympha,
Eucomonympha). У других родов между гаметоцитом
и бесполой клеткой существуют столь резкие мор-
фологические различия, что можно было бы отнести их
к разным семействам (у видов рода Leptospironym-
pha). Гаметы также могут быть у одних родов ди-
морфными (Trichonympha, Eucomonympha), у других —
морфологически неразличимыми (Barbulanympha, Mac-
rospironympha).
В эугамном периоде копуляция гамет у всех гипер-
мастигид происходит в свободноподвижном состоянии,
но морфологическая картина весьма разнообразна.
Так, например, у рода Trichonympha гаметы диморф-
ные. У других родов гипермастигид гаметы морфоло-
гически неразличимы или отличаются только разме-
рами; женская гамета не имеет специального места
для внедрения мужской гаметы; слияние гамет проис-
ходит сбоку и мужской пронуклеус мигрирует к ядру
женской гаметы.
Метагамный период у большинства гипермастигид
характеризуется тем, что в зиготе происходит частич-
ная или полная реорганизация всех клеточных орга-
нелл.
6) В ходе полового процесса у всех гипермастигид
формируются цисты или псевдоцисты, образование
которых по времени совпадает с началом линьки та-
ракана. Они предназначены для защиты паразитов от
неблагоприятных условий при смене хитиновой вы-
стилки задней кишки и для их вывода наружу в окру-
жающую среду, где происходит заражение молоди и
других личинок таракана. Инцистирование у разных
родов гипермастигид наблюдается на различных ста-
диях полового процесса. У Trichonympha и Macrospi-
ronympha инцистируется гаметоцит и под защитой
оболочки цисты идет гаметогенез, у Leptospironympha
еирога — зигота. У Urinympha, Rhynchonympha и
Barbulanympha настоящих цист нет; неблагоприятные
условия во время линьки хозяина их зиготы перено-
сят в состоянии псевдоцисты, т. е. клетка округляет-
ся, кутикула уплотняется, жгутиковые зоны остаются
внутриклеточными, клетка неподвижна.
7) Продолжительность полового процесса значи-
тельно варьирует у разных родов. У одних родов он
растягивается на значительный срок и включает по-
коящиеся стадии, когда никаких изменений не про-
исходит. Так, у Barbulanympha половой процесс на-
чинается за 50 дней до линьки таракана, но на ста-
дии зиготы приостанавливается, и около 35 дней ни-
каких изменений в клетке не наблюдается; за 3 дня
до линьки происходит реорганизация клеточных ор-
ганелл и образование псевдоцисты; завершение по-
лового процесса — мейоз происходит спустя 1 день
после линьки таракана. Покоящиеся стадии наблю-
даются также у Eucomonympha и Rhynchonympha. У
других родов развитие стадий полового процесса не-
прерывное и относительно быстрое. Так, у Tricho-
nympha и Macrospironympha половой процесс начи-
нается за 6—8 дней до начала линьки хозяина и
протекает непрерывно; гаметогенез происходит в
цистах; эксцистирование гамет и последующие ста-
дии полового процесса осуществляются в кишечнике
перелинявшего таракана в течение первых 2 дней
после линьки.
У всех гипермастигид половой процесс заканчи-
вается спустя 1—2 дня после окончания линьки хо-
зяина.
Жизненный цикл жгутиконосцев
из термитов и тараканов и связь
его со способом заражения хозяев
Оксимонады и гипермастигиды, населяющие ки-
шечник низших термитов и тараканов рода Crypto-
cercus, близки между собой в морфофизиологическом
отношении. Однако в ходе исторического становле-
ния системы паразит—хозяин у этих жгутиконосцев
в каждой из групп хозяев сложился свой жизненный
цикл; по-разному происходит и их трансмиссия сре-
ди насекомых.
Жизненный цикл жгутиконосцев из тараканов
Cryptocercus включает вегетативную фазу, которая
осуществляется в нелиняющем хозяине и состоит из
длинного ряда бесполых поколений, и половой про-
цесс, который отмечен у жгутиконосцев только в
период линьки хозяина (Cleveland, 1957). В ходе
полового процесса образуются цисты и псевдоцис-
ты — стадии, предназначенные для выведения жгути-
коносцев из кишечника хозяина в момент его линь-
ки и последующего заражения ими других членов
колонии. У самого Cryptocercus линька личинок и
вылупление молоди из яиц строго приурочены друг
к другу. Тараканы линяют 1 раз в год (кроме нимф
младшего возраста, которые линяют дважды в год).
Заражение жгутиконосцами молоди и личинок
разных возрастов Cryptocercus происходит в момент
линьки последних, когда вместе с экскрементами и
огрызками древесины они заглатывают цисты и псев-
доцисты простейших. Механизм смены хитиновой
выстилки кишечника у линяющей личинки таракана
таков, что личинка в это время не лишается всей
своей протистофауны — часть цист и псевдоцист не
выводится наружу, а остается в кишечнике и после
линьки заканчивает половой процесс. Это обстоя-
тельство способствует быстрому восстановлению
численности жгутиковых в кишечнике хозяина, ко-
торому они так необходимы для нормальной жизне-
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
385
деятельности. Таким образом, у жгутиконосцев из
Ciyptocercus способ трансмиссии жгутиковых от хо-
зяина к хозяину тесно связан с половым процессом.
Жизненный цикл жгутиконосцев из низших тер-
митов включает только вегетативную фазу, состо-
ящую из длинного ряда бесполых поколений, ко-
торый прерывается линькой хозяина и вновь вос-
станавливается после линьки. При каждой линьке
личинка термита теряет всю свою жгутиковую фауну
и затем вновь приобретает ее. Личинки низших тер-
митов линяют не менее 1 раза в месяц. Единствен-
ным способом заражения молоди и перенесших
линьку личинок от личинок, не линяющих в данный
момент, является заражение посредством проктоде-
ального питания, характерного для термитов. Оно
заключается в том, что личинки слизывают друг у
друга выделения из порошицы, которые наряду с
питательными веществами для вскармливания моло-
ди содержат жгутиковых. При этом жгутиконосцы в
выделениях находятся в «мумифицированном» со-
стоянии (т. е. имеют плотную кутикулу, а жгутики
плотно прижаты к телу) (Догель, 1956). Это состо-
яние — удивительное приспособление жгутиковых
для успешного заражения ими нового хозяина. Быс-
трое воспроизведение численности жгутиковых в ки-
шечнике перенесшей линьку личинки обеспечивает-
ся массовыми вспышками деления вновь приобре-
тенных жгутиконосцев (Andrew, Light, 1929).
Несмотря на тщательные поиски многих исследо-
вателей, половой процесс у подавляющего большин-
ства жгутиконосцев, населяющих кишечник низших
термитов, не найден. Однако Кливленду удалось на-
блюдать отдельные случаи полового процесса у одно-
го из видов Trichonympha и некоторых Pseudotricho-
nympha (Cleveland, 1965а, 1965b), а также у жгути-
коносцев из примитивного австралийского термита
Mastotermesdarwiniensis (Cleveland, 1966с). Как сле-
дует из наблюдений Кливленда, половой процесс в
этих редких случаях не является обязательным в жиз-
ненном цикле и не связан со способом трансмиссии
жгутиконосцев от хозяина к хозяину.
Существование регулярного и обязательного по-
лового процесса в жизненном цикле жгутиконосцев
из Cryptocercus и отсутствие его у близких видов
из термитов позволяет поставить вопрос о том, что
первично — явление сексуальности или асексуаль-
ности в этой группе простейших. Приведенные дан-
ные о различных способах трансмиссии жгутиконос-
цев в 2 группах хозяев и о распространении и роли
полового процесса в жизненном цикле простейших
из тех и других хозяев свидетельствуют в пользу
предположения некоторых исследователей об утрате
полового процесса жгутиконосцами из термитов. Ве-
роятно, выпадение полового процесса из жизненного
цикла жгутиконосцев из термитов — результат фор-
мирования в ходе эволюции в данной системе пара-
зит—хозяин иного способа их трансмиссии, чем у
жгутиконосцев из Cryptocercus. Особенности биоло-
гии хозяина, связанные с частыми линьками, стиму-
лировали развитие способа трансмиссии, исключаю-
щего потерю жгутиковой фауны, без которой хозяин
не в состоянии нормально существовать. Надежный
способ передачи жгутиковой инфекции от одного
25 Протесты, часть 1
термита к другому посредством проктодеального пи-
тания исключил из жизненного цикла жгутиконос-
цев половой процесс как необходимое звено для за-
вершения цикла развития симбионтов. Вероятно,
редкие случаи полового процесса, наблюдаемые у
жгутиконосцев из термитов, отражают предковую
способность жгутиконосцев реагировать на гормон
линьки хозяина. Поскольку тараканы рода Crypto-
cercus предположительно являются современными
потомками тех палеозойских тараканов, от которых
дивергировали термиты (McKittrick, 1965), то, веро-
ятно, явление сексуальности среди оксимонад и ги-
пермастигид, паразитирующих в Cryptocercus, пер-
вично, асексуальность же у жгутиконосцев из ни-
зших, термитов вторична.
Культивирование
В последние годы удалось наладить культивиро-
вание жгутиконосцев, населяющих кишечник Crypto-
cercus, in vitro (Ritter, 1974; Ritter et al., 1978). Ha
основании изучения физико-химических свойств со-
держимого задней кишки Cryptocercus была создана
синтетическая Среда, свободная от целлюлозы, на ко-
торой жгутиконосцы живут и размножаются неогра-
ниченно долгое время. Инокуляция, культивирование
и микроскопирование жгутиконосцев осуществляется
в особой камере, где поддерживается газовый режим,
имитирующий анаэробные условия существования
простейших in vivo (95 % азота и 5 % углекислого
газа при свободном кислороде в пределах 0.8—20 цм
при 13 °C).
Экология
Гипермастигиды и оксимонады вместе с трихомо-
надами, грибами и бактериями составляют большое
сообщество организмов, населяющих кишечник низ-
ших термитов и тараканов рода Cryptocercus. Боль-
шая часть жгутиконосцев, входящих в состав фауны
кишечника, является облигатными симбионтами, ко-
торые играют важную биологическую роль в жизни
хозяев. Сложившиеся симбиотические отношения
между жгутиконосцами и их хозяевами носят харак-
тер мутуалистических взаимоотношений. Еще в 30-х гг.
нашего столетия Кливлендом было показано, что
присутствие жгутиконосцев в кишечнике необходи-
мо для нормального пищеварения и существования
хозяев; искусственно освобожденные от жгутиконос-
цев насекомые погибают через несколько дней, не-
смотря на обильную пищу (Cleveland, 1926; Cleve-
land et al., 1934). Выяснилось, что низшие термиты
и Cryptocercus являются насекомыми-ксилофагами,
т. е. питаются исключительно древесиной, но сами
они не в состоянии ее переваривать. Эту функцию
берут на себя жгутиконосцы, которые, обладая набо-
ром ферментов, расщепляющих целлюлозу, перево-
дят частицы древесины в легко усваиваемые вещест-
ва, главным образом глюкозу и ацетаты (см.: Honig-
berg, 1970). Эти субстраты абсорбируются из кишки
насекомого-ксилофага и далее используются ими в
386
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
процессах биосинтеза. В 70-х гг. было обнаружено,
что жгутиконосцы, имеющие в пищевых вакуолях
кусочки древесины, сами содержат многочисленные
эндобиотические бактерии (Bloodgood, Fitzharris, 1976),
и высказано предположение, что некоторые целлю-
лазы, принимающие участие в расщеплении древеси-
ны в фагоцитарных вакуолях жгутиконосцев, воз-
можно, имеют бактериальное происхождение. Одна-
ко вскоре на аксенической культуре Trichonympha
sphaerica экспериментально было доказано, что цел-
люлазная активность исходит от самих жгутиконос-
цев (Yamin, 1981). При этом жгутиконосцы были
предварительно освобождены от эндобиотических
бактерий, для чего использовали антибиотики. В
культуральную среду вводили меченную по углероду
целлюлозу и затем определяли конечные продукты
ее внутриклеточного метаболизма. Оказалось, что они
содержали метку. Таким образом, четко было по-
казано, что целлюлазная активность трихонимф не
зависит от эндобиотических бактерий. Несомненно,
что жгутиконосцы обеспечивают питание продуктами
разложения древесины не только себе, но и своим
хозяевам. Однако многие биохимические аспекты
физиологической стороны симбиоза жгутиконосцев
и насекомых-ксилофагов еще до сих пор остаются
неясными. Еще менее ясны симбиотические отноше-
ния жгутиконосцев и их внутриклеточных бактерий.
Эволюция
Эволюция гипермастигид
В настоящее время несомненно, что Trichomona-
dida и Hypermastigida тесно связаны между собой.
Практически все крупные исследователи этих жгути-
коносцев на светомикроскопическом уровне отмеча-
ли сходство в их организации (наличие парабазаль-
ного аппарата, сходный тип митоза) (Kirby, 1949;
Grasse, 1952; Honigberg, 1973). Электронно-микро-
скопическое изучение трихомонадид и гипермасти-
гид выявило их сходство в организации корешковой
системы (Hollande, Carruette-Valentin, 1971, 1972).
На основании этих данных Trichomonadida и Hyper-
mastigida были объединены в один надотряд — Ра-
rabasalidea (Honigberg, 1973; Levine et al., 1980). По
современным воззрениям, предками гипермастигид
были трихомонадоподобные жгутиконосцы. Если это
так, то они обитали в тех древних тараканах, от
которых эволюировали термиты. Однако в современ-
ных Cryptocercus, которые являются реликтами тре-
тичного периода (Бей-Биенко, 1950) и рассматрива-
ются в качестве предков современных термитов, три-
хомонады не найдены. Можно предположить, что
трихомонады были утрачены Cryptocercus по мере
стабилизации механизма трансмиссии между члена-
ми колонии, т. е. вторично.
По мнению большинства исследователей, среди
гипермастигид четко просматриваются 2 параллель-
ные эволюционные линии — Lophomonadina и Tri-
chonymphina — от общего трихомонадоподобного
предка (Grassi, 1911; Догель, 1922; Kirby, 1949; Ho-
nigberg, 1963b). Ветвь Lophomonadina состоит из
наиболее примитивных Lophomonadidae, затем идут
Joeniidae и, наконец, Kofoidiidae (Kirby, 1949). Эво-
люция лофомонадин происходила главным образом
среди низших термитов. В кишечной фауне Crypto-
cercus присутствует лишь 1 представитель этой вет-
ви — Prolophomonas.
Филогенетические связи внутри ветви Trichonym-
phina просматриваются менее ясно. Вероятно, среди
них в начале филогенетического ряда идут Hoplo-
nymphidae и Staurojoeninidae, а затем Eucomonymp-
hidae, Trichonymphidae и Spirotrichosomatidae. Се-
мейства Spirotrichonymphidae и Holomastigotoididae,
вероятно, отделяются от трихонимфин в самом нача-
ле этой ветви, либо представляют собой третью
самостоятельную ветвь, начинающуюся непосредст-
венно от трихомонадоподобного предка (Grasse,
Hollande, 1942; Hollande, Carruette-Valentin, 1971).
Trichonymphina широко эволюировали в обеих груп-
пах хозяев. Общими родами, широко распространен-
ными среди низших термитов и Cryptocercus, явля-
ются Trichonympha и Leptospironympha.
Роды Deltotrichonympha и Koruga, населяющие
кишечник австралийского термита Mastotermes dar-
winiensis — наиболее примитивного из ныне живущих,
сочетают в своей организации черты Lophomonadina
и Trichonymphina (Hollande, 1986). Нам представляется
возможным рассматривать их как филогенетического
предка тех древних гипермастигид, от которых эво-
люировали Lophomonadina и Trichonymphina, либо
как отдельную слепую эволюционную ветвь от три-
хомонадоподобных предков.
Сопряженная эволюция
жгутиконосцев отрядов
Oxymonadida и Hypermastigida
и их хозяев
Жгутиконосцы отрядов Oxymonadida и Hypermas-
tigida — это весьма специализированные паразиты,
которые в своем распространении ограничены лишь
2 группами хозяев — низшими термитами и тарака-
нами рода Cryptocercus (исключение составляют: среди
Oxymonadida род Monocercomonoides, имеющий ши-
рокий круг хозяев, включая других насекомых и раз-
личных позвоночных; среди Hypermastigida род Lo-
phomonas, паразитирующий в тараканах родов Blatta,
Blattella и Periplaneta). Высшие термиты обладают
иной кишечной протофауной, чем низшие, и она
сравнима с протозойной фауной других насекомых.
Кишечная фауна других таракановых (кроме
Cryptocercus) также не содержит Oxymonadida и Hy-
permastigida. Эта группа жгутиконосцев объединяет
приблизительно 45 родов и 250 видов. Подавляющее
большинство родов жгутиконосцев являются специа-
лизированными либо для термитов, либо для Cryptocer-
cus, но существует 3 рода жгутиконосцев, представите-
ли которых распространены среди обеих групп хозя-
ев, — Oxymonas, Trichonympha и Leptospironympha.
Данные о современном распространении жгутико-
носцев среди их хозяев, об отсутствии перекрестной
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
387
инфекции между хозяевами симбионтов и о морфо-
физиологической близости жгутиконосцев привели
исследователей к выводу, что эти жгутиконосцы
могли быть приобретены обеими группами хозяев
только от общих предков (Cleveland et al., 1934;
Kirby, 1937, 1949; Grasse, 1952). Учитывая современ-
ные данные о филогении отрядов Isoptera и Blattodea
(McKittrick, 1965), можно утверждать, что первичны-
ми хозяевами анцестральных оксимонад и гипермас-
тигид были палеозойские тараканы, от которых в
позднем палеозое или раннем мезозое дивергировали
термиты. (Вероятно, современные реликтовые тара-
каны рода Cryptocercus являются теми дожившими до
наших дней представителями таракановых, от кото-
рых дивергировали термиты.)
Гипермастигиды являются наиболее древней груп-
пой жгутиконосцев, которые большую часть своей
эволюции претерпели в палеозойских тараканах до
дивергенции от них термитов. Об этом свидетельст-
вует широкое распространение многих Hypermastigi-
da в Cryptocercus и в филогенетически далеких груп-
пах низших термитов. Oxymonadida — также давно
сложившаяся группа жгутиконосцев, которая была
представлена в общем Сгур/осегси^-подобном пред-
ке. В настоящее время Oxymonadida известны среди
термитов только в сем. Calotermitidae, но наличие
этих жгутиконосцев в Cryptocercus позволяет пред-
положить их более широкое распространение в про-
шлом.
За период эволюционного развития в низших тер-
митах оксимонады и гипермастигиды почти не пре-
терпели или претерпели весьма ограниченные эволю-
ционные изменения. Такой вывод позволяет сделать
анализ современного распространения этих жгутико-
носцев в их хозяевах (Kirby, 1937, 1949). Строгая
специфичность к хозяину у них наблюдается сравни-
тельно редко. В большинстве случаев один вид жгу-
тиконосца встречается в разных видах одного рода
или даже в разных родах хозяина, часто географичес-
ки весьма удаленных друг от друга.
Современное распространение жгутиковых среди
термитов отражает филогенетические взаимоотноше-
ния между ними. Возможно, эволюционный путь жгу-
тиконосцев из термитов был связан со стабильностью
и устойчивостью видов во времени, что является
следствием утраты полового процесса и перехода к
исключительно агамному размножению.
Иной эволюционный путь проделали Oxymonadi-
da и Hypermastigida, паразитирующие в кишечнике
Cryptocercus, за время становления 2 его видов. Со-
временные виды Cryptocercus — С. punctulatus (Се-
верная Америка) и С. relictus (Дальний Восток, Рос-
сия) — это реликты третичного периода. Они сфор-
мировались как самостоятельные виды в условиях
полной географической изоляции в результате изме-
нения условий существования их предка — какого-
то третичного вида Cryptocercus (Бей-Биенко, 1950).
2 современных вида тараканов, разделенных геогра-
фическим барьером с третичного периода, обладают
в настоящее время строго специфичной для каждого
из них паразитофауной (Cleveland et al., 1934;
Бобылёва, 1975). Параллельно с формированием се-
вероамериканского и дальневосточного видов Cryp-
tocercus шло обособление строго специфичных для
каждого хозяина видов жгутиконосцев, т. е. в эволю-
ции паразитов и хозяев наблюдался филогенетичес-
кий параллелизм.
Таким образом, жгутиконосцы из термитов и
жгутиконосцы из Cryptocercus за один и тот же ис-
торический период прошли разные эволюционные
пути. Первые проделали только ограниченные эво-
люционные изменения и не создали видоспецифич-
ных для каждого хозяина комплексов, а вторые до-
стигли такой специфичности, проделав путь дивер-
гентного видообразования. Возможно, это различие
обусловлено сохранением полового процесса в жиз-
ненном цикле жгутиконосцев из Cryptocercus и
большими эволюционными возможностями, связан-
ными с рекомбинацией генного фонда.
Систематический обзор
Подотряд Lophomonadina Light, 1927
Гипермастигиды, у которых жгутики сосредоточе-
ны в одной зоне, занимающей апикальное положе-
ние. Расположение 4 «привилегированных» кинето-
сом справа от остальных соответствует таковому у
трихомонадид. Корешковая система как у трихомо-
надид. Контакт поперечно исчерченных корешковых
фибрилл с «привилегированными» кинетосомами со-
храняется и в интерфазе. Цитоскелет образован в ос-
новном структурами пельта-аксостилярного комплек-
са. Зачаток веретена между атрактофорами присутст-
вует на протяжении интерфазы. При делении старые
структуры резорбируются и дочерние органеллы об-
разуются de novo.
Включает 4 семейства.
Сем. Joeniidae Janicki, 1915
1 апикальная жгутиковая зона разнообразной
формы; внутри зоны жгутики располагаются про-
дольными рядами; массивный аксостиль пронизыва-
ет все тело; 2 лентовидные парабазальные ламеллы;
многочисленные парабазальные тела обвивают аксо-
стиль; атрактофоры являются придатками параба-
зальных ламелл.
Паразиты низших термитов. Включает 6 родов.
Род Joenia Grassi, 1885 (рис. 320, 321)
Треугольная по форме жгутиковая зона. Аксо-
стиль имеет расширение передней части в виде
чаши («capitulum»), где помещается ядро. Параба-
зальные филаменты и парабазальные тела образуют
сложную перистую структуру вокруг ядра. Хорошо
изучен электронно-микроскопически (Hollande, Va-
lentin, 1969). Известно 3 вида — J. annectens, J, du-
boscqui и . J. clevelandi из термитов рода Calotermes
и Neotermes.
388
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Joenina Grassi, 1919
Жгутиковая зона имеет форму полукруга. 2 пара-
базальных филамента закручены вокруг ядра и аксо-
стиля.
1 вид — J. pulchella из Porotermes adamsoni.
Род Joenopsis Cutler, 192СГ
Подковообразная жгутиковая зона. 2 парабазаль-
ных филамента и 2 парабазальных тела колбасовид-
ной формы.
Известно 2 вида — J. polytricha и J. cephalotricha
из термитов рода Archotermopsis.
Род Parajoenopsis Saleem, 1969
Недостаточно изучен.
Род Projoenia Lavette, 1969
Парабазальный аппарат подобен таковому у De-
vescovina (Trichomonadida).
1 вид — Р. sawayai.
Род Placojoenia Radek et Hausmann,
1994
Подковообразная жгутиковая зона. Парабазальные
тела в 5—7 рядов по спирали обвивают аксостиль.
Свободные диктйосомы разбросаны в цитоплазме.
Характерный признак — многочисленные цитоплаз-
матические пластинки. Хорошо изучен электронно-
микроскопически (Radek, Hausmann, 1994а).
Известен 1 вид — Р. sinaica из термита Kaloter-
mes sinaicus.
Сем. Lophomonadidae Kent, 1880
Расположение жгутиков на апикальной части
клетки напоминает корону; аксостиль проходит
через все тело; парабазальные тела располагаются по
кругу. Комплекс «привилегированных» кинетосом и
корешковых фибрилл как у трихомонадид.
Включает 3 рода.
Род Lophomonas Stein, 1860 (рис. 317—319)
Хорошо изучен электронно-микроскопически
(Hollande, Carruette-Valentin, 1972). Известно 2 вида.
L. blattarum имеет овальную или грушевидную форму
тела размером до 100 мкм. Мощный аксостиль в виде
слуховой трубки. Ядро локализовано внутри расши-
рения передней части аксостиля «Capitulum». Обна-
ружено чрезвычайное усложнение наружной мемб-
раны ядра. L. striata имеет веретенообразную форму
тела, величина которого может составлять до 57 мкм.
На поверхности тела находятся бактерии рода Fusi-
formis. Известны цисты.
Паразитируют в домашних и полудомашних тара-
канах (Blatta, Periplaneta, Blattella).
Род Prolophomonas Cleveland, Hall,
Sanders et Collier, 1934
Известен 1 вид — P. tocopola. Паразитирует в дре-
весиноядном таракане Cryptocercus. Жгутиковая зона
насчитывает 10 —15 жгутиков, аксостиль трубчатый,
форма парабазальных тел неизвестна. Prolophomonas
рассматривается в качестве связующего звена между
паразитофаунами Cryptocercus и другими таракано-
выми.
Род Microjoenia Grassi, 1892
Мелкие жгутиконосцы (не более 20 мкм). Форма
тела грушевидная. Около 20 жгутиков расположены
по кругу. Парабазальные тела лежат наподобие оже-
релья. Аксостиль проходит через все тело.
2 вида из термитов рода Reticulitermes.
Сем. Kofoidiidae Light, 1927
Известен 1 вид — Kofoidia loriculata из Рагапео-
termes. Мало изучен.
Сем. Rhyzonymphidae Grasse et
Hollande, 1951
Известен 1 вид — Rhyzonympha jahieri из Anacan-
thotermes ochraceus. Мало изучен.
Подотряд Trichonymphina Poche, 1913
Гипермастигиды с 2 или 4 билатерально-симмет-
ричными жгутиковыми зонами. Парабазальные ла-
меллы обычно образуют ростральную трубку, кото-
рая является центральной осевой структурой в клет-
ке, поэтому билатеральная симметрия в организации
сочетается с радиальной. Связь поперечно исчерчен-
ных корешковых фибрилл с «привилегированными»
кинетосомами опосредована морфоплазмой. Жгути-
ковые зоны занимают или передний отдел тела, или
большую часть тела и подразделяются на ростраль-
ный и постростральный районы; либо автономные.
В постростральном районе жгутики MOiyr быть ор-
ганизованы в продольные и меридиональные ряды,
концентрические круги, в 2 леотропно закрученные
спирали. В формировании цитоскелета принимают
участие как парабазальные ламеллы, так и структу-
ры пельта-аксостилярного комплекса. Обычно рав-
ное разделение систем при делении, с общим или
частичным сохранением старых структур, когда
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA. ОТРЯД HYPERMASTIGIDA
389
новые системы формируются. В основе классифи-
кации на семейства лежит организация жгутиков в
постростральном районе. Включает 8 семейств.
Сем. Hoplonymphidae Light,
1926
Передний конец тела прикрыт наружной шапоч-
кой. Имеется 2 билатерально-симметричные, тре-
угольные по форме жгутиковые зоны, которые занима-
ют переднюю треть тела и разделены узкой зоной
поверхности, свободной от жгутиков. В пределах
каждой из зон жгутики располагаются продольными
рядами. Парабазальные ламеллы лежат свободно, не
образуют ростральную трубку. Типичный множе-
ственный парабазальный аппарат. Хорошо развит
пельта-аксостилярный комплекс. Ядро заключено
в ядерный мешок. В интерфазе 2 лентовидных ат-
рактофора с атрактосферами. Поверхность тела по-
крыта симбиотическими бактериями. Включает
4 рода.
Род Barbulanympha Cleveland, Hall,
. Sanders et Collier, 1934 (рис. 322)
Крупные жгутиконосцы (примерно от 100 до
300 мкм). Форма тела напоминает желудь. Передний
конец прикрыт шапочкой. На основании рострума
больше жгутиковых рядов, чем на вершине. Имеют
своеобразный аксостилярный аппарат.
Известно 4 вида из тараканов рода Cryptocer-
cus.
Род Urinympha Cleveland, Hall,
Sanders et Collier, 1934 (рис. 323)
Крупные жгутиконосцы (примерно от 75 до
300 мкм), имеют длинное и узкое тело. Есть наруж-
ная шапочка. Жгутиковые зоны имеют одинаковое
число рядов как на вершине, так и на основании.
Характерная черта — при движении все жгутики од-
ной жгутиковой зоны действуют как единое целое.
Существует 2 вегетативные формы в зависимости от
клеточного цикла: грушевидная в состоянии перед
делением клетки и длинная, узкая — между деле-
ниями. Поверхность тела покрыта палочковидными
бактериями.
Известно 2 вида из тараканов рода Cryptocer-
cus.
Род Rhynchonympha Cleveland, Hall,
Sanders et Collier, 1934
Жгутиконосцы внешне весьма похожи на Urinym-
pha, но более крупные. Возможно, синоним Urinym-
pha (Hollande, Carruette-Valentin, 1971).
Известен 1 вид — Rh. tarda из Cryptocercus punc-
tulatus.
Род Hoplonympha Light, 1926
Тело сигарообразное. По характеру движения по-
хож на Urinympha. Недостаточно изучен.
Известен 1 вид — Н. natator из Paraneotermes
simplicicornis.
Сем. Staurojoeninidae Grassi, 1917
Гипермастигиды, у которых билатеральная сим-
метрия сочетается с четвертичной радиальной сим-
метрией. Наружная шапочка отсутствует. 4 жгутико-
вые зоны разделены между собой свободными от жгу-
тиков клеточными поверхностями. Жгутиковые зоны
маленькие относительно остальной части тела. В
пределах зоны жгутики располагаются продольными
рядами. Парабазальные ламеллы свободные, не обра-
зуют ростральную трубку. Типичный множественный
парабазальный аппарат. Хорошо развит аксостиляр-
ный аппарат. Ядро на основании рострума. 2 либо 4
атрактофора. На поверхности тела симбиотические
бактерии. Включает 2 рода.
Род Staurojoenina Grassi, 1917
Крупные жгутиконосцы (около 200 мкм). Тело по
форме напоминает амфору, горлышко которой увенча-
но 4 чередующимися жгутиковыми зонами и лопастями
поверхности, свободной от жгутиков. 4 лентовидных ат-
рактофора, из которых лишь 2 длинных контактируют
с ядром, а 2 коротких отходят от парабазальных ламелл.
Известно 3 вида из термитов сем. Calotermitidae и 2 вида
из термитов рода Bifiditermes.
Род Idionympha Cleveland, Hall, Sanders et
Collier, 1934
Известен 1 вид — I. perissa из Cryptocercus punc-
tulatus. Отличительная черта — присутствие зачатка
веретена в интерфазе.
Сем. Trichonymphidae Kent, 1880
Гипермастигиды, у которых билатеральная симмет-
рия сочетается с радиальной. Передний конец тела
имеет наружную шапочку. Жгутиковая зона занимает
от V3 до 2/3 тела, состоит из продольных рядов, расхо-
дящихся радиально из-под наружной шапочки. Ряды
жгутиков прерываются круговой щелью, разделяющей
тело на рострум и построструм. На уровне круговой
щели наблюдается удвоение числа жгутиковых рядов
построструма. Эктоплазма имеет дольчатое строение,
ряды жгутиков проходят в глубоких бороздках между
эктоплазматическими гребнями. Парабазальные ла-
меллы образуют ростральную трубку конической фор-
мы. Типичный множественный парабазальный аппарат.
Аксостилей нет. В интерфазе 1 атрактофор в просвете
ростральной трубки. Ядро хромосомного типа.
Включает 2 рода.
390
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Trichonympha Leidy, 1877
(рис. 324—326)
Относительно крупные гипермастигиды от 45
(Т. lutea из Cryptocercus relictus) до 360 мкм в
длину (Г. collaris из Zootermopsis angusticollis).
Форма тела в основном удлиненная, веретеновидная,
но встречается и грушевидная. У трихонимф, насе-
ляющих кишечник тараканов рода Cryptocercus, яд-
ро заключено в ядерный мешок. Известно более 30 видов
из тараканов рода Cryptocercus и 3 семейств низших
термитов — Calotermitidae, Hodotermitidae, Rhinoter-
mitidae.
Род Protrichonympha Saleem, 1952
Мало изучен. Известен 1 вид — Р. pristina из Аг-
chotermopsis wronghtoni.
Сем. Eucomonymphidae Cleveland,
Hall, Sanders et Collier, 1934
Гипермастигиды, у которых билатеральная сим-
метрия сочетается с радиальной. Передний конец
тела прикрыт шапочкой. Жгутики покрывают все
тело. В роструме жгутики располагаются продольны-
ми рядами, в построструме — меридиональными ря-
дами со слабым леотропным закручиванием. Число
рядов в роструме и построструме одинаково. Эктоп-
лазма рострума имеет дольчатое строение. Параба-
зальные ламеллы образуют ростральную трубку. Па-
рабазальные филаменты идут в построструме под
каждым жгутиковым рядом, к ним в цитоплазме при-
легают диктиосомы. Аксостили свободные, иногда
собраны в пучок. Атрактофоры появляются только
во время митоза.
Включает 2рода.
Род Eucomonympha Cleveland, Hall, Sanders
et Collier, 1934 (рис. 328, вкл.; 329)
Жгутиконосцы почти округлой формы тела; около
100 мкм. Ростральная трубка в виде кувшина. Ядро
лежит при основании рострума. Существуют в 2 мор-
фоформах: свободноживущие округлые формы в про-
свете кишечника и удлиненные формы, прикрепляю-
щиеся к хитиновой выстилке кишки.
Известно 2 вида из тараканов рода Cryptocercus.
Род Pseudotrichonympha Grassi
et Fpa, 1911
Форма тела вытянутая. Крупные жгутиконосцы —
до 500 мкм. Ядро лежит в середине тела.
Известно более 15 видов из низших термитов сем.
Rhinotermitidae.
Сем. Teratonymphidae Koidzumi,
1921
Крупные гипермастигиды, около 300 мкм, соче-
тающие в своей организации билатеральную симмет-
рию с радиальной. Все тело покрыто жгутиками. В
роструме жгутиковые ряды идут продольно; в пост-
роструме жгутики располагаются концентрически-
ми кругами, опоясывающими все тело. Ростральная
трубка, парабазальный аппарат как у Trichonympha.
Многочисленные аксостили. В интерфазе 1 атракто-
фор в просвете ростральной трубки.
Включает 1 род.
Род Teratonympha Koidzumi, 1921
(рис. 327)
Известен 1 вид — Т. mirabilis из термитов рода
Reticulitermes.
Сем. Spirotrichosomatidae Hollande
et Carruette-Valentin, 1971
Гипермастигиды, у которых билатеральная сим-
метрия сочетается с радиальной. Наружная шапочка
присутствует. В роструме 2 жгутиковые ленты рас-
полагаются бок о бок, почти параллельно друг другу,
в построструме они идут как леотропные спирали,
охватывающие тело жгутиконосца. Число оборотов
жгутиковых лент в построструме значительно варь-
ирует. Между рострумом и построструмом существу-
ет короткая безжгутиковая зона. В некоторых родах
присутствуют жгутиковые ленты 2 типов — первич-
ные, о которых сказано выше, и вторичные, которые
также несут жгутики, но берут свое начало от пер-
вичных. Как в первичных, так и во вторичных лен-
тах жгутики объединены в короткие ряды, распола-
гающиеся поперек ленты. Во многих родах кине-
тосомы жгутиков одного ряда сливаются в общий
пучок (fasciculus), который идет в толще эктоплаз-
мы, и лишь при выходе на поверхность тела жгутики
вновь становятся свободными. Парабазальные ламел-
лы образуют ростральную трубку. Парабазальный ап-
парат в виде парабазальных филаментов, подстилаю-
щих жгутиковые ленты в построструме, и диктио-
сом, следующих по ходу спиралей жгутиковых лент.
Аксостили тонкие, длинные и могут быть свободны-
ми либо группируются в пучок. Атрактофоры в виде
тонких длинных нитей, иногда с атрактосферами.
Ядро обычно при основании рострума и заключено
в ядерный мешок; при делении нередко мигрирует в
задний конец клетки. Первичные жгутиковые ленты
во время деления раскручиваются, а вторичные лен-
ты и их жгутики исчезают.
Включает 6 родов, паразитирующих в тараканах
рода Cryptocercus и термитах рода Stolotermes.
ТИП POLYMASTIGOTA. КЛАСС PARABASALEA, ОТРЯД HYPERMASTIGIDA
391
Род Apospironympha Cleveland et Day, 1958
2 типа жгутиковых лент. Многочисленные вто-
ричные жгутиковые ленты возникают из ростраль-
ных либо передних постростральных частей первич-
ных жгутиковых лент и идут до конца тела. Они
слабоспиральные.
Известно 4 вида из термита Stolotermes africanus.
Род Bispironympha Bobyleva, 1969 (рис. 330)
Только первичные жгутиковые ленты, которые в
построструме делают всего 1 оборот по леотропной
спирали. Жгутиковые спирали идут до конца тела.
Ядро при основании рострума.
Известен 1 вид — В. unica — из таракана Crypto-
cercus relictus.
Род Colospironympha Cleveland et Day, 1958
2 типа жгутиковых лент. Как у Apospironympha,
первичные жгутиковые ленты располагаются в ростру-
ме и передней части построструма, где делают не-
сколько оборотов по леотройной спирали. Вторичные
жгутиковые ленты берут свое начало от первичных, но в
отличие от Apospironympha жгутики, возникающие в
них, располагаются через определенные интервалы.
Известно 3 вида из термита Stolotermes africanus.
Род Leptospironympha Cleveland, Hall, Sanders
et Collier, 1934 (рис. 331)
Имеются только первичные жгутиковые ленты.
Форма тела удлиненная, расширяющаяся кзади.
Жгутиковые ленты делают в построструме от 2 до
10 оборотов. Ядро при основании рострума. Извест-
но более 10 видов из тараканов рода Cryptocercus и
термитов рода Stolotermes.
Род Macrospironympha Cleveland, Hall,
Sanders et Collier, 1934
Крупные гипермастигиды. Форма тела почти ша-
ровидная. Имеются только первичные жгутиковые
ленты. Очень короткий рострум. 2 широкие (3—
5 мкм) первичные жгутиковые ленты, делающие 5—
6 оборотов в построструме, подстилаются такими же
по ширине внутренними лентами. Во время деления
ядро мигрирует кзади.
Известен 1 вид — М. xylopletha — из таракана
Cryptocercus punctulatus.
Род Spirotrichosoma Sutherland, 1933
(рис. 332)
Форма тела удлиненная, расширяющаяся кзади. 2
типа жгутиковых лент. Первичные ленты располага-
ются в роструме и передней части построструма, где де-
лают несколько оборотов по леотропной спирали. На
остальной части тела располагаются вторичные
ленты, которые являются продолжением первичных,
но более тонкие; иногда ветвистые, несут много fasci-
culi.
Известно более 10 видов из термитов рода Stolo-
termes.
Сем. Spirotrichonymphidae Grassi, 1917
Жгутиконосцы, сочетающие черты билатеральной
и радиальной симметрии. На переднем конце тела
имеется гиалиновая шапочка. Жгутиковые ленты на-
чинаются на вершине рострума и идут левозакручен-
ными спиралями до конца тела. Передний конец тела
несколько удлинен, и есть ростральный район тела.
Парабазальные ламеллы подстилают жгутиковые лен-
ты. Парабазальный аппарат в виде диктиосом по ходу
жгутиковых лент. Аксостиль трубчатый или состоит
из длийных фибрилл. Ядро прикрепляется к ростру-
му посредством ядерного мешка. Атрактофоры в
виде тонких, длинных нитей, которые следуют за
жгутиковыми лентами, иногда на значительном рас-
стоянии. Во время деления идет процесс раскручива-
ния жгутиковых лент.
Населяют кишечник низших термитов родов Reti-
culitermes, Anacanthotermes, Neotermes, Coptotermes,
Scheadorhinotermes, Leucotermes, Porotermes.
Известно 3 рода.
Род Spirotrichonympha Grassi et Foa, 1911
(рис. 333)
Средние и крупные жгутиконосцы. Форма тела
веретеновидная. В роструме жгутиковые ленты рас-
положены очень тесно и идут в глубине цитоплазмы.
Состоит из 2 подродов — Spirotrichonympha Grassi,
1911 и Spironympha Grassi, 1911. Для первого подро-
да характерен трубчатый аксостиль, для второго —
аксостиль из длинных фибрилл.
В подроде Spirotrichonympha известно более 30
видов, в подроде Spironympha — 5.
Род Spirotrichonymphella Grassi,
1911
Известно 2 вида. Мало изучен.
Сем. Holomastigotoididae Janicki,
1915
Крупные и средние гипермастигиды, обладающие
билатеральной и радиальной симметрией. Жгутико-
вые ленты (их число варьирует от 2 до 60) начинают-
ся на вершине тела и идут до конца тела в виде пра-
возакрученных спиралей. Нет рострального района
тела. Каждая жгутиковая лента состоит из 1 ряда ки-
нетосом и фиброзной ленты, прилегающей к базаль-
ным телам снаружи. Парабазальные пластинки рас-
392
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
полагаются под жгутиковыми лентами в передней
трети тела. Ядро находится в верхней части тела. Ат-
рактофоры являются придатками парабазальных ла-
мелл. Дистальные концы атрактофоров в интерфа-
зе соединены зачатком веретена. «Привилегирован-
ными» кинетосомами являются первые кинетосомы
каждого ряда. Парабазальный аппарат в виде дик-
тиосом по ходу спиралей. Аксостили связаны с каждой
жгутиковой лентой. Населяют кишечник низших тер-
митов семейств Psammotermitidae и Hodotermitidae.
Включает 3 рода.
Род Holomastigotoides Grassi et Foa, 1911
Крупные жгутиконосцы. Форма тела коническая.
Число лент варьирует от 2 до 60. Аксостиль в виде тон-
ких длинных и свободных нитей. Я. tusitala изучен
электронно-микроскопически (Lingle, Salisbury, 1995).
Известно около 40 видов из термитов родов Prorhi-
notermes, Coptotermes, Heterotermes, Psammotermes.
Род Holomastigotes Grassi, 1892
Известно 3 вида из термитов родов Hodotermes и Re-
ticulitermes из которых наиболее изучен Я. elongatum.
Род Rostronympha Duboscq, GrassG et Rose,
1937
Медузоидная форма тела. Отличительная черта —
наличие хоботка на переднем конце тела. Хоботок
сократимый, легко отпадающий.
Известен 1 вид — R. magna из Anacanthotermes
ochraceus.
Кливлендом (Cleveland, 1966а, 1966b) была опи-
сана фауна кишечника австралийского термита Mas-
totermes darwiniensis. Это 2 вида рода Deltotricho-
nympha Cleveland, 1966 — D. operculata и D. па-
па — и 1 вид рода Koruga Cleveland, 1966 — К. bonita.
Эти морфологически близкие виды позднее были ис-
следованы под электронным микроскопом (Tamm,
Tamm, 1973; Hollande, 1986). Оказалось, что органи-
зация этих гипермастигид сочетает в себе черты как
Trichonymphina (имеют рострум, целиком покрытый
жгутиками, 2 интерфазных атрактофора фибрилляр-
ной структуры), так и Lophomonadina (нет ростраль-
ной трубки, во время деления ростральные струк-
туры разрушаются и вновь образуются в дочерних
клетках). Авторы рассматривают жгутиконосцев Del-
totrichonympha и Koruga как связующее звено меж-
ду двумя эволюционными линиями гипермастигид —
Lophomonadina и Trichonymphina. Вероятно, сле-
дует объединить эти роды в отдельное сем. Delto-
trichonymphidae и создать самостоятельный подот-
ряд.
Hypermastigida incertae
sedis
1. Cyclojoenia australis из Stolotermes ruficeps среди Lophomo-
nadina.
2. Виды рода Holomastigotes Grassi, 1911 (6 видов) из терми-
тов родов Anacanthotermes и Reticulitermes.
3. Micromastigotes grassei из термитов рода Postelectrotermes.
THE PHYLUM POLYMASTIGOTA BUTSCHLI, 1884
SUMMARY
The main features characteristic of the phylum are the following:
all species of Polymastigota are heterotrophic, colourless, mainly com-
mensals and parasitic flagellates of various internal organs of animals
and man. The specific unit of kinetic apparatus is mastigont or 2—4
kinetosomes with complex flagella roots, characteristic of each phy-
lum. In all classes the karyomastigont is present and the polymerisati-
on of kinetid takes place. Mitochondria are absent in all species. Many
species have hydrogenosomes.
The phylum includes:
Class Diplomonadea Wenyon, 1926
Order Retortamonadida Grass6, 1952
Order Diplomonadida Wenyon, 1926
Class Oxymonadea Grasse, 1952
Class Parabasalea Honigberg, 1973
Order Trichomonadida Kirby, 1947
Order Hypermastigida Grassi et Fod, 1911
THE CLASS DIPLOMONADEA WENYON, 1926
Diplomonads are rather small (about 20 pm long) free-living and
symbiotic flagellates. Many of them possess axial symmetry, owing to
the binary complex of cell organelles. Most of the species have 2
karyomastigonts located in the front part of the cell. The karyomasti-
gont includes 1—4 flagella, their kinetosomes, microtubules, lamellae,
fibrillar components, and one nucleus. Mitochondria, Golgi apparatus,
and axostyl are absent.
Food is ingested by phagocytosis and pinocytosis over cell surface
or through a special structure — cytostome. The life cycle is simple,
no sexual phenomena have been observed. Trophozoites multiply by
binary fission. In certain species the resting cysts are formed.
Only few species are pathogenic, and all of them belong to the
order Diplomonadida. The great majority of the diplomonads are har-
mless commensals. Free-living diplomonads inhabit freshwater reser-
voirs with anaerobic conditions.
THE ORDER RETORTAMONADIDA GRASSE, 1952
This order includes one family Retortamonadidae Wenrich, 1932
with 2 genera: Retortamonas and Chilomastix. The genera are repre-
sented by commensals of the digestive tract of man, vertebrate and
invertebrate animals. The trophozoits possess 2—4 flagella, one ka-
ryomastigont, a cytostome located ventrally. The recurrent flagellum
passes through the cytostomal groove. The life cycle comprises two
stages: the motile trophozoite, multiplying in the host intestine by
binary fission, and a cyst covered by thick wall, and serves to disse-
minate the species. About 30 species of Retortomonadida are known
today.
THE ORDER DIPLOMONADIDA WENYON, 1926
The order includes two families: Enteromonadidae and Hexamiti-
dae. There are three genera of enteromonads: Caviomonas with 2 fla-
gella, Trimitus with 3 flagella, and Enteromonas with 4 flagella. The
family Hexamitidae is divided into 2 subfamilies: Hexamitinae Kent,
1880 and Giardininae Kulda, Nohynkova, 1978.
Hexamitinae: Hexamita, Spironucleus, Trepomonas, Trigonomo-
nas.
ТИП POLYMASTIGOTA. SUMMARY
393
I Giardininae: Giardia, Octomitus.
The genera Trepomonas, Trigonomonas include free-living flagel-
lates. The genus Hexamita consists of free-living and parasitic species.
Two genera — Giardia and Octomitus are exclusively parasitic. Giar-
dia intestinalis is a parasite of man, Hexamita meleagridis — a para-
site of birds, fish and oysters, Spironucleus meleagridis is a pathoge-
nic to the turkey.
Trophozoites of the family Hexamitidae are characterized by a
2-fold rotational symmetry. They have 8 flagella, 2 karyomastigonts
and a double set of cytoplasmic organelles. Each mastigont has 4 fla-
gella: one of them is recurrent.
Flagellates of the genus Giardia differ from other diplomonads
with the adhesive disk. The organelle is situated on the flattened ven-
tral surface of the body, and represents the structure of attachment.
Species of Diplomonadida have cysts. The cysts are oval bodies
covered with rather thick wall. Ripe cysts have 4 nuclei, intercytoplas-
mic axonemes of flagella, polysaccharides in the cytoplasm. Giardia
intestinalis is transmitted by cysts, which are resistant to freezing, to
hot temperature, to water and other environmental;factors.
Trophozoites multiply by longitudinal binary fissuon. Nuclear di-
vision is a semiopen mitosis. Binary nuclear division occurs during
the early stages of encystment. Plasmotomy in Giardia follows excyst-
ment of the doubled cell.
THE CLASS OXYMONADEA GRASSE, 1952
Oxymonadids are parasitic flagellates inhabiting the hindgut of
the lower termites and cockroaches of the genus Cryptocercus (except
for several species of Monocercomonoides and Polymastix, which live
in other insect groups as well as in vertebrates). Together with Hyper-
mastigida, they compose a large community of microorganisms that
play a important biological role in the life of their xylophagous hosts,
as they take an active part in splitting of cellulose. Close symbiotic
interrelations are formed between these microorganisms and their
hosts that it is a form of mutualism. Oxymonadids occur in the intes-
tinal lumen either in the free state or attached to the chitin lining,
using a special attaching organelle; The life cycle in some Oxymona-
dida includes only the free, stage, while in others, an alternation of the
free and attached stages is observed. The organization of Oxymonadi-
da is based on the karyomastigont with two pairs of flagella intercon-
nected with a particular structure, preaxostyle, from which the axosty-
le emerges. Oxymonadida are predominantly mononuclear organisms,
however, among them there are genera with both temporary and per-
manent polykaryomastigont organization.
This order includes three families: Polymastigidae, Oxymonadi-
dae, and Pyrsonymphidae.
Flagellar apparatus. The ventral kinetosomes are oriented at
an acute angle to each other, while dorsal ones have an orthogonal
orientation to each other. The flagella are of the acronematic type. The
transitional flagellar zone is short and presented by a transverse plate
with no additional structures. There are three types of flagellar orien-
tation: all flagella are free (Polymastix, Oxymonas)', one or two flagel-
las are bent backwards and are connected to the cell membrane at
some distance (Monocercomonoides)', all flagella are recurrent and run
in deep grooves of the plasma membrane (Pyrsonympha).
Preaxostyle and pelta-axostyle complex. The preaxo-
style is a paracrystalline structure connecting the ventral and dorsal
kinetosome pairs and serves as the MTOC. The axostyle is the main
cytoskeleton element and passes along the longitudinal body axis. It is
a multilayered structure composed of strictly oriented microtubule
rows. The anterior microtubule rows are embedded into the preaxo-
style. Individual microtubules in each row are interconnected with
bridges; less close connections also exist between the rows. In many
oxymonadids the axostyle is able to contract and works as an active
locomotor organelle. The axostyle contraction is based, on the sliding
of microtubular rows along each other due to dinein arms activity.
However, the fine mechanism of this process is different in different
species. In many oxymonadids there are additional microtubular struc-
tures, such as paraxostyle and pseudoaxostyle (Oxymonas, Pyrsonym-
pha, Sauromonas). Pelta and funis (a band lining the recurrent flagel-
lum in Monocercomonoides) also have the microtubular organization.
Nucleus and mitosis. The nucleus is usually vesicular, with
one or several nucleoli. Its structure and division are studied predomi-
nantly at the light microscopical level. Mitosis is characterized by
complete retention of the nuclear envelope throughout the mitosis and
by an intranuclear position fo the entire mitotic apparatus, both the
spindle and the chromosomes (closed intranuclear pleuromitosis). The
ultrastructural study of Pyrsonympha shows that the centriolar plate
that is located on the inner side of the nuclear envelope represents the
MTOC; the process of formation of two half-spindles that gradually
distance from each other to form the central spindle and chromosomal
filaments.
Cytoplasm. Oxymonadids have no morphologically distinguis-
hable mitochondria. Dense microbodies similar to hydrogenosomes
have been described in Pyrsonympha. However, their cytochemical
nature and relation to enzymes of the anaerobic energy complex is not
proved.
The Oxymonadida have no typical Golgi apparatus. However, the
presence of numerous small electron-opaque vacuoles has been noti-
ced, which, according to the current concepts, may be considered as
individual elements functioning in the cell as the Golgi apparatus.
Nutrition. The majority of oxymonads engulf wood particles or
bacteria, which are abundant in the intestine of their xylophagous
hosts. The nutrition occurs by means of phagocytosis. The small oxy-
monadids have osmotrophic nutrition by pinocytosis.
Symbionts. The body surface of oxymonadids is covered by
symbiotic bacteria (predominantly by spirochetes). As shown by
means of electron microscope, specialized structures are formed in the
site of attachment of the prokaryotic symbiont to the cell membrane
by both the flagellate and the symbiont. These membrane structures
provide a strong anchoring of the symbiont to the body surface.
Polymerization of karyomastigonts. In the order Oxy-
monadida a tendency has been observed of a transition in the cell
organization from the forms with one karyomastigont to the tempo-
rary polykaryomastigont state (Opisthomitus, Oxymonas, Sauromo-
nas) or to the permanent polykaryomastigont organization (Probosci-
diella, Barroella). In a way of polymerization of the karyomasti-
gonts/ the preaxostyle decreases markedly, pelta and paraxostyle are
reduced, however, the axostyle is preserved, and usually, the number
of axostyles corresponds to the number of karyomastigonts in the
cell.
Reproduction. The division in all Oxymonadida is longitudi-
nal. Mitosis can occur both in the free state and at the attachment
stage. In some Oxymonadida, during the division, migration of the
nucleus to the posterior end of the cell and degeneration of the axo-
style have been observed. Plasmotomy can lead to the free or to the
attachment stage in the life cycle.
The oxymonads from the intestine of the lower termites reproduce
exclusively asexually. Oxymonadids parasitizing in the cockroach
Cryptocercus reproduce asexually in the period between moults of the
host as well. However, in the moulting period of Cryptocercus, Oxy-
monadida necessarily have the sexual process.
Sexual process. The sexual process (gamete copulation or
autogamy) occurs in the genera Oxymonas, Notila and Saccinobaccu-
lus under the effect of the moult hormone of Cryptocercus. During the
gamete fusion, not only nuclei fuse but also axostyles. The nuclei of
Oxymonadida enter one-step meiosis accompanied by a partial or
complete reorganization of all cell organelles. The meiosis has not
been studied at the ultrastructural level nor with cytophotometry.
Transmission. The mode of transmission of oxymonads from\
host to host is similar to transmission of hypermastigids. Flagellates
of lower termites are transmitted via proctodeal feeding typical of
termites. The nymphs of various age lick off anal excreta containing
mummified flagellates. Transmission of oxymonads from Cryptocer-
cus is strictly connected with the sexual process. Nymphs of Crypto-
cercus become infected with flagellates through eating cysts and
pseudocysts which are formed during the sexual process and shed
from the host intestine.
Evolution. Phylogenetic connections of Oxymonadida with
other flagellates are not clear. The closest to Oxymonadida in their
morphologic cell organization are Retortamonadida and Diplomonadir
da. Among Oxymonadida, the most primitive family undoubtedly is
Polymastigiidae. Two lines can be traced in evolution of the family
Oxymonadidae. Thus, in the subfamily Oxymonadinae, the evolution
was based on the process of polymerization of karyomastigonts, while
in the subfamily Saccinobaculinae, the evolution is associated with
perfection of the axostyle as an active locomotor organelle.
A detailed systematic analysis of families and genera of the order
Oxymonadida is presented.
THE CLASS PARABASALEA HONIGBERG, 1973
Parabasalea are commensals and endoparasitic flagellates of ani-
mals and man. The protists possess a specific parabasal complex of
organelles, which consists of Golgi apparatus (from one to many dic-
tyosomes) and parabasal filaments (from one to many in different
species). The number of flagella varies in different species. The undu-
lating membrane is characteristic of Trichomonadida. The flagellar
394
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
i rootlet system consists of fibrillar elements, which initiate the cyto-
skeletal microtubules of pelta-axostylar complex. The kinetid as a
whole is presented by 4 kinetosomes, which are strongly connected to
the nucleus forming a karyomastigont. Mitochondria, are absent. Hyd-
rogenosomes are the obligatory organelles of the cell.
Sexual process is known in some genera. The multiplication
is agamic by means of binary fission of cells. The nucleus division
is closed pleuromitosis with extranuclear mitotic spindle. During the
division chromosomes attached to the interior membrane of the nucle-
us.
More than 300 species of the class Parabasalea are placed in two
orders: Trichomonadida Kirby, 1947 and Hypermastigida Grassi et
Бой, 1911.
THE ORDER TRICHOMONADIDA KIRBY, 1947
This order includes commensals and endoparasitic flagellates,
which inhabit digestive tract, uro-genital organs of man, some mam-
malians, and other internal organs of many groups of invertebrate and
vertebrate animals. Trichomonads have 4—6 smooth flagella. The re-
current flagellum forms the undulating membrane. The most species
have one karyomastigont and one parabasal apparatus. The complex of
flagellar rootlets consists of small fibrillar bands and a costa. The
axostyl is well developed. The hydrogenosomes are numerous in cy-
toplasm. The nuclear division is typical of Trichomonadea and presen-
ted with closed intranuclear mitosis with, extranuclear spindle.
The life cycles of trichomonads are studied insufficiently. The
most data concern the trophozoites inhabiting the internal organs and
tissues of a host. The existence of cysts has not been proven yet. The
life cycle of Trichomonas vaginalis, the species pathogenic to man, is
studied in detail. The infection of man is induced by trophozoites. The
same way of infection is characteristic of Tritrichomonas foetus,
T. suis, T. caviae, and other parasites of animals.
The order Trichomonadida includes approximately 200 species,
which are distributed everywhere. The order comprises 4 families.
Monocercomonadidae (10 genera) are parasites of fish, amphibi-
ans, reptiles, birds, guinea pigs, man.
Devescovinidae (8 genera) are commensals of the intestine of
termites.
Calonymphidae (5 genera) are commensals of intestine of termi-
tes.
Trichomonadidae (8 genera) are parasites of man and animals.
THE ORDER HYPERMASTIGIDA GRASSI ET FOA, 1911
Hypermastigids are parasitic flagellates inhabiting the intestine of
lower termites and cockroaches of the genus Cryptocercus (with the
exception of Lophomonas that parasitizes in cockroaches Blatta, Blat-
tella, and Periplaneta). This group of organisms achieves the highest
level of cellular organization among all flagellates. Most authors iden-
tify two major evolutionry branches of Hypermastigida: lower (Lo-
phomonadina) and higher (Trichonymphina).
Flagellar apparatus. Numerous flagella (from tens to several
thousands) are located in one zone (Lophomonadina) or several zones
(Trichonymphina). Inside the zone the flagella are arranged in a cer-
tain pattern: a complete or partial circle, one or several plates, longi-
tudinal or spiral rows. The flagella have unusually long kinetosomes
in many species and a complex system of kinetosomal fibrils connec-
ting all kinetosomes to each other. The flagellar zone and flagella-as-
sociated organelles form the mastigont system. Lophomonadina have
one mastigont system, while Trichonymphina have two, or sometimes
four. As this occurs the cell organization shows bilateral symmetry
combined with the radial one.
Among the kinetosomes, a group of «privileged» kinetosomes is
identified; their functional role is evident in cell morphogenesis in the
course of cell division. In Lophomonadina this group was shown to
consist of four kinetosomes located on the right of the rest of the flagel-
lar zone. It produces rootlet structures: the cross-striated fibrils of para-
basal filaments, atractophores and preaxostyle fibrils forming the pelta-
axostyle complex. The rootlet structures in the lower hypermastigids are
similar to those of trichomonads. In the higher Hypermastigida the num-
ber and arrangement of the «privileged» kinetosomes vary significantly:
Unlike Lophomonadina, in Trichonymphina the direct connection of the
«privileged» kinetosomes and rootlet filaments is very short-lived and is
revealed only in dividing cells. Early in the course of morphogenesis,
their connection is split and is masked by an electron-dense cytoplasm
(the morphoplasm). The morphoplasm is thought to act as an informati-
on carrier from the «privileged» kinetosomes to the rootlet structures
during the vegetative part of the life cycle.
Cytoskeleton. In Hypermastigida the cytoskeleton is formed
by two types of structures: parabasal lamellae composed of densely
packed cross-striated fibrils (with periodicity of 42—47 nm) and
microtubules. Each of the cross-striated fibrils is identical, in their size
and structure, to the parabasal fibril of trichomonads. In Lophomona-
dina the cytoskeleton is formed mainly by microtubular structures
such as the thick axostyle running throughout the entire body, and the
pelta; the cross-striated fibrils are poorly developed. In Trichonym-
phina the cytoskeleton consists predominantly of parabasal lamellae
located just beneath the kinetosomes. The microtubular structures take
part in formation of the nuclear sleeve and the external cap.
Parabasal apparatus. Hypermastigida have numerous argen-
tophilic Janicki-type parabasal apparatuses consisting of the parabasal
filament and parabasal body. The parabasal filament is composed of one
or several elementary cross-striated fibrils, which are continuations of
distal ends of the parabasal lamellae. The parabasal body, whose shape
in Hypermastigida varies significantly, according to its ultrastructural
and cytochemical organization, is the Golgi apparatus. The parabasal
filament is always oriented parallel to the pile of Golgi cistemae.
Nuclear and mitotic apparatuses. Hypermastigids have
one nucleus. Data on the nuclear organization are mainly based on
light microscopy; while the ultrastructure of the nucleus has been stu-
died in a limited number of species. One of the observed features of
the nuclear structural organization is a complicated nuclear envelope,
outgrowths of the outer membrane in some lower Hypermastigida
(Lophomonas and Placojoenia). In many trichonymphines, interphase
chromosomes remain morphologically well formed, although somew-
hat looser as compared with mitotic chromosomes. The chromosomes
are fixed with their kinetochores to the inner nuclear envelope memb-
rane. Almost nothing is known about the ultrastructural morphology
of the chromosomes.
Atractophores are organelles characteristic of Hypermastigida and
predominantly have cross-striated structure. During the mitosis they
form and apply the division spindle to the nucleus. They are formed
and act in contact with the «privileged» kinetosomes and are conside-
red as elements of the rootlet system. In some hypermastigids the
atractophores are permanent structures, in others they appear only du-
ring mitosis. The mitosis in Hypermastigida has been studied in detail,
particularly at the ultrastructural level and is classified as a closed
extranuclear pleuromitosis. This type of mitosis is characterized by
1) a complete conservation of the nuclear envelope throughout the
mitosis; 2) an extranuclear position of the entire spindle apparatus,
including the MTOCs, central spindle and chromosomal fibres; only
the chromosomes remain inside the nuclear envelope; 3) the stage of
the chromosome equatorial plate is not pronounced, and the prophase
is directly followed by anaphase.
Hydrogenosomes. Characteristic of the Hypermastigida living
under anaerobic conditions are hydrogenosomes, functional analogues
of mitochondria. In hydrogenosomes, all enzymes responsible for the
anaerobic metabolic pathways have been found. Hydrogenosomes
often are merely scattered in the cytoplasm, occasionally they are lo-
cated among kinetosomes along the flagellar bands.
Nutrition. Hypermastigids eat predominantly small pieces of
wood that are abundant in the hindgut of the xylophage host. The food
particles are captured in the posterior, non-structurized end of the
body. The nutrition process has been shown to be of the mixed type,
with both phagocytosis and active pinocytosis. Processes of defecation
in Hypermastigida are poorly studied yet.
Symbionts. Hypermastigida are characterized by the presence
of various prokaryotic symbionts both on the surface of the cell mem-
brane and in the cytoplasm. However, the character of the symbiotic
relations between flagellates and their bacteria are not known.
Asexual reproduction and transmission. In the period
between moults of the host, Hypermastigida that inhabit its intestine
reproduce only asexually. Both mitosis and cell divisional morphoge-
nesis have been mainly studied in detail at the light microscopy level.
In the moulting period of the host, most of its protistofauna is lost and
then is restored after the moult. Hypermastigid transmission among
the lower termites which have the moult once a month, occurs via the
proctodeal feeding typical of these insects. In the moment of the trans-
mission, the flagellates are found in excretions in mummified state.
The fast increase of the number of the flagellates after the moult of
the host is due to frequent divisions of the flagellates newly acquired
from non-molting nymphs. Unlike the hypermastigids parasitizing in
lower termites, the flagellates of Cryptocercus necessarily have a
sexual process that is intimately connected with transmission of hyper-
mastigids from one host specimen to another. Cysts and pseudocysts
that are formed in the course of the sexual process and are shed into
the surrounding environment are the stages of the transmission of
ТИП POLYMASTIGOTA. ЛИТЕРАТУРА
395
flagellates among nymphs of different ages in the cockroach colony. Unlike
the lower termite larvae, Cryptocercus nymphs moult only once a year.
Sexual process. The sexual process in all Hypermastigida is
initiated by the moult hormone (ecdyzone); however, its start, durati-
on, and ending strongly vary in different Hypermastigida genera. Rat-
her varying morphological patterns have been observed in different
flagellate genera when comparing individual periods (progamic^ euga-
mic, and metagamic) of the sexual process and the ways of its perfor-
mance (copulation of gametes or autogamy). An extremely wide vari-
ability is seen in various Hypermastigida genera with respect to the
haploid or diploid state of the nucleus at the beginning of sexual
process. Finally, the chromosomal reduction can occur by one-step or
two-step meiosis in different genera. It should be noted that the cyto-
logical pattern of the sexual process in Cryptocercus flagellates is
studied in detail only at the light microscopy level, whereas neither
electron microscopic nor cytophotometric data have been presented on
the DNA content in nuclei at different meiosis stages.
Biological role in the life of hosts. Hypermastigida
are obligatory symbionts taking an active part in decomposing the
wood, which is the food of the lower termites and cockroaches of the
genus Cryptocercus. The cellulose activity has been experimentally
proven to emanate from the flagellates. However, some biochemical
aspects of physiology of the symbiosis of the flagellates and their
insect xylophagous hosts remain unclear.
Evolution. According to the current concept, ancestors of Hy-
permastigida were trichomonad-like flagellates assumed to live in the
gut of the ancient cockroaches from which the termites evolved. Pro-
bably, the predominant part of evolution of Hypermastigida occurred
in the cockroaches prior to divergence of the termites from them. In
Hypermastigida, two parallel evolutionary lines, Lophomonadina and
Trichonymphina, appeared from their common trichomonad-like an-
cestor. However, there are no Trichomonades in the present-day Cryp-
tocercus. Subsequently, the evolution of Lophomonadina occurred
predominantly in the lower termites. In the present-day Cryptocercus,
only one genus, Prolophomonas, is known. The Trichonymphina evol-
ved in both the lower termites and cockroaches. Common to both host
groups are genera Trichonympha and Leptospironympha. The sugges-
tions on the evolutionary interrelations of Hypermastigida could be
clarified by application of the molecular-biological analysis of DNA
or RNA.
A detailed systematic analysis of families and genera of the order
Hypermastigida is presented.
ЛИТЕРАТУРА21
Бей-Биенко Г. Я. Насекомые, таракановые И Фауна СССР.
Нов. сер. М.; Л.: Изд-во АН СССР, 1950. Т. 40.
(Бобылёва Н. Н.) Bobyleva N. N. Morphology and evolution of
intestinal parasitic flagellates of the far-eastern roach Cryptocercus
relictus // Acta Protozool. 1975. Vol. 14. P. 109—160.
Гинецианская T. А., Добровольский А. А. Частная паразито-
логия. Паразитические простейшие и плоские черви. М., 1978.
Догель В. А. Исследования над паразитическими простейшими
из кишечника термитов. 1. Tetramitidae // Зоол. вести. 1916. ТЛ,
вып. 1. С. 1—54.
(Догель В. A.) Dogiel V. A. Untersuchungen an parasitischen Рго-
tozoen aus dem Darmkanal der Termites. III. Trichonymphidae //
Arch. rus. Protist. Obschch. 1922. Bd 1. S. 172—234.
Догель В. А. Общая протистология. M., 1951.
Догель В. А. Протозойные симбионты термитов и их общее
биологическое значение в жизни хозяев И Инфекционные и прото-
зойные болезни полезных и вредных насекомых. М.: Сельхозпгз,
1956.
Догель В. А. Зоология беспозвоночных. М., 1981.
Дубинин В. Б. Новый вид Trichomonas из кишечника средне-
азиатского дикобраза II Зоол. журн. 1938. Т. 17, № 1. С. 187—
190.
Жуков Б. Ф., Мыльников А. П. Культивирование свободножи-
вущих бесцветных жгутиконосцев из сооружений биологической
очистки# Простейшие активного ила. 1983. Т.8. С. 142—-152.
Карапетян А. Е. Методика культивирования лямблий И Цито-
логия. 1960. Т. 2, № 3. С. 379—384.
Карапетян А. Е. О патогенности Lamblia intestinalis для белых
мышей И Мед. паразитол. и паразитарн; болезни. 1962. Т. 31, № 5.
С. 697—701.
Падченко И. К. О цикле развития трихомонад # Вести, зоол.
1975. Т. 5, № 5. С. 52—56.
Пак С. М. Трихомонады кишечника краснохвостой песчан-
ки И Тр. Ин-та зоол. АН Каз. ССР. 1960. Т. 12. С. 82—85,
Райков И. Б. Ядро простейших. Происхождение и эволюция.
Л., 1978.
(Райков И. Б.) Raikov I. В. The protozoon nucleus. Morphology
and evolution # Wien; New York: Springer, 1982.
(Райков И. Б.) Raikov I. В. The diversity of forms of mitosis in
Protozoa: a comparative review # Eur. J. Protistol. 1994. Vol. 30.
P. 253—269.
Селюкайте 3. Б. Трихомонады грызунов: морфология, жизнен-
ный цикл, некоторые биологические особенности И Паразитоло-
гия. 1977. Т. 11, № 1. С. 65—71.
Серавин Л. Н. Эукариоты, лишенные важнейших клеточных
органелл (жгутиков, аппарата Гольджи, митохондрий) и главная
задача органеллологии И Цитология. 1992. Т. 34. С, 3—33.
Соловьёв М. М. Изучение процесса деления Lamblia duodena-
lis в культуре И Мед. паразитол. и паразитарн. болезни. 1963. Т. 32,
№1. С. 96—101.
21 Литература к разделу «Тип Polymastigota».
Соловьёв М. М., Ченцов Ю. С. Электронно-микроскопичес-
кое изучение Lamblia muris в связи с особенностями экологии па-
разита И Мет. паразитол. и паразитарн. болезни. 1966. Т. 35, № 6.
С. 667—672.
Соловьёв М. М., Ченцов Ю. С. Ультраструктура цист Lamblia
muris И Паразитология. 1970. Т. 4, № 6. С. 510—514.
Соловьёв М. М., Ченцов Ю. С. Изучение трофозоитов Lamb-
lia intestinalis из культуры методом растровой электронной мик-
роскопии # Мед. паразитол. и паразитарн. болезни. 1976. Т. 45,
№6. С. 650—654.
Терас Ю.Х. Изменчивость осарсол-чувствительности Tricho-
monas vaginalis. I. Осарсол-резистентность in vitro И Трихомоноз
урогенитального тракта. Таллин. 1963. С. 118—132.
Тимофеев Б. А., Петровский В. В. Трихомонады и трихомоно-
зы сельскохозяйственных животных. М., 1967.
Шарапов М. Е., Соловьёв М. М. Изучение роста Lamblia in-
terstinalis в культуре с целью совершенствования методики культи-
вирования И Мед. паразитол. и паразитарн. болезни. 1976. Т. 45,
№6.0 665—660.
Andrew В. J., Light S. F. Natural and artificial production of so-
called «mitotic flares» in the intestinal flagellates of Termopsis angu-
sticollis II Univ. Calif. (Berkeley) Publ. Zool. 1929. Vol. 31. P.433—
440.
Beams H. W., Sekhon S. S. Further studies on the fine structure
of Lophomonas blattarum with special reference to the so-called
calyx, axial filament and parabasal body # J. Ultrastr. Res. 1969.
Vol. 26. P. 296—315.
Beams H. W., Tahmisian T. N., Anderson E., Wright B. Studies
on the fine structure of Lophomonas blattarum with special reference
to the so-called parabasal apparatus # J. Ultrastr. Res. 1961. Vol. 5.
P. 166—183.
Benchimol M., Kachar B., de Souza W. The structural organiza-
tion of the pathogenic protozoan Tri trichomonas foetus as seen in
replicas of quick frozen, freeze-fractured and deep etched cells # Biol.
Cell. 1993. Vol. 77. P. 289—295.
Benchimol M., Johnson P. J., de Souza W. Morphogenesis of
the hydrogenosome, an ultrastructural study # Biol. Cell. 1996.
Vol. 87, N 3. P. 197—205.
Bishop A. Observations upon Embadomonas intestinalis in cultu-
re # Parasitology. 1934. Vol. 26, N 1. P. 17—25.
Bloodgood R. A., Fitzharris T. P. Specific associations of proka-
ryotes with symbiotic flagellate protozoa from the hindgut of termite
Reticulitermes and the wood-eating roach Cryptocercus # Cytobios.
1976. Vol. 17. P. 103—122.
Bloodgood R. A., Miller K. R., Fitzharris T. P., McIntosh J. R.
The ultrastructure of Pyrsonympha and its associated microorga-
nisms # J. Morphol. 1974. Vol. 143. P. 77—105.
Boeck W. C., Drobohlav J. The cultivation of Entamoeba histo-
lytica II Amer. J. Hyg. 1925. Vol 25, № 3. P. 371—407.
Brugerolle G. Etude ultrastructurale du trophozoite rt du kyste
chez le geme Chilomastix Alexeieff, 1910 (Zoomastigophorea, Retor-
/
396
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
tomonadina Grassi, 1952) И J. Protozool. 1973a. Vol. 20, N 4.
P. 574—585.
Brugerolle G. Sur Г existence de vrais kystes chez les Trichomo-
nadines intestinales. Ultrastructure des kystes de Trichomitus bat-
rachorum Perty, 1852; Trichomitus sanguisugae Alexeieff, 1911 et
Monocercomonas tipulae Mackinnen, 1910 // C. R. Acad. Sci. Paris.
Ser. D. 1973b. T. 277. P. 2193—2196.
Brugerolle G. А Г etude cytologique et phyletique des Diplizoai-
res (Zoomastigophorea, Diplozoa Dangeard, 1910) // Protistologica.
1974. T. 10, N 1. P. 83—90.
Brugerolle G. Contribution a Г etude cytologique et phyletique
des DipTozoaires (Zoomastigophorea, Doplozoa Dangeard, 1910).
V. Nouvelle interpretation de 1’organization cellulaire de Giardia //
Protistologica. 1975a. T. 11, N 1. P. 99—109.
Brugerolle G. Contribution a 1’ltude cytologique et phyletique
des Diplozoaires (Zoomastigophorea, Doplozoa Dangeard, 1910).
Vol. 1. Caracteres generaux des Diplozoaires // Protistologica. 1975b.
T. 11, N 1. P. Ill—118.
Brugerolle G. Etude de la cryptopleuromitose et la morphogenese
de division chez Trichomonas vaginalis et chez plusieurs gemes de
Trichomonas primitives // Protistologica. 1975b. T. 11, N 4. P. 457—468.
Brugerolle G. Ultrastructure du gerne Retortomonas Grassi, 1879
(Zoomastigophorea, Retortomonadida Wenrich, 1931) // Protistologi-
ca. 1977. T. 13, N 2. P. 233—240.
Brugerolle G. Etude ultrastructurale du flagelle parasite Polymas-
tix melolonthae (Oxymonadida) // Protistologica. 1981. Vol. 17.
P. 139—145.
Brugerolle G. Structural diversity of Trichomonads at the basis
for systematic and evolutionary consideration // Acta Univ. Carolina.
Biol. 1986. Vol. 30. P. 199—200.
Brugerolle G., Joyon L. Sur la structure et la position systemati-
que du genre Monocercomonoides (Travis, 1932) 7/ Protistologica.
1973. Vol. 9. P. 71—80.
Brugerolle G., Kbnig H. Ultrastructure of organization of the cy-
tosceleton in Oxymonas an intestinal flagellate of termites // J. Euk.
Microbiol. 1997. Vol. 44. P. 305—313.
Brugerolle G., Joyon L., Oktem N. Contribution a 1’ltude cyto-
logique et phyletique des Diplozoaires (Zoomastigophorea, Diplozoa
Dangeard, 1910). 1. Etude ultrastructurale de gerne Trepomonas (Du-
jardin, 1841) // Protistologica. 1973a. T. 9, N 3. P. 339—348.
Brugerolle G., Joyon L., Oktem N. Contribution a 1’ltude cyto-
logique des Diplozoaires (Zoomastigophorea, Diplozoa Dangeard,
1910. II. Etude ultrastructurale du gerne Spironucleus Lavier, 1936 //
Protistologica. 1973b. T. 9, N 4. P. 495—502.
Brugerolle G., Joyon L., Oktem N. Contribution a 1’ltude cyto-
logique et phyletique des Diplozoaires (Zoomastigophorea, Diplozoa
Dangeard, 1910). IV. Etude ultrastructurale du gerne Octomitus (Pro-
wazek, 1904) // Protistologica. 1974. T. 10, N 4. P. 457—463.
Camp R. R., Mattern C. F. T., Honigberg Bi M. A study of Di-
entamoeda fragilis Jepps, Dobell. I. Electronmicroscopic observations
of the binucleate stages; II. Taxonomic position and revision of the
genus 7/ J. Protozool. 1974. Vol. 21, N 1. P. 69—82.
Cleveland L. R. Symbiosis among animals with special reference
to termites and their intestinal flagellates // Quart. Rev. Biol. 1926.
Vol. 1. P. 51—60.
Cleveland L. R. Morphology and mitosis of Teranympha 7/ Arch.
Protistenk. 1938. Vol. 91. P. 442—451.
Cleveland L. R. Brief accounts of the sexual cycles of the flagel-
lates of Cryptocercus /7 J. Protozool. 1956. Vol. 3. P. 161—180.
Cleveland L. R. Correlation between the molting period of Cryp-
tocercus and sexuality in its Protozoa U J. Protozool. 1957. Vol. 4.
P. 168—175.
Cleveland L. R. Movement of chromosomes in Spirotrichonym-
pha to centrioles instead of the ends of central spindle 7/ J. Protozool.
1958. Vol. 5. P. 63—68.
Cleveland L. R. The centrioles of Trichonympha from termites
and their functions in reproduction 7/ J. Protozool. 1960. Vol. 7.
P. 326—341.
Cleveland L. R. Fertilization in Trichonympha from termites 7/
Arch. Protistenk. 1965a. Vol. 108. P. 1—5.
Cleveland L. R. Fertilization in Pseudotrichonympha 7/ Arch.
Protistenk. 1965b. Vol. 108. P. 6—7.
Cleveland L. R. General features of the flagellate and ameboid
stuges of Deltotrichonympha operculata and D. nana, sp. nov. 7/ Arch.
Protistenk. 1966a. Vol. 109. P. 1—7.
Cleveland L. R. Reproduction in Deltotrichonympha 7/ Arch.
Protistenk. 1966b. Vol. 109. P. 8—14.
Cleveland L. R. General features and reproduction in Koruga bo-
nita, gen. et sp. nov. 7/ Arch. Protistenk. 1966c. Vol. 109. P. 18—23.
Cleveland L. R., Day M. Spirotrichonymphidae of Stolotermes /7
Arch. Protistenk. 1958. Vol. 103. P. 1—53.
Cleveland L. R., Hall S. K., Sanders E. P., Collier J. The wood-
feeding roach Cryptocercus, its protozoa and die symbiosis between
protozoa and roach // Mem. amer. Acad. Arts, Sci. 1934. Vol. 17.
P. 185—342.
Cochrane S., Smith H., Buhse H., Scammel J. Structure of the
attached stage of Pyrsonympha in termite Reticulitermes flavipes Kol-
lar /7 Protistologica. 1979. Vol. 15. P. 259—270.
Cross J.B. The flagellates subfamily Oxymonadinae 7/ Univ.
Calif. Publ. Zool. 1946. Vol. 53. P. 67—162.
da Silva N. S., Dias Filho В. P., de Souza W. Identification and
localization of an adhesin on the surface of Tritrichomonas foetus //
Parasitol. Res. 1999. Vol. 85. P. 984—992.
Diamond L. S. The establishment of various trichomonads of ani-
mals and man in axenic cultures 7/ J. Parasitol. 1975. Vol. 43, N 3.
P. 488—490.
Dobell О. C., Leidlaw P. P. On the cultivation of Entamoeba his-
tolytica and some other entozoic amoebae /7 Parasitology. 1926.
Vol. 18, N 2. P. 283—318.
Doflein F. Lehrbuch der Protozoenkunde. Jena, 1953.
Gibbons I. R., Grimstone A. V. On flagellar structure in certain
flagellates 7/ J. Biophys., Biochem. Cytol. 1960. Vol. 7. P. 697—716.
Grassi P. -P. Traitl de Zoologie. Anatomie, systematique, biolo-
gie. T. I. Phylogenie Protozoaires: generalites Flagelles. Fasc. 1. Paris:
Masson, 1952.
Grassi P.-Р., Hollande A. Les affmites et 1’evolution des Tricho-
nymphines 7/ C. R. Acad. Sci., Paris. 1942. Vol. 215. P. 35—37.
Hilber C.P., Hammond D.M., Caskey F.H., Johnson A.E.,
Fizgerald P. R. The morphology and incidence of trichomonas of
swine, Trichomonas suis (Gruby, Delafond), Tritrichomonas rotunda
n. sp. and Trichomonas buttreyi n. sp. 7/ J. Protozool. 1960. Vol. 7,
N 2. P. 159—171.
Hollande A. Morphologie, ultrastructure et affmites de Deltotri-
chonympha operculata Sutherland (1953) symbionte du termite austra-
lien Mastotermes darwiniensis Frogatt. 7/ Protistologica. 1986. Vol. 22.
P. 415—439.
Hollande A., Carruette-Valentin J. La lignee des Pyrsonymphi-
nes et les caracteres infrastructuraux communs aux genre Opisthomi-
tus, Oxymonas, Saccinobacculus, Pyrsonympha et Streblomastix /7
C. R. Acad. Sci. Ser. D. 1970a. Vol. 270. P. 1587—1590.
Hollande A., Carruette-Valentin J. Appariement chromosomique
et complexes synaptonematiques dans les noy aux en cours des depo-
lyploidisation chez Pyrsonympha flagellata: le cycle evolutif des Pyr-
sonymphines symbiontes de Reticulitermes lucifugus /7 C. R. Acad.
Sci. Ser. D. 1970b. Vol. 270. P. 2550—2553.
Hollande A., Carruette-Valentin J. Les atractophores, reduc-
tion du fuseau et la division cellulaire chez les Hypermastigines. Etude
infrastructurale et revision systematique des Trichonymphines et des
Spirotrichonymphines // Protistologica. 1971. Vol. 7. P. 5—100.
Hollande A., Carruette-Valentin J. Le probleme du centrosome
et la cryptopleuromitose atractophorienne chez Lophomonas striata //
Protistologica. 1972. Vol. 8. P. 267—278.
Hollande A., Enjument M. Morphologie et reproduction de Tri-
chomonas aulacodi nov. sp. — parasite de Trionomus (Aulacodus)
swinderianus Temming. Etude du cycle chromosomatique № Ann. Sci.
Natur. Zool. Biol. Anim. 1953. T. 15, N 3—4. P. 439—448.
Hollande A., Valentin J. Morphologie et infrastructure du genres
Barbulanympha, hypermastigine symbiontique de Cryptocercus punc-
tulatus Scudder. 7/ Protistologica. 1967. Vol. 3. P. 257—268.
Hollande A., Valentin J. Appareil de Golgi, pinocytose, lysoso-
mes, mitochondries, bacteries symbiontiques, atractophores et pleuro-
mitose chez les Hypermastigines du genre Joenia et Affinites entre
Joeniides et Trichonymphines 7/ Protistologica. 1969. Vol. 5. P. 39—
86.
Honigberg В. M. A contribution to systematics of the non-pig-
mented flagellates 7/ Progress in Protozoology (Prague, 1961). New
York: Acad. Press, 1963a. P. 68—69.
Honigberg В. M. Evolutionary and systematic relationship in the
flagellate order Trichomonadida Kirby 7/ J. Protozool. 1963b. Vol. 10,
N1.P.20—63.
Honigberg В. M. Protozoa associated with termites and their role
in digestion 7/ Biology of Termites 7 Eds. K. Krishna, F. M. Weesner.
New York; London: Acad. Press, 1970. Vol. 2. P. 1—36.
Honigberg В. M. Fine structure of the mastigont system of Tri-
chomonas foetus (Riedmuller) /7 J. Protozool. 1971. Vol. 18, N 1.
P. 183—198.
Honigberg В. M. Remarks upon trichomonad affinities of certain
parasitic protozoa /7 Progress in Protozoology: Abstract of papers read
ТИП POLYMASTIGOTA. ЛИТЕРАТУРА
397
at 4th Int. Congr. Protozool. Sept. 1973. Univ. Clermout. Clermout :
Ferrand, 1973. P. 187.
Honigberg В. M. Trichomonads of veterinary impotrance. New
York etc., 1978a. P. 164—258.
Honigberg В. M. Trichomonads of importance in human medici-
ne. New York etc., 1978b. P. 276—424.
Honigberg В. M., Bennet C. J. Lightmicroscopie observations on
structure and division of Histomonas meleagridis (Smith) // J. Proto-
zool. 1971. Vol. 18, N 4. P. 687—697.
Honigberg В. M., Kuldova J. Structure of a Nonpathogenic His-
tomonad from the cecum of Galliform birds and revision of the tri-
chomonad family Monocercomonadinae Kirby // J. Protozool. 1969.
Vol. 16, N 3. P. 526—535.
Hughes W. F., Zander D. V. Isolation and culture of Hexamita
free of bacteria // Poult. Sci. 1954. Vol. 33, N 7. P. 810—815.
Hungate R. E. Mutualistic intestinal protozoa // Biochem. Physi-
ol. Protozoa. 1955. Vol. 2. P. 159—199.
Jouon L., Mignot J. P., Kattar M. P., Brugerolle G. Comple-
ments a l’6tude des Trichomonadida et plus particulement de cineti-
de // Protistologica. 1969. T. 5, N 2. P. 309—326.
Kazubski S. L. Structure and evolution of Trichomonadida //
Wiad Parazitol. 1990. Vol. 36. P. 175—180.
Kirby H. A species of Proboscidiella from Kalotermes (Crypto-
termes) dudleyi Banks a termite of Central America, with remarks on
the oxymonad flagellates // Quart. J. Microscop. Sci. 1928. Vol. 72.
P. 355—386.
Kirby H. Flagellates of the genus Trichonympha in termites //
Univ. Calif. Publ. Zool. 1932. Vol. 37. P. 349—476.
Kirby H. Host-parasite relations in the distribution of Protozoa in
Termites // Univ. Calif. Publ. Zool. 1937. Vol. 41. P. 189—212.
Kirby H. Devescovinid flagellates of termites. I. The genus De-
vescovina // Univ. Calif. Publ. Zool. 1941. Vol. 45. P. 1—92.
Kirby H. Flagellate and host relationships of Trichomonad flagel-
letes // J. Parasitol. 1947. Vol. 33. P. 214—228.
Kirby H. Systematic differentiation and evolution of flagellates in
Termites // Rev. Soc. Mexic. Hist. Nat. 1949. Vol. 10. P. 57—79.
Kubai D. F. Unorthodox mitosis in Trichonympha agilis: kine-
tochore differentiation and chromosome movement // J. Cell Sci. 1973.
Vol. 13. P. 511—522.
Kudo R. R. Protozoology. Springfields; Illinois, 1966.
Kulda J., Nohynkova E. Parasitic Protozoa I Ed. J. P. Kreier.
New York; London: Acad. Press, 1978. Vol. 2. P. 1—138.
Kuldova J., Kulda J. Division of Tritrichomonas foetus in cultu-
re II J. Protozool. 1974. Vol. 21, N 3. P. 153.
Lahgford G. M., Inoue S. Motility of the microtubular axostyle
in Pyrsonympha // J. Cell Biol. 1979. Vol. 80. P. 521—538.
Levine N. D. et al. A newly revised classification of the Proto-
zoa // J. Protozool. 1980. Vol. 27. P. 37—58.
Lingle W. L., Salisbury J. L. Ultrastructure of the parabasalid pro-
tist Holomastigotoides // J. Euk. Microbiol. 1995. Vol. 42. P. 490—509.
Mattern C.F.T., Honigberg B.M. The mastigont system of
Trichomonas gallinae (Rivolta) as by electron microscope // J. Proto-
zool. 1967. Vol. 14, N 2. P. 320—329.
McKittrick F. A. A contribution to understanding of cockro-
ach-termite affinities II Ann. Entomol. Soci 1965. Vol. 58. P. 18—22.
Monteiro-Leal L. H., da Cuhna-e-Silva N. L., Benchimol M., de
Souza W. Isolation and biochemical characterization of the costa of
Tritrichomonas foetus // Europ. Cell Biol. 1993. Vol. 60. P. 235—242.
MUller M. Peroxisomes and hidrogenosomes in Protozoa // J. His-
tochem., Cytochem. 1973. Vol. 21, N 11. P. 955—957.
Muller M. The hydrogenosome // J. Gen. Microbiol. 1993.
Vol. 139. P. 2879—2889.
Nie D. Morphology and taxonomy of the intestinal Protozoa of
the guinea-pig, Cavia porcella // J. Morphol. 1950. Vol. 86, N 3.
P. 381—494.
Nielson H. M., Ludvik J., Nielson R. On the ultrastructure of
Trichomonas vaginalis Donne // J. Microscop. 1966. Vol. 5, N 2.
P. 220—250.
Ohkuma M., Noda S., Horikoshi K., Kudo T. Phylogeny of sym-
biotic methanogens in the gut of the termite Reticulitermes speratus //
FEMS Microbiol. Let. 1995. Vol. 134. P. 45—50.
Pitelka D. R., Schooley C. N. The fine structure of the flagellar
apparatus in Trichonympha // J. Morphol., 1958. Vol. 102. P. 199—
246.
Puytorac P., de, Grain J., Mignot J. Precis de Protistologie.
Paris, 1987.
Radek R., Hausmann K. Placojoenia sinaica n. g., n. sp., a sym-
biotic flagellate from the termite Kalotermes sinaicus // Eur. J. Protis-
tol. 1994a. Vol. 30. P. 25—37.
Radek R., Hausmann K. Endocytosis, digestion and defecation in
Flagellates // Acta Protozool. 1994b. Vol. 33. P. 127—147.
Ritter H. A fluid system for the cultivation, light microscope
examination and manipulation of obligate anaerobes // J. Protozool.
1974. Vol. 21. P. 565—568.
Ritter H., Jnoue S., Kubai D. Mitosis in Barbulanympha. I. Spin-
dle structure, formation and kinetochore engagement; II. Dynamics of
two-stage anaphase, nuclear morphogenesis and cytokinesis // J. Cell
Biol. 1978. Vol. 77. P. 638—684.
Tamm S., Tamm S. L. The fine structure of the centriolar ap-
paratus and associated structures in the flagellates Deltotrichonym-
pha and Koruga. I. Interphase # J. Protozool. 1973. Vol. 20. P. 230—
245.
Wenrich D.H. Culture experiments on intestinal flagellates.
III. Species from amphibiens and reptiles // Parasitology. 1947. Vol. 33,
N 1. P. 62—70.
Wenyon С. M. Histological observations on the possible pathoge-
nicity of Trichomonas intestinalis and Chilomastix mesnili, with a
note on Endolimax nana // J. Trop. Med. 1920. Vol. 23, N 1. P. 125—
127.
Wenyon C.M. Protozoology. London, 1926.
Wenyon С. M. Protozoology. A manual for medical men, veteri-
narians and zoologists. London, 1965.
Woodrum D. T., Linck R. W. Structural basis of motility in the
microtubular axostyle: implication for cytoplasmic microtubule struc-
ture and function // J. Cell Biol. 1980. Vol. 87. P. 404—414.
Yamin M. A. Flagellates of the orders Trichomonadida Kirby,
Oxymonadida Grass6 and Hypermastigida Grassi a. Foa. reported
from lower termites (Isoptera families: M asto ter mi tidae, Kalotermiti-
dae, Hodotermitidae, Termopsidae, Rhinotermitidae and Serritermiti-
dae) and from the wood-feeding roach Cryptocercus (Dictyoptera:
Cryptocercidae) // Socibiology, 1979. Vol. 4. P. 1—119.
Yamin M. A. Cellulose metabolism by the flagellate Trichonym-
pha from a termite is independent of endosy mbiotic bacteria // Scien-
ce. 1981. Vol. 211. P. 58—59.
Yemilohun P. F. Isolation and characterization of flagella from
Trichomonas vaginalis // Parasitol. Res. 1998. Vol. 84. P. 800—805.
Тип PLASMODIOPHORA Zopf, 1884 —
ПЛАЗМОДИОФОРИДЫ
Плазмодиофориды — очень небольшая по количеству видов и
весьма своеобразная группа облигатных внутриклеточных парази-
тов различных водорослей, водных и наземных сосудистых расте-
ний, а также грибов. Заражение плазмодиофоридами нередко при-
водит к появлению у хозяев патологических новообразований. Ос-
новная вегетативная (трофическая) стадия жизненного цикла
плазмодиофорид — многоядёрный организм, лишенный клеточной
стенки и обычно именуемый в литературе плазмодием. Основной
тип питания осмотрофный; какие-либо специализированные
структуры или временные образования, обеспечивающие питание,
отсутствуют. В жизненном цикле плазмодиофорид имеются жгути-
ковые стации и цисты; показано наличие мейоза (точнее, присут-
ствие синаптонемального комплекса) при формировании цист (по-
коящихся спор).
История изучения
Плазмодиофориды известны уже более сотни лет.
В первую очередь их изучение связано с тем, что
многие из них являются паразитами культурных рас-
тений, причем таких распространенных, как карто-
фель, капуста, томаты и т. д. Научная литература,
посвященная плазмодиофоридам, довольно обширна
и включает немало обзоров и монографий (см.,
напр.: Соок, 1933; Karling, 1942, 1968; Sparrow,
1960; Olive, 1975; Мазин, Проценко, 1976; Miller,
Dylewski, 1983; Buczacki, 1983; Lee et al., 1985; Dy-
lewski, 1990; Braselton, 1995). Однако несмотря на
все сказанное, целый ряд важнейших вопросов, ка-
сающихся биологии, систематики и филогенетичес-
ких взаимосвязей плазмодиофорид с другими макро-
таксонами низших эукариот, остаются до сих пор
совершенно неясными. Более того, среди специалис-
тов по сей день нет единого мнения о природе этих
организмов и соответственно о том, к какому царст-
ву живых существ их следует относить.
Классификация
Некоторые исследователи считают, что плазмо-
диофорид следует рассматривать в качестве класса в
составе грибов (Fungi), другие включают их в число
миксомицетов (Myxomycetes, Mycetozoa), третьи от-
водят им место среди представителей простейших
(Protozoa) (De Вагу, 1887; Alexopoulos, 1962; Honig-
berg et al., 1964; Karling, 1968; Ainsworth, 1973;
Olive, 1975; Levine et al., 1980; Buczacki, 1983; Lee
et al., 1985; Page, 1987; Dylewski, 1990; Braselton,
1995).
Мы будем рассматривать эти организмы как са-
мостоятельный тип Plasmodiophora Zopf, 1884 (=Plas-
modiophoromycota) в составе протистов (Protista).
Тип включает единственный класс Plasmodiopho-
rea Cook, 1928 с 1 отрядом Plasmodiophorida Cook,
1928 и 1 сем. Plasmodiophoridae Zopf, 1884.
Морфология и физиология
Вегетативный плазмодий
Многоядерные вегетативные плазмодии двух ви-
дов плазмодиофорид (Tetrату ха parasitica и Spon-
gospora subterranea) изображены на рис. 345, А и
345, В. Электронно-микроскопические исследования,
выполненные на Plasmodiophora brassicae, Sorospha-
era veronicae, Spongospora subterranea, Polymyxa be-
tae, P. graminis, Woronina pythii, Ligniera verrucosa и
Membranosorus heterantherae, выявляют значительное
сходство ультраструктурной организации вегетатив-
ных плазмодиев всех изученных видов плазмодиофо-
рид (Williams, McNabola, 1967, 1970; Williams,
Yukawa, 1967; Miller, Dylewski, 1983; Miller et al.,
1985). Вегетативные плазмодии, целиком погружен-
ные в цитоплазму клетки-хозяина, лишены какой-
либо специализированной оболочки и окружены
обычной плазматической мембраной. В них имеется
хорошо развитый шероховатый ЭПР и много сво-
бодных рибосом; многочисленные тельца Гольджи
(диктиосомы) часто обнаруживаются в непосредст-
венной близости от ядер. Плазмодии содержат также
митохондрии со сравнительно небольшим числом
трубчатых крист; весьма характерно наличие боль-
шого количества липидных включений от 0.1 до
0.8 мкм в диаметре.
ТИП PLASMODIOPHORA
399
Рис. 345. Различные стадии жизненного цикла некоторых плазмодиофорид. (По: Karling, 1942, 1968, модифицировано).
А — восьмиядерный плазмодий Tetramyxa parasitica, окружающий ядро клетки-хозяина; Б — крупный спорангиосорус Woronina glomerata в септирован-
ном филаменте Vaucheria; В — гипертрофированные клетки растения, содержащие многоядерные плазмодии Spongospora subterranea; Г — цистосорусы
S. subterranea в оболочке клетки-хозяина; Д — цисты Plasmodiophora brassicae; Е — цистосорусы Sorosphaera veronica’, Ж — зрелые зооспорангии Poly-
myxa graminis с выводными трубками, проходящими через прилежащий слой клеток.
Fig. 345. Select stages of the life cycle of some plasmodiophorids.
A — octonucleate plasmodium of Tetramyxa parasitica, close about the nucleus of host cell; Б — large sporangiosorus of Woronina glomerata in Vaucheria; В —
hypertrophied plant cells with multinucleated plasmodia of Spongospora subterranea; Г — cystosori of 5. subterranea in the host-cell envelope; Д — cysts of
Plasmodiophora brassicae; E — cystosori of Sorosphaera veronica; Ж — mature zoosporangia of Polymyxa graminis.
Наиболее хорошо изучена ультраструктура вто-
ричного вегетативного плазмодия (см.: «Жизненный
цикл»), где особый интерес представляет зона так
называемого интерфейса между мембраной паразита
и цитоплазмой хозяина, поскольку она во многом
отражает характер паразито:хозяинных взаимоотно-
шений. При этом тонкая организация интерфейса
существенно различается, хотя, как уже было отме-
чено выше, ультраструктура плазмодия весьма сход-
на у разных видов плазмодиофорид. У Plasmodiopho-
ra brassicae паразит отделен от цитоплазмы хозяина
семислойной структурой толщиной около 23 нм, ко-
торая состоит из 2 тесно сближенных унитарных
мембран. Уильямс с соавт. (Williams, Yukawa, 1967;
Williams, McNabola, 1967, 1970) обозначают это об-
разование как «оболочка плазмодия», однако спе-
циальные исследования позволяют заключить, что
наружная мембрана «оболочки» формируется вслед-
ствие инвагинации плазмалеммы клетки-хозяина, ко-
торая наблюдается еще в начале процесса проникно-
вения паразита в клетку (иными словами, представ-
ляет собой паразитофорную вакуоль). Следует отме-
тить, что самообразование паразитофорной вакуоли
было показано лишь для Р. brassicae (Miller et al.,
1985). Интерфейс между плазмодием Sorosphaera ve-
ronicae и цитоплазмой клетки Veronica persica пред-
ставлен 1 трехслойной плазмалеммой паразита тол-
щиной 9—12 нм. Аналогично устроена зона кон-
такта паразита и хозяина у Polymyxa betae и Beta
vulgaris. Оболочка плазмодия Woronina pythii состо-
ит из единой электронно-плотной структуры, в кото-
рой не удается различить отдельные слои. Толщина
этой оболочки составляет 19—24 нм на ранних ста-
диях развития плазмодия и 8—9 нм перед его деле-
нием. Подобная же редукция толщины оболочки
наблюдается у Plasmodiophora brassicae в период
прекращения роста плазмодия (Williams, McNabola,
1967). У данного вида плазмодиофорид это является
результатом разрушения мембраны паразитофорной
вакуоли (Buczacki, 1983).
400
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
У клетки растения, инфицированной Plasmodio-
phora brassicae, ядро и ядрышки оказываются значи-
тельно крупнее, чем у соседних неинфицированных.
Ее цитоплазма обогащена рибосомами, амилопласта-
ми, митохондриями, диктиосомами и шероховатым
ЭПР, причем наблюдается тенденция к концентра-
ции этих органелл вблизи паразита (Williams, McNa-
bola, 1967; Williams, Yukawa, 1967). Крупная цент-
ральная вакуоль клетки-хозяина постепенно фраг-
ментируется и в конце концов исчезает совсем.
Параллельно с уменьшением ее объема происходит
увеличение объемов цитоплазмы хозяина и плазмо-
дия. Неинфицйрованные клетки растения, прилежа-
щие к зараженным и даже связанные с ними цитоп-
лазматическими мостиками (плазмодесмами), не об-
наруживают никаких изменений в своей организации
и метаболизме, что свидетельствует об очень тесных
взаимоотношениях паразита и хозяина. Значитель-
ное увеличение размеров растительной клетки после
ее заражения имеет место и в случае других предста-
вителей плазмодиофорид (рис. 345, В).
К концу вегетативного роста плазмодии почти
полностью заполняют собой клетку-хозяина. Пере-
ход к цистогенезу Р. brassicae сопровождается разру-
шением наружной мембраны двухслойной «оболоч-
ки» (паразитофорной вакуоли) плазмодия, и паразит
оказывается отделенным от цитоплазмы хозяина
только собственной плазмалеммой. В это время ядра
всех плазмодиев, находящихся в одной клетке, син-
хронно делятся (Williams, McNabola, 1967). Органи-
зация цитоплазмы хозяина нарушается, и клетка
очень быстро дегенерирует. На этой стадии были
обнаружены глубокие инвагинации плазматической
мембраны плазмодиев, захватывающие окружающую
цитоплазму растительной клетки, включая такие ор-
ганеллы, как пластиды и митохондрии. Однако про-
исходит ли при этом образование пищеварительных
вакуолей, т. е. имеет ли здесь место фагоцитоз, пока
неясно. В процессе цистогенеза в цитоплазме плаз-
модия появляются многочисленные мелкие везикулы
0.1—0.3 мкм в диаметре, окруженные унитарной
мембраной и содержащие тонкофибриллярный мат-
рикс. Такие везикулы отсутствуют во время вегета-
тивного роста. После последнего ядерного деления
эти везикулы постепенно выстраиваются вокруг каж-
дого индивидуального ядра с прилежащей к нему
цитоплазмой с органоидами. Наконец, мембраны ве-
зикул сливаются в одну, окружающую одноядерные
участки, на которые таким образом разбивается вся
протоплазматическая масса плазмодия. Фибрилляр-
ный матрикс, который содержится внутри везикул,
оказывается локализованным на наружной поверх-
ности плазматических мембран развивающихся цист
и участвует в формировании их оболочек.
Ядро и митоз
Ядерные деления в вегетативном плазмодии по
классификации Райкова (1978, 1986) следует отнес-
ти к одной из разновидностей полузакрытого орто-
митоза. Единственное центральное ядрышко, сохра-
няющееся на протяжении всего деления, в метафазе
становится вытянутым в направлении полюсов вере-
тена, а конденсированные хромосомы выстраиваются
в отчетливую экваториальную пластинку в виде коль-
ца. Если ядро на этой стадии рассматривать сбоку,
то расположенные перпендикулярно друг другу вытя-
нутое ядрышко и экваториальное хроматиновое
кольцо образуют очень характерную крестообразную
фигуру, благодаря чему митоз у плазмодиофорид
получил наименование крестовидного ядерного деле-
ния (cruciform nuclear division) (Cook, 1928, 1933;
Karling, 1942, 1968; Sparrow, 1960; Olive, 1975;
Мазин, Проценко, 1976; Dylewski, 1990; Braselton,
1995).
Электронно-микроскопические исследования со-
матических митозов в растущих вегетативных плаз-
модиях (спорангиальных и цистогенных) Polymyxa
betae, Sorosphaera veronicae, Plasmodiophora brassi-
cae и Woronina pythii выявили принципиальное сход-
ство ультраструктурной организации аппарата
крестовидного ядерного деления у различных видов
плазмодиофорид (Braselton et al., 1975; Dylewski,
Miller, 1983b, 1984a; Dylewski, 1990; Braselton,
1995). Каждое интерфазное ядро, как правило име-
ющее более или менее правильную округлую форму,
содержит единственное отчетливое центральное яд-
рышко, состоящее из плотного фиброгранулярного
материала (рис. 346, А). В непосредственной близос-
ти от ядра были найдены пары центриолей, лежащие
с двух его противоположных сторон. От них ради-
ально расходятся микротрубочки, часть из которых
огибает наружную поверхность ядерной оболочки
(рис. 346, А). Центриоли в каждой паре находятся
под углом 180° друг к другу («конец к концу») и
сохраняют такое расположение на протяжении всех
стадий митоза в вегетативном плазмодии. Отличия
профазных ядер от интерфазных заключаются в по-
явлении мелких, неправильной формы хроматино-
вых агрегатов, которые к концу этой стадии увели-
чиваются в размерах и становятся более электрон-
но-плотными.
Метафаза характеризуется формированием четкой
экваториальной пластинки, которая располагается
перпендикулярно продолжающему сохраняться яд-
рышку, несколько вытянутому вдоль оси веретена
(рис. 346, Б). Ядерная оболочка остается интактной,
за исключением полюсов, где она перфорируется и
частично разрушается проникающими в ядро микро-
трубочками, идущими от центроконусов, содержа-
щих центриоли. Помимо полярных микротрубочек к
ортомитотическому аппарату относятся также мик-
ротрубочки, отходящие от участков хромосом, кото-
рые предположительно соответствуют кинетохорам.
В метафазе и анафазе появляется большое количес-
тво внутриядерных мембранных везикул, отпочковы-
вающихся от внутренней мембраны исходной ядер-
ной оболочки. Ядрышко разделяется на 2 равных
фрагмента, синхронно расходящихся к полюсам
вместе с дочерними хромосомным пластинками. В
телофазе происходит окончательное формирование
новых ядерных оболочек в результате слияния мем-
бранных пузырьков внутри оболочки материнского
ядра. Полярные «окна» закрываются значительно
раньше — еще в поздней метафазе.
ТИП PLASMODIOPHORA
401
Рис. 346. Схемы организации интерфазных и митотических ядер плазмодиофорид.
А — интерфазное ядро спорангиального плазмодия Woronina pythiix Б — метафаза крестовидного ядерного деления в цистогенном плазмодии Sorosphaera
veronicae; В — метафаза спорангиального митоза у Woronina pythii. вв — внутриядерные везикулы, даг — диктиосомы аппарата Гольджи, мт — микро-
трубочки, хр — хроматин, ц — центриоли, эпр — цистерны шероховатого ЭПР, яд — ядрышко, яо — ядерная оболочка. (А — по: Dylewski, Miller,
1983b; Б — по: Braselton et al., 1975; В — no: Dylewski, Miller, 1984b).
Fig. 346. Schematic drawing of an interphase and mitotic nuclei of plasmodiophorids.
A — interphase nucleus of sporangiogenous plasmodium of Woronina pythii; Б — metaphase of cruciform nuclear division in cystogenous plasmodium of Soro-
sphaera veronicae; В — metaphase of sporangial mitosis in Woronina pythii. ее — intranuclear vesicles, даг — dictyosomes (Golgi bodies), мт — microtubules,
xp — chromatin, ц — centrioles, эпр — rough endoplasmic reticulum, яд — nucleolus, яо — nuclear envelope.
Общий план организации митотического аппарата
и способ формирования дочерних ядерных оболочек
являются принципиально сходными для всех ядерных
делений, наблюдаемых в жизненном цикле каждого
отдельного вида и всех плазмодиофорид вообще. Од-
нако, как было хорошо показано на примере W. pyt-
hii (Dylewski, Miller, 1984b), имеются определенные
отличия в морфологической картине спорангиальных
митозов (рис. 346, В) и ядерного деления, предшест-
вующего образованию спорангиев (так называемого
26 Протисты, часть 1
транзитного, или промежуточного, митоза) у плазмо-
диофорид. Основное отличие митозов, протекающих
в спорангиях, заключается в полной резорбции яд-
рышка еще до начала деления; поэтому эта фаза жиз-
ненного цикла даже выделялась ранее некоторыми
исследователями как акариотная стадия. Таким обра-
зом, в морфологическом отношении спорангиальный
митоз не является крестовидным ядерным делением.
Другой его отличительной чертой является изменение
исходной ориентации центриолей в каждой паре
402
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
(180°): они расходятся от концов друг друга и распо-
лагаются под углом 30—35°. Следует отметить, что
переход к новой морфологической картине ядерного
деления осуществляется постепенно. Так, транзит-
ный митоз был назван еще и промежуточным не
только из-за своего места в жизненном цикле парази-
та. В начале этого деления ядрышко начинает дис-
пергироваться, однако его фрагменты еще продолжа-
ют присутствовать в течение профазы, «метафазы и
анафазы; они полностью элиминируются лишь к
концу телофазы.
Тем не менее несмотря на существование морфо-
логических различий между тремя формами вегета-
тивного ядерного деления (т. е. крестовидного, тран-
зитного и спорангиального), все они обладают фунда-
ментальным сходством в организации митотического
аппарата у всех изученных видов плазмодиофорид.
К этому следует добавить, что архитектура веретена
принципиально такая же и при мейозе, только пер-
форации на полюсах в исходной ядерной оболочке,
пожалуй, несколько шире, чем при митозе (Braselton,
Miller, 1973; Dylewski, Miller, 1984b). Наконец, спо-
соб образования оболочек дочерних ядер во всех слу-
чаях один и тот же.
Очень интересное явление было обнаружено у
W. pythii в процессе трансформации зрелого вегета-
тивного плазмодия в цистосорусы (Dylewski, Miller,
1983е, 1984b). Все ядра внутри каждого плазмодия
одновременно входят в профазу редукционного деления,
обнаруживая отчетливые синаптонемальные комп-
лексы. Однако приблизительно 16 % этих пахитен-
ных ядер мигрируют к периферической части плаз-
модия, где образуют агрегат, отделяющийся от ос-
новной части цитоплазмы специальной мембраной.
Затем все агрегированные ядра сливаются, формируя
массивное плеоморфное ядро. В то время как ос-
тальные ядра в плазмодии завершают первое мейоти-
ческое деление, наблюдается дегенерация массивно-
го, слившегося ядра и окружающего его небольшого
участка цитоплазмы. Предполагается, что происхо-
дящая таким образом элиминация части ядер цисто-
генного плазмодия вызвана необходимостью поддер-
жания определенной ядерно-цитоплазматической
пропорции в каждой образующейся одноядерной
цисте.
Зооспоры и цисты
Исследования зооспор плазмодиофорид под элек-
тронным микроскопом четко показали, что оба жгути-
ка, имеющие типичную (9 + 2) конфигурацию фиб-
рилл аксонемы, лишены мастигонем. Кинетосомы,
находящиеся на переднем конце зооспоры, распола-
гаются под острым углом по отношению друг к другу
(от 20 до 45° у разных видов). Направленный назад
жгутик обычно в 3—3.5 раза длиннее переднего и
имеет отчетливо сужающийся концевой участок (Ко-
le, Gielink, 1961; Aist, Williams, 1971; Miller et al.,
1985).
Весьма существенным моментом в жизненном
цикле большинства облигатных внутриклеточных па-
разитических простейших является их проникнове-
ние к месту своей локализации в организме хозяина.
В особенности этот вопрос актуален для плазмодио-
форид, поскольку толстая и прочная целлюлозная
оболочка растительных клеток представляет серьез-
ную преграду на пути заражения. Длительное время
считалось, что зооспоры проникают прямо через кле-
точную оболочку, оказывая на нее какое-то воздейст-
вие химической природы. Однако на представителях
разных видов и родов плазмодиофорид — VT. polycys-
tis, W. pythii, Sorodiscus cokeri, Polymyxa betae и дру-
гих — было установлено наличие иного, очень свое-
образного механизма, с помощью которого паразиты
инъецируют клетки хозяина.
Детально, с применением электронно-микроско-
пической техники этот вопрос был исследован у
Plasmodiophora brassicae (Aist, Williams, 1971). Под-
вижные зооспоры прикрепляются своим задним концом
к стенке растительной клетки, вскоре после чего их
жгутики прекращают свою работу. Они закручива-
ются вокруг зооспоры, плотно прилегая к ее повер-
хности, а затем менее чем за 1 мин втягиваются
внутрь тела. Вскоре зооспоры совершенно округля-
ются и инцйстируются. Наружная оболочка форми-
руется, по всей видимости, путем вывода на поверх-
ность тела содержимого многочисленных мелких ве-
зикул, которые в это время появляются в цитоплазме
и затем сливаются с плазматической мембраной
цисты. В процессе созревания такой зооспоровой
цисты в ее цитоплазме возникает и увеличивается в
размерах окруженная унитарной мембраной вакуоль.
В каждой цисте имеются также липидные включе-
ния и 1 хорошо развитое тельце Гольджи, ассоции-
рованное с шероховатым ЭПР.
Однако самой главной особенностью такой цис-
ты является развитие в ней своеобразного аппарата
инвазии, формирующегося приблизительно через 2 ч
после того, как зооспора прикрепляется к стенке
растительной клетки (рис. 347; 348, А). Он пред-
ставляет собой длинную трубку, образованную путем
впячивания (инвагинации) внутрь цитоплазмы плаз-
матической мембраны цисты (Rohr). Наружный ко-
нец трубки всегда ориентирован в направлении кле-
точной стенки растения-хозяина и закрыт пробочкой
из материала незначительной электронной плотности.
Центральное пространство этого образования запол-
нено осмиофильным фибриллярным материалом,
очевидно адгезивной природы. В глубине трубки
лежит электронно-плотный, заостренный на одном
конце, палочковидный шип (Stachel) длиной более
700 нм. Наружная оболочка этого шипа состоит из
продольно расположенных, тесно сближенных фиб-
рилл толщиной 3.8 ± 1.1 нм. Тупой конец шипа на-
ходится почти у основания трубки, которая в этом
месте резко сужается и образует загибающийся в сто-
рону узкий рукав (Schlauch) длиной 1 мкм и более,
заполненный гомогенным электронно-светлым мате-
риалом; предполагается, что его конец остается откры-
тым прямо в цитоплазме цисты (рис. 347; 348, А).
Вскоре после того как в зрелой зооспоровой цис-
те завершится образование аппарата инвазии, с его
помощью осуществляется проникновение паразита в
клетку растения (рис. 348). Это происходит через
2.5—3.5 ч после инцистированйя зооспор. Прибли-
ТИП PLASMODIOPHORA
403
Рис. 347. Схемы двух серийных срезов через зооспоровую цисту Plasmodiophora brassicae с развитым аппаратом инвазии. (По: Aist, Wil-
liams, 1971).
в — вакуоль, лв — липидные включения, п — «пробочка», пмц — плазматическая мембрана цисты, я — ядро, т — «трубка» (Rohr), к — «копье» (Sta-
chel).
Fig. 347. Schematic drawing of two serial sections of zoospore cyst of Plasmodiophora brassicae with the invasion apparatus.
в — cyst vacuole, ле — lipid droplets, n — Rohr plug, пмц — plasma membrane of the cyst, я — nucleus, m — Rohr, к — Stachel.
зительно за 15 мин до начала этого процесса наблю-
дается сильное увеличение размеров вакуоли, кото-
рая заполняет около половины объема цисты. В ре-
зультате выпячивания (эвагинации) трубки нахо-
дящийся в ней фибриллярный адгезивный материал
выходит наружу и плотно прилегает (приклеивается)
к стенке растительной клетки, менее чем за 1 мин
формируя так называемый адгезориум (рис. 348, Б),
Дальнейшее выворачивание трубки приводит к тому,
что находившийся в ее глубине плотный шип своим
заостренным концом вонзается в оболочку клетки-
хозяина и прокалывает ее насквозь (рис. 348, В). За-
тем, практически за 1 с через канал в адгезориуме в
образовавшееся отверстие впрыскивается одноядер-
Рис. 348. Схема процесса проникновения паразита (Plasmodiophora brassicae) в клетку корневого волоска растения-хозяина. (По: Aist,
Williams, 1971).
А — зрелая зооспоровая циста на поверхности стенки растения-хозяина; Б — увеличение объема вакуоли, эвагинация трубки и начало образования адгезо-
риума; В— шип прокалывает стенку растительной клетки; Г— аппарат инвазии сразу после инъекции амебоидного зародыша в растительную клетку.
ад — адгезориум, в — вакуоль, ле — липидные включения, п — «пробочка», скх — стенка клетки-хозяина, ст — стенка трубки, сц — стенка цисты,
' фам — фибриллярный адгезивный материал, ш — «шланг» (Schlauch), к — «копье» (Stachel).
Fig. 348. Diagrammic representation of zoospore (Plasmodiophora brassicae) penetration of the host cell wall.
A — encystment of zoospore on host cell wall, Б — expansion of zoospore vacuole and development of adhesorium, В — penetration of host wall by Stachel,
Г -г- continued expansion of zoospore vacuole presumably resulted in the passage of parasite cytoplasm into host cell, ad — adhesorium, в — cyst vacuole, лв —
lipid droplets, n — Rohr plug, скх — host cell wall, cm — Rohr, сц — cyst wall, фам — adhesive material, at — Schlauch, к — Stachel.
404
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ный амебоидный зародыш паразита. Осуществление
этого процесса становится возможным в результате
резкого увеличения размеров вакуоли, которая за-
полняет собой весь внутренний объем цисты, благо-
даря чему зародыш быстро проталкивается в клетку-
хозяина (рис. 348, Г). Таким образом, инвазия про-
исходит путем инъекции паразита сквозь прокол в
стенке растительной клетки, а пустая оболочка
цисты остается прикрепленной снаружи. Время от
начала формирования адгезориума до момента про-
никновения амебоидного зародыша в клетку состав-
ляет приблизительно 1 мин. Предполагается, что та-
кой способ инвазии присущ и первичным и вторич-
ным зооспорам плазмодиофорид (Aist, Williams,
1971; Dylewski, 1990). Причем часть вторичных зоо-
спор у Р. brassicae после своего образования может
не покидать оболочку клетки-хозяина, а внедряться
непосредственно в соседние клетки, лежащие в бо-
лее глубоких слоях ткани растения. Считается, что
длина шипа вполне достаточна, чтобы проколоть 2
прилежащие друг к другу клеточные стенки, общая
толщина которых составляет около 150—450 нм.
Формирование еще одного типа специализиро-
ванных структур, возникающих на заключительном
этапе дифференциации вегетативного плазмодия в
спорангии, известно у представителей некоторых ро-
дов. Так, у Polymyxa спорангиогенез сопровождается
образованием особых выводных трубок, через ко-
торые осуществляется выход вторичных зооспор за
пределы оболочки клетки-хозяина (рис. 345, Ж).
Электронно-микроскопическое исследование (D’Amb-
ra, Mutto, 1977) показало, что они формируются из
специальных одноядерных участков, отделенных от
собственно зооспорангия перегородкой. В дальней-
шем, после разрушения этой перегородки, зрелые
зооспоры выходят из спорангиев через отверстие
возникшего трубковидного образования. Подобного
рода приспособления, хотя и менее выраженные
морфологически, имеются также у представителей
рода Ligniera и, возможно, некоторых других плаз-
модиофорид (Miller et al., 1985).
Морфология и тонкая организация зооспоровых
цист и других стадий жизненного цикла плазмодио-
форид свидетельствует о крайней степени специали-
зации и большой древности этой группы организмов,
прошедших длительный эволюционный путь адапта-
ции к паразитическому образу жизни и развитию в
клетках растений и грибов.
Жизненный цикл
Наибольшее препятствие для детального изучения
жизненных циклов плазмодиофорид представляет со-
бой их облигатный внутриклеточный паразитизм. В
большинстве ранних работ исследователи неизбежно
сталкивались с неконтролируемой контаминацией ис-
следуемого материала посторонними почвенными ор-
ганизмами, в первую очередь простейшими (голыми
амебами и жгутиконосцами). Только после разработки
методики длительного содержания паразитов в куль-
туре каллусной ткани растения-хозяина в аксеничных
условиях на синтетической среде (Ingram, 1969; Wil-
liams et al., 1969) стало возможным существенно при-
близиться к расшифровке жизненных циклов плазмо-
диофорид.
На рис. 349 изображена обобщенная схема веро-
ятного жизненного цикла плазмодиофорид, основан-
ная на данных по наиболее исследованным видам
(Plasmodiophora brassicae, Sorosphaera veronicae,
Spongospora subterranea и ряда др.). Одноядерные
цисты (покоящиеся споры) наземных видов могут
длительное время находиться в почве, сохраняя свою
жизнеспособность (например, у Plasmodiophora bras-
sicae до 3—8 лет), но могут прорастать и без пе-
риода покоя, причем их прорастанию способствуют
корневые выделения растения-хозяина (Мазин, Про-
ценко, 1976; Власов, 1986). Через отверстие в обо-
лочке цисты выходит первичная зооспора. Она
имеет 2 апикальных жгутика неравной длины. При
плавании короткий жгутик направлен вперед и со-
вершает вращательные движения, а более длинный,
направленный назад жгутик двигается волнообразно.
Первичные зооспоры проникают в клетки растения-
хозяина (в случае наземных хозяев чаще всего в
корневые волоски) и локализуются там в виде од-
ноядерных амебоидных зародышей, которые затем
дают начало первичным, или спорангиальным, веге-
тативным плазмодиям (рис. 349).
При инфицировании растения-хозяина первичны-
ми зооспорами в каждой зараженной клетке, как
правило, оказывается сразу много одноядерных аме-
боидов паразита. Не исключено, что некоторые из
них могут сливаться друг с другом сразу по несколь-
ку штук, давая начало молодым многоядерным плаз-
модиям, которые далее продолжают расти, проходя
последовательные синхронные ядерные деления
(Tommerup, Ingram, 1971).
Зрелый первичный плазмодий распадается на од-
ноядерные сегменты — спорангии (зооспорангии),
которые часто объединены в группы — спорангиосо-
русы (рис. 345, Б). В развивающихся зооспорангиях
происходит ряд митотических делений, после чего из
них выходят одноядерные вторичные зооспоры (от
4 до нескольких десятков из каждого спорангия у
разных видов), по внешнему виду сходные с первич-
ными. Выход осуществляется либо через отверстие в
тонкой стенке спорангия, либо, как у представителей
рода Polymyxa. через специальные выводные трубки
(рис. 345, Ж).
Вторичные зооспоры вновь инфицируют клетки
растения-хозяина, давая начало развитию вторично-
го, или цистогенного, плазмодия, но, как отмечают
многие авторы, перед этим происходит их попарное
слияние (Cook, 1933; Kole, 1954; Tommerup, Ingram,
1971; Ingram, Tommerup, 1972, и др.). Однако это
слияние лишь имитирует копуляцию гамет, так как
вслед за ним не происходит кариогамии, и оба ядра
сохраняют свою индивидуальность, т. е. имеет место
вегетативное (Keskin, 1964, — цит. по: Olive, 1970),
или агамное (Серавин, Гудков, 1984), слияние кле-
ток. Отмечается также, что молодые вторичные аме-
боиды Moiyr сливаться друг с другом внутри заражен-
ной ими клетки (Sparrow, 1960; Tommerup, Ingram,
1971). Именно на стадии развития вторичного ве-
гетативного плазмодия многие виды плазмодиофорид
ТИП PLASMODIOPHORA
405
Рис. 349. Обобщенная схема жизненного цикла плазмодиофорид. (По: Karling, 1968; Ingram, Tommerup, 1972; Olive, 1975, модифициро-
вано).
1 — циста (покоящаяся спора); 2 — прорастающая циста; 3 — первичная зооспора; 4 — молодой первичный (спорангиальный) плазмодий; 5 — кресто-
видные ядерные деления; 6 — расчленение плазмодия на одноядерные спорангии; 7 — группа спорангиев (спорангиосорус); 8 — отдельный зрелый спо-
рангий с четырьмя зооспорами; 9 — вторичные зооспоры; 10 — слияние (плазмогамия) вторичных зооспор; 11 — двухъядерный продукт плазмогамии (мо-
лодой вторичный плазмодий); 12 — вторичный (цистогенный) плазмодий; 13 — крестовидные ядерные деления; 14 — кариогамия (попарное слияние
ядер) во вторичном плазмодии; 15 — «акариотная стадия» (профаза первого мейотического деления); 16а, 166 — двухступенчатый мейоз; 17 — расчлене-
ние плазмодия на одноядерные цисты; 18 — группа цист (цистосорус).
Fig. 349. Diagram of the plasmodiophorid life cycle.
1 — cyst (resting spore); 2 — cyst germination; 3 — primary zoospore; 4 — young primary (sporangiogenous) plasmodium; 5 — cruciform nuclear divisions;
6 — cleavage of plasmodium into uninucleate sporangia; 7 — sporangiosorus; 8 — mature sporangium with four zoospores; 9 — secondary zoospore; 10 — fusi-
on (plasmogamy) of the secondary zoospores; 11 — binucleate product of plasmogamy (young secondary plasmodium); 12 — secondary (cystogenous) plasmodi-
um; 13 — cruciform nuclear divisions; 14 — karyogamy in the cystogenous plasmodium; 15 — «akaryotic» stage (prophase of the first meiotic division); 16a,
166 — two-step meiosis; 17 — cleavage of plasmodium into uninucleate cysts; 18 — cystosorus.
вызывают патологические новообразования у расте-
ния-хозяина (рис. 350). Причина заключается в том,
что паразиты, находясь внутри клетки, побуждают ее
к ненормально усиленному росту (гипертрофии) и де-
лению (гиперплазии), причем сами они оказываются
включенными в каждую дочернюю клетку хозяина.
Экспериментально было также показано, что зара-
женные клетки каллусной ткани растений в культуре
растут и делятся быстрее, чем незараженные (Ingram,
1969). Численность паразитов может увеличиваться
за счет фрагментации (множественной плазматомии)
растущих вторичных плазмодиев внутри клетки-хозя-
ина (Sparrow, 1960; Karling, 1968). Параллельно с
увеличением размеров паразита его ядра претерпева-
ют последовательные синхронные крестовидные де-
ления.
Развитие вторичного плазмодия обычно заверша-
ется 2 последовательными ядерными делениями, на
протяжении которых отсутствуют ядрышки (Cook,
1928, 1933; Karling, 1942, 1968). Томмерап и Ингрэм
406
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 350. Патологические новообразования у растений, вызываемые заражением плазмодиофоридами. (По: Karling, 1942, 1968).
А — корни капусты, инфицированной Plasmodiophora brassicae; Б — здоровый и больной экземпляры морской травы Zostera папа, пораженный Plasmo-
diophora bicaudata; В — клубень картофеля, пораженный Spongospora solani.
Fig. 350. Plant diseases caused with plasmodiophorids.
A — cabbage roots infected with Plasmodiophora brassicae; Б — Zostera папа, intact and infected with Plasmodiophora bicaudata; В — potato tuber infected
with Spongospora solani.
(Tommerup, Ingram, 1971), изучая жизненный цикл
Plasmodiophora brassicae в культуре каллусной ткани,
пришли к выводу, что ядра зрелого вторичного плаз-
модия сливаются попарно перед первым из этих 2
ядерных делений, которые, по их мнению, представ-
ляют собой двухступенчатый мейоз. Это согласуется с
результатами других исследований, выполненных на
Р. brassicae, Spongospora subterranea, Octomyxa ас-
hyae и других (см.: Buczacki, Moxham, 1980; Dylewski,
1990), в которых сообщается о наличии кариогамии и
следующего сразу вслед за ней мейоза. Таким образом,
если правы авторы упомянутых выше работ, то дипло-
фаза у плазмодиофорид чрезвычайно кратковременна.
Тем не менее четкие документальные доказательства
кариогамии получены не были, а наличие мейоза на
этой стадии жизненного цикла подтверждается лишь
присутствием синаптонемальных комплексов в про-
фазе первого ядерного деления. Эти комплексы были
обнаружены, в частности, у Sorosphaera veronicae,
Plasmodiophora brassicae, Polymyxa betae, Woronina
pythii и других (Braselton, Miller, 1973^ Miller et al.,
1985; Dylewski, 1990).
Завершение вегетативного роста вторичного плаз-
модия внешне проявляется в наступлении так назы-
ваемой акариотной стадии (буквально — безъядер-
ной). Такое наименование было дано в ранних рабо-
тах, поскольку ядра в живом материале становятся
менее отчетливыми вследствие диссоциации в них
единственного центрального ядрышка. Представляет-
ся наиболее вероятным, что эта стадия соответствует
профазе первого деления мейоза.
Наконец, после завершения второго мейотического
деления вокруг каждого образовавшегося ядра обо-
собляется участок цитоплазмы и плазмодий расчле-
няется на одноядерные гаплоидные цисты (покоящи-
еся споры); клетка-хозяин к этому времени уже по-
гибает. Цисты либо свободно располагаются внутри
оболочки растительной клетки, либо объединены в
цистосорусы в виде небольших групп полых или губ-
чатых сфер, уплощенных дисковидных структур (ха-
рактер их организации является важным системати-
ческим признаком). В дальнейшем оболочка клетки-
хозяина разрывается и цисты попадают во внешнюю
среду. Распространению наземных видов плазмодио-
форид способствуют различные почвенные обитатели
(дождевые черви, насекомые), токи воды, а также де7
ятельность человека, связанная с выращиванием
сельскохозяйственных культур.
ТИП PLASMODIOPHORA
407
Продолжительность одного жизненного цикла у
плазмодиофорид очень невелика. Так, Власов (1986)
в работе с искусственно выращенными растениями
сахарной свеклы проследил прохождение всех ос-
новных стадий развития Polymyxa betae в течение
20 сут. Приблизительно столько же времени требу-
ется Plasmodiophora brassicae для завершения жиз-
ненного цикла в культуре каллусной ткани (Tommerup,
Ingram, 1971); от момента заражения до образования
заметных вздутий на корешках всходов капусты про-
ходит всего 9—11 дней, а продолжительность всего
жизненного цикла паразита в этих условиях состав-
ляет около полутора месяцев (Мазин, Проценко,
1976).
Питание
Следует отметить еще одну очень важную, с
нашей точки зрения, сторону биологии плазмодио-
форид. Принято считать, что эта группа организмов
исходно и исключительно характеризуется осмо-
трофным типом питания. Дилевский (Dylewski, 1990)
специально подчеркивает, что у вегетативных плаз-
модиев отсутствует способность к амебоидной под-
вижности и фагоцитозу. Однако некоторые наблюде-
ния, выполненные на отдельных видах, позволяют
подозревать, что первичным способом питания, ве-
роятно, все же была фаготрофия. Так, согласно
Цопфу (Zopf, 1894, — цит. по: Karling, 1968), моло-
дые амебоидные зародыши и плазмодии Woronina
glomerata, паразитирующие в талломе Vaucheria,
способны формировать пищеварительные вакуоли,
включающие участки протопласта водоросли с хро-
матофорами. Были описаны также и другие, правда
достаточно сомнительные, случаи фаготрофного пи-
тания у Octomyxa, Polymyxa graminis и Ligniera junci
(см.: Karling, 1968; Olive, 1975). Некоторые сви-
детельства в пользу того, что вегетативные плазмо-
дии Plasmodiophora brassicae способны захватывать
участки цитоплазмы клетки-хозяина, имеются в ра-
боте Уильямса и Мак-Наболы (Williams, McNabola,
1967). Тем не менее существование настоящего фа-
гоцитоза у плазмодиофорид по-прежнему нуждается
в проверке.
Специфичность паразитов
к хозяину и практическое
значение
Некоторые плазмодиофориды способны инфици-
ровать лишь несколько или даже какой-то один вид
растения, тогда как другие имеют широкий круг раз-
личных хозяев. Так, например, представитель рода
Ligniera — L. betae — известен только из корней
обычной свеклы (Beta vulgaris), а другой вид этого
же рода — L. junci — был обнаружен в корневых
волосках и корнях значительного числа сосудистых
растений (Karling, 1968). Возбудитель весьма вредо-
носного заболевания капусты, известного под назва-
нием килы, — Plasmodiophora brassicae — может
развиваться в клетках еще около 200 видов хозяев,
принадлежащих к тому же семейству крестоцветных
(Karling, 1968). Woronina polycystis паразитирует в
гифах целого ряда различных водных грибов из числа
сапролегниевых (Sparrow, 1960; Karling, 1968), в то же
время W. pythii инфицирует исключительно предста-
вителей одного рода Pythium (Dylewski, Miller, 1983а).
Таким образом, плазмодиофориды обладают разной
степенью специфичности к хозяину.
Plasmodiophora brassicae Woron., 1877 — это
возбудитель наиболее вредоносной болезни капусты
и других растений семейства крестоцветных. На
корнях пораженного растения развиваются уродли-
вые опухолевые разрастания (рис. 350, А), которые,
достигая определенного размера, разрушаются при
участии почвенных микроорганизмов. Эта болезнь,
имеющая в настоящее время всесветное распростра-
нение, издавна была известна и наносила серьезный
ущерб овощному хозяйству, уничтожая в иные годы
до половины всего урожая капусты. В 1872 г. Рос-
сийское общество садоводов объявило конкурс с
премией за выяснение причин болезни и разработку
методов борьбы с ней. Первое предварительное со-
общение о возбудителе этого заболевания было сде-
лано основоположником отечественной фитопатоло-
гии М. С. Ворониным на заседании Санкт-Петербург-
ского Общества естествоиспытателей в феврале 1874 г.,
а в 1877 г. появилась его статья «Plasmodiophora
brassicae. Организм, причиняющий капустным рас-
тениям болезнь, известную под названием килы». В
этой работе в главных чертах был описан цикл раз-
вития патогена, и, как пишут Мазин и Проценко
(1976), «приходится лишь изумляться правильности
наблюдений и выводов, сделанных М. С. Ворониным
с помощью самых скромных средств». Почти все
намеченные им методы борьбы с этим заболеванием
сохраняют свою актуальность и до сих пор. Работа
Воронина (1877) о киле капусты вошла в мировой
фонд классических исследований по болезням расте-
ний. Так, например, в 1934 г. она была опублико-
вана в США на ^английском языке в серии «Класси-
ки фитопатологии».
Известен еще один представитель плазмодиофо-
рид, имеющий большое практическое значение, —
Spongospora solani Brunchorst, 1887 (син.: S. subter-
rane а (Wall.) Lagerch., 1891). Этот вид поражает
клубни и корни картофеля (рис. 350, В), томаты и
другие растения семейства пасленовых. Цистогенные
плазмодии спонгоспоры, развивающиеся в перифери-
ческих клетках клубня картофеля, образуют характер-
ной формы губчатые комочки покоящихся спор —
цистосорусы (рис. 345, Г). Это заболевание встреча-
ется почти во всех странах умеренного пояса. Другой
широко распространенный вид плазмодиофорид,
принадлежащий к роду Polymyxa, — Р. betae Keskin,
1964 — развивается в поверхностных клетках эпи-
дермиса корней свеклы обыкновенной (круг его рас-
тений-хозяев ограничен сем. Chenopodiaceae). Осо-
бенно активное изучение этого паразита началось
после того, как в 70-е гг. была показана его роль в
передаче вируса некротического пожелтения жилок
свеклы (см.: Власов, 1986). Подобные данные были
получены также и в отношении Р. graminis.
408
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Заметки о филогении
и положении в системе
низших эукариот
Тот факт, что в одном жизненном цикле имеются
стадии развития, сходные с таковыми у миксомице-
тов, грибов-фикомицетов и некоторых саркодовых,
затрудняет решение вопроса о положении плазмодио-
форид в системе низших эукариот, и поэтому, как мы
уже отмечали ранее, среди исследователей до сих пор
нет единого мнения по данному вопросу: одни счита-
ют их родственными грибам (прежде всего хитридие-
вым), другие включают в число миксомицетов и, на-
конец, третьи отводят им место среди представителей
простейших (De Вагу, 1887; Alexopoulos, 1962; Ho-
nigberg et al., 1964; Olive, 1970, 1975; Ainsworth, 1973;
Levine et al., 1980; Lee et al., 1985; Page, 1987; Dy-
lewski, 1990; Braselton, 1995).
Рассматривая вопрос о возможных предковых
формах плазмодиофорид, Карлинг (Karling, 1968), а
вслед за ним Оляйв (Olive, 1970, 1975) высказывают
предположение, что это были какие-то протеомик-
сиды (Proteomyxida), ранее включаемые в качестве
самостоятельного таксона в состав Protozoa (Honig-
berg et al., 1964). Среди протеомиксид имеется не-
мало видов, которые проникают внутрь водорослевых
клеток и питаются, фагоцитируя участки их цитоп-
лазмы. Считается, что некоторые из них могут обра-
зовывать плазмодиоподобные амебоидные формы
(возможно, многоядерные, хотя их цитологическое
изучение не проводилось), которые затем распадают-
ся на сегменты, дающие начало зооспорангиям (зоо-
цистам). Отмечались зооспоры, несущие 2 жгутика
неравной длины. В некоторых случаях наблюдалось
формирование покоящихся спор (цист). Однако из-
за крайне слабой изученности этой группы организ-
мов сколько-нибудь связные данные об их жизнен-
ных циклах отсутствуют. Карлинг (Karling, 1944)
описал новый фаготрофный организм — Phagomyxa
algarum, встречающийся в клетках морских водоро-
слей. Плазмодии поглощают участки водорослевого
протопласта; в них были отмечены синхронные
ядерные деления (однако свидетельства крестовид-
ных делений отсутствуют). Наблюдалось формирова-
ние спорангиев и зооспор, сходных по виду с та-
ковыми у плазмодиофорид, но покоящиеся споры
найдены не были. Карлинг (Karling, 1944, 1968) рас-
сматривает этот организм как промежуточную фор-
му между Proteomyxa и Plasmodiophorea.
Идея о близости протеомиксид и плазмодио-
форид была высказана еще Ланкестером (Lankester,
1885), который установил новый класс Proteomyxa в
составе группы «монад» Ценковского (Monadineae
Cienkowski, 1865) и 2 известных к тому времени
родов Plasmodiophora и Tetramyxa. Однако в насто-
ящее время хорошо известно, что так называемые
протеомиксиды — сборная группа организмов, при-
надлежащих к разным классам и даже типам про-
стейших, поэтому, уже начиная с системы Левайна с
соавт. (Levine et al., 1980), Proteomyxida как конк-
ретный систематический таксон больше не сущест-
вует.
Гипотеза Карлинга о происхождении плазмодио-
форид от каких-то факультативных паразитических
амебоидных простейших, которые обладали фаго-
трофным способом питания и проникали внутрь рас-
тительных клеток, а также были способны к транс-
формации в жгутиковую стадию, кажется достаточно
убедительной. Их эволюция шла по пути адаптации
к облигатному внутриклеточному паразитизму, что
привело к усложнению их жизненного цикла и по-
явлению специального аппарата инвазии. Следует от-
метить, что если голозойное питание у плазмодиофо-
ровых действительно имеется, то, очевидно, их зна-
чимость как возможной промежуточной группы
между грибами и их предками возрастает.
Систематический обзор
Сем. Plasmodiophoridae Zopf,
1884
Развернутый диагноз, составленный на основе оп-
ределений, даваемых в упомянутых выше работах и
переосмысленных в свете новых данных, которые
были получены с помощью электронно-микроскопи-
ческих и других исследований, можно сформулиро-
вать следующим образом.
Облигатные внутриклеточные паразиты растений
со сложным жизненным циклом, включающим поко-
ящиеся цисты, первичные зооспоры, спорангиальный
плазмодий, спорангии, вторичные зооспоры, цисто-
генный плазмодий; основной тип питания осмотроф-
ный; вегетативная плазмодиальная стадия лишена
клеточной стенки, какие-либо специализированные
структуры или временные образования, обеспечива-
ющие питание, отсутствуют; увеличение числа ядер
за счет их последовательных синхронных крестовид-
ных делений; плазмодий может подвергаться плазмо-
томии (шизогонии) на несколько дочерних; зооспо-
ры с 2 гладкими (без мастигонем) апикальными жгу-
тиками неравной длины; для ряда видов доказано
формирование зооспоровых цист со сложным аппа-
ратом инвазии; вторичные зооспоры (и вегетативные
плазмодии) способны сливаться попарно без после-
дующей за этим кариогамии; непосредственно перед
формированием покоящихся цист, по всей видимос-
ти, происходит кариогамия и мейоз; цисты лежат
свободно, или агрегированы в небольшие 1руппы,
или формируют цистосорусы; спорангии отдельные
или объединены в спорангиосорусы.
Вся систематика этого семейства исходно постро-
ена на том, каким способом организованы зрелые
цисты: они могут быть одиночные и лежать в виде
рыхлой массы внутри оболочки клетки хозяина, но
могут быть объединены в цистосорусы различных
типов. Характер, форма и размеры спорангиев также
имеют определенную таксономическую ценность.
Тем не менее Карлинг (Karling, 1942, 1968) и другие
авторы неоднократно отмечали, что эти признаки
недостаточны для четкого разграничения родов и
видов. Так, было показано, что форма и размеры
ТИП PLASMODIOPHORA. ЛИТЕРАТУРА
409
цистосорусов и спорангиев Woronina pythii (паразита
водных грибов из рода Pythium) могут заметно варь-
ировать и в значительной степени зависят от ви-
довой принадлежности хозяина (Dylewski, Miller,
1983а). Определенная зависимость формы и разме-
ров цистосорусов от того, в клетках какой ткани
растения-хозяина они развиваются, была продемон-
стрирована у Ligniera veronicae (Miller et al., 1985)
и ряда других плазмодиофорид. Однако для нахож-
дения более адекватных систематических признаков
требуются дальнейшие исследования.
Всего к настоящему времени известно немногим
более 3 десятков видов плазмодиофорид, принадле-
жащих к родам Plasmodiophora, Tetramyxa, Octomyxa,
Sorosphaera, Spongospora, Sorodiscus, Ligniera, Woro-
nina и Polymyxa\ некоторые авторы поддерживают вы-
деление еще одного самостоятельного рода — МетЬ-
ranosorus (для М. heteranterae Ostenfeld et Petersen,
1930), другие же склоняются к тому, что его следует
признать синонимом Sorosphaera или Sorodiscus (Каг-
ling, 1968; Miller et al., 1985; Dylewski, 1990).
Род Plasmodiophora отличается от всех остальных
родов отсутствием цистосорусов. Цисты (рис. 345, Д)
лежат свободно, не соединяясь друг с другом. Спо-
рангии мелкие, лежащие свободно или объединенные
в спорангиосорусы. Этот род включает несколько
видов — паразитов наземных и водных высших рас-
тений.
У родов Tetramyxa и Octomyxa цисты соединяются
в небольшие группы: по 2 или 4 у первого и по 8 у
представителей второго рода.
Род Sorosphaera преимущественно имеет сфери-
ческие или эллипсоидные цистосорусы (рис. 345, Е).
Для рода Sorodiscus характерны уплощенные, диско-
видной формы цистосорусы. У рода Spongospora
(рис. 345, В) формируются губчатые, пронизанные
каналами и бороздками цистосорусы, часто овальной
или сферической формы (рис. 345, Г).
Представители остальных родов, имеющие цисто-
сорусы неопределенной формы и строения, различа-
ются по размерам и характеру организации споранги-
ев. Так, у Ligniera спорангии обычно мелкие, соеди-
ненные в рыхлые спорангиосорусы. У Woronina
спорангии более крупные, чем у предыдущего рода, и
встречаются по отдельности, хотя могут и образовы-
вать спорангиосорусы (рис. 345, Б). Спорангии Poly-
myxa крупные, удлиненной формы, дольчатые, с
длинными выводными трубками (рис. 345, Ж).
THE PHYLUM PLASMODIOPHORA ZOPF, 1994
SUMMARY
The phylum Plasmodiophora is a very specific group of obligato-
ry endoparasites of terrestrial and aquatic plants, algae, and fungi.
Different species of plasmodiophorids diverse in specificity to host
organisms (some of them are capable to infect a rather wide variety of
hosts, while there are plasmodiophorids arranged for some or even
any species of host), and their distribution follows that of their hosts,
accordingly.
First known species of plasmodiophorids — Plasmodiophora
brassicae — was described by Woronin in 1877. At the present time,
there is rather extensive literature, devoted to these group of protists,
especially to species, which caused serious diseases of food plants
(such as potatoes, cabbage, etc.). However, the number of questions
concerning life cycles and some other aspects of their biology still
remain unclear. The possible phylogenetic relationships of plasmodio-
phorids are quite doubtful also.
All plasmodiophorids have a complex life cycle. The main trophic
stage of plasmodiophorids is a multinucleate, unwalled plasmodial
protoplast (plasmodium), and it is reproduced in the life cycle twice.
Uninucleate thick-walled cysts (resting spores) germinate into primary
zoospores. The zoospores bear two flagella of unequal length without
mastigonemes. They penetrate into host cells and give rise to young
primary plasmodia, which feed and enlarge; the number of nuclei in-
creases by multiple synchronous mitoses. Mature primary (sporangio-
genous) plasmodia cleave into uninucleate sporangia (zoosporangia),
which are grouped into sporangiosori in many species of plasmodio-
phorids. Nuclei in sporangia undergo divisions during zoosporogene-
sis, and a number of uninucleate secondary zoospores are released at
the end of this process. Secondary zoospores infect the host’s cells,
and their protoplasts give rise to young secondary (cystogenous) plas-
modia. The growth of plasmodia is accompanied by multiple synchro-
nous mitotic nuclear divisions. At the end of development of seconda-
ry plasmodia meiotic nuclear divisions occur, which is preceded, as
considered, by binary karyogamy (but the existence and timing of
karyogamy in the life cycle of plasmodiophorids has not been convin-
cingly documented). At any rate, the diplophase in these protists is
quite transit. Then plasmodium cleaves into uninucleate haploid cysts
(resting spores). The specific aggregations of resting spores (cystoso-
ri) are characteristic feature of many species of plasmodiophorids.
Resting spores are released into the environment after the cell wall of
the host cell breaks off. They may remain viable up to several years
in some species.
Plasmodiophorids develop a very specific and complex infection
apparatus within the zoospores encysted prior to penetration through
the host cell wall. This apparatus is organised on the principle of
syringe, which inoculates the uninucleate protoplast of parasite into
the host cell cytoplasm. This fact and other features of plasmodiopho-
rides give evidence to an extreme degree of specialization and evolu-
tionary antiquity of this group of organisms.
The phylum Plasmodiophora is a comparatively homogenous
taxon. It includes the single class Plasmodiophorea with the order
Plasmodiophorida comprising a single family Plasmodiophoridae.
More than thirty species of plasmodiophorids are known to the present
time only. They are classified into nine genera: Plasmodiophora, Tet-
ramyxa, Octomyxa, Sorosphaera, Spongospora, Sorodiscus, Ligniera,
Woronina, Polymyxa. Some authors distinguish also Membranosorus
as one more taxonomically valid genus of plasmodiophorids.
ЛИТЕРАТУРА
Власов Д. Ю. Изучение цикла развития Polymyxa betae Kes-
kin И Микобиология и фитопатология. 1986. Т. 20. С. 350—353.
Воронин М. С. Plasmodiophora brassicae. Организм, причиняю-
щий капустным растениям болезнь, известную под названием килы //
Тр. СПб. О-ва естествоиспытателей. 1877. Т. 8. С. 171—201.
Мазин В. В., Проценко Е. П. Возбудитель килы крестоцвет-
ных — Plasmodiophora brassicae Woron. М., 1976.
Райков И. Б. Ядро простейших, морфология и эволюция. Л.,
1978.
Райков И. Б. Пути эволюции митотического аппарата у низ-
ших эукариот // Систематика простейших и их филогенетические
связи с низшими эукариотами. Л., 1986. С. 26—56. (Тр. ЗИН АН
СССР; Т. 144.)
Серавин Л. Н., Гудков А. В. Основные типы и формы агамных
слияний клеток у простейших И Цитология. 1984. Т. 26. С. 123—
131.
Ainsworth G. С. Introduction and keys to higher taxa // The fun-
gi. Vol. 4B. New York, 1973. P. 635—648.
410
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Aist J, R., Williams P. H. The cytology and kinetics of cabbage
rot hair penetration by Plasmodiophora brassicae // Can. J. Bot. 1971.
Vol. 49. P. 2023—2034. "
Alexopoulos C. J. Introductory mycology (2nd ed.). New York,
1962.
Braselton J. P., Miller С. E. Centrioles in Sorosphaera // Mycolo-
gia. 1973. Vol. 65. P. 220—226.
Braselton J. P., Miller С. E., Pechak D. G. The ultrastructure of
cruciform nuclear division in Sorosphaera veronicae (Plasmodiopho-
romycete) // Amer J. Bot. 1975. Vol. 62. P. 349—358.
Buczacki S. T. Plasmodiophora. An inter-relationship between
biological and practical problems // Zoosporic plant pathogenes. A
modern perspective. London; New York: Acad. Press, 1983. P. 161—
191. ’
Buczacki S. T., Moxham S. E. Karyogamy in Plasmodiophora
brassicae // Trans. Brit. Mycol. Soc. 1980. Vol. 75. P. 439—444.
Cook W. R. I. The methods of nuclear division in the Plasmodio-
phorales // Ann. Bot. 1928. Vol. 62. P. 347—377.
Cook W. R. I. A monograph of the Plasmodiophorales // Arch.
Protistenk. 1933. Bd 80. S. 179—254.
D’Ambra V., Mutto S. The ultrastucture of Polymyxa betae zoo-
spore exit-tube differentiation // Can. J. Bot. 1977. Vol. 55. P. 831—
839.
De Вагу A. Comparative morphology and biology of the Fungi,
Mycetozoa and Bacteria (English trans.). London, 1887.
Dylewski D. P. Phylum Plasmodiophoromycota // Handbook of
Protoctista. Boston. Jones and Burlett Publishers. 199Q. P. 399—416.
Dylewski D. P., Miller С. E. Systematic and host range studies
of Woronina pythii (PlasmodiOphoromycetes) and host, Pythium
species, from axenic culture // Mycologia. 1983a. Vol. 75. P. 412—
422.
Dylewski D. P., Miller С. E; Cruciform nuclear division in Woro-
nina pythii (PlasmodiOphoromycetes) // Amer. J. Bot. 1983b. Vol. 70.
P. 1323—1339.
Dylewski D. P., Miller С. E. Nuclear elimination during cyst for-
mation in Woronina pythii (PlasmodiOphoromycetes) // Mycologia.
1983c. Vol. 75. P. 271—279.
Dylewski D. P., Miller С. E. The ulrastructure of mitosis during
sporangiogenesis in Woronina pythii (PlasmodiOphoromycetes) // Pro-
toplasma. 1984a. Vol. 121. P. 42—53.
Dylewski D. P., Miller C. E. The ultrastructure of meiosis in Wo-
ronina pythii (PlasmodiOphoromycetes) // Mycologia. 1984b. Vol. 76.
P. 1075—1088.
Honigberg В. M., Balamuth W. Bovee E. C. et al. A revised clas-
sification of the phylum Protozoa 7/ J. Protozool. 1964. VoL ll.
P.7—20.
Ingram D.S. Growth of Plasmodiophora brassicae in host cal-
lus II J. Gen. Microbiol. 1969. Vol. 55. P. 9—18.
Ingram D. S., Tommerup 1. C. The life history of Plasmodiopho-
ra brassicae Woron. // Proc. Roy. Soc. London. Ser. B. 1972.
Vol. 180. P. 103—112.
Karling J. S. The Plasmodiophorales. New York, 1942.
Karling J. S. Phagomyxa algarum n. gen., n. sp., an unusual para-
site with plasmodiophoralan and proteomyxean characteristics //
Amer. J. Bot. 1944. Vol. 31. P. 38—52.
Karling J. S. The Plasmodiophorales. New York; London, 1968.
Kole A.; P. A contributios to the knowledge of Spongospora sub-
terranea (Wall.) Lagerh,, the cause of powdery scab of potatoes //
Tijdschr. Platenziekten. 1954. Bd 60. S. 1—65.
Kble A. P., Gielina A. J. Electron microscope observations of the
flagella of the zoosporarigial zoospores of Plasmodiophora brassicae
and Spongospora subterranea // Proc. Kon. Ned. Acad. Wetersch. Ser. C.
196L Vol. 64. P. 157—161.
- Lankester E. R. Protozoa // Encycl. Brit. (9th ed.). 1885. Vol. 19.
P. 830—866.
Lee J. J., Hutner S. H., Bovee E. C. (Eds). An illustrated guide
to the Protozoa. Lawrence, 1985.
Levine N. D., Corliss J. O., Cox F. E. G. et al. A newly revised
classification of the Protozoa // J. Protozool. 1980. Vol. 27. P. 37—58.
Miller С. E., Dylewski D. P. Zoosporic fungal pathogenes of lower
plants U Zoosporic plant pathogenes. A modern perspective. London;
New York: Acad. Press, 1983. P. 249—283.
Miller С. E., Martin R. W., Dylewski D. P. The ultrastructure of
plasmodia, sporangia and cystosori of Ligniera verrucosa (PlasmodiO-
phoromycetes) // Can. J. Bot. 1985. Vol. 63. P. 263—275.
Olive L.S. The Mycetozoa: a revised classification // Bot. Rev.
1970. Vol. 36. P. 59—89.
Olive L. S. The Mycetozoans. New York ect., 1975.
Page F. C. The classification of «naked» amoebae (phylum Rhizo-
poda) // Arch. Protistenk. 1987. Bd 133. S. 199—217.
Sparrow F. K. Aquatic Phy corny cetes. Michigan, 1960.
Tommerup L, Ingram D. S. The life-cycle of Plasmodiophora
brassicae Woron. in Brassica tissue cultures and in intact roots // New
Phytol. 1971. Vol. 70. P. 327—332.
Williams P. H., McNabola S. S. Fine structure of Plasmodiophora
brassicae in sporogenesis // Can. J. Bot. 1967. Vol. 45. P. 1665—
1669.
Williams P. H., McNabola S. S. Fine structure of the host-parasite
interface of Plasmodiophora brassicae in cabbage // Phytopathology.
1970. Vol. 60. P. 1557—1561.
Williams P. H., Yukawa J. B. Ultrastructural studies on the host-
parasite relations of Plasmodiophora brassicae // Phytopathology.
1967. Vol. 57. P. 628—687.
Williams P. H., Reddy M. N., Strandberg J. O. Growth of nonin-
fected and Plasmodiophora brassicae infected cabbage callus in cultu-
re // Can. J. Bot. 1969. Vol. 47. P. 1217—1221.
Тип MYCETOZOA de Вагу, 1859 — МИКСОМИЦЕТЫ
К типу относятся собственно миксомицеты и церкомонады, име-
ющие множество общих черт в строении жгутиковых клеток (Кар-
пов, 1990). Жгутиковые клетки представлены вегетативными осо-
бями или зооспорами с 1—2 апикальными жгутиками. Корешковая
система жгутиков однообразно устроена в пределах типа и представ-
лена характерным набором латеральных микротрубочковых корешков
и 1 коротким фибриллярным корешком. Митохондрии с трубчаты-
ми или пузырьковидными кристами. В жизненном цикле обычно
есть плазмодиальная и амебоидная стадии, формирующие фило-
подии или субпсевдоподии филозного типа.
Сапрофитные и фаготрофные, водные и почвенные организмы.
В типе 2 класса: Cercomonadea и Eumycetozoea.
Класс Cercomonadea Mylnikov,
1986 — Церкомонады
К классу относятся миксомицеты, не образующие
плодовых тел.
Отряд Cercomonadida (Vickerman)
Mylnikov, 1986
Амебоидные организмы с 2 гладкими гетеродинамичными
жгутиками. В амебоидном состоянии жгутики сохраняются. Псев-
доподии типа лобоподий или филозных субпсевдоподий образу-
ются в любом участке, но чаще на заднем конце. В жизненном
цикле нескольких видов обнаружено образование многоядерных
плазмодиев, а также цисты. При передвижении клетки ползают
по субстрату, реже плавают. Питаются бактериями и мелкими
водорослями, захватываемыми псевдоподиями. Хищничества не
отмечено. Цитостома нет. В клетках найдены стрекательные орга-
неллы типа микротоксицист, трихоцист и кинетоцист. Один из
микротрубочковых корешков образует конус, направленный к яд-
ру. Кремниевых структур не отмечено. Митохондрии с трубчаты-
ми или везикулярными кристами.
Свободноживущие. Обитают на дне морских и пресных водо-
емов, в очистных сооружениях, обильны в почве.
История изучения
Дюжарден в начале века (Dujardin, 1841) описал
амебоидных жгутиконосцев рода Cercotnonas, имею-
щих 1 жгутик. Затем в 1886 г. И. Красильщик опи-
сал новый род Cercobodo, который отличался от
Cercomonas дополнительным жгутиком, направлен-
ным назад. Большинство систематиков и исследова-
телей тонкой структуры этих организмов считают
оба названия синонимами (Patterson, Zolffel, 1991),
поскольку в действительности второй жгутик у Cer-
comonas часто малозаметен. В начале XX в. церко-
монад относили к отряду Rhizomastigida Doflein,
1916. Затем Холланд (Hollande, 1952) поместил их в
подотряд Bodonina. Наиболее полный определи-
тель данной группы жгутиконосцев составил Жу-
ков (1971).
В настоящее время после изучения ультраструк-
туры церкомонад стало ясно, что они в значитель-
ной мере отличаются как от бодонид, так и от
других групп зоофлагеллат (Мыльников, 1986). Ка-
валье-Смит (Cavalier-Smith, 1993) объединил церко-
монад и тауматомонад вместе с некоторыми аме-
боидными жгутиконосцами в класс Heteromitea Ca-
valier-Smith, 1993 и отнес их к типу Opalozoa
Cavalier-Smith, 1991. Затем после изучения последо-
вательности нуклеотидов генов 185 рРНК церкомо-
нады были помещены в класс Monadofilosa Cavalier-
Smith, 1996, который относится к типу Rhizopoda
Dujardin, 1835 emend. Cavalier-Smith, 1996. В насто-
ящее время их сближают с миксомицетами (протос-
телидами) и помещают в качестве самостоятельного
класса в тип Myxomycetozoa (Карпов, 1997; наст,
издание).
Классификация
В класс Cercomonadea входит 1 отряд Cercomona-
dida. В нем выделяют 2 семейства (различаются по
наличию жгутикового кармана и длине переднего
жгутика) с 6 родами и более чем 60 видами.
Класс Cercomonadea Mylnikov, 1986
Отряд Cercomonadida Mylnikov, 1986
Сем Cercomonadidae Kent, 1880
Сем. Heteromiiidae nom nov
412
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 351. Морфология церкомонад.
А, Б — Cercomonas longicauda: А — трофонт; Б — подвижная форма; В, Г — трофонт С. metabolicus: Д — С. plasmodialis: Е — С. cometa: Ж — фраг-
ментация плазмодия С. longicauda. (А, Б, Ж — по: Мыльников, 1985а; В, Г — по: Жуков, Мыльников, 1987; Д — по: Мыльников, 19856; Е ориг.
А. П. Мыльникова).
Fig. 351. Cercomonad morphology.
А, Б — Cercomonas longicauda: А — trophozoit, Б — moving cell; В, Г — trophozoit of C. metabolicus: Д — C. plasmodialis: Е — С. cometa: Ж — plasmo-
dium fragmentation of C. longicauda.
Морфология и физиология
В момент активного ползания по субстрату или
плавания большинство церкомонад имеет овальную
форму тела (рис. 351). Ядро расположено в передней
части клетки, вблизи базальных тел жгутиков. Двига-
тельный жгутик направлен вперед и совершает греб-
ные движения, рулевой тянется назад под телом жгу-
тиконосца. Псевдоподии различной формы и разме-
ров образуются на любом участке тела, но чаще на
заднем конце. Форма псевдоподий сходна с таковой
у амеб Leptomyxa, Acanthamoeba, Vanella. У 2 видов
псевдоподии ветвятся. Толщина псевдоподий состав-
ляет 0.2—1.0 мкм.
Клетки ограничены унитарной мембраной. У 2 ви-
дов обнаружен уплотненный слой гликокаликса. На-
личие каких-либо плотных покровов или чешуек не
отмечено (рис. 352, вкл.). Жгутиковый карман най-
ден у представителей рода Heteromita (Bodomorpha).
Рулевой жгутик может проходить в продольной бо-
роздке, или внутри канала, или свободно. Мастигоне-
мы не обнаружены. В амебоидном состоянии, в плаз-
модии и при делении клетки жгутики сохраняются.
Этим церкомонады отличаются от миксомицетов и
шизопиренид (см. с. 417 и 485). Аксонема жгутиков
устроена обычным образом (по формуле 9 + 2). Цен-
тральные микротрубочки аксонемы упираются в не-
большую аксосому. Поперечная пластинка незначи-
тельно приподнята над поверхностью клетки. От са-
ТИП MYCETOZOA. КЛАСС CERCOMONADEA
413
Рис. 354. Схема строения клетки церкомонад. (Ориг. А. П. Мыль-
никова).
ик — исчерченный фибриллярный корешок, л — липидные капли, лм —
латеральные микротрубочковые корешки, мк — микротрубочковый конус,
пж — передний жгутик, рж — рекуррентный жгутик, св — сократитель-
ная вакуоль, э — экструсомы. Остальные обозначения как на рис. 352.
Fig. 354. Scheme of cercomonad cell structure.
ик — striated fibrillar rootlet, л — lipid droplets, лм — lateral microtubular
rootlets, мк — microtubular cone, пж — anterior flagellum, рж — recurrent
flagellum, ce — contractile vacuole, э — extrusomes. Other abbreviations are
the same as on fig. 352.
теллитов кинетосом отходят: микротрубочковый ко-
нус (не у всех клеток), охватывающий ядро, и 2 или
3 ленты, состоящие из 3—11 микротрубочек. От-
дельные микротрубочки проходят в стенке жгутико-
вого кармана или канала и редко в псевдоподиях. По-
перечно исчерченные структуры малозаметны.
Ядро пузырьковидного типа, обычно яйцевидной
формы, содержит 1 центрально расположенное яд-
рышко. Размер ядра у мелких видов достигает 1—2,
у крупных 2—5 мкм. Деление клеток продольное.
Оболочка ядра во время митоза исчезает, от центрио-
лей отходят микротрубочки веретена.
Вблизи ядра располагается 1 крупная митохонд-
рия (рис. 353, вкл.) (Heteromita) или несколько мел-
ких (рис. 352). Кристы митохондрии трубчатые или
везикулярные. Микротельца, ограниченные мембра-
ной, с мелкозернистым содержимым, лежат около
ядра или в других частях клетки. У Heteromita (Во-
domorpha) reniformis микротельце, митохондрия и яд-
ро тесно примыкают друг к другу и синхронно де-
лятся. Сократительная вакуоль у пресноводных видов
имеет обычное строение. Аппарат Гольджи представлен
у мелких видов 1 диктиосомой, состоящей из не-
скольких цистерн, у крупных — несколькими диктио-
сомами. ЭПР развит незначительно. Обобщенная схема
строения церкомонад представлена на рис. 354.
Для церкомонад характерны стрекательные орга-
неллы 3 типов: микротоксицисты, трихоцисты и ки-
Рис. 355. Экструсомы церкомонад. (Ориг. А. П. Мыльникова).
А, Б — микротоксицисты: покоящаяся (А) и выстрелившая (F); В — кине-
тоциста; Г — осмиофильное тело; Д, Е — трихоцистоподобная структура
на поперечном (Д) и продольном (£) срезах.
Fig. 355. Cercomonad extrusomes.
А, К — microtoxicysts: intact (A), after extrusion (К); В — kinetocyst; Г —
osmiophylic body; Д, E — trichocyst-like structure in cross (Д) and longitudi-
nal (E) sections.
нетоцисты (рис. 355). Первые образуются в аппарате
Гольджи, а остальные — в лакунах ЭПР. Микроток-
сицисты при выстреливании впрыскивают свое со-
держимое в бактерию и лизируют ее. Функции других
экструсом неясны. Кроме того, в клетках часто встре-
чаются субмембранные осмиофильные тела, участ-
вующие в скольжении жгутиконосца по субстрату.
Церкомонады питаются мелкими бактериями, ре-
же мелкими водорослями, захватываемыми псевдопо-
диями. В культуре лишь небольшое число клеток
беспорядочно плавает в толще среды, остальные ак-
тивно ползают по субстрату. При остановке жгутико-
носец расплющивается и начинает активно питаться.
При этом работа жгутиков замедляется. У некоторых
видов во время ползания рулевой жгутик совершает
волнообразные движения. Участие псевдоподий в пе-
редвижении клеток не установлено.
Жизненный цикл
Наиболее детально жизненный цикл изучен у пред-
ставителей сем. Cercomonadidae. Он состоит из мо-
надной, амебоидной, плазмодиальной стадий и ста-
дии цисты. Переход из монадной стадии в амебоид-
ную происходит в течение нескольких секунд.
Цисты появляются при старении культуры. У одних
видов оболочка цист складчатая (рис. 356), у дру-
гих — гладкая. Пробочка не обнаружена. Внутри
цист находятся ядро, уплотненная цитоплазма с ос-
миофильными гранулами (полисахариды) и капли
липидов. Жгутики отсутствуют. Рибосомы распола-
гаются плотно упакованными рядами. При эксцисти-
ровании оболочка цисты разрушается.
Рис. 356. Внешний вид цисты. (Ориг. А. П. Мыльникова).
Fig. 356. General view of cyst.
414
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 357. Строение плазмодия.
(Ориг. А. П. Мыльникова).
Рис. 358. Внешний вид
трофонта Cercomonas lon-
gicauda.
(Ориг. А. П. Мыльникова).
Fig. 358. General view of
Cercomonas longicauda
trophozoit.
Fig. 357. Plasmodium structure.
Некоторые виды в культуре регулярно образуют
многоядерные разветвленные плазмодии — клеточ-
ные образования, содержащие увеличенное число
клеточных органелл: ядер, жгутиков, сократительных
вакуолей и других компонентов клетки (рис. 357; 363,
вкл.). Плазмодии, по некоторым данным, появляются
в результате многократной) митоза и нарушений ци-
тотомии. Плазмодий отпочковывает отдельные клет-
ки и постепенно переходит в одноядерное состоя-
ние. Плодовых тел и слияния клеток не отмечено.
Половой процесс достоверно неизвестен.22
Практическое значение
Несколько видов церкомонад являются надежны-
ми индикаторами сапробности вод (SladeCek, 1973).
Они могут служить также показателем качества ра-
боты очистных сооружений. Имея сходный с бодо-
нидами ареал, церкомонады играют существенную роль
в круговороте органического вещества в морских и
пресных водоемах, в первую очередь в составе бен-
тоса и перифитона, где они достигают значительной
численности (Larsen, Patterson, 1990; V0rs, 1992).
Это необходимо учитывать при выяснении роли про-
стейших в природных экосистемах.
Филогения
Некоторые авторы относили церкомонад к бес-
цветным представителям хризофитовых водорослей
(Starmach, 1980). Сходство отмечалось в гетеродина-
22 Только у Sainouron предполагается копуляция (Жуков, 1971;
Patterson, Zolffel, 1991).
Рис. 359. Внешний вид трофонта Cercomonas granulifera. (Ориг.
А. П. Мыльникова).
А — подвижная особь, Б — трофонт.
Fig. 359. General view of Cercomonas granulifera trophozoit.
A — moving cell, Б — trophozoit.
мичности жгутиков и амебоидности особей. Однако
структура корневой системы жгутиков, строение
цист, покровы и жизненные циклы в этих группах
существенно различаются. На основании изучения
последовательности нуклеотидов генов 185 рРНК пока-
зано, что Cercomonas является предковой группой для
Heteromita и, возможно, Thaumatomonadida, а также
Testaceafilosia (Cavalier-Smith, 1996). Большое сход-
ство наблюдается у церкомонад со слизистыми грибами,
особенно с протостел идами (Protostelida). Их объеди-
няет наличие мйкротрубочкового конуса, способ рас-
положения кинетосом, яйцевидная форма ядра, спо-
соб питания, наличие микротелец, стадия плазмодия.
Однако церкомонады не развиваются на твердых сре-
дах и не образуют плодовых тел (наиболее характерный
признак слизистых грибов). Церкомонад можно рас-
сматривать как предковую группу протостелид (Schus-
ter, Pollack, 1978; Карпов, 1990).
Систематический обзор
Сем. Cercomonadidae Kent, 1880
(Син.: Cercobodonidae Hollande, 1942)
Клетки амебоидные, ползающие, с 2 голыми гете-
родинамичными жгутиками. Микротрубочковый конус
обычно охватывает ядро. Псевдоподии широкие, раз-
личной формы, в том числе разветвленные. Образуют
плавающие клетки, многоядерные плазмодии и цис-
ты. Стрекательные органеллы типа микротоксицист
и трихоцист. Обитают в почве, морских и пресных
водоемах.
В состав семейства входит 3 рода.
ТИП MYCETOZOA. КЛАСС CERCOMONADEA
415
Рис. 360. Cercomonas соте-
ta на стадии бляшки.
(Ориг. A. IL Мыльникова).
Fig. 360. Aggregation of Cer-
comonas cometa.
Рис. 361. Внешний вид трофонта Helkesimas-
tix faecicola. (Ориг. А. П. Мыльникова).
А — в профиль, Б — со спинной стороны.
Fig. 361. General view of Helkesimastixfaeci-
cola trophozoit.
A — profile, Б — from dorsal side.
Род Cercomonas Dujardin, 1841
(Син.: Cercobodo Krassiltschick, 1886;
Dimastigamoeba Blochmann, 1895).
Клетки ползающие, активно метаболирующие, с
2 гетеродинамичными жгутиками. Длинный, перед-
ний жгутик совершает гребные движения. Задний
жгутик проходит под вентральной стороной тела.
Псевдоподии различной формы, чаще образуются на
заднем конце тела. Ядро обычно в передней поло-
вине тела. Вблизи базальных тел жгутиков начинает-
ся микротрубочковый конус, который охватывает яд-
ро. Цисты круглые, с тонкой или утолщенной оболоч-
кой.
Типичный представитель — Cercomonas longica-
uda — имеет яйцевидное или веретеновидное тело.
Длина 18—36, ширина 9—14 мкм (рис. 358). Боль-
шая псевдоподия образуется на заднем конце тела. У
трофонтов псевдоподии окаймляют периметр тела.
Сократительная вакуоль обычно находится сзади. Жгу-
тики равной длины. В момент остановки движения
образует простые или разветвленные псевдоподии, ко-
торые активно поглощают бактерии. Порошица в за-
днем конце тела. Предполагается, что в перемещении
клеток по субстрату активную роль играет рулевой
жгутик, совершающий ундулирующие движения. Раз-
множается продольным делением надвое. Образует
шаровидные цисты диаметром 6—7 мкм. Клетки
крупного жгутиконосца С. granulifera наполнены боль-
шим количеством гранул запасного вещества. В ста-
дии трофонта форма тела практически не отличает-
ся от амебы (рис. 359).
Другой представитель — С. cometa образует раз-
ветвленные псевдоподии и выделяет в стадии тро-
фонта обильную слизь. Слой трофонтов образует
крупную бляшку, заметную невооруженным глазом
(рис. 360). После окончания формирования бляшки
клетки расползаются.
Род Massisteria Larsen et Patterson,
1990
Клетки малоподвижны, уплощены и прижаты к
субстрату. 2 коротких жгутика неактивны. Клетки
образуют очень тонкие разветвленные псевдоподии,
содержащие мелкие стрекательные органеллы и
пучки микротрубочек. В жизненном цикле описаны
плавающие клетки и многоядерные особи, напомина-
ющие плазмодии. Цисты неизвестны.
1 вид — М. marina Larsen et Patterson, 1990.
Диаметр трофонтов равен 2.5—5.0 мкм (рис. 362,
В, Г)- 2 неактивных жгутика отходят на дорсальной
стороне клетки. Псевдоподии ветвятся и иногда анас-
томозируют. Дочерние особи образуют бляшки (клас-
тер), окруженный слизью (мукоидным материалом).
Плавающие особи не образуют псевдоподий, не пита-
ются и несут 2 гетеродинамичных жгутика. Обитают
в солоноватых и морских водах.
Род Helkesimastix Woodcock et Lapage,
1914
Клетки ползающие, сильно метаболирующие,
продолговатые, с небольшим рострумом. Передний
жгутик сильно редуцирован, задний тянется назад и
равен по длине телу. Задний жгутик сопровождается
двойным рядом гранул. Ядро в передней части тела,
сократительная вакуоль в задней. Небольшая псевдо-
подия образуется только на заднем конце тела.
Обитают в почве и пресных водоемах.
Представители рода Helkesimastix в целом напо-
минают Heteromitidae, но отличаются от последних
интенсивной метаболией тела и способом ползания
по субстрату. 1 вид — И faecicola (рис. 361). По не-
которым данным (Жуков, 1971), отмечен половой
процесс (гологамная копуляция). Встречается в со-
ставе бентоса и в очистных сооружениях.
Сем. Heteromitidae nom. nov.
(Син.: Bodomorphidae Hollande, 1952;
non Heteromitidea Cavalier-Smith, 1993)
Диагноз. Двухжгутиковые ползающие протесты
с ригидным телом и 2 гетеродинамичными жгутика-
ми. Микротрубочковый конус отсутствует. Передний
конец образует небольшой рострум. Жгутики выхо-
дят из небольшого углубления. Задний конец тела
приподнят над субстратом. Передний жгутик редуци-
рован и совершает частые гребные движения. Задний
жгутик соприкасается с субстратом. Задний конец
тела постоянно раскачивается. Экструсомы типа ки-
нетоцист. Плазмодии не образуются. Обитают на дне
пресных и морских водоемов, а также в почве.
К сем. Heteromitidae относятся род Heteromita и,
возможно, роды Allantion и Sainouron.
Имя рода Bodomorpha было признано младшим
синонимом Heteromita (Patterson, Zolffel, 1991). По-
416
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 362. Внешний вид некоторых церкомонад.
А, Б — трофонт Heteromita reniformis: А — в профиль, Б — со спинной стороны; В, Г — Massisteria marina: В — неподвижный трофонт со жгутика-
ми (ж), Г — плавающая клетка (мигрант); Д — Allantion tachyploon: Е — Sainouron mikroteron. (А, Б — ориг. А. П. Мыльникова; В, Г — по: Patterson,
Fenchel, 1990; Д — по: V0rs, 1992; Е — по: Жуков, 1971).
Fig. 362. General view of some cercomonads.
А, Б — trophozoit of Heteromita reniformis: A — profile, Б — from dorsal side; В, Г — Massisteria marina: В — trophozoit with flagella (ж), Г — swimming
cell; Д — Allantion tachyploon: E — Sainouron mikroteron.
этому новое название семейства (Heteromitidae вмес-
то Bodomorphidae) дано по имени рода Heteromita.
Род Heteromita Dujardin, 1841
(Син.: Bodomorpha Hollande, 1952)
По форме тела и характеру движения представи-
тели Heteromita напоминают бодонид (см. с. 212).
Клетки овальные или в форме запятой с небольшим
рострумом. Ниже рострума из небольшого углубле-
ния отходят 2 жгутика. Передний жгутик редуциро-
ван, совершает быстрые машущие движения и на-
правлен в сторону. Задний жгутик контактирует с
субстратом. При движении задний конец тела раска-
чивается. Небольшие псевдоподии отходят чаще от
переднего конца клетки. Цисты известны.
Обитают в почве, пресных и морских водах.
Heteromitidae incertae sedis
Из почвы были описаны 2 рода мелких ползаю-
щих жгутиконосцев (Sandon, 1924). По наличию не-
большого апикального углубления (жгутикового карма-
на) и редукции переднего жгутика, который незаметен,
напоминают Heteromita. Уточнение их системати-
ческого положения возможно после изучения ульт-
раструктуры данных жгутиконосцев.
Род Allantion Sandon, 1924
(рис. 362, Д)
Клетки овальные, немного изогнутые, ригидные.
Задний жгутик, контактирующий с субстратом, отхо-
дит из небольшого субапикального углубления. Перед-
ний жгутик мало заметен. Задний конец тела содер-
жит несколько светопреломляющих гранул; во время
движения он раскачивается и не соприкасается с суб-
стратом. Цисты известны.
Обитают в почве, пресных и морских водах.
Единственный вид — A. tachyploon Sandon, 1924.
Длина клеток 7—14 мкм.
Род Sainouron Sandon, 1924
(рис. 362, Е)
Клетки овальные или бобовидные, слабометабо-
лирующие, ползающие. Задний жгутик свободный,
соприкасается с субстратом. Задний конец клетки
приподнят над субстратом. Передний жгутик незаме-
тен, но предполагается. Сократительная вакуоль не
описана. Движение быстрое, вибрирующее. Отмечена
копуляция. Цисты известны.
Обитают в почве и пресных водах.
Типовой вид — S. mikroteron Sandon, 1924.
Средняя длина клеток 3—5 мкм, реже встречаются
особи до 14 мкм.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
417
THE PHYLUM MYCETOZOA DE BARY, 1859
THE CLASS CERCOMONADEA MYLNIKOV
SUMMARY
The cercomonads are a group of free-living widespread amoeboid
flagellates with two smooth heterodynamic flagella, which remain on
amoeboid stage. The cells are limited by a plasmalemma. The
dense envelope or scales are not marked. The flagella are present du-
ring all life stages besides cysts. These organisms produce lobopodia
or filose subpseudopodia in any part of the cell, but more often at the
posterior end. The ovoid nucleus is located in the anterior part of a
cell. The nuclear envelope disappears during mitosis and mitotic spin-
dle is connected with centrioles.
Anterior flagellum is directed ahead and makes flapping movement;
the recurrent flagellum runs back under the ventral cell side being loca-
ted in longitudinal groove, in cytoplasmic tube, or freely. Flagellar root-
lets are arranged in the same mode as in zoospores of myxomycetes. In
some species the single microtubules can be found in pseudopodia.
Microbodies with granular contents, limited by a membrane, often
apply to the nucleus or occur in other parts of a cell. Fresh-water
species have contractile vacuoles of usual structure. One or more usual
dictyosomes occur in cytoplasm. Cells normally contain one type of
extrusomes (microtoxicysts, trichocysts or kinetocysts). Mitochondria
with tubular or vesicular cristae.
The life cycle includes formation of multinuclear plasmodia with
multiplied cell organelles, and cysts. When ageing the plasmodium
buds single cells. Fruiting bodies are not known. Cysts are produced
in ageing culture. The plug has not been found in the cyst. The sexual
process is authentically not known.
Trophonts feed on bacteria and small algae, which are captured by
pseudopodia. Cytostome is absent. Predation is not known.
Cercomonads widely occur in fresh, brackish, marine and sewage
waters, and often in soil. Some cercomonads are used as the indicators
of saprobolity of waters. They play an essential role in circulation of
organic matter in marine and fresh waters.
The class Cercomonadea contains one order Cercomonadida, con-
sisting of two families, which have more than 60 species.
THE FAMILY CERCOMONADIDAE KENT, 1880
(SYN.: CERCOBODONIDAE HOLLANDE, 1942)
Cells are gliding amoeboid with two long heterodynamic flagella.
Microtubular cone usually covers a nucleus. Pseudopodia are of various
forms, branching included. Life cycle consists of monad, amoeboid,
plasmodial and cyst stages. The transformation from monad stage in
amoeboid one appears in a few seconds. Cytoplasm contains
microtoxicysts and trichocysts. Occurrence in marine and fresh waters,
and soil.
The family contains 3 genera.
Genus Cercomonas Dujardin, 1841
Genus Massisteria Larsen et Patterson, 1990
Genus Helkesimastix Woodcock et Lapage, 1914
The representatives of the genus Helkesimastix resemble Hetero-
mitidae and differ from the latter by intensive metaboly of the body
and the movement mode.
THE FAMILY HETEROMITIDAE NOM. NOV.
(SYN.: BODOMORPHIDAE HOLLANDE, 1952;
NON HETEROMITIDEA CAVALIER-SMITH, 1993)
Biflagellar gliding protists with rigid body and two heterody-
namic flagella. Microtubular cone is absent. The cell has a small ros-
trum at the anterior end. Flagella arise from a small pocket. Anterior
flagellum is shortened and makes frequent sweeps. The posterior end of
the body is shaking during the movement. Cells contain kinetocysts.
Plasmodia are not formed. Occurrence in fresh and marine waters, also
in soil.
This family contains the genus Heteromita Dujardin, 1841 and,
probably, two more genera: Allantion Sandon, 1924 and Sainouron
Sandon, 1924, which have been described from soil. The latter orga-
nisms have one or two flagella and rigid body. The anterior flagellum
is reduced considerably or absent. During the gliding movement the
posterior cell end is directed away from the substratum.
The systematic position of Allantion and Sainouron can be revised
after a study of their ultrastructure.
Phylogeny. On the basis of study SSU rRNA genes it is shown
that Cercomonas is an ancestral group for Heteromita and, probably,
Testaceafilosea. Cercomonads seem to demonstrate some similarity
with mycetozoans, especially with Protostelida. They have analogous
microtubular cone, arrangement of kinetosomes, ovoid nucleus, mode
of feeding, microbodies, plasmodial stage bf life cycle. However cer-
comonads are not able to form the fruiting bodies as mycetozoans. It
is possible to consider cercomonads as an ancestral group for protos-
telids and for myxomycetes as a whole.
Класс Eumycetozoea Zopf, 1884 — Эумицетозои
(слизевики, миксомицеты)
Широко распространенная группа свободноживущих назем-
ных организмов, обитающих преимущественно на остатках расти-
тельного, реже животного и грибного происхождения. Обладают
голозойным (фаготрофным) типом питания, поглощают разнооб-
разные микроорганизмы, простейших, клетки водорослей и гри-
бов. Имеют сложный жизненный цикл. Образуют плодовые те-
ла — спорокарпы.
История изучения
История изучения этих организмов насчитывает
уже около 200 лет. В последние десятилетия интерес
к ним стимулировался многочисленными цитологичес-
кими, биохимическими, биофизическими и генетичес-
кими исследованиями, в которых слизевики выступали
в роли модельных объектов. Кроме того, изучение
этой своеобразной группы организмов имеет большое
значение для эволюционных и филогенетических пос-
троений, а также систематики грибов и простейших.
Слизевикам были посвящены работы многих вы-
дающихся микологов и протистологов (de Вагу, 1859;
Rostafinski, 1875; Lister, 1894, 1911, 1925; Ячевский,
1907; Martin, 1960; Alexopoulos, 1969; Jahn et al.,
1974; Olive, 1975, и др.).
Де Бари (de Вагу, 1887) впервые убедительно по-
казал, что между слизевиками и грибами существует
лишь внешнее сходство. Он отнес слизевиков к цар-
ству животных и предложил для них новое название
Mycetozoa (животные-грибы), тем самым обратив
внимание на их двойственную природу. Тип (отдел)
Mycetozoa включал 2 класса: Acrasiees и Myxomyce-
tes. В первый вошли слизевики, миксамебы которых
агрегируют, не сливаясь, и образуют псевдоплазмо-
дий. Класс Myxomycetes был разделен на Endospo-
геае и Exosporeae в зависимости от типа спороноше-
ния. В 1875 г. Ростафинский опубликовал первую
специальную монографию по слизевикам (Rostafihs-
ki, 1875). В его систему также были включены акра-
зиевые, но исключен род Ceratiomyxa.
В 1970 г. американский миколог и протозоолог
Олайв возродил концепцию де Бари о близости мик-
27 Протисты, часть 1
418
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
сомицетов к животным (Olive, 1970). В значитель-
ной мере это было связано с открытием своеоб-
разной группы — протостелид. Олайв предложил
рассматривать слизевиков в типе Protozoa, указав
на то, что в силу традиций, а также отсутствия оп-
ределенности в оценке «таксономического расстоя-
ния», отделяющего слизевиков от других низших
эукариот, не существует единой точки зрения на
положение слизевиков в системе низших эукариот.
Следствием двойственности биологии этих организ-
мов является одновременное существование 2 систем
миксомицетов, причем в одной рассматриваются
только миксогастриды и цератиомиксовые (Martin,
Alexopoulos, 1969), а в другой особое внимание уде-
ляется протостелидам, которые проявляют наиболь-
шее сходство с простейшими (Olive, 1967, 1969,
1975).
Классификация
Слизевиков относят либо к грибам (отдел Мухо-
mycota — Hawksworth et al., 1995), либо к простей-
шим (класс Eumycetozoea — Olive, 1975). Причем
если в первой системе отдел Myxomycota включает
2 класса — Myxomycetes и Protosteliomycetes, то в
системе Олайва к миксомицетам (Eumycetozoea) от-
носят 3 подкласса — Protostelia, Dictyostelia и Myxo-
gastria. Подклассы Protostelia, Dictyostelia и Myxogas-
tria ниже рассматриваются раздельно.
Положение Dictyostelia в рамках системы миксо-
мицетов наиболее спорно (Кусакин, Дроздов, 1998;
Anderson, 1998), однако в силу традиций в данном
издании они рассматриваются совместно со слизеви-
ками (Карпов, 1997). Сокращения фамилий авторов-
микологов даны в соответствии с принятыми в ми-
кологической литературе (Kirk, Ansell, 1992). Названия
таксонов, используемые в микологической литературе,
приведены в скобках после знака «=», если объем
таксона в ботанической классификации соответству-
ет соответствующему таксону в зоологической клас-
сификации. В последнее время появились данные о
сходстве слизевиков и церкомонад (Карпов, 1997). В
качестве связующего звена между этими двумя груп-
пами протистов рассматривают Hyperamoeba flagel-
lata (Карпов, Мыльников, 1997).
Морфология
и физиология
Большинство миксомицетов имеет довольно слож-
ный жизненный цикл. Вегетативная стадия может
быть представлена мелкими, одноядерными или со-
держащими несколько ядер амебоидными клетками,
формирующими тонкие, нитевидные псевдоподии
(филоподии), либо многоядерными плазмодиями, до-
стигающими в отдельных случаях макроскопических
(десятки сантиметров) размеров. Многие слизевики
образуют жгутиковые стадии, имеющие, как прави-
ло, 2 апикальных (обычно неравной длины) жгутика,
лишенных мастигонем. Последние три десятилетия
ознаменовались применением электронного микро-
скопа в изучении морфологии и систематики этих
организмов. Особенно много новых данных было
получено об ультраструктуре зооспор (Jacobson,
Adelman, 1975; Nelson, Scheets, 1975, 1976; Ishigami,
1977; Spiegel et al., 1995). Однако эти данные каса-
ются очень небольшого числа видов, что не позволя-
ет полноценно использовать их для систематическо-
го анализа в таксономии.
На определенном этапе своего развития слизеви-
ки формируют различного рода плодовые тела, со-
держащие споры. Споруляция осуществляется одним
из трех возможных способов.
1. Дифференцировка одиночных одноядерных
или содержащих несколько ядер амебоидных клеток
(в ряде случаев возникающих в результате предвари-
тельной сегментации многоядерного плазмодия) в
простые стебельчатые спорокарпы, несущие 1 или
несколько спор (протостелиды),
2. Агрегация отдельных одноядерных амеб с по-
следующим формированием многоклеточного псев-
доплазмодия, дающего, в свою очередь, начало сте-
бельчатому многоспрровому спорокарпу (диктиосте-
лиды).
3. Развитие многоспоровых плодовых тел (сидя-
чих или на ножках) непосредственно из многоядер-
ного плазмодия (миксогастриды).
, У многих миксомицетов в жизненном цикле име-
ется половой процесс. Кроме того, у целого ряда
видов обнаружен феномен слияния вегетативных
особей. Серавин и Гудков (1984а), обобщая имев-
шийся к тому времени фактический материал по
различным группам протистов, выделили несколько
основных типов агамных слияний клеток, из ко-
торых три — псевдоконъюгация, псевдокопуляция
и плазмодизация — встречаются у представителей
класса Eumycetozoea. Псевдоконъюгация представ-
ляет собой частичное (с помощью цитоплазматиче-
ских мостиков), временное объединение партне-
ров. При псевдокопуляции происходит полное (хотя
и временное) слияние вегетативных особей, внешне-
имитирующее копуляцию гамет. Наконец, плазмоди-
зацйя — это полное агамное слияние клеток, при-
водящее к формированию из амебоидов или жгу-
тиконосцев постоянных или временных плазмодиев.
Г еографическое
распространение и экология
Большинство видов слизевиков имеют широкое
распространение, многие рассматриваются как кос-
мополиты. Однако некоторые виды имеют довольно
ограниченные ареалы, приуроченные к определен-
ным природно-климатическим зонам. Экология слизе-
виков изучена явно недостаточно. Кислотность суб-
страта — наиболее важный абиотический фактор,
влияющий на распространение слизевиков в природе
(Stephenson, 1988). Слизевики встречаются обычно в
гнилой древесине и под корой отмерших деревьев, в
листовом опаде, в почве, в помете растительноядных
животных, как правило, во влажных затенённых
местах. Однако даже в местах с очень низкой влаж-
ностью слизевики часто присутствуют в виде покоя-
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
419
щихся стадий или спор, что подтверждается наблю-
дениями при проращивании их на природных субст-
ратах во влажных камерах. Большинство изученных
почв из разных районов — от пустынь до лесов и
окультуренных зон — содержат споры или цисты
слизевиков, причем их количество колеблется от
10 до 10 000 на 1 г почвы (Madelin, 1984; Feest,
1987).
Подкласс Protostelia L. S. Olive, 1970 — Прото стел иды
Морфология и физиология
Трофическая стадия
Эта фаза протостелид представлена мелкими од-
ноядерными или содержащими несколько ядер аме-
боидными клетками. У некоторых форм развивается
многоядерный сетчатый плазмодий (рис. 364, А). В от-
личие от подобных образований у типичных плазмо-
диальных миксомицетов (представителей подкласса
Myxogastria) в плазмодиях протостелид отсутствует ха-
рактерный ритмический «челночный» ток цито-
плазмы. Для всех видов этого подкласса, как и в це-
лом для Eumycetozoea, свойственно формирование
филозных псевдоподий, выступающих из участков ги-
алиновой цитоплазмы. Однако у ряда форм при опре-
деленных условиях может наблюдаться также образова-
ние лобоподий (Olive, Stoianovitch, 1969; Olive, 1975).
Питание
Питаются протостелиды преимущественно бакте-
риальными организмами, но, кроме того, многие спо-
собны фагоцитировать клетки дрожжей и споры раз-
личных грибов. В некоторых случаях отмечается кан-
нибализм. Своеобразный способ питания был описан
у Ceratiomyxella tahitie ns is, Nematostelium gracile и
Schizoplasmodium cavostelioides. Эти протостелиды
вызывают образование перфораций в стенках некото-
рых дрожжевых и водорослевых клеток. Выпуская в
образовавшееся отверстие псевдоподиальный вырост,
они с его помощью начинают поглощать участки кле-
точного протопласта, включая их в относительно
крупные пищевые вакуоли. Затем псевдоподия втяги-
вается обратно, оставляя от подвергшейся нападению
клетки лишь пустую оболочку (Whitney, Bennett, 1984).
Аналогичный способ питания известен лишь у ряда
представителей класса Filosea.
Жгутики
Многие виды протостелид способны образовы-
вать жгутики. Обычно присутствуют 2 близкоассо-
циированных, лишенных мастигонем жгутика: длин-
ный, направленный вперед, и расположенный у его
основания короткий, повернутый в противополож-
ную сторону. Нередко имеются и дополнительные
пары жгутиков (Olive, 1975). У ряда видов встреча-
ются также одиночные жгутики. Можно выделить
две главные морфологические группы жгутиковых
клеток у протостелид (Spiegel, 1981а, 1981b; Spiegel
et aL, 1986, 1995). Первая группа включает предста-
вителей таких родов, как Ceratiomyxa, Ceratiomyxella,
Protosporangium, Cavostelium и других, клетки кото-
рых способны к активному плаванию и перемещают-
ся в толще воды, двигаясь по спиральной траек-
тории. Они имеют так называемую зооспороподоб-
ную форму тела: конусовидный передний конец,
несущий длинный и короткий жгутики (последний
может отсутствовать), и цилиндрическую, расширя-
ющуюся на заднем конце остальную часть тела. Дру-
гая группа включает такие формы, как Planoprotos-
telium, например Р. aurantium, клетки которых яв-
ляются амебоидными по своей морфологии и не
способны к активному плаванию. Биение жгутиков
вызывает лишь незначительные хаотическйе переме-
щения организма.
Особенности ультраструктурной
организации
Митохондрии всех изученных видов имеют труб-
чатые или иногда везикулярные кристы. В клетках
трофических стадий протостелид присутствуют ше-
роховатый ЭПР, диктиосомы аппарата Гольджи, со-
кратительные и пищеварительные вакуоли. Гиали-
новые участки цитоплазмы на периферии амебоидных
особей содержат разрозненную сеть микрофиламен-
тов актинового типа. В дискретных псевдоподи-
альных выростах были также найдены организован-
ные пучки микрофиламентов (Spiegel et al., 1979;
Рис. 364. Nematostelium sp. (По: Olive, 1975).
A — плазмодий, Б — спорокарп.
Fig. 364. Nematostelium sp.
A — plasmodium, Б — sporocarp.
420
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Spiegel, 1982а; Whitney, Bennett, 1984; Bennett,
1986).
Жгутиковый аппарат Cavostelium bisporum, Се-
ratiomyxella tahitiensis, Protosporangium articulatum,
Ceratiomyxa fruticulosa и других видов, клетки кото-
рых способны к активному плаванию и могут при-
нимать «зооспороподобную» форму тела, связан с
ядром специальной системой корешковых микро-
трубочек и состоит из 2 кинетосом, расположенных
приблизительно под прямым углом по отношению
друг к другу (Furtado, Olive, 1971b; Nelson, Scheets,
1975; Olive, 1975; Spiegel, 1981a, 1981b, 1982b;
Spiegel et al., 1986). Помимо конусовидного мик-
ротрубочкового деривата, направленного в сторону
ядра, с ними ассоциирован еще целый набор оп-
ределенным образом организованных корешковых
структур. От передней кинетосомы отходит длин-
ный, направленный вперед жгутик; вторая кинето-
сома либо лежит свободно, либо дает начало корот-
кой, повернутой назад ундулиподии. Дополнительные
жгутики, как например у Protosporangium articulatum,
выступают чаще всего ближе к заднему концу клет-
ки и имеют такой же состав и организацию ба-
зального аппарата (включая всю систему кореш-
ковых дериватов кинетосом), но не образуют связи
с ядерной оболочкой и не способны генерировать
направленное перемещение клетки (Spiegel et al.,
1986).
Амебоидные по своей морфологии особи Plano-
protostelium aurantium имеют 1 или несколько раз-
розненных жгутиков без какой-либо определенной
их локализации на поверхности тела. В основании
жгутика располагается единственная кинетосома с
системой корешковых микротрубочек, которые сво-
бодно заканчиваются в цитоплазме, не образуя связи
с ядерной оболочкой (Spiegel, 1982а).
Наружная поверхность трофических клеток про-
тостелид представляет собой разнообразно органи-
зованный тонкофибриллярный гликокаликс (Nelson,
Scheets, 1975; Dykstra, Aldrich, 1978; Spiegel et al.,
1986). На поверхности жгутиковых особей Ceratio-
myxella tahitiensis были обнаружены овальные че-
шуйки, формирующиеся в везикулах аппарата Голь-
джи (Furtado, Olive, 1971Ь). Своеобразный клеточный
покров имеется у представителей рода Endostelium,
Е. zonatum и Е. amerosporum, — безжгутиковых
форм протостелид. Он представляет собой довольно
сложное образование 0.25—2.0 мкм толщиной, ос-
нову которого составляет тонкофибриллярный мате-
риал, местами организованный в виде пучков, ори-
ентированных параллельно клеточной поверхности.
Этот покров одевает клетку сверху и сзади, тогда
как передний, гиалиновый фронт цитоплазмы с фи-
лозными субпсевдоподиями выступает за его грани-
цы (Bennett, 1986). Детали ультраструктурной орга-
низации и общая морфология (включая организацию
клеточного покрова) этих протостелид очень сход-
ны с таковыми у амеб рода Gocevia (G.fonbrunei и
G. placopus) (Pussard et al., 1977; Page, Willumsen,
1980; Bennett, 1986), принадлежащих к сем. Cochli-
opodidae подкласса Testacealobosia, класса Lobosea
(Page, 1987).
Ядра, размножение, цисты
Ядра протостелид в основном везикулярного
типа, с единственным, обычно занимающим цент-
ральное положение ядрышком. Во всех случаях,
когда ядерное деление подробно изучалось на ульт-
раструктурном уровне (в основном у форм, способ-
ных формировать жхугиковые клетки), было уста-
новлено, что оно, согласно классификации Райкова
(1986), представляет собой открытый ортомитоз.
Перед началом деления ундулиподии жгутиков втя-
гиваются в цитоплазму, аксонемы и корешковые де-
риваты кинетосом резорбируются; ядрышко и ядер-
ная оболочка отсутствуют. На полюсах веретена рас-
полагаются центриоли, причем у некоторых видов
сразу в количестве нескольких пар (Spiegel, 1982b;
Spiegel et al., 1986). У безжгутиковых форм протос-
телид какие-либо отчетливые внеядерные ЦОМТы
пока обнаружены не были (Heath, 1981). Непосред-
ственно после кариокинеза в каждом дочернем ядре
появляется несколько мелких «проядрышек», кото-
рые затем сливаются в одно крупное центральное
ядрышко. У одноядерных форм сразу за делением
ядра следует цитокинез.
Особи, содержащие несколько ядер, способны
подвергаться плазматомии, в результате которой в от-
личие от обычного цитокинеза могут обособляться
неравные по объему и количеству ядер (если их ис-
ходно было более 2) клетки. Плазматомия не связана
с завершением процесса ядерного деления и происхо-
дит спонтанно, при этом втягивания и резорбции
жгутиков не происходит (Olive, 1975).
Амебоидные и плазмодиальные особи протосте-
лид могут инцистироваться. Это наблюдается обычно
в стареющих культурах и связано, по всей видимости,
с истощением пищевых ресурсов, изменением pH и
накоплением в среде продуктов обмена. Эксцистиро-
вание происходит сразу, как только цисты помеща-
ются в свежую среду.
Жизненные циклы
У одноядерных форм протостелид любая из трофи-
ческих особей может непосредственно становиться
преспоровой клеткой. У плазмодиальных видов нача-
лу образования плодовых тел предшествует сегмента-
ция плазмодиев на одноядерные или содержащие не-
сколько ядер фрагменты, выполняющие функцию пре-
споровых клеток. Процесс споруляции схематически
изображен на рис. 365. Уплощенная преспоровая клет-
ка (рис. 365, А) становится полусферической, так как ее
цитоплазма постепенно концентрируется в центре,
покидая все более утончающуюся краевую зону
(рис. 365, Б). На поверхности клетки появляется тон-
кая предохранительная оболочка, препятствующая
высыханию, которая покрывает спорокарп (плодовое
тело протостелид) в течение всего процесса спороге-
неза. В то же время в базальной части преспоровой
клетки дифференцируется участок цитоплазмы, полу-
чивший название «стелиоген» (рис. 365, В, Г), кото-
рый начинает секретировать трубчатую (полую внут-
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
421
ри) ножку. По мере ее роста протопласт с ядром под-
нимается над поверхностью субстрата (рис. 365, Д). Когда
ножка достигает окончательной длины, он окружается
плотной оболочкой и становится спорой (рис. 364, Б\
365, Е). Весь процесс спорогенеза обычно занимает
30—60 мин.
Спорокарпы протостелид обычно несут одну-
единственную спору — одноядерную или содержа-
щую несколько ядер, хотя некоторые виды имеют
2 или даже 4 (редко больше) споры (рис. 364, Б).
Форма спор может быть различной: округлая, оваль-
ная, продолговатая или грушевидная (систематичес-
кий признак). Стенка спор обычно гладкая, но у
ряда видов (например, у представителей рода Schi-
zoplasmodiopsis) она определенным образом структу-
рирована (Dykstra, 1978). У многих протостелид зре-
лые споры просто отпадают от конца ножки. Scfti-
zoplasmodium cavbstelioides имеет уникальное среди
Eumycetozoea приспособление для рассеивания спор.
Вскоре после созревания между стенкой споры и
оболочкой спорокарпа у этого вида появляется газо-
вый пузырек, который увеличивается в объеме, до-
стигая иногда размеров самой споры. Затем проис-
ходит разрыв наружной оболочки, в результате чего
спора (балистоспора) высвобождается, вылетая из
спорокарпа в горизонтальном направлении (Olive,
1975). Споры подавляющего большинства протосте-
лид способны прорастать без какого-либо периода
покоя, продуцируя амебоидные или жгутиковые тро-
фические клетки.
Итак, последовательную смену стадий развития у
протостелид можно обобщенно представить в следу-
ющем виде:
X I
трофическая стадия -» спорогенез спора
циста
Эта принципиальная схема протекания жизненно-
го цикла имеет довольно разнообразные варианты ре-
ализации в онтогенезе конкретных видов. Рассмот-
рим для иллюстрации 2 наиболее характерных в этом
отношении примера.
Protosporangium bisporum (рис. 366). Спорокар-
пы этого вида обычно содержат 2 двухъядерные не-
отпадающие споры (Olive, Stoianovitch, 1972). Вско-
ре после прорастания происходит синхронное деле-
ние их ядер и последующее обособление в общей
сложности 8 одноядерных жгутиковых клеток, даю-
щих начало амебоидным трофическим особям
(рис. 366, Б—£). Амебы становятся многоядерными
(сетчатые плазмодии не образуются), и их количест-
во увеличивается в результате плазматомии (рис. 366,
Затем происходит постепенная фрагментация
амебоидных особей на одноядерные преспоровые
клетки (рис. 366, 3, И), приступающие к спорогенезу
(рис. 366, К). Одноядерное состояние сохраняется до
заключительного этапа формирования спор, перед
которым происходит 2 последовательных митоти-
ческих деления (рис. 366, Л).
Ceratiomyxella tahitiensis (рис. 367). Этот вид име-
ет очень своеобразный жизненный цикл (Olive, 1970,
1975). Отпадающие, содержащие несколько ядер спо-
ры прорастают (рис. 367, A, F), и образующиеся при
этом амебоидные клетки вскоре инцистируются,
формируя зооцисты (рис. 367, В). Все ядра в зооцисте,
исключая одно-единственное, дегенерируют. Остав-
шееся ядро обычно претерпевает 3 последователь-
ных митотических деления, сопровождаемых цитоки-
незом, в результате чего образуется 8 одноядерных
Ж1угиковых особей (рис. 367, Г—Ж). В дальнейшем
каждая из них способна давать начало многоядерно-
му сетчатому плазмодию (рис. 367, 3-—М). Митоти-
ческие деления в плазмодии имеют тенденцию быть
синхронными (по крайней мере, в отдельных его
участках). Перед началом споруляции плазмодий
становится дольчатым и затем сегментируется на со-
держащие по нескольку ядер преспоровые клетки
(рис. 367, Н), которые далее развиваются в споро-
карпы (рис. 367, О, ГГ).
Наличие полового процесса у протостелид (за ис-
ключением одного случая у Ceratiomyxa fruticulosa)
не установлено. В то же время агамные слияния
особей — достаточно распространенное явление у
представителей этой группы протистов (Olive, 1970,
1975). Присутствие псевдоконъюгации и псевдоко-
пуляции предполагает существование у протостелид
Рис. 365. Спорогенез у Nematostelium sp. (пояснения в тексте).
(По: Olive, 1975).
Fig. 365. Diagrammatic illustration of sporogenesis in Nematostelium sp.
(see comments in the text).
A
422
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 366. Схема жизненного цикла Protosporangium bisporum. (По: Olive, Stoianovitch, 1972).
А — спорокарп; Б — прорастание; В — митотические деления ядер; Г — сегментация спорового протопласта; Д — обособление одноядерных жгутико-
вых клеток; Е — амебоидные трофические особи; Ж — многоядерные амебоидные протопласты; 3 — фрагментация многоядерных протопластов; И — об-
разование одноядерных преспоровых клеток; К, Л — спорогенез.
Fig. 366. Life cycle of protostelid Protosporangium bisporum.
A — sporocarp; Б — spore germination; В — mitotic division of the nuclei; Г — segmentation of spore protoplast; Д — flagellate cells formation; E — amoebo-
id cells; Ж — multinuclear amoeboid cells; 3 — fragmentation of multinuclear amoeboid cells; И — prespore cell formation; К,Л — sporogenesis
различных форм агамных генетических взаимоотно-
шений (Серавин, Гудков, 19846), однако этот вопрос
остается пока еще совершенно неизученным.
В ядрах сегментирующихся преспоровых клеток
у С. fruticulosa были обнаружены синаптонемальные
комплексы хромосом (Furtado, Olive, 1971а), что
подтверждает наличие полового процесса у этого ви-
да. Однако на каком именно этапе жизненного цик-
ла цератиомиксы происходит восстановление дипло-
идного состояния, пока нельзя сказать достаточно
определенно.
Г еографическое
распространение и экология
О географическом распространении и экологии
протостелид известно очень мало. Большинство ви-
дов, найденных в зоне умеренного климата, отмече-
ны также в тропиках (Baker, 1975; Moore, Spiegel,
1995; Stephenson et al., 1999). К протостелидам с
очень широким ареалом можно отнести вид С. fruti-
culosa, который встречается практически во всех
природно-климатических зонах. Однако некоторые
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
423
ж
Рис. 367. Схема жизненного цикла Ceratiomyxella tahitiensis. (По: Olive, 1975).
А — спора; Б — прорастание споры; В — зооциста; Г—Ж — последовательные стадии формирования и обособления одноядерных жгутиковых особей;
3—М — развитие многоядерного сетчатого плазмодия; Н — сегментация плазмодия и образование многоядерных преспоровых клеток; О, П — развитие
спорокарпа.
Fig. 367. Life cycle of Ceratiomyxella tahitiensis.
A — spore; Б — spore germination; В — cyst formation; Г—Ж — flagellate cells formation; 3—M — early stages in plasmodial development by nuclear division
without cell division; H — mature reticulate plasmodium and prespore cell formation; О, П — sporogenesis.
тропические виды имеют ограниченные ареалы, как
например С. morchella и С. spaerosperma. Выявлены
виды, встречающиеся в определенных микроместообита-
ниях, причем их спектр у одних и тех же видов может
меняться в различных природно-климатических зонах.
Например, Nematostelium gracile и N. ovatum, обита-
ющие в зоне умеренного климата только в листовой
подстилке, в тропиках заселяют преимущественно воз-
душный опад (омертвевшие части эпифитов и листья
деревьев). Protostelium mycophaga, найденный на воз-
душном опаде в умеренной зоне, в тропиках отмечен
только в листовой подстилке (Moore et al., 1996).
424
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Систематический обзор
В подкласс Protostelia входит 1 отряд Protosteliida
с 3 семействами. 2 из них — Cavosteliidae и Ceratio-
myxidae — включают виды, имеющие в жизненном
цикле жгутиковые стадии; в сем. Protosteliidae включа-
ют только безжгутиковые формы (Olive, 1970, 1975).
Сем. Cavosteliidae L. S. Olive, 1970
(= Cavosteliaceae L.S.Olive, 1964)
Среди его представителей имеются, пожалуй,
наиболее просто организованные виды протостелид.
Описано 4 рода — Cavostelium L. S. Olive, 1964, Се-
ratiomyxella L. S. Olive et Stoian., 1971, Planoproto-
stelium L.S. Olive et Stoian., 1971, Protosporangium
L.S. Olive et Stoian, 1972, различающихся по харак-
теру организации трофических стадий, строению и
размерам плодовых тел, количеству спор, их форме
и т. д. Жизненные циклы Protosporangium bisporum
и Ceratiomyxella tahitiensis были рассмотрены ранее
(см.: рис. 366; 367). Трофические стадии видов рода
Cavostelium и Planoprotostelium aurantium в основ-
ном представлены одноядерными формами.
Сем. Ceratiomyxidae L. S. Olive, 1970
(= Ceratiomyxaceae J.Schrdt., 1889)
Этот таксон был перенесен в подкласс Protostelia
из состава миксогастрид, куда он включался ранее
(Olive, 1970). Основанием для этого послужило боль-
шее сходство организации плазмодиев и плодовых
тел представителей этого семейства с таковыми у
протостелид. Семейство включает 1 род.
Род Ceratiomyxa J. Schrot., 1889
Род объединяет 3 вида. Один вид — С. fruticulosa
является наиболее хорошо изученным и широко рас-
пространенным в природе (Martin, Alexopoulos, 1969).
Перед плодоношением плазмодий, погруженный в
слизистый матрикс, поднимается над поверхностью
субстрата либо в виде ветвящихся бело-молочных
вертикальных выростов, либо в виде более компакт-
ного образования, несколько напоминающего по
внешнему виду пчелиные соты. Затем сетчатая плаз-
модиальная масса, расположенная на поверхности
этих структур, сегментируется на одноядерные пре-
споровые клетки, которые формируют спорокарпы
на тонких полых ножках (Olive, 1975). Спорогенез
у С. fruticulosa был впервые детально описан Фа-
минциным и Ворониным в 1873 г. Как уже упомина-
лось ранее, в преспоровых клетках были найдены
ядра, находившиеся в профазе первого мейотическо-
го деления (Furtado, Olive, 1971а), Исходно одно-
ядерная спора становится четырехъядерной, а сразу
после прорастания — восьмиядерной. Затем следует
цитокинез и образуется 8 одноядерных жгутиковых
клеток. Некоторые авторы наблюдали плазмагамию
таких особей, но следует ли кариогамия непосредст-
венно за слиянием их цитоплазмы, остается пока
неясным (Olive, 1975).
Сем. Protosteliidae L. S. Olive et
Stoian., 1966
(= Protosteliaceae L. S. Olive, 1962)
Семейство объединяет 9 родов: Clastostelium
L. S. Olive et Stoian., 1977; Endostelium L.S. Olive,
W. E. Benn, et Deasey, 1984; Microglomus L. S. Olive
et Stoian., 1977; Protostelium L. S. Olive et Stoian.,
1960; Protosteliopsis L. S. Olive et Stoian., 1966; Ne-
matostelium L. S. Olive et Stoian., 1970; Schizoplasmo-
diopsis L. S. Olive, 1967; Tychosporium Spiegel., 1995
и Schizoplasmodium L. S. Olive et Stoian., 1966, ха-
рактеризующихся отсутствием жгутиковых стадий. Без-
жгутиковые протостелиды, вероятно, неоднократно
возникали в процессе эволюции от жгутиковых форм.
Так, например, и трофическая и плодовая стадии
жизненного цикла Protostelium mycophaga чрезвы-
чайно сходны с таковыми у Planoprotostelium auranti-
um. Их различие заключается главным образом в от-
сутствии жгутиков у первого из этих 2 видов (Olive,
1975; Spiegel, 1982а). Трофические стадии Protos-
telium, Protosteliopsis и Endostelium в основном
представлены одноядерными (иногда содержащими
несколько ядер) амебами, тогда как Schizoplasmodi-
um, Nematostelium и Schizoplasmodiopsis образуют мно-
гоядерные сетчатые плазмодии (Olive, 1975; Olive et al.,
1984).
Подкласс Dictyostelia L. S. Olive, 1970 (= Dictyostelidae Raper,
1984) — Диктиостелиды
Морфология и физиология Трофическая стадия
Dictyostelium discoideum, впервые описанный в
1935 г. Рэпером (Raper, 1935), в настоящее время наи-
более хорошо изученный вид диктиостелид (хотя он и
не самый широко распространенный в природе пред-
ставитель этой группы протестов). Поэтому описание
некоторых стадий развития клеточных слизевиков
будет дано преимущественно на основе данных, полу-
ченных при исследованиях этого вида.
Эта фаза жизненного цикла представлена одно-
ядерными амебоидными клетками. При активном пе-
ремещении по субстрату на переднем конце амеб
имеется зона прозрачной гиалиновой цитоплазмы, из
которой время от времени выступают филоподиаль-
ные отростки, однако могут формироваться и псев-
доподии лобозного типа.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
425
Питание
Питаются диктиостелиды главным образом бакте-
риальными организмами, но некоторые штаммы ряда
видов (например, Polysphondilium pallidum и Dictyos-
telium discoideum) удается культивировать в лабора-
тории в аксеничных условиях (Hohl, Raper, 1963;
Watts, Asworth, 1970). Встречаются if такие формы,
как D. caveatum, которые способны поглощать не
только бактерии, но и миксамеб других миксомице-
тов, нападая даже на клетки более крупные, чем они
сами, и «откусывая» от них «кусочки» (Waddell, 1982;
Waddell, Vogel, 1985). Среди видов диктиостелид
нередки случаи каннибализма (Olive, 1975; Waddell,
Duffy, 1986).
Ядра, размножение
и жизненные циклы
Интерфазные ядра амеб большинства диктиосте-
лид имеют несколько (от 2 до 5) мелких перифери-
ческих ядрышек, хотя у представителей рода Acyto-
stelium часто содержится лишь 1 эксцентрично рас-
положенное ядрышко (Hohl et al., 1968; Olive, 1970).
У Polysphondilium и Dictyostelium вблизи ядра имеют-
ся дискретные ЦОМТы (Roos, 1975; Moens, 1976;
Omura, Fukui, 1985). Во время митоза осевое бипо-
лярное веретено располагается почти целиком внутри
ядра, оболочка которого остается интактной, за ис-
ключением полярных отверстий, через которые про-
ходят микротрубочки, связанные с расположенными
на вершинах веретена ЦОМТами (полузакрытый ор-
томитоз). После завершения митоза следует цитоки-
нез (см. схему жизненного цикла D. discoideum, —
рис. 368, А—В). В условиях, неблагоприятных для
развития дальнейших стадий жизненного цикла, тро-
фические особи диктиостелид способны переходить
в состояние покоя, выделяя вокруг себя плотную обо-
лочку и превращаясь в так называемые микроцисты
(Bonner, 1978, 1982).
Истощение пищевых запасов и начало голодания
стимулируют переход к следующему («социальному»)
этапу жизненного цикла.
Преагрегация
(рис. 368, Г)
В течение кратковременного (4—8 ч) преагрега-
ционного периода осуществляются серьезные преоб-
разования морфологии и физиологии трофических
амеб, характеризующие первую ступень их диффе-
ренцировки. Клетки прекращают делиться и несколь-
Рис. 368. Схема жизненного цикла Dictyostelium discoideum (пояснения в тексте). (По: Olive, 1975).
Fig. 368. Life cycle of Dictyostelium discoideum (see comments in the text).
426
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 369. Последовательные стадии жизненного цикла Dictyostelium discoideum. (По: Bonner, 1971).
А — отдельные трофические особи, Б — последовательные этапы агрегации, В — стадия мигрирующего псевдоплазмодия, Г — формирование плодовых
тел.
Fig. 369. Diagrammatic illustration of sporogenesis in Dictyostelium discoideum.
A — vegetative myxamoebae, Б — cell aggregation, В — the migrating pseudoplasmodium, Г — frutification.
ко уменьшаются в размерах. Изменяются адгезивные
и другие свойства поверхностной мембраны амеб.
Происходит быстрое снижение фагоцитарной актив-
ности, вплоть до полной потери способности к фаго-
цитозу. Наблюдается экзоцитоз имеющегося содер-
жимого пищеварительных вакуолей, которые вскоре
исчезают совсем. В конце этой и непосредственно в
начале следующей стадии жизненного цикла в амебах
появляются аутофагические вакуоли — самоперева-
ривание, по всей видимости, становится основным
источником энергии в течение всего дальнейшего
развития этих организмов (Maeda, Takeuchi, 1969;
Yamamoto et al., 1981). Экспериментально показано,
что если клетки D, discoideum, находящиеся на более
поздних этапах дифференцировки, «вернуть в исход-
ные условия», то они еще довольно длительный пери-
од оказываются неспособными к фагоцитозу (Takeu-
chi et al., 1983).
Агрегация (рис. 368, Д; 369, Б)
Первым видимым свидетельством окончания пре-
агрегации и начала следующей стадии развития слу-
жит появление центров агрегации — небольших
групп или отдельных клеток, которые привлекают к
себе других амеб. Вскоре движущиеся по направле-
нию к этим центрам особи формируют сплошные
Рис. 370. Агрегация у Dictyostelium discoideum. (По: Olive, 1975).
Fig. 370. Cell aggregation in Dictyostelium discoideum.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
427
клеточные «потоки» (рис. 370). Агрегация осуществ-
ляется благодаря выделению в среду специального
привлекающего амеб вещества — аттрактанта, пер-
воначально названного «акразином» (Bonner, 1947).
Для целого ряда видов (таких, как D. mucoroides,
D. purpureum, D. rosarium, D. discoideum) эту функ-
цию выполняет цАМФ. Было установлено также, что
цАМФ вызывает положительный хемотаксис у амеб
D. discoideum в очень незначительной концентра-
ции — 10"6—10"5 М (Konijn et al., 1967, 1969; Ко-
nijn, 1973). Следует; отметить, что трофические
особи практически не привлекаются этим вещест-
вом, а только амебы, прошедшие дифференцировку
на стадии преагрегации. Экспериментальные иссле-
дования показали, что клетки способны восприни-
мать пространственные градиенты аттрактанта и дви-
жутся к его источнику более или менее прямоли-
нейно. Специальные рецепторы, воспринимающие
цАМФ, располагаются равномерно по всей поверх-
ности амеб. Местное повышение концентрации
цАМФ индуцирует образование псевдоподии в сти-
мулируемом участке, т. е. локомоция происходит в
направлении максимальной концентрации комплек-
сов цАМФ—рецептор (Gerisch et al., 1975).
Однако цАМФ не является универсальным хемо-
аттрактантом для всех видов диктиостелид. D. minu-
tum, D. lacteum, D. polycephalum, D. vinaceo-fuscum и
другие имеют иные, причем, по всей видимости, раз-
личные «акразины» (Konijn, 1973; Bonner, 1982). У
Polysphondilium1 violaceumn Р. pallidum эту функцию
выполняет олигопептид с молекулярной массой око-
ло 1500.
Псевдоплазмодий
(рис. 368; 369; 371)
В конце агрегации формируется вертикально рас-
положенная конусовидная клеточная масса, вокруг
которой секретируется общая слизистая оболочка.
Это многоклеточное образование, которое в даль-
нейшем ведет себя как единый организм, носит на-
звание псевдоплазмодия. Обычно псевдоплазмодий
принимает горизонтальное положение и начинает
перемещаться по субстрату. На выбор направления
миграции оказывает влияние целый комплекс факто-
ров окружающей среды. Псевдоплазмодий обладает
фото-, термо- и хемотаксисом, причем его передний
конец более чувствителен к определенным стимулам,
чем остальные участки тела (Raper, 1940, 1951; Bon-
ner et al., 1950; Bonner, 1982). Разные виды демон-
стрируют различную реакцию на воздействие одного
и того же фактора. Например, Dictyostelium discoide-
um, D. mucoroides, D. purpureum и Polysphondilium
violaceum весьма чувствительны к степени освещен-
ности, тогда как Dictyostelium polycephalum оказыва-
ется практически индифферентным в этом отноше-
нии.
У D, discoideum и некоторых других видов кле-
точных слизевиков уже в начале этой стадии жиз-
ненного цикла наблюдаются признаки дальнейшей
дифференцировки клеток, усиливающиеся по ме-
ре приближения заключительного этапа развития.
Рис. 371. Сорогенез у различных видов диктиостелид. (По: Olive,
1975).
А — Dictyostelium discoideum, Б — D. mucoroides and D. purpureum, В —
Polysphondilium sp.
Fig. 371. Frutification in the dictyostelids.
A — Dictyostelium discoideum, Б — D. mucoroides and D. purpureum, В —
Polysphondilium sp. .
Даже с помощью простых цитологических методов
окраски можно видеть, что мигрирующий псев-
доплазмодий D. discoideum отчетливо подразделяется
на два основных участка — передний и задний,
в соответствии с распределением 2 типов клеток
(рис. 372, А) — предножковых и преспоровых (Bon-
ner et al., 1955; MacWilliams, Bonner, 1979). Клетки
преспорового участка характеризуются появлением в
них специализированных, так называемых преспоро-
вых, вакуолей (Hohl, Yamamoto, 1969; Maeda, Take-
uchi, 1969), окруженных унитарной мембраной со
слоем плотного материала с внутренней стороны и
содержащих тонкофибриллярный матрикс. В даль-
нейшем эти структуры принимают участие в форми-
ровании оболочки спор, сливаясь, по всей видимос-
ти, с плазматической мембраной преспоровой клет-
ки (Ikeda, Takeuchi, 1971). В предножковых клетках
такие вакуоли отсутствуют, однако в них в большом
количестве имеются аутофагические вакуоли, содер-
жащие кислую фосфатазу (Maeda, Takeuchi, 1969;
Yamamoto et al., 1981; Yamamoto, Takeuchi, 1983).
Клетки преспорового и предножкового участков
псевдоплазмодия хорошо отличаются друг от друга
по подвижности (Kitami, 1983а), степени адгезивное-
428
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Л 5 В
Рис. 372. Схема процесса сорогенеза у Dictyostelium discoideum
(пояснения в тексте). (По: Olive, 1975).
Fig. 372. Frutification in Dictyostelium discoideum (see comments
in the text).
ти своей поверхности (Yabuno, 1971) и целому ряду
других (биохимических, морфологических и ультра-
структурных) признаков.
Соотношение клеток 2 типов в псевдоплазмодии
является величиной более или менее постоянной
(MacWilliams, Bonner, 1979). После дезагрегации
псевдоплазмодия D, discoideum и последующего цен-
трифугирования образовавшейся клеточной суспензии
удалось получить 2 фракции, содержащие раздельно
преспоровые и предножковые клетки. Их количест-
венное соотношение в среднем соответственно равно
2:1. При фракционировании указанным способом
только задних частей псевдоплазмодиев было уста-
новлено, что наряду с преспоровыми в них присут-
ствует в среднем около 19% клеток, соответствующих
предшественникам ножки (Ratner, Wayne, 1983).
Несмотря на наличие у целого ряда видов дик-
тиостелид очень ранних признаков такой специали-
зации клеток, их дифференцировка все же является
достаточно лабильной, так как допускает взаимную
трансформацию одного клеточного типа в другой.
Еще Рэпер (Raper, 1940) показал, что если псевдо-
плазмодий D. discoideum разрезать перпендикулярно
его продольной оси на несколько частей, то каждая
из них через несколько часов восстанавливает орга-
низацию интактного псевдоплазмодия (с нормальной
пропорцией и распределением преспоровых и пред-
ножковых клеток). Было отмечено, что конверсия
клеток-предшественников ножки в преспоры осу-
ществляется дольше, чем обратное превращение
(Gregg, 1965; Sakai, 1973).
Согласно имеющимся данным, судьба амеб в ис-
ходно гомогенной массе трофических особей опре-
деляется тем, на какой стадии ядерного цикла их
застала стимуляция к началу «коллективного» этапа
развития. Амебы, находящиеся в S и G2 фазе, диф-
ференцируются в преспоровые клетки, тогда как пред-
ножковые формируются из особей в фазе G{ (McDo-
nald, Durston, 1984; Sharpe, Watts, 1985; Gomer, Fir-
tel, 1987). Вместе с тем было показано, что
существует ряд факторов химической природы, име-
ющих непосредственное отношение к клеточной
специализации. Одним из наиболее существенных
морфогенов у диктиостелид является цАМФ (в том
числе у видов, не использующих это вещество в ка-
честве «акразина») (Kay et al., 1979; Kitami, 1983а,
1983b; Schaap, Spek, 1984; Gerisch, 1987). Кроме
того, образование предножковых клеток связывают с
присутствием некоего низкомолекулярного вещества,
в то время как спорообразованию способствует
NH3 (Kay et al., 1979, 1984). Очевидно, помимо ис-
ходной, зависящей от фазы ядерного цикла клеточ-
ной «предопределенности» существуют дополнитель-
ные механизмы химического контроля специализации
клеток в виде веществ-регуляторов. Тип клеточной
дифференцировки может быть изменен в ходе онто-
генеза.
Феномен миграции псевдоплазмодия еще не
получил достаточно исчерпывающего объяснения.
Однако ясно, что его перемещение по субстрату
обусловлено индивидуальной подвижностью состав-
ляющих его клеток, которые обладают различной
двигательной активностью (Kitami, 1983а, 1983b).
Было установлено наличие определенных хемотак-
сисов у отдельных клеток и существование внутри
псевдоплазмодия переднезаднего и радиального гра-
диентов концентрации веществ-аттрактантов (Корас-
hik, 1982; Odell, Bonner, 1986), что по крайней мере
в какой-то степени должно обеспечивать необходи-
мые условия для координированного поведения кле-
точной массы как единого многоклеточного организма.
Кульминация и спорообразование
(рис. 368, Ж, 3; 369, Г; 371, Б;
372, Б, Б).
После некоторого периода направленного пере-
мещения по субстрату (продолжительность которого
в значительной степени определяется условиями ок-
ружающей среды) псевдоплазмодий прекращает дви-
жение и начинает формировать плодовое тело (со-
рокарп). Псевдоплазмодий (теперь — сороген) зани-
мает вертикальное положение, и клеточная масса
вступает в фазу своей окончательной дифференци-
ровки. Группа клеток, появляющаяся перед этим на
заднем конце слизевика и проявляющая все призна-
ки, свойственные предшественникам ножки, оказы-
вается лежащей на субстрате в основании сорогена
(рис. 372, Б). Из них начинает формироваться ба-
зальный диск будущего сорофора (ножки сорокар-
па). Первые клетки ножки появляются в апикальной
папилловидной части сорогена (рис. 372, £*). Они хо-
рошо отличаются от дающих им начало предножко-
вых клеток благодаря своей сильной вакуолизации.
Новые клетки ножки продолжают пристраиваться
сверху, тогда как первые погружаются вниз, проходя
через центр сорогена. Постепенно они выделяют во-
круг себя толстую целлюлозную стенку и погибают.
Вокруг сердцевины из плотно упакованных оболочек
мертвых клеток секретируется двухслойная целлю-
лозная трубка, покрытая снаружи слизистой плен-
кой. Наконец, первые ножковые клетки достигают
основания сорогена и соединяются с базальным дис-
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
429
ком, клетки которого к этому времени также образуют
целлюлозные стенки. С этого момента сороген начи-
нает подниматься над субстратом и ножка становит-
ся видна снаружи (рис. 372, В).
Все новые и новые предножковые клетки надстра-
иваются сверху, претерпевая соответствующую транс-
формацию, до тех пор пока полностью не истощается
весь их запас. Внешне это проявляется в исчезнове-
нии апикального папилловидного выступа сорогена
(рис. 371). На конце ножки, поднимающейся у D. dis-
coideum на высоту до 5 мм над поверхностью субстрата,
остается только масса преспоровых клеток, которые
к этому времени уже заканчивают свою дифференци-
ровку в споры. Споры одеты толстой целлюлозной
оболочкой и устойчивы к воздействию неблагоприят-
ных условий окружающей среды. Из спор при про-
растании вновь выходят амебоидные трофические
особи (рис. 369, А).
Агамные клеточные
взаимодействия
В жизненном цикле диктиостелид при образова-
нии клеточных контактов на стадии преагрегации и
ранних этапах агрегации неоднократно наблюдались
случаи псевдоконъюгации и псевдокопуляции (Huff-
man et al., 1962; Huffman, Olive, 1964; Sinha, Ash-
worth, 1969; Olive, 1975). Наличие псевдоконъюга-
ции в образующихся центрах агрегации было также
показано с помощью электронного микроскопа (Kirk
et al., 1971). Более того, существование цитоплазма-
тических мостиков между клетками (и обмен содер-
жимым их цитоплазмы) удалось обнаружить и на
более поздних стадиях развития у D. discoideum (Ono
et al., 1972). Интересно отметить, что поиски в псев-
доплазмодиях каких-либо специализированных меж-
клеточных контактов (типа щелевых), присутствие
которых считалось необходимым для координации
процессов дифференцировки клеток, не дали поло-
жительного результата (Johnson et al., 1977). Появление
рекомбинантных фенотипов у D. discoideum (при от-
сутствии, как мы видели, полового процесса) известно
уже давно (см.: Olive, 1975). Были найдены и цито-
логические доказательства существования «класси-
ческого» парасексуального процесса, при котором
возникшие диплоидные ядра (2п =14) претерпевают
вегетативную гаплоидизацию (Sinha, Ashworth, 1969;
Brody, Williams, 1974; Olive, 1975). Очевидно, у этих
организмов должны существовать и иные формы па-
расексуального, а также пресексуального процессов
(Серавин, Гудков, 19846).
Альтернативный путь развития
(макроцисты)
При определенных условиях, неблагоприятных для
образования плодовых тел (повышенная влажность,
темнота, измененный состав среды и др.), некоторые
виды диктиостелид способны вступать на альтерна-
тивный путь протекания жизненного цикла, ведущий
к формированию так называемых макроцист. Найден-
ные у D. mucoroides, D. discoideum, D. purpureum, D. mi-
nutum, D, giganteum, Polysphondilium violaceum и
P. pallidum макроцисты представляют собой половую
фазу развития этих клеточных слизевиков, среди кото-
рых есть как гомо-, так и гетероталлические формы
(Blaskovics, Raper, 1957; Erdos et al., 1972, 1973; Filo-
sa, Dengler, 1972; Collins, 1979; Szabo et al., 1982, и др.)
При смешивании в одной культуре штаммов проти-
воположных типов спаривания вскоре наблюдается
появление двухъядерных клеток. Внешне они ничем
(даже размерами) не отличаются от обычных трофи-
ческих форм, хотя и возникают в результате попарно-
го слияния амебоидных особей. Лишь спустя некото-
рое время продукты слияния становятся морфологи-
чески хорошо отличимы благодаря значительному
увеличению объема их цитоплазмы. Параллельно с
этим происходит сначала «набухание», а затем и сли-
яние обоих ядер, в результате чего возникает так назы-
ваемая гигантская клетка зиготы (O’Day, 1979; Chag-
la et al., 1980). Было подтверждено высказывавшееся
предположение (Szabo et al., 1982), что одноядерные
амебоидные особи, дающие в результате слияния на-
чало двухъядерным клеткам, представляют собой спе-
циализированные гаметы (O’Day et al., 1987а, 1987b).
Образовавшиеся таким образом гигантские клет-
ки начинают привлекать к себе одноядерных амеб,
выступая фактически в роли центров агрегации. В
ряде случаев было показано, что при этом использу-
ется тот же цАМФ-опосредованный клеточный хе-
мотаксис (O’Day, 1979; Abe et al., 1984). После
окончательного сформирования плотного агрегата
(вокруг которого образуется так называемая первич-
ная оболочка, которая напоминает слизистую плен-
ку, окружающую мигрирующий псевдоплазмодий),
гигантская клетка приступает к фагоцитозу окружа-
ющих ее амебоидных особей (Erdos et al., 1972,
1973; Filosa, Dengler, 1972). Этот процесс продолжа-
ется до тех пор, пока не будут съедены все агреги-
рованные амебы, после чего гигантская клетка окру-
жается вторичной, а затем и третичной оболочкой,
окончательно превращаясь в макроцисту.
Приблизительно на стадии формирования вторич-
ной оболочки в синкарионе были обнаружены синап-
тонемальные комплексы хромосом (Erdos et al., 1972).
Наличие мейоза подтверждается также появлением
при последующем прорастании макроцист рекомби-
нантных фенотипов (Macinnes, Francis, 1974). Перед
окончательным вступлением в период покоя макро-
циста становится многоядерной.
В процессе прорастания цитоплазма макроцисты
сначала распадается на сравнительно крупные одно-
ядерные «проамебы», которые, в свою очередь, де-
лятся, продуцируя множество трофических амебоид-
ных особей нормального размера, выходящих в окру-
жающую среду.
Г еографическое
распространение и экология
Диктиостелиды длительное время рассматрива-
лись как преимущественно копрофильные (обитаю-
щие на помете животных) организмы с очень широ-
430
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ким ареалом (Brefeld, 1869; Raper, 1984). Однако в
последнее время благодаря систематическим иссле-
дованиям ряда авторов было выявлено много новых
видов диктиостелид, встречающихся в разнообразных
микроместообитаниях (Cavender, 1973; Hagiwara,
1996). Диктиостелиды найдены в почвах различного
типа, помете животных, гниющих растениях и грибах
и в ризосфере многих растений. Наибольшее число
видов обитает в листовой подстилке и почвах листо-
падных лесов в зоне умеренного климата. Среди них
наиболее часто встречается Dictyostellium mucoroi-
des. Основным фактором, влияющим на встречае-
мость и распространение диктиостелид, служит нали-
чие пищевых ресурсов в виде почвенных бактерий.
Так, например, значительное увеличение численнос-
ти почвенных бактерий ведет к увеличению числа
клеток D. mucoroides и, наоборот, снижение числен-
ности популяции бактерий ведет к снижению числен-
ности клеток диктиостелид (Kuserk, 1980).
Систематический обзор
Диктиостелиды характеризуются способностью
формйровать хорошо развитую недифференцирован-
ную ножку на стадии образования сорокарпа. Един-
ственный отряд Dictyosteliida объёдиняет 2 семейст-
ва: Acytosteliidae и Dictyosteliidae (Raper, 1984).
Сем. Acytosteliidae
(= Acytosteliaceae Raper ex Raper
et Quinlan, 1958)
Включает 1 род с 4 видами, различающимися глав-
ным образом по форме и размерам спор (Olive, 1975).
Род Acytostelium Raper, 1956
Первым из описанных и наиболее хорошо изу-
ченных представителей этого таксона является
A. leptosomum (см;: Raper, Quinlan, 1958). Из одного
клеточного агрегата у A. leptosomum обычно возни-
кает несколько папилловидных сорогенов, каждый
из которых дает начало сорокарпу, состоящему из
терминального соруса спор на длинной тонкой
полой ножке. У представителей этого рода отсут-
ствуют специализированные предножковые клетки и
все клетки сорокарпа можно считать преспоровыми
(так как по окончании кульминационной стадии раз-
вития они все превращаются в споры). Ножка у
видов рода Acytostelium, в отличие от остальных
Dictyostelia не имеет клеточного строения и син-
тезируется, по всей видимости, самими преспоровы-
ми клетками. Она представляет собой тонкую полую
трубку, чрезвычайно напоминающую спорокарпы
протостелид. Ее стенки содержат параллельно рас-
положенные и ориентированные в продольном на-
правлении целлюлозные фибриллы (Hohl et al.,
1968).
Хотя обычно у Acytostelium образуются много-
споровые сорокарпы, имеются данные, что при оп-
ределенных условиях культивирования могут проду-
цироваться двуспоровые и даже односпоровые пло-
довые тела, фактически неотличимые от протосте-
лидных. Эти факты, по мнению некоторых авторов
(Olive, 1967, 1970, 1975), свидетельствуют о прямой
филогенетической линии развития от Protostelia к
Dictyostelia, где Acytostelium занимает промежуточ-
ное положение.
Сем. Dictyosteliidae
(= Dictyosteliaceae Rostaf., 1875)
Включает 3 рода — Dictyostelium Bref., 1870; Po-
lysphondilium Bref., 1884 и Coenonia Tiegh., 1884,
различающиеся по характеру организации плодовых
тел (сорокарпов).
Род Dictyostelium Bref., 1870
Род самый обширный среди клеточных слизеви-
ков (известно 35 видов). Первый представитель этого
таксона — D. mucoroides — был обнаружен и описан
Брефельдом в 1869 г. (Brefeld, 1869). Виды диктио-
стелиумов различаются главным образом по форме,
размерам и окраске сорокарпов и спор. Кроме того,
у D. minutum, D. deminutivum, D. lacteum и ряда дру-
гих форм в жизненном цикле отсутствует миграцион-
ная фаза и псевдоплазмодии приступают к спорогене-
зу сразу после своего образования. Формирование
ножки внутри клеточной массы у D. mucoroides начи-
нается практически сразу по окончании процесса аг-
регации, после чего псевдоплазмодий принимает го-
ризонтальное положение и мигрирует по субстрату
(рис. 371, Б). Аналогичная ситуация имеет место и у
D. purpureum.
Очень интересные особенности имеет в своем раз-
витии D. polycephalum. У этого вида каждый образо-
вавшийся клеточный агрегат подразделяется на неко-
торое количество отдельных, очень тонких и длин-
ных (до 1 см) псевдоплазмодиев, у которых в период
миграции отсутствует дифференцировка на презумп-
тивные споры и клетки ножки. Перед началом куль-
минации эти псевдоплазмодии в свою очередь под-
разделяются на несколько частей, каждая из которых
самостоятельно дает начало плодовому телу (Olive,
1975).
Сорокарпы D. rosarium кроме обычного крупного
терминального соруса спор имеют еще и небольшие
дополнительные сорусы, расположенные по всей
длине ножки и сидящие непосредственно на ее по-
верхности.
Род Polysphondilium Bref., 1884
Этот род характеризуется, как и Dictyostelium
mucoroides, выступающим сзади продолжением внут-
реннего участка ножки на стадии мигрирующего
псевдоплазмодия (рис. 371, В). В псевдоплазмодиях
отсутствует дифференцировка на преспоровые и
предножковые клетки. В процессе удлинения ножки
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
431
во время кульминационной фазы развития на за-
днем конце поднимающегося над субстратом соро-
гена происходит отшнуровка небольших клеточных
масс. Эти отделившиеся участки последовательно,
начиная в самых нижних, дифференцируются на не-
сколько мелких вторичных сорогенов, развиваю-
щихся затем в отдельные сорусы на ножках. Это
приводит к образованию серии мутовок из несколь-
ких вторичных ветвей, расположенных по всей
длине основного ствола, на конце которого распо-
лагается 1 более крупный терминальный сорус
(рис. 371, В). 2 наиболее распространенных вида —
Р. violaceum и Р. pallidum — хорошо различается
по окраске спор: фиолетовой и белой соответст-
венно.
Род Coenonia Tiegh., 1884
Представлен единственным известным видом С. den-
ticulata.
Ножка этого клеточного слизевика расширяется
сверху в виде чашевидного образования с зубчатыми
краями, в котором находится сорус округлых желто-
ватых спор. Сорокарпы иногда имеют мутовки вто-
ричных сорусов.
Подкласс Myxogast|*ia L. S. Olive, 1970 (= Myxogastres Fr., 1829) —
Миксогастриды
Myxogastria — самый крупный подкласс в классе
Eumycetozoea. Сейчас известно около 800 видов, от-
носящихся к 5 отрядам (порядкам, по ботанической
классификации).
Отличительный признак миксогастрид — свобод-
ножйвущий многоядерный плазмодий, обладающий
амебоидной активностью и ритмичным (челночным)
током цитоплазмы. При определенных условиях из
плазмодия образуется спорокарп (спорофор), в кото-
ром формируются споры.
История изучения
Внешнее сходство спорокарпов миксогастрид с
плодовыми телами некоторых гастеромицетов послу-
жило причиной первоначального их отнесения к
классу грибов Gasteromycetes (Persoon, 1801). Фриз
(Fries, 1829), основываясь на наблюдениях за ста-
дией плазмодия, предложил выделить их в отдель-
ный подкласс Myxogastres, оставляя их тем не менее
в пределах класса Gasteromycetes. Вальрот (Wallroth,
1833) заменил название Myxogastres на Myxomyce-
tes; в этот таксон кроме миксомицетов вошли так-
же отдельные роды гастеромицетов. В том же году
Линк (Link, 1833) разделил Myxomycetes и Gastero-
mycetes на 2 отдельных класса, исключив из первого
все роды гастеромицетов.
В 1960 г. Мартин (Martin, 1960) предложил раз-
делить отдел Mycota на 2 подотдела: Myxomycotina
с единственным классом Myxomycetes и Eumycotina,
куда вошли настоящие грибы. В свою очередь, класс
Myxomycetes был разделен на 2 подкласса: Сега-
tiomyxomycetidae с 1 порядком Ceratiomyxales и
Myxogastromycetidae, состоящий из 5 порядков: Li-
ceales, Trichiales, Echinosteliales, Stemonitales и Phy-
sarales. В дальнейшем эта система была детально
разработана в монографии Мартина и Алексопулоса
(Martin, Alexopoulos, 1969). Объем и состав этих
порядков практически полностью совпадает с объ-
емом и составом соответствующих отрядов в класси-
фикации Олайва (Olive, 1975), которая использована
в настоящем издании.
Морфология и физиология
Плазмодии
Как уже отмечалось, отличительным признаком
миксогастрид служит свободноживущий многоядерный
плазмодий. Отсутствие клеточной стенки у плазмо-
дия создает условия для фагоцитоза и пиноцитоза (и,
следовательно, внутриклеточного пищеварения), т. е.
голозойного способа питания (Reyter, Chastellier,
1977; Madeline 1984). Плазмодий активно фагоцити-
рует бактерии, простейших, водоросли, мелких бес-
позвоночных, споры грибов и различные твердые
пищевые частицы. Тем не менее ряд видов удается
поддерживать в лабораторной культуре на питатель-
ной среде в аксеничных условиях (Gray, Alexopou-
los, 1968).
Плазмодий может активно мигрировать и облада-
ет различными положительными и отрицательными
таксисами (Konijn, Koevenig, 1971; Hader, Schrecken-
bach, 1984). Способ движения аналогичен способу
передвижения некоторых саркодовых. Из плазмодия
миксомицетов выделен сократительный белок актин,
участвующий в организации движения (Hatano et al.,
1980).
Организация плазмодия имеет существенное зна-
чение для систематики миксомицетов, особенно на
уровне отряда. На основании наблюдений за плазмо-
диями разных видов в условиях культуры было вы-
делено 3 основных их типа: протоплазмодий, афано-
плазмодий, фанероплазмодий (Alexopoulos, 1960).
Все они на ранних этапах развития имеют значи-
тельное морфологическое сходство, исчезающее на
более поздних стадиях.
Протоплазмодий имеет микроскопические раз-
меры и характеризуется отсутствием циркулирующих
токов протоплазмы. Этот тип плазмодия характерен
для представителей отрядов Echinosteliida и Liceida.
Однако если протоплазмодии эхиностелиевых не об-
ладают способностью к слиянию (Haskins,1978), то
протоплазмодии лициевых способны сливаться друг с
другом (Wollman, Alexopoulos, 1967).
432
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 373. Участок фанероплазмодия Physarum polycephalum. (По:
Olive, 1975). \
Fig. 373. Phaneroplasmodium of Physarum polycephalum.
Афаноплазмодий характерен для видов отря-
да Stemonitida. Он имеет тонкий, веерообразно раз-
вернутый передний край, за которым располагается
плохо различимая сетчатая протоплазматическая
масса. Афаноплазмодий отличает слабое развитие
или полное отсутствие слизевого чехла, имеющегося
у фанероплазмодия, а также наличие особой стадии
«коралла» перед началом формирования спорофо-
ров; кроме того, этот тип плазмодия менее устойчив
к обезвоживанию, чем остальные (Collins, 1979).
Фанероплазмодий наиболее обычен для видов
отряда Physarida (Rammeloo, 1976), однако он име-
ется и у некоторых видов отряда Liceida (McManus,
1966). Этот тип плазмодия подобен афаноплазмо-
дию, но значительно толще, с отчетливой сетью не-
одинаковых по толщине сосудов и развитым слизе-
вым чехлом (рис. 373), часто окрашен в различные
оттенки красного, желтого, розового, коричневого и
черного цветов. Имеются сведения о достаточной
стабильности окраски плазмодия в условиях культу-
ры (Collins, 1979). Однако в целом таксономичес-
кая ценность этого признака низкая. Так же как и у
афаноплазмодия, отличительной чертой зрелого фа-
нероплазмодия является так называемый челночный
ток цитоплазмы, отсутствующий только у протоплаз-
модиальных форм. В отличие от афаноплазмодия
фанероплазмодий, так же как и протоплазмодий,
может обитать в менее влажной среде. Стадия «ко-
ралла» также отсутствует* а при образовании споро-
форов фанероплазмодий, как и протоплазмодий эхи-
ностелиевых, может претерпевать плазматомию.
Весьма вероятно, что это сходство может быть свя-
зано с филогенетической близостью отрядов Echi-
nosteliida и Physarida.
Некоторые авторы выделяют четвертый тип плаз-
модия — плазмодий видов отряда Trichiida, занимающий
по своей морфологии промежуточное положение
между афаноплазмодием и фанероплазмодием. Он
характеризуется наличием токов протоплазмы, но от-
личается от последнего меньшими размерами и сла-
бой пигментацией (Indira, 1964; Rammeloo, 1976).
Спорокарпы
(плодовые тела, спорофоры)
Для миксогастрид наряду с наличием многоядер-
ного плазмодия характерен особый тип спороноше-
ния, при котором плазмодий превращается в споро-
карп. Структуры этой стадии обладают наиболее
важными таксономическими признаками. К ним от-
носятся гипоталлус, или подслоек, который обра-
зуется как основание спорокарпа (рис. 375, Л, Р);
перидий, покрывающий споровую массу и имею-
щий вид либо тонкой, прозрачной или непрозрачной
мембраны, либо плотной оболочки, он может быть
одно- или многослойным; колонка (колумелла) —
стерильная часть спорангия, может быть непосред-
ственным продолжением ножки или дериватом ни-
жней части спорокарпа (рис. 375, Ж, 3, М, С);
ложная колонка (псевдоколумелла) — агрегат
извести или узелков капиллиция в полости споро-
карпа; капиллиций — система нитей, трубочек,
пленок, служащая для рассеивания спор. Кроме этих
признаков в таксономии миксогастрид используется
еще ряд структур спорокарпов (рис. 375, Д, Е, М, Н, Т).
Типы спорокарпов. Спорокарпы миксогастрид
отличаются большим разнообразием (рис. 374, А—Д,
вкл.; 375; 376). При переходе в генеративную фазу
плазмодий превращается в спорокарп одного из
4 типов: плазмодиокарп, эталий, псевдоэталий, спо-
рангий. Встречаются виды, обладающие спорокарпа-
ми промежуточного строения, а также спорокарпами
нескольких типов в одной колонии.
Плазмодиокарп — наиболее простой тип спо-
рокарпа, когда весь плазмодий покрывается оболоч-
кой и, не меняя формы, целиком преобразуется в
спорокарп (рис. 375, О). Вследствие этого плаз-
модиокарпы всегда сидячие (без ножки), среди них
встречаются ветвящиеся, сетчатые, повторяющие
форму плазмодия в период превращения в спорокарп.
Плазмодиокарпы имеются у представителей отрядов
Liceida, Trichiida, Physarida и не найдены у Stemoni-
tida и Echinosteliida.
Спорангии — самостоятельные, сидячие или на
ножках спорокарпы, формирующиеся из отдельных
частей плазмодия (рис. 375, Б, Д, Ж, К, Л, П, Р). При
этом каждая из них развивается в отдельный споро-
карп с характерными формой, цветом и структурой.
У некоторых видов число спорокарпов, развившихся
из одного плазмодия, может достигать нескольких де-
сятков и даже сотен. Этот тип спорокарпа встречает-
ся во всех отрядах и семействах класса.
Этали и — подушковидные образования, форми-
рующиеся из ветвящихся плазмодиев, у которых внут-
ренние ответвления образуют спорангии, в то время
как протоплазма внешних (наружных) ответвлений
разрушается и преобразуется в кортекс, окружающий
весь эталий (рис. 376). Остатки боковых стенок спо-
рангиев иногда сохраняются в виде пленок или пере-
городок (псевдокапиллиция). Характерно, что виды,
имеющие эталии, как правило, никогда не образуют
других типов спорокарпов. Эталии встречаются толь-
ко у видов отрядов Liceida, Physarida и Stemonitida и
не встречаются в отрядах Trichiida и Echinosteliida.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
433
Рис. 375. Строение спорофоров миксогастрид.
А-—2> — Cribraria argillacea, спорангии; В — споры; Г — деталь узелка псевдокапиллиция с диктидиновыми гранулами; Д — Arcyria ferruginea, споран-
гии; Е — споры, нити капиллиция и перидий с сетчатой орнаментацией; Ж — Comatricha tenerrima, спорангий; 3 — участок капиллиция и колонки; И —
деталь капиллиция и спора; К — колония спорангиев; Л — Lamproderma scintillans, 2 спорангия; М — капиллиций и колонка; Н — споры и деталь ка-
пиллиция; О — Didymium dubium, плазмодиокарп; П — D. minus, спорангии; Р — D. nigripes, спорангии; С — деталь ножки и колонки спорангия; Т —
нить капиллиция и споры. (А—Г, 3—Т — по: Nannenga-Bremekamp, 1979; Д—Е — по: Lister, 1925).
Fig. 375. Various structures of fruiting bodies of myxogastrids.
А, Б — Cribraria argillacea, sporangia; В — spores; Г— detail of pseudocapillitium and dictydine granules; Д — Arcyria ferruginea, sporangium; E — spores,
capillitium threads and detail of peridium; Ж — Comatricha tenerrima, sporangium; 3 — part of capillitium and columella; И — capillitium and spore; К — spo-
rangia; Л — Lamproderma scintillans, two sporangia; M — capillitium and columella; H — spores and thread of capillitium; О — Didymium dubium, plasmodio-
carp; П — D. minus, sporangia; P — D. nigripes, sporangia; C — parts of stalk and columella; T — thread of capillitium and spores.
28 Протисты, часть 1
434
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 376. Эталии Lycogala epidendrum. (По: Горленко, 1981).
Fig. 376. Aethalia of Lycogala epidendrum.
Псевдоэталии состоят из множества споранги-
ев, плотно прилегающих друг к другу, которые, одна-
ко, не теряют индйвидуальности благодаря сохранив-
шимся боковым стенкам, по крайней мере на ранних
стадиях развития. Псевдоэталии могут быть, так же
как и спорангии, сидячими или на ножках и встреча-
ются в отрядах Liceida, Physarida, реже Trichiida, Ste-
monitida.
Таким образом, если рассматривать класс в целом,
то тип спорокарпа образует непрерывную гамму пе-
реходов от одной формы к другой. Наибольшим раз-
нообразием в этом смысле обладает отряд Physarida,
где встречаются практически все типы спорокарпов.
Однако в некоторых случаях этот признак достаточ-
но стабилен на уровне семейств. Так, представители
сем. Reticulariidae (отр. Liceida) имеют преимуществен-
но эталии, а виды сем. Liceidae этого же отряда —
спорангии и редко плазмодиокарпы.
Спорангии могут быть сидячими или на ножке,
шаровидными, цилиндрическими, подушковидными,
яйцевидными, дисковидными, многолопастными,
сжатыми с боков, конусовидными и т. д. В пределах
каждой из перечисленных основных форм можно
выделить множество отклонений. Например, развет-
вленные спорангии могут иметь различную форму
ветвления, сферические спорангии часто бывают не-
сколько вытянуты или приплюснуты, что придает
спорангию в целом другой вид. Форма спорангия,
несмотря на сильную изменчивость даже в пределах
вида, — признак, имеющий большое таксономичес-
кое значение. Предел его варьирования ограничен
рамками одного вида, и поэтому он может быть ис-
пользован как критерий видового уровня в сочета-
нии с другими признаками.
Окраска спорангиев также сильно варьирует в за-
висимости от внешних условий. Значение этого при-
знака в пределах высших таксонов различно. Напри-
мер, в сем. Trichiidae преобладает красная, желтая и
коричневая окраска. Красный цвет различных оттен-
ков встречается чаще всего у видов рода Arcyria, у
одного вида Trichia — Т. floriformis — и у монотип-
ного рода Metatrichia (М. vesparium). Для спорангиев
родов Trichia и Hemitrichia наиболее характерный
цвет — желтый. В сем. Trichiidae цвет спорокарпа
имеет только видовое значение, причем в пределах
одного вида этот признак варьирует незначительно. В
сем. Stemonitidae преобладают черный и коричневый
цвета спорокарпа, что может служить как родовым,
так и видовым признаком. Например, для представи-
телей рода Lamproderma (рис. 375, Л—Н) характерен
металлический или радужный оттенок в окраске спо-
рангия, а для видов родов Comatricha и Stemonitis —
коричневый. В сем. Physaridae цвет — сильно варь-
ирующий признак даже на уровне вида. Это связано
прежде всего с различием в количестве извести и ее
распределением по поверхности перидия, что, в свою
очередь, зависит от условий окружающей среды. Так,
например, у широко распространенного вида Physa-
rum viride окраска спорангия находится в зависимос-
ти от качества откладываемых крупинок извести. Она
может быть оранжево-красной, желтой, зеленоватой
или даже серой с металлическим блеском. Эта гамма
переходов может быть представлена у спорангиев од-
ной колонии.
Таким образом, цвет спорокарпов — видовой ди-
агностический признак. Его можно использовать
только с учетом всех возможных вариаций, коли-
чество которых неодинаково в разных таксонах. Из-
за сильной изменчивости этот признак можно при-
менять только наряду с другими признаками на уров-
не вида.
Морфогенез, структура, окраска и форма ножки
спорокарпа. На основании наблюдений за развитием
ножки спорокарпа некоторые авторы разделяют мик-
согастрид на 2 подкласса (Ross, 1957, 1973). Для пер-
вого характерен эпигипоталлический тип развития
спорокарпа, а для второго — субгипоталлический.
При эпигипоталлическом развитии ножка представ-
ляет собой вырост гипоталлуса, а при субгипоталли-
ческом развитии она формируется из тех же участков
протоплазмы плазмодия, что и перидий.
Структура ножки — хороший систематический
признак на уровне рода. Например, у видов Comatricha
она заполнена фибриллярным материалом, у Macbri-
deola — полая, а у Stemonitis — полая или заполне-
на аморфным материалом. В отряде Trichiida этот
признак может использоваться на видовом уровне.
Окраска ножки может иметь значение как на уровне
рода, так и вида. Форма ножки, а также ее отсут-
ствие или наличие могут служить диагностическим
признаком на уровне рода или вида, хотя из-за ва-
риабельности этого признака к нему нужно отно-
ситься с осторожностью. Например, у Trichia varia
имеется вся гамма переходов от спорангиев с хоро-
шо выраженной ножкой до сидячих.
Перидий зрелого спорокарпа вскрывается (рис. 374,
В, Д), и споры свободно рассеиваются, причем этому
способствуют специальные структуры — капиллиций
и псевдокапиллиций (рис. 375, А, Д).
Наиболее простой способ раскрытия спорокарпа
выражается в неупорядоченном растрескивании пе-
ридия. Этот способ часто встречается у видов с
плазмодиокарпом и значительно реже у видов со
спорангиями. Более специализированный тип раст-
рескивания — разрушение только апикального
конца спорокарпа, при этом растрескивание может
осуществляться продольной щелью (Licea biforis),
кольцевой щелью (L. parasitica), кольцевой щелью с
образованием крышечки (L. kleistobolus). Наконец, у
некоторых видов Licea растрескивание происходит
вдоль специальных швов на перидии.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
435
Споры (рис. 374, Е, Ж;
375, В, Е, И, Н. Т; 377, Б, вкл.)
У большинства миксомицетов споры сферичес-
кие, реже яйцевидные, эллипсоидальные. Они быва-
ют свободными или объединенными в комплексы, в
последнем случае они обычно полиэдрической или
неправильной формы. Форма и размер спор недораз-
вившихся спорофоров, а также спорофоров, сформи-
ровавшихся в неблагоприятных условиях, также могут
сильно отличаться. Цвет споровой массы и отдельных
спор — один из важнейших диагностических призна-
ков. Окраска спор в массе часто не совпадает с цве-
том отдельных спор, который можно наблюдать на
препаратах в световом микроскопе. Указанные при-
знаки у некоторых видов могут сильно варьировать.
У видов рода Echinostelium цвет споровой массы бе-
лый, бежевый, розоватый. Под микроскопам споры
выглядят практически бесцветными (прозрачными).
У ряда видов, например Ысеа, толщина оболочки не-
равномерная, из-за чего некоторая ее часть (обычно
примерно треть) выглядит более светлой, чем осталь-
ная.
Структура экзоспория у миксомицетов имеет важ-
ное значение при разграничении видов. В большин-
стве монографий и определителей используются данные
о строении экзоспория, полученные с применением
иммерсии. В последнее время появились работы по
изучению тонкой структуры экзоспория с использо-
ванием сканирующего электронного микроскопа
(Schoknecht, Small, 1972; Rammeloo, 1974, 1975,
1990; Новожилов, 1993). Основными элементами эк-
зоспория являются бородавочка и шип. Главное раз-
личие этих структур заключается в том, что бородавоч-
ка имеет закругленную вершину, а шип — заостренную.
Бородавочки могут быть одно- или многовершинны-
ми или булавовидными. В последнем случае диаметр
вершины больше диаметра основания (рис. 374, Ж).
Сетчатый тип экзоспория имеет несколько моди-
фикаций: сетчато-бородавчатый — с ребрами ячеек,
образованными слившимися бородавочками, как на-
пример у Comatricha irregularis, Lamproderma atros-
porum, сетчато-складчатый — с ребрами, уплощен-
ными настолько, что они становятся похожи на
складки, например у Dianema depressum, Protophysa-
rum phloiogenum или Lamproderma cribrarioides
(рис. 374, E). Следует отметить, что подобная диф-
ференциация сетчатых типов экзоспория возможна
только после изучения в сканирующем электронном
микроскопе; в световом микроскопе, даже при ис-
пользовании иммерсионного объектива, эти типы эк-
зоспория, как правило, неразличимы.
Капиллиций и псевдокапиллиций
Настоящий капиллиций присутствует во всех по-
рядках эндоспоровых миксомицетов (рис. 374, Д;
375, Е, 3, М; 377, А, Б; 378). Он образуется как сис-
тема вакуолей различной формы в протоплазме плаз-
модия в период превращения последнего в спорокарп
(Gray, Alexopoulos, 1968; Ellis et al., 1973; Gaither,
1974). У некоторых родов стемонитовых — Comat-
Рис. 378. Строение капиллиция. (По: Горленко, 1981).
А — отдельная нить капиллиция Trichia sp., Б — общий вид капиллиция
Stemonitis sp.
Fig. 378. Structure of capillitium.
A — capillitium thread of Trichia sp., Б — capillitium of Stemonitis sp.
richa, Stemonitis, — капиллиций формируется путем
образования нитей, отходящих от ножки или колон-
ки (Ross, 1973).
Псевдокапиллиций встречается у большинства
видов со спорокарпами в виде эталиев или псевдоэта-
лиев и, как правило, является остатком перидия
боковых стенок редуцированных спорангиев. Так,
псевдокапиллиций имеется у видов Enteridium
(отр. Liceida), а также у Fuligo, Mucilago (отр. Physarida).
Псевдокапиллиций обычно представлен тяжами или
перфорированными пластинками (Reticularia), скульп-
турированными или гладкими трубочками неправиль-
ной формы (Lycogala), щетинками (Tubifera) или
утолщенными частями боковых стенок спорангия,
которые свешиваются с краев апикальных концов
спорангиев в виде нитей и образуют каркас псевдо-
эталия (Dictydiaetalium). Сетчатый перидий Dictydi-
шп и Cribraria, очевидно, гомологичен псевдокапил-
лицию указанных родов.
Изучение структуры капиллиция привело к изме-
нению взглядов на положение некоторых родов в сис-
теме. Так, электронно-микроскопические исследова-
ния показали, что капиллиций рода Minakatella по-
лый, вследствие чего этот род логично перевести из
436
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
одного семейства в другое (Keller et al., 1973); так же
обстоит дело и с родом Prototrichia (Ellis et al., 1973).
У стемонитовых форма сети капиллиция, особенно
его периферической части, может служить ключевым
признаком на уровне рода для Comatricha и Stemonitis
или видовым признаком для большинства представи-
телей родов Lamproderma и Diachea. Орнаментация
капиллиция часто играет важную роль как 'критерий
видового уровня в сем. Trichiidae, так же как и узелки
с известью капиллиция в сем. Physaridae.
Среди миксомицетов широко распространено яв-
ление выведения кальция из цитоплазмы в виде солей
на стадии перехода плазмодия в спорокарп (рис. 377,
В, Г). В частности, углекислый кальций откладывает-
ся в ножке спорангия, в капиллиции, в перидии или
на нем у видов отр. Physarida. По своей структуре из-
весть может быть либо в форме аморфных гранул
(представители сем. Physaridae), либо в виде кристал-
лов (Didymium) или даже в форме конгломератов
кристаллов (Lepidoderma). Из других таксонов каль-
ций найден в спорокарпах некоторых видов — Tri-
chia contorta, Calomyxa metallica, Metatrichia vespari-
um\ у последнего он входит в состав органического
соединения (Nelson et al., 1977). Кальций обнаружен
в ножке спорангия у Diachea leucopodia и в перидии
основания спорангия Lepidoderma iridescens (Stemo-
nitida), а также в диктидиновых гранулах Cribraria
(Liceida) (Schoknecht, 1975). Однако следует отме-
тить, что у Perichaena кальций присутствует в форме
оксалата, тогда как у представителей отр. Physari-
da — в виде углекислого кальция и фосфата. Кроме
того, кальций у видов отр. Physarida концентрируется
в основном между мембранами перидия, а у Pericha-
ena — на его поверхности. Кроме кальция в состав
отложений у видов Perichaena входит кремний.
Жизненный цикл
Несмотря на значительное разнообразие внешне-
го вида трофических стадий и плодовых тел, плазмо-
диальные слизевики демонстрируют принципиальную
общность в организации своего жизненного цикла.
Полный жизненный цикл в лабораторной культу-
ре занимает от нескольких дней до нескольких не-
дель в зависимости от того, к какому конкретно виду
относится данный организм. Кроме того, на его про-
должительность существенное влияние оказывают так-
же условия культивирования. Схема типичного жиз-
ненного цикла плазмодиальных слизевиков представ-
лена на рис. 379.
Трофическая гаплофаза. Каждая зрелая спора у
большинства представителей подкласса содержит, как
правило, 1 гаплоидное ядро. При прорастании через
пору или щель в споровой оболочке выходит един-
ственная одноядерная клетка. При недостаточной
влажности окружающей среды из споры высвобожда-
ется амебоидная особь (миксамеба), тогда как в при-
сутствии воды — жгутиковая клетка (рис. 379, Б). В
случае изменения соответствующих условий может
легко происходить их взаимная трансформация. У
некоторых видов плазмодиальных слизевиков в ре-
зультате митоза и цитокинеза, происходящих непо-
средственно перед началом прорастания, из споры
выходит не 1, а сразу несколько одноядерных особей
одновременно (например, у Badhamia affinis их число
достигает 8).
И амебоидные и жгутиковые клетки содержат од-
ноядрышковое ядро и обычный набор цитоплазмати-
ческих органелл: аппарат Гольджи, митохондрии с
трубчатыми кристами, шероховатый ЭПР, сократи-
тельную вакуоль. Жгутиконосец имеет продолгова-
тую («зооспороподобную») форму тела. Наиболее
типично присутствие единственной пары близкоас-
социированных жгутиков, расположенной на пере-
днем конце клетки, — 1 длинного, направленного
вперед, и 1 короткого («рудиментарного»), лежаще-
го у основания первого и повернутого в обратную
сторону (этот жгутик часто практически неразличим
на светооптическом уровне) (Olive, 1975). Иногда
оба жгутика могут иметь одинаковую длину; в ряде
случаев отмечалось присутствие дополнительных
(«сверхштатных») жгутиков (Olive, Stoianovitch,
1971; Mir et al., 1984). Жгутики лишены мастиго-
нем.
Электронно-микроскопическое изучение строе-
ния базальной части жгутикового аппарата у целого
ряда видов плазмодиальных слизевиков (Physarum
flavicomum, Ph. polycephalum, Ph. cinereum, Echinos-
telium minutum, Stemonitis pallida, Didymium iridis и
др.) выявило принципиальное единообразие его ор-
ганизации (Schuster, 1965; Aldrich, 1968; Ishigami,
1977; Haskins, 1978; Wright et al., 1979; Mir et al.,
1982, и др.). С 2 кинетосомами, расположенными
непосредственно на переднем конце тела и ориенти-
рованными под углом друг к другу, ассоциирован
определенный набор цитоплазматических микротру-
бочковых дериватов. Среди них, в частности, сущест-
венное развитие получает конусовидная структура
(состоящая из микротрубочек, которые берут начало
от дискретного электронно-плотного ЦОМТа, лежа-
щего недалеко от основания кинетосомы переднего
жгутика), которая связывает базальную часть жгути-
ков с ядерной оболочкой. Внутрь этого конуса ока-
зывается включенной диктиосома аппарата Гольджи.
В целом, как можно видеть, общая организация жгу-
тикового аппарата клеток у плазмодиальных слизе-
виков весьма сходна с таковой у многих жгутиковых
протостелид (Spiegel, 1981а, 1981b; Spiegel et al.,
1986).
При изучении ультраструктуры миксамеб выясни-
лось, что в них присутствует развитый микротрубоч-
ковый цитоскелет, включающий ядерно-центриоляр-
ный комплекс, который соответствует по своему со-
ставу и организации базальному аппарату жгутиков
(Aldrich, 1968; Wright et al., 1980; Roobol et al.,
1982; Clayton et al., 1983; Mir et al., 1982). Отличие
заключается главным образом лишь в редуцирован-
ном состоянии некоторых микротрубочковых структур.
Процесс трансформации миксамебы в жгутиконосца
включает перемещение всего комплекса к перифе-
рии клетки, формирование аксонем ундулиподий и
удлинение соответствующих цитоплазматических де-
риватов (Schuster, 1965; Aldrich, 1968; Haskins, 1978;
Wright et al., 1980; Uyeda, Furuya, 1985). Предпола-
гается, что в регуляции взаимопревращений амебоид-
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
437
Рис. 379. Схема типичного жизненного цикла миксогастрид (пояснения в тексте). (По: Olive, 1975).
А — выход миксамеб или зооспор из спор, Б—Г — слияние гаплоидных клеток и образование зиготы, S' — образование микроцист, Д — образование
плазмодия, Е — рост плазмодия, F — формирование склероция (макроцисты), Ж — формирование плодовых тел (спорофоров) и спорогенез, 3 — сфор-
мировавшийся спорофор.
Fig. 379. General life cycle of the myxogasrids.
A — germinating spore, Б—Г — swarm cells fusing and zygote formating, S' — microcysts, Д — plasmodium formating, E — mature fan-shaped plasmodium,
E' — sclerotium (macrocysts), Ж<— developing sporophores and sporogenesis^ 3 — sporophore.
ной и жгутиковой форм клеток у Physarum polycep-
halum принимают участие ионы Са2+ (Uyeda, Furuya,
1986).
Клетки этой стадии жизненного цикла плазмоди-
альных слизевиков активно питаются, фагоцитируя
различные пищевые организмы (бактерии, дрожжи),
и делятся. Собственно жгутиконосцы не способны к
мультипликации, поэтому непосредственно перед на-
чалом каждого нового деления происходит втягива-
ние жгутиков внутрь клетки и резорбция их аксонем.
При наступлении неблагоприятных условий миксаме-
бы могут переходить к состоянию покоя, секретируя
вокруг себя плотную оболочку и превращаясь в сфе-
рической формы микроцисты (рис. 379, Б'). В них
сохраняется ядерно-центриолярный комплекс, что
свидетельствует об их способности продуцировать
жгутиковую клетку непосредственно в процессе про-
растания (Schuster, 1965; Olive, 1975).
Инициация диплофазы (рис. 379, Б, В). Гаплоид-
ные особи плазмодиальных слизевиков сливаются по-
парно. Показано, что у гетероталличных видов слия-
ние может происходить как между 2 жгутиконосцами
или между 2 миксамебами, так и между жгутиковой и
амебоидной клетками (Olive, 1975). Для приобрете-
ния способности (физиологической компетентности)
к слиянию клеткам требуется некоторый период, со-
ставляющий, например, у Perichaena vermiculdris 12—
18, а у Didymium iridis 10—12 ч (Ross, 1967а, 1967b).
Никаких морфологических отличий этих клеток от
некомпетентных особей не наблюдается. Индукция
компетентности происходит при достижении опреде-
ленной критической плотности популяции и связана
с синтезом и выделением в среду специфического
растворимого фактора (Shipley, Holt, 1980). Пуско-
вой механизм, «включающий» синтез этого фактора,
связан, по всей видимости, с клеточными контактами
и не зависит от изменений запасов пищи в окружаю-
щей среде (Pallotta et al., 1983).
Непосредственно за плазмагамией следует слияние
ядер обоих партнеров, что хорошо было продемонст-
рировано на примере Physarum gyrosum (см.: Olive,
1975). У Didymium iridis кариогамия наблюдается че-
рез 4—20 мин после образования двухъядерной клет-
ки (Ross, 1967а). Возникшая таким образом зигота
дает начало диплоидной плазмодиальной стадии жиз-
ненного цикла.
438
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Кроме того, известно немало случаев, когда разви-
тие плазмодиальной фазы осуществляется без предва-
рительного слияния гаплоидных клеток. Так, напри-
мер, описан штамм D. nigripes, который формирует
плазмодии без осуществления плазмо- и кариогамии
одноядерных особей (Кегг, 1967, 1968, 1970; Kerr,
Kerr, 1967). Возможно, что диплоидное состояние
ядер достигается в этом случае в результате эндоми-
тоза. Было показано, что у этого организма и микс-
амебы, и плазмодии содержат ядра, варьирующие по
плоидности. Есть также свидетельства того, что из
гаплоидных миксамеб могут развиваться гаплоидные
плазмодии. Подобные вариации плоидности наблюда-
лись и у ряда других видов (Ross, 1966). Некоторые
авторы рассматривают формы, у которых отсутствуют
гаметы (но не мейоз), как апогамные (Stosch, 1935;
Olive, 1975). Апогамный способ развития плазмодиев
может наблюдаться и у таких видов, как например
Physarum polycephalum (Anderson et al., 1976; Pallotta
et aL, 1983).
Плазмодий (рис. 379, E), Это основная вегетатив-
ная стадия жизненного цикла миксогастрид. Один из
наиболее широко распространенных в лабораторных
культурах и достаточно хорошо изученных предста-
вителей этой группы протистов — Ph, polycepha-
lum — имеет типичный фанероплазмодий с отчетли-
выми главными сосудами и развитой сетью более
мелких сосудов, вливающихся в сплошной, вееро-
образно развернутый протоплазматический передний
край (рис. 373).
Формирование плазмодиев осуществляется следу-
ющим образом. Одновременно с увеличением объема
цитоплазмы происходят последовательные синхрон-
ные деления исходного диплоидного ядра, в результа-
те чего образуются содержащие некоторое количе-
ство ядер амебоидные микроплазмодии (рис. 379, Д),
На примере Ph, polycephalum и Didymium iridis было
показано, что одноядерные миксамебы, микроплаз-
модии и уже достаточно крупные многоядерные плаз-
модии могут сливаться, образуя еще более крупные
плазмодиальные стадии (Olive, 1975). Однако плаз-
модий может развиваться и без предварительного
объединения мелких форм. Характерный челночный
ток цитоплазмы появляется в плазмодиях, когда они
имеют еще микроскопические размеры. У Perichaena
vermicularis, например, он начинается приблизитель-
но на двадцатиядерной стадии.
В природе при наступлении неблагоприятных ус-
ловий (понижение влажности и температуры, исто-
щение пищевых запасов и т. д.) плазмодии могут пе-
реходить в неактивное, своего рода инцистированное
состояние, формируя так называемые склероции
(рис. 379, Е'). В лабораторной культуре синхронное
развитие склероциев у Physarum polycephalum может
быть получено, например, простым помещением мик-
роплазмодиев в непитательную солевую среду. У не-
которых видов с крупным фанероплазмодием склеро-
ций сохраняет его форму, у большинства же видов
образуются маленькие склероции (макроцисты, или
«сферулы»), так как плазмодий предварительно дро-
бится путем расчленения протоплазмы на большое
число отделенных друг от друга участков (Stewart,
Stewart, 1961; Alexopoulos, 1966; Stiemerling, 1971).
Ядра остаются на стадии Gx ядерного цикла и входят
в фазу Sj только в начале процесса прорастания (Hut-
terman, 1973). Стенки склероция, так же как и слизе-
вой чехол плазмодия, состоят из фибриллярного мат-
рикса, упакованного более плотно в склероции и
менее плотно в плазмодии. Однако слизевой чехол
состоит в основном из галактозы, тогда как стенки
склероция — преимущественно из галактозамина
(McCormick et aL, 1970а, 1970b). Склероции в основ-
ном сохраняют свою жизнеспособность от 1 года до
3 лет. Есть сведения, что у Badhamia utricularis этот
срок может достигать 7 лет.
Афаноплазмодий и протоплазмодий не образуют
склероций, а только комплекс макроцист, покрытый
общим чехлом (Aldrich, Blackwell, 1976).
В плазмодиях имеется обычный набор цитоплаз-
матических органелл (многочисленные диктиосомы
аппарата Гольджи, митохондрии с трубчатыми крис-
тами, гладкий и шероховатый ЭПР, различные типы
вакуолей и везикул), а также присутствует большое
число разнообразно организованных микрофиламен-
тозных структур, обеспечивающих как подвижность
организма в целом, так и характерные движения его
цитоплазмы. Как показали иммунофлюоресцентные
исследования с применением специфических анти-
тел к актину, а также электронно-микроскопические
работы, выполненные на Physarum polycephalum,
Clastoderma debaryanum, Fuligo septica, Perichaena
vermicularis и других видах истинных слизевиков,
микрофиламенты в плазмодиях могут располагаться
в виде полигональной сети или компактных пучков,
формирующих продольные, кольцевые или спираль-
но закрученные фибриллы (Wohlfarth-Botterman,
1964; McManus, Roth, 1965; Rhea, 1966; Usui, 1971;
Gawlitta et aL, 1980; Naib-Majani et aL, 1982, 1983,
и др.). Во многих случаях наблюдалась непосредст-
венная связь микрофиламентов с плазматической
мембраной. Концентрация актина и характер его ор-
ганизации четко коррелируют с дифференциальной
локомоторной полярностью тела плазмодия.
Электронно-микроскопические и иммунофлюо-
ресцентные исследования с применением специфи-
ческих антител к тубулину показали, что в плазмодиях
Physarum polycephalum в отличие от одноядерной аме-
боидной стадии совершенно отсутствуют цитоплазма-
тические микротрубочки (Wohlfarth-Botterman, 1964;
Rhea, 1966; Havercroft, Gull, 1983; Diggins, Dove,
1987, и др.). В то же время фанероплазмодий Ph.mel-
leum и протоплазмодии Clastoderma debaryanum со-
держат цитоплазматические микротрубочки, которые
часто организованы в плотные пучки (McManus, Roth,
1967). Более того, у Ph.melleum были обнаружены до-
вольно крупные, ассоциированные с микротрубочка-
ми электронно-плотные тельца, очевидно представля-
ющие собой цитоплазматические ЦОМТы.
Имеются сведения (Indira, 1964), что плазмодии
Stemonitis herbatica, Physarum compressum и Arcyria
cinerea при помещении их в стерильную воду распа-
даются (путем плазматомии) на мелкие, очевидно од-
ноядерные, жгутиковые клетки. Эти факты (прини-
мая во внимание отсутствие в плазмодиях центрио-
лей) свидетельствует о том, что базальный аппарат
жгутиков может довольно быстро формироваться
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
439
de novo. У неидентифицированного до вида предста-
вителя рода Physarum было описано образование из
плазмодиев одноядерных амебоидных особей (Ross,
Cummings, 1967). К сожалению, из-за очень малого
количества подобного рода наблюдений и слабой
изученности даже известных примеров, интерпрета-
ция этих данных является весьма затруднительной.
Не исключено, что данный феномен представляет
собой достаточно редкую в условиях лабораторной
культуры форму бесполого размножения плазмоди-
альных слизевиков.
Особенности организации митозов на амебоид-
ной и плазмодиальной стадиях жизненного цикла.
Многочисленные свето- и электронно-микроскопи-
ческие исследования, выполненные на Ph. flavico-
тит, Ph. polycephalum, Didymium nigripes, Arcyria
cinerea, Echinostelium minutum и других видах, четко
продемонстрировали, что вегетативные ядерные де-
ления на плазмодиальной и амебоидной стадиях
жизненного цикла истинных слизевиков представля-
ют собой 2 разных типа митозов (Kerr, 1967; Al-
drich, 1969; Mims, 1972а; Hinchee, Haskins, 1980a,
1980b; Havercroft, Gull, 1983, и др.). Так, в плазмо-
диях в ходе митоза сохраняется интактная ядерная
оболочка. Центриоли отсутствуют; веретено форми-
руется внутриядерно. ЦОМТом веретена у Physarum
polycephalum служит небольшое, не связанное с обо-
лочкой ядра аморфное тельце, возникающее в конце
интерфазы; в профазе оно дает отростки пучков
микротрубочек (Tanaka, 1973). Веретено деления об-
ладает осевой симметрией. Таким образом, в соот-
ветствии с классификацией Райкова (1986) в плаз-
модиях имеет место закрытый внутриядерный орто-
митоз. В то же время у миксамеб оболочка ядра во
время деления полностью разрушается уже к позд-
ней профазе и отсутствует вплоть до телофазы. Би-
полярное осевое веретено формируется внеядерно.
Следовательно, в данном случае наблюдается типич-
ный открытый ортомитоз (Райков, 1986). Вдобавок
ко всему, на полюсах веретена присутствуют центри-
оли.
Наличие в одном жизненном цикле 2 типов веге-
тативных ядерных делений является характерной
особенностью представителей подкласса Myxogastria
(Olive, 1975). Следует помнить, что миксамебы явля-
ются гаплоидными организмами и за кариокинезом у
них следует цитокинез. Ядра же плазмодиев диплоид-
ные, и митозы приводят лишь к увеличению их коли-
чества в цитоплазме.
Формирование плодовых тел и спорогенез
(рис. 379, Ж). Выход плазмодиев на поверхность
субстрата перед началом образования плодовых тел
обусловлен целым комплексом факторов окружаю-
щей среды. В экспериментах с лабораторными куль-
турами установлено, что к их числу в первую оче-
редь относится истощение пищевых ресурсов, изме-
нение pH и температуры. Собственно плодоношение
стимулируется значительным увеличением степени
освещенности плазмодиев.
Наиболее мелкие и просто организованные споро-
карпы встречаются у таких протоплазмодиальных
форм, как представители родов Licea и Echinostelium,
причем у последних они во многих отношениях на-
поминают спорокарпы протостелид (Olive, 1975). У
некоторых видов при развитии спорангиев происхо-
дит распадение плазмодия на множество отдельных
многоядерных участков. Каждый из них в дальней-
шем самостоятельно дифференцируется в стебельча-
тый или сидящий на поверхности субстрата споран-
гий.
Спорогенез в плодовом теле протекает следую-
щим образом. Плазмодиальный протопласт последо-
вательно расчленяется на мелкие одноядерные фраг-
менты, представляющие собой преспоровые клетки.
Гаплоидизация осуществляется в процессе созрева-
ния спор (Carroll, Dykstra, 1966; Aldrich, 1967, 1970;
Aldrich, Mims, 1970; Haskins et al., 1971, и др.).
Профаза первого мейотического деления обычно
становится различимой спустя приблизительно 20 ч
после разделения цитоплазмы на преспоровые фраг-
менты (благодаря появлению отчетливых синаптоне-
мальных комплексов). По крайней мере первое из
2 делений мейоза протекает с формированием внут-
риядерного веретена и сохранением ядерной оболоч-
ки (за исключением «полярных окон»); центриоли
отсутствуют. 3 из 4 образующихся в конечном итоге
гаплоидных ядра дегенерируют. Приблизительно в
это же время в каждой споре можно обнаружить
появление аппарата Гольджи и пары центриолей
(Aldrich, 1967; Mims, 1972b).
Г еографическое
распространение и экология
Большинство видов миксогастрид — космополи-
ты. Среди них можно назвать такие виды, как Arcy-
ria cinerea, A. dunudata, Comatricha typhoides, Fuligo
septica, Lycogala epidendrum, Stemonitis fusca, встре-
чающиеся на всех материках в различных природно-
климатических зонах. Однако некоторые виды име-
ют более узкие ареалы. Так, например, Hemitrichia
clavata отмечен исключительно в зоне умеренного и
холодного климата, тогда как Craterium paraguayen-
se и Tubifera bombarda отмечены только в тропиках
и субтропиках (Stephenson et al., 1994). Предста-
вители рода Cribraria обитают преимущественно в
хвойных лесах на гнилой древесине хвойных с кис-
лым значением pH (Stephenson, 1988).
Наиболее хорошо исследованы миксогастриды
лесных сообществ тропиков и зон с умеренным кли-
матом (Maimoni-Rodella, Gottsberger,1980; Stephen-
son, 1988, 1989; Eliasson, 1991; Новожилов, 1993;
Stephenson, Stempen, 1993; Ing, 1994), тогда как све-
дений о видах, обитающих преимущественно в пус-
тынях (Blackwell, Gilbertson, 1980; Новожилов, Го-
лубева, 1986) и тундрах (Ing, Smith, 1980; G0tzsche,
1984, 1989, 1990; Stephenson, Laursen 1993; Новожи-
лов и др., 1998а, 19986), еще недостаточно. Миксо-
гастриды встречаются обычно в гнилой древесине и
под корой отмерших деревьев, в листовом опаде, в
почве, обычно во влажных затененных местах. Од-
нако даже в местах с очень низкой влажностью сли-
зевики часто присутствуют в виде покоящихся ста-
дий или спор, что подтверждается наблюдениями
при проращивании их на природных субстратах во
440
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
влажных камерах. Среди миксогастрид имеется груп-
па криофильных альпийских и субальпийских видов,
способных образовывать спорофоры рядом с таю-
щим снегом (Meylan, 1931; Kowalski, 1970). Выделе-
ны экологические группы на коре живых деревьев
(Brooks et al., 1977; Frederick, Pendergrass, 1990),
помете растительноядных животных (Eliasson, Lun-
dqvist, 1979; Сох, 1981; Eliasson et al., 1991), в лис-
товой подстилке (Harkonen, 1981), на мхах и пече-
ночниках (Stephenson, Studlar, 1985). Эти примеры
показывают, что экологические ниши, занимаемые
миксомицетами, гораздо разнообразнее, чем счита-
лось до последнего времени. Плодовые тела крупных
миксогастрид могут служить источником пищи и
убежищем для некоторых грибов (Rogerson, Stephen-
son, 1993) и насекомых (Stephenson et al., 1994);
последние, в свою очередь, участвуют в распростра-
нении спор миксомицетов.
Систематический обзор
В составе подкласса Myxogastria обычно выделя-
ют 5 отрядов: Echinosteliida (= Echinqsteliales), Tri-
chiida (= Trichiales), Stemonitida (= Stemonitales),
Physarida (= Physarales) и Liceida (= Liceales) (Mar-
tin, Alexopoulos, 1969; Olive, 1970, 1975; Levine et
al., 1980; Новожилов, 1993).
Ранее к Myxogastria относили род Ceratiomyxa, од-
нако, как уже упоминалось выше, в последнее время,
прежде всего благодаря электронной микроскопии,
были получены данные, указывающие на близость Cera-
tiomyxa к протостелидам, причем экзогенные споры Ce-
ratiomyxa, видимо, можно рассматривать как гомолог
односпорового спорофора протостелид (Olive, 1970).
Некоторые авторы относят к Myxogastria еще 1 от-
ряд — Echinosteliopsida L. S. Olive, 1970 с 1 семей-
ством — Echinosteliopsidiidae L. S. Olive, 1970, в ко-
торое входят 2 рода: Echinosteliopsis Reinchardt et
L. S. Olive, 1967 и Bursula Sorok., 1876. Представитель
последнего — В. crystallina Sorok., 1876 — известен
только на территории России. Этот вид рассматрива-
ется некоторыми авторами в качестве связующего
звена между вампиреллидами и цератиомиксовыми
(Гоби, 1884). Однако Ячевский (1907) считает, что
этот вид нельзя включать в цератиомиксовые. Необ-
ходимы дальнейшие исследования жизненного цикла
и ультраструктуры этого малоизученного миксомице-
та для более ясного понимания его положения в сис-
теме. Пока же преждевременно относить его к Myx-
ogastria.
Отряд Echinosteliida L. S. Olive, 1970
(= Echinosteliales W. Martin, 1961)
Спорокарпы представляют собой спорангии, ску-
ченные или рассеянные, на ножках, до 1.5 мм высо-
той, обычно меньше. Ножка прозрачная в проходя-
щем свете, заполнена гранулярным материалом. Ко-
лонка имеется у большинства видов. Капиллиций у
некоторых видов отсутствует, у большинства имеется
в виде одиночной нити или системы нитей, отходя-
щих от окончания колонки или ножки; нити капил-
лиция ветвящиеся, прямые, иногда образуют сеточку.
Споры в массе белые, коричневые, бурые, бежевые,
розоватые; оболочки спор равной толщины или с
дисковидными утолщениями, гладкие или орнаменти-
рованные шипиками. Количество спор в зрелых спо-
рокарпах бывает очень мало. Например, спорангии
Е. lunatum содержат всего 4—8, редко 14 спор. Спо-
рокарп развивается по субгипоталлическому типу.
Плазмодий — протоплазмодий.
В настоящее время в отряде выделяют 2 семейст-
ва—- Echinosteliidae и Clastodermatidae.
Сем. Echinosteliidae L. S. Olive, 1970
(= Echinosteliaceae Rostaf., 1873)
В семействе имеется единственный род Echino-
stelium de Вагу, 1873. В мире известно около
20 видов этого рода. Его представители обладают
самыми маленькими спорокарпами среди миксогаст-
рид. Этот признак, а также сходство одноядерных
миксамеб протостелид (род Cavostelium)^ прото-
плазмодия эхиностелиевых рассматриваются некото-
рыми авторами как подтверждение их филогейети-
ческой близости (Olive, Stoianovitch, 1971). Однако
строение корешкового аппарата жгутиков зооспор
Cavostelium (С. apophysatum) и Echinostelium (Е. mi-
nutum, Е . lunatum, Е. bisporum) различно. У Cavoste-
lium apophysatum каждый жгутик имеет индивидуаль-
ное базальное тело, не связанное с другими базаль-
ными телами, тогда как базальные тела жгутиков
Echinostelium bisporum, Е. minutum и Е. lunatum объ-
единены попарно (Furtado, Olive, 1970; Spiegel,
1981а, 1981b; Whitney et al., 1982). Стенка спор у
Echinostelium и Cavostelium также имеет различия. У
первого она состоит из электронно-прозрачного
внутреннего и электронно-плотного наружных слоев,
пронизанных каналами, у второго имеется только
1 электронно-плотный слой (Whitney et al., 1982;
Haskins, McGuinness, 1986).
Echinostelium рассматривается как связующее зве-
но между протостелиевыми и миксогастриевыми
(Blackwell, Alexopoulos, 1975). Для сбора видов Ec-
hinostelium наиболее приемлемым можно считать ме-
тод влажной камеры. Первые спорангии обычно по-
являются на 2—7-е сутки после помещения субстра-
та в чашку Петри. Образцы спорангиев Echinostelium
часто теряют споры, из-за чего рекомендуется делать
постоянные препараты, а также микрофотографии
или рисунки.
Сем. Clastodermidae L. S. Olive, 1975
(= Clastodermataceae Alexop.
et T. E. Brooks, 1971)
Спорокарпы — спорангии на ножках, шаровид-
ные, темноокрашенные. Перидий после созревания
спорангия исчезает или сохраняется в виде лепестко-
видных долек, объединенных в основании спорангия
в чашечку. Капиллиций в виде простых или ветвя-
щихся нитей, отходящих от основания спорангия или
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA
441
колонки. Окончания нитей связаны с лепестковидны-
ми остатками перидия или образуют расширения, на-
поминающие чешуйкоподобные остатки перидия.
Споры в массе темно-коричневые.
В семействе 2 рода: Clastoderma A. Blytt, 1880 и
Barbeyella Meyl., 1914. Представители обоих родов
имеют трофическую стадию — протоплазмодий — и
субгипоталлический тип развития спорокарпа, что
позволяет выделить оба рода из отряда Stemonitida и
поместить их в отдельное семейство отряда Echino-
steliida.
Отряд Liceida L. S. Olive, 1970
(= Liceales Е. Jahn, 1928)
Спорокарпы в виде спорангиев, псевдоэталиев
или эталиев, обычно больше 200 мкм в диаметре.
Капиллиций отсутствует. Колонка отсутствует. Псев-
докапиллиций имеется или отсутствует, иногда хо-
рошо развит. Споры в массе светлые или яркоокра-
шенные, в проходящем свете бесцветные или окра-
шены в желтый, коричневый, красновато-коричневый
цвета.
Различные представители отряда имеют как про-
топлазмодиальный, так и фанероплазмодиальный ха-
рактер организации диплоидной трофической ста-
дии — плазмодия.
Отряд включает 3 семейства: Liceidae, Reticularii-
dae и Cribrariidae.
Сем. Liceidae L.S.Olive, 1970
(=Liceaceae Rostaf., 1873)
В семействе 1 род — Licea Schrad., 1797. Описа-
но около 30 видов. Большинство видов — эпифиты,
а также обитатели гниющих растительных остатков,
несколько видов — копрофилы. Наиболее характер-
ный признак рода — отсутствие в спорокарпах как
капиллиция, так и псевдокапиллиция. Фанероплазмо-
дий отмечен только у 2 видов: Lretiformis и L.varia-
bilis.
В настоящее время отсутствуют четкие представ-
ления о родственных связях Licea с другими таксо-
нами миксогастрид. Некоторые виды обладают сход-
ными признаками с Perichaena (Trichiidae) и Listerel-
la (Dianemidae) ( Eliasson, Keller, 1990; Eliasson et al.,
1991).
Сем. Reticulariidae L.S.Olive, 1970
(= Enteridiaceae Farr, 1982)
В семействе 4 рода: Tubifera J. F. GmeL, 1792; En-
teridium Ehrenb., 1819; Lycogala Pers., 1794; Dictydia-
ethalium Rostaf., 1873.
Tubifera объединяет виды, у которых боковые
стенки спорангиев сохраняются после созревания
спорокарпа.
У Enteridium псевдокапиллиций состоит из нитей,
перфорированных пленок или их комбинаций.
Виды рода Lycogala, среди которых наиболее из-
вестный L. epidendrum; формируют округлые эталии
до 2 см в диаметре (рис. 376), на ранних стадиях
развития имеющих ярко-красную или розовую окрас-
ку; псевдокапиллиций состоит из сморщенных, вет-
вящихся или простых трубочек.
Виды Dictydiaethalium образуют эталии, при-
чем боковые стенки спорангиев, формирующих эта-
лий, сохраняются частично в виде нитевидных ру-
диментов, отходящих от верхних стенок споран-
гиев.
Сем. Cribrariidae L. S. Olive, 1970
(= Cribrariaceae Rostaf., 1873)
В семействе 2 рода: Cribraria Pers., 1794 и Lind-
bladia Fr., 1849. Спорокарпы у представителей се-
мейства — спорангии на ножках (рис. 375, А), иногда
сидячие, псевдоэталии или эталии. В структурах спо-
рокарпа имеются так называемые диктидиновые гра-
нулы (рис. 375, Г). Перидий (за исключением Lind-
bladia) разрушается на большей части поверхности
после созревания спорокарпа и сохраняется только в
виде нитей. Нити образуют сеточку на всей или толь-
ко апикальной части спорокарпа, в последнем случае
в основании спорокарпа из остатков перидия образу-
ется чашечка (рис. 375, А). Псевдокапиллиций отсут-
ствует. Многие виды очень трудно отличить друг от
друга из-за чрезвычайной изменчивости признаков
спорокарпов под влиянием факторов окружающей
среды. Наиболее стабильными и имеющими наиболь-
шую диагностическую ценность следует считать раз-
мер и форму узелков сеточки перидия. Большинство
видов не удалось пока культивировать в лаборатор-
ных условиях.
Отряд Trichiida L.S.Olive, 1970
(= Trichiales T. Macbr., 1922)
Спорокарпы — спорангии или плазмодиокарпы,
иногда псевдоэталии, обычно больше 200 мкм в ди-
аметре. Колонка отсутствует. Капиллиций имеется в
виде нитей или трубочек с различной орнамента-
цией. Нити капиллиция свободные или связаны с
внутренними стенками спорокарпа. Споры в массе
светлые, красновато-коричневые (у Listerella рага-
doxa черные). В отряде 2 семейства: Dianemidae и
Trichiidae.
Сем. Dianemidae L. S. Olive, 1970
(= Dianemaceae Т. Macbr., 1899)
Спорокарпы у видов этого семейства — плазмо-
диокарпы или спорангии. Перидий обычно однослой-
ный, реже двухслойный, с наружным гранулярным
слоем. Капиллиций состоит из сплошных гладких,
ровных или четковидных нитей, отходящих от осно-
вания спорокарпа. Нити простые, слабо ветвящиеся,
никогда не образуют сеть.
В семействе 4 рода: Calomyxa Nieuwl, 1916; Dia-
пета Rex, 1891; Listerella E. Jahn, 1906; Minakatella
G. Lister, 1921. Виды Calomyxa имеют слабоорна-
ментированные нити и свободные споры, тогда как
442
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
у представителей Minakatella нити капиллиция глад-
кие, а споры объединены в комплексы. Наиболее
оригинальное строение имеет капиллиций Listerella
paradoxa, обладающий четковидными кольцевидны-
ми утолщениями. Положение Listerella в системе
миксогастрид остается спорным. На основании таких
признаков, как темная споровая масса и наличие
протоплазмодия, этот род относят также к отряду
Liceida (Kowalski, 1967).
Сем. Trichiidae LS. Olive, 1970
(= Trichiaceae Rostaf, 1873)
Характерным отличительным признаком семейст-
ва можно считать структуру капиллиция, который
состоит из простых или ветвящихся, полых трубо-
чек — элатер, свободных или объединенных в сеть.
Трубочки капиллиция имеют различную орнамента-
цию (рис. 374, А, вкл.; 377, А, Б, вкл.).
Известно 10 родов: Arcyria F. Н. Wigg., 1780; Агсу-
odes О. F. Cook, 1902; Calonema Morgan, 1893; Cornu-
via Rostaf., 1873; Hemitrichia Rostaf., 1873; Metatrichia
Ing, 1964; Oligonema Rostaf, 1875; Perichaena Fr.,
1817; Prototrichia Rostaf, 1876; Trichia Haller, 1768.
При их разграничении используются признаки капил-
лиция и перидия. Границы между родами не всегда
четко определены, так же как и их положение в сис-
теме.
У представителей сем. Trichiidae нити капилли-
ция полые. Отсутствие четко выраженного протока
в нитях капиллиция у видов Prototrichia позволяет
ставить вопрос о правомочности положения этого
рода в сем. Trichiidae (Ellis et al., 1973). В пределах
этого семейства можно проследить пути, по которым
шло усложнение капиллиция. Наиболее просто ус-
троенный тип капиллиция, представленный редко
ветвящимися трубочками, имеется у Perichaena. Ус-
ложнение капиллиция происходило по 2 основным
направлениям: во-первых, за счет усложнения систе-
мы в целом и, во-вторых, путем усложнения отдель-
ных элементов системы. Следующим этапом на пути
усложнения капиллиция, видимо, следует считать ка-
пиллиций Oligonema, который состоит из отдельных
слабоорнаментированных элатер, не связанных с пе-
ридием. У видов Trichia заметно усложнение орна-
ментации элементов капиллиция за счет образования
шипиков и спиральных утолщений на нитях капил-
лиция. Для родов Arcyria и Hemitrichia характерны
связь капиллиция с перидием и образование сети
нитей, а также их способность вытягиваться при со-
зревании спорангия и изменении влажности, что
приводит к растрескиванию перидия (рис. 375, Д).
Основное отличие между указанными родами заклю-
чается в орнаментации отдельных элементов капил-
лиция (рис. 375, Е).
Отряд Stemonitida L. S. Olive, 1970
(= Stemonitales Т. Macbr., 1922)
Спорокарпы в виде спорангиев, иногда эталиев
или псевдоэталиев. При созревании перидий сохра-
няется или исчезает. Споры прорастают одной-двумя
(реже больше) зооспорами или миксамебами. Споро-
карп развивается эпигипоталлически, т. е. гипотал-
лус расположен на субстрате, а протоплазма концен-
трируется над ним и преобразуется в спорангий.
Ножка формируется внутри зачатка спорокарпа и
связана с гипоталлусом, но не с перидием. Ножка
полая или заполнена фибриллярным материалом.
Известь отсутствует (изредка имеется), иногда при-
сутствует желатиновидное или восковидное вещест-
во. Капиллиций обычно в виде системы нитей, не
связанных с перидием и исходящих от колонки или
непосредственно от основания спорокарпа (если спо-
рокарп — эталий) (рис. 375, Ж—Н). Псевдокапилли-
ций отсутствует. Споры в массе коричневые или
черные, в проходящем свете окрашены так же, как
в массе, или темно-коричневые. Ассимилятивная
(трофическая) стадия — афаноплазмодий.
В отряде 1 семейство — Stemonitiidae.
Сем. Stemonitidae
(= Stemonitidaceae Fr., 1832)
Семейство насчитывает 14 родов. В пределах се-
мейства имеются роды с весьма размытыми граница-
ми. Это Amaurochaete Rostaf., 1873; Comatricha Pre-
uss, 1851; Stemonitis Gled, 1753. В результате реви-
зии как самого семейства, так и отдельных его родов
(Ing, Nannenga-Bremekamp, 1967; Nannenga-Breme-
kamp, 1967, 1974; Nannenga-Bremekamp et al., 1984)
были выделены такие роды, как Collaria Nann.-Bre-
mek., 1967; Paradiacheopsis Hertel, 1954; Stemonaria
Nann.-Bremek., R. Sharma et Y. Yamam., 1984; Ste-
monitopsis (Nann.-Bremek.) Nann.-Bremek., 1975;
Symphytocarpus Ing et Nann.-Bremek., 1967. Один из
самых известных родов миксомицетов — Stemoni-
tis — имеет рассеянные или скученные, цилиндри-
ческие спорангии на ножках. Гипоталлус мембрано-
видный, обычно хорошо развит. Перидий исчезает
после созревания спорангия. Ножка переходит в спо-
рангий как колонка, капиллиций отходит от колон-
ки в виде ветвящихся нитей, окончания которых об-
разуют сплошную сеточку под перидием (рис. 378,
Б). Споры в массе черные, коричневые или бурые, в
проходящем свете фиолетово-коричневые.
Отряд Physarida L. S. Olive, 1970
(= Physarales Т. Macbr., 1922)
Самый обширный по числу таксонов отряд,
включающий 2 семейства. Спорокарпы всех типов,
сидячие или на ножках (рис. 375, О—ГГ). Часто спо-
рангии сочетаются в одной группе с плазмодиокар-
пами. Известь обычно входит в состав перидия, ка-
пиллиция или одновременно во все структуры спо-
рокарпа в виде аморфной (гранулированной) или
кристаллической формы (рис. 378, В, Г). Капилли-
ций состоит из трубочек различной формы, в местах
ветвления которых могут образовываться узелки с
известью. Споры в массе черные или коричневые.
Ассимилятивная стадия — фанероплазмодий.
ТИП MYCETOZOA. КЛАСС EUMYCETOZOEA. SUMMARY
443
Сем. Physaridae LS. Olive, 1970
(= Physaraceae Chevall., 1826)
Семейство включает 10 родов: Badhamia Berk.,
1853; Badhamiopsis T.E. Brooks et H.W. Keller, 1976;
Cienkowskia Rostaf., 1873; Craterium Trentep., 1797;
Erionema Penz., 1898; Fuligo Haller, 1768; Leocarpus
Link, 1809; Physarum Pers., 1794; Physarella Peck,
1882; Protophysarum M. Blackw. et Alexop./ 1975.
Они объединяют около 200 видов. Для представите-
лей этого семейства характерно наличие аморфной
(гранулярной) формы извести в нитях капиллиция
(за исключением Protophysarum).
Наиболее крупный род Physarum насчитывает
свыше 100 видов. Один из видов — Ph. polycepha-
lum известен как модельный объект во многих ге-
нетических, физиологических и биохимических ис-
следованиях и является одним из наиболее хорошо
изученных организмов. Другой представитель семей-
ства — Fuligo septica — имеет один из самых круп-
ных плазмодиев, из которого может сформироваться
эталий до 20 см в длину. Один из монотипных ро-
дов — Protophysarum — рассматривается некоторы-
ми авторами как связующее звено между Echinosteli-
ida и Physarida (Collins, 1979). Protophysarum phloio-
genum — единственный вид семейства, в капиллиции
которого не найдены гранулы извести. Внешне споро-
карпы Р. phloiogenum напоминают спорокарпы Lamp-
roderma (Stemoniida), однако наличие фанероплазмо-
дия и субгипоталлический тип развития спорокарпа
(Blackwell, 1974) позволяют отнести этот таксон к
Physarida.
Сем. Didymiidae L.S. Olive, 1970
(= Didymiaceae Rostaf. ex Cooke, 1877)
Семейство включает 7 родов: Diderma Pers, 1794;
Didymium Schrad., 1797; Lepidoderma de Вагу, 1873;
Mucilago Battarra, 1755; Physarina Hohn, 1909; Squa-
muloderma Kowalski, 1973; Trabrooksia H.W. Keller,
1980. Они объединяют около 130 видов. Отличи-
тельным признаком семейства является наличие пре-
имущественно кристаллической извести в перидии
(рис. 377, Г) и отсутствие таковой в капиллиции.
Известь наружного слоя перидия различной формы:
аморфная, гранулированная, кристаллическая, иног-
да образует скорлуповидный слой или откладывается
в виде чешуек различных размера и формы (рис. 377,
В). Как и в сем. Physaridae, здесь имеется исключе-
ние — монотипный род Trabrooksia, основанный на
Т, applanata Keller, 1980, наиболее характерным
признаком которого является отсутствие извести в
структурах спорокарпа.
THE PHYLUM MYCETOZOA DE BARY, 1859
THE CLASS EUMYCETOZOEA ZOPF, 1884
SUMMARY
The class Eumycetozoea (myxomycetes) includes three subclas-
ses: Protostelia (protostelids), Dyctiostelia (cellular slime molds) and
Myxogastria (plasmodial slime molds), which are phagotrophic bacte-
rivorous amoebae-like terrestrial protists. They typically occur in soil,
litter, decaying wood and other vegetable materials, in which their
swarm cells, myxamoebae and plasmodia feed and grow. Only few
species can grow in axenic or monoxenic laboratory culture. About
1000 species of plasmodial slime molds have been described in the
world.
Myxomycetes are known primarily as a mesophylic group of orga-
nisms. Many species are cosmopolitan, the great majority occur in tem-
perate regions. Some tropical and desert species have limited areas of
distribution. A few species have been reported only from alpine habitats.
Data of moist chambers experiments show that myxomycetes are often
present in the different types of substrata as dormant stages (microcysts
or sclerotium) even in the unsuitable microhabitats. For this reason and
due to the fact that sporangia often form far from the site of main
plasmodium feeding, it is very difficult to recognize the ecological role
of myxomycetes in terrestrial communities. Some species inhabit only
definite microhabitats, occupying various substrates, i. e. coarse wood
debris, litter, bark of living trees and shrubs, canopy of tropical forest,
dung of herbivorous animals etc. Some species form sporocarps near
melting snow on litter, plant debris, and on living plants, others form
bryophyte-myxomycete associations. Myxomycetes are known to offer
food, shelter, and a breeding place to various species of insects and
mites. In general, it must be stated that there is a great insufficiency of
information concerning the ecology of Myxomycetes.
Life cycle of the Myxomycetes includes two different assimilative
stages. The assimilative phase of the Myxomycetes can be presented
by small uninucleate or multinucleate ameboid cells with filopodia,
and by multinucleate plasmodia. The plasmodium becomes converted
into one or more fruiting bodies (sporocarps), which produce spores.
There are four types of myxomycete sporocarps: sporangium, aethali-
um, pseudoaethalium, and plasmodiocarp. Sporocarps are usually
composed of 6 parts: hypothallus, stalk, columella, peridium, capilli-
tium and spores. In some fruiting bodies a pseudocolumella or a pseu-
docapillitium may be present. Not all of these components are present
in all fruiting body types. Most plasmodial myxomycetes, except pro-
tostelids, form multispored sporocarps. Protostelids produce microsco-
pic fruiting bodies that usually bear a single spore at the tip of a
delicate stalk. Germinating spores give rise from one to four and oc-
casionally more myxamoebae or swarm (flagellated) cells.
Both phases grow, feed, and divide forming rather considerable
population in the substrate. Some species have heterothallic, sexual
cycle, which includes meiosis at the spore producing stage, some have
apomictic cycle without meiosis. Both «versions» of the life cycle
may be realized in one and the same species. Myxamoebae and swarm
cells may transform one to another, sometimes under the control of
availability of free water or food resources. Typical swarm cell is
equipped with one or two (sometimes more pairs) unequal flagella
without mastigonemes.
Although the flagellar apparatus of myxomycete swarms in gene-
ral has a similar organization, significant variability of details of fla-
gellar rootlets structure is found in different orders and genera. These
differences concern the number of microtubules and their placement.
These variations are expected to have not only taxonomic, but also the
phylogenetic value. Usually myxamoebae or swarm cells are uninuc-
leate and are regarded as haploid. Most myxomycetes are heterothallic
species with sexual process (gametes may be a swarm of cells or
myxamoebae). Some species have homothallic life cycle. Agamic cell
fusions were also reported.
The vegetative stage of cellular clime molds (Dyctiostelia) repre-
sented with small amoeboid cells. Amoebae assimilate about 1000
bacterial cells in a period of 3—4 hours under 22 °C, and strongly
influence the number and composition of soil bacteria. They stream
together in great numbers, forming compact, multicellular association,
or pseudoplasmodia. Aggregation process is stimulated with excretion
of some chemical compounds («acrasins») by amoebae, such as cAMP
444
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
(Dictyostelium discoideum). Probably different kinds of acrasins are
excreted by different species of dictyostelids. Pseudoplasmodia sub-
sequently develop fructifications, or sorocarps that consist of suppor-
tive stalks and unwalled sori containing propogative spores. Someti-
mes (under unfavorable conditions) dictyostelids may produce macro-
cysts instead of sporocarps.
Mycetozoa incertae sedis
Hyperamoeba flagellata Alexeieff,
1923
Вид впервые был выделен Алексеевым из фекалий
лошади в 1923 г. (Алексеев, 1923) и изучен на свето-
оптическом уровне. Недавно Н. flagellata была найде-
на на поверхности льда пресного водоема, выделена в
клональную культуру и исследована в световом и элек-
тронном микроскопах (Карпов, Мыльников, 1997).
Жизненный цикл этого протиста включает 3 стадии:
амебоидную, жгутиковую и стадию цисты (рис. 380).
Причем трофонтами являются только амебоидные
клетки, которые могут также и размножаться путем
деления надвое. При увеличении плотности клеток в
культуре у трофонтов вырастает 1 направленный вперед
жгутик, клетка превращается в монаду, которая уже не
питается и не делится, а служит для расселения.
Жгутиконосцы имеют вытянутое тело с заострен-
ным передним концом, из которого выходит единст-
венный жгутик. Длина клетки 12—17 мкм, а жгутик
примерно равен по длине телу (рис. 380). Время
жизни таких амебофлагеллат невелико. Они теряют
жгутик и превращаются в амеб, а чаще всего — в
цисты. Цисты образуются при неблагоприятных усло-
виях непосредственно из амебоидных клеток. Они
имеют шаровидную форму и гладкую толстую стенку.
Изучение ультраструктуры жгутиковой формы по-
казало, что наиболее информативной структурой яв-
ляется жгутиковый аппарат (рис. 381). Его строение
почти идентично таковому зооспор протостелиды
Cavostelium bisporum (Карпов, Мыльников, 1997).
Для тех и других характерен конус из микротрубо-
чек, идущий к ядру от фибриллярной ножки (коре-
шок 1, — рис. 381), а также набор латеральных
микротрубочковых корешков, обозначаемых обычно
под номерами 2—5 (Spiegel, 1981а) (рис. 381; 382).
Вместе с тем в жизненном цикле Hyperamoeba не об-
разуются плодовые тела (Карпов, Мыльников, 1997),
наличие которых характеризует всех протостелие-
вых. Таким образом, Hyperamoeba, отличается от
протостелиевых отсутствием плодовых тел и поэтому
не может быть отнесена к ним.
Среди церкомонад фактически не встречаются
виды, у которых, подобно Hyperamoeba, жгутиковая
стадия не являлась бы трофонтом. Большинство из
них сочетают амебоидную и жгутиковую формы в
одной трофической стадии. Пожалуй, только у Mas-
sisteria происходит разделение амебо-жгутиковой ста-
дии на амебоидную трофическую (причем жгутики со-
храняются, но не активны) и жгутиковую расселитель-
ную (Patterson, Fenchel, 1990). У Hyperamoeba
flagellata это разделение более четкое: жгутиковая
стадия кратковременна в жизни вида и имеет в отли-
чие от церкомонад не 2 жгутика, а только 1. Отличия
жгутикового аппарата Н. flagellata и представителей
церкомонад также весьма существенны (рис. 382), по-
этому объединение этих простейших в один таксон не
представляется возможным.
Вместе с тем по особенностям жизненного цикла
и строению кинетиды Hyperamoeba обладает сходст-
, 10 мкм ,
Рис. 380. Основные жизненные формы Hyperamoeba flagellata в культуре. (Ориг. А. П. Мыльникова).
А — жгутиковая стадия, Б — трансформация жгутиковой стадии в амебоидную, В—Е — различные амебоидные формы, Ж — циста.
Fig. 380. Stages of life cycle in the culture of Hyperamoeba flagellata.
A — flagellate, Б — flagellate transformation into amoeboid form, В—E — different amoeboid forms, Ж — cyst.
ТИП MYCETOZOA. ЛИТЕРАТУРА
445
Рис. 381. Схема строения жгутикового аппарата Hyperamoeba fla-
gellata. Вид с вентральной стороны клетки.
бк — безжгутиковая кинетосома, ж — жгутик, жк — жгутиковая кинетосо-
ма, фс — фибриллярная связка. 7—5 — номера корешков.
Fig. 381. Scheme of flagellar apparatus structure of Hyperamoeba fla-
gellata. Ventral view.
бк — nonflagellar basal body, ж — flagellum, жк — flagellar basal body,
фс — fibrillar connection. 7—5 — rootlet numbers.
bom и с церкомонадами, и с протостейидами (Карпов,
1997).
На рис. 382 схематически изображены основные
элементы корешковой системы жгутиков (вид с вент-
ральной стороны клетки) представителей двух семейств
церкомонад (Cercomonas и Heteromita), Hyperamoeba
и протостелиды Cavostelium, Сходная ориентация ки-
нетосдм, расположение и строение корешков свиде-
тельствуют о том, что корешки, отмеченные одинако-
выми номерами, могут считаться гомологичными. Зо-
оспоры С. bisporum содержат наиболее полный набор
Рис. 382. Схемы строения жгутиковых аппаратов. Вид с вентраль-
ной стороны клетки.
А — Cercomonas, Б — Heteromita, В — Hyperamoeba, Г — Cavostelium.
кп — кинетосома переднего жгутика, кз — кинетосома заднего жгутика.
7—5 — номера гомологичных корешков.
Fig. 382. Schemes of flagellar apparatus structure in Cercomonas
(A), Heteromita (Б), Hyperamoeba (B) and Cavostelium (Г). Ventral
view.
кп — basal body of anterior flagellum, кз — basal body of posterior flagel-
lum. 7—5 — numbers of homologous rootlets.
корешков среди протостелид, у которых строение
жгутикового аппарата очень консервативно (Spiegel,
1981а). Кроме того, этот вид может представлять и
образец строения корешковой системы у зооспор
миксогастриевых, которые являются по отношению
к протостелидам сестринской группой (Spiegel et al.,
1995).
Следовательно, в филогенетическом смысле Hyper-
amoeba представляет собой переходное звено, связы-
вающее церкомонад и миксомицетов в одну, по-види-
мому монофилетическую, группу протистов (Карпов, 1997).
MYCETOZOA INCERTAE SEDIS
HYPERAMOEBA FLAGELLATA ALEXEIEFF, 1923
SUMMARY
Hyperamoeba flagellata is a fresh-water protist of uncertain
taxonomic position. It exists in three living forms: as an amoeba, a
flagellate, and a cyst. The flagellated stage of H. flagellata has only
one long smooth flagellum, which is directed anteriorly. It emerges
from the flagellar basal body (kinetosome) located at the apical end
of the cell. Two cones of microtubules extend from the flagellar base
towards the nucleus. The outer cone originates from the lateral side
of the flagellar kinetosome, whereas the inner cone arises from a
MTOC associated with the flagellar kinetosome by a short fibrillar
rootlet. The second (nonflagellar) kinetosome lies ventrally and is
directed to the right side of the cell. It has two microtubular rootlets,
which pass laterally in the same direction. The microtubular cones
overlap the anterior end of the nucleus. A dictyosome lies bet-
ween the basal bodies and the nucleus. Mitochondria with tubular
cristae have an electron dense core which is oriented along the
longitudinal axis. The structure of the flagellated stage of H. flagel-
lata is compared with flagellated cells of protostelids and cercomo-
nads.
It is proposed that this organism may represent a phylogenetic
link between protostelids and cercomonads.
ЛИТЕРАТУРА23
Гоби X. Я. О группе Ameboideae И Тр. С.-Петербург, о-ва ес-
тествоиспытателей. СПб., 1884. Т. 15. С. 1—36.
Горленко М. В. (ред.). Жизнь растений. М., 1976. Т. 2. Грибы.
23 Литература к разделу «Тип Mycetozoa».
Горленко М. В. (ред.). Курс низших растений. М„ 1981.
(Карпов С. A.) Karpov S. A. Cercomonads and Their Relationship
to the Myxomycetes // Arch. Protistenk. 1990. Bd 148. S. 297—307.
(Карпов С. А., Мыльников А. П.) Karpov S. A., Mylni-
kov A. P. The ultrathin structure of colourless flagellate Hyperamo-
446
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
eba flagellata with special reference to the flagellar apparatus // Eur.
J. Protistol. 1997. Vol. 33. P. 349—355.
(Карпов С. А., Новожилов Ю. К., Чистякова Л. Е.) Karpov
S. A., Novozhilov Yu. К., Chistiakova L. E. The comparative study
of zoospore cytoskeleton in Symphytocarpus confluens, Arcyria cine-
rea and Lycogala epidendrum (Eumycetozoa) // Проблемы ботаники
на рубеже XX—XXI веков: Тез. докл. II (X) съезда Рус. Ботан.
О-ва. СПб.: БИН РАН, 1998.
Красильщик И. Материалы к естественной истории и систе-
матике флагеллат. О Cercobodo laciniaegerens nov. gen. et nov.
sp. // Зап. Новорос. об-ва естест. 1886. T. 2, вып. 1. С. 211—245.
Кусакин О. Г., Дроздов А. Л. Филема органического мира.
Часть 2: Prokaryotes, Eukaryotes: Microsporobiontes, Archemonado-
biontes, Euglenobiontes, Myxobiontes, Rhodobiontes, Alveolates, He-
terokontes. СПб.: Наука, 1998.
Мыльников А. П. Особенности тонкого строения жгутико-
носца Bodomorpha reniformis // Цитология, 1984. Т. 26, N 11.
С. 1308—1310.
Мыльников А. П. Новый вид Cercobodo amoebinus (класс Zo-
omastigophorea Calkins). // Биология внутренних вод. Информ,
бюл., 1985 a. N 65. С. 22—25.
Мыльников А. П. Бесцветный жгутиконосец Cercobodo plas-
modialis sp. n. // Биология внутренних вод. Информ, бюл. 1985 б.
N 68. С. 29—31.
Мыльников А. П. Биология и ультраструктура амебоидных
жгутиконосцев Cercomonadida ord. nov. И Зоол. журн. 1986. Т. 65.
С. 683—692.
Новожилов Ю. К. Определитель грибов России. Отдел Мух-
omycota. Вып. 1. Класс Myxomycetes. СПб.: Наука, 1993.
Новожилов Ю. К., Голубева О. Г. Эпифитные слизевики из
Монгольского Алтая и пустыни Гоби И Микология и фитопатоло-
гия. 1986. Т. 20. С. 368—374.
(Новожилов Ю. Е., Шнитлер М., Стефенсон С. Л.) Novozhi-
lov Y. К., Schnittler М., Stephenson S. L. The myxomycetes of Rus-
sian Subarctic and Arctic areas // Микология и фитопатология.
1998a. Vol. 32, N 1. P. 18—29.
(Новожилов Ю. E., Шнитлер M., Стефенсон С. Л.) Novozhi-
lov Y. K., Schnittler M., Stephenson S. L. Analysis of myxomycete
diversity of Russian Subarctic and Arctic areas II Микология и фито-
патология. 19986. Vol. 32, N 2. P. 27—33.
Райков И. Б. Пути эволюции митотического аппарата у ни-
зших эукариот // Систематика простейших и их филогенетические
связи с низшими эукариотами). Л., 1986. С. 26—56. (Тр. ЗИН АН
СССР; Т. 144).
Серавин Л. Н., Гудков А. В. Основные типы и формы агам-
ных слияний клеток у простейших // Цитология. 1984а. Т. 26. № 2.
С. 123—131.
Серавин Л. Н., Гудков А. В. Возможные формы агамных гене-
тических взаимоотношений у протистов и пути становления поло-
вого процесса // Цитология. 19846. Т. 26. № 11. С. 1224—1236.
(Фаминцин А., Воронин М.) Famintzin A., Woronin М. Uber
zwei neue Formen von Schleimpilzen: Ceratium hydnoides und Cera-
tium poroides // Mem. Acad. Imp. Sci. St. Petersburg. 1873. Ser. 7,
Bd 20. S. 1—16.
Ячевский А. А. Микологическая флора Европейской и Азиат-
ской России. Слизевики. М., Изд. В. Рихтер, 1907. Т. 2.
Abe К., Orii Н., Okada Y., Saga Y., Yanagisawa К. A novel
cyclic AMP metabolism exhibited by giant celle and its possible role
in the sexual development of Dictyostelium discoideum // Devlop.
Biol. 1984. Vol. 104. P. 477—483.
Ainsworth G. C., Bisby G. Dictionary of the fungi. Ed. 7. Kew,
Surrey, 1983. 443 p.
Ainsworth G. C. Introduction and keys to higher taxa // The
Fungi. Vol. 48. New York, 1973. P. 635—648.
Aldrich H. C. Ultrastructure of meiosis in three species of Physa-
rum // Mycologia. 1967. Vol. 59. P. 127—148.
Aldrich H. C. The development of flagella in swarm cells of the
myxomycete Physarum flavicomum // J. Gen. Microbiol. 1968.
Vol. 50. P. 217—222.
Aldrich H. C. The ultrastructure of mitosis in myxamoebae and
plasmodia of Physarum flavicomum // Amer. J. Bot. 1969. Vol. 56.
P. 290—299.
Aldrich H.C. Pre- and postmeiotic events in spores of the myx-
omycete Didymium ridis // J. Cell Biol. 1970. Vol. 47. P. 4a (Abstr.).
Aldrich H. C., Blackwell M. Resistant structures in the Myxomy-
cetes // Weber D. J., Hess W. M. The fungal spore. New York etc.,
1976. P. 414—461.
Aldrich H. C., Mims C. W. Synaptonemal complexes and meid-
sis in myxomycetes I I Amer. J. Bot. 1970. Vol. 57. P. 935—941.
Alexopoulos C. J. Gross morphology of the plasmodium and its
possible significance in the relationschips among Myxomycetes // My-
cologia. 1960. Vol. 52. P. 1—20.
Alexopoulos C. J. Introductory mycology (2nd ed.). New York,
1962.
Alexopoulos C. J. Morphogenesis in the Myxomycetes // The
Fungi (Ainsworth and Sussman, eds.). Acad. Press: N. Y., 1966.
Vol. 2. P. 211—234.
Alexopoulos C. J. The experimental approach to the taxonomy of
the Myxomycetes // Mycologia. 1969. Vol. 61. P. 219—239.
Anderson C. L. Phylogenetic relationships of the Myxozoa // In:
Evolutionary Relationships Among Protozoa I Eds. G. H. Coombs et
al. London: Kluwer, 1998. P. 341—350.
Anderson R. W., Coocke D. J., Dee J. Apogamic development of
plasmodia in the myxomycete Physarum polycephalum: a cinematog-
raphic analysis // Protoplasma. 1976. Vol. 89. P. 29—40.
Baker J. G. Annotated checklist of the simpler mycetozoans of
North Carolina 11 J. Elisha Mitchell Sci. Soc. 1975. Vol. 90. P. 135—
136.
Bell R., Achenbach F. Reactivation of cytoplasmic actomyosin in
Physarum plasmodia extracted with glycerol and dimethylsulphox-
ide II J. Cell Sci. 1987. Vol. 87. P. 231—239.
Bennett W. E. Fine structure of the trophic stage of Endostelium
olive Bennett et Deasey, 1984 (Eumycetozoea, Protosteliia) I I Protis-
tologica. 1986. T. 22. P. 205—212.
Blackwell M. A study of sporophore development in the myx-
omycete Protophysarum phloiogenum // Arch. Mikrobiol. 1974.
Bd 99. S. 331—344.
Blackwell M., Alexopoulos C. J. Taxonomic studies in the Myx-
omycetes. IV. Protophysarum phloiogenum, a new genus and species
of Physaraceae // Mycologia. 1975. Vol. 67. P. 32—37.
Blackwell M., Gilbertson R. L. Sonoran Desert Myxomycetes //
Mycotaxon. 1980. Vol. 11. P. 139—149.
Blaskovics J. S., Raper К. B. Encystment stages of Dictyosteli-
um // Biol. Bull. 1957. Vol. 113. P. 58—88.
Bonner J. T. Evidence for the formation of cell aggregates by
chemotaxis in the development of the slime mold Dictyostelium dis-
coideum // J. Exp. Zool. 1947. Vol. 106. P. 1—26.
Bonner J. T. The life cycle of cellular slime molds // Nat. Hist.
1978. Vol. 87. P. 70—79.
Bonner J. T. Evolutionary strategies and developmental constra-
ints in the cellular slime molds // Amer. Natur. 1982. Vol. 119.
P. 530—552.
Bonner J. T., Clark W. W., Neely C. L., Slifkin M. K. The ori-
entation to light and the extremely sensitive orientation to temperature
gradients in the slime mold Dictyostelium discoideum // J. Cell Comp.
Physiol. 1950. Vol. 36. P. 149—158.
Bonner J. T., Chiquoine A. D., Kolderie M. Q. A histochemical
study of differentiation in the cellular slime molds // J. Exp. Zool.
1955. Vol. 130. P. 133—158.
Brefeld O. Dictyostelium mucoroides. Ein neuer Organismus und
der Verwandschaft der Myxomyceten // Abh. Senckenberg. natur-
forsch. Ges. 1869. Bd 7. S. 85—107.
Brody T., Williams K. L. Cytological analysis of the parasexual
cycle in Dictyostelium discoideum // J. Gen. Microbiol. 1974.
Vol. 82. P. 371—383.
Brooks T. E., Keller H. W., Chassain M. Corticolous myxomy-
cetes VI. A new species of Diderma I I Mycologia. 1977. Vol. 69.
P. 179—184.
Carroll G. C., Dykstra R. Synaptonemal comlexes in Dydimium
iridis // Mycologia. 1966. Vol. 58. P. 166—169.
Cavalier-Smith T. The protozoan phylum Opalozoa I I J. Euk.
Microbiol. 1993. Vol. 40. N 5. P. 609—615.
Cavender J. C. Geographical distribution of Acrasieae // Myco-
logia. 1973. Vol. 65. P. 1044—1054.
Chagla A. H., Lewis К. E., O’Day D. H. Ca2+ and cell fusion
during sexual development in liquid cultures of Dictyostelium discoi-
deum // Exp. Cell Res. 1980. Vol. 126. P. 501—505.
Charvat I., Ross I. K., Cronshaw J. Ultrastructure of the plasmo-
dial slime mold Perichaena vermicularis. I. Plasmodium // Protoplas-
ma. 1973. Vol. 76. P. 333—351.
Clayton L., Pogson С. I., Gull K. Ultrastructural and biochemical
characterisation of the cytoskeleton of Physarum polycephalum myx-
amoebae//Protoplasma. 1983. Vol. 118. P. 181—191.
Collins N. R. Myxomycete biosystematics: some recent develop-
ment and future research opportunites I I Bot. Rev. 1979. Vol. 45.
P. 145—201.
ТИП MYCETOZOA. ЛИТЕРАТУРА
447
Сох J. J. Notes on coprophilous myxomycetes from the Western
United States // Mycologia. 1981. Vol. 73. P. 741—747.
de Bary A. Die Mvcetozoen. Ein Beitrag zur Kenntnis der Nie-
dersten Organismen. 2nfl ed. Leipzig; Engelman, 1859.
de Bary A. Comparative morphology and biology of the Fungi,
Mycetozoa and Bacteria. Oxford, 1887.
Diggins M. A'., Dove W. F. Distribution of acetylated alpha-tubu-
lin in Physarum polycephalum // J. Cell Biol. 1987. Vol. 104.
P. 303—309.
Dykstra M. J. Ultrastructure of the genus Schizoplasmodiopsis
(Protostelia) // J. Protozool. 1978. Vol. 25. P. 44—49.
Dykstra M. J., Aldrich H. C. Successful demonstration of an elu-
sive cell coat in amoebae // J. Protozool. 1978. Vol. 25. P. 38—41.
Eliasson U. The myxomycete biota of the Hawaiian Islands //
Mycol. Res. 1991. Vol. 95. P. 257—267.
Eliasson U. H., Keller H. W. Relationships in the Liceales // Fo-
urth Intern. Mycol. Congr. IMC 4, Regensburg, Germany 28 august—
3 sept. 1990, Abstr. / Ed. A. Reisinger, A. Bresinsky. Regensburg,
1990. P. 18.
Eliasson U. H., Lundqvist N. Fimicolous Myxomycetes // Bot.
notiser. 1979. Bd 132. S. 551—568.
Eliasson U. H., Keller H. W., Schoknecht J. D. Kelleromyxa, a
new generic name for Licea fimicola (Myxomycetes) // Mycol. Res.
1991. Vol. 95. P. 1201—1207.
Ellis T. T., Scheetz R. W., Alexopoulos C. J. Ultrastructural ob-
servations on capillitial types in the Trichiales (Myxomycetes) //
Trans. Amer. Microsc. Soc. 1973. Vol. 92. P. 65—79.
Erdos G. W., Nickerson A. W., Raper К. B. Fine structure of
macrocyst in Polysphondilium violaceum // Cytobiologie. 1972.
Vol. 6. P. 351—366.
Erdos G. W., Raper К. B., Vogen L. Mating types and macrocyst
formation in Dictyostelium discoideum // Proc. Nat. Acad. Sci. U. S.
1973. Vol. 29. P. 1828—1830.
Feest A. The quantitative ecology of soil Mycetozoa // Prog. Pro-
tis. 1987. Vol. 2. P. 331—361.
Filosa M. F., Dengler R. E. Ultrastructure of macrocyst formati-
on in the cellular slime mold, Dictyostelium mucoroides: extensive
phagocytosis of amoebae by specialized cell // Develop. Biol. 1972.
Vol. 29. P. 1—16.
Frederick L., Pendergrass L. Corticolous Myxomycetes: bark of
living trees as a distinct ecological niche // Fourth Intern. My cologo
Congr. IMC 4, Regensburg, Germany 28 August—3 Sept. 1990,
Abstr. / Ed. A. Reisinger, A. Bresinsky. Regensburg, 1990. P. 19.
Fries E. M. Myxogastres // Systema Mycologicum. Gryphiswal-
diae, 1829. Vol 3. P. 67—199.
Furtado J. S., Olive L. S. Ultrastructural studies of protoste-
lids: the amoebo-flagelate stage // Cytologia. 1970. Vol. 2. P. 200—
219.
Furtado J. S. Olive L. S. Ultrastructural evidence of meiosis in
Ceratiomyxa fruticulosa // Mycologia. 1971a. Vol. 63. P. 413—416.
Furtado J. S., Olive L. S. Ultrastructure of the protostelid Cerati-
omyxella tahitiensis, including scale formation // Nova Hedwigia.
1971b. Vol. 21. P. 537—576.
Gaither T. W. Ultrastructure of the capillitium and spores of
Physarum polycephalum // Bull. Torrey Bot. Club. 1974. Vol. 101.
P. 86—190.
Gawlitta W., Wolf К. V., Hoffmann H.-U., Stockem W. Studies
on microplasmodia of Physarum polycephalum. I. Classification
and locomotion behavior I I Cell and Tiss. Res. 1980. Vol. 29. P. 71—
86.
Gerisch G. Cyclic-АМР and other signals controlling cell deve-
lopment and differentiation in Dictyostelium // Ann. Rev. Biochem.
1987. Vol. 56. P. 853—879.
Gerisch G., Hulser D., Malchow D., Wick U. Cell communicati-
on by periodic cyclic-АМР pulses // Phil. Trans. Roy. Soc. London.
1975. B. 272. P. 181—192.
Gomer R. H., Firtel R. A. Cell-autonomous determination of
cell-type choice in Dictyostelium development by cell-cycle phase //
Science. 1987. Vol. 237. P. 758—762.
Goetzsche H. F. Contributions to the myxomycete flora of Ice-
land // Acta Bot. Isl. 1984. Vol. 7. P. 13—26.
Goetzsche H. F. Myxomycetes from Greenland // Opera Bot.
1989. Vol. 100. P. 93—103.
Goetzsche H. F. Notes on Icelandic Myxomycetes // Acta Bot.
Isl. 1990. N 10. P. 3—21.
Gray W. D., Alexopoulos C. J. Biology of Myxomycetes. New
York: Roland Press Comp. 1968.
Gregg J. H. Regulation in the cellular slime molds // Develop.
Biol. 1965. Vol. 12. P. 377—393.
Hader D. P., Schreckenbach T. Phototactic orientation in plas-
modia of the acellular slime mold, Physarum polycephalum // Plant
and Cell Physiol. 1984. Vol. 25. P. 55—61.
Hagiwara H. Dictyostelids in Japan. X. Two new species of Dic-
tyostelium, D. pseudo-brefeldianum and D. robustum // Bull. Nation.
Sci. Mus., Tokyo, Ser. B. 1996. Vol. 22, N 2. P. 47—54.
Harkbnen M. Myxomycetes developed on litter of common Fin-
nish trees in moist chamber cultures // Nordic J. Bot. 1981. Vol. 1.
P. 791—794.
Haskins E. F. A study of the amoebo-flagellate transformation in
the slime mold Echinostelium minutum de Bary // Protoplasma. 1978.
Vol. 94. P. 193—206.
Haskins E. F., Hinchee A. A., Cloney R. A. The occurrence of
synaptonemal complexes in the slime mold Echinostelium minutum de
Bary //J. Cell. Biol. 1971. Vol. 51. P. 898—903.
Haskins Б. F., McGuinness M. D. Comparative ultrastructural
observations of spore wall structure in six species of Echinostelium
and three species of Eumycetozoa // Mycologia. 1986. Vol. 78.
P. 613—618.
Hatano S., Owaribe K., Matsumura F., Hasegawa T., Takaha-
shi I. Characterization of actin and myosin isolated from Physarum //
Canad. J. Bot. 1980. Vol. 58. P. 750—759.
Havercroft J. C., Gull K. Demonstration of different patterns of
microtubule organization in Physarum polycephalum myxamoebae
and plasmodia using immunofluorescent microscopy I I Eur. J. Cell
Biol. 1983. Vol. 32. P. 67—74.
Hawksworth D. L., Kirk P. M., Sutton В. C., Pegler D. N. Ain-
sworth and Bisby’s Dictionary of the Fungi. 8th edit. CAB Internat.
1995.
Heath I. B. Nucleus-associated organelles in fungi // Int. Rev.
Cytol. 1981. Vol. 69. P. 191—221.
Hinchee A. A., Haskins E. F. Open spindle nuclear division in
the amoebal phase of the acellular slime mold Echinostelium minutum
with chromosomal movement related to the pronounced rearrangement
of spindle microtubules // Protoplasma. 1980a. Vol. 102. P. 117—
130.
Hinchee A. A., Haskins E. F. Closed spindle nuclear division in
the plasmodial phase of the acellular slime mold Echinostelium minu-
tum // Protoplasma. 1980b. Vol. 102. P. 235—252.
Hohl H. R., Hamamoto S. T. Ultrastructure of spore differentia-
tion in Dictyostelium: the prespore vacuole // J. Ultrastruct. Res. 1969.
Vol. 26. P. 442—453.
Hohl H. R., Raper К. B. Nutrition of cellular slime molds. II.
Growth of Polysphondilium pallidum in axenic culture // J. Bacteriol.
1963. Vol. 85. P. 199—206.
Hohl H. R., Hamamoto S. T., Hemmes D. E. Ultrastructural as-
pects of cell elongation, cellulose synthesis and spore differentiation
in Acytostelium leptosomum, a cellular slime mold I I Amer. J. Bot.
1968. Vol. 55. P. 783—796.
Huffman D. M., Olive L. S. Engulfment and anastomosis in the
cellular slime molds (Acrasiales) // Amer. J. Bot. 1964. Vol. 51.
P. 465—471.
Huffman D. M., Kahn A. J., Olive L. S. Anastomosis and fusi-
ons in Dictyostelium // Proc. Nat. Acad. Sci. U. S. 1962. Vol. 48.
P. 1160—1164.
Hutterman A. Biochemical events during spherule formation of
Physarum polycephalum // Berlin Dsch. Bot. Ges. 1973. Vol. 86.
P. 55—76, 172—174.
Ikeda T., Takeuchi I. Isolation and characterization of a prespore
specific structure of the cellular slime mold, Dictyostelium discoide-
um // Develop. Growth. Differ. 1971. Vol. 13. P. 221—229.
Indira P. U. Swarmer formation from plasmodia of myxomyce-
tes // Trans. Brit. Mycol. Soc. 1964. Vol. 47. P. 531—533.
Ing B. The phytosociology of myxomycetes // The New Phytolo-
gist. 1994. Vol. 126. P. 175—202.
Ing B., Nannenga-Bremekamp N. E. Notes on Myxomycetes.
XIII. Symphytocarpus nov. gen. Stemonitacerum I I Proc. Kon. Ned.
Akad. Wetensch. Ser. C. 1967. Vol. 70. P. 217—233.
Ing B., Smith L. R. I. Two myxomycetes from South Georgia //
Brit. Antarct. Surv. Bull. 1980. N 50. P. 118—120.
Ishigami M. A light and electron microscopy study of the flagel-
late to amoeba conversion in the myxomycete Stemonitis pallida //
Protoplasma. 1977. Vol. 91. P. 31—54.
Ishigami M., Motomochi K. Cytoplasmic streaming and distribu-
tion of actomyosin fibrils in the aphanoplasmodium of Stemonitis her-
batica // Protoplasma. 1988. Suppl. 1. P. 35—41.
Jacobson D. N., Adelman M. R. Amoebo-flagellate transformati-
on in Physarum polycephalum // J. Cell Biol. 1975. Vol. 47. P. 97—
105.
448
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Jahn Т. L., Bovee Е. С., Griffith D. L. Taxonomy and evolution
of the Sarcodina: a reclassification // Taxon. 1974. Vol. 23. P. 483—
496.
Jermyn K. A., Duffy К. T. J., Williams J. G. A new anatomy of
the prestalk zone in Dictyostelium // Nature. 1989. Vol. 340. P. 144—
146.
Johnson G., Johnson R., Miller M., Borysenko J., Revel J.P. Do
cellular slime molds form intercellular junctions? // Science. 1977.
Vol. 197. P. 1300.
Kay R. R., Town C. D., Gross J. D. Cell differentiation in Dicty-
ostelium discoideum // Differentiation. 1979. Vol. 13. <P. 7—14.
Kay R., Gross J., Peacey M., Jermyn K., Dhokia B., Kopachik
W., Brookman J., Pogge R. Dictyostelium morphogenes and how
they might act // J. Embryol., Exp. Morphol. 1984. Vol. 82. Suppl.
P. 173.
Keller H. W., Aldrich H. C., Brooks T. E. Corticolous Myxomy-
cetes. II: Notes on Minakatella longifila with ultrastructural evidence
for its transfer to the Trichiaceae // Mycologia 1973. Vol. 65.
P. 768—778.
Kerr S. J. A comparative study of mitosis in amoebae and plas-
modia of the true slime mold Didymium nigripes // J. Protozool. 1967.
Vol. 14. P. 439—445.
Kerr S. J. Ploidy level in the true slime mold Didymium nigri-
pes // J. Gen. Microbiol. 1968. Vol. 53. P. 9—15.
Kerr S. J. Nuclear size in plasmodia of the true slime mold Di-
dymium nigripes I I J. Gen. Microbiol. 1970. Vol. 63. P. 347—356.
Kerr N. S., Kerr S. J. A new hypothesis concerning plasmodium
formation in the true slime mold, Didymium nigripes // Amer. Zool.
1967. Vol. 7. P. 230.
Kirk D., McKeen W. E., Smith R. Cytoplasmic connections be-
tween Dictyostelium discoideum cells // Canad. J. Bot. 1971. Vol. 49.
P. 19—20.
Kirk P. M., Ansell A. E. Authors of fungal names. Edit. Internat.
Mycolog. Inst, and Inst, of С. A. B. Internat. 1992.
Kitami M. Notes on behaviour of prestolk and prespore cells of
Dictyostelium at the culmination stage // Sci. Repts. Yokohama Nat.
Univ. 1983a. Sec.l. P. 37—45.
Kitami M. Preliminary findings on the cell movement in the mig-
rating pseudoplasmodium of Dictyostelium discoideum /Z^Sci. Repts.
Yokohama Nat. Univ. 1983b . Sec.2. P. 19—27.
Konijn T. M. Chemotaxis and aggregation in slime molds // Be-
havior of microorganisms. London; New York, 1973. P. 48—61.
Konijn T. M., Koevenig J. L. Chemotaxis in Myxomycetes or
true slime molds // Mycologia. 1971. Vol. 63. P. 901—906.
Konijn T. M., Van de Meene J. G. C., Bonner J. T., Barkley
D. S. The acrasin activity of adenosine-3’, 5’-cyclic phosphate// Proc.
Nat. Acad. Sci. U. S. 1967. Vol. 58. P. 1152—1154.
Konijn T. M., Chang Y.-Y., Bonner J. T. Synthesis of cyclic
AMP in Dictyostelium discoideum and Polysphondilium pallidum //
Nature. 1969. Vol. 224. P. 1211—1212.
Kopachik W. Orientation of cells during slug formation in Dicty-
ostelium // W. Roux’s Arch. Develop. Biol. 1982. Vol. 191. P. 348—
354.
Kowalski D. T. Observation on the Dianemaceae // Mycologia.
1967. Vol. 59. P. 1075—1084.
Kowalski D. T. The species of Lamproderma // Mycologia. 1970.
Vol. 62. P. 621—672.
Kuserk F. T. The relationship between cellular slime molds and
bacteria in forest soil // Ecology. 1980. Vol. 6, № 6. P. 1474—1485.
Larsen J., Patterson D. J. Some flagellates (Protista) from tropi-
cal marine sediments // J. Natur. Hist. 1990. Vol. 24. P. 801—937.
Levine N. D., Corliss J.O., Cox F. E. G. et al. A newly revised
classification of the Protozoa H J. Protozool. 1980. Vol. 27. P. 37—
58.
Link J. H. E. Myxomycetes // Handbuch zur Erkennung der nut-
zbarsten und am haufigsten vorkommenden Gewachse. 3. Ordo Fungi,
Subordo 6. Myxomycetes. Berlin, 1833. Bd 3. S. 405—422, 432—
433.
Lister A. A monograph of the Mycetozoa. 3rd ed. I Revised by
G. Lister; British Museum (Nat. Hist.). London, 1925.
Macinnes M. A., Francis D. Meiosis in Dictyostelium mucoroi-
des П Nature. 1974. Vol. 251. P. 321—324.
MacWilliams H. K,, Bonner J. T. The prestolk-prespore-pattern
in cellular slime molds // Differentiation. 1979. Vol. 14. P. 1—22.
Madelin M. F. Myxomycete data of ecological significance //
Trans. Brit. Mycol. Soc. 1984. Vol. 83. P. 1—19.
Maeda Y., Takeuchi I. Cell differentiation and fine structures in
the development of the cellular slime mold, Dictyostelium discoide-
um // Develop. Growth, Differ. 1969. Vol. 13. P. 221—229.
Maimoni-Rodella R. C. S., Gottsberger G. Myxomycetes from
the forest and the cerrado vegetation in Botucatu, Brazil: A compara-
tive ecological study // Nova Hedwigia. 1980. VoL 34. P. 207—246.
Martin G. W. The systematic position of the Myxomycetes //
Mycologia. 1960. Vol. 52. P. 119—129.
Martin G. W., Alexopoulos C. J. The Myxomycetes. Iowa City:
Univ. Iowa Press, 1969.
McCormic J. J., Blomquist J., Rusch H. P. Isolation and charac-
terization of an extracellular polysaccharide from Physarum polycep-
halum // J. Bacteriol. 1970a. Vol. 104. P. 1110—1118.
McCormic J. J., Blomquist J., Rusch H. P. Isolation and char-
acterization of a galactosamine wall from spores and spherules of
Physarum polycephalum 7/ J. Bacteriol. 1970b. Vol. 104. P. 1119—
1125.
McDonald S. A., Durston A. J. The cell cycle and behaviour in
Dictyostelium discoideum // J. Cell Sci. 1984. Vol. 66. P. 195—204.
Mclnnes M. A., Francis D. Meiosis In Dictyostelium mucoroi-
des H Nature. 1974. Vol. 251. P. 321—323.
McManus M. A. Civilisation on angar of the plasmodia of Licea
biforis, Licea variabilis, and Cribraria violacea // Micologia. 1966.
Vol. 58. P. 479-483.
McManus M. A., Roth L. E. Fibrillar differentiation in myxomy-
cete plasmodia И J. Cell Biol. 1965. Vol. 25. P. 303—318.
McManus M. A., Roth L. E. Microtubular structure in myxomy-
cete plasmodia // J. Ultrastr. Res. 1967. Vol. 20. P. 260—266.
Meylan Ch. Contribution a la connaissance des myxomycetes du
Jura et des Alpes H Bull. Soc. Vaud. Sci. Nat. 1931. Vol. 57: P. 301—
307.
Mims C. W. An ultrastructural study of precleavage mitosis in
the myxomycete Arcyria cinerea // J. Gen. Microbiol. 1972a. Vol. 71.
P. 53—62.
Mims C. W. Centrioles and Golgi apparatus in postmeiotic spores
of the myxomycete Stemonitis virginiensis // Mycologia. 1972b.
Vol. 64. P. 452—456.
Mir L., Moisand A., Wright M. Relationships between the ante-
rior kinetosome and microtubule organizing center I in amoebae of the
myxomycete Physarum polycephalum // Biol. Cell. 1982. Vol. 45.
P. 268.
Mir L., Wright M., Moisand A. Variations in the number of
centrioles, the number of microtubular organizing centers I and the
percentage of mitotic abnormalities in Physarum polycephalum amoe-
bae I I Protoplasma. 1984. Vol. 120. P. 20—35.
Moens P. B. Spindle and kinetochore morphology of Dictyosteli-
um discoideum H J. Cell Biol. 1976. Vol. 68. P. 113—122.
Moore D., Spiegel F. W. A new technique for sampling protoste-
lids // Mycologia. 1995. Vol. 87. P. 414—418.
Moore D., Stephenson S. L., Spiegel F. W. Protostelids from tro-
pical forests of Costa Rica and Puerto Rico // Inoculum. 1996.
Vol. 47. P. 22.
Naib-Majani W., Stockem W., Wohlfarth-Botterman K.-E., Os-
born M., Weber K. Immunocytochemistry of the acellular slime mold
Physarum polycephalum. II. Spatial organization of cytoplasmic
actin И Eur. J. Cell Biol. 1982. Vol. 28. P. 103—114.
Naib-Majani W., Stockem W., Weber K., Wehland J., Wohl-
farth-Botterman K.-E. Cytoplasmic actin patterns in Physarum as re-
vealed by NBD-phallacidin staining // Cell Biol. Int. Rept. 1983.
Vol. 7. P. 637—640.
Nannenga-Bremekamp N. E. A revision of the Stemonitales //
Proc. Kon. Ned. Akad., Wetensch. Ser.C. 1967. Vol. 70. P. 201—216.
Nannenga-Bremekamp N. E. De Nederlandse Myxomyceten.
Zutphen, 1974.
Nannenga-Bremekamp N. E., Jamamoto J., Scharma R. Stemo-
naria, a new genus in the Stemonitaceae and two new species of Ste-
monitis (Myxomycetes) // Proc. Kon. Ned., Akad., Wetensch. Ser. C.
1984. Vol. 87. P. 449—469.
Nelson R. K., Scheets R. W. Swarm cell ultrastructure in Cerati-
omyxa fruticulosa // Mycologia. 1975. Vol. 67. P. 733—740.
Nelson R. K., Scheets R. W. Thread phase ultrastructure in Cera-
tiomyxa fruticulosa // Mycologia. 1976. Vol. 68. P. 144—151.
Nelson R. K., Scheets R. W., Alexopoulos C. J. Elemental com-
position of Metatrichia vesparium sporangia // Mycologia. 1977.
Vol. 5. P. 365—375.
O’Day D. H. Aggregation during sexual development in Dictyos-
telium discoideum // Canad. J. Microbiol. 1979. Vol. 25. P. 1416—
1426.
O’Day D. H., McConachie D. R., Rivera J. Appearance and de-
velopmental kinetics of unique cell type in Dictyostelium discoideum.
is it the gamete phase of sexual development? // J. Exp. Zool. 1987a.
Vol. 242. P. 153—159.
ТИП MYCETOZOA. ЛИТЕРАТУРА
449
O’Day D. H., Rama R. A., Lydan M. A. Gamete formation ref-
lects the sexual pheromone hierarchy of Dictyostelium giganteum //
Experientia. 1987b. Vol. 43. P. 619—621.
Odell G. M., Bonner J. T. How the Dictyostelium discoideum
grex crawls // Phil. Trans. Roy. Soc. London. 1986. Vol. 312.
P. 487—525.
Olive L. S. The Protostelida — a new order of the Mycetozoa //
Mycologia. 1967. Vol. 59. P. 1—29.
Olive L. S. Reassignment of Gymnomycota 11 Science. 1969.
Vol. 164. P. 857.
Olive L. S. The Mycetozoa. A revised classification // Bot. Rev.
1970. Vol. 36. P. 59—89.
Olive L. S. The Mycetozoans. New York etc.: Acad. Press, 1975.
Olive L. S., Stoianovitch C. Monograph of the genus Protosteli-
um // Amer. J. Bot. 1969. Vol. 56. P. 979—988.
Olive L. S., Stoianovitch C. A minute new Echinostelium with
protostelid affinities // Mycologia. 1971. Vol. 63. P. 1051—1062.
Olive L. S., Stoianovitch C. Protosporangium: a new genus of
Protostelids // J. Protozool. 1972. Vol. 19. P. 563—571.
Olive L. S., Bennett W. E., Deasey M. C. The new protostelid
genus Endostelium I I Mycoiogia. 1984. Vol. 76. P. 884—891.
Omura F., Fukui Y. Dictyostelium MTOC: structure and linkage
to the nucleus // Protoplasma. 1985. Vol. 127. P. 211—221.
Ono H., Kobayashi S., Yanagisawa K. Cell fusion in the cellular
slime mold, Dictyostelium discoideum // Cell Biol. 1972. Vol. 54.
P. 665—666.
Page F. C. The classification of «naked» amoebae (Phylum Rhi-
zopoda) // Arch. Protistenk. 1987. Bd 133. S. 199—217.
Page F. C., Willumsen N. B. S. Some observations of Gocevia
placopus (Hulsmann, 1974), an amoeba with a flexible test, and on
Gocevia-like organisms from Denmark, with comments on the gene-
ra Gocevia and Hyalodiscus // J. Natl. Hist. 1980. Vol. 14. P. 413—
431.
Pallotta D., Blanchard S., Larue H. Food deprivation is not a
prerequisite for the amoebal to plasmodial transition in Physarum po-
lycephalum // Develop. Genet. 1983. Vol. 4. P. 117—127.
Persoon С. H. Synopsis methodica fungorum. Gottingae, 1801.
Pussard M., Scnaud J., Pons R. Observations ultrastructurales sur
Gocevia fonbrunei Pussard, 1965 (Protozoa, Rhizopoda) // Protistolo-
gica. 1977. T. 13. P. 265—285.
Rammeloo J. Structure of the epispore in the Trichiaceae (Trichi-
ales, Myxomycetes) as seen with scanning electron microscope // Bull.
Soc. Roy. Bot. Belg. 1974. Vol. 107. P. 353—359.
Rammeloo J. Structure of the epispore in the Stemonitales (Myx-
omycetes) as seen with scanning electron microscope // Bull. Jard.
Bot. Nat. Belg. 1975. Vol. 45. P. 3—4.
Rammeloo J. Notes concerting the morphology of some myx-
omycete plasmodia cultured in vitro // Bull. Soc. Roy. Bot. Belg.
1976. Vol. 109. P. 195—207.
Rammeloo J. The contribution of scanning electron microscopy
in the study of Myxomycetes // Fourth Intern. Mycological Congr.
IMC 4, Regensburg, Germany 28 August—3 Sept. 1990, Abstr. / Ed.
by A. Reisinger, A. Bresinsky. Regensburg. 1990. P. 18.
Raper К. B. Dictyostelium discoideum, a new species of slime
mold from decaying forest leaves // J. Agr. Res. 1935. Vol. 50.
P. 135—147.
Raper К. B. Pseudoplasmodium formation and organization in
Dictyostelium discoideum // J. Elisha Mitchell Sci. Soc. 1940.
Vol. 56. P. 241—282.
Raper К. B. Isolation, cultivation, and conservation of simple
slime molds // Quart: Rev. Biol. 1951. Vol. 26. P. 169—190.
Raper К. B. The Dictyostelids. Princeton-New Jersey: Princeton
Univ. Press. 1984.
Raper К. B., Quinlan M. S. Acytostelium leptosomum: a unique
cellular slime mold with an acellular stalk // J. Gen. Microbiol. 1958.
Vol. 18. P. 16—32.
Ratner D., Wayne B. Comparison of differentiating Dictyosteli-
um discoideum cells types separated by an improved method of den-
sity gradient centrifugation // Exp. Cell Res. 1983. Vol. 143. P. 1—
13.
Reyter A., Chastellier C., de. Morphometric and cytochemical
studies of Dictyostelium discoideum in vegetative phase. Digestive
system and membrane turnover // J. Cell Biol. 1977. Vol. 75.
P. 200—217.
Rhea R. P. Electron microscopic observations on the slime mold
Physarum polycephalum with specific reference to fibrilla structures И
J. Ultrastruct. Res. 1966. Vol. 15. P. 349—379.
Rogerson С. T., Stephenson S. L. Myxomyceticolous fungi 11
Mycologia. 1993. Vol. 85. P. 456—469.
29 Протисты, часть 1
Roobol A., Havercroff J. C., Gull K. Microtubule nucleation by
the isolated microtubule-organizing center of Physarum polycephalum
myxamoebae // J. Cell. Sci. 1982. Vol. 55. P. 365—381.
Roos U. P. Mitosis in the cellular slime mold Polysphondilium
violaceum I I J. Cell Biol. 1975. Vol. 64. P. 480—491.
Ross I. K. Capillitial formation in the Stemonitaceae // Mycolo-
gia. 1957. Vol. 49. P. 621—627.
Ross I. K. Chromosome numbers in pure and gross cultures of
myxomycetes // Amer. J. Bot. 1966. Vol. 53. P. 712—718.
Ross I. K. Syngamy and plasmodium formation in the myxomy-
cete Didymium iridis // Protoplasma. 1967a. Vol. 64. P. 104—119.
Ross I. K. Growth and development of the myxomycete Pericha-
ena vermicularis. II. Chromosome numbers and nuclear cycles //
Amer. J. Bot. 1967b. Vol. 54. P. 1231—1236.
Ross I. K. The Stemonitomycetidae, a new subclass of Myxomy-
cetes II Mycologia. 1973. Vol. 65. P. 477—485.
Ross I. K., Cummings R. J. Formation of amoeboid cells from
the plasmodium of myxomycete // Mycologia. 1967. Vol. 59.
P. 725—732.
Rostafinski J. Sluzowce (Mycetozoa). Monografia. Pariz, 1875.
Sakai Y. Cell type conversion in isolated prestolk and prespore
fragments of the cellular slime mold Dictyostelium discoideum // De-
velop. Grows., Diff. 1973. Vol. 15. P. 11—19.
Schaap P., Spek W. Cyclic-АМР binding to the cell surface du-
ring development of Dictyostelium discoideum // Differentiation.
1984. Vol. 27. P. 83—87.
Schoknecht J. D. SEM and x-ray microanalysis of calcareous de-
posits in Myxomycete fructification // Trans. Amer. Microsc. Soc.
1975. Vol. 94. P. 216—223.
Schoknecht J. D., Small E. B. Scanning electron microscopy of
the acellular slime moulds(Mycetozoa — Myxomycetes) and the tax-
onomic significance of surface morphology of spores and acces-
sory structures // Trans. Amer. Microsc. Soc. 1972. Vol. 91. P. 30—
410.
Schuster F. L. Ultrastructure and morphogenesis of solitary sta-
ges of the true slime molds // Protistologica. 1965. T. 1. P. 49—62.
Sharpe P. T., Watts D. J. The role of the cell cycle in differenti-
ation of the cellular slime mold Dictyostelium discoideum // Mol. and
Cell Biochem. 1985. Vol. 67. P. 3—9.
Shipley G. L., Holt С. E. Fusion competence: an inducible state
necessary for zygote formation in the myxomycete Didymium iridis //
Eur. J. Cell Biol. 1980. Vol. 22. P. 225.
Sinha U., Ashworth J. M. Evidence for the existence of elements
of a parasexual cycle in the cellular slime mold Dictyostelium discoi-
deum // Proc. Roy. Soc. 1969. Ser. B. Vol. 173. P. 531—540.
Spiegel F. W. Phylogenetic significance of the flagellar apparatus
in protostelids (Eumycetozoea) // BioSystems. 1981a. Vol. 14.
P. 491—499.
Spiegel F. W. Ultrastructure of the flagellate protostelid Cavoste-
lium apophysatum phylogenetic significance // MSA Newsletter.
1981b. Vol. 32. P. 48.
Spiegel F. W. The ultrastructure of the trophic cells of the protos-
telid Planoprotostelium aurantium // Protoplasma. 1982a. Vol. 113.
P. 165—177.
Spiegel F. W. Mitosis in the protostelid Planoprotostelium auran-
tium // Protoplasma. 1982b. Vol. 113. P. 178—188.
Spiegel F. W., Olive L. S., Brown R. M. Roles of actin during
sporocarp culmination in the simple mycetozoan Planoprotostelium
aurantium // Proc. Nat. Acad. Sci. U. S. 1979. Vol. 76. P. 2333—
2339.
Spiegel F. W., Feldman J., Bennett W. E. Ultrastructure and
development of the amoebo-flagellate cells of the protostelid Proto-
sporangium articulatum // Protoplasma. 1986. Vol. 132. P. 115—128.
Spiegel F. W., Lee S. B., Rusk S. A. Eumycetozoans and molecu-
lar systematics // Canad. J. Bot. 1995. Vol. 73. P. 738—746.
Stephenson S. L. Distribution and ecology of Myxomycetes in
temperate forests. I. Patterns of occurrence in the upland forests of
south-western Virginia // Canad. J. Bot. 1988. Vol. 66. P. 2187—2207.
Stephenson S. L. Distribution and ecology of Myxomycetes in
temperate forests. II. Patterns of occurrence on bark surface of living
trees, leaf litter, and dung // Mycologia. 1989. Vol. 81. P. 608—621.
Stephenson S. L., Laursen G. A. A preliminary report on the dis-
tribution and ecology of Myxomycetes in Alaskan tundra // Bibliothe-
ca Mycologia. 1993. Vol. 150. P. 251—257.
Stephenson S. L., Stempen H. Myxomycetes. A hadbook of
slime molds. Portland; Oregon: Timber Press, 1994.
Stephenson S. L., Studlar S. M. Myxomycetes fruiting upon bry-
ophytes: coincidence or preference? // J. Bryology. 1985. Vol. 13.
P. 537—548.
450
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Stephenson S. L., Kalyanasundaram I., Lakhanpal T. N. A
comparative biogeographical study of myxomycetes in the mod-Appa-
lachians of eastern North America and two regions of India // J. Bio-
geography. 1993. Vol. 20. P. 645—657.
Stephenson S. L., Wheeler Q. D., McHugh J. V., Fraissinet
P. R. New North American associations of Coleoptera with Myxomy-
cetes // J. Nat. Hist. 1994. Vol. 28. P. 921—936.
Stephenson S. L., Landolt J. C., Moore D. L. Protostelids, dicty-
ostelids, and myxomycetes in the litter microhabitat of the Luquillo
Experimental Forest, Puerto Rico I I Mycol. Res. 1999. Vol. 103, N 2.
P. 209—214.
Stewart P. A., Stewart В. T. Membrane formation during sclero-
tization of Physarum polycephalum plasmodia // Exp. Cell Res. 1961.
Vol. 23. P. 471-478.
Stiemerling R. Die Sklerotisation von Physarum confertum H Cy-
tobiologie. 1971. Vol. 3. P. 127—136.
Stosch H. A., von. Untersuchungen uber die Entwicklungsge-
schichte der Myxomyceten. Sexualitat und Apogamie bei Didymiace-
en // Planta. 1935. Bd 23. S. 623—658.
Szabo S. P., O’Day H. D., Chagla A. H. Cell fusion, nuclear fu-
sion, and zygote differentiation during sexual development of Dicty-
ostelium discoideum // Develop. Biol. 1982. Vol. 90. P. 375—382.
Takeuchi I., Ishida S., Amagai A. Phagocytosis and exocytosis by
differentiating Dictyostelium discoideum cells // Plant. Cell Physiol.
1983. Vol. 24. P. 395—402.
Tanaka K. Intranuclear microtubule organizing center in early
prophase nuclei in the plasmodium of the slime mold, Physarum po-
lycephalum // J. Cell Biol. 1973. Vol. 57. P. 220—224.
Usui N. Fibrillar differentiation in a microplasmodium of the
slime mold Physarum polycephalum // Develop. Growth, Diff. 1971.
Vol. 13. P. 241—255.
Uyeda T. G. P., Furuya M. Cytoskeletal changes visualized by
fluorescence microscopy during amoeba-to-flagellate and flagellate-
to-amoeba transformations in Physarum polycephalum // Protoplasma.
1985. Vol. 126. P. 221—232.
Uyeda T. G. P., Furuya M. Effects of low temperature and calci-
um on microfilament structure in flagellates of Physarum polycepha-
lum // Exp. Cell Res. 1986. Vol. 165. P. 461—472.
Uyeda T. G. P., Furuya M. Evidence for active intractions be-
tween microfilaments and microtubules in myxomycete flagellates // J.
Cell Biol. 1989. Vol. 108. P. 1727—1735.
Waddell D. R. A predatory slime mold // Nature. 1982. Vol. 298.
P. 464—466.
Waddell D. R. Duffy К. T. I. Breakdown of self/nonself recogni-
tion in cannibalistic strains of the predatory slime mold, Dictyostelium
caveatum // J. Cell Biol. 1986. Vol. 102. P. 298—305.
Waddell D. R., Vogel G. Phagocytic behavior of the predatory
slime mold, Dictyostelium caveatum. Cell nibbling // Exp. Cell Res.
1985. Vol. 159. P. 323—334.
Wallroth G. W. F. Flora cryptogamia Germaniae. Norimbergae,
1833. Vol. 2.
Watts D. J., Ashworth J. M. Growth of myxamoebae of the cel-
lular slime mold Dictyostelium discoideum in axenic culture // Bio-
chem. J. 1970. Vol. 119. P. 171—174.
Whitney K. D., Bennett W. E. An ultrastructural study of fee-
ding techniques in three protostelids // Canad. J. Bot. 1984. Vol. 62.
P. 1750—1755.
Whitney K. D., Bennett W. E., Olive L. S. Observation on Echi-
nostelium bisporum II Mycologia. 1982. Vol. 74. P. 677—680.
Wohlfarth-Botterman К. E. Differentiations of the ground cy-
toplasm and their significance for the motive force of amoeboid mo-
vement // Primitive Motile Systems in Cell Biology. New York, 1964.
P. 79—109.
Wollman C., Alexopoulos C. J. The plasmodium of Licea biforis
in agar culture // Mycologia. 1967. Vol. 59. P. 423—430.
Wright M., Moisand A., Mir L. The structure of the flagellar
apparatus of the swarm cells of Physarum polycephalum // Protoplas-
ma. 1979. Vol. 100. P. 231—250.
Wright M., Mir L., Moisand A. The structure of the profla-
gellar apparatus of the amoebae of Physarum polycephalum: relation-
ships to the flagellar apparatus // Protoplasma. 1980. Vol. 103.
P. 69—81.
Yabuno K. Changes in cellular adhesiveness during the develop-
ment of the slime mold Dictyostelium discoideum // Develop. Growth,
Diff. 1971. Vol. 15. P. 181—190.
Yamamoto A., Takeuchi I. Vital staining of autophagic vacuoles
in differentiating cells of Dictyostelium discoideum // Differentiation.
1983. Vol. 24. P. 83—87.
Yamamoto A., Maeda Y., Takeuchi I. Development of an
autophagic system in differentiating cells of the cellular slime mold
Dictyostelium discoideum // Protoplasma. 1981. Vol. 108. P. 55—
69.
Тип RHIZOPODA Siebold, 1845 — РИЗОПОДЫ
Гетеротрофные, одноклеточные или плазмодиальные протисты
с лобоподиями, филоподиями или ретикулоподиями, обычно обла-
дают амебоидной подвижностью. Жгутиковые стадии кратковре-
менны и весьма различны по строению, встречаются редко. Не-
которые классы не имеют жгутиковых форм. Кристы в мито-
хондриях различной формы. Многие имеют домик или раковинку.
Свободноживущие, редко паразитические. Преимущественно фа-
готрофы.
Тип весьма гетерогенен по составу. Довольно большая группа
ризопод имеет неопределенное таксономическое положение в
этом типе. В частности, в Rhizopoda incertae sedis помещены
аталамиды и моноталамиды, обычно включаемые вместе с фора-
миниферами в класс Granuloreticulosea. Однако исследования уль-
траструктуры аталамйд Penardia и Gymnophris (см.: «Отряд Atha-
lamida») показали, что не все аталамиды могут быть отнесены к
гранулоретикулозным амебам. Более того, многие представители
аталамйд и моноталамид не имеют гранул и двунаправленных
токов цитоплазмы в ретикулоподиях, т. е. не могут быть отнесены
по этим признакам к гранулоретикулозным амебам. Между тем
четкая очерченность и большой объем класса фораминифер, ди-
агноз которого полностью совпадает с диагнозом гранулоретику-
лозей, фактически вынуждает упразднить таксон Granuloreticulosa
de Saedeleer, 1934 как младший синоним Foraminifera D’Orbigny,
1826. Фораминиферы, таким образом, исключаются из ризопод
и рассматриваются в этом издании в качестве самостоятельного
типа.
В тип Rhizopoda включены классы Lobosea, Heterolobosea, Fi-
losea, Peloflagellatea (Pelomyxa palustris) и Xenophyophorea.
Классификация
Тип Rhizopoda Siebold, 1845
Класс Lobosea Carpenter, 1861
Подкласс Gymnamoebia Haeckel, 1862
Отряд Euamoebida (Lepsi, 1960) Page, 1987
Отряд Leptomyxida (Pussard et Pons, 1976) Page, 1987
Отряд Loboreticulatida Page, 1987
Отряд Acanthopodida Page, 1976
Подкласс Testacealobosia de Saedeleer, 1934
Отряд Arcellinida Kent, 1880
Отряд Himatismenida, Page, 1987
Отряд Trichosida Moebius, 1889
Класс Heterolobosea Page et Blanton, 1985
Отряд Schizopyrenida Singh, 1952
Отряд Acrasida (Schroeter, 1886) Page et Blanton, 1985
Класс Filosea Leidy, 1879
Подкласс Aconchulinia de Saedeleer, 1934
Отряд Cristidiscoidida Page, 1987
Отр^д Cristivesiculatida Page, 1987
Подкласс Testaceafilosia de Saedeleer, 1934
Отряд Gromiida Claparede et Lachmann, 1859
Класс Peloflagellatea Goodkov et Seravin, 1991
Отряд Pelobiontida (Page, 1976)
Класс Xenophyophorea Schulze, 1904
Отряд Psamminida Poche, 1913
Отряд Stannomida Tendal, 1972
Класс Lobosea Carpenter, 1861 — Л обозные амебы
Амебоидные организмы, образующие псевдоподии лобозного
типа и (иногда) субпсевдоподии различной формы. Токи цитоплаз-
мы при перемещении клеток плавные, равномерные. Кристы мито-
хондрий трубчатого типа. Представители класса не имеют в жиз-
ненном цикле жгутиковые стадии и не образуют «плодовые тела».
Половой процесс обнаружен только у 1 вида из подкласса Testacea-
lobosia и еще у 1 — представителя подкласса Gymnamoebia —
предполагается его наличие. Пресноводные, морские и почвенные
организмы. Распространены всесветно. Таксон весьма гетероген-
ный по своему составу, включает 2 подкласса: Gymnamoebia
(голые лобозные амебы) и Testacealobosia (раковинные лобозные
амебы).
Подкласс Gymnamoebia Haeckel,
1862 — Голые лобозные амебы
Представители подкласса передвигаются посредством амебо-
идного движения и не имеют внеклеточных покровных структур
типа раковины или тектума. Половой процесс достоверно неизвес-
тен, лишь у 1 вида предполагается его наличие.
Таксон несомненно сборный; представители 4 входящих в
него отрядов — Euamoebida, Leptomyxida, Loboreticulatida и Acan-
thopodida — сильно различаются по морфологии и особенностям
биологии, поэтому в настоящем изданйи отряды рассматриваются
по отдельности.
Отряд Euamoebida (Lepsi, 1960) Page,
1987
Центральный отряд подкласса Gymnamoebia, представителей
которого традиционно и обозначают словом «амеба». Для боль-
шинства видов характерно наличие сложного, высокодифференци-
рованного надмембранного комплекса плазмалемма—гликокаликс.
Многие виды (но не все) образуют цисты покоя. Свободноживу-
452
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
щие (морские, пресноводные, почвенные) и паразитические орга-
низмы. У одного вида (Sappinia diploidea) предполагается наличие
полового процесса.
История изучения
Считается общепризнанным, что Резель фон Ро-
зенхоф был первым, кто наблюдал живую амебу в
1755 г. Он дал этому организму название «маленький
протей» («der kleine Proteus»), Интересно отметить,
что проведенный впоследствии тщательный сравни-
тельный анализ описаний и рисунков, сделанных Ро-
зенхофом (Kudo, 1959), позволил заключить, что в
действительности он, скорее всего, наблюдал очень
своеобразный организм, подробно описанный спустя
более ста лет Греефом (Greeff, 1874) под названием
Pelomyxa palustris, который в настоящее время ис-
ключен как из состава подкласса Gymnamoebia, так и
вообще из числа лобозных амеб. (Гудков, Серавин,
1991).
Линней (1758 г.) сначала назвал «протея» Розен-
хофа Volvox chaos, а позднее (1767 г.) изменил это
название на Chaos protheus и тем самым установил
род Chaos, В 1822 г. Винсент ввел новое название —
«амеба». Amoeba proteus — «классический» предста-
витель рассматриваемой группы простейших — была
впервые обнаружена Палласом в 1766 г. и названа им
Volvox proteus. Однако лишь в работе Лейди 1878 г.
этот вид получил свое более или менее адекватное
морфологическое описание (и современное наимено-
вание), впоследствии дополненное и уточненное Ша-
фером (Schaeffer, 1916). Собственно говоря, начиная
с подробной монографии Лейди (Leidy, 1879), напи-
санной по результатам его исследований амеб Север-
ной Америки, а также работ Пенара (Penard, 1890,
1902) и ряда других авторов, выполненных в Европе,
и начинается настоящая история изучения разнообра-
зия свободноживущих представителей этой группы
простейших. Существенную роль в этом отношении
сыграла также более поздняя работа Шафера (Scha-
effer, 1926), в которой автор установил четкие диаг-
нозы целого ряда новых, а также уже известных к
тому времени видов амеб и, кроме того, определил
границы некоторых таксонов более высокого таксо-
номического уровня.
В 1849 г. Грос в Москве описал первую паразити-
ческую амебу из полости рта человека — Entamoeba
gingivalis, а в 1875 г. Леш в Петербурге открыл воз-
будителя амебиаза человека. Непатогенную амебу че-
ловека описал Льюис в 1870 г., а в 1903 г. Шаудинн
оставил для этого непатогенного вида название Ле-
ша — Amoeba (Entamoeba) coli Loech. Патогенной
же амебе он дал новое видовое название Entamoeba
histolytica. Существенный вклад в изучение паразити-
ческих амеб человека и животных внесли работы Эп-
штейна (см.: Эпштейн, 1941). Сейчас уже накоплено
значительное количество данных по самым различ-
ным видам амебоидных простейших, как факульта-
тивных, так и облигатных паразитов (Гинецинская,
Добровольский, 1978; Крылов, 1994), однако систе-
матика этих организмов до настоящего времени оста-
ется практически неразработанной.
Классификация
За период с 1980 по 1995 г., т. е. после опублико-
вания системы, принятой Международной Комис-
сией протозоологов (Levine et al., 1980), классифика-
ция амебоидных протистов претерпела существенные
изменения: был установлен тип Rhizopoda, выделен
новый класс Heterolobosea, соответственно установ-
лены новые отряды и семейства, а также выделены
17 новых родов. В основе используемой в настоящем
разделе системы амебоидных протистов лежит клас-
сификация, предложенная Пэйджем (Page, 1987).
Класс Lobosea Carpenter, 1861
Подкласс Gymnamoebia Haeckel, 1866
Отряд Euamoebida (Lepsi, 1960) Page, 1987
Сем. Amoebidae (Ehrenberg, 1838) Page, 1987
Сем. Thecamoebidae (Schaeffer, 1926) Page, 1987
Сем. Hartmannellidae (Volkonsky, 1931) Page, 1974
Сем. Paramoebidae (Poche, 1913) Page, 1987
Сем. Vexilliferidae Page, 1987
Сем. Vannellidae (Bovee, 1970) Page, 1987
Incertae sedis:
Сем. Echinamoebidae Page, 1975
Сем. Entamoebidae Chatton, 1925
Род Dinamoeba Leidy, 1874
Род Phreatamoeba Chaves, Balamuth et Gong, 1986
Морфология и движение
Форма тела
Представители подкласса Gymnamoebia не имеют
постоянной формы тела и перемещаются посредст-
вом так называемого амебоидного движения. Амебы,
неподвижно сидящие на субстрате (рис. 383), а также
особи, осуществляющие незначительные, хаотические
перемещения (рис. 384), могут иметь самую разнооб-
разную форму. (Это дало основание некоторым ран-
ним авторам утверждать, что амебы вообще не имеют
определенной формы.) Однако активно, направленно
передвигающаяся клетка обычно принимает харак-
терную локомоторную форму тела (рис. 385), которая
у многих видов обладает динамически стабильными,
хорошо выраженными определенными морфологи-
ческими особенностями (Schaeffer, 1926; Bovee,
1953; Page, 1988). В современной литературе, говоря
о форме амебы, всегда имеют в виду именно локомо-
торную форму.
При движении на заднем (уроидном) конце тела
простейшего происходит постоянная интернализация
плазматической мембраны. Это часто сопряжено с
образованием характерных морфологических картин
организации уроида, основные типы строения кото-
рого представлены на рис. 386.
Амеба может открепляться от субстрата и свобод-
но парить, пассивно перемещаясь с токами воды.
При этом она обычно принимает своеобразную фло-
тирующую форму (рис. 387).
Цитоплазма клетки подразделяется на оптически
прозрачную гиалоплазму, не содержащую включений,
и заполненную различного рода включениями гра-
нулоплазму (рис. 388). В гиалоплазме сосредоточена
ТИП RHIZOPODAJCnACC LOBOSEA. ПОДКЛАСС GYMNAMOEBIA
453
Рис. 383. Неперемещающая-
ся Paradermamoeba valamo.
Fig. 383. Non-moving Para-
dermamoeba valamo.
Рис. 384. Ненаправленно (хаотич- .
но) перемещающаяся Paraderm-
amoeba valamo.
Fig. 384. Non-directionally (chaoti-
cally) moving Paradermamoeba va-
lamo.
Рис. 385. Локомоторная форма
Paradermamoeba valamo.
Fig. 385. Locomotive form of Pa
radermamoeba valamo.
Рис. 386. Основные типы организации уроида у голых амеб.
А — бульбовидный, Б — морулярный, В — складчатый, Г — виллозно-бульбовидный, Д — «кустовидный».
Fig. 386. Main types of uroid organization in naked amoebae.
A — bulbous, Б — morular, В — plicate, Г — villous-bulbous, Д — fasciculate.
Рис. 387. Разнообразие флотирующих форм голых амеб.
Fig. 387. Diversity of floating forms in naked amoebae.
Рис. 388. Организация клетки голых амеб (схема).
1 — гиалоплазма, 2 — гранулоплазма. пс — псевдоподии, сп — субпсев-
доподии, л — гиалиновые лопасти.
Fig. 388. Cell organization of a naked amoeba (scheme).
7 — hyaloplasm, 2 — granuloplasm. nc — pseudopodia, cn — subpseudopo-
dia, л — hyaline lobes.
454
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
основная часть цитоскелетных элементов клетки, а в
гранулоплазме — ядро, органеллы, вакуоли и различ-
ные цитоплазматические включения.
Большая часть амеб способна образовывать раз-
личного рода клеточные отростки, временные вы-
росты — псевдоподии. Многие современные авторы
помимо термина «псевдоподии» используют также
термин «субпсевдоподии», предложенный Пэйджем.
Псевдоподиями обычно называют выросты цитоплазмы
клетки, включающие в свой состав как гиалоплазму,
так и гранулоплазму; субпсевдоподии (рис. 388) со-
стоят только из гиалоплазмы и не принимают учас-
тия в перемещении основной массы цитоплазмы
клетки.
Морфотипы голых амеб
Совокупность особенностей морфологической ор-
ганизации локомоторной формы амебы легко опи-
сывается через понятие «морфотип» — комплексную
характеристику организации локомоторной формы
амебоидного организма (Смирнов, Гудков, 1999).
Морфотипом называется совокупность признаков,
описывающих динамически стабильную организацию
локомоторной формы амебы. Все разнообразие локо-
моторных форм в пределах отряда Euamoebida сво-
дится к 13 основным морфотипам (Смирнов, Гудков,
1999).
Ниже дана классификация морфотипов голых ло-
бозных амеб (рис. 389).
1. Политактический (рис. 389, А) — полипо-
диальная форма с несколькими четко дифференциро-
ванными, различного размера псевдоподиями, образу-
ющимися в передней части клетки. Пример: Amoeba
proteus при своем обычном перемещении по субстра-
ту.
2. Ортотактический (рис. 389, Б) — ортотак-
тическая форма (см.: Grebecki, Grebecka, 1978); тело
клетки сильно вытянуто и несколько сплющено с бо-
ков, несет многочисленные латеральные складки,
дискретные псевдоподии на переднем конце тела от-
сутствуют. Пример: Amoeba proteus при быстрой не-
прерывной локомоции.
3. Пальмелоидный (рис. 389, В) — полипо-
диальная форма с многочисленными псевдоподиями
примерно одинаковой длины. Пример: Polychaos fas-
ciculatum при своем обычном перемещении по субст-
рату.
4. Монотактический (рис. 389, Г) — моно-
подиальная форма; тело клетки практически округ-
лое в поперечном сечении. Адгезивный уроид отсут-
ствует. Пример: Trichamoeba sinuosa.
5. Стриатный (рис. 389, Д) — уплощенная,
овальная или вытянутая форма с несколькими прак-
тически параллельными дорсальными гребнями. При-
мер: Thecamoeba striata.
6. Ругозный (рис. 389, Е) — уплощенная, про-
дольно вытянутая форма с многочисленными беспо-
рядочными дорсальными гребнями и складками. При-
мер: Thecamoeba sphaeronucleolus.
7. Ланцетовидный (рис. 389, Ж) — уплощен-
ная ланцетовидная форма с распластанными по суб-
страту боковыми участками тела и гладкой дорсаль-
ной поверхностью. Пример: Paradermamoeba valamo.
8. Языковидный (рис. 389, 3) — уплощенная
вытянутая форма с гладкой дорсальной поверх-
ностью. Пример: Dermamoeba granifera.
9. Вееровидный (рис. 389, Я) — уплощенная,
вееровидная, лопатовидная или полукруглая форма с
широкой серповидной фронтальной зоной гиалоплазмы.
Передняя кромка тела ровная; субпсевдоподий нет.
Пример: Vannella simplex.
10. Майорельный (рис. 389, К) — уплощен-
ная, неправильно-треугольная или вытянутая форма с
четко выраженной фронтальной гиалиновой каймой
разной ширины. Клетка может образовывать широ-
кие треугольные субпсевдоподии. Пример: Mayorelfa
cantabrigiensis.
11. Дактилоподиальный (рис. 389, Л) — уп-
лощенная варьирующая форма с широкой фронталь-
ной гиалиновой каймой, из которой образуются паль-
цевидные дактилоподии. Пример: Korotnevella bulla.
12. Парамебный (рис. 389, М) — уплощенная
неправильно-вееровидная форма с четко выраженной
фронтальной зоной гиалоплазмы, из которой обра-
зуются небольшие пальцевидные субпсевдоподии.
Часто эти субпсевдоподии продолжаются в непра-
вильной формы дорсальные гребни, идущие по по-
верхности тела клетки до ее заднего конца. Пример:
Neoparamoeba aestuarina.
13. Вексиллиферный (рис. 389, Н) — упло-
щенная неправильно-треугольная форма тела с мно-
гочисленными длинными и тонкими субпсевдоподия-
ми, расположенными по направлению движения.
Пример: Vexillifera bacillipedes.
Движение
Как можно видеть, характерная форма тела амеб
непосредственным образом связана с их движением.
Поэтому нет ничего удивительного в том, что попыт-
ки объяснить феномен амебоидного движения пред-
принимались уже очень давно.
Исторически так сложилось, что наиболее интен-
сивно механизмы амебоидного движения изучались
на примере представителей сем. Amoebidae (Amoeba
proteus и Chaos carolinense). При наблюдении за рас-
тущей псевдоподией (или всей движущейся клеткой)
на ее переднем конце хорошо видно гиалиновую ша-
почку, которая переходит в очень узкую гиалиновую
кайму, окружающую всю клетку. При движении впе-
ред цитоплазма течет по осевой части псевдоподии и
кажется очень жидкой (принято говорить, что в этом
случае она находится в состоянии золя и носит назва-
ние эндоплазмы). В кортикальной области она, на-
против, очень вязкая (находится в состоянии геля и
называется эктоплазмой).
Буквально до недавнего времени широко обсужда-
лись 2 наиболее распространенные теории амебоид-
ного движения — теория Маста, или теория потока
под давлением, и теория Аллена, или теория сокра-
щения фронтальной зоны. Основные положения пер-
вой теории были высказаны Мастом еще в 1926 г. и
в дальнейшем дополнялись и развивались самим ав-
ТИП RHIZOPODA, КЛАСС LOBOSEA, ПОДКЛАСС GYMNAMOEBIA
455
Рис. 389. Морфотипы представителей отряда Euamoebida.
А — политактический, Б — ортотактический, В — пальмелоидный, Г — монотактический, Д — стриатный, Е — ругозный, Ж — ланцетовидный, 3 —
языковидный, И — вееровидный, К — майорельный, Л — дактилоподиальный, М — парамебный, Н — вексиллиферный. Для Д—3 приведены также по-
перечные сечения клеток.
Fig. 389. Morphotypes of representatives of order Euamoebida.
A — polytactic, Б — orthotactic, В — palmelloid, Г — monotactic, Д — striate, E — rugose, Ж — lancet-like, 3 — spatulate, И — fan-shaped, К — mayorel-
la-like, Л — dactylopodian, M — paramoebian, H — vexilliferian. For Д—3 transverse sections of the cells are also given.
456
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 390. Механизмы амебоидного движения по Масту (А) и Ал-
лену (Б).
Fig. 390. Mechanisms of amoeboid movement according to Mast
(A) and Allen (Б).
тором, а также рядом других исследователей (Mast,
1926, 1931; Goldacre, Lorch, 1950; Wehland, Weber,
1979). Суть ее заключается в том (рис. 390, А), что в
результате сокращения эктоплазмы на заднем конце
клетки создается избыточное давление. Из-за этого
возникает ток эндоплазмы в направлении переднего
конца клетки. Когда золеобразная эндоплазма дости-
гает гиалиновой шапочки, она растекается наподобие
струй фонтана (так называемое фонтанирующее дви-
жение эндоплазмы) и, достигая кортикальной зоны
клетки, превращается в эктоплазму, имеющую геле-
образную консистенцию. На заднем конце амебы (в
уроиде) при этом происходит обратное превращение
эктоплазмы в эндоплазму. Эти процессы происходят
так быстро, что создается впечатление плавного не-
прерывного тока цитоплазмы. В результате описан-
ных процессов клетка продвигается вперед. Главное
отличие теории Аллена (Allen, 1961, 1973) заключа-
ется в том, что он считал зону сокращения эктоплаз-
мы локализованной не на заднем, а на переднем
конце клетки амебы. Согласно Аллену (рис. 390, Б),
золеобразная эндоплазма на переднем конце тела
амебы переходит в гелеобразное состояние (экто-
плазму) в процессе сокращения; при этом она подтя-
гивает за собой следующую порцию эндоплазмы. Эк-
топлазма остается сокращенной до тех пор, пока
снова не перейдет в уроиде в разжиженное состояние
золя.
Мы не будем сейчас подробно останавливаться на
критике этих теорий амебоидного движения — об-
стоятельный анализ их недостатков был проделан в
свое время в работах Серавина (1967, 1977). Главное,
что отмечает этот автор, «теория Аллена, как и тео-
рия Маста, отражает лишь некоторые стороны, всего
лишь некоторые аспекты амебоидного движения, но
не дает приемлемой картины в целом» (Серавин,
1977, с. 125). Серавин выдвинул свою гипотезу, кото-
рая действительно объясняет все существующее раз-
нообразие локомоторных форм амебоидных организ-
мов, — «гипотезу о множественных механизмах,
обеспечивающих амебоидное движение». Она вклю-
чает следующие основные положения.
1. Общим для всех типов амебоидных организмов
и клеток является то, что в основе формирования и
функционирования их двигательных систем, как бы
они внешне не различались, лежит одинаковая сово-
купность механизмов.
2. У разных типов амебоидных организмов и кле-
ток степень участия одних и тех же механизмов в
организации нормального движения неодинакова, что
отражается на форме тела амебоидных организмов и
клеток и характере их движения.
3. Степень развития и способ организации кле-
точных структур, принимающих участие в амебоид-
ном движении, у разных видов амебоидных организ-
мов и клеток могут быть весьма различными, и, более
того, они могут изменяться даже у одной и той же
амебы в зависимости от условий среды, что сказыва-
ется на форме тела и особенностях движения клеток.
«Благодаря всем этим обстоятельствам, — пишет
Серавин (1977, с. 131), — и возникает то большое
разнообразие амебоидных организмов и клеток, силь-
но различающихся по строению, форме тела и харак-
теру движения».
Многие из механизмов и процессов, о которых
говорил в своих работах Серавин, в настоящее время
определены и изучены благодаря использованию со-
временных методов световой и электронно-микрос-
копической техники, а также иммуноцитохимии и
флуоресцентно-аналоговой цитохимии (см. обзоры:
Grebecki, 1982; Stockem, Klopocka, 1988). Показано,
в частности, что у амеб имеется высокодинамичная
контрактильная система, связанная с плазматической
мембраной клетки и представленная дифференци-
ально организованным кортикальным слоем актино-
вых и миозиновых филаментов, который обладает
Рис. 391. Схема организации эктоплазматического кортекса Amo-
eba proteus.
А — распределение толщины микрофиламентозного слоя вдоль продоль-
ной оси тела движущейся амебы, Б — организация кортекса в передней
части тела, В — организация кортекса в задней части тела амебы.
Fig. 391. Organization of ectoplasmic cortex in Amoeba proteus
(scheme).
A — distribution of microfilament layer thickness along the longer axis of the
moving amoeba, Б — cortex organization in the frontal part of the cell, В —
cortex organization in the posterior part of the cell.
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYMNAMOEBIA
457
Рис. 392. Направление движения эктоплазматического кортекса
Amoeba proteus в процессе движения амебы по субстрату.
А — прикрепление клетки к субстрату в зоне уроида, Б — прикрепление
в средней части тела, В — прикрепление передним концом тела.
Fig. 392. Direction of ectoplasmic cortex movement in Amoeba pro-
teus during its locomotion on substrate.
A — attachment of the cell to the substrate in uroid region, Б — attachment
in the middle part of the cell, В — attachment by the frontal part of the cell.
логической и физиологической полярностью. Так,
толщина слоя актиновых филаментов минимальна
на переднем конце клетки, в центральной части дви-
жущейся амебы его толщина увеличивается, а в рай-
оне уроида уменьшается (рис. 391, А). Толщина слоя
миозина также минимальна на переднем конце и
увеличивается к середине, однако в отличие от ак-
тинового миозиновый слой в клетке равномерно
утолщается по направлению к уроидной зоне, и мак-
симальная его толщина наблюдается на заднем кон-
це клетки. Организация микрофиламентозного кор-
текса также различна вдоль оси тела движущейся
клетки. В передней трети тела амебы актиновые фи-
ламенты располагаются в виде продольных рядов,
связанных между собой, а также с интегральными
белками плазматической мембраны (рис. 391, Б).
Цитоскелет задней части тела устроен иначе — ак-
тиновые филаменты образуют сложную трехмерную
сеть, в которой залегают толстые миозиновые фила-
менты (рис. 391, В),
На основании этих и других данных Гребецким
(Grebecki, 1982) была предложена новая теория аме-
боидного движения — теория генерализованного
кортикального сокращения. Суть ее сводится к сле-
дующему. Весь кортикальный цилиндр амебы являет-
ся сократимым. Трехмерное сокращение акто-миози-
нового кортекса приводит к сжатию эндоплазмы, на-
ходящейся в состоянии золя, в результате чего она в
соответствии с градиентом давления устремляется к
переднему концу амебы. Вместе с ее током в пере-
дний конец тела перемещаются молекулы глобулярного
актина (G-актина), которые постоянно образуются на
заднем конце клетки путем деполимеризации актино-
вых микрофиламентов, входящих в состав кортекса.
Эктоплазматический слой амебы, сокращаясь, дви-
жется относительно субстрата, и это движение про-
исходит в направлении участка прикрепления клетки
к субстрату (рис. 392). Таким образом выглядит рабо-
та механизма, создающего движущую силу при локо-
моции амебы с лобоподиями.
Слой микрофиламентов на переднем конце тела
амебы открепляется от плазмалеммы. Он непроница-
ем для любых включений в эндоплазме, но молекулы
G-актина свободно проникают сквозь него, как
сквозь сито (рис. 393). Процесс фильтрации эндо-
плазмы, текущей вперед в соответствии с градиентом
давления, приводит к увеличению размеров гиалино-
вой шапочки. На внутренней поверхности плазма-
леммы на переднем конце клетки имеются центры
полимеризации актина, и проникший в гиалиновую
шапочку G-актин начинает полимеризоваться, фор-
мируя новый кортикальный слой микрофиламентов,
который связан с плазматической мембраной. Сразу
же после окончания этого процесса начинается со-
кращение образовавшегося слоя микрофиламентов.
Поскольку его связь с мембраной сохраняется, он
оказывает на цитоплазму давление, направленное
назад. Вследствие этого рост лобоподии прекращает-
ся. В это же время происходит постепенная деполи-
меризация старого, ранее открепившегося от плазма-
леммы участка микрофиламентозного кортекса. На
следующем этапе новый слой актиновых филаментов
также теряет контакт с мембраной. Он продолжает
сокращение, двигаясь назад, а мембрана движется в
противоположную сторону под напором новой пор-
ив - гель Ш • золь G-актин
Рис. 393. Схема работы системы микрофиламентов на переднем
конце растущей лобоподии. (По: Grebecki, 1990).
1—4 — последовательные стадии изменения организации микрофиламен-
тов. Стрелки — направление движения тока эндоплазмы и поверхностной
мембраны лобоподии соответственно.
Fig. 393. The work of microfilament system on the tip of growing
lobopodium.
7—4 — successive stages of microfilament organization. Arrows indicate di-
rection of movement of endoplasm flow and lobopodium cell membrane res-
pectively.
458
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ции эндоплазмы, фильтрующейся через него впе-
ред, — происходит дальнейший рост гиалиновой
шапочки и т. д. Таким образом осуществляется рост
лидирующей псевдоподии.
Теория генерализованного кортикального сокра-
щения (Grebecki, 1982, 1994) в совокупности с дан-
ными по механизмам создания, изменения, регуля-
ции и координации градиентов вязкостно-эластиче-
ских свойств эндоплазмы (Taylor, Condeelis, 1979)
хорошо объясняет многие, хотя и не все аспекты
локомоции амеб «протеусного» типа. Однако уже из
сказанного можно видеть, что в обеспечении амебо-
идного движения даже какого-то одного морфоди-
намического типа клеток задействованы не один, а
целая совокупность относительно дискретных меха-
низмов. Наконец, экспериментально показано, что
любые воздействия на клетку, которые прямо или
опосредованно влияют на работу того или иного ме-
ханизма, участвующего в организации амебоидного
движения, приводят к изменению характера движе-
ния и формы тела простейшего (иными словами, при
определенном комплексе экспериментальных воздей-
ствий — в рамках физиологических условий —
можно искусственно изменить амебный морфотип).
Все это однозначно свидетельствует в пользу гипоте-
зы Серавина.
Клеточный покров, ядро
и органеллы
Представители подкласса Gymnamoebia в боль-
шинстве своем имеют хорошо дифференцированный
надмембранный комплекс плазмалеммы. Хотя у не-
которых форм клеточный покров представляет со-
бой просто тонкий слой аморфного гликокаликса, у
многих видов он состоит из структур, хорошо за-
метных на электронно-микроскопическом уровне
(рис. 394, вкл.). В современной классификации амеб
строение гликокаликса является одним из важней-
ших систематических признаков (Page, 1988).
Большинство амеб одноядерные, некоторые —
двухъядерные, но известно также несколько многоя-
дерных видов (Page, 1988, 1991). Встречается 2 ос-
новных морфологических типа ядер (Райков, 1982):
везикулярные (с 1 крупным ядрышком, занимающим
более или менее центральное положение в ядре) и
гранулярные (с большим количеством относительно
мелких ядрышек, располагающихся преимуществен-
но по периферии ядра). В ядрах некоторых видов
присутствует так называемая внутриядерная пластин-
ка — фибриллярный слой, прилежащий к внутрен-
ней мембране ядерной оболочки. Это образование
может быть достаточно сильно развитым и сложно
организованным, например наподобие «сотового
слоя» Amoeba proteus (рис. 399, Д, вкл.) и ряда дру-
гих представителей сем. Amoebidae (Pappas, 1959;
Daniels, 1973; Page, 1986; Смирнов, Гудков, 1997).
Представители родов Paramoeba и Neoparamoeba
имеют парасому — ДНК-содержащую самореплици-
рующуюся органеллу, которая находится обычно
около ядра. При делении ядра парасома делится неза-
висимо, но синхронно с ним (Grell, Benwitz, 1970).
Некоторые авторы (Hollande* 1980) полагают, что па-
расома — очень древний кинетопластидоподобный
симбионт, высокоинтегрированный с хозяином.
Как и все представители класса Lobosea, голые
амебы имеют митохондрии с трубчатыми анастомози-
рующими кристами (Page, 1987). У некоторых видов
(представителей сем. Amoebidae) обнаружено явление,
которое мы обозначаем как митохондриальный гете-
роморфизм — одновременное присутствие в цитоп-
лазме одной и той же клетки митохондрий, морфоло-
гически хорошо отличающихся друг от друга, например
по плотности митохондриального матрикса (Flickin-
ger, 1974; Смирнов, Гудков, 1997).
Аппарат Гольджи всегда организован в виде дик-
тиосом, число и размеры которых у различных ви-
дов варьируют (Page, 1986, 1987, 1988). Большинст-
во пресноводных видов имеет сократительные ва-
куоли, окруженные спонгиомом трубчатого или
везикулярного типа. У многих видов амеб в цитоп-
лазме присутствуют эндобиотические бактерии, либо
лежащие свободно в цитоплазме клетки, либо за-
ключенные в симбионтофорную вакуоль (Page, 1986;
Смирнов и др., 1995). Автотрофные эндоцитобионты
(«зоохлореллы»), напротив, встречаются у амеб
крайне редко, хотя потенциальная возможность ус-
тановления такого рода ассоциаций в ряде случаев
была экспериментально доказана (Гудков и др.,
1993; Карпов, 1993). В числе прочих включений в
цитоплазме амеб следует упомянуть специфические
кристаллы, которые нередко являются существенным
компонентом цитоплазмы этих простейших. Они за-
ключены в специальные вакуоли и у разных видов
могут иметь различные строение и размеры (Bovee,
Jahn, 1973; Stockem, 1978; Page, 1986).
Питание
Амебы питаются путем эндоцитоза, поглощая
оформленные пищевые частицы посредством фа-
гоцитоза, а_жидкости, содержащие растворенные ор-
ганические вещества, — с помощью пиноцитоза
(рис. 395). Механизмы процессов эндоцитоза в на-
стоящее время достаточно хорошо изучены (см.:
Stockem, Klopocka, 1988; Christofidou-Solomidou et
al., 1989). Эндоцитоз может осуществляться в любом
участке клетки. Никаких специализированных для
питания участков тела у амеб не выявлено.
Фагоцитоз происходит при участии микрофила-
ментозной системы клетки, которая локально диф-
ференцируется непосредственно под плазмалеммой в
местах формирования фагосомы (Jeon, Jeon, 1983).
На электронограммах часто заметен плотный «во-
ротничок» из микрофиламентов, расположенный во-
круг образующейся пищевой чашечки (рис. 396, вкл.).
Подавляющее большинство амеб — полифаги.
Они не имеют строгой пищевой специализации и фа-
гоцитируют бактерии, одноклеточные водоросли,
других простейших и их цисты. Многие представите-
ли сем. Amoebidae, которое включает преимущест-
венно крупные виды амеб (например, Amoeba proteus
и Chaos carolinense). способны активно охотиться на
такую высокоподвижную добычу, как инфузории Ра-
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYMNAMOEBIA
459
Рис. 395. Эндоцитоз (схема).
А — пиноцитоз, Б — фагоцитоз.
Fig. 395. Endocytosis (scheme).
А — pinocytosis, Б — phagocytosis.
ramecium и Tetrahymena. В пищеварительных вакуо-
лях амебы Chaos illinoisense неоднократно были от-
мечены даже мелкие многоклеточные животные (ко-
ловратки). В то же время среди амеб встречаются не-
которые виды, для которых свойственна в той или
иной степени выраженная пищевая специализация.
Так, представители рода Trichamoeba питаются пре-
имущественно одноклеточными водорослями, а поч-
венные виды рода Deuteramoeba — почти исключи-
тельно конидиями некоторых грибов. Вероятно, в пи-
тании самых мелких амеб, у которых практически не
обнаружены фагосомы (они сами по размеру сравни-
мы с крупными бактериями), существенную роль иг-
рает пиноцитоз растворенных органических веществ
из окружающей среды.
Размножение
и жизненные циклы
Голые амебы размножаются простым делением
клетки, непосредственно перед которым обычно про-
исходит митотическое деление ядра. У многоядерных
видов ядра, как правило, делятся синхронно. Основ-
ным типом деления ядра у амеб является ортомитоз;
при этом встречаются все его разновидности — от-
крытый, полузакрытый и закрытый (Райков, 1982;
Page, 1988).
Размножение амеб чаще всего происходит следую-
щим образом. Непосредственно перед делением амеба
прекращает передвигаться и распластывается по суб-
страту (многие виды при этом принимают весьма ха-
рактерную розетковидную форму) (рис. 397). Проис-
ходит резорбция сократительных вакуолей и диктио-
сом, которые впоследствии формируются de novo в
каждой из дочерних клеток. Митохондрии делятся са-
мостоятельно и распределяются более или менее по-
ровну по обеим дочерним клеткам. Вначале делится
ядро, затем следует цитокинез.
Частота клеточных делений у мелких амеб очень
высока и может в лабораторных условиях достигать
3—4 раз в сутки. У крупных видов она существенно
ниже — клетки делятся 1 раз в течение 2—3 сут. В
то же время крупные амебы в лабораторных услови-
ях и при строго ограниченном количестве доступной
пищи могут существовать, не делясь, до 70 сут.
В состав подкласса Gymnamoebia входят только
агамные организмы; половой процесс ни у одного из
представителей этого таксона достоверно не описан
(Page, 1988, 1991), хотя его наличие предполагается
у Sappinia diploidea (Goodfellow et al., 1974). У этого
вида при инцистировании 2 клетки тесно прижима-
ются друг к другу и покрываются одной общей обо-
лочкой, внутри которой, по-видимому, происходит
объединение их цитоплазмы. Из цисты выходит 1
клетка, однако до сих пор остается неясным, что
происходит в цисте с ядрами после слияния цито-
плазмы обоих партнеров.
Единственной стадией жизненного цикла у боль-
шинства амеб является амебоидный трофозоит, одна-
ко некоторые виды также способны образовывать по-
коящиеся стадии — цисты. Циста может иметь од-
нослойную или двухслойную оболочку (рис. 398); она
способна переживать высушивание и сохранять жиз-
неспособность в течение длительного времени (иног-
да до нескольких лет).
При инцистировании у амеб происходят очень
серьезные морфологические изменения, затрагиваю-
щие практически все клеточные органеллы; умень-
Рис. 397. Схема последовательных стадий (А—Г) бинарного деления амеб. (А — форма, называемая «розетка деления»).
Fig. 397. A scheme of successive stages (A—Г) of amoebae binary fission. (A — a form called «division rosette»).
460
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 398. Цисты амеб с двухслойной (А) и однослойной (5) стен-
кой.
Fig. 398. Amoebae cysts with two-layered (A) and one-layered (Б)
wall.
шается содержание воды в цитоплазме клетки, падает
ее объем. Клетка округляется и секретирует на своей
поверхности плотную оболочку — стенку цисты. Ма-
териал для ее формирования синтезируется при участии
аппарата Гольджи. Таким образом образуются цисты
с однослойной стенкой. В некоторых случаях после
секреции наружной стенки цисты, которую также на-
зывают эктоцистой, клетка продолжает сжиматься,
теряя воду. Цитоплазма ее отделяется от стенки экто-
цисты. После окончания сжатия клетка секретирует
еще одну, внутреннюю стенку цисты — эндоцисту.
Так образуется циста с двухслойной стенкой. При эк-
сцистировании амеба перфорирует стенку цисты и
выходит наружу через образовавшееся отверстие.
По-видимому, практически все амебы, не образую-
щие цист, при наступлении неблагоприятных условий
могут переходить в состояние псевдоцисты. При этом
клетка округляется, токи цитоплазмы в ней практи-
чески прекращаются, однако никакой специфической
оболочки на поверхности клетки не формируется.
Систематический обзор
Сем. Amoebidae (Ehrenberg, 1838)
Page, 1987 (рис. 399, вкл.)
Это семейство в числе прочих включает и самые
крупные виды, относящиеся к подклассу Gymnamoe-
bia. Так, например, Chaos carolinense может дости-
гать в длину 3, а иногда и 5 мм (средние размеры
амеб этого вида составляют около 1 мм). Около
0.5 мм составляют средние размеры амеб Amoeba
proteus. Большинство представителей семейства ха-
рактеризуются полиподиальным или пальмелоидным
морфотипами при обычном перемещении по субстра-
ту и ортотактическим или моноподиальным морфо-
типами — при активной непрерывной локомоции.
Все виды, входящие в состав семейства, одноядерные,
за исключением представителей родов Chaos и Pa-
rachaos, включающих только многоядерные формы.
Встречаются практически все формы организации
гранулярного типа ядер (с периферическими ядрыш-
ками, с лентой ядрышкового материала и т. д.). Лишь
немногие виды способны образовывать цисты. У
большинства видов клеточный покров представлен
разными формами филаментозного гликокаликса,
у некоторых гликокаликс аморфный. Многие виды
имеют хорошо развитую внутриядерную пластинку,
часто организованную в виде «сотового слоя». Все
представители семейства — пресноводные или поч-
венные организмы. Наконец, целый ряд видов этого
семейства — Amoeba proteus, Chaos carolinense, Po-
lychaos dubium — уже давно стали «классическими»
лабораторными модельными объектами клеточной
биологии.
Роды: Amoeba Вогу de St. Vincent, 1822; Chaos
(Linnaeus, 1767) Page, 1991; Deuteramoeba Page, 1987;
Hydramoeba Reynolds et Looper, 1928; Parachaos Wil-
lumsen, Siemensma et Suhr-Jessen, 1973; Polychaos
Schaeffer, 1926; Trichamoeba Fromentel, 1874.
Сем. Thecamoebidae (Schaeffer, 1926)
Page, 1987 (рис. 400, вкл.)
Амебы, входящие в состав этого семейства, не об-
разуют дискретных псевдоподий. При перемещении
вся клетка движется, как единое целое, и лишь при
смене направления движения может на короткое
время образовывать короткие широкие псевдоподии.
Представители семейства характеризуются стриат-
ным, ругозным, языковидным и ланцетовидным мор-
фотипами. Большинство видов имеют 1 ядро везику-
лярного типа с компактным или состоящим из не-
скольких частей центральным ядрышком; некоторые
виды имеют ядро с периферическими ядрышками.
Представители рода Thecochaos, не найденные совре-
менными исследователями в живом виде и известные
только по постоянным (тотальным) препаратам, —
многоядерные. За исключением одного вида — Sap-
pinia diploidea, ни один вид из этого семейства не
образует цист. Практически все представители этого
семейства имеют толстый, плотный, как правило,
аморфный гликокаликс, хотя в пределах некоторых
родов (Dermamoeba, Paradermamoeba) он сложно диф-
ференцирован и достигает толщины 500 нм (Смирнов,
Гудков, 1994). В состав этого семейства входят пресно-
водные, морские, а также почвенные виды.
Особо следует выделить род Pseudothecamoeba,
который резко отличается от всех остальных родов
этого семейства. В него входят полиподиальные
амебы, имеющие гранулярные ядра и филаментозный
гликокаликс. Многие авторы отмечали, что Pseudo-
thecamoeba значительно ближе к представителям сем.
Amoebidae, нежели сем. Thecamoebidae. Вероятно, в
будущем систематическое положение этого рода бу-
дет пересмотрено.
Роды: Dermamoeba Page et Blakey, 1979; Para-
dermamoeba Smirnov et Goodkov, 1993; Parvamoeba
Rogerson, 1993; Pseudothecamoeba Page, 1988; Sappi-
nia Dangeard, 1896; Thecamoeba Fromentel, 1874; The-
cochaos Page, 1981.
Сем. Hartmannellidae (Volkonsky,
1931) Page, 1974 (рис. 401, вкл.)
Все представители этого семейства характеризу-
ются моноподиальным морфотипом и имеют 1 ядро
везикулярного типа. У некоторых видов стенка ядра
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYM NAM О ЕВ IА
461
изнутри армирована слоем филаментозного материа-
ла. Большинство видов образуют цисты с однослой-
ной стенкой и без пор. Клеточный покров представ-
лен тонким слоем аморфного гликокаликса, у не-
которых видов в его состав входят пятиугольные
чашковидные структуры. В составе семейства имеют-
ся пресноводные, морские и почвенные виды.
Роды: Cashia Page, 1974; Glaeseria Volkonsky,
1931; Hartmannella (Alexeieff, 1912) Page, 1974; Sacc-
amoeba (Frenzel, 1892) Bovee, 1972.
Сем. Paramoebidae (Poche, 1913)
Page, 1987 (рис. 402, вкл.)
Представители этого семейства характеризуются
дактилоподиальным, майорельным и парамебным
морфотипами. Одноядерные виды; ядра везикулярно-
го типа. У представителей родов Paramoeba и Janic-
kina имеется парасома. Клеточный покров представ-
лен толстым, стратифицированным гликокаликсом
(род Mayorella) и чешуйками различной формы
(роды Paramoeba и Korotnevella) (Гудков, 1988).
Представители рода Janickina — паразиты морских
беспозвоночных — имеют тонкий аморфный глико-
каликс. Ни один известный вид, входящий в состав
семейства, по-видимому, не образует цист. В составе
семейства преимущественно морские виды, однако есть
также пресноводные и почвенные организмы.
Роды: Janickina Chatton, 1953; Korotnevella (Pa-
ge, 1981) Goodkov, 1988; Mayorella Schaeffer, 1926;
Paramoeba Schaudinn, 1896.
Сем. Vexilliferidae Page, 1987
(рис. 403, вкл.)
Представители этого семейства характеризуются
вексилиферным и парамебным морфотипами. Все
они одноядерные и имеют ядра везикулярного типа;
у представителей рода Neoparamoeba около ядра на-
ходится 1 или несколько парасом. Клеточный пок-
ров сложно дифференцирован: у Vexillifera в составе
гликокаликса присутствуют цилиндрические гексаго-
нальные гликостили, у Pseudoparamoeba — корот-
кие куполообразные, также гексагональные в осно-
вании гЯикостили, у Neoparamoeba — пентагональ-
ные гликостили, между которыми располагаются
длинные, тонкие филаменты. Цисты у представите-
лей этого семейства не формируются. Известен лишь
1 пресноводный вид (Vexillifera bacillipedes), осталь-
ные — морские.
Роды: Neoparamoeba Page, 1987; Pseudoparamoe-
ba Page, 1979; Vexillifera Schaeffer, 1926.
Сем. Vannellidae (Bovee, 1970) Page,
1987 (рис. 404, вкл.)
Представители этого семейства характеризуются
вееровидным или языковидным морфотипами. Одно-
ядерные, ядра везикулярного или гранулярного типа.
Клеточный покров представлен относительно длин-
ными пентагональными гликостилями (роды Vannella
и Pessonella) или слоем коротких призматических
гликостилей, расположенных в плотном аморфном
гликокаликсе (род Platyamoeba). За редким исключе-
нием, не образуют цист. Морские, пресноводные и
почвенные виды.
Роды: Pessonella Pussard, 1973; Platyamoeba Pa-
ge, 1969; Vannella Bovee, 1965.
Gymnamoebia incertae sedis
Сем. Echinamoebida Page, 1975
Объединяет мелких уплощенных амеб, обра-
зующих заостренные, очень тонкие субпсевдопо-
дии — эхиноподии. Пресноводные и почвенные
виды.
Роды: Comandonia Pernin et Pussard, 1979; Echi-
namoeba Page, 1975; Filamoeba Page, 1967.
Сем. Entamoebidae Chatton, 1925
Все представители семейства, за исключением
Entamoeba moshkovskii, являются паразитами различ-
ных животных и человека. Характерной особен-
ностью большинства представителей семейства явля-
ется деление ядер в цистах. Количество образующих-
ся ядер в цистах постоянно у каждого конкретного
вида (например, Entamoeba histolytica — 4, Е. coli —
8); встречаются также виды с одноядерными циста-
ми.
Роды: Entamoeba Casagrandi et Barbagallo, 1895;
Endolimax\ lodamoeba.
Род Dinamoeba Leidy, 1874
Амебы при движении имеют грушевидную форму;
на поверхности тела присутствуют многочисленные
мелкие выросты цитоплазмы, напоминающие тупоко-
нические гиалиновые субпсевдоподии; предполагает-
ся, что единственный до сих пор описанный вид
этого рода — D. mirabilis, — двухъядерный. Вид
описан и изучен в начале века и до настоящего вре-
мени не был реизолирован и переисследован.*
Род Phreatamoeba Chaves, Balamuth
et Gong, 1986
Амебы уплощенные, с очень немногочисленными
фронтальными, конической формы субпсевдоподия-
ми или субцилиндрические при активной локомоции.
* Некоторые исследователи приходят к заключению (Simpson
et al., 1997), что род Phreatamoeba следует признать синонимом
Mastigamoeba и отнести, таким образом, Ph, balamuthi к мастиг-
амебам — группе простейших, также не имеющей определенного
положения в системе низших эукариот. К числу мастигамеб пред-
лагается относить и род Dinamoeba (Griffin, 1988; Simpson et al.,
1997).
462
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Преимущественно многоядерные, хотя встречаются од-
ноядерные формы. Имеется временная одноядерная од-
ножгутиковая стадия. Образуют цисты покоя. Един-
ственный известный вид — Ph. balamuthi — анаэроб-
ный организм, лишенный митохондрий.
Экология и условия обитания
Голые амебы населяют практически все сущест-
вующие водные и наземные экотопы. Представители
этой группы были обнаружены даже в почвах и озе-
рах Антарктиды и в грунтах термальных источников
Новой Зеландии. Можно достаточно уверенно гово-
рить о существовании специфической солоноводной
и пресноводной фаун амеб, ареалы которых практи-
чески не перекрываются между собой (Page, 1983;
1988). Солоноводные виды населяют исключительно
морские местообитания, а пресноводные встречают-
ся в пресных водах, почве, на поверхности растений,
коре деревьев и т. д. Среди пресноводных амеб (фак-
тически под ними понимаются виды, способные раз-
множаться в минеральной среде с соленостью 1—
2 %о) есть преимущественно или исключительно
почвенные виды, облигатные пресноводные виды и
виды, встречающиеся в обоих местообитаниях. В
частности, исключительно пресноводными являются
все виды родов Amoeba и Chaos, аЧточвенными —
представители рода Deuteramoeba. Необходимо, од-
нако, учитывать, что почвенные амебы на самом де-
ле обитают в слое капиллярной воды, заполняющей
пространство между частицами грунта (т. е. реально
также являются водными организмами). Многие поч-
венные виды образуют цисты и при пересыхании
грунта инцистируются.
И у морских и у пресноводных видов нет четкого
разделения на планктонных и бентосных амеб. В
планктоне могут быть обнаружены флотирующие
формы практически любых видов. В то же время
многие мелкие виды амеб обитают в макроагрегатах
водорослей, хлопьях детрита, бактериальной слизи и
на других субстратах, «парящих» в толще воды. Они
могут достигать значительной численности — до
68 тыс. экз./мл, а в эвтрофированных озерах — до
380 тыс. экз./мл. В макроагрегатах водорослей обна-
руживали до 12 млн экз./мл амеб (Arndt, 1993).
Приведенные цифры с очевидностью показывают,
что голые амебы являются важным компонентом эко-
систем. Их численность в бентосе пресноводных озер
может достигать 69.7 млн экз./мл и в некоторых слу-
чаях существенно превышать численность других
групп протистов. В почвах насчитывается до 2 млн
экз. амеб на 1 грамм сухой массы почвы, что состав-
ляет существенную часть общей численности почвен-
ных протистов. В лабораторных опытах было показа-
но, что амебы являются важным компонентом круго-
ворота углерода, азота и фосфора в почвах.
Подавляющее большинство свободноживущих
представителей подкласса Gymnamoebia — аэроб-
ные или микроаэрофильные формы. Однако сущест-
вует и практически не изучена фауна анаэробных
амеб, из которой известно пока лишь всего несколь-
ко видов, в частности Flamella citriensis и Vannella
peregrinia (Bovee, 1956; Смирнов, Фенхел, 1996;
Смирнов, 1997).
Биогеография амеб изучена крайне слабо. По-ви-
димому, подавляющее большинство видов — космо-
политы, однако их распределение в локальных мес-
тообитаниях очень гетерогенно, что существенно за-
трудняет фаунистические исследования (Page, 1976).
Одни и те же виды могут быть обнаружены и в
Европе, и в Северной Америке, в то же время фауна
двух соседних прудов в пригороде европейского го-
рода может сильно отличаться.
Отряд Leptomyxida (Pussard
et Pons, 1976) Page, 1987
Отряд объединяет уплощенных амеб, как правило, имеющих
уроид адгезивного типа (рис. 405). Некоторые представители этого
таксона существуют в виде индивидуальных амеб, а некоторые —
в виде крупных, Многоядерных плазмодиев. Половой процесс неиз-
вестен; отмечены случаи агамных слияний особей. Для ряда видов
характерно образование так называемых цист пищеварения, пред-
назначенных для переваривания фагоцитированного материала.
Классификация
Современная классификация отряда, разработан-
ная в публикации Пэйджа (Page, 1987) и используе-
мая нами в настоящем разделе, в основе своей при-
держивается системы, предложенной ранее Пуссаром
и Поном (Pussard, Pons, 1976).
Подотряд Rhizoflabellina Page, 1987
Сем. Flabellulidae Bovee, 1970
Сем. Leptomyxidae (Pussard et Pons, 1976) Page, 1987
Подотряд Leptoramosina Page, 1987
Сем. Gephyramoebidae Pussard et Pons, 1976
Сем. Stereomyxidae Grell, 1966
Морфология и движение
Общий план строения клетки Leptomyxida сходен
с таковым Euamoebida. Отличительной чертой этой
группы является прежде всего наличие у всех ее
представителей развитого адгезивного уроида. Поли-
морфизм амеб обычно более высок, нежели у Euamo-
ebida, — большинство представителей этой группы
способно при быстром перемещении по субстрату
принимать моноподиальную форму, тогда как для их
обычной, локомоторной-фцрмы характерны 4 основ-
ные морфотипа (рис. 406).
1. Ризомонотактический (рис.406, А) —
моноподиальная форма с адгезивным уроидом. При-
мер: Rhizamoeba saxonica.
2. Флабеллульный (рис.406, Б) — уплощен-
ная форма с обширной фронтальной зоной гиало-
плазмы и неровным, изрезанным передним краем, но
без субпсевдоподий. Уроид адгезивного типа, часто с
длинными филаментами. Пример: Flabellula citata.
3. Парафлабеллульный (рис. 406, В) — уп-
лощенная форма с обширной фронтальной зоной ги-
алоплазмы, из которой образуется множество корот-
ких субпсевдоподий. Уроид адгезивного типа. При-
мер: Paraflabellula reniformis.
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYMNAMOEBIA
463
Рис. 405. Адгезивный уроид (схема).
Fig. 405. Adhesive uroid (scheme).
4. Сетчатый (рис. 406, Г) — многоядерные
плазмодии с полиаксиальным током цитоплазмы. В
передней части движущегося плазмодия образуются
лоборетикулоподии (анастомозирующие лобоподии),
а вдоль его заднего края — многочисленные адгезив-
ные филаменты. Пример: Leptomyxa reticulata.
В отряде имеются как одноядерные, так и много-
ядерные виды. Ядра, как правило, везикулярного ти-
па. Сократительные вакуоли имеются у пресновод-
ных видов и отсутствуют у морских.
Практически все представители отряда имеют тон-
кий аморфный, недифференцированный гликокаликс.
У одного вида — Rhizamoeba saxonica — имеются суб-
мембранные поверхностные структуры («коллосо-
мы»), представляющие собой ре!улярно расположен-
ные тяжи из электронно-плотного материала (Page,
1986). У представителей сем. Stereomyxidae в цитоп-
лазме присутствуют своеобразно устроенные центры
организации микротрубочек — центриолеподобные
тельца (подобного рода образования известны еще
только у представителей отряда Acanthamoebida), ас-
социированные с диктиосомами аппарата Гольджи
(Benwitz, Grell, 1971а, 1971b).
Механизмы амебоидного движения у представите-
лей этого отряда практически не изучались. В преде-
лах группы наблюдается несколько различных мор-
фологических картин локомоции.
1. Клетки, обладающие ризомонотактическим
морфотипом, движутся при помощи аксиального
тока цитоплазмы. Он в значительно меньшей степени
равномерный, чем у представителей отряда Euamoe-
bida; часто наблюдаются кратковременные толчки и
выбросы гиалоплазмы в передней части тела локомо-
торной формы. Характерной особенностью движения
представителей рода Rhizamoeba являются кратковре-
менные выбросы гиалоплазмы из переднего конца клет-
ки в направлении, противоположном движению. При
этом клетка в передней части как бы складывается попо-
лам, но направление движения не изменяется (рис. 407).
2. У амеб, обладающих флабеллульным и пара-
флабеллульным морфотипами, при передвижении
непрерывно происходит «развертывание» вееровид-
ной фронтальной гиалоплазмы от центра тела к его
краям. При этом ее краевые участки отделяют-
ся от центральной зоны глубокими бороздами. Эти от-
деленные участки смещаются при движении назад вдоль
краев тела амебы и, достигнув его заднего края, входят
в состав адгезивного уроида клетки (рис. 408).
3. У амеб, обладающих сетчатым морфотипом,
ток цитоплазмы называют полиаксиальным. При
этом направленные токи цитоплазмы можно наблю-
дать одновременно в разных участках плазмодия, и
их направления не совпадают — обычно даже нет ка-
кого-либо одного генерального направления тока ци-
топлазмы. Плазмодий перемещается медленно, хао-
тично, часто меняя направление движения (рис. 409).
Рис. 406. Морфотипы представителей отряда Leptomyxida.
А — ризамебный, Б — флабеллульный, В — парафлабеллульный, Г — сетчатый (2 варианта организации).
Fig. 406. Morphotypes of representatives of the order Leptomyxida.
A — rhizamoeboid, Б — flabellulate, В — paraflabellulate, Г — reticulate (two variants of organization).
464
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 407. «Выброс» цитоплазмы в передней части тела движу-
щейся Rhizamoeba sp.
Fig. 407. «Eruption» of cytoplasm in the frontal part of the cell of
locomoting Rhizamoeba sp.
При механическом раздражении, уменьшении со-
держания кислорода в среде или воздействии иных
неблагоприятных факторов все представители отряда
теряют свою обычную морфологическую организа-
цию и образуют моноподиальную локомоторную
форму, которая быстро перемещается по субстрату.
Ток цитоплазмы у такой амебы аксиальный, неравно-
мерный. Попав в благоприятные для нее условия,
клетка перестает активно перемещаться,, теряет мо-
ноподиальную форму и принимает характерную для
нее, обычную локомоторную форму.
Размножение и жизненные
циклы
Практически у всех представителей отряда отме-
чено как бинарное, так и множественное деление.
Процесс деления клетки протекает по одному плану
с таковым у Euamoebida; ядра у всех видов делятся
митотически. Крупные плазмодиальные формы (на-
пример, Leptomyxa reticulata) часто размножаются
фрагментацией.
Многие виды способны образовывать цисты
покоя, обладающие двухслойной стенкой. У некото-
рых видов внутри цист может происходить деление
клетки. Иногда после образования внешней стенки
цисты — эктоцисты клетка внутри нее фрагмен-
тируется на несколько частей и каждая их них об-
разует свою внутреннюю стенку цисты. Таким обра-
зом, внутри одной оболочки оказывается несколько
отдельных эндоцист, из которых при эксцистиро-
вании выходят отдельные, самостоятельные плазмо-
дии.
У одного вида — Flabellula baltica отмечены би-
нарные слияния клеток в культуре. Продукт слияния
сохраняет все черты организации обычной локомо-
торной формы. Последующую обратную фрагмента-
цию не наблюдали. Возможно, здесь имеет место
конгрегация — одна из известных форм агамных
слияний клеток протистов (см.: Серавин, Гудков,
1984, 1987).
Рис. 409. Направление «по-
лиаксиального» тока ци-
топлазмы в плазмодии Lep-
tomyxa reticulata.
Fig. 409. Directions of «ро-
lyaxial» cytoplasm flow in
plasmodium of Leptomyxa
reticulata.
Рис. 408. Последовательный этапы «развертывания» фронтальной зоны гиалоплазмы у Fla-
bellula (А—Е).
Стрелками показано направление перемещения отдельных участков цитоплазмы.
Fig. 408. Successive stages of «unfolding» of the hyaloplasm frontal zone in Flabellula (A—E).
Direction of movement of different parts of the cytoplasm is indicated by arrows.
ТИП RKIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYMNAMOEBIA
465
Питание, экология и условия
обитания
Все представители отряда — полифаги. Пита-
ются они путем фагоцитоза, захватывая подходящие
по размеру пищевые частицы любой природы. Пред-
ставители сем. Leptomyxidae после некоторого пери-
ода активного фагоцитирования пищевого материала
инцистируются, образуя цисты пищеварения, в кото-
рых происходит переваривание заглоченного мате-
риала.
Представители отряда населяют пресноводные,
морские и почвенные местообитания. В морских
местообитаниях они зачастую превосходят по своему
разнообразию представителей отряда Euamoebida и,
вероятно, играют важную роль в этих сообществах.
Подавляющая часть водных форм — бентосные ор-
ганизмы.
Систематический обзор
Подотряд Rhizoflabellina Page,
1987
Этот таксон включает 2 семейства, которые объ-
единяют как одноядерные, так и многоядерные фор-
мы протистов, обладающих флабеллульным, пара-
флабеллульным, ризамебным и сетчатым морфоти-
пами.
Сем. Flabellulidae Bovee, 1970
(рис. 410, вкл.)
Для представителей этого семейства характерны
флабеллульный или парафлабеллульный морфотипы.
Преимущественно одноядерные организмы, хотя у
ряда видов наблюдается тенденция к появлению
многоядерных форм. Цисты неизвестны. Клеточный
покров образован аморфным гликокаликсом. Пре-
имущественно морские виды.
По всей видимости, к этому же семейству следует
отнести также род Flamella Schaeffer, 1926, хотя
Пэйдж (Page, 1976, 1988) относит его к сем. Hyalo-
discidae Poche, 1913. Систематическое положение
этого семейства пока остается неясным, однако
большинство современных авторов склоняются к
тому, что род Hyalodiscus Hertwig et Josser,
1874 следует относить к классу Filosea (отряд Асоп-
chulinia). Фишбек и Бови (Fishbeck, Bovee, 1993)
предположили, что род Flamella может относиться к
сем. Flabellulidae. Различия между Flamella и Hyalo-
discus достаточно очевидны, и мы принимаем оба
эти вполне обоснованные предположения. Сем. Ну-
alodiscidae мы не включаем в состав класса Lobosea.
Роды: Flabellula Schaeffer, 1926; Flamella (Scha-
effer, 1926) Michel et Smirnov, 1999; Paraflabellula
Page et Willumsen, 1983.
Сем. Leptomyxidae (Pussard et Pons,
1976) Page, 1987 (рис. 411, вкл.;
412, вкл.)
Представители этого семейства характеризуются
ризамебным и/или сетчатым морфотипами (многие
виды обладают значительным полиморфизмом).
Встречаются либо одноядерные амебы с отчетливо
выраженной тенденцией к приобретению многоядер-
ности, либо многоядерные микроплазмодии; некото-
рые виды способны к агамным слияниям. Представи-
тели рода Rhizamoeba часто активно перемещаются
по субстрату, тогда как представители 2 других
родов, входящих в состав семейства, преимущест-
венно существуют в виде малоподвижного плазмодия
и лишь при сильном механическом или ином разд-
ражении способны принимать моноподиальную ло-
комоторную форму. В числе особенностей биологии
лептомиксид следует упомянуть их способность к
образованию цист пищеварения после фагоцитиро-
вания пищи. Для большинства видов известны и
цисты покоя. Клеточный покров у большинства со-
стоит из тонкого аморфного гликокаликса; у неко-
торых представителей рода Rhizamoeba имеются суб-
мембранные электронно-плотные структуры —
«коллосомы». Представители рода Balamuthia — па-
разиты и патогенны для обезьян. Пресноводные,
морские и почвенные виды.
Роды: Rhizamoeba Page, 1972; Leptomyxa Goo-
dey, 1914; Balamuthia Visvesvara, Schuster et Marti-
nez, 1993.
Подотряд Leptoramosina Page, 1987
Таксон включает 2 семейства, объединяющих од-
ноядерных протистов, основная трофическая форма
которых обладает сетчатым морфотипом.
Сем. Gephyramoebidae Pussard et
Pons, 1976
Монотипический таксон с 1 родом и видом Ge-
phyramoeba delicatula Goodey, 1914. Среди одноядерных
особей изредка встречаются формы, содержащие не-
сколько ядер. Цисты с 1 оболочкой. Почвенный вид.
Сем. Stereomyxidae Grell, 1966
В состав семейства входит единственный род Ste-
reomyxa Grell, 1966, объединяющий очень своеобраз-
но организованных, исключительно морских протис-
тов. Длинные ветвящиеся, относительно ригидные,
лобозного типа псевдоподии не анастомозируют и не
формируют настоящие сетчатые плазмодии. В цито-
плазме присутствуют специфические ЦОМТы (Веп-
witz, Grell, 1971а, 1971b) — центриолеподобные
тельца — подобные тем, что имеются у Corallomyxa
chattoni (отряд Loboreticulatida) и всех представите-
30 Протесты, часть 1
466
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
лей отряда Acanthopodida (Page, 1987). Эта группа
организмов еще очень плохо изучена, и помещение
их в отряд Leptomyxida является в достаточной сте-
пени провизорным.
В состав сем. Stereomyxidae до недавнего времени
включали также род Stygamoeba Sawyer, 1975, одна-
ко, как показали недавние ультраструктурные иссле-
дования (Смирнов, 1996), этот род должен быть вооб-
ще исключен из класса Lobosea в силу особенностей
своего тонкого строения.
Отряд Loboreticulatida Раде, 1987
Многоядерные, с сетчатым морфотипом, ригидные плазмоди-
альные организмы с анастомозирующими псевдоподиями лобозно-
го типа. Все известные виды — морские. От основного тела пери-
одически отрываются отдельные, направленные вверх «ветви», кото-
рые разносятся токами воды и дают начало новым организмам.
Единственным способом размножения является фрагментация. Ульт-
раструктурное исследование одного вида — Corallomyxa chattoni —
показало, в частности, присутствие центриолеподобного ЦОМТа,
как и у представителей рода Stereomyxa (Grell, Benwitz, 1978).
К этому отряду, созданному Пэйджем в 1987 г.,
относится 1 сем. Corallomyxidae Page, 1987 с единствен-
ным родом Corallomyxa Grell, 1966, который до этого
входил в состав сем. Stereomyxidae (Grell, 1966).
Отряд Acanthopodida Page, 1976
Весьма гомогенный в систематическом отношении таксон.
Объединяет более или менее уплощенных амеб, которые обладают
короткими заостренными субпсевдоподиями, образующимися из
фронтальной зоны гиалоплазмы. Эти так называемые акантоподии
имеют осевой цитоскелет, образованный плотным аксиальным
пучком микрофиламентов. Кроме того, акантоподии вблизи от
своего основания могут разветвляться надвое. Все известные виды
одноядерные, цистообразующие.
Классификация
В состав отряда входит 1 семейство — Acanth-
amoebidae Sawyer et Griffin, 1975, включающее 2
рода — Acanthamoeba Volkonsky, 1931 и Protacanth-
amoeba Page, 1981.
Морфология, размножение,
питание, экология
Все представители отряда характеризуются акант-
амебным морфотипом. Организация клеток в целом не
отличается от таковой других голых лобозных амеб.
Все амебы одноядерные, ядра везикулярного типа. Су-
щественной ультраструктурной особенностью аканта-
меб является наличие специфически устроенных
ЦОМТов — центриолеподобных телец (сходных по
своему строению с таковыми представителей родов
Stereomyxa и Corallomyxa). которые лежат в непосред-
ственной близости к ядру и ассоциированы с диктио-
сомами аппарата Гольджи. Клеточный покров пред-
ставлен тонким аморфным гликокаликсом.
Клетки размножаются бинарным делением, ядра
делятся митотически. Все представители отряда об-
разуют цисты покоя, при этом у Acanthamoeba эндо-
циста часто имеет звездообразную форму. В местах
контакта эктоцисты и эндоцисты находятся цистные
поры — специализированные участки стенки цисты,
через которые клетка выходит наружу при эксцисти-
ровании. Представители рода Protacanthamoeba цис-
тных пор не имеют, эктоциста и эндоциста у них
гладкие, округлой формы.
Все виды, входящие в состав отряда, полифаги.
Питаются преимущественно бактериями. Населяют
практически все пресноводные, морские и почвенные
местообитания и, вероятно, являются одними из
самых широко распространенных амеб (Page, 1988).
Некоторые представители рода Acanthamoeba (на-
пример, A. castellanii, A. culbertsoni) могут стано-
виться факультативными паразитами и патогенны для
человека и животных — часто они служат причиной
возникновения у них тяжелых заболеваний, в част-
ности амебного менингоэнцефалита.
THE PHYLUM RHIZOPODA SIEBOLD, 1875
THE CLASS LOBOSEA CARPENTER, 1861
THE SUBCLASS GYMNAMOEBIA HAECKEL, 1862
SUMMARY
The class Lobosea unites phagotrophic amoeboid organisms
(amoebae) with lobose type of pseudopodia (when more or less
filiform pseudopodia are present, they are produced from broader
hyaline lobopodium; the «subpseudopodia»). Amoebae move by ste-
ady flow of cytoplasm. Mitochondrial cristae are of tubular or,
rarely, vesicular type (these organelles are absent from obligate pa-
rasites). There are neither flagellate stages nor fruiting bodies in life
cycles.
Free-living (marine, soil, freshwater) and parasitic (facultative and
obligate) organisms, distributed worldwide.
There are two subclasses: Gymnamoebia (naked lobose amoebae)
and Testacealobosia (testate lobose amoebae).
THE SUBCLASS GYMNAMOEBIA HAECKEL, 1862
The subclass includes lobose amoebae without such extracellular
structures as test or tectum. This is apparently a heterogeneous taxon.
Representatives of this group differ essentially by their morphology
and biology. There are four orders in the subclass: Euamoebida, Lep-
tomyxida, Loboreticulatida and Acanthopodida.
The order Euamoebida includes «classical» naked amoeboid pro-
tists, which are traditionally called «amoebae», because the notion
«amoeba» was first applied by Vincent in 1822. Most amoebae are
uninucleate, rarely binucleate, sometimes multinucleate. There are two
main morphological types of nuclei — vesicular and granular. There
are complex differentiations of glycocalyx in most species.
Life cycles are very simple and include only amoeboid trophozoites,
which multiply by binary fission, preceeded by mitotic nuclear division
(mainly by typical orthomitosis). Cyst stage is presented in some species.
There is only one species (Sappinia diploidea), which is thought to pos-
sess a sexual process (however this statement needs verification). There
are six families in the order: Amoebidae, Thecamoebidae, Harmannelli-
dae, Vannellidae, Paramoebidae and Vexilliferidae.
The order Leptomyxida includes both uni-and multinucleate flat-
tened amoeboid organisms, sometimes large multinucleate plasmodial
forms. The high level of morphological polymorphism is characteristic
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС GYMNAMOEBIA. ЛИТЕРАТУРА
467
<?f this group. Nuclei are usually of vesicular type. Glycocalyx thin,
amorphous. Adhesive uroid in some species. Representatives of the
family Stereomyxidae possess peculiar mucrotubule organizing centre.
Sexual process is unknown, but agamic cell fusions are common. The
order comprises two suborders: Rhizoflabellina (families Flabellulidae
and Leptomyxidae) and Leptoramosina (families Stereomyxidae and
Gephyramoebidae).
The order Loboreticulatida includes only one family Corallomyxi-
dae with the single genus Corallomyxa. Multinucleate, rigid plasmo-
dial protists with anastomosing lobose pseudopodia. Fragmentation is
the only way of reproduction. Only marine species known.
The order Acanthopodida includes the single family Acanth-
amoebidae. These are small flattened amoebae with specific short
sharp subpseudopodia («acanthopodia») with axial microfilamentous
core, forming from the frontal hyaloplasm of the cell. Glycocalyx thin,
amorphous. Single nucleus of vesicular type. There are peculiar «cen-
triole-like bodies» associated with Golgi complex. Multiplication
only by binary fission. Cysts in all species. Some species are pathoge-
nic.
Free-living gymnamoebae inhabit a great variety of ecotopes.
There seem to exist specific marine and freshwater faunas, which
hardly coincide. Amoebae can reach great numbers and are an impor-
tant component of ecosystems. The majority of free-living gymnamoe-
bia are aerobic and microaerophilic, but there is a peculiar anaerobic
fauna, which is yet poorly known. Most gymnamoebae seem to be
cosmopolitan.
ТУРА24
Allen R. D. A new theory of amoeboid movement and endoplas-
mic streaming // Exp. Cell Res. 1961. Vol. 8. P. 17—31.
Allen R. D. Biophysical aspects of pseudopodium formation and
retraction // The Biology of Amoeba. New York: Acad. Press, 1973.
P. 202—245.
Arndt H. A critical review of the importance of rhizopods (naked
and testate amoebae) and actinopods (heliozoa) in lake plankton //
Mar. Microb. Food Webs. 1993. Vol. 7. P. 3—29.
Benwitz G., Grell K. G. Ultrastruktur mariner Amoeben. II. Ste-
reomyxa ramosa. // Arch. Protistenk. 1971a. Vol. 113. P. 51—67.
Benwitz G., Grell K. G. Ultrastruktur mariner Amoeben. III. Ste-
reomyxa angulosa П Arch. Protistenk. 1971b. Vol. 113. P. 68—79.
Bovee E. C. Morphological identification of free-living amoebi-
da И Proc. Iowa Acad. Sci. 1953. Vol. 60. P. 599—615.
Bovee E. C. Some observation on the morphology and activities
of a new ameba from citrus wastes, Flamella citriensis n. sp. // J.
Protozool. 1956. Vol. 3. P. 151—153.
Bovee E. C. The lobose amebas. I. A key to the suborder Cono-
podina Bovee and Jahn, 1966 and descriptions of thirteen new and
little known Mayorella species // Arch. Protistenk. 1970. Vol. 112. P.
178—227.
Bovee E. C., Jahn T. L. Taxonomy and phylogeny // The Biology
of Amoeba I Ed. K. W. Jeon. New York: Acad. Press, 1973. P. 38—82.
Christofidou-Solomidou M., Brix K., Stockem W. Induced pino-
cytosis and cytoskeletal organization in Amoeba proteus — a combi-
ned fluorescence and electron microscopic study // Eur. J. Protistol.
1989. Vol. 24. P. 336—345.
Daniels E. Ultrastructure // The Biology of Amoeba / Ed.
K. W. Jeon. New York: Acad. Press, 1973. P. 125—169.
Fishbeck D. W., Bovee E. C. Two new amoebae, Striamoeba spa-
rolata n. sp. and Flamella tiara n. sp., from fresh water // Ohio J. Sci.
1993. Vol. 93. P. 134—139.
Flickinger C. J. The fine structure of four «species» of Amoeba И
J. Protozool. 1974. Vol. 21. P. 59—68.
Goldacre R. J., Lorch I. J. Folding and unfolding of protein mo-
lecules in relation to cytoplasmic streaming, amoeboid movement and
osmotic work // Nature. 1950. Vol. 166. P. 487—499.
Goodfellow L. P., Belcher J. H., Page F. C. A light- and electron-
microscopical study of Sappinia diploidea, a sexual amoeba // Protis-
tologica. 1974. Vol. 10. P. 207—216.
Grebecki A. Supramolecular aspects of amoeboid movement //
Progress in Protozoology. Warszawa, 1982. P. 117—130.
Grebecki A. Membrane and cytoskeleton flow in the motile cells
with emphasis on the contribution of free-living amoebae I I Int. Rev.
Cytol. 1994. Vol. 148. P. 37—80.
Grebecki A., Grebecka L. Morphodynamic types of Amoeba
proteus: a terminological proposal // Protistologica. 1978. Vol. 14. P.
349—358.
Greeff R. Pelomyxa palustris (Pelobius), ein amoebenartiger Or-
ganismus des sussen Wassers 11 Arch. Microscop. Anat. 1874. Bd 10.
S. 53—73.
Grell K. G. Amoben der familie Stereomyxidae I I Arch. Protis-
tenk. 1966. Vol. 109. P. 147—154.
Grell K. G., Benwitz G. Ultrastruktur mariner amoben. I. Para-
moeba eilhardi Schaudinn 11 Arch. Protistenk. 1970. Vol. 112.
P. 119—137.
Grell K. G., Benwitz G. Ultrastruktur mariner Amoben. IV. Corallo-
myxa chattoni n. sp. // Arch. Protistenk. 1978. Vol. 120. P. 287—300.
Griffin J. L. Fine structure and taxonomic position of giant
amoeboid flagellate Pelomyxa palustris И J. Protozool. 1988. Vol. 35.
P. 300—315.
ЛИТЕРА
Гинецинская T. А., Добровольский А. А. Частная паразитология.
Паразитические простейшие и плоские черви. М.: Высш, шк., 1978.
Гудков А. В. Korotnevella nom. п. (Gymnamoebia, Paramoebi-
dae) — новое родовое название для майорелла-подобных амеб,
имеющих чешуйки в составе клеточного покрова. И Зоол. журн.
1988. Т. 67. С. 1728—1730.
Гудков А. В., Серавин Л. Н. Новое представление о природе
«гигантской амебы» Pelomyxa palustris', положение этого организ-
ма в системе низших эукариот (Peloflagellata classis n.) // Зоол.
журн. 1991. Т. 70. С. 5—16.
Гудков А. В., Карпов А. С., Малышева Л. В. Электронно-мик-
роскопическое исследование искусственно полученной (новообра-
зованной) симбиотической ассоциации амеб с хлорофитовыми во-
дорослями И Цитология. 1993. Т. 35. С. 127—133
Карпов А. С. Некоторые свойства искусственно полученных
симбиотических ассоциаций амеб с хлорофитовыми водорослями
Chlorella sp. И Цитология. 1993. Т. 35. С. 115—126.
Крылов М. В. Возбудители протозойных болезней домашних
животных и человека. Т. 1. СПб.: ЗИН РАН, 1994.
(Райков И. Б.) Raikov I. В. The protozoan nucleus. Morphology
and evolution. Wien: Springer, 1982.
Серавин Л.Н. Двигательные системы простейших. Л.: Наука, 1967.
Серавин Л. Н. Амебоидное движение И Движение немышеч-
ных клеток и их компонентов. Л.: Наука, 1977. С. 121—132.
Серавин Л. Н., Гудков А. В. Основные типы и формы агамных
слияний клеток у простейших И Цитология. 1984. Т. 26. С. 123—131.
(Серавин Л. М., Гудков А. В.) SeravinL. N., Goodkov А. V.
Euhyperamoeba fallax Seravin et Goodkov, 1982 (Lobosea, Gymna-
moebia) — multinucleate marine limax amoeba — morphology, bio-
logical peculiarities and systematic position // Acta Protozool. 1987.
Vol. 26. P. 267—284.
(Смирнов A. B.) Smirnov A. V. Stygamoeba regulata n. sp. (Rhi-
zopoda) — a marine amoeba with an unusual combination of light-
microscopical and ultrastructural features // Arch. Protistenk. 1996.
Vol. 146. P. 299—307.
(Смирнов A. B.) Smirnov A. V. An illustrated survey of gymn-
amoebae isolated from anaerobic sediments of the Niva Bay (the
Sound) (Rhizopoda, Lobosea) И Ophelia. 1999. Vol. 50. P. 113—148.
(Смирнов А. В., Гудков A. B.) Smirnov A. V., Goodkov A. V.
Freshwater gymnamoebae with a new type of surface structure Para-
dermamoeba valamo and P. levis sp. n. (Thecamoebidae), and notes
on the diagnosis of the family // Acta Protozool. 1994. Vol. 33.
P. 109—115.
(Смирнов А. В., Гудков A. B.) Smirnov A. V., Goodkov A. V.
Description of the large multinucleate lobose amoeba Chaos glabrum
sp. n. (Lobosea, Amoebidae), with notes on the diagnosis of the genus
Chaos II Acta Protozool. 1997. Vol. 36. P. 227—233.
(Смирнов А. В., Гудков A. B.) Smirnov A. V., Goodkov A. V. A
new approach to the description of gymnamoebae: illustrated classifica-
tion of the basic morphotypes // Protistology. 1999. Vol. 1. P. 20—29.
(Смирнов А. В., Фенхел T.) Smirnov A. V., Fenchel T. Vahl-
kampfia anaerobica n. sp. and Vannella peregrinia n. sp. (Rhizopo-
da) — anaerobic amoebae from a marine sediment // Arch. Protistenk.
1996. Vol. 147. P. 189—198. \
Смирнов А. В., Осипов Д. В., Раутиан M. С. Эндоцитобионты
у представителей двух отрядов амеб — Euamoebida и Leptomyx-
ida И Цитология. 1995. Т. 37. С. 403—414.
Эпштейн Г. В. Паразитические амебы. М.; Л.: Медгиз, 1941.
24 Цитируемая и рекомендуемая литература к разделу «Тип Rhi-
zopoda. Класс Lobosea. Подкласс Gymnamoebia».
468
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Hollande A. Identification du parasome (Nebenkem) de Janickina
pigmentifera a un symbionte (Perkinsiella amoebae nov. gen. nov. sp.)
apparente aux flagelles kinetoplastides I I Protistologica. 1980. T. 16.
P. 613—625.
Jeon K. W., Jeon M. S. Generation of mechanical forces in pha-
gocytosing amoebae: light and electron microscopic study // J. Proto-
zool. 1983. Vol. 30. P. 536—538.
Kudo R. R. Pelomyxa and related organisms // Ann. N. Y. Acad.
Sci. 1959. Vol. 78. P. 474—486.
Leidy J. Freshwater rhizopods of North America // Rept. U. S. Geol.
Surv. Territ. 1879. Vol. 12. P. 1—324.
Lepsi I. Fauna Republicii Populare Romine. Protozoa.
Vol. 1. Rhizopoda. F. 2. Euamoebidea. Budapest: Acad. Republ.
Popul. Romine. 1960.
Levine N. D., Corliss J. O., Cox F. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R., Lom I. J., Lynn D.,
Merinfeld E. G., Page F. C., Poljansky G., Spraque V., Vavra J.,
Wallace F. G. A new revised classification of the Protozoa // J. Proto-
zool. 1980. Vol. 27. P. 37—58. ..
Mast S. O. Structure, movement, locomotion and stimulation in
Amoeba // J. Morphol., Physiol. 1926. Vol. 41. P. 347—425.
Mast S. O. Locomotion in Amoeba proteus (Leidy) // Protoplas-
ma. 1931. Vol. 14. P. 321—330.
Page F. C. A further study of taxonomic criteria for Umax amoe-
bae, with description of new species and a key to genera // Arch.
Protistenk. 1974. Vol. 116. P. 149—184.
Page F. C. An illustrated key to freshwater and soil amoebae.
Ambleside: Freshwater Biol. Ass., 1976.
Page F. C. Marine Gymnamoebae. Cambridge: Inst. Terr. Ecol.,
1983.
Page F. C. The genera and possible relationships of the family
Amoebidae, with special attention to comparative ultrastructure 11 Pro-
tistologica. 1986. Vol. 22. P. 301—316.
Page F. C. The classification of «naked» amoebae (Phylum RhiT
zopoda) П Arch. Protistenk. 1987. Vol. 133. P. 199—217.
Page F. C. A new key to freshwater and soil Gymnamoebae I I
Ambleside: Freshwater Biol. Ass., 1988.
Page F. C. Nakte Rizopoda. Nakte Rizopoda und Heliozoea. Stutt-
gart; New York: Fischer, 1991. (Protozoenfauna; Bd 2).
Pappas G.D. Electron microscope studies on amoebae // Ann.
N. Y. Acad. Sci. 1959. Vol. 78. P. 448—473.
Penard E. Etudes sur les rhizopodes d’eau douce // Mem. Soc.
Phys. Hist. Nat. Geneve. 1890. T. 31. P. 1—230.
Penard E. Faune Rhizopodique du bassin du Leman. Geneve:
Kundig, 1902.
Pussard M., Pons R. Etude des genres Leptomyxa et Gephruamo-
eba (Protozoa, Sarcodina). I. Leptomyxa reticulata Goodey, 1915 И
Protistologica. 1976. Vol. 12. P. 151—168.
Schaeffer A. A. Notes on the specific and other characters of
Amoeba proteus Pallas (Leidy), A. discoides spec. nov. I I and A.
dubia spec. nov. Arch. Protistenk. 1916. Vol. 37. P. 204—228.
Schaeffer A. A. Taxonomy of the Amebas. Pap. Dept. Mar. Biol.,
Carnegie Inst, of Washington. 1926. Vol. 24. P. 1—116.
Simpson A. G. B., Bernard C., Fenchel T., Patterson D. J. The
organization of Mastigamoeba schizophrenia n. sp. more evidence of
ultrastructural idiosyncrasy and simplicity in pelobiont protists I I Eur.
J. Protistol. 1997. Vol. 33. P. 87—98.
Stockem W. Fine structure of cytoplasmic crystals in Amoeba
proteus I I Cytobiologie. 1978. Vol. 17. P. 301—306.
Stockem W., Klopocka W. Ameboid movement and related phe-
nomena H Int. Rev. Cytol. 1988. Vol. 112. P. 137—183.
Taylor D. L., Condeelis J. S. Cytoplasmic structure and contracti-
lity in amoeboid cells I I Int. Rev. Cytol, 1979. Vol. 56. P. 57—144.
Wehland J., Weber K. Effects of the actin-binding protein
DNAse I on cytoplasmic streaming and ultrastructure of Amoeba pro-
teus II Cell Tiss. Res. 1979. Vol. 199. P. 353—372.
Подкласс Testacealobosia de Saedeleer,
1934 — Раковинные лобозные амебы
Подкласс объединяет амебоидные организмы, которые пере-
двигаются посредством образования лобоподий и строят однока-
мерную «раковину» или имеют плотные, отделенные от цитоплаз-
мы покровные структуры. Одноядерные, двухъядерные и многоя-
дерные формы. Гетероморфизма ядер нет ни на одной стадии
жизненного цикла. Размножение бесполое. У одного вида (Arcella
vulgaris) описан половой процесс (автогамия), протекающий в осо-
бых, мейотических цистах. Пресноводные, бриофильные и почвен-
ные организмы; описано небольшое число морских интерстици-
альных видов.
Включает 3 отряда, в состав которых входит около 1000 видов.
Классификация
Традиционно выделявшаяся группа раковинные
амебы (Testacea) в настоящее время разделена на
4 отряда, относимые по типу псевдоподий к разным
классам типа Rhizopoda. Это отряды Arcellinida, Hi-
matismenida и Trichosida (класс Lobosea) и отряд
Gromiida (класс Filosea).
В настоящем издании система подкласса Testacea-
lobosia принимается в следующем виде:
Отряд Arcellinida Kent, 1880
Сем. Microcoryciidae de Saedeleer, 1934
Сем. Microchlamyidae Ogden, 1985
Сем. Arcellidae Ehrenberg, 1832
Сем. Centropyxidae Deflandre, 1953 /
Сем. Plagiopyxidae Bonnet, 1959 /
Сем. Lamtopyxidae Bonnet, 1974 1
Сем. Distomatopyxidae Bonnet, 1979
Сем. Difflugiidae Wallich, 1864
Сем. Hyalospheniidae Schulze, 1877
Сем. Lesquereusiidae Ogden, 1979
Сем. Paraquadrulidae Loeblich et Tappan, 1961
Сем. Mississippiellidae Huddleston et Haman, 1985
Сем. Cryptodifflugiidae Jung, 1942
Сем. Phryganellidae Jung, 1942
Отряд Himatismenida Page, 1987
Сем. Cochliopodiidae (de Saedeleer, 1934) Page, 1987
Отряд Trichosida Moebius, 1889
Род Trichosphaerium Schneider, 1878
Отряд Arcellinida Kent, 1880
Морфология и физиология
Строение клетки
У раковинных амеб собственно амебоидная клет-
ка заключена в раковину («домик»), которая состоит
из агглютинированного клеткой экзогенного матери-
ала или из секретированных ею частиц (рис. 413).
Тело амебы, как правило, не заполняет полость рако-
вины целиком, а закреплено в ней при помощи эпи-
подий (рис. 416, Д, вкл.) — выростов цитоплазмы,
прикрепленных изнутри к стенке раковины. Иногда
цитоплазматическое тело амебы соединяется с внут-
ренней поверхностью раковины лишь в области устья,
по его периферии.
Снаружи клетка покрыта плазмалеммой, в состав
которой входит тонкий слой недифференцированного
гликокаликса. В цитоплазме находится ядро (ядра) и
набор органелл, типичный для аэробных корненожек.
Ядра раковинных амеб обычно везикулярного ти-
па, с более или менее компактным ядрышком,
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
469
реже — гранулярного типа. Большинство представи-
телей отряда одноядерные, но для некоторых харак-
терно наличие 2 ядер (многие представители рода Ar-
cella, — рис. 416, £); известны также многоядерные
виды (Arcella megastoma). У изученных представите-
лей родов Arcella, Hyalosphenia, Nebela, Lesquereusia
оболочка ядра изнутри армирована слоем фибрил-
лярного материала. Деление ядра на ультраструктур-
ном уровне изучено только у Arcella vulgaris, у кото-
рой оно носит характер закрытого ортомитоза.
Митохондрии имеют тубулярные кристы, кото-
рые могут анастомозировать. Матрикс митохондрий
может быть электронно-плотным (у изученных пред-
ставителей родов Cryptodifflugia, Hyalosphenia, Cent-
ropyxis) или электронно-светлым (Lesquereusia, Net-
zelia). У разных представителей одного и того же
рода митохондрии могут иметь или плотный, или
светлый матрикс, однако гетероморфизм митохонд-
рий у раковинных амеб неизвестен.
Для Testacealobosia характерно наличие в цито-
плазме многочисленных диктиосом аппарата Гольд-
жи. Он принимает участие в синтезе цементных ве-
зикул, содержимое которых скрепляет частицы, вхо-
дящие в состав стенки раковины. У видов, имеющих
секретированную раковину, аппарат Гольджи участ-
вует в синтезе идиосом для ее построения.
В клетке раковинных амеб обычно имеется 1 или
2 сократительные вакуоли; известны также виды, у
которых их число достигает нескольких десятков.
Вакуоли опорожняются либо в полость раковины,
либо наружу.
Шероховатая эндоплазматическая сеть (ШЭПС) у
многих видов образует плотные скопления в цито-
плазме, которые находятся около ядра, а иногда —
окружают его. Эти скопления эндоплазматической
сети прокрашиваются гематоксилином и другими
красителями, окрашивающими рибонуклеопротеиды,
и иногда совершенно маскируют ядро. В литературе
начала века их часто описывали под названием «хро-
мидии» и рассматривали даже как одну из альтерна-
тивных форм существования ядра клетки у раковин-
ных амеб.
У видов, которые синтезируют для построения
стенки раковины кремниевые чешуйки, известна мо-
дификация ШЭПС — силикалемма. В ее цистернах
происходит синтез идиосом. Некоторые раковинные
амебы способны покрывать тонким слоем кремния
фагоцитированные частицы, например бактерии.
Впоследствии эти частицы встраиваются в стенку ра-
ковины.
В составе цитоскелета раковинных амеб выявле-
ны микрофиламенты и, по крайней мере у некоторых
видов, микротрубочки. Пучки микрофиламентов об-
наружены в псевдоподиях движущихся амеб; их скоп-
ления имеются в различных участках цитоплазмы.
Микротрубочки в псевдоподиях не отмечены, однако
у некоторых видов рода Arcella они ассоциированы с
поровыми комплексами и располагаются вокруг ядра.
В цитоплазме клеток находится большое количест-
во различных включений, которые представляют
собой кристаллы (в основном оксалаты), липидные
глобулы, цементные везикулы и материал для постро-
ения дочерней раковины. В цитоплазме большинства
Рис. 413. Схема организации клетки раковинной амебы.
а — раковина, б — эпиподии, в — ядро, г — сократительная вакуоль, д —
шероховатый ЭПР, е — диктиосома, ж — митохондрия, з — ксеносома в
вакуоли, и — пищеварительная вакуоль, к — псевдоподии.
Fig. 413. Organization of testate amoebae cell.
a — shell, 6 — epipodia, в — nucleus, г — contractile vacuole, d — rough
endoplasmatic reticulum, e — dictyosome, ж — mitochondrion, з — xenoso-
me in a vacuole, и — digestive vacuole, к — pseudopodia.
изученных видов наряду с митохондриями находятся
микротельца, окруженные одинарной мембраной.
Многие раковинные амебы факультативно или об-
лигатно содержат в цитоплазме различные эндоцито-
биотические организмы (например, Difflugia gramen,
D. lobostoma, Hyalosphenia papilio, Cucurbitella mes-
piliformis). Как правило, это зоохлореллы — одно-
клеточные зеленые водоросли из рода Chlorella.
Иногда в цитоплазме амеб отмечали наличие водоро-
слей из родов Protococcus и Scenedesmus. Симбиоз с
хлореллами у некоторых видов носит, видимо, обли-
гатный характер, в то время как другие могут обхо-
диться и без симбионтов. Показано, что виды, содер-
жащие хлореллы, гораздо дольше других раковинных
амеб могут обходиться без пищи (некоторые крупные
виды диффлюгий — до 3 мес). При помещении их в
темноту водоросли постепенно исчезают из цито-
плазмы и амебы вскоре гибнут. Бактерии в цитоплаз-
ме пока обнаружены только у Difflugia geosphaera.
Они лежат свободно в цитоплазме клетки, однако ха-
рактер взаимоотношений амеб с бактериями неясен.
Наличие раковины, придающей более или менее
стабильную форму цитоплазматическому телу клетки,
создает предпосылки для компартментализации ци-
топлазмы — разделения ее на различные в функцио-
нальном отношении участки (рис. 413). Это разделение
хорошо выражено у Testacealobosia, имеющих округ-
лую или грушевидную раковину (представители рода
Difflugia и морфологически схожих с ним родов), и
практически отсутствует у амеб с уплощенной ракови-
ной (типа раковин Arcella spp. и Centropyxis spp.).
У амеб с вытянутой или округлой раковиной ядро
всегда располагается в задней (аборальной) ее части.
470
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Вокруг него обычно находится плотное скопление
ШЭПС. Этот участок цитоплазмы иногда называют
«зоной плотной ЭПС». На периферии этой части
клетки располагаются диктиосомы, иногда — запасные
идиосомы и сократительные вакуоли. Никаких дру-
гих органелл и пищеварительных вакуолей здесь нет.
Ближе к устью (в оральной части цитоплазмы)
располагаются все митохондрии, вакуоли, кристал-
лы, цементные везикулы и частицы, фагоцитирован-
ные клеткой для построения дочерней раковины.
Псевдоподии начинают формироваться несколько
глубже границы устья раковины, и поэтому ближай-
шая к нему часть цитоплазмы представляет собой
сеть переплетающихся и анастомозирующих тяжей.
Раковина
Подавляющее большинство Testacealobosia имеют
жесткую раковину, которая не способна к какой-
либо конформации. Исключением является предста-
витель рода Microchlamys — М. patella, раковина ко-
торого представляет собой находящийся на спинной
стороне тела амебы мягкий щиток, который может
«складываться». Этот щиток образован одним слоем
гексагональных субъединиц — альвеол. Цитоплазма-
тическое тело этой амебы покрыто тонким мембран-
ным мешком, который прикреплен к внутренней по-
верхности щитка и имеет центральное отверстие для
выхода псевдоподий. Такая организация является ис-
ключением, у всех остальных раковинных амеб подо-
бного мешка нет.
Форма раковин чрезвычайно разнообразна — они
могут быть дисковидными, колпачковидными, оваль-
ными, грушевидными, полусферическими или почти
сферическими. Края латерально сжатых или упло-
щенных раковин бывают окружены килем разной
ширины.
Если раковина уплощена, то часть ее, на которой
находится устье, называется брюшной поверхностью,
а противоположная — спинной. У раковин, удлинен-
ных по оси, проходящей через центр устья, различа-
ются передняя (устьевая) и задняя части. Подобное
же разделение раковин на приустьевую, переднюю и
заднюю части применяется и в отношении раковинных
амеб, имеющих уплощенную брюшную поверхность
и эксцентрично расположенное устье (рис. 436; 437).
При этом если задняя часть раковины вздута, то она
называется брюшком, если же небольшое вздутие
имеется и в передней части раковины, то для его
обозначения употребляется термин «козырек».
Раковины Arcellinida однокамерные, хотя внутри
них могут встречаться структуры, подразделяющие
их полость. Внутри раковины Pontigulasia проходит
узкий мостик, построенный из органического мате-
риала. У близких родов Zivcovicia и Lagenodifflugia
уже имеется плотная диафрагма, являющаяся продол-
жением внутренней стенки раковины и отделяющая
полость основной части раковины от шейки; для вы-
хода псевдоподий в этом случае служат 1 или 2 от-
верстия в диафрагме. Диафрагма имеется и у Lesque-
reusia spiralis. У многих почвенных раковинных амеб
(например, у Plagiopyxis) вентральная часть ракови-
ны глубоко вдается в ее полость, отделяя брюшко от
ограниченной козырьком камеры; в этом случае устье
лежит в глубине раковины.
Форма и расположение устья у раковинных амеб
чрезвычайно широко варьируют. Устье может быть
круглым, эллипсоидным, трех-, четырех- й многоло-
пастным, зубчатым, щелевидным, иметь форму вось-
мерки и т. д. Устье может располагаться в нижнем
конце вытянутой раковины (Difflugia), в центре
брюшной поверхности (Arcella, Cyclopyxis) или экс-
центрично (Centropyxis, Plagiopyxis). Оно может на-
ходиться непосредственно на поверхности раковины
(Phryganella), либо на конце так называемой усть-
евой воронки — впячивания вентральной поверхнос-
ти, вдающегося внутрь раковины (Cyclopyxis, Centro-
pyxis, Arcella). У некоторых видов рода Arcella устье
располагается на конце устьевой трубки, направлен-
ной вниз от конца устьевой воронки. Иногда устье
открывается в камеру, ограниченную козырьком.
Встречаются раковинные амебы с крупным устьем
(его диаметр может составлять больше половины ди-
аметра раковины) так называемые эвристомные
формы, и микростомные раковинные амебы, у кото-
рых округлое или радиально-симметричное устье
оказывается значительно суженным. Уменьшение
эксцентрично расположенного устья приводит к
формированию щелевидного устьевого отверстия. У
представителей сем. Mississippiellidae устье образо-
вано серией из 10—18 мелких овальных отверстий,
располагающихся дуговидными рядами. Устье может
быть окружено валиками, воротничками, края устья
могут утолщаться или заходить один на другой, об-
разуя «губы»; приустьевая часть может вытягиваться
в удлиненную шейку.
На раковинах Arcellinida могут встречаться мно-
гочисленные выросты. Различают шипы (выросты, в
которые заходит полость раковины) и не имеющие
внутренней полости иглы.
По общим чертам строения можно выделить сле-
дующие морфологические типы раковин раковинных
амеб.
1. Акростомный — устье терминальное, ось
симметрии раковины проходит через центр устья. В
этом типе различаются простые акростомные (ради-
ально-симметричные, — рис. 414, А), уплощенные ак-
ростомные (рис. 414, Б) и изогнутые акростомные ра-
ковины, характеризующиеся билатеральной симметрией.
2. Трахелостомный — с устьем на шейке,
либо располагающейся на продольной оси раковины
(рис. 414, В), либо изгибающейся (рис. 414, Г) и от-
деленной сужением от расширенного брюшка.
3. Аксиальный — раковина в виде части сфе-
ры с уплощенной вентральной поверхностью, центр
которой Занят устьем. Устье может быть округлым,
треугольным, зазубренным. Иногда имеется устьевая
воронка, вдающаяся в глубь раковины (рис. 414, Д, Е).
4. Эллипсостомный — эллипсоидное устье
располагается в центре уплощенной вентральной по-
верхности раковины, имеющей овоидные очертания.
Ось устья совпадает с продольной осью вентральной
поверхности раковины (рис. 414, Ж, 3).
5. Пропилостомный — устьевая воронка как
бы образует переднюю камеру, открывающуюся сна-
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
471
ружи на вентральной поверхности раковины, а внутри
заканчивающуюся собственно устьем (рис. 414, И).
6. Диплостомный — имеется обширная вес-
тибулярная полость, ограниченная со стороны вент-
ральной подошвы диафрагмой с 2 почковидными от-
верстиями. Собственно устье располагается в верхней
части вестибулярной полости. Имеется скелетная
система, укрепляющая эту полость (рис. 414, К).
7. Плагиостомный — с эксцентричным устьем,
лежащим на вентральной подошве, что определяет би-
латеральную симметрию раковины (рис. 414, Л, М).
Возможно развитие устьевой воронки, иногда с «уз-
дечками», соединяющими ее с дорсальной стенкой ра-
ковины.
8. Плагиостомный с козырьком — устье от-
крывается в камеру, ограниченную козырьком. От-
верстие этой камеры располагается на подошве рако-
вины (рис. 414, Н, О).
9. Криптостомный — сходный с плагиостом-
ным, но устье имеет резко выраженную тенденцию к
уменьшению размеров и обычно принимает вид узкой
поперечной щели (рис. 414, 77, Р).
10. Криптостомный с козырьком — отлича-
ется от плагиостомного с козырьком крайней редук-
цией дистального отверстия ограниченной козырь-
ком камеры до линейной или полулунной щели
(рис. 414, С, Г).
У некоторых видов Arcella устье окружено пора-
ми. Поры могут находиться и в апикальной части
раковины (Hyalosphenia и Nebela). Их функциональ-
ное значение остается неясным. Во время эксцисти-
рования у Nebela tincta наблюдали выход псевдопо-
дий через латеральные поры.
По способу построения раковины подразделяются
на агглютинированные и состоящие из гомогенного
материала. Стенка агглютинированных раковин со-
стоит из частиц, скрепленных органическим цемен-
том (матриксом), который синтезируется клеткой.
Частицы могут быть фагоцитированы амебой из ок-
ружающей среды (ксеносомы) или секретированы са-
мой клеткой (идиосомы и протеинозные альвеолы).
Иногда в состав стенки раковины входят как ксено-
сомы, так и частицы, синтезированные клеткой. Ра-
ковины, состоящие из гомогенного материала, обыч-
но образованы несколькими органическими или ми-
неральными слоями.
Амебы, строящие агглютинированные раковины,
зависят от имеющегося в окружающей среде строи-
тельного материала и поэтому в разных условиях
могут иметь различную их структуру. Как правило,
в качестве ксеносом используются песчинки и пан-
цири диатомовых водорослей (рис. 415, А—Г, вкл.).
Преобладающими минеральными компонентами та-
ких раковин являются кремнезем и кварц. Многие
виды Testacealobosia, видимо, обладают способно-
стью к выбору определенных частиц для построения
раковины: одни виды рода Difflugia строят гладкие
раковины из небольших уплощенных кварцевых час-
тиц, в то время как другие формируют шероховатые
раковины из относительно крупных частиц. Некото-
рые виды в качестве материала для построения ра-
ковины используют преимущественно панцири диа-
томовых водорослей.
Рис. 414. Морфологические типы раковин раковинных амеб.
(Ориг).
А — простая акростомная раковина; Б — упрощенная акростомная ракови-
на; В, Г — трахелостомные раковины; Д, Е — аксиальная раковина с усть-
евой воронкой; Ж, 3 — эллипсостомная раковина; И — пропилостомная
раковина; К — диплостомная раковина; Л, М — плагиостомная раковина;
Я, О — плагиостомная раковина с козырьком; Я, Р — криптостомная рако-
вина; С, Г — криптостомная раковина с козырьком. Е, 3, Л, Н, П, С — вид
с вентральной стороны, Д, Ж, И, К, М, О, Р,Т — вид сбоку, у — устье.
Fig. 414. Morphological types of testate amoebae shells.
A — simple acrostomic shell; Б — flattened acrostomic shell; В, Г — trachelos-
tomic shells; Д, E — axial shell with aperture funnel; Ж, 3 — ellipsostomic
shell; И — propolostomic shell; К — diplostomic shell; Л, M — plagiostomic
shell; Я, О — plagiostomic shell with a «peak»; Я, P — cryptostomic shell; C,
T — cryptostomic shell with a «peak». E, 3, Л, H, П, C — ventral view, Д, Ж,
И, К, M, O, P,T — lateral view, у — aperture.
У ряда раковинных амеб (например, у Heleopera)
преобладающим компонентом раковины являются че-
шуйки фагоцитированных ими мелких филозных ра-
ковинных амеб, в основном из рода Euglypha. При
культивировании в условиях, исключающих возмож-
ность питания этими амебами, Heleopera spp. строит
раковину только из органического материала. У не-
которых раковинных амеб (например, Centropyxis
spp.) встречаются все переходы от органических ра-
ковин к агглютинированным. Экзогенными частица-
ми у них часто покрыта только спинная сторона ра-
ковины (рис. 415, Ж).
Эндогенные минеральные частицы (идиосомы)
представляют собой кремнеземные или известковые
образования различной формы. У Nebela они имеют
вид чешуек различной формы (рис. 415, Д, Е). У Les-
quereusia spiralis они палочковидные, слегка изогну-
тые (рис. 416, А, В, вкл.), а у Netzelia представлены
небольшими округлыми частицами (рис. 416, В, О- У
Paraquadrula идиосомы состоят из извести и имеют
472
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
вид пластинок, а у Mississippiella они образованы
ромбоидными кристаллами кальцита.
Раковину из протеинозных альвеол, скрепленных
матриксом, имеют Arcella (рис. 416, 3), Centropyxis,
Microchlamys и некоторые другие роды. Альвеолы —
это полые пентагональные образования (рис. 416, Ж,
И). В качестве основного компонента в их состав
входит вещество белковой природы, близкое к кера-
тину.
Показано, что раковина Arcella содержит ами-
нокислоты: глицин, аланин, лейцин, аргинин, валин,
фенилаланин, цистеин, тирозин и глутаминовую кис-
лоту. В старых раковинах альвеолы обычно заполне-
ны неорганическими соединениями, например соля-
ми железа. Наружная поверхность может быть пок-
рыта дополнительным слоем матрикса.
Псевдоподии и движение
Перемещение и захват различных частиц осуще-
ствляются при помощи псевдоподий. Иногда выделя-
ют 2 их типа: экзолобоподии, состоящие только из
гиалоплазмы (особой их формой являются ретикуло-
( лобоподии — заостренные на концах и разветвляю-
щиеся с образованием анастомозов), и эндолобопо-
дии, в состав которых входит и гиало- и гранулоп-
лазма.
Мелкие и средние раковинные амебы при пе-
редвижении обычно поднимают домик вертикально.
Цитоплазма при таком движении растекается по суб-
страту, образуя вентральный валик, на котором ползет
амеба. Подобное перемещение очень характерно для
Centropyxis, Arcella, мелких диффлюгий (рис. 417).
Амебы, имеющие крупные агглютинированные
домики, очень редко отрывают их от субстрата. Как
правило, при движении такая амеба образует длин-
ные псевдоподии, концы которых прикрепляются к
субстрату. Потом псевдоподии втягиваются, подтас-
кивая раковину (рис. 418).
Скорость движения раковинных амеб различна, но
в целом это малоподвижные организмы. Скорость пе-
ремещения Arcella vulgaris 51—90 мкм/мин, т. е. 3—
5.5 мм/ч; Difflugia crenulata перемещается со ско-
ростью 60 мкм/мйн; Quadrulella symmetrica —
32 мкм/мин.
Рис. 417. Перемещение арцелл.
вв — вентральный валик цитоплазмы. Стрелкой показано направление пе-
ремещения.
Fig. 417. Movement of Arcella species.
вв — ventral «plate». The direction of movement is indicated by an arrow.
Питание и осморегуляция
Большинство раковинных амеб — полифаги. Они
питаются бактериями, водорослями, спорами грибов,
жгутиконосцами, инфузориями или даже коловратка-
ми. В пищеварительных вакуолях почвенных рако-
винных амеб часто обнаруживаются частицы отмер-
ших растений. Неоднократно описывалась способ-
ность раковинных амеб к пробуравливанию стенок
водорослей, вслед за чем в образовавшееся отверстие
клетка просовывает псевдоподии и водоросль «вы-
едается» изнутри. Некоторые виды (Arcella arenaria,
A, polypora) — облигатные фитофаги.
Отмечены случаи питания некоторых раковинных
амеб другими раковинными амебами. Так, например,
Heleopera и Nebela фагоцитируют Euglypha и Trine-
та (класс Filosea) и в дальнейшем встраивают идио-
сомы заглоченных раковин в собственную раковину.
Мелкая Hyalosphenia platystoma прикрепляется к ра-
ковинам крупных раковинных амеб, проделывает
сбоку отверстие и опустошает их.
Интенсивность питания достаточно высока — ин-
тервал между заглатыванием отдельных пищевых
частиц у некоторых видов составляет 4—7 мин. Про-
должительность переваривания пищи составляет
30—60 мин, фаза дефекации длится 2—3 мин. При
дефекации псевдоподии втягиваются, вакуоль с непе-
реваренными остатками подходит к мембране в об-
ласти устья й выбрасывает свое содержимое во внеш-
нюю среду.
Сократительные вакуолй у раковинных амеб ра-
ботают с разной интенсивностью в зависимости от
условий окружающей среды, но обычно интервал
между систолами составляет 20—60 с.
Размножение и жизненные
циклы
Размножение происходит путем бинарного деле-
ния. Детали этого процесса отличаются у представи-
телей различных родов, однако в целом можно выде-
лить 2 наиболее характерных типа морфогенеза до-
черней особи.
У Arcella и других раковинных амеб, имеющих
органическую альвеолярную раковину, при подготов-
ке к делению в цитоплазме накапливаются так назы-
ваемые текагенные гранулы — везикулы, содержа-
щие в упакованном виде альвеолы для построения
дочерней раковины. У Arcella диаметр этих гра-
нул около 500 нм, а высота до 900 нм; их общий
объем может составлять до 7 % объема цитоплазмы
клетки.
У Arcella vulgaris процесс деления начинается с
того, что часть цитоплазмы выпячивается из устья
материнской раковины, образуя текагенную почку
(рис. 419). По размеру она примерно равна материн-
ской особи. Текагенные гранулы перемещаются к
поверхности этой почки и располагаются под ней в
1 слой. На этом этапе клетка секретирует на повер-
хность почки слой гомогенного вещества. Вслед за
этим происходит синхронный выброс содержимого
ТИП RHIZOPODA. КЛАСС LOBOSEA, ПОДКЛАСС TESTACEALOBOSIA
473
Рис. 418. Перемещение крупных диффлюгий.
а—г — последовательные стадии. А — вид сверху, Б — вид сбоку.
Fig. 418. Movement of large Difflugia species.
a—г — successive stages. A — view from above, Б — lateral view.
гранул. Оказавшись на поверхности, альвеолы увели-
чиваются в размерах и сливаются друг с другом. Ви-
димо, первоначальное скрепление альвеол происхо-
дит за счет ранее секретированного вещества.
Затем амеба образует куполообразную псевдоподию,
которая окружает только что образовавшуюся раковину.
Внутреннее пространство этого купола определяет
окончательную форму й размеры дочерней раковины.
Увеличение размеров дочерней раковины происходит за
счет движений цитоплазмы текагенной почки, которая
«расталкивает» ее изнутри. Затем цитоплазматический
купол быстро втягивается. Весь процесс формирования
дочерней раковины занимает около 10 мин.
В результате взаимного сжатия альвеол формиру-
ется гексагональная структура стенки дочерней рако-
вины. В местах, где соприкасаются 3 альвеолы, обра-
зуются микроскопические поры. Системой таких пор
оказывается окружена каждая альвеола.
На последних этапах деления дочерняя и мате-
ринская раковины соединены друг с другом специ-
альным замком из содержимого текагенных гранул.
Он имеет вид муфты, проходящей через устья рако-
вин и «зацепленной» за края устий изнутри. Когда
завершается распределение цитоплазмы между клет-
ками, замок разрушается и происходит окончатель-
ное деление клеток.
В цитоплазму дочерней клетки мигрируют мито-
хондрии и другие органеллы. Происходит митотичес-
кое деление^ядра, и одно из ядер перемещается в до-
чернюю клетку. Разрушается замок, происходит
окончательное разделение цитоплазмы, и дочерняя
клетка переходит к самостоятельному существова-
нию. После достройки раковины цитоплазма отделя-
ется от ее стенок и образуются эпиподии, на которых
подвешено тело взрослой особи.
Подобный ход морфогенеза характерен для всех
раковинных амеб, строящих раковину из органичес-
ких альвеол. Однако формирование цитоплазмати-
ческого купола при морфогенезе подробно прослеже-
но только у Arcella.
У молодых раковин каждая альвеола имеет внут-
реннюю полость, с возрастом просвет альвеол часто
заполняется неорганическим веществом: у арцелл это
железо; у Centropyxis hirsuta —марганец. С накопле-
нием неорганических веществ связана более темная
окраска старых раковин. ..
У раковинных амеб, строящих агглютинирован-
ные раковины, делению предшествует период актив-
ного поглощения ксеносом. В начале деления из
устья материнской раковины выпячивается участок
цитоплазмы (рис. 420). Он быстро увеличивается в
размерах и принимает форму, примерно соответст-
вующую форме будущей раковины. К поверхности
этого выпячивания перемещаются вакуоли с ксено-
сомами и с органическим цементом. Иногда амеба
образует пальцевидные, весьма подвижные псевдопо-
дии, которые могут захватывать частицы из внешней
среды. Происходит экзоцитоз содержимого вакуо-
лей, и ксеносомы скрепляются органическим цемен-
том. Крупные частицы из внешней среды могут про-
сто «прилипать» к стенке дочерней раковины. Ядро
претерпевает деление, часть органелл и одно из ядер
474
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 419. Схема морфогенеза Arcella spp. (Ориг.).
А — клетка, готовая к делению; Б, В — формирование выроста цитоплазмы и перемещение текагенных гранул; Г — образование цитоплазматического ку-
пола (к) и начальное формирование раковины; Д — материнская и дочерняя раковины, скрепленные замком (з) в области устья; Ё — клетки после деле-
ния. тг — текагенные гранулы, я — ядра.
Fig. 419. Morphogenesis of Arcella spp.
A — a cell ready for division; Б, В — cytoplasmic bud formation and movement of thecagenous granules; Г — cytoplasmic dome (к) formation and first stages
of shell formation; Д — daughter cells connected by a «coupling» (з) in the aperture area; E — cells after division, тг — thecagenous granules, я — nuclei.
мигрируют в дочернюю клетку. Вслед за этим про-
исходит окончательное разделение цитоплазмы кле-
ток. Дочерняя клетка еще некоторое время достраи-
вает раковину, фагоцитируя ксеносомы и включая
их в стенку раковины. Изнутри раковина выстилает-
ся слоем органического цемента; за счет него также
формируется органическая каемка вокруг устья и за-
полняются промежутки между частицами. У взрос-
лой особи цитоплазма.отделяется от стенок ракови-
ны и образуются эпиподии.
У амеб, которые включают в состав раковины на-
ряду с ксеносомами еще и идиосомы, морфогенез ра-
ковины проходит по той же схеме, хотя его детали
могут варьировать у разных видов.
В литературе имеются описания процесса, назван-
ного экзувиацйей. У Hyalosphenia papilio иногда из
устья раковины выпячивается цитоплазма. Объем
этого выроста растет, в то время как объем цитоплаз-
мы в раковине уменьшается. На поверхности этого
выпячивания формируется раковина, после чего в нее
перетекает вся цитоплазма клетки. Ядро переходит в
новую раковину без каких-либо изменений; деления
не происходит. Старая раковина остается пустой. Не-
которые современные исследователи подвергают со-
мнению существование процесса экзувиации.
Часто можно встретить описание конгрегации —
объединения 2 раковинных амеб, слияния их цито-
плазматических тел и последующего инцистирования
продукта слияния. Однако ни одному исследователю
не удалось пока достоверно наблюдать эксцистирова-
ние после подобного процесса.
Покоящиеся стадии раковинные амебы образуют
при наступлении неблагоприятных условий (измене-
ние температуры, понижение влажности). При этом
псевдоподии втягиваются, сократительная вакуоль
прекращает свою деятельность. Устье раковины за-
крывается эпифрагмой, представляющей собой про-
зрачную органическую мембрану, или пробкой из
ксеносом или идиосом. Внутри такой раковины часто
находится «предцистная» форма амебы (псевдоцис-
та), которая отличается отсутствием цистной оболоч-
ки. Истинная циста покрыта плотной защитной обо-
лочкой, в состав которой могут входить и ксеносомы
(рис. 421). Как правило, циста сферическая или слег-
ТИП RHIZOPODA, КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
475
Рис. 420. Схема морфогенеза Difflugia spp. (Ориг.).
А — клетка, готовая к делению; Б — образование цитоплазматического выроста; В, Г — начальные этапы формирования дочерней раковины; Д — завер-
шение формирования дочерней раковины, эпиподии еще не образовались; Е — клетки после деления, к — ксеносомы; п — псевдоподии, агглютинирую-
щие экзогенный материал; я — ядро.
Fig. 420. Morphogenesis of Difflugia spp.
A — a cell ready for division; Б — cytoplasmic bud formation; В, Г— first stages of daughter shell formation; Д — completion of daughter shell formation,
epipodia not formed yet; E — cells after division, n pseudopodia capturing exogenous material, к — xenosomes, я — nucleus.
ка уплощенная. Эксцистирование происходит при
попадании цисты в благоприятные условия.
Arcella vulgaris помимо обычных, покоящихся
цист образует особые мейотические цисты, в кото-
рых происходит половой процесс, протекающий по
типу автогамии. Мейотическое деление ядра при
этом прослежено на ультраструктурном уровне. Из
8 образующихся ядер 4 пикнотизируются и перевари-
ваются, а оставшиеся сливаются попарно, восстанав-
ливая исходное двухъядерное состояние клетки. До-
стоверных наблюдений полового процесса у других
раковинных амеб пока нет.
Жизненный цикл Testacealobosia, как правило,
представляет собой серии бинарных делений, кото-
рые прерываются инцистированием, происходящим
при наступлении неблагоприятных условий. У Arcel-
la vulgaris в него включается половой процесс, одна-
ко происходит он регулярно или нет, пока неясно.
По-видимому, у некоторых видов в него могут нере-
гулярно включаться конгрегация и экзувиация, одна-
476
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 421. Циста раковинной амебы (схема).
а — агглютинированная «пробка», закрывающая устье раковины; б — ра-
ковинка; в — агглютинированная дополнительная стенка цисты; г — сек-
ретированная стенка цисты; д — цитоплазма клетки.
Fig. 421. Testate amoeba cyst.
a — agglutinated «plug» closing shell aperture; 6 — shell; в — agglutinated
supplementary cyst wall; г — secreted cyst wall; d — cell cytoplasm.
ко все эти процессы нуждаются в более подробном
описании.
Для одного из видов раковинных амеб — Difflugia
limnetica, характеризующегося чередованием в жиз-
ненном цикле бентической и планктонной фаз, было
описано явление цикломорфоза. Во время бентичес-
кого периода жизни раковина состоит из мелких пес-
чинок и воротничок у нее развит слабо, а в эпилим-
нионе в состав раковины входят панцири диатомей и
имеется развитый воротничок.
Экология и распространение
Условия обитания
Отряд Arcellinida составляют преимущественно
пресноводные организмы, лишь небольшая их часть
обитает в интерстициали морских побережий. Мож-
но утверждать, что раковинные амебы встречаются
почти всюду, где имеется доступная для них вла-
га, — от крупных озер и рек до пространств между
частицами почвы.
В крупных пресноводных водоемах можно го-
ворить об отдельных сообществах раковинных амеб,
характерных для фитали и профундали. В фитали
преобладают раковинные амебы с мелкими раковинами
(Paraquadrula, Cryptodifflugia, Phryganella, мелкие ви-
ды рода Difflugia), формы с раковинами из орга-
нического материала, с идиосомами или с редкими
мелкими инкрустирующими частицами. В этой зоне
часто встречаются формы с уплощенными в дорсо-
вентральном направлении раковинами; относительно
большой процент видов имеет шипы или иглы.
С увеличением глубины и с переходом к обита-
нию на поверхности донных осадков увеличивается
количество видов с крупными раковинами, появляет-
ся большое число форм с крупными инкрустирующи-
ми частицами, преобладают шаровидные или округ-
лые в сечении раковины без шипов и игл. В зоне,
лишенной растительности, население раковинных
амеб донных осадков слабо зависит от глубины водо-
ема. В профундали обычно доминирует род Difflugia,
наряду с которым могут встречаться представители
родов Pontigulasia, Lesquereusia, Heleopera, Phryga-
nella.
В настоящее время доказано, что существует 3
основных сообщества пресноводных раковинных
амеб, которые позволяют судить о характере водо-
ема и о типе его осадков. Эти сообщества характер-
ны для типичных олиготрофных озер с богатыми
кислородом придонными слоями, эвторфных озер
с отложениями гиттии и недостатком кислорода в
придонных слоях и для небольших озер с низким
значением pH воды и с отложениями дью на дне.
Кроме этих 3 встречаются и смешанные сообщества.
Следует отметить, что большая часть пресноводных
раковинных амеб представлена так называемыми ин-
дифферентными видами, не характерными для како-
го-либо определенного типа водоемов.
В болотах раковинные амебы бывают связаны
преимущественно со сфагнумом, причем заселяют в
основном пазухи листочков мха. Имеются торфоби-
онтные виды, часто встречающиеся в болотах, и тор-
фоксенные виды, обнаруживаемые там лишь изредка.
К типичным торфобионтным видам относятся Difflu-
gia bacillifera, Hyalosphenia papilio, H. elegans, Nebe-
la tenella. Для группы болотных раковинных амеб
также показано наличие комплексов видов, прису-
щих определенным типам болот. Широко известна
классификация сообществ раковинных амеб, предло-
женная Харнишем (Harnisch, 1927), который выделя-
ет 5 таких комплексов, встречающихся в различных
типах или участках болот. Основным фактором, оп-
ределяющим характер этих сообществ, является
влажность среды. В пределах одного болота раковин-
ные амебы обычно распределяются в прямой зависи-
мости от влажности мха, причем по мере уменьше-
ния влажности увеличивается количество микростом-
ных форм.
Еще более четко выражена зависимость от влаж-
ности среды сообществ раковинных амеб, обитаю-
щих в лиственных мхах. И в этом случае можно
проследить связь микростомии раковинных амеб с
уменьшением влажности окружающей среды.
Исследование вертикального распределения рако-
винных амеб в пределах моховой подушки показы-
вает, что можно выделить 3 зоны: верхний, зеленый
слой мха, практически не заселяемый раковинными
амебами; слой с отмирающими листочками, в кото-
ром встречается большое количество преимущест-
венно уплощенных плагиостомных видов, среди ко-
торых редки формы, строящие раковины из ксеносом;
слой почвы, пронизанной ризоидами мха, в котором
преобладают крупные сфероидные формы с ракови-
нами, состоящими из большого количества ксено-
сом.
В почве обитает большое количество видов ра-
ковинных амеб, образующих там иногда довольно
плотные популяции. Наиболее густо бывает заселена
подстилка, в которой благодаря наличию системы
чрезвычайно тесных полостей (между отдельными
листьями в опаде) преобладают раковинные амебы с
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
477
уплощенными раковинами. Для фауны раковинных
амеб подстилки характерно преобладание видов с
органическими раковинами либо с раковинами, пос-
троенными из идиосом. При переходе к гумусовому
горизонту полости между отдельными почвенными
частицами становятся значительно больше, вследст-
вие чего в этом слое преобладают раковинные амебы
с относительно крупными, объемистыми, часто сфе-
рическими или близкими к этому типу раковинами.
В фауне данного горизонта преобладают раковинные
амебы с раковинами, построенными Из ксеносом,
причем здесь встречается большое количество спе-
цифических терробионтных видов, для которых ха-
рактерна отчетливо выраженная тенденция к крип-
тостомии. Только для почв характерны многие виды
рода Plagiopyxis, Centropyxis cryptosoma, С, vandeli и
другие виды.
В интерстициальной фауне отмечаются некото-
рые эврибионтные представители отряда Arcellinida.
Единственный характерный только для интерстициа-
ли род отряда — Pomoriella (сем. Hyalospheniidae).
Очень часто в пробах песка, взятых на морских побе-
режьях, встречается Centropyxis constricta, рассматри-
ваемый на этом основании как псаммофильный вид.
Малые размеры раковин амеб и их способность к
цистообразованию обеспечивают возможность их
легкого распространения. Распространение раковин-
ных амеб при помощи ветра было доказано их отсут-
ствием в свежевыпавшем снегу и постоянным присут-
ствием пустых раковин или цист в снегу, лежавшем
длительное время в горах и на ледниках. Возможно
распространение раковинных амеб животными. Ин-
цистированные раковинные амебы неоднократно об-
наруживались в загрязнениях перьевого покрова птиц
и шерстного покрова млекопитающих. Отмечены
случаи переноса раковинных амеб панцирными кле-
щами.
Географическое распространение
Вопрос о географическом распространении рако-
винных амеб не является окончательно решенным в
первую очередь из-за неравномерного изучения их
фауны в разных регионах земного шара. Все основ-
ные данные в этой области были получены до при-
знания самостоятельности групп раковинных амеб с
лобоподиями и с филоподиями, и поэтому отмечен-
ные закономерности в равной степени относятся как
к подклассу Testacealobosia (класс Lobosea), так и к
подклассу Testaceafilosia (класс Filosea).
Из 1630 видов и подвидов раковинных амеб обо-
их подклассов, описанных до 1975 г., 1032 были об-
наружены в Европе, 684 в Африке, 428 в Южной
Америке, 424 в Северной и Центральной Америке,
229 в Азии, 89 в Антарктиде.
Неоднократно выдвигавшаяся гипотеза об абсо-
лютно космополитном распространении раковинных
амеб в последние десятилетия подвергается сомнению.
Можно утверждать, что не все раковинные амебы
являются космополитами, но тенденция к космопо-
литизму выражена у этой группы значительно более
резко, чем у многих других групп животных.
Среди раковинных амеб встречаются убиквисты с
космополитным распространением. Чрезвычайно ши-
роко распространены космополиты, связанные с оп-
ределенными типами среды, так что в большинстве
случаев видовой состав раковинных амеб скорее оп-
ределяется типом биотопа, чем географическим по-
ложением территории. Отмечены виды с рассеянным
распространением; есть основания полагать, что су-
ществуют отдельные виды, ограниченные в своем
распространении определенными географическими
зонами. Неоднократно делались попытки выявления
комплексов видов, характерных для отдельных геог-
рафических зон, однако результаты таких работ не
могут считаться убедительными, до тех пор пока все
эти зоны не будут исследованы хотя бы с примерно
одинаковой степенью тщательности.
Была выдвинута гипотеза, объясняющая законо-
мерности распространения некоторых раковинных
амеб с позиций теории континентального дрейфа.
Ван Ойе (Van Оуе, 1956) удалось выделить комплекс
видов, характерный для участков суши, составлявших
Гондвану (Южная Америка, Новая Зеландия, Австра-
лия, Антарктида). На примере рода Nebela он пока-
зал наличие общих видов в Новой Зеландии и Юж-
ной Америке и различие видового состава этого рода
в фауне Новой Зеландии и Палеарктики. Проведен-
ный недавно анализ распространения бриофильных
видов также позволил выделить 2 крупных комплекса
видов, один из которых по своему распространению
соответствует границам Гондваны, а другой встреча-
ется в Северной Америке, Европе и Азии, что соот-
ветствует Лавразии.
Если рассматривать данные по распространению
отдельных экологических группировок раковинных
амеб, то можно найти определенные закономернос-
ти. Считается, что фауна водных раковинных амеб
состоит в основном из широко распространенных
видов, хотя некоторые авторы допускают наличие
местных фаун. Неоднократно подчеркивалась ориги-
нальность тропической фауны. Фауна раковинных
амеб мхов и примитивных почв считается космопо-
литной. Исключения составляют Nebela cartesi, N.
cockayini и Certesella martiali, отмеченные только в
южном полушарии, и Nebela vas, встречающаяся в
южном полушарии и приэкваториальных зонах се-
верного полушария. В почвах имеются, как правило,
2 фракции: субвоздушная космополитная, близкая к
сообществу раковинных амеб мхов, и космополитная
эдафическая. К этому комплексу в некоторых слу-
чаях добавляются почвенные виды с ограниченным
распространением: либо почвенный тропический
комплекс, характерный для тропической зоны Афри-
ки, Южной Америки, Мексики, Непала, либо поч-
венный комплекс умеренных и холодно-умеренных
зон Канады, высокогорий Мексики и Японии. Фауна
почвенных амеб лесного гумуса Европы состоит
только из широко распространенных видов.
Раковинные амебы часто встречаются в очень
больших количествах и в некоторых биотопах могут
являться доминирующей в количественном отноше-
нии группой. Максимальная их численность в зо-
не фиталй олиготрофных озер может достигать 6, в
почве — 800 млн/м2. Биомасса раковинных амеб в
478
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
почве может, по некоторым данным, доходить до
800 мг/м2, что соответствует 8 кг/га. Тодовая про-
дуктивность раковинных амеб в 40—100 раз выше
их биомассы, что связано с небольшой (3—8 дней)
продолжительностью жизни одного поколения.
Практическое значение
Можно предположить, что организмы, встречаю-
щиеся в таких количествах, как Arcellinida, могут иг-
рать весьма существенную биологическую роль. Рако-
винные амебы, питающиеся бактериями, водорослями
и некоторыми простейшими, сами могут являться до-
бычей некоторых инфузорий, малощетинковых червей,
тихоходок, нематод и мальков рыб. Весьма вероятно,
что почвенные раковинные амебы участвуют в процессе
разложения опада. Неоднократно высказывалось пред-
положение, что эти организмы, отмирая, могут обога-
щать почву азотистыми соединениями.
Выделение фаунистических комплексов раковин-
ных амеб, характерных для определенных условий
обитания, делает возможным использование этой
группы протистов, довольно хорошо сохраняющихся
в субфоссильном состоянии, для историографическо-
го анализа изменений этих условий.
Исследование видового состава раковинных амеб
в колонках торфа в первую очередь позволяет выде-
лять горизонты, соответствующие различной степени
обводнения. Вместе с пыльцевым анализом такой
анализ может быть использован для датировки пери-
одов обводнения и осушения в послеледниковый пе-
риод. Раковинные амебы могут быть использованы и
в палеолимнологическом анализе, так как выявление
определенных комплексов видов в озерных отложе-
ниях дает возможность судить о характере водоема
(олиго-, мезо- или эвтрофный), его глубине и наличии
или отсутствии плавающей или погруженной расти-
тельности в момент образования соответствующего слоя
отложений. В последнее время делаются попытки
использования раковинных амеб в биодиагностике
почв — их видовой состав и соотношение различных
морфологических типов раковин являются доста-
точно чуткими индикаторами физико-химических
свойств исследуемой почвы.
Систематический обзор
Отряд Arcellinida Kent, 1880
Корненожки, имеющие раковины или иные пок-
ровные структуры с 1 устьем, построенные из органи-
ческого или органического и неорганического мате-
риала. Пресноводные.
Сем. Microcoryciidae de Saedeleer,
1934
Цитоплазматическое тело покрыто полужесткой
или жесткой оболочкой, которая обладает некоторой
способностью к изменению формы, особенно в об-
ласти устья. В состав оболочки могут быть включены
инородные частицы. Оболочка часто образует склад-
ки вокруг устья. Одно- и многоядерные формы.
Роды: Amphizonella Greef, 1866; Diplochlamys Greef,
1866; Microcorycia Cockerell, 1911; Parmulina Penard,
1902 (рис. 422); Penardochlamys Deflandre, 1953; Zo-
nomyxa Nusslin, 1884.
Сем. Microchlamyidae Ogden, 1985
Органическая, способная к изгибанию или ригидная
раковина. Цитоплазматическое тело окружено при-
крепляющейся к раковине мембраной с единственным
отверстием. Ядро занимает центральное положение.
Единственный род: Microchlamys Cockerell, 1911
(рис. 423).
Сем. Arcellidae Ehrenberg, 1832
Раковинка плотная, органическая, с выраженной
альвеолярной структурой (иногда альвеолы чрезвы-
чайно мелки), дисковидная или колпачковидная.
Устье центральное, расположено на впяченной
внутрь вентральной поверхности раковины. Одно-,
двух- и многоядерные формы.
Роды: Antarcella Deflandre, 1928; Arcella Ehren-
berg, 1832 (рис. 424); Pyxidicula Ehrenberg, 1834
(рис. 425).
Примечание: Многими авторами отрицается самостоятель-
ность рода Antarcella, отличающегося от рода Arcella одноядер-
ностью.
Сем. Centropyxidae Deflandre, 1953
Раковина либо органическая (альвеолярная струк-
тура при помощи светового микроскопа не выявляет-
ся), без ксеносом, либо агглютинированная, причем
количество ксеносом и их характер различны. Устье
центрально расположенное или эксцентричное, почти
всегда лежит на впячивании вентральной поверхнос-
ти, образующей уплощенную подошву. Раковина в
виде части сферы, дисковидная или билатерально-
симметричная. Ядро 1.
Роды: Centropyxis Stein, 1857 (рис.426—428);
Collaropyxidia Zivcovic, 1975; Cyclopyxis Deflandre,
1929; Ellipsopyxella Bonnet, 1975; Ellipsopyxis Bon-
net, 1965 (рис. 429); Geopyxella Bonnet et Thomas,
1955 (рис. 430); Gillardella Stepanek, 1952; Paracen-
tropyxis Bonnet, 1959 (рис. 431); Planhoogenraadia
Bonnet, 1977; Proplagopyxis Schonbom, 1964; Proto-
plagopyxis Bonnet, 1959; Pseudawerintzewia Bonnet,
1959; Trigonopyxis Penard, 1911 (рис. 432).
Примечание: Леблих и Таппан (Loeblich, Tappan, 1964) вы-
деляют роды Trigonopyxis и Ciclopyxis в самостоятельное сем. Tri-
gonopyxiadae, которое характеризуется органической радиально-
симметричной раковиной с ксеносомами, с уплощенной или вогну-
той вентральной поверхностью без наружной шейки. Приведенных
признаков явно недостаточно для выделения обладающих ими ор-
ганизмов из сем. Centropyxidae.
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
479
Сем. Plagiopyxidae Bonnet,
1979
Раковина агглютинированная, билатерально-сим-
метричная, характеризуется криптостомным типом
строения. Устье часто открывается в камеру, ограни-
ченную козырьком. Ядро 1.
Роды: Bullinularia Penard, 1911 (рис. 433^; Geo-
plagiopyxis Chardez, 1965; Hoogenraadia Gauthier-Li-
evre et Thomas, 1958; Oopyxis Jung, 1942 (рис. 434);
Plagiopyxis Penard, 1910 (рис. 435).
Сем. Lamtopyxidae Bonnet, 1974
Раковина агглютинированная, с вентральной по-
дошвой и внутренней билатеральной симметрией,
которая замаскирована внешней радиальной симмет-
рией. Пропилостомные формы — имеется вестибу-
лярная полость, которая открывается на уровне по-
дошвы окруженным зубцами отверстием, а внутри
заканчивается эллиптическим устьем. Раковина со-
стоит из 2 камер — брюшка и вестибулюма.
Единственный род: Lamtopyxis Bonnet, 1974
(рис. 436).
Сем. Distomatopyxidae Bonnet, 1979
Раковина агглютинированная, с внешней радиаль-
ной симметрией. Имеется вестибулярная полость, ха-
рактеризующаяся билатеральной симметрией. На
уровне вентральной подошвы раковины располагает-
ся дистальное отверстие вестибулярной полости, за-
крытое диафрагмой с имеющимися в ней отверстия-
ми. Собственно устье располагается в верхней части
вестибулярной полости.
Единственный род: Distomatopyxis Bonnet, 1979
(рис. 437).
Сем. Difflugiidae Wallich, 1864
Раковина агглютинированная, с осевой симмет-
рией и терминально расположенным устьем. Устье
округлое, лопастное, окруженное зубцами или эллип-
соидное. Одно- и многоядерные формы.
Роды: Cucurbitella Penard, 1902 (рис. 439); Dif-
flugia Leclerc, 1815 (рис. 438); Lagenodifflugia Medi-
oli et Scott, 1983; Maghrebia Gauthier-Lievre et Tho-
mas, 1960 (рис. 440); Pentagonia Gauthier-Lievre
et Thomas, 1958; Pontigulasia Rhumbler, 1895
(рис. 441); Protocucurbitella Gauthier-Lievre et Tho-
mas, 1958 (рис. 442); Pseudocucurbitella Gauthier-Li-
evre et Thomas, 1958; Pseudopontigulasia van Oye,
1956; Schwabia Jung, 1942; Sexangularia Awerintzew,
1906; Suiadifftugia Green, 1975; Zivcovicia Ogden,
1987.
Примечание: Юнг (Jung, 1942a, 1942b) выделяет из рода
Difflugia роды Globonota, Acipyxis, Planodifflugia, Pycnochila, Lo-
boforamina, Eustoma и Cingodifflugia. Это деление не принимается
в современной литературе.
Сем. Hyalospheniidae Schulze, 1877
(=Nebelidae Taranek, 1882)
Раковина органическая, с идиосомами или с соче-
танием идиосом и ксеносом. Идиосомы не образуют
черепицеобразного покрова. Раковина в большинстве
случаев вытянута по продольной оси, часто сжата с
боков. Устье эллипсоидное или щелевидное, реже
круглое.
Роды: Ampullatoria van Oye, 1949; ApolimiaKor-
ganova, 1987; Awerintzewia Schouteden, 1906; Certe-
sella Loeblich et Tappan, 1961; Geamphorella Bonnet,
1959; Heleopera Leidy, 1879 (рис. 444); Heleoporella
Couteaux, 1978; Hyalosphenia Stein, 1857 (рис. 443);
Jungia van Oye, 1949; Leptochlamys West, 1901; Meta-
heleopera Bartos, 1954; Nebela Leidy, 1875 (рис. 445);
Paranebela Jung, 1942; Pomoriella Golemansky, 1970
(рис. 446); Pseudonebela Gauthier-Lievre, 1954.
Примечание: Юнг (Jung, 1942a, 1942b) выделяет из рода
Nebela роды Leidyella, Argynnia, Deflandria, Penardiella, Porosia,
Apodera, Alocodera, Schaudinnia, Pterygia и Umbonaria. Это деле-
ние не принято в современной литературе.
Сем. Lesquereusiidae Ogden, 1979
Раковина построена из идиосом, иногда из идио-
сом и ксеносом. Идиосомы представлены кремнезем-
ными палочками, пластинками или имеют неправиль-
ную форму. Устье круглое, овальное или удлиненное.
Роды: Lesquereusia Schlumberger, 1845 (рис. 447;
448); Netzelia Ogden, 1979; Quadrulella Cockerell,
1909 (рис. 449).
Сем. Paraquadrulidae Deflandre, 1953
Раковина образована четырехугольными известко-
выми пластинками. Устье овальное или щелевидное.
Одноядерные формы.
Роды: Lamtoquadrula Bonnet, 1979; Paraquadrula
Deflandre, 1932.
Сем. Mississippiellidae Huddleston et
Haman, 1985
Раковина состоит почти целиком из кальцита без
следов кремнезема, покрыта не пластинками, а пра-
вильными ромбическими кристаллами кальцита. Устье
образовано серией овальных отверстий (10—18), об-
разующих дуговидные ряды.
Единственный род: Mississippiella Haman, 1982.
Сем. Cryptodifflugiidae Jung, 1942
Характерно наличие ретикулолобоподий (заост-
ренных или ветвящихся эктоплазматических псевдо-
подий, способных к образованию анастомозов). Ра-
ковина органическая, прозрачная, лишь изредка с
отдельными ксеносомами.
480
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
426
429, Д
437
4J5, Б
Рис. 422—452, Раковины представителей разных родов Testacealobosia. (Ориг.).
422 — Parmulina cyathus: А —- вид сбоку, Б — вид сверху. 423 — Microchlamys patella (по: Penard, 1902). 424 — Arcella gibbosa. 425 — Pyxidicula
operculata. 426 — Centropyxis plagiostoma: A — вид с вентральной стороны; Б — вид сбоку. 427 — Centropyxis sylvatica. 428 — Centropyxis puteus: A — вид
сбоку, Б — вид с вентральной стороны. 429 — Ellipsopyxis lamottei, А — вид с вентральной стороны, Б — вид сбоку. 430 — Geopyxella sylvatica var.
globulosa: A — вид с вентральной стороны, Б — вид сбоку. 431 — Parantropyxis sp., вид сбоку (схематично). 432 — Trigonopyxis arcula. 433 — Bullinularia
indica. 434 — Oopyxis cophostoma. 435 — Plagiopyxls oblonga: A — вид с вентральной стороны, Б — вид сбоку (схематично). 436 — Lamtopyxis callistoma
(по: Bonnet, 1974): А — вид сбоку (схематично); Б — устье снизу. 437 — Distomatopyxis sp. (по: Bonnet, 1970): вид сбоку (схематично). 438 — Difflugia
oblonga. 439 — Cucurbitella mespiliformis. 440 — Maghrebia spatula. 441 — Pontigylasia Compressa. 442 — Protocucurbitella cOroniformis: A — общий вид,
Б — часть устья. 443 — Hyalosphenia papilio (по: Defiandre, 1953). 444 — Heleopera petricola. 445 — Nebela lageniformis (no: Deflandre, 1953). 446 —
ТИП RHIZOPODA, КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA
481
450
451
Рис. 422—452 (продолжение).
Pomoriella valkanovi (по: Golemansky, 1970). 447 — Lesquereusia modesta. 448 — Lesquereusia spiralis.
449 — Quadrule Ila symmetrica. 450 — Cryptodifflugia compressa (no: Penard, 1902). 451 — Cryptodifflugia
oviformis: схема псевдоподиальной сети (no: Hedley, Ogden, 1979). 452 — Phryganella acropodia.
Fig. 422—452. Shells of Testacealobosia from different genera.
422 Parmulina cyathus: A — lateral view, Б — view from above. 423 — Microchlamys patella. 424 — Arcella gibbosa. 425 — Pyxidicula operculata. 426 —
Centropyxis plagiostoma: A — ventral view, Б — lateral view. 427 — Centropyxis sylvatica. 428 — Centropyxis puteus: A — lateral view, Б — ventral view.
429 — EllipSopyxis lamottei: A — ventral view, Б — lateral view. 430 — Geopyxella sylvatica var. globulosa: A — ventral view, Б — lateral view. 431 —
Paracentropyxis sp., lateral view (scheme). 432 — Trigonopyxis arcula. 433 — Bullinularia indica. 434 — Oopyxis cophostoma. 435 — Plagiopyxis oblonga: A —
ventral view, Б — lateral view (scheme). 436 — Lamtopyxis callistoma: A — lateral view (scheme), Б — aperture, view from below. 437 — Distomatopyxis sp.
lateral view (scheme). 438 — Difflugia oblonga. 439 — Cucurbitella mespilliformis. 440 — Maghrebia spatula. 441 — Pontigulasia compressa. 442 — Protocu-
curbitella coroniformis: A — general view, Б — a part of aperture. 443 — Hyalosphenia papilio. 444 — Heleopera petricola. 445 — Nebela lageniformis. 446 —
Pomoriella valkanova. 447 — Lesquereusia modesta. 448 — Lesquereusia spiralis. 449 — Quadrulella symmetrica. 450 — Cryptodifflugia compressa. 451 —
Cryptodifflugia oviformis: a scheme of pseudopodial network. 452 — Phryganella acropodia.
31 Протесты, часть 1
482
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Роды: Cryptodifflugia Penard, 1890 (рис. 450; 451);
Difflugiella Cash, 1904; Petelopella Loeblich et Tappan,
1961; Wailesella Defiandre, 1928.
Сем. Phrygandllidae Jung, 1942
Характерно наличие ретикулолобоподий. Ракови-
на агглютинированная, с большим количеством ксе-
носом. Одноядерные формы.
Единственный род: Phryganella Penard, 1902
(рис. 452).
Отряд Himatismenida Page, 1987
Представители этого отряда существенно отлича-
ются по своей организации от Arcellinida. Многие
авторы (Ogden, 1985; Pussard et al., 1977) относят их
к голым амебам (Gymnamoebia) или рассматривают
как переходную группу между раковинными и голы-
ми амебами. Размножение бесполое, половой про-
цесс неизвестен.
Отряд включает 1 семейство — Cochliopodiidae
(de Saedeleer, 1934) Page, 1987 с 3 родами: Cochlio-
podium, Gocevia и Paragocevia. Морские и пресно-
водные виды.
Сем. Cochliopodiidae
(de Saedeleer, 1934) Page, 1987
Род Cochliopodium (Hertwig
et Lesser, 1874) Bark, 1973
Представители этого рода (рис. 415, 5) с дорсаль-
ной стороны покрыты гибким щитком (тектумом),
состоящим из одного слоя чешуек (см.: Bark, 1973),
который прилегает к цитоплазме клетки (рис. 453).
Эпиподий нет. Тектум не заходит на вентральную
сторону клетки и не имеет дифференцированного
устья.
При перемещении по субстрату амеба образует
псевдоподии, которые часто высовываются из-под пе-
реднего края тектума (см.: Page, 1988). Тело амебы
вместе с тектумом может очень сильно конформиро-
ваться, при этом тектум повторяет очертания цитоп-
лазматического тела клетки. Чешуйки, составляющие
Рис. 453. Организация клетки Cochliopodium sp., вид сбоку.
т — тектум. Стрелкой показано направление перемещения.
Fig. 453. Cell organization of Cochliopodium sp., lateral view.
m — tectum. The direction of movement is indicated by an arrow.
Рис. 454. Флотирующая форма Cochliopodium sp.
m — тектум.
Fig. 454. Rotating form of Cochliopodium sp.
m — tectum.
тектум, синтезируются при участии аппарата Гольд-
жи (Yamaoka et al., 1984) и транспортируются к по-
верхности клетки, где и включаются в состав текту-
ма. Фагоцитоз происходит только на вентральной
стороне тела, свободной от тектума.
Свободно плавающая в воде амеба часто прини-
мает характерную колоколовидную форму (рис. 454).
При инцистировании клетка округляется, цитоплаз-
ма отделяется от тектума и покрывается слоем фиб-
риллярного материала. Таким образом, фибриллярная
стенка цисты снаружи оказывается покрыта чешуй-
ками, ранее составлявшими тектум. В ходе эксцис-
тирования чешуйки синтезируются в аппарате Голь-
джи и новый тектум формируется под стенкой цис-
ты. Клетка покидает цисту через отверстие в ее
стенке; от цисты остается пустая фибриллярная обо-
лочка, покрытая чешуйками, которые составляли
тектум до инцистирования амебы. Фактически тек-
тум формируется заново при каждом цикле инцисти-
рование—эксцистирование.
Ультраструктура клетки типична для амебоидных
организмов; компартментализации цитоплазмы нет.
При делении клетки тектум распределяется поровну
между 2 дочерними особями; в отличие от представи-
телей отряда Arcellinida у Cochliopodium нет никако-
го специфического процесса морфогенеза тектума
дочерней особи.
Род Gocevia Valkanov, 1932
Этот род объединяет амеб, организация которых
(рис. 455) в целом схожа с организацией представи-
телей рода Cochliopodium, но которые вместо текту-
ма, состоящего из чешуек, покрыты сверху толстой
фиброзной оболочкой (кутикулой) (Page, Willumsen,
1980). У некоторых видов кутикула покрыта слоем
посторонних частиц различных формы и размера.
Одноядерные; 1 вид (G. binucleata) двухъядерный.
При локомоции образуют несколько коротких лобо-
подий, которые могут высовываться из-под переднего
края кутикулы.
Ультраструктурные исследования показали на-
личие у G. fonbrunei ЦОМТа в цитоплазме и микрот-
рубочек в составе цитоскелета амебы (Pussard et al.,
ТИП RHIZOPODA. КЛАСС LOBOSEA. ПОДКЛАСС TESTACEALOBOSIA. SUMMARY
483
Рис. 455. Организация клетки Gocevia sp., вид сбоку.
к — кутикула. Стрелкой показано направление перемещения.
Fig. 455. Cell organization of Gocevia sp., lateral view.
к — cuticle. The direction of movement is indicated by an arrow.
1977). Так же как и у Cochliopodium, при делении у
Gocevia кутикула материнской клетки распределяет-
ся поровну между дочерними особями.
Род Paragocevia (Hulsmann, 1974)
Представители рода Paragocevia отличаются тем,
что образуют при локомоции множество (до сотни)
коротких заостренных псевдоподий, направленных
вперед. Кутикула прилегает к поверхностной мемб-
ране клетки и при конформации амебы повторяет
форму цитоплазматического тела клетки. Эпиподий
нет.
Отряд Trichosida Moebius, 1889
Многоядерные амебоидные организмы, имеющие
тонкую (около 1 мкм) гибкую раковину, которая со-
стоит из слоя фиброзного материала. На некоторых
стадиях жизненного цикла раковина покрыта слоем
эндогенных спикул из кальцита. 1 род — Trichospha-
erium Schneider, 1878.
Раковина плотно прилегает к телу амебы и пол-
ностью окружает его; короткие конические гиалино-
вые псевдоподии (дактилоподии) выходят через мно-
гочисленные устья диаметром 4-—7 мкм. Некоторые
наблюдения (Scheehan, Banner, 1973) свидетельству-
ют о том, что устья затянуты тонкой пленкой пред-
положительно полисахаридной природы. Способ
прохода дактилоподий сквозь нее непонятен.
Раковина очень пластичная и меняет форму при
движении организма. Перемещающаяся амеба образу-
ет широкую фронтальную гиалиновую псевдоподию,
к поверхности которой плотно прилегает раковина.
При движении амебы клеточная мембрана и ракови-
на перемещаются по поверхности субстрата, «пере-
матываясь» подобно гусенице трактора (Polne-Fuller,
1987). Дактилоподии не принимают участия в движе-
нии.
Размеры особей варьируют от 10 до 1000 мкм.
Клетка многоядерная, крупные особи могут содер-
жать сотни ядер. Ядра делятся митотически, несин-
хронно. Митоз закрытый. Размножение происходит
делением пополам, крупные особи могут последо-
вательно претерпевать серию бинарных делений без
достижения исходного размера. У одного вида описано
множественное деление (Polne-Fuller, 1987). При этом
материнская клетка диаметром около 60 мкм делит-
ся на 10, а клетка диаметром 1200 мкм — более
чем на 260 дочерних особей.
Жизненный цикл трихосфериума до сих пор окон-
чательно не выяснен. На одних стадиях цикла ра-
ковина покрыта спикулами, а на других спикулы
отсутствуют. Бесспикульная форма трихосфериума
напоминает голых амеб и была даже описана как
самостоятельный вид — Pointifex maximus Schaeffer,
1926 (подробнее см.: Page, 1983). Шаудинн (Schau-
dinn, 1899) предположил, что раковину покрытую
спикулами, имеет шизонт, а гладкую — гамонт. При
дальнейшем изучении этих организмов не удалось
достоверно наблюдать ни шизогонию, ни гаметого-
нию (Angell, 1976; Schuster, 1976), однако в литера-
туре продолжают употреблять оба эти термина.
Морские организмы; обычны в прибрежных водах
и эстуариях. Питаются преимущественно водорос-
лями.
THE PHYLUM RHIZOPODA SIEBOLD, 1845
THE CLASS LOBOSEA CARPENTER, 1861
THE SUBCLASS TESTACEALOBOSIA DE SAEDELEER, 1934
SUMMARY
This subclass comprises lobose amoebae, which possess «shell»
(or «test») — a rigid structure outside the cell — and do not exhibit
nuclear heteromorphism. Three orders are considered within the sub-
class — Arcellinida, Himatismenida and Trichosida. The validity of
placing the latter two orders into this subcluss is discussed.
The chapter includes summarised data and illustrations on shell
morphology and structure, cell organisation, movement, division and
encystment, habitats and distribution of representatives of this group.
Detailed systematic account includes diagnoses and generic composition of
families. Illustrations of representatives of most genera are presented. Refer-
ence list includes 58 essential monographs and papers on this group.
The shell of the representatives of the order Arcellinidae («typi-
cal» lobose testate amoebae) has one opening (aperture) through
which pseudopodia emerge. Shell and aperture shape varies greatly.
The shell is usually single-chambered though structures dividing it
(«bridge», «diaphragm») are known for representatives of several
genera. The shell may bear spines or protuberances. Morphological
classification of shells is presented.
The shell may consist either of homogeneous layer(s) of mineral
or organic material or of particles glued together with organic matrix
(agglutinated shells). The latter case occurs much more often. The
origin of particles may be different: they may be either secreted within
the cell (idiosomes and proteinaceous alveioli) or be phagocyted from
the environment (xenosomes). Shell composition is usually charac-
teristic of species and genera. At the same time combination of idio-
somes and xenosomes, as well as of xenosomes and proteinaceous
alveoli is possible in one and the same shell.
Idiosomes are usually siliceous, less often calcareous. Proteina-
ceous alveoili are organic. A great variety of particles may be used as
xenosomes: sand granules, diatom frustles, idiosomes of engulfed tes-
tate amoebae. The degree of selectivity in choosing xenosomes for
shell formation is yet obscure.
The cell is usually attached to the inside of the shell wall by
means of epipodia. It has thin undifferentiated glycocalyx. The nucle-
us is vesicular, rarely granular. There is usually one nucleus in the
cell, but two- and multinuclear species are also known. Mitochondria
484
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
have tubular cristae. Numerous dictyosomes of the Golgi complex are
characteristic of many species. They play an important role in secre-
ting material for the building of the shell. Rough EPR is also well-de-
veloped. A modification of rough EPR — silicalemma — has been
described for testate amoebae, secreting siliceous particles. Presence
of endobiotic algae is typical of many species.
Cytoplasm compartmentalisation is charactristic of testate amoe-
bae. It is especially well-pronounced in species with large ovoid or
pyriform shells. Aboral region of the cell usually contains nucleus
surrounded by rough EPR, dictyosomes and sometimes contractile va-
cuole. Mitochondria, food vacuoles,, cement vesicles, ^vacuoles with
particles for shell formation are all located in the part of the cell
adjoining the aperture. Formation of pseudopodia starts slightly inside
the cell, the cytoplasm in the aperture region appears foamy.
Division is by binary fission followed by morphogenesis of the
daughter shell. The main stages of division process are described for
amoebae with agglutinate shells with xenosomes and with proteina-
ceous alveoli. General pattern of the process is as follows: the amoeba
stops feeding, its cytoplasm volume increases, a part of cytoplasm is
protruded from the aperture. Into it cement vesicles and vaculoles with
particles for shell formation are transported and then arranged in a
layer below the membrane of the cytoplasmic bud. The contents of
cement vesicles and vaculoles with particles are then excretred onto
the surface of the bud and the particles are «glued» together with the
cement. The nucleus divides, the daughter nucleus and some organel-
les are also transported to the future daughter cell. Cell division takes
place and epipodia are formed in the daughter cell. Some changes in
the construction of the newly formed shell may take place after the
division, but soon it hardens and further changes become impossible.
Classification within the group is traditionally performed almost
exclusively on the basis of shell morphology. However, intraspecific
variability of shell shape and material is still poorly studied. There are
reasons to think that it may be considerable, but this knowledge is yet
to be applied in systematics.
There are two main ways of movement of lobose testate amoebae.
Amoebae with small or medium shells often rise the shell, vertically
when moving on substrate. Cytoplasm outside the shell forms a kind
of ventral plate. Amoebae with large agglutinate shells move without
rising the shell by means of long pseudopodia. Their ends attach to the
substrate and then pseudopodia contract, «dragging» the cell ahead.
The majority of testate amoebae is polyphagous. They feed on
bacteria, algae, fungi spores, ciliates, etc. Penetration of algae walls
and the shells of other testate amoebae has been observed in some
species. Obligatory feeding on other testate amoebae and subsequent
use of their shell plates for the construction of the daughter shell has
been described.
Lobose testate amoebae are mainly freshwater organisms. They
inhabit soils and mosses, lakes and streams. In the lakes they dwell in
plankton, benthos and periphyton of macrophytes. Some main trends
of their distibution in these habitats are presented. Some species can
live in brackish water, some were found in interstitial marine sands.
Data on geographical distribution of testate amoebae are presen-
ted. They are shown to be mainly cosmopolitan, though existence of
specific local faunas has been described.
Practical role of testate amoebae is discussed. It is shown that
testate amoebae may serve as paleolimnological and soil diagnosis
indicators.
Representatives of the order Himatismenida differ from Arcellini-
da essentially. This order comprises one family only — Cochliopodi-
dae — including three genera. Cochliopodium species possess the
«tectum» — a flexible covering consisting of scales — which is not
separated from the cell. Tectum covers «dorsal» side only and has no
differentiated aperture. Substrate-moving amoeba forms pseudopodia
emerging from the «front» edge of the tectum. Cell body together with
tectum can change its shape considerably. Scales of tectum are syn-
thetised in the dictyosomes of Golgi complex. Rotating amoeba often
acquires bell-like shape. Division is by binary fission; scales of the
tectum are divided between parental and daughter amoebae without
any particular «morphogenesis». The covering of representatives of
two other genera is called «cuticle». It is does not consist of scales but
is fibrous. It is also flexible and is equally divided between the divi-
ding cells.
Flexible nature of the covering and the lack of morphogenesis
suggest .that the order Himatismenida should be excluded from Testa-
cealobosia.
Representatives of the order Trichosida (one genus only) possess
thin flexible shell with numerous apertures. On certain stages of the
life cycle the shell can be covered by calcareous spicules. Details of
biology and life cycle are poorly known. For ascertainig the systema-
tic position of this order further study is needed.
ЛИТЕРАТУРА25
Аверинцев С. В. Rhizopoda пресных вод. СПб., 1906.
Гельцер Ю.Г., Корганова Г. А., Алексеев Д. А. Почвенные
раковинные амебы и методы их изучения. М., 1985а.
Гельцер Ю. Г., Корганова Г. А., Алексеев Д. А. Практическое
руководство по идентификации почвенных тестаций. М., 19856.
Angell R. W. Observations on Trichosphaerium platyxyrum sp.
n. // J. Protozool. 1976. Vol. 23. P. 357—364.
Bark A. W. A study of the genus Cochliopodium Hertwig et Les-
ser, 1874 // Protistologica. 1973 // T. 9. P. 119—138.
Bartos E. Korenonozce radu Testacea. Bratislava, 1954.
Bonnet L. Le peuplement thecamoebien des sols // Rev. Ecol.
Biol. 1964. T. 1. P. 123—408.
Bonnet L. Les Distomatopyxidae fam. nov. et al structure diplo-
some chez les Thecamoebiens (Rhizopoda, Testacea) // C. R. Acad.
Sci. 1970. T. 217. P. 1189—1191.
Bonnet L. Les Lamtopyxidae fam. nov. et al structure propylos-
tome chez les Thecamoebiens (Rhizopoda, Testacea) // C. R. Acad.
Sci. Ser. D. 1974. T. 278. P. 2935—2937.
Cash J., Hopkinson J. The British freshwater Rhizopoda and He-
liozoa. Vol. 2. London, 1909.
Chardez D. Ecologie generale des Thecamoebiens (Rhizopoda,
Testacea) // Bull. Inst. Agron. et Stat. Rech. 1965. T. 3. P. 307—341.
Chardez D. Histoire naturelie des Protozoaires Thecamoebiens //
Naturalistes Belg. 1967. T. 48. P. 484—576.
Decloitre L. Le genre Euglypha Dujardin // Arch. Protistenk.
1962. Bd 106. S. 51—100.
Decloitre L. Le genre Euglypha (Complements a jour du 31 de-
cembre 1974, de la Monographie du genre parue en 1962) // Arch.
Protistenk. 1976. Bd 118. S. 18—33.
25 Цитируемая и рекомендуемая литература к разделу «Тип
Rhizopoda. класс Lobosea. Подкласс Testacealobosia».
Decloitre L. Le genre Cyclopyxis (Complements a jour du 31
decembre 1974, de la Monographie du genre parue en 1929) // Arch.
Protistenk. 1977a. Bd 119. S. 31—53.
Decloitre L. Le genre Nebela (Complements a jour du 31 decem-
bre 1974, de la Monographie du genre parue en 1936) // Arch. Protis-
tenk. 1977b. Bd 119. S. 325—352.
Decloitre L. Le genre Centropyxis (I complements a jour du 31
decembre 1974, de la Monographie du genre parue en 1929) // Arch.
Protistenk. 1978. Bd 120. S. 63—85.
Decloitre L. Le genre Centropyxis (II complements a jour du 31
decembre 1974, de la Monographie du genre parue en 1929) // Arch.
Protistenk. 1979. Bd 121. S. 162—192.
Deflandre G. Le genre Arcella Ehrenberg // Arch. Protistenk.
1928. Bd 64. S. 152—282.
Deflandre G. Le genre Centropyxis Stein // Arch. Protistenk.
1929. Bd 67. S. 322—375.
Deflandre G. Etude morphologique sur le genre Nebela Leidy
(Rhizopoda, Testacea) // Ann. Protistol. 1936. T. 5. P. 201—322.
Deflandre G. Ordre des Thecamoebiens H Traite de Zoologie.
1953. T. 1, fasc. 2. P. 97—148.
Gauthier-Lievre L., Thomas R. Les genres Difflugia, Pentago-
nia, Maghrebia et Hopgenraadia (Rhizopodes testaces) en Afrique //
Arch. Protistenk. 1958. Bd 103. S. 241—370.
Golemansky V. Rhizopodes nouveaux du psammon littoral de la
men Noir // Protistologica. 1970. Vol. 6. P. 365—371.
Golemansky V. Psammonobiotidae fam. noy. Une nouvelle famil-
le de Thecamoebiens du psammon supralittoral des mers П Acta Pro-
tozool. 1974. T. 13. P 137—142.
Golemansky V. Thecamoebiens psammoriobiontes du supralitto-
ral vietnamen de la Mer Chinoise et description de Cryptodifflugia
brevicolla sp. n. (Rhizopoda: Arcellinida) // Acta Protozool. 1979.
T. 18. P. 285—282.
ТИП RHIZOPODA. КЛАСС HETEROLOBOSEA
485
Golemansky V. Description de trois thecamoebiens (Protozoa:
Rhizopodea) nouveaux des eaux souterraines littorales des mers П
Acta protozool. 1981. T. 20. P. 115—119.
Golemansky V. Thecamoebiens interstitielies: etat actuelle de nos
connaissances sur leur taxonomie, ecologie et repartition geographi-
que//Ann. Stat. Biol. Besse-en-Chandesse. 1983. T. 17. P. 335—351.
Grebecki A. Dynamix of the contractile system in the pseudopo-
dial tips of normally locomoting amoebae, demonstrated in vivo by
video-enhancement // Protoplasms. 1990. Vol. 154. P. 98—111.
Grospietsch T. Wechseltierchen (Rhizopoda). Stuttgart, 1958.
Grospietsch T. Monographische Studie der Gattung Hyalosphae-
nia Stein // Hydrobiologie. 1965. Bd 26. S. 211—241.
Harnisch O. Einige Daten nur rezenten und fossilen Testaceen-
Rhizopodenfauna der Sphagnen // Arch. Hydrobiol. 1927. Bd 18.
S. 345—360.
Hedley R. H., Ogden C. G. Biology and fine structure of Euglyp
ha rotunda (Testacea, Protozoa) // Bull. Brit. Mus. Nat. Hist. (Zool.).
1973. Vol. 25. P. 121—137.
Hedley R. H., Ogden C. G. Reticulopodia in testate amoebae
(Rhizopoda, Protozoa) // Bull. Brit. Mus. Nat. Hist. (Zool.). 1979.
Vol. 37. P. 1—6.
Hedley R. H., Ogden C. G.Kraft J. I. Observation on clonal
cultures of Euglypha acantophora and E. strigosa (Testacea: Proto-
za) // Bull. Brit. Mus. Nat. Hist. (ZooE). 1974. Vol. 27. P. 103, 111.
Jung W. Sudchilienische Thekamoben // Arch. Protistenk. 1942a.
Bd 95. S. 253—356.
Jung W.-Illustrierte Thekamoeben-Bestimmungstabellen. I. Die
Systematik der Nebelinen // Arch. Protistenk. 1942b. Bd 95. S. 357—
390.
Leidy J. Freshwater rhizopods of North America. Washington,
1879.
Loeblich A. R., Tappan H. Suprageneric classification of the Rhi-
zopodea II J. Paleontology. 1961. T. 35. P. 245—330.
Loeblich A. R., Jr., Tappan H. Protista. 2. Sarcodina, chiefly
«Thecamoebians» and Foraminiferida // Treatise on Invertebrate Pale-
ontology, Pt., C / Ed. R. C. Moore. Lawrence, Kansas: Geol. Soc.
Amer.; Univ. Kansas Press, 1964.
Mignot J. P., Raikov I. B. New ultrastructural data on the morp-
hogenesis of the test in the testaceen Arcella vulgaris // Eur. J. Protis-
tol. 1990. T. 26. P. 132—141.
Mignot J.P., Raikov LB.Evidence for meiosis in the testate
amoeba Arcella // J. Protozool. 1992. T. 39. P. 287—289
Netzel H. Die Gehausebildung bei der Thekamoben-Gattung
Arcella (Rhizopoda, Testacea) // Cytobiologie. 1971. Bd 3. S. 89—
92.
Netzel H. Die Bildung der Gehausewand bei der Thekamobe Cen-
tropyxis discoides (Rhizopoda, Testacea) // Ztschr. Zellforsch. 1972a.
Bd 135. S. 45—54.
Netzel H. Die Schalenbildung bei Difflugia oviformis (Rhizopo-
da, Testacea) // Ztschr. Zellforsch. 1972b. Bd 135. S. 55*7-61.
Netzel H. Morphogenese des Gehauses von Euglypha rotunda
(Rhizopoda, Testacea) II Ztschr. Zellforsch. 1972c. Bd 135. S. 63—
69.
Ogden C. G. The flexible shell of the freshwater amoeba Microc-
hlamys patella (Claparede et Lachmann, 1859) (Rhizopoda: Arcellini-
da) // Protistologica. 1985. T. 21. P. 141—152.
Ogden C. G., Hedley R. H. An atlas of freshwater testate amoe-
bae. S. 1: Brit. Mus. (Nat. Hist.); Oxford Univ. Press, 1980.
Page F. C. Marine gymnamoebae / Inst. Terr. Ecol. 1983.
Page F. C. The classification of «naked» amoebae (Phylum Rhi-
zopoda) // Arch. Protistenk. 1987. Bd 133. S. 199—217.
Page F. C. A new key to freshwater and soil Gymnamoebia. Am-
bleside, 1988.
Page F. C., Willumsen N. B. S. Some observations on Gocevia
placopus (Huelsmann, 1974), an amoeba with a flexible test, and on
Gocevia-like organisms from Denmark, with comments on the gene-
ra Gocevia and Hyalodiscus // J. Nat. Hist. 1980. Vol. 14. P. 413—
431.
Penard E. Faune rhizopodique du bassin du Leman. Geneve,
1902.
Polne-Fuller M. A multinucleate marine amoeba which digests
seaweeds II J. Protozool. 1987. Vol. 34. P. 159—165.
Pussard M., Senaude J., Pous R. Observations ultrastructurales
sur Gocevia vonbrunnei Pus sard, 1965 (Protozoa, Rhizopoda) // Pro-
tistologica. 1977. T. 13. P. 265—285.
Schaudinn F. Untersuchungen fiber den Generations-wechsel von
Trichosphaerium sieboldi Schn И Abh. Preuss. Akad. Wiss. Phys.-
Math., Berlin (Suppl.). 1899. P. 1—93.
Scheehan R., Banner F. T. Trichosphaerium — an extraordinary
testae rhizopod from coastal waters // Est. Coast, mar. Sci. 1973.
Vol. 1. P. 245—260.
Schonborn W. Studien fiber die Gattung Difflugiella Cash (Rhi-
zopoda, Testacea) // Limnologica. 1965. T. 3. P. 315—328.
Schonborn W. Beschalte Amoben (Testacea). Wittenberg, 1966a.
Schonborn W. Testaceen als Bioindikatoren im System der See-
typen. Untersuchungen in Masurischen Seen und im Suwalki-Gebeit
(Polen) II Limnologica. 1966b. T. 4. P. 1—11.
Schuster F. L. Fine structure of the schizont stage of the testate
marine amoeba, Trichosphaerium sp. // J. Protozool. 1976. Vol. 23.
P. 86—93.
Sudzuki M. Marine Testacea of Japan // Sesoko Mar. Sci. Lab.,
Tech. Rep. 1979. Vol. 6. P. 51—61.
Thomas R. Le genre Plagiopyxis // Hydrobiologie. 1958. Bd 10.
S. 198—214.
Van Oye P. On the thecamoebian fauna of New Zealand with
description of four species and biogeographical discussion // Hydrobi-
ologie. 1956. Bd 8. S. 16—37.
Yamaoka I., Kawamura N., Mizuno M., Nagatini Y. Scale for-
mation in an amoeba, Cochliopodium sp. // J. Protozool. 1984.
Vol. 31. P. 267—272 .
Класс Heterolobosea Page et Blanton, 1985
В состав класса входят амебы, для которых характерны так
называемое эруптивное движение цитоплазмы при локомоции и
уплощенные, обычно дискоидные кристы митохондрий. Многие
виды способны к жгутиковой трансформации (Page, Blanton,
1985; Page, 1987а). Пресноводные, морские и почвенные организ-
мы. Распространены всесветно. Класс несомненно сборный, весь-
ма гетерогенный в систематическом отношении.
Краткая история изучения
В состав этого таксона, выделенного в 1985 г.
Пэйджем и Блантоном (Page, Blanton, 1985), вошли
организмы, длительное время считавшиеся принадле-
жащими к разным классам протистов. Так, шизо-
пирениды входили в качестве отряда Schizopirenida
Singh, 1952 в состав класса Lobosea Carpenter, 1861,
а акразиевые имели статус самостоятельного класса
Acrasea Schroeter, 1886 (Levine et al., 1980). Правда,
предположение об их возможной близости высказы-
валось и ранее (Page, 1978). Первоначально Пэйдж
и Блантон включили в свой новый класс только сем.
Acrasidae из числа бывших представителей класса
Acrasea, однако позднее Пэйдж (Page, 1987а) счел
необходимым включить в него также сем. Guttuli-
nopsidae. Кроме того, в состав класса вошли некото-
рые амебоидные организмы, обладающие уплощен-
ными кристами митохондрий, которые ранее имели
неясный систематический статус. Собственно исто-
рия изучения этого таксона в его нынешнем составе
началась более века назад — в 1873 г., когда Цен-
ковский в России открыл первую акразиду Guttulina
rosea (в настоящее время ее относят к роду Pochei-
па). Типовой вид рода Acrasis — A. granulata был
описан Ван Тиегемом в 1880 г. во Франции и с тех
пор не был ни разу реизолирован. Типовой вид рода
Guttulinopsis — G. vulgaris был открыт Олайвом в
1901 г. Эти и родственные им организмы из-за спо-
собности образовывать плодовые тела длительное
486
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
время рассматривали в составе Mycetozoa независи-
мо от того, какой систематический ранг был присво-
ен этому таксону в той или иной классификации
низших эукариот (Olive, 1975; Levine et aL, 1980).
Что же касается шизопиренид, которые традицион-
но рассматривались в числе голых лобозных амеб
(Page, 1976), то 2 основных рода — Vahlkampfia
(под названием Schizopyrenus) и Naegleria также из-
вестны с начала нашего века.
Классификация
Отряд Schizopyrenida Singh, 1952
Сем. Vahlkampfiidae (Jollos, 1917) Page et Blanton, 1985
Сем. Gruberellidae Page et Blanton, 1985
Отряд Acrasida (Schroeter, 1886) Page et Blanton, 1985
Сем. Acrasidae (Van Tieghem, 1880) Olive, 1970
Сем. Guttulinopsidae Olive, 1970
формации, показали, что у амебоидных трофозоитов
отсутствуют не только центриоли (кинетосомы) и
какие-либо ЦОМТы, но и вообще цитоплазматиче-
ские микротрубочки. Иными словами, кинетосомы,
формирующие жгутики, и корешковый микротру-
бочковый аппарат возникают у этих организмов de
novo в процессе трансформации, причем очень быс-
тро (Schuster, 1963; Walsh, 1984; Fulton, 1993).
Большинство видов в составе класса (за исклю-
чением родов Gruberella и Euhyperamoeba) — одно-
ядерные организмы. Ядра, как правило, везикулярно-
го типа. Клеточный покров всех представителей
класса представлен тонким аморфным гликокалик-
сом. Кристы митохондрий уплощенные, дискоидной
формы. Диктиосом аппарата Гольджи, как правило,
нет; вместо них имеются скопления мелких, морфо-
логически недифференцированных везикул, принад-
лежность которых к аппарату Гольджи может быть
выявлена лишь на цитохимическом уровне.
Морфология и движение
Биология и размножение
Амебоидные трофозоиты большинства предста-
вителей класса Heterolobosea организованы по одно-
му плану и представляют собой мелких лимаксных
амеб, демонстрирующих при локомоции четко выра-
женное эруптивное движение (рис. 456). Направлен-
ный вперед ток цитоплазмы неравномерный, проис-
ходит отдельными толчками. Каждый такой толчок
приводит к образованию новой гиалиновой полусфе-
ры (отчетливого выступа) в передней части тела аме-
бы — таким образом организм продвигается вперед.
Аналогично происходит и образование псевдоподий •
при изменении направления движения простейшего
(рис. 459). Исключение составляют представители
семейств Euhyperamoebidae и Gruberellidae, локомо-
ция которых имеет промежуточный характер между
таковой у Lobosea (со свойственным им равномер-
ным течением цитоплазмы при локомоции) и Hete-
rolobosea. Stachyamoeba и Rosculus в норме имеют
уплощенную локомоторную форму (рис. 457) и лишь
при очень быстром перемещении могут принимать
форму лимаксного типа.
В составе класса имеются виды, способные к
трансформации в жгутиконосцев. Организация жгу-
тиконосцев варьирует — они могут иметь или не
иметь цитостом (т. е. в ряде случаев жгутиконосец
не способен питаться); количество жгутиков (рис. 458),
как и строение жгутикового аппарата, варьирует
у представителей разных родов (Page, 1988, 1991).
Ультраструктурные и иммуноцитохимические иссле-
дования ряда видов, способных к жгутиковой транс-
Жизненный цикл Heterolobosea включает в себя
стадии трофозоита и цисты покоя. Трофозоиты мно-
гих видов способны трансформироваться в жгутико-
носцев, и, таким образом, в их жизненном цикле по-
является третья стадия — жгутиковая (рис. 460). Как
инцистирование, так и жгутиковая трансформация
происходят нерегулярно, и причины, их вызываю-
щие, еще во многом не изучены.
Организация жгутиконосцев варьирует у разных
видов, различен и их удельный вес в жизненном
цикле протистов (Fenchel, Patterson, 1986; Page, 1988,
1991; Broers et al., 1990). У некоторых видов жгути-
ковая трансформация происходит лишь изредка, при
этом жгутиконосец не способен питаться и делиться.
В других случаях жгутиконосец имеет цитостом и
питается так же, как и амебоидный трофозоит; де-
ление может осуществляться как на амебоидной, так
и на жгутиковой стадиях цикла. И наконец, у ря-
да видов жгутиковая стадия является доминирую-
щей. Жгутиконосец питается и делится, а изредка
образующаяся амебоидная стадия к делению не спо-
собна.
Циста покоя у Heterolobosea, как правило, имеет
однослойную стенку; у некоторых видов имеются
цистные поры — специализированные участки стен-
ки цисты, прорывая которые, амеба при эксцистиро-
вании выходит наружу.
Весьма своеобразно протекает инцистирование
у многих представителей отряда Acrasida (Olive,
1975; Page, 1988). Непосредственно перед инцисти-
Рис. 456. Эруптивное движение лимаксной амебы. Схема.
Fig. 456. Eruptive movement of a limax amoeba. Scheme.
ТИП RHIZOPODA. КЛАСС HETEROLOBOSEA
487
Рис. 457. Stachyamoeba lipophora (A), Rosculus ithacus (Б).
Fig. 457. Stachyamoeba lipophora (A), Rosculus ithacus (Б).
Рис. 458. Разнообразие жгутиковых стадий Heterolobosea.
Fig. 458. Diversity of flagellate stages in Heterolobosea.
Рис. 459. Образование эруптивных псевдоподий при изменении
направления движения (показано стрелками) клетки.
Fig. 459. Eruptive pseudopodia formation in the course of changing
the direction of cell movement (indicated by arrows).
рованием амебы объединяются в крупные агрега-
ты, после чего происходит образование цист (или
спор, как их обычно называют в данном случае).
Эти агрегаты, которые часто имеют весьма слож-
ную, древовидную форму, называют плодовыми те-
лами, или сорокарпами, по аналогии с миксомице-
тами.
Рис. 460. Жизненные циклы Heterolobosea.
А — без жгутиковой стадии, Б — со жгутиковой стадией. Пояснения в
тексте.
Fig. 460. Life cycles of Heterolobosea.
A — flagellate stage absent, Б — flagellate stage present. Explanations in the
text.
Следует отметить также своеобразие биологии
Euhyperamoeba fallax — единственного достоверно
описанного представителя сем. Euhyperamoebidae.
Эти амебы способны к попарным и множественным
агамным слияниям. «Продукты слияния» через неко-
торое время снова распадаются на индивидуальные
особи (Серавин, Гудков, 1987).
488
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Питание
Все представители класса питаются посредством
эндоцитоза. Основной пищей являются бактерии, но
амебы могут фагоцитировать и другие подходящие
по размеру пищевые частицы. Naegleria fowleri об-
разует для захвата пищевых частиц специализиро-
ванные трубчатые псевдоподии, которые ряд авторов
называют амебостомами (см.: John et al., 1985). Эти
псевдоподии не имеют постоянной локализации и
могут образовываться в любом месте клетки, не яв-
ляясь, таким образом, постоянными клеточными
структурами. Отдельно следует упомянуть питание
Euhyperamoeba fallax — амебы, основной пищей ко-
торой служат нитчатые синезеленые водоросли. Клет-
ка фагоцитирует нить водоросли с одного конца,
сматывая ее внутри себя в клубок. Для фагоцитиро-
вания больших агрегатов водорослей отдельные аме-
бы могут объединяться во временные плазмодии.
После захвата и переваривания пищи такой плазмо-
дий вновь распадается на отдельные особи (Серавин,
Гудков, 1987).
Экология
и распространение
Представители класса Heterolobosea распростра-
нены всесветно и населяют практически все экото-
пы — от тропических озер и морей до почв и вод
Антарктиды. Подавляющее большинство видов обли-
гатно свободноживущие, однако есть и опасные фа-
культативные паразиты человека и млекопитающих,
например Naegleria fowleri и N. australiensis, вызы-
вающие у человека амебный менингоэнцефалит. Эти
2 пресноводных вида очень широко распространены
в природе, однако в норме они не патогенны. Лишь
при повышении температуры воды до 30 °C и выше
они переходят в патогенную форму, способную про-
никать в организм человека через слизистые оболоч-
ки носоглотки. Эти виды активно изучаются, осо-
бенно актуальными их исследования стали после
того, как было выяснено, что N. fowleri способна
достигать очень высокой численности в подогретой
воде плавательных бассейнов и охладительных пру-
дах электростанций.
Heterolobosea играют важную роль в потоках ве-
щества и энергии в пресноводных, почвенных и мор-
ских экосистемах. Их численность, достигающая в
лесных почвах 6100—10000 экз. на 1 г по^вы, срав-
нима с численностью всех других групп протистов,
а иногда и превосходит ее. Биогеография Heterolo-
bosea изучена крайне слабо и, по-видимому, подчи-
няется тем же закономерностям, что имеют место и
в случае Lobosea. Макропространственная (в пред-
елах крупных территорий) гетерогенность практи-
чески незаметна, а микропространственная (донные
грунты озера, почва небольшого луга) может быть
очень велика.
Систематический обзор
Отряд Schizopyrenida Singh, 1952
Представители отряда не образуют плодовых тел.
Сем. Vahlkampfiidae (Jollos, 1917)
Page et Blanton, 1985
Семейство включает моноподиальных амеб с от-
четливо выраженным эруптивным движением. Яд-
рышки сохраняются в процессе митоза (закрытый
ортомитоз или, как его называли раньше, — промитоз),
формируя так называемые полярные массы. Факти-
чески все виды, не способные к жгутиковой транс-
формации, объединены в род Vahlkampfia, тогда как
остальные входят в состав других родов, представи-
тели которых способны формировать жгутиконосцев
различной организации. Пресноводные и морские
организмы.
Роды: Adelphamoeba Napolitano, Wall et Canz,
1970; Heteramoeba Droop, 1962; Naegleria (Alexeieff,
1912) Calkins, 1913; Paratetramitus Darbyshire, Page
et Goodfellow, 1976; Pemina El Kadiri, Joyon et Pus-
sard, 1992; Singhamoeba (Singh et Hanumiah, 1979)
Sawyer, Nerad et Munson, 1992; Tetramastigamoeba
Singh et Hanumaian, 1977; Tetramitus Perty, 1852;
Vahlkampfia Chatton et Lalung-Bonnaire, 1912; Willa-
ertia De Jonckheere, Dive, Pussard et Vickerman,
1984.
Сем. Gruberellidae Page et Blanton,
1985
В состав этого явно гетерогенного семейства вхо-
дят 2 монотипических рода амеб — Stachyamoeba
Page, 1975 и Gruberella Page, 1983, которых объеди-
няет лишь формальное сходство в картине деления
ядра (закрытый ортомитоз с дезинтеграцией ядрыш-
ка). Жгутиковые стадии отсутствуют. Stachyamoeba
lipophora при локомоции обычно уплощенная, вее-
ровидная и лишь при очень быстром перемещении
способна принимать моноподиальную форму тела,
демонстрируя при этом эруптивный ток цитоплазмы
(Page, 1987а). Gruberella flavescens — морская мно-
гоядерная амеба — при перемещении постоянно
имеет моноподиальную форму (Page, 1984), однако
характер ее амебоидного движения сильно отличает-
ся от типичного эруптивного движения представите-
лей Vahlkampfiidae.
Отряд Acrasida (Schroeter, 1866)
Page et Blanton, 1985
Все представители отряда, за исключением одного
рода, формируют плодовые тела.
ТИП RHIZOPODA. КЛАСС HETEROLOBOSEA. SUMMARY
489
Сем. Acrasidae (Van Tieghem, 1880)
Olive, 1970
Трофозоиты представляют собой моноподиальных
амеб с эруптивным током цитоплазмы и морфологиче-
ски неотличимы от таковых, входящих в состав сем.
Vahlkampfiidae. Особенностью этих организмов явля-
ется агрегация трофозоитов перед инцистированием и
образование ими дифференцированных агрегатов цист
(спор) — сорокарпов (Olive, 1975; Page, 1988). У од-
ного вида в жизненном цикле известна жгутиковая ста-
дия (Olive et al., 1983).
Роды: Acrasis (Van Tieghem, 1880) Olive, 1970;
Pocheina Loeblich et Tappan, 1961.
Сем. Guttulinopsidae Olive, 1970
Трофозоиты уплощенные, часто неправильно-тре-
угольной или вееровидной формы, с обширной фрон-
тальной зоной гиалоплазмы. Представители рода Gut-
tulinopsis образуют сорокарпы (Olive, 1975; Raper et al.,
1977), тогда как Rosculus ithacus — единственный вид
рода Rosculus. будучи морфологически совершенно не-
отличимым от амебоидных трофозоитов Guttulinopsis spp.,
не формирует плодовых тел (Page, 1974, 1988).
Роды: Guttulinopsis Olive, 1901; Rosculus Hawes,
1963. ;
Heterolobosea incertae sedis
Сем. Euhyperamoebidae Seravin et
Goodkov, 1982
Монотипический таксон. Единственный достоверно описан-
ный к настоящему времени вид — Euhyperamoeba fallax — мор-
ской многоядерный организм. Амебы перемещаются, устойчиво
сохраняя моноподиальную форму тела, при этом наблюдается
равномерный ток цитоплазмы. Токи цитоплазмы, имеющие эруп-
тивный характер, также наблюдаются у этого простейшего, одна-
ко они не принимают непосредственного участия в организации
локомоции (Серавин, Гудков, 1987). Цисты неизвестны. Жгутико-
вая стадия в жизненном цикле отсутствует. Фактически основная
причина того, что Е. fallax помещена в класс Heterolobosea, —
плоские кристы митохондрий. В целом тонкая организация этих
амеб весьма сходна с таковой Gruberella flavescens. Euhyperamo-
eba fallax характеризуется способностью к некоторым типам
агамных слияний клеток — бинарной и множественной псевдоко-
пуляции, а также временной плазмодизации (в случае «коллектив-
ного питания» скоплениями нитчатых цианобактерий) (Серавин,
Гудков, 1984, 1987).
Род Percolomonas Fenchel
et Patterson, 1986
Организмы, для которых известна лишь жгутиковая ста-
дия. Имеет 4 жгутика и общую организацию клетки, очень сход-
ную с таковой представителей рода Tetramitus (Fenchel, Patterson,
1986).
Род Psalteriomonas Broers, Stumm
et Vogels, 1990
Анаэробные организмы, не имеющие нормальных митохонд-
рий. Доминирующей стадией в жизненном цикле является жгути-
ковая. Четырехъядерные жгутиконосцы имеют 16 жгутиков оди-
наковой длины со сложно организованной корешковой системой.
Лимаксного типа амебоидные клетки образуются лишь в присут-
ствии небольших количеств кислорода в среде И имеют только 1
ядро. Деление клеток возможно на обеих стадиях жизненного
цикла, причем имеет место закрытый ортомитоз с сохранением
ядрышка и формированием полярных масс (типичный промитоз)
(Broers et al., 1990).
THE PHYLUM RHIZOPODA SIEBOLD, 1845
THE CLASS HETEROLOBOSEA PAGE ET BLANTON, 1985
SUMMARY
The class Heterolobosea was established in 1985 by F. C. Page
and R. L. Blanton to unify amoeboid organisms, which had been for-
merly considered to belong to different classes of protists. This taxon
is apparently systematically heterogeneous and should be considered
only as a nominal unification of quite different organisms, which are
amoeboid at all or some stages of their life cycle.
Freshwater, marine and soil organisms.
Trophic stages of heteroloboseans usually are naked amoeboid
organisms, often with alternate, transitory (with rare exception) flagel-
late stage. Resting cysts afe common. Amoebae demonstrate «erupti-
ve» movement during locomotion. Stachyamoeba and Rosculus^ nor-
mally have a flattened locomotive form and acquire limax form with
eruptive locomotion only in very fast advance. Locomotion of Euhy-
peramoeba and Gruberella spp. can be considered intermediate be-
tween that of Heterolobosea and Lobosea due to steady flow of their
cytoplasm. Mitochondrial cristae are flattened, discoid. Organisms are
usually uninucleate, sometimes multihucleate. Nucleus of vesicular
type. Closed intranuclear orthomitosis is characteristic, often with bi-
nary division of nucleolus («promitosis»). Dictyosomes of the Golgi
complex are usually lacking; instead there are small morphologically
undifferentiated vesicles. Flagellates are nearly always formed by
transformation of amoebae. Organisation of flagellates and their role
in the life cycle varies. There are bi- and quadriflagellate species. In
some species flagellates are incapable to feeding (have no cytostome)
and cell division. In other cases feeding and reproduction can take
place on both amoeboid and flagellate stages. And, finally, there are
some species considered as heteroloboseans, which have dominant fla-
gellate stage in their life cycle. Asexual organisms. Different forms of
agamic cell fusions are common for some species (Euhyperamoeba
fallax). Cell aggregation with subsequently fruiting bodies formation
is characteristic of representatives of the order Acrasida.
Heteroloboseans are distributed worldwide and inhabit a great va-
riety of ecotopes. They play an important role in sea water, marine
and soil ecosystems. Most of them are free-living, but some species are
capable to give facultative parasitic forms, which cause dangerous dise-
ases, such as amoeboid menningoencephalithus in man (some Naegleria
spp.).
The class Heterolobosea includes two orders: Schizopyrenida (do
not form fruiting bodies) and Acrasida (fruiting bodies formed).
The family Euhyperamoebidae Seravin et Goodkov, 1982, genera Per-
colomonas Fenchel et Patterson, 1986 and Psalieriomorids Broers, Stumm
et Vogels, 1990 are considered as Heterolobosea incertae sedis.
490
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ЛИТЕРАТУРА26
Серавин Л. Н., Гудков А. В. Основные типы и формы агамных
слияний клеток у простейших // Цитология 1984. Т. 26. С. 123—131.
(Серавин Л. Н., Гудков А. В.) Seravin L. Na, Goodkov А. V.
Euhyperamoeba fallax Seravin et Goodkov, 1982 (Lobosea, Gymn-
amoebia) — multinucleate marine limax amoebpa morphology, biolo-
gical peculiarities and systematic position И Acta Protozool. 1987.
Vol. 26. P. 267—284.
Broers С. A. M., Stumm С. K., Vogels G. D., Brugerolle G. Psal-
teriomonas lanterna gen. nov., sp. nov., a free-living amoeboflagellate
isolated from freshwater anaerobic sediments // Eur. J. Protistol. 1990.
Vol. 25. P. 369—380.
Fenchel T., Patterson D. J. Percolomonas cosmopolitus (Ruinen)
n. gen., a new type of filter feeding flagellate from marine plankton //
J. Mar. Biol. Ass. U. K. 1986. Vol. 66. P. 465—482.
Fulton C. Naegleria: a research partner for cell and developmental
biology H J. Euk. Microbiol. 1993. Vol. 40. P. 520—532.
John D. T., Cole T. B., Bruner R. A. Amoebostomes of Naegleria
fowleri П J. Protozool. 1985. Vol. 32. P. 12—19.
Levine N. D., Corliss J. O., Cox F. E. G., Deroux G., Grain J.,
Honigberg В. M. et al. A new revised classification of the Protozoa //
J. Protozool. 1980. Vol. 27. P. 37—58.
Olive L. S. The mycetozoans. New York etc.: Acad. Press, 1975.
Olive L. S., Stoianovitch C., Bennett W. E. Descriptions of acra-
sid cellular slime molds: Pocheina rosea and a new species, Pocheina
flagellata П Mycologia. 1983. Vol. 75. P. 1019—1029.
26 Литература к разделу «Тип Rhizopoda. Класс Heterolobosea».
Page F. C. Rosculus ithacus Hawes, 1963 (Amoebida, Flabelluli-
dae), and the amphizoic tendency in amoebae I I Acta Protozool. 1974.
Vol. 13. P. 143—154.
Page F. C. A revised classification of the Gymnamoebia (Proto-
zoa, Sarcodina) // Zool. J. Linn. Soc. 1976. Vol. 58. P. 61—77.
Page F. C. Acrasis rosea and the possible relationship between Acra-
sida and Schizopyrenida I I Arch. Protistenk. 1978. Vol. 120. P. 169—181.
Page F. C. Gruberella flavescens (Gruber, 1889), a multinucleate
lobose marine amoeba (Gymnamoebia) // J. Mar. Biol. Ass. U. K.
1984. Vol. 64. P. 303—316.
Page F. C. The classification of «naked» amoebae (Phylum Rhi-
zopoda) II Arch. Protistenk. 1987a. Vol. 133. P. 199—217.
Page F. C. Transfer of Stachyamoeba lipophora to the class Hete-
rolobosea // Arch. Protistenk. 1987b. Vol. ГЗЗ. P. 191—197.
Page F. C. A new key to Freshwater and soil Gymnamoebae. Am-
bleside: Freshwater Biol. Ass., 1988.
Page F. C. Nakte Rhizopoda. Nakte Rhizopoda und Heliozoea
(Protozoenfauna, Bd 2). Stuttgart; New York: Fisher, 1991.
Page F. C., Blanton L. The Heterolobosea (Sarcodina: Rhizopo-
da), a new class uniting the Schizopyrenida and the Acrasidae (Acra-
sida) II Protistologica. 1985. Vol. 21. P. 121—132.
Raper К. B., Worley A. C., Kessler D. Observations on Guttuli-
nopsis vulgaris and Guttulinopsis nivea // Mycologia. 1977. Vol. 69.
P. 1016—1030.
Schuster F. An electrone microscope study of the amoeba-flagel-
late, Naegleria gruberi (Schardinger). I. The amoeboid and flagellate
stages // J. Protozool. 1963. Vol. 10. P. 297—313.
Walsh C. Synthesis and assembly of the cytoskeleton of Naegleria
gruberi flagellates // J. Cell. Biol. 1984. Vol. 98. P. 449—456.
Класс F i I о s e a Leidy, 1879 — Филозные амебы
Амебоидные организмы, обладающие филозными псевдоподи-
ями (филоподиями), — длинными, тонкими, заостренными, дихо-
томически ветвящимися и лишь изредка анастомозирующими вы-
ростами гиалоплазмы, не содержащими микротрубочкового цито-
скелета. Одноядерные и многоядерные виды; гетероморфизма
ядер нет ни на одной стадии жизненного цикла. У 2 видов описан
мейоз, у всех остальных половой процесс неизвестен. Класс не-
сомненно сборный, в котором таксоны объединены только по
формальным признакам организации псевдоподий. В его составе
выделяют 2 подкласса --- Aconchulinia (голые филозные амебы) и
Testaceafilosia (раковинные филозные амебы).
Подкласс Aconchulinia de Saedeleer, 1934 — Голые филозные
амебы
Филозные амебы, тело которых не заключено в раковину.
Сборная группа; представители разных семейств этого подкласса
очень сильно различаются по строению и биологии.
История изучения
Существенные морфологические различия между
лобоподиями и филоподиями были отмечены в конце
прошлого века; они нашли свое отражение в соответ-
ствующем разделении корненожек на 2 крупные
группы (см.: Аверинцев, 1906). В пределах обеих
групп встречаются как голые амебоидные организмы,
так и формы, имеющие [^а^овину. Некоторые авторы
при построении системы рассматривали наличие/от-
сутствие раковины как признак более высокого ранга,
чем тип псевдоподий, однако большинство исследо-
вателей придавало существенное значение органи-
зации псевдоподий. Именно последняя точка зрения
отражена и в современной систематике амебоидных
протистов (Levine et al., 1980; Page, 1987).
Длительное время голыми филозными амебами
называли только представителей 2 современных се-
мейств — Nucleariidae и Vampyrellidae. Путаница в
определении понятия «филоподии» привела к тому,
что в состав подкласса в разное время входили роды
и семейства, которые современные авторы (см., напри-
мер, Siemensma, 1991) относят к типу Foraminifera.
Ультраструктурные исследования выявили прин-
ципиальные различия в строении гранулоретикуло-
подий и филоподий (наличие/отсутствие микротру-
бочек), и их результаты являются базой современного
разграничения этих 2 таксонов. В ряде ультраструк-
турных исследований (см.: Patterson, 1985) было
показано, что представители отдельных родов про-
тистов, которых по морфологической организации
относят к солнечникам (Heliozoa), по строению
псевдоподий и организации цитоскелета схожи с фи-
лозными амебами и резко отличаются от солнечников.
В 1987 г. они были включены в состав класса Filo-
sea в ранге сем. Pompholyxophryidae (Page, 1987).
Классификация
Общепринятая классификация голых филозных
амеб до сих пор не разработана — среди авторов нет
единства в вопросе о границах подкласса. В настоя-
щее время в его состав включают 4 семейства, объ-
ТИП RHIZOPODA. КЛАСС FILOSEA. ПОДКЛАСС ACONCHULINIA
491
единенных в 2 отряда. Принятая в этом разделе сис-
тема в основном соответствует таковой, разработан-
ной Пэйджем (Page, 1987), однако в нее внесены не-
которые изменения и дополнения. В состав семейств
включены только хорошо изученные роды, которые
современные авторы (Siemensma, 1991, и др.) с уве-
ренностью относят к этим семействам. Видовой со-
став рода Nuclearia приводится в соответствии с пос-
ледней ревизией рода (Patterson, 1984).
Подкласс Aconchulinia de Saedeleer, 1934
Отряд Cristidiscoidida Page, 1987
Сем. Nucleariidae Cann et Page, 1979
Сем. Pompholyxophryidae Page, 1987
Отряд Cristivesiculatida Page, 1987
Сем. Vampyrellidae Zopf, 1885
Сем. Arachnulidae Page, 1987
Морфология
Строение клетки
По организации клетки можно довольно четко
различить 2 жизненные формы филозных амеб —
распластанную (субстратную) и радиальную (флоти-
рующую). В первом случае клетка распластана по
субстрату, она образует псевдоподии и активно пере-
мещается, во втором — округлая, с направленными в
разные стороны филоподиями. Такая клетка пассив-
но парит в толще воды и перемещается с ее потока-
ми. Представители ряда таксонов могут образовывать
обе формы; для некоторых описана только флотиру-
ющая форма или только субстратная.
Общий план строения клетки не имеет каких-либо
черт, необычных для протистов, однако по ряду при-
знаков представители различных таксонов голых фи-
лозных амеб сильно отличаются друг от друга, и по-
этому мы рассматриваем их отдельно по семействам.
Сем. Nucleariidae
Cann et Page, 1979
Среди голых филозных амеб наиболее детально
исследованы представители рода Nuclearia (рис. 461—
463). Все виды, входящие в этот род, имеют обе жиз-
ненные формы и легко переходят от флотирования к
перемещению по субстрату и наоборот. Флотирую-
щая форма округлая, с длинными филоподиями, а пе-
ремещающаяся по субстрату — обычно продольно
вытянута. У многих видов клетки имеют многослой-
ную или однослойную мукополисахаридную оболоч-
ку толщиной до 7 мкм, однако никакого дифферен-
цированного экзоскелета у них не обнаружено. У не-
которых видов отмечено дихотомическое ветвление
филоподий.
При движении N. simplex (рис. 461) уплощена,
продольно вытянута и обычно образует 2 пучка фи-
лоподий, направленных вперед, по ходу движения, и
назад. Напротив, N. delicatula (рис. 462) при переме-
щении обычно имеет округлую форму и не расплас-
тывается по субстрату. N. radians (рис. 463), для ко-
торой описана только флотирующая форма, никогда
Рис. 461. Nuclearia simplex. (По: Pernin,
1976).
А — субстратная форма с двумя пучками филопо-
дий, Б — с одним (передним) пучком филоподий,
В — флотирующая форма.
Fig. 461. Nuclearia simplex.
А — substrate form with two bundles of filopodia,
Б — with one (frontal) bundle of filopodia, В — floa-
ting form.
Рис. 462. Nuclearia delicatula. (По: Cann,
1986).
A — субстратная форма, Б — флотирующая
форма. Виден толстый гликокаликс (а), к которо-
му прикрепились бактерии (б).
Fig. 462. Nuclearia delicatula.
А — substrate form, Б — floating form. Thick glyco-
calyx (a) with attached bacteria (6) can be seen.
492
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 463. Nuclearia radians, флотирующая форма. (По: Greef,
1866).
а — гликокаликс, б — цитоплазма клетки.
Fig. 463. Nuclearia radians, floating form.
a — glycocalyx, 6 — cell cytoplasm.
не перемещается активно по субстрату, а лишь пас-
сивно парит с токами воды.
Большинство видов рода одноядерные, однако
N. delicatula имеет до 50 везикулярных ядер в одной
клетке. Гликокаликс у большинства видов представ-
лен толстым слоем фиброзного мукополисахаридно-
го материала (Сапп, 1986). У ряда видов (N. simplex,
N. radians) обнаружена капсула из фибриллярного
материала, заключающая в себе ядро (Mignot, Sa-
voie, 1979). Было показано также наличие кор-
тикального слоя микрофиламентов под мембраной
клетки и в филоподиях (Mignot, SaVoie, 1979). У N.
radians в цитоплазме находятся единичные микрот-
рубочки, и похоже, что перинуклеарная капсула иг-
рает определенную роль в их организации (Сапп,
Page, 1979). Вокруг ядра обычно расположены мно-
гочисленные диктиосомы. Сократительная вакуоль,
как правило, одна, но известны и виды с множест-
венными сократительными вакуолями.
Единственный детально изученный вид рода Vam-
pyrellidium — V. perforans (рис. 464) (см.: Surek,
Melkonian, 1980) — по организации и светомикрос-
копической морфологии весьма схож с Nuclearia
simplex. В филоподиях у него отмечены аксиальные
пучки микрофиламентов, а в цитоплазме — отдель-
ные неупорядоченные микротрубочки (Patterson et
al., 1987). Ядро заключено в капсулу из фибрилляр-
ного материала и результаты ультраструктурного ис-
следования показывают, что она, вероятно, играет
определенную роль в организации микротрубочек. В
состав гликокаликса входят длинные тонкие фила-
менты, расположенные перпендикулярно мембране.
Представитель рода Leptophrys — L. elegans
(рис. 465) — изучен слабо, ультраструктура его
неизвестна. Флотирующая форма у этого вида не
описана, субстратная — распластанная, неправиль-
ная, часто разветвленная. Характерно присутствие
нескольких зон гиалоплазмы, из которых образуются
филоподии. Клетка содержит несколько ядер и име-
ет множественные сократительные вакуоли.
Рис. 464. Vampyrellidium perforans. (По: Surek, Melkonian, 1980; рисунок выполнен Б. В. Громовым).
А—Е — последовательные стадии оседания флотирующей формы.
Fig. 464. Vampyrellidium perforans.
А—Е — successive stages of floating form discent.
ТИП RHIZOPODA, КЛАСС FILOSEA. ПОДКЛАСС ACONCHULINIA
493
Рис. 465. Leptophrys elegans. (По: Hertwig, Lesser, 1874).
Fig. 465. Leptophrys elegans.
Сем. Pompholyxophryidae Page,
1987
В состав семейства входят организмы, по своей
морфологической организации напоминающие сол-
нечников. Эти амебы (рис. 466) никогда не переме-
щаются активно по субстрату, а только флотируют.
Клетка округлая, с радиальными филоподиями; име-
ется экзоскелет, состоящий из слоя кремниевых че-
шуек или кремниевых сфер. Обычно у одного орга-
низма имеется несколько морфологических типов
чешуек. Цитоскелет псевдоподий представлен акто-
миозиновыми филаментами; в его составе нет мик-
ротрубочек (Patterson, 1985). В центральной части
клетки всегда находится ядро, окруженное диктиосо-
мами, но никакой более сложной дифференцировки
цитоплазмы не отмечено. Чешуйки (сферы) синтези-
руются клеткой, однако этот процесс подробно не
прослежен. Паттерсон (Patterson, 1985) отметил в
цитоплазме Pompholyxophrys punicea многочислен-
ные вакуоли, содержащие кремниевые сферы на раз-
ных стадиях синтеза.
Сем. Vampyrellidae Zopf,
1885
Представители рода Vampyrella (рис. 467) не
имеют каких-либо ярко выраженных особенностей
организации, и их тонкое строение типично для аме-
боидных организмов. Многие виды способны к агам-
ным слияниям. Одноядерные клетки Lateromyxa gab
Иса обязательно сливаются при контакте друг с дру-
гом и способны образовывать крупные (до 800 мкм)
синцитии. Иногда внешний (кортикальный) слой ци-
топлазмы синцития сильно вакуолизован. У этого
вида ядро заключено в уплощенную цистерну ЭПР,
а внутри ядра имеется ламина — тонкий электрон-
но-плотный слой, прилежащий изнутри к стенке ядра
(Huelsmann, 1993).
Сем. Arachnulidae Page,
1987
Единственный детально изученный вид — Arach-
nula impatiens (рис. 468) — резко отличается по ор-
ганизации от всех остальных представителей под-
класса. Это очень крупный (до 1 мм) организм.
Флотирующей формы в жизненном цикле нет; суб-
стратная — распластанная, часто разветвленная или
сетчатая. Многочисленные филоподии образуются из
периферической гиалиновой каймы или из лопастей
гиалоплазмы; они ветвятся и иногда анастомозируют.
Клетка содержит большое количество ядер.
Рис. 466. Pompholyxophrys degrooti.
А— внешний вид, Б----разрез (схема), а — чешуйки двух типов, б — цитоплазма клетки.
Fig. 466. Pompolyxophrys degrooti.
А — general view, Б — section (scheme), a — scales of two types, 6 — cell cytoplasm.
494
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 467. Vampyrella lateritia.
А — локомоторная форма, Б — флотирующая форма, В — питающаяся амеба (а) на клетке нитчатой водоросли (б). (А — по: Cienkowski, 1865; Я, В —
по: Roepstorf et al., 1994).
Fig. 467. Vampyrella lateritia.
A — locomotive form, Б — floating form, В — feeding amoeba (a) on the cell of filamentous alga (6).
Рис. 468. Arachnula impatiens. (По: Cienkowski, 1876, с изменениями).
A — субстратная форма, Б — A. impatiens (а), питающаяся конидиями Cochliobolus sativus (6).
Fig. 468. Arachnula impatiens.
A — substrate form, Б — feeding on conidia of Cochliobolus sativus.
Псевдоподии и движение
Флотирующие формы голых филозных амеб пере-
мещаются пассивно, с токами воды, и могут исполь-
зовать псевдоподии для прикрепления к субстрату
или к пищевым объектам. Субстратные формы актив-
но перемещаются при помощи филоподий.
Данные о строении и функционировании псевдо-
подий Filosea отрывочны, а механизм функциониро-
вания филозных псевдоподий никогда не был объек-
том пристального внимания исследователей. Основ-
ной способ перемещения клетки — прикрепление
конца филоподии к субстрату с последующим сокра-
щением филоподии и подтягиванием клетки. Свое-
образным феноменом является движение путем
«складывания» филоподий, характерное для Nuclea-
ria. При этом псевдоподия прикрепляется к субстра-
ту концом и перегибается посередине. В месте пере-
гиба образуется капля гиалоплазмы, в которую с
обоих концов втягиваются остатки псевдоподии.
Клетка при этом подтягивается вперед, и образовав-
шаяся капля гиалоплазмы сливается с цитоплазмой
клетки. Образуется новая филоподия, и процесс пов-
торяется.
Описан весьма своеобразный механизм сокраще-
ния псевдоподий у флотирующей формы Nuclearia
(Mignot, Savoie, 1979; Patterson, 1983). По-видимому,
сокращение псевдоподии происходит за счет инваги-
наций ее мембраны/ которые быстро отшнуровыва-
ются в виде мелких везикул и транспортируются в
цитоплазму клетки.
В цитоскелете филоподий практически нет мик-
ротрубочек. В центральной части филоподии обычно
имеется развитый аксиальный пучок микрофиламен-
тов (скорее всего, акто-миозиновых), однако пока не-
ясно, каким образом цитоскелет участвует в движе-
ниях филоподии.
Питание
Для всех Aconchulinia характерно голозойное пи-
тание. Представители разных семейств имеют свои
особенности реализации механизмов поиска пище-
вых объектов, их захвата и фагоцитоза.
ТИП RHIZOPODA. КЛАСС FILOSEA. ПОДКЛАСС ACONCHULINIA
495
Семейства Nucleariidae
и Pompholyxophryidae
Представители этих двух семейств чаще всего яв-
ляются полифагами и фагоцитируют мелкие пище-
вые объекты — детрит, бактерии, одноклеточные
водоросли. Помимо обычного фагоцитоза Nuclearia
delicatula имеет и другой тип питания — она может
«выедать» содержимое мертвых или поврежденных
клеток водорослей (Cienkowski, 1865). При этом об-
разуется псевдоподия, которая проникает внутрь
клетки и там ветвится, фагоцитируя ее содержимое.
Здоровые клетки водорослей этот вид не атакует
(Сапп, 1986). Представитель рода Vampyrellidium —
V. perforans — наряду с фагоцитозом целых клеток
может продырявливать оболочки клеток водорослей
и при помощи проникающей внутрь клетки псевдо-
подии фагоцитировать ее содержимое (Surek, Melko-
nian, 1980). По характеру питания V, perforans боль-
ше похож на представителей сем. Vampyrellidae.
Сем. Vampyrellidae
Представители семейства отличаются своеобраз-
ным способом питания. Они атакуют интактные
клетки водорослей, продырявливают их стенку и
проникают внутрь, выедая содержимое клетки. Дета-
ли этого процесса могут различаться у разных видов,
однако само нападение на клетку у всех видов рода
Vampyrella протекает сходным образом. Амеба плот-
но прикрепляется к поверхности клетки водоросли,
распластывается по ней и принимает «подушкооб-
разную» форму. По описаниям Гоби (1915), в теле
амебы вблизи стенки клетки водоросли возникает
вакуоль, которая быстро увеличивается в размерах.
Вскоре в клеточной стенке водоросли образуется от-
верстие, а протопласт водоросли сжимается, теряет
контакт со стенками клетки и «всасывается» внутрь
этой вакуоли. Многие исследователи (Cienkowski,
1865; Klein, 1882; Zopf, 1885) наблюдали, как Vam-
pyrella прикрепляются к клетке при помощи не-
скольких филоподий, а затем образуют более толс-
тую псевдоподию, которая плотно прилегает к стенке
водоросли, пробуравливает ее и проникает внутрь,
фагоцитируя протопласт водоросли. (Аналогичным
образом, как уже было отмечено, может питаться и
Vampyrellidium perforans из сем. Nucleariidae.) Late-
romyxa gallica после прикрепления к стенке водоро-
сли покрывается снаружи плотной оболочкой, про-
буравливает стенку клетки водоросли, а затем про-
никает целиком внутрь клетки и фагоцитирует
протопласт водоросли. В случае нападения на нитчатую
водоросль амеба проникает!^ в соседние клетки,
пробуравливая поперечные перегородки между ними.
Сем. Arachnulidae
Arachnula impatiens может фагоцитировать раз-
личные мелкие пищевые частицы в разных участках
своего разветвленного тела. Специфической пищей
этого вида являются конидии почвенных грибов, ко-
торые она пробуравливает, образуя небольшое (1—
2 мкм) круглое отверстие в стенке конидии. Основ-
ная часть содержимого конидии высасывается (подобно
тому, как это делают Vampyrella), затем внутрь ко-
нидии проникает псевдоподия, при помощи которой
фагоцитируется остаток содержимого (см.: Old, Dar-
byshire, 1978, 1980).
Размножение
и жизненные циклы
Биология представителей различных семейств в
пределах этого подкласса принципиально различает-
ся. У многих видов она тесно связана с особенностя-
ми пищевого поведения (при этом обычно формиру-
ются сложные жизненные циклы).
Семейства Nucleariidae
и Pompholyxophryidae
Представители этих двух семейств обладают про-
стыми жизненными циклами, в которых последова-
тельно чередуются стадии питания и роста и бинарные
деления особей. Некоторые виды способны инцисти-
роваться. Клетки Vampyrellidium perforans способны
сливаться, формируя небольшие плазмодии, дальнейшая
судьба которых остается неясной; для этого рода
также описано формирование плазмодиев при кол-
лективной атаке амебами большого пищевого объек-
та. После поглощения пищи эти плазмодии быстро
распадаются на отдельные особи (Surek, Melkonian,
1980).
Сем. Vampyrellidae
Жизненные циклы представителей этого семейст-
ва — наиболее сложные среди голых филозных амеб.
Часто жизненный цикл тесно связан с особенностями
питания вида; иногда некоторые его стадии проходят
внутри клеток водорослей.
В наиболее простом случае {Vampyrella lateri-
tia, — рис. 469) цикл (см.: Roepstorf et al., 1994)
включает в себя нападение на клетку водоросли, вы-
сасывание ее содержимого и инцистирование амебы
на поверхности клетки водоросли с образованием
цисты пищеварения. После переваривания пищи про-
исходит кариокинез и из цисты выходит несколько
дочерних клеток. В неблагоприятных условиях мно-
гие виды образуют цисты покоя, способные сохра-
няться длительное время.
Lateromyxa gallica имеет весьма сложный жизнен-
ный цикл (рис. 470; 471), Этот вид детально изучен
как на светомикроскопическом, так и на ультра-
структурном уровне, и тем не менее биология его ос-
тается до конца не выясненной. Нападающая на клет-
ку водоросли амеба обычно представляет собой мно-
гоядерный синцитий. После прикрепления к клетке
водоросли амеба покрывается снаружи плотной обо-
лочкой и проникает внутрь водоросли (при этом пус-
тая оболочка остается снаружи, на поверхности водо-
496
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 469. Схема жизненного цикла Vampyrella lateritia. (По: Roepstorf et al., 1994, с изменениями).
А — нападение амеб на клетку водоросли, Б — циста пищеварения, В — множественное деление в цисте, Г — выход дочерних особей из оболочки
цисты.
Fig. 469. Life cycle of Vampyrella lateritia (scheme).
A — amoebae attacking alga cell and phagocyting algal protoplast, Б — digestion cyst, В — multiple division inside the cyst, Г— daughter cells coming out of
the cyst envelope.
в
Рис. 470. Lateroxyma gallica: начальные стадии жизненного цикла. (По: Huelsmann, 1993).
А — циста заражения; Б — фрагментирующийся синцитий фагоцитирует клетки водоросли; В — цисты пищеварения.
Fig. 470. Leptomyxa gallica: first stages of the life cycle.
A — infection cyst, Б — fragmentating syncytium phagocyting alga cells, В — digestion cysts.
ТИП RHIZOPODA, КЛАСС FILOSEA. ПОДКЛАСС ACONCHULINIA
497
Рис. 471. Схема жизненного цикла Late-
romyxa gallica. (По: Roepstorf et al.,
1993, с изменениями).
А — нападение на клетку водоросли и фаго-
цитоз протопласта водоросли; Б, В — цисты
пищеварения; Г — деление и выход дочер-
них особей во внешнюю среду; Д — циста
покоя (необязательная стадия, показана пре-
рывистыми стрелками).
Fig. 471. Life cycle of Lateromyxa gallica (scheme).
A — amoebae attacking alga cell and phagocyting algal protoplast; £, В — infection cysts; Г — division and daughter cells coming out of the cyst; Д — resting
cyst (non-obligatory stage, shown by broken line).
росли). По аналогии с терминологией, используемой
при описании других паразитов водорослей (см.: Гро-
мов, 1976), эта стадия может быть названа цистой за-
ражения. Проникший в клетку нитчатки синцитий
фагоцитирует протопласт водоросли и может разде-
литься на несколько особей в ходе проникновения в
соседние клетки водоросли через поперечные перего-
родки. Амеба, которая фагоцитировала достаточное
количество пищевого материала, покрывается плот-
ной оболочкой и образует цисту пищеварения. Циста
всегда образуется внутри выеденной клетки водоросли,
а если синцитий фрагментировался по мере роста и
перемещения внутри нитчатки, то образуется цепоч-
ка цист, находящихся на разных стадиях развития.
Непереваренные остатки пищи собираются в круп-
ной вакуоли в центре клетки. Стадия цисты пищева-
рения плавно переходит в стадию репродуктивной
цисты. Происходит синхронное митотическое деле-
ние ядер, а впоследствии — цитокинез. Обычно из
цисты выходят 3—4 одноядерные амебы, которые
покидают оболочку клетки водоросли и поражают
новые клетки.
При неблагоприятных условиях цист^ пищеваре-
ния трансформируется в цисту покоя. При этом
объем ее цитоплазмы уменьшается и внутри прежней
цистной оболочки формируется небольшая круглая
циста. Непереваренные остатки пищи выбрасываются
в пространство между стенками цисты. Эксцистиро-
вание происходит в подходящих условиях (в лабора-
32 Протисты, часть 1
тории — при переносе цист в свежую питательную
среду).
Кариологические исследования L. gallica (Roeps-
torf et al., 1993) показали, что в цистах покоя происхо-
дит мейотическое деление ядра. Судьба продуктов мей-
оза не прослежена, и этот вопрос требует пристального
изучения. Ясно, однако, что (как и в случае лобозных
раковинных амеб) филозные амебы не могут больше
рассматриваться как облигатно агамные формы.
Систематический обзор
Отряд Cristidiscoidida Раде, 1987
Кристы митохондрий уплощенные.
Сем. Nucleariidae Cann et Page, 1979
Одноядерные или многоядерные формы, преиму-
щественно субстратные или преимущественно фло-
тирующие в зависимости от вида. Характерно пере-
движение путем сокращения филоподий с последую-
щим подтягиванием клетки; у флотирующих форм
имеется своеобразный механизм формирования фи-
лоподий. Полифаги, пищевые частицы обычно фаго-
цитируют. Цисты размножения неизвестны.
498
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Определение, описания видов, ревизии, ссылки на
литературу см.: Pernin, 1976; Patterson, 1984; Cann,
1986; Siemensma, 1991.
Роды: Gobiella Cienkowski, 1865; Leptophrys Her-
twig et Lesser, 1874; Nuclearia Cienkowski, 1865;
Vampyrellidium Zopf, 1887.
Сем. Pompholyxophryidae Page, 1987
Округлые организмы с радиальными филоподия-
ми, покрытые слоем чешуек. Единственная жизнен-
ная форма — флотирующая. По морфологической
организации сходны с солнечниками, однако не
имеют экструсом и микротрубочковой аксонемы
внутри псевдоподий.
Изображения амеб, ревизии, описания видов, их
определение и ссылки на литературу см.: Roijackers,
Siemensma, 1988.
Роды: Pinaciophora Greef, 1873; Pompholyxophrys
(Archer, 1869) Roijackers et Siemensma, 1988; Rhabdi-
ophrys Rainer, 1968.
Отряд Cristivesiculatida Page, 1987
Кристы митохондрий трубчатого или везикуляр-
ного типа.
Сем. Vampyrellidae Zopf, 1885
Уплощенные одноядерные амебы, активно пере-
мещающиеся и питающиеся водорослями. Крупные
виды фагоцитируют водоросли целиком, мелкие про-
буравливают стенки клеток водорослей, проникают
внутрь и выедают их содержимое. Для некоторых
видов характерны сложные жизненные циклы, опи-
саны цисты размножения. Определение видов рода
Vampyrella см.: Siemensma (1991); ультраструкту-
ру — Roepstorf et al., 1993, 1994; род Lateromyxa
подробно описан Хюльсман (Huelsmann, 1993).
Роды: Lateromyxa Huelsmann, 1993; Vampyrella
Cienkowski, 1865.
Примечание. Некоторые авторы включают в состав семей-
ства род Hyalodiscus Hertwig et Lesser, 1874 и более десятка дру-
гих родов. Большинство из них известно только по первоописани-
ям. Представители всех этих родов должны быть реизолированы и
детально исследованы, прежде чем можно будет уверенно опреде-
лить их систематическое положение. Современные данные см:
Page, 1988 — для Hyalodiscus', Roepstoff et al., 1994 — для Gobiel-
la и некоторых других родов. Хюльсман (Hiilsmann, 1993) отмеча-
ет, что разными авторами в семейство включалось до 30 видов (в
составе разных родов), для большинства из которых имеются лишь
отрывочные и недостоверные сведения. Большое количество аме-
боидных паразитов водорослей, обладающих сложными жизненны-
ми циклами (иногда с включением жгутиковых стадий), было опи-
сано в начале нашего века (см.: Громов, 1976).
Сем. Arachnulidae Раде, 1987
Очень крупные (до 1 мм), малоподвижные, силь-
но разветвленные многоядерные амебоидные орга-
низмы. Питаются конидиями грибов (в частности,
Cochliobolus sativus) путем пенетрации их стенок с
последующим фагоцитозом содержимого. Жизнен-
ный цикл сложный, включает цисты пищеварения и
цисты размножения. Литературу и другие сведения
по этому семейству см.: Old, 1977; Old, Darbyshire,
1980; Siemensma, 1991.
Единственный род: Arachnula Cienkowski, 1876.
THE PHYLUM RHIZOPODA SIEBOLD, 1845
THE CLASS FILOSEA LEIDY, 1879
THE SUBCLASS ACONCHULINIA DE SAEDELEER, 1934
SUMMARY
The class Filosea Leidy, 1879 includes amoeboid organisms with
filose pseudopodia (filopodia) — long, thin, dichotomically branching
and rarely anastomosing. They consist of hyaloplasm and lack micro-
tubular cytoskelet. Uni- and multinulear forms, no nuclear heteromor-
phism. Sexual process known for two species only. Two subclasses —
Aconchulinia (naked filose amoebae) and Testaceafilosia (testate lobo-
se amoebae).
The subclass Aconchulinia unites filose amoebae without shells.
The taxon is apparently heterogeneous.
The chapter includes information on the history of study, mor-
phology and movement, biology and reproduction, feeding, life cycles
and distribution of the representatives of Aconchulinia. Four families
within this subclass differ significantly and are treated separately. Special
attention is paid to the algivorous amoebae and to the evolution of their
life cycles related with the food objects. The chapter is provided with
illustrations of representatives of all families of naked filose amoebae.
The detailed systematic account includes diagnoses of families of
filose testate amoebae; generic composition of each family is listed.
Classification of naked filose amoebae is still to be elaborated.
The classification used in this paper in general corresponds to that of
Page (1987), with some amendments. Only those genera are listed,
which are with certainty attributed to the filosid families by modern
authors (Siemensma, 1991, etc.). Specific composition of Nuclearia
genus is given according to the last revision of the genus (Patterson,
1984).
In the course of history of amoebae studies rhizopods were some-
times divided into testate and naked, each group comprising both lo-
bose and filose amoebae. Nowadays the type of pseudopodia is consi-
dered as a feature of higher systematic rank than presence/absehce of
the test.
For a long time, the name «naked filose amoebae» was applied
only to the representatives of two modern families — Nucleariidae
and Vampyrellidae. A certain confusion existed about the term «filo-
podia», and some representatives of modern Granuloreticulosea were
sometimes treated within Filosea. Now these two taxa are clearly de-
fined on the basis of presence/absence of microtubules in pseudopo-
dia. The group, previously treated as heliozoans, was shown to be
close to filose amoebae and is now considered as the family Pompho-
lyxophryidae within Filosea.
Among naked filose amoebae two «life forms» are common —
«flattened» («substrate») and «radiate» («floating»). In the former
case the cell forms pseudopodia and moves, in the latter it is carried
with the water currents. Some amoebae have both, for some only one
of them was described.
Representatives of taxa within Aconchulinia differ from each
other by* the cell organization.
ТИП RHIZOPODA. КЛАСС FILOSEA. ПОДКЛАСС ACONCHULINIA. ЛИТЕРАТУРА
499
The Family Nucleariidae. This is the best studied group of this
subclass. All species have both life forms. Floating form is rounded,
substrate form is usually elongated. Dichotomic branching of pseudo-
podia occurs in some species.
Most species are uninuclear, but Nuclearia delicatula may have
up to 50 vesicular nuclei in the cell. Glycocalyx consists of a thick
layer of fibrose material, Nucleus of some species is enclosed in a
capsule of fibrillar material.
The family Pompholyxophryidae. Representatives of this family
morphologically resemble heliozoans. They exist only in floating
form. The cell is rounded, with radial pseudopodia. There is an exo-
skeleton of silicious scales or spheres. One organism may have scales
of different types. Pseudopodial cytoskeleton consists of actin-miosin
filaments; there are no microtubules.
The family Vampyrellidae. For the representatives of this family
agamic fusions were observed. Sometimes the cortical layer of syncy-
tium is vacuolised.
The family Arachnulidae. The only well-studied species of this
family — Arachnula impatiens — differs greatly from other repre-
sentatives of the subclass. It is a large organism (up to 1 mm), there
is no floating form, the substrate form is often branched or net-like.
Numerous pseudopodia branch and anastomose. The cell contains
many nuclei.
Substrate forms of naked filose amoebae move by means of filo-
podia. There is a central axial bundle of microfilaments in filopodia
and practically no microtubules. Data on their structure and functio-
ning are scarce. In most cases the end of filopodium is attached to the
substrate, then filopodium contracts and the cell is «dragged» forward.
For Nuclearia a peculiar «folding» of pseudopodia was observed. Fi-
lopodium attaches to the substrate and folds iri the middle. A drop of
hyaloplasm is formed, into which both «halves» of filopodium are
drawn. The cell is dragged forward and hyaloplasmic drop merges
with the cytoplasm of the cell.
Representatives of different filosid families have peculiar means
of searching, capturing and phagocyting food objects. Members of
Nucleariidae and Pompholyxophryidae are polyphagous and feed on
small objects — bacteria, unicellular algae, etc. Nuclearia delicatula
may devour the contents of damaged algal cells. Vampyrellidium per-
forate is capable of penetrating whole algal cells. Its feeding resem-
bles that of members of family Vampyrellidae, which attack intact
algal cells, perforate their walls and devour the contents. This process
is described in some detail.
Arachnula impatiens (family Arachnulidae) feed almost exclusely
on soil fungi conidia. It penetrates the conidia wall and devours the
contents in a way similar to that of Vanipyrella. Members of Nuclea-
riidae and Pompholyxophryidae have rather simple life cycles. Some
of them form cysts, some are capable of forming small plasmodia.
Some representatives of Aconchulina have complicated life cycles,
connected with their feeding behaviour. The most complicated ones
are characteristic of the represenatives of the family Vampyrellidae. In
the most simple case it consists of attacking an algal cell, devouring
of its contents and formation of a «cyst of digestion» on the surface
of the algal cell. Inside the cyst division occurs and several daughter
cells leave the cyst. Resting cysts are also common. Lateromyxa gal-
lica is shown to possess a very sophisticated life cycle, differing from
that described above for other Vampyrellidae.
ЛИТЕРАТУРА27
Аверинцев C. Rhizopoda пресных вод И Тр. Имп. СПб. О-ва
Естествоисп. 1906. Т. 36. С. 120—336.
Гоби X. Я. Монография семейства Vampyrellaceae // Ботан.
зап. СПб ун-та. 1915. Т. 16.
Громов Б. В. Микроорганизмы — паразиты водорослей. Л.:
Изд-во ЛГУ, 1976.
Cann J. Р. The feeding behavior and structure of Nuclearia deli-
catula (Filosea, Aconchulinia) // J. Protozool. 1986. Vol. 33. C. 392—
396.
Cann J. P., Page F. C. Nucleosphaerium tuckeri nov. gen. nov.
sp. — a new freshwater amoeba without motile form in a new family
Nucleariidae (Filoseay Aconchulinia) feeding by ingestion only //
Arch. Protistenk. 1979. Vol. 122. P. 226—24b.
Cienkowski L. Beitrage zur Kenntnis der Monaden // Arch. Mik-
rosk. Anat. 1865. Bd 1. S. 203—232.
Greef R. Ober einige in der Erde lebende Amoeben und andere
Rhizopoden // Arch. Mikrosk. Anat. 1866. Bd 2. S. 299—331.
Hertwig R., Lesser E. Ober Rhizopoden und nahestehende Orga-
nismen // Arch. Mikrosk. Anat. 1874. Bd 10. S. 67—78.
Huelsmann N. Lateromyxa gallica n. g, n. sp. (Vampyrellidae): a
filopodial amoeboid protist with a novell life cycle and conspicuos
ultrastructural characters // J. Euk. Microbiol. 1993. Vol. 40. P. 141—
149.
Klein J. Ueber Vampyrella // Bot. Ztschr. 1882.
Levine N. D., Corliss J. O., Cox F. E. G., Deroux G., Grain J.,
Honigberg B.M., Leedale G.F., Loeblich A. R., Lom J., Lynn D.,
Meisterfeld E. G., Page F. C., Poljansky G., Sprague V., Varra J.,
Wallas F. G. A new revised classification of the Protozoa // J. Proto-
zool. 1980. Vol. 27. P. 37—58.
Mignot J.-P., Savoie A. Observations ultrastructurales sur Nucle-
aria simplex Cienkowski (Protozoa, Rhizopoda, Filosea) // Protistolo-
gica. 1979. T. 15. P. 23—32.
Old К. M. Giant soil amoeba cause perforation of conidai of Coch-
liobolus sativus // Trans. Brit, mycol. Soc. 1977. Vol. 68. P. 277—281.
Old К. M., Darbyshire J. F. Soil fungi as food for giant amoe-
bae II Soil. Biol. Biochem. 1978, Vol. 10. P. 93—100.
Old К. M., Darbyshire J. F. Arachnula impatiens Cienk., a my-
cophagous giant amoeba from soil // Protistologica. 1980. T. 16.
P. 277—287.
Page F. C. The classification of ’naked’ amoebae (Phylum Rhizo-
poda) // Arch. Protistenk. 1987. Vol. 133. P. 199—217.
Page F. C. A new key to fresh-water Gymnamoebia. Ambleside,
1988.
Patterson D. J. On the organization of the naked filose amoeba,
Nuclearia moebiusi Frenzel, 1897 (Sarcodina, Filosea) and its implica-
tions // J. Protozool. 1983. Vol. 30. P. 301—307.
Patterson D. J. The genus Nuclearia (Sarcodina, Filosea): species
composition and characteristics of the taxa // Arch. Protistenk. 1984.
Vol. 128. P. 127—139.
Patterson D. J. On the organization and affinities of the amoeba
Pompholyxophrys punicea Archer, based on ultrastructural examinati-
on of individual cells from the wild material // J. Protozool. 1985.
Vol. 32. P. 241—246.
Patterson D. J., Surek B., Melkonian M. The ultrastructure of
Vampyrellidium perforance Surek et Melkonian and its taxonomic po-
sition among the naked filose amoebae // J. Protozool. 1987. Vol. 34.
P. 63—67.
Pernin P. Etude sur le vivant d’une amibe a pseudopodes filosa:
Nuclearia simplex Cienkowski, 1865 (Protozoa, Rhizopodea, Filosia,
Aconchulinida) // Protistologica. 1976. T. 12. P. 555—562.
Roepstorf P., Huelsmann N., Hausmann K. Karyological inves-
tigation of the Vampyrellid filose amoeba Lateromyxa gallica Huels-
mann, 1993 // Eur. J. Protistol. 1993. Vol. 29. P. 302—310.
Roepstorf P., Huelsmann N., Hausmann K. Comparative fine
structural investigations of interphase and mitotic nuclei of vampyrel-
lid filose amoebae // J. Euk. Microbiol. 1994. Vol. 41. P. 18—30.
Roijackers R. M. M., Siemensma F. J. A study of cristidiscoidid
amoebae (Rhizopoda, Filosea), with descriptions of new species and
key to genera and species // Arch. Protistenk. 1988. Vol. 135. P. 237—
253.
Siemensma F.J. Nackte Rhizopoda. Klass Filosea Leidy, 1879
(Protozoenfauna, Bd 2). Stuttgart; New York: Fischer, 1991. S. 132—
170.
Surek B., Melkonian M. The filose amoeba Vampyrellidium per-
forans nov. sp. (Vampyrellidae, Aconchulinida): axenic culture, fee-
ding behaviour and host range specificity // Arch. Protistenk. 1980.
Vol. 123. P. 166—191.
Zopf W. Die Pilzthiere oder Schleimpilze // Breslau, 1885.
27 Литература к разделу «Тип Rhizopoda. Класс Filosea. Под-
класс Aconchulinia».
500
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подкласс Testaceafilosia de Saedeleer, 1934 — Раковинные
филозные амебы
Корненожки, тело которых заключено в раковину с 1 или 2
отверстиями (устьями) для выхода псевдоподий (филоподий). Гете-
роморфизма ядер нет ни на одной стадии жизненного цикла. Оби-
тают в пресных водах, почве, мхах, а также в интерстициали побе-
режий морей и океанов.
Во многих исследованиях раковинные филозные амебы рас-
сматриваются вместе с представителями подкласса Testacealobosia
(раковинные лобозные амебы) как единая группа «раковинные
амебы» (Testacea).
Классификация
Общепринятой классификации раковинных фи-
лозных амеб не существует. В ходе дальнейшего из-
ложения мы будем придерживаться классификации,
чаще всего соответствующей принятой Бови (Bovee,
1985). Синонимика родов приведена в основном по
Хаману (Haman, 1988).
Подкласс Testaceafilosia de Saedeleer, 1934
Отряд Gromiida Claparede et Lachmann, 1859
Подотряд Gromiina Delage et Herouard, 1896
Сем. Amphitrematidae Pochfc, 1913
Сем. Chlamydophryidae de Saedeleer, 1934
Сем. Gromiidae Reuss, 1862
Сем. Psammonobiotidae Golemansky, 1974
Сем. Pseudodifflugiidae de Saedeleer, 1934
Сем. Volutellidae Sudzuki, 1979
Подотряд Euglyphina Bovee, 1985
Сем. Cyphoderiidae de Saedeleer, 1934
Сем. Euglyphidae Wallich, 1864
Сем. Paulinellidae de Saedeleer, 1934
Сем. Trinematidae Hoogenraad et de Groot, 1940
Морфология и физиология
Строение клетки
Клетка заключена в раковину. Цитоплазма ее
покрыта плазмалеммой, в состав которой входит
тонкий слой недифференцированного гликокаликса.
В цитоплазме находится ядро (ядра) и набор орга-
нелл, типичный для аэробных корненожек. У мел-
ких видов клетка заполняет полость раковины почти
целиком и прикрепляется к раковине в районе устья,
а у крупных она обычно занимает только часть ра-
ковины и закреплена в ней при помощи эпипо-
дий — выростов цитоплазмы, присоединяющихся
изнутри к стенке раковины (рис. 472).
Одноядерные или многоядерные формы. Ядро
обычно везикулярного типа, с более или менее ком-
пактным ядрышком, реже — гранулярного типа.
Ядерная ламина (слой фибриллярного материала,
прилегающий изнутри к стенке ядра) отсутствует.
Деление ядра на ультраструктурном уровне изучено
только для 2 видов рода Euglypha, у которых оно
носит характер закрытого ортомитоза (Ogden, 1979).
Митохондрии имеют тубулярные анастомозирую-
щие или везикулярные кристы. Матрикс митохондрий,
как правило, электронно-плотный; гетероморфизм
митохондрий неизвестен. В цитоплазме находятся
многочисленные диктиосомы аппарата Гольджи. В
них синтезируются идиосомы и цементные везикулы,
содержимое которых скрепляет идиосомы, входящие
в состав стенки раковины. Шероховатый ЭПР обра-
зует плотные скопления в цитоплазме, обычно окру-
жающие ядро. В клетке имеется 1 или 2 сократи-
тельные вакуоли.
В составе цитоскелета раковинных амеб выявлены
микрофиламенты (по-видимому, акто-миозиновые)
и, по крайней мере у некоторых видов, микротру-
бочки. Пучки микрофиламентов обнаружены в фи-
лоподиях движущихся амеб, их скопления также
имеются в периферических участках цитоплазмы
клетки.
Наличие раковины, придающей стабильную фор-
му цитоплазматическому телу клетки, приводит к
разделению цитоплазмы клетки на разные в функци-
ональном отношении участки (компартментализация
цитоплазмы, — рис. 472). Как правило, ядро распо-
лагается в задней (аборальной) части клетки. Вокруг
него обычно находится плотное скопление шерохо-
ватого ЭПР. Этот участок цитоплазмы иногда назы-
вают зоной плотной ЭПС. На периферии этой части
клетки располагаются диктиосомы и сократительные
Рис. 472, Схема строения клетки раковинной филозной амебы.
а — раковина; б — ядро; в — ядрышко: г — шероховатый ЭПР; д — со-
кратительная вакуоль; е — эпиподии; ж — диктиосомы; з — запасные че-
шуйки в цитоплазме; и — митохондрии; к — пищеварительные вакуоли,
цементные везикулы и другие включения; л — филоподии.
Fig. 472. Organization of filose testate amoeba cell.
a — shell; 6 — nucleus; в — nucleolus; г — rough endoplasmic reticulum;
d — contractile vacuole; e — epipodia; ж — dictyosomes; з — platelets sto-
red in cytoplasm; и — mitochondria; к — digestive vacuoles, cement vesicles
and other inclusions; л — filopodia.
ТИП RHIZOPODA. КЛАСС FILOSEA. ПОДКЛАСС TESTACEAFILOSIA
501
вакуоли. Никаких других органелл и пищеваритель-
ных вакуолей здесь нет. В периферической части
цитоплазмы расположены запасные идиосомы, кото-
рые иногда собраны в стопки по 3—4 штуки.
Ближе к устью (в оральной части цитоплазмы)
располагаются все митохондрии, вакуоли, кристаллы,
цементные везикулы и другие включения. Псевдопо-
дии начинают формироваться несколько глубже гра-
ницы устья раковины, и поэтому ближайшая'к нему
часть цитоплазмы представляет собой сеть перепле-
тающихся и анастомозирующих тяжей.
Раковина
Форма раковин разнообразна. Встречаются почти
все типы раковин, перечисленные в разделе «Testace-
alobosia». Представители 2 подотрядов принципиаль-
но различаются по строению и природе раковин.
Подотряд Gromiina
У видов этого подотряда раковина органическая,
секретированная. Иногда в нее вкраплены посторон-
ние частицы (ксеносомы). Детали строения раковины
известны лишь для некоторых видов.
Раковина Gromia секретированная, двухслойная.
Оба слоя состоят из гомогенного органического мате-
риала. Более толстый, наружный слой раковины пер-
форирован, а тонкий, внутренний не имеет перфора-
ции (рис. 473 а, б). В состав стенки раковины входит
Б
Рис. 473. Строение стенки раковины Gromia oviformis. (По: Hed-
ley, I960, с изменениями).
А — схема поперечного среза через стенку раковины, Б — перфорирован-
ная структура наружного слоя стенки раковины, а — внешний, перфориро-
ванный слой; б — внутренний, гомогенный слой; в — клеточная мембрана;
г — цитоплазма клетки.
Fig. 473. The structure of shell wall in Gromia oviformis.
A — transverse section of shell wall (scheme), Б — perforated structure of the
outer layer of shell wall, a — outer, perforated layer of shell wall; 6 — inner
homogenous layer of shell wall; в — cell membrane; г — cell cytoplasm.
Рис. 474. Схема организации устья раковины Gromia oviformis.
(По: Hedley, 1960, с изменениями).
А — устье закрыто, Б — устье наполовину открыто, В — устье открыто.
а — лепестки оральной капсулы, б — раковина, в — цитоплазма.
Fig. 474. Organization of aperture of Gromia oviformis shell (sche-
me).
A — aperture closed; Б — aperture half-open; В — aperture open, a —
«lobes» of the oral capsule; 6 — shell; в — cytoplasm.
большое количество железа. У G. oviformis своеоб-
разно организовано устье раковины (рис. 474). Име-
ется оральная капсула, состоящая из нескольких
(обычно около 9) мукополисахаридных лепестков
(Hedley, 1960). При движении цитоплазмы через
устье раковины лепестки могут либо почти пол-
ностью перекрывать просвет устья (когда псевдопо-
дии втянуты), или же разворачиваться, открывая
устье раковины (когда псевдоподии выпущены)
(рис. 474).
Органическая раковина Amphitrema имеет два
устья, находящихся на ее противоположных концах
(рис. 478). Стенка раковины трехслойная. Наружный
слой ее гранулярный, средний образован 2—3 слоя-
ми фибрилл, а внутренний состоит из плотно пере-
плетенных фибрилл. Изнутри раковина выстлана му-
коидным матриксом, а снаружи покрыта слоем ве-
щества полисахаридной природы.
Подотряд Euglyphina
Представители этого подотряда строят раковину
из идиосом, скрепленных органическим матриксом.
Идиосомы обычно кремниевой природы и имеют вид
пластинок (чешуек) различной формы, иногда с зуб-
цами или выростами.
Расположение идиосом в раковине в большинстве
случаев упорядочено. Они могут быть уложены
вплотную друг к другу или перекрываться. Иногда
(Assulina) все идиосомы одинаковые. У многих видов,
однако, в состав раковины одновременно входят иди-
осомы нескольких типов, различающиеся по форме и
502
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
размеру. У некоторых представителей (Trinema) устье
образовано более мелкими чешуйками, чем тело ра-
ковины. Представители рода Euglypha имеют ха-
рактерные зубцы на чешуйках, окружающих устье
(рис. 498). Многие виды формируют чешуйки с игло-
видными выростами. Вырост может располагаться на
краю (Е. acanthophora, Е. brachiata) или посередине
чешуйки (Е. cristata). У некоторых видов (Placocista
spinosa) иглы представляют собой тонкйе удлинен-
ные идиосомы, вставленные между обычными чешуй-
ками.
Устье может быть окружено воротничком из орга-
нического матрикса; воротничок может быть ровным
(Sphenoderia lenta) или зубчатым (Tracheleuglypha
dent ata).
Псевдоподии и движение
Филозные раковинные амебы, подобно голым фи-
лозным амебам (подкласс Aconchulinia), передвига-
ются посредством образования филоподий (рис. 472).
По организации псевдоподий представители рода
Gromia существенно отличаются от остальных Testa-
ceafilosia.
При движении клетка образует 1 или несколько
филоподий, которые прикрепляются концами к суб-
страту и сокращаются, подтягивая клетку. У некото-
рых видов можно наблюдать «складывание» филопо-
дии при ее сокращении. При этом филоподия пере-
гибается примерно посередине, а в месте перегиба
образуется капля гиалоплазмы, в которую филоподия
втягивается одновременно с обоих концов. Капля
гиалоплазмы при этом разрастается до размеров не-
большого гиалинового островка, который «вливает-
ся» в цитоплазму клетки.
Псевдоподии Gromia представляют собой сеть
разветвляющихся и анастомозирующих цитоплаз-
матических нитей (рис. 483). В местах анастомозов
могут образовываться островки гиалоплазмы, иногда
довольно большого размера.
У многих представителей обоих подотрядов при
движении часть гиалоплазмы выходит через устье ра-
ковины и распластывается по субстрату. Образуется
гиалиновый диск неправильной формы, от которого
берут начало филоподии.
Питание
Относительно небольшие размеры Testaceafilosia
позволяют им фагоцитировать лишь самых мелких
протистов, бактерии и детрит. Некоторые виды мо-
гут фагоцитировать мелкие одноклеточные водоро-
сли. Известны симбионтсодержащие виды, среди ко-
торых одним из наиболее интересных является Раи-
linella chromatophora. Эта амеба облигатно содержит
в цитоплазме 2 крупные цианобактерии. Механизм
передачи симбионтов при делении клетки не изучен.
Интересно, что изначально эта амеба была описана
как одноклеточная водоросль Cyanospira (Chodat,
1920).
Размножение и жизненные
циклы
Размножение происходит путем бинарного деле-
ния и сопровождается морфогенезом дочерней рако-
вины.
У представителей подотряда Euglyphina идиосомы
в составе стенки раковины скреплены органическим
матриксом. Идиосомы синтезируются в аппарате
Гольджи и транспортируются в мембранных оболоч-
ках в район ядра клетки, где накапливаются на пери-
ферии зоны шероховатого ЭПР. Там же накаплива-
ются многочисленные цементные везикулы.
Рис. 475. Схема деления и морфогенеза дочерней раковины Eu-
glypha sp. (По: Ogden, 1991).
А, Б — выпячивание участка цитоплазмы, миграция идиосом и начало фор-
мирования дочерней раковины; В, Г — увеличение объема цитоплазмы до-
черней особи за счет ее вакуоляризации и окончательное формирование
дочерней раковины; Д — деление ядра и миграция одного из ядер в цито-
плазму дочерней особи; Е — отделение дочерней особи.
Fig. 475. Division and daughter shell morphogenesis in Euglypha
sp. (scheme).
А, Б — protrusion of cytoplasmic bud, idiosome migration and first stages of
daughter shell formation; В, Г — increase of cytoplasm volume of the daughter
cell by means of vacuolarization and completion of daughter shell formation;
Д — nucleus division and migration of one of the daughter nuclei to the cyto-
plasm of daughter cell; E — separation of the daughter cell.
ТИП RHIZOPODA, КЛАСС FILOSEA. ПОДКЛАСС TESTACEAFILOSIA
503
В начале деления из устья материнской раковины
выпячивается участок цитоплазмы (рис. 475). Он
быстро увеличивается в размерах и принимает
форму, примерно соответствующую форме будущей
раковины. К поверхности этого выпячивания пере-
мещаются вакуоли с идиосомами и с органическим
цементом. Экскретированные чешуйки упорядочен-
но располагаются на поверхности цитоплазмы, при
этом сначала занимают свои места околоустьевые
чешуйки, а потом ряд за рядом все остальные. На
этой стадии чешуйки удерживаются пучками микро-
филаментов на концах пальцевидных цитоплазмати-
ческих выростов и могут перемещаться по поверх-
ности будущей дочерней клетки. Впоследствии иди-
осомы скрепляются органическим цементом, образуя
стенку дочерней раковины. Ядро, претерпевает деле-
ние. Часть органелл и одно из ядер мигрируют в
дочернюю клетку. Вслед за этим происходит оконча-
тельное разделение цитоплазмы клеток. Дочерняя
клетка еще некоторое время достраивает раковину,
выстилая ее изнутри слоем органического цемента.
У взрослой особи цитоплазма отделяется от стенок
раковины и образуются эпиподии.
Деление у представителей подотряда Gromiina
протекает аналогичным образом, но изучено оно
значительно менее детально. Гомогенная раковина
дочерней клетки формируется за счет перемещения
к ее поверхности цитоплазмы вакуолей, содержащих
материал стенки раковины, и их последующей экск-
реции. Для некоторых видов, имеющих относительно
гибкую раковину, отмечали «прямое» деление клет-
ки путем образования продольной перетяжки на ра-
ковине. В этом случае деление не сопровождается
классическим морфогенезом дочерней раковины; од-
нако эти наблюдения нуждаются в проверке.
Жизненный цикл представителей рода Gromia ос-
тается не до конца понятным. Некоторые авторы
(Hedley, Wakefield, 1969) описывали образование
громией одножгутиковых гамет путем множественно-
го дробления клетки (гамонтогамии, по терминоло-
гии этих авторов). Неясно, происходит ли при этом
мейотическое деление ядра.
Рис. 476. Схема строения цисты Euglypha sp.
А — циста без «стебелька», Б — циста со «стебельком», а — стенка рако-
вины, б — стенка цисты, в — цитоплазма клетки, г — пробка в устье ра-
ковины, д — «стебелек».
Fig. 476. Organization of cyst in Euglypha sp. (scheme).
A — cyst without a «stalk», Б — cyst with a «stalk», a — shell wall, 6 —
cyst wall, в — cell cytoplasm, г — «plug» in shell aperture, d — «stalk».
Рис. 477. Множественная псевдоконъюгация Euglypha rotunda.
(По: Серавин, Гудков, 1984).
Fig. 477. Multiple pseudoconjugation in Euglypha rotunda.
Большинство видов способно инцистироваться.
При этом клетка перестает образовывать псевдопо-
дии, объем цитоплазмы существенно уменьшается за
счет удаления из нее воды. Сократительные вакуоли
перестают функционировать. Некоторые виды обра-
зуют в устье раковины пробку из экскретированных
идиосом и экзогенных частиц, скрепленных органи-
ческим матриксом. Клетка округляется и покрывает-
ся стенкой из органического материала, в которую
могут быть включены идиосомы. В некоторых случа-
ях циста подвешена внутри раковины на стебельке
(рис. 476).
Половой процесс у раковинных филозных амеб не
описан, однако для некоторых видов (Euglypha rotun-
da) известен процесс бинарной или множественной
псевдоконъюгации (Hedley, Ogden, 1973). При бинар-
ной псевдоконъюгации отдельные филоподии 2 осо-
бей сливаются, образуя цитоплазматический мостик.
При множественной псевдоконъюгации происходит
частичное слияние филоподий нескольких особей.
При этом за счет характерного расположения рако-
вин объединившихся организмов (рис. 477) образуют-
ся так называемые розетки (Серавин, Гудков, 1984).
В цитоплазматических мостиках, соединяющих клет-
ки, встречаются пищеварительные вакуоли и другие
включения, однако ни в одном случае не отмечено
перехода ядер из клетки в клетку.
Экология и распространение
Большинство представителей Testaceafilosia оби-
тают в пресноводных водоемах, почвах и мхах. Пред-
ставители подотряда Gromiina населяют преимущест-
венно пресноводные экотопы (кроме интерстициаль-
ных видов), в то время как Euglyphina встречаются во
всех перечисленных экотопах, кроме интерстициали.
Особую группу среди Testaceafilosia составляют
организмы, обитающие в интерстициали морей и
океанов. Эта интерстициальная фауна была открыта
Големанским (Golemansky, 1976) и интенсивно изу-
чается в течение последних лет. В состав группы вхо-
дит около 90 видов, часть из которых представлена
облигатными псаммобионтами.
Супралитораль как среда обитания характеризует-
ся значительными колебаниями солености (от 0.1 до
35 %о) и содержания кислорода. В связи с этим мно-
гие интерстициальные виды являются эвригалинными
(например, Psammonobiotus communis встречается в
504
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
пресных водах и при солености до 18 %о). Видовое
разнообразие облигатных псаммобионтов снижается
по мере удаления от моря, и на расстоянии 12—20 м
от уреза воды они начинают вытесняться факуль-
тативными псаммобионтами, часть из которых широ-
ко распространена в пресноводных водоемах (Trine-
та lineare, Cyphoderia ampulla).
Все псаммобионтные виды, по данным Големанс-
кого, являются космополитами, при этом одни и те
же виды обнаруживаются в зонах, резко различаю-
щихся по солености (например, в супралиторали гви-
нейского побережья Атлантики и в супралиторали
Черного и Балтийского морей). Как правило, видовое
разнообразие больше на супралиторали внутренних
морей, чем на океанических побережьях.
Систематический обзор
Подотряд Gromiina Delage et
Herouard, 1896
Сем. Amphitrematidae Pochfe,
1913
Раковина органическая, иногда с ксеносомами. На
противоположных концах раковины располагается 2
устья.
Роды: Amphitrema Archer, 1867 (рис. 478, 479);
Archerella Loeblich et Tappan, 1961 (-Ditrema Archer,
1877).
Примечание. В род Archerella включают формы, имеющие
раковину без ксеносом, с устьями, не окруженными воротничками.
Некоторые исследователи считают такое разделение рода нецеле-
сообразным.
Сем. Chlamydophryidae
de Saedeleer, 1934
Раковина органическая или из кремниевых идио-
сом, округлая или вытянутая.
Роды: Capsellina Penard, 1909; Chlamydophrys Ci-
enkowski, 1876; Lecythium Hertwig et Lesser, 1874
(рис. 480); Penardeugenia Deflandre et Deflandre-Riga-
ud, 1958 (-Eugenia Averintzev, 1906); Rhogostoma
Belar, 1921.
Примечание. В состав семейства иногда включают роды La-
genidiopsis Golemansky, 1974 (рис. 481; 482) (-Lageniopsis Char-
dez, 1977); Lagunculina Rhumbler, 1904 (=Ovulina Gruber, 1884);
Pseudolagenidiopsis Sudzuki, 1979. Согласно некоторым современ-
ным авторам, эти роды должны быть отнесены к форамйниферам
(см.: Medioli, Scott, 1983; Orgen, Couteaux, 1989).
Сем. Gromiidae Reuss,
1862
Раковина чисто органическая или с небольшим
количеством ксеносом, постоянной формы или спо-
собная к незначительным изменениям формы. Име-
ется дифференцированное околоустьевое образова-
ние — оральная капсула. Ветвящиеся и анастомози-
рующие псевдоподии образуют сеть (в отличие от
других представителей отряда).
Единственный род: Gromia Dujardin, 1835
(рис. 483).
Сем. Psammonobiotidae Golemansky,
1974
Раковина билатерально-симметричная, органичес-
кая или агглютинированная, с ксеносомами или иди-
осомами разнообразной формы, округлая в сечении
или сжатая в дорсовентральном направлении. Устье
располагается под углом к продольной оси тела и
находится у большинства видов в центре ворон-
ковидного расширения раковины. 1 ядро. Псаммоби-
онтные формы.
РоДы: Alepiella Golemansky, 1970 (рис. 484);
Centropixiella Valkanov, 1970 (рис. 485); Chardezia
Golemansky, 1970 (рис. 486); Corythionelloides Sud-
zuki, 1979; Messemvriella Golemansky, 1973
(рис. 487); Micramphora Valkanov, 1970 (рис. 488);
Micramphoraeopsis Sudzuki, 1979; Micropsammella
Golemansky, 1970 (рис. 489); Micropsammelloides
Sudzuki, 1979; Ogdeniella Golemansky, 1982 (=Am-
phorellopsis Golemansky, 1970); Psammonobiotus Go-
lemansky, 1967 (рис. 490); Pseudocorythion Valkanov,
1970 (рис. 491).
Сем. Pseudodifflugiidae de Saedeleer,
1934
Раковина вытянутая, агглютинированная, с ксено-
сомами, иногда с идиосомами.
Роды: Clyopelina Penard, 1902; Diaphorodon Ar-
cher, 1869; Frenzelina Penard, 1902; Maroltia Haman,
1988 (=Baileya Averintzev in Schouteden, 1906;
-Pamphagus Bailey, 1853); Nadinella Penard, 1899;
Plagiophrys Claparede et Lachmann, 1859; Pseudodif-
flugia Schlumberger, 1845; Rhumbleriella Golemansky,
1970.
Сем. Volutellidae Sudzuki,
1979
Раковина органическая, трубчатая, с улитковид-
ным завитком, с темными пластинками, без удлинен-
ной устьевой зоны; с септой, делящей раковину на
камеры, или без нее. Устье округлое. Псаммобионт-
ные формы.
Роды: Chardezella Haman, 1988 (=Voluta Chardez,
1972; =Volutella Chardez, 1972) (рис. 492); Pseudovo-
lutella Sudzuki, 1979.
Примечание. Систематическое положение родов, составля-
ющих семейство, требует уточнения. Возможно, они должны быть
отнесены к фораминиферам (см.: Golemansky, 1992).
ТИП RHIZOPODA. КЛАСС FILOSEA. ПОДКЛАСС TESTACEAFILOSIA
505
Рис. 478—506. Раковины представителей разных видов Testaceafilosia.
478. Amphitrema flavum. (По: Defiandre, 1953). 479. Amphitema stenostoma. (По: Defiandre, 1953). 480. Lecythium hyalinum. 481. Lagenidiopsis elegans. (По:
Golemansky, 1976). 482. Lagenidiopsis valkanovi. (По: Golemansky, 1979). 483. Gromia oviformis. 484. Alepiella tricornuta. (По: Golemansky, 1970). 485.
Centropyxiella platystoma. (По: Golemansky, 1981). 486. Chardezia caudata. (По: Golemansky, 1974). 487. Messemvriella filosa. (По: Golemansky, 1974). 488.
Micramphora tokioensis. (По: Sudzuki, 1979). 489. Micropsammella retorta. (По: Golemansky, 1974). 490. Psammonobiotus communis. (По: Golemansky, 1974).
506
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
499
Q
506
505
Рис. 478—506 (продолжение).
491. Pseudocorythion wailesi. (По: Sudzuki, 1979). 492. Chardezella cochlea. (По: Sudzuki, 1979). 493. Campascus interstitialis. (По: Golemansky, 1981). 494.
Corythionella minima. 495. Cyphoderia ampulla. 496. Pseudocyphoderia orientalis. (По: Sudzuki, 1979). 497. Assulina seminulum. 498. Euglypha acanthophora.
(По: Hedley et al., 1974). 499. Euglyphidion enigmaticum. 500. Placocista spinosa. 501. Playfairina caudata. 502. Sphenoderia lenta. 503. Tracheleuglypha acolla.
504. Paulinella chromatophora. 505. Corythion dubium. 506. Trinema enchelys. Рис. 484, 485, 487, 489, 490, 493, 496: A — вид с брюшной стороны, Б — вид
сбоку. Рис. 486: А — вид с брюшной стороны: Б, В — вид сбоку. Рис. 491: А — вид сверху, Б — вид сбоку. Рис. 498: А — общий вид, Б — вид устья, В —
приустьевая пластинка (увеличена).
Fig. 478—506. Shells of the representatives of different species of Testaceafilosia.
Fig. 484, 485, 487, 489, 490, 493, 496: A — ventral view, Б — lateral view. Fig. 486: A — ventral view; Б, В — lateral view. Fig. 491: A — view from above,
Б — lateral view. Fig. 498: A — general view, Б — aperture view, В — aperture funnel (enlarged).
ТИП RHIZOPODA. КЛАСС Fl LOSEA. ПОДКЛАСС TESTACEAFILOSIA. SUMMARY
507
Gromiina incertae sedis
Edaphonobiotus Schonborn et al., 1983; Microquad-
rula Golemansky, 1968.
Примечание. Систематическое положение большинства ин-
терстициальных раковинных амеб неясна. Многие виды первона-
чально описаны исключительно по пустым раковинам. Более позд-
ние наблюдения показали, что некоторые из таких видов имеют
гранулоретикулоподии. Вследствие этого роды, к которым отно-
сятся эти виды, были включены в различные семейства форамини-
фер.
Судзуки (Sudzuki, 1979) сгруппировал известные к тому вре-
мени роды в 4 семейства. Современные исследователи (Bovee,
1985) включают роды, отнесенные Судзуки к семействам Micfam-
phoridae и Pseudocorythionidae, в состав сем. Psammonobidtidae
Golemansky, 1974. Роды, отнесенные Судзуки к сем. Lagenidiopsi-
dae, относят либо к сем. Chlamydophryidae de Saedeleer, 1934
(Bovee, 1985), либо причисляют к форамийиферам (см.Г Medioli,
Scott, 1983; Ogden, Couteaux, 1989). Роды, Отнесенные Судзуки к
сем. Volutellidae, иногда рассматривают как фораминифер (Gole-
mansky, 1992).
Подотряд Euglyphina Bovee,
1985
Сем. Cyphoderiidae de Saedeleer,
1934
Раковина удлиненная, обычно изогнутая у перед-
него конца, ретортовидная, покрыта мелкими округ-
лыми или овальными, кремнеземными идиосомами,
иногда налегающими друг на друга, так что создает-
ся впечатление гексагональной структуры. Иногда на
раковине могут встречаться отдельные ксеносомы.
Устье простое или с тонким воротничком.
Роды: Campascus Leidy, 1879 (рис. 493)^ Corythi-
onella Golemansky, 1970 (рис. 494);; Cyphoderia
Schlumberger, 1845 (рис. 495); Pseudocyphoderia Sud-
zuki, 1979 (рис. 496); Pseudowailesella Sudzuki, 1979;
Schaudinnula Averintzev, 1906.
Сем. Euglyphidae Wallich, 1864
Раковина состоит из овальных кремниевых плас-
тинок (идиосом), удлиненная, акростомная. Устье ок-
руглое или эллипсоидное.
Роды: Ampullatoria van Оуе, 1956; Assulina Eh-
renberg, 1872 (рис. 497); Euglypha Dujardin, 1840
(рис. 498); Euglyphidion Bonnet, 1960 (рис. 499); He-
teroglypha Thomas et Gauthier-Lievre, 1959; Pareug-
lypha Penard, 1902; Placocista Leidy, 1879 (рис. 500);
Playfairina Thomas, 1961 (рис. 501); Sphenoderia
Schlumberger, 1845 (=Hologlypha Ehrenberg, 1872)
(рис. 502); Tracheleuglypha Defiandre, 1928 (рис. 503);
Valkanovia Tappan, 1966 (=Euglyphella Valkanov,
1962).
Сем. Paulinellidae de Saedeleer, 1934
Прозрачная раковина из крупных удлиненных
пластинок с закругленными краями, которые, нале-
гая друг на друга, создают впечатление гексагональ-
ной структуры. В цитоплазме имеется 2 крупные
симбиотические цианобактерии. Ядро 1.
Единственный род: Paulinella Lauterborn, 1895
(рис. 504).
Примечание. Этот род также был описан как одноклеточ-
ная водоросль Cyanospira Chodat, 1920 (Loeblich, Tappan, 1964;
Tappan, 1980).
Сем. Trinematidae Hoogenraad et de
Groot, 1940
Раковина из округлых или овальных идиосом, уп-
лощенная в дорсовентральном направлении. Устье
эксцентричное.
Роды: Corythion Taranek, 1882 (рис. 505); Trine-
та Dujardin, 1841 (рис. 506).
THE PHYLUM RHIZOPODA SIEBOLD, 1845
THE CLASS FILOSEA LEIDY, 1879
THE SUBCLASS TESTACEAFILOSIA DE SAEDELE£R, 1934
SUMMARY
This subclass comprises filose amoebae possessing «shell» (or
«test») —- a rigid structure outside the cell. Earlier this group was
often considered together with lobose testate amoebae within the order
Testacea, due to the presence in both of more or less similarly orga-
nised «shell». Nowadays the former «Testacea» are divided, according
to the nature of pseudopodia, into subclasses Testacealobosia and Tes-
taceafilosia and are classified within different rhizopod classes.
The subclass Testaceafilosia comprises one order, Gromiida,
which is divided into two Suborders: Gromiina and Euglyphina. In
general, the systematics of the group is poorly developed and many of
its taxa are probably not homogeneous.
In the chapter light-microscopical and ultrastructural data on the
representatives of Testaceafilosia are considered and summarised. The
chapter includes data arid illustrations oh shell structure, cell organisa-
tion, movement, division and encystment, habitats and distribution of
representatives of this group. Detailed systematic account includes di-
agnoses and generic composition of families. Synonyms for each
genus are listed. Illustrations of representatives of most genera are
presented.
The classification used in this paper follows that of Bovee (1985).
Generic synonymy is given according to Haman (1988).
The cell of representatives of Testaceafilosia is enclosed in a shell
(«test»). It has one or, rarely, two openings (apertures) through which
pseudopodia (filopodia) emerge. The cell may fill entire shell cavity
or part of it, in which case it is attached to the inside of the shell wall
by means of epipodia.
The cell has thin undifferentiated glycocalyx. The nucleus is
vesicular, rarely granular. Both uninuclear and multinuclear species
are known. Mitochondria have tubular anastomosing or vesicular
cristae. Numerous dictyosomes of the Golgi complex are charac-
teristic of many species. They play an important role in secreting
material for the building of the shell. Rough EPR is also well-deve-
loped. Compartmentalisation of cytoplasm is characteristic of species
with large shell.
508
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Shell shape varies greatly. All morphological types of shells
(discussed in «Testacealobosia») are found in Testaceafilosia as
well.
Two suborders within Gromiida differ by the nature of shell. The
shells of Gromiina are made of secreted organic layers in which xeno-
somes are sometimes embedded. Shells of Euglyphina are agglutinated
and consist of siliceous flattened idiosomes («scales») glued with or-
ganic matrix. Shape of scales varies, indentations and spines are com-
mon.
Movement of filose testate amoebae by means of filopodia is des-
cribed. Representatives of Gromia genus are shown to differ from the
rest of the group by the pseudopodial organisation. They form a net
of branching and anastomosing pseudopodia. Division is by binary
fission followed by morphogenesis of die daughter shell. The main
stages of division process of representatives of Euglyphina are descri-
bed. Division in Gromiina is poorly known.
Testaceafilosia live in freshwater, soil and moss habitats. Several
groups of Gromiina inhabit marine interstitial sands.
ЛИТЕРАТУРА28
Серавин Л. H., Гудков А. В. Основные типы и формы агамных
слияний у простейших // Цитология. 1984. Т. 26, № 2. С. 123—130.
Berrend R.E. Filopodial movement in Cyphoderia ampulla
(Ehr.) // Primitive motile systems in cell biology / Eds R. D. Allen,
N. Kamiya. New York: Acad. Press, 1964. P. 433—443.
Bovee E. C. An illustrated guide to protozoa. Kansas, 1985.
Chardez D. Thecamoebiens du mesopsammon des plages de la
Mer du Nord // Rev. Verviet. Hist. Nat, 1977. T. 34. P. 18—34.
Chodat R. Algues de la region du Grand St. Bernard // Soc. Bot.
Geneve Bull. 1920. T. 2, N 12. P. 293—305.
Deflandre G. Orders des Testacealobosa, Testaceafilosa, Thala-
mia ou Thecamoebiens (Rhizopoda Testacea) /-Traite de Zoologie.
Paris, 1953. Vol. 1, fasc. 2. P. 97—148.
Golemansky V. Rhizopodes nouveaux du psammon littoral de la
mer Noire // Protistologica. 1970. Vol. 6. P. 365—371.
Golemansky V. Psammonobiotidae fam. nov. Une nouvelle famil-
le de Thdcamoebiens du psammon supralittoral des mers // Acta Pro-
tozool. 1974. T. 13. P. 137—142.
Golemansky V. Rhizopodes psammobiontes (Protozoa, Rhizopo-
da) du psammal supralittoral des cotes de 1’Atlantique // Acta Zool.
Bulgar. 1976. T. 4. P. 23—29.
Golemansky V. Description de trois thecamoebiens (Protozoa:
Rhizopodea) nouveaux des eaux souterraines littorales des mers //
Acta protozool. 1981. T. 20. P. 115—119.
Golemansky V. Thecamoebiens interstitiels (Rhizopoda: Arcelli-
nida, Gromida et Monothalamida) du supralittoral francais de
1’Atlantique dans la region du Roscoff (Bai de Morlaix) // Acta Zool.
Bulgar. 1992. T. 45. P. 3—15.
Haman D. Annotated lexicon of testate lobosid and filosid amoe-
bean genera, subgenera and type species (Sarcqdina: Protozoa: «The-
28 Цитируемая и рекомендуемая литература к разделу «Тип Rhi-
zopoda. Класс Filosea. Подкласс Testaceafilosia».
camoebians»)// Rev. Esp. Micropaleontol. 1988. Vol. 20, N 2.
P. 205—244.
Hedley R.H. The iron-containing shell of Gromia oviformis
(Rhizopoda) // Quart. J. Microscop. Sci. 1960. Vol. 101, N 3.
P. 279—293.
Hedley R. H., Ogden C. G. Biology and fine structure of Euglyp-
ha rotunda (Testacea, Protozoa) // Bull. Brit. Mus. nat. Hist. (Zool.)
1973. Vol. 25. P. 121—137.
Hedley R. H., Wakefield J. St. J. Fine structure of Gromia ovifor-
mis (Rhizopodea, Protozoa) // Bull. Brit. Mus. nat. Hist. (Zool.) 1969.
Vol. 18. P. 69—89.
Hedley R.H., Ogden C.G., Kraft J. I. Observations on clonal
cultures of Euglypha acantophora and E. strigosa (Testacea: Pro-
tozoa) // Bull. Brit. Mus. Nat. Hist. (Zool.). 1974. Vol. 27. P. 103,
111.
Loeblich A. R., Tappan H. Protista. 2. Sarcodina, chiefly «Theca-
moebians» and Foraminiferida // Treatise on Invertebrate Paleontolo-
gy, Pt., C / Ed. R. C. Moore. Lawrence; Kansas: Geol. Soc. Amer.;
Univ. Kansas Press, 1964.
Medioli F. S., Scott D. B. Holocene Arcellacea (Thecamoebians)
from Eastern Canada // Cushman Found. Foram. Res. Spec. Public.
1983. Vol. 21. P. 1—63.
Ogden C. G. An ultrastructural study of division in Euglypha
(Protozoa: Rhizopoda) // Protistologica. 1979. Vol. 15. P. 541—556.
Ogden C. G. Observations of clonal cultures of Euglyphidae
(Rhizopoda, Protozoa) // Bull. Brit. Mus. nat. Hist. (Zool.) 1991.
Vol. 41, N 4. P. 137—151.
Ogden C. G., Couteaux M.-M. Interstitial marine Rhizopods
(Protozoa) from littoral sands on the East Coast of England // Eur. J.
Protistol. 1989. Vol. 24. P. 281—290.
Sudzuki M. Marine Testacea of Japan // Sesoko Mar. Sci. Lab.
Tech. Rept. 1979. Vol. 6. P. 51—62.
Tappan H. The paleobiology of plant protists. San Francisco: Fre-
eman et Co., 1980.
Класс Peloflagellatea Goodkov et Seravin, 1991 —
Пелофлагеллаты (Pelomyxa palustris)
Pelomyxa palustris (Greeff, 1874) — единственный вид в соста-
ве класса — пресноводный многоядерный амебоидный протист,
имеющий жгутики, которые не принимают участие в обеспечении
клеточной локомоции.
Этот организм обычно встречается в небольших пресноводных
озерах, прудах, застойных заводях рек и тому подобных местооби-
таниях, богатых разлагающимся органическим материалом расти-
тельного происхождения (Dejdar, 1931; Leiner et al., 1954; Kudo,
1957; Daniels et al., 1966; Andresen et al., 1968). Иловые отложения
на дне такого рода водоемов (сапропель) характеризуются практи-
чески полным отсутствием кислорода или очень незначительным
его содержанием (Brand, 1946; Van Bruggen et al., 1983; Fenchel,
Finlay, 1991). Приуроченность P. palustris именно к таким услови-
ям окружающей среды была подтверждена экспериментально (An-
dresen et al., 1968; Chapman-Andresen, Hamburger, 1981; Van Brug-
gen et al., 1988).
P, palustris обнаружена в самых различных частях
Европы, Азии и Америки: Англия и Гебридские ос-
трова (Archer, 1877; Gulliver, 1888; Veley, 1905;
West, 1905; Whatley, 1976), Франция (Schirch, 1914;
Hollande, 1945), Швейцария (Penard, 1893, 1902), Че-
хословакия (Stolz, 1900; Dejdar, 1931; Fortner, 1934),
Германия (Greeff, 1874; Bott, 1906; Okada, 1930; Lei-
ner et al., 1954), Дания (Andresen et al., 1968), Нидер-
ланды (Van Bruggen et al., 1988), Россия (Аверинцев,
1906; Серавин, Гудков, 1987а), Индия (Bourne, 1891),
США (Leidy, 1879; Mast, 1934; Kudo, 1957; Daniels et
al., 1966). Кроме того, Кудо (Kudo, 1957) сообщает,
что у него имеются сведения о находках Р, palustris
на территории Ирландии и Италии. Наконец, Андре-
сен (Andresen, 1973) упоминает о том, что пеломиксу
находили также в северной части Африканского кон-
тинента. Таким образом, Р. palustris имеет чрезвы-
чайно широкое, практически всесветное распростра-
нение.
ТИП RHIZOPODA. КЛАСС PELOFLAGELLATEA
509
История изучения
Среди протистов, пожалуй, нет больше такого
вида, который, подобно Р. palustris, имел бы столь
богатую «приключениями» историю изучения и
представления о природе которого и положении в
системе низших эукариот менялись столь часто и
радикально. Можно сказать, что уже самое начало
изучения этого организма было не совсем обычным.
Название Pelomyxa palustris и ее подробное описа-
ние были даны Рихардом Греефом (Richard Greeff) в
1874 г., хотя, как сообщает этот автор (Greeff,
1874), впервые он обнаружил пеломиксу в 1867 г. й
присвоил ей название Pelobius, сделав краткое опи-
сание своей находки в 1870 г. Однако выяснилось,
что название Pelobius уже тогда было преаккупиро-
ванным, причем дважды: Эрихсон (Erichson) еще в
1832 г. использовал это наименование для предста-
вителя водных жесткокрылых (отряд Coleoptera), а
Фитзингер (Fitzinger) в 1843 г. обозначил им одного
из земноводных (класс Amphibia).
Причина того, что жук, амфибия и простейшее
независимо получили одно и то же наименование,
обусловлена сходством в условиях обитания этих ор-
ганизмов, отраженных в смысловом значений слова
pelobius'. корень «ре!» берет начало от греческого
pelos («ил», «грязь»), а греч. bios, как известно, оз-
начает «жизнь». Поэтому позднее Грееф (Greeff,
1874) был вынужден дать ему новое родовое наиме-
нование — он заменил окончание -bius на -туха (от
греч. тиха — слизь); тогда же он присвоил ему и
видовой эпитет «palustris».
Таким образом, датой неформального открытия
пеломиксы является 1867 год. Тем не менее инте-
ресно отметить следующее. Как известно, считается
общепризнанным, что первым, кто наблюдал живую
амебу, был Резель фон Розенхоф (R6sel von Rosen-
hof). Обнаруженный им в 1755 г. амебоидный орга-
низм, который он назвал «der kleine Proteus», Лин-
ней впоследствии обозначил сначала как Volvox
chaos (1758), а затем как Chaos protheus
тогда как Мюллер (Muller) в 1786 г. использовал для
него название Proteus diffluens (ей.: Lorch, 1973).
Как показал тщательный сравнительный анализ име-
ющихся данных (Kudo, 1959), организм, увиденный
Розенхофом, в действительности более всего напо-
минает Pelomyxa palustris.
Последовательность изменений научных представ-
лений о природе Р. palustris и положении этого орга-
низма в системе живого будет подробно рассмотрена
нами далее.
Классификация
Класс Peloflagellatea Goodkov et Seravin, 1991
включает единственный отряд Pelobiontida (Page, 1976)
с единственным сем. Pelomyxidae (Schulze, 1877)' и
единственным монотипическим родом Pelomyxa
(Greeff, 1874).
Морфология, особенности
организации,
систематический обзор
Внешняя морфология, способ
движения и «традиционное»
представление о природе
и систематическом положении
Р. palustris
Пеломикса перемещается в пространстве посред-
ством амебоидного движения. Движущиеся особи
имеют вид толстого цилиндра; на заднем конце тела,
как правило, присутствует полусферической формы
утолщение или вздутие, отделяющееся от остальной
части тела более или менее выраженной перетяж-
кой, — уроид (рис. 507, вкл.). На поверхности урои-
да, как правило, присутствуют многочисленные,
иногда довольно крупные гиалиновые выросты эктоп-
лазмы (рис. 508, вкл.) — вилли (villi). При движении
наблюдается равномерный, направленный вперед ак-
сиальный ток цитоплазмы с отчетливым ее фонтани-
рованием на переднем конце тела (Okada, 1930; For-
tner, 1934; Mast, 1934; Holland, 1945; Leiner et al.,
1954; Kudo, 1957; Серавин, 1967; Andresen, 1973). В
числе особенностей этого вида следует отметить его
весьма значительные размеры: в среднем они состав-
ляют 600—800 мкм, однако многие особи имеют в
длину до 1 мм, а отдельные экземпляры могут дости-
гать даже до 5 мм (Greeff, 1874; Penard, 1902; Fort-
ner, 1934; Kudo, 1957; Bovee, 1985). P. palustris —
многоядерный организм; крупные и средних разме-
ров особи содержат, как правило, несколько сотен
(подсчитано было до 400) ядер, однако имеются све-
дения, что некоторые мелкие экземпляры содержали
всего лишь несколько ядер (Leiner et al., 1954; Kudo,
1957; Daniels et al., 1966; Daniels, Breyer, 1967; An-
dresen, 1973; Daniels, 1973; Chapman-Andresen, 1978,
1982, 1985). Характерными особенностями P. palust-
ris является также присутствие большого количества
оптически пустых, называемых структурными, вакуо-
лей, которые, по некоторым подсчетам, занимают до
75 % всего объема цитоплазмы (цитоплазма выглядит
под световым микроскопом как бы вспененной), на-
личие гликогеновых тел и отсутствие обычного ком-
плекса сократительной вакуоли (Greeff, 1874; Leidy,
1879; Korotneff, 1879; Gulliver, 1888; Stolz, 1900; Pe-
nard, 1902; Fortner, 1934; Leiner et al., 1954; Kudo,
1957; Daniels et al., 1966; Andresen et al., 1968; Chap-
man-Andresen, 1971; Whatley, Whatley, 1983). Нако-
нец, в цитоплазме пеломиксы всегда присутствует не-
сколько форм облигатных эндобиотических бакте-
рий (рис. 509, А, Б, вкл.) (Veley, 1905; Holland, 1945;
Leiner et al., 1954; Kudo, 1957; Daniels et al., 1966;
Andresen et al., 1968; Chapman-Andresen, 1971; What-
ley, 1976; Whatley, Whatley, 1980, 1983; Van Bruggen
et al., 1983; Vogels et al., 1984). Насколько этот при-
знак является характерным свойством Р. palustris,
свидетельствует тот факт, что об этом довольно под-
робно писали еще старые авторы. Так, Пенар (Ре-
510
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
nard, 1902) считал, что он первым высказал сообра-
жение о присутствии у пеломиксы эндобионтов, од-
нако еще Грубер (Gruber, 1885) предполагал их нали-
чие в цитоплазме P.villosa, которую сам же Пенар
(Penard, 1902) свел в синоним Р. palustris. Бурн (Bo-
urne, 1891), давая описание Р. viridis, впоследствии
также сведенной в синонимР. palustris (Kudo, Р957),
прямо указал на присутствие симбиотических бакте-
рий у этого организма. •
На базе имевшегося до второй половины нашего
столетия фактического материала об организации
Р. palustris систематическая принадлежность этого
организма к амебам практически ни у кого не вызы-
вала сомнений. Правда, в качестве курьеза можно
упомянуть о взглядах Галливера (Gulliver, 1888), ко-
торого размеры и сильная вакуолизированность ци-
топлазмы пеломиксы натолкнули на мысль, что она
представляет собой организм, состоящий из множес-
тва отдельных мелких одноядерных клеток.
Морфология локомоторной формы Р. palustris и
ее способ перемещения характерны для представите-
лей настоящих лобозных амеб (Rhizopoda, Lobosea),
на основании чего Р. palustris и относили к числу
представителей этого таксона, нередко из-за ее раз-
меров именуя «гигантской амебой» (см.: Penard,
1902; Doflein, Reichenow, 1953; Kudo, 1957; Page,
1976; Крылов и др., 1980; Levine et al., 1980). Даже
после того как были произведены электронно-мик-
роскопические исследования пеломиксы и получило
развитие совершенно иное представление о природе
этого организма, некоторые исследователи продол-
жали относить Р. palustris к числу Rhizopoda либо в
составе таксона Lobosea, либо наряду с ним (Bovee,
1985; Page, 1987, 1988, 1991; Sleigh, 1989; Карпов,
1990).
Представление о Р. palustris как
организме, родственном
проэукариотам
Данные, полученные в результате ранних элект-
ронно-микроскопических исследований Р. palustris
(см.: Daniels et al., 1966; Daniels, Breyer, 1967; An-
dresen et al., 1968; Andresen, 1973; Chapman-Andre-
sen, 1973; Daniels, 1973; Whatley, 1976), послужили
для многих исследователей основой для радикально-
го изменения представлений о природе этого орга-
низма. Было сделано заключение, что пеломикса
представляет собой крайне примитивный организм,
живой реликт, близкий по своей организации к
предковым формам современных протистов (Jahn et
al., 1974; Margulis, 1974, 1981; Whatley, 1976; What-
ley, Whatley, 1980, 1983; Margulis, Schwartz, 1982;
Corliss, 1984). Основанием для такого вывода послу-
жило то обстоятельство, что у нее не были найдены
митохондрии, тельца Гольджи, ЭПР, микротельца,
центриоли, ЦОМТы, и лишь наличие ядерной обо-
лочки, как отмечает Маргелис (Margulis, 1974,
1981), дает формальное право относить Р. palustris к
числу эукариот. В соответствии с этими взглядами
на природу пеломиксы она была выделена в особый
тип Caryoblastea (Karyoblastea)29(Margulis, 1974; см.
также: Margulis, 1981; Corliss^ 1984; de Puytorac et
al., 1987). Считалось, что кариобластеи «наиболее
примитивные из всех живущих эукариот» (Margulis,
Schwartz, 1982, с. 72). Следует отметить, что иссле-
дователи, признающие пеломиксу родственной про-
эукариотным организмам, как правило, придержива-
ются симбиогенетической теории происхождения
эукариотной клетки. Р. palustris использовалась
рядом авторов в качестве возможной иллюстрации
своих представлений о возникновении митохондрий
из эндобиотических бактерий (Whatley, 1976; What-
ley, Whatley, 1980, 1983, и др.). «Не исключено, —
пишет Маргелис, — что Pelomyxa... является пря-
мым связующим звеном с теми организмами, кото-
рые когда-то приобрели протомитохондрии в качес-
тве эндобионтов» (Margulis, 1983. С. 184).
Идею о примитивности Р. palustris поддержал и
развил в своих работах Кавалье-Смит (Cavalier-
Smith, 1983, 1987, 1991, 1993, 1995). Более подроб-
но аргументацию этого исследователя мы рассмот-
рим несколько позже, сейчас же отметим, что он
разработал свою, отличную от остальных систему
протистов, в которой поместил Р. palustris наряду с
другими, по его мнению примитивными, первично-
безмитохондриальными амебоидными организмами в
тип Archamoeba, входящий в отдельное царство Ar-
chezoa, противопоставив его тем самым остальным
простейшим, составляющим царство Protozoa (см.
также: Corliss, 1994).
Новое представление об
организации и природе Р. palustris
Электронно-микроскопические исследования, вы-
полненные в отличие от предыдущих работ с учетом
того, что Р. palustris обитает в условиях дефицита
кислорода, позволили выявить совершенно иную
картину тонкой организации этого организма, нежели
представлялось ранее. Обнаружено, что у пеломиксы
имеются многочисленные мелкие тельца Гольджи,
разбросанные по всей эндоплазме клетки (Серавин,
Гудков, 1987а; Гудков, Серавин, 1991а). Они являют-
ся производными мембран мощно развитой вакуо-
лярной системы пеломиксы и функционально про-
должают быть тесно с ней связанными, поскольку
часть продуцируемых этими структурами везикул и
микровакуолей поступает непосредственно в эндоп-
лазматические вакуоли. В целом вакуолярная систе-
ма Р. palustris представляет собой единый, весьма
пластичный транспортно-распределительный аппа-
рат клетки; иными словами, она соответствует по
своей природе мембранам гладкого ЭПР (Гудков,
Серавин, 1991а), и, таким образом, вакуоли пело-
миксы нельзя считать просто «структурными вакуо-
лями». Сравнительно-морфологический анализ пока-
зывает, что чрезвычайно сходное развитие вакуолярной
системы свойственно также ряду других пресновод-
ных протистов (инфузорий, амеб), у которых, как и
29 Корлисс (Corliss, 1984) справедливо предложил заменить на-
чальную букву «С» в исходном наименовании данного таксона на
букву «К» (от греч. karyon — ядро).
ТИП RHIZOPODA. КЛАСС PELOFLAGELLATEA
511
у пеломиксы, отсутствует специализированный ком-
плекс сократительной вакуоли (Penard, 1902; Mas-
hansky et al., 1963; Page, 1978, 1988). Например,
цитоплазма голой лобозной амебы Pseudothecamoeba
proteoides структурно организована очень сходно с
таковой Pelomyxa palustris, а мембраны ее вакуоляр-
ной системы представляют собой гипертрофирован-
ный ЭПР (Page, 1978).
Интересно отметить, что еще Лейнер с соавт.
(Leiner et al., 1954), изучая Р. palustris на светоопти-
ческом уровне, высказали сомнение относительно
присутствия митохондрий у этого вида, что впослед-
ствии было подтверждено электронно-микроскопи-
ческими данными (Daniels et al., 1966; Daniels, Brey-
er, 1967; Andresen et al., 1968; Andresen, 1973; Chap-
man-Andresen, 1973; Daniels, 1973; Whatley, 1976).
Однако отсутствие этих органелл у пеломиксы —
вполне закономерное явление. Сейчас хорошо извес-
тно, что многие не только паразитические, но и сво-
бодноживущие протисты, обитающие, как и пело-
микса, в условиях дефицита кислорода (в сапропе-
ле), могут быть лишены митохондрий (Fenchel et al.,
1977; Finlay, Fenchel, 1989; Fenchel, Finlay, 1991;
Серавин, 1992). Ныне известно несколько сот видов
таких протистов, причем вторичность отсутствия
митохондрий у них просто не вызывает сомнений
(см.: Серавин, 1992).
Во многих случаях, хотя и не всегда, у таких про-
тистов появляются особые микротельца — гидроге-
носомы, содержащие специфический, отличный от
других типов микротелец набор ферментов, но мор-
фологически часто довольно сходные с ними (Muller,
1975; De Duve, 1983; Finlay, Fenchel, 1989; Fenchel,
Finlay, 1991). Правда, в отношении поверхностной
мембраны гидрогеносом среди исследователей нет
единого мнения. Более того, показано, что в ряде
случаев она унитарная, а иногда имеется 2 тесно
сближенные мембраны, часто спаянные воедино
электронно-плотным веществом; наконец, обычная
мембранная оболочка может вообще отсутствовать
(см.: Серавин, 1992).
В цитоплазме Р. palustris уже давно были найдены
небольшие гранулы или, как их называли некоторые
авторы, «сфероидальные тельца» (Daniels et al., 1966;
Daniels, Breyer, 1967; Andresen et al., 1968; Andresen,
1973; Daniels, 1973), весьма напоминающие микро-
тельца и, возможно, представляющие собой гидроге-
носомы (Серавин, Гудков, 19876). Правда, Чапман-
Андресен и Гамбургер (Chapman-Andresen, Hambur-
ger, 1981) посчитали их пероксисомами, однако, по
данным самой Чапман-Андресен (Chapman-Andresen,
1981), у пеломиксы практически отсутствует каталаз-
ная активность цитоплазмы, тогда как для пероксисом
характерно наличие этого фермента (Muller, 1975;
De Duve, 1983).
Следует особо отметить, что анаэробные и мик-
роаэробные простейшие из разных, иногда далеко
отстоящих друг от друга систематических групп
(т. е. потерявших митохондрии независимо друг от
друга и соответственно независимо сформировавших
свои гидрогеносомы в тех случаях, когда они имеют-
ся), часто вступают в тесную симбиотическую ассо-
циацию с метаногенными архебактериями (Fenchel
et al., 1977; Van Bruggen et al., 1983, 1984, 1986;
Vogels et al., 1984; Finlay, Fenchel, 1989; Broers et
al., 1990; Fenchel, Finlay, 1991, 1992; Esteban et al.,
1993). Эти симбионты метаболически тесно связаны
с гидрогеносомами, утилизируя появляющийся в них
молекулярный водород и образуя метан. Специаль-
ные исследования показали, что 2 вида эндобиоти-
ческих бактерий, характерных для Р. palustris, пред-
ставляют собой именно метаногенные формы (Van
Bruggen et al., 1983, 1988; Vogels et al., 1984). Более
того, у столь различных пресноводных организмов^
обитающих в сапропеле, как Р. palustris, инфузории
Metopus striatus и Trimyema compressum, а также
амебофлагеллата Mastigella sp., обычным эндобион-
том является один й тот же вид архебактерйи —
Methanobacterium formicicum (Van Bruggen et al.,
1984, 1985, 1988; Goosen et al., 1990). Все эти
факты свидетельствуют о глубочайших параллелиз-
мах в эволюции представителей различных групп
протистов, перешедших 'к существованию в анаэроб-
ных и микроаэробных условиях, и естественным об-
разом объясняют вторичность отсутствия Митохонд-
рий у Р. palustris.
Очень важное значение для понимания истинной
природы Р. palustris имеют данные по организации ее
цитоскелета, а также тот факт, что у нее имеются
настоящие жгутики.
Электронно-микроскопические исследования по-
казывают, что весьма существенным компонентом
опорно-двигательного аппарата пеломиксы являются
тубулиновые структуры (Серавин, Гудков, 1987в;
Griffin, 4988; Гудков, 1989), причем в кортикальной
зоне клетки они получают наибольшее развитие и
структурную организованность (рис. 510, вкл.); мощ-
ные пучки микротрубочек часто пронизывают в про-
дольном направлении гиалйновые билли уроида.
Присутствие такого микротрубочкового цитоскелета
совершенно не характерно для лобозных амеб, опор-
но-двигательный аппарат которых представлен глав-
ным образом кортикальной системой микрофила-
ментов (Taylor, Gondeelis, 1979; Stockem et al., 1982;
Stockem, Klopockaj 1988, и др.), хотя цитоплазмати-
ческие микротрубочки и были найдены у ряда пред-
ставителей этой группы простейших (Bowers, Korn,
1968; Page, 1979, 1980, 1981; Preston, 1985; Christi-
ani et al., 1986).
В эктоплазме пеломиксы также были обнаружены
рудиментарные кинетосомы, обладающие нетипич-
ным строением и поэтому названные субцентриолями
(Гудков, 1989). Каждая субцентриоль ассоциирована
с ЦОМТом и имеет цитоплазматический микротру-
бочковый дериват, принимающий непосредственное
участие в организации эктоплазматического микро-
трубочкового цитоскелета пеломиксы.
Уже ранние исследователи отмечали, что на по-
верхности тела движущихся амебоидным способом
Р. palustris присутствуют относительно многочислен-
ные тонкие неподвижные «щетинки» («bristles»)
(Blochmann/ 1894; Okada, 1930; Daniels, 1973;
Bovee,* 1985; Page, 1987, 1988). В действительности,
как уже было сказано, они представляют собой на-
стоящие, хотя и неподвижные жгутики (рис. 511)
(Серавин, Гудков, 1987г; Griffin, 1988; Гудков,
512
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
1989). Контрактильная система ундулиподий не
только отклоняется от стандартного микротрубочко-
вого паттерна (9 + 2), но и существенно варьирует
даже у одной и той же особи. Были найдены жгутики
с формулой строения аксонемы 8 + 1,9 +0, 9+1,
10 + 1, 10 + 5 и даже 14 + 3. Весьма значительным
изменениям подвергаются также кинетосомы (Гуд-
ков, 1989), хотя корешковая система в виде орга-
низованного пучка микротрубочек, часто идущего
вдоль поверхности тела (или аксиально пронизываю-
щего гиалиновые уроидные вилли), сохраняется
(рис. 511, вкл.). В целом жгутиковый аппарат Р. pa-
lustris явно находится в сильно редуцированном со-
стоянии.
Как известно, виды, способные к амебоидному
движению, независимо возникают в различных, не-
посредственно не связанных филогенетически, круп-
ных таксонах жгутиконосцев (Масюк, 1981, 1985;
Серавин, 1989), при этом у них очень часто наблюда-
ется в той или иной степени выраженная редукция
жгутикового аппарата. Это объясняется утратой ис-
ходного функционального значения жгутиков в связи
с переходом к принципиально иному типу клеточной
локомоции (Серавин, 1985, 1989). «Классическим»
примером такого рода может служить безжгутиковая
трихомонада Dientamoeba fragilis, длительное время
считавшаяся настоящей лобозной амебой (Dobell,
1940; Эпштейн, 1941; Doflein, Reichenow, 1953;
Camp et al., 1974, и др.). Подобная ситуация имеет
место и в случае Р. palustris, которая, очевидно, на-
ходится на одном из этапов эволюционного перехода
от жгутикового типа клеточной организации к амебо-
идному (Гудков, Серавин, 1991).
Теперь, рассматривая место Р. palustris в системе
низших эукариот, можно однозначно утверждать,
что она не является «karyoblastean» и что тип Кагу-
oblastea sensu Margulis, 1974 в действительности не
существует.
Кавалье-Смит (Cavalier-Smith, 1983) создал осо-
бое подцарство Archezoa в царстве Protozoa, в ко-
торое включил 4 типа, по его мнению наиболее
примитивных, первично-безмитохондриальных про-
тистов: Archamoeba, Metamonada, Parabasalia и Mic-
rosporidia. К первому из них он отнес, в частности,
такие амебоидные организмы, как Pelomyxa и Enta-
moeba (Cavalier-Smith, 1983). Позднее этот исследо-
ватель (Cavalier-Smith, 1987) вывел Archezoa из со-
става Protozoa, придав этим таксонам равнозначный
систематический уровень царств. Дальнейшую ис-
торию развития представлений о составе Archezoa
можно назвать историей последовательных исключе-
ний. Представители Archezoa, по мнению Кавалье-
Смита, первично лишены митохондрий, пероксисом,
гидрогеносом, диктиосом аппарата Гольджи и имеют
70S рибосомы, подобно бактериям, а не 80S, как у
большинства других эукариот. На основании того,
что Parabasalia обладают хорошо развитыми диктио-
сомами и имеют гидрогеносомы, они были удалены
из состава Archezoa и перенесены в царство Protozoa
(Cavalier-Smith, 1987). Таким образом, в царстве Ar-
chezoa оставалось 3 типа: Archamoeba, Metamonada
и Microsporidia. Archamoeba были представлены ро-
дами Mastigamoeba, Mastigella, Mastigina, Pelomyxa,
Entamoeba, Phreatamoeba. Позднее этот исследова-
тель (Cavalier-Smith, 1991, 1993) пришел к заключе-
нию, что Entamoeba, скорее всего, вторично лишены
митохондрий, т. е. потеряли их в процессе эволю-
ции, и должны быть исключены из числа архезоев.
Соответственно он перенес их в царство Protozoa, а
остальных архамеб объединил в класс Pelobiontea (с
диагнозом типа Archamoeba) с двумя отрядами Mas-
tigamoebida (Mastigamoeba, Mastigella, Mastigina, Pe-
lomyxa) и Phreatamoebida (Phreatamoeba).
Следует сразу отметить, что включение Phreata-
moeba в архамеб совершенно непонятно, поскольку
ранее уже было показано (Chavez et al., 1986), что в
цитоплазме Ph. balamuthi — единственного известно-
го вида в отряде Phreatamoebida — присутствуют ор-
ганеллы, которые ультраструктурно подобны мито-
хондриям протистов, живущих при низком содержа-
нии кислорода в среде. Кроме того, кинетосома
единственного жгутика у этого организма соседствует
с безжгутиковой кинетосомой («barren-kinetosome»),
что не соответствует диагнозу типа Archamoeba. На-
конец, по своим молекулярно-биохимическим свой-
ствам Ph. balamuthi значительно более сходна с выс-
шими эукариотами, нежели с теми, что находятся
ближе к основанию филогенетического дерева (см.:
Morin, Mignot, 1995). Поэтому не удивительно, что
Cavalier-Smith (1995) опять был вынужден вывести
этот вид из состава архамеб и отнести к числу Proto-
zoa. (Ранее, в разделе, посвященном голым лобозным
амебам (Rhizopoda, Lobosea, Gymnamoebia), уже от-
мечалось, что некоторые исследователи (Simpson et
al., 1997) сводят род Phreatamoeba в синоним Mas-
tigamoeba.)
В свою очередь, изложенные здесь, а также дру-
гие30 данные по Pelomyxa palustris, однозначно пока-
зывают, что этот организм также не может более рас-
сматриваться как «archezoan» и должен быть удален
из типа Archamoeba; соответственно класс Pelobion-
tea прекращает свое существование.
Ясно также, что Р. palustris нельзя отнести ни к
числу лобозных амеб, ни вообще к какому-либо так-
сону в составе Rhizopoda von Siebold 1845, emend.
Page, 1987, поскольку ее организация свидетельству-
ет о непосредственной связи со жгутиконосцами. Во
всяком случае Р. palustris сохраняет значительно
больше сходств с предковыми жгутиконосцами, чем,
например, Dientamoeba fragilis, которую тем не
менее относят к числу трихомонад (отряд Trichomo-
nadida) в составе типа Parabasalia (Camp et al., 1974;
Sleigh, 1989; Brugerolle, 1991a, 1991b).
На основании всего вышесказанного P. palustris
была выделена в самостоятельный класс Peloflagella-
tea (Гудков, Серавин, 19916).
В заключение этого раздела представляется необ-
ходимым сказать следующее. Формально в типе Ar-
chamoeba остался лишь отряд Mastigamoebida в со-
ставе родов Mastigamoeba, Mastigella и Mastigina. В
действительности же он представляет собой прови-
30 Филогенетический анализ результатов по секвенированию
«тяжелой» рРНК Pelomyxa sp. показывает, что данный организм по
этому признаку является весьма эволюционно продвинутым и дале-
ко отстоит от форм, близких к основанию филогенетического де-
рева (Morin, Mignot, 1995).
ТИП RHIZOPODA. КЛАСС PELOFLAGELLATEA
513
зорный таксон, объединяющий группу организмов не-
ясного систематического положения. Считающаяся
характерной для мастигамеб организация корешко-
вой системы жгутика — «unikont» с «nucleus connec-
tor» (Cavalier-Smith, 1991* Brugerolle, 1991a, 1991b),
с одной стороны, встречается у некоторых типичных
простейших (Protozoa), таких как Cercomonadida и
Protostelida (см.: Карпов, 1990), а с другой стороны,
ее нет у Mastigella sp. (Van Bruggen et al., 1985)и. Тот
факт, что во всех названных таксонах отсутствует
набор основных клеточных органелл (на основании
чего они были помещены в царство Archezoa), может
быть результатом их параллельной эволюции в связи
с переходом к обитанию в условиях дефицита кисло-
рода. Более того, даже одновременное отсутствие ми-
тохондрий, диктиосом аппарата Гольджи и микроте-
лец не есть признаки, характерные только для Arche-
zoa. Все эти органеллы отсутствуют у Entamoebidae
или, например, у анаэробной инфузории Balanthidium
coli, у которых их утрата в процессе эволюции не
вызывает сомнений (см.: Серавин, 1992). Константы
седиментации рибосом мастигамеб до сих пор неиз-
вестны, однако следует напомнить, что рибосомы Ра-
rabasalia, обладающих гидрогеносомами и хорошо
развитыми диктиосомами, имеют константу седимен-
тации 70S (Champney et al., 1992). Наконец, почему-
то в литературе по Archezoa не упоминается тот
факт, что в цитоплазме инцистирующейся метамона-
ды Giardia lamblia появляются настоящие диктиосо-
мы (Reiner et al., 1990; Gillin et al., 1991). Иными
словами, есть все основания полагать, что мастигаме-
бы и некоторые другие архезои не являются эволюци-
онно примитивными первично-безмитохондриальны-
ми организмами* и поэтому существование такого
таксона, как Archezoa, представляется более чем со-
мнительным.
Краткая ревизия рода Pelomyxa
С тех пор как в 1874 г. Р. Грееф установил род
Pelomyxa, многие исследователи неоднократно вклю-
чали в него те или иные виды. Длительное время,
как справедливо отмечает Пэйдж (Page, 1983), среди
систематиков вообще установилась тенденция счи-
тать чуть ли не всех крупных лобозных многоядер-
ных амеб принадлежащими к роду Pelomyxa, Начало
этому фактически было положено Вильсоном, кото-
рый отнес обнаруженный им многоядерный амебо-
идный организм к роду Pelomyxa, выделив его в са-
мостоятельный вид Р. carolinensis (Wilson, 1900); за
ним последовали и многие другие авторы (Mast,
1938; Rice, 1945; Kudo, 1946, 1951, 1959; Daniels,
1973; Bovee, Sawyer, 1979; Bovee, 1985, и др.). В
общей сложности было описано около 20 различ-
ных видов пеломикс. Правда, еще Пенар (Penard,
1902) возражал против такого подхода. Шаффер
(Schaeffer, 1926), восстановив линнеевский род
* Некоторые более поздние авторы по-прежнему сближают
Р. palustris с мастигамебами, объединяя их в один отряд Pelobionti-
da Page, 1976 emend. Griffin, 1988, и продолжают развивать идею
об их эволюционной примитивности относительно других групп
жгутиковых простейших (Simpson et al., 1997).
33 Протисты, часть 1
Chaos, перенес в него Pelomyxa carolinensis Wilson,
считая его более поздним синонимом Chaos chaos L.
В роде Pelomyxa он оставил единственный, по его
мнению, валидный вид Р. palustris Greeff. Андресен
(Andresen et al., 1968; Andresen, 1973), Пэйдж (Page,
1976, 1981, 1983) и ряд других исследователей про-
должали развивать идеи Шаффера о необходимости
удаления из рода Pelomyxa всех многоядерных форм,
не имеющих других сходств с типовым видом. В
конечном итоге (после исключения из рода Chaos
одноядерных видов и некоторых уточнений в диаг-
нозе рода Pelomyxa} это стало общепризнанным
(см.: Levine et al., 1980; Page, 1987, 1988, 1991).
В конце прошлого и начале этого века был опи-
сан целый ряд организмов, действительно очень сход-
ных с Р. palustris, которые были помещены в род Pe-
lomyxa в качестве самостоятельных видов. Блохман
(Blochmann, 1894) на основании различий в окраске
пеломикс выделял 3 их вида: Р. palustris, Р. greejfi и
Р. villosa. Бурн (Bourne, 1891) описал Р. viridis, име-
ющую зеленый цвет. Существенное значение окраске
придавал также Пенар (Penard, 1902), когда обосно-
вывал выделение Р, belewskii в самостоятельный вид.
Однако, как показали исследования многих авторов,
использование этого признака для разделения видов
пеломикс не является правомочным, поскольку ок-
раска этих организмов определяется характером за-
глоченного пищевого материала (Okada, 1930; Hol-
lande, 1945; Kudo, 1957; Andresen et al., 1968).
Лэйди (Leidy, 1879) описал P. villosa, основыва-
ясь в первую очередь на таком признаке, как при-
сутствие у движущихся форм развитых уроидных
билли. Однако эти структуры имеются и у Р. palust-
ris, что было отмечено Греефом (Greeff, 1874) в
первоописании вида. Поэтому, хотя некоторые авто-
ры поддержали выделение Р. villosa (Gruber, 1885;
Blochmann, 1894), Пенар (Penard, 1902) высказал
справедливое сомнение относительно его валид-
ности, а впоследствии он был окончательно сведен
в синоним Р. palustris (Kudo, 1957; Bovee, Jahn,
1973).
Пенар в ряде работ (Penard, 1893, 1902, 1904)
описал несколько новых, по его мнению, представи-
телей рода Pelomyxa, а также перенес в него некото-
рые виды, описанные другими авторами под иными
родовыми названиями: Р. belewskii Penard, 1885,
Р. fragilis Penard, 1904, Р. beryllifera Penard, 1902,
P. vivipara Penard, 1902, P. binucleata (Gruber, 1885),
P. paradoxa Penard, 1902, P.prima (Gruber, 1885),
P.tertia (Gruber, 1885). Однако еще Голд (Gould,
1894), сравнивая организацию Р. palustris и Р. belew-
skii, пришла к выводу, что они, по сути, ничем не
отличаются друг от друга. Сравнительно-морфологи-
ческий анализ имеющихся данных, а также сведения,
полученные при изучении жизненного цикла Р. palus-
tris, позволили окончательно установить, что Р. be-
lewskii, Р. fragilis, Р. vivipara и Р. binucleata в дейст-
вительности являются синонимами Р. palustris, тогда
как реальность существования остальных пенаровс-
ких видов Pelomyxa весьма сомнительна (Chapman-
Andresen, 1978, 1982; Page, 1981).
Что касается Р. parvialveolata, описанной Корот-
яевым (Korotneff, 1879), то тот же Пенар (Penard,
514
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
1902) вполне справедливо заключил, что имеющиеся
данные не позволяют ни идентифицировать этот ор-
ганизм с Р. palustris, ни рассматривать его в качестве
самостоятельного вида в этом роде. Наконец, принад-
лежность Р, caulleryi Hollande, 1945 к роду Pelomyxa
вообще очень сомнительна (Andresen et al., 1968),
хотя Чапмен-Андресен (Chapman-Andresen, 1978) не
исключает, что эта форма может представлять собой
одну из фаз жизненного цикла Р. palustris.
Подводя итог изложенному, можно заключить,
что Р, palustris Greeff, 1874 является пока единствен-
ным валидным видом в этом роде.
THE PHVLUM RHIZOPODA SIEBOLD, 1845
THE CLASS PELOFLAGELLATEA GOODKOV ET SERAVIN, 1991
(PELOMYXA PALUSTRIS)
SUMMARY
The class Peloflagellatea was established by Goodkov and Seravin
in 1991 as a monotypical taxon for adopting the systematic position
of the unusual protist Pelomyxa palustris in the system of the lower
eukaryotes. There is reason to consider other organisms in this chapter
(see below).
P. palustris is a free-living, multinucleate amitochondrial amoebo-
id organism. It has extremely wide, nearly worldwide distribution. The
locomoting specimen resembles a typical monopodial lobose amoeba,
with a steady axial flow of cytoplasm and its fountaining streaming in
the frontal end of the body. The average size of the body is 600—
800 pm, however many specimens reach up to 1 mm in length, and
some of them can reach even up to 5 mm. The number of nuclei can
vary, accordingly, and reach as many as 2000 per cell. Pelomyxa in-
habit in small freshwater backwater lakes, ponds, stagnant river wa-
ters, and other similar habitats, rich in decaying organic material (sap-
ropel), and characterised by anaerobic or microaerobic conditions.
Thus, in the previous studies it was frequently given the trivial name
«giant backwater amoeba».
P. palustris was discovered in 1867 and briefly described (under
the name Pelobius) in 1870 by Greeff. A more detailed description of
this organism under the present name was published by Greeff in
1874. Since Greeff’s works, it was studied by many authors during
one and a half centuries, and there is extensive literature, devoted to
this organism. However, its phylogenetic relationships, life cycle, and
many other essential biological features still remain unclear.
P. palustris is an exceptional case among all protists, because the
conception of its nature and position in the system of living organisms
varied repeatedly and quite radically. According to the traditional
point of view, P. palustris was considered to be a lobose amoeba and
classified among other rhizopods. It is also considered as an evolutio-
nary extremely primitive, primarily amitochondrial organism, and de-
fined as a special phylum Karyoblastea. The idea of primary primiti-
veness of P. palustris was supported and developed by some authors.
Thus, it was placed in the phylum Archamoeba within the kingdom
Archezoa, opposed to the majority of the other protists, which are
typical eukaryotes and assembled into other kingdoms (there are some
later modifications of this system, but without essential changes of the
conception view on Pelomyxa*s nature). At last, some other authors
considered P. palustris as aquatic fungi.
The recent investigations suggest that the absence of mitochondria
in P. palustris is not a primary, but secondary, and accounts for adap-
tation to life under low oxygen conditions. It has hydrogenosome-like
bodies and obligatory methanogenic endocytobionts. All these features
are common also for a number of different amitochondrial sapropelic
protists. It may explain the deep morpho-physiologycal parallelisms in
the evolution of quite different low eukaryotic organisms, which are
adapted to anaerobic and microaerobic conditions.
P. palustris possesses true, but nonmotile flagella, and well deve-
loped microtubular cytoskeleton with polar organisation. The contrac-
tile system of undulipodia deviates from the standard microtubular
pattern (9 + 2) and varies even in the same specimen (axonemes with
8 + 1, 9 + 0, 9 + 1, 10 + 1, 10 + 5 and even 14 + 3 microtubular
patterns were found). As a whole, the flagellar apparatus of P. palus-
tris is obviously strongly reduced. This may be explained by the loss
of locomotive function of flagella, which is connected with die radical
evolutionary change of the main type of cell locomotion. Apparently,
Pelomyxa is at one of the stages of the evolutionary transformation
from die flagellate type of cellular organisation to the amoeboid one.
It should be considered as flagellate, but it is not possible at the pre-
sent time to designate its taxonomic and phylogenetic relationships
with any known phylum of flagellates.
The complete life cycle of P. palustris still remains unclear. There
is only fragmentary and poorly documented evidence on the presence
of several stages, such as cysts and small uni- or binucleate amoeboid
forms. The plasmotomy is considered as the main way of cellular
multiplication, but the method of nuclei division is not yet satisfacto-
rily established.
More than twenty species of Pelomyxa have been described by
different authors. However, some of them are now recognised as sy-
nonyms of polymorphic species P. palustris, and the others are either
considered invalid or are included in different genera, which are not
related to Pelomyxa.
Various authors suggest close relation of Pelomyxa with the gene-
ra Mastigamoeba, Mastigella, Mastigina and Phreatamoeba (the latter
was synonymised with Mastigamoeba by some investigators). Altho-
ugh such a relation seems to be doubtful in many respects, we include
Mastigamoeba, Mastigella and Mastigina in Peloflagellatea as genera
incertae sedis.
ЛИТЕРАТУРА31
(Аверинцев C.) Averintzev S. Freshwater Rhizopoda. St. Peters-
burg: Merkushev, 1906.
Гудков А. В. Ультраструктура гигантской амебы Pelomyxa pa-
lustris. I. Цитоплазматические микротрубочки, субцентриоли и
жгутики. Сравнительно-морфологический анализ организации //
Цитология. 1989. Т. 31. С. 371—379.
Гудков А. В., Серавин Л. Н. Ультраструктура гигантской
амебы Pelomyxa palustris. III. Вакуолярная система: ее природа,
организация и функциональное значение И Цитология. 1991а.
Т. 33. С. 17—25.
Гудков А. В., Серавин Л. Н. Новое представление о природе
«гигантской амебы» Pelomyxa palustris; положение этого организ-
ма в системе низших эукариот (Peloflagellata classis n.) И Зоол.
журн. 19916. Т. 70. С. 5—16.
Карпов С. А. Система протистов. Омск, 1990.
31 Литература к разделу «Класс Peloflagellatea».
Масюк Н.П. Морфологический параллелизм и систематика
водорослей И Проблемы эволюционной морфологии и биохимии в
систематике и филогении растений. Киев: Наукова думка, 1981.
С. 176—188.
Масюк Н. П. О типах морфологической структуры тела водо-
рослей и основных напрвлениях их эволюции // Ботан. журн. 1985.
Т. 70. С. 1009—1018.
Серавин Л. Н. Двигательные системы простейших. Л.: Наука,
1967.
Серавин Л. Н. Вариации в строений аксонемы жгутика и
происхождение этой органеллы И Цитология. 1985. Т. 9. С. 971—
985.
Серавин Л. Н. Существует ли в действительности такой так-
сон — подцарство Protozoa? // Биол. науки. 1989. Т. 7. С. 4—13.
Серавин Л. Н. Эукариоты, лишенные важнейших клеточных
органелл (жгутиков, аппарата Гольджи, митохондрий), и главная
задача органеллологии // Цитология. 1992. Т. 5. С. 3—33.
ТИП RHIZOPODA. КЛАСС PELOFLAGELLATEA. ЛИТЕРАТУРА
515
Серавин Л. Н., Гудков А. В. Аппарат Гольджи у амебы Pelo-
myxa palustris И Докл. АН СССР. 1987а. Т. 296. С. 249—250.
Серавин Л. Н.» Гудков А. В. Цитоплазматические гранулы, по-
добные микротельцам, у амебы Pelomyxa palustris И Цитология.
19876. Т. 29. С. 600—603.
Серавин Л. Н., Гудков А. В. Особенности организации цито-
скелета у амебы Pelomyxa palustris И Цитология. 1987в. Т. 29.
С. 835—837.
Серавин Л. Н., Гудков А. В. Жгутики пресноводной амебы Pe-
lomyxa palustris И Цитология. 1987г. Т. 29. С. 721—724.
Andresen N. General morphology И The Biology of Amoeba I
Ed. K. W. Jeon. New York; London : Acad. Press, 1973. P. 99—123.
Andresen N., Chapman-Andresen C., Nilsson J. R. The fine
structure of Pelomyxa palustris // Compt. Rend. Trav. Labor. Carls-
berg, Ser. chim. 1968. Vol. 36. P. 285—320.
Archer W. Resume of recent contributions to our knowledge of
freshwater Rhizopoda. Pt 4 // Quart. Jorn. Microscop. Sci. 1877.
Vol. 68. P. 330—353.
Blochmann F. Kleine Mitteilungen uber Protozoen // Biol. Zent-
ralbl. 1894. Bd 14. P. 82—91.
Bott K. Uber die Fortpflanzung von Pelomyxa palustris nebst
Mitteilungen uber ihren Bau // Arch. Protistenk. 1906. Bd 8. P. 120—
158.
Bourne A. G. On Pelomyxa viridis sp.n. and on the vesicular na-
ture of protoplasm // Quart. J. Microscop. Sci. 1891. Vol. 32.
P. 257—374.
Bovee E.C. Class Lobosea Carpenter, 1861 // An Illustrated
Guide to the Protozoa. Lawrence: Allen Press. 1985. P. 158—211.
Bovee E. C., Jahn Th. L. Taxonomy and phylogeny // The Biolo-
gy of Amoeba / Ed. K. W. Jeon. New York; London: Acad. Press,
1973. P. 37—82.
Bovee E. C., Sawer T. K. Marine flora and fauna of the north-
eastern United States. Protozoa: Sarcodina: Amoebae // NOAA Techn.
Rept. NMFS. Circ. 419. 1979.
Brand Th. Anaerobiosis in Invertebrates. 1946.
Broers С. A. M., Stumm С. K., Vogels G. D. Psalteriomonas lan-
terna gen. nov., sp. nov., a free-living amoeboflagellate isolated from
freshwater anaerobic sediments // Eur. J. Protistol. 1990. Vol. 25.
P. 369—380.
Brugerolle G. Flagella and cytoskeletal systems in amitochondrial
flagellates: Archamoeba, Metamonada amd Parabasalia // Protoplasma.
1991a. Vol. 164. P. 70—90.
Brugerolle G. Cell organization in free-living amitochondriate he-
terotrophic flagellates // The Biology of Free-living Heterotrophic Fla-
gellates, eds D. J. Patterson, J. Larsen. Oxford: Clarendon Press,
1991b. P. 133—148.
Camp R. R., Mattern C. F. N., Honigberg В. M. Study of Dien-
tamoeba fragilis Jepps et Dobell. 1. Electronmicroscopic observations
of the binucleate stage. 2. Taxonomic position and revision of the
genus // J. Protozool. 1974. Vol. 21. P. 69—82.
Cavalier-Smith T. A 6-kingdom classification and a unified phy-
logeny // Endocytobiology. II Eds W. Schwemmler, H. E. A. Schenk.
Berlin: Gruyter, 1983. P. 1027—1034.
Cavalier-Smith T. The origin of cells: a symbiosis between
genes, catalysis, and membranes H Cold Spring Harbor Symp. Quant.
Biol. 1987. Vol. 52. P. 805—824.
Cavalier-Smith T. Archamoebae: the ancestral eukaryotes? //
BioSystems. 1991. Vol. 25. P. 25—38.
Cavalier-Smith T. Kingdom Protozoa and its 18 phyla // Micro-
biol. Rev. 1993. Vol. 57. P. 953—994.
Cavalier-Smith T. Cell cycles, diplokaryosis and the archezoan
origin of sex // Arch. Protistenk. 1995. Vol. 145. P. 189—207.
Champney W. S., Chittum H. S., Samuels R. Ribosomes from
trichomonad protozoa have prokaryotic characteristics // Int. J. Bioc-
hem. 1992. Vol. 24. P. 1125—1133.
Chapman-Andresen C. Biology of the large amoebae // Ann.
Rev. Microbiol. 1971. Vol. 25. P. 27—48.
Chapman-Andresen C. Endocytic process // The Biology of
Amoeba ZEd. K. W. Jeon. New York; London: Acad. Press, 1973.
P. 319—348.
Chapman-Andresen C. The life cycle of Pelomyxa palustris // J.
Protozool. 1978. Vol. 25. P. 42A.
Chapman-Andresen C. Peroxidase activity in large freshwater
amoebae // Abstr. 6th Int. Congr. Protozool. Warszawa. 1981.
Vol. 53.
Chapman-Andresen C. Identification of Pelomyxa binucleata as
a stege in the life cycle of P. palustris // J. Protozool. 1982. Vol. 29.
P. 499—500.
Chapman-Andresen C. Nuclear characteristics of Pelomyxa pa-
lustris II Abstr. 7th Int. Congr. Protozool. Nairoby. 1985. P. 293.
Chapman-Andresen C., Hamburger K. Respiratory studies of
the giant amoeba Pelomyxa palustris // J. Protozool. 1981. Vol. 28.
P. 433—440.
Chavez L. A., Balamuth W., Gong T. A light and electron mic-
roscopical study of a new, polymorphic free-living amoeba, Phreata-
moeba balamuthi n. g., n. sp. // J. Protozool. 1986. Vol. 33. P. 397—
404.
Corliss J. O. The kingdom Protista and its 45 phyla // BioSys-
tems. 1984. Vol. 17. P. 87—126.
Corliss J. O. An interim utilitarian («user-friendly») hierarchical
classification and characterization of the protists // Acta Protozool.
1994. Vol. 33. P. 1—51.
Daniels E. W. Ultrastructure // The Biology of Amoeba I Ed.
K. W. Jeon. New York; London: Acad. Press, 1973. P. 125—169.
Daniels E. W., Breyer E. P. Ultrastructure of the giant amoeba
Pelomyxa palustris // J. Protozool. 1967. Vol. 14. P. 167—179.
Daniels E. W., Breyer E. P., Kudo R. R. Pelomyxa palustris Gre-
eff. 2. Its ultrastructure // Ztschr. Zellforsch. 1966. Bd 73. P. 367—
383.
De Duve C. Microbodies in the living cells // Sci. Amer. 1983.
Vol. 248. P. 52—62.
De Puytorac P., Grain J. and Mignot J.-P. Precis de protistolo-
gie. Peris. 1987.
Dejdar E. Analyse der naturlichen Standortsbedingungen von Pe-
lomyxa palustris Greeff // Arch. Protistenk. 1931. Vol. 74. P. 198—
206.
Doflein F., Reichenow E. Lehrbuch der Protozoenkunde. Jena:
Fischer, 1953.
Esteban G., Guhl В. E., Clarke K. J., Embley T. M., Finlay B. J.
Cyclidium porcatum n. sp.: a free-living anaerobic scuticociliate con-
taining a stable complex of hydrogenosomes, eubacteria and archaeo-
bacteria // Eur. J. Protistol. 1993. Vol. 29. P. 262—270.
Fenchel T., Finlay B. J. The biology of free-living anaerobic cili-
ates // Eur. J. Protistol. 1991. Vol. 26. P. 201—215.
Fenchel T., Finlay B. J. Production of methane and hydrogen by
anaerobic ciliates containing symbiotic methanogenes // Arch. Micro-
biol. 1992. Vol. 157. P. 475—480.
Fenchel T., Perry T., Thane A. Anaerobiosis and symbiosis with
bacteria in free-living ciliates // J. Protozool. 1977. Vol. 24. P. 154—
163.
Finlay B. J., Fenchel T. Hydrogenosomes in some anaerobic pro-
tozoa resemble mitochondria // FEMS Microbiol. Lett. 1989. Vol. 65.
P. 311—314.
Fortner H. Untersuchungen an Pelomyxa palustris Greeff. Studi-
en zur Biologie und Physiologie des Tieres. 1. Teil // Arch. Protistenk.
1934. Vol. 83. P. 381—464.
Gillin F. D., Reiner D. S., McCaffery M. Organelles of protein
transport in Giardia lamblia // Parasitology Today. 1991. Vol. 7.
P. 113—116.
Goosen N. K., Wagener S., Stumm С. K. A comparison of two
strains of the anaeribic ciliate Trimiema compressum // Arch. Micro-
biol. 1990. Vol. 153. P. 187—192.
Gould L. J. Notes on the minute structure of Pelomyxa palustris
(Greeff) II Quart. J. Microsc. Sci. 1894. Vol. 36. P. 295—296.
Greeff R. Pelomyxa palustris (Pelobius), ein amoebenartiger Or-
ganismus des sussen Wassers // Arch. Microscop. Anat. 1874. Bd 10.
P. 53—73.
Griffin J. L. Fine structure and taxonomic position of the giant
amoeboid flagellate Pelomyxa palustris // J. Protozool. 1988. Vol. 35.
P. 300—315.
Gruber A. Studien uber Amoben // Zeitschr. Wissenschaft. Zool;
1885. Bd 41. P. 186—225.
Gulliver G. Note on the minute structure of Pelomyxa palustris //
J. Roy. Mier. Soc. (London). 1888. Vol. 1. P. 11—12.
Hollande A. Biologie et reproduction des rhizopodes des genres
Pelomyxa et Amoeba et cycle evolutif de l’Amoeboiphilus destructor
nov. gen. nov. spec., Chytridinee (?) parasite de Pelomyxa palustris
Greeff II Bull. Biol. France et Belg. 1945. Vol. 79. P. 31—66.
Jahn T. L., Bovee E. C., Griffith D. L. Taxonomy and evolution
of the Sarcodina: a reclassification // Taxon. 1974. Vol. 23. P. 483—
496.
Korotneff A. Etudes sur les Rhizopodes // Arch. Zool. Exp. et
Gen. 1879. Vol. 8. P. 467—482.
Kudo R. R. Pelomyxa carolinensis Wilson. I. General observation
on the Illinois stock H J. Morphol. 1946. Vol. 78. P. 317—351.
Kudo R. R. Observations on Pelomyxa illinoisensis // J. Morphol.
1951. Vol. 88. P. 145—184.
516
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Kudo R. R. Pelomyxa palustris Greeff. I. Cultivation and general
observations // J. Protozool. 1957. Vol. 4. P. 154—164.
Kudo R. R. Pelomyxa and related organisms // Ann. N. Y. Acad.
Sci. 1959. Vol. 78. P. 474-486.
Leidy J. Freshwater rhizopods of North America // Rept.
U. S. Geol. Surv. Territ. 1879. Vol. 12. P. 1—324.
Leiner M., Wohlfeil M., Schmidt D. Pelomyxa palustris Greeff //
Ann. Sci. Natur. (Zool.), Ser. 1954. Vol. 16. P. 537—594.
Levine N. D., Corliss J. O., Cox F. E. G., Deroux G., Grain J.,
Honigberg В. M., Leedale G. F., Loeblich A. R. Ill, Lom J., Lynn
D., Merinfeld E. G., Page F. C., Poljansky G., Sprague V., Vavra J.,
Wallace F. G. A newly revised classification of the Protozoa // J. Pro-
tozool. 1980. Vol. 27. P. 37—58.
Lorch I. J. Some historical aspects of amoeba studies // The Bio-
logy of Amoeba I Ed. K. W. Jeon. New York; London: Acad. Press,
1973. P. 1—36.
Margulis L. Five-kingdom classification and the origin and evo-
lution of cells II Evolution Biology Vol. 7. New York; London: Ple-
num Press, 1974. P. 45—78.
Margulis L. Symbiosis in cell evolution. Life and its environment
on the early Earth. San Francisco: Freeman, 1981.
Margulis L., Schwartz К. C. Five kingdoms: an illustrated guide
to the phyla of life on Earth. San Francisco: Freeman, 1982.
Mashansky V. F., Seravin L. N., Vinnichenko L.N. Ultra-
structure of the ciliate Loxodes rostrum (O. F. M.) in relation to the
mode of its water transport // Acta. Protozool. 1963. Vol. 1.
P. 403-410.
Mast S. O. A amoeboid movement in Pelomyxa palustris (Gre-
eff) H Physiol. Zool. 1934. Vol. 7. P. 470—478.
Mast S. O. Amoeba and Pelomyxa vs. Chaos // Turtox News.
1938. Vol. 16. P. 56—57.
Morin L., Mignot J.-P. Are Archamoeba true Archezoa? The
phylogenetic position of Pelomyxa sp. // 2nd Eur. Congr. of Protisto-
logy, 21—26 July, Clermont-Ferran, 1995. P. 70. (Abstr. 174).
Muller M. Biochemistry of protozoan microbodies, peroxyso-
mes, — glycerophosphate oxydase bodies, hydrogenosomes // Ann.
Rev. Microbiol. 1975. Vol. 29. P. 467—483.
Okada J. K. Uber den Bau und die Bewegungsweise von Pelo-
myxa II Arch. Protistenk. 1930. Vol. 70. P. 131—154.
Page F. C. A revised classification of the Gymnamoebia (Proto-
zoa: Sarcodina) // Zool. J. Linn. Soc. 1976. Vol. 58. P. 61—77.
Page F. C. An electron-microscopical study of Thecamoeba pro-
teoides (Gymnamoebia), intermediate between Thacamoebidae and
Amoebidae // Protistologica. 1978. Vol. 14. P. 77—85.
Page F. C. Eugene Penard’s slides of Gymnamoebia: re-examina-
tion and taxonomic evaluation // Bull. Brit. Mus. Nat. Hist. (Zool.).
1981. Vol. 40. P. 1—32.
Page F. C. Marine Gymnamoebia. Cambridge, 1983.
Page F. C. The classification of «naked» amoebae (phylum Rhy-
zopoda) II Arch. Protistenk. 1987. Vol. 133. P. 199—217.
Page F. C. A new key to freshwater and soil gymnamoebae with
instructions for culture. Ambleside, 1988.
Page F. C. Nackte Rhizopoda // Protozoenfauna. Bd 2. Nackte
Rhizopoda und Heliozoea. Stuttgart; New York: Fischer. 1991. P. 7—
170.
Penard E. Pelomyxa palustris et quelques autres organisms inte-
rieurs II Arch. Sci. Phys, et Nat. 1893. Vol. 29. P. Г65—184.
Penard E. Faune rhizopodique du bassin du Leman. Geneva;
Kundig et Fils. 1902.
Penard E. Quelques nouveaux Phizopodes d’ean douce // Arch.
Protistenk. 1904. Vol. 3. P. 391-422.
Reiner D. S., McCaffery M., Gillin F. D. Sorting of cyst wall pro-
teins to a regulated secretory pathway during differentiation of the
primitive eukaryote, Giardia lamblia // Eur. J. Cell. Biol. 1990.
Vol. 53. P. 142—153.
Rice N. E. Pelomyxa carolinensis (Wilson) or Chaos chaos (Lin-
naeus)? II Biol. Bull. 1945. Vol. 88. P. 139—143.
Schaeffer A. A. Taxonomy of the amebas // Pap. dep. Mar. Biol.
Carnegie Inst. Wash. 1926. Vol. 24. P. 1—116.
Schirch P. Beitrage zur Kenntnis des Lebenscyclus von Arcella
vulgaris Ehrbg. und Pelomyxa palustris Greeff // Arch. Protistenk.
1914. Vol. 33. P. 247—271.
Sleigh M. A. Protozoa and other protists. Cambridge: Univ.
Press. 1989.
Stolz A. Beobachtungen und Versuche uber die Verdaung und
Bildung der Kohlenhydrate in einem amoebenartigen Organismus, Pelo-
myxa palustris Greeff // Ztschr. wiss. Zool. 1900. Vol. 68. P. 625—668.
Van Bruggen J. J. A., Stumm С. K., Vogels G. D. Symbiosis of
methanogenic bacteria and sapropelic protozoa H Arch. Microbiol.
1983. Vol. 136. P. 89—96.
Van Bruggen J. J. A., Zwart К. B., Van Assema R. M., Stumm
С. K., Vogels G. D. Methanobacterium formicicum, en endosymbiont
of the anaerobic ciliate Metopus striatus McMurrich // Arch. Microbi-
ol. 1984. Vol. 139. P. 1—7.
Van Bruggen J. J. A., Stumm С. K., Zwart K.B., Vogels
G. D. Endosymbiotic methanogenic bacteria of the sapropelic amoeba
Mastigella // FEMS Microb. Ecol. 1985. Vol. 31. P. 187—192.
Van Bruggen J. J. A., Zwart К. B., Hermans J. G. F., Van Hove
E. M., Stumm С. K., Vogels G. D. Isolation and characterization of
Methanoplanus endosymbioticus sp. nov., an endosymbiont of the ma-
rine sapropelic ciliate Metopus contortus Quennerstedt // Arch. Micro-
biol. 1986. Vol. 144. P. 367—374.
Van Bruggen J. J. A., Van Rens G. L. M., Geertman E. J. M.,
Zwart К. B., Stumm С. K., Vogels G. D. Isolation of methanogenic
endosymbiont of the sapropelic amoeba Pelomyxa palustris Greeff //
J. Protozool. 1988. Vol. 35. P. 20—23.
Veley L. J. A further contribution to the study of Pelomyxa palus-
tris (Greeff) II J. Linn. Soc. (Zool.). 1905. Vol. 29. P. 374—395.
Vogels G.D., Van der Drift C., Stumm C.K., Keltjens J. T. M.,
Zwart К. B. Methanogenesis: surprising molecules, microorganisms and
systems // Antonie Van Leeuwenhoch. 1984. VoL 50. P. 557—567.
West G. S. Observations on freshwater rhizopods, with some re-
marks on their classification // J. Linn. Soc. (Zool.). 1905. Vol. 29.
P. 108—117.
Whatley F. R., Whatley J. M. The establishment of organelles //
Endocytobiology. I. Berlin; New York: de Gruyter, 1980. P. 849—
865.
Whatley F. R., Whatley J. M. Pelomyxa palustris // Endocytobi-
ology. II. Berlin; New York: de Gruyter, 1983. P. 413—426.
Whatley J. M. Bacteria and nuclei in Pelomyxa palustris: com-
ments on the theory of serial endosymbiosis // New Phytol. 1976.
Vol. 76. P. 111—120.
Wilson H. V. Notes on a species of Pelomyxa // Amer. Natur.
1900. Vol. 34. P. 535—550.
Класс Xenophyophorea Schulze, 1904 —
Уникальные морские глубоководные простейшие, обитающие
в основном в батиальной и абиссальной зонах Мирового океана.
Тело ксенофиофорий представляет собой многоядерный плазмо-
дий, заключенный в систему ветвящихся трубочек органического
происхождения (гранелляр). В цитоплазме обнаружено большое
количество кристаллов сульфата бария (гранелл). Покровы тела
состоят из ксенофий — инородных тел, склеенных веществом му-
кополисахаридного типа. Ксенофиями служат раковины форами-
нифер, спикулы губок, скелеты радиолярий, панцири диатомовых
водорослей, минеральные частицы донного осадка. Наряду с ксе-
нофиями в стенках раковин ксенофиофорий расположены тяжи
фекальных пеллет (стеркомары), состоящие из отдельных пеллет
(стеркомат, или стерком) и ксантосом — небольших округлых
телец, которым приписывается экскреторная функция. Дополни-
тельными опорными структурами раковин у ряда форм являются
Ксенофиофорий
линеллы — волокнистые нити, оплетающие раковину снаружи или
внутри и проникающие между ксенофиями. Ксенофйофории —
самые крупные морские простейшие, достигающие размеров 25 см.
Они являются важным компонентом глубоководного бентоса.
История изучения
Эти необычные простейшие всегда представляли
большую трудность для диагностики. Их часто отно-
сили к другим группам беспозвоночных или рассмат-
ривали как «явно неорганические образования». Оп-
ТИП RHIZOPODA, КЛАСС XENOPHYOPHOREA 517
ределение осложнялось хрупкостью ксенофиофорий
и присутствием в пробах в виде фрагментов. Недоста-
точная степень изученности этих организмов объяс-
нялась также ограниченностью материала, что связано
с их преимущественно глубоководным образом жизни.
Впервые ксенофиофории были описаны как при-
митивные фораминиферы по материалам экспедиции
1882 г. на «Тритоне» в северную Атлантику (Brady,
1883). Обрабатывая сборы экспедиции на «Челленд-
жере», Геккель выделил загадочные организмы в
группу глубоководных губок (Haeckel, 1895). Гез
описал крупные листовидные организмы, пойманные
в восточной части Тихого океана во время экспеди-
ции на «Альбатросе», как специфический тип аг-
глютинирующих фораминифер (Goes, 1892). Сборы
экспедиций на «Вальдивии», «Альбатросе», «Сибоге»
и «Челленджере» позволили Шульцу классифициро-
вать непонятных животных как гигантские корне-
ножки. Он же назвал их ксенофиофориями (Schulze,
1904, 1907а, 1907b). В 1912 г. русский зоолог Ще-
потьев (1912а, 19126) собрал и определил мелковод-
ных ксенофиофорий с коралловых рифов у Цейлона.
Основной вклад в изучение систематики этой
группы сделали датский исследователь Тендал и ан-
гличанин Гуди (Tendal, 1972, 1979, 1985, 1989, 1994,
1995—1996; Tendal, Gooday, 1981; Tendal et al.,
1982; Gooday, Tendal, 1988; Gooday, 1991, 1996).
Многочисленные сборы ксенофиофорий в пос-
леднее время позволили оценить их роль в глубоко-
водном бентосе и описать ряд новых таксонов (Ка-
менская, 1987, 1988, 1998; Levin et al., 1986; Levin,
Thomas, 1988; Каменская, Саидова, 1998).
До настоящего времени нет единства мнений о
ранге таксона и его положении в системе Protista.
Первоначально ксенофиофории наряду с форамини-
ферами были выделены в самостоятельный отряд
(Schulze, 1904). Листер относил их к фораминифе-
рам в ранге семейства (Lister, 1909). Тендал рассмат-
ривал ксенофиофорий в ранге подкласса (Tendal,
1972). Самый высокий ранг они занимали в класси-
фикации простейших Маргулис (см.: Tendal, 1989).
Там ксенофиофории были выделены в тип Xenophy-
ophora. Мы, следуя принятой в настоящем руковод-
стве системе, рассматриваем ксенофиофорий в ранге
класса Xenophyophorea Schulze, 1904 в типе Rhizo-
poda Siebold, 1845.
Классификация
Класс Xenophyophorea Schulze, 1904
Отряд Psamminida Poche, 1913
Сем. Psammettidae Tendal, 1972
Сем. Psamminidae Haeckel, 1889
Сем. Syringamminidae Tendal, 1972
Сем. Cerelasmidae Tendal, 1972
Отряд Stannomida Tendal, 1972
Сем. Stannomidae Haeckel, 1889
Морфология и физиология
Ксенофиофории характеризуются большим раз-
нообразием формы тела. Она может быть сферичес-
кой, дисковидной, древовидной, листовидной или
напоминать рыхлую губку. Размеры ксенофиофорий
колеблются от 1. мм у некоторых видов рода Psam-
metta до 200—250 мм у видов рода Stannophyllum. В
среднем размеры корненожек не превышают 20—
70 мм.
Окраска простейших определяется цветом агглю-
тинированных частиц (ксенофий). Если ксенофии
отсутствуют или прозрачны как скелеты радиолярий,
то ксенофиофории имеют коричневатую окраску.
Покровы тела Xenophyophorea устроены довольно
сложно. Важным элементом, выполняющим защит-
ную и опорную функции, являются ксенофии — ино-
родные включения, представляющие собой спикулы
губок, раковины фораминифер, скелеты радиолярий,
минеральные частицы донного осадка. Обычно каж-
дый вид характеризуется определенным типом ксено-
фий, однако отмечен ряд исключений: у Psammetta
arenocentrum и Р. globosa тип ксенофий меняется с
возрастом. Молодь первого вида использует в качестве
ксенофий минеральные частицы осадка, молодь вто-
рого агглютинирует раковины фораминифер. Взрослые
особи встраивают в стенку раковины спикулы губок.
Возможно также использование простейшими 2 ти-
пов ксенофий в различных частях раковины. Так, у
Stannophyllum concretum покровы состоят из раковин
фораминифер, а внутренняя часть заполнена скелетами
радиолярий (Tendal, 1972). Ксенофии между собой
скрепляются цементирующим веществом мукополи-
сахаридного типа.
Наряду с ксенофиями в стенках раковины распо-
ложены стеркомары — длинные темные ветвящиеся
тяжи диаметром 35—540 мкм. У разных видов они
бывают различной формы и длины. Стеркомары
плохо развиты в роде Maudammina. Чрезвычайно
сильное развитие стеркомар характеризует виды ро-
дов Cerelasma и Stannophyllum.
Стеркомары состоят из стерком и ксантосом (Ten-
dal, 1972).
Стеркомы (или стеркоматы) представляют собой
фекальные пеллеты корненожек. Они бывают 4 ти-
пов: 1) эллипсоидные темно-коричневые тела разме-
ром 8—60 мкм, в состав которых входят различные
мелкие частички, гранеллы и черные кристаллы не-
известного происхождения; 2) продолговатые обра-
зования желтого цвета, состоящие из сферических
элементов диаметром 1—4 мкм, никогда не содержат
гранелл; 3) сферические стеркомы размером около
10 мкм, содержащие исключительно гранеллы; 4) редко
встречающиеся образования из складчатых мембран.
Ксантосомы у ксенофиофорий представляют со-
бой округлые красноватые тела размером 1—7 мкм.
Им приписывается экскреторная функция. Обычно
хаотично разбросаны среди стерком. И стеркомы, и
ксантосомы присущи не только ксенофиофориям, но
и некоторым другим группам ризопод.
Основным элементом органической части ракови-
ны ксенофиофорий являются тонкие, дихотомически
ветвящиеся трубочки, содержащие цитоплазму. Тру-
бочки цилиндрической формы, толщина стенки ме-
нее 0.5 мкм. Родам Maudammina, Galatheammina и
Reticulammina свойственна слабо развитая трубчатая
система. В родах Psammetta, Cerelasma, Syringammi-
па и Stannoma она развита хорошо (Tendal, 1972).
518
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
В гомогенной цитоплазме расположены ядра и
гранеллы.
Ксенофиофории имеют ядра 3 типов: 1) сфери-
ческой формы, 2—10 мкм в диаметре, с явно выра-
женным ядрышком; 2) эллипсоидной формы, 4—9 мкм
в длину, с диффузной хроматиновой сетью; 3) мел-
кие, 0.5—1 мкм, круглые ядра. Наиболее часто встре-
чаются ядра первого типа, вторые отмечены реже,
третьи — очень редко. По данным Шульца и Ще-
потьева (Schulze, 1904, 1907а; Щепотьев, 1912а), ядра
первого и второго типов представляют собой стадии
развития одного ядра, ядра третьего типа рассмат-
риваются как образующиеся гаметы. Считая ксено-
фиофорий гетерокариотными организмами, Тендал
(Tendal, 1972) приписывает ядрам первого типа со-
матические функции, а ядрам третьего типа — гене-
ративные.
Уникальной особенностью ксенофиофорий мож-
но считать наличие в цитоплазме гранелл — много-
численных кристаллических образований размером
2—5 мкм, с высоким содержанием сульфата бария.
Неизвестно, образуются ли гранеллы в цитоплазме
или выбираются псевдоподиями из донного осадка.
Функция их также неизвестна.
Псевдоподии у фиксированных ксенофиофорий
описаны не были, однако в качестве их остатков
рассматривали капли безъядерной цитоплазмы на от-
крытых концах органических трубочек (Schulze,
1907а, 1912). На современных фотографиях глубоко-
водных участков дна с живыми ксенофиофориями
отчетливо видны радиально расходящиеся от тела
простейших длинные тонкие тяжи, интерпретируе-
мые как псевдоподии (Lemche et al., 1976; Tendal,
Gooday, 1981). Длина их в несколько раз превосхо-
дит диаметр тела и в некоторых случаях может до-
стигать 6—12 см.
Своеобразными опорными структурами органи-
ческого происхождения являются линеллы. Они пред-
ставляют собой волокнистые нити до нескольких
миллиметров в длину. Уплощенными концами ли-
неллы прикрепляются к ксенофиям, стенкам органи-
ческих трубочек и стеркомар, другим линеллам. Бы-
вают 3 видов: 1) одиночные, 2) скрепленные между
собой цементом, 3) анастомозные и ветвящиеся. Ли-
неллы найдены только у видов отряда Stannomida и
за его пределами не встречаются. Линеллы располо-
жены в основном внутри раковины, однако у неко-
торых видов рода Stannophyllum образуют густую по-
верхностную сеть.
Питание
Исходя из того что в цитоплазме ксенофиофорий
не обнаружены пищеварительные вакуоли с пищевы-
ми частицами, а последние аккумулируются вне плаз-
мосодержащих трубочек, у этих простейших предпо-
лагается наличие внеклеточного (или лакунарного)
пищеварения (Tendal, 1972, 1979).
По-видимому, при помощи псевдоподий корне-
ножки выбирают из поверхностного слоя осадка
представляющие пищевую ценность частички. Они
собираются в компактные массы в пространствах
между органическими трубочками и стенками рако-
вин. В этих лакунах происходит пищеварение, пита-
тельные вещества поступают в клеткуу, а неперева-
ренные остатки склеиваются в стеркомы и служат для
образования новых стеркомар (Tendal, 1972). Подо-
бный тип питания обнаружен у некоторых форами-
нифер и не является уникальным среди Protista.
Размножение и развитие
Данные о размножении ксенофиофорий отрывоч-
ны (Schulze, 1907а; Щепотьев, 1912а). Тем не менее
к настоящему времени можно считать доказанным,
что 1) ксенофиофории размножаются гаметами, при-
чем каждая особь имеет несколько периодов размно-
жения; 2) гаметы несут 2 жгутика; 3) в развитии при-
сутствуют 1 или 2 амебоидные стадии; 4) существует
различие между ювенильными и взрослыми особями
(Tendal, 1972).
Экология
и распространение
Ксенофиофории — важный компонент глубоко-
водной фауны. Основная масса этих гигантских кор-
неножек обитает в абиссальной и батиальной зонах
океана. На мелководье и в ультраабиссали они из-
вестны пока только по единичным находкам (Tendal,
1972; Каменская, Саидова, 1998). Изучение ко-
личественного распределения простейших на ложе
океана выявило приуроченность группы преимущес-
твенно к эвтрофным областям, где биомасса ксе-
нофиофорий (вместе с раковиной) составляла 1—
2 г/м2, а доля в общей биомассе макробентоса варь-
ировала в районах от 10 до 100 % (Каменская, 1987,
1988).
Ксенофиофории служат пищей многим донным
беспозвоночным. Так, фрагменты Stannophyllum zo-
narium обнаружены в кишечнике моноплакофоры
Neopilina (Tendal, 1985), остатки ксенофиофорий
неоднократно отмечались в пищеварительном тракте
некоторых глубоководных изопод, голотурий и морс-
ких звезд (Соколова, 1986).
Географическое распространение ксенофиофорий
изучено недостаточно. Оба отряда представлены в
Тихом, Индийском и Атлантическом океанах. Наи-
более широко распространены роды Cerelasma,
Stannophyllum и Galatheammina. Только в Индийс-
ком океане обнаружен род Maudammina, только в
Тихом — Cerelpemma. Распространение родов Syrin-
gammina и Reticulammina ограничено Тихим и Ат-
лантическим океанами.
Эврибатные роды Psammetta, Cerelasma и Stan-
nophyllum обнаружены от литорали до абиссали, а
виды последнего встречаются в ультраабиссали.
Роды Cerelpemma и Stannoma эндемичны для абиссали,
а роды Occultammina и Psammina найдены в абисса-
ли и ультраабиссали. Все остальные роды приуро-
чены к батиальной и абиссальной зонам Мирового
океана (Tendal, 1972; Tendal et al., 1982; Gooday,
1991; Каменская, Саидова, 1998).
ТИП RHIZOPODA. КЛАСС XENOPHYOPHOREA
519
Положение в системе
протистов
Наличие таких признаков, как двухслойная (труб-
чатая органическая и агглютинированная) раковина,
разделение ядер на соматические и генеративные, при-
сутствие стерком, ксантосом, лакунарное пищеварение,
размножение при помощи двухжгутиковых гамет,
сближает ксенофиофорий с фораминиферами, а
также с комокиидами. Однако организация тела в
виде многоядерного плазмодия и некоторые особен-
ности размножения близки у Xenophyophorea с
Eumycetozoea. По-видимому, ксенофиофории зани-
мают промежуточное положение между форамини-
ферами и Eumycetozoea (Tendal, 1972).
Рис. 512. Представители класса Xenophyophorea.
А — Reticulammina sp., Б — Stannophyllum sp.
Fig. 512. General view of some xenophyophores. A — Reticulammi-
na sp., Б — Stannophyllum sp.
Систематический обзор
Отряд Psamminida Poche, 1913
Ксенофиофории не имеют опорных структур в
виде линелл. Раковина различной формы, хрупкая,
негибкая.
Сем. Psammettidae Tendal, 1972
Раковина массивная, грубая. Форма тела шаро-
видная, дисковидная или неопределенных очертаний.
Специфический поверхностный слой и полости в
раковине отсутствуют. Ксенофии расположены бес-
системно и скреплены только в местах соприкосно-
вений. Состав ксенофий различен: спикулы губок,
раковины фораминифер или крупнозернистый дон-
ный осадок.
Роды: Homogammina Gooday et Tendal, 1988; Ma-
udammina Tendal, 1972; Psammetta Schulze, 1906.
Сем. Psamminidae Haeckel, 1889
Обычно раковина твердая, может быть очень
хрупкой. Тело шаровидной, дисковидной или неоп-
ределенной формы, иногда с большими лопастями.
Внешние ксенофии собраны в поверхностный слой,
иногда слоев может быть несколько. Ксенофии со-
стоят из крупных зерен песка, скелетов радиолярий
или раковин планктонных фораминифер.
Роды: Cerelpemma Laubenfels, 1936; Galatheam-
mina Tendal, 1972; Psammina Haeckel, 1889; Reticul-
ammina Tendal, 1972 (рис. 512, A); Semipsammina
Tendal, 1975.
Сем. Syringamminidae Tendal, 1972
Раковина хрупкая, пористая. Сцементированные
ксенофии образуют трубчатую систему, внутри кото-
рой находятся органические компоненты клетки и
стеркомары. Ксенофии состоят из раковин секреци-
онных фораминифер или гранул кварца.
Роды: Aschemonella Brady, 1879; Occultammina
Tendal, Swinbanks et Shirayama,1982; Syringammina
Brady, 1883.
Сем. Cerelasmidae Tendal,
1972
Раковина относительно мягкая. Тело неправиль-
ной закругленной формы, с большими полостями.
Число ксенофий обычно уступает количеству цемен-
тирующего вещества, которое наряду со стеркомара-
ми выполняет опорную функцию. У одних видов ксе-
нофии отсутствуют, у других имеют различное проис-
хождение и могут состоять из минеральных частиц,
спикул губок, раковин фораминифер или скелетов
радиолярий.
Род: Cerelasma Haeckel, 1889.
Отряд Stannomida Tendal,
1972
Ксенофиофории имеют линеллы. Раковина гибкая.
Сем. Stannomidae Haeckel,
1889
Раковина относительно мягкая, хрупкая, иногда
древовидная, иногда представляет собой рыхлую не-
оформленную массу или тонкую пластинку. Ксено-
фии самые разнообразные.
Роды: Stannoma Haeckel, 1889; Stannophyllum Ha-
eckel, 1889 (рис. 512, Я).
В настоящее время класс Xenophyophorea вклю-
чает 2 отряда, 5 семейств, 14 родов и 57 видов.
520
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
THE PHYLUM RHIZOPODA SIEBOLD, 1845
THE CLASS XENOPHYOPHOREA SCHULZE, 1904
SUMMARY
The xenophyophores are marine rhizopod plasmodial protozoans,
constructing agglutinated tests. The test is composed of foreign partic-
les («xenophyae») held together with organic cement. The protoplasm
is organized as a branched multinucleate plasmodium surrounded by
organic sheath. An exceptional feature is the accumulation in the cy-
toplasm of crystals of BaSO4. The test ranges from a few millimeters
to 25 cm in size.
The class comprises 2 orders, 5 families, 14 genera, and 57 spe-
cies. Xenophyophores occur from bathyal to hadal zones of the oce-
ans.
ЛИТЕРАТУРА32
Каменская О. E. Ксенофиофории и комокиации в трофичес-
ких цепях глубоководного бентоса // Питание морских беспозво-
ночных и его роль в формировании сообществ: М., 1987. С. 15—
22.
Каменская О. Е. Количественное распределение комокиаций
и ксенофиофорий в южной части Атлантического океана И Струк-
турно-функциональные исследования морского бентоса. М.: Изд.
ИОАН, 1988. С. 15—20.
Каменская О. Е. Ксенофиофории (Protozoa, Xenophyophorea)
из сборов 16-го рейса НИС «Дмитрий Менделеев». I. Новый вид
рода Reticulammina И Бентос высокоширотных районов. М.: Изд.
ВНИРО, 1998. С. 121—124.
Каменская О. Е., Саидова X. М. Переописание вида Psammina
planata (Saidova, 1970) — ультраабиссальной ксенофиофории из
Курило-Камчатского желоба И Бентос высокоширотных районов.
М.: Изд. ВНИРО, 1998. С. 125—128.
Соколова М. Н. Питание и структура глубоководного макро-
бентоса. М., 1986.
(Щепотьев A) Schepotieff A. Untersuchungen uber niedere Or-
ganismen. IL Die Xenophyophoren des Indischen Oceans // Zool. Jb.
Anat. Ontog. Tiere. 1912a. Bd 32. S. 245—286.
(Щепотьев A.) Schepotieff A. Rhizopodenstudien // Arch. Protis-
tenk. 19126. Suppl. 15. S. 219—242.
Brady H. B. Notes on some of the Reticularian Rhizopoda of the
«Challendger» Expedition. Part I. On new or little known arenaceous
types II Q. J. Microscop. Sci. 1879. 19. P. 20—63.
Brady H. B. Syringammina, a new type of arenaceous Rhizopo-
da //Proc. Roy. Soc. 1883. Vol. 35. P. 155—161.
Goes A. On a peculiar type of arenaceous foraminifera from the
American tropical Pacific, Neusina agassizi // Bui. Mus. comp. Zool.
Harv. 1892. Vol. 23. P. 195—198.
Gooday A. J. Xenophyophores (Protista, Rhizopoda) in box-core
samples from the abyssal northeast Atlantic Ocean (Biotrans area):
their taxonomy, morphology and ecology // J. Foram. Res. 1991.
Vol. 21, N 3. P. 197—212.
Gooday A. J. Xenophyophores (Protista), including two new spe-
cies from two abyssal sites in the northeast Atlantic // J. Foram. Res.
1996. Vol. 26. P. 193—208.
Gooday A. J., Tendal O. S. New xenophyophores (Protista) from
the bathyal and abyssal northeast Atlantic Ocean // J. Nat Hist. 1988.
Vol. 22. P. 413—434.
Haeckel E. Report on the Deep-Sea Keratoza // Rep. Challenger.
1889. Vol. 32. P. 1—92.
Haeckel E. Account of E. Haeckel’s work on «the Deep-Sea Ke-
ratoza» H Nat. Sci. 1895. Vol. 7. P. 35.
32 Литература к разделу «Класс Xenophyophorea».
Honigberg B.M., Balamuth W. Subphylum Sarcomastigophora
nom. nov. to embrace the flagellate and amoeboid assemblages of
protozoans II J. Protozool. 1963. Suppl. 10. P. 27.
Lemche H., Hansen B., Madsen E. J., Tendal O.S., Wolff T.
Hadal life as analyzed from photographs // Vidensk. Medd. Dan. Na-
turhist. Foren. Kpbenhavn. 1976. Vol. 139. P. 263—336.
Levin L. A., Thomas C. L. The ecology of xenophyophores (Pro-
tista) on eastern Pacific seamounts // Deep Sea Res. 1988. Vol. 35,
N 12. P. 2003—2027.
Levin L. A., DeMaster D. J., McCann L. D., Thomas C. L. Ef-
fects of giant protozoans (class: Xenophyophorea) on deep-seamount
benthos H Mar. Ecol. Prog. Ser. 1986. Vol. 29. P. 99—104.
Lister J. J. The Xenophyophoridae // Treatise on Zoology I Ed.
R. Lancaster. S. 1., 1909. P. 284—286.
Poche F. Das System der Protozoa // Arch. Protistenk. 1913.
Bd 30. S. 125—321.
Schulze F. E. Die Xenophyophoren der «Siboga»-Expedition //
Siboga-Exped. Monogr. IV. 1904. P. 1—18.
Schulze F. E. Die Xenophyophoren, eine besondere Gruppe der
Rhizopoden I Wiss. Ergeb. Tiefsee-Exped. 1907a. Bd 2. S. 1—35.
Schulze F.E. Die Xenophyopora / Bull. Mus. comp. Zool. Harv.
1907b. Vol. 51. P. 205—229.
Schulze F. E. Xenophyophora // Zool. Anz. 1912. Bd 39. S. 38—
43.
Tendal O. S. A monograph of the Xenophyophoria (Rhizopoda,
Protozoa) // Galathea Rep. 1972. Vol. 12. P. 7—99.
Tendal O. S. Aspects of biology of Komokiacea and Xenophyop?
horia II Sarsia. 1979. Vol. 64, N 1. P. 13—17.
Tendal O. S. A preliminary account of the Komokiacea and
Xenophyophorea //Peuplements profonds du golf de Gascogne I Eds
L. Laubier, Cl. Monniot. 1985. P. 263—265.
Tendal O. S. Phylum Xenophyophora // Handbook of the Protoc-
tista / Eds L. Margulis et al. Boston, 1989. P. 135—138.
Tendal O. S. Protozoa Xenophyophorea Granuloreticulosa: Psam-
mina zonaria sp. nov. from the West Pacific and some aspects of the
growth of xenophyophores // Mem. Mus. Hist. Nat. 1994. Vol. 161.
P. 49—54. (Resultats des Campagnes Musorstom; Vol. 12).
Tendal O. S. Synoptic checklist and bibliography of the Xenophy-
ophorea (Protista), with a zoogeographical survay of the group // Ga-
lathea Rep. 1995—1996. P. 79—101.
Tendal O. S., Gooday A. Xenophyophoria (Rhizopoda, Protozoa)
in bottom photographs from the bathyal and abyssal NE Atlantic //
Ocean. Acta. 1981. Vol. 4. P. 415—422.
Tendal O. S., Swimbanks D. D., Shirayama J. A new infaunal
xenophyophore (Xenophyophorea, Protozoa) with notes on its ecology
and possible trace fossial analogues // Ocean. Acta. 1982. Vol. 5, N 3.
P. 325—329.
Rhizopoda incertae sedis
Отряд Athalamida Haeckel, 1862
Наиболее полное описание этой группы было
сделано Бови (Bovee, 1985а), поэтому в настоящем
издании характеристика группы приводится преиму-
щественно на основании его работы. Род Arachnula
здесь не рассматривается, так как позднее он был
переведен в класс Filosea (Hausmann, Hulsmann,
1996).
Аталамиды — гетерогенная группа ризопод, пред-
ставленная ретикулоподиальными формами без рако-
вины. Одни из них сильно напоминают форамини-
фер, лишенных раковин {Rhizoplasma kaiseri, Ponto-
туха flava), другие морфологически более сходны с
RHIZOPODA INCERTAE SEDIS. ОТРЯД ATHALAMIDA
521
Рис. 513. Biomyxa vagans. (По: Bovee, 1985).
A — особь с сократившимися ретикулоподиями, Б — подвижная особь в
расправленном состоянии, р — ретикулоподии.
Fig. 513. Biomyxa vagans.
А — cell with involved reticulopodia, Б — moving cell with extended reticu-
lopodia. p — reticulopodia.
миксомицетами (Reticulomyxa filosa}. Остальные ви-
ды помещены в этот отряд только потому, что их не-
возможно отнести к каким-либо другим известным
таксонам. Отсутствует типичный представитель отря-
да. Все это позволило рассматривать отряд Athalami-
da в качестве таксона неопределенного таксономическо-
го положения в типе ризопод.
Аталамиды обитают в различных биотопах от прес-
ных вод и почв до морей, некоторые представлены
комменсалами. В отряде принято выделять сем. Bio-
myxidae, в которое входят виды с непостоянной,
быстро меняющейся формой тела, и сем. Chlamydo-
myxidae, представители которого имеют более или
менее определенную, чаще округлую форму тела.
Однако, учитывая неопределенный объем отряда, в
который разные авторы включают разное количество
родов, мы опустим деление на семейства и приведем
наиболее известных представителей.
Род Biomyxa Leidy, 1879 (рис. 513)
Тело клетки не имеет определенной формы. Ре-
тикулоподии ветвящиеся, анастомозирующие, грану-
лярные, иногда с двунаправленным движением гра-
нул вдоль них. Размеры клетки составляют более
100 мкм в диаметре. Обитают во мхах и копрофиль-
ной фауне.
Род Pontomyxa Topsent, 1892
Тело клетки более или менее округлой формы,
желтого цвета. Является комменсалом туникат. Раз-
меры тела в расправленном состоянии достигают
10 см. Многоядерные.
Род Reticulomyxa Nauss, 1949 (рис. 515)
Тело клетки более или менее округлой формы,
белого цвета. Периферические ретикулоподии пред-
ставляют собой подвижную сеточку. Встречаются в
мокром мусоре. Размеры тела в расправленном состо-
янии (с ретикулоподиями) достигают 60 мм. Многоя-
дерные.
Род Rhizoplasma Fervorn, 1896 (рис. 514)
Тело клетки более или менее определенной, часто
округлой формы, оранжевого или желтого цвета. Пе-
Рис. 514. Rhizoplasma kaiseri. (По: Bovee, 1985).
Подвижная особь в расправленном состоянии, р — ретикулоподии.
Fig. 514. Rhizoplasma kaiseri.
Moving cell with extended reticulopodia. p — reticulopodia.
рис. 515. Reticulomyxa filosa. (По: Bovee, 1985).
Подвижная особь в расправленном состоянии, р — ретикулоподии.
г Fig. 515. Reticulomyxa filosa.
Moving cell with extended reticulopodia. p — reticulopodia.
522
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 516. Схема строения клетки Penardia cometa. (По: Микрюков, Мыльников, 1995).
аг — диктиосома, б — бактерия, гт — грибовидное тело, к — кинетосома, л — липидная капля, м — митохондрия, мт — микротельце, мтб — микро-
трубочки, on — окаймленные пузырьки, пс — псевдоподия, св — сократительная вакуоль, э — экструсома, эпр — эндоплазматический ретикулум, я —
ядро.
Fig. 516. Cell structure of Penardia cometa.
аг — Golgi apparatus, 6 — bacterium, гт — mushroom-like body, к — basal body, л — lipid droplet, м — mitochondria, мт — microbody, мтб — microtubu-
les, on — coated vesicles, nc — pseudopodium, ce — contractile vacuole, э — extrusome, эпр — endoplasmic reticulum, я — nucleus.
риферические ретикулоподии представляют собой
подвижную сеточку. Организмы морские, обычно
крупные (до 10 мм, включая псевдоподии).
Кроме описанных родов к аталамйдам относят
также роды Gymnophris Cienkowsky, 1876 и Penar-
dia Cach, 1904. Исследования пресноводных Gym-
nophris cometa и Penardia cometa (Микрюков, Мыль-
ников, 1996) показали, что в жизненном цикле Gym-
nophris cometa имеется как минимум 3 стадии:
трофическая подвижная амеба с ретикулоподиями,
передвигающаяся, однако, при помощи лобоподий;
двухжгутиковая зооспора; шаровидная циста. Жиз-
ненный цикл Penardia cometa характеризуется нали-
чием только амебоидной трофической стадии, кото-
рая обычно формирует агрегации из нескольких кле-
ток.
Ультратонкое строение амебоидных стадий обоих
видов практически идентично (рис. 516). Покровы
представлены плазмалеммой с тонким слоем глико-
каликса. Ядро крупное, пузырьковидное, расположе-
но в центре клетки. С дорсальной стороны клетки
неподалеку от ядра находится пара кинетосом и ап-
парат Гольджи. Митохондрии имеют пластинчатые
кристы. Экструсомы о0ои}( видов представлены мик-
ротоксицистами, которые, однако, различаются по
строению. Микротрубочки ретикулоподий, по-видимо-
му, не имеют морфологически выраженного ЦОМТа.
Обычно аталамид относят к классу Granuloreticu-
losea, поэтому для них должны быть характерны пу-
зырьковидные или трубчйтые кристы в митохондри-
ях. Пластинчатые кристы в митохондриях обоих
видов резко выделяют их из группы гранулоретику-
лозных амеб, поэтому авторы справедливо помещают
оба рода в отдельное сем. Gymnophryidae Mikrjukov
et Mylnikov, 1996 с неопределенным таксономиче-
ским положением в системе протистов.
Некоторые авторы включают в отряд еще 2 ро-
да — Chlamydomyxa Archer, 1875 (Bovee, 1985а) и
Theratromyxa (Микрюков, Мыльников, 1996).
Отряд Monothalamida Haeckel, 1862
Основной признак группы — органическая одно-
камерная раковина, часто дополнительно кальцини-
рованная, в которую может включаться и инородный
материал. Ретикулоподии обычно выходят из отвер-
стий раковины — псевдостомов. Смена поколений
неизвестна. Встречаются в морских и пресных водо-
емах, а также в наземных условиях обитания. Опи-
сание моноталамид приведено по работе Бови (Bo-
vee, 1985b).
Сем. Amphitremidae Poche,
1913
Раковина имеет 2 и более псевдостома, из кото-
рых выходят ретикулоподии.
Род Amphitrema Archer,
1869 (рис. 517)
Раковина овальная, с 2 псевдостомами, инкрусти-
рована снаружи инородными частицами. Ретикулопо-
дии выходят из 2 полюсов раковины, размеры кото-
рой около 70 мкм в длину.
Род Diplophrys Barker,
1868 (рис. 518)
Раковина тонкая, органическая, размером до
20 мкм, округлая; с 2 псевдостомами на противопо-
ложных полюсах.
Род Ditrema Archer, 1877
В раковине накапливаются соли железа; она ко-
ричневого цвета, с толстыми стенками. По размерам
и форме близка к раковине представителей рода Am-
phitrema.
RHIZOPODA INCERTAE SEDIS. ОТРЯД MONOTHALAMIDA
523
Рис. 517. Amphitrema wrigtianum.
(По: Bovee, 1985).
Рис. 518. Diplophrys archeri. (По: Bovee,
1985).
Рис. 519. Microgromia haeckeliana.
(По: Bovee, 1985).
pa — раковина.
A — одиночная особь, Б — группа клеток.
pa — раковина.
Fig. 517. Amphitrema wrigtianum.
pa — test.
Fig. 518. Diplophrys archeri.
A — single cell, Б — group of the cells.
Fig. 519. Microgromia haeckeliana.
pa — test.
Род Microcometes Cienkowsky, 1876
Раковина прозрачная или желтого цвета; с более
чем 2 псевдостомами; обычно их 3—5, из них выхо-
дят ретикулоподии.
Сем. Lieberkuehnidae de Saedeleer,
1934
Раковина большая; с 1 псевдостомом; не сплю-
щенная с одной стороны; пластичная или твердая.
Род Allelogromia de Saedeleer, 1934
Раковина более или менее твердая, с налипшими
частицами; однослойная.
Род Diplogromia (Rhumbler) de Saedeleer,
1934
Раковина твердая, с налипшими частицами; двух-
слойная.
Род Lieberkuehnia Claparede et Lachmann,
1859 (рис. 520)
Раковина гладкая и пластичная.
Сем. Microgromiidae de Saedeleer,
1934
Раковины с 1 псевдостомом, мелкие (менее 20 мкм);
с септой или без септы в шейке раковины; пресноводные.
Род Apogromia de Saedeleer, 1934
В шейке ракцвины нет септы, сократительная ва-
куоль 1.
Род Belaria de Saedeleer, 1934
В шейке раковины имеется симметрично располо-
женная септа.
Род Heterogromia de Saedeleer,
1934
В шейке раковины нет септы, сократительных ва-
куолей несколько.
Род Microgromia (Hertwig et Lesser)
de Saedeleer, 1934 (рис. 519)
В шейке раковины имеется асимметрично распо-
ложенная септа.
Оба эти отряда обычно объединяют вместе с фо-
раминифераМи в класс или тип Granuloreticulosa de
Saedeleer, 193:4 со следующими признаками: ветвя-
щиеся и анастомозирующие между собой псевдопо-
дии, внутри которых проходят одиночные или неупо-
рядоченные группы микротрубочек; внутри псевдо-
подий имеет место одновременное двунаправленное
Рис. 520. Lieberkuehnia wagnerella. (По: Bovee, 1985).
pa — раковина.
Fig. 520. Lieberkuehnia wagnerella.
pa — test.
524
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
движение практически всех органелл, за исключени-
ем ядер; клетки содержат 1 или несколько ядер, ко-
торые расположены в центральном утолщении цито-
плазмы и могут быть заключены в раковину; кристы
в митохондриях трубчатые или пузырьковидные.
Между тем не все представители аталамид и мо-
ноталамид имеют двунаправленное движение цито-
плазмы и гранулы в ретикулоподиях. Кроме того,
первые электронно-микроскопические исследования
показали гетерогенность этой группы на ультра-
структурном уровне.
Пример исследования Penardia cometa (Микрю-
ков, Мыльников, 1995, 1996) показывает, что по
крайней мере не все аталамиды могут быть отнесены
к гранулоретикулозным амебам. Более того, другие
представители этой группы не изучены электронно-
микроскопическими методами и могут оказаться со-
вершенно отличными от гимнофриид. В связи с этим
мы поместили аталамид как таксон с неопределен-
ным положением в рамках типа Rhizopoda.
Между тем четкая очерченность и большой объем
класса фораминифер, диагноз которого полностью
совпадает с диагнозом гранулоретикулозей, факти-
чески вынуждают упразднить таксон Granuloreticulo-
sa de Saedeleer, 1934 как младший синоним Foramini-
fera d’Orbigny, 1826.
RHIZOPODA INCERTAE SEDIS
SUMMARY
THE ORDER ATHALAMIDA HAECKEL, 1862
This heterogenous group is represented by naked rhizopods with
reticulopodia. Some of them are similar to naked foraminiferans, whe-
reas others are more similar to myxomycetes. The typical repre-
sentative is absent.
Predominantly freshwater organisms, but occur also in moist soil
and on marine bottom. Their internal structure and biology are very
little known.
Because of taxonomic uncertainty and insufficient knowledge we
did not divide this order into families. There are four genera in this
order: Biomyxa Leidy, 1879; Rhizoplasma Fervom, 1896; Reticulo-
myxa Nauss, 1949; Pontomyxa Topsent, 1892.
The representatives of two other genera, which were considered
to belong to this order (Gymnophris Cienkowsky, 1876 and Penardia
Cach, 1904) have been investigated recently by Mikrjukov and Myl-
nikov (1996). These authors have shown that G. cometa has at least
three stages in their life cycle: trophic amoeba with reticulopodia,
which moves nprmally using the lobopodia, biflagellate zoospore, and
rounded cyst. P. cometa has only one amoeboid trophic stage, which
often forms aggregations of several cells. The ultrathin structure of
amoeboid cells in both species is almost identical. They have vesicular
nucleus, dictyosome, two basal bodies, microtoxicysts, and mitochon-
dria withlamelliform cristae. As far as granuloreticulous amoebae
have to have vesicular, or tubular cristae in mitochondria, these aut-
hors excluded both genera from Athalamida and established a new
family Gymnophryidae Mikrjukov et Mylnikov, 1996 with uncertain
taxonomic position among protista.
Some other aythors include in the order Athalamida 2 more gene-
ra: Chlamydomyxa and Theratromyxa.
The description of each genus is presented in the chapter.
THE ORDER MONOTHALAMIDA HAECKEL, 1862
The order includes reticulous amoebae having organic test
with one chamber. They are predominantly freshwater organisms,
but also occur in marine habitats. These organisms belong to 3
families.
Amphitremidae Poche, 1913, which includes the genera Amphit-
rema, Ditrema, Diplophrys and Microcometes.
Lieberkuehnidae de Saedeleer, 1934, including the genera Lie-
berkuehnia, Allelogromia andDiplogromia.
Microgromiidae de Saedeleer, 1934, including the genera Apog-
romia, Heterogromia, Microgromia and Belaria.
The description of the order and genera has been made after
Bovee, 1985.
ЛИТЕРАТУРА33
(Микрюков К. А., Мыльников А. П.) Mikrjukov К. A., Mylnikov
A. P. Fine structure of an unusual rhizopod Penardia cometa, containing
extrusomes and kinetosomes // Eur. J. Protistol. 1995. Vol. 31. P. 90—96.
Микрюков К. А., Мыльников А. П. Новые данные о строении
и жизненном цикле аталамидных амеб (Protista, Athalamida) //
Зоол. журн. 1996. Т. 75, вып. 9. С. 1283—1293.
33 Литература к разделу «Rhizopoda incertae sedis. Отряд Atha-
lamida. Отряд Monothalamida».
Bovee E. C. Order Athalamida Haeckel, 1862 // An illustrated
guide to the Protozoa. Lawrence: Soc. Protozool. USA, 1985a.
P. 246—248.
Bovee E.C. Order Monothalamida Haeckel, 1862 // An illustra-
ted guide to the Protozoa. Lawrehce : Soc. Protozool. USA, 1985b.
P. 248—252.
Hausmann K., Hiilsmann N. Protozoology. Stuttgart; New York :
Georg Thieme, 1996. 338 p.
Отряд Komokiida Kamenskaya,
1992 — Комокииды
Агглютинирующие простейшие, раковина которых представ-
ляет собой систему ветвящихся трубочек равного диаметра. Стен-
ки раковины простые, двухслойные, агглютинированный слой со-
стоит в основном из частичек ила, иногда с вкраплениями облом-
ков фораминифер. Трубочки наполнены цитоплазмой и большим
количеством фекального материала в виде пеллет, которые назы-
ваются стеркоматами. Глубоководные, донные организмы, обитаю-
щие в Тихом, Индийском и Атлантическом океанах. В централь-
ных районах океанов комокиации являются одной из основных
групп глубоководного бентоса.
История изучения
По материалам, собранным на нескольких глубо-
ководных станциях в рейсах НИС «Витязь», «Акаде-
мик Курчатов», «Галатея», «Метеор» и «Таласса», в
1977 г. была описана новая группа простейших. Тен-
RHIZOPODA INCERTAE SEDIS. ОТРЯД KOMOKIIDA
525
дал и Хесслер дали ей название Komokiacea (от рус-
ского слова «комочки») и с определенными оговор-
ками отнесли ее к фораминиферам (Tendal, Hessler,
1977). В системе фораминифер комокиации были
отнесены к подотряду Textulariina (Tendal, Hessler,
1977; Tappan, Loeblich, 1982; Loeblich, Tappan,
1988).
Столь недавнее описание группы объясняется тем,
что из-за необычного строения тела комокиаций до-
лгое время не считали живыми организмами, а при-
нимали за слипшиеся частицы донного осадка или
останки умерших животных. Глубоководный образ
жизни, чрезвычайная хрупкость и разрушение при
стандартных методах размывки бентосных проб так-
же ограничивали возможность изучения этих про-
стейших.
Основная заслуга в исследовании морфологии,
разработке систематики, анализе географического
распространения комокиид принадлежит датскому
исследователю Оле Тендалу (Tendal, Hessler, 1977;
Tendal, 1979, 1985). Сейчас группа находится в ста-
дии интенсивного изучения. Исследуются как сами
простейшие, так и их роль в глубоководных донных
сообществах (Gooday, Cook, 1984; Каменская, 1987,
1989, 1990, 1993; Mullineaux, 1988; Schroder et al.,
1989).
Большую дискуссию вызывает систематическое
положение комокиид. В системе фораминифер ко-
мокииды были отнесены к подотряду Textulariina
Delage et Herouard, 1896 и рассматривались там в
ранге надсем. Komokiacea Tendal et Hessler, 1977
(или Komokioidea Tappan et Loeblich, 1982). В пос-
ледней классификации форамифер (Loeblich, Tap-
pan, 1988) комокииды также рассматривались как
фораминиферы подотряда Textulariina. Однако не-
обычная форма тела простейших и ограниченные
сведения об их биологии заставляли других исследо-
вателей с осторожностью говорить о принадлежнос-
ти комокиид к фораминиферам или вовсе отрицать
ее (Саидова, 1981). Поэтому позднее комокииды бы-
ли выделены в самостоятельный отряд неясного так-
сономического положения в классе Granuloreticulo-
sea (Каменская, 1992). В последнее время некоторые
исследователи также считают комокиид своеобраз-
ной и независимой от фораминифер группой про-
тистов. Мы рассматриваем комокиид в качестве от-
ряда incertae sedis в типе Rhizopoda.
Морфология и физиология
Отличительной чертой комокиид является мно-
гообразие форм тела. Основной элемент морфоло-
гии — цилиндрические ветвящиеся трубочки. В
большинстве случаев ветвление дихотомическое. Все
многообразие форм тела у комокиид можно свести
к двум основным типам: с центром роста, когда
можно говорить о центральных и периферических
ветвях, и без центра организации, когда простейшие
напоминают моток спутанных нитей. Толщина тру-
бочек обычно постоянна по всей длине, однако су-
ществует ряд исключений, когда наблюдается разли-
чие диаметра центрального ствола и боковых ответ-
влений или постепенное уменьшение толщины тру-
бочки с каждым последующим ветвлением. У одних
видов трубочки внутри цилиндрические, у других
они разделены по всей длине поперечными септами
на камеры. Довольно часто наблюдается срастание
трубочек (анастомозы). Кроме того, внешний вид
многих комокиид зависит не только от характера
ветвления трубочек, но и от наличия донного осад-
ка, заполняющего пространство между ними (Tendal,
Hessler, 1977).
Трубочки двухслойные, внутренний органический
слой обычно тонкий, не более 0.5 мкм. Агглютини-
рованный слой различной толщины, в среднем
около 10 мкм. Комокииды, как правило, агглютини-
руют частицы пелитовой фракции донного осадка,
но у некоторых мелководных видов во внешнем не-
органическом слое отмечены алевритовые компонен-
ты донного осадка. Отсутствие видимых пор в рако-
вине объясняется или их временным характером, или
деформацией стенки при фиксации (Tendal, Hessler,
1977). Окраска комокиид зависит от количества и
характера агглютинированных частиц. Обычно она
рыжевато-коричневая. Если агглютинированный слой
тонкий, то просвечивают темные стеркоматы, прида-
вая комокиидам сероватый оттенок. Размеры комо-
киид колеблются от 1 мм до нескольких сантимет-
ров.
Основным содержимым трубочек являются ци-
топлазма и стеркоматы. Цитоплазма может быть или
сильно разжижена, или иметь вид тонких тяжей. На
поверхности некоторых раковин были обнаружены
обрывки цитоплазмы, которые, по-видимому, пред-
ставляют собой остатки псевдоподиальной системы.
В цитоплазме расположены ядра сферической или
овальной формы, с гранулированным содержимым,
размером от 4 до 10 мкм у разных видов и стерко-
маты — темно-коричневые тела размером 10—
30 мкм, состоящие из массы мелких частиц, склеен-
ных вместе веществом мукополисахаридного типа.
Стеркоматы обнаружены также в просветах между
цитоплазмой и стенкой раковины (Tendal, Hessler,
1977; Tendal, 1979).
Питание
По-видимому, процесс питания у комокиид про-
исходит следующим образом: псевдоподии захваты-
вают пищевые частицы из окружающего донного
осадка. Они транспортируются внутрь раковины, где
перевариваются, а непереваренные остатки склеива-
ются в стеркоматы. В отличие от большинства других
ризопод у комокиид эти фекальные пеллеты не вы-
брасываются из раковины, а аккумулируются внутри
трубочек. Весьма вероятно, но требует дальнейшего
изучения предположение Тендала о наличии у комо-
киид и лакунарного пищеварения (Tendal, 1979), ко-
торое было найдено у Allogromia laticollaris. Частицы
пищи перевариваются не в пищеварительных вакуо-
лях, а в просветах между стенкой раковины и цито-
плазмой (Lengsfeld, 1960). Это позволяет небольшо-
му объему цитоплазмы справляться с относительно
большими массами пищевых частиц. Аккумулирован-
526
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 521. Представители отряда Komokiida.
А — Septuma sp., Б — Edgertonia sp., В — Normania sp., Г — Lana sp.
Fig. 521. General view of some komokiids.
ные фекальные пеллеты, по мнению Тендала, могут
служить субстратом для развития микрофлоры и,
обогащенные таким образом, перевариваться комо-
киидами повторно, причем этот процесс может пов-
торяться несколько раз.
Экология и распространение
Комокииды являются преимущественно глубо-
ководными организмами. Основная масса нахожде-
ний этих простейших приурочена к глубинам более
3000 м. На абцссальных и ультраабиссальных глуби-
нах Мирового океана частота встречаемости Komo-
kiida в бентосных пробах составляет более 70 %
(Каменская, 1987, 1990).
Изучение количественного распределения этих
Rhizopoda показало, что на ложе океана они являют-
ся одной из ведущих групп макробентоса. В среднем
по океану биомасса комокиид (включая раковину)
составляет 50—100 мг/м2, в центральных районах
океанов она увеличивается до 100—500 мг/м2 . (Ка-
менская, 1987, 1993). Для комокиид не отмечено
четкой зависимости между их обилием и троф-
ностью района обитания (Tendal, Hessler, 1977; Ка-
менская, 1987). В эвтрофных океанических и оли-
готрофных областях они могут быть единственными
представителями макробентоса. В целом наблюдается
закономерное увеличение доли комокиаций в общей
биомассе бентоса с увеличением глубины и по на-
правлению от прибрежных районов океана к его
центральной части от долей процента до 50—100 %.
Комокииды обитают в верхних слоях донного осад-
ка, обычно не погружаясь в него глубже 5—10 см.
Большая часть видов приурочена к верхнему санти-
метровому слою грунта (Tendal, Hessler, 1977). Здесь
RHIZOPODA INCERTAE SEDIS. ОТРЯД KOMOKIIDA. ЛИТЕРАТУРА
527
они служат доступной пищей для некоторых бентос-
ных беспозвоночных: голотурий, эхиурид, морских
звезд (Соколова, 1986, 1998).
Анализ вертикального распределения комокиид
выявил эврибатные роды Lana, Septuma и Edgertonia,
встречающиеся от батиали до ультраабиссали. Комо-
киации обнаружены в Филиппинском, Идзу-Бонинс-
ком, Рюкю, Волкано, Тонга, Палау, Яп и Марианс-
ком ультраабиссальных желобах Тихого океана (Ка-
менская, 1989). Основная масса видов приурочена к
абиссальным глубинам 3000—6000 м. Единственный
мелководный род Globipelorhiza обитает на глубинах
60—720 м у побережья Швеции и Норвегии. Наибо-
лее широко распространены в Мировом океане виды
родов Septuma, Lana и Edgertonia (рис. 521).
Систематический обзор
В настоящее время отряд Komokiida включает
три семейства: Komokiidae, Rhizamminidae и Bacu-
lellidae, в которых насчитывается 14 родов и около
30 видов.
Сем. Komokiidae Tendal et Hessler,
1977
Раковины различной формы: древовидные, кусто-
видные или напоминающие моток спутанных нитей.
Трубочки цилиндрические, иногда с перетяжками.
Расстояние между последовательными разветвления-
ми трубочек значительно больше их диаметра.
Роды: Cerebrum Schroder, Medioli et Scott, 1989;
Globipelorhiza Cedhagen et Mattson, 1991; Ipoa Tendal
et Hessler, 1977; Komokia Tendal et Hessler, 1977;
Lana Tendal et Hessler, 1977; Normanina Cushman,
1928; Reticulum Schroder, Mediolo et Scott, 1989; Sep-
tuma Tendal et Hessler, 1977.
Сем. Rhizamminidae Wiesner, 1931
Раковина крупная, трубчатая, ветвящаяся. Диа-
метр более 100 мкм, длина до нескольких сантиметров.
Агглютинированные трубочки выстилаются внутри
органическим слоем. Просветы их заполнены цито-
плазмой и массой стеркомат.
Род: Rhizammina Brady, 1879 с единственным ви-
дом Rh. algaeformis Brady, 1879.
Сем. Baculellidae Tendal
et Hessler, 1977
Форма тела разнообразная за счет коротких боко-
вых отростков. Внутренние полости также заполне-
ны стеркоматами.
Роды: Arbor Schroder, Medioli et Scott, 1989; Ba-
culella Tendal et Hessler, 1977; Catena Schroder, Me-
dioli et Scott, 1989; Chondrodapis Mullineaux, 1988;
Edgertonia Tendal et Hessler, 1977.
RHIZOPODA INCERTAE SEDIS
ORDER KOMOKIIDA KAMENSKAYA, 1992
SUMMARY
The komokiids are agglutinated protozoa. The test consists of a
system of branched tubules of even diameter. The test wall simple, con-
sisting of an inner organic layer and a thicker agglutinated outer layer.
Stercomata accumulate within the tubules. The test of various shapes:
tree-, bush-, tube-like or forming loose clump of anastomosing tubules.
The order comprises 3 families, 14 genera and approximately
30 species. Komokiida are marine organisms. They occur in the
Atlantic, Pacific and Indian Oceans mainly at depths of more than
3000 m.
ЛИТЕРАТУРА34
Каменская О. E. Ксенофиофорий и комокиации в трофичес-
ких цепях глубоководного бентоса// Питание морских беспозво-
ночных и его роль в формировании сообществ. М.: Изд. ИОАН,
1987. С. 15—22.
Каменская О. Е. Количественное распределение комокиаций
и ксенофиофорий в южной части Атлантического океана И
Структурно-функциональные исследования морского бентоса. М.:
Изд. ИОАН, 1988. С. 15—20.
Каменская О. Е. Особенности вертикального распределения ко-
мокиаций в Тихом океане И Тр. ИОАН. М., 1989. Т. 123. С. 55—58.
Каменская О. Е. Предварительные данные о комокиациях (Fo-
raminifera, Komokioidea) с трансатлантического разреза по 31—
36° ю. ш. И Тр. ИОАН. М., 1990. Т. 126. С. 62—66.
Каменская О. Е. О положении комокиид в классе Гранулоре-
тикулозей (Granuloreticulosea de Saedeleer, 1934) И Питание морс-
ких беспозвоночных в разных вертикальных и широтных зонах.
М.: Изд. ИОАН, 1992. f
Каменская О. Е. Комокиации из района подводной горы Валь-
дивия (юго-восточная Атлантика) И Тр. ИОАН. М., 1993. Т. 127.
С. 72—81.
Саидова X. М. О современном состоянии системы надвидовых
таксонов кайнозойских бентосных фораминифер. М.: Изд. ИОАН,
1981.
Соколова М. Н. Питание и структура глубоководного макро-
бентоса. М., 1986.
Соколова М. Н. Характеристика питания морских звезд и кре-
веток в батиали Норвежского моря И Бентос высокоширотных
районов. М.: Изд. ВНИРО, 1998. С. 22—33.
Brady Н. В. Notes on some of the reticularian Rhizopoda of the
«Challenger Expedition». 1. On new or little known arenaceous
types Л Quart. J. Microsc. Sci. 1879. Vol. 19. P. 20—63.
Cedhagen T., Mattson S. Globipelorhiza sublittoralis gen. et sp.
nov., a komokiacean (Protozoa: Foraminifera) from the Scandinavian
sublittoral II Sarsia. 1991. Vol. 76, N 3. P. 209—213.
34 Цитируемая и рекомендуемая литература к разделу «Отряд
Komokiida».
528
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Delage V., Herouard Е. Traitd de Zoologie Concrete. La Cellule
et les Protozoaires. Paris, 1896. T. 1.
De Saedeleer H. Beitrag zur Kenntnis. der Rhizopoden, morpho-
logische und sysiematische Untersuchungen und ein Klassifikations-
versuch // Mus. Roy. Hist. Nat. Belg. 1934. Mem. 60.
Gooday A., Cook P. An association between komokiacea forami-
nifera (Protozoa) and Palludicellina ctenostomes (Biyozoa) from the abys-
sal north-east Atlantic // J. Natur. Hist. 1984. Vol. 18, N 5. P. 765—784.
Lengsfeld A. M. Nahrungsaufnahme und Verdauung bei der Fora-
minifere Allogromia laticollaris // Helgolander wiss. Meersunnters,
1969. Vol. 19. P. 385—400.
Loeblich A. R., Jr., Tappan H. Foraminiferal genera and their
classification. New York, 1988. Vol. 1. P. 40—42.
Mullineaux L. S. Taxonomic notes on large agglutinated forami-
nifers encrusting manganese nodules, including the description of a
new genus, Chondrodapis (Komokiacea) // J. Foram. Res. 1988.
Vol. 18. P. 46—53.
Schroder C. J., Medioli F. S., Scott D. B. Fragil abyssal fora-
minifera (including new Komokiaces) from the Nares Abyssal Plai-
ne // Micropaleontology. 1989. Vol. 35. P. 10—48.
Tappan H., Loeblich A. R., Jr. Granuloreticulosa // Synopsis and
Classification of Living Organisms / Ed. S. P. Parker. Vol. 1. New
York : McGrow-Hill Comp., 1982. P. 527—552.
Tendal O. S. Aspects of the biology of Komokiacea and Xeno-
phyophoria II Sarsia. 1979. Vol. 1. P. 13—17.
Tendal O. S. A preliminary account of the Komokiacea and
Xenophyophorea // Peuplements profonds du golf de Gascogne / Eds
L. Lauber, Cl. Monniot. S. 1., 1985. P. 263—265.
Tendal O. S., Hessler R. R. An introduction of the biology and
systematics of Komokiacea (Textulariina, Foraminifera) // Galath.
Rep. 1977. Vol. 14. P. 165—194.
Wiesner H. Die Foraminiferen der deutschen Sudpolar Expedi-
tion 1901—1903 // Deutsche Sudpolar Expedition 1901—03 / Hera-
usg. von E. Drygalski. S. 1., 1931. Vol. 12. P. 53—165.
Отряд Blastocystida Zierdt, 1988
emend. Belova, 1992 — Бластоцисты
Своеобразная группа паразитов, заселяющих кишечник неко-
торых беспозвоночных и позвоночных, в том числе и человека.
Тело овальной, эллипсоидной, реже амебоидной формы; централь-
но обычно располагается электронно-прозрачная центральная ва-
куоль, в ряде случаев разделенная цитоплазматическими мембрана-
ми на отдельные компартменты. Много митохондрий, содержащих
кристы трубчатого типа. Гликокаликс структурированный, хорошо
выражен; органеллы, похожие на центриоли, не обнаружены. Цис-
терны аппарата Гольджи собраны в стопку. Жгутики отсутствуют.
Анаэробы.
История изучения
История изучения бластоцист начинается с пер-
вых рисунков Каннингхема (Canningham, 1881). Но
по-видимому, бластоцист находили задолго до Кан-
нингхема. Об очень похожих организмах упоминал в
1849 г. в своей рукописной работе Леш (Loech).
Бриттен (Brittan, 1849) описал бластоцисты под на-
званием аннулярные тельца. Каннингхем назвал об-
наруженных им паразитов зооспорами. Он предпо-
лагал, что в кишечнике зооспоры развиваются, как
амебы, а во внешней среде формируют жгутики, как
трихомонады. В более поздних работах (Ucke, 1908;
Bensen, 1909) бластоцист часто описывали как одну
из стадий развития трихомонад. В работах Боне и Про-
вачека (Bohne, Prowazek, 1908; Prowazek, 1911, 1912)
на рисунках изображены типичные бластоцисты, кото-
рые определены как цисты Trichomonas intestinalis.
Сомнение в принадлежности бластоцист к трихо-
монадам высказал Добель (Dobell, 1908), а позднее
Веньон (Wenyon, 1910). В 1911 г. появляется иссле-
дование Алексеева (Alexeieff, 1911) по так называе-
мым цистам трихомонад, выполненное в основном на
паразитах из тритонов: Triturus cristatus (гребенча-
тый тритон) и Т. marmoratus (мраморный тритон).
Первоначально Алексеев был склонен считать цисто-
подобные образования стадиями развития Entamoeba
ranarum, но затем изменил свою точку зрения и
решил, что цисты трихомонад относятся к грибам, и
дал этим организмам название Blastocystis enterocola
Alexeieff, 1911. Брумпт (Brumpt, 1912) описал В. ho-
minis из человека и под влиянием работ Алексеева
также отнес этот вид к грибам.
Впервые бластоцисты были достаточно обосно-
ванно определены как Protozoa Циердтом с соавт.
(Zierdt et al., 1967); позднее эта точка зрения была
подтверждена (Белова, 1992).
Классификация
Система бластоцист в настоящее время представ-
ляется следующим образом.
Отряд Blastocystida Zierdt, 1988 emend. Belova, 1992
Сем. Blastocystidae Zierdt, 1988
Род Blastocystis Alexeieff, 1911
Морфология и физиология
Форма тела бластоцист чаще всего круглая
(рис. 522, А, Б*, Д) или овальная (рис. 522, Л), реже
амебоидная (рис. 522, В). Лобоподии формируются
нечасто и довольно медленно, этим и объясняется
сравнительно редкое появление амебоидных стадий.
При бинарном делении бластоцисты напоминают по
форме гантели (рис. 522, Г). Стадии бластоцист из
кишечника хозяев и в культуре существенно разли-
чаются по размерам. Стадии, обитающие в кишечни-
ке, мельче культуральных.
Аналогичные различия отмечены в числе ядер у
стадий бластоцист, развивающихся в кишечнике и в
культуре. У кишечных стадий число ядер варьирует
от 1 до 4 (редко 8—32), в культуре — от 1 до 64
(реже более 100). Ядра по Романовскому—Гимза ок-
рашиваются в рубиновый цвет и локализуются в ци-
топлазме, которая чаще всего тонким слоем распола-
гается по периферии тела бластоцист. В двухъядер-
ных и четырехъядерных стадиях бластоцист ядра
локализуются обычно полярно по отношению друг к
другу (рис. 522, 3, И). Центральную часть тела пара-
зита занимает центральная вакуоль, которая не окра-
шивается по Романовскому—Гимза или очень слабо
красится в серо-синий цвет. Цитоплазма красится по
Романовскому—Гимза в голубой цвет различной ин-
тенсивности у всех видов бластоцист.
RHIZOPODA INCERTAE SEDIS. ОТРЯД BLASTOCYSTIDA
529
Рис. 522. Виды бластоцист из домашних животных. (По: Белова, 1996).
А — Blastocystis anatis из кишечника, Б — В. anatis из культуры, В — В. anseri из культуры, Г — В. galli из кишечника, Д — В. galli из культуры, Е —
В. numidae из кишечника, Ж — В. meleagridis из кишечника, 3 — В. egui из кишечника, И — В. suis из кишечника, К — В. suis из культуры, Л —
В. Bovis из кишечника, М — В. ovis из кишечника, Н — В. ovis из культуры.
Fig. 522. Species of blastocysts from domestic animals.
A — Blastocystis anatis from intestine, Б — B. anatis in culture, В — В. anseri in culture, Г — В. galli from intestine, Д — B. galli in culture, E — B. numidae
from intestine, Ж — В. meleagridis from intestine, 3 — B. equi from intestine, И — В. suis from intestine, К — В. suis in culture, Л — B. bovis from intestine,
M — B. ovis from intestine, H — B. ovis in culture.
При изучении бластоцист in vivo с использовани-
ем светового фазово-контрастного микроскопа они
обычно выглядят как малоподвижные организмы
круглой формы, с большой центральной вакуолью.
Центральная вакуоль может занимать 3/4—4/5 всего
тела паразита. По периферии располагается тонкий
слой цитоплазмы с включенными в нее ядрами.
В электронном микроскопе у всех стадий бласто-
цист, развивающихся в кишечнике и культуре, выяв-
лен хорошо выраженный гликокаликс (рис. 523,
вкл.). Под гликокаликсом расположена типичная
плазматическая, или наружная, мембраца. На ней
имеются «карманы», представляющие собой неболь-
шие конические углубления с электронно-плотными
34 Протисты, часть 1
краями. В цитоплазме содержится большое число
рибосом и мелких вакуолей. Часто вакуоли с наруж-
ной стороны окружены рибосомами. Митохондрии с
электронно-плотным матриксом содержат кристы
трубчатого типа (рис. 523). Митохондрии располага-
ются либо на некотором расстоянии от ядра, либо
на разных полюсах рядом с ядром. Аппарат Гольджи
представлен несколькими цистернами, собранными в
стопку, и располагается рядом с ядром на разных
полюсах. Ядро окружено 2 мембранами, содержит
глыбки хроматина, расположенные на периферии яд-
ра. Между оболочкой и электронно-плотной частью
ядра имеется электронно-прозрачное пространство.
Ядро содержит ядрышко.
530
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 524. Схема жизненного цикла бластоцист. (По: Белова, 1992).
А — циста второго порядка (расселительная стадия); Б—Д — бинарное агамное деление, повторяющееся многократно; Е, Ж — гамонты (?); 3, И — гаме-
ты (?); К, Л — зигота (?); М, Н — циста первого порядка.
Fig. 524. The scheme of blastocyst life cycle.
A — cyst of the second generation (motile stage); Б—Д — agamic cell division; E, Ж — gamonts (?); 3, И — gametes (?); К, Л — zygote (?); My H — cyst of
the first generation.
Центральная вакуоль ограничена мембраной, эту
мембрану еще называют внутренней. Центральная ва-
куоль занимает большую часть клетки. Она разделена
мембраной на отдельные участки разной величины.
Между отдельными компартментами центральной ва-
куоли часто содержится небольшое количество ци-
топлазмы с включенными в нее митохондриями и ри-
босомами. При размножении в центральной вакуоли
формируется большое число (до нескольких десятков)
особей. В таких клетках цитоплазма занимает лишь
узкое пространство между плазматической мембра-
ной и мембраной центральной вакуоли (рис. 523).
Жизненный цикл
Жизненный цикл бластоцист в настоящее время
можно представить следующим образом. Рассели-
тельные стадии (вторичные цисты) представлены не-
большими амебками с относительно толстой оболоч-
кой и крупным ядром (рис. 524, А). Расселительные
стадии формируются в результате деления много-
ядерной стадии с крупными ядрами (первичные
цисты) (рис. 524, Н). Процесс формирования цисто-
подобных стадий мы называем цистцгонией. Рассе-
лительные стадии в организме хозяина приступают к
бинарному делению, повторяющемуся многократно
(рис. 524, Б—Д). Этот процесс мы называем мерого-
нией, а продукты деления — мерозоитами; размеры
ядер в мерозоитах такие же, как и у расселительных
стадий. Затем часть мерозоитов превращается в мно-
гоядерные стадии с мелкими ядрами — предположи-
тельно гамонты (рис. 524, Е, Ж). Гамонты дают на-
чало мелким (2.5 х 2.5 мкм) голым амебкам с мелкими
ядрами — предположительно гаметам (рис. 524, 3, И).
Этот процесс мы называем гаметогонией. Процесс
кариогамии не удалось наблюдать, хотя встречались
двухъядерные особи с мелкими, очень близко распо-
ложенными, в ряде случаев соприкасающимися друг
с другом ядрами. Предположительно, кариогамия за-
вершается формированием зиготы (рис. 524, К, Л).
Зигота трансформируется в первичную цисту, содер-
жащую большое число (до 32) одноядерных рассели-
тельных стадий — вторичных цист (рис. 524, М, Н).
Практическое значение
Практическое значение бластоцист определяется
их патогенностью для хозяев. Имеется ряд сообще-
ний, подтверждающих этиологическую роль Blasto-
cystis hominis при острых и хронических гастроэнте-
ритах у людей. У иммунодефицитных людей В. ho-
minis может вызывать даже ревматоидные артриты,
RHIZOPODA INCERTAE SEDIS. ОТРЯД BLASTOCYSTIDA
531
локализуясь в суставной жидкости. Аналогичные кли-
нические признаки наблюдаются у домашних живот-
ных, зараженных бластоцистами.
Систематический обзор
Отряд Blastocystida Zierdt, 1988
emend. Belova, 1992
Сем. Blastocystidae Zierdt, 1988
Характеристика семейства совпадает с характе-
ристикой отряда (см. с. 528).
Род Blastocystis Alexeieff, 1911
Характеристика рода совпадает с характеристи-
кой отряда и семейства. В настоящее время в роде
Blastocystis насчитывается 23 вида.
Ниже приводятся описания наиболее важных в
практическом отношении видов, которые были об-
наружены у человека и домашних животных.
Blastocystis hominis Brumpt, 1912.
Хозяева. Homo sapiens (человек); Pan troglody-
tes (шимпанзе) — предположительно; Macaca nemestri-
па (свинохвостый макак) — предположительно; Cavia
porcellus (морская свинка) — экспериментальное заражение.
Локализация. Слепой отросток толстого ки-
шечника.
Распространение. Всесветное.
Описание. Форма тела круглая, овальная, реже
амебоидная. Различают 3 стадии: вакуолярную, гра-
нулярную и амебоидную. Размах изменчивости раз-
меров стадий, обнаруживаемых в фекалиях, состав-
ляет 2—50 мкм; чаще встречаются стадии от 5 до
20 мкм в диаметре. Число ядер в одной особи варь-
ирует от 1 до 4, чаще всего встречаются двухъядер-
ные стадии. Стадии, развивающиеся в культуре на
аксеничной питательной среде, имеют большее мор-
фологическое разнообразие. Число ядер в культу-
ральных стадиях может достигать нескольких десят-
ков, а максимальный диаметр сферических форм —
порядка 100 мкм.
Blastocystis anatis Belova, 1991 (рис. 522, А, Б).
Хозяин. Anas platyrhynchos (домашняя утка).
Локализация. Слепые отростки толстого кишечника.
Распространение. Всесветное.
Описание. Бластоцисты имеют овальную, круг-
лую, реже амебоидную форму. Длина 6.2—у40.2, ши-
рина 5—32.5 мкм. Число ядер в одной особи колеб-
лется от 1 до 4. Стадии бластоцист, развивающиеся
в культуре, значительно разнообразнее по размерам
и числу ядер. Размеры В, anatis в культуре варьируют
в пределах 2.5—56.2 х 2.5—56.2 мкм.
Blastocystis anseri Belova, 1992 (рис. 522, В).
Хозяин. Anser anser (домашний гусь).
Локализация. Слепые отростки толстого ки-
шечника.
Распространение. Всесветное.
Описание. Формы культуральных стадий варь-
ируют от круглой и овальной до эллипсоидной и
амебоидной. Размеры колеблются в пределах 7.5—
46.2 х 7.5—46.2 мкм. Число ядер в одной особи ча-
ще всего варьирует от 1 до 8; редко встречались
экземпляры, содержащие более 20 ядер. Цитоплазма
у В. anseri красится по Романовскому—Гимза в
бледно-голубой цвет. Пелликула часто имеет кон-
центрически расположенные складки.
Blastocystis galli Belova et Kostenko, 1990 (рис. 522,
Г,Д).
Хозяин. Gallus gallus (домашняя курица).
Локализация. Слепые отростки толстого ки-
шечника.
Распространение. Всесветное.
Описание. Форма тела бластоцист варьирует от
круглой и овальной до эллипсоидной и гантелевид-
ной. Размеры стадий, развивающихся в слепых от-
ростках кишечника, варьируют в пределах 2.5—
47.5 х 2.5—46.3 мкм. Число ядер в кишечных стади-
ях бластоцист обычно колеблется от 1 до 4, реже
встречаются особи с 8—32 ядрами. Стадии, развива-
ющиеся в культуре, многообразнее. Размеры их варь-
ируют в пределах 2.5—110x2.5—110 мкм, число
ядер обычно колеблется от 1 до 32, реже встречают-
ся особи, содержащие до 64—100 ядер.
Blastocystis numidae Belova, 1992 (рис. 522, Е).
Хозяин. Numida meleagris (домашняя цесарка).
Локализация. Слепые отростки толстого ки-
шечника.
Распространение. Всесветное.
Описание. Форма тела бластоцист варьирует от
круглой и овальной до гантелевидной и амебоидной.
Размеры колеблются в пределах 5—40 х 5—33.8 мкм.
Число ядер в одной особи варьирует от 1 до 4.
Blastocystis meleagridis Belova, 1992 (рис. 522, Ж).
Хозяин. Meleagris galldpavo (домашняя индейка).
Локализация. Слепые отростки толстого кишеч-
ника.
Распространение. Европа, Азия, предположи-
тельно всесветное.
Описание. Форма тела круглая, овальная, ци-
линдрическая, амебоидная. Размеры варьируют в пре-
делах 2.5—55.1 х51.3 мкм. Число ядер в одной осо-
би колеблется от 1 до 4.
Blastocystis equi Belova, 1992 (рис. 522, 3).
Хозяин. Equus caballus (лошадь).
Локализация. Слепая кишка.
Распространение. Европа.
Описание. Форма тела овальная или круглая.
Размеры 12.3—37.5 мкм. Число ядер в одной особи
варьирует от 1 до 4. По Романовскому—Гимза цен-
тральная вакуоль окрашивается в серо-голубой цвет,
а цитоплазма — от бледно-голубого до темно-сине-
го, неравномерно, ячеисто.
Blastocystis suis Belova, 1992 (рис. 522, Я, К),
Хозяева. Sus scrofa domestica (домашняя свинья);
Sus scrofa (кабан).
532
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Локализация. Слепая кишка.
Распространение. Европа.
Описание. Формы стадий, развивающихся в ки-
шечнике, варьируют от круглой и овальной до ганте-
леобразной и амебоидной. Число ядер в одной особи
колеблется от 1 до 2, размеры 5—16.3 х 6.25—17.3 мкм.
Стадии, развивающиеся в культуре, крупнее — 8.8—
50 х 8.8—50 мкм, обычно круглой и овальной фор-
мы, реже амебоидной. Число ядер в одной особи ко-
леблется от 1 до 30.
Blastocystis bovis Belova, 1992 (рис. 522, Л).
Хозяин. Bos taurus (крупный рогатый скот).
Локализация. Слепая кишка.
Распространение. Европа, Азия; предположи-
тельно всесветное.
Описание. Вид описывается только по стадиям
из слепой кишки. Форма этих стадий круглая, оваль-
ная или амебоидная. Размеры варьируют в пределах
5—20 х 5—20 мкм. Число ядер в одной особи колеб-
лется от 1 до 4. Ядра обычно располагаются полярно.
Blastocystis ovis Belova, 1992 (рис. 522, М, Я).
Хозяин. Ovis aries (овца).
Локализация. Слепая кишка.
Распространение. Европа, Азия; предположи-
тельно всесветное.
Описание. Форма бластоцист варьирует от
круглой и овальной до амебоидной. Размеры колеб-
лются в пределах 3.8—16.3 х 5—17.5 мкм. Число ядер
в одной особи от 1 до 2, размножение осуществля-
ется бинарным делением. Растут бластоцисты на пита-
тельной среде плохо. Стадии, развивающиеся в куль-
туре, овальные и амебоидные. Число ядер 1—2, мно-
гоядерные стадии не обнаружены. Размеры стадий,
развивающихся в культуре, незначительно отличают-
ся от размеров стадий, паразитирующих в кишечнике,
и колеблются в пределах 3.8—18.8x5—18.8 мкм.
RHIZOPODA INCERTAE SEDIS
THE ORDER BLASTOCYSTIDA ZIERDT, 1988, EMEND. BELOVA, 1992
SUMMARY
Blastocysts are parasites inhabiting intestine of some invertebrates
and vertebrates, humans included. Blastocysts have been found in ne-
arly all species of domestic animals: ducks, geese, hens, fowl, turkeys,
cats, horses, pigs, cattle, sheep, camels and lamas. They normally have
rounded or oval body shape, and amoeboid forms also occur.
Blastocysts reproduce by binary and polytomous division. In case of
binary division blastocysts reminiscent the dumb-bells. Number of nuclei
on the stage of blastocysts reproducing my multiple diyisipn fluctuates
from 2 to 100 and more depending on the species of blastocysts. Stages
developing in culture contain the maximum number of nuclei.
An electron microscopy study has revealed a pronounced glyco-
calyx in all stages of blastocysts. A typical plasmic membrane is situ-
ated below the glycocalyx. Situtated on the plasmic membrane are
«pockets» representing small conical holes with electron dense mar-
gins. The cytoplasm contains a large number of ribosomes and small
vacuoles. Mitochondria with electron dense matrix contain tubular
cristae. Golgi apparatus is represented by several dictyosomes. The
central vacuole is restricted by cytoplasmic membrane. It occupies a
large part of the cell and is divided by thin threads of cytoplasm into
separate parts of different size. Up to several tens of individuals are
formed in the central vacuole during reproduction.
The following system of blastocysts has been accepted:
ORDER BLASTOCYSTIDA ZIERDT, 1988, EMEND. BELOVA, 1992
Parasitic organisms of oval, ellipsoid, less frequently of amoeboid
shape; electron transparent central vacuole is situated in the centre, in
several cases divided by cytoplasmatic membranes into different com-
partments; mitochondria with tubular cristae; anaerobes; glycocalyx
pronounced; flagella and centriole-like structures have not been found.
FAMILY BLASTOCYSTIDAE ZIERDT, 1988
GENUS BLASTOCYSTIS ALEXEIEFF, 1911
The genus includes 23 species.
ЛИТЕРАТУРА35
Белова Л. M. Мировая фауна и морфофункциональная организация
бластоцист (Protista, Rhizopoda). СПб., 1992. (Тр. ЗИН РАН; Т. 244).
Крылов М. В. Определитель паразитических простейших. СПб.,
1996.
Alexeieff A. Sur la nature des formations dites «kystes de Tricho-
monas intestinalis»// Comp. Rend. £ean. Soc. Biol. 1911. T. 71.
P. 296—298.
Bensen W. Untersuchungen uber Trichomonas intestinalis und va-
ginalis des Menschen // Arch. Protistenk. 1909. Bd 18. S. 115—127.
Bohne A., Prowazek S., von. Zur Frage der Flagellatendysente-
rie // Arch. Protistenk. 1908. Bd 12. S. 1—8.
Boreham P. F. L., Stenzel D. J. Blastocystis in humans and ani-
mals: morphology, biology, and epizootiology // Adv. Parasitol. 1993.
Vol. 32. P. 1—70.
Brittan F. Report of series of microscopical investigations on
the pathology of cholera // London Med. Gaz. 1849. Vol. 9. P. 530—
542.
Brumpt E. Colite a Tetramitus mesnili (Wenyon, 1910) et colite
a Trichomonas intestinajis Leuchart, 1879 — Blastocystis hominis
35 Цитируемая и рекомендуемая литература к разделу «Отряд
Blastocystida».
n. sp. et formes voisines // Bull. Soc. Pathol. Exot. 1912. T. 5.
P. 725—730.
Canningham D. D. On the development of certain microscopic
organisms occuring in the intestinal canal // Quart. J. Microsc. Sci.
1881. Vol. 21. P. 234.
Dobell C. Some remarks upon the «autogomy» of Bodo lacertae
(Grassi) // Biol. Cbl. 1908. Bd 28. S. 548.
Losch F. A. Massenhafte Entwickelung von Amoeben im Dick-
darn // Arch. Pathol. Anat. Phis. Klin. Med. 1875. Bd 65. S. 196—211.
Prowazek S., von. Zur Kenntnis der Flagellaten des Darmtrak-
tus // Arch. Protistenk. 1911. Bd 23. S. 96.
Prowazek S., von. Beitrage zur Protozoen und verwandter Orga-
nismen von Sumatra (Deli) // Arch. Protistenk. 1912. Bd 26. S. 250.
Ucke R. Trichomonaden und Megastomen im Menschendarm //
Cbl. Bacteriol. 1. Abt. Orig. 1908. Bd 4. S. 231.
Wenyon С. M. A new flagellate Macrostoma mesnili sp. n. from
the human intestine, with sonie remarks on the supposed cyst of Tric-
homonas II Parasitology. 1910. Vol. 3. P. 210—216.
Zierdt С. H. Blastocystis hominis, a long misunderstood intestinal
parasite // Parasitol. Today. 1988. Vol. 4. P. 15—17.
Zierdt С. H., Rude W. S., Bull B. S. Protozoan characteristics of
Blastocystis hominis // Amer. J. Clin. Pathol. 1967. Vol. 48. P. 495—
501.
Тип FORAMINIFERA d’Orbigny, 1826 — ФОРАМИНИФЕРЫ
Одноклеточные организмы, тело которых защищено органи-
ческой, агглютинированной или полностью секреционной ракови-
ной, которая может быть однокамерной, псевдоколониальной,
псевдокамерной и многокамерной (число камер обычно 6—15(30),
реже до нескольких сотен, в последнем случае камеры могут быть
дифференцированы). Камеры связаны между собой фораменами
(отверстиями), у более сложно устроенных — интегрирующими
системами: столонами, внутренними устьевыми структурами, тун-
нелями, каналами; раковины обычно имеют 1, 2 или несколько
наружных отверстий — устьев, служащих местом выхода грануло-
ретикулоподий (=ретикулоподий), образующих вокруг раковины
ретикулум. Жизненные циклы с чередованием полового и беспо-
лого поколений, с промежуточной редукцией. Гаметы двух- и трех-
жгутиковые или амебоидные. Одно из поколений (чаще половое)
может отсутствовать. Ядерный аппарат со сложными изменениями
в течение жизненного цикла, лишь у низших форм часть цикла
одноядерна, а у остальных многоядерность сохраняется в течение
всего жизненного цикла (кроме зиготы), при этом у наиболее про-
двинутых форм возникает ядерный дуализм. Мейоз двухступенча-
тый, митоз происходит по типу закрытого плевромитоза. Кристы
митохондрий трубчатые, у планктонных форм имеются криптосо-
мы и микровилли. Преимущественно морские донные свободножи-
вущие организмы, но есть и планктонные формы. Изредка наблю-
дается переход к паразитизму. Ископаемые формы найдены в от-
ложениях от кембрия до голоцена.
Общая характеристика
Фораминиферы представляют собой естествен-
ную, хорошо очерченную группу организмов, одну
из самых больших, высокоорганизованных и своеоб-
разных среди простейших. Они включают более
4.5 тыс. родов, около 40 тыс. видов, большая часть
которых (свыше 30 тыс.) — вымершие. Число со-
временных видов, по оценке разных авторов, состав-
ляет от 5 до 10 тыс. Полное перечисление всех пер-
воописаний (без исключения синонимов) в Каталоге
фораминифер (США) достигает 200 000 (Hottinger,
1982). Фораминиферы представляют одну треть всех
известных протистов и примерно 2.5 % всех извест-
ных на Земле организмов.
Одной из самых характерных черт этой группы
является наличие раковины — в простейших случа-
ях органической, у более эволюционно продвинутых
видов — агглютинированной (посторонние частицы,
чаще всего песчинки, скреплены органическим или
известковым цементом), у высших форм — полностью
секреционной, известковой. Ультраструктура извест-
ковой стенки различна у наиболее продвинутых
представителей разных классов, представляющих
особые филогенетические линии развития, и исполь-
зуется как диагностический признак высокого таксо-
номического ранга. Раковина фораминифер обтека-
ется снаружи слоем цитоплазмы, так что их скелет
в строгом смысле не внешний, а внутренний (рис. 539,
2, 3; 541, 5).
Еще более важным признаком, на котором по-
строена вся система фораминифер, служит строение
раковины и ее камер. В простейшем случае ракови-
ны одно- или двухкамерные; у большинства много-
камерных видов число камер не превышает 5—30,
но есть высокоорганизованные формы с числом ка-
мер, достигающим нескольких сотен. В этих случаях
камеры могут быть дифференцированы по форме и
функции, что является высокой ступенью организа-
ции для простейших. Многообразие жизненных
форм раковин фораминифер, типов их симметрии
настолько велико, что делает эти организмы исклю-
чительной моделью для исследования основных прин-
ципов морфогенеза. У многокамерных форм возни-
кают системы интеграции, позволяющие преодолеть
разобщенность отдельных подразделений, — это
столоны, внутренние устьевые системы, туннели и
каналы. Последние имеют собственные известковые
стенки и отличаются особой сложностью. В них
циркулирует эктоплазма. Функцию каналов можно
сравнить с таковой кровеносной системы, сосуды ко-
торой имеют твердые известковые стенки. Это уни-
кальная организменная система высокого уровня, встре-
чающаяся среди протистов только у фораминифер.
Цитоплазматическое тело фораминифер находит-
ся в основном внутри раковины; наружу через спе-
циальные отверстия — устья выходит сеть тонких,
анастомозирующих между собой гранулоретикулопо-
дий, главные функции которых — передвижение и
захват пищи. Ретикулоподии резко отличаются от
широких лобоподий амеб; а тем более от жгутиков
и ресничек. Они служат диагностическим признаком
ранга типа и хорошо отличают фораминифер от дру-
гих протистов.
Ядерный аппарат фораминифер также несет чер-
ты высокой организации и своеобразия: у них име-
534
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ется многоядерность и ядерный дуализм, выражаю-
щийся в том, что ядра дифференцированы по стро-
ению и функции на вегетативные и генеративные.
Ядра фораминифер претерпевают сложные измене-
ния в процессе жизненного цикла, который характе-
ризуется гетерофазным чередованием поколений.
При этом гаплоидная и диплоидная фазы цикла со-
ставляют почти равные части. Классический двух-
фазный жизненный цикл фораминифер занимает
обычно 1 год, а иногда и 2. Лишь у представителей
низшего рода Allogromia он завершается за 8 ч
(Grell, 1973; Lee et al., 1979). Известны случаи,
когда после выхода молоди родительская особь еще
продолжала существование. Вероятно, длительность
годичных жизненных циклов фораминифер адапти-
рована к годовой цикличности обилия питательных
веществ, цветения водорослей. Долгие периоды веге-
тативного существования требуют активности и жиз-
ненной стратегии, сравнимой с таковой многокле-
точных организмов. В этом отношении фораминифе-
ры — не очень типичные простейшие. Присущий им
темп роста очень мал, а продолжительность жизни
необычайно велика по сравнению с другими протес-
тами.
Гаметы подавляющего большинства фораминифер
имеют 2, редко 3 жгутика; иногда встречаются амебо-
идные, по-видимому утратившие жгутик. Митохонд-
рии взрослых особей и гамет имеют трубчатые кристы.
Средний размер большинства видов 0.5 мм, масса
отдельного организма 0.01—0.5 мг. Среди роталиел-
лид и глабрателлид есть группы, отличающиеся более
мелкими размерами — до 0.1 мм (Pawlowski, Lee,
1991). Большинство фораминифер едва различимы
невооруженным глазом, но среди как вымерших, так
и современных форм встречаются виды размером с
монету (2—3 см) или даже с ладонь (12—16 см) (ис-
копаемые нуммулитесы, современные корнуспирои-
десы, палочковидные батисифоны). Это настоящие
гиганты в мире простейших. Механизм функциони-
рования таких крупных клеток, их ядерный аппарат
до сих пор не изучены и представляют огромный ин-
терес.
Подавляющее большинство видов форамини-
фер — донные формы, передвигающиеся по столо-
нам водорослей, гидроидам, грунту, зарывающиеся в
грунт на 2—10, иногда до 30 см (Goldstein, Harben,
1993), чаще свободноживущие, иногда прикреплен-
ные, ложноколониальные. Прикрепленные формы
разных, не родственных групп приобретают конвер-
гентное сходство, приспосабливаясь к прикреплению
и распространению по субстрату. Среди форамини-
фер известны комменсалы и очень редко — парази-
ты. Небольшая часть видов перешла к парению в
верхней толще воды (преимущественно до 200 м) и
приобрела глубокие адаптации к пелагическому об-
разу жизни.
Фораминиферы практически всеядны, употребля-
ют в пищу бактерии, дрожжи, водоросли, других про-
тистов, мелких многоклеточных животных, их яйца и
личинки, детрит, растворенную в воде органику. Но
сами они редко становятся пищей для других орга-
низмов и их участие в пищевой цепи пока сложно
оценить. В немногих известных случаях (заглатыва-
ние рыбами, иглокожими) фораминиферы использо-
вались, скорее, как источник извести.
Фораминиферы — почти исключительно морские
организмы, составляющие одну из наиболее важных
групп океанического бентоса во всех районах Миро-
вого океана — от литорали до абиссальных впадин.
Небольшое число видов обитает в опресненных во-
дах устьев рек, а также в грунтовых водах повышен-
ной солености — в пустынях Африки и Средней
Азии (Бродский, 1928; Gauthier-Lievre, 1935; Янков-
ская, Михалевич, 1972; Михалевич, 1976). Биомасса
донных фораминифер может составлять от 20 до
80 %, иногда превышая массу макробентоса. Их
может быть так много, что они напоминают «живой
песок». Фораминиферы, занимающие огромные про-
странства морского дна, представляют весьма значи-
тельный резервуар запасной энергий. Многие виды
фораминифер служат индикаторами состояния вод-
ных масс — они могут указывать на загрязнение, а
также на богатство питательными веществами, что, в
свою очередь, может быть использовано для опреде-
ленных районов перспективного рыбного промысла.
У донных фораминифер-эпибионтов обнаружена спе-
циальная защитная система (Yankb et al., 1994). Ее
состояние может использоваться как очень чувстви-
тельный биомаркер для предсказания экологических
последствий в районах антропогенного загрязнения.
Отмершие раковины фораминифер участвуют в
образовании морских осадков, составляя важнейший
компонент толщи осадочного чехла Мирового океана
на всех этапах его формирования. В осадках отлага-
ются не только донные, но и планктонные формы,
отмершие раковины которых падают «дождем» на
дно сквозь толщу морской воды со средней ско-
ростью 2 см/с, образуя так называемые голубые, или
глобигериновые, илы. Они занимают в Мировом оке-
ане примерно треть всего дна (около 120 млн км2);
их толщина местами доходит до нескольких метров.
В результате химических и тектонических процессов
эти илы со временем превратились в известняки.
Следовательно, фораминиферы являются их породо-
образующим элементом; входят они наряду с кокко-
литофоридами и в состав мела. В меловом периоде
фораминиферы благодаря своей активности стали ос-
новой известковых отложений в условиях недостатка
СО2. Из-за этого воды океанов мира оказались насы-
щенными растворенным карбонатом кальция, который
является активным поглотителем углекислого газа:
Н2О
СаСОз + СО2 —Са(НСОз)2.
Бикарбонат, Са(НСО3)2, — хорошо растворимая
соль, способная накапливаться в океане бесконечно
долго, тем самым спасая планету от парникового эф-
фекта и перегрева. Такова биосферная роль форами-
нифер, осуществляющих карбонат-бикарбонатное
равновесие в природе (Михалевич, 1999).
Раковины фораминифер известны с кембрия; име-
ются отдельные сообщения о более ранних находках.
Фораминиферы — единственная группа животных,
ископаемые остатки которых столь полно представ-
лены и изучены на протяжении всей геологической
истории Земли. Это обстоятельство наряду с особен-
ТИП FORAMINIFERA. ИСТОРИЯ ИЗУЧЕНИЯ
535
ностями строения их раковин (в онтогенезе в основ-
ном последовательно повторяются черты строения
предков) дает уникальный материал для изучения их
филогении и построения их естественной системы.
Комплексы пустых раковин на дне океанов (та-
натоценозы) отражают условия обитания живых
комплексов и используются при палеореконструкци-
ях прежних экологических условий, восстановлении
очертаний палеоконтинентов. Фораминиферы неза-
менимы при определении возраста осадочных пород,
в том числе нефтегазоносных. В связи с их огромной
ролью при поиске энергетических полезных ископа-
емых в течение двух веков изучались преимущест-
венно их раковины, а основные сведения о строении
их живой клетки и биологические сведения получе-
ны лишь в конце нашего века.
История изучения
Первые упоминания о фораминиферах относятся
к 5 в. до н. э., когда Геродот заметил в известняках
египетских пирамид округлые, напоминающие моне-
ты образования, позднее названные нуммулитами. В
России в 18—19 вв. такие известняки называли «че-
чевичными» камнями, а известняки с удлиненными
раковинами фузулинид, похожими на зерна, —- «хлеб-
ными» камнями. Леонардо да Винчи изображал в
своих орнаментах мелких фораминифер, которых он
рассматривал с помощью трубки с двумя увеличи-
тельными стеклами на концах — задолго до изобре-
тения микроскопа. Сохранилось несколько изобра-
жений фораминифер, сделанных в 18 в. (Beccarius,
1731; Janus Plancus, 1739, — цит. по: Loeblich, Tappan,
1988), и более позднего времени, когда их рассмат-
ривали уже как живые организмы. В конце 18—на-
чале 19 в. появляются первые научные работы, по-
священные этой группе (Batsch, 1791; Soldani, 1789—
1798; Fichtel, Moll, 1798; Lamarck, 1801; Montfort,
1808; Фишер, 1829, — цит. no: Loeblich, Tappan,
1988). Первые классификации фораминифер, осно-
ванные на строении их раковин, даны в начале 19 в.
(de Blainville, 1825; d’Orbigny, 1826, 1839, — цит.
no: Loeblich, Tappan, 1988). Впервые название «фо-
раминиферы» — несущие отверстия (форамены) —
было введено д’Орбиньи. Он рассматривал их как
класс, состоящий из 10 семейств, 5 из которых ва-
лидны до сих пор. Во всех вышеупомянутых работах
фораминифер считали моллюсками, вплоть до работ
Дюжардена (Dujardin, 1835, — цит. по: Loeblich,
Tappan, 1988), отнесшего их к простейшим, к груп-
пе Rhizopoda, на основании наличия у них нитевид-
ных разветвляющихся псевдоподий.
Благодаря целому ряду последующих работ, по-
священных морфологии и классификации форамини-
фер (Schultze, 1854; Carpenter, Parker, Jones, 1862;
Reiss, 1862; Jones, 1875; Schwager, 1877, — цит. no:
Loeblich, Tappan, 1988), было расширено представле-
ние об их составе и введено много новых таксонов.
Швагер, Райс и особенно Шульце (Schultze, 1854;
Reiss, 1862; Schwager, 1877, — цит. по: Loeblich,
Tappan, 1988) уделяли большое внимание строению
стенки раковин. Шульце придавал этому признаку
даже больший диагностический вес, чем строению
самой раковины; им же было положено начало изуче-
нию живой клетки этих простейших.
Новый этап в изучении группы ознаменовали ра-
боты английского исследователя Брэди (Brady,
1884, — цит. по: Loeblich, Tappan, 1988), основанные
на обширных сборах современных форм на судне
«Челленджер». Атлас рисунков Брэди с новыми оп-
ределениями Баркера (Barker, 1964, — цит. по: Lo-
eblich, Tappan, 1988) не утратил своего значения и в
настоящее время, несмотря на то что объем видов
Брэди понимал очень широко. Последующие класси-
фикации конца 19—начала 20 в. (Neymayr, 1887,
1889; Delage, Herouard, 1.896; Eimer, Fickert, 1899;
Lister, 1903; Rhumbler, 1911; Schubert, 1921, — цит.
no: Loeblich, Tappan, 1988) развивали систему, пред-
, ложенную Брэди. Среди них наиболее оригинальны
работы Румблера (Rhumbler, 1911) и Листера (Lister,
1903, — цит. по: Loeblich, Tappan, 1988); последний
много внимания уделял изучению диморфизма ра-
ковин, которое связано с чередованием поколений,
открытым ранее Мюнье-Шальма (Munier-Chalmas,
1880).
В первой половине 20 в. вышли одна за другой 3
классификации Кушмана (Cushman, 1925, 1927, —
цит. по: Loeblich, Tappan, 1988; Cushman, 1948), мо-
дифицировавшего систему Брэди. Он довел число се-
мейств фораминифер до 50, что вызвало бурную нега-
тивную реакцию европейских специалистов, назы-
вавших его систему «сумасшедшей классификацией».
Тем не менее в течение нескольких десятилетий она
служила основой всех систематических работ по этой
группе и была более популярна, чем другие (Galloway,
1933; Chapman, Parr, 1936; Glaessner, 1945; Hofker,
1951; Sigal, 1952; Pokorny, 1958, — цит. no: Loeblich,
Tappan, 1988). Из перечисленных работ наиболее ори-
гинальным подходом отличались 2: в одной из них в
основу выделения таксонов положено строение устья
(Hofker, 1951), в другой — количество камер (Sigal,
1952). Число включенных в них семейств составляло
соответственно 23 и 62.
Первая попытка обосновать систему форамини-
фер филогенетически принадлежит Неймайру (Ney-
mayr, 1887, 1889, — цит. по: Loeblich, Tappan, 1988).
Начиная с работ Кушмана, авторы предлагаемых сис-
тем уделяли этому вопросу значительное внимание.
Но наиболее последовательный и глубокий филоге-
нетический эволюционный подход, основанный не
только на морфологии, но и на данных геологичес-
кой летописи, характеризует систему, данную Фур-
сенко и Раузер-Черноусовой (1959) в «Основах пале-
онтологии». К сожалению, она не охватывала всего
многообразия форм, поскольку включала лишь груп-
пы, которые были найдены на территории бывшего
СССР.
Вторая половина 20 в. характеризуется началом и
затем бурным расцветом электронно-микроскопичес-
ких исследований стенки раковин фораминифер. Ряд
работ в этой области стали классическими (Reiss,
1957, 1959, 1960; Reiss, Merling, 1958, — цит. по:
Loeblich, Tappan, 1988; Towe, Cifelli, 1967; Hansen,
Reiss, 1971a, 1971b; Bellemo, 1974a, 1974b; Hemleben
et al., 1977; Leutenegger, Hansen, 1979; Be et al., 1980,
536
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
и мн. др.). Первые шаги были сделаны Райсом (Reiss,
1957, 1959, — цит. по: Loeblich, Tappan, 1988), пере-
строившим на основании полученных данных систе-
му роталиацей (Reiss, 1963). Ему принадлежит заслу-
га открытия биламеллярной стенки. Обнаружив у не-
которых роталиацей биламеллярную стенку, он выделил
их в особые таксоны, оторвав от других близких по
морфологии групп. Этот же подход сохранился и в
двухтомной классификации Лёблика и Таппан «Trea-
tise» (Loeblich, Tappan, 1964), ставшей на четверть
века настольной книгой всех фораминиферологов. Позже
было доказано, что биламеллярную стенку имеют все
роталиацей, а первоначально ее не удалось обнару-
жить у части из них вследствие методических труд-
ностей. Разорванные первоначально близкородствен-
ные формы дискорбид и кассидулинид были вновь
воссоединены. Хотя увлечение новыми данными уль-
тратонкого строения стенки повлекло за собой и в
ряде других случаев необоснованные таксономические
изменения (например, разрыв форм с микрозернис-
той и гиалиновой стенкой), в целом оно дало новый
импульс развитию систематики группы и привело к
резкому возрастанию числа выделенных новых таксонов
различного ранга. В «Treatise» вошло 127 семейств, а
к моменту переиздания этой классификации (Loeb-
lich, Tappan, 1988) было описано еще 104 семейства,
207 подсемейств (из них только после 1980 г. —
40 семейств, 66 подсемейств, 173 рода).
Наряду с электронно-микроскопическими иссле-
дованиями продолжались исследования морфологии
раковин в световом микроскопе. Работы Хоттингера
(Hottinger, 1978, 1982) поражают своей ювелирной
точностью и глубиной морфофункционального ана-
лиза. В классификациях второй половины нашего
века сохранялись 2 направления: в первом придается
первостепенное значение составу и структуре стенки
раковин, во втором — плану строения раковин.
Первое направление продолжали развивать Лёблик и
Таппан ^Loeblich, Tappan, 1988) и их последователи.
Хотя с некоторыми положениями их классификации
1988 г. нельзя согласиться, тем не'менее этот фун-
даментальный двухтомный труд, включающий все
известные роды фораминифер, прекрасно иллюстри-
рованный, с полной ревизией и синонимией всех
родовых и надродовых таксонов остается до сего дня
непревзойденным и значение его трудно переоце-
нить. Однако, как справедливо полагает Хэйнс (Ha-
ynes, 1990), использование этой классификации в
качестве стандарта и успехи прикладной палеонто-
логии иногда противостоят появлению новых вари-
антов системы, что, в свою очередь, может привести
к ошибочным выводам в стратиграфии. Он приводит
яркие примеры, когда разделение глобигеринид на
Globogerinidae и Globorotaliidae помогло обнаружить
резкие переломы в горизонтах мелового и четвертич-
ного периодов. Сходная картина наблюдалась при раз-
делении орбитальных фораминифер. «Перемены такого
рода — яркий показатель здоровья науки, и жалобы
по этому поводу напоминают таковые по поводу то-
го, что Коперник разрушил стабильность птоломее-
вой системы» (Haynes, 1990).
Системы, предложенные отечественными иссле-
дователями (Подобина, 1978, 1998; Субботина и др.,
1981; Саидова, 1981; Басов, 1990; Маслакова, 1990;
Михалевич, 1992, 1998; Григелис, 1995; Раузер-Чер-
ноусова и др., 1996), а также рядом зарубежных авто-
ров (Hohenegger, Piller, 1975; Brasier, 1980, — цит.
по: Lee, 1990; Hansen, 1979; Hohenegger, 1981, 1990;
Jones, 1984, 1994), представляют второе направление.
К сожалению, все они уступают по полноте охвата
материала классификации Лёблика и Таппан.
Большая часть всех известных работ по форами-
ниферам (до 70-х гг. нашего столетия) посвящена
изучению их скелетов, чаще ископаемых. Библиог-
рафия только по работам, касающимся систематики,
настолько огромна (свыше 6000 названий), что одно
упоминание авторов, внесших существенный вклад в
изучение группы, составляет солидный список (см.:
Loeblich, Tappan, 1988). Современная литература по
группе почти необъятна. С начала 80-х гг. (см.: Hay-
nes, 1981) и поныне ежегодно выходит более 500 ра-
бот. Поэтому особое значение приобретают катало-
ги. Наиболее известный из них начат еще в 1940 г.
(Ellis, Messina, 1940, — цит. по: Loeblich, Tappan,
1988) и состоит к настоящему времени из 82 объем-
ных томов. Кроме того, имеется «Индекс родов и
видов фораминифер» (Sherborn, 1888, — цит. по:
Loeblich, Tappan, 1988), переизданный Тальманом
(Thalmann, 1960), и каталог планктонных форамини-
фер (Saito et al., 1976, — цит. по: Loeblich, Tappan,
1988).
В России микропалеонтологические исследова-
ния начаты петербургским профессором Меллером
(1878, 1880, — цит. по: Фурсенко, Раузер-Черноусова,
1959). В 30—70-е гг. нашего века сложилась круп-
ная школа отечественной микропалеонтологии, дол-
гое время возглавляемая А. В. Фурсенко и Д. М. Ра-
узер-Черноусовой, которая недавно отметила свой
столетний юбилей выходом «Справочника по систе-
матике мелких фораминифер палеозоя» (под ред.
Вдовенко и др., 1993). Работы других крупных фо-
раминиферологов этого периода (О. С. Вялова,
А. М. Волошиной, А. А. Крашенинникова, М. С. Ба-
раша, Т. Н. Горбачик, М. Н. Соловьевой, Е. А. Рейт-
лингер, Л. Ф. Плотниковой, В. Н. Беньямовского,
О. Богуш и др.) внесли огромный вклад в изучение
систематики и стратиграфии ископаемых форамини-
фер различных регионов бывшего СССР и дна Ми-
рового океана. Крупный центр этой школы сложил-
ся в Петербурге, во ВНИГРИ, выпустившем ряд мо-
нографий в серии «Ископаемые фораминиферы
СССР» и «Микрофауна СССР» (Н. Н. Субботина,
А. К. Богданович, Н. А. Волошинова, В. П. Василенко,
Л. Г. Дайн, Е. В. Быкова, Е. В. Мятлюк), а также в
«Трудах» ВНИГРИ, Геологического и Палеонтологи-
ческого институтов АН СССР и ВСЕГЕИ. Эти ис-
следования продолжают микропалеонтологи следующе-
го поколения — С. П. Яковлева, В. А. Басов, А. Я. Аз-
бель, Л. А. Алексейчик-Мицкевич, Э. М. Бугрова,
Г. П. Пронина (к сожалению, эта школа в . настоящее
время переживает большие трудности).
Стратиграфическое распространение крупных
таксонов фораминифер и этапы их исторического
развития освещены в работах как отечественных,
так и зарубежных авторов (Фурсенко, Раузер-Черно-
усова 1959; Loeblich, Tappan, 1964; Раузер-Черно-
ТИПFORAMINIFERA.КЛАССИФИКАЦИЯ
537
усова, 1972, — цит. по: Loeblich, Tappan, 1988; Но-
henegger, Piller, 1975; Ross, Haman, 1989).
В 20 в. получили широкое развитие фаунистичес-
кие, экологические и зоогеографические исследования
современных фораминифер. В России это школы
Зоологического института РАН, Санкт-Петербург
(3. Г. Щедрина, В. И. Михалевич, Т. Г. Лукина), Инсти-
тута океанологии РАН, Москва (М. С. Бараш, X. М. Са-
идова, Н. В. Беляева, И. И. Бурмистрова, Т. А. Хусид)
и некоторые другие (И. А. Басов, Л. В. Поляк — Ин-
ститут Арктики и Антарктики, Санкт-Петербург).
За рубежом наиболее значительны работы Бэнди
(Bandy et al., 1964, 1965, — цит. по: Debenay et al.,
1996), Болтовского (Boltovskoy, 1953—1984, — цит.
no: Loeblich, Tappan, 1988), Би (Be, 1957—1982, —
цит; no: Debenay et al., 1996), Флегера (Phleger, 1951,
1960, — цит. no: Debenay et al., 1996), Мюррея (Mur-
ray, 1963—1996, —: цит. no: Debenay et al., 1996),
Кеннета (Kennet, 1976, — цит. no: Lee, Anderson,
1991), Дебенея с соавт. (Debenay, 1985—1996, —
цит. no: Debenay et al., 1990—1998), а также Де
Лака (De Laca et al., 1980) и Липса (Lipps, 1983).
Позже были начаты (Lee et al., 1971; Seiglie, 1971)
и успешно продолжены (Nagy, Alve, 1987; Alve, 1991;
Yanko et aL, 1994, 1998; Kaminski et al., 1995) ис-
следования влияния загрязнений на фауну форами-
нифер прибрежных морей. Работа Бреслера и Янко
(Bresler, Yanko, 1995) положила начало изучению
химической экологии фораминифер.
Как уже указывалось, преимущественное внимание
уделялось изучению скелета фораминифер, даже у
ныне живущих форм. Их цитология, биология, жиз-
ненные циклы до сих пор изучены недостаточно.
После первых шагов в этом направлении (Schultze,
1854; Munier-Chalmas, 1880; Lister, 1895; Rhumbler,
1911; Munier-Chalmas, Schlumberger, 1883, 1885;
Schaudinn, 1894—1903, — цит. no: Loeblich, Tappan,
1988) в начале нашего века были выяснены более
подробно жизненные циклы нескольких видов (Winter,
1907; Myers, 1935, 1936, 1940; Foyn, 1936; Jepps,
1942; Le Calyez, 1938, 1946, 1950, :—цит. no: Loeb-
lich, Tappan, 1988). Важный вклад внесли классичес-
кие работы Грелля и его учеников и ряд более позд-
них работ (Arnold, 1982, 1984; Lee et al., 1991; Gol-
dstein, Moodley, 1993; Roettger et al., 1998; Stouff et
al., 1999a).
Наиболее полно изучен на световом и электрон-
но-микроскопическом уровне жизненный цикл Cibi-
cides lobatulus (Воронова, 1976, 1978а, 19786, 1979)
и Saccammina alba (Goldstein, Т988). Первые цито-
фотометрические исследования ядер были проведе-
ны Вороновой и Селивановой (1976) и Цех (Zech,
1964). Электронно-микроскопические исследования
жизненных циклов других видов пока фрагментарны
(Dahlgren, 1967а, 1967b; Schwab, 1969, 1973, 1976;
Angell, 1971; Cesana, 1971, 1972, 1975, 1978; Bert-
hold, 1977; McEnery, Lee, 1981; Arnold, 1982; Paw-
lowski et al., 1995b; Goldstein, 1997).
Появился ряд работ и по жизненным циклам
планктонных форм (Lee et al., 1965; Be et al., 1976;
Spindler et al., 1978; Be, 1980). Конец века ознаме-
новался Серьезным изучением цитоплазмы форами-
нифер на^ электронно-микроскопическом уровне.
Обобщающие сведения приведены в работах Андер-
сона и Би (Anderson, Be, 1978), Андерсона (Ander-
son, 1988), Ли (Lee, 1990, 1993) и Ли и Андерсона
(Lee, Anderson, 1991). В результате всех этих иссле-
дований картина строения цитоплазмы форамини-
фер и особенностей их жизненных циклов (Ander-
son, 1984; Воронова, Михалевич, 1985; Lee et al.,
1991) становится в общих чертах ясна.
Кроме того, имеется ряд работ, в той или иной
мере посвященных биологии (Lipps, Ronan, 1974;
Hallock, 1981а, 1986; Hickman, Lipps, 1983; Faber,
1991; Lee, Anderson, 1991, и др.) и питанию форами-
нифер (Muller, Lee, 1969; Murray, 1973; Lipps, 1975;
Boltovskoy, Wright, 1976; Lee, 1980; Haynes, 1981;
Brand, Lipps, 1982; De Laca, 1982; Alexander, De
Laca, 1987; Faber, Lee, 1991a) и их взаимоотношени-
ям с симбионтами (Hallock 1981b, 1982, 1984; Lee,
Anderson, 1991). Особенно большой вклад в изучение
проблем симбиоза внесли работы Ли с соавт. (см. ра-
боты 1963—1991 гг.), Андерсона и Би (Anderson, Be,
1976а, 1976b), Хоттингера (Hottinger, 1982), Леуте-
неггер (Leutenegger, 1977, 1983, 1984). Наиболее ши-
роко все направления изучения фораминифер (цито-
логия, симбиоз, экология, биология, физиология, пи-
тание, систематика и даже характеристика ДНК)
представлены в работах школы Ли.
Исследование группы становится все более углуб-
ленным и всесторонним, начато изучение последова-
тельностей нуклеотидов ДНК, протеинов, ассоции-
рованных с минеральными элементами в стенке рако-
вин (King, Hare, 1972; Schroeder, Bada, 1977; Langer
et al., 1993; Lee, 1993, 1995; Wray et al., 1993; Stat-
hoplos, Tuross, 1994; Pawlowski et al., 1995a), имеет-
ся попытка применения данных по аминокислотному
составу нескольких современных видов и родов в
таксономии (Haugen et al., 1989; Pawlowski et al.,
1995a, 1997; Fahrni et al., 1997), появились первые
биохимические работы (Schwab, Schlobach, 1973;
Anderson, Be, 1976a; Schroeder, Bada, 1977; Laatsch,
Schwab, 1984; Faber, Lee, 1991b; Bresler, Yanko,
1995). На излюбленном классическом объекте — Al-
logromia laticollaris начаты физиологические иссле-
дования (Lee, Pierce, 1963; Schwab, Hofer, 1979).
Однако фораминиферы все еще представляют об-
ширное поле для исследования: неизвестны внутрен-
нее строение многих раковин, ультраструктура стен-
ки некоторых крупных таксонов, жизненные циклы и
строение цитоплазмы и ядер огромного большинства
видов, особенно у нодозариат и текстуляриан; даже
инвентаризация видов и родов фораминифер еще да-
лека от завершения.
Классификация
В тип Foraminifera входят следующие таксоны.36
Класс Astrorhizata Saidova, 1981
Подкласс Lagynana Mikhalevich, 1980
Отряд Lagynida Mikhalevich, 1980
36 Звездочкой (*) отмечены таксоны, положение которых
сильно изменилось по сравнению с их прежним положением в сис-
теме. В настоящем издании система фораминифер приводится по:
Михалевич, 1999.
538
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подкласс Ammoscalariana Mikhalevich, 1980
Отряд Ammoscalariida Mikhalevich, 1980
Подкласс Astrorhizana Saidova, 1981 (=Astrorhizicae Saidova,
1981, part.)
Отряд Astrorhizida Haeckel, 1894
Отряд Dendrophryida Haeckel, 1894
Надсем. Dendrophryoidea Haeckel, 1894
Надсем. Schizamminoidea Ndrvang, 1961
Отряд Saccamminida Lankester, 1885
Подотряд Saccamminina Lankester, 1885
Надсем. Saccamminoidea Brady, 1884
Надсем. Crithioninoidea Goes, 1894
Подотряд Psammosphaerina Haeckel, 1894
Отряд *Parathuramminida Mikhalevich, 1980
Надсем. Parathuramminoidea E. V. Bykova, 1955
Надсем. Mafginarioidea Loeblich et Tappan, 1986
Отряд Hippocrepinida Saidova, 1981
Надсем. Hippocrepinoidea Rhumbler, 1895
Надсем. Botellinoidea Chapman et Parr, 1936
(включая *Caligelloidea Reytlinger, 1959)
Класс Spirillinata Maslakova, 1990
Подкласс Ammodiscana Mikhalevich, 1980
Отряд Ammodiscida Mikhalevich, 1980
Отряд Ammovertellida Mikhalevich, 1998
Отряд Tournayellida Hohenegger et Piller, 1973
Подкласс Spirillinana Maslakova, 1990
Надотряд Archaediscoida Pojarkov, 1980,
Отряд Archaediscida Pojarkov, 1980
Отряд Lasiodiscida Mikhalevich, 1993
Надотряд Involutinoida Hohenegger et Piller, 1977
Отряд Involutinida Hohenegger et Piller, 1977
Надсем. Involutinoidea Butschli, 1880
Надсем. Triasinoidea Loeblich et Tappan, 1986
Отряд Hottingerellida Mikhalevich, 1993
Надотряд Spirillinoida Hohenegger et Piller, 1975
Отряд Spirillinida Hohenegger et Piller, 1975
Подотряд Spirillinina Hohenegger et Piller, 1975
Подотряд Spirotrocholiniria Mikhalevich, 1993
Отряд Patellinida Mikhalevich, 1992
Надсем. Hergottellbidea Loeblich et Tappan, 1984
Надсем. Patellinoidea Rhumbler, 1906
Отряд Seabrookinida Mikhalevich, 1980
Класс Miliolata Saidova, 1981
Подкласс Schlumbergerinana Mikhalevich, 1992
Отряд Schlumbergerinida Mikhalevich, 1980
Подкласс Miliolana Saidova, 1981
Надотряд Squamulinoida Mikhalevich, 1988
Отряд Squamulinida Mikhalevich, 1988
Надотряд Cornuspiroida Jirovec, 1953 .
Отряд Cornuspirida Jirovec, 1953 (=отряд Cornuspirida
Mikhalevich, 1980 (part.), подотряд Cyclogyrina Saido-
va, 1981)
Подотряд Comuspirina Jirovec, 1953
Подотряд Hemigordiopsina Mikhalevich, 1986
Надсем. Hemigordiopsidea Nikitina, 1969
Надсем. Shanitoidea Mikhalevich, 1986
Надотряд Costiferoida Mikhalevich, 1988
Отряд Costiferida Mikhalevich, 1988
Надотряд Milioloida Delage et Herouard, 1896
Отряд Nubeculariida Jones, 1875
Надсем. Nubeculariidea Johes, 1875
Надсем. Discospirinoidea Wiesner, 1931
Отряд Miliolida Delage et Herouard, 1896
Подотряд Miliolina Delage et Herouard, 1896
Надсем. Quinqueloculinoidea Cushman, 1917
Надсем. Milioloidea Ehrenberg, 1839
Надсем. Austrotrillinoidea Mikhalevich, 1986
Подотряд Alveolinina Mikhalevich, 1980 (=Alveolini-
dae Loeblich, Tappan, 1964; Alveolinellinina Saido-
va, 1981)
Отряд Soritida Schultze, 1854 (=Orbitolinida Ehrenberg,
1839; Orbitolitida Wedekind, 1937)
Надсем. Peneroplidea Schultze, 1854
Надсем. Meandropsinoidea Henson, 1848
Надсем. Soritidoidea Ehrenberg, 1839
KnacCfNodosariata Mikhalevich, 1992
Подкласс Hormosinana Mikhalevich, 1992
Отряд Hormosinida Mikhalevich, 1980
Надсем. Hormosinoidea Haeckel, 1894
Надсем. Cribratinoidea Loeblich et Tappan, 1984
Отряд Nouriida Mikhalevich, 1980
Отряд Pseudopalmulida Mikhalevich, 1992
Подкласс Nodosariana Mikhalevich, 1980
Отряд Lagenida Delage et Herouard, 1896
Отряд Nodosariida Calkins, 1926
Подотряд Syzraniina Mikhalevich, 1992
Подотряд Nodosariina Calkins, 1926
Надсем. Nodosarioidea Ehrenberg, 1838
Надсем. Colanielloidea Fursenko, 1959
Отряд Polymorphinida Mikhalevich, 1980
Подотряд Polymorphinina Mikhalevich, 1980
Подотряд *Pleurostomellina Mikhalevich, 1993 (=над-
сем. Rotaliacea в системе Loeblich, Tappan, 1988)
Подотряд Delosinina Revets, 1989
Отряд Vaginulinida Mikhalevich, 1992
Класс Rotaliata Mikhalevich, 1980
Подкласс Textulariana Mikhalevich, 1980
Надотряд Dusenburyinoida Mikhalevich, 1998
Отряд Dusenburyinida Mikhalevich, 1998
Надотряд Lituoloida Calkins, 1909
Отряд Haplophragmiida Mikhalevich, 1992
Надсем. Ammosphaeroidinoidea Cushman, 1927
Надсем. Haplophragmioidea Eimer et Fichert, 1899
Надсем. Labyrinthidomatidoidea Loeblich et Tap-
. pan, 1988
Отряд Lituolida Lankester, 1885
Надсем. Lituolinoidea de Blainville, 1827
Надсем. Biokovinoidea Gusczic, 1977
Надсем. Loftusioidea Brady, 1884
Надсем. Coscinophragmatoidea Thalmann, 1951
Отряд Cyclolinida Mikhalevich, 1992
Надсем. Cyclolinoidea Loeblich et Tappan, 1964
Надсем. Cyclopsinelloidea Loeblich et Tappan, 1984
Отряд Spiroplectamminida Mikhalevich, 1992
Надсем. Spiroplectamminoidea Cushman, 1927
Надсем. Spirotextularioidea Saidova, 1975
Отряд Textulariida Delage et Herouard, 1896
Надсем. Textularioidea Ehrenberg, 1938
Надсем. Septotextularoidea Loeblich et Tappan,
1985
Надотряд Ataxophragmioida Fursenko, 1958
Отряд Ataxophragmiida Fursenko, 1958
Подотряд Ataxophragmiina Fursenko, 1958
Надсем. Ataxophramoidea Schwager, 1877
Надсем. Textularielloidea Groenhagen et Luterba-
cher, 1966
Надотряд Trochamminoida Saidova, 1981
Отряд Trochamminida Saidova, 1981
Подотряд Trochamminina Saidova, 1981
Надсем. Trochamminoidea Schwager, 1877
Надсем. Remaneicioidea Loeblich et Tappan, 1964
Подотряд Carterinina Mikhalevich, 1980
Подкласс Rotaliana Mikhalevich, 1980
Надотряд Buliminoida Saidova, 1981
Отряд Stilostomellida Saidova, 1981
Отряд Bolivinitida Saidova, 1981
Подотряд Spirobolivinina Mikhalevich, 1998
Подотряд Bolivinitina Saidova, 1981
Надсем. Loxostomatoidea Loeblich et Tappan, 1962
Надсем. Bolivinitoidea Cushman, 1927
Надсем. Parabrizalininoidea Revets, 1996
Надсем. Eouvigerinoidea Cushman, 1927
Надсем. Pavoninoidea Eimer et Fickert, 1899
Надсем. Milettioidea Saidova, 1981
Отряд Cassidulinida d’Orbigny, 1839
Отряд Buliminida Saidova, 1981
Подотряд Turrilinina Saidova, 1981
Подотряд Buliminina Saidova, 1981
Подотряд Tubulogenerinina Mikhalevich, 1998
Подотряд Caucasinina Mikhalevich, 1998
Надотряд Robertinoida Mikhalevich, 1980
Отряд Robertinida Mikhalevich, 1980
Подотряд Caratobuliminina Mikhalevich, 1980
Подотряд Robertinina Mikhalevich, 1980
Надотряд Discorboida Ehrenberg, 1838
Отряд Rosalinida Delage et Herouard, 1896
Надсем. Rosalinoidea Reiss, 1963
Надсем. Discorbinelloidea Sigal, 1952
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
539
Надсем. Sphaeroidinoidea Cushman, 1927
Надсем. Siphoninidoidea Cushman, 1927
Отряд Discorbida Ehrenberg, 1838
Отряд Glabratellida Mikhalevich, 1994
Отряд Planorbulinida Mikhalevich, 1992
Подотряд Planorbulinina Mikhalevich, 1992
Подотряд Acervulinina Schultze, 1854
Отряд Asterigerinida d’Orbigny, 1839
Надсем. Asterigerindidea d’Orbigny, 1839
Надсем. Clypeorbioidea Sigal, 1952
Отряд Rotaliida Fursenko, 1958
?Отряд Chilostomellida Haeckel, 1894
Надотряд Nonionoida Saidova, 1981
Отряд Nonionida Saidova, 1981
Подотряд I^onionina Saidova, 1981
Надсем. Nonionoidea Schultze, 1854
Надсем. Almaenoidea Myatlyuk, 1959
Подотряд Astrononioniha Saidova, 1981
Отряд Elphidiida Saidova, 1981
Отряд Orbitoidida Copeland, 1956
Отряд Nummulitida Carpenter, Parker et Jones, 1862
Класс Globigerinata Mikhalevich, 1980
Отряд Heterochelicida Mikhalevich, 1992
Отряд Globorotaliida Mikhalevich, 1980
Отряд Globigerinida Carpenter, Parker et Jones, 1862
Морфология и физиология*
Особенности строения скелета
фораминифер
Морфология раковины
Фораминиферы имеют однокамерную или много-
камерную раковину, обтекаемую снаружи слоем ци-
топлазмы, так что скелет их в строгом смысле не
внешний, а внутренний (рис. 539, 2, 5; 541, 5). Типы
строения раковин чрезвычайно многообразны, имен-
но они придают такое своеобразие всей группе. Ос-
новные типы скелета следующие (рис. 525):
1) ветвящиеся, изогнутые или иной непра-
вильной формы; 2) одноосные (камеры располо-
жены в 1 или в 2—5 параллельных рядов); 3) не-
правильно- клубковидные (обороты навивают-
ся беспорядочно) и правильно клубковидные
(обороты располагаются под правильными углами друг
к другу, угол зависит от числа камер в обороте и их
расположения); 4) спирально свернутые (спи-
раль может быть плоской — плоскоспиральные,
конической — трохоидные или еще более высо-
кой вытянутой формы, спирально-винтовые).
Свернутые раковины могут быть эволютными
(снаружи видны все обороты спирали) и инволют-
ными (последний оборот или даже одна последняя
камера охватывает все предыдущие). Инволютность
может быть частичной (снаружи видна лишь часть
оборотов, часто одна сторона раковины инволютная,
другая — эволютная (у трохоидных форм)).
Этими морфологическими типами не исчерпы-
вается все морфологическое разнообразие раковин.
Существуют гетероморфные типы, когда в процессе
онтогенеза раковина начинает свое развитие по од-
* Данные по сфоению скелета и цитоплазмы фораминифер и
их жизненным циклам приводятся по: Михалевич, 1999.
ному типу, а заканчивает — по другому, в результате
чего она состоит из отделов разного строения. На-
пример, начальный отдел может быть плоскоспи-
ральным, поздний — выпрямленным (Ammobaculites,
Ammotium, Lingulina, Amphicoryna, — рис. 525, 13,
A—Д, 14, Б; 547, III, 7, 15, 20) или начальный —
трохоидный, поздний — 3-, 2- или 1-рядный (многие
Ataxophragmiida, — рис. 547, V, 5, 7). У милиолид
часто начальный отдел клубковидный, поздний —
спирально-плоскостной или выпрямленный (рис. 527,
А, Б; 547, III, 14, 15). Во многих не родственных
группах последние камеры часто располагаются цик-
лически (рис. 525, 13, Д; 547, III, 8, 21, V, 13, 31).
Иногда в одной раковине последовательно сменяют-
ся 3 разных типа строения.
Гетероморфность может распространяться не
только на способы расположения камер, но и на их
форму, размеры, строение устья (рис. 525, 13\ 532;
547, III, 8, V, 2, 25). Тип строения раковин (наряду
с другими чертами их морфологии — строение устья,
камер) мы считаем ведущим таксономическим при-
знаком ранга класса; более частные детали их стро-
ения характеризуют отряды.
В случае двухкамерной или многокамерной ра-
ковины начальная округлая камера называется про-
локулусом. У многих милиолят за пролокулусом сле-
дует вторая узкая трубчатая камера, образующая
вокруг него полный или неполный оборот, — флек-
состиль (рис. 527). Форма камер фораминифер чрез-
вычайно разнообразна — она может быть округлой,
овальной, трубчатой, веерообразной, серповидной,
трапециевидной, звездчатой, V-образной или иной
(рис. 525; 547). Как правило, она характеризует
виды и роды, но в отдельных случаях специфична
для таксонов более высокого ранга. Так, у милиолят
преобладают трубчатые камеры различной длины; у
спириллинат — длинные трубчатые камеры, образу-
ющие несколько оборотов; у астроризат обычны ка-
меры ветвящейся или иной неправильной формы; у
нодозариат встречаются камеры редкой V-образной
формы. Внутренность камер может быть усложнена
дополнительными структурами — сетчатыми или
ячеистыми образованиями, столбиками, неполными
перегородками, клапанами (рис. 531, 5, 6, А, Б*, 533,
4; 534, 3). Число камер чаще от 4—5 до 20—30, но
может доходить до нескольких десятков и даже со-
тен (рис. 531, 7; 532; 547, III, 16, 21, V, 31). Камеры
отделены друг от друга септами, при пересече-
нии которых со стенкой раковины образуется шов
(рис. 526). Швы могут быть на одном уровне с по-
верхностью раковины либо углубленными, выпуклы-
ми, натечными; по форме они могут быть прямыми,
косыми, в различной степени изогнутыми, иногда
зигзагообразными (рис. 525; 547).
Устье
Сообщение внутренней цитоплазмы раковин с на-
ружной средой осуществляется через специальные
отверстия — устья (рис. 528). Внутренние устья
камер, предшествующих последней, обычно называ-
ют фораменами. В некоторых группах первичные фо-
рамены зарастают, но во внутренней перегородке
540
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
541
Рис. 526. Обозначение частей раковины (на примере трохоид-
ной многокамерной раковины Globorotalia).
А — спиральная сторона, Б — пупочная сторона, В — боковая сторо-
на. к — камеры, кл — киль, п — пупок, пк — периферический край,
ш — швы.
Fig. 526. Designation of the shell parts (illustrated by the troc-
hospiral multichambered shell of Globorotalia).
A — spiral side, Б — umbilical side, В — lateral side, к — chambers,
ui — sutures, n — umbilicus, пк — peripheral edge, кл — keel.
Рис. 527. Шлифы раковин милиолят.
А — Sarmatiella; Б, В — Nubeculinella (за круглым пролокулусом сле-
дует трубчатая камера — флексостиль, затем последующие камеры).
Fig. 527. Shell sections of some Miliolata.
A — Sarmatiella', Б, В — Nubeculinella (rounded proloculus followed
by the second tubular chamber — flexostyle, then by the subsequent
chambers).
ных и некоторых плоскоспиральных форм), реже на
периферических концах камер (рис. 529; 547, V, 2,
12, 20). Форма устьев разнообразна — округлые,
овальные, петлевидные, щелевидные, ситовидные,
древовидные, лучистые. Они могут быть усложнены
дополнительными скелетными образованиями — как
наружными (шейка, отворот, губа), так и внутренни-
ми (ребра, пластинки, зубы, желобки, трубки, усть-
евые камерки). Ребра, пластинки и зубы выполняют
основную опорную функцию, так как расположены
по ходу сильного тока цитоплазмы и наиболее разви-
конвергентно образуются новые отверстия в другом
месте, исполняющие ту же функцию, дейтерофораме-
ны (Ceratobuliminidae). Устья могут быть единичны-
ми или множественными, занимать различное поло-
жение по отношению к последней камере (у ее ос-
нования или на вершине) и ко всей раковине (на
пупочной или спиральной стороне, периферическом
крае (редко), — рис. 525, 9, Б). У вытянутых форм
устье может быть конечным. У эволюционно продви-
нутых родов могут быть дополнительные устья, рас-
положенные в швах на пупочной стороне (у трохоид-
Рис. 525. Строение раковины.
1 — Plagiophrys (Lagynana), однокамерная органическая раковина: п — подостиль. 2 — Haliphysema (Astrorhizana), однокамерная агглютинированная вет-
вящаяся, укрепленная в грунте раковина с устьем, включающим иглы губок. 3 — Psammosphaera (Astrorhizana), однокамерная агглютинирванная раковина
без определенного устья, с торчащей спикулой губки. 4 — Lagena (Nodosariata, Nodosariana), однокамерная известковая раковина со скульптурой. 5 —
Grigelis (Nodosariata, Nodosariana), однорядная раковинах псевдокамерами, отделенными друг от друга пережимами. 6 — Frondicularia (Nodosariata, Nodo-
sariana), однорядная раковина с V-образными камерами. 7 — Glomospira (Spirillinata, Ammodiscana), неправильно-клубковидная раковина, обороты которой
образованы длинной трубчатой камерой. 8 — Quinqueloculina (Miliolata, Miliolana), правильно-клубковидная раковина с 2 трубчатыми камерами в каждом
обороте: А—В — внешний вид, Г — схема расположения камер на поперечном разрезе. 9 — спирально-плоскостные раковины: А — Spirillina (Spirillinata,
Spirillinana), эволютная раковина с длинной трубчатой второй камерой; Б — Almaena (Retaliate, Rotaliana), частично эволютная раковина с дополнительными
устьями на периферических концах камер; В, Г — Elphidium (Retaliate, Rotaliana), инволютная раковина с открывающимися в швах отростками каналов.
10 — трохоидные раковины: А — Neoglabratella (Rotaliata, Rotaliana), сизигий (по: Myers, 1940); Б — Tretomphalus (Retaliate, Rotaliana) с шаровидной
пловучей камерой; В — Candeina (Globigerinata) с шовными устьями; Г — Globigerinita (Globigerinata) с пупочной буллой (дополнительной скелетной
пластинкой). 11 — Bolivina (Rotaliata, Rotaliana), двухрядная раковина. 12 — спирально-винтовые раковины: А — Polymorphina (Nodosariata, Nodosariana),
сквозь стенку последней камеры просвечивает энтозолениевая трубка; Б — Euuvigerina (Rotaliata, Rotaliana) с наружной скульптурой. 13 — гетероморфные
раковины: А — Vulvulina (Rotaliata, Textulariana); Б — Fissotextularia (Rotaliata, Textulariana) с дополнительными устьями на периферических концах камер;
В — Pavonina (Retaliate, Rotaliana); Г, Д — Vandenbroeckia (Miliolata, Miliolana): Г — метало-, Д — микросферическая формы. 14 — Lingulina (Nodosariata,
Nodosariana): А — однорядная мегалосферическая, Б — свернутая микросферическая формы.
Fig. 525. Structure of the shell.
I — unilocular organic shell (p) of Plagiophrys (Lagynana), n — podostyle. 2 — branching unilocular agglutinated shell bored in the soil — Haliphysema
(Astrorhizata, Astrorhizana), using Sponge spicules. 3 — Psammosphaera (Astrorhizata, Astrorhizana), unilocular agglutinated shell without definite aperture, with
the Sponge spicule produced. 4 — Lagena (Nodosariata, Nodosariana), unilocular lime secreted shell with the sculpture. 5 — Grigelis (Nodosariata, Nodosariana),
uniserial shell with the pseudochambers separated from each other by the shell wall constrictions. 6 — Frondicularia (Nodosariata, Nodosariana), uniserial shell with
V-shaped chambers. 7 — Glomospira (Spirillinata, Ammodiscana), irregular-glomerate shell with the volutions formed by the long tubular chamber. 8 — Quinque-
loculina (Miliolata, Miliolana), regular-glomerate shell with the two tubular chambers per volution (A—В — outward appearance, Г — transverse section). 9 —
Planospiral shells: A — Spirillina (Spirillinata, Spirillinana), evolute shell with the long tubular second chamber; Б — Almaena (Rotaliata, Rotaliana), partly evolute
shell with additional apertures at the peripheral chamber ends; В — Elphidium (Rotaliata, Rotaliana), involute shell with the canal processes opened in the sutures.
10 — Trochospiral shells: A — Neoglabratella (Rotaliata, Rotaliana), syzigy, Б — Tretomphalus (Rotaliata, Rotaliana) with the floating chamber, В — Candeina
(Globigerinata) with the sutural apertures, Г— Globigerinita (Globigerinata) with bulla (additional skeletal plate). 11 — Bolivina (Rotaliata, Rotaliana), biserial shell.
12 — High trochospiral shells: A — Polymorphina (Nodosariata, Nodosariana), entosolenian tube is seen through the shell wall of the last chamber, Б — Euuvigerina
(Rotaliata, Rotaliana) with external structure. 13 — Heteromorphic shells: A — Vulvulina (Rotaliata, Textulariana); Б — Fissotextularia (Rotaliata, Textulariana) with
the additional apertures at the peripheral ends of the chambers, В — Pavonina (Rotaliata, Rotaliana); Г, Д — Vandenbroeckia (Miliolata, Miliolana): Г — megalo-,
Д — microspheric forms. 14 — Lingulina (Nodosariata, Nodosariana): A — uniserial megalosperic form, Б — coiled microspheric form.
542
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 528. Строение устья.
1 — положение устья: а — устье в основании септальной поверхности, б—е — приподнятое над основанием септальной поверхности (б — единичное,
в—э — множественное, г — древовидное). 2 — наружные элементы строения устья: а — выступ; б — шейка; в — шейка с отворотом; г — шейка с губой;
д, е — пластинчатый зуб (а—д — конечное устье). 3, 4 — внутренние скелетные образования устья (3: а, б — лучистое (радиальное) устье нодозариат;
в—н — устьевые зубы милиолят, простые (в, л) и сложные. 4: а—г — внутренние зубы, пластинки и желобки роталиат; д, е — энтозолениевые трубки
нодозариат).
Fig. 528. Structure of the aperture.
1 — position of the aperture: a — aperture at the base of the septal surface, 6—e — elevated above the base of the septal surface (6 — singular, в — d — multiple,
e — dendrithic). 2 — external elements of apertural structure: a — projection; 6 — neck; в — neck with a collar; г — neck with a lip; d, e — plate-like tooth
(a—d — terminal aperture). 3, 4 — inner skeletal formations of the aperture (3: a, 6 — radial aperture of Nodosariata, в—н — apertural teeth of Miliolata, simple
(в, л) and complex: 4: a—г — inner teeth, plates and grooves of Rotaliata; d, e — entosolenian tubes of Nodosariata).
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
543
Рис. 529. Дополнительные устья.
1 — Polysegmentina (Miliolana), в начальном отделе трубчатые камеры, клубкообразно (сигмоидально) навитые, без дополнительных устьев, в позднем отделе
камеры широкие, с дополнительными шовными устьями, расположены плоскоспирально. 2 — Trocholinopsis (Spirillinana) с дополнительными шовными
устьями по спиральному шву между оборотами длинной трубчатой камеры. 3—6 — дополнительные устья у Textulariana: 3, 4 — дополнительные устья на
периферических концах камер (3 — Norvanganina, 4 — Plotnikovina); 5, 6 — дополнительные пупочные устья (5 — Polystomammina, 6 — Rotaliammina).
7—9 — дополнительные устья у Rotaliana: 7 — Pseudoeponides (на спиральной стороне, параллельно спиральному шву), 8 — Cribroelphidium (шовные устья),
9 — Strebloides (шовные устья на пупочной стороне). 10 — дополнительные шовные устья у Globigerinoides (Glibigerinata).
Fig. 529. Additional apertures.
1 — Polysegmentina (Miliolana), tubular chambers of the initial part glomerately (sigmoidally) coiled, lacking additional apertures, later chambers broad, planospi-
rally coiled, having additional apertures. 2 — Trocholinopsis (Spirillinana) — additional sutural apertures in the spiral suture between the volutions of the long tubular
chamber. 3—6 — additional apertures of Textulariana: 3, 4 — additional apertures at the peripheral chamber margins (3 — Norvanganina, 4 — Plotnikovina)', 5,
6 — umbilical additional apertures (5 — in Polystomammina, 6 — in Rotaliammina). 7—9 — additional apertures of Rotaliana: 7 — in Pseudoeponides (on the
spiral side, in parallel to the spiral suture), 8 — in Cribroelphidium (sutural apertures ), 9 — in Strebloides (sutural apertures on the umbilical side). 10 — additional
sutural apertures of Globigerinata (Globigerinoides).
ты у вытянутых, удлиненных форм, достигая особен-
ной сложности у милиолят, булиминид (роталиата),
нодозариат. Так называемый пластинчатый зуб мили-
олят (рис. 528, 2, д\ 549) в отличие от их внутренних
зубов (рис. 528, 3, в—н) относится к наружным обра-
зованиям, поэтому мы предложили назвать его усть-
евой лопастью (Михалевич, 1981). Строение устья
может меняться в процессе онтогенеза (например, от
единичного к множественному). Зуб милиолят может
меняться от простого к сложному (рис. 528, 3, в—н*
530). Вообще внутренние зубы милиолят уникальны
и достигают большой сложности и разнообразия. У
агглютинированных форм, стенка которых в отличие
от секреционной стенки не позволяет сформировать
дополнительные опорные структуры, нередко опор-
ную функцию несут включенные в состав приусть-
евой стенки спикулы губок (рис. 525, 2; 547, 7, As-
trorhizana, 3). Все многообразие дополнительных
образований устья связано с их большой и разноооб-
разной функциональной нагрузкой в связи с усилен-
ным движением цитоплазмы в устьевой области. У неко-
торых примитивных однокамерных форм с органичес-
кой или агглютинированной раковиной (Lagynana, As-
trorhizana) оформленное устье отсутствует (рис. 525, 3).
Различные черты строения устья имеют разный
таксономический вес. Так, внутренние зубы милиолят
характерны только для этого класса. Энтосолениевые
трубки и радиальное устье нодозариат (рис. 528, 3, а,
б, 4, д, е; 547, IV), имеющиеся у их агглютинированных
(Nodosinum) и секреционных форм (Dentalina), — так-
же важный диагностический признак ранга класса.
Для представителей класса Rotaliata характерны силь-
но развитые, сложные зубные пластинки, желобки,
клапаны, не встречающиеся ни в одном другом классе
(рис. 528, 4, а—г).
У планктонных форм (класс Globigerinata) устья
сильно увеличены в размерах (рис. 539, 7). Положение
устья по отношению к последней камере в большин-
стве классов фораминифер конечное. Только в клас-
544
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 530. Изменения строения устья и зуба на разных возрастных стадиях у Flintina Ungulata.
1 — внешний вид раковин (А, Б — с боковых сторон; В — со стороны устья); 2—6 — вид раковин и устья с боковой стороны на разных возрастных ста-
диях (по: Михалевич, 1983).
Fig. 530. Growth stages of the apertural structures in Flintina Ungulata.
1 — outward appearance of the shell (А, Б — from the lateral sides, В — from the apertural side); 2—6 — lateral view at the different growth stages.
cax Rotaliata и Globigerinata оно изначально базаль-
ное — в основании устьевой поверхности последней
камеры (рис. 539, 7; 547, V). Положение устья по от-
ношению к раковине часто используется в разных
классах как признак семейственного ранга, а более
мелкие особенности строения устья — в качестве ро-
довых признаков. Так, многие роды в подклассах тек-
стуляриана и роталиана и в классе глобигерината были
выделены на основании наличия у них дополнитель-
ных устьев (рис. 525, 10, В, 13, 529; 539, 1; 547, V,
20) . В то же время способность образовывать до-
полнительные устья характерна лишь для некоторых
классов.
Интегрирующие системы
У особей с большим числом камер и оборотов
сообщение цитоплазмы отдельных камер между со-
бой и с внешней средой становится затруднитель-
ным. Это противоречие снимается возникновением
интегрирующих систем различной сложности (рис. 531,
7—4).
1. Столоны — у многокамерных форм система
взаимосвязанных внутренних устьев большой протя-
женности (в классе Miliolata — у альвеолинид; в под-
классах Textulariana — у высших литуолид, орбито-
линид, Rotaliana — у орбитоидид, — рис. 531, 4).
2. Туннели — система просветов между внут-
ренними перегородками (рис. 531, 7; 533, 5) (в под-
классах Ammodiscana — у фузулинид, Miliolana — у
альвеолинид). У первых они проходят между отложе-
ниями внутреннего скелета (хоматами) и резорбиро-
ванными отверстиями в нижней части септ, у вто-
рых — это система просветов, которые составляют
спиральные перегородки, не доходящие до межка-
мерных септ. В обоих случаях они расположены
между элементами внутреннего скелета (эндогенное
происхождение), имеют органическую выстилку и, по
существу, не имеют собственных стенок. Сходные
происхождение и функцию имеют проходы между
последовательными, расположенными одна над дру-
гой камерами конечного отдела у Rhapydionina (Mili-
olana). Токи цитоплазмы проходят в просветы между
множественными устьевыми отверстиями и столбика-
ми этих последовательных камер. Эти проходы не-
верно назывались в литературе каналами, но в отли-
чие от последних они не имеют собственных стенок.
3. Внутренние устьевые системы — внут-
ренние образования устья; могут быть сильно разви-
ты и проходить через всю камеру от устья к предыду-
щему форамену, соединяя последовательно между
собой форамены всех предшествующих камер. Они
получили большое развитие у Rotaliata, особенно в
подклассе Rotaliana (рис. 528, 4, в, г), встречаются у
моноаксиальных Globigerinata, изредка — у Nodosa-
riata (рис. 528, 4, д) (Михалевич, 1993а, с. 7).
4. К а н а л ы, — отличаются от столонов и туннелей
морфологически (они экзогенного, а не эндогенного
происхождения, проходят не внутри камер, а между
ними) (Hottinger, 1978) и функционально — в них
циркулирует не внутренняя, а наружная цитоплазма,
которая доставляет к внутренним частям клетки кис-
лород, осуществляет связь с внешней средой и отделе-
на от внутренней цитоплазмы с помощью специаль-
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
545
Рис. 531. Детали внутреннего строения раковин и структуры стенки.
1 — Praefusulina (Ammodiscana, Fusulinoida), спирально-свернутая инволютная раковина со сложным внутренним строением: км — камерки, са — септаль-
ные арки, се — септы, сп — септальные поры, ту — туннель, ус — устье. 2, 3 — фрагменты системы каналов роталиан: 2 — Baculogypsina, х 240; 3 —
Rotalia, х 240. 4 — схема системы столонов фораминифер: ор — ось раковины, ос — оси. столонов, пс — плоскость столонов. 5 — ячеистая стенка
Cyclammina (шлиф). 6 — столбики на внутренней стенке милиолян: А — Shanita, Б — Archaias. 7 — бифонтинальная стенка роталиан с двуслойными
септами. 8 — поперечный срез стенки некоторых текстуляриан: ос — органический слой, п — псевдопоровые канальцы. 9 — иглы планктонного вида
Prosphaeroidinella, свободно сочлененные со стенкой раковины. (1 — по: Фурсенко, Раузер-Черноусова, 1959; 2—4 — по: Hottinger, 1978; 5 — по: Brady,
1884; 6: А — по: Loeblich, Tappan, 1988; Б — по: Seiglie et al., 1977; 7, 8 — по: Loeblich, Tappan, 1964; 9 — Hofker, 1971).
Fig. 531. Details of the inner shell structure and wall ultrastructure.
1 — Praefusulina (Ammodiscana, Fusulinoida), spirally coiled involute shell with complex inner structure: yc — aperture, ce —. septa, cn — septal pores, ca —
septal archs, my — tunnel, km — chamberlets. 2, 3 — fragments of Rotaliata’s canal system: 2 — Baculogypsina, x 240; 3 — Rotalia, x 240. 4 — the scheme of
the Foraminiferal stolon system: op — shell axis, oc — stolon axes, nc — plane of stolons. 5 — cancellate wall of Cyclammina (section). 6 — pillars of the inner
shell wall of Miliolana: A — Shanita, Б — Archaias. 7 — bifontinal Rotaliana’s wall with bilamellar septa. 8 — transverse section of the wall of some Textulariana:
oc — organic layer, n — pseudopore canaliculi. 9 — spines of the planktonic species Prosphaeroidinella, loosely conjuncted (set into hole) with the chamber wall.
35 Протисты, часть 1
546
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
зк
Рис. 532. Дифференциация камер у многокамерных фораминифер.
1 — Orbitolina (Textulariana), с крупной зародышевой камерой (зк), начальная часть трохоидная, поздние камеры дисковидные, затем циклические, с различ-
ной внутренней структурой: имз — инфрамаргинальная зона с камерками; мз — маргинальная зона с прямоугольными камерами, подразделенными вторич-
ными перегородками на ряд более мелких ячей (от 4 до 16); рд — радиальные перегородки радиальной зоны с радиальными проходами (рх); ц — центральная
сетчатая зона из разветвляющихся и соединяющихся перегородок; я — ячеистый слой стенки краевой зоны. 2 — Biplanispira (Pellatispiridae, Rotaliana): А —
поперечное сечение, Б — продольное сечение. В центре — крупные зародышевые камеры (зк), в следующем обороте — малочисленные камеры средних
размеров (кер), в последующих оборотах — многочисленные мелкие камеры (мк). 3 — Heterostegina (Nummulitidae, Rotaliana). Камеры начальных оборотов
серповидные (ек), неподразделенные, в последующих оборотах подразделены на камерки (пк); у камерок, расположенных ближе к периферическому краю,
могут быть межсептальные столбики (кс). 1 — по: Douglass, 1960, — цит. по: Loeblich, Tappan, 1964; 2 — по: Cole, 1957, — цит. по: Loeblich, Tappan, 1988;
3 — по: Eames et al., 1968 из: Loeblich, Tappan, 1988.
Fig. 532. Differentiation of the chambers in the multichambered Foraminifera.
1 — Orbitolina (Textulariana) (with large embrional chamber — зк), earlier part trochospiral, later chambers discoidal, than cyclical, with different internal
structure: ц central cancellate zone formed by ramificating and coalescing partitions, pd — radial partititions of the radial zone with the radial passages (рх), мз —
marginal zone with the rectangular chambers, subdivided by the second order beams into smaller cellules (from 4 to 16), имз — inframarginal zone with the
chamberlets, л — alveolar wall layer of the marginal zone. 2 — Biplanispira (Pellatispiridae, Rotaliana): A — transverse section, Б — longitudinal section. Large
embrional chambers in the center (зк), middle size chambers small in number in the next whorl (кер), numerous small chambers in the successive whorls (мк). 3 —
Heterostegina (Nummulitidae, Rotaliana). Chambers of the initial whorls crescentiform (ck), riot subdivided, in subsequent whorls chambers subdivided into cham-
berlets (пк), in the chamberlets situated nearer to the peripheral margin, the interseptal pillars may be present (кс).
Рис. 533. Микрогранулярная стенка фораминифер.
1 — схема ультраструктуры монофонтинальной микрогранулярной стенки (вое — внутренний органический слой). 2 — Clavulina (Textularioida, Textularia-
na): А — внешний вид раковины, Б — ультраструктура ее микрогранулярной стенки, х-2000. 3 — ультраструктура микрогранулярной стенки Rugosofusulina
(Fusulinoida, Ammodiscana), х 1800. 4 — схема усложнений внутренней полости камер (поперечные и радиальные перегородки и столбики) у Abrardia
(Orbitolinidae? Textulariana). 5 — усложнения внутренней стенки и полости камер у Schwagerina (Fusulinoida, Ammodiscana): дополнительные отложения
скелетного вещества (хоматы) (хо) и проходящий между ними туннель (т)', пк — канальцы псевдопор. 6 — схема поперечного среза стенки Fusulinoida с
ампулообразными канальцами псевдопор: т — тектум, к — кериотека. 7 — аксиальное сечение Mesorbitolina (Textularioida) с явственным наружным эпи-
дермальным слоем (светлый) (эс), х 50. 8 — поперечный срез стенки Simplorbitolina (Textularioida, Textulariana): видны ампулообразные канальцы псевдопор,
х 100. 9 — аксиальное сечецие Sumatrina (Neoschwagerinidae, Fusulinoida, Ammodiscana), на котором видна толстая кериотека (к), тонкий слой тектума (т)
и ампулообразные канальцы псевдопор, х 300.1 — по: Loeblich, Tappan, 1964; 3 — по: Т. Н. Исаковой, устн. сообщ.; 4 — по: Neumann, Damotte, 1960; 5 —
схема по; Thompson, 1961 (фрагмент) из: Loeblich, Tappan, 1964; 6 — схема по: Фурсенко, Раузер-Черноусова, 1959; 7 — схема по: Douglass, 1960; 8 —
схема по: Douglass, 1964 из: Loeblich, Tappan, 1988; 9 — схема по: Ozawa, 1927.
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
S47
3
8
Fig. 533. Microgranular wall of Foraminifera.
1 — scheme of the ultrastructure of the mohofontinal microgranular wall (eoc — inner oranic lining). 2 — Clavulina (Textularigid^ T^xJuMana): A external
view of the shell, Б — ultrastructure of its microgranular wall, x 2000. 3 — ultrastructure of the microgranular wall of Rugosofusulin^ (F4§yjinoid^ Ammodiscana),
xl800. 4 — scheme of the complication of the inner chamber lumen (transversal and radial partititions and pillars) in Abrardia (Oryt^liaidae, Textulariana). 5 —
complication of the inner wall and chamber lumen in Schwagerina (Fusulinoida, Ammodiscana): additional deposition of the skeletal matter (ehomata) (xo), tunnel
(m) passing between them, пк — pseudopore canaliculi. 6 — scheme of the transverse wall section of Fusulinoida with the атроц1е-ДШе canaliculi of the pseudo-
pores: m — tectum, к — keriotheca. 7 — axial section of Mesorbitolina (Textularioida) with the distinct outer epidermal layer (fight one) (^c), x 50. 8 — transverse
wall section of Simplorbitolina (Textularioida, Textulariana): the ampoule-like pseudopore canaliculi are seen, xlOO. 9 — axial section of Sumatrina (Neoschwage-
rinidae, Fusulinoida, Ammodiscana), thick keriotheca (к), thin layer of tectum (m) and ampoule-like pseudopore canaliculi are seen, x 300.
548
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ных образований, не имеющих, по современным дан-
ным, внутренней органической выстилки. При этом
достигается высокая степень дифференциации ци-
топлазмы. Каналы можно рассматривать как сложный
устьевой аппарат. Подобной уникальной системы нет
ни в одной другой группе протистов. Системы кана-
лов, развитые в различной степени, давно известны во
многих семействах роталиат и могут рассматриваться
как пример параллелизма в близкородственных груп-
пах. Обычно у них имеются спиральные и радиальные
каналы с множеством более мелких разветвлений
(рис. 531,2,5). Недавно описанные каналы у предста-
вителей подкласса Spirillinana являют пример парал-
лелизма в более далеких таксонах. Они не достигают
такой сложности, как каналы роталиан, в простейшем
случае представляя систему щелей (например, Lasio-
discidae) (рис. 547, II, 4—6). У более сложно органи-
зованных Hottingerellidae имеются спиральный канал
и щелевидные проходы, связывающие его с полостью
камер (рис. 547, II, 10, б). Поскольку наличие каналов
рассматривается как диагностический признак ранга
отряда, среди спириллинан также были выделены со-
ответствующие отряды (Михалевич, 19936, рис. 1—3;
рис. 547, II, 5, 10, б\
Описанные выше интегрирующие системы — это
системы нового, организменного уровня. Они позво-
ляют снять разобщенность, нарастающую по мере
увеличения числа камер, оборотов спирали и общих
размеров раковин, ведут к более глубокой частичной
или полной компартментализации и дифференциа-
ции цитоплазмы, повышают уровень организации
клеточной системы.
Стенка раковин
Химический состав
Органическая (тектиновая) стенка низших фора-
минифер (лагинаны) и органическая основа стенки
всех остальных групп состоит в основном из мукопо-
лисахаридов и склеропротеинов. Наиболее детально
ее состав изучен у Heterostegina depressa (Rotaliana)
(Weiner, Erez, 1984). На долю протеинов приходится
около 0.5 % ее массы. Составляющие стенку макро-
молекулы относятся к 2 основным классам веществ:
в одном преобладают протеины, богатые аспарагино-
вой кислотой, в другом — протеины, богатые серином
и глицином и, вероятно, связанные с полисахаридами.
Биохимические свойства макромолекул матриц, обна-
руженных у гетеростегины, подобны свойствам орга-
нических матриц других типов. Мукополисахаридная осно-
ва стенки (гликокаликс) играет барьерную роль, защищая
организм от влияния загрязнений среды, что прослежено на
биохимическом уровне (Bresler, Yanko, 1995).
В состав агглютинированной раковины входят
песчинки, спикулы губок и другой посторонний ма-
териал, скрепляемый вначале органическим, а затем
минеральным цементом, который секретируется ор-
ганизмом (Bender, Hemleben, 1988). Появление сек-
реционных известковых элементов можно объяснить
экзоцитозом вакуолей, в которых известковые грану-
лы образуются в результате карбоангидразозависимо-
го транспорта НСО3~ в сочетании с переносом Са++ (в
вакуоль) в обмен на Н+ (в цитоплазму). В процессе
участвует Са++/Н+ АТФ-аза, поскольку условием каль-
цинации является защелачивание вакуолярной среды
(Лабас, Михалевич, 1987); вакуоли поставляются ап-
паратом Гольджи.
У подавляющего большинства фораминифер ске-
лет известковый, чаще кальцитовый, реже арагони-
товый, который, как правило, в силу особенностей
своей кристаллической решетки более богат строн-
цием. Арагонитовый скелет известен для крупной
группы робертинид, для ряда других групп — инво-
лютинид, некоторых планктонных фораминифер
(фавузеллиды) (Григелис, Горбачик, 1980; Горбачик,
1983, — цит. по: Loeblich, Tappan, 1988). Наличие
арагонитового скелета, на наш взгляд, не является
таксономическим признаком высокого ранга. У Trocho-
lina (Involutinida) один из видов имеет кальцитовую
раковину, другой — арагонитовую. У некоторых фо-
раминифер наблюдались прослои кальцита и араго-
нита в одной раковине (Archambault-Guezou, Lefev-
re, 1981). Кроме того, в ископаемом состоянии ара-
гонит легко переходит в кальцит. Поэтому мы
сохраняем робертинид как самостоятельный таксон
не потому, что стенка их раковин арагонитовая, а на
основании морфологии их раковин. Известковый ске-
лет включает небольшие примеси соединений Mg,
Мп, Fe, Al, Sr и некоторых других элементов. Состав
их изменчив, изучен недостаточно, однако высокая
магнезиальность известковых игл милиолят может
служить дополнительной характеристикой подклас-
са, а преобладание железистого цемента характерно
для некоторых агглютинирующих видов и родов.
Среди фораминифер описан ряд агглютинирую-
щих родов и 1 секреционный род Silicoluculinina с
кремнеземной стенкой. Поскольку минеральный со-
став скелета определяется органической матрицей, а
состав ее аминокислот у организмов с известковым
и кремнеземным скелетом различен и строго специ-
фичен для таксонов высокого ранга (фораминиферы,
радиолярии, акантарии, хризофитовые, гаптофито-
вые, диатомовые, губки и другие, — King, 1977),
такие формы заслуживают выделения в самостоя-
тельный таксон не ниже класса. Однако в связи с
тем что секреторное происхождение кремнезема в
стенке фораминифер, несмотря на работы Хансена
и Ханзликовой (Hansen, Hanzlikova, 1974), все же не
доказано окончательно, положение таких форм в
системе остается проблематичным. Скорее всего, ок-
ремнение стенки раковин — результат вторичных
процессов, идущих при фоссилизации. Как показал
Голубев (1981), при разрушении органических пле-
нок, окружающих кристаллы, последние становятся
плохо растворимыми. Это имеет место не только
при фоссилизации, но и при обжиге. Если рядом
имеется кремнезем (например, прокаливание проис-
ходит на стекле), — кристаллы окремняются.
Ультраструктура стенки раковин
За два последних десятилетия появились новые
электронно-микроскопические исследования ультра-
структуры стенки фораминифер (см. раздел «Исто-
рия изучения»).
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
549
Тектиновая стенка
Ультраструктура тектиновой стенки изучена в ос-
новном у однокамерных представителей из классов
Lagynata (целиком органическая раковина — у А1-
logromia и др.) и Astrorhizata (агглютинированная
раковина — у Iridia) (Hedley, 1963; Lengsfeld, 1969;
Hedley et al., 1973, Pierce, Nathanson, 1974; C6sana,
1981; Schwab, Plapp, 1983; Cartwright et аЦ 1989).
Она состоит из внутреннего фибриллярного и на-
ружного электронно-плотного слоев. Иногда может
присутствовать еще и наружный гранулярный слой.
Толщина фибриллярного слоя различна не только у
разных видов, но и у одного и того же животного и
зависит от формы его раковины или физиологичес-
кого состояния. Особенности строения этой стенки
не удалось использовать в систематике, и, видимо,
поэтому ее изучение не привлекло большого внима-
ния исследователей. Часто тектиновая стенка покры-
та редкими песчинками, а некоторые ее участки
могут быть полностью агглютинированными (напри-
мер, у Amphifenestrella),
Агглютинированная стенка
Органический слой толщиной 0.5 мкм подстила-
ет стенку и агглютинированных и секреционных ра-
ковин. Это внутренний органический слой (ВОС)
(рис. 531, S; 533, 7; 534, 7; 535, 7, 3), отделяющий
цитоплазму от стенки раковин.
Агглютинированная стенка (рис. 525, 7—3, 13, А,
К; 531, 5, S; 545, 7—4, S; 546, 2, А, В, Е, М; 547, 7,
77, 7, 2, 777, 7—3, 7V, 7—5, V, 7—73) в простейшем
случае состоит из зерен кварца, скрепленных тем же
органическим веществом, что и подстилающий стен-
ку слой. Тончайший слой этого вещества обволаки-
вает каждую песчинку и может образовывать тонкие
расслаивающиеся прослои между рядами песчинок.
Эти прослои носят случайный, нерегулярный харак-
тер. Кроме песчинок в состав стенки фораминифер
могут входить пустые раковины других организмов (в
том числе и фораминифер), частицы слюды, спику-
лы губок и иной посторонний материал. Некоторые
роды (JTechnitella) строят раковину исключительно из
спикул губок. В условиях культуры, при отсутствии
природного грунта, они используют любой доступ-
ный материал (осколки стекла, угля). Песчинки в
стенке раковин могут располагаться в один или не-
сколько слоев, быть сцементированы слабо, рыхло
или твердо, прочно. Многие виды агглютинирующих
фораминифер способны отсортировывать частицы по
размерам (грубопесчанистая, тонкопесчанистая стен-
ка), иногда в стенке раковин одного вида имеются
слои, состоящие из частиц разного размера, разной
степени сцементированности (например, у Rhabdam-
mina parabyssorum внутренний слой твердый, наруж-
ный — рыхлый). Обычно более тонкопесчанистые
слои находятся у внутренней части стенки и возле
устья; наружные слои более грубые.
Наличие агглютинированной стенки мы исполь-
зуем как таксономический признак ранга подкласса;
частные особенности ее строения могут служить ро-
довыми и видовыми признаками.
Секреционная известковая стенка
Секреционная известковая стенка тоже выстлана
изнутри органическим слоем — такого же состава,
как у органических и агглютинированных раковин
(Angell, 1967; Banner et al., 1973). Эта органическая
выстилка сохраняется, если растворить известковую
раковину в кислоте. Она обычно хорошо видна как
тонкая пленка у пустых растворенных раковин, по-
вторяя очертание последних.
Все многообразие описанных типов известковой
секреционной стенки можно свести к нескольким ос-
новным типам.
1. Микрогранулярная стенка (рис. 533, 7—9)
(термин Раузер-Черноусовой и Герке, — см.: Рау-
зер-Черноусова, Герке, 1971) — над ВОС беспоря-
дочно и рыхло (не ориентированно) расположены
довольно крупные (1—5 мкм) известковые зерна
округлой, овальной, угловатой или иной формы
(рис. 533, 7—3). «Субангулярная, равнозернистая,
разнозернистая» и т. д. типы стенок, описанные в
литературе, — ее частные случаи. Каждое из зерен
окружено тонкой органической оболочкой; более
толстые расслаивающиеся прослои того же органи-
ческого вещества встречаются беспорядочно и пре-
рывисто между группами зерен, как и в случае аг-
глютинированной стенки. Иногда зерна отсортирова-
ны по размерам. В световом микроскопе эта стенка
выглядит шероховатой, сахаристой. Она встречается
наряду с агглютинированной в подклассах Astrorhi-
zana, Ammodiscana, Hormosinana, Textulariana. Меж-
ду агглютинированной и микрогранулярной стенками
нет резкой границы — часто встречаются формы со
смешанным типом. Среди 142 агглютинированных
видов, изученных Бендером (Bender, 1995), формы с
известковым цементом составляли 21%. Поскольку
некоторые агглютинированные виды могут сами сек-
ретировать известковые зерна (Михалевич, 1995;
рис. 533, 2), можно предположить, что переход в
эволюции от агглютинированной стенки к микрогра-
нулярной происходил постепенно. Поэтому мы не
придаем микрогранулярной стенке такого высокого
таксономического веса, как это принято в системах
Дёблика и Таппан (Loeblich, Tappan, 1964; 1988) и
их последователей.
Агглютинированная и микрогранулярная стенки
во многом сходны. Очень часто они массивные, тол-
стые, тяжелые. Дополнительные элементы скелета,
усложнения стенки возникают при этих типах почти
исключительно внутри раковин (сетчатые, ячеистые,
губчатые, альвеолярные слои, столбики, группы сли-
вающихся столбиков, холмики, хоматы, — рис. 531,
5, 6, А, Е; 533, 4, 5).
2. Игольчатая (фарфоровидная) стенка милиолят
(рис. 534) — над ВОС расположен известковый слой
удлиненных кристаллов 3 различных способов уклад-
ки: толстый рыхлый срединный слой из беспорядоч-
но пересекающихся длинных игл (иногда в виде ко-
ротких цилиндрических кристаллов) либо планок,
облицованный изнутри и с поверхности плоскими
плитчатыми кристаллами, уложенными параллельно
поверхности наподобие паркета. Именно наружному
«паркетному» слою обязаны высшие милиоляты
550
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 534. Игольчатая (фарфоровидная) стенка фораминифер.
1 — схема ультраструктуры монофонтинальной игольчатой (фарфоровидной) стенки {вое — внутренний органический слой). 2 — Pyrgo (Miliolana): А —
внешний вид, Б — ультраструктура его стенки, х 39000. 3 — столбики во внутренней полости камер Fusarchaias (Miliolana). 4, 5 — отложения дополни-
тельного раковинного вещества на поверхности раковин некоторых видов Quinqueloculina (Miliolana): 4, А; 5, А, Б — вид с боковых сторон. 4, К; 5, В — вид
со стороны устья. 2 — по: Михалевич, 1988; 3 — по: Reichel, 1952; 4, 5 — по: Михалевич, 1983.
Fig. 534. Porcelaneous (composed from needles) Foraminiferal wall.
1 — scheme of the ultrastructure of monofontinal porcelaneous wall {eoc — inner organic layer). 2 — Pyrgo (Miliolana), A — outer view, Б — its shell ultrast-
ructure x 39 000. 3 — pillars in the inner chamber lumen of Fusarchaias (Miliolana). 4, 5 — deposition of the additional shell material on the shell surface of
some species of Quinqueloculina (Miliolana) (4, A, 5, А, Б — side views, 4, Б, 5, В — apertural views).
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
551
Рис. 535. Монофонтинальная стекловато-лучистая (радиальная) и бифонтинальная стенка фораминифер.
1 — схема строения монофонтинальной стекловато-лучистой (радиальной) стенки (вое — внутренний органический слой). 2 — Lingulina (Nodosariata),
продольный разрез, видны слои нарастания, х 42. 3 — схема строения бифонтинальной стенки у вновь образованной камеры: А — 2 секреционные пластины
по обе стороны сом, образованные одновременно; Б — схема строения стенки и порового канальца на примере донной Amphistegina (Rotaliana) у более
ранней камеры: по обе стороны от сом верхний и нижний известковый слои, образованные при начале формирования этой камеры, нижний известковый
слой отделяется от цитоплазмы (ци) внутренним органическим слоем (вое), последующие слои нарастания известковой стенки отделены друг от друга
органическими слоями (ос), дпп — дополнительная поровая пластинка, ми — митохондрии, мп — микропоры, нос — наружный органический слой, п —
пора, пк — поровый каналец, пп — поровая пластинка. 4 — ультраструктура стенки и пор Globorotalia (Globigerinata) на поперечном разрезе стенки: А —
у молодой камеры, х 4000 (в верхней части поры начинают расширяться воронкообразно); Б — у более ранней камеры, х 1600, видны слои нарастания и
краст (к), сужающий отверстия пор снаружи. 2 — по: Ellis, Messina, 1940; 3 — по: Leutenegger, 1977; 4 — схема по: Hemleben et al., 1977.
Fig. 535. Monofontinal hyaline-radial (radial) and bifontinal wall of Foraminifera.
I — schematic structure of monofontinal hyaline-radial (radial) wall (вое — inner organic lining). 2 — Lingulina (Nodosariata), x 42, longitudinal section. 3 —
schematic structure of bifontinal wall in the newly formed chamber: A — two secreted lamella at the both sides of the com (middle (primary) organic membrane),
formed simultaneously, Б — schematic structure of the wall and pore canaliculi illustrated by the earlier chamber of the bottom Amphistegina (Rotaliana): upper and
lower lime layers on the both sides of com, which are formed at the very beginning of chamber formation, lower lime layer is separated from the cytoplasm (ци) by
the inner organic lining (вое), subsequent layers of growth of lime wall are separated from each other by the organic layers (oc), hoc — outer organic layer, n —
pore, пк — pore canaliculi, nn — pore plate, мп — micropores, mu — mitochondria. 4 — shell and pore ultrastructure in Globorotalia (Globigerinata) (transverse
section of the wall): A — in the young chamber x 4000 (mark funnel-like pore widening in their upper part), Б — in the more younger chamber, x 1600, layers of
- growth and crust (к) narrowing the pore openings from the outside are seen.
фарфоровидным блеском в световом микроскопе. Он
нередко бывает разрушен, и тогда стенка теряет, свой
блеск, становясь тусклой, шероховатой. Каждый из
кристаллов, как и в предыдущем типе стенки, имеет
тонкую органическую оболочку. Электронно-мик-
роскопические исследования низших представителей
милиолят — шлумбергеринан с агглютированной
стенкой (Михалевич и др., 1986) также позволили
обнаружить у них известковые секретированные эле-
менты, характерные для стенки высших милиолят
(длинные иглы на внутренней стороне стенки и зуба,
короткие толстые палочки в срединном слое стен-
ки, — рис. 536, 7, вкл.). Этот факт наряду с полным
морфологическим сходством (квинквелокулиновый
тип строения раковин, строение устья и зуба) свиде-
тельствует о родстве этих групп, обоснованности
552
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
выведения их из состава текстуляриат (Саидова, 1981),
а также о том, что, как и в случае с микрогрануляр-
ной стенкой, переход от агглютинированного типа
стенки к секреционному совершался в эволюции
этих групп постепенно. Полное сходство строения
этих раковин ранее рассматривалось как конвергент-
ное, в действительности они представляют разные
этапы в эволюции группы.
В отличие от микрогранулярной Стенки при
игольчатой стенке на наружной стороне раковин
может возникать разнообразная скульптура (ячеи,
ребра, шипики, — рис. 534, 4, 5; 547, III, 7, 10, 12,
15). Нередки при этом типе стенки и внутренние
столбики (например, у Shanita, Archaias, — рис. 531,
6; 534, 3).
3. Стекловато-лучистая, или радиальная
(но не часто употребляемый термин «радиально-лу-
чистая»!), стенка нодозариат (рис. 535, 1, 2) — рас-
положенные над ВОС удлиненные столбчатые крис-
таллы (скорее всего, образованные более мелкими
кристаллитами), которые окружены тончайшей орга-
нической мембраной и ориентированы перпендику-
лярно поверхности стенки; между ними остаются от-
верстия размером 1—1.5 мкм (псевдопоры) (рис. 535,
1; 536, 3). У начальных камер раковин четко выраже-
ны слои нарастания (рис. 535, 2) . В световом мик-
роскопе стенка выглядит стекловатой (лягены, нодо-
зарии, лентикулины). Этот тип стенки исследован
недостаточно.
У современных нодозариат описана моноламел-
лярная стенка; у некоторых древних форм (пермь,
триас) стенка также моноламеллярная, у ряда других
(Eonodosaria, Eogeinitzina) описан внутренний мик-
рогранулярный слой й наружный радиальный (Гу-
бенко, 1989). Возможно, первый является следстви-
ем перекристаллизации. Во всяком случае, в процессе
исторического развития он исчезает. У современных
фораминифер сочетание микрогранулярного и ради-
ального слоев не отмечалось. Процесс возникнове-
ния третьего типа стенки менее ясен, однако она
отличается от микрогранулярной упорядоченным
ориентированным расположением столбчатых крис-
таллов и существенно отличается от биламеллярной
стенки роталиат, описанной ниже. При этом типе
стенки встречены почти исключительно наружные
дополнительные скелетные образования (ребра, шиш-
ки, шипы, — рис. 525, 4; 538, Е—К; 547, IV, 6\ 548,
7); внутренние септы только у вымерших Colani-
ellidae (рис. 546, 2, Л).
4. Уникальный тип секреционной известковой
стенки представляет монокристаллическая сек-
реционная стенка высших спириллинан. Она иссле-
дована недостаточно. По-видимому, кристаллы рас-
положены очень тесно, упорядоченно, строго пер-
пендикулярно ВОС и, вероятно, в один слой, в
результате чего могут вести себя оптически как еди-
ный кристалл или 2—3 кристалла. Слои нарастания
не обнаружены. Как видно, у Patellina corrugata
(Berthold, 1976) она покрыта не только изнутри, но
и снаружи тонким органическим слоем. У ископае-
мых форм нередко отмечен внутренний гранулярный
известковый слой (например, у Archaediscus), воз-
можно, возникший в результате диагенеза. У совре-
менных спириллинан такой слой отсутствует. В све-
товом микроскопе этот тип стенки выглядит стекло-
ватым, часто скульптурирован снаружи. Он обладает
высокими структурными возможностями, так как у
некоторых представителей этой группы (например, у
Lasiodiscida, Hottingerellida) обнаружена даже систе-
ма каналов (рис. 547, II, 4—6, 10), хотя и довольно
примитивная (Hottinger, 1976; Азбель, 1986).
Перечисленные выше 4 типа секреционной из-
вестковой стенки образуются по одну сторону от
ВОС, во всяком случае, у слабо изученных 3-го и
4-го типов СОМ (см. ниже) пока не обнаружена.
Оба этих типа проявляют во многом сходные свой-
ства. Так, кристаллы наружных ребер Nodosaria af-
finis вели себя как несколько крупных кристаллов
(Hay et al., 1963). В обоих таксонах сильно развита
способность к образованию ламелл, многократно ок-
ружающих всю раковину с каждым этапом роста
(ортол амеллярность).
5. Принципиально иное строение имеет била-
меллярная (=бифонтинальная, — см. ниже) стен-
ка роталиан и глобигеринат (535, 3, 4; 536, 4, 5;
537). Часто в русскоязычной литературе ее называют
двухслойной. Наиболее наглядно ее строение изо-
бражено в классической работе Хемлебена с соавт.
(Hemleben et al., 1977). Кроме подстилающего рако-
вину снизу ВОС у них имеется второй срединный
органический слой (мембрана) — COM (primary or-
ganic membrane) (рис. 535, 3, 4; 537). По обе сторо-
ны от нее одновременно закладываются и затем фор-
мируются внутренний и внешний известковые слои.
При этом считалось, что функция матрицы перехо-
дит от ВОС к СОМ. Внутренние септы также сразу
закладываются двойными (рис. 531, 7; 537). Внут-
ренний известковый слой ниже СОМ сохраняет пос-
тоянную толщину на протяжении жизни особи. По
мере роста раковины кнаружи от СОМ количество
нарастающих дополнительно слоев может быть раз-
личным, толщина их неодинакова. Они также разг-
раничены дополнительными органическими мембра-
нами (органическими слоями — ос). Мы предлагаем
закрепить за ними как термин выражение «слои на-
растания». Они формируются при многократном учас-
тии обтекающего раковину наружного слоя цито-
плазмы. Вновь образованные слои простираются над
всеми предыдущими камерами, в результате стенка
предшествующих камер толще, чем у последующих
(в особенности, чем у самой последней). Наиболь-
шего развития эти дополнительные слои достигают у
планктонных форм, образуя мощную корку («crust»)
(рис. 535, 4; 536, 4, Б). Этот тип стенки имеет тон-
кую единообразную упорядоченную структуру: мель-
чайшие кристаллиты, окруженные тончайшим слоем
органического вещества, объединены в небольшие
плитки (plates), уложенные друг над другом в верти-
кальные столбики перпендикулярно поверхности
стенки. В световом микроскопе стенка выглядит
прозрачной, гиалиновой, в отличие от бесцветной
стенки нодозариат она часто слабо-желтоватая или
розоватая, по-видимому благодаря большему присут-
ствию органических прослоев. Если столбики на-
правлены не перпендикулярно к поверхности рако-
вины, а под косым углом (35°, 45°), причем направ-
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
553
Рис. 537. Схема строения бифонтинальной стенки, пор и цитоплазмы донных роталиан. (По: Leutenegger, 1977; Leutenegger, Hansen,
1979 — объединенная схема).
в — вакуоль, вое — внутренний органический слой, г — аппарат Гольджи, дпп — дополнительная поровая пластинка, л — липиды, м — митохондрии,
мп — микропора, ос — органический слой, п —пора, пе— пероксисомы, пк — поровый канал, пп — поровая пластинка, р — рибосомы, с — стенка, се —
септа, си — симбиотические водоросли, сом — срединная органическая мембрана, хр хроматин, ци— цитоплазма, я — ядро, яп — ядерные поры.
Fig. 537. Schematic structure of bifontinal wall, pores and cytoplasm of the bottom Rotaliana.
n — роге, мп — micropores, nn — pore plate, dnn— additional pore plate, пк — pore canal, com — middle (primary) organic membrane, oc — organic layer,
eoc — inner organic lining, ce — septa, c — wall, ци — cytoplasm, я — nucleus, яп — nuclear pores, xp — chromatin, г — Golgi apparatus, м — mitochond-
ria, ne — peroxisomes, p — ribosomes, в — vacuole, л — lipids, cu — symbiont algae.
ление угла наклона меняется то в одну, то в другую
сторону, стенка теряет блеск и прозрачность и ста-
новится матовой.37 (Зернистая и гиалиновая стенки
часто встречаются у видов одного рода, например у
Turrilina (Hansen, 1972)). Такую стенку называли
микрозернистой. Выделение таксонов высокого ран-
га на основании наличия микрозернистой стенки
привело к разрыву в системе близкородственных
групп (см. раздел «История изучения»). Иногда в
стенке одной и той же раковины имеются участки с
перпендикулярными и косыми столбиками, напри-
мер у Globorotalia (Маслакова, 1978).
Дополнительные скелетные образования при этом
типе стенки достигают особого разнообразия и слож-
37 Такую же картину можно наблюдать на раковинах моллюс-
ков: блестящие перламутровые слои образованы перпендикулярно
расположенными кристаллами, а тусклые, сахаристые — косыми,
разнонаправленными.
ности — внутренние клапаны, пластинки, желобки,
системы каналов у роталиан (рис. 528, 4, а—г; 531,
2, 3), наружные ребра, шишки, шипы, иглы, а также
прикрывающие пупок или швы дополнительные ске-
летные пластинки роталиат и глобигеринат (рис. 525,
70, Г, 72, Я; 531, 9; 538, А—Д\ 539, 7, 2; 547, V,
24, Б, 26, Б), не встречающиеся в других классах.
Может быть, внешняя скульптура раковин действи-
тельно иногда придает стенке большую прочность,
как считалось ранее, однако чаще всего ее располо-
жение и направление указывают на связь с интенсив-
ными токами цитоплазмы в том же направлении
(Hemleben, 1975; Hemleben et al., 1977), а бугорки и
кили могут служить местом прикрепления ретикуло-
подий. Длинные толстые сплошные шипы некоторых
донных форм (рис. 538, Е, И) не гомологичны внеш-
не похожим на них шипам Calcarina, Baculogypsina и
Baculogypsinoides (рис. 547, V, 30), внутри которых
554
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 538. Наружные дополнительные скелетные образования у Rotaliata (Rotaliana) (А—Д) и у Nodosariata (Nodosariana) (Е—К).
А ребра Brizalina; Б, В — гранулы и натечные швы у Planulina, вид со спиральной и боковой стороны; Г, Д — дополнительные скелетные пластинки
(Г — у Lamellodiscorbis, Д — у Astrononion); Е — шипы по периферическому краю у Lenticulina; Ж — периферический киль и ячеистые структуры в
верхней и базальной частях раковины у Lagena; 3—К — продольные ребра у Amphicoryna (3) и Marginulina (И — вид с боковой стороны, К — вид с пери-
ферического края).
Fig. 538. Outer additional skeletal formations in Rotaliata (Rotaliana) (А—Д) and Nodosariata (Nodosariana) (E—K).
Rotaliana: A — costae of Brizalina; Б, В — granules and thickened sutures of Planulina, views from the spiral and lateral sides; Г, Д — additional skeletal plates
(Г — Lamellodiscorbis, Д — Astrononion). Nodosariata (Nodosariana): E — peripheral spines in Lenticulina; Ж — peripheral keel and cancellate structures of the
upper and basal parts of Lagena; 3—К — longitudinal costae of Amphicoryna (3) and Marginulina (И — lateral view, К — peripheral view).
Рис. 539. Особенности планктонных и донных форм.
1 — раковина планктонного Globigerinoides с иглами и последней выводковой камерой. 2 — раковина планктонной Hastigerina с иглами, нитевидными
псевдоподиями и цитоплазматической пузыристой капсулой (пк). 3 — органическая раковина донного вида Myxotheca (Lagynata) с ретикулоподиями. 4 —
микротрубочки в псевдоподиях планктонной Globigerina (Globigerinata): А — продольный срез, х 35000; Б — поперечный срез, х 38000. м — митохондрии;
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
555
❖Л 5
мт — микротрубочки {стрелками указаны фибриллярные мостики
между микротрубочками); ст — стереоплазма (пучок продольных мик-
ротрубочек). 5 — микровилли в цитоплазме Globigerina (Globigerinata),
х 24 000 (фрагмент внутреннего содержимого вакуоли). 6 — Globigerina
(Globigerinata) (х 600): крипт — криптосомы, расположенные в цитоп-
лазме камер, сохраняющих форму после растворения раковины в резуль-
тате специальной обработки. 3 — по: Loeblich, Tappan, 1988; 4 — схема
по: Febvre-Chevalier, 1971; 5 — схема по: Leutenegger, 1977; 6 — схема
по: Lee et al., 1965.
Fig. 539. Peculiar structure of some planktonic and bottom forms.
1 — shell of planktonic Globigerinoides with the spines and brood chamber (the last one). 2 — shell of planktonic Hastigerina with spines, filose pseudopodia and
cytoplasmic bubble capsule {пк). 3 — organic shell of the bottom species Myxotheca (Lagynana) with reticulopodia. 4 — microtubules in pseudopodia of planktonic
Globigerina (Globigerinata): A — longitudinal section, x 35 000; Б — transverse section, x 38 000. cm — stereoplasm (bundle of longitudinal microtubules), мт —
microtubules, м — mitochondria, fibrillar bridges between microtubules are showed by arrows. 5 — microvillies in the cytoplasm of Globigerina (Globigerinata),
x 24 000 (fragment of the inner content of vacuole). 6 — Globigerina (Globigerinata), x 600: крипт — cryptosomes, situated in the cytoplasm of the chambers,
preserving their form after special dissolution of the shell.
556
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
проходят каналы. Не гомологичны они и иглам планк-
тонных форм (рис. 539, 7, 2), служащим опорой рети-
кулоподиям и растянутой на иглах цитоплазме, приспо-
собленной к парению (Hastigerina) (рис. 539, 2). Еще
более специализированное образование — иглы Globi-
gerina, которые могут быть закреплены подвижно на-
подобие сустава в выступах, имеющих углубление в
центре (рис. 531, 9; 539, 7), и при необходимости могут
поворачиваться либо сбрасываться во врегёя опускания
раковин на дно. Внешние концы этих игл слабо либо
вовсе не кальцифицированы, благодаря чему они ста-
новятся гибкими и не ломаются.
Пятый тип стенки возникает в результате появле-
ния центров секреции по обе стороны от СОМ в виде
одновременно закладывающихся 2 пластин. Мы на-
зываем такую стенку бифонтинальной (Михале-
вич, 1992, 1995, 1999), что позволяет противопоста-
вить ее первым четырем типам стенок, которые обра-
зуются в виде одного слоя над ВОС и относятся к
монофонтинальному типу (прежнее название «одно-
слойная» приводило к смешению понятий, так как в
монофонтинальной стенке могут присутствовать слои
разного генезиса, например слои с зернами, отсорти-
рованными по размерам, слои нарастания).
Существует еще один, особый тип стенки — у
картерин. В ней крупные уплощенные овальные и ве-
ретеновидные известковые зерна уложены в 1 слой
параллельно поверхности раковины и скреплены ор-
ганическим цементом с примесью мельчайшего аг-
глютинированного материала. Поскольку пока неясно,
секретируются ли эти зерна самим организмом или
взяты извне, систематическое положение этого выде-
ленного нами отряда (Михалевич, 1980, 1999) окон-
чательно не установлено.
Между основными перечисленными типами стен-
ки имеются различия в способе секреции извести.
Стенка милиолят образуется в результате гетероген-
ной нуклеаций кальцита (редчайшее явление среди
всех живых организмов). Стенка пателлин и, по-ви-
димому, остальных спириллинат образуется по осо-
бому, безматричному типу как следствие постепенно-
го разрастания на конце трубчатой камеры и по на-
ружному краю раковины, а не с поверхности, как у
других групп с известковым скелетом. Микрограну-
лярная, нодозариевая радиальная и бифонтинальная
стенки возникают по эпитаксиальному типу (на орга-
нической матрице) в результате поляризованной сек-
реции. При этом если переход от органической стен-
ки к агглютинированной и от нее к микрогрануляр-
ной и милиолятной мог совершаться постепенно (см.
с. 549, 552) и, по-видимому, также постепенно мог
произойти переход от микрогранулярной к радиаль-
ной стенке (по сообщению Исаковой, у фузулинид
наблюдались округлые зерна, уложенные в столбики,
перпендикулярные поверхности раковины, — Миха-
левич, 1992, 1999), то бифонтинальная стенка рота-
лиат могла возникнуть лишь в результате резкого,
скачкообразного изменения характера секреции —
появления второго центра секреции и одновременной
закладки 2 пластин (Лабас, Михалевич, 1987).
Из всех перечисленных выше типов стенки наибо-
лее древними являются тектиновая и агглютиниро-
ванная, возникшие еще в докембрии. Известковая
микрогранулярная стенка, а также стенка нодозариан
и спириллинан возникли в девоне, карбоне, милио-
лян — в карбоне, перми. Наиболее поздно сформи-
ровалась стенка роталиан — в триасе, юре; расцвет
организмов с этим типом стенки начался с мелового
периода. Фораминиферы представляют собой уни-
кальный объект для изучения процессов биоминера-
лизации, эволюции скелета на протяжении геологи-
ческой истории.
Строение пор
Предложенное еще в прошлом веке деление фора-
минифер на перфората и имперфората (Carpenter et
al., 1862) многими авторами принимается до сих пор
и входит в диагнозы многих таксонов. Тем не менее
стенка всех фораминифер в той или иной мере про-
ницаема и имеет разнообразные мелкие отверстия.
Их можно обнаружить даже у раковин с органичес-
кой стенкой. Характер поровых отверстий тесно свя-
зан с типом стенки. У агглютинированных и микро-
гранулярных раковин это либо случайные неоформ-
ленные просветы между зернами агглютината или
секретированной извести, либо тонкие, нередко изви-
тые канальцы с органической выстилкой (у некото-
рых текстулярий), либо ветвящиеся или ампулообраз-
ные, более крупные полости й канальцы в стенке фу-
зулинид и высших атаксофрагмиид, либо прямые
короткие тонкие канальцы («canaliculi») некоторых
астроризат (Masonella, — 547,1, Astrorhizana, 7), ли-
туолид и атаксофрагмиид (рис. 533, 7, 5, 6, 8, 9).
Собственно, всю систему щелей и альвеол в толс-
той губчатой микрогранулярной стенке (рис. 531, 5)
можно рассматривать как способствующую обмену с
внешней средой. Все эти полости чаще замкнуты с
какой-либо одной стороны, но проницаемость стенки
обеспечивается просветами между частицами и про-
ницаемостью самих органических мембран. Все эти
детали строения используются в систематике фора-
минифер, однако мы (Михалевич, 1980, 1981, 1992,
1999) придаем им меньший таксономический вес,
чем Лёблик и Таппан (Loeblich, Tappan, 1988), кото-
рые выделили раковины одинакового строения, но с
наличием или отсутствием небольших поровых ка-
нальцев в микрогранулярной и агглютинированной
стенках в крупные, далеко отстоящие друг от друга
таксоны (например, гомеоморфные Prolixoplectidae и
Eggerellidae). Мы рассматриваем эти особенности
строения как тесно связанные с особенностями мик-
рогранулярной стенки в целом.
Прямые, очень узкие поровые канальцы без до-
полнительных поровых пластинок (ситовидная поро-
вая пластинка в основании отмечена) существуют и
при стекловато-лучистом (радиальном) типе стенки
(рис. 535, 7). Органическая внутренняя выстилка в
канальцах при этом типе стенки пока не обнаружена.
Поровые отверстия спириллинан исследованы недо-
статочно. Наиболее подробно они изучены у Patellina
corrugata (Berthold, 1976). Это довольно широкие,
воронкообразные углубления, снаружи напоминающие
ямки. Их боковые стенки выстланы тонким органичес-
ким слоем, снизу они замкнуты ситовидными плас-
тинками с очень мелкими микропорами (0.01 мкм),
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
557
их внутренняя полость заполнена гранулированным
органическим веществом.
Стенка милиолят описывается как неперфориро-
ванная. Однако мы наблюдали отдельные, неравно-
мерно разбросанные, неправильной формы отверстия
на поверхности раковины Borelis pulchra (рис. 536,
2), а также у некоторых агглютинированных милиолят
(Михалевич, 1983; Михалевич и др., 1986). У пене-
роплиса даже отмечены неправильные и разветвляю-
щиеся поровые канальцы. Известны поры и у некото-
рых триасовых и верхнепермских милиолят (Milioli-
porinae, Kamuraninae). У одного из видов Nubeculina
из голоцена Кларк (Clark, 1993) наблюдал регуляр-
ные поровые отверстия на внутренней, более тонкой
и плотной части стенок, слепо заканчивающиеся в
более толстом рыхлом наружном слое, имеющем ти-
пичное для милиолят строение. Мы сталкиваемся здесь
с таким же явлением, как у многих астроризан и тек-
стуляриан с толстой стенкой: поровые канальцы сфор-
мированы лишь частично, в остальной части стенки
проницаемость обеспечивается более рыхлым распо-
ложением ее структурных единиц. У большинства
милиолян функцию пор заменяют просветы между
рыхлыми кристаллами.
Во всех описанных выше случаях мы имеем дело
с псевдопорами. Собственно поры имеются только в
бифонтинальной стенке (рис. 535, 5, 4; 536,. 4, 5;
537) — обычно это равномерно расположенные, ци-
линдрические неветвящиеся поровые канальцы с соб-
ственной органической выстилкой вдоль стенок,
толстой губчатой поровой пластинкой с ситовидны-
ми отверстиями (микропорами) на уровне СОМ и
более тонкими пластинками на уровне последующих
органических слоев (Hemleben et al., 1977; Leuteneg-
ger, 1977; Spindler, 1978). Число поровых пластинок
и микропор пока не может быть использовано в
диагностических целях в силу неполноты данных.
Среди роталиан наиболее крупные поры отмечены у
дискорбид (у Discorbis vesicularis — 2—2.5 мкм), а
также у Robulus midwayensis (0.5) и Bulimina margi-
nata (0.25—0.5 мкм). У планктонных форм (Globige-
rinata) размеры пор обычно крупные (у Globigerina
bulloides размер воронкообразных пор 0.5 мкм в ниж-
ней части, 2 мкм — в верхней, расширенной части).
Псевдопоры нодозариан более мелкие (рис. 536, 5):
размер овальных пор у Nodosaria affinis — 0.16 мкм
в ширину, 0.33 мкм в длину. Размеры пор в разных
отрядах нодозариан и роталиан служат лишь вспо-
могательным диагностическим признаком. У планк-
тонных форм наружная часть пор резорбируется, ис-
чезают вторичные поровые пластинки, входное от-
верстие расширяется до воронкообразного (рис. 535,
4, А), напоминая «щели» в капсулярной мембране
радиолярий (конвергентное сходство структур у
планктонных организмов). Перед процессом гамето-
генеза поры многих планктонных фораминифер за-
печатываются вторичными отложениями извести и
перестают функционировать (Be et at, 1980). Осо-
бое строение пор у глобигеринат (рис, 535, 4; 536,
4, Б) — таксономический признак ранга класса.
В настоящее время известно, что поровые плас-
тинки проницаемы для О2, СО2, ионов солей, раство-
ренных низкомолекулярных органических веществ,
благодаря чему поры играют важную роль в процес-
сах дыхания, осморегуляции, поддержания ионного
равновесия клетки с окружающей средой и в некото-
рых других процессах метаболизма. Они избавляют
клетку от излишней герметизации. У многих донных
видов (Nonionella Stella, Buliminella tenuata, Bolivina
argentea, Cassidulina cornuta и др.) возле пор обнару-
жены скопления митохондрий (рис. 535, 3, Б; 537) и
симбиотических водорослей (Leutenegger, Hansen,
1979; Hottinger, 1982), что подтверждает дыхатель-
ную функцию пор. Поры играют и защитную роль,
препятствуя проникновению бактерий.
До работ Бертольда (Berthold, 1976) о функции
пор судили главным образом на основании их морфо-
логии. Бертольд провел оригинальный эксперимент:
через отверстие тонкой аллюминиевой пластинки в
морскую воду была опущена раковина Patellina cor-
rugata вершиной конуса вниз. Более широкое осно-
вание возвышалось над пластинкой; находящееся на
нем устье было запечатано силиконовым маслом и
парафином. Единственными отверстиями раковины
оставались погруженные в морскую воду поры. Раст-
воренный в воде краситель нейтральный красный ак-
тивно проникал через поры; спустя некоторое время
его концентрация в раковине намного превысила та-
ковую в воде. Сквозь прозрачную стенку раковины
были видны циркулирующие потоки цитоплазмы,
свидетельствующие о том, что клетка жива.
Наблюдения в световом микроскопе (Myers, 1935)
позволили сделать вывод о том, что поры, как и
устье, служат местом выхода ретикулоподий. Однако
уже Ле Кальве (Le Calvez, 1947) и Арнольд (Arnold,
1954), а впоследствии и другие ученые (Towe, Wright,
1963; Sliter, 1974; Berthold, 1976; Leutenegger, 1977)
показали, что поры закрыты изнутри либо органичес-
ким слоем, либо ситовидной пластинкой и никак не
могут служить местом выхода псевдоподий. Послед-
ние могут выходить лишь через устья и отверстия ка-
налов. У некоторых планктонных фораминифер (Ог-
bulina), не имеющих одного главного устья, в стенке
раковины находятся мелкие поровые отверстия и
между ними более крупные, обеспечивающие выход
псевдоподий. Последние в отличие от пор на самом
деле являются устьями (Berthold, 1976) (рис. 546, 2, Н).
Цитология и физиология
Новые данные по ультраструктуре клетки фора-
минифер уже могут быть частично использованы в
систематике.
У живых особей цитоплазма обычно ярко окраше-
на: желтовато-зеленоватая, коричневатая, бурая, оран-
жевая, бордовая, что зависит от содержащихся в ней
различных включений и водорослей. В период разм-
ножения она очищается и становится прозрачной или
молочно-белой. Резкие механические, химические, тем-
пературные и световые воздействия заставляют жи-
вотное быстро втянуть свою цитоплазму внутрь рако-
вины. При этом освободившиеся последние камеры
заполняются жидкостью из сжавшихся вакуолей, ко-
торая, возможно, играет защитную, буферную роль
между организмом и окружающей средой. У боль-
558
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
шинства фораминифер нет постоянной четкой гра-
ницы между эндо- и эктоплазмой. Лишь у высших
форм, имеющих систему каналов, в которых цирку-
лирует эктоплазма, это разделение становится посто-
янным. Наружный слой цитоплазмы обтекает тонким
слоем раковину донных форм, у планктонных он об-
разует более мощный слой, а у Hastigerina — специ-
ализированную пузыристую капсулу, напоминающую
калимму радиолярий (рис. 539, 2).
Ретикулоподии
От наружного слоя цитоплазмы отходит сеть тон-
ких, обычно ветвящихся и анастомозирующих между
собой нитей — ретикулоподий (псевдоподий) (рис. 525,
7; 539, 2, 3; см. также: Михалевич, 1999, рис. 28, 7).
Эта сеть — ретикулум — у донных форм может быть
приподнята или растекается по субстрату веерообраз-
но, у сидячих форм может распространяться широ-
кой полосой (ламеллоподии), у планктонных она от-
ходит радиально во все стороны, окутывая раковину
«облаком». Диаметр ризоподиальных нитей от 30 нм
до 1—2 мкм (у планктонных тоньше), длина их в не-
сколько и даже в десятки раз превышает размеры ра-
ковин. В вытянутых ретикулоподиях (рис. 539, 4, А)
наружная часть их цитоплазмы — реоплазма — со-
вершает быстрый ток в двух противоположных на-
правлениях (Allen, 1964) со скоростью до И мкм/с
(Samuel et al., 1984). В ней содержатся мелкие грану-
лы (0.5 х 5 мкм), придающие ретикулоподиям зер-
нистый вид (из-за чего их часто называют гранулоре-
тикулоподиями), плотные темные частицы (возмож-
но, источник фосфолипидов) и ряд еще более мелких
частиц, среди которых имеются частицы, организую-
щие движение (MOV — motility organizing vesicles).
Они окружены фибриллярным материалом (Bowser,
Rieder, 1985; Bowser et al., 1985). Могут встречаться
в реоплазме и митохондрии, и лизосомы. Внутрен-
няя, осевая часть ретикулоподий — стереоплазма —
более темная и плотная, состоит из пучка микротрубо-
чек, протянувшихся вдоль ретикулоподий (рис. 539,
4, А). Микротрубочки могут иметь 1—2 коротких
поперечных отростка (рис. 539, 4, Б) (Febvre-Cheva-
lier, 1971; Travis, Bowser, 1986), связывающих их друг
с другом, и не могут образовывать таких сложных
пространственных структур, как мультивалентные
микротрубочки в аксоподиях радиолярий. Попереч-
ные отростки (мостики) связывают микротрубочки и
с другими окруженными мембраной органеллами, и с
клеточной мембраной. Механизм движения (Travis,
Bowser, 1991, — цит. по: Lee, Anderson, 1991) осу-
ществляется следующим образом: динеин превращает
свободную энергию гидролизата АТФ в механиче-
скую работу, позволяя микротрубочкам вращаться
друг за другом подобно храповику; от вращающихся
микротрубочек движение передается связанной с ними
цитоплазме. Этот сложнейший процесс нуждается еще
в дополнительных исследованиях.
У планктонных фораминифер на поздних стадиях
онтогенеза ретикулоподии становятся жесткими, пе-
рестают анастомозировать, приобретая некоторое
конвергентное сходство с аксоподиями других план-
ктонных протистов (солнечников, радиолярий, акан-
тарий). Внутрь их врастают известковые иглы. Фора-
миниферы и здесь пошли по своему особому пути: во
всех случаях, когда возникает необходимость в жест-
ких структурах, у них формируются не органические,
а минеральные известковые образования (иглы, рако-
вины).
Ретикулоподии — важный систематический при-
знак типа фораминифер, резко отличающий их от
жгутиковых, инфузорий, амеб. Функции ретикулопо-
дий многообразны: передвижение, захват пищи, ося-
зание, образование новых камер, цист, оболочки во-
круг сизигиев (см. раздел «Общая характеритика»).
Комплекс псевдоподий в устье раковины называют
подостилем (рис. 525, 7). Вытянутые концы ретику-
лоподий прилипают к субстрату. В результате сокра-
щения ретикулоподиальных нитей раковина подтяги-
вается к месту прикрепления. Нужна немалая сила,
чтобы подтянуть таким образом крупные раковины
или удержаться на субстрате, водорослях, не быть
снесенным током воды. При передвижении необхо-
дима способность к ориентации: упавшие с водоро-
слей животные могут находить обратный путь на то
же место (Zmiri et al., 1974). У некоторых мелковод-
ных видов наблюдался положительный термотаксис и
отрицательный фототаксис. У форм, имеющих кана-
лы, устье обычно зарастает (открываясь вновь только
перед процессом образования новой камеры) и рети-
кулоподии выходят наружу через отверстия кана-
лов — через «заднюю дверь», по образному выраже-
нию Хоттингера (Hottinger, 1982). Это дает животно-
му возможность сохранять мобильность и при
втянутой цитоплазме, уходить от вредных химичес-
ких воздействий, ставить раковину на киль, повора-
чивая к свету боковые поверхности (что важно в слу-
чае наличия симбионтов): Мы наблюдали, как донная
Massilina secans и другие виды милиолид прикрепля-
ются ретикулоподиями к поверхностной пленке воды
в чашке Петри, устьем кверху, и «плавают» в таком
положении иногда по нескольку дней в поисках
пищи. У некоторых видов раковина движется вперед,
увлекаемая сильным током цитоплазмы. У М. secans
она движется в сторону, противоположную той, куда
направлены ретикулоподии, — как бы пятится задом.
С помощью ретикулоподий фораминиферы могут пе-
редвигаться по плоскому субстрату, проходя 11—
188 мм в день (Wetmore, 1988). Laticarinina paupera-
ta проходила 12—16 мм в день по поверхности грун-
та и 5 мм в день, передвигаясь в грунте (Weinberg,
1991). По нашим наблюдениям, Massilina secant про-
двинулась за 1 мин на 100 мкм, затем за 15 мин одо-
лела расстояние длиной 215 мкм. При своем продви-
жении она оставляла хорошо заметные следы светло-
го органического вещества на дне чашки Петри. Во
время передвижений по водорослям у фораминифер-
эпифитов Textularia bocki и Quinqueloculina ungeria-
na по всему пути их следования ретикулоподии ос-
тавляли следы из секретируемого ими органического
вещества (предположительно, сульфатсодержащих
гликозаминоглюканов), служащего питательной сре-
дой бактериям и грибам (Langer, Gehring, 1993). Пос-
ледние, в свою очередь, служат пищей названным фо-
раминиферам. Целенаправленное повторное возвра-
щение фораминифер-эпифитов к своим «фермам»
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
559
возможно на основе хемотаксиса. Таким образом,
взаимоотношения фораминифер-эпифитов и водоро-
слей приносят двустороннюю выгоду: водоросли за-
висят от организмов, очищающих их поверхность, а
фораминиферы используют эту поверхность как ис-
точник пищи. Вещества, выделяемые отмершими раз-
лагающимися водорослями, уменьшают вредное вли-
яние загрязнений (в том числе тяжелыми металлами)
на фораминифер (Bresler, Yanko, 1995).
Кроме функций передвижения (до 2 см в день) и
прикрепления ретикулоподии способны осуществлять
захват мелких частиц или отдельных организмов. Фа-
бер и Ли (Faber, Lee, 1991а) даже называют их «ап-
паратом для захвата пищи». Ретикулоподии разветв-
ляются по дну, проникая в интерстициальные прост-
ранства между частицами грунта на глубину 2—3 мм
либо образуя в толще воды приподнятую над рако-
виной сеть. В последнем случае они нуждаются в
поддержке. Эту функцию у сидячих донных видов
осуществляют приподнятые разветвления раковин,
иногда расположенные возле устья иглы губок
(рис. 525, 2); у многих планктонных видов — это
длинные скелетные иглы (см. раздел «Секреционная
известковая стенка») (рис. 539, 1,2).
Выбор, захват и перемещение пищи могут вклю-
чать сложные манипуляции (Anderson, 1988) — вра-
щение, передачу от одной ретикулоподии к другой,
поднятие на вершину псевдоподиальных нитей. При-
касаясь к добыче снаружи, ретикулоподии увеличива-
ют давление, и поверхность жертвы (рачки, домики
диатомей) лопается в наиболее слабых местах, подобно
орешку. Тогда отдельные нити проникают внутрь и
отделяют мягкие внутренние части, вылущивая содер-
жимое из панциря таким образом, что создается впе-
чатление точных и сложных осмысленных действий.
Затем они переносят кусочки пищи в пищеваритель-
ные вакуоли, находящиеся внутри раковины или в ре-
тикулуме. Обратным током цитоплазмы непереварен-
ные частицы, имеющие вид серовато-коричневых ша-
риков, выносятся к вершинам псевдоподий и
выбрасываются во внешнюю среду. Избирательное
отношение к пищевым частицам при кратком сопри-
косновении, вероятно, обеспечивается хемосенсорны-
ми процессами (Anderson, 1988). У Massilina secans в
культуре мы наблюдали, как убитые клетки Chlorella
втягивались животным в ретикулоподии. В тонких нитях
эти включения выглядели как вздутые узелки и переме-
щались цитоплазматическим током в сторону устья с
большой скоростью — примерно 20—25 мкм/с. Воз-
ле устья они собирались в комок, превышающий ди-
аметр устья (Михалевич, 1999, рис. 28, 2).
Имеются сведения (Schultze, 1854; Christiansen,
1971), что ретикулоподии некоторых видов форами-
нифер (например, Elphidium) способны выделять па-
рализующее жертву вещество: когда пойманное жи-
вотное (быстро движущиеся копеподы, капреллиды,
кумацеа или медленно движущиеся иглокожие, аме-
бы) прилипает к нитям ретикулоподий, оно долго
борется, судорожно пытается вырваться, но посте-
пенно слабеет, окончательно запутывается в липких
нитях и погибает. Картина, заснятая на пленку, вы-
глядит драматично. Некоторым животным удается
вырваться, но они падают на грунт и беспомощно
шевелят лапками, неспособные к прежнему движе-
нию.
Своеобразно добывание пищи агглютинирован-
ной Pilulina argentea. Она выстраивает в глубине
грунта домики (Christiansen, 1971) — полости, стен-
ки которых выложены изнутри крупными песчинка-
ми, а сверху, на уровне поверхности грунта, закрыты
«крышей» из ила и грязи. Домики в 3—4 раза круп-
нее самой раковины животного. Пилулина выставля-
ет сквозь «крышу» ретикулоподии и поджидает до-
бычу. Схватив какого-нибудь мелкого рачка, неосто-
рожно подплывшего к колеблющимся нитям, она
затаскивает его внутрь домика. Когда домики вскры-
вали, в них обнаруживали протиста, крепко держа-
щего свою жертву. Есть предположение, что ретику-
лоподии фораминифер выделяют вещества, возможно,
привлекающие либо парализующие животных. Так,
Dunaliella прекращала движение, достигнув ретику-
лоподиальной сети аммонии (Rotaliana) (Lee et al.,
1961).
Фораминиферы могут быть хищниками и поедать
других фораминифер. Floresina высококонической
многокамерной раковиной (Rotaliana) заползала на
превосходящие ее по размерам линзовидные ракови-
ны 3 видов Amphistegina (Rotaliana), проделывала до
10 отверстий в их стенке и извлекала цитоплазму из
нескольких камер. Через 3—7 дней жертвы обычно
погибали, но некоторые из них, после того как фло-
резина откреплялась, восстанавливались. Иногда
Amphistegina удавалось отразить нападение хищника
(Hallock, Talge, 1994). Флорезины нападали только
на взрослые раковины, на поверхности которых они
затем размножались.
Отдельные сведения, накопленные по питанию
фораминифер в культуре, и полевые наблюдения (см.
с. 534, 537) показывают, что питание фораминифер
разнообразно. Они могут использовать растворенные
в воде аминокислоты и другие органические вещест-
ва (например, Notodendrodes antarcticus — De Laca et
al., 1981; Pararotalia spinigera и Rosalina macropo-
ra, — Bresler, Yanko, 1995), могут быть детритофага-
ми, бактериофагами (обычно ниже освещенной зо-
ны), поедать экскременты, водоросли (преимущест-
венно диатомовые, но также дуналиеллу, хлореллу и
др.), дрожжи, различных простейших, яйца, личинок,
мелких рачков (например, копеподы Calanoida —
главный компонент в питании глобигеринид). Неко-
торые виды питаются избирательно, но большинство,
по-видимому, всеядны. Известны случаи каннибализ-
ма — поедания собственных зигот у Glabratella огпа-
tissima (Myers, 1940), Patellina corrugata, Heterotheca
lobata (Grell, 1988).
Каждый из способов питания может осуществ-
ляться активно (передвижение, увеличение токов
воды движением псевдоподий) или пассивно. Пассив-
ный тип питания особенно характерен для сидячих
форм либо укрепившихся, «укоренившихся» в грунте
(рис. 525, 2; 547, /, Astrorhizana, 9). Приподнятые над
поверхностью дна формы улавливают детрит, оседаю-
щий из толщи воды. Раковины могут лежать на по-
верхности грунта (Astrorhiza) либо передвигаться в
его поверхностном слое, оставляя в нем ходы, чаще
на глубине 1—2 см. В последнем случае раковины
560
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
обычно удлиненной формы, при которой грунт ока-
зывает наименьшее сопротивление (рабдаммины, ре-
офаксы, многие милиолиды).
Распределение пищи влияет на многие стороны
жизни фораминифер: наступление полового размно-
жения, размеры, распространение. Так, планктонные
виды с иглами поглощают преимущественно зо-
опланктон, а без игл — фитопланктон (Hemleben,
Spindler, 1983). Последние точно повторяют в своем
движении смещение максимума хлорофилла вверх и
вниз между 0 и 95 м, а также уменьшение и исчез-
новение фитопланктона в высоких широтах. Эта
картина четко отражается в современных и ископа-
емых осадках. Сезонность чередования поколений у
донных фораминифер тоже нередко связана с оби-
лием пищи, цветением водорослей.
Фораминиферы могут быть эпифитами на водоро-
слях (чаще всего на Enteromorpha, Zostera, Zaniche-
На, Ulva, Polysiphonia, Laminaria, редко на Fucus и
Codium, — Lee et aL, 1969; на Sargassum natans, —
Spindler, 1980), эпибионтами на раковинах моллюс-
ков, панцирях рачков, гидроидах (Dobson, Haynes,
1973), полихетах (мы наблюдали Cibicides grossepun-
ctatus на элитрах антарктической полихеты Picnogo-
пит sp.), других фораминиферах. На створках Chla-
mys opercularis в Ирландском море был обнаружен
51 вид фораминифер-эпибионтов, причем часть из
них были нехарактерны для этого местообитания
(Harward, Haynes, 1976). Иногда фораминиферы са-
ми служат подвижным субстратом для других живот-
ных, как например Buccella frigida для некоторых
турбеллярий (Zapata, Varela, 1981).
Кроме того, фораминиферы могут быть коммен-
салами. Cibicides refulgens соскребает с раковины
гребешка водоросли и бактерии. В то же время осо-
би этого вида образуют над собой агглютинирован-
ные трубки и выпускают через них ретикулоподии,
питаясь как фильтратор (Михалевич, Воронова, 1985);
кроме того, растворяя раковину моллюска, С. reful-
gens паразитически использует его аминокислоты
(Alexander, De Laca, 1987). Случаи паразитизма
среди фораминифер единичны. Planorbulina может
быть эндопаразитом других фораминифер (Banner,
1971), Rosalina паразитирует на двустворчатых мол-
люсках (Todd, 1965). Все эти паразитические формы
относятся к подклассу Rotaliana. Известны еще 2
случая паразитирования фораминифер из подкласса
Nodosariana: Fissurina и Lagena (первый выкусывает
гранулки в ретикулоподиях Discorbis mediterranen-
sis, — Le Calvez, 1947).
Многие животные (рыбы, иглокожие) заглаты-
вают фораминифер (Daniels, Lipps, 1978). Мелким
видам гастропод (Walker, 1971) и полихет (Lipps,
Ronan, 1974) и их молодым стадиям фораминиферы
могут служить пищей. Они, скорее всего, использу-
ются не как источник калорий, а как источник
СаСО3, но один из видов скафопод может вытяги-
вать цитоплазму фораминифер из раковин наружу.
Нематоды пробуравливают раковины фораминифер,
их популяции быстро растут на фораминиферовой
культуре. Некоторые гастроподы, являясь избира-
тельными хищниками, могут существенно влиять на
численность поедаемого вида (например, на числен-
ность Islandiella islandica). Другая гастропода, Litto-
rina littorea, также пробуравливает раковины фора-
минифер (Walker, 1971). Нередки ископаемые рако-
вины со следами хищников (округлые отверстия в
стенке раковин).
Ретикулоподии принимают также активное учас-
тие в формировании новых камер. Этот процесс изу-
чен у Discorbis (Le Calvez, 1938), Rosalina floridana
(Angell, 1979), Clavulina (Coleman, 1980), Trochammi-
na inflata (Angell, 1990), Textularia candeiana (Bender,
1992). У последней он протекает следующим обра-
зом. Перед образованием новой камеры организм
усиленно питается. Последующий процесс можно
разделить на ряд стадий. >
1. Выход веера ретикулоподий через устье, послед-
ней камеры, который затем несколько укорачивается
и формирует вокруг себя цисту из плотно упакован-
ных частиц детрита, не скрепленных цементом, из-за
чего циста может быть легко разрушена.
2. Формирование внутри цисты, перед устьем пос-
ледней камеры, полости примерно такого же разме-
ра, как будущая камера, которая определяет прост-
ранство для ее построения.
3. Отбор частиц грунта для построения новой ка-
меры и распределение их по всему фронту выступа в
направлении от предпоследней камеры к устью с од-
новременным цементированием их органическим ве-
ществом и образованием наружного органического
слоя (=оболочки — envelope).
4. Отложение биоминерализованной извести, в на-
правлении от дистальной к проксимальной части стен-
ки камеры. Ориентированное расположение кристал-
лов, формирующее цилиндрические полости поровой
системы.
5. Органическая выстилка пор.
6. Формирование тонкого органического слоя с
внутренней стороны камеры после того, как живот-
ное покидает цисту; затем наращивание этого слоя в
несколько рядов в течение последующего роста.
Таким образом, у многокамерных агглютиниро-
ванных раковин текстуляриан, а также хормозинан
слои нарастания имеются, но только в органической
части стенки (Bender, 1995). Весь процесс длится
24 ч.
Textularia относятся к подклассу Textulariana клас-
са Rotaliata. У высших представителей этого класса
в подклассе Rotaliana процесс формирования новой
камеры наиболее подробно на современном уровне
изучен у Rosalina floridana (Angell, 1979). Он тоже
длится 24 ч. Начальные стадии — образование веера
ретикулоподий, цитоплазматического выступа, со-
впадающего по очертаниям с формой будущей каме-
ры, и цисты протекают так же, как у Textularia.
Затем формируется «мембранный комплекс» цито-
плазматического выступа, по-видимому, соответству-
ющий наружной органической оболочке (envelope) у
Textularia. Формирование цитоплазматического вы-
ступа завершается образованием мембраны. Она, по-
видимому, и представляет собой СОМ, поскольку в
отличие от Textularia у Rosalina floridana, как и у
всех представителей с бифонтинальной стенкой (см,
с, 552, 556), кристаллы извести откладываются по
обе стороны от мембраны (рис. 535, 3; 537).
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
561
Начало и завершение процесса кальцификации
прослежено Анжел (Angell, 1979) наиболее подроб-
но. Сначала мелкие кристаллиты видны как мириа-
ды мелких темных точек, затем — как более круп-
ные и светлые центры кристаллизации. В световом
микроскопе на этой стадии на поверхности образу-
ющейся камеры видны разбросанные белые крапин-
ки. Примерно через час после их появления стано-
вится видимым и устье будущей камеры. Постепенно
центры кристаллизации увеличиваются в размерах,
края их сливаются. Но между этими заполненными
известью пространствами остаются незаполненными
поровые полости. По-видимому, на поверхности ци-
топлазматического выступа предсуществует химичес-
кая «разметка» плазматической мембраны на зоны,
участвующие в образовании извести, и «островки», в
пределах которых это образование отсутствует, —
будущие поры (Лабас, Михалевич, 1987). Равномер-
ность расположения и упорядоченность центров сек-
реции определяется, по-видимому, аппаратом Гольд-
жи. «Пятнистый» характер известковой секреции, ее
отсутствие в области пор можно предположительно
объяснить паттерном зон экзоцитоза везикул, в мем-
бране которых функционирует карбоангидраза и
Ca+VH* АТФ-аза. В работе Анжел дальнейший про-
цесс не прослежен, однако, по-видимому, в данном
случае, как и у Textularia, формируется органичес-
кая выстилка пор и ВОС. Раковинообразующая сек-
реция извести и мукополисахаридная секреция орга-
нических элементов разделены во времени, по оче-
реди сменяя друг друга (Лабас, Михалевич, 1987).
Наконец, ретикулоподии выходят из устья новой ка-
меры, разрушая цисту. Процесс образования ВОС
над цитоплазматическим содержимым заканчивается
в каждой камере позже, когда уже идет образование
следующей (Spindler, 1978). Таким образом, распо-
ложение новых камер определяется положением фо-
раменов, отражающих направление цитоплазмати-
ческого потока, а их форма контролируется длиной
и расположением ретикулоподий, которые формиру-
ют первую наружную органическую оболочку (enve-
lope) — матрицу последующей кальцификации. Вы-
ход цитоплазмы за пределы камеры и образование
новой камеры осуществляются циклически, с опре-
деленной периодичностью, у молодых чаще и быст-
рее, чем у взрослых. Одновременно помимо форми-
рования последней камеры появляются новые орга-
нический и известковый слои на предшествующих
камерах. Так возникают слои нарастания (см. раздел
«Секреционная известковая стенка»). При этом в об-
ласти пор на уровне новых органических слоев у
донных бифонтинальных форм формируются допол-
нительные поровые пластинки (рис. 535, 3, б\ 537).
Возникновение новых камер у планктонных ви-
дов (глобигеринат) имеет некоторые особенности
(Hemleben, 1969; Towe, 1971; Spindler, Roettger, 1973).
Они не строят цисту, у игольчатых форм первичная
органическая мембрана натягивается, как тент, меж-
ду известковыми иглами. Затем по обе стороны от
нее происходит отложение извести, как у донных
роталиат.
Формирование новых камер в классе Miliolata,
прослеженное у Spiroloculina hyalina (Angell, 1980),
36 Протисты, часть 1
принципиально иное. Хотя сначала перед появлением
новой камеры их ретикулоподии тоже строят цисту,
она, однако, окружает всю раковину, а не только
будущую камеру. После образования цисты в ней
остаются ретикулоподии и через устье предыдущей
камеры выходит небольшое количество цитоплазмы.
С этого момента и начинается построение новой ка-
меры. В отличие от Rosalina и Textularia у Spirolo-
culina до начала секреции новой камеры не форми-
руется ни цитоплазматический выступ, ни окружающяя
его оболочка (мембрана — envelope). Цитоплазма
спиролокулины поставляет к месту образования стенки
два рода вакуолей: с органическим веществом (иног-
да наблюдались уплощенные фибриллы) и с удли-
ненными кристаллами извести. Как те, так и другие
вакуоли (пузырьки) открываются и одновременно
опорожняются в пространство, образующееся между
тонким слоем цитоплазмы и ретикулоподиями. Так
формируется довольно толстая органическая матри-
ца, в которую внедряются известковые кристаллы (у
роталиат кристаллы извести отлагаются на мембра-
не, а не внутри нее). Тонкий слой цитоплазмы фор-
мирует наружную часть стенки, затем ретикулоподии
достраивают ее изнутри, тоже поставляя органичес-
кие и известковые компоненты. Причем камера фор-
мируется не сразу вся целиком, как это имеет место
у роталиат, а сегментами. Сначала движущийся по-
перек предыдущей камеры тонкий слой цитоплазмы
формирует часть камеры, внедряя иглы беспорядоч-
но, затем наращивает ее вперед; процесс продолжа-
ется вплоть до завершения построения камеры и
формирования зуба. Псевдоподии добавляют новые
элементы изнутри также последовательно, после
того как тонкий участок стенки уже сформирован
цитоплазмой снаружи. На этом этапе кристаллы ук-
ладываются ориентированно — параллельно поверх-
ности стенки. Стенка постепенно утолщается. Далее
ретикулоподии завершают построение каждого пос-
ледовательно сформированного сегмента стенки по-
явлением изнутри толстой мембраны (органического
слоя, подстилающего стенку), отличающейся по со-
ставу и плотности от органического матрикса, в ко-
торый внедрены кристаллы извести. Каждый крис-
талл имеет тонкую органическую оболочку, сохраня-
ющую форму кристалла и хорошо различимую после
его растворения. Известковые иглы имеют 0.2 мкм
в длину и 0.01 мкм в ширину.
Во время образования последней камеры к ранее
сформированным камерам продолжают добавляться
новые порции органического и известкового матери-
ала, в результате чего начальные камеры становятся
толще последующих, но слои нарастания не образу-
ются. В ретикулоподиях, достраивающих стенку ра-
ковины, не просматривается почти никакой структу-
ры — только пузырьки с органическим и известко-
вым веществом. Ретикулоподии, прикрепляющие
раковину к цисте, имеют множество продольных
микротрубочек и филаментов. С их помощью после
завершения формирования новой камеры раковина
поворачивается в цисте и выходит из нее. На по-
строение цисты уходит несколько часов; от момента
выхода цитоплазмы из устья камеры до полного по-
строения новой камеры проходит еще 6 ч.
562
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
По-видимому, у разных видов и родов милиолид
этот процесс несколько варьирует. По нашим наблю-
дениям, циста при построении новой камеры у моло-
дых мелких милиолид окружала всю раковину, а у
крупных массилин обычно закрывала раковину лишь
частично. Иногда животное выходило из цисты рань-
ше, чем завершалось построение новой камеры, и
окончание процесса было хорошо видно: бесфор-
менная масса цитоплазмы в месте построения нового
участка клубилась подвижным «облаком» (Михале-
вич, 1999, рис. 28, 3); активность псевдоподий в этот
период была велика, и раковина часто меняла по-
ложение — буквально поворачивалась с боку на бок.
Вновь выстроенная камера тонкая и прозрачная (Ми-
халевич, 1999, рис. 28, 4). Пока построение новой ка-
меры не завершено, границы последовательно сфор-
мированных сегментов отчетливо различимы, так как
наиболее молодые участки отличаются и наибольшей
тонкостью и прозрачностью. Построение камеры под
цистой происходило за 8 ч, без цисты процесс иногда
длился до 2 дней.*
У представителей класса Spirillinata (на примере
Patellina corrugata) процесс роста стенки камеры
также совершается при участии ретикулоподий; об-
разование цисты не наблюдается, рост ограничивает-
ся краями стенки трубчатой камеры; началу процес-
са кальцификации не предшествует образование ци-
топлазматического выступа. Морфогенетические
процессы и в этой группе фораминифер глубоко от-
личаются от таковых у роталиат. Первыми формиру-
ются ближайшие к устью структуры, а не удаленные
от него, строительный материал добавляется по
краю уже имеющегося, как при строении обычной
стены в высоту. Но и в этом случае, по-видимому,
должен действовать механизм секреции, благодаря
которому происходит поочередное отложение то из-
вести, то органического вещества, образующего внут-
ренний и внешний слои раковины и выстилку пор.
Таким образом, у рассмотренных представителей,
относящихся, по нашей системе, к 3 разным классам
(Rotaliata, Miliolata, Spirillinata), механизм роста и
образования новых камер имеет глубокие различия и
характеризует особые эволюционные линии развития
(Михалевич, 1999). Мы считаем, что сходство про-
цессов формирования новой камеры у Textulariana и
Rotaliana подтверждает родство этих групп и опро-
вергает общепринятые взгляды об их конвергентном
происхождении, так как различий (формирование
СОМ и откладывание извести по обе стороны от нее
у роталиан) у них меньше, чем сходства (построение
цисты только вокруг вновь строящейся камеры; фор-
мирование цитоплазматического выступа и окружаю-
щего его органического слоя; поочередное, а не од-
новременное, как у милиолят, откладывание извести
* У Archaias angulatus (Soritida, Mitiolata) (Wetmore, 1999),
имеющего широкие камеры, при формировании новой камеры сна-
чала образуется цитоплазматический выступ. Детально процессы
образования органического матрикса и кальцификации в этой ра-
боте не прослежены. Вряд ли есть основания предполагать, что
типичная монофонтинальная фарфоровидная стенка A. angulatus
формируется подобно бифонтинальной стенке роталиат со слож-
ными порами. По-видимому, сходство ограничивается только обра-
зованием цитоплазматического выступа, в остальном же процесс
кальцификации Происходит у этого вида по типу других милиолят.
органического вещества; формирование новой ка-
меры сразу целиком и не от устья, а навстречу ему).
Механизм роста раковин и камер может нару-
шаться в экстремальных условиях: при сильных ме-
ханических воздействиях, при крайних, почти ле-
тальных значениях температур, солености. Напри-
мер, дисковидные соритиды в этих условиях могут
становиться цилиндрическими или выстраивать по-
лудиск, перпендикулярный основному; камеры пра-
вильно-клубковидных милиолид начинают нарастать
беспорядочно, принимать уродливую, изогнутую фор-
му — т. е. теряется тип симметрии, свойственный
данному виду. Причиной этого является дезорганизация
цитоплазматического тока. Но иногда разрушенная с
одной стороны раковина достраивается до первона-
чальной формы. Все это заставляет предположить
наличие механизма контроля роста, регулирующего
формирование камер по оси симметрии и закреплен-
ного генетически. По-видимому, это регулирование
определяется биохимическим градиентом первичных
мембран, покрывающих цитоплазму и ретикулопо-
дии.
Цитоплазматические органеллы
Основные цитоплазматические органеллы пред-
ставлены на рис^ 537. Митохондрии (рис. 537, 539,
4, Б\ 541, 2, 3), которые у фораминифер бывают
до 0.5 мкм в диаметре и 1—5 мкм в длину, име-
ют трубчатые кристы. Пероксисомы (микротельца)
обычно заполнены темной гранулярной массой
(рис. 537) и отмечены как у донных, так и у планк-
тонных фораминифер. Диктиосомы аппарата Гольд-
жи (рис. 537) выделяют карбогидразу, а также слизь,
которая транспортируется в пузырьках к поверхнос-
ти раковины и к ретикулоподиям и служит для при-
крепления к различным поверхностям, а иногда для
удержания жертвы. Вакуоли с кристаллами извести
для построения скелета, энзимы, в том числе пище-
варительные, лизосомы (пищеварительные вакуоли)
также секретируются в аппарате Гольджи. Перевари-
вание пищи происходит, по-видимому, не только в
этих вакуолях, но и непосредственно в цитоплазме
(лакунах) (Leutenegger, 1977). Основные запасные
питательные вещества в клетке фораминифер нахо-
дятся в виде жировых капель. Обнаруженные у не-
которых видов (Archaias angulatus, Cyclorbiculina
compressa) зерна крахмала являются результатом де-
ятельности симбиотических водорослей. ЭПР у фо-
раминифер хорошо развит. Кроме того, в цитоплаз-
ме фораминифер встречаются ксантосомы (пигмен-
тированные гранулы), кольцевые мембраны (скорее
всего, резервный материал для построения ядерных
мембран в процессе размножения, — Spindler, Неш-
leben, 1982). Есть органеллы, которые встречаются
только у планктонных фораминифер, — это крипто-
сомы (Lee et al., 1965) и микровилли (Anderson, Be,
1976а, 1976b; Leutenegger, 1977). Первые имеют ок-
руглую или неправильную форму (рис. 539, 6), раз-
мером 15—30 мкм; иногда занимают более полови-
ны камеры; ассоциируются с углеводами. Роль их
неясна. Система микровиллей (фибриллярный, или
везикулярный, ретикулум) — уникальный механизм
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
563
плавучести. Это изогнутые, разветвленные, плотно
упакованные в вакуолярном пространстве трубки до
10 мкм в диаметре (рис. 539, 5). По-видимому, заме-
на тяжелых ионов легкими меняет удельную массу
микровиллей, обеспечивая раковинам медленное
вертикальное движение (Hansen, 1975).
Симбиоз
У многих крупных, секретирующих известь дон-
ных фораминифер, обитающих на глубине 10—70 м
(Alveolinidae, Soritidae — Miliolana; Amphisteginidae,
Calcarinidae, Nummulitidae — Rotaliana), и планктон-
ных фораминифер эуфотической зоны (Globigerini-
dae, Hastigerinidae, Globorotaliidae) в цитоплазме на-
ходят клетки водорослей (рис. 537) (преимущественно
динофлагелляты и диатомовые, но также хризофиты,
родофиты, хлорофиты и, по-видимому, гаптофиты). У
представителей Nonionida и Elphidiida найдены от-
дельные хлоропласты (Lopez, 1979). Часть этих водо-
рослей — комменсалы и паразиты, но, как правило,
какой-либо один вид является симбионтом для оп-
ределенного вида фораминифер. В настоящее время
симбионты обнаружены более чем в 20 семействах в 3
разных классах этих протистов. Многие группы близ-
кородственных фораминифер связаны с одними и
теми же или близкородственными симбионтами. У ро-
талиат симбионты принадлежат только к диатомовым,
у милиолят (Peneroplidae, Archaiasinidae) — к родофи-
товым и хлорофитовым. Сходство симбионтов нуммули-
тид (роталиаты) и альвеолинид (милиоляты) рассмат-
ривается как исключение (Leutenegger, 1984). Потреб-
ности симбионтов к различной освещенности влияют
на распределение их хозяев по глубинам. Во время бес-
полого размножения дочерние особи наследуют сим-
бионтов от родительской клетки, после полового про-
цесса выбирают их из окружающей среды. У Hastige-
rina depressa симбионты менялись по сезонам. Клетка
хозяина предоставляет симбионтам постоянную среду
обитания и продукты питания, например азотистые
соединения. Внутриклеточные диатомовые по сравне-
нию со свободноживущими содержали повышенные
количества глюкозы и глицерина (Kremer et al., 1980).
Хозяин, в свою очередь, получает продукты фотосинте-
за и питательные вещества (углеводороды, некоторые
аминокислоты, например глютамин, аланин). Такие
взаимоотношения особенно выгодны в среде, бедной
пищевыми продуктами (Lee, 1980). Кроме того, вся
система симбионт—хозяин получает дополнительную
энергию. Возможно, различные организмы канализи-
руют эту энергию по-разному. У исследованных фора-
минифер она резко увеличивает скорость кальцина-
ции и, вероятно, производство органических мембран.
По данным Хэллок (Hallock, 1981b), скорость кальци-
нации у симбионтсодержащих фораминифер возрастает
вдвое. Радиоуглеродным методом (Kuile, Erez, 1987)
доказано использование НСО3-симбионта при построе-
нии скелета фораминифер. Гипотетически результат
этих процессов можно выразить следующей формулой:
Са++ + 2НС0з---> СаСОз + СО2 + Н2О.
кальцификация
фотосинтез
Amphistegina lobifera может иметь запас углерода,
полуденного на свету, откладывать его в специаль-
ных вместилищах и затем расходовать постепенно.
Милиоляты таких запасов не делают и в отличие от
симбионтзависимой амфистегины используют полу-
ченный углерод немедленно. В клетке хозяина сим-
бионты обычно окружены тонкой мембраной, меша-
ющей вторжению лизосом (рис. 537), но в условиях
голода, особенно в темноте, фораминиферы перева-
ривают их. У родофициевых — симбионтов пене-
роплиса такая мембрана отсутствует; они свободно
лежат в цитоплазме хозяина, что может рассматри-
ваться как более продвинутая фаза симбиотических
отношений (Lee, 1990).
Днем симбионты находятся в наружной цитоплаз-
ме, ночью— внутри раковин. Удаление симбионтов
вызывает тот же эффект, что и темнота: сокращение
продолжительности жизни и уменьшение размеров
раковин.
Симбиотические отношения сложились у фора-
минифер в процессе длительной эволюции и повли-
яли на размеры и форму раковин, их внутреннюю
морфологию (усложнение, появление большого чис-
ла камер и камерок), строение стенки (появление
более тонких, прозрачных участков — «окон», час-
тичная резорбция органического вещества). Эти из-
менения способствовали более удобному размеще-
нию симбионтов, наибольшей светопроницаемости.
Сходные морфологические изменения раковин иско-
паемых форм дают основание предполагать также и
у них наличие симбионтов (фузулиниды).
Положение симбионтов внутри протиста зависит
от формы раковины хозяина. У билатерально-сим-
метричных форм, стоящих на периферическом крае
раковины и поворачивающих боковые стороны к
свету под большим или меньшим углом (Amphistegi-
na, Elphidium, Heterostegina), симбионты находятся
на обеих сторонах раковины, у плоских форм, лежа-
щих одной стороной на субстрате (Sorites), — на
верхней стороне раковины. В обоих случаях симби-
онты располагаются в периферической части камер;
часто их наблюдали возле пор.
Интересно размещение симбионтов у некоторых
планктонных видов, имеющих иглы: ночью симбион-
ты (динофлагелляты) находятся внутри раковин, но с
рассветом покидают ее и размещаются вдоль игл в
псевдоподиальной сети хозяина (Be et al., 1977).
Присутствие симбионтов влияет на изотопный со-
став раковин (фракционирование О18 и С13).
Кроме симбионтов в цитоплазме фораминифер
найдены паразитические Sporozoa (Nyholm, 1962).
Ядро
Размеры ядер фораминифер варьируют от очень
мелких (5—12 мкм) до более крупных (40—100 мкм)
и очень крупных (0.6 мм у гигантских раковин Bathy-
siphon). Число хромосом известно для небольшого
числа видов, оно невелико — 2, 3, 9, 18. Ядра имеют
типичное для эукариот строение (рис. 537; 540; 541,
1—3). Количество и размер ядерных пор варьируют у
разных видов и, кроме того, зависят от физиологи-
ческого состояния клетки. У многих однокамерных
564
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
3
*
ТИП FORAMINIFERA. МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
565
агглютинированных видов ядерная мембрана утолще-
на снаружи или изнутри фиброзным или губчатым
(сотовым, сетчатым) слоями (рис. 540, 7) (например,
у Boderia albicollaris), более жесткая. Более тонкая и
эластичная оболочка ядер многих высших секреци-
онных фораминифер позволяет изменять форму ядра
и не препятствует их проникновению через узкие
проходы из камеры в камеру (рис. 540, 5). Их ядра
чаще бывают не только округлой, но и лопастной,
четковидной, амебоидной (рис. 540, 2) (термин см.:
МсЕпегу, Lee, 1976), неправильной, причудливой
формы. Ядро молодого гамонта Elphidium ponticum
имело округлые очертания, зрелого — лопастные
(Райков и др., 1998; Михалевич, 1999). У низших од-
нокамерных фораминифер также наблюдались ядра
гамонта амебоидной формы (Allogromia laticollaris,
Myxotheca arenite ga,— Schwab, 1971). По-видимому,
такая форма возникает у них из предшествующей ок-
руглой в процессе гаметогенетических изменений яд-
ра, как это было прослежено у Allogromia laticollaris
(МсЕпегу, Lee, 1976) и у Saccammina alba (Goldstein,
1988).
В длинной трубчатой камере Quinqueloculina sp.
мы наблюдали удлиненное, вытянутое вдоль камеры
крупное ядро (Михалевич, 1999) (рис. 542, 2). У не-
которых видов оболочка ядра с внешней стороны
образует выросты (ворсинки), складки, извилины
(Ovammina ораса, Globigerinoides sacculifer), что, по
мнению Дальгрена (Dahlgren, 1967а), ведет к увели-
чению поверхности ядра и усилению его функций.
Гистохимические исследования ядер фораминифер по-
ка немногочисленны (Lee et al., 1965; Dahlgren, 1967b;
Schwab, 1971; Воронова, 1976, Spindler et al., 1978;
Райков и др., 1998, и некоторые др.). Они позволили
обнаружить внутри ядер скопления хроматинового
вещества (ДНК) и нуклеолы — обычно более темно
окрашенные и крупные гранулы, содержащие белок
и РНК. Скопления ее молекул часто видны снаружи
от ядерных пор. Нуклеолы могут быть разбросаны по
всему ядру (рис. 540, 2; 541, 7) или скапливаться на
его периферии (рис. 537). Видимо, их расположение
и размеры меняются при изменении физиологиче-
ского состояния ядра. Хроматин в неделящихся ядрах
обычно деконденсирован.
С тех пор как были впервые описаны 2 типа ядер
у фораминифер (Grell, 1956) — вегетативные (или
соматические, или Ма) и генеративные (или А/u), ко-
личество известных видов с ядерным диморфизмом
значительно увеличилось (см.: Lee et al., 1965; Воро-
нова, 1976, 1978а; Muller-Merz, Lee, 1976; Leuteneg-
ger, 1977; Grell, 1979; и мн. др.). В настоящее время
стало ясно, что в процессе эволюции фораминифер
произошла полимеризация ядер и их дифференци-
ровка на соматические (рис. 537; 540, 7—3; 542, 2,
5), контролирующие функции обмена, и генератив-
ные, дающие начало новому поколению (рис. 539, 6;
540, 4; 553, 3) (по классификации Райкова (1978), —
большие пузырьковидные и малые пузырьковидные ядра).
В соматических ядрах в 3 раза больше РНК и проте-
ина, чем в генеративных, а количество ДНК в тех и
других одинаково. Дифференцировка соматических
ядер необратима. Генеративные ядра могут превращаться
в соматические в случае утраты последнего.
Все обнаруженные к настоящему моменту виды с
ядерным дуализмом относятся, по принятой нами
классификации, к 2 родственным классам Rotaliata и
Globigerinata и к своеобразному по строению рако-
вин и ультраструктуре их стенки классу Miliolata.
Данные по последнему классу весьма скудны, но об-
наружение соматических и генеративных ядер у эво-
люционно продвинутого рода Sorites (S. margina-
lise •— Mtiller-Merz, Lee, 1976; S. orbicularis, — Le-
utenegger, 1977), а также у более просто устроенной
Quinqueloculina sp. (рис. 542) свидетельствует о том,
что в эволюции этой группы тоже возник ядерный
дуализм.
У низших однокамерных лягинан и астроризан,
так же как и у своеобразной группы спириллинат,
ядерный дуализм не обнаружен. Первые исследова-
ния жизненных циклов и ядер многокамерных агглю-
тинированных трохаммин (низшие роталианы) (Sala-
mi, 1976; Angell, 1990; Pawlowski et al., 1995) показали,
что ядра их агамонтов тоже не дифференцированы.
Гомокариотность может использоваться как допол-
нительный таксономический признак этих таксонов.
Ядерный аппарат нодозариат практически не иссле-
дован.
Ядерный дуализм — одна из самых характерных
особенностей фораминифер. Кроме того, у них про-
изошла полимеризация ядер. Например, число гене-
ративных ядер может достигать от нескольких десят-
ков до нескольких тысяч и даже десятков тысяч; у
некоторых видов произошла полимеризация и сома-
тических ядер (4—6 у Rosalina, Glabratella, Globige-
rinoides; 40—60 у Sorites), Для фораминифер извес-
тен и второй путь увеличения ядерного вещества —
полиплоидия. Цитофотометрические исследования,
показавшие возрастание плоидности в ядре в 20—
30 раз, проводились для Patellina corrugata, Rotaliel-
la heterocariotica (Zech, 1964), Cibicides lobatulus
Рис. 540. Ядра фораминифер.
1 — ядро Boderia (Astrorhizata), x 1500, с сотовым слоем (с); в — вакуоль. 2 — соматическое ядро Orbulina (Globigerinata) «амебоидной» формы, х 2900.
3 — мигрирующее соматическое ядро (Ма) Cibicides lobatulus (Rotaliana) четковидной формы, протянувшееся вдоль четырех камер; видны сужения (су) в
местах его прохождения из камеры в камеру через форамены; часто Ма не имеет четких очертаний, дегенерирует; цитоплазма сохраняет форму камер, хотя
стенка раковины растворена (ее прежнее положение — спиральный и межкамерный швы (сил, мил)), х 1200. 4 — Ми агамонта С. lobatulus в процессе мейоза;
белое пространство — растворенный спиральный шов (сил) раковины, х 120Ю. 1 — схема по: Schwab, 1977; 2 — схема по: Be et al., 1977; 3,4 — по:
Воронова, Михалевич, 1985.
Fig. 540. Foraminiferal nuclei.
1 — nucleus of Boderia (Astrorhizata), x 1500, with honeycomb layer (с); в — vacuole. 2 — somatic nucleus of Orbulina (Globigerinata) «amoeboid» in form,
x2900. 3 — rosary-like migrating somatic nucleus (macronucleus — Ma) of Cibicides lobatulus (Rotaliana), stretched out along the four chambers, in the places
of its passing through foramens from chamber to chamber — the constrictions (cy) are seen, part of the macronucleus has not clear outline as it is degenerating;
cytoplasm preserves the form of the dissolved chambers, its previous position is marked by the spiral and interchamber sutures (сил, мил), x 1200. 4 — Generative
nuclei (Mu) of the C. lobatulus agamont in the process of meiosis (white space — the dissolved spiral suture (сил) of the shell), x 1200.
566
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТИП FORAMINIFERA. ЖИЗНЕННЫЕ ЦИКЛЫ
567
Рис. 542. Ядра Quinqueloculina sp.
7 — внешний вид раковины, 2 — схема последовательных поперечных сре-
зов последней камеры, 3 — вид ядер на поперечных срезах. Ма — сомати-
ческое ядро, Ми — генеративные ядра; х 120.
Fig. 542. Nuclei of Quinqueloculina sp.
1 — outer view of the shell, 2 — scheme of the successive transver-
se sections of the last chamber, 3 — view of the transverse sectio-
ned nuclei. Ma — somatic nucleus, Mu — generative nuclei; xl20.
(Воронова, Селиванова, 1976). Полигеномное ядро от-
мечено также для Psammophaga simplora (Arnold,
1984) и Saccammina alba (Goldstein, 1988). Механизмом
создания полиплоидного ядра служит, вероятно, не до-
веденное до конца деление. Дифференцировку ядер и
их последующую полимеризацию и полиплоидизацию
можно рассматривать как важный шаг в эволюции ядер-
ного аппарата, ведущий к интенсификации его функций
и повышению метаболической активности.
Гетероморфизм ядер протистов встречается еще
только у инфузорий, однако у них он имеет сущест-
венные отличия (см. раздел «Место фораминифер в
системе протистов»).
Митозы фораминифер происходят по типу закры-
того внутриядерного плевромитоза (Myers, 1935;
Zech, 1964; Воронова, 19786; Райков, 1978), но могут
иметь и некоторые черты ортомитоза (Cesana, 1978).
Во время гаплоидных митозов у Cibicides lobatulus
(Воронова, 1976; Воронова, Михалевич, 1985) и у
других видов (Le Calvez, 1938; Schwab, 1969, 1973,
1976) отмечены центросомы (рис. 541, 1) ицентрио-
ли (рис. 543, 7). Интересно, что у жгутиковых дочер-
них клеток гамонтов Myxotheca arenilega центросомы
с центриолями, а у безжгутиковых агамет — без них.
Клеточные центры деления наиболее полно изучены
у Iridia lucida (Cesana, 1978; рис. 541, 7, 2). Центро-
сомы с центриолями, как бы охваченные снизу плот-
ной акроплазматической чашечкой, образуются сна-
ружи от ядерной оболочки. В этом месте на ядерной
оболочке появляются довольно крупные окна. Затем
напротив центросом, внутри от ядерной оболочки, по-
является слой плотной плазмы, который служит цент-
ром организации микротрубочек веретена (ЦОМТ),
идущих к кинетохорам хромосом (полуверетена). На
поздних стадиях митоза полуверетёна расходятся к
противоположным полюсам ядра. Часть микротрубо-
чек соединяет оба ЦОМТа между собой, образуя
внутриядерное центральное (латеральное) веретено,
лежащее параллельно ядерной оболочке. Мейоз фо-
раминифер изучен наиболее подробно Вороновой
(19786) у ядер агамонта Cibicides lobatulus (рис. 543,
2). Ею установлена предмейотическая дупликация ДНК,
двухступенчатый характер мейоза; на стадии дипло-
тены (рис. 543, 2, Е) наблюдались хиазмы. Хиазмы во
время мейоза отмечены также у Patellina corrugata
(Berthold, 1977). В общих чертах мейоз фораминифер
протекает, как у большинства простейших и много-
клеточных, но как у Cibicides lobatulus, так и у других
видов, у которых исследовался мейоз {Patellina corru-
gata, Iridia lucida), не выражена экваториальная плас-
тинка и практически отсутствует выраженная метафаза.
Ядерный аппарат фораминифер претерпевает силь-
ные изменения на протяжении их сложного жизнен-
ного цикла.
Жизненные циклы
Среди разных групп эукариот, у которых имеется
смена ядерных фаз, фораминиферы занимают срав-
Рис. 541. Строение делящихся ядер и гамет фораминифер.
7 — генеративное ядро донной Iridia lucida (Astrorhizata), х 30000, в начале деления, хр — хроматин, ц — центросомы. 2 — дальнейший процесс деления
ми того же вида, х 64000. м — митохондрия, не — нити веретена, хр — хроматин, цомт — центр деления, яп — ядерные поры. 3 — гамета Iridia lucida
(Astrorhizata), х 41000, продольное сечение, в — вакуоль, xi — поперечный срез инкорпорированного жгутика, Ж2 — продольный срез жгутика, м —
митохондрия, мт — микротрубочки, ц — центриоль, я — ядро. 4 — гамета планктонной Hastigerina pelagica (Globigerinata), х 4500. 5 — участок стенки
молодой камеры Heterostegina (Rotaliana), х 154000. с — стенка, слева внутренняя цитоплазма, справа — наружная, сом — срединная органическая мемб-
рана. 7, 2 — схема по: Cesana, 1978; 3 — схема по: Cesana, 1972; 4, 5 — по: Spindler et al., 1978 (5 — схема).
Fig. 541. Structure of regenerating Foraminiferal nuclei and gametes.
7 — generative nucleus of Iridia lucida (Astrorhizata), x 30 000 at the beginning of division, ц — centrosomes, xp — chromatin. 2 — further process of development
of generative nucleus of the same species, x 64 000. цомт — center of division, xp — chromatin, яп — nuclear pores, не — spindle threads, м — mitochondria.
3 — gamete of Iridia lucida (Astrorhizata), x 41 000, longitudinal section, ц — centrioles, xi — transverse section of the flagellum incorporated, X2 — longitudinal
section of flagellum, м — mitichondria, я — nucleus, мт — microtubules, в — vacuoles. 4 — gamete of planktonic Hastigerina pelagica (Globigerinata), x 4500.
5 — part of the shell wall of the young chamber of Heterostegina, x 154 000. c — wall, inner cytoplasm from the left, outer cytoplasm — from the right, com —
middle (primary) organic membrane.
568
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
2
ТИП FORAMINIFERA. ЖИЗНЕННЫЕ ЦИКЛЫ
569
нительно примитивное место. Их жизненные циклы
относятся к гетерофазным, т. е. гапло- и диплофазы
составляют примерно равные части цикла (антитети-
ческая смена поколений) с некоторым превалиро-
ванием гаплофазы. Между первым агамным поколени-
ем (Aj) и гамогонией (В) у некоторых видов могут
«вклиниваться» повторные агамные поколения (А2, А3).
Таким образом, схему чередования поколений изо-
бражают как А—В—А—В либо как Aj—А2—А3—В.
Дополнительные агамные поколения известны у мно-
гих, низших однокамерных форм (например, у Allog-
romid) и у псевдодвухкамерной Spirillina vivipara.
Грелль (Grell, 1979) не наблюдал подобного повторе-
ния у высших роталиат, однако аналогичное явление
отмечено у Discorbis orbicularis и Discorinopsis agua-
gui', у Ammonia tepida агамные поколения многократно
повторялись, особенно в культуре (Goldstein, Mood-
ley, 1993). В культуре же у некоторых видов (напри-
мер, у Elphidium crispum, — Hofker, 1930) происхо-
дит полное выпадение стадии гамонта и наблюдается
длительное агамное размножение. Повторные агам-
ные поколения были отмечены также для милиолят
(Cornuspira, Calcituba, Spiroloculina). Жизненные
циклы с повторными агамными поколениями были
названы апогамными (МсЕпегу, Lee, 1976).
У одного из планктонных видов, напротив, наблю-
далась только половая часть цикла (Be, 1982). Воз-
можно, несмотря на интенсивное изучение этого ви-
да, его агамонты пока просто не обнаружены.
Фораминиферы — почти единственная группа
животных, произошедших от жгутиконосцев, кото-
рая сохранила в своей эволюции такое соотношение
гапло- и диплофаз. Среди низших растений анало-
гичная форма жизненного цикла широко распрост-
ранена. Описана она также у миксоспоридий (Успен-
ская, 1982). Сохранение в длительной эволюции фо-
раминифер гетерофазной смены поколений можно
рассматривать как дополнительное свидетельство их
происхождения от жгутиковых.
К настоящему-времени примерно у 100 видов
фораминифер в той или иной степени изучены жиз-
ненные циклы, в том числе у 26 из них при помощи
электронного микроскопа. Полные жизненные цик-
лы установлены у 20 видов. Лишь для 6 из них были
Рис. 543. Процессы деления в ядрах Cibicides lobatulus (Rotalia-
na).
1 — митоз ядер агамет С. lobatulus (рисунки с препарата): А — появление
центросомы (ц) и начало конденсации хромосом, Б — начало деления (про-
фаза) (ц — центриоли, не — нити веретена). 2 — мейоз ядер агамонта
С. lobatulus: А — пролептотена; Б — лептотена; В — зиготена; Г, Д — па-
хитена; Е — диплотена, 3 хромосомы с хиазмами, в центре ядра простое
пересечение хромосом; Ж — диакинез; 3 — метафаза I; И — анафаза I;
К — телофаза I, Л — профаза II, М — анафаза II; Н — телофаза II. 1 —
по: Воронова, Михалевич, 1985; 2 — по: Воронова, 19786.
Fig. 543. Processes of division in the nuclei of the Cibicides lobatu-
lus (Rotaliana).
7 — mitose of the agametes nuclei of C. lobatulus (drawings from the prepara-
tion): A — appearing of the centrosome (ц) and the beginning of chromosome
condensation, Б — beginning of the division (prophase) (ц — centrioles, не —
spindle threads). 2 — meiosis of the agamont nucleus of the C. lobatulus: A —
proleptotena, Б — leptotena, В — zigotena, Г, Д — pachitena, E — diplotena,
three chromosomes with chiasmata, in the center of the nucleus the mere cros-
sing of the chromosomes, Ж — diakinesis, 3 — metaphase I, И — anaphase I,
К — telophase I, Л — prophase II, M — anaphase II, H — telophase II.
проведены электронно-микроскопические исследова-
ния ядер (Cibicides lobatulus, — Воронова, 1976,
19786; Allogromia laticollaris, — Schwab, 1971; McE-
nery, Lee, 1976; Myxotheca arenilega, — Foeyn, 1936;
Schwab, 1969; Ovammina opaca, — Dahlgren, 1964,
1967a; Saccammina alba, — Goldstein, 1988, Tro-
chammina sp., — Pawlowski et al., 1995b).
Жизненный цикл фораминифер чаще всего годич-
ный, а его стадии обычно — сезонны. Зимой, весной
и летом, как правило, наблюдается бесполое размно-
жение — агамогония, в результате чего из агамет об-
разуются гаплоидные гамонты (рис. 544, 9—13). Га-
могония у большинства изученных фораминифер на-
чинается осенью, с понижением температуры, но
может встречаться и летом; длительность ее состав-
ляет 24—30 ч. Гамогония — половой процесс, при
котором гамонт образует гаметы, которые затем сли-
ваются и дают начало агамонту (или шизонту, или
диплонту). При этом у некоторых видов (Patellina
corrugata, многие высшие роталиаты) родительские
гамонты объединяются попарно пупочными* Сторона-
ми в сизигий (рис. 525, 10, А), стенка ракойины меж-
ду ними растворяется, и гаметы копулируют в замк-
нутом пространстве. Такой способ размножения
называют гамонтогамным, или пластогамным (напри-
мер, у Discorbis patelliformis, Rosalina leei, Glabratel-
la sulcata, у разных видов Bolivinella). Связывающая
материнские раковины мембрана в отличие от обыч-
ных раковинных мембран состоит из не содержащих
сульфаты мукополисахаридов (Lipps, Erskian, 1969).
Снаружи сизигий окружается плотной органической
мембраной. У Patellina corrugata могут образовывать-
ся агрегаты из нескольких раковин. У ряда плас-
тогамных видов в ходе эволюции возникла аутога-
мия — копуляция гамет, образующихся у одной
особи (Rotaliella heterocariotica, R. roscoffensis). Про-
цесс образования гамет изучен у гамонтогамных
видов более полно, чем у гаметогамных, когда гаметы
копулируют в воде (Schwab, 1969, 1973; Воронова,
1976, 1978а, 19786; Cesana, 1978; Arnold, 1982, 1984;
Grell, 1988). Число гамет в этом случае достигает не-
скольких десятков тысяч и в десятки, а иногда и бо-
лее раз превосходит их количество у пластогамных
видов.38 У одного из гаметогамных видов (Myxotheca
arenilega) также доказана аутогамия. У Hastigerina
pelagica (Spindler et al., 1978) иногда после множест-
венного деления и образования гамет остатки цито-
плазмы родительских особей регенерируют, восста-
навливают растворенные камеры и продолжают су-
ществование и даже рост. Такие же наблюдения
имеются и для других видов (Spiroloculina hyalina, —
Arnold, 1955; Bolivina doniezi, — Sliter, 1970). Bee
это свидетельствует о высокой регенеративной спо-
собности фораминифер.
У многих низших форм с тектиновой раковиной
(Lagynana) наряду с обычным жизненным циклом
встречается еще и почкование, и цитотомия (множес-
твенное почкование — например у Allogromia), и про-
стое деление надвое. Агамогонию можно считать сле-
дующим этапом бесполого размножения (множествен-
38 Особенно велико число гамет у представителей пелагических
родов.
570
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТИП FORAMINIFERA. ЖИЗНЕННЫЕ ЦИКЛЫ
571
ное деление). В процессе эволюции произошли прогрес-
сивные изменения и в половой части цикла — от изога-
мии (Glabratella patelliformis) к анизогамии {Rubratella),
иногда с избирательной сексуальной активностью гамет.
Так, у Rubratella intermedia гамонты разных, а не оди-
наковых размеров. Гаметы их тоже анизогамные: по-
томство от более мелких гамонтов имеет более мелкие
гаметы с более мелкими ядрами. Сексуальная диффе-
ренциация изучена экспериментально у Metarotaliella
parva (Weber, 1965). Молодые гамонты не могут спа-
риваться друг с другом, а только с более взрослыми и
зрелыми. Взрослые гамонты могут друг с другом спа-
риваться, но их реактивность по отношению друг к другу
стимулируется более молодыми гамонтами (Lee, 1990).
Рассмотрим жизненный цикл на примере наибо-
лее полно изученного гаметогамного вида Cibicides
lobatulus. Начнем с агамогонии (рис. 544, 7, 2).
После слияния двухжгутиковых гамет образуется
диплоидная зигота, вырастающая в молодой агамонт,
ядро которого сразу приступает к диплоидным мито-
зам. В результате первого деления образуется 2 ядра
(рис. 544, 3); одно из них становится соматическим,
или вегетативным, — Ма. Оно увеличивается в раз-
мерах, обогащается кариолимфой, в нем появляются
богатые РНК ядрышки (нуклеолы). Из второго ядра
в результате последующих делений возникает снача-
ла 2, а затем до нескольких десятков генеративных
ядер — Ми. В процессе агамогонии Ма увеличивает-
ся в размерах и продвигается из камеры в камеру по
направлению к центру, а Ми мигрируют от центра к
периферии, к последним камерам, где и находятся до
конца. На конечной стадии они проходят 1-е и 2-е
редукционные деления (двухступенчатый мейоз), в
результате которых образуются гаплоидные ядра.
Интересно отметить, что в результате асинхрон-
ности мейозов в клетке одновременно существуют и
диплоидные и гаплоидные ядра. Возможно, это явле-
ние широко распространено у многоядерных про-
стейших. В мейозе участвуют только Ми. Хотя в
некоторых случаях в Ма можно наблюдать как бы
попытки совершить мейоз, он не бывает доведен до
конца. При этом Ма либо только увеличйвается в
размерах, либо, как у Rotaliella roscoffensis и неко-
торых других видов (Grell, 1979), в нем происходит
конденсация хромосом и образование внутриядерно-
го веретена. Хромосомы остаются унивалентными.
После разрушения ядерной оболочки они выходят в
цитоплазму, становятся пикнотическими и исчезают.
После мейоза в Ми могут происходить еще пов-
торные гаплоидные митозы, обеспечивающие огром-
ное количество молоди у гаметогамных видов. К
концу массовых делений микронуклеусов, обычно
после повторных митозов, Ма дегенерирует — теря-
ет отчетливые очертания, а затем распадается на от-
дельные фрагменты, которые растворяются в цито-
плазме. Вокруг вновь образовавшихся гаплоидных
Ми обособляется цитоплазма и формируется ракови-
на — это агаметы, дающие начало гамонту — по-
ловому поколению, которое и вступает в процесс
гамогонии: ядра взрослых гамонтов делятся (гапло-
идные митозы) и дают начало гаметам.
Таков жизненный цикл этого вида, в общих чер-
тах отражающий жизненные циклы других высших
фораминифер. Он отличается от последних только
тем, что на стадии гамогонии у него, так же как и на
стадии агамогонии, образуется Ма (иногда на стадии
формирования первой камеры агаметы, но чаще на
стадии 4—5 камер). Во время серии последователь-
ных гаплоидных митозов гамогонии у генеративных
ядер Ма становится лентообразным, четковщщым (рис. 540,
3) и продвигается к 5-й и 6-й камерам, увлеченный
токами цитоплазмы, при этом части его могут отры-
ваться. К концу митозов гамогонии он дегенерирует.
Агамонты всех изученных видов роталиат, глобиге-
ринат и милиолят многоядерные и гетерокариотные.
Ядерный дуализм гамонтов наблюдался только у Cibi-
cides lobatulus. Наличие его подтверждено цитофото-
метрически, и от ядерного дуализма агамонтов он от-
личается тем, что возникает на гаплоидной основе.
Ввиду чрезвычайной сложности исследования
ядерных процессов у фораминифер и интерпретации
полученных цитологических данных, выполненных
на разных уровнях исследования, ряд ценных работ
по цитологии и жизненным циклам фораминифер
(особенно проведенных на световом уровне, но иногда
даже и на электронно-микроскопическом) не дает
полного представления о сущности происходящих
ядерных процессов. Поэтому относительно некото-
рых видов мы не можем точно сказать, гетерокари-
отны они или гомокариотны и на какой стадии жиз-
ненного цикла. Образование ядер будущих гамет у
гамонта из единственного ядра в большинстве работ
не прослежено, что объясняется значительными ме-
тодическими трудностями: до гамогонии цитоплазма
еще не очищена от включений и Ма, занимающий
не более одной миллионной объема особи, трудно
отличить от ядер инородных организмов (остатков
пищи, комменсалов, симбионтов); в популяции мо-
гут встречаться особи с разрушенными Ми и амик-
ронуклеарные; вообще в цитоплазме нередко отме-
чается то или иное количество дегенерировавших
ядер разного происхождения; окраска метиловым зе-
леным пиронином и железным гематоксилином мас-
кирует ядра.
При морфологическом сходстве агамонтов и га-
монтов многоядерные особи могут быть приняты за
Рис. 544. Жизненный цикл Cibicides lobatulus (Rotaliana).
1 — гаметы (л); 2 — зигота (2л); 3, 4 — однокамерные агамонты (2л) (5 — двухъядерный, 4 — трехъядерный); 5—8 — растущие многокамерные агамон-
ты с Ма и множественными мелкими Ми. мигрирующими от центральных камер к периферии (R\. Ri — 1-е и 2-е редукционные деления, Ма — дегенери-
рует), ц — циста вокруг агамонта (л); 9 — агаметы (л), вышедшие из цисты; 10—13 — растущие гамонты (л).
Fig. 544. Life cycle of the Cibicides lobatulus (Rotaliana).
1 — gametes (л); 2 — zygote (2л); 3, 4 — unilocular agamonts (2л) (3 — binucleate, 4 — threenucleate), 5—8 — growing multinucleate agamonts with the
macronucleus (Ma) and multiple small micronuclei migrating from the central chambers to the periphery (R\, Ri — 1-st and 2-nd reductional (meiotic) divisions,
Ma is degenerating), ц — cyst around the agamont (л); 9 — agametes (л), having leaving the cyst; 10—73 — growing gamont (л).
572
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
агамонтов. Ми агамонтов ярче и легче идентифициру-
ются. Лишь окраска по Фёльгену полной серии до-
статочно тонких срезов дает возможность выявить все
ядра, а цитофотометрия позволяет подтвердить их
принадлежность к тому или иному поколению. Но
твердые включения в цитоплазме фораминифер за-
трудняют получение полной серии срезов, а цитофо-
тойетрические исследования пока единичны, поэто-
му некоторые цитологические наблюдения могут в
дальнейшем не подтвердиться, как не подтвердилась
«хромидиальная» теория (Le Calvez, 1938).
Первоначально считалось, что генеративные ядра
формируются из так называемых хромидиев, сущест-
вующих наряду с главным ядром. При определенных
способах приготовления ядерных срезов ядерный ап-
парат С. lobatulus предстает в виде сочетания Ма и
«хромидия». Последний на самом деле содержит по
крайней мере 3 компонента: микронуклеусы, остатки
дегенерировавших ядер, остатки ядерного материала
инородных организмов (Воронова, Михалевич, 1985).
В толстых срезах эти компоненты накладываются
друг на друга. В настоящее время можно утверждать,
что в ряде случаев хромидиальная сеть представ-
ляет цитоплазматическую РНК (рибосомальная сеть).
Плотные «хромидии», состоящие из ДНК, по-види-
мому, и есть генеративные ядра, так же как и «сфе-
рические тельца», описанные у Polistomella (=Elphi-
dium) (Lister, 1895). Все вышеизложенное дает осно-
вание предполагать, что ядерный дуализм гамонтов
фораминифер, обнаруженный у Cibicides lobatulus,
на самом деле, возможно, распространен в этой груп-
пе простейших гораздо шире.
Исследование ядер и жизненных циклов не изу-
ченных пока в этом отношении представителей клас-
са Nodosariata, обладающих своеобразными типами
строения раковин и ультраструктурой ее стенки, яв-
ляется первоочередной задачей, поскольку сущест-
венно дополнит общую картину и, вероятно, сможет
пролить свет на филогенетические связи этой группы.
Представители однокамерных Astrorhizata с орга-
нической или агглютинированной раковиной и свое-
образной примитивной по строению группы Spirilli-
nata относятся к гомокариотным фораминиферам. В
их жизненном цикле обычен одноядерный гамонт,
дающий ядра многочисленных гамет, и многоядерныи
агамонт с однородными ядрами без дифференциации
Ма и Ми. У Iridia lucida (Le Calvez, 1936, 1953;
Cesana, 1972, 1975, 1978) агамонт остается одноядер-
ным вплоть до начала образования агамет. У других
агамонтов Lagynana и Astrorhizana одноядерный пе-
риод в жизни агамонта хотя и короткое время, но все
же существует в отличие от изученных роталиат, где
практически одноядерна лишь зигота (Воронова, Ми-
халевич, 1985). Гольдштейн (Goldstein, 1988) отмечает,
что из 40—70 ядер агамонта образуется лишь 3—
30 агамет, остальные ядра погибают. Гольдштейн ос-
торожно называет их негенеративными; никаких дру-
гих различий между ними и Ми она не обнаружила.
По-видимому, здесь мы встречаемся с явлением, от-
меченным и у других фораминифер (Allogromia lati-
collaris, — Schwab, 1976; Patellina corrugata, — Ber-
thold, 1971; Cibicides lobatulus, — Воронова, Миха-
левич, 1985; Михалевич, 1999), когда часть ядерного
и цитоплазматического материала погибает и не ис-
пользуется непосредственно при развитии потомства.
У планктонной Hastigerina pelagica эти остатки со-
держали вакуоли, несколько ядер и жгутики и об-
разовывали сферические тельца, способные двигаться
(Spindler et al., 1978).
Арнольд (Arnold, 1984) подробно исследовал свое-
образие формирования Ми у гамонта Psammophaga
simplora. Сравнение этой стадии жизненного цикла
данного вида с аналогичной фазой гетерокариотного
гамонта Cibicides lobatulus, а также с гетерокариотны-
ми агамонтами других видов показывает, что гамого-
нию Psammophaga simplora все же нельзя отнести к
гетерокариотному типу, поскольку (как и указывает
автор) Ми, образующиеся в первичном ядре, последо-
вательно сменяют его, а не существуют одновременно.
Большую часть жизни у гамонта Р. simplora имеется 1
ядро. После выхода генеративных ядер из-под оболоч-
ки первичного ядра в последнем практически не оста-
ется хроматинового материала и оно существует еще
некоторое время не как полноценное ядро, а как его
остаток. Здесь мы опять сталкиваемся с неполным ис-
пользованием ядерного вещества при формировании по-
томства — как бы с потерей части его. При истинной
гетерокариотности на протяжении основной части су-
ществования агамонта (у Cibicides lobatulus и гамонта)
Ма и Ми сосуществуют одновременно.
Исследования Арнольда еще раз показали слож-
ность и многообразие ядерных изменений в процес-
се жизненного цикла фораминифер и необходимость
дополнительных исследований. И все же анализ име-
ющихся на сегодняшний день сведений по жизнен-
ным циклам примитивных однокамерных фораминифер
не позволил пока обнаружить у них гетерокариот-
ность; была четко установлена лишь многоядерность
агамонтов. Исключение среди Lagynana представляет
Phthanotrochus arcanus (Arnold, 1978), у которого с
некоторой долей сомнения автор называет более
крупные дегенерирующие ядра (8—14 мкм) Ма. Они
существуют одновременно с множеством более мел-
ких (1.6—2.4 мкм) Ми.
Ph. arcanus в отличие от остальных Lagynana
имеет хотя и органическую, но многокамерную тро-
хоидную раковину роталоидного типа. Если ядерный
дуализм у данного вида действительно существует, то
это подтверждает его родство с другими морфологи-
чески сходными известковыми секреционными рако-
винами роталиат и проливает свет на происхождение
последних.
Систематическое положение Ph. arcanus в этом
случае должно быть изменено (ближе к низшим пред-
Рис. 545. Различия в строении микро- (А) и мегалосферических (5) раковин у одного и того же вида.
Д А, Б — у Hormosinana (Reophax)', 2—4 — у Textulariana (2, А, Б — Textilina, 3, А, Б — Norvanganina, 4, А, Б — Textella)', 5—6 — у Nodosariana (5, А,
Б — Dimorphina, б, А, Б — Amphicoryna); 7—8 — у Rotaliana (7, А, Б — Rectobolivina, 8, А, Б — Jadammina).
ТИП FORAMINIFERA. ЖИЗНЕННЫЕ ЦИКЛЫ
573
Fig. 545. Differences in the structure of the micro- (A) and megalospheric (Б) shells of the same species.
1, А, Б — in Hormosinana {Reophax), 2—4 — in Textulariana (2, А, Б — Textilina, 3, А, Б — Norvanganina, 4, А, Б — Textella), 5—6 — in Nodosariana (5,
А, Б — Dimorphina, 6, А, Б — Amphicoryna}, 7—8 — in Rotaliana (7, А, Б — Rectobolivina, 8, А, Б — Jadammind).
574
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ставителям роталиат с агглютинированной ракови-
ной — к подклассу Textulariana). Предположение
Арнольда (Arnold, 1979) о происхождении высших
секреционных форм от многокамерных органических
раковин сходного строения, развиваемое им на при-
мере Periptigma и Miliolata, получит в этом случае
еще одно подтверждение в параллельной группе фо-
раминифер.
Обзор всех известных в настоящее время данных
по жизненным циклам фораминифер позволяет раз-
бить их на 3 группы (Воронова, Михалевич, 1985;
Михалевич, 1999).
1. С гомокариотными многоядерными агамонтами
и одноядерными гамонтами (Lagynana, Astrorhizana,
Spirillinata); исключение составляет Iridia lucida с од-
ноядерным агамонтом.
2. С гетерокариотными агамонтами и одноядерны-
ми гамонтами (Miliolata, большинство известных Ro-
taliata, Globigerinata).
3. С ядерным дуализмом как агамонтов, так и га-
монтов (некоторые Rotaliata: Cibicides lobatulus', воз-
можно, Elphidium).
Ясно, что первый из них является исходным
типом, что подтверждается работами Грелля и его
школы (Weber, 1965; Grelle, Bardele, 1977; Grell,
1979, 1988), в которых прослежены разные этапы
дифференцировки соматических ядер: от более при-
митивных, когда Ма во время мейоза не только уве-
личивается в размерах, но в нем происходит конден-
сация хромосом (хотя и остающихся унивалентными)
и даже формируется веретено (Rotaliella roscoffen-
sis), как бы не до конца повторяя процессы, проис-
ходящие в микронуклеусах, до простого увеличения
в размерах с увеличением количества ДНК (7?. hete-
rocariotica). Дифференциация ядер у Cibicides loba-
tulus зашла достаточно далеко (Воронова, Михале-
вич, 1985; Михалевич, 1999). Вероятно, именно
кратное увеличение ДНК Ма, происходящее парал-
лельно с его увеличением в микронуклеусах, при не
доведенном до конца процессе деления в первых в
отличие от вторых и объясняет механизм создания
полиплоидного ядра.
В гаметогенезе разных групп фораминифер име-
ются свои особенности. Ряд видов, приступая к раз-
множению, формирует цисты из слабо сцементиро-
ванного постороннего материала (Cibicides lobatulus,
Rosalina globularis) — такие же как у Rosalina flori-
dana при построении новых камер. У агглютиниро-
ванной однокамерной Cribrothalamina alba в стенке
раковины образуются 10—15 крупных отверстий —
«пор», дающих выход гаметам (Goldstein, Barker, 1988).
У донных роталиат гаметы могут выходить как через
расширенные резорбцией поры (Cibicides lobatulus),
так и через устье (Ammonia tepida), у планктонных
форм — только через широкие устья. Пелагические
формы имеют еще ряд особенностей. Выход гамет у
них может быть связан с лунными циклами (напри-
мер, у Hastigerina pelagica, — Spindler, 1979).
При гаметогенезе происходят большие изменения
и в скелете, — в результате вторичной гаметогене-
тической кальцификации ячеистая у многих планк-
тонных фораминифер ультраструктура поверхности
сглаживается (535, 4, Б; 536, 4, Б), стенка становится
толще, поры забиваются известью почти полностью,
раковина сбрасывает подвижно сочлененные с нею
иглы (рис. 531, 9; 539, 7) и погружается на дно (Be et
al., 1976; Be, 1980). Есть виды, которые образуют
последнюю репродуктивную камеру особой формы
(Globigerinoides sacculifer, — рис. 539, 7). Нечто по-
добное можно встретить и у донных форм (Sorites
orbicularis, — Kloss, 1980). Среди донных имеются
виды с кратковременной пелагической стадией в их
жизненном цикле. Наиболее яркий пример — плаву-
чие гамонты Tretomphalus (рис. 525, 10, Б) и Cymba-
loporetta с шарообразной камерой внизу раковины.
Их агамонтов без такой камеры относили к роду Ro-
salina. В период гаметообразования гамонт Tretom-
phalus bulloides инцистируется. Перед самым выходом
гамет он формирует большую флотирующую, напол-
ненную газом камеру снизу трохоидной раковины,
освобождается из цисты и всплывает к поверхности
воды. Там двухжгутиковые гаметы покидают материнс-
кую раковину и копулируют в воде. Зигота снова
опускается на дно. На этом пелагическая часть цикла
заканчивается. Пелагические стадии донных форм спо-
собствуют более широкому их расселению (Михалевич,
1983; Воронова, Михалевич, 1985).
Наличие 2 ядерных фаз в жизненном цикле фо-
раминифер проявляется не только в цитологических
особенностях, но также и в строении скелета. Как и
у других организмов с подобной сменой поколений,
она может быть изоморфной и гетероморфной, так
как обе формы эволюируют в значительной степени
автономно. Агамонты большинства фораминифер
имеют микросферическую, гамонтьг— мегалосфери-
ческую раковину (у спириллинат соотношение об-
ратное). В простейшем случае они отличаются лишь
размерами начальной камеры — пролокулуса (что и
отражено в их названии) и общими размерами: мик-
росферические агамонты крупнее макросферических
гамонтов. Примером могут служить Cibicides loba-
tulus, Rotaliella heterocariotica, Ammonia beccarii, Ja-
dammina zernovi (рис. 545, 8). Если агамное поколе-
ние повторяется дважды, появляются микросфери-
ческие агамонты (А{) и макросферические агамонты
(А2) (например, y^Elphidium crispum, Heterostegina
depresso). У ряда высокоорганизованных групп фо-
раминифер с очень большим числом камер (напри-
мер, у некоторых Nummulitacea подкласса Rotalia-
na — Biplanispira) 2 начальные камеры мегалосфе-
рического гамонта (пролокулус и дейтероконх) очень
крупные, иногда крупнее остальных (рис. 532, 1, 2,
А, Б). Обе они окружены общей неперфорированной
стенкой, отличающейся от стенки остальной ракови-
ны, имеют 1 центральную крупную округлую пору и
ряды пор в основании септы. Их называют зароды-
шевыми камерами, илц эмбрионами.
Как у агглютинирующих, так и у секретирующих
видов наблюдается и более резкое проявление гетеро-
морфизма (Воронова, Михалевич, 1985; Михалевич,
1999; рис. 525, 75; 545, 7—7; 547, III, 17, IV, 13, V, 2).
Раковины разных генераций могут отличаться друг от
друга очертаниями, общими пропорциями, соотноше-
нием разных отделов, наличием какого-нибудь отдела
лишь у одной из них. Например, часто у выпрямлен-
ных форм в разных таксонах начальный плоскоспи-
ТИП FORAMINIFERA. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
575
ральный трохоидный или клубковидный отдел имеет-
ся у агамонтов, но отсутствует у гамонтов. При этом
бесполое поколение с микросферической раковиной
часто повторяет более примитивные предковые чер-
ты. Таким образом, по онтогенезу раковин можно су-
дить о предшествующих этапах филогенетического
развития группы.
Строение гамет
Гаметы подавляющего большинства форамини-
фер двухжгугиковые, с жгутиками различной длины
(рис. 541, 3, 4\ 544, 7), содержат ядро, жировые
капли, митохондрии, осмиофильные гранулы, микро-
тельца, диктиосомы аппарата Гольджи; у видов с
избирательной сексуальной активностью — акросо-
му (Cesana, 1972).
У Glabratella и Nummulites venosus гаметы трех-
жгугиковые. Гаметы с таким числом жгутиков не
встречаются у саркодовых и вообще чрезвычайно
редки у простейших (у некоторых Sporozoa). У аме-
боидных гамет {Patellina corrugata, Spirillina vivipa-
ra, Metarotaliella simplex, ряда Rotaliella) отсутствие
жгутика вторично. Он утрачен, так как в нем нет
необходимости при гамонтогамном способе размно-
жения. Гаметы некоторых милиолид имеют образо-
вание, внешне сходное с гаптонемой, — прямой,
торчащий вперёд аксостиль; ультраструктура его до
сих пор не исследована. Жгутики же фораминифер
имеют на поперечном срезе обычное для всех жгу-
тиков строение (2 центральные и 9 пар перифери-
ческих фибрилл) (рис. 79). У Boderia turneri (Hedley
et al., 1968) и Operculina cf. complanata (Roettger et
al., 1998) один из жгутиков имеет поверхностные
волоски — мастигонемы.
Основные направления
эволюции
Несмотря на многочисленные классические ис-
следования (Догель, 1929; Полянский, 1970; Полян-
ский, Райков, 1977), морфологические закономернос-
ти эволюцйи на уровне одноклеточных разработаны да-
леко не так основательно, как у многоклеточных.
Скелет фораминифер отражает основной план стро-
ения одноклеточного организма; в процессе эволю-
ции группы он подвергся наиболее сильным изме-
нениям. Принципиальное значение для понимания
эволюции группы и построения ее системы имеет во-
прос о том, какие признаки считать первичными,
какие — возникшими параллельно и независимо в
разных группах, а какие — конвергентно; при этом
мы исходим из положений Шмальгаузена (1969).
Изменения скелета фораминифер происходили в 2
главных направлениях — формы раковины и ее стен-
ки. Это — два взаимосвязанных процесса. Для удоб-
ства изложения рассмотрим каждый из них в отдель-
ности.
Одна из основных особенностей эволюции скеле-
та фораминифер — возникновение многокамернос-
ти, отражающей периодический, неравномерный ха-
рактер роста. Первые проявления этой тенденции
встречаются даже у раковин с органической стенкой.
В каждой из основных линий развития этот процесс
получил дальнейшее развитие. Его можно рассматри-
вать как процесс полимеризации, столь распростра-
ненный у одноклеточных. В эволюционном плане
многокамерность фораминифер можно сравнить с
возникновением многоклеточности, когда повыше-
ние сложности организма идет за счет полимериза-
ции внутриклеточных компонентов, а не за счет по-
лимеризации клеток (Михалевич, 1999).
Из 4 типов полимеризации (Подлипаев и др.,
1974) у фораминифер мы наблюдали 2 (рис. 546, 7):
1) фиссацию (разделение первоначально имев-
шихся образований, — например, у Calligella, Рага-
thikinella, Ammoscalarid)\
2) дополнительную полимеризацию (добавление
новых гомологичных единиц, — как типичные при-
меры — реофаксы, квинквелокулины, нодозарии, но-
нионы, дискорбисы). Подавляющее большинство фо-
раминифер пошли по второму пути; первый характерен
для примитивных групп (подклассы Laginana, Astror-
hizana и Ammodiscana). Возникновение многокамер-
ное™ по способу фиссации при непрерывном росте
(в отличие от прерывистого, достроечного роста) мы
называем ложной многокамерностью (псевдомного-
камерностью); сочленение камер в обоих случаях
различное (Михалевич, 1981, рис. 2; рис. 546) и
может рассматриваться как признак высокого таксо-
номического ранга.
Возможно сочетание двух способов (многоэтаж-
ная полимеризация): подразделение на камерки внут-
ри сформированных камер в группах средней слож-
ности организации (Spirocyclinidae из литуолид, Phe-
nacophragma из эндотирид, своеобразные группы
Carterina, Patellina) и у наиболее высокоразвитых
многокамерных форм в линиях с различным типом
стенки — орбитолин (Textulariana, — рис. 532, 7),
фузулинид (Ammodiscana, — рис. 531, 7), милиолят
(вертикальные и тангенциальные септы альвеоли-
нид, — рис. 547, III, 16), высших роталиат (Hetero-
stegina, Cycloclypeus), где этот способ дополнитель-
ный, но не ведущий. Переход от камер, одинаковых
по форме и размерам (Saccamminopsis, Hormosinella,
Grigelis, — рис. 546, 2, А, Б; последний род выделен
на основании способа сочленения камер и их равной
величины, — Михалевич, 1981), к неодинаковым
(рис. 546, 2, В—Н), как это имеет место у большин-
ства видов, — следующий этап дифференциации на
базе предшествующей полимеризации. Его конеч-
ный результат — уменьшение числа камер в послед-
них оборотах, вплоть до 1 (рис. 546, 2, 3—Н), также
проходил параллельно в разных группах (Ammospha-
erulina из Литуолид, Periloculina из милиолят, Lage-
noglandulina из нодозариат, Orbulina из глобигери-
нат). Такого рода дифференциация ведет к умень-
шению автономности камер, подчинению их целому
организму, усложнению организменной структуры.
У высших многокамерных текстуляриан (орбитолина)
и роталиан (нуммулитес, бипланиспира, гетеросте-
гина), где число камер особенно велико, происходит
дальнейшая глубокая дифференциация камер, когда
начальные зародышевые и следующие за ними меди-
576
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
2
ТИП FORAMINIFERA. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
577
анные, экваториальные и периферические слои ка-
мер имеют различное строение (рис. 532).
Широко распространено мнение, что многокамер-
ные формы произошли от двухкамерных со второй
трубчатой камерой, например литуолиды от аммодис-
цид. Мы считаем, что хотя такой путь развития дей-
ствительно имел место, он представляет собой лишь
второстепенный, побочный путь (Михалевич, 1999).
Подобные формы возникали параллельно в разных фи-
логенетических ветвях. Они характеризуются рядом
примитивных черт, особым сочленением камер, со-
хранением трубчатой камеры на ранних стадиях раз-
вития (Toumayellidae, Lituotubidae, Patellinidae; многие
Miliolidae, — рис. 527). У большинства многокамерных
форм (нодозариаты, роталиаты) последующие камеры на-
растают сразу же вслед за округлым пролокулусом.
Анализ встречаемости разных типов раковин в
крупных таксонах (классах, подклассах, — Михале-
вич, 1981, 1999) показал, что, несмотря на парал-
лельное возникновение морфологических типов во
многих таксонах, их распределение и богатство не-
одинаковы в каждой ветви (рис. 547, I—V)- Наимень-
шее их число — у примитивных тектиновых и аг-
глютинированных форм (Astrorhizata — в основном
однокамерные), своеобразной боковой ветви сек-
реционных форм (Spirillinata), специализированных
планктонных форм (Globigerinata), наибольшее — в
подклассах Textulariana и Rotaliana, где почти каж-
дому морфологическому типу агглютинированных и
микрогранулярных раковин соответствует изоморф-
ный аналог с бифонтинальной стенкой (рис. 547, V).
В этих же таксонах получил наибольшее развитие в
отличие от спирально-плоскостных асимметричный
трохоидный тип раковин, обеспечивающий кратчай-
шее расстояние между начальными и последними ка-
мерами и создающий наибольшие филогенетические
возможности, особенно в случае возникновения до-
полнительных отверстий в пупочной области, которые
связывают цитоплазму с внешней средой. Трохо-
идные раковины встречаются и в классе спирилли-
ната. Однорядные раковины у роталиан — исключе-
ние (Stilostomella)-, они, скорее всего, являются вто-
рично-однорядными. Во всяком случае, по строению
устья и зуба эти животные хорошо отличаются от
однорядных нодозариат с радиальным или простым
устьем. Кроме обилия и разнообразия однорядных
форм (рис. 547, IV, 1—3, 8—12, 14) у нодозариат
встречаются особые полиморфиноидный (рис. 547,
IV, 19, 20) и плектофрондикуляриевый типы с шев-
ронообразно изогнутыми камерами (рис. 547, IV, 5,
16, 18). Из спирально свернутых раковин у нодо-
зариат представлен только плоскоспиральный тип
(рис. 538, Е; 547, IV, 21). Последний легко отличить
от плоскоспиральных роталиат по строению и пол-
ожению устья (у них оно конечное, а не в основании
устьевой поверхности).
Примитивные однокамерные раковины встреча-
ются лишь в низших таксонах и у низших пред-
ставителей милиолят и нодозариат (мы не считаем
ляген вторично однокамерными). У милиолят преоб-
ладают клубковидные типы, не представленные у
высших роталиат, трохоидный тип для милиолят
(рис. 547, III, 18, 19) — исключение. (В связи с этим
мы изменили место Fisherinidae в системе — пере-
вели их из Cornuspiracea в Peneroplidea). Исследова-
ние ультраструктуры стенки фишеринид (Михале-
вич, 1988) подтвердило их принадлежность к мили-
олятам.
У прикрепленных видов самых разных таксонов
(вплоть до высших) раковины приобретают непра-
вильную форму конвергентно (Haddonia, Manorella
из Placopsilinidae, Textulariana; Cornuspiramia, Nube-
culinopsis из Miliolata; Bullopora из Nodosarioida, No-
dosariana; Cibicidella, Vagocibicides, Miniacina из Ro-
taliata). Большое число параллелизмов дает развитие
устья и его структур (рис. 528; 547) в разных груп-
пах — от единичного устья к множественному и си-
товидному, от простых внешних и внутренних зубов
до сложных внутренних пластинок и трубок (Миха-
левич, 1981, 1999). В развитии устья также четко
прослеживаются процессы полимеризации и дальней-
шей олигомеризации. Множественное устье с нефик-
сированным положением в виде не имеющих четкой
формы отверстий у многих астроризат мы считаем
первично-множественным, а появление у них одного
фиксированного, морфологически оформленного
устья более крупного размера рассматривается как
прогрессивный процесс олигомеризации (Михалевич,
1995). Появление нескольких отверстий на устьевой
поверхности вместо одного у высших многокамерных
форм — новая ступень полимеризации. В этом слу-
чае возникает вторично-множественное устье.
Примеры постепенного усложнения устья в этом
направлении можно найти почти во всех крупных
Рис. 546. Процессы полимеризации, дифференциации и олигомеризации в эволюции фораминифер.
1 — способы полимеризации: фиссация (путем внедрения стенки во внутреннюю полость камер или пережимов стенки): А — Caligella, Б — Paratikhinella,
В — Ammoscalaria; дополнительная полимеризация (при достроечном росте): Г — Discammina, Д — Haplophragmoides, Е — Earlandinita, Ж — Dusenbury-
ina, 3 — Nodosaria. 2 — дифференциация камер и их последующая олигомеризация в разных филогенетических линиях (классах) фораминифер: А, Б —
камеры не дифференцированы по размерам (А — Saccamminopsis (Hormosinana), Б — Grigelis (Nodosariana)); В—Н — камеры дифференцированы по раз-
мерам (В — Reophax (Hormosinana); Г — Lunucammina (Nodosariana); Д — Saracenaria (Nodosariana); E — Trochammina (Textulariana); Ж — Neoconorbina
(Rotaliana); 3, И — Idalina (Miliolana); К — Daucinoides (Nodosariana); JI — Colaniella (Nodosariana); M — Ammosphaerulina (Textulariana); H — Orbulina
(Globigerinata). Ж—Н — олигомеризация камер на конечной стадии (вплоть до объемлющей последней камеры)). 1 — по: Михалевич, 1981; 2 — по: Ми-
халевич, 1999.
Fig. 546. Processes of polymerization, differentiation and oligomerization in the Foraminiferal evolution.
1 — modes of polymerization: fission: by the way of wall intrusions into inner chamber lumen or by the wall constrictions — subdivision of previously formed unit
(A — Caligella, Б — Paratikhinella, В — Ammoscalaria), additional polymerization (growth by the addition of the new units), Г — Discammina, Д — Haploph-
ragmoides, E — Earlandinita, Ж — Dusenburyina, 3 — Nodosaria, 2 — differentiation of the chambers and their subsequent oligomerization in different Forami-
niferal phylogenetic lineages (classes): А, Б — chambers not differentiated in dimensions (A — Saccamminopsis (Hormosinana), Б — Grigelis (Nodosariana)), В—
H — chambers differentiated in dimensions (B — Reophax (Hormosinana), Г — Lunucammina (Nodosariana), Д — Saracenaria (Nodosariana), E — Trochammina
( Textulariana), Ж — Neoconorbina (Rotaliana), 3, И — Idalina (Miliolana), К — Daucinoides (Nodosariana), JI — Colaniella (Nodosariana), M — Ammosp-
haerulina (Textulariana), H — Orbulina (Globigerinata); Ж—Н — oligomerization of the chambers at the final stage (up to the fully embracing last chamber)).
37 Протисты, часть 1
578
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
таксонах. Особенно много их среди Textulariana
(почти в каждом семействе литуолид, текстуляриид,
трохамминид, атаксофрагмиид можно составить та-
кие параллельные ряды), у многих Rotaliana и Milio-
lana, некоторых псевдомногокамерных и многока-
мерных Spirillinata (торнайеллиды, эндотириды, фузу-
линиды). У Nodosariana множественное устье образуется
очень редко (Cribrorobulina), что можно связать с пре-
обладанием у них лучистого устья (рис. 528, 3, а, б;
547, IV, 3, 11, 16, 21) — достаточно большого и уже
подразделенного радиальными ребрами. Интересно
отметить, что среди Rotaliana множественное и ситовидное
устье встречаются, как правило, у более примитивных
форм, не имеющих системы каналов (Cribrogloborotalia,
Poroeponides). При наличии этой системы устьевые
отверстия иногда совсем исчезают (некоторые эльфи-
дииды).
Процесс образования связанных с устьем же-
лобков и трубок привел к частичному или полному
обособлению цитоплазмы и ее дальнейшей глубокой
дифференциации. У роталиат можно проследить раз-
витие внутренних скелетных образований устья — от
коротких пластинок и язычков (Turrilinoidea, Episto-
minoides, Ammonia, Eponidella, Globorotalites) до же-
лобообразных пластинок (Siphogerinoides, Eouvigeri-
na, Orthokarstenia) и трубок и сложных зубных плас-
тинок (Bolivina, Brizalina, Uvigerina и особенно
Bulimina, Stainforthia), все более удлиняющихся и об-
разующих, как и у Osangularia и Colettes, сплошную
систему, которая соединяет все камеры. Эта система
взаимосвязанных внутренних устьевых элементов
функционально сходна с системой каналов, но не го-
мологична им. Устьевые трубки роталиат и нодозари-
ат (эллипсолагены, полиморфиниды) различаются по
строению и могут рассматриваться как конвергент-
ные. Устьевой зуб также возникает параллельно и не-
зависимо в нескольких группах. У Schlumbergerinana
он представлен примитивным, часто плохо заметным
образованием. Наибольшее развитие устьевой зуб по-
лучил у Milioloida, где у родов Cribrolinoides, Torto-
nella, Flintina (Михалевич, 1976, 1999; рис. 528, 3, г,
ж—и, м; 530) он достигает такой сложности, что мо-
жет быть назван зубной пластинкой (иного проис-
хождения, чем у Bulimina (Rotaliata)). Устьевые зубы
милиолид нередко множественны (Dentostomina, неко-
торые Spiroloculina, — полимеризация), могут быть
неодинаковых размеров и формы (Dentostomina —
дифференциация); изредка они сливаются в единое
образование (Tortonella, Cribrolinoides — дальнейшая
олигомеризация) (рис. 528, 3, г, ж, м). Морфологиче-
ское богатство скелетных структур устья связано с их
большой и разнообразной функциональной нагруз-
кой. Дополнительные шовные устья часты у низших
и особенно у высших Rotaliata (рис. 529, 3—6, 7—9;
547, V, 20); особенно большое развитие они полу-
чили у планктонных форм (рис. 525, 10, В; 529, 10;
546, 2, Н); встречаются у некоторых спириллинат
(рис. 529, 2; 547, II, 5), у милиолят — лишь в виде
исключения (рис. 529, 1); у Nodosariata не развиты
(кроме Delosina). Появление дополнительных усть-
ев — важный прогрессивный признак в развитии
группы, которому ранее не уделялось должного вни-
мания.
В эволюции протистов значительную роль играет
прогрессивное увеличение размеров тела. Однако
самые крупные особи — гиганты в мире протистов
(от 1 до 18 см) встречаются среди примитивных од-
нокамерных и двухкамерных форм. У многокамер-
ных форм рост сдерживается разобщенностью камер.
Системы, достигшие высокой степени полимеризации
и дифференциации, для обеспечения существования
организма нуждаются в интеграции отдельных час-
тей. Интегрирующие системы (столоны, внутренние
устьевые системы, туннели, каналы) фораминифер
описаны выше (см. раздел «Интегрирующие систе-
мы»), они возникают конвергентно в разных ветвях
в наиболее продвинутых группах и обеспечивают
связь начальных камер с поздними. Зачатки прими-
тивной системы каналов имеются у палеозойских ла-
зиодисцид, инволютинид (у вымерших в мелу Hottin-
gerellida), у юрских спириллинид (Spirotrocholina, —
Азбель, 1986) (рис. 547, II, 4—6, 10, Б). Характерно,
что у всех этих форм она возникла лишь при трохо-
идном типе раковин, создающем, как отмечалось
выше, дополнительные структурные возможности.
Каналы отличаются от столонов и туннелей мор-
фологией, происхождением и функцией. Самая со-
вершенная система каналов имеется у высших рота-
лиат (рис. 531, 2, 3; 547, V, 27, 30). Совокупность
каналов представляет собой уникальную систему ор-
ганизменного уровня, сравнимую по функции с кро-
веносной системой высших многоклеточных, но
функционирующую в одной клетке.
Большинство групп с развитой системой тунне-
лей вымерло (фузулиниды из класса спириллина-
та — в перми; орбитолины из подкласса текстуляри-
ана — в эоцене). Alveolinina (милиоляна) приобрели
систему туннелей в мелу, некоторые формы сохра-
нились в тропиках до сих пор. Среди спириллинат,
развивших примитивную систему каналов (Михале-
вич, 19936, 1999), дожили до настоящего времени
лишь формы без каналов (спириллиниды, пателли-
ниды, аннулопателлиниды). Rotaliida и Nummulitida
с каналами существуют с мела по настоящее время,
некоторые из них (Elphidiidae) сумели приспосо-
биться к жизни в холодных водах. <
Часть фораминифер пошла по иному пути в ус-
транении противоречия между многокамерностью и
единством всего организма: у взрослых особей про-
исходит резорбция стенок предшествующих камер и
как бы возврат к однокамерному состоянию (Pegidii-
dae — Rotalioida, Ellipsolingulina, Lagenoglandulina,
Bombulina (рис. 548) — Nodosarioida, Orbulina —
Globigerinidae). Этот путь имеет меньшее прогрес-
сивное значение и не получил широкого развития.
Стенка раковин фораминифер менялась от орга-
нической до агглютинированной и затем до секреци-
онной. Наибольшие изменения произошли в разви-
тии секреционных раковин (Михалевич, 1999). Ради-
альная стенка нодозариат и стенка спириллинат
отличаются от микрогранулярной большей упорядо-
ченностью укладки кристаллов и органических про-
слоев. Возникновение бифонтинальной стенки можно
считать изменением типа ароморфоза. Она отличается
наиболее высокой степенью упорядоченности всех
ее элементов: строго ориентированной укладкой
ТИП FORAMINIFERA. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
579
кристаллов, четкой разграниченностью органических
и известковых слоев, не только четкой разметкой
пространства порами, но и возникновением поро-
вых канальцев со сложной внутренней структурой
(рис. 535, 5, Б\ 537). Эти особенности связаны с
более равномерным, густым и упорядоченным распо-
ложением центров секреции. Их пространственная
организация определяется, по-видимому, аппаратом
Гольджи. При упорядоченной укладке правильных
кристаллов стенка достигает наибольшей прочности
при наименьшей ее толщине. У представителей с
агглютинированной и микрогранулярной стенкой
развиты внутренние ее утолщения (рис. 531, 5; 547,
7, Astrorhizana, 6, 2>, 7) — отмеченные раньше аль-
веолярные, губчатые слои, которые изредка встреча-
ются у астроризат (Daitrona, Masonella) и получили
особое развитие у разных отрядов текстуляриан, а
также у фузулинид. Отложения внутренних скелет-
ных образований (столбики орбитолин, бьекови-
нид — текстуляриана, меандропсинид — милиоляна,
хоматы фузулинид, — рис. 531, 6, А, Б; 533, 4, 5;
534, 3) особенно развиты при микрогранулярной и
фарфоровидной стенке. Наиболее полное развитие
они получили у Fusulinoida. Эти структурные осо-
бенности характеризуют надсемейства и отряды. Не-
которое конвергентное сходство прослеживается и в
строении псевдопор в микрогранулярной стенке, и в
единичных случаях их возникновения в фарфоровид-
ной стенке (см. раздел «Строение пор»). И те и
другие могут быть извилистыми, ветвистыми, окан-
чиваться слепо. Возможность внешних образований
на наружной стенке возникла лишь при полностью
секреционной стенке (ребра, шипы, иглы, ячеи, гра-
нулы у милиолян, нодозариан, спириллинан, ротали-
ан, глобигеринат, — рис. 525, 72, Б\ 526, Б\ 531, 9;
538; 539, 7, 2; 547, 777, 7, 72, 75, 7V, 6, 78, V, 77, 23).
Детали их строения могут служить родовыми и ви-
довыми признаками. Дополнительные скелетные
пластинки, прикрывающие пупок или швы, получи-
ли широкое развитие лишь у роталиат и глобигери-
нат. Наиболее развиты они у высших роталиат в
подклассе Rotaliana и у Globigerinata (рис. 525, 70, г;
538, Г, Д; 547, V, 24, Б, 26, Б, 28, Б), но в некоторых
случаях известны и в низшем подклассе этого клас-
са, у Textulariana (Asterotrochammina, Dobrogelina,
Arenagula (рис. 547, V, 9—77). У спириллинат, вы-
сших милиолят и нодозариат дополнительные ске-
летные пластинки не возникли. Их наличие можно
рассматривать как признак групп высокого таксоно-
мического ранга, их форма и положение характери-
зуют более низкие таксоны.
Рассмотренные типы секреционной известковой
стенки сложились в процессе эволюционного разви-
тия фораминифер (рис. 550) и характеризуют опреде-
ленные филогенетические линии развития (в приве-
денной ниже системе — классы; см. также: Михале-
вич, 1999). У высших представителей этих классов
(их высших подклассов — Spirillinana, Miliolana, No-
dosariana, Rotaliana) возник свой особый тип ультра-
структуры стенки. Разные типы укладки кристаллов в
известковой стенке у милиойян, спирилинан, нодоза-
риан, роталиан — пример дивергенции, но лучистое
расположение столбиков кристаллов, перпендику-
Рис. 548. Bombulina (Nodosariana).
1 — внешний вид раковины; 2 — продольное сечение, видна частично ре-
зорбированная первая камера. (По: Михалевич, 1983).
Fig. 548. Bombulina (Nodosariana).
1 — outer view of the shell; 2 — longitudinal section, partly resorbed first
chamber is seen.
лярных поверхности раковины, в монофонтинальной
стенке спириллинан и нодозариан и в бифонтиналь-
ной стенке роталиан возникло параллельно. Тенден-
ция к смене агглютинированной стенки секрецион-
ной известковой при одном и том же типе строения
раковин, как показали морфологические и экспери-
ментальные данные (Михалевич, 1992, 1995, 1998,
1999; см. также раздел «Секреционная известковая
стенка») и анализ их геологической истории, также
возникала параллельно в каждой линии.
Проведенный выше анализ ультраструктуры стенки
современных форм заставляет нас по-новому взгля-
нуть на сборную группу палеозойских Fusulinacea. В
ее состав входят формы с различными типами рако-
вины и стенки. Количество параллелизмов, и так до-
статочно большое среди фораминифер, еще увеличи-
вается существованием параллельной системы фузу-
линацей.
Сравнение слоев наиболее сложной стенки фузу-
линид с таковой высших текстуляриан (орбитолина)
показывает их большое сходство (Михалевич, 1999Ъ
наружный, органический слой соответствует текту-
му, срединный, микрогранулярный — кериотеке; ам-
пулообразные альвеолы подобны таковым неошваге-
рин (ср.: pl. 307, 2—4 и 169, 7; 177, 7 и 182, 5 в
работе Лёблика и Таппан — Loeblich, Tappan, 1988;
см. также рис. 533, 6—9). Микрогранулярная стенка
фузулинид, возможно, отличалась большей упорядо-
ченностью зерен (Т. Н. Исакова, устн. сообщ.), что
объясняет появление волокнистости при диагенезе.
Фиброзный волокнистый слой возникает при диаге-
незе и радиальной и микрогранулярной стенки (Ho-
henegger, Piller, 1975). Отсутствие принципиальных
различий в строении стенки текстуляриан и палео-
зойских фузулиноид показывает, что ее строение не
может служить надежным диагностическим призна-
ком для этих таксонов. Поэтому при ревизии сбор-
ной группы прежних фузулинацей (в понимании
Лёблика и Таппан, — Loeblich, Tappan, 1988) мы
опирались главным образом на морфологические
признаки раковин и устья, особенно на начальных
580
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
стадиях развития. Низшие фузулинацеи, в понима-
нии Лёблика и Таппан (Loeblich, Tappan, 1988), по-
падают в предлагаемой нами системе в различные
классы (см. раздел «Систематический обзор») в за-
висимости от формы раковины, а также появления
лучистого слоя стенки.
Некоторые особенности ядер и их изменения в
процессе жизненного цикла в таксонах разной степе-
ни продвинутое™ были отмечены ранее ;(см. разделы
«Ядро», «Жизненные циклы»; см. также: Михалевич,
1999). У низших лягинан наряду с обычным жиз-
ненным циклом встречается простое деление надвое.
Агамогонию можно считать следующим этапом бес-
полого размножения (множественное деление). В
процессе эволюции произошли прогрессивные изме-
нения и в половой части цикла — от изогамии (Glab-
ratella patelliformis) к анизогамии (Rubratella), иногда
с сексуальной активностью гамет. Как показал ана-
лиз современных данных по цитологии и жизненным
циклам, направленность основных эволюционных из-
менений этих структур и циклов у фораминифер идет
параллельно тенденциям прогрессивного развития их
скелетов (Михалевич, 1980, 1981, 1999). Гетерокари-
отность возникла в наиболее эволюционно продви-
нутых группах (см. разделы «Ядро», «Жизненные
циклы») — у глобигеринат, высших милиолят и вы-
сших роталиат. У низших роталиат (текстуляриан)
агамонты гомокариотны. Во всяком случае, Павловс-
кому с соавт. (Pawlowski et al., 1995b) не удалось об-
наружить у агамонта трохаммины ядерного дуализма.
Скорее всего, такими же окажутся и агамонты ни-
зших милиолят (шлумбергеринаны). Не исключено,
что такая же картина будет обнаружена у высших и
низших нодозариат, данные по которым пока отсут-
ствуют, так как ранее их включали в роталиат и не
проявляли к ним специального интереса. У более
примитивных спириллинат, у которых агамонты даже
высших представителей гомокариотны, тем более
трудно ожидать обнаружения диморфизма у низших
форм (аммодискан). Таким образом, определенный
систематический подход помогает наметить на буду-
щее стратегию исследования ядерного аппарата.
Нельзя не отметить и такое своеобразное направ-
ление развития фораминифер, как их совместная эво-
люция с симбионтами. У многих секреционных фора-
минифер (в продвинутых группах роталиат — Amphis-
tegina, Calcarina, Nummulites и милиолят — Alveolina,
Sorites) сложились с симбионтами длительные отно-
шения (Amphistegina даже стала симбионтзависимой).
Это повлияло на размеры и форму раковин, их внут-
реннюю морфологию (усложнение, появление боль-
шого числа камер и камерок), строение стенки (час-
тичная резорбция органического вещества, появление
более тонких, прозрачных участков, обеспечивающих
доступ света к симбионтам). Вся система симбионт—
хозяин получает дополнительную энергию. У исследо-
ванных видов такие взаимоотношения резко увеличи-
вают скорость кальцинации и, вероятно, производство
органических мембран и ведут к увеличению общих
размеров особей. В различных филогенетических ли-
ниях развития фораминифер формы, эволюировав-
шие совместно с симбионтами, достигли наибольшей
сложности организации.
Как видно из вышеизложенного, фораминиферы
дают поразительное число примеров параллелизма в
строении раковины и устья, что можно объяснить их
широким современным и геологическим распростра-
нением, разнообразными условиями существования,
огромным многообразием форм. Можно сказать, что
они используют почти все варианты комбинаций из
заданного природой набора признаков, составляю-
щих универсальное пространство возможностей.
Зная сочетание различных черт строения у форами-
нифер и тенденции их развития, можно предсказать
нахождение определенных форм (например, различ-
ные типы строения устья милиолид — наличие усть-
евой лопасти, простого или сложного зуба, множест-
венного устья при различных способах навивания —
квинквелокулиновое, трилокулиновое, билокулино-
вое, сигмоилиновое, — рис. 549; см. также: Миха-
левич, 1999). Так, долгое время у сигмоилиновых ра-
ковин не были описаны формы с множественным
устьем, у спиролокулиновых — со сложным зубом;
позднее они были найдены (Longiapertina Seiglie,
1979; Bidentina Mikhalevich, 1988). Таким образом,
современная система форминифер, несмотря на мно-
гие еще нерешенные вопросы, обладает и прогности-
ческой ценностью.
Наибольшее число параллелизмов в пределах
всего подтипа фораминифер охватывает первые сту-
пени их усложнения (Михалевич, 1981, 1999): воз-
никновение многокамерное™, способы образования
камер, наиболее простые случаи усложнения устья и
появления дополнительных устьев. Эти тенденции
развиваются до тех пор, пока дивергенция не исчер-
пывает всех возможностей старых структур (столоны
и туннели сменяются каналами, агглютинированная
стенка — известковой). Как мы видели выше, агглю-
тинированный тип стенки обладает ограниченными
структурными возможностями. При известковой сек-
реционной стенке значительное число запретов, по-
видимому, оказывается снятым. Все это дает нам
основание относить возникновение известкового сек-
реционного скелета, особенно бифонтинальной стен-
ки, к новообразованиям типа ароморфозов, сущест-
венно изменяющих план строения организма, его ос-
новные свойства.
Проведенный анализ показал, что изменения ус-
тьев, интегрирующих систем и типов стенки проис-
ходили параллельно у раковин разного типа строе-
ния, но в каждой из рассмотренных филогенетических
ветвей тенденции развития реализованы в неодина-
ковой степени, достигают разного уровня совершенства
и имеют своеобразные черты. Поэтому при рассмот-
рении классов, мы берем за основу план строения
раковин, способ навивания, тип симметрии, а так-
же учитываем комплекс признаков других структур
и ультраструктуру стенки (последняя характеризует
подклассы, иногда отряды).
Качественное своеобразие эволюции наиболее
бедно в примитивных подклассах Lagynana и Astro-
rhizana. Они обладают преимущественно однокамер-
ными, часто неправильными раковинами, простым
устьем и, как правило, простой стенкой, органичес-
кой или агглютинированной, реже микрогрануляр-
ной. Их агамонты долго сохраняют 1 ядро; разделе-
ТИП FORAMINIFERA. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
581
Строение устья
Рис. 549. Строение устья у милиолят с разным типом навивания камер. (По: Михалевич, 1999).
Fig. 549. Apertural structures in Miliolana with the different types of chamber arrangement.
ние ядер на соматические и генеративные у них от-
сутствует; нет четкой дифференциации на экто- и
эндоплазму. Можно предположить, что после цито-^
логических исследований многие из них окажутся
принадлежащими к другим типам протистов, как это
произошло с Pelosina, — после проведенного иссле-
дования мы относим ее к ксенофиофориям (Михале-
вич, Воронова, 1999).
Гомокариотность характерна и для своеобразной
древней примитивной ветви — современных спирил-
линат, обладающих двухкамерной раковиной с длин-
ной второй трубчатой камерой (или произошедших
582
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Рис. 550. Схема развития разных классов фораминифер в геологическом прошлом. (По: Михалевич, 1999).
1 — подкласс Lagynana; 2 — подкласс Astrorhizana; 3 — класс Miliolata; 4—9 — класс Spirillinata (4—7 — подкласс Spirillinana: 4 — надотряд Spirillinoida;
5 — надотряд Involutinoida; 6, 7 — надотряд Arcchaediscoida (6 — отряд Lasiodiscida, 7 — отряд Archaediscida); 8, 9 — подкласс Ammodiscana (8 —
псевдодвухкамерные трубчатые формы, 9 — псевдомногокамерные Taumayellida и многокамерные Fusulinoida)); 10, 11 — класс Nodosariata (10 — подкласс
Hormosinana, 11 — подкласс Nodosariana); 72, 73 — класс Rotaliata (72 — подкласс Textulariana, 73 — подкласс Rotaliana); 14 — класс Globigerinata.
Fig. 550. Scheme of the development of different Foraminiferal classes in geological history.
7 — subclass Lagynana; 2 — subclass Astrorhizana; 3 — class Miliolata; 4—9 — class Spirillinata (4—7 — subclass Spirillinana: 4 — superorder Spirillinoida; 5 —
superorder Involutinoida; 6, 7 — superorder Archaediscoida (6 — order Lasiodiscida, 7 — order Archaediscida); 8, 9 — subclass Ammodiscana (8 — pseudotwo-
chambered tubular forms, 9 — pseudomultichambered Toumayellida and multichambered Fusulinoida)), 10, 1,1 — класс Nodosariata (10 — subclass Hormosinana,
77 — subclass Nodosariana); 72—73 — class Rotaliata (72 — subclass Textulariana, 73 — subclass Rotaliana); 14 — class Globigerinata.
от них), с простым устьем, но с упорядоченной
структурой секреционной стенки у их высших форм.
Их древние формы сочетают ряд плезиоморфных
черт (наличие трубчатой камеры у многих из них
хотя бы на ранней стадии, псевдокамерность, при-
митивные септы, простое устье, клубковидное на-
вивание) с апоморфными (наличие туннелей и кана-
лов, дополнительных устьев, трохоидной формы рако-
вин).
У милиолят и нодозариат типы строения раковин
довольно многообразны (у первых преобладают не-
правильно- и правильно-клубковидные, у вторых —
одноосные, спирально-винтовые и инволютные плос-
коспиральные, отличающиеся от плоскоспиральных
раковин роталиат конечным положением устья).
Каждый из этих классов обладает особыми типами
усложнения устьев (у милиолят — с зубами, у нодо-
зариат — радиальные либо с трубкой), в обоих клас-
сах не развиты дополнительные устья, дополнитель-
ные скелетные пластинки. Интегрирующие системы
первых представлены столонами и туннелями; у вто-
рых эти системы почти не развиты. Стенка у высших
подклассов обоих классов монофонтинальная, раз-
личной укладки. Ядерный диморфизм обнаружен у
милиолят (рис. 542) (по нодозариатам нет данных).
Какое бы из рассмотренных выше эволюционных
направлений мы не взяли — строение ядер, жизнен-
ные циклы с ядерным дуализмом, типы строения ра-
ковин (наиболее многообразны, с преобладанием
плоскоспиральных и особенно трохоидных), устья
(сложные зубные пластинки, клапаны и трубки, до-
полнительные устья), дополнительные наружные ске-
летные пластинки, взаимосвязанные внутренние усть-
евые системы, сложные интегрирующие системы ка-
налов — все они наиболее разнообразно и сложно
представлены у высших роталиат. Появление бифон-
тинальной стенки, обладающей структурой, которая
обеспечивает легкость и прочность раковин при наи-
меньшей толщине, сделало для роталиат возможны-
ми ароморфные преобразования, резкое возрастание
темпов эволюции, увеличение числа высших и низ-
ших таксонов, адаптивную радиацию, появление
планктонных форм. Происхождение последних от
донных роталиан типа дискорбисов или нонионов до-
казывается морфологическим сходством и наличием
донной стадии в их жизненном цикле. Арнольд (Аг-
ТИП FORAMINIFERA. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ
583
nold, 1978) предполагает, что милиоляты произошли
от предковых форм, подобных Periptigma (Lagynana).
Мы считаем (Михалевич, 1992, 1999) вероятным про-
исхождение и других классов фораминифер также от
лагинан (рис. 551), среди которых можно найти при-
митивные прототипы основных морфологических
форм высших подклассов (рис. 547, /, Ammoscalaria-
па, 5, 6). Предположение о возможности происхож-
дения роталиат от Phthanotrochus arcanus (Lagynana)
(Loeblich, Tappan, 1988, pl. 10, fig. 7, 9) подкрепля-
ется данными по их жизненным циклам и ядерному
аппарату. Это единственный вид лагинан, у которо-
го, возможно, существует ядерный дуализм (Arnold,
1978) (см. с. 572).
Оба примера лишний раз подтверждают, что пер-
воначально складывался основной тип раковин, а
затем, в процессе эволюции, происходило изменение
состава и ультраструктуры раковинной стенки. В ре-
зультате в классах Spirillinata, Miliolata, Nodosariata,
Rotaliata почти каждому агглютинированному или
микрогранулярному типу соответствует изоморфный
аналог с секреционной стенкой (рис. 547, II, 1, 9.
III, 1, 11. IV, 1, 10; 3, 11; 4, 14; 5, 16. V, 1, 14; 3,
16; 6, 18; 7, 22; 9, 24; 10, 26; 11, 28; 12, 29), а
низшим подклассам — высшие. В предшествующих
системах такие гомеоморфные раковины рассматри-
вались как возникшие конвергентно. Мы же рас-
сматриваем их как родственные формы, последова-
тельные этапы развития в каждой из этих групп, что
особенно наглядно подтверждается при сравнении
текстуляриан и роталиан: сходство процессов обра-
зования новых камер (см. раздел: «Ретикулоподии»),
морфологическое сходство в строении не только ра-
ковины, камер, устья, но и деталей строения пупоч-
ной области (Михалевич, 1972а, 1999), пупочных
концов камер, величины и положения дополнитель-
ных устьев, дополнительных скелетных пластинок в
пупочной области (рис. 547). Мало вероятно, чтобы
абсолютно полное совпадение до мельчайших под-
робностей у таких сложных форм (пары Tiphotroc-
ha—Neoconorbina, Arenagula—Neocribrella, Asterot-
rochammina—Discorbis, Dobrogelina—Sestronophora,
Spiroplectinata—Euvigerina, Minouxia—Sporobulimina,
Toretammina—Oridorsalis и мн. др.) (рис. 547, V, 9,
24; 10, 26; И, 28; 12, 29) могло столько раз возни-
кать конвергентно, представляя зеркальный вариант
больших групп, отличающихся друг от друга лишь
составом и строением стенки. Причем совпадают не
только основные структуры, но и основные тенденции
их развития, стадии онтогенеза. Яркие примеры та-
кого совпадения можно найти и у нодозариат: столь
характерное только для этой группы лучистое и
асимметрично-щелевидное устье возникло еще у их
агглютинированных форм (Nodosinum, Cuneata)
(рис. 547, IV, 3; см. также: Михалевич, 19986). О
возможности постепенного перехода от агглютиниро-
ванной стенки к секреционной известковой уже го-
ворилось выше (см. раздел: «Секреционная известковая
стенка», а также: Михалевич, 1995, 1999). Поэто-
му прежних текстуляриан, объединявшихся только
на основании наличия у них агглютинированной
стенки, мы рассматриваем как сборную группу. Из
их состава мы переводим псевдодвухкамерные труб-
Рис. 551. Схема происхождения разных классов фораминифер.
Fig. 551. Scheme of the origin of different Foraminiferal classes.
чатые раковины, клубковидные многокамерные ра-
ковины одного из милиолятных типов строения и
раковины с нодозариатными типами строения в со-
ответствующие классы — Spirillinata, Miliolata, No-
dosariata в качестве их более примитивных подклас-
сов. Выявление среди инволютных литуолид и среди
атаксофрагмиид других агглютинированных предста-
вителей класса Nodosariata требует дополнительного
морфологического анализа.
Выяснение за последние годы закономерностей
процессов образования новых камер у представите-
лей классов Spirillinata и Miliolata (см. раздел: «Ре-
тикулоподии») подтверждает глубокие отличия их от
роталиат. В результате в подклассе Textulariana ос-
таются только представители с роталиатными типа-
ми строения. Такой подход позволил сформулиро-
вать более четкие диагнозы и устранить значитель-
ное число параллелизмов, возникавших в прежних
системах.39 В класс Astrorhizata вошли только при-
митивные однокамерные и псевдомногокамерные
формы с тектиновой, агглютйнированной и микро-
гранулярной стенкой. Наиболее примитивным и, по-
видимому, сборным является подкласс Lagynana,
нуждающийся в дополнительных исследованиях. Вы-
сокоорганизованные специализированные планктон-
ные формы входят в класс Globigerinata./—
Скелет отражает план строения организма и от-
личается большой консервативностью. Вероятно, ос-
новные типы раковин сложились во времена кемб-
рия—карбона (рис. 550), далее происходили измене-
ния строения стенки, внутренних структур раковин,
39 Кроме того, по-видимому, многие параллельно существовав-
шие таксоны были созданы исследователями, изучавшими группы
какого-либо отдельного исторического периода изолированно, вне
связи со всей системой в целом. Данные Фарни с соавт. (Fahrni et
al., 1995, 1997) и Павловского с соавт. (Pawlovski et al., 1994, 1995,
1997) подтвердили на молекулярном уровне близость подкласса
Schlumbergerinana с подклассом Miliolana, подкласса Textulariana с
подклассом Rotaliana, а также показали, что число основных фило-
генетических трупп фораминифер ближе к 5, а не к 12, как это
принято в большинстве современных классификаций.
584
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
их устья. Фораминиферы представляют уникальную
группу, поскольку их филогенетическая история,
таксономия, распространение в прошлом имеют под-
робную и объективно контролируемую историческую
основу. Представленная ниже система фораминифер
не противоречит данным исторической геологии.
Наиболее древними, но сохранившимися до наших
дней являются Lagynana, Astrorhizana и Spirillinata,
известные с кембрия. В ордовике появились древ-
ние Nodosariata и, по-видимому, Miliolata, потомки
которых процветают и ныне. Давшие пышный рас-
цвет в девоне и карбоне Fusulinoida со сложно ус-
троенной массивной раковиной целиком вымерли. У
них не появилось ни значительных усложнений
строения устья, ни наружной скульптуры стенки. Их
интегрирующие системы — туннели — более при-
митивные структуры по сравнению с системой кана-
лов роталиан. Именно массивность и несовершенст-
во микрогранулярной стенки могли стать причиной,
закрывшей этой высокоразвитой группе дальнейший
путь эволюционных преобразований. Агглютиниро-
ванные формы роталиат (Textulariana) существуют с
карбона до наших дней. С триаса стали заметны, а
в мелу достигли пышного расцвета современные вы-
сшие роталиаты — подкласс Rotaliana и несколько
позже, с Юры, — произошедшие от них планктон-
ные Globigerinata. И те и другие представляют вы-
сшую ступень в развитии группы, как млекопитаю-
щие среди остальных позвоночных. Они же наибо-
лее богато представлены в современных морях.
Место в системе протистов
Представление о протистах как о типе карди-
нально изменилось с развитием электронной микрос-
копии. Это была сборная, скорее экологическая, чем
систематическая, группа. Преимущественно однокле-
точные эукариотные организмы в настоящее время
объединяются в группу протистов с различным чис-
лом типов (от 18 до 45) (Крылов и др., 1980; Corliss,
1984). Наметилась также тенденция к увеличению
числа царств и объединению их в доминионы (Ку-
сакин, Дроздов, 1998). Сравнение фораминифер с
близкими группами прежних саркодовых (Старобо-
гатов, Михалевич, 1985; Михалевич, 1999) показыва-
ет их глубокие различия. Между фораминиферами
и радиоляриями больше различий (ядерный аппарат,
псевдоподии, состав и форма скелета, жизненные
циклы и образ жизни), чем сходства (трубчатые
кристы митохондрий, двухжгутиковые гаметы и за-
крытый внутриядерный плевромитоз). Если сравнить
ядерный аппарат фораминифер с таковым радиоля-
рий (1 высокополиплоидное ядро) и акантарий (мно-
гоядерность, по-видимому без полиплоидии), то ока-
жется, что фораминиферы в отличие от этих групп
использовали в своей эволюции не один, а оба воз-
можных пути. Ядерный диморфизм среди протистов
встречается еще только у инфузорий, но у них име-
ются чаще 1, а не множество Ми и иные жизненные
циклы. Обычный путь эволюции генома — перво-
начальная полимеризация ядер, затем полимеризация
геномов (полиплоидизация) и последующая олигоме-
ризация ядер. На пути полимеризации геномов и
олигомеризации ядер фораминиферы продвинуты в
меньшей степени, чем инфузории. Судьба соматичес-
кого ядра у них иная, чем у радиолярий и инфузо-
рий, — оно не наследуется дочерними клетками, а
разрушается и у бесполой стадии. Инфузории давно
рассматриваются как тип. Сходство жгутика Boderia
turneri (Lagynana) (см. раздел: «Строение гамет») со
жгутиком хризомонадовых позволило рассматривать
эти группы как близкие (Loeblich, 1974). Недавно
жгутики с мастигонемами обнаружены еще у одного
вида фораминифер из высших роталиат — у Nummu-
lites venosus (Roettger et al., 1998). Однако сближе-
нию фораминифер с хризомонадовыми противоречит
наличие у хризомонад кремнеземного скелета. На-
личие у фораминифер трехжгутиковых форм и спо-
собность к откладыванию извести скорее позволяет
сближать их с Haptophyta (Prymnesiomonada) (Ми-
халевич, 1980, 1999; Старобогатов, Михалевич, 1985).
Особенности ядерного аппарата фораминифер,
черты его высокой специализации и большой про-
двинутое™, а также своеобразие морфофизиологи-
ческого направления эволюции скелета форамини-
фер, создавших новую уникальную интегрирующую
структуру организменного уровня — каналы, осо-
бенности строения их гранулоретикулоподий, гамет
и жгутиков, а также необычность жизненных циклов
(антитетическая смена поколений с промежуточной
редукцией) позволили нам поднять их ранг до под-
типа (Михалевич, 1980), а затем до типа (Миха-
левич, 1999). Анализ последовательностей нуклеоти-
дов ДНК у фораминифер показал их несходство со
всеми другими изученными последовательностями из-
вестных групп прокариот и эукариот и подтвердил
уникальность этой группы протистов (Langer et al.,
1993).
Систематический обзор"
Принципы построения
макросистемы фораминифер
(критерии выделения таксонов)
При выделении таксонов мы опирались на комп-
лекс следующих признаков: количество камер (1—2
или много), форма раковины и тип навивания, ульт-
раструктура стенки, наружные и внутренние допол-
нительные скелетные образования, строение устья,
наличие дополнительных устьев, интегрирующих
систем, форма камер и другие более мелкие призна-
ки, а также тенденции их развития. Основными из
перечисленных признаков мы считаем 3 первыхГБ
паре признаков форма раковины—структура стенки
мы отдаем приоритет первому. Такой подход харак-
терен для школы Фурсенко (Фурсенко, Раузер-Чер-
ноусова, 1959; Подобина, 1978; Соловьева, 1978;
Яковлева, 1979; Саидова, 1981; Субботина и др.,
1981; Басов, 1990; Маслакова, 1990; Раузер-Черноу-
сова, 1990; Вдовенко и др., 1993; Григелис, 1995).
* Система публикуется по: Михалевич, 1999.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
585
Многие особенности в усложнении внутренней
структуры камер, их стенки, а также в строении
устья мы рассматриваем как возникшие параллель-
но у разных типов раковин с неодинаковым планом
строения (Михалевич, 1999). Поэтому план строения
раковин положен в основу выделения классов. В
классах Spirillinata и Nodosariata, так же как это
было ранее в классах Miliolata (Саидова, 1981) и
Rotaliata (Михалевич, 1980), агглютинированные
формы, аналогичные по строению высшим секреци-
онным формам, относятся соответственно к более
примитивным подклассам Ammodiscana и Hormosi-
nana. В подклассах Lagynana и Astrorhizana известны
только виды с более примитивным типом стенки.
Таксоны выделялись с использованием категорий
признаков единого уровня (например, надсемейства
выделялись на основании усложнения внутренней
полости камер и внутреннего усложнения стенки).
В тех случаях, когда морфологический признак еще
не устоялся (например, способ навивания у неко-
торых примитивных форм варьирует, на ранних эта-
пах становления многокамерности тип строения ра-
ковин не выражен отчетливо), за ним признавался
меньший таксономический вес, чем в случае его ста-
билизации.
Палеозойские формы рассмотрены в единой сис-
теме с современными, что позволило устранить зна-
чительное число параллелизмов предшествующих
систем. Крупные, целиком вымершие таксоны рас-
смотрены кратко. Таксоны, положение которых
сильно изменились по сравнению с их прежним мес-
том в системе, отмечены звездочкой (*).
Класс Ast го rhizat a Saidova, 1981 (рис. 547, /)
Раковина однокамерная, иногда псевдоколониаль-
ная или псевдокамерная, сферической, удлиненной,
трубчатой, звездчатой, ветвящейся или иной непра-
вильной формы; стенка монофонтинальная, иногда с
заметными псевдопорами, органическая у Lagynana и
Ammoscalariana, агглютинированная или микрогра-
нулярная у Astrorhizana, простая или частично под-
разделенная неполными перегородками, может быть
утолщенной изнутри (губчатой, лабиринтовой); устье
может отсутствовать, может быть первично-множес-
твенным (с отверстиями неправильной формы, не
фиксированными по положению, иногда располо-
женными на концах трубок) или единичным у более
эволюционно продвинутых форм (обычно с фикси-
рованной формой и положением, может быть на
приподнятой шейке, иметь небольшую губу); агамон-
ты одноядерны, ядерный дуализм не обнаружен; га-
меты двухжгутиковые или амебоидные; свободножи-
вущие или (очень часто) прикрепленные. Кембрий—
голоцен.
Примечание. В виде исключения, в этот класс включены
некоторые формы с полными органическими септами либо с неус-
тойчиво многокамерной органической раковиной.
Подкласс Lagynana Mikhalevich, 1980
Раковины с одним из астроризатных типов строе-
ния раковины и устья; стенка раковины органичес-
кая, часто сильно ожелезненная, характерного корич-
невого или ржаво-красного цвета, обычно простая;
псевдоподии ветвящиеся или нитевидные; кроме
обычных жизненных циклов часты почкование, цито-
томия, простое деление надвое, гаметы двухжгутико-
вые или амебоидные; свободноживущие и прикреп-
ленные донные формы, часто в пустых раковинах
фораминифер и трубках червей; морские, но встреча-
ются пресноводные роды, 2 рода перешли к парази-
тизму (Rhynchosaccus, Thalamophagd). Кембрий—го-
лоцен.
Примечание. У 2 древних родов Labyrinthochitinia и Chiti-
nolagena (ордовик) отмечена лабиринтовая стенка, у первого из
них, а также у современных Belaria и Cystophrys имеются внутрен-
ние подразделения стенки, однако они неустойчивы и недостаточ-
но исследованы.
В этот слабо изученный подкласс входят наибо-
лее примитивные представители фораминифер. Ряд
особенностей строения раковины и цитоплазмы про-
ходит в этой группе первые этапы своего становле-
ния. У многих из них раковина заполнена цитоплаз-
мой лишь частично, значительная ее часть выходит
наружу. Ядро 1, обычно крупное (40—79 мкм), ок-
руглое или овальное, с довольно жесткой ядерной
оболочкой. У представителей многих родов отмече-
ны формы с несколькими мелкими ядрами (до 10,
реже до 150), возможно, находящиеся в процессе
размножения. Цитоплазма лагинан не дифференци-
рована на экто- и эндоплазму. У Allogromia протеи-
новая основа раковины образуется эндоплазмой на
цитоплазматической мембране, а не эктоплазмой ре-
тикулоподий. У Shepheardella тонкий слой цитоплаз-
мы покрывает раковину снаружи (как у высших фо-
раминифер), у других лагинан такого слоя не наблю-
далось. Для Saedeleria отмечена стенка из 2 слоев.
Представители одного из родов лагинид (Rhumble-
rinella) секретируют известковые иглы. У многих
форм наблюдается переход к псевдо- и многокамер-
ности. Такие формы в соответствии с нашей класси-
фикацией мы перевели из Allogromiida в отряд Аш-
moscalariida (подкласс Ammoscalariana).
Отряд Lagynida Mikhalevich,
1980
Раковина округлой, овальной, удлиненной или не-
правильной разветвляющейся формы; стенка голая
или с редкими мелкими песчинками, разбросанными
по поверхности, может быть с дополнительными
586
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
внутренними неполными септами; устьев 1, несколь-
ко, либо оно отсутствует, у некоторых родов устье на конце
вытянутой шейки; гаметы двухжгутиковые. Голоцен.
Примечание. У Rhumblerinella на поверхности раковины из-
вестковые иглы, плотно уложенные, но не скрепленные цементом.
Семейства: Lagynidae Schultze, 1854 (с подсем.
Belariinae Mikhalevich, 1999, с внутренними септами;
роды Belaria и Cystophrys); Maylisoriida^ Е. V. Byko-
va, 1961.
Отряд Allogromiida Fursenko, 1958
Раковина округлая, овальная, удлиненная, палоч-
ковидная; стенка голая либо может быть покрыта
песчинками полностью или на отдельных участках,
но при этом сохраняет эластичность; устьев 1, не-
сколько, либо оно отсутствует, может быть округлой
или неправильной формы, часто с вытянутой шей-
кой, с отворотом губы или с энтосолениевой труб-
кой; гаметы (там, где они известны) амебоидные. Ор-
довик—голоцен.
Примечание. Род Rhynchosaccus может паразитировать на
более крупных фораминиферах.
Семейства: Allogromiidae Rhumbler, 1904;
Shepheardellidae Loeblich et Tappan, 1948; Labyrin-
thochytinidae Mikhalevich, 1999 (стенка лабиринто-
вая, может быть с внутренними подразделениями —
роды Labyrinthochitinia и Chitinolagena).
Подкласс Ammoscalariana Mikhalevich, 1980
Раковина с тенденцией к многокамерности, но
камеры сформированы в результате разделения труб-
чатой камеры органическими септами (Ammosca-
lariidae) или имеют неустойчивый характер; стенка
органическая или частично агглютинированная; га-
меты (там, где они известны — Phthanotrochus) аме-
боидные; свободноживущие формы. Эоцен—голо-
цен.
Примечание. Сборная группа, нуждающаяся в дополни-
тельном изучении. Входящие в этот подкласс формы имеют раз-
ный тип раковин (однорядный — Hospitellidae, трохоидный —
Phthanotrochus, милиолятный — Periptigma, плоскоспиральный —
Ammoscalariidae). У высших фораминифер такие различия рас-
сматриваются как признаки, характеризующие отряды. Но поскольку
становление многокамерности здесь только начинается (например,
у Periptigma камеры нестойкие, часто отваливаются; трохоидная
спираль у Phthanotrochus может выпрямляться, становиться не-
правильной) и формы эти изучены недостаточно, они пока остав-
лены в одном таксоне.
Если подтвердится, что Periptigma — предковая форма мили-
олят, a Phthanotrochus — роталиат, и их систематическое положе-
ние будет изменено, в отряде Ammoscalariida останутся только
псевдомногокамерные формы.
Отряд Ammoscalariida Mikhalevich,
1980
Раковина удлиненной, чашечковидной, плоскоспи-
ральной, примитивной трохоидной или примитивной
милиолятной формы, у прикрепленных может быть
неправильной, разветвляющейся; у Ammoscalariidae
и Hospitellidae — псевдомногокамерная, у Phthanot-
rochidae — неустойчиво многокамерная; стенка го-
лая или с песчинками, которые могут образовывать
сплошной покров, но внутренние септы при этом
только тектиновые; устье 1, конечное. Эоцен—голо-
цен.
Примечание. Род Thalamophaga (Hospitellidae) — паразит
многокамерных известковых фораминифер.
Семейства: Phthanotrochidae Arnold, 1978 (вклю-
чая Periptigma); Ammoscalariidae Mikhalevich, 1982;
Hospitellidae Loeblich, Tappan, 1984 (включая свобод-
ные однорядные формы типа Nodellum в качестве са-
мостоятельного подсемейства).
Подкласс Astrorhizana Saidova, 1981
(= Astrorhizicae Saidova, 1981, part.)
Раковины с одним из астроризатных типов стро-
ения камеры и устья; стенка агглютинированная или
микрогранулярная; соматические ядра крупные,
часто с жесткой оболочкой, укрепленной сотовым
или иным дополнительным слоем; гаметы двухжгути-
ковые (у Boderia один из жгутиков с мастигонема-
ми); свободноживущие либо (очень часто) прикреп-
ленные. Нижний кембрий—голоцен.
Примечание. От однокамерных представителей подкласса
Lagynana отличаются агглютинированной или микрогранулярной,
а не голой органической стенкой, от всех остальных классов —
однокамерностью и неправильной формой раковин, от однокамер-
ных Lagenida (Nodosariata) — агглютинированной или микрогра-
нулярной неупорядоченной структурой частиц стенки.
Отряд Astrorhizida Haeckel,
1894
Раковина свободная, одиночная, трубчатая или с
трубками, отходящими от центрального тела; стенка
агглютинированная, простая; устья — открытые
концы трубок — 2 или несколько. Нижний кемб-
рий—голоцен.
Семейства: Astrorhizidae Brady, 1881; Bathy-
siphonidae Avnimelech, 1952; Silicotubidae Vyalov,
1968; Vanhoeffenellidae Saidova,. 1980; Hippoj^epinel-
lidae Loeblich et Tappan, 1984. * '
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
587
Отряд Dendrophryida Haeckel, 1894
Раковины прикрепленные или свободные, оди-
ночные или ложноколониальные, неправильной, час-
то причудливой формы, сильно ветвящиеся; стенка
агглютинированная, простая или усложненная внут-
ренними перегородками, или лабиринтовая, с раз-
личным количеством цемента, чаще рыхлая, эластич-
ная; устье — открытые концы разветвлений, или оно
нефиксированное, в виде перфораций в стенке.
Надсем. Dendrophryoidea Haeckel,
1894
Стенка раковины простая. Верхний карбон—го-
лоцен.
Семейства: Dendrophryidae Haeckel, 1894; No-
todendrodidae de Laca, Lipps et Hessler, 1980; Dryorhi-
zopsidae Loeblich et Tappan, 1984.
Надсем. Schizamminoidea Norvang,
1961
Раковина с внутренними подразделениями полос-
ти камер либо с лабиринтовой стенкой. Ордовик—
голоцен.
Примечание. Большинство родов современные, лишь среди
диффузилинид имеется 2 древних рода.
Семейства: Diffusilinidae Loeblich et Tappan, 1961;
Schizamminidae Norvang, 1961; Halyphysemidae Loeb-
lich et Tappan, 1984.
Отряд Saccamminida Lankester, 1885
Раковина округлая, овальная, слабо удлиненная или
полусферическая, свободная или прикрепленная, оди-
ночная или ложноколониальная; стенка агглютиниро-
ванная, простая или усложненная (лабиринтовая, яче-
истая, с внутренними перегородками); устьев 1, много,
либо оно отсутствует. Ордовик—голоцен.
Подотряд Saccamminina Lankester,
1885
Раковина одиночная. Ордовик—голоцен.
Надсем. Saccamminoidea Brady, 1884
Раковина с простой стенкой. Ордовик—голоцен.
Семейства: Saccamminidae Brady, 1884; Steg-
namminidae Moreman, 1930.
(JJryctodermidae), или с полостью камер, подразде-
ленной внутренними септулами (Daitronidae, Crithio-
nidae). Ордовик—голоцен.
Семейства: Crithionidae Goes, 1894; Oryctoder-
midae Saidova, 1981; Daitronidae Mikhalevich, 1995.
Подотряд Psammosphaerina Haeckel,
1894
Раковина ложноколониальная или ложномногока-
мерная. Ордовик—голоцен.
Семейства: Psammosphaeridae Haeckel, 1894; Ро-
lysaccamminidae Loeblich et Tappan, 1984; Lacustrinel-
lidae Mikhalevich, 1995.
Отряд ‘Parathuramminida Mikhalevich,
1980
Раковина свободная или прикрепленная, одиноч-
ная или ложноколониальная, округлой, удлиненной
или неправильной формы; стенка микрогранулярная,
простая или усложненная, перфорированная или не
перфорированная; устьев 1, много, либо оно отсутст-
вует. Силур—пермь.
Примечание. Данный отряд отличается от Saccamminida
только строением стенки. Мы полагаем, что черты морфологичес-
кого сходства обоих отрядов не конвергентные, но свидетельству-
ют о родстве этих групп. Поэтому отряд Parathuramminida перене-
сен нами из прежних Fusulinacea в класс Astrorhizata. Микрогра-
нулярную стенку мы рассматриваем как этап формирования
стенки, следующей за агглютинированной. Плохая сохранность
этих древних форм (нижний силур—пермь) и неясность многих
деталей строения пока препятствуют проведению более глубокой
ревизии.
Надсем. Parathuramminoidea
E.V. Bykova, 1955
Раковина с простой стенкой, с неподразделенной
внутренней полостью. Верхний силур—нижний кар-
бон.
Семейства: Parathuramminidae Е. V. Bykova, 1955;
Archaesphaeridae Malakhova, 1956.
Надсем. Marginarioidea Loeblich
et Tappan, 1986
Раковина с усложненной стенкой: лабиринтовой,
либо крупноперфорированной, либо пронизанной ра-
диальными канальцами, ведущими наружу. Нижний
силур—верхняя пермь.
Семейства: Tuberitinidae A. D. Miklukho-Maklay,
1958; Auroriidae Loeblich et Tappan, 1986; Eovolutini-
dae Loeblich et Tappan, 1986; Marginaridae Loeblich et
Tappan, 1986.
Надсем. Crithioninoidea Goes, 1894
Раковина co стенкой, пронизанной правильны-
ми или неправильными альвеолярными канальцами
Отряд Hippocrepinida Saidova, 1981
Раковина свободная или прикрепленная, псевдо-
двухкамерная, реже псевдомногокамерная, как пра-
588
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
вило, удлиненная, от округлого, явственно обособ-
ленного или почти не выраженного пролокулуса отходит
вверх вторая трубчатая псевдокамера, которая может
быть прямой, разветвленной на конце, изогнутой, слабос-
вернутой или в виде конуса или воронки, она может
быть неподразделенной, или с неполными внутренними
септами, либо с наружными или внутренними пережима-
ми; стенка агглютинированная или микрогранулярная;
устье конечное, округлое или овальное — открытый, ши-
рокий или суженный конец трубки: у разветвленной
раковины соответственно 2 устья. Силур—голоцен.
Надоем. Hippocrepinoidea Rhumbler,
1895
Раковина с неподразделенной внутренней поло-
стью. Силур—голоцен.
Семейства: Hippocrepinidae Rhumbler, 1895;
Hyperamminidae Eimer et Fickert, 1899; Ammovolumi-
nidae Chernykh, 1967.
Надоем. Botellinoidea Chapman
et Parr, 1936 (включая ‘Caligelloidea
Reytlinger, 1959)
Раковина псевдомногокамерная, с псевдокамера-
ми, образованными неполными септами или пережи-
мами стенки. Верхний силур—голоцен.
Семейства: Botellinidae Chapman et Parr, 1936;
Paratikhinellidae Loeblich et Tappan, 1984; Hormosi-
nellidae Rauser et Reytlinger, 1986.
Класс Spirillinata Maslakova, 1990 (рис. 547, //)
Раковины псевдодвухкамерные (с длинной второй
трубчатой камерой, навитой вокруг пролокулуса)
или произошедшие от них (псевдомногокамерные,
многокамерные у высших представителей, но обыч-
но с сохранением трубчатой части на протяжении
1—2 оборотов), с различными типами навивания и
расположения камер: клубковидным (очень часто,
особенно в начальном отделе), плоскоспиральным
(более часто в конечном отделе, после предшеству-
ющего клубковидного), иногда двухрядным (обычно
в конечном отделе), встречается и трохоспиральный
тип, у высших супермногокамерных форм раковины
могут становиться сферическими или фузулиноидны-
ми; псевдокамеры возникают в результате пережимов
наружной стенки, а также образований внутреннего
скелета или коротких септул, камеры многокамер-
ных форм обычно сохраняют примитивные черты
происхождения от предшествующих трубчатых (бу-
дучи не совсем правильными, с короткими прямыми
или лишь слабо изогнутыми септами, на концах ко-
торых отсутствуют небольшие отвороты губы, так
характерные для септ Rotaliata, волнистые септы
встречаются только в этой группе), у высших пред-
ставителей камеры приобретают более правильную
форму, иногда могут быть подразделены на камерки
(в обоих подклассах); интегративные системы — в
виде туннелей у высших супермногокамерных форм,
а также примитивные системы каналов у эволюцион-
но продвинутых трубчатых форм; стенка изучена не-
достаточно, монофонтинальная, агглютинированная
(часто с железистым цементом) или микрогрануляр-
ная у низших форм (Ammodiscana), у их многока-
мерных форм иногда очень толстая, с несколькими
слоями, не отличающимися принципиально от тако-
вых у Textulariana, но с особыми дополнительными
внутренними отложениями (валики, гребни, крючки,
хоматы) на дне камер, не встречающимися в других
группах, стенка у представителей высшего подкласса
(Spirillinana) полностью секреционная, радиальная,
может быть с регулярными псевдопорами в виде
ямок (согласно литературным данным), слои нарас-
тания не образуются, у древних форм обычны 2 слоя
в стенке — микрогранулярный и радиальный, у 1п-
volutinida стенка, как правило, радиальная, у совре-
менных спириллинид радиальные группы кристалли-
тов ведут себя оптически как один кристалл, наруж-
ная скульптура, а также внутренние столбики и
септы хорошо развиты; рост трубчатой камеры не-
прерывный, происходит постепенным приращением
стенки по ее краю, у многокамерных форм процесс
роста не исследован; устье конечное или произошед-
шее от него (у трохоидных форм имеет тенденцию
смещаться на брюшную сторону), всегда простое, без
внутренних структур, единичное или (редко) мно-
жественное (у некоторых эволюционно продвинутых
многокамерных форм), у многокамерных трохоид-
ных форм иногда с наружной лопастью или пластин-
кой Т-образной или S-образной формы, встречаются
дополнительные устья; агамонты современных спи-
риллинат гомокариотны (их пролокулус крупнее,
чем у гамонтов, — во всех других классах соотно-
шение обратное), гаметы (там, где они известны)
амебоидные; донные, преимущественно свободножи-
вущие формы. Кембрий—голоцен.
Примечание. Этот класс — одна из наиболее древних и
недостаточно изученных групп фораминифер, имеющих некоторое
конвергентное сходство с Miliolata (происхождение от трубчатых
форм с сохранением начальной трубчатой части у многих много-
камерных представителей, конечное положение устья, образование
раковины фузулиноидной формы у некоторых супермногокамерных
раковин, наличие туннелей) и отчасти с Rotaliata (у многокамер-
ных плоско- и трохоспиральных форм). Однако его представители
отличаются от представителей Miliolata не только отсутствием
внутренних устьевых структур (их характерного зубного аппарата),
формой камер (у большинства многокамерных форм утерявших
трубчатый характер) и ультраструктурой секреционной стенки у
высших подклассов, но также и тенденциями их развития: форми-
рованием у многокамерных представителей преимущественно не-
трубчатых камер, скорее напоминающих камеры представителей
Rotaliata, преобладанием плоскоспиральных и трохоидных (несвой-
ственных милиолятам) раковин, развитием дополнительных устьев
и системы каналов. От представителей Rotaliata они отличаются
прежде всего трубчатой псевдодвухкамерной или псевдомногока-
мерной раковиной их низших форм и происхождением (роталиат
мы рассматриваем как произошедших скорее от древних многока-
мерных лягинан типа Phthanotrochus, когда и начальные камеры
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
589
образуются в результате дополнительной полимеризации, а не
фиссации), конечным положением устья и отсутствием внутренних
устьевых структур (в противоположность базальному устью рота-
лиат со сложными внутренними зубными пластинками), характе-
ром их камер и септ (так же, как и ультраструктурой монофонти-
нальной известковой стенки у высших представителей). Конечное
положение устья спириллинан отчетливо видно не только у их
псевдодвухкамерных трубчатых форм, но также у их псевдомного-
камерных и многокамерных раковин. Достаточно взглянуть на
такие их типичные формы, как Toumayella, Endothyranella, с их
округлым устьем — как на конце трубчатой камеры, или Endothy-
randpsis, Endothyra, Globoendothyra с полукруглым устьем — как
на конце полуцилиндрической трубки (см.: Loeblich, Tappan, 1988,
pl. 232, 6—14’, 244, 1—6\ 245, 5; 248, 7—11), Сходство некоторых
из них (например, Tournayella) с Fisherina и Fisherinella (Miliola-
ta) поразительно. Это конечное (а не базальное — вопреки указан-
ному в диагнозах) положение их широкого открытого устья отра-
жается на характере их коротких внутренних септ, разделяющих
камеры. При действительно базальном устье, которое имеется у
роталиат, септы устроены иначе.
Некоторые роды, ранее помещавшиеся в Textulariana, также
имеют конечное округлое или полукруглое устье и некоторые осо-
бенности камер и септ, указывающие на их происхождение от
предковых форм с трубчатыми камерами. В соответствии с нашей
классификацией они должны быть переведены в Spirillinata; Lituo-
tuba и все сем. Lituotubidae, некоторые Charentiinae (Melathrokerb
on, Karaisella), Hottingeritidae, Buccicrenatinae, Kutsevella из Am-
momarginulininae, возможно, род Budashevella и некоторые дру-
гие. Многие плоскоспиральные литуолиды нуждаются в ревизии,
так как характер устья их ранних стадий не изучен.
Как было показано выше, стенка палеозойских форм (преж-
ние Fusulinacea) принципиально не отличается от агглютиниро-
ванной и микрогранулярной стенки текстуляриан, но первые пред-
ставляют гетерогенную группу, нуждающуюся в ревизии.
Исходя из принципа приоритета морфологии раковины, мы пере-
вели некоторые их таксоны в другие группы в соответствии с планом
их строения (например, Parathuramminida, Paratikhinellidae — в Astror-
hizata; Colaniellida, Pseudopalmulinidae — в Nodosariata; Biplanatidae,
Nezzazatidae — в Textulariana). Эту ревизию нельзя считать завершен-
ной.
В подклассе Ammodiscana оставлены только псевдодвухкамер-
ные трубчатые формы или формы, чье происхождение от таких
трубчатых предшественников не вызывает сомнений. Палеозойс-
кие формы рассмотрены менее подробно, чем современные.
Подкласс Ammodiscana Mikhalevich, 1980
Раковины с агглютинированной или микрограну-
лярной стенкой. Кембрий—голоцен.
Следующие палеозойские отряды не рассматрива-
ются здесь подробно: Palaeospiroplectamminida Mik-
halevich, 1999 (начальная часть как у Toumayellida,
поздняя двухрядная — Palaeospiroplectamminidae);
Koskinobigenerinida Mikhalevich, 1999 (раковина це-
ликом двухрядная, трубчатая, или псевдомногокамер-
ная свернутая начальная часть отсутствует; стенка
агглютинированная и микрогранулярная (часть преж-
них Palaeotextulariacea — Koskinobigenerinidae, ?Se-
mitextulariidae без Pseudopalmulinidae и, вероятно, без
Palaeotextulariidae — перенесены в Nodosariata)), от-
ряды Biseriamminida, Tetrataxida, Endothyrida (вклю-
чая роды Melathrokerion и Karaisella из Charentiinae,
перенесенные из Textulariana), Fusulinida, Ozawainellida.
Отряд Ammodiscida Mikhalevich,
1980
Раковина co второй трубчатой камерой, навитой
плоскоспирально, иногда слаботрохоидной у свобод-
ноживущих форм, у прикрепленных — раскручиваю-
щейся, неправильно навитой.
Семейства: Ammodiscidae Reuss, 1862; Tolypam-
minidae Cushman, 1928; *Pseudoammodiscidae Conil
et Lys, 1970; *Pseudolituotubidae Conil et Longerstaey,
1980.
Отряд Ammovertellida Mikhalevich,
1999
Раковина co второй трубчатой камерой, навитой
клубковидно по крайней мере в начальном отделе
в отличие от плоскоспирально навитых аммодис-
цид.
Семейства: Usbekistaniidae Vyalov, 1968; Am-
movertellidae Saidova, 1981.
Отряд Toumayellida Hohenegger
et Piller, 1973
Раковина co второй трубчатой камерой вокруг
пролокулуса, делающей 1 или несколько оборотов в
одной плоскости или стрептоспирально, иногда по
высокой спирали (Plagioraphe), или с нерегулярным
чередованием типов навивания, с последующим под-
разделением на псевдокамеры, иногда в конечных
оборотах — на камеры, обычно несколько непра-
вильной формы и неодинаковых размеров, послед-
ние камеры могут быть расположены в 1 ряд; стенка
агглютинированная (Lituotubidae, Plagioraphidae) или
микрогранулярная (Toumayellidae, Moravamminidae);
устье в виде округлого или полукруглого отверстия
на конце последней камеры. Средний девон—голо-
цен.
Примечание. Сохранившийся на значительном протяже-
нии отрезок трубчатой камеры, характер его сочленения с проло-
кулусом, расположение камер и их неодинаковая форма, конечное
устье отличают эти формы от настоящих литуолид и свидетельст-
вуют об их сходстве и родстве с аммодисцидами.
Семейства: Moravamminidae Pokorny, 1951, То-
urnayellidae Dain, 1953; *Lituotubidae Loeblich et
Tappan, 1984; Plagioraphidae Mikhalevich, 1999 (ра-
ковина высококоническая или цилиндрическая, за
пролокулусом следуют трубчатые камеры, каждая из
которых длиною в 1 оборот или несколько более
одного).
590
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подкласс Spirillinana Maslakova, 1990
Раковина с известковой секреционной стенкой,
радиальной целиком или только в наружном слое, с
богато представленными дополнительными скелет-
ными образованиями на наружной стенке раковины,
в пупочной области, у многокамерных форм иногда с
подразделением камер на камерки, с дополнительны-
ми устьями и примитивно устроенной системой кана-
лов у высших форм. Карбон—голоцен.
Примечание. Клубковидные формы с радиальной стен-
кой — тупиковая ветвь, трохоидные формы образовали примитив-
ную систему каналов. Агглютинированные формы встречаются на-
чиная с кембрия и до наших дней, микрогранулярные вымерли в
карбоне и перми, в развитии форм с радиальной стенкой наблюда-
ется преемственность: архедисциды — карбон, лазиодисциды —
карбон—пермь, инволютиниды — пермь—мел, спириллиниды —
триас—голоцен. Мы рассматриваем их как разные этапы развития
одной филогенетической ветви.
Надотряд Archaediscoida Pojarkov,
1980
Раковина псевдодвухкамерная, со стенкой, состо-
ящей из наружного радиального и внутреннего мик-
рогранулярного слоев извести, поровые отверстия не
описаны.
Отряд Archaediscida Pojarkov,
1980
Раковина овальная или линзовидная, со второй
трубчатой полуцилиндрической камерой, навитой
инволютно, обороты навивания могут быть непра-
вильно-клубкообразными либо слабосигмоидными,
поздние обороты могут быть плоскоспиральными, у
продвинутых форм полость камер с подразделением
на псевдокамеры, вся раковина может быть покрыта
вторичными мощными слоями утолщения, особенно
развитыми в пупочной области. Нижний и верхний
карбон.
Примечание. У некоторых родов микрогранулярный слой
отсутствует и вся стенка радиальная. По-видимому, этот слой об-
разуется на внутренней части стенки в результате процессов диаге-
неза.
Семейства: Archaediscidae Cushman, 1928; As-
teroarchaediscidae A. D. Miklukho-Maklay, 1957; Pla-
noarchaediscidae Gubenko, 1989; Glomodiscidae Mik-
halevich, 1999 (внутренняя полость трубчатой камеры
подразделена внутренними скелетными образования-
ми; роды Glomodiscus, Uralodiscus, Tournarchaedis-
cus, Permodiscus).
Отряд Lasiodiscida Mikhalevich, 1993
Раковина от слаботрохоидной, почти плоскоспи-
ральной до высококонической, со второй трубчатой
полуцилиндрической камерой, навитой эволютно, пу-
почная область заполнена радиально-фиброзной мас-
сой, с бугорками, столбиками, у продвинутых родов
она пронизана щелями каналов, вдоль спирального
шва (перпендикулярно ему) с наружной стороны ра-
ковины могут быть шовные устья, снабженные труб-
чатыми выростами у специализированных форм. Ни-
жний карбон—пермь.
Примечание. Трубчатые выросты (мостики) ведут в нави-
тую трубчатую камеру через дополнительные устья и вместе со
щелями в пупочной области образуют примитивную систему кана-
лов. Lasiodiscida отличается от Archaediscida формой раковины,
правильным эволютным навиванием оборотов, наличием прими-
тивной системы каналов.
Семейства: Lasiodiscidae Reytlinger, 1956; Мо-
notaxinoididae Mikhalevich, 1999 (слаботрохоидная
раковина, без дополнительных устьев и боковых мос-
тиков).
Надотряд Involutinoida Hohenegger
et Piller, 1977
Раковина псевдодвухкамерная, со стенкой из 1
слоя радиально расположенных кристаллов, с псев-
допоровыми отверстиями, арагонитовая.
Примечание. В результате перекристаллизации стенка
часто становится полностью микрогранулярной.
Отряд Involutinida Hohenegger et Piller,
1977
Раковина без системы каналов. Нижняя пермь—
верхний мел.
Примечание. Сем. Ventrolamminidae, представители кото-
рого имеют многокамерную трохоидную раковину и иное строение
устья, выведено нами из состава отряда.
Надсем. Involutinoidea Butschli,
1880
Внутренняя полость камер без дополнительных
образований.
Семейства: Involutinidae Butschli, 1880; Planis-
pirillinidae Piller, 1978; Aulotortidae Zaninetti, 1984;
Triadodiscidae Zaninetti, 1984; Hirsutospirellidae Zani-
netti, Ciarapica, Cirilli et Cadet, 1985.
Надсем. Triasinoidea Loeblich et
Tappan, 1986
Полость трубчатой камеры подразделена столбиками.
Семейство Triasinidae Loeblich et Tappan, 1986.
Отряд Hottingerellida Mikhalevich, 1993
Пупочная область, заполненная пластинчатым ра-
ковинным веществом, пронизана системой каналов,
которая включает спиральный канал, параллельный
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
591
внутреннему краю трубчатой камеры, и систему ще-
левидных проходов, связывающих его с камерой.
Юра?—нижний мел.
Семейство Hottingerellidae Mikhalevich, 1993.
Надотряд Spirillinoida Hohenegger et
Piller, 1975
Раковина псевдодвухкамерная или многокамер-
ная, стенка из 1 кристалла извести или мозаики не-
скольких кристаллов; псевдопоры заполнены органи-
ческим гранулированным веществом, согласно лите-
ратурным данным, имеют в основании ситовидную
пластинку с микропорами.
Отряд Spirillinida Hohenegger et Piller,
1975
Раковина спиральноплоскостная или трохоидная,
псевдодвухкамерная. Верхний триас—голоцен.
Подотряд Spirillinina Hohenegger et
Piller, 1975
Раковины без системы каналов. Верхний триас—
голоцен.
Семейства: Spirillinidae Reuss et Fritsch, 1861;
Conicospirillinidae Mikhalevich, 1999 (с трохоидной
раковиной и пупочной областью, заполненной вто-
ричным раковинным веществом).
Подотряд Spirotrocholinina
Mikhalevich, 1993
Раковина с пластинами в пупочной области, иду-
щими от одного внутреннего края камеры к другому,
с проходящим между ними спиральным каналом.
Семейство Spirotrocholinidae Mikhalevich, 1993.
Отряд Patellinida Mikhalevich, 1992
Раковина трохоидная, с начальным отрезком
трубчатой камеры, составляющим часть оборота или
несколько оборотов, иногда совсем выпадающим, и
с более поздним многокамерным отделом, обороты
которого образованы 2, реже 3 узкими полулунными
камерами или 1 циклической камерой; камеры мо-
гут быть подразделены на камерки дополнительными
септами или радиальными трубочками. Нижняя юра—
верхний мел—голоцен.
Надсем. Hergottelloidea Loeblich et
Tappan, 1984
Раковина с простыми камерами, не подразделен-
ными на камерки.
Примечание. По-видимому, в надсемейство следует вклю-
чить сем. Chilostomellidae из роталиат, роды которого (Chilosto-
mella, Chilostomellus) имеют необычную для роталиат раковину с 2
камерами в обороте и конечное, а не базальное устье. Однако
отряд Chilostomellida нуждается в предварительной ревизии.
Семейства: Ungulatellidae Seiglie, 1964 (рако-
вина Ungulatella — типового рода семейства — близ-
ка по строению агглютирированной раковине Plagio-
raphe); Placentulinidae G. К. Kasimova, Poroshina et
Geodakchan, 1980 (без Ashbrookininae, но, по-види-
мому, включая род Metapatellina из Ungulatellidae);
Hergottellidae Loeblich et Tappan, 1984; Pannellainidae
Loeblich et Tappan, 1984 (возможно, сюда же следует
включить близкий по строению род Heronallenita —
устье неясно).
Надсем. Patellinoidea Rhumbler, 1906
Раковина с камерами, подразделенными на камер-
ки полусептами или радиальными трубочками.
Семейства: Patellinidae Rhumbler, 1906; *An-
nulopatellinidae Loeblich et Tappan, 1964; Paleopatel-
linidae Mikhalevich, 1999 (раковина сходна по стро-
ению с раковинами Patellinidae, даже внутренняя
колумелла может присутствовать, но трубчатая не-
подразделенная камера вокруг пролокулуса отсутст-
вует).
Отряд *Seabrookinida Mikhalevich,
1980
Раковина уплощенная, неправильно-овальная в
очертаниях, на ранней стадии с пролокулусом и 2 ка-
мерами по обе стороны от него, последующие каме-
ры полностью охватывают предыдущие, чередуются,
располагаясь под углом 180° друг к другу (билокули-
новый тип строения); устье конечное, овальное или
щелевидное, обычно окружено губой, соединенной
внутренней трубкой с предыдущей камерой. Верхний
мел—голоцен.
Примечание. Необычное для роталиат строение раковины
и конечное положение устья заставляют нас вывести отряд из Ro-
taliata и (хотя и с некоторой долей сомнения) поместить его в
данном классе.
Монотипический таксон, имеющий некоторое конвергентное
сходство с билокулинидами (Miliolana), но отличающийся от него
ультраструктурой стенки.
Семейство Seabrookinidae Cushman, 1927.
Класс Miliolata Saidova, 1981 (рис. 547,///)
Раковины, однокамерные (редка), псевдодвухка-
мерные, псевдомногокамерные, многокамерные, пре-
имущественный тип навивания — неправильно- и
правильно-клубковидный, плоскоспиральный (чаще
в сочетании с клубковидным), трохоидный — лишь
в виде исключения; у высших представителей рако-
592
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
вины могут быть циклическими и фузулиноидными;
может сохраняться флексостиль; камеры преимущес-
твенно трубчатые, чаще по 2, реже 3 в обороте (у
Soritida и высших Alveolinida они широкие и их
число в обороте значительно больше), у высших
представителей их может быть несколько тысяч,
полость камер может быть подразделена септами
(нередко нескольких порядков), столбиками, у про-
двинутых форм имеются интегрирующие системы
в виде столонов и туннелей; стенка у низших форм
агглютинированная, у высших — секреционная, мо-
нофонтинальная, игольчатая (фарфоровидная), с псев-
допорами (между рыхло расположенными игольчатыми
кристаллами), рост новой камеры происходит посте-
пенно, участок за участком, в направлении от осно-
вания к устью, органическая и известковая секреция
происходит одновременно; устье конечное, у низших
форм — простое, у высших — с наружным или
внутренним зубом различного строения, ситовидное
(вторично-множественное), дополнительные устья в
виде исключения; ядерный аппарат исследован
слабо, у высших секреционных форм он гетерокари-
отный, у агамонта Quinqueloculina мы наблюдали уд-
линенный Ма и множество Ми\ у Sorites известна
полимеризация соматических ядер; у Triloculina га-
меты с 2 жгутиками и аксостилем; донные, преиму-
щественно свободноживущие формы, но встречаются
и прикрепленные. Карбон—голоцен.
Подкласс Schlumbergerinana Mikhalevich, 1992
Раковина многокамерная, одного из милиолятных
типов строения; стенка агглютинированная или сме-
шанного типа (известковые иглы с большим количес-
твом агглютинированного материала). Мел—голоцен.
Примечание. Представители этого подкласса не достигают
такого многообразия и такой сложности строения раковины и
устьевого аппарата, как представители высшего подкласса милио-
лят — мил ноля ны.
Отряд Schlumbergerinida Mikhalevich,
1980
Раковина с правильно-клубковидным типом нави-
вания в 3, 5 и более плоскостях либо становящаяся
плоскоспиральной, последняя камера может быть
сильно объемлющей, камеры трубчатые, по 2, реже
по 3 в обороте; устье конечное, в виде округлого или
овального отверстия, с 1 или несколькими зубами или
без зубов, иногда множественное (с трематофором).
Мел—голоцен.
Примечание. В состав этого отряда входят некоторые
роды, стенка которых считается состоящей из кремнезема. Однако
секреционное происхождение кремнезема в стенке раковины этих
представителей пока не доказано (см. раздел «Химический со-
став»).
Семейства: Rzehakinidae Cushman, 1933; Sig-
moilopsidae Vella, 1957; Siphonapertinidae Saidova,
1975; Schlumbergerinidae Mikhalevich, 1980; Spiroglu-
tinidae Mikhalevich, 1983.
Подкласс Miliolana Saidova, 1981
Раковины псевдомногокамерные, псевдодвухка-
мерные или многокамерные (однокамерные в виде
исключения — Squamulinida), клубковидные (ред-
ко — однорядные, трохоидные) или плоскоспираль-
ные (чаще в сочетании с клубковидной частью), с
2 трубчатыми камерами в обороте и флексостилем
либо у некоторых плоскоспиральных (Alveolinida,
Soritida) с большим количеством широких камер
в обороте; устье простое, или с зубом различного
строения, либо ситовидное, дополнительные устья,
как правило, не развиты, у высших форм имеется
система внутренних перегородок и камерок, сообща-
ющихся фораменами, столонами, туннелями, система
каналов отсутствует; стенка секреционно-известко-
вая, фарфоровидная, монофонтинальная, с рыхлым
срединным слоем беспорядочных удлиненных
(игольчатых или палочковидных) кристаллов, выст-
ланным изнутри и (как правило) снаружи облицо-
вочным слоем кристаллов (удлиненной, плитчатой
или иной формы), уложенных горизонтально (парал-
лельно наружной стенке) и более плотно, со случай-
ными, неправильной формы отверстиями между
ними на наружной поверхности стенки; поровые от-
верстия на поверхности постэмбриональных камер
нерегулярные, неправильной формы, видимые только
на электронном уровне, оформленные поровые ка-
нальцы, характерные для нодозариан и роталиан, от-
сутствуют. Карбон—настоящее время.
Примечание. Внутренний органический слой стенки со-
стоит из фибриллярного слоя и пересекающихся трубчатых эле-
ментов (Leutenegger, 1977). У Nubeculina обнаружены поровые от-
верстия на внутреннем слое стенки, ведущие вглубь до середины
толщины стенки, у Peneroplis — изогнутые, ветвящиеся канальцы
в рыхлом срединном слое стенки. И те и другие замкнуты слепо,
выходы на поверхность не оформлены.
Надотряд Squamulinoida Mikhalevich,
1988
Раковина однокамерная. Верхний мел—голоцен.
Отряд Squamulinida Mikhalevich,
1988
Раковина однокамерная. Верхний мел—голдцен.
Семейство Squan^ulinidae Reuss et Fritsch, 1861.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
593
Надотряд Cornuspiroida Jirovec, 1953
Раковина псевдодвухкамерная, без флексостиля.
Отряд Cornuspirida Jirovec, 1953
(=Order Cornuspirida Mikhalevich, 1980 (part.),
suborder Cyclogyrina Saidova, 1981)
Раковина свободная либо прикрепленная, псев-
додвухкамерная, с округлым пролокулусом и длинной
второй трубчатой камерой, навитой плоскоспираль-
но, клубковидно, зигзагообразно либо неправильно,
может быть распрямленной; устье простое; стенка
простая. Карбон—голоцен.
Подотряд Cornuspirina Jirovec, 1953
Раковина свободная или прикрепленная, псевдо-
двухкамерная, вторая камера навита плоскоспирально
по крайней мере в начальном отделе свободноживу-
щих форм, у прикрепленных форм может быть сильно
измененной, слегка ветвящейся. Карбон—голоцен.
Семейство Cornuspiridae Cushman, 1919 (с 4
подсемействами).
Подотряд Hemigordiopsina
Mikhalevich, 1986
Раковина свободная или прикрепленная, вторая
трубчатая камера навита неправильно-клубкообразно
по крайней мере в начальном отделе. Пермь—голо-
цен.
Надсем. Hemigordiopsidea Nikitina,
1969
Стенка раковины простая. Пермь—голоцен.
Семейство Hemigordiopsidae Nikitina, 1969 (с
4 подсемействами).
Надсем. Shanitoidea Mikhalevich, 1986
Раковина с внутренними выростами стенки. Ниж-
няя пермь—верхний триас.
Семейства: Baisalinidae Loeblich et Tappan, 1986
(с 2 подсемействами); Shanitidae Mikhalevich, 1986 (с
1 подсемейством).
Надотряд Costiferoida Mikhalevich,
1988
Раковина без флексостиля, с камерами, располо-
женными в 1 ряд — прямой или изогнутый, камеры
неправильно-грушевидные или широкоовальные; ус-
тье простое, без зуба, может быть на конце вытяну-
той шейки, с отворотом. Верхний триас.
38 Протисты, часть 1
Отряд Costiferida Mikhalevich, 1988
Характеристика отряда совпадает с таковой под-
отряда.
Семейство Costiferidae Senowbar-Daryan et Za-
ninetti, 1986.
Надотряд Milioloida Delage
et Herouard, 1896
Раковина многокамерная, с флексостилем, каме-
ры изначально трубчатые, по 2, реже 2.5—3 в обо-
роте.
Отряд Nubeculariida Jones, 1875
Раковина свободная или прикрепленная, плоско-
спиральная, эволютная, камеры изначально трубча-
тые, по 2, реже по 3 в обороте, у продвинутых форм
могут становиться циклическими; устье простое, мо-
жет быть с шейкой и губой; стенка простая или с
внутренними неполными перегородками. Средний три-
ас—голоцен.
Надсем. Nubeculariidea Jones, 1875
Внутренняя стенка камер простая. Средний три-
ас—голоцен.
Семейства: Nubeculariidae Jones> 1875; Ophtal-
midiidae Wiesner, 1920; Meandroloculinidae Bogdano-
vich, 1981; Nodobaculariellidae Bogdanovich, 1981;
Wiesnerellidae Saidova, 1981; Spiriamphorellidae Se-
nowbary-Daryan et Zaninetti, 1986; Stellarticulinidae
Mikhalevich, 1988.
Надсем. Discospirinoidea Wiesner, 1931
Внутренняя стенка камер усложнена неполными
перегородками, последние камеры располагаются
циклически. Верхний мел—голоцен.
Семейство Discospirinidae Wiesner, 1931.
Отряд Miliolida Delage et Herouard,
1896
Раковина в подавляющем большинстве случаев
свободная, с изначально клубковидным навиванием,
с вторичными переходами к плоскоспиральному рас-
положению камер у некоторых родов, в большинстве
случаев с инволютными оборотами, часто с инво-
лютными 1—2 последними камерами у продвинутых
групп (альвеолиниды) одного из «фузулиноидных»
типов, камеры чаще трубчатые, иногда последние
сильно уплощенные (Hauerinidae); устье простое
лишь у небольшой группы более примитивных форм,
у всех остальных усложненное — с наружным плас-
тинчатым зубом (устьевой лопастью), или внутрен-
ним зубом различной степени сложности, либо сито-
594 СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
видное; у 2 родов известны дополнительные шовные
устья (Hauerinidae); стенка простая, или с внутрен-
ними неполными перегородками, либо со сложной
системой внутренних перегородок, полностью деля-
щих камеру на камерки, с дополнительной системой
внутренних подразделений. Нижний мел—голоцен.
Подотряд Miliolina Delage et Herouard,
1896
Раковина в основном сохраняет один из милио-
лидных типов строения; стенка камер простая, или
усложненная, либо с неполными перегородками. Ниж-
ний мел—голоцен.
Надсем. Quinqueloculinoidea Cushman,
1917
Стенка простая, камеры неподразделенные. Ниж-
ний мел—голоцен.
Примечание. Семейства этого таксона выделены на основе
строения раковин и расположения их камер по одному из милио-
лятных типов (квинквелокулиновому; сигмоилиновому; трилокули-
новому; билокулиновому — например, Pyrgo, с последними
объемлющими камерами; спиролокулиновому) либо их различных
сочетаний, иногда с наличием выпрямленного отдела. Подсемей-
ства, которые здесь не приводятся, выделены на основании особен-
ностей строения устьевого аппарата.
Семейства: Hauerinidae Schwager, 1876; Tubi-
nellidae Rhumbler, 1906; Quinqueloculinidae Cushman,
1917; Spiroloculinidae Wiesner, 1920; Massilinidae
Thalmann, 1941; Sigmoilinitidae Luczkowska, 1974;
Glomulinidae Saidova, 1981; Triloculinidae Bogdano-
vich, 1981; Zoyaellidae Saidova, 1981; Pyrgoidae Mik-
halevich, 1983; Miliolechnidae Zaninetti, Ciarapica, Ci-
rilli et Cadet, 1985; Labalinidae Mikhalevich, 1988.
Надсем, Milioloidea Ehrenberg,
1839
Раковина co сложной стенкой, с многочисленны-
ми псевдопорами на поверхности, полость камер не
подразделена. Средний эоцен—голоцен.
Семейства: Miliolidae Ehrenberg, 1839; Neagui-
tinidae Andersen, 1984.
Надсем. Austrotrillinoidea Mikhalevich,
1986
Внутренняя полость частично подразделена не-
полными перегородками. Верхний мел—голоцен.
Примечание. Надсемейство занимает промежуточное поло-
жение между подотрядами Miliolina (с простыми камерами и стен-
кой) и Alveolinina (с камерами, полностью поделенными на камер-
ки усложненной системой перегородок и столбиков).
Семейства: Riveroididae Saidova, 1981; Aust-
rotrillinidae Mikhalevich, 1986; Brebinidae Mikhale-
vich, 1986.
Подотряд Alveolinina Mikhalevich, 1980
(= Alveolinidae Loeblich et Tappan, 1964;
Alveolinellinina Saidova, 1981)
Раковина многокамерная, в начальном отделе у
микросферических форм клубковидная (квинквело-
кулиновая, трилокулиновая), очень небольшая, у ме-
галосферических — билокулиновая (за пролокулу-
сом еще 1 камера), все остальные обороты обеих
генераций инволютные, плоскоспиральные у Fabula-
riidae и Alveolinidae либо клубкообразные у Кега-
mosphaeridae, раковина по форме шаровидная, вере-
теновидная (фузулиноидная), с очень большим чис-
лом камер, полностью поделенных на камерки
усложненной системой перегородок и столбиков
(септальные и тангенциальные перегородки могут де-
лить камеру на 2 и более этажей), когда вторичные
спиральные перегородки не доходят до ограничива-
ющих камеру септ, образуются пресептальный и пос-
тсептальный туннели; устье множественное — сито-
видное или в виде многочисленных отверстий на
септальной поверхности раковины (всегда выше ос-
нования септальной поверхности). Нижний мел—го-
лоцен.
Примечание. У Alveolinella наблюдались поры у пролоку-
луса и первых оборотов, напоминающих поры Peneroplis (Hofker,
1971).
Семейства: Alveolinidae Ehrenberg, 1839; Fabu-
lariidae Ehrenberg, 1839; Keramosphaeridae Brady, 1884.
Отряд Soritida Schultze, 1854
(=Orbitolinida Ehrenberg, 1839;
Orbitolitida Wedekind, 1937)
Раковина многокамерная, с 3 и более обычно ши-
рокими камерами в обороте, флексостиль обнаружен
у одной из генераций не у всех представителей, на-
вивание камер плоскоспиральное (в виде исключе-
ния — трохоидное), чаще эволютное, реже инволют-
ное, раковины с тенденцией к веерообразному и
циклическому расположению последних камер, реже
выпрямленному; стенка простая у низших представи-
телей, у ранних камер с ямками или прободенная, у
высших представителей — со сложным внутренним
скелетом из пластинок и столбиков, с резко возрас-
тающим числом камер, у высокоорганизованных
представителей могут быть столоны; устье — без
зуба, простое либо сложное, множественное, всегда
выше основания септальной поверхности. Верхний
мел—голоцен.
Надсем. Peneroplidea Schultze,
1854
Стенка камер простая, каморы немногочислен-
ные, не подразделенные. Верхний мел—голоцен.
Семейства: Peneroplidae Schultze, 1854; Fishe-
rinidae Millett, 1899; Fisherinellida^ Saidova, 1981.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
595
Надсем. Meandropsinoidea Henson,
1848
Раковина плоскоспиральная на ранней стадии, поз-
же может быть дискоидальной, веерообразной, ци-
линдрической или конической, маргинальная зона
камер с интерсептальными столбиками и септулами,
подразделяющими камеры не до конца; устье мно-
жественное, редко простое, одиночное. Верхний
мел—голоцен.
Примечание. Сем. Rhapydionidae перенесено из альвеоли-
нид на основании строения начальной части раковины — плоско-
спиральной, с несколькими широкими камерами в обороте.
Семейства: Meandropsinidae Henson, 1848; Rha-
pydionidae Keijer, 1945; Fusarchaiasidae Saidova, 1981;
Hottingerinidae Mikhalevich, 1988.
Надсем. Soritidoidea Ehrenberg, 1839
Раковина с полостью камер, разделенной на ка-
мерки, сообщающиеся с помощью столонов, которые
связывают камерки предыдущего и последующего ря-
дов, но не камерки одного ряда. Верхний мел—голоцен.
Семейства: Soritidae Ehrenberg, 1839; Archaia-
sidae Cushman, 1927; Praerhapydionidae Hamaoui et
Fourcade, 1973; Cycledomiidae Mikhalevich, 1988.
Класс Nodosariata Mikhalevich, 1992 (рис. 547, IV)
Раковины однокамерные и многокамерные (псев-
домногокамерные в виде исключения), преимуще-
ственный тип раковин одноосный, спирально-винто-
вой (в виде особой полиморфиноидной спирали, не
встречающейся в других таксонах), плоскоспираль-
ный (почти исключительно инволютный), трохоид-
ный тип не представлен; камеры часто округлые,
треугольные, V-образные, пальмовидные, дополни-
тельные внутренние перегородки камер лишь у 1
вымершего таксона; наружная скульптура у секреци-
онных форм хорошо развита, но наружные дополни-
тельные скелетные пластинки и интегрирующие системы
(столоны, туннели, каналы) отсутствуют (за исклю-
чением встречающихся изредка внутренних устьевых
структур — взаимосвязанных энтосолениевых трубок);
стенка монофонтинальная, у низших представителей
(Hormosinana) агглютинированная, может быть с из-
вестковым цементом и микрогранулярной, у высших
(Nodosariana) — радиальная, с регулярными псевдо-
порами; рост прерывистый, способ образования но-
вых камер не изучен; устье конечное даже у сверну-
тых форм, единичное или множественное, простое
(округлое, овальное, щелевидное, трирадиальное) ли-
бо радиальное, может быть с внутренней (энтосоле-
ниевой) трубкой (не аналогичной трубкам роталиат),
щелевидное устье может быть асимметричным, с на-
висающим колпачком, 3 последних типа устья (ради-
альное, с энтосолениевой трубкой и асимметричное)
уникальны для данного таксона также, как и имею-
щаяся у некоторых родов устьевая камерка, допол-
нительные устья не развиты; данные по строению
ядер и гамет отсутствуют; преимущественно свобод-
ноживущие, донные, описан 1 паразитирующий вид.
Ордовик—голоцен.
Примечание. По-видимому, сюда же следует перенести из
прежних Fusulinina сем. Palaeotextulariidae Galloway, 1933, в стен-
ке которого имеется слой радиальных кристаллов. Эти древние
формы (карбон—пермь) нуждаются в дополнительных исследова-
ниях и здесь подробно не рассматриваются. Д. М. Раузер-Черно-
усова (1990) отмечает преемственность палеозойских и мезозой?
ских нодозариат, постепенное исчезновение микрогранулярного
слоя стенки в процессе исторического развития. У монотипическо-
го подотряда Delosinina отмечена своеобразная система каналов,
открывающихся отверстиями в швах, однако она изучена недоста-
точно (Revets, 1989).
Подкласс Hormosinana Mikhalevich, 1992
Раковина с одним из нодозариатных типов распо-
ложения камер и строения устья; стенка монофонти-
нальная, агглютинированная или микрогранулярная,
может быть усложнена изнутри, псевдопоры не изу-
чены. Ордовик—голоцен.
Примечание. Вероятно, не только Pseudopalmulidae, но и
ряд других форм прежних Palaeotextulariacea (исключая Koskinobi-
generininae) будут переведены в данный класс после дополнитель-
ных исследований. Для выведения родов, принадлежащих к под-
классу Hormosinana, из состава подкласса Textulariana требуется
дополнительное изучение строения их устья на начальных стадиях,
особенно в семействах Mayncinidae и Charentinidae. Роды Lamina,
Triplasia, Flabellammina, Flabellamminopsis, Ammopalmula имеют
один из нодозариатных типов строения и, по-видимому, должны
быть переведены в подкласс Hormosinana.
Отряд Hormosinida Mikhalevich,
1980
Раковина представляет собой прямой или изогну-
тый ряд увеличивающихся в размерах камер; стенка
агглютинированная, скрепленная органическим или
известковым цементом, часто сильно ожелезненным,
или микрогранулярная, может быть пронизанной ка-
нальцами или альвеолярной; устье конечное, про-
стое, округлое (может быть с приподнятой шейкой и
губой) или щелевидное, иногда ситовидное, у одного
рода (Nodosinum) с внутренними радиальными ребра-
ми, у форм с разветвленной последней камерой усть-
ев 2. Ордовик—голоцен.
596
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Примечание. Однорядные агглютинированные формы с
внутренним зубом (Dusenburyinidae), относимые ранее к хормози-
нидам; мы включаем их в подкласс Textulariana (Михалевич,
19986).
Надсем. Hormosinoidea Haeckel,
1894
Раковины с простой стенкой.
Семейства: Hormosinidae Haeckel, 1894; Reo-
phacidae Cushman, 1910; *Oxinoxisidae Vyalov, 1968;
*Glaiicoamminidae Saidova, 1981; Cuneatidae Loeblich et
Tappan, 1984; *Earlandinitidae Loeblich et Tappan, 1984.
Надсем. Cribratinoidea Loeblich
et Tappan, 1984
Раковина с усложненной внутренней структурой
стенки (с канальцами или альвеолярная).
Семейства: Cribratinidae Loeblich et Tappan,
1984; Thomasinellidae Loeblich et Tappan, 1984.
Отряд Nouriida Mikhalevich, 1980
Раковина с камерами, расположенными по поли-
морфиноидному типу; стенка агглютинированная. Мел—
голоцен.
Семейство Nouriidae Chapman et Рагг, 1936.
Отряд Pseudopalmulida Mikhalevich,
1992
Раковина двухрядная по крайней мере в началь-
ном отделе, с характерными камерами V-образной
формы; стенка микрогранулярная. Девон.
Семейство Pseudopalmulidae Е. V. Bykova, 1959
(по-видимому, без рода Paratextularia).
Подкласс Nodosariana Mikhalevich, 1980
Раковина с одним из нодозариатных типов распо-
ложения камер и строения устья; стенка секрецион-
ная, известковая, монофонтинальная, стекловато-лу-
чистая (радиальная), с псевдопорами, имеющими очень
узкие канальцы (до 0.33 мкм в диаметре) и ситовидную
поровую пластинку в основании, но, согласно литера-
турным данным, без дополнительных ситовидных плас-
тинок; с хорошо развитыми слоями нарастания и на-
ружной скульптурой (ребра, шипы, иглы, бугорки).
Примечание. У некоторых древних форм в стенке имеется
микрогранулярный слой, появившийся, скорее всего, в результате
перекристаллизации. В системе Лёблика и Таппан (Loeblich, Tap-
pah, 1984) многие близкородственные формы сходного строения
помещены в разные, удаленные друг от друга таксоны из-за осо-
бенностей строения их стенки (например, Robuloididae с «одно-
слойной стенкой» и Lenticulinidae с «двухслойной»). Но этот при-
знак у нодозариан «не работает». Даже сами авторы были вынуж-
дены включить в Ichtyolariidae также формы с двойной стенкой,
так как и та и другая стенки встречаются иногда у разных предста-
вителей одного рода. Хотя в диагнозах «Классификации» 1988 г.
(Loeblich, Tappan, 1988) и говорится о «двухслойной» (биламел-
лярной) стенке, по-видимому, все же картину искажают слои на-
растания, так как при электронно-микроскопических исследова-
ниях СОМ у нодозариан пока не обнаружена, и, следовательно, их
стенку нельзя отнести к биламеллярному (бифонтинальному) типу.
Отряд Lagenida Delage et Herouard,
1896
Раковина однокамерная; устье множественное или
(чаще) единичное, округлое (простое или радиаль-
ное, может быть на вытянутой шейке, с отворотом)
или щелевидное (может быть асимметричным, с на-
висающим колпачком), как округлое, так и щелевид-
ное устья могут быть с энтосолениевой трубкой. Верх-
ний силур—юра—голоцен.
Примечание. У древнего рода Eolagena из силура наруж-
ный слой стенки микрогранулярный, скорее всего перекристалли-
зованный.
Семейства: Lagenidae Reuss, 1862 (с под-
семействами Lageninae Reuss, 1862, Cribrolageninae
Mikhalevich, 1993); Ellipsojagenidae A. Silvestri, 1923
(с подсемействами Ellipsolageninae A. Silvestri. 1923;
Oolininae Loeblich et Tappan, 1961; Parafissurininae
R.W. Jones, 1984).
Отряд Nodosariida Galkins, 1926
Раковина многокамерная или псевдомногокамер-
ная, однорядная. Силур—голоцен.
Подотряд Syzraniina Mikhalevich,
1992
Раковина псевдомногокамерная (псевдокамеры
образованы пережимами стенки), в поперечном сече-
нии округлая или квадратная, с несколько вдавлен-
ными боковыми сторонами; у некоторых древних
родов внутренний слой стенки микрогранулярный;
устье простое, округлое. Силур—девон—карбон—
пермь, голоцен.
Семейство Syzraniidae Vachard, 1981 (включая
современный род Grigelis Mikhalevich, 1981).
Подотряд Nodosariina Calkins, 1926
Раковина многокамерная, иногда с объемлющими
камерами, в поперечном сечении округлая, овальная,
уплощенная, треугольная или субквадратная; стенка
у современных форм только радиальная, у древних
может быть с микрогранулярным слоем; устье округ-
лое (простое или радиальное), щелевидное, иногда
множественное. Триас—голоцен. /
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
597
Примечание. Формы с двух- и трехрядной раковиной
(хотя бы только в начальном отделе) исключены из состава отряда.
Если у Textulariana (см. ниже) двухрядные раковины происходят
от плоскоспиральных, то у Nodosariana они ведут свое происхож-
дение как от плоскоспиральных (Palmulidae), так и от спирально-
винтовых форм (Plectofrondiculariidae, Partizaniidae) и попадают
соответственно в разные отряды. Формы с V-образными (пальмо-
видными) камерами могут иметь место в обеих этих линиях, а
также среди целиком однорядных форм.
Надсем. Nodosarioidea Ehrenberg,
1838
Раковина с неподразделенной полостью камер.
Семейства: Nodosariidae Ehrenberg, 1838; Fron-
diculariidae Reuss, 1860; Nodosinellidae Rhumber,
1895; Ptychocladiidae Elias, 1950; Geinitzinidae Bozor-
gnia, 1973; Pachyphloiidae Loeblich et Tappan, 1984;
Ichtyolariidae Loeblich et Tappan, 1986; Daucinoididae
Mikhalevich, 1993.
Надсем. Colanielloidea Fursenko,
1959
Раковина с полостью камер, подразделенной вер-
тикальными радиальными септами.
Семейство Colaniellidae Fursenko, 1959.
Отряд Polymorphinida Mikhalevich,
1980
Раковина многокамерная, спирально-винтовая или
произошедшая от нее (двухрядная, с однорядным от-
делом или даже целиком вторично-однорядная), ка-
меры, как правило, удлиненные; стенка радиальная
(изредка описывают микрогранулярную стенку);
устье округлое (радиальное или простое) или щеле-
видное, в обоих случаях может быть с энтосоление-
вой трубкой.
Подотряд Polymorphinina Mikhalevich,
1980
Раковина с округлым, радиальным или щелевид-
ным устьем, с энтосолениевой трубкой или без нее,
может иметь дополнительную устьевую камерку.
Пермь, триас—юра, голоцен.
Семейства: Polymorphinidae d’Orbigny, 1839;
Glandulinidae Reuss, 1860; Plectofrondiculariidae
Cushman, 1927; Partizaniidae Loeblich et Tappan, 1984.
Подотряд ‘Pleurostomellina
Mikhalevich, 1993
Раковина с устьем, нависающим в виде капюшона,
энтосолениевые трубки последовательных камер со-
единяются в одну систему, их расположение у одно-
рядных раковин отражает двухрядный характер пред-
ковых форм. Мел—голоцен.
Примечание. 2 рода, не имеющие устьевой трубки, исклю-
чены из состава плевростомеллид. У Ellipsolingulina трубка корот-
кая, вероятно вследствие того, что внутренние стенки камер у
взрослой особи растворяются, образуя общую полость раковины.
Стенка плевростомеллид исследована недостаточно, но отмечены
случаи микрогранулярной стенки, хотя их и включали ранее в под-
класс Rotaliana (=надсем. Rotaliacea в системе Лёблика и Тап-
пан, — Loeblich, Tappan, 1988).
Семейство Pleurostomellidae Reuss, 1860.
Подотряд Delosinina Revets, 1989
Раковина высокоспиральная, трехрядная, со взду-
тыми удлиненными сильно объемлющими камерами,
быстро увеличивающимися в размерах; устье отсутст-
вует, от основания септальной поверхности отходит
явственно очерченная аркообразная область, прони-
занная мелкими порами, крупные дополнительные
шовные устья ведут в каналы, расположенные под
швами, которые, в свою очередь, ведут к пронизан-
ной порами устьевой области. Голоцен.
Примечание. У единственного рода этого монотипического
семейства, ранее относимого к роталиатам, бифонтинальная стен-
ка не обнаружена (Revets, 1989). Строение его раковины также
сходно с раковинами полиморфинид.
Семейство Delosinidae Рагг, 1950.
Отряд Vaginulinida Mikhalevich, 1992
Раковина многокамерная, плоскоспиральная, по
крайней мере в начальном отделе или у одной из ге-
нераций (бесполой), у микросферических форм поз-
дний отдел может быть развернутым, либо мегало-
сферическая раковина полового поколения может
быть целиком однорядной (Spirolingulinidae, Palmuli-
dae), как у Lingulina; обороты, за исключением Turk-
menella, инволютные; стенка радиальная; устье еди-
ничное (щелевидное или радиальное) или множест-
венное. Пермь, триас, юра, мел, голоцен.
Семейства: Vaginulinidae Reuss, 1860; Lenticu-
linidae Chapman, Parr et Collins, 1934; Marginulinidae
Wedekind, 1937; Robuloididae Reiss, 1963; Palmulidae Sa-
idova, 1981; Spirolingulinidae Loeblich et Tappan, 1986.
Класс Rotaliata Mikhalevich, 1980 (рис. 547, V)
Раковины только многокамерные, широко пред-
ставлены все правильные типы строения раковин,
клубковидный тип практически отсутствует, преиму-
щественными типами являются трохоидные и плос-
коспиральные, а также произошедшие от тех и дру-
гих двух- и трехрядные (редко однорядные), у про-
двинутых форм нередки циклические раковины,
внутренние усложнения полости камер чрезвычайно
598
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
развиты: от выростов, пластинок, столбиков (у Tex-
tulariana) до септ и септул нескольких порядков у
высших представителей обоих подклассов, у кото-
рых число камер может достигать нескольких тысяч,
причем у Orbitolina и Nummulites центральные, меди-
анные и периферические камеры дифференцированы
по структуре; часто имеются дополнительные ске-
летные пластинки (над швами, над пупочной или
устьевой областью), особенно у высших роталиат;
интегрирующие системы сильно развиты (столоны,
туннели и взаимосвязанные внутренние устьевые
структуры у Textulariana; столоны, взаимосвязанные
внутренние устьевые структуры и сложные системы
каналов у Rotaliana); стенка у текстуляриан агглюти-
нированная и микрогранулярная, монофонтинальная,
с псевдопорами, часто усложненная изнутри — яче-
истая, альвеолярная, тубчатая, у роталиан — бифон-
тинальная, с порами, имеющими цилиндрические по-
ровые канальцы с несколькими поперечными поро-
выми пластинками, вновь формирующаяся последняя
камера образуется сразу целиком, в направлении от
устья к основанию, известковая и органическая сек-
реция разделены во времени; устье обычно базаль-
ное — в основании септальной поверхности как у
свернутых, так и у некоторых двух- и трехрядных
форм, но также может быть ареальным и ситовид-
ным, у вытянутых форм может становиться конечным;
внутренние структуры устья чрезвычайно развиты в
обоих подклассах, но достигают особой сложности у
высших роталиат, представлены зубными пластинка-
ми различной длины, формы и сложности (иногда с
боковыми отростками), язычками, желобками, сифо-
нами, клапанами, часто образующими единые взаи-
мосвязанные внутренние устьевые системы; дополни-
тельные устья имеются у обоих подклассов (шовные,
пупочные, на концах камер); известные агамонты
текстуляриан многоядерные, гомокариотные, агамонты
высших роталиат многоядерные, гетерокариотные, га-
меты двухжгутиковые, реже трехжгутиковые (у Glabra-
tella, Nummulites venosus)', преимущественно свободно-
живущие, донные, редко прикрепленные, известно
несколько паразитических видов. Карбон—голоцен.
Подкласс Textulariana Mikhalevich, 1980
Раковина с одним из роталиатных типов располо-
жения камер и строения устья; стенка монофонти-
нальная, агглютинированная или микрогранулярная,
может быть усложнена изнутри (альвеолярная, лаби-
ринтовая, губчатая, с дополнительными неполными пе-
регородками, столбиками и т. д.). У многих отмечены
псевдопоры и псевдопоровые канальцы, иногда развет-
вляющиеся, наружная скульптура отсутствует, но до-
полнительные скелетные пластинки в пупочной облас-
ти могут быть; у продвинутых форм с большим числом
камер интегрирующие системы в виде столонов и тун-
нелей, внутренние устьевые структуры подобны тако-
вым роталиан, но менее развиты. Карбон—голоцен.
Надотряд Dusenburyinoida
Mikhalevich, 1998
Раковина однорядная; устье с внутренним зубом.
Голоцен.
Отряд Dusenburyinida Mikhalevich,
1998
Характеристика отряда совпадает с таковой над-
отряда.
Примечание. Среди однорядных многокамерных раковин с
агглютинированной стенкой большинство представителей отно-
сится к подклассу Hormosinana. Определить принадлежность этих
форм к подклассу Hormosinana или Textulariana помогает строение
устья (как, например, у Nodosinum, Cuneata), форма раковины и
камер (например, у Polychasmia, Adelungia). Устье с внутренним
зубом у Desenburyina неоспоримо свидетельствует о ее принадлеж-
ности к Textulariana (Михалевич, 19986).
Семейство Dusenburyinidae Loeblich et Tap-
pan, 1984.
Надотряд Lituoloida Calkins, 1909
Раковина линзовидная, плоская, округлая, удли-
ненная, с плоскоспиральным или стрептоспираль-
ным навиванием по крайней мере на ранних стадиях,
свернутая целиком или на поздних стадиях с вы-
прямленным однорядным отделом цилиндрической
или веерообразной формы, либо камеры могут ста-
новиться циклическими, выпрямленный отдел обыч-
но меньше свернутого. Карбон—голоцен.
Примечание. У высших таксонов всех 3 входящих сюда от-
рядов чаще имеется микрогранулярная стенка и усложненная внут-
ренняя структура, большинство из них вымерло в юре и в мелу.
Отряд Haplophragmiida Mikhalevich,
1992
Раковина со стрептоспиральным навиванием
камер по крайней мере в начальном отделе, эволют-
ная или инволютная, поздний отдел может быть вы-
прямленным; устье в основании септальной поверх-
ности или над нею, единичное или множественное,
у выпрямленных форм — конечное. Триас—голо-
цен.
Надсем. Ammosphaeroidinoidea
Cushman, 1927
Раковина с простой стенкой. Средняя юра—голо-
цен.
Семейства: Ammosphaeroidinidae Cushman,
1927; Recurvoididae Alekseychik-Mitskevich, 1973;
Ammobaculinidae Saidova, 1981. ,
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
599
Надсем. Haplophragmioidea Eimer
et Fichert, 1899
Раковина с альвеолярной стенкой. Средний три-
ас—голоцен.
Семейства: Haplophragmiidae Eimer et Fichert,
1899; ?Mesoendothyridae Voloshinova, 1958.
Надсем. Labyrinthidomatidoidea
Loeblich et Tappan, 1988
Раковина с полостью камер, подразделенной внут-
ренними септами. Нижний и верхний мел.
Семейства: Barkerinidae Smout, 1956; Labyrint-
hidomatidae Loeblich et Tappan, 1988.
Отряд Lituolida Lankester, 1885
Раковина с плоскоспиральным навиванием камер
по крайней мере в начальном отделе, эволютная или
инволютная, поздний отдел может быть выпрямлен-
ным; устье в основании септальной поверхности или
ареальное, единичное или множественное, у выпрям-
ленных форм — конечное. Нижний карбон—голоцен,
но большинство таксонов с юры.
Примечание. Семейства выделяются на основании эволют-
ной или инволютной раковины, наличия или отсутствия выпрям-
ленного отдела, формы раковины, подсемейства — на основании
особенностей строения устья.
Надсем. Lituolinoidea de Blainville,
1827
Раковины целиком свернутые или с выпрямлен-
ным отделом, с простой стенкой.
Примечание. Вероятно, среди Mayncinidae, Flabellammini-
dae и Ammomarginulinidae есть формы, давшие начало высшим
нодозариатам, а не роталиатам, но для их выявления нужен тща-
тельный морфологический анализ, особенно выяснение строения
устья у камер начальных оборотов.
Семейства: Lituolidae de Blainville, 1827; Pla-
copsilinidae Rhumbler, 1913; Sphaeramminidae Cushman,
1933; Haplophragmoididae Mayne, 1952; ?FIabellam-
minidae Podobina, 1978; Ammomarginulinidae Podobi-
na, 1978; Ammobaculitidae Alekseychik-Mitskevich, in
Subbotina, 1981; Mayncinidae Loeblich et Tappan, 1985;
Nautiloculinidae Loeblich et Tappan, 1985.
Надсем. Biokovinoidea Gusczic,
1977
Раковина целиком свернутая или с коротким вы-
прямленным отделом, утолщенной стенкой, услож-
ненной внутренней структурой — псевдоальвеоляр-
ной (с канальцами, обычно не доходящими до внут-
ренней или наружной поверхности, но иногда
открывающимися на одну из них или на обе, напри-
мер у Lituoliporidae).
Примечание. У Biokovinidae кроме открывающихся нару-
жу канальцев ниже них находится слой, напоминающий альвео-
лярный, имеются межсептальные столбики, имеются также слои,
похожие по положению и форме на фенотеку и кериотеку фузули-
нид, у Charentinidae основание септ утолщено наподобие хомат.
Стенка Biokovinidae по сложности как бы переходная между стен-
кой Biokovinidae и Loftusioidea, граница между этими труппами
нечеткая. Сем. Hottingeritidae перенесено нами в подкласс Ат-
modiscana (класс Spirillinata), так как для его типового рода ти-
пично не базальное, а конечное устье — широкий открытый
конец последней камеры. Большое число камер в обороте, имею-
щих несколько неправильную форму, также более характерно для
представителей псевдокамерных и многокамерных аммодискан.
Подсем. Buccicrenatinae (сем. Cyclamminidae) тоже перенесено
нами в подкласс Ammodiscana, так как строение его устья, камер
и способ их сочленения, а также подобные хоматам внутренние
утолщения стенки свидетельствуют о его сходстве с торнайеллида-
ми и эндотиридами, произошедшими от трубчатых форм.
Семейства: Biokovinidae Gusczic, 1977; Lituoli-
poridae Gusczic et Velic, 1978; Charentiidae Loeblich
et Tappan, 1985.
Надсем. Loftusioidea Brady, 1884
Раковина целиком свернутая или с выпрямлен-
ным отделом, цилиндрическим или расширяющимся
до веерообразного; стенка утолщенная, с отдельными
септулами (Discamminidae, Spiropsammidae) или с
хорошо развитым альвеолярным слоем, иногда со
столбиками.
Примечание. Сем. Mesoendothyridae со стрептоспираль-
ным навиванием оборотов переведено нами в отряд Haplophragmi-
ida, а сем. Spirocyclinidae с циклическими камерами, с внутренни-
ми столбиками и септами -4- в отряд Cyclolinida. Сем. Spiropsam-
midae перенесено из отряда Spiroplectamminida, поскольку имеет
сильно развитые плоскоспиральный и узкий однорядный, а не
двухрядный выпрямленный отделы. Самым примитивным в этой
группе является сем. Discamminidae с органическими септами, но
с толстой стенкой с глубокими, частично изолированными полос-
тями. У Loftusiida кроме альвеолярного слоя имеются столбики.
Семейства: Loftusiidae Brady, 1884; Cyclammi-
nidae Marie, 1941 (без Buccicrenatinae), Choffatellidae
Mayne, 1958; Discamminidae Mikhalevich, 1980; Spi-
ropsammidae Seiglie et Baker, 1984; Pseudochoffatelli-
dae Loeblich et Tappan, 1985; Labyrinthinidae Mik-
halevich, 1992 (включая Biplanata из Nezzazatidae c
внутренними дополнительными септами, зубчатыми по
краю).
Надсем. Coscinophragmatoidea
Thalmann, 1951
Раковины прикрепленные, в начальном отделе
спирально свернутые, в позднем отделе неправиль-
ные, могут быть разветвленными, с утолщенной, аль-
веолярной стенкой.
Семейства: Coscinophragmatidae Thalmann, 1951;
Haddonidae Saidova, 1981.
Отряд Cyclolinida Mikhalevich, 1992
Начальный отдел раковины плоскоспиральный
или с колеблющейся осью навивания, поздний — ве-
600
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ерообразный и кольцевой; устье множественное, в
виде округлых отверстий по периферическому краю
раковины.ЛОра—эоцен.
Надсем. Cyclolinoidea Loeblich
et Tappan, 1964
Раковина с простой стенкой и неподразделенной
полостью камер.
Семейство Cyclolinidae Loeblich et Tappan,
1964.
Надсем. Cyclopsinelloidea Loeblich
et Tappan, 1984
Раковина с полостью камер, подразделенной стол-
биками, или септами, или и тем и другим, у наиболее
продвинутых семейств имеются столоны.
Семейства: Spirocyclinidae Munier-Chalmas,
1887 (перенесено из Loftusioidea, поскольку имеются
столбики и септы); Ilerdorbidae Hottinger et Cans,
1982 (с внутренними септами первого и второго по-
рядка, столонами); Orbitopsellidae Hottinger et Cans,
1982 (co столбиками и столонами; исключая Laby-
rinthinidae Mikhalevich, 1992 — с начальным плос-
костным и поздним выпрямленным цилиндрическим
отделом, простой стенкой, но с внутренними перего-
родками и столбиками); Cyclopsinellidae Loeblich et
Tappan, 1984 (с внутренними столбиками и столона-
ми).
Отряд Spiroplectamminida Mikhalevich,
1992
Раковина удлиненная, может быть расширена в
верхней части, образована начальным плоскоспи-
ральным отделом и последующим выпрямленным
двухрядным отделом, изредка переходящим в одно-
рядный; устье как в спирально свернутом отделе, так
и в двухрядном чаще расположено в основании сеп-
тальной поверхности, реже приподнято над основа-
нием, в позднем отделе может быть на вершине пос-
ледней камеры, единичное или множественное. Кар-
бон—голоцен.
Примечание. Плоскоспиральный отдел спироплектамми-
нид составляет значительную часть раковины, гораздо большую,
чем у текстулярий, но все же меньшую, чем двухрядный выпрям-
ленный отдел. Последний по ширине может быть равен свернуто-
му отделу, но чаще расширен, особенно в верхней части. У Textu-
lariopsidae и Pseudobolivinidae раковина целиком двухрядная, спи-
ральный отдел редуцирован.
Надсем. Spiroplectamminoidea
Cushman, 1927
Раковина с простой стенкой и неподразделенной
плоскостью камер.
Семейство Spiroplectamminidae Cushman, 1927.
Надсем. Spirotextularioidea Saidova,
1975
Раковина с внутренними септами, подразделяющи-
ми полость камер, или с альвеолярной стенкой.
Мел—голоцен.
Семейства: Spirotextulariidae Saidova, 1975 (внут-
ренние септы у периферических концов камер); No-
valesiidae Loeblich et Tappan, 1984 (с радиальными
септами); Ecougellidae Loeblich et Tappan, 1985 (пе-
ренесено из Loftusiacea, так как выпрямленный отдел
двухрядный).
Отряд Textulariida Delage et Herouard,
1896
Раковина удлиненная, двухрядная, чаще уплощен-
ная, может быть с конечным однорядным отделом,
начальный отдел часто «псевдотрехрядный» — с 1
дополнительной камерой; устье в основании септаль-
ной поверхности либо приподнятое, единичное или
множественное, Moiyr быть дополнительные устья на
периферических концах камер. Юра, миоцен, палео-
цен, голоцен.
Примечание. По-видимому, большинство родов текстуля-
риид произошло от плоскоспиральных форм, во всяком случае,
имеются виды (например, Textularia occidentalis Mikhalevich,
1978, — см.: Михалевич, 1978а, рис. 53—55); Fissotextularia flori-
dana (Cushman, 1922, — см. рис. 545, 4, а, б\ 547, V, 2, а, 2, б), у
которых одна из генераций целиком двухрядная, а другая имеет
рудиментарный плоскоспиральный отдел. Иногда остается только
1 рудиментарная камера, которая может быть остатком либо плос-
коспирального отдела, либо трохоидного. Но так как для боль-
шинства видов изучена лишь одна из генераций и раковины с ру-
диментарным трохоидным отделом ни в одной из них пока не об-
наружены, мы оставляем текстуляриид в составе Lituoloida, а не
Axatophragmioida, куда их перевели Лёблик и Таппан (Loeblich,
Tappan, 1988).
Большинство представителей отряда — кайнозойские формы,
к вымершим относятся лишь жившие в юре и мелу Textulariopsi-
dae.
Надсем. Textularioidea Ehrenberg,
1938
Раковины с простой стенкой и неподразделенной
полостью камер.
Семейства: Pseudobolivinidae Wiesner, 1931; Тех-
tulariidae Ehrenberg, 1938; Bigenerinidae Saidova,
1981; Textulariopsidae Loeblich et Tappan, 1982; Tex-
tularioididae Loeblich et Tappan, 1984 (прикреплен-
ные формы).
Надсем. Septotextularoidea Loeblich
et Tappan, 1985
Раковины с полостью камер, подразделенной сеп-
тами.
Семейства: Tawitawiidae Loeblich et Tappan, 1961;
Septotextulariidae Loeblich et Tappan, 1985.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
601
Надотряд Ataxophragmioida Fursenko,
1958 ,
Раковина высокотрохоидная или спирально-вин-
товая, целиком или только в начальном отделе, позд-
ний отдел может быть выпрямленным (многорядным,
двухрядным, однорядным, смешанным) или цик-
лическим, циклические обороты могут составлять
большую часть раковины; устье единичное (в осно-
вании септальной поверхности или конечное, иногда
с зубом) или множественное, у циклических форм
имеются многочисленные округлые отверстия по пе-
риферическому краю раковины; могут быть дополни-
тельные устья на периферических концах камер. Ни-
жний карбон—голоцен.
Отряд Ataxophragmiida Fursenko,
1958
Характеристика отряда совпадает с таковой надотряда.
Примечание. Кроме рассматриваемого ниже подотряда
Ataxophragmiina в отряд Ataxophragmiida входят вымершие по-
дотряды, которые здесь подробно не рассматриваются: Dicyclinina
Munier-Chalmas, 1857 (нижний и верхний мел), раковина которого
состоит в основном из циклических камер с очень небольшим,
едва возвышающимся конусом посередине, и Orbitolinina Copeland,
1956 (юра—олигоцен) с высококонической или низкоконической
раковиной. Формы, входящие в эти подотряды, имели сложное
внутреннее строение. Наибольшей сложности достигли представи-
тели подотряда Orbitolinina (рис. 532, 7). У них имеются эмбрион,
состоящий их крупных протероконха и дейтероконха, периэмбри-
ональная зона из более крупных камер, центральная, радиальная и
маргинальная зоны, в каждой из которых имеются сложные под-
разделения камер (нередко первого и второго порядка), камеры
могут быть разной формы. Кроме того, могут встречаться ряды
камер без подразделений внутренней полости, либо с альвеоляр-
ной структурой или столбиками. Такой же сложной дифференци-
ации камер, начиная с эмбриональных, достигают высшие предста-
вители подкласса Rotaliana — Nummulitida (рис. 532, 2, 5), возник-
шие позже (палеоцен). Более совершенная бифонтинальная стенка
и новая организменная система — каналы позволили им занять
прочную экологическую нишу в современном океане.
Подотряд Ataxophragmiina Fursenko,
1958
Раковина спирально-винтовая или чаще высоко-
трохоидная, с удлиненным прямым конечным отде-
лом, обычно с уменьшающимся числом камер (4—
3—2—1), либо число камер в конечном отделе мо-
жет возрастать (Valvulinidae); устье в основании септальной
поверхности, или конечное, иногда с зубом или мно-
жественное на вершине последней камеры, изредка
устье бывает прикрыто дополнительной скелетной
пластинкой с устьевыми отверстиями. Верхний кар-
бон—голоцен.
Примечание. Расцвет подотряда приходится на юрское и
меловое время, но его представители, в том числе высокоорганизо-
ванные, со сложной внутренней структурой, многочисленны и в
современных морях. Семейства выделяются в основном по соотно-
шению отделов с разным числом камер в начальных оборотах и в
конечном отделе. Раковины с поровыми канальцами и без них, но
имеющие сходную морфологию, мы рассматриваем как близкород-
ственные формы.
Надсем. Ataxophragmoidea Schwager,
1877
Раковины с неподразделенной полостью камер,
простой стенкой, но поровые канальцы могут присут-
ствовать; могут быть дополнительные устья на пери-
ферических концах камер.
Семейства: Ataxophragmiidae Schwager, 1877;
Valvulinidae Berthelin, 1880; Verneuiliiiidae Cushman,
1911; Globotextulariidae Cushman, 1927; Spiroplecti-
natidae Cushman, 1928; Eggerellidae Cushman, 1937;
Verneuilinoididae Suleymanov, 1973; Tritaxidae Plotni-
kova, 1979; Prolixoplectidae Loeblich et Tappan, 1985;
*Pseudogaudryinidae Loeblich et Tappan, 1985.
Надсем. Textularielloidea Groenhagen
et Luterbacher, 1966
Раковина с полостью камер, подразделенной раз-
личным образом (септами, столбиками); стенка может
быть с ретикулярным внутренним слоем, изредка с по-
ровыми канальцами; устьевая область может быть при-
крыта дополнительной скелетной пластинкой с мно-
гочисленными отверстиями.
Семейства: Liebusellidae Cushman, 1933; Pfen-
derinidae Smout et Sugden, 1962 (без рода Dobrogeli-
na с низкой трохоидной раковиной, который мы пе-
ревели в Trochamminoida); Coskinolinidae Moulade,
1965; Textulariellidae Groenhagen et Luterbacher, 1966;
*Chrysalidinidae Neagu, 1968; Cuneolidae Saidova, 1981;
Pemerinidae Loeblich et Tappan, 1984.
Надотряд Trochamminoida Saidova,
1981
Раковина низкотрохоидная, часто временно при-
крепляющаяся, обычно с выпуклой спиральной сто-
роной и уплощенной или вогнутой пупочной, у не-
скольких родов спиральная сторона плоская, а пу-
почная сильно выпуклая (как у Cibicides), тенденция
к развертыванию раковины и образованию выпрям-
ленного отдела слабо выражена, пупочные концы
камер часто особой конфигурации (удлиненные, изо-
гнутые, лопастные); устье в основании септальной
поверхности обычно на пупочной стороне, реже на
периферическом крае, изредка ареальное, приподня-
тое над основанием, — в этом случае может быть
множественным (Jadammina, Arenoparrella); очень
часто имеются дополнительные устья на пупочной
стороне, в швах, или весь пупок открытый, могут
быть дополнительные скелетные пластинки в пупоч-
ной области (Asterotrochammina, Dobrogelina). Кар-
бон—голоцен.
Примечание. Иногда (например, у Jadammina) имеется од-
новременно базальное устье и над ним ареальное (рис. 545, 5, а, б)^
Если первое зарастает, — остается лишь ареальное, которое в
таком случае можно рассматривать как вторичное.
602
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отряд Trochamminida Saidova,
1981
Раковины с агглютинированной стенкой, прочной
или очень тонкой, эластичной, с внутренними септа-
ми или без них.
Подотряд Trochamminina Saidova,
1981
Характеристика подотряда совпадает с таковой
отряда.
Надсем. Trochamminoidea Schwager,
1877
Раковина с простой стенкой и неподразделенной
полостью камер.
Семейства: Trochamminidae Schwager, 1877;
Conotrochamminidae Saidova, 1981; Rotaliamminidae
Saidova, 1981; Vialovinidae Suleymanov, 1983; Valvu-
lamminidae Loeblich et Tappan, 1986 (с подсем. Dob-
rogelininae Mikhalevich, 1992 с дополнительной npo-
боденной скелетной пластинкой, прикрывающей пу-
почную область).
Надсем. Remaneicioidea Loeblich
et Tappan, 1964
Раковины с полостью камер, подразделенных до-
полнительными септулами или складками стенки.
Семейства: Remaneicidae Loeblich et Tappan,
1964; *Nezzazatidae Hamaoui et Sant-Marc, 1970 (пе-
ренесено из Fusulinoida); Asterotrochamminidae Bron-
nimann, Zaninetti et Whittaker, 1983; *Dictyopsellidae
Bronnimann, Zaninetti et Whittaker, 1983 (перенесено
из Ataxophragmiida); Pyreninidae Mikhalevich, 1999 (c
выпрямленным расширенным отделом).
Подотряд Carterinina Mikhalevich,
1980
Раковина co стенкой из удлиненных, суженных
на концах известковых спикул, уложенных парал-
лельно поверхности и скрепленных органическим
цементом с небольшим количеством тонкопесчани-
стого материала; начальные камеры простые, бо-
лее поздние подразделены септулами; последние ка-
меры могут располагаться неправильно или цикли-
чески.
Семейство Carterinidae Loeblich et Tappan,
1955.
Подкласс Rotaliana Mikhalevich, 1980
Раковина с одним из роталиатных типов располо-
жения камер и строения устья; стенка секреционная,
известковая, бифонтинальная, с порами и нескольки-
ми ситовидными поровыми пластинками в поровом
канальце (размер пор в среднем 0.25—0.5 мкм, но
бывают более мелкие или более крупные, до
2.5 мкм), с сильно развитой наружной скульптурой,
у многих родов с дополнительными скелетными
пластинками в пупочной области, над устьем и над
швами, с внутренними структурами большой слож-
ности (кроме дополнительных септ имеются клапа-
ны, желобки, пластины, язычки в устьевой и при-
устьевой области, иногда связанные между собой в
единую систему); интегрирующие системы в виде
столонов, внутренних устьевых систем и хорошо
развитой сложной системы каналов. Триас—голо-
цен.
Примечание. Система подкласса нуждается в ревизии, в
основу которой должно быть положено изучение строения внут-
ренних устьевых структур, известных далеко не для всех родов.
Такую ревизию проводят в последнее время Хансен и Ревете (Han-
sen, Revets, 1992; Revets, 1996), но поскольку она еще не заверше-
на, дать новую систему всего подкласса пока не представляется
возможным.
Надотряд Buliminoida Saidova, 1981
Раковина удлиненная; одно-, двух- (может быть с
начальным плоскоспиральным отделом, а в послед-
нем отделе становится однорядной) либо трехрядная,
трохоидная, спирально-винтовая, в трех последних
случаях обычно с переходом к двух- и однорядной,
раковина также может быть округлой, из 2 рядов
спирально свернутых камер (кассидулиновый тип), с
тенденцией к разворачиванию; полость камер обычно
(кроме 1 семейства) не подразделена; у 2 родов име^-
ется примитивная система каналов; устье петлевид-
ное, отходящее от основания септальной поверхнос-
ти либо конечное, единичное, реже множественное, у
более примитивных форм без зуба, у эволюционно
продвинутых — с зубными пластинками различной
формы и сложности (в виде пластинок, язычков, же-
лобков, полуцилиндрических сифонов), пластинки
могут быть изогнутыми, складчатыми, прикрепляю-
щимися к стенке предыдущей камеры, могут иметь
свободные отростки (иногда зубчатые), могут высту-
пать над краем устья, зубные пластинки последова-
тельных камер часто связаны между собой и образу-
ют единую внутреннюю устьевую систему; изредка
встречаются дополнительные шовные устья. Средняя
юра—голоцен.
Примечание. Как и в подклассе Textulariana, двух- и одно-
рядные формы произошли от плоскоспиральных (отряд Boliviniti-
da) и от трохоидных (отряд Buliminida) форм. Среди B*olivinitida
много представителей с раковиной, гомеоморфной раковинам спи-
роплектамминид и текстуляриид, среди Buliminida — раковинам
атаксофрагмиид, среди Cassidulinida — раковинам плекторекурво-
идид. В усложнении строения устья и зубных пластинок у Bolivi-
nitida и Buliminida много параллелизмов. Зубная пластинка фор-
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
603
мируется за счет внутренних слоев стенки. Среди изученных родов
исключение составляет только род Stainforthia, чья сложная зубная
пластинка образована и внутренним и наружным слоем, как и вся
стенка раковины. На этом основании Ревете (Revets, 1996) отно-
сит этот род к incertae sedis.
Отряд Stilostomellida Saidova, 1981
Раковина однорядная, прямая или слегка изогну-
тая; устье конечное, округлое, часто с зубом. Верх-
ний мел—голоцен.
Примечание. Возможно, вторично-однорядные. Строение
устья (наряду с ультраструктурой стенки раковин) позволяет легко
отличать эти однорядные раковины от представителей нодозариат.
Семейство Stilostomellidae Finlay, 1947.
Отряд Bolivinitida Saidova, 1981
Раковина обычно уплощенная, двухрядная цели-
ком или имеет плоскоспиральный, начальный отдел,
камеры чаще широкие, уплощенные, последние не-
редко с тенденцией к однорядности, могут стано-
виться веерообразными, полость камер может быть с
дополнительными подразделениями, система каналов
отсутствует; устье в основании септальной поверх-
ности, приподнято над ней или конечное, чаще пет-
левидное или щелевидное, реже округлое, у эво-
люционно продвинутых родов со сложной зубной
пластинкой, может быть множественным. Верхний
мел—голоцен.
Подотряд Spirobolivinina
Mikhalevich, 1999
Раковина с плоскоспиральным начальным отде-
лом, поздний отдел — двухрядный, выпрямленный.
Примечание. От подотряда Bolivinitina данный подотряд
отличается наличием хорошо выраженной начальной спиральной
части раковины. В поперечном сечении раковины уплощенные
или субцилиндрические, полость камер без внутренних подразде-
лений, устье петлевидное, у большинства известных родов с систе-
мой зубных пластинок. Лёблик и Таппан (Loeblich, Tappan, 1988)
свели сем. Spirobolivinidae Saidova, 1981 в синоним Lacosteinidae,
а подсем. Sigmavirgulininae — в синоним Fursenkoinidae. Мы вос-
станавливаем оба таксона, так как Spirobolivinidae отличается от
Lacosteinidae (характеризуется цилиндрической раковиной и взду-
той удлиненной последней камерой) уплощенной раковиной и ка-
мерами, близкими к Bolivina, a Sigmavirgulininae — отличается от
Fursenkoinidae не только уплощенными камерами, формой камер и
устья, но и наличием плоскоспирального отдела.
Семейства Lacosteinidae Sigal, 1952; Spiroboli-
vinidae Saidova, 1981 (с подсем. Sigmavirgulininae Sa-
idova, 1981).
Подотряд Bolivinitina Saidova, 1981
Раковина двухрядная целиком или с переходом к
однорядности в последнем отделе.
Примечание. Надсемейства различаются главным образом
строением устья и его внутренних структур, а также на основании
наличия или отсутствия вторичного подразделения камер.
Надсем. Loxostomatoidea Loeblich
et Tappan, 1962
Раковина двухрядная целиком, конечные каме-
ры с тенденцией к однорядности, полость камер без
внутренних подразделений; устье простое, без зуб-
ной пластинки. Верхний мел—голоцен.
Семейства: Loxostomatidae Loeblich et Tappan,
1962; Bolivinellidae Hayward, 1980; Tortoplectidae Loe-
blich et Tappan, 1985.
Надсем. Bolivinitoidea Cushman,
1927
Раковина двухрядная целиком, последние камеры
могут быть с тенденцией к однорядности, полость ка-
мер без внутренних подразделений; устье с хорошо
развитым зубным аппаратом: петлевидное устье окру-
жено полностью или частично приподнятым ободком,
нижний край которого уходит внутрь, образуя зубную
пластинку, состоящую из 2 частей — прикрепленной,
уходящей внутрь к форамену предыдущей камеры, и
свободной, обычно возвышающейся над устьем, но
иногда не заметной снаружи. Верхний мел—голоцен.
Примечание. Свободная часть зубной пластинки может
быть не развита.
Семейства: Bolivinitidae Cushman, 1927; Boli-
vinidae Glaessner, 1937 (с родами Coryphostoma и
Cassidella, перенесенными из Fursenkoidae); Bolivi-
noididae Loeblich et Tappan, 1984.
Надсем. Parabrizalininoidea Revets,
1996
Раковина двухрядная целиком или с переходом к
однорядной, уплощенная; камеры без вторичных под-
разделений; устье обычно петлевидное, горизонталь-
ное, с зубной пластинкой в виде полуцилиндрическо-
го сифона, сифоны последовательных камер образу-
ют единую систему.
Семейство Parabrizalinidae Revets, 1996.
Надсем. Eouvigerinoidea Cushman,
1927
Раковина двухрядная с тенденцией к одноряднос-
ти или однорядная в конечном отделе, толстая с уп-
лощенными боковыми сторонами или цилиндричес-
кая, камеры без вторичных подразделений; устье пет-
левидное или конечное, может быть на приподнятой
шейке, зубная пластинка в виде полуцилиндрическо-
го желобка (сифона), сифоны последовательных
камер связаны между собой, образуя единую систему.
Нижний мел—голоцен.
Примечание. Из последнего семейства мы исключаем формы с
трохоидной (трехрядной) начальной частью (подсем. Tubulogenerini-
пае). Ранее включавшееся в это надсемейство вымершее сем. Lacostei-
nidae (Loeblich, Tappan, 1988) включено нами в подотряд Spirobolivi-
nina, так как его типовой род имеет плоскоспиральный начальный отдел.
Семейства: Eouvigerinidae Cushman, 1927; Sip-
hogenerinoididae Saidova, 1981.
604
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Надсем. Pavoninoidea Eimer et Fickert,
1899
Раковина двухрядная, с конечным веерообразным
однорядным отделом, полость камер без вторичных
подразделений; устье конечное, в виде длинного ряда
отверстий на устьевой поверхности. Миоцен—голо-
цен.
Примечание. Сем. Pavoninidae перенесено из Buliminacea,
так как его представители характеризуются наличием уплощенной
двухрядной раковины и отсутствием устьевого зуба.
Семейство Pavoninidae Eimer et Fickert, 1899.
Надсем. Milettioidea Saidova,
1981
Раковина удлиненная, двухрядная целиком или
только в начальном отделе (с последующим одноряд-
ным), в поперечном сечении округлая или овальная,
полость камер подразделена радиальными перегород-
ками или перегородками, идущими в двух направле-
ниях — продольно и горизонтально, в результате
чего в месте их соприкосновения с наружной стен-
кой видны гексагональные ячеи; устье петлевидное,
отходящее от основания септальной поверхности
либо конечное, округлое, окруженное воротничком
с ободком, зубная пластинка сложная, как у Bolivina
(Afrobolivina), либо желобообразная, у начальных
камер может отсутствовать. Голоцен.
Семейство Milettiidae Saidova, 1981.
Отряд Cassidultnida d’Orbigny,
1839
Раковина округлая, линзовидная, овальная или уп-
лощенная, расположенные двухрядно, чередующиеся
камеры свернуты в спираль, обычно плоскую, изред-
ка трохоидную (Cassidulinita), конечный отдел может
быть выпрямленным в плоскости навивания или пер-
пендикулярно к этой плоскости (Ehrenberginidae);
устье овальное, петлевидное или в виде щели, часто
изогнутой, расположено параллельно основанию сеп-
тальной поверхности или под углом к ней, у одного
рода конечное, может быть со вторым дополнитель-
ным отверстием выше первого, у многих родов при-
крыто устьевой пластинкой с задней стороны стенки,
на передней стороне стенки может быть небольшой
зуб в виде язычка или зазубренный. Палеоцен—го-
лоцен.
Семейства: Cassidulinidae d’Orbigny, 1839; Eh-
renberginidae Cushman, 1927; Cassidulinitidae Saidova,
1981; Orthoplectidae Loeblich et Tappan, 1984.
Отряд Buliminida Saidova,
1981
Раковина трохоидная, спиральновинтовая или
трехрядная, в конечном отделе часто с переходом к
однорядности, в поперечном сечении обычно округлая,
овальная, треугольная, камеры чаще вздутые, шаро-
образные или удлиненные; без дополнительных под-
разделений на камерки, у двух родов имеется прими-
тивная система каналов (у малого числа видов);
устье чаще петлевидное, отходящее от основания
септальной поверхности, может быть конечным, на
удлиненной шейке, может быть множественным (си-
товидное устье); внутренние структуры устья — от
простых до сложно устроенных зубных пластинок,
полуцилиндрических трубок, которые могут образо-
вывать единую систему; у нескольких родов встреча-
ются дополнительные шовные устья. Средняя юра—
голоцен.
Подотряд Turrrliniпа Saidova, 1981
Раковина трохоидная, спиральновинтовая или
трехрядная, в поперечном сечении округлая, оваль-
ная или треугольная, камеры быстро увеличивающи-
еся в размерах, обычно сильно выпуклые, округлые
или удлиненные; устье петлевидное в основании сеп-
тальной поверхности или множественное, без зуба
или с маленькой простой зубной пластинкой, могут
быть дополнительные шовные устья. Средняя юра—
голоцен.
Семейства: Turrilinidae Cushman, 1927 (с под-
сем. Sporobulimininae Mikhalevich, 1999 — с множес-
твенным устьем, роды Sporobulimina и Sporobulimi-
nella); Tosaiidae Saidova, 1981; Virgulinellidae Loeb-
lich et Tappan, 1984.
Подотряд Buliminina Saidova,
1981
Раковина как у Turrilinacea, у некоторых родов с
сильно объемлющими последними камерами; усть-
евое отверстие как у Turrilinacea (округлое конеч-
ное, на вытянутой шейке, петлевидное, множествен-
ное), но зубная пластинка присутствует всегда и
имеет сложное строение: стержень ее желобообраз-
ный (может изгибаться, расширяться, быть склад-
чатым или иметь другие особенности строения), со-
единяет край устья со стенкой предыдущей камеры
выше форамена, возвышающийся свободный край
пластинки может выступать наружу, он может быть
зазубренным (crista — гребешок). Палеоцен—голо-
цен.
Семейства: Buliminidae Jones, 1875; Uvigerini-
dae Haeckel, 1894; Reussellidae Cushman, 1933; Buli-
minellidae Hofker, 1951; Fursenkoidae Loeblich et
Tappan, 1961 (гомеоморфные раковины с «зернис-
той» стенкой, но без родов Coryphostoma и Cassidel-
la); Stainforthiidae Reiss, 1963; Trimosinidae Saidova,
1981.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
605
Подотряд Tubulogenerinina
Mikhalevich, 1999
Раковина с трехрядным начальным отделом, пере-
ходящим в двух- и однорядный, в поперечном сече-
нии округлая или овальная; устье обычно конечное,
часто на конце небольшой шейки, зубная пластинка
в виде полуцилиндрического желоба (сифона), си-
фоны последовательных камер связаны между собой.
Верхний мел—голоцен.
Семейство Tubulogenerinidae Saidova, 1981 (ве-
роятно, с родом Rectobulimlna, перенесенным из Tur-
rilinidae).
Подотряд Caucasinina Mikhalevich,
1998
Раковина с начальным низкотрохоидным широко-
округлым, состоящим из нескольких оборотов, и позд-
ним высокотрохоидным выпрямленным трех- или
двухрядным отделами, в поперечном сечении округ-
лая; устье петлевидное, отходящее от основания сеп-
тальной поверхности, зубная пластинка обычно кла-
панообразная, известна не у всех родов.
Семейство Caucasinidae N. К. Bykova, 1959 (без
рода Tremachora; с подсем. Bagatellinae N. К. Byko-
va, 1959).
Надотряд Robertinoida Mikhalevich,
1980
Раковина трохоидная и высокотрохоидная, каме-
ры подразделены перегородками различного положе-
ния и направления в разных таксонах, подразделяю-
щими камеру частично или отгораживающими от
нее отдельные камерки, чаще в пупочной области;
стенка арагонитовая; устье на септальной поверхнос-
ти обычно и внутрикраевое и петлевидное, реже
устье ареальное, может также быть на перифериче-
ских краях камер, могут быть дополнительные шов-
ные устья, основное устье у предшествующих камер
зарастает полностью или частично: в первом случае
межкамерный форамен образуется в результате ре-
зорбции, во втором он гомологичен лишь части пре-
жнего устья. Средний триас—голоцен.
Примечание. 2 вымерших таксона, которые в классифика-
ции Лёблика и Таппан (Loeblich, Tappan, 1988) рассматриваются
как подсемейства и соответствуют в нашей системе подотрядам
(вымершие в юре Duostominina и меловые Conorboidina), здесь не
рассматриваются.
Отряд Robertinida Mikhalevich,
1980
Характеристика отряда совпадает с таковой надо-
тряда.
Подотряд Ceratobuliminina
Mikhalevich, 1980
Раковина низкотрохоидная; устье в основании
септальной поверхности внутрикраевое и отходящее
от нее вверх в виде петли (у Ceratobuliminidae) или
аркообразного отверстия (Reinholdellidae), у Episto-
minidae устье на периферическом крае камер или
возле него, у предшествующих камер устье зарастает,
внутренний форамен (дейтерофорамен) образуется в
результате резорбции над внутренней перегородкой,
которая может быть короткой, отграничивающей
часть камеры, либо замыкать камерки, в результате
чего с наружной стороны стенки раковина выглядит
двухрядной или со звездчатой структурой вокруг
пупка. Юра—голоцен.
Примечание. Семейства выделяются на основании различ-
ного строения и положения устьев и внутренних перегородок —
прямых или изогнутых, идущих в различных направлениях у раз-
ных представителей, иногда прилегающих к наружной стенке
только на одной стороне раковины.
Семейства: Ceratobuliminidae Cushman, 1927;
Epistominidae Wedekind, 1937; Reinholdellidae Seiglie
et Bermudez, 1965; Epistominoididae Saidova, 1981.
Подотряд RObertinina Mikhalevich,
1980
Раковина трохоидная, сильно уплощенная, с пе-
реходом к плоскоспиральной (Alliatinidae) или вы-
сокая, асимметричная, спирально-винтовая (Roberti-
nidae); камеры подразделены двойной поперечной
перегородкой, образованной вдавлением стенки, в
результате чего формируются маленькие дополни-
тельные камерки, составляющие отчетливые серии;
устье в основании септальной поверхности внутри-
краевое и поднимающееся от нее вверх в виде петли,
изредка оно бывает ареальным, септальный межка-
мерный форамен гомологичен части устья; могут
встречаться дополнительные шовные устья. Палео-
цен—голоцен.
Семейства: Robertinidae Reuss, 1850; Alliatini-
dae McGowran, 1966.
Надотряд Discorboida Ehrenberg,
1838
Раковина трохоидная; стенка кальцитовая; фора-
мен обычно гомологичен прежнему устью.
Примечание. Отряды выделяются на основании строения
раковины и степени усложнения внутренней полости камер и устья
(Михалевич, 1992, 1997а, 1998а) (с учетом последних данных по
внутренним структурам Discorbida, Rosalinida и Rotaliida Хансена
и Реветса,— Hansen, Revets, 1992).
Так, отчетливо видна филогенетическая линия от Rosalinida с
неподразделенной полостью камер и простым устьем к Discorbida
С клапаном (кроющей пластинкой), делящим устье надвое, но не
образующим дополнительных камерок, затем к Asterigerinida с
более сложным внутренним подразделением камер, образованием
дополнительных камерок у наиболее продвинутых форм с очень
большим числом камер и появлением интегрирующей системы
столонов и, наконец, к Rotaliida, у которых кроме устьевого кла-
606
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
пана имеется примитивная система каналов в виде щелей и высо-
коразвитая система каналов, состоящая из спирального и сети от-
ветвляющихся каналов (иногда имеется сочетание системы щелей
и каналов). Внешнее строение раковин у представителей этих от-
рядов иногда полностью совпадает.
Остальные отряды надотряда отличаются в основном особен-
ностями морфологии их раковин.
Отряд Rosalinida Delage et Herouard,
1896
Раковина низкотрохоидная, обычно с выпуклой
спиральной стороной и слабовыпуклой, плоской или
вдавленной пупочной, изредка раковина двояковы-
пуклая (Bronnimaniidae) или приближается к шаро-
образной — с объемлющими оборотами или сильно-
объемлющей последней камерой (Sphaeroidinoidea),
у некоторых родов спираль может развертываться,
пупок открытый или закрытый, пупочные концы ка-
мер могут быть с расширенным клапаном (некото-
рые Bagginidae), синусоидным вырезом (Rosalinidae,
Discorbinellidae), маленьким^ зубчиками (Rotalielli-
dae); внутренность камер простая, неподраздеденная;
устье в основании септальной поверхности (внутри-
краевое — на пупочной стороне, или протягиваю-
щееся от периферического края до пупка, или
центрально-краевое — экваториальное), или на сеп-
тальной поверхности — ареальное, ситовидное, от-
ходящее вверх от основания септальной поверхности
в виде высокой щели, которая расположена парал-
лельно периферическому краю и лежит в углублении
септальной поверхности, или, в виде исключения,
конечное, с небольшим отворотом (Siphoninidoidea);
дополнительные устья чаще на пупочной стороне (у
внутреннего края камер или в швах), но изредка
встречаются и на спиральной. Нижний мел—голо-
цен.
Надсем. Rosalinoidea Reiss,
1963
Раковина низкотрохоидная, обычно с выпуклой
спиральной стороной и слабовыпуклой, плоской или
вдавленной пупочной, спиральная сторона эволют-
ная, пупочная — инволютная, изредка раковина дво-
яковыпуклая (Bronnimaniidae, у Rectoeponides с вы-
прямленным отделом), пупок открытый или за-
крытый, пупочные концы камер могут быть с
расширенным клапаном (некоторые Bagginidae), си-
нусоидным вырезом (Rosalinidae), маленькими зубчи-
ками (Rotaliellidae); устье в основании септальной
поверхности внутрикраевое (на пупочной стороне;
может доходить до периферического края или оги-
бать клапанообразный пупочный конец камеры и за-
ходить в шов между последней и предпоследней ка-
мерами), изредка ситовидное (некоторые Eponididae,
у них же устье может быть прикрыто изрешеченной
пластинкой); дополнительные устья чаще на пупоч-
ной стороне (в пупочной области или шовные), но
изредка встречаются и на спиральной стороне (Hele-
ninidae). Нижний мел—голоцен.
Семейства: Bagginidae Cushman, 1927; Eponi-
didae Hofker, 1951; Rosalinidae Reiss, 1963; Patellinel-
lidae Bugrova, 1990; Rotaliellidae Loeblich et Tappan,
1984.
Надсем. Discorbinelloidea Sigal,
1952
Надсем. Discorbinelloidea отличается от надсем.
Rosalinoidea строением устья. Для этого таксона ха-
рактерно центрально-внутрикраевое (экваториальное)
или ареальное устье, либо устье, отходящее от осно-
вания септальной поверхности в виде вертикальной
щели, идущей параллельно периферическому краю и
расположенной в нише, которая образована вдавле-
нием септальной поверхности. Палеоцен—голоцен.
Семейства: Discorbinellidae Sigal, 1952; Pseu-
doparrellidae Voloshinova, 1952; Parrelloididae Hofker,
1956; Planulinoididae Saidova, 1981; Svratkinidae Bug-
rova, 1990.
Надсем. Sphaeroidinoidea Cushman,
1927
Раковина трохоидная, с сильно выпуклыми каме-
рами, с уплощенной пупочной стороной или близкая
к шарообразной, с объемлющими последними оборо-
тами или объемлющей последней камерой; устье в виде
отверстия в центре пупочной области (Bueningiidae),
либо в виде серии трубок, пронизывающих пупочную
область (Pegidiidae), либо в виде щели, приподнятой
над основанием септальной поверхности, может быть
с клапаном (Sphaeroidinidae); встречаются дополни-
тельные шовные устья на спиральной стороне (Тге-
machoridae). Эоцен—голоцен.
Примечание. Гетерогенное надсемейство, включает свое-
образные формы, филогенетические связи которых с остальными
розалинидами неясны. У Pegidiidae начальные камеры по мере рос-
та резорбируются. У Tremachoridae (миоцен) бифонтинальная
стенка не обнаружена (Revets, 1991): либо она перекристаллизова-
на, либо это своеобразные формы среди роталиан с более прими-
тивной стенкой.
Семейства: Sphaeroidinidae Cushman, 1927; Pe-
gidiidae Heron-Allen et Earland, 1928; Tremachoridae
Lipps et Lipps, 1967; Bueningiidae Saidova, 1981.
Надсем. Siphoninidoidea Cushman,
1927
Раковина трохоидная целиком, с закрытым пуп-
ком, либо трохоидная только на ранней стадии, на
поздней может становиться почти плоскоспираль-
ной, стрептоспиральной, выпрямленной — двух- или
однорядной; устье даже на ранней, свернутой стадии
конечное, с отворотом. Верхний мел—голоцен.
Семейство Siphoninidae Cushman, 1927.
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
607
Отряд Discorbida Ehrenberg, 1838
Раковина сходная по строению с раковинами Ro-
salinidae, но с подразделенной полостью камер: близ-
ко к пупочной области стенка камеры образует про-
ксимальные неперфорированные кроющие пластин-
ки (клапаны), делящие устье надвое и частично
прикрывающие его, эти пластинки могут прирастать
или не прирастать к стенке раковины, вследствие
этого, в пупочной области, под клапанообразными
концами камер появляются воронкообразные расши-
рения, которые в совокупности образуют непрерыв-
ную структуру и сообщаются с пупочной областью,
но отделены от остальной части камеры. Средний
эоцен—голоцен.
Семейство Discorbidae Ehrenberg, 1838.
Отряд Glabratellida Mikhalevich,
1994
Раковина трохоспиральная, чаще высокая (высо-
котрохоидная или даже спирально-винтовая), реже
слабовыпуклая, обычно с небольшим числом оборо-
тов и камер в обороте (4—5, редко 7), пупочная
сторона уплощенная, вогнутая шги слабовыпуклая,
чаще с закрытым пупком, с наружной скульптурой
стенки на пупочной стороне в виде радиальных ре-
бер или шишек, расположенных по ходу цитоплазмы
и, по-видимому, облегчающих соединение 2 роди-
тельских раковин при пластогамном размножении,
внутренняя полость камер без подразделений и без
проксимальной пластинки, прилегающей к устью;
устье в основании септальной поверхности послед-
ней камеры чаще в виде высокой арки или петли на
пупочной стороне, доходящее до пупка, реже щеле-
видное, ближе к периферическому краю, либо це-
ликом пупочное; дополнительные устья не развиты;
у Heronallenidae устье в области пупка прикрыто
позже сформированными пластинками; размноже-
ние по способу пластогамии (гамонтогамии), гаметы
трехжгутиковые. Нижний эоцен—голоцен.
Примечание. Трехжгутиковые гаметы среди фораминифер
известны только у глабрателлид и у Nummulites venosus.
Семейства: Glabratellidae Loeblich et Tappan,
1964; Buliminoididae Seiglie, 1970; Heronallenidae
Loeblich et Tappan, 1986.
Отряд Planorbulinida Mikhalevich,
1992
Раковина чаще прикрепленная, трохоидная цели-
ком или только на ранней стадии, обычно с эволют-
ной уплощенной спиральной и выпуклой инволют-
ной пупочной стороной, поздний отдел может быть
выпрямленным, циклическим, коническим, ветвя-
щимся или в виде неправильной массы, состоящей
из очейь большого числа камер, часто утративших
правильную форму; внутренность камер неподразде-
ленная; стенка обычно с очень крупными порами;
устье у целиком трохоидных форм в основании сеп-
тальной поверхности экваториальное или протягива-
ющееся на пупочную или спиральную сторону, иног-
да на обе, на спиральной стороне может простирать-
ся вдоль внутренних концов нескольких последних
камер, часто оторочено губой, могут быть дополни-
тельные пупочные и шовные устья на пупочной сто-
роне, у выпрямленных форм устье конечное, у форм,
образующих неправильную массу камер, — множес-
твенное: в виде округлых отверстий, напоминающих
поры («устьевые поры») на поверхности стенки
камер, или многочисленных аркообразных отверстий
на одной из сторон или на периферическом крае в
швах между камерами. Палеоцен—голоцен.
Примечание. Представители Planorbulinida имеют специа-
лизированные видоизмененные раковины, приспособленные к сй-
дячему образу жизни. Среди них также много комменсалов и
встречаются паразитические формы (см. с. 560).
Подотряд Planorbulinina
Mikhalevich, 1992
Раковина свободноживущая или прикрепленная,
трохоидная целиком или только на ранней стадии,
начальный трохоидный отдел обычно с эволютной
уплощенной спиральной и инволютной выпуклой
пупочной сторонами, реже частично эволютный на
обеих сторонах (Planulinidae), конечный отдел мо-
жет быть выпрямленным (одно- или двухрядным),
циклическим, в виде беспорядочно нагромождаю-
щейся массы, в которой камеры расположены в 1
слой или образуют слабо приподнятый конус; устье
единичное, у трохоидных форм оно расположено в
основании септальной поверхности, экваториальное
или протягивающееся на одну из сторон или на обе,
на спиральной стороне может проходить под внут-
ренними концами нескольких последних камер, час-
то оторочено губой, могут быть дополнительные
устья на пупочной стороне, у выпрямленных форм
устье конечное, у циклических и распространяю-
щихся неправильной массой — множественное (ок-
руглые устьевые поры на поверхности камер или
аркообразные отверстия в швах). Верхний эоцен—
голоцен.
Примечание. Представители сем. Fabianiidae Deloffre et
Hamaoui, 1973 имеют камеры, подразделенные внутренними вер-
тикальными и горизонтальными перегородками, и заслуживают
ранга подотряда. Составляющие его роды (верхний палеоцен—
верхний эоцен) вымерли, и поэтому этот таксон здесь не рассмат-
ривается.
Семейства: Planorbulinidae Schwager, 1877; Ci-
bicididae Cushman, 1927; Cymbaloporidae Cushman,
1927; Victoriellidae Chapman et Crespin, 1930; Planu-
linidae Bermudez, 1952; Annulocibicididae Saidova,
1981; Stichocibicididae Saidova, 1981; Stensioinidae
Bugrova, 1990.
608
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Подотряд Acervulinina Schultze, 1854
Раковина прикрепленная, толстая, почти диско-
видная, с выпуклой верхней поверхностью или не-
правильной формы, часто древовидной, с очень
большим количеством мелких камерок округлой или
полигональной формы, расположенных многочис-
ленными слоями; стенка даже у ископаемых форм
ярко окрашена в розовый, бордовый, оранжевый
цвета (возможно, из-за распада хлорофилловых пиг-
ментов эндобионтов), поверхность выпуклых округ-
лых камер может напоминать ягоду малины, иногда
камеры снаружи уплощены и даже вдавлены, закры-
ты пластинкой с порами, у древовидных форм (Но-
motrematidae) внутри от основания вверх отходят
полые неперфорированные цилиндрические трубки с
двойной стенкой, которые могут разветвляться, во-
круг них в несколько слоев располагаются мелкие
камеры; устья — многочисленные округлые отверс-
тия в стенке камер или в стенке углубленных плас-
тинок, закрывающих вход в камеры, намного круп-
нее соседних пор (в литературе описывались как
«крупные» поры). Палеоцен—голоцен.
Примечание. Трехкамерная молодая форма мегалосфери-
ческой раковины Miniacina ведет свободный образ жизни, здтем
оседает на дно, от третьей камеры отрастают трубчатые выросты,
и вокруг них нарастают округлые камеры. Микросферическая ра-
ковина оседает на стадии 5—9 камер, образующих отчетливую
трохоидную спираль. Взрослые раковины хомотрематид достигают
7 мм в высоту. Они настолько не похожи на остальных секрецион-
ных фораминифер, что долгое время их принимали за кораллы.
Сидячий образ жизни привел к большому разнообразию в нараста-
нии поздних камер и к преобразованию конечного устья в этой
группе фораминифер.
Семейства: Acervulinidae Schultze, 1854; Но-
motrematidae Cushman, 1927.
Отряд Asterigerinida d’Orbigny, 1839
Раковина трохоидная целиком (у малокамерных
форм) или только на ранних стадиях (у форм с боль-
шим числом камер и камерок), полость камер под-
разделена вторичными септами, которые либо не до-
ходят до противоположной стенки и в этом случае не
образуют камерки, либо прикрепляются к стенке ра-
ковины и формируют дополнительные камерки, у
многокамерных форм имеются столоны, каналы от-
сутствуют. Нижний мел—голоцен.
Надоем. Asterigerinoidea d’Orbigny,
1839
Раковина трохоспиральная, двояковыпуклая или
уплощенная, обычно с одной инволютной стороной
(пупочной), реже инволютны обе стороны (у Palme-
rinella, Amphistegina), количество камер сравнитель-
но невелико, пупок почти всегда закрытый, пупоч-
ные концы камер обычно простые, без лопастей и
вырезов, полость камер подразделена вторичными
септами, последние либо не доходят до противопо-
ложной стенки и в этом случае не образуют камерки
(Eponidellinae) или, прикрепляясь к стенке раковины
в пупочной области, формируют вторичные камерки,
которые частс^ имеют вид звездообразных структур в
центре раковины на пупочной стороне (Alfredinidae,
Asterigerinatidae, Asterigerinidae, Amphisteginidae —
у последних имеются еще и межсептальные столби-
ки), столоны и каналы отсутствуют; устье в основа-
нии септальной поверхности последней камеры, эк-
ваториальное или на пупочной стороне, часто закан-
чивается в складке септы возле периферического
края (Nuttalides), может подниматься вверх от осно-
вания этой поверхности и при этом быть поделен-
ным вдоль вертикально (Palmerinella из Epistomariidae)
или может быть ареальным, ситовидным, у некото-
рых представителей (Eponidellinae из Epistomariidae)
с образованием новых камер часть устья зарастает,
другая его часть служит межкамерным фораменом;
дополнительные устья могут быть связаны со швами
(Epistomaroides, Monspielensina) либо с дополнитель-
ными внутренними перегородками (в этом случае они
располагаются параллельно периферическому краю —
Epis tomaria, Pseudoeponides). Нижний мел—голо-
цен.
Семейства: Asterigerinidae d’Orbigny, 1839;
Epistomariidae Hofker, 1954; Gavelinellidae Hofker,
1956; Asterigerinatidae Reiss, 1963; Amphisteginidae
Reiss, 1963; Osangulariidae Loeblich et Tappan, 1964;
Heterolepidae Gonzales-Donoso, 1969; Alfredinidae
S. N. Singh et Kalia, 1972; Karreriidae Saidova, 1981;
Quadrimorphinidae Saidova, 1981; Trichohyalidae Sai-
dova, 1981; Coleitidae Loeblich et Tappan, 1984; Glo-
borotalitidae Loeblich et Tappan, 1984 (вымерли в
мелу); Oridorsalidae Loeblich et Tappan, 1984.
Надсем. Clypeorbioidea Sigal, 1952
Раковина обычно трохоидная только на ранней
стадии или у одной из генераций, поздние обороты
могут располагаться в одной плоскости, могут быть
объемлющими, образуя раковину фузулинового типа
или почти сферическую, эмбриональные камеры
очень крупные, последующие — мелкие, многочис-
ленные; развита система столонов. Верхний мел—
верхний эоцен.
Семейства: *Clypeorbidae Sigal, 1952; Boreloi-
didae Reiss, 1963.
Отряд Rotaliida Fursenko, 1958
Раковина трохоидная по крайней мере на ранней
стадии развития, на поздней может становиться плос-
коспиральной, в некоторых случаях камеры распола-
гаются кольцеобразно, число камер чаще обычное
(5—15), но может быть и велико, вытянутые пупоч-
ные концы камер с клапанами, пупок закрытый или
с вторичным раковинным веществом, образующим
вертикальные столбики (могут быть расположены в
несколько ярусов), между столбиками находятся ще-
левидные отверстия — примитивные каналы, кроме
пупочных и околопупочных полостей и щелей могут
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
609
быть внуТрисептальные щели, кроме того, у боль-
шинства родов имеется хорошо развитая система ка-
налов — спиральный канал и сообщающиеся с ним
септальные, в свою очередь разветвляющиеся на
более мелкие; устье в основании септальной поверх-
ности (экваториальное или на пупочной стороне, ще-
левидное или в виде ряда округлых отверстий), часто
зарастает, и в таком случае функционирует только
система каналов; дополнительные устья пупочные и
шовные. Верхний мел—голоцен.
Примечание. Роталииды — самые высокоорганизованные
и сложно устроенные представители секреционных известковых
фораминифер, вершина развития группы. Возникнув в мелу, они
быстро заняли широкую экологическую нишу. Семейства выделя-
ются с учетом особенностей системы каналов. К настоящему вре-
мени ряд высокоорганизованных семейств вымер, в современных
океанах известны лишь раковины нотороталиид и калькаринид в
тропических мелководных морях, а также и раковины аммониид,
особенно мелководного рода Ammonia, широко распространенного
в теплых и умеренных водах краевых морей всех океанов.
Семейства: Rotaliidae Ehrenberg, 1839; Calcari-
nidae Schwager, 1876; Chapmaninidae Thalmann, 1938;
Notorotaliidae Hornibrook, 1961; Cuvillierinidae Loeb-
lich et Tappan, 1964; Ammoniidae Saidova, 1981; Dic-
tioconoidessidae Bugrova, 1990.
?Отряд Chilostomellida Haeckel,
1894
Раковина трохоидная целиком или только на ран-
ней стадии, округлой, обтекаемой формы, слабо
сдавлена по оси навивания или перпендикулярно ей,
с сильнообъемлющими оборотами и небольшим чис-
лом камер в обороте (3—4, иногда в последних обо-
ротах по 2 камеры), камеры вздутые, быстро увели-
чивающиеся в размерах, иногда последняя камера
сильно объемлющая, полость камер неподразделена;
устье в основании септальной поверхности, узкое,
щелевидное, с губой, нависающей козырьком. Верх-
ний мел—голоцен.
Примечание. Положение отряда проблематично. Возмож-
но, после ревизии он будет переведен в класс Spirillinata, надсем.
Hergotelloidea.
Семейства: Chilostomellidae Brady, 1881; Chi-
lostominidae Finger et Gaponoff, 1986.
Надотряд Nonionoida Saidova, 1981
Раковина плоскоспиральная.
Отряд Nonionida Saidova, 1981
Раковина плоскоспиральная, двусторонне-сим-
метричная или несколько асимметричная, инволют-
ная или полуинволютная, уплощенная или почти ша-
рообразная с относительно небольшим числом
камер, пупок открытый или заполнен скелетным ма-
териалом, может быть прикрыт лопастью, могут быть
дополнительные скелетные пластинки над швами и
над пупочной областью, под которыми образуются
39 Протисты, часть 1
дополнительные полости, пупочная область может
быть также пронизана щелями, служащими для вы-
хода цитоплазмы (Protelphidium), но настоящей сис-
темы каналов нет; устье в основании септальной по-
верхности, экваториальное или заходит на пупочную
сторону, изредка может быть множественным, пери-
ферическим и конечным; у некоторых родов имеют-
ся дополнительные шовные или пупочные устья.
Верхний мел—голоцен.
Подотряд Nonionina Saidova,
1981
Раковина без дополнительных скелетных пласти-
нок.
Надоем. Nonionoidea Schultze,
1854
Раковина с устьем в основании септальной поверх-
ности.
Семейства: Nonionidae Schultze, 1854; Nonio-
nellidae Voloshinova, 1958; Spirotectinidae Saidova,
1981.
Надсем. Almaenoidea Myatlyuk,
1959
Раковина с дополнительными щелевидными усть-
ями, расположенными вдоль периферического края
камеры и окруженными отворотом, на последней ка-
мере устье может приподниматься на вытянутой
шейке, экваториальное устье в основании септаль-
ной поверхности имеет тенденцию смещаться на
одну из сторон. Средний эоцен—голоцен,
Семейства: Almaenidae Myatlyuk, 1959 (без
рода Saraswati Kalia, 1982, который на основании
трохоидной раковины и дополнительных устьев, па-
раллельных периферическому краю, перенесен в Не-
leninidae (Rosalinida)).
Подотряд Astrononionina Saidova,
1981
Раковина с дополнительными скелетными плас-
тинками над швами и пупочной областью, под кото-
рыми образуются дополнительные полости, открыва-
ющиеся дополнительными пупочными устьями в
швах или над пупком. Средний эоцен—голоцен.
Семейства: Astrononionidae Saidova, 1981; *Bis-
saciidae Loeblich et Tappan, 1988.
Отряд Elphidiida Saidova, 1981
Раковина плоскоспиральная, симметричная, инво-
лютная, у одного рода с выпрямленным отделом,
число камер относительно невелико; пупок закрытый,
хорошо развита система каналов, состоящая из 2 сим-
610
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
метричных спиральных каналов, отходящих от них
подшовных каналов, открывающихся 1 или 2 рядами
отверстий мелких септальных канальцев, могут также
быть развиты вертикальные пупочные каналы, откры-
вающиеся в пупочной области; мелкие округлые усть-
евые отверстия в основании септальной поверхно-
сти или над нею могут зарастать и открываться только
перед образованием новой камеры. Палеоцен—голо-
цен.
Семейства: Elphidiidae Galloway, 1933; Cribro-
elphidiidae Voloshinova, 1958.
Отряд Orbitoidida Copeland,
1956
Раковина дисковидная или линзовидная, с боль-
шим числом камер, подразделенных на камерки, с
системой столонов, системы каналов нет. Верхний
мел—верхний эоцен.
Семейства: Orbitoididae Schwager, 1876; Lepi-
torbitoididae Vaughan, 1933 (без Clypeorbinae Sigal,
1952); Linderinidae Loeblich et Tappan, 1984.
Отряд Nummulitida Carpenter,
Parker et Jones, 1862
Раковина плоскоспиральная, эволютная или инво-
лютная, линзовидная, дискоидальная, округлая, с очень
большим числом камер, которые могут быть подразде-
лены на камерки, экваториальные и конечные камеры
могут быть дифференцированы, последние могут быть
кольцевыми, может присутствовать септальный кла-
пан, как у роталиид; имеется сложная система кана-
лов: спиральный маргинальный корд, система внутрен-
них спиральных каналов и подшовные каналы у более
ранних форм, у более поздних и эволюционно продви-
нутых родов каналы модифицированы или замещены
интерсептальными каналами, часто присутствует и
система столонов. Палеоцен—голоцен.
Примечание. Нуммулитиды — наиболее сложная и совер-
шенная группа фораминифер. Большинство ее семейств вымерли к
эоцену, но в сем. Nummulitidae много современных представите-
лей. Красивые крупные раковины Assilina, Cycloclypeus и особен-
но Heterocyclina, Heterostegina, Operculina и Nummulites обычны в
мелководных тропических морях.
Семейства: Nummulitidae de Blainville, 1827;
Discocyclinidae Galloway, 1928; Pellatispiridae Hanza-
wa, 1937; Asterocyclinidae Bronnimann, 1951.
Класс Globigerinata Mikhalevich, 1980
Раковины многокамерные, трохоидные и плос-
коспиральные (у Heterochelicida — с зачаточным
спиральным и хорошо развитым выпрямленным от-
делами), с небольшим числом камер в обороте, про-
грессивным увеличением размеров пупочной облас-
ти, разворачиванием спирали и неплотным сочлене-
нием камер, окруженных и удерживаемых широким
слоем цитоплазмы или пузыристой капсулой (у про-
двинутых специализированных форм); камеры силь-
но уплощенные у плоских форм, у остальных шаро-
видные или выпукло-удлиненные; часто развиты до-
полнительные скелетные пластинки над пупком или
швами; стенка секреционная, бифонтинальная, с
мощным утолщением наружных слоев во время гаме-
тогенеза (краст), гладкая либо шиповатая и ячеистая,
может быть с длинными радиальными иглами, непо-
движными либо сидящими свободно в углублениях
наподобие суставов, поровые канальцы воронкооб-
разные, с 1 срединной, поровой пластинкой (диаметр
наружного порового отверстия до 2 мкм), нараста-
ние новых камер сходно с таковым Rotaliata; устье
базальное, в основании последней камеры, занимает
значительную часть раковин; часто имеются допол-
нительные устья; ретикулоподии радиальные, не
анастомозирующие; агамонты многоядерные, гетеро-
кариотные, у Globigerina bulloides и Globigerinoides
ruber с полимеризацией также и соматических ядер,
имеются уникальные органеллы — криптосомы и
микровилли, гаметы двухжгугиковые, их число во
много раз больше, чем у донных; свободноживущие,
пелагические. Юра—голоцен.
Примечание. 3 вымершие в мелу группы — Planomalinida,
Rotaliporida и Globotruncanida, а также Hantkeninida, вымершие в
миоцене, ниже не рассматриваются.
Отряд Heterochelicida Mikhalevich,
1992
Раковина на ранней стадии спирально навитая
(трохоидная или плоскоспиральная), впоследствии
двух- и трехрядная (или сразу двух- и трехрядная —
спиральный отдел может отсутствовать по крайней
мере у одного из поколений), с тенденцией увеличе-
ния числа камер на поздней стадии до образования
многорядной раковины, реже с уменьшением их
числа до образования однорядной стадии, камеры
сильно выпуклые или почти шаровидные; стенка
ребристая, реже шиповатая; устье — аркообразное
отверстие в основании последней камеры или конеч-
ное, у Laterostomella с внутренней зубной пластин-
кой; могут быть дополнительные шовные устья. Ни-
жний мел—голоцен.
Семейства: Heterochelicidae Cushman, 1927;
Guembelitriidae Montanaro Gallitelli, 1957; Chiloguem-
belinidae Reiss, 1963.
Отряд Globorotaliida Mikhalevich, 1980
Раковина трохоспиральная, уплощенная, кониче-
ская, с килем или равномерно раздутая, с широким
периферическим краем, пупок обычно закрытый;
ТИП FORAMINIFERA. СИСТЕМАТИЧЕСКИЙ ОБЗОР
611
стенка с толстым наружным известковым слоем, но
гладкая или в бугорках и ямках, без игл; устье —
щелевидное или аркообразное отверстие в основа-
нии септальной поверхности на пупочной стороне,
дополнительные пластинки отсутствуют; дополни-
тельные устья обычно не развиты. Нижний палео-
цен—голоцен.
Семейства: Globorotaliidae Cushman, 1927;
Pulleniatinidae Bronnimann et Bermudez, 1953; Trun-
corotaloididae Loeblich et Tappan, 1961.
Отряд Globigerinida Carpenter, Parker
et Jones, 1862
Раковина трохоидная, иногда только на ранней ста-
дии, последние камеры могут быть с тенденцией к
плоскоспиральному расположению, или одна последняя
камера полностью объемлет все предыдущие, пупок
обычно широкий, зияющий или прикрыт дополни-
тельной пластинкой, камеры шарообразные, иногда
слабо связанные друг с другом, без цитоплазмы легко
отламываются (Boliella); стенка с толстым известко-
вым ячеистым слоем (краст), с крупными порами и мо-
жет быть покрыта тонкими длинными монокристал-
лическими иглами, сидящими свободно в углублениях
на приподнятых сосочках (Globigerinidae) или закреп-
ленными неподвижно, в поперечном сечении иглы
могут быть округлыми, трехгранными, трирадиальны-
ми, могут располагаться по всей поверхности ракови-
ны или только на ее периферических концах; в других
случаях стенка гладкая (Candeinidae), может быть с
ямками или сетчатой (Catapsydracidae), часто имеются
дополнительные скелетные пластинки над устьем и
швами; устье обычно огромное, зияющее, в пупочной
области, куда открываются и устья предшествующих
камер, может быть множественным у раковин с объ-
емлющей последней камерой (многочисленные округ-
лые отверстия крупнее пор); дополнительные устья в
швах и в пупочной области хорошо развиты во многих
таксонах. Средний эоцен—голоцен.
Семейства: Orbulinidae Schultze, 1854; Globige-
rinidae Carpenter, Parker et Jones, 1862; Candeinidae
Cushman, 1927; Catapsydracidae Bolli, Loeblich et
Tappan, 1957; Hastigerinidae Bolli, Loeblich et Tap-
pan, 1957; Globigerinitidae Bermudez, 1961.
THE PHYLUM
SUM
Foraminifera are unicellular organisms whose body is protected
by the shell. In primitive forms the shell is organic (mucopolysacha-
rids), in more advanced it is agglutinated, then with partly secretions
cement and finally secretious throughout; the last one may be mono-
fontinal (microgranular, porcelaneous, hyaline-radial) with the pseu-
dopores, or bifontinal with the complex pore canaliculi. The shell may
be unilocular, pseudocolonial, pseudochambered and multichambered
(with chamber number usually 6—15, 30, more rarely up to several
hundreds (in the latter case the chambers can be strongly differentia-
ted). Chambers are connected with each other by the foramina, in the
advanced groups by more complex integrative systems: stolons, tun-
nels, inner apertural systems and canals.
The shells have one, two or several external openings — apertu-
res (in the most primitive forms aperture may be absent). In advanced
groups the inner apertural structures (teeth, ribs, tooth-plates of diffe-
rent complexity) and the additional apertures arise. The aperture ser-
ves for the protruding of granuloreticulopodia, usually branching and
anastomosing, forming the reticulum around the shell. Life cycles with
the alternation of the sexual and asexual generations, with the interme-
diate reduction. Gametes mostly biflagellate, heterocontous, more ra-
rely threeflagellate or amoeboid. One of the generations (more often
sexual one) may be absent. Nuclear apparatus with the complex chan-
ges during the life cycle, only in the lower forms part of the life cycle
is mononuclear, in the rest ones it is multinucliate during the whole
life cycle (except zigote), in the most advanced forms the nuclear
dimorphism arises. Meiosis is twostepped, mitoses goes as a closed
pleuromitoses. Mitochondrial cristae are tubular. Planktonic forms
have cryptosomes and microvilli. Mostly sea bottom free-living orga-
nisms, but planktonic group also exists. Parasitic forms very rare.
Cm—Holocene.
THE CLASS ASTRORHIZATA SAIDOVA, 1981
Shells unilocular, sometimes pseudocolonial or pseudochambered,
of subsphaerical, elongated, tubular, stellate, branching or other irre-
gular form; wall monofontinal, sometimes with the visible pseudopo-
res, tectinous in Lagynana and Ammoscalariana, agglutinated or mic-
rogranular in Astrorhizana, may be simple or partly subdivided by the
partitions in the Lagynana and Astrorhizana, may be thickened from
the inside (spongy, labyrinthic); aperture may be entirely absent, may
be primarily multiple (openings often irregular in form and position or
FORAMINIFERA
MARY
situated at the end of the tubes), or single in more advanced forms
(usually of definite form and position, may be slightly elevated on the
neck, may have a small lip); agamonts uninuclear, nuclear dualism
unknown, gametes biflagellate or amoeboid (in some Lagynana); free-
living or (very often) attached (among Lagynana freshwater forms are
also met, two genera — Rhynchosaccus and Thalamophaga — are pa-
rasitic). Cm—Holocene.
Subclasses: Lagynana Mikhalevich, 1980; Ammoscalariana Mik-
halevich, 1980 (insufficiently studied); Astrorhizana Saidova, 1981.
THE CLASS SPIRILLINATA MASLAKOVA, 1990
Shells pseudotwochambered (with long tubular second chamber
around proloculus) or derived from it (pseudomultichambered, multi-
chambered in higher representatives, but often with the initial tubular
part preserved during 1—2 volutions), with different coiling types —
glomerate (very often, especially at the initial part), pianospiral (more
often at the adult stage, being initially streptospiral), sometimes bise-
rial in the last part, trochospiral one also occur, in higher supermulti-
chambered forms shell may become spherical or fusiform; pseudo-
chambers formed by constrictions of the outer shell wall, by the for-
mation of endoskeletal thickenings or by the short septula, chambers
in multichambered forms usually bearing the primitive features, sho-
wing their origin from the tubular one (being somewhat irregular, with
short right or only slightly curved septa lacking characteristic for
Rotaliata’s septa lip at the septal ends), fluted septa occur only in this
group, chambers in advanced groups becoming more regular; integra-
tive systems — tunnels in advanced supermultichambered forms, pri-
mitive canal systems in some advanced tubular forms; wall insuffici-
ently studied, monofontinal, agglutinated (often with femigenous ce-
ment) or microgranular in lower forms (Ammodiscana), in
multichambered forms sometimes very thick, with different layers, not
differing in principle from those of the thick walled Textulariana, but
with special additional inner shell depositions (nods, nodosites, hooks,
chomata) on the chamber floor not met in other groups, in higher
subclass (Spirillinana) completely secretious, radial, may be pitted Бу
regular pseudopores (according to the literature), layers of growth are
not formed, in ancient forms often two layers are marked — microg-
ranular and radial, in Involutinida — usually radial, in recent Spirilli-
nids — radial crystal units act optically as a single crystal, outer scul-
pture and inner pillars well developed; growth of tubular part uninter-
612
СИСТЕМАТИЧЕСКИЙ ОБЗОР
rupted, occurs by lateral accretion along the edge of the chamber, in
multichambered forms unclear; aperture terminal or derived from it (in
trochoid forms tending to displace on the umbilical side), always sim-
ple, never forming inner apertural structures, single or (rarely) multip-
le (m advanced multichambered forms), in multichambered trochoid
forms outer apertural structures sometimes present (valvular flap, T-
shaped or S-shaped plate), additional apertures often occur; agamonts
of recent Spirillinana homokariotic (their proloculus larger then that of
the gamonts — the reverse correlation), gametes (if known) amoebo-
id; bottom, mostly free living forms. Cm—Holocene.
Remarks. As it was shown above the wall of the palaeozoic
forms (former Fusulinacea) does not differ in principle from the ag-
glutinated and microgranular textulariana’s wall, but it is heterogenous
group and needs the profound revision. Based on priority of the shell
structure rather than wall ultrastructure we moved some of their taxa
to the other groups according to shell organisation plan (for instance
Parathuramminida, Paratikhinellidae — to Astrorhizata; Colaniellida,
Pseudopalmulinidae — to Nodosariata; Biplanatidae, Nezzazatidae —
to Textulariana). This revision could not be considered as finished.
Left in the subclass Ammodiscana are only pseudotwochambered tu-
bular forms or the forms whose origin from these ones is doubtless.
The palaeozoic forms are regarded here in less detail than the recent
ones.
Subclasses: Ammodiscana Mikhalevich, 1980; Spirillinana Masla-
kova, 1990.
THE CLASS MILIOLATA SAIDOVA, 1981
Shells unicellular (very seldom), may be pseudotwo-chambered,
pseudomultichambered, but mostly multichambered, the predominant
type of coiling — irregular- and regular glomerate, planospiral more
often combined with one of the glomerate ones (trochospiral only as
an exception); shells of the higher representatives could be cyclical
and fusulinoid in form; the presence of flexostyle is typical of the
majority of the groups; chambers predominantly tubular, usually two
per volution (only in Soritida broad and more numerous per volution),
in higher representatives chamber number increases to several thou-
sands, chamber lumen could be subdivided by the septa (sometimes by
the primary and the secondary ones), by pillars, advanced forms may
have integrative systems (stolons, tunnels), interconnected inner aper-
tural systems not developed, canal system absent; shell wall — mono-
fontinal, agglutinated in the lower forms (subclass Schlumbergerina-
na), in the higher ones (subclass Miliolana) — porcellaneous, with the
irregular not put into shape pseudopores between irregular randomly
oriented needles of crystal units, chamber growth occurs by the gra-
dual addition of the wall part by part, organic and secretious elements
are laid simultaneously; aperture terminal in position (even in Alveo-
linids and Peneroplids — above the base of apertural face), simple in
the most primitive forms, in more complex ones — with the outer
(flap) or inner tooth of a special structure, sometimes aperture secon-
darily multiple, additional apertures non developed (only as an excep-
tion); nuclear apparatus heterokariotic in higher secretious forms, so-
metimes with the polymerization of somatic nuclei (Sorites); in Trilo-
culina gametes are known as biflagellate (and having the axostyle?);
bottom forms, mostly free living, more rarely attached. Carbon—Ho-
locene.
Subclasses: Schlumbergerinana Mikhalevich, 1992; Miliolana Sa-
idova, 1981.
THE CLASS NODOSARIATA MIKHALEVICH, 1992
Shells unilocular and multichambered (pseudomultichambered as
an exception), predominant shell types — uniserial, high-trochospiral
(in a special polymorphinoid spire, absent in any other taxa), pianos-
piral (nearly exceptionally involute), typical trochospiral type is absent
completely; chambers often subcircular, triangular, V-shaped, palma-
te; inner additional skeletal chamber septa present only in one fossil
taxon (Collaniellidae), outer shell sculpture well developed in secfe-
tious forms, but outer additional skeletal plates are absent, integrative
systems poorly developed (stolons and tunnels absent, inner apertural
systems occur rarely, canal system marked only in Delosinida, purely
studied); wall monofontinal, agglutinated or microgranular in the
lower representatives (Hormosinana), in the higher ones (Nodosaria-
na) — hyaline-radial, with regular pseudopores having cylindrical
pseudopore canaliculi but without additional sieve-plates (according to
the literature basal sieve-plate present), growth interrupted, process of
forming of the new chambers has not been studied; aperture terminal
even in the coiled forms, single or multiple, simple or radial, slit-like,
asymmetrically-fissured, hooded, all the apertural types may be with
entosolenian tube (non analogues to the apertural structures of Rotali-
ata), (radial, asymmetrically fissured and hooded aperture, as well as
entosolenian tube are unique and are found only in this class), additi-
onal apertures not developed; data on nuclear apparatus and gamete
structure absent; mostly free-living, bottom forms, one parasitic speci-
es is described. Ordovician—Holocene.
Subclasses: Hormosinana Mikhalevich, 1992; Nodosariana Mik-
halevich, 1992.
THE CLASS ROTALIATA MIKHALEVICH, 1980
Shells multichambered only, all regular types of chamber arrange-
ment well presented (glomerate type practically absent), mostly
trochospiral, pianospiral and derived from both of them biserial and
triserial ones (rarely secondarily uniserial), in advanced forms often
cyclical; inner complications of chamber lumen are strongly develo-
ped: from thickened layers of the wall of different structure (labyrin-
thic, spongy, alveolar) and pillars in Textulariana up to septa and
septula (of die first and second order) in higher representatives of both
subclasses; in advanced forms of both subclasses chamber number
could increase to several hundreds and thousands, and in this case they
are often differentiated in their structure into central, median and pe-
ripheral layers (Qrbitolina, Nummulites)', additional skeletal plates
above the sutures and umbilicus are common, especially in higher
Rotaliata; integrative systems strongly developed (stolons, tunnels and
the interconnected inner apertural systems in Textulariana, stolons,
inner interconnected apertural systems and very complex canal sys-
tems in Rotaliana); wall in Textulariana monofontinal, agglutinated
and microgranular, with pseudopores, with various complication of the
inner wall, which could be cancellate, alveolar, spongy, in Rotalia-
na — bifontinal, with true regular pores, having cylindrical pore cana-
liculi with some transverse sieve plates besides the basal one; growth
interrupted, the contours of every new chamber forming entirely at
once, organic and lime secretion are divided in time, alternating; aper-
ture usually at the base of the apertural face (basal) both in coiled and
in some biserial and triserial forms (in the last ones could become
secondarily final), may also be areal and sieve-like, may have various
inner structures (especially developed in Rotaliana, but occurring in
Textulariana also): tooth-plates, subcylindrical or closed cylindrical
plates, tongues, valves, which could be interconnected and could form
a single system, additional apertures also well developed in both sub-
classes (sutural, umbilical, peripheral); agamonts of the higher Rotali-
ana (in Textulariana insufficiently studied) multinucleate, heterokario-
tic, gametes biflagellate, more rarely threeflagellate (Glabratella,
Nummulites venosus). Mostly free living, bottom forms, rarely attac-
hed, some parasitic species are known. Devonian—Holocene.
Subclasses: Textulariana Mikhalevich, 1980; Rotaliana Mikhale-
vich, 1980.
THE CLASS GLOBIGERINATA MICHALEVICH, 1980
Shells multichambered, trochospiral and planospiral, more rarely
derived from them multiserial to uniserial (Heterochelicida), with a
small number of chambers per volution, with the progressive enlar-
ging of the sizes of umbilical area up to nearly uncoiled spire and
loose conjunction of the chambers, encircled and supported by the
wide cytoplasmic layer or by the vesicular capsula (in advanced spe-
cialized forms (Hastigerina); chambers strongly flattened in flat forms,
in the rest ones — subsphaerical or inflated and elongated; additional
sutural and umbilical skeletal plates well developed; wall lime secre-
tious, bifontinal, with the great thickening of die outer wall layers
during the gametogenesis (crust), smooth or hispid and alveolar from
the surface, may have long radial needles, immobile or setting loosely
in the holes, having the opportunity to turn like in joints, pores regu-
lar, usually large, pore canaliculi funnel shaped, with one middle
sieve-plate, growth of the new chambers like in Rotaliana; aperture
situated at the base of the chamber (basal), usually large, occupying
significant part of the shell, additional apertures very often; pseudopo-
dia radial, non anastomosing; agamonts multinucleate, heterokariotic,
in Globigerina bulloides and Globigerinoides ruber with the polyme-
rization of the somatic nuclei also, the unique organellas — criptoso-
mes and microvilli — are present; gametes biflagellate, their Humber
many times larger then in the bottom species; free-living pelagic
forms. Jurassic—Holocene.
Remarks. The representatives of the three fossil groups (Plano-
malinida, Rotaliporida, Globotruncanida) and also Hantkeninida are
not regarded here.
ТИП FORAMINIFERA. SUMMARY
613
Foraminifera are one of the largest highly organized and well
outlined groups of Protists, known from the praecambrian till recent
time. The comparison of the Foraminifera with the other groups of the
former sarcodines showed sharp differences between them (in the
structure of nuclear apparatus, pseudopodia, in the life cycles, form
and composition of the skeleton) (Mikhalevich, 1999).
The features of the foraminiferal nuclear apparatus (polymerizati-
on of nuclei, genoms, the nuclear dimorphism), the arising of the
multichambered and deeply differentiated shell with the unique integ-
rative structure of the organism level (the canal system), along with
the features of their granuloreticulopodia, gametes and flagella, their
special life cycles with the antithetic alternation of generations (as
well as the originality of the motpho-functidnal direction of their evo-
lution) — all these characters permitted us to raise the rank of this
group up to the phylum. The presence in the majority of the species
of the multichambered test is one of the most characteristic features of
the Foraminifera.
The profound differentiation and integration of the skeletal and
cytological structures of this group is amazing for the organisms of
the unicellular level. Based on the study of morphological features of
their skeletons and cytoplasm, on the ultrastructure of their shell wall
as well as on the development of all these features during the geolo-
gical history a new conception of the foraminiferal evolution and
macrosystem is presented: the phylum Foraminifera consists of 6 clas-
ses, 11 subclassesr 66 orders. The separation of taxa within the phy-
lum is based on the complex of features: test morphology and the type
of coiling, number of chambers (one, two or many), modes of cham-
ber formation (true or pseudomultichamberedness) and chamber con-
junction, shell wall composition and its ultrastructure, inner and outer
additional shell skeletal structures, structure of the aperture, presence
of the additional apertures and the integrative systems, chamber form
and some other minor features along with their development tenden-
cies.
Special attention has been paid to the evolutionary significant fe-
atures and to the degree of their development, reflecting the level of
differentiation and integration of the skeletal and cytoplasmic structu-
res (Mikhalevich, 1992, 1999). These especially include such apomor-
phic features as the multichamberedness, additional apertures, integra-
tive systems (not only stolons, tunnels and canals but also intercon-
nected inner apertural structures, creating the compartmentalisation
and the profound differentiation of the cytoplasm), as well as the
differentiation of the nuclear apparatus.
Some of these features and cytoplasmic features were not used pre-
viously for the characterization of the highest Foraminiferal taxa. In spite
of the insufficient cytological data now it is possible to use some of them
in taxonomy, firstly the features of the nuclear apparatus.
Within the Foraminifera the nuclear dimorphism is characteristic
of the highest subclasses of the classes Miliolata and Rotaliata and for
the class Globigerinata.
In Spirillinata even the representatives of their higher subclass —
Spirillinana — are homokariotic, as well as the representatives of the
primitive subclasses Lagynana and Astrorhizana, and evidently of the
subclass Textulariana. It is possible to suppose that in all the other
lower subclasses (Ammodiscana, Schlumbergerinana, Hormosinana)
the nuclear dimorphism is also absent.
The differentiation between endo- and ectoplasm is not marked
clearly in more primitive subclasses (Lagynana, Astrorhizana). It is
more pronounced in the other groups of the bottom Foraminifera,
especially in the organisms having canals, and is very distinct in the
planktonic forms, where the ectoplasm is situated around the shell like
vide layer, sometimes even forming calimma-like vesicular layer, con-
vergent to calimma of Radiolaria.
The presence of the two cytoplasmic organellas — microvilli and
cryptosomes — could also be used as a; taxonomic feature of the class
Globigerinata.
Foraminiferal taxonomy is based mainly on the features of their
skeleton. To describe the complete and precise diagnoses of high taxa
one not only needs to make comparative analysis and to generalize the
known data, but also to watch die development of the basic features
in different groups and to weigh the taxonomic value of the traditio-
nally used and newly introduced features.
In the pair of features: test morphology — structure of the
wall — we give the paramount importance to the first one, conside-
ring the second one as having an essential, but co-ordinate importance
in the classification. This approach is in line with the findings of the
Fursenko’s school (Rauser-Chernousova, Subbotina, Maslakova, Sai-
dova and others), but is opposite to the commonly used classification
of Loeblich and Tappan, 1988, whose remarkable labour creates the
basis of the new opportunities for further taxonomic studies.
The accumulation of electron-microscopic data on the shell wall
ultrastructure has led to the exaggeration of its taxonomic value and
has caused the greatest disagreement in evaluation of this feature. We
consider: it of secondarily significance, being only of subclass but not
of class character. Summarising the most recent data on the secretious
lime wall the following main types could be separated:
1. Microgranular wall — formed by disorderly, randomly situa-
ted grains of different form and dimensions above the inner organic
lining. This wall may be very thick and may form complex inner
structures — alveolar, labyrinthic, sponge-like, honey-combed, stolo-
niferous layers, pillars, chomata. So far as there is no clear cut bound-
ary between the agglutinated and microgranular wall, as it was shown
by many authors and by our own data, hence historically the transition
from the first to the second one could take place gradually. That is
why the difference between the microgranular and the agglutinated
wall is not of high taxonomic value. The microgranular wall, along
with the agglutinated one, is characteristic of the subclass Astrorhiza-
na and of the subclasses Ammodiscana, Schlumbergerinana, Hormosi-
nana, Textulariana.
The comparison of the most complex Fusulinid wall with that of
the advanced Textulariana (Orbitolina for instance) shows their great
resemblance: microgranular character, outer organic layer seen in Me-
sorbitolina (Textulariana) corresponding to the tectum, middle microg-
ranular one — to keryotheka, the ampoule-like alveoli in Simplorbito-
lina (Textulariana) and Sumatrina (Fusulinida) are also similar. Cho-
mata (Schwagerina) show the example of the inner complication of the
wall. The absence of principle differences in the wall ultrastructure of
both of the named groups gives the basis to revise the heterogenous
palaeozoic Fusulinid taxa according to the test morphology of their
different groups.
2. Miliolate porcelaneous wall — its needle-like crystal units in
the middle layer (except of the inner and outer layers) are situated
loosely, randomly and are disoriented. This type can also form the
inner pillars (Fusarchaias) and besides external sculpture. It has been
shown earlier (Mikhalevich, Rodionova and Sinjakova, 1986) that
Dentostomina (Schlumbergerinana) with the agglutinated wall also has
long lime secretious needles on the surface of the inner wall, of the
aperture and of the tooth surface typical of the porcellaneous miliolate
wall. This fact along with the full resemblance of the shell structure
confirms their relationship with the Miliolata and gives the ground for
excluding such forms from the previous Textulariacea. Earlier the test
similarity of agglutinated Schlumbergerinana and porcellaneous Mili-
olana was regarded as convergent. The transition from the first to the
second wall type of these groups in the geological history could have
also taken place gradually.
3a. Hyaline-radial (or radial) wall — elongated pillar-like units
of crystals set compactly and oriented perpendicularly to the surface
above the inner organic layer. This type has no inner complication of
the wall, but may form the outer sculpture. Layers of growth (secon-
darily layers) could clearly be seen. The type is characteristic of No-
dosariana.
3b. The radial wall ultrastructure of the higher Spirillinata is
similar in packing of the crystal units to Nodosariana’s one. The pa-
cking is so regular that the crystal units often act optically as mono-
crystal. Apparently both types appeared independently. Spirillinana’s
wall does not form secondary layers of growth but sometimes forms
the inner structures and has some other peculiar features (in pore char-
acter, in the mode of growth).
In the ancient forms of both subclasses many species have a mic-
rogranular layer of the wall. It is unknown whether the two layers
were laid simultaneously or the microgranular one is a result of the
process of diagenesis. But it is possible to mark the gradual disappea-
rance of the microgranular layer during the process of historical deve-
lopment in the later forms, may be due to the better preservation. In
all the types, discussed above, the shell matter is situated on one side
of the inner organic lining.
4. The bilamellar wall of higher Rotaliata and Globigerinata has,
in principle, another structure (Hemleben et al., 1977; Angell, 1979,
. and oth.). Besides the Inner Organic Lining (IOL), there is a Primary
Organic Membrane (POM), and the very first two layers are formed
at once on both its sides — the inner and the outer lime layers.
Since in Russian the bilamellar wall is usually called as two-lay-
ered one, and as the layers could have different genesis in the different
groups — in order to escape any confusion and to oppose it to all
others — we named it as a bifontinal one and the previous types —
as monofontinal. The plate-like crystal units of bifontinal wall are
packed compactly in compact vertical pillars (columns) presenting a
uniform regular structure. When these pillars are set at the angle of
35—45 degrees — the wall looses its transparency. Primarily the
614
СИСТЕМАТИЧЕСКИЙ ОБЗОР
overstatement of this feature had lead to the break of some relative
groups of the higher Rotaliata. Later it became clear that the pillars of
plate crytallits could be situated perpendicularly or be inclined, not
only in the shells of one species, but even in the same shell. Their
position could also be changed during the process of diagenesis. In
this type4 of wall structure the secondary lamina (layers of growth) are
also pronounced, especially in some Globigerinata where they form
the crust. Bifontinal type provides the shell with the most durability,
resistance along with the least thickness and lightness.
It also gives wide structural opportunities to the development of
both the outer (spicules, striae, tuberculi, additional skeletal plates)
and inner shell structures (apertural tongues, plates, grooves, valves).
Unlike the previous types, this type can arise as a result of a sudden
change in the mode of the test secretion and of the appearance of two
centres of the secretion simultaneously on the both sides of the POM
and then the synchronous laying of the two lamellae. The openings in
the wall permitting communication of the cytoplasm with the environ-
ment present in any kind of the shell wall ultrastructure. But only in
the bifontinal type of the wall they have a complex structure with the
additional sieve plates and may be called true pores. Their character
could be used as a taxonomic feature of the subclass Rotaliana and
class Globigerinata.
All the openings of the monofontinal wall could be denoted as
pseudopores. These are irregular openings of the organic, agglutina-
ted, microgranular and porcellaneous wall and also (according to the
literature) regular, but less complex than in Rotaliana openings of
Nodosariana and of Spirillinana. The latter have some peculiarities as
compared with the Nodosariana. The pseudopores of this two subclas-
ses are characteristic of each of them.
The enumerated types of the secretious wall differ also in the
mechanism of the physiological process of growth and chamber for-
mation. The comparison of this process in Textulariana and Rotaliana
shows that they have much in common (Angell, 1979, 1990; Bender,
1992): the protuberance of the reticulopodian veneer and formation of
the envelope around it, outlining the new chamber just at once, shell
matter is laid on the membrane, not inside it, the organic and mineral
secretion are separated in time. These facts also confirm the unity of
both subclasses in the same class.
In Miliolata the growth of the new chamber goes gradually, seg-
ment by segment, organic and mineral secretions are going simulta-
neously, calcareous crystals are intruded into the organic layer.* Anot-
her peculiarity of the Miliolate wall is that it is formed as a result of
the heterogenous nucleation of calcite opposite to the majority of Fo-
raminifera and of other animals. In Spirillinata the growth is also
going gradually, by the accretion of the end of the chamber (Berthold,
1976). Data on the Nodosariata growth are absent.
A special attention has been paid to the evolution of main skeletal
structures in different foraminiferal groups. Their skeleton reflects
their basic plan of structure, connected with the functions of the whole
organism. The multichambered tests are unique among Protista. They
represent the result of the polymerization (multiplication of the homo-
logous elements). The multichamberedness could be regarded to some
extent as an analogue of multicellularness.
Among the different modes of polymerization foraminiferans used
just two (Mikhalevich, 1981, 1999):
1. Fission — subdivisions of the chambers already formed under
uninterrupting growth. It occurs in more primitive classes (Astrorhiza-
ta, Spirillinata, occasionally in Nodosariata and Miliolata).
2. Additional polymerization: addition of new units — goes under
interrupted growth and has been found in more advanced groups (No-
dosariata, Miliolata and especially Rotaliata and Globigerinata).
Chamber conjunction differs in both cases.
In the first case pseudochambers are formed, in the second one —
true chambers. This feature could also be used as a taxonomic one of
the high rank groups. In the higher groups of Miliolata and Rotaliata,
two modes may be combined: the multilevel polymerization, when
chambers are divided into chamberlets by the partitions.
* In Archaias angulatus (Soritida, Miliolata) (Wetmore, 1999)
with the broad chambers the anlage is formed before the building of
the new camera. The process of formation of organic matrix and the
process of calcification are not searched in this work in details. But it
is hardly possible to suppose that the processes of formation of the
typical monofontinal pocelaneous wall of A. angulatus could be simi-
lar to those of Retaliate bifontinal wall with the complex pores. Ob-
viously the similarity is seen only in the formation of the anlage and
the rest of the processes of calcification are going in this species as in
other Miliolata.
The transition from the chambers equal in form and dimensions
(in some representatives of Nodosariata — Saccamminopsis, Grigelis)
to the unequal ones (in the majority of Miliolata and Nodosariata, in
all Rotaliata and Globigerinata) presents further differentiation on the
basis of the preceding polymerization. It is especially clearly seen in
the heteromorphous tests (Vulvulina, Fissotextularia (Textulariana),
Semitextularia (Hormosinana), Vandenbroeckia (Miliolana)). It often
goes along with the oligomerization — the decreasing of the chamber
number in the last volutions, sometimes to a single one. We can see
such parallel examples in different classes (Idalina (Miliolana), Cola-
niella (Nodosariana), Ammosphaerulina (Textulariana), Orbulina
(Globigerinata)). Such feature may be used as a characteristic feature
of the subfamily rank, for instance, we used it for the separation of the
subfamily Involvohauerininae. The further profound differentiation
takes place in higher Miliolata (Alveolina, Raadshoovenia) and espe-
cially in higher Textulariana and Rotaliana showing similar tendenci-
es, when in supermultichambered forms the number of chambers are
hundreds and thousands and they are presented by the embryonal,
medial, equatorial and peripheral layers of different chamber structure
(Orbitolina (Textulariana), Biplanispira, Heterostegina (Rotaliana)). In
some cases the number of chambers could be a feature characteristic
of classes, i.e. Astrorhizata are unilocular or pseudochambered, Spiril-
linata are mostly pseudotwochambered with the second tubular cham-
ber, which is often preserved even when the last volutions (in their
higher representatives) are subdivided into chamberlets or form some
additional chambers. The rest of the classes are mostly (as Miliolata,
Nodosariata) or exclusively multichambered (as Rotaliata, Globigeri-
nata).
The next structure important in taxonomy is an aperture, which
also changes strongly during the evolutionary process. Its significance
is caused by its large functional role. Its development in different
groups shows many examples of parallelism: in die more primitive
class (Astrorhizata) it develops from primarily multiple aperture not
strictly fixed in position and form (Cribratina, Microcometes — Lagy-
nana, Orbulinelloides — Astrorhizana) to singular one with definite
form and position (Heterogromia — Lagynana, Lagenammina, Piluli-
na — Astrorhizana), in more advanced classes (Miliolata, Nodosaria-
ta, Rotaliata) from primarily singular to the secondary multiple ones,
from the simple outer structure to the inner ones. The inner apertural
structures could be regarded as apomorphic feature as they not only
support the strong cytoplasmic streaming but also lead to the partial
compartmentalisation or to the complete isolation of the cytoplasmic
flows in the apertural area and to the profound differentiation of the
cytoplasm.
At the same time, the position of the aperture and its specific
structures are original in each class in consideration, presenting sepa-
rate phylogenetic branches (Mikhalevich, 1999). This structures could
be used as their diagnostic features: the inner teeth of Miliolata (with
their interesting ontogenesis, repeating phylogenesis), the Nodosariata
radial aperture, met only in this group, as well as entosolenian tubes
and also the hooded and asymmetrically fissured aperture. The inner
apertural complication of Rotaliata achieves the most complexity and
diversity — the inner tooth-plates and grooves, growing longer and
longer, and finally becoming connected in the continuous system
(inner apertural system), functionally similar to some extent to the
canal system, but not analogous to it. The system of inner apertural
valves (or paries proximus) in Discorbidae plays the same role. Glo-
bigerinata apertures are usually strongly enlarged, becoming high ar-
ched, yawning as one of the ways of adaptations to the planktonic
mode of life. In more primitive classes Astrorhizata and Spirillinata
the aperture is simple, only in the most advanced higher multicham-
bered Spirillinana the special external T-shaped or S-shaped plate is
developed, not found in any other group. In the classes having lower
subclasses the peculiarities of the inner apertural structures are of the
same character in both lower and higher ones: simple and complex
teeth or trematophore in Schlumbergerinana (Miliammina, Dentosto-
mina, Schlumbergerina) and Miliolana (Quinqueloculina, Tortonella,
Pseudohauerina), radial and asymmetrically fissured aperture in Hor-
mosinana (Nodosinum, Cuneata) and Nodosariana (Nodosaria, Fissu-
rina), inner apertural systems of interconnected tooth-plates in Textu-
lariana (Clavulina) and Rotaliana (Bulimina). The same could be said
about the position of the main aperture: it is terminal at the end of the
tubular chamber in both subclasses of Spirillinata and of Miliolata
(even in the miliolate forms with broad chambers, such as Alveolini-
dae and Peneroplidae the aperture is never connected with the base of
apertural surface). In both subclasses of Nodosariata it is also termi-
nal, even in the coiled forms (Lenticulina). Opposite to the previous
classes in the coiled forms (both pianospiral and trochospiral ones) of
the both subclasses of Rotaliata and in Globigerinata the aperture is
ТИП FORAMINIFERA. SUMMARY
615
basal — it is situated at the base of the apertural face of the last
chamber, at least initially. Even in the elongated monoaxial retaliate
forms derived from the coiled ones the aperture often preserves its
basal position being connected with the base of the apertural face not
only in the young stages but at the adult ones also (Textilina, Norvan-
ganina (Textulariana), Bolivina, ^Brizalina, Stainforthia (Rotaliana)).
The change of position of Rotaliata’s aperture is connected with the
profound change of the plan of their shell structure, of the whole
organism. As to the additional apertures appearing in this group (su-
tural, umbilical, peripheral ones), we consider them to be an important
apomorphic feature. There was paid too little attention to them in the
diagnoses of the high rank taxa. Additional apertures are strongly de-
veloped in both subclasses of Rotaliata, and especially in the class
Globigerinata. They are rather well developed in Spirillinata. In Mili-
olata they occur only as an exception. That is why we separated such
genera into special subfamilies. In Nodosariata the additional apertu-
res are nearly fully absent (only in Delosina).
Taking into account their great functional and evolutionary signi-
ficance, we consider the presence of additional apertures as a feature
of high taxonomic value.
Great numbers of chambers and their volutions makes difficult the
communication between them and the environment. This contradiction
is removed owing to arising of integrative systems: more primitive
stolons and tunnels and more complicated canal system. Their presen-
ce could be undoubtedly regarded as apomorphic feature. Integrative
systems arise in the more advanced groups of the different phyletic
lineages and in different periods: the stolons of the fossil and recent
Textulariana and Rotaliana, the tunnels of Paleozoic Fusulinids, of the
Cretaceous and recent Alveolinids. The primitive canal system was
recently discovered by Hottinger (1976) and by Azbel (1986) in Pa-
laeozoic Lasiodiscida and Involutinida, in Jurassic Spirillinids. This
fact became the basis for the separation of the new taxa of the order
level (Hottingerellida, Lasiodiscida, Spirotrocholinina). The canal sys-
tem of the subclass Rotaliana achieves the highest development and
complexity as was shown by Hottinger and his co-authors (Hottinger,
1978; Bilman et al., 1980) in their scrupulous works. The pseudopo-
dial ectoplasm is circulating in canals, carrying oxygen and the other
components. Their function could be compared (to some extent) with
the blood system of the higher multicellular animals, but having the
skeletal origin and functioning in one cell. It is also an example of the
profound differentiation of the inner and outer cytoplasm, unusual for
the unicellular level.
So far as the type of shell construction reflects the plan of the
foraminiferal organism, and taking into account also the conservative
state of the skeletal structures, we consider them to have the highest
taxonomic value. The analysis of the occurrence of the different shell
types in the different foraminiferal branches shows that their diversity
and richness are not the same in each branch. Their minimal number
is met in the primitive tectinous forms (Lagynana) and in Astrorhiza-
na, which usually have simple subsphaerical, tubular or irregular uni-
locular test, sometimes pseudochambered or pseudocolonial. Together
with the primitive skeleton, they have some more primitive features:
more primitive nuclear apparatus and non differentiated cytoplasm as
mentioned above. They also often preserve the ability to reproduce
with the simple division or budding.
There is no high diversity in the tubular pseudo-two-chambered
Spirillinata or in the multichambered, strongly specialised planktonic
forms.
The multichambered Miliolata (with the predominance of the irre-
gular glomerate forms and derived from them regular glomerate ones)
are clearly more varied. Planospiral coiling is also met in this group,
mostly combined with the glomerate beginning. The trochospiral shell,
however, occurs in Miliolata as an exception only. That is why we
have checked the ultrastructure of Fisherinella wall with trochospiral
test to confirm its belonging to Miliolata (we moved it in Soritida). In
Miliolata shells with a few tubular chambers per volution and with the
flexostyle prevail.
The trochoid type is absent completely in Nodosariata, where pre-
dominant uniserial and less common biserial forms exist. A special
polymorphinoid and plectofrondicularian types not found in any other
class are also present. Nodosariata planospiral shells are (except in one
case) evolute and are clearly distinguished from those of Rotaliata, not
only by their shell-wall ultrastructure in higher subclasses, but also by
the position of the aperture at the end of the shell, and never at the
base of the last chamber.
The highest diversity of the shell types is in Rotaliata, here the
trochospiral forms prevail and derived from them secondarily mono-
axial, planospiral ones are also numerous, the glomerate types are
practically absent. It is clearly seen that nearly every morphological
type of the agglutinated or microgranular Textulariana has its isomor-
phous analogue with the bifontinal wall: Montsechiana — Eulinderina
(with annular chambers), Arenagula — Neocribrella and Minouxia —
Sporobulimina (both pairs with additional apertural seave plates),
Dobrogelina — Sestronophora (with umbilical seave plates), Toretam-
mina — Oridorsalis (with additional apertures on the spiral side).
Only these two subclasses have additional outer skeletal plates, aper-
tural and umbilical sieve plates, well-developed additional apertures,
the structural peculiarities of the umbilical region and the form of the
chamber umbilical ends in the representatives of both subclasses coin-
cide sometimes to the slightest details (for example in some Troch-
amminids (Textulariana) and Discorbids (Rotaliana)). It is important
that not only all the basic structures of the shell and of the main and
additional apertures, as well as of their inner apertural structures are
similar in the representatives of the both subclasses, but also the ten-
dencies of their ontogenetic and evolutionary development. The pro-
cess of formation of the new chambers also goes similarly in Troch-
ammina, Clavulina, Textularia (Textulariana) and Rosalina (Rotalia-
na), differing from the gradual accretion of the Spirillinata’s tubes and
from the process of the new chamber formation in Miliolata.
It is unlikely that such a coincidence in such complex forms could
arise convergently many times, producing mirror variants of the large
groups, differing from each other only by shell composition and shell-
wall structure. Excellent examples of such coincidence of the aggluti-
nated and secreted shells are also clearly seen in other phyletic branc-
hes, not only in the primitive Ammodiscana with Spirillinana but also in
the complex forms of Nodosariata and Miliolata. The nodosariate radial
aperture (characteristic of this whole class and often observed in higher
Nodosariana) arose already in their agglutinated genus Nodosinum (Hor-
mosinana), as well as the other types of Nodosariata aperture, not found
in any other class (for example asymmetrically-fissured aperture of Cu-
neata (Hormosinana) and of Fissurina (Nodosariana)).
In Miliolata not only the chamber form, mode of coiling and
apertural structures coincide in both subclasses (Schlumbergerinana
and Miliolana), but also the details of their development during onto-
genesis (for instance, the stages of multiple and complex teeth deve-
lopment).
Such profound similarity in the shell and apertural construction
and in the ontogenetic development processes of the isomorhpic rep-
resentatives with the agglutinated (lower subclasses) and lime secreted
(higher subclasses) wall in several lineages witnesses their belonging
to several phyletic branches. Hence the transition from the agglutina-
ted to the lime secreted wall was going in each of this branches inde-
pendently and in parallel. Only the representatives of the higher sub-
classes of each of the phylogenetic branches (classes) in consideration
have a special type of their lime secreted wall ultrastructure. The re-
gularity of its ultrastructural elements enlarges from disorderly packed
Miliolana’s units of crystals to more regularly packed ones in
Spirillinana’s and Nodosariana’s radial wall and to the most perfect
and regular bifontial Rotaliana’s and Globigerinata’s wall.
All their lowest subclasses have the similar agglutinated wall and
represent the earlier stage of the evolutionary development of the cor-
responding classes. However in each of these phyletic branches (clas-
ses) the tendencies of development are realised to a different degree.
They achieve a different level of perfection and have their own featu-
res. That is why we put the plan of the shell structure, mode of coi-
ling, type of symmetry and apertural structure into the base of the
classes, bearing in mind also the complexes of the features of the other
structures and the wall ultrastructure.
Following this principle we unite such forms with the analogous
test morphology in one class. Thus in 4 classes — Spirillinata, Milio-
lata, Nodosariata, Rotaliata — there are lower subclasses with the ag-
glutinated and microgranular wall and higher ones with a special ul-
trastructure of a fully secretious wall. We regard this lower subclasses
as earlier steps in the evolution of the co-ordinate classes, while in the
previous systems they were regarded as convergent forms to their
secretious isomorphic forms. The close affinity of Schlumbergerinana
to Miliolana within the class Miliolata and of the Textulariana and
Rotaliana within the class Rotaliata is now confirmed by the works at
the molecular level (Fahmi et al., 1995, 1997). Only the higher sub-
classes differ in their wall ultrastructure. All this lower subclasses are
agglutinated. Thus a shell-wall ultrastructure could be used as a diag-
nostic feature of the subclass level only. Such an approach permits
formulation of a more distinct diagnosis and elimination of the con-
siderable number of parallelisms of previous systems.
Eariier these lower subclasses (Ammodiscana, Schlumbergerina-
na, Hormosinana, Textulariana) and also subclass Astrorhizana, with
quite a different plan of their shell structure, were united only at the
basis of one feature — an agglutinated wall — in a heterogenous
616
СИСТЕМАТИЧЕСКИЙ ОБЗОР
taxon Textulariacea (or Textulariina). We moved from it (Mikhale-
vich, 1992, 1999) the unilocular forms, the pseudotwochambered
forms, multichambered forms with one of Miliolate or Nodossariate
types to different classes, accordingly to their test morphology. We
also removed such forms from the previous Fusulinacea (Fusulinina).
The radial layer in the wall of some of them confirms this change. The
classification given here does not contradict the data of their geologi-
cal history, which gives an objectively controlled historical backgro-
und. Probably the basic skeletal types were formed in the periods
Cm—C, then the changes in the aperture, the inner complications of
the chamber lumen and chamber differentiation along with the chan-
ges in shell-wall structure took place (in parallel in each phyletic
branch). Astrorhizata and Spirillinata — are the most ancient groups
but existing until recently. Nodosariata (Ord) and Miliolata (evidently
Ord) appeared later. Fusulinoida, after its magnificent bloom, died out
completely. They did not give significant complication neither of the
aperture, nor of the external wall structure. Their integrative systems
(tunnels) were more primitive then the canals of Rotaliana. May be the
thickness, massiveness and the imperfectness of their microgranular
wall had closed for them the possibility of further transformations.
The agglutinated Rotaliats (Textulariana) have been in existence
from the Carboniferous till present, recent higher Rotaliata achieved
their magnificent bloom since Cretaceous. The rise of their bifontinal
wall we regard as aromorphosis (promising evolutionary change),
which led to their highest diversity and progressive complexity of all
their morphological structures, including inner apertural structures and
canal systems, leading to the profound compartmentalisation and dif-
ferentiation of the cytoplasm. It also gives them the possibility of the
adaptive radiation, exploded enlarging of the number of the new taxa
and occupying a new ecological niche, giving planktonic Globigerina-
ta. Both groups represent the highest step in the foraminiferal deve-
lopment and in this sense could be compared with Mammalia among
the other vertebrates.
Recent Foraminifera inhabit all the depths of the World Ocean
from littoral to abyssal. Some of them also occur in the salt lakes, in
the underground salt waters of the deserts, in the kelp. Their biomass
sometimes exceeds that of macrobenthos, achieving from 20 to 80 %.
It was considered for a long time that most of them were widely
distributed or cosmopolitan. However the investigations of the last
decades have clearly shown that their distribution is strictly subject to
bathymetric and zoogeographical zonation patterns as in the other ani-
mal groups. Many foraminiferal species could be used as indicators of
the definite water masses, of the currents and of the upwelling sensi-
tively reflecting the changes in temperature and salinity, the concent-
ration of the oxygen and of the other chemical components. The bot-
tom Foraminifera usually needs the limited quantities of the oxygen
and can exist in hypoxide environment. Some of them (Ammonia bec-
carii, Elphidium excavatum, Quinqueloculina seminulum) can become
the facultative anaerobes for a short time. In such conditions their
mitochondria have the tendency to regroup. Forms living deeper in the
sediments are abandoned to the less oxygen quantities than the super-
ficial ones. The light is also an important factor influencing foramini-
feral distribution, especially of the symbiont bearing forms. Above the
abiotic factors the biotic ones are also of great importance, the-nutri-
tional sources being the first of them. Just owing to the richness of the
organic matter the silty sediments are often more favourable for them.
The general model combining the influence of both biotic and abiotic
factors of the foraminiferal distribution was offered by Hottinger
(1988). The microhabitats of Foraminifera are mostly connected with
the amount of food available, with the character of the sediment, of
hydrodynamism, with the abundance of algae, especially Corallina and
Ulva. The different types of pollution also influence foraminiferal po-
pulations and sometimes causes morphological abnormalities of their
shells. The role of the Foraminifera in the trophic chains is not abso-
lutely clear yet, but some new data are represented as well as some
cases of predation and parasitism (both of the Foraminifera and on
them). There-is also evidence showing the role of Foraminifera in
bioerosion. Owing to the processes of the calcification both the bottom
and planktonic Foraminifera are one of the most significant compo-
nents of the sediment cover of the World Ocean. The Globigerine
oozes occupy nearly */з of the Ocean floor sometimes reaching several
meters in thickness. The fossil foraminiferal sediments reflecting the
previous ecological environments provide the keys to the under-
standing of the geological history of the Earth.
ЛИТЕРАТУРА40
Азбель А. Я. Новые фораминиферы Оксфорда Русской плат-
формы // Палеонтол. журн. 1986. № 1. С. 27—32.
Бараш М. С. Планктонные фораминиферы в осадках северной
части Атлантики. М.: Наука, 1970.
Басов В. А. Классификация и фиилогенез лягенид // Тез. XI
Всес. микропалеонтол. совещ. М., 1990. С. 21.
Бродский А. Л. Foraminifera (Polythalamia) в колодцах пусты-
ни Кара-Кум // Тр. Среднеаз. гос. ун-та. Сер. 8а. Зоол. 1928.
Вып. 5. С. 1—16.
Брынов О. П. Кристаллические единицы в структуре стенки
раковины четвертичных фораминифер семейства Cassidulinidae и
Islandiellidae // Вести. Моск, ун-та. Сер. 4. Геол. 1990. № 1. С. 87—
90.
Бугрова Э. М. Новое в системе роталоидных фораминифер //
Проблемы современной микропалеонтологии. Л.: Наука, 1990.
С. 22—36.
Вдовенко М. В., Раузер-Черноусова Д. М., Рейтлингер Е. А.,
Сабиров А. А. (с участием Гроздиловой Л. П.). Справочник по
систематике мелких фораминифер палеозоя (за исключением эн-
дотироидей и пермских многокамерных лагеноидей). М., 1993.
Воронова М. Н. О полиплоидии соматических ядер форами-
ниферы Cibicides lobatulus // Цитология. 1976. Т. 18, № 4.
С. 509—512.
Воронова М. Н. Ядерный дуализм гамонтов фораминиферы
Cibicides lobatulus И Цитология. 1978а. Т. 20, № 8. С. 859—867.
Воронова М. Н. Мейоз у фораминиферы Cibicides lobatulus И
Там же. 19786. № И. С. 1328—1331.
Воронова М. Н. Жизненный цикл фораминиферы Cibicides lo-
batulus: Автореф. дис. ... канд. биол. наук. 1979.
40 Цитируемая и рекомендуемая литература к разделу «Тип Fo-
raminifera».
Поскольку в монографии Леблика и Таппан (Loeblich, Tappan,
1988) дана полная библиография предшествующих работ по систе-
матике и морфологии раковин, данный список включает по пре-
имуществу работы по цитологии и биологии фораминифер, а
также работы, изданные после 1988 г.
Воронова М. Н., Михалевич В. И. Современные представле-
ния о жизненных циклах фораминифер // Жизненные циклы про-
стейших. Л., 1985. С. 48—66, табл. 1—12. (Тр. Зоол. ин-та АН
СССР; Т. 129).
Воронова М. Н., Селиванова Г. В. Исследование генератив-
ных ядер фораминиферы Cibicides lobatulus методом цитофото-
метрии // Протозоология. 1976. Вып. 1. С. 144—150.
Голубев С. Н. Реальные кристаллы в скелетах кокколитофо-
рид. М.: Наука, 1981.
Григелис А. А. Фораминиферы юрских отложений Юго-За-
падной Прибалтики. Вильнюс, 1995.
Губенко Т. А. Типы строения стенки раковин раннекаменно-
угольных фораминифер: Автореф. дис. ... канд. геол.-минерал,
наук. Владивосток, 1989.
(Догель В. A.) Dogiel V. A. Polymerization als ein Prinzip der
progressiven Entwicklung bei Protozoa // Biol. Zbl. 1929. Bd 49, N 8.
S. 451—459.
Карпов С. А. Система протистов. Омск, 1990. (Омск. пед. ин-т.
Биол. сер.; Вып. 1).
Крашенинников В. В. Микроструктура стенки некоторых
кайнозойских фораминифер и методика ее изучения в поляризо-
ванном свете // Вопр. микропалеонтол. 1956. Т. 1. С. 37—48.
Крылов М. В., Добровольский А. А. Макросистема и филоге-
ния споровиков И Принципы построения макросистемы одноклеточ-
ных животных. Л., 1980. С. 62—74. (Тр. Зоол. ин-та АН СССР; Т. 94).
Крылов М. В., Добровольский А. А., Исси И. В., Михале-
вич В. И., Подлипаев С. А., Решетник В. В., Серавин Л. Н., Старо-
богатов Я. И., Шульман С. С., Янковский А. В. Новые представле-
ния о системе одноклеточных животных И Там же. С. 122—132.
Кусакин О. Г., Дроздов А. Л. Филема органического мира.
СПб.: Наука, 1998. 358 с.
Лабас Ю. А., Михалевич В. И. Раковинообразование у Textu-
lariata и Rotaliata (Foraminifera): Тез. докл. и сообщ. IV съезда
ВОПР. Л.: Наука, 1987. С. 11.
Манцуроба В. Н., Горбачик Т. Н. Новые данные о строении
раковин трохолин // Вопр. микропалентол. 1982. Т. 25. С. 116—
129.
ТИП FORAMINIFERA. ЛИТЕРАТУРА
617
Маслакова Н. И. Глоботрунканиды юга европейской части
СССР. М.: Наука, 1978.
Маслакова Н. И. Критерии выделения высших таксонов фо-
раминифер // Систематика и филогения беспозвоночных. Крите-
рии выделения высших таксонов. М.: Наука, 1990. С. 22—27.
Маслакова Н. И., Горбачик Т. Н. Класс Foraminifera. Фора-
миниферы // Микропалеонтология. М.: Изд. Моск, ун-та, 1995.
С. 13—111.
Михалевич В. И. Возрастная изменчивость антарктических
видов рода Trochammina Parker et Jones и ее значение для систе-
матики подсемейства Trochammininae (Foraminifera) // Результаты
биологических исследований советских антарктических экспеди-
ций. Л., 1972а. С. 5—40. (Исследование фауны морей; Т. 11 (19),
вып. 5).
Михалевич В. И. Донные фораминиферы материковой отмели
западно-экваториальной Африки и вопросы их зоогеографии //
Океанология. 19726. Т. 12, вып. 3. С. 520—526.
(Михалевич В. И.) Mikhalevich V. L. New data on the forami-
nifera of the Ground waters of the Middle Asia // Int. J. Speleol. 1976.
N 8. P. 167—175.
Михалевич В. И. Зоогеография фораминифер шельфов тро-
пической Атлантики // Фауна и систематика одноклеточных жи-
вотных. Л., 1978а. С. 59—96. (Тр. Зоол. ин-та АН СССР;
Т. 78).
Михалевич В. И. Особенности распределения трех подотря-
дов фораминифер (Textulariina, Miliolina, Rotaliina) // Океаноло-
гия. 19786. Т. 133. С. 48—60.
Михалевич В. И. Вертикальное распределение и особенности
экологии донных фораминифер шельфов тропической Атланти-
ки И Биология моря. 1979. № 3. С. 3—10.
Михалевич В. И. Систематика и эволюция фораминифер
в свете новых данных по их цитологии и ультраструктуре // Прин-
ципы построения макросистемы одноклеточных животных. Л.,
1980. С. 42—61. (Тр. Зоол. ин-та АН СССР; Т. 94).
Михалевич В. И. Параллелизм и конвергенция в эволюции
скелетов фораминифер. Л., 1981. С. 19—41, табл. 1. (Тр. Зоол.
ин-та АН СССР; Т. 107).
Михалевич В. И. Донные фораминиферы шельфов тропичес-
кой Атлантики. Л., 1983. (Зоол. ин-т АН СССР).
Михалевич В. И. Система подкласса Miliolata (Foraminifera) И
Систематика, экология и стратиграфия милиолят (Фораминифе-
ры) / Под ред. В. И. Михалевич. Л., 1988. С. 77—110. (Тр. Зоол.
ин-та АН СССР; Т. 184).
Михалевич В. И. Ультраструктура фораминифер // Проблемы
современной микопалеонтологии. Л.: Наука, 1990. С. 14—22.
Михалевич В. И. Фораминиферы (Foraminifera) залива Рыбий
хвост. СПб. : Гидрометеоиздат, 1991. С. 27—38. (Информ. Бюл.
Сов. Ант. Эксп.; № 116).
Михалевич В. И. Макросистема фораминифер: Автореф. дис.
... докт. биол. наук. СПб., Зоол. ин-т РАН, 1992.
(Михалевич В. И.) Mikhalevich V. I. New higher taxa of the
subclass Nodosariata (Foramimifera) // Zoosystematica Rossica.
1993a. N 2. P. 5—8.
(Михалевич В. И.) Mikhalevich V. I. New higher taxa of the
class Spirillinata (Foraminifera) // Ibid. 19936. P. 9—11.
Михалевич В. И. Новый род и вид семейства Ammoscalariidae
(Foraminifera) И Зоол. журн. 1994. Т. 73, вып. 6. С. 18—21.
(Михалевич В. И.) Mikhalevich V. I. A new classification of the
class Astrorhizata (Foraminifera) II Zoosystematica Rossica. 1995.
N 3 (2). P. 161—174.
(Михалевич В. И.) Mikhalevich V. I. The new conception of the
foraminiferal macrosystem // Appl. micropaleontol. Environm. Sci.
Tel Aviv Univ., 1997a. P. 88—89.
(Михалевич В. И.) Mikhalevich V. I. Main directions of the
foraminiferal evolution. Evolution of the life on the Earth. Tomsk
Univ. (Russia), Oregona Univ. (U, S. A.), 19976. P. 44—45.
Михалевич В. И. Макросистема фораминифер // Изв. РАН.
Сер. биол. 1998а. № 2. С. 266—271.
Михалевич В. И. О систематическом положении семейства
Dusenburyinidae (Foraminifera) // Зоол. журн. 19986. Т. 77, № 8.
С. 976—978.
Михалевич В. И. Система и филогения фораминифер. СПб.;
Омск, 1999. 185 с.
Михалевич В. И., Воронова М. Н. Гаметогенез у гаметогам-
ных Фораминифер // Материалы 3-го съезда ВОПР: Тез. докл.
Вильнюс, 1982.
Михалевич В. И,, Воронова М. Н. Арктические цибицидесы.
Проблемы четвертичной палеоэкологии и палеографии Баренцева
и Белого морей // Тез. Всес. конф. Мурманск, 1985. С. 94—95.
40 Протисты, часть 1
Михалевич В. И., Воронова М. Н. О систематическом поло-
жении рода Pelosina (Xenophyophoria Protista, Ins. sedis) // Зоол.
журн. 1999. T. 78, № 2. С. 133—141.
Михалевич В. И., Дебеней Ж.-П. Параллелизм и конверген-
ция в развитии устья Фораминифер // Полевые и эксперименталь-
ные биологические исследования. СПб., Омск, 1998. С. 3—31. (Рос.
гос. пед. ун-т им. А. И. Герцена, Омский гос. пед. ун-т; Биол.
сер. Вып. 4).
Михалевич В. И., Родионова М. К., Синякова Г. Н. Ультра-
структура стенки двух агглютинированных родов фораминифер
квинквелокулинового типа строения // Систематика простейших и
их филогенетические связи с низшими эукариотами. Л., 1986.
С. 66—71. (Тр. Зоол ин-та АН СССР; Т. 144).
Плотникова Л. Ф., Егорова С. И. Особенности строения
стенки раковин Атаксофрагмиид // Палеонтология и стратигра-
фия мезозоя Украины. Киев: Ин-т геол, наук, 1982. С. 24—25.
Подлипаев С. А., Наумов А. Д., Боркин Л. Я. К определению
понятий полимеризации и олигомеризации // Журн. общ. биол.
1974. Т. 35, № 1. С. 100—113.
Подобина В. М. Систематика и филогения Гаплофрагмиидей.
Томск: Изд. Томск, гос. ун-та, 1978.
Подобина В. М. Фораминиферы и биостратиграфия палеогена
Западной Сибири. Томск: Изд. Томск, гос. ун-та, 1998.
Полянский Ю. И. О морфологических закономерностях эво-
люции Простейших И Зоол. журн. 1970. Т. 49, вып. 4. С. 560—
569.
Полянский Ю. И., Райков И. Б. Полимеризация и олигомери-
зация в эволюции Простейших И Значение процессов полимериза-
ции и олигомеризации в эволюции. Л., 1977. С. 29—32.
Пронина В. М. Позднепермские милиоляты Закавказья // Сис-
тематика, экология и стратиграфия Милиолят (Фораминиферы).
Л., 1988. С. 49—54. (Тр. Зоол. ин-та АН СССР; Т. 184).
Райков И. Б. Ядро простейших. Морфология и эволюция. Л.:
Наука, 1978.
(Райков И. Б., Караджан Б. П., Михалевич В. И.) Rai-
kov I. В., Karajan В. Р., Mikhalevich V. I. Ultrastructure of the ga-
mont Shell and Nucleus in the Polythalamous Foraminifera Elphidium
ponticum II Europ. J. Protistol. 1998. T. 34. P. 153—161.
Раузер-Черноусова Д. M. О систематическом положении Ho-
дозариидей и о составе надотряда Lagenoida (Фораминиферы) //
Тез. XI Всес. микропалеонтол. совещ. М., 1990. С. 109.
Раузер-Черноусова Д. М., Герке А. А. Терминологический
справочник по стенкам раковин фораминифер. М.: Наука, 1971.
Раузер-Черноусова Д. М., Рейтлингер Е. А. Систематика над-
родовых таксонов отряда Hormosinida (Foraminifera) // Палеонтол.
журн. 1986. Т. 20, № 4. С. 11—16.
Раузер-Черноусова Д. М., Бенш Ф. Р., Вдовенко М. В., Гиб-
шман Н. Б., Левен Э. Я., Липина О. А., Рейтлингер Е. А., Со-
ловьева М. Н., Чедия И. О. Справочник по систематике форами-
нифер палеозоя (Эндотириды, Фузулиноиды). М.: Наука, 1996.
Саидова X. М. Бентосные фораминиферы Мирового океана.
М. : Наука, 1976.
Саидова X. М. О современном состоянии надвидовых таксо-
нов кайнозойских бентосных фораминифер. М.: Изд. Ин-та океа-
нол. АН СССР им. П. П. Ширшова, 1981.
Соловьева М. Н. Интегративные системы фораминифер И
Вопр. микропалеонтол. 1978. Вып. 21. С. 3—16.
(Старобогатов Я., Михалевич A.) Starobogatov Ja., Mikhale-
vich A. On the heterogeneyty of the Sarcodina II Abstr. 7th Intern.
Congress Protozool. Nairobi. Kenia. 1985. N 189. P. 189.
Субботина H. H., Волошинова H. А., Азбель А. Я. (ред.). Вве-
дение в изучение фораминифер. Классификация мелких форами-
нифер мезо-кайнозоя. Л.: Изд. Всес. науч.-исслед. геологоразвед.
ин-та (ВНИГРИ), 1981.
Успенская А. В. Новые данные о жизненных циклах и биоло-
гии миксоспоридий (Protista) // Докл. АН СССР. 1982. Т. 262.
С. 503—507.
Фурсенко А. В. Введение в изучение фораминифер. Новоси-
бирск: Наука, 1978.
Фурсенко А. В., Раузер-Черноусова Д. М. Простейшие. Под-
класс Foraminifera. Фораминиферы И Основы палеонтологии. М.:
Изд. АН СССР. 1959. С. 115—346.
Шмальгаузен И. И. Проблемы дарвинизма. Л.: Наука, 1969.
Яковлева С. П. К систематике юрских литуолин // Палеонтол.
журн. 1979. № 1. С. 12—211.
Янковская А. И., Михалевич В. И. Фораминиферы озера
Иссык-Куль и грунтовых вод Средней Азии // Докл. АН СССР.
1972. № 4. С. 1005—1W8.
618
СИСТЕМАТИЧЕСКИЙ ОБЗОР
Aldredge A, L., Jones В. М Hastigerina pelagica: Foraminiferal
habitat for planktonic dinoflagellates // Mar. Biol. 1973. Vol. 22, N 2.
P. 131—135.
Alexander S. P., De Laca T. E. Feeding adaptations of the forami-
niferan Cibicides refulgens living epizoically in the antarctic scallop
Adamussium colbecki // Biol. Bull. 1987. Vol. 173, N 1. P. 136—159.
Allen R. D. Cytoplasmic streaming and locomotion in marine fo-
raminifera II R. D. Allen, N. Kamiya. Primitive Motile Systems in
Cell Biology, New York: Acad. Press, 1964. P. 407—433.
Alterbach A. V., Heeger T., Linke P., Spindler M., Thies A.
Miliolinella subrotundl (Montagu), a miliolid foraminifer building
large detritic tubes for a temporary epibenthic lifestyle H Mar. Micro-
paleontol. 1993. Vol. 20. P. 293—301.
Alve E. Benthic Foraminifera in sediment cores reflecting heavy
metal pollution in Sorfiord, Western Norway // J. Foram. Res. 1991.
Vol. 21, N 1. P. 1—19.
Alve E. Benthic foraminiferal responses to estuarine pollution: a
review // Ibid. 1995, Vol. 25, N 3. P. 190—204.
Alve E,, Olsgard F. Benthic foraminiferal colonization in experi-
ments with copper-contaminated sediments // J. Foram. Res. 1999.
Vol. 29, N 3, P. 186—196.
Anderson O. R. Cellular specialization and reproduction in plan-
ktonic Foraminifera and Radiolaria // Marine Plankton Life Cycle
Strategies. Boca Raton, Florida, 1984. P. 35—66.
Anderson O. R. Comparative Protozoology — ecology, physiolo-
gy, life history. New York: Springer, 1988.
Anderson O. R., Be A. W. H. A cytochemical fine structure of
phagotrophy in a planktonic foraminifer, Hastigerina pelagica (d’Or-
bigny) // Biol. Bull. 1976s. Vol. 151, N 3. P. 437—449.
Anderson O. R., Be A. W. H. The ultrastructure of a planktonic
foraminifer, Globigerinoides sacculifer (Brady), and its symbiotic di-
noflagellates // J. Foram. Res. 1976b. Vol. 6, N 1. P. 1—21.
Anderson O. R., Be A. W. H. Recent advances in foraminiferal
fine structure Researches // Foraminifera I Eds R. H. Hedley,
C. G. Adams. London: Acad. Press, 1978. Vol. 3. P. 121—203.
Anderson O. R., Timtivate-Choy S. Cytochemical evidence for
peroxisomes in planktonic Foraminifera // J. Foram. Res. 1984.
Vol. 14, N 3. P. 203—206.
Angell R. W. The text structure and composition of the Forami-
nifer Rosalina floridana // J, Protozool. 1967. Vol. 14. P. 299—307.
Angell R. W. Observations on gametogenesis in the foraminifer
Myxotheca // J. Foram. Res, 1971. Vol. 1, N 1. P. 39—42.
Angell R. W. Calcification during chamber development in Rosa-
lina floridana // Ibid. 1979. Vol. 9, N 4. P. 341—353.
Angell R. W. Test morphogenesis (chamber formation) in the fo-
raminifer Spiroloculina hyalina Schultze // Ibid. 1980. Vol. 10, N 2.
P. 89—101.
Angell R. W. Observations on reproduction and juvenile test bu-
ilding in the foraminifer Trochammina inflata // Ibid. 1990. Vol. 20,
N 3. P. 246—247.
Archambault-Guezou T., Lefevre R. Some applications of ion
microscopy to biomineralization involved in sedimentary rocks //
Scanning Electron Microscopy. 1981. Vol. 1. P. 503—514.
Arnold Z. M. A note on foraminiferan sieve-plates // Contr.
Cushm. Found. Foram. Res. 1954. Vol. 5, N 2. P. 77.
Arnold Z. M. An unusual feature of Miliolid reproduction // Ibid.
1955. Vol. 6. P. 94—96.
Arnold Z. M. Biological observatins on the foraminifer Spirolo-
culina hyalina Schulze // Univ. Calif. Publ. Zool. 1964. Vol. 72.
Arnold Z. M. An allogromiid ancestor of the miliolidean Forami-
nifera // J. Foram. Res. 1978a. Vol. 8, N 2. P. 83—97.
Arnold Z. M. Biological evidence for the origin of polythalamy
in Foraminifera // Ibid. 1978b. Vol. 8, N 2. P. 147—166.
Arnold Z. M. Biological clues to the origin of miliolidean fora-
minifera // Ibid. 1979. Vol. 9, N 4. P. 302—321.
Arnold Z. M. Psammophaga simplora n. gen., n. sp., a polygeno-
mic Californian saccamminid // Ibid. 1982. Vol. 12, N 1. P. 72—79.
Arnold Z. M. The gamontic cariology of the saccamminid fora-
minifer Psammophaga simplora Arnold 7/ Ibid. 1984. Vol. 14, N 3.
P. 171—187.
Arnold Z. J., d’Escrivan F., Parker W. C. Predation and avoi-
dance responses in the Foraminifera of the Galapagos hydrotermal
mounds // Ibid. 1985. Vol. 15, N 1. P. 38—42.
Banner F. T. A new genus of the Planorbulinidae an endoparasite
of another foraminifer // Rev. Esp. Micropaleontol. 1971. Vol. 3, N 2.
P. 113—128.
Banner F. T., Sheehan R., Williams E. The organic skeletons of
Rotaline Foraminifera: a review // J. Foram. Res. 1973. Vol. 3, N 1.
P. 30—42.
Barker R. W. Taxonomic notes on the species figured by
H. B. Brady in his Report on the Foraminifera dredged by H. M. S.
«Challenger» during the years 1873—1876 // Soc. Econ. Paleontol.
and Mineral. Spec. Publ. 1960. Vol. 9. P. 1—238.
Barmawidiaja D. M., Jorissen F. J., Puskaric S., Van der
Zwaan G. J. Microhabitat selection by benthic foraminifera in the
Northern Adriatic Sea // Ibid. 1992. Vol. 22, N 4. P. 297—317.
Be A. W. H. Gametogenic calcification in a spinose planktonic
foraminifer Globigerinoides sacculifer (Brady) // Mar. Micropaleontol.
1980. Vol. 5, N 3. P. 283—310.
Be A. W. H. Biology of planktonic foraminifera // Foraminifera:
Notes for a short Course. Studies in Geol. Vol. 6. Univ. Tenessee,
Khoxville, 1982. P. 51—92.
Be A. W. H., Anderson O. R., Roger O. Gametogenesis in plan-
ktonic foraminifera // Science. 1976. Vol. 192, N 4242. P. 890—892.
Be A. W. H., Hemleben C., Anderson O. R., Spindler M., Ha-
cunda J., Tuntivate-Choy S. Laboratory and field observations of li-
ving planktonic Foraminifera // Micropaleontology. 1977. Vol. 23,
N 2. P. 155—179.
Be A. W. H., Hemleben C., Anderson O. R., Spindler M. Cham-
ber formation in planktonic Foraminifera // Ibid. 1979. Vol. 25, N 3.
P. 294—307.
Be A. W. H., Hemleben C., Anderson O. R., Spindler M. Pore
structure in planktonic Foraminifera // J. Foram. Res. 1980. Vol. 10,
N 2. P. 117—128.
Be A. W. H., Spero H. T., Anderson O. R. Effects of symbiont
elimination and reinfection on the life processes of the planktonic
foraminifer Globigerinoides sacculifer // Mar. Biol. 1982. Vol. 70,
N 1. P. 73—86.
Bellemo S. Studies on the ultrastructures of calcareous Foramini-
fera with hyaline perforate walls // Paleontol. Inst. Univ. Uppsala.
1974a. N 153. 14 p.
Bellemo S. Ultrastructure in the foraminifer Cibicides floridanus
(Cushman) // Ibid. 1974b. N 154. 11 p.
Bender H. Chamber formation and biomineralization in Textula-
ria candeiana d’Orbigny (Sarcodina: Textulariina) // J. Foram. Res.
1992. Vol. 22, N 3. P. 229—242.
Bender H. Test structure and classification in agglutinated fora-
minifera // Proc. Fourth Intern. Workshop on Agglutin. Foram. Kra-
kow, Poland. Sept. 12—19, 1993 / Eds M. A. Kaminsky et al. Kra-
kow, 1995. P. 27—70. (Grzybowski Found. Sp. Publ.; N 3).
Bender H., Hemleben C. Calcitic cement secreted by agglutina-
ted foraminifers grown in laboratory culture // J. Foram. Res. 1988.
Vol. 18, N 1. P. 42—46.
Bernhard J. M., Alve E. Survival, ATP pool, and ultrastructural
characterization of benthic foraminifera from Drammensfjord (Norway):
response to anoxia // Mar. Micropaleontol. 1996. Vol. 28. P. 5—17.
Berthold W.-U. Untersuchungen uber die sexuelle Differenzie-
rung der Foraminifere Patellina corrugata Williamson mit einen Beit-
rag zum Entwicklungsgang und Shalenbau // Arch. Protistenk. 1971.
Bd 113, N 3. P. 147—184.
Berthold W.-U. Ultrastructure and function of wall perforations
in Patellina corrugata Williamson, Foraminifera // J. Foram. Res.
1976. Vol. 6, N 1. P. 22—29.
Berthold W.-U. Synaptinemale komplexe bei Patellina corrugata
Williamson (Protozoa, Foraminifera) // Cytobiologie. 1977. Vol. 14.
P. 253—258.
Berthold W.-U. Ultrastructure analysis of the endoplasmic algae
in Amphistegina lessonii d’Orbigny, Foraminifera (Protozoa) and their
systematic position (in German with English summary) // Arch. Pro-
tistenk. 1978. Bd 120. S. 16—62.
Billman H., Hottinger L., Oesterle H. Neogene recent Rotaliid
Foraminifera from the Indopacific Ocean; their canal system, their
classification and their stratigraphic use // Schweiz. Palaeontol. Abh.
1980. Vol. 101. P. 71—113.
Boltovskoy E^ Wright R. Recent Foraminifera. Hague: W. Junk, 1976.
Bowser S. S., Rieder C. L. Evidence that cell surface motility
in Allogrmia is mediated by cytoplasmic microtubules // Can. J. Bio-
chem. Cell Biol. 1985. Vol. 73. P. 608—620.
Bowser S. S., McGee-Russell S. M., Rieder C. L. Digestion of
pray in Foraminifera is not anomalous: A correlation of light micros-
copic, cytochemical and HVEM technics to study phagotrophy in two
allogromiids // Tiss. Cell. 1985. Vol. 17. P. 823—839.
Brand T. E., Lipps J. H. Foraminifera in the trophic structure of
shallow-water communities // J. Foram. Res. 1982. Vol. 12, N 2.
P. 96—105.
Bresler V., Yanko V. Chemical ecology: a new approach to the
study of living benthic epiphytic foraminifera // Ibid. 1995. Vol. 25,
N 3. P. 267—280.
ТИП FORAMINIFERA. ЛИТЕРАТУРА
619
Brummer G.-J., Hemleben C., Spindler M. Ontogeny of extant
spinose planktonic foraminifera (Globigerinidae): A concept exampli-
fied by Globigerinoides sacculifer (Brady) and G. ruber (d’Orbigny) //
Mar. Micropaleontol. 1987. Vol. 12, N 4. P. 357—381.
Carpenter W. B., Parker W. K., Jones T. R. Introduction to the
study of Foraminifera. Roy. Soc. London, 1862.
Cartwright N. G., Gooday A. J., Jones A. R. The morphology,
internal organization, and taxonomic position of Rhizammina algae-
formis Brady, a large agglutinated deep-sea foraminifer // J. Foram.
Res. 1989. Vol. 19, N 2. P. 115—125.
Cesana D. Le mecanisme de lb mitose gamogonie chez le Fora-
minifere Iridia lucida // C. R. Acad. Sci. Paris. Ser. D. 1971.
Vol. 272. P. 3057—3060.
Cesana D. Ultrastructure des gametes chez un Foraminifere Iridia
lucida Le Calvez // Ibid. 1972. Vol. 274. P. 1044—1047.
Cesana D. Les stades initaux de la gamogonie chez Iridia lucida
Le Calvez (Foraminifera. Lagynidae) // Ibid. 1975. Vol. 281, N 4.
P. 263—266.
Cesana D. La mitose gamogonique chez Iridia lucida (Foramini-
fera, Lagynidae) // Ann. Sci. Natur. Ser. 12. 1978. T. 20, N 3.
P. 287—320.
Cesana D. Real structure and new interpretation of Foraminiferal
test // C. R. Acad. Sci. Paris. 1981. Ser. 3. Vol. 292, N 2. P. 197—200.
Christiansen O. Notes on the biology of Foraminifera // Vie et
milieu. Suppl. 22. 1971. Vol. 2. P. 465—478.
Clark F. E. Morphological details of the genus Nubeculina Cus-
hman, 1924 // J. Foram. Res. 1993. Vol. 23. P. 201—204.
Coleman A. R. Test structure and function of the agglutinated fora-
minifera Clavulina // J. Foram. Res. 1980. Vol. 10, N 2. P. 143—152.
Colom G. Foraminiferos Ibericos. Introduction al estudio de las
especies bentonicas recientes // Investigac. Pesquera. Barcelona, 1974.
T. 38, N 1.
Corliss J. O. The kingdom of Protista and its 45 phyla // BioSys-
tems. 1984. Vol. 17. P. 87—126.
Culver S. J. Foraminifera // J. H. Lipps. Fossil Prokariotes and
Protists. Blockwell; Boston, 1993. P. 203—247.
Culver S. J., Buzas M. A. The effects of anthropogenic habitat
disturbance, habitat destruction, and global warming on shallow mari-
ne benthic foraminifera // J. Foram. Res. 1995. Vol. 25, N 3. P. 204—
212.
Cushman J. A. Foraminifera. Their classification and Economic
use. 4th ed. Cambridge; Mass.: Harvard Univ. Press, 1948.
Dahlgren L. On nuclear cytology and reproduction in the mono-
thalamous foraminifer Ovammina opaca Dahlgren // Zool. Bidr. Up-
psala, 1964. Bd 36. H. 3. S. 315—334.
Dahlgren L. On the ultrastructure of the gamontic nucleus and the
adjacent cytoplasm of the monothalamous foraminifer Ovammina
opaca Dahlgren // Ibid. 1967a. Bd 37. H. 2. S. 77—112.
Dahlgren L. On the nuclear distribution of RNA and DNA and
on the ultrastructure of nuclei and adjancent cytoplasm of the Forami-
nifers Hipocrepinella alba. Heron-Allen and Earland and Globobulimi-
na turgida (Bailey) // Ibid. 1967b. Bd 37. H. 2. S. 113—138.
Daniels R. A., Lipps J. H. Predation on Foraminifera by Antarctic
fish // J. Foram. Res. 1978. Vol. 8, N 2. P. 110—113.
Debenay J.-P., Redois F. Distribution of the twenty seven domi-
nant species of the shelfbenthic foraminifers on the continental shelf,
north of Dakar (Senegal) // Marine Micropaleontol. 1997. Vol. 29.
P. 237—255.
Debenay J.-P., Pawlowski J., Decrouez D. Les Foraminiferes ac-
tuels. Masson, Paris. Collect. 3 Cycl. et Recherche. Paris, 1996a.
Debenay J.-P., Guillou J.-J., Lesourd M. Cristallogenese de la
calcite d’origine colloidale dans les tests des foraminiferes // C. R.
Acad. Sci. Paris. 1996b. T. 323, ser. Ila. P. 171—178.
Debenay J.-P., Eichler B., Guillou J.-J., Eichler-Coelho P., Co-
elho C., Porto-Filho E. Comportement des peuplements de foramini-
feres et comparaison avec 1’avifaune dans une lagune fortement stra-
tifiee: La Lagoa da Conceicao (SC, Brasil) // Rev. Paleobiol. Geneve.
1997. Vol. 16, N 1. P. 55—75.
De Laca T. E. Use of dissolved amino acids by the foraminifer
Notodendrodes antarctikos // Amer. Zoologist. 1982. Vol. 22, N 3.
P. 683—690.
De Laca T. E., Lipps J. H., Hessler R. R. The morphology and
ecology of a new large agglutinated Antarctic foraminifera (Textulariina:
Notodendrodidae nov.) // Zool. J. Linn. Soc. 1980. N 69. P. 205—224.
De Laca T. E., Karl D. M., Lipps J. H. Direct use of dissolved
organic carbon by agglutinated benthic Foraminifera // Nature. 1981.
Vol. 289. P. 287—289.
Deutsch S., Lipps J. H. Test structure of the foraminifer Carteri-
na//J. Paleontol. 1976. Vol. 50. P. 312—317.
Dietz-Elbraechter G. Untersuchungen iiber die zooxanthella der
Foraminifere Heterostegina depressa d’Orbigny 1826 // «Meteor» For-
schergeben (c) b. 1971. S. 41—47.
Dobson M., Haynes J. Association of foraminifera with hydroids on
the deep shelf// Micropaleontology. 1973. Vol. 19, N 1. P. 78—90.
Pouglas R. G., Woodruff F. Deep-Sea benthonic foraminiferen //
The Sea. Vol. 7. Wiley Interscience. I Ed. Emiliani. New York, 1981.
P. 1233—1327.
Ellison R. L., Broome R., Ogilvie R. Foraminiferal response to
trace metal contamination in the Potapsco River and Baltimore Harbo-
ur, Maryland // Mar. Pollut. Bull. 1986. Vol. 17, N 9. P. 419—423.
Faber W. W., Jr. Distribution and substrate preferance of Pene-
roplis planatus and P. arietinus from the Halophila Meadow near Wad:
Taba, Eilat, Israel // J. Foram. Res. 1991. Vol. 21. N 3. P. 218—221.
Faber W. W., Jr., Lee J. J. Feeding and growth of Peneroplis
planatus (Fichtel and Moll) # Montfort. Symbiosis. 1991a. N 10.
P. 63—82.
Faber W. W., Jr., Lee J. J. Histochemical evidence for digestion
in Heterostegina depressa and Operculina ammonoides (Foraminife-
ra) II Endocytobiol. Cell. Res. 1991b. N 8. P. 53—59.
Fahrni J. F., Pawlowski J. Identification of actins in Foraminife-
ra: Phylogenetic perspectives Я Europ. J. Protistol. 1995. Vol. 31.
P. 161—166.
Fahrni J. F., Pawlowski J., Richardson S., Debenay J.-P., Zani-
netti L. Actin Suggests Miliammina fusca (Brady) is related to porcel-
laneous rather than to agglutinated foraminifera // Micropaleontology.
1997. Vol. 43, N 2. P. 211—214.
Febvre-Chevalier C. Constitution ultrastructurale de Globigerina
bulloides d’Orbigny 1826 (Rhizopoda — Foraminifera) // Protistologi-
ca. 1971. T. 7, fasc. 3. P. 311—324.
Foeyn B. Foraminiferen Studien. 1. Der Lebenzyklus von Discor-
bina vilardeboana d’Orbigny Я Bergens Mus. Arbok. Naturvidens ka-
peling rekke. 1936. N 2. P. 1—22.
Gauthier-Lievre L. Sur une des singularites de 1’oued Rhir: des
Foraminiferes Thalassoides vivants dans les eaux sahariennes // Bull.
Soc. H. N. Afr. du Nord. 1935. T. 26. P. 142—147.
Geslin E., Debenay J.-P., Lesourd M. Abnormal wall textures
and test deformation in Ammonia (hyaline foraminifer) // J. Foram.
Res. 1998. Vol. 28, N 2. P. 148—157.
Goldstein S. T. On the life cycle of Saccammina alba Hedley,
1962 II J. Foram. Res. 1988. Vol. 18, N 4. P. 313—327.
Goldstein S. T. Gametogenesis and the antiquity of the reproduc-
tive pattern in the Foraminiferida // Ibid. 1997. Vol. 27, N 4. P. 319—
328.
Goldstein S., Barker W. W. Test ultrastructure and taphonomy
of the moonothalamous agglutinated foraminifer Cribrothalamina
n. gen., alba (Heron-Allen and Earland) // Ibid. 1988. Vol. 18, N 2.
P. 130—136.
Goldstein S. T., Barker W. W. Gametogenesis in the monothala-
mous agglutinated foraminifer Cribrothalammina alba // J. Protozool.
1990. Vol. 37, N 1. P. 20—27.
Goldstein S. T., Corliss В. H. Deposit feeding in selected deep
sea and shallow-water benthic foraminifera // Deep: Sea Res. 1994.
Vol. 41. P. 229—241.
Goldstein S. T., Harben E. B. Taphofacies implications of infau-
nal foraminiiferal assemblages in a Georgia salt marsh, Sapelo Is-
land // Micropaleontology. 1993. Vol. 39, N 1. P. 53—62.
Goldstein S., Moodley L. Gametogenesis and life cycle of the
foraminifer Ammonia beccarii (Linne) forma tepida (Cushman) //
J. Foram. Res. 1993. Vol. 23, N 4. P. 213—221.
Grell K. G. Der Generationswechsel der polythalamen Foramini-
fere Rotaliella heterocariotica // Arch. Protistenk. 1954. Bd 100, N 2.
S. 268—286.
Grell K. G. Der Generationswechsel der polythalamen Foraminifere
Rotaliella heterocaryotica // Ibid. 1956. Bd 100, N 2. S. 211—235,
Grell K. G. Protozoology. Berlin etc.: Springer, 1973.
Grell K. G. Cytogenetic systems and evolution in Foraminifera //
J. Foram. Res. 1979. Vol. 9, N 1. P. 1—14.
Grell K. G. The life-cycle of the monothalamous foraminifer He-
terotheca lobata, n. gen., n. sp. // Ibid, 1988. Vol. 18, N 1. P. 54—74.
Grell K. G., Bardele C. F. Light and electron microscopical stu-
dies of the foraminiferen Rotaliella heterocariotica Я Abstr. Fifth In-
tern. Congr. Protozool., New ¥prk. New York, 1977. P. 369.
Hallock P. Trends in test shape with depth in large, symbiont-be-
aring foraminifera // J. Foram. Res. 1979. Vol. 9, N 1. P. 61—69.
Hallock P. Light dependence in Amphistegina Я Ibid. 1981a.
Vol. 11, N 1. P. 40—46.
Hallock P. Algal symbiosis: a mathematical analysis Я Mar. Biol.
1981b. Vol. 62, N 4. P. 249—255.
620
СИСТЕМАТИЧЕСКИЙ ОБЗОР
Hallock Р. Evolution and extinction in larger Foraminifera //
Third North Amer. Paleontol. Conv., Proc. 1982. Vol. 1. P. 221—225.
Hallock P. Distribution of selected species of living algal symbi-
ont-bearing Foraminifera on two Pacific coral reefs // J. Foram. Res.
1984. Vol. 14, N 4. P. 250—261.
Hallock P., Talge H. K. A predatory foraminifer Floresina am-
phiphaga n. sp., from the Florida Keys // Ibid. 1994. Vol. 24, N 4.
P. 210—213.
Hallock P., Cottey T. L., Forward L. B., Halas J. Population
biology and sediment production of Archaias angulatus (Foraminiferi-
da) in Largo Sound, Florida // J. Ibid. 1986. Vol. 16, К L P. 1—8.
Hallock P., Talge H. К., Cockey E. M., Muller R, D. A new
disease in reef-dwelling foraminifera: implications for coastal sedi-
mentation // Ibid. 1995. Vol. 25, N 3. P. 280—286.
Hansen H. J. Two species of Foraminifera of the genus Turrilina
with different wall structure // Lethaia. 1972. Vol. 5. P. 39—45.
Hansen H. J. On feeding and supposed buoyancy mechanism in
four recent Globigerinid Foraminifera from the Gulf of Elat, Israel //
Rev. Esp. Micropaleontol. 1975. N 2. P. 325—341.
Hansen H. J. Test structure and evolution in the Foraminifera //
Lethaia. 1979. Vol. 12, N 2. P. 173—182.
Hansen H. J., Hanzlikova E. Ultrastructure of some silicieous Fo-
raminiferida // Rev. Esp. Micropaleontol. 1974. Vol. 6, N 3. P. 447—466.
Hansen H. J., Reiss Z. Electron microscopy of Rotaliacean wall
structures // Bull. Geol. Soc. Denmark. 1971a. Vol. 20. P. 329—346.
Hansen H. J., Reiss Z. Scanning electrone microscopy of wall
structures in some benthic and planktonic Foraminiferida // Rev. Esp.
Micropaleontol. 1971b. Vol. 4, N 2. P. 169—181.
Hansen H. J., Revets S. A revision and reclassification of the
Discorbidae, Rosalinidae and Rotaliidae // J. Foram. Res. 1992.
Vol. 22, N 2. P. 166—181.
Hansen H. J., Reiss Z., Schneiderman N. Ultrastructure of bila-
mellar walls in Foraminiferida // Ibid. 1969. Vol. 1, N 3. P. 293—316.
Harney J. N., Hallock P., Talge H. K. Observations on a trimor-
phic life cycle in Amphistegina gibbosa populations from the Florida
Keys II J. Foram. Res. 1998. Vol. 28, N 2. P. 141—148.
Harward N. J. B., Haynes J. R. Chlamys opercularis (Linn-
aeus) as a mobile substrate for Foraminifera // Ibid. 1976. Vol. 6, N 1.
P. 30—36.
Haugen J.-E., Sejrup H.-Р., Vogt N. B. Chemotaxonomy of Qu-
aternary benthic Foraminifera using amino acids // Ibid. 1989. Vol. 19,
N 1. P. 38—52.
Hay W. W., Towe К. M., Wright R. Ultrastructure of some se-
lected foraminiferal tests // Micropaleontology. 1963. Vol. 9, N 2.
P. 171—196.
Haynes J. R. Foraminifera. New York: Wiley and Sons, 1981.
Haynes J. R. The classification of the Foraminifera — a review
of historical and philosophical perspectives // Paleontology. 1990.
Vol. 33, N 3. P. 503—528.
Hayward B. W., Brasier R. C. Taxonomy and distribution of
present day Bolivinella // J. Foram. Res. 1980. Vol. 10, N 2. P. 102—
116.
Hayward B. W., Grenfell H. R., Reid С. M., Hayward K. A. Re-
cent New Zealand shallow-water benthic Foraminifera: taxonomy, eco-
logic distribution, biogeography, and use in paleoenvironmental asses-
sment I Institute of geological & nuclear sciences. Monograph N 21.
(New Zealand Geological Survey; paleontol. bull. N 75). Lower Hutt;
New Zealand, 1999. 258 p.
Hedley R. H. The significance of an «inner chitinous lining» in
Saccamminid organization with special reference to a new species of
Saccammina (Foraminifera) from New Zealand // New Zealand J. Sci.
1962. N 3. P. 375—389. (New Zealand Oceanograph. Inst. Contrib.;
N 127).
Hedley R. H. Cement and iron jn the arenaceous Foraminifera //
Micropaleontology. 1963. Vol. 9, N 4. P. 433—441.
Hedley R. H. The biology of Foraminifera // Intern. Rev. Gener.
Experim. Zook / Eds W. J. L. Felts, R. J. Harrison. New York; Lon-
don, 1964. P. 1—48.
Hedley R. H., Parry D. M., Wakefield J. St. J. Reproduction in
Boderia turneri. (Foraminifera) // J. Nat. Hist. 1968. N 2. P. 147—
151.
Hedley R. H., Ogden C. G., Wakefield J. St. J. Shell ultrastruc-
ture in allogromiid Foraminifera (Protozoa) // Bull. Brit. Mus. Nat.
Hist. Zool. 1973. Vol. 24, N 9. P. 467—474.
Hemleben C. Zur Morphogenese planktonischer Foraminiferen //
Zitteliana. 1969. Vol. 1. P. 91—133.
Hemleben C. Spine and pustule relationships in some Recent
planktonic Foraminifera H Micropaleontology. 1975. Vol. 21, N 3.
P. 334—341.
Hemleben C., Spindler M. Recent advances in research on living
planktonic Foraminifera II Reconstruction of marine paleoenviron-
ments / Ed. J. E. Meulenkamp. 1983. P. 141—170. (Utrecht Micro-
pal. Bull.; Vol. 30).
Hemleben C.^ Be A. W. H., Anderson O. R., Tuntivate S. Test
morphology, organic layers and chamber formation of planktonic fo-
raminifer Globorotalia menardii (d’Orbigny) // J. Foram. Res. 1977.
Vol. 7, N 1. P. 1—25.
Hemleben C., Spindler M., Anderson O. R. Modem Planktonic
Foraminifera. New York: Springer, 1989.
Hickman C. S., Lipps J. H. Foraminiferivory: selective ingestion
of Foraminifera and test alternations produced by the neogastropod
Olivella // J. Foram. Res. 1983. Vol. 13, N 2. P. 108—114.
Hofker J. Der Generationswechsel von Rotalia beccarii var. fle-
vensis, nov. var. II Ztschr. Zellforsch. Mier. Anat. 1930. Bd 10.
S. 756—768.
Hofker J. The toothplate Foraminifera // Arch. Neerland. Zool.
1951. Bd 8. S. 353—373.
Hofker J. Studies of Foraminifera. Pt Ш. Systematic problems //
Publ. Naturhist. Genootschap Limburg. 1971. Reeks 21, Afl. 1—3.
Hohenegger J. Ichtiolaria denticostata n. sp., eine charakteristis-
che Foraminifere des Unteren Lias Mitteleuropa // Stuttgarter Beitr.
Naturkunde. Ser. B. 1981. N 74.
Hohenegger J. On the way to optimal suprageneric classification
of agglutinated Foraminifera // Paleoecology, Biostratigraphy, Paleo-
ceanography and Taxonomy of Agglutinated Foraminitera / Eds
S. Hemleben et al. Netherlands.: Kluwer Acad. Publ., 1990. P. 77—
105.
Hohenegger J. Determination of Upper Triassic and Lower Juras-
sic Ichtyolarias using morphogenetic programs // Micropaleontology.
1993. Vol. 39, N 3. P. 233—262.
Hohenegger J., Piller W. Wandstrukturen und Grossgliederung
der Foraminiferen // Oster. Acad. Wiss. Sitzungsberichte. 1975.
Abt. 1, Bd 184, Bis. 1, H. 5. S. 67—98.
Hohenegger J., Piller W., Baal C. Horizontal and vertical spatial
microdistribution of Foraminifers in the shallow subtidal Gulf of Trieste,
North Adriatic Sea // J. Foram. Res. 1993. Vol. 23, N 2. P. 79—102.
Hottinger L. An Early Umbilical Canal System in Trocholina
schouberti n. sp. from the Lower Cretaceous of North Eastern Moroc-
co H Eclog. Geol. Helvetiae. 1976. Vol. 69, N 3. P. 815—820.
Hottinger L. Comparative anatomy of elementary shell structures
in selected larger Foraminifera // Foraminifera / Eds R. H. Hedley,
C. G. Adams. London, 1978. Pt 3. P. 203—206.
Hottinger L. Larger Foraminifera, Giant Cells with a historical
background // Naturwissenschaften. 1982. Bd 69. S. 361—371.
Hottinger L., Dreher D. Differentiation of Protoplasm in Num-
mulitidae (Foraminifera) from Elat, Red Sea // Mar. Biol. 1974.
Vol. 25. P. 41—61.
Jones R. W. A revised classification of the unilocular Nodosariida
and Buliminida (Foraminifera) // Rev. Esp. Micropaleontol. 1984.
Vol. 16. P. 91—160.
Jones R. W. The Challenger Foraminifera. Oxford: Oxford Univ.
Press, 1994.
Kaminski M. A., Kuhnt W. Tubular agglutinated Foraminifera as
indicators of organic carbon flux // Proc. Fourth Intern. Workshop
Aggl. Foram. Krakow, Poland, Sept. 12—19, 1993.1 Eds M. A. Ka-
minski et al. Krakow, 1995. P. 141—144. (Grzybowski Found. Spec.
Publ.; N 3).
Kaminski M. A., Boersma A., Tyszka J,, Holbourn A. E. L.
Response of deep-water agglutinated Foraminifera to dyoxic conditi-
ons in the California Borderland Basins // Ibid. P. 131—140.
King K., Jr. Amino acid survey of recent calcareous and siliceous
deepsea microfossils // Micropaleontology. 1977. Vol. 23, N 2.
P. 180—193.
King K., Jr., Hare P. E. Amino acid compposition of planktonic
Foraminifera: A paleobiochemical approach to evolution // Science.
1972. Vol. 175, N 4029. P. 1461—1463.
Kloos D. P. Studies on the foraminifer Sorites orbicularis // Geol.
Mijubouw. 1980. Vol. 59, N 4. P. 375—383.
Kremer В. P., Schmaljohann R., Roettger R. Features and nut-
ritional significance of photosyntheites produced by unicellular algae
symbiotic with larger Foraminifera // Mar. Ecol, Progr. Ser. 1980.
Vol. 2, N 3. P. 225—228.
Kuile B., Erez J. Uptake of inorganic carbon and internal carbon
cycling in symbiont-bearing benthonic Foraminifera // Mar. Biol.
1987. Vol. 94, N 4. P. 499—509.
Laatsch H., Schwab D. Identification and quantification of caro-
tenoids and carotenoid esters from the foraminiferen Allogromia lati-
collaris // J. Foram. Res. 1984. Vol. 14. N 1. P. 36—49.
ТИП FORAMINIFERA. ЛИТЕРАТУРА
621
Langer М. R. Biosynthesis of glycosaminoglicans in foraminife-
ra: a review // Mar. Micropaleontol. 1992. Vol. 19. P. 245—255.
Langer M. R., Gehring C. A. Bacteria fanning: a possible fee-
ding strategy of some smaller motile Foraminifera // J. Foram. Res.
1993. Vol. 23, N 1. P. 40—47.
Langer M. R., Lipps J. H., Piller W. E. Molecular paleobiology
of protists: amplification and direct sequencing of foraminiferal
DNA // Micropaieontology. 1993. Vol. 39, N 1. P. 63—68.
Le Calvez J. Recherches sur les Foraminiferes. I. Development et
Reproduction // Arch. Zool. Experm. Gen. 1938. T. 80. P. 163—333.
Le Calvez J. Entosolenia marginata, foraminifere apogamique ec-
toparasite d’un autre foraminifere Discorbis vilardeboanus // Comt.
Rendu Hebdomadaire Seanc. Acad. Sci. Paris. 1947. Vol. 224.
P. 1448—1450.
Le Calvez J. Recherches sur les Foraminiferes. II. Place de mei-
ose et sexualite // Arch. Zool. Experim. Gen. 1950. T. 87, Pt 4.
P. 211—243.
Le Calvez J. Ordre des foraminiferes // Traite de Zoologie / Ed.
P. Grasse. 1953. Vol. 1 (2). P. 149—265.
Lee J. J. Nutrition and Physiology of the Foraminifera // Bio-
chemistry and physiology of Protozoa. 2nd ed. London: Acad. Press,
1980. Vol. 3. P. 43—66.
Lee J. J. Phylum Granuloreticulosa (Foraminifera) // J. O. Cor-
liss. Handbook of protoctista. Boston, 1990.
Lee J. J. On a Piece of Chalk // Updated. J. Euk. Microbiol. 1993.
Vol. 40. N 4. P. 395—409.
Lee J. J., Anderson O. R. (Eds) Biology of Foraminifera. Lon-
don, 1991.
Lee J. J., Bock D. The importance of feeding in two species of
Soritid Foraminifera With algal symbionts // Bull. Mar. Sci. 1976.
Vol. 26, N 4. P. 530—537.
Lee J. J., Hallock P. Algal symbiosis as the driving force in the
evolution of larger Foraminifera // Eds J. J. Lee, J. F. Frederik. Endo-
cytobiology. Ill // Ann. N. Y. Acad. Sci. 1987. Vol. 503. P. 330—
347.
Lee J. J., Muller W. A. Trophic dynamics and niches of salt
marsh Foraminifera // Amer. Zool. 1973. Vol. 13. P. 215—223.
Lee J. J., Pierce S. Growth and physiology of Foraminifera in the
laboratory. IV. Monoxenic culture of an allogromiid with notes on its
morphology II J. Protozool. 1963. Vol. 10. P. 404—411.
Lee J. J., Zucker W. Algal symbiosis in the foraminifer Archai-
as // Ibid. 1969. Vol. 16, N 1. P. 71—81.
Lee J. J., Pierce S., Tentchoff M., McLaughlin J. J. A. Growth
and physiology of foraminifers in the laboratory. Prt 1. Collection and
maintenance // Micropaleontology. 1961. Vol. 7, N 4. P. 461—466.
Lee J. J., Freudental H. D., Muller W. A., Pierce S., Gross-
man R. Growth and physiology of Foraminifera in the laboratory.
Pt 3. Initial studies of Rosalina floridana (Cushman) // Micropaleonto-
logy. 1963. Vol. 9, N 4. P. 449—466.
Lee J. J., Freudental H. G., Kossoy V., Be A. W. H. Cytological
observations on two planktonic Foraminifera, Globigerina bulloides
d’Orbigny, 1826 and Globigerinoides ruber (d’Orbigny, 1839) Cush-
man, 1927 // J. Protozool. 1965. Vol. 12, N 4. P. 531—542.
Lee J. J., McEnery M.? Pierce S., Freudental H. D., Mul-
ler W. A. Tracer experiments in feeding littoral Foraminifera // J. Pro-
tozool. 1966. Vol. 13, N 4. P. 659—670.
Lee J. J., McEnery M. E., Rubin H. Quantitative Studies on the
growth of Allogromia laticollaris (Foraminifera) // Ibid. 1969a.
Vol. 16, N 2. P. 377—395.
Lee J. J., Muller W. A., Stone R. J., McEnery M. E., Zucker W.
Standing crop of Foraminifera in sublittoral epiphytic communities of
a Long Island salt marsh // Mar. Biol. 1969b. Vol. 4. P. 44—61.
Lee J. J., Tietjen J. H., Stone R. J., Muller W. A., McEnery M.,
Saks N., Mastropaolo C., Kennedy E. Effects on environmental
stress on the community structure, productivity, energy flow, and mi-
neral cycling in salt marsh epyphytic communities // Proc, third Nati-
on. Symp. Radioecol. 1971. Vol. 2. P. 816—828.
Lee J. J., McEnery M. E., Kahn E., Shuster F. Symbiosis and
the evolution of larger Foraminifera // Micropaleontology. 1979.
Vol. 25, N 2. P. 118—140.
Lee J. J., McEnery M. E., Garrison J. R. Experimental studies
of larger Foraminifera and their symbionts from the Gulf of Elat on
the Red Sea // J. Foram. Res. 1980a. Vol. 10, N 1. P. 31—47.
Lee J. J., McEnery M. E., Lee M. J., Reidy J. J., Garrison J. R.,
Roettger R. Algal symbionts in larger Foraminifera // Endocytobiol.,
Endosymb. Cell Biol. 1980b. P. 113—124.
Lee J. J., Reimer Ch. W., McEnery M. E. Diatoms endosymbi-
onts from larger Foraminifera of the Red Sea // Botan. Mar. 1980.
Vol. 23. P. 41—48.
Lee J. J., Saks N. M., Kapiotou F., Wilen S. M., Shilo M. Effects
of host cell extracts on cultures of endosymbiotic diatoms from larger
Foraminifera // Mar. Biol. 1984. Vol. 82, N 2. P. 113—120.
Lee J. J., Hunter S. H., Bovee E. C. An illustrated guide to the
Protozoa. Lawrence: Kansas. Soc. Protozool., 1985a. P. 16—17.
Lee J. J., Lee M. J., Weis D. S. Possible adaptive value of endo-
symbionts to their protozoan hosts // J. Protozool. 1985b. Vol. 32,
N 3. P. 380—382.
Lee J. J., Anderson R. O., Karim B., Beri J. Additional insight
into the structure and biology of Abyssotrema pacifica (Broennimann,
Van Dover and Whittaker) from the East Pacific Rise // Micropaleon-
tology. 1991. Vol. 37, N 3. P. 303—312.
Lena H. Cytological studies in Allogromia flexibilis (Wiesner)
(Foraminifera) // Int. Rev. Hydrobiol. 1972. Bd 57, H 4. S. 637—644.
Lena H., Freire F. Estudios citologicos en Allogromia laticollaris
Arnold (Protozoa, Foraminifera) // Physis Sec. A. 1974. Vol. 33,
N 86. P. 123—133.
Lengsfeld A. M. Zum Feinbau der Foraminifere Allogromia lati-
collaris. I. Mutt.: Zellen mit ausgestreckten und eingezogenen Rhizo-
podien // Helgoland Wiss. Meeresunters. 1969. Bd 19, N 2. S. 230—
261.
Leutenegger S. Ultrastructures de Foraminiferes perfores et im-
perfores ainsi que de leurs symbiotes // Cahiers Micropaleontol. 1977.
N 3. P. 1—52.
Leutenegger S. Specific host-symbiont relationship in larger Fo-
raminifera // Micropaleontology. 1983. Vol. 29, N 2. P. Ill—125.
Leutenegger S. Symbiosis in benthic Foraminifera: specificity
and host adaptations // J. Foram. Res. 1984. Vol. 14, N 1. P. 16—35.
Leutenegger S., Hansen H. J. Ultrastructural and radiotracer stu-
dies of pore function in Foraminifera // Mar. Biol. 1979. Vol. 54, N 1.
P. 11—16.
Levy A., Mathieu R., Poignant A., Rosset-Moulinier M. Discor-
bidae and Rotaliidae: a classification to be revised // J. Foram. Res.
1986. Vol. 16, N 1. P. 63—71.
Lewis К. B. A key to the recent genera of the Foraminiferida //
New Zealand Dept. Sci. Industr. Res. Bull. 1970. N 196. (New Zea-
land Oceanogr. Inst. Mem.; N 45).
Lipps J. H. Wall structure systematics, and phylogeny studies of
Cenozoic planktonic Foraminifera // J. Paleontol. 1966. Vol. 35. P. 245—330.
Lipps J. H. Feeding strategies and test function in foraminifer //
Proc. «Benthonics 75». Halifax; Nova Scotia, 1975. P. 26.
Lipps J. H. Biotic interactions in benthic Foraminifera // Bio-
tic interactions in Recent and fossil benthic communities / Eds
M. J. S. Tevesz, P. L. McCall. New York: Plenum Publ., Corp. 1983.
P. 331—376.
Lipps J. H. Prokaryotes and Protista // Fossil prokariotes and Pro-
tists I Ed. J. H. Lipps. Tenessee, Vol. 18. P. 1—19.
Lipps J. H., Erskian M. G. Plastogamy in Foraminifera Glabra-
tella ornatissima // J. Protozool. 1969. Vol. 16, N 3. P. 422—453.
Lipps J. H., Ronan T. E. Predation on Foraminifera by the Po-
lychaete worm, Diopatra // J. Foram. Res. 1974. Vol. 4, N 3. P. 139—
143.
Lister J. J. Contributions to the life history of the Foraminifera //
Roy. Soc. London Phil. Trans. Ser. B. 1895. Vol. 186. P. 401—453.
Loeblich A. R., Jr. Protistan Phylogeny as indicated by the fossil
record // Taxon. 1974. Vol. 23, N 2/3. P. 277—290.
Loeblich A. R., Jr., Tappan H. Treatise on Invertabrate Paleonto-
logy. Pt C. Protista. 2. Sarcodina, chiefly «Thecamoebians» and JFora-
miniferida. Ed. R. C. Moore. Lawrence: Geol. Soc. Amer, and Univ.
Kansas Press, 1964. Vol. 1, 2.
Loeblich A. R., Jr., Tappan H. Foraminiferal genera and their
classification Los Angeles: Van Nostraand Comp. (New York), 1988.
Vol. 1, 2.
Loeblich A. R., Jr., Tappan H. Present status of foraminiferal
classification. Studies in Benthic Foraminifera // Proc. Fourth Intern.
Sympos. on Benthic Foraminifera. Benthos ’90. Sandai, 1990. Tokai
Univ. Press, 1992. P. 93—102.
Loeblich A. R., Jr., Tappan H. Foraminifera of the Sahul
Shelf and Timor sea. 1994. (Cushm. Found. Foram. Res. Spec.
Publ.; N 3).
Lopez E. Algal chloroplasts in the protoplasm of three species of
benthic Foraminifera: taxonomic affinity, variability, and persistence //
Mar. Biol. 1979. Vol. 53. P. 201—211.
Lutze G. F., Thiel H. Epibenthic Foraminifera from elevated mic-
rohabitats: Cibicidoides wuellerstorfi and Planulina ariminensis //
J. Foram. Res. 1989. Vol. 19, N 2. P. 153—159.
Lutze G. F., Weber G. Habitat and asexual reproduction of Cyc-
lorbiculina compressa (d’Orbigny), Soritidae // Ibid. 1980. Vol. 10,
N 4. P. 251—260.
622
СИСТЕМАТИЧЕСКИЙ ОБЗОР
Marshalek D. S. Observations on Iridia diaphana, a marine fora-
minifer // J. Protozool. 1969. Vol. 16, N 4. P. 599—612.
Matsushita S., Kitazato H. Seasonality in the benthic foramini-
feral community and the life history of Trochammina hadai Uchio
in Hamana Lake, Japan // Paleoecology, Biostratigraphy, Paleoceano-
graphy and Taxonomy of Agglutinated Foraminifera I Eds C. Hemle-
ben et al. Netherlands: Kluwer Acad. Publ., 1990. P. 695—715.
МсЕпегу M. E., Lee J. J. Allogromia laticollaris: a foraminifer
with unusual appogamic metagenic life cycle // J. Protozool. 1976.
Vol. 23, N 1. P. 94—108.
МсЕпегу M. E., Lee J. J. Cytological and fine structural studies
of three species of symbiont bearing larger Foraminifera from the
Sea // Micropaleontology. 1981. Vol. 27, N 1. P. 71—83.
Muller W. A. Competition for food and other Niche-Related stu-
dies of three species of salt-marsh Foraminifera // Mar. Biol. 1975.
Vol. 31. P. 339—351.
Muller W. A., Lee J. J. Apparent indespensability of bacteria in
foraminiferan nutrition // J. Protozool. 1969. Vol. 16, N 3. P. 471—478.
Muller-Merz E., Lee J. J. Symbiosis in the larger foraminiferan
Sorities marginalis (with notes on Archaias sp.) // Ibid. 1976. Vol. 23,
N 3. P. 390—396.
Mullineaux L. S. Taxonomic notes on larger agglutinated forami-
nifers encrusting manganese nodules, including the description of a
new genus, Chondrodapis (Komokiacea) // J. Foram. Res. 1988.
Vol. 18, N 1. P. 46—53.
Munier-Chalmas E. Sur le dimorphisme des Nummulites // Bull.
Soc. Geol. France. Ser. 3. 1880. T. 8. P. 300—301.
Murray J. W. Wall structure of some agglutinated Foraminiferi-
da // Paleontology. 1973. Vol. 16, pt 4. P. 777—786.
Myers E. H. The life history of Patellina corrugata. Williamson,
a foraminifer // Bull. Scripps. Inst. Oceanogr. Univ. California. Techn.
ser. 1935. Vol. 3, N 15. P. 355—391.
Myers E. H. The life-cycle of Spirillina vivipara Ehrenberg with
notes on morphogenesis, systematics and distribution of the Foramini-
fera II J. Roy. Microsc. Soc. 1936. Vol. 56. P. 120—146.
Myers E. H. Observations on the origin and fate of flagellate
gametes in Multiple tests of Discorbis (Foraminifera) // J. Mar. Biol.
Ass. 1940. Vol. 24, N 1. P. 201—226.
Myers E. H. Biology, ecology and morphogenesis of a pelagic
Foraminifera// Stanford Univ. Publ. Biol. Sci. Ser. 9. 1943. N 1.
P. 1—30.
Nagy J., Alve E. Temporal changes in Foraminiferal faunas and
impact on pollution in Sandebukta, Oslo Fiord // Mar. Micropaleontol.
1987. Vol. 12, N 2. P. 109—128.
Neymayr M. Die Staemme des Thierreiches. Wirbellose Thiere.
Bd 1. Wien : F. Tempsky, 1889. S. 153—206.
Nyholm K. G. A study of the foraminiferan Gypsina // Zool. Bidr.
Uppsala. 1962. Vol. 33. P. 201—206.
Nyholm K. G., Nyholm P. G. Ultrastructure of monotjalamous
Foraminifera // Zoon. 1975a. Vol. 3, N 2. P. 141—154.
Nyholm K. G., Nyholm P. G. On the microtubules of some mo-
nothalamous Foraminifera, especially Cylindrogullmia alba // Ibid.
1975b. Vol. 3, N 2. P. 151—154.
d’Orbigny A. Tableau methodique de la classe des Cephalopo-
des II Ann. Sci. Nat. Paris. 1826. Ser. 1, T. 7. P. 245—314.
d’Orbigny A. Foraminiferes des Iles Canaries // Barker-Webb et
Berthelot. «Hist. Nat. Iles Canaries» Zool. Paris, 1839. T. 2, pt 2.
P. 119—146.
Pawlowski J., Lee J. Taxonomic notes on some tiny shallow
water Foraminifera from the Northern Gulf of Elat (Red Sea) // Mic-
ropaleontology. 1991. Vol. 37, N 2. P. 149—163.
Pawlowski J., Lee J. J. The life cycle of Rotaliella elatiana
n. sp. — a tiny macroalgavorous foraminifer from the Gulf of Eilat //
J. Protozool. 1992. Vol. 39, N 1. P. 131—143.
Pawlowski J., Bolivar I., Fahrni J., Zaninetti L. Taxonomic
identification of Foraminifera using ribosomal DNA sequenses // Mic-
ropaleontology. 1994. Vol. 40, N 4. P. 373—378.
Pawlowski J., Bolivar I., Fahrni J., Zaninetti L., Kitazato H.
Partial LSUrDNA sequences of Trochammina sp. (Foraminiferida) //
Proc. Fourth Intern. Workshop Aggl. Foram. Krakow, Poland.
Sept. 12—19, 1993 I Eds M. A. Kaminski et al. Krakow, 1995a.
P. .227—232. (Grzybowski Found. Spec. Publ.; N 3).
Pawlowski J., Swiderski Z., Lee J. J. Observations on the ultra-
structure and reproduction of Trochammina sp. (Foraminiferida) //
Ibid. 1995b. P. 233—237.
Pawlowski J., Bolivar I., Fahrni J., Vargas C., Gouy M., Zaninet-
ti L. Extreme differences in rates of molecular evolution of Foraminifera
Revealed by Comparison of Ribosomal DNA Sequences and the Fossil
Record // Mol. Biol. Evol. 1997. Vol. 14, N 5. P. 498—505.
Pierce S., Nathanson M. E. Electron microscopical investigations
on surface-related structures in Allogromia (Protozoa: Foraminiferi-
da) // Trans. Amer. Mier. Soc. 1974. Vol. 93, N 2. P. 170—180.
Piller W. E. Wall structures of Palaeotextulariid Foraminifers and
discussion of microgranular test walls // Palaeoecology, Biostratigra-
phy, Palaeooceanography and Taxonomy of Agglutinated Foraminife-
ra // Eds C. Hemleben et al. S. 1., 1990. P. 25—35.
Reiss Z. Reclassification of perforate Foraminifera // Bull. Geol.
Surv. Israel. 1963. Vol. 35. P. 1—111.
Reiss Z., Hottinger L. The Gulf of Aqaba. Ecological Micropale-
ontology // Ecological Studies, Analysis and Synthesis. Berlin etc.:
Springer, 1984. Vol. 50.
Revets S. A. Structure and taxonomy of the genus Delosina Wi-
esner, 1931 (Protozoa: Foraminifera) // Bull. Brit. Mus. (Nat. Hist.)
Zool. Ser. 1989. Vol. 55. P. 1—9.
Revets S. A. The reclassification of Tremachora Lipps and Lipps,
1967 in the suborder Delosinina // J. Foram. Res. 1991. Vol. 21, N 3.
P. 252—255.
Revets S. A. The generic Revision of five families of Rotaline
Foraminifera. Pt I. The Bolivinitidae; Pt II. The Anomalinidae, Alaba-
minidae, Cancrisidae and Gavelinellidae. 1996. (Cushm. Found.
Foram. Res. Spec. Publ.; N 34).
Rhumbler L. Die Foraminiferen (Thalamophoren) der Plankton-
Expedition. Ergebnissen. Plankton-Exped. Humboldt-Stifftung. Kiel;
Leipzig, 1911. Bd 3. S. 1—476.
Roettger R. Larger Foraminifera: reproduction and early stages of
development in Heterostegina depressa // Mar. Biol. 1974. Vol. 26,
N 1. P. 5—12.
Roettger R. Unusual multiple fission in the gamont of the larger
foraminiferen Heterostegina depressa // J. Protozool. 1978. Vol. 25,
N 1. P. 41—44.
Roettger R., Hallock P. Shape trends in Heterostegina depressa (Pro-
tozoa, Foraminifera) // J. Foram. Res. 1982. Vol. 12, N 3. P. 197—204.
Roettger R., Dettmering C., Kruger R., Schmaljohann R., Ho-
henegger J. Gametes in nummulitids (Foraminifera) // J. Foram. Res.
1998. Vol. 28, N 4. P. 345—349.
Ross C. A., Haman D. Suprageneric ranges of Foraminiferids //
Ibid. 1989. Vol. 19, N 1. P. 72—84.
Rupp G., Bowser S. S., Manella C. A., Rieder C. L. Naturally
occuring Tubulin-containing paracrystals in Allogromia: Immunocy-
tochemical identification and functional significance // Cell Motil. Cy-
toskel. 1986. Vol. 6. P. 363—375.
Salami M. B. Biology of Trochammina cf. T. quadriloba Hoeg-
lund (1947), an agglutinated foraminifer // J. Foram. Res. 1976.
Vol. 6. P. 142—153.
Samuel S., Bowser H., Israel A., Samuel M., Russel M. G., Rei-
der C. L. Surface transport properties of reiculopodia: do intra cellu-
lar and extracellular motility share a common mechanism? // Cell.
Biol. Intern. Rep. 1984. Vol. 8, N 12. P. 1051—1063.
Schaudinn F. Untersuchungen an Foraminiferen. I. Calcituba po-
lymorpha // Ztschr. Wiss. Zool. 1895. Bd 59. S. 191—232.
Schroeder R. A., Bada J. L. Kinetics and mechanism of the
epimerization and decomposition of threonine in fossil Foraminifera //
Geochim. et Cosmochim. Acta. 1977. Vol. 41, N 8. P. 1087—1095.
Schultze M. S. Ueber der Organismus der Polythalamien (Forami-
niferen) nebst Bemerkungen ueber die Rhizopoden im Allgemeinen.
Leipzig: Engelmann, 1854.
Schwab D. Electronen microscopische Untersuchungen an der Fo-
raminifere Myxotheca arenilega Schaudinn // Ztschr. Zellforsch. Mic-
rosk. Anat. 1969. Bd 96. S. 295—324.
Schwab D. Elektronenmikroskopische Untersuchungen intracellu-
lar lebenden Einzellem in Foraminiferen // Ztschr. Naturforsch. 1971.
Bd 26b, H. 12. S. 1341—1344.
Schwab D. Electron microscopic studies on the foraminifer Allog-
romia laticollaris Arnold. Mitosis in agamonts // Protoplasma. 1972.
Bd 75. S. 79—89.
Schwab D. Centrosomal bodies during meiosis in the foraminifer
Myxotheca arenilega Schaudinn // Ibid. 1973. Bd 78. S. 339—341.
Schwab D. Gametogenesis in Allogromia laticollaris // J. Foram.
Res. 1976. Vol. 6, N 4. P. 251—257.
Schwab D., Hofer H. W. Metabolism in protozoan Allogromia
laticollaris Arnold // Ztschr. mikroskop.-anat. Forschung. 1979. Bd 93,
H. 4. S. 715—727.
Schwab D., Plapp R. Quantitative chemical analysis of the shell
of the monothalamous foraminifer Allogromia laticollaris Arnold //
J. Foram. Res. 1983. Vol. 13, N 1. P. 69—72.
Schwab D., Schlobach H. Carotene wax pigments in the forami-
niferan Allogromia laticollaris Arnold // J. Protozool. 1973. Vol. 20,
N 2. P. 339—341.
ТИП FORAMINIFERA. ЛИТЕРАТУРА
623
Schwab D., Schlobach H. Pigments in monothalamous Foramini-
fera // J. Foram. Res. 1979. Vol. 9, N 2. P. 141—146.
Schwab D., Schwab-Stey H. Hexagonal structures at the plasma
membrane of the foraminifer Allogromia laticollaris Arnold // Ibid.
1981. Vol. 11, N 3. P. 212—216.
Schwab-Stey H., Schwab D. The transformation of Cytoplasmic
Microtubules into Helices and Paracrystals by Halothane in the Fora-
minifer Allogromia laticollaris Arnold // Ztschr. mikroskop.-anat. For-
schung. 1979. Bd 93. H. 4. S. 751—762.
Schwager C. Quadro del proposto sistema de classificazione dei
foraminiferi con guscio // R. Comitato Geol. Italia, Bull. 1877., Vol. 8,
N 1—2. P. 18—27.
Seiglie G. A. A preliminary note on the relationships between
Foraminifers and pollution in two Puerto Rican Bays // Carib. J. Sci.
1971. Vol. 11, N 2. P. 93—98.
Sen-Gupta В. K., Machain-Castillo M. L. Benthic Foraminifera
in Oxygen pure habitats // Mar. Micropaleontol. 1993. Vol. 20. P. 182—
201.
Sigal J. Ordre des Foraminifera. Traite de Paleontol. T. 1. Paris.
1952. P. 133—301.
Sliter W. V. Bolivina doniezi Cushman et Wickenden in clone
Culture II Contr. Cushm. Found. Foram. Res. 1970. V. 21. P. 87—99.
Sliter W. V. Test ultrastructure of some living benthic Foramini-
fers II Lethaia. 1974. Vol. 7. P. 5—16.
Spindler M. The development of the organic lining in Heteroste-
gina depressa (Nummulitidae: Foraminifera) // J. Foram. Res. 1978.
Vol. 8, N 3. P. 258—261.
Spindler M. Lunar periodicity of reproduction in the planktonic
foraminifer Hastigerina pelagica // Mar. ecol. Progr. Ser. 1979. Vol. 1,
N 1. P. 61—64.
Spindler M. The pelagic gulfweed Sargassum natans as a habi-
tat for the benthhic Foraminifera Planorbulina acervalis and Rosalina
globularis // Neues Jb. Geol. Paleontol. Monatschr. 1980. H. 9. S. 569—
580.
Spindler M., Hemleben Ch. Formation and possible function of
annulate lamelie in a planktonic Foraminifer // J. Ultrasstructure Res.
1982. Vol. 81. P. 341—350.
Spindler M., Roettger R. Der Kammerbauvorgang der Grossfo-
raminifere Heterostegina depressa (Nummulitidae) II Mar. Biol. 1973.
Vol. 18. P. 146—159.
Spindler M., Anderson O. R., Hemleben C., Be A. W. H. Light
and electron microscopic observations of gametogenesis in Hastigeri-
na pelagica (Foraminifere) // J. Protozool. 1978. Vol. 25, N 4.
P. 427—433.
Stathoplos L., Tuross N. Proteins and DNA from modern plank-
tonic Foraminifera // J. Foram. Res. 1994. Vol. 24, N 1. P. 49—59.
Stouff V., Lesourd M., Debenay J.-P. Laboratory observations of
asexual reproduction (schizogony) and ontogeny of Ammonia tepida
with the comments on the cycle II J. Foram. Res. 1999a. Vol. 29, N 1.
P. 75—85.
Stouff V., Geslin E., Debenay J.-P., Lesourd M. Origin of morpho-
logical abnormalities in Ammonia (Foraminifera): studies in laboratory
and natural environments // J. Foram. Res. 1999b. Vol. 29, N 2. P. 152—
171.
Tendal O. S., Hessler R. R. An introduction to the biology and
systematics of Komokiacea (Textulariana, Foraminiferida). Scand. Sci.
Press, 1977. P. 165—194.
Thalmann H. E. An index to the genera and species of forami-
nifera. 1890—1950. Calif. G. Vanderbilt Found. Stanford Univ.,
1960.
Todd R. A new Rosalina (Foraminifera) parasitic on a bivalve //
Deep Sea Res. 1965. Vol. 12. P. 831—837.
Toksvad T., Hansen H. J. A study of calcareous cement in agglu-
tinated Foraminifera // Publ. Contin. Shelf Inst. 1983. Vol. 108.
P. 159—169.
Towe К. M. Lamellar wall construction in planktonic foraminife-
ra // Proc. Second Planktonic Conf. 1971. Vol. 2. P. 1213—1218.
Towe К. M., Cifelli R. Wall ultrastructure in the calcareous Fora-
minifera: Crystallographic aspects and a model for calcification //
J. Paleontol. 1967. Vol. 41, N 3. P. 742—762.
Towe К. M., Wright R. C. Ultramicrostructure of some selected
foraminiferal tests. 1963. Vol. 9. P. 171—195.
Towe К. M., Berthold W.-U., Appleman D. E. The crystal-
lography of Patellina corrugata Williamson: a-axis preffered orientati-
on II J. Foram. Res. 1977. Vol. 7, N 1. P. 58—62.
Travis J. L., Bowser S. S. A new model of reticulopodial motility
and shape: evidence for a microtubule-based motor and an actin skele-
ton // Cell Motility and the Cytoskeleton. 1986. Vol. 6. P. 2—14.
Tyszka J., Kaminski M. A. Factors controlling the distribution of
agglutinated Foraminifera in Aalenia-Bajocian dysoxic facies (Pieniny
Klippen Belt, Poland) // Proc. Fourth Intern. Workshop Aggl. Foram.,
Krakow, Poland, Sept. 12—19, 1993 I Eds M. A. Kaminski et al. Kra-
kow, 1995. P. 271—291. (Grzybowski Found. Spec. Publ.; N 3).
Walker D. A. Etching of the test surface of benthonic foramini-
fers due to ingestion by the gastropod Littorina littorea Linne // Canad.
J. Earth Sci. 1971. Vol. 8, N 11. P. 1487—1491.
Weber F. W. Ueber die Paarung der Gamonten und der Kerndu-
alismus der Foraminifere Metarotaliella parva Grell // Arch. Protis-
tenk. 1965. Bd 108. S. 217—270.
Wefer G., Berger W. H. Stable isotopes in benthic Foraminifera:
seasonal variation in large tropical species // Science. 1980. Vol. 209,
N 4458. P. 803—805.
Weinberg J. R. Rates of movement and sedimentary traces of
deep-sea Foraminifera and mollusca in the laboratory // J. Foram. Res.
1991. Vol. 21, N 3. P. 213—217.
Weiner S., Erez J. Organic matrix of the shell of the foraminifer
Heterostegina depressa // Ibid. 1984. Vol. 14, N 3. P. 206—212.
Wetmore R. A. Burrowing and sediment movement by benthic
Foraminifera, as shown by time-lapse cinematography // Rev. Paleobi-
ol. 1988. Vol. 2. P. 921—927.
Wetmore R. A. Chamberformation in Archaias angulatus //
J. Foram. 1999. Vol. 29, N 1. P. 69—75.
Williamson D. F., Roettger R., Schmaljohann R., Keigwin L.
Oxygen and carbon isotopic fractination and algal symbiosis in the
benthic foraminiferan Heterostegina depressa // Palaeogeogr., Palaeo-
climatol., Palaeoecol. 1981. Vol. 33. P. 231—251.
Wray C. G., Lee J. J., De Salle R. Extraction and enzymatic
characterization of foraminiferal DNA // Micropaleontology. 1993.
Vol. 39, N 1. P. 69—74.
Yanko V., Kronfeld J., Flexer A. Response of Benthic Foramini-
fera to various pollution sources: implications for pollution monito-
ring // J. Foram. Res. 1994. Vol. 24, N 1. P. 1—18.
Yanko V., Ahmad M., Kaminski M. Morphological deformities
of benthic foraminiferal tests in response to pollution by heave metals:
implications for pollution monitoring // J. Foram. Res. 1998. Vol. 28,
N 3. P. 177—201.
Zapata M. A., Varela M. S. Epibiosis de Turbelarios sobre Buc-
cella frigida (Cushman) (Foraminifera, Protozoa) // Bol. Soc. Biol.
Concepcion, Chile. 1981. Vol. 52. 245—246.
Zech L. Zytochemische Messungen an den Zellkernen der Fora-
miniferen Patellina corrugata und Rotaliella heterocariotica // Arch.
Protistenk. 1964. Bd 107. S. 295—330.
Zmiri A., Kahan D., Reiss Z. Aspects of the thermotactic respon-
se in Foraminifera // J. lYotozool. (Suppl.). 1972. Vol. 19. P. 56.
Zmiri A., Kahan D., Hochstein S., Reiss Z. Phototaxis and ther-
motaxis in some species of Amphistegina (Foraminifera) // Ibid. 1974.
Vol. 21, N 1. P. 133—139.
Zucker W. H. Fine structure of planktonic Foraminifera and
their endosymbiotic algae: Ph. D. Thes., City Univ, of New York,
1973.
624
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Тип FORAMINIFERA d’Orbigni, 1826 — ФОРАМИНИФЕРЫ
Экология и зоогеография41
Экология
Фораминиферы — обширная группа простейших,
представленная на всех широтах Мирового океана
от литорали до ультраабиссали. Среди них встреча-
ются свободноживущие донные фораминиферы, при-
крепленные и планктонные организмы. Свободножи-
вущие донные фораминиферы обитают не только на
поверхности морского дна, но и на глубине несколь-
ких десятков сантиметров в мягких осадках (Buzas,
1965; Boltovskoy, 1966b; Wollenburg, Mackensen,
1998), при этом численность живых представителей
всех видов более или менее монотонно убывает с
глубиной (Басов, Хусид, 1983; Bernhard, 1989; Мо-
odley, 1990). В то же время иногда отмечали появ-
ление пиков численности этих организмов в осадках
на глубине нескольких сантиметров. В хрошо аэри-
рованных осадках фораминиферы опускаются на
большую глубину (Corliss, Emerson, 1990), в грунтах
же с высоким содержанием органического вещества
и малой концентрацией кислорода они сосредоточи-
ваются в поверхностном горизонте и вблизи ходов
роющего макробентоса (Langer et al., 1989). На рас-
пространение группы литоральных фораминифер в
значительной степени влияет воздействие на них мор-
ских волн, генерируемых ветром того или иного пре-
обладающего направления (Преображенская, Троицкая,
1991). Корсун с соавт. (1994) отмечают, что наблюде-
ние за культурами, фотографирование фораминифер и
следов их движения опровергли бытовавшее ранее
мнение об их малой подвижности внутри осадков.
У живых фораминифер раковины ориентированы
определенным образом. Крупные трубчатые и ветвя-
щиеся раковины располагаются перпендикулярно по-
верхности грунта (De Laca, 1986; Cedhagen, 1988).
Уплощенные мелкие формы перемещаются главным
образом в пределах поверхностного слоя осадков (Ki-
tazato, 1988; Langer et al., 1989). В то же время на-
блюдения в аквариальной культуре показали, что ра-
ковины Bolivina pseudopunctata располагались пер-
пендикулярно поверхности стенки аквариума устьем
вниз, образуя сплошную сеть из ризоподиальных
нитей, которая занимала очень большую площадь.
Прикрепленные фораминиферы в качестве субст-
рата используют камни, донную растительность, же-
лезо-марганцевые конкреции, покровы донных жи-
вотных, спикулы губок и другие объекты, включая
планктонные организмы. Распространены они также
повсеместно на всех широтах и глубинах Мирового
океана.
Планктонные фораминиферы для успешного па-
рения в толще воды строят раковину или небольшого
41 При написании этого раздела была использована книга Бол-
товского и Райте (Boltovskoy, Wright, 1976), а в основу классифи-
кации положена система, предложенная Лёбликом и Таппан (Loeb-
lich, Tappan, 1987). .
размера, или тонкостенную, украшенную значитель-
ным количеством больших и малых пор и игл. У от-
дельных видов вокруг раковины можно наблюдать
обрастание в виде эктоплазматического студенистого
вещества, также позволяющего увеличить плавучесть
организма.
Встречаясь повсеместно, фораминиферы в то же
время чутко реагируют на воздействие окружающей
среды. Такие факторы, как температура, соленость,
характер грунта, пищевой фактор, а также глубина и
связанные с ними давление, освещенность, величины
pH, содержание в воде растворенного кислорода и
карбоната кальция и многие другие показатели, ока-
зывают решающее влияние на существование, раз-
множение и в конечном счете на распространение и
географию отдельных видов фораминифер.
Влияние температуры
Температура, без сомнения, оказывает сильное
воздействие на жизнедеятельность и существование
фораминифер. Лабораторные опыты наглядно пока-
зывают, что различные виды этих организмов пред-
почитают или вообще могут жить только в опре-
деленном температурном диапазоне. Так, например,
Buccella frigida, Resigella moniliforme, Eggerelloides
scabra в культуре живут и размножаются при 15—
20 °C, Cribroelphidium excavatum — при 10—15, Mi-
liammina fusca — при 10—20 °C (Murray, 1968).
Buliminella elegantissima, Epistominella exigua могут
жить в более широком температурном диапазоне —
от 3.8 до 14.5 °C (Boltovskoy, Lena, 1969). В то же
время Elphidium selseyense, Turbinulina depresura,
Ammonia batavus могут выдерживать в культуре тем-
пературу до -1.8 °C, а размножаться при 24 °C (Vo-
orthuysen, 1960). У вида Allogromia laticularis отме-
чен рост раковины при 15—30 °C (Lee, McEnery,
1970). Размножение же у Spirillina vivipara описано
при 18—21, а рост раковины — при 26 °C (Brads-
haw, 1961).
Приуроченность отдельных групп видов форами-
нифер к определенному диапазону температур (сте-
нотермность) в природных условиях отмечается во
многих работах. Конечно, температура находится в
непосредственной зависимости от широтного и бати-
метрического факторов, и их влияние трудно разгра-
ничить. Но все же можно выделить виды, живущие в
условиях постоянного низкого температурного режи-
ма, или виды, обитающие при температуре порядка
20—30 °C, или формы литоральной зоны, выдержи-
вающие сильные суточные и сезонные колебания
температуры, особенно в умеренных поясах.
Приуроченность планктонных видов форамини-
фер к определенным широтным поясам еще более
четко выражена. Так, например, обитатели холодных
ТИП FORAMINIFERA. ЭКОЛОГИЯ И ЗООГЕОГРАФИЯ
625
и умеренных вод Neogloboquadrina pachyderma вы-
держивают довольно большие температурные перепа-
ды поверхностного слоя воды: от -1.4 до 24 °C, Glo-
bigerina bulloides от -0.7 до 27, Globirotalia inflata от
-0.4 до 27, Globigirinita glutinata от 0.3 до 30 °C. В
то же время формы тропической зоны океана, напри-
мер Globigerinoides trilobus, G. sacculifer, Globigerina
conglobata, Globorotalia menardii, живут в поверхнос-
тном слое воды при 15—30 °C. Виды же Candeina
nitida и Globoquadrina hexagona в Атлантическом и
Индийском океанах обитают при 25—30 °C (Be, Tol-
derlund, 1971).
Температура изменяется с глубиной и, следова-
тельно, оказывает влияние и на вертикальное рас-
пространение фораминифер. При этом в большей за-
висимости от глубины находятся фораминиферы с
известковой раковиной: высокое давление и низкая
температура приводят к низкому содержанию в воде
карбоната кальция, а это угрожает растворением ра-
ковин.
Кроме того, в зависимости от температуры нахо-
дятся такие морфологические признаки, как строе-
ние, орнаментация (дополнительные скелетные обра-
зования) и размеры раковин. Было показано, что ра-
ковины широко распространенных видов в холодных
водах имеют большие размеры (Rhumbler, 1911) и
менее орнаментированы (Lutze, 1962), хотя есть и
противоположные мнения (Boltovskoy, 1962, 1966а;
Вё, 1972, и др.). Бе (Be, 1968) отмечает, что планк-
тонные виды в тропических широтах имеют более
перфорированную раковину, чем в холодных и уме-
ренных водах. Болтовской (Boltovskoy, 1962, 1966а)
указывает, что виды Globigerinoides trilobus, G. ruber
в холодных водах утрачивают вторичные устья.
Влияние солености
С соленостью морской воды несомненно прямо
связаны жизненная активность организма, его физио-
логические функции. Однако ее влияние на жизнь и
размножение фораминифер все же не столь значи-
тельно, как влияние других факторов. Конечно, по-
давляющее большинство фораминифер предпочитает
нормальную морскую соленость порядка 30—40%о,
но они могут выдерживать иногда значительное
опреснение (7—10%©) или гиперосолонение (50—
90 %©) воды, хотя процесс воспроизводства возможен
лишь в условиях, близких к нормальной солености.
Бентосные виды фораминифер более приспособ-
лены к пониженной солености, ее сезонным и суточ-
ным колебаниям. В лабораторных условиях типичные
морские виды из подотряда роталииды, помещенные
в пресную воду, оставались живы после 5 недель эк-
сперимента (Schultze, 1854).
В то же время можно назвать группу видов —
Ammotium cassis, Jadammina macrescens, Miliammina
fusca, Verneuilinulla advena, Trochammina inflata,
Cribroelphidium longipontis, R. excavatum — типич-
ных обитателей литорали северных и умеренных
широт, на жизнедеятельность которых помимо силь-
ного перепада температур, малой глубины, постоян-
ного воздействия приливно-отливного течения су-
щественное влияние оказывает и периодическое оп-
реснение воды (5—15 %©).
В заливах, бухтах и лагунах, где наряду с другими
факторами изменчив и фактор солености, фауна фо-
раминифер представлена богаче как в качественном,
так и в количественном отношении по сравнению с
открытым океаном.
Планктонные виды более чувствительны к изме-
нению степени солености. Обычно они живут при со-
лености, равной 34—36 %©, но могут выдерживать
колебания от 33 до 40 %©. Нижним же пределом вы-
живания разных видов некоторые авторы считают со-
леность, равную 27 (Waller, Polski, 1959) или даже
31—33.5 %© (Boltovskoy, 1966b).
Соленость оказывает также влияние на морфологию,
размер и орнаментацию раковин. Известно, что фора-
миниферы, живущие в условиях пониженной солености,
имеют более тонкие, хрупкие, слабо орнаментирован-
ные раковины по сравнению с теми же видами, обита-
ющими при нормальной морской солености (Le Cal-
vez, Le Calvez, 1951; Kurc, 1961; Brodniewicz, 1965;
Wright, 1968, и др.). Фурсенко (1959) указывал, что у
вида Ammonia beccarii, найденного в Белом море (со-
леность 18—20 %©), раковины были меньше, тоньше,
с меньшим числом камер, без орнаментации и пупоч-
ной шишки по сравнению с раковинами того же вида
из Средиземного моря (соленость 37—39 %©). Увели-
чение солености выше нормы также, видимо, оказыва-
ет влияние на строение раковины, но этот вопрос пока
еще недостаточно изучен.
Пищевой фактор
Несомненно, пищевой фактор является определя-
ющим не только при распределении фораминифер,
но оказывает значительное влияние на их числен-
ность. Чем больше пищи, тем богаче фауна. При
этом ризоподиальная сеть наряду с функциями дви-
жения, роста, размножения, прикрепления, газооб-
мена и вывода метаболитов играет главенствующую
роль в процессе питания. Ризоподии образуют ог-
ромную биологически активную поверхность, кото-
рая покрывает большую площадь дна. Столь обшир-
ная ловчая сеть с успехом обеспечивает трофичес-
кие потребности фораминифер (Корсун и др., 1994).
Пищей для них служат в первую очередь фитоплан-
ктон (диатомовые, низшие водоросли, споры), а
также бактерии, органический детрит, инфузории,
личинки иглокожих, ракообразных, нематоды. Отме-
чена избирательная способность у некоторых видов
фораминифер к определенной пище. Так, Шаудин
(Schaudinn, 1895) отмечает, что пищей для Patellina
и Discorbis служат инфузории и личинки копепод.
Корсун с соавт. (1994) отмечают, что цикл работ,
проведенных в проливе Мак-Мердо, выявил не толь-
ко способность фораминифер к абсорбции РОВ, но
и то, что РОВ иногда составляют преобладающую
долю рациона (De Laca, 1985).
Фораминиферы нередко поселяются на покровах
морских беспозвоночных. В ряде случаев отмечено
травление выемок в карбонатных раковинах и био-
кластах (De Laca, Lipps, 1972; Lipps, 1983; Smyth,
626
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
1988; Venec-Peyre, 1988). Тонкое цитологическое
исследование Cibicides refulgens показало, что псев-
доподии этого факультативного эктопаразита пенет-
рируют ракушку моллюска-хозяина и по тончайшим
каналам проникают в его мантийную полость, выса-
сывая плазму; поглощение и транспорт свободных
аминокислот прослежены с помощью радиактивной
метки (Alexander, De Laca, 1987). В свою очередь
эти организмы оказываются захваченными более круп-
ными животными, которые «пасутся» на грунте, или
же илоедами, пропускающими грунт с фораминифе-
рами через свой кишечный тракт. Наряду с ними
найдены стенофаги, питающиеся также и форамини-
ферами (Lipps, 1983). Если крупные донные живот-
ные заглатывают фораминифер целиком, то мелкие
проделывают в раковине отверстия (Baumfalk et al.,
1982; Sharma, Kumar, 1987).
Пищевой фактор влияет как на горизонтальное, так
и на вертикальное распространение фораминифер, а
также на их размеры. Так, например, в сублиторали,
где пищи больше, количество фораминифер возраста-
ет, при этом они обычно крупнее, чем на литорали.
Без сомнения, существует прямая зависимость
между богатством фауны фораминифер и общей вы-
сокой биологической продуктивностью района, на-
пример зоны апвелинга (Басов, 1978, 1979).
Выявлена зависимость роста раковин форамини-
фер от сезонных и широтных изменений количества
пищи. Например, у Elphidium crispum в высоких ши-
ротах в «голодные» зимние месяцы нарастающие но-
вые камеры намного мельче, короче, с меньшей ор-
наментацией, чем выросшие в летнее время (Myers,
1943). Это подтверждается и лабораторными опыта-
ми (Murray, 1963). Недостаток пищи может, видимо,
вызвать появление неправильных, уродливых камер
и даже раковин.
Обладая высокой численностью и соответственно
довольно большой биомассой, особенно в высокоши-
ротных экосистемах, где отмечено возрастание доли
мейобентоса при общем уменьшении биомассы дон-
ной фауны (Голиков, Аверинцев, 1977), форамини-
феры играют в биоценозе значительную роль и, без
сомнения, являются важным промежуточным трофи-
ческим звеном для всей донной фауны. z
Влияние грунта и химизма среды
Характер грунта, его состав, размеры, степень за-
иленности, содержание в нем СаСО3 влияют на жиз-
недеятельность фораминифер. Известно, что виды с
агглютинированной раковиной обладают избиратель-
ной способностью при ее построении и, следова-
тельно, находятся в большей зависимости от грунта,
чем формы с секреционной раковиной. Некоторые
авторы отмечают предпочтительное расселение и
для ряда видов с известковой раковиной на опреде-
ленных субстратах. Так, например, Verneuilinulla ad-
vena (Leslie, 1965; Vilks, 1968), Cribroelphidium lon-
gipontis (Myers, 1943; Atkinson, 1971), Cibicides re-
fulgens (Morishima, 1948; Pujos, 1972), Angulogerina
angulosa (Pujos, 1972) встречены преимущественно
на крупнопесчанистых грунтах. Виды же Buliminella
elegantissima, Islandiella norcrossi (Leslie, 1965), Eg-
gerelloides scabra, Uvigerina peregrina (Bouysee, Le
Calvez, 1967) обитают на глинистых или илистых
субстратах. В то же время Дигас (1980) в Баренце-
вом море отмечает максимальную концентрацию фо-
раминифер с известковой раковиной на песках и
илистых песках, тогда как чистые илы содержат ма-
лое количество видов с секреционной раковиной. На
строение, величину, толщину известковых раковин и
на их распространение в большой степени влияет
количество карбоната кальция в грунте и концентра-
ция его в придонном слое воды. На больших глуби-
нах (более 3000 м), где содержание растворенного
СаСО3 невелико, а содержание углекислоты повы-
шено, происходит растворение раковин планктон-
ных фораминифер, выпавших в осадок (Беляева,
1969; Саидова, 1969). Там же иногда обнаруживают-
ся живые бентосные формы с известковой ракови-
ной. Дигас (1980) показала, что в Баренцевом море
численность известковых раковин бентосных фора-
минифер значительно возрастает с востока на запад
в десятки и сотни тысяч раз. Численность же только
живых фораминифер лишь в 2.5 раза превышает
аналогичные показатели для восточной части моря
(Корсун и др., 1994), что можно объяснить высокой
скоростью растворения пустых известковых раковин
в восточной части Баренцева моря и низкой интен-
сивностью этого процесса на западе. Процесс раст-
ворения карбонатного материала наиболее интен-
сивен в поверхностном, окисленном слое осад-
ков, поэтому вышележащая водная толща пересыщена
по кальциту, а нижележащие осадки благоприятны
для сохранения известковых скелетов (Корсун и др.,
1994).
Кроме того, от величины частиц грунта, его за-
иленности зависит характер и количество пищи, при-
годной для фораминифер.
Величина pH, содержание растворенного в воде
кислорода, микроэлементов, остатков на грунте орга-
нических веществ, освещенность, взмученность вод-
ной толщи и многие другие факторы оказывают су-
щественное воздействие на распространение и чис-
ленность фораминифер.
Помимо этого, горизонтальные и вертикальные
течения оказывают огромное влияние на расселение
бентосных (перенос гамет) и планктонных видов.
Глубина обитания
С увеличением глубины снижается температура,
увеличивается давление, уменьшается освещенность
а следовательно, отчасти и количество пищи, кисло-
рода, карбоната кальция. Все это довольно подробно
было рассмотрено ’ выше. Следовательно, можно за-
ключить, что глубина оказывает влияние на рост,
размножение, горизонтальное и вертикальное рас-
пространение фораминифер. Кроме того, некоторые
авторы утверждают, что с увеличением глубины на-
блюдается тенденция к округлению раковины, увели-
чению ее размеров и орнаментации (Bandy, 1960,
1963, 1964). Однако существуют и противоположные
мнения, суть которых сводится к тому, что с увели-
ТИП FORAMINIFERA. ЭКОЛОГИЯ И ЗООГЕОГРАФИЯ
627
чением глубины происходит уменьшение раковины и
исчезновение орнаментации (Frerichs, 1970).
Количество видов с известковой раковиной на глу-
бинах свыше 4000—5000 м уменьшается, так как со-
кращается содержание в воде растворенного карбоната
кальция и увеличивается количество углекислоты. Но
все же в Тихом океане на глубинах порядка 5000 м,
т. е. ниже критических, нам удалось обнаружить из-
вестковые раковины Pyrgo serrata, Fissurina alveolata,
F. cucullata, Oolina longispina и других (Лукина,
1980).
Несомненно, что на больших глубинах возможен
процесс кремнезацйи (замещения кальция кремни-
ем). Так, еще Брэди (Brady, 1884) утверждал, что на
больших глубинах виды рода Miliolina имеют крем-
ниевую раковину, а на малых те же виды выделяют
известковый скелет. Ныне живущие виды форамини-
фер с кремниевой раковиной отмечены только на
больших глубинах.
Зоогеография
При составлении зоогеографической характе-
ристики фораминифер — этой наиболее массовой и
всесветно распространенной группы морских про-
стейших — было использовано батиметрическое де-
ление океана на следующие фаунистические зо-
ны: супралитораль, литораль, сублитораль, батиаль,
абиссаль и ультраабиссаль (Виноградова, 1977).
Супралитораль — зона на границе моря и
суши, располагается выше уровня максимального
прилива. Она только эпизодически покрывается
водой при ветровых нагонах или же лишь заплески-
вается морским прибоем. Это зона неблагоприятна
для жизни фораминифер, поскольку они не могут
выносить постоянного пересыхания.
Литораль — приливно-отливная зона моря, на-
ходится между уровнем максимального прилива и от-
лива. Эта зона также подвергается сильному суточно-
му и сезонному колебанию температуры и постоянно
находится под сильным влиянием приливно-отлив-
ных и ветровых течений, часто с пониженной соле-
ностью (из-за впадения рек или таяния льдов). В ли-
тературе существуют данные о том, что некоторые
виды фораминифер могут переносить частичное пе-
ресыхание в течение 2—4 ч (например, виды родов
Peneroplis, Cabcarina, Baculogypsina, — Fischer,
1966). Постоянно изменяющиеся условия литорали,
резко различающиеся в разных широтных поясах, оп-
ределяют своеобразие фауны, состоящей из сравни-
тельно небольшого числа видов, но с довольно высо-
ким процентом эндемичных форм.
В северных умеренных широтах из фораминифер
на литорали можно встретить Reophax curtus, Атто-
tium cassis, Verneuilinulla advena, Cribroelphidium lon-
gipontis, Miliammina fusca, Jadammina macrescens
(Лукина, 1985a, 19856, 1988, 1990).
Сублитораль — зона морского дна, соответ-
ствующая шельфу, или материковой отмели, — от
нижней границы литорали до глубины 200 м (грани-
ца шельфа). В Антарктиде эта зона опускается до
глубины 500—800 м, хотя максимум видового разно-
образия приходится на глубину 300—400 м. Эта ба-
тиметрическая зона неоднородна и подразделена на
верхний, средний и нижний отделы.
В верхнем отделе сублиторали, границу которого
разные авторы проводят в районе отметки 20—25 м
(Phleger, 1960, 1965; Murray, 1973, и др.), еще чувст-
вуется влияние сильного перемешивания воды. Фауна
фораминифер верхнего отдела сублиторали представ-
лена группой литоральных видов, обогащенная теми
формами, которые не могут жить в неблагоприятных
и изменчивых условиях литорали. Например, в рай-
оне архипелага Земли Франца-Иосифа можно встре-
тить не только Reophax curtus, Ammotium cassis, но и
Lobatula lobatula, Astrononion gallowayi, Cribroelphi-
dium bartletti, C. hollandense, C. clavatum и других
(Лукина, 1977).
Кроме того, по наблюдениям ряда авторов (Фур-
сенко, Фурсенко, 1970; Лукина, 1985а), расселение
фораминифер в акваториях с сильным перемешива-
нием водных масс в значительной степени зависит от
характера грунта, и особенно от того или иного водо-
рослевого субстрата, на котором живут фораминифе-
ры (ризоиды Laminaria, Cystoseira, Ulva, Rhodomella,
Ahnfeltia, Zostera) (Goldstein, 1999).
Границу между средним и нижним отделами суб-
литорали разные авторы проводят на основании из-
менения видового состава и количества форамини-
фер на глубинах 50, 70—125 м (Phleger, 1960); 35,
55, 75—85, 120, 180 м (Phleger, 1965); 80, 180 и
350 м (Murray, 1973).
Фауна фораминифер среднего отдела шельфа
наиболее богата, так как глубины 50—100—150 м
обычно в высшей степени благоприятны для всех
процессов жизнедеятельности фораминифер (тем-
пература, соленость, освещенность, кислород, пища,
pH, содержание карбоната кальция и т. д.). Здесь
можно встретить представителей родов Textularia,
Trochammina, Saccammina, Haplophragmoides, Spi-
roplectammina, Triloculina и многих других. Кроме
того, в донных осадках можно найти и раковины
планктонных видов, поскольку глубины сублиторали
обычно приурочены уже к открытым участкам мо-
рей.
Глубже, в пределах нижнего отдела сублиторали,
появляются виды родов Cassidulina, Uvigerina, Marti-
notiella, Recurvoides, Glomospira, Triloculina и другие.
Батиаль — зона морского дна, соответствую-
щая континентальному склону — от границы кон-
тинентального шельфа (200 м) до глубины 3000 м.
Переходная зона между сублиторалью и батиалью
располагается от 200 до 500—1000 м. На границе
шельфа и склона проходит четкая фаунистическая
граница, приуроченная к материковому склону. Для
склона характерен больший угол падения (средний
наклон 6°, а на шельфе 0.2—1°). Это зона муть-
евых потоков, с мозаично расположенными грунта-
ми, перемешиванием грунтов, при этом отложение
мягких осадков происходит только в защищенных
впадинах, у подножий хребтов. Для батиали харак-
терны хорошая аэрация, слабые изменения темпера-
туры и солености, что способствует многообразию
фауны как макро-, так и мейобентоса. Батиаль так-
же подразделяется на 2 зоны; граница одной из них
628
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
проходит от глубины 300—400 до 1100 м, а дру-
гой — от 1100 до 1700 и.
Для батиальной зоны отмечены специфические
виды как фораминифер (Хусид, 1973), так и пред-
ставителей других групп морских организмов. В цен-
тральной части Тихого океана в пределах батиали
встречены фораминиферы родов Pyrgo, Amphicory-
na, Marginulina, Ceratobulimina, Cymbaloporella,
Cymbaloporetta, Bolivina, Gyroidina, Pulleftia и мно-
гие другие (Лукина, 1980).
Абиссаль — зона морского дна, соответствующая
глубинам океанического ложа (от 3000 до 6000 м).
На глубинах 3000 и 4500 м происходит самая резкая
смена фаун и не только на уровне видов, но и на
уровне более крупных таксонов. Для абиссальной
зоны характерны большая монотонность условий су-
ществования, ограниченность пищевых ресурсов, ма-
лое число видов макробентоса при общей количе-
ственной бедности. При этом фауна фораминифер
представлена довольно большим числом видов. Так,
в Тихом океане на глубинах 3000—5000 м отмечено
100 видов, относящихся к родам Oolina, Fissurina,
Pavonina, Thurammina, Rhabdammina, Astrorhiza,
Cyclammina, Technitella, Psammosphaera и другим
(Лукина, 1980). В абиссальной зоне Атлантического,
Тихого и Индийского океанов обнаружен прибли-
зительно один и тот же комплекс глубоководных
видов. При этом интересно отметить, что виды,
встреченные на глубинах батиали и абиссали в
Тихом и Атлантическом океанах, в Арктике и Ан-
тарктике, как правило, обитают на меньших глуби-
нах, что, видимо, связано с достаточно низкими тем-
пературами этих относительно мелководных участков,
которые соответствуют глубоководным температур-
ным показателям в Атлантическом и Тихом океанах.
Глубже 5000 м видовой состав фауны форамини-
фер заметно обедняется, особенно за счет массового
растворения раковин известковых видов. Граница
критической глубины проходит приблизительно на
уровне 3200 м на широте 40° северного и южного
полушарий и по изобате 4800 м — в тропиках. Это
критическая точка, глубже которой СаСО3 в услови-
ях низкой температуры, высокого давления и повы-
шенного содержания углекислоты уже не может су-
ществовать в виде нерастворенного вещества (Беля-
ева, 1969). Хотя отмечены случаи нахождения в
тропической части Тихого океана большого коли-
чества пустых раковин планктонных фораминифер
хорошей сохранности вплоть до глубины 5000 м
(Лукина, 1980).
Ультраабиссаль — зона наибольших океани-
ческих глубин (6000—И 000 м). Для этой зоны ха-
рактерно общее обеднение фауны как макро-, так и
мейобентоса. Среди фораминифер на этих глубинах
в Тихом океане отмечено лишь 4 вида с агглютини-
рованной раковиной — Hyperammina friabilis, Н. по-
vaezealandiae, Nodosinum gaussicum, Alveolophragmi-
um pussilum (Лукина, 1980).
Количество донной фауны зависит от состава и
распределения донных осадков, макро- и микрорель-
ефа дна, а также от системы придонных и даже по-
верхностных течений, влияющих на распределение
жизни во всей толще воды (Филатова, 1977). Между
продуктивностью поверхностных вод океана и количест-
вом органического вещества в донных осадках, служа-
щих пищей донной фауне, существует прямая связь.
В разных географических зонах и глубинных диа-
пазонах Мирового океана можно проследить не толь-
ко изменение видового состава фауны фораминифер,
но и численность отдельных видов, и, как следствие
этого, изменение биомассы, которую составляют они
в общей системе биоценоза.
В литоральной зоне, как отмечалось выше, обыч-
но встречается ограниченное количество видов при
небольшом проценте живых особей, но численность
доминирующей формы достигает, как правило, очень
большой величины — 500 экз./10 см2, а биомасса —
0.8 г/м2 у берегов Мурмана (Корсун и др., 1994).
На глубинах шельфа во всех широтах Мирового
океана можно встретить наибольшее число видов
фораминифер, которые к тому же представлены и
наибольшим числом экземпляров, то есть образуют
наибольшую плотность поселения.42 По данным мно-
гих авторов (Phleger, 1951; Walton, 1955; Uchio,
1960), наибольший по сравнению с другими глубин-
ными зонами процент живых особей фораминифер
отмечен в пределах шельфа. Так, в верхнем отделе
шельфа наших арктических морей и архипелага
Земли Франца-Иосифа наиболее массовые виды —
Lobatula lobatula, Elphidiella frigida, Cribroelphidium
clavatum и другие достигают плотности поселения от
1 тыс. до 600 тыс. экз. в пересчете на 1 м2 поверх-
ности дна. При этом их биомасса равна 4—29 г/м2
(Лукина, 1977, 19856, 1988, 1990). У восточных бе-
регов Камчатки биомасса наиболее часто встречаю-
щихся видов составляет 20—40 г/м2. Шельфовая
зона тропических областей заселена также очень
большим количеством видов фораминифер, плот-
ность поселения и величина биомассы которых
очень велики.
В северо-западной части Индийского океана у
восточных берегов Индостана в сублиторальной зоне
биомасса фораминифер изменяется от долей граммов
до 17 г/м2 (Desai, Kutti, 1967), а наиболее высокая
численность живых бентосных фораминифер отмече-
на в этом районе в пределах глубин 2610—4400 м
(Хусид, 1988). При этом многие авторы указывают
на мозаичное расселение раковин фораминифер в
поверхностном слое осадка (Дигас* 1970; Троицкая,
1970; Olsson, Eriksson, 1974; Кириенко, 1977). В на-
стоящее время четкого объяснения этому явлению
еще нет, но, возможно, оно связано с характером
грунта и пищей. Величина «пятен мозаики» может
достигать 1—2 м в диаметре.
Количество раковин фораминифер на глубинах
батиали и абиссали в разных широтах и районах
Мирового океана неодинаково и зависит от многих
факторов. При относительно постоянных величинах
температуры, солености, высокого давления, pH, со-
держания кислорода и пищи на величину плотности
поселения батиальных и абиссальных видов влияют
течения, рельеф дна, скорость образования осадков
42 При подсчете фораминифер и их биомассы можно пользо-
ваться только данными о раковинах, содержащих плазму (т. е. про-
водить предварительное подкрашивание материала бенгальской
розовой краской).
ТИП FORAMINIFERA. ЭКОЛОГИЯ И ЗООГЕОГРАФИЯ
629
и содержание карбоната кальция. Обычно процент
живых особей фораминифер на глубинах 1000—
6000 м невелик. И объяснить это явление можно
«разбавляющим» эффектом быстрого процесса на-
копления осадков, когда на дно океана падает «дождь»
из оседающих взвешенных в воде частиц, в том числе
из пустых раковин планктонных фораминифер, насе-
ляющих в огромном количестве толщу водной массы
(50—1000 м). В тропических областях образуется тол-
стый слой осадков из глобигериновых раковин.
Придонные течения переносят с места на место
песчаный грунт с раковинами фораминифер, концен-
трируя его у подножий водных хребтов и материко-
вых склонов.
Абиссальная зона характеризуется ограничен-
ностью пищевых ресурсов и соответственно малым
количеством видов. Биомасса макробентоса в откры-
тых районах океана в умеренных широтах на глуби-
нах 3000—4000 м колеблется от 0.1 до 1 г/м2. На
фоне общего обеднения, фауны с увеличением глуби-
ны значительно возрастает роль фораминифер в со-
здании биомассы глубоководного бентоса (Зенкевич,
1967; Саидова, 1970). В Курило-Камчатском желобе,
по расчетам X. М. Саидовой, фораминиферы в сред-
нем содержат 2—3 г живой протоплазмы в пересчете
на 1 м2 при численности 18—56 тыс. экз./м2 живых
раковин. Это вполне сопоставимо с биомассой мно-
гоклеточных организмов.
Изучение большого фактического материала из
различных частей Тихого и Индийского океанов поз-
волило установить, что в зоне апвелинга у берегов
Перу и Чили биомасса фораминифер на отдельных
участках шельфа может достигать 100 г/м2, на конти-
нентальном склоне 5.8—70.3 г/м2, а в абиссальной
зоне этого района средняя величина биомассы снижа-
ется до 13 г/м2 (Хусид, 1977, 1978, 1988).
Сопоставление величин биомассы фораминифер и
макробентоса показало, что биомасса фораминифер
иногда превышает биомассу макробентоса. Особенно
значительна роль фораминифер в приконтинентальных
районах океана. У берегов Южной Америки (Тихий океан)
в сублиторали на долю фораминифер приходится до
50 % общей биомассы бентоса, в батиальной зоне до
70—90, в абиссальной до 20—80, а в ультраабиссаль-
ной до 50 % общей биомассы (Хусид, 1988).
Зоогеографическое районирование, проведенное
на основе анализа распространения многих групп
животных, неоднородности качественного состава
фауны фораминифер в разных широтах Мирового
океана, позволило выделить комплексы видов фора-
минифер, приуроченные к следующим поясам: ар-
ктическому и антарктическому, бореальному и анти-
бореальному, северному и южному умеренно-тепло-
водным и тропическому (Peres, 1961). Различия
между ними особенно наглядно проявляются на не-
больших глубинах. Так, на шельфе Арктики у бере-
гов Земли Франца-Иосифа и у побережья Кольского
полуострова среди массовых арктическо-бореальных
видов можно назвать Lobatula lobatula, Elphidiella
frigida, E. arctica, Cribroelphidium bartbetti, C, clava-
tum, Astrononion gallowayi (Лукина, 1977). В вос-
точном секторе Арктики на шельфе Восточно-Си-
бирского моря в группу массовых арктическо-боре-
альных видов входят Psammosphaera fusca, Reophax
subfusiformis, Ammotium cassis, Haynesina orbiculare
(Todd, Low, 1966; Лукина, 1978, 1994).
В пределах шельфа Берингова моря в бореальном
поясе уже иной состав группы доминирующих по
численности видов фораминифер — Cuneata arctica,
Verneuilinulla advena, Ammotium cassis, Spiroplectam-
mina biformis, Adercotryma glomerata, Islandiella is-
landica (Саидова, 1994).
Еще южнее, у южных берегов о-ва Сахалин и
южных Курильских островов, можно наблюдать сме-
шивание группы бореальных фораминифер с тепло-
водными формами. Наряду с бореальными видами
Miliammina fusca, Verneuilinulla advena, Trochammina
inflata можно встретить такие тепловодные формы,
как Textularia conica, Cribroelphidium fax, C. reti-
culosum, E. jensoni, Pararotalia nipponica, Ammonia
baccari (Лукина, 1975).
На шельфе заливов Желтого моря среди наиболее
широко распространенных видов на долю бореаль-
ных форм (Verneuilinulla advena, Ammobaculites ех-
iguus) приходится небольшой процент. В подавляю-
щем же большинстве фауна состоит из тепловодных
видов — Trochammina asiatica, Tiphotrocha convexa-
concava, Lagena perlucida, L. semilineatq (Щедрина,
Лукина, 1984).
Особенно резкое изменение в составе фауны фо-
раминифер можно наблюдать при переходе к тропи-
ческому поясу. На малых глубинах в Тихом океане в
районе экватора в массовом количестве обитают Ver-
tebralina striata, Peneroplis planatus, Marginopora ver-
tebralis, M. plicata, Alveolinella guoyi, Tinoporus spen-
gleri (Лукина, 1979).
Таким образом, приблизительно на одних и тех
же глубинах шельфа в разных широтах можно про-
следить постепенное изменение состава фауны от
арктической, арктическо-бореальной до тропической.
С увеличением глубины уменьшаются различия фи-
зическйх характеристик водных масс и видовой со-
став фауны фораминифер склона и ложа во всех
океанах вне зависимости от географических зон имеет
много общих форм. Следовательно, с уменьшением
глубин увеличивается эндемизм, а космополитизм же
характерен для глубоководных видов фораминифер.
Среди общих закономерностей горизонтального
распространения фораминифер следует отметить уве-
личение количества видов по направлению к экватору,
богатство и разнообразие фауны Антарктики и своеоб-
разие видового состава районов коралловых рифов.
Изучение видового состава современных форами-
нифер и закономерностей их распространения очень
важно при составлении палеокомплексов, так как по-
могает выяснить глубины, температуру обитания
предков современных видов, точнее установить воз-
раст отложений. Кроме того, фораминиферы, являясь
весьма многочисленной группой простейших, заселя-
ют все биологические ниши Мирового океана, а в не-
которых районах Арктики и Антарктики их можно
рассматривать в качестве главных компонентов мейо-
бентоса (фауны, средний размер особей которой не
превышает 1 мм); по своей роли в биоэнергетике
донных экосистем иногда они могут превосходить
организмы макробентоса.
630
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ЛИТЕРАТУРА43
Басов И. А. Связь количественного распределения бентосных
фораминифер на западном шельфе Африки и условия их обита-
ния // 2-я Всесоюз. конф, по биологии шельфа. Ч. 1. Киев, 1978.
С. 16—17.
Басов И. А. Экология бентосных фораминифер в зоне апвел-
линга у Юго-Западной Африки // Вопр. микропалеонтол. / ГИН
АН СССР. 1979. Вып. 22. С. 135—146.
Басов И. А., Хусид Т. А. О биомассе бентосных фораминифер
в осадках Охотского моря И Океанология. 1983. °Т. 23, вып. 4.
С. 648—655.
Беляева Н. В. Планктонные фораминиферы в поверхностном
слое осадков Тихого океана // Микрофлора и микрофауна в совре-
менных осадках Тихого океана. М., 1969. С. 73—119.
Виноградова Н. Г. Фауна шельфа, материкового склона и
абиссали // Биология океана. Т. 1. Биологическая структура океа-
на. Океанология. 1977. С. 178—198.
(Голиков А. Н., Аверинцев В. Г.) Golikov A. N., Averin-
cev V. G. Distribution patterns of benthic and ice biocoenoses in the
high latitudes of the Polar Basin and their part in the biological struc-
ture of the World Ocean // Polar Oceane / Ed. M. J. Dwinvebar. Cal-
gary, 1977. P. 331—363.
Дигас Л. А. Зоогеографическое районирование Баренцева мо-
ря по фораминиферам // Вопросы геологии Южного Урала и По-
волжья. Вып. 6. Ч. II. Кайнозой. Саратов, 1970. С. 127—142.
Дигас Л. А. Количественное распределение известковых бен-
тосных фораминифер в современных осадках Баренцева моря И
Палеонтологические исследования на Украине: Материалы 1-й
годич. сес. Украин. палеонтол. о-ва. Киев, 1980. С. 66—77.
Зенкевич Л. А. 39-й рейс (НИС «Витязь») И Природа. 1967.
№ 2.
Кириенко Е. А. О комплексах фораминифер в донных отло-
жениях Белого моря // Стратиграфия и палеогеография четвертич-
ного периода Севера Европейской части СССР. Петрозаводск,
1977. С. 40—47.
Корсун С. А., Погодина И. А., Тарасов Г. А., Матишов Г. Г. Фо-
раминиферы Баренцева моря (гидробиология и четвертичная па-
леоэкология) Мурм. Морск. биол. ин-т. Кольск. науч, центр. Апа-
титы, 1994.
Лукина Т. Г. О фауне фораминифер малых глубин Южного
Сахалина // Образ жизни и закономерности расселения современ-
ной и ископаемой микрофауны. М., 1975. С. 85—89.
Лукина Т. Г. Фораминиферы верхних отделов шельфа у архи-
пелага Земля Франца-Иосифа // Исслед. фауны морей СССР. 1977.
Т. 14 (22). С. 72—105.
Лукина Т. Г. Фораминиферы Новосибирского мелководья //
2-я Всес. конф, по биологии шельфа. Ч. 2. Киев, 1978. С. 66—67.
Лукина Т. Г. Зависимость распределения фораминифер от глу-
бины в различных географических зонах Тихого океана // Матери-
алы совещ. Хабаровск, 1979. С. 86—87.
Лукина Т. Г. Глубоководные фораминиферы центральной части
Тихого океана // Исслед. фауны морей СССР. 1980. Т. 24 (32).
С. 3—203.
Лукина Т. Г. Фораминиферы верхнего отдела шельфа губы
Чупа (Кандалакшский залив, Белое море) И Исслед. фауны морей
СССР. 1985а. Т. 31 (39). С. 109—119.
Лукина Т. Г. Фораминиферы Онежского залива и сопредель-
ного бассейна Белого моря И Исслед. фауны морей СССР. 19856.
Т. 33 (41). С. 113—126.
Лукина Т. Г. Фораминиферы из района Сонострова (Канда-
лакшский залив, Белое море) // Исслед. фауны морей СССР. 1988.
Т. 38 (46). С. 4—30.
Лукина Т. Г. Фораминиферы Новосибирского мелководья //
Исслед. фауны морей СССР. 1990. Т. 37 (45). С. 105—129.
Лукина Т. Г. Фораминиферы Чаунской губы Восточно-Сибир-
ского моря // Экосистемы и фауна Чаунской губы и сопредельных
вод Восточно-Сибирского моря // Исслед. фауны морей. 1994.
Т. 48 (56). С. 98—115.
Преображенская Т. В., Троицкая Т. С. Литоральные форами-
ниферы как показатель гидродинамики среды И Биология моря.
1991. № 4. С. 17—26.
Саидова X. М. Распространение и условия обитания современ-
ных бентосных фораминифер в Тихом океане И Микрофлора и
43 Литература к разделу «Тип Foraminifera» Экология и зоогео-
графия фораминифер».
микрофауна в современных осадках Тихого океана. М., 1969.
С. 120—193.
Саидова X. М. Бентосные фораминиферы района Курило-
Камчатского желоба по материалам 39-го рейса НИС «Витязь» //
Тр. ИОАН СССР. М., 1970. Т. 86.
Саидова X. М. Экология шельфовых сообществ фораминифер
и палеосфера голоцена Берингова и Чукотского морей И ИОРАН.
М., 1994. С. 93.
Троицкая Т. С. Условия обитания и распределение форами-
нифер в Японском море (семейства Elphidiidae, Cassidulinidae и
Islandiellidae) // Тр. ИГиГ СО АН СССР. 1970. Вып. 7. С. 136—
161.
Филатова 3. А. Глубоководная донная фауна И Биология океа-
на. Т. 1. Биологическая структура океана. Океанология. 1977.
С. 88—96.
Фурсенко А. В. Подкласс Фораминиферы. Общая часть // Ос-
новы палеонтологии. Справочник для палеонтологов и геологов
СССР. Т. 1. Простейшие. М.: Изд-во АН СССР, 1959. С. 115—
168.
Фурсенко А. В., Фурсенко К. Б. О фораминиферах лагуны
Буссе и условиях их существования // Тр. ИГиГ СО АН СССР.
1970. Вып. 71. С. 114—135.
Хусид Т. И. Распределение бентосных фораминифер в заливе
Аляски и восточной части Алеутского желоба // Тр. ИОАН СССР.
1973. Т. 91.
Хусид Т. А. Биоценозы бентосных фораминифер в Перуанско-
Чилийском желобе // Тр. ИОАН СССР. 1977. Т. 108. С. 25—36.
Хусид Т. И. Биомасса и количественное распределение бен-
тосных фораминифер в северо-западной части Индийского океа-
на И Морская микропалеонтология. М.: Наука, 1978. С. 185—191.
Хусид Т. А. Роль фораминифер в бентосе океана И Морская
стратиграфия и микропалеонтология. М.: Наука, 1988. С. 101—
114. <
Щедрина 3. Г., Лукина Т. Г. Фораминиферы Тихоокеанского
шельфа юго-восточной Азии / ЗИН АН СССР. Л., 1984.
Alexander S. F., De Laca Т. Е. Feeding adaptations of the fora-
miniferan Cibicides refulgens living epizoically and parasitically on
the antarctic scallop Adamussium colbecki // Biol. Bull. 1987.
Vol. 173. P. 136—159.
Atkinson K. The relationship of Recent foraminifera to the sedi-
mentary facies in the turbulent zone Cardigan Bay, Waler // J. Nat.
Hist. 1971. Vol. 5, N 4. P. 385—439.
Bandy O.L. General correlation of foraminiferal structure with
environment // Inter. Geol. Congr., 21st. Copenhagen, 1960. Vol. 22.
P. 7—19.
Bandy O. L. Dominant paralic Foraminifera of Southern Califor-
nia and the Gulf of California // Contr. Cushman Found. Foram. Res.
1963. Vol. 14, pt 4, N 269. P. 127—134.
Bandy O. L. Foraminiferal trends associated with deep-water
sands. San Pedro and Santa Monica basins, California // J. Paleontol.
1964. Vol. 38. P. 138—148.
Baumfalk Y. A., Fortuin A. R., Mok R. Ph. Talpinella cunicula-
ria n. gen. n. sp., a possible foraminiferal parasite of late cretaceous
Orbitoides // J. Foram. Res. 1982. Vol. 12. N 3. P. 185—196.
Be A. W. H. Shell porosity of Recent planktonic foraminifera as a
climatic index // Science. 1968. Vol. 161. P. 881—884.
Be A.W. H. Orbulina universa d’Orbigny-foraminiferal indicator
species of latitudinal occurences in the Indian Ocean // Inter. Geol.
Congr., 24th. Montreal, 1972. Vol. 7. P. 4.
Be A. W. H., Tolderlund D. S. Distribution and ecology of living
planktonic foraminifera in surface waters of the Atlantic and Indian
Oceans // The Micropaleontology of the Oceans. Cambridge: Univ.
Press, 1971. P. 105—149.
Bernhard J. M. The distribution of bentic Foraminifera with res-
pect to oxygen concentration and organic carbon levels in shallow
water Antarctic sedimenta // Limnol. and Oceanogr. 1989. Vol. 34,
N 6. P. 1131—1141.
Boltovskoy E. Planktonic Foraminifera as indicators of different
water masses in the South Atlantic // Micropaleontology. 1962. Vol. 8,
N 3. P. 403—408.
Boltovskoy E. Depth at which Foraminifera can survive in sediments //
Contr. Cushman Found. Foram. Res. 1966a. Vol. 17, pt 2. P. 43—45.
Boltovskoy E. The subtropical/subantarctic zone of convergence
(Atlantic Ocean, western part) // Serv. Hidrogr. Nat., Argentina.
1966b. H. 640. P. 1—69.
ТИП FORAMINIFERA. ЭКОЛОГИЯ И ЗООГЕОГРАФИЯ. ЛИТЕРАТУРА
631
Boltovskoy Е., Lena Н. Microdistribution des Foraminiferes ben-
thoniques vivants // Micropaleontol. 1969. Vol. 12, N 3. P. 177—185.
Boltovskoy E., Wright R. Recent Foraminifera. The Hague, 1976.
Bouysee Ph., Le Calvez Y. Etude des fonds marins compris entre
Penmarch et Groix (Sud-Finistere) // Bull. Bur. rech. geol et. Minteres.
1967. Vol. 2. P. 39—73.
Bradshaw J. S. Laboratory experiments on ecology of Foramini-
fera H CCFFR. 1961. Vol. 12, N 3. P. 87—106.
Brady H. B. Report on the Foraminifera collected by H. M. S. Chal-
lenger during the years 1873—1876 // Rep. Sci. Res. Exp. Voyage
H. M. S. Challenger. Zool. 1884. Vol. 9. P. 1—814.
Brodniewicz I. Recent and certain Holocene Foraminifera of the
Southern Baltic Sea // Acta Paleontol. Polon. 1965. Vol. 10, N 2.
P. 131—236.
Buzas M. A. The distribution and abundance of Foraminifera in
Long Island Sound // Smithsonian Inst. Misc. Colin. 1965. Vol. 149,
pt 1. P. 1—87.
Cedhagen T. Phoxichilidium tubulariae, en Daligt kand avenak
Havsspindel // Goteborge Naturhistoriska Musei Arstryck. 1988.
P. 35—37.
Corliss В. H., Emerson S. Distribution of Rose Bengal stained
benthic Foraminifera from Nova Scotian continental margin and Gulf
of Maine // Deep-Sea Res. 1990. Vol. 37, N За. P. 381—400.
De Laca T. E. Trophic position of benthic rhizopoda in McMurdo
Sounds // Antarct. J. U. S. 1985. Vol. 19, N 5. P. 147—149.
De Laca T. E. The morhpology and ecology of Astrammina ra-
ra // J. Foram. Res. 1986. Vol. 16, N 3. P. 216—223.
De Laca T. E., Lipps J. H. The mechanism and adaptive signifi-
cance of attachment and substrate pitting in the foraminiferan Rosalina
globularis d’Orbigny // J. Foram. Res. 1972. Vol. 2, N 2. P. 68—72.
Desai B. N., Kutti M. Studies on the benthic fauna of Cochin
back-water// Proc. Ind. Acad. Sci. Ser. B. 1967. Vol. 46, N 4.
P. 123—142.
Fischer P. H. Ecologie de certain foraminiferes du haut niveau sur
les cotes du Pacifique // Bull. zool. France. 1966. Vol. 91, N 3.
P. 295—300.
Frerichs W. E. Distribution and ecology of benthonic foraminife-
ra in the sediments of the Andaman Sea // Cushman Found. Foram.
Res. Contr. 1970. Vol. 21, N 4. P. 123—147.
Goldstein S.T. Foraminifera: A biological overview // Modern
Foraminifera. Dordrecth etc.: Kluwer Acad. Publ., 1999. P. 37—
55.
Kitazato H. Locomotion of some benthic foraminifera in and on
sediments II J. Foram. Res. 1988. Vol. 18, N 4. P. 344—349.
Kurc G. Foraminiferes et Ostracodes de L’etange de Thau // Inst.
Peches Marit., Rev. Trav. 1961. Vol. 25. P. 134—248.
Langer M., Hottinger L., Huber B. Functional morphology
in low-diverse benthic foraminiferal assemblages from tidal flats on
the North Sea // Senckenberg. marit. 1989. Vol. 20, N 3—4. P. 81—
99.
Le Calvez J., Le Calvez Y. Contribution a 1’etude des Foramini-
feres des eaux saumatres. I. Etange de Canet et des Salses // Vie et
Milieu. 1951. P. 237—254.
Lee J. J., МсЕпегу M.E. Autogamy in Allogromia laticollaris
(Foraminifera) // J. Protozool. 1970. Vol. 17, N 2. P. 184—195.
Leslie R. J. Ecology and paleoecology of Hudson Bay Foramini-
fera // Bedford Inst. Oceanogr. Rept. 1965. Vol. 65, N 6. P. 192.
Lipps J. H. Biotic interactions in benthic foraminifera // Biotic In-
teractions in Recent and Fossil Benthic Communities I Eds
M. J. J. Tevez, P. L. McCall. New York, 1983. P. 331—376.
Loeblich A. R., Jr., Tappan H. Forminiferal genera and their
classification. New York, 1987. Vol. 2.
Lutze G. F. Variationsstatistik und Okologie bei rezenten Forami-
niferen // Paleontol. Zt. Sch. 1962. Bd 36. S. 252—264.
Moodley L. Southern North Sea seafloor and subsurface distribu-
tion of living benthic foraminifera // Netherlands J. Sea Res. 1990.
Vol. 27, N 1. P. 57—71.
Morishima M. Foraminiferal thanatocoenosis of Ago Bay, Kii
Peninsula, Japan // Rep. Comm. Treat. Mar. Ecol. Paleoecol. 1948.
Vol. 7. P. 89—91.
Murray J. W. Ecological experiments on Foraminiferida // J. Mar.
Biol. Assoc. U. K. 1963. Vol. 43, N 3. P. 621—642.
Murray J. W. Living foraminifera of lagoons and estuaries //
Micropaleontology. 1968. Vol. 14, N 4. P. 435—455.
Murray J. W. Distribution and ecology of living benthic forami-
niferids. New York, 1973.
Myers E. H. Life activities of Foraminifera in relation to marine
ecology // Amer. Phil. Soc. Proc. 1943. Vol. 86, N 3. P. 439-458.
Olsson J. I., Eriksson B. Horizontal distribution of meiofauna
within a small area, with special reference to Foraminifera // Zoon.
1974. Vol. 2. P. 67—84.
Peres J. M. Oceanographie biologique et biologie marine. Tomo
1. La vie benthique. Paris: Press. Univ. France, 1961.
Phleger F.B. Foraminifera distribution. Ecology of Foraminifera
northwest Gulf of Mexico // Geol. Soc. Amer. Mem. 1951. Vol. 46. P. 1—
88.
Phleger F. B. Ecology and distribution of recent Foraminifera.
Baltimore, 1960.
Phleger F.B.Depth patterns of benthonic Foraminifera in the
Eastern Pacific // Progr. Oceanogr. 1965. Vol. 3. P. 273—287.
Pujos M. Repartition des biocoenoses de Foraminiferes benthiqu-
es sur le plateau continental du Golfe de Gascogne a 1’ouest de
1’embouchure de la Gironde // Rev. Espan. Micropaleontol. 1972.
Vol. 4, N 2. P. 141—156.
Rhumbler L. Die Foraminiferen (Thalamophoren) der Plankton
Expedition. Ergebnisse // Plankton-Exped. Humboldt-Stifftung. 1911.
Bd 3. S. 1—476.
Schaudinn F. Untersuchungen an Foraminiferen. I. Calcituba po-
lymorpha // Ztschr. Wiss. Zool. 1895. Bd 59. S. 191—232.
Schultze M. S. Uber den Organismus der Polythalamien (Forami-
niferen) nebst Bemerkungen uber die Rhizopoden in Allgemeinen.
Leipzig: Engelmann, 1854.
Sharma V., Kumar R. Significance of benthic foraminiferal pre-
dation in species diversity // Curr. Sci. (India). 1987. N 18. P. 948—
949.
Smyth M.J. The foraminifer Cymboloporella tabellaeformis
(Brady) bores into gastropod shells // J. Foram. Res. 1988. Vol. 18,
N 4. P. 277—285.
Todd R., Low D. Foraminifera from the Arctic ocean off the
Eastern Siberian coast // Geol. Sur. Res. U. S. Geol. Sur. Prof. Pap.
1966. Vol. 550-C. P. 79—85.
Uchio T. Ecology of living benthonic Foraminifera from the San
Diego area // Cushman Found. Foram. Res. Spec. Pap. 1960. Vol. 5.
P. 1—72.
Venec-Peyre M. T. Two new species of bioeroding Trochammini-
dae (Foraminiferida) from French Polynesia // J. Foram. Res. 1988.
Vol. 18, N 1. P. 1—5.
Vilks G. Foraminiferal study of the Magdalen Shallows, Gulf of
St. Lawrence // Maritime Sediments. 1968. Vol. 4, N 1. P. 14—21.
Voorthuysen J. H. Die Foraminiferen des Dollart-Ems-Estuari-
um // Verh. Kon. Ned. Geol. Minjb. Gen., Geol. 1960. Bd 19.
S. 237—269.
Waller H. O., Polski W. Planktonic Foraminifera of the Asiatic
shelf // Cushman Found. Foram. Res. Contr. 1959. Vol. 10, N 4.
P. 123—126.
Walton W. R. Ecology of living benthonic Foraminifera, Todos
Santos Bay, Baja, California // J. Paleontol. 1955. Vol. 29. P. 952—
1018.
Wollenburg J. E., Mackensen A. On the vertical distribution of
living (Rose Bengal stained) benthic Foraminiferes in the Arctic
Ocean // J. Foram. Res. 1998. Vol. 28, N 4.
Wright R. C. Miliolidae (Foraminiferos) recientes del estuarid del
Rio Quequen Grande (Prov. de Bs. As.) // Rev. Mus. Argentine Cienc.
Nat., Hidrobiol. 1968. Vol. 2, N 7. P. 225—256.
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ*
акантамебы 466
акантарии 170, 548, 558, 584
акантоециды 321, 332
акантоцефалы 116
акразиды 485
акразиевые 417, 485
акритархи 141
аксолотль 36—37
акулы 68, 78, 89, 94
алвеолаты 135
алыча 31, 116
альвеолиниды 544, 563, 575, 595
амбистомы 37
амебы 24, 123, 126, 130—131, 133, 136, 139—141, 143, 145—
146, 148, 154, 157, 159, 163—164, 166—167, 171, 173,
176—178, 180—184, 186—188, 269, 340, 404, 412, 415, 418,
420—421, 424—429, 444, 451—452, 454, 456—478, 482—
491, 493—495, 497—498, 500, 502—503, 507, 509—515,
522, 524, 528, 533, 558—559
аммодисканы 580, 599
аммодисциды 577, 589
амниоты 51
амфибии 36, 42, 47, 50—52, 65, 81—82, 89, 92, 187, 212, 221,
226, 229—230, 232, 239, 247—248, 297, 301—302, 307—
309, 311—312, 341, 355, 358, 363, 365, 368, 509
амфилиниды 62
амфистегины 563
аннелиды 37, 49—50, 53, 74, 93, 98, 102, 105, 107—109, 237
антилопы 248, 250
аппендикулярии 37
аптериготы 37
апузомонады 147, 164
артроподы 247
архамебы 128—129, 133, 142, 512
архебактерии 126, 142, 511
архезои 129, 133, 142, 512—513
археоциаты 76
архицеломаты 108
арцеллы 473
астазии 188, 202, 204, 206
астроризаты 539, 556, 577, 579
асцидии 20, 37
атаксофрагмииды 556, 578, 583, 602
аталамиды 136, 451, 520—522, 524
бабочки 32, 91
бактерии 32, 64, 71, 78—79, 89, 94, 138, 142, 154, 157, 167,
171, 173, 176—180, 183—186, 195, 207, 220, 226, 234, 242,
* Все указатели приведены в авторской редакции.
266, 268—269, 293, 314, 316, 321, 323, 325, 339, 344, 346,
348, 353, 356—357, 364, 379—380, 385—386, 388—389,
411, 413, 415, 425, 430—431, 437, 458-459, 466, 469, 472,
478, 488, 495, 502, 509—512, 534, 557—558, 560, 625
батисифоны 534
безлегочные 65
бесхвостые 36, 50, 247, 301, 307, 309, 358
бесчелюстные И, 43, 76
бикозоециды 135, 149, 173—174, 258, 262, 264—269, 316,
318—319
билатерии 108
блохи 249
бобры 28
бодониды 173, 176, 211—220, 222—223, 225—227, 236, 238—
241, 314, 317, 411, 416
бокоплавы 25
бородавочники 250
брахиаты ИЗ
брахиоподы 115
броненосцы 231
брюхоногие 26, 36, 41, 43, 54, 60, 65, 70, 76, 115, 227
булиминиды 543
бьековиниды 579
вакуолярии 285, 288
вампиреллиды 440
вампиры 235
верблюды 212, 249—250
веслоногие 59, 201, 209, 272
вирусы 94, 180, 188, 407
водомерки 243, 253
водоросли 24, 71, 74, 76, 123—125, 128, 135, 139—140, 146,
161—163, 166, 184—186, 207—208, 210, 226, 266, 270,
274—275, 281, 289—290, 295, 316—317, 327, 330, 407, 411,
413, 431, 458—459, 467, 469, 472, 478, 483, 488, 495, 497—
498, 502, 507, 534, 558—560, 625
вольвоциды 152
воротничковые жгутиконосцы 35, 49, 128, 134—135, 145—146,
158, 173, 186, 314, 321—323, 326—329, 331,. 336
вторичнополостные 21, 111
вторичноротые 53, 98, 100, 103, 108—109, 113, 115
высшие ракообразные 38
высшие термиты 386
гады 16
галлообразующие клещи 26
гаптомонады 317
гаптофиты 128, 134—135, 276—277, 280—283, 317, 548, 563
гастеромицеты 431
гастреады 100
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
633
гастроподы 227, 241, 560
гелиофлагеллаты 316
гельминты 25
гемогрегарины 56
гемоспоридии 56
гетеронематины 202
гетеростегины 548
гиббоны 93
гидра 91
гидроидные 25, 73, 534, 560
гимнофрииды 524
гипермастигиды 156, 162, 357—358, 368—369, 372, 375—381,
383—392
гипермастигины 154, 180
гирокотилиды 62
гистомонады 363—364
гифохитридиевые 135
глабрателлиды 534, 607
глобигеринаты 172, 552—553, 557, 561, 571, 575, 579—580
глобигериниды 536, 559
глоссины 233
гнатостомулиды 111
головастики 308, 310—311
головоногие 18, 26, 38, 45, 62, 76, 89, 111
голотурии 60, 518, 527
гоминиды 44
горбуша 94
грамположительные бактерии 226
гранулоретикулозеи 451, 524, 527
гранулоретикулозные амебы 136, 451, 522, 524
граптолиты 76
гребешки 560
гребневики 19, 45, 103, 106, 111
грегарины 56, 166
грибы 24, 79, 93, 123—125, 128, 130, 134—135, 142, 146, 154,
158, 164, 172, 176, 184, 290—291, 385, 398, 404, 407-409,
414, 417—419, 431, 440, 445—446, 459, 472, 495, 498, 528,
558
громия 503
грызуны 23, 46, 52, 66, 114, 230, 245—247, 250, 358, 361, 363,
368, 395
губки 19, 24—25, 45, 49, 76, 100, 103—104, 110, 115, 134, 321,
328—329, 336, 516—517, 519, 543, 548—549, 559, 624
гуси 14, 531
гусиные 363
двоякодышащие 70
двукрылые 26, 232—233, 235, 240, 243, 245, 247
двупарноногие 37
двустворчатые моллюски 46, 76, 560
девесковиниды 363, 366—367
дейтеростомии 101, 104—105
десятиногие 93
диатомовые 135, 145, 166, 173, 281, 471, 476, 516, 548, 559,
563, 625
динозавры 83
диномонады 155
динофилиды 37, 112
динофитовые 147—148, 158, 162, 169, 172—173, 180, 184
динофлагеллиды 149
динофлагелляты 219, 563
дипломонадеи 337—338, 341, 346
дипломонадиды 183, 337—338, 340—341, 347
дипломонады 126, 128, 130—131, 133, 139—141, 143, 154, 176,
341, 344—349
дискорбиды 536, 557
дискорбис 582
диффлюгии 173, 469, 472
дождевые черви 406
дрожжи 167, 300, 346, 419, 437, 534, 559
41 Протисты, часть 1
дрозды 12
дрозофилы 30—31
дуналиеллы 559
единороги 14
ежи 26
жаброногие 93
жабы 308
жаворонки 46
жгутиконосцы 24, 35, 49, 67, 123, 128—129, 134—135, 138,
140—141, 143, 145—146, 148—150, 152, 154, 158—159,
161—162, 167, 171, 173, 178—184, 186—187, 202, 204,
209—217, 219—221, 223—224, 226—236, 238, 240—243,
245, 247—248, 251—253, 260, 262—264, 266—269, 273—
277, 280—288, 290, 298—299, 301, 310—314, 316—317,
319, 321—323, 325—330, 332, 335—338, 340—341, 343—
351, 353—355, 357—360, 364, 366—369, 372—373, 378—
382, 384—392, 395, 404, 411—413, 416, 418, 436-437, 444,
446, 472, 486, 488—489, 512, 569
желто-зеленые водоросли 186
жесткокрылые 26, 509
жиардии 347
живородящие 62
жуки 97, 509
жуки-плавунцы 91
зайцеобразные 52
зеленые водоросли 72—73, 76, 125, 149, 155—156, 159, 161—
162, 195
земляные лягушки 308
земноводные 11, 26, 34, 50, 82, 114, 509
змеи 59, 62, 248, 312
золотистые водоросли 1, 275, 289
зооспоровые грибы 123—124, 135, 146, 290, 402, 404, 408
зоофиты 98
зоофлагеллаты 266, 316, 321, 411
зоохлорелла 458, 469
зубастые карповые рыбы 52
зубры 28, 46
иглокожие 18—19, 26, 35, 37, 48, 60, 75—76, 98, 534, 559—
560, 625
изоподы 518
иксодовые 26, 232, 248
инволютиниды 548, 578, 590
индейки 348, 366, 531
инфузории 18, 24, 35, 56, 59—60, 64, 67, 70, 123, 125, 128,
131, 140, 142, 145—148, 150—152, 154, 157—159, 161—
162, 167, 169—171, 173—174, 176—188, 231, 243, 321, 329,
458, 472, 478, 510—511, 513, 558, 567, 584, 625
ихтиозавры 40
кабаны 531
капреллиды 26, 559
кариобластеи 510
кассава 245
кассидулиниды 536
каулерпа 140
кета 94
кинетопластиды 24, 123, 125, 128, 149, 173—174, 180, 201—
202, 208, 211—223, 225—226, 229, 236, 238, 240—241, 243,
252, 297, 314, 316, 319
кистеперые 21, 38, 48, 50—51, 81
китообразные 27, 38, 40, 59, 79
кишечнодышащие 19, 105
кишечнополостные 18—19, 21, 35, 53, 59, 74—75, 91, 98,
100—101, 107—109, 111
клещи 11, 26, 112, 231—232, 235, 248, 477
клопы 66, 229, 233—235, 240, 243, 245, 249, 252—253
634
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
клопы-слепняки 245
книдарии 37, 40, 45, 59, 74—75, 103—104, 106, 111, 148
коалы 36
кобылки 59
козы 249—250
кокколитофориды 172, 276—277, 280—282, 534
коккосфериды 277, 281
кокосовые пальмы 245
кокцидии 56, 179—180
коловратки 16—17, 37, 48, 140, 231, 272, 459, 472
кольчатые черви 18—19, 25, 53, 66, 73, 75, 111, 341
комары 180, 243, 247—248
комокиации 520, 524—527
комокииды 519, 524—527
копеподы 559, 625
копытные 20, 27, 41, 61, 80
кораллы 25, 75, 432, 608
корнеголовые 36
корненожки 24, 185, 468, 478, 490, 500, 517—518
корнуспироидесы 534
костистые рыбы 56
краб-разбойник 54
кремневые губки 76
крестоцветные 407, 409
криптобии 179, 211—220, 222—229, 236, 238—239, 241—242
криптомонады 1, 128, 149, 155—156, 180, 194—198
криптофитовые 1, 134—135, 147, 158, 176, 184
кровососущие 232, 235—236, 238, 247—250
крокодилы 96, 231, 236, 248
кругоресничные инфузории 56, 59, 145, 171, 174, 176, 179, 188
крыложаберные 101
ксенофиофорий 516—520, 527, 581
ктенофорообразные 108
кузнечики 66
кумовые 26, 559
лабиринтодонты 50
лабиринтоморфы 290—291
лабиринтулы 145, 166
лазиодисциды 578
ланцетники 43
ластоногие 15, 27, 59
латимерия 21
лейшмании 157, 212, 215—216, 222, 224, 230, 233—235, 238—
239, 245—246, 252
ленивцы 36, 230—231, 246—247
ленточные черви 25, 62, 64
лептомиксиды 465
лептомонады 243, 251—252
летучие мыши 97, 232, 235, 249—250
лигеиды 245
литуолиды 544, 556, 575, 577—578, 583, 589
лофомонадины 368—369, 372, 375—378, 386
лошадиные 42, 81
лучистые 17—18, 541
лягинаты 552, 577
лягушки 89, 308
лямблии 180, 338, 346—347, 395
магнолия 66
мадрепоровые 75
малломонас 272
малощетинковые черви 25
мамонты 12, 81
маниот 245
масличные пальмы 245
мастигамебы 513
мастигофоры 150
мастодонты 38, 81
меандропсиниды 579
мегасфероморфы 73
медузы 18—19, 73
меритерий 81
меротрихи 285, 288
метазоа 117, 140, 144, 188, 321
метамонады 128—129, 141—142, 513
мечехвосты 38, 48, 50, 76
мизостомиды 112
микроспоридии 128, 131, 134—135, 138—139, 145—146, 154
миксамебы 417, 425, 436—440, 442
миксогастриды 424, 431—432, 434, 438—442
миксогастриевые 440, 445
миксомицеты 125, 135—136, 140, 157, 398, 408, 411—412,
417—419, 425, 431, 435—436, 440, 442, 445, 487, 521
миксоспоридии 24, 135, 188, 569
милиолиды 539, 558, 560, 562, 575, 578, 580
милиоляны 556—557, 579
милиоляты 539, 543, 548—549, 551, 556—557, 562—563, 569,
571, 575, 577—580, 582—583, 586, 588, 592
млекопитающие И, 16—17, 27, 34, 36, 38, 40—44, 46, 50—52,
57, 59, 61, 68, 76, 78—79, 82, 88, 91, 93, 97, 179, 212, 216,
221, 230—232, 237—239, 245—247, 249—250, 297, 337,
340—341, 346, 355, 364, 477, 488, 584
многоклеточные 24, 30, 35, 49, 56, 67, 73, 76, 79, 98, ПО, 115,
123—126, 133—134, 144, 146, 152, 156—157, 162, 164, 167,
170, 173, 182, 185—186, 202, 321—322, 328—329, 336, 459,
534, 575, 578, 629
многоножки 37—38, 62, 355—356
мокрецы 248
моллюски 11, 15—18, 20, 26, 36, 38, 40—41, 43, 45—46, 48,
53—55, 60, 62, 65, 68, 76, 79, 89, 93—94, 98, 111, 115, 143,
183, 212, 227, 231, 241, 243, 281, 294—295, 301, 535, 560,
626
моноплакофоры 20, 518
моноталамиды 451, 522, 524
моноцеркомонадиды 365
морские свинки 301, 341, 366, 531
москиты 226, 230, 233—234, 245, 247—248, 252
муравьеды 36, 231, 246
муравьи 36
мухи 12, 212, 233—235, 237, 243, 248
муха це-це 233—235, 237
мухи-кровососки 248
мушки 30, 46, 97
мхи 440, 476, 500, 503, 521
мшанки 25, 50, 101, 105—107, 115
наездники 59
насекомые 11, 16—18, 21, 23—24, 26—27, 30—32, 36—38, 43,
45, 51, 59, 61, 76, 85, 88, 93—94, 97, 114—115, 145, 186,
212—213, 215—217, 226, 229, 231—238, 240, 243, 245, 247,
252, 272, 341, 348, 355—356, 365—366, 368, 384—386, 395,
406, 440
насекомые-ксилофаги 385—386
насекомые-хозяева 234—235, 243
наутилоидеи 76
нематоды 16, 25, 38, 48, 88, 117, 231, 243, 364, 478, 560, 625
немертинообразные 108
немертины 25, 53, 102, 108
неопилины 20
неошвагерины 579
непарнокопытные 79
неполнозубые 246
нодозарианы 556—557, 579, 592, 596
нодозариаты 537, 539, 543, 552, 565, 575, 577—580, 582—583,
595, 599, 603
нодозарии 552, 575
нонионы 582
носороги 12
нотонейрон 105 \
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
635
нуммулитесы 534, 575
нуммулитиды 563
оболочники 19
овении 115
оксимонадиды 162, 341
оксимонадовые 337, 348—351, 354—355
оксимонады 133, 154, 180, 337, 348—350, 352—357, 384—385, 387
олигохеты 93
олистодискусы 285
онихофоры 50, 76, 89
оомицеты 135, 290
опалинаты 297, 301—302, 310—311
опалиниды 152, 298, 303—306, 308—311
опалины 135—136, 146, 162, 302, 307—310, 312
опистомастиготы 214, 243
опоссумы 247, 249
орбитоидиды 544
орбитолиниды 544
орбитолины 575, 578—579
ортонектиды 25
остракоды 76
офиуры 62, 65, 117
охромонады 269
пальмовый вор 54—55
пальмы 245
панцирные жгутиконосцы 67, 148—149
панцирные клещи 477
парабазалии 128—129, 154, 156, 180, 337, 349—350, 357
парастазии 206
парнокопытные 179, 249—250
пателлиниды 578
пателлины 556
пауки 26, 57, 69—70, 115
паукообразные 11, 18, 26, 38, 40, 50, 76
пединелловые водоросли 135
пединелломорфиды 135
пелагомонадацеи 273
пелагомонадиды 273
пелагомонасы 273—274
пелагофитовые 258, 273
пеломиксы 142, 508—511, 513
пенероплисы 557, 563
пентастомиды 112
первичнополостные 99—100, 111, 115
первичноротые 49, 98, 100, 102, 106, 108—109
первоптицы 44, 49, 82—83
переносчики-пиявки 232, 235
перепончатокрылые 26, 61
петалонамы 74
пилулины 559
пингвины 44—45, 61
пироплазмиды 56, 142, 157, 179—180
пирсонимфы 357
пиявки 25, 62, 93, 115, 227—229, 231—232, 235, 237—239,
241, 247—248, 341
плазмодиофориды 398т—402, 404, 406—409
плазмодиофоровые 408
планарии 227, 241
пластинчатожаберные 26
плацентарные 97
плеченогие 16, 48, 50, 76, 89
плоские черви 98, 111, 194—195, 282, 317, 335, 395,467,489, 549
погонофоры 20, 48, 53, 64, 75, 89, 93, 101, 109, ИЗ, 115
позвоночные 13, 17, 38, 40,42, 45, 76, 98, 231—232, 235—236, 251
покрытосеменные 187
поликладиды 68
полимастигины 133, 152, 314, 348
полимастиготы 337
полиморфиниды 578
полипиды 113
полипы 18, 73, 100, 108
полихеты 25, 37, 53, 66, 75, 93, 105, 112, 115, 560
полужесткокрылые 26, 233, 243, 252
полухордовые 98
пресмыкающиеся 11, 26, 34, 40, 82, 114, 248
приапулиды 53
приматы 44, 79, 93, 249
примнезиофитовые 276
проамебы 429
прокариоты 45, 71, 73, 123—126, 128, 138—139, 142, 144, 161,
180, 186, 226, 348, 353, 356, 379, 584
промастиготы 214, 224, 226, 230, 243, 245, 247, 253
простейшие 16—22, 24, 35, 40, 43, 45, 52, 55—56, 60, 64, 67,
70, 76, 100, 116, 123—125, 134, 140—141, 143—146, 148,
157—162, 166—167, 169—171, 179—182, 184—188, 202,
207, 210, 217, 229, 236, 243, 251—252, 275, 284, 289, 291,
312—313, 321, 323—324, 337, 347, 358, 362—363, 377, 381,
384—385, 395, 398, 402, 404, 408—409, 414, 417—418, 431,
444, 446, 452, 458, 467, 478, 486, 489—490, 508—511,
513—514, 516—518, 524—526, 532—535, 559, 567, 571—
572, 574—575, 624, 627, 629—630
протей 452
протеомиксиды 408
протеромонады 135—136, 146, 297—300, 311
протисты 21, 24, 49, 87, 116, 121—136, 138—184, 186—188,
190, 192, 194—196, 201, 206, 210, 218—219, 222, 225—226,
229, 238, 251, 270, 272, 275, 284—285, 287—288, 290, 293,
297, 301—302, 304, 312, 316, 336, 349—350, 378, 384, 398,
415, 418, 421, 424, 438, /144—446, 451—452, 462, 464—465,
478, 485—486, 488, 490—491, 502, 508—512, 514, 519, 522,
525, 533—534, 548, 558—559, 563, 567, 578, 581, 584
протобионты 71—72
протоктисты 123
протоопалины 310—311
проторнея 105
протостелиды 136, 411, 414, 418—422, 424, 430, 436, 439—
440, 444—445
протостелиевые 440, 444
протостомий 101, 104, 108
протоцилиата 312
проэукариоты 510
прямокрылые 26
псевдодендромонады 136, 266—267, 269, 316—319
псевдозухии 36, 83, 96
птицы 11—12, 15—17, 20, 25—27, 32, 34, 36, 38, 42, 49, 52,
60, 62, 82—83, 88, 95—97, 230, 232, 247—248, 251, 341,
358, 363—364, 366, 477
пятиустки 112
рабдаммины 560
рабдомонадины 202
равноногие 26
равноресничные 35, 56
радиолярии 19, 76, 135—136, 143, 157, 170, 172—173, 179,
184, 516—517, 519, 548, 557—558, 584
раки 25—26, 54—55
раки-отшельники 54—55
раковинные амебы 24, 173, 176, 178, 184—186, 451, 468 —470,
472, 474, 476—478, 482, 484, 490, 500, 502
ракообразные И, 18, 26, 36, 38, 43, 50, 53—54, 56, 58, 61, 70,
76, 87, 89, 93, 112, 114, 173, 207, 227, 242, 266, 625
ракушковые 93
рафидофитовые 285—287
рачки 201, 206, 209, 272, 559—560
реофаксы 560, 575
рептилии 36, 38, 42, 49—52, 56, 59, 61—62, 82—83, 92» 96,
227, 230—232, 235, 238—239, 245, 248, 252, 297» 301, 341,
355, 358, 366
636
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
ресничные черви 25, 37, 44, 60, 68, 140, 184
ретортамонадиды 339—341
ретортамонады 133, 140, 338, 349
родофитовые 563
родофициевые 563
роталианы 548, 552—553, 556—557, 562, 565, 575, 577, 579,
582—584, 592, 598, 606
роталиаты 172, 543, 548, 552—553, 556, 561—563, 569, 571—
572, 574—575, 577—580, 582—584, 586, 588—589, 591,
595, 597—599
роталиацеи 536
роталиеллиды 534
роталииды 609, 625
ротоногие 58, 93
рукокрылые 27
ручейники 26, 363
рыбообразные 11, 26, 56, 61
рыбы 11, 15—17, 21, 26, 34, 38, 40, 42—43, 48, 50—52, 54,
56—57, 59, 61—62, 65, 70, 76, 79, 81, 116, 173, 179, 187,
208, 219—220, 222, 226—232, 237—239, 241, 248, 281—
282, 297, 301, 312—313, 348, 358, 365, 478, 534, 560
сабеллитиды 116
сайгаки 28
саламандры 65
сапролегниевые 407
сапролегнии 290
саранчовые 66
сарган 54, 56
саркодовые 178, 181, 184, 408, 431, 574—575, 584
сеймурии 82
синезеленые водоросли 72—73, 76, 176, 184, 195, 488
синуровые 270—271
синурофитовые 258, 270—273
синуры 272
сипункулиды 112, 116
сифоновые водоросли 140
сифонофоры 19, 25
скафоподы 560
сколециды 100—101, 108, 111
скорпионы 26, 57, 76, 89, 243
скребни 43, 111, 116
слепни 235, 248—249
слизевики 128, 417—419, 424, 427-^31, 436—439, 446
слоны 38, 59, 79, 81, 250
соколиные 52
солнечники 123, 135—136, 145, 148, 157, 161—164, 166, 170,
176, 178, 180, 184, 490, 493, 498, 558
соритиды 562
сосальщики 18, 25
сосущие инфузории 56, 60, 64, 148, 159, 167, 184
спириллинаны 548, 552, 556, 579, 589
спириллинаты 539, 556, 565, 574, 578—581, 588
спириллиниды 578, 588
спиролокулиновые 580
спиролокулины 561
спирохеты 180, 348, 353, 357, 379
спонгомонады 314, 316
споровики 24, 52, 56, 123, 128, 141—142, 146—147, 159, 161—
162, 185
споровые 201, 210
стегоцефалы 38, 50, 81—82
стеллерова корова 17
стемонитовые 435—436
стопоходящие 61
стрекающие 111
строматопораты 76
сувойки 327.
суктории 56, 148
сумчатые 230—231
сфагновые 184, 263, 287
сфагнум 476
сфеномонадины 202
сфероморфиды 116
сцифоидные 18, 25, 73
табуляты 76
тайнобрачные 85
таракановые 26, 386—388, 395
тараканы 161—162, 348—350, 353—357, 366, 368—369, 377,
380—391
тауматомонады 148, 411
текстулярианы 537, 560, 575, 579—580, 583, 589, 598
текстуляриата 552
текстулярии 556, 600
текстулярииды 578, 600, 602
тентакуляты 113
термиты 36, 179, 341, 348—350, 353—357, 363, 366, 368—369,
377, 380—381, 384—392, 395
типулиды 363
тли 17, 26
траустохитридиевые 135
трахеломонасы 207
трематоды 25
триатомины 235
трилобитообразные 50, 73
трилобиты 49—50, 58, 75—76
трипаноплазмы 236
трипаносоматиды 24, 149, 211—226, 229—239, 243, 245, 247,
252—253
трипаносомы 179, 212—213, 215—217, 220—226, 228—238,
240, 243, 245, 247—251
трипомастиготы 214, 229—232, 245, 247
тритоны 232, 517, 528
трихомонадиды 156, 358, 362—365, 367, 376—378, 386—388
трихомонады 130—131, 148—149, 154, 183, 340, 348, 350,
357—358, 361—364, 367—368, 385—386, 395, 512, 528
трихонимфины 368—369, 372, 374—378, 386
трохамминиды 578
трохаммины 565
туникаты 521
турбелляриеобразные 111
турбеллярии 19, 25, 48, 53, 70, 75, 108, 140, 560
тутовый шелкопряд 31, 114
угри 62
улитки 89, 219
усоногие 18, 87, 93
фавузеллиды 548
факусы 207
фикомицеты 408
филоподиальные амебы 424
фитомоносы 245
фишериниды 577
флагеллаты 187, 446
фораминиферы 18, 76, 123, 136, 140, 143, 145, 161—162, 167,
172—173, 179, 184, 187, 451, 504, 507, 516—520, 523—525,
527, 533—537, 539, 543, 548—549, 552, 556—560/ 562—
563, 565, 567, 569, 571—572, 574—575, 578—580, 583—
586, 588, 607—610, 624—630
форель 94, 229
форониды 106—107, 115
фузулинацеи 579
фузулиниды 535, 544, 556, 563, 575, 578—579, 599
хвостатые амфибии 36, 50, 65, 301
хелицеровые 26, 38, 50
хиолитиды 76
хиолиты 76
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
637
хитоны 70
хитридиевые 142, 154, 408
хищные 27, 141, 148
хлореллы 469, 559
хлоромонадовые 285
хлорофиты 467, 563
хоаномастиготы 214
хоанофлагеллаты 149, 314, 316, 321—329
хоботные 42, 53, 81
хордовые 19—20, 37, 57, 65, 98, 103
хормозинаны 560
хризомонадиды 148
хризомонады 146, 152, 155, 180, 187, 262, 265—267, 270, 272,
281, 298, 301, 314, 316, 319, 584
хризофитовые 123, 125, 149, 173—174, 176, 180, 184, 258—
260, 262—264, 266, 270—274, 276, 287, 290, 317—318, 414,
548
хризофиты 263, 266, 281, 563
хромисты 128, 135—136, 281, 290, 311
хромобионты 135
хромофитовые 135, 274
хромофиты 135—136
хромулиниды 270
хромулины 263
хрящевые 57, 65, 94, 246—247
целоматы 108—109
цепедеи 310
цератиомиксовые 418, 440
цератиомиксы 422
церкомонадиды 148, 176, 187
церкомонады 136, 174, 411—414, 418, 444—445
цестоды 25, 38
цианобактерии 123, 156, 489, 502, 507
цикадовые 26
цилиаты 142, 154
цинодонты 51
человекообразные 79
червеобразные 53, 73, 100
черви 11, 18—19, 22, 25, 35, 37—38, 40, 43-44, 53, 60, 62, 64,
66, 68, 70, 73, 75, 88, 98, 100—103, 105, 108, 111, 114—116,
140, 236, 341, 395, 406, 467, 478, 585
черепахи 44, 94, 248, 301
четырехлегочные пауки 69
четырехлучевые кораллы 108
чешуекрылые 26
членистоногие 17, 22, 25, 35, 38, 40—42, 48, 50, 52, 57—61,
66, 68—69, 73—76, 93, 95, 98, 103, 112, 114—115, 117, 145,
251
шаттонеллы 285
шелкопряды 31, 114
шизопирениды 412, 485—486
шимпанзе 79, 531
шиншиллы 346
щетинкочелюстные 113
щитковые 76
щупальцевые 53, 113
эвглениды 148, 155, 158, 173—174, 176, 184, 201—202, 205—
209, 211
эвгленовые 24, 123, 125, 128, 146, 149, 156—157, 166, 173,
185, 201—210, 213, 219, 236, 275, 289
эвгленозои 201
эвгленоидеи 201
эвглены 185, 188, 202—205, 207
эвтрептины 202
эвтрепции 202, 204
эллипсолагены 578
эльфидииды 578
энантиозои 110
эндотириды 575
энтамебы 144
энтеромонадиды 347
эогиппусы 42
эпимастиготы 214, 224, 231—232, 236, 243, 247
эукариоты 25, 123—126, 128—129, 133—136, 138—139, 141—
145, 157—158, 164, 180, 187, 196, 213, 222, 224, 236, 238,
251—252, 290, 316, 395, 398, 408—409, 418, 446, 467, 486,
509—510, 512, 514, 563, 567, 584
эуметазои 110
эумицетозои 417
эуриптериды 38
эуфаузиевые 26
эхиностелиевые 431—432, 440
эхиуриды 53, 111, 527
явнобрачные 85
ящерицы 56, 62, 248, 300—301, 348
ящерохвостые 82
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Acalephae 100
Acanthamoeba 139, 145, 158, 412, 466
Acanthamoebidae 466—467
Acanthobdella 62
Acanthocephala 99—101, 103, 105—106, 111
Acanthocorbis 332, 335
Acanthoeca 332, 336
Acanthoecida 137
Acanthoecidae 322, 325, 328, 332, 336
Acanthoecopsis 335
acanthophora, Euglypha 502
Acanthopodida 451, 466—467
Acari 40, 88
Acervulinina 608
achyae, Octomyxa 406
Acnidaria 106
Acoela 53, 60, 69
Acoelomata 98, 100, 102—103, 106, 108—111, 118
Aconchulinea 135
Aconchulinia 451, 465, 490—491, 493—495, 497—499, 502
Aconchulinida 499
acosta, Hypotrichomonas 366
Acotylea 68
Acrania 104, 109
Acrasea 485
Acrasida 135, 141, 143—144, 451, 486, 488—490
Acrasidae 485—486, 489—490
Acrasis 485, 489—490
Acrida 59
Acridiella 59
Acritarcha 141
Actinobdella 232
Actinotrochozoa 49, 106—107
acus, Belong 54
acus, Cyclidiopsis 205, 207
acus, Euglena 204—205
acus, Heteronema 202
Acytosteliidae 430
Acytostelium 425, 430, 447, 449
adamsoni, Porotermes 388
Adamussium 630
Adelphamoeba 488
Adelungia 598
Adercotryma 629
adleri, Leishmania 245
advena, Verneuilinulla 625—627, 629
aegyptum, Trypanosoma 250
Aerania 118
* Курсивом выделены названия видов и родов.
Aerobacter 346
aerogenes, Klebsiella 346
aestuarina, Neoparamoeba 454
aethiopica, Leishmania 245—246
affinis, Badhamia 436
affinis, Nodosaria 552, 557
Afrenulata 88—89
africana, Chroomonas 196
africanus, Stolotermes 391
agamae, Leishmania 245
agilis, Rhynchobodo 241
agilis, Trepomonas 346
Agnatha 76
Ahnfeltia 627
akashiwo, Heterosigma 288—289
alba, Cribrothalamina 574
alba, Saccammina 537, 565, 567, 569
albicollaris, Boderia 565
Alepiella 504
algaeformis, Rhizammina 527
algarum, Phagomyxa 408, 410
algeriensis, Labyrinthula 294—296
Allantion 415—417
Allelogromia 523—524
Allogromia 525, 528, 534, 537, 549, 565, 569, 572, 585, 624, 631
Allogromiida 586
Almaenoidea 609
Alveolata 128, 137
Alveolina 580
Alveolinella 594, 629
Alveolinidae 538, 563, 594
Alveolinina 538, 578, 594
Alveolophragmium 628
Alytes 36
Amaurochaete 442
amazoensis, Leishmania mexicana 247
Ambistoma 36, 52
Amera 99
americana, Leishmania 246
americana, Reclinomonas 275
americanum, Trypanosoma 248
amerosporum, Endostelium 420
Ammobaculites 539, 629
Ammodiscana 538, 544, 549, 575, 585, 588—589, 599, 611
Ammodiscida 589
Ammonia 569, 574, 578, 609, 624—625, 629
Ammoscalaria 575
Ammoscalariana 538, 583, 585, 611
Ammoscalariida 586
Ammoscalariidae 586
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
639
Ammosphaeroidinoidea 598
Ammosphaerulina 575
Ammotium 539, 625, 627, 629
Ammovertellida 589
Amniota 42—43
Amoeba 164, 178, 181, 184, 187, 445, 447, 449, 452, 454, 458,
460, 462, 466—468, 484—485, 515—516
Amoebidae 452, 454, 458, 460, 466—468, 516
amoebinus, Cercobodo 446
Amoenoscopa 332
Amphibia 48, 509
Amphifenestrella 549
Amphileptus 171, 184
Amphilinida 114
Amphinomidae 75
Amphistegina 559, 563, 580
Amphisteginidae 563, 608
Amphitrema 501, 504, 522, 524
Amphitrematidae 500, 504
Amphitremidae 522, 524
Amphizonella 478
Amphorellopsis 504
ampulla, Cyphoderia 504, 508
Ampullatoria 479, 507
Anacanthotermes 388, 391—392
anaerobica, Vahlkampfia 467
Anas 531
anatis, Blastocystis 531
Anchitherium 80—81
andrejvi, Phlebotomus 246
anduzei, Lutzomyia 246
angulatus, Archaias 562
angulosa, Angulogerina 626
angulosa, Stereomyxa 467
angustatus, Bodo 240
angusticollis, Zootermopsis 357, 390, 395
Animalcula 17
Animalia 110, 113, 134, 136
Anisonema 207, 209
Ankylochrysidaceae 270
annamense, Trypanosoma 250
annectens, Joenia 376—377, 379, 387
Annelida 42—43, 53, 74, 99—100, 102—105, 110—112, 117
Anoplasma 230
Anoplura 232
Anser 531
anser, Anser 531
anseri, Blastocystis 531
Antarcella 478
antarcticus, Notodendrodes 559
Antennata 50, 103—104
Anthomedusae 59
Anthozoa 25, 100
Antimhinum 117
Antracotherrium 115
Anura 50, 231, 253, 308, 312
apheles, Chrysochromulina 277, 284
Apheloecion 332
Apicomplexa 56, 110, 139, 141
Aplacophora 99
Aplonis 119
Apocyanaceae 245
Apodotrocha 37
Apogromia 523—524
Apolimia 479
apophysatum, Cavostelium 440, 449
Apospironympha 390—391
Appendicularia 99, 103
applanata, Trabrooksia 443
Apsilis 116
apterus, Pyrrhocoris 235
Apusomonadida 186
Apusomonas 186
Arachnactis 117
Arachnida 41, 69, 99
Arachnula 493, 495, 498—499, 520
Arachnulidae 491, 493, 495, 498—499
Aranei 40, 69
Araneomorphae 69
Arbor 527
Arborea 74
arcanus, Phthanotrochus 572, 583
Arcella 176, 178, 468—473, 475, 478, 484—485, 516
Arcellacea 508
Arcellidae 468, 478
Arcellinida 451, 468, 470, 476—478, 482-485, 508
Archaediscida 538, 590
Archaediscidae 590
Archaediscoida 538, 590
Archaediscus 552
Archaeocyatha 106—107, 110
Archaeopteryx 82, 119
Archaeornis 44, 49, 82
Archaeomithes 82—83
Archaias 552, 562
Archaiasinidae 563
Archamoeba 142, 188, 510, 512, 514—516
Archamoebae 125, 128, 133, 137, 515
Archelon 44
Archerella 504
Archezoa 125, 128—129, 131, 133—134, 137, 142, 188, 510,
512—514, 516
Archiannelida 115
archibaldi, Leishmania 246
Archibilateria 108—109
Archicoelomata 104, 108
Archotermopsis 388, 390
arctica, Elphidiella 629
Arcyodes 442
Arcyria 434, 438—439, 442, 446, 448
Arecaceae 245
Arenagula 579, 583
arenaria, Arcella 472
Arenicola 93
arenilega, Myxotheca 565, 567, 569
arenocentrum, Psammetta 517
argentea, Bolivina 557
argentea, Pilulina 559
argentipes, Phlebotomus 246
ariarii, Trypanosoma 249
ariasi, Phlebotomus 246
aries, Ovis 532
armata, Rhynchobodo 241
oroides, Histiona 275 F
Arthropoda 43, 48, 50, 57, 69, 99—100, 102—106, 110, 112,
119—120
Articulata 103—106, 116
articulatum, Protbsporangium 420, 449
Arvicola 46
Ascetospora 144
Aschelminthes 102—105, 111, 119—120
Ascidia 103
Ascidiacea 99
Asclepiadaceae 245
Ascothoracida 89
asiatica, Trochammina 629
Assulina 501, 507
Astasia 201, 205, 207—209
Asterigerinida 608
Asterigerinoidea 608
640
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Asteroarchaediscidae 590
Asterotrochammina 579, 583, 601
Astomata 187
Astrammina 631
Astrononion 627, 629
Astrononionina 609
Astrorhiza 559, 628
Astrorhizata 537, 549, 572, 577, 583, 585, 587, 589, 611
Astrorhizida 586
Astrorhizidae 586
Astrosiga 330
Ataxophragmiida 538—539, 601—602
Ataxophragmiidae 601
Ataxophragmiina 538, 601
Ataxophragmioida 601
Ataxophragmoidea 601
Atelocerata 106
Athalamia 135, 137
Athalamida 186, 451, 520—521, 524
atrosporum, Lamproderma 435
attenuates, Parabodo 241
auerswaldae, Xenusion 75
augusta, Tritrichomonas 358, 368
aulacodi, Tritrichomonas 368
aulastomi, Chilomastix 340
Aulomonas 328, 331
aurantium, Planoprotostelium 419—420, 424, 449
aurelia, Paramecium 185
austeni, Glossina 250
australiensis, Naegleria 488
australis, Cyclojoenia 392
Austrotrillinoidea 594
avicularis, Opisthomitus 356
avium, Trypanosoma 230, 248
Awerintzewia 479
Axobranchia 100
Axolotl 119
axonucleata, Protoopalina 310—311
baccari, Ammonia 629
bacillata, Merotricha 288
bacillifera, Difflugia 476
bacillipedes, Vexillifera 454, 461
backeri, Trypanosoma avium 230
Baculella 527
Baculellidae 527
Baculogypsina 553, 627
Baculogypsinoides 553
Badhamia 436, 438, 443
Baileya 504
balamuthi, Phreatamoeba 462, 512, 515
Balamuthia 465
Balanoglossi 100
Balantidium 179
baltica, Flabellula 464
baltica, Macoma 296
barbari, Trypanosoma 232, 254
Barbeyella 441
Barbulanympha 377—379, 383—384, 389, 396—397
Barroella 348, 353, 355—356, 393
bartletti, Cribroelphidium 627, 629
batavus, Ammonia 624
Bathysiphon 563
batrachorum, Hexamastix 366
batrachorum, Trichomitus 358, 363
Batrachosauria 114
beccarii, Ammonia 574, 625
beckeri, Cryptobia 228
Belaria 523—524, 585—586
belewskii, Pelomyxa 513
Belone 54
Benthogastraea 108
berhei, Trypanosoma 250
Berthelinia 41
beryllifera, Pelomyxa 513
Bicoecales 275
Bicosoeca 264—266, 268, 275, 319
Bicosoecaceae 268, 275
Bicosoecales 135, 137, 189, 264—265, 267, 269, 274—275,
318—319
Bicosoecida 189, 275, 318
Bicosta 332
Bidentina 580
biforis, Licea 434
Bilateria 35, 48—50, 53, 75, 93, 98—111, 113, 118
Biliphyta 137
binucleata, Gocevia 482
binucleata, Pelomyxa 513, 515
Biokovinoidea 599
Biomyxa 521, 524
Biomyxidae 521
Biplanatidae 589
Biplanispira 574
Birgus 54—55
Bison 46
Bispironympha 391
bisporum, Cavostelium 420, 440, 444—445
bisporum, Echinostelium 450
bisporum, Protosporangium 421, 424
Bivalvia 41, 46, 99
Blastocrithidia 213—215, 220, 223—224, 226, 232—236, 243,
251—256
Blastocystida 528—529, 531—532
Blastocystidae 528, 531—532
Blastocystis 528, 530—532
Blatta 368, 386, 388, 394
blattarum, Lophomonas 378, 388, 395
Blattella 368, 386, 388, 394
bocki, Textularia 558
bocki, Xenoturbella 120
Boderia 565, 575, 584, 586
Bodo 173, 178, 212—213, 216, 223, 225—227, 240—241, 252—
253, 301, 532
Bodomorpha 412, 416, 446
Bodomorphidae 415—417
Bodonida 213, 240, 251—252
Bodonidae 213, 219, 227, 240, 253
Bodonina 253, 255—256, 411
boipevae, Zelleriella 312—313
Bolivina 557, 569, 578, 603—604, 624, 628
Bolivinella 569
Bolivinitida 603
Bolivinitina 603
Bolivinitoidea 603
bombarda, Tubifera 439
Bombulina 578
bonita, Koruga 392, 396
borealis, Spirorbis 93
Borelis 557
Borkovia 136
borreli, Cryptobia 216, 219, 222, 224—225, 228, 241
Bos 532
botrytis, Codonosiga 321—322, 336
bouffardi, Trypanosoma 230
bovis, Blastocystis 532
bovis, Giardia 347
bovis, Trypanosoma 249
Box 313
Brachiata 105, 110, 113, 115
Brachiopoda 49—50, 89, 99—100, 102—103, 106—107, 113
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
641
Braconidae 59
branchialis, Cryptobia 216, 219, 228, 241, 254
Branchiostoma 43
Branchiotremata 99
Branchiura 112
Brassica 115, 410
brassicae, Plasmodiophora 398—400, 402, 404, 406—407, 409—
410
braziliensis, Leishmania 245—247, 253
brevicollis, Calcaritermes 356
brevicula, Wallaceina 222, 245, 252
brevipalpis, Glossina 250—251
Brizalina 578
brucei, Trypanosoma 212, 219, 229, 231, 233—235, 238, 250,
255—256
Bryozoa 40, 50, 99—100, 102, 105—107, 112—113, 528
Buccella 560, 624
Buccicrenatinae 589, 599
Budashevella 589
Bufo 255, 313
Bufonidae 309
bufonis, Karotomorpha 297, 301
Bulimina 557, 578
Buliminella 557, 624, 626
Buliminida 604
Buliminina 604
Buliminoida 602
bulla, Korotnevella 454
Bullinularia 479
bullocki, Cryptobia 228
bulloides, Globigerina 557, 610, 625
Bullopora 577
Bursula 440
caballus, Equus 531
Cafeteria 189, 264, 266—267, 275, 318
Cafeteriaceae 267, 275
Calanoida 559
Calcarina 553, 580
Calcarinidae 563, 609
Calcaritermes 356
calceolata, Pelagomonas 273, 275
Calcituba 569, 631
Calliacantha 332
Calliobdella 228
calmettei, Trypanosoma 248
Calocoris 245, 252
Calomyxa 436, 441
Calonema 442
Calonympha 367
Calonymphidae 358, 365—366, 394
Calotermes 355—356, 387—388
Calotermitidae 355—356, 387, 389—390
Calotheca 332
cameli, Trypanosoma 250
Campanoeca 335
Campascus 507
Campyloacantha 332
Campylomonas 195, 197—198, 200
candeiana, Textularia 560
Candeina 625
Candida 300, 346
cantabrigiensis, May&rella 454
Capitellidae 93
Capsellina 504
carassii, Cryptobia 241
carassii, Trypanosoma 248
Carinaria 241
carinariae, Cryptobia 227, 241
Cariosphaeroides 73
Carnivora 46
carolinense, Chaos 454, 458, 460
carolinensis, Pelomyxa 513, 515—516
carterae, Hymenomonas 280
Carterina 575
Carterinina 602
cartesi, Nebela 477
Caryoblastea 133, 135, 137, 510
Cashia 461
Cassidulina 557, 627
Cassidulinida 538, 602, 604
Cassidulinidae 604, 630
cassis, Ammotium 625, 627, 629
Castellanella 251
castellani, Acanthamoeba 139, 145, 158
catastomi, Cryptobia 228
Catena 527
Catenulida 53
cati, Giardia 347
catostomi, Cryptobia 228
caucasicus, Phlebotomus 246
Caucasinina 605
caudata, Hexamita 346
caudata, Mallomonas 272
caudata, Protoopalina 309
caudatus, Bodo 225—227, 240, 253
Caudofoveata 89, 115
Caulerpa 140
caulleryi, Pelomyxa 514
caveatum, Dictyostelium 425
Cavia 313, 397, 531
cavia, Monocercomonoides 349
caviae, Chilomitus 366
caviae, Tritrichomonas 358, 363, 394
Caviomonas 341, 347, 392
Cavosteliidae 424
cavostelioides, Schizoplasmodium 419, 421
Cavostelium 419—420, 424, 440, 444-—445, 449
cebus, Trypanosoma 249
celiae, Phlebotomus 246
Celloniella 270
Centropyxidae 468, 478
Centropyxis 176, 178, 469—473, 477—478, 484—485
Cepedea 302, 306, 309, 311—313
Cephalocarida 89
Cephalochordata 99—100, 103, 105
Cephalopoda 62, 99
Cephalorhyncha 111, 115
Cephalothamnion 241
Cephalothamnium 213, 227, 241, 254
cephalotricha, Joenopsis 388
ceramodactyli, Leishmania 245
Ceratiomyxa 417, 419—421, 424, 440, 447—448
Ceratiomyxales 431
Ceratiomyxella 419—421, 424, 447
Ceratiomyxidae 424
Ceratiomyxomycetidae 431
Ceratobulimina 628
Ceratobuliminidae 541, 605
Ceratobuliminina 605
Cercobodo 411, 415, 446
Cercomonadea 135—137, 411, 413, 415, 417
Cercomonadida 187, 411, 417, 446, 513
Cercomonadidae 411, 413—414, 417
Cercomonas 338, 411, 414—415, 417, 445
Cerebrum 527
Cerelasma 517—519
Cerelasmidae 517, 519
Cerelpemma 518—519
cerevisiae, Saccharomyces 346
642
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Certesella 477, 479
cervi, Trypanosoma 248
Cestoda 99, 103
Cestodaria 99, 103
Chaetognatha 42, 49, 99—100, 102—107, 109—110, 113
chagasi, Leishmania 245, 247, 256
chalumnae, Latimeria 48
Chaos 452
Chardezella 504
Chardezia 504
Charentiinae 589
Charnia 74
Charniodiscus 74
charnwoodensis, Pseudovendia 75, 117
Chattonella 285—289
chattoni, Corallomyxa 465—467
Chelicerata 50, 99, 103—104, 106, 112
chelodina, Trypanosoma 248
Chenopodiaceae 407
chensiensis, Rana 309
cheopis, Xenopsylla 249
Chilomastix 337, 339—341, 392, 395, 397
Chilomitinae 358, 365
Chilomitus 365—366
Chilomonas 170, 176, 181, 189, 195—197, 200
Chilopoda 104
Chilostomellida 609
chinensis, Phlebotomus 246
Chironomus 93
chiton, Chrysochromulina 280, 284
Chlamydomyxa 522, 524
Chlamydomyxidae 521
Chlamydophryidae 500, 504, 507
Chlamydophrys 504
Chlamys 560
Chlorarachnion 284
Chlorarachniophyceae 284
Chlorarachniophyta 284
Chlorella 467, 469, 559
Chlorodesmos 273
Chloromonadida 285
Chloromonadinae 289
Chloromonadophyceae 285
Chloromonadophyta 285
Chlorophyta 140, 143—144
Choanoeca 323, 325, 331, 336
Choanoflagellata 35, 321
Choanoflagellida 322, 328, 336
Choanomonada 135, 137, 321—323, 325, 327, 329, 331, 333,
335—336
Choanomonadea 135, 137, 322, 336
Chondrodapis 527—528
Chondrostei 94
Chordata 57, 99—100, 102—106, 110, 113, 117—118
chromatophora, Paulinella 502
Chromista 125, 134—135, 137, 290, 314—315, 317, 319—320
Chromulina 263, 269
Chromulinaceae 269
Chromulinales 268—269, 274
Chroomonas 194—196, 198, 200
Chrysamoeba 189, 262, 269
Chrysamoebaceae 269
Chrysocapsaceae 269
Chrysochromulina 277, 280—281, 284
Chrysococcus 269
Chrysodidymus 273
Chrysolepidomonadaceae 269
Chrysomeridiales 270, 274
Chrysomonadida 275
Chrysomonadina 99
Chrysomonadinae 275
Chrysophyceae 135, 137, 140, 143—144, 189, 258—259, 261,
263—265, 267—270, 274—275, 284
Chrysophyta 135—137, 258—259, 261, 263—265, 267—271,
273—275, 284, 289
Chrysosphaera 270
Chrysosphaeraceae 269
Chrysothallaceae 269
Chrysotila 277
Chuaria 73
Chytridiomycetes 137
Chytridiomycota 142, 144
Cibicidella 577
Cibicides 537, 560, 565, 567, 569, 571—572, 574, 601, 626,
630
Cibicididae 607
Ciliata 104, 148, 186, 189—190
ciliatorum, Leptomonas 243
Ciliophora 110, 139, 142—144, 161, 185
Cimex 249
cinerea, Arcyria 438—439, 446, 448
cinerea, Nepa 235, 243, 252, 255
cinereum, Physarum 436
citriensis, Flamella 462, 467
Cladocera 59
Cladospongia 330
claparedi, Amphileptus 171
Clastoderma 438, 441
Clastodermataceae 440
Clastodermatidae 440
clavata, Hemitrichia 439
clavatum, Cribroelphidium 627—629
Clavulina 560
clevelandi, Coronympha 365
clevelandi, Joenia 387
Clyopelina 504
Clypeorbioidea 608
Cnemidophorus 52
Cnidaria 49, 102—104, 106, 110—111, 119—120, 336
Cnidosporidia 52, 104, 110
cobitis, Trypanosoma 226, 254
Coccolithophoridae 284
Coccolithus 281—282
Coccosphaerales 276—277, 281
Cochliobolus 498—499
Cochliopodidae 420, 484
Cochliopodium 482—485
cockayini, Nebela 477
Codium 560
Codomonas 264
Codonocladium 330
Codonosiga 321—322, 326, 328, 330, 336
Codonosigopsis 330—331
Coelenterata 53, 99—102, 106, 111
Coelomata 35, 42, 48, 52—53, 98—104, 106—111, 113, 115,
119—120
Coelomocoela 99, 119
Coelurosauria 83
Coenobita 54—55
Coenobitidae 54
coenocystis, Labyrinthula 296
Coenonia 430—431
Colacium 201, 207, 209—210
Colanielloidea 597
colbecki, Adamussium 630
Coleites 578
Coleoptera 118, 450, 509
coli, Amoeba (Entamoeba) 452
coli, Balanthidium 179, 513
coli, Entamoeba 461
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
643
Collaria 442
collaris, Trichonympha 390
Collaropyxidia 478
Colospironympha 391
Colponema 148
colubrorum, Monocercomonas 365
Comandonia 461
Comatricha 434—436, 439, 442
compressa, Cyclorbiculina 562
compression, Physarum 438
compression, Trimyema 511, 515
confusum, Trypanosoma 250
congolense, Trypanosoma 234, 250, 254, 256
conica, Bicosoeca 266
conica, Textularia 629
Conion 332
Conopodina 467
conorhini, Trypanosoma 229, 255
consociatum, Phalansterium 316
constricta, Centropyxis 477
contorta, Trichia 436
Copelata 104
Copepoda 59, 207, 210—211
Coptotermes 391—392
Corallomyxa 465—467
Corallomyxidae 466—467
Cornuspira 569
Cornuspiramia 577
Cornuspirida 593
Comuspirina 593
Comuspiroida 593
cornuta, Cassidulina 557
Cornuvia 442
coronaris, Barroella 356
Coronympha 365
corrugata, Patellina 552, 556—557, 559, 562, 565, 567, 569, 572,
575
corrugatus, Spirorbis 93
Corythion 178, 507
Corythionella 507
Corythionelloides 504
Coscinophragmatoidea 599
Cosmoeca 332
Costia 241
Costiferida 593
Costiferoida 593
Cotylea 69
Craterium 439
crenulata, Difflugia 472
Cribraria 435—436, 439, 441, 448
Cribrariaceae 441
Cribrariidae 441
cribrarioides, Lamproderma 435
Cribratinoidea 596
Cribroelphidiidae 610
Cribroelphidium 624—629
Cribrolinoides 578
Cribrorobulina 578
Cribrothalamina 574
Cricosphaera 284
Crinolina 333
crispum, Elphidium 569, 574, 626
cristallina, Bicosoeca 275
cristata, Euglypha 502
cristata, Galerida 46
cristatus, Triturus 528
Cristidiscoidida 451, 491, 497
Cristivesiculatida 451, 491, 498
Crithidia 213, 222, 224, 226, 234, 236, 243, 248, 251—255
Crossopterygii 48
crucifer, Cyathobodo 320
Crucispina 333
Crustacea 50, 69, 99, 104, 106, 114
cruzi, Trypanosoma 215, 224—225, 229, 231, 233—235, 238,
240, 249, 253—255
Cryptobia 213, 216, 219—220, 222, 224—225, 227—229, 236,
241, 253—256
Cryptobiidae 213, 219, 227, 241—242
Cryptocellides 68
Cryptocercus 348, 354—357, 366, 368, 377, 380—381, 383—
391, 393—397
Cryptodifflugia 469, 476, 482, 484
Cryptodifflugiidae 468, 479
Cryptomonada 200
Cryptomonadida 194, 200
Cryptomonadidae 197, 200
Cryptomonas 196, 198, 200
Cryptophyceae 135, 137, 194, 200
Cryptophyta 128, 135, 137, 148, 194—195, 197, 199—200
cryptosoma, Centropyxis 477
Cryptotermes 397
crystallina, Bursula 440
Crystallolithus 282
Ctenaria 103
Ctenophora 25, 49, 59, 88, 99—100, 102, 104—105, 110—111,
118, 120, 336
ctenostomes, PalludiceIlina 528
cucullata, Fissurina 627
Cucurbitella 469, 479
culbertsoni, Acanthamoeba 466
Culex 247
Culicidae 243
culicis, Blastocrithidia 226, 253
Cuneata 583, 598
cunicularia, Talpinella 630
curtus, Reophax 627
curvifilus, Bodo 223, 225—226, 240, 252
cyanoguttatum, Herichthys 228
Cyanophyta 72—73
Cyanospira 502, 507
Cyathobodo 317—320
Cyclammina 628
Cyclidiopsis 204—207
Cycloclypeus 575, 610
Cyclojoenia 392
Cyclolinida 599
Cyclolinoidea 600
Cyclopida 210
Cyclopsinelloidea 600
cyclopum, Cephalothamnium 227, 242, 254
Cyclopyxis 176, 470, 478, 484
Cyclorbiculina 562
Cymbaloporella 628
Cymbaloporetta 574, 628
Cynodontia 51, 82
Cyphoderia 176, 504, 507—508
Cyphoderiidae 500, 507
cyprini, Cryptobia 241
Cyrtotreta 105
Cystoseira 627
Cystotrypanosoma 248
Cytomorpha 102
dahlii, Cryptobia 228, 241
Daitrona 579
danica, Ochromonas 262, 275
danilevskyi, Trypanosoma 221—222, 224, 248, 250, 252, 255
darwiniensis, Mastotermes 385—386, 392, 396
Dasy tricha 190
davidi, Phytomonas 245
644
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
deanei, Crithidia 226
deanei, Trypanosoma 249
debaryanum, Clastoderma 438
Decapoda 54
delicatula, Gephyramoeba 465
delicatula, Nuclearia 491—492, 495, 499
Delosinina 597
Deltotrichonympha 386, 392, 396—397
Deltotrichonymphidae 392
deminutivum, Dictyostelium 430
dendrocoeli, Cryptobia 227, 241
Dendrocoelum 227
Dendromonas 317
Dendrophryida 587
Dendrophryoidea 587
Dentalina 543
dentata, Tracheleuglypha 502
Dentostomina 578
depressa, Heterostegina 548, 563, 574
depression, Dianema 435
depresura, Turbinulina 624
Dermamoeba 454, 460
Dermanyssus 248
Dermocystidium 296
Deropyxis 281
Desenburyina 598
designis, Bodo 226, 240, 253
Desmarella 322, 326, 328, 330, 336
Desmodes 235, 250
desouzai, Crithidia 226
Deutencephala 102
Deuteramoeba 459—460, 462
Deuterostomia 49, 53, 93, 98—110, 113, 115
Devescovina 366, 388, 397
Devescovinidae 358, 365—366, 394
Devescovininae 358
Diachea 436
Diacronema 280—281, 283
Dianema 435, 441
Dianemaceae 441, 448
Dianemidae 441
Diaphanoeca 332—333, 335—336
Diaphorodon 504
Dickinsonia 75
dicoelobius, Paedophoropus 60
Dicrateria 277
Dictydiaethalium 441
Dictydium 435
Dictyocha 135, 137, 189
Dictyochophyceae 135, 137, 189
Dictyostelia 135, 137, 144, 418, 424
Dictyosteliida 430
Dictyosteliidae 424, 430
Dictyostelium 424—425, 427, 430, 444, 446—450
Dicyemida 111
Didacna 79
Didinium 70, 184
Didymiidae 443
Didymium 436—439, 443, 446, 448—449
Dientamoeba 141, 188, 364—366, 512, 515
Dientamoebinae 358
diffluens, Proteus 509
Difflugia 173, 176, 178, 469—472, 476, 479, 484-485
Difflugiella 482, 485
Difflugiidae 468, 479
diffusa, Leishmania 247
digitatum, Phalansterium 314, 316, 319—320
Dimastigella 213, 227, 242, 252—253, 256
dimidiata, Cepedea 302, 306, 309, 313
diminuta, Malmiana 228
dimorphon, Trypanosoma 250
Dinamoeba 452, 461
Dinenympha 348, 351, 357
Dinobriaceae 269
Dinoflagellata 99
Dinophilida 37, 112
Dinophilidae 105, 120
Dinophylus 112
Dinophyta 143
Dipleurozoa 75, 118
Diploblastica 100, 103, 110—111
Diplochlamys 478
diplocostata, Stephanoeca 325, 336
Diploeca 331
Diplogromia 523—524
diploidea, Sappinia 452, 459—460, 466—467
Diplomonadea 126, 135, 137, 140, 337—339, 341, 343, 345, 347,
392
Diplomonadia 337—338, 341, 343, 345—347
Diplomonadida 129, 137, 189, 337—338, 344—345, 347—348,
355, 392—393
Diplonema 211
Diplophrys 522, 524
Diplosiga 330—331
Diplosigopsis 331
Diplotheca 334
Diplozoa 188, 337, 396
Dipnoi 70
Diptera 232, 235, 243, 250, 255—256
Discocelis 264, 268, 275
Discocyclinidae 610
discoides, Arcella 468, 485
discoideum, Dictyostelium 424—429, 444, 446—450
Discorbida 607
Discorbidae 607
Discorbinellidae 606
Discorbinelloidea 538, 606
Discorbis 557, 560, 569, 583, 625
Discorboida 605
Discorinopsis 569
Discospirinoidea 593
Distigma 201, 204, 209
Distomatidae 337
Distomatopyxidae 468, 479, 484
Distomatopyxis 479
Ditrema 504, 522, 524
Dityscus 91
dobelli, Hegneriella 306, 313
Dobrogelina 579, 583, 601
dogieli, Par enteroxenos 115
domestica, Sus scrofa 531
doniezi, Bolivina 569
donovani, Leishmania 245—246, 252—253
Dorvilleidae 37
Drosophila 46, 97, 117, 120, 255
Drosophilidae 97
dubium, Poly chaos 460
duboscqi, Joenia 387
duboscqi, Phlebotomus 246
Dunaliella 559
duodenalis, Giardia 345—346
Dusenburyinida 598
Dusenburyinoida 598
Duttonella 248—250
Ebriadina 99
eburneense, Trypanosoma 249
Echinamoeba 461
Echinamoebida 461
Echinamoebidae 452
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
645
Echinoderida 100
Echinodermata 53, 99—100, 102—106, 109—110, 113, 336
Echinosteliaceae 440
Echinosteliales 431, 440
Echinosteliida 431—432, 440—441, 443
Echinosteliidae 440
Echinosteliopsida 440
Echinosteliopsidiidae 440
Echinosteliopsis 440
Echinostelium 435—436, 439—440, 447, 449—450
Echiura 105, 117
Echiurida 42, 89, 100, 103, 105, 110
Echiuroidea 102
Echiurus 119
Ecterocoelida 108
Ectoprocta 103, 105
Edaphonobiotus 507
Edentata 230, 247
Edgertonia 527
Eggerellidae 556, 601
ehrenbergii, Euglena 211
elegans, Hyalosphenia 476
elegans, Leptophrys 492
elegantissima, Buliminella 624, 626
Ellipsolingulina 578, 597
Ellipsopyxella 478
Ellipsopyxis 478
elmassiani, Trypanosoma 250
elongatum, Holomastigotes 392
Elphidiella 628—629
Elphidiida 539, 563, 609
Elphidiidae 578, 610, 630
Elphidium 559, 563, 565, 569, 572, 574, 624, 626
Embadomonadidae 337—338
Embadomonas 341, 395
Emiliania 284
Enantiozoa 106, 110
encomeli, Trypanosoma 249
Endolimax 397, 461
Endoprocta 99, 103
Endosporeae 417
Endostelium 420, 424, 446, 449
Endothyra 589
Endothyranella 589
Endothyranopsis 589
Endothyrida 589
Endotrypanum 213, 220, 224—225, 230—231, 240, 247, 253
enriettii, Leishmania 245
Entamoeba 131, 179, 189, 395—396, 452, 461, 512, 528
Entamoebidae 142, 452, 461, 513
Entelegyne 69
enterica, Giardia 338
enterica, Megastoma 338
Enteridiaceae 441
Enteridium 435, 441
Enterocoela 99, 102, 108, 119
Enterocoelida 108
enterocola, Blastocystis 528
Enteromonadida 337—338, 341, 347
Enteromonadidae 338, 345, 347, 392
Enteromonas 338, 346—347, 392
Enteromorpha 560
Enteropneusta 88—89, 99—100, 103, 105
Enterozoa 106, 110
Entoconchidae 36
Entodiniomorpha 35, 140, 142
Entomosigma 288
Entoprocta 102, 105, 118
Entosiphon 202, 209, 211
Eogeinitzina 552
Eohippus 80
Eonodosaria 552
Eouvigerina 578
Eouvigerinoidea 603
Ephelota 60
Ephydatia 329, 336
epidendrum, Lycogala 439, 441, 446
Epihippus 81
Epineura 98, 100, 103, 108
Epipyxis 188
Epistominella 624
Epistominoides 578
Epistylis 321
Epithelioneura 103
Epithelioneuria 103
equinum, Trypanosoma 250
equiperdum, Trypanosoma (Trypanozoon) 223, 229, 231, 235,
251
Equus 80, 531
Euamoebida 451—452, 454, 462—467
Euamoebidea 468
Eucoelomata 102
Eucomonympha 372, 383—384, 390
Eucomonymphidae 369, 372, 375, 377—378, 386, 390
Eudorinidae 35
Eugenia 504
Euglena 176, 181, 183, 187—188, 201—202, 204—205, 207,
209—211
Euglenales 201, 211
Euglenida 202, 209—211, 253
Eugleninae 210
Euglenoidea 135, 137, 143, 201—203, 205, 207, 209—210
Euglenoidina 211
Euglenophyceae 210
Euglenophyta 139, 143, 201, 210, 254, 275
Euglenozoa 125, 131, 135, 137, 141, 201, 203, 205, 207, 209—211,
213, 215, 217, 219, 221, 223, 225, 227, 229, 231, 233, 235, 237,
239, 241, 243, 245, 247, 249, 251, 253, 255, 263, 265
Euglypha 176, 178, 471—472, 484—485, 500, 502—503, 507—508
Euglyphella 507
Euglyphidae 500, 507—508
Euglyphidion 507
Euglyphina 500—503, 507—508
Euhyperamoeba 140, 467, 486—490
Euhyperamoebidae 486, 489
Eumetazoa 99, 102—103, 110—111
Eumycetozoa 446—447
Eumycetozoea 135, 137, 140, 411, 417—419, 421, 423, 425, 427,
429, 431, 433, 435, 437, 439, 441, 443, 446, 449, 519
Eumycotina 431
Eunicidae 37
Euphorbiaceae 245
eupora, Leptospironympha 384
Eupyrgus 60
Euryalae 62
Eutreptia 204, 209
Eutreptiida 202, 209—210
Euvigerina 583
evansi, Trypanosoma 212, 223, 235, 250
excavatum, Cribroelphidium 624
exigua, Epistominella 624
faecicola, Helkesimastix 415
faecicola, Proleptomonas 212—213
Falcomonas 199—200
Falconiformes 52
fallax, Euhyperamoeba 140, 187, 487—490
fallisi, Trypanosoma 225—226, 255
falshawi, Trypanosoma 248
familiaris, Blastocrithidia 243, 256
646
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
fasciatus, Nosopsyllus 249, 255
fasciculata, Crithidia 222, 224, 243, 253—255
fasciculatum, Polychaos 454
fennica, Parastasia 205—206, 210
Fibrocapsa 285, 288—289
Filamoeba 461
filosa, Reticulomyxa 521
Filosea 135, 137, 176, 419, 451, 465, 468, 472, 477, 490—491,
493—495, 497—501, 503, 505, 507, 520
Filosia 499
Fisherinella 589
Fisherinidae 577, 594
fissa, Hexamita 346
Fissotextularia 600
Fissurina 560, 627—628
Flabellula 462, 464-Л65
Flabellulidae 462, 465, 467, 490
Flagellata 100
flagellata, Hyperamoeba 135, 137, 444—446
flaundersi, Spriggina 74
flava, Pontomyxa 520
flavescens, Gruberella 488—490
flavicollis, Calotermes 356
flavicomum, Physarum 436, 439, 446
flavipes, Reticulitermes 348, 351, 357, 396
flaviscutellata, Lutzomyia 247
Flintina 578
Floresina 559
floridana, Fissotextularia 600
floridana, Rosalina 560, 574
floriformis, Trichia 434
fluviatilis, Ephydatia 329, 336
foetus, Tritrichomonas 180, 358, 362—364, 367, 394—397
fonbrunei, Gocevia 420, 449, 482
Foraminifera 135, 137, 140, 143—144, 451, 490, 520, 524, 527—
528, 533, 535, 537, 539, 541, 543, 545, 547, 549, 551, 553,
555, 557, 559, 561, 563, 565, 567, 569, 571, 573—575, 577,
579, 581, 583—585, 587, 589, 591, 593, 595, 597, 599, 601,
603, 605, 607, 609, 611, 624—625, 627, 629—631
formicicum, Methanobacterium 142, 180, 188, 190, 511, 516
fowleri, Naegleria 488, 490
fragilis, Dientamoeba 141, 188, 364—366, 396, 512—513, 515
francai, Phytomonas 245
Frenulata 89
Frenzelina 504
friabilis, Hyperammina 628
frigida, Виссе Ila 560, 624
frigida, Elphidiella 628—629
frobeniusi, Trypanosoma 250
fruticulosa, Ceratiomyxa 420—422, 424, 447—448
Fucus 560
Fuligo 140, 435, 438—439, 443
Fungi 134, 136—137, 139, 141, 144, 188, 296, 398, 409—410,
446—449, 499
Fungia 75
fusca, Glossina 250
fusca, Milliammina 624—625, 627, 629
fusca, Psammosphaera 629
fusca, Stemonitis 439
fuscipes, Glossina 249—250
fuscipleuris, Glossina 250
Fusiformis 353, 356, 388
fusiformis, Karotomorpha 301
Fusulinacea 579, 587, 589, 612
Fusulinida 589
Fusulinoida 579, 584, 602
galatheae, Neopilina 48
Galatheammina 517—519
galbana, Isochrysis 281
Galerida 46
galli, Blastocystis 531
gallica, Lateromyxa 493, 495, 497, 499
gallinae, Dermanyssus 248
gallinae, Trichomonas 363—364
gallinarum, Trypanosoma 248
gallopavo, Meleagris 531
gallowayi, Astrononion 627, 629
Gallus 531
gallus, Gallus 531
gambiense, Trypanosoma 250
Gasteromycetes 431
Gastroneuralia 105—106
Gastropoda 99
Gastrotricha 89, 99—101, 103, 105, 111
gaussicum, Nodosinum 628
Geamphorella 479
Geminigera 199—200
Geoplagiopyxis 479
Geopyxella 478
geosphaera, Difflugia 469
Gephyramoeba 465
Gephyramoebidae 462, 465, 467
Gephyrea 100
gerridis, Blastocrithidia 243, 254, 256
Giardia 129, 138, 140, 142, 179, 189, 338, 341, 343—348, 393,
396, 513, 515—516
Giardiidae 338, 348
giganteum, Dictyostelium 429, 449
giganteus, Megaloceras 38
Gigantomonadinae 358
Gigantomonas 366
Gillardella 478
gingivalis, Entamoeba 452
Glabratella 559, 565, 569, 571, 574—575, 580, 598
Glabratellida 607
Glaeseria 461
Globigerina 556—557, 610, 625
Globigerinata 539, 543—544, 557, 565, 574, 577, 579, 583—584,
610
Globigerinida 611
Globigerinidae 563, 578, 611
Globigerinoides 565, 574, 610, 625
Globipelorhiza 527
Globoendothyra 589
Globoquadrina 625
Globorotalia 553, 625
Globorotaliida 610
Globorotaliidae 536, 563, 611
globosa, Psammetta 517
globularis, Rosalina 574, 631
globus, Monocercomonoides 355
glomerata, Adercotryma 629
glomerata, Woronina 407
Glomodiscidae 590
Glomospira 627
Glossina, 212, 238, 247—251, 254
Glossiphonidae 247
glutinata, Globigirinita 625
Glyptotermes 356
Gnathobdellaea 62
Gnathostomulida 89, 104—105, 111, 120
Gobiella 498
Gocevia 420, 449, 482—483, 485
gomezi, Lutzomyia 246
Goniomonadidae 194, 197, 200
Goniomonas 195—197, 200
Gonyostomum 285—287, 289
gorbuscha, Oncorhynchus 94
Gordiacea 100, 111, 115
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
647
gracile, Nematostelium 419, 423
gracilis, Euglena 181, 183, 205, 207—208
gracilis, Prinia 46
gracilis, Trilobius 243
gramen, Difflugia 469
graminis, Polymyxa 398, 407
grandis, Trachelomonas 336
granifera, Dermamoeba 454
granulata, Acrasis 485
granulifera, Cercomonas 415
Granuloreticulosa 451, 520, 523—524, 528
Granuloreticulosea 136—137, 451, 498, 522, 525, 527
Graptolitha 100, 103
Graptolitoidea 99
grassei, Micromastigotes 392
grayi, Trypanosoma 248
greeffi, Pelomyxa 513
Gregarinea 56
Gregarinida 141
Grigelis 575, 596
Gromia 501—504, 508
Gromiida 451, 468, 500, 507—508
Gromiidae 500, 504
Gromiina 500—501, 503—504, 507—508
Gruberella 486, 488—490
Gruberellidae 486, 488
guatemalense, Trypanosoma 249
Guillardia 199—200
gulikae, Leishmania 245
guoyi, Alveolinella 629
Guttulina 485
Guttulinopsidae 485—486, 489
Guttulinopsis 485, 489—490
guyanense, Trypanosoma 249
guyanensis, Leishmania braziliensis 246
Gymnamoebae 467—468, 490, 516
Gymnamoebia 135, 451—453, 455, 457—463, 465—467, 482,
485, 490, 499, 512, 516
gymnodactyli, Leishmania 245
Gymnophryidae 522, 524
Gymnophrys 137
gyrans, Pavlova 283
Gyraulus 79
Gyroidina 628
Gyropaigne 209
gyrosum, Physarum 437
habilis, Homo 76
Habrobracon 59
Haddonia 577
Haematomonas 250
haematopotae, Crithidia 248
Haemopis 241
Haemosporidia 56
Halisarca 120
Haplogyne 69
Haplophragmiida 598
Haplophragmioidea 599
Haplophragmoides 627
Haptophyceae 275—276, 281, 284
Haptophyta 135—137, 143—144, 276—277, 279, 281, 283—284,
584
Hartmannella 461
Hartmannellidae 452, 460
Hastigerina 556, 558, 563, 569, 572, 574
Hastigerinidae 563, 611
Hegneriella 305—306, 309—313
Heleopera 471—472, 476, 479
Heleoporella 479
helicis, Cryptobia 216, 219, 227, 241
Heliozoa 137, 467, 484, 490
Heliozoea 468, 490, 516
Helix 89, 289, 312
Helkesimastix 415, 417
Hemichordata 53, 99—100, 102, 104, 106, 109—110, 113
Hemigordiopsidea 593
Hemigordiopsina 593
Hemiptera 232, 243, 245, 252—254, 256
Hemirhamphus 54
Hemiselmididae 194, 197, 200
Hemiselmis 197, 200
Hemistasia 213, 241, 253
Hemitrichia 434, 439, 442
herbatica, Stemonitis 438, 447
Hergottelloidea 591
Herichthys 228
herkulea, Gigantomonas 366
Herpetomonas 213—215, 221—222, 236, 243, 248—250, 254—
255
Herpetosoma 229—230, 235, 247, 249
hertigi, Leishmania 226, 245, 253
Heterakis 364
Heteramoeba 488
heteranterae, Membranosorus 409
heterocariotica, Rotaliella 569, 574
Heterochelicida 610
Heterocyclina 610
Heteroglypha 507
Heterogromia 523—524
Heterolobosea 135, 137, 141, 143—144, 451—452, 485—490
Heteromita'lM, 412—417, 445
Heteronema 202, 209
Heteronematida 202, 209—210
Heterosigma 285—286, 288—289
Heterostegina 548, 563, 574—575, 610
Heterotermes 392
Heterotheca 559
hexactis, Ophionotus 62
hexagona, Globoquadrina 625
Hexamastix 365—366
Hexamita 129, 183, 338, 346—348, 392—393, 397
Hexamitidae 338, 341, 345, 347—348, 392—393
Hibberdia 270
Hibberdiaceae 270
Hibberdiales 270, 274
Himatismenida 451, 468, 482—484
Hipparion 81
hippicum, Trypanosoma 250
hipposideri, Trypanosoma 249
hirsuta, Centropyxis 473
Hirudinea 99, 103, 238
Hirudinidae 247
Hirudo 241
Histiona 275
histolytica, Entamoeba 179, 395—396, 452, 461
Histomonas 363—366, 397
Hodotermitidae 377, 390, 392, 397
Hologlypha 507
Holomastigotes 392
Holomastigotoides 376—378, 392, 397
Holomastigotoididae 369, 372, 375, 377, 386, 391
hominis, Blastocystis 528, 530—531
hominis, Pentatrichomonas 363, 365, 367
Homoptera 232
Hoogenraadia 479, 484
Hoplonympha 389
Hoplonymphidae 369, 372, 374—375, 377—378, 386, 389
Hormosinana 538, 549, 585, 595, 598
Hormosinella 575
Hormosihida 538, 595
648
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Hormosinoidea 538, 596
Hornellia 289
Hospitellidae 586
Hottingerellida 538, 552, 578, 590
Hottingerellidae 548, 591
Hottingeritidae 589, 599
howa, Postelectrotermes 356
hyalina, Spiroloculina 561, 569
Hyalodiscidae 465
Hyalodiscus 449, 465, 485, 498
Hyalomma 235, 255
hyalommae, Blastocrithidia 232, 235, 255
Hyalosphenia 469, 471—472, 474, 476, 479
Hyalospheniidae 468, 477, 479
Hydramoeba 460
Hydrozoa 100, 120
Hydruraceae 270
Hydrurales 270, 274
Hydrurus 270
Hyla 313
Hylidae 309
Hymenomonas 280, 284
Hymenoptera 61, 232
hymenostoma, Oxytricha 171, 187
Hyopsodus 79—80
Hyperammina 628
Hyperamoeba 135—137, 186—187, 418, 444-^145
Hypermastigida 129, 135, 137, 140, 179, 353, 355, 358, 368—
369, 371, 373, 375, 377, 379, 381, 383, 385—387, 389, 391—
397
Hyponeura 98, 100
Hypopylaria 245
Hypotricha 35, 64, 70
Hypotrichomonadinae 358, 365
Hypotrichomonas 365—366
Hyracotherium 42, 80
Ichthyobdellidae 247
Ichthyobodo 213, 227, 241
Ichthyostegalia 81
Idionympha 389
ignotum, Trypanosoma 250
illinoisense, Chaos 459
Imantonia Til
indica, Cryptobia 241
indicum, Trypanosoma 248
infantum, Leishmania 245—246
inflata, Globorotalia 625
inflata, Hexamita 346
inflata, Microrhopalodina 352
inflata, Trochammina 560, 625, 629
Infusoria 100
inopinatum, Trypanosoma 231, 248
Insecta 41, 50, 69, 99, 104, 232—233
intermedia, Lutzomyia 246
intermedia, Rubratella 571
intermedia, Zelleriella 306
intermedius, Phlebotomus 245
intestinalis, Chilomastix 313
intestinalis, Cryptobia 216, 219—220, 222, 228, 241
intestinalis, Giardia 179, 338, 340, 346—347
intestinalis, Lamblia 338
intestinalis, Protoopalina 306, 309
intestinalis, Trichomonas 528
Invertebrata 98, 119
Involutinida 590 .
Involutinoida 590
Involutinoidea 590
lodamoeba 461
iridescens, Lepidoderma 436
Iridia 549, 567, 572, 574
iridis, Didymium 436—438, 446, 449
irregularis, Comatricha 435
irridipennis, Glyptotermes 356
islandica, Islandiella 560, 629
Islandiella 560, 626, 629
Islandiellidae 630
Isochrysidaceae 276
Isochrysidales 276, 281, 317, 320
Isochrysis 277, 281
Isonema 211
Isoptera 387, 397
Isotricha 190
ithacus, Rosculus 489
jaculans, Pleuromonas 186, 240, 251
jaculum, Leptomonas 235, 243, 252
Jadammina 574, 601, 625, 627
jaegeri, Zelleriella 312
jahieri, Rhyzonympha 388
Janickina 461, 468
japonica, Fibrocapsa 289
japonica, Opalina 302
jensoni, Elphidium 629
Joenia 369, 376—380, 387, 396
Joeniidae 369, 386—387
Joeniides 396
Joenina 369, 388
Joeninia 378
Joenopsis 388
jubilans, Cryptobia 216, 219, 222, 228
junci, Ligniera 407
Jungia 479
kaiseri, Rhizoplasma 520
Какоеса 334
Kalotermes 397
Kalotermitidae 397
Kamptozoa 99—100, 104—106, 112—113, 117
Kamuraninae 557
Karaisella 589
Karotomorpha 297—298, 301, 311—313
Karyoblastea 510, 512, 514
karyophilus, Leptomonas 243
Karyorelictida 70
Kentrosiga 330, 336
Khawkinea 205—206
Kinetoplastida 143, 186, 213, 240, 251—256, 299
Kinetoplastidea 135, 137, 143, 211, 213, 215, 217, 219, 221, 223,
225, 227, 229, 231, 233, 235, 237, 239, 241, 243, 245, 247,
249, 251—253, 255, 263, 265
Kinetoplastmonada 252
Kinorhyncha 99—101, 103, 105, 111, 115
Klebsiella 346
kleistobolus, Licea 434
Kofoidiidae 369, 386, 388
kohl-yakimovi, Tripanosoma ninae 250
Котта 199—200
Komokia 527
Komokiida 135, 137, 524—527
Komokiidae 527
Komokioidea 525, 527
Korotnevella 454, 461, 467
Koruga 386, 392, 396—397
Kutsevella 589
Labyrinthidomatidoidea 599
Labyrinthodyction 296
Labyrinthomorpha 144, 291, 294, 296
Labyrinthomorphea 135, 137, 290—291, 293, 295—296
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
649
Labyrinthomyxa 290, 293, 295—296
Labyrinthula 290—291, 293—296
Labyrinthulea 291
Labyrinthulida 135, 137, 291, 295—296
Labyrinthulidae 290—291, 295—296
Lacerta 52, 56, 300
lacertae, Trichomitus 363, 532
lacteum, Dendrocoelum 227, 241, 427, 430
Lagena 560, 629
Lagenida 596
Lagenidae 596
Lagenidiopsis 504
Lagenodifflugia 470, 479
Lagenoeca 331
Lagenoglandulina 575, 578
Lagomorpha 52
Lagottrix 93
Lagynana 537, 543, 569, 572, 574, 580, 583—586, 611
Lagynata 549
Lagynida 585
Lagynidae 586
Lamblia 67, 138, 179, 338, 341, 345—346, 395, 513, 515—516
lamblia, Giardia 138, 140, 142, 338
Lamellisabella 119
Laminaria 296, 560, 627
Lamproderma 434—436, 443, 448
Lamtopyxidae 468, 479, 484
Lamtopyxis 479
Lamtoquadrula 479
Lana 527
Lasiodiscida 538, 552, 590
Lasiodiscidae 548, 590
lateritia, Vampyrella 495
Lateromyxa 493, 495, 498—499
Laticarinina 558
laticollaris, Allogromia 525, 528, 537, 565, 569, 572, 631
Latimeria 48
latro, Birgus 54
latum, Gonyostomum 288
Lecithoepitheliata 60
lectularis, Cimex 249
Lecythium 504
Leda 46
leei, Rosalina 569
legeri, Protrichomonas 365
Leishmania 212—213, 215—216, 220, 224—226, 230, 235—236,
240, 245—247, 252—256
lenta, Sphenoderia 502
Lepidoderma 436, 443
Lepidoptera 91, 232
Lepocinclis 205
Leptochlamys 479
Leptodactylidae 309
Leptodiscus 172
Leptomonas 213, 215, 220—221, 223, 231, 235—236, 243, 248,
252, 254, 256
Leptomyxa 412, 463—465, 468
Leptomyxida 451, 462, 466—467
Leptomyxidae 462, 465, 467
Leptophrys 492, 498
Leptoramosina 462, 465, 467
leptosomum, Acytostelium 430, 447, 449
Leptospironympha 383—384, 386, 391, 395
leptovasorum, Phytomonas 245
Lesquereusia 469—471, 476, 479
Lesquereusiidae 468, 479
leucopodia, Diachea 436
Leucotermes 391
lewisi, Trypanosoma 212, 229—230, 235, 249—250, 253, 255
Lewisonella 249
Licea 434—435, 439, 441, 447—448, 450
Liceaceae 441
Liceales 431, 440—441, 447
Liceida 431—432, 434—436, 440—442
Liceidae 434, 441
Lieberkuehnia 523—524
Lieberkuehnidae 523—524
Ligniera 398, 404, 407, 409—410
Lima 46
Umax, Berthelinia 41
Umax, Tamanovalva 119
limnetica, Difflugia 476
Limulus 115
Lincus 245
Lindbladia 441
lineare, Trinema 504
lingardi, Trypanosoma 248
Lingula 48
Lingulina 539, 597
Liphistiomorphae 69
lipophora, Stachyamoeba 488, 490
Listerella 441—442
littorea, Littorina 560
Littorina 560
Lituolida 599
Lituolinoidea 599
Lituoloida 598
Lituotubidae 577, 589
lobata, Heterotheca 559
lobatulus, Cibicides 537, 565, 567, 569, 571—572, 574
lobifera, Amphistegina 563
Loboreticulatida 451, 465—467
Lobosea 135, 137, 176, 420, 451—453, 455, 457—459, 461, 463,
465—469, 471, 473, 475, 477, 479, 481, 483, 485—486,
488—490, 510, 512, 515
lobostoma, Difflugia 469
Loftusioidea 599
logocaudense, Trypanosoma 249
longa, Astasia 205, 207—208
Longiapertina 580
longipalpis, Glossina 249—250
longipalpis, Lutzomyia 247, 256
longipes, Phlebotomus 246
longipontis, Cribroelphidium 625—627
longispina, Oolina 627
Lopadorhynchus 119
Lophomonadidae 369, 386, 388
Lophomonadina 369, 386—387, 392, 394—395
Lophomonas 368—369, 377—378, 386, 388, 394—396
Lophophorata 103, 113
Loricifera 105, 111, 119
loriculata, Kofoidia 388
Loxodes 70, 148, 516
Loxodonta 59
Loxostomatoidea 603
lucida, Iridia 567, 572, 574
lucifugus, Reticulitermes 357, 396
lucilia, Crithidia 243
lunatum, Echinostelium 440
luridus, Troilus 245, 252
Lurus 44
lutea, Trichonympha 390
luteus, Olisthodiscus 288—289
Lutzomyia 246—247, 256
Lycogala 435, 439, 441, 446
Lygaeidae 245, 249, 254, 256
Lygaeus 256
m’baikiensis, Sauromonas 356
Macaca 256, 531
42 Протисты, часть 1
650
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Macbrideola 434
Machairodus 38
Масота 296
macrescens, Jadammina 625, 627
macropora, Rosalina 559
Macrospironympha 383—384, 391
Macrostoma 341, 532
Maghrebia 479, 484
magna, Hibberdia 270
magna, Trichonympha 380
major, Leishmania 246
Malakostraca 61
Mallomonadaceae 270, 273
Mallomonas 270—273
Malmiana 228
Mammalia 48, 61, 82
Mandibulata 99, 103—104
Manorella 577
mantis, Squilla 58
marginalis, Sorites 565
marginata, Bulimina 557
marginata, Cepedea 313
marginatus, Dityscus 91
Marginopora 629
Marginulina 628
mariagerensis, Postgaardi 205, 211
marina, Massisteria 415
marmoratus, Scorpaeniehthys 228
marmoratus, Triturus 528
marocanum, Trypanosoma 250
Maroltia 504
Marsupialia 97
martiali, Certesella 477
martini, Phlebotomus 246
Martinotiella 627
Masonella 556, 579
Massisteria 415, 417, 444
Mastigamoeba 133, 137, 468, 512, 514
Mastigamoebida 128, 512
Mastigella 128, 511—514, 516
Mastigina 128, 512, 514
Mastigophora 56, 67, 190, 251—255, >13
Mastotermes 385—386, 392, 396
Maudammina 517—519
maximus, Trichosphaerium 483
Mayorella 176, 454, 461, 467
Mecoptera 232
medicinalis, Hirudo 241
mediterranensis, Discorbis 560
Megaloceras 38
Megastoma 338, 469
Megatrypanum 230, 247—248
Melathrokerion 589
meleagridis, Blastocystis 531
meleagridis, Hexamita 393
meleagridis, Histomonas 363—366
meleagridis, Spironucleus 346, 348
Meleagris 531
meleagris, Numida 531
melleum, Physarum 438
melolonthae, Monocercomonoides 352, 396
melophagium, Trypanosoma (Megatrypanum) 248
Melophagus 248
Membranosorus 409
menardii, Globorotalia 625
menneri, Vendomia 75
Menoidium 209
Merostomata 99
Merotricha 285—286, 288
Meryhippus 80—81
mesnili, Chilomastix 341, 397, 532
Mesohippus 80—81
Mesozoa 88, 99—100, 102—103, 110—111
mespiliformis, Cucurbitella 469
Messemvriella 504
Metaheleopera 479
Metamonada 125, 128—129, 133, 141—142, 144, 188, 512, 515
Metaphyta 49, 76
Metarotaliella 571, 575
Metatrichia 434, 436, 442, 448
Metazoa 35—36, 43, 48—49, 52, 60, 67, 71, 73, 76, 99—104,
106, 110—111, 116—120, 128, 130, 133—134, 137—141,
144, 328
Methanobacterium 142, 180, 188, 190, 511, 516
Metopus 177, 183, 190, 511, 516
mexicana, Leishmania 245—247
Micramphora 504
Micramphoraeopsis 504
Microchlamyidae 468, 478
Microchlamys 470, 472, 478, 485
Microcometes 523—524
Microcorycia 478
Microcoryciidae 468, 478
Microgromia 523—524
Microgromiidae 523—524
Microjoenia 388
Micromastigotes 392
Micropsammella 504
Micropsammelloides 504
Microquadrula 507
Microrhopalodina 348, 352—353, 356
Microsporidia 52, 110, 123, 125, 128, 137—138, 141, 188, 190,
512
Microsportella 335
midwayensis, Robulus 557
migonei, Phlebotomus 245
Milettioidea 604
Miliolana 538, 544, 563, 578—579, 591—592
Miliolata 538, 544, 561—562, 565, 574, 577, 583—585, 588—
589, 591
Miliolida 593
Miliolidae 577, 594, 631
Miliolina 538, 594, 627
Milioliporinae 557
Milioloida 538, 578, 593
Milioloidea 538, 594
milliaris, Spirorbis 93
mimosa, Dimastigella 242, 252—253
Minakatella 435, 441—442, 448
Miniacina 577, 608
Minouxia 583
minutum, Dictyostelium 427, 430
minutum, Echinostelium 436, 439—440
Miohippus 42, 80
mirabilis, Dinamoeba 461
mirabilis, Teratonympha 390
Miridae 252
miridarum, Blastocrithidia 243, 252
Mississippiella 472, 479
Mississippiellidae 468, 470, 479
mitra, Bicosoeca 266
mobilis, Caviomonas 341
Modiolus 46
Moeritherium 81
Mollusca 42, 99—100, 102—106, 110, 119
Molluscoidea 100
Monas 173, 187
mongolensis, Phlebotomus 246
moniliforme, Resigella 624
moniliformis, Desmarella 322, 326
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
651
Monocercomonadidae 358, 365, 394
Monocercomonadinae 358, 365, 397
Monocercomonas 363, 365, 396
Monocercomonoides 348—352, 355, 386, 393, 396
Monocercomonoididae 349
Monocosta 334
Monogenea 88, 99, 103
Monoplacophora 20, 48, 88—89, 99
Monosiga 325—328, 330, 336
Monosigida 137
Monosigidae 322, 328—329, 336
Monotaxinoididae 590
Monothalamia 135, 137
Monothalamida 508, 522—524
Monotremata 51
monterogei, Endotrypanum 247^ 253
Moravamminidae 589
moshkovskii, Entamoeba 461
mossoense, Trypanosoma 250
Mucilago 435
mucoroides, Dictyostelium 427, 429—430, 446—448
multiforme, Trypanosoma 250
multilineata, Triadopsis 219, 253
Multituberculate 51, 76
muralis, Lacerta 56
muris, Tritrichomonas 358, 361—364, 368, 395
murium, Trypanomonas 249
murmanense, Trypanosoma 248
muscarum, Herpetomonas 222, 243
musculi, Trypanosoma 235
mutabilis, Hexamita 346
Mycetozoa 100, 135, 137, 141, 398, 410—411, 413, 415, 417,
419, 421, 423, 425, 427, 429, 431, 433, 435, 437, 439, 441,
443—445, 447—449, 451, 486
Mycota 431
Mygalomorphae 69
Myriapoda 41, 50, 62, 99
Mystacocarida 89
Myxochrysidaceae 269
Myxochrysis 262, 269
Myxogastres 431, 447
Myxogastria 135, 137, 418—419, 431, 439—440, 443
Myxogastromycetidae 431
Myxomycetes 186, 398, 417—418, 431, 443, 445—450, 524
Myxomycota 418, 446
Myxomycotina 431
Myxosporidia 123
Myxotheca 565, 567, 569
Myxozoa 137, 143, 188, 446
Myzostomida 89, 99, 112
Myzostomidae 117
nabicula, Leptomonas 220
Nabidae 245, 252
Nadinella 504
Naegleria 486, 488—490
Naegrelia 141
Nannoeca 334
Nannomonas 248, 250
nanum, Trypanosoma 250
nasuta, Rhynchomonas 241
natans, Sargassum 560
natator, Hoplonympha 389
Naucoris 66
Nautiloidea 89
Nautilus 48
Nebela 178, 469, 471—472, 476-477, 479, 484
Nebelidae 479
necator, Ichthyobodo 227, 241
necatrix, Costia 241
necatrix, Vairimorpha 138
nelsoni, Hexamita 346, 348
Nemathelminthes 98—103, 106, 110—111, 118
Nematochrysopsidaceae 270
Nematoda 88, 99—101, 103, 105, 111
Nematodes 99
Nematomorpha 99, 101, 103, 111
Nematorhyncha 103, 111
Nematostelium 419, 423—424
Nemertinea 102
Nemertini 53, 98—100, 102—106, 110, 119
Nemertodermatida 53
nemestrina, Macaca 531
Neoconorbina 583
Neocribrella 583
Neogloboquadrina 625
Neoparamoeba 454, 458, 461
Neopilina 48, 518
neopolitana, Ochrosphaera 282
Neorhabdocoela 44
Neotermes 356, 387, 391
Nepa 66, 235, 243, 252, 255
Nephroselmis 189
Netzelia 469, 471, 479
nevadensis, Zootermopsis 357
Nezzazatidae 589, 599, 602
nigripes, Didymium 438—439, 448
nipponica, Pararotalia 629
nitida, Candeina 625
nitrophilus, Parabodo 240—241, 2S2
nivarletti, Rhinoestrus 235
nodosa, Stegophiura 62
Nodosaria 552, 557
Nodosariana 538, 560, 577—579, 595—597
Nodosariata 538, 544, 571, 578, 583—586, 589, 595
Nodosariida 596
Nodosariina 538, 596
Nodosarioida 577—578
Nodosarioidea 597
Nodosinum 543, 583, 595, 598, 628
Nonionella 557
Nonionida 539, 563, 609
Nonionina 609
Nonionoida 609
Nonionoidea 609
norcrossi, Islandiella 626
nordicus, Phytomonas 234, 245, 252
Normanina 527
norrisii, Stephanoeca 335
norvegicus, Rattus 249
Nosopsyllus 249, 255
Notila 351—352, 354—355, 357, 393
Notodendrodes 559
Notoneuralia 105—106
Notoneuron 105
Notorotaliidae 609
Notosolenus 209
Nouriida 596
novaezealandiae, Hyperammina 628
Nubeculariida 593
Nubeculariidea 593
Nubeculina 55.7, 592
Nubeculinopsis 577
Nucellosphaeridium 73
Nuclearia 491—492, 494—495, 498—499
Nucleariidae 490—491, 495, 497—499
Nucleosphaerium 499
Nucula 46
Numida 531
numidae, Blastocystis 531
652
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
numidae, Trypanosoma 248
Nummulitacea 574
Nummulites 610
Nummulitida 539, 578, 601, 610
Nummulitidae 563, 610
obtrigonoidea, Opalina 302, 313
occidentalis, Textularia 600
Occultammina 518—519
ochraceus, Anacanthotermes 388, 392
Ochromonadales 269
Ochromonas 173, 188, 262, 269, 275
Ochrosphaera 280, 282
Octomitus 344—345, 347—348, 393, 396
Octomyxa 406—407, 409
Oestridae 235
Ogdeniella 504
Oligochaeta 99, 103, 112
Oligomera 99
Oligonema 442
Olisthodiscus 285—286, 288—289
olmeca, Lutzomyia 247
Olynthus 100
oncopelti, Crithidia 226, 243, 244
oncopelti, Leptomonas 223, 227
Oncorhynchus 94
Onega 75
Onychophora 88, 99—100, 103—106, 110, 112, 119
Oolina 627—628
Oomycetales 290
Oomycetes 135—137, 144, 290
Oopyxis 479
opaca, Ovammina 565, 569
Opalina 170, 302—303, 305—306, 308—309, 311—313
Opalinata 135—137, 297, 299, 301, 303, 305, 307, 309, 311—313
Opalinatea 135, 137, 301, 303, 305, 307, 309, 311—312
Opalinida 303, 312
Opalinidae 303, 311—313
Opalozoa 188, 411, 446
opercularis, Chlamys 560
operculata, Deltotrichonympha 392
Operculina 575, 610
Ophionotus 62
Ophryoscolecidae 64
Opisthomitus 353, 355—356, 393, 396
Opistomitus 351
orbiculare, Haynesina 629
orbicularis, Sorites 565, 569, 574
Orbitoides 630
Orbitoidida 610
Orbitolina 598
Orbulina 557, 575, 578, 630
orculaeformis, Parvicorbicula 334
orculaeformis, Pleurasiga 333
Oridorsalis 583
orientalis, Phlebotomus 246
ornatissima, Glabratella 559
Orohippus 80
Orthokarstenia 578
Orthonectida 99, 103, 111
Osangularia 578
Osteolepiformes 50, 81
Ostracodermi 76
Ostracodes 631
Ostraea 46
ovalis, Karotomorpha 301
Ovammina 565, 569
ovata, Monosiga 325—327
ovatum, Gonyostomum 288
ovatum, Nematostelium 423
oviformis, Gromia 501, 508
oviformis, Cryptodifflugia 481
ovinus, Melophagus 248
Ovis 532
ovis, Blastocystis 532
Oweniidae 105, 114—115
Oxymonadea 135, 137, 337, 341, 348—349, 351, 353, 355, 392—
393
Oxymonadida 129, 137, 179, 188, 341, 344, 347, 349, 355, 386—
387, 393, 396—397
Oxymonadidae 349, 355—356, 393
Oxymonadina 348
Oxymonadinae 350—352, 354—356, 393, 396
Oxymonas 348, 351—353, 355—356, 386, 393, 396
Oxytricha 171, 187
Ozawainellida 589
pachyderma, Neogloboquadrina 625
Paedophoropus 60, 65
Paguridae 54—55
Palaeodictyoptera 38
Palaeospiroplectamminida 589
pallida, Stemonitis 436, 447
pallidipes, Glossina 249—250
Palludicellina 528
palpalis, Glossina 249—-250
palustris, Pelomyxa 128, 142, 176, 183, 187, 190, 451—452, 467,
508—516
Pamphagus 177, 504
Pan 531
panamensis, Lutzomyia 246
pangonia, Crithidia 248
pangoniae, Leptomonas 248
Pantopoda 69, 99, 106, 112
papatasi, Phlebotomus 246
Paphia 46
papilio, Hyalosphenia 469, 474, 476
Parabasalea Г35, 137, 140—141, 337, 357—359, 359, 361, 363,
365, 367—369, 371, 373, 375, 377, 379, 381, 383, 385, 387,
389, 391—394
Parabasalia 512—513, 515
Parabasalidea 386
Parabodo 213, 240, 252
Parabrizalininoidea 603
parabyssorum, Rhabdammina 549
Paracentropyxis 478
Parachaos 460
Paradermamoeba 454, 460, 467
Paradiacheopsis 442
paradoxa, Listerella 441
paradoxa, Myxochrysis 262, 263, 269
paradoxa, Pelomyxa 513
paraensis, Lutzomyia 246
Paraflabellula 462, 465
Paragocevia 483
paraguayense, Craterium 439
Parahistomonas 366
Parajoenopsis 388
Paramecium 170, 181—182, 184—187, 313, 458
Paramoeba 458, 461, 467
Paramoebidae 452, 461, 466—467
Paranebela 479
Paraneotermes 388—389
Parannelida 100
Paranotila 357
Paraphysomonadaceae 269
Paraphysomonas 173, 180, 260
Paraquadrula 4П\, 476, 479
Paraquadrulidae 468, 479
Pararotalia 559, 629
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
653
Pararthropoda 103
parasitica, Licea 434
Parastasia 204—207, 209—210
Paratetramitus 488
Paratikhinellidae 588—589
Paratrichomonas 365
Parazoa 99, 101—103, 110, 115
Parenchymia 105
Parenteroxenos 36, 115
Pareuglypha 507
parisii, Pentatrichomastix 365
Parmulina 478
parva, Chrysochromulina 277
parva, Hexamita 348
parva, Metarotaliella 571
Parvamoeba 460
parvialveolata, Pelomyxa 513
Parvicorbicula 334
parvum, Prymnesium 280, 282, 284
pascheri, Woloszynskia 180, 190
Pashysoeca 331
patella, Microchlamys 470, 485
patelliformis, Discorbis 569, 571
patelliformis, Glabratella 580
Patellina 552, 556—557, 559, 562, 565, 567, 569, 572, 575, 625
Patellinida 591
Patellinidae 577, 591
Patellinoidea 591
Paulinella 502, 507
Paulinellidae 500, 507
pauperata, Laticarinina 558
Pavlova 277, 280—281, 283—284
Pavlovales 276—277, 282, 284
Pavlovophycidae 276
Pavonina 628
Pavoninoidea 604
pecaudi, Trypanosoma 250
pecorum, Duttonella 250
pecorum, Trypanosoma 250
pedicellata, Diaphanoeca 332, 334
pedifer, Phlebotomus 246
Pedinellales 187
Pedinellomorpha 136—137
Pedinomonas 186
Pegidiidae 578, 606
pelagica, Hastigerina 569, 572, 574
pelagicus, Coccolithus 282
Pelagomonadaceae 273
Pelagomonadales 273
Pelagomonas 273, 275
Pelagophyceae 135, 137, 258, 273—275
Pellatispiridae 610
Pelobates 36
Pelobatidae 309
Pelobiontea 512
Pelobiontida 128, 451, 509
Pelobius 467, 509, 514—515
Peloflagellata 133, 137, 467, 514
Peloflagellatea 135, 451, 508—509, 511—515
Pelomyxa 128, 142, 176—177, 183, 187, 190, 451—452, 467—
468, 508—516
Pelomyxidae 509
pelophila, Phanerobia 242
Pelosina 581
Peltogaster 36
Penardeugenia 504
Penardia 136—137, 451, 522, 524
penardii, Vacuolaria 288
Penardochlamys 478
Peneroplidae 563, 594
Peneroplidea 538, 577, 594
Peneroplis 592, 594, 627, 629
Pennatulida 74
Pentagonia 479, 484
Pentastomida 99—100, 103—104, 106, 110, 112, 119
Pentatomidae 245, 252, 254
Pentatrichomastix 365
Pentatrichomonadinae 365
Pentatrichomonas 363, 365, 367
Peranema 176, 184, 202, 207, 209
percae, Trypanosoma 248
Percolomonas 141, 143, 489—490
peregrina, Uvigerina 626
peregrinia, Vannella 462, 467
perforans, Vampyrellidium 492, 495, 499
Perichaena 436—438, 441—442, 446, 449
Periloculina 575
Periplaneta 368, 386, 388, 394
Periptigma 574, 583, 586
Peripylaria 245
perissa, Idionympha 389
Peritricha 56, 59, 176, 178, 185, 190
Perkinsiella 468
perlucida, Lagena 629
Pernina 488
perplexa, Choanoeca 323, 325, 336
persica, Veronica 399
peruensis, Lutzomyia 246
pessoai, Lutzomyia 246
pessoai, Phlebotomus 245
Pessonella 461
Petalomonas 207, 209
Petalonamae 74
Petelopella 482
Phacus 176, 205—206, 209—210
Phaeocystis 276—277, 280
Phaeodermatium 270
Phaeophyta 144
Phaeoplacaceae 269
Phaeosacciaceae 270
Phaeothamniaceae 269
Phagocytellozoa 110
Phagomyxa 408, 410
Phalansteriida 186, 314, 319—320
Phalansteriidae 314, 316, 319
Phalansterium 314, 316, 319—320
Phanerobia 242
Phenacophragma 575
Phlebotomus 245—246
phloiogenum, Protophysarum 435, 443, 446
Phoronida 53, 88—89, 99—100, 102—103, 105—107, 112—113,
118
Phoronoidea 106
Phoxichilidium 631
Phreatamoeba 133, 137, 452, 461, 512, 514—515
Phreatamoebida 512
Phryganella 470, 476, 482
Phryganellidae 468, 482
Phthanotrochidae 586
Phthanotrochus 572, 583, 586, 588
Phyllomitus 241
phyllostomae, Trypanosoma 249
Physarales 431, 440, 442
Physarida 432, 434—436, 440, 442—443
Physaridae 434, 436, 443
Physarum 434, 436—439, 443, 446—450
Phytomastigophorea 276, 285
Phytomonadida 35
Phytomonadina 99
Phytomonas 213, 216, 220, 225, 229, 233—236, 245, 252—254
654
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Picnogonum 560
pifanoi, Leishmania 247
Pilulina 559
Pinaciophora 498
Pipidae 309
Piroplasma 246
Piroplasmida 56
Piscicola 62
piscicola, Zelleriella 312—313
pisciformis, Siredon 36
Placentalia 51, 97
Placocista 502, 507
Placojoenia 369, 372, 376—379, 388, 394, 397
Placopsilinidae 577
placopus, Gocevia 420, 449, 485
Placozoa 88, 104, 110, 118
Plagiophrys 504
Plagiopyxidae 468, 479
Plagiopyxis 470, 477, 479, 485
planatus, Peneroplis 629
planctonica, Bicosoeca 265
Planctosphaerea 88—89
Plancus 535
Planhoogenraadia 478
Planoarchaediscidae 590
Planoprotostelium 419—420, 424, 449
Planorbis 93
Planorbulina 560
Planorbulinida 607
Planorbulinidae 607
Planorbulinina 607
Plantae 134, 136, 138—139, 144
Planuloidea 99
Plasmodiophora 135, 137, 398—410
Plasmodiophorales 410
Plasmodiophorea 398, 408—409
Plasmodiophorida 398, 409
Plasmodiophoridae 398, 408—409
PlasmodiOphoromycetes 410
Plathelminthes 49, 98—100, 102—106, 108, 110—111, 114,
119
Platodes 99
Platyamoeba 461
Platyhelminthes 120
Platypleura 332, 334
platyrhynchos, Anas 531
platystoma, Hyalosphenia 472
Playfairina 507
Plethodontidae 65
Pleurasiga 334
Pieurochrysis 277, 284
Pleurocystis 284
Pleuromonas 186, 240, 251
Pleurostomellina 597
plicata, Marginopora 629
Pliohippus 42, 80
Pocheina 485, 489—490
pochmannii, Chroomonas 195
Podaxonia 106, 113
Poecilia 52
Poeciliopsis 52
Pogonophora 40, 42, 53, 88—89, 99—100, 102—106, 108—110,
113, 115, 120
Polistomella 572
polycephalum, Dictyostelium 427
polycephalum, Physarum 436—439, 443
Polychaeta 40, 99, 103, 105, 112—115, 186
Polychaos 454, 460
Polycladida 60, 75, 105
polycystis, Woronina 402, 407
Polyfibula 334—335
Polykrikos 67
Polymastigida 67, 337
Polymastigidae 348—349, 355, 393
Polymastigina 348, 358
Polymastigota 133, 135, 137—138, 337—339, 341, 343, 345,
347—349, 351, 353, 355, 357—359, 359, 361, 363, 365,
367—369, 371, 373, 375, 377, 379, 381, 383, 385, 387, 389,
391—393, 395, 397, 399
Polymastix 348—349, 351, 353, 355, 393, 396
Polymera 99
Polymorphinida 597
Polymorphinina 597
Polymyxa 398—400, 402, 404, 406—407, 409—410
Polyoeca 334—335
Polyplacophora 70, 99
Polypodium 25
poly рога, Arcella 472
Polyposthia 68
Polyposthides 68
Polyposthiidae 68
Polysiphonia 560
Polysphondilium 425, 427, 429—430, 447—449
Polytoma 161
polytricha, Joenopsis 388
Polyzoa 102
pomatia, Helix 89, 91
Pomoriella 477, 479
Pompholyxophryidae 490—491, 493, 495, 498—499
Pompholyxophrys 493, 498—499
ponticum, Elphidium 565
Pontigulasia 470, 476, 479
Pontomyxa 520—521, 524
porcellus, Cavia 531
porci, Trypanosoma 250
Porifera 49, 99, 102, 104, 106, 110, 115, 119
Poroeponides 578
Porolepiformes 50
Porotermes 388, 391
Porphyra 139, 188
Postelectrotermes 356, 392
Postgaardi 205, 211
Poteriodendron 264
pouchetii, Phaeocystis 277, 280
Prasinophyceae 148, 189
Priapulida 88—89, 99—100, 103, 105, 111, 115
Priapuloidea 102
Prinia 46
pristina, Protrichonympha 390
Proboscidea 186
Proboscidiella 355—356, 393, 397
Prochordata 98, 100
Procoelomata 108—109
Procryptobia 213, 227, 242, 256
Progoneata 104
Projoenia 388
Prolecithophora 53
Proleptomonas 212, 236
prolifera, Caulerpa 140
Prolixoplectidae 556, 601
Prolophomonas 386, 388, 395
Proplagopyxis 478
Prorhinotermes 392
Prosopyga 106
Prosopygia 113
Protacanthamoeba 466
Protencephala 102, 108
proteoides, Pseudothecamoeba 511, 516
Proteomonas 196, 200, 243, 252
Proteomyxa 408
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
655
Proteomyxida 296, 408
Proteomyxidea 296
Proteromonadea 135, 137, 297, 299, 312
Proteromonadida 313
Proteromonadidae 297, 301, 312—313
Proteromonas 297—298, 300—301, 311—313
Proterospongia 329, 336
Proteus 509
proteus, Amoeba 164, 181, 184, 452, 454, 458, 460
proteus, Notila 357
proteus, Volvox 452
Protheroteridae 41
protheus, Chaos 452, 509
Protiosphargis 44
Protococcus 469
Protocucurbitella 479
Protodonta 76
Protomonadida 337
Protomonadina 254
Protoopalina 302—303, 306, 309—313
Protophysarum 435, 443, 446
Protoplagopyxis 478
Protomaea 105
Protospongia 328—330
Protosporangium 419—421, 424, 449
Protostelia 135, 137, 418—419, 424, 430, 443, 447
Protostelida 141, 414, 417, 449, 513
Protosteliidae 424
Protosteliomycetes 418
Protosteliopsis 424
Protostelium 423—424, 449
Protostomia 49, 93, 98—99, 101—110, 113, 115
Prototrichia 436, 442
Protozoa 59—60, 67, 99—100, 102—104, 106, 110—111, 118,
123, 134, 136—137, 140, 143, 186—190, 201, 211, 252—255,
275, 284, 289, 296, 313, 395—398, 408, 410, 418, 446, 448—
449, 468, 485, 490, 499, 508, 510, 512—516, 520, 524, 527—
528
Protrichomonadinae 358
Protrichomonas 365
Protrichonympha 390
Protrochozoa 106—107
Prowazekella 301
Prowazekia 240, 301
proximo, Euglena 207
Prymnesiales 276—277, 282
Prymnesiida 276, 284
Pfyriihesiomonada 584
Prymnesiophyceae 275—276, 284
Prymnesiophycidae 276
Prymnesium 280—282, 284
Psalteriomonas 154, 180, 489—490, 515
Psammetta 517—519
Psammettidae 517, 519
Psammina 518—520
Psamminida 451, 517, 519
Psamminidae 517, 519
psammobia, Chromulina 269
Psammonobiotidae 484, 500, 504, 507—508
Psammonobiotus 503—504
Psammophaga 567, 572
Psammosphaera 628—629
Psammotermes 392
Psammotermitidae 392
Pseudawerintzewia 478
Pseudobodo 173, 264, 268—269, 275, 318
Pseudocoelomata 102, 108
Pseudocorythion 504
Pseudocucurbitella 479
Pseudocyphoderia 507
Pseudodendromonada 135, 137
Pseudodendromonadida 275, 317, 320, 325
Pseudodendromonadidae 317
Pseudodendromonas 317, 319—320
Pseudodevescovina 365
Pseudodifflugia 504
Pseudodiffhigiidae 500, 504
Pseudoeponides 608
Pseudomicrosportella 335
Pseudonebela 479
Pseudopalmulida 596
Pseudoparamoeba 461
Pseudopontigulasia 479
pseudopunctata, Bolivina 624
Pseudothecamoeba 460, 511
Pseudotrichomonas 365
Pseudotrichonympha 372, 385, 390, 396
Pseudovendia 75, 117
Pseudovolutella 504
Pseudowailesella 507
Psychodopygys 246
Pteria 46
Pteridinium 74
Pteridomonas 187
Pterobranchia 40, 89, 99—100, 103, 105, 108
pulchella, Joenina 388
pulchra, Borelis 284, 557
Pullenia 628
Pulmonata 241
punctulatus, Cryptocercus 355, 357, 368, 387, 389, 391,
396
punicea, Pompholyxophrys 493, 499
purpureum, Dictyostelium 427, 429—430
pussilum, Alveolophragmium 628
Pycnomonas 248, 251
Pyrenomonas 199—200
Pyrgo 594, 627—628
pyriformis, Tetrahymena 189
Pyrrhocoris 235
pyrrhocoris, Leptomonas 235, 252
Pyrrophyta 1
Pyrsonympha 348, 350—353, 357, 393, 395—397
Pyrsonymphidae 348—350, 355, 357, 393
Pyrsonymphina 348
pythii, Woronina 398—402, 406—407, 409—410
Pythium 407, 409—410
Pyxidicula 478
Quadrostiatae 69
Quadrulella 472, 479
quartana, Khawkinea 205
quilliermondi, Candida 346
Quinqueloculina 558, 565, 592
Quinqueloculinoidea 594
radians, Chrysamoeba 262, 269
radians, Nuclearia 491—492
Radiata 18, 35, 75, 99, 101—102, 106, 110—111
Radiolaria 56, 137, 143—144
rajae, Trypanosoma 248
Rana 52, 309, 312—313
ranarum, Entamoeba 528
ranarum, Opalina 302—303, 305—309, 311
Rangea 74
rangeli, Trypanosoma 229, 233—235, 249, 253—254, 256
Ranidae 309
Raphanus 115
Raphidomonadida 285, 289
Raphidophyceae 135, 287, 289
Raphidophyta 135—137, 285, 287, 289
656
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
rattorum, Trypanosoma 249
Rattus 249
rattus, Rattus 249
Reclinomonas 275
Rectoeponides 606
Recurvoides 627
Reduviidae 249, 255
relictus, Cryptocercus 357, 368, 387, 390—391
Remaneicioidea 602
reniformis, Bodomorpha 413, 462
reniformis, Heteromita 413
reniformis, Paraflabellula 462
Reophax 627, 629
Reptilia 48, 238, 248
Resigella 624
Reticulammina 517—520
Reticularia 435
Reticulariidae 434, 441
reticulata, Leptomyxa 463—464, 468
Reticulitermes 348, 351, 357, 388, 390—392, 395—397
Reticulomyxa 521, 524
reticulosum, Cribroelphidium 629
retiformis, Licea 441
Retortamonadia 337—339
Retortamonadida 129, 137, 338, 341, 355, 392—393
Retortamonadidae 337—338, 392
Retortamonas 337, 339—341, 392
Rhabdammina 549, 628
Rhabdiophrys 498
Rhabdomonadida 202, 209—210
Rhabdomonas 204, 209
Rhabdospira 209
rhesii, Trypanosoma 249
rhinoestri, Leptomonas 235
Rhinoestrus 235
Rhinomonas 194
Rhinotermitidae 355, 357, 390, 397
Rhipidodendron 314, 316, 319—320
Rhizammina 527
Rhizamoeba 462—463, 465
Rhizochromulina 135, 137, 189
Rhizoflabellina 462, 465, 467
Rhizoflagellata 99
Rhizomastigida 411
Rhizoplasma 520—521, 524
Rhizopoda 100, 104, 135—137, 139, 143, 185, 189, 410—411,
449, 451—453, 455, 457, 459, 461, 463, 465—469, 471,
473, 475, 477, 479, 481, 483—485, 487, 489—491, 493,
495, 497—501, 503, 505, 507—517, 519—529, 531—532,
535, 631
Rhizopodea 296, 485, 499, 508
rhodesiense, Trypanosoma 224, 250
Rhodomella 627
Rhodomonas 199—200
Rhodophyta 139, 141, 144, 196
Rhogostoma 504
Rhombozoa 111
Rhumbleriella 504
Rhynchobdellaea 62
Rhynchobodo 213, 216, 241
Rhynchocala 118
Rhynchocoela 102
Rhynchoidomonas 236
Rhynchomonas 213, 216, 241, 253
Rhynchonympha 383—384, 389
Rhyzonymphidae 369, 388
rigidus, Leptomonas 223, 252
Robertinida 605
Robertinina 605
Robertinoida 605
Robulus 557
Rodentia 46, 52
rodhaini, Trypanosoma 250
roenbergensis, Cafeteria 189, 275
Rosalina 559—561, 565, 569, 574, 631
Rosalinida 538, 605—606, 609
Rosalinidae 606—607
Rosalinoidea 538, 606
rosanoffii, Chromulina 263
rosarium, Dictyostelium 427, 430
roscoffensis, Rotaliella 569, 571, 574
Rosculus 486, 489—490
rosea, Guttulina 485, 490
Rostronympha 392
Rotaliana 538, 544, 548, 559—560, 562—563, 574, 577—579,
584, 597—598, 601—602
Rotaliata 538, 543—544, 560, 562, 565, 574, 577—578, 583, 585,
588, 591,597, 610—611
Rotaliella 565, 569, 571, 574—575
Rotaliida 539, 578, 605, 608
Rotaliidae 609
Rotalioida 578
rotans, Trepomonas 346
Rotatoria 99, 101
rotatorium, Trypanosoma 221, 224, 231, 247—248, 253
Rotifera 25, 99—100, 103, 105, 111
rotunda, Euglypha 396, 485, 503, 508
rotundus, Desmodes 235, 250
rougeti, Trypanosoma 251
ruande, Trypanosoma 250
ruber, Globigerinoides 610, 625
Rubiaceae 245
rubra, Euglena 205
Rubratella 571, 580
rufescens, Chrysococcus 269
ruficeps, Stolotermes 392
Saccammina 537, 565, 567, 569, 627
Saccamminopsis 575
Saccamoeba 461
Saccharomyces 346
Saccinobacculinae 348, 355—357
Saccinobacculus 351—355, 357, 393, 396
sacculifer, Globigerinoides 565, 574, 625
Sacculina 36
Saepicula 334
Sainouron 415—417
saleuta, Discocelis 264, 268, 275
Salientia 311
Salivaria 220, 225, 235, 237, 247, 249
Salmo 94, 256
salmonis, Hexamita 346, 348
salmositica, Cryptobia 216, 219, 225, 228—229, 241, 253, 255—
256
Salpingoeca 328, 331—332
Salpingoecida 137
Salpingoecidae 322, 328, 331, 336
salpinx, Cyathobodo 317, 320
saltans, Bodo 227, 240, 253
sanguisuga, Haemopis 241
sapiens, Homo 44, 76, 79, 531
Sappinia 452, 459-460, 466-467
Saprolegnea 135, 137
Saprolegnia 135, 137, 290—291, 293, 295—296
Saprolegniales 290
Sarcinochrysidaceae 270
Sarcinochrysidales 270, 274
Sarcodina 189, 448, 468, 485, 490, 499, 508, 515—516
Sarcomastigophora 110, 313, 520
Sargassum 560
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
657
Saroeca 334
sativus, Cochliobolus 498—499
sativus, Raphanus 115
saturnalis, Protoopalina 302—303, 306, 313
Sauromonas 352—353, 355—356, 393
Savillea 334
sawayai, Projoenia 388
saxonica, Rhizamoeba 462—463
scabra, Eggerelloides 624, 626
Scaphopoda 99
Scenedesmus 469
schaudinni, Endotrypanum 247
Schaudinnula 507
Scheadorhinotermes 391
scheini, Trypanosoma 248
Schizamminoidea 587
Schizobodo 301
Schizochytrium 291, 295—296
Schizocoela 102, 108
Schizophyta 379
Schizoplasmodiopsis 421, 424, 447
Schizoplasmodium 419, 421, 424
Schizopyrenida 135, 451, 486, 488—490
Schizopyrenus 486
Schizotrypanum 249
Schlumbergerinana 538, 578, 592
Schlumbergerinida 592
Schwabia 479
Scolecida 49, 98, 100—101, 103—104, 106—111, 113
Scombresocidae 54
Scorpaenichthys 228
scrofa, Sus 531
Scyphozoa 120
Seabrookinida 591
Sedentaria 40
Selenococcidium 254
selseyense, Elphidium 624
semen, Gonyostomum 287—289
semilineata, Lagena 629
senegalensis, Leishmania 245
septica, Fuligo 140, 438—439, 443
Septotextularoidea 600
Septuma 527
sergenti, Phlebotomus 245
serpens, Phytomonas 216, 229, 234, 254
Serpula 93
serrata, Pyrgo 627
Sestronophora 583
Sexangularia 479
sexguttatus, Calocoris 245
Seymouriamorpha 82, 92
Shanitoidea 593
Shepheardella 585
Shisotrypanum 220
Silicoflagellata 99
Silicoluculinina 548
simiae, Trypanosoma 250
simius, Rhynchobodo 241
simplex, Cyathobodo 320
simplex, Metarotaleilla 575
simplex, Vannella 454, 491—492
simplicicornis, Paraneotermes 389
simplora, Psammophaga 567, 572
sinaica, Placojoenia 378, 388, 397
sinaicus, Kalotermes 355
Singhamoeba 488
sinuosa, Trochamoeba 454
Siphogerinoides 578
Siphonaptera 232
Siphoninidoidea 606
Siphonophyceae 140, 185
Sipuncula 105, 117
Sipunculata 104
Sipunculida 89, 99—100, 103, 105, 110
Sipunculoidea 102, 106
Siredon 36
Slopalinata 136
sokolovi, Vendia 75
solani, Spongospora 407
Solenogastres 89
Solifugae 40
somaliense, Trypanosoma 250
Sorites 565, 574
Soritida 538, 592, 594
Soritidae 563, 595
Soritidoidea 595
Sorodiscus 402, 409
Sorosphaera 398—400, 404, 406, 409—410
soudanense, Trypanosoma 250
Spathidium, 60
spatuliger, Lincus 245
spengleri, Tinoporus 629
Spermatozopsis 189
sphaerica, Trichonympha 386
Sphaerodendron 328, 330
Sphaeroeca 328, 330, 336
Sphaeroidinoidea 606
sphaeronucleolus, Thecamoeba 454
Sphenoderia 502, 507
Sphenomonadida 202, 209—210
Sphenomonas 209
sphyrelata, Poly fibula 335
spinigera, Pararotalia 559
spinosa, Placocista 502
Spinther 75
Spiralia 50, 105, 107—110, 115
spiralis, Lesquereusia 470—471
Spiraloecion 335
Spirillina 569, 575, 624
Spirillinana 538, 548, 579, 588, 590, 611—612
Spirillinata 538, 562, 572, 574, 577—578, 583—585, 588—589,
599, 609, 611
Spirillinida 591
Spirillinina 591
Spirillinoida 591
Spirobolivinina 603
Spirochaeta 250
Spirocyclinidae 575, 599—600
Spiroloculina 561, 569, 578
Spiromonas 167
Spironucleus 345—348, 392—393, 396
Spironympha 391
Spiroplectammina 627, 629
Spiroplectamminida 600
Spiroplectamminoidea 600
Spiroplectinata 583
Spirorbis 93
Spirotextularioidea 600
Spirotrichonympha 372, 378, 391, 396
Spirotrichonymphella 391
Spirotrichonymphidae 369, 372, 375, 377, 386, 391, 396
Spirotrichosoma 391
Spirotrichosomatidae 369, 372, 375, 377—378, 386, 390
Spirotrocholina 578
Spirotrocholinina 591
splendidum, Rhipidodendron 314, 319—320
Spongia 144
Spongiaria 99—100, 102—103
Spongomonada 135, 137
Spongomonadida 186, 314—315, 319—320, 325
658
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Spongomonadidae 314, 316, 319
Spongomonas 314, 316, 319—320
Spongospora 398, 404, 406—407, 409—410
Sporobulimina 583, 604
Sporozoa 52, 56, 100, 104, 115, 123, 563, 575
Sportelloeca 335
Spriggina 49, 74—76, 117
Spumella 173, 178, 260
Squamata 313
Squamulinida 538, 592
Squamulinidae 592
Squamulinoida 538, 592
Squidae 119
Squilla 58
Stachyamoeba 486, 488—490
staheli, Phytomonas 245
Stannoma 517—519
Stannomida 451, 517—519
Stannomidae 517, 519
Stannophyllum 517—519
Staurojoenia 378
Staurojoenina 378—379, 389
Staurojoeninidae 369, 372, 374—375, 377, 386, 389
Stegophiura 62
Stelexomonas 328, 332
Stella, Nonionella 557
Stemonaria 442, 448
Stemoniida 443
Stemonitales 431, 440, 442, 448—449
Stemonitida 432, 434, 436, 440—442
Stemonitidae 434, 442
Stemonitis 434—436, 438—439, 442, 447—448
Stemonitbpsis 442
Stentor 184—186
stepanovi, Onega 75
Stephanacantha 332, 335
Stephanocodon 264
Stephanoeca 325, 332, 334—336
Stercoraria 234, 237—238, 247—248
Stereomyxa 465—467
Stereomyxidae 462—463, 465—467
Stichogloeaceae 269
stilbia, Cryptobia 228
Stilostomella 577
Stilostomellida 603
stipitatus, Cyathobodo 317
Stolotermes 390—392, 396
Stomochordata 99, 103
Stomoxys 249
Storeatula 199—200
Streblomastix 348, 351, 357, 396
Striamoeba 467
striata, Lophomonas 388
striata, Thecamoeba 454
striata, Vertebralina 629
striatus, Metopus 190, 511, 516
Strigiiformes 52
strix, Streblomastix 357
Strombomonas 204, 209—210
Stygamoeba 466—467
Stylonychia 185
subsalsa, Chattonella 287—289
subterranea, Spongospora 398, 404, 406—407, 410
subulata, Crithidia 248
Suctoria 56, 60, 64, 176
Suiadifflugia 479
suis, Blastocystis 358, 363—364, 367, 394, 531
suis, Tritrichomonas 358, 363—364, 367, 394
suis, Trypanosoma 251
sulcata, Glabratella 569
sulcata, Proteomonas 196, 200
sulcatus, Entosiphon 202
Suprapylaria 245
swynnertoni, Glossina 249—250
symmetrica, Quadrulella 472
Symphytocarpus 442, 446—447
Syndetophyllum 335
Synura 270—273, 275
Synuraceae 270, 273
Synurales 270, 273
Synurophyceae 135, 137, 258, 270—271, 273—275
Syracosphaera 284
Syringammina 517—520
Syringamminidae 517, 519
Syzraniina 596
Tabanida 249
Tabanidae 235, 247—249
tabaniformis, Glossina 250
tachinoides, Glossina 249—250
taeniata, Rhynchobodo 241
tahitiensis, Ceratiomyxella 419—421, 424, 447
Talpinella 630
Tamanovalva 119
tarda, Urinympha 389
Tardigrada 99—100, 103—104, 106, 110, 112, 119
tarentolae, Leishmania 245, 253
taurus, Bos 532
Technitella 549, 628
Temnocephala 99
temporaria, Rana 309, 313
tenax, Trichomonas 358, 363, 365, 367
tenella, Nebela 476
Tentaculata 42, 53, 99, 101, 103—110, 113
tenuata, Buliminella 557
tepida, Ammonia 569, 574
Teratonympha 390
Teratonymphidae 369, 377, 390
termitidis, Sauromonas 356
termitis, Monocercomonoides 355
Termopsidae 355, 357, 397
Termopsis 395
terrestris, Arvicola 46
terrestris, Rana 309
territans, Culex 247
tertia, Pelomyxa 513
Testacea 185—186, 468, 484—485, 500, 507—508
Testaceafilosia 135, 414, 451, 477, 490, 498, 500—503, 505,
507—508
Testacealobosia 135, 420, 451, 466, 468—471, 473, 475* 477,
479, 481» 483—484, 500—501, 507—508
Tetradendracea 66
Tetrahymena 149, 181, 184, 189, 459
Tetramastigamoeba 488
Tetramastix 301
Tetramitus 301* 488—489, 532
Tetramyxa 398, 408—409
Tetrapoda 38, 61
Textularia 558, 560—561, 600, 627, 629
Textulariana 538, 544, 549, 560, 562* 574—575, 577—579,
583—584, 588—589, 595—598, 602* 611—612
Textulariellidae 601
Textularielloidea 538, 601
Textulariida 600
Textulariidae 600
Textulariina 525, 528
Textularioidae 600
Textularioididae 600
Textulariopsidae 600
Thaliacea 99, 103
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
659
Thaumatomonadida 186, 320, 414
Thaumatomonas 186
Thecamoeba 454, 460, 516
Thecamoebidae 452, 460, 466—467
Thecochaos 460
theileri, Trypanosoma 225, 230, 232, 235, 248
Theileria 230
Theratromyxa 522, 524
Theriodontia 52
thienemanni, Desmarella 322, 336
Thraustochytrida 135, 137, 291, 295—296
Thraustochytriidae 291, 295
Thraustochytrium 291, 293, 295—296
Thurammina 628
tigrinum, Ambistoma 36
tincta, Nebela 471
Tinoporus 629
Tiphotrocha 583, 629
tipulae, Monocercomonas 363, 396
tocopola, Prolophomonas 388
Toretammina 583
torosus, Triturus 232
Tortonella 578
tortuosa, Trigomonas 346
Tournayella 589
Toumayellida 589
Toumayellidae 577, 589
Trabrooksia 443
Tracheleuglypha 502, 507
Trachelomonas 176, 201, 204, 209—210
Trachelonema 70
tragelaphi, Trypanosoma 248
transvaaliense, Trypanosoma 248
trapidoi, Lutzomyia 246
Trematoda 99, 103
tremulans, Pseudobodo 269
Trepomonas 343—348, 392—393, 396
Tretomphalus 574
Triadopsis 219, 253
Triasinoidea 590
Triatoma 234, 253—255
triatomae, Blastocrithidia 224, 233—234, 243
triatomae, Trypanosoma 249
Tribrachium 75
Tricercomonas 338, 347
Trichamoeba 454, 459—460
Trichia 434, 436, 442
Trichiaceae 442, 448—449
Trichiales 431, 440—441, 447, 449
Trichiida 432, 434, 440—441
Trichiidae 434, 436, 441—442
Trichomitus 358—359, 363, 396
Trichomonadida 129, 135, 137, 141, 179, 313, 344, 357—359,
361, 363, 365, 367, 386, 388, 392—394, 396—397, 512
Trichomonadidae 358, 365, 367, 394
Trichomonadinae 358, 365, 367
Trichomonas 129, 180, 189, 249, 358—359, 362—365, 367,
394—397, 528, 532
Trichonympha 372, 378—386, 390, 395—397
Trichonymphidae 369, 372, 374, 377—378, 386, 389, 395
Trichonymphina 369, 386, 388, 392, 394—395
trichophorum, Peranema 202
Trichoplax 110, 115, 118
Trichoptera 232
Trichosida 451, 468, 483—484
Trichosphaerium 468, 483—485
Tricladida 75
Trigomonas 346
Trigonopyxis 478
Trilobita 50, 99, 106
Trilobitoidea 106
Trilobitomorpha 99, 103—104
Trilobius 243
trilobus, Globigerinoides 625
Triloculina 592, 627
Trimitus 347, 392
Trimyema 511
Trinema 178, 472, 502, 504, 507
Trinematidae 500, 507
Triploblastica 100—101, 103, 110—111
Triton 119
Tritrichomonadinae 358, 365 *
Tritrichomonas 358, 361—364, 367, 394—397
Triturus 232, 528
Trochaeozoa 105
Trochammina 560, 569, 625, 627, 629
Trochamminida 538, 602
Trochamminidae 602, 631
Trochamminina 538, 602
Trochamminoida 538, 601
Trochamminoidea 538, 602
Trochelminthes 100
Trochodendracea 66
Trochozoa 49—50, 102, 106—113
troglodytes, Pan 531
Troilus 245, 252
tropica, Leishmania 245
trutta, Salmo 94
trypaniformis, Dimastigella 227, 242, 253, 256
Trypanomonas 249
Trypanoplasma 241, 252, 254
Trypanosoma 212—213, 215, 219, 221—222, 214—226, 229—
238, 247—256
Trypanosomatida 213, 243, 251
Trypanosomatidae 212—213, 219* 236, 243, 251—256
Trypanosomatina 252
Trypanozoon 223, 248—251, 255
Tubifera 435, 439, 441
Tunicata 99—100, 103—104, 118
Turbellaria 44, 53, 99, 103
Turbinulina 624
turneri, Boderia 575, 584
Turrilina 553
Turrilinina 604
tusitala, Holomastigotoides 376—378, 392
Ulva 560, 627
umbilicalis, Porphyra 139
umbratilis, Lutzomyia 246
undulata, Chilomastix 341
undulata, Diaphanoeca 332, 334
ungeriana, Quinqueloculina 558
unguiculata, Acanthocorbis 335
unica, Bispironympha 391
Uniramia 50
Urceolus 176, 207, 209
Urinympha 378—379, 381—384, 389
Urochordata 105
Urodela 50
Urodelomorpha 50
urundiense, Trypanosoma 250
ussuriensis, Notilia proteus 357
utricularis, Badhamia 438
uvella, Polytoma 161
uvella, Spongomonas 314
uvella, Synura 270, 272
Uvigerina 578, 626—627
Vacuolaria 285—289
vaginalis, Cryptobia 216, 219, 241
660
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
vaginalis, Trichomonas 180, 358, 362—364, 367
Vaginulinida 597
Vagocibicides 577
Vahlkampfia 467, 486, 488
Vahlkampfiidae 486, 488—489
Vairimorpha 138
valamo, Paradermamoeba 454, 467
Valkanovia 507
Vampyrella 493, 495, 498—499
Vampyrellaceae 499
Vampyrellidae 490—491, 493, 495, 498—499
Vampyrellidium 492, 495, 498—499
vandeli, Centropyxis 4TT
Vanhoeffenellidae 586
vanhoffi, Glossina 251
Vannella 454, 461—462, 467
Vannellidae 452, 461, 466
Varela 560
varia, Trichia 434
variabilis, Licea 441, 448
vas, Nebela 477
Vaucheria 407
Vendia 75
Vendomia 75
venezuelense, Trypanosoma 250
Vermes 99—100, 102
vermicularis, Perichaena 437—438
Vermiculites 73
Vermidea 100
Veronica 399
veronicae, Ligniera 409
veronicae, Sorosphaera 398—400, 404, 406
verrucarum, Lutzomyia 246
verrucosa, Ligniera 398, 410
Vertebralina 629
vertebralis, Marginopora 629
Vertebrata 43, 47, 98—100, 103—105, 236
ver tens, Pyrsonympha 350, 352—353, 357
vesicularis, Discorbis 557
vesparium, Metatrichia 434, 436, 448
vespertilionis, Trypanosoma 249
vestita, Paraphysomonas 180
Vexillifera 454, 461
Vexilliferidae 452, 461, 466
vickersae, Trypanosoma 249
villosa, Pelomyxa 510, 513
vinaceo-fuscum, Dictyostelium 427
Viola 117
violaceum, Polysphondilium 427, 429, 431, 447, 449
virescens, Vacuolaria 286—289
virguloidea, Opalina 302—303, 307
viride, Physarum 434
Viridiplantae 125
viridis, Euglena 207
viridis, Lacerta 300
viridis, Pelomyxa 510, 513
viridis, Proteromonas lacertae 297, 300—301, 312—313
viridis, Vacuolaria 288
vivax, Trypanosoma 249
vivida, Calliobdella 228
vivipara, Pelomyxa 513
vivipara, Spirillina 569, 575, 624
Viviparus 79
vlkianum, Diacronema 280—281, 283
vlkii, Pseudodendromonas 320
Voluta 504
Volutella 504
Volutellidae 500, 504, 507
Volvocidae 35
Volvox 452, 509
volvox, Sphaeroeca 336
vorax, Procryptobia 242
Vorticella 184
vulgaris, Arcella 469, 472, 475
vulgaris, Beta 399
vulgaris, Guttulinopsis 485
Wailesella 482
Wallaceina 213, 215—216, 221—222, 224, 243
Waskia 341
wellcomei, Psychodopygys 246
wenrichi, Parahistomonas 366
wentrichi, Paratrichomonas 365
whitmani, Phlebotomus 245
Willaertia 488
Woloszynskia 180, 190
woodcocki, Trypanosoma 248
Woronina 398—400, 406—407, 409—410
wrightii, Leishmania 246
wronghtoni, Archotermopsis 390
wrublewskii, Trypanosoma 248
Xanthomonadina 99
Xanthophyceae 137, 140, 143—144
Xanthophyta 186
Xenophyophora 517, 520
Xenophyophorea 135, 137, 140, 451, 516—517, 519—520, 528
Xenophyophoria 520, 528
Xenophyophoridae 520
Xenoprorhynchus 60
Xenopsylla 249
Xenoturbella 120
Xenoturbellida 89
Xenusion 75
Xiphosura 48
xylopletha, Macrospironympha 391
ylephiletrix, Lutzomyia 246
zachsi, Lamellisabella 119
Zanichelia 560
Zelleriella 306, 309, 311—313
zernovi, Jadammina 574
zeteki, Barroella 356
Zivcovicia 470, 479
Zoa 110
zonarium, Stannophyllum 518
zonatum, Endostelium 420
Zonomyxa 478
Zooflagellida 99
Zoomastigophora 253
Zoomastigophorea 186, 188, 251, 255, 275, 313, 395—396,
446
Zootermopsis 357, 390
Zoothamnium 190
Zostera 296, 560, 627
УКАЗАТЕЛЬ ТЕРМИНОВ
абиссаль 20, 516, 518, 526, 534, 627—630
автогамия 161—162, 354—355, 382—384, 468, 475
автогенез 19, 125
автоинвазия 245
авторадиография 305
автотрофы 64, 73, 124, 152, 167, 169, 195, 266, 458
агамета 163, 567, 569, 571—572
агамогония 159, 569, 571, 580
агамонт 565, 567, 569, 571—572, 574—575, 580, 585, 588, 592,
598, 610
агломерация 229, 232
адаптация 20, 29—36, 38—39, 44, 48—50, 53, 61, 66, 94—95,
115—116, 129, 172, 180—188, 207, 216, 222, 230, 236, 239,
252, 295, 302—303, 308—309, 313, 337, 341, 358, 404, 408,
534, 582
адгезориум 403—404
аденозинмонофосфат 234
акантоподии 466
акантосомы 225—226
акариомастигонт 367
акариотный 401, 406
акарология 11
акинетопластия 223
акклиматизация 12, 27
аккумуляция 231, 235
акразин 427—428
акрогастеры 105
акронема 317, 350
акросома 574—575
аксонема 149—150, 211, 218—220, 265, 274, 297, 300, 314,
323, 325, 343—345, 348, 359, 361, 402, 412, 420, 498, 512,
514
аксоподия 135, 143, 145, 147—148, 166, 172, 558
аксосома 218—219, 412
аксостиль 337, 344—345, 348—359, 362—363, 365—368,
375—377, 382, 387—392, 575, 592
актин 129, 146, 148, 271, 322, 419, 431, 438, 456—457
акто-миозин 457, 493—494, 500
акцептор 154
аланин 183, 472, 563
аллантоис 51
аллогенез 38, 41—43, 47
алломорфоз 35—36, 38
аллопатрическое видообразование 84
аллотетраплоидия 31
альвеола 147, 470—473, 478, 556, 579
амастигота 215, 223, 226, 230, 243, 245, 247
амбулакральная система 35, 48
амебиаз 452
амебоидизация 138, 143
амебоидность 143
амебостом 488
амебофлагеллаты 128, 365, 444, 511
амилопласт 400
амилоплектин 157
аминокислота 78, 144, 183—185, 472, 537, 548, 559—560, 563,
626
амитоз 158—159
амплификация 157
амфиплоидность 124
амфистегин 563
анаболия 54, 56—57
анастомоз 166, 172, 222—223, 291, 415, 458, 463, 465—466,
469—470, 472, 479, 490, 493, 501—502, 504, 518, 521, 523,
525, 533, 558, 610
анафаза 224, 262, 295, 300, 306, 325, 346, 362, 380, 400, 402
анаэробы 128—129, 142, 337, 346, 348, 528
андрогенез 24
анемия 228
анизогаметы 297
анизогамия 161, 571, 580
антераксантин 270
антераксин 286
антибиотики 31—32, 226, 346, 364, 386
антиген 358, 364, 367
антропогенез 19, 116
анус 106, 112, 140, 366
арагонит 277, 281, 548, 590, 605
арахнология 11
аргинин 472
ареал 30, 42, 87, 235, 249, 414, 418, 423, 439, 462
арогенез 34, 38, 42—44
ароморфоз 21, 34—36, 38, 116, 160, 578, 580
артроподизация 50
археи 71
археотрох 104
архитектоника 13
архицеломаты 104, 108
асексуальность 385
астаксантин 262
АТФ 142, 152, 183, 379, 558
АТФ-аза 548, 561
ауксотрофы 226
аутогамия 569
аутофагия 426=—427
аутэкология 27
афаноплазмодий 431—432, 438, 442
ацентрическое веретено 306
аэрация 624,627
базальное тело жгутика 149—150, 323, 330, 375, 378, 412, 415,
440, 544, 589, 591, 598—599, 601, 610
662
УКАЗАТЕЛЬ ТЕРМИНОВ
базальный комплекс жгутика 265, 273, 315, 323, 391, 420, 428,
436, 589
бактериофаги 184, 321, 559
бактериофагия 226
батиаль 516, 518, 527, 627—629
батиметрия 624, 627
батисифон 534
батрахология 11
батрахоморфия 50
бентос 173, 176, 321, 327, 414—415, 462, 516—517, 520, 524,
526—527, 534, 629—630
беспозвоночные 11—12, 16, 18—21, 23—25, 27, 35—36, 38,
40, 43, 49, 53, 62, 66, 73, 76, 88—89, 95, 98, 101—102, 108,
111, 114—115, 117, 139, 173, 183, 186, 211—212, 227—229,
231—232, 235—236, 241, 243, 337—338, 341, 358, 365, 395,
431, 461, 516, 518, 520, 527—528, 625
биваленты 162, 295, 308
биогенетический закон 19, 53
биогеография 462, 488
биогеоценоз 32—33
биокласты 625
биомаркер 534
биоминерализация 556, 560
биопродукция 263
биосинтез 183, 386
биосфера 21, 43, 84, 115, 145, 534
биота 178, 186
биотоп 177, 180, 183—185, 207, 477, 521
биоценоз 12, 20, 185, 626, 628, 630
биоценология 27
биоэнергетика 629
бифонтинальная стенка 172, 556—557, 560—561, 577—580,
582, 597—598, 601—602, 606, 610
бластомер 104, 113
бластопор 98, 102, 104—106
борьба за существование 19
брадителия 46—47
бродяжка 56
брызгальце 56, 61
бурса 62, 65
валин 472
вегетация 207
венд 71, 73—76, 116—117, 144, 188
вестибулюм 479
вещества-аттрактанты 428
вещества-регуляторы 428
видообразование 21, 30—33, 46, 52, 84, 88, 114, 133, 188, 387
видоспецифичность 387
вирулентность 249, 364
вирус 94, 180, 188, 226, 407
внеклеточная цитоплазматическая сеть 290—291, 295
внеядерное веретено деления 362—363, 380
внутриядерное веретено деления 300, 339, 353, 567
— пространство 211, 223, 290, 297, 306, 341, 439, 567, 571
внутриядерный интердесмоз 158, 346
— митоз 128, 158, 201, 358, 439, 584
— плевромитоз 211, 348, 354
воротничок 105, 314, 316, 321—325, 329—331, 333, 458, 470,
476, 502, 504, 507, 604
ворсинки 565
выводковая сумка 61
галлоцианин 305—306
гальванотаксис 170
гамета 49, 141, 149, 161—163, 169, 308, 354—355, 382—384,
404, 418, 503, 518—519, 530, 534, 569, 571—572, 574—575,
580, 584, 595, 626
гаметогамия 569, 571
гаметогенез 49, 161, 308, 311, 382, 384, 557, 565, 574, 610
гаметоцит 161, 354—355, 382, 384
гамогония 56, 159, 569, 571—572
гамонт 163, 307—308, 483, 530, 565, 567, 569, 571—572, 574—
575, 588
гамонтогамия 503, 569, 575, 607
гамонтоциста 302, 307—308
гамоны 169
гаплоидизация 166, 429, 439
гаплонт 354—355, 382—384
гаплофаза 436, 569
гаптонема 147, 276—277, 281—283, 317, 575
гастральная полость 107
гастрея 19, 53, 98, 104
гастродерма 111
гастронейрон 104—105
гастротрох 104
гастроцель 100
гастроэнтерит 530
гаструла 49, 104
гельминтоз 116
гематофагия 240
гемоглобин 78, 93, 126
гемолимфа 98, 232—234, 238, 243, 245
гемоцель 234
ген 32, 57, 71
генеалогия 33, 78—79, 88, 92, 95—96
генерация 52, 96, 262, 574, 594, 597, 600, 608
геном 30—33, 46, 54, 57, 78, 116—117, 124—126, 141, 160—
163, 196, 224, 584
генотип 30—32, 42, 48, 57, 162, 183
генофонд 30—32, 87
географическая раса 87
геотаксис 170
герпетология 11, 27
гетеробатмия 48, 66—67
гетеродинамичность 414
гетеродинамные жгутики 201—202, 209, 211, 217, 219, 240—
241, 411, 415
гетерозигота 30, 32
гетерокариотность 518, 572, 574, 580
гетероклеточность 144
гетероконтность 135, 152, 201, 258, 264, 266—271, 276, 283,
285, 287—288, 290, 335
гетероксенность 224, 228, 232, 236, 240—241, 245, 247
гетероморфизм 124, 215, 458, 468—469, 490, 500, 539, 567,
574
гетерономность 22, 24, 50, 115—116
гетеростегины 548
гетероталлия 429, 437
гетеротопия 54
гетеротрофия 72, 123—124, 135, 141, 155—156, 162, 166—167,
169, 171—174, 176—179, 181, 183—187, 195—197, 201—
202, 205, 207—211, 217, 222, 238—239, 251, 266, 297,
301—302, 311, 321, 337, 451
гетерохроматин 223—224, 323
гетерохрония 45, 54
гиалоплазма 452, 454, 462—463, 466, 472, 489—490, 492—494,
502
гибрид 31, 52, 87, 94, 114—116, 162, 251
гибридизация 31, 33, 78—79, 87, 94, 116
гидрогеносомы 125, 128, 142, 152, 154—155, 180, 183, 337,
350, 357—358, 362, 368, 378—379, 511—513
гидроцель 116
гиперконденсация 223
гиперморфоз 38
гиперплазия 405
гипертрофия 38, 405, 511
гипогенез 36—37, 42, 45
гипоморфоз 36—37
гипоталлус 434, 442
УКАЗАТЕЛЬ ТЕРМИНОВ
663
гипофаринкс 234
гипофиз 308
гистоны 157
глазок 43, 195, 202, 295
гликоген 93, 157, 205, 266, 305, 317, 362, 377, 379, 509
гликозаминоглюканы 558
гликокаликс 146, 204, 215—217, 241, 260, 321, 353, 379, 412,
420, 451, 458, 460—461, 465—466, 468, 486, 492, 500, 522,
528—529, 548
гликолиз 226, 379
гликопротеины 215—216, 351
гликосомы 225—226
гликостили 146, 461
глицин 183, 472, 548
глутаминовая кислота 472
глюкан 205
глюкоза 226, 385, 563
головастик 308, 310—311
голофиты 201
Гольджи аппарат 129, 141—142, 148, 152, 156, 187, 195, 205,
225, 262, 265, 271, 274, 277, 280, 287—288, 292, 294, 297—
301, 305, 315, 317—318, 323, 337—338, 341, 345, 348, 350,
357, 363, 365, 377, 395, 398, 402, 413, 419, 436, 438—439,
458, 460, 463, 466, 469, 482, 486, 500, 502, 510, 512—515,
522, 528—529, 548, 561—562, 574—575, 579
гомеоморфность 556, 583, 604
гомеостаз 42
гомозигота 32
гомокариотность 305, 565, 572, 574, 581
гомоквадрантное дробление 113
гомология 11, 13, 22, 42, 44, 48—49, 51, 53, 56—57, 65, 67—
70, 78, 92—95, 104, 107, 109—110, 113—114, 125—126,
129, 138—140, 162, 164, 186, 202, 208, 220—222, 224, 236,
238, 266, 290, 295, 311, 322, 347, 365, 435, 445, 553, 556,
575, 578, 605
гомономность 67
гонада 109, 364
гонадотропин 308
гормон 308, 384—385
гравитаксис 170
гранеллы 516—518
гранулоплазма 452, 454, 472
гранулоретикулоподии 490, 533, 584
граны 155
гуанин 287
гумус 477
дактилоподии 454, 461, 483
дарвинизм 19—21, 29—33, 51, 84, 88, 92, 94, 114—117, 188
девиация 56—57
девон 47—48, 76, 81, 97, 556, 584, 589, 596
дегенерация 21, 43, 348, 354—355, 382, 400, 402, 421, 439,
571—572
деградация 31, 36, 43, 100, 227, 379
дедукция 15
дезагрегация 428, 488
дезорганелл изация 141, 143
деконденсация 224
дендробионты 41
дентин 56
деполимеризация 377, 457
десмосомы 217
детрит 167, 174, 176—177, 183—184, 266, 462, 495, 502, 534,
559—560, 625
детритофаги 76, 559
дефекация 167, 233, 235, 380, 472
диагенез 552, 579, 590
диадиноксантин 205, 260, 270, 274
диапауза 26
диатоксантин 260, 270, 274
диафрагма 470
дизруптивный отбор 32—33
диктиосома 125, 128—129, 142—143, 156, 195, 205, 225, 262,
265, 274, 280, 286, 289, 292, 305, 315, 317, 323, 327, 337—
339, 341, 357, 376—377, 388, 390—392, 398, 400, 413, 419,
436, 438, 458—459, 463, 466, 469—470, 486, 492—493, 500,
512—513, 562, 574—575
динеин 149—150, 219, 351, 558
диплеурула 37
диплоидизация 138, 143—144
диплонт 355, 382—384, 569
диплотена 567
диплофаза 162, 406, 437, 569
дискинетопластия 223
дискоболоцисты 148, 262
диссимиляция 73
домены 138
домик 35, 53, 89, 146, 201, 204—205, 258, 260, 264—269, 319,
321—322, 325—326, 328—332, 451, 468, 472, 559
доминантность 32
дрейф генов 31
дробление 49—50, 98, 108, 113, 115, 438, 503
дублеты 149, 218—220, 323
дупликация 229, 363, 567
естественный отбор 19, 30, 32, 44, 58
жабры 18, 34, 36, 41—42, 57, 82, 179, 219, 227—228
жгутик 24, 35, 49, 124—125, 128—129, 135, 138, 141—143,
147—152, 159, 166—167, 172—174, 176, 178, 180, 184, 187,
194—197, 201—206, 208—209, 211, 213—223, 225—228,
232—234, 236, 240—243, 248, 258—260, 262, 264—271,
273—274, 276—277, 280—283, 285—291, 293—295, 297—
298, 300—304, 308, 310—311, 314, 316—319, 321—325,
329—331, 334, 336—341, 343—351, 354—359, 361—369,
372, 374—380, 382, 384—385, 387—392, 395, 398, 402, 404,
408, 411—416, 418—421, 424, 436—438, 440, 444—445,
451, 462, 485—486, 488—489, 498, 508, 511—515, 518, 528,
533—534, 558, 567, 569, 572, 574—575, 584, 586, 592
железа 59—61, 69, 146, 205, 229—230, 232—235, 245, 247—
248, 266, 269, 314, 472, 501, 522, 548
желточник 70
желудок 66, 232, 235, 366
желудочек 34
жир 157, 179, 186, 205, 211, 281, 562, 574—575
загрязнение 145, 196, 248, 477, 534, 537, 548, 559
запасные питательные вещества 156—157, 258, 266, 290, 317,
323, 339, 415, 470, 501, 534, 562
зародыш 13, 16, 32, 43, 53—54, 62, 65, 98, 104—105, 404, 407
зачаток 35, 44, 54, 57, 67, 70, 363, 378, 387, 389, 392, 442, 578
зигота 32, 161—163, 170—171, 272, 280, 282, 308, 310—311,
355, 382, 384, 429, 437, 530, 533, 559, 571—572, 574
зигоциста 300, 302, 308, 310
зоб 235
золь 454, 456—457
зооид 326
зоопланктон 321
зооспора 135, 149, 262, 269—270, 290—291, 293—295, 300,
402, 404, 408, 411, 418, 440, 442, 444—445, 522, 528
зооспорангий 404, 408
зооциста 408, 421
цдиоадаптация 21, 33—35, 38, 43, 47
идиогенез 34
идиоплазма 42
идиосома 469—472, 474, 476—477, 479, 500—504, 507
известняк 71, 143, 534—535
изменчивость 18—19, 29—32, 45, 84, 88, 95, 114, 160—161, 164,
172, 183, 187—188, 210, 303, 309, 364, 383, 395, 434, 441, 531
664
УКАЗАТЕЛЬ ТЕРМИНОВ
изобата 628
изогамета 272, 280, 282, 294
изогамия 161, 262, 276, 571, 580
изолят 31, 213
изоляция 45
изотоп 235, 563
изоформа 219
имаго 243, 366
иммобилизация 60
иммунитет 32, 231
иммунодиагностика 367
иммуноцитохимия 456, 486
импрегнация 266
инвагинация 104, 199, 217, 221, 234, 293, 343, 379, 399—400,
402, 494
инвазия 228, 230—231, 233—235, 346, 363—364, 402, 404, 408
инволютная раковина 539, 598—599, 606, 609—610
инволютность 539
индикация 145, 178
индукция 234, 308, 427, 437
инициация 94, 437
инокуляция 229—230, 235, 381, 385
интердесмосома 346
интеркинез 354, 356
интернализация 452
интерстициаль 476—477, 500, 503
интерстициальный вид 173, 183, 468, 477, 503, 507, 559
интерфаза 157, 194, 205, 223, 252—253, 262, 286, 292, 295, 306,
308, 345, 352, 377—378, 387, 389—390, 392, 400, 425, 439
интрогрессия 33
интродукция 12
инфекция 212, 228, 235, 245, 339, 385, 387, 395, 400, 404, 407
инфильтрат 249
инцистирование 17, 156, 170—171, 183, 217, 222—223, 225,
272, 281, 293—294, 300, 307—308, 310, 339, 345—346, 363,
384, 402, 420-421, 438, 459, 462, 465, 474—475, 477, 482,
486, 489, 495, 503, 513, 574
К-тело 290
кайнозой 76, 527, 630
кала-азар 246—247
калории 560
кальцинация 522, 548, 563, 580
кальцификация 276, 556, 561—563, 574
каннибализм 185, 228, 235, 240, 419, 425, 559
капиллиций 432, 434—436, 440—443
карбоангидраза 561—562
карбон 38, 97, 583—584, 587, 590, 592—593, 595—596, 598—
601
кариогамия 162, 383, 404, 406, 408, 424, 437—438, 530
кариокинез 196, 420, 439, 495
кариолимфа 157, 378, 571
кариология 11, 24, 92, 210, 497
кариомастигонт 67, 152, 337—338, 341, 344, 346—350, 352—
353, 355—359, 365—367
кариоплазма 223
кариотип 87
карцинология 11
катагенез 36, 38, 42, 47
катаморфоз 36—37, 41
квинквелокулиновый тип строения раковин 551, 580, 594
кДНК 222—223
кембрий 48—49, 75—76, 89, 97, 116, 133, 188, 281, 533—534,
583—586, 588—590
кериотека 579
кинета 67, 70, 303—304, 306, 308, 312
кинетида 49, 67, 141—142, 151—152, 186, 311, 325, 444
кинетом 152, 304
кинетопласт 152, 211, 213—215, 222—223, 225, 229, 236,
240—241, 243, 247—248, 251—252, 297, 299, 319
кинетопластия 223
кинетосома 35, 67, 70, 128—129, 141, 148—152, 195—196,
202, 211, 215—219, 222—223, 258, 260, 262, 265—266,
268—271, 273—274, 277, 280, 283, 286, 288, 293—295, 297,
300, 304, 314, 316—317, 321, 323, 325, 337—341, 343—346,
350—352, 357, 359, 362—363, 365, 368, 372—378, 380—
381, 387—388, 390—392, 402, 413—414, 420, 436, 445, 486,
511—512, 522
кинетохоры 224—225, 262, 287, 295, 354, 362, 378, 380, 400,
567
кинетоциста 148, 411, 413, 415
кинобласт 35
киноцит 35
кладизм 95—97
клетка-хозяин 158, 180, 398—400, 403—407
клон 87, 202, 364, 444
книдоциста 148
кокколиты 276—277, 280—282
коксальные поры 69
коллосомы 463, 465
колониальность 25, 35, 67, 144, 171, 174, 176, 201, 209, 211,
259, 264, 266, 268, 270, 273, 276—277, 290, 295, 314, 321—
323, 325—330, 332
колония 19, 35, 73, 86, 144—145, 159, 198, 207, 227, 241—242,
262, 270—273, 280, 282, 290—295, 314, 316—317, 319,
322—323, 326, 328—330, 334, 384, 386, 432, 434
комиссура 105
комменсализм 179, 211, 228, 241, 297, 301, 308—309, 311, 337,
347—348, 357—358, 366—367, 521, 534, 560, 563, 571, 607
компартмент 195—196, 528, 530
компартментализация 469, 482, 548
конвергенция 21, 39—42, 92, 95, 108—109, 160, 534, 541, 552,
557—558, 562, 575, 577—579, 583, 587—588, 591
конверсия 428
конгломерат 436
конгрегация 464, 475
конидии 459, 495, 498
конкреции 624
конкуренция 29, 31, 38, 84, 89
консументы 173
контаминация 234—236, 245, 248, 404
конъюгация 161—162, 164, 169, 325
конъюктива 231
копрофил 241, 441, 521
копуляция 60, 161, 163, 169, 294, 300, 308, 310, 354—355, 364,
382—384, 404, 415—416, 418, 569, 574
кора 418, 439
корешок 125, 129, 150—151, 195—196, 201—202, 216, 218—
219, 221, 260, 264—265, 267—271, 273, 277, 286, 288, 290,
293, 297, 300—301, 304, 314—315, 317—318, 321, 323, 325,
337—338, 341, 344, 355, 357—358, 362, 407, 411, /144—445
кормус 86
кортекс 35, 159, 432, 457
космополитизм 418, 439, 462, 477, 504, 629
коста 51—52, 59—60, 80, 82, 246, 358, 362—363, 365—367
кофактор 226
Кребса цикл 305
кремнезация 627
криопротектор 364
криофилы 440
криптобиаз 228
криптосома 533, 562, 610
криптостомия 477, 479
кристаллиты 552, 561
кристы 124—125, 129, 135—136, 152, 154, 187, 195, 201, 205,
222, 227, 252, 262, 264—265, 268—270, 274, 281, 286, 290,
292, 294, 297, 299, 301—302, 305, 311, 314—319, 321, 323,
329, 398, 411, 413, 419, 436, 438, 451, 458, 469, 485—486,
489, 497—498, 500, 522, 524, 528—529, 533—534, 562, 584
кровепаразит 211, 216, 226, 228, 231, 238—239, 241, 247—248
УКАЗАТЕЛЬ ТЕРМИНОВ
665
кровепаразитизм 216, 228—229, 236, 239
кровообращение 15, 34
кроссинговер 160, 162, 164
ксантосома 516—517, 519, 562
’ ксантофиллы 156, 260
ксеносома 471, 473^474, 476—479, 482, 501, 504, 507
ксилофаги 366, 385—386
куколка 91
культивирование 181—182, 186—187, 201, 208, 243, 252, 291,
300, 337—338, 340, 346, 358, 364, 367, 381, 385, 395, 425,
430, 436, 441, 471
кутикула 35, 52, 58, 67, 105, 109, 146, 166, 201, 204, 350, 382,
384—385, 482—483
лакуна 62, 413, 518, 562
ламелла 201, 205, 260, 270, 272, 274, 280, 286—287, 289, 344,
368, 375—378, 380, 387—392, 552
ламеллоподия 558
ламина 493, 500
латекс 74
лейкозин 262, 281
лейкопласт 141, 155, 157, 196, 198, 262, 290
лейцин 472
леотропный 351, 357, 372, 388, 390—391
лизис 413
лизосома 152, 156, 167, 184, 202, 299, 305, 313, 350, 558, 562—
563
линеллы 516, 518—519
линька 55, 354—355, 366, 380—385
липиды 157, 184, 195, 201, 203, 209—210, 262, 274, 280, 282—
283, 285-287, 305, 323, 398, 402, 413, 469
литораль 518, 534, 624—627
личинка 19—20, 36—37, 43, 49, 55, 57, 62, 75, 82, 91, 93,
Ю4—106, 111—113, 115—116, 173, 234, 243, 272, 281, 363,
384—385, 534, 559, 625
лобоподия 143, 166, 363, 411, 419, 451, 457, 463, 468, 477, 482,
490, 522, 528, 533
лоборетикулоподия 463
локомоция 427, 454, 457—458, 460—461, 463, 482—483,
485—486; 488—489, 508, 512
лорика 322, 328, 332—335
лютеин 286
лямблиоз 338, 346—347
макробентос 518, 520, 526—527, 534, 624, 628—629
макромутант 30
макронуклеус 35, 70, 231, 243
макросистема ПО, 187, 252, 284, 584
макротаксон 143—144
макрофаги 228, 230
макроциста 429, 438
макроэволюция 21, 30, 44, 116, 187—188
максиллы 57
малакология 11
малаты 379
мальпигиевы сосуды 52, 234, 243, 245
мандибулы 50, 57
мантийная полость 626
мантийный комплекс 70
мантия 60
мастигонемы 125, 129, 135, 149, 194—195, 197, 201—202,
240—241, 259, 262, 264—271, 273, 277, 281, 285, 287—288,
290, 293, 298, 317—319, 323, 358, 402, 408, 412, 418—419,
436, 575, 584, 586
мастигонт 215, 346, 351, 359, 362—363, 367—368, 380—381
мацерация 71
мезобласты 98
мезодерма 35, 57, 66, 98, 112
мезозой 21, 36, 38, 40, 51, 76, 311, 387, 595
мезосома 105, 108
мейобентос 626—629
мейоз 124—125, 143, 157, 161—164, 171, 280, 282, 290, 294—
295, 297, 300, 302, 308, 354—355, 382, 384, 398, 402, 406,
408, 429, 438—439, 490, 533, 567, 571, 574
меланизм 31
мембранеллы 70, 184
менингоэнцефалит 249
мерогамия 161, 382
мерогония 56, 159
мерозоиты 530
метаболия 166, 187, 204, 241, 415
метамерия 22, 24, 50, 53, 62, 65—66, 69—70, 75, 107, 112
метаморфоз 26, 57, 82, 112, 308
метанефридий 66, 142, 154, 180, 511
метасома 105, 109
метатрохофора 112
метафаза 158, 224, 262, 280, 287, 295, 300, 306, 325, 346, 362,
400, 402, 567
механорецепторы 234
механотаксис 169
механохимия 187
миграция 26, 55, 162, 206, 229, 232—234, 245, 262, 271, 281,
287, 295, 300, 325, 348, 354, 356, 364, 380, 384, 390—391,
402, 427—428, 430—431, 473—474, 503, 571
микология 123, 417—418
микробентос 176, 207
микровилли 147, 172, 321, 533, 562—563, 610
микроворсинки 219, 232—234, 344, 347
микрогамета 161
микронуклеус 157, 571—572, 574
микроплазмодий 438, 465
микропланктон 76, 186, 207
микропопуляция 231
микропора 556—557, 591
микростомия 476
микротельце 159, 187, 225, 268—269, 345, 350, 362, 378, 413—
414, 469, 510—511, 513, 515, 562, 574—575
микротоксицисты 148, 411, 413—414, 522
микротрихии 64
микротрубочка 49, 128, 146—150, 152, 159, 166, 169, 195, 202,
204, 211, 215—226, 243, 260, 262, 264—266, 269—271, 274,
277, 280, 282—283, 286—287, 293—295, 297—298, 300,
303—304, 306, 311, 314—315, 317, 323, 325, 338—339, 344,
347, 350—354, 357, 362, 377—378, 380, 400, 412—413, 415,
420, 425, 436, 438—439, 444, 463, 469, 482, 486, 490, 492—
494, 500, 511—512, 514, 522—523, 558, 561, 567
микрофагоцитоз 308
микрофауна 536, 630
микрофибрилла 233, 344, 378
микрофиламенты 146—150, 166—167, 169, 219, 271, 286, 305,
322—323, 327, 338, 351, 363, 419, 438, 457—458, 466, 469,
492, 494, 500, 503, 511
микрофлора 71, 73, 526, 630
микрофоссилии 73, 188
микроциста 425, 437
микроэволюция 21, 30—31, 33, 44, 52, 116, 210
микроэлементы 626
мимикрия 35
минерализация 178, 207
минеральный скелет 76, 114, 179, 208, 328, 364, 519, 548
миозин 457
миоцен 44, 46, 600, 604, 606, 610
митоз 125, 128—129, 151, 157—159, 162, 164, 196, 201, 208,
223—225, 229, 252—253, 262, 266, 272, 280, 287, 295,
300—302, 306, 325, 339—341, 346, 348, 353—354, 362—
363, 368, 377—378, 380, 386, 390, 400-^02, 413—414, 425,
436, 439, 483, 488, 533, 567, 571
митохондрион 222—223, 226—227, 241—242
митохондрия 73, 124—126, 128—129, 131, 133, 135—136, 138,
141—143, 152, 154—155, 159, 180, 183, 187, 195, 197, 201,
43 Протисты, часть 1
666
УКАЗАТЕЛЬ ТЕРМИНОВ
205, 211, 222—223, 252, 262, 264—265, 268—270, 273—
274, 281, 286, 289—290, 292, 294, 297, 299—301, 305—306,
311, 314—319, 321, 323, 325, 327, 329, 337—339, 341, 345,
348, 350, 357, 362, 378, 395, 398, 400, 411, 413, 419, 436,
438, 451, 458—459, 462, 469—470, 473, 485—486, 489,
497—498, 500—501, 510—514, 522, 524, 528—530, 533,
557—558, 562, 574—575, 584
мицелий 290
млечники 245
многокамерность 533, 539, 544, 559—560, 565, 572, 574—575,
577—578, 580, 583, 585—586, 588—592, 595, 597—599,
608, 610
мозг 37, 43—44, 50, 53—54, 57, 105, 111, 140, 179, 230, 246
монада 140, 149, 258, 408, 444
монотомия 56, 262, 273, 363
монофилия 41, 48—49, 52, 115, 117, 134
морфогенез 25, 54, 159, 362, 374—375, 377, 380—381, 434,
472—474, 482, 502—503, 533
морфогены 428
морфоплазма 372, 374—376, 378, 388
морфотип 154, 454, 458, 460—463, 465—466
морфофизиологический прогресс 34, 42—43
мукополисахариды 350—351, 548, 569
мукоциста 148, 280, 282—283, 285—286, 288—289, 316
мумификация 366, 385
мускулатура 35, 54, 59, 62, 65
мутант 30, 33
мутация 29—33, 45, 48, 84, 96, 126, 160
мутовка 431
наследственность 95
нематоциста 25, 148
неотения 36—37, 112
неотрох 106
нефридий 70, 112
номинализм 95
номогенез 42, 114
нотонейрон 105
нуклеиновая кислота 35, 71, 93—94, 114, 134
нуклеоид 222—223
нуклеола 565, 571
нуклеоморф 155, 158, 195—197, 199
нуклеоплазма 262
нуклеосома 157
обрастания 12, 187, 321, 624
однополость 42
одноступенчатый мейоз 162, 354—355, 384
окаймленные пузырьки 225
— ямки 167
окремнение 548
оледенение 76
олигомеризация 22, 67—70, 111, 114—116, 186—187, 577—
578, 584
олигопептиды 427
олигоцен 44, 46, 80, 601
онтогенез 11, 16, 19, 21—22, 32—33, 36—38, 43, 45, 50, 53—
57, 78, 84, 102, 115—116, 310, 322, 368, 421, 428, 535, 539,
543, 558, 575, 583
оогамия 161, 290
оогонии 364
ооциты 364
опалесценция 303
опистомастигота 214, 243
оплодотворение 17, 162, 170, 382
опоясывающая ламелла 260, 270, 272, 274, 280
опреснение 534, 625
опущенный жгутик 135, 267—268
органеллология 142—143, 187, 395, 514
организмы-химеры 196
органогенез 59, 64
ордовик 48, 76, 188, 584
ортогенез 32, 42, 44
ортогон 105, 111
ортоламеллярность 552
ортомитоз 158, 205, 262, 270, 276, 280, 285, 287, 297, 306, 325,
337, 345, 400, 420, 425, 439, 459, 469, 488—489, 500, 567
ортоселекция 32, 42
осеменение 41, 52, 364
осморегуляция 35, 166—167, 172, 472, 557
остии 50, 69
палеозой 21, 38, 48, 73, 75—76, 107, 116, 385, 387, 536, 578—
579, 585, 589, 595
палеокомплексы 629
палеоконтиненты 535
палеолит 12
палеореконструкция 535
палеосфера 630
палеоцен 600—601, 604—608, 610—611
палингенез 38, 53—54
палинтомия 159, 171, 205, 307—308
панкинетопластия 223
панцирь 44, 76, 258, 272, 281, 471, 476, 516, 559—560
парадесмос 346, 362
паразит—хозяин 227, 229, 237, 399
паразитизм 11, 18, 22, 24—26, 36, 43, 56, 60, 62, 67, 111, 114—
116, 131, 138, 141—142, 145, 149, 154, 159, 167, 170—171,
179—180, 187, 201, 211—214, 216—217, 219—222, 226—
228, 230—232, 235, 238—239, 241, 243, 245, 247—249, 252,
290, 295, 337, 341, 346—349, 354, 358, 363—364, 368,
380—381, 385—388, 390, 395, 402, 404, 407—408, 451—
452, 467, 511, 532—533, 560, 563, 585—586, 595, 598, 607
паразитофауна 22, 387—388
паразитофорная вакуоль 234, 399—400
паразитоценоз 22
паразиты И, 20—22, 24—25, 29, 35—36, 45, 56, 88, 91, 128—
129, 142, 145, 171, 179—180, 187, 211—213, 215—216,
227—236, 238—241, 243, 245, 247—250, 252, 290, 295, 309,
313, 337—338, 355—357, 362, 367—368, 382, 384—387,
395, 398—400, 402, 404—405, 407—409, 452, 461, 465—
466, 488, 497—499, 528—529, 534, 563, 586
параксиальная пластинка 218—219
— структура 219, 236, 314
параксиальный тяж 149, 201—202, 211, 218—220, 226, 273—
274
параксостиль 352—353, 356—357
парамебный 454, 461
парамилии 205
парамилон 157, 201, 205, 207, 209
парамитоз 158
параподии 50, 66
парасома 458, 461
паренхима 53, 105, 107, 113
паренхимула 49, 111
партеногенез 17, 24, 87, 114
патоген 407
патогенность 179—180, 228, 234, 238, 243, 249—250, 301,
337—338, 341, 346—348, 357—358, 363—364, 367, 395,
452, 465—466, 488, 530
пахитена 402
педицел 332—335
педогенез 37
педоморфоз 37
пектин 280
пеллеты 516—517, 524—526
пелликула 64, 147, 344, 531
пельта 350, 352—353, 355, 357—359, 362—363, 365, 367—368,
375, 377, 387—389
пептон 346, 364
УКАЗАТЕЛЬ ТЕРМИНОВ
667
перидий 434—436, 440—443
перилемма 146
перинуклеарная оболочка 196
— капсула 492
перинуклеарное пространство 155, 157, 180, 260, 262, 265,
345.378
перипласт 147, 194, 196—199
перипластидная цитоплазма 199
перипластидное пространство 195
перипластидный геном 196
перистальтика 166, 204, 233
перифитон 24, 414
пермь 47, 82, 97, 552, 556, 578, 587, 590, 593, 595—597
пероксисомы 125, 128, 1.52, 154—155, 225, 511—512, 562
перфорация 400, 402, 419, 435, 441, 460, 501, 587, 625
пилорус 245
пиноцитоз 167, 184, 202, 221—222, 266, 293, 297, 299, 301,
304—305, 308, 337, 344, 347—348, 353, 379, 431, 458—459
пиреноид 67, 156, 195, 197, 199, 201, 205, 207, 209, 260, 270,
274, 280, 282—283, 286, 289
пиримидиновый синтез 226
пировиноградная кислота 154
пиронин 305—306, 571
пируваты 142, 154, 379
пищеварение 117, 167, 171, 186, 233—234, 337, 385, 431, 462,
465, 495, 497—498, 518—519, 525
плавники 59, 62, 81—82
плагиоаксония 107
плагиостомия 476
плазмагамия 437
плазмалемма 140, 146—150, 167, 184, 194—195, 199, 202—
204, 215—221, 225, 233—234, 241, 243, 258, 260, 268,
270—271, 274, 277, 282—283, 285, 288, 291, 304, 315, 317,
327, 360, 362, 399—400, 451, 457—458, 468, 500, 522
плазматомия 405, 420—421, 432, 438
плазмиды 32
плазмодесмы 400
плазмодизация 143, 164, 418, 489
плазмодии 140—141, 143, 145—146, 164, 179—180, 258, 262,
269, 289—291, 294, 398—400, 402, 404—408, 411—415,
418—421, 424, 431—432, 434—441, 443, 451, 462—466,
488, 495, 516, 519
плазмодиокарп 432, 434, 441—442
плазмотомия 140, 354, 408
плаколиты 281
планктон 55, 73, 141, 172—173, 183—184, 187, 196, 207, 209,
258, 263, 266, 268—269, 272—273, 276, 280—283, 287, 321,
327, 330—332, 462, 476, 519, 533—534, 536—537, 543, 548,
552, 556—563, 569, 572, 574, 577—578, 582—584, 624—630
планула 37, 49, 106
пластиды 73, 141—143, 152, 155, 196—198, 205, 266, 271, 290,400
плевромитоз 128, 158, 211, 290, 294, 337—338, 348, 354, 357—
358, 362, 380, 533, 567, 584
плексус 53, 105, 115, 230
плеоморфизм 230—232, 402
плоидность 225, 306, 438, 565
подостиль 558
покровы 34, 41—42, 57—58, 64, 70, 83, 116, 125, 135, 146—
149, 186, 194, 196, 207, 211, 215—217, 235, 238—239, 241,
271—273, 277, 281—283, 288, 290—291, 297—298, 301,
303, 312, 315—316, 338, 355, 412, 414, 420, 451, 458, 460—
461, 465—468, 477—479, 486, 516—517, 522, 586, 624—625
полигенисты 48, 50, 52
поликдриомастигонт 348, 352—353, 355
поликинетопластия 223, 242
полимеризация 22, 35, 59, 67, 115—116, 138, 140, 187, 225,
233, 310—311, 337, 347, 350, 353, 355, 365, 457, 565, 567,
575, 577, 584, 589, 592
полиморфизм 29—32, 207, 210, 251, 309, 312, 363, 462, 465
полиморфиниды 577—578
полиплоидизация 87, 567, 584
полиплоидия 31, 565, 584
полипы 18, 73, 100, 108
полифилия 21, 48—50, 52, 117
полицеллюляризация 144
полуверетено 223—224, 345—346, 353—354
полудесмосома 232
популяция 21—22, 27, 29—33, 40, 42, 44—45, 84—85, 87—88,
92, 114, 119, 160, 166, 172—174, 176, 179—183, 185, 188,
196, 207—208, 214, 223, 227, 231—232, 235, 243, 272, 281,
300, 307—308, 363, 366, 430, 437, 476, 560, 571
порошица 35, 385, 415
поры 88, 148, 157, 195, 205, 281—282, 292, 323, 345, 436, 466,
469, 471, 473, 486, 556—557, 560—561, 574, 579, 590, 592,
594, 597—598, 601—602, 607—608, 610—611
построструм 389—391
почкование 56, 159, 229, 569, 585
Пратта среда 346
преагрегация 425—427, 429
преадаптация 84
преаксостиль 348—353, 375, 377
предциста 474
преспоры 420—422, 424, 427—430, 439
преформизм 16—17
привилегированные кинетосомы 368, 372, 374—378, 380—381,
387—388, 392
присоски 219, 350, 357
прогамонты 307—308
прогенез 36—37
пролокулюс 539, 577, 589, 591, 593—594
промитоз 488—489
пронуклеус 162, 355, 382, 384
протерозой 71, 73—74, 76, 89, 116
протистофауна 384
протомитохондрии 510
протонефридии 113
протоплазма 431—432, 434—435, 438, 442, 629
протоплазмодий 431—432, 438—442
протопласт 226, 407—408, 419, 421, 439, 495, 497
протостомии 101, 104, 108
прототрох 104
протофауна 386
протрофозоиты 308, 310
профаза 162, 196, 224, 262, 280, 287, 295, 300, 306, 325, 340,
354, 356, 363, 380, 382, 400, 402, 406, 424, 439
профундаль 476
проядрышки 420
псаммобионты 503—504
псевдоаксостиль 352, 356
псевдозвездчатая структура 202
псевдокапиллиций 432, 434—435, 441—442
псевдоконъюгация 164, 418, 421, 429, 503
псевдокопуляция 163, 418, 421, 429, 489
псевдоплазмодий 144, 417—418, 427—430
псевдоподия 124, 136, 141, 143, 145, 166, 184, 258, 269, 291,
293, 324—325, 363, 365, 379, 411—416, 418—419, 424, 427,
451, 454, 458, 460, 465—466, 468-^74, 479, 482—483, 486,
488, 490—491, 493—495, 498, 500—504, 518, 522—523,
525, 535, 557—559, 561—562, 584—585, 626
псевдопоры 552, 557, 579, 585, 588, 591, 594—596, 598
псевдостом 522—523
псевдоциста 231, 363, 380—382, 384, 460, 474
псевдоэталий 432, 434—435, 441—442
радиация 33, 35, 38—39, 41, 45, 49—51, 61, 76, 133, 626
радиоуглеродный метод 563
раковина 41, 54—56, 59—60, 76, 89, 91, 143, 172, 247, 451, 468—
474, 476—479, 482—483, 490, 500—504, 507, 516—520, 522—
527, 533—537, 539, 541, 543—544, 548—549, 551—553, 556—
563, 565, 569, 571—572, 574—575, 577—611, 624—629
668
УКАЗАТЕЛЬ ТЕРМИНОВ
раковинообразование 561
раса 87
растение-хозяин 402, 404—405, 407, 409
реактивность 571
регенерация 17, 25, 569
регресс 36, 43, 59, 116, 129
редукционизм 89, 115
редукционная олигомеризация 70
редукционное деление 402, 571
редукция 36, 41—42, 44, 50—51, 60, 65—66, 68—70, 80, 125,
141, 143, 150, 159—164, 166, 196, 222, 233, 268, 314, 316,
347, 384, 399, 416, 471, 512, 533, 584
резистентность 32, 182, 250
резонаторы 59
резорбция 162, 170, 207, 280, 295, 300, 339—340, 346, 363, 382,
387, 401, 420, 437, 459, 544, 557, 563, 574, 578, 580, 605--606
рекапитуляция 53—57, 104
рекомбинация 161—162, 166, 387, 429
ректальные железы 230
ректум 234, 243, 245
рекуррентный жгутик 149, 211, 217—219, 240—242, 260, 269,
339, 349—351, 355
реликты 48, 88—89, 108, 112, 116, 231, 368, 386—387
реоплазма 558
реотаксис 169
реофаксы 560, 575
репликация 158—159, 196
реснички 35, 40, 56, 59—60, 67, 70, 124, 147—148, 152, 166,
184, 297, 301, 304, 533
ретикулолобоподии 472, 479, 482
ретикулоподии 143, 166, 172, 451, 520—524, 533, 553, 556—
562, 583, 585, 610
ретикулум 172, 196, 225, 533, 558—559, 562
ретикулярный митохондрион 222—223, 241
рецепторы 152, 216, 427
рецессивность 29—30, 32, 160
рибонуклеопротеиды 469
рибосомы 125—126, 128, 133, 138—139, 152, 155, 157, 195,
225, 299, 305, 323, 344—345, 378—379, 398, 400, 413,
512—513, 529—530
ризоиды 140, 476, 627
ризомы 140
ризопласт 260, 262, 266, 270—272
ризоподии 166—167, 260, 269, 558, 624—625
ризостиль 195, 288
рифей 73, 76
РНК 126, 133, 138, 157, 182, 196, 305—306, 565, 571—572
РОВ 166—167, 625
роговица 40
родамин 222
родопсин 203
ростеллум 352, 356
рострум 167, 240—242, 375, 380, 382, 389—392, 415—416
рРНК 94, 126, 128—129, 133, 138—139, 196, 213, 226, 236,
274, 290, 411, 414
рудименты 222, 314, 441
рулевой жгутик 149, 152, 211, 287, 339, 412—413, 415
сальтационизм 84, 116
самозарождение 17
самооплодотворение 87, 162
самопереваривание 426
саморазвитие 43
сапробность 207, 211, 414
сапропель 174, 177, 508, 511
сапрофиты 124, 184, 207, 209, 290, 295, 411
сателлиты 218, 323, 412
светособирающий каротиноид, комплекс 270
свободноживущие 22, 25, 88, 117, 128, 141—142, 145, 149, 154,
157, 159, 167, 169—172, 177, 181, 183, 185—188, 201,
206—207, 209, 211—214, 216—220, 222—223, 225—227,
236, 238—239, 242, 252—253, 268, 270, 276, 301, 311, 314,
317, 321, 337, 341, 346—348, 390, 395, 411, 417, 431, 451—
452, 462, 488, 511, 533—534, 563, 585—586, 588—589, 592,
595, 598, 610, 624
свободноплавающие 56, 176, 178, 184, 227, 241, 264, 266, 269,
276—277, 280, 290, 317, 319, 321, 326, 328—329, 331
сегментация 32, 42—43, 49—50, 57, 60—62, 67, 69, 74—75, 98,
100, 109, 111—112, 115—116. 217, 281, 404, 408, 418,
420—422, 424, 561—562
сегрегация 224
седиментация 125, 138—139, 513
сезонность 114, 181, 187, 302, 560, 624—627
секреция 225, 227, 233—234, 241, 299, 420, 427-428, 437,
460, 468—469, 471—473, 501, 519, 533, 543, 548—549,
551—552, 556, 558—559, 561—563, 565, 572, 574, 577—
580, 582—583, 585, 588, 590, 592, 596, 598, 602, 608—610,
626
селекция 186
семенные мешки 227, 241
— сумки 60, 69
септы 50, 195, 504, 523, 525, 539, 544, 552, 574—575, 582,
585—586, 588—589, 591—592, 594, 597—602, 608
сердечно-сосудистая недостаточность 249
серодиагностика 93
сестринские группы 97, 133, 445
— линии 96
— хромосомы 164
— ядра 164, 354, 380
сизигий 558, 569
сиквенсы 213, 226, 236
силикалемма 469
силур 46, 76, 596
симбиогенез 33, 73, 123, 510
симбиоз 35, 64, 73, 89, 115, 119, 123, 154, 157—158, 161—162, 172,
179—181, 195—196, 201, 226, 266, 314, 323, 348, 353, 355—357,
368, 379—381, 385—387, 389, 395,458,467,469, 502, 507, 510—
511, 537, 557—558, 562—563, 571, 580
симпатрическое видообразование 84
симпласты 67, 146
синаптонемальные комплексы 162, 295, 402, 406, 422, 439
сингамия 49, 302, 308, 310
синглеты 219
синкарион 162, 300, 429
синура 272
синцитий 124, 493, 495, 497
систола 472
склеропротеины 548
склероций 438
скрещивание 16, 30, 42, 45, 52, 87
слоевище 140—141
соленость 45, 172, 178, 180, 182, 188, 226, 328, 462, 503—504,
534, 562, 624—625, 627—628
сомателлы 140—141, 305, 363, 365
соматонемы 146, 298—299, 301
сорокарпы 430—431, 487, 489
сорусы 430—431
сотовый слой 458, 460
спаривание 59, 429, 571
сперматека 219
сперматозоид 16, 161, 364
спермин 112, 157
спикулы 483, 516—517, 519, 543, 548—549, 602, 624
спленомегалия 228
спонгиом 458
спонгоспоры 407
спора 19, 163, 212, 266, 294, 325, 398, 404, 406-408, 418—419,
421,424,427,429—432,434-436,439—442,472,487,489,625
спорангии 281, 290, 294—295, 401, 404, 408—409, 432, 434—
436, 439—442
УКАЗАТЕЛЬ ТЕРМИНОВ
669
спорангиогенез 404
спорангиосорус 404, 408—409
спорический мейоз 124—125, 162
споровая масса 432, 435—436
спорогенез 420—421, 424, 430, 439
спорогония 56
спорокарп 417—418,420-421, 424, 430-432, 434—436, 439—443
спороношение 417, 432
спорообразование 428
спорофор 431—432, 435, 440
споруляция 418, 420—421
стабилизирующий отбор 31—32, 45, 48, 87, 187
статоцист 44
стелиоген 420
стенотермность 624
стенофаги 626
стереоплазма 558
стеркомары 516—-519
стеркоматы 516—517, 524—525, 527
стеркомы 516—519
стигма 195, 201—203, 207, 209, 260, 262, 270—271, 274, 276,
280, 283, 285, 288, 290, 293—294
стилет 60, 356
столоны 533—534, 544, 578, 580, 582, 592, 594—595, 598, 600,
602, 605, 608, 610
стоматоцисты 270
страминопилы 128, 135
стратиграфия 117, 536, 630
стратификация 461
строма 199, 280, 286
строматолиты 71—72, 76
сублитораль 626—629
субпсевдоподии 411, 420, 451, 454, 461—462, 466
субституция 20, 62, 64—66, 117
субцентриоль 511
суккуленты 65
сукцессии 24, 185
сукцинатдегидрогеназа 305
сульфаты 569
супралитораль 503—504, 627
сурры болезнь 212
сферомастигота 215, 231
сферулы 71, 316, 438
схизоцель 98, 107—108
Т-соединение 332, 334
тагмы 60
таксисы 169, 431
таллом 140, 146, 407
танатоценозы 535
тахителия 46, 48
тека 147, 273, 322, 328, 331
тектум 451, 482, 579
телеология 13, 19, 44
телогенез 38, 42, 47
теломорфоз 35—36, 41
телофаза 224, 262, 280, 287, 295, 300, 325, 340, 354, 362, 400,
402, 439
тентакулы 321—323, 325—326, 329—330, 333
тергиты 69
терморезистентность 182, 309
термотаксис 170, 558
терробионты 477
тетразолий 222
тетрапл оидность 171
тигмотаксис 169
тилакоиды 155—156, 195, 197, 199, 201, 205, 260, 270, 272,
274, 280, 286—287, 289
тирозин 472
токсицисты 148
томиты 159
томонт 159
торфобионты 476
трансдукция 33
трансмиссия 22, 212, 235—236, 241, 368, 384—386
трансовариальная передача 232, 235
транспозоны 32
трахеи 41—42, 52, 58, 66
третичные млекопитающие 79
трехчленные мастигонемы 265, 267, 298
триас 44, 47—48, 51, 76, 82, 552, 556—557, 584, 590—591,
593, 596—599, 602, 605
триптиказа 364
триптозы 364
трихиты 202
трихомоноз 358, 363—364, 367, 395
трихоцисты 64, 148, 184, 199, 285—288, 411, 413^—414
трофозоит 181, 202, 206, 272, 297, 299—301, 303—312, 337—
341, 345—349, 363—366, 395, 459, 486, 489
трофонт 415, 444
трофосома 64
Трохеи гипотеза 104—106, 115
трохофора 37, 104—105, 108, 111, 113, 116
тубулемма 146, 211, 215—217, 219, 243, 297—299, 301, 303—
304, 311, 338
тубулин 148, 219, 438, 511
У-соединение 332
убиквисты 196
ультраабиссаль 518, 520, 527, 624, 627, 629
ундулиподии 148—149, 202, 208, 219, 420, 436, 512
ундулирующая мембрана 149, 214, 217, 225, 236, 337, 357—
363, 365—367, 415
уроид 452, 454, 456—457, 462—463, 509, 511—513
устьице 194
фагосомы 184, 458—459
фаготрофия 143, 207, 209, 280, 316, 407—408, 411, 417, 451
фагоцитобласт 35
фагоцитоз 156, 167, 184, 266, 293, 344, 350, 353, 379, 400,
407-408, 419, 426, 429, 431, 437, 458, 462, 465, 469-^72,
474, 482, 488, 494-^95, 497—498, 502
фагоциты 35, 53, 230
фактор эволюции 19, 30
фалькс 303—304, 308
фанерозой 71, 89
фанероплазмодий 431—432, 438, 441—443
фенилаланин 472
фенотип 30—32, 57, 84, 429
ферменты 89, 91, 93, 154—156, 167, 179, 183, 195, 222, 226,
233, 347, 350, 362, 366, 385, 511
фетализация 37
фибриллы 93, 149—150, 157, 195, 199, 223—224, 265, 268,
276—277, 280, 299, 304, 319, 323, 329, 341, 344, 346, 349—
350, 352, 355—356, 362, 368, 372, 374—378, 380, 387—388,
391, 402, 430, 438, 501, 561, 575
фикобилины 135, 156, 195
фикоцианин 195, 198—199
фикоэритрин 195, 197, 199
филамент 146, 148—149, 154, 219, 259, 271, 273, 283, 321, 323,
329, 352, 357, 361—363, 365—366, 372, 375—377, 387—
388, 390, 456—457, 461—463, 492—493, 561
филема 123, 186, 446
филлодии 65
филлокладии 65
филогенез 19, 21—22, 33—34, 38, 42, 50, 52—55, 57—58, 67,
70, 92, 94, 96, 104, 114—116, 160, 187, 310
филоподии 143, 166, 411, 418, 451, 477, 490—495, 497—498,
500, 502—503
фильтраторы 76, 560
670
УКАЗАТЕЛЬ ТЕРМИНОВ
филэмбриогенез 30, 33, 38, 54—57, 115
фиссация 575, 589
фиталь 476—477
фитогельминтология 116
фитолиты 76
фитопланктон 187, 560, 625
фитофаги 245, 472
флексостиль 539, 592—594
флотирование 452, 462, 491—494, 497—498, 574
фонтанирование цитоплазмы 456, 509
форамены 533, 535, 539, 544, 561, 592
фоссилизация 548
фосфатаза 299, 305, 353, 427
фосфолипиды 558
фотопериодизм 27
фотопериодичность 230
фоторецептор 152, 201—202, 209, 270—271, 273, 295
фотосинтез 65, 71, 73, 179, 183, 185, 195, 201—202, 205, 207—
209, 260, 266, 273, 280, 285—286, 563
фототаксис 169, 558
фототрофы 125, 135, 141, 155—156, 173, 194—196, 198, 201—
202, 206—207, 270, 276, 280—282, 285—287
фукоксантин 258, 270, 272, 274, 280, 286
хемоаттрактант 427
хеморецепция 559
хемотаксис 169, 427—429, 559
хиазмы 344, 567
хиатусы 84, 87
хитин 35, 41—42, 50, 52, 57—58, 75, 89, 93, 109, 124, 128, 146,
348, 384, 390
хищник 27, 29, 38, 45—46, 52, 79, 141, 148, 167, 176, 181,
184—185, 187, 202, 226, 240, 245—247, 252, 559—560
хищничество 31, 184, 226, 228, 235, 411
хлоропласт 123—126, 128—129, 135, 141—142, 155—157, 159,
166, 180, 184, 195—199, 201, 205, 207, 209, 258, 260, 262,
269—272, 274, 276—277, 280—283, 285—290, 563
хлорофилл 124—125, 128, 135, 141, 156, 195, 201, 205, 260,
270, 272, 274, 276, 280—281, 286—287, 290, 560
хоаномастигота 214
хоаноциты 49, 328—329, 336
хоаны 81
хоботок 53, 228, 230, 232, 241, 247, 277, 392
хозяин-ксилофаг 353, 379
холодоустойчивость 26, 181, 187
хоматы 544, 549, 579, 588
хондриом 213
хорда 20, 54, 57, 62, 65—66
хризоламинарин 258, 262, 274, 280, 282
хроматиды 306, 354, 378, 380
хроматин 157—158, 223, 225, 262, 286, 352, 357, 378, 400; 518,
529, 565, 572
хроматофоры 67, 407
хромидии 469, 572
хромосомы 21, 30—31, 46, 91, 148, 157—158, 161—162, 164,
182, 194, 196, 201, 205, 211, 223—225, 252, 262, 280, 287,
295, 299—300, 306, 308, 325, 340, 346, 352—354, 362—363,
378, 380, 382—384, 389, 400, 422, 429, 563, 567, 571, 574
цАМФ 427—429
целлюлазы 380, 386
целлюлоза 124, 146—147, 179, 205, 280, 290, 322, 331, 366,
379, 381, 385—386, 402, 428—430
целом 35, 53, 58, 62, 65, 93, 98, 102, 107—110, 112—113, 116,
213, 245, 428, 535, 575
целомодукты 66—67
ценогенез 43, 53—54
центриоль 138, 141, 158, 295, 323, 377, 400—401, 420, 438—
439, 486, 510, 528, 567
центродесмос 346, 362
центроконус 400
центросомы 567
цефализация 42—43, 50, 57
циклоз 35
цикломерия 98
цикломорфоз 476
циста 17, 142, 148, 170—171, 176, 180—181, 183, 185, 187,
196, 201—202, 206—207, 209, 214—215, 217, 223, 227, 236,
240, 242—243, 262—263, 272, 282, 290, 293—294, 299—
300, 308, 316, 321, 326—327, 337—341, 345—348, 355—
356, 363, 366, 368, 380—382, 384, 388, 395, 398, 400, 402—
404, 406, 408—409, 411, 413—416, 419—421, 444, 451,
458—462, 464—466, 468, 474—475, 477, 482, 486—487,
489, 495, 497—498, 503, 522, 528, 530, 558, 560—562, 574
цистеин 364, 472
цистиды 113
цистогенез 400, 402, 404, 407—408
цистообразование 227, 466, 477
цистосорусы 402, 406—409
цитозин 287
цитоид 141, 144
цитоидизация 138—139, 141
цитокинез 158—159, 196, 206, 222, 227—229, 295, 300, 306,
310, 325, 339—340, 346—347, 354—356, 363, 380, 420—
421, 424—425, 436, 439, 459, 497
цитопиг 167
цитоплазма 35, 67, 140—142, 146—148, 154—156, 158—159,
161—164, 167, 170, 172, 180, 183—185, 195, 199,204—205,
207, 216—217, 219—221, 225—227, 234, 260, 262, 265—
266, 271—272, 274, 280, 282, 285—289, 291—294, 297,
299—300, 304—305, 308, 311, 323, 325, 337, 339, 343—358,
362—363, 366—367, 372, 374, 376—380, 382, 388, 390—
391, 398—400, 402, 406—408, 413, 419—420, 424, 429,
431—432, 436, 438—439, 451—452, 454, 456—461, 463—
465, 468—470, 472—474, 482, 485—486, 488—489, 492—
494, 497, 500—503, 507, 509—513, 516—518, 524—525,
527—531, 533, 537, 539, 541, 543—544, 548—549, 552—
553, 556—563, 569, 571—572, 577—578, 585, 607, 609—611
цитоскелет 125, 129, 136, 146—148, 150, 159, 167, 169, 194—
195, 220, 236, 316, 323, 339, 344, 351—352, 375, 377, 387—
388, 436, 454, 457, 466, 469, 482, 490, 493—494, 500, 511,
515
цитостом 151, 167, 184, 201—202, 208—209, 211, 214, 216,
220—222, 226, 240—243, 264, 266—269, 297, 299, 301, 308,
317—319, 338—341, 344, 347—348, 411, 486
цитотомия 414, 569, 585
цитофаринкс 211, 220—222, 318—319, 338
цитофотометрия 537, 565, 571—572
ЦОМТ 196, 225, 272, 314, 350, 352, 354, 362, 378, 380, 420,
425, 436, 438—4-39, 465—466, 482, 486, 510—511, 522, 567
Чагаса болезнь 249
чагома 249
чешуйки 146—147, 149, 194—195, 199, 259—260, 262, 266—
267, 269—272, 276—277, 280—283, 290—291, 317—319,
. 412, 420, 443, 461, 467, 469, 471, 482, 493, 498, 501—503
чешуя 52, 56—57, 68, 70
Шаудинна окраска 452, 483, 625
шельф 206, 209, 263, 280, 627—630
шизогония 159, 408
шизонт 483, 569
штамм 32, 79, 207, 223, 226, 231, 234, 249, 425, 429, 438
щеточная каемка эпителия 232—234, 344, 347
щиток 470
щупальца 40, 48, 53, 59—60, 108, 112, 167, 184, 327
эвагинация 403
эвгленоидное движение 201, 203—204
УКАЗАТЕЛЬ ТЕРМИНОВ
671
эволютная раковина 539, 593, 598—599, 606, 610
эволюционизм 29—31
эволюционисты 31, 83
эволюция И, 13, 17—25, 27—73, 75—79, 81—84, 87—89, 91—
97, 101—105, 108, 110—112, 114—120, 123—124, 126,
128—129, 133, 138—144, 159—162, 164, 179—180, 184—
188, 196, 201, 208, 210, 213, 217, 219, 221—222, 231, 236—
240, 252, 273, 275, 281, 284, 289, 301, 309, 311—312, 319,
328, 338, 341, 347, 355, 358, 365, 368—369, 385—387, 392,
395, 404, 408—409, 417, 424, 446, 511—514, 533, 535, 541,
549, 552, 556, 562—563, 565, 567, 569, 571, 574—575,
577—585, 579, 588, 602—603, 610
эврибатные виды 518, 527
эврибионты 174, 477
эвригалинные виды 503
эвристомные формы 470
эвтрофные водоемы 172, 187, 207, 209, 287, 462, 476, 518,
526
эжектисомы 148, 194, 196—197, 199
эйдос 12
экзолобоподии 472
экзоскелет 491, 493
экзоспория 435
экзофтальмия 228
экзоциста 170
экзоцитоз 426, 473, 548, 561
экзувиация 474—475
экология 11, 17—18, 20—27, 29—30, 32, 34—35, 41, 44, 48,
51, 55, 61, 70, 79, 83—85, 87—88, 91, 94—95, 108—109,
112, 143, 171—174, 177, 179—180, 183—188, 196, 202, 206,
210, 212, 232, 239, 251, 263, 266, 272, 280, 287, 291, 295,
302, 308—309, 327, 346, 385, 395, 418, 422, 429, 439—440,
462, 465—466, 476—477, 488, 503, 518, 526, 534—535, 537,
584, 601, 609, 624—625, 627, 629—631
экосистемы 31, 84, 172, 180, 276, 414, 462, 488, 626, 629—
630
экотопы 462, 488, 503
эксконъюганты 162
экскреция 35, 167, 225—226, 503, 516—517
экструзия 148
экструсомы 147—148, 184, 187, 209, 241, 262, 268—269, 273—
274, 280, 285, 413, 415, 498, 522
эксцистирование 171, 183, 308, 310, 346, 382, 384, 413, 420,
460, 464, 466, 471, 474—475, 482, 486, 497
эктодерма 105
эктокомменсалы 241—242
эктопаразиты 179, 227—228, 241, 626
эктоплазма 147, 289, 291, 304—305, 372, 379, 389—390, 454,
456-457, 479, 509, 511, 533, 558, 585, 624
эктосимбионты 353, 356
эктоциста 460, 464, 466
элатеры 442
элитры 560
эллипсолагены 578
эмбриогенез 18, 24—25, 49, 54, 56—57, 126
эмбрион 53—54, 57, 61, 346, 574
эндемизм 26, 518, 627, 629
эндобионты 180, 183, 185, 196, 379—380, 386, 458, 511
эндокомменсалы 228
эндолобоподии 472
эндомастигота 215—216, 230—231, 243, 245, 247, 252
эндомитоз 383, 438
эндопаразиты 36, 38, 115, 179—180, 183, 187, 201, 206—207,
209—210, 216, 251, 337—338, 344—348, 357—358, 560
эндоплазма 35, 147, 152, 156, 225, 277, 288, 293, 305—306,
377—378, 454, 456—458, 469, 510, 581, 585
эндосимбиоз 24, 142, 196, 226, 356—357
эндосома 349
эндоциста 170, 460, 464, 466
эндоцитобионты 458, 467, 469
эндоцитоз 167, 180, 338, 379, 458, 488
энергида 67
энзимы 379, 562
энтероциты 228, 232—234, 243
энтодерма 111, 232
эоцен 46, 79, 81, 578, 586, 594, 600, 606—611
эпибионты 174, 183, 201, 207, 209, 327, 534, 560
эпигенез 16
эпидерма 111
эпидермис 53, 64, 235, 407
эпизоотии 228
эпилимнион 476
эпимастигота 214, 224, 231—232, 236, 243, 247
эпиподии 468, 473—474, 482—483, 500, 503
эпителизация 53, 107
эпителий 52—53, 64, 103—104, 179, 219, 227—229, 232—235,
241, 344, 347, 364, 366
эпифиты 423, 441, 558—560
эпифрагма 474
эруптивное движение 485—486, 488
эстеразы 93
эстерон 308
эталии 432, 434—435, 441—443
эугамный 384
эукариотизация 125, 138—139
эукинетопластия 223, 241—242
эумитоз 158
эуцит 141 )
эхинеон 260
эхиноподии 461
юра 44, 49, 82, 281, 556, 578, 584, 591, 596—602, 604—605,
610
ядро 35, 67, 73, 84, 100, 125, 128, 140—141, 143, 146—148,
152, 155—159, 161—164, 166, 171, 187, 194—196, 199, 201,
205—206, 208, 210—211, 214—215, 223—225, 243, 247,
252—253, 260, 262, 265—276, 280—295, 297—300, 302,
305—308, 310—312, 314—315, 317, 321, 323, 325, 327,
337—341, 343—350, 352, 354—358, 362—363, 365, 367—
368, 375—378, 380, 382—384, 387—392, 395, 398, 400, 402,
404—406, 408—409, 411—415, 418—422, 424—425, 429,
436, 438—439, 444, 454, 458—461, 463—466, 468—469,
473-475, 478—479, 483, 486, 488—490, 492—493, 497,
500, 502—504, 507, 509, 518—519, 522, 524—525, 528—
532, 534, 537, 563, 565, 567, 569, 571—572, 574—575,
580—582, 584—586, 592, 595, 610
ядрышко 157—158, 194, 196, 201, 205, 223—224, 262, 280,
286, 292, 300, 305—306, 315, 323, 345, 352, 355—357, 363,
378, 400—402, 405—406, 413, 420, 425, 458, 460, 468,
488—489, 500, 518, 529, 571
яичники 70, 228, 364
яйцеклад 61
яйцеклетки 161, 170
СПИСОК СОКРАЩЕНИИ
БИН
БиНИИ СПбГУ
ВИЗР
ВИНИТИ
ВНЙГРИ
ВНИРО
воз
ВСЕГЕИ
гин
ГОСНИОРХ
ЗИН
ИБВВ
ИБМ
ИБР
ИгиГ СО РАН
ИОАН
ИОРАН
ИЭВБ РАН
ИПЭЭ
ЛГПИ
ЛГУ
МГУ
ММБИ
РАН
СНГ
СПбГУ
цин
— Ботанический институт РАН (Санкт-Петербург)
— Биологический научно-исследовательский институт СПбГУ
— Всероссийский институт защиты растений (Санкт-Петербург)
— Всероссийский институт научной и технической информации (Москва)
— Всероссийский научно-исследовательский геолого-разведывательный институт (Санкт-Пе-
тербург)
— Всероссийский научно-исследовательский институт рыбного хозяйства и океанологии
(Москва)
— Всемирная организация здравоохранения
— Всероссийский научно-исследовательский геологический институт (Санкт-Петербург)
— Геологический институт РАН (Москва)
— Государственный научно-исследовательский институт озерного и речного рыбного хозяй-
ства (Санкт-Петербург)
— Зоологический институт РАН (Санкт-Петербург)
— Институт биологии внутренних вод РАН им. И, Д. Папанина (Борок)
— Институт биологии моря РАН (Владивосток)
— Институт биологии развития РАН (Москва)
— Институт геологии и геофизики Сибирского отделения РАН (Новосибирск)
— бывший Институт океанологии АН СССР (Москва)
— Институт океанологии РАН (Москва)
— Институт экологии Волжского бассейна РАН (Тольятти)
— Институт проблем экологии и эволюции РАН им. А. Н. Северцова (Москва)
— бывший Ленинградский государственный педагогический институт им. А. И. Герцена
— бывший Ленинградский государственный университет им. А. А. Жданова
— Московский государственный университет им. М. В. Ломоносова
— Мурманский морской биологический институт РАН
— Российская академия наук
— Союз независимых государств
— Санкт-Петербургский государственный университет им. Д. И. Менделеева
— Институт цитологии РАН (Санкт-Петербург)
АТФ — аденозинтрифосфат РНК — рибонуклеиновая кислота
ВОС — внутренний органический слой РОВ — растворенные органические вещества
ДМСО — диметилсульфоксид рРНК — рибосомальная РНК
днк — дезоксирибонуклеиновая кислота СМТ — субмембранные микротрубочки
иРНК — информационная РНК СОМ — срединная органическая мембрана
Ма — макронуклеус СТЭ — синтетическая теория эволюции
мДНК — митохондриальная ДНК ФАС — S-образная лента микротрубочек
Ми — микронуклеус ЦМТ — цитостомальные микротрубочки
МТР — микротрубочки, подстилающие стенки жгутикового ЦОМТ — центр организации микротрубочек
кармана и ротового аппарата ЭПР — эндоплазматический ретикулум
НП — нуклеотидные последовательности
СВЕДЕНИЯ ОБ АВТОРАХ
БЕЛОВА Лариса Михайловна, Зоологический институт РАН.
199034 Санкт-Петербург, Университетская наб., 1.
Научные интересы: фауна, систематика, распростране-
ние, морфофункциональная организация и жизненные циклы пара-
зитических простейших.
Основные публикации
Белова Л. М., Крылов М. В. Множественные формы малатде-
гидрогеназы (К.Ф. 1.1.1.37) у Crithidia asclepii и С. oncopelti (Mas-
tigophora, Trypanosomatidae) И Паразитология. 1984. Т. 28, вып. 6.
С. 425—429.
Крылов М. В., Белова Л. М. Полиморфизм малатдегидрогеназы
у критидий (Protozoa, Mastigophora) // Докл. АН СССР. 7985.
Т. 280, № 6. С. 1426—1428.
Белова Л. М. Мировая фауна и морфофункциональная органи-
зация бластоцист (Protista, Rhizopoda) И Тр. ЗИН РАН. 7992.
Т. 244. 58 с.
Belova L. М. Blastocystis galli (Rhizopoda, Lobosea) — a parasite
or a commensal? // Arch. Protistenk. 1992. Bd 141. S. 215—218.
Белова Л. M. Фауна Blastocystis // Паразитология. 1995. T. 29,
вып. 3. C. 208—213.
БОБЫЛЁВА Нина Николаевна, Институт цитологии РАН.
194064 Санкт-Петербург, Тихорецкий пр., 4.
E-mail: vasilek@cit.ras.spb.su
Научные интересы: клеточная биология протистов.
Основные публикации
Bobyleva N. N. Morphology and evolution of intestival parasitic
flagellates of the far-eastern roach Cryptocercus relictus // Acta Proto-
zool. 1975. Vol. 14. P. 109—160.
Bobyleva N. N., Kudrjavtsev B. N., Raikov I. B. Changes of the DNA
content of differentiating and adult macronuclei of the ciliate Loxodes
magnus (Karyofelictida) // J. Cell Sci. 1980. Vol. 44. P. 375—394.
Бобылёва H. H., Сергеева Г. И. Структурная организация хро-
матина ядер, неспособных к делению, на примере макронуклеу-
сов инфузории Loxodes magnus // Цитология. 7997. Т. 33. С. 14—
19.
Бобылёва Н. Н., Скарлато С. О. Структурная организация не-
активного хроматина жгутиконосцев-трипаносоматид рода Crithi-
dia // Докл. РАН. 1994. Т. 339. С. 551—554.
Sergejeva G. /., Bobyleva N. N. Polynemic structures in the diffe-
rentiated macronucleus of the ciliate Biirsaria ovata Beers 1952 // Acta
Protozool. 7995. Vol. 35. P. 115—124.
ГАВРИЛОВА Ольга Владимировна, Санкт-Петербургский
государственный университет, Биологический институт.
198904 Санкт-Петербург, Старый Петергоф, Ораниенбаумское
шоссе, 2.
E-mail: Olga.Gavrilova@paloma.spbu .ru
Научные интересы: цитология и филогения эукариоти-
ческих микроорганизмов.
Основные публикации
Громов Б. В., Гаврилова О. В. Ультраструктура зооспор одно-
клеточной водоросли Bracteacoccus minor (Chlorophyta, Chlorococ-
cales) // Ботан. журн. 1986. T. 71. С. 66—71.
Громов Б. В., Гаврилова О. В. Особенности ультраструктуры
зооспор одноклеточной зеленой водоросли Neochloris alveolaris //
Цитология. 1987. Т. 29. С. 637—641.
Гаврилова О. В., Громов Б. В. Строение зооспор зеленой водо-
росли Chlorotetraedron bitridens (Palmellaceae) И Ботан. журн. 7988.
Т. 73. С. 486—497.
ГРОМОВ Борис Васильевич, Санкт-Петербургский государ-
ственный университет, Биолого-почвенный факультет:
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: Вoris.Gromov @ paloma. spbu .ru
Научные интересы: строение микроорганизмов, парази-
тизм, симбиоз, биологически активные вещества автотрофных
микроорганизмов.
Основные публикации
Громов Б. В. Строение бактерий. Л.: Изд-во ЛГУ, 7985.
ГУДКОВ Андрей Владимирович, Санкт-Петербургский госу-
дарственный университет, Биолого-почвенный факультет.
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: good@good.usr.pu.ru
Научные интересы: биология, эволюционная морфология,
филогения и систематика протистов.
Основные публикации
GoodkovА. V., Seravin L.N. Pelomyxa palustris: amoeba, caryo-
blastean, archezoan, or peloflagellatan? II Cytology (Russia). St. Peter-
sburg, 7995. Vol. 37. P. 1053—1063.
Серавин Л. H., Гудков А. В. Агамные слияния протистов и про-
исхождение полового процесса. СПб.; Омск, 7999.
ЖУКОВ Борис Федорович, Институт биологии внутренних
вод РАН.
157742 п/о Борок Ярославской обл.
E-mail: zhukov@ibiw.yaroslavl.su
Научные интересы: протозоология (протистология), гете-
ротрофные жгутиконосцы, систематика, филогения, экология.
Основные публикации
Жуков Б. Ф. Определитель бесцветных свободноживущих жгу-
тиконосцев подотряда Bodonina Hollande И Биология и продуктив-
ность пресноводных организмов. Л.: Наука, 7977. С. 241—284.
Жуков Б. Ф. Определитель бесцветных жгутиконосцев отряда
Bicosoecida Grasse et Deflandre (Zoomastigophorea, Protozoa) //
Биология и систематика низших организмов. Л.: Наука, 1978.
С. 3—28.
KarpoffS. A., Zhukov В. F. Ultrastructure and taxonomic position
of Apusomonas proboscidea Alexeieff // Arch. Protistenk. 1986.
Bdl31. S. 13—26.
Жуков Б. Ф., Карпов С. А. Пресноводные воротничковые жгу-
тиконосцы. Л.: Наука, 1985. 120 с.
Жуков Б. Ф. Атлас пресноводных гетеротрофных жгутиконос-
цев (биология, экология и систематика). Рыбинск, 1994. 160 с.
ИВАНОВ Артемий Васильевич (1906—1992), профессор,
академик РАН, лауреат Ленинской премии.
Зоолог-эволюционист, автор классических исследований по
сравнительной анатомии, филогенетике и систематике паразити-
ческих брюхоногих моллюсков, ресничных червей и погонофор
(включая вестиментифер, открытых в глубоководных гидротер-
мах). Разрабатывал общие проблемы эволюционной морфологии,
филогенетики и систематики, а также проблемы происхождения и
674
сведения об авторах
построения системы многоклеточных животных. Автор ряда мо-
нографий и учебных пособий. Работал в Ленинградском (ныне
Санкт-Петербургском) государственном университете и в Зоологи-
ческом институте РАН.
Основные публикации
Иванов А. В. Морфологические адаптации к паразитическому
образу жизни И Проблемы общей паразитологии // Учен. зап. ЛГУ.
Сер. биол. 1937. Т. 3, вып. 4. С. 53—94.
Иванов А. В. Бескишечные турбеллярии (Acoela) южного по-
бережья Сахалина И Тр. ЗИН АН СССР. 1952. Т. 12. С. 40—132.
Иванов А. В. Погонофоры. М.; Л.: Наука, 1960. 271 с. (Фауна
СССР. Нов. сер.; Вып. 73).
Ivanov А. V. Pogonophora. London: Acad. Press, 1963. 479 p.
Иванов А. В. Происхождение многоклеточных животных. Фи-
логенетические очерки. Л.: Наука. 1968. 288 с.
Иванов А. В., Мамкаев Ю. В. Ресничные черви, их происхож-
дение и эволюция. Л.: Наука, 1973. 222 с.
Ivanov А. V. Analysis of the embryonic development of Pogono-
phora in connection with problems of phylogenetics // Ztschr. zool.
Sist. und Evolutionsforsch. 1988. Bd 26. S. 161—185.
Ivanov A. V. On the systematic position of Vestimentifera // Zool.
Jb. Syst. 1994. H.4. S. 410—456.
КАМЕНСКАЯ Ольга Евгеньевна, Институт океанологии
им. П. П. Ширшова РАН.
117218 Москва, ул. Красикова, 23.
E-mail: kamenska@bentos.ioras.msk.ru
Научные интересы: глубоководная мейо- и макрофауна.
Основные публикации
Виноградова Н. Г, Галкин С. В., Каменская О. Е., Левен-
штейнР.Я., Романов В. Н. Распределение глубоководной фауны
на широтном трансокеаническом разрезе в Южной Атлантике
(примерно по 31 градусу ю. ш.). М.: Наука, 1990. С. 7—19. (Тр.
ИОАН; Т. 126).
Каменская О. Е. Комокиации из района подводной горы Валь-
дивия (юго-восточная Атлантика). М.: Наука, 1993. С. 2—81. (Тр.
ИОАН; Т. 127).
Каменская О. Е. Конспект системы класса Xenophyophorea
(Protozoa) # Бентос северных морей Евразии 1 ВНИРО. 1997.
С. 43—56.
КАРПОВ Сергей Алексеевич, Санкт-Петербургский государ-
ственный университет, Биолого-почвенный факультет.
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: karpov@skl027.spb.edu .
Научные интересы: протистология, симбионтология, кле-
точная биология, систематика.
Основные публикации
Жуков Б. Ф., Карпове. А. Пресноводные воротничковые жгу-
тиконосцы. Л.: Наука, 1985. 120 с.
Карпов С. А. Система протистов. Омск, 1990. 195 с.
KarpovS. A. (ed.). The biology of free-living heterotrophic flagel-
lates II Cytology (Russia). St. Petersburg, 1995. Vol. 37.
КОЛЧИНСКИЙ Эдуард Израилевич, Институт истории ес-
тествознания и техники РАН, СПб. филиал.
199034 Санкт-Петербург, Университетская наб., 5/2.
Научные интересы: история теории эволюционного уче-
ния, детерминация органической эволюции и методы ее познания,
эволюция эволюций, эволюция биосферы, особенности эволюци-
онного процесса в современных условиях, история русско-немец-
ких научных связей.
Основные публикации
Завадский К. М., КолчинскийЭ. И. Эволюция эволюций: исто-
рико-критические очерки проблемы. Л.: Наука, 1977.
Колчинский Э. И. (ред.-сост.). Развитие эволюционной теории
в СССР, 1917—1970-е гг. Л.: Наука, /977.
Колчинский Э. И. Эволюция биосферы: историко-критические
очерки исследований в СССР. Л., 1990.
Колчинский Э. И. В поисках советского «союза» философии и
биологии. Дискуссии и репрессии в 20-х—начале 30-х годов.
СПб.: Дом Буланина, 7999.
Kolschinsky Е. I. Zur Rolle Deutscher Wissenschaften bei der Ent-
wicklung der Biologie in Russland im 18. Jahrhundert // Europa in der
Friihen Neuzeit. Koln etc., 1997. Bd 4. S. 289—307.
ЛЕНЦМАН Наталья Валерьевна, Санкт-Петербургский госу-
дарственный университет, Биолого-почвенный факультет.
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: arcella@nvl. stud.pu.ru
Научные интересы: протистология, амебоидные протис-
ты, ультраструктура, систематика раковинных амеб.
Основные публикации
Smirnov А. V., LentsmanN. V., Nasonova Е. S. Vertical distributi-
on of naked and testate amoebae (Lobosea: Gymnamoebia, Testacea-
lobosia) in the bottom sediments of a freshwater lake // Proc. St. Pe-
tersburg nat. Soc. Ser. 1. 1998. Vol. 92. P. 74—81.
ЛУКИНА Татьяна Григорьевна, Зоологический институт
РАН.
199034 Санкт-Петербург, Университетская наб., 1.
E-mail: LTG@aster.zin.ras.spb.ru
Научные интересы: современные бентосные форамини-
феры Арктического бассейна и Тихого океана, фаунистика, рас-
пространение, зоогеография.
Основные публикации
Лукина Т. Г. Фораминиферы верхнего отдела шельфа у архи-
пелага Земля Франца-Иосифа // Биоценозы шельфа Земли Фран-
ца-Иосифа и фауна сопредельных акваторий. Л., 1977. С. 72—105.
(Исслед. фауны морей; Т. 14 (22)).
Лукина Т. Г. Глубоководные фораминиферы центральной
части Тихого океана. Л., 1980. 204 с. (Исслед. фауны морей; Т. 24
(32)).
Лукина Т. Г. Фораминиферы Новосибирского мелководья //
Экосистемы Новосибирского мелководья и фауна моря Лаптевых
и сопредельных вод. Л., 1990. С. 105—129. (Исслед. фауны морей;
Т. 37 (45)).
Лукина Т. Г., Гальцова В. В., Владимиров М. А. Мейобентос
Чаунской губы Восточно-Сибирского моря // Экосистемы и фауна
Чаунской 1убы и сопредельных вод Восточно-Сибирского моря.
СПб., 1994. С. 67—97. (Исслед. фауны морей; Т. 45 (56)).
Лукина Т. Г. Фораминиферы материкового склона Курильской
островной гряды И Фауна материкового склона Курильской ос-
тровной гряды. СПб., 1994. С. 45—76. (Исслед. фауны морей; Т. 46
(54)).
МИХАЛЕВИЧ Валерия Иосифовна, Зоологический институт
РАН.
199034 Санкт-Петербург, Университетская наб., 1.
E-mail: MVI@zisp.spb.su
Научные интересы: фораминиферы: морфология, фауна,
систематика, эволюция, зоогеография.
Основные публикации
Михалевич В. И. Отряд фораминиферы — Foraminifera d’Or-
bigni, 1826 // Определитель фауны Черного и Азовского морей.
Киев: Наукова думка, 1968. С. 9—21.
Михалевич В. И. Параллелизм и конвергенция в эволюции ске-
летов фораминифер И Тр. ЗИН АН СССР. Л.: Наука, 1981. Т. 107.
С. 19—41.
Михалевич В. И. Донные фораминиферы шельфов тропической
Атлантики II Тр. ЗИН АН СССР. Л.: Наука, 1983. С. 1—47.
Михалевич В. И. Система подкласса Miliolata (Foraminifera) //
Систематика, экология и стратиграфия милиолят (Фораминифе-
ры) / Под ред. В. И. Михалевич. Л.: Наука, 1988. С. 77—110. (Тр.
ЗЙН АН СССР; Т. 184).
Михалевич В. И. Макросистема фораминифер // Изв. РАН.
Сер. биол. 1998. №2. С. 266—271.
Михалевич В. И. Система и филогения фораминифер. СПб.;
Омск, 1999. 185 с.
МЫЛЬНИКОВ Александр Петрович, Институт биологии
внутренних вод РАН.
157742 п/о Борок Ярославской обл.
E-mail: mylnikov@ibiw.yaroslavl.su
Научные интересы: протозоология (протистология), гете-
ротрофные жгутиконосцы, ультраструктура, систематика, филоге-
ния, экология.
Основные публикации
Мыльников А. П. Анаэробные поли- и ризомастигины некото-
рых полисапробных зон И Биология низших организмов / Под ред.
М. М. Камшилова и др. Рыбинск, 1978. С. 128—136.
Мыльников А. П. Определитель свободноживущих жгутиконос-
цев отряда Diplomonadida (Wenyon) Brugerolle И Водные сообщес-
тва и биология гидробионтов / Под ред. В. Н. Яковлева. Л.: Наука,
1985. С. 174—198.
Мыльников А. П. Биология и ультраструктура амебоидных
жгутиконосцев Cercomonadida ord. n. И Зоол. журн. 1986. Т. 65,
вып. 5. С. 683—692.
СВЕДЕНИЯ ОБ АВТОРАХ
675
Мыльников А. П. Некоторые таксономические признаки церко-
монадид И Биология внутренних вод. Информ, бюл. 1992. Ъ 93;
С. 35—39.
Микрюков К. А., Мыльников А. П. Новые данные о строении и
жизненном цикле аталамидных амеб (Protista: Athalamida) // Зоол.
журн. 1996. Т. 75, вып. 9. С. 1284—1294.
НОВОЖИЛОВ Юрий Капитонович, Ботанический институт
им. В. Л. Комарова РАН.
197376 Санкт-Петербург, ул. проф. Попова, 2.
E-mail: mixus@YN1091.spb.edu
Научные интересы: систематика, экология, биогеография
грибов и слизевиков.
Основные публикации
Новожилов Ю. К. Определитель грибов России. Отдел Мухо-
mycota. Вып. 1. Класс Myxomycetes. СПб.: Наука, 1993. 288 с.
Novozhilov Y. К., Schnittler М. Nivicole Myxomycetes of the Khi-
bine Mountains (Kola peninsula // Nordic J. Bot. 1997. Vol. 16.
P. 549—561.
Novozhilov Y. K., Schnittler M., StephensonS. The myxomycete
biota of Russian subarctic and arctic areas (preliminary report) // Mi-
cologia i Fitopatologia. 1998. Vol. 32, N 1. P. 18—29.
Каратыгин И. В., Нездойминого Э. Л., Новожилов Ю. К., Жур-
бенко М.П. Грибы Российской Арктики. СПб.: Изд-во СПб. гос.
хим.-фарм. акад., 1999. 212 с.
ПОЛЯНСКИЙ Юрий Иванович (1904—1993), профессор,
чл.-кор. РАН.
Первый президент Всесоюзного, а позже Всероссийского об-
щества протозоологов. Автор более 300 научных публикаций,
главным образом в области протозоологии, в том числе по эво-
люции и генетике простейших. Один из авторов «Большого прак-
тикума по зоологии беспозвоночных», учебника для средней
школы «Общая биология» (29 изданий на русском языке, пере-
веден на языки бывших союзных республик, а также на француз-
ский, испанский и португалький языки). Велика его роль в пере-
издании тематических монографий В. А. Догеля и в публикаци-
ях их на английском и немецком языках. Возглавлял кафедру
Зоологии беспозвоночных Ленинградского (ныне Санкт-Петер-
бургского) государственного университета, активно участвовал в
создании Института цитологии АН, в рамках которого организо-
вал и возглавил Лабораторию цитологии одноклеточных организ-
мов.
Публикации последних лет
Полянский Ю. И. О закономерностях микро- и макроэволюции
у одноклеточных эукариот // Проблемы генетики и теории эволю-
ции. Новосибирск, 1991. С. 229—241.
Polansky Yu. I. Protozoology and the problem of species // J. Pro-
tozool. 1992. N 1. P. 177—180.
СЕРАВИН Лев Николаевич, Санкт-Петербургский государст-
венный университет, Биолого-почвенный факультет.
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: invert@ontil.bio.spbu.ru
Научные интересы: клеточная биология, протистология,
этология и физиология простейших, а также многоклеточных ор-
ганизмов.
Основные публикации
Серавин Л. Н. Двигательные системы простейших (строение,
механохимия и физиология). Л.: Наука, 1967. 332 с.
Серавин Л. Н. Теория информациии с точки зрения биолога.
Л.: Изд-во ЛГУ, 1973. 160 с.
Серавин Л. Н. Простейшие... Что это такое? Л.: Наука, 1984.
174 с.
СеравинЛ. Н. Ctenophora — Гребневики. СПб.; Омск, 1998.
84 с.
Серавин Л. Н, Гудков А. В. Агамные слияния простейших и
происхождение полового процесса. СПб.; Омск, 1999. 154 с.
СМИРНОВ Алексей Валериевич, Санкт-Петербургский госу-
дарственный университет, Биолого-почвенный факультет.
199034 Санкт-Петербург, Университетская наб., 7/9.
E-mail: smirnov@as2187.spb.edu
Научные интересы: протистология, амебоидные протес-
ты, экология, ультраструктура, систематика голых амеб.
Основные публикации
Smirnov А. V. Stygamoeba regulata n. sp. (Rhizopoda) — a mari-
ne amoeba with an unusual combination of light-microscopical
and ultrastructural features // Arch. Protistenk. 1995. Bd 146. S. 299—
307.
Smirnov A. V., Fenchel T. Vahlkampfia anaerobica n. sp. and Van-
nella peregrinia n. sp. (Rhizopoda) — anaerobic amoebae from a ma-
rine sediment // Arch. Protistenk. 1996. Bd 147. S. 189—198.
Smirnov A. V., Goodkov A. V. Freshwater gymnamoebae with a
new type of surface structure Paradermamoeba valamo and P. levis
sp. n. (Thecamoebidae), and notes on the diagnosis of the family //
Acta Protozool. 1994. Vol. 33. P. 109—115.
Smirnov A. V., Goodkov A. V. Study of Polychaos annulatum Pe-
nard, 1902 comb. Nov. (Gymnamoebia, Amoebidae) with taxonomic
analysis of Polychaos fasciculatum-like species // Eur. J. Protistol.
1998. Vol. 34. P. 1—9.
Smirnov A. V., Goodkov A. V., GoobanovD. L. An illustrated guide
to gymnamoebae. Part I. The family Thecamoebidae. WWW-homepa-
ge. http: // rem.ifmo.ru/dox/amoebae.htm
СУХАНОВА Ксения Мироновна, Российский государствен-
ный университет им. А. И. Герцена, факультет биологии.
191186 Санкт-Петербург, наб. р. Мойки, 48.
Научные интересы: протозоология, морфология, физио-
логия, экология, эволюция, жизненные циклы протистов.
Основные публикации
Суханова К. М. Опалиниды и инфузории пищеварительного
тракта бесхвостых амфибий (Anura) Каневского заповедника /
Материалы по изучению истории и природы Каневского заповед-
ника. Киев, 1962. С. 98—117.
Суханова К. М. Температурные адаптации у простейших. Л.:
Наука, 1968. 268 с.
Суханова К. М. Адаптации простейших к факторам внешней
среды: полиморфизм вида и популяции по признаку теплоустойчи-
вости И Вопросы экологии простейших. Л.: Наука, 1978. С. 26—
36.
Суханова К. М. Растительные жгутиконосцы // Фауна аэротен-
ков. Атлас. Л.: Наука, 1984. С. 40—82.
Суханова К. М. Свободноживущие инфузории — индикаторы
термического режима пресных водоемов // Инфузории в биотести-
ровании. СПб., 1997. С. 104—109.
ФРОЛОВ Александр Олегович, Зоологический институт
РАН.
199034 Санкт-Петербург, Университетская наб., 1.
E-mail: frol@neva.spb.ru
Научные интересы: фауна, сравнительная морфология,
происхождение и эволюция паразитических простейших.
Основные публикации
Фролов А. О. Мировая фауна грегарин. Семейство Monocysti-
dae И Тр. ЗИН АН СССР. Л., 1991. 126 с;
Фролов А. О. Происхождение трипаносоматид И Паразитоло-
гия. 1993. Т. 27, вып. 2. С. 97—107.
Frolov А. О., KarpovS. A. Comparative morphology of kineto-
plastids // Cytology (Russia). St. Petersburg, 1995. T. 37. P. 1072—
1096.
Фролов А. О., СкарлатоС.О. Тонкое строение и механизмы
адаптации низших трипаносоматид в полужесткокрылых насеко-
мых И Цитология. 1995. Т. 37. С. 539—560.
Frolov A., SkarlatoS. Unusual pattern of mitosis in free—living
flagellate Dimastigella mimosa (Kinetoplastida) // Protoplasma. 1998.
Vol. 201. P. 110—114.
ЧИБИСОВА Ольга Ильинична, Московский государствен-
ный университет им. М. В. Ломоносова, Биологический факультет.
119899 Москва, Ленинские горы.
Научные интересы: почвенные простейшие.
Основные публикации
Чибисова О. И., Лепинис А. К., Гельцер Ю. Г., Гептнер В. А.
Определитель Protozoa почв Европейской части СССР. Вильнюс,
1973. 170 с.
СОДЕРЖАНИЕ
Предисловие.................................................................................. 5
Preface .................................................................................... 7
ОБЩАЯ ЧАСТЬ .................................................................................. 9
Краткая история зоологии (Ю. И. Полянский).................................................... 11
Summary ....................... ч........................................................... 29
Пути и закономерности эволюции (А. В. Иванов, Э, И. Колчинский) ............................. 29
Summary ..................................................................................... 85
Систематика и ее задачи (А. В. Иванов) ...................................................... 85
Summary ................................................................................ 97
Система животного мира (А. В, Иванов) ................................................ . 97
Summary ..........................................................,.......................... ИЗ
Литература ................................................................................. 114
ПРОТИСТЫ. ОБЩИЕ ВОПРОСЫ .................................................................. 121
Система протистов и проблемы их мегасистематики (С. А. Карпов).............................. 123
Summary ................................................................................. 136
Пути эволюции протистов (Л. Н. Серавин) .................................................... 138
Summary .................................................................................... 145
Общая характеристика протистов (Ю. И. Полянский, К. М. Суханова, С. А. Карпов) ............. 145
Summary ................................................................................. 185
Литература................................................................................. 185
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ ..................................................................... 191
Тип Cryptophyta (Б. В. Громов, О. В. Гаврилова) .. ...................................... 193
Summary .................................................................................. 200
Литература ............................................................................... 200
Тип Euglenozoa ............................................................................. 201
Класс Euglenoidea (К. М, Суханова) .................................................. 201
Summary .................................................................................... 210
Литература............................................................................... 210
Класс Kinetoplastidea (А. О. Фролов) ...................................................... 211
Summary ................................................................................. 251
Литература............................................................................... 251
Тип Chrysophyta ......................................................................... 257
Класс Chrysophyceae (К. М. Суханова, С. А. Карпов)....................................... 258
Отряд Bicosoecales (С. А. Карпов) .......................................................... 264
Класс Synurophyceae (К М. Суханова, С. А. Карпов) .......................................... 270
Класс Pelagophyceae (К. М. Суханова, С. А. Карпов) ...................................... 273
Summary ................................................................................. 274
Литература............................................................................... 275
Тип Haptophyta (К. М. Суханова) ............................................................ 276
Summary ................................................................................. 283
Литература .............................................................................. 284
Тип Raphidophyta (К. М. Суханова)........................................................ 285
Summary ................................................................................. 289
Литература............................................................................... 289
Тип Saprolegnia ......................................................................... 290
Класс Labyrinthomorphea (К. М. Суханова) ................................................ 290
Summary ................................................................................. 296
Литература ................................................................................. 296
Тип Opalinata ........................................................................... 297
Класс Proteromonadea (К. М. Суханова) ................................................... 297
Класс Opalinatea (К М. Суханова) ........................................................ 301
Summary ................................................................................. 312
Литература.................................................................................. 312
«Chromista» incertae sedis ................................................................ 314
Отряд Spongomonadida (С. А. Карпов) . . ................................................... 314
Отряд Pseudodendromonadida (С. А. Карпов) ............................................. 317
Summary ................................................................................. 319
Литература .............................................................................. 320
Тип Choanomonada (С. А. Карпов, Б. Ф. Жуков)............................................. 321
Summary ................................................................................. 335
Литература .............................................................................. 336
СОДЕРЖАНИЕ
677
Тип Polymastigota .............................................................................. 337
Класс Diplomonadea (К. М. Суханова) ............................................................ 337
Класс Oxymonadea (Н. И. Бобылёва) .............................................................. 348
Класс Parabasalea .............................................................................. 357
Отряд Trichomonadida (К. М. Суханова) .......................................................... 358
Отряд Hypermastigida (Н. Н. Бобылёва)........................................................... 368
Summary ........................................................................................ 392
Литература...................................................................................... 395
Тип Plasmodiophora (А. В. Гудков) .............................................................. 398
Summary ........................................................................................ 409
Литература..................................................................................... 409
Тип Mycetozoa................................................................................... 411
Класс Cercomonadea (А. П. Мыльников) ........................................................... 411
Summary ........................................................................................ 417
Класс Eumycetozoea (JO. К. Новожилов, А. В. Гудков) ............................................ 417
Summary ........................................................................................ 443
Mycetozoa incertae sedis ....................................................................... 444
Hyperamoeba flagellata (С. А. Карпов) .................................................. 444
Summary ...................................................................................... 445
Литература...................................................................................... 445
Тип Rhizopoda................................................................................... 451
Класс Lobosea ................................................................................ 451
Подкласс Gymnamoebia (А. В. Смирнов, А. В. Гудков) ............................................. 451
Summary ........................................................................................ 466
Литература ..................................................................................... 467
Подкласс Testacealobosia (О. И. Чибисова, А. В. Смирнов, Н. В. Ленцман)......................... 468
Summary ........................................................................................ 483
Литература...................................................................................... 484
Класс Heterolobosea (А. В. Смирнов, А. В. Гудков) .............................................. 485
Summary ....................................................................................... 489
Литература..................................................................................... 490
Класс Filosea .................................................................................. 490
Подкласс Aconchulinia (А. В. Смирнов)........................................................... 490
Summary ........................................................................................ 498
Литература...................................................................................... 499
Подкласс Testaceafilosia (Я. В. Ленцман, А. В. Смирнов, О. И. Чибисова) ........................ 500
Summary ........................................................................................ 507
Литература...................................................................................... 508
Класс Peloflagellatea (А. В. Гудков, Л. Н. Серавин)............................................. 508
Summary ........................................................................................ 514
Литература..................................................................................... 514
Класс Xenophyophorea (О. Е. Каменская) ......................................................... 516
Summary ........................................................................................ 520
Литература ..................................................................................... 520
Rhizopoda incertae sedis ....................................................................... 520
Отряд Athalamida (С. А. Карпов) ................................................................ 520
Отряд Monothalamida (С. А. Карпов).............................................................. 522
Summary ........................................................................................ 524
Литература...................................................................................... 524
Отряд Komokiida (О. Е. Каменская) .............................................................. 524
Summary ........................................................................................ 527
Литература...................................................................................... 527
Отряд Blastocystida (Л. М. Белова) ............................................................. 528
Summary ........................................................................................ 532
Литература .................................................................................... 532
Тип Foraminifera (В. И. Михалевич) ............................................................. 533
Общая характеристика (533). — История изучения (535). — Классификация (537). — Морфология и
физиология (539). — Жизненные циклы (567). — Основные направления эволюции (575). — Место в
системе протистов (584). — Систематический обзор (584). — Summary (611). — Литература (616).
Тип Foraminifera (Т. Г. Лукина) ................................................................ 628
Экология и зоогеография (624). — Литература (630).
Указатель русских названий..................................................................... 632
Указатель латинских названий.................................................................... 638
Указатель терминов.............................................................................. 661
Список сокращений ............................................................................ 672
Сведения об авторах .......................................................................... 673
CONTENTS
Preface ......................................................................................... 5
GENERAL ZOOLOGY. Introduction to the whole edition .............................................. 9
Essey on the history of zoology (G. I. Poljansky)................................................ 11
Summary ....................................................................................... 29
Pathways and principles of the evolution (A. V. Ivanov, E. I. Kolchinsky)........................ 29
Summary ......................................................................................... 85
Systematics and its purposes (A. V. Ivanov) ..................................................... 85
Summary ......................................................................................... 97
Classification of the animal world (A. V. Ivanov) .............................................. 97
Summary ......................................................................................... 113
References..................................................................................... 114
PROTISTS. GENERAL PART .......................................................................... 121
Classification of protists and problems of their megasystematics (S. A. Karpov) ................ 123
Summary ......................................................................................... 136
Trends of evolution in protists (L. N. Seravin) ................................................ 138
Summary ......................................................................................... 145
General characterization of protists (G. I. Poljansky, К. M. Sukhanova, S. A. Karpov)........... 145
Summary ........................................................................................ 185
References...................................................................................... 185
PROTISTS. SPECIAL PART (Taxa review of protists)............................................... 191
Phylum Cryptophyta (В. V. Gromov, О. V. Gavrilova) ........................ 193
Summary ........................................................................................ 200
References...................................................................................... 200
Phylum Euglenozoa............................................................................... 201
Class Euglenoidea (К. M. Sukhanova) ............................................................ 201
Summary ........................................................................................ 210
References...................................................................................... 210
Class Kinetoplastidea (A. O. Frolov) ........................................................... 211
Summary ....................................................................................... 251
References...................................................................................... 251
Phylum Chrysophyta.............................................................................. 257
Class Chrysophyceae (К. M. Sukhanova, S. A. Karpov) ........................................ 258
Order Bicosoecales (S. A. Karpov) .............................................................. 264
Class Synurophyceae (К. M. Sukhanova, S. A. Karpov)........................ 270
Class Pelagophyceae (К. M. Sukhanova, S. A. Karpov) ........................ 273
Summary ........................................................................................ 274
References...................................................................................... 275
Phylum Haptophyta (К. M. Sukhanova) .............................. 276
Summary ................................................................................. . . 283
References.................................................................................... 284
Phylum Raphidophyta (К. M. Sukhanova) ............................................... , 285
Summary ........................................................................................ 289
References..................................................................................... 289
Phylum Saprolegnia ............................................................................. 290
Class Labyrinthomorphea (К. M. Sukhanova) ...................................................... 290
Summary ........................................................................................ 296
References...................................................................................... 296
Phylum Opalinata................................................................................ 297
Class Proteromonadea (К M. Sukhanova) .....................................................• 297
Class Opalinatea (К. M. Sukhanova) .......................................................... 301
Summary ....................................................................................... 312
References.................................................................................... 312
«Chromista» incertae sedis ..................................................................... 314
Order Spongomonadida (S. A. Karpov) ..................................................... . 314
Order Pseudodendromonadida (S. A. Karpov) ................................................... 317
Summary ........................................................................................ 319
References...................................................................................... 320
Phylum Choanomonada (S. A. Karpov, B. F. Zhukov)................................................ 321
Summary ...................................................................................... 335
References.................................................................................... 336
Phylum Polymastigota ........................................................................... 337
СОДЕРЖАНИЕ
679
Class Diplomonadea (К. M. Sukhanova)............................................................... 337
Class Oxymonadea (N. N. Bobyleva) ......... ....................................................... 348
Class Parabasalea.................................................................................. 357
Order Trichomonadida (К. M. Sukhanova) ............................................................ 358
Order Hypermastigida (N. N. Bobyleva).............................................................. 368
Summary ........................................................................................... 392
References......................................................................................... 395
Phylum Plasmodiophora (A. V. Goodkov).............................................................. 398
Summary ........................................................................................... 409
References......................................................................................... 409
Phylum Mycetozoa .................................................................................. 411
Class Cercomonadea (A. P. Mylnikov) ............................................................... 411
Summary ........................................................................................... 417
Class Eumycetozoea (Yu. K. Novozhilov, A. V. Goodkov) ............................................. 417
Summary ........................................................................................... 443
Mycetozoa incertae sedis .......................................................................... 444
Hyperamoeba flagellata (S. A. Karpov) ............................................................. 444
Summary ........................................................................................... 445
References......................................................................................... 445
Phylum Rhizopoda ...................................................... ....................... 451
Class Lobosea ................................................................................. 451
Subclass Gymnamoebia (A. V. Smirnov, A. V. Goodkov)................................... 451
Summary ........................................................................................... 466
References......................................................................................... 467
Subclass Testacealobosia (О. I. Chibisova, A. V. Smirnov, N. V. Lentsman) ......................... 468
Summary ........................................................................................... 483
References......................................................................................... 484
Class Heterolobosea (A. V. Smirnov, A. V. Goodkov) ................................................ 485
Summary ........................................................................................... 489
References......................................................................................... 490
Class Filosea...................................................................................... 490
Subclass Aconchulinia (A. V. Smirnov) ............................................................. 490
Summary .......................................................................................... 498
References......................................................................................... 499
Subclass Testaceafilosia (N. V. Lentsman, A. V. Smirnov, О. I. Chibisova)......................... 500
Summary ........................................................................................... 507
References......................................................................................... 508
Class Peloflagellatea (A. V. Goodkov, L. N. Seravin) .............................................. 508
Summary ........................................................................................... 514
References......................................................................................... 514
Class Xenophyophorea (О. E. Kamenskaya).......................................................... 516
Summary ............................,............................................................. 520
References......................................................................................... 520
Rhizopoda incertae sedis .......................................................................... 520
Order Athalamida (S. A. Karpov) ................................................................... 520
Order Monothalamida (5. A. Karpov) ........................................ ....................... 522
Summary ........................................................................................... 524
References......................................................................................... 524
Order Komokiida (О. E. Kamenskaya)................................................................. 524
Summary ........................................................................................... 527
References......................................................................................... 527
Order Blastocystida (L. M. Belova} ................................................................ 528
Summary ........................................................................................... 532
References......................................................................................... 532
Phylum Foraminifera (V. I. Mikhalevich) ........................................................... 533
General description (533). — Historical sketch (535). — Classification (537). — Morphology and phisiolo-
gy (539). — Life cycles (567). — Main trends of evolution (575). — Place in the classification of pro-
tists (584). — Review of taxa (584). — Summary (611). — References (616).
Phylum Foraminifera (T. G. Lukina) ................................................................ 628
Ecology and zoogeography (624). — References (630).
Index of Russian names ............................................................................ 632
Index of Latin names .............................................................................. 638
Index of terms .................................................................................... 661
List of abbreviation .............................................................................. 672
List of authors.................................................................................... 673
Научное издание
ПРОТИСТЫ:
РУКОВОДСТВО ПО ЗООЛОГИИ.
ЧАСТЬ 1
Утверждено к печати
Зоологическим институтом РАН
Редактор издательства Е. И. Васьковская
Художник Е. И. Кудина
Технический редактор Е. В. Траскевич, В. В. Шиханова
Корректоры О. М. Бобылева, Ю. Б. Григорьева, И. А. Крайнева и А. X. Салтанаева
Компьютерная верстка Е. М. Сальниковой, Л. Н. Напольской
Лицензия ИД № 02980 от 06 октября 2000 г.
Сдано в набор 16.06.1999. Подписано к печати 09.12.2000.
Формат 60x90 Vg. Бумага офсетная. Гарнитура Таймс. Печать офсетная.
Усл. печ. л. 85. Уч.-изд. л. 105.2. Тираж 1000 экз. Тип. зак. № 3284. С 233
Санкт-Петербургская издательская фирма «Наука» РАН
199034, Санкт-Петербург, Менделеевская лин., 1
nauka @ aspid. nw .ru
Санкт-Петербургская типография «Наука» РАН
199034, Санкт-Петербург, 9 лин., 12
---------------Entamoeba histolytica
—------Dictyostelium discoideum
---Physarum polycephalum
----Arabidopsis thaliana
----Reticulomyxa filosa
P-----Trypanosoma cruzi
।—Chlamydomonas reinhardtii
— Naegleria gruberi
«.-тубулин
I—Tetrahymena pyriformis
-Ji--Plasmodium falciparum
L ।— Homo sapiens
tHomo sapiens
j—-------------Schizophyllum commune
l_j-------Schizosaccharomyces pombe
—-----““Neurospora crassa
— Entamoeba histolytica
------- Reticulomyxa filosa
-------Chondrus crispus
Porphyra purpurea
Reticulomyxa filosa
Physarum polycephalum
— Dictyostelium discoideum
—;----—Trichomonas vaginalis
Giardia lamblia
—Hexamita sp.
— Caenorhabditis elegans
__r- Caenorhabditis elegans
‘-Caenorhabditis elegans
r— Caenorhabditis briggsae
. p Drosophila melanogaster
Ip Drosophila melanogaster
|p Homo sapiens
4 [--Drosophila erecta
Hj—Gallus gallus
l—Gallus gallus
------Schizophyllum commune
-----Encephalitozoon hellem
I-------Pneumocystis carinii
J --------— Schizosaccharomyces pombe
L pi-------------Geotrichum candidum
। i----Saccaromyces cerevisiae
‘----------Candida albicans
i---Collectotrichum graminicola
j—l ~ 1— Collectotrichum gloeosporioides
0
rt
’---Trichoderma viride
— Trichoderma viride
f-Penicillium digitalum
Neurospora crassa
Emericella nidulans
Acremonium chrysogcnum
-Physarum polycephalum
r-Chlamydomonas reinhardtii
r Oryza sativa
p npZea mays
Arabidopsis thaliana
_rArabidopsis thaliana
I ‘-Arabidopsis thaliana
pOryza sativa
|i— Zea mays
|_p Arabidopsis thaliana
•-Arabidopsis thaliana
r Porphyra purpurea
П----Porphyra purpurea
"h—Achlya klebsiana
’-----Ectocarpus variabilis
rEuplotes crassus
‘•Euplotes octocarinatus
- p Stylonychia lemnae
h Paramecium tetraurelia
L ’Tetrahvmena thermonhila
'Tetrahymena thermophila
j—Babesia bovis
1-Plasmodium falciparum
r- Eimeria tenella
^-Toxoplasma gondii
г Naegleria gruberi
4i—Euglena gracilis
I rTrynanoso
Trypanosoma brucei
Trypanosoma cruzi *
г Leishmania mexicana
*- Leishmania major
Rhodobionta
Mycetozoa
Trichomonadida
Diplomonadida
Metazoa
Fungi
Microsporidia
Fungi
Mycetozoa
Chlorobionta
Rhodobionta
Stramenopiles
Ciliophora
Sporozoa
Percolozoa
Euglenozoa
-----------------------Saccharomyces cerevisiae
Pneumocystis carinii
-------Neurospora crassa
--------------Drosophila melanogaster
~~ ।—Homo sapiens
1------Homo sapiens beta
--------Giardia lamblia
----------;-----Euplotes crassus
-Trichomonas vaeinalis
----Oxytricha nova
----Plasmodium falciparum
।---Leishmania major
----------1 | Trypanosoma cruzi
LpTrypanosoma brucei
[Trypanosoma brucei
------Tetrahymena termophila
-I---Plasmodium falciparum
L—Toxoplasma gondii
— Cryptosporidium parvum
r~Fucus distichus
l-Fucus vesiculosus
центрактин
Diplomonadida
Ciliophora
Trichomonadida
Ciliophora
Sporozoa
Euglenozoa
Ciliophora
Sporozoa .
Stramenopiles
Percolozoa
Rhodobionta
Chlorobionta
Rhodobionta
_________pPhytophthora infestans
*—Phytophthora megasperma
‘ j- Achlya bisexualis
ULPythium irregulare
*-Phytophthora infestans
----Naegleria fowleri
______ Entamoeba histolytica
-----------------------------Chondrus crispus
— Acetabularia cliftonii
Cyanidioschyzon merolae
______p Chlamydomonas reinhardtii
L Vol vox carteri
г- Solanum tuberosum
__ r Arabidopsis thaliana
’—Pisum sativum
j- Arabidopsis thaliana
1-------Solanum tuberosum
LI— Oryza sativa
ir Arabidopsis thaliana
Hr Solanum tuberosum
Ir-Arabidopsis thaliana
^-Solanum tuberosum
----Pneumocystis carinii
—Absidia glauca
r-Histoplasma capsulatum
|—P“" Emencella nidulans
L ------Thermomyces lanuginosus
___(Saccharomyces cerevisiae
'—Kluyveromyces lactis
[---Schizosaccharomyces pombe
L|_j—Puccinia graminis
Ц----Filobasidiella neoformans
'—Phaffia rhodozima
r— Schistosoma mansoni
'—Diphyllobothrium dendriticum
г Homo sapiens
^-Schistosoma mansoni
.p Drosophila melanogaster
___rHomo sapiens
I LHomo sapiens
[r-Drosophila melanogaster
. L—Drosophila melanogaster
rHydra attenuata
П-Podocqryne
l Pmiliani
Chlorobionta
Fungi
Metazoa
ч ‘-rodocoryne carnea
1----Emihania huxleyi
‘Acanthamoeba castellanii
rPhysarum polycephalum
ITDictyostehum discoideum
i Dictyostelium discoideum 8
i—Dictyostelium discoideum A3-SI
LrDictyostelium discoideum 12
^•Dictyostelium discoideum
"Rhizopoda"