






































































































































































































































































































































































































































































































































Автор: Грант В.Г.
Теги: экология растений этология растение и окружающая среда биологические науки биология ботаника
Год: 1984




Текст
В. Грант.
Видообразование
у растений
Plant Speciation
Verne Grant
Second Edition
Columbia University Press
New York 1981
В. Грант
Видообразование
у растений
Перевод с английского
Н. О. Фоминой
под редакцией
акад. А. Л. Тахтаджяиа
Москва «Мир» 1984
ББК 285 + 28.57
Г 77
УДК 581.5
Грант В.
Г 77 Видообразование v растений: Пер. с англ.— М_: Мир.
1984,—528 с, ил.
Автор иотграфиА — ewenH* аввртвж.нН пчвВ. )*' 1МА0ИМЙ сомт-
ввг.твлп. во аага аг.аалавяия орговк,оов> |М: Хар. т»| НАСТОашм
амага пасами»** имеймин дестажшмим в оЛлагга aaamiioiMl ботаинаи в
шгрпав онм^еяага juhrim ежевамх оЛ»»п«л. Pauerr;m<> зпяпс аи)п»в-
вмддаоА ивмаанмаостм. utwraiocni та<тулж«.«1 ж гапп>пфп«»«аиа par змвмм
ПйОрдемми. мгататамог» мШВМКевМ. ышйажжвм* с япмлйдам ««кам. •
такжа других фак-прла • «и^>!?жмм«мя* у ржетсямИ. В еаесс-ж вряясрт ирн-
МАМЫ домы* ОО 4оа<* «*м .300 андам рагтааяй
Предмаажааеш зла 4о<апмпв. спсцылясгоя а ввлаегж иаолпцкж я гметсмя*
тика раегеаиА. для геяегиноа
?004000000— 454
Ml(tl>-м
НО-84
ББК MJ-2B.57
S84-S81.4
Виаоврамжааиа у расгам!
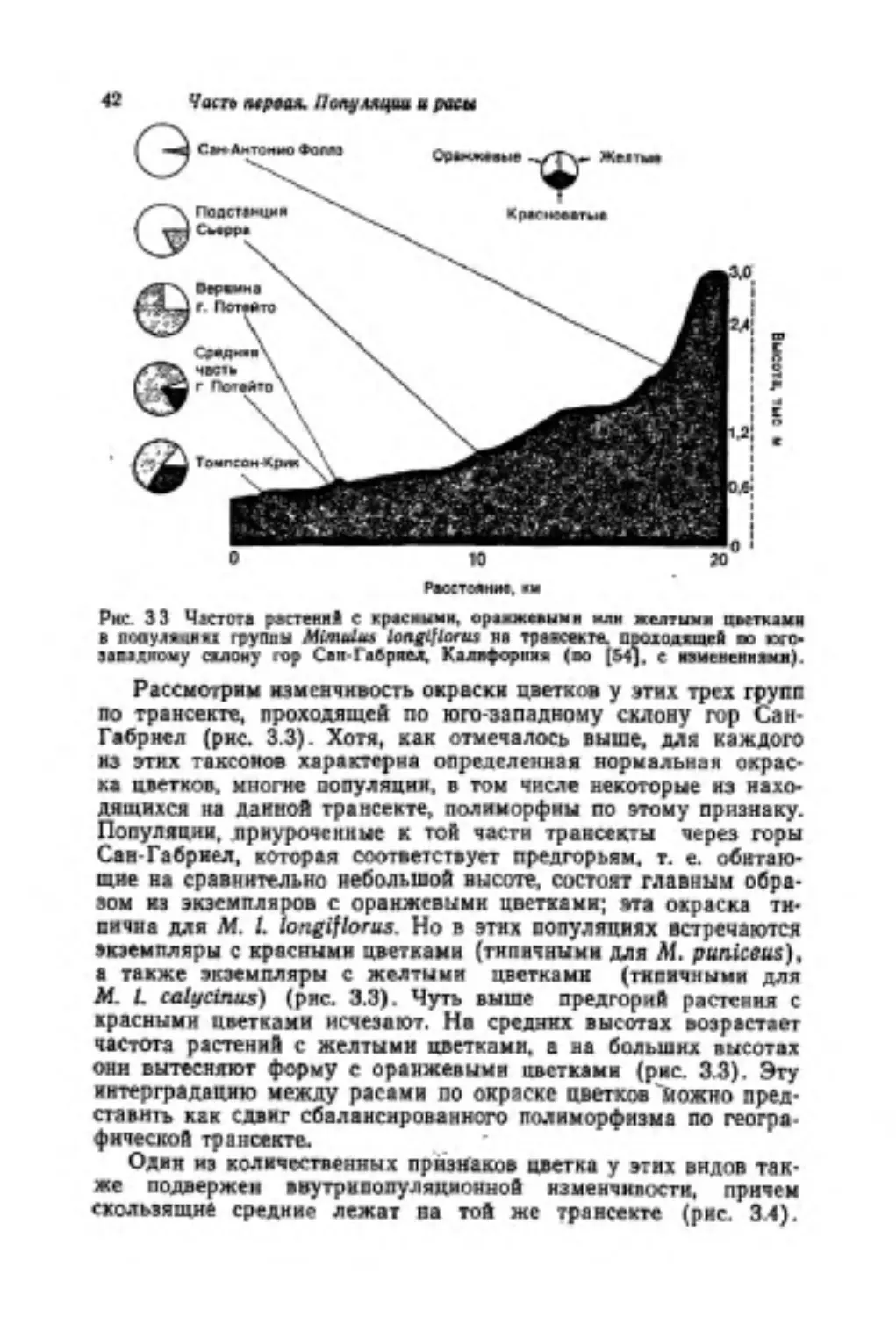
Науяеый радмтор М. Ь Микялыы. .Мл парчи радакгор 3. В Сддлирииисын
Хуаржмк В. А. Мешмаок Дуло*<г»««вш1 рсяавпм А. Я Мции
ТжхиячеиапЯ рддактар ГТ Спивал Каррактоу U И Каеаяаы
ИЬ М 3IIT
Слава а набор I4XA4. ПэЛШиСааС а оеяап 03 ЮМ Формат вОХЭД'/д
В>магп тммервфеяаа .4 I. Га&ЯМйУря лп7«раммаа Пеытн аыеоииЯ Об wet Ю80 <ум я-
Ут. кр. отт. 3]. Усл. веч. а. М Уа-йид. л. Й Ид*. ,ч Твудк КЮО »м Здк. О.
Ц<ы I рчм €0 ма.
ИЗДАТЕЛЬСТВО «МИР». 12М». Мосжяа, Н ПО. ГСП. 1*4 FUtcnM м« 2
Мюсаоасхаи ?игк>гр(<)ма М II Саю>пал|гр*фпргма пр» Г=сувирс-г»»мневи юзиватитг СССР
о яевам «матеднт». в-мигоИчн м внмжнсД таогвелп
Мпгиы, II.1I0S, Нпгатаяекая JM-. *- I-
Редакция литературы по биололги
1971, 1961 Columbia University Press
£) Перенос на pyccul иш, «Мир», 1984
Предисловие
редактора перевода
Предлагаемая вниманию читателя книга «Видообразование у
растений» принадлежит перу видного американского ботаника,
профессора Техасского университета в городе Остине и члена
Национальной академии США Верна Гранта. Грант —автор
восьми книг, в том числе «Естественная истории семейства поле-
мониевых» (1959 г.), «Происхождение адаптаций» (1963 г.),
«Архитектура зародышевой плазмы» (1964 г.), «Колибри к
опыляемые ими метки» (1968 г., совместно с Карен Грант).
«Генетика цветковых растений» (1975 г.) и «Эволюция организ-
мов» (1977 г.), вышедшая в 1980 г. в русском переводе. Многие
его работы посвящены генетическому и таксономическому изу-
чению большого американского рода Gilia из семейства Polcmo-
niaoeae.
Книга Гранта посвящена видообразованию у высших расте-
ний. преимущественно цветковых. Хотя Грант касается почти
всех аспектов видообразования, главное внимание он уделяет
генетике (особенно цитогенетике) и репродуктивной биологии,
В книге Гранта читатель найдет летальное и достаточно совре-
менное освещение таких вопросов, как генетические н экологи-
ческие различия видовых особенностей, роль хромосомных пере-
строек в эволюции, роль гибридизации (в том числе интрогрес-
сии) в видообразовании, эволюционная роль полиплоидии и
анеупловлии н, наконец, рол» различных специализированных
генетических систем, включая апомиксис. Все эти разделы со-
держат много интересных и часто свежих данных и представ-
ляют собой очень ценный обзор проблемы видообразования у
ВЫСШИХ растений
Естественно, что в книге под таким названием автор не мог
обойти общую теорию вида, или эйдологию. Но именно в этой
части книга Гранта является наиболее спорной. Как и в преж-
них своих работах Грант придерживается концепции так назы-
ваемого «биологического вида». Как известно, ата концепция
формировалась в 40 х годах, главным образом под влиянием
6
ПргЛамвие редитора перевод»
популяцкоино генетических идей Ф. Г. Добржаиского, изложен-
ных и первом издании его классической книги «Генетика и про-
исхождение видов» (New York, 1937). Она стала чуть ли не не-
отъемлемой составной частью синтетической теории эволюции
и особенно пропагандировалась Эрнстом Майром. Концепцию
«биологического вида» приняли многие эоологи (в основном
специалисты по наземным позвоночным и энтомологи) и часть
ботаников. В ботанике она имела некоторый успех лишь в 40-х
и отчасти в 50-х годах (в ее пользу высказывался и автор этих
строк). Но постепенно стали выявляться все изъяны этой кон-
цепция и против вое выступил ряд крупных биологов (преиму-
щественно ботаников) как советских, так и зарубежных В рус-
ской литературе очень убедительную ее критику дал К. М. За-
вадский в своей книге «Вид и видообразование» (Л.: Наум,
1968). Средн зарубежных ученых, выступивших против концеп-
ции «биологического вада», следует упомянуть таких крупных
ботаников, как П. Рейвн (1976) и А. Кронквня (1978).
Против концепции «биологического вида», определяемого
как репродуктивно изолированные скрещивающиеся между со-
бой популяции, можно сделать следующие основные возраже-
ния. Прежде всего, а отличие от таксономической категории
вида, категория биологического вида не универсальна и приме-
нима только к «бнпа ректальным видам» (термин, употребляе-
мый Грантом), что признает и Грант. Но она не универсальна
даже для бипареитальяых видов. Основной критерий «биологи-
ческого вида» — репродуктивная изоляция, вескрешиваемость
с другими; даже близкими видами,— в той или иной мере при-
меним только к симпатрическям видам. Что же касается алло-
патрических видов, то здесь репродуктивная изоляция не имеет
строго облигатного характера. Общеизвестно, что во многих
случаях «хорошие» виды растений, разделенные в течение дли-
тельного времени, исчисляемого миллионами лет, непреодоли-
мыми географическими барьерами, при совместном произраста-
нии в культуре успешно скрещиваются. Таких примеров очень
много. Так, два географически очень удаленных вида плата-
на— Platanut orientals (Восточное Средиземноморье и Запад-
ная Азия) и Р. occidenlalis (восточные районы США) — при
совместном произрастании легко гибридизируют я дают впол-
не плодовитый гибрид с нормально протекающим мейозом.
Однако, несмотря на легкую скрещиваемость этих двух плата-
нов, ни один систематик не согласится объединить их в одни
вид. Подобным же образом легко скрещиваются китайский вид
Catalpa ovata и восточно-североамериканский вид С. bignonioi-
des. Как и гибрид платана, гибрид катальпы вполне плодовит
и широко культивируется. С такой же легкостью скрещиваются
многие аллопатрические внды многих других растений, что хо-
Прдмкми рШааора яерекда 1
рошо известно садоводам Таким образом, последовательно*
применение критерии яеекрешиваемости должно привести к
объединению многих бесспорных аллопатрических видов
Ограничивая применение концепция «биологического апла»
только бипарентальныни («дву родительскими») организмами.
Грант применяет совершенно иные критерии к уинпаренталь-
ным («однородитсльскнм») группам. К ним он применяет поня-
тие «микровида» (жорданона), принципиально отличное от по-
нятия «биологического вида». Под мнкровиломи Грант пони-
мает однотипные популяции в преимущественно унипареиталь-
иых группах растений, слегка отличающиеся морфологически
друг пт друга. Он различает клональные микровиды (размно-
жающиеся вегетативно), агамоспермные микроинды, гибридные
по конституции гетерогамные микровнды (например. Rosa cant-
по) н. наконец, автогамные микровиды. Грант прав в том, что
с таксономической точки зрения эти группы растений известны
своей сложностью, и он, конечно, прав, что концепция биологи-
ческого вида к ним неприменима. Он считает, что зти группы
представляют собой «смесь», состоящую обычно из нескольких
первоначальных «биологических пидои» и многочисленных
производных «микровндон» По его мнению, ята «смесь» ис мо-
жет быт разложена на составляющие ее элементы «обычными
1аксономическимн методами». Такое утверждение — следствие
неправильного представления автора как о принятой в совре-
менной систематике концепция вида, так и о современных ме-
тодах систематики
Грант определяет таксономический вид как «группу морфо-
логически сходных индивидуумов» и считает его (в отличие от
«биологического вида») «искусственной категорией», Мало кто
из современных систематиков согласится с определением таксо-
номического вида как группы, сходной лишь морфологически.
Прежде всего современный систематик старается не ограничи-
ваться лишь внешней морфологией, по асе чаще использует
даипые внутренней морфологии, включая ультраструктуру, на-
пример ультраструктуру экзины пыльцевых зерен Но. кроме
того, для современной систематики характерно возрастающее
стремление опираться также на данные биохимии, генетики, ци-
тогенетики и даже молекулярной биологии. Наконец, для совре-
менной систематики вил есть отнюдь ис чисто морфологическое,
но прежде всего морфолого-экоиого-географичест-ое понятие,
аксономнческий вид- совершенно реальная категория, без
'’РоЯ “ожст обойтись ни одна область теоретической н
р ладной биологии, сельского хозяйства, лесоподствв н т. л,
.Т™** °’₽а"а природы. Совершенно очевидно, что любой био
й- ' o6wK^ с которым имеет дело исследователь или
> Должен быть определен до вида.
8 Превшими» (мбвстори мргаойа
Вид — это основная категория таксономической иерархии и
одна на самых фундаментальных категорий биологии. Его мож-
но определить как обособившуюся в процессе эволюции систему
клонов или популяций, объединенную общими признаками (йор-
фологяческими, экологическими, биохимическими, генетически-
ми, цитологическими и другими), общим происхождением и об-
щим географическим ареалом и достаточно четко отделенную
от близких видов как совокупностью своих признаков, так я раз-
личными (у разных видов разными) изолирующими барьерами.
Это самое общее определение виза, применимое как к бинарен-
тальным, так и к уннпарентальиым организмам, как к популя-
ционным, так и к клональным образованиям. Для систематики
такие категории, как «биологический вид», «мякроякд» (жорда-
нон) или «полувида излишни и в таксономической иерархии им
нет места. Следует также помнить, что такие понятия, как
«сяигамеоя», «экотипа, «клнматип», «биотипа, «изореагента и
прочие, используемые в генетически! и генэкологических иссле-
дованиях, также нельзя считать таксономическими категориями
Несмотря иа хорошо известную неравноценность видов у раз-
ных групп организмов, их разную структуру н разные спосо-
бы размножения, категория вила в современной биологии явля-
ется универсальной. Наибольшие сомнения вызывают виды у
облигатяо-самоопыляющихся и агами ых растений. Всем язве
стиы неимоверные трудности, с которыми сталкиваются оасте-
матнки, изучающие виды ястребиики (Hieracium) или одуван-
чика (Taraxacum). Но трудности здесь скорее технические, чем
принципиальные. Используя различные современные методы си-
стематики, включая молекул ярко-биологические, н изучая объ-
екты в природе и на экспериментальных делянках, опытный
систематик почти всегда установит естественные границы вида,
его объем и связи с родственными вилам». Поэтому, вопреки
мнению Гранта (и многих других авторов), здесь вполне воз-
можно объединить близкородственные «жорданоны» в настоя-
щие линнссвские виды
Несколько слов об употреблении Грантом таксономической
номенклатуры. Он не учитывает одного важного различия двух
международных кодексов номенклатуры —ботанического и зоо-
логического. Последний регламентирует название только одной
категории таксонов рангом ниже вида — название подвида, со-
стоящее из трех слов. При этом триноминальное название пи-
шется без предшествующего третьему члену слова, обозначаю-
щего ранг (например, подвид домовой мыши —,Мих muse ulus
domestlcus). В ботанической номенклатуре признается несколь-
ко таксономических категорий рангом ниже вида (подвил раз-
новидность, форма, подформа), требующих указания их раита.
Грант не следует этому обязательному правилу и постоянно
ПрсЛл-ЛМШ рс&юара .пгрнчРа
9
употребляет триноминальные названия без указания ранга внут-
ривидового таксона (например, GiUa сапа speciota). Неясно, ка-
ков таксономический ранг третьего члена.
Последняя глава книги Гранта «Вид и иакроэволюиия» со-
держит ряд положений, с которыми я не могу полностью согла-
ситься. Но так как предисловие — не самое подходящее место
для полемики, то я позволю себе отослать читателя, интересую-
щегося втими вопросами, к моей статье о хакроэволкщии в мире
растений, опубликованной в 1983 г. («Макроэполюцииниые
процессы в история растительного мира». — Бот жури., 1983,
т. 68, № 12, с. 1593—1603).
В заключение я хотел бы сказать несколько слов о почти
полном отсутствии ссылок на русскую литературу в книге Гран-
та. Давно прошли те времена, когда западному ученому можно
было более или менее обойтись без ссылок на русскую литера-
туру. Положение резко изменилось уже в первые десятилетия
нашего века, а в настоящее время любой ученый просто обязан
знать важнейшие советские работы по своей специальности. Неко-
торые видные западные ученые, например крупнейшие амери-
канские ботаники Л. Дж. Стеббинс и А. Кронквнст. давно это
поняли и успешно пользуются русской литературой, добросо-
вестно ссылаясь на важнейшие работы. В этом отношении мо-
жет служить примером последняя книга А. Кронквиста «Инте-
грированная система классификации цветковых растений»
(A. Cro'nquist. An integrated system of classification of flowering
plants. Columbia University Press, New York, 1981), в которой
после каждого раздела приводится обширный список русской
литературы, набранный кириллицей. Поэтому вызывает некото-
рое удивление отсутствие у Гранта ссылок на книгу В. Л. Кома-
рова «Учение о виде у растений» (М.— Л., 1940). статью
Н. И. Вавялова «Лимнеевскнй вид как система» (1931), книгу
М. А. Розановой «Экспериментальные основы систематики расте-
ний» (М. — Л.. 1946) и книгу К. М. Завадского «Вид н видооб-
разование» (Л.: Наука, 1968). классическую работу А. С. Семе-
пова-Тяньшакского «Таксономические границы вида и его под-
разделения» (Зап. Ими. Акад, наук, 1910, т. 25, 1, с. 1—29), из-
данную в том же году на немецким языке в Германки. Не упо-
минаются также цитогенетические работы Г. А. Леаитского.
И все же. несмотря на эти, а возможно, и некоторые другие
недостатки, выход в свет книги Гранта представляет собой
крупное событие в ботанической литературе. Я уверен, что пуб-
ликация на русском языке насыщенной фактами и оригиналь-
ной по построению книги Гранта принесет большую пользу бо-
таникам-систематикам, экологам, цитогенетикам, а также расте-
ниеводам и селекционерам н стимулирует исследования и этих
направлениях.
А. Л. Тахтиджян
Предисловие
к русскому изданию
Перке английское издание «Вндообразованвя у растений» выш-
ло а свет в 1971 г., а переработанное второе издание — в 1981 г.
Я очень рад. что моя книга выходит в русском переводе и что
тем самым содержащийся в ней материал станет доступным
более широкому кругу читателей в Советском Союзе.
В «Видообразования у растений» рассматривается эволюция
рас. видов и групп видов у наземных растений и в особен кости
у цветковых растений. Обсуждается природа этих биологиче-
ских единиц и порождающие их эволюционные процессы.
Вид представляет собой основную единицу органической
эволюции, но, к сожалению, об этом иногда забывают. Некото-
рые биологи пытаются объяснить эволюцию как результат всего
двух процессов —мутационного процесса и отбора, ие прини-
мая во внимание вид н видообразование. Разумеется, сочета-
нием мутационного процесса н отбора действительно можно
объяснить молекулярную эволюцию, в значительной степени —
эволюцию бактерий и простые изменения, происходя-
щие в процессе эволюции эукариотических организмов.
Однако этими двумя процессами нельзя адекватно объяснить
эволюцию самых сложных эукариотических организмов. Эволю-
ционные изменения, происходящие в сложных организмах, дол-
жны быть пропущены через вид. Как показано в этой книге,
эго не только теоретическая необходимость, но в факт, установ-
ленный наблюдениями.
Связь между видом и эволюцией была незыблемо закрепле-
на Дарвином в заглавии его классического труда «О происхож-
дении видов» и многократно подчеркивалась многими эволю-
ционистами на всем протяжении нынешнего века.
Высшие растения — это, безусловно, сложные эукариотиче-
ские организмы, и в процессе эволюции они проходят через ря-
ды последовательно сменяющих друг друга вадов. Многие за-
коны эволюции, установленные на дрозофиле, птицах и млеко-
питающих, вполне приложимы к высшим растениям. Но в ре-
Предисловие
11
продуктивном поведении и видовой оргаиизацни высших расте-
ний есть также некоторые особенности, присущие им одних, что
и определяет характер их эволюционного развития, отличный
от эволюции большинства животных. Эти особые черты эво-
люции растений описаны а моей книге.
Переходи or общих принципов и широко распространенных
моделей к отдельным примерам, нельзя нс заметить, что до-
вольно многие иркводпмые в книге примеры относятся к флоре
западных районов Северной Америки. Такой выбор отражает
особенности научной биографии автора. Западная часть Север-
ной Америки — это. во-первых, огромная лаборатории, предо-
ставленная нам самой природой для изучения эволюции расте-
ний. а во-вторых, это лаборатория, ь которой я жил и в кото-
рой протекала ббльшзн часть моей научно-исследовательской
работы
Я очень хорошо понимаю, что русская ботаническая и биоло-
гическая литература отражена в настоящей книге крайне недос-
таточно. В этом повинен языковый барьер Большое значение
переводов с русского на английский и е английского на русский
состоит в том. что они позволяют преодолевать этот барьер
Я хочу воспользоваться предоставившейся мне возможностью
высказать свою признательность акад. А. Л Тахтаджяву и
проф. А. В. Яблокопу, рехоиеидовавшим «Видообразование у
растений» к переводу, Н О. Фоминой, сделавшей перевод, и
акад А. Л. Тахтаджяиу, отредактировавшему его. а также из-
дательству «Мир». Книге очень повезла, что редактором рус-
ского издания оказался акад. Тахтаджян.
Остии. Техас, США Верн Гране
Предисловие
Эта книга посвящена явлениям видообразования у высших рас-
тений Видообразование рассматривается в пей как процесс,
имеющий много различных аспектов н подверженный влиянию
многих факторов. Мы начнем е описания популяций и рас
(Часть первая). Затем перейдем к обсуждению природы видов
И их поведения (частя вторая и третья). Далее рассматрива-
ются первичная дивергенция видов (часть четвертая), естест-
“н“’Г"в₽и“3‘“и" и |И^РИД|,ОС видообразование (часть пя-
т я). Полиплоидия, агамоспермия и другие слецналнэирован-
c"Ctcmw. связанные с гибридяостыо, а также
комплсксы, в основе которых лежат такие генетичс-
истемы, описаны в частях с шестой по восьмую вклю-
12 Лрейаслмш
чктельно. Последняя глада (часть девятая) содержит краткий
очерк макроэаолкщии и свете исследований, проводимых иа
видовом уровне.
Короче говоря, в этой книге всесторонне трактуется вволю
дня растений на уровне видов и групп видов, главным обратом
применительно к высшим растениям.
Автор исходит из предпосылки, что естественный, или эво-
люцмонирующнй, вид — одна из основных единиц организации
живой материк. Эволюционирующий вид, подобно весы другим
биологическим единицам, обладает рядом особых общих
свойств, которые можно выявить н объяснить. Эти свойства не
удается обнаружить, если рассматривать видовые единицы все-
го лишь как бесконечный ряд отдельных сущностей, имеющих
свои специфические особенности, т. е. так, как они рассматри-
ваются в чисто описательной таксономии, хотя подобный под-
ход поаволяет заложить необходимый фундамент. Их можно
выявить, применяя те методы анализа и синтеза, которые успеш-
но используются в других разделах теоретической биологии.
Первое издание этой книги, опубликованное в 1971 г., широ-
ко используется. Однако за десять лет, прошедшие со времени
выхода в свет первого издания, в той области биологии, которая
занимается изучением растительных вндои. произошло много
событий. В течение этого времени не только появлялось мно-
жество новых исследований, ио и развивались мои собственные
представления об этой области биологии. Настало время для
коренного пересмотра текста.
Во втором издании несколько изменено распределение ма-
териала по главам. Добавлен ряд новых глав. Весь текст попол-
нен новыми данными. Обсуждение некоторых концепций дове-
дено до современного уровня.
Несмотря на то, что в новом издании библиография была
значительно расширена, она все еще остается далеко не полной.
Литература по втиы проблемам столь обширна, что при состав-
лении обзоров приходится производить некоторый отбор. К со-
жалению, в процессе такого отбора многие хорошие работы не-
избежно выпадают из поля зрения.
Карин А. Грант, критически прочитавшая всю рукопись при
подготовке первого издания книги, прочитала все новые главы,
написанные для этого издания. Ока взяла на себя также ос-
новной труд по чтению корректур.
Перепечатка рукописи была выполнена Дугласом Дьюком и
Маргарет Буленджер. Я выражаю им свою благодарность
Д-р Викки П. Реберн и другие редакторы издательства Ко-
лумбийского университета, как всегда, оказывали мне всяческую
помощь н содействие.
Отамаяич богини
Ухнмрситет штата Техас
Январь 1Я81
В. Г.
Часть первая
Популяции и расы
1 Размножение растений
Одно из основных свойств живой материв — ее способность к
размножению. Этой способностью наделены организованные
единицы самых разнообразных размеров и сложности — от мак-
ромолекул ДНК. хромосом в клеток до отдельных организмов
и скрещивающихся популяций. В згой книге нас интересует
одна из важнейших размножающихся единиц живого мира, а
именно вид. Концепция вила как размножающейся единицы
имеет вековую давность и кратко сформулирована и известном
положении: «подобное рождает подобное».
У большинства животных и у многих растений размножение
неразрывно связано с полом. Но высшие растения произошли в
процессе эволюции от одноклеточных форм, у которых половой
процесс и процесс размножения не только обособлены, но даже
антагонистичны. Размножение у простейших происходит путем
деления: одна клетка делится на две. Половой процесс у этих
организмов происходит путем конъюгации две клетки сливают-
ся а одну. Таким образом, весь жизненный цикл простейших
слагается из чередования фаз клеточного деления, ведущего к
увеличению численности популяции, и полового процесса, ве-
дущего к уменьшению числа особей.
Это глубокое различие между половым процессом и размно-
жением сохраняется в скрытой форме у высших растений и
животных. Наличие у высших организмов полового процесса
означает, что для осуществления размножения необходимо уча-
стие двух особей, тогда как в отсутствие Этого процесса размно-
жение обеспечивает одна особь с помощью того или иного спо-
соба почкования.
У высших растений обособленность этих двух процессов
можно продемонстрировать как на физиологическом, так и иа
популяционном уровне. У многих или во всяком случае у боль-
шинства многолетних растений наряду с половым наблюдается
и вегетативное размножение. Как половое, так и вегетативное
размножение приводит к истощению пищевых запасов роди-
тельских растений; оба способа требуют определенных затрат
и
Чат мрачи Пичглииш и расы
энергхи. А поскольку запасы энергии у родительских растений
ограничены. такое столкновение потребностей полового и веге-
тативного размножения разрешается у многих растений с по-
мощью отрицательной корреляции между этими противополож-
ными способами размножения (703, 705]; см. также более позд-
ние работы, например [351]. Обильное образование семян
нередко сочетается со слабым вегетативным размножением, и
наоборот, мощное вегетативное размножение часто сопровож-
дается уменьшением количества семян.
Так, у некоторых сортов земляники удаление усов вызывает
заметное увеличение количества плодов, тогда как другие ре-
монтантные сорта выбрасывают мало усов или не образуют ик
вовсе [380, 703]. Барвинок (Vinca major') обычно широко рас-
пространяется с помощью длинных ползучих побегов, но обра-
зует мало плодов; если, однако, обрезать эти ползучие побеги
и at допускать образования новых, то растения начинают завя-
зывать больше семян [703].
Важное значение размножения как такового достаточно оче-
видно. Это — необходимое условие для продолжения существо-
вания популяции или вида во времена. И это единственный спо-
соб увеличения числа особей и заселения новых территорий.
Функция полового процесса, оправдывающая его включение
в жизненный цикл. несмотря на то что он (вменяет эффектив-
ность размножения популяции или айда, меиее очевидна и все
еше остается предметом дискуссий. Принято считать, что поло-
вой процесс, связанный с чередованием перекрестного оплодо-
творения и мейоэа, служит механизмом, обусловливающим ре-
комбинацию генов. Рекомбинация генов в свою очередь пред,
стазляет собой главный источник наследственной изменчивости
у видон с половым размножением. А наследственная изменчи-
вость обеспечивает соответствующие реакции видов на неодно-
родные или изменчивые условия среды.
Отсюда следует, что вегетативное а другие формы бесполого
размножения обладают преимуществом в более или менее по-
стоянной среде, к которой данный вид уже хорошо адаптирован,
поскольку эти формы размножения препятствуют образованию
слабо адаптированных рекомбинационных типов или сводит его
к минимуму, тогда как образование проростков в результате
полового процесса создает преимущество в изменяющейся или
неоднородной среде [280, 709, 759, 785]. Существуют среды и
того и другого типа, в необходимы оба способа размножения.
Очевидно, что в системе размножения вида, процветающего на
протяжении длительного периода времени, должны сочетаться
половой и бесполый процессы.
0 самом деле, системы размножения растений сочетают в
себе и половые, и бесполые способы размножения. Каждый из
Г Л. /. PaAMWXtnt* рос гг**Д
15
этих способов выполняет свою функцию в жнзви данного вида.
Однако каждый нз них оказывает при этом свое особое вл канне
на природу данного вида н его структуру.
Отклонения от случайного перекрестного
оплодотворения
При проведении анализа удобно постулировать идеальное соче-
тание условия для полового размножения в некой гипотетиче-
ской популяции, а затем рассмогреть различные отклонении от
этого идеального состояния, наблюдаемые в реально сущест-
вующих популяциях.
Допустим, что для нашей гипотетической популяции харак-
терно свободное скрещивание, или панмиксия. Она не только
размножается исключительно путем перекрестного оплодотво-
рения. но при этом перекрестное оплодотворение происходит в
таких условиях, что гаметы объединяются в пары чисто случай-
ным образом. Предполагается, что любая женская гамета, воз-
никшая в этой популяции, с равной вероятностью может быть
оплодотворена любой мужской гаметой. Отсюда следует, что
при панмиксии особи, составляющие поколение потомков в дан-
ной популяции, представляют собой продукты разных лар га
мет, И.1ЯТЫХ случайным образом нз фонда гамет, созданного ро-
дительским поколением.
У реально существующих ендов растений отклонения от пан-
миксин обычно выражаются в следующих четырех формах
I) перекрестное оплодотворение происходит преимущественно
между соседними растениями (вицинизм) и приводит к скре-
щиванию между родственными особями, или инбридингу;
размножение происходит путем самооплодотворения (автога-
мии)— теснейшая форма иибридиига; 3) новые растения раз-
виваются из почек, образуемых родительским растением (веге-
гагнпное размножение); 4) новое поколение растений возникает
из ссмин. развивающихся на материнском растении без овлодо-
твор-иня (агамоспермия)
Первые два из перечисленных здесь способов размножения —
вицинизм н автогамия — слагаются из оплодотворения и следую-
щего за ним мейозп и представляют собой, твкнм образом, фор-
мы патового размножении. Дла последних способа — вегета-
тивное размножение и агамоспермия — обходятся без оплодо-
творения и представляют собой, следовательно.формы бесполо-
го или апомиктического размножения.
Всех особей, возникших из одной родительской особи в ре-
зультате митотических делений, называют членами одного кло-
• гакнм образом, вегетативное размножение и агамоспермия
•едут к образованию клона или ряда клонов.
18 Част* лераая- и расы
Существует также важное различие между первым из пере-
численных выше способов размножения и тремя остальными.
Только этот первый способ требует перекрестного оплодотворе-
ния. В трех других случаях оплодотворения либо не происходят
вовсе, либо в нем участвуют гаметы, образованные одним роди-
тельским растением. Соответственно в первом случае размно-
жение осуществляется при участии двух родительских особей
(бипареитальиое размножение), а в трех остальных — лишь од-
ной (унипареитальиое размножение).
Совершенно очевидно, что популяция, состоящая из особей,
возникших в результате унипарентальною размножения, будь
то клоны или икбрелкые линии, обладает одним важным отли-
чием от настоящей скрещивающейся популяции, членьа которой
возникают в результате настоящего полового размножения.
Случайное скрещивание в обширной популяции — идеали-
зированная ситуация, полезная в качестве некоего исходного
стандарта. Возможно. что в действительности не существует ни
одного широко распространенного айда растений, достаточно
близкого к этому состоянию. У известных кам видов растений,
размножающихся половым путем, наблюдается та или иная
степень вица к иа на. Однако у некоторых реальных раститель-
ных видов обнаружено крайне резкое отклонение от случайного
скрещивания в сторону строго унипарентальиого разм ноженки.
В некоторых группах растений бкпаректальное размножение
может быть полностью заменено вегетативным размножением,
агамоспермией и, возможно, автогамией.
Гораздо чащи, однако, встречается некое сочетание унипа-
ректального я бипарентальиого размножения у одного и того
же растения. Такие растения занимают промежуточное поло-
жение между растениями со строго унипарентальным и строго
бвларентальным размножением. В сущности, как отмечает
Густафсон [317], между истинно половым размножением и
различными способами унипареитальноги размножения имеется
ряд переходных типов.
Вегетативное размножение
Существует много разданных способов вегетативного размноже-
ния: при помощи наземных столонов н ползучих побегов, под.
земных корневищ к клубней, укорененных отводкой, клубнелу-
ковиц и луковиц, придаточных почек на срезанных стеблях или
опавших листьях и выводковых почек, образующихся в цветке
или соцветии. Последний способ называют живорождением, бо-
лее правильно называть его ложным живорождением [639].
Вегетативное размножение широко распространено у много-
летних покрытосеменных, встречаясь во всех основных группах.
Гл 1. Разлмхекие рмгеииЛ IT
Среди многолетников виды, не способные размножаться вегета-
тивным путем, крайне редки. Густафсон [317J приводит данные,
свидетельствующие о том. что в некоторых скандинавских фло-
рах примерно 80% всех видов покрытосеменных способны к
той или иной форме неге готнвного размножения, а примерно у
50% многолетних покрытосеменных различные формы вегета-
тивного размножения достигли такого развития, что дают им
возможность очень быстро расселяться.
Секвойя вечнозеленая (Sequoia sempervirens) способна да-
вать корневую поросль, представляя собой в этом отношении
исключение среди хвойных |40й]. Когда старое дерево отмирает,
из корневой поросли вырастают молодые деревца, которые в
конце кондов превращаются в большие деревья, окружающие
кольцом исходный родительский лень. Со временем вокруг от-
дельных деревьев, образующих такое вторичное кольцо, могут
образоваться третичные кольца. Поскольку секвойя может до-
живать до 1300 лет ([406]; см. также (352]). продолжительность,
жизни этих клопов приходится исчислять в тысячелетиях
Согласно оценкам Джепсона [405], в лесу примерно 80% зре-
лых деревьев секвойи происходят из корневой поросли н при-
мерно 20%—из семян. По нашим наблюдениям, деревья, воз-
никшие в результате полового и бесполого размножения, рас-
пределены по территории рошв в вокруг нее далеко не случай-
ным образом. В густой центральной части некоторых рощ раз-
множение происходит почти исключительно за счет корневой
поросли, тогда как проросткам удается выжить только на от-
крытых периферических участках.
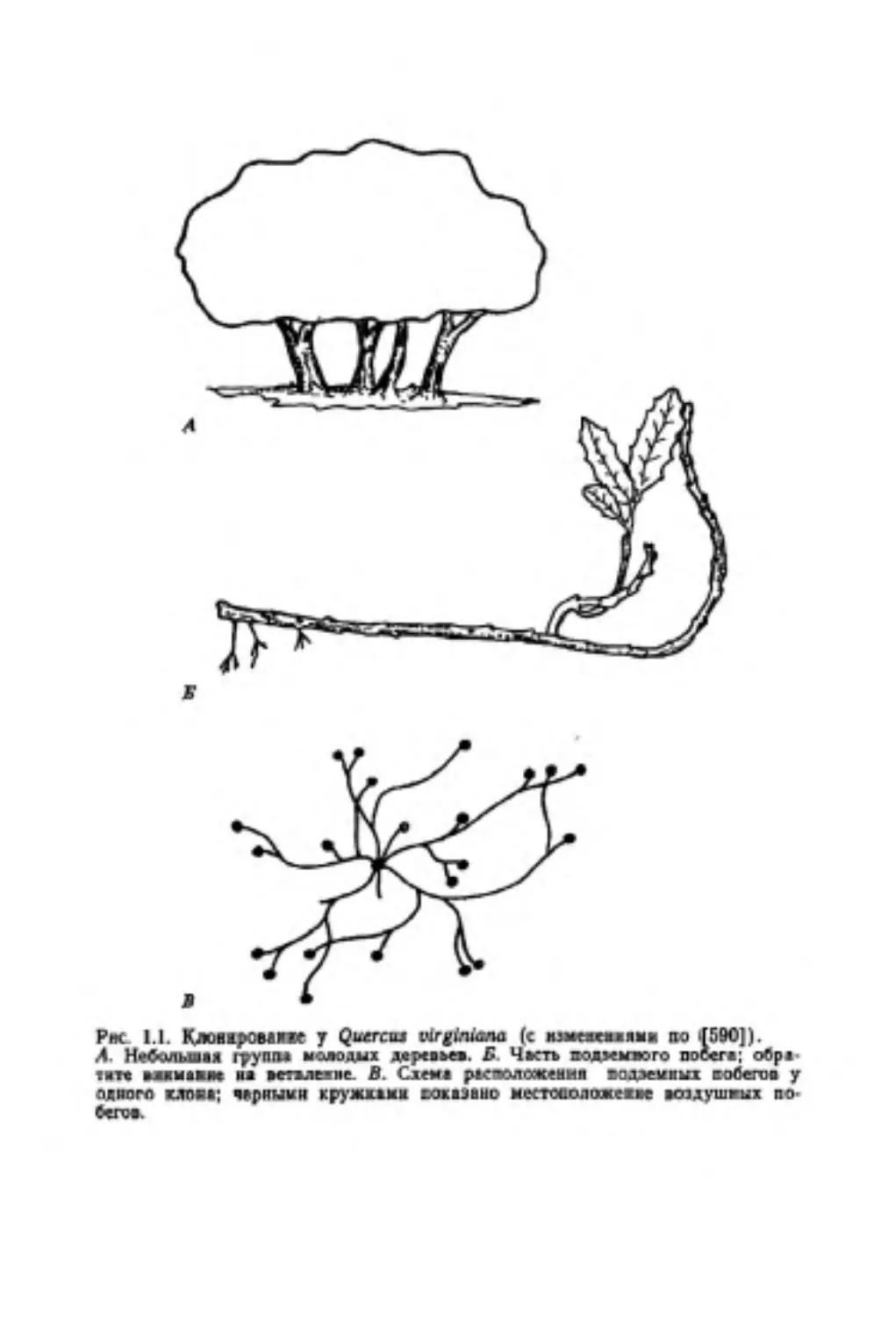
У некоторых видов дубов имеются стелющиеся подземные
побеги, которые в дальнейшем могут дать начало новым расте-
ниям Например, по данным наблюдений в Техасе, молодые по-
бега Quercus vtrginiana иногда бывают соединены с переплете-
нием подземных побегов, расходящихся от расположенного в.
центре дерева (рис. 1.1), Зрелые деревья этого вида часто об-
разуют островки или клональные колонии [5<Ю].
Осины Populus tremuloides и Р. grandidentata дают мощ-
ные корневые отпрыски от пия. Впоследствии соединения между
корневыми отпрысками и родительским растением отмирают,
так что возникают обособленные молодые деревья. В природе
осина, как правило, встречается в виде клонов, в которые мо-
.^^1.ВХ0ДИть от нескольких деревьев до большого их числа
[403].
некоторые клоны Populus tremuloides в шт. Юта достигают
значительных размеров. Один такой клон занимал площадь в
га п состоял из 15000 деревьев; другой занимал более 40 га,
8П гК.ОГп’ЫХ ®“л0 “•сч«7а«о 47000 деревьев, а третий —около
в восточной части Северной Америки клоны Р. tremuloi-
2—43
Рве 1.1. Кловжромв» у Querciu virgimana (с кзавеякваявв во 4590]).
Л. Небольшая группа молодых деревьев. Б Часть подэемяого побега; обра
1вте влнмавм ва ветвление В. Схема расположения подземных побегов у
одного сапна; черными кружками хаки в нс местосолажекк вепдушгых по.
бегов.
Гл. I. Гтял-клпм (мтглиЛ
I»
des ие столь велики. Самый большой клон. обнаруженный а
этом районе, занимал чуть больше 1,5 га 1430).
Обширные клопы осины должны быть довольно старыми, и
некоторые из них, возможно, возникли еще в позднем плейсто-
цене В гор» к шт Юта Populus iremuloides образует много жиз-
неспособных семян, во венком случае в отдельные годы, но в
современных климатических условиях, когда в начале лета
осадков выпадает мало, доля прорастающих семян ничтожна.
В этой области осина размножается, по-видимому, только веге-
тативным способом. Поскольку климат, для которого типичны
летние засухи, сложился п Юте уже 8000 лет назад, возможно,
что и возраст некоторых клонов осины также достигает
8000 лет. На протяжении столетий или тысячелетий вегетатив-
ного размножения особой расе осины, рано покрывающейся
листвой, удалось заселить вершины гор в штате Юта. которые
прежде находились подо льдом и снегом [125).
Festuca rubra представляет собой многолетний злак, размно-
жающийся как вегетативно, при помощи корневищ, так и поло-
вым путем и являющийся ветроопыляемым и самонесовмести-
мым видом. Харбера [338] исследовал состав популяций этого
злака к Шотландии, основываясь на выборках, взятых с обшир-
ных квадратов. Растения, обладающие различными фенотипи-
ческими признаками и плодовитые при перекрестном опылении,
рассматривались как принадлежащие к разным генотипам.
О'.-: кино несовместимые растения С <МШ1Ма1111111 фНМНШИМ-
сними признаками рассматривались как разные клоиы одного
н того же генотипа.
В результате изучения фенотипических признаков выборок и
нх поведения при скрещивании Харбера [338] пришел к выводу,
что в квадрате площадью 80 м! представлено относительно не-
много разных генотипов, но что многие растения принадлежат
к одному и тому же клону. Один из генотипов был распростра-
нен на участке диаметром около 220 я. По мнению Харберда,
Для того чтобы какой либо клон мог покрыть участок такой пло-
щади, возраст его должен достигать нескольких сотен, а может
быть, и тысячи лет. Некоторые другие клоны были небольшими.
Таким образом, изучение выборок нз популяций Festuca rubra
показало, что они состоят а основном из нескольких генотипов,
которые распространились путем клонирования по большим
участкам, а также из нескольких других генотипов, образующих
лишь небольшие клопы. Перекрестное опыление при помощи
ветра у самоиесовмсстимого злака с такой клональной структу-
рой популяции должно обычно приводить к несовместимым ком-
бинациям [338].
У родственного Festuca rubra другого многолетнего вида —
г. оо/пн —наблюдается иная картина. F ovina расселяется го-
2*
SO Чат перво* Пеяумцаи а расы
раздо медленнее чем F. rubra. У F. ovina Харберд (339] обнару-
жил многочисленные генотипы на квадрате размером около 9 и*.
В этом случае в отличие от F. rubra соотношение между бес-
полым и половым размножением, по-видимому, сдвинуто в сто-
рону полового размножения. *
Прибрежиоморскому злаку Spartina patens на восточной час-
ти Северной Америки также свойственны ауткроссинг, веге
татнвное размножение корневищами в образование клонов. От-
дельные генотипы можно идеитнфицировагь благодаря сущест-
вующим между ними различиям по изоферментам и составить
карту их мнкрогеографичесхого распространения В том районе
Северной Каролины, где проводилось изучение, на маршруте
длиной 200 и был обнаружен 101 генотип. Некоторые генотипы
образуют довольно обширные клоны, тогда как другие пред-
ставлены всего несколькими отдельными растениями [724].
Примером исключительно вегетативного размножения слу-
жит хорошо известный случай с Elodca canadensis (Hydrocha-
rltaceac) в Европе, В восточной части Северной Америки, отку-
да происходит это двудомное растение, засоряющее водоемы,
оно размножается как половым путем (семенами), так и веге-
тативно, выбрасывая побеги и образуя зимние почки. Женские
растения были завезены аз Северной Америки на Британские
острова примерно в 1840 г. Ввиду отсутствия на чужой для него
территории мужских растений этот канадский сорняк мог раз-
множаться только вегетативно. Тем нс менее за период с 1840
по 1880 г. ему удалось распространиться почти по всем внут-
ренним водоемам Европы [317].
Агамоспермия
При агамоспермия отдельное растение образует жизнеспособ-
ные семена, которые содержат зародыши, возникшие без опло-
дотворения. Особенности развития таких зародышей сложны и
варьируют от случая к случаю (см. гл. 31). Новый зародыш в
семени может развиваться из иеоилодотоорекиой яйцеклетки
(партеногенез), из какой либо другой клетки или ядра зароды-
шевого мешка (апогамия) или же из какой-нибудь соматиче-
ской клетки семязачатка (адвентивная эмбриокия). Агамосперм-
ное образование семян может происходить без опыления или же
С опылением (псевдогамия). В последнем случае пыльца сти-
мулирует рост эндосперма, необходимого для нормального раз-
вития семени и зародыша [317].
Различные пути эмбрионального развития сходны в том, что
во всех случаях клеточные лилии, ведущие к образованию Ново-
го зародыша, обходятся как без мейоза, так и без оплодотворе-
ния. В результате, если не считать некоторых исключительных
Гл. I. РазмлшлллилpuL/tnsi
31
процессов, происходит образование семян, содержащих зароды-
ши, которые генетически идентичны материнскому растению.
Агамоспермия широко распространена среди высших расте-
ний Она встречается у многих представителей семейств Grami-
neae. Composite и Rosaceae, а также п ряде более мелких се-
мейств Наиболее хорошо известные примеры относятся к Шега-
cium. Taraxacum, Crepis, Citrus н Роа. Агамоспериные растения
обычно бывают многолетними. Их размножающиеся половым
путем родичи если и известны, го неизменно принадлежат к пе-
рекрестиоопыляющимся видам, у которых перекрестное опыле-
ние обеспечивается самонссовместнмостью. двудомностью нли
какой-либо другой системой размножения по типу ауткроссин-
га; это позволяет считать, что для предков агаиоспериных рас-
тений было характерно исключительно перекрестное оплодо-
творение (317].
В некоторых группах растений агамоспермия полностью вы-
теснила половое размножение (облигатная агамоспермия).
У других растений часть семян образуется путем агамоспермии,
а часть — в результате полового процесса (факультативная ага
моспермня). Как и в случае вегетативного размножения, четкой
границы между половым н бесполым размножением провести
нельзя; эти два способа размножения связаны между собой ря-
дом переходных состояний [317].
При факультативной агамоспермии характер изменчивости
популяции меняется популяция распадается на ряд групп, ехм:-
гоящнх из идентичных особей, которые отличаются друг от дру-
га по второстепенным признакам. Такая популяция состоит из
скоплений более или менее обособленных агамоспермпых мни-
ровидоя.
Автогамия
Для гермафродитных покрытосеменных характерен широкий
«венгр систем скрещивания —от облигатного ауткроссинга на
одном конце до фактически пат ной автогамии —на другом. Гер-
мафродитные цветки могут быть полностью самонесовместимы
и не завязывают семян после самоопылении, как у Gilia capita-
ta capitata (Polemoniaceac). или же само несовместимость мо-
жет быть неполной, как у G. capitata tomentosa, у которой мно-
гие самоопылявшиеся цветки образуют всего лишь по несколь-
ку семян [266]. Неполной и частичной может быть также само-
совместимость. Cheiranthus cheiri (Crucifcrae) полностью само-
совмсстнм при искусственном самоопылении, если же подверг-
"уть его в одинаковой степени самоопылению и перекрестному
опылению, то большая часть (92%) завязавшихся семян оказы-
вается результатом перекрестного опыления [46].
22 Часть aepaat Пати/мирш и роем
Полностью самосовыестимые покрытосеменные растения
варьируют либо в отношении деталей строения цветка, создаю-
щих преимущества для ауткроссиига или самоопыления, либо
и отношении того и другого вместе. В природных популяциях
Clarkta unguiculata (Onagraceae), имеющей в высокой степени
прогакдрические цветки с выступающими наружу репродуктив-
ными органами, частота ауткроссикга, определяемая по испыта-
ниям потомства, составляет 96% [857). В отличие от этого строе-
ние цветка С. ecilia допускает значительное спонтанное само-
опыление, и в двух популяциях этого лида ауткроссинг состав-
лял 43 и 45% [856. 858).
У многих самосовместнмых растений тычинки и рыльца од-
ного и того же цветка расположены очень близко друг от друга
и созревают одновременно, так что обычно самоопыление про-
исходит автоматически. Исследователи, изучающие автогамные
растения, единодушно считают, что автогамия обычно не бы-
вает настолько полной, чтобы исключить некоторый ауткроссинг.
Самоопыление может преобладать, однако время от времени
происходит перекрестное опыление, осуществляемое ветром или
насекомыми; поэтому рассматриваемую систему размножения
правильнее называть преимущественной автогамией [5. 425).
Произведенные сравнительно недавно опенки частоты аут-
кроссинга у нескольких видов преимущественно автогамных зла-
ков дала следующие результаты:
Hordeam lubatura — 1% (32)
Hordeum cutgara — I % H 4% |B, 308)
Avena barbate — 2% [398]
4t>«wa fatua — от 1 no 12% н 3% (394. 398(
Bromar mallil —4% (398)
Можно было ожидать, что существуют примеры полной или
почти полной автогамии, и в некоторых случаях такое ожида-
ние действительно оправдалось. По имеющимся оценкам часто-
та ауткроссикга у Ftstuca microitachyi (Gramineae) составляет
0% [6]. Мелкие цветки Polemonium micranlhum (Polemoniaceae)
самоопыляются еще в бутонах, а затем, уже после опыления,
раскрываются всего на несколько часов, так что никакой оче-
видной возможности для опыления их насекомыми нет. Попыт-
ки искусственного перекрестного опыления этих растений оказа-
лись безуспешными; псе потомство, полученное при этих искус-
ственных скрещиваниях, было результатом самоопыления.
Имеющиеся данные позволяют считать, что некоторые популя-
ции Р. micranlhum действительно полностью ннбредны (301).
Автогамия нли, точнее, преимущественная автогамия — обыч-
ный и широко распространенный способ размножения у покры-
Гл. I. Гимшиииш рш-rtnuA
23
тосеменных, особенно у однолетних травянистых растений [317,
780). В сем. Polemoniaccae это наиболее часто встречающийся
способ опыления, установленный я опытах по скрещиванию
примерно у 45 видов н предсказываемый на основании полевых
наблюдений еще у 30 видов. Автогамия а атом семействе встре-
чается. насколько это известно, только у многолетних его пред
ставителей [301].
Система размножения сказывается на характере изменчивос-
ти данной популяции. Если два нли несколько гомозиготных ин-
дивидуумов, различающихся ло двум или большему числу ге-
нов. размножаются путем ауткроссипга. то при этом п резуль-
тате рекомбинации сильно возрастает индивидуальная изменчи-
вость и большинство рекомбинантов остаются гетерозиготными
по крайней мере по одному гену. Непрерывное самоопыление
приводит к противоположным результатам. Если исходная по-
пуляция состоит из индивидуумов, гетерозиготных во двум нли
большему числу генов, то с каждым поколением самоопыления
доля гетерозиготных индивидуумов уменьшается с равномерной
скоростью, а первоначальный ряд гетерозиготных и гомозигот-
имх рекомбинантов сводится к меньшему числу гомозиготных
типов Следует ожидать, что строго автогамная популяция дол-
жна состоять главным образом из ряда истинно ннбредных чне
тых линий.
В реальных популяциях автогамных растений эти ожидае-
мые результаты могуч действительно наблюдаться нли не наб-
людаться в зависимости от других факторов Прежде всего
автогамия, как мы уже видели, обычно не бывает облигатной;
время от времени в преимущественно автогамной популяции
происходят скрещивания между различными биотипами, служа-
щие источником новой изменчивости [280. 345. 782]. Кроме того,
непрерывное самоопыление нс приведет к снижению в популя-
ции доли гетероэигот с ожидаемой скоростью, если эти гетеро
зиготы обладают каким либо Селективным преимуществом по
сравнению с гомозиготами [Зов, 401].
Недавними исследованиями Алларда и его сотрудников было
установлено наличие у некоторых преимущественно автогамных
растений постоянной гстерозиготности, обусловленной преиму-
ществом гетерозигот. Это было обнаружено у Secale cereale [5,
*01, 403); Phaseolus lunalus [7. 344]; Avena fatua [394. 404] и
Avena barbate 1100].
Фактические состав популяций у многих автогамных видов
растений оказывается, таким образом, гораздо сложнее, чем
следовало бы ожидать на основании экстраполяций от концеп-
ции чистых линий; это в последние годы подчеркивают многие
исследователи. Не следует, однако, думать, что простые модели
нереалистичны во всех случаях. Популяции некоторых автогам-
24 Чат первая. Лалрм^ш и рот
них аилов приближаются к простому составу, слагаясь из одно-
го или нескольких гомозиготных биотипов, Популяции других
видов содержат больший запас изменчивости, и частота ауткрос-
синга в анх выше, так что они приближаются в этих отношениях
к популяциям видов, у которых регулярно происходит пере-
крестное опыление.
Рекомбинационные системы
В размножении растений, как было ясно показано в предыду-
щем параграфе, участвуют как процессы, создающие изменчи-
вость. так и процессы, обеспечивающие воспроизведение геноти-
пов. Равновесие между процессами этих двух типов создается
той частью генетической системы, которая известна под иахва-
нием рекомбинационно*. Рекомбинационная система устанав-
ливает различные точки равновесия для разных популяций и
видов. В диапазоне от максимальной рекомбинации до макси-
мальной репликации полезно различать три модальных состоя-
ния: открытые, ограниченные закрытые рекомбинационные
системы. Для них характерны соответственно широкий хуткрос-
сииг, преимущественная автогамия и агамоспермия.
На рекомбинационную систему оказывают влияние многие
факторы, перечисленные в табл. 1.1. Эти факторы обсуждаются
в обзорах Гранта [294] и Солбрнга |759]. Важно отметить, что
некая определенная степень регулирования рекомбинации мо-
жет быть достигнута в результате различных сочетаний этих
регулирующих факторов.
Как указывает Сол бриг [759], некоторые факторы, регули-
рующие рекомбинацию, непосредственно контролируются отбо-
Таблкца 1.1. Компоненты рекомбинационной системы растени*
([280], с изменениями)
I Факторы. регулирующие количество реконвнншои их I ооколнхе; ре-
гуляния осунзествляетея во время мейоха
1 . Число хромосом
2 Частота кроссагнгомра
3 CtepBHMKCTb -яОрнхоп
II. Факторы. регулмруиицяе количество режомбяяацвя на 1 повеление; ре-
гуляши осуществляется во время оплодотворения
4. Система г-крещнмяжя
S. Система омылен»
6. Потенциал рвеселеняи
7. Число «ми н характер их прорастания
а Величина популяции
9. барьеры для серащивани в внешне наыируюше механизмы
lit. Фактор, регулирующий выевчоство расомбажаци в еаипицу временя
10 Прсдолжтгельясгтр «еаграциа
Гл. I. Ралмшхжгчме pocTvntfA
»
ром, тогда как на другие такие факторы отбор же влияет. Ком-
поненты рекомбинационной системы, непосредственно контроли-
руемые отбором, соответствую? факторам I. 2, 4, 5, 6 и 9
(табл. 1.1). Факторы 7 и 10 контролируются силами отбора, ре-
гулирующими иные характеристики, нежели соотношение, реком-
бинации и регуляции как таковое. Л факторы 3 и 8 не подвер-
жены, согласно Солбригу [759]. прямому действию отбора.
Солбрмг [756, 759] вы двинул идею о том, что рекомбинаци-
онные системы сопряжены с определенными «расходами» и что
открытые рекомбинационные системы требуют больших расхо-
дов, чем ограниченные или закрытые системы. Открытая реком-
бинация связана с генетическими или так называемыми мейотн-
чесхимн расходами, измеряемыми числом возникающих плохо
адаптированных рекомбинантов, а также с энергетическими
расходами, связанными с приспособлениями цветков для привле-
чения опылителей и образованием избытка пыльцы для пере-
крестного опыления. Очевидно, что открытые рекомбинацмои
мыс системы обладают преимуществами, перекрывающими ути
расходы.
Среднее число пыльцевых зерен на семязачаток действи-
тельно гораздо выше у перекрестиоопыляющихся растений, чем
у самоопылителей. Число пыльцевых зерен на семязачаток у
видов с о алл ичиим и системами скрещивания составляет в сред-
нем [ 132):
У облигатных пергкрестнсюамлителсЛ —5М9
у фжкультативаых гм-рифдегноолылмпмей — 797
у прс1пгузд«сп»«я»о автогзмямх рдел««ий — 16В
у облигатно автогпмких растений — Л
Im pat tens capensis образует клейстогамные и хазмогамные
цветни и семена, развивающиеся в результате как самоопыле-
ния. гак и перекрестного опыления. Энергетические расходы на
семема этих двух типов составляют, согласно оценкам. 135 кал
для семян из хазмогамных цветков и 65 кал для семян из клей-
стог амии х цветков [576]
Экология рекомбинационных систем
Причину разнообразия рекомбинационных систем у растения
следует искать в неодинаковости требований, предъявляемых к
размножению местообитаниями разного типа. Здесь вступает в
действие erne один спектр —от открытых местообитаний до
замкнутых сообществ.
Открытые местообитания, будь они постоянно открытыми,
как п пустыне, млн периодически открытыми, как на первых
стадиях экологической сукцессии, или искусственными, как на
X Чап» пераоя. Популацш| а расы
возделываемых полях, благоприятны для заселения. Заселению
и свою очередь способствуют ограниченные и закрытые реком-
бинационные системы. Такне рекомбннационньЛ системы обес-
печивают сохранение постоянства при размножении. Популя-
ция, адаптированная к данному открытому местообитанию и
обладающая ограниченной или закрытой рекомбинационной
системой, может воспроизводить свой адаптивный генотип(ы) и
быстро увеличивать свою численность в пределах имеющегося
местообитания. Экспериментирование с новыми типами реком-
бинантов, которые могли бы понизить успех-размножения, сво-
дится к минимуму.
Виды растений с ограниченными и закрытыми рекомбина-
ционными системами действительно встречаются обычно в от-
крытых местообитаниях и иа ранних стадиях экологической
сукцессии; например, однолетники пустынь, однолетники Среди
эемноморсхой области, сорняки, травянистые растения первых
стадий экологической сукцессии и агамоспсрмные растения в
областях, обнажившихся после окончания плейстоценового оле-
денения.
Растения с открытыми рекомбинационными системами, на-
против, встречаются в закрытых растительных сообществах; к
ими относятся, например, деревья и кустарники лесов умерен-
ной эоны, многие злаки степей и равнин и многие многолетние
травянистые растения подлеска.
В закрытых растительных сообществах вероятность выжи-
вания проростков очень мала, что связано с конкуренцией. Рас-
тительная популяция год за годом приносит урожай семян, но
новым проросткам удается сохраниться лишь в тех очень ред-
ких случаях, когда в сообществе освобождается подходящее
место. По имеющимся оценкам, за три последние столетия в
девственном лесу в Нью-Гемпшире нз всех проростков бука
Fagus grandifolui сохранялось по одному на 0,04 га в 10 лет
(см. обзор (1201). В случаях, подобных этому, когда раститель-
ная популяция обладает огромной избыточной плодовитостью,
она может позволить себе иметь открытую рекомбинационную
систему и. следовательно, множество плохо адаптированных
рекомбинационных зигот.
Существуют две точки зрения относительно сил отбора, при-
ведших к развитию открытых рекомбинационных систем.
Мое первоначальное мнение [280] сводилось к тому, что вы-
сокая избыточная плодовитость растений в закрытых сообщест-
вах дает нм возможность использовать долговременное преиму-
щество открытой рекомбинационной системы, а именно созда-
ние изменчивости, которая позволит справиться с будущими
изменениями условий среды, Если рассматривать это с другой
точки зрения, то можно сказать, что многим существующим в
Гл I. Разляпжеши р<хтекий
27
настоящее время в природе видам растений с открытыми ре-
комбинационными системами удалось сохраниться на протяже-
нии длительного времени и дожить до наших дней благодаря
таком рекомбинационной системе Наблюдаемое во многих со-
временных сообществах скопление подобных видов может быть
результатом меж де нового или межвидового отбора
Более простая альтернативная гипотеза была выдвинута не-
давно Левином [472] и Солбригом [756. 759]. Растения в закры-
тых сообществах находятся в условиях непрерывно изменяю-
щейся .биотической среды. Постоянно возникают новые генотипы
патогенных организмов, растительноядных животных и расте-
ний конкурентов. Открытая рекомбинационная система порож-
дает новые паризиты, необходимые растительной популяции для
того, чтобы соответствовать изменяющимся элементам окру-
жающей ее среды. В этом случае открытые рекомбинационные
системы могли развиться в результате индивидуального отбора.
Такая гипотеза весьма привлекательна, по требует дальнейшей
проверки.
Остается вкратце коснуться некоторых терминологических
вопросов. Выражения «возникший в результате г-отбор а» и
«возникший я результате К отбора», используемые в популя-
ционной экологии, соответствуют заселяющему виду и нилу
закрытого сообщества, однако они не содержат никаких указа-
ний относительно типа рекомбинационной системы. Тем не ме-
нее различные авторы применяют эти термины в более широ-
ком смысле, распространяя их и на другие альтернативные ре-
комбинации ииые системы
Некоторые авторы для обозначения противоположных эко-
логических условий, связанных с рекомбинационными система
ми разного типа, пользуются терминами «стабильные—неста-
бильные» или «предсказуемые—непредсказуемые». Однако эти
термины могут привести к некоторой путанице. Так, среда, бла-
гоприятствующая К-отбору. считается более предсказуемой, чем
среда, способствующая г-отбору [399, 638]. Однако, как мы
только что видели, закрытое сообщество, содержащие типы, воз-
никшие в результате АС-отбора, находится в более непредска-
зуемой биотической среде, чем если бы оно находилось в от-
крытом местообитании. Этой путаницы можно избежать, ис-
пользуя простые описательные термины — открытое местооби-
тание и закрытое сообщество.
2 Локальные расы
Расы —иго популяция или системы популяций какого-либо ви-
да, статистически отличающиеся по составу своих генофондов
и по своим генетически детерминированным фенотипическим
признакам от других популяций или систем популяций, относя-
щихся к тому же виду. Это определение охватывает широкий
спектр различных популяционных единиц —от локальных скре-
щивающихся популяций до обширных географических рас. При
обсуждении зтих единиц в зволюцноином плане удобно разли-
чать два уровня расовой изменчивости: локальные расы и геог-
рафические расы. Первым посвящена данная глава, а вторые
будут рассмотрены в гл. 3.
Мы подходим к проблеме дифференциации локальных рас
с теоретической точки зрения. Будут рассмотрены такие факто-
ры, регулирующие «тот процесс, как поток генов н создаваемые
им эффекты, взаимодействие потока генов с естественный отбо-
ром и взаимодействие между отбором и дрейфом генов. Раэбор
каждой группы регулирующих факторов сопровождается изло-
жением соответствующих эмпирических данных.
Расселение и поток генов
Растения, будучи постоянно прикреплены к одному месту, спо-
собны расселяться на двух стадиях своего жизненного цикла:
пыльца перекосится с помощью ветра, насекомых, птиц или
других факторов; перенос плодов или семян осуществляют раз-
личные физические или биотические факторы. Каждый ботаник,
которому приходилось наблюдать распространение семян в при-
роде. всиоре обнаруживает, что оно следует некой общей моде-
ли: большая часть производимых данным материнским расте-
нием семян, которые не были съедены животными, локализует-
ся поблизости от материнского растения, тогда как небольшое
число семян разносится на более далекие расстояния.
Такого рода общие наблюдения подкреплены в некоторых
случаях количественными данными. Солсбери (704] подсчитал
число семян Verbascum thapsus, унесенных ветром из разные
расстояния от родительского растения. Основная масса семян
упала на землю на расстоянии примерно 3,5 м от родительско-
го растеяия, однако за пределами этого 3,5-метрового радиуса
число семян быстро сокращалось с увеличением расстояния.
Аналогичная картина наблюдается для распространяемых вет-
ром плодов Senecio jacobaea (704].
Гл 1. Лаллмми росы
»
Такая картина распространения оказалась типичной для ря-
да растений и животных. Кривая распределения расстояний по
частоте положительно экспессиииа н асимметрична. Значитель-
ная доля единиц расселения остается вблизи родительской осо-
би. Затем с увеличением расстояния от этой исходной точки
кривая частот быстро снижается.
Приведем один конкретный пример, хорошо иллюстрирую-
щий это характерное распределение. Колуэлл (117] выбрасывал
Рас. 2 1. Рвспроетрваеаве сосноаоД пмльиы ветрам по двум радиусам и»
Тахмые рвсстопния от нсточпнкд пылыш (1171
— в направлении ветра; II — под углом 4 5> к ваоравлемню астра
* воздух помеченную радлоактипиими изотопами пыльцу сосны
Pinus coulteri и прослеживал за ее распространением по двух
радиусам от исходной точки. На рис. 2.1 представлена кривая
распределения сосновой пыльцы, переносимой ветром. Большая
часть пыльцы упала на землю недалеко от источника, пролетев
по ветру от 3 до 10 м. А дальше количество пыльцы быстро
уменьшалось. На расстоянии от 45 до 120 м от источника хот»
и было найдено некоторое количество пыльцы, но оно было
очень невелико |117|.
Для изучения распространения пыльцы пчелами только что.
раскрывшиеся цветки хлопчатника (Gossypium kirsutum и
G. arbor сит) обрызгивали синей краской и прослеживали за
последующим распространением краски по хлопковому пол»
(804]. Посте посещений пчел частицы краски были обнаружены
иа расстоянии 25 м от обработанных цветков. Однако по мере
удаления от места нанесения краски количество меченой пыльцы
30 Часть лсраал. Псяулсциа к расы
равномерно убывало; »то позволяет считать, что активность
пчел была сосредоточена на растениях в пределах тонного со-
седства [804].
Если описанный выше характер распространения семян про-
должается на протяжении ряда последовательных поколений,
то следует ожидать, что родственные друг другу растения ока-
жутся ближайшими соседями. Если же таким образом будет
распространяться пыльца, то перекрестное опыление будет про-
исходить преимущественно между родственными растениями,
растущими друг подле друга, В результате в популяции с пере-
крестным опылением возникнет значительная степень инбри-
динга. Однако, поскольку в норме в такой популяции происхо-
дит также распространение семян и пыльцы на большие рас-
стояния, инбридинг будет дополняться небольшой, но, вероят-
но, значимой степенью свободного перекрестного опыления [280,
2981
Следует отметить, что хотя характер расселения описывается
-общей для разных организмов положительно эксцессявной и
асимметричной кривой, расстояния, в пределах которых оно
происходит, могут сильно различаться. Короче говоря, речные
типы организмов обладают различной способностью к расселе-
нию
Расселение представляет собой физическую основу потока
генов. Сам по себе поток генов наблюдать удается редко;
обычно о нем судят иа основе данных по расселению. Однако
при изучении межсортовых засорений у сельскохозяйственных
культур поток генов можно наблюдать и измерить.
Такне исследования были проведены на многочисленных ви-
дах культурных растений. По степени засорения между двумя
-сортами какого либо перекрестно опыляющегося вила культур-
ных растений, растущих па соседних экспериментальных участ-
ках опытного поля в практически одинаковых условиях среды
и завязывающих семена после свободного опыления при помо-
щи ветра или пчел, можно судить о величине потока генов меж-
ду ними при данном разделяющем их расстоянии. Как показы-
вают результаты этих исследований, характер потока генов
сходен с характером расселения.
Хорошим примером служит люцерна (Mcdlcago satioa).
Бред кер и др [64] использовали две линия люцерны, различаю-
щиеся по окраске цветков, детерминируемой одним геном. Ли-
нии с белыми цветками гомозиготна по рецессивному гену, а
нормальная окраска — доминантный признак. Две эти линии
выращивали па отдельных экспериментальных участках при
различных расстояниях между участками, к они свободно опы
лились местными перепончатокрылыми (Bomb ил и ВетЫг)
Процент засорения среди проростков следующего поколения па
Гл. i. .Чокаллпыг расы
3
Таблица 2.1. Засорение линки Mcdicago
saliva с белыми цветками генами линии е
окрашенными цветками при различных
расстояниях между экспериментальными
участками [64]
Расстояние между Здсореаае. хуиломевнеегв мрсиоачжтоарылыым, Ч
1*2 г IMJ г
ГЛ 42,9 19.6
101 30.9 9.7
201 ю.о .1.3
4Q3 6.» 6,5
«15 5.2 4.4
I2M 9.3 0.0
1616 «.» 0.0
участках линии с белыми цветками служит показателем потока
домниантмото аллеля (табл. 2.1).
Как показывает таблица, поток доминантного маркирующего
аллеля очень велик при небольших расстояниях (50 м). но с
увеличением расстояния снижается. Следует, однако, отмстить,
что даже при расстояниях от 800 до 1600 м был обнаружен
довольно значительный поток генов [64].
Обычно при небольших расстояниях между эксперименталь-
ными участками с разными сортами степень засорения одного
сорта другим невелика. На экспериментальных участках сред-
него размера с опыляемой ветром кукурузой (Zea mays), раз-
деленных расстоянием в 15 м, засорение составляло всего 1%
((46); см. также [635]). У опыляемых насекомыми репы (Bras-
sica гара) и редьки (Raphanus salivas) при таком же расстоя-
ния между экспериментальными участками межсортовое засо-
рение также снижалось до небольшого уровня [46]. У перекрест-
но опыляющейся редьки межсортовое засорение при расстоянии
между участками 4.5 м составляло всего 2% [128].
Панмиктические единицы и соседства
В большой непрерывкой популяции какого-либо бнпарептально-
го вида панмиктическая единица—это свободно скрещиваю-
щаяся группа, в пределах которой гаметы объединяются слу-
чайным образом [909]. У организмов, у которых свободное скре-
щивание иосит локальный характер, панмиктическая единица
представляет собой, в сущности, субпопуляцию. Если же сво-
32 Veer» нервен Ловцицш в расы
водное скретиваиие, что часто случается, не является локаль-
ным, то следует различать такую единицу, как соседство [911].
В панмиктической единице с числом особей Л'» происходит
некоторая доля (1/Х.) инбридинга. Коэффициент 1/N. исполь-
.ауют для вычисления величины соседства (Х«), в котором про-
исходит неслучайное скретиваиие. Таким образом, величину
соседства вычисляют, вводя некую чисто гипотетическую пан-
миктическую единицу, в которой имеются отклонения от панмкк-
-сив [911]. Концепция соседства имеет более широкое примене-
ние. чем панмиктическая единица, а поэтому она более полезна
в популяционной биологии растений.
Область, занимаемая соседством в непрерывной популяция,
распределенной в двухмерном пространстве, представляет собой
окружность радиусом 2а, где о — среднее квадратичное откло-
нение расстояний, на которые происходит расселение. В этой
области сосредоточено 86,5% родителей особей, находящихся
в центре окружности. Эта оценка представляет собой матема-
тическое следствие описанного выше выведения величины со-
седства из гипотетической панмиктической единицы [911]. Вели-
чина соседства оценивается числом размножающихся особей в
области, занимаемой этим соседством, т. е. в окружности с ра-
диусом 2а.
Величина панмиктической единицы или соседства зависит
от расстояний, на которые происходит расселение, и от скорости
потока генов Если индивидуумы, принадлежащие к большой
непрерывной популяция, обычно мигрируют на далекие расстоя-
ния, то панмиктическая единица или соседство может прибли-
жаться по величине к популяции в целом. Если же индивиду-
умы. составляющие большую непрерывную популяцию, ведут
сидячий образ жизни и расселяются а предела! небольшой об-
ласти, то скрещивания будут происходить исключительно между
соседними индивидуумами. В этом случае панмиктическая еди-
ница илн соседство представляет собой небольшую локальную
труппу внутри популяции в целом. Если же расстояния, на ко-
торые происходят миграции, варьируют, так что имеет место
значительная миграция на небольшие и некоторая миграция
на большие расстояния, то панмиктические единицы и соседст-
ва будут иметь промежуточную величину (909).
Оценки величины соседства получены лишь для нескольких
видов растений н животных. По недавний оценкам величины
соседств у Drosophila pseudoobscura V,=3229 и 6479 для двух
местностей в Колорадо и V,—4922 и 6436—для двух местностей
в Калифорнии [132]. Соответствующие оценки для Drosophila
nigrospiracula из трех местностей в Аризоне составили N,—
“21 256; 55483 и 145780 [415] Такая величина скрещивающей-
Гл 2. Локальнее рисы
33
ся группы будет препятствовать дифференцированию локаль-
ных рас.
У тех нескольких видов растений, которые до сих пор изу-
чались с точки зрения величины соседств, скрещивающиеся
группы во всех случаях малы, и в этом они резко отличаются
от дрозофилы. Оценки величины соседств у четырех видов рас-
тений в Иллинойсе дали следующие результаты-
Phlox pHosa (PoLetnuniBceae). //,—75—282 [CBOj
LioJi'is otptra (Campofetac), Д',—30—191 [481]
Uolris r.ffilftdracea (CoaposItoeL N>a 166 [712]
Lifkospermum ca'olimense (Boraginaceae), Ak—4 [432]
Кроме того, для Linanthus parryae (Polemonlaceae) в Кали-
форнии также, вероятно, характерны небольшие соседства, од-
нако надежных количественных данных по этому виду ист
[203, 204, 207].
Поток генов и расовая дифференциация
Величина панмиктической единицы млн соседства оказывает
влияние па распределение генетических вариаций в пределах
популяции в целом. Если не учитывать действия естественного
отбора, то можно считать, что эти генетические вариации име-
ют тенденцию к равномерному распространению в результате
широкого перекрестного опыления. И наоборот, если перекрест-
ное плылеяне происходит внутри отдельных небольших субпопу-
ляцнй, то это способствует локальной дифференциации.
Райт [909] на основе математических соображений показал,
что если панмиктическая единица состоит примерно из 10 раз-
множающихся особей, то в обширной непрерывной популяции
может возникнуть значительная локальная расовая дифферен-
циация независимо от действия естественного отбора. Если чис-
ло размножающихся особей в панмиктической единице дости-
гает 100, то следует ожидать, что расовая дифференциация бу-
дет происходить не в локальных, я в региональных масштабах.
Если же свободно скрещивающаяся группа состоит из 1000 и
более особей, то расовая дифференциация невелика и происхо-
дит на значительных расстояниях, поскольку распределение ва-
риаций зависит от системы скрещивания у данного вида. Не-
значительное расселение на далекие расстояния задерживает
региональную дифференциацию в обширной непрерывной попу-
ляции [909].
Величина соседства у перечисленных выше видов растений
v*VaeT си-’’ЪЧ0€ отклонение от широкой панмиксии [478].
a Phlox рИоза, Liairis aspera и L cylindracea величина мог-
34 Чосп переил. Псмуляцш и расы
ла вы допускать региональную расовую дифференциацию на
основе одной лишь структуры популяции. Локальная расовая
дифференциация у Lithospermum carolinicnse и вероятно, у
Linanthus parryae может, no-вкдимому. быть результатом одной
лишь структуры по типу соседств.
В популяциях lananthus parryae * l-iatris cylindracea дейст-
вительно происходит морфологическая дифференциация в ло-
кальных масштабах [203. 204, 712]. Эта дифференциация не свя-
зана пи с эдафическими факторами, ни с какими-либо иными
известными факторами среды.
Llnanthui parryae (Polemoniaceae)—самонееовместимое
однолетнее растение, опыляемое насекомыми и произрастающее
в пустыне Мохаве и на примыкающих к ней горах. Популяции
в западной частя пустыни Мохаве полиморфны по окраске цвет-
кои. причем в одних районах формы с голубыми и белыми цвет-
ками встречаются вперемешку, а в других они разделены.
Иногда колонии с чисто голубыми цветками растут вблизи ко-
лоний с исключительно белыми цветками или непосредственно
примыкают к яим, и такая ситуация сохраняется неизменной на
протяжении многих лет (203, 204]. Обе цветовые формы растут
на одной н той же песчаной равнине, и никаких различий в ус-
ловиях кх обитания ие замечено. Вполне возможно, что эти ло-
кальные расы, различающиеся по окраске цветков, —результат
ограниченного потока генов и величины соседств и что отбор,
определяемый условиями среды, ие играл никакой роли в их
возникновения, однако подобный вывод ни в коей мере нельзя
считать бесспорным.
Взаимодействие между потоком генов и
отбором
Как мы видели, большая непрерывная свободно скрещивающая-
ся популяция теоретически может дифференцироваться из ряд
локальных рас на основании одного лишь характера скрещива-
ния и без участия отбора, если область расселения ограничена
и панмиктические единицы или соседства невелики [909]. Этот
теоретический вывод подтверждается некоторыми наблюдения-
ми, которые были приведены выше. В природе, однако, субпо-
пуляция, обитающие в разных частях какой-лнбо области, не-
редко оказываются в различных условиях среды и, следовательно,
подвергаются разным давлениям отбора. Естественный от-
бор. таким образом, действует в сочетании с системой скрещи-
вания данного вида.
В условиях свободного скрещивания, происходящего в широ
них масштабах в большой популяции, влияния отбора, направ-
ленного на адаптацию к той или ивой конкретной локальной
Гл. J. Локампы» роен
»
среде, сводятся на нет непрерывным притоком генов из других
местностей. Давления отбора умеренной интенсивности иогут
обеспечить сохранение обширных региональных, но не обособ-
ленных локальных рас. И наоборот, лишь очень интенсивный
отбор может обеспечить сохранение локалышл рас в условиях
мощного потока генов
Если, однако, популяция при такой же обшей величине ока-
зывается вследствие своей системы скрещивания подразделен-
ной на мелкие панмиктические единицы или соседства, то мате-
риалом для отбора может служить генетическая изменчивость
в каждом локальном участке без вмешательства потока генов
из других участков. Подобное положение вещей благоприятно
для (формирования и поддержания под действием отбора ряда
локальных рас, адаптированных к соответствующим средам.
Такал ситуация благоприятствует эволюции вида а целом, по-
скольку любая панмиктическая единица' или соседство, обла-
дающие признаками, имеющими общую адаптивную ценность,
а не только локальное значение, может распространяться за
счет других скрещивающихся единиц, наделенных менее благо-
приятными признаками [909, 912].
Следует подчеркнуть, что именно равновесие между интен-
сивностью отбора и скоростью потока генов определяет, будет
лк происходить локальная расовая дифференциация или нет.
По своей интенсивности отбор может варьировать от слабого
ДО сильного, а скорость расселения — от низкой до высокой;
поэтому следует ожидать, что в природе должны встречаться
самые разнообразные сочетания эти двух факторов. Однако
некоторые акторы склонны выделять в качестве веной нормы
какое-либо один Сочетание, низкую скорость расселения и ин-
тенсивность отбора от умеренной до высокой [196] или высокую
скорость расселения и очень интенсивный отбор [23). В приводе
схшсствует много возможных точек равновесия н много норм.
Некоторые из них способствуют локальной расовой дифферен-
циации. другие же— ист.
Метафора «дисперсности (зернистости) средня (environmen-
tal цтатп) позволяет взглянуть па эту проблему несколько ина-
че. Среда обит-шня данной популяции кли системы популяций
обычно пространственно неоднородна. Однако эта пространст-
венная неоднородность может быть «тонкодисперсной» или
♦трубоднспсрсной» в зависимости от величины организмов и
радиуса их расселения [486]. Приведем в качестве примера са-
ванну, где участки леса перемежаются с участками лугов Пти-
цы одного из видов могут летать над всей саванной и корннть-
ся повсюду, не испытывая давлений отбора ни со стороны лес-
ных. пи со стороны луговых участков. Для этих пгнц данная
среда тпккоднепереная Популяция прикрепленных к субстрату
У
36 Часть пераал Полцляцлш и расы
однолетних растений, расселяющихся на небольшие расстояния,
будет, однако, подвергаться днзрупгнвным давлениям отбора
лесной и луговой среды по отдельности. Для такиУ растений та
же самая среда оказывается грубоднсперсной.
Микрогсографическая расовая
дифференциация
Среди перекрестно опыляющихся растений известны многочис-
ленные случаи, когда различные локальные расы встречаются
в соседних местностях, разделенных расстоянием примерно
100 м или менее, к, несмотря на это. сохраняют свои отличи-
тельные расовые признаки из поколения в поколение. В таких
случаях экологические условия в соседних колониях заметно
различаются. Возможно поэтому, что подобная локализованная
расовая дифференциация обусловлена действием дизруптмвиого
отбора в отдельных маленьких скрещивающихся группах. Оче-
видно, что в этих случаях днзруптивный отбор превалирует над
потоком генов.
Локальная расовая дифференциация, коррелированная с
различиями в условиях среды, наблюдалась средн прочих у
следующих перекрестноопыляюшихся видов (перечисляются в
хронологическом порядке):
Hlaracutm tunbetfalum [840]
GlUa eapitata [2Й6, 267]
Ctarkia xanttana [491]
Gottorn ршяйит [186]
Pinui nvinJifota <Ску«ллас и Бжмхам [766] сообщала
о хллячкя рмздых рмляеий. wo
теперь оан представляются фежо
PoltitfUia gLuulutasa
Agroshs tenuis
HeiiunJhu* annuae X bolandtri
Diptaeas longiflofut
EschxcJuitixia caitfomica
CMlnxxa heisfcphglla
Pituu aibicatdix
Pinas mac/готова
Дн/Днхап/Аи.т» tfdoraium
Prucalypiut olnlnallt
тмш«ескиш1 моднфякщяямя [Ь|])
[W]
(«]
[790]
[54]
[M]
(121)
l«U
[97]
(24. Б40)
(457)
Гл 2 Локалкьме pact*
37
В примере с Agrostis tenuis выявляются некоторые сущест-
венные черты. Этот свободно скрещивающийся ветроопыляемый
злак имеет более или менее непрерывное распространение на
волнистых холмах в Уэльсе, где такие факторы среды, как поч-
ва. высота, климат и интенсивность выпаса, варьируют. В соот-
ветствии с этими различиями в среде в выборках из популяций
Agrostis tenuis обнаруживаются наследственные или эком- М*
ские различия. Следовательно» популяции реагируют на отбор,
происходящий я локальной среде, несмотря на непрерывность
распространения и перекрестное опыление с помощью ветра
Эта локальная дифференциация в некоторых случаях обуслов-
лена. вероятно, слабым или умеренным лизруитнвным отбором
в сочетании с распространением пыльцы на относительно не-
большие расстояния [651
3 некоторых других местностях на популяции Agrostis te-
nuis действует иная комбинация сил.'В районах свинцовых руд-
ников в Уэльсе этот злак растет ма почвах с высоким содержа-
нием свинца, я также на прилегающих к ним почвах, не содср
жащнх свинец в токсичных количествах. На почвах этих двух
классов существуют экотипы, нс переносящие свинец, и экотипы,
толерантные к нему. В некоторых местах эти две расы очень тес-
но соседствуют и. уж во всяком случае, находятся достаточно
близко одна от другой, чтобы было возможно их перекрестное
опыление при помощи ветра. В этом случае расовые различия,
очевидно, сохраняются благодаря дизруптнвному отбору, кото-
рый достаточно интенсиве и, чтобы перекрыть эффекты межра-
ссеых скрещиваний [402. 422, 540].
Какие факторы отбора участвуют □ микрогсографической
расовой дифференциации растений? Климатические факторы
обычно изменяются постепенно по макрогеографическим тран-
ссктам н поэтому вряд ли могут бить причиной резко выражен-
ной локальной расовой дифференциации. Исключения встреча-
ются на высоких горах в умеренной зоне, где интенсивность та-
кого фактора, как ветер, может изменяться при небольших из-
менениях высоты, и эти изменения коррелируют с различными
экотипами данного вида. Примерами служат Pinus atbicaulis.
Р тиггауапа и несколько других высокогорных древесных рас
тений (97J.
В отличие от этого эдафнчеекяе факторы часто резко изме
няются на небольших расстояниях. По этой причине, а также
потому, что почва представляет собой чрезвычайно важный ком
понент среды для семян и проростков, элифические факторы
пссбенио хорошо соответствуют рати факторов отбора, приводи
Щепо к дифференциации микрогеографических рве у растений.
> большинства растений, перечисленных в приведенном выше
списке. — от классического турсссоиовскою примера с Hieracium
3S Чат «рвал. Z7«w«4“> расы
я до изученного недавно эвкалипта, а также у подробно иссле-
дованной полевицы (Agroitis) и других маков в Уэльсе —ло-
кальная расовая дифференциация коррелирует с изменениями
Мафических факторов среды.
Сочетание отбора с генетическим дрейфом
В популяционной системе колониального типа, состоящей ид
маленьких изолированных иля полуиэолвроваиных популяций,
пространственное рааделеяие снижает интенсивность потока ге-
нов и благоприятствует действию генетического дрейфа. Если,
как это часто случается, нзо.гяроваиные популяции живут в рав-
ных средах, то отбор может действовать также в сочетании с
дрейфом генов. Это приводит к образованию ряда локальных
рас. специфически адаптированных к своим локальным средам.
Расовая изменчивость в таких случаях не образует посте-
пенных географических градиентов или клик, а носит несколько
беспорядочный характер. Каждая колония или группа соседних
колоний обладают собственными отличительными чертами.
Предсказанный характер расовой изменчивости наблюдается
у ряда видов растений е популяционными системами колониаль-
ного типа в областях с разнообразными условиями среды. На-
пример. различные вады Cupressua а Калифорнии часто обра-
зуют ряд изолированных рошип: некоторые из этих рощиц об-
ладают достаточно четко выраженными отличительными приз-
наками. так что специалист по кипарисам может по одному
лишь внешнему виду данного экземпляра, ие обращаясь к гер-
барной этикетке, определить, откуда ои происходит [904]. Вто-
ростепенная расовая изменчивость хорошо развита также у
различных травянистых растений е колониальной структурой
популяционных систем. Примерами служат CUia achdledoha
в Калифорнии, секция Chelranthus рода Erysimum и Nigella de-
genii на островах Эгейского моря [273, 749. 811]
3 Географические расы
Локальная расовая изменчивость постепенно переходит в геог-
рафическую изменчивость. Различие между этими двумя видами
изменчивости достаточно произвольно, но полезно с практике
ской точки зрения. Локальные расы занимают центральное мес-
то в популяпнонно-бнплогнчеекях исглглованиях, тогда как бо-
лее широкие расовые группировки находятся в сфере интересов
систематиков, и их обычно рассматривают как подвиды, а иног-
да как разновидности, различаемые в формальной таксономии.
Характер внутривидовой географической н.змсичивоетн у
растений чрезвычайно разнообразен. В сущности, каждый вид
растений а неком описательном смысле неповторим. Тем не ме
нее ясе многообразие типов изменчивости' можно объединять в
несколько общих категорий. Крупные (например, нелокальные)
расы можно разбить ва трв категории, непрерывные географи-
ческие расы, дизъюнктивные географические расы и экологиче-
ские расы.
Непрерывная географическая изменчивость
В случае непрерывной географической изменчивости локаль-
ные популяции обычно полиморфны, что является результатом
потока генов и уравновешивающего отбора; по по географиче-
ской трансекте можно вилеть постепенный сдвиг частоты дан-
ного варианта. Полиморфизм постепенно переходит в локаль-
ные расовые различия, которые в свою очередь постепенно об-
разуют различия на уровне географических рас.
Генетически детерминированные фенотипические признаки,
подверженные внутривидовой изменчивости, образуют, следова-
тельно, градиенты вдоль географических траисскт. Подобные
радненты признаков носят название клип. Клина —это посте-
пенное изменение какого-либо признака в ряду популяций, ле-
жащих ня некой географической трансекте (теоретическое об-
суждение клик см. [201J)
Клииальная, нли непрерывная, географическая изменчивость
наблюдается у видов, обладающих более или менее иепрерып
ной популяционной системой и занимающих область, в которой
наблюдается постепенный переход от одних условий среды и
Другим. Примерами служат виды равнинных злаков и деревьев
лесов умеренной зоны, имеющие широкое распространение.
luntptrui virginiana в восточной части США варьирует по
воему биохимическому составу, а также по морфологическим
ризнакам. Флейк. Раллоф н Тернер (215], используя метод га-
«о
Чапе нервах. Пацллкии и раем
Рас. 31 Общий «МЛ раствижЛ и форма апети у трех представителей группы
Miawliu kmttflvtu ([MJ: моиеимлатура измене»).
Л — М рамтееш Б — М I'vyplhrm fangiflorut В — М. luagtflufus culy-
anus
эорой хроматографии в сочетании со специальными методами
нумеричесхой таксономии, проанализировали соединения из
группы терпеноидов в ряде популяций, распределенных по
траисекте, идущей от г. Вашингтона (федеральный округ Ко-
лумбия) до Техаса, на расстоянии около 240 км друг от друга.
Эти авторы обнаружили, что популяции различаются между
собой по содержанию разных терпеноидных компонентов и что,
кроме того, существует постепенная, или клика льна я. изменчи-
вость по этому признаку вдоль траисекты, идущей с северо-
востока на юго-запад [215]. У этого же вида наблюдаются сход-
ные постепенные изменения в содержании летучих масел по
другой длинной траисекте —от реки Миссисипи до Небраски
Группа Mimulus longiflorus (Scrophulariaceae) состоит из
четырех таксонов кустарниковых растений, растущих в южной
Калифорнии. Три из тех форм, которые мы здесь рассмотрим,
таксономически известны как .М. longiflorus longiflorus, М. I
calycinus и Л4 punictus, и принадлежат к секции Diplacus рода
Mimulus [607]; их рассматривают также как виды выделенного
из Mimulus рода Diplacus [54]. Географическую изменчивость
этой группы изучал Бикс [54]. Нас здесь интересует, во-первых,
китертрадацвя между М longiflorus longiflorus и М I, calycinus.
а во-вторых, иитерградапия между М. I. longiflorus я не пол-
ностью изолированным М punictus
Общий вид растений этих трех форм и их цветки показаны
на рис. 3.1. а их распространение — на рис. 3.2. Mimulus puni
Гл. J. Гтеафтесии ркы
41
eras —высокий прямостоячий кустарник с красными цветка и я,
сидящими на длинный цветоножках и имеющими короткие
трубки; ои растет па прибрежных холмах н равнинах вблизи
океана. Mimulut long if lor us longiflorui — pacKHAHcnaii кустар-
ник средней высоты с розонато-ораижевымн цветками проме-
жуточной длины; он встречается на выжженных солнцем пред-
горьях в чапаральной зоне удаленных от берега гор на юге Ка-
лифорнии. Mimuius long if lor us calycinus растет на каменистых
участках »тях гор на больших высотах и в районах, гранича-
щих с пустыней Мохдпе. Это — небольшое, низкорослое, разрас-
тающееся в стороны растение; цветки у него желтые, с длинной
трубкой и короткой цветоножкой. Интерградацня наблюдается
между М. L longiflorus и М. I. calycinus на промежуточных вы-
сотах на удаленных от моря горах, а также между М L longif-
lorus и М. punieeus в предгорьях прибрежной полосы (54].
v?2 г'“ТаЙвзесюе распростриевае групвы MlnuUlu iiMgifloriu я юж-
ВОЯ Каляфорют (ю [54|_ с „змеяенижии).
42 Чат мдмл в расы
Рис 33 Частота раггениЛ С краевыми, орамжаамми или желтыми цветками
в пооулмии tpynnw Mimda» lotittfionu пн трэтсекте одохамак! по «о-
закатному склону гор Сап Габриел. Калпрорпиа (по (54). е «змевелиааиз),
Рассмотрим изменчивость окраски цветков у утих трех групп
по трансекте, проходящей по юго-западному склону гор Саи-
Габриел (рис. 3.3). Хотя, как отмечалось выше, для каждого
из утих таксонов характерна определенная нормальная окрас-
ка цветков, многие популяции, в том числе некоторые из нахо-
дящихся на данной трансекте, полиморфны по этому признаку.
Популяция, приуроченные к той части трансекты через горы
Сав-Габрнел, которая соответствует предгорьям, т. е. обитаю-
щие на сравнительно небольшой высоте, состоят главным обра-
зом из экземпляров с оранжевыми цветками; эта окраска ти-
пична для М. I. longiflorus. Но в этих популяциях встречаются
экземпляры с красными цветками (типичными для М. punictus),
а также экземпляры с желтыми цветками (типичными для
М. /. calycinus) (рис. 3.3). Чуть выше предгорий растения с
красными цветказги исчезают. На средних высотах возрастает
частота растений с желтыми цветками, а иа больших высотах
они вытесняют форму с оранжевыми цветками (рис. 33). Эту
интерградацню между расами по окраске цветков Можно пред-
ставить как сдвиг сбалансированного полиморфизма по геогра-
фической трансекте.
Один из количественных признаков цветка у этих видов так-
же подвержен ввутрнаопуляционной изменчивости, причем
скользящие средние лежат на той же трансекте (рис. 3.4).
Гл г Гюгрсфгиыш расы
43
В этом случае сдвиг происходит в определенно# золе трансекты
на средних высотах. Ряд популяций типичных М. longtflorus
tonglftorus на небольших высотах, т. е. а предгорьях, почти од-
нороден по атому количественному признаку; такая же одно-
родность наблюдается у ряда популяций М. I. calycinus в раз-
ных местах на больших высотах (рис. 3.4). По существу такие
же ступенчатые сдвиги были обнаружены при оценке различ-
ных других признаков цветка и вегетативных органов у этих
популяций, а затем все эти признаки были выражены в значе-
ниях гибридного индекса [54].
Между М. longijtorus longtflorus и М. puniceus наблюдается
также иатергдодация по различным траисектам, идущим от
удаленных от моря предгорий к прибрежным холмам н равни-
нам [54]. Если ограничиться этими данными, то М. puniceus
Мяягноки
№•*4 нм»'
ТмлОмп. 19М
ТолпХхып. 1Я60
Уо-мр-веи
Сам Амтомн&ФйАМ
£**«>•
Вмриинд г Лотыто
Сравним масть г ЛоееЛю
Гер» ГИ>г«4*те
Сл**4ж«м»ые Томпсона
Т««пеоиС4ймик
ПяППДВЛЬГМ, 1К&
Ляг-лДиплом law
I I____________I______L—______4______I___________
•90 *90 -10 0 Ю 20 30
। Огмосмзыьмая длина tpvOui ммчяи а иаатеномди
Рас. 3.4. Внутри- я иежполуаяаномяая яяняячявоеп. по одному ю количе-
Ственвых пржзшгпв пытка у группы Mimulux longifloru» на дзук трансепта*
прояомщнх по удами nun ОТ побережья горам в пжвоА Квлжфоржяя (по
J»4], с иыдеттмя).
(Мегявалась pa”"®a “Т-жау девой труСжя вепчихй в длмвой Двегояожхн
Ддя каж*°в Noey"®»" представлены в графкчесжой ферые средам
ПВЧЕНИ». стаадартяОФ откжэпгпйс дцааа»ои яяигкзквссти.
<4 Чаете «рева. ЛопрлЯ1НШ u расы
можно рассматривать как одну из географических рас М. tongl-
florus. Во взаимоотношениях между этими популяционными
системами имеется, однако, ряд интересных осложмнющкх мо-
ментон, которые мы рассмотрим в одной из последующих глав.
Дизъюнктивные географические расы
Непрерывная географическая изменчивость наблюдается в тех
случаях, когда сама популяционная система непрерывна или
почти непрерывна я достаточно велика. А это возможно при
условии пространственной непрерывности или полунепрерывно
сти ее местообитания Противоположная ситуация наблюдается
в тех случаях, когда местообитание данного вида раэбито на
относительно небольшие области, распределенные в пространст-
ве с перерывами (например, ряд изолированных горных вершин
в случае какого-либо альпийского вида). В таких случаях из-
менчивость носит совершенно иной характер.
Популяционная система такого вида относится к колониаль-
ному типу, т. с. состоит нз изолированных или полуиэолироваи-
ных популяций, занимающих обособленные местообитания.
Каждая изолированная колония становится специфически адап-
тированной к условиям своей локальной среды. Отдельные ко-
лонии. обитающие в различных средах, дивергируют. Всему это-
му способствует отбор в сочетании с пониженным обменом ге-
нами между колониями. И если величина колонии постоянно
или периодически бывает достаточно мала для того, чтобы
дрейф генов мог оказаться эффективным фактором, то эти коло-
нии будут также приближаться к мономорфизму.
Вследствие этого характер изменчивости какого-либо вида,
населяющего экологические островки, резко отличается от ха-
рактера изменчивости вида с обширной и непрерывной популя-
ционной системой. Географическая изменчивость такого видя
прерывистая, географические расы разобщены, и в зависимости
от величины популяции степень полиморфизма может сильно
понизиться. И если в большой непрерывной популяции какого-
либо вида границы между географическими расами проводятся
более или менее произвольно, то у вид» с популяционной сис-
темой островного типа географические расы дискретны и раз-
граничены не произвольно.
Эти противоположные типы географической изменчивости
можно наблюдать у двух видов Gilia — и. latiflora и О. leplant-
ha (Polemoniaceae). Указанные виды принадлежат к одной и
той же секции Arachnion данного рода; оба они однолетние,
оба диплоидные (2л—18), размножаются преимущественно пу-
тем перекрестного опыления и произрастают в общем в одной
и той же области в южной части Калифорнии. Оба вида поли-
Гл. 3. Ггыра^плслиг ролы
43
типические, с 4-5 географическими расами, которые рассматрива-
ются систематиками как подвиды. Как показали зксперимеп
тельные исследования 1299]. конспецифичные расы иятерфер-
тильиы у обоих видов. Однако на этом нх сходство кончается
GUia latlflora встречается на песчаных равянпах а пустыне
Мохаве и примыкающих к ней долинах. Это местообитание не
прерывно, как и местообитание G. latiflora. Географические
расы этого вида в большинстве случаев интерградируют на об-
ширных пространствах, и границы между ними устанавливают-
ся в значительной мере произвольно [300].
GUia leptantha произрастает па открытых песчаных полянах
в сосновых лесах на горах южной Калифорнии. Ее местооби-
тания разбросаны по ряду изолированных горных хребтов. Та-
кне системы изолированных популяций образуют разобщенные
географические расы, сходные по фенотипическим признакам,
но разделенные хорошо выраженной прерывистостью в характе-
ре измени на ости (300).
Экологические расы
Географические расы аллопатричны, они занимают различные
географические области Пространственные азнимоотиоше|и<я
между расами могут также носить характер смежно* симпат-
рнн раса Л и раса Б занимают разные местообитания в одной
и той же области, скрещиваются между собой и интерградируют
в многочисленных эонах коятакта. но каждая из них в своем
собствеппом местообитании сохраняет отличительные расовые
признаки.
Такой тип расовой изменчивости создается в тех областях,
где два или несколько местообитаний, подходящих для предста-
вителей рлиого и того же вила, распределены п пространстве в
виде пятен или в шахматном порядке. Примером служит саван-
на, в которой участия леса чередуются с участками лугов. Че-
редующиеся участки с разными типами почвы или с разными
типами материнской породы также могут быть заняты различ-
ными экологическими расами, то же самое наблюдается в го-
ристой местности па соседних горных склонах, обращенных
» разные стороны
GUia achilteilolia (Polemoniaceae) встречается в одной нз
частей Береговых хребтов (Калифорния), в саванне с участка-
ми виргинского дуба. Популяции этого вила отличаются силь-
ной изменчивостью. Однако его различные лохальпые варианты
можно объединить в две большие группы или экологические
расы, известные в систематике под названиями G. a, achilltifo-
Чан G a multlcaulis.
<в Чат креол. Покцлпрш «
GiUa achillaijolia achUleifolta — более обычная и широко
распространенная форма — встречается на открытых солнеч-
ных поляках. Ее крупные сине-фиолетовые цветки собраны в
головки и опыляются пчелами. G. a. multicaulis растет иа полу-
затененных участках редколесья в тех же областях. У нее мел-
кие бледно окрашенные и самоопыляющиеся цветки Эти два
экологические расы при экспериментальных скрещиваниях дают
плодовитых гибридов (во всяком случае, в некоторых сочета-
ниях) и иитерградируют в природе в тех местах, где открытые
поля соприкасаются с участками леса [2731.
По всей области обитания Cilia achuleifolia лесные и от-
крытые полевые местообитания чередуются в шахматном поряд-
Ге S reaioatwctue овсы
47
не. Это отражено и в характере распространения мух эколо-
гических рас данного вида (рис. 3 5).
Конспецифичные высотные расы воаиикают у видов, насе-
ляющих гористые области. Будут ли эти высотные расы по
своим пространственным взаимоотношения и экологическими
или географическими расами, зависит от рельефа местности.
Сплошные горные массивы с относительно пологими склоками
благоприятны для развития географических рас благодаря на-
личию обширных непрерывных пространств на разных высо-
тах. В отличие от этого в пересеченных горных массивах с
крутыми склонами те же высотные зоны представляют собой
ряд обособленных мелких пятен, нередко смежпо-симпатричс-
ского типа, что благоприятствует развитию экологических рас.
Разумеется, между этими двумя основными типами рельефа су-
ществует ряд промежуточных типов.
Аналогичным образом элафическне расы могут представ
лить собой экологические расы нли мелкие локальные расы или.
наконец, нечто промежуточное в зависимости от величины тер-
ритории, занимаемой дайной расой, и от ее характера.
Факторы отбора, участвующие в формировании
рас
Много лет назад Турессон (840) установил адаптивную природу
многих расовых признаков, наблюдаемых у растений. Его выво
Ды были основаны иа трех группах данных. Во-перкых, отличи-
тельные расовые признаки часто представляют собой адаптации
к условиям среды, преобладающим а местообитании данной ра-
сы Так, раса Atripicx litorale, обитающая на защищенном от
ветров южном побережье Швеции, представлена высокими пря-
мостоячими растениями, тогда как раса, населяющая открытое
ветрам западное побережье, — низкорослая и разрастающаяся
вширь (840). Во-Ъторых, параллельные расовые различия наблю-
даются у других видов, встречающихся в тех же местообита-
ниях. например у Alriplex sarcophyllum и Л. ргаесох. И нако-
нец. в-третьих. у одной расы .4 literal» при выращивании ее в
среде, типичной для другой расы, в результате фенотипической
модификации развиваются признаки зтой другой расы [840,
Среда, в которой обитает то или иное растение, характери-
аусгся рядом аспектов, и все эти аспекты влияют на формиро-
вание расовых признаков. Наибольшее внимание уделялось
такому важному аспекту, как климат. Преобладающее большин-
ство экотипов, изучавшихся Турессоиоы (840, 841. 843] у Atri-
Plex, Geum. Caltha, Achillea н многих других родов, относятся
именно к климатическим экотипам. Классические исследования.
a
Част» мрете Популяции и расы
проведенные Клаузеном, Кеком и Хнси [101, 103] ив Potentilla
glandulosa. Achillea millefolium н других видах, также касаются
климатических рас.
Селективное воздействие климата ясно выступает а расовых
различиях по срокам цветения. В северных районах Швеции
вегетационный период короче, чем в южных. В соответствии с
этими климатическими различиями северные линки Fragaria
vesca, Campanula rolundifolia, Oeum rivale и £ alt ha palustris
зацветают раньше, чем южные линии тех же видов при совмест-
ном выращивании на однородных экспериментальных участиях
[МО, М3], Подобным же образом расы Potentilla glandulosa и
комплекса Achillea millefolium из альпийской эоны гор Сьерра-
Невада в Калифорнии зацветают иа экспериментальных участ-
ках раньше, чем расы, обитающие в тех же горах на средних
высотах [101. 103].
В формировании рас у некоторых видов явно участвуют эда-
фичеекме факторы. Во многих случаях образование эндемичных
рас (и видов) коррелирует с отложениями гипса или выходами
известняка [847]. В гл. 2 наряду с другими примерами упомина-
лись эдафические расы Agrostis tenais на почвах, содержащих
свинец, н на обычных почвах.
В некоторых случаях оказалось возможным продемонстри-
ровать существование зависящих от эдафических условий расо-
вых различий внутри одной климатической расы. Популяции
АсЛШеа borealis. Cilia capitata capitata и Streplanlhus glundu-
losus, состоящие каждая из одной климатической расы, встре-
чаются в Калифорнии как иа серпентинитовых, так и иа несер-
пемтнннтовых почвах. Как показал Круккеберг [446], нежду по-
пуляциями, обитающими на тех и других почвах, имеются ра-
совые различия в отношении физиологической выносливости к
серпентинитовым почвам. Эдафическая расовая дифференциа-
ция у многих видов накладывается на климатическую расовую
дифференциацию [446. 452].
Снискал [733] описала у Brassica nigra биотические расы.
В Малой Азии В. nigra встречается как в естественных усло-
виях, так и в посевах в. campestris. Эти две расы В. nigra раз-
личаются по комбинации вегетативных признаков, особенностям
шегков и семян. Более специализирована та раса, распростра-
нение которой ограничивается полями, засеянными В. campest-
ris; ее отличительные признаки дают ей возможность расти
средн растений В. campestris. а семенам — вызревать и распро-
страняться вместе с семенами последней [733]. Подобным же
образом некоторые расы Camrlina satrira (СгисИегае) относятся
к неспециализированным сорнякам (С. a. saliva), тогда как
другие расы (С. з. linicola) — к специализированным сорнякам
льняных полей [780, 824].
Гл 3. Г<кире^<е<ес«ье рат
4»
Распял я дифференциация в отношении животных-опылите-
лей обнаружена у нескольких видов Polcmonlceae [301]. Раз-
линия в строении цветка между разными расами Gtlia splendent
коррелируют с опылением их преимущественно жужжалвми
(Bombylius). Euloruhus (Cyrtidael с нх длинными хоботками н
колибри. У Eriastrum densifolium и OUta achilleifotia есть расы
с широким зевом венчика, произрастающие в Калифорнии вдоль
береговой линии и опыляемые крупными видами пчел, а есть
расы с маленьким зевом, опыляемые мелкими видами имел н
произрастающие на удаленных от моря горных хребтах. К слож-
ному переплетению расовой дифференциации могут добавиться
у многих видов растений расы, различия между которыми свя-
заны с опылением (301].
Количественных данных о скорости образования рас немно-
го. Очень интересны в этом плане исследования по расам Mcmu-
lus guttatus (Scrophulariaceae) в бассейне озера Бонневиль в
шт. Юта [860]. Бассейн этого древнего озера в течение плейсто-
ценового и голоценового периодов из территории, покрытой во-
дой и льдом, превратился а сушу, причем время этих измене-
ний можно установить.
Широко распространенный Mtmulus guttatus заселял многие
из обнажившихся при этом участков суши и образовал дивер-
гентные расы. Возраст этих популяций можно определить на ос-
новании известных возрастов нх местообитаний. Таким образом
можно установить, что за 4000—14 000 дет имел место средний
уровень расовой дифференциации в отношении количественных
признаков и скрещиваемости (507. 860]. Это по осторожной оцен-
ке; на самом деле эволюционные изменения замяли, возможно,
лишь часть этого времени.
Часть вторая
Природа видов
4 Биологический вид
Биологические вид —основная единица организации бинарен-
тальвых живых существ, представляющая собой систему скре-
щивающихся популяций, репродуктивно изолированную ог дру-
гих таких систем Иными словами, это общая сумма скрещи-
вающихся между собой особей, а следовательно— самая обшир-
ная единица нормального бипаренталького размножения. К био-
логической концепции вида приходят различными путями.
Первоначальная концепция вида возникла на основе прямых
наблюдений в природе. Первым естествоиспытателям, так же
как и нх современным последователям, занимающимся популя-
ционной биологией, было очевидно, что живые организмы не об-
разуют континуум, а распадаются на более нлн менее дискрет-
ные скопления скрещивающихся между собой особей. Напри-
мер, члены семейства кошачьих делятся на такие дискретные
группы, как львы, тигры, рыся, домашние кошки и т. п. Каждая
группа размножается в чистоте, или, как выражались первые
естествоиспытатели, порождает себе подобных, а гибридов меж-
ду группами либо не существует вовсе, либо они столь редки,
что заслуживают особого внимания. Эта прерывистая картина
взаимоотношений и изменчивости наблюдается во многих се-
мействах животных и средн некоторых высших растений. Обо-
собленные размножающиеся внутри себя группы и есть виды.
Что касается видов в царстве растений, то лучше всего обра-
титься к тому, что писали об этом Чезалызиио (Cesalpino). в
De plantls librl (1583). Джон Рэй (Ray) в Hiatoria plantarura
(1686). а также Линней в СтШса botanica 15121 н Philosophia
botanica (1761; (513]).
Чезальпиио утверждал, имея в виду виды, что подобное
всегда порождает подобное (см. [702]). Джоя Р»й пошел даль-
ше, утверждая, что растения, выросшие нз одного семени и
вновь продуцирующие себе подобные через семена, принадле-
жат к одному н тому же виду [702]. <Не существует более на-
дежного критерия внла, — писал ом, — чем то, что о« разиио-
Гл. 4. Бмлыичссгшй »*-Э
51
ждется в чистоте, развиваясь из семени и ие выходя га прнсу*
дне ему пределы» (см. (140]).
Линнеи продолжил эти высказывания, подчеркнув различия
между видами и разновидностями. «Творен Природы, создавая
виды, наложил на слон создания вечный закон плодиться и
размножаться, воспроизводя себе подобных. Правда, во многих
случаях ои наделил их правом На некоторые отклонения во
внешнем виде, но не на превращение одного вида в другой»
(Critica botanica; ср. [Ь53]). Совсем иначе обстоит дело с раз-
новидностями. «Существует столько разновидностей, сколько
существует растений, разнившихся из семян данного вида»
(Philoxophia botanica; ср (653): см. также [419])
За недостатком места, мы не можем сколько-нибудь подроб-
но изложить здесь дальнейшую историю концепции вада (см.
(245, 280. 568]). Достаточно сказать, что в XIX и ь начале XX ев.
многие естествоиспытатели высказывали соображения, предвос-
хитившие биологическую концепцию вида. Современная биоло-
гическая концепция вида была сформулирована Рспшсм. Дю-
Рие. Майром, Добржаиским и другими в период примерно с
1929 по 1957 г. Соответственно сами скрешипаюшнеся группы
получили название биологических видов.
Если биологические вилы представляют собой обскобленные
я репродуктивном отношении группы, то очевидно, что между
ними должны существовать барьеры, препятствующие гибриди-
зация; обнаружение таких барьеров послужило бы вторым до-
водом в пользу концепции биологического вида Оказалось, что
зтн постулированные барьеры, или репродуктивные изолирую-
щие механизмы, действительно существуют. Стерильность мула
вошла в легенды, н первые селекционеры иногда называли по-
лучаемые ими стерильные гибриды растительными мулами.
Ьюффоп в 1749 г., а вслед за ним Кант в 1775 г. н многие дру-
гие ученые использовали стерильность потомков п качестве глав-
ного объективного критерии для разграничения отдельных ви-
дов. Однако вес обстоит не так просто.
Скрещивания между биологическими видами предотвраща-
ются разнообразными изолирующими механизмами. Некоторые
пары видов действительио дают стерилыше гибриды, однако от
других сочетаний видов можно искусственный путем получить
плодовитые гибриды, хотя я природе зтн виды обычно не скре-
щиваются Друг с другом вследствие существования различных
внешних барьеров, таких, как отвергание партнера чужого вида
или механические преграды, делающие скрещивание невозмож-
Рме. 4 2 Cilia e.apitata (PofanOBaKM*).
Изобрзжешое здесь ржтеиме прживд-
зезогт м поавмзу Gc capiiata.
ft 4 £41оде/11чдоша eud W
Пример
Cilia capitata и G. tricolor — родственные виды, принадлежащие
it одной и той же сети» рода Gilia (Polemoniaceae). Это одно-
летние травянистые растения с диплоидным набором хромосом
(2л—18), размножающиеся преимущественно в результате пере-
крестного опыления.
Gilia tricolor произрастает в долинах и предгорьях централь-
ной и северной Калифорнии. G. capitata встречается в тех же
районах, ио, кроме того, заходит дальше на север, юг и запад,
доходя до береговой линии (рис. 4.1). Во многих районах от-
дельные растения обоих видов растут в непосредственной бли-
зости друг от друга. Эти два вида симоатричны.
Морфологические признаки типичных представителей этих
двух видов изображены на рис. 4.2 н 4.3. Растения Gilia capUa-
Рж. 43. О ilia tricolor
64 Чат «теряя. Лрарода впОол
ta крупные я высокие, а О. tricolor — относительно мелкие.
У G capitata оонпетяе образует плотную головку, а у О. trico-
lor око рыхлое. У G. capitata на чашелистиках имеются узкие
средние жилки, а у G. tricolor — широкие зеленые полосы. Эти
два вида различаются также по форме венчика. Кроме того,
у G. capitata венчик одноцветный, сине-фиолетовый, а у G. tri-
color ои трехцветный, с пурпурными пятками иа сине-фиолето-
вом фоне и оранжевой трубкой.
Ни тот ни другой вид нельзя, однако, назвать однородным.
О. capitata подвержен значительной географической изменчиво-
сти. Признаки цветков, коробочек и семян варьируют у разных
рас по траисекте, идущей от прибрежных песчаных дюи до гор,
расположенных дальше от берега, в области, окружающей за-
лив Сан-Франциско (рис. 4.4). Между этими и другими расами
0. capitata наблюдается полная интерградация. Географическая
изменчивость наблюдается и у G. tricolor, но у этого вида она
менее выражена. При этом каи у О. capitata, так и у G. tricolor
Рас 4.4. Цветка, аоробочак и саыеиа трех рас OUta capitata, проюрастяижях
м траисекте. ксуые* от ривр<*ж»<1Й .тлим лалава Сян-Фрвшмско (Каха-
6прмая| к горам, расположая имм иа сеаар от наго
Л G(Ua capitata chamtitonis Б — О.с. tomentoia, В — G.c. глр#о/а.
Гл. 1. Вшлоитсхий «ив
М
географическая изменчивость не выхолит за пределы данного
аила. Промежуточные формы между этими двумя видами неиз-
вестны; ни одного гибрида между ними обнаружено не было.
Внутри каждого вида Gllta и между ними были проведены
экспериментальные скрещивания Скрещивали семь линий Оз-
lia capitata, принадлежащих к пяти географическим расам. Эта
расы скрещиваются друг с другом во всех сочетаниях, причем
в большинстве сочетаний скрещиваются очень легко Межрвсо-
вые гибриды F| обладают высокой или частичной плодови-
тостью. образуя от 50 до 90% полноценной пыльцы и обильные
семема. Развивающиеся из них растения F} обычно жизнеспо-
собны. но небольшая доля растений обладает пониженной жиз-
неспособностью [266. 268]. Северная и южная расы G. tricolor
также оказались кнтерфертильными [269].
В отличие от этого многократные попытки провести скрещи-
вания между GUia capitata и С. tricolor всегда оказывались
безуспешными. Перекрестное опыление между этими видами
привадит к образованию недоразвитых семян, ио не дает ни
нормальных семян, ни гибридов Fi. Кроме того, установлено,
что G. tricolor не скрещивается ни с одним из других родствен-
ных видов GUia G capitata с тоудом удается искусственно скре-
стить с каким-нибудь другим близким видом, ио получающиеся
при этом гибриды высоко стерильны [269. 2751-
Таким образом, G tricolor и G capitata являются самостоя-
тельными биологическими видами. Между представителями ви-
да G. tricolor происходит половое размножение, приводящее к
образованию интерградирующих вариаций. Подобное же скре-
щивание с аналогичными результатами происходит между пред-
ставителями G. capilata. Однако G. tricolor и G. capitata ие мо-
гут скрещиваться и действительно никогда не скрещиваются
между собой, так как этому препятствуют очень эффективные
репродуктивные изолирующие механизмы.
Биологический вид как естественное следствие
полового размножения в биотическом
сообществе
К концепции биологического вида можно прийти и третьим
путем. Можно показать, кто половой процесс, происходящий
в неоднородной среде, неизбежно должен сопровождаться орга-
низацией скрещивающихся внутри себя популяций в обособлен-
ные биологические виды Предпосылки к подобным взглядам
излагались Добржаиским [165] и Кэйном [79]. Кэйн писал:
5€ Чат «горал Природа ялшса
«Доств точно лжшь представить послодстма я«ограюге«кяо41 гмС^вДиза-
цаж иажду ве«мн ими* жмаущимм жиаотжымв, тгсби оосжнтк краАиюю важ-
ность npoaccej мдообраюини. Ecu бы act межамдоные барьеры вдруг
икмм то чукоы были устраяекы, то ато врмвело бы к ужасаажему сголпо-
тво ренте гибридов со эсезозможнмим сотетаипиа признаков Ни один ан-
дивадуум н« быв бы ЛоДЖжМы пЛрвлеы ядаптнрамн и тому ила поыу об-
разу жизиж, a Msorw првммки каждого на hhi могла бы «жаватвся адапти-
ромняымя для пома иесовмс<тныых целеЯя
А Добржаиский [165] утверждал, «то «продолжение жизни
возможно лишь и том случае, если генные комплексы, согласо-
ванность которых проверена естественным отбором, будут пре-
дохранены от разрушения, неизбежного при неограниченной
гибридизации». Мы построим наши дальнейшие рассуждения,
использовав эти высказывания в качестве основы.
Любая пригодная для обитания локальная область на по-
верхности земного шара обычно содержит разнообразные эко-
логические ниши, занимаемые различными енмпагричеекяии
популяциями организмов. Каждая популяция адаптирована к
своей особо* нише в биотическом сообществе благодаря нали-
чию у нее определенной комбинации физиологических н морфо-
логических признаков. Каждая комбинация признаков основана
в свою очередь па определенной комбинации генов.
Половое размножение представляет собой механизм, обус-
ловливающий рекомбинацию генов. Рекомбинацию генов нель-
зя считать абсолютным благом: в одних случаях она приводит
к благоприятным результатам, а в других —к неблагоприятным.
Адаптивные свойства различных возможных комбинаций генов
широко варьируют в зависимости от входящих в кил гемов и
взаимодействий между ними.
Особи, принадлежащие к данной скрещивающейся популя-
ции, могут варьировать по нескольким наследственным призна-
кам. но тем не менее они способны скрещиваться друг с другом,
давая жизнеспособное плодовитое потомство. Такая популяция
полиморфна по ряду неадаптированных генов. Поясним это на
простом примере. Геи А представлен в популяции двумя алле-
лями, Л, н А,, а другой геи В точно так же представлен двумя
формами. В, к Вг. Девять диплоидных генотипов, которые мо-
гут получиться в результате полового процесса из этих двух
полиморфных генов (А^В^, АА|В,В,. AjAjSjB:), обла-
дают индивидуальными различиями; тем не менее все они —
нормальные представители своей популяции, адаптированные к
жизни и более или менее успешному размножению в зкологиче-
ской эоне, занимаемой их популяцией. Кроме того, хорошие
адаптивные признаки нескольких рекомбинационных типов —
»то не просто счастливое совпадение, а результат естественного
отбора, который через предшествующие поколения оказывал
Га. 4. Биолоатский аид
В?
свое действие на данную скрещивающуюся популяцию, сохра-
няя благоприятные аллелн с хорошими комбинационными спо-
собностями.
Другая популяция, занимающая другую экологическую ни-
шу в том же сообществе, обладает, подобно первой популяции,
определенной адаптивной комбинацией признаков, которая под-
вержена изменчивости в известных пределах Эта популяция,
однако, полиморфна по другому набору коадапгириваиных те-
аов. Допустим, чти оиа содержит аллели А Я А И Я, и ко
торые, комбинируясь друг с другом, да юг девять диплоидных
генотипов АгЛввдВг АА^А^ВпВ^ Все эти генотипы,
несмотря на индивидуальные различим, приспособлены к жизни
в нише, занимаемой их популяцией
Последствия скрещивания между членами этих двух популя-
ции весьма различны, если рассматривать их в плане адагттиро
ванн ости по то и ков Рассмотрим, например, девять типов потом-
ков F2. получающихся я результате скрещивания генотипов
А|Л| В\В\ и Два родительских типа, яновь появляю-
щиеся в поколении несомненно, жизнеспособны; жизнеспо-
собным может оказаться и тип F». Остальные шесть генотипов,
представляющие собой неиспытанные рекомбинации родитель-
ских генов, могут оказаться плохо адаптированными как к раз-
личным средам одной н другой роиггелъеких особей, так н к
любой другой доступной нм среде Если гены А и В не сцепле-
ны между собой. го можно ожидать, что такие жизнеспособные
или субвитя.тъиые рекомбинационные типы составят 10 из 16 зи
гот F?.
Согласно правилам‘Менделя. при других родительских парат
из агах двух популяций, например A,АДАХи т д.,
доля рекомбинантов в F,. не относящихся ни к Р типу. ни к
Fi-типу. будет такой же. а именно 62,5% Поэтому в целом по
томкл, получающиеся в результате гибридизации между двумя
популяциями, различающимися по одной простой адаптивной
комбинации генов. —это преимущественно рекомбинанты с по-
ниженной приспособленностью.
Доля субвитальных рекомбинантов быстро повышается с
увеличением числа ггнимх различий между родительскими по-
пуляциями. Допустим, что две популяции содержат различные
аллели пяти песцегтленмых между собой генов, от А до Е, опре
дечякниих различные адаптивные типы в каждой популяции
В таким случае скрещивание между AiAt...EiEt и А* А.. £,£,
ласт зиготы Fj, из которых только 3,3% будут генотипически
£i0^*JN с родительскими особями или с особями F|. Остальные
96,7% зигот Ft составят рекомбинанты, жизнеспособность ко-
торых п большей или меньшей степени понижена Если роди-
тельские популяции различаются по адаптивной комбинации те-
58
9КГ4 tTOfat Прироса
вое, которую образуют десять независимых генов, то такие суб-
витальные рекомбинанты составят 99,9% всех зигот Fa.
Иными словами, 966797 из каждого миллиона зигот Fj, воз-
никших в результате гибридизации между популяциями, разли-
чающимися по адаптивной комбинации генов, состоящей из пя-
тя иесцеплениых между собой генов, будут обладать плохо адап-
тированной генетической конституцией. А иа каждый миллион
зигот Fa, образовавшихся в результате гибридизации между по-
пуляциями. которые различаются по адаптивной комбинвпин
признаков, определяемой десятью независимыми генами. 999 022,
по все* вероятности, не смогут выжить ни в какой из доступных
им сред,
Коиечио, реальные популяции будут так илц, иначе откло-
няться от зтих упрощенных моделей. Некоторые факторы, на-
пример сцепление, несколько уменьшают генетический груз,
создаваемый гибридизацией. Другие факторы, такие, как слож-
ность генетических различий между симпвтрическими видами,
напротив, увеличивают этот груз, причем последние факторы,
возможно, перевешивают первые. Таким образом, вытекающие
из наших численных примеров оценки потенциального груза,
создаваемого гибридизацией между физиологически дифферен-
цированными сим патричес кими популяциями, вероятно, не сле-
дует считать преувеличенными.
Если при скрещиваниях между дифференцированными сим-
патрическими популяциями наблюдаются огромные потери ре-
продуктивного потенциала, то сирещнваиия внутри популяции
обычно бывают благоприятными. Особи, скрещивающиеся С
другими членами своей популяции, обычно оставляют гораздо
больше потомков, чем особи, скрещивающиеся с членами чужих
популяций. В таких условиях любые генетически детерминиро-
ванные отвергания членов чужих популяций н качестве брачных
партнеров или любые барьеры, препятствующие скрещиванию,
будут подхвачены отбором и будут распространяться в каждой
симпатричесиой популяции.
Таким образом, мы приходим к выводу, что половое размно-
жение, если оно выходит за пределы отдельных популяций дан-
ного биотического сообщества, приводит к распаду аавптквных
комбинаций генов внутри каждой популяции. Угроза продолже-
нию существования таких симпатрических популяций тем боль-
ше, чем более свободно они могут скрещиваться друг с другом.
И наоборот, те биотические сообщества, которые состоят из
репродуктивно изолированных скрещивающихся внутри себя
популяций, или. иными словами, биологических видов, остаются
стабильными.
Гл, 4, лиЛ
59
Концепция генофонда вида
Концепцию биологического вида можно выразить, связав ее с
представлением о генофонде. Концепции вида, основанная на
генофонде, хорошо сформулирована Добржа неким |168].
«Мспаелеягкая популяция — vrn ряпрохунтнаял» оппе<псетпп piiwnnxtnuiHic-
ся половым путем скрещивающихся между собой шмидмкж o6.iaa.x-
юшг общий геиофоилом ЬнаскигичегяиА пн.< — его самая обшнргдя к са-
мая емкая меядемвекая жжулшнх Падамдоааае группировки. такие, как
юп|и»ды роли it в, J* ожидают общими пяофинхаии, а зоэгочу дита-
ны ГипывгнчесдеА реймыюстн. прмсрпгА менде.чшсиим пооу.тикмм Одлахо
виды шфф«р«мин|юммы нм комплексы подчиненных мендележких полтля-
жий, которые можвго называть еодвндами. расами или локальными копуле
ииммн>
Существенное возражение, выдвигаемое рядом авторов [195.
196. 471] против такого определения, состоит в том. что пере
крестное оплодотворение и обмен генами происходят не на ви-
довой уровне, а внутри локальных популяций и субпопуляцнй.
Что касается самих этих фактов, то они верны, и мы нх будем
обсуждать в следующем разделе, однако при их истолковании
упускается из виду самое главное.
Имеющиеся в данном генофонде гены доступны всем парам
способных к скрещиванию индивидуумов. Образуются ли все та
кие пары в действительности или нет, значения не имеет. Кон-
цепция генофонда содержит некий элемент потенциальности
Две особи, находящиеся на противоположных сторонах одной
и той же обширном видовой популяции, могут объединить свои
гены; их потомки черпают гены из двух удалениых одна от дру-
гой частей обширного генофонда. Однако особь, принадлежащая
к другому, репродуктивно изолированному виду, не может чер-
пать гены из генофонда первого вида, даже если она живет с
ним па одной территории Следовательно, мы имеем полное
право представлять себе биологические виды как популяции, об
лвдающке обширными генофондами, которые доступны для всех
входящих в их состав особей.
Причины и следствия, вытекающие из концепции
Какова основа генотипического н фенотипического сходства
между особями, принадлежащими к одному и тому же биологи
тесному виду? Согласно традиционной точке зрения, поток ге-
нов обладает значительным гомогенизирующим и интегрирую-
шим действием в обширных и свободно скрещппвкнипхея попу-
ляционных системах на уровне видов и региональных подразде
n'roi*' в“дов- Такой точки зрения придерживался еще Дарвин
1152]. «Перекрестное оплодотворение играет очень важную роль
• природе, обеспечивая сохранение особей, принадлежащих к
0° Част» втсрад. Природа видов
одному и тому же виду алн к одной и той же разновидности,
в чистоте и их единообразие».
В наши дни эти же взгляды усиленно пропагандирует Майр
[565. 569— 571J Приведем некоторые цитаты из его работ.
(НсеогвивеДсше оопуляжш Tanystpfera gslaiea находятся] в центре не*
ферыисго погожа генов, ждущего в сбоях нз гр явлениях через весь втог
остромо* материк (564|
Устойчивый кмтенгннный аратов гедев. обусловлю нм й процессом лб-
и«»а гсиаиа. служит главным фактором, обеспечя** кишем генетическую имтч-
трвроеамносп между повуллияями дммго вада [6691.
У всех аироко распространенных ершгветающкх видов отяоентчлыю во
д»мжжых оргаякэмов, оо-анднмому, происходят доетаточмо нитеяснвим* об-
мен гевами. обеспечжвающжя амажую одвородносп геиофсидои всех досаль*
них вопуляилД [570].
«Отмечая ээразжтелхмую аюрфологкческую однородность болмкннстоа
широко распространенных аалов t. высоко* чмезмаобгыо. в в... своей етатм
(19М| особен во полччркмвад выравнивающее действие потока генов» (571).
Изложенная выше точка зрения была довольно широко рас-
пространена средн эволюционистов, включая и меня самого
[281. 293]. Однако аа последнее время ряд исследователей вы
сказались и против нее,
Эрлих и Рейвн [196] утверждают, чго «ие следует представ-
лять себе вид как эволюционную единицу, объединяемую свя-
зующими силами потока генов, (потому что] в природе поток
генов гораздо более ограничен, чем обычно принято считать...»
На это указывают и другие авторы [195. 197, 660. 753].
Эти авторы основывают свои возражения против традицион-
ной точки зрении иа данных об относительно ограниченном по-
токе генов и зачастую небольших размерах соседств, о которых
мы говорили и гл. 2. Эта груши данных, несомненно, имеет
отношение к рассматриваемому вопросу.
Обратившись вновь к этой проблеме в 1980 г. [298], я при-
шел к выводу, что ив приверженцы традиционной точки эреиня,
яя их нынешние противники нс привели достаточно убедитель-
ных данных в пользу своих высказываний. Сторонники тради-
цжжиых взглядов сделали несколько правдоподобных, ио до-
вольно смелых допущений, которые необходимо подкрепить
более твердыми данными. Традиционная точка зрения и в са-
мом деле уязвима.
В спою очередь в аргументации, приводимой критиками тра-
диционных взглядов, есть два слабых места. Во-первых, эти
критики рассматривают процесс расселения, ограничиваясь рам-
ками лишь одного поколения. Между тем, изучая поток генов в
масштабах географических рас и видов, необходимо рассматри-
вать более обширный промежуток времени, охватывающий мно-
го поколений Во-вторых, цитированные выше авторы придают
слишком большое значение расселению на небольшие расстоя-
ния, которое, безусловно, представляет собой наиболее эероят-
Гл. 4.
6t
ное течение событий; однако они преуменьшают роль событий»
сопровождающих расселение па далекие расстояния, которые
также могут иметь биологическое значение
Как же обстоит дело в настоящее время? Приходится при-
знать, что окончательного решения пока нет. Имеющиеся дан-
ные можно истолковать по разному. Я наложу здесь краткие
итоги проведенного мною недавно анализа этой проблемы [298].
Рассмотрим сначала изменение во времени расстояния, на
которое происходит расселение, для организмов с противопо-
ложными способностями к расселению. Это изменение зависит
от двух факторов, среднего расстояния, на которое происходит
расселение за 1 поколение и продолжительности генерации
Расстояние, на которое происходит расселение у Drosophila
pseudoobscura и сс близких родичей в Калифорнии, составляет
в среднем 361 м. В природе у этих мух появляется примерно по
7 поколений я год С учетом этих двух факторов можно счи-
тать обоснованной оценкой расселение на 2527 км за 1000 лет.
Это расстояние сопоставимо с длиной или шириной области рас-
пространения этого вида в западной части Северной Америки
Сравните эту величину с величиной расстояния, на которое
способно расселяться какое-либо многолетнее травянистое рас-
тение Liat г is aspera расселяется в среднем за одно поколение
па 3.5 ы» а продолжительность генерация этого растения состав-
ляет 5 лет [481]. Соответственно кумулятивное расстояние, на
которое происходит расселение, составит всего 0,7 км за
1000 лет. или 7 км за 10000 дет. Это расстояние соответствует
лишь очень незначительной части ареала вида. Продолжнтоль-
иость существования вила может оказаться недостаточной для
его широкого расселения.
Приведенные выше основные параметры расселения типичны
дли этих двух групп организмов; сходные оценки имеются дли
других видом дрозофилы и для других многолетних травянистых
растений (см. [298]). Эти два организма представляют дзе
крайности: подвижные мухи с высокой способностью к расселе-
нию и прикрепленные к субстрату многолетние растения с низ-
кой способностью к расселению. В одном случае за период в
1000 или 10000 лет может произойти расселение на большие
расстояния; в другом случае это. по-видимому, невозможно.
Хотя расселение представляет собой физическую основу по-
тока генов, эти процессы не идентичны, к их не следует смеши-
вать. Пас здесь интересует поток генов во времени нли, точнее,
то. что мы называем генетически эффективным потоком генов
во времени. Это обмен генами между популяциями, различаю-
щимися по частоте аллелей, который происходит на протяжении
двух или большего числа поколений н п котором участвуют трн
или более популяции.
52 Часть поры. Природа айда*
Между расселением во времени и генетически эффективным
потоком генов есть одно важное различие. В первом случае
-суммарное расстояние. па которое происходит расселение, ли-
нейно возрастает со временем. Но во втором случае частота
иммигрирующих генов быстро снижается на каждом этапе и в
каждом поколении, пока в конце концов процесс генетически
эффективного обмена генами не Прекращается вовсе |2?8].
Рассмотрим гипотетически# пример. Допустим, что популя-
ции I, II, III и IV распределены по трансекте, пересекающей
.ареал данного вида с запада на восток. В популяции I имеется
новый селективно нейтральный аллель Ля. частота которого
равна 100%. Популяции II, III и IV содержат прежний аллель
Л, с частотой 100%. Обмен генами между соседними популяция-
ми происходит в обоих направлениях со скоростью т—0.10.
Ъ таком случае частота («) аллеля А* в этих четырех популя-
циях по прошествии трех поколений составит:
Популяцяи I 9.-0,755
Популяция П о,-ОД 10
Популяция III 9,-0.025
Популяция IV 9,-0.001
Частота нового аллеля резко снижается на каждом этапе
-короткого ступенчатого процесса иммиграции. На каком-то
этапе этого Процесса аллель А, становится настолько редким,
-что вероятность его включения в следующую выборку мигрантов
ничтожна. На этом этапе процесс генетически эффективного
обмена генами прекращается. Расселение как таковое продол-
жается. но генетически эффективный поток генов иссякает
1т<ли бы генетически эффективный обмен генами продолжал-
ся на протяжении очень многих этапов, то очевидно, что ему
была бы необходима мощная поддержка со стороны отбора.
В таком случае яам следовало бы изменить свое допущение,
предположив, что аллель А, селективно ие нейтрален, а сохра-
няется отбором. При этом частота аллеля Аз могла бы позрас.
тать на каждом этапе миграционного процесса, таи что вероят-
ность его включения в следующую выборку мигрантов была бы
достаточно велика. Но повышение частоты того или иного алле-
ля под действием отборе требует времени. На протяжении
.ряда последовательных поколений генетически эффективный об-
мен генами, создаваемый совместным действием миграции и
-отбора, будет протекать гораздо медленнее, чем расселение как
таковое.
Гл 4. БиологткклЛ вид
4»
Рассмотрим вновь пример с дрозофилой в свете этих сооб-
ражений. Как мы уже видели, такой высоко подвижный орга-
низм с коротким временем генерации, как дрозофила, может
Расселиться во области, соответствующей видовому ареалу. за
ООО лет; ио для того чтобы генетически аффективный поток
генов распространился по той же области с учетом действия от-
бора на каждом этапе, вполне может потребоваться в 50 или
даже в 100 раз больше времени. Такие периоды времени при-
ближаются к историческому возрасту данного вида
Вплоть до этого момента все ваши рассуждения свидетель-
ствуют в пользу аититрялиинониой точки зрения. Существуют
серьезные сомнения относительно того, может ли лоток генов
оказывав гомогенизирующее воздействие на внутривидовую из-
менчивость и широко распространенных видовых популяциях в
течение периодов времени порядка 10000 лет. Эффективный
лоток генов в областях, соответствующих видовому ареалу, стал-
кивается с затруднениями даже у высокоподвижиых видов с
коротким временем генерации Но особенно остро стоит эта
проблема для прикрепленных к субстрату многолетних растений
с длительным временем генераций и ограниченной способностью
к расселению.
Однако, прежде чем так легко отбросить традиционную точ-
ку зрения, следует обратить внимание на другую группу дан-
ных. свидетельствующих о том, что поток генов оказывает влня-
нис иа характер видовой изменчивости. В популяционных систе-
мах с бнларектальиым размножением, состоящих из множества
географических рас, наблюдается большая однородность в пре-
делах обширных областей при непрерывной структуре популя-
ииоииой системы, чем при разорванной (см. гл. 3). Это разли-
чие коррелирует с наличием или отсутствием открытых кана-
лов для обмена генами. Кроме того, в бтптарентальпых популя-
ционных системах однородность внутривидовой изменчивости
гораздо выше, чем в унипарептальиых (см. гл. 1 и 5) И снопа
это различие коррелирует с иаличиеи или отсутствием эффек-
тивного механизма для обмена генами.
Причины несоответствия между двумя группами данных не-
известны. Быть может, было бы полезно вновь рассмотреть дан-
ные по расселению. Методы, используемые при количественных
исследованиях расселения, применимы лишь на ограниченных
участках. Эти методы не позволяют выявить события, связан-
ные с расселением иа большие расстояния, что. возможно, ведет
к недооценке средних расстояний, па которые происходит рас-
селение. Внесение в существующие оценки поправок, учитываю-
ших некоторое расселение на большие расстояния, могло бы
способствовать примирению этих двух групп фактических
данных
<4 Часть сгорая, ПрироР* «ьйол
Виды-двойники
Наиболее существенная черта биологических видов, как мы ви-
дели. связана с их взаимоотношениями при скрещивания. Спо-
собность особе» успешно обмениваться генами, т. е. свободно
скрещиваться между собой и производить плодовитое и жизне-
способное потомство, характеризует их как представителей
одного и того же биологического вида, тогда как неспособность
к свободному н успешному обмену генами — признак обособ-
ленности биологических видов. Все остальное, в том числе и
морфологические различия. — это надстройка.
Родственные биологические виды действительно отличаются
друг от друга по внешним морфологическим признакам, равно
как и по лежащим в их основе физиологическим признакам, по-
скольку многие морфологические признаки входят в состав
адаптивных комбинаций признаков соответствующих видов. Три
дициоиные таксономические методы классификации и определе-
ния основаны на этой обычной ассоциации внешних морфологи-
ческих признаков с физиологическими особенностями.
Но такая корреляция не всегда выдерживается. Известны
случаи, когда хорошие биологические вилы морфологически аб-
солхпяо неразличимы. Такие виды получили название видов-
двойников.
Классическим примером видоп-даойинков служат Drosophi-
la pseudoobscura и О perslmilis. Прежде считалось, что эти мор-
фологически сходные мухи принадлежат к одному виду—
D. pscudoobscura, одиако в дальнейшем при некоторых «внутри-
видовых» скрещиваниях обнаружилось, что скрещиванию меж-
ду ними препятствует стерильность гибридов Затем оказалось,
что эти интерстерильные группы образуют снмпатрическхе по-
пуляции на обширных территориях в западной части Северной
Америки, не гибридизируя между собой, н в соответствии с этим
онн были признаны отдельными видами.
После поисков, предпринятых рядом исследователей, были
в конце концов обнаружены небольшие морфологические раз-
личия в строении половых органов и крыльев у самцов, а также
некоторые поведенческие, физиологические и цитологические
различия (обзор см. [569]). Эти различия очень невелики, в их
трудно выявить обычными таксономическими методами.
Между тем мухи легко узнают друг друга. Когда самцов и
самок Drosophila pseudoobscura и D perstmilis помещают вмес-
те в популяционный ящик в лаборатории, скрещивания проис-
ходят исключительно или преимущественно между особями,
принадлежащими к одному и тому же виду; и лишь в редких
случаях в обычных лабораторных условиях самки и самцы раз-
ных видов копулируют друг с другом. Подобным же образом
Гл. 4. Биологический вид ©>
в природе, где очень много возможностей для гибридизации,
особи, принадлежащие к разным видам, спяривлются крайне
редко, и межвидовые гибриды неизвестны 1170). Очевидно, дро-
зофилы умеют различать виды D pxttuioobstura и D perximitit
независимо от того, умеют ли это делать систематики
Виды двойники существуют и в других комплексах видов
дрозофилы, и во многих других группах животных (обзор см.
[569]). Внды-даийникн встречаются также во многих родах пыс-
ишх растений {279J, причем у последних это явление часто со-
четается с полиплоидией.
Тетра плои дни й вид е геномом ЛА SB морфологически сходен
и может интерградировать с предковым диплоидным видом АА
по двум причинам. Во первых, что самое главное, тетраплоид-
нын вид содержит Л-геном Во-вторых. тетрвплоид. вероятно,
будет мы теп л ять некоторое число особей родительского типа 4,
особенно если это сегментный аллотетраплопд. По тем же при-
чинам этот тетраплоид будет сходен со своим вторым диплоид-
ным родительским видом ВВ. Вследствие различим в строении
хромосом диплоидные виды АА и ВВ образуют при скрещива-
нии более или мепее стерильных гибридов F|, гибриды между
тетраплондыым видом ААВВ н любым нз диплоидных роди-
тельских видов, будучи трнплондамп, также стерильны. Таким
образом, популяция. которую по морфологическим признакам
можно было бы принять за непрерывно пнтерградирующую по-
пуляцию и которую систематик мог бы рассматривать как еди-
ный вид, представляет собой на самом деле группу из трех ин-
терстер пльпых видов-двойникои [283].
В качестве иллюстрации приведем группу видов Gilia Irons-
montana. которую тщательно проанализировала Дэй ||5В).
В эту группу входят мелкие однолетние травянистые растения,
встречающиеся в пустыне Мохаве п на прилегающих к ней тер-
риториях в западной части Северной Америки Это преимущест-
венно автогамные растения, однако у пих происходит некоторое,
во тем не менее биологически значимое перекрестное опыление
при помощи насекомых. Общий вид этих растений показан на
рве. 4.5 и 4.6.
Группа Gilia transmonfana состоит не из трех, а из пяти род-
ственных диплоидных и тетраплопдных видов. Озим из тетра-
плоидиых видов — G trans топ tana происходит от двух дип-
лоидов — G, minor и G clolwyi, а другой тетраплоид — G. та
Ног — от днплондов G. minor н G aiiquanta {158]. Таким обра-
зом, у обоих тетраплоидпых видов есть одни общий диплоидный
предок (О Mitor). Все эти пять видов в ранних таксономиче-
ских работах рассматривались как разные формы одного вида.
Результаты опытов по искусственной гибридизации свиде-
тельствуют о том, что эти пять видов, сами по себе вполне
5--4S
« Чисг» агирил. Природа «Лч
плодовитые, высоко ввтерстерильны в любых комбинациях.
В природе они симпатричны н растут в самых различных соче-
таниях. не скрещиваясь между собой [158. 283. 30CJ.
Дэй [158] выращивала и сравнивала ряд линий, представ-
ляющих известный диапазон изменчивости каждого вида, в од-
нородных условиях среды в вегетационном домике. Она обнару-
жила. что эти виды различаются, не очень сильно, ио различия
между ними постоянны и затрагивают почти все части расте-
ния (см. рис. 4.5—4,7). Таи, Cilia allquanta отличается своими
несколько более крупными цветками, но у остальных четырех
видов цветки о дика косо мелкие (рис. 4.6). У С. minor семена
мельче, чем у других видов (рис. 4.7). Листья, окраска венчика,
коробочки и другие признаки характеризуются незначительны-
ми межвидовыми различиями.
Одиако эти внешние фенотипические различия нс дают воз-
можности проводить идентификацию во всех случаях. Вернем-
ся еще раз к величине семян. У Cilia minor вес семян колеблет-
Рве. 4 5. Форма роста у трупы валов-лвоЗкикоа рада GUia [158].
Гл 4. БчшапюаЛ гид
St
Рж 4 в Цвет»» пят» апдо»-ис4анм>в СЛо, <и«Лраж»«>ш и рве. 4 5
415*1
А — G. eJoAepf. Б — О. transmonlana. В —О minor Г— 6 malior Д — С. <Ш-
4М>п/а
Рис 47 Семена те» же пят» «клсж-лвоАнюпв GtUa (IS*).
Л — G. eto*e»it Б — G Irmunontana. В —0. minor. Г—О. malior. Д-GaU-
«впив
ся от 10 до 20 мт, у G. transmontana—от 20 до 30 иг, у б do-
bryi—от 30 до 85 мт. Эти три вида различаются по средней ве-
личине семян (рис. 4.7). во образуют непрерывный ряд, если
рассматривать диапазон изменчивости нескольких видов (1581.
В общем GUia transmonlana заполняет морфологический
разрыв между своими диплоидными предками О. minor и
О cloieyi, так что ясно выраженные рааличия между этими
тремя видами найти трудно. Аналогичным образом G. malior
яитерградирует с G. minor И б. allquanta. А поскольку в
G transmoniana, и б malior содержат геном G. minor, оба
5»
Ь8 Чапа ргорая Природа оидоо
тстраплондиых вида имеют много общих морфологических
признаков.
Проблемы таксономической идентификации в группе Gilia
transmontana не имеют, однако, отношения к вопросу о биоло-
гическом составе sroh группы. Cilla transmonlana, G. malior,
О minor, G. cioktyi H G. ahquania образуют пять отдельных
скрещивающихся групп и предс1авллют собой, таким образом,
хорошие биологические вилы независимо от того, можно ли во
всех случаях идентифицировать их но внешним морфологиче-
ским признакам или пет.
5 Видовые единицы
у унипарентальных растений
Концепция биологического вида относится к бимаренгальным
организмам. Опп нс распространяется иа у пипа ректальные ор
ганнзмы. не образующие Скрещивающихся групп Между тем
уннпаректальное размножение обычно у растений, а также в
разных группах, принадлежащих к другим царствам. поэтому
необходима разработать параллельные пли альтернативные
концепции вила, приложимые к единицам изменчивпетн п этих
унипарентальных группах.
Системы унипарентальных популяций
Уннпаренталъные организмы образуют популяции и обширные
системы популяций. Популяции состоят из генотипически сход-
ных особей. Генотипическое сходство обусловлено происхожде-
нием от общих предковых линий и усиливается иод действием
стабилизирующего отбора в данной нише или данном местооби-
тании. Эти два фактора — общие предки и стабилизирующий
отбор — приводят к тому, что родственные особи в своей среде
определенным образом интегрируются, образуя популяции (574,
4
Итак, между унипарептальнымн и бнпарептааьнимн организ-
мами существуют как черты .сходства, так и различия в отно-
шении характера их популяций и популяционных систем.
И у упнпареигальиых, и у бипарентальпых организмов наблюла*
ются следующие процессы- I) наследование одинаковых генов
пт общих предков. 2) распространение определенных генов но
всей папуллиионпой системе или виду в результате миграции
и естественного отбора, 3) подавление распространения утих
генов в другие виды, обусловлениос нх неспособностью мигри-
ровать в другую среду или воспроизводиться в ней. Под дейст-
вием всех этих процессов родственные особи, несущие одинако-
вые гены, объединяются в популяции, обитающие в одной и той
же среде, независимо от того, размножаются тги особи половым
Путем или нет (574. 732].
Пппулянии организмов, размнпжяющкхся половым путем н
при участии двух родительских особей, связаны между собой
двумя дополнительными процессами, отсутствующими н строго
унниарентадьных группах. Один из этих процессов — скрещива-
ние между особями, облегчающее распространение генов в по-
пуляционной системе. Другой процесс репродуктивная нзоля
пня препятствующая распространению этих генов в другие ви-
лы [732].
70 ¥«т» «терм. Природа «иди
Степени интеграции
Существомшие сил, интегрирующих популяции и присущих
только бяпареитальиым организмам, заставляет думать, что
внелогические виды представляют собой или могут представлять
более интегрированные единицы, чем популяционные системы
уиипарентальных организмов. Можно сказать, что особи, при-
надлежащие к одно* и той же популяции и одному н тому же
виду, у уиипарентальных организмов связаны узами происхож-
дения от общей родительской особи, а у бкпаректальных — уза-
ми общих родителей, а также узами скрещивания. Сцепления,
возникающие в результате перекрестного оплодотворения, слу-
жат фактором интеграции у популяций и видов бипарситальиых
организмов, отсутствующим в уиипарентальных группах.
Фактически дискретные группировки видов обычно различи-
мы у организмов с половым размножением (примером служит
большинство родов к семейств у млекопитающих и ппш), но
явно отсутствуют во многих уиипарентальных группах расте-
ний, примером которых служат крупные группы видов таких ро-
дов, как Rubus и Hieracium.
Кроме того, как было показано в гл. I, проводя различия
между бипарентальным и уинпаректальным размножением, мы
не отражаем в полной мере существующее положение вещей.
У известных нам видов растений наблюдается целый спектр
систем размножения —от широкого аутярогемига на одном кон-
це спектра через всевозможные промежуточные состояния до
строго унипареитальиого размножения иа другом его конце. От-
сюда следует, что степень интеграции, достигаемая у этих видов,
также варьирует, в притом коррелятивно: от максимальной у
свободно скрещивающихся видов до относительно рыхлой струк-
туры в преимущественно уиипареитяльямх группах.
Иными словами, если, следуя Мегличу (574] и Симпсону
1732). считать, что видовая структура существует как у бипареи-
тальных, так и у унипареитальиых организмов, то возникает
необходимость различать разные уровни интеграция у видов с
различной биологией размножения. Хорошо интегрированные
биологические виды — одно из состояний, наблюдаемых в при-
роде. От этого состоянии имеется постепенный переход к раз-
личным промежуточным уровням интеграции, существующим у
многих видов высших растений с частично, преимущественно
или исключительно уиипарентальиым размножением.
В системах популяций бипаректальных организмов изменчи-
вость имеет иерархическую структуру: группы локальных рас
объединяются в географические расы, а группы географических
рас —в биологический вид. В системах популяций унипарен-
тальяых растений аналогичной иерархической структуры измен-
Гл. 5 Budjtuf ^ынаци д уныларекпмчых ратчиЛ 7»
чнвости кс наблюдается. Вместо этого у них обычно можно ви-
деть ряды морфологически различных единиц. или микровидов,
одного п того же ранга. Эти микровидн часто группируются
вокруг одного или нескольких предковых видов с половым
размножением.
Микровиды
Термином михровид (или жорданон) обозначают в ботанике
любую однородную популяцию, которая морфологически слегка
отличается от родственных ей однородных популяций. В такой
трактовке этот термин приложим к обширному кругу популя-
ций. оказываясь синонимом чистых линий, иекоторых локальных
рас, а иногда видов-двойников нт п , и его действительно ис-
пользуют в этих н других случаях. Поскольку этот термин прп
менялся широко и достаточно свободно, он. естественно, пере-
стал быть точным и его заменили другими, более строгими тер-
минами.
Однако в практике применения термина мнкровнд есть одна
особенность, ради которой его следует сохранить. Ботаники
обычно используют этот термин для обозначения слаболнффе-
ре и цирова иных популяций, относящихся к группам растении с
унипвренгальным размножением.
Так, микровиды описаны в автогамной группе Erophila verna
и в агамоспермией секции Moriferus рола киЬих. Eropltlla verna
н секция Moriferus рода Rubus, конечно, очень сильно различа-
ются по систематическим связям, жизненным формам и систе-
ме размиожении. На фоне таких очевидных различий между
этими двумя группами сходство их ио характеру изменчивости,
достаточное, чтобы оправдать применение к ним обшей концеп-
ции михрпнилпв, прн<5бретает важное значение. Кроме того,
этот общий элемент в характере изменчивости, к которому vwk-
но приложить термин мнкровнд коррелирует с двумя apyiUMK
общими свойствами, а именно с естественной гибридизацией я
упнпарентл.'тьиым размножением.
Не все ботаники признают эти общие свойства или хотя бы
знают о них Это особенно справедливо в отношении тех ис-
следователей, которые впервые стали применять термин микро-
вид: использование ими этого термина служит примером выде-
ления, в значительной мере интуитивного, некой популяцион-
ной единицы в ряду изменчивости известного типа Тем самым
была создана предпосылка для формулировки более опрсде
ленной концепции микровида.
Микровиды определяются здесь как популяции, принадле-
жащие к преимущественно уинларппальным группам растений.
« обшей довольно однородным и лишь-незначнтелыю отличаю-
Л HaiT» вторая ПрароЬа виЗов
шямся одна от другой морфологически. Их распространение не-
редко бывает ограничено относительно небольшой географичес-
кой областью, хотя »то н не обязательно. Они часто имеют гиб-
ридное происхождение (прямо иди косвенно) и часто навсегда
сохраняют гибридную конституцию.
В зависимости от способа воспроизведения микровиды де-
лятся на четыре главных класса: 1) клональные мнкровнды. вос-
производящиеся путем вегетативного размножении, например
Opunlia. 2) агамоспермные мнкровнды. размножающиеся при
помощи агамоспермии, например секция Moriferus рода Rub us.
3) гетерогамные мнкровнды, воспроизводящиеся при поиоши
таких генетических систем, как у Oenothera biennis или Rosa
canlna; 4) автогамные микровнды, которые преимущественно
автогамны н гомозиготны по хромосомам, как у Erophila.
Следует указать, что микровиды классов 1 к 2 размножа-
ются бесполым путем, а классов 3 и 4 — половым.
Автогамные мнкровнды в большинстве случаев не исключи-
тельно автогамны; частично у них наблюдается ауткроссинг и,
кроме того, нормальный половой цикл. Следовательно, такие
автогамные михровяды представляют собой один из типов био-
логических видов. Концепции автогамных микроандов, к которой
мы здесь подошли с одной стороны, в значительной мере сов-
падает с концепцией биологических видов двойников, изложен-
ной в гл. 4. Дальнейшее обсуждение видов-двойников в авто-
гамных группах растений дано в гл. 11.
Проблема вида у клональных н агамосперм кых микровкдов.
не способных к половому размножению и не обладающих свой-
ствами биологических видов, осложняется в еше большей сте-
пени. чем у автогамных микровидов. У гетерогамных мнкрови
дов, также обладающих гибридной конституцией, хотя они и
размножаются половым путем, также возникают сложности в
связи с проблемой вида. Примеры клональных, агамоспермных
и гетерогамных микроандов описаны в соответствующих главах
восьмой части.
Гибридность и микровнды
Гибридность обычно ассоциирована с микровидами Растения,
принадлежащие к агамоспериным и гетерогамным мнкровилам.
представляют собой перманентные гибриды. Они обычно высо-
ко гетерозиготны и в случае клональных михровидов могут быть
межвидовыми гибридами. Автогамные мнкровнды в отличие от
микровидов трек остальных классов не сохраняют гибридной
конституции н могут быть полностью или почти полностью го-
мозиготными: однако. подобно другим микровидам, оин часто
возникают хак продукты гибридизации.
Гл. S. Видовые е&лшуы ц ржи-хий 73
В случае гибридных микровидов гетерозиготные индиаклу-
умм создают однородные популяции в результате апомиксиса
или гетерогамии В процессе своего возникновении мнкроекд
проходит через стадию, когда его распространение сильно огра-
ничено; а случае же успешной эволюции микровкд Достигает
стадии широкого географического распространения. Процесс
роста популяции ог гибридного индивидуума через небольшой
K3IOII и эндемичный микровид до широко распространенного ми
кроаида протекает непрерывно. В некий нс поддающийся опре-
делению момент иа поздних стадиях этого постепенного про-
цесса клональный, агамоспермный или гетерогамный мнкровнд
приобретает право рассматриваться как таксономически само-
стоятельная единица.
Естественная гибридизация между двумя или несколькими
видами, прежде размножавшимися половым путем, за которой
следует уикпареитальное размножение образовавшихся гибри-
дов и жизнеспособных вариантов, вышепляющнхея в дальней-
ших поколениях, приводят к формированию многочисленных
михроендов. В группах растений, обладающих этим сочетанием
эволюционных факторов, обычно можно обнаружить ряд как
широко распространенных, так н эндемичных микровидов, ко-
торые в процессе филогенеза наложились на исходные биоло-
гические виды, затушевывая существующую между ними пре-
рывистость.
В таких группах растений изменчивость не ограничена опре-
деленными рамками, соответствующими границам вида, по ох-
ватывает два или несколько родительских видов и все возник
шне от них мнкровиды. Подобные группы растений создают
огромные таксономические трудности Биологическая коицепипк
вила иепрнложима к такой группе как целому и непригодна
дли нее. Альтернативные таксономические концепции вида на
практике также оказываются песостоятельнымн. Причина этих
неудач заключается в том, что рассматриваемые группы расте-
ний—это смесь, состоящая обычно из нескольких исходных
биологических видов и многочисленных происходящих от них
мнкровидов.
Кроме того, это смесь, которую нельзя с помощью обычных
гаксоаоммчеекнх методов разделить на составные элементы.
Примеры таких гибридных комплексов, состоящих из биологи-
ческих видов н мнкровндов, будут приведены в главах, отно-
сящихся к восьмо! части книги.
Пересмотр вопроса о гомогенности вида
Унипаренталъмые растения могут пролить некоторый свет на
•опрос, обсуждение которого не было доведено до конца в пре-
74 Часть сгорал. Прлрода радов
дыдушей главе, л именно на вопрос о роли потока генов я скре-
щивания в усилении гомогенности. Как уже говорилось ранее
(см. гл. 4), известные нам скорости потока генов слишком ма-
лы, чтобы этот поток мог оказывать существенное гомогенизи-
руюшее воздействие на изменчивость многолетних растений со
свободным неродственным скрещиванием на больших площадях
в течение периодов в 10000 лет.
У уннпареитальных растений уровень как обмена генами, так
и скрещивания, конечно, гораздо ниже, чем у растений со сво-
бодным неродственным скрещиванием. Тем не менее гомоген-
ность наблюдается и у мнкроикдов с обширным ареалом. Что
же определяет гомогенность у уиипареитальиых растений?
Сходство индивидуумов в пределах отдельных видов не обя-
зательно приписывать воздействию потока генов и скрещива-
ния. Существует другое объяснение, которое, несмотря на его
очевидность, часто упускают из виду, а именно рост численно-
сти индивидуумов, происходящих от общего предка.
Рост численности популяции происходит как у бнпареяталь-
кых, так н у унипареитальиых организмов. Отличить эффекты
роста численности от эффектов, создаваемых потоком генов у
бипареитальных растений, трудно. Но у уяиварсктальных расте-
ний сходство между индивидуумами должно быть обусловлено
главным образом происхождением всей популяции от одного
общего предка (в отличие от моих прежних взглядов; см. [293]).
Репликативный характер воспроизведения, быть может, в зна-
чительной мере обусловливает гомогенность видовой популяции
также и у бнпарентальных растений, хотя у них роль этого
фактора по сравнению с ролью потока генов остается неизвест-
ной [298].
6 Вопрос о виде у растений
У высших растений наблюдается чрезвычайно широкий диапа-
зон типов организации родственных особей в популяционные
системы. В царстве растений, как и в царстве животных суще-
ствуют хорошо интегрированные биологические виды, однако
это не единственная форма организации популяций у растении.
У них имеются также большие и разнообразные популяционные
системы иа уровне, таксономически соответствующем секции
или подроду, причем эти системы не подразделяются на четко
обособленные виды. Кроме того, у растений встречаются виды,
занимающие по уровню интеграции промежуточное положение
между этими двумя крайними ситуациями.
Далее в этой главе мы займемся следующими вопросамн с
какой частотой встречаются у растений эти различные типы
организации вида? Какие факторы определяют их возникиове
ине?
Для того чтобы найти подход к решению этих вопросов, не-
обходимо установить некий эталон для сравнения, В качестве
такого эталона удобно использовать ряд дискретных к хорошо
интегрированных биологических видов, наблюдаемых у многих
ттли даже у большинства высших животных, например в семей
стве кошачьих, после чего можно рассматривать сходства с этим
Эталоном или отклонения от исто, обнаруживаемые у растений
У растений отклонения от организации, типичной для хорошо
развитого биологического вида, принимают обычно две формы.
У бипарентальных растений между двумя или несколькими
скрещивающимися группами может существовать репродуктив-
ная изоляция, занимающая промежуточное положение между
уровнями изоляции, характерными для биологических видов п
рас. Млн же бнпареитальное размножение может быть заме-
щено каким-либо способом укнпарепталыюго размножения.
В том и другом случаях отклонения могут колебаться от уме-
ренных до чрезвычайно резких. И соответственно этим двум
формам отклонений от принятого нами эталона в их крайних
выражениях мы различаем два главных типа <невидоо>. а имен-
но — полувиды и мнкровиды. Мнкровнды были рассмотрены в
предыдущей главе, а здесь мы рассмотрим полупили.
Как будет показано в дальнейшем, такие <нсвиды» (потире
sic») не столь уж необычны, во всяком случае в некоторых фло-
рах, по сравнению с биологическими видами. Их весьма широ-
кое распространение приписывают отчасти природе процессов
видообразования, а отчасти основным характеристикам самих
растений.
П Часть вторая Прирова шбое
Полувиды как результат постепенного
видообразования
Обычный способ видообразования у растений и животных с пе-
рекрестный оплодотворением Состоит в постепенной диверген-
ции двух или нескольких популяционных систем. На ранней
стадии дивергенции вти популяционные системы ведут себя как
географические или экогеографические расы. Они занимают
соседние территории, свободно скрещиваясь друг с другом и
интерграднруя в зовах контакта. На поздней стадии диверген-
ции взаимоотношения, между этими популяционными системами
приобретают характер взаимоотношений между снмпатрнческн-
мн видами, обитающими на одной н той же территории, ио не
скрещивающимися друг с другом. Между ранней и поздней
стадиями дивергенции и репродуктивной изоляции наблюдают-
ся различные промежуточные стадии, когда популяции не пред-
ставляют собой нн хороших рас, ни хороших видов, ио связаны
Друг с другом благодаря незначительному скрещиванию и сла-
бому обмену генами. Популяционные системы, находящиеся на
таких промежуточных стадиях дивергенции, были удачно назва-
ны полувидами (обзор и библиографию см. [282]).
Процесс географического видообразования будет рассмотрен
в гл. 13. Здесь же нас интересует один из результатов этого
процесса. Постеленная н непрерывная эволюционная диверген-
ция от географической расы до симпатрического вида неизбеж-
но приводит к тому, что в любой данный момент можно обна-
ружить несколько популяционных систем, находящихся на про-
межуточной стадии полувидов.
Во многих родах растений мы находим дизъюнктивные алло-
патрические популяции, которые занимают, каждая в пределах
своего ареала, сходные экологические местообитания к морфо-
логически дифференцированы до такой степени, что системати-
ки присваивают им разные видовые названия, ио которые при
экспериментальных скрещиваниях оказываются илтерфертиль-
ными, Примером служат Catalpa bienonuMes из восточной час-
ти США и С. ovata из Китая [778f Другой пример — РШапиз
ractmosa и Р. vrighlii, произрастающие соответственно в оро-
шаемых горными потоками долинах Калифорнии в в каньонах
Аризоны. Arbutus nenzieiii и A. aritoniea, произрастающие в ле-
систых районах Калифорнии и Аризоны, разделенных обширной
пустыней, служат третьим примером; в данном случае неизвест-
но, дают ли они при скрещиваниях плодовитое потомство нлн
нет. В этих и других аналогичных случаях отдельные популя-
ционные системы уже нельзя считать' расами, ио еще нельзя
рассматривать как биологические виды. Они находятся на по-
лупидооом уровне дивергенции.
Г» б Вопрос о тбе у ратнвЛ
Т7
Интересную промежуточную ситуацию представляют собой
перекрывающиеся кольца рас. Такая ситуация возникает в тех
случаях, когда ряд рас иитертрадирует по какой-либо географи-
ческой трансекте. Терминальные расы такого ряда, различаю-
щиеся евльпее всего, сосуществуют симпатричсски, ие скрещи
ваясь одна с другой и сколько-нибудь значительной степени.
Таким обратом, у одной и той же популяционной системы в
разных частях ее ареала проявляются черты, свойственные ра-
се или виду. Наиболее хорошо изученные примеры кольцевого
перекрывания рас относятся к птицам и другим животным (об-
зор см. (569J). Среда растений, принадлежащих к флоре запад-
ных районов США, подобные примеры обнаружены, в частно-
сти. в следующих группах видов: Gilia capttala. Ipomopsis aggre-
gate Aquilcgia. chrysantha, Mimuius longijlorus в Pinus ponaero-
sa ([54. 266, 268, 270. 334], а также Grant, неопубликованные
данные).
Mimuius puniceus (Scrophulariaceae) встречается в прибреж-
ной зоне на юге Калифорнии, а М. tongiflorus— в удалей них
от побережья горах (рис. 3.2). В южной части Калифорнии, иа
обширном пространстве от береговой линии до расположенных
дальше от моря гор, между этими двумя единицами происходит
полная интерградации (см. гл. 3). Однако в тех частях гор
Санта-Ана, где местообитания, пригодные для М. puniceus и
М. longijlorus находятся очень близко друг от друга, популя-
ции этих двух видов сосуществуют, не интерграднруя ,54]. Таким
образом. М puniceus и М. longi/torus ведут себя во многих об-
ластях как расы, а в некоторых местах как снмпатричесхие
виды.
Полувиды как результат естественной
гибридизации
Процесс эволюционной дивергенция ие обязательно неуклонно
идет лишь в одну сторону. Он обратим в довольно втирохих пре-
делах, поэтому полувиды могут возникать ие только как еди-
ницы, находящиеся на пути превращения в виды, по также и
как продукты гибридизации между видами, которые прежде
были хорошо изолированы.
Несмотря на то что биологические виды интерстеркльны и
хорошо изолированы в других отношениях, между ттими могут
происходить естественная гибридизация и обмен генами, до тех
пор лона эффективность барьеров, препятствующих скрещива-
нию, ие достигнет 100%. Эти барьеры между родственными
видами растений, принадлежащих к одному и тому же роду
или секции, в сущности, часто могут быть до известкой степени
78
Част» порах ПрипСа saioo
разрушены. Нередко можно встретить два симпатрических ва-
да. различающихся и не скрещивающихся между собой иа боль-
ших пространствах, ио образующих гибриды в одной нли не-
скольких местностях.
Межвидовая гибридизация — потенциальная угроза целост-
ности родительских видов как отдельных скрещивающихся
групп. Превратится ли эта потенциальная угроза в свершив-
шийся факт или иет — вопрос Другой. Межвидовая гибридиза-
ция может привести к разнообразным конечным результата»
(см. часть пятую). Некоторые из иих результатов не нарушают
обособленность участвующих а гибридизации видов, а поэтому
в данный момент они нас не интересуют.
Здесь мы рассмотрим образование путем гибридизации но-
вых популяций, позволяющих преодолеть барьеры к скрещияа-
нию. морфологический разрыв и экологическую дифференциа-
цию между первоначально обособленными родитепьскнмн вида-
ми. Такне связующие популяции гибридного происхождения де-
лятся на два класса: скопления гибридов (hybrid swarms) в
интрогрессивные популяции. Их образование поворачивает про-
цесс видообразования вспять и, если гибридизация происходит
в достаточно широких масштабах, превращает виды, бывшие
прежде самостоятельными, п полувиды.
Скопление гибридов представляет собой сложную смесь ро-
дительских форм, гибридов Fi, особей, возникших в результате
возвратных скрещиваний и расщепления. В таких скоплениях
наблюдается, как и следует ожидать, очень высокая степень
индивидуальной изменчивости В литературе описано много при-
меров скоплений гибридов, но мы приведем здесь лишь некото-
рые из них.
Скопления гибридов между Geum rivals и G. urbanum встре-
чаются во многих местах в Англии и Европе [230. 557]. Aquil»-
gia formosa и A. pubtscsns образуют скопления гибридов в раз-
личных зонах контакта этих двух видов в горах Сьерра-Невада
в Калифорнии [96. 270]. Колонии, образуемые тремя или четырь-
мя видами Baptisia (Leguininosae) и их гибридами, встречаются
иа берегу Мексиканского залива в Техасе [9—11].
При многократных возвратных скрещиваниях гибридов с
одним из родительских видов возникает интрогрессивная попу-
ляция, сходная с данным родительским видом, но проявляющая
изменчивость п сторону другого родительского пкда [15, 16].
Если интрогрессия заходит достаточно далеко, то она может
Привести к стиранию морфологических и экологических разли-
чий между исходными видами, которые в результате будут низ-
ведены до уровня полувидов.
В некоторые колонии Iris hexagona gigantiaurulra в дельте
Миссисипи иа юге шт. Луизиана иитрогрессирует Iris fulva. так
Г* в. Вопрос о пВс у ротяаа
79
что морфологический разрыв между нормальными незасорсн-
пыми формами этих двух видов заполняется их диапазоном из-
менчивости [686]. Хотя эти два вида Iris в большинстве облас-
тей, где они сиипатркчиы, обособлены один от другого, эго
обособление не полное и в некоторых зовах гибридизации они
иитергралиругот, соединяясь через ряд промежуточных форм. Та-
ким образом, они представляют собой полувиды гибридного
происхождения.
Определение биологического вида как наиболее инклюзив-
ной скрещивающейся группы оказывается несостоятельным в
случае естествен по гибридизирующих видов и полувидов Там.
где происходит ограниченный обмен генами между полувидами,
а остальном изолированными друг от друга, наиболее инклю-
зивной скрещивающейся единицей является не единый биологи-
ческий вид. а совокупность полувидов. Такую совокупность на-
зывают спигамсоиом. Сннгамеоп — самая обширная скрещиваю-
щаяся система в группе гибридизирующих видов. Сннгамеоп
ведет себя иа своей внешней границе как хорошо изолирован-
ный биологический вид, но отличается от него более сложной
внутренней структурой.
Сиигамсоны обычны у разных родов растений: Plans, Quer-
cus, Geum, Apuilegla, Iris н др. Примеры будут описаны в гл. 18.
Масштабы проблемы вида
У бипареитальных растений полувиды гибридного происхожде-
ния достаточно обычны, во всяком случае в некоторых флорах.
Например, в результате изучения различных родов растений со
свободным неродствеипым скрещиванием во флоре западной
части США у меня создалось впечатление, что еннгамеоиы,
состоящие на гибридных полувидов, часто более обычны, чем
хорошо выраженные биологические виды Это наблюдается у
Quprcus, Ccanothus, Diplacus, Iris и у свободно скрещивающих-
ся представителей Gilia в западных областях США. У других
родов свободно скрещивающихся растений из тех же областей,
например Finns. Aquilegla и Ipomopsis, гибридные полувиды
составляют устойчивое меньшинство.
Гибридные комплексы, содержащие многочисленные уиипа-
реитальные микровиды, столь же обычны. Комплексы агамо-
спермных микронидов обнаружены у таких северных родов, как
Antennaria, Crepis, Hieracium, Taraxacum. Роа. Ranunculus, Ru-
bus и Potentilla. Густафсон [317] перечислил 73 рода растений
Из всех частей земного шара, содержащих агамоспермные мик-
ровиды. Некоторые другие роды, в том числе Allium. Festuca и
Saxifraga. содержат унипареитальные гибридные формы, раз-
множающиеся путем ложного живорождения.
во
Часть вторая. ПраааОа ььОов
Отклонения от эталона хороших биологических видов пред-
ставляют собой, таким образом, обычное явление у высших
растений; достаточно обычны и крайне сильные отклонения, о
чем свидетельствуют случаи распада видовой организации. Во
всех досконально изученных примерах участвует гибридизация
в сочетании либо с уиипарентальным, либо с бипарентальным
размножением. В пом заключается одно из самых важных раз-
личий между растениями и многими группами высших живот-
ных, у которых хорошие виды являются правилом, а сингамео-
ны или агамные комплексы — исключением.
Причины и следствия, вытекающие из концепции
вида у растений
Сравнивая хорошо изученные группы высших растений со мио-
гимн группами птиц, млекопитающих и таких родов насекомых,
как Drosophila, мы обнаруживаем, что у растений дискретные
биологические виды в обшем встречаются реже и что для них
более характерны иные ситуации. Отсюда следует, что причин-
ные факторы, определяющие различные типы организации айда
в двух царствах живой природы, также должны различаться.
Поэтому сравнительный подход к проблеме вида может пролить
некоторый свет на те факторы, от которых зависит организация
вида у высших растений.
Очевидное и коренное различие между растениями и живот-
ными—это степень сложности индивидуального организма.
Развитию относительно простого растительного организма со-
ответствует открытая система роста с серийным образованием
новых частей. Тело животного имеет гораздо более сложное и
топко сбалансированное строение, а кроме того, оно развивается
как единое целое а результате закрытой системы роста. Логично
допустить, что организмы этих двух типов различаются также
по сложности и ивтегрироианности генетических систем, конт-
ролирующих их рост и развитие.
Одна эта предпосылка позволяет понять два главных факто-
ра. способствующих частичному или полному распаду видовой
организации во многих группах растений. Эти дня фактора —
частая межвидовая гибридизация и уннпареиталыюе размно-
жение.
Виду, обладающему высокоинтегрированным и точно сбалан-
сированным генотипом, межвидовая гибридизация нанесет отно-
сительно больший вред, чем виду с более простым и менее стро-
го координированным набором генов. Относительно простая
физиолого-морфологическая организация тела растения, отра-
жающая соответственно олюентельно более простой генотип,
Гл. Вопрос о у растопил 6 b
рпзмэжио. делает растения более устойчивыми к межвидовым
обменам генами, чем большинство групп животных (279]. Сле-
довательно, из открытой системы роста растений вытекает воз-
можность разнообразных форм вегетативного и агамоспермиого
размножения, в результате чего численность гибридов и их по-
томков будет увеличиваться.
Как писал Густафсон [317]: «Виды явнобрачных растений
снова и снова оказываются жертвой своих собственных черт:
своей свободной системы роста, своего вегетативного распрост-
ранения. Эти свойства’непрерывно способствуют возобновлению
популяции, ее изменению в направлении развития полнплонд-
ности и апомиксиса».
Эти основные свойства растений не только сказываются иа
характере растительных видов, но и оказывают глубокое влия-
ние на макроэволюцию растений. Как мы увидим в дальнейшем,
в процессе филогенеза растений неоднократно имело место об-
разование анастомозов между линиями, которые прежде были
обособленными. Если у животных видообразование обычного
типа приводит к формированию филогенетического древа, то у
растений око нередко ведет к образованию филогенетической
сети.
6—45
7 Разные типы видов
Общий термин «вид» используется в самый разных значениях.
Поэтому, для того чтобы уменьшить число возможных недора-
зумений. желательно в каждом конкретном случае с помощью
прилагательных иля приставок указывать, какой именно смысл
а него вкладывается.
Многочисленные авторы, занимавшиеся этой проблемой,
различают разные типы видов, давая им разные названия [см.
53, 79, 81, 157, 244. 279, 293, 564, 570. 732, 737, 752, 855].
Принятая здесь классификация типов видов вобрала в себя
некоторые черты более ранних систем, в частности систем Кэй-
иа [79] и Симпсона [732], но распределяются они иным образом.
Настоящая система отличается, кроме того, от большинства
прежних тем, что в вей последовательно используется единст-
венный критерий — характер классифицируемых биологических
объектов.
Типы видов
Как доказывает опыт, биологам необходимо различать по край-
ней мере четыре типа видов, чтобы охватить все разнообразие
биологических объектов. Иными словами, существует не менее
четырех различных тылов видовых единиц, для которых в бота-
нике и зоология необходимо иметь четыре различные концепции
вида. В биологии растений полезно иметь еще одну, пятую кон-
цепцию вида.
Пять типов видов, различаемых в этой книге, перечислены
ниже с указанием некоторых часто употребляемых синонимов:
1. Таксономический вид (морфологический вид, фенети-
четкий вид)
2. Биологический вид (генетический вид)
3. Мнкроцид (агамиый вид)
4. Последовательные (successional) виды (палеонтологи-
ческие виды)
5. Биосистематическнй вид (эковид, ценоонл)
Характеристики видов этих пятя типов приведены в табл. 7.1.
Следует указать, что каждая из перечисленных выше коицеп
Пий вила имеет силу в своей собственной сфере. Однако яри
использовании той или иной концепции вида за пределами этой
сферы возникают недоразумения. Как уже отмечалось, недора-
зумения могут возникнуть также в тех случаях, когда термин
«видь используют би соответствующих пояснений, что бывает
достаточно часто.
Гл 7. rttntt rtJoJthr
Таблица 7.1. Характеристики видом разных типе»
Тип 1л*пгп>«иирг«яые >лгаиы Сфер» яминки
TdHL'<HII)MH4KKH11 Тайгоны; группы мир- рилагичесхя сходных индивид уумоа Группы с морфологическими раз- личии ин, используемыми в фор- мально* классификации
&аалогический Системы шмулядий. разкноааюшнАсЯ Ви- .томи вутем Организмы с оловаам рлгчмс1ж«- хисм, принадлежащие к одному й(н-ы*»1н«'»му уровню
.Чпжроипа Популяции унналрем ПЛМЫХ организмов У ныаарснтвлм1ыс организмы, в том числе беслолые и парасексу- адьмые организмы
Послсдомгелыше ва । к Фнлепачсг&ме линии Биологические виды н мнжриеядм •о времени
Ьноснстема тжче 1 рупяы, плодовитые Групаы раслсни* с половый раз-
скпй мл при енрешинамжнх мможенвем. исжусстпен iki niflp*- днзировлиные па жслеримгпталь- хых участках
Биологические виды, микровилы и последовательные виды в
отличие от двух других типов —естественные единицы в том
смысле, что оии существуют в природе, а не просто категории,
созданные человеком. Эти три типа можно объединить в слож-
ную единицу — эволюционный вид Симпсона (732], который мы
обсудим в одним нз дальнейших разделов этой главы.
Таксономический вид
ТакетюмичеекнА вид. предстааднющий собой основную едини-
цу формальной классификации, — универсальная категория,
встречающаяся у всех форм живой материн, как это предусма-
тривается кодексами ботанической и зоологической номенклату-
ры. Кодексы номенклатуры определяют также положение вида
в иерархии таксономических категорий В ботанике такую
иерархию составляют (в восходящем порядке). форма, разно-
видность, подвид, вид, ряд, секция, подрод, род (при атом опу-
скаются различные возможные промежуточные категории, обра-
зуемые при помощи приставок над* или под-)-
Таксономический вид ОСПОМн на морфологическом сходстве
и различии и представляет собой совокупность морфологически
сходных индивидуумов, отличающуюся от других таких совокуп-
ностей. Полагаться на внешние морфологические признаки прак-
тически необходимо, поскольку морфологические признаки, в
том числе окраска, — единственные признаки, которые можно
постоянно использовать при повседневной работе с любой груп-
пой организмов. Таксономический вид должен быть единицей.
6’
*4 Voctb вторая Природа видов
которую было бы удобно применять в практической работе по
классификации и определению.
Недостаток критерия морфологического сходства и различия
состоят в том. .что с его помощью невозможно объективно уста-
новить уровень различия, заслуживающий видового ранга. Нет
способа, который бы позволял объективно реиигтъ, представляет
ли собой данный таксон вид. разновидность нли ряд. Поэтому
разные систематики могут создавать н создают различные си-
стемы классификации для одной и той же группы организмов.
Несом ней но, что прерывистости в ряд)- изменчивости, раз-
граничивающие биологические виды, можно использовать также
для разграничения таксономических видов. В тех случаях, ког-
да это возможно, следует отождествлять таксономический вид
с видом биологическим. Однако такая возможность имеется не
всегда. Во всяком случае, это нельзя считать существенным нли
обязательным элементом концепции таксономического вида.
Субъективный характер таксономического вида отражен в
высказываниях равных ученых. Приведем прежде всего одну
цитату нз Дарвина [152, 154]:
•Н* подлежит сомксаню, что ди настоящего времеял не удалось праве
ста ясной осраянчно! черты между видами м оохвндвмя. г. е формами,
которые, по ииенпо некоторых натуралистов, приближаются к видам, но м
вполне достигают итого ранга. иля между поднайми и хорошо выражении-
ми разновидностями, или, жаяоме®, между ими* четко выражешымм рюяо-
еядностямя и илдиа к дуальными различиями. Эти различия плавно оережодгт
ОДИ1 в другие, слипнись в одни непрерывный ряд—
Из всего сказанного ново, <гто термин *пнд> я считаю прсж)в*жьпым. грн-
дума иным рахи удобства для обовжвчемия груип особая, близко между со-
бой схожих, и существенно не отличающимся от термина гравво—даоствт.
который обозначают формы, имея равно различающееся я более колеблю-
щиеся [в своих прививках]*
Эта точка зрения, доведенная до своего логического конца,
$ “л | сформулирована Р яга ком следующим образом (цнт. do
«Вид — »то сообщество (или весимвио блвако рохствеииих сообтест»',
отличи талы ые морфологические л риз наш ко троса, го мхехию компетаят-
яого систем а пгжа. достатово четко выражаю, чтобы ояраадата приевосяаа
«му (зли им) видового нвлваяия».
Недавно Кронквкст [131] дал разумное определение таксо-
номического вида:
«Виды—это самые медике гр углы, постоянио ж пхледоввтельяо сохра-
няющве свою обособленность и различимые обычными способами»
Дополняя это определение. Кронкввсг [131] пишет: «после-
довательно сохраняющие обособленность означает, что все или
очень большая доля рассматриваемых особей явно принадле-
жат к одной или к другой группе, а не находятся где-то посере-
дине между ними-. Слом «обычными способами» подчеркивают,
Гл 7. Рляиле Тиам ак>м
86
•что следует применять традиционные и практичные методы.
Так, дли различения видов грибов принято (и необходимо) ис-
пользовать микроскоп, тогда как при определении иидоа семей-
ных растений обычно пользуются простой или препаровальной
лупой.
Практическая применимость для определения и классифика-
ции представляет гобой, таким образом, один нз важнейших
критериев таксономического вада. А отсюда следует, что так-
сономический вид —искусственная категория, которая во мно-
гих случаях не совпадает с естественными единицами. Ибо, как
удачно выразился Бодри [53]: «Природа... развивалась, совер-
шенно ие заботясь о тех практических проблемах, с которыми
придется столкнуться ее нсследователям>.
Биологический вид как одна нз стадий процесса
дивергенции
Биологический вид рассматривался в гл. 4 в качестве одной из
ЧОЦНИИИШИШ единиц. Однако му единицу МОЖНО рассматри-
вать также как одну из стадий более или менее непрерывного
ряда уровней дивергенции популяционных систем. Узловым точ-
кам в таком ряду соответствуют локальные расы, аллопатриче-
ские полувиды, аллопатрические виды и снмпатрвческие виды
Рис. 7.1. Псклехователькые стадха процесса мхтелеияой перяичяоН хиьер-
пекнин
86 Чкти «тори. Природа видов
(рис. 7.1). При гибридизации между двумя биологическими ви-
дами эти единицы вновь обретают статус снмпатрвчесхих полу-
видов и образуют новую сборную единицу — сипгамеоп.
Хота теоретически иерархия популяционных систем отлича-
ется от иерархии таксономических категорий, на практике меж-
ду ними существует определенное соответствие, когда дело ка-
сается групп растений с обычным половым размножением. Для
популяционных систем приняты следующие таксономические
эквиваленты:
Локальная раса — ие различается таксономически
или рассматривается как раз-
новидность
Географическая раса — подвид; некоторые авторы счи-
тают се разновидностью
Аллопатрический, полу- — часто рассматривается как
вид таксономический вид
Симпатрический полу- — таксономический вид
вид гибридного проис-
хождения
Аллопатрический био- — таксономический вид
логический вид
Симпатрический бноло- — таксономический вид
гический вид
Группа видов —ряд
Сиигамвои • — ряд. секция
Последовательные виды
Поскольку биологический вид реагирует иа изменяющиеся усло-
вия среды, весьма вероятно, что с течением временя ом под-
вергнется изменениям. Изменяющийся биологический вид, рас-
сматриваемый в перспективе геологического времени, превра-
щается в пару последовательных видов. Другими славами,
последовательные вилы—это члены одной и той же эволюцион-
ной линия, существующие в разные периоды времени в обла-
дающее различными признаками. Вернемся вновь к рис. 7.1;
А и В ва одном определенном уровне схемы — это два биологи-
ческих вида; тогда как А и В в разных точках одной линии —
два последовательных вила
Филетическая эволюция — процесс непрерывный, а поэтому
два последовательных вида связаны, по крайней мере теорети-
чески. рядом интерградирующих форм, Таким образом, решение
о том, где провести границу между ними, неизбежно становится
произвольным. На практике принятие такого решения часто об-
легчается наличием разрывов в палеонтологической летописи.
Гл. 7. Разные тмил видов
Для многих современных видов хвойных известны третичные
предки. Ниже перечислены некоторые пары видов, относящиеся
к сем. Pinacete и произрастающие на западе Северной Амери-
ки. Слева указаны виды, произраставшие в районе Большого
Бассейна в миоцене, а справа —нх современные эквиваленты,
встречающиеся в хвойных лесах .западных районов Америки
131). Члены каждой пары очень сходны морфологически м. по-
видимому, представляют собой родственные последовательные
виды.
Мвоцея
Pinus ahordensls
Р Jlorist&nii
Р. puingmjolia
Picea lahonhiw
Р. зовотвшй
Ttuga m»'.'ti-KsL4ldts
Т. sotuvnensii
Pseudo!suga sonotwuls
Abies ahwdsitslt
A concotoeoidri
A tlamatHensis
A. latiearpus
A sonarmen sis
A. taxirlous
Соерсмежнвя anoxa
P. cont aria
P ponderosa
P nonlicofa
P. engelmannii
P Ьг^мсНалл
T. mertetuiana
T. kcterophyUa
P. menzlesil
A rtcuroata
A concotor
A- shastensh
A magiufica
A grandis
A. amabilit
Другие примеры последовательных видов рода Pinu$ можно
наблюдать у третичных и современных сосен с закрытыми шиш-
ками в прибрежных районах Калифорнии [30].
Эволюционный вид
Соединив биологический, уимпарентальный и последователь-
ные виды, можно получить комбинированную концепцию эво-
люционного вида.
Эволюционный вид по определению Симпсона (731], пред-
ставляет собой популяцноииую систему, обладающую следую-
щими характеристиками: I) это линия, состоящая из последо-
вательных популяций предков-потомков, существующая в прост-
ранстве и во времени; 2) эволюция этой линии обособлена от
эволюции других таких линий, т. е. от других видов; 3) эта ли-
ния обладает собственной «единственной а своем роде эволю-
ционной ролью». т. е. соответствует своей особой экологической
нише в биологическом сообществе, 4) ока имеет собственные
эволюционные тенденция, будучи способной изменять свою эво-
люционную роль на протяжении своей истории [732].
м
Чва» Природа «к!»
Такими характеристиками обладают как унипарентальные,
так и биларентальны» организмы. Следовательно, концепция
эволюционного вида охватывает более разнообразные системы
скрешиааиия и носит более широкий характер, чем концепция
биологического вида в бипарентальных группах.
Концепция эволюционного вида придает перспективность
проблеме гибридизации к интегрированности видов. Естествен-
ная гибридизация неизбежно понижает интегрированность уча-
ствующих в ней видов. Однако в свете концепции эволюционно-
го вида главный вопрос состоит не в том, происходит ли между
двумя данными видами гибридизация, а в том. утрачивают ли
два гибридизирующих вида свои различные экологические и
эволюционные роли нли иет. Если, несмотря иа некоторую гиб-
ридизацию. они не сливаются, то, следовательно, в эволюцией
иой перспективе они остаются отдельными видами [732J.
Концепция эволюционного вида проливает также свет на
причины гомогенности видов. Мы уже обсуждали эту проблему
в связи с биологическими и унвпаренгальныии видами (см.
гл. 4 и 5) и имели основания усомниться в том, что поток генов
может служить главным фактором, определяющим гомогенность
во многих группах растений. Рассматривая те же самые при-
меры с точки зрении концепции эволюционного вида, мы вводим
в обсуждение исторический фактор, а вместе с ним — удовлетво-
рительное альтернативное объяснение: гомогенность видовой по-
пуляции может в значительной мере определяться происхожде-
нием от общего предка.
Биосистематические категории
Выделение бноснстематкчсских категорий основано на плодо-
витости при скрещиваниях, определяемой в экспериментах по
искусственной гибридизации. Хорошо известна следующая
иерархия этих категорий (в восходящем порядке): экотип, эко-
вид, ценовид, компариум [96, 100, 102, 136, 840, 842).
Экотип слагается из «всех членов данного вида, приспособ-
ленных к выживанию в среде определенного типа в пределах
общего ареала распространении данного вида». Два или не-
сколько экотипов «способны свободно обмениваться генами без
снижения плодовитости или мощности потомков» [102].
Эковид представляет собой совокупность экотипов. Два или
несколько эковидов разделяются барьерами неполной стериль-
ности: например, между ними возможна искусственная гибриди-
зация, причем гибриды F, могут обладать значительной, но не
полной стерильностью.
Ценовиды — это совокупности родственных Йковидов. Раз-
личные ценоекды можно скрещивать друг с другом — обычно
Гл. 7. Разные типы виды
С трудгми, и получающиеся гибриды F| высоко или полностью
стерильны. Поэтому обыск генами между цснопндами очень ог-
раничен или не происходит вовсе
Компарнум слагается из группы родственных цемовидов.
Различные ком парку мы нс удастся искусственно скрещивать
друг с другом, так как этому препятствует очень сильно вира*
женная несии честим ость.
Представленная здесь система популяционных единиц носит
особый характер. Она основана на одной, но при этом очень
важном аспекте изоляции, а именно на внутренней репродуктив-
ной изоляции. Биоснстсматические категории дают возможность
систематизировать (в тех случаях, когда они известны) взаимо-
отношения между группами по плодовитости
В «нормальной» группе растений бпосистематические кате-
гории можно приравнять к соответствующим популяционным
единицам примерно следующим образом >котип=локальная
раса; экоеид и ценовнд=биологический вид: компарнум-био-
логический вид. сингамеон или группа пилон. Корреляции эти
не слишком точны н «типичных» группах и еще менее точны во
всех остальных.
Причина этого заключается я тон, что преграды, обуслов-
ленные стерильностью, представляют собой лишь одну группу
Изолирующих механизмов. Существуют, кроме того, внешние
изолирующие механизмы, которые могут быть весьма эффектип
ны в природных условиях В некоторых группах растений нме.
ются хорошо развитые внешние изолирующие механизмы, но
слабо развитые внутренние механизмы. Поэтому биосистсмаги-
ясские категории имеют более непосредственное отношение к
ситуациям, возникающим на экспериментальных участках, чем
к тому, что происходит в природе.
Единицы классификации культурных растений
Виды и группы видов, в которые входят культурные растения,
содержат больше отклоняющихся форм, чем аналогичные груп-
пы тех же размеров у дикорастущих растений, а использование
географических критериев затруднено. ЧТО связано С условиями
возделыяаиия. Как указывают Харлан и де Вст [347]: «Методы
формальной таксономии не очень пригодны для классификации
культурных растеиийк. Для этих целей необходим какой-то иной
способ классификации.
Харлан и де Вет [347] предложили иерархию категорий, в
основе которой лежит генофонд. На видовом тройне и выше они
различают три категории генофондов GP-I,’GP-2 и GP-3.
Первичный генофонд (GP1) эквивалентен биологическому
виду Хорошо изолированные виды относят к разным катего-
90
Уотт» вторая. Природа видов
ринм GP-1. Если растения, принадлежащие к одной к той же
группе, рвзличаютен по плоидности. то их обычно отворят к раз-
ным GP-i.
Вторичный генофонд (GP-2) представляет собой группу био-
логических видов, которые могут быть скрещены с культурный
растением. Барьеры в виде стерильности или пониженной жиз-
неспособности гибридов существуют, но они могут быть преодо-
лены. Категория GP-2 примерно соответствует ценовнду в био-
систематике.
Третичный генофонд (GP-3) — наиболее обширная из раз-
личаемых в этой иерархии единиц, она соответствует наивыс-
шей скрещиваемости и наивысшему потенциально возможному
переносу генов. Гибриды между разными GP3 получить труд-
но, к они нежизнеспособны нли полностью стерильны Обмен
генами между разными GP-3 достигается только с помощью
специальных методов, таких, как выращивание зародышей в
культуре для получения гибридов или удвоение хромосом для
песетаипвления плодовитости.
У Triticam к GP-1 принадлежат виды Т. топососсшп, Т lur-
gidum, Т. tlmopheevi и Т. aestivum. В единицу GP2, к которой
принадлежит TritUum, входят также роды Aegilope, Secate,
llaynaldia и три вила Agropyron. Единица GP-3, в которую вхо-
дят эти виды пшеницы и родственные ей роды, содержит, кро-
ме того, несколько других видов Agropyron и несколько видов
£/ртшгз [347].
Некий внд GP-1, относящийся к какой-либо группе культур-
ных растений, обычно содержит не только возделываемые, но
и спонтанные формы (либо сорняки, либо дикорастущие, либо
и те и другие вместе). Харлан и де Вет [347] предлагают ис-
пользовать для классификации этих альтернативных форм кате-
горию подвида. Вид GP-I делится на два подвида, Л и В. К под-
виду В относят сорняк и дикорастущую форму, т. е. спонтанные
расы. Подвид А сохраняют для культурных сортов.
Вид GP-1 н его подвиды А в В имеют формальный таксоно-
мический статус, и им присваиваются латинские названия.
Существует также ряд категорий рангом ниже подвида, не
имеющих формального статуса. Их можно представить в сле-
дующем виде:
Подвид В
Раса
Подраса
Подвил А
Раса
Подраса
Культивар
Ливня, клон иля генотип
Гл. 7. ’«ял «идо»
91
Биологический вид или таксономический вид?
Разногласия по проблеме вида сосредоточены на взаимоотно-
шениях между таксономическими и биологическими видами.
Между этими двумя типами видо^ имеются как сходства, так
я различия. В ранний период истории концепции вида говорили
преимущественно о сходстве, тогда как ближе к нашему времени
стали уделять больше пни мания различиям
Натуралисты раннего периода—Джон Рэй |61>н]. Линней
[512—515], Бюффои [74] и другие — занимались высшими жи-
вотными и растениями н пределах местной фауны и флоры. Сле-
довательно. они имели дело С симпатрнческими популяциями
организмов, размножающихся половым путем и различающихся
по хорошо выраженным морфологическим признакам. Виды,
которые они выделяли, можно было одинаково хорошо опреде-
лить как по взаимоотношения и при скрещивании, так и по пре-
рывистости в изменчивости морфологических признаков. Самая
ник-тюзиапая единица бипарскталыюго размножения н основ-
ная таксономическая единица были для них синонимами Ины-
ми словами, между биологическими и таксономическими вида-
ми никакого различия не делалось, ла в нужды в ней не было.
Для многих групп организмов, изучаемых современными
методами, также наблюдается хорошее соответствие между кри-
терием. основанным на способности к скрещиванию, и крите-
рием морфологической прерывистости. Прерывистости в измен-
чивости. разграничивающие биологические виды, делают также
возможным распознавание таксономических видов В таких
группах таксономический вид остается синонимом биологиче-
ского вида.
Однако критерии, основанные на скрещиваемости и на мор-
фологических различиях, не всегда совпадают. Расширение си-
стематических биологических исследований со времен первых
«л тура л истов выявило многочисленные примеры, в которых эти
два критерия не совпадают, что положило начало проблеме вида.
Существуют три часто встречающиеся ситуации, в которых
ис удается или не представляется возможным приравнять таксо-
номический вид к биологическому виду. Это касается аидоп-
двпйкиков, полувидов и видов с уинпарентальныи размноже-
нием. Каждая из этих ситуаций широко распространена у выс-
ших растений.
Система таксономических категорий уииверса.тьпя. Все орга-
низмы следует распределить по родам и видам, и каждому иа
них следует присваивать отдельное иаавакие. На не псе орга-
низмы размножаются половым путем. А в бесполых группах
биологических видов не существует.
91
Чаете вторая Природа видов
Кроме того, таксономическая иерархия отличается жест-
костью. В вей есть две категории — вид и подвид. Каждое рас-
тение, согласно правилам ботанической номенклатуры, относит-
ся к определенному виду. Полувидов в таксономической иерар-
хии не существует Но некоторые популяционные системы в-
природе находятся на полувидоаом уровне дивергенции. Беле
угодно, их можно считать таксономическими видами,ио они не
являются биологическими видами.
В ссязи с этим уместно напомнить, что иерархия таксоно-
мических единиц была соадана до появления теории эволюции.
Лииясевский вид был отлит в креационистскую форму. Линией
писал в 1764 г. (цит. по [768)1: «Мы различаем столько видов,
сколько различных форм было сотворено изначально». Со вре-
мени Линнея никаких серьезных изменений в таксономическую
иерархию внесено не было. Но естественные популяционные
системы эволюционируют. Их не всегда удается втиснуть в ядо-
эволюционный» набор категорий. Семена, из которых выросла
проблема вида, были посеяны давно.
Третий случай, в котором критерии скрещиваемости и мор-
фологических различий вступают в противоречие, это виды-
двойники. Такие виды представляют собой настоящие биологи-
ческие аиды, но нх очень трудно различить по морфологическим
признакам, вследствие чего и нельзя считать хорошими таксоно-
мическими видами. Эго ие означает, что родственные виды-
двойники невозможно различить по фенотипическим призна-
кам. Фактически такое различие можно провести в любой груп-
пе достаточно полно изученных видов-двойников. Однако для
того чтобы произвести идентификацию в этих группах, необхо-
димо приложить гораздо больше усилий, чем это требуется в
обычной таксономической практике. В обычной практике разде-
ление биологических видов может оказаться неудобный или да-
же неосуществимым, поэтому за единицу формальной классифи-
кации приходится принимать весь комплекс видов-двойников
Такое положение возникает обычно в связи с аллополиплои-
дией. Рассмотрим группу трех родственных видов (двух диплои-
дов и одного тетраплоида) с геномами АЛ, ВВ и ЛАВВ. Одни
лишь внешние морфологические признаки могут оказаться не-
достаточно надежными в качестве критериев для различения
этих видон. Однако если помимо морфологических данных рас-
полагать сведениями о числе хромосом, то этого обычно бывает
достаточно для идентификации. Два диплоидных вида АА я
ВВ — морфологически крайние типы в пределах данной группы,
и их, как правило, можно отличить одни от другого по внешним
признакам. Морфологически промежуточный вид ЛАВВ надеж-
но отличается от обоях диплоидов даже по одному лишь числу
хромосом. Таким образом, виды АА. ВВ и ЛАВВ можно плен-
Гл. 7. Разнит типы тиЯ&л
&Э
ткфнпнрпвать ио совокупности наружных н внутренних феноти-
пических признаков Однако первый этап идентификации —
определение числа хромосом, для чего необходимо исследовать
живой растительный материал под микроскопом — процедура,
выходящая далеко за пределы методов, которые можно считать
стандартными для определения видов у висшнх растений
Здесь мы сталкиваемся с новой формой старого расхожде-
ния между естественной и искусственной системами классифн
канни. Первая отражает генетические связи, но может оказать-
ся неудобной для применения, вторая полезна для практическо-
го использования и таксономии, но не всегда отражает биологи-
ческое родство Расхождение между этими подходами к класси-
фикации обычно затрагивает высшие категории. Однако она
вновь возникает на видовом уровне во всех тех случаях, когда
таксономические виды, различаемые в повседневной практике,
пеэкпнваленткы биологическим видам (283].
Таким образом, концепция вида применительно к соврсмеп
иым оргапизмам дуалнетичиа, поскольку она основывается как
иа морфологических критериях, так и на критерии скрещивае-
мости. Такой дуализм не создает никакой проблемы вида в
группах, в которых жги два критерия совпадают Но пе во всех
группах дело обстоит так благополучно. Какому же критерию
следует отдавать предпочтение? Среди биологов нет единодуш-
ного мнения по этому вопросу. Представители различных школ
отвечают на него по-разному, и их ответы отражают различную
практику, которой они придерживаются
Развитие концепции вида, начавшееся во времена ранних
натуралистов от общей исходной точки, в дальнейшем пошло
□о разным направлениям Одно из таких направлений привело
к развитию концепция вида, принятой в таксономии Другое
частично независимое направление привело к созданию совре-
менной биологической концепции вида. В результате при ис-
пользования термина вид в таксономии и в популяционной био-
логии в него вкладывают различный смысл.
Во многих дискуссиях по проблеме вида не проводится раз-
личия между этими двумя употреблениями термина. Нередко
биологическую концепцию вида критикуют с позиций таксоно-
мической концепции, не признавая различий в точках зрения.
Надо ли объяснять, что подобные дискуссии лишь способствуют
сохранению путаницы.
Делались также попытки упорядочить положение, ограничив
применение термина вид и используя его лишь в одном из его
традиционных значений. К сожалению, разные авторы расхо-
дятся во мнениях о том, какое именно из этих значений следует
выбрать. Одни авторы (131, 244, 762 н др.] считают, что термин
вид следует закрепить за формальной таксономией, а для ре-
5М Часть вторая. Природа аидс-а
продуктивно изолированных скрещивающихся групп создать
новые термины. Другие [100. 279 (но не [282]). 525 и др] пред-
почитают сохранить термин вид для изолированных скрещиваю-
щихся групп. Ни одно из этих противоположных предложений
принято не было, н вряд ли то или другое из них найдет широ-
кое признание, поскольку обе школы исследователей пренебре-
гают исторически сложившимися традициями в применении
термина вид.
Выход на этого семантического тупика, позволяющий достичь
желанной целя — внести ясность в обсуждения, избежав при
этом опасности пойти наперекор издавна установившимся тра-
дициям в использовании термина, — состоит в том, чтобы при-
нять термины «биологический вида и «таксономический пил»,
вкладывая в них различный смысл {79, 157. 282, 652). Целесо-
образность разграничения этих двух типов видов била доказана
при изложении и обсуждении проблемы в этой и в предшест-
вующих главах. Задумайтесь еще раз над видамн-двойннкамн
в сеете этого обсуждения. Каждый кз видов-двойников по от-
дельности представляет собой хороший биологический вид н
может быть признан за таковой исследователями, интересую-
щимися его генетическими или экологическими характеристика-
мн. В то же время группу видов-двойников н целом при обычной
'идентификации можно рассматривать как один таксономический
вид.
Критика биологической концепции вида
Многие авторы предлагали концепции вида, совершенно ие сов-
местимые с биологической концепцией (см., например, Беннет
156]. Бурна [76]; Гейтс [236]? Дэвидсон ПБ5); Кроикэиет [130].
Левин [475]; Мейсон [558]; Мосхуин 15871; Рейвн [660. 661]; Со-
ках. Кровелло [754]; Эрлих. Холм [195]; Эрлих, Рейвн [196].
Некоторые кз этих работ открыто направлены против биологи-
ческой концепции вида (Бспиет, Кронквмст, Москунн, Рейвн.
Сокал и Кровелло, Эрлих). Кроме того, некоторые работы, на-
правленные против биологической концепции вида, часто с одоб-
рением цитируют другие авторы (см., например. Орндаф [631]
о работе Сокала и Кровелло [754] и многие другие).
Перечисленные выше авторы • большинстве своем ботаники;
одни (Бурна) — зоолог, публикующий свои работы а ботаниче-
ском журнале; два (Эрлих, Сокал) —зоологи, сотрудничающие
с соавторами боташшаии. Несогласие с биологической концеп-
цией вида или возражения против нес высказывают, как показы-
вает этот перечень, в основном ботаники, ио также и отдельные
зоологи.
Гл 7. Ралпж типы видов
96
В этом разделе мы рассмотрим некоторые нз основных плз-
рожсний против биологической концепции вида и попытаемся
ответить на них.
Авторы старшего поколения (Бурив. Гейтс. Дэвидсон. Мей-
сон) отстаивают номиналистическую концепцию видя’ реальны
лишь индивидуумы. а виды это просто скопления индивиду-
умов. Подобная точка зрения лишена здравого смысла приме
нптельно к бнпаргитнльнмм организмам, у которых скрещиваю-
щиеся популяции действительно представляю? собой реально
существующие репродуктивные единицы и группируются в более
обширные популяционные системы
Авторы более поздних работ (Рейви, Сок ал и Кровелло.
Эрлих н Холм, Эрлих и Рейв») подчеркивают важность скре-
щивающейся популяции, но нс принимают биологический вид.
Эти авторы правы по первому вопросу. Но на границе между
популяционными системами действительно существуют барь-
еры. препятствующие скрещиванию между этими популяциями.
Точки зрения упомянутых выше авторов можно придерживаться
только в гоя случае, если пренебречь этим вторым аспектом пли
принизить его значение
Сокал и Кровелло [754] рассмотрели поэтапно весь процесс,
необходимый для распознавания биологического вида, подверг-
нув его логическому анализу. Оки приходят к выводу, что био-
логической концепцией вида трудно пользоваться па практике.
Совершенно верно, что для выделения биологического вида в
любой данной группе необходимо провести много полевых и экс-
периментальных исследований, а также проделать большую ра-
боту' с музейным» коллекциями. Для некоторых п>упп растений
такая работа проведена Соклл и Кровелло (754], подчеркивая
эти трудности, совершенно умалчивают о том, что на самом де-
ле все необходимые операции в ряде случаев были проведены.
Лсвм [475]. Рейви [660], Сокал [753]. Эрлих и Ренан [190]
считают, что интеграция обширных популяционных систем видо-
вого масштаба не может быть результатом обмена генами, кпх
полагают другие ученые, поскольку этот процесс иосит локали-
зованный характер. Вполне возможно, что это справедливо дли
некоторых групп организмов, в том числе для многих растений
(обсуждение этого вопроса см. в гл. 4). Мн здесь будем исхо-
дить из допущения, что оно справедливо Затем названные выше
авторы делают заключение, что недостаточная интегрирован-
ность. обусловленная слишком обширным потоком гемов, подры-
вает статус биологического вида как эволюционной единицы.
Мы можем согласиться с предпосылкой, но не с выводом.
Биологический вид состоит пз действительно или потенциально
скрещивающихся между собой индивидуумов, репродуктивно
изолированных от других таких видов. Широкое скрещивание в
«8
Часть сгорая. ПрароОа mdoo
пределах данной популяционной системы не представляет со-
вой существенной особенности биологического вила, существен-
ное значение имеет его нескрещипаемость с другими популяци-
онными системами.
Многие ботаники полагают, что. согласно биологической кон-
цепции вида, все или почти все растения должны быть сгруп-
пированы в дискретные видовые единицы. Это не реализуется
но многих группах растений и а разных флорах, и такое несоот-
ветствие между наблюдением и теорией вызывает глубокое не-
довольство биологической концепцией вида. Подобное отноше-
ние к «той концепция не высказывается прямо в публикуемых
работах, и поэтому его нельзя документально подтвердить, как
это обычно принято. Тем не менее оно существует; оио прояв-
ляется при неофициальных обсуждениях и подразумевается в
некоторых опубликованных материалах [130, 660, 754, 756].
Вот еще один пример того, как в биологическую концепцию
вида вкладывают большее значение, чем это нужно. Биологиче-
ский вид представляет собой одну из стадий процесса днверген
цин. как уже отмечалось в этой главе; существуют также и
другие стадии незавершенного видообразования и вторичного
слияния видов. Следовательно, ряд популяционных систем а
любой данный момент времени состоит как на биологических
видов, так и из полувидов. В сущности, скорее всего следует
ожидать, что в каждый данный момент своей истории не все
популяции будут сгруппированы в дискретные биологические
виды. Важно, что в каждый момент существует некоторое чис-
ло биологических видов.
Одна из характерных черт биологических единиц вообще со-
стоит в том. что процессы их размножения постепенны и тре-
буют значительного времени. Деление одной хромосомы, клетки
иля особи иа две дочерние хромосомы, клетки или особи прохо-
дит черев ряд промежуточных стадий. Тем не менее никто не
считает хромосомы, клетки и особей, после того как они образо-
вались, произвольно выделенными единицами организации. Точ-
но так же биологический вид в процессе подразделения на два
или несколько новых видов может проходить через промежуточ-
ные стадии, и это отнюдь не должно наносить ущерб статусу
дочерних видов как реально существующих единиц на их ко-
нечных и хорошо развитых стадиях.
Общая особенность всех работ, перечисленных в начале это-
го раздела, — поиски единой универсальной концепции вида.
В иих обсуждаются <виды> без всяких дальнейших уточнений.
Авторы работ, направленных против биологической концепции
вида, пытаются использовать эту концепцию в качестве удов-
летворительного универсального критерия вила я обнаружи-
вают. что она для этого не подходит Однако, как мы отмечали
• Гл 7. Ркты! ты видов 97
ранее, единственный критерий вида, имеющий универсальное
применение, — это практичность н удобство для ндентнфикаиин
и классификации, и этот критерий дает нам таксономическую
единицу, которая в своей основе искусственна и часто яе соот-
ветствует естественным единицам.
Для того чтобы сделать возможным полезный критический
диалог о виде, необходимо проводить различие между сущнос-
тями, называемыми здесь таксономическими видами и биологи-
чески ин видами. Эти сущности перекрываются, но они в корне
различны. Однако в некоторых статьях, направленных против
биологической концепции вида, открыто отвергается как рас-
хождение между таксономическим и биологическим видами, так
и сами эти термины (см., например, |76)). в других же расхож-
дение просто игнорируется, а термин <вид> используется попе-
ременно в разных значениях.
Я мог бы привести многочисленные примеры работ, в кото-
рых термин «вида без прилагательных используется в смысле
таксономического вида в одной части предложения или абзаца
и в смысле биологического вида в другой части того же самого
абзаца. Такие колебания между двумя различными концепция-
ми вила во многих случаях, по-вихимому, не осознаны, но они
всегда служат источником недоразумений. Всякая логически
обоснованная критика биологической концепции вида должна
признавать различие между биологическим и таксономическим
видом.
7—as
Часть третья
Генетические и экологические
аспекты вида
8 Генетическая основа
различий между видами
Изучение наследственности при межвидовых в межрасоеых
скрещиваниях проводится многими исследователями во многих
группах растений. Как показали результаты этих работ, близ-
кие виды или расы дифференцированы в отношении систем ге-
нов. различных типов хромосомных перестроек и систем сцеп-
ления. Поскольку обзор соответствующих данных уже опубли-
кован нами в другой книге [294], здесь мы ограничимся лишь
кратким их изложением.
Генные системы
Генные системы, участвующие в дифференциации видов у рас-
тений. можно классифицировать следующим образом. По систе-
мам каждого из перечисленных ниже типов имеются эмпириче-
ские данные (см. [294]).
1. Простые меиделееские системы.
2. Сериальные генные системы, состоящие из двух или
нескольких генов, контролирующих последовательные
этапы какого-либо метаболического процесса или про-
цесса развития.
3. Системы множественных генов, состоящие из генов с
аддитивным действием.
4. Системы оппозиционных генов, состоящие из множест-
венных генов и (или) сериальных генов в сочетании с
контролирующими геиаыи, обладающими негативным
действием.
5. Системы модификаторов, состоящие из главных генов
и генов-модификаторов.
6. Цитоплазматические факторы.
7. Системы, состоящие из взаимодействующих друг с дру-
гом цитоплазматических и хромосомных генов.
Простые меиделевские системы встречаются не очень часто,
ио они существуют. Хорошими примерами служат плодовитые
Гл 8 Ггнеггяеспал основа paaon-iuA между ыдлми ОТ
межвидовые скрещивания у Aquitegia [648]. Цветки большинст-
ва видов имеют шпоры, но у примитивного вида A. ecalcarata
шпоры отсутствуют. Признак наличия или отсутствия шпор на-
следуется как различие, определяемое одним геном, причем на-
личие шпор — доминантный признак при скрещиваниях между
A. ecalcarata п A. vulgaris, а также между A. ecalcarata и
A. canadensis. Различие по тому же признаку наследуется как
определяемое двумя генами н при скрещиваниях A. ecalcarata х
ХА. chrysantha и Л. ecalcaralaxA. alpina дает в Fj соотнпше-
ине 15:1. Различие в ориентации цветка также определяется
одним геном, и при скрещиваниях A. ecalcarataxA. chrysanlha
поникающие цветки доминируют над прямостоячими (648).
Преобладающая часть различий при межрасовых и межви-
довых скрещиваниях у растений наследуется как признаки, обус-
ловленные множественными генами (т. е. соответствует типам
2—5 из приведенного выше перечня). Так обстоит дело у Agui-
legia У этой группы растений, как и у большинства других
групп, окраска цветной наследуется очень сложным образом.
Л межвидовые различия по высоте растений, форме листьев,
длине шпор, величине лепестков к т. п. детерминируются у
Aquilegia множественными генами [648).
В простой системе множественных генов, контролирующих
какой-либо количественный признак, гены обладают аддитивных
действием к доминирование отсутствует. Это приводит к возник
иовеиию промежуточного фенотипа в Fi. непрерывкой изменчи-
вости в широком диапазоне и промежуточной моды в Fa и не-
прерывной изменчивости в более узких пределах в последующих
поколениях. Классическим примером такой системы служит
наследование различий в длине венчика при скрещиваниях меж-
ду разновидностями Nlcvtiana longijlora (182) (рис. 8 1).
Вряд ли следует говорить, что взаимодействия между гена-
ми обычно гораздо сложнее, чем в приведенном примере. Гак,
компоненты какой-либо системы множественных генов у данной
расы или вида могут обладать не только одним лишь аддитив-
ным действием на тот или иной количественный признак; один
компоненты системы могут оказывать на развитие данного при-
знака аддитивные эффекты, а другие —негативные. Когда две
такие внутренне сбалансированные оппозиционные системы ге-
нов сочетаются в результате межрасовой или межвидовой гиб-
ридизации. то в поколениях Fa, а при иекоторык условиях и у
гибридов Fi. наблюдается трансгрессивное растепление, причем
количественное выражение данногп признака у гибридов выхо-
дит за пределы его изменчивости у обоих родительских типов.
В сериальных генных системах отдельные гены детермини-
руют различные этапы какого-либо процесса в развитии. Такне
системы играют важную роль в наследовании окраски цветков.
?•
Члсп г утл. Гснтмскаа u асомеияескыг аспекты гада
При изучении генетической основы окраски цветков обычно об-
наруживается. что ее наследование определяется комплементар-
ными генами, т. е. один геи создает условия, необходимые для
действия другого гена, контролирующего следующий этап. В на-
следовании окраски цветков нередко участвуют также ингиби-
торы. роль которых состоит в оппозиционном взаимодействии с
сериальной генной системой.
Рассмотрим, например, различие между двумя близкими ви-
дами Vio/e, выражающееся в наличии или отсутствии одной из
особенностей в окраске цветка: для цветков V. tricolor харак-
терно темное пятно, обычно отсутствующее у V. aroensls Кла-
узен [95, 96] показал, что у обоих видок имеются комплементар-
ные темы, определяющие образование этого пятна. Но V. arven-
Гл. 6 Генетическая ос/сзеа различив жжбд еи^ам 10|
я* содержит, кроме того, один или два геил ингибитора, ползи-
ляющне образование пятна. У потомков от скрещивания между
этими видами происходит сложное расщепление по этому при*
знаку рЛ5].
Системы оппозиционных генов, состоящие из генов е про-
тивоположными. положительными и отрицательными воздейст-
виями на развитие данного признака, встречаются часто. О ши-
роте их распространения можно судить по результатам изуче-
ния наследования 16 многофакторных признаков у межрасовых
гибридов Potenlilla glandulosa. Семь или даже восемь из этих
признаков контролируются системами оппозиционных генов
[9BJ.
Классический пример межвндоиых различай и системах мо-
дификаторов наблюдается у аилов Gossupium hirsutum и G bar-
badense. Многие признаки этих двух близкородственных видов
определяются гомологичными генами. Наличие нли отсутствие
пятен на лепестках, например, контролируется у каждого из
этих видов парой гомологичных аллелей гена S. При скрещива-
ниях внутри каждого пила наличие пятна доминирует над его
отсутствием, и и F, эти признаки расщепляются в отношении
3:1. Но при межвидовых скрещиваниях между растениями с
питиями н без пятен потомки F, носят промежуточный харак-
тер. а в F. наблюдается непрерывная изменчивость Объясня-
ется это тем. что хотя главные гены, определяющие наличие или
отсутствие пятна на лепестках (и различные другие признаки),
у Этих двух видов хлопчатника гомологичны, они несут различ-
ные наборы генов модификаторов. Система модификаторов у
G. barbadense усиливает проявление признака пятна, а сиогвет-
стпующая система у G. hirsutum подавляет его |349).
Интересный тип взаимодействия генов наблюдается а тех
случаях, когда у потомков межвидовых гибридов появляются
совершенно новые и непредсказуемые признаки. Для Antirrhi-
num ma/us и A. glutinosum характерны типичные двугубые цвет-
ки; такие же цветки имеются у гибридов F, между этими вида-
ми и у большинства потомков F,. однако у некоторых индивиду-
умов F, цветки больше похожи иа цветки их далекого родича,
относящегося к роду Rhinanlhus [519J. Точно так же у Digitalis
при одном из межвидовых скрещиваний у некоторых потомков
появляется новый признак цветка, а именно шпоры, отсутствую-
щие у родительских видов [715].
Новые фенотипы наблюдаются также у гибридных потомков
Ряда других групп растений, а не только у Antirrhinum и Digita-
lis. Следовательно, это явление носит общий характер, оно по-
лучило название макрорекомбииации (краткий обзор с библио-
графическими ссылками см. [294]). Макрорекомбннапии имеют
эволюционное значение, поскольку они показывают, что меж-
in
Hum rftru. Гмтгисж u молижмеи» «смет» >«я
видовая гибридизация может порождать не только промежуточ-
ное формы, во также формы, не похожие ни на один из роди-
тельских видов.
Гены, кодирующие ферменты
Электрофорез позволяет выявить те генетические различия в
ферментах, которые никак не проявляются фенотипически Эти
различия могут иметь место на любом уровне организации: иа
уровне индивидуума у гетерозигот или полиплоидов; между от-
дельными особями; в пределах одного вида, между видами и
между таксонами более высокого ранга. Гены, определяющие
электрофоретическую изменчивость, отличаются от генов, кото-
рые определяют большинство внешних фенотипических призна-
ков.
В последние годы методы электрофореза используются чрез-
вычайно широко, в связи с чем накопилось огромное количество
данных по электрофоретической изменчивости как у животных,
так и у растений. Здесь мы можем лишь в общем вкде коснуть-
ся этой большой проблемы. Появилось несколько новых обзоров
по электрофоретической изменчивости у растений [255, 335, 412];
есть также другие ценные обзоры, посвященные главным обра-
зом, но не исключительно, животным [28, 508].
При исследовании 13 ферментов в й популяциях Tragopogon
dutnus, 3 популяциях Т. porrtfolius к 3 популяциях Т. pratensis
полиморфизм по этим ферментам был обнаружен у первых двух
видов. У всех видов наблюдалось большое сходство между по-
пуляциями, л чем свидетельствуют показатели генетической
идентичности, разные 0,97—1,00. Вместе с тем между этими
тремя вндами но всех попарных сочетаниях существуют четко
выраженные различия, а показатели генетической идентичности
колеблются от 0.50 до 0.62 [694].
У очень близкородственных видов Clarkla btloba и Г. Ungu-
lata показатель генетической идентичности высокий (/=0,88).
В отличие от этого у менее близкородственных видов С. гиЫсип-
da а С. franciscana показатель генетической идентичности ниже
(/-0,28) [262—255].
При сравнении Phaseolus vulgaris и Ph. cocclnsus по двум
ферментным системам — эстеразы и лейцинамвнопелтядазы —
методом электрофореза между ними были обнаружены значи-
тельные различия в положения полос этих ферментов. У алло-
полиплоидного гибрида, полученного в результате скрещивания
этих видов, были обнаружены все полосы, образуемые родитель-
скими видами, что свидетельствует об аддитивности в наследо-
вании этих ферментов [884]. Такая аддитивность у аллополвлло-
идов гибридного происхождения — явление обычное. Одиако в
Гл. В Глнл7пялскаа осмола различий между подали 103
некоторых случаях аддитивности не наблюдается на электро
фореграммс аллополиплоида отсутствуют полосы. имеющиеся
у родительских видов, или, напротив, появляются новые полосы,
хак, например, у искусственных аллополкплоидов Nicotiana
(669). В таких случаях у межвидовых гибридов и у аллополи
плоидов, по-видимому, имеют место те или иные сложные взаи-
модействия между генами.
Замечания относительно регуляции
В периодической литературе довольно широко обсуждается
вопрос о теиах-регуляторах и регуляции развития как ключевых
элементах в днффсреиинаннн видов я в процессе видообразова-
ния (см., например, (77, 440, 896]).
Регуляция развития, несомненно, играет важную роль в соз-
дании фенотипических различий между видами. Это давно приз-
нано. и не об этом здесь речь.
Обсуждение рассматриваемого вопроса в приведенных выше
и в других недавно опубликованных статьях носит особый ха-
рактер В них регуляцию относят за счет действия геиое-регу-
ляторов и побочных эффектов хромосомных перестроек. Здесь
нас интересуют гены регуляторы, а хромосомные перестройки
будут рассмотрены в этом аспекте в гл. 14. Гецы регуляторы
противопоставляют структурным генам, а структурный ген —
это, в сущности, то же самое, что и классический меиделевский
геи, рассматриваемый в несколько ином плане. Выдвигаемое
авторами упомянутых работ положение состоит в том. что эво-
люция «зависит главным образом от мутаций генов-регуляторов
н лишь во вторую очередь — от изменений структуриых генов»
[8961.
Разграничение структуриых и регуляторных генон возникло
а результате изучении бактерий, и его стали широко применять
также к высшим организмам, где оно неуместно У высших рас-
тений нет ни одного класса генов-регуляторов. Регуляция обес-
печивается гетерогенной совокупностью контролирующих генов.
В »ту совокупность входят темы-ингибиторы, мииус-модифика-
торы, системы оппозиционных генов, гетерохроматиновые блоки
и контролирующие элементы (которые, возможно, представляют
собой гетерохроматиновые блоки) (см. |294|). Кроме того, у
высших растений разграничение между структурными и регуля-
торными генами оказывается несостоятельным: некоторые типы
генон-регулнторов па самом деле представляют собой структур-
ные темы, как »то хорошо видно на примере генов ингибиторов.
Итак, изменения в характере развития играют важную роль
в соэдаиин межвидовых различий у высших растений. Но эти
1<М Уот грта. Гнплжш а жоммчсскхы ослглгы лшЪ
изменения нельзя связать с генетическими элемента мн какого-
либо одного типа. Они представляют собой результат действия
генов и генных взаимодействий многих типов.
Хромосомные перестройки
Структурные перестройки хромосом возникают в результате
разрывов хромосом в последующего воссоединения разорванных
кондов каким-либо иным способом. В зависимости от кисла раз-
рывов, их местоположения к способа воссоединении различают
несколько типов перестроек, главные из которых представлены
иа рис. 8 2.
Во многих группах растений при цитологическом научении
близких видов и их гибридов F, обнаруживается, кто эти виды
различаются по инверсиям, транслокациям и другим структур-
ным перестройкам. В качестве иллюстрации здесь будут описа-
ны два классических примера.
Эвери {25] провел исчерпывающий цитогенетический анализ
гибридного треугольника Nicotians, состоящего из видов N. ala-
ta, N. langsdorffii и N. bonariensis, У этих видов имеется по
9 пар хромосом, относящихся к разным классам в зависимости
от величины и морфологии в пределах данного набора и до не-
которой степени—между наборами, принадлежащими разным ви-
дам. В мейозе у гибридов Fi от скрещивания N. alata и N. langs-
darflil хромосомы Ль В, « D образуют депочку, что указывает
на существование между родительскими вилами различий по
двум последовательным транслокациям в этих хромосомах.
Картина конъюгации при скрещивания N. alata я N. bonariin-
ait показывает, что эти виды различаются по реципрокной
транслокации в хромосомах Л| к В, в по двум последователь-
ным транслокациям в хромосомах Ль Вг и £>. На основании
этих данных можно предсказать, что N. langsdorflii и N. bona-
litnsis дифференцированы по ряду транслокаций, затрагиваю-
щих все хромосомы в вх наборах, за исключением хромосом С
и Ег. Каргины конъюгации, наблюдаемые у гибридов, получен-
ных в результате скрещивания N. langsdorffii н N. bonariensis,
подтверждают это предсказание (25].
Столь же остроумный цитогенетический анализ видов l.ilium
martagon. L. kansonii н гибрида Fi между ними провел Ричард-
сов [682]. У этих двух видов лилий по 12 пар хромосом. Карти-
на конъюгации хромосом у гибрида показывает, что родитель-
ские виды различаются по небольшим парацентрическим ин-
версиям в шести из этих двенадцати хромосом [682|.
Это относительно простые и ясные примеры В большинстве
случаен ВИДЫ,различаются по многочисленным невыявлеиным
1.Т*р«инал»нев н»н'м
Парачм*рямееям жеирсиа
Ct. ABCD EFQHi Cr. ABCD _ EFGHI
«м. ABCD У > EFQ ИМ ABCD t ' tFGMI
Сг. A8CD S F ОИ£ Сг ABCD EFGHI
*о» ABCD t 1 Им. t и t ABGFE _ DCHI
ААг>миицмя б Тмкпоанцив
Сг. ABCD PFOHI Ст. ABCD EFGHI
Нш. ABCD ll EFGMGHI ИМ, 1 t AD EF BCGMI
7 Доимая Мфгроанаа траносжаци»
Cr ABCD EFGHI MNQP QRST
ИЖ ABCD t EFGMBT MNOF 1 . QRI
атхапсгкяаим riwna
Ct ABCD EFGHI -MHOP ^MT
И*в. ABCD - СЯДТ MNOP ’l EFGHI
ВПоемдомт«»ы<ы* траяапожации
ABCD FFGHI MNOP ORST uvw xvz
тммсвеиацм* A BCD
QRST MNOP r— Г EFGHI DVW xvz
1 ЛОЖКАМИ ABCD ORST MNQP ? . XV2 UVW ГГСНГ
Рис. 8.2. Типы хромолшвыт перестроек [283].
В каждом случае стандартная хромосома (ст) сравкиваетсв с хромосомой,
измелившейся а резулвтате перестройки falx). Стрелками показаны точки
риврым ва стандартной хромосома, срн которых может возникнуть даыяяи
перестройка. Центромера мюбрамеиа в мае овале, располагающегося в се-
редине или пусти в середжяе к рокотом и
106 Чет rpms. Геигтмекм и гмшмаы аслпти «ада
перестройкам, вызывающим аберрации при конъюгации в
мейозе.
Во многих родах растений были обнаружены межвидовые
различия, обусловленные структурными перестройками хромо-
сом. Такие перестройки часто встречаются в группах травяни-
стых растений, особенно у однолетних. Хорошо известные при-
меры наблюдаются у Brassica. Clarkia. Crepis, Elymus. Galeop-
sis, Gossypium, GiUa, Layia, Madia н Nico liana.
Системы сцепления
Генные системы нередко характеризуются особыми типами
сцеплении. Следовательно, межвидовые различия —это разли-
чия ие только в генных системах, но также н в наборах сцеп-
ленных генов или в системах сцепления. Известны системы
сцепления разных типов. Помимо обычного сцепления генов в
стандартных хромосомах существуют сцепления, затрагивающие
лишь определенные типы генных систем; примерами служат:
многофакторное сцепление, сцепление M—V, сцепленные функ-
циональные генные системы, сцепленные адаптивные генные
комбинации и сбалансированные полигеиные системы.
Две из этих систем имеют отношение к эволюционным про-
цессам. которые будут обсуждаться в последующих главах. Эти
системы кратко рассматриваются ниже. Обзор систем саепле
ния в целом можно найти у Гранта [294].
Хромосомные перестройки тесно связаны с системами сцеп-
лении. Хромосомы — это носители обычных для данного вида
групп сцепления. Структурные перестройки — источник измене-
ний обычного сцепления. Некоторые перестройки в гомозиготном
или гетерозиготном состоянии служат носителями систем сцеп-
ления определенных типов,
Многофакторное сцепление
У покрытосеменных растений модальное гаплоидное число хро-
мосом п обычно равно 7—14. Следовательно, если множествен-
ные факторы, определяющие два признака, по которым разли-
чаются две расы или два вида, распределены по набору хроно»
сом более или менее случайным образом, то некоторые яэ ге
нов для каждого из «тех признаков должны находиться в одной
и той же хромосоме (или хромосомах). Короче говоря, сцепле-
ние между этими двумя признаками должно быть частичным
или многофакторным. Весьма вероятно, что с одним из двух
таких признаков или с ними обоими частично связан таким же
образом третий количественный признак. Продолжая рассуж-
дать в том же направлении, можно ожидать, что все признаки,
Гл. t. Глтичгслая огком риывчШ! яехдл пЛиш 107
находящиеся под многофакторным контролем, по которым раз-
личаются две расы или два вида, связаны, прямо или косвенно,
системой переплетающихся между собой слабых сцеплений. По-
добная система сцепления не предотвращает, но сильно ограни-
чивает рекомбинацию при межрасовой и межвидовой гибриди-
зации [98. 99, 294].
Эти ожидаемые многофакторные сцепления обнаружены в
нескольких экспериментах по гибрндизации Хорошие данные
получили Клаузен и Хиси [99] при межрасовом скрещивании у
Potentilla glandulosa (2л—14). Они изучали корреляции между
14 признаками в обширной расщепляющейся популяции F»; бы-
ло известно, что II на этих признаков детерминируются двумя
или несколькими генами. Из 91 пары признаков у 67 наблюда-
лась слабая, но статистически значимая корреляция. Ни один
из признаков ие расщеплялся независимо от всех других; на-
против, каждый признак коррелировал со многими или с боль-
шинством других 13 признаков. Корреляции между определен
ними парами признаков, такими, как длина лепестков и длина
чашелистиков, возможно, обусловлены плейотропным действием
генов, одиако многие другие сочетания признаков, морфогенетм-
ческн пс связанных друг с другом, как, например, опушенность
растения и вес ссмяи, также наследовались совместно, и в этих
случаях следует предполагать наличке многофакторного сцеп-
ления [99).
Аналогичные результаты были получены позднее для ряда
количественных признаков при межвидовом скрещивании Mimu-
ius levisii и М. cardinalis [377J.
Многофакторное сцепление обусловливает корреляцию меж-
ду признаками в пределах расы или вида, получившую название
генетической связности [99], или связности признаков.
Сцепление М—V
При изучении межвидовых гибридов в нескольких родах рас-
тений были получены данные, свидетельствующие о наличии
сцепления между генами, детерминирующими раххичиые морфо-
логические признаки, и генами жизнеспособности. Это широко
распространенное, а быть может, и всеобщее явление называют
сцеплением М—V (292). Термин «морфологические гены» отно-
сится в данном контексте к любым генам, Определяющим види-
мые фенотипические признаки, в том числе окраску разных
частей растений Пол генами жизнеспособности (viability genes)
имеются в ваду любые гены, оказывающие влияние на рост и
мощность
Гены жизнеспособности могут быть плейогропными, оказы-
вая действие также ив морфологические признаки, и наоборот
10*1 Часть трать л Гапетиьаст.иа м хплагщчл-лиг лтлекты мьаа
как в случае альбиносных проростков Нам хотелось бы иметь
возможность исключить подобные случаи плейотропин. Поэтому
мы ограничимся рассмотрением таких фенотипических призна-
ков, которые затрагивают цветки или плоды, ио ке могут быть
выявлены иа стадии вегетативного роста и которые коррелиру
ют с различиями в жизнеспособности. Наблюдаемые в таких
случаях корреляции позволяют предположить скорее наличие
сцепления между отдельными морфологическими генами н ге-
мами жизнеспособности, а не эффекты плейотропин.
Таким образом, мы здесь имеем дело с данными об аллель-
ных различиях между видами или расами растений по сцеплен-
ным генам М и V. Самая простая из возможных ситуаций —
сцепленная пара генов М—V без существенного взаимодейст-
вия между генами Один из родительских видов имеет при атом
генотип M.Va/MaVa. а другое — генотип MtVt/MiVb. Допустим,
что генам V, и V» соответствуют различные оптимальные для
роста условия среды. В таком случае у потомков гибрида, выра-
щиваемых в среде, благоприятствующей одному из аллелей V,
соотношение фенотипов при многофакторном расщеплении по
морфологическому маркеру М будет отклоняться от нормально-
го. Различные белее сложные примеры с участием частичного
или многофакторного сцепления между двумя нлв несколькими
генами V и двумя или несколькими генами М дают сходные ре-
зультаты [292].
Иктерфертильиые виды Mimulus cardinaiis и М. lewis и
(Scrophularlaceae) встречаются соответственно на небольших
и значительных высотах в горах Калифорнии. Проростки одной
из растущих иа побережье линий М. cardinaiis хорошо выжи-
вают в саду на низменности (Ставфорд), во в'альпинарии
(Тимберлейн) смертность среди ннх очень велика. Сеянцы пы
сокогорной расы М. lewisii, напротив, хорошо выживают а Тим
берлайие. но в Станфорде средн анх наблюдается высокая
смертность [615].
Эти два вида различаются также по окраске цветков — розо-
вой у Mimulus lamisii и оранжевой у М. cardinaiis. Окраска
цветков наследуется как различие по одному гену, причем розо-
вая окраска доминирует. Нойс я Хисн [616] обнаружили, что
среди потомков F» от межвидового скрещивания при выцащива-
нни в условиях, в общем благоприятных для них, соотношение
растений с розовыми и оранжевыми цветками составляло при-
мерно 3:1.
Однако при выращивания а Тимберлайне соотношение розо-
вых и оранжевых цветков в F> составило 8,7 к I, а в одной пов-
тор кости Fi. выращенной летом в Станфорде, было получено со-
отношение 1,7 к I. Затем Нобс н Хиси сравнивали соотношения,
полученные в одном и том же саду в Станфорде зимой, т. е. в
Гл. 8 Генетическая ос.нсаа различи* доеду витали 10®
прохладную погоду, и жарким летом. В заимей популяции F»
соотношение розовых и оранжевых цветков составило 4,4 к 1,0.
а в летней — 1,7 к 1,0 (616).
Такие измененные соотношения фен«ггнпоп сопровождались
в каждом случае высокой гибелью проростков. Смертность, оче-
видно, была вызвана избирательной элиминацией типов cardina-
Its в альпийской зоне или в прибрежной полосе зимой и типов
Ifwisii летом у побережья. Кроме того, гены определяющие об-
щую физиологическую адаптацию проростков к условиям кли-
мата. должны быть сцеплены с геном окраски цветков, для то-
го чтобы получались наблюдаемые отклонения в соотношениях
фенотипом [616].
Сходные данные были получены при межрасовом скрещива-
нии у Potentilla glandulosa и при межвидовых скрещиваниях у
Gossypium, Lycoptfticon, Phastolus и Oryza Эти данные позво-
ляю! считать, что изучавшиеся расы и виды несут разные алле-
ли сцепленных генов М и V [98. 292. 447. 448. 683. 70b. 800. 801].
Во многих группах растений удается определить гены.содер
жвшиеси в гомологичных хромосомах у интерфертильных видов
иля рас, однако нам почти ничего не известно о генетических
факторах, относящихся к тем затронутым перестройками уча-
сткам. по которым различактгея хромосомы ннгсрстернльных
видов В самом деле, к изучен ню этой проблемы обращались
редко венду того, что наложение на генетический Анализ ос
ложняюшил эффектов гибридной стерильности делало ее почти
неразрешимой Тем не менее для нескольких групп растений мы
теперь располагаем некоторыми предварительными сведениями
относительно того, какие гены локализованы в перестроенных
участках хромосом [2Я7].
Эти сведения получены в результате экспериментов по гиб-
ридизации между парами видов Ctarkia (329. 378. 379]. Gassy-
pium 1238, 241. 680, 800. 801), Tfiticum [219. 534, 716, 718]. Zea
(551, 829], Gilia [287] и Melilofus (70S]. В каждом нз этих слу-
чаев данные носят косвенный характер, и анализ нх сложен
Здесь можно подвести итоги только по главным моментам; бо-
лее полный обзор содержится в работах Гранта |280, 2Я7].
Полученные данные свидетельствуют о том, что в каждой из
этих пяти групп растений морфологические гены и гены жнзие
способности лежат в участках, затронутых перестройкой. Рас-
смотрим в качестве примера результаты, полученные при гиОрн
лизании видов Gilia.
Gilia malior и G. madocensis различаются uo многим струк-
турным перестройкам, чем и объясняется в значительной мере
высокая стерильность их гибридов F.. Эти виды различаются
также по генам жизнеспособности, что ведет к значительному
разрушению гибридов n F,—F., и по многим морфологических
по
Чает» гретая. Глетпчетхш а ято.юлкетявв аспекты «тЛт
признакам. Отбор среди гибридных потомков на жизнеспособ-
ность в условиях среды, благоприятных для типов modoeensu.
и отбор на плодовитость привели к возникновению трех мощ-
ных и плодовитых лиииП, из которых две гомологичны по струк-
турным перестройкам и плодовиты при скрещиваниях с С. лго-
doctiuis. Объяснить этот результат очень трудно, если не пред-
положить, что отдельные гены жизнеспособности находятся в
некоторых из участков, затронутых перестройками. Полученные
таким образом жизнеспособные ланки были сходны с С. modo
crnsis по большинству своих морфологических признаков, а »то,
по-видимому, означает, что некоторые морфологические гены
сцеплены с генами жизнеспособности в одних и тех же пере-
строенных участках [288J.
Таким образом, мы располагаем данными о том. что в не
скольких группах растений в затронутых перестройкой участках
хромосом локализованы сцепленные между собой морфологиче-
ские гены и гены жизнеспособности. Вероятно, между система-
ми сцепления M—V у видов, гомологичных по хромосомам, и
у видов, различающихся по структурным перестройкам, нет ка-
ких-либо глубоких различий.
Интегрированный генотип
Как мы убедились, каждая часть данного организма или любой
из его количественных признаков — результат морфогенетичес-
ких процессов, контролируемых системой генов. Отсюда сле-
дует. что Для нормального развития данного органа или призна-
ка необходимо гармоничное взаимодействие отдельных компо
пеитов такой системы. Кроме того, для успешного функциониро-
вания организма в целом необходима координация между его
отдельными частями н признаками. А поэтому различные систе-
мы генов, принадлежащие данному генотипу, подвергаются от-
бору на гармоиичное взаимодействие друг с другом.
Функциональное и селективное значение гармоничных взаи-
модействий между генами понимали многие исследователи
(Bridges. [68] и другие). «Природные виды слагаются из инди-
видуумов, гены которых находятся в состоянии внутреннего
равновесия, с тем чтобы обеспечивать гармоничное физиологи
четкое и морфологическое развитие одного поколения за дру-
гим» [102]. Каждый вил обладает не только виутреиним генети-
ческим равновесием, ио и собственной, отличной от другой, адап-
тивной комбинацией генов. Как внутреннее генетическое равно-
весие, так и адаптивная комбинация генов могут быть
разрушены в результате межвидовой гибридизации.
В Fs и в последующих поколениях потомков межвидовых
схрещлвапвй обычно бывает немало карликовых и стерильных
Гл I. Гелтпеаал оскала рамикив мвхду лидами 111
индивидуумов, индивидуумов с пониженной жизнеспособностью
или с какими-либо иными отклонениями. Растения GUia malior.
G. modocensU и их гибриды F, вполне жизнеспособны. Но в по-
коления! Fs —Fa 42% проростков были слабыми, чахлыми или
полулегальными, можно, кроме того, добавить к этому значи-
тельную гибель вследствие неспособности семян к прорастанию
[289]. Дисгармоничные взаимодействия между генами разных
видов могут также проявляться и Ft. Хорошим примером служат
красиилистиыс карликовые гибриды, обнаруженные среди жиз-
неспособных растений в Fi от скрещивания Gossypium arbor turn
и G. htrsutum. Такое полулегальное состояние обусловлено
комплементарным действием двух гемов. Rt, a R,», имеющихся
у некоторых линий родительских пндоп [239].
Иными словами, виды различаются по своим системам гене-
тического равновесия и по своим адаптивным комбинациям ге-
нов.
9 Изолирующие механизмы
Основные формы изоляции
Гибридизация и обмен генами между видами предотвращаются
или интенсивность этих процессов снижается и результате су-
ществования многочисленных и разнообразных изолирующих
механизмов. Эти механизмы можно разделить иа три основных
класса: 1) пространственные, 2) средовые, 3) репродуктивные.
Классификация изолирующих механизмов представлена в
табл. 9.1. Изоляция у растений довольно подробно рассматри-
вается в ряде обзоров [282. 472, 780].
Пространственная, или географическая, изоляция сущест-
вует между любыми двумя аллопатрическими видами растений,
географические ареалы которых разделены разрывами, превы-
шающими обычный радиус распространения нх пыльцы или се-
мян, Так, например, обстоит дело с Platanus occidentalis и
Р. orientaUs, которые распространены соответственно в восточ-
ное части США и в восточной части Средиземноморской области
и которые, несмотря иа интерфертильность, очевидно, не могут
скрещиваться между собой в природных условиях [780].
Средовая, или экологическая. изоляция возникает в резуль-
тате дифференциации видов по их экологическим предпочте-
ниям Благодаря этой дифференциации виды, сосуществующие
а одной и той же географической области, занимают в ней раз-
личные местообитании. Если между игами видами происходит
гибридизации и если нх местообитания обособлены, то большая
часть их гибридных потомков не сможет выжить ввиду отсутст-
вия среды промежуточного или рекомбинационного типа [14].
Так, Salvia apiana и S. iMlUJera. которые экологически диффе-
ренцированы, ио симпатрнчиы иа юге Калифорнии я которые
способны в ограниченной степени скрещиваться друг с другом,
не гибридизируют в ненарушенной природной среде, хотя иногда
образуют скопления гибридов в районах, где природные усло-
вия были искусственно изменены [17, 202, .101).
Репродуктивная изоляция создается в результате блокиро-
вания обмена генами между популяциями, в блокирование бы-
вает обусловлено генетическими различиями в строении репро-
дуктивных органов и в биологии размножения или во взаимо-
отношениях по плодовитости. Барьеры, препятствующие скре-
щиванию, могут быть внешними, действующими до перекрестного
опылении, как в случае временнбй. механической или этоло-
гической изоляции; или же они могут бьегь внутренними, дейст-
вующими после перекрестного опыления, как в случае несов-
местимости, нежизнеспособности, стерильности и разрушения
гибридов Барьеры, обусловленные несовместимостью между
Гл. 9. Икмирующм мсхапизны
113
Таблица 9.1. Классификация изолирующих механизме®
у растений
I Простржяствсмные механизмы
1 Гсопмфичсская нжоляцяя
!!. Срехавые ммьннзмы
2 Эжилогичесхжя жюляижж
111 Репродуктивные мвхаякзмы
А. Вмешмяе
3 . Вре*е*«»«йя жаояяцжя
«) ссзожиая
б| СУТОЧЖАЯ
4 . Механическая нэоляжжя
5 . ^голэппеагам изо/ниин
6 Изоляция. обусловлен »•« аатссаиивй
Б Внуаряикмв
? Несовиветныпгп.
а| ао оплел от ооуеяжя
Г>| после гАЛодотэоремя
8. Нежизя*еж>соЛжхп> гибридов
9. Стерильность гибридов
10 Разрушение гнбрждоа
Цветковая
ИЗОЛЯЦИЯ
родительскими видами, можно разделить на действующие до и
после оплодотворения.
Само собой разумеется, что а реальных ситуациях три ОС-
поеные формы изоляции могут сочетаться различным образом.
Различные виды репродуктивных изолирующих механизмов
также обычно действуют в разнообразных комбинациях.
Внешняя репродуктивная изоляция
Временная изоляция между двумя видами растений создается
в тех случаях, когда их периоды цветения н опыления приходят-
ся иа разное время года или суток. Эта изоляция может быть
частичной, если пики цветения двух видов разделены достаточ-
но большим временным интервалом, ио периоды цветения пере-
крываются. или же полной, если периоды цветения двух видов
совершенно не перекрываются. Соответствуют не примеры мож-
но найти у Pinas, Phlox, Salvia и во многих других группах рас-
тений.
Oenothera brevipes и Ое. davaeformis (Ona grace ас), напри-
мер, растут вместе в пустыне на западе США. цветут одновре-
менно и опыляются одиночными пчелами, главным образом
андреиами, однако гибриды между ними редки. Оказалось, что
Цветки Ое. brevipes раскрываются до восхода солнца и их по
сещают пчелы, активные ранним утром, тогда как цветки
Ое. davaeformis раскрываются в позднее послеполуденное вре-
мя в их посещают пчелы, летающие в конце дня (657].
1 Н Часть трепа. Гекетткш а тюлосичесаие шмат и аава
Механическая изоляция создается результате структурных
различий между цветками двух или нескольких видов, препят-
ствующих перекрестному опылению между этими видами или
нарушающих его. Различия в строении цветков, приводящие к
механической изоляции, встречаются главным образом у тех
растений, у которых цветки имеют сложную структуру (напри-
мер. в семействах Orchidaceae, Papilionaceae. Labiatae. Scrophu-
lariaoeae и Asclepiadaceae).
Так. Pedicularis groenlandica и P. attollens растут вместе в
горах Сьерра-Невада в Калифорнии, цветут одновременно и
опыляются одними и теми же видами шмеле*, ио не гибридизи-
руют. Отчасти это обусловлено различиями в структуре цветков
у этих двух видов. Пыльца Р. groenlandiea переносится к рыль-
цу на спинке шмелей, а пыльца Р. attollens—на передне* поверх-
ности головы [540, 765J. Другие способы механической изоля-
ции выработались между видами Stanhopea. Ophrgs. Aquitegia.
Asclepias, Penstcmon, Mimuius. Salvia. Phlox и Cordia (обзор
см. [474J).
Этологическая изоляция, обеспечиваемая особенностями по-
ведения, благодаря которым индивидуумы предпочитают выбк
рать в качестве брачных партнеров представителей собственного
вида, играет очень важную роль к репродуктивной изоляции
у высших животных. Предполагалось, что у растений подобная
изоляция совершенно отсутствует. Тем ие менее она встречается
у некоторых растений в качестве вторичного эффекта видоспе-
цифичного поведения насекомых-опылителей, посещающих цвет-
ки этих растений. Пчелы, бражники и в меньшей степени неко-
торые другие насекомые, посещающие цветки, способны к сен-
сорным восприятиям, которые дают им возможность отличать
друг от друга разные виды цветков по запаху, форме и окрас-
ке; кроме тога, они обладают инстинктом постоянства, застав-
ляющим их кормиться преимущественно на цветках одного вида
при ряде последовательных посещений Это постоянство в посе-
щении цветков ограничивает перекрестное опыление, так что
оно не выходит, в основном или всецело, за пределы одного вша
из скмпатрической смеси видов растений, различающихся по
опознавательным знакам цветков [266, 267].
Этологическая изоляция родственных видов, обусловленная
приверженностью пчел к посещению цветков одного вида, на-
блюдается у Pedicularis [765; Grant, неопубликованные данные],
Stanhopea [177]. Lythrum [471], Rhinanlhus [456], Cercidium
(417) и у некоторых других родов. Недавно была ппнеана ото-
логическая изоляция двух видов Anigozanthos (Haemodoraceae)
в западной часта Австралии, обеспечиваемая медососами (Me-
hphagldae) [382].
Гл. 9. Изолирующие мехапазшл
115
У однолетних растений часто встречаются симпатрическне
смеси вндо», размножающихся путем неродственного скрещива-
ния и автогамии, или же двух нли нескольких преимущественно
авгогамиых видов. Автогамная система размножения создает
а таких случаях барьер, препятствующий перекрестному опыле-
нию между разными видами. Левин (474J рассматривает это
как изолирующий механизм, п мы следуем его точке зрения
здесь н в табл. 9.1.
Концепция репродуктивной изоляции приложима главным
образом к организмам, для которых характерны половое раз-
множение и ауткроссннг. Ее можно расширить, с тем чтобы рас-
пространить ее на смеси автогамных видов растений с видами,
размножающимися путем ауткроссипга, а также на смеси двух
или нескольких преимущественно автогамных видов, как мы зто
сделали выше; однако она лишена смысла в приложевии к
строго унипарентальным группам растений и не должна исполь-
зоваться для таких групп.
Внутренняя репродуктивная изоляция
Несовместимость при скрещивания» создается разного рода
барьерами, препятствующими развитию жизнеспособного гиб-
ридного зародыша Некоторые из этих барьеров действуют до
оплодотворения, а другое — после него.
В некоторых случаях пыльцевое зерно не прорастает па
рыльце чужого вида, как при скрещивании Datura meteloides
и Л stramonium, ил к же пыльцевая трубка не может достаточно
успешно расти в чужеродном столбике, как при скрешиаапик
Iris terms и I. tenuis (25, 710). Или же, если происходит оплодо
творение и формируется гибридный зародыш, .тот зародыш мо-
жет преждевременно погибнуть вследствие дегенерации гибрид-
ного зплосперма, как это бывает при некоторых скрещиваниях
у Dalura. Iris. Gossypium и других родов.
Межвидовые гибриды F, часто обладают нормальной мощ-
ностью, однако иногда у них наблюдаются различные конститу-
Ц1юналь>1ые дефекты, известные под общим названием нежизпе
способности гибридов, которан блокирует обмен генами между
видами в вегетативной фазе поколения F,. Некоторые линии
Crepis tectorum при скрещивании с С. capillans дают нежизне-
способные гибриды F, (381). В F, от некоторых межвидовых
скрещиваний у Gossypium, GUia н Papaver появляются красные
карликовые растения (239, 273—275. 459, 538]. При многих меж-
видовых скрещиваниях с участием Nicotiana langsdorffii и дру-
гих родительских типол, относящихся к роду Nicotianu, у гобрн
дов F, развиваются генетически детерминированные опухоли
(7431.
116 Часп трвпл. Гечеппеекш а амхмлыеоам аспекты е»?а
Гибридная стерильность, как и несовместимость, охватывает
широкий крут разнообразных явлений. В зависимости от того,
на какой стадии развития проявляется действие барьера, гиб
рядную стерильность можно разделить на днол ОНТ иую стериль-
ность и стерильность гамет. Явления стерильности можно также
классифицировать в зависимости от факторов, которые их конт-
ролируют, иа генную, хромосомную и цитоплазматическую сте-
рильность. В реальных случаях гибридной стерильности наблю-
даются разнообразные комбинации различных стадий и факто-
ров.
Взаимодействия между генами растений разных видов часто
приводят к абортивному развитию пыльников или других важ-
ных органов у гибрида. Дисгармоничные взаимодействии ядер-
иых генов с чужеродной цитоплазмой вызывают мужскую сте-
рильность у некоторых гибридов Zea, Epilobium, Streptocarpus.
Nicotiana, Capsicum. Solatium и и других группах растений [89,
107. 183. 312, 386, 416. 377. 619, 637, 651J.
Неблагоприятные генные взаимодействия иногда проявля-
ются также на стадиях мейоэа и гаметогенеза, тем или иным
образом нарушая эти тонко координированные процессы. Гене-
тически детерминированная стерильность гамет обнаружена у
гибридов Nicotiana, Geum, Cilia, Bromus и других родов [229,
476, 309, 878J.
Виды растений часто различаются по хромосомным пере-
стройкам, как это отмечалось в предыдущей главе. Перестройки
могут приводить к нарушению у гибрида образования нормаль-
ных бивалентов и(влн) нормального распределения хромосом а
анафазе мейоза. При этом у части продуктов мейоза возникают
дупликации или нехватки определенных участков хромосом, и
в результате из них не развиваются жизнеспособные пыльцевые
зерна н зародышевые мешки. Доля таких нежизнеспособных
гаметофитов быстро возрастает е увеличением числа структур-
ных перестроек. Например, у гибрида, гетерозиготного всего по
шести независимым транслокациям, ожидаемая фертильность
пыльцы составляет лишь 1,5%, что сравнимо по уровню сте-
рильности с высокостерильныии межвидовыми гибридами.
Хорошо известно, что иа поведение хромосом при мейоэе
оказывают воздействие как генные факторы, так и структурные
различия в хромосомах [138. 407, 670]. Во многих случаях эти
две причины, вызывающие нарушения мейоза, а следовательно,
стерильность гамет, встречаются одновременно. В сущности
хромосомную стерильность, наблюдаемую особенно часто у on-
рсдсленных жизненных фору растений, можно рассматривать
хак состояние, накладывающееся на более широко распростра-
ненное и. вероятно, играющее главную роль явление генной сте-
рильности.
Гл. 9. Июшмюаре лсхачидик
117
Если гибрид а конечном итоге дает какое-то число потомков
Fj или В|, то среди них, по все# вероятности. будет некоторая
доля слабых или неполноценных индивидуумов. Как уже гово-
рилось и гл. 8, и поколениях F> —Р« от скрещиваний Gilia та-
lior и G. modocensii примерно 42% потомков обладали понижен-
ной жизнеспособностью. В F» от скрещиваний Laifia eailtardtoi-
des и L. hieracioides (Compositae) пониженная жизнеспособ-
ность наблюдалась у 80% индивидуумов [96].
Выводы
Ценные в адаптивном отношении генные комбинации, имеющие
си у отдельных видов, при гибридизации могут распадаться.
Разрушение гибридов — проявление распада адаптивных генных
комбинаций. Другие репродуктивные изолирующие механизмы
служат для защиты элях адаптивных генных комбинаций от
распада.
Репродуктивные изолирующие механизмы могут развиваться
как побочные продукты эволюционной дивергенции. Разрушение
гибридов, несомненно, представляет собой такой побочный про-
дукт. Эволюционная дивергенция может служить и обычно слу-
жит непосредственной причиной различных других форм репро-
дуктивной изоляции. Так, механические и сезонные различия
между Salvia теШ/ега и S. apiana связаны главным образом с
различием в признаках их нормальных опылителей — мелких
пчел, вылетающих весной, и крупных летних пчел соответствен-
но, и нх воздействия на репродуктивную изоляцию представля-
ют собой, возможно, побочные продукты дифференциации.
Однако это не единственный способ возникновения репро-
дуктивной изоляции. Как мы увидим в дальнейшем, механизмы,
действующие в родительском поколении, могут в некоторых слу-
чаях создаваться в результате отбора па изоляцию как тако-
вую Адаптивные генные комбинации у таких видов предохра-
няются от распада механизмами, возникающими в результате
прямого отбора, направленного на создание репродуктивной изо-
ляции.
10 Экологические взаимодействия
Между различными видами, обитающими на одной и той же
территории, происходят генетические и энологические взаимо-
действия. Генетические взаимодействия выражаются в свобод-
ном скрещивании, ограниченном скрещивании или полном от-
сутствии скрещивания в зависимости от степени и способа ре-
продуктивно* иаоляцни. Что касается экологических взаимодей-
ствии, то такие виды могут вступать или не вступать в конку-
ренцию в зависимости от различных факторов; если же между
ними существует конкуренция, то она может привести к разно-
образным последствиям. Кроме тога, экологические взаимодей-
ствия между видами связаны е их репродуктивными взаимодей-
ствиями.
Конечно, экология — слишком обширная область, чтобы мож-
но было достаточно подробно изложить здесь ее принципы, да
в этом в нет нужды, поскольку существует ряд хороших совре-
менных руководств по этому предмету; вместе с тем она столь
тесно связана с излагаемыми здесь проблемами, что совсем не
коснуться ее мы иа можем. Поэтому нам пришлось пойти на
некоторый компромисс: в этой главе дается краткий очерк из-
бранных аспектов экологии, особенно важных для понимания
проблемы видообразования у растений. В соответствии с такой
задачей мы ограничимся замечаниями о некоторых типах взаи-
модействия между родственными видами.
Экология всегда отличалась богатством терминологии Со-
временная экология в этом отношении также не составляет иск-
лючения; единственное отличие заключается в том, что терми-
нологии ее стала новее. Так. например, в настоящее время при
обсуждении экологической ниши используются такие термины,
как ширина киши, перекрывание ниш. гиперобъем ниши, фун-
даментальная ниша, реализованная ниша, поведенческая киша
т. п. Во многих случаях эти термины — просто новые названия
для существовавших ранее понятий. Для тех целей, которые мы
себе ставим в данной главе, эти термины не нужны, а поэтому
они либо не используются вовсе, либо упоминаются лишь мимо-
ходом, что дает нам возможность сосредоточить внимание иа
самих экологических явлениях и процессах.
Сосуществование
Термины аллопатрия и симпатрия полезны для описания про-
странственных взаимоотношений между популяциями (564].
Аллопатрия иаблюдАтся в тех случаях, когда популяции
обитают в разных областях. Следует различать две возможно-
Га tO. 3*<Motawatr
119
ста; а) облает разрешены, б) области соприкасаются (пара-
натрия).
Симпатрня наблюдается тогда, когда популяции обитают на
одной и той же территории, так что особи одной популяция на-
ходятся в пределах нормальной облает я расселения другой по-
пуляции. н наоборот. Здесь опять-таки существуют две возмож-
ности [282). а) Смежная симпатрня: популяции занимают раз-
ные ниши в одном и том же местообитании, например в саван-
не. где участки лесной растительности чередуются с открытыми
грасслсндамк; однако при этом особи, принадлежащие к разным
популяциям, обычно не живут в тесном соседстве. Эти популя-
ции симпатричиы в генетическом смысле, коль скоро гаметы нли
зиготы одной из них находятся в пределах нормальной области
расселения другой, и наоборот, по не вступают в непосредствен-
ный контакт в зколошческоы смысле, б) Биотическая симпат-
рия популяции обитают в одной и той же нише, и принадлежа-
щие к пим особи часто живут в тесном соседстве: таким обра-
зом. они сямватрнчиы не только генетически, ио и экологически
Близкородственные внды растений будут занимать сходные
или одни н те же географические области и обладать сходными
экологическими предпочтепиямн просто вследствие тесных фи-
логенетических связей между ними. Они могут быть аллопатри-
ческими. смежно- иля биотически-снмпатрическнмн в зависимо-
сти от различных факторов.
Биотическая симпатрня —самая тесная форма сосущество-
вания. Она может служить отправной точкой в процессе межви
левого отбора, ведущего к разнообразным конечным результатам.
Эффекты межвидового отбора
Сосуществование родственных видов может привести и нередко
приводит к конкуренции между видами за ресурсы, необходимые
обоим видам, ио имеющиеся в ограниченном количестве. Если
в этой конкуренции одни внд превосходит другой, то вступает в
действие межвидовой отбор; затем извилистыми путями этот
отбор может привести к различным конечным результатам.
Один из результатов межвидового отбора —замещение ви-
дов. Это весьма обычный процесс в истории жизни. Другой ре-
зультат— развитие у одного вила механизмов, наносящих вред
виду-конкуренту (например, аллелопатия [592]). Третий воз
кожный результат —экологическая дифференциация между
двумя конкурирующими видами, позволяющая понизить уровень
примой конкуренции между ними.
Этот последний процесс можно проиллюстрировать с по
мощью простой модели. Допустим, что два экологически сход-
ных и симпатричссхнх вида X и У конкурируют в гетерогенной
130
Часп трапа. Гттлкляг а гкиюмпкыа aaumi аяЛа
среде (£). имеющей три грани, ил* пиши (£>, Ег, Ее). Каждый
ид полиморфен и содержит ряд различных типов (Х>, Л» и
т. д), адаптированных к этим трем нишам. Полиморфный тип
Xi обладает превосходством в нише Е\, а тип Уз —в нише Е*.
В начале процесса межвидового отбора виды распределены
следующим образом:
Среда Е, Е, Е»
Вид X Xi (обладает пре- X» Ха
имуществом)
Вид Y Г| >а Г» (обладает пре-
имуществом)
К концу процесса отбора получается следующая картина:
Среда £i £» £з
Вид X Х| Ха
Вид У У» Уа
Каждый вид стал более узко специализированным к той ни-
ше, в которой он Обладает селективным преимуществом. Оба
вида продолжают занимать общую нишу, но у каждого из них
есть теперь и отдельная ниша. Степень перекрывания ниш и
прямая конкуренция между двумя видами снизились.
Межвидовой отбор, действующий таким образом, приводит
в конечном счете к повышению специализации у родственных
симпатрических видов и к смещению признаков [73J в области
симпатрин у частично симпатрических видов.
Сосуществование без межвидового отбора
или замещение видов
Сосуществование ие обязательно влечет за собой межвидовую
конкуренцию. Два вида могут сосуществовать, ие вступая в
ведущую к отбору конкуренцию, если их численности снижают-
ся под действием какого-либо иного фактора, например сильно-
го хищничества. В таком случае процесс межвидового отбора
не развивается.
Возможна и такая ситуация, когда два сосуществующих вида
с одинаковыми потребностями конкурируют за какой-либо огра-
ниченный ресурс лишь в небольшой степени, потому что оин
нуждаются в незначительных количествах этого ресурса. В та-
ких случаях процесс межвидового отбора протекает медленнее,
и он. возможно, менее эффективен по сравнению с другими
стохастическими факторами, воздействующими па обилие bit-
д. в
Возможны также случаи, когда сначала возникает межвидо-
вая коикуреиция. а затем следует межвидовой отбор; однако
процесс отбора не лоехдгает такого уровня, чтобы привести к за-
мещению того или другого вида или к экологической дифферен-
Гл 10. Эюшаним лмимодгЛстыл
121
циации. Межвидовой отбор — процесс длительный, я за то пре
ня, пока он происходит, направляющий его фактор может изме-
ниться. А это может привести к реверсии относительных селек-
тивных преимуществ конкурирующих видов иля разных форм
(при полиморфизме)
Если изменения факторов среды носят циклический харак
тер. причем каждый цикл занимает меньше времени, чем процесс
полного замещения одного вила другим, то между двумя сим-
патрнческимн видами, находящимися в состоянии конкуренции,
может установиться неустойчивое равновесие я они будут сосу-
ществовать постоянно.
Согласно принципу конкурентного исключения (340], посто-
янное сосуществование между двумя полными конкурентами
невозможно. Этот принцип в общем виде иереи как теоретиче-
ски. так и вмпкрически, однако ни в одном конкретном случае
его не следует применять некритически. Как было показано вы-
ше, сосуществование нс обязательно должно сопровождаться
межвидовой конкуренцией, а межвидовая конкуренция не всег-
да несовместима с сосуществованием
Эффекты экологических требований
Организмы разных типов предъявляют очень различные требова-
ния к ресурсам среды, в том числе и к ограничивающим ресур-
сам У часто встречающихся видов крупных организмов, как
правило, требования бывают более высокие, чем у часто встре-
чающихся вндоп мелких организмов. Вследствие этого межви-
довая конкуренция за ограничивающие ресурсы, имеющие ре-
шающее значение, обычно бывает более острой среди крупных
организмов, чем среди мелких.
Можно предсказать, что эти различия перейдут в различия
в интенсивности межвидового отбора. У крупных организмов
межвидовой отбор, вероятно, быстро приведет к замещению ви-
дов У мелких организмов, у которых межвидовая конкуренция
слабее или даже пренебрежимо мала, процесс межвидового от-
бора протекает медленнее и может не дойти до полного завер-
шения, г. е. до замещеякя видов, в пределах данной территории.
У таких организмов достижение симпатрия и ее сохряпеннс осу-
ществляются легче (282, 895].
Некоторые эмпирические данные подтверждают правильность
этих предсказаний Во флоре западных областей США близкие
виды таких древесных растений, хак Pinos и Quercus. и таких
многолетних травянистых растении, как Polomonium и Calothor-
tus. обычно занимают различные экологические эоны па з*о-
географических трансептах Сиипатрнчсскне контакты между
родственными видами в этих группах обычно ограничиваются
122 Чат грт*. Гыпгжш и эгожмхмиаи огпжты акда
областями перекрывания их местообитания. В »то# флоре био-
тическн-снмг.агрнчсскне скопления близких видов встречаются
обычно, и почти исключительно, среди однолетних травянистых
растения, таких, как Clarkla и Gllia. Например, в природе не-
редко можно встретить два, три и даже больше видов Gilia.
растущих рядом (282, 283].
Сходные корреляции наблюдаются у животных. Симпатркче-
ские скопления близких видов существуют у таких мелхях на-
секомых, как дрозофилы, комары и цикадки E/ythroneura. Про-
тивоположную крайность представляют крупные млекопитаю-
щие, у которых близкие виды обычно аллопатричиы или пара-
патрнчкы. У чешуекрылых, пищевые потребности которых иа
стадии имаго очень малы, взрослые особи, принадлежащие к
разным видам, часто занимают широко перекрывающиеся пи-
щевые ниши, тогда как усиленно питающиеся гусеницы тех же
самых видов занимают обособленные узкие пищевые ниши [282,
695L
Экологические взаимодействия и изолирующие механизмы
близких видов связаны между собой. В результате симпатрнче-
схих контактов возникают ситуации, вызывающие разного рода
реакции. В некоторых группах растений эта реакция выража-
ете» » экологической аивергенцяи и обособлении, влекущих за
собой более сильную экологическую изоляцию, тогда как дру-
гие группы реагируют усилением репродуктивной изолинии в
области симпатрии. Возможно, что во многих группах эти две
реакции объединены тем или иным образом. Важным фактором,
определяющим тип реакции у данного вида растений, является
то, в какой степени — высокой, умеренной или низкой —ок нуж-
дается в ресурсах среды.
Биотическая симпатрия, сохраняющаяся иа протяжении дли-
тельного времени, зависит, по-видимому, от совместного дейст-
вия двух факторов: слабой или неполной межвидовой конкурен-
ции и сильной репродуктивной изоляции.
11 Формы взаимоотношении
по плодовитости
Селекционеры дакни поняли, что разные группы растений резко
различаются по той легкости, с которой происходит скрещива-
ние между видами, и по плодовитости гибридов. В некоторых
родах близкие виды скрещиваются с трудом, а гибриды между
ними, если их удается получить, обычно стерильны. Другую
крайность составляют группы растений, у которых возможны
широкие скрещивания между морфологически различными вида-
ми, секциями или даже родами, и гибриды, полученные от таких
скрещиваний, плодовиты.
Результаты различных экспериментов по гибридизации в
очень многих случаях непредсказуемы. Однако при сравиитель
ном изучении достаточно большой выборам групп растений вы-
являются некоторые общие закономерности.
Основная часть данной главы будет посвящена обзору ряда
групп растений, главным образом из северного умеренного поя-
са, которые изучались в генетико-таксономическом плаке раз-
ными исслсдоэателями. Эти растения сгруппированы здесь по
жизненным формам и системам скрещивания. В заключитель-
ных параграфах очерчены общие закономерности и указаны не-
которые корреляции.
Древесные растения
Ceanothus
Североамериканский род Ceanothus (Rhamnaceae) занимает
область от южной части Канады до Гватемалы с центром рас-
пространения на тихоокеанском побережье. Из 55 видов этого
рода 44 эндемичны дли Калифорнии. Это кустарники с мелки-
ми душистыми цветками Опыление производят разнообразные
насекомые, обеспечииаюшис облигатное перекрестное оплодо-
творение у самонесовмеетммых видов и преимущественное пере-
крестное оплодотворение у самосовместимых видов Число хро-
мосом 2л = 24 у всех известных видов [614].
Виды этой группы делятся иа дм хорошо выраженные сек-
ции Ceanothus и Cerastes. Характер взаимоотношений между
видами у калифорнийских представителей Cerastes и между
Cerastes и Ceanothus установил Нове [614], сочетавший полевые
и экспериментальные исследования.
Виды секции Cerastes встречаются в самых разнообразных
экологических Местообитаниях на тихоокеанском побережье.
Они дифференцированы как морфологически, так и экологиче-
124 Часть,гргтьа. Гечетюмекме ч мимогижлы аспгкги euda
спи. В большинстве комбинаций эти виды (или полувиды) раз-
делены географически. В остальных сочетаниях их ареалы пере-
крываются. и оии встречаются на разных, но привыкающих
друг к другу типах почв в одной и той же области [614].
Ареал Ceanothus cuneatus перекрывается с ареалами по
крайней мере 5 других видов секции Cerastes в области, рас-
положенной к северу от бухты Сан-Франциско в Калифорнии.
Ceanothus cuneatus широко распространен на песчаниках, а в
некоторых районах, где па поверхность выходят серпентиниты,
образуя отдельные пятна среди окружающего песчаника, он
встречается вместе с двумя эндемичными для серпентинитов
видами — С. jepsonii в С. pumilus. В некоторых из таких зон
контакта естественной гибридизации не происходит или почти
ие происходит, тогда как в других имеет место значительная
гибридизация между С. cuneatus и видами или полувидами,
растущими иа выходах серпентинитов [614].
Нобс скрещивал друг с другом 12 видов секции Orasfes;
Домдоы ДЭ F, вм пет«рм мыяаостм
F, жвымкпоосвнс, с«ми« «мямАются дерамльмо
— Зшмквммие пгодре м мг«мвс%
Рже. 11,1, Плодовитость (Зрелое ери агрнявввкжв между рваными видвми
«явим Claries роде Ceanothut (613].
Гл II Фермы ллшиыхтвшма ло плоОсмгосп
125
результаты представлены на рис. 11.1. Все »ти вилы легко скре-
щиваются и дают в F, жизнеспособные гибриды. Гибриды об-
разуют жизнеспособные семена и пыльцу; фертильная пыльца
составляет от 88 до 97% всей пыльцы. Мейоэ у гибридов также
протекает нормально с регулярным формированием 12 бивален-
тов (614].
Члены секций Cerastes и Ceanothus симпатричны на обшир-
ной территории, и виды, принадлежащие к этим двум секциям,
обычно растут в тесном соседстве в одном и том же раститель-
ном сообществе и цветут одновременно. Естественные межсек-
ционные гибриды очень редка, несмотря на обширность области
контакта [614].
В свете приведенных данных интересно было обнаружить,
что между представителями этих двух секций существует реяро-
СЕКЦИЯ CERASTES
СЕКЦИЯ ЕОСЕЛМОТИиЗ
Лолуммем с/См* !*»*.» мы г мчОри^ы
Сввщванл не MMWWHMCb
Рж 11 2 Плодовитость шС-ридав гри гтрпцжмнмех между член л ин двух
секинЛ рода CeaMtkttt [613]. '
126 Чат rpms. Гемтимааи а пииояпгтк асяыты вам
дуктпвная изоляция, обеспечиваемая эффективными внутренне
ми барьерами Попытки Нобса произвести межсскцяонные скре-
щивании в большинстве случаев оказались совершенно безус-
пешными; гибриды не давали семян (рис. 11.2). В тех редких
случаях, когда удавалось получить несколько гибридных семян
и проростков, они обычно оказывались нежизнеспособными и
погибали на ранних стадиях. Еще реже межсекциоиные гибриды
достигали зрелости; при этом они были стернльиымн (фертиль-
ная пыльца составляла от 0 до 8%), и течение мейоза было
нарушено. При конъюгации хромосом у спонтанного гибрида
между С. masonii и С- foliosus число бивалентов госта ал ял о от
2 до 11, в среднем 7 [614].
Quercus
К обширному и широко распространенному роду Quercus (Fa-
gaceae) принадлежат многочисленные виды деревьев и кустар-
ников. Все они диплоидны, причем во всех случаях, в которых
был проведен цитологический анализ, оказалось, что диплоид-
ный набор (2л) составляют 24 хромосомы. Дли этих растений
характерно перекрестное встроопылеиие Все виды распределе-
ны между двумя четко разграниченными подродами — Quercus
и Erythrobalunus. Данных о взаимоотношениях между видами
дубов довольно много, ио они фрагментарны и разбросаны по
разным источникам; поэтому мы в своем изложении следуем
обзорам, составленным для некоторых географических областей
{58,780).
Виды или полувиды, принадлежащие к одному и тому же
подроду, обычно распределены по различным, но часто заходя-
щим одна в другую, подобно переплетенным пальцам, климати-
ческим или эдафическим зонам одной и той же области. Так,
среди калифорнийских представителей подрода Quercus вид
Quercus lobata произрастает на дне долин, a Q. douglasii — в
сухих предгорьях; Q. durata — ея выходах серпентинитов, а
Q durnosa — иа окружающей территория, где серпентиниты
отсутствуют. В Техасе Q moh'iana встречается на известняках,
Q. HavardU — ns песках, a Q. grisea— на выходах изверженных
иород [589].
Там, где виды или полувиды одного к того же подрода при-
ходят в контакт, они часто гибридизируют; в результате обра-
зуются обширные сикгамеоны, как эго будет описано в одной из
последующих глав. Естественные гибриды между видами, при-
надлежащими к одному подроду, по-видимому, более или меиее
плодовиты, и мейоз протекает у них нормально [780]. Такер
(839] обнаружил, что у гибридов между Quercus garryana и
Q. durata (весьма отдаленное скрещивание в морфологическом
Гл II Форни взаиттоиимиЛ по плодогяиып 1ST
смысле) Образуется от 93 до 97% фертильной пыльцы и иейоэ
протекает нормально, одна ко семена этого гибрида стерильны;
стерильными оказались также семена гибрида между Q garrya-
па и Q dumosa.
Дубы, принадлежа тис к подродам Qucrcus н Erythrobalanus,
довольно часто контактируют также и в природе. Тем не менее
гибриды между подродами в природе не обнаружены, а при
искусственных скрещиваниях возникают редко [770].
Ribts, Pinus. Eucalyptus. Prosopis
К роду Ribcs (Saxifragaceae) относятся свыше 150 видов кустар-
ников. Те из них, которые были изучены цитологически, оказа-
лись диплонлами (2п = 16). Этот род делится на четыре хорошо
различающихся подрода и на мноточисленные секции.
Кип 1429] к другие исследователи проводили множество скре-
щиваний между разными видами Ribes. Если деление иа секции
привести в соответствие с данными по скрещиваниям, то выяв-
ляются следующие закономерности. Во всех испытанных комби-
нациях вилы, принадлежащие к одной и той же секции, легко
скрещиваются друг с другом и лают плодовитых гибридов.
Скрещивания между представителями разных секций одного и
того же подрода бывают успешными не во всех случаях, меж-
секциоивые гибриды, если их удастся получить, обладают час-
тичной плодовитостью. Главный барьер, препятствующий скре-
щиванию у рола Ribas, существует между разными подродами,
которые хороню различаются также и морфологически. Репро-
дуктивная изоляция между подродами создается резко выра-
женной несовместимостью. В тех редких случаях, когда втот
барьер преодолевается, возникающие гибриды F, стерильны
[429].
В общем сходная карткпа наблюдается у рода Pinus
(2л-24), который делится на два хорошо выраженных подро-
да — Pinus (—Diploxyton) и Strobus (^Haptoxylon) Виды под-
рода Pinus успешно скрещиваются во многих комбинациях.
Успешными обычно оказываются скрещивания в пределах ка-
иейлибо группы видов, таких, как группа Pinus pondcrosa или
группа Р. radiala Некоторые из таких межвидовых гибридов
плодовиты. Подобным же образом представителей подрода
Strobus можно искусственно скрещивать в некоторых комбина-
циях. Однако нм одного успешного скрещивания между членами
разных подродов провести не удалось вследствие существующе-
го меяеду ними эффективного барьера несовместимости [129.
180, 579. 685. 906, 907].
Подобные же взаимоотношения обнаружены между видами
рода Eucalyptus (Myrtaceae) в Австралии. Этот род делится на
1№ Чмп r pm г Гмтлмти и мололиеоам шлегтм аш»а
пять подродов с многочисленными видами, у большинствч из
которых число хромосом 2п=22. Прайор [650] сообщает, что ви-
ды, принадлежащие к одному и тому же подроду, обычно зани-
мают экологически различные местообитании и в ряде извест-
ных случаев шперфертнльны. В отличие от этого виды, отно-
сящиеся к разным подродам, часто встречаются вместе, а скре-
щивание между ними предотвращается высоко эффективными
внутренними барьерами [660].
Аналогичная ситуация была описана сравнительно недавно
для Prosopis (Legwninoaae) [729].
Многолетние травянистые растения
с резко выраженными различиями
в механизме цветка
Aquilegia
Aquilegia (Raminculaceae)—один из родов многолетиях тра-
вянистых растений с яркими цветками, число хромосом 2п—14.
Этот род широко распространен в Евразии и Северной Амери-
ке. По всей этой области произрастают многочисленные имено-
ванные виды, многие из которых, по-видимому, представляют
собой географические расы. Разнообразные формы, относящие
ся к этому роду, естественно, распадаются на четыре или, быть
может, пять главных групп видов [270, 646].
1) Самый примитивный вид —эго Aquilegia ecalcarata из
Гималаев с белыми или пурпуровыми цветками, лишенными
шпор. Ои близок к родственному роду Isopyrum (2л—14). имею-
щему сходные цветки. 2) По всей Евразии и в северной части
Северной Америки распространено большое число форм, в том
числе группа видов A. vulgaris с поникающими синими или пур-
пуровыми цветками, снабженными длинными шпорами. Эта
группа делится на две подгруппы: подгруппа собственно A. vul-
garis с загнутыми шпорами н другая подгруппа A. alpina с пря-
мыми шпорами. 3) По всему умеренному поясу Северной Аме-
рики распространена группа A. canadensis, для которой харак-
терны помикающие красные цветки с толстыми шпорами.
4) В западных районах Северной Америки произрастают члены
группы A. chrisaalha с прямостоячими желтыми цветками, не-
сущими длинные тонкие шпоры.
В различных районах Саперной Америки трн группы biuob
распространены симпатрнчеекк. В некоторых местностях они об-
разуют естественные гибриды, сохраняя обособленность в дру-
гих районах, где они снмпатрическн перекрываются [270].
Первые работы, касающиеся взаимоотношений между вида-
ми Aquilegia по плодовитости, проведенные Андерсоном и Ша-
Г А II. Фирмы взоимоотмашспый по плодовитости 12Э
фером [21]. были продолжены Празмо (646—648). Как показали
эти исследования, члены четырех описанных выше групп видов
легко скрещиваю!си между собой в любых комбинациях и дают
плодовитые или полу плодовитые гибриды F( с нормальным те-
чением мейоза.
Так. Fi от скрещиваний двух даликнх в пределах этого рода
видов — Aquilegia ecalca/aia и /1. cu/farts — дает 42*% фертиль-
ной пыльцы. 19% фертильных семян и большое число intiomkob
Fj [645]. Fi от скрещивания .4. ecalcarala и Д chrysantha даст
62% фертильной пыльцы и 8% фертильных семян |647]. Потом-
ки Fi or скрещивания A. ccalrarata н .4 canadensis также лают
полуфертнльную пыльцу н семена |6491 А и Fi от скрещиваний
между членами групп Л vulgaris, A. enrysaniha и A canadensis
получаются полуплодовитые гибриды.
Таким образом, скрещиванию между группами видов у это
го рода препятствуют, очевидно, главным образом внешние
барьеры. У североамериканских водосборов эти барьеры созда-
ются экологическими, механическими и этологическими изоли-
рующими механизмами. Виды, принадлежащие к группе Aqui
legia vulgaris, обычно опыляюкя шмелями, виды группы A ca-
nadensis— колибри, я группы 4 chrysanlha — бражниками (94,
2701
Скрещивания между Aquilegia и isopyrum приводит к со-
вершенно иным результатам, нежели скрещивания внутри рода
Aquilegia Скалинскя [734] пыталась скрестить Aquilegia ecatca-
rala с Isopyrum thatief routes; при этом она обнаружила несов-
местимость па разных стадиях до в после оплодотворения.
Пыльцевые трубки Isupqtum прорастают в столбике Aquilegia
на незначительное расстояние Некоторые яйцеклетки все же
оплодотворяются Но затем, п течение двух первых педель эмб
риопвльного развития, либо происходит дегенерация эндоспер-
ма, либо ие ра лвмпаегся сам зародыш, так что и конечном иго-
те никаких гибридных проростков ие образуется (732).
Орхидные
Хорошо известно, что в группе орхидных можно проилипдить
скрещивании между очень далекими формами Виды рода Кал
da успешно скрещиваются с представителями девяти других р<г
Лов Представ и гслсА десяти родов подтрибы Laelieac (Cal/lcua.
Epidendfum. Laelia и другие) скрещивали примерно в 20 раз-
личных межродовых комбинациях Несколько родов подтрибы
Oncidieae успешно скрещиваются с родами, принадлежащими к
трем другим подтрибам — Coch liodcac, Comparctticac и Tricho-
centreac [463).
9-45
130 Чет гртя Гпгтичеож и жомонкюг «пмгм »ьЛа
К роду орхидных Ophrys в Средиземноморской области от-
оеится ряд видов, которые при искусственных скрещиваниях
яруг с другом дают гибриды, производящие фертильную пыль-
цу. Эти виды, встречаясь в природе в различных симпатриче-
скнх комбинациях, не гибридизируют в сколько-нибудь значи-
тельной степени. Их изоляция в природных условиях обеспечи-
вается главным образом различиями в строении цветков,
создающими этологические и механические барьеры, препятст-
вующие межвидовому опылению, которое производят пчелы
[7911
В одной из секций орхидных рода Vanda у межвидовых гиб-
ридов происходит нормальный мейоз с полной конъюгацией
хромосом, но у гибридов между видами, относящимися к дру-
гой секции того же рода, мейоз нарушен [424]. Нормальное те-
чение мейоза наблюдается также у гибридов между некоторыми
вшами Dendrobium [721].
Секция Oncocyclus рода Iris
Секция Oncocyclus рода iris (Iridaceae) — разнообразная груп-
па ирисов, произрастающих в юго-западных областях Азин.
Крупные цветки этих растений снабжены специализированными
механизмами, различающимися у главных групп видов; опыле-
ние производят крупные пчелиные (Xytocopa, Bombas, Antho-
phora). Изоляция разных видов в природе создастся географи-
ческими. экологическими, временными я механическими барь-
ерами. Никаких внутренних репродуктивных изолирующих
механизмов у этих видов не возникло. При искусственных меж-
видовых скрещиваниях, производимых во многих комбинациях,
снижения способности к скрещиванию или плодовитости гибри-
дов по сравнению с тем, что наблюдается при внутривидовых
скрещиваниях, нет или почти нет. Почтя у всех гибридов F, фер-
тильность пыльцы была близка к 90% [27].
Antirrhinum, Pens tenon, Epiphyllum
Рассмотрим кратко три другие группы многолетних растений с
яркими цветками, принадлежащие к этим группам вндм иитер-
фертильиы в широких пределах.
Большинство видов рода Antirrhinum (Scrophulariaceae). от-
носящихся к секции Anlirrhinaslrum (2п—16), более или менее
легко скрещиваются друг с другом и дают плодовитых гибри-
ден. Изоляцию между этими видами создают главным образом
экологические и этологические барьеры. Но виды секции Antir-
rhinastrum изолированы от Antirrhinum orontium и от секции
Гл IL Форлт еэвььоо’иоыиииа iw плодовитости 131
«ли выделенного в качестве самоего»тельного (сегрегатного)
рола Дклплл барьерам» иссоп мести мости [49, 560].
Три вида Penslcmon секции PeUanthera (Scrophulariaceae)
обитают в разных экологических зонах, ио произрастают енмпа-
трическн в различных комбинациях на юте Калифорнии, к этим
видам относятся Penslemon grinneilii, Р. spectabilis и Р centran-
thifolius Они резко различаются по строению и окраске цветков:
у Р. grinnelli венчик двугубы#, с широким зевом, у Р speciabilis
цветки Синеватые, с широкой трубкой, а у Р centranthifoliut —
красные, вороиковидные.
При скрещиваниях на экспериментальных участках все три
вида интерфертнльиы. В природе они гибридизируют в некото-
рых областях, где встречаются совместно, оставаясь изолирован-
ными в других областях. Изоляция создается главным образом
механическими и этологическими факторами. Penstemon grinnel-
Hi обычно опыляется пчелами-камеищицами (Xylocopa). Р. spec-
tabilis — осами псеваомазоридами и средних размеров пчелины-
ми, а Р. centranthilolius — колибри [808].
Виды Epiphyltum (Cactaceae) также сильно различаются ио
своим ярким цветкам. В пределах рода Epiphyllum, а также
между видами, которые иногда помешают в разные выделенные
из него роды, возможны отдаленные скрещивания. Так, можно
получить жизнеспособные гибриды, скрещивая между собой
вид с красными цветками, опыляемый колибри, и формы с бе-
лыми цветками, опыляемые бабочками [353].
Многолетние травянистые растения
с ярко выраженными межвидовыми различиями
в механизме цветка
Geum
Весьма обширный род Geum (Rosaceae). к которому относятся
примерно 56 видов, широко распространен по всему северному
полушарию, а также в умеренных областях южного полушария.
Эго многолетние травянистые растения с перекрестным опыле-
нием Основное число хромосом в трнбе, к которой относится
Geum. х—7. В роде Geum не известно ни одного диплонда, все
его члены — полиплоиды (от 4* до 12х) с числом хромосом от
2л = 28 до 2п - 84 [230. 231 j.
ГаеВски [230] делит род Geum на II подродов. Большая
часть генетико-таксономических исследований проводилась на
двух нз этих подродов, а именно — Oreogeum и Geum. К подроду
Oreogeum относятся три вида, распространение которых ограни-
чено горными районами Европы Два хорошо известных вида
в-
132 Чат грета* Гемеппесеее « тгмтеааи ошетх ««во
подрода Oreogeum — О. monlanum (4х) и G. replans (6х). Бо-
лее обширны* и широко распространенны* подрод Geum содер-
жит 25 видов, в том числе следующие (все они гексаплоилы);
G rivale, О. urbanum. G. macrophyllum. G. molle, G. coccineum.
G. hlspldum и G. eanadense [230].
В природе изоляция между этими видами создается глав-
ным образом экологическими и сезонными барьерами. Там. где
эти барьеры разрушаются, между видами происходит естествен-
ная гибридизация. Geum rivale и G. urbanum гибридизируют во
многих зонах контакта по всей Европе [228, 230].
На экспериментальных участках эти виды легко скрещива-
ются друг с другом. Любые два вила подрода Geum можно
скрещивать независимо от различий в уровне плондиости. Пред-
ставителя подрода Geum успешно скрещиваются с видами Oreo-
geum и четырех других подродов Но между группами более
высокого ранга — родом Geum и родственными родами в'аИ-
steinia и Coluria — вступают в действие очень эффективные
барьеры несовместимости, и все попытки провести межродовые
скрещивания оказываются безуспешными [230].
Жизнеспособность искусственных межвидовых гибридов
варьировала от гетерозисной до сублетальпой при разных ком-
бинациях скрещиваемых форм. В пределах подрода Geum гиб-
риды Fi от скрещиваний Geum macrophyllum с G. rivale. G. топ-
tanum или G. eanadense обладали нормальной мощностью, ио
рост гибридов между G. macrophyllum и О. urbanum. G. molle
или G. hispidum был сильно подавлен. Большая часть гибридов
от скрещиваний между членами разных подродов была жизне-
способной, ио гибриды F] от скрещиваний Geum и РГогопспвйт
никогда не цвели [230].
Стерильность гибридов также варьировала в широких преде-
лах— от почти нормальной плодовитости до полной стерильно-
сти. Так, |Дпример, в подроде Geum фертильность пыльцы гиб-
ридов F, от скрещивания Geum rivale и О, urbanum составляла
78. а семян — 72%; высокая плодовитость ияблюдялась также
в Fi от скрещиваний Geum rivale с G. coccineum, G. molle и
G. hispidum. Но гибриды G. rivale и G macrophyllum в Fi были
высокостерильны: фертильная пыльца я завязавшиеся семена
составляли всего 1%; очень высокой стерильностью отличались
и многие другие гибриды в подроде Geum. В подроде Oreogeum
гибриды между О. monlanum и G reptans производили высоко-
стерильную пыльцу и совершенно стерильные семена [230].
Некоторые скрещивания представителей подрода Geum и
Oreogeum друг с другом давали полуплодовигых гибридов Fj;
например, у гибридов G monlanum с G rivale фертильная
пыльца составляла 34%, а фертильные семена — 15% При др у
тих комбинациях •представителей подрода Geum и Oreogeum,
Г Я И. Фермы езаимллятммвнмй ял плоЛовпасги 1X3
например G. rivatexG. replant. получались высокостерилъиые
гибриды Все прочие скрещивания между «йенами разных под-
родов также приводили к высокой стерильности [230].
У гибридов между членами группы видов. в которую входят
Geum rioale. G. urbanum, G. nolle и G hisptdun, наблюдалась
довольно высокая способность хромосом к кон питании В раз-
ных клетках этих гибридов образовывалось от 17 до 21 бивален-
та. и их хромосомы были гомологичны пли почти гомологичны.
Но в клетках стерилыгых гибридов от скрещивании G macro-
phyllum с G rival? конъюгация сильно нарушается, число би-
валентов варьирует от 0 до 7, а уиивалептм многочисленны
Между двумя из трех геномов G, rival* (бх. подрод Geum) и
двумя геномами G. monlanum (4х, подрод Огео^еит) наблюда-
ется хорошая гомология. У гибридов между членами подрода
Geum и других подродов способность хромосом к конъюгации
была понижена [230].
Судя по поведению спонтанного нм фи пл он да от Geum г i vale
(6х) и G. macrophyllum (бг), эти виды различаются по хромо-
сомным перестройкам. У гибридов F> конъюгация нарушена,
число бивалентов равно 0—7, и гибриды эти, как уже говори-
лось, высокоетсрильпы От них были получены два растении
F* с удвоенным числом хромосом, т. е. 12х; в клетках этих ам-
ф илл он лов при мейозс формировалось до 42 бивалентов» и они
были плодовиты (229).
Есть, однако, другое фолы, ушмммм на то» что у гиб-
ридов Geun fiiiale и G macrophy Uum имеются нарушения в
конъю|зини хромосом, контролируемые генами. Если скрестить
G. rlvate или G. macrophyllum с третьим гексаплоидным видом,
G alepptcum, то происходит полная конъюгация с образованием
21 бивалента, иными словами, хромосомы G. rivale н G. macro-
phyllum более гомологичны, чем можно было бы думать, судя по
особенностям мейпэа у гибрида между этими видами 1229). Та
кнм образом, стерильность этого и других гибридов между вида-
ми рода Geum обусловлена. вероятно, пе только хромосомными,
но и генными различиями [229—231].
Solatium
К секции Tubcrarium рода Solomon (Solanaceae) относится
группа клубиеобразуюшнх видов со свободным неродственным
ек реши ван нем, которые образуют полиплоидный ряд от 2х до
6х 12). Эти виды легко скрещиваются в большинстве ком-
бинаций. Некоторые межвидовые гибриды плодовиты, и все их
хромосомы образуют биваленты, другие стерильны и способ-
ность хромосом к конъюгации у них понижена, но в общем все
эти виды цитогенетически весьма близки. Имеются данные, евм-
*33 Чат гртя. Гпчгаиаж и моюшшмш аатты аиОа
детельствующие о том, «то некоторые виды различаются по не-
большим структурным перестройкам [123, 311, 354, 542, 813,
-8UJ.
Секция Apogon рода Iris
Одна из естественных групп среди безбородых ирисов (секция
Apogon рода Iris) — ряд Californicae, или тихоокеанские ири-
сы, состоящая ю 11 полувидов, распространенных по тихоокеан-
скому побережью США, от шт. Вашингтон до юга Калифорнии.
К этой группе относятся свободно опыляющиеся многолетние
травянистые растения с числом хромосом 2л —40. В окраске и
форме их ярких цветков наблюдаются межвидовые различия,
во эти различия невелики, и, кроме того, они связаны интергра-
дацией [461. 462].
В пределах обшей области распространения тихоокеанских
ирисов отдельные полувиды занимают различные экологические
зовы. Например. Iris douglaslana растет вдоль береговой линии;
I. macrosiphon — дальше от берега, в предгорьях, а /. hartwe-
gii —в горах Сьерра-Невада, в эоне сосен подрода Diploxylon.
Местообитания этих в других полувидов во многих случаях не-
прерывно переходят одни в другие, что создает возможность для
краевых или смежно-симпатрических контактов между ними; а
таких случаях между подвидами происходит естественная гиб-
ридизация в многочисленных комбинациях [461, 462). Образуе-
мый ими сингамеон будет описан в гл. 18.
Смит и Кларксон [741], а также Ленц [462] искусственно
скрещивали тихоокеанские ирисы друг с другом в разнооб-
разных комбинациях. Эти полувиды легко скрещиваются. Гиб
ряды Fi вполне жизнеспособны, плодовиты и производят от 84
до 99% фертильной пыльцы. Мейоз у гибридов протекает нор-
мально с образованием 20 бивалентов.
Самые близкие родичи тихоокеанских ирисов — группа евра-
зийских видов с 40 хромосомами, принадлежащая к ряду SiM-
ticae. Члены этих двух рядов легко скрещиваются друг с другом,
образуя жизнеспособные гибриды, которые, однако, стерильны
1золее далекий родич тихоокеанских ирисов — Iris tenuis
(2л-28). относящийся к ряду Evansia [462]. I. tenuis растет в
Орегоне, где он симпатричен с такими члеиами ряда Calilorni-
еае. как /. tenax. Естественной гибридизации между /. tenuis и
1. tenax ие происходит; как установили Смит и Кларксон [741],
между ними существует эффективный барьер несовместимости.
Этот барьер создается блоками, действующим и на нескольких
Гл. 11. Формы паилиы пошгмиа ло плодооагости 135
стадиях. Пыльцевые трубки в пестиках чужого вида растут мед-
ленно и нередко разрываются. Оплодотворение либо пе происхо-
дит вовсе, либо задерживается. А развитие гибридных зароды-
шей в тех редких случаях. когда они образуются, прекращается
иа стадии 12 клеток. Искусственных гибридов между /. tenuis
и I. tenax полупить не удается.
Stlene и др.
К роду Silent (Caryophyllaccae) относятся многочисленные ви-
лы многолетних травянистых растений и несколько однолетни-
ков. Их основное число хромосом х=12; у этого рода известно
много диплоидных видов (2л—24), особенно в Европе, ио до-
вольно обычны и полиплоиды.
В западных районах Северной Америки произрастают 33 ви-
да травянистых многолетников с различной плотностью — от
2х до 8х; в большинстве случаев это тетраплоиды. ио есть среди
них также диплоиды, гексаплоилы н октоплоиды [447. 450]. Эти
заладноамгриканскис виды обладают резко выраженными мор-
фологическими различиями [451].
Крукгбсрг [448. 451] обнаружил, что западиоамерикаиские
виды легко скрещиваются между Собой в большинстве комбина-
ций и получающиеся при этом гибриды жизнеспособны, ио, за
немногими исключениями, высокостерильны. Конъюгация хро-
мосом у гибридов очень ограничена; наблюдаются у них также
и другие нарушения мейоза [448.451].
Обширные исследования, проведенные Викери [860] на М|‘-
mulus (Scrophulariaccae). показывают, что закономерности, об-
наруженные у Geum. наблюдаются также у многолетних пред-
ставителей секции Simiolus рода Mimulus, среди которых цент-
ральное место занимают группы М guttatus и М. giabralus. Эти
же закономерности выявликпея у Vernonia (Compoxitae) [418].
Однолетние травянистые растения
GUia
Одним из естественных подразделений гетерогенного рода GUia
(Polemoniaceae) является Секция GUia, или облнстиениостебсль-
вые гилин. К этой секции относятся десять видов однолетних
травянистых растений Два вида гилин — GUia capitata я G tri-
color— мы уже рассматривали в гл. 4. во в связи с другой
проблемой
Облнствеиностебельные гилин произрастают на тихоокеан
IX
Чат гатя. Гаятчкиа а ажожнмчегхи ооттм падл
сном побережье Северной Америки н в умеренном поясе Южной
Америки, где ареалы разных видов перекрываются и где они
часто встречаются в тесном соседстве в различных комбинациях.
Система размножения варьирует у разных членов группы от
свободного перекрестного опылеиия до автогамии. У шести ви-
дов число хромосом 2п = 18, а у трех тетраплондов —2п=36.
Еще один, десятый, вид не изучался ни цитологически, ни гене-
тически [274, 286).
Все эти виды в большинстве комбинаций разделены мощ-
ными барьерами несовместимости. Если и удается получить
гибриды Fi. то они высокостерильны. Мейоа у гибридов нару-
шен, причем способность хромосом к конъюгации настолько по-
нижена, что в некоторых случаях образуется всего по одному
биваленту на клетку [268).
Низкая способность к конъюгации, наблюдаемая у гибридов,
обусловлена главным образом различиями между родительски-
мм видами по структурным перестройкам, о чем свидетельст-
вуют особенности искусственных амфнплоидов. У стерильных
гибридов с нарушенным мейоза», полученных нз эксперимен-
тальных участках путем скрещивания разных видов в четырех
различных комбинациях, произошло спонтанное удвоение хро-
мосом; в результате возникли амфнплондиыс потомки, хромосо-
мы которых нормально конъюгировали с образованием бивален-
тов, а плодовитость колебалась от средней до высокой [268,
286). В первых поколениях амфнплоидных потомков наблюда-
лось расщепление как по поведению хромосом, так и по плодо-
витости; это служит веским доказательством того, что в данном
случае, помимо хромосомных перестроек, имеют также место
нарушения мейоза. контролируемые генами [271].
Подобная же картина наблюдается в близкой к предыдущей
секции Arachnion, состоящей также нз многочисленных видов
однолетних травянистых растений, которые часто встречаются
в природе снмпатрнчески. Виды эти обычно разделены эффек-
тивными барьерами несовместимости, а гибриды между ними
в тех случаях, когда их удается получить, высокостерильиы.
Стерильность гибридов имеет в основном хромосомную, а час-
тично— генную природу [158, 285, 300].
Layla
К роду Layla (Compositae) принадлежит IS видов однолетних
трав, произрастающих в предгорьях н долинах Калифорнии и
Цветущих весной. В этой области тесная симпатрия 2 — 3 ви-
дов—обычное явление. По числу хромосом эти виды можно
разбить на три серии: у шести диплоидных видов число хромо-
rt II. Формы ахшжятг>ваше*ша па плоОолигооли
137
Рис ПЛ Плодовитость гнСрид-лв при скреишвааиях между рвмиии внда-
нж Madia [96].
сом 2п-14, у восьми диплоидных видов — 2п—16 к у ОДНОГО
тетраллоидного— 2п=32 [428].
Виды, имеющие по 7 пар хромосом, успешно скрещиваются
между собой. Большинство видов с 8 парами хромосом также
способны скрещиваться друг с другом. Однако между видами с
7 и видами с 8 парами хромосом, а также между некоторыми
видами с 8 парами существуют эффективные барьеры несовме-
стимости [96, 102].
1Э* Часп трегьл. ГtntruviMU и rwaaoMtcvu аникты гида
Гибриды между некоторыми видами, например между Layla
gaillardioidet н L. hetarotricha, нежизнеспособны, а такие скре-
щивания, как L galllardioidesXL. hitracioldta, сопровождаются
разрушением гибридов. При многих межвидовых скрещиваниях
наблюдается стерильность гибридов, сопровождающаяся пару
шеннямя мейоза {96. 428].
Способность хромосом к конъюгации у межвидовых гибри-
дов сильно различается в разных комбинациях скрещиваемых
видов. При скрещивании диплоидных видов, принадлежащих к
одной и той же группе, у потомков F, конъюгируют все или поч-
ти все хромосомы. В отличие от этого у гибридов от скрещива-
ний между членами различных групп видон способность хромо-
сом к конъюгации понижена. Так, у гибридов между Layla hate-
rotricha (л—8) и L glandulota (л=8) образуется от 0 до 2 би-
валентов, а у гибридов между L. heterotricha (п=8) и L. pla-
tyglossa (п=7) —от 0 до 3 бивалентов [96, 428].
Клаузен [96] полагает, что снижение способности хромосом
к конъюгации у межвидовых гибридов Layla обусловлено ген-
ной дисгармонией между родительскими вилами, а ие структур-
ными перестройками в их хромосомах. Одиако имеются данные,
свидетельствующие о том. что стерильность у Layla вызывается
как хромосомными, так к генными различиями. У гибридов F,
от скрещивания Layla platygloua (п=-7) и L pmtachaata
(л-8) образуется от 0 до 3 бивалентов. Эти гибриды путем
удвоения хромосом дали некоторое количество тетраплоидиых
потомков с числом хромосом 2л—30. У тетраплоидных растений
хромосомы нормально конъюгировали, образуя 14—15 бивален-
тов, мультввалситов н вторичных ассоциаций, по-видимому, не
было. Последние стадии мейоза были, однако, нарушены, и эти
тетраплоиды оказались стерильными (102, 428].
Сходную картину Клаузен и его сотрудники [96) обнаружили
у родственного Layla многолетника Madia (СошрозНае)
(рис. 11.3).
Clarkia в др.
Род Clarkia (Onagraceae) содержит много видов однолетних
трав, которые часто встречаются в природе симпатрическп. Как
и в случаях с GUia и Layla, жги виды изолированы мощными
барьерами несовместимости. При некоторых скрещиваниях гиб-
риды оказываются нежизнеспособными. Если же гибриды дости-
гают зрелости, то они бывают более нли менее стерильными.
Стерильность в значительной степени обусловлев» хромосом-
ными различиями [496].
В сущности такая же картина наблюдается у многих других
родов однолетних растений, например у Brassica (СгисИегае,
Гл. II. Формы влнмыогатеытеД те «xedoeuromi 139
|916]), Spergutaria (Caryophyllaceae, [6561), в группе Alriplex
triangularis (Chenopodiaceae, [320]), в однолетних секциях Ni-
cotiana (Solanaceae. [26. 250]), Collinsia (Scrophulariaceae. (232,
233, 235]). Microseris (Compositae, [92]), Helianthus (Cooiposi-
tae, [374]), Slephanomcria (Compositae, [251]), в секции Cera-
tochloa рода Bromus (Gramineae [332. 794, 795]), Avena (Gra-
mineae. [653]).
Автогамные однолетники
Cilia inconspicua
Комплекс Cilia inconspicua образует группа низкорослых авто-
гамных видов с мелкими цветками, относящихся к секции Агас-
hnion рода Gi/in (Polemoniaceae). Известно 25 видов, принад-
лежащих к этому комплексу. Они широко распространены по
всем аридным оЛзастям западной части Северной Америки, где
встречаются 23 вида, а также в умеренном поясе Южной Аме-
рики. Основное число хромосом у них х—9 Это обычно диплои-
лы и тетраплоиды; известей также одни октоплокд [283].
Виды, входящие в »тот комплекс, сходны, хотя и неидентич-
ны по своим морфологическим признакам, н нх трудно опреде-
лять обычными таксономическими способами. В гл. 4 были рас-
смотрены и проиллюстрированы черты сходства между пятью
видами комплекса Cilia inconspicua, а именно G fransmonta-
па, С. malior, G. minor, G. aliquanta и G. clokeqi. Кроме того,
эти растения часто образуют симпптрнческие скопления из 2,
3 или 4 видов, вследствие чего некоторые из ранних системати-
ков пришли к логическому заключению, что все зтн формы от-
носятся к одному изменчивому виду кли двум котерградирую-
щнм видам. В настоящее время на основе данных, полученных
в полевых и экспериментальных исследованиях, установлено,
что группа G. inconspicua представляет собой комплекс видои-
мойиикое [283].
Между всеми этими видами почти в любых комбинациях
действуют эффективные барьеры несовместимости (рис. 114).
Если удается получить межвидовые гибриды, то они почти во
всех случаях высоко стерильны (ряс 11.4); конъюгация хромо-
сом у гибридов выражена слабо. Несколько групп данных сви-
детельствуют о том, что виды этой группы различаются по хро-
мосомным перестройкам, т. е. о том, что стерильность гибридов
имеет хромосомную природу; однако имеются и другие данные,
говорящие о наличии также и геннпй стерильности [158, 283,
140
Часть третья Гечетиюъые и «ь:л«т« вис>а
- с«9мм«*ми» еечскмию «• >д*поск
— Г •«во»** выоо«*осг«р««пы*ы о - 9% фертильное л мшим)
Гивря** г->трс'«*ии*мм (12 — MS ферпиьмеа пыл»цы|
Рве. 11.4. Фертильность гмйрвдев ком скрещшанааж м«жду видамв-доАкв*
кама на группы OUia <п4ош?*с*и [283]
INT — О. interior; AUO — О. auatrcn'cddentalie; JAC —О jacaiu; MIN —
О. minor; BRE — G. breccierum, DIEG — G dtsgenus; TWE — G twdyl;
INC —G. incoiupicaa вела Str; TRA —0. fraasmonJona; MAL — 0. mattor;
FLA—0 ftotocincia; OPH —G. ophthalmotdes; S It — G. unuata tt; S I —
0. xinun/a 1; MOD —G modnwuts; TET — 0. tetrakreccia; С 4ж — О craisi-
*0йа 4x. C 8x G cfastifoUa 8r. OCH — G. oc.kroieuca; MEX — G mericaia;
CLO — 0. BRA — G. ebradbury»; ALI — G alt^uania.
Erophlla, Elymus н др.
Комплекс Efophila verna (Crueiferae)—группа морфологически
сходных автогамных однолетних трав, произрастающих в Евро-
пе. Основное число хромосом к—7. В этом комплексе есть дкп-
лоиды (2n~U). полиплоиды и пазличпые аиеуплоиды с числом
хромосом вплоть до 2л64 [901J.
Всю эту группу было принято рассматривать как один таксо-
номический вид — Draba vtrna. или Erophlla verna. Ее гскети-
Гл. 11. Формы ялтмоотматвкий по плодовитости HI
ко-тэксонлмичсский состав анализировали несколько исследова-
телей, а в особенности Винге [901]. который считает, «по она
состоит из четырех видов-двойников. Эти пиан встречаются в
Европе симпатрнчески в разных попарных комбинациях. Винге
1901 ] показал, что между видами группы Erophlla oerna су шест
вует барьер стерильности, создаваемый несовместимостью и
различиями в структуре хромосом.
Как показывают экспериментальные данные, симлатрнческие
виды-двойники, разделенные барьером хромосомной стерильно
стн, существуют также у автогамного многолетнего злака Ehj-
tnus glaucm |750, 751 ].
Судя по цитотаксоиомнчсскнм данным, такие же симнатрн-
ческие виды-двоими кн имеются по крайней мере в трех других
группах автогамных однолетников- rfcit satiua (begun- О
(337J, Camtssonia migrant ha {Onagraceae, [658]) и Мм/деЯа
albicauhx (Loasaceae).
Общие модели
Наша следующая задача заключается в том, чтобы обобщить
все данные, представленные а этой главе При этом можно вы
явить следующие общие модели {к качестве примеров приведе-
ны тс таксоны, которые подробно рассматривались о предыду-
щих разделах главы),
1. Модель Ceanothus Древесные растения со свободным ис-
родс-тиеиным скрещиванием и более иди менее неслеппалнзнро-
ванными цветками. Разные виды совместимы друг с другом,
плодовиты при скрещннамнях, и нх хромосомы гомологичны в
широких пределах. В природе виды изолированы или полунзо-
лированы посредством экологических и других внешних факто-
ров Между разными секциями или подродами существует силь-
ный барьер несовместимости. Ceanolhus, Quercus, Ribes, Pinus,
Prosopis, Eucalyptus
2. Модель Aquilegia. Многолетние травянистые растения со
свободным неродственным скрещиванием и специализированны-
мн цветками. Разные виды совместимы Друг с другом, плодов и
ты при скрещиваниях, и их хромосомы гомологичны в широких
пределах. Изоляция между видами создается механическими и
этологическими барьерами, а также экологическими и другими
внешними факторами. Между разными секциями, подродами или
родами существует сильная несовместимость. Aquilegia, Antir-
rhinum, Penstemnn. Epiphyllum. секция Oncoeyclus рода Iris,
орхидеи.
3. Модель (jeum Многолетние травы со свободным нерод-
ственным скрещиванием и умеренными межвидовыми различия-
ми в строении цветков. Разные виды совместимы друг с другом
в широких пределах, но между секциями или подродами суще-
ствуют эффективные барьеры несовместимости. Виды, принад-
лежащие к одной я той же группе видов, иитерфертклъиы и их
хромосомы гомологичны; гибриды между видами, относящимися
к разным группам, характеризуются хромосомной или генной
стерильностью. Родственные виды изолированы в природе эко-
логическими или другими внешними факторами. Изоляция сход-
на с наблюдаемой у тина 2. но она в большей мере обусловлена
хромосомной н генной стерильностью, чем механизмами, созда-
ваемыми строением цветка. Geum, Sotanum, секция Apogon рода
Iris, Silent секция Slmiolus рода Mimulus, Vernonia.
4. Модель Madia. Однолетние травы с разнообразными систе-
мами размножения —от свободного неродепккного скрещива-
ния до инбридинга. Близкие виды обычно изолированы барьера-
ми несовместимости, а также хромосомной и генной стерильно-
сти. Brassica, Spergularia. Atriplex, Clarkia, Gilia. Nicotiana,
Collinsia, Layia, Madia, Microseris. Helianthus. Stephanomeria,
Bromus, Avena.
5. Модель Gilla inconspicua. Однолетние или, реже, много-
летние травы с автогамной системой размножения. Отдельный
таксономический вил состоит кз нескольких или многих видов-
двойников. изолированных барьерами несовместимости, а также
хромосомной и генной стерильностью. Gillo Inconspicua, Erophi-
la verna. Elymus glaucus, Vida saliva. Camissonia micrantha.
Mentzetia alblcautis.
Большая часть генетико-таксономических исследований про-
водилась иа группах растений, встречающихся в северном уме-
ренном поясе. О прироХе барьеров, создаваемых стерильностью,
и их распространении среди растений южного умеренного и тро-
пического поясов известно гораздо меньше. Это отразилось и
иа подборе родов, рассмотренных в данной главе. Поэтому те
типы, которые здесь были выделены, отнюдь ие исчерпывают все
возможности, и в будущем число их, несомненно, возрастет в
результате дальнейших генетико-таксономических исследований.
Тропический американский древесный род Theobroma (Ster-
culiaceae) отличается высокой межвидовой несовместимостью,
подобно умеренным однолетним родам, и значительной нежиз-
неспособностью гибридов [2].
В Австралии некоторые группы древесных растений, такие,
как Podocarpus. Proteaceae, некоторые Myrtaeeae и некоторые
Epacridaceae, претерпели дифференциацию а отношении струк-
туры хромосом, сравнимую с наблюдаемой во многих группах
травянистых растений северных флор (325. 413. 414 . 746. 747).
Цитологические и морфологические данные позволяют считать,
что австралийской флоре, возможно, присуще пскоторое сносов-
разие, заключающееся в наличия а пей древесных растений со
специализированными цветками, опыляемыми животными, н с
дифференциацией по структуре хромосом внутри родов и меж-
ду ними.
Другая интересная и своеобразная черта австралийской и
новозеландской флоры связана с родом Epilobium (Onagraceae),
к которому относятся автогамные многолетние травы. Плодови-
тость при скрещиваниях и гомологичность хромосом — общее
правило для большинства межвидовых комбинаций в этой груп-
пе [ббб].
Рассматривая модели, которые мы идентифицировали и об-
судили выше, следует отметить ряд общих черт в генетических
системах филогенетически неродственных друг другу родов.
Черты сходства между Ceanothus, Quercus и Pinus или между
Madia, Clarkia и Gilia, а также коллективные различия между
зтимн группами родов выражены очень резко. Выделенные ти-
пы, по-андимому, отражают подлинные конвергенции между
членами разных семейств растений.
Общие корреляции
Для того чтобы создать более широкую основу для наших обоб-
щений, можно использовать более обширный набор групп рас-
тений, чем тот. который мы рассматривали до сих пор в этой
главе Несколько лет назад я просмотрел литературу по 7'2 груп-
пам (родам, секциям или подтрибам) растений, которые изуча-
лись в геиетико-таксономическом аспекте. Я распределил эти
группы растений по разным категориям на основе наличия или
отсутствия несовместимости и барьеров стерильности между
родственными группами Эти же самые филт>ды можно также
сгруппирован, в три класса в соответствии с жизненными
фирмами
Корреляция между жизненной формой и типом взаимоотно-
шений по плодовитости представлена в табл 111. Рассмотрим
скачала распределение барьеров несовместимости. Виды, при
надлежащие к одной и той же секции или группе видов, обычно
легко скрещиваются между собой у древесных растений и мно-
голетних трап, ио быняют разделены барьерами несовместимости
у однолетних трав (табл. 11.1).
В распространении барьеров стерильности выявляется та же
самая корреляция с жизненной формой Гибриды Fi. получен-
ные при скрещиваниях между родственными видами, обычно
плодовиты или частично плодовиты у древесных растений и
многолетних трав, ио у однолетних трав они а большинстве слу-
чаев стерильны (табл II I).
К подобным же выводах пришли н другие авторы (ИЗ, 144.
144 Чат rper»a Гмзпшсш и жолоогчсин аспекте еж>в
Таблица 11.1. Распространение барьеров несовместимости и стерильности в группах растений.
относящихся к различным [280] с нэп жизненным формам (по енеииямн)
Чтиво фммд. которш :«р|щп1И4 между влкМсШВ
видами £4>м»ио и^мв>9кгт а глсдуюскжм р^вультвтвм
Жммсаиаа форма
с*реший*ейкст* с’трмвмеост» гв4ри««
•аассзадм знака в меовли МЯ1ЫВ
Деревья и кустарника 19 2 II 2
V Ммпгт.мстнне траяы 2» 0 15 6
Однолетние трааы 3 19 3 19
194, 783], основываясь на нехавнсниом изучении имеющихся
данных.
В нескольких случаях оказалось возможным сравнивать раз-
ные секции одного и того же рода в отношении корреляции
между жизненными формами и эффективности барьеров сте-
рильности. У рода Helianthus многолетние виды, состаплякицне
группу Н. giganteus, оказались интерфертнльмыми, тогда как
однолетники, относящиеся к группе Я. оллниз. иитерсте-
рнлькы (374].
У Crepis (Compositae) члены трех секций однолетних трав,
а именно Яostia, Phytodesia и Nemauchenes, при скрещивании
друг с другом дают стерильные гибриды. В отличие от этого
скрещивания между многолетними видами секции Lepidoseris
в большинстве случаев дают плодовитые гибриды; однако п
другой секции многолетников (Berinia) межвидовые гибриды во
многих случаях стерильны 133].
Однолетние виды рода Knautia (Dlpsacaceae). относящиеся
к секциям Tricheroides и Knautia, хороню различаются по споим
геномам и изолированы друг от друга эффективными внешними
и внутренними репродуктивными барьерами. Многолетние же
виды, принадлежащие к секции Trichera, напротив, сходны по
своим геномам, в репродуктивная изоляция между ними выра-
жена слабо (190].
Так случилось, что большинство деревьев, кустарников и
многолетних трав, включенных п каш обзор, размножаются пу-
тем свободного неродственного скрещивания, тогда как многие
из рассмотренных групп однолетников содержат частично илн
преимущественно самоопыляющиеся формы. Возможно поэтому,
что наблюдаемая в табл. 11.1 корреляция между стерильностью
и жизненной формой отражает на самом деле более глубокую
зависимость между типом системы размножения и образованием
барьера стерильности.
1л ll Форма, «лаалюогтеошгик^ пи плоЛмш.'иои
14-S
Выводы
Многие группы многолетних растений северной умеренной зоны
характеризуются своеобразным типом изоляции: близкие вилы
обычно плодовиты при скрещиваниях друг с другом, и их изоля-
ция в природе обеспечивается главным образом экологическими
и другими внешними факторами; но изоляция между более да-
лекими видами, принадлежащими к разным секциям, подродам
или родам, обеспечивается внутренними барьерами, блокирую-
щими скрещивание или же успешное развитие гибридных за-
родышей.
Эти барьеры, создаваемые несовместимостью и нежизнеспо-
собностью и контролируемые генами, возникают в таксономиче-
ской структуре данной группы растений внезапно. Внутренние
каналы, по которым происходит обмен генами между видами,
открыты в довольно широких таксономических пределах, но .за
этими пределами они становятся закрытыми.
Для многих однолетних и некоторых многолетних трав уме-
ренной зоны северного полушария характерен иной тип изоля-
ции. У них между близкими видами обычно действуют внутрен-
ние изолирующие механизмы, создающие несовместимость и
гибридную стерильность. Кроме того, в этих группах преобла-
дает стерильность, обусловленная структурой хромосом. Неред-
ко у них существуют также внешние изолирующие механизмы
и генная стерильность, однако эти последние играют лишь вто-
ростепенную роль.
Можно считать, что группы видов, плодовитые при скрещи-
ваниях, и группы видов, стерильные при скрещиваниях, облада-
ют некоторыми общими для тех и других основными механизма-
ми изоляции. Экологическая изоляция, репродуктивная изоля-
ция, создаваемая внешними механизмами, и различные барьеры,
контролируемые генами, имеются и у тех и у других групп расте-
ний. Одно из главных различий между ними — интерполяция
хромосомной стерильности в пределах некоторого круга родст-
венных форм в одном случае и отсутствие этого—в другом (280).
10—«*.
о
Часть четвертая
Эволюционная дивергенция
12 Способы видообразования
Виды различаются, как мы видели в гл. 8, по адаптивным приз-
накам, контролируемым сложными комбинациями генов. Сле-
довательно. видообразование сопряжено с возникновением но-
вых к различных комбинаций генов в обособленных популяциях.
Этот аспект процесса видообразования относится к эволюцией
иым видам вообще. Что же касается биологических видов, со-
стоящих из перекрестно оплодотворяющихся организмов, то у
них видообразование связано, кроме того, с развитием репродук-
тивных изолирующих механизмов.
При обсуждении видообразования удобно рассматривать
упомянутые выше события — развитие новых генных комбина-
ций и изолирующих механизмов — как завершение процесса ви-
дообразования. Теперь мы можем перейти к рассмотрению сил.
обусловливающих процесс видообразования, н Нолей, на кото-
рых протекает «тот процесс.
Силы и поля
Эволюционные силы, участвующие в видообразовании, можно
разделить на две группы; силы, создающие изменчивость, и си-
лы, закрепляющие возникшие изменения в дочерних популя-
циях. Ниже перечислены силы, создающие изменчивость и иг-
рающие определенную роль в процессе видообразования; каж
дой из этих сил дано буквенное обозначение, которым мы будем
пользоваться в дальнейшем.
Изменчивость, существующая в обычных условиях (е)
Гибрилизация (Л)
Новые мутации (и)
Силы, которые закрепляют изменчивость и значение которых
для видсюбразопэияя либо установлено, либо предполагается
некоторыми учеными, сводятся к следующим.
Отбор с широким свободным неродственным скрещива-
нием (а)
I Л. 12 Способы »U$wf>pain*&*U1
I4T
Инбридинг в небольших популяциях (и отбор) (^)
Инбридинг в результате самооплодотворения (и отбор)
Or)
Инбридинг в результате ассортативного сирешимиия
„ «•>
Силы, создающие изменчивость, и силы, закрепляющие эту
паиенчиоость. могут комбинироваться самыми разнообразными
способами, в результате чего возникают различные двустадий-
ные процессы, например a—s. u— i, и т. п.
Поля, на которых могут происходить процессы видообразо-
вания, бывают аллопатрическими, смежно-симпатрическими и
биотическн-симпатрическиин. В результате сочетания этих трех
полей с различными комбинациями сил возникают многочислен-
ные способы видообразования.
Способы видообразования
Число возможных комбинаций ИЗ 3 ТИПОВ СИЛ, СОЗДАЮЩИХ 113-
менчньостъ, 4 типов сил, закрепляющих эту измеичивостъ, к
3 тнпоп полей, конечно, достаточно велико, по многие из этих
возможных способов нереалистичны и ими можно пренебречь.
Остается ряд способов, из которых одни известны, другие веро-
ятны, но существование нх не установлено, а третьи хотя н спе-
кулятивны, но часто упоминаются в текущей литературе.
Ниже перечислены, с краткими замечаниями, известные, ве-
роятные или серьезно обсуждаемые способы видообразования.
1. Аллопатрическое поле, о—г Географическое видооб-
разование. Хорошо известно.
2. Аллопатрическое поле, и—ip. Квантовое видообразо-
вание. Известно.
3 Аллопатрическое поле, v—if. Аллопатрическое видо-
образование, связанное с самооплодотворением. Сход*
но с квантовым видообразованием Известно у расте-
ний.
4. Смежио-снмпатрнческое поле, о— <й Смежно-симпат*
ричесьое видообразование, Весьма вероятно у насеко-
мых со свободным неродственным скрещиванием и у
других беспозвоночных, занимающих разные узкие
нищи и сосуществующих в одном и том же местооби-
тании.
5 Бнотнчески-епмпатричссхое поле, А—к—г Аллопо-
лмпломдпое видообразование. Хорошо известно у рас-
тений.
б. Внотяческн-симпатрнческое поле, Л—i Рекомбинаци-
онное видообразование и гибридное видообразование
4U Частл ытмртял. Эаожочтнмиил Лт»»рт««|ри
с сегрегацией внешних барьеров. Весьма вероятно у
растений
7. Биотическисимпатрическое поле, о—«/. Биотически-
симпатрнческое видообразование у самооплодотво-
ряютцнхся организмов Вероятно для некоторых рас-
тений. беспозвоночных и протестов.
в. Биотически-симпатрическое поле, о—s или о—<«.
Биотическисимпатрическое видообразование у сво-
бодно скрсщивающяхся организмов при действии ляэ-
Й-ппаввого отбора или ассортативного скрещивания.
екоторые ученые рассматривают этот способ как
реально возможный.
Обсуждение
Описанная выше система классификации способов видообразо-
вания на основе поля и действующих сил взята нз моей более
ранней работы [282J. Эта система носит общий характер и при-
ложима к животным, растениям и протестам. Со времени ее
первого опубликования в 1963 г. появились три другие общие
классификации способов видообразования [77, 894. 913].
Система Буша [77] основана на географическом поле; систе-
ма Уайта [894]— на поле и наличии или отсутствии хромосом-
ных перестроек; система Райта [913] —на поле, а также на
том. является ли дивергенция первичной или она связана с гиб
риднзацией. Кроме того, между этими системами имеются па-
раллелизмы, перекрывания в различия. так что подробное срав-
нение их завело бы нас слишком далеко. И все же здесь сле-
дует сделать несколько частных замечаний.
Олив из выделяемых Бушем [77] способов видообразова-
ния—это парапатрическое видообразование. Я не пользуюсь
этим термином, потому что парапатрик, в сущности, представ-
ляет собой одну из форм аллопагрни. и. следовательно, в этом
термине пет необходимости. Парапатрическое видообразование
в том смысле, какой в пего вкладывает Буш [771 по-видимому,
соответствует моему «аллопатрическому видообразованию у си-
дячих организмов» (см. [282]), а это один из подтипов географи-
ческого видообразования. В системе Уайта [894], претендующей
на приложимость ко всем высшим организмам, упущено не-
сколько способов видообразования, играющих важную роль у
высших растений, а поэтому она ие совсем применима при изу-
чении эволюции растений.
Уайт [894] выделяет особый способ видообразования, кото-
рый он называет стасипатричсским видообразованием. При ста-
сипатрнческом видообразовании в облаете, занимаемой данным
видом, появляется новая хромосомная перестройка, которая об-
Гл IS- Способы авдообраооплпил 149
разует а этой области новую гомозиготу по структурной пере
отройке. Этот способ видообразования Уайт противопоставляет
аллопатрическим способам видообразования Я считаю данные,
иа основе который выделяется стасипагрическое видообразова-
ние. неубедительными, а сам способ гипотетическим, хотя Уайт,
очевидно, со мной ие согласится. По крайней мере некоторые
из приводимых им примеров можно так же хорошо или даже
лучше объяснить как случаи квантового видообразования, свя-
занные с хромосомными перестройками.
Наша собственная система, изложенная выше, дает приемле-
мую основу для обсуждения способов видообразования у расте-
ний. Обратите внимание иа то, что способы 5 и 6 в нашем пе-
речне представляют собой типы гибридного видообразования
у растений, Они будут обсуждаться в дальнейших главах, в пя-
той частя книги Другие способы — это аспекты первичной эво-
люционной дивергенции, которые рассматриваются в следую-
щей главе лол названием первичного видообразования.
13 Первичное видообразование
Пол первичным видообразованием здесь имеется в виду про-
цесс эволюционно# дивергенции между популяциями до видом?
го уровня. Мы относим к первичному видообразованию все явле-
ния. обугловливаюшие первичную эволюционную дивергенцию,
и исключаем из него явления гибридного видообразования н
усиления изоляции, которые обсуждаются отдельно в последую-
щих главах.
Процессу видообразования в качестве начального условия
необходим запас генетической изменчивости. Такая изменчи-
вость представляет собой всеобщее явление в органическом ми-
ре. Существует три уровня изменчивости, важных для видообра-
зования; полиморфизм, локальная расовая изменчивость н геог-
рафическая изменчивость. Эти типы изменчивости уже были
описаны в гл. 2 и 3.
Соотношение между разными уровнями изменчивости опре-
деляет путь первичного видообразования. Наиболее хорошо
известен и документирован путь от полиморфиэмдлевез локаль-
ные, а затем географические пасы до видов. Этот путь хорошо
описывается географической теорией видообразования, которую
мы рассмотрим более подробно в следующем разделе этой гла-
вы. Следует также рассмотреть альтернативные сокращенные
пути возникновеиня новых дддоя Путь, при котором выпадает
формирование географических рас, носит наааавма .аплитового
видообразовании. В таких случаях новый вид развивается, цд
какбГлибб локальной расы. Путь биологически симпатрическо-
П> видообр аэовання~ббхСДЯт как лока льные, та к н геогсаФмче-
схие тисы Предполагается. что новый вид развивается__нсдо-
средственно из какой-лвбо уариациудшДорфйого вида Пер-
вичное видообразование в биотически-симпатричесхом поле —
дискуссионная проблема, которую мы попытаемся разобрать в
одном из дальнейших разделов этой главы.
Географическое видообразование
Географическая теория видообразования имеет давнюю исто-
рию. Ее основные концепции неоднократно формулировались с
разными дополнениями и разъяснениями рядом зоологов, в том
числе такими выдающимися учеными, как М. Вагнер (863],
К. Джордан (421]. Д. С. Иордаи [420], Рекш [676], Добржая-
схий [166] и Май|Г[5б4]. Обзор этой проблемы в историческом
плане представлен в работе Майра [569].
Гл. 13 Первично? 1S1
Теория географического видообразования в ее современной
форме утверждает, что географические расы — эго предшествен-
ники видов в непрерывном процессе эволюционной дивергенции
(см рис. 7.1). Пространственная изоляция популяций на расо-
вой сталии дивергенции создает возможность для развития и
поддержании у отдельных популяций комбинаций генов, опреде-
ляющих отличительные морфологические н физиологические
признаки этих популяций На этой же сталии начинает закла-
дываться репродуктивная изоляция. А формирование видов
представляет собой дальнейшее развитие указанных процессов.
Репродуктивные изолирующие механизмы, развивающиеся в
течение периода пространственной изоляции двух или несколь-
ких популяций, позволяют этим популяциям сосуществовать
друг с другом, нс скрещиваясь в тех случаях (еслу такопые воз
инкаю]). когда в результате расширения своих ареалов эти по-
пуляции оказываются в одной и той же области.
Данные в пользу згой теории можно вкратце суммировать,
следуя Майру [564]: 1) между признаками, разделяющими геог-
рафические расы, н признаками, разделяющими виды, нет ка-
чественных различий; 2) во многих группах обычны системы
популяций, занимающие промежуточное положение между геог-
рафическими расами и видами, т. е. полувиды; 3) известны слу-
чаи, когда в одной части ареала данного вида рид интергра-
диру ющих рас замещают друг друга географически, а в другой
его части симпатрически перекрываются; 4) между географиче-
скими расами одного вида часто можно обнаружить частичную
репродуктивную изоляцию.
Примеры промежуточных стадий между географическими
расами и видами, о которых говорится в пунктах 2 и 3, приво-
дились в гл 6 Пункт 4 обсуждается более подробно далее в
настоящей главе Здесь мы опишем пример, в котором все ста-
дии дивергенции, требуемые географической теорией видообра-
зования, Можно обнаружить в пределах одной естественной
группы растений
Этим примером служит группа родственных между собой ряс
и видов диплоидных однолетних растений, распространенных в
пустынных и гористых районах на юго-западе США принад-
лежащих к более обширной секции Arachnion рода GUia Мор-
фологические, экологические, географические и цитогенетиче-
ские взаимоотношения между этими расами и видами рас-
смотрены Грантом и Грант [263. 300].
Наиболее ярко выраженные географические расы GUia lafi-
flora хорошо дифференцированы морфологически Эти расы ин
т^рфергильны и непрерывны, а поэтому пни связаны и природе
рядом ннтерградирующнх форм (рис 13 1. А) Близкий к
(ifiofa вид G teptantha состоит из ряда географических рас, ко-
a ееЛяГвисл
Я M'M’tft*
я a.
О 150
Рис 13.1. Пять стадий
дивергенции у ниже та-
лмА свинки ДлзсАдеп
(PolemoniicM*) в юго-
мзадиой части Север
вой Америк в.
А. Непрерывные геогм-
фвяескне рисы. Б. Дпзъ-
юпктивкыс географнче-
сияе расы. Я. Маргиналь-
но -п»мпитрнеескне полу-
виды Г Аллопдтрнио.кве
мды Д Слмпатрыческне
ДД¥
Гл. 13 ntptutnot
153
торые также различаются морфологически и китерфартильны,
ио занимают разобщенные области, в связи с чем между ними
нет непрерывной интерградапки (рис. 13.1. Б)
Для Gilia latiflora, G. leptantha и родственного нм третьего
полувида G tenutflora характерно в значительной степени алло-
патрическое распространение с перекрыванием в пограничных
районах (рис 13.1. В). Морфологические и экологические раз
личин между этими тремя полувидами более выражены, чем
между расами в каждом из них. Способность к скрещиванию
между разными подвидами и плодовитость образующихся при
этом гибридов также поиижеиы по сравнению с тем. что наблю-
дается при межрасовых скрещиваниях Теи не менее пути для
обмена генами остаются открытыми. В некоторых областях сим-
патрнческого контакта эти полувиды гибридизируют; я других
областях енмпатрни пин остаются обособленными.
Gilta ocluoleuca и G. mexicana занимают совершенно отдель-
ные области (рис. 13 1, Г) Эти два вила разделены мощным
барьером стерильности и сильно выраженной прерывистостью
морфологических признаков
Gilia ochroleuca симпатрична на юге Калифорнии с еннга-
иеоном G tmuiflora—leplanlha—taliftora. и особи всех этих
форм во многих местах растут вместе и цветут одновременно
(рис. 13.1,Д). Морфологическая прерывистость между этими
двумя популяционными системами сохраняется на протяжении
всей обширной зоны симпатрии. Преграды, препятствующие
скрещиваниям между этими двумя формами, и стерильность
гибридов выражены в значительной степени.
Квантовое видообразование
Квантовое видообразование лаключаетск в том, что от предко-
вого вида какого-либо свободно скрещивающегося организма
отпочковывается новый и дивергентный вид, приходя через про-
межуточную стадию географически изолированной или патуизо-
.лированиой периферической популяции Силами, закрепляющи-
ми изменчивость, служат отбор н дрейф геиоа, действующие
совместно в маленькой периферической дочерней популяции.
Такое определение процесса квантового видообразования соот-
ветствует определению, данному ранее Грантом [282]
Желательно помнить это первоначальное определение кван-
тового видообразовапня. поскольку за последнее время в лиге
ратуре появилось несколько совершенно ошибочных определений
этого процесса [29. 892). Ошибки возникли из-за того, что Уайт
[891] не читал работу Гранта [281), а Айала и Валентайн [29]
нс читали книгу Добржанского и др [171].
IM
Часть четвертая. Эьолю^мля дичрмчцил
^вантовое вндообралпванне было впервые описано Майрон
(565] в его оригинальной и важной статье. Формулировке этого
способа предшествовали две белее ранние концепции Майра, а
именно — концепция аберрантных признаков в периферических
популяциях и принцип основателя [564]. а также его новая в то
время концепция генетических революций [565].
Обсуждая способы видообразования в работе 1963 г. (282].
я принял модель Майра [5651. придавая ей очень большое зна-
чение, но присоединил к ней также две другие концепции, со-
вершенно очевидно имеющие самое непосредственное отношение
к этому способу видообразования Эти ранние концепции, кото-
рым Майр [565] не уделил должного внимания, —концепция
Райта [906] о взаимодействии отбора с дрейфом генов я кванто-
вая эволюция Симпсона [730]. Обе концепции относились к мел-
ким изолированным популяциям и обе предсказывали, что в
таких популяциях создаются чрезвычайно благоприятные усло-
вия для быстрых и коренных эволюционных изменений. При
создания этих концепций их никак ие связывали с процессом
видообразования, однако они были вполне совместимы с теория-
ми видообразования. Концепция взаимодействия отбора с дрей-
фом генов предусматривала движущую силу, необходимую для
эволюционной дивергенции. Теркин квантовое видообразование,
как мие казалось, соответствует комбинации участвующих в
этом процессе факторов, поэтому а 1963 Г.. обсуждая указанный
процесс в своей книге, я впервые ввел этот термин [282].
Квантовое видообразование можно сравнить с географиче-
ским видообразованием. Но если географическое видообразова-
ние— процесс постеленный и консервативный, то квантовое ви-
дообразование вызывает быстрые и коренные изменения гено-
типа или фенотипа илв же одновременно и того н другого.
Мы исходим из того, что поток генов в более или менее неп-
рерывных центральных популяциях какого-либо перекрестно-
опыляющегося вида оказывает поглощающее действие на новые
вариации. Кроме того, эти условия благоприятствуют генам, об-
ладающим хорошей комбинационной способностью, что позво
ляет им смешиваться с чужими генами, поступающими с непре-
рывным потоком генов. Если же несколько особей из такой
популяции оказываются в стороне от основного русла потока
генов в виде полунзолированной периферической популяция (на-
пример, вновь основанной колонии), го это ведет к инбридингу
с сопровождающими его резко выряженными генетическими и
фенотипическими последствиями В большинстве случаев такие
резкие изменения скорее будут иметь низкую адаптивную цен-
ность, что повлечет за собой вымирание згой популяции. Однако
а одиой или нескольких изолированных периферических попу-
ляциях могут произойти радикальные изменения, которые при-
Гл 13 Пе/халпог виЗоовгямкишс
155
ведут к визкикиовевню нового хорошо адаптированного состоя-
ния. и в результате подобная популяция даст начало отклоняю-
щемуся дочернему виду [565, 569).
Этот новый дочерний вид, если ои сможет выжить в природе,
будег разрастаться, и по мере увеличения его численности нач-
нет возрастать запас его изменчивости Со временем ои дает
начало новому поколению дочерних видов, которые будут воз-
икать путем квантового видообразования.
Характер изменчивости, наблюдаемой у многих видов или
групп видов, свидетельствует в пользу индигового вндообраао-
вакич В основной части такого вида, занимающей центральную
область ареала, наблюдается географическая изменчивость кли-
нального и консервативного типа, Но по периферии ареала
предкового вида в географически изолированных местах встре-
чаются расы и виды с отклоняющимися признаками. Майр
[564, 565, 569] приводит в качестве примера ракетохвостых зи-
мородков (Tanysiptera galatea), обитающих на Новой Гвинее
и прилежащих островах, и дронго (Dicrurus hollentoius).
Позднее Карсон [83—86] получил цитогенетические и био-
географические данные о квантовом видообразовании у Drosop-
hila и. и частности. у дрозофил Гавайских островов.
Квантовое видообразование у Clarkia
В результате биосистематнческях и цитогенетических исследо-
ваний однолетнего рода Clarkia (Onagrarcac). проведенных
Люисом и его сотрудниками [493, 494, 496. 497. 499, 859]. было
обнаружено несколько пар видов, взаимоотношения между ко-
торыми явно относятся к типу родители — потомки. Так, совер-
шенно очевидно, что Clarkia biloba и С. lingulata филогенети-
чески представляют собой соответственно предковый и дочер-
ний виды [499]. То же самое отннснтся к таким парам видов,
как С. rubicurida н С. franciscana; С. unguiculata и С exilis',
С. mildrediae и С. virgala; С. mildrediae и С. stellala [493, 494.
8591
Дочерпни вид отличается от предкового по морфологическим
признакам, экологическим предпочтениям, расположению участ-
ков и хромосомах или же по всем трем характеристикам одно-
временно и нередко занимает ограниченную географическую об-
ласть, расположенную на периферии ареала предкового нила
[493, 499].
Лгоис [493] отмечает, что периферические популяция в эко-
логически краевых местообитаниях данного вида а климатиче-
ски неблагоприятные годы бывают обречены на полное или поч-
ти ладное вымирание. Снижение численности, близкое к выми-
ранию, с последующей регенерацией популяции из одной или
IM leers fmriniu. Уголоционнап Оимримци
нескольких генотипически абепраитных выживших особей иног-
да непосредственно ведет к быстрому образованию дивергент-
ного дочернего вида (493). Иными слонами, быстрое видообра-
зование бывает иногда результатом резких колебаний числен-
ности популяций в периферических областях. Люис [493] припи-
сывает такие быстрые эволюционные изменения действию край-
не сильного, или «катастрофической», отбора, однако правиль-
нее рассматривать эти изменении как результат совместного
действия отбора и дрейфа генов
Очень часто можно обнаружить, что в пределах одного вида
Clarkia периферические популяции отклоняются по морфологи-
ческим и хромосомным признакам. Такие полуиэолированные
периферические локальные расы служат примером популяций,
которые иногда могут дать начало новому дочернему виду с
отклоняющимися признаками [493, 859|.
Хромосомные перестройки, по которым различаются разные
виды Clarkia. ие похожи иа те, по которым различаются геогра-
фические расы одного и того же вида И »то eHie раз свидетель-
ствует о том. что многие новые виды Clarkia возникли не из
географических рас, а из отклоняющихся периферических попу-
ляций (493).
Пример Clarkia biloba и С. Ungulata был подробно проана-
лизирован Люксом и Робертсом [499]. Это самосовместимые. ио
преимущественно свободно скрещивающиеся однолетние травы.
Они очень близки морфологически и генетически, но разделены
барьером хромосомной стерильности. Главное морфологическое
отличие касается формы лепестков, которые у С. biloba сердце-
видные. а у С. Ungulata в виде язычков. Число хромосом у
С. biloba 2 л—16. а у С. Ungulata 2л—18 [499]. Эти виды очень
сходны также по электрофоретическим характеристикам [254].
На филогенетическую взаимосвязь между этими двумя ви-
дами указывают число хромосом и экологические
предпочтения. У Clarkia biloba п = 8. что близко к основному
числу х=7 для рода Clarkia. тогда как у эволюционно более
продвинутого С. Ungulata п—9. С. biloba занимает более влаж-
ные местообитания, чем С. Ungulata, к, таким образом, она н
экологически ближе к предковой форме Цктотаксономнчсские
и экологические исследования предоставляют ням достаточно
веские данные, позволяющие считать, что вид С. Ungulata про-
изошел от С. biloba [499].
Географические взаимоотношения между эти мн видами поз-
воляют сделать дальнейшие заключения относительно того, как
происходило их образование в филогенезе. Clarkia biloba зани-
мает обширную область в северной части Сьерра-Невады к при-
лежащих j ней районах Калифорнии. С. Ungulata, напротив,
занимает очень небольшую область в одной нэ речных долин на
Г Я. 13. ritjMwtfW* въджбриюаам/е
IKT
южной оконечности «реала С biloba Такое ограниченное н пе-
риферическое распространение С. Ungulata позволяет предпо-
лагать. что этот вид возник от предкового вила С. btloba очень
быстро и совсем недавно [499].
Цитогенетический анализ межвидовых гибридов выявляет
характер и степень хромосомных изменении, обусловливающих
дифференциацию Cla/kia Ungulata от С btloba. Четыре нз
хромосом, входящих в хромосомные наборы этих двух видов,
гомологичны и образуют при мейозе биваленты. Две другие
хромосомы каждого набора образуют у гибрида кольцо из че-
тырех хромосом, что указывает на существующие у этих двух
ВИДОВ различия по реципрокной транслокации именно в этих
хромосомах [499].
Остальные хромосомы образуют у гибрида цепь из пяти хро-
мосом. Дополнительная девятая хромосома Clafbia Ungulata
располагается в середнпе цепи: это свидетельствует о том. что
она представляет собон третичный 1 рисомн к. Таким образом,
у предков С. Ungulata должна была произойти втирал и нсза
виенмая транслокации, и одна нз участвующих в ней хромосом
сохранилась в гомозиготном состоянии, т. е. в виде третичного
тетрасомнка, в результате чего у современной С. Ungulata по-
лучился набор из девяти хромосом. Кроме того, лишняя девятая
хромосома С. Ungulata, очевидно, несет гены, детерминирующие
характерную для этого вида форму жтсисов [499].
Итак, в генетической дифференциации Clarkla btloba к
С. Ungulata важную роль играют две отдельные транслокации.
Их совместное действие и создаст в основном барьер стериль-
ности между этими двумя видами. Та транслокация, кагорам
сохранилась в виде третичного тетрасомнка, вызвала анеуплонд-
ное увеличение числа хромосом от л=8 у С. btloba до я«9 у
С. Ungulata. Лишняя девятая хромосома у С. Ungulata обуслов-
ливает основ к or морфологическое различие между этим видом
и предковым видом С. btloba [499].
Солбрнг [7S5] также постулировал происхождение некоторых
видов Gutierrena (Coniposilac) в результате квантового видо-
образования.
Аллопатрическое видообразование, связанное
с самооплодотворением
У многих са несовместимых н преимущественно свободно скре-
щивающихся растений гем не менее в какой-то степени проис-
ходит самооплодотворение. У таких растений эффекты инбри-
динга в дочерних популяциях могут быть обусловлены системой
размножения, а ие величиной популяции. Результатом является
1» Чвсг» vnepraL 9кмоцмпа> Oumpwmfn»
видообразование (способ 3; см. гл. 12), очень сходное с кванто-
вым видообразованием (способ 2). Очевидное различие между
этими способами состоит в сочетании сил: а — 0 вместо о — 4-
Кроме того, у растений, способных к самоопылению, аффекты
инбридинга выражены обычно менее редко, чем у растений С
облигатным перекрестным опылением.
Во многих случаях прохождение популяции через «бутылоч-
ное горлышко» низкой численности и самоопыление могут про-
исходить одновременно и совместно воздействовать из изменчи-
вость в дочерней популяции. На периферии предкового вида в
результате закрепления изменчивости вследствие инбридинга
или ограниченных размеров популяции или того и другого мо-
жет возникнуть новая локальная раса или узко эндемичный вид.
Clarkia biloba и С. lingulata самосовместимы, и вполне возмож-
но. что в процессе формирования С. Ungulata наряду с ограни-
чением размеров популяции происходило некоторое самоопыле-
ние.
Аллопатрическое видообразование у растений, связанное с
-самоопылением, также очень близко к симпатрнческому вилооб
разеванию с самоопылением (способы 3 и 7; см. гл. 12). В неко-
торых случаях имеющихся данных недостаточно, для того чтобы
провести различие между этими двумя способами.
Из этих двух возможностей в большинстве случаев более ве
роятио видообразование иа каком-либо периферическом
участке за пределами основного ареала предкового вида, Если
дочерняя популяция в какой-то степени способна к свободному
скрещиванию, как это характерно для большей части автогам-
ных растений, то в случае симпвтрии с предковым видом ее от-
клоняющиеся вариации будут поглощаться, а в случае аллопат-
рин они могут сохраниться и закрепиться Кроме того, в неко-
торых случаях биогеографические данные свидетельствуют в
пользу какого-либо аллопатрического периферического участка.
Вполне возможно, однако, что существуют примеры действи-
тельно сямпатрического видообразования при участии самоопы-
ления и следует вести поиски таких примеров.
Вероятным примером способа 3 служат виды GUia tenuiflora,
С. ausfrooccidenialis и G. jacens. Эти три диплоидных вида
встречаются в южной части Береговых хребтов в Калифорнии
(рис. 13.2); они очень близки друг другу, ио морфологически
хорошо различимы. Во всех трех возможных комбинациях они
даюг полуплодовнтые гибриды Ft со слегка нарушенной конъюга-
цией хромосом [300]. По сравнению с преимущественно свобод-
но скрещивающимся видом G. tenui/lora два других вида не-
сколько упрощены как морфологически, так и в отношении си-
стемы размножении, представляя собой небольшие автогамные
однолетники. Кроме того, они занимают более аридные месго-
Гл. 13. Перечне* виЛюврв»*»**
1б->
Рис. 13.2. Географн'нккс* распростратнае G. 1мш]1<*а и двум дочерних еа*
1 огам вых ваше» а юж но! Ка.ъформим.
обитания, чем G. tenuiflora. Следовательно, эволюция, вероятно,
шла от некой предковой формы, сходной с G. tenuiflora. к
G. austrooccidentalls и G. jacens в качестве дочерних форм.
Географическое распространение этих аилов показано на
рис. 13.2. Как можно видеть. Gilia austrooccidentalls и G. jacens
занимают ограниченные н более илк менее изолированные обла-
сти к югу от более обширного ареала G. tenuiflora. Принимая
во внимание все изложенные адесь обстоятельства, следует сде-
лать вывод, что G. austrooccidentalls и G. jacens дивергнровали
от предкового вида G. tenuiflora в качестве видов, приспособлен-
ных к более аридной среде, и что отправными точками диверген-
.ции послужили какяе-то краевые в географическом и экологи-
ческом смысле популяции этого предкового вида.
Другой возможный пример - различающаяся по хромосомам
пара видов Clarkia rubicunda и С franciscana. С. franciscana
представляет собой узко андемичиый автогамный вид. область
распространения которого приминает к области распростране-
ния его вероятного предка С. rubicunda в северной части Кали-
форнии (497]. Хоти электрофоретические данные и не подтверж-
дают прямо такое происхождение [253], они н нс исключают его.
Еще одним примером служат диплоидные автогамные одно-
летние представители Vicia (Leguminosae) [576], Mimulus
160 Чат чгхртаа. Эяолочшжлшл доаерянцая
(Scrophulariaccae) [435] и Stephanomarla (Compoaitae) [253,
255. 257]. Mimulus nasutus считается автогамным дочерним вя-
лом свободно скрещивающегося М gultaius [435]. A Stephanome-
ria malheurensis—автогамный вид. происходящий от широко
распространенного н свободно скрещивающегося S. exigua сого-
лапа [253. 256. 257].
Готлиб [253] высказывает мнение, что Stephanomeria mat-
Jieurensis — автогамный дочерний вид симнатричсского проис-
хождения. Действительно, вид S. malheurtnsis известен только
из одной местности на северной окраине ареала S. exigua сото-
naria в юго-восточной части шт. Орегон. Необходимо обследо-
вать обширную область к северу и востоку от этой местности,
поскольку вполне вероятна что там будет обнаружен S. mat-
hturensis. Это изменило би всю биогеографическую картину.
Возможно, что S. malheurensis зародился в какой-то симпатри-
ческой области, однако более вероятно (по приведенным выше
причинам), что он возник аллолатрически где-либо на перифе-
рии ареала предкового видя, став впоследствии маргииалыю-
еимпагрическим с S. exigua.
Биотически-симпатрическое видообразование
Стоящая перед нами проблема заключается в том, чтобы уста-
повить, может ли первичная дивергенция на видоном уровне
происходить в пределах одной скрещивающейся популяции. Это
давний, много обсуждавшийся и все еще не решенный вопрос.
Добржанскнй [168] в Мазер [561] отмечают, что полимор-
физм и симпатричсские виды — это проявление альтернативных
реакций живых организмов на среду, в которой на обшей тер-
ритории имеются разнообразные ниши. Далее Мазер (во не
Добржанский) высказал мнение, что виды могут возникать не-
посредственно из полиморфных типов при наличии некоторых
определенных условий отбора. Тодэй и другие исследователи
развивали эту точку зрения, получив некоторые эксперимен-
тальные данные, подтверждающие ее [572, 752. 810, 825, 827).
Майр [565, 569, 570], напротив, возражает против возможности
первичного видообразования в биотнческн-сямпатрической сре-
де. Все высказывания за и против продолжаются на протяжении
многих лет. ио проблема до сих пор остается нерешенной.
Попытаемся проанализировать все, что с нею связано, и нач-
нем ваш анализ с перечисления и определения круга нерешен-
ных вопросов
Пер яичное симпатрнческое видообразование ведет от поли-
морфизма к видам по кратчайшему пути, иа котором нет проме-
жуточной стадии географической и микрогеографичеекой изоля-
о
Гл. 13. /.'ехеичлог
101
пик. Поэтому, во-первых, следует выяснить, возможен ли теоре-
тически такой пуп. эволюции, во-вторых, вероятен ли он теоре-
тически; и, наконец, в-третьих, действительно ли он существует
в природе. Тодэй и его сотрудники, а также их исследователи
занимались Двумя первыми вопросами, а Майр — третьим.
Теоретические затруднения, связанные с первичным симпа-
трическии видообразованием, наиболее велики в том случае,
когда популяция основателей состоит из перекрестно оплодотво-
ряющихся особей В этом случае процессу дивергенции непре-
рывно противостоит скрещивание между дивергентными поли-
морфными типами. В условиях строгого самооплодотворения эти
трудности в значительной мере сглаживаются Следовательно,
обсуждение теоретической возможности первичного видообразо-
вания следует начинать с рассмотрения этой проблемы п том
виде, а каком она существует в системе, размножающейся при
помощи перекрестного оплодотворения.
Дивергенция между разными полиморфными типами а пе-
рекрестиооплодотворяхицейся популяции усиливаете» дизруп-
тинным отбором [561. 570. 810, 825. 827]. Этот тип отбора дей-
ствует в условиях, когда один аспект гетерогенной среды благо-
приятен для одного полиморфного типа, а другой ее аспект —
для другого. Некоторые эксперименты, проведенные на Droso-
phila melanogaster. показали, что сильный днэруптивный отбор
ведет к дивергенции, несмотря на случайное скрещивание [579,
810. 825, 827]. В проведенных в конце шестидесятых и начале
семидесятых годов экспериментах на комнатной мухе (Musca
domestlca) были получены сходные результаты [640. 752].
Напротив, п перекрестпооплодотворяющейся. но панмиктиче-
ской популяции воздействие днзруптявного отбора может про-
должаться все время благодаря системе ассортативного скре-
щивания Тенденция генетически сходных индивидуумов скре-
щиваться друг с другом усиливает дизруптивный отбор.
Дли проявления действия дизруптив.чого отбора в условиях
случайного скрещивания необходимы очень сильные давления
отбора. Давления отбора, применявшиеся а экспериментах на
дрозофиле, варьировали от очень сильных [810] до слабых [242],
Которые, одиако, в природных условиях все еще рассматрива-
лись как сильные.
Длн того чтобы сопоставить результаты экспериментов по
Ди эруптивному отбору г. ситуацией в панмиктических природ-
ных популяциях, следует выяснить, происходит ли сильный от-
бор также и в природе. Данные о существовании а природных
популяциях селективных ценностей порядка 20% или выше бы-
ли получены Фордом [218]. который пришел к выводу, что такая
высокая селективная ценность—явление достаточно общее и
обычное
I га Чосг» «егасргаа. Змлжволжи
Мне кажется, что вывод Форд» основах на ряде примеров,
выбранных для такого специального исследования именно по-
тому, что в них особенно заметно проявляются реакции на дей-
ствие естественного отбора. В природе высокая селективная
ценность — скорее исключение, чем правило. Но как бы то ни
было, сильный отбор преобладает по крайне мере в некоторых
природных популяциях, и в таких случаях диаруптквный отбор
может оказывать свое действие, несмотря на случайное скре-
щивание.
Мы упомянули об ассортатшп|ом скрещивании в качестве
возможного вспомогательно фактора. Ассортативное скрещива-
ние имеет потенциально важное значение, потому что оно лает
дизруптивному отбору возможность вызвать эволюционную ди-
вергенцию при более низких иитенснииостях отбора, чем это
необходимо в условиях панмиксии. Комбинация ассортативного
скрещивания с дизруптивиым отбором открывает более широкие
возможности для енмпатркческой дивергенции в природе.
Быть может, нам следовало бы. однако, подойти к проблеме
симпатрического видообразования с другой точки зрения. Мы
вправе усомниться в допустимости экстраполяции от полимор-
физма к видообразованию Эти дне формы сямпатрической ге-
нетической дифференпнации. несомненно, имеют некоторые об-
щие черти. Но между ними существуют также качественные
различия.
Полиморфизм представляет собой изменчивость такого типа,
при котором различии между видами обусловлены различиями
в сложных комбинациях генов. Скрещивания между двумя хо-
рошо адаптированными полиморфными формами, по всей веро-
ятности. дадут некоторое количество менее хорошо адаптиро-
ванных потомков. Однако при скрещивании между двумя хоро-
шо адаптированными видами доля плохо адаптированных потом-
ков будет несравненно выше. Отсюда следует, что плата за диз-
руптивный отбор, оцениваемая по генетической смерти, при ис-
тинном снмпатрическом видообразовании должна быть гораздо
выше, чем при поддержании в усилении полиморфной системы,
и вполне может оказаться чрезмерно высокой для большинства
природных популяций.
Итак, мы не располагаем биогеографическими данными, ко-
торые свидетельствовали бы о существовании у перекрестио-
оплодотворяюшнхея организмов биотически-симпатричссхото
Видообразования, хотя для географического и квантового видо-
образования такие данные имеются. Кроме того, постулируя
видообразование путем дизруптввиого отбора в перекрестно-
оплодотворяющейся популяции, мы сталкиваемся с очень боль-
шими затруднениями теоретического характера, которые сгла-
живаются при рассмотрении самооплодотворяющсйся популя-
Гл IS. Перличчо* лид<юбразовлчие
1»
ции. Первичное биотически симпатрмческое видообразование у
перекрестнооплодотворяюшихся организмов не было ни доказа-
но. ни исключено, в связи с чем по этому вопросу желательно
собрать больше данных.
Репродуктивная изоляция как побочный продукт
дивергенции
Следует ожидать, что образование различных генных комбина-
ций, детерминирующих различные комбинации адаптивных при-
знаком и днвергирующих популяциях, будет во многих случаях
в порядке побочного действия снижать возможность скрещива-
ний между такими популяциями. Короче говоря, можио ожидать
возникновения репродуктивной изоляции как побочного продук-
та эволюционной дивергенции. В пользу такого способа возник-
новения репродуктивной изоляции, на который впервые указал
Дарвин [IS2, 153], а впоследствии и многие другие авторы, сви-
детельствуют многочисленные данные как по растениям, гак и
но животным.
Между разными расами одного вида обычно существует час-
тичная репродуктивная изоляция. Ранее в этой книге мы рас-
сматривали примеры расовой дифференциации в огмошеним
климата, агентов опыления и других факторов внешней средн.
Частичная сезонная изоляция нередко наблюдается между раз-
ными климатическими расами, принадлежащими к одному виду.
А между расами, для которых характерны разные агенты опы-
лении, может возникнуть частичная механическая и зтолиги-
ческая изоляция.
Saluia meUifera и 5 apiana относятся к отдельным енмпат-
рнческнм видам, обитающим в южной частя Калифорнии. В их
способности сохранять свои отличительные признаки в областях
сосуществования главную роль играет механическая изоляция.
Эти два вида очень сильно различаются по строению цветков,
приспособленных к опылеяию разными пчелами: цветки $. те/-
Jijera опыляют такие формы, как Chloralictus н Anthophora,
имеющие средние размеры, а цветки 5. apiana опыляют круп-
ные пчелы-каментииы рода Xyloropa [264]. Сезонная изоляция
иозннкла, возможно, как побочный продукт этих рам ячий в спо-
собе опыления. Salvia mallifera цветет ранней весной, в период
пика активности своих опылителей, a S. apiana — в конце весны
н в Начале лета, в период вылета пчел-камеящиц. Этологнче-
Полиция "" дР>тоЛ "обочиый эффект тех же самых разли-
" В строа,ин ипет*°®, поскольку пчелы научаются посещать
*ко тг цвегкн, строение которых обеспечивает им возмож-
ность успешно выполнить свои функции. Наконец, некоторые
1Г
164
9am «пясртш». Эплюцваиял дккр:ечцы»
данные указывают иа существование между этими видами барь-
ера частичной несовместимости, которая может быть также
сакзана с различиями в строении их цветков [264].
Длина столбика по отношению к длине пыльцевой трубки—
фактор, играющий важную роль н совместимости при перекрест-
ном опылении у цветковых растений. В пределах данной расы
рост пыльцевой трубки координирован с нормальной для этой
расы длиной столбика. Но две расы, различающиеся по вели-
чине цветков, нередко бывают разделены, кроме того, барьером
частичкой несовместимости. Так, при искусственном перекрест-
ном опылении между двумя расами Gilia ochroleuca с крупны-
ми пветками у них образуются многочисленные коробочки и
семена; ио между расами с крупными и расами с мелкими
цветками этого же вили существует ясно выраженный барьер,
препятствующий их успешному скрещиванию. Расы G. ochw
leuca с крупными и мелкими цветками различаются также по
легкости скрещивания с чужеродным видом 0. сапа [300].
Большое число искусственных перекрестных опылений было
проведено у диплоидных видов секции Arachnion рода Gilia
(300]. Полученные результаты, сгруппированные по уровням
таксономической дивергенции, представлены в табл. 13.1. Как
видно из таблицы, у растений этой группы перекрестная совме-
стимость снижается е увеличением эволюционной дивергенции.
Виды рода Picea а одних комбинациях успешно скрещива-
ются друг е другом я дают гибриды, однако в других случаях
скрещивания оказываются безуспешными. Райт [905] обнару-
жил, что не удаются в большинстве случаев скрещивания меж-
ду видами, ареалы которых сильно удалены один от другого и
которые хорошо дифференцированы морфологически. Напротив,
Таблица 13.1. Эффективность барьеров, создаваемых несовмести-
мостью, на разных уровнях дивергенции среди диплоидных
представителей секции Arachnion рода Gilia [300]
Tan екфеядамава Часло яепу «асм» Засл* гиврвд- лалжйШжхсл мых ижлжив-^
MUI МММ > :3а [|| <
Между членами одной популяции 4 17.8 23
М«жлу римыин географим»гкним расами одного м того же вида 26 15.3 12
Между разными кмдамм. отмосжщж- ммса к одвой в той же секция 103 3.7 3
Между гвлмяма секции •I'uc’urJon ДМЕЛЛиНЫШ! ВИДАМИ друГИХ CWK- мй —• — — — 41 0,004 0.038
Г1 13 Пвряпим видом р^ова-.ur
1|Ж
вады Picea, занимающие соседние ареалы, а большинстве слу-
чаев скрещиваются довольно легко Такая корреляция позво-
ляет считать, что барьеры несовместимости возникли, вероятно,
в процессе дивергенции [905).
Генные комбинации, лежащие я основе межрасовых разли-
чий. создаются в процессе отбора на основании своих гармонич-
ных и сбалансированных воздействий на развитие в пределах
каждой расы. Однако многие межрасовые гибриды получат по
наследству генотипы, лишенные внутреннего равновесен.
И в самом деле, у межрасовых гибридов многих видов растений
в той влн иной степени наблюдаются пониженная жизнеспособ-
ность, частичная плодовитость и разрушение.
Крукеберг (449| сравнивал плодовитость гибридов Fi. полу-
ченных при скрещиваниях между разными популяциями Slrtp-
tanlhus glantfulosus (Cruelferae) к 334 различных комбинациях.
При зюи он обнаружил статистически значимую отрицатель-
ную корреляцию между плодовитостью гибридов и географиче-
ским расстоянием, разделяющим родительские популяции Пло-
довитость гибридов понижается с увеличением расстояния меж-
ду родительским и расами, а после того, как зго расстояние
превысит 80—120 км. она резко падает (449].
Все способы репродуктивной изоляции к изоляции, созда-
ваемой средовыми факторами, существующие между хорошими
биологическими видами, существуют также, во всяком случае
в зарождающейся форме, между расами одного вида. Для того
чтобы произошло усиление изоляции от уровня, типичного для
рас, до уровня, типичного для видон, необходимо лишь продол*
жение процесса дивергенции.
Выводы
В гл 11 были рассмотрены некоторые результаты процесса ви-
дообразования В данной главе мы рассмотрели одну из фаз
этого процесса — фазу, обозначаемую как первичное видообра-
зование. Попытаемся теперь связать то, что было сказано в
этой главе, с материалом гл. 11. Какие группы явлений видооб-
разования из описанных я гл. II можно считать результатом
пепничнигп видообразования?
‘У древесных растений с беспорядочной системой опыления,
примером которых служат Ceanothus, Querrus. Pinus и другие
ролы, наблюдаются взаимоотношения, указывающие иа посте-
пенную дивергенцию, связанную с длшельно существующими
экологическими условиями. В результате такой дивергенции
образую! ся расы, полувиды и. в конечном итоге, группы видов,
между которыми существуют внутренние изолирующие механиз-
мы и которые рассматриваются таксономически хак секции ион
IS1 Чат waepraa. Эаомоааопнал ЛлаертцаЛ
пвдроды. Естественная гибридизация часто проке холит на ран-
не» я промежуточной стадиях дивергенции. Для того чтобы
объяснил, тип взаимоотношений, существующих между валами
Ceonothut, нет нужды постулировать какие-либо иные процессы,
кроме процессом первичного видообразования и простой естест-
венной гибридизации, рассматриваемой как процесс, обратный
первичной дивергенции.
У многолетних трав со специализированными системами опы-
ления, подобных Aquilegia, Antirrhinum, Penstenum (частично)
и многим орхидным, наблюдаются в общем сходные взаимоот-
ношения. У них также выявляется постепенная дивергенции
через ряд стадий до состояния полной внутренней изоляции,
прячем иа ранних и средних стадиях нередко происходит ревер-
сия, вызываемая естественной гибридизацией. Дивергенция
обусловлена различиями в агентах опыления, а также длитель-
но существующими экологическими условиями. Тип диверген-
ция, наблюдаемый у Aquilegia, так же как я тип. характерный
для Ceanotlua, можно удовлетворительно объяснить как ре-
зультат первичного видообразования.
У таких многолетних травянистых растений, как Geum и
виды секции Apogon рода Irin, у которых нет сильно выражен-
ных межвидовых различий в системе опыления, также имеются
некоторые общие черты с типами Aquilegia и Ceanothua, ио при
этом у инх имеется и ряд отличий, как, например, хромосомная
стерильность между группами видов. Логично приписать общие
черты некой общей причине в процессе первичного видообразо-
вания. Что же касается отличительных явлений, характерных
для типа Geum, в частности наличия различных хромосомных
перестроек у разных групп видов, то они требуют иного объяс-
нения н будут дополнительно обсуждаться в одной нэ дальней-
ших глав.
Тип Madia, обнаруженный у многих родов однолетних трав,
еще сильнее отличается от типов Cranothus и Aquilegia. так как
у принадлежащих к этому типу растений наблюдается хромо-
сомная стерильность между близкими видами Эту черту нель-
зя отнести полностью за счет сил. способствующих диверген-
ции как таковой н обсуждавшихся в настоящей главе. Однако
у типа Madia есть другие черты, указывающие на действие про-
цессов первичного видообразования
14 Структурные изменения
хромосом при видообразовании
На протяжении XX в. дифференциация «илов растений и жи-
вотных отношении структуры хромосом изучалась с различ-
ных точек зрения. В первые десятилетия были описаны межви-
довые различия в структуре хромосом у таких ролов растений,
как Nicotiana. Triticum и Viola. Исследовались различные типы
перестроек и их поведение при конъюгации. Была установлена
зависимость между поведением структуриых гетерозигот и про-
цессе мейоза и стерильностью межвидовых гибридов, а также
генетическим повелением аллонолиплоидов и других форм гиб-
ридного происхождения. Результаты исследований этого перио-
да суммированы Дарлингтоном [138, 139].
На следующем этапе стало возможным попытаться понять
смысл обнаруженных ранее цитогенетических явлений, задав-
шись следующим вопросом, какова функциональная роль видо-
специфичного кариотипа в жизни самого вида? Поиски ответа
на этот вопрос велись в различных направлениях. Структурные
перестройки связывали с благоприятными эффектами положе-
ния [248], благоприятными системами сцепления генов [150. 790.
795] или с обоями этими факторами [277).
С расширением иигигеиетичсскнх исследований и вовлече-
нием в пих большего числа родов растений стало также воз-
можным установить, в каких таксономических группах встреча-
ется изменение последовательности участков в хромосомах. Дар-
лингтон [143. 144, 150]. Стеббинс (SO, 794], Грант [280]. Эрсн-
дорфер [191] и Люнс [490] выявили некоторые обшпе тенденции
и корреляции у высших растений. Основные выводы, сделан-
ные этими авторами, были излажены в гл. 11
Позднее интерес сосредоточился на процессах видообразо-
вания, ведущих к формированию видослсцнфичных кариотипов.
Этой проблемой занимались исследователи нескольких школ.
В 50—60 х голах она обсуждалась применительно к растениям
Люксом [493, 494, 497] и мною [284], а применительно к иасс-
?0“ым— байтом [890—892], Карсоном [84], Джоном и Лютом
[406, 4071. Обсуждения продолжались и в 70-х годах (см., на-
пример. [85, 87, 293. 483, 611, 645. 894]).
Статьи, в которых рассматриваются эволюционные аспекты
изменения последовательности участков в хромосомах, очень
различны по своей научной цен костя Многие авторы, по-види-
«ому, занимались этой проблемой, ие будучи знакомы со взгля-
“"«н Других исследователей, высказанными ранее. Во многих
„„Э” Удастся внимание лишь какой-либо одной стороне
*6в Ч»т нпсрпя. Экмацшима* дмидимци
В этой главе мы обсудим два взаимосвязанных вопроса. Эго.
"во-первых.— функциональная роль видоспеиифичиой структуры
хромосом и, во-вторых, способ установления нового кариотипа
а промессе видообразования. Конечно, первую проблему можно
было бы обсуждать, не связывая ее со второй А некоторые ав-
торы пытаются обсуждать вторую проблему, почти или совершен-
но ие касаясь первой Однако мы в своем изложении исходим
из того, что проблема создания новой последовательности уча-
стков в хромосоме неотделима от проблемы се функциональ-
ного значения.
Теория эффекта последовательности участков
в хромосомах
Фенотипическое выражение данного гена нередко зависит от
его пространственных взаимоотношений с соседними генами в
хромосоме. Эго явление хорошо известно под названием эффек-
та положения гема. Гольдшмидт [247—249] расширил концеп-
цию аффекта положения, распространив ее на определенную
последовательность участков хромосом в геноме в целом. Каж-
дая последовательность участков обладает собственным инди-
видуальным «эффектом последовательности» (pattern effect) и
создает свой собственный характерный фенотип [248. 249]. От-
сюда следует, что изменение этой последовательности служит
источником новых эффектов последовательности, а тем самым
средством для создания новых комбинаций признаков
[248. 249).
Затем Гольдшмидт установил корреляцию различий между
видами во физиологическим и морфологическим признакам с их
различиями во взаиморасположении хромосомных участков.
Между этими двумя типами межвидовой дифференциации су-
ществует эволюционная связь. Формирование нового вида на-
чинается с крупномасштабного мутационного события в хромо-
сомах— с «системной мутации», которая перемешивает и пере-
распределяет хромосомные участки. В результате такого пере-
распределения (если оно ие оказывается гибельным) возникает
новый вид (248, 249].
Ряд эволюционистов (обзор см. [282]) выдвинули серьезные
возражения против гальашммдтовскоА теории видообразования.
Системные мутации должны возникать вначале в гетерозигот-
ном состоянии, и несущие их особи, будучи полными структур-
ными гетерозиготами. окажутся стерильными. Кроме того, си-
стемные мутанты — в гомозиготном или гетерозиготном состоя
пик —скорее всего будут нежизнеспособны. Даже если данный
системный мутант сможет преодолеть «бутылочное горлышко»
начальной стерильности и нежизнеспособности, что весьма сом-
Гл 14 Струауркме хромкам при юйообраэомишм
16»
цитель но, то как он сможет утвердиться в популяции, состоящей
из немутангных особей?
Взгляды Гольдшмидта в нх первоначальной форме не встре-
тили широкой поддержки си стороны ЭВОЛЮЦИОНИСТОВ. Онн бы-
ли приняты, одни ко, с существенными исправлениями некото-
рыми учеными, занимающимися видообразованием у растений.
К их числу относятся Люис и Рейвн (493, 494, 497]
.Пюис и Рейвн постулировали ряд хромосомных пере-
строек— не столь многочисленных, чтобы вызвать полную сте-
рильность у гетеразигот, или с не столь резко выраженными
фенотипическими эффектами, чтобы привести к нежизнеспособ-
ности,—и тем самым устранили одно из главных возражений
против теории Гольдшмидта Они высказали мнение, что какой-
то фактор — быть может, гены-мутаторы (493, 497] или вынуж-
денный инбридинг (494] — в определенные периоды в истории
растительной популяции может вызвать разрывы хромосом я
структурные перестройки. Некоторые из новых геномов могут
обладать такими эффектами последовательности. которые по-
рождают новые физиологические свойства и дают своим носи-
телям возможность адаптироваться к новым экологическим
местообитаниям (497]. Такие ценные в адаптивном отношении
гено** «огут закрепиться н гомозиготном состоянии в какой-
большой дочерней популяции, а затем раелристрз
•ревратитьея в новым дочерний вид (494. 497].
была предложена еше одна модификация теории
а Уилсон и его сотрудники [440, 645, 896| высказа-
но хромосомные перестройки служат источником
•арактере генной рсгулацп. по которым различа-
ть
оч
от
Ул
они I
Тедр^
Даже осяо.
эффекта последовлтельностн участков обладает
таинствами с теоретической точки зрения. Име
• количество данных, свидетельствующих о том,.
<н, так и у животных функционально связан-
гсиы нередко локализованы в хромосомах
ъии, структурные перестройки разрывают
новые пространственные взаимоотношения
"казывастся на фенотипе. С эмпирической
ельства в пользу того, что эффект после-
важную роль в эволюции, менее убедн-
ожепия гена у растений наблюдаются
оомосомпые перестройки в большинстве
гаметпых фенотипических изменений,
ффеиты положения встречаются чаше.
-и вредными.
.а последовательности в качестве полного влч
-мго объяснения межвидовых различий в кариотипе
I7V Чат тыртал. Жмоциотаа дааармцаа
у растеняИ имеет, как я уже отмечал в других работах (284.
293). одни фатальный недостаток. У всех жизненных форм рас-
тений дквергирующие виды характернзуются различными ком-
бинациями адаптивны! признаков. Если бы эти межвидовые
различия адаптивных свойств определились эффектом последо-
вательности, то можно было бы ожидать, что Им будут сопут-
ствовать соответствующие межвидовые различия кариотипов во
всех группах растений. Однако на самом деле этого нет. Как
мы убедились в гл. II, в некоторых таксономических группах
и у некоторых жизненных форм растений изменение последо-
вательности участков в хромосомах связано с видообразованием,
а в других — нет. Поэтому главное эволюционное значеиве из-
менения последовательности участков в хромосомах следует
искать ве в прямых эффектах последовательности. а в чем-то
ином.
Теория, основанная на сцеплении генов
Другое основное объяснение значения изменения последователь-
ности участков в хромосомах для видообразования состоит в
том, что происходящие при этом перестройки создают возмож-
ность для сцепления генов. На эту мысль наводит прежде всего
установленный цитогенетикой факт, что инверсии и транслока-
ции в гетерозиготном состоянии служат механизмами, позво-
ляющими выключить некоторые гены из процесса эффективной
рекомбинация. Логично было приписать сходную функцию сцеп-
зелия перестройкам, закрепившимся в гомозиготном состоянии
в процессе видообразования, как это и сделали Дарлингтон
(142]. Дарлингтон и Мазер [ISO], Стеббинс (790) и Грант [277].
ч Теория, объясняющая изменения последовательности участ-
ков в хромосомах на основании сцепления генов, подтверждает-
ся также данными о том. что этот процесс постоянно сопровож-
дает видообразование в тех группах растений, особенно одно-
летних и некоторых многолетних, у которых сильно ограничен-
ная рекомбинация, no-видимому, обладает высокой селективной
ценностью подругам причинам.
Как указывает Стеббинс [780, 782]. травянистым растениям,
первыми заселяющим новые местообггта ния, особевностя од-
нолетним. необходима относительно высокая однородность при
размножении семенами, для того чтобы быстро увеличить чис-
ленность любого данного генотипа, доказывающего свою адап-
тивную ценность; такое требование репродуктивного постоянства
обычно обеспечивается у однолетников небольшим числом хро-
мосом или низкой частотой хиазм, а также самоопылением. Пе-
речень факторов, ограничивающих рекомбинацию у однолетних
расзеиий. был впоследствии расширен за счет включения в него
Га 14 Струюррпме шдетмшм хромосом при 9*)юбрлмамш 171
также и межвидовых различий в последователь мости участков
[277, 280, 785).
На основе данных, под ученных в некоторых экспериментах по
гибридизации, считалось, что различия между видами растений
в последовательности хромосомных участков, обусловленные пе-
рестройками, не зависят от морфологических и физиологических
различий между родительскими видами [780). С таких позиций
было трудно усмотреть какую-либо генетическую связь между
изменениями последовательности участков в хромосомах и меж-
видовыми различиями по адаптивным прнзпакям Однако п
дальнейшем были получены данные [287, 707), указывающие на
то, что в нескольких группах растений сцепленные группы ге-
мов, определяющих цепные в адаптивном отношении морфоло-
гические н физиологические признаки, локализуются в участках,
подвергшихся перестройкам (см. гл. 8). Эти данные устраняют
существовавшее прежде теоретическое препятствие.
Дополнительные данные в пользу теории изменении после-
довательности участков, основанной на сцеплении генов, были
получены Бэбкоком и его школой в результате изучения редук-
ции основного числа хромосом у Crepis в процессе филогенеза.
Бэбкок и его сотрудники (33, 34, 37. 793J путем сравнительных
морфологических и цитологических исследований установили,
что в филогенезе рода Crcpis преобладающим направлением был
переход от многолетних травянистых растений с основным чис-
лом хромосом Х"6 к однолетникам с х-3. Стеббинс [780] вы-
сказал правдоподобное предположение, что адаптивная ценность
низкого основного числа хромосом состояла в усилении сиеплс-
ння генов у возникших в результате однолешмх видов.
Далее стило возможным показать, что в механизме умень-
шения числа хромосом у С г ери,г участвовали неравные реципрок-
ные транслокации [33, 610, 834). Тем самым была установлена
корреляция между изменением последовательности участков в
хромосомах и изменениями в группах сцепления.
Закрепление новых структуриых гомозигот
Закрепление хромосомных перестроек у структурных гомозигот
в дочерней популяции сопровождается в качестве побочного
эффекта образованием барьеров хромоеоммой стерильности
между новой и старой популяциями, Процесс этот представляет
собой, таким образом, особый случай первичного видообразова-
ния, Это один из аспектов видообразования, который все еше
остается малопонятным.
Если принять теорию изменения последовательности участ-
ков в хромосомах, основанную ив сцеплении генов, то становит-
ся логически необходимым постулировать для родительской по-
173 Чат чегасргал. Эволюционная дияяроянция
сулицни некое предковое состояние генного и связанного с
ним хромосомного полиморфизма. Такой генный и структурный
полиморфизм мог быть либо сбалансированным я длительным,
либо переходным н кратковременным.
Другой необходимый постулат состоит в том, что одна ИЗ
возможных гомозиготных комбинаций генов, встречающаяся в
полиморфной родительской популяции в качестве одного кз ва-
риантов. имеет наивысшую адаптивную ценность в каком-либо
новом местообитании, доступном для этой популяции. В таком
случае выщепеицы, гомозиготные по этому сочетанию генов,
могут заселять повое местообитание я успешно размножаться
в нем Образующаяся в результате дочерняя популяция будет
отличаться от основной частя родительской популяции как по
своим генам, так и по структуре хромосом [284. 894].
Конечно, этот процесс может происходить независимо » раз-
ных инвертирующих линиях. Два или несколько различных но-
вых местообитаний, доступных для данной популяции, могут
Оказаться местом рождения двух или нескольких генных или
структурных гомозигот, происходящих от одной и той же измен-
чивой родительской популяции В результате возникнут адаптив-
ные различия я связанные с ними изменения последовательяо-
сти участков в хромосомах у трех или большего числа попу-
ляций.
У дрозофилы, прямокрылых и Clarkia (Onagraceae) типы
хромосомных перестроек, по которым различаются близкие ви-
лы, ае похожи иа те. которые встречаются в пределах вида в
качестве обычных полиморфных или расовых вариантов [493,
497, 770, 875, 892]. У дрозофилы, кроме того, полиморфизм по
строению хромосом лучше всего развит в нейтральных популя-
внях данного вила, тогда хак периферические популяции часто
бывают мономорфным! в отношении последовательности генов
183, 84, 88). Эти факты свидетельствуют о том, что виды, у кото-
рых сейчас наблюдается широко распространенный хромосом-
ный полиморфизм, когда-то в прошлом были мономорфными
J84, 88, 408, 770, 891],
Какие способы видообразования обусловливают закрепление
новой последовательности участков в хромосомах? Имеющиеся
«ведения носят косвенный характер, н их нельзя считать окон-
чательными. Некоторые исследователи отдают предпочтение
симлатричесхому способу видообразования (252, 253, 894].
Однако следует ожидать, что сохранение и размножение особей
с новыми последовательностями в области, завитой особями с
последовательностями предкового типа, будет затруднено. Дру-
гие исследователи, в том числе и я, считают, что новый вид,
как правило, возникает из какой-либо изолированной моно-
морфной периферической популяции предкового вида [86. 87.
Гл М. Стметрркие аажиечи хдвхосяж пив ачйообраы-оача» ПЭ
Выводы
Как было показано к гл. 11. дифференциация близких видов
по структуре хромосом — одна из ярко выраженных особенно-
стей однолетних растений, относящихся к типам Madia и Cilia
inconspicua Процессы первичного видообразования ча основе
генной изменчивости не позволяют объяснить угу особенность.
Но теория, основанная на сцеплении генов, дает удовлетвори-
тельное объяснение преобладанию у однолетних растений из
мекенных последовательностей участков п хромосомах.
Однолетние растения обычно играют в своих сообществах
роль пионеров. Эта экологическая роль благоприятствует рас-
тениям с ограниченными рекомбинационными системами, а хро-
мосомные перестройки служат аффективными факторами, огра-
ничивающими рекомбинацию. Близкие виды однолетников, пер-
выми осваивающие новые местообитания, обладают различными
наборами сцепленных адаптивных генных комбинаций, связан-
ных с различной последовательностью участков в их хромосо-
мах
Тип Geum, как это было также показано в гл. 11. занимает
промежуточное положение между типами Madia и GUia incon-
spicua. с одной стороны, и типами Ceanothus и Aquitegia — с
другой. Близкие виды многолетних трав, относящиеся к типу
Geum, генетически дифференцированы и репродуктивно изоли-
рованы при помаши внешних механизмов, как у типов Ceanot-
Itus ii Aquilegia. и эти общие черты можно в целом объяснить
первичной дивергенцией на основе генной изменчивости. Но хро-
мосомная стерильность обнаружена между группами видов в
ролах, относящихся к типу иеит. Таким образом, здесь тоже
имело место изменение последовательности участков в хромосо-
мах. но на более высоком таксономическом уровне, чем у родов,
относящихся к типам Madia к GUia inconspicua.
Можно сделать вывод, что процесс дивергенции в отношении
расположения участков в хромосомах, наблюдающийся в со-
временный период у быстро эполюцновирукицих однолетних
растений, происходил в каком-то более раннем цикле видообра-
зования у многолетних растений, принадлежащих к типу Geum.
Иными главами, у Geum и у подобных ему родов самой недав-
ней Сталин видобразования, не связанной с изменением после-
довательности участков в хромосомах, предшествовала сталия,
сопровождающаяся изменениями этой последовательности Быть
может, в более ранний период видообразования такие многолет-
ние роды играли в сообществах роль пионеров, сравнимую с
ролью Однолетников в некоторых недавних сообществах
Древесные растения северного умеренного пояса, очевидно,
претерпели изменения последовательности участков в хромосо-
174 Чат чтааргаа Эчитрпчяаа iuatpmipia
мах в еще более ранние периоды своей эволюционной истории.
Целые роды или семейства в настоящее время цитологически
стабильны; все известные виды, которые к ним относятся, имеют
одно и то же основное число хромосом, отличаясь, однако, от
родственных родов нля семейств но этому числу, Так. у Betula-
сеае х=14. у Fagaceae х=12, а у Corylaceae л=14, 11 и 8 [151 ).
В австралийской флоре также известны группы древесных рас-
тений, как, например, Proteaceae и Myrtaceae, у которых, судя
по имеющимся данным, в далеком прошлом имело место изме-
нение последовательности участков в хромосомах [747J.
В заключение следует отметить, что нам известно достаточ-
но много относительно изменения последовательности участков
в хромосомах и его роли в видообразовании, ирднако целый ряд
проблем все еще остается невыясненным.
15 Отбор на репродуктивную
изоляцию
Репродуктивная изоляции в ее разных формах может разви-
ваться и развивается как побочный продукт эволюционной ди-
вергенции между популяциями, о чем уже говорилось и гл. 13.
На этот основной процесс накладывается другой, комплементар
иый, способ возникновения изоляции. Некоторые типы механиз-
мов репродуктивной изоляции создаются в результате прямого
отбора на изоляцию как таковую в случае, если это состояние
выгодно для «х или иных конкретных популяций. Предполага-
ется, что те особи из двух симлатрических популяций, которые
производят нежизнеспособные или стерильные гибриды внесут
меиьший вклад в будущие поколения, чем сестринские особи
из тех же родительских популяций, которые не гибридизируют.
Следовательно, у каждого из диух видов частота генетических
факторов, детерминирующих какое-либо препятствие или а вер-
сию к межвидовой гибридизации, будет из поколения в поколе,
иие повышаться. Происходящий в этик направлениях естествен-
ный отбор приведет, таким образом, к усилению репродуктив-
ной изоляции, первоначально возникшей как побочный резуль-
тат дивергенции.
Особые условия
На способность отбора усиливать за рождающуюся репродук-
тивную изоляцию оказывают влияние разные условия. Некото-
рые из этих условий необходимы, а другие — благоприятны для
отбора иа изоляции.
Совершенно очевидно, во первых, что этот особый процесс
отбора возможен лишь в том случае, если участвующие в нем
виды снмлатрическис или по меньшей мере маргипа-тыю-симпат
ричсские
Во-вторых, результаты гибридизации должны быть вредны-
ми. Некоторые или все гибриды в F, или в последующих поко-
лениях должны быть стерильными или нежизнеспособными. Та-
кая ситуация обычно наблюдается при межвидовой гибридиза-
ции у растений, как мы уже отмечали в предшествующих гла-
вах; однахо она не распространяется иа псе случаи гибрндиза
Нии у растений.
Какой-либо гибридный генотип в F, или в последующих по-
колениях может быть стерильным, но обладать в том местооби-
тании, в котором он встречается, более высокой вегетативной
мощностью, чем любой из родительских видов. Если хорошо
адаптированное гибридное растение способно к вегетативному
176
4»m чгпргм. Эхит^юти Лиерммчиж
или агамоспермкому размножению, то оно может затеи рас-
пространиться и повысить свою численность по сравнению с ро-
дительскими видами а среде, обычной для этих последних. В ра-
де групп растений наблюдались реальные примеры гибридов,
численность которых превышала численность родительских форм
в определенных местообитаниях. Отбор на изолинию происходит
в обычной комплементарной ситуации, когда в результате меж-
видовой гибридизации действительно значительно снижается
число хорошо адаптированных потомков, производимых гибриди-
зирующими родительскими особями.
Еще одно условие, способствующее эффективному действию
отбора ва репродуктивную изоляцию, состоит в том, что утра-
та репродуктивного потенциала, происходящая при гибридиза-
ции, иесьма невыгодна для участвующих в гибридизации орга-
низмов . Селективная невыгодность снижения репродуктивного
потенциала сильно варьирует у организмов разных классов. На
одном конце находятся организмы с короткой продолжитель-
ностью жизни, такие, как поденки а однолетние растения, особи
которых имеют возможность размножаться лишь одни раз в
жизни. А ва противоположном конце находятся долгоживущие
древесные или травянистые растения замкнутых сообществ, ко-
торые обычно кз года в год, десятилетие за десятилетием произ-
водят огромный избыток семян И, разумеется, множество рас-
тений и животных занимают промежуточно* положение между
этими двумя крайностями.
Селективная невыгодность утраты репродуктивного потен-
циала велика в случае однолетних растений и других зфемер-
ных организмов, ко для долгоживущих растений замкнутых
сообществ она пренебрежимо мала (780]. Отсюда следует, что
отбор на репродуктивную изоляцию может быть особенно силь-
ным у однолетников и относительно неэффективным у многолет-
них растений с большой продолжительностью жизни [169, 785].
Некоторые многолетние травы, возможно, приближаются к 0Д-
иолетипам по своей способности реагировать иа отбор, направ-
ленный на создание изоляции (479].
Далее, отбор на репродуктивную изоляцию не обязательно
ведет к полному предотвращению гибридизации, как указал
Эрмаи [198]. Репродуктивная изоляция будет усиливаться до
уровня, необходимого для того, чтобы предохранять популяции
от суммарных вредных эффектов гибридизации. Этот уровень
зависит от толерантности популяций к селективной элиминации.
Эрмаи [198] считает, что минимально необходимый уровень ре-
продуктивной изоляции — это уровень, при котором «обмен ге-
нами между видовыми популяциями ис превышает той скоро-
сти, с которой рассеянные гены могут быть элиминированы есте-
ственным отбором».
Гл. 15. ОгОор ы рлпроруягимдю изоляцию ITT
И, наконец, отбор на создание репродуктивно* изоляции мо-
жет действовать только на изолирующие механизмы, сущест-
ву юшке в родительском поколении. Особенно эффективно отбор-
усиливает этологические барьеры и барьеры несовместимости.
При этом такой действующий ло оплодотворения барьер, как.
этологическая изоляция, сохраняет репродуктивный потенциал
родительских видов более успешно, чем какой-либо барьер, всту-
пающий в действие, подобно несовместимости, после оплодотво-
рения.
Важная роль этологической изоляции у животных может
быть частично или в значительной хере приписана отбору, на-
правленному на создание изоляции У растений же одной это-
логической изоляции обычно недостаточно, чтобы предотвра-
тить гибридизацию, потому что приверженность насекомых-
опылителей к одним и гем же цветкам время от времени нару-
шается. и в таких случаях несовместимость может оката ibeir
чрезнычайио существенной В связи с этим следует ожидать,
что у растений отбор на изоляцию создает не только этологи-
ческие барьеры, но и барьер несовместимости
Барьеры несовместимости состоят у растений из полого комп-
лекса блокирующих механизмов, действующих до и после опло-
дотворения Механизмы, действующие до оплодотворения, имеют
высокую селективную ценность, так как сохраняют яйцеклетки
для внутривидовых скрещиваний. Однако механизмы, действую-
щие после оплодотворения и препятствующие образованию зре-
лых гибридных семян, также обладают' селективным преимуще-
ством по сравнению с отсутствием таких механизмов, поскольку
они позволяют сохранить питательные вещества, количество ко-
торых ограничено, для запасания в негибридных семенах [2&Ц.
Эксперименты по искусственному отбору
Гипотеза о том. что отбор способен усиливать репродуктивную
изоляцию, била подвергнута проверке в экспериментах с искус-
овсяным отбором.
Первые такие эксперименты были проведены на дрозофиле.
Смешанные культуры, содержавшие два вила (Drosophila pscu-
aoobscura и D. persimiHs) или две линии одного и того же вида
(D melanogasier) подвергались па протяжении ряда поколений
отбору, направленному протии гибридов В процессе такого
искусственного отбора этологические барьеры между видами
или линиями усиливались [443, 445, 8741.
У растений аналогичные эксперименты по отбору провел
Патерниапи [634). Патериками использовал два сорта Zea mays.
различающиеся по окраске зерен, формы с белыми и желтыми
зернами высевали вперемешку, создавая им возможность для
12-43
IT* Чат штвфтаа Эаолвчиоянва Рамр^пциа
перекрестного опылен»» .Чатем проводил» искусственный отбор,
сохраняя початка, содержавшие главным образом иегибрид-
иые семена. После такого отбора на протяжении пяти поколе-
ний уровень спонтанной межсортовой гибридизации упал с 47
до 3% у сорта е желтыми семемами и е 36 до 5% у сорта с бе-
лыми семенами. Усиление репродуктивной изоляции было час-
тично обусловлено большими различиями между сортами по
срокам цветения, а частично — развитием барьеров несовмести-
мости, действующих да оплодотворения.
Данные по природным популяциям
Как предсказывает теория, отбор, направленный на создание
изоляции, действительно происходит, о чем свидетельствуют »кс-
леримеиталькые данные. Следовательно, наша задача состоит
а том. чтобы оцепить роль этого процесса в природе. Для групп
видов характерны три типа географического распространения,
позволяющие сравнивать родственные аллопатрические и сим-
патрнчсскме популяции по относительной степени нх изоляции
и родительском поколении (рис. 15.1).
В простом случае ареалы двух близких видов перекрывают-
ся, так что некоторые расы одного вида аллопатричиы по от-
ношению к другому виду, а другие—симпатричны (рис, 15.1_А).
Аллопатрические и снмпатрические виды можно сравнивать по
степени изоляции между ними.
В самых первых работах по изучению отбора на изоляцию
а природе была использована именно такая ситуация. Исследо-
вания проводились главным образом на парах видов плодовой
мушки и лягушек и касались этологической изоляции. Оказа-
лось, что в большинстпе случаев этологическая изоляция выра-
жена сильнее между симпатрическими расами какой-либо пары
илов, чем между аллопатрическими расами тех же самых двух
• •
£
Vac 131 Тря типа тгроВмяясяогь ряепрсетряаяння я групп, ендов. в ко-
торой воэвожмо сраьжяаать относительный уровень ртяр.иуктяеяой азоля-
TU41 между аллеюатрвчеекяма а саапатрачесаамв потух шля и и (объяснения
•см. в тесте).
Гл. IS. Отбор па рряродртлк/ю иголырю
I?»
видов (см. (199, 200] и приведенную в этих работах библиогра-
фию).
Другим источником данных послужили сравнения между ви-
дами в целом в группах видов, которые состоят из нескольких
симпатрических видон, занимающих центральную часть ареала,
я находящегося иа периферии географически изолированного
вида (рис. 15.1, Б),
Сравнения степени этологической изоляции в группах видов
с распространением подобного типа проводились у разных жи-
вотных Приведем а качестве примерив группы видов Drosophi-
la guarani [4391. D. wUllstoni [172], Erebla tyndarus [518] и Oam-
busia all inis [385]. В каждой из этих групп видов этологическая
изоляция была выражена относительно сильно между сиипатри-
necxHMH видами и слабее между периферическим видом и одним
из нейтральных видов.
В подобной ситуации (рис. 15.1.Б) по крайней мере одни
вид в каждой паре видов оказывался в условиях сиипатрик. а
следовательно, подвергался отбору па усиление изоляции. Это
осложняет картину. Более четкие данные можно получить, срав-
нивая эффективность барьеров между двумя или несколькими
симпатрическими видами с барьерами между двумя или не-
сколькими аллопатрическими видами (рис. 15.1,6). Такие срав-
нительные исследования мощности барьеров иесовиестимостя
проводились на облпстаепиостебелы1ых гмлиях, о чем пойдет
речь в следующем разделе.
Барьеры несовместимости у Gilia
V растений барьеры несовместимости могут возникнуть и воз-
никают как побочные эффекты эволюционной дивергенции На
ато указывает наличие у многих видов явной, хоти и слабо вы-
раженной несовместимости между расами. Данные о том, что
такая перекрестиям несовместимость у видов облиствениосте-
бельвых гилий является побочным эффектом дивергенции, были
приведены в гл. 13. Но перекрестная несовместимость может
создаваться также как механизм, усиливающий изоляцию
Облиственностебельиые гилии — естественная группа одно-
летних растений, произрастающих на тихоокеанском побережье
Северной Америки и в умеренном поясе Южной Америки (см.
М. II) Девять из десяти видов, составляющих эту группу, изу-
чались с цитотаксоиомической, генетической, а также экологи-
ческой точек зрения. Все Полученные межвидовые гибриды вы-
«жостермльны. и мейоз у них нарушен. Имеются косвенные дан-
ные, указывающие на то, что все девять видов окажутся иитер-
стерильными во всех возможных комбинациях. Между иекоторы-
12*
«80
Чат wpraa. Эюлтрошт&тргтчаа
ми видами существуют достаточно мощные барьеры несовмести-
мости; между другими эти барьеры менее эффективны [274,
2861
Облиствениостебельиые гилии представляют собой однолет-
ние травянистые растения, которые цветут весной, а в коше
весны или начале лета, образовав семена, отмирают. Поэтому
снижение продукции семян в результате возинкноееиия стериль-
ных гибридов крайне невыгодно этим растениям в селективном
отношении. В некоторых популяциях GUia обнаружен полимор-
физм по скрещиваемости с чужим видом, что создает возмож-
ность для действия отбора [290].
Девять видов облнствеияостебельиых гилий, изученных гене-
тически, можно разделить иа две подгруппы на основе их геог-
рафического распространения.
Пять видов — Oilia capitata. G. tricolor и другие — встреча-
ются в предгорьях и долинах Калифорнии н прилежащих райо-
нах. Ареалы этих видов сильно перекрываются (рис. 4.1). Особи,
принадлежащие к двум или нескольким видам этой подгруппы,
растут и цветут в тесном соседстве во многих естественных мес-
тообитаниях (рис. 15.2).
пгедгорныг и долинных аилы
пеимоьскне виды
Рас 1S2 Пары млов обяпствепоспОельпи гадай, растущее а природе а
трсаьи еоседотае [Й90].
Гл. ti. Отбор на рслроЛцКТилпро изоляцию IfiI
Остальные четыре вида — Cilia millefoliaia, G. Mvinii и дру-
сие — встречаются на морском берегу в четырех далеко спетой-
щнх одна от другой местностях в Северной и Южной Америке.
Эти приморские виды лапистые аллолатричны по отношению
друг к другу. Два из них имеют редкие снмпатрические контак-
ты с некоторыми предгорными и долинными видами, а ареалы
двух других целиком лежат за пределами области раса рос । ра-
ке иия видов первой подгруппы (рис. 15.2).
Эти две географические подгруппы облиственностебельных
гнлнй заметно различаются по эффективности барьеров несов-
местимости. Эффективность барьера несовместимости можно
оцепити количественно по среднему числу гибридных семян, про-
изводимых па одни цветок при перекрестном опылении чужим
лидом в контролируемых условиях.
Снмпатрические предгорные и долинные виды разделены
очень мощными барьерами несовместимости У этих видов про-
дукция гибридных семян на один цветок колебалась от 0,0 до
1.2 при средней 0,2 для многочисленных скрещиваний между
разными пилами В отличие от этого приморские виды легко
скрещиваются между собой. Скрещивания между разными при-
морскими вилами давали от 7.7 до 24Л гибридных семян на одни
цветок, а средняя для межвидовых скрещиваний во всех комби-
нациях составила 18,1 семян иа цветок [290|.
Как отмечалось выше, в качестве побочного эффекта дивер-
генции следует ожидать некоторой несовместимости при скрещи-
ваниях Но уровень дивергенции между видами в обеих геогра-
фических подгруппах этих гнлий примерно одинаков. Различия
в скрещиваемости, наблюдаемые между двумя географическими
подгруппами видов, нельзя объяснить на основе одной лишь
эволюционной дивергенции. Высокую степень несовместимости
прн скрещиваниях между симлатрмческнми видами предгорий
н долин наиболее разумно отнести за счет отбора, направлен-
ного на создание изоляции [290].
Подтверждение этому дает изучение гнлий, принадлежащих
к секции Arachnion В этой группе нельзя проводить сравнения
третьего типа (рис. 15.1.fl), так как в ней пет подгруппы алло-
патрических видов, но сравнения второго типа (рис. 15.1, Б)
провести можно. В результате многочисленных скрещиваний
между снмпатрическими видами в этой группе была выявлена
сильная несовместимость, выразившаяся в том. что среднее чис-
ло гибридных семян иа цветок составило 3.4 Охнако прн скре-
щиваниях между центральными м периферическими видами не-
совместимость слабее и среднее число гибридных семян на цве-
ток равно 7.6 [290J.
U2 Часто teraepraa. Эклюцкаппая Зипрмяаа»
Опознавательные знаки цветков
Пчелы различные другие насекомые-опылители обладают ин-
стинктом постоянства. в силу которого они всякийра з посещают
преимущественно цветки растений одного вида. Этот инстинкт,
как уже было сказано в гл. 9, лежит в основе этологической
изолинии между разными пилами растений, которые опыляются
пчелами или другими насекомыми.
Классическими экспериментами Фриша было показано, что
медоносные пчелы способны распознавать определенные цвета,
формы и аапахи, по которым различаются цветки разных видов
во многих группах растений [221, 222]. Такие же способности
обнаружены у других пчел и насекомых, посещающих цветки
[206, 253]. Облетая цветки и постоянно выбирая цветки одного
вида, насекомое руководствуется их отличительвой окраской (в
частности, ее узором), формой в(или) запахом.
Несколько лет назад был проведен обзор мяогочнелеишх
ролов покрытосеменных с целью оценить относительное значение
признаков цветков в систематике растений с различной систе-
мой опыления. Оказалось, что 38% признаков, используемых
систематиками для различения близких видов растений, опы-
ляемых пчелами, относятся к лепесткам и другим внутренним
частям цветка. Примерно такой же процент признаков цветка
используется в систематике тех видов растений, которые опы-
ляются другими насекомыми, также неизменно посещающими
один и те же цветки. Что же касается тех растений, которые
беспорядочно опыляются многими видами неспециализирован
иых насекомых, то у них лишь 15% признаков, используемых
для различения близких видов, относятся к цветку (265].
При этих подсчетах межвидовые различия по тем признакам
цветка, которые связаны с механической изоляцией, ие отделяли
от различий по признакам, связанным с этологической изоля-
цией. Тем не менее можно с достаточной уверенностью допус-
тить, что какая-то часть этой внушительной массы признаков
цветка у покрытосеменных, постоянно опыляемых одними и те-
ми же видами насекомых, служат опознавательными знаками
для опылителей. В подтверждение такого допущения можно
привести результаты, полученные при более подробном изучения
некоторых частных случаен.
Так, например. Gilia capitata capitata и G. с. chamitsonit
различаются помимо прочих признаков по форме лепестков и по
запаху цветков. На экспериментальном участке, где высеяны
вперемешку эти две формы гилии и где их посещают пчелы, от-
дельные насекомые обычно сохраняют верность одной или дру-
гой форме Такое поведение пчел позволяет считать, что при вьь-
Гл. 15. Отбор не рвпрсОрлтиомрю изолларио
1U
боре цветков они руководствуются формой лепестков, запахом
цветков нли одновременно обоими этими признаками [265. 267].
Дополнительные данные, свидетельствующие о возможном
адаптивном значении опознавательных знаков цветков для соз-
дания этологической изоляции, были получены позднее Левином
и Керстером [478], а также Левином и Шаалем [484] на флок-
сах. Распространение этих видов соответствовало типу, изобра-
женному на рис. 15.1, А.
В восточной части США ареалы Phlox pilosa и Р. glaberrima
перекрываются, и во многих местах эти виды биотически енм-
иатричиы. Эти два вида несовместимы, но между ними нет дос-
таточно надежной механической нли сезонной изоляции. Их
опыляют одни и те же виды чешуекрылых, что приводит в сме-
шанных колониях к довольно значительному опылению одного
вида другим (478. 479(.
И у Phlox pilosa. и у Р glaberrima цветки обычно розовые.
У Р. pilosa встречаются также формы с белыми цветками в ка-
честве полиморфных вариантов В большинстве полиморфных
популяций Р pilosa форма с белыми цветками встречается ред-
ко, но в некоторых популяциях она преобладает.
Географическое распространеиие популяций Phlox pilosa с
преимущественно белыми цветками представляет большой инте-
рес. Эти популяции всегда симпатричны с популяциями Р. gla-
berrima, у которого цветки розовые Такая корреляция позволяет
«читать, что белая окраска цветков в смешанных колпкнях по-
могает бабочкам-опылителям распознавать разные виды этого
растения [478].
Бабочки, перелетающие от Phlox glaberrima к Р. pilosa в
смешанных к<хао<шях, явно предпочитают Р. pilosa с розовыми
цветками и избегают форму с белыми цветками. Подсчет пыль-
девых зерен Р. glaberrinia па рыльцах Р. pilosa показывает, что
опылители переносят в пять раз больше пыльцевых зерен этого
вила па розовые цветки Р. pilosa. чем на такое же число белых
цветков Р. pilosa [479]. Таким образом, в смешанных колониях
фирма Р. pilosa с белыми цветками, очевидно, обладает селек-
тивным преимуществом по сравнению с формой с розовыми цвет-
ками. Этим, по-видимому, объясняется высокая частота, дости-
гаемая формой Р. pilosa с белыми цветками в области перскры
шшия с Р glaberrima [479].
У нескольких групп растений, произрастающих в Скалистых
горят в штате Колорад» и опыляемых одними и теми же насе-
комыми. наибольшая дивергенция по призпаху высоты соцветия
наблюдается в условиях еимпатрии. Высота, на которой нахо-
дится соцветие, оказывает влияние на постоянство посещения
опылителей. Дивергенция по высоте соцветия представляет со-
бой. возможно, результат отбора на изоляцию [862].
1Ы
Часть *t тле ртах. Эволюциокчал
Исторические аспекты
Первые высказывания по теории отбора, направленного на соз-
дание вэоляции. в новейший период эволюционных исследований
принадлежат Фишеру [213], Добржа некому (167} и Хаксли
[392] Их формулировки были краткими. но весьма многозначи-
тельными и послужили стимулом для последующих исследова-
ний.
Одна кз задач, возникших вслед за этим, состояла в более
точном указ а инн условий, в которых можно ожидать действия
предполагаемого процесса отбора. Другая задача заключалась
в получении убедительных доказательств того, что процесс этот
действительно происходит в природе. В шестидесятых годах на-
шего века эти задачи были выполнены. Дальнейшие исследова-
ния, проводившиеся в семидесятых годах, еще больше укрепили
представление о роли отбора в усилении изолирующих механиз-
мов. м в настоящее время оно завоевало всеобщее призвание.
С исторической точки зрения интересно отметить, что и Дар-
вин, и Уоллес еще в XIX в. рассматривали возможность отбора
на создание релродуитявной изоляции. Дарвин [153] писал:
«Можчо Дслустнтъ... что мрожхаюгцвмуея виду было бы выгодно, если
бы он окмалгя в какой-то небольшой степе» я бесллэд«мм орн ек решив*
яки € родительской формой или с кавой-лебо Другой рийовддаоегью, по-
тому что таким обратом возндмо бы мевыое шгпкммх и ухудшенных потом-
ков, ыогушик затеи примешивать пос крон к новому виду, каюдяшешуги
в процессе форыироважяя Но тот, кто даст себе труд порвзмыслжтъ -ад те-
ма стадиями, про годя мерея которые эта началах* степень бесплодия мо-
жет уевлмаатьса под действами естеегмивого отборе. _ убедите* а чрмымаЯ*
ной сложности угой проблемы. По яремом раем махаем мп мне кажется. что
это в« могло быть достигнуто путем естественного отбора».
Подобный же взгляд Дарвии высказал в «Происхождении
видов» [1541
Уоллес [873], однако, считал, что естественный отбор мог
создавать преграды скрещиванию между диверпгрующими сим-
патрнчсскими видами. В статье, озаглавленной «Влияние есте-
ственного отбора на стерильность и плодовитость», Уоллес [873]
писал:
«Многим может паказвтъея, что •♦осхолъяу бестподле должно быть ВЫ-
ГОДНО зарождающими нндан. когда омм эонвиадгт одну к ту же иди со-
седнее облаете, так как оно ячвтралвауат аффекты межвидовых схрчшва-
нм1, тп око должно усалмыться под действием «сгестымвпт отбора. его
покажется ехде Солее вероятным, если допустить, н Н ТО Ш № убеля-
лись. есть основания, что ьводояитость бывает подвержен изменяямости,
пожалуй, столь же часто, «ы другие орнпакв- Особ«яво следует огме-
ТМ-Ъ. сто подобный эффект воэнижжег ве в результате Сйхраиамнм бесплод-
ию ыриамтс* по арнчиие их беспл.тджя, во п риультвтс пгпалппцег поста
гибридных потомков, которые более иалочвелеквы, менее способам обсела-
Гл. 15. Отбор па рспродуктилчую изоляцию IHB
четь npoA<M««we сушестьомння ооА расы к менее ЦНЯКОАМШ к лен-
jiuw услоожяы среды, чем любая м> чистых форм Имение эта неоолмоиея-
и ость гибридных потомков и ест главжж н постальму число таких тибря*
лов будет всегда жиже там, где баешкмм будет мли&лаьшим. постольку те
части двух форм, в которых бс^плсдие постигнет наиболхтеА стеке», будут
обладать преимуществом м в деиздюм счете вмжжаут в борьба за суиаест-
воианм».
Уоллес считал также, что различным симпатрнчсскии видам
растений, опыляемых насекомыми, постоянно посесцахмаими
цветки одного к того же вида, выгодно иметь на цветках разные
опознавательные знаки, гак как подобные, знаки снижают часто*
ту опыления пыльцой чужого вида. Приведем еще одну цитату
1873].
*Бытъ может, и«г*иж> дли тоги, чтобы гомочъ ньечкомым писезать вся*
мА ри лишь одам ц,аатои. что врелиммЯно важно для сохрлигяия дачного
вила, у растения, ми встающих а одно и то же время я емешавжых колена-
ял. цветки обычно сильно различаются по форы? и окраска Эту особенность
лекального распределения (жрхсхи цветкза можно сравнить а отношении м»
мммчшми с Оюошамтслыюй окраской у животжыд То-сво так же как ок*
раоса у этих еккледнэх лает ьозможвостъ скобам разного таза узнавать
Друг лругж тс*» самым ивбегать гтернлмых союдо мгжду размыми вила-
ми. форма и омрвска цветков у каждого вида растена*. о тли чающая их от
цветке* другая видов, обычно растущая вокруг. помогает иасекимыш-опыли-
телмм кзСмгать переноса выльем одного нитка на рылыи чужого айда»
Взгляды Уоллеса огн-релнли его время, поэтому его идея не
привлекла внимания, я затем была забыта В основном та же
мысль, по сформулированная в генетических терминах, была
вновь высказана Фишером н другими в начале современного пе-
риода изучения эволюции, о чем уже говорилось выше Я счи-
тал уместным выразить запоздалое признание гипотезе Уоллеса,
назвав отбор, направленный на создание репродуктивной изоля-
ции. эффектом Уоллеса (290].
После того как я в 1966 г. поставил вопрос относительно
обоснованности модели Уоллеса о происхождении стерильности,
возникли некоторые проблемы. Отбор на создание репродуктив-
ной изоляции не может оказывать мня иве па гибридную сте-
рильность. Все зависит от истолкования того, какой смысл вкла-
дывал Уоллес в термин стерильность. Уоллес и Дарвин применя-
ли этот термин нс в современном его смысле, т. е. для них ои
означал не только стерильность гибридов, но и то. что мы те-
перь называем несовместимостью. Так, Дарвин (152, гл. 9]. рас-
сматривая беечлодяе, проводит различие между «бесплодием
Двух видов прн первом скрещивании» и «бесплодней происшед-
ших от них гибридов»; Уоллес [8731 также проводил это раз-
личие
Я расцениваю весь этот абзац Уоллеса (873, с. 173—186] сле-
дующим образом Уоллес иногда имел в виду гибридную сте-
lee VatTB wraeproa. Экитгият дмркяция
рилыюсть, н в этих случаях ои был неправ, но иногда он имел в
виду несовместимость при скрещиваниях, а в таких случаях он
был на верном пути. Я склонен делать упор на этих последних
случаях, пренебрегая первыми, и отдавать должное Уоллесу зь
его первые шаги в этой трудной и сложной проблеме.
Эффект Уоллеса и смещение признаков
Эффект Уоллеса к смещение признаков создают сходную карти-
ну изменчивости в Группах видов с географическим распростра-
нением типа Л (рас. 15.1), а именно—максимальную дифферен-
циацию признаков н зоне симпатрии. В связи с этим высказы-
вались мнения, что аффект Уоллеса «может быть включен в бо-
лее широкую концепцию смешения признаков» (474].
Однако здесь возможны различные толкования. Я рассмат-
риваю эффект Уоллеса и смешение признаков не как концеп-
ции, одна из которых включает в себя другую, а как две парал-
лельные концепция. Эффект Уоллеса возникает в результате ре-
продуктивных взаимодействий между видами, а смешение приз-
наков— в результате экологических взаимодействий. Эффект
Уоллеса — продукт отбора, направленного на снижение межви-
довой гибридизации, смешение признаков — результат отбора,
направленного ва снижение межвидовой конкуренции.
В классической работа Брауна и Уилсона {73] о смешении
признаков все внимание сосредоточено почти исключительно на
различиях в признаках, не имеющих отношения к размножению,
н на экологических взаимодействиях Авторы лишь вкратце ка-
саются различий по признакам, участвующим в создании ре-
продуктивной изоляции Концепция отбора, направленного иа
создание репродуктивной изоляции, возникла в результате рас-
суждений в совершенно ином направлении, нежели то, которое
развивается в качестве основной темы в статье Брауна и Уил-
сона.
Недавно термин смещение репродуктивных признаков был
использован Валевом (8861 в связи с некоторыми примерами, от-
носящимися к растениям. Это как бы перекинуло мостнк между
концепциями эффекта Уоллеса и смещения признаков. Выше я
отметил, что эти дм концепции параллельны, однако можно
также сказать, что в некоторых случаях они сливаются.
Один из примеров Валена (886] касается величины цветков
у пар видов Solanum в Мексике. Цветки S. Iwnholtzianum и
3. grayi имеют одинаковую величину в аллопатрических частях
своих ареалов, но в симпатрическях областях они заметно раз-
личаются по величине. То же самое наблюдается у другой пары
видов — 5. citrulUJolium и 5 rostratum. Величина цветков у этих
растений коррелирует с размерами опыляющих их пчел. Эго
\ Гл 15. Ог5о/> м> ртцкитуктнлфо иоллца» 187
смещение првзкакоа объясняют тем, что в областях симпатрии
оно снижает конкуренцию за пчел, относящихся к одному виду.
Такая дифференциация, по-видимому, ведет к развитию механи-
ческой кзоляцни между видами [886J.
Я согласен с тем. что это — пример смещения признаков и
что он удачно назван смещением репродуктивных признаков,
поскольку дифференциация, очевидно, возникает в результате
конкуренции за жизненно важный фактор, в данном случае—эа
опылителей. Репродуктивная изоляция при этом, несомненно,
возникает, однако она, по видимому, не является продуктом от-
бора иа создание изоляции как таковой.
Параллельный случай, относящийся к окраске цветков я дру-
гим их признакам, описан для пары видов fuchsia parviflora и
F. incliandra [87]. Здесь также различия в признаках цветков,
наблюдаемые между двумя видами в области симпатрии, связа-
ны с более выраженными различиями между их опылителями
в этой области, чем в аллопатрических областях.
Во многих реальных случаях, касающихся цветковых расте-
ний н нх опылителей, два селективных процесса — отбор на сни-
жение конкуренции за опылителей н отбор на изоляцию цвет-
ков, — вероятно, переплетаются друг с другом.
Часть пятая
Естественная гибридизация
и ее результаты
16 Естественная гибридизация
Проблема естественное гибридизации в животном и раститель-
ном царствах всегда привлекала особое внимание человека.
Гречесхая мифология изобилует самыми причудливыми и неве-
роятными примерами гибридов между животными, составители
средневековых травников описали несколько подлинных гибри-
дов в европейское флоре. Возникновение научного подхода к
вопросим гибридизации датируется серединой XVIII в.
Появление трудов, принадлежащих школам ^ёльрейтера
Линнея, которые придерживались противоположных вз: лидов,
следует считать началом научного исследования проблемы гиб-
ридизации у растений. <VorlluflEC Nachricht» Кёльрейтера [444]
публиковалась с 1761 по 1766 г. Труд Линнея eDisqutsitio de se-
xu plantarum» [513] впервые увидел свет в 1760 г. В 1751 г. одни
из учеников Лнннся, придерживавшийся его взглядов. — Хаарт-
маи [321]—опубликовал свой труд cPIaniae Hybridae».
После Кёльрейтера и Лнннся вопрос о гибридизации расте-
ний в природе обсуждался многими авторами иа всем протяже-
нии XIX н XX вв., что привело к накоплению обширнейшей лите-
ратуры по данному вопросу. Объем литературы столь велик, что
дать ее полный обзор не представляется возможным; поэтому
мы н нс пытаемся это сделать.
Ьбльшая часть литературы по гибридизации в природе носит
описательный характер и состоит из сообщений о гибридах, об-
наруженных в той нли иной группе растений. Подобного рода
специальные сведения занимают надлежащее нм место в архи-
вах систематики. Биолога-эволюциониста больше интересуют об-
щие выводы, подкрепляемые частными данными. В соответствии
с этим в нашем изложении основное внимание уделяется общим
принципам.
На протяжении длительной истории изучения естественной
гибридизации у растений рассматривались две общие проблемы:
1) являются ли естественные гибриды уродцами нли представ-
ляют собой нормальное явление в какова частота их появления
Гл И. бгтктаеипвя
19»
в разных группах расгеиий нли флорах? 2) приводит ли есте-
ственная гибридизация к каким-либо значительным эволюцион-
ным последствиям’ Иными словами, возникают ли иногда из.
естественных гибридов новые расы или новые виды’ Эти проб-
лемы обсужлалнсь многими авторами, принадлежавшими к раз-
ным поколениям, причем нередко разные авторы, основываясь
на одних и тех же данных, приходили к различным выводам.
В этой и в последующих главах мы попытаемся разобрать две
сформулированные здесь обширные проблемы.
В настоящей главе рассматривается образование спонтанных
гибридов в природе. Гл. 17 посвящена эволюционным продук-
там таких гибридов, вплоть до расового уровня. В гл. 19 обсуж-
дается возникновение новых видов в результате гибридизации.
Многим читателям будет полезно, если мы перечислим здесь
некоторые из наиболее значительных обзоров. Очень некныи
справочным изданием может служить книга Робертса «Гибри-
дизация растений до .Менделя» [688], в которой собраны выдерж-
ки из трудов ранних исследователей — от Линнея и Кельрейтера
до Фоккс. Современные обзоры по естественной гибридизации у
растений с соответствующими литературными ссылками принад-
лежат Хейзеру [365, 372], Андерсону (15. 16]. Стеббинсу [780,
786), Бейкеру [39). Стейсу [767] н Левину [476). Сводки по
естественной гибридизации у животных опубликованы в числе
прочих Майром [569. 570] и Ремингтоном [672].
Гибридизация как процесс, обратный
дивергенции
Образование двух или нескольких репродуктивно изолирован-
ных популяций из общей предковой скрещивающейся популя-
ции—процесс чрезвычайно длительный. Дивергенция между от-
дельными лиинямн может в конце концов достигнуть уровня
полной репродуктивной изоляции, когда скрещивание между ни-
ми становится уже совершенно невозможным. У высших расте-
ний такая стадия абсолютной репродуктивной изоляции, вероят-
но, почти по всех случаях бывает обусловлена развитием полной
несовместимости между линиями, поскольку всегда существуют
спогхзбы обойти полную стерильность гибридов, а внешние изо-
лирующие механизмы бывают совершенными лишь п редких
случаях, а быть может, и никогда.
Между начальной и конечной стадия ян видообразования
происходит неуклонное снижение интенсивности скрещивания
дивергируюшнх популяций и. напротив, прогрессивное развитие
изоляции, создаваемой средовыми факторами, и репродуктивной
изоляпии. Спонтанное скрещивание между популяциями, кото-
рые и прошлом ливергнровплн до уровня дизъюнктивных рас.
4Ю Уисте «жпи. Есткппыя « те рпцмтаты
полувидов или видов и которые разделены частично* экологиче-
ской или репродуктивной изоляцией или как той. так и другой,
представляет собой естественную гибридизацию.
Термин гибрид используется чрезвычайно широко, в том чис-
ле для обоаиачения продуктов скрещивания между особями с
различными генотипами, принадлежащими к одной и той же по-
пуляция, и продуктов прививок, в которых привои и подвои при-
надлежат к разным видам или родам. Для того чтобы можно
было плодотворно обсуждать естественную гибридизацию с эво-
люционной точки зрения, необходимо несколько ограничить при-
менение этого термина. Стеббинс [780] определяет гибридизацию
как «скрещивание между индивидуумами, принадлежащими к
отдельным популяциям, которые обладают различными адаптнв
иыми нормами». В определении Стеббинса подразумевается
имевшая место в прошлом эволюционная дивергенция, а в оп-
ределении. которое мы сформулировали выше, о не* говорится
прямо
Гибридизация в этом смысле представляет собой процесс,
-обратный эволюционной дивергенции. Процесс этот может быть
локальным или распространяться более широко в зависимости
от того, сколь велика охватываемая гибридизацией географиче-
ская область. Гибридизация может происходить между сямпа-
трическамн видами «ли между расами, которые были прежде
разобщены я дискретны, ио затем носстаиовили контакт по ка-
кому-либо новому географическому фронту. В последнем случае
результатом будет полоса изменчивых промежуточных типов,
известная под названием эоны вторичной ннтерградацни, или
гибридной зоны (см. [672]). В конечном счете результаты есте-
стаей ио* гибридизации могут быть очень кратковременными
или, как мы увидим п дальнейшем, длительными.
Гибриды и скопления гибридов
Встречающиеся в природе межвидовые гибриды (по-видииому,
ггнбрнды первого поколения) обнаружены во всех крупных груп-
пах растений и во всех хорошо изученных флорах. Хаартман
{321] в свое* книге «Plantae Hybridae», опубликованной в 1751 г,
• описал ряд естественных гибридов, а также спонтанные гибри-
ды, возникшие в садах. Одним из них был естественный гибрид
между Trifolium герепз и Т. pratensc. встречающийся в разных
частях Швеции (си. [688]).
К 1891 г. Кернер (431) имел возможность привести список
тветоечающихся в природе гибридов европейской флоры, в кото-
рый дошло свыше 1000 межвидовых гибридов, относящихся ко
многим группам растений — от мхов, папоротников и хвощей до
двойных к сережкоцветных деревьев я травянистых покрытосе-
Гл. 16 Сттиля гиОрчдихщчл
19г
мениых. Болес новые списки естественных гибридов опубликова-
ны Алленом [4], Хейзером [365] и Стейсом [767].
Ботаники уделяют особенно большое внимание деревьям и
кустарникам ТОГО или иного региона, а поэтому у крупных дре-
весных растений гибридные индивидуумы, даже если это еди-
ничные зкэемпляры, обычно бывают выявлены, и сообщения о-
них публикуются довольно часто. Кернер [4311 обнаружил иа
Дунае гибридную иву. вероятно имевшую конституцию Salit
incanaxdaphnoides. в 1852 г., т. е. в то время, когда существо-
вание естественных гибридов все еще оспаривалось ботаниками.
Конечно, и после 1852 г. в природе было обнаружено множест-
во гибридных ив, возникших в результате самых разнообразных
межвидовых комбинаций (ср. [73б]|.
Многие авторы описали естественные гибриды дубов, проис-
ходящие от разных родительских видов (см., например, [57. 122.
343. 633, 839, 866. 903]). В Калифорнии изредка встречаются
гибриды между Pinus cvulteri и Р jellreyi. а в шт. Миннесота —
между Picea mariana и Р. glauta [516. 917]. У Ceanothus доволь-
но обычны внутрисекцвонные гибриды; кроме того, в природе
встречаются отдельные межсекционные гибриды [537, 614]. Спи-
сок естественных гибридов древесных и травянистых растений
можно продолжать бесконечно.
Гибрид F; может быть плодовнгым, частично плодовитым,
высокостерильным или полностью стерильным. Во всех случаях,
за исключением последнего, он производит некоторое количест-
во потомков последующих поколений Различие между высокой
степенью гибридной стерильности и полной стерильностью ни-
жет быть очень невелико, составляя доли процента, по, по мне-
нию многих ученых, имеет при этом важное биологическое зна-
чение.
Стеббинс [756] приводит один численный пример, который
тем не менее достаточно реалистичен Допустим, что у нас есть
гибрид многолетнего травянистого растения, образующего корне-
вища, например Elpmus. и что этот гибрид высокостерплеи —
фертильность его семяи равна всего 0.001% Со временем такой
гибридный индивидуум может дать начало клону, выбрасываю-
щему ежегодно по 1000 цветоносных побегов с 200 цветков на
каждом. Это составит а общей сложности 200000 цветков, а сле-
довательно. 200 000 потенциальных семязачатков к год А посту-
лированная 0,001 %-ная фертильность семян означает, что этот
гибридный клон может дать два семени в год или 200 семян за
столетие [786].
Частично плодовитые гибриды R способны размножаться
половым путем в результате самоопыления, родственных скре-
щиваний с сестрииснимн гибридными растениями или возврат-
ных скрещиваний с одним или обоими родительскими видами.
« И Чат юта. Есткпетал те/вЛазшгш и м ргцмптм
Получающиеся при этом потомки F» могут затем продолжать
скрещиваться друг с другом или с исходными родительскими
растениями. В результате образуется скопление гибридов —
•крайне изменчивая смесь видов, гибридов, беккроссов и реком-
-бинлнтов. возникающих в более поздних поколениях.
Скопления гибридов известны во многих группах растений.
Для наших целей достаточно перечислять несколько типичных
примеров. У древесных растений скопления гибридов описаны
в числе прочих у luniperus [209. 697], Quercus (59, 343]. Aescu-
Jus [341. 342], Eucalyptus [115] и Opunlia [304]. Средн травяни-
стых покрытосеменных аналогичными примерами служат Carex
1179), Viola (66, 700], Geum [230, 557]. Aquilegla [94, 270], Mi-
.mulus секция Diplacus [54], EpUobium [666] и Helianthus [366].
Среди папоротников скопления гибридов можно обнаружить у
.Pieris, исследованного Уокером [870].
Полный список естественных гибридов и скоплений гибридов
у растений начинался бы с Achillea и кончался бы Zinnia. Сяу-
чайная или спорадическая естественная гибридизация —нор-
мальное явление в царстве растений, возникающее в тех слу-
чаях, когда два нли несколько родственных видов приходят в
.контакт Но явление это не универсально. Привлекают внимание
те исключительные случаи, в которых два иля несколько близ-
ких видов произрастают аа одной и той же территории, не гиб-
ридизируя (насколько об этом можно судить по данным поле-
вых исследований).
GUia tricolor не гибридизирует со своими симпатричссхими
родичами в Калифорния [269]. В Великобритании Ranunculus
acres, R. repens и R. bulbosus часто встречаются вместе, однако
гибриды между ними неизвестны [350]. Papaoer dubium и Р. 1е-
coquil также сосуществуют на Британских островах, не гибри-
дизируя [539]. Lotus corniculatus и близкие ему виды сохраняют
обособленность в Калифорнии и на севере Европы, ио. по-види-
мому. гибридизируют в Средиземноморской области 1366]. Семь
видов LeaventPorttUa симпагричиы иа юго-востоке США. ко не
образуют гибриды [692]. У Sedum п центральных районах Мек-
•сикн и у Silene на западе Северной Америки гибриды очень ред-
ки [116. 451]. luniperus virginlana и 1 ashei не гибридизируют
в различных зонах симпатрического контакта на плато Озарк
на юге центральной части США, хотя в более ранних работах
и сообщалось о существовании гибридов между ними (1, 215].
Внутренний контроль над гибридизацией
Самая важная причина отсутствия гибридизации между родст-
веяными симпатрическимв видами это несовместимость в ши
роком смысле, т. е. наличие ие только внутренних барьеров, лей-
Гл 16 Еснстветшл айяк1пза<»м
193
ствующих до оплодотворения, но также барьеров, действующих
на ранних стадиях после оплодотворения.
Сели говорить о перечисленных в предыдущем разделе слу-
чаях отсутствия гибридизации, то имеются экспериментальные
данные, указывающие на существование очень мощных барьеров
несовместимостии между видами групп Gilia tricolor, Ranunculus
acris. Lotus corniculatus н Leavenworthia |269. 306, 350. 692].
Естественные гибриды между членами разных секций редки
у С eanol fins к не известны у Quercus. Кроме того, как показали
экспериментальные скрещивания, у этих родов существуют очень
мощные барьеры кесовместкмосгм между видами, принадлежа-
щими к разным секциям, о чем уже говорилось в гл. 11 (см.
]614]1
Однако барьеры несовместимости — это лишь часть пробле-
мы Papaecr dubium н Р. lecoquii легко скрещиваются на экспе-
риментальных участках, по. как ми уже отмечали, в Великобрп
тапни в естественных условиях они не гибридизируют [638]. Кру-
кенбергу [451] удалось добиться успеха почти во всех скрещива-
ниях, которые он пытался произвести между 28 эанадкоамсри
канскими видами Sllene. по в природе эти виды сохраняют обо
соблеиность и лишь в редких случаях образуют гибриды.
Внешний контроль над гибридизацией
Известно много примеров, когда данная пара родственных видов
растений или животных не гибридизирует в природе, но легко
образует гибриды в ботаническом салу или зоопарке Кельрей
тер писал об этом еще в 1761 г. |444. 688]:
«[Маловерок 1шх чтобы] пн том уворяаоченном устройстве раствтельяо
то шргтм. которое создаю природа, могло воднииуп гибридно. расттяиа
Прирока которая пест да. даже в случаях кажущегося ве.тмпвНшсго Осело-
рядка. привержен* к самому великолепному порядку, яр л асе исключила эту
путаижду (Ojuuko) а ботигичгских гадах, где в тесним пространстве собрв*
гм ект»1к1»мож1чые растения со всех частей яечиого шаря. гнбрмдвма рагтенмк
могут моти ничуть, огобепчо сели разместить их в соответствии с их ежтемв-
тячесжим положением, а следовательно. п«жестеть радом рвстмшп, облит-
этшнс канбо'шшнш сходством По краАмеВ мер» лдссь человек d ивметвоы
Семем арсюстаазаот растения и ту возможность. которую он дает своим
ЖН5ОТИМ», примемикмм ю очежь удаленных одна ох другой частей земного
шара, которых ои попрека природе держит к плеву я зоопарке алн в etae
бо.те огранпеняом пространстве».
Позднее Кернер |431 ] сформулировал положение о pain мес
^обитания в ограничении естественной гибридизации более кон
креню.
«Еслв данный мд м некой либо местности благоденствует, предгтавлов
больuiHM числом инднпяхуалышх растений и воэоСаюоляетгп в потомках,
которые в осипяном не изменены, то можно допустить» чти 0£Гамк)ЭвНЯ зто-
гп пила соответствует почве и климату дайной местности П отсутствие н*
1Э_лч
1Й4 Чат лягая. Етсгмпная шбраримцал в аа раармтаты
кой гаримноиой СОЯМ) ие ножжг быть и реки о ороцмеянин ьам; накроем,
он рано или ведано должен вымереть. Это соответствье климата м почем ор
ганяаенхх рктетка пропмюшеел его внешней форме должно сущест-
вовать также а сля вновь вооняжшего гибрида, с тем ттобы те несколько
раетенмй. которые аолвлккттся в каком-либо нести, котлы выжить в свояк
аарвовачаяьамт местообитаниях и »ть начало иногочнслеажын котомкан
Иногда таков соответствие существует, а кжхда его нет. В ооежедием слу-
чая (жбржд ееяоаает. «лак поя«>ш№нгь> на свет. Цо если даже «го оргакюа-
выв шпшроаева к how и планету того наста, гм он ВОМП, <му пред-
стоит еще выдержать борьбу с теми задави. которые уже обосновалась я
этой меетаоетн, а а ссобеаяогтж со своими собствеяными рахительскянн ев-
дама. Если ап* ооеладака вышно растут а равной местности и чжленность
ал велика, то новой форме нелегко тввиаветв территорнейв
Кернер приходит к выводу, ото гибридные растения могут
обосноваться в местообитании своих предков только в том слу-
чае, веля они в адаптивном смысле превосходят родительские
ияды или если они могут найти подходящее свободное местооби-
тание. не та пятое этими последними. Подобные условия встреча-
ются редко. Кернер [431] приводит в качестве примеров Rhode
dendron. Salvia и Nuphar, гибридные формы которых успешно аа-
селилн новые местообитания, отличные от местообитаний роди-
тельских видов
В начале современного периода изучения естественной гиб-
ридизации важное значение факторов среды в предотвращении
или допущении естественной гибридизации подчеркивали Дю
Рие 1181]. Вига ид [895]. Эплннг [202]. Андерсон [14. 15]. Хейзер
[3651. Стеббинс [780] и многие другие исследователи.
Виганд [895] указывал на то, что естественная гибридизация
между разными видами растений нередко бывает Ограничена
определенными местностями, в которых среда была нарушена в
результате вмешательства человека. На Ньюфаундленде я в
шт. М»и. например, гибридные формы Amelanvhier и Rubus
встречаются в нарушенных местообитаниях вдоль железнодо-
рожных путей, тогда как в нетронутых лесах пли а лесах, гра-
ничащих с этими путями, произрастают родительские виды в
чистом виде [895]-
Это наблюдение было распространено на многие другие груп-
пы растений У ирисов естественная межвидовая гибридизация
происходит главным образом в местообитаниях, нарушенных в
результате деятельности человека — земледелия, постройки до-
рог. лесных разработок я т. П„— как в дельте Миссисипи, так
н на тихоокеанском побережье [15, 462. 655, 686. 860]. Salvia
apiana и 5. mellifera. которые сямпатрнчны на обширной терри-
тории в южной части Калифорнии, остаются обособленными там,
где нх полывно-кустариичковос сообщество находится в слоем
естественном состоянии, по в тех местах, где среда была искус-
ственно нарушена, между ними наблюдается локальная гибри-
дизация (17. 202. 264].
Гл. 16. f«TKTn»uuu aullpiuWaw
IM
В 1891 г Кернер дал объяснение корреляции между гибри-
дизацией и разрушением местообитаний, которое в несколько
расширенном виде сейчас принимается большинством нсслсдо-
вателей. В замкнутом стабильном сообществе для образующих
ся время от времени гибридных зигот подходящих местообита-
ний нет. Стабилизирующий отбор элиминирует их почти сразу
же после нх появления. Но в тех местах, где естественное со-
общество нарушается в результате строительства дороги, пере
выпаса и т. п. что приводит к созданию новых свободных место-
обитаний, гибриды могут утвердиться и это действительно про
всходит. Иными словами, изоляция, создаваемая средой, подав-
ляет гибридизацию между совместимыми друг с другом вилами
о стабильном замкнутом сообществе, но в открытом местообита-
нии она частично утрачивает свою эффективность.
Гибридизация местообитания
Андерсом [14, 15] расширил концепцию контроля над гнбрилнзп
дней со стороны среды, распространив ее на второе и последую-
щие поколения гибридов Он указывал на существование тако-
го же расщепления по физиологическим различиям, как и по
различиям морфологическим. Если FI носит промежуточный ха
рактер и обладает единообразием в отношении экологических
предпочтений, то Fj и последующие поколения отличаются боль
шим разнообразием рекомбинантов по физиологическим призна-
кам Если родительские виды различаются по нескольким неза
висимым физиологическим признакам, то последующим поколе-
ниям их гибридных потомков потребуются десятки или сотни
экологических ниш. Андерсон [14. 15] приходит к выводу, что
потомки Fi могут удовольствоваться промежуточным местооби-
танием, но что для выжиааиии сколько-нибудь представитель-
ной выборки поколения F, потребуется разнообразный набор
экологических ниш —своего рода «гибридизация местообита
ния>.
Как писал Андерсон [15, гл. 17].
«Хороша теетм, что гибридам, диы того чтобы выжить. исобхолими
прочел)точи», а мостообигами» Не учитывалось, адиако. то, кто дли сохра-
нения не только первого тибри тиого пожолеиия. но и дальнейших поколений
пеобяо.исми мс просто промежуточные местообигаиня. а местообитания,
предоставлявши» тохже все иоаможяис сочетания ковтрастирутощвт разлв-
ini! ntpBouau.ibHWi местообитаний Толыю путем гибридизации иестооСвта-
имя метут сохраниться в природ» гибрн.шм» рекомОинаьии»
Промежуточные местообитания встрсчаклсм в природе не
слишком редко. Но местообитания, которые в достаточной степе-
ни гибридизированы, чтобы допустит* относительно свободно**
196
‘lam. чтя Есттлгкяая ечбрнбизация и ее ргацягч-
воспроизведение межвидового растительного гибрида, гораздо
менее обычны. И такие гибридизированные местообитания соз-
даются в современном мире главным образом в результате дея-
тельности человека, например строительства дорог, сельскохо-
зяйственных работ и лесоразработок [15].
Замечу мимоходом, что необходимо проводить различие меж-
ду гибридизированными и свободными местообитаниями. Второе
и последующие поколения межвидовых гибридов могут выжить
как в тех. так и в других местообитаниях, но по разным причи-
нам. в открытом местообитания — вследствие ослабления меж-
видовой конкуренции и стабилизирующего отбора, а а гибриди-
зированном местообитании — вследствие разнообразного набора
новых ниш. Можно представить себе скопление гибридов, произ-
растающих либо в однородном открытом местообитании, либо
в смешанном, но более или менее замкнутом местообитании
Следовательно, гибридизации местообитания представляет со-
бой ие единственное условие, которое создаст возможность для
обоснования расщепляющихся гибридных потомков
Основной аффект деятельности человека, упоминавшейся вы-
ше, состоит в частичном удалении природной растительности
и создании тем самым открытых местообитаний. Часто »ти дей-
ствия сопровождаются искусственной гибридизацией местооби-
тания вследствие смешения почвы разных типов, создания новых
градиентов затененности или влажности и т. п. А в тех местах,
где такай гибридизации происходит, создаваемые ею возможно-
сти для воспроизведения гибридов играют, вероятно, второсте-
пенную роль по сравнению с теми, которые создает наличие сво-
бодных участков.
Открытые местообитания наряду с гибридизацией местооби-
таний или без нее — обычный результат деятельности человека
в современном мире. Но иа протяжении всей историк земли,
вплоть до современности и включая ее. многократно происхо-
дили разрушения биотических сообществ и возникновение но-
вых местообитаний под действием чисто природных факторов.
Обвалы, наводнения, извержения вулканов н пожары, вызван-
ные ударом молнии, создавали новые открытые местообитания
локального масштаба. Процессы горообразования, поднятия су-
ши из моря, ледниковые периоды и периоды засухи оказывали
на природные сообщества воздействия крупного масштаба. Да-
леко идущие последствия имели также биотические факторы,
подобные иммиграции новых крупных растительноядных орга-
низмов или новых возбудителей заболеваний.
Климатические и геологические изменения обусловливают
миграции растений, приводящие к новым симпатрическим кон-
тактам между видами. В то же самое время, как полагают
Андерсов [14. 16] и другие авторы, эти физические изменения
Гл 16 Есттлегшая либридяятлил
197
или нэмепившиесн биотические факторы или тс и другие могут
открыть новые незанятые местообитания. которые будут слу-
жить местом размножения для естественных гибридов
Сопоставление высших растений с высшими
животными
Естественные гибриды обнаружены в разных крупных группах
высших животных Соответствующие списки для птиц состав-
лены Кокремом 1116] н Грэем [3081, а для млекопитающих —
Грэем [307]. Ценные обзоры естественной гибридизации у живот-
ных принадлежат Хэббсу [386]. Мвйру (569, 570] и Моору (.584].
Подробное обсуждение сложной проблемы гибридизации у
животных и оспещекие различных связанных с ней частных во-
просов читатель найдет в этих сводках и в приведенных в них
обширных библиографиях Здесь же нас интересует нс гибриди-
зация у животных как таковая, а широкое сравнение между жи-
вотными и растениями в отношепин частоты гибридизации и ее
эволюционной роли Эти два царства действительно различают-
ся по указанным параметрам
Установлено, что естественная гибридизация оказывает влия-
ние на характер изменчивости в отдельных группах животных,
в частности у некоторых бабочек, цикадок, рыб и птиц [383. 386.
387, 672. 696. 722. 723]. Перечет, примеров можно было бы про-
должить. Вместе с тем во многих других группах животных, та-
ких, как двукрылые или млекопитающие, естественная межви-
довая гибридизация, очевидно, встречается редко и не влечет
за собой никаких последствий.
Но увеличение числа тех или других примеров не поможет
нам глубже понять роль гибридизации в эволюции животных.
Как было показано в этой главе, естестве иная гибридизация
наблюдается в одних группах растений, по отсутствует в дру-
гих Широкие сравнения между царствами растений и живот-
ных в отношении важности гибридизации пока еще нельзя про-
водить на удовлетворительной количественной основе, и поэто-
му приходится искать другие подходы к этой проблеме
Несколько поколений ботапикоп считали, что гибридизация
играет важную роль в эволюции растений (см., например, [14
16, 163. 181, 205, 317, 365. 431. 519. 609. 780J). А несколько ппко-
Ленин зоологов пришли к выводу, что гибридизация не играет
важной общей роли в эволюции животных (см , например, 1165,
214. 564, 569. 882. 889])
Я привожу здесь высказывания различных авторов нс для
того, чтобы решить научный вопрос подсчетом голосов или апел-
ляцией к авторитетам Суть дела состоит в том. что авторы-бо-
таники и авторы-зоологи выражают свои взгляды, основываясь
198 Чат мпл Епктоем* гявриЛ^цая а м рггцмтты
на больших запасе знаииЯ, который не следует легко отбрасы
вить.
С этим связан другой вопрос, мнения do которому также до-
вольно единодушны, а именно вопрос об относительной частоте
хороших биологических видов в царстве животных и царстве
растений. Как отмечалось в гл. 6, отклонения от идеального
стандарта дискретных биологических видов очень обычны у
высших растений, но гораздо менее обычны у высших животных.
Эти отклонения от организации биологического вида в случае
высших растений обычно сопровождаются естественной гибри-
дизацией.
Другой подход состоит в сравнении частоты полиплоидии в
двух царствах. Хорошо известно, что примерно половила видов
высших растений — полиплоиды, тогда как среди животных по-
липлоидия известна лишь в нескольких группах. Кроме того,
большинство встречающихся в природе полиплоидных расте-
ний — аллополиплонды {102]. Это служит явным указанием на
важное значение гибридизации в эволюции растений; аналогич-
ные указания в отношении царства животных отсутствуют.
Этологические барьеры, препятствующие гибридизации, у
высших животных развиты гораздо сильнее, чем у растений.
У животных эти барьеры, препятствующие скрещиванию, игра-
ют главную роль и высоко эффективны, если ие считать тех слу-
чаев, когда создаются аномальные условия. В большинстве
групп растений подобные этологические барьеры совершенно от-
сутствуют. А в тех группах, в которых они существуют. — в эво-
люционно наиболее продвинутых семействах цветковых расте-
ний— они носят вторичный характер, находясь в зависимости
от поведения посещающих цветки насекомых, и поэтому меиее
эффективны, чем непосредственные предпочтения при выборе
брачного партнера, существующие у животных.
Хэббс [386] и Майр [569] отмечают, что среди животных гиб-
ридизация обычна главным образом в группах с наружным
оплодотворением, в частности у рыб и у некоторых амфибий, но
ис у рептилий, птиц, млекопитающих и насекомых, для которых
характерно внутреннее оплодотворение. Правда, и здесь не об
ходится без редких исключений. Царство растений в целом срав-
нимо с той частью царства животных, которой свойственно на-
ружное оплодотворение, поскольку у всех растений перекрестное
оплодотворение обеспечивается внешними агентами. С этим
коррелирует также сходство в отношении частоты гибридиза-
ции у растений и у животных с наружным оплодотворением.
17 Интрогрессия
Воспрон-чведеиие естественных гибридов в очень многих слу-
чаях следует по определенному пути, известному под названием
интрогрессии Интрогрессивная .гибридизация, по определению
Днд^ргпмя^Tr. jinwtnpiiu^ полираi мне скрезцнванш! _еГгЯ?Т-
лдд Обеими родительскими популяци*
ми» Она приводит к переносу генов от одного вида нли полувида
к другому, что сопровождается преодоленной барьеров, препят-
ствующих скрещиванию {15, 16. 19].
Рамине работы Дю Рис [181] н Марсден-Джонса [557] по-
могли понять явление интрогрессии, хотя эти авторы п не ис-
пользовали данный термин [293, 365]. Марсден-Джонс [557] про
вел генетическое и полевое исследование Geum urbanum. G. riva-
ls и их гибрида. О’, intermedium, я Англии. Родительские виды
широко распространены в Англии и экологически разделены;
гибриды также часто встречаются в некоторых местообитаниях.
Марсден-Джонс [557] обнаружил два места, в которых формы,
получающиеся в результате возвратных скрещиваний G. rivaled
у б. intermedium, замешали чяетых О rtieto Он утверждает,
что это «пример возникновения дикой популяции нового типа в
результате межвидовой гибридизации» [557].
Дю Рие провел популяционные исследования групп видов у
Draccphyttum. Coprusma и Salix в новозеландской и скандинав-
ской флорах. По его данным, члены какой-либо игры видов гиб-
ридизируют в определенных местах, а кроме того, у популяций
тех же родительских видов наблюдается конвергенция по мор-
фологическим признакам в зонах снмпатричсскоги перекрыла
ння Дю Рис объяснял эту конвергенцию гибридизацией, за ко-
торой следуют возвратные скрещивании. В результате такого
процесса один вид. как он выразился, «засоряется» определенны-
ми генами другого вида [161].
Факторы, способствующие интрогрессии
Рассмотрим колонию, состоящую из одного млн двух видов If нх
гибридов F|. В такой колонии действуют генетические и эколо-
гические факторы, которые способствуют возвратным скрешияэ
нияи, а тем самым направляют воспроизведение гибридов по
пути интрогрессии. Рассмотрим сначала погтуляцнонно-генети-
ческие факторы.
Естественный гибрид обычно бывает представлен одним или
несколькими растениями, находящимися в окружении обширной
Популяции одного или обоих родительских видов Далее, гмбрн-
ЭХ) Чат ««гол. Етегл'ямдл тбридимцая в га ретультотн
ди Fi обычно бывают по крайней мере полустерильными. а час-
то и высокостерильиыми. В таких условиях они вносят в гамет-
иый фонд колония очень небольшое число жизнеспособных га-
мет по сравнению с вкладом родительских видов. Вследствие
этого в очень большой часта случаев эффективных оплодотворе-
ний, в которых участвует какой-либо гибрид, вторым партнером
окажется растение одного иа родительских видов, что приведет
к появлению потомков от возвратных скрещиваний, т. е бекжрос-
сов [780J.
Экологические факторы, способствующие интрогрессии, пред-
ставляют собой естественный результат условий в местообита-
ниях, которые были описаны в гл 16. Каждый родительский вид
хорошо адаптирован к той экологической нише, которую он
обычно занимает. В открытом или промежуточном местообита-
нии могут поселиться гибриды Fi и некоторые потомки Ft про-
межуточного типа. Открытое местообитание или местообитание,
до некоторой степени гибридизированное, может дать приют не-
которым рекомбинантам, возникшим к результате естественной
гибридизация Однако большую часть возможностей для за-
крепления гибридных потомков в обычном месте встречи между
двумя видами или полувидами получают те генотипы, которые по
своим физиологическим признакам приближаются к одной или
другой родительской форме Необходимыми экологическим и пред-
почтениями скорее всего будут обладать потомки от возвратных
скрещиваний, для которых существующие в этом месте условии
среды, по всей вероятности, окажутся благоприятными [15, 19].
Бэтке и Олстон [38] проанализировали состав одной гибрид-
ной колонии Baptisia я Техасе. В качестве критериев они ис-
пользовали морфологические и биохимические признаки. На ос-
нове этих критериев они установили, что колония имеет следую-
щий состав.
Ba/Uiaia 1еисорЛма <70 растений
вярТГМа ipUe'ora'pe SCI раепкве
Г«бриты Fi М распева
Продукты пптпрятяых скрещиваний с В Ituco- 37 ряспвитй
рЧоев
Продукты возаратвых скрещиваний с В. грЛмго- 19 растений
Таким образом, в естествспных условиях растения, происходя-
щие от гибридов, распадаются иа два класса реципрокных бек-
кроссов [38).
Описанные выше популяционно-генетические и экологические
факторы продолжают действовать в последующих поколениях
гибридов и ведут к повторным возвратным скрещиваниям. В ре-
Га. 17.
201
вульгате некоторые гены или хромосомные участки одного вида
проникают в генофонд другого вила. А вследствие этого полу-
чившие их популяции вида реципиента конвергируют в наврав
пенни вида-данора по разным морфологическим, биохимическим
н физиологическим признакам [15. 16, 19. 181]. .
Методы
Дня самых старых и наиболее широко распространенных мето-
ла изучения интрогрессии основаны на ислолковании гибридно-
го индекса и «многомсрных> диаграмм разброса (pictorializcd
scatter diagrams). Получающиеся прн атом графики двух ти-
пов дают наглядное представление о том, что происходит в ин-
трогрессивных популяциях; таким образом, эти графики служат
как дли анализа, так и для описания процесса интрогрессии.
Вначале Андерсон и его сотрудники пользовались гибридным
индексом. Многомерные диаграммы разброса — существенное
усовершенствование, введенное ими позднее [15, 18].
Основная причина, обусловившая использование этих стан-
дартных методов и их последующее усовершенствование, связа-
на с многофакторным сцеплением. Как уже говорилось в гл 8.
оио представляет собой частичное н переплетающееся сцепление
между различными количественными признаками. Miюгофактор-
ное сцепление ведет к слабой ассоциации признаков, или связ-
ности признаков, у расщепляющихся гибридных потомков.
Андерсон (13. 15, 16] показал, что многофакторное сцепление и
связность признаков создают основу для обнаружения н коли-
чественном оценки результатов интрогрессии.
Следует ожидать, что в результате возвратных скрещива-
ний и интрогрессии возникнет изменчивая популяция, прибли-
жающаяся по своим фенотипическим признакам к одному из
родительских видов Гипотезу о том, что такое сочетание измен-
чивости и конвергенции фактически обусловлено интрогрессией,
можно проверить, использовав критерий ассоциации признаков.
Этот критерий применим в случае признаков, не связанных меж-
ду гобой морфогенетмческп, как, например, опушениостъ стебля
и окраска цветков; on неприменим к таким п.тейотрплиым приз-
накам, как форма листьев и форма чашелистиков.
Если какие-либо гемы, определяющие один признак, вмести
от вила донора в интрогрессивную популяцию другого вида, то
тогда в результате многофакторного сцепления рекуррентная
родительская форма получит вместе с ними другие генные снс-
т«мы и детерминируемые ими признаки Поэтому интрогрессив-
ная популяция будет походить на рекуррентную родительскую
форму. но будет варьировать в направлении вида доюра не
2U3 Чат мгв». Етстаоишя гибрадаяпрар и м раумтат
только по одному количественному признаку, а по многим или
по кем таким признакам [15. 16].
Метод гибридного индекса предполагает рассмотрение опре-
деленного числа, например десяти, отдельных признаков, разли-
чающихся у двух предполагаемых родительских видов. Прила-
гаем экстремальному состоянию каждого признака, обнаружен-
ного у вида А, по произвольной шиле оценку 0, противопо-
ложному его состоянию у вида Б —оценку 2, а промежуточному
состоянию у известных иля предполагаемых гибридов F|—
опенку 1. Или же некоторым морфологическим признакам при-
дается относительно больший нес., чем другим, для того чтобы
отразить их относительную полезность в качестве показателей
лежащих а их основе генетических детерминант. В первом слу-
чае общая опенка, или гибридный индекс, отдельного растения,
принадлежащего к виду А и измеренного по десяти признакам,
будет равна нулю, а значение гибридного индекса для растения,
принадлежащего к виду Б, будет равно 20. Эти противополож-
ные оценки служат эталоном для определения промежуточности
н изменчивости гибридной популяции.
Далее следует определить значения гибридного индекса для
каждого отдельного растения в выборке, взятой из гибридной
колонии, и представить распределение значений индекса по
частоте п виде гистограммы. Колония, состоящая из родитель-
ских видов и гибридов Fi. представлена на гистограмме рядом
пиков в экстремальных и промежуточных областях. В отличие
от этого популяция, подверженная сальной интрогрессии, пред-
ставлена значениями гибридного индекса в диапазоне от зна-
чений. характерных для гибридов Ft, до значений, характерных
для одной кз родительских форм. Наконец, у популяции, подвер-
женной слабой интрогрессии, выявляется заметный сдвиг моды
от одного из экстремальных состояний к промежуточному, сопро-
вождаемый усилением индивидуальной изменчивости.
В следующем разделе будет описано применение метода гиб-
Йидных индексов к реальным примерам интрогрессии у Iris и
'elianthus (рис. 17.2 м 17.5).
Метод с использованием гибридного индекса объединяет ре-
зультаты измерения отдельных признаков. Два растения могут
иметь одинаковые промежуточные оценки вследствие разных,
ио компенсаторных отклонений от некой экстремальной комби-
нации признаков, в связи с чем с помощью этого метода анали-
за сведений относительно корреляции признаков получить почти
не удается. Напротив, при использовании метода многомерных
диаграмм разброса эти сведения сохраняются, так что во многих
случаях предпочтение отдают последнему методу.
Обычная диаграмма разброса, в которой два количественных
признака представлены осями координат, а каждое отдельное
Гл. 17 Нюроцхюм
3»
растение —точкой, отражает изменчивость данной популяции по
двум признакам. В многомерной диаграмме разброса дополни-
тельные признаки представлены длиной усиков, отходящих от
этих точек. Подобная диаграмма почти с одного взгляда позво-
ляет охватить всю картину изменчивости в популяции, варьиру-
ющей одновременно ио трем нли большему числу признаков
Затем можно перейти к построению ряда многомерных диаг-
рамм разброса для разных популяций предполагаемых роди-
тельских видов и популяций, происходящих от них в результате
интрогрессии. Такие ряды дают возможность непосредственно
сравнивать несколько популяций во диапазону и коррелирован-
ное™ их изменчивости В следующем разделе это показано на
примере I г it. Использование коэффициентов корреляции для пар
изменчивых признаков в гибридных популяциях, казалось бы,
должно давать определенное преимущество, поскольку эти ко-
эффициенты количественно оценивают меру связности призна-
ков. Я пользовался этим методом [270), однако ом так и не по-
лучил широкого распространения
Сравнительно недавно несколько исследователей, хорошо
знакомых со статистикой нли математикой, внесли ряд усовер-
шенствований в метод гибридных индексов, используя класспфн
цируюшне функции [216. 608]. различные метрические методы
[608, 882], метод главных компонент [60S, 629]. многомерный
статистический анализ [6291 и кластерный анализ [126]. Все эти
методы довольно сложны. Ьолее подробное нх описание можно
найти в перечисленных здесь работах.
В ранних работах по исследованию интрогрессии использо-
вались морфологические признаки Еще одно относительно пс-
давкее новшество — использование биохимических признаков,
выявляемых методами электрофореза, хроматографии и други-
ми. Включение биохимических признаков, конечно, позволило
расширить ту фактическую основу, иа которую опирается изу-
чение интрогрессии.
Среди ранних исследований. использующих биохимические
данные, следует назвать работы Тернера и его сотрудников иа
Bapttsia (Lcguminusac) [9, 38. 845). Вслед за этими работами
были опубликованы первые общие обзоры по этой проблеме
[8, 9]. В настоящее время использование биохимических призна-
ков вошло в повседневную практику. В следу кинем разделе мы
опишем исследование интрогрессии у Juniper их; в этом исследо-
вании были использованы как биохимические., так и другие
сложнейшие методы анализа.
ЛИ Чатл*гая.Етп»тммаЛридмт^ла№1мг11Мпаы
Примеры
Из многочисленных хороших примеров интрогрессии мы рас-
смотрим здесь только четыре. Выбранные нами примеры иллю-
стрируют различные аспекты этого процесса н разные подходы
к его изучеиию.
Iris
Ирисы, растущие и дельте Миссисипи в южной части шт. Луи-
зиана, изучали Райли [686] и другие. В ненарушенных природ-
ных местообитаниях в этой области Iris futoa и I. hexagona gi-
gant icaerutea' симпатричиы и ие гибридизируют. I. hexagona gi-
ganticacrulea (—/. giganlicaerulea) растет на заволоченных поч-
вах, а /. /ulva — на более сухих соседних участках и в лесах.
Там, где природные местообитания разрушаются человеком и
вместо них возникают пастбища и канавы, между двумя этими
нрисамн происходит гибридизация и появляются интрогрессии-
иые популяции [686].
Райли выбрал четыре колонии н подробно изучил их. Коло-
ния О — это чистая популяции Iris hexagona glgantuaerulea,
растущая иа нетронутых болотах. Колонии Н-1 и Н-2—более
или меиее гибридизированные популяции, растущие на пастби-
ще в 150 м от колонии О Колония чистая популяция /. Jul-
па, растущая в дубовом лесу в нескольких километрах от коло-
нии О. Незаверенные популяции (G и F) служат эталонами для
сопоставления гибридизированных популяций (HI и H-2J.
Из каждой колонии Райли собрал по 23 растения; для каж-
дого растения были проведены измерения по 7 признакам, отно-
сящимся к цветку (рис. 17.1). Затем результаты измерений бы-
ли оценены по произвольной шкале, по которой экстремальное
fiiltia-подобиое состояние получает оценку 0, a giganticaerulea-
подобное состояние — опенки 2, 3 или 4. Оценки по всем семи
признакам для каждого растения объединяли затем в значения
гибридного индекса, колебавшиеся от 0 до 17. Распределения
значений индекса по частоте для каждой колонии представляли
в виде гистограмм (рис. 17.2),
• В егтлмчне от «Международного кодекса мхуюппискоА номсяхлатуры»
«Международный кодекс ботдкмчеаюй ноиекклатури» крнлиагт кссмошжо
тамсоаюинческжх категорий рампам ниже вида Поэтому после вашния мда
еоАясиипго указывать таксоиомкческкй pair- мутротидиксго такгожа, если
ок пркэодитдм (см. Ч. Джеффри. Бвологммеская аимкшиитура, Изд-во
«Мир», М. I960, стр 27—28}. Так, после вдмакия frit hexagona должно
следом?» «зиЬзр», «van иди «forma» в мвнсимостн от того, кадей ранг
сридзет:» жймиию gigani.cnfraiea — Прим.
Гл. П. HnTfoiftKU»
ЭД
Кок показывают эти гистограмм», колония Н-2 по своим
морфологическим признакам и значительной мере сходна с
Iris giganiicarrulta. ио содержит несколько гибридных растений.
Колония Н-1, однако, представляет собой интрогрессивную по
[гулянию, и ее диапазон изменчивости в значительной степени
Рис, П I Цветя» и чяпмлвгтики (уеалнчмы) tris fut'.'a (А) и I krragona
Maniirmruha (Б) [15]
заполняет разрыв в морфологических признаках между I. gl-
ganticaerulea и / fuloa [6861-
Позднее Андерсом [15] представил эти же данные Райли
[*>86] в виде многомерных диаграмм разброса (рис. 17,3 и 17.Ч).
эти диаграммы выявляют корреляцию признаков в родительских
‘и гибридных популяциях.
Спустя еще некоторое время Рэндолф и его сотрудники [655]
вновь изучили этот случай Проведя исчерпывающий анализ из-
менчивости по 13 признакам у многочисленных аллопатрических
и симпатрических популяций »тих двух видов, они пришли к
выводу, что эффекты интрогрессии локализованы в нарушенных
местообитаниях в пределах эоны симпатрии. Насколько можно
было установить, каждый из видов Iris в значительной части
своего ареала не испытывал воздействия генов другого вида.
Часть пятая. Есгсстыьмая пиритизация и ле риу.итигм
Рве. 17.2. Р»с®ределенве по частоте значений пебрвдмых иадексоа для
23МСТМИ1 а каждой и четырех колоний ирисов в южной части Л)хшиы
г —Me fuloa. G. — l. haxagoaa gtgaatlracrulia, tt-1 — пип у ляп ня, подверг-
шаяся сильной иитрофмсмя. М-2 — популяция. подвергшаяся слабой китро-
грвосмм.
Helianthus
Наш следующий пример — виды подсолнечника Helianthus
annuus и Н bolanderi, изученные Хейзером [366]. Масличный
подсолнечник (Н. annuus) растет как сорняк ло всему умерен-
ному поясу Северной Америки В Калифорнии Н. annuus не яв-
ляется одним из элементов местной растительности; вероятно,
он сравнительно недавно был интродуцирован индейцами.
Н. bolandert встречается н Калифорнии и Орегоне как в есте-
ственных местообитаниях, так и в зарослях сорняков. Местная
раса Н. bolanderi растет в этих двух штатах на выходах сер-
Рис. 17 3 Мкг1гпмср>гме дмгр«миы разброс* /rif /ufoa. / hriuigona gigun
Нелегала и двух гибридных килоняЛ (на (15], по ядмннм (686)).
Т* же самые мнулвцм, чп> и и* рис. 112
зов
Часть ur« &т»гтохмлал .•а«р1йии<<и» и ее рщмтагч
Ряс 17.4 ООьясивж еяполоа. исполыовашл » рас. 173
пситинитов в предгорьях, тогда как сорная раса распростране-
на в центральной долине Калифорнии [366].
Эти два вида подсолнечника различаются по многим иорфо
логическим признакам. Melianthus annuus — высокое растение
с овальными листьями, а Н. bolandert — растения от низкой до
средней высоты с ланцетовидными листьями. Кроме того, они
различаются также по опушеиности, обертке соцветия, централь-
ной части цветоложа, язычковым цветкам и. семянкам. Такие
морфологические различия служат маркерами, позволяющими
производить оценку отдельных растений и анализировать попу-
ляции методом гибридных индексов (366).
Как показали эксперименты по гибридизации. Melianthus
annuus и Н. bolandert легко скрещиваются между собой в обоих
направлениях. Гибриды F> промежуточны по морфологический
признакам, конъюгация хромосом п мейоэе у них нарушена, а
пыльца и семена высокостернльны [366].
Melianthus annuus и Н. bolanderi приходят в контакт в раз-
ных районах Калифорнии. Периоды цветения у них перекрыва-
ются, и в ряде мест они гибридизируют. На рис. 17.5.Д, пока-
зана картина изменчивости в одном скоплении гибридов в до-
лине р. Сакраменто в Калифории^ На основе морфологических
14—45
ГиОрмдный индекс
Рис. 17Л. Частота распределен*! зиачеиий гибридного илдекеа в лиги волу-
ляиииз Hckafitkus спя и и» к Н. bvtenderl [385].
Л. Раса Н. Manderi с серпенткнхтсвых мча предгорий в Калвфорами.
Б. Pica Н. annuus их штата У в: сур и В Сорвав раса Н. boi&ndtri из делим
Каз»Форпми Г Н. annuut из должны Сан-Хоакин Д. Сжопленае гибридов нз
долины реки Сакраменто.
210 Часп алгм. Стпчим mCpuSuxupis и «« ра/мгты
признаков и коррелированной с ними фертильности пыльцы был
сделан вывод, что ага гибридная колония состоит из двух роди-
тельских вило*, нескольких гибридов Fi и двух типов реципрок-
ных беккроссов (.366].
Признаки неиитрогрессированных популяций Helianthus из
Сент-Луиса (шт. Миссури) и Н bolanderi. произрастающих на
выходах серпентинитов в северной части Калифорнии, показаны
на рис. 17.5. А и fi. На рис. 17Д Г и Д изображены для сравис
НИИ популяции тех же двух видов кз центральной долины Кали-
форнии. где они растут как сорняки. Можно видеть, что в каж-
дой нэ двух последних популяций изменчивость направлена в
сторону другой. Диапазоны изменчивости этих популяций нахо-
дятся в пределах изменчивости беккроссов из скоплений гибри-
дов; поэтому был сделан логический вывод, что они представ-
ляют собой продукты интрогрессивной гибридизации [366].
Распространение расы Helianthus bolanderi, экстремальной
по морфологическим признакам, ограничено серпентинитовыми
почвами предгорий, тогда как интрогрессивная раса того же ви-
да встречается каи сорняк в центральной долине вместе с
Н. annuus. Такое экогеографнческое распространение с доста-
точным основанием позволяет предположить, что вслед за ин-
тродукцией, вероятно индейцами, Н. annuus и Калифорнию на-
чалась гибридизация между этой формой и местной дикорасту-
щей формой Н. bolanderi [366]. Интрогрессии от Н. annuus к
Н. bolanderi привела затем к образованию долинной расы Н. bo-
landeri, которая смогла распространиться по всем возникшим
в это же время сорным местообитаниям. По всей вероятности,
Н. annuus я nein-ральлых районах Калифорнии испытал в свою
очередь воздействие интрогрессии со стороны Н bolanderi Воз-
можно, что возраст интрогрессивной долинной расы Н. bolande-
ri составляет всего несколько сотен -лет [366].
Оливьери и Джейн [629] еще раз проанализировали этот слу-
чай. использовав более обширный набор признаков, в том числе
и биохимических, и проведи более сложный статистический ана-
лиз имеющихся данных. Полученные ими гистограммы, как и
следовало ожидать, слегка отличаются от гистограмм Хейзера
(рис. 17.5), но. по-видимому, соответствуют его первоначальной
иитеплретацки.
Helianthus annuus перекрывается и гибридизирует е други-
ми однолетними видами подсолнечника в других районах США.
например с Н. debiiis и Н. argophyllus в Техасе и с Н. petiolaris
иа Великих равнинах и на юго-западе. Н. annuus дал начало
интрогрессивным расам, изменчивость которых направлена в
сторону Н. debibs в Техасе и Н. petiolaris — в центральных райо-
нах США. Н. petiolaris существует также в качестве интрогрес-
сивной расы (364. 368—371. 373].
Гл 17 Интрогрессия
211
/uniper us
Juniper us virgiruana широки распространен на востоке США, а
/. scopulorum — на западе Их ареалы » основном аллопатрнчны
и переходят один и другой с несколькими небольшими участка-
ми контакта на границах (рве 17 6). Фассет [209] сообщает о
скоплениях гибридов, выявленных им иа основе изучения морфо-
логических признаков
Сравнительно недавно иееколькимм исследователями [216]
был проведен анализ популяции этих можжевельников иа длим
«ой траткекте от восточной части Миссури до западной части
Монтаны (риг 17.6) с использованием биохимических прилив
ков -43 содержащихся в листьях соединений, входящих в со-
став летучих масел. Полученные данные были подвергнуты дис-
криминантному анализу
На основе оценок, полученных в результате такого анализа,
авторы згой работы построили шкалу гибридиости, которую
можно было использовать для пострення гистограмм Характер
изменчивости в ряде гистограмм — по одной для каждой попу-
ляции на трансекте от А до Z — показывает, что интрогрессия
происходит от Juniper us scopulorum к 1. virglniana в северо-
западной части области, занимаемой чтим последним [216). Че-
тыре такие гистограммы представлены на рис. 17.7.
Pinas
Интересный пример интрогрессии в рамках исторического вре-
мени, основанный на налеоботаиичесжнх данных, описан Мейсо-
ном [558]. Этот пример относится к двум вилам сосны. Pinus
remorata и Р. muricata, произраставшим в прибрежной части
Калифорнии в период от плейстоцена до современной эпохи.
В плейстоцене популяции Pinus remoraia па острове Санта-
Крус морфологически сильно отличались от родственной Р mu-
ricata, которая в то время была распространена на материке
в Калифорнии. В позднем плейстоцене или в постплейгтоиспо-
вос время Р. muricata мигрировала иа остров Санта-Крус. Меж-
ду этими двумя пилами происходила естественная гибридизация,
продолжавшаяся и в современную эпоху, что привело к изме-
нению признаков у некоторых популяций Р. remoraia. Ископае-
мые данные, собранные в Калифорнии, свидетельствуют о про-
исходившей в плейстоцене интрогрессии в обратном направле
нии —or Р remarata к Р muricata-, этот процесс продолжается
н в настоящее время (457),
14»
Ряс. П.е Геогр«фжчегжпе расп ростр* мешав luni^nu Virginians и J всори(огвл [216].
Буквама nowzMMc иестсл вхождение изучавшихся ппиуляки*.
Гл. 17. Нкгрчмркям
213
Рж- 17.7. Гжтогрвммы гиб рядных шкал для ооалляций Junt perns иа трансеп-
те, идущей от шт. Мжсуря до шт Момтэяв (216]
Вяличниы для построения гибридной шкалы получены • результате джкри*
мнняятяол) анллизн «3 летучих масел, содержишися в листьях и стеблях.
D — популяции / Qi>fuUaiM и» Миссури м юго-воетошой части
Неброски X—маудмня J. scopalorum ю Модтяны. Н и V — £вс итттро-
гмяпмме попудяцмл кз ааграяичиай johu между >тммм двумя видами
1 сложение на карте ятях и других популяций на даеноА трансекте показано
на рас. 17 6.
. Связность признаков как критерий гибридностн
Андерсон (13, 15, 16] рассматривал связность признаков — сла-
бую ассоциацию между разными признаками и изменчивой по-
пуляции— как диагностический признак гибридностн, о чем уже
говорилось выше. Это заключение было осиовако как иа теоре-
тических, так и на эмпирических данных. Связность признаков—
ожидаемый результат многофакторного сцепления в любой рас-
щепляющейся гибридной популяции. Эго было подтверждено
экспериментально на искусственных гибридных популяциях.
814 Часп пятая. Lmcnenna» гибриЛтуия а tr рпумти
Обычно связность признаков наблюдается также в природных
гибридных популяциях.
Заключение Андерсона иа протяжении многих лет пользо-
валось широким признанием. Однако недавно было показано,
что такое слишком широкое обобщение неоправданно [297].
Рассмотрим сначала эмпирические данные, относящиеся к
природным гибридным популяциям. Эти данные касаются почти
исключительно многолетних растений, обычно травянистых.
У таких растений особи одного или обоих родительских расте-
ний сохраняются достаточно долго, чтобы иметь возможность
смешиваться е гибридами Fi и последующими поколениями.
Если они попадают в выборки на популяций, как это часто бы-
вало в ранних исследованиях, то они искажают результаты в
пользу связности признаков.
При Повторном анализе данных по нескольким старым при-
мерам — Iris Aquilegia, Ozytropis и другим группам растений,
у которых была описана связность Признаков, — в каждой вы-
борке были обнаружены особи родительского типа. После иск-
лючения этих особей кз выборки и проведения нового анализа
одной только гибридной субооауляцни связность признаков за-
метно снижалась [297].
Уровень связности признаков можно измерить по коэффи-
циентам корреляции для разных пар признаков. А степень сни-
жения связности признаков в гибридной субпопуляции по срав-
нению с популяцией в целом можно измерить по различию меж-
ду коэффициентами корреляции для одной в той же лары приз-
наков в выборках из той и другой.
Дальнейшими исследованиями в том же направлении было
установлено, что в разных случаях степень снижения связности
сильно варьирует. Таи. например, в одной гибридной популяции
Opuntia при анализе популяции в целом наблюдалась статисти-
чески эначииая корреляция между признаками почтя во всех
попарных комбинациях, одиако в гибридной субполуляции такая
корреляция была обнаружена лишь для одной пары признаков.
Данные о саязиосш признаков в этой гибридной популяции —
артефакт, обусловленный способом Взятия выборки. В отличие
от этого в гибридной популяции Iris уровень статистически вы-
являемой связности признаков хотя и ниже, чем предполагалось
прежде, но все же достаточно высок (297].
Итак, связность признаков — обычная, но ие всеобщая чер-
та. характеризующая природные гибридные популяции расте-
ний. Ока проявляется, например, у некоторых гибридных попу-
ляций Iris и Aquilegia при правильном взятии выборок. ио не
наблюдается у некоторых гибридных популяций Opuntia. Ozyt-
ropis я Gilia [297]. Первоначальное обобщение претендовало иа
слишком большую универсальность.
Гл. 17. Интрогрессия
216
Для тогп чтобы понять это, пая придется вновь вернуться
к теоретическим допущениям, на которых было основано преж-
нее обобщение Некоторые ня этих допущение относятся к инеш-
ннм факторам, определяющим связность признаков, а другие —
к внутренним.
Самый важный внешний фактор — естественный отбор.
Предполагалось, что отбор в природных гибридных популяциях
всегда будет благоприятствовать генотипам, приближающимся
к родительским типам Это допущение, несомненно, состоятельно
во многих или даже в большинстве случаев. Но нет никаких
причин, по которым можно было бы считать, что опо всегда
справедливо. В тех случаях, когда имеются новые и разнообраз-
ные среды, доступные дли заселения, отбор будет благоприятст-
вовать также новым рекомбинантным типам.
Что касается внутренних факторов, то реальной силой сле-
дует считать многофакторное сцепление, и оно способствует
связности признаков; ио рекомбинация генов —также реальная
сила, которая действует в противоположном направлении, вызы-
вая рекомбинации признаков. В результате любая данная гиб-
ридная популяция представляет собой некое равновесие между
этими противоположными тенденциями Положение точки рав-
новесия в каждом отдельном случае в значительной мере за ни-
сит от рекомбинационной системы данного растения (см. гл. 1).
В общем следует ожидать, что у растений с высоким основным
числом хромосом связность признаков в гибридных популяциях
будет слабее, чем у растений с низким основным числом-, кро-
ме того, у однолетних растений связность слабее, чем у много-
летних
Экологическая сегрегация морфологических
типов'
В гибридных популяциях происходит расщепление не только по
морфологическим и биохимическим признакам, но также и по
физиологическим признакам, определяющим экологические пред-
почтения Однако в отличие от первых двух точно оценить фи-
апологические признаки трудно и сведения о иих публикуются
редко. Тем не Meow полевые наблюдения свидетельствуют о том,
что в гибридных популяциях происходит сегрегация по экологи-
ческим предпочтениям.
Интрогрессивная популяция, или скопление гибридов, часто •
растет в неоднородных или до некоторой степени гибрнднзиро- 1
ванных местообитаниях, расположенных между местообитания- *
ми популяций родительских видов. В таких случаях микрогеог-
рафнческое распределение беккроссов резвых типов бывает ие-
21в Час Го ллтсл, £<гкгммшм тбрадизауся s « ри^.отатн
случайным. Напротив. как и следовало ожидать, особи, прибли-
жающиеся по своим морфологическим признакам к одному из
родительских видов, обычно имеют тенденцию скапливаться в
тех участках гибридного местообитания, которые больше всего
сходны е нормальной для «того вида средой. И наоборот, гиб-
ридные выщепекцы противоположного типа и беккроссы зани-
мают главным образом ниши, приближающиеся к местообита-
ниям другого родительского вида. Я наблюдал такую экологи-
ческую сегрегацию различных морфологических форм в гибрид-
ных популяциях Aquilegia, Mimulus sect Diptacus, Ipomopsis я
Gilia. Несомненно, другие ботяинки наблюдали то же самое и у
других растений. Это явление было описано Бенсоном, Филлип-
сом и Уайлдером [59] в гибридной популяции Quercus.
В этом случае родительскими полувидами были Quercus
douglasii и Q. turbinella subsp. califomica. Q. douglasil—листо-
падное дерево с сизо голубоватыми листьями, растущее в пред-
горьях центральной и северной Калифорнии на противополож-
ной от моря стороне. Quercus turbinella subsp. califomica — веч-
нозеленый кустарник, растущий иа пустынных и полупустынных
горных склонах в центральной и южной Калифорнии. Эти два
полувида встречаются вместе и гибридизируют в разных местах
в предгорьях южной и центральной Калифорнии. Популяция,
подробно проанализированная Бенсоном. Филлипсом н Уайлде-
ром [59]. произрастает в этой зоне контакта, иа перевале Тихон.
Техокская популяция занимает экологически разнообразную
область, где склоны холмов обращены к разным сторонам све-
та. Жаркие сукне склоны, обращенные иа юг и юго-восток, боль-
ше всего соответствуют обычному местообитанию Quercus tur-
binella subsp. califomica. Склоны, обращенные на север и севе-
ро-восток, также сухие, но получают меньше солнечных лучей
и приближаются по своим условиям к обычной среде более се-
верного вида Q douglasii.
Бенсон. Филлипс и Уайлдер [59] провели измерения по 11 ве-
гетативным признакам в выборке из техоиской популяции и
оценили их по методу гибридных индексов. При этом оказалось,
что гибридная популяция в целом содержит большое число бек-
, кроссов, приближающихся к Quercus douglasil. а также бек-
кроссое. приближающихся к Q. turbinella subsp. califomica, при-
чем относительные количества тех и других примерно одинако-
вы. Однако при повторном анализе данных с учетом микрогеог-
рафнческого местоположения отдельных растений вмяпилась
другая картина. На склонах, обращенных на север и на северо-
восток, растения морфологически напоминали Q. douglasii.
В отличие от этого иа склонах, обращенных на юг и юго-восток,
было много растений, приближавшихся по своим гибридным
индексам к Q. turbinella subsp califomica [59].
Гл, 17, Игетрогреаия
217
Сцепление М—V в связи с интрогрессией
Гены, детерминирующие у высших растений, различные морфо
логические признаки. в том числе те. которые раньше других
выявляются у цветков, плодов или но поздних стадиях вегма»
ТНВ1ЮГО развития, обычно сцеплены с генами, воздействующими
на роет н мощность иа ранних стадиях развития. Близкие виды
растений часто несут аллели сцепленных друг с другом морфо-
логических генов и генов жизнеспособности. Примером такого
сцепления служит сцепление .4— V. которое мы вкратце обсуж-
дали » гл 8 Кроме того, имеются данные о существовании сис-
тем сцепления *1—1' в перестроенных участках, по который
различаются хромосомы видов растений, стерильных при скре-
щиваниях, а также в гомологичных участках у рас или видов,
плодовитых при скрещиваниях (см. гл 8).
Сиеп.теине M—V, несомненно, широко распространенное, а
быть может, и универсальное явление в мире растений Здесь мы
обсудим отношение этого широко распространенного типа сцеп-
лении к анализу интрогрессии. Допустим, что мы обнаружили
популяцию растений, характеризующуюся следующими черта-
ми: 1) популяция подвержена изменчивости: 2) растения по
комбинации морфологических признаков приближаются к неко-
ему виду А, по изменчивы в паправленин другого вида Б: 3) они
приближаются также к виду А по своим экологическим пред-
почтениям: 4) они плодовиты яри скрещивании с видом А, но
их гибриды с видом Б обладают хромосомной стерильностью.
Используя стандартные методы анализа интрогрессии, мы
пришли бы к выводу, что ваша гипотетическая популяция пред-
ставляет собой продукт интрогрессии определенных генов от ви-
да Б к виду А Мы постулировали бы имевшую место в прош-
лом гибридизацию АхБ. за которой следовали возвратные скре-
щивания с родительский видом А, сопровождавшиеся отбором
беккроссов, хорошо соответствовавших среде вида А
Интрогрессивная гибридизация, несомненно, будет приво-
дить к описанным выше эффектам Однако следует указать, что
такие же конечные результаты могут быть получепы при другом
способе гибридизации Если гены, определяющие морфологиче
ские признаки, обычно сцеплены с генами жизнеспособности и
если виды растений различаются ио аллелям генов, входящих
» эти системы спеплення М—V. то межвидовая гибридизация,
за которой следуют непрерывный инбридинг и отбор инбредиых
продуктов, приведет к комбинации признаков, перечисленных а
одиом из предыдущих абзацев. Следовательно, такую комбина-
цию признаков нельзя считать абсолютным критерием интро-
грессии |292).
218 Чат кмил Есгаепыт mtpwWwutu» и аа релуытагн
Я наблюдал в эксперименте с Gilia проявление эффектов, по-
добных тем, которые возникают при интрогрессии, но в отсут-
ствие последней. Роднге.тмкиии вилами служили Gilia modocen
sis н 6. malior. Они представляют собой автогамные тетрапло-
идные виды, принадлежащие к комплексу G. inconspieua, с ко-
торым мы уже поаиакоыилнсь в гл. 11. Здесь пас интересуют
некоторые инбредные линии, происходящие от гибридов F», по
лученных при скрещивании G malior и modocensis. в частности
линии, обозначенные на рис. 17.8 хак Ветвь II [288, 292].
Гибриды F, от скрещивания между Gilia malior и С. model-
ceruis высоко, но ие полностью стерильны, а конъюгация хромо-
сом у них очень сильно нарушена. Помимо нарушения коиыога-
пии у гибридов, имеются и другие цитогенетические данные,
указывающие на то, что родительские виды различаются по
многочисленным независимым перестройкам в разных участках
[285. 289].
Эти гибриды Fi дали некоторое число потомков без измене-
ния плоидвости. которых размножали путем инбридинга иа про-
тяжении ряда поколений. Полученные таким образом инбредные
линии подвергли отбору на плодовитость и мощность в среде,
благоприятной для типов, сходных е Gilia modocensis. Морфо-
логические признаки как таковые при отборе ие учитывались.
Рис 17 а Два интерстерлльнмх вкла ОИл и <ыж* на гнбрклов между па
ми (2921
Этот гибехл был пс-тучти в ряттлыхтв иибрклхжтв и отбора ив жпнвеечо
собпость По морфологии а плозовитостн пр» скрчшяшяяях ои банлон к
одаому ВТ рмаительохнл вялое.
Гл. 17. ИктрОфгиня
219
Растения, получившиеся после отбора в ряде поколений, были
плодовитыми и мощными [283* 289].
Одна группа родственных линий, происходивших пт одной и
той же родительской особи из F2 и названная Ветвью II, оказа-
лась морфологически очень близкой к Gilia modocensls
(рис 17 8) В ней, однако, наблюдалась изменчивость в сторону
б. malior
Плодовитые растения из Ветви II исппльэовллн для возврат-
пых скрещиваний с обоими родительскими видами. Гибриды, по-
лученные от скрещивания между гибридом из Ветви II и Gilia
modocensis. были плодовиты, и все их хромосомы конъюгирова-
ли с образованием бивалентов, тогда как при воэиратпом скре-
щивании с (?. malior получались высокостерильные гибриды с
на ру шейной конъюгацией |288|.
Допустим теперь, что растения Ветви II были бы обнаруже-
ны в природе н изучены с морфологической, экологической и ци-
тогенетической точек зрении Очевидно, их определили бы как
отклоняющуюся расу Gilia modocensis. получившую некоторые
гены от G malior в результате интрогрессии п обладающую по-
этому соответствующими этим генам морфологическими призна-
ка мн. Однако в данном случае нам известна вен родословная
Ветви Л, и мы можем утверждать, что никакого возвратного
скрешпвамия у се предков не происходило [289, 292}.
Таким образом, как ИЗ экспериментальных исследований, так
и из теоретических рассуждений следует, что эффекты, ие отли-
чимые от эффектов интрогрессии, могут возникать и иным путем
Существующие методы анализа ннгрогрессин не дают возмож-
ности различать: а) возвратное скрещивание и отбор и б) отбор
во взаимодействии со сцеплением Al— V |292|.
Учитывая вес это, становится необходимым пересмотреть кри-
терии, ия основе которых определяют интрогрессию, а также не-
которые примеры, ндентнфниирпваппые при помощи этих крите-
риев Быть может, иаилучшим критерием иа данном уровне на-
ших знаний служит система скрещивании растений, о которых
идет речь. У преимущественно алтигамимх растений размноже
ннс гибрида путем возвратных скрещиваний крайне маловеро-
ятно Относить наблюдаемую у таких растений изменчивость,
сходную с Епирогрессией. за счет инцкнресснк недопустимо
У некоторых гибридизирующих автогамных растении, в част-
ности у группы Elymus glaocus и группы Aegiiops variabilis
[751, 781. 919], изменчивость, сходная с наблюдаемой при интро-
грессии. п самом деле была истолкована как интрогрессия
Однако в свете того, что нам известно о сцеплении .М— V. при
пятые н этих случаях интерпретации следует, возможно, пере,
смотреть
220 Чат лхпя. Бепсттиишл гибридизация в « дгаулгоп*
В отличие от этого в группах растений, раэмиожающихся при
помощи ауткроссннга, наиболее вероятный способ размножения
гибридов — возвратные скрещивания. Это прежде всего следст-
вие самой системы размножения. Кроме того, это обусловлено
соотношением числа гамет, поставляемых в гвметиый фона мио
гочислениымн плодовитыми растениями родительских видов н
редкими стерильными гибридными растениями, соответственно,
о нем уже говорилось выше в этой главе.
Если, обнаружив у свободвоопылнющихся растений картину
изменчивости, сходную с интрогрессией, сделать вывод о нали-
чии у них интрогрессии, то такой вывод скорее всего окажется
правильным, соответствуя точке зрения Андерсона [15. 15].
Стеббинса [780] и других авторов Все примеры интрогрессии,
выбранные в качестве иллюстраций для данной главы (Zrte, Не-
lianthus, Juniper us к т. п.), относятся к перекрестиоогылякицвм-
ся растениям.
Следует ожидать, что в группах растений с системой раз-
множения. промежуточной между ауткроссингом и инбридингом,
размножение гибридов происходит разнообразными смешанны-
ми способами. Судя по данным Пази и Цохари [636|, так, воз
можно, обстоит дело в некоторых частях группы Aegtlops папа-
bilis.
Трансгрессия барьеров хромосомной
стерильности
Интрогрессия между видами растений, различающимися по
структурным особенностям кариотипов, должна быть связана с
прохождением генов через барьер хромосомной стерильности.
В этом разделе нам предстоит рассмотреть механизм, с по-
мощью которого происходит трансгрессия барьеров хромосом-
ной стерильности. Мы изложим здесь в пересмотренном и рас-
ширенном виде материалы, публиковавшиеся ранее [280. 282).
Начнем с рассмотрения некоторых предварительных усло-
вий. Представим себе некую гибридную популяцию, состоящую
из особей двух видов и гибридов между ними. Растения размно-
жаются при помощи ауткроссннга. Хромосомы родительских ви-
дов различаются по структурным перестройкам, ведущим к сни-
жению конъюгации или несбалансированности гамет у гибри-
дов нли же одновременно к тому и другому, в результате чего
у гибридов наблюдается частичная или полная хромосомная
стерильность. Некоторые гены, которые у одного из родитель-
ских видов были локализованы в этих хромосомах, попав в ге-
нотип другого родительского вида, образуют рекомбинанты, бо-
лее адаптированные к среде этого другого вида. Ихзожеиные
Гл. 17. Интрогрессия
221
здесь условия, по всей вероятности, часто встречаются в при-
роде.
Процесс трансгрессии стерильности, происходящий, согласно
ожиданиям, при этих условиях, сложен. Он слагается ия несколь-
ких отдельных субпроцессов. действующих одновременно. Во-
первых, растения, составляющие гибридную популяцию, размпо
жаются при помошн агткроссинга Во вторых, гомсо логичные
хромосомы структуриых гетерознгот образуют биваленты, и по
крайней мере в некоторых из материнских меток пыльны к ма-
теринских клеток мегаспор наблюдается кросгинговср Отбор
гамег происходит так. чтобы элиминировать образовавшиеся
при менозс метки с нехватками или дупликациями н сохранить
гсиетнчески сбалансированные гаметофиты. В то же время есте-
ственный отбор среди членов данной популяции благоприятст-
вует сохранению некоторых межвидовых рекомбинационных ге-
нотипов в среде одного из родительских видов
Мы рассмотрим сначала процесс трансгрессии стсрклыюстя
Применительно к одной определенной модели В этой модели
имеются вид-рсинпнент (/?) и вид-донор (О), вводящий некото-
рые их своих геноа я первый вил путем интрогрессии Рекомби-
национные генотипы, содержащие некоторые гены вида О на ге-
нетическом фоне вида R. имею, высокую адаптивную ценность
я среде вида R Адаптивная пег ioctb таких межвидовых реком-
би ил циомных типов в этой сред- выше, чем ценность комбина-
ции соответствующих аллелей у нснитрогресснроаа иного ви-
да R.
Гены, входящие в обладающую адаптивной ценностью ком-
бипанкю. находятся я хромосомах, различающихся у этих двух
видов по структурным перестройкам
Для того чтобы упростил, обсуждение, мы сосредоточим вни-
мание па одной хромосоме (/), присутствующей v родительских
видом я двух разных структурных формах (/, и /л) Хромосомы
Л и /,< отличаются по одной транспозиции, показанной на
рис 17 9, А, В процессе филогенетической истории хромосомы
/е один нз участков и левом плече поменялся местами с одним
из участков в правом плече, но а хромосоме /, эта перестройка
ие имела места, п результате чего хромосомы этих двух типов
различаются по своей структуре, как это показано на рис 17.9, А.
Центральные, вли интерстициальные, участки хромосом h и
1,1 гомологичны (рис I7 9. Л) Мы постулируем, что гены вида
D, образующие с генами вила R ценную в адаптивном смысле
рекомбинацию, локализованы в этих интерстициальных участ-
ках хромосомы Zrf. Геиы вида R, входящие в ту же самую цен-
ную комбинацию гемов находятся в соответствующих гомоло-
гичных участках хромосомы /, или же в этой, а также п в дру-
гих хромосомах данного набора
•«A
Рве 17.9. Цитогяяетнчвсхое поведанне жары хромосом, различавшийся по
своей структур#, и гнбрнлж!Я популаан-
Родитвлыгхив ниды ft a D ncryt в ааодаг в данную вопуляцио дм частично
гомологичные фермы >то! хромосомы (обоаначеавые соответственно Л В /<),
различающееся во одной тряиспоягамн А Роиительосне формы. Б. Продук-
ты мвйоэа в F, turn у гебряссв, волученвых при всмрвтнык скрешяваиинх,
с нроссвяговфом между сдечдми. В Продукты мейоза в Fi иди у пгёрнков,
•овучеиимх при возвратных ежрекхивамивх, с лвоЯным кроссмиговером
между участкхмн. nuaaepi шнмжя всрестрсйле
Гл. 17. //мтросресгал
223
У гибриде ж RxD конъюгация хромосом I может быть нару-
шена в различной степени вследствие отсутствия полной гомоло-
гии между ними Следует ожидать, что у^бипален гое t,h в тех
случаях, котла они образуются, будет довольно регулярно про-
исходить кроссинговер в гомологичных интерстициальных участ-
ках (рис. 17 9.6). При таком кроссииговсре половина клеток,
образующихся в результате мейоза. несет нехвагки и дуплика-
ции по смешенным участкам (рис. 17.9, Б), и их них развиваются
нож из неспособные гамегофнты. что обуслошММС! uaiycicpnab-
кость гибридов Иными словами, когда в результате кроссинго-
вера смешенный участок понадает в гомологичную хромосому,
пн. создавая мех натку п дупликацию. пыстуиает в роли фактора
стерильности.
Хромосомы с нехватками к дупликациями будут выпадать нз
фонда гамет в каждом поколении. То же самое будет происхо-
дить с генами вида D. локализованными в интерсгпцнальиых
участках этих хромосом
Функциональными окажутся лишь те гаметы, которые несут
иекроссовсрныс хромосомы /, или /а (рис 179,5). Следователь-
но, гибрид F| после скрещивания со своими сибсами лает по-
томков Fs. расщепляющихся на лил родительских и одни гиб-
ридный класс {/,/,. Шы. bit), а при возвратном скрещивании с
видом R происходит расщепление иа одни родительский л идин
гибридны А классы (LL, /J.r) Генотипы hl л пол у стерильны в
Fa. н Bt. так же как и в Fi В отличие от *того среди потом-
ков последующих поколений плодовиты лишь те. которые обла-
дают родительскими Генотипами — Ыг и 1Л1Л.
Таким образом, прн обычном кроссинговере в хромосоме /
(рнс 17.9,5) гибридная популяция расщепляете»: но морфоло-
гическим н физиологическим признакам, которые определяются
генами, локализованными в згон хромосоме, иа родительские
типы и тип Fi Интрогрессия генов, лежащих н хромосоме /. от
вида D к виду R блокирована, так как эти гены сцеплены г фак-
торами стерильноеiи. находящимися в этой же самой хромо-
соме
Если, однако, кроссинговер в структурной гстерозиготе про-
исходит d точках, отделяющих смещенные участки от гомоло-
гичных интерстициальных участков, то генетические результаты
гибридизации окажутся совершенно иными. Двойной кроссииго-
вер. который приведет к такому отделению, схематически изоб-
ражен на рис 17.9.5. Такого рода кроссинговер может происхо-
дить или не происходить в поколении F»; если он ие произойдет
в Fi. то он может произойти в любой структурной гетерознготе
в любом последующем поколении ог возвратного скрещивания.
В результате такого двойного кроссинговера возникнут крос-
соверные хромосомы типа /м (рис. 17.9, В) Зги хромосомы ге-
22ч Уисте |шш. естгсгепишл riM/Mdu-vw и ег ргг/штаты
нетяческп сбалансированы, в связи с чем содержащие ях гаме-
ты жизнеспособны. По свое* структуре они гомологичны хромо-
сомам /„ поэтому способны образовать фертильные гетерозиго-
ты 1,1, а и тем самым включаться в хромосомны* фонд вида R
Сбалансированные хромосомы /«несут также в своих нейт-
ральных интерстициальных участках’обширные блоки генов ви-
да D. Поэтому хромосомы 1,4. которые внутренне сбалансиро-
ваны н одновременно гомологичны хромосомам I,. могут служить
мостиком для перехода генов вида D к виду R. Вслед за этим
в результате естественного отбора может произойти увеличение
частоты новых хромосом /« или их частей в среде вида R.
Мы рассмотрели процесс трансгрессии стерильности иа при-
мере упрошенной модели с одной транспозицией. Можно пока-
зать. что этот процесс будет происходить таким же образом при
наличии перестроек других типов, в частности транслокаций и
реннверсий. и соответствующих кросеннговеров Кроме того, что
весьма примечательно, этот процесс будет действовать при нали-
чии большего числа перестроек в отдельных хромосомах или
плечах хромосом, с тем чтобы обходить как мощные, так и сла-
бые барьеры хромосомной стерильности.
Естест венный отбор, благоприятствуюшнй сохранению новой
межвидовой комбинации генов, протаскивает важные гены ви
да-донора через барьер стерильности, состоящий из хромосом-
ных перестроек, число и типы которых могут быть различны.
Тахой отбор может привести к созданию новой расы, сходной
по своим морфологическим и физиологическим признакам с ви-
дом-донором, но в отношении плодовитости принадлежащей к
группе вида-реципиента.
Остается рассмотреть роль генов, локализованных в пере-
строенных участках, в процессе интрогрессии через барьер хро-
мосомной стерильности. Эти гены ограничивают упомянутую
выше конвергенцию.
Те определяющие морфологические признаки гены, которые
локализованы в перестроенных участках вида D. будут элими-
нированы вместе с саиими этими участками в результате отбо
ра гамет в гибридной популяции. Если гены морфологических
признаков вида D сцеплены с генами жизнеспособности этого
вида в тех же участках, то они будут мнивинровагься также и
естественным отбором в среде вида R. Сцепления двух типов —
сцепление между морфологическими генами и генами хромосом-
ной стерильности и сиеплепае генов Л!—V в перестроенных
участках —препятствуют слишком сильному сближению интро-
грессивной расы с видом-донором по морфологическим призма
кам. Иными словами, перестроенные участки ограничивают инт-
рогрессию чужих генов и играют роль тормозов в процессе кон
вергенции морфологических и физиологических признаков
Гл. 17. Нктрырсссня
Z25
Интрогрессия и расообразованне
Во многих группах видов путем интрогрессии образуются отлезь
яые расы Показательным примером служит группа Helianlhus
annuus, о которой мы уже говорили
Расы интрогрессивного происхождения описаны п числе про-
чих у следующих родов Dracophyllum [181]. Coprosma (1811.
Abies [563] (обзор си [780]); Тradeseantia (II9|; Cist us [137];
Helianlhus [366]; GUia {Ж 300]; Pinus [334] и Purshia [786].
Во многих случаях расы интрогрессивного происхождения сям-
патричпы, по крайней мере частично, с видом- или полувидом-
донором.
Образование рас путем интрогрессии весьма часто происхо-
дит у южнокалифорнийских диплоидных гнлий с крупными
цветками, принадлежащих к секции Arachnion. Виды и полуви-
ды этих гнлнй встречаются симпатрически во многих разных
комбинациях, занимая образовавшиеся недавно аридные и по-
луаридные местообитания Расы интрогрессивного происхожде-
вин. по-видимому, хорошо адаптируются к некоторым из этих
местообитаний. Судя по морфологическим, зкологическим н ци-
тогенетическим данным [300], перечисленные ниже географиче-
ские расы, получившие подвидовые названия, происходят от сле-
дующих родительских форм-
С laUJ/ora eleven
G tepTimffea /гспкчгза
G сапа грпбпэла
G nfhtoteitra ctiotda
G latiflara интрогрессаи ст G. t cruder а
G leptantba ieptaUha интрогрессия CT
Q laliflofa
G сала интрогрессия от G. teplanfha par-
fiuiii
G odtr^euca tnxonaia интрогрессия от
G. letrtMiha Sfptdniha
Интрогрессивная гибридизация— фактор, вызывающий кон-
вергенцию по морфологическим признакам и зкологическим
предпочтениям в зоне симаатрнческого контакта между видами
или полувидами. Другие факторы действуют в противоположном
м 1ммеимя
При наличии подходящих условий симпятрическне контакты
могут также привести к отбору на репродуктивную изоляцию
и (или) смешение признаков, как было отмечено в гл 15. В та-
ких случаях в снмлатрическоА зоне усиливается дивергенция
между видами Симпатрня — эго состояние, вызывающее разно-
образные реакции
Смешение признаков, хорошо известное у животных, у рас-
тений олнеанп лишь в 01 дельных случаях По мнению некоторых
ботаников, растения в этом отношении не изучались достаточно
15 -45
226
Чып литы. Естяптоошя говрвЛиачяя и « цпвлтати
тщательно. Возможно, что это отчасти справедливо. Мне, одна-
ко, хотелось вы подчеркнуть роль интрогрессии; конвергенция
по морфологическим и физиологическим признакам, обусловлен-
ная интрогрессией, встречаете* у растений часто и, возможно,
преобладает над смещением признаков во многих или даже в
большинстве случаев.
Выводы
Естественная гибридизация между симпатрнческнми видами или
полувидамн растений происходит довольно часто при наличии
подходящих экологических условий. Имеется много описатель-
ных, но ие количественных данных, свидетельствующих о том.
что естественная гибридизация чаще встречается у растений, чем
у высших животных с внутренним оплодотворением.
Гибридизация у растений недостаточно коррелирует с эффек-
тивностью внутренних репродуктивных барьеров между роди-
тельскими видами, вплоть до предельной точки, т. е. абсолют-
ной перекрестной несовместимости. Вместе с тем наблюдается
тесная корреляция между гибридизацией и наличием открытых
местообитаний, в которых могли бы обосноваться гибриды и их
потомки В современном мире такие открытые местообитания
создаются в широких масштабах в результате деятельности че-
ловека, ко они возникали также на протяжения всей геологиче-
ской история и результате различных природных процессов.
Воспроизведение естественных гибридов в группах растений,
размножающихся при помощи ауткроссянга, обычно происходит
путем возвратных скрещиваний и интрогрессии. Инфильтрация
генов от вида-донора к вкду-реципиеиту гораздо сильнее ограни-
чена в тех случаях, когда родительские виды различаются по
хромосомным перестройкам, а не несут совершенно гомологич-
ные хромосомы.
В группах автогамных растений гибридизация может при-
вести к конечным результатам, очень сходным с теми, которые
обычно считают указанием иа интрогрессию. Альтернативный
путь слагается из инбридинга (а не возвратного скрещивания),
расщепления и отбора.
Интрогрессивная гибридизация играет ключевую роль в фор-
мировании рас у многих преимущественно своболноопыляюшях-
ся групп растений. Расы интрогрессивного происхождения об-
наружены в группах видов, принадлежащих к родам Pinus,
Abies. Quercus, Purshia, Cislus, Coprosma. Dracophyllum, Helian-
thus, Gilia. Tradescantia и другим.
18 Сингамеон
Локализованная естественная гибридизация и ограниченный об-
мен генами между полувидами, которые обычно бывают репро-
дуктивно изолированы, приводят к объединению этих полувидов
в более объемлющую (inclusive) скрещивающуюся единицу.
Эта более крупная единица представляет собой сингамеон.
Подобна» ситуация была описана Лотси [520. 52IJ для одной
из групп рода Betula На европейском континенте произрастает
много форм Betula различного и к тому же спорного таксоно-
мического ранга, и все эти формы гибридизируют в широких
масштабах. Лотси [5201 пришел к выводу, что *Betula пред-
ставляет собой очень большое скрещивающееся сообщество —
единый сингамеон»
Дю Рие [181] дал обзор многочисленных аналогичных слу-
чаен у Nothofagus на Новой Зеландни, Geum — в Европе. Me-
landrium в Швеции, Salix — в северной части Европы н т. п.
Все эти группы, по его мнению, представляют собой «большие
и высо копил и морфные «гибрид» синга мео и ы. в которых виды
более или менее утратили свою самостоятельность » (с 384)
Позднее рядом исследователей была разработана концепция
сингамеона [53, 134. 279|. В современных терминах сингамеон
можно определить как «общую сумму видов или полувидов, ко-
торые связаны между собой частой или эпизодической гибриди-
зацией. происходящей в природе; [таким образом] это любая
гибридизирующая группа вндоо„> [279. с. 67].
Сингамеон — самая «объемлющая» скрещивающаяся едини-
ца в гибридизирующей группе видов. Ом может иметь относи-
тельно простую внутреннюю структуру и состоять из двух-трех
полувидов, но иногда ок бывает и очень сложных. Образовав-
шиеся в некоторых группах растений снигамеопы представляют
собой обширные сети полувидов, состоящие из многом келейных
таксономических видая и занимающие ареалы, сравнимые по
своей протяженности с целыми континентами.
По виешнему виду и по репродуктивным взаимодействиям с
другими популяционными системами сингамеон сходен с биоло-
гическим видом, т е. оц является репродуктивно изолированной
единицей; вместе с тем он отличается от биологи веского вида
своей более сложной внутренней структурой Следует отметить,
что основные составные части сипгамеонов в формальной систс
матмке рассматриваются как виды, и это совершенно правиль-
но; они обычно действительно представляют собой хорошие так*
сономнческис виды.
15’
Часть лягал. Бггепявпмал > г. ил л>гат-
228
Сиигамеоаы существуют но многих родах растения Выше
уже были перечислены некоторые давно известные примеры из
taopw Европы к Новой Зеландии. В западной части Северной
мерики много сингамеоиов обнаружено у Pinus, Juniper us,
Quercus. Mimulus. sect Diplacus, Iris. Aquilegia, GUia sect.
Arachnion и у других родов. В предшествующих главах уже бы-
ли описаны некоторые аспекты этих сингамеоиов. Здесь мы рас-
смотрим структуру сингамеоиов в целом на примере двух групп
растений.
Тихоокеанские ирисы
Самый сложный сиигамеон, тщательно изученный одним иссле-
дователем. — ато. вероятно. ирисы, произрастающие на тихооке-
анском побережье Северной Америки (462].
Ряд Californicae секции Apogoп рода Iris представляет собой
естественную группу, состоящую из II таксономических видов,
которые распространены от южной Калифорнии до шт. Вашинг-
тон. Их взаимоотношения по плодовитости обсуждались в гл. II.
Между ними не существует сильной внутренней изоляции.
Рас 1&1. Структура епгаымна у ирисов тяхоокикхого побережья (4621.
Толстые линии, соеджвяюакве рашые вилы, означают высокую стеомъ гиб-
ридизации; тиикме липки — умереяиую гибридизацию, прерывистые ли*нн —
ие.»нвчит«лыиую гн&рядя^мию. Буквы в кружках — сокрицеиные названия
равный полувидов рода JrU BRA — / bracif^la; CHR — / скгумрЬиЬ
U. DOU —/. touriatianA. FER —/. HAR — I. hurlvgit: INN —
/ 1п.чглп4плг<я. MAC — / таегаирЛйл. MUN — /. mtauu; PUR — /. purdjp;
TEN — /. Ifпал. TMA — I. tawtstifna; XTH — I. thoapsonil-l. doaf/asii]-
до x 1. 1ллсрп1пиз1а.
ГA. IS. Cwr-fOMf ОЯ
219
Эти виды или полувиды занимают разнообразные экологии?
ские эоны в пределах их общего ареала. Так. Iris dnugtasiana
встречается идол», берега, / macrosiphon—в предгорьях г т. л.
Местообитания этих и других полувидов часто переходят одно
в другое, создавая возможность для маргинальных или сисжно-
симпатричсских контактов Кроме того, во многих местах жато-
|ическне барьеры разрушаются в результате лесных разработок,
прокладки дорог и других нарушений среды, вызванных деятель-
ностью человека. Как следствие этого между полувидами начи-
нается естественная гибридизация в многочисленных комбина-
циях [461. 462].
Все эти таксономические виды, за исключением / munzii.
связаны между собой в результате происходящей между ними
естественной гибридизации. Иными словами, из 11 таксономи-
ческих видов лишь одни — I. munzii—представляет собой изо
лированный вил. а 10 остальных — это связанные друг с другом
полувиды, образующие еннгамеоны. как показано па рис. 18.1
Некоторые полувиды, как, например. / douglastana. зани-
маю! в сингамеоне центральное положение, будучи связаны со
многими другими полувидами, тогда как другие, подобно / hart-
vegii, являются периферическими и евазакы лишь с одним по-
лувидом Некоторые пары полувидов гибридизируют в очень зна-
чительной степени, другие —в умеренной, а третьи — в очень
слабой (рис. 181).
Белые дубы
В обоих подродах луба — Quercus (белые дубы) и Erg-
throbalanut (черные дубы) — снигамеоны образовались неза-
инсимо Между этими двумя сиигамсонами существует сильная
репродуктивная изоляция Сингамеон белых дубов наиболее об-
ширен кик географически, таи и таксономически.
Белые дубы в США научались многими ботаниками. Лите-
ратура по естественной гибридизации этих дубов очень велика
н разбросана по разным паданиям Пальмер [633] собрал све-
дении ио всем известным для Северной Америки гибридным
комбинациям, однако после выхода его обзора (1948 г.) появи
лось много новых статей на эту тему, а других общих обзоров
сделано нс было. Я попытаюсь обрисовать здесь в основных
чертах сингамеон белых дубов в США Прн этом будут прнве
лены ссылки на ключевые работы, поскольку дать полный об
зор литературы не представляется возможный.
Для удобства изложения разделим сингамеон белых дубов
на региональные группы Мы начнем наш обзор с калифорний
230 .wax f стествмям мбрлйжция а «* ркумтяти
Ржг. J85. Структура еингамаовл белых вубо» (полрад Qu</c«i) в К*-
ляфорнп.
ской группы, а затем проследуем через всю страну к восточ-
ным штатам.
Ареалы семи видов белых дубов, встречающихся в Калифор-
нии, перекрываются в различных комбинациях. В некоторых
частях перекрывающихся облягте*, где экологическая изоляция
нарушена, они гибридизируют. На рис. 18.2 эти полувиды рас-
положены в порядке, примерно соответствующем их географи-
ческому распространению с севера иа юг, а соединяющие их
лниин отражают степень гибридизации между ними.
КалифорииПская группа этого сингамеона интересна тем.
его она объединяет дубы, резко различающиеся по своим мор
фологическим и экологическим признакам. Quercus garryana —
лесное дерево с большими рассеченными листьями, распростра
иеииое в северо-западно* части тихоокеанского побережья США
Q. dumosa и Q turbirulla — низкорослые кустарники с мелкими
колючими листьями, растущие соответственно в чапаральных
и пустынных фитоценоза». Несмотря на такие резкие различия,
эти формы связаны либо прямо, гибридизируя друг с другом,
либо косвенно, через промежуточные популяции.
В определенных местах иа севере Калифорнии лсаюс дере-
во Q. garryana гибридизирует с формой засушливых предго-
рий, Q. douglasii [780), и с вилами кустарниковых зарослей,
Гл. 18 Сангамгоп
231
Q. dumosa и Q. du rata [837]. На юге Калифорнии <?. douglasii
гибридизирует с чапаральным Q. dumosa. a Q. dumom—( пус-
тынным Q. lurbinella [57, 836]. Эти и другие комбинации пока-
заны на рис. 18.2.
Судя по морфологическим признакам, одной из родительских
форм единственного гибридного дуба, растущего иа пустынных
горах в южной Калифорнии, был Quercus Inbata [838]. Q. toba-
la, распространенный в предгорных долинах, в настоящее время
не встречается вблизи того места, где растет гибрид; возможно,
однако, что он встречался там во время одной нз плювиальных
фаз плейстоцена Раньше считалось, что другой родительской
формой был обильный в этой местности Q. turbiiuUa [838]. Прав-
да. основываясь на данных более поздних исследований, можно
думать, что другой родительской формой был новый юго-запад-
ный вил. названный Q. cormliut muileri (К. Nixon, личное сооб-
щение; см. также Madrono).
Область распространения сиигамеона белых дубов тянется
дальше на юго-запад, от Аризоны в Юты до западной частя
Техаса, где он представлен формами, показанными на рис. 18.3.
Эти полувиды расположены иа схеме также в соответствии с их
географическим распространением. Гибридизация между пус-
тынной формой Q turbineUa н растущей в Скалистых горах
формой Q. gambelii в настоящее время происходит иа севере
Аризоны и происходила в Юге в прошлом, о чем свидетельству-
ют старые изолированные гибридные клоны [125]
Рас. 183. Структура скигамюиа белых дубов Inoapo* Queroir) a wo
эапахяой чает» США
Я32 Чат яягы. Дгтуетеаннм шбридазяцая > т» рмумгагы
Хотя в настоящее время Quercus gambelii Скалнцтых гор в
крупноплодный дув (Q. тасгосагра) восточной частя США ге-
ографически изолированы, в прошлом, вероятно в плейстоцене,
они были связаны гибридизацией, о чем свидетельствует нали-
чие промежуточных признаков у некоторых аберрантных попу-
ляций [573].
На юго-западе Quercus turbinella гибридизирует также с
Q. grisea, который в свою очередь гибридизирует с Q. mohriana,
а этот последний — е Q havardl [589] (рис. 18.3).
В Техасе Quercus stellata занимает центральное положение
в скнганеоне белых дубов. Здесь он гибридизирует с Q. havardi.
Q. mvhriana. Q. virginiana. Q. minima, Q. oteoides, Q. margaretta,
Q. alba. Q. lyraia, Q. тасгосагра. Q. prinus и Q. sinuata. Некото-
рые из этих полувидов гибридизируют также друг с другом, на-
пример Q. virginiana с Q minima (589. 593].
Кроме того, иа востоке США Quercus stellata гибридизирует
с этими же полувидами, а также с другими, например с Q. bico-
lor в Q prtnoides [343, 632].
В восточной части Северной Америки белый дуб Quercus alba
занимает центральное положение в сингамеоие. гибридизируя
f fН ft
О ГжОямды О NcMff
♦ WV н
а аМ ГиОраздь. Q pnnui
Рис 184. Листья Qiurcus atba и его джух гмбрихоя [343]
Гл 18. Синголюл
2»
Рис IB 5 Структура ошгаиеока белых лувое (поарол Quercus) * ао-
crwMPfi мает» США [3431
здесь с 11 другими полувидами' Q. miehauxH. Q. muehlenbergii,
Q. prinoldes. Q. lyrata. Q macrocarpa, Q stellaia, Q. bicolor.
Q. austrina. Q. prlruis, Q. margaretta, и Q robur (343]. Фариа
листьев к ее изменчивость у двух ну этих гибридных комбина
ций покапаны иа рис. 18.4.
Хардин [343] провел прекрасное исследование снигамеона
белых дубов в восточной части Северной Америки, уделив особое
внимание Quercus alba. Многие растущие здесь виды имеют ши-
рокое распространение, причем области нх распространения в
значительной степени перекрываются с областями распростране-
ния других видов В зонах симпатрии время от времени проис-
ходит естественная гибридизация Структура образовавшегося
в результате снигамеона схематически показана иа рис. 18.6.
234 Чят лягая Еспсгяяяшя гявридмацая ж w рщмпгы
Как можно жалеть. 14 из 16 различных вадов — члены этого
сяигамеока [343].
Черсшчатый дуб (Quercus robur) выращивают в восточной
части Северной Америки, где он спонтанно гибридизирует в не-
скольких местах с Q. alba (343). На Британских островах, иа
своей родине, Q. robur гибридизирует с Q. petraea [126, 767].
Таким образом, свигамеои белых дубов перекидывает мост че-
рез Атлантический океан и продолжается в Европе.
19 Гибридное видообразование
К числу важнейших проблем эволюции растений относится
проблема, связанная с ролью гибридизации в видообразовавии,
которую обсуждало нс одно поколение исследователей, Служит
ли естественная гибридизация одним из механизмов возникно-
вения новых видов растений?
Некоторые исследователи раннего периода отвечали на этот
вопрос утвердительно. Однако, как мы увидим далее, эти же
самые авторы упускали нз виду некоторые аспекты проблемы,
в особенности расщепление в гибридном потомстве, и их утпер
дизельные ответы были поэтому преждевременными и в значи-
тельной мерс основывались на интуиции. Тем ие мспсс эти пер-
вые обсуждения поддерживали интерес к проблеме ло тех пор,
пока у современных исследователей не появилась возможность
подойти к ней с позиций генетики.
В одной из первых глав было показано, что новые расы воз-
никают иногда в результате естественной гибридизации Это
процесс гибридного образования рас, но необязательно — про-
цесс гибридного видообразования Новая раса, образовавшаяся
в результате процессов, включающих гибридизацию, может в
дальнейшем продолжать ливер тировать до видового уровни в
условиях географической изоляции от исходной предковой расы.
Это особый случай географического видообразования, прн кото
ром гибридизация спадала необходимую изменчивость иа одной
из промежуточных стадий. Дальнейшее обсуждение этого слу-
чая нас здесь ие интересует
Под гибридным видообразованием мы имеем в виду возник-
новение нового вила непосредственно из какого-либо естествен-
ного гибрида Такое определение сразу выдвигает на первый
план самую суть проблемы. ясно, что тот самый патовой про-
цесс. который привел к образованию естественного гибрида, вы-
зовет также разрушение содержащейся в нем комбинации генов
в результате расщепления в последующих поколениях. Следова-
тельно. существенную часть мехаинзма гибридного видообразо-
вания составляет стабилизация репродуктивного поведения гиб-
ридов
Стабилизация воспроизведения гибридов
Расщепление в F, пли в любом из последующих поколений, воз-
никших в результате межвидового скрещивания, может быть
полностью подавлено или сильно ограничено Известно много
3M
Часть мгал. Етсгеанлая гиОрид^жщаь а м prignrant
разных механизмов, с помощью которых это достигается: 1) ве-
гетативное размножение; 2) агамоспермия. 3) перманентная
гетерозиготность по транслокациям. 4) перманентная нечетная
полиплоидия; 5) амфиплоидия; 6) рекомбинационное видообра-
зование. 7) выщелление нового типа, изолированного внешними
барьерами.
Способы 1 и 2. объединяемые под общим названием апомик-
сиса, вкратце описаны в одном из последующих разделов этой
главы и более подробно рассматриваются в части восьмой Спо-
собы 3 и 4 также рассматриваются в части восьмой на примере
генетических систем группы Oenothera biennis и группы Коза
canina соответственно- Способ 5, амфиплоидия или аллополи-
плондия, описан в общих чертах в этой главе, я затем подробно
разбирается в части шестой.
Способы 6 к 7 в отличие от способа 5 связаны с рекомбина-
цией иа гомоплоиаиом уровне Способ 6. названный рекомбина-
ционным видообразованием, заключается в образовании нового
рекомбинанта, гомозиготного по факторам хромосомной стериль-
ности. Общее представление о нем дается в этой главе, а в
гл. 20 ои рассмотрен более подробно. Способ 7 заключается в
образовании нового рекомбинанта, изолированного внешними
барьерами. Мы называем этот способ гибридным видообразова-
нием при наличии внешних барьеров и вкратце рассмотрим его
водном из последующих разделов данной главы.
Все перечисленные здесь способы создают возможность для
размножения любого данного генотипа гибридного происхожде-
ния. Это может быть генотип либо самого гибрида F,, либо одно-
го из происходящих от него последующих поколений, и ои мо-
жет бить гетерозиготным или гоиознготным. -
При способах 6 и 7 рекомбинант гибридного происхождения
достигает состояния, при котором ои размножается в чистоте,
благодаря тому что он. по крайней мере иа время и по крайней
мере частично, становится гомозиготным. При способах 1—4 ге-
нотип гибридного происхождения размножается в чистоте, со-
храняя свое высокогетерозиготное состояние. Способ 5 представ-
ляет собой некое компромиссное решение, приводящее к комби
нации признаков, присущих гомозиготности, с признаками гете-
розиготиостн. При этом способе форма гибридного происхожде-
ния гомозиготна в том, что касается мейоза, и поэтому
размножается в чистоте, но сохраняет противоположные генные
системы двух родительских видов.
В любом случае конкретная генная комбинация гибридного
происхождения, если она обладает адаптивной ценностью в ка-
кой-либо доступной для нее среде, способна увеличить свою чис-
ленность и образовать собственную популяцию в местности со-
ответствующей этой благоприятной среде. Такая производная
Г я 19 Гибридное ридообразоРание
131
поп).шипя может представлять собой новый вид или микровид
гибридного происхождения
Способ стабилизации воспроизведения гибрида определяет
тип вида, возникающий в результате гибридизации. Вегетагив-
иое размножение и агамоспермия — бесполые способы воспро-
изведения В результате этих способов унипареитального раз-
множения может возникнуть клональный или агамоспсрмный
микровид. Перманентная структурная или мумерическая гиб-
рндиоегь (способы 3 и 4) также приводят к возникновению уни-
нарентальиых микровидов В отличие от этого амфиплоилия,
рекомбинационное видообразование и гибридное вилообратона
иие при наличии внешних барьеров представляют собой способы
стабилизации, действующие в условиях полового размножения.
Следовательно, оии приводят к образованию новых биологиче-
ских видов
Согласно концепциям, рассмотренным п гл. 7, новые виды,
возникающие от гибридов или от форм гибридного происхожде-
ния,—это во всех случаях эволюционные виды Но лишь в том
случае, если половое размножение и свободное неродственное
скрещивание сохраняются иа всем протяжении критических ста-
дий формирования, можно говорить о гибридном происхождении
новых биологических видов. Трудности гибридного ПИДООбраЗО'
ввния достигают наивысшей степени в этом последнем случае,
где половое размножение остается в силе. Возиикиовение нового
биологического вида в результате гибридизации— теоретически
самый сложный, но вместе с тем и самый интересный процесс
История вопроса
В доменделеяский период возможность гибридного происхожде
ния новых видов растений обсуждали Линией а 1744 и 1760 гг.
Уильям Герберт в 1820 г. Шарль Нодзн — п 1863 г и Антой
Кернер в 1891 г. В начале послемендслевского периода эту ли-
нию рассуждений продолжили Лотси, Хайата и другие. Взгляды
перечисленных исследователей нашли отражение в их работах
Линией (514]. Герберт [376]. Нодэн (609]. Кернер (4311. Лотси
(5191 и Хайата [355], обзор см, Дю Рис ] 181].
Линней и его ученик Рулберг впервые высказали мысль о
гибридном происхождении йовых видов растений в 1744 г в свя-
зи с Peloria — одной из форм Linaria, характеризующейся ра
диальио-симметричными енетками В uDisquisitm de Seeu Plan-
latum» 1514] Ли к ней вновь говорит о гибридном видообразова-
нии «невозможно сомневаться в том. чго существуют иовые
виды, возникшие в результате гибридизации».
Кернер (431] основывал свои рассуждения иа известных фак-
тах о том, что скрещивание порождает новые формы и что во
23в Часто нагая. tcrecnmnae иерийыачм и ее реецмтагы
многих группах растетги* встречаются естественные межвидо-
вые гибриды; из этого он делал вывод, что «гибриды могут ди-
вить начало новым видам*. Конечно, в природе образуется мно-
го гибридов, но лишь немногим из них удается стать новыми
видами. По мнению Кернера, важным ограничивающим факто-
ром в данном случае является среда.
Только в тех местах, где имеется подходящее и открытое
местообитание, ие занятое родительскими видами, гибридные
растения могут закрепиться и увеличить свою численность.
Одна ко тайпе открытые местообитания встречаются в природе
повсюду, так же. как и виды гибридного происхождеиня. В ка-
честве примеров Кернер приводит Rhododendron, Salvia и Nu-
phar а Европе.
Так, Rhododendron intermedium представляет собой продукт
гибридизации между R lerrugineum и Я. hlrsutum. В некоторых
местах в Альпах R intermedium изредка встречается вместе с
родительскими видами, но в других местах он образует обшир-
ные собственные популяции и значительно превосходит но чис-
ленности родительские типы В таких местах R. intermedium
достигает статуса хорошего вида [431].
Линией и Кернер были выдающимися ранними сторонника-
ми идеи о том. что естественные гибриды могут служить отправ-
ными точками для возникновения новых видов Анализ их рас-
суждений показывает, что оба они пе уделяли должного внима-
ния двум важным аспектам этой проблемы, без учета которых
невозможно создать удовлетворительную гипотезу гибридного
видообразования, а имеиио —стерильности и расщеплению
Линнею и Кернеру не было известно явление расщепления.
Кернер, собственно, постулирует, что «гибриды передают свою
форму потомкам в неизменном виде... при условии исключения
пыльцы от других видов*. Линией проводил мало эксперимен-
тальных исследований по гибридизация растений, а эксперимен-
тальные исследования Кернера касались других аспектов раз-
множения растений. Поэтому ни тот, ни другой не могли себе
представить, что при переходе от естественного гибрида к дочер-
нему виду в случае плодовитых межвидовых скрещиваний воз-
никают какие-то проблемы, связанные с передачей наследствен-
ных признаков.
Синеок первых авторов, занимавшихся гибридизацией расте-
ний, которые знали о существовании явления расщепления, но
не обсуждали формирование видов в результате гибридизации,
достаточно велик: от Кёльрейтера [441] до Менделя, Дарвина и
Фокке [2171.
Большой интерес представляет статья Надэна [609]. посколь-
ку в ней обсуждается и расщепление, и гибридное видообразо-
вание. Нодэн рассматривал первое как установленный факт, а
Гл 19. ГuGpudttoe валЗссбмзование
239
второе — как некую возможность Он понимал, что явление рас-
щепления создает трудности для теории гибридного видообразо-
вания Тем не менее он допускал возможность исключительных
случаев, в которых гибридные признаки закреплялись к после-
дующих поколениях, и считал, что такие случаи могут принести
к возникновению новых видов (609].
Осложнение, создаваемое стерильностью, в ранний период
изучения гибридного видообразования также отбрасывалось
Было известно, что некоторые пары видов при скрещиваниях
дают стерильных гибридов, а другие —плодовитых; поэтому
считалось, чго гибридное видообразование возможно лишь п тех
случаях, когда гибриды плодовиты (431]. Таким образом, вни-
мание было сосредоточено только па одном особом аспекте осей
проблемы, а проблема гибридного видообразования и целом ка-
залась проще, чем опа есть на самом деле.
Создание теории современной генетики в первые десятиле-
тия нашего века дало, наконец, возможность правильно поста-
вить старую проблему гибридного видообразования, учитывая
при этом пс только морфологические и физиологические призна-
ки, но также барьеры, создаваемые стерильностью гибридов, и
расщепление. Для того чтобы решить эту проблему, необходимо
было найти некий механизм, с помощью которого из межвидо-
вого гибрида мог бы ьозиикмуть новый, внутренне изолирован-
uuii константный вид без утраты способности к половому раз-
множению
Первым крупным достижением в этом направлении была ги-
потеза Винге [898. 899], касающаяся амфиплоидин (сам этот
термин появился гораздо позднее). Согласно этой гипотезе, но-
вый константный вид может возникнуть от гибрида между дву-
мя существовавшими ранее видами в результате удвоения у
него числа хромосом. Гипотеза Винге была вскоре подтверждена
экспериментальными данными, полученными другими исследо-
вателпми для NicoUana, Raphanus-Bt asslca, Galeopsis и других
растений (105, 417. 596, 597|.
Амфиплоидия — зто способ видообразования, связанный с
однонаправленным увеличением числа хромосомных наборов,
тогда как эволюции многих о ядов растений происходила, оче-
видно. на диплоидном уровне, н поэтому оставался неясным во-
прос о том. возможно лн гибридное видообразование без изме-
нения плоидиостн
Сам Винге вернулся к этому вопросу в своих более поздних
работах, проводившихся на Tragopogon и Erophila (900, 901]. а
Лампрехт (458] —в работе на Phaseolus Эксперименты на Тга-
gopogon, Erophila и Phaseolus проводились со специальной
целью; установить, возможно ли возиикнпвеиие новых плодови-
тых видов нли микровидов в результате гибридизации, не со-
г-М) Чат лятм Стсгвгнчая .--uJpwsMua» а «• ргм/мгагн
правождаюшеЖся амфиплоидным удвоением Эти эксперименты
проложили путь другим ученым, исследовавшим иной, отличны*
от амфиллоидии процесс гибридного видообразования, извест-
ный теперь под названием рекомбинационного видообразования.
Введение в амфиплоидию
Хромосомная стерильность создается в результате структурных
перестроек между родительскими геномами, что приводит к на-
рушениям, видимым или невидимым, течения мейоза у гибрида:
гомеологичные хромосомы либо не конъюгируют нормально с
образованием бивалентов и ие расходятся, как положено, к по-
люсам в анафазе, либо конъюгируют, но расщепляются, образуя
прн этом дочерние ядра, несущие нехватки и дупликации. В лю-
бом случае какая-то часть продуктов мейоза оказывается несба-
лансированной и не лает полноценные споры и гаметы Эта
часть нежизнеспособных спор, а тем самым и степень стериль-
ности. быстро возрастает с увеличением числа гетерозиготных
перестроек (см. гл. 8 и 9).
Допустим, что у гибридов, обладающих хромосомно* стериль-
ностью. произошло удвоение числа хромосом. В таком случае,
если отвлечься на время от других возможных осложняющих
факторов, мейоз у этих гибридов будет протекать нормально. с
образованием фертильных гамет.
Такое восстановление фертильности иа новом полиплоидном
уровне наиболее ясно выражено в первом из указанных выше
случаев, в котором хромосомы диплоидного гибрида не обра-
зуют нормальных бивалентен Обозначим хорошо различающие-
ся по своей структуре геномы родительских видов буквами А и
В. Два диплоидных вида имеют при этом конституцию АА и
ВВ. а гибрид между ними — конституцию АВ. V хромосом, при-
надлежащих к набору А, нет гомологичных партнеров, с кото-
рыми они могли бы конъюгировать при меЯозе у гибрида, нет
гомологичных партнеров и у хромосом из набора В Однако у
образующегося при этом аллотетраплоидиого гибрида (или ам-
фидиплоада) с геномом ДАВ В положение совершенно иное.
У него имеется по гомологичному партнеру для каждой хромо-
сомы каждого генома, поэтому мейоз и образование гамет могут
протекать нормально.
Во втором из приведенных выше случаев гомеологичные хро-
мосомы гибрида образуют биваленты, но расщепляются и дают
генетически несбалансированные продукты мейоза. Диплоидные
родительские виды содержат различные субгеиомы— Л, и А,.
Происходящий от них гибрид Г, имеет конституцию A,At и ха-
рактеризуется хромосомно* стерильностью при явно нормаль-
ном меЯозе. В этом случае также у тетра плоимого производного
ГА. 19. Гибридно* л»3оо<5ри»*ани<
21Г
АЛ .ЛА* плодовитость по всей вероятности, восстановите*, по
крайней мере частично, в результате тот. что Дарлингтон |138|
назвал дифференциальным сродством (differential affinity).
Каждая Л.-хромосома и каждая А. хромосома у этого тетра
плоила имеет полностью гомологичного партнера Такие томо-
логичные хромосомы предпочтительно КО1ГЪЮГНруЮТ Друг с дру-
гом и расходятся к противоположным полюсам, образуя струк-
турно и генетически сбалансированные продукты мейозд.
Тетрадлондная форма, происходящая ОТ гибрида. обЛйДйКН
шего хромосомной стерильностью, незапнеимо от того, имеет ли
она геном ЛАВВ или АЛЛМг. не только плодовита, но и ре-
продуктивно изолирована от своих диплоидных родительских
видов Скрещиванию между тетра плоиднымн и диплоидными
растениями нередко препятствуют барьеры несовместимости, и
гибриды, если они образуются, представляют собой триплоплы.
а поэтому обычно стерильны
Классические примеры Primula ke&ensis, Raphanobrassica и
Galeopsis letrahit иллюстрируют различного типа геномы, встре-
чающиеся у экспериментальных амфиплоидов
Гибрид Fi от скрещивания между Primula floribunda
(2л —18) н Р. veriiallata (2л-18). широко известный под назва-
нием Р heui/rntis. — диплоидное многолетнее травянистое расте-
ние. подобно своим родительским видам У этого гибрида про-
исходила конъюгации с образованней нормальных бивалентов,
1м> пи был аысокистсрилем. В три разных гола это п остальном
стерильное гибридное рз стен не спонтанно выбрасывало фер-
тильные побеги, которые, как показало цитологическое месте-
дованнс. представляют собой тпранлонды (2л = 36) и при конъ-
югации хромосом у них образуются преимущественно бива-
ленты.
Очевидно, у- этих тстраплондных побегов пред почтительно
конъюгировали полностью гомологичные хромосомы Эти фер-
тильные побеги возникали в результате удвоении хромосом в
сома i ячеек нх клетках почки или какой либо части почки. Они
давали сечена, нз которых развивались плодовитые и вполне
однородные потомки Fj (612 649)
Противоположная ситуация была обнаружена у Raphano-
bfassica — эмфнплонда. полученного в результате скрещивания
Raphanus satious и Brassica aleracea. Оба родительских вид» —
лнилонды с 2л — 18 хромосом. У гибридов К| конъюгация хромо-
сом полностью нарушен», и own высоко стерильны Эти гибри-
ды дали некоторое количеств» «редуцированных диплоидных
гамм От слияния таких гамет были получены тегрвплоидиые
(2л = 36) растения F* у которых мейол протекал бгт нарушений
с образованием нормальных бивалентов и которые по большей
части были вполне плодовитыми, давая морфологически одно-
1в—45
W Часть алгол. Есгкгкюшл гибр^имцяя > « рармтагы
(вдвое поколение F». Эта нова» плодовита а размножающаяся а
чистоте линия изолирована барьером стерильности от диплоид-
ных родительских видов [4171.
daleopsit letrahit (2л —32) в отличие от Primula kewensis и
Raphanobrasslca представляет собой тетравлондиый вид. встре-
чающийся в природных условиях в северной части Европы и в
Лани. Мюитцинг [596, 597] показал, что это однолетнее травя-
нистое растение является аифкплондом, возникшим в Европе
от других родственных диплоидных видов: G. pubescens (2л—16)
и О. speciosa (2л—16).
У гибридов Fi. полученных в результате искусственного скре-
щивания между Oaleopsis pubescent и О. speciosa, мейоз кару
шеи, и они высоко стерильны; фертильная пыльца составляет у
них всего 8%, а число образующихся бивалентов — от 5 до 8
От гибридов Fi, несмотря на их высокую стерильность, было
получено поколение Fi. состоявшее на нескольких стерильных
диплондов и одного стерильного триплонда. Последний возник
в результате слияния одной вередуцврованной и одной редуци-
рованной гаметы F|. Этот триплонд при возвратном скрещива-
нии с О. pubescent образовал одно семя, из которого в F» раз-
вилось тетрапломдное растение. Вероятно, оно возникло в ре
вульгате оплодотворения нередуцироваииой Зп-яйцеклетхи нор-
мальным 1л-слермием. У этого тетраплоидного растения F> в
метафазе мейоза образовалось 16 бивалентов, а фертильная
пыльца составила 70%. Оно дало плодовитое тетраплоидное по-
томство F< [596, 597].
Этот искусственный аллотетраплоид сходен С возникшим в
природе Galeopsis tetrahit как по морфологическим признакам,
так и по числу хромосом. Кроме того, подобно дикорастущему
G. tetrahit, он изолирован от С. pubescens и G. speciosa барь-
ером несовместимости Его можно было бы рассматривать как
синтетическую форму О. tetrahit. Чтобы проверить это предполо-
жение, Мюитцинг скрестил искусственный тетраплоид с природ-
ным G. tetrahit. Скрещивание прошло без затруднений, а расте-
ния Fi были плодовиты, и конъюгация хромосом протекала у
них нормально. Это был первый пример экспериментального
создания существующего в природе амфиплоидкого вида [596,
Введение в рекомбинационное видообразование
Рекомбинационное видообразование — термин, принятый для
обозначения одной из наименее известных форм гибридного
видообразования [291]. Этот процесс происходит в рамках систе-
мы полового воспроизведения и ведет к образованию дочернего
вида, изолированного от родительских видов, как и при амфи-
Гл 19. Гибридпал жюдоо^рал.’вйчиг
2U-
пл он ди к. барьером хромосомной стерильности. Однако в этом
случае в отличие от амфиплондни дочерний вид остается на том
же уровне пломдностц. что и родительские формы
Такой результат достигается благодари возникновению и за-
креплению в последующих поколениях гибридов новой рекомби-
национной формы, гомозиготной по разным независимым хро-
мосомным перестройкам, по которым различались родительские
виды Такая новая гомозиготная рекомбинационная форма пло-
довита при скрещиваниях внутри себя, по стерильна при скре-
щиваниях с любым из родительских видов.
Процесс рекомбинационного видообразования может привес-
ти к созданию дочерних видов, окруженных барьерами стериль-
ности, различающимися по своей эффективности. Степень изо-
ляции дочернего вида прямо пропорциональна степени стериль-
ности при скрещиваниях между родительскими видами.
Основную идею, на которой построена гипотеза рекомбина-
ционного видообразования, высказал Мюнтцинг (595, 598. 601].
Согласно предложенной км концепции, отдельные пары факто-
ров, обусловливающих хромосомную стерильность. могут реком-
бинироваться, а результате чего образуется новая плодовитая
форма; однако дальнейшей разработкой этой концепции Мюнт-
цииг не занимался. Герасимова (237] подтвердила правильность
концепции Мюитциигл в экспериментальных работах по трансло-
кациям у См pis. Стеббинс [776, 777, 786] рассматривал в общем
плане рекомбинацию факторов стерильности в связи с гибрид-
ным видообразованием. Позднее Стеббинс [782] и Грант [280]
предложили конкретную генетическую модель рекомбинатюн-
пого видообразования. Еше позднее, в 1963 г, я опубликовал
обзор по этой проблеме, собрав все имевшиеся к тому времени
фактические данные [282].
Итак, мы вкратце ознакомили читателя с рекомбинационным
видообразованием, с тем чтобы сопоставить этот способ с дру-
гими способами гибридного видообразования В следующей гла-
ве оно будет рассмотрено более подробно
Отметим мимоходом, что процесс рекомбинации способен
создавать новые барьеры генной стерильности из прсдсушество-
вавших барьеров совершенно с тем же успехом, с каким он соз-
дает барьеры хромосомной стерильности. Имеются некоторые
данные, укалывающие иа образование путем рекомбинации но-
вых барьеров генной стерильности у Phascvlui и новых контро-
лируемых генами барьеров несовместимости у Gilia [273. 458].
Об этой стороне проблемы известно, однако, слишком мало, что-
бы она заслуживала специального обсуждения в этой книге.
В ибшем этот процесс в значительной мере сходен с процессом,
в котором участвуют факторы хромосомной стерильности и ко-
торый будет описан в дальнейшем.
16*
244 Voces клгал. Естесгвгыая .•utfuSuatUfUt и м рпумтагы
Гибридное видообразование при наличии
внешних барьеров
Гибридизация между двумя плодовитыми при скрещивании
видами растений может привести к образованию разнообразны»
рекомбинантов с разными комбинациями морфологических и
физиологических признаков, по которым различались родитель-
ские формы Некоторые из этих новых комбинаций признаков
могут обусловить создание внешней изоляции того или иного
типа между гибридным потомством и родительскими видами.
Если такие рекомбинанты, изолированные внешними барьерами,
хорошо адаптированы в какой-либо доступной им среде, то они
могут затем увеличивать свою численность на территории, зани-
маемой родительскими видами.
В одном из первых разделов злой главы мы уже приводили
исторический пример возможного гибридного видообразования
у Rhododendron, изучавшийся Кернером (431]. Кернер постули-
ровал. что R. intermedium произошел от гибрида между R. fer-
ruginsum и R hirsutum и сумел обосноваться в некоторых мес-
тах в Альпах. Этот дочерний вид гибридного происхождения
отличается от родительских видов по окраске нветкои и до не-
которой степени — по предпочитаемым им почвам. На основе
того, как излагает этот пример Кернер, следует считать, что
/?. intermedium частично изолирован от R. ferrugineum и R hir-
sutum поведением опыляющих эти растения пчел, а, возможно,
также и здафическими факторами Более новое описание гибри-
да R. intermedium и его родительских видов дал Хеги {359].
Delphinium recurvatum, D, hesperium к D. gypsopnilum со-
ставляют группу плодовитых при скрещивании друг с другом
диплоидных видов, распространенных в предгорьях и долинах
Калифорнии. Судя по морфологическим и экологическим приз-
накам, D. gypsophilum возник в результате выщепления из
скопления гибридов от скрещивания между D. recurvatum и
D. hesperium. По экологическим предпочтениям и морфологиче-
ским признакам D gypsophilum занимает промежуточное поло-
жение между этими предполагаемыми родительскими вилами.
Среди потомков Fa, полученных при искусственном скрещива-
нии между D. recurvatum и D. hesperium, некоторые экземпля-
ры морфологически сходны с D. gypsophilum, В природе D. gyp-
sophilum сохраняет обособленность От двух других видов глав-
ным образом благодаря экологической изоляции [495]
В Калифорнии обитают три родственных диплоидных
(2п=1б) вида Penstemor. (Р. centranttutolius, Р. grinnellii и
Р spectabilis), до некоторой степени плодовитые при скрещива-
ниях друг с другом. Они различаются по строению цветков, спо-
Гл 19 Гибрид, мг в«ккл?б(М1Х>мм>ы
245
собам опыления II экологическим особенностям Сопоставление
морфологических признаков этих грех видов, дачные экологи
ческнм исследований и природных условиях и признаки естест-
венных 1нбридип между мини позволяют считать, что Р. specta-
bills возник в результате гибридизации между Р. centrantijolius
и Р ghnnellil Нельзя, однако, исключить возможность того, что
•ил Р. spec tab ilis возник в процессе первичного видообразова-
ния В природе эти три вида изолированы др • меха-
нический и. этологическими и экологическими барьерами [808,
809].
Стеббинс и Ферлан [791| постулировали гибридное нромс-
хождепие Oohrys murbeckct от скрещивания между О. fusca и
О. lutea в Средиземноморской области. В атом случае обособ-
ленность предполагаемых дочернего к родительских видов под-
держивается главным образом этологическими и экологическими
изолирующими факторами [791].
Такер и Сауэр [839] описали ряд диплоидных поп
Amaranlhus в дельте р. Сакраменто в Калифорнии, где в настоя-
щее время, возможно, происходит гибридное образование
диплоидных видов. В этом регионе обнаружено несколько хоро-
шо выраженных видов (Л. powellii, A. caudatus, A retroflex из)
и одновременно несколько отклоняющихся локальных популя-
ций, у которых наблюдаются п разных комбинациях признаки
названных выше видов. Эти отклоняющиеся популяции нли не-
которые из них, по видимому, представляют собой различные
диплоидные гибридные производные уже существующих милов
Лотагопйде (8.39J-
На Аляске Carex rostrata обычно встречается п пресноводных
озерах, а С roiundafa — на торфяных болотах. В переходных зо-
нах между этими двумя местообитаниями обнаружены скопле-
ния гибридов, образовавшихся в результате скрещиваний меж-
ду названными выше видами В некоторых местах из скоплений
гибридов вышеилнется новая, морфологически отличающаяся
плодившая популяция, и численность ее по сравнению с числен-
ностью родительских видов возрастает Эта новая форма рас-
сматривается как дочерний вид гибридного происхождения п
получила название С. paludiuagans [179]
Новая Зеландия — один нз центров видообразования для ро-
да Epilobium с 37 местными видами, занимающими широкий
спектр местообитаний Это многолетние травы, для которых ха-
рактерны вегетативное размножение, мелкие цветки и преиму-
щественно автогамная система размножения Epilobium — гомо-
плонды с 2н-ЗЬ Продукты скрещивания между разными вида-
ми плодовиты п большинстве комбинаций Симпатрня представ-
ляет собой обычное явление. при этом репродуктивная изоляция
246 Чисто лктох Егтгстаеклэт оиблиЛмечил » •» pnyrtrau
обеспечивается автогамией, а также экологической дифферен-
циацией [666].
Однако иногда репродуктивные барьеры разрушаются, так
что гибриды и скопления гибридов хорошо известны. Обнаруже-
но также несколько однородных популяций гибридного проис-
хождения. Некоторые иа них находятся ия расовом, а другие —
на видовом уровне Так, предполагается, что Epilobium purpura-
turn на острове Южный — результат скрещивания между Е. eras-
sum и Е. porphyrium. Трудно сказать что-либо определенное от-
носительно этого н некоторых других случаев, ио в обшем пред-
ставляется вероятным, что разнообразие видов Epilobium на
Новой Зеландии увеличилось путем гибридного видообразова-
ния (666].
Недавно был описан интересный пример гибридного видооб-
разования у древовидных папоротников (Cyatheaceae) в Пуэрто-
Рико (1I8J. Виды Alsophila bryophila, A. dryopteroides и Nephe-
lea porioricensis (у всех 2л «69 II) гибридизируют во всех воз-
можных комбинациях. Образующиеся при этом гибриды плодо-
виты н дают в F» гомоплоидных выщепенцев, часть которых со-
храняется в природе.
Одно из гомоплоидных гибридных производных от скрещива-
ния между Alsophila dryopteroides и Nephelea porioricensis обра-
зует однородную популяцию, превосходящую по численности по-
пуляции родительских видов в той же местности. Эта гибридная
популяция размножается половым путем. Никаких данных о ве-
гетативном размножении нет [118].
Размножение в чистоте у этой формы гибридного происхож-
дения обеспечивается, по-видимому, следующей системой вос-
произведения. Гибридный папоротник F, производит рекомби-
наитные споры, из которых развиваются обоеполые гаметофиты.
Мужские и женские гаметы развиваются иа гаметофите в ре-
зультате мвтотических делений, а поэтому они генетически идеи
тичны. В результате самооплодотворения внутри гаметофита об-
разуется гомозиготный спорофит, несущий такую же генную ком-
бинацию, как и та рекомбинантная спора, из которой этот гаме-
тофит развился. Такой спорофит дает начало спорам, вз кото-
рых развиваются новые гаметофиты, так что в конце концов
возникает колония идентичных индивидуумов данного нового
типа [118].
Изолированность такой популяции гибридного происхожде-
ния обеспечивается внешними этологическими барьерами и са-
мооплодотворением (118].
Гл. 19. ГибрийпЫ 24?
Введение в апомиктическое видообразование
Иногда оказывается, что один из межвидовых гибридов n F>
нлн п каком-либо из последующих ранних поколений обладает
исключительном адаптивной цепкостью Его высокогетсроэигог
нын генотип в гаком случае может быть наделен гетерозисным
з4>4>ектом или гомеостазом или одновременно и тем и другим.
Подобный гибрид потенциально заслуживает воспроизведения
и увеличения численности Но размножение межвидового гибри-
да, превосходящего по своей адаптивной ценности родительские
пяди, сопряжено е двумя очевидными затруднениями. Первое из
них — гибридная стерильность, второе — неспособность размно-
житься в чистоте. Существует, однако, несколько способов, поз-
воляющих обойти эти затруднения, ня что уже давно указал
Дарл ннн он (138, гл 16J.
Любая гетсрозигота, произошедшая в резу.тыате мсжвндсъ
вого скрещивания, может преодолеть внутренне присущие ей
стерильность и расщепление, затрудпикмцне моспронзнедепие, и
обеспечить размножение своего особого гетерозиготного геноти-
па при помощи нескольких способов стабилизации, перечислен-
ных в начале этой главы (способы I—5). Здесь в связи с гиб-
ридным видообразованием мы упомянем и двух первых спосо-
бах, объединяемых под общим названием апомиксис В этом
разделе дается лишь общее npeflciявление об апомиктическом
видообразовании, а более подробно оно будет рассматриваться
в части восьмой
О punt ia fulgida и О sptnosior — два различных няда какту-
сов. растущих в пустынях на юго-западе США и в Мексике. Они
мрем я от времени скрещиваются, давая гибриды с высоко сте-
рильными семенами. Несмотря на стерильность семян, гибрид-
ные растения достигли большого обилия вдоль реки Хила в
Аризоне благодаря своей способности к вегетативному размно-
жению опавшими члениками стебля [304],
Европейские ежевики, принадлежащие к секции Monferus
рода Rubus. образуют большой комплекс гибридизирующих ви-
дов, размножающихся половым путем, и их апомиктических
производных. Их гибриды размножаются как путем агамоспер-
мии, так и нет-ативно. Гибриды каждого типа могут размно-
жаться и распространяться при помощи семян, образующихся
бесполым путем, нли в результате укоренения опавших кусочков
стебля, многие мнкровнды возникли именно такими способами
обладающий хорошо выраженными морфологически-
ми признаками и экологическими предпочтениями. может таким
обратом распространиться аломиктически и создать свою соб-
24в 7<кг» ллгоя. Еспсткляал лвридизацш и к (мзуиьгегы
ствекиую популяцию. Таили популяция. состоящая из одинако-
вых гибридных индивидуумов апомиктического происхождения,
если она недостаточно хорошо обособлена морфологически иля
узко эндемична, или же характеризуется одновременно и тем и
другим, представляет собой клональный или агамоспермны*
вид. Если же гибридные производные обладают комбинацией
признаков, легко распознаваемой в обычной таксономической
практике, и если этой комбинации признаков удалось распрост-
раниться путем апомиксиса по определенной географической об-
ласти, то образовавшаяся в результате популяция достигает ста-
туса таксономического вида.
Выводы
Полезно кратко охарактеризовать известные способы гибридно-
го видообразования у растений.
I. Гибридное видообразование в сочетании с половым вос-
произведением; новый биологический вил возникнет непосредст-
венно кз межвидового гибрида.
I. Гибридное видообразование при наличии внешних
барьеров; формирование нового рекомбинационного типа
в потомстве гибрида, отделенного от родительских видов
внешцими изолирующими механизмами.
2. Рекомбинационное видообразование; гибрид, характе-
ризующийся хромосомной стерильностью или полусте-
рнльностью, лает начало гомопловлным или неудвоившим-
ся производным, обладающим новой гомозиготной реком-
бинацией различных независимых хромосомных пере-
строек. по которым различаются родительские виды; но-
вая плодовитая линия характеризуется собственной ком-
бинацией признаков и стерильна при скрещиваниях с ро-
дительскими видами.
3. Амфиплоидия; гибрид, обладающий хромосомной сте-
рильностью, путем удвоения числа хромосомных наборов
дает начало плодовитой полиплоидной форме, которая
изолирована от родительских ендов барьером, создавае-
мым хромосомной стерильностью.
II. Гибридное видообразование в сочетании с бесполым или
почти бесполым размножением; размножение гибрида при помо-
щи механизмов, обходящих нормальный половой или хромосом-
ный цикл, и распространение возникшей гибридной формы по
определенной географической области, так что по величине и
составу, популяции она достигает уровня унипареитального мнх-
ровида или вида. . _ .. •
Гл It. Гпвепдяог ааЛюСжвмшше 249
4. Апомиктическое видообразование: новый клональный
или агамоспермиый микровид развивается соответственно
из межвидового гибрида путем вегетативного раэмиоже-
кия или агамоспермиого образования семян (часть вось-
мая).
5. Перманентная структурная или нумеряяеская гибрнл-
иость; какая-либо гетерозигота по транслокация размно-
жается по генетической системе типа Oenothera biennis
или численность какого-либо нечетного полиплоида уве
личкввется по генетической системе Rosa сапта, в ре-
зультате чего образуется популяция, соответствующая
но величине гетерогамному микровиду (часть восьмая).
20 Рекомбинационное
видообразование
Рекомбинационное видообразование — это формирование и за-
крепление в потомстве гибрида, характеризующегося хромосом-
ной стерильностью или полустерильностью, нового гомозиготно-
го рекомбинанта по двум или нескольким независимым пере-
стройкам, которыми различаются родительские виды
В результате процесса рекомбинационного видообразования
могут возникнуть дочерние виды, изолированные слабыми или
сильными барьерами стерильности. Дать объяснение гибридному
происхождению новой гомоолоидиой линии, которая сама пло-
довита, но высокостерильна при скрещиваниях с другими пред-
ставителями своего роди, несомненно, труднее Однако, если мы
сможем найти удовлетворительное объяснение атому случаю, у
нас окажется все необходимое, для того чтобы понять гене-
тически более простые случаи, в которых функционируют слабые
барьеры стерильностм.
В этой главе мы опишем гипотетический механизм рекомби-
национного видообразования, дадим обзор соответствующих экс-
периментальных данных и попытаемся понять возможную роль
этого процесса в природе.
Гипотеза
Довольно четкую модель, описывающую интересный с теорети-
ческой точки зрения случай рекомбинационного видообраэова-
ння с возникновением сильных барьеров стерильности, предло-
жили Стеббинс [782] и Грант [280]. Эта модель предусматривает
возникновение барьера хромосомной стерильности между двумя
гибридизирующими родительскими видами в результате незави-
симых друг от друга структурных перестроек. Мозель основана
нз более ранних работах Мюитцинга и других авторов, о чеы го-
ворилось при вводном обсуждении агой проблемы в гл. 19.
Допустим, что два родительских вида изолированы друг от
друга барьером хромосомной стерильности, создаваемым двумя
или иесколькими независимыми хромосомными перестройками
Гибрид между этими двумя видами, обладающий хромосомной
стерильностью, хотя и неполной, может дать начало одному или
большему числу новых гомозиготных реиомбинантов по струк-
турным перестройкам. Сами эти новые формы плодовиты, но
проявляют хромосомную стерильность при скрещивании друг
с другом и с родительскими видами [280. 282, 780, 782].
Такой процесс рекомбинации можно проиллюстрировать на
простом примере. Допустим, что два родительских вида ралли-
251
Г А. К. Ptwriwiavionnix лидосб fAAOtan**
ГОИЗД’ГОГЫ Pt
AA M CC DO
AA M C,C, D.O,
а.в. ec do
A*Afc 1.1. D.D.
Рис. 20.1. Пивелепяе пр« скресхмваник гибрида, гетауюавготаого по лауш не-
зависимым тражлокаиям.
Перми траксл оваций затрагивает участки а в В в хромосомах А и В, а вто*
рай—участки с м А хромосомах С и D. У ралктелидого вада Р. послед о
вательипеть учветмоя отййдартмдя во всех четырех хромосомах, тогда как
вкп Р? несет обе трляслскаиня. Гибриды F. образуют гамети 16 классов.
У г в мет 12 классов жмеютеж нехватки и дтпликашги по участкам в — b, уча-
сткам с — d нли сбехм парам участков. Гаметы четырех других классов сба-
лаиенроманы и ж из «способны. Этж четыре класса фуккнмоиалвных гамет
продуцируют в F, различные тпш гетероанют по травелонлцяяы (на схеам
ие показаны) и четыре типа структурных гомозигот (онаааны), Дальнейшие
объяснения см. в тексте.
чаются по даун транслокациям » четырех хромосом». Они со-
держат хромосомные наборы ЛЯ BBCCDD и A>AfBtB,CtCaDfDt
соответственно (см. рис. 20.1). Гибрид Fi от скрещивания
между ними имеет конституцию AAtBB,CCaDDc, гетерозиготен
252 Тосте «гос Еаесткнма гивршЗкэоч». » " р^ч^пт
по двум независимым транслокациям, и поэтому 25% его гамет,
как н следовяло ожидать, фертильвы (см. рис, 20.1).
В число функционирующих гамет входят гаметы двух роди-
тельских типов, которые нас здесь ис интересуют, и гаметы двух
рекомбинационных типов (ABCaDc и Л»В.СО). которые интере-
суют пас, поскольку они могут дать в Fs начало новым гомо-
зиготным типам АЛ ВВ Ctfit D.D, и AtAtBJhCCDD (рис. 20.1).
Эти новые гомозиготные рекомбинанты полностью плодовиты
при скрещиваниях внутри себя, но частично стерильны прн скре-
щиваниях друг с другом и с родительскими вилами.
Разумеется, если число независимых транслокаций больше,
то они создают более мощный барьер хромосомной стерильности.
Так. следует ожидать, что гетерозигота по шести независимым
транслокациям даст всего 1.56% фертильных гамет. Межвидо-
вые гибриды растений, прпнаводяшне примерно такие количест-
ва фертильной пыльцы, считаются высокосгеркльиыми. В об-
щем доля фергнлышх гамет, производимых гетерооиготамн по
транслокациям, составляет 1/2". где л —число независимых
транслокаций.
Кроме того, по мере усиления барьера, создаваемого хромо-
сомной стерильностью, число новых фертильных типов гомози-
гот. которые могут возникнуть п Fi ялв в последующих поколе-
ниях, также возрастает. И эти новые, гомозиготные по структур-
ным перестройкам рекомбинанты или большинство из них будут
изолированы от родительских видов мощными барьерами сте-
рильности.
Вернемся вновь к примеру гибркда, гетерозиготного по шес-
ти независимым транслокациям. Этот гибрид может дать в пос-
ледующих поколениях 62 новых плодовитых гомозиготных ре-
комбинанта. Такие рекомбинанты при возвратных скрещиваниях
с родительскими видами дадут гибридов, производящих от 3 до
50% фертильных гамет. Прн атом гибриды, полученные от воз-
вратных скрещиваний рекомбинанта, отличающегося самой вы-
сокой частотой, с любым из родительских андон, дадут 12.5%
фертильных гамет.
Структурные перестройки иных типов помимо транслокаций,
в частности транспозиции н реянверенн, также создают возмож-
ность для усиления барьеров хромосомной стерильности путем
рекомбинации. Транспозицией называют обмен участками в пре-
делах одной хромосомы, как это показано на рис. 8.2. Реннпер
сия представляет собой результат двух последовательных инвер-
сий в одной и той же области хромосомы, причем одна из ин-
версий заключена в другой. Если ABCDEF — последователь-
ность генов в стандартной хромосоме, то в хромосоме с ренивер-
сией оиа имеет вид AECDBF. У структурных гетерозигот крос-
еннговер между транспозициями или в пределах реинверсии при-
Гл 20 Рско.ч^илацаоччое бнОообрвювапие
253
водит к образованию хромосом с нехватками и дупликациями.
Поэтом)* множественные независимые транспозиции или реки-
версии п гетерозиготном состоянии могут создать высокую cic-
netib хромосомной стерильности при условии, что кроссипгопер
всегда происходит между транспозициями или внутри реп и вер-
сий Функциональные гаметы представляют собой нскроссовср-
ные типы по транспозициям или реииверсням
Средн функциональных гамет будет несколько таких, у кото-
рых отдельные транспозиции или реинвсрсии, затронувшие раз-
ные хромосомы, рекомбинированы. Эти гаметы могут дать в Ft
или в более поздних поколениях новые плодовитые типы струк-
турных гомозигот, которые будут изолированы oi родительских
видов барьерами стерильности Транспозиции и реипверспи. по-
добно транслокациям, могут рекомбинироваться, давай новые
плодовитые изолированные липни.
В процессе рскоибпиаипоиного видообразования важную
роль играет система скрещивания участвующих в нем растений.
Система скрещивания, существующая в гибридной популяции,
будь то свободное Неродствен нос скрещивай не или самооплодо-
творение. оказывает решающее влияние иа образование этой
гибридной популяцией новых плодовитых видов, изолированных
внутренними барьерами Инбридинг способствует рекомбинаци-
онному видообразованию [280].
Непосредственный конечный результат рекомбинационного
видообразования - возникновение структурной гомозиготы ново
го типа Структурная гомозиготность достигается быстрее всего
потомками какой либо структурной гетерознготы в условиях ни-
бридинга Новая гомозиготнан форма, коль скоро она образова-
лась. также лучше всего закрепляется и сохраняется системой
инбридинга. Свободное неродственное скрещивияяс в гибридной
популяции, напротив, сильно снижает вероятность как возник-
новения новой структурно-гомозиготной формы, так и после-
дующего сохранения такой формы я качестве самостоятельной
линии, размножающейся в чистоте [280, 282].
Поучительно сопоставить легкость закрепления новой струк-
турной гомозиготы в простом гипотетическом случае при про-
тивоположных системах скрещивания Допустим, что родитель
скис виды различаются по двум независимым перестройкам, как
иа рис 20 1 В результате гибридизации между этими видами
возникнет смешанная колония, состоящая, например, из двух
растений F| и четырех растений Р(. Допустим далее, что вое
произведение гибридов ласт ряд форм F?, которые будут одина-
ково жизнеспособны и одинаково плодовиты Какова же будет
частота одной определенной новой структурной гомозиготы (т е
ЯЛ ВВ CrfCj D,DC по рис 20 1) в гибридном покоэенин F> при
самооплодотворении н при случайном скрешнванни7
25* Часть аяпя Дстеггвгньая швриЛч»!»» к м ,мз,жгаты
При самооплодотворении зта гомозиготная форма составит
11% поколения F». Если, однако. растения размножаются путем
свободного скрещивания и если гибриды Fi случайно скрещива-
ются друг с другом и с растениями Pi в гибридной колонии, то
результаты будут совершенно иные В этих условиях частота той
же самой новой гомозиготы среди гибридов Fi составит от О
до 5% (а зависимости от того, какие гибриды Fs будут воспро-
изводиться), с модой в левой части этого диапазона.
При свободном скрсщнваани в типичной гибридной колонии,
состоящей из смеси растений Fi и растений родительских типов,
возникновение определенной новой структурной гомозиготы воз-
можно. Но вероятность появления такого генотипа невелика, а
кроме того, если он и возникнет, то ои не будет размножаться
в чистоте. В гибридной колонии такого же состава самооплодо-
творение во много раз более аффективно в смысле создания и
закрепления нового вида.
Нам осталось вкратие обсудить судьбу различий по феноти-
пическим признакам между родительскими видами в процессе
рекомбинационного видообразования.
При рекомбинации участков хромосом рекомбинируют также
локализованные в них гены. И когда новая рекомбинация хро-
мосомным перестроек закрепляется в гомозиготном состоянии,
то закрепляются и входящие в нее гены. В результате дочерний
вид может унаследовать новую комбинацию морфологических и
физиологических признаков родительских видов, и эта новая
комбинация признаков будет размножаться в чистоте.
Экспериментальный подход
Гипотеза рекомбинационного видообразования создавалась на
протяжении 30 лет В течение этого времени иа представителях
ряда групп растений проводились эксперименты, относящиеся к
рекомбинационному видообразованию Самые первые экспери-
менты принадлежали Герасимовой [237], Винге [901] в Лампрех-
ту (458).
Эти первые эксперименты ставились не для проверки отдель-
ных аспектов гипотезы рекомбинационного видообразования, ко.
торые были разработаны позднее. По сути дела, некоторые из
ятях первых фактических данных были получены в эксперимен-
тах, которые и не были настоящими экспериментами в гои, что
касается гипотезы рекомбинации. Взаимоотношения между тео
рией рекомбинационного видообразования и экспериментом не
всегда складывались так. как это должно было быть, если исхо-
дить аз гой картины развития науки, какую обычно описывают
в учебниках. Тем ке менее со временем накопилось довольно
много экспериментальных данных, если понимать термин экспе-
Г* Ю. Ргжолбаиочиожч!» выЭообримокве 25*
римект в широком смысле и если учитывать как первоначальные
фактические данные различных исследователе*, так и выведен-
ные ими заключение
Экспериментальные данные по рекомбинационному видооб-
разованию были получены в исследованиях на Crepis [2371.
Erophila [9011, Nicotiana [742, 744], Elymus [782], Collinsia [162,
233] и Cilia [291]. Все эти дачные подтверждают правильность
по крайне* мере одного аспекта гипотезы рекомбинационного
видообразования, К числу более полных экспериментов, в кото-
рых исследовалось одновременно несколько аспектов гипотезы,
относятся эксперименты на Nicotiana, Elymus и Cilia. Большин-
ство проведенных исследования имело дело со слабыми барьера-
ми стерильности. Только в экспериментах с Elymus и Cilia были
предприняты попытки проверить гипотезу рскомбинацноипого
видообразования в условиях очень мощных барьеров хромосом-
ной стерильности.
Ранние эксперименты
Герасимова (237| проводила свои исследования на четырех ли-
ниях Crepis tectorum (2л—81, различавшихся по гомозиготным
транслокациям в одной или в обеих парах хромосом. Нормаль-
ная линия С tectorum характеризуется стандартной последова-
тельностью участков во всех четырех хромосомах- A.B.C.D.
У мутантпой липни I имеется гомозиготная транслокация, затра-
гивающая хромосомы Л и D, а у мутантной линии II — .хромо-
сомы В и С. Эти две липин, плодовитые сами по себе, оказались
частично стерильными при скрещиваниях друг е другом, чего и
следовало ожидать. Полустерильный гибрид F: от скрещивания
между линиями I и II. которые были гетерозиготны по двум не-
зависимым транслокациям, дал поколение Ft, содержавшее це-
лый ряд гетерозигот и гомозигот по транслокациям
Одно из растений Ft, названное линией nova. содержало обе
транслокации в гомозиготном состоянии и было плодовитым.
Линия nova отличается от линий 1 и II одной транслокацией,
поэтому можно было предположить, что при скрещивании со
своими непосредственными родительскими формами опа даст
частично стерильных гибридов. От нормальной липни Crepis tee-
totum линия nova отличается двумя независимыми транслока-
циями. а поэтому следует ожидать, что скрещивания между эти-
ми линиями будут еще более стерильными. Герасимова [237]
подтвердила »тх>. получив гибрид Ft между линяпми teetotum я
nova, который оказался полугтерильиыи; мейоз у него был на-
рушен, а фертильные семена составляли 30%.
2S6 Уотт» алгол. Есгкгмпмя иЛроЭлмялл '* регул» гать
Это был первый случай экспериментального получении новой
изолированной плодовитой формы путем рекомбинации незави-
симых факторов, создающих хромосомную стерильность. Барьер
стерильности в атом случае выл слабым к фенотипически линии
ие различались.
Группа Erophlla verna (х-7) представляет собой комплекс
автогамных однолетних видов-двойников с различным, полипло-
идным или ансуплоидным, числом хромосом (901], о чем уже
говорилось в гл. 11. Винге скрещивал две следующие формы
£ чета: Е. conlertilolia (л=15) и Е. ololacra-petiolata (п — 32).
У гибридов конъюгация хромосом была сильно нарушена и от-
мечалась высокая стерильность как пыльцы, так и семян; фер-
тильность семян составляла всего 3% нормальной (901].
Поколение F, состояло из 451 растения, средн которых наб-
людалось расщепление по морфологическим признакам, плодо-
витости и числу хромосом. Четыре растения F> послужили осно-
вателями для четырех семей F> н нх иибредиых потомков вплоть
до поколения Гт. Ливии продолжали расщепляться, достигая
единообразия в разное время — от поколения Fi до F». Восемь
линий, которые были подвергнуты цитологическому анализу,
достигли в Fi—F, стабильности по числу хромосом, которое во
всех случаях отличалось от исходного и было различный в раз-
ных линиях. От скрещивания (л-15) X (п-32) произошли ли-
нии с п—22. 23, 25, 29. 31 и 34. Эти линии были плодовитыми,
морфологически единообразными и представляли собой новые
ансуплоидвые типы [961|.
Эксперименты Винте показали, что гибридизация между ро-
дительеннми видами, находящимися иа разных уровнях плоид-
ности, может привести к возникновению новых плодовитых кон-
стантных аиеуплоидных типов. Предполагается, хотя соответст-
вующих возвратных скрещиваний не производилось, что гибри-
ды от скрещиваний этих анеуплоилов с родительскими видами
будут до некоторой степени стерильны. Этот эксперимент остав-
ляет открытым вопрос о том, может ли гибридюацна давать так-
же начало новым микровидам на гомоплоидном уровне
Последующие эксперименты при существовании
слабых барьеров стерильности
Следующая группа экспериментов, которую нам предстоит рас-
смотреть. проводилась па Nlcotlana, CoUuuia, Elpmus и Gilia-.
они отличаются от первых экспериментов тем. что ко времени
нх проведения формулировка гипотезы рекомбинационного видо-
образования уже стала вырисовываться более четко Это позво-
ляло планировать эксперименты таким образом, чтобы с их по-
мощью можно было действительио подвергнуть проверке гипо-
Гл 20 PftovflUMatfUonttiy ли&эобр^оа&^ие
257
тезу или ее наиболее существенные части. В этом разделе мы
рассмотрим экслер имен гы, приводившиеся с Nicotiana н Collin-
sia в условиях сущее 1 новация слабых барьеров стерильности
Смит и его сотрудники [742, 744] провели длительный с еле к
цноимый эксперимент на гимоплондных инбредных линиях, про-
исходящих ог скрещивания между разными видами Nicotiana.
Оба родительских вида. .V. langtdorjii и A' sanderae. — дкплон-
ды с 2л=18. Они различаются по величине цветков (у Л\ lang-
sdorfii трубка венчика короткая, а у Л' sandcrae — длинная), а
также по вегетативным и другим признакам.
У гибрида Ft пыльца была полустерилкман (число фертиль-
ных пыльцевых зерен сое iявляло в среднем 55%), а семена —
довольно ферт ильным и Мейоз был несколько иа рушен. Большая
часть хромосом конъюгировала, образуя биваленты, по некото-
рые из пт соединялись в цепочку или оставались одиночными;
у таких хромосом наблюдалось отставание в анафазе [744J.
Снижение фертильности пыльцы, верою но, обусловлено главным
образом этими нарушениями конъюгации
В F; были заложены три селекционные линии, которые про-
латжл.1И вести путем самоопылении силе и.пых родительских рас-
тений вплоть ли Fio Искусственный отбор производили по дли-
не трубки венчика В одной липни вели отбор на длинную -руб
К), как у родительского вида Nicotiana sanaerae, в другой--на
короткую трубку, как у Лг. langidorjii, а в третьей — на трубку
промежуточной длины Все три линии реагировали на отбор ио
величине цветков, но при этом стали различаться также по дру-
гим признакам цветка и веггтатпяпым признакам, не подвергав-
шимся отбору. Они стали отличаться морфологически как друг
от дриа, так и от родительских видов (744).
Отобранные таким образом линии оставались диплом им ы мн,
подобно родительским видам Наибольший иитсрес для нас
представляю! диплоидные производные с короткими венчиками
и другими связанными с этим признаками, так как у них а по-
колениях F< — Fo фертнль;юсть пыльцы повысилась поч.и до
нормального уровня, тогда как две другие линии в поздних по-
колениях оставались или становились лол усгер ильными {7-14].
Формы с мелкими цветками при скрещивании с обеими ро-
дительскими линиями давали папустсрмлкных гибридов. Фер-
тильность пыльцы у тибрилов. полученных при возвратомскре-
uiKBaiiiin с Nicotiana langsdorfii, состай?1плн 46—58%. а при воз-
вратном скрещивании с .V. sandcrac — 60—61%. Как у тех, так
п у труни гибридов отмечались некоторые пткломсиня в мейо-
зс Таким образом, линия с мелкими цветками была стерильна
при скрещивании с яюбой из родительских линий, так же как и
эти линии при скрещивании одна с другой (742, 744J.
Г'-
25в Част» мш. Есткгмш» :ибрю)имцая rt praytnm
Гарбер и его сотрудники изучали гибридов и гибридных по-
томков от скрещиваний Collinsia corymbmax С. tinctorla н С. со-
rymbosaxC. multicolor [162. 233[. Об» родительских вида ди-
плоидны (2л—14) и различаются по ряду морфологических
признаков
Межвидовые гибриды F> содержали траислокациоиные коль-
ца иа 6 хромосом и характеризовались высоко* плодовитостью.
У растений F> и Fi меиоэ протекал нормально, с образованием
бивалентов. Большинство растений, составлявших поколения F:
и Fa. имели геномы родительских типов. Однако в этих поко-
лениях были три линии, представляющие каждое иа двух перво-
начальных скрещиваний, которые обладали новыми рекомбина-
ционными геномами, отличавшимися от родительских по транс-
локациям. Кроме того, эти линии отличались новыми комбина-
циями морфологических признаков [162, 233].
Более поздние эксперименты при существовании
мощных барьеров стерильности
Первый эксперимент по рекомбинационному видообразованию
при наличии очень мощного барьера хромосомной стерильности
был проведен на гибридном травянистом растении на комплек-
са Elymus glaucus. Родительскими видами этого гибрида были
Elymus glaucus и SUanion jubalum; у обоих видов число хро-
мосом 2п=28. Этот эксперимент, поставленный Стеббинсом, опи-
сан в его кратком сообщении [783].
У этого межвидового гибрида происходит конъюгация хро-
мосом с образованием 11—14 бивалентов, но он высоко стери-
лен: количество фертильной пыльцы не достигает 1%, а число
семян равно примерно 1 на 10000 цветков. Цитогенетически*
анализ искусственных аллополиплошюв, происходящих от этого
гибрида, показывает, что стерильность его имеет частично хро-
мосомную природу я обусловлена мелкими перестройками, ие
очень сильно нарушающими образование бивалентов, а частич-
но-генного характера (783, 786).
Гибриды Fi. происходящие от указанных выше родительских
видов, получали несколько раз от скрещиваний между разными
микроаилаии группы Elymus glaucus и разными линиями Sita-
шоп jubalum или родственного ему S. hist г ух. Затем проводили
возвратные скрещивания растений F,. полученных от различных
гибридных комбинаций, с £. glaucus; доля семян и растений В.
была при этом очень невелика: в результате нескольких скрещи-
ваний было получено в общей сложности 173000 цветков, кото-
рые принесли всего 15 семян В1 и II неумоеиных беккроссов.
Растения В, в большинстве случаев представляли собой по мор-
Гн 29 /’екозеввявчвоиисе апЗ«>5|»эоваи®е
239
фологки и плодовитости реверсии к Е. gtaucus, а полому они
нас здесь не интересуют |783].
Одно из растений В, обладало, однако, морфологическими
признаками как гибрида F|, так и Etymus gtaucus, послуживше-
го отцовским растением, и било полустернльпым. При самоопыле-
нии оно дало в поколениях lt и Ь мощное и полностью плодо-
витое потомство. Эти ннбредиые семьи характеризовались рас-
шеплспием по морфологическим признакам с некоторой измен-
чивостью в направлении Silanion tubatum [783].
Одно растение из поколения 1г скрестили затем с исходной
родительской линией Etymus gtaucus. чтобы выяснить нх вза-
имоотношения по фсргилытостн Гибриды Fi. полученные в ре-
зультате такого тестирующего скрещивания, оказались высоко
стерильными, дао всего 0—3% фертильной пыльны |783].
Работа, проведенная иа Etymus. продвинула еще иа одни
шаг экспериментальное доказательство рекомбинационного ви-
дообразования. Она подтверждает эту гипотезу на важном при-
мере с очень мощным барьером стерильности, имеющим, по
крайней мере частично, хромосомную природу. Изолированное
гоз'оплоидное производное было получено, однако, путем воз-
врат исхо скрещивания в поколении Fi, а ие путем прямого ин-
бридинга. При такой постановке эксперимента снижается его
применимость к тем реально существующим в природе ситуа-
циям, в которых скорее всего можно ожидать рекомбинационно-
го видообразования
Следующий эксперимент, который пам предстоит рассмот-
рен., был поставлен с парой видов из труппы Gilia inconspicua
и их гибридными производными [2911. Родительские вилы G. та-
liar и G. modocensis— автогамные однолетние растения с чис-
лом хромосом 2л —36. Гибриды F> от скрещивания между ними
пысокосгерилыты фертильная пыльца у них составляет всего
2%. а всхожесть семян—менее l-Ь Конъюгация хромосом у
гибрида сводится к образованию 1 — 10 бивалентов Имеются не-
зависимые данные, свидетельствующие о том. что такое нару-
шение конъюгации а значительной степени обусловлено струк-
турными перестройками между родительскими видами, так что
гибридная стерильность имеет главным образом хромосомную
природу [289. 291].
Этот гибрид, несмотря их свою стерильность, дал некоторое
чисто потомков F; и последующих поколений иа гомоплондиом
уровне Значительно больше ппловниы растений в поколениях
Гз и Fe характеризовались пониженной жизнеспособностью или
стерильностью или же пдиовречеиттп и тем и другим. Растения
F. были использованы дтя закладки ряда пибредиых линий, в
которых на протяжении последующих поколений проводился от-
бор на мощность и плодовитость в сочетании с нормальным те-
17>
ЯП Vacn ши Естественны skSpwJhwir*» « •• гячмы-ягы
чениеи мейоза. Этот отбор оказался успешным в нескольких ли-
ниях, которые в конечном итоге достигли в последующих поко-
лениях полной мощности и плодовитости при нормальном тече-
нии мейоза и без изменения плоидности (289. 291К
Плодовитые гомоплоидиые растения, полученные от стериль-
ных межвидовых гибридов, скрещивали затем с родительскими
видами и между собой, для того чтобы выяснить их генетиче-
ские взаимоотношения Одна группе линий, известная под наз-
ванием «Ветвь II», оказалась как бы восстановленной формой
Cilia modocensis, соответствуя ей как по своим фенотипическим
признакам, так и по плодовитости при скрещиваниях. Ветвь II
ке служит подтверждением гипотезы рекомбинационного видо-
образования. *
Другое плодовитое гомоплондное производное, названное
«Ветвь III», унаследовало морфологические признаки родитель-
ских пилон в повой комбинации. Ветвь III была сходна с GUia
modocensis по четырем признакам, с G. malior — по трем и с
промежуточной, подобно гибридам Fi — по четырем другим при-
знакам (291). Характер ветвления и форма листьев у этой груп-
пы линий и родительских видов показаны на рнс. 20.2.
При скрещиваниях Ветви III с GUia malior. G. modocensis к
Ветвью II наблюдалась хромосомная стерильность У гибридов
Ft от скрещиваний G. malior с Ветвью III, например, конъюга-
ция хромосом была нарушена (в среднем образовывалось 13 би-
валентов). а фертильная пыльца составляла 4—18% Всхожесть
семян была ниже 1%. Таким образом Ветвь III изолирована от
родительских и сестринских форм мощным барьером стериль-
ности (291),
Поведение Ветви III по всем существенным аТпектам соот-
ветствует тому, чего следовало ожидать на основании гипотезы
рекомбинационного видообразования. Она была получена от
межвидового гибрида е высоиой хромосомной стерильностью пу-
тем чистого инбридинга. Шаг за шагам в результате отбора в
ряде последовательных поколений опа достигла полной плодо-
витости в сочетании с нормальным мейозом и без изменения
плоидности. При скрещиваниях с родительскими видами
Ветвь III дает стерильных гибридов. Она обладает собственным
особым сочетанием морфологических признаков родительских
видов, в также их факторов хромосомной стерильности.
Диалогичные экспериментальные исследования проводятся в
настоящее время на видах рода Огуга. У О saliva к у О. glaber-
rima число хромосом 2п—24. Гибриды Ft между ними высоко-
стерильны. Путем возвратных скрещиваний этих гибридов с ро-
дительскими видами можно получить дальнейшие поколения
потомков. Среди ннбредиых потомков от этих последующих
скрещиваний было несколько форм с новыми рекомбинациями
Гл №. РгыаЛшшчоаняол шОыврахмшаа **'
Рис 20.2 Характер »етв.тени! к ферма листьев у двух иигерстерильимх вм
лов Oilia в у Ветом П1 — плодовито* изолирован»»! гомовлоидмой фермы,
лронслолявей от гибрида между нишв [291]
См ’«же рис 17 в, oTaortiuiiii<B к тем же родительским видам и происхо-
дите* от ап Ветви II.
признаков родительских видов. Это позволяет предполагать ро-
ком б низ сноннос видообразование Особенности утих новых лк*
кий в отношении плодовитости пока еще не выяснены [708].
Роль блочного наследования в рекомбинационном
видообразовании
Некоторые экспериментальные данные, относящиеся к Gossy-
pium, Clarkia и Cilia, позволяют предположить, что перестроен-
ные участки хромосом у близких видов несут различные аллели
генов, оказывающих влияние на рост и мощность. У пар видов,
принадлежащих к этим и другим родам, в перестроенных участ-
ках хромосом можно установить локализацию генов, детермини-
262 ¥агуь пятая. Естестыняал габрг^изаф/ч и ее рг<у".та?ы
рующих мор4>ОЛйГНЧСС,<ъс признаки. Фактические данные, ка-
саюшнеся блочного наследования систем генов М—V, были из-
ложены в друга# моей работе [283], а краткий обзор этих дан-
ных приведен в гл. 8.
Здесь нам предстоит рассмотреть влияние на рекомбинаци-
онное видообразование генов М— V, локализованных в пере
строенных участках, по которым различаются родительские ви-
ды. Допустим, что между двумя видами, которые различаются
по блокам генов М—V, имеющим такую же протяженность, как
и структурные перестройки, происходит естественная гибриди-
зация. Допустим далее, что гибридное потомство растет и раз-
множается в местообитания одного нз родительских видок (Pi),
ио не в отличающемся по экологическим условиям местообита-
нии другого вида (Р>).
Очевидно, что в такой ситуации отбор под действием среды
будет благоприятствовать гибридным вышепенпам. несущим
аллели жизнеспособности вида Р, Но, кроме того, можно ожи-
дать, что в этой гибридной популяции действие отбора будет
направлено на сохранение целых блоков генов или. что то же
самое, перестроенных участков хромосом вида Pi в ущерб соот-
ветствующим участкам вида Р,. Таким образом, можно ожидать,
что в данном случае гомоплоидныс гибридные производные по
своей зависящей от хромосом плодовитости, критическим эколо-
гическим предпочтениям и связанным С ними морфологическим
признакам будут ревертнровать к генотипу одного из родитель-
ских видов (Pi).
Ииеино такой результат наблюдался при контролируемых
экспериментальных условиях в потомстве гибрида от екрешн-
вання Gilia malior и О. modocensis, обладавшего хромосомной
стерильностью Мы имеем в виду гомоплоидные линии, назван
ные Ветвью II. Эти линии в условиях среды, типичной для
G. modocensis. подвергали отбору на мощность, но ие на мор-
фологическое сходство с О. modocensis иля плодовитость при
скрещиваниях с последним. Тем не меиее в результате такого
отбора была получена форма, гомологичная по своим хромосо-
мам G. modocensis, плодовитая прн скрещивании с этим роди-
тельским видом и морфологически очень сходная с ним [288].
Пример с Ветвью II был рассмотрен в гл. 17 и проиллюстриро-
ван нв рис 17.8.
Из всего сказанного можно сделать вывод, что гены, опреде-
ляющие жизнеспособность и локализованные в участках, за-
тронутых перестройками, потенциально способны тормозить или
сдерживать процесс рекомбинационного видообразования. Этот
потснииа.ты1ый тормоз может начать действовать, если гибрид-
ная популяция растет в местообитании одного из родительских
видов
Га 2V wnMjWOMniir
26Т
Наиболее подходящим местом для рекомбинационного видо-
образования служит, вероятно, какое инбудь новое, измененное
или «рекомбинационное», местообитание, находящееся поблизо-
сти от гибридной популяции Такая среда может оказаться не-
обходимой в качестве местообитания для индивидуумов с новой
комбинацией перестроенных участков и может послужить фак
тором отбора, способствующим сохранению этих участков, если
ппн несут блоки генов, обладающие адаптивными свойствами.
Таким образом, следует ожидать, что рекомбинационное видо-
образование будет происходить главным образом а тех место-
обитаниях, которые по Своим экологическим характеристикам
отличаются от местообитаний родительских видов.
О роли рекомбинационного видообразования
в природе
Конечная цель проведения экспериментов по рекомбинационно-
му видообразованию заключается в том, чтобы установить осу-
ществимость этого процесса и его роль в эволюции растений
Попытаемся теперь выяснить роль рекомбинационного впдообра
зевании в природе в сэетс имеющихся экспериментальных дан
иых.
Теоретически рекомбинационное видообразование можно
сравнить с явлением амфиплоидия, о котором известно гораздо
больше Эти два процесса представляют собой два альтернатив-
ных пути от гибрида с хромосомной стерильностью к новому
птодовигому изолированному дочернему виду Как показывают
многочисленные экспериментальные и цлтотаксоночнческне дан-
ные. амфип.тондный путь наблюдается в эволюции растений
очень часто. Логично предположить, что рекомбинационное аи-
дсюбразованне также представляет собой один из обычных про-
цессов, происходящих в природе; оно. быть может примерно
стать же обычно, как и амфиплоидия, но в отличие от последней
ис оставляет следов, которые можно было бы выявить лх-тем
цнготаксономических исследований.
Эксперименты, проводившиеся на GUia.— e.-.i тег венные. в
которых формы, происходящие от высокоетерн.тытого гибрида,
были получены путем чистого инбридинга.—могут преподать
нам урок в этом отношении. В первых поколениях поптмкоп от
скрещивания G. malior н G modoc.en.sis наблюдалось значитель-
пое разрушение гибридов Эти лицин едва удавалось сохранить
в благоириятпих искусственных условиях; что же касается их
?->чо»Н,,^ИНЯ С1 ссг&-г,|*иных условиях, то оно еще менее вероятно
|2Я9). у некоторых из этих линий, которым удалось преодолеть
барьер, создаваемый разрушением гибридов, происходил, кроме
тога, полира 1 к одному из родительских видов [288|. В отличие
ЭМ Часть пятил рстесптялая шйри^имция и п л».1у=гатн
от них единственный новый дочерний вид, которому удалось воз-
никнуть, на протяжении своей долгой истории в условиях экспе-
риментальной среды оставался конституционно слабый 1291].
Эксперименты с Gilia показали, что при гибридизации между
высоко интерстеркльиыми и хорошо дифференцированными ви-
дами преодолеть барьер, создаваемый разрушением гибридов,
крайне трудно.
В прежних формулировках гипотезы рекомбинационного ви-
дообразования эффективность барьера, обусловленного разруше-
нием гибридов, не учитывалась. В теоретических и эксперимен-
тальных исследованиях этого способа видообразования все вни-
мание было сосредоточено на одном аспекте проблемы — гиб
ридной стерильности. Амфкплохдия представляет собой один
из способов, позволяющих избежать хромосомной стерильности
у межвидового гибрида, а рекомбинационное видообразование
можно рассматривать как альтернативный выход из того же
тупика. Однако при этом оставалось недостаточно ясным, поче-
му амфнплондня даст также возможность преодолеть и барьер,
создаваемый разрушением гибридов, тогда как рекомбинацион-
ное видообразование такой возможности лишено.
Наиболее подходящий вывод, к которому мы можем прийти,
исходя из имеющихся в нашем распоряжении эксперименталь-
ных данных, состоит в том, что рекомбинационное видообразо-
вание— гораздо менее обычный способ видообразования у рас-
тений, чем амфиплондия.
Экспериментальные исследования показали, однако, что в
искусственных условиях рекомбинационное видообразование
имеет место. Это в свою очередь позволяет допустить, что такой
процесс может иногда происходить и в природе.
Примером естественного вида, который, вероятно, возник и
результате рекомбинационного видообразования, служит Gilia
achilleifolia' (2л = 18). Этот диплоидный вил встречается в Ка-
лифорнии. и южной части Берегового хребта Считается, что ои
возник в результате гвбридиэации между какими-то древними
представителями групп диплоидных видов G capitata и G. ange-
lensis (273].
В пользу этой филогенетической гипотезы имеются достаточ-
но убедительпые данные. Прежде всего растения GUia achillei-
folia по морфологическим признакам, относящимся ко всем их
частям, занимают промежуточное положение между двумя ро-
дительскими видами: кроме того, окн крайне изменчивы по сво-
им признакам, и такая изменчивость выражается в форме диф-
ференциации на локальные расы. Некоторые расы G achitttifo-
lia морфологически близки к G. capitata. тогда хак другие близ-
ки к G angelensis; систематики иногда даже путали' первых с
G. capitata, а вторых с G angtlmsis [273].
t л. 20 РгммОамациопкт aupoo6fnuo«anuo 265
Два предполагаемых родительских вида в настоящее время
гибридизируют в природе » некоторых районах на юге Калифор-
нии Весьма важно, что возникшие и результате згой гибриди-
зации интрогрессивные формы морфологически сходны с Gilia
achillei/oUa (273].
Известно, что часть хромосом Gilia achilleifolla гомологич-
на. а часть иегомологичнз хромосомам членов групп видов
G capitala и О. angtUnsis [274]. Соотношение геномов всех этих
форм совместимо с гипотезой рекомбинационного пронсхожде
нкя G. aeMllti/olia. Кроме того, G. achiileifolia самосовместима.
а некоторые се расы преимущественно автогамны. Таким обра-
зом, система размножения этой формы благоприятствует закреп-
лению нового генома рекомбинационного типа.
Необходимо провести дополнительные генстико таксономиче-
ские исследования с целью увеличить число вероятных приме-
ров. Необходимы также более чистые экспериментальные ис-
следования по рекомбинационному видообразованию. Мы еше
очень далеки от того, чтобы оценить, сколь велика роль реком-
бинационного видообразования в эволюции растений.
21 Гибридные комплексы
У многих групп видов, принадлежащих к разным родом и раз-
ным семействам растений, наблюдается сходный характер из-
менчивости. Этому общему типу изменчивости присущи следую-
щие черты. Группа видов содержит многочисленные и разнооб-
разные формы, иитерграднруюшие по морфологическим призна-
кам и экологическим предпочтениям. Ввиду такой интерграда-
ции эти разные формы можно было бы считать 'иенами одного
полнтипического вида, но диапазон морфологической и экологи-
ческой изменчивости группы в целом сильно превышает ее диа-
пазоны у обычных политнпическнх видов, относящихся к тому
же роду. Кроме того, некоторые из этих форм в тех или иных
зонах симнатрического контакта остаются более или менее обо-
собленными. Интерградация между ними не похожа на ннтер-
градацкю между расами.
Таксономически такая группа является «критической» и
«грудкой». Ни одна система классификации, созданная на осно-
ве общепринятых гербарных и палевых методов, не дает воз-
можности удовлетворительно произвести обычное определение.
Если же проанализировать данную группу с применением тех
же рутинных морфологических и географических методов, ио не
с таксономической, а с популяционно-биологической точки зре-
ния, то выясняется, что она представляет собой комплекс отно-
сительно слабо выраженных видов, полувидов н{илн) мнкрови-
лов. Генетико-таксономические данные, если их удается полу-
чить. нередко подтверждают концепцию о группе взаимосвязан-
ных видов, поскольку при этом обнаруживается существование
барьеров стерильности или других репродуктивных барьеров
между некоторыми морфологически различными и енмпатриче
скнми формам».
У групп растений, обладающих такой характерной изменчи-
востью. нет ни одной общей для всех генетической системы. Они
могут состоять из диплоидных или полиплоидных популяций
или же одновременно нз тех н других. Размножение может быть
половым, почти половым (субсексуальиым) или бесполым. Раз-
личные группы растений, создающие затруднения для система-
тиков, сходны в общих чертах по характеру изменчивости, но
отнюдь не вее они сходим по типу своих генетических систем.
Существует, однако, и другой фактор, общий для всех групп
растений, обладающих описанным выше типом изменчивости.
Этот фактор — широко распространенная естественная гибриди-
зация в пределах данной группы и закрепление значительного
числа форм гибридного происхождения. В соответствии с этям
такие группы можно объединить под общим названием гибрид-
/ 4. 2/ l ийрийные ьом.ыс*
267
пых комплексов. Гибридный комплекс —это группа видов, со-
стоящая из трех или большего числа исходных предковых видов
и ил гибридов нли потомков этих гибридов, в которой в резуль-
тате естественной гибридизации морфологическая прерывистость
между исходными формами оказалась стертой [272|.
Классификация
Гибридные комплексы можно классифицировать в зависимости
от способа стабилизации воспроизведения естественных гибри-
дов млн нх потомков [272]. В одной их предыдущих глав мы пе-
речислили известные способы гибридного видообразования;
здесь мы вновь приведем этот перечень, с тем чтобы на него бы
л о удобнее ссылаться.
Известны следующие способы стабилизирующего размноже-
ния гибридов- 1) вегетативное размножение, 2) агамоспермия.
3) перманентная гетерозиготность по транслокациям. 4) перма-
нентная нечетная полиплоидия; о) амфиплоидия 6) рскомбнна-
цнопнос видообразование, 7) гибридное видообразование при
наличии внешних барьеров Эти способы создают возможность
для размножения и чистите высокогетерозиготного генотипа (1 —
5) или гомозиготного выщеленца (6—7) з условиях бесполого
(1—2) или полового (3—7) размножении.
Если процесс гибридного видообразования независимо повто-
ряется в разных линиях, так что из трех нли более исходных
видов образуется два или более гибридных вида, то возникает
гибридный комплекс. Тип гибридного комплекса соответствует
способу стабилизации воспроизведения гибридов.
1. Гомогамный комплекс Формы гибридного происхождения
размножаются половым путем с нормальным течением мейоти-
ческиго цикла. Дочерние виды гибридного происхождения ди-
плоидпы или по меньшей мере неулвоившиесм и гомиилоидные
по плюшем и ю к исходным родительским видам _
2. Полиплоидный комплекс. Формы, возникшие в результате
естественной гибридизации, представляют собой четные поди
илоиды (обычно амфнплоиды], размножающнсся_половым пу-
усы --------------------------------- --------------- ~~
3. АгампыЙ комплекс Увеличение численности гибридов пли
форм гибридного происхождения полностью или частично осу-
ществляется при помощи пеоплодотворенных семяи. Эту концеп-
цию можно расширить, с тем чтобы включить т» нее бесполые
папоротники у которых гибриды размножаются путем апога
метни.
4 Клональный комплекс Численность гибридов пли форм
гибридного происхождения увеличивается бесполым путем, при
помощи различных способов вегетативного размножения.
ЭИ Чат алгол Еиитасмаа шбралиацил и и резумгаги
5. Гетерогамный комплекс. Гибриды или формы гибридного
происхождения представляют собой перманентные гетероэиготы
по транслокациям или перманентные нечетные полиплоиды.
В гомогамных комплексах дочерние вилы возникают от роди-
тельских видов в результате рекомбинационного видообразова-
ния или гибридного видообразования при наличии внешних
барьеров В соответствии с этим гомогамный комплекс может
состоять из видов, разделенных барьером хромосомной стериль-
ности. или же из видов, плодовитых при скрещиваниях между
собой.
У двух генетических систем — перманентной гетерозиготностя
по транслокациям и перманентной нечетной полиплоидии —
имеется несколько общих черт, а именно продолжающаяся гете-
розиготность по хромосомам, комплементарные геномы в муж-
ских и женских гаметах и половое размножение, нередко само-
оплодотворение. Структурная или нуиерическая гибрндность
поддерживается благодаря половому размножению, нередко са-
мооплодотворению. из поколения в поколение. Эта две генетиче-
ские системы также дают начало гибридным комплексам сход-
ных типов. Вследствие такого сходства по ряду признаков для
обозначения создаваемых этими системами эволюционных про-
дуктов (гетерогамный микровил и гетерогамный комплекс)
можно использовать общую терминологию. Тем не менее гете-
рогамные комплексы делятся на два подтипа: комплексы, со-
держащие диплоидные гетерозяготы по транслокациям, н комп
лсксы, состоящие из нечетных полиплоидов.
Приведенная выше классификация гибридных комплексов
носит аналитический характер. Реальные гибридные комплексы,
развивающиеся в той или иной группе растений, могут содер-
жать смесь разных типов. Многие высокие и некоторые низкие
полиплоиды являются агамоснермиыми. Поэтому агамные комп-
лексы нередко накладываются иа полиплоидные комплексы,
комбинируясь с вини. Подобным же образом в разных ветвях
одной и гой же группы растений может иметь место как реком
билационпое видообразование, так и амфиплоидия. В таких слу-
чаях возникает частично гомогамный, а частично полиплоидный
гибридный комплекс. В реально существующих клональных
комплексах наряду с полиплоидией, агамоспермией или поло-
вым размножением гибридов нередко встречается вегетативное
размножение.
Роль системы размножения
Естественные гибриды образуются между видами растений, об-
ладающими любой системой размножения. Однако результат
естественной гибридизации в очень сильной степени зависит от
Гл 21 Гибриднл*с ьомпмкси
2Г>Ч
системы размножения участвующих в ней растений, и влияние
этой системы может сказаться на типе разминающегося в конеч-
ном итоге гибридного комплекса
У однолетних растений, гибриды которых располагают лишь
коротким периодом времени для размножения, гибридизация в
условиях свободного скрещивания приводит к интрогрессии, в в
условиях самооплодотворения — к амфиплондии [273]. Эти ожи-
даемые результаты очень четко подтверждаются у однолетних
гн ли А, у которых формы с перекрестным оплодотворением ха*
рактермзуются сильно выраженной интрогрессией, тогда как
автогамные формы дают амфинлондов. Таким образом в гибри-
дизирующих группах однолетних растений автогамная система
размножения способствует образованию полиплоидных комплек-
сов.
У многолетних цветковых растений, способных к ваетатив-
ному размножению, полиплоидия. как мы увидим в части шес-
той. встречается относительно часто, н. разумеется, такие расте-
ния также образуют дочерние клоны Поэтому следует ожидать,
что широкая гибридизация в группе многолетних растений, раз
мпожакнцнхея вегетативным способом, приведет к образованию
полиплоидных и клональных комплексов
У папоротников, которые нвляю»ся многолетними растения-
ми, полиплоидия гораздо чаше встречается в равнослоровых
группах. чем в разноспоровых [442]. По-видимому, у раинпспо-
ровых значительное место занимает симооплодотворснне [442|.
Таким образом, следует ожидать, что равноспоровозть и само
оплодотворение способствуют развитию полиплоидных комплек-
сов в гибридизирующих группах папоротников.
Как было птмечсно в предыдущей главе, самооплодотворение
у однолетних и у миоголеттгх растений служит фактором, спо-
собствующим рекомбинационному видообразованию. Таким об-
разом, следует ожидать, что гомогамные комплексы, состоящие
пз видов н микровндов. характеризующихся хромосомной сте-
рильностью. при наличии подходящих местообитаний будут раз-
виваться в гибридизирующие группы преимущественно а итогам
пых растений.
У большинства агамоспермпых растений их предковые фор
мы. размножающиеся половым путем, бывают двудомными, од.
иодпмными или самоиесовмсстнмымн [317. 318]. Гусифсоп [317,
318] отметил, что растения, обладающие той или*иной из >гих
систем размножения, в особеняостм гибридные растении, бхдут
иметь очень небольшие шансы продолжить свое существование
путем полового размножения в изолированных насаждениях,
тогда как агамоспермия и веге га тинное размножение — относи-
тельно легкие пути, позволяющие выйти пз тупика стерильности.
Облигатное перекрестное опыление у гибридов представляет го-
tTO Чисть шияя. Етсгаеялал м ft рпумтагы
бой, таким образом, фактор, благоприятствующий разлитию
агамкых и клональных комплексов.
Автогамия преобладает у перманентных гетерозигот по транс-
локациям группы Oenothera biennie и, по-видимому, у перма-
нентных нечетных полиплоидов группы Rosa canina. Каждая из
этих групп возникла от предковых форм, для которых были ха-
рактерны нормальный мейоз и свободное опыление. В каждой
группе самооплодотворение составляет один иэ компонентов ге-
нетической системы, обеспечивающей поддержание перманент-
ной гибрндиостк. В экологическом отношении формы гибридного
происхождения особенно успешно развиваются в сорных или
окультуренных местообитаниях; так обстоит дело, в частности,
в группе Oenothera.
Не исключено, что предковые популяции обладали совершен-
но необходимыми им гетерозисными свойствами и поддерживали
эти свойства при помощи непрерывного перекрестного опыления.
В таком случае при заселении нового местообитания, где не бы-
ло подходящих опылителей, у них развилась в качестве компен-
сации система перманентной гетероэиготиости, основанная ил
самооплодотворении при участии неодинаковых гамет [144, 146].
Согласно этой гипотезе, главным стимулом, вызвавшим точки
цню по крайней мере некоторых гетерогамных комплексон, была
продолжавшаяся зависимость популяций от гетерозиса на фоне
обеднения среды опылителями, происходившее одновременно из-
менение системы размножения — от свободного опыления к са-
мооплодотворению — явилось частью механизма, обеспечиваю-
щего подгонку всей генетической системы к новым условиям су-
ществования.
Гомогамный комплекс
Гомогамный комплекс представляет собой гибридный комплекс,
в котором новые производные виды возникли в результате про-
цессов гибридного видообразования, не сопряженных с резки-
ми изменениями в генетической системе. Нам уже известны про-
цессы рекомбинационного видообразования прн наличии барь-
еров хромосомной стерильности и гибридного видообразования
при наличии внешних барьеров. Третий возможный процесс, ко-
торым следует заняться в будущих исследованиях, —формиро-
вание новых, контролируемых генами барьеров стерильности и
несовместимости в результате рекомбинации. В зависимости от
того, какой из этих различных процессов ответствен за видооб-
разование, возникающие виды могут быть изолированы Один от
другого и от родительских видов барьерами хромосомной сте-
рильности. внешними барьерами, геняой стерильностью и кссов-
Гл Я ГибрыОпыг «ожалдеж
271
местнмостъю или же одновременно и внутренними л внешними
барьерами.
В любом случае производные виды остаются на том же уров-
не плоидности. что и предковые виды. Кроме того, у гибридных
производных сохраняются нормальный половой цикл и нормаль-
ное течение мейоза Если родительские виды представляют со-
бой диплондов, размножающихся половым путем, то нх дочерние
виды —также дипломам с половым размножением.
В гибридных комплексах всех других типов производные ви-
ды можно отличать от родительских ио их цитогене immcckiim приз-
накам или системе размножения. Однако и гомогамных комп-
лексах это невозможна. поскольку гибридные производные со-
храняют исходную генетическую систему предковых видов.
Вследствие этого анализировать гомогамные комплексы очень
трудно, хотя и возможно. Гомогамный комплекс легко может
остаться незамеченным, и многие из ннх, вероятно, так и нс бы-
ли распознаны. Этим, вероятно, и объясняется немногочислен-
ность известных примеров.
Гомогамный комплекс облиствейиостебел иных
гнлий
Облнсгвенносгебельные гмлии (секция Cilia рода Cilia) группа
стерильных при скрещиваниях друг с другом однолетних травя-
нистых растений, шнооко изучавшихся как таксономически, так
и цитогенетически Большинство видов этой группы диплом
дны (2л-18). а некоторые — геграплондны (2*«3б) Вес виды
размножаются половым путем, а системы размножения варьнру
ют от облигатного перекрестного опыления до преобладающего
инбридинга (272, 274].
Взаимоотношения между Cilia capitata и G tricolor в этой
группе были описаны в гл 4 как пример пары хороших биологи-
ческих видов, В гл. 9 был дан обзор плодовитости при скрещи-
ваниях в пределах згой секции в целом, чтобы пронллюстриро
вать некий обший тип взаимоотношений, обычных для одиолст*
них растений, —тип Madia. Здесь будет описана сетчатая фило-
гения этой группы как пример гомогамного комплекса
Мы рассмотрим виды облнственностебельиыч гилпй, произ-
растающих п Калифорнии— центре распространения и вероят-
ном центре происхождения этой секции, а точнее вилы, произ-
растающие в материковой Калифорнии, без близлежащих остро-
вов Эти виды перечислены на рис 21.1 Все инн диплоидны, за
исключением G cliiwrum (2n-4x-36)
Облиствешюстебельные гилин деЛ1Гтся на три группы видов,
различающиеся по морфологическим признакам и экологическим
предпочтениям- J) группа GUia tricolor, в которую входят G. tri-
Г2
Часть пятая Елтесгмтал а к результаты
Pile 21 I Филогения сблпствеяностебельных тихий а Кзлнф?рн/л (272).
Спаошянми ливнями вокамны основные навравлсявя лнвергянцян; преры-
в«1ы«« — вым в аоптляцяи шбрндного вржхожаскня. В« пн формы —
лиялонды. м меключвовем твтрааловянаго G еНоогшп.
color к G. angetensis, 2) группа G mlllefoliaia и родственные
формы: 3) группа G. capitata. Зги три группы ендов отражают
главные ветви в филогении рассматриваемой секции (рис. 21.1).
Крайние члены группы — G. capitata и G. tricolor, противополож-
ные признаки которых были описаны в гл 4 (см. рис 4.2 и 4.3).
Giiie capitata представляет собой сложный полнтипический
вид, состоящий из необычайно разнообразных и многочисленных
форм, которые распадаются на две крайние группы и гибридов
между ними, занимающих промежуточное положение. Крайние
формы G. capttata дивергяровали в прошлом до видового уровня
и все еще сохраняют, подобно видан, способность оставаться
обособленными в некоторых зонах симпатричесжого контакта.
В других областях, однако, крайние формы связаны между со-
бой расами гибридного происхождения [266, 263].
Gilia acliilleifolia по морфологическим и экологическим приз-
накам запинает промежуточное положение между определенны-
ми ветвями групп G. capitata и О. tricolor. Кроме того, у этого
вида наблюдается необычайно сильная изменчивость в направ-
лении обеих предполагаемых родительских групп. Некоторые из
Гл 21 Гвбеайпы ииишкы
гп
таких вариантов соответствуют естественным гибридам, извест-
ным между этими двумя группами видов. Все имеющиеся дан-
ные свидетельствуют о гибридном происхождении G achillrifo-
На. показанном на рве. 21.1 [274].
Цитогенетические и морфологические данные с достаточным
основанием позволяют предположить, что Cilla clivorum (2п—
= 36)—амфиплонл. происходящий от G. achilleilolia и G. mil-
ItfoluUa. как это показано на рис. 21.1 (271, 274(.
Популяции, промежуточные по морфологическим признакам
между Gilia capitata и G. angelcnsis, встречаются в каньоне
Сайта Ана, на юге Калифорнии, а популяции, морфологически
промежуточные между G. capitata и G. miUefoliata. — на мысе
Мсндоснпо, на севере Калифорнии. Таксономический статус этих
популяций неясен, и на рис. 21.1 они просто зафиксированы как
таковые
Рис. 21.1 позволяет понять, как в результате гибридизации
ветвящаяся филогенетическая схема превращайся в сетчатую,
без изменения генетических систем у.форм гибридного происхож-
дения. На ней показаны также последовательные уровни гибри-
дизации: виды, которые сами возникли в результате гибридиза-
ции, становятся предками более позднего поколения видов и по-
пуляций гибридного происхождения.
Еще иснэучениые гомогамные комплексы, несомненно, суще-
ствуют у Pinus, Qutrcut к у многих других групп растений, в ко-
торых имеются диплоидные или гомоплондпые виды, иногда
гибридизирующие друг с другом. Филогенетический анализ дру-
гих примеров таких комплексов заслуживает внимания в качест-
ве предмета дальнейших исследований.
1Й-45
Часть шестая
Полиплоидия
22 Полиплоидия:
распространение и частота
Исторический очерк
Полиплоидией называют особое арифметическое соотношение
между числом хромосом у родственных организмов, различаю-
щихся по этому числу. В простейшем случае у полиплоидного
организма число хромосом вдвое больше, чем у какой-либо род-
ственной формы. В других случаях число хромосом у полиплоида
увеличено ие вдвое по сравнению с гаплоидным их числом
у одной из родственных форм, а в большее число раз, например
в 3 или 6 (Зх или 6х). Или, наконец, число хромосом у поли-
плоида равно сумме нх чисел у двух родственных форм, обла-
дающих меньшим числом хромосом, как в ряде 2Л-10, 12, 22.
Во всех этих случаях у полиплоидного организма наблюдается
более высокое число хромосом в результате прибавления доба-
вочных целых хромосомных наборов, имеющихся у одного или
нескольких предковых организмов. Короче говоря, полиплои-
дия—это наличие у одного организма трех или большего числа
хромосомных наборов
Явление полиплоидия было открыто в начале нашего иека.
когда цитогенетика и цитотаксономия растений находились на
стадии первых ранних поисков. Главную роль в его открытии
сыграли Винклер [902], предложивший термин полиплоидия, и
Винге [898], частично объяснивший сущность этого явления.
Винклер [902] изучал вегетативные прививки и химеры у
Solatium. Он обнаружил, что из каллуса, развивающегося на
стебле на поверхности разреза, может регенерировать новый
побег, причем из стеблей диплоидных S. nigrum развивались
таким образом тетраплоидные растения. Винклер предположил,
что такие тетрапл с идиые растения возникают в результате со-
матического удвоения в первоначально диплоидной ткани.
Винте [898] сравнивал родственные виды, принадлежащие к
одному и тому же ролу, по числу хромосом. Он использовал ре-
зультаты собственных подсчетов хромосом у Chenopodium, а
также опубликованные ранее данные подсчетов Тахары [820] на
Chrysanthemum При этом Вииге обнаружил, что число хромо-
Гл п Полиплоидия распространю* и частота Гб
сом у разных видов образует упорядоченные ряды. У разных
видов Chenopodium 2п-18 и 36, а у разных видов Chrysanthe-
mum 2л-18, 36. М, 72 и 90. Таким образом, у близких видов
число хромосом оказалось кратным некоторому общему основ-
ному числу,
Для объяснения этих наблюдении Винге [698] предложил
оказавшуюся крайне плодотворной гипотезу удвоения числа хро-
мосом у межвидовых гибридов. По его мнению, полиплоидный
ряд возникает в результате следующей последовательности со-
бытий. Межвидовая |ибридизаиия дает начало гибридным рас-
тениям. которые стерильны вследствие нарушения у них конъ-
югации хромосом. Иногда у гибрида нлн у его потомков проис-
ходит спонтанное удвоение числа хромосом, и они становятся
плодовитыми Затем такие плодовитые полиплоидные растении
могут размножиться н превратиться в новый константный вид.
Таким образом, два диплоидных вида путем гибридизации мо-
гут лать качало новому кграплоидному виду; в результате гиб-
ридизации между диплоиюм и тетраглон.'лм может возникнуть
гекеавлоидпый вид. а при гибридизации между другими форма-
ми — полиплоиды с более высокой плоидпостью (898J.
Гипотеза Винге вскоре была подтверждена экспериментально
путем искусственной межвидовой гибридизации у Nicotiana,
Rophanohrassica н Gateopsis (106. 127, 596]. Классические приме-
ры Raphanohrassica и Galeopsis были описаны в гл 19
Совершенно очевидно, что между яяг1спиямн полиплоидии, с
которыми имел дело Винклер и которыми занимались Винге и
его последователи, имеются существенные различия В керном
случае зто внутривидовая негибридкаи поллплоидия. а во вто-
ром — полиплоидия, связанная с межвидовой гибридизацией
Полиплоидия первого типа возникает в результате умножения
одного и тот же отбора хромосом, а полиплоидия второго ти-
па— к результате удвоения структурно несходных хромосомных
наборов у межвидового гибрида Эти различия стали яснее бла-
годаря терминам автополиплондня и алл ополи пл он дня. введен-
ным Кихарой и Они (438]. Термин амфиллоидия — удобный см
мои им термина аллополмплоидия [
Масштабы проблемы и ее суть
Изучение полиплоидии необычайно расширилось со времени
первых исследований, и эти обусловило появление огромного
количества литературы Анализ плонднпстн играет важнупо роль
в таких разнообразных областях исследований, как цитогенетн
на, физиология, селекция растений, иитотансомомия н биогеогра-
фия Совершенно очевидно, что рассмотреть здесь проблему но-
пиплоиднн в ее полком объеме невозможно
ЙГ6 Часто шестая. Полиплоид ил
Хорошие обзоры различных аспектов полиплоидии составили
Мюитцинг (599. 600. 602], Дарлингтон 1139. 148], Клаузен. Кек
и Хиси [102], Деве Л. и Лёве Д. [528, 5321 Стеббинс [780. 787].
Тышлер (831]. Ригер [684]. Готтшалк [258] и Люкс [505].
В этой книге основное впимаиие уделено эволюционным ас-
пектам полиплоидия у высших растений. Для того чтобы выпол-
нить эту задачу, необходимы лишь некоторые данные, относя-
щиеся к различным областям науки, в которых занимаются
полиплоидией. Поэтому мы изложим здесь выборочные цитогак-
сономичесхне, цитогенетические я биогеографические данные.
При отборе этих данных мы руководствовались их важностью
для стоящего перед нами вопроса.
В ближайших разделах этой главы мы рассмотрим данные,
которые показывают, что полиплоидия действительно играла
очень важную роль в эволюции высших растений, особенно по-
крытосеменных и папоротников. В пользу такого заключения сви-
детельствуют две группы фактов: значительная часть видев выс-
ших растений — полиплоиды; различные роды достигают высо-
ких уровней плоидности- Совершенно очевидно, что в связи с
этим возникает вопрос: чем объяснить наблюдаемые факты?
В чем причины такого успеха полиплоидного состояния п эволю-
ции растений? Мм попытаемся выяснить этот вопрос в после-
дующих главах.
Полиплоидные ряды
В целях иллюстрации мы перечислим несколько хорошо извест-
ных примеров полиплоидии у покрытосеменных. Примеры эти
сгруппированы по типу полиплоидных рядов.
Простейший случай — род, секция или группа видов, в кото-
рую входят диплонды и тетраплоиды. Классическими примера-
ми служат Dahlia. Gossypium и группа Nicotiana tabacum.
У нескольких диплоидных Mnot'Dahlia 2п-32. тогда как са-
довая, или изменчивая, георгина (D vaiiabilis) — тетрэплоид
(2П-64) (подробности и библиографию см. (151J). Многочис-
ленные дикорастущие виды Gossypium и выращиваемые в Ста-
ром Свете G. arboteum и G. keroaceum — диплоиды. тогда как
выращиваемые в Новом Свете хлопчатники, принадлежащие к
видам О. hirsuium и G. barbadense,—тетраплоиды. Число хро-
мосом равно у ннх соответственно 2л=26 и 2п—52, а основное
число х=13 [391]. Группа Nicotiana tabacum (х= 12) содержит
ряд диплоидных видов (2п-24). в том числе N. glutinosa. а так-
же тетраплондный (2п=48) настоящий табак. У. tabacum [250].
Следующий тип — просто продолжение предыдущего У чле-
«оп полиплоидных рядов этого типа основное число хромосом
Гл 22 Полиплоидия; распространения « мостста 277
Рис 221 Гкммиоадше р*ды У САгяа»и*лпшп (г-9) (6О?|
П«мая метафаза чеПозя а млтерп:|сжн\ клегкая пильаы Л — С mankinol
fix. и—9). В —С. iadkum Их. л—18) B — C /а^иили (fix. a-JT). Г—
C огла/ия (fix. л—3fi) Д — C. nxxomwa (IOi. n—15). E - C. patificum (lOx.
e-45).
увеличивается в четкое число раз, т. е. равно 2х. 4х. 6х. 8л.
Примером такого ряда, достигающего гексаплокдиого уровня,
служат вилы рода TrUicum с числом хромосом 2л—14, 28 и 42
при основном числе х—7. Более длинный полиплоидный ряд об-
наружен у Chrysanthemum (х=9), что уже отмечалось ранее.
В этом роде встречаются формы с числами хромосом, ранными
2л. 4х. 6х. 8х и 10х (рис. 22,1). и даже более высокими Еще
длиннее полиплоидный ряд у Potentilla (х = 7). содержащий фор-
мы с числами хромосом, кратными всем четным числам — от 2х
до 16х (фактические данные к библиографию см. [181]).
Полиплоидные ряды, в которые входят формы с числами хро-
мосом, кратными как четным, так и мечетным числам, можно
проиллюстрировать па примерах Rosa и Crepis У диплоидных
роз 2п—14. В группе Rosa canina есть, кроме того, тетраплоид
иые. пеитаплокдные и гексаилоидиые формы с числом хромо-
сом 2п = 28, 35 и 42 (см [151]) В группе Crepis occidentals
(х=|1) полиплоидный ряд длиннее: в исто входят формы 2х,
Зх, 4х, 5х, 7х я 8х [36]. Нечетные полиплоидные виды к этих и
S.’B Часто шатал. ПолшыачЛи
других подобных группах размножаются бесполым иля субеек-
суальчыми способами.
У дибазических полиплоидов число хромосом равно сумме
двух анеуплоидных диплоидных чисел [144]. Примером, относя-
щимся к роду Brassica, служит тетра плои дный ви д В. парит
(2л=38), происходящий от В. oleracea (2л=18) и В. campestris
(2л—20) [102. 848]. Другой дибазическнй полиплоидный ряд об-
наружен в одной ил групп рода Clarkia с гаиетным числом хро-
мосом л=8, 9, 17 н 26. С. prostrata (2л=6х-52), вероятно, про-
изошел от С. daoyi (2п=4х—34) и С speciosa (2л-2х-18):
С. davyi в свою очередь — тетраплоидиое производное двух
диплоидных видов с 2л—16 и 2л—18 соответственно [496].
Ядра полиплоидов содержат много дуплицированного гене-
тического материала, поэтому они часто могут выдержать поте-
рю одной или нескольких пар хромосом. В результате получа-
ются модифицированные полиплоидные ряды, приводящие в ко-
нечном итоге к тому, что Дарлингтон назвал полиплоидным
сбросом (polyploid drop). Такого рода измененный ряд обнару-
жен у Hmperis (СгосИгтае), где у разных видов гаметпое чис-
ло п-7, 14, 13 и 12 (144. 148. 151].
У папоротников встречаются полиплоидные ряды, в которых
основное число хромосом х=37, 41 или другим столь же высо-
ким числам. Например, в группе Polypodium outpace (х—37)
есть диплоидные, тетраплоидиые и гекса плоили ые формы с
2п-74. 148 к 222. В основе другого полиплоидного ряда у Polys-
lichum лежит х=41 В таких случаях может оказаться, что ос-
новное число, выведенное из существующего полиплоидного ря-
да, иа самом деле не соответствует истинному основному числу
в филогенетическом смысле. Существующие в настоящее время
диплоидные члены ряда представляют собой, возможно, давно
возникшие полиплоиды из какого-либо более раннего эволюцион-
ного цикла [554].
Аналогичные рассуждения справедливы и в отношении неко-
торых сравнительно высоких основных чисел, обнаруженных у
других покрытосеменных. В трибе Pomoideae' сем. Rosaceae
основное число х=|7. В других трибах этого семейства основ-
ные числа хромосои не выходят за пределы, обычные для дни-
лендов: Rosoidcae (л—7), Prunoideae (х—8) и Spiraeoideae
(х—9). На основе этих данных Дарлингтон высказал предполо-
жение, что триба Pomoideae представляет собой модифицирован-
ное тетраплоидиое производное более примитивных представите-
лей Rosaceae с 7 парами хромосом [138]. Позднее Стеббинс
[780]. исходя из морфологических и цитологических признаков,
предположил, что Pomoideae (х=17)—это, вполне возможно, ал-
1 Приильнее Maloddeae. — Прия, pop.
Г.» 22 Полиплоидия (i^kdol гранение в чаггота
279
лотетра ллоиды. возникшие в результате гибридизации Prunoi*
deae (г—в) и Spiraeoideac (х —9)1.
Возможно, что число хромосом х—13 у Gossypium — вторич-
ное основное число. первоначальным основным числом у этого
рода было л —7. Такое основное число характерно и для многих
других родов Malvaceae. От него в результате удвоения и поли-
плоидного сброса могли произойти состоящие из 13 пар хромо-
сомные наборы ныне существующих диплоидных видов Gossy-
pium [832].
Распространение полиплоидии
в разных таксономических группах
Полиплоидия широко распространена среди покрытосеменных.
Ока часто встречается у представителей Ro^aceae, Compos Нас.
Rubiaceac, Iridaceae. Gramineac» и многих других семейств.
Определенно, хотя и относительно редко, она всгрсчастся в та-
ких семействах, как Cacsalpinaceac, Paswfloraceae и Po'.cmonia-
сеас. В ряде других семейств, например у Fagaceae и Berberida-
сеае, полиплоидные формы существуют, но они редки.
Стеббинс [780] обратил внимание на межродовые различия
частоты полиплоидия в нескольких семействах покрытосемен-
ных Так. в сем. Salicaceae полиплоидия обычна у Salix, но ред-
ка встречается у Populus. В сем Betulaceae полиплоидные фар
мы обычны у Belula, ни ОТМХтЛЫЮ редки у Дйш Полиплои-
дия часто встречается у Thaltetrum, но редко —у Aquilegia (сем.
Ranunculaceae), часто — у Ttdipa, но редко — у I.ilium (Lilia-
ceae) [780].
У голосеменных полиплоидия возникает редко и спорадиче-
ски [433, 831]. У гинкго и саговников она неизвестна. Сообща-
лось о тетранлонлки у Gnctum и Welwitsthia. но в настоящее
время ш сообщения считаются сомнительными [159]. У Ephed-
ra, однако, тетраплопдия встречается часто.
Средн хвойных один из очень немногих подлинных естествен-
ных полиплоидов — эти Sequoia semperuirens (Taxodiaceae)
<2я=бх—66); близкий к нему род Metasequoia — диплонт
(2ч — 22) Другой пример — FUzroya cupressoides (CupresSflCCae)
с 2л-4л —44 [326]. У hmipents известны триплопдиые и тетра
плоили ые культивары (см. гл. 34).
У Pteridopbyta в отличие от голосеменных полиплоидия рас-
пространена очень широко и достигает самых высоких уровней,
* Имей точим ipcMvt грмлегжижм.-п.ч В II Гладком (см ее статью
«О происхождении похссггЬгтпа МакНЗеае, Ботангх. журя, 1972, S7, 42—49).
По мнению Л. Кр^нквжтг |А CronqulsL eAn Intererated Svetin о! ClasjiH
cation of Flnwcrinff Plan’s». Columbia Univ. Pres*. NY. 1981). гномы Глаз-
ковой ближе к ястим — ре*}.
180 Часть метал. Полиплоидия
известных в царстве растение. Большинство Polypodiaceae—вы-
сокие полиплоиды. Полиплоидия обнаружена у обоих родов Psi-
lotalei — Psilotum и Tmesipleris. Вилы Equisetum гомоплондиы
с очень высохни числом хромосом (2пя216). что свидетельст-
вует о давно возникшей полиплоидии. Среди Lycopodia les высо-
кий уровень плоидиости встречается у Lycopodium к Isoetes, по
Selaginella' сохраняет низкое диплоидное число хромосом
(2л-18). Хорошие сводки данных по числу хромосом у Ptery-
dophyta составили Ментон (554] и Лёве и др. [533].
Средн Bryophyta полиплоидии очень часто встречается у
мхов, ио довольно редко—у печеночников. Полиплоидия также
хорошо известна у зеленых водорослей и хароаых, но необычна
у бурых и красных водорослей (505].
В царстве грибов и в царстве животных полиплоиды крайне
!>еши. Обзор данных по грибам содержится в работе Люиса
505]. Обзоры по полиплоидии у животных см. Бунгенберг [75].
'аЛт (893, 894 ] и Люис [505].
Средн беспозвоночных возможные случаи полиплоидии обна-
ружены у Lumbridus н других дождевых черней [75, 588). Наи-
более хорошо известные примеры относятся к членистоногим-
Artrmia salina (Crustacea), Trichoniscus elicabethae (Isopoda),
Otiorrhunchus app. (Curculionidae, Coleoplera), Solenobia triquet-
rella (Psychidae, Lepidoptera) и OMhiph.Ua polystigma (Cha-
maemylldae, Diptera) [76, 517, 769].
Среди позвоночных полиплоидия обнаружена у некоторых
рыб (713, 714). Что касается полиплоидии у других групп рыб,
например Salmonldae, то единого мнения по этому вопросу не
существует [713. 714, 888J. Несколько случаев полиплоидии из-
вестно у амфибий в ящериц, в частности у ящериц рода Chemi-
dophoras [63)
Таким образом, полиплоидия спорадически встречается в
царствах животных и грибов, но характерна она для царства
растений.
Диапазон хромосомных чисел
Число хромосом у покрытосеменных колеблется от 2п—4 до
Ип зв 500 И то и другое крайние значения обнаружены у дву-
дольных: низкое число у naplopappus gracilis (2л —4). а высо-
кое—у одного иа видов Kalanchoe (2 л as 500) (40. 395]. Другой
высокий полиплоид среди двудольных — Buddleja coloilei, вид.
' В современных системах эти три рсха откосят к трем ииостоятельаыи
лоркляам — Lpeopaiiales. SelaqineUales a Iwiaies. — Приял ред.
Гл ?? Полипмидия- /нклдотрояспгмг а частота
у которого 2н«300 (CM. (loll). У ОДНОДОЛЬНЫХ' число хромо*
сом колеблется от 2л*- 6 до 2л=22б.
Часто i а распределения видов покрытосеменных с разным
гаплоидным числом хромосом показана на рис. 22 2 Приведен-
ные но этом рисунке кривые построены на осмоое выборки из
17 138 видео, перечкелелных в «Хромосомном атласе» [149. 151].
Анализ этой выборки и сведение полученных данных в таблицы
было проведено д-ром Хдуордом Латимером и мною в 1951 г.
В настоящее время эти таблицы можно было бы значительно
расширить за счет многочисленных новых подсчетов хромосом,
проведенных начиная с середины пятидесятых готов и собран-
ных воедино в сводках Федорова [210] н Мура [583]. Однако вы-
борка. пл которой построены кривые на рис 22 2, достаючно
велика и репрезентативна. так что зтот рисунок хороши отра-
жает главные статистические особенности числа хромосом у
цветковых растений
Как показывает рис. 22.2, существуют тысячи видов покры-
тосеменных с 14—1о и более парами хромосом. Болес высокие
числа образуют почти непрерывный анеуплондный ряд от л —14
Ло н=85. это1 ряд тянется далее с многочисленными разрывами
до п-154. а затем каблюдастся скачок к п — 250 У небплъгипго
числа видов, как и можно было ожижать, обнаружены еще бо-
лее высокие хромосомные числа.
Напяысшее число хромосом в царстве растений характерно
для папоротников и близких к ним форм [521, 522, 533, 554]. Со-
матические клетки Tmesipteris tannensis содержат свыше 400
хромосом [5541. Еще выше число хромосом у Ophioglossum По
диплоидный ряд у этого рода начинается с‘2л»240. характер-
ного для 7 видов У О vulgotum и О thermal? 2п —4Й0: у О. аю-
ricum 2л ••720 (рнс.22 3). У О nipponicum 2п —960. а наивысшее
число хромосом. 2л—1260, обнаружено у О reticuiatum (резуль-
таты некоторых новых подсчетов я ссылки на прежние публика-
ции по Ophiogtosium см [521. 522J)
Диапазон уровней плоидности
Обращаясь от обсуждения диапазона по хромосомным числам
к обсуждению диапазона го уровням плоидности, мы нередко
мемся с сгр •. осложнением. Хромосомы можно
сосчитать. Что же касается уровня плоидьпсти. го во многих
группах растений его можно точно определив метолом сравне-
1 г г»гхи опплопмпк ндпч» ч чг пиело \poMMvM (2л—4) обвару«ено
у №Ье/?Ге1Л|£*ы (Uacwn Н Н Ьпхоаскнч 3 В. Бетаин* «урн, ВО,
V 9 I9fir»l н v Cn/jicrf.-iih, 1-<т-,'.оГслг (Соьллсасчм А П. Пробатпоа II С.,
bojiiHH* жури . 12 , АЬ 2 1Ч7Г) —
282 Часть метал Полиплоиды»
мня, однако во многих других группах его нельзя определить не-
посредственно, а можно лишь оценить, исходя из некоего гипо-
тетического основного числа [1.39, 8.32].
Если род состоит из видов с числом хромосом 2п = 14, 28, 42
и 56. то нетрудно установить, что он содержат тетраплоиды. тек-
саплоиды октоплоиды. Сомнения возникают в том случае, если
род содержит только формы с 2л-28 и 2п = 56. Казалось бы.
носителей такого числа хромосом следует считать соответствен-
но шплондами н тетраплоидамн, но подобное заключение мо-
жет быть ошибочным; более вероятно, что это тетраплоиды н
окголлоиды. ио и такой вывод остается всего лишь гипотезой.
Гл 22 Помиипидил цвлп/олграченм в частота 2*3
Тем ие менее и качестве основы для определения уровня плоид-
иости или частоты полиплоидов в данной группе растений гипо-
теза. которая, возможно, верив, лучик, чем факт, который, воз-
можно. неверен.
По данным Ментон [554], вес 13 видов Equisetum, принад-
лежащие к двум секциям этого рода, имеют 2л яг 216 Решив
иа основе одного этого факта, что у Equisetum полиплоидия от-
сутствует. мы допустили бы ошибку. На самом деле все виды
Equtsttum находятся. по видимому, на одном и том же высоком
уровне плотности. Надеяться иа то, что когда-нибудь будет
установлено основное число хромосом у этого таксономически
идол крова иного и гомоплондиого рода, мы ие можем и. следо-
вательно. не можем оценить этот уровень плоидностн [354].
Каких уровней достигает полиплоидия у покрытосеменных?
Начнем рассмотрение этого вопроса с перечисления некоторых
родов покрытосеменных, содержащих высокие полиплоиды. у
которых основное число хромосом известно по непосредствен-
ным цитотаксономнческим данным, и поэтому верхний уровень
плоидностн можно определить с достаточной степенью достовер-
ности.
Чаелб
Рис. 222 Частота расврехслеьИЯ 17134 яплоо Двудольных по гаплояхному
числу хромосом [282]
Лаб Распрехелв-не в диапазоне от в-2 хо п-BI в « Г Распрсхе-телп
в диапазоне от л-37 до о—170 Еще у четмрсх ьихот, которыа на графиках
ж показаны. «вело хроиотоя лежит и пределах от п-|32 до л-750 4 я
В — олиохольаме, £ к Г — дмулоть'чзе
Vfirr, wKfiw Полиплоидия
Рис 223 Мкт,р»<нг,йя клетка споры OfMogtouum aoortrum примерно е
ЭКО Оиоаленпми [г,22|
Полиплоидный ряд у Potentilla (л—7) имеет диапазон от
2п-14 до 2п—112 Potentilla haematochroa, у которой 2га—112,
представляет собой 16-плокл. Полиплоидный ряд Chrysanthe-
mum (л=9) содержит формы с числом хромосом, кратным всем
четным числам от 2х до 10х, а затем, после некоторого разры-
ва. — формы с такими высокими числами, как 2л—160 и 198.
Ch. lacustre, у которого 2п=198, по-видимом у, представляет со-
бой 22-плокл. В одной из групп рода Seneao с основным числом
хромосом х=5 имеются виды, находящиеся на нижнем я сред-
нем уровнях плоидиости—от 2х до 16х. У 5. roberti-friesii, от-
носящегося к этой группе, 2п%180, т. е. он представляет собой
примерно 36-плонд (фактические данные н библиографию см.
По сообщению Хзйра н Бейцеяберга [328]. у Роа litorosa из
Новой Зеландии 2/1=263—265 Известно, что у Роа основное
число т-7. Следовательно, Р. litorosa — 38-плонд. у которого
Гл 21 По шплочОил яыплжтявхлше а шготтп 28i
произошел полиплоидный сброс Это ттаттвысшттй уровень плоил
ocni. известил у однодольных [328].
Buddltia I Loganiaceac) и Katanchoe (Craxsjlaceae) служат
примерами родов покрытосеменных с очень высоким числом хро-
мосом, у которых основное число, а следовательно, н уровень
плоидности можно определить лишь косвенным путем. В роле
Buddleja есть виды с числом хромосом 2х. 4х и Gx. если считать
основным числом х —19. В таком случае В colvilel с 2naj300 —
примерно 16-плопд. Возможно, однако, что х—19 у Buddleja и
близких к иему родов представляет собой вторичное основное
число давнего тетраплоидного происхождения. Тогда В colvilei
уже не 16-илоид, а 32-нлоид. У представителей рода Kalanchor
(Crassulaccae) преобладающее основное число х—17. Прн этом
виды Kulanchoe с $й»500 следует считать примерно 30-пловда-
мп с анеу плондными отклонения и я. Но сели, что представляется
вероятным, х = 17 потучеко от одной из старых тстраилоилвых
фирм, то приведенную оценку уровня плоттдпости следует
удвоить. Kalanchoe, по-акдимому, достигает наивысшего уровня
плоидности— примерно 6Ох (фактические данные и библиогра-
фию см. [151]).
У папоротников м близких к нам групп фактические или иэ-
вестпые основные числа как уже отмечалось, в общем высокие
Ментон [554] Приводят следующие основные числа для четырех
родов Polypodiaceac Лгр/етит (X —36), Polypodium (х—37).
Dryoptrris (х=4|). Poli/itichum (х—41). В каждом из этих ро.
дов есть полиплоидный ряд с указанным основным числом.
Так, например, в группе Polypodium vulgarc (х—37) имеются
диплонды, тетраплоиды и тскеаплоиды; у последних 2п-222
Однако нынешние основные числа почти наверное соответствую г
полиплоидным числам далекого прошлого Исходные. или пер-
воначальные, основные числа у этих родов папоропшкои оста-
ются неизвестными Можно, однако, без большого риска допус-
тить. что современные полиплоидные папоротники находятся на
достаточно высоких уровнгх лдоттдтюетн [534].
Ophioglossum (Opliiog ossaceae) образует полиплоидный ряд.
начипающиПся с 2х-2к —240 и доходящий через 1х. 6х и 6х до
более чем 10х у О reticulation. у которого 2л —1260 Было выска
заио правдоподобное предположение, что у Opkioglvssum пекти-
нов, тми первоначальное, основное число 15. Из этого сле-
дует. чтп современные «диплоидные» виды — 16 илоиды. а
О reticulatum — 84 п-топд [521, 613].
Леван [467J изучат жпзнеспособность различных ттскуссгтки
иых полиплоидных форм Phleum pratense (х = 7) Он установил,
что II-плотные фхтрзты т-птофеевки. содержащие 77 хромосом,
жизнеспособны, тогда как у 12 плоидов с 84 хромосомами жиз-
неспособность сильно понижена В этом случае пороговое для
2ВС Часть жвстая. Поляплоадия
жизнеспособности число хромосом лежит в пределах между 77
84. У растений с 12х и 13л митоз был нарушен; это позволяет
вред|юложять, что течение митоза играет важную роль в опре-
делении этого порога (467].
Со сходными цитологическими я физиологическими затрудне-
ниям» встречаются, вероятно, высокие полиплоиды, возникаю-
щие в природе. Но все же уровни полиплоидии, обнаруженные
в природе, значительно превосходят верхние пределы, дости-
гаемые а экспериментальных условиях. Несомненно, что высокие
полиплоиды, существующие в природе в настоящее время, — со-
храненная отбором выборка из древних линий, которые успеш-
но н постепенно преодолевали разнообразные физиологические
затруднения.
В большинстве случаев полиплоидии, известных у животных,
она остается на низких уровнях плоидности, лишь изредка дости-
гая умеренно высоких уровней. Тах, полиплоидные формы у
Cntmuiophorui и у других япцерни —трнплоиды; у амфибий —
по большей части триялоиды и тетраплоиды, хотя у трех видов
известны гекса плонды и октоцлонды [63]. У долгоносиков (Cur-
culionidae) большинство полиплоидов также трнплоиды, хотя
известно несколько тетра-, пента- н гексаплоидов. Такой же
диапазон уровней плоидности характерен для других семейств
насекомых, в которых встречаются полиплоиды [517].
Частота
Наша следующая задача состоит в том, чтобы оценить долю
полиплоидных видов средн высших растений Дать точную оцен-
ку частоты полиплоидии так же трудно, как и оценить ее уров-
ни и по тем же причинам. Если, желая оставаться на твердой
почве фактических данных, вычислять процент полиплоидных
форм по данным цятотаксояоыическото анализа полиплоидных
рядов, то мы неизбежно упустим из виду многочисленные древ-
ние Полиплоидные виды и тем самым серьезно занизим частоту
полиплоидии. Единственный способ избежать такого искаже-
ния — принять некоторые допущения относительно исходных,
или первоначальных, основных чисел
Среди ранних оценок частоты полиплоидии у покрытосемен-
ных укажем на оценки, приведенные Мюитцингом. Дарлингто-
ном и Стеббинсом. Мюнтцинг (599, 600] и Дарлингтон [139]
привели численные данные, свидетельствующие о том, что при-
мерно половина видов покрытосеменных — полиплоиды. По
приближенной оценке Стеббинса [780]. полиплоиды составляют
30—35% всех видов покрытосеменных.
Позднее я попытался подойти к «той проблеме, сопоставив
имеющиеся данные Во частоте распределения видов поирытосе-
Гл 22 Полиплоидия: распространи* и «итога 'Л?
мен мых с различным числом хромосом (рис. 22.2) с правдопо-
добным и широка распространенным допущением о том. что для
этом группы первичное основное число х—7, 8 и (или) 9 (282].
В гаком случае гаплоидное число хромосом л-14 может слу-
жить удобным пограничным пунктом виды с 14 ларами хромо
сом и более при всех статистических расчетах относят к поли-
плоидам, я виды с меньшим числом пар хромосом —к липло-
идам.
Такой подход га к же нс свободен от ошибок, что связано с
осложнения и н. создаваемыми энеуплаидией. Средн покрытосе-
менных имеется несколько тетраплоидных видов, у которых
2н —12. 10 или даже меньшему числу пар; эти виды происходят
от нисходящего акеуплоидного ряда Некоторые другие виды с
2н—14 или большим числом пар, напротив, представляют собой
диплоидные продукты восходящей анеуплпидин. Мы не знаем,
сколько видов в нашей выборке принадлежат к каждой из этих
категорий. Следует, однако, отметить, что эти два источника
ошибок компенсируют друг друга.
При помощи вычислений, произведенных описанным выше
методом, были получены следующие оценки относительной час-
тоты полиплоидных видов у покрытосеменных (282]'
Дэудс/ь^ые <3%
Одедолмиве 58*»
Поарытаг.-меиные и «глав 47%
Недавно Голдблат [246] вновь вычислил частоту полиплоидов у
однодольных, воспользовавшись билее полными данными, при-
веденными в сводках Федорова [210] и Мура [583], н получил
оценку 55% (вместо 58%).
Голдблат [246] считает, что я провожу пограничную линию
слишком высоко (между и -13 и 14) и поэтому ноя опечка час-
тоты полиплоидии слишком занижена. На самом деле при пер-
вом анализе этого вопроса я провел эту границу ниже и получил
другие оценки, но публиковать их не стал. Если постулировать,
что формы, у коюрых п —13 пли выше. — полиплоиды, то на ос-
нове тех же самых фактических данных получаются следующие
оценки относительной дачи полиплоидных видов:
Двудсмьнм* 49%
OsnoKUMiwc 60%
Покрытссечемнме в целом о2%
Я нс решился бы однако, провести границу ниже, чем на уровне
п-13, так как считаю, что восходящая аиеуплоидня игра,ч
важную роль вплоть до л = 12, а иа уровнях п-13, 14 и т. д
встречается е довольно значительной частотой Бить может, нс
ЗШ Чат щегол. ПолшиооЛш*
тайная частота полиплоидных видов у покрытой ценных нахо-
дится приблизительно посередине между двумя приведенными
выше оценками,
В отличие от покрытосеменных у хвойных только 1,5% поли-
плоидных видов, а у саговниковых не обнаружено ни одного по-
липлоидного вида [4X1].
Для Pteridophyta подходящей выборкой может служить пе-
речень хромосомных чисел, составленный Ментон |554]. В этом
перечне таксономические виды, содержащие формы двух или
нескольких уровней плоидности, рассматриваются для интере-
сующих нас здесь целей как два или несколько видов, а гибри-
ды в расчет не принимаются. Составленный таким образом пе-
речень содержит 103 вида, и в нем представлены все крупные
группы Pteridophyta. Только пять из этих видов, в том числе три
вида Selaglnella с 2я —18, — первичные днплоиды Полученная
при »том опенка частоты полиплоидов у Pteridophyta равна 95%
23 Типы полиплоидов
Разработка классификации
Между аютопслинлаидамя и аллополиллсилами (или амфнллон-
дамп) существует фундаментальное различие. Кнхара и Ойо
[438) первоначально определили это различие в общем виде:
авгополиплоидия в отличие от аллополи пл он дни является ре-
зультатом удвоения одного и того же набора хромосом.
К первом издании «Современных успехов цитологии» Дар-
лингтон [138] сформулировал это несколько иначе: автополип-
лоид представляет собой полиплоид, возникший путем удиоеиии
или сложения хромосомных наборов какой-либо структурной
гомозиготы: следовательно, это организм, содержащий три или
большее число наборов гомологичных хромосом В отличие от
этого аллополнплоид возникает в результате удвоения числа
хромосом у межвидового гибрида; следовательно, это полиплоид,
содержащий отдельные наборы негомологнчиых хромосом |136).
Тем, кто начинал изучение этой проблемы, было ясно, что
между гомологичными и пегомологмчиыым хромосомами, между
структурными гомозиготами и структурными гетероз и гота мн и
между гомозиготными индивидуумами н межвидовыми гибрида-
ми существует полная пмтертрадации Таким образом, а пт ппо-
ли и л он дня н аллополиплоидия, согласно приведенному выше
определению,— крайние звенья непрерывного ряда, члены кото-
рого постепенно переходят друг в друга. Во мисмих случаях мо-
жет оказаться полезным дальнейшее подразделение этих проти-
воположных классов и выделение некоторых нромежуточиых ти-
пов.
Мюнтцииг [599], рассмотрев различные определения а в топ о-
лнп л опдоо. предложил следующую концепцию. Авгополнплоиды
содержат множественные хромосомные наборы, которые иден-
тичны н структурном отношении, но могут различаться по ген-
ному составу В дальнейшем Клаузен. Кек и Хиси [102] выдели-
ли два основных типа а от ополи пл он,щв: I) а влипал иплоиды.
образующиеся из плодовитых мсжрасовыч гибрилов. и 2) авти-
полнплопды, образующиеся на обычных негибридиых индиви-
дуумов
Дарлингтон (138] различает три основных типа вллотстраплон-
дии в зависимости ог степени структурной дифференциации
между входящими в их состав геномами Примерами этих трех
типов служат Raphanobrassicu, Primula btwinsit и Crepis rubra-
feetida
Наиболее важные данные относительно Raphanobrassica и
Primula kewensis были приведены в гл. 19. в которой читатель
найдет интересующие его подробности.
19-46
290
Чат шастал. ПалыиаеРия
В экстремальном случае, как у RaphanobrastUa, структурные
различия между геномами очень велики, поэтому у диплоидного
гибрида хромосомы нс конъюгируют, однако у тетрапловдов.
происходящих от этих гибридов, они образуют биваленты. Для
промежуточного состояния, примером которого служит Primula
kewnsis, характерна несколько менее сильная структурная диф-
ференциация хромосом, так что они конъюгируют и образуют
биваленты и у диплоидного гибрида, и у его тетраплоидного
производного. Слабая структурная дифференциация между ро-
дительскими геномами, таках, как между Crepis rubra и С. foe-
tida. делает возможным образование бивалентов у диплоидного
гибрида и усиленное образование квадривалектов у происходя-
щего от него тетраплоида. Следовательно, аллополиплоиды это-
го третьего типа по своим цитогенетическим признакам прибли-
жаются к автополколоидам [138].
Последующие авторы продолжали разработку этой обшей
темы. Клаузен, Кек и Хисн [102] классифицируют амфкплонды
на основе двух взаимосвязанных признаков, а именно эффектив-
ности барьера стерильности между родительскими видами и
степени конъюгации между геномами родительских форм. По их
системе Raphanobrasslca представляет собой иитерненоспсци-
фнчный амфнплоид с неконъюгируюшими геномами; Primula
kevensis — нктерэкиспеинфнчный амфиплонд с конъюгацией
между геномами. Между этими двумя типами существуют раз-
личные промежуточные комбинации. Для обозначения амфнпло-
идов типа Р kewensis и Raphanobrasslca соответственно Стеб-
бинс [778, 780] ввел полезные термины сегментный аллополи-
плоид и геномный аллополиплоид,
На гексаплоидном, октоплоидяом или более высоких уров-
иях плоидности растение может быть одновременно и авто- и
аллополиплоидом. Полиплоиды такого типа были удачно назва-
ны автоаллооолнплоидамн. Они возникают от гибридов между
двумя родительскими формами, принадлежащими к разным аи-
лам н имеющими различные геномы, одна из которых или обе —
автогюлнплошы. Так, в результате гибридизации между авто-
тетраплондои и отличающимся от него по геному диплоидом об-
разуется триплоидный гибрид, который при удвоении дает гек-
саплоид, содержащий четыре генома одного вида и два генома
другого вида.
В заключение этого раздела перечислим главные типы поли-
плоидов с указанием их генетической конституции. В соответст-
вии с принятой практикой каждый геном обозначен одной нз
букв алфавита. Между геномами А, В и т. д. существуют боль-
шие структурные различия, поэтому у межвидовых гибридов
конъюгация хромосом обычно бывает нарушена. Геномы или
субгеиомы A,, At и т. д. по своей структурной дифференциации
Гл 23 Типы
291
занимают промежуточное положение, вследствие чего как у ди-
плоидного гибрида, так и у его амфиплоидного производного
наблюдается частичная или полная конъюгация хромосом. Ди-
плоидные виды, которые самостоятельно или в комбинации друг
с другом образуют разного рода полиплоиды, имеют геномы 44,
4f4r. ВВ и т. х
I Автополиплонды
I. Истинный автополиплоид ЛЛАЛ
2. Межрасовый а в тополи пл он д ЛЛЛЛ
П. Амфиплонды
3. Сегментный аллополнплоид 4*4«4гАг
4 Геномный аллополиплоид ААВВ
5. Автоаллополиплоид ААААВВ
Обратите внимание па то, что типы 2, 3,4 и 5 являются гиб-
ридными полиплоидами и только тип 1 негнбрндного происхож-
дения
Цитологические критерии
Основными критериями, по которым различаются автополнплои-
ды и амфиплонды, служат поведение хромосом, плодовитость,
соотношения при расщеплении и морфологмя. В отдельных слу-
чаях все эти критерии могут оказаться несостоятельными. Не-
сомненные амфиплонды часто проявляют цитогенетические свой-
ства звтополиплоидов и наоборот
Наиболее очевидная особенность поведения хромосом, по ко-
торой, как следует ожидать, должны различаться аяте пол иллои-
ды н амфиплонды, — это наличие или отсутствие мультивален-
•JOK при мейозе.
Образование мультнвалентов ограничивается, однако, часто-
той хиазм, которая в свою очередь зависят от величины хромо-
сом и генетических условий. Вследствие этих ограничивающих
факторов у автотетраплондов число ьвндривалентов почти всег-
да бывает ниже максимально иозможпого Так. у автотетрапло
идиой ржи (Secflte с ere ale. х-7) образуется от 0 до 6 квадрива
лентов на клетку, тогда как максимальное их число равно 7 IV,
а средняя частота кплдрнвалгитов варьирует от 2 до 4 IV у раз
иых линий (397, 604].
Вместе с тем у сегментных аллополиплоидов возможно и
часто наблюдается образование некоторого числа мультнвален-
тов. Например, тетр а плоил Primula kewensis, как уже говори
лось выше, обычно образует биваленты, но время от времени у
него появляются квадривалеиты. Возпикковемия квапривален-
тов у этого и других сегментных аллополиплоидовследует ожи-
дать, поскольку 'между хромосомами, принадлежащими к раз-
личным субгеномам, существует частичная гомолог
19*
2*2 Чат шестая. Полиплоидия
Если эти гомеологячиые хромосомы могут конъюгировать у
диплоидного гибрида, то они могут иногда конъюгировать н у
его амфиплоидкого производного. Иными словами, избиратель-
ная конъюгация может оказаться ие вполне эффективной Такая
межгеномиая (или так называемая гетерогеиетическая) конъ-
югация может просто привести к образованию бивалентов. Но
если гетерогенетическая конъюгация сочетается с нормальной
гомогенетической конъюгацией с участием тех же партнеров, то
образуются ассоциации более высокого порядка —тривалелты
и хвадривалекгы. У сегментного аллотетраплонда, имеющего ге-
ном A,AtAsAt, четыре хромосомы, принадлежащие к любому
данному хромосомному типу 1, могут время от времени конъюги-
ровать, образуя квадрнвалент 1А,-1А,-1Аг14>-
Генетические критерии
Что касается плодовитости, то Дарлингтон [138] сформулировал
следующее общее правило' между плодовитостью диплоидного
гибрида и плодовитостью его тетраплоидного производного су-
ществует отрицательная корреляция. Плодовитые гибриды дают
стерильных тетраплоидов, стерильные гибриды — плодовитых
тетраплоилоо, а гибриды, обладающие промежуточной плодо-
витостью. дают тетраплоидов с промежуточной плодовитостью.
В таком случае автополнплоидов можно отличить от аифиплон-
дон по стерильности первых и плодовитости вторых.
Это обобщение логически вытекает из ожидаемых различий
о поведении хромосом между автополиплоидами и амфиплонда-
ми. о чем говорилось в предыдущих разделах. У автополиплоида
неравномерное расщепление по гомологичным хромосомам, по-
лученным от мультивалента, ведет к стерильности гамет В про-
тивоположность этому у амфиплоидоп гомологичные хромосомы,
полученные от бивалентов, расщепляются равномерно, в резуль-
тате чего образуются фертильные гаметы
Мы видели, однако, что на самом деле поведение хромосом
у полиплоидов нередко Отклоняется от ожидаемого. Автополи-
плоиды, у которых конъюгация бивалентов генетически детерми-
нирована. могут быть высокоплодовитыми, а амфнплоиды могут
быть стерильными по разным причинам. У амфиплоида, обра-
зующего мультиаалеиты, может происходить неравномерное рас-
щепление хромосом, как у типичного автополиплоида, что при-
водит к формированию несбалансированных и нежизнеспособных
гамет. Или же у амфиплоида наблюдается нормальная — в цито-
логическом отношении—конъюгация, ио образующиеся била-
лепты гстерогеистичны; иными словами, конъюгировавшие хро-
мосомы расщепляются равномерно в количественном отношении,
Гл 23. Типы лмштлашТсы
253
ни образуют генетически несбалансированные продукты реком-
бинации. Амфиплокд снова дает стерильные гаметы. Или. на*
конец. генетическая сгернльносгь может сочетаться у диплоид-
ного гибрида с хромосомной стерильностью м передаваться его
амфиплондным потомкам.
Экспериментальные данные по плодовитости полиплоидов
разных типов согласуются с реальным положением вещей. Об-
щее правило о наличии отрицательной корреляции по плодови-
тости справедливо во многих случаях, однако из него имеются
также многочисленные исключения Несомненно, известно много
примеров авто тетра плондов с пониженной плодовитостью и
аллотетраплоидов с повышенной плодовитостью Но существует
также и много противоположных примеров.
Сирс [716] создал ряд диплоидных гибридов и производных
аллотетраплоидов между десятью диплоидными видами Trilb
cum, Aegilops и Haynakila (га — 7) В 16 гибридных комбинациях
ему удалось сравнить соответствующие Fi и аллотетраплопды в
отношении конъюгации хромосом и плодовитости. У разных
диплоидных гибридов уровень образования бивалентов варьиро-
вал от высокого до низкого. У аллотетраплоидов плодовитость
колебалась от 100 до 0% Однако никакой постоянной зависимо-
сти между степенью конъюгации у диплом лов Fi н степенью
плодовитости их тетраллондных производных обнаружено нс бы-
ло Гибриды F> с низким уровнем конъюгации в некоторых еду
чаях производят плодовитых амфиллондов. а и других —сте-
рильных в зависимое! и от прочих фдморов [716].
Ожидаемое различие между автополнплондамн п амфиплои-
дэми по поведению при скрещиваниях также выводится из раз-
личных экстремальны\ типов поведения хромосом, описанных
выше У любого автотетр а плоила следует ожила 1ь тстрасомною
наследования тех при знаков, во которым он гетерозиготен, тогда
как амфнллоид должен сохранять постоянство и размножаться
в чистоте.
Теоретически выделенные для полиплоидов соотношения при
расщеплении сильно отличаются от соотношений у дяплоидов
Рассмотрим гетерозиготный автотстраплоид с генетической кон-
стнгуцией TTtt по гену 7, локализованному вблизи центромеры
в четырех хромосомах' которые распределяются по гаметам слу-
чайным образом. Следует ожидать, что соотношение зигот в F2
от этой ггтсрозмготы составит 1/777:34 гетеро? иготы • ]itti
Если ген 7 находится иа расстояния 50 единиц хромосомной
карты от центромеры, то теоретическое соотношение зигот в F?
должно быть 17777 >9.8 гетерозиготы Uttt Соотношения, на-
блюдаемые в экспериментах, лежат между этими двумя теорети-
ческими оценками [511. 580]. В любом случае частота класса
гомозигот по рецессивному гену a F? гораздо ниже при тетра-
Частя шастая. ЛолшиоаДй
сонном наследования (между 1/22 я 1/36), чем при диеомном
наследовании (1/4).
В отличие т автополкплоидов у аллотетрапловда, несущего
гены ТТ в одной паре хромосом в гевоме А и гены tt в другой
паре хромосом в геноме В. вообще не следует ожидать расщеп-
ления по гену Т.
Тетр асом ное наследование, однако, не всегда происходит у
всех автотетраплондое и не обязательно отсутствует у алло-
тетраплоидов, Оно может наблюдаться у сегментного аллотетра-
пловда в случае образования у него мультнвалентов. В свою
очередь у автотетраплопда, в особенности если он образовался
давно, могла произойти генетическая дивергенция, в результате
которой прежде гомологичные аллелн превратились в отдельные
пары дулляхатиых генов.
У амфиплоида может происходить некоторое расщепление,
но не в таких соотношениях, как при тетрасомии. Допустим, что
у вас есть сегментный аллотетраплоид, несущий пару аллс
лей ТТ в одной паре хромосом одного субгенома и пару аллелей
tt в другом субгеиоые. Такой сегментный аллотетраплоид обычно
образует гаметы Tt и зиготы ТТН. Однако иногда у него образу-
ются гстерогенетические пары (Tit и Т/t} с последующим неза-
висимым распределением аллелей (; в результате возникает не-
которое число гамет tt. которые, объединяясь, лают некоторое
число зигот tttt. Таким образом, сегментный аллотетраплоид из.
редка дает вышепенцев, соответствующих диплоидной родитель-
ской форме tt.
Герстель и Филлипс [240] провели ряд остроумных экспери-
ментов с синтетическими амфнплоидами Gossypium и Nicotiana.
которые дали им возможность установить корреляцию между
соотношениями при расщеплении и близостью геномов. У неко-
торых амфиплоядов, происходящих от видов, не связанных близ-
ким родством, расщепления не наблюдается. У других амфипло-
ядоа, происходящих от близкородственных видов, как, например,
Gossypium arboreum и С. herbaceum. наблюдается расщепление
в соотношениях, характерных для тетрасомиков. Третьи амфн-
плонды, различающиеся не полностью, но в достаточной степени,
изредка расщепляются по некоторым генам-маркерам [240].
Морфологические критерии
Часто считают, что по морфологическим признакам автополн-
плоид должен походить на диплоидный родительский вид. тогда
как амфиплоид должен быть сходен с межвидовым гибридом.
Однако экспериментальные автополиплоиды не всегда сходны с
диплоидными родительскими видами. Ясно, кроме того, что если
какой-либо сегментный аллополвпловд может расщепляться в
Г* 33. Тили помлмидов
Таблица 23 1 Анфнплоиды. которых путали с одной или с
обеими родительскими формами [102]
Праш-мля м
Sparlina toans^ndii Phlaum pratense Pw еплна Iris verncolor Lriogonujn fasticulatu/n tsp. fotiolo- ШП Earner actiostUa ffratteca napus ffafoopsia ietrahit Pcnstem'in neoteticus .Madia L^figracilii Artemisia doaglariana all emit lor a 5 stricta P nttiCAU.r: P. cxiHs в P mpina t virgintia F. f 1 у pie am я £. f. poii folium P angtocospas я ft taojafohus fl oitnacea r. B. COBpeilrit G speaosa и G. pttbescens P lanas и P. acureus iM g> air Jit Л за/adorjU я A ludovictana
направлении одного диплоидного родительского вида по одному
гену, то он может таким же образом расщепляться и по другим
гемам и в конечном счете окажется очень сходным с диплома
ным видом как морфологически, так и экологически. Автоалло-
полиплоиды также часто проявляют морфологическое сходство
с определенным диплоидным видом.
У природных амфиплоидиых видов и в самом деле обычно
наблюдается изменчивость в сторону одного или обоих диплоид-
ных родительских видов. Клаузен, Кек и Хиси в 1945 г. [102]
составили список амфиплоидое, которых систематики ошибочно
принималн за одну из родительских форм или как за одну, так
и за другую; мы приводим этот список в табл. 23.1. Позднее
было обнаружено много других аналогичных примеров, и мож-
но считать, что изменчивость подобного рода достаточно обычна
(см. [283]). Это показывает, что сильное морфологическое сход-
ство между данным полиплоидом и данным днплондом не мо-
жет служить надежным критерием нвтополиплоидин
Относительная частота автополнплоидов
и амфнплоилов в природе
Определить, что данный природный полиплоид представляет
собой автополиплоид или амфкплоид, —отнюдь не легкая зада-
ча, поскольку различные критерии, которыми мы располагаем,
в пограничных случаях оказываются ненадежными. Сегментные
алло полиплоиды и автоаллополнплоиды по морфологическим
признакам и по поведению хромосом могут быть похожи на ав-
тополнплоидо». Даже после проведения значительного числа
39S Часть tuecraa. ПэлиалонЗил
экспериментов в онтогенетических исследгми1ний не всегда уда-
ется дать окончательный ответ на этот вопрос. Тем не менее
проблему эту ве следует считать неразрешимой. Если при изу-
чении данного полиплоида принять во внимание все критерии
и ресинтезнровать его экспериментально из его предполагаемых
родительскиI форм, то можно определить природу и происхож-
дение этого полиплоида достаточно удовлетворительво.
Сейчас, оглядываясь назад, можно сказать, что в первых
работах по естественной полиплоидии проблему эту чрезмерно
упрощали. И Дарлингтон [138, 139], и Мюйтцииг ]599] считали,
что автополиплоиды часто встречаются в природе и играют важ-
ную роль в эволюции растений. Такое заключение было основа-
но главным образом иа широком распространении у многих так-
сономических видов растений морфологически сходных диплоид-
ных и полиплоидных форм [138, 139, 599]. В его пользу свиде-
тельствовало также образование мультивалевтое у многих из
тех же полиплоидных форм (599]. Однако в настоншее время эти
две группы данных нельзя рассматривать как достаточно веские
аргументы в пользу подобного заключения
Клаузен. Кек и Хисн [102] опубликовали критический обзор
данных примерно по 28 тщательно изученным группам расте-
ний. содержащим природные полиплоиды. Здесь мы наложим их
заключения (102, гл. 6—8] в обобщенном и упрошенном виде.
Из 28 примеров природных полиплоидов только три признаны
несомненными и три других — возможными автополиплоидами.
Несомненные автополиолоиды —это Galax aphylla, Biscutelta
laevigata и Zea perennis; возможные автополиплонды — Vacci-
nlam uliginosum. Eragrostis pallescens. Galium motlugo н 0. oe
rum. Таким образом, преобладающее большинство природных
полиплоидов в этой выборке хорошо изученных и подвергнутых
повторному критическому анализу случаев являются амфиплон-
ламн [102].
Позднее Стеббинс [778] еще раз проанализировал все эти
примеры и перевел некоторые из возможных случаев в категорию
сомнительных Единственным несомненным автополиплондом
Стеббинс считает Galax aphylla.
По данным Ментон [554], у папоротников и близких к ним
групп широко распространены гибридизация и амфиплоидия.
бдиако, по ее мнению, естественная автополиплоидия у Ptertdo-
phyla отсутствует, за одним возможным исключением — Psilo-
tum.
Справедливости ради следует добавить, что некоторые из бо-
лее поздних авторов все еще продолжают считать, что автопо-
липлиидня играет или могла бы играть более важную роль в
природе, чем это вытекает из рассмотренных выше исследований
[143, 146, 543]. За последние годы перечень известных амфипло-
Гл. 23. Тшш аолаплоа^ол ИГ
идов сально увеличился, тогда как список возможных природ
иых автополиплоидои увеличился лишь веэначктельио.
Наиболее обоснованный вывод, который можно сделать из
имеющихся фактических данных во сосудистым растениям, со-
стоит в том, что амфввлоидия встречается гоохздо чаше и рас-
пространена гораздо шире, чем автополнплондия. Кроме того,
из немногочисленных твердо установленных природных автопо-
липлоидов по крайней мере некоторые представляют собой меж-
расовых автоплоидов, как, например, Biscutella laevigata и So-
latium tuberosum. Тахам образом, в полиплоидии у растений
тем или иным способом бывает обычно замешана гибридизация
24 Факторы, способствующие
возникновению полиплоидии
Вопрос о том. почему полиплоидия играет столь важную роль
в эволюции растения, очень сложен. Его многочисленные и раз-
нообразные аспекты широко обсуждались многими авторами
Были попытки установить корреляции между полиплоидией и
климатом, географической широтой, высотой над уровнем мо-
ря. типом местообитания, жизненной формой, системой размно-
жения, гибридным происхождением, величиной клеток, величи-
ной хромосом, их структурой, характером и поведением половых
хромосом, генотипом и другими факторами. Некоторые из этих
факторов рассматриваются в настоящей главе
По-видимому, не существует какой-то одной причины, опре-
деляющей эволюционный успех полиплоидии. Вместе с тем раз-
личные факторы, способствующие полиплоидии, пе равноценны
по своему значению.
В предпринятых мною попытках дать здесь анализ явления
полиплоидии в эволюционном плайе оказалось полезным раз-
делить регулирующие его факторы на три категории: 1) перво-
степенные. или фундаментальные, факторы; 2) второстепенные,
или факторы, играющие значительную роль; 3) третьестепенные,
или относительно маловажные факторы. Именно а этом поряд-
ке в последующих разделах этой главы рассматриваются усло-
вия. благоприятствующие полиплоидии. Такой подход к пробле-
ме позволяет сформулировать ряд объяснений.
Преимущества, внутренне присущие
амфиплоидному состоянию
Хорошо известно, что гибриды между вполне дифференцирован-
ными расами или видами часто обладают повышенной мощ-
ностью или жизнеспособностью или же повышенным физиологи-
ческим гомеостазом. Генетические механизмы, лежащие в основе
гетерозиса, усиленно изучались и обсуждались. Предполага-
лось, что у гибридов диапазон ферментативной активности ши-
ре. чем у их родительских форм, и что они более разнообразны
в биохимическом отношения. Нас здесь интересуют, однако, не
генетические причины, а фенотипические результаты гибридно-
сти. Гетерозис и физиологический гомеостаз — признаки, кото-
рые можно наблюдать у многих гибридов в царствах животных
и грибов, а также и в растительном царстве. Гетерозисные и го-
меостатические свойства многих гибридов, по всей вероятности,
имеют адаптивную ценность для своих носителей.
Гл 2 f Фолгорн, innate гвумщис возмиллоогдию /юяиллоьЛиа 299
Прн нормальном половом размножения гибриды не могут
вое про изводит г. в чистоте свое гетерозиготное состояние. Если
гибрид стерилен, то он вообще ие способен к половому размно-
жению. Если же он плодовит, то в последующих половых поко-
лениях ои утрачивает свои цеииые свойства — гетерозис и го-
меостаз. Однако аыфиплондня представляет собой такую гене-
тическую систему, которая позволяет выйти из тупика (1381.
Амфнплоид восстанавливает утраченную плодовитость и может
размножаться половым путем, сохраняя способность воспроиз-
водить в чистоте свой высокогетерозиготный генотип. Он обла-
дает преимуществами гибридностн в сочетании с преимущест-
вами. которые дает размножение в чистоте.
Поэтому в тех случаях, когда данный природный гибрид об-
ладает гетерозисными н гомеостатическими свойствами, амфп-
плоидмя дает возможность воспроизводить эти ценные в адап-
тивном отношении свойства в ряду последую И IX поко-
лений. Кроме тоги, амфиплоидня не препятствует полностью
расщеплению и рекомбинации, если ие считать случаев крайне
резко выраженных геномных аллополвпловдов, ио допускает
появление некоторых новых вариантов, создавая тем самым воз-
можность для возникновения более тонких адаптивных призна-
ков и их закрепления в последующих поколениях
Высокая частота иол кп ломаных видов у высших растений,
преобладающее бояьшипстоо которых —а мфиплокды, а осталь-
ные— главным образок межрасовые авгололиллоиды, свиде-
тельствует о том, что данная генетическая система успепгно
справляется с воспроизведением адаптивных гибридных гено-
типов в рамках патового размножения.
Другая черта полиплоидов, которая может оказаться выгод-
ной во многих ситуациях,— это большая забуференпостъ их ге-
нотипов по сравнению с дпп.топдамн, обеспечиваемая наличием
многочисленных дупликаций Полиплоидный индивидуум лучше
забуферен в физиологическом отношении, чем соответствующий
диплома И он забуферен лучше, чем диплома в отношении раз-
множения. гак как благодаря гетрасомному наследованию он
медленнее расходует имеющиеся у пего запасы изменчивости.
Такими буферными свойствами обладают как автопат и пл он ды,
так н амфиплоиды.
Способы образования
Существует два основных способа возникновения полиплоидов:
соматическое удвоение при митозе и отсутствие редукции хромо-
сом при иейозе [362].
300
Часп чисты Пошлмчды
При соматическом удвоении хромосомы во время митоза де-
лятся. но не расходятся к полюсам и оказываются все в одном
дочернем ядре В результате соматическая клетка с числом хро-
мосом 2л дает начало дочерней клетке с числом хромосом 4л.
Это соматическое удвоение может затронул» некоторые вегета-
тивные ткани, не имеющие отношения к развитию цветков, на-
пример корневые клубеньки; такие случаи иве здесь не интере-
суют. Но если удвоение произойдет в зиготе нли в зародыше,
находящемся иа ракинх стадиях развития, то ово приведет к
изменению всего растения. Илн же ово может произойти в одной
из почек к, следовательно, оказать влияние на одну из ветвей
растения. Тетраплоид Primula kewensis, о котором мы уже го-
ворили в гл. 19, возник в результате соматического удвоения,
происшедшего у некоторых цветоносных ветвей диплоидного
гибридного растения.
Отсутствие редукции связано с тем, что при мейозе не про-
исходит образования клеточной стеики. Если зто имеет место в
диплоидных материнских клетках пыльцы и материнских клет-
ках зародышевого мешка, то образуются диплоидные споры и
гаметы. В таком случае после слияния двух иередуцироваиных
гамет могут сразу образоваться тетраплондпые зиготы. Именно
так возникла Raphanobraasica, как было отмечено в гл. 19.
Иногда отсутствие редукция может затронуть лишь одну ли-
нию половых клеток, что приведет к развитию либо диплоидных
пыльцевых зерен, либо диплоидных яйцеклеток. В таких слу-
чаях образование нового тетраплоидвого индивидуума происхо-
дит в два этапа. На первом этапе в результате слияния 2п- и
1л гамет образуется Зл-зиготв, а в следующем поколении — на
втором этапе —нсредуцнроваиаая Зл-гамета, сливаясь с нор-
мальной In-гаметой, дает тстрвплонд. Искусственный аллотетра-
плоид Oaleoptis tatrahil был получен именно в результате таких
последовательных этапов (см. гл. 19).
Менее обычный способ возникновения полиплоидии связан с
полиспермией, т. е. оплодотворением яйцеклетки двумя мужски-
ми ядрами. Этот процесс описан у Ustera ovata и у некоторых
других орхидей [324].
Амфиплонды обычно образуются из диплоидных гибридов,
как в случаях Primula kewensls и Raphanobrassica. Высказыва-
лись также предположения, что в некоторых группах, в частно-
сти у Zauschneria и Galium, амфиплонды возникли в результате
гибридизации между автополиплондами. принадлежащими к
разным видам [102]. Если эта интерпретация верна, то в подоб-
ных случаях сначала происходил процесс удвоения, а гибриди-
зация имела место позднее.
Г.1 24 Факторы, спосоЬс1»^ющи« паамлжмм^ш
301
Полиплоидия и жизненная форма
Мюигиинг [599] провел анализ числа хромосом у 4К родов тра-
вянистых растений, относящихся к покрытосеменным, ссели ко-
торых были как многолетние, так и однолетние виды Он уста-
новил наличие корреляции между числом хромосом и характе-
ром роста Как правило, у многолетних видов данного рола чис-
ло хромосом выше, чем у однолетних его представителей. Во
многих случаях более высокое число хромосом у многолетних
видов служит мпным свидетельством полиплоидного происхож-
дения. Так, в родах Helianthus, Euchlaezea, Sorghum и других
имеются многолетние полиплоиды и однолетние ди плои ды [599].
Стеббинс [775] подтвердил заключения Мюнтцинга н допол-
нил его обзор, добавив к нему 202 рода двудольных Он соста-
вил таблицы числа хромосом для растений, относящихся к каж-
дому из трех классов жизненных форм, а именно к древесным
растениям, многолетним травянистым растениям н однолетним
или двулетним травянистым растениям. Высокая частота поли-
плоидных видов обнаружена у 52% родов травянистых п у 36%
древесных ролов растений Таким образом тенденция к поли-
плоидии сильнее выражена у травянистых двудольных Если же
выборку травянистых родин разбить на под групп и многолетних
и однолетних, то станет ясно, что полиплоидия чаще встречает-
ся у многолетних родов [775].
Густафсон также установил, что на севере Европы частота по-
липлоидии статистически выше у многолетних Gramineae, Ra-
nunculaceae. Crucilcrae, Lepuminosae и Compusilae. чем у одно-
enuu представителей этих семейств [3171. Следует ОТМСТЯТ»,
что Pteridophyta, у которых Ментон [554] обнаружила такую
высокую частоту полиплоидны, — преимущественно многолетние
травимнегые растения
Мюптиинг [399] пытался объяснить корреляцию, наблюдае-
мую между потпплопдиеЛ н многолетним образом жизни, до-
пустив, что полип.тоидиость оказывпет влияйот но жизненную
форму. Он указал, что у экспериментальных авгополштлоидов
скорость роста в общем ниже, чем у диплоидных родительских
форм. Такое замедление роста могло бы во многих случаях
принести к превращению одни.к-тнего растения в многолетнее
при удвоении числя хромосом [559. 654].
Полое поздние ан юры предпочли представши эту чавнеи-
мостъ, отменяв местами причину и следствие. По их мнению,
мн of иле гни ft образ жизни с его возможное™ мн для всгстатнз
КОГО размножения благоприятствует полиплоидии [143. 146. 317.
318. 775]. Веским доводом в пользу такой юиерпрстацин факти-
ческих данных служит корреляция, подмеченная Дарлингтоном
Один । • о»-ем». немногих полгнктпдоа срети хвойных, Sequoia
302
Част» шкта*. ПояшмлЗы
strnparvirens. представляет собой также один из крайне редких
видов хвойных, способных регенерировать из корневых отпрыс-
ков [из].
Каким образом вегетативное размножение благоприятствует
полиплоидии? На этот вопрос разные авторы ответ а ют по-раз-
ному. По мнению Дарлингтона (146]. вегетативное размножение
благоприятствует выживанию уже возникших полиплоидов. По
мнению Же Густафсона [317], клональное размножение снижает
нормальную рекомбинацию, но облегчает функционирование
иередуцироеаиных гамет.
Более простое и, по-видимому. более общее объяснение со-
стоит в том. что большая продолжительность жизни многолет-
них трав, а в особенности тех, которые способны к вегетативно-
му размножению, сильно повышает вероятность возникновения
соматического удвоенна у гибридного растения [276. 775]. Мно-
голетний травянистый гибрид Primula kewensis в первый год
своего существования не образует фертильных тетраплоидных
ветвей, однако в последующие годы это наблюдалось в трех
отдельных случаях. следовательно, мы вправе допустить, что
этот гибрид, возможно, никогда не дал бы начала плодовитым
амфнплоидам. если бы продолжительность его жизни была ко-
роткой.
Факторы, имеющее первостепенное значение
Относительно природы и распространения полиплоидии сделано
три важных обобщения. Во-первых, полиплоидия представляет
собой характерную особенность царства растений и только этого
царства, причем и здесь она встречается главным образом у
многолетних растений с большой продолжительностью жизни.
Во-вторых, полиплоидия у растений обычно носит форму амфи-
плондин. И в-третьих, амфиплоидия представляет собой генети-
ческую систему, дающую возможность воспроизводить при по-
мощи полового размножения гибридный генотип, обладающий
ценными адаптивными свойствами. В сущности, это, вероятно,
наилучшая генетическая система для выполнения столь сложной
задачи. Теперь, руководствуясь этими обобщениями, можно
определить основные факторы, способствующие возникновению
полиплоидии.
Полиплоидия способствует сочетание трех основных факто-
ров: 1) большая продолжительность жизни и наличие возмож-
ностей к вегетативному размножению; 2) первичное видообразо-
вание, сопровождающееся хромосомными перестройками; 3) вы-
сокая частота естественной межвидовой гибридизации. В преды-
дущих главах книги эти три фактора рассматривались как
самостоятельные явления вне связи с полиплоидией.
Гл 21 Факторы, ттююбттедкхцие ааииккмекшо млаялоидш 2ПЛ
Важно понимать, что полиплоидия, как весьма частое и ши-
роко распространенное вволюционное событие, зависит от сов-
местного действия всех трех факторов. Отсутствие любого иэ
этих фаьторов в данной группе организмов сильно задерживает
или полностью исключает формирование полиплоидных видов.
Второй н третий факторы играют особо важную роль. Первый
фактор важен, но ire столь необходим; как мы увидим ниже, у
однолетних травянистых растений его отсутствие может компен-
сироваться другими второстепенными факторами.
Перечисленные три основных фактора особенно характерцы
для высших растений И именно у высших растений часто воз-
никают полиплоидные виды, достигающие пышного развития.
В царстве животных, где полиплоидия — явление чрезвычайно
редкое, эти три фактора обычно отсутствуют или же встречают-
ся как исключения. Таким образом, как это указывает Густаф-
сон [317]. глубокие различия, существующие между высшими
растениями и высшими животными, привели к глубоким разли-
чиям в характере их эволюции, и одно из этих различий — име-
ние полиплоидии.
В царстве растений полиплоидия распространена неравно-
мерно, и по многих случаях ее наличие или отсутствие коррели-
рует с наличием или отсутствием одною из основных факторов.
В тех группах растений, у которых видообразование происхо-
дит по Типу Ceanothus, полиплоидия в основном отсутствует.
В гл. 11 были описаны примеры, относящиеся к Quercus, Ribes.
Ceanothus и другим родам покрытосеменных. Кхошу [433] ука-
зывал иа многочисленность нсполнплондиых видов среди хвой-
ных. Все эго—древесные растения с большой П|х>должнтель-
костью жизни отдельных индивидуумов, у которых часто наблю-
даются вегетативное размножение н естественная гибридизация.
Но близкие гибридизирующие виды ие различаются но хромо-
сомным перестройкам (33]. Короче говоря, факторы I и 3 нали-
цо. по фактор 2 отсутствует.
Мюитцинг [599] показал, что частота полиплоидии определен-
но ниже у однолетних травянистых растений по сравнению с
многолетними, принадлежащими к тому же роду. Хейзер [367]
устанопи.1, что полиплоидия наблюдается относительно редко
средн сорной флоры штата Индиана, и показал, что *то корре-
лирует с преобладанием среди процветающих сорняков расте-
ний с однолетним жизненным циклом. Густафсон (317, 318] так-
же обращает внимание иа то, что среди однолетних растений
вообще и в сорняковой флоре, состоящей главным образом из
однолетних видов, в частости процент полиплоидных видов ни-
зок
Между однолггтими травянистыми растениями, относящими-
ся к роду Madia, схыесгауют межвидовые различия по структу-
Э04 Veer» амстол. Полиялсадил
ре хромосом. Естесгвенная гибридизация происходят у них час*
то, но индивидуальная продолжительность жизни невелика и
возможности для вегетативного размножения отсутствуют. Та-
ким образом, у них наличествуют факторы 2 и 3, но ист фак-
тора 1.
Широта и высота местности
Давно известно, что частота полиплоидии у высших растений
коррелирует с географической широтой. Процент полиплоидных
видов во флоре данной области закономерно возрастает в на-
правлении от низких широт к высоким. Такай тенденция впер-
вые была замечена Хагерупом {323] н Тышлером [830]. В ранних
данных оо этому вопросу имелись разного рода ошибки, которые
были устранены в более поздних исследованиях, однако общие
выводы оставались прежними (833].
Новые исследования оказались также значительно более
обширными по сравнению с ранними обзорами. Региональные
флоры, упоминавшиеся в этих ранних работах, подверглись бо-
лее полному повторному изучению, а, кроне того, к их числу
добавились новые флоры. Оригинальные исследования слишком
многочисленны, чтобы нх можно было здесь перечислить; обзо-
ры. содержащие обширные библиографии, опубликовали Лёве А.
и Леве Д (528, 530] и Хансльт [336]. Более новые данные под-
тверждают сделанные ранее обобщения.
В табл. 24.1 приведен процент полиплоидных покрытосемен-
ных во флорах ряда областей, лежащих от 36 до 84° с ш. в
Евразии и Арктике. Данные взяты из работ Лёве А. в Лёве Д.
[530], а также Хаиельта [336] и несколько обобщены; для выяс-
нения дальнейших подробностей читатель может обратиться к
этим обзорам. Все перечисление в табл. 24.1 региональные фло-
ры были подвергнуты исчерпывающему цитотаксон омическому
изучению и расположены в порядке возрастания широты мест-
ности Можно видеть, что частота полиплоидных видов возрас-
тает с 37% в Средиземноморской области до 74% иа Шпицбер-
гене и что она вообще выше в арктической и субарктической
зонах [336, 524, 528, 530, 8331
Одна из последних оценок частоты полиплоидных видов сре-
ди сосудистых растений арктического региона в целом состав-
ляет 60% в низких и 70% в высоких северных широтах [532].
На тихоокеанском побережье Северной Америки частота по-
.-И1ЛСН..:>В также упеличнмпся с МараВМИММ широты ВТ Ка-
лифорнии до Аляски [387]. Имеется ряд данных, указывающих
иа го. что подобная же тенденция к увеличению доли полиплои-
дов в высоких широтах наблюдается также в Южном полуша-
рии [325, 336].
Гл. 34. Флкпр», спокЪсткукщал мтшштдеаекше iummjoo^ioi Мб
Таблица 24.1. Частота полиплоидных видов покрытосеменных в
разных широтных зонах в Евразии и Арктике (обобщено по
данным Лёве А. а Лёве Д. (530] и Ханельта [336])
ООлает*
Сицилия
Венгр»»*
Пардубпп* (ЧССР1
[Дежтржльпоя Ея^кпи
Шлезвиг- ГолыитеЯш
Дания
А и.«Я
Юго-западная паста Грекиандми
Фарерски* о ва
Исландия
Шисцк*
Финляндия
Иоролгмя
Северо-тападяая «сть Аляпгя
О-в Девой
Шнв-пС^р ел
Земля Фраша-Иосифа
Земле Пирк
Гр.Д С.*,. Пшшыонды. %
38—38 37,0
44-47 46.а
ГС—1» дй.«
50 52.3
46—55 50,7
54 —го 54.5
54-5И 53.5
50-61 52.8
60-62 74.0
62 п.о
63-66 71.2
W-69 56.9
СО—70 57.3
58—71 57,6
сз 59.3
75 Т6.0
77-61 74.0
ВО-82 75.0
52-64 В5.9
Частота полиплоидных видов отиосктелыю велика в высо-
ких горах. По данным Ханелыа [336], процент полиплоидии во
флорах различных высоких гор Евразии, ?\мерики и Новой Гви-
неи составляет от 45 до 85, Иногда частота полиплоидов бывает
выше на высоких горах, чем иа близлежащих низменностях, но
это случается не всегда [336].
Лёве А. и Лёве Д. [531] сравнивали частоту полиплоидии в
соседних альпийских и лесных флорах на г Вашингтон н в го-
рах Уайт-Мауитиис в шт. Нью-Гэмпшир. В альпийской эоне,
выше верхней границы лесной раетительпостя, 63,6% таксонов
сосудистых растений — полиплоиды, тогда как среди соседней
лесной растительности полиплоиды составляют только 45%. Та-
ким образом, частога полиплоидии в альпийской эоне сущест-
венно выше [631].
Эти факты достаточно определенны Но что они означают?
Им давали различные интерпретации Чтобы показать все раз-
нообразие этих интерпретаций, я вкратце изложу их здесь.
Подробно они обсуждаются Леве А. и Лёве Д. [528. 531], Стеб-
бинсом [780. 787]. Джонсоном, Пакером и Ризе [410]. а также
Ханельтом [336.
1. В суровом холодном климате полиплоиды образуются под
действием темпера турных шоков [323].
20—35
ЗОВ Чат шкпя, Похоллоадля
2. Полиплоиды лучше адаптированы к холодному климату
я к другим связанным с этим климатом условиям среды, чем
дипломам (524, 52В, 531].
3. Корреляция между полиплоидией и широтой местности
или высотой над уровнем моря носит косвенный характер. По-
липлоидия чаще всего встречается у многолетних травянистых
растений, размножающихся вегетативным путем, н эта жизнен-
ная форма преобладает в арктических и альпийских флорах
1317. 318].
4. Изменения климата в плейстоцене, в том числе продвиже-
ние и отступление ледяных щитов и ледников, вызывали изме-
нения в распространении существующих видов. Возникали но-
вые контакты, происходила естественная гибридизация и обра-
зовывались гибридные полиплоиды (554].
5. Изменения климата и рельефа местности в плейстоцене
способствовали естественной гибридизации в новых комбина-
циях, как было сказано выше. С отступлением льдов открылись
новые местообитания, которые можно было заселять. Амфиплок-
ды особенно хорошо подходят для заселения этих новых свобод-
ных местообитаний [192, 780, 787].
Первое из этих объяснений теперь уже считается несостоя-
тельным. Решающей стадией является не образование поли-
плоидов. а их закрепление в статистически значимых соотно-
шениях, выявляемое при сравнении флор в целом. Третье объяс-
нение также можно отбросить. Корреляция между полиплоидией
н географической широтой действует в пределах гауппы много-
летних травянистых растений в разных флорах [528].
Все остальные объяснения достаточно правдоподобны, одна-
ко иа основе тех статистических к флористических данных, ко-
торыми мы располагаем в настоящее время, им нельзя дать
окончательную оценку. Необходимы более подробные исследо-
вания.
Экологические факторы
Одни из подходов к очерченной выше проблеме состоит в том,
чтобы изучить частоту полиплоидов в различных экологических
группах данной северней флоры.
Джонсон и Пэхер [409] собрали данные о частоте полиплоид-
ных покрытосеменных в разных частях одной из долин в север-
ной части Аляски на 68* с.ш. Частота полиплоидов в пределах
этой долины сильно различалась в разных местообита явях, по-
вышаясь в нарушенных местообитаниях с влажной почвой н
неглубоким залеганием вечной мерзлоты и понижаясь в более
стабильных местообитаниях с более сухой почвой и более глу-
боким залеганием мерзлоты.
Гл 21. Факгоры, глэгобггвтйлцм# *ммдемл«£м4*к 307
Эрендорфер [193] проанализировал различные растительные
сообщества во флоре низменных районов Австрии с точки эре*
нкя частоты полиплоидов. Самый высокий процент полиплоидов
был обнаружен на лугах, недавно созданных в этой области
человеком, тогда как в лесных сообществах частота полиплои-
дов была ниже.
Другой подход к этой проблеме состоит в том. чтобы подроб-
но изучить географическое к экологическое распространение
родственных диплоидных н полиплоидных видов з отдельных
группах растений.
Как у покрытосеменных, так и у папоротников наблюдается
следующая общая картина полиплоидные представители данной
группы имеют широкое распространение, тогда как нх диплоид-
ные родичи распространи.in менее широко (534. 780]. У папорот-
ников. относящихся к родам Cystopteris, Polypodium, Dryopteris
и Asptemuffi, полиплоиды многочисленны, а дипломам либо ред-
ки, либо представляют собой реликтовые или вымершие формы
[55*1]. Подобное же различие между дипломдамм и полиплоида-
ми наблюдается в группе biscutella laevigata (Cruci ferae) (552,
553].
В комплекс Gilia inconspicua (Polcmoniaceae) вводят много-
численные диплоидные и тетраплондные вилы, произрастающие
в пуегыннх и горах западной части Северной Америки В этой
группе и географнчссксе, н экологическое распространение д.ш-
лондных аилов в общем ограничено по сравнению с их амфн-
плондными приизводиыми. Многие нз диплоидных индии опреде-
ленно представляют собой релимы. В Оглнчис от дмплоидоп
полиплоиды имеют широкое распространение и очень обильны
на обширных и разнообразных по своим условиям территориях
(283]. Тетра клонимые гнлнм прн выращивании их на экспери-
ментальных участках отличаются также от своих диплоидных
родичей более высокой мощностью.
Данные, полученные при использован ни описанных здесь
двух подходов, свидетельствуют в пользу объяснений 4 н 3.
Однако эти данные нельзя считать решающими, и проблема все
еще остается открытой.
По моему мнению, подтверждаемому результатами моих соб-
ственных исследований иа нескольких группах полиплоидов и
днплондов хак в природе, так н па экспериментальных участ-
иях, амфиплоиды и в самом деле часто характеризуются более
высокой мощностью, гомеостатическими буферными свойствами
и способностью и аднптаппн пп сравнению сп своими лппзопл-
ными родичами. Вследствие этого амфнп.юнлм в ItpttpOM не-
-«бладают адаптивным превосходством ПйД предковыми
диплоидными форма ин. Появление на территории предковых
видон новых пригодных для заселения местообитаний, кпннкаю-
2СЖ
306 Часп ыссгая. Полшитгды
щих в результате кзмененнй климата, разрушений, причиняе-
мых деятельностью человека, или других нарушений среды, соз-
дает те экологические предпосылки, при которых амфиплонды
могут воспользоваться своими преимуществами.
Полиплоидия и система размножения
Амфиплонды у однолетних растений возникают главным обра-
зом в результате отсутствия редукции и последующего слияния
вередуцироваииых гамет [102]. Если однолетнее гибридное рас-
тение размножается при помощи ауткросснига, то вероятность
соединения двух керелуцироваккых гамет очень невелика. Та-
кой гибрид может начать интрогрессию, но вряд лн ои за свою
короткую жизнь сумеет произвести амфнплокда,- Однако если
однолетнее гибридное растение размножается при помощи авто-
гамии, то вероятность соединении двух иередуцнроваииых гамет
значительно повышается [278].
Две однолетние секция рода GUia — GUia к Aracknion
(х-9) — насчитывают вместе 39 видов; все эти виды, за исклю-
чением одного, были изучены цитотаксономически и генетичес-
ки. Что касается плоидности, то среди 38 цитологически изучен-
ных видов обнаружено 23 диплоида, 14 тетраплоидов и 1 окто-
плоид [281, 283, 286]. В отношении размножения у этих же ви-
дов иаблюдается весь диапазон систем — от облигатного
ауткросснига до автогамного инбридинга. Весьма примечатель-
но, что все 15 полиплоидных видов автогамные [278]. У одного
в целом автогамного тетраплоидиого вида имеется раса, размно-
жающаяся при помощи ауткроссиига.
На ряде примеров была прослежена родословная однолетних
амфиллоидиых видов GUia и установлено нх происхождение от
определенных автогамных диплоидных видов. Оказалось, что от
автогамных диплоидных видов GUia происходит относительно
больше амфиплоидов, чем от диплоидных GUia с перекрестным
Оплодотворением.
Такая же корреляция между автогамией и возникновением
полиплоидов обнаружена и у других родов, в секциях, состоя-
щих из однолетних видов. Большинство полиплоидных видов
среди однолетних представителей родов Galeopsls. Madia. Mio
raserti, Clarkia, Menttella. Amsinckia и Esehscholtila — автогам
кые формы [278]. Наблюдаемая корреляция согласуется с пред-
ставлением о том, что автогамия является одним из второсте-
пенных факторов, способствующих возникновению полиплоидии,
который в известной пепени компенсирует короткую продолжи-
тельность жизни каждого индивидуального растения у однолет-
них видов.
Гл. 24 факторы, способсгаующис ло.пгяломды* 309
Многолетние травянистые растения, у которых жмиплпидиш
широко распространена, обычно размножаются при помпши
перекрестного опыления и часто бывают самонссовмсспгчимн
Корреляцию между тремя факторами — характером роста, по-
липлоидией и системой размножения — можно в этом случае
объяснить несколькими разными способами
Густафсон (317] вслед за Люксом [489] счшает, что у сами
несовместимых дни Лондон в столбике создаются условия, кото-
рые благоприятствуют функционированию иерсдуцдроваыных
пыльцевых черен, образуемых этим же самым растением; это
приводит к формированию тегрэплонднык или по меньшей мере
тркпломдиых зигот По моему мнению, совершенно достаточна
более простое и более прямое объяснение. Многолетние гибрид*
ные растения располагают достаточным временем. а следова-
тельно, и большими возможностями, чтобы претерпеть полипло-
ид мое удвоение, обычно путем соматического удвоения, по иног-
да также путем соединения нерсдуцированиых гамет. Таким об-
разом. по Густафсону, причинная связь существует между сяыо-
несовчестимостью и полиплоидией. а многодетность —коррели-
рованный с ними фактор. тогда как. с моей точки зрения при-
чинная связь существует между Miioro.neniocibio и налипло-
ид не fi. а коррелированный фактор — свободное скрешнпание.
На эту проблему можно взглянуть и под иным углом зрения
У растений, обладающих системой самонеспвместимости гамето-
фиги ого типа или типа Nicotiana, полпплоидизацня превращает
самонссов честимый диплоид в самосовместииый тетраплоид
|488 494] У Solanum и Nicotiana (сем Solanaceac) и у некото-
рых представителей сем. Scrophulariaccac и Lcpunnnosae дипло-
идные виды обычно гампмегаяместимы, а природные полиплоид-
ные виды самооовместн-им [494].
Мёллер [594] высказал мнение, что установление полиплоид-
ного состояния должно быть особенно затруднено у раздельно-
полых организмов. Предполагалось, что главная причина этого
заключается в нарушении механизма определения иола у тетра-
плопдов с половым:! хромосомами XXV Y Меллер |5бМ] сделал
такое предположение, надеясь, что пнп послужит объяснением
редкости полиплоидии у Ж1гво1иых, в большинстве своем раз-
дельнополых, и се широкого рпспрпстрямепия у растений, кото-
рые по большей части гермафродитом.
Полученные в дальнейшем фактические данные частично под-
тверждают. а частично противоречат гипотезе Меллера. Твер-
до установленные примеры полиплоидии у животных действи-
тельно обнаружены либо в группах, размножающихся партено-
генетически (Дг/стпйг satora. равноногие раки, чсхлоноскп, турец-
кий скосарь. Ochthiphtla). либо у гермафродитов (дождевые
черви) (75, 769]. Вместе с тем полиплоидия возникла и у некого-
810
Чаете шесты. Пслаамм^ы
рых двудомных растений хак в природе (Salix, Rumex, Galium),
так и в экспериментальных условиях (Mtlandrium) [161, 522,
780, 885)
Влияние величины хромосом
Дарлингтон давно высказал предположение, что механика де-
ления хромосом и механика клеточного деления, возможно, иг-
рают роль факторов, ограничивающих полиплоидию Между ве-
личиной клеток, величиной хромосом и полиплоидным числом
хромосом существуют реципрокные зависимости (13Я)
Известно, что скорость роста у автооолиплоилов часто бы-
вает ниже, чем у родственных им диплоидоа; это обусловлено,
по крайней мере частично, задержкой клеточного деления [599).
На некотором уровне плоидности, различном для разных групп
растений, искусственный полиплоид переступает некий порог
жизнеспособности, который опять-таки частично определяется
особенностями митоза (см, гл. 22).
Клетки, образующие питательные ткани в зародышевых меш-
ках и пыльниках, часто или обычно содержат эндополиплоидиые
ядра. Такая клетка — продукт многократных эндомитозов; она
не претерпевает клеточных делений. Эти эндополиплоидиые
клетки достигают гораздо более высоких уровней плоидности,
чем растения в целом. У некоторых представителей Ranuncula-
ceae. Papaveraceae и Compositae находящиеся в зародышевом
мешке клетки-антиподы могут достигать 64-пломдмостн [602].
Наивысшая степень плоидности обнаружена у Phaseolus cocci-
мм ( = Р. muitiflorus), зародышевый мешок которых содержит
4096-плоидные ядра [147].
Отдельные клетки, которые не должны делиться, явно могут
функционировать при гораздо более высоких уровнях плоидио-
сти, чем целые растения, рост которых осуществляется путем
клеточных делений. Это различие между терминальными члена-
ми клеточных линий и растущими растительными организмами
в смысле толерантности к высоким уровням плоидности согла-
суется с гипотезой Дарлингтона.
Отсутствие или редкость полиплоидии у некоторых Liliaceae
и у некоторых древесных растений объясняются, возможно, тре-
бованиями. предъявляемыми клеточной механикой. У Lilium.
FritiUaria и у некоторых других родов лилейных с крупными
хромосомами полиплоиды либо не достигают высоких уровней
плоидности, либо отсутствуют вообще. Быть может, в таких слу-
чаях хромосомы просто слишком велики, так что прн высокой
плоидности клеточное деление становится невозможным [138,
139].
Гл 24 факгорк, спасоОстеуюсиие oommou/ovumo пимллоийии 311
У древесных двудольных камбиальные клетки узкие, н это,
ло-виднмому, ограничивает как число хромосом, так и нх вели-
чину. Относительно низкая частота полиплоидии у древесных
двудольных по сравнению с многолетними травянистыми дву-
дольными и полное ес отсутствие в некоторых ролах и семейст-
вах двудольных, возможно, обусловлены именно этой особен-
ностью камбиальных клеток [139. 775].
Влияние генотипа
Нарушение конъюгации хромосом у межвидового гибрида соз-
дает необходимые условия, делающие возможным отсутствие
редукции и образование амфиллоидов. Нарушение конъюга-
ции— результат различий между родительскими геномами по
хромосомным перестройкам. Однако, как показали многочислен-
ные исследования [139, 502, 670, 687, 868], поведение хромосом
во время ыейоэа регулируется также определенными генами.
Отсюда следует, что на частоту отсутствии редукции, в тем са-
мым в иа возникновение полиплоидов у ряда гибридов с хромо-
сомной стерильностью, могут оказывать влияние генотипические
различия по тем или иным генам, контролирующим мейоэ [286,
816. 817).
Гены, детерминирующие различные отклонения в мейозе. на-
пример асниапсис нлн формирование аномального веретена, из-
вестны у кукурузы, дрозофилы и других генетически хорошо нау-
ченных организмов. Нас здесь особенно интересуют те из генов,
регулирующих мейоэ, которые вызывают нарушение второго
мейогнческого деления, в результате чего образуются нерелуци-
рованные гаметы; такие гены были обнаружены у Datura stra-
monium и Zea mays [25, 679, 709).
У Datura stramonium имеется рецессивный мутантный геи,
известный пол названием dyad, который вызывает образование
к концу иейоза диад нерелуцнроваппых спор как в мужской, так
и в женской линиях. Диплоидное растение Datura, гомозиготное
по гену dyad, при самооплодотворении дало начало некоторому
числу тетра плотных потомков [25. 709). Растении Zea mays.
несущие рецессивный аллель гена elongate в гомозиготном сос-
тоянии, дают некоторое число нерсдуцировапиых яйцеклеток, по
нормальную гаплоидную пыльцу. Поэтому первое поколение по-
томков растений elongate состоит главным образом из трипло-
ндоя. В последующих поколениях эти триплонды могут давать
тетрпплопдов и более высоких полиплоидов [679].
У диплоидных гибридов Fi от скрещивания Rumcx tkyrsiflo-
rus и R acetosa наблюдается ипдивндуальная изменчивость в
Э13 Часть мистол. Помплож>ил
отношении мейоза. У некоторых растений F, мсйоз протекает
нормально, и они образуют тетрады; у других второе иейотн-
ческое деление отсутствует, и они образуют диады иередуниро-
ванных гамет. Выщеплеиае форм, образующих диады, происхо-
дит также в поколекиях F> и Bt Потомки F, от гибридных инди-
видуумов с нормальным мейозом представляют собой дипломов
(2п—14). Но потомки Ft от гибридных растений, образующих
диады, —тетраплоиды или почти тетраплоиды (2л«25) [816,
8171
Восемь видов секции GUia рода GUia скрещивали искусст-
венно в различных комбинациях; было получено 15 гибридов,
достигших стадии цветения. У четырех из втих 15 типов межви-
довых гибридов произошло спонтанное удвоение числа хромо-
сом путем соединения иередуцироваииых гамет, в результате че-
го было получено плодовитое или пол у плодовитое амфнп.тоид-
ное поколение Ft- Все эти четыре гибрида, давшие амфнплоид-
ных потомков, имели в качестве одного нли обоих родительских
видов GUia millefoliata и (или) G. oaldiviensu. G. millefoliala и
G. valdivierwis — два близкородственных диплоидных (2л—18)
вида, принадлежащих к одной и той же геномной группе, Таким
образом, выборка искусственных спонтанных амфиплоидое сек-
ции GUia рода GUia отличается необычно высокой концентра-
цией геномов milleloliala [286].
У одного из гибридов между GUia millefoliala и G. achillelfo-
На, выращиваемых на экспериментальных участках, заложенных
в трех повторностях, наблюдалось удвоение хромосом. В отли-
чие от этого иа участках, которые также были заложены в не-
скольких повторностях, но на которых были высеяны гибриды,
не имевшие в качестве родительских форм ни G. millefoliata, ни
G. oaUtidensis, амфиплонды не появлялись [286]. В свете этих
данных предстааинется знаменательным, что из трех известных
природных тетраплоилных видов, относящихся к секции GUia
рода GUia, два несут геном millefoliata [286].
Левин [470] обнаружил аналогичное явление у рода Phlox
в восточной части Северной Америки В этой области встреча-
ются пять аллотсграплондных видов Phlox. Четыре из этих пяти
тетраплоидоя несут геном диплоидного вида Phlox pilosa [470].
Таким образом, создается впечатление, что родительские ви-
ды генотипически различаются по способности производить ам-
фнплоидных потомке» как в природе, так и на эксперименталь-
ных участках при более или менее одинаковых прочих условиях.
Возможно, что гены, входящие в состав различных геномов,
представляют собой один из третьестепенных факторов, воздей-
ствующих на распределение полиплоидов в пределах данного
рода или секции, в состав которых входят разные симпатричес-
кие виды.
Гл а. Факюры, * гтснгл-м #йгмиллОдМи10 «ОЗвялоядня 3|3
Общие выводи
Полиплоидия предстамляет собой генетическую систему» часто
встречающуюся и широко распространенную среди высших рас-
тений. В разных таксономических группах сосудистых растений
полиплоидия распределена неравномерно, и распространение ее
зависит от Мпнмата, жизненной формы растении, его системы
размножения, структуры хромосом, их величины и от других
факторов. Причины эволюционного успеха полиплоидии у выс-
ших растений кроются в этих разнообразных корреляциях, и их
можно выявить, проведя соответствующий анализ.
Огромное болыинистио истречающихся в природе полиплоид*
пых видон растений, подвергнутых генетико-таксономическому
анализу, оказались амфмллоидлмн. Дмфнплоидмя представляет
собой определенного типа перманентную гнбрндность. Эта гене-
тическая система дает возможность межвидовому гибриду пре-
одолеть барьер стерильности и воспроизводить в чистоте свою
гибридную оаепгушп Кроме того, она позволяет растениям
испильтоаать преимущества, присущие высокогетерозиготному
у» в частности гетерозис и физиологический ГОНГОМ I
многочисленных гибридных комбинациях, которые содержат эти
выгодные свойства
Способность какой либо группы растений испапьзимагь »•
своем эволюционном развитии преимущества, предоставляемые
вмфкплоидиой генетической системой, зависит от разнообразных
факторов, различающихся также по своему' значению. Один фак-
торы оказывают влияние на возникновение нового полиплоида, а
другие—иа последующее создание нм природной популяции.
Как в возникновении, так и в закреплении нового полиплоида
участвуют разного рода внутренние и имгшине факторы
Первостепенное н фундаментальное значение имеет сочета-
ние трех условий Первое наличие диплоидных в и дож, несу-
щих различные геномы или субгеноыы. Второе —естественная
гибридизация между этими видами Третье—большая продол-
жительность жизни многолетних растений, попншающая вероят-
ность соматического удвоения, или» и качестве частичной компен-
сации у короткоживущих однолетних видлв, — автогамная си-
стема размножения, повышающая вероятность соединения перс-
аудированных гамет. В случае отсутствия у данной группы рас-
тений любого из этих трех условий следует ожидать, что поли-
плоидов в есй нет или они редки Изучение распространения по-
липлоидии в .зависимости от хромосомных перестроек, гнбрид-
яосгн и многолетнего образа жизни (или автогамии. заменяю-
щей его у однолетников) показало, что полиплоидия и в самом
деле часто встречается только в тех группах растений, которые
обладают этими признаками.
314 Чает» шспа. Поыллоива*
Не во всех группах растений, отвечающих этим трем глав-
ным условиям, полиплоидия распространена одинаково равно-
мерно или встречается одинаково часто. Неравномерное рас-
пространение полиплоидии среди разных групп растений, срав-
нимых в отношении факторов первостепенного значения, свиде-
тельствует о существовании второстепенных и третьестепенных
факторов.
Нарушение местообитания и стрессовые условия среды, часто
в холодном климате, принадлежат к числу внешних факторов
второстепенного значения, благоприятствующих возникновению
полиплоидии. К второстепенным внутренним факторам, препятст-
вующим возникновению полиплоидии, относятся в числе прочих
хромосомы, имеющие слишком крупные по сравнению с клеткой
размеры, и клетки, величина которых мала по сравнению с раз-
мерами н числом содержащихся в них хромосом. Наличие или
отсутствие генов, вызывающих нарушение второго деления мейо-
за, а следовательно обусловливающих образование вередуииро-
ваниых гамет, —третьестезеяный фактор, который, возможно,
позволяет объяснить относительно неравномерное участие раз-
личных диплоидных видов в создании амфиплондов в некоторых
родах.
25 Полиплоидный комплекс
Нередко те самые факторы, которые способствуют образова-
нию одного нового тетраплоидното вида, способствуют также
возникновению других новых тетраплондов кз других диплокдов.
относящихся к згой же группе видов. Эти же факторы могут
затем привести к образованию тстраплоидамн гекса пл оидаь и
октоллондов. В результате такой непрерывной полиялондизации
ни разным ветвящимся направлениям получается полиплоидный
комплекс.
Одна из простейших возможных структур полиплоидного
комплекса представлена на рис. 25 I. Изображенный на этом
рисунке комплекс состоит из трех диплоидных видов, из двух
возможных аллотетраплэндпых производных и одного аллогек-
са плопда. Вместе они представляют сабой шесть видав двой-
ников, изолированных барьерами хромосомной стерильности, а
возможно, и другими, связанными с последними, барьерами;
однако эти виды содержат много одинакового генетического ма-
териала, и поэтому диапазоны изменчивости по морфологичес-
ким признакам, н экологическим предпочтениям у них перекры-
ваются.
Другие возможные типы этого простого полнплондпого комп-
лекса могут состоять из автотстраплопдов к вллотстраплсндов,
вптоаллогсксапдоидов и геномных аилогсксаплоидов или алло-
октоплоидов и гексаплоидов. При наличии достаточно обширного
ряда диплоидных видов в качестве основного фонда эволюция
путем полиплоидии может создать очень большой полиплоидный
комплекс, содержащий многочисленные тетра ллоадиыс и гекса-
плондпыо виды, а некоторые потомки этих последних достигают
в нем еще более высоких уровней плоидности
Методы анализа
Полиплоидный комплекс можно идентифицировать при помощи
цитотаксонсм ячеек их методов Филогенетические взаимоотно-
шения между входящими в него видами-двойниками можно
установить при помаши цитогенетических и гекетнко-такеоилми-
веских методов Филогенетический анализ полиплоидного комп-
лекса можно довести до любой степени полноты. Для наших
целей полезно разбить анализ полиплоидного комплекса на че-
тыре главные стадии.
Первая стадия — это пдептифнквцня полиплоидного комп
лскса как такового. Перед нами большая и таксономически
трудная группа растительных видов Судя по морфологическим
31«
Vorra шатал ПолшмоиЛм
Рис. 25.1. Структура простого полиплмдяаго хамплскса.
признакам и по географическому распространению, нх можно
считать видами-двойниками, но таксономические границы этих
видов, их ареалы и генетическая природа вевзвестны, пептону
следует применить цитотаксснюмические методы. Проведение
ряда предварительных подсчетов числа хромосом может выявить
наличие в этой группе растеяиЯ полиплоидного ряда, что послу-
жит первым указанием иа то. что она представляет собой поли-
плоидный комплекс.
Расширение цитотаксононвческих исследований позволит нам
далее выяснить многие детали и завершить картину. Затем сле-
дует перейти ко второй стадии анализа, иа которой иногда мож-
но обнаружить, что разные растения, обладающие сходными
морфологическими признаками и встречающиеся вместе в одной
и той же местности, имеют различные четные числа хромосом.
Эти данные позволяют предположить, что между видами-двой-
никами, находящимися на разных уровнях плоидности. сущест-
вует тесная филогенетическая связь. Или же можно обнаружить
ряд форм с разными и иеиитерграднрующими морфологически-
ми признаками, находящихся ип одном и том же основном уров-
не плоидности и составляющих, таким образом, вероятную ос-
нову изучаемого полиплоидного комплекса. По мере накопле-
ния данных по подсчету хромосом в природных популяциях
можно начать устанавливать корреляции между тем или иным
уровнем плоидности и определенными морфологическими приз-
наками, биохимическими компонентами или величиной пыльце-
вых зерен и устьиц. Установив такие корреляция, можно путем
экстраполяции в известных пределах предсказывать таксономи-
ческие и географические границы некоторых видов-двойников.
Теперь на основе обширных цхтотаксононических данных мож-
но описать в общих чертах наш полиплоидный комплекс.
Для третьей стадии необходимо прибегнуть к эксперимен-
тальным методам, хотя и в несколько предварительном плане.
Гл 25, Полыплонйн&д ксм.ыъс
317
Для этого проводятся скрещивания между некоторыми видлми-
двойниками и определяется плодовитость получении* гибридов.
Эта третья, нли бносистсматичеекая, стадия анализа необходи-
ма дли juju, чтобы убедиться в правильности некоторых родст-
венных отношений, предсказанных ранее на основе цитогаксомо
мических данных Ьиосистсматическне методы дают возмож-
ность определить некоторые филогенетические связи внутри по-
липлоидного комплекса.
Четвертая стадия состоит в приложении экспериментальных
генети ко-таксономических методов кп всем членам изучаемого
комплекса. При по мош к обычных методов искусственной гибри-
дизации. анализа геномов и в тех случаях, когда это возмож-
но, — рсснитсза прослеживав «си родословная одного тстраилонд-
иого вида от его диплоидных предков Используй те же методы,
следует далее усыновить родословные других тетра плоили ых
членов комплекса Наконец, этими же методами устанавлива-
ются родословные тех его членов, которые обладают бодсс вы
со кой плоид|гостыо. Теперь ст» носится возможным проследить
филогенетическое происхождение определенных полиплоидных
видов от определенных диллоидиых видов. Постулированную та-
ким образом филогению можно подкрепить данными цнтогенс-
гичсского анализа искусе;венных межвидовых гибридов, полу-
ченных при с у друг с другом разных полиплоидны!
видов Филогенетический анализ полиплоидного комплекса за-
горней
Здесь полезно провести аналогию с химическими методами
анализа Мы можем сказать, что тайное неизвестное соединение
представляв! собой белок, установив, что в его состав входят
такие зле менты, как азот н сера. Затем, применяя более тонкие
аналитические методы, можно определить его химическую фор
мулу. При помощи ешс более сложных и трудоемких методов
можно вывести его структурную формулу, отражающую химиче-
ские связи между спсгаалнющимп его атомами Химические фор-
мулы известны для многих белков, однако лишь для очень пе-
мпоснх ил них полностью установлена структурная формула.
Так обстоит дело н с изучением полиплоидного комплекса
Идентифицировать ту или иную группу растений хак полиплоид-
ный комплекс (стадия I) относительно легко Достаточно серь-
езный иитотаксомомнческнй анализ (стадия 2) провести значи-
тельно труднее, и он требует многих лет работы. Еще более
трудную задачу представляет бипсистематическнй анализ (ста-
дия 3). который едва ли можно осуществить в одиночку. Излиш-
не говорить, что полный генептко таксономический анализ одно-
го полиплоидного комплекса (стадия 4) — чрезвычайно трудоем-
кая задача, поглощающая массу промечи.
318 Часп шесто*. Полиплоа&й»
Заслуживают упоминания некоторые часто допускаемые ме-
тодологические ошибки. Специалисты по цитотаксономии иног-
да переоценивают надежность описательных цитотаксоном иче-
ских методов в применения к полиплоидному комплексу, недо-
оценивая в то же самое время важность гекетико таксономиче-
ского подхода. Разница между цито- и гекетико-таксоиомиче-
скими методами по затрате труда и времени очень велика; одна-
ко для сколько-нибудь надежного филогенетического анализа
полиплоидного комплекса дополнительные усилия, связанные с
применением экспериментального генетико-таксономического
подходя, абсолютно необходимы
Многие цитотаксоиомнеты неверно интерпретируют получен-
ные ими результаты, рассматривая их в свете таксономической,
а не биологической или эволюционной концепции вида. Цитотак-
совомнст обнаруживает группу диплоидов к тетраплоидов со
сходными морфологическими признаками; он рассматривает их
как различные «хромосомные расы» одного и того же вида
Другая группа диплоидов и тетраплоидов, сходных между собой,
ио отличающихся по морфологии от первой группы, рассматри-
вается как другой вид, состоящий нз двух ^хромосомных рас».
В результате получается таксономическая система, в которой час-
то бывают смешаны естественные родственные связи и взаимо-
отношения по плодовитости. Морфологически различные тетра-
ллондиые типы могут быть на самом деле противоположными
вышепекцами одного и того же сегментного аллотетраплоида
(ЛзАИИт) и в качестве таковых должны рассматриваться как
относящиеся к одному и тому же виду. В таких случаях погра-
ничную черту, разделяющую разные виды, следует проводить не
а еередиие комплекса A^A.A,Ai. а между этим комплексом в це-
лом и диплоидными видами А,А, и ЛЛ>.
Многие группы растений были идентифицированы как поли-
плоидные комплексы. Довольно большое число полиплоидных
комплексов уже подверглось детальному цитотаксоиомическому
анализу, а некоторые из них — н предварительному биосисгема-
тичесхому анализу. И лишь для нескольких полиплоидных комп-
лексон был проведен обширный, хотя и неполный генетико-так-
соиомический анализ.
В двух следующих разделах рассматриваются главные осо-
бенности некоторых полиплоидных комплексов, изученных цито-
таксоаомнческя к биоеястематнчески. Далее будут рассмотрены
полиплоидные комплексы Triticum—Aegllops, Bromus и Gilia,
по которым были проведены обширные генетико-таксономиче-
ские исследования.
Га 25 ПллшиовРпьй комплекс
31
Цитотаксономнческие примеры
Эрендорфер и его сотрудники провели цитотаксономлческ’1й яиа-
ini полиплоидных комплексов в нескольких ветвях рола Galium
iRubiaceae) Такие полиплоидные комплексы описаны в группе
G pumilum в Европе [184], в группе G. anisophyllum также в
Европе [191]. в группе G. Incanum в Турпин и соседних с ней об*
ластах [185] и в группе G. multiflorum в западной части Север
гой Америки [161, 189].
Гр)ппа Galium pumilum (х—II) в Европе содержит поли-
плоидный ряд пт диплонллэ до охтоплоидпз Диплондний член
комплекса, G au^frutcum. встречается как реликт ледникового
времени в разобщенных районах Альп, где входящие я этот
комплекс полиплоиды занимают обширные и непрерывные арса
ты 1184].
Полиплоидный комплекс в группе Galium anisophytlum я
Европе содержит псе четные полиплоиды от 2л до Юл Здесь
снова липлояды встречаются в ряде убежищ, разбросанных по
горам Европы, а тетраплоиды занимают непрерывный п обшир-
I ый ареал. Распространение видов с высоким уровнем МОЙА*
гости показано па рис. 25 2 |191].
San/cwfa ctassicaulis и родственные ей фооиы (Unibell I Ге гае)
образуют еще озим полиплоидный комплекс с основным числом
хромосом х-8 [55! В таксономический вид S crasslcaulis вхо
дят тетраплоиды. гекса плоили и октоплоилы, занимающие об-
мирную область вдоль тихоокеанского побережья Северной
Америки н Чили Полагают, что диплоидные виды 5. laciniata п
5 hoffmannil, встречаюшиеся в прибрежных районах Ка.тифор
пни. служат вероятными предками тениплоидиого 5. crassicau’
lit Другой диплоидный вил, S bipinnatifida. суля по его морфо
логическим признакам, возможно, был одной из предковых форм
некоторых октоплоидпыч членов этот комплекса [55].
Группа папоротников, принадлежащих к роду Asplenium
(Polypodiaccac) и распространенных в восточной части Север
пой Америки, образует еще один полиплоидный комплекс при
основном числе хромосом x~3G [86-11. В этом комплексе есть
диплоидные виды, обладающие крайним* морфологическими
ппизияками. — A. montanum. A. platyneuron и A. rhizophyllum
(2л—72) В некоторых случаях промежуточные по морфологи-
ческим признакам формы являются стерильными диплоидными
или триплоидными гибридами. Другие—плодовитые— враче
жуточяые формы предо™ вл яют собой тетраплоиды (2n— 144)
Таким образом, тетраплоид A pinnatilidum зяньмлет по морфа
логическим признакам промежуточное положение между A mon-
tanum и A. rhizophyllum и. вероятно, является амфкплондным
производным этих двух диплоидных видов [864].
"(161] wn;jX\fdcfM> tunfjoQ anXdj ж «птвжаггпк
-eda 'яоГиагпшгоа и «огмогамт MHaMtdrjcdrocd эохэжхф«Ьоод Л<1
Гл 25 По.шллотЭчиА комплекс
321
Родственные взаимоотношения в этой группе Asphtnium,
установленные Вагнером [864] прн помощи цитотаксоиомичсских
методов, были затем проверены с применением хемотаксоном и*
ческнх методов (740). Методом хроматографии на бумаге было
исследовано содержание определенных химических соединений
у разных видов. Оказалось, что естественные гибриды и алло-
тетраплонды содержат в разных комбинациях вещества, имею*
шисся у нх предполагаемых диплоидных родительских видов
[740]. Хемотаксонсмкческис данные подтвердил и филогению,
разработанную на основе цшатоксопомичесхих давших.
Другие полиплоидные комплексы, изученные ниготаксоними-
чески ми методами. — это группа Polypodium vulgar* (Polypo-
diaceae) в Европе и Северной Америке и группа Trillium cams ch-
tatetnse (Ldiaceae) в Японии (454. 554 556]. Среди миогочис
ленных полиплоидных комплексов в сем Gramineae, исследо-
ванных цитотаксоиомнчсскимн методами, укажем группу Огу-
zopsis-Stipa в Северном полушарии, циркумполярную группу
Paa alpina и группу Bouteloua curttpendula североамериканских
равнин [262, 411, 627].
Бмосистематнчсские примеры
Группа Phacelia httirophylla состоит из свободно скрещиваю
щнхеп многолетних и двулетних травянистых растений, широко
распространенных пи всей западной части Северной Америки и
южной части Южной Америки. Эта группа сложна в такс о пом и
ясском отношении, так как содержит много разных, по ингер-
градирующих морфологических форм. Хекард [338] установил,
что данная группа представляет собой полиплоидный комплекс
(х— 11), состоящий нз шести известных диплоидных аилов, мно-
гочисленных тетра пл он лов и нескольких гекса илпидоа
Как показывают результаты искусственном гибридизации,
между диплоидными видами существуют хорошо развитые барь-
еры, препятствующие скрещиванию, и барьеры, создаваемые
стерильностью гибридов. В отличие от диплоидов различные
тетраплоидные формы в бпльшнпствс случаев довольно легко
скрещиваются друг с другом, давал полуплодовнгых или плодо-
витых гибридов. Эти различия в степени внутренней изоляции
коррелируют с различиями в характере измениниосги На лип
лоидном уровне таксоны обычно разделены разрывам н в мор-
фологических признаках, тогда как на тетр а пл о идиом уровне
таксоны часто образуют непрерывный ряд благодари ннтергра-
дацни по этим признакам [358].
Полиплоидные представители группы Phacdia hderophylla,
вероятии, представляют собой амфиплоидов, возникших в ре-
зультате гибридизации между диплоидными видами в различ-
21-45
^22 Часть шестая. Полиллоя^ил
иых комбинациях; такое толкование послужило бы дальнейшим
объяснением их нитерградаиин по морфологическим признакам.
У нас пока нет никаких данных относительно происхождения
различных тетраплоидных и гекса пл ом дм ых форм.
Хскард [358] объединил морфологически близкие диплоидные
и тетраолокдиые формы в один вид. Следовательно, единицы,
которым были даны видовые названия, являлись во многих слу-
чаях таксономическими видами, но отнюдь нс биологическими
или эволюционными видами. Полученные Хекардом генетические
данные иногда не согласуются с морфологическими данным и,
чего и следовало ожидать Так. диплоидны А вид Phacelia nemo-
ralis не удалось искусственно скрестить с тетраплоидным Ph. пе-
moralis, однако тетраплоидиый Ph. nemoralis скрещивается с
теграплондиым Ph. californica, образуя высокоплодовитые гиб-
риды. Дальнейшие этапы анализа этого интересного полиплоид-
ного комплекса должны состоять в том, чтобы выделить вчерне
биологические виды на тетраплон дном и гекса лломдком уровнях
и установить их родословную.
Род Clarkia (Onagraceae) в западной части Северной Амери-
ки н в умеренной эоне Южной Америки содержит более обшир-
ный полиплоидный комплекс [496]. У диплоидных видом 2п —10,
14, 16 и 18 хромосомам при первоначальном основном числе
х»7. Эти диплоидные виды распадаются иа несколько ясно вы-
раженных групп видов, которые по праву возведены в ранг от-
дельных родов. Godetia, Euchardium и Clarkia sensu stricto.
Однако те филогенетические линки, которые на диплоидном
уровне дивергнруют, на тстраплондвои и гексаплоидном уров-
нях сливаются я различных комбинациях Такне анастомозы
служат основанием для того, чтобы рассматривать всю эту со-
вокупность как один род Clarkia sensu Into (196].
Постулированная сетчатая филогения рода Clarkia так, как
ее представляли я 1955 г., схематически показана на рис. 25.3.
После 1955 г, Люнс и его сотрудники получили много дополни-
тельных данных. На рис. 25.3 изображена вероятная родослов-
ная различных полиплоидных видов Clarkia. построенная на ос-
нове морфологических и цитологических данных [496].
Группа Achillea millefolium (Compositae) — полиплоидный
комплекс (л—9), широко распространенный в Северном полу-
шарии [102, 188] Собственно A. millefolium в Старом Свете н
A. borealis в Новом Свете являются гексаплоидами; A. colllna
и A. virescens в Старом Свете и североамериканский 4. lanulo-
за — тетра пл силами. В Европе встречаются два европейских
диплоидных вида—A. asplerufolia (на влажных лугах) и A. хе-
tacea (в сухих степях) (102, 189].
Ареалы диплоидных видов Achillea asplenifolia и Л. set ас га
перекрываются, и, несмотря на сильные внутренние изолирую-
324 Чист* шегтол. ПолиллоиОм
вше барьеры, эти виды гибридизируют в природе, вот» и в ог-
раниченной степени. В Альпах происходит интрогрессия от
A. setacsa к A. asplenifolia на диплоидном уровне. Тетраплокд
A. collina — амфиплонд, происходящий от этих двух диплоиден.
Затем A. collina гибридизирует с другим тетраплондным видом
A. virescens. В некоторых сочетаниях естественная гибридизация
происходит также между тетраплоядными и амфиплоидными
видами: A. collina (4х)ХА. millefolium (бх) и A. lanulosa (4х)Х
ХА. borealis (бх); при этом образуются аиеуплондные потомки,
что еще больше осложняет характер изменчивости всей этой
группы [189].
Цохари и Фельдман [918] обнаружили сходную ситуацию в
полиплоидном комплексе, в который входят Triticum и Aegilopt
(Gramineac). Здесь так же, как и у Achillea. разные аллотетра-
плоидные виды с различной геномной конституиней гибридизи-
руют в природе, что еще больше усложняет н без того сложную
изменчивость этой группы (918].
Комплекс Gilia inconspicua
Группа GlUa Inconspicua (х=9)— это часть секции Arachnion
рола Gilia, которая состоит из. автогамных однолетников. Эта
группа широко распространена в западной части Северном
Америки и на юге Южной Америки. В Северной Америке изве-
стно 23 вида этого комплекса, а в Южной — 2 вида; есть веские
основания считать, что в каждой из этих областей существуют
и другие еще неизвестные виды. Из 23 североамериканских видов
13 диплоидны (2л = 18) и 19 тетравлоидны (2п=36). Эти тетра-
пломды представляют собой амфиплоидов, и в большинстве слу-
чаев их предки установлены
В гл. II группа Gilia Inconspicua была приведена в качестве
примера одного нз типов межвидовых взаимоотношений, обыч-
ных среди автогамных однолетних растений. Часть этой группы,
к которой относятся О. transmontana и родственные ей виды,
уже рассматривалась в гл. 4 в связи с проблемой видов двойни
ков (см. ряс. 4.5). Здесь мы ввовь рассмотрим эту группу как
полиплоидный комплекс, который подвергся широкому генети-
ко-таксономическому изучению, но все еще не проанализирован
до конца.
Диплоидные виды этой группы распадаются на 5 геномных
групп. Родственные вады, принадлежащие к одной и той же ге-
номной группе, обычно различаются по субгеномам [300].
Гибриды Р|, полученные от 17 скрещиваний между тетра-
плоимыми видами в разных комбинациях, были подвергнуты
цитогенетическому анализу. У большей части этих гибридов
конъюгация хромосом в мейозе протекала на среднем уровне.
Гл. К. Полиплоидный пояпмкх 325
Так, я Fi от скрещивания GUia malior и G. transmonlana наблю-
далось в среднем ло У бивалентов при диапазоне от 7 до 15 11,
тогда как прн полной конъюгации образуется 18 бивалентов.
Уровень конъюгации хромосом у этого н у сходных с ним гиб-
ридов позволяет предположить, что два участвовавших в гибри-
дизации тетраплоидных вида имеют одни общий геном, по раз-
личаются по второму геному [285].
Для некоторых пар тетра плои д ныл видов это предположение
подтвердилось; общий диплоидный геном идентифицировали,
получив соответствующие триплондные гибриды и установив, что
конъюгация хромосом происходит, как и предполагалось, по ти-
пу Drosera. Следовало ожидать, что у тркплондных гибридов,
полученных в результате скрещивании между тетраплоидпими
и диплоидными видами, принадлежащими к одной из линяй
предков —потомков, конъюгация будет происходить по ти-
пу >9I1+<1SL
Именно такой тип конъюгации и был обнаружен у гриплонд*
пых гибридов GUia sinuala (4x)xG. tat if lor a (ii) и G. modocen-
гй (4x)xG latiflcra (2x) [285] Следовательно, геном G tati(lo-
ra. или геном T, имеется как у G. smuata. так и у G modo*
censis Ila основе морфологических и экологических данных мож-
но сделать вывод, что именно G latlflora. а не какой либо дру-
гой член Т геномной группы послужил одним из диплоидных ро-
дительских видов и для G. sinuata, и для G. modocensis. Что же
касается видов, сыгравших роль второй родительской формы
для этих двух тетрапломдкых видов, то они различи млея как по
споим геномам, так и морфологически.
Дэй [153| провела подробное таксономическое исследование
двух тетр а пл оидпых видев и нх предполагаемых диплоидных ро-
дительских видов в подгруппе GUia transmontana. Морфологи-
ческие. экологические, цитологические н цитогенетические дан-
ные свидетельствовали п том. что родословная этих двух тегра-
плондов, G transmontana к G. mahor, носи г сетчатый характер
и что они происходят от трех диплоидных видов, как эго пока-
зано на рис. 25 4. Дэй затем подтвердила правильность такой
филогении, искусственно рееннтезировав G transmontana и
G malior из нх диплоидных родительских форм к показав, что
каждый искусственный амфлплонд дает плодовитых потомков
при скрещивании с соответствующим природным тегрэплоид-
иым видом (158].
Сходные данные были получены для родословной теграпло-
ндниги вида, известного под названием GUia ophrhalmoides 111.
Считалось, что этот вид происходит от скрещивания между
G. clokeyi (2х) и G. тепгачд (2*). Сломинные тетраплондмыс
потомки, полученные от этого диплоидного межвидового гибри-
да, оказались хорошими G. optuhalmoides, чти подтвердило пред-
ад
Чат шестая. Лашыоидал
Рис 20.4. Филогении двух частей полниондного комплекса а группе Gltia
(acaaspicua (по дажпым [158, 283. 300])
полагавшуюся филогению ({300]. а также неопубликованные
данные V. A. Grant). Имеются и другие данные, указывающие
на частично общую, а частично отличающуюся родослоеную
близкого к G. ophthalmoiiies 111 тетраплоидного вида —G. орМ-
halmoidcs I.
Эту сетчатую систему родственных связей можно расширить,
включив в нее Qilia sinuala. G. modocensis и другие тетраплоид-
иые вады. Сетчатая родословная двух частей этого полиплоид-
ного комплекса изображена на рис. 25 4
Triticum и Aegilops
Хорошо известный полиплоидный ряд пшеницы составляет часть
более крупного полиплоидного комплекса, в который входит
Aegilops. Triticum и Aegilops связаны между собой на полипло-
идном уровне Разграничение старых родовых единиц — пшени-
цы и вгнлолса,— основанное иа различиях в признаках колоса
и колосков, оказывается, таким образом, несостоятельным. Оче-
видно, Triticum и Aegilops составляют одни род, и специалисты
по генетике пшеницы склонны перенести виды, относившиеся до
сих пор к Aegilops. в род Triticum. Однако па таксономическая
и номенклатурная реорганизация еще не закончена, и пока что
Гл. iS. Полиплоидный МЯЛЛ1КЛ
32Г
Таблица 25 1 Система классификации и геномная конституция
пшениц (Triticum) (основана на данных Фельдмана (211);
данные о распространении по Харлану н Цохарн (348]. а также
по Цохарн и др. (919])
I. Т monococcim Охкохрпянка. 2х. А<4
а) Дмсрастущая раса Т. « ЬогоГйжт
Ближний Всего» or Турами до Иряз* и южная частя Вымнет
00.1JOC1 роза*
б) Кудьтумая раса Г m ляолос&ссил!
2 Т i/trgiaum Культурная двузернянка н твердая п-пепяиа 4х А *8 В
а) Д иорастущая раса Т t dicocc-jutes
Ближний Босток or Турция и Израиля до Ирана
6) К.>льгур*4И плптт»1 раса Г f duotcum
В) Культурные ю.эиэеря'ые расы. Т t durum, turgidum. poionue®.
carthkcum
3 Т limopheinii и AAGG
а| Дикорьстрцажа paca 7. t агат at i curt
На юг* СССР, в Грузии х Армении*
б) Культурная раса
4 Т wtivum ( — Г (Ndeore). Мягкая. или обелмовсмияя, пиаенякя, бл
AABBDD
а) Культурные пленчатые расы Т ar tprita. нядсЛа. меЦоеи
GJ Культурные голозерные расы 7. о*. udtiivum. c^/fupactum, арЛшго*
едегизн
1 Г «оганлнч агт*ям«гя такие I Южном Кры«<-. а ыжнгн «зет* 3*1 и.» ore Ла-
амы*ав я • Южжы » иосг«нмзм 3* к* maim ытяжм! анз. 1 агоЛл. актмммгся Afr-
мскм и • ствт|»_^•остьим* «астм Т«рмм |Ца«ам Н Н .ос ell» — Лрим рей1
4 Г areraiuuw нортжаекч также к смн|и-впст<»«»вЛ «ueiK Т>уа»и и в Сгмргоы
Право — Лдкм р«<>
мы имеем дело с двумя перекрывающимися наборами родовых
и видовых названий.
Triticum—Aegilops — группа однолетних злаков, принадле-
жащих к подтрибг Tnricinae трибы Hordeae’ Как у Triticum
Aegilops, так и у трибы в целом основное число хромосом ж—7.
В отношении рода Triticum в прежнем узком смысле в пос-
ледние годы наблюдается тенденция к низведению многих групп,
прежде считавшихся видами, до ранга подвидов, или культива-
ров. что упрощает систематику пшениц. В табл. 25.1 представ-
лена система пшениц, основанная иа работе Фельдмана [211].
Геномный анализ пшениц проводили Кнхара [436. 437]. Сирс
(718. 719] и многие другие. Полиплоидная пшеница получила
геном А от днплоидмого вида Triticum monococcum Источник
генома Я у тетраплондиой Т. turgidum неизвестен; мы вновь вер-
немся к этому вопросу чуть ниже. Геном D у гексап.юидноА
Т. aestivum происходит от диплоидного вида Aegilops squarro-
* Согласно ормилу орнорггега, эту трибу следует называть TfRIceae
(см Цы-лев Н. Н. Злаки СС<_Р. И»д-«о «Наук*», 1976, с. 105). — Прим
м
Част» шестая. ПомтлоиОия
sa(—T. /шзсЛи)1, запимаютего обширный ареал в западной
часта Азии и на юге Средней Азии.
Геном Я имеется как у Triticum turgidum. так и у Г. aesli-
vum. Попытки найти диплоидный вид с конституцией ВВ оказа-
лись безуспешными. Одно время полагали, что Aegilops speltoi-
des( — TriUeum speltoides) является носителем генома В, однако
цитогенетические и электрофоретические данные противоречат
Лой гипотезе, и в настоящее время она а своей первоиачальиой
форме отвергнута. Теперь считается, что Ле. speltoides содержит
геном SS. Что касается необнаруженного диплоида ВВ, то воз-
можно. что он вымер. Другая возможность состоит в том, что
диплоида ВВ никогда и ие существовало. Геном Я, имеющийся
у Г. turgidum, мог возникнуть у тетраплоидной Т. turgidum в
результате дивергенция от какого-нибудь Другого родственного
генома, возможно S [211).
Филогению полиплоидного ряда, ведущего к мягкой пшени-
це, можно представить следующим образом: 1) гибридизация
между дикорастущей однозернянкой (АА) и иеидентифициро-
ваикым диплондом, несущим геном Я, или каким то предшгст-
венииком Я, с образованием амфиллоидной дикорастущей дву
асрнянки (ААЯВ); 2) превращение дикорастущей двузернянки
в культурную пленчатую, а затем в культурную твердую пшени-
цу в результате изменений, вызванных селекцией а условиях
доместикации; 3) гибридизация между культурной двузернянкой
и Aegilops squarrosa (DD) с образованием амфиллоидиой
мягкой пшеницы (AABBDD); 4) селективные изменения при
культивировании, приведшие к превращению пленчатой пшени-
цы а голозерную мягкую пшеницу.
Эти изменения происходили в западных и юго-западных
районах Азии. При археологических раскопках в этих районах
в ряде мест найдены карбонизированные зерна Triticum rnono-
coccum. Т. turgidum и Г. aestivum. Возраст древнейших найден-
ных зерен этих трех видов соответствует 9000 лет до в. э. (211J.
Дикорастущая двузернянка встречается в настоя шее время
на Ближнем Востоке от Израиля и Ливана до реки Тигр и Кас-
пийского моря [348]. Твердые пшеницы выращивают главным
образом в Средиземноморской области, где климат относитель-
но мягкий [919]. Aegilops squarrosa распространен от прикаспий-
ского региона до среднеазиатских степей и доходит на востоке
до Ташкента и Алма-Аты [919]. Этот вид. очевидно, представлен
рядом экотипов, в том числе экотипами, адаптированными к
1 И Н. Цмлп (toe сК) считает Aegilops squarrosa сшвопвмом A Irian-
ciatis tobip iftaniiaUs распространенного от Средиземноморья до Ср* дней
Аля*. • Де (азмсАй ом прюжмт едипетсетмьным вхмм и дзясе относят к
другой сеяния родл A egilops Прим ред
Гл 2S По.шиижЬша ломялеч
зй
резко континентальному климату. Экологические адаптации
Ае squareosa составляют самый важный вклад итога вида в
формирование мягких пшениц. Налипнем у гексаллоидной мяг-
кой пшеницы генома О объясняется се широкое экологическое
разнообразие и ее способность расти в холодном и сухом степ-
ном климате ([919]; Харлан, личное сообщение)
Интересно отметить, что этот процесс продолжается у искус-
ственно синтезированного аллооктоплоила TritUale — амфипло
идного производного Triticum aestivum (6х) XSecale cereate (2х),
который можно выращивать в северных широтах н в горных
раПопах
Triticum timopheeuii обладает геномом AAGG (табл. 25.1)
и представляет собой, таким образом, еще одно тетраплоидиое
производное Т. monococcum. Другой диплоидной родительской
формой мог быть Aegilops speltoides (SS) или такой близкий
к нему вид, как Ае. bicornis (S‘S‘) Геном S близок х геному
G. Эта филогенетическая гипотеза допускает, что геном S прев-
ратился в геном G у тетраплоида Г. timopheecii [211]
Среди большого числа видов, относимых к старому роду
Aegilops, встречаются диплоиды, тетраплоиды в гексаплоилы.
У Aegilops различают пять геномов и еще большее число субте-
номпе Пить геномных групп, имеющихся у диплоидных видов,
во большей части совладают с таксономически ни секциями это-
го рода. Главные геномы, субтеноыы и их диплоидные носители
перечислены в табл 25 2 Некоторые субгеномы (М1', D!,S' и
т п). известные только у полиплоидов, в табл. 25 2 ис вклю-
чены.
Геномы полиплоидных видов Aegilops приведены в табл. 25.3,
из которой видно перекрешиваиие филогений этих видов. Так,
Таблица 25.2. Геномы н нх диплоидные носители у
Aegilops [437. 918]
Гспм Cydrui5u Дмп.кмдкйЛ мл
Mt Ar ruuiifei •
S S’ Ar NeomU
5 At iftaroiwute. loKglsuxma
5 Ле iptUntdei UgMtica
D Ae rjuaf'ata
С с At Caudata
с- ,4* n.wbrKafa
.м м At corrtoiu
м« At iMiaHxJala
I тыгн« • ик'мии* аымзютса • crwund моно
„mull рел АгаЫулдоиа lew Uiewbll Н op сП. С — Рэм ргд.
330 Часть шестая. ПолаллоиНия
Таблица 25.3. Геномная конституция
полиплоидных видов Aegilops (918)
Пздип*зяд*м> *»A
Г«ОИ
Ле с/аиа (♦*)
А», спим (6х)
A* iwenalit
Ав. ьвШечляа
Ac cyUnd/fCA
Ac oafiabUu в Ал. Aotschpl
Ас МшиЫ/в
Ас со/шплаеи
Ав bitinaalfs
Ac iriarislaia (4г)
Ал tnariflala (fix)
Ac ocata
DDM-M”
DDOWM^M"
DDCOAUM'
DDCC
COSO
OC»CC
CX>MW
С-С-ЛРМ*
ОЫШ
COM'M'MW
OC'MW
DDM*M*
два диплоидных вида, ведущих геномы DD и ММ. дали начало
тетравлоидам с конституцией DDMM. Один из этих диплоидов
(DD) я другой днплоад (СС) дали тетраплоид DDCC и т. д.
Разные тетраплоидные виды нередко имеют по одному общему
диплоидному предку и различаются по другому. Процесс рекой-
бинацни геномов продолжается на уровне гексаплоидов.
. Сетчатый характер взаимоотношений в группе Triticum—
Aegilops представлен здесь хак сложение различных геномов
в результате амфка-тондин Однако это лишь часть проблемы
Имеются данные, как прямые, так и косвенные, свидетельствую-
щие о естественной гибридизации и интрогрессии между некого
рыми участвующими во всех описанных выше процессах видами.
В результате такой гибридизации изменялись морфологические
признаки, а. возможно, и геномы этих видов (£11, 636, 918, 919)
Старые полиплоидные комплексы
С течением времени диплоидные виды, входящие в данный
полнялондный комплекс, могут вымереть. Могут вымереть так-
же и низкие полиплоиды, так что единственными живыми пред-
ставителями комплекса останутся формы с более высокой пло
ид костью.
Группа Bromus carinatis (Gratnineae) служит примером по-
липлоидного комплекса, далеко продвинувшегося в своем разви-
тии. В этой группе есть формы, достигшие высоких уровней пло-
идиостя. например В. arttonicus (12л). однако многие ее дипло-
идные и тетраплоидные представители вымерли Стеббинс и его
сотрудники (779. 781. 794. 795] установили филогению 12х- и
8х видов этой группы.
Гл 2S. Полиплоидный колпмкс
331
Комплекс Bromus carinatus принадлежит к секции Cerato-
Моа рода Bromus, ио входящие в пего виды содержат геномы
двух близких секций, Bromopsis и Nsohromus. Все эти расте-
ния— многолетние злаки, нередко рыхлокустовые. произрастаю-
щие а Северной и Южной Америке. Основное число хромосом
х—1.
В секции CeratoMoa рода Bromus" диплоидные или тетра-
влондиые виды неизвестны. В. catharticus и его родичи, произ-
растающие в Южной Америке,— аллогексаплоиды, несущие
геномы, происходящие от диплоидных видов, которые вымерли
(рис 25 5).
Bromus carinalus и В. marginatus, растущие в Северной Аме-
рике. — аллооктоплоиды. Три из имеющихся у них четырех ге-
номов гомологичны геномам южноамериканских гексаплойдоа.
Четвертый геном L, судя ио цитологическим и морфологическим
данным, происходит от какого то североамериканского диплоид-
ного вида, относящегося к секции Bromopsis (рис. 25.5). Эти
октоплоиды амфиплоидпого происхождении возникли, вероятно,
в западной части Северной Америки в конце третичного перио-
да, возможно в плиоцене. Приходится допустить, что в прош-
лом в Северной Америке существовал и какие-то гсксаплоидкые
виды, гомологичные современным южноамериканским гексан.>о-
идам (779, 794].
Bromus anzonicus — амфиилоид с числом хромосом 12х. Он
также гомологичен южноамериканским гексаплоидам по трем
из своих шести геномов (рис 255). Остальные три генома
В. arisonicus происходят, по-вндимому. от какого то аллогекса-
илонда, принадлежащего к родственной секции Ntobromus
(рас 25.5) [795].
Двекадцатиплоидиый вид Bromus arizonicus — обычный и
агрессивный сорняк иа большей части своего ареала в Аризоне
и Калифорнии. Его диплоидные и тстрацлондные предковые
формы вымерли Его гексаплпидный предок, относившийся к
секции Ceralothloa, в Северной Америке вымер, возможно в пе-
риод неблагоприятных климатических изменений в плиоцене
плейстоцене, по дожил до наших диен в более умеренных кли-
матических условиях Южной Америки Между тем 12-плоидпое
производное этих вымерших ди плои лов и низких полиплоидов —
агрессивный вид, процветающий в настоящее время к Демонст-
рирующий. как непрерывное амфпплоидиое удвоение может
вдохнуть новую жизнь в старый полиплоидный комплекс [779,
781].
* Некоторые спстекатвии, в тон числе Н Н Шелев Сое eit. стр 207).
с«п>»> с, ко.ли CrrttoMou ролл Bromus еычеитоагвзьным родин Csruto
<Ыоа — Прим род
ЗЮ
Часть восстал. Полиплоидия
Рис. 255 Филэгенетнческл малмоотиовеихя мкокооолиоловдмы! налов
Bromiu {78IJ
Днплзкдгшс или тетрволс'ядпые иосхгтелж хежсторих нсхсдпых геномов не-
Jвестиы
Остатки древних полиплоидных комплексов
Стеббинс (787, 789] отметил наличие обратной корреляции меж-
ду степенью морфологической спспкалноацни и уровнем плоид-
ноети разных семействах покрытосеменных Оказалось, что в
пределах данного семейства растений для морфологически более
примитивных родов характерно яоливлоидаое основное число
Гл 25 Полиплаж^чыА комлмкс
333
хромосом Напротив, у родов с более специализированными
морфологическими признаками основные числа — диплоидные
Приведем несколько примеров [789):
LiliKeee
М' । filer ксае
Pctanoniiceac
ilydraphyllace*e
Сагарам tae
Морфологически
прммхтканые виды, ПО-
лмпжжАные
основные ЧИСЛА
TojltUiA
DtiHUfi
С ant на, Cobata
WigarzdiA
Heliantbeie
Морфологически
слеииАЛИЗирсммныг ви-
ды. ДЯВЛОЩШМе OCHOA-
иыс имел»
Trillium, S cilia
Таилалл1а
Gilia, РМок
Nemnphila
Cidwieae
Эволюционно продвинутые роды в этих семействах сохра-
нили днплоидность своих предков Сохранение дхплоидиостн
играло, возможно, важную роль в нд способности достигнуть
высоких ступеней эволюционного прогресса. Вместе с тем при-
надлежащие к тем же семействам роды с высокими хромосом-
ными числами — давние полиплоиды, нередко остатки старых
полиплоидных комплексов, сохранившиеся в неизменном состоя-
нии (787, 789].
В некоторых семействах древесных двудольных все предста-
вители отличаются высоким основным числом хромосом, неко-
торые авторы [780, 787, 832] рассматривают это как сохранение
давнего полиплоидного состояния, что кажется правдоподобным.
Magnoliaceie Х-Г9
Tr<xhn<lrn<3rer**« *-19
€?rodwphyj|ica>e ж -19
SalkactM *=19
Hippoc«»tM«ceae *-20
Rlatanaceai ж-21
У большинства главных групп Pteridophyla основные хро-
мосомные числа лежат а полиплоидно* диапазоне, а у некото-
рых групп —в верхней его части [533, 554]. Например:
Pviypvdiwn. ek (Polypodiac«e) х=37
PsfMtm (PtiloUnae) x-SO, S2
Гпхеп'ргягаи (Pailolaceax) a—19»
ЕдЫмТип (Equisdaceae) х=|ОЙ
Ofhlogtwutm (OphlOElassareae) x—120
334 Часть аштая. Похилясидия
Наиболее вероятный вывод, к которому можно прийти на
основе этих данных, состоит в ток, что многие виды современ-
ных PteridophyU — дожившие до наших дней остатки теологи-
чески древних полиплоидных комплексов [554].
Необратимость
Эволюция, направленная от диллонда к тетраплоиду и поли-
плоиду более высокого уровня, в основном необратима. Она мо-
жет частично повернуть в обратную сторону в результате поли-
плоидного сброса (см. гл. 22 и 27). Так, у Htsptrit, у которого
х-7, число хромосом в гаметах тетра илоядных видов п=14,
13 и 12.
Некоторые исследователи высказывали мнение о возможно-
сти резкого и полного возврата из тетраплоидиого в днплонд-
иое состояние посредством образования полигаплоидов [160.
667]. Гаплоидные или политаплондные потомки от тетраплоид-
иых родительских форм, безусловно, будут днплондиымн. Такие
полнгаплонлы иногда действительно возникают в полиплоидных
популяциях растений. Однако полигаплоид, происходящий от
аллотетраплоида. и по определению, и по результатам наблю-
дений представляет собой диплоидный гибрид, характеризую-
щийся хромосомной стерильностью, к его главная возможность
размножаться половым путем — это вторичное удвоение Поли-
гаплоиды вряд ли могут повернуть вспять эволюцию, направ-
ленную в сторону полкплоиднзацнк.
Подавление изменчивости
Полиплоидное состояние по сравнению с диплоидным несколько
ограничивает воэмнкпояение мутационной и рекомбинационной
изменчивости, и эти ограничения становятся более жесткими с
повышением уровня плоидности. Дупликация генов и полисом-
ное наследование подавляют выражение новых мутаций. Эти
факторы, а также избирательное спаривание ограничивают об-
разование новых рекомбинантов.
Тышлер [831] отмечает, что мутантные проростки, обычные у
Triticum monococcum (2х), у Г. durum (4х) встречаются реже,
а у 7. aestluum (6х) крайне редки.
Полиплоидное состояние ограничивает возникновение измен-
чивости, ио не исключает ее полностью. Сегментные аллополи-
плонды могут расщепляться до некоторой степени по межвидо-
вым генным различиям У полиплоидов изредка происходит
вышеплеиис мутантов 1 полнсомкых соотношениях. Тем не ме-
нее у полиплоидов по сравнению с аиплоидамн создающий из-
Гл. 25. Полыплс»й*ый комплекс
33$
мспчивость механизм, связанный с половым процессом, подав*
лен.
Следует таким образом ожидать» что у полиплоидных видов
способность реагировать на изменяющиеся условия среды выра-
жена слабее, чем у диплоидных видов Безусловно, новые поли
плоили часто развиваются очень успешно, более успешно, чем
их диплоидные родичи, в местообитаниях разных типов (см.
гл. 24). Их успех объясняется наличием у них генетической сис-
темы. допускающей размножение определенного |ибрндного ге-
нотипа в чистоте. Но эта генетическая система одновременно
налагает ограничения на образование новых вариаций и гем са-
мым устанавливает пределы потенциальным зналюиионным воз-
можностям данной полиплоидной линии.
По мерс того как данная филстическая линия достигает (в
основном необратимо) высших уровней плоидностн, эти ограни-
чения эволюционного потенциала возрастают. Полиплоидная
система, квк утверждает Стеббинс (780), способна создавать
многочисленные родственные виды или даже роды, обладающие
я обшем сходными признаками, однако опа, очевидно, не прини-
мала никакого участия в возникновении новых семейств или
порядков.
Диплоилизапня
Давно возникшие голнплонды обычно больше похожи на днп-
лоидоя» чем вновь образовавшиеся. Это явление известно под
названием дмпломдкэацкм. Днплоидизация влияет как на цито-
логические признаки полиплоидов, так и па их генетическую
Полагают, что для кисть образовавшегося, или «сырого»,
полиплоида, если это авто полиплоид, должна быть характерна
высокая частота мультняалентов, а если это сегментный алло-
полиплонд. то образование некоторого их числа. Мультнвалеиты
снижают плодовитость полиплоида Естественный или искусст-
венный отбор на плодовитость будет благоприятствовать генам
(если такие гены имеются в генофонде), способствующим обра-
зованию бнпялситот» за счет мулъгивалеитов
Джайлс и Рэндолф [243] сравнивали частоту мультивалеп-
топ и одной автотс-траплоидной линии кукурузы (Zeo mays.
х--10) в начале и в конце 10-летнего периода отбора на плодо-
витость У шесть образовавшихся автотетраплендиых растений
Я—Ю IV содержались в 89% материнских клеток пыльцы, а у
Их потомков 10-го поколении —только в 52% этих клеток Час-
тота образования бивалентов возрастала по мере снижения час-
тоты квадрквалентов. Искусственный отбор на плодовитость,
вызвавший это изменение, яозможио, сопровождался бессозиа-
356 Уотте ur.ru. ПолиллоиЛил
тельным отбором на генетические факторы, влияющие на способ
конъюгации хромосом при мейозе.
Отбор генов, оказывающих влияние на мейоз, вероятно, сыг-
рал важную роль в эволюция многих естественных амфиплоя-
дое. У гексаплоидиоА пшеницы Triticum aestivum наблюдается
значительная гомология между разными геномами. Однако го-
меологичные хромосомы обычно не конъюгируют, а образуют
биваленты, состоящие из гомологичных хромосом. Такое пове-
дение в мейозе обусловлено одним геном, локализованным в
длинном плече пятой хромосомы пшеницы. Прн отсутствии этого
гема образуются мультивалеиты. Если этот геи (ген 5В) имеет-
ся, то ои подавляет образование мультнвалентов н способствует
конъюгации гомологичных бивалентов (686].
Дупликатные факторы достаточно часто встречаются у новых
амфиллоидов и обычны у старых. Дупликатные локусы являют-
ся следствием сложения родственных геномов Дупликатные
факторы могут оказаться ненужными для функционировании
организма; в таком случае функция одного из локусов может
отклониться от первоначальной функция. Если такой днвергнро-
вавший ген начнет выполнять какую-либо другую близкую
функцию, выгодную для данного организма, то он будет сохра-
няться отбором.
Интересный пример дивергенции между факторами, которые
прежде были дупликатиыки, обнаружен у аллотетраплоидных
хлопчатников Gostypium hiriutum и О. barbadtnst Их геномы
имеют конституцию A ADD, Диплоиды АА — хлопчатники, выра-
щиваемые в Старом Свете, а диплоиды DD — дикорастущие аме-
риканские виды
Группа сцеплении, в которую входит ген R. детерминирую-
щий пигмент антоциан, и ген С1. детерминирующий скученность
соцветия, содержится как в геномах А а D. так и в геноме АЛ
тетранлондов. У диплоидое АА н DD ген Я представлен в одном
локусе, но у тсграпловдов A ADD —в двух локусах (R, и Я»).
Подобным же образом ген CI представлен в одном локусе у
дипломов АА, а возможно, и у диплондов DD, ио у теграпло-
илов AADD- в двух локусах (С1\ и Ch). В геноме тетрапло-
идея эти гены располагаются следующим образом: —С1а —
в геноме A. a R, — CI, — в геноме D (802. 8C3J
Прн возникновении тетраплондного хлопчатника гены Ri и
R] и С/| и С1г были, несомненно, дуплякатнымн факторами.
Однако у современного тетраплондяого хлопчатника они уже не
являются дупликатяыми; гены R, и R, взаимодействуют как па-
ра комплементарных факторов; то же самое можно сказать и
относительно генов С/, и Ch [802. 803].
Эволюция в направлении более высоких уровней плоидности
усложняет мейоэ и создает более жесткие ограничения прн рае-
Гл. 25. килпллкс
'ОТ
шеплении к рекомбинации. Процесс диплонднэацни противодей-
ствует этим тенденциям, давая возможность формам с высокой
или средней илоилкостыо вести себя в теистическом отношении
как более или менее настоящие диплоиды.
Можно только предполагать, что диплоидюация — это фак-
тор, лающий возможность некоторым полиплоидным комплек-
сам эволюционировать до высоких уровней плоидности. Кроме
того, этот фактор, вероятно, обусловливает способность некото-
рых родов с высокими основными числами (например, Epilo-
Oium, х=18; Equisctum, х—108) к усиленному видообразованию
на старых уровнях плоидности. Короче говоря, диплоилязацик
может вдохнуть а давно возникшие полиплоиды новую жизнь.
а «
26 Природные автополиплоиды
Основными критериями, по которым у растений а природе мож-
но отличать автополнплондов от амфиплоидоа, служат, хак уже
было отмечено в гл. 23, следующие: образование мулыквалеи-
тов, стерильность, тетрасомкое наследование и морфологическое
сходство с родственными диплоидами. Эти критерии полезны,
ио их нельзя считать диагностическими, и в отдельных случаях
они легко могут привести к ошибочным заключениям.
Было время, когда иа основе этих критериев считали, что
автополиплоидия широко распространена у растений в природе
Критическая переоценка фактических данных, предпринятая
Клаузеном, Кеком и Хисн [102]. заставила изменить это звклю
некие. По сравнению с амфиплоидией автополипломдия в при-
роде — явление, необычное у высших растений, однако оно
встречается в ряде групп.
Тем не менее в литературе продолжают появляться вообще
пня об ввтополиплондии. Слишком часто такие Сообщения бы-
вают основаны на недостаточных данных. Например, утвержде-
ние об автокодиплоядком происхождении тетраплоидкых пред-
ставителей группы EpUobium angustilolium основано на цитоге-
нетических данных, весьма обширных и наводящих на размыш-
ления, но не вполне убедительных [586, 587].
К яакболее хорошо документированным примерам природной
авгополиплоидин у растений откосятся те, в которых можно с
достаточной уверенностью назвать родительские виды полипло-
идов и в которых известны также плодовитость при скрещива-
ниях и взаимоотношения между диплоидными родительскими
популяциями. Для получения таких данных необходимо, конеч-
но. комбинировать полевые и экспериментальные методы, по
своей разрешающей способности значительно превосходящие
обычные цнтотаксояомяческие методы.
В этой главе мы рассмотрим три подлинных случая природ-
ной автополиплоидии к вкратце упомянем еще о двух примерах.
Недостаток места ие аает нам возможности рассмотреть раз-
личные недавние сообщения, в которых заключение об автопо-
липлоидии делается без достаточных на то оснований.
Группа Biscutella laevigata
Biscutella laevigata к близкие к ней виды (Cruciferae) — это
группа многолетних травянистых растений, встречающихся в
гористых местностях Европы. Классическими исследованиями
Гл 26 Природное автмолч/LiQvdci 339
Ментом [ц52, 553] было установлено, что средн них есть формы
с числом хромосом 2х. 4х и 6х (х=9) Гексаплоилы известны
только из одной местности и Испании. То, что нами известно па
сегодняшний день об этом группе, откосится главным образом
к диплоидам и тетра галоида и.
Тетраплоидная форма Biscutella laevigata sensti ttrieto имеет
непрерывный ареал, встречаясь в Альпах на всем нх протяже
нин. Дипломам, известные под общим названием В coronopifo-
На. образуют ряд разобщенных популяций на низменностях к
северу н к востоку от Альп. В виде исключения один диплоид-
ный вил встречается в горах а окрестностях Вены.
Диплоиды из отдельных районов различаются морфологиче-
ски. и прежде их описывали как разные виды. Однако Мг»гтои
[5S3J. скрестив нх между собой, обнаружила, что гибриды Р|
плодовиты и что у НИХ происходит нормальная конъюгация хро
носом Следовательно, эти лип л он ды, по-видимому, п|
ляют собой расы, принадлежащие к одному виду, а их тстрапло-
кдное производное, В. laevigata, — межрасовый амогетраплоид.
У Biscutella laevigata иногда встречаются квадрива ленты,
что наводит на мысль об автотезраплондии, но не может слу-
жить решающим доказательством Волге убедительным доводом
представляется обнаружение тривалентов у искусственных три
плоидннх гибридов, полученных при скрещивании В laevigata
с различными диплоидным и расами [553].
Географическое и экологическое распространение этпт ди
плоидов м тетра пл он дев и сочетании с тем, что нам известно
о геологической истории области, где они встречаются, позво-
ляет считать, что диплоидные популяции низменностей являют-
ся реликтами межледниковой флоры Что же хасается альпий-
ского тетраплоида Biscutella laevigata, то ои распространен в
области, которая во время последнего оледенения находилось
подо льдом, и поэтом) он. вероятно, пмнгрировал сюда в пос-
леледниковый период 1552. 553]
Межрмсоные комбинации генов оказались, по эндныому. цеп-
ными в адаптивном отношсиип для этлго автотетр а плоила
В связи с >тлм можно । pe.iположить, что ллпллмлная ряса,
встречающаяся в горах вблизи Вены, участвовала е качестве
одной из родительских форм и меж расовом скрещивании или
скрешиввпиях. давших начало удачному альпийскому' автотегра-
плонду [102, 552. 553].
Группа Solatium tuberosum
Счожиая группа Solatium tuberosum очень тшиелыю изучена
рядом всследооателсй и представляет собой иибочее хорошо
проанализированный пример природной авгогаолиплсигдин
И*
•34» Часть метал Иолиплоидил
Начнем с того, что обрисуем таксономическое толожение
этой группы. Секция Taber arium рода .Sofanum представляет со-
бой большую секцию, состоящую из 32 рядов, распространенных
в Северной и Южной Америке (123]. К ряду Tuberose относят-
ся 6 видов, в том числе S fu^erosum sensu lato, встречающийся
в Перу. Чили и Аргентине Пять из этих видов — дикорастущие
растения, а шестой. S. tuberosum, содержит дикорастущие расы
ю Патагимнм и Чили, а также многочисленные культурные расы,
выращиваемые в районе Анд (по [123}; другой таксономический
подход к ряду Tuberosa см. [354р.
Это свободно скрещивающиеся многолетние травянистые рас-
тения. образующие клубки. Диплоидные формы самонесовмес-
тимы и перекрестно опыляются шмелями н другими пчелиными;
помимо размножения семенами у них происходит вегетативное
размножение прн помощи клубней [175]. У полиплоидов. выри
щиваемых в культуре, к пыльца, и семена обычно стерильны, и
их размножают вегетативно (354. 812. 814].
Solarium tuberosum sensu lato — сложный таксономический
вид. Он содержит не только ряд дикорастущих н культурных
форм, о чем уже упоминалось, но также полиплоидный ряд. со-
стоящий кз форм 2х, Зх. 4х и 5х при х-12 [814].
В соответствии с классификацией культурного картофеля,
созданной Доддсом [174]. существует семь главных форм Sola-
num tuberosum sensu lato. различающихся no морфологам, рас-
пространению и плоидиости. Для этих таксонов Доддс ясиоль*
дует неформальную категорию «группа»; его группы соответст-
вуют культурным расам по другим классификациям Ниже пе-
речислены эти семь групп с указанием их плондности и некото-
рых других черт.
1. Stenotomum. 2х. Высокие плато в Андах.
2. Phureja. 2х. Далины в Андах.
3. Andigena. 4х. Картофель, выращиваемый в Андах.
4 Tuberosum (—S. tuberosum sensu stricto) 4x Обычный
культурный картофель, Европа и Северная Америка..
5. Chaucha. Зх. Стерильные семена.
в Juzepczukii- Зх. Стерильные семена
7. Curtitobum. Sx. Стерильные семена.
Филогенетические взаимоотношения между этими таксонами
схематически изображены на рис. 26 1 (по данным [174, 175])
Кроме них на згой схеме указан вид Solanum ataule, принадле-
жащий к ряду Acaulia. Выращиваемый в Андах картофель andi-
gena считается тетраплоидным производным меж расовых скре-
щиваний между stenotomum (2х) и phureja (2х> Тетраплоид,
очевидно, возник спонтанно в условиях первобытного земледе-
лии. Следовательно, обыкновенный культурный картофель, так-
Гл 26. Приробчи« мтоломилм^м
Ml
Рис. 2$ I Филогения основных культурных рис картофеля Sctanum iubtro-
лит [17«]
Объяснения си в тексте
же тетраплоидный. предстаэлсят собой продукт селекция этого
более примитивного андийского картофеля.
Обыкновенный картофель, на основе достаточно веских ци-
тогенетических данных, давно считали автотетраплоидом. У не-
го наблюдается высокая частота квадривалситов при мейоэе.
Наибольшей частоты — 9 IV на клетку —они достигают у под-
разновидности (subvariety) Chippewa [812]. У некоторых под-
разновидностей средняя частота хвадривалектов составляет
4.4 IV [584]. Такому цитологическому прививку соответствует
тстрасомия по некоторым генам, наблюдаемая у культурных
рас картофеля [78].
Данные из других источников окончательно решают вопрос
в пользу автогеграплоидии Искусственный триплоид, получек
иый в результате гибридизации между тетраплондиой расой tu-
berosum к диплоидной stenotomum. содержит в среднем 8,2 III
иа клетку. Такая высокая частота тривалеятов свидетельствует
342 Часп шестая Полиплоидия
о том, «по культурный картофель — автогетраплоид. содержащий
хромосомы stenotomum (384J.
Дальнейшие доказательства дает редуцированная иля поли-
гаплоидная форма Solatium tuberosum sensu strlcto, у которой
2п«-24 вместо нормальных 2н —48. У некоторых полигаплонлое
наблюдается 12 11. т. е„ иными словами, полная конъюгация в
большинстве шГёток и высокая фертильность пыльцы. Кроме
того, эти полигаплоиды при скрещивании с некоторыми днпло-
идными культиварами дают плодовитых гибридов [384, 812].
Наконец, диплоидные расы очень близки генетически. Между
ними нет никаких барьеров, препятствующих скрещиванию.
У межрасовых гибридов Fi происходи? нормальная конъюгация
С образованием бивалентов. В одних сочетаниях возникают пло-
довитые гибриды, тогда как в других сочетаниях образующиеся
гибриды имею? стерильную пыльцу [175, 815].
В этом разделе внимание было сосредоточено иа автотетрэ-
плоидни в группе Solanum tuberosum. Чтобы завершить карти-
ну. следует отметить, иго среди дикорастущих видов секции
Tuberarium амфнллоидия довольно обычна. Так, Solanum acaule
(ряд Acaulla) — аллотетраплонд, a S. demissum (ряд Demis-
3fl) — аллогекса плонд [384, 812].
Группа Dactylis gio тс г at а
Ежа сборная — Dactylis gtomerata (Gramineae), 2п»14 к 28,—
самонесовместимое свободною: ылякнцееся растение, способное
также к вегетативном)' размножению. Эта группа широко рас-
пространена в Евразии1, и к ней относится обширный ряд мор-
фологических и экологических вариантов на двух уровнях пло-
идиостн [797].
На диплоидном уровне Стеббинс и Цохари [797] различают
11 подвидов; 4 из них представляют собой крайние варианты в
отношении морфологии. Они встречаются среди растительности
старых типов и имеют прерывистые ареалы, тянущиеся от Ги-
малаев до Канарских островов и островов Зеленого Мыса. Эти
4 географические расы, по-видимому, образовались в результа-
те Давней первичной дивергенции на диплоидиом уровне
Остальные 7 подвидов занимают промежуточное положение
между четырьмя крайними типами и связаны с более совре-
менными типами растительности, в том числе с пастбищными
субклнмаксамн я сорными местами. Они, возможно, представля-
ют собой результат скрещиваний и ннтерградации между пер-
вичными расами [797].
' А также о М*ка?оие1ти. Сеаврээй Африке я Азам. — Прим, ptd
Гл 26- Природные аетохслыллоиды
343
Диплоидные расы ингерфсртильны. Фертильность пыльцы у
гибридов Fi колеблется от лолустерилъной до полностью фер-
тильной, а семена фертнльны [797).
Тетраплоиды по сравнению с липлоидамн встречаются чаше,
распространены более широко и крайне изменчивы Морфологи
ческне и экологические данные позволяют считать, что тетра-
плоиды возникли от различных межрасовых гибридов Таким
образом, они являются мсжрасовыми амфнллоидамн [797].
Скрещивания между тетраплоидиой и диплоидной расами
ежи сборной удаются с трудом, а их гибриды Ft высокостериль
вы [797]. Daciylis giomtraia в настоящее время представляет со*
бой, таким образок, таксономическим вид, состоящий на двух
биологических видов.
Ьаръер стерильности между Амилоидами и тетраплондамм
эффективен, по не является абсолютным. Тринлопдные гибриды
при опылении пыльной тетраллоидов завязывают некоторое ко-
личество семян. Наиболее мощные потомки от возвратных скрс
щиваннй — теграплонды. Поэтому возможна интрогрессия от
диплом дон к тетраплоидным популяциям через промежуточных
три плои дои. Такая интрогрессия - еще один фактор, вносящий
вклад п изменчивость тетраплоиднои популяционной системы
[797].
Другие примеры
Мы приведем здесь еще два примера природных эвюгимипло-
идов с указаниями па литературу, в котором можно найти даль-
нейшие подробное! К
Medicaga saliva, 2п —16 и 32 Диплоидная форма Af saliva
( — .М coerulea)— дикорастущее растение, распространенное от
Турции до Ирана и Каспийского моря. Тстраплопдная форма
(М. даГйчт sensu stricto) — культурное растение, возникшее, не-
видимому, спонтанно в области распространения диплоида. а
впоследствии распространившееся в качестве культурной формы
в Европе [465].
Galax aphylla (Diapenslaceac). 2п=12 и 21. Единственный
таксономический вид монотипического рода, распространенный
и юго-восточной части США. Во многих местах по всему ареалу
этого таксономического вида можно встретить растущие рядом
диплоидные и тетра плои д и ыс индивидуумы [41]. Такая картина
сильно склоняет к предположению об автотетраллоидии [102,
778. 779. 789],
Часть седьмая
Анеуплоидия
27 Анеуплоидия
Различия между индивидуумами или популяциями (или клетка-
ми) по числу хромосом выражаются в трех основных формах:
полиплоидия, анеуплоидия и агматоплоилия. Если полиплоидия
представляет собой различия по числу целых хромосомных на-
боров. то анеуплоидней называют различия по числу отдельных
хромосом, а агматоплоидией — различия по числу хромосомных
участков.
К анеуплондной изменчивости относится очень разнородная
группа явлений. Отклонения от стандартного числа хромосом
могут быть как в сторону увеличения, так и в сторону уменьше-
ния. Эти численные отклонения могут затрагивать одну хромо-
сому (полясомия, моносомия), одну пару хромосом или несколь-
ко их пар. Такие лишние хромосомы могут быть гомологами
членов нормального комплемента или ие иметь к ним отношения
(добавочные, сверхкомплектные, или В-хроносомы). Кроме того,
анеуплоидия может возникать на диплоидном или полиплоид-
ном уровнях.
Аиеуплоклия иа полиплоидном уровне включает несколько
различных явлений. Различные попарные комбинации разных
диплоидных основных чисел создают аиеуплоидиую изменчи-
вость среди аллотетраплоидкых производных. Так. два вида с
8 парами хромосом каждый дают аллотетраплоид с 16 парами
хромосом, тогда хак скрещивание между видом с 8 и видом с
9 парами даст аллотетраплоид с 17 парами хромосом. У многих
полиплоидов в результате нарушений мейоза возникают сверх-
комплектные пары хромосом, Кроме того, у полиплоидов возмо-
жен пилналоидный сброс, т. е. нисходящая (descending) анеу-
плоидия.
У одного и того же отдельного растения или в одной и той
же популяции можно встретить одновременно различные типы
аиеуплоидкой изменчивости D данной группе растений может
наблюдаться анеупловдия и иа диплоидном, и иа полиплоидном
уровнях. В одной и той же группе растений анеуплоидия может
также встречаться одновременно с агматонолиплоидней.
Гл 27 АчгдодеьЭде
345
В этой главе мы рассмотрим несколько примеров обычной
анеуилондмн на полиплоидном к диплоидном уровнях. Полисо
мин и В-хромосом мы здесь касаться не будем (см. 1.434]).
Нисходящая анеуплоидия на диплоидном уропие
Нисходящие аисуилоидные ряды мд диплоидном уровне идее
етны у доппльно большого числа групп растений Стеббинс
(780] перечислил 25 таких групп. В гл. 14 мы привели несколько
примеров нисходящей аксуплоидии наряду с другими относя
шкмнея к этому явлению сведениями Ниже представлено не-
сколько других примеров; цифры к этом списке соответствуют
гаплоидному числу хромосом (и), а скобками ( ) обозначены
необнаруженные промежуточные числа
Crepis (Composite), 6-5-4-3 [33]
Youngia (Cocipositac). 8-( ) о [36]
Eriophyllum и Pseudobaiiia (Compositac), 8-7 ( )-4-3 [82]
Nicoliana (Solanaceae), !2-( J-10-9 |250)
Phacella (HydrophyПассас). I1-10-9-8-7 [90. 91]
Clarbia (Or.agrace-ac). 745-5 [492]
Knaulia (Dipsacaceae), 10-( )-8 [191]
Ccphalana (Dipeacaccae) 9-( )-5(l9l]
Lesquerella (Cruciferaei' 8-7-6 5 (693]
Persoonia и родственные роды (Proteaeeae) 7-( )-5 [413. 411)
IrUiltaria (Liliaccac) 12-( )-9(144, 148]
Иногда нисходящие ааеуилокдпые ряди бывают образованы
родами, принадлежащими к какому-либо одному семейству нэп
трнбе Ограничимте двумя примерами В трибе Poletnonieae сем
Polemomaceae наблюдается ряд 9 8 7 6. а и сем LoranthaceBe—
ряд 12 11-10-9 8 (42. 281].
Среди покрытосеменных у 783 видов диплоидное число хро-
мосом низкое — от п-2 до л-6 Распределение видна по числу
хромосом выглядит следующим образом [282]:
л=6 у 408 видов
|»=Л » 225 »
я-4 а 134 >
л-3 > 15 »
п-2 » I вида
Все эти виды представляют собой результат редукции основного
числа хромосом в грицессе филогенеза
Механизм анеуплондной редукции на диплоидном уровне
связан с неравными реципрокными трдислокацнямн [33. 610,
8341. В предковом хромосомном наборе хромосомы А и В обме-
За 6
Уесть сс-Зь лол. Л
пинаются участками, очень неравными по длине, так что весь
генетически активный материал хромосомы А переносится в хро-
мосому В, тогда как центромера хромосомы А получает в об-
мен лишь небольшой инертный участок. Затем ставшая инерт-
ной хромосома А утрачивается а процессе одного из клеточных
делений. В результате получается хромосомный набор, содержа-
щий иа одну пару хромосом меньше.
Классическое описание этого механизма дал Тогби (834] на
основе цитологического аналиаа Crepis neglecta (2л-8) и С. fu-
liginosa (2п-6). Последний вид с редуцированным числом хро-
мосом происходит от предкового вида, близкого х С. neglecta.
Отдельные хромосомы у этих видов можно идентифицировать в
соматических и в мейотических клетках по величине и морфоло-
гии. Четыре хромосомы в наборе С. neglecta обозначены Л, В.
С. D. а три хромосомы С. fuliginosa — А, В. D (рис. 27.1).
л
Рнс. 37.1. Хромосомные наборы лгут ькиов Crept! и гибрида между ни-
ми №34).
Л. гаплоидных набор озиатинчскиа храмом» С. neglects.
£. Гаплоидный набор сомлтимотнх хромосом С. Isligieoea
В Диплоидный набор соиатичкких хромосом тибрим Н
Г Коигмогандх меВотнчееккк хромосом п матафом 1 у гибрида Fl.
Гл 27. Лгидллоягдия
W
Конфигурации пар, получающихся при конъюгации хромо-
сом у межвидовых гибридов, показывают, что эти два вида раз-
личаются по трем инверсиям, по одной транслокации в хромо-
сомах Л к О и по другой транслокации в хромосомах В и С.
Особенно существенна зга вторая транслокация' одна часть
хромосомы В вида С. fuliginosa конъюгирует с хромосомой В
ила С negtecta. а другая ее часть — с хромосомой С этого по-
следнего (рис 27 1) Это показывает, что хромосома В вида
С. luliginosa содержит участки, гомологичные хромосоме С ви-
да С negtecta.
В наборе С. fuiiginosa нет хромосомы С как таковой, и соот-
ветственно основное число хромосом у этого вида меньше; одна-
ко генетический материал, имевшийся у предковых форм хро-
мосомы С, содержится у С. luliginosa в хромосоме В Это про-
изошло в процессе филогенеза в результате неравной реципрок-
ной транслокации между предковыми хромосомами В и С, ко-
торая привела к образованию новой составной хромосомы В у
С. luliginosa [834].
Восходящая анеуплоидни на диплоидном уровне
У ряда групп растений обнаружена также восходящая аиеупло-
идия иа диплоидном уровне Ниже перечислено несколько при-
меров: как к п предыдущем перечне, цифры соответствуют чис-
лам л, а недостающие числа обозначены скобками ( ).
Frltillarla (I iliaceae), 12 13 [144. 148]
Narcissus (Ainarv)lidaceae), 7-( )-10-ll [212]
Crocus (Indaccae), 12-13-14 15 (69|
Phacrlia (Hydrophyllaceae), 11-12-13 [90. 91. 119]
Clarkia (Onagraceae), 7-8-9 [492]
Cephalaria и Succisia (Dipsacaceae). 9-10 [191]
Неудивительно, что существуют анеуплондные ряды, кото
рые один исследователи считают восходящими, а другие —нис-
ходящими. У Podocarpus (Podocarpaecac) с гаплоидными чис-
лами 10-11-12-13-( 1-I7-18-19. по мнению Кхошу [433], в про-
цессе филогенеза имела меею восходящая анеуплоидня, а по
мнению Хейра и Бейцеиберга [327],— нисходящая анеуплоидня.
В грибе Astereae (Compositae) есть формы с гаплоидными чис
лани 4-5-6-( )-8-9 Согласно одной школе исследователей, ос-
новное число хромосом у предковых форм х—5 (или 4) [844.
846]. согласно другой школе, исходное основное число х—9
[720. 760].
Одним из способов возникновения анеуолоидпого увеличения
числа хромосом может быть неправильное разделение центроме-
ры. Будучи сложной структурой, центромера редко делится по-
34в
Чапа седалая. Ляа^плсяаил
I 2
С.МОЫ ажжв
3 4
I 2 3 4
с л>«м» вменяв । и >
4
Ряс. 27.2. Гомологии некоторых хромосом Ctarlua btloia it С. tingalala > яон-
фигтраиля, обрахукицявсх яря ах копыоеоикя [199].
перечло с образованием двух половинок, каждая из которых со-
храняет способность к координации с веретеном. В таких слу-
чаях одна метацентрическая (или субметацмгтричяекая) хромо-
сома дает начало двум телоцеятрическим (или субтелоцеитри-
чесхим) хромосомам (144, 148].
У Frit Mana основное число х-12 и хромосомный набор со-
стоит нз 10 телоцевтрических и 2 метацентрических хромосом.
Fritillaria pudica. у которой п —13, содержит 12 телоцеитриче-
сккх и 1 метацентрическую хромосому. Следовательно, анеупло-
ядиое увеличение от п—12 до п—13 произошло, вероятно, в ре-
зультате превращения одной метацентрической хромосомы в аве
телоцентрические [144, 148). Подобным же увеличением числа
телоцентрических хромосом обусловлено аиеуплоидиое увеличе-
ние у narcissus и Crocus [69, 212].
Другой механизм аиеуплоидиого увеличения числа хромосом
состоит во включении в нормальный хромосомный набор сверх-
комплектной пары хромосом в результате отставания во время
клеточного деления; ато приводит к образованию тетрасомнка
(2л-|-2). Сверхкомплектные или тетрасомиые хромосомы могут
затем дквергировать от своих гомологов путем взаимообменов
с другими иегомологичными хромосомами и в результате дивер-
генций в функциях генов.
Исходное основное число у Clarkia (Onagraceae) х-7, а ви-
ды с 8 и 9 парами хромосом возникли позднее [492]. Анеуплоид-
иое увеличение у Clarkia, по крайней мере в некоторых случаях,
Гл. 27. Л/вп>ллоидил
34»
произошло путем включения и набор тетрасомиых хромосом,
претерпевших транслокации.
Люис и Робертс (499] провели подробное патогенетическое
исследование пары близкородственных видов — Clarkia biloba
(п=8) и С. Ungulata (n=B). Sth авторы установили, что второй
из указанных видов произошел от первого Конъюгации хромо
сом у гибрида между этими видами свидетельствует о том. что
их геномы различаются по двум транслокациям и одной инвер-
сии, но в остальном гомологичны. Одна на транслокаций нахо-
дится в девятой хромосоме С Ungulata
Девятая хромосома С. Ungulata вместе с двумя другими хро-
мосомами этого вида и двумя хромосомами С biloba образует
у гибрида цепь из 5 хромосом Конфигурация этой цепи н ее
состав представлены на рис 27.2. Очевидно, девятая хромосома
возникла а результате обмена плечами между двумя хромосо-
мами С. biloba с терминальными участками 1-2 и 3-4. что при-
вело к образованию новой хромосомы 1-4 Затем две исходные
и одна новая хромосома закрепились у предка С. Ungulata Та-
ким образом, в наборе этого последнего па одну пару хромо-
сом больше, чем у предкового С biloba [499].
Полиплоидный сброс
Анеуллоидпая редукция па полиплоидном уровне была удачно
названа Дарлингтоном полиплоидным сбросом (polyploid drop
[144, 146, 148)). Полиплоид часто может выдержать утрату од-
ной или нескольких пар хромосом ввиду наличия у него дуплн-
катных факторов в гомологичных пли гомеологичпых хромосо-
мах
Дарлингтон [148] приводит примеры полиплоидного сброса
у представителей 20 групп растений. Некоторые из них перечне,
лены ниже; числа п дамы в следующем порядке: дпплоиды, по-
липлоиды и редуцированные ансуплоиды.
Ilesperls (Crucifcrac), 7-14-13-12
Linum (Linaceae), (8. постулировано) 16-15
Veronica (Scrophulanaceae), 7-21-20
Mai/deae (Qramineae), 5-10-9
У Proteaceae анпестральное основное число x = 7: полипло-
идное удвоение до х-14 произошло в ранний период истории
этого семейства (в примитивном подсемействе Peraoonioideae)
Это давнишнее тстрал-имшяоо число хромосом встречается так-
же в производных подсемействах Protcoideac и Grevilleoideae,
В подсемействе Proteoideae имеется, кроме того, нисходящий
ряд х-14-13-12 11-10. a v Grevilleoideae ряд х-14-13-11 10
[414].
3S0
¥лег» седьмом Лмемомои^ия
Анеуплоидное увеличение у полиплоидных
гибридов
Межвидовые гибриды е уинваленткымн хромосомами, не имею-
щими гомолога при мейозе. представляют собой источник аиеу-
плоидной изменчивости в экспериментальных условиях и в
меньше* степени — в природных популяциях, Анеуплоидия обыч-
но бывает возрастающей, поскольку приобретение сверхкомп-
лектной хромосомы переносится легче, чем утрата членов ос-
новного набора. Эту форму анеуплоидия чаше можно обнару-
жить на полиплоидном уровне, чем на диплоидном, вследствие
более высоких буферных свойств полиплоидных генотипов.
Хорошим примером служит скрещивание между гибридом и
«го потомками в группе Erophlla verna (Cruciferae) [901J. Этот
пример уже был рассмотрен в гл. 20; здесь мы упомянем о нем
лишь вкратце в связи с акеуплоидней.
Гибрид был получен в результате скрещивания между фор-
мой с 15 парами н формой с 32 парами хромосом. У гибридов
F, наблюдались многочисленные нсконъюгировавшие хромосо-
мы. В Fj произошло расщепление по числу хромосом и по дру-
гим признакам.
В последующих инбредиых поколениях выщепенцы выдели-
лись в различные стабильные линии Как показал цитологиче-
ский анализ восьми таких стабильных линий, они содержали
следующие анеуплоидкые наборы хромосом: л = 22, 23. 25, 29. 31
н 34. Анеуплоидия встречается также в природных популяциях
группы Erophlla чета в Европе, я вполне возможно, что она
возникла у них таким же образом [901].
Два тетраплоидных вида Cilia (G. malior н G modoctnais)
« одинаковым числом хромосом (2л—36) дали при скрещивании
гибрид F,. у которого при мейозе наблюдалось много веконъю-
тировавших хромосом. Этот гибрид в свою очередь дал несколь-
ко гомоплоидов, а также один стабильный анеуплоид с 2л —
-50 (287[.
Аиеуплоидныс числа хромосом гибридного происхождения у
полиплоидов иногда закрепляются благодаря агамоспермии На-
пример. анеуплонднвя изменчивость в группе Bouteloua curtipen-
dula (Gramincac) обусловлена сочетанием полиплоидии, гибри-
дизации и агамоспермии [261, 262). Эти и другие сходные при-
меры будут рассмотрены в главах, посвященных агамоспермии
(см. часть восьмую).
Carex
Род Carex привлекает внимание своим длинным и почти непре-
рывным авеуплондным рядом—от л =6 до л—56,— который со-
держит подряд все гаметные числа от л-12 до л-43 (156, 361,
Гл, 77. Ллл^п,юи^ил
351
380] Короткие последовательные аиеуплоилиые ряды (sensu la-
te) обнаружены в нескольких секциях Carex. Примерами слу-
жат Dittanies (п—28—37). Extensae (п—30 35). Acutae
(л=34 42) и группа Carex lariophyllea (п=3|—34) [156].
У Carex известны основные числа х=6. 7. 8 и 9: предполага-
ется. что у этого рола встречается также «»5. которое, возмож-
но, было первоначальным основным числом [371, 534, 867].
Кроме того, у Carex существуют полиплоиды с основным чи-
слом х=5 и другими вторичными основными числами Так, к
группе С. siderosticta есп, формы с п«6 и п= 12, в секции Capil-
tares (х—9) есть диплоидные, тетраплоидпае и гексап.тоидные
виды, а в группе С slenarMa— формы с и —17 и л-3-1. У Ca-
rex описаны полиплоидные ряды, основанные па х=л, 6. 7. 8 и
9 [534. 867). Таким образом, часть наблюдаемой у Carex айву-
п.1оид1юй иэмсичипоети можно отнести за счет различных по-
бочных аффектов полиплоидии [534, 867]. Монобазнчсская поли-
плоидии с различными основными числами даст сходные, но
разные производные числа, а днбазнческая полиплоидия запол-
нит разрывы Так, дпплоиды с 7 и 8 парами хромосом и разных
Комбинациях дадут ряд тетраплоидов с п—14, 15 н 16. Этот ряд
может продолжаться в обоих направлениях за счет полиплоид-
ных сбросов и приобретений
Кроме того, V рода Carex происходит сстеспю|П|ая гибриди-
зация Друри [1/9] сообщает, что ив Аляске почти все виды сек-
ции Vesieariae участвуют в гибридизации. В природе известим
гибриды между Carex rostrata, с одной стороны, и С. rotuadata.
С. memhranacea и С physocarpa— с другой Из скопления гиб-
ридов. возникшего в результате скрещиваний между С rostrata
к С. rolundata в одной из частей Аляски, выщсплкстся новая
константная н плодовитая форма, численность которой возрас-
тает и которая развивается в новый вид гибридного проиехпж-
леячя, получивший название С. paludtaagans [179].
Ббльшая часть аиеуплоидной изменчивости у Carex обуслов
лена гибридизацией и полиплоидией, однако у Carex, по-видимо-
му. встречается также агматоклоидня, о чем будет сказа-
но в гл. 28.
Claytonia virgirtica
Claytonia oirgmica (Portnlacaceae) — небольшое многолетнее
травянистое клубнелуковичиос растение, широко распространен-
ное в центральных и восточных областях Северной Америки.
В этой группе имеется обширный аисуплопдный ряд. Розуэлл
(698) и Люмс [503] обнаружили в ней формы со следующими
значениями 2п ; 12. 14, 16, 17, 18, 19. 20, 22, 24, 26. 28, 30. 31, 32,
^2
Часть Л
mtft fc. jR/f
2л = 12 2л =14 2л=16
w:» ИдьЛ
2п«17 2л» в 2п-19
ПЙц41 *':•
2л=20 2/1=26 2о=2Й
•mW -И
2л =30 2л=32
2Л’34
Рас. 27.3 Лиетплоклиаа юмснчааскть у ClatfiOKia virglnica (по >(69в) с
мсистиям а).
Хромссыам по большей части «ткмякя в метафаи I мейота Неямыогя-
роепшне хромоссыы бьтые, нуаьтявааепты выхелтсм течками.
-34, 36, 41, 48, 72. Некоторые из соответствующих этим числам
хромосомных наборов изображены на рис. 27.3.
Первоначальным основным числом в этой группе было, ве-
роятно, а-8 [698]. Изменчивость по числу хромосом можно све-
сти к нескольким компонентам диплоидной и полиплоидной
анеуплоидии. Прежде всего в ней имеется нисходящая анеупло-
идия на диплоидной основе от х—8 к 7 и к 6 Затем в резуль-
тате разных попарных комбинаций этих днплондов возникает
ряд теграилондов с числами л, равными 16, 15, И к, возможно,
12, Число л, равное 12, обнаружено также у триплоидов (х=8,
2л = 3х — 24), а форма с я=|1—также гриллоид. Большие чис-
Гjl ?7. Л-гуллоиЗил
Ш
ла соответствуют полиплоидам более высокого уровня, 48-хро-
мосом мая форма— это 6х. а 72-хромосомная— это, вероятно. 9х
Наконец. наблюдается некоторая амсуплоидкая измеичнвосп.
вблизи четных полиплоидных уровней; примером служит п—17
н 16 (698].
Анеуплондная ММСтивость иногда наблюдается в пределах
разных популяций В одной из популяций в Картндже (шт Те-
хас) число хромосом колебалось от 2л-24 до 2л-41; чаще все-
го встречались числа 2п~ 21—36 н 28—32. Оказалось, что в этой
популяции модальные числа различны в разные годы, а я по-
пуляции Сент-Луиса— ранней весной и летом [503. 504]. Имог
да наблюдаются различия в числе хромосом я разных частях
одного и того же растения. Например, у одного индивидуума в
популяции Картмджа в клетках стебля и в материнских клетках
пыльцы 2п = 30. а в клетках корня 2п —28 (506].
В восточной части Северной Америки широко распростране-
ны днплонды (х—8), тетраплоиды (ж —14. 16, 16) и триплоиды
(х= 11. 12). причем они в значительной мере симиатричны Дил-
ломды, у которых х-7 и 6, имеют более ограниченное распрост-
ранение |307]. Эти флрмы рассмэтринаюгся как «цнтотипы> и
«рзсы> одного вида (507]. С эволюционной точки зрения, одна-
ко, Claytonia virginica следует рассматривать как нелроанвлнзп-
роваиую полиплоидную группу видов, на которую наложилась
анеуплоидия.
Высокие основные числа у покрытосеменных
У многих родов и некоторых семейств древесных покрытосемен-
ных основные числа высокие и лежат в интервале от х—12 до
ж—21. Вопрос о происхождении таких высоких основных чисел
обсуждается чрезвычайно широко В качестве одного из воз-
можных объяснений нередко предлагали давно возникшую по-
лю-.юн ЫН»
Чтобы облегчить анализ этой проблемы, удобно разбить вы-
сокие основные ЧИСМ иа две подгруппы Довольно высокие чис-
ла (х-19 -21) встречаемся у Magnoliaceae. Trochodenckareae,
Hippocastanaccae. Platanaceae и др Умеренно высокие числа
(х = 12—16) обнаружены в некоторых других семействах древес-
ных растений, например у Fagaccae (х=12), Асегасеас (х- 13).
Betulaceae (х—14). Ulmacrae (х=14). Juglandaceae
(х« 16) и др.
По мнению ряда авторов (144, 780, 832], очень высокие основ-
ные числа первой подгруппы (ж-19-21) — результат Армией
полиплоидии. Я согласен’ что в данном случае это наиболее ве-
роятное заключение (см. гл. 22 п 25).
23-45
354 Часть седьмая. Але^ллоидия
Рассмотрим далее подгруппу с умеренно высокими основны-
ми числами <х— 12—16). Стеббинс [775. 778. 780. 787] считает,
что это также древние полиплоидные числа. Альтернативное
объяснение, которым пренебрегают, состоит в том. что такие
числа — результат восходящей энеуплондим [293].
Это подводит нас к вопросу о первоначальном основном чис-
ле у покрытосеменных Высказывалось предположение (J91. 282,
283). что оно лежит в пределах ж —7—9. У многих семейств по-
крытосеменных, в том числе у некоторых примитивных, таких,
как Аппопасеас [869], встречаются одновременно гаплоидные
числа п -7, 8 и 9. которые, по-видимому, и представляют собой
основные числа для этого класса
Многие исследователи [143. 150. 191. 194. 325. 659, 664, 665,
869] считают, что г—7 является первоначальным основным чис-
лом. Я, однако, не вижу никаких серьезных причин, по которым,
следовало бы выбрать именно х=7. Сколько-нибудь надежным
заключением, вытекающим из имеющихся данных, предсравня-
ется все же допущение о неком первоначальном основном числе
в пределах л=7—9".
Приняв л-7-9 в качестве исходной точки, можно легко
представить себе, что такие умеренно высокие числа, как л=12.
13 и 14. могли быть древнею тетраплоидного происхождения в
результате полиплоидного сброса или без него. Но их с равным
успехом можно считать и продуктами восходящей анеуплоидии.
Существует ряд причин, по которым следует ожидать, что
восходящая анеуплоидия имеет место во многих группах лесных
деревьев и кустарников. Высокое диплоидное число хромо-
сом— одни из способов максимизировать генерацию рекомби-
национной изменчивости в популяции растений, а экологические
условия, существующие в стабильных лесных сообществах, бла-
гоприятствуют открытым рекомбинационным системам. Поэто-
му доминантные и су б доминантные лесные вилы часто подвер-
гаются отбору на эиеуплоидное увеличение. Следовательно,
можно ожидать, что многие древесине растении с высокими чис-
лами хромосом — настоящие дипломы.
’• А. Тахтаджяаом (Прсмехожлеиве и расселение икткоя1л рястсииа.
«Наука», 1970, с 33) было ямаиэмо крыпажмкакне, что ванбали вероят-
нее осмовное число хромосоя у раиммх цветковых мстсмиВ было 6. По Стаб-
бжису (Stebbitts G. L., Flowering pl*nts, evolutioa above the sptciot level. 1974.
Harvard University Press, Gambndg*. Massachusetts, p 206) яерюначалмме
основные числа мегюг быть 6. 7 или в — Прим. ред.
28 Агматоплоидия
Анеуплоидня (sensu stricto) означает изменение числа целых
хромосом. В отличие от этого агматоплондией называют раз-
личия по числу независимо образующихся пар хромосомных
фрагментов в данной группе (534» 550).
Эти два типа изменения числа хромосом коррелируют с ти-
пом центромеры. У бол ыии метка растений м животных плечи
хромосом прикреплены к локализованным центромерам, вос-
производящим себя и свои хромосомы на протяжении ряда кле-
точных поколений. Приобретение или утрата одной или несколь-
ких пар хромосом данным набором зависит от приобретения или
утраты соответствующих центромер Точно так же плечи или
фрагменты хромосом могут сохраниться в данной клеточной ли-
нии или популяции только в той случае, если омн прикреплены
К центромере Изменения числа хромосом бывают анеуилонл-
лыми (sensu AtTicto> нлн ПОЛИПЛОИДНЫМИ
У небольшого числа групп растений и животных центромеры
диффузные. Активность таких центромер ие локализована в ка-
ком-то одном участке, а распространяется по всей длине данной
хромосомы Хромосомы, содержащие диффузные центромеры,
получили название полицентрических [61. 5О9|. фрагменты, об-
разующиеся при поперечных делениях полицентрической хромо-
сомы. могут воспроизводиться на протяжении ряда клеточных
делений, если их разорванные копим заживляются; таким обра-
зом. они могут стать постоянными компонентами данной попу-
ляции. Диффузные центромеры создают возможность для воз
никнивеним агматоплондин [509, .534]-
Атматопломдиыс ряды нельзя омнчнть or собственно апсу-
плотных рядов, если распили ап. лишь простым перечнем хро-
мосомных чисел Как в том. так н в другом случае можно встре-
тить. _'•< -10. 12. II
Таким образом, термин анеуплоидня в сто общем значении
все еше остается полезным в описательных целях, когда мы
имеем дело с агзмемчнвостыо числа хромосом, ис связанной с
полиплоидией. Явление, известное ныне под пазваинсм агмато-
плоидни, прежде относили к анеуплоидии, однако после уста-
новления его природы и его особых черт оно было выделено как
самостоятельное явление [5S0J. В результате такого терминоло-
гического выделенки анеуплоидня, сохраняя снос первоначаль-
ное широкое значение, приобрела вместе с тем и более узкий
смысл Н зтой главе нам придется использовать термин аиеупло-
илпя как в узком, tan п в общем его значении, указывая всякий
раз. чтд именно имеетен в виду
S3*
3S6
Част» tetouex. ЛжичаотЗил
Из всего того, что будет сказано в этой главе, станет ясно,
что наши представления об агматоплоидии пока еще находятся
на сталии первых поисков. Цитогенетические данные во многих
случаях остаются спорными. Связь между агматоплоидвей и ви-
дообразованием неясна. Мы ие знаем, как достигается репродук-
тивная изоляция между разными агматоплоидныыи популяция-
ми. И нам непонятно, в чем заключается адаптивная ценность
агматоплоидиыл изменений
Полицентрические хромосомы
В царстве животных хромосомы с диффузными центромерами
обнаружены у некоторых Hemiptcra. или клопов, у Coccidae,
Aphldae и Heteroptera. а также у клещей [5C9J. Среди высших
растений диффузные центромеры встречаются у Juncaceae (Lu-
zuta) и Cypcraceae (Eleocharix, Scirpus, Carex); предполагают,
что они имеются также у Zinglberaceae и Мозасеае (обзор и ли-
тературу см. [534]).
Полицентрические хромосомы, содержащие диффузные цент-
ромеры, отличаются по морфологии и поведению от хромосом
обычного типа с локализованной центромерой. Вспомним, что
у обычных хромосом в метафазе митоза наблюдается ясно вы-
раженная центромерная перетяжка н что эти центромеры вы-
страиваются на метафазной пластинке, а плечи хромосом сво-
бодно располагаются п разных положениях. У полицентрических
хромосом таких перетяжек нет, а вдаль метафазной пластинки
выстраиваются целые хромосомы. Когда в анафазе обычные
хромосомы перемещаются к полюсам, то впереди находится
центромера, которая тянет за собой плечи хромосомы. Поли-
центрические хромосомы, напротив, при перемещении к полюсу
бывают расположены примерно в одной плоскости, а иногда нх
концы даже опережают среднюю часть [534].
На рис. 28 1 можно видеть упомянутые выше особенности
полицевгрических хромосом на примере делящихся клеток
Luzuta campestris (Juncaceae) [72].
Возникал вопрос о том, распределяется ли активность цент-
ромеры по длине полицентрической хромосомы непрерывно нля
прерывисто? Иными словами, является лн днффуэность центро-
меры полной или неполной? Данные по Luzuta purpurea свиде-
тельствуют в пользу второй возможности. Хромосомы этого ви-
да. по-видимому, содержат многочисленные центромерные уча-
стки [61].
Хорошо известно, что ацентрические фрагменты обычных
хромосом имеют тенденцию к отставанию во время клеточного
деления и рано млн поздно утрачиваются жизнеспособными кле-
точными линиями. У организмов с диффузными центромерами
можно ожидать, однако, что фрагменты хромосом завершат нор-
Гл 28 Аллагоплсидил
357
малыше метафалные и анафазные движения. Хока несли инду-
цировал у Eleocharis pal us tris (Cypcraccac) разрывы хромосом
прн гюмошн рентгеновских лучей. Он обнаружил, как и ожила*
лось, что в метафазе фрагменты хромосом, даже мелкие, рас*
полагались по экваториальной пластинке, а а анафазе квправля*
лвсь к полюсам [330. 331}.
Риг 2Й1 Митоз у Luzeda rampBtris (2л-12) [72]
Вил с!кжу; видно располсжсиие хромосом в метафазе и рапгтсП апвфаэе.
Слота иные разрывы хромосом у организмов с полицентриче-
скими хромосомами могут, следовательно, привести к возникло
пению восходящих агматоплоидных рядов. У дайной группы
видов может наблюдаться значительная изменчивость по числу
хромосомных пар. а наборы, состоящие из большого числа хро-
мосом, будут содержать по крайней кере несколько более мел-
ких членов [330. 534].
Это не означает, что агматоплоидня валяется неизбежным
результатом фрагментации полицентрических хромосом В ее
возникновении, несомненно, участвуют и другие факторы, как
цитологические, так и селективные Уайт [889, 800] обратил вни-
мание на один, возможно важный, цитологический фактор, а
именно на роль теломеры, или терминального участка, хромосо-
мы. Если хромосомные фрагменты не имеют нормально зажив-
ленных концов, то они ие могут долго оставаться жизнеспособ-
ными (889. 890].
Фактически анеуплоидия (yensu lato) встречается не во всех
группах организмов, обладающих полицентрическими хромого-
зм
Часть седьмая. ЛягуплоиЗня
нами. Средн Heteroptera с диффузными центромерами у неко-
торых семейств (Pentatomidae и Corixidw) число хромосом от-
«чается большим постоянством, тогда как в других семействах
оно колеблется в довольно широких пределах [ЯЙ9]. Разные ви-
ды Luzula, Eleocharix и Carex сохранили низкое основное число
хромосом (889]. У Eleocharis анеуплоидня (sensu lato) встреча-
ется относительно редко.
Carex
У рода Carex, как уже отмечалось в гл. 27. анеуплоидня (sensu
stride) распространена достаточно широко, однако есть данные
о том, что у этого рода встречается также и агматоплондия.
В хромосомах Carex нет ясно различимой локализованной
центромеры. Судя по морфологии хромосом, они. по-видимому,
обладают диффузной центромерой. Кроме' того, в анафазе при
движении к полюсам эти хромосомы часто бывают направлены
концами вперед. Все это указывает кв полмиентричность хромо-
сом и позволяет предполагать, что наблюдаемую изменчивость
по числу хромосом, возможно, частично обусловливает фрагмен-
тация (156, 5341.
Хейльбори [360, 361] сравнивал Carex pilulifcra и С. ericeto-
гшп в отношении обшей длины н числа соматических хромосом.
Он обнаружил, что у вида с более высоким числом хромосом —
С. ericetorum (л—15) — хромосомы короче, чем у вида с баке
низким их числом — С. pilulifera (л—9). Число длинных хромо
сом в наборе снижается по мере увеличения общего числа хро-
мосом (360. 361J. Более поздние исследователи подтвердили это
наблюдение на других видах группы Carex. У видов с боле* вы-
сокими числами хромосом хромосомы обычно короче, чем у
близких им видов с низким числом хромосом (156, 5341.
Обратная зависимость между числом хромосом и их длиной
может быть обусловлена помимо фрагментации и рядом других
факторов. Следовательно, эта группа данных ие может служить
доказательством того, что фрагментация и агиагоплоидия уча
снижали в наблюдаемой у Carex изменчивости числа хромосом.
Однако зависимость между величиной н числом хромосом в со-
четании с наличием полицентрических хромосом и необычайно
широким развитием амеуплоидии (sensu lato) сильно склоняет
к мнению о том, что в видообразовании у рода Carex участвова-
ла агматоплоядия.
Luzula
Как показали Малейрос и др. 1549]. Норденшельд [619. 622]
и Браун [72], хромосомы Luzula (Juncaceae) обладают диффуз-
ными центромерами. Эти и другие авторы обсуждали воэмож-
зае
масть того, что фрагментация хромосом участвовала в создании
различий по числу хромосом у этого рода
Число хромосом у Lutula колеблется от 2л —6 у £ purpurea
до 2л-66 и 72 у £. pitona Число 2«- 12 обнаружено у многих
видов и представляет собой основное число для данного рола.
У этол» рода есть обыкновенный полиплоидный ряд. основанный
на х«6, достигающий октоилоилиого уровня [619|.
У Luzula наблюдается также аисуплоидня (sensu lato). Так,
у L. spicata обнаружены смежные числа п — 6 и7«1 у A orcst-
га — л—10 и И В каждом случае набор с более высоким чис-
лом хромосом содержит две маленькие хромосомы, соответству-
ющие одной большой хромосоме в наборе с меньшим числом
хромосом Таким образом, анеуплонднос увеличение, вероятно,
обусловлено фрагментацией н аг.м атоллом дне А |619. 620|.
Кроме того. Норденшельд [619) обнаружил у этой группы
полиплоидные ряды особого типа, в которых общая длина всех
хромосом остается примерно постоянной Отдельные хромосомы
данного тетраплимда примерно вдвое меньше хромосом диплои-
дов, а хромосомы октоглоида примерно вдвое меньше, чем у тс-
трапломдов. Такне соотношения наблюдаются в группах £ cam-
pt sir is и spicata. Норденшельд объясняет эти одновременные
изменения числа и величины фрагментацией всех хромосом
предкового набора. Численные ccxirHOUieiiiiH между видами но-
сят полиплоидный характер, однако удвоение происходит, воз-
можно, путем фрагментации, а не умножения, и возникающие
прн этом ряды представляют собой агматоплоидию особого зи-
ла (616—621J.
Эта гипотеза относительно апи атолл ом дин подтверждается
поведением крупных и мелких хромосом при нх конъюгации
друг с другом у межвидовых гибридов, подученных прн скрещи-
вании видов, различающихся по уровню плотности. Lutula сат-
pestris н £ paHesccns содержат по G пар больших хромосом,
а у £. iudelua 24 пары мелких яроМОСОМ; следовательно, но
числу хромосом этот вид формально можно считать октопло-
ндом. При мейозе у межвидовых гибридов каждая нз шести
больших хромосом, принадлежащих диплоидному родительскому
виду, конъюгирует с несколькими мелкими хромосомами, при-
надлежащими £ sudetica (2^ 24) [619—621].
Сурсгасеае и Juncaceae
Сем. Сурсгасеае характеризуется несколькими необычными и
хорошо внражепгыии цитоло! кческими и эмбриологическими
особенностями. На эти особенности указывали Хейльборп [360,
361] и многие болег гпхдине исследователи.
260 Часть седьмая. Апеумопдил
Во первых, у некоторых родов Сурегасеае имеются полицент-
рические хромосомы. Как уже было сказано, у Eltochans и Ca-
ret центромеры диффузные. В отличие от этого у Fimbrietylit
в у некоторых видов Cyperus центромеры локализованные.
У Bulbostylit, по-видимому. встречаются центромеры как одно-
го. так н другого типа [634].
Во-вторых, у Сурегасеае в качестве коррелированного состоя-
ния встречается также агматополиплоидия Довольно обширные
и непрерывные апеуплондные ряды (sensu lato) наблюдаются у
Caret. Некоторая анеуплоидия обнаружена у Eleocharis. Одна-
ко виды Fimbristylis образуют четный нли почти четный поли-
плоидный ряд.
В-третьих, у различных представителей Сурегасеае два мейо-
ткческнх деления происходят в порядке, противоположном тому,
в котором они происходят у других растений. Конъюгировавшие
хромосомы расходятся не при первом, а при втором делении
мейоза. В отличие от обычной последовательности первое деле-
ние эквациоииое, а второе — редукционное [156, 867].
Девис [156] отмечает, что у покрытосеменных и у насекомых
постредукциоиный мейоз тесно коррелирует с диффузным со-
стоянием центромер Причины такой корреляции неясны.
В-четвертых, для Сурегасеае характерен особый способ об-
разования пыльцы. В каждой тетраде микроспор три дегеиери
руют и только одна развивается в функциональное пыльцевое
зерно (156. 361. 867].
Интересно, что большинство этих необычных цитологических
и эмбриологических признаков проявляются вновь в сем. Зии-
сасеае.
Диффузные центромеры обнаружены у Liuula, иак уже го-
ворилось ранее н этой главе, но они, iio-вилямому, не встреча-
ются у June иг. В соответствии с этим у Luiuta наблюдается зна-
чительная анеуплоидия (sensu lato). включая некоторую агма-
топлоидию, тогда как у luruus анеуплоидия почти отсутствует
Согласно Норденшельлу [622], у Juncaceae происходит постре-
дукционный мейоз; у них образуются все четыре микроспоры, но
они остаются связанными в тетраде, что напоминает картину,
наблюдаемую у Сурегасеае [644, 914].
Перечисленные ныше черты сходства по цитологическим и
онтогенетическим признакам между Juncaceae и Сурегасеае
вместе со сходством пи различным морфологическим и анато-
мическим признакам указывают на значительно более тесное
родство между этими двумя семействами, чем допускалось в ста-
рых системах классификации, и которыхJuncaceaeсчитали род-
ственными лилиям, а Сурегасеае—макам (130, 330, 544, 914].
Часть восьмая
Специализированные
генетические системы
29 Перманентная
гетерозиготность
по транслокациям
Перманентная тетероэиготность по транслокациям представляет
собой своеобразную н высокоспецналнзнрованную генетическую
систему, в основе которой лежат последовательные транслока-
ции и которая выражается в образовании больших хромосом-
ных колец. Во многих группах растений довольно обычны реци-
прокные транслокации. В природных популяциях таких групп
небольшие кольца (состоящие из 4 или 6 хромосом) встречают-
ся как полиморфные варианты наряду с соответствующими
структурными гомозиготами, образующими биваленты В неко-
торых случаях растения, для которых характерны небольшие
кольца, достигают я популяциях большой частоты вследствие
какого-либо селективного преимущества гетерознтот. В несколь-
ких случаях благодаря преимуществу гегерозитот в популяциях
сохраняется высокая частота растений, у которых образуются
большие кольца (обзор см. |294]). Эти случаи близки к состоя-
нию перманентной гетерозигогности по транслокациям, но не
соответствуют ему полностью
Перманентные тетероэнготы по транслокациям гетерозиготны
По последовательным транслокациям, в которых участвуют по
1Ш1ЫМА мере несколько, е обычно большая часть или все хро-
мосомы данного набора В соответствии с этим у них при мейозе
образуются большие или полные хромосомные кольца. Геноти-
пически ало — диллонды. состоящие из двух геномов особого
типа, известных под названием комплексов Реннера.
Перманентные гетерозиготы по транслокациям прн половом
размножении воспроизводят в чистоте свою структурную гете-
розиготность с выгцеплением либо небольшого числа гомозигот-
ных индивидуумов, либо вообще без выщепления. Обычно это
достигается путем передачи одного комплекса Рейнера через
яйцеклетки, а другого —через пыльцу Односторонняя передача
осуществляется при помощи системы сбалансированных дета-
лей При оплодотворении (как правило, при самооплодотворе-
нии) два комплекса Рейнера воссоединяются, и. таким образом.
3G2 Чаете ооеемал. Смциамшцхман>ше имешжлив систблы
данное гетерозиготное состояние передается от одного полового
поколения следующему.
Итак, перманентная гетерозиготность по транслокациям —
это генетическая система, для которой характерно1 сочетание
следующих признаков, послелога ильные транслокации, хромо
сомные кольца от средних до больших, системы сбалансирован*
мыл деталей, половое разы ножек нс. обычно путем самооплодо-
творения, воспроизведение в чистоте определенного состояния
структурной гетерозиготностя. , s .
Перманентная гетерознгота по транслокациям, обладающая
пенной в адаптивном отношении генетической конституцией, мо-
жет создать популяцию идентичных индивидуумов и распростра-
ниться по территории с благоприятными условиями. Она может
превратиться в гетерогамный мнкроинд.
в свою очередь гетерогамный микро в ид в результате даль-
нейшего развития может стать составной частью более крупной
совокупности — гетерогамного комплекса. Гетерогамный комп-
лекс представляет собой гибридный комплекс, состоящий из ге-
терогамных мнкровидов и нх предковых видов или полувидов,
у которых мейоз протекает нормально с образованием бивален-
тов.
Генетическая система у Oenothera
Многочисленные. геирогамные микровиды, принадлежащие к
группе Oenothera biennis в Северной Америке и Европе, —тра-
вянистые диплоиды (2л-14), размножающиеся преимуществен-
но прн помощи самоопыления. Они являются перманентными
гетерознготамн по транслокациям, причем у многих мнкровидов
образуются полные кольца лэ всех 14 хромосом, а у некого
рых— более мелкие кольца. Растение ОелоМега. образующее
кольца, после самооплодотворения дает исключительно кольце-
образующих потомков: классы структурных гомозигот, которых
следовало бы ожидать, среди этих потомков отсутствуют. Гете-
роэнготы по транслокациям воспроизводят свою гибридную кон
етитуцию в чистоте прн помощи полового размножения.
Цитогенетика Oenothera разрабатывалась рядом исследовате-
лей. Классическими считаются работы де Фрила, открывшего
аномальное поведение этих растений при скрещиваниях, и ра-
боты Реннера, давшего объяснение такому поведению. Затем на
протяжении более 40 лет анализом этой проблемы усиленно
занимались Клеланд я его ученики. Хорошие обзоры ранних
исследований составили Клеланд [109] н Дарлингтон [139].
Большое количество фактических данных собрано в моиогра
фни Клеланда с соавторами, опубликован мой в 1950 г. Обзор
со ссылками на миогочислемяые литературные источники дал
Гл 2D ntpuvwina* trttfOJuMHHtxTb по трслс.юклцилч 363
Клеланд (112]. Краткий обзор современного состояния проблемы
принадлежит Гранту J294K
11 целях удобства мы опишем здесь генетическую систему
перманентных гетерознгш по транслокациям, используя терми-
ны, относящиеся к классическому объекту, из котором было
изучено >то явление, тек Oenothera МнпагсЬшпа |рпс 291).
Этот вид широко распространен r Европе и Северной Америке
|6<М. 606], при конъюгации и мебозс у него образуются кольцо
из 12 хромосом и 1 бивалент (рис 29 2). Этот вид, ткни обра
зом, гетерозиготен г»о последовательным транслокациям в 12 из
имеющимся у него 14 хромосом.
IW 29 1 O/rtoxlr/d lAihafJ.iana (—Or ег$ •кгокраЛа. Ое giazioctana) [604].
ЭМ Часгг 0се»жал, Свгчиалиэирсвалчьее гыспгчлслли еиспмы
Взаиморасположение герминальных участков хромосом
Oenothera lama'Ckiana можно описать, сопоставляя его с нх рас-
положением у близкого структур ио-гомозиготного вида Ое. hoo-
keri, произвольно принятого за стандартное. В табл. 29.1 указа-
ны последовательные номера, присвоенные 14 плечам 7 хромо-
сом стандартной расы Ое hookeri. У большинства индивидуумов
Ое. hookeri два гаплоидных набора хромосом структурно гомо-
логичны, поэтому при мейозе они образуют нормальные бива-
ленты. Гаплоидные наборы Ое. lamarckiana отличаются от аиа
логичных наборов стандартной, или дефрнзовской, расы Ое. hoo-
keri по нескольким транслокациям (табл. 29.1). Кроме того, два
набора у Ое. lamarckuuia отличаются один от другого по после-
довательпым транслокациям во всех хромосомах, за исключе-
нием хромосом с терминальными участками 1—2 (табл. 29.1).
Таким образом, наблюдаемую кольцеобразную фигуру иа 12
хромосом и 1 бивалент можно объяснить конъюгацией гомоло-
гичных плеч при мейозе (рис. 29,2. 6).
В метафазе I хромосомы, образующие кольцо, выстраиваются
на веретене зигзагообразно, чтобы затем разойтись к разным
полюсам (рис. 29-2, Б). В анафазе I соседние хромосомы кольца
отходят к противоположным полюсам; иными сломами, хромо-
сомы. лежащие в кольце через одну, отходит к одному и тому
же полюсу. Таким образом, к концу мейоза два дифференциро-
ванных генома вновь воссоединяются а интактном виде [109,
1121.
Геномы особого типа, обнаруженные у Oenothera, известны
ПОД общим названием комплексов Реннера, к каждый из инх
имеет собственное название. Два комплекса Реннера, имею-
щиеся у Ое. tamarcklana, носят названия velans и gaudene
(табл. 29.1). Следовательно, Ое. tamarcklana имеет гетерозигот-
ную геномную конституцию velansfgaude/is. Описанный выше
тип расхождения в мейозе кольцеобразующих хромосом право-
Таблица 29.1. Сравнение стандартного расположения терминаль-
ных участков хромосом в комплексах Реннера с расположением
этих участков у Oenothera lamarckiana [111, 111]
Ь-д Распа» вшит
Реймера a iimamuimm
Oe. hookeri, раса де Фри- Зв standard hooken 1—1 3—4 S—6 7—8 9—10 11— <2 13—14
Ое. lamarckis- velans 1-2 3—4 >-8 7—6 9-10 11—1J 13-14
па ftaudens 1—2 3—12 5-5 7—11 Я-4 8-14 13—IB
365
Гл 29 Перканемгчая tticpv зиготность ло трачслокацояж
А
vetonj
9*udrns
Б
Рнс 29 2 Кончю'хш и |1йсм:мд«ние хромосом у Оепо/Лего lamardhcna
<2*-14) (200)
Этот из арс^ывжллет собой trtepu.iKOTy (Ntou gardens Л Дикемеэ. вид-
но кольцо iu 12 чроуосом м I бммлент л Метафиз 1, кольцо раихлюжено
па вгрстенг т.тьпм оОрзтзч. чтобы п анифах хроятосо'чи мог.та расходиться
я pj’iiuv милюсам через сдмт (черед) ющееся рэсхождеиае)
дит к точу, что в одно дочернее ядро переходит целиком комп*
леке velons, а в другое —комплекс gawfens (рис. 2У 2. Б).
Иными Glonn мн, гетсрозигога veians'gaudens обычно образует
лишь два класса гамет зл'ш и gardens, если иметь в виду
только последовательность участков в хромосомах
Следовало бы ожидать, чго при самооплодотворении «ли
при скрещивании сибсов Oenothera lamartkiana злет потомков
трех типов velans'velans. velans'gaudens и gaudenx/gauden* в
менделсвском соотношении 1:2:1 Вместо этого, как уже гово-
рилось. образуются только гетсрознготы Отсутствие ожидав-
шихся классов структурных гомозигот обусловлено действием
системы гбалансированных легален (109. 112, 673].
Такая генная система считается из комплементарных деталь
ных факторов, сбалансированных в гетерозшопюм состоянии
Двойная гегерозигота Jr^) жизнеспособна. гомозиготные ре-
комбинации « (т?Н нс:*|,:>иес,,осо^пм Каждый набор
комплементарных летальных факторов сцеплен с одним комп-
лексом Реннера. С гчеточ сбалансированных лет злей геном
Oenothera lamarcktana можно записать в виде - Тн-
^вПОГЯ!
иы oeians/velans н gaudenslgaudens гомоэнготны по легальным
факторам и гибну г кг стадии .штаты
Чвст» есч»и«. Омцымидареави/ыы .Cne/wwTew пктеми
зев
С различными комплексами Реннера сцеплены не только ле-
тальные факторы, но также гены, детерминирующие те или иные
фенотипические признаки (см. (109, 1121 а также сжатый об-
зор Гранта (294|). Кроме того, гены, находящиеся в разных хро-
мосомах. принадлежащих к одному и тому же комплексу Ренне-
ра, передаются в составе одной группы сцепления. Поэтому гиб-
риды по транслокациям воспроизводят в чистоте не только свое
особое состояние структурной гетерозиготное™, ио также, и
притом одновременно, особую совокупность фенотипических
признаков
Несколько компонентов генетической системы перманентной
гетерозиготное™ по транслокациям варьируют в зависимости
от вида в пределах группы Oenothera lamarckiana. Одни из та-
ких изменчивых компонентов — величина кольца. Кольцо из
12 хромосом, имеющееся у Ое. lamarckiana. не типично для груп-
пы в целом. У одной популяции О». irrigua (*0е. hooker! hewet-
Ш) в Нью-Мексико постоянно образуется кольцо из 8 хромо-
сом + 3 бивалента. Чаще всего, однако, у микровидов, населяю-
щих всю центральную и восточную части США, образуется коль-
цо из 14 хромосом (111, 114].
Система сбалансированных деталей также подвержена из-
менчивости. У представителей рассматриваемой группы, оби-
тающих ив западе США. эти летали отсутствуют или встреча
ются спорадически. Сбалансированные летали характерны, од-
нако, для гетерогамных микровидоа. распространенных в цент-
ральных и восточных областях Северной Америки и в Европе.
Существуют системы сбалансированных деталей двух типов
(рис. 29 3). В системах первого типа, примером которых служит
Oenothera lamarckiana, действие деталей проявляется на стадии
зиготы; в системах второго типа, широко раслростраиснпых в
Северной Америке, оно сказывается на гаметофитах. Соответ-
ственно растения энотеры с зиготными деталями производят
полустерильиые семена, а растения с гаметофитнымн деталя-
ми — полустерихьиую пыльцу [109, 111, 112, 114). Еще ОДНО след-
рис 29 3 Дм типа систем сбиансипованных деталей у ОеяоСЬега [113]
Слева. Система е участием зятстных деталей. Сарова Система с участием
гамето0итимх леталсЛ [113].
(л 29. txt хромистость ло гра-*.лакццмях JQ7
сгоне системы гамстпфптных деталей состоит в том, чти данный
гетерогамный ымкровид постоянно передаст один из своих комп-
лексов Реннера через пыльцу, а другой — через яйцеклетки.
Источники изменчивости
Причины наследственной изменчивости у кольцеобразующих
энотер разнообразны. Одним нз важных источников новых форм
служит происходящий время от времени ауткросенмг между
микрюикдами. у которых обычно наблюдается инбридинг. Дру-
гие процессы создают новые вариации внутри самой кибрелиий
линии Хотя Oenothera tamarcklana к родственные ей формы
после самоопыления обычно производят единообразное потонет
ДО, тем не менее у них возникают с некоторой низкий частотой
новые формы, в тон числе дефризовскис нутации.
Прежде чем обсуждать частоту мутаций у виотеры, необхо-
димо уяснить себе, что некоторые гены сцеплены в кольцеобра-
эуюшей гстерознготе с комплексом Реннера, а другие нс сцеп
лены с ним Последние расщепляются обычным образом У та-
кой неполной гетерознготы по транслокациям, как Oenothera
lumurckiaita, растепление по хромосомам, образующим бивален-
ты, происходит независимо от расщепления по хромосомам, об-
разующим кольцо. поэтому гены, локал нэпе а нны». в fin вален i пых
хромосомах, не входят в комплексы Реннера. Гемы, находящиеся
в герминальных конъюгирующих участках кольцеоб разу тощих
хромосом, могут более или менее свободно переходить у комп-
лексных гетерозигот из одного комплекса Реннера в другой. Та-
кие гены также не зависит от какого-то одного из этих комплек-
сов (см. [112]). Разумеется, эти свободно рекомбинирующиеся
гены ие Дают начали новым «мутатам» у Oenothera
Гелы, которые детерминируют характерные фенотипические
признаки, наследуемые э виде единого блока через любой данный
комплекс Реннера и перс-Тамар мыс своим потомкам любой дан-
ной комплексной гстсрозиизтой, должны располагаться на не-
кой ьюгируюшмх дифференциальных участках хромосом с транс-
локациями (112, 139}. Эти участки .хромосом являются прокси-
мальными тг находятся по соседству с транс локационными раз-
рывами. участвующие в этом хромосомные механизмы рассмот-
рены Грантом (2SM). Сбалансированные легальные факторы
должны быть также локализованы в этих дифференциальных
участках Таким образом, гетерознготы по транслокациям вос-
производят я чистою определенную гетерозиготную ком би на
иию генов в этой части генома при иомпщи rcueiичсской систе-
мы, описанной в и ли идущем рлздслс
Однако изредка, хотя и с равномерными интервалами, про-
исходят цитогенетические события, которые отделяют один или
ЗД Voct* kk>*u. Сл«ч—ьыы^мшии— Mivnmuw сасгмм
несколько генов, локализованных в дифференциальных участ-
ках, от соответствующих комплексов Реннера и дают им воз-
можность проявиться в новых гомозиготных сочетаниях. Одно
из таких событий — происходящий изредка кроссинговер и об-
мен генами между комплексами Рейнера. При этом какой-либо
ген может иногда оказаться в противоположном комплексе Рен-
нера. Затем в результате самоопыления возникает несколько
потомков, которые гомозиготны по этому гену и появляются в
качестве новых стабильных мутантных типов.
Другое событие —возникновение новых реципрокных транс-
локаций в данном комплексе Реннера. Такие транслокации мо-
гут разорвать большое кольцо комплексной гетероэнготы на
меньшие кольца н биваленты Затем после самоопыления целые
хромосомы н блоки генов могут образовать гомозиготных выше-
пеицев Возникающие при помощи описанных выше и других
способов редкие гомозиготные выщепенцы представляют собой
диплоидных мутантов энотеры (см. [112, 114. 674]).
У хромосом, образующих кольца, при разделении в анафа-
зе 1 наблюдаются разного рола нарушения. Время от времени
возникает иераехождеяне, в результате чего образуется некото-
рое число гамет с 8 хромосомами. Такне гаметы в свою очередь
дают несколько трисомиков с измененными фенотипами. Боль-
шинство мутантов у Oenothera lamarckiana оказались трисоми-
ками с 2л —15 [112, 114).
Второй главный источник новых вариаций у энотеры — гиб-
ридизация между различными комплексными гетерозиготами.
Два гетерогамных микроиида. каждый нз которых при самооп-
лодотворении дает обычно потомков одного типа, нередко дают
в первом гибридном поколении четыре класса потомков. Так,
при скрещивании Oenothera lamarckiana (velanslgaudens) и
Ое. strigosa (deprimens/stringens) получаются все четыре воз-
можные диплоидные комбинации velansldeprlmens. velanslstrin-
gens, gaudensldeprimens, gaudensistnngens [112. 139].
Многие из этих новых комбинаций геномов, возникающих при
указанном выше и других гибридных скрещиваниях, содержат
больше бивалентов или мелких колец, чем любой из родитель-
ских видов [112, 139]. Некоторые лары генов, сцепленные у ро-
дительских микровкдое с комплексами Реннера и сохраняющие-
ся у этих микровидов в состоянии перманентной гетероэвготио-
ста, у межмикровидовых гибридов будут распределяться неза-
висимо. В последующих поколениях такие гены могут начать
выщепляться в качестве новых гомозиготных типов. Если эти
гомозиготы окажутся жизнеспособными, то новые вышепсниы
станут постоянным добавлением к фонду изменчивости данного
гибридного комплекса.
Гл. И. Пе/мачечпая гетерозит аасп но гралслсклция* ,еу
У некоторых гибридов первого поколения, полученных от
скрещивания между двумя гетерогамными микровндами, име-
ются полные или почти полные кольца, ассоциированные с по-
вой комбинацией сбалансированных летальных факторов Та-
кие гибриды будут размножаться в чистоте. Они представляют
собой потенциальные зачатки новых гибридных микровидов,
происходящих непосредственно от скрещиваний между сущест-
вующими гибридными микровидами 1112].
Гетерогамный комплекс в группе
Oenothera hookeri-biennis
Североамериканские виды подрода Oenothera рода Oenothe-
га —высокие многолетние или однолетние травянистые расте-
ния, часто сорные, с желтыми цветками, распускающимися ве-
чером. Все они- диялопды с 2п=14 Эта группа усиленно
изучалась с цитогенетической точки зрения Клеландом и его
школой [110—114] и в таксономическом плане —Мюнцем [604—
606]. Североамериканские энотеры — таксономически трудная
группа вследствие характерного для пих широкого диапазона
изменчивости и отсутствия ясно выраженных границ между ви-
дами.
При ревизии этой группы Мюнц (606] выделил десять таксо-
номических ВИДОВ. МНОГИХ ИЗ которых ЛОЛН1ИЛИЧССКНС. с одним
или несколькими подвидами. Клеланд 1112—114] различает
сходный ряд таксономических видов или «групп рас», как зи их
называет Системы Мюнца и Клеланда, хотя они и не идентич-
ны. сходны и хорошо согласуются одна с другой (та б- 29 2)
После того как была написана эта глава, Рсйвн, Дитрих н
Штуббе (662] опубликовали новую работу по таксономии подро-
да Oenothera Прннятаг ими система существенно отличается от
системы Мюнца Все эти три системы представлены н сопостав-
лены и табл 29 2.
В большинстве генетических исследований используется но-
менклатура Клеланда Во избежание путаницы мы также будем
придерживаться этой номенклатуры в нашем обсуждении Так-
сономические эквиваленты единиц, различаемых Клеландом,
можно найти в табл 29 2
Вид Oenothera lamartklana ролом из Европы был случайно
завезен в Северную .Америку Нас здесь интересуют главным
образом остальные девять видов, возникшие в Северной Амери-
ке и составляющие группу Ое hooheri-hiennis
Географическое распространение девяти местных видов по-
казано па рис. 29.4 и 29 3. Ни Клеланд, нм Мюнц не опубл ико-
на.то хороших карт географического распространения этих ви-
24-45
tn>
Чет «осэзша. Славна м гоуо«дяпы» ммтиакк» пктг«м
Таблица 29 2. Североамериканские таксоны Oenothera поХрода
Oenothera, различаемые Мюицем [606J. Клеландом {114] и Рев-
аком и др. [662]
Cnevnta Mieuua Ma*u««a гм КлелйПУ Намаяна io PcImp и X9
Ое. hooheri (9 подвило*) Of. hooitffi Off. eiaiu (hockffi M.UO WM • eriaia\
Or longissima (2 под- вид» > Of haokeri Oe tongistima
Ол jamesli Oe kocken Oe
Ое elafa Oc etata Of. elala (более шгра- ия кокиепция)
— — Or ww'pi (яыде/ен hj hooUrt)
Oe. argillUuia Oe afgitbr-ata Oe. araillicda
Or grandiflofa L'Heri* tlcr (не de Vrtea> Oe. partlflosa Of graa^ifiora Oe. g fMiIi/lor a
tubsp. narvifiora Oe rwviltora II Oe оаАгу^ла
subep аядоййдйаа Oe parr i f lor a [ Oe. pars-(flora
Of- bsetvus e
subsp caeciarum Of bifnnit II Of biennis
SMbip. Lentrah^ Or biennis 1 Or Inennit
50fc*p (Ш31ГО1ПОЛ- tana Oc biennis 111 Oe. aurJronwn/ana
Oe singota (3 подвжд1| Oe. eryikrofcpala (амжлж Oe. sfng&ra Ои tMnfeQ
Oe lamarckiana Oe giaaaniana
мяткэироеан • Север nil Амер! м| de Vries
доз Поэтому ареалы, изображенные на наших каргах, лишь
приблизительные, хоти и основаны на опубликованных • этими
авторами сведениях. Как указывал Клеланд (111—114). рас-
сматриваемые таксономические виды распадаются на два ряда
по типу генетической системы. Одна подгруппа обладает нор-
мальней диплоидной генетической системой. У входящих в нее
растений крупные, свободно опыляющиеся цветки; при мейозе
у них образуются (за некоторыми исключениями) биваленты, и
они не содержат сбалансированных деталей. Другая подгруппа
состоит из перманентных гетерезигот по транслокациям. У вхо-
дящих в нее растений образуются крупные хромосомные кольца
(по большей части из 14 хромосом), они размножаются при по-
мощи самоопыления и содержат сбалансированны* летали.
Биваленты образуются у следующих видов: Oenothera Лоо-
ken. Ое. argilticola. Ое. grandi/lora и Ое. data. От них происхо-
дят хольцеобразующае виды: О*, parviflora, Ое. biennis. Ое. stri-
gose и Ое. lamarckiana. В некоторых популяциях Ое. boohed,
Ое. jamesu и Ое. tongissima образуются кольца небольших или
Рис 294. Геигрвфмческсе расгростр*иенте в Северной Ашрмке вмхов
lhera подрода Oenothera, образующих бивалелы (по давным MMU |606]
н Клславда [114|).
Гл 29 Пея*®*»е<*чо * гммро-шлэг*аст» яо г ране лп*е^ик *
373
средних размеров; иногда эти популяции несут также сбаланси-
рованные летали, занимая, таким образом, промежуточное по-
ложение между двумя основными подгруппами (112, 114. 604.
606|.
Филогенетические корни видов, у которых образуются коль-
ца. явно следует искать средн аидов, формирующих биваленты
Последние, несомненно, обладают нормальной половой генетиче-
ской системой. Они предпочитают умеренно влажный климат,
что считается в данной группе растений примитивным призна-
ком. Некоторые из видов, образующих биваленты, разобщены •«
носят реликтовый характер, чти указывает па их достаточно
древний возраст В противоположность этому кильиеобразуюшпе
виды с их производной генетической системой агрессивны и час-
то являются соринками Они заселяют распаханные поля на
Среднем Западе и проникают в северные облает, недавно нахо-
дившиеся подо льдом
Очень знаменательно поэтому, что одни комплекс Реннера,
известный как hookcri Johansen, обычек н широко распростра-
нен у Oenothera hookeri и встречается также у Ое grandiflora и
Or. Qfglllicoia. В комплексе hooberi Johansen терминальные уча-
стки располагаются следующим сбраээм 1—2 3—4 5—6 7—10
9—8 II —12 13—14. Этот комплекс Реннера — не единственный,
обнаруженный у Ое kookerl. но он вс-рспется чаше других
Кроме того, отдельные хромосомы из комплект hooJteri Johan-
sen часто встречаются в других комплексах По я им причинам
комплекс hookeri Johansen считается комплексом с пред копим
расположением тернннагьмых участков в данной группе [114,
606, 798. 799].
Три или четыре таксономических вида кольиеобрнзуюшнх
Oenothera плохо различимы и очень изменчивы как фенотипи-
чески, так к генотипически. Фенотипически о»'и варьируют от
мезофитов с широкими тонкими листьями до ксерофитов с уз-
кими толстыми листьями. Чтп касается геномов. то у Oenothera
parent lor a. Ое biennit и Ое tirigoza известно примерка 130 комп-
лексов Реннера (это число получено в результате подсчета раз-
личных комплексов, перечисленных Клеландом (114, см прило-
жение]) Эти комплексы, соединяясь попарно. образуют бесчис-
ленное множество локальных или широко распространенных ге-
терогамных мнкровндэв.
Вследствие этого филогенетические взаимоотношения между
производными кольнеобразукипимп видами и предковыми вщп«
ми. образующими бнвиленты. весьма сложны и до сих пор не
выяснены, Некоторые филогенетические связи можно предста-
вить себе п общих чертах, однако еше остаются большие пробе-
лы. В целом можно сказать, что филогения этой группы поскт
сетчатый характер, так как кдлшеобразующпе таксономические
374
Часть аасыюл. Сптциллили^а^ые тшетыческме системы
Рис- 296. Постулиро*лни1« фмшгмня мсстжых североа мерин дютих внаав
подрзда Oenothera (на [113, 114] с яеволмпиив шмеяениями)
виды я мнкровиды предел авляют собой продукты гибридизация
между разными парами родительских видов.
Реконструкция филогении, созданная Клеландом [113, 114].
основана на данных по морфологии, географии, распределению
концевых участков хромосом, пластидам и 5 факторам, или фак*
торам самояесовместм мости, которые обусловливают появление
пыльцевых деталей в системах сбалансированных деталей. Дан-
ные, полученные в разных областях исследования, не всегда сов-
падают, к это вызывает сомнения в правильности некоторых час-
тей филогенетической схемы. Суть филогенетической гипотезы
Клеланда отражена в сжатом виде на рис. 29.6 и в нижеследую-
щих дополнительных замечаниях
По мнению Клеланда, центр возникновения группы Ое. Лпо-
ЙуМЙНЦ|В находится в Мексике и Центральной Америке. Из
этого центра примитивные формы мигрировали в умеренные об-
ласти Северной Америки; пять последовательных волн миграции
далн начало пяти видам или полувидам, получившим соответ-
ственно названии популяций 1. II, III. IV и V. Популяция 1—*
непосредственный предок Ое. aegdlicola, популяция II—•
Ое grandiflo/a, а популяции III и IV - Ое. hookeri. Популяции
III и IV — гипотетические. Популяции 1—IV являются предками
кольисобразующих Oenothera. Как показано на рнс. 29.6, оив
гибридизировали в разных комбинациях, дав начало колъцеоб-
Га 29. Перманентная {стерохиотность по тр®*мокя«»шд 375
разу ютим типам. Кроме того, с течением времени в разных ли-
ниях 1’акап.'1ивэлнсь транслокации.
Принятые» в этой гипотезе допущения относительно центра
происхождения и волн миграции необязательны Кроме того,
собственно Ое hookai, согласно этим допущениям, нс участво-
вала в создании производных типов, что представляется, однако,
маловероятным, млн принять но внимание обширность области,
в которой опа енмлатрпчио с Ос. strigasa. Следовательно, было
бы, возможио, лучше си казаться от некоторых из этих допуще-
ний п просто постулировать, что в Северной Америке произошла
дивергенция на несколько предковых видов, из которых одни из
лестны, а другие неизвестны. Теперь мы можем перейти к рас-
смотрению взаимоотношений кпльцеобразующих типов, по кою-
рым имеется больше данных Верхняя nacib филеи енсшчсскоп
сети (рис 29 6) известна лучше, чем ее лсиоваиис
У комплекса Oenothera biennis, передаваемого через яйце-
клетку (или у тал называемого альфа комплекса), расположение
терм .шальных участков и двух известных популяциях относится
к типу hookeri Johanson; во многих других популяциях, зани-
мающих обширную территорию, наблюдается иной тип располо-
жения отлнчаюшийся от hookeri Johanson на одну транслока-
цию, наряду с другими типами, различающимися сильнее
Oenothera biennis 1 и II очень сходны фенотип и чески, их
предковые липин пардлдеэды и пропс» m же
двух родительских видом Оли им из этих видав был Ое g ranch-
flora, от которого Ос. biennis 1 (но не Ое biennis II) получила
комплекс hooker i Johansen. С другой стороны, Ое biennis I н II
имеют общих предков с Ос sirigosa Но з/п^озл-подобпыс приз-
наки локализованы в передаваемом через пыльцу (бета) комп-
лексе у Ое. biennis I и в женском (альфа) комплексе у Ое. bien-
nis I! У Ое. biennis I и II альфа- и бета комплексы ревертиро*
папы. Ос biennis III происходит от скрещивания между Ос bien-
nis I 2 и Ое Ыегзлгх II d {114] (рис 296)
Комплексы Реиьери у Oenothera parviflora, Ое strlgosa и
Ос. biennis (за исключением упомянутого выше альфа-комллскса
Ое. biennis 1) силы™ отличаются or предкового комплекса hooke-
ri Johansen, будучи уда темы от него па три нли более трансло-
кации, Диа комплекса Реннера у Ое parviflora I. очень далеки
от Комплексов нх предполагаемых предков — Ое argillicola и
Ое, grandiflora. Ое. parviflora II также далек от своего предпо-
лагаемого предка Ое argillicola по своему геному
Oenothera parviflora I имеет общих предков с Ое. biennis по
своим альфа-комплексам Ое parviflora II в отличие от Ое par-
viflora I песет в своих альфа комплексах slrigosa-подобные при-
знаки и, по видимому, имеет общую родословную с Ое sttigosa
(рис. 20 6)
376
V«rt аосьнал. Спгтюмзарошми ижтичаоим сиспгш
Гетерогамные мнкровиды в группе
Oenothera biennis
У хольцеобраэуюшкх видов существует ряд разнообразных
комплексов Реннера, передаваемых как через яйцеклетку, так
и через пыльцу. Число альфа- и бета-комплексов, перечисленных
Клеландом [114, см. приложение], указано в табл. 29.3, а в
табл. 29.4 приведены некоторые примеры. Возможное число
диплоидных комбинаций этих альфа- и бета-комплексов в каж-
дом кольпеобр азующем таксоне достигает нескольких десятков
или сотен.
Таблица 29.3. Число известных комплексов
Реннера у Oenothera parvijlora и Ое. strigo-
se (по данным о числе комплексов, приве-
денным Клеландом [114])
Tieece Чмсап мафа ЧК14 6«T»- к:«(Ц/«сиа
Ое partijltjfs I 9 2
Ое parwlltra I] 6 4
Oa birrtnu ' 14 27
Ое Ыептя 11 12 7
Oe biennis 111 4
Oe. stngota 2? 11
Любая конкретная диплоидная комбинация двух комплексов
Рейнера определяет конкретный гетерогамный микровид. Гете
рогамные микровиды представляют собой основные популяци-
онные единицы гетерогамных комплексов, оправдывающие су-
ществование генетической системы энотеры, поскольку они явля-
ются размножающимися в чистоте гибридами, гетерозиготными
по группе последовательных транслокаций и по генам, локали-
зованным в хромосомах с транслокациями Гетерозигота по то-
му или иному гену, если она обладает адаптивной ценностью,
может размножать свою гетерозиготную конституцию при помо-
щи генетической системы, свойственной энотере, и распростра-
ниться а качестве гетерогамного вида в локальном иля регио-
нальном масштабе.
Клеланд не состаинл списка комплексных гетерозигот или
гетерогамных микроаидов как таковых однако их можно рекон-
струировать на основе его данных, скомбинировав альфа- н бе-
та-комплексы. зарегистрированные по отдельности в одной и
той же местности [114; см. приложение]. Каждый кольцеобра-
эующий вид насчитывает следующее число михровидов: Oenot-
Таблиц» 29 4 Избранные комплексы Реннера в группе Oenothe-
ra hookert-biermis [114]
Ban Рас«о»жямав *г*мв*аам«ых учаоюо* Немалое вомлеае* мяв м*етяас*ь
Ot. toottri 1—2 3—4 5—5 7—14 9-8 11-12 13-14 HooJtffi Johansen
Or hoakrrt 1-2 3-4 »-e 7-8 *-1» 11-12 13-14 de Vriea
Or. etats 1—4 3—2 5-6 7—1» 11-12 13-3 Чааульгвмк. Макеты
Of parviftora l i—i a 3-7 5—14 4-10 9-8 11-12 2-0 Киифтои-Форж. Виргиния
Oil Шиин» I 1-2 3-4 5-14 7-ie 9-8 i 11-12 13—C Средний Зилад
Of brnrnt I 1—9 3—10 S—11 7-4 3-8 11—12 13-6 Р*чаюм, Виргиния
Of Singrid 1—7 3-4 5-8 2-10 »—tl 6—12 13—14 Келоридс-Слриагс. Колорадо
Oe. slngvia 1-7 3—4 b-9 6-e 10-2 11-12 13-14 Сгтерлеяд. Небрасха
Таблица 29.5. Хромосомная констггуция пяти гетерогамных мим-
роеидоа а группе Oenothera biennis (на основе данных Клеланда
ЦП])
Во Ммхмоегь Кмаагкс Р|ГЯУМЖ«1«Г
Or Ммжтгрм, Колорадо Лета хл|фа 1-5 1-И п 6—14 5-5 7—8 7-12 9—10 0-5 11-12 Л-12 15-в 1Э-М
Or. rlrlg&ia Кллр<с. бега альфа 1—4 L—13 п 3—10 S—в 7-6 7-в 9-Ь 9-11 11-12 10—12 13-14 2-14
Or. bltanls 1 Bipina ai. Индиана б «ГТ* альфа 1-2 II и 7-8 7—10 5-12 9—13 11—10 11-12 13—И
Or bitfnnis 1! Сгшфо восток бета лифа 1-2 3—12 3-2 U 7-10 4-10 п 11-13 11-12 13—14
Or parvijlors 1 Клифтом Форм. Вирги НИИ бета альфа 1-4 1-19 3-2 3—7 S-1J 5-М 7-10 4-10 II 11-8 И-12 13—14 2-6
Гл 29 Перлитсмтал по грапсмк&цахм 379
hera par atftor a — 4 ммкровида; Or. biennis — 90 мн кров и дов и
Ое strigosa - 41 микроенд.
В табл. 29.5 показана в целях иллюстрации геномной консти-
туция нескольких гетерогамных мнкроиидов Хотя протяжен-
ность ареалов этих и большинства других мнкровилов в данной
группе неизвестна, многие из них, вероятно, имеют локальное
распространение; некоторые же виды распространены широко.
Например, микровнд Oenothera biennis И, обозначенный в
табл 29 5 как североеосточный, обнаружен в риле мест в
шт. Пенсильвания, Нью-Йорк, проа Онтарио, Нью Брансуик и
Квебек; этот мнкрояид, очевидно, представляет собой удачную
гетерозиготную мпабшаамю
За последние годы Левин и его сотрудники стали применять
для анализа мн крив и дов группы ОеноГйега biennis методы элек-
трофореза
На одном из полей в шт. Коннектикут популяция Oenothera
biennis была исследована ги 19 ферментным локусам Пять из
этих 19 локусов были гетерозиготными, т. е средний уровень
гетероз Hi отности был очень высок составляя 26% (5/19). Кроме
того, у всех, индивидуумов в этой популяции оказался один и
тот же гетерозиготный генотип по этим ферментным локусам.
Иными словами, уровень полиморфизма у них низок Эту попу-
ляцию, а возможно, и ряд популяций на всем протяжении одной
обширной территории в шт. Коннектикут характеризует один
определенный гетерогамный микровмд (4/21.
Левин |472] пег л ело вал 44 популяции Oenothera biennis I в
пп Иллинойс по 20 ферментным локусам Средняя гетерозигот-
ногти составляла 4.5% локусов Уровни полиморфизма в этих
популяциях были следующими
I генотип у 59% популяций
2 генотипа у 27% >
3—5 генотипов у 16% »
Основным типом изменчивости в данном случае является не
внутрилопулмиионная, а ыежпопуляанонная изменчивость
Средняя гегерслиготногти по ферментным локусам в более
обширном ряде популяции группы Oenothera biennis составляет-
9Д% у собстэгнно Ое biennis, 143% у Ое pan^tura и 2,8% у
Ое strigose Эти данные сравнимы с данными по образующей
биваленты Ос argilliurta. у которой уровень гетсролпготностм
составляет 8 0% |4вЗ. 487].
Утверждалось, что генная гетерознготпасть в ipynne Ое. bien-
nis ниже, чем можно было бы ожидать у перманентных гетеро*
зигот |472]. Возможно, однако, что функция генетической систе-
мы Oenothera состоит те в поддержании высоких уровней reie-
розиготпости, а в том. чтобы обеспечивать репликацию опреде-
линией гстеролиготной комбинации гемов.
380 Часть somnas. Сгицшиюиробахгии сештачеоом системе
Гетерогамные микровнды и комплексы
в других группах растений
Подрод Oenothera рода Oenothera не является уроженцем Евро-
пы; он был запеки сюда из Северной Америки, акклиматизи-
ровался и эволюционировал на протяженна двух или трех
веков. Различные линии энотеры, завезенные в Европу
(обычно на кораблях, в балласте), гибридизировали с образо-
ванием ряда новых местных гетерогамных микровидов. Реннер
[675] различал в Европе 18 кольцеобраэующнх микровидпе под-
рода Oenothera.
Некоторые из них, например европейский вид Oenothera Ыеп-
nit. по-видимому, возникли в Северной Америке, но большинство
из зтих 18 европейских видов возникло de novo в результате гиб-
ридизации. происходившей в Европе. Самый известный из них —
Ое lamardtiana (velanstgaudens). Комлекс veiana близок к
Ое hookeri, а комплекс gaudens лежит в пределах диапазона
изменчивости Ое. biennis. Гибридизация между Ое. biennis и
Ое. hookeri происходила в Европе, и продукт этой гибридизации.
Ое. lamarckiana. распространился по всей Европе, а позднее
проник в Северную Америку. К другим хорошо известным евро-
пейским видам относятся Ое. suaoeolens (albicans/flavens),
Ое mollis (simulant Iplanans) и Oe. rubrieaulis (lingens/rubens)
' ll Южной Америке Oenothera (подрод Oenothera) представ-
лен секцией или подсекцией Mumia. параллельной собственно
энотерам Северной Америки. Изучение Oenothera-Mumia, пред-
принятое недавно Дитрихом [164]. показало, что эта группа со-
стоит из 57 таксонов, из которых 20 образуют биваленты, а
37 — кольца. Среди кольцеобразующих таксонов встречаются
растения с большими кольцами (из 14 и 12 хромосом) и расте-
ния с мелкими кольцами. Таким образом, Oenothera Mumia
представляет собой еше один хороню развитый гетерогамный
комплекс [164].
Кольца из 14 хромосом обнаружены у представителей дру-
гих подродов Oenothera я у других родов сем Onagraceae, род-
ственных Oenothera. Ниже перечислены такие таксоны; псе
они — обитатели Северной Америки
Oenothera подрод Raimannia (3561
Oenothera подрод liartmannia [322]
Oenothera полрод Laoauzia [322]
Oenothera полрод Knei/fia; Oe. perennis [807]
Gayophylum heterozygum [497, 560)
Gaura biennis и G. tnangulata [663]
Calylophus serrulalus [835]
Гл 29 Перюиалпа» гегеромгопосте по тххислжшщкл 341
Мелкие транслокациоиные кольца часто встречаются в более
обширной группе родов, родственных Oenothera, а именно п
большой трибе Onagrrae, а также в маленькой трибе Наоуеае.
Кариотип у представителей утих родов характеризуется двумя
существенными особенностями— хромосомами более нлн менее
одинаковой длины с медианными пеитромсрамн и ранными пле-
чами Кроме того, во крайней мере у подрода Oenothera (рода
Oenothera) проксимальные участки вблизи центромеры гетеро-
хромат нчпы Эти особенности облегчают возникновение н ус-
пешное функционирование реципрокных транслокаций. Затем
на основе полиморфизма по мелким кольцам могут образоваться
полные гетерозиготы ио транслокациям (114, 459).
Центральноамериканский вид fthoeo spaihacea (2л—12, Com-
melinaceae), прежде известный под названием R discolor, яв-
ляется паяной гетерозиготой по транслокациям с кольцом из 12
хромосом и, возможно, с зиготными деталями. У близкой к нему
формы в Британском Гондурасе, Я. spathacea concolor (2л=12),
происходит нормальное образование бивалентов. Геном /? а. сол-
color гомологичен одному из двух геномов кольцеобразукицего
R spaihacea spathacea-, источник другого генома этого послед-
него вида неизвестен [20, 711. 725, 807J.
Некоторые из указанных выше гетерогамных групп имеют
простую (Rhoeo spaihacea) яти относительно простую (Gayop-
hytum heteroeygom) таксономическую структуру, тогда как у
других она сложная (Oenofhera-Muneia). Таксономическая
сложность той нлн иной гетерогамной группы, вероятно, частич-
но определяется числом способных гибридизировать предковых
видов, а частично—диапазоном доступных для заселения эколо-
гических местообитаний. Такая группа, как североамерикан-
ские виды подрода Oenothera рода Oenothera имеющая пример
ио четыре предковых вила и занимающая обширную и разнооб-
разную территорию, в процессе последовательных циклов тибри-
лизацня может дать начало многочисленным и разнообразным
микровидам.
30 Перманентная
нечетная полиплоидия
Термин перманентная нечетная полиплоидия (permanent odd
polyploidy) используется здесь для обозначения генетической
системы особого типа, а не в буквальном значении входящих в
него слов. Пентаплондное агамоспермное растение Crepis или
трнплоидное живородящее растение Allium — это в буквальном
смысле «перманентные нечетные полиплоиды», однако они выхо-
дят за рамки этой концепции в принятом нами определении
Под перманентной нечетной полиплоидией мы имеем в виду яв-
ление. когда растение воспроизводит в чистоте некое состояние
нечетной эуплоидиости при помощи полового размножения. Это
осуществляется путем передачи одного генома через пыльцу и
двух или нескольких геномов через яйцеклетки.
Такая генетическая система обнаружена в группе Rosa cani-
ne (Rosaceae) и l.rucopogon juniperinus (Epacridaceae); к ней
приближается генетическая система группы Cardaminr. атаса
(Cruciferae).
Таксономическая изменчивость в одной известной группе
перманентных нечетных полиплоидов принимает форму ряда ге-
терогамных микровидов, связанных в один гетерогамный комп-
лекс.
Генетическая система Rosa canina
Большая часть микровидпе группы Rosa canina в Европе — пей-
та пл он ды (х=7, 2п—35); в этой группе имеется также несколько
тетраплоидных и гексаплоидных форм. Мы рассмотрим репро-
дуктивный цикл типичной певтаплоилиой формы
При мейозе у пеитаплондной розы собачьей 14 из ее 35 хро
мосом конъюгируют с образованием 7 бивалентов, а остальные
(21 хромосома) представляют собой унявалеиты. То же самое
наблюдается при конъюгации хромосом в материнских клетках
пыльцы н материнских клетках зародышевого мешка (7 114-211).
Но распределение хромосом в этих двух клеточных линиях про-
исходит совершенно оо-разному {139. 819]. Схематически это
показано па рис. 30.1.
В метафазе 1 в материнских клетках зародышевого мешка
хромосомы 7114-211 распределяются по двум группам: 7 бива-
лентов выстраиваются по экваториальной пластинке, а 21 не-
конъюгировавшая хромосома собирается на микропилярном
конце. Хромосомы, образующие биваленты, разделяются и во
время анафазы 1 расходятся к противоположным полюсам пере
тена. В телофазе 7 конъюгировавших хромосом соединяются с
Га Л) П^ммснтавя нечегчая лоАыплоиогая
Рис 30 I Прпцсо; Mrirtia п слнмне гамет компжм<?нтйр*нх гмаап в групп»
Нм сап ни [207],
Комьям артижнс хроугкомы черные, а > ншзлгтшс — белые Покампы
только 2 Оивалеггаге и С снижалейтмил хромосом.
2J меконъю тиров я вшей хромосомой. Таким образом, к концу
первого мейотического деления 28 хромосом обычно бывают рас*
положены на мн кропил ярмом конце клетки, а 7 — на другом ее
конце. Второе мейотическое деление протекает нормально и
приводит к образованию линейной тетрады, состоящей из двух
больших к дв}л маленьких ядер. Одно из крупных 28-хромосом-
пых ядер, находящихся па мнкропилярпом конце, превращается
в функциональный зародышевый мешок (рис 30 1) [J39, 390,
819).
В материнских клетках пыльцы унмеаленты и биваленты
располагаются в экваториальной плоскости. Во время первого
деления хромосомы, образующие биваленты, разделяются к пер-
выми направляются к полюсам, а затем делятся и следуют ха
ними к полюсам унивалентяые хромосомы. Во время второго де*
пения ксигыогнрующке хромосомы делятся и расходятся к новым
полюсам Некоиъюггруюшис хромосомы при втором делении
также разделяются, хотя и с опозданием, я дочерние хроыпгомы
начинают пе|К?мец1втъся к полюсам^ ио большая их часть до по-
люсов не доходит [J39. 819].
ЗМ Часп пикая. С.здчввлиьирсадаиь* гамеяииспм милы
В результате мейоза я мужской липин образуются дочерние
ядра, содержащие основное комплемент из 7 конъюгирующих
хромосом и варьирующего числа неконъюгирующих хромосом.
В одной выборке из 63 микроспор, в которых подсчитывали чис-
ло хромосом, 9 клеток содержали по 7 хромосом, а остальные
54 — от 8 до 22 хромосом. Микроспоры различались по величине
и форме и в своем большинстве были абортивными. Функцио-
нальными были пыльцевые зерна с набором нз 7 хромосом из
конъюгирующего генома (рис. 30.1) [139, 207,819J.
У тетраплондов при конъюгации образуется 7П +14 I, а у
гехсаплоидов— 7 П+281. Эти типы, таким образом, сходны с
пентаплоидвмн, содержащими 711+211, в том отношении, что
они содержат хак конъюгирующие, так и иекоиъюгнрующне ге-
номы Кроме того, подобно пентаплоидам. они передают унвва-
лентные хромосомы через яйцеклетки [819J.
Ббльшая часть мнкровидов группы Rosa canina несет, как
отмечалось выше. 2п-5х-3б хромосом (149, 151, 819). Сначала
полагали, что у розы собачьей это постоянное пентвплокдаое
состояние поддерживается прн помощи агамоспермии [390. 819],
однако позднее было показано, что они размножаются пазовым
путем [207. 316]. Ряд авторов утверждает, что эти растения в
значительной мере самоопыляются (139, 146. 602, 780), ио поми-
мо этого у них в некоторой степени происходят также ауткрос-
сииг и гибридизация Желательно получить дополнительные ко-
личественные данные о частоте инбридинга и ауткроссинга у
этих растений.
При самооплодотворении у пеитаплоидиого растения Rosa
canina происходит слияние женской гаметы с 28 хромосомами
и мужской гаметы с 7 хромосомами. В результате этого события
восстанавливается пемтаплоидиое состояние (2п=35), а кроме
того, образуется потомство, сходное с родительским растением
По своему геному (7 П+211). Перекрестное опыление между
сестринскими пентаплоидиыми растениями приводит к таким же
результатам. Следовательно, роза собачья воспроизводит в чис-
тоте состояние нечеткой полиплоидии при половом размножении.
Гетерогамный комплекс в группе Rosa canina
Группа Rosa canina составляет отдельную секцию (секция Cani-
пае) рода Rosa. К этой секции относятся дикорастущие колючие
кустарники с цветками от мелких до средних или, в редких слу-
чаях, крупных размеров (рис. 30.2); встречаются они в мелко
лесье н на полях по всей Европе н в соседних с ней частях се-
верной Африки и на Ближнем Востоке.
К секции Caninae относятся многочисленные микровилы, из
которых одни расароетранейы широко, а другие — локальна
Гл. 30 Лгрламантиал нагнал лоим» доа^ия
Рис 302 Мнкровглы -»уппе Яма canina (по дьнным [339] I.
А — Я. dumctotum (5л) 5 — R. loamMie (5*) д— Я. agrtUn (5л). Г —
R miarAntha (5л) Д — R. гиЪгЦоЧа (4л).
причем между ними происходит доводьио сложная иптергр аха-
ния Вследствие этого разные авторы представляют себе таксо-
номию этой группы по разному, как это обычно бывает с гибрид-
ными комплексами Тэкхолм [819] различает 34 таксономичес-
ких вида, объединяя их а 5 групп видов. Хеги [359] низводит
некоторые из этих таксономических видов до ранга подвидов, ко
различает в каждой вядпвой или подвидовой единице многом не-
25-45
Звв Чат восьмая. Слсмиаяизмровсмчьи гытвяясяы сотым
веяные разновидности н формы. Так, Коза canina sensu stricto
и R dumetorum он считает подвидами R. canina sensu lato. Но
R canina sensu stricto имеет при этом 7 разновидностей и
12 форм, a R. dumetorum —Ь разновидностей и 10 форм [3S9],
В связи с этим принято считать, что рассматриваемая группа
таксономически разнообразия и сложна.
На основе морфологических признаков секцию Caninae мож-
но считать тесно спаянной единицей, хорошо отделенной от дру-
гих секций рода Rota (рис. 30 2), поскольку. с одной стороны.
Caninae морфологически достаточно ясно отличается от других
секций [819], а с другой — входящие в эту секцию мнкровиды
иитерградируют в многочисленных комбинациях. Так, R. canina
sensu stricto интерградврует с R dumetorum, R. stylosa, R pou-
zini и R chavini, R. dumetorum в свою очередь интерграднрует с
R. stylosa, а кроме того, — с R. corii/olio и R. lomerUelia [359].
Основное число хромосом в группе Rosa canina как я
в других секциях этого рода. Однако в сепии Caninae не изве-
стно ин одного диплоида. Большинство входящих в нее микро-
видов —пентаплоиды; есть также несколько тетраплощов или
гексаплоидов [819].
Для рассмотрения частоты распределения разных уровней
плоидности в группе Rosa canina мы примем за основу состав-
левиый Твкхолмом [819] список таксономических видов и снабдим
его данными о числе хромосом в соответствии со снедениями, опуб-
ликованными самим Тэкхолыом [819] и последующими автора-
ми (библиографию см. [149]). Из 34 таксономических видов 15 —
пентаплоиды, 3—тетраплоиды, 3 — гекеаплонды, 4 содержат как
пситаплондные, так и гексаплоидные формы, а по 9 видам под-
счетов не производилось.
К пеятаплоидам (2п~35) относятся: Rosa canina sensu stri-
ct, R. dumetorum, R. micrantha, R tomenlosa и R. stylosa (час-
тично). К тетраплоидаи (2л -28)— R. rubrijolla, R. pomifera и
R mollis К гексаплондам (2n-42) — R. junzillii. R. pouzini,
R. chavini и R. stylosa (частично).
Представления Тэкхолма о геномной конституции пентапло-
идиого микровида, основанные на наблюдениях за конъюгацией
хромосом, можно выразить формулой AABCD Исходя из ха-
рактера поведения при мейозе некоторых искусственных гибри-
дов между микровидамн, ое полагал, что у разных ми кровил ое
из группы Rasa canina одни геном общий, а по другому геному
они различаются. Так, три пентапловдвых микровнда могут
иметь формулы AABCD, AACDE я AADEF соответственно.
Тэкхоли [819] считал пентаплоидные мнкровиды непосредствен-
ными продуктами гибридизации между одним предковым дип-
лоидным видом и различными октоплондными видами Поль-
Гл. 30. Перманентная нечетная лоляъюиЗия ЗВ?
дуясь геномными формулами, микропид. образующийся в ре-
зультате скрещивания между АА и AABRCCDD. можно обозна-
чить AABCD; скрещивание AAxAACCDDEE даст микровид
AACDE и т. д.
Последующие цитогенетические исследования заставили мо-
дифицировать эти заключения в некоторых важных отношениях.
Густафсон и Х^кэнссон [319] скрещивали три микроенда Rosa
canina (5х и бх) с диплоидным видом R. rugosa (2п — 711), при-
надлежащим к другой секции. У этих искусственных гибридов
при иеПозе образовывалось по 14 бивалентов. Это свидетельст-
вует о том, что полиплоидные R. canina гомологичны не по двум,
а но трем геномам. Они аетотриплоидмы по одному из своих 5
или б хромосомных наборов [316, 319|. Фагерлиид (2OBJ пошел
дальше, указывая на иекоторую гомологию между всеми гено-
мами полиплоидных R. canina.
На основе шгтогеметических данных полученных Густафсо-
ном и Хёканссоном [319}. Густафсон (316] предложил следующие
формулы геномов: дли Rosa canina sens.i slricto — Д*Лр4|СА>,
а для родственного микронида R rubiginosa — BBBCF. Геном С
обнаружен у R rugosa, лпюсящейся к другой секции, и, возмож-
но, распространен у данного рода более широко 1139]. Основы-
ваясь нз этих и ряде дополнительны к данных, Фагерлиид [208]
низвел бы некоторые из обозначенных буквами геномов до еуб-
геиомов {см. также (62, 915]). Во всяком случае, мнкровиды
R. canina, по-видимому, представляют собой аггоаллополипло-
иды, геномы которых в разной степени различаются по своей
структуре
Геномная конституция полиплоидных Rosa canina имеет важ-
ное значение для проблемы происхождевия новых микровидов.
Классическая точка зрении Тэкхолма. считавшего, что различ-
ные микровнды R. canina — строгие аллополиплоиды и поэтому
размножаются в чистоте, привела к заключению, что этн мик-
ро виды — арнмые продукты гибридизации между разными ком-
бината । [Him г чьеннх видов (819]. Согласно згой точке зре-
ния, ряд потенция л ьпо возможных мнировидов ограничивается
числом гибридизирующих предковых видов.
Если же исходить из того, что микровнды Rosa canina но
своей конституции валяются автоаллополиглопдами. то тогда
один высокогстерозиготным .мнкровна *ожет непосредственно
дать начало ряду новых ми крови доз в результате расщепления
по генным различиям между геномами. Так, у пен гаплоида с
конституцией A^AtAiCD хромосомы At, которые обычно обра-
зуют унивалеиты, могут н:«огда включаться в конъюгирующие
наборы, и тогда они будут в дальнейшем передавать своя гены
следующему поколению как через мужские, так н через женские
гаметы [319].
25*
3S8 Часп еопмал С/кциалемроааинмс гсчсгачсскис сиспше
Короче говоря, одна автоаллополиплоидиый микровил мото-
ры* обычно размножается а чистоте, может иногда выщепить
один или несколько новых микровидов. размножающихся в чис-
тоте. Этот процесс позволяет объяснить возникновение очень
большого числа микровнлов Rosa canina, реально существую-
щих в природе [319].
Правдоподобную картину филогенеза комплекса Rosa canina
воссоздал Тзкхолм [819]. Нам неизвестно ни одного диплоидного
вида н нн одного полиплоида с нормально протекающим исйо-
зом, который обладал бы морфологическими признаками, ха-
рактерными для секции Caninae. Все современные формы обла-
дают производной генетической системой. Следовательно. эта
группа, вероятно, довольно древняя. Кроме того, современные
производные микрпвмды занимают в Европе территории, на
которые оказал сильное воздействие ледниковый период. Дип-
лоидные предки группы Rosa canina, по-видимому, вымерли в
плейстоцене, а существующие в наши дни гибридные комплексы
возникли в плейстоценовое и постллейстоцековос время [819J.
Leucopogon funiperinus
Перманентная нечетная полиплоидия была обнаружена также у
Leucopogon juniperinus (Epacridaceae) в Австралии [745]. Этот
вид — трнплоид, размножающийся половым путем, с числом
хромосом 2п-Зх-12. В процессе мейоза два гаплоидных набора
образуют биваленты, тогда как третий гаплоидный набор не
проявляет никакой тенденции к конъюгации Эго растение ведет
Матвримсдав «*•»*• пыпь^
и пыльцевое зерно
Матеонмсяое клетм аасюдыайввого ыввкв
И )аМАММШ«мь.й ы«де>«
Р»с. 30.3 Формирование гамет аомплежятавиых типов ыужско<* и
ежой линях у Leucofiogon jenifcriAitA (4li-*-4i) {747].
Зародышевый иешж г«ы»ржит четыре уымвллеята. ио а пыльцевых Мрвах
их кт
Гл Л7. Пбрмйн/лТмлв мгмтмде ЗвЭ
себя. таким образом, как аллотриплоид с геномной конститу-
цией .4АВ (745, 746].
В первой метафизе материнские клетки как зароднтсвого
мешка, так и ПЫЛЫ1М конъюгилуют с образованием 411+41-
В анафазе I в материнских клетках обоих типов все уннваленты
отходят к одному полюсу В материнских клетках зародышево-
го мешка кпя полюс —функциональный, а в материнских клет-
ках пыльны — гсфупкциоиа.тькый В результате ядра зароды-
шевого мешка к яйцеклетки несут по 8 хромосом, принадлежа-
щих к геномам Л и В. тогда как функциональные пыльцевые
зерна несут по четыре хромосомы генома А (рис. 30.3). При
оплодотворении эосстаиавлпваютсн соматическое число хромо-
сом 2п= 12 и геномная конституция 4ЛВ [745. 746].
Одно из условий, облегчавших возникновение сбалансиро-
ванной системы элимпиацни комплементарных гамет у Leucopo-
gon junipennus, — своеобразный способ развития пыльцы у этих
растений У Leucopogon, как и у других представителей трибы
Styphellcae сем Epacridaceae, три микроспоры в каждой тетра-
де абортивные ц только одна развивается в зрелое пыльцевое
зерно Это функционирующее пыльцевое зерно представляет со-
бой одно из двух 4-хромосомных продуктов мейоза у L. juniperi-
nus (рис. 303). У этого вида возможно существование системы
элиминации гамет с мужской стороны без всякого сокращения
общего выхода фертильной пылким [746, 747].
Leucopogon junipennus морфоло- ически и цитологически еди-
нообразен на всем протяжении обширной территории, занимае-
мой им в Австралии. Это единый аллотриплоидный вил ,746]
В этой группе растений не произошло образования гетерогамно-
го комплекса
Cardamine insuela
Между Cardamine am nr а я С rioalaris (Cruciferae) в Швейца-
рии изредка происходит естественная гибридизация. Оба роди-
тельских вида — днплояды (2п=16), а их естественные гибри-
ды—триплоиды (2л=3х=24). Добавочный хромосомный набор
они получают от С rivularis и имеют геномную конституцию
RRA [852].
Гибриды высокостерильпы и дают всего 2—3% фертильной
пыльцы Тем не менее они хорошо воспроизводятся и распрост-
ранились локально 1и> площади 16 га, гас ОНИ преобладают над
родительскими видами. Размножаются они как вегетативным,
так и половым путем Гибридные растения, получившие назва-
ние С. insuela. размножаются половым путем, представляющим
собой некую модификацию схемы размножения Rosa canina
|852. 853].'
ВО Чат Соажа. Сшчшишврооакчме «-«rwooM итж
Тршиоидные растения Cardamine tnsueta испытывались по
потомству в трех сериях прн разных условиях. Потомство три-
плоила, выращивавшегося в открытом поле. состояло нз 21 рас-
тения; все они выли триплоидами (Зх—24). Потомство тркпло-
ида в контролируемых сестринских скрещиваниях состояло из
5 растений; 4 из анх были тричлоидаиа, а один — тетраплоидом
с геномом RRRA. Потомство от возвратных скрещиваний трипло-
ида с С. rlvularis состояло из форм 2х, Зх, 4х и 5*. Таким обра-
зом. насколько это известно, в природных условиях тршиоидные
расгення производят исключительно триолоикые растеши, а
прн искусственных близкородственных скрещиваниях дают пре-
имущественно. но не исключительно триплондное потомство
(8521.
Обычная конфигурация, получающаяся прн конъюгации в
метафазе I у природного триплоида. — 8 II-f-8 I; предположи-
тельный геном этих форм RR I1-I-A I. Расхождение хромосом в
анафазе 1 варьирует в различных клетках. Часто встречается
соотношение 8:16, обнаруженное у 29% проанализированных
клеток. Таким образом, в этом случае выявляется тенденция к
поляризованному расхождению хромосом. У 7% клеток были
обнаружены реституционные ядра с 24 хромосомами. Нередко
наблюдается также неравномерное распределение хромосом,
что приводит к образованию аиеуплоидных продуктов мейоза
Cardamine inaueta производит функциональные 8- и 16-хро-
мосомиые яйцеклетки и 8- к 16-хромосомиыс пыльцевые зерна.
Ои отличается в этом отношении от Rosa canina, у которой жен-
ские гамегы полиплоидные, а мужские гаплоидные.
Следовало ожидать, что у Cardaminc insueta при соединении
8- и 16-хромосомных пыльцевых зерен с 8- я 16-хромосомными
яйцеклетками образуется потомство, принадлежащее к полипло-
идным классам 2х, Зх и 4я. Кроме того, следовало бы ожидать,
что нередуцированные гаметы из 24-хромосомных реституцион-
ных ядер дадут начало некоторому числу потомков 5х (а также
индивидуумам других уровней плоидности). И в самом деле,
такие полиплоиды (2х—5х) были обнаружены в потомстве
С. insucta при искусственных возвратных скрещиваниях, о чем
уже говорилось выше. Однако потомки С. insueta, произрастав,
шие в открытом поле, не образовывали полный ряд таких поли-
плоидных форм; здесь обнаружены только триплоиды, о чем
уже говорилось ранее [852].
Можно было также ожидать, что среди этих потомков будет
некоторое число аиеуплоидных форм, однако нх не было ни в
природных, ин в экспериментальных популяциях. У Cardamine
insueta проявляется определенная тенденции к воспроизведению
в чистоте своей триплоидиой конституции в природных уело-
Гл. 30. Лерлюпектая нелегкая полиплоидия 39!
виях. Механизмы, обусловливающие иной характер размноже-
ния. еще не вполне понятии [8.52].
Cardamine атаеа и С. rivularis дали также плодовитый авто-
аллогексаплокд (2л —48. RRRRAA), известный иод названием
С. schuhii (852]. Здесь мы имеем дело со случаем, когда одна
пара диплоидных родительских видов даст начало формам, на-
ходящимся на двух различных уровнях плоидиости н размно-
жающимся в чистоте. Урбаиска продолжает свои интересные
исследования на С. insueta и С. schuleU, и, по-видимому. в буду-
щем нам станет более понятной биология размножения этих
видов.
Эволюционный потенциал гетерогамных
комплексов
Дарлингтон (139] обратил внимание на некоторые интересные
сходные черты в цитогенетике размножающихся в чистоте не-
четных полиплоидов группы Rosa cantna н размножающихся в
чистоте гетерозигот по транслокациям группы Oenothera biennia.
У каждой группы есть несколько регулярно конъюгирующих
хромосом пли участков хромосом: это 7 бивалентов у Rosa ca-
nina и терминальные конъюгирующие участки у Oenothera bien-
nis. V каждой группы есть также несколько нскотгыогируюших
хромосом или участков - упиваленты у Rosa и дифференциаль-
ные участки у Oenothera [139] (см. также [7Я0]) В хромосомном
наборе каждой группы наблюдается разделение функций между
конъюгирующими и неконъюгнрующнмн участками Некоторые
хромосомы или их участки, регулярно совершающие конъюга-
цию и подвергающиеся кроссипговсру, обеспечивают изменчи-
вость. Другие хромосомы или участки хромосом, которые пе
конъюгируют, несут гены, детерминирующие постоянные приз-
наки разных микроаиаов [139].
Гибридизация между диплоидными вилами, образующими
биваленты, лает в каждом случае ряд иумерических или струк-
турных гибридов. Эти гибриды передают одну часть своего хро-
мосомного набора через пыльцу, а другую —через яйцеклетки,
и гаметы двух комплементарных типов часто соединяются в
результате самооплодотворения. Таким образом, численность
гибридов увеличивается, и они развиваются в гетерогамные мик-
ровиды. Гибридные микровнды заполняют рлтрыви между чред-
ковыми видами и дают начало гетерогамному комплексу в его
первичиой стадии. Затем микровнды гибридизируют друг с дру-
гом, давая начало новым вторичным микрювидам. которые еще
больше обогащают изменчивость гетерогамного комплекса на
его поздних стадиях.
393 Часть восьмая. Слгпиа-аоьроваыив cewrwucKUt системы
Как у Rosa canina, так и у Oenothera biennis значительные
части генотипа «заперты» (locked up) в состоянии перманентной
гетерознготиоств а не конъюгирующих участках хромосом, поэто-
му эти растения обычно размножаются в чистоте по таким ге-
терозиготным комбинациям генов. Однако происходящие в виде
исключений в этих же учвстках конъюгации или кроссикговери
приводят время от времени к выщеплению новых константных
типов. Таких редких выщепенцев можно рассматривать как
запоздалые продукты гибридизации родительских видов, кото-
рые обычно размножаются в чистоте, ко высокогетероэиготны.
Эти константные гибридные выщепенцы. если они окажутся
ценными в адаптивном отношении, могут превратиться в новые
мнкровиды и тем самым еще больше увеличить общую измен-
чивость гетерогамного комплекса.
Гетерогамный комплекс в стадии зрелости содержит десятки
или сотни микровидов, происхождение которых прямо или кос-
венно связано с гибридизацией и которые заселили территории,
сравнимые по площади с материками. Такие гетерогамные комп-
лексы, примером которых служат группы Rosa canina и Oenot-
hera biennis, несомненно, вволюционнруют очень успешно.
И все же остается фактом, что у гетерогамных мнкровкаоа
значительные части генотипа «заперты». Рекомбинация у иих
очень сильно ограничена тем же самым хромосомным механиз-
мом. который обеспечивает кратковременный эволюционный
успех. Можно ожидать, что в конечном итоге эти ограничения
сильно препятствуют появлению новых вариаций и загоняют
гетерогамный комплекс в молюциоиный тупик [141, 780].
31 Агамоспермия
Агамоспермией называют образование семян без оплодотворе-
ния Это одна из основных форм апомиксиса, или размножения
без оплодотворения, у высших растений Само агамоспермиое
размножение включает в себя очень разнообразные и сложные
явления, которые будут описаны в дальнейшем Агамоспермия
широко распространена у цветковых растений, о кем свидетель-
ствует перечень, представленный в табл. 31 I В этой главе
дается обзор эмбриологических и цитогенетических явлений как
необходимой основы для понимания происхождения изменчиво-
сти у агамоспермиых растений Опираясь на эти данные, мы
сможем затем перейти в следующей главе к рассмотрению »во-
люцноивого развития агамиых комплексов.
Болес подробное изложение этих проблем читатель найдет
в классической монографии Густафсона «Апомиксис у высших
растений» [317]. Многие дополнительные данные, полученные
позднее, содержатся в обзорах Стеббинса [780]. Нюгреиа
]625]. Фряксслла [223]. Батталья [48] я Рутисхэуэера [701].
Эмбриологические пути
Различные пути эмбрионального развития, ведущие к агамо-
спсрмному образованию семян, показаны па рис 31 I В этой
схеме использованы классификация к термппологиг. Густафсона
[317] и сложные явления представлены в .несколько упрошенной
форме, с тем чтобы сделать возможными широкие сопоставле-
ния.
Нормальный жизненный цикл семейного растения, являю-
щийся предковым лтя разного типа агамоспермиых жизненных
циклов, слагается, как известно, из чередования диплоидного
спороф1Гтпого и гаплоидного гакстофитного поколений; эти по-
коления разделены такими событиями, как мейоэ и оплодотво-
рение, которые СОответстиепио сокращают и восстанавливают
соматическое число хромосом (рис. 31.1,4). При агамоспермиых
жизненных циклах, которые обеспечивают сохранение более или
менее постоянного числа хромосом у ряда поколений растений,
развивающихся из семян, образование зародыша происходит
без мейоза н без оплодотворения.
Этот результат достигается двумя способами, соответствую-
щими двум основным типам агамоспермии — гаметофитному
апомиксису и адвентивной эмбрноняи При гаметофитном апо-
миксисе гаметофит как морфологическое образование сущест-
вует. ио редукции .хромосом не происходит (рис. 31.1,£). В от-
А НОРМАЛЬНЫЙ UHKft ЛоЛОВО<*0 РАЗМНОЖрМАИ
L ГАМГОеиГМыИ АПОМИКСИС
Д АДМНТИвНАЯ ЭМБРИОНИЯ
Рве. 311. Слособы аг«-м-п*рм«сго рммяожвлжя (два ернтм привелся жиэмевнмЛ мнм рахгтежмя с «ормвдышм во-
ловик раэмиожемаем).
Гл. 31. Аслнослерлил
Рис 31 2 Авхаория у Crepis accident alls (во [792] с измеяеиичмн),
А. Делггхаяся матери-си я <лгткд хародышпиго мешка (1), в под ней ув»-
лкчшаюшаигй спчатнмесяая клетка (i) Б Лияе*м*я тетрада (3). а псд'ией
сонлтяческвя клетка. развивающаяся в апоспоркчегккй мродмшезый ме-
шок (4) В .Чатеркгскля клетка ззроды^теязто мешка (J), aerenepii зрения
при нервен зеленям, а вод wed одпоклеточаий i поспорив, и й 1вреды»«шВ
мешок (4).
Рис. 31 3 Дшызосмрмя у ЛпГепдалв portitdU. матерннскея к.тетха мр<ип»
шевого из кптпрой эаааяваегся псге.туяирспаиаыЛ ирошшевый ме-
ик« [850].
личие от этого при адвентивной эмбрионпи гэгегэфита нет, и
с(»тветс1мекио выгадает чередование покатекиЙ (рис 31.1.В).
Как показано на рис. 31.1.6. при гаметофитиом апомиксисе
возможны два альтернативных пути, ведушче от материнского
спорофита к нередуинрованжлгу гаметефнту, и еще два пути»
ЭК Чат аопжи Смцагмаи/юаыяиг шмтичгсш састглы
ведущие от этого гаметофит» к новому зародышу. При апоспо-
рии женский гаметофит образуется иа соматических клеток ну-
целлуса или халазы семязачатка (рис. 31.2). При диплоспории
женский гаметофит образуется непосредственно из одной мате-
ринской клетки зародышевого мешка, не претерпевшей мейоза
(рис. 31.3).
В любом случае женский гаметофит содержит яйцеклетку с
нсредуцироеанным числом хромосом и разного рода дополни-
тельные клетки. При диплоидном партеногенезе из такой яйце-
клетки без оплодотворения развивается новый зародыш. При
апогаметии какая-либо другая (нс яйцеклетка) клетка женско-
го гаметофита образует зародыш (рис. 31.1,5).
Алогамиое развитие зародышей описано у ряда покрытосе-
менных. Густафсон приводит примеры развития зародышей из
синергид у Taraxacum. Hitracium, AlchamiUa. Alnus и Poa и
примеры развития их из антиподов у Hleracium. Etaiostema и
Allium (317). Но. как отмечает Густафсон, в природе это явле-
ние, вероятно, не имеет большого значения для покрытосемен-
ных.
Остальные процессы, сочетаясь друг с другом, образуют две
полные линии развития, которые можно назвать диплоспория—
партеногенез и апоспория—партеногенез. Оба эти последова-
тельности встречаются в четко выраженной форме у цветковых
растений (табл. 31.1). Оки встречаются также в смешанной
форме у некоторых покрытосеменных, у которых развиваются
и днплоспоровые, и апослоровые зародышевые мешки
(табл 31.1).
Наконец, при адвентивной эмбрионни новый зародыш разви-
вается непосредственно из нуцеллуса или интегумента семяза-
чатка на материнском растения н. таиим образом, совершенно
обходит зародышевый мешок. Адвентивная эмбриоиия представ-
ляет собой тот тип агамоспермии, который в наибольшей степе-
ни приближается к живорождению как одной пз форм вегета-
тивного размножения (или к ложному живорождению, согласие
ваи дер Пэйлу (639|). В обоих случаях новый индивидуум обра-
зуется бесполым путем в соцветие материнского растения. Но
при адвентивной эмбриопни дочернее растение начинает разви-
ваться как зародыш, находящийся в семени, тогда как при
псеодоживорожленин оно развивается из воздушной луковнчкн,
возникающей внутри цветка или на его месте.
У некоторых агамоспермиых покрытосеменных для апомик-
тического развития семени необходимо опыление. Такое стиму-
лирующее воздействие опыления, необходимое апомиктическому
цветковому растению в качестве предпосылки для образования
семени, известно под названием псевдогамии; при этом опыление
служит либо общим стимулом, инициирующим развитие зеро-
Г.’ J! А. а ткпсрмл
397
дыша, либо, что более обычно. за опылением следует слияние
ядра спермин с ядрами эндосперма. Этот процесс оплодотворе-
ния инициирует развитие эндосперма, необходимого для нормаль-
ного роста зародыша. Сам зародыш, однако, не получает ника-
кого |енеткчсского материала ог растения, поставляющего пыль-
цу. и по своему генотипу полностью сходен с материнской фор-
мой.
Агамоспермное растение может производить семена исклю-
чительно бесполым путей или же может время от времени да-
вать потомков а результате полового процесса В первом случае
имеет место облигатный, а во втором — факультативный апо-
МИКС1К.
Одно и то же факультативно агамоспермное растение может
я разные годы давать в разных соотношениях семена, образую-
щиеся Пановым и бесполым путем, или даже давать те н .трутне
семена в урожае данного года Иными словами, половой путь
развития зародыша,схематически представленный на рис. 31 1,Л,
может действовать в одной линки развития семян, тогда как в
другое время или в другом месте на том же самом растении
развитие семян может происходить альтернативным атампсперм-
ным путем (рис. 31.1. Б и В).
Hicracium aurantiacum в Карпатах (Польша) образует жиз-
неспособные семязачатки двух типов в одних и техжесоиветия.х.
Некоторые семязачатки содержат редуцированные зародыше-
вые ыешкн, образовавшиеся в результате нормального мейоза,
тогда как другие содержат керсдукированные зародышевые
мешки, возникшие в результате апоспории. В последних заро-
дыш н эндосперм развиваются автономно, тогда как в первых
им необходимо оплодотворение как предпосылка к дальнейше-
му развитию. Вследствие этого в одном соцветии семянки обра-
зуются как половым, так и бесполым путем. Соотношение семя-
нок этих двух типов различно у разных растений |73].
У папоротников, где агамоспермия, разумеется, отсутствует
то определению аналогичную рать играет апогаметия. пред-
ставляющая собой преобладающий способ беспалого образова-
ния зародышей у этой группы У впогамяых папоротников пос-
ледовательность событий начинается с образования в спорая-
гнях материнского растения переду иированиых спор. Эти споры
дают начало гаметофитам, несущим нсрсдуиипиваииое число
хромосом. Затем иэ вегетативных клеток гаметофита без опло-
дотворения развивается новый спорофит [554]
Таксономическое распределение
В табл 31.1 представлен список атамоспсэмиых цве-ковых рас-
тений Этот перечень составлен на осноэс работ Густафсона
]317] и Нюгрена [625]. в которых можно найти ссылки на ориги-
*6 Чат eonnas Слечиамвироаамне мштнчжпм шстмм
нальные иссиедования. Следует поикать, кто многие таксономи-
ческие виды, перечисленные табл. 31.1. содержат как половые,
так и агамоспермные формы. Виды сгруппированы а этой таб-
лице, во-первых, по типу агамоспермии и, во-вторых, по принад-
лежности к тому или иному семейству растений.
Этот список неполон, во дает достаточно хорошее представ-
ление о распространении агамоспермии у покрытосеменных.
Можно видеть, что каждая ил основных форм агамоспермии
встречается у представителей многих семейств. Так, днцлоспо-
рия— партеногенез встречается у многочисленных представите-
лей сложноцветных, а также во многих других семействах. Апо-
спория — партеногенез встречается у разных представителей ро-
зоцветных и в других семействах. Адвентивная эмбриоиия наб-
людается довольно часто у представителей семейств лилейных,
рутовых и других.
Для Parthenium argentatum характерны одновременно и дип-
лоспория—партеногенез, и апоспория—партеногенез; такая
ситуация, когда обе системы агамоспермнего размножения
встречаются вместе у одного и того же вида, обнаружена и у не-
которых других растений (табл. 31.1). Эти две близкие формы
гаметофитиого апомиксиса наблюдаются у разных видов Роа
в разных породах Hieraeium.
Псевдогамия широко распространена у агамоспермных по-
крытосеменных, в том числе у видов Роа. НиЬиз. Potentllta, Ra-
nunculus, Hypericum. Parthenium, Citrus и Allium. К группам,
у которых псевдогамия не наблюдается, относятся Hieraeium.
Tarasacum, Anlcnnaria, Crepis и Caiamagrostis [317].
Если рассматривать представленный в табл. 31.1 список ви-
дов с точки зрения их географического и климатического рас-
пространения. то можно выявить некоторые тенденции. Многие
примеры адвентивной змбрионии относятся к тропическим и
субтропическим растениям, тогда как гаметофнтный апомиксис
чаще встречается у растений северных регионов с более холод-
ным климатом.
Что касается числа хромосом, то в выборке агамоспермных
цветковых растений очень велика доля полиплоидов [317]. Мно-
гие из них—высокие полиплоиды, как, например, Anlcnnaria
alplna с 2л—84. Другие — мечетные полиплоиды, как триплоиды
Crepis и Matus. У третьих наблюдается широкий диапазон аиеу-
плондиых чисел хромосом, варьирующих вблизи некоторой по-
лнплондиой моды. Таким образом, полиплоидна в той или иной
форме обычно бывает связана с агамоспермией. Тем не менее
в родах Citrus. Nothoscordum. Potentilla и других имеется неко-
торое число агамоспермных диплоидов.
Густафсон [317] и Стеббинс [780] указывают, что бесполые-
диплоиды встречаются чаще в группах с адвентивной эмбрио-
Таблица 31.1 Избранные примеры агамоспермиых покрытосе-
менных (по [317] и [625]; данные по ClMia [543])
I ДиПЛ1КЬ0рЖ>~«1рТ11ЮГвЯ«
Dicotyledcoieae
Bajincphu жеае
Ва1лли>рИага глрогпеа (2л-94—IIX); Я globosa (большое чжж>)
CompotitaH
Ля/г.".па.ч'а ulpiua (7я=84) и 13 други видов
Cbond'Ula осзл!Ьо1ер1я (2/1—15) я 8 других видов (2я—15, 20)
Erlgtron аплши (2л—27) н 2 других вида (2л-а7. 32—ОД
fapaiartom glandtdatum (2л-51)
Jhc'actwn подрод EufiUractum, М1югочжсзсккые виды (2л-18,
27, 36)
/лги dealaia (2л—21)
£нв^л-Д;л cef»?a'rtua (2л = 76) и 5 других видов (2л=7б)
Taraxacum. мио г очН слеши ые вады (2л-24. 32, 40, 43)
Crweileree
Лгаби hoUocUti (2л-14» 21)
PLumtxf.a*MfB*
Slattce otaeefolia cottjusa (2л—27)
баигигвеем
tfaiuiarpn/a cardaia (2л=94—104)
ТМулшНммем
ITiks.Vof<!4u2 vlri&jtAa (2л—27)
Urlieacoe
£juriMT<flK» гйгЛрпсбжт (2ft-52) и 3 других вида (2л-60, 80)
Monoco<yle4oneae
Amaryllidsceac
Соовта pt-lunc alata (2л- 48)
Zephyr atuhea (лллпа (2л—24)
Burrnanniauae
Burmannia corteitu (2n-64—72)
Grarameae
Caiamagroriis cansdmns (2л—42—68) и 6 других видев (2ft—42—
112)
Рол giauci (2л-42—70) и 4 других вила
<1. Лтк.гпприя—пвртсяогеме.)
DtcotytedorHoe
СзлкювИае
Crecis acuminata (2п-ЗД 44) и 8 лругпх влдоп (2л—22—88)
llitrazium подрод Pilonlla, многочнелехиые ыцы (2л—27. 36.
<2, 46)
GutLkrae
Hypencuffi ptfisrafMi (2я—32)
Polygonoceae
Atrapiwria (rtaeutM (2л-58?)
Rarur<i I я севе
Hananculiu au'teomug (2n=l6, 32. 40. 48)
Rosa<cae
Alchemilta ar.'eatis (2л—48) и 2 других вида
Mains ЛирвАллли (2п—5»)
Poieniilia argenlea (2л-14, 28 35, 42. 56); Р ргаесох (2л—42)
Rirbas секция Atorrjerw». мжгхшнглениые виды (2л«21—49)
5огЬыв сАдтделкхрЬм (2л—68); S hgbnda (2л -68)
•UibcaceBB
Etaiailema смилило/«ля (?л-5О) ; Е latifolium (2л-60)
М од ocoCyled опеле
Gram! оме
Paspatum diUlptum (2л—40)
Pm ample (2л-63) я 5 друпа аядса
III. Ддшлоспорм»— аартемоггаеэ к апоспория—(ыртеиогенед
Cocapoatae
Anienfaria car pat lea (2л—40—42)
Partkenitun afgenlatUM (2л—54—144); Я (лгалшл (2л-54—90)
RttuetM
Pottntttla wna (2л—42, 49. 84)
IV Ад»«лтш<вя амЛряояня
Dicctykdoncae
ABHCJfJiKeae
Mangifera wSica (2л-4О. ~52)
Belulaceae
rugpsa (2л-98)
BuAiccte
Sarcococca pruniformti (2л—28) друпю гады
CicUceK!
Opuntla ficus-indica (2л—88) а другие мдм
Cappandaceae
Capparis frondasa
CeJaatraceae
Ctlasirus scandent (2л—46}
£мэлул1ьд laiifoliiLi л друлм ащш
CloaUcaM
Cluua rcsta; C nu’rvx
Eupborblaceae
Euphorbia dulas (2л-12. 28)
Guttiferee
GsrdnLa тлпрал1&ла (2д—7B)
Malahiftaeaaa
Hlptagt nuidablota
Myrtaccae
Eugenia tnalazc^nsis (2л—22) н другие вады
ОДшкеае
ОсЛла se/rulata (2л-35)
R utaсеве
Ciirtu, ы«ого«1К'-ле<иньае вады (2л—18, 27. 36)
Triphuta вмгая11о(а
Хаги Колу turn inutgti
MooocolykdoBeae
Аг а севе
SpathipapOwn patinii (2л—18)
Lllltceae
AUium odonun (2л-16. 32)
Hasta catrulfa (2a-60)
Noltwscordum fragrau (2л—16—19, 24)
Smtadna racemosa
Tedipa gesncriana (2л-24, 36)
Orehidaceae
NLgritMa nigra (2л-64)
Zeuxtae iulcoia (2ч - 44)
Zygopglaiujn macitapl (2л-40)
Гл У J Агаиоинржл ч
401
пней, подобно HothoscorcUum fragrans н многим видам Citrus,
чем в группах с гаметофмтным апомиксисом. Единственными из-
вестными знплоидами с гаметофмтныи апомиксисом являются,
по видимому, некоторые члены комплексом Potent ilia argenlea,
Ranunculus auricomus, Hieraeium umbellatum н Arabis hotbocl-
Ш[3\7, 625, 780).
Мкс не известно ин одного документированного случая ага-
моспермии у голосеменных. Не содержится таких примеров и в
сводном обзоре Фриксслла [223).
Ментон [э541 перечисляет следующие примеры а йогам и ых
папоротников (Polypodiaceac) с указанием в тех случаях, когда
это известно, уровней полиплоидии
borrori (2л. Зх, 4х, 5х). D remeta (За), D atrate (Зиг)
ЯйеодМел* p»i ypod.cites
Cyrtomiufn lolcaf*"» I lx). C jerftmei (3r),C cwpoHdrum (3л)
Jiptowcm mananthti (3x)
Prttoee af'OfiU'fiurta (lr)
Pttris eretica (2x. 3x. 4x)
У всех этих апотамных папоротников высокие хромосомные
числа— от 2н — 58 у одной формы Pieris cretica до 2н — 205 у
одной из форм Dryopteris berreri. Следует заметить, что боль-
шингтво вв них — нечетные полиглоты, главным образом
трнплсиди, хотя иногда встречаются и пеитаплоиды [5541 О мно
гик случаях апогамгтни у Ptetis сообщил также Уокер [87 !|
Клековски (4411 отмечает, что у папоротников генетическая
система коррелирует с продолжительностью жизни гаметофитно-
го поколения. Большая продолжительность жизни таметофитно-
го поколения обычно сочетается с головым размножением и
ауткроссннгом. Апсиаметмя же часто наблюдается у растений
с короткой продолжительностью жизни гаыстофитнэго поко-
ления.
Система размножения
Густафсон [317] иссяедоваг 35 родов цветковых растений, у
которых имеются как агамоспермные виды, так и вилы, размно-
жающиеся половым путем. Цель этого исследования состояла в
том. чтобы определить тип системы размножения у половых ро-
дичей агамоспермных растений Полученные результаты можно
сгруппировать следующим образом В 10 из частично агамо-
спермпых родов половые виды двудомные и (или) однодомные.
У 16 родов полови# формы самонесовместпмы. У 7 из этих ро-
дов механизм цветка способствует ауткроссингу. Лишь у одно-
го нлн, возможно, у двух из 35 родов, входящих в эту выборку,
обнаружены самосплодотворяющисся цветки [317).
26-48
402 Veer» mm Слгциалияирс^аммыл ггмгпмдеш* с*ст*м*
Примеры агамослерммых растений с двудонными или одно-
домиыыя родичами имеются в родах Е latest etna, Balanophora,
Wikatroemia, Antenna ria, Euphorbia м Alnus. Самонесовмсстимые
родичи агамоспермиых растений встречаются у Calamagrostis.
Zephyranlhes, Rub из, Parthenium. Taraxacum и Hieracium. Bur-
mannia— один из исключительных родов, содержащих самооп-
лодогворяющиеся виды, родственные агамоспермным видам
(3I7J.
Самояесовместимость оказалась наиболее обычной системой
размножения в выборке частично агамоспермиых ролов. Дву-
домность к однодомность. которые сраакимаьио редки у покры-
тосеменных а пелом, у частично агамоспермных родов встреча-
ются с непропорционально высокой частотой. Из этот вытекает
чрезвычайно интересное общее заключение, что ауткроссннг,
осуществляемый прн помощи той нли иной системы размноже-
ния, преобладает у половых родичей почти всех агамоспермиых
цветковых растений [317].
Гибридность и агамоспермия
Тесная связь между агамоспермией и гибридизацией отмечалась
многими исследователями. занимавшимися этой проблемой пос-
ле Эрнста [205). Все агамоспермные цветковые растения, кото-
рые изучались генетически, оказались высокогетерозиготными.
Большинство из них — межвидовые гибриды идя потомке гиб-
ридов. Болес ранние обзоры относящихся к этой проблеме дан-
ных принадлежат Густафсону (317] и Стеббинсу [780].
Первая группа данных, приведшая к этому заключению, как
указывали Эрнст [205] м последующие авторы, вытекает из кар-
тины изменчивости большинства групп агамоспермных видов.
Такие группы часто содержат очень большое число слабо выра-
женных мхиронндов. Различные мнкровнды столь многочислен-
ны и столь взаимосвязаны морфологически, что удовлетвори-
тельная таксономическая трактовка этих групп становится фак-
тически невозможной. Подобная группа в целом по характеру
изменчивости сходна с сильно увеличеаиыи скоплением гибри-
дов.
Эта характерная картина изменчивости уже давно известна
у комплексов агамоспермных видов Hurracium, Taraxacum.
Antennarla, Crepis. Rubus, Crataegus, Poienlilla, Alchemilla. Poa.
Calamagrostis н Citrus.
Для нескольких комплексов видов, в которые входят как
половые, так и агамоспермные популяции, было отмечено, что
агамоспермные формы содержат различные комбинация и ре-
комбинации морфологических признаков, имеющихся у двух или
несиольяих половых видов. Такие данные, свидетельствующие
ГЛ. 31. Асалссперяия 407
о гибридном происхождении агамоспсрмкых видов, получены
UU Antennaria (773. 774). для группы Crepis occidental is J34>).
подрода Eubatus рода Rubus (315], Pateniilla (см. [317]). Part-
henium (689] и Calamagroslis [626].
Прямые данные, свидетельствующие о естественной гибриди-
зации. получены для Aniennaria [773, 774], Parthenium (689] и
Hypericum (см [317]). Искусственные гибриды соответствуют
некоторым природным агамоспермным формам у Ал/еллаг/а,
Potentate н Rubus.
Как отмечалось рамсе и как показывает табл. 31.1, преобла-
дающее большинство агамоспермных цветковых растений — по-
липлоиды В одной нз предыдущих глав мы узнали, что у выс-
ших растений 6oui.au а я часть встречающихся в природе поли-
плоидов— амф|шлоиды. Цитогенетические и морфологические
данные, свидетельствующие об а мфи плоидности. получены для
следующих зганпспермиых полиплоидов- Hieracium. crepis, Ru-
utenlilte, Роа и Calamagrostis (см. [31Zjj.
Многие агамоспермвые формы подобны межвидовым гнбри
дам: пыльца у них стерильная или полустеркльиая, мейоз нару-
шен или пыльники абортлвны Агамоспермпые формы с муж-
ской стерильностью встречаются, например» у Taraxacum. Сгс*
pis, Hietacium, Ranunculus, Hipericum. Crataegus, Potenlilla, Ru-
?!<.. ii ДЙМВ [317].
Данные о мужской стерильности у пепсевдсгамных апомнк
юн. подобных Taraxacum и Crepis. у которых пыльца не прини-
мает участия в агаиоспсрмпом образовании семеня, можно ин-
терпретировать по-разиому. Стабилизирующий отбор на муж-
скую фертильность у этих форм ошаблск. а следовательно, от-
сутствие фертильной пыльцы не обязательно свидетельствует с
гибридном происхождении (139. 317]. Однако у таких псевлогам-
ных апомиктов, как Potentate, у которых для формирования се-
мян необходимо некоторое количество фертильной пыльны, про-
дуцирование бос-.ьшого количества стерильной пыльны после
нарушенного мейоза, несомненно, служит веским доказательст-
вом 1ибридиости.
Факультативные аломикты, у которых эпизодически проис-
ходит половое размножение, иногда дают в Ft очень широкий
спектр вьппепенцев (см. рис. 31 4). Масштабы расщепления я
потомстве Ft фзкульгатияиого авомикта можно сравнить с рас-
щеплением в поколении F? |ибрпда, вмвккшего я результате
межвидового схрешиваплм в той же группе э астений Такого
рода широкое растепление, наблюдаемое у Potentate, Rubus,
Sorbus и Citrus, вскрывает гибридную конституцию факульта-
тивно ж «но глтсриисього уастсшм Короче говоря
агпмоспермпос растение ведет себя при размиожаил как меж-
видовой гибрид.
26*
*04 4«rr t<x**a* Смцшмл»иро^ам>ам wennrvwe системы
Образование зародыша бесполым путем связано с гнбрнд-
костью также и у папоротников. Почти все апогамиые папорот-
ники, изученные Ментон (554). ииын гибридное происхождение.
Возникновение гаметофитного апомиксиса
Эмбриологические пути прн гаметофнтиом апомиксисе проходят
вблизи тех мест, где обычно протекают мейоз и оплодотворение,
ио обходят оба этих процесса. Всякая постоянная и устойчивая
агамоспермная линия должна обходиться и без мейоза, и без
оплодотворения Между ген эти два процесса не зависят один
От другого. Интересно поэтому найти такие случаи, в которых
имеет место только одна из этих двух последовательных фаз
гаметофитного апомиксиса
Примеры одной только апоспории обнаружены у Leonlodon
hispldus, Pieris hieracioides и у некоторых других видов сложно-
цветных, размножающихся половым путем. Зародышевые меш-
ки с ясно выраженными яйцеклетками или без них возникают
у этих видов из соматических клеток семязачатка. Однако эти
зародышевые мешки не дают начала зародышам. За апоспорией
не следует партеногенетического развития яйцеклетки или апо-
га много развития ее клеток-спутннкоэ [60. 317]. Подобным же
образом в одной линии Parthsnium argentatum нередуцирэван-
иые яйцеклетки образуются в результате апоспории или дни-
доспорив, но затем эти клегии не претерпевают партеногенети-
ческого развития [644].
Комплементарную ситуацию представляет собой гаплоидный
партеногенез, или развитие из редуцированного и иеоплодотво-
ренного яйца гаплоидного зародыша. Это наблюдал Хагеруп
[324] у нескольких датских видов орхидей. Хагеруп [324] при-
водит иллюстрации партеногенетических гаплоидных зародышей
Listera Ooata (n—17), Orchis strictifolius (л-20). Platanthera
chlorantha (n—21), Cephalanthera longifolia (n—16) и C. dama-
sonium (n=)6). Эти гаплоидные индивидуумы должны быть
стерильными, и по агой, а также по другим причинам должны
представлять некое прекодяшее состояние в природе
В царстве животных и у некоторых растений гаплоидный
партеногенез хорошо известен как лабораторное явление; но у
животных в отличие от высших растений ок стал нормальным
процессом размножения в нескольких группах. Например, у всех
Hymenoptera и у некоторых Coleoptera, Hemiptera и Acarina
самцы развиваются в рез)ыьтате гаплоидного партеногенеза
(764. 887. 893].
Как указал Густафсон [317], наличия лишь одной фазы из
двухфазного процесса гаметофитного апомиксиса недостаточно
для того, чтобы агамослермяое размножение могло стать перма-
Гл. 31 Алдахлер.чая
405
иентным процессом. Однако с) шее в нонан нс каждой из этих изо
лированных фаз по отдельности свидетельствует о тенденции в
направлении агамоспермии [317). Агаыосперыная систем я раз
множения может возникнуть в том случае, когда две комплсмск*
тарные тенденции— выпадение мейоза и диплоидный партено-
генез — встречаются одновременно у одною и того же орга-
низма
Чю касается детерминации на генном уровне, то можно до-
пустить. что эти две тенденции —развитие в обход мейоза и
ишй парте i f I', мм рымрукися гаыышмв гена-
ми или генными системами. В таком случае гамстофитиый апо-
миксис можно рассматривать как выражение комплементарного
взаимодействия между этими отдельными юлами
Аллели отдельных тенов или юнных систем, контролирую-
шк.\ апомиксис, присутствовали, возможно, в предковых поло-
вых популяциях в виде рецессивных факторов Случайное скре
щивэиие между носителями противоположных факторов могло
привести к образованию рецессивного коиплснектарного ге»ю-
тнпа. который вел себя как агамоспермпая форма. Сбалансиро-
ванная комбинация генов, детерминирующих агямоспермное
воспроизведение такого генотипа, может, однако, легко разру-
шится я потомстве этого нового а гам ослер много растения в слу
час аут кросс и ига [317. 644].
При адаеиШВ1ОЙ эмбрномин, когда новый зародыш развива-
ется на соматических клеток семязачатка, одного-единстпепиого
изменения в эмбриологическом пути оказывается достаточно,
чтобы обойти н мскоз. и оплодотворение. Следааательно, вот-
ки км овение адвентивной эмбрионии из предкового полового цик-
ла и эмбриологи тескон, а возможно, и в генетическом аспекте
проще, чем возникновение гаметофнтного апомиксиса.
Поведение при скрещиваниях
Наследование гамет офи г кого апомиксиса можно изучать на по-
томках о1 скрещиваний между агамоспеэмеы»и; фирмами. Такие
СКрешнвагия производились в ряде групп растений. Большая
часть опубликованных данных носит предварительный характер,
и они неубедительны. Результаты некоторых экспериментов по
гибридизации, проведенных на Potentate, Роа, Calamagroitii и
Rubus, представляют, однако, значительный интерес и будут из-
вожены в этом разделе
У Poientitla можно сравнить результаты внутри видовых скре-
тираний между вгамоспермнымн формами с результатами меж-
видовых скрещиваний. Внутривидовые скрещивания Potentilia
vemaxP. оегпа и Р. tabernaccnontanaxP. tahernaemontana лают
гибридов Fi, которые агамсспермны подобие двум своим роди-
«Зв
Часть аосьяьал. Сясциалширолапши ыштиискил аитлшл
тел ьс хим формам. В отличие от атого при межвидовых скрещм-
ваииях между Р. recta и Р. adscharica. а также Р. canescens к
Р. иегпа поколение F> состоит главным образом или целиком ив
половых гибридов. Следовательно, у Poientilla агамоспермное
состояние разрушается при межвидовой, но не при внутривидо-
вой гибридизации (ср. [317]).
Клаузен и др. [104] скрещивали шесть агамоспермных видов
Роа в восьми различных сочетании. Агамоспермными родитель-
скими формами были Р. pratensls, Р. ampla, Р. сотргезза,
Р. canbyi, Р. scabrella и Р arida. Потомства разных индивиду-
умов нз Fi выращивали по отдельности и получили семьи Р2.
Некоторые из индивидуумов F2 были отобраны в качестве ро-
дительских особей для получения отдельных семей К2. Семьи
F'i и F> обследовали для выяснения наличия или отсутствия рас-
щепления. Семьи, в которых наблюдалось расщепление, расце-
нивались как результат полового размножения родительских ин-
дивидуумов. тогда как семьи, все члены которых были одно-
родными и походили на материнское растение, расценивались
как результат агамоспермного размножения (1041.
От скрещивания Роа ampla с Р. prat ens is было получено
35 растений F,. каждое кз которых анализировали по потомст-
ву. получив 35 семей F2. Из этих семей Ft 20 были признаны
агамоспермными, а 15—половыми. Затем от ряда растений Рь
взятых кз семей обей категорий, было получено в г» 13 агамо-
слермвых и 9 половых семей. Другие межвидовые гибридные
комбинации агамоспермных Роа подобным же образом расщеп-
лялись в F: и Fj на агамоспермные и половые потомства [ 104].
В сущности, многие нз агамоспермных семей не были совер-
шенно однородными и матроклиниыми. но содержали некоторое
число половых выщепенцев. Доля половых индивидуумов в
семьях Fa. в остальном агамоспермных, колебалась от 10 до
90%. Отсюда следует, что некоторые агамоспермные индивиду-
умы F, вькокоагамоспермиы. тогда как другие агамоспермиы
лишь в слабой степени [104].
Самый же главный момент состоит в том, что расщепление
по агамоспермии у гибридов между агамоспермиыми видами
Рол носит сложный характер. В F3 н в F> иногда происходят ре-
версии от агамоспермии к половому размножению. И наоборот,
некоторые половые особи Fa лают начало агамослермиым г>.
Ряд скрещиваний между агамоспсрмами был опнсаи у еже-
вики. Скрещивание между Rubus plicatus и R. cassias дало
крайне изменчивое поколение F,, У гибрида F, от скрещивания
Rubus polyanthemus с R. insularU пыльца была фертильной, и
от него было получено поколение F>, которое очень сильно рас-
щеплялось. Подобная же изменчивость наблюдалась в Fi н F»
от скрещиваний между другими агамоспермными видами Rubus.
Гл. 31. Агалде/крлшл
40?
И в данном случае гибридизация приводит к разрушению ага-
моспермии и частичному восстановлению полового размножения
|3151
У Calamagrostis скрещивание между двумя половыми вида»
ни, С. epigeios и С. arundiruicea, даст гибридов F| и Fj, которые
образуют некоторое число длплоспоровых зародышевых мешков
[624, 626]. Гены, детерминирующие агамоспермию, содержатся,
следовательно, у полового вида н могут соединиться при гибри-
дизации. Эго важный факт, поскольку он показывает, каким
образом такой агамоспермный вил Calamagrosti*. как С purpu-
rea. о котором известно, что он представляет собой гибридное
производное С. epigeios и другого лалового вида, может возник-
нуть в природе [624. 626].
Эксиеримгн 1альиыс данные свидетельству ют о том, что гл-
метофитный апомиксис наследуется, как и следовало ожидать,
сложным образом. Гибридизация между половыми видами мо-
жет иногда привести к сборке генной комбинации, детермини-
рующей агамоспермию. И наоборот, гибридизация между двумя
агамоспермиями видами нлн же между одним агамоспермным
в одним половым родительским видом может разрушить эту же
самую генную комбинацию и восстановить половое размножение
у части потомства. Эти половые типы могут затем воспроизво-
дить 1.ОВЫЙ набор агамоспермных форы в следующем поко-
лении.
Расщепление
В патовом потомстве факультативных атамоспермных форм
обычно наблюдается высокая степень и I. ди и и дуальной изменчи-
вости. Особенно заметно расщепление по морфологическим приз-
накам, оно затрагивает также число хромосом и филнологнчес-
кую мощность.
На рис. 31.4 показана индивидуальная изменчивость призна-
ков плода у полового потомства Ft грейпфрута Citrus parodist:
родительским растением, давшим пыльцу, был лимон (С. ftmon).
Факультативно агамсспсрмные формы секции Mortfrrttf рода
Rubus после ауткросскигв во многих гибридных комбинациях
давали очень изменчивое потомство. Некоторые из этих комби-
наций. такие, как Rubus pltcatusxR. cues ms, дающие расщеп-
ляющееся поколем не F|, упоминались в предыдущем разделе.
Ауткроссннги между R plicatus или другими агамоспсриными
видами, с одной стороны, н половым днплондом R. tomenlosus —
с другой, дают поколения Ft, которые по морфологии н плодо-
витости сходны с расщепляющимся поколением Г> (315).
Гг.ппа Rubus ас и f us представляет собой сложную совокуп-
кисть лглыислермпых мнкровидпв ня Скандинавском полуостро-
Varrft лекалам, Слгфмьшларовамкмг .rwrruw«icw системы
A
Б В
Г
Д
E
Рис 31.4 Рисаиммгии* по пржакакам плоха а полоаом потомстве факужта-
тнвжо пгамогпер'Ююго растеимя Citrus (аз Frost по (717]).
Л Жепсаое pojumaacKoe растение — грейпфрут С parodist Б. Мужское ро-
азпвзпаиж растение — пином С fimon. В—Е. Четыре ратных индивидуума F1.
ве. Многие из этих микровидов — гибриды (либо гибриды F|,
либо вышелеипы F>), образовавшиеся п результате скрещива-
ний между Я. caesius (<x)i различными предсуществовавшими
видами группы Я. acutus Эти микровнды различаются по приз-
накам, характеризующим листья, стебель, колючки, опушенностъ
и т. п.
Лилфорсс (см. (315J) обнаружил в Скания (Швеция) гиб-
ридное растение, которое он счел гибридом F< от скрещивания
Rubus gothicus и Я. caesius (Я. gothicus— один из микровидов
группы Я. acutus). Пыльна этого растения была полностью фер-
тильной. Его пересадили в Ботанический сад в Лунде, где оно
дало многочисленное поколение Fi, а затем несколько потомков
F, и F.,
Поколение Fa от скрещивания Rubus gothicus и Я. caesius
представляло большой интерес, поскольку оно содержало ряд
выщепенцев, сходных е различными, имеющими собственные
названия инкровидами группы Я. acutus. Так. некоторые инди-
видуумы были сходны с Я. pruinosus— у них были тайне же
листья с семью прилистниками и прямые колючки Другие рас
тения Fj походили иа Я- acutus, Я. permlxtus, Я. waMbergd,
R. gothicus, R. polycarpus, Я. ruedensis или Я. caesius. Как и сле-
довало ожидать, в Fa были и такие растения, которые не походи-
ли ни иа один из именованных микровкдов, встречающихся в
природе [315].
Гл 31 Агамы игр ми я
409
Нарушения мейоза у факмльтлткано агамоспермпого вида
могут привести к образованию функциональных гамет г различ-
ным числом хромосом В таких случаях нозиикаюг зиготы с чис-
лами хромосом, образующими анеуялоидлым ряд. Такая вису*
плондная изменчивость наблюдалась у потомков Роа и других
злаковых
Граи [310} определял число хромосом у отдельных растений
в поколениях Fi от нескольких межвидовых скрещиваний между
факультативно атамислернными представителями Роа. Одно из
таких скрещиваний tP. scabreUaxP pralcnsis}, к котором число
хромосом у родительских растений было 2п-83 н 68 соответст-
венно дало п F; потом коп. которые по числу хромосом обрати
вали следующий ряд. 2л —65. 66. 67. 68. 68. 70. 70, 71.71, 71. 74.
75. 75, 79 Прн другом скрещнпапнн между Р ampla (2п — 64) и
Р pratensii (2л = 68) число хромосом у гибридов F| варьирова-
ло от 2л—60 до 2г!— 117, Подобная апеуплоидная изменчивость
была обнаружена в поколениях Fi от скрещиваний разных форм
Роа примерно в 16 гибридных комбинациях [310]
Клаузен и его сотрудники изучали физиологическую мощность
потомков от межвидовых скрещиваний между агамоспсрмнымн
Роа. В обшеы гибриды Fi были жизнеспособными, так же как и
ббльшая часть нх агамоспермных потомков Г?, Однако среди
половых потомков F«. полученных от тех же самых межвидовых
скрещиваний. наряду с мощными растениями было много сла-
бых в нежизнеспособных индивидуумов Такое расщепление н
рекомбинация по мощности средн половых потомков шбрпдоо
Рои представляет собой обычное разрушение гибридов Аглмп-
слермиые потомки тех же самых гибридов в значительной мере
избегают этого [104]
Характер изменчивости
Основным источником новых вариаций в группе агамоспермных
растений служи? гибридизация. Самыми обычными гибридными
комбинациями являются1 половая форма X факультативно ага-
мослсрмная форма и факультативно агамислермная форма X
Хфамлътатнвно агамоспермпая форма. В гибридизации в ка-
честве отцовской формы могут участвовать лаже облигатные
агамоспермы. ери условии, что они способны продуцировать хо-
тя бы незначительное количество фертильном пыльны. Гнбридн
аирунотис родительские растения могут находиться иа любом
уровне теистической дифференциации—от малых биотипов до
видов, связанных лишь отдаленным родством.
Вторым источником изменчивости служит автосегретация.
АвтоссгрсгаиисА называют спонтанные изменения в генотипе
410
Тест» ескьмч. Сгициамииро»атм мптплпик системы
яйцеклеток, развивающихся без оплодотворения. Так, у Taraxa-
cum мейоэ в материнской клетке зародышевого мешка может
быть нарушен, что приводят к образованию яйцеклетки, у кото-
рой число хромосом больше или меньше, чем у материнского
растения. Или же гетерогеиетическая конъюгация и кросскнго-
вер в материнской клетке зародышевого мешка могут привести
к образованию генотипически измененной яйцеклетки. Такие
яйцеклетки, развиваясь партеногеистичесни, могут дать расте-
ния, отклоняющиеся по фенотипу, Автосегрегацня может проис-
ходить как у облигатно, так и у факультативно агамоспермных
форм [317].
Третьим источником изменчивости служат почковые мутации,
приводящие к развитию фенотипически измененных побегов. Та-
кие вегетативные спорты были обнаружены несколько раз у
агамоспермией ежевики [315, 317].
Отклоняющиеся индивидуумы, независимо от способа своего
возникновения, могут затем размножаться бесполым путем.
Агамоспермиая система обеспечивает создание изменчивости
различными способами и чередующееся с втими вспышками из-
менчивости точное воспроизведение любых благоприятных новых
вариантов путем бесполого размножения.
В предыдущем разделе было описано, как одво-едимстпен-
иое гибридное растение иэ группы Rubus aculus произвело по-
ловых потомков F*. сходных с различными встречающимися в
природе агамоспермными микровидами, относящимися к той же
самой группе видов. Разных выщепепдев Fi можно было отнести
к R. aculus sensu stricto, R. pruinosus, R. polycarpus и г. д. При-
родные мнкровиды, вероятно, возникли таким же образом, т. е.
как половые вышепсипы гибридов, а затем достигли статуса
мнкровидов в результате увеличения численности путем агамо-
спермии и расширения занимаемой области [315].
В предыдущем разделе было также показано, что а группе
Роа pratensis у половых потомков гибридов существует авеупло-
кдная изменчивость. И в этом случае было обнаружено анало-
гичное состояние анеуплокдии в природе.
Лёве [523] подсчитал число хромосом у отдельных растений
одного вз членов группы Рао pratensis, известного под назва-
нием Р. irrigata в растущего на участке диаметром 10 м в
Исландия. Нв этой небольшой площади Лёве нашел растения
с 2л=82, М. 87, 91, 95. 98. 105, 108, 111, 112, 113 н 119 [523].
Таксономический вид Р pralensis в целом на протяжении об-
ширной занимаемой нм цнркумбореальной области образует
почти непрерывный анеуЛлоидный ряд от 2л = 22 до 2п=147
[98, 310, 523].
Гл, 3t, Азалосперлия
41!
Преимущества агамоспермии
Агамоспермия — это одна мз генетических систем, которая осно-
вана на гнбридности и которая делает возможным воспроизве-
дение гибридного генотипа. Уместно спросить: в чем же состоят
особые адаптивные преимущества агамоспермией генетической
системы’ В 30-х годах Дарлингтон высказал (но не развил) две
точки зрения, послужившие отправными в рассуждениях после-
дующих авторов. В первом издании «Современных успехов ци-
тологии» [138] Дарлингтон рассматривал агамоспермию под руб-
рикой «стабилизации гябрндностм». В последующих работах
Дарлингтона [140. 141. 145] концепция стабилизации гнбрндно-
сти уже не выдвигается. Вместо этого в «Эволюции генетиче-
ских систем* |141, 145] агамоспермия представляется как <из-
<и । ание стерильности».
Необходимо заметить в скобках, что Дарлингтон в назван-
ных выше работах на самом деле пользуется термином апомик-
сис. Однако в данной книге термин апомиксис используется в
том смысле, какой в него вкладывал Густафсон [317], т. е. для
обозначения более широкого круга процессов бесполого разм-
ножения. чем апомиксис Дарлингтона Апомиксис в понимании
Дарлингтона— это по существу то же самое, что в данной кни-
ге обол качается термином агамоспермия.
Представление об агамоспермии как о способе размножения,
позволяющем избежать стерильности, икеет широкое примене-
ние, Ми и гне агамоспермные формы д различных родах расте-
ний — стери.и мыс или полустернльвые гибриды, как отмечалось
ранее в этой главе. Нечетные полиплоиды. в особенности три-
плонды, обычно бывают стерильными н часто встречаются сре-
ди агамоспермных растений.
Существуют, однако, плодовитые агамоспермные гибриды.
X ним относятся гибриды между вихаыи рода Crfrur мейоэ у
них притекает нормально, и они плодовиты [677. 880]. Но при
этом кгогис гибридные формы Citrus размножаются путем
адвективной змбриопип. В данном случае агамоспермия не поз-
воляет избежать стернльиостн.
Таким образом, коинепиия стабилизации гибридное™ при-
ложима к известным случаям агамоспермии более широко и в
более общей форме, чем концепция избегания стерильности.
Представим себе, что п результате естественной гибридиза-
ции возникает некий вмсокогстсрозн! отныА генотип, обладаю-
щий цепными в адаптивном отношении гетерозисными свойства-
ми Эю1 iciiOTiin может представлять собой гибрид Ft или отяо
ситься к одному из Солее поздних поколении. ом может быть
стерильным или плодовитым. В любом случае данный гетеро-
аиготмый генотип э-.-ого гибрида будет разрушен в ре
412 Часть восьмая Са<цмм(.лр0«а^<ые секетическме системы
нормального полового размножения. Однако агамоспермия дает
возможность этому самому гибриду воспроизводиться в частоте
при помощи семян. Агамоспермиый гибрид может воспроизво-
дить точные копки своего ценного в адаптивном отношении ге-
нотипа в многочисленных растениях, развившихся иэ продуци-
руемых мм семян.
Тот же самый процесс копирования, который мы только что
описали применительно к агамоспермии, может осуществляться,
н иногда осуществляется, путем вегетативного размножения.
Однако для семян обычно характерны некоторые адаптивные
свойства, а именно состояние покоя, устойчивость к различным
воздействиям и в особенности способность к распространению,
которыми ие наделены в такой же степени органы вегетативного
размножения растений того же вида. Агамоспермия как генети-
ческая система унаследовала эти особые преимущества, прису-
щие размножению семенами
Амфнплондия — другая генетическая система, которая мо-
жет создавать и часто создает копии гибридного генотипа.
В природе возникают, однако, определенные типы ну мернческих
гибридов, которые не могут сохраняться при помощи амфиплои-
лми, но могут поддерживаться при помощи агамоспермии.
Нечетные полиплоиды — трип.юнды, пентаплоиды н т. д..—
если они обладают собственной адаптивной ценностью, могут
сохраняться при помощи последовательных поколений расте-
ний, разнявшихся из семян, которые образовались в результате
агамоспермии, но не в результате амфннлондии. Естественные
гибриды между родительскими видами с высокими полиплоид-
ными числами хромосом могут оказаться ие способными удваи-
вать число своих хромосом, йе выходя за пределы, являющиеся
пороговыми для упорядоченного клеточного деления В угли
случае опять таки агамоспермный механизм размножения поз-
воляет выйти из тупика. Наблюдаемую средн агамоспермиых
растений высокую частоту полиплоидов Двух типов — нечетных
полиплоидов и высоких полиплоидов — можно интерпретировать
как аргумент в пользу представления о том, что агамоспермия
способна привести к успеху в тех ситуациях, в которых амфи-
плоидия терпит поражение.
Как подчеркивают Густафсон [317] н другие авторы, агамо-
спермия не обязательно означает полный отказ от полового раз-
множения. В агамоспермных линиях время от времени может
происходить половое размножение, давая начало новым гибрид-
ным генотипам. Новые гибриды, образовавшиеся таким обра-
зом, могут затем размножаться в процессе еще одного цикла
агамоспермии. В результате внутри данной агамоспсрмной груп-
пы могут время от времени возникать новые адаптивные гетеро-
зиготы, которые за гем воспроизводятся в свою очередь в массо-
вых количествах при помощи агамосоермного механизма.
Гл. 9J. Агллос&рмнл 413
Агамоспермия, следовательно, обладает известными собст-
венными преимуществами по сравнению с другими конкретными
генетическими системами. Агамоспермия делает возможным
размножение редкого гомоллоядного гетерозиготного выше вен-
ца в гнбрнднов популяции растений, размножающихся при по-
мэгци ауткроссинга, тогда как половое размножение обеспечить
этого не может. Она сохраняет нечетные полиплоиды и высокие
н-орнчные анеуплоиды, что невозможно при амфнпломдим. Она
делает возможным размножение гибридов с высокими поли-
плоидными числами хромосом, когда дальнейшее удвоение числа
хромосом и амфиплоидия были бы безрезультатными. Она обес-
печивает бесполое размножение семенами, на что неспособно
вегетативное размножение.
32 Агамный комплекс
-Сочетание процессов естественной гибридизации и агамосперм-
ного размножения привело с течением времени ж возникновению
ряда микровидов, несущих в различных рекомбинациях призна-
ки родительских видов. Образующаяся а результате структура
представляет собой агамный комплекс, в котором агамосперм-
ные гибридные производные в процессе филогенеза наложились
иа первоначальные половые виды.
Структура простого агамного комплекса схематически пока-
зана на рис. 32.1. На этом рисунке можно видеть многочислен-
ные агамоспермные производные, заполняющие разрывы и мас-
кирующие Прерывистости в морфологических признаках между
диплоидными половыми видами. Изменчивость в пгамяых комп-
лексах — это наиболее сложные картины изменчивости, извест-
ные у растений. Для форм, входящих в агамиые комплексы
Rubus и Hieraeium, были опубликованы тысячи латинских наз-
ваний видового или ннфравилового ранга.
Проблема вида в том или ином агвмном комплексе частично
разрешима. Диплоидные половые вады, если они сохранились
-ДО настоящего времени, можно обнаружить, дать им названия
н нанести на карту их ареалы. Отдельные агамоспермные произ-
водные, если они обладают достаточно однородной в ясно вы-
раженной комбинацией признаков в пределах данной географи
ческой области, также следует признать таксономическими ви-
дами и дать нм названия
Однако в некоторых частях агамиого комплекса проблема
вида неразрешима. Между диплоидными половыми видами и
достигшими наибольшего успеха и более широко распростра-
нившимися агамоспермными видами находится множество ага-
ыоспермиых вариантов более низкого ранга — от малых б моти-
вов до локализованных агамоспермиых микровидов. Какие из
них следует выделять таксономически и в каком таксовомкчес
ком ранге, зависит от точки зрения и соображений удобства, в
на этот счет мнения исследователей часто расходятся.
Филогенетический анализ данного агамного комплекса со-
пряжен, кроме того, со многими трудностями, вытекающими из
большой сложности этой проблемы. Стадии анализа, обрисован
ные выше применительно к полиплоидным комплексам, вполне
приложимы к атаыиым комплексам. Некоторые агамиые комп-
лексы. такие, как группа Crepis occUtentaUs, подвергались де-
тальному цитотаксономаческому анализу, а другие, подобно сек-
ции Moriferus рода Rubus, — детальному биосистематическому
диализу, но ин один большой агамный комплекс не был исчерпы-
еваюшям образом изучен а генетико-таксономическом плане.
Гл, 93. АглмчыЛ 41S
Комплекс Crepis occidentalis
Группа Crepis occidentalis (л—11)—совокупность однолетних
травянистых растений. растущих и яападных областях Северной
Америки. Эти растения встречаются на сухих склонах гор: центр
их распространения находится на Колумбийском плато, а ареал
тянется от Тихоокеанского побережья до Скалистых гор. В эту
группу входят половые диплоиды и многочисленные агаиоспсрм-
н ые полиплоиды [36].
Бэбкок и Стеббинс [36] проанализировали группу Crepis
occidentalis ва ссиове гербарных материалов, полевых исследо-
ваний, иитотакоономических исследований и наблюдений за жи-
выми растениями, пересаженными и» природных местообитаний.
Гибридизацию в экслсрнмситалъиых условиях провести ие уда-
лось. поэтому прямые данные относительно теистических взаи-
моотношений между видами отсутствуют,
Бэбхои и Стеббинс разбили вариации в группе Crepis occiden-
talis иа 9 таксономических видов: С- acuminata, С. bakeri.
Рее 33 1 Структура аростэто лгаииого комплекса (ЭВ].
416 Часге «схожая Смииамоирэеамие «емтичесюк систем*
С. barbigera, С. елз/й, С. intermedia, С. modocensis. С. monticota,
С. occidentals и С. pleurocarра. В большинстве случаен эти так-
сономические виды содержат диплоидную половую популяцию
и различные агамоспермные формы, которые морфологи чески
более или менее сходны с диплондом. Агамоспермные формы,
если они вообще имеют названия, рассматриваются как трино-
минальные категории forma apomictica. Таким образом, в группе
С. occldentalis именованные таксономические виды представляют
собой сложные и искусственные единицы.
В эту группу входят семь известных половых диплоидных
видов. У следующих таксономических видов — Crepis acuminata.
С. hokeri, С. exllls, С. modocensis, С. monticola. С. occldentalis
к С. pleurocarpa — они рассматриваются как ивфравидовые еди-
ницы, В группе С occldentalis половые диплоидные популяции
составляют основные биологические виды. Эти семь диплоидных
нов хорошо различаются по морфологическим признакам
Лишь один из нит (С. acuminata 2х) имеет широкое географиче-
ское распространение; ареалы остальных шести видов относи-
тельно ограничены и перекрываются снмпатрически н различных
местах на северо-западе Тихоокеанского побережья 136].
Все агамоспермные члены группы Crepis occldentalis — поли-
плоиды (основное число X" 11). достигающие октоплонляого
уровня. Среди них есть все возможные нечетные полиплоиды
(Зх, 5х, 7х). Морфологически некоторые из агамоспермиых ти-
пов очень сходны с каким-либо одним диплоидным видом, но у
большинства из иих комбинируются признаки двух или несколь-
ких диплоидных видов. Именно в этой суперструктуре полипло-
идных агамоспермных форм можно обнаружягь интерградению
между различными крайними комбинациями морфологических
ф^изнаков [36]
Агамоспермные популяции отличаются друг от друга по
признакам листьев, стеблей, соцветий, цветков и семянок Неко-
торые кз иих имеют локальное, а другие широкое географичес-
кое распространение. Те популяции, которые распространили
свою отличительную совокупность прнзпаков по сколько-нибудь
значительной географической области, получили неформальное
таксономическое признание. Им присвоены триноминальные ла-
тинские названия неформальной категории forma apomictica
(apm). каи уже упоминалось выше. На рис. 32.2 изображены
соцветия (корзинки) девяти получивших названия агамосперм-
ных микровидов из группы Crepis occldentalis. Примером хоро-
шо выраженного агамоспермпого микровида служит С. pteuro-
carpa apm. grayi. форма встречающаяся в северо-восточной час-
ти Калифорнии. Это. возможно, пентаплонд. На основе морфо-
логических признаков его считают гибридным производный по-
ловых диплоидных форм С. pleurocarpa и С. occldentalis Другой
Е
Ж 3 В
A
S
в
д
Рис 31 i Союпяч лепят агамоаершшт ипромлое Спри оаМопМи
(по (36))
А—с.о. арт. twmilitr. Калифорния 6—С. о арпт. лшсоЛи. Ватикггап.
В — С.о, орт. calycuictp, Колорадо Г —Со. арт. Юп. Д — Сл. tpm
eUlpUca. гемрпо» часть Кынфораяи. £ —С. о. арт. daUaWaa. АльСерта.
Ж — Со. apoi rpdbt/gU, Моатааа. 3— С.о. open hamUtonamt. cesepaaa
часть Калпфоркп. И — Со арт. стшаа, Втштггав.
27—46
TIB Част» аосалая. Сллциамаироваимм вснегмскм сасплы
агамоспермммй вид. распространенный в горах северо-восточно!
части Калифорнии, — С acuminata арпт. sierras, вероятно пред-
ставляющий собой трнплокл.
Всего в монографии Бабкока и Стеббинса приводится 113 та-
ких форм, получивших название апомиктических. Это лишь не-
большая доля всего множества обособленных агамоспермных
поцуляшгй Crepis в западней частя Северной Америки
Число агамоспермных микровидов относвтелиьо велико в об-
ластях, находящихся вблизи цсктра распространения половых
популяций. Половые и факультативно апомиктические популя-
ции являются непрерывным источником возникновения новых
агамоспермных микровилов в центральных частях ареала. По
периферии области распространения всего комплекса, напротив,
число мнкровилов меньше, ио каждый из них имеет более об-
ширный ареал. Здесь некоторым отдельным агамоспермным
формам, очевидно, благоприятствовал естественный отбор, по-
скольку они были лучше приспособлены к региональным усло-
виям среды, и эти формы размножились, заселив обширные тер-
ритории (36)
Филогенетическую историю комплекса Crepis occidentalis
можно воспроизвести п общих чертах по косвенным данным.
Современное географическое распространение каждого из по-
ловых диплоидных видов коррелирует с распространением опре-
деленных климатических и эдафичесхих провинций, сложивших-
ся в плиоцене и в раннем плейстоцене. Следовательно, ареалы
диплоидных видов, вероятно, установились в этот же период, от
раннего плиоцена до раннего плейстоцена, в ответ иа установ-
ление более сухого климата и излияния лав [36].
Диплоидный вил. после того как ом обосновался, гибридизи-
ровал в различных комбинациях, давая аллотетраплоидных про-
изводных. Процесс гибридизации продолжался, возможно, до
недавнего времени н привел к возникновению агамоспермных
никровкдов, представляющих собой вторичных и третичных гиб-
ридов. так как некоторые из э™х никровкдов имеют высокие
полиплоидные числа хромосом и связаны сложными родственны-
ми взаимоотношениями. Позднее, в течение плейстоцена, поли-
плоидные агамоспермиые формы широко распространились, вы-
теснив диплоидные половые формы, которые оказались ограни-
ченными двумя центрами распространения в северо-западной
части тихоокеанского побережья Северной Америки.
Европейские ежевики
Обширный род Rubus (х—7) делится на четыре подрода; самый
сложный из ких — подрод Rubus. к которому относятся ежевики.
Подрод Rubus образовал разнообразные совокупности видов и
Гл 93 Аым**Д немилые
419
ыикровидов п ряде умеренных областей земного гилра в Европе.
Азии Средн зем поморской области, в восточной части Северной
Америки, на тихоокеанском побережье Северной Америки it п
районе Кордильер в Центральной и Южной Америке.
Ежевики Евразии и восточной части Северной Америки об
разуют одну подгруппу Rubus, формально выделяемую как сек-
ции МогОггш Флора европейских ежевик— наиболее хорошо
изученная часть этом секции, и служит основным предметом
Обсуждения в данном разделе Опа делится иа две естественные
ншегтные под названиями Moriferi Veri н Rubi Corylijo-
/м. Однако европейские ежевики не представляют собой само-
стоятельной естественной единицы, обособленной от членов еек
НИИ Morifcrus, произрастающих в восточной части Северной
Америки и в Азин н некоторые виды ежевики ил двух послед-
них областей, несомненно, филогенетически связаны со своими
европейскими родичами (314, .315, 317]
Ежевики (подрод Rubus) и европейские ежевики в частно-
сти (часть секции Afori/erus) изучались рядом систематиков
и ; енетнков — Фокке. Лидфорссох. Густафсоном и другими
Побудительной причиной таких исследований, как такгоиомнче-
еккх, так и генетических, была «почти неисчерпаемая изменчи-
вость этих ежезтк» [317] Пытаясь отразить эту изменчивость в
таксономическом плане, различные литры ив протяжении ряда
.чет опубликовали около 4 -5 тыс. латинских названий Эти наз-
вапия относятся к единицах' самого разного уровня — от широко
распространенных видов до узко локализованных форм.
Для того <пи0ц проиллюстрировать масштабы и • .
у еврокйсш ГустЦюоа и итожил вкратце систему их
классификации созданною бочее ранним исследователем этой
траппы, Судре, Судре ралли чает м группе Moriferi Verl 691
ммьровнд; псе эти микрппи.ты он объединяет в 109 таксономиче-
ских видов В одной лишь Европе существует около 1930 имено-
ванных взрнететов Morifen veri [317] Другая основная ветвь
европейских ежевик, Ruh CorylRoIii, содержит один таксономи-
ческий вид (Rubus caesius), по бесчисленное множество микро-
вндов [315]
Европейские ежевики образуют полиплоидный ряд (х = 7).
К нем представлены следующие уровня плоидности 2х. Зх, 4х.
5х, Ьл и. возможно, 7х. Кроме того, в этой группе имеется не-
сколько вторичных амеуплондов с числом хромосом, отклоняю-
щимся от полиплоидною. Большая часть этих мнкровндов —
тетраплонды. Несколько трнплокдных н пеитаилондны.х микро*
видов занимают обширные ареалы [315. 317]
Ежевики восточной части Северной Америки также образуют
полиплоидный ряд с основным числом хромосом х=7; среди них
есть триплоидм. пантаплоиды и гексаплоилы. а также несколько
4ЭТ Чат wvsjbuk. эвчтсчлгкае системы
диплоидных видов [315J. В отличие от этого все формы, произ-
растающие на тихоокеанском побережье. — высокие полиплоиды
с уровнем плоидности от 6х до 12х [71, 102].
Пшиплоидиые европейские ежевики валяются по большей
части факультативно агамоспермиыки формами. Установлено,
что многие нз них — гибриды или гибридные производные, о чем
свидетельствуют в разных случаях морфологические признаки,
стерильность пыльцы, состояние печетпов полиплоидное™ н(нли)
высокий уровень растепления прн возврате к половому размно-
жению. Было также обнаружено, что и у восточноамериканских
ежевик существует агамоспермия. Помимо образования агамо-
спермией семян ежевика распространяется, как вто хорошо из-
вестно, при помощи вегетативного размножения [315, 317].
В Европе большинство агамоспермных форм образуют ге-
ографически локализованные мнкровиды. Некоторые из этих
локальных мнкровидов достаточно обособлены; другие же гиб-
ридизируют и иитерградкруют с родственными микровндами.
Среди европейских агамоспермных форм некоторые образовали
географически широко распространенные популяции, превра-
тившись. таким образом, в агаиоспермные виды
Филогенетическую основу агамиого комплекса у европейских
ежевих образуют диплоидные половые вады. В западной часта
Евразии известно пять диплоидных вадов: Rubus ballet. R. тса-
nescens. R. moschus, R. iomenlosus и R ulmifolius (2n = 14). По-
лагают, что и некоторые другие вилы, такие, как R. alnicola,—
диплоиды. Возможно также, что в эволюционном развитии евро-
пейского агамного комплекса сыграли некую роль и другие по-
ловые диплонды. произрастающие в восточной части Северной
Америки, например R. allegheniensis (315, 317].
Все диплоидные половые виды европейских ежевих припал
лежат к группе Mori/eri Veri. Они хорошо отличаются от агамо-
спермных микровидов и видов, а также друг от друга. Некото-
рые из них имеют широкое, а другие —ограниченное распростра-
нение.
Rubus Iomenlosus и R. и/лы/о^из характеризуются обширными
ареалами, тянущимися от южной части Европы до Малой Азии;
R. moschus встречается в горах Кавказа; R. incanescens — на
западе Средиземноморской области, a R. botlel — ua Канарских
островах. Возможный диплоцд R. alnicola распространен в Пи-
ренеях и горах Юры. В тех местах, где эти виды встречаются в
природе, они часто гибридизируют с образованием стерильных
гибридов. Многие иэ агамоспермных михровидов Moriferi Veri—
давнишние или недавние производные таких гибридов [315, 317].
В целом диапазон изменчивости агамоспермных Moriferi Ve-
rt в Европе сильно выходит за пределы крайних отклонений а
морфологических признаках, наблюдаемых у европейских ди-
Гл. S2. .4лхмчы4 комплекс
♦21
пдонлов. Но некоторые европейские агамоспермы морфологпче-
скн очень близки к некоторым восточное меряна псин и дкпло-
идвм. таким, как Rubus alleghemensis Из этого можно сделать
вывод, что среди диплоидных лаловых предков агамного комп-
лекса Европы были виды, распространение которых и настоящее
время ограничено восточной частью Северной Америки, а также
различными хребтами в вММЛЮб части Евразии (315, 317]
В группе Rubi СогуЩоШ ДИПЛОМЫ неизвестны. В эту группу
входит, однако. сизая ежевика (Rubus caesius, 2п-28)—алло-
тетраплоил. происходящий от диплоидных видов, которые. ве-
роятно, уже вымерли. R caesius представляет собой фякульта*
тимно агамослермкыЙ вид. Он свободно гибрмдилирует с други-
ми видами п микровндамя ежевики, но изолирован от них таки-
ми барьерами, как стерильность и псжизиесппсобнпгть гибридов.
Подобно диплоидам в группе Monfen Veh. он образ;.», т
ленный естественный вид (315. 317].
Rubus caesius i ибрилизирует с ннплондами R. ulmifolius и
R lomentosus. а также с другими диплоидными н полиплоидны*
ми вилам»- ежевики Многочисленные агамоспермные микровнды
Rubi CoryUfniti образовались в результате расщепления среди
потомков от происходивших ранее гибридизаций между Rubus
caesius и различным»» предстаимтглнми Moriferi Vari. Дру»»и»
агамоспермные Rubi Corylifohi продукты вторичных возврат*
пых скрещиваний между Rubus caesius и другими членами груп-
пы Rubi Contitfolii Так. естественные гибриды между R caesius
и R gothicus (Corylifolii) в результате расщеплении дают нача-
ло формам. сходных» с другими нменсваиными мнкгювидаыи
Rubi Corylifolii, такими, как R acutus, R trahlbetgii и R pruino-
sus (314. 315. 317].
Обыкновенная малина Rubus iclaeus (2л = 14). принадлежа-
щая к другому полроду, Idaeobafus, гибридизирует с R caesius
(2л = 28). Триплондные гибриды стервлыш. Однако иногда из
нередуиировзгпых гамет образуются плодовитые пли oo.ijiltq-
довитые тетраплоидные и гексэплсидмые гибриды Эти гибриды
в результате возвратных скрещиваний с К caesius вносит в
группу R caesius отдельные гены малины Некоторые агзмо-
сперчныс формы Rubi Corylifolii пл-пилимому, представляю» со-
бой продукты гибридизации R idaeus и R caesius Таким обра-
кш, обогащению изменчивости и комплексе европейских ежевнк
способствовал также один диплоидны Л вил, не принадлежащий
к подроду Rubus (317]
Диплоидные половые виды ежевики в Европе в настоящее
время встречаются в ниде реликтов на юге Средиземноморской
области. Некоторые из них изолированы географически. В наши
дни американские днплолды. конечно, изолированы от европей-
ских диплоидоа Возможно, однако, что в доледниковое или
*В УотИОГЫШЙ. ММППМЛПМ састиш
межледниковое время (иля же яви в то. так и в другое) эти
диплоидные виды имели более обширные ареалы и сильнее пе-
рекрывалось друг с другом. Быть может, именно тогда и возник-
ла первая волна аллотетрапловдое и агамоспермных производ-
ных [314].
Эта межледниковая флора ежевкк вынуждена была отступить
во время последнего оледенения, но в послеледниковое время с
улучшением климатических условна она. вероятно, вновь рас-
пространилась и дала новые поколения агамоспермных гибрид-
ных производных. В настоящее время агамоспермные ежевики
в отличие от своих диплоидных половых предков занимают об-
ширный ареал по всей северной и восточной Европе. Агамо-
спермвые мнкровиды гибридного происхождения успешно засе-
лили холодные области этого материка, которые еще недавно
находились подо льдом (314. 315]
Taraxacum
Широко распространенный род Taraxacum встречается как в
Старом, так и в Новом Свете, как в Северном, так и в Южном
полушариях. Центр его распространения находится в Евразии,
где ой на востоке доходит до Японии, а иа севере проникает в
Заполярье. Многочисленные варианты сгруппированы в 32 сек-
ции. Основное число хромосом *—8. Известны формы 2х, Зх. 4х.
5х и 6х, а также несколько авеуплокдов.
У этого рода различают 28 половых диплоидных видов. Хо-
рошо известным примером служит Taraxacum kok-saghyz (2п=
— 16) из Средней Азии. В Австрии обнаружена диплоидная по-
ловая форма Т officinal? (224]. Большинство половых дипло-
ндов самоиесовместимы. Многие иэ них относятся к нескольким
примитивным секциям этого рода [681] Некоторые — узко виде-
мичиые или реликтовые формы. Так. Т. pieninicum — реликт
ледникового времени, встречающийся в горах в Польше, а
Т. cylleneum эндемичен для одной горы в Греции [225, 227, 545].
Искусственные гибриды между диплоидными половыми ви-
дами высокостерильны при одних комбинациях родительских
форм [224. 317, 547] и плодовиты прн других [681].
Полиплоидные представителя Taraxacum являются агамо-
спермами, а большинстве случаев облигатными, но иногда фа
культативными. Агамоспермиые таксоны заходят в своем рас-
пространении в арктическую область, я в других флорах не ре л
ко растут как сорняки [148, 317. 681. 763]. Фюрнкранц [226] об-
наружил. что в Австрии полиплоиды обычно встречаются в сме-
шанных популяциях с диплоидамн.
Гл J? АгалмыЛ комплекс
42J
Наиболее обычный уроне ин плондносги у этого рода — трн-
плсииня. наблюдаемая а 114 из 218 таксонов, в которых лроИЗ
водились подсчеты (681] Например, одуванчик лекарственный
(Гога.хясат о//«йя/е)— это, как правило, агамоепгрмный три-
плонд (2« = 24). хотя у этого вида известна и диплоидная фор-
ма Часто встречаются тстраплоидные формы Taraxacum
(60 таксонов); известны также пситаплоидиые формы (15 так-
сонов) и одна гсксаплопдная форма [681].
Дтя полиплоидных агаиоспермяых форм характерен нару-
шенный мейоз с низкий уровнем конъюгации. Некоторые из них
лают стерильную пыльцу. а другие ие образуют ее вовсе. При
ат амоспермном размножении материнская клетка зародышевого
мешка делится с образованием диады Нередупироеаиная хала
зальная клетка этой диады образует зародышевый мешок. За-
тем из нсредушрованиой яйцеклетки партенитенетичеекк разви-
вается зародыш [546, 681).
У агамоспермных Taraxacum имеется множество мккровидов.
По данным Густафсона (317]. в Скандинавии насчитывается
450 микровидов. Олтард (630] недавно описал 26 новых «виловэ
(читай «микровидон») в одной только Дании. Согласно Ричард-
су [681]. у этого рода о целом описано почти 2000 агамосперм
иых чпкровидов. Микровилы распадаются иа скопления, кото-
рые соответствуют примерно 26 таксономическим секциям [681].
Эти мнкровиды довольно однородны, хотя даже в отдельных
популяциях существует некоторая индивидуальная генетическая
изменчивость., как это было показано для Taraxacum officinale
1757]
Taraxacum произрастает в основном в умеренном поясе.
с центром распространения к центральных и западных районах
Евразии Ричардс [6£1] считает, что это—древний род. пропс-
ходящий из западных частей Гималаев; оттуда наиболее про-
двинутые фор» ы распространились в Европу, в арктические и
другие регионы Изменения климата и плейстоцене способство-
вал и koi тактам между раэаыкя видами, гибридизации и обра-
лияаиию в этот период многочислен пых агамоспермных микро-
видов.
Филогении этою рода сложна и не вполне понятна Ричардс
[6*1] ncxiaiaeT. что при гибридна в пнях в результате которых
возникли многие из нынешних агамоспермных мнкровндов.
одним из родительских видов были представители секций Сега-
topho/a и rVc/ica Так, секция Bulgaria представляет собой ре-
зультат скрещивания Ceratophorax Р). а секция AIpiпа — ре-
зультат скрещивания ArcticaXTaraxaium ("'секции Vulgarta).
Для того чтобы подтвердить эго предположение, необходимо,
однако, провести более подробные исследования.
424
4acf Смфимймм/ммаммм* сигт«лы
Агамные комплексы типа Rubus Mori/erus
Агамные комплексы Rubus. Crepis и Taraxacum принадлежат к
определенному типу, наблюдаемому также у рада других групп
растений северной умеренной области. Этот достаточно часто
встречающийся тип удобно назвать типом Rubus Moriferus.
поскольку он наиболее подробно изучен на примере этой
группы.
Наиболее существенные черты агамных комплексов типа Ru-
bus Morijerus сводятся к следующим. Эти комплексы насчиты-
вают сотни или тысячи взаимосвязанных агамоспермиых и над о
внлов. бее эти микровиды гибридного происхождения. Они про-
дуцируют семена путем гаметофитиого апомиксиса и нередко
также до некоторой степени — половым путем. Агамоспермные
формы являются полиплоидами, четными пли нечетными, причем
нечетные более обычны. Полиплоидные агамоспермные формы
филогенетически и таксономически представляют собой над-
стройку над рядом ясно выраженных половых диплоидных ви-
дон, существующих в настоящее время. Половые диплоиды. од-
нако. в большинстве случаев имеют ограниченные или даже ре-
ликтовые ареалы, тогда как их агамоспермные производные бо-
лее агрессивны и имеют более широкое распространение
Помимо комплексов Rubus. Crepis и Taraxacum к типу Rubus
Moriferus относятся некоторые другие агамные комплексы, пере-
численные ниже; обзоры по большей части этих комплексов
можно найти у Густафсона [317], а комплексы Cotoneaster,
Townsendia и Hieracium pilosella рассмотрены Саксом [710], Би-
меной [52] и Скалинска [735].
Hieracium подрод Pilosella (а=7) в Евразии
Hutracium подрод Hieracium (х=7) в Северном полуша-
рии
Alchemilla подрод Alchemilla (х—8) в Северном полу-
шарии
Alchemilla подрод Alchemilla (х—8) в Евразии, Новом
Свете и Австралии
Townsendia (х—9) в западных районах Северной Аме-
рики
Hypericum (х=8) в Евразии
Parihenium (х=9) в южных районах Северной Америки
Sorbus (х—17) в Евразии
Cotoneaster (х—17) в Евразии
An/ennaria (х=7) в Северном полушарии
Calamagroslis (ж = 7) в Евразии
Вес эти примеры, разумеется, различаются в деталях. Так,
подрод Hieracium содержит много диплоидных половых видов,
Гл 33 •«.’вжялгЛ КЧНПЛЛКС
435
Рк 32 X Раелртхгряясгжо ВоиМоад eurtlpiKduta (262)
Три приведение 3JV4K рлтнгокляоетя г-.держ1т следу ошее wejeo хромо
«м В с cw^Upeniuio — 2п-40. 41—54; В. с carspUcta -2л-&5—К».
И с ’«ж'»-2ч-20. 40-41
а у AIrhemilla опи редки. У Sorbus и у Coloneaster также хоро-
шо представлены половые диплоиды. но здесь это старые днп-
лои локированные гллотстрап лонды (х>-17). У Antenna ria
(х-*.7), Parthenium (л-9) и Calamagrostis (к-7) предковые
половые виды были, по-аиднмому, тетр аплой да ми
Интересно, что большинство перечисленных здесь агамных
комплексов типа Rubus Moriferus относится к сем. Compositae
и Rosace а с, очень успешно развивающимся в Северном полуша-
рии. К сложноцветным принадлежат Crepis, Taraxacum, Hie ra-
dium. Townsendia. Parthenium и Antennaria; к розоцветным —
Rubui, Atchemilia, Sorb us и Coloneaster Кроме того, среди при-
веденных примеров есть один представитель сем. Gramincae —
Calamagrostis и один представитель сеи Hypericaceae — Нуре-
rteum.
426 Часть восьмая. Слс^иалимрсмкмь^ /тнспнаскм системы
Комплекс Bouteloua curtipendula
Гранины трааы. образующие комплекс Bouteloua curtipendula.
широко распространены и грасслеядах Северной Америки—от
Канады до Мексики, а также встречаются в других разобщен-
ных областях Центральной и Южной Америки. Гулд и Кападиа
1262), произведя ревизию этой группы, выделила в ней 12 андон.
Один из них, собственно В curtipendula, имеет очень обширный
ареал, пересекающий всю центральную часть США и Мексики
в виде широкого пояса (рис. 32.3). Остальные 11 видов имеют
более ограиичеииое распространение (рис. 32.4) (2621.
Число хромосом у этих видов варьирует от 2л—20 до 2л—103
(табл. 32.1) при основном числе х=10. Среди них есть дипло-
иды, тетраплоиды, гексаплоилы и более высокие полиплоиды
вплоть до I Ох. а также многочисленные аисуплонды [261, 262]
Таблица 32.1. Число хромосом у разных членов комплекса
Bouteloua curtipendula (л —10) (260—262]
В «А Арьлл Числд 9л
В. Iriama В ялт&а В. imtflora В. varnocMi В. refleta Южная Мексика От ш*ятрадьаа8 каста Мексики ю Г сидураса От Юти до северный гаДоно» а«гтра.’¥яг-й части Мексики От заодчоео Тыка ю Ныо- МексжкО От КдемфоржиВского * ал ив а до 20 20 20 (30 у одиого пгбри- 21 jk П. 25. 23, ЗА 40 20
В pedldUata В. diiticha Центральная Медали От троакесяов Мексики до Вероятно, ДММЯД (СУ- ДЯ по величине пыль- це® ыа зерен) То же
В. vanfvdenii В. /мдеа А. dtatau В. purpurea В. CMftlfiMBttta var. /«лам var. curlipendula t/и. ca ел pitot a П*РУ _ Остром Kapntfcucca Cacretax То же Оахака Це!ггр*льиаи Меиздта Мексика От «>жно* части Каиды до Темса От юго'завидно* иаспг США до пвм1 частя Ммгсша. а также в Южной Америке Не известно, возможно, ДЯЖЛОНД То же 40 20. 40.42 40. 41, 42. 43, 44. 45, 46. 43, 50, 51. 52. 53, 54. 55. 56, 58. 59. 60, 62, 68. 60. 62, 64. 60. 72, 77. 78, 80. 82. М. Й5. «, 87. 88. 90. 91. 92, 94. 96. 90. 100, 102. 103
Гл 32 4л>члшВ кохял.'кс
427
Высокие полиплоиды и апсуплоиды группы Bouteloua curti-
pend и {а предо являют собой агамоспермиыс формы, размножаю-
щиеся путем апоспории и псевдогамии; у них часто наблюда-
ются нарушенный мейоз и низкая степень конъюгации хромо-
сом. Половые гекса пл оиды или половые аисугшоиды близкие к
гексаплондному уровню, в этой группе не известны. Днплоиды.
тетраплопды и анеуадоиды невысокого уровня размножаются,
насколько »то известно, половым путем. Мейоз протекает нор-
мально у диплоидов и некоторых тетраплокдов. а у других те-
траплоптов и анеуплоилоп с низким числом хромосом — с уме-
ренными нарушениями [220. 259. 346, 561].
Произведя подсчет хромосом в одной нз выборок, Фрстср и
Браун |220| обнаружили, что граница между половым размно-
жением и агамоспермией проходит в этой ipynne па уровне
2п =52 Все растения, у которых число хромосом зыше 52. раз-
множаются при помощи агамоспермии, а те. у которых оно равно
52 или ниже, во всех известных случаях размножаются половым
путем. Возможна, однако, что в дальнейшем будут обнаружены
агамоспермных фичмы < HH3KIM ЧИСЛОМ хромосо* ДОО
Диплоидные подовые виды, такие, как Bouleloiui unifiora,
В Iriacno, В. media и другие, встречаются главным образом от
субтропических до тропических широт и и^еют относ-исшио ог«
ракнченмые ареалы. То же самое относится к половым тетра-
ДОШЧ В purpurea и частично—к В (рис. 32.4).
Половые диплоидные и тетрапломдиые формы В. curlipendula
tenuis также встречаются в Мексике (рис. 3z-3).
В 07Л1ГЧЖ о? диплоидов представите.! г. атой группы с высо-
кими хромосомными числами, а именно Bovtetoua curtipendula
Liifhpendulii (2n-4O—64) и В. с. caespHosa <2п-53—103), име-
ют очень обширные ареалы, расположенные севернее п тяну-
щиеся мере) равнины до южных частей Камалы (рис. 32 3) Как
отмечалось ранее, растения с более высокими числами хромо-
сом — в д| глазом примерно 5г— 10-г являются а:амосперм-
ными. На карте, представленной на рис 32.5, можно видеть,
что эти агамоспепмиие формы В curtipcndula занимают об-
ширную область на юго-западе США и на севере Мексики
[2S9J.
Еще одна черта, характерная для видов Boulrlnua с высоки-
ми хромосомными числами. — наличие у них обширных апсупло-
идцых рядов. В таб.1. 32.1 можно видеть почти непрерывные
аиеуплоилные ряды .-ишь с несколькими очень небольшими раз-
рыпаки. хак. напри нор. у В rurlioenduta curtipetuluta (2л = от40
до 64) и В с caespilosa (2п«от 58 до 103) При подсчетах хро-
мосом в выборках эуплондпые числа встречаются чаше всего,
а различные аиеуклоидные числа представляют собой отклоне-
ния от этих зуллоидныч yawn (261. 262(
М-
Рж- 324. Раиростраиеаве 9 oaomai аилов (лплояммх » тетраааотлиыт) коиптееси ЯояМочш nrtlpendata [2621
На аорте аоизанн титло южыя часть ареала В иЫЦога я «асрмя часть ареш В <в«1с*а.
Гл. 32. AtaMtttd л0млмлс
«»
Рис 325 Ркл|><хт|ння» половил и «ганосоарииых форм Boultlma сил-
hprrubpa [259].
Такой обширный анеуплоядиый ряд возникает в результате
комбинации процессов естественной гибридизации, нарушенного
мейоза и атамоспсрмиого размножения
Естественная гибридизация между популяциями с разным
числом хромосом ведет к выщепленйю аисуплоидных форм в
последующих поколениях. Вероятно, в этом — причина возникно-
вения простых аисуллоидвых рядов, наблюдаемых у полового
вида Bouteloua u-arnockii (см. табл. 32.1). В. irarnotkii (2х, 4х)
гибридизирует в западной части Техаса с В. curtipendula caespi-
tosa — видом, характеризующимся высоким хромосомным чис-
лом (2С>2| Полагают, что наиболее обширный аиеуплоидный ряд
в пределах самого вида В. curtiptnduta возник в результате бо-
лее сложных гибридизаций внутри этого таксономического вида,
а также между ним и различными другими видами данной груп-
пы [261, 426).
430
Vacr» fooAOJt. Специализированные «еягтиймекде сютемм
Рх 3?6 Первая акафвэв в додвацрйс* матврнипо* «лгвср пыльцы самого
рк-гемяя Bcuteloua curtipendula caespitcui с висоюш явслом хромосом;
обрдпгге вншмакме м очмь мерла номерную сегрегацию хромосом [2f>2]
У растений вида Bouteloua curtipendula с высокими хромо-
сомными числами мейоз часто бывает нарушит, о чем уже гово-
рилось выше. Наиболее интересная для нас особенность мейоза
заключается в очень неравномерном расщеплен кв в анафазе I:
ббльшая часть хромосом направляется к одному полюсу и лишь
несколько — к другому (рис. 32.6). Клетки, получившие боль-
шое число хромосом, по-видимому, дают жизнеспособные пыль-
цевые верна, которые в свою очередь могут дать аиеуплоидных
потомков. Различные анеуплокдиые формы, после того как <ии
образовались, мотуг затем воспроизводиться прн помощи агамо-
спермии {261, 262].
Подводя итоги, можно сказать, что группа Bouteloua curti-
pendula представляет собой половой полиплоидный комплекс на
тстраплондиом уровне, происходящий от В. uniflora, В. curlipen-
duia 2х и различных других диплоидных видов. От итого полово-
го полиплоидного комплекса возник агамиый комплекс с диапа-
зоном плоидности от 5* до 10х. А вти ауплондиые агамосперы-
иые формы в свою очередь дали начало обширному и почти не-
прерывному ряду вторичных анеуплоидов.
Цнркумбореальныс группы Роа pratensie и Р. palustris—
агамные комплексы, сходные в общих чертах с группой Boute-
loua curtipendula. В группе Роа половые формы — низкие полип-
Гл 33. Лмшшл яоммкг
«I
лопды или, что реже, они диплоидны (х—7). Формы бо-
лее высокой плондиости являются агамоспериными или живо-
родящими Эуплоидиые узлы соединены почти непрерыв-
ным рядом вторичных анеуплоидов. В гл. 31, посвященной
агамоспермии, уже говорилось о том, что у таксономического
вида Р. praiensis число хромосом варьирует от 2п=22 да
2» = 147.
У агамоспсрмного комплекса Potentilta gracilis (к-Т) иа за-
паде Северной Америки также преобладает вторичная акеупло-
идия. Число хромосом у итого комплекса варьирует от 2л» 56
до 2ns 109 (100].
Тип агамоспермных комплексов, характерный для Bouteloua,
Роа и Potentilla. можно рассматривать как одни из вариантов
более обычного типа Robust Moriferus. у которого вторичная
анеуплоидня развевалась в широких масштабах.
Citrus
Род Citrus принадлежит к полтрнбе Cltrmae трибы Citreae под-
семейства Aurantioideae, сен. Rutaceae Подтриба Citrinae—
группа из 13 родов вечнозеленых иля — в редких случаях —
листопадных деревьев и кустарников, приносящих плоды с соч-
ной мякотью. Большинство представителей Citrinae — уроженцы
Юго-Восточной Азии Малайзии и Австралии; некоторые члены
группы встречаются в Африке [818].
Citrus принадлежит к группе из шести родов Citrinae. свя-
занных близким родством; все они приносят плоды типа апель-
синов или лимонов. Помимо самого Citrus в эту группу входят ро-
ды Fortunella, Eremocitrus, Poncirus, Clymenia и Microcilrus.
Представители всех этих родов могут скрещиваться между со-
бой. Они произрастают в Индии и Китае и по пепи островов в
Тихом океане доходят до Австралии (818].
Самый большой и наиболее широко распространенный род в
этой группе — Citrus с 16 вилами и двумя подродами.
В табл. 32.2 представлена в сжатом виде система рола Citrus
по Свинглю [818]
К подроду Papeda относятся дикорастущие виды с мелкими
щитками и несъедобными плодами. Соковые мешочки мякоти
плодов у Papeda содержат капельки едкого масла. Для подрода
Citrus, напротив, характерны крупные душистые цветки и
съедобные плоды, соковые мешочки мякоти которых нс содер-
жат едкого масла С indica и С. tachibana, по-видимому, дико-
растущие виды. Остальные виды подрода Citrus (виды 9—16 в
табл. 32.2) культивируются, и большинство из них, в частности
С. sinensis и С. parodist, дикорастущем состоянии неизвестны.
432
Часть восьмая. гечетичккм системы
Таблица 32.2 Система рода Citrus (по [818])
I. Подрод Ра ped а
I. С. iOuMgemis — нчдаг вапеда Китай
2 С- talipes — кхкл памела. Ивана и Бирма
3. С л|«г<мГА|а мелкоовепговав папедд Фиипсмиы
4. С- ctieMca иелебессмая памела. Сулавеси Филиппины
5. С яыкгор<егй — меламемАсква пааеда Юго-Востсянаа .Ъ«я и Поли*
ияаня
6 С. kyurU —маврикийская памда. Юго-Восточмая Дам* и Ма>
лайаяя
II Подрод Citrus
Г. С indlcus — tnunritacub сихтй апельсин. Восточная ость ГMa-
В. С. (всбД-апд — тдчиблнгхкй аэельсяж. Япония
S. С. medico — цитрон. Восплмаи часть Амг. в настоящее время
широко выплакивается в культуре
10. С. Limon — лимон. Юго-Восточная Азы; широко выращивается
II. С. auranttfoha лайм Восточная Даня я Малайзия; игжроко вы-
ращивается в культуре
IX С auranfium — померанец. Юго-Восточная Азия, широко выраши-
паетсч в культуре
IX С. sinensis — сладкий амльсин Китай я Индокитай; широко вы*
риЕивяскя в культуре
14 С. reticulata — матдарим Юго-Восточная Алия и Филиппины; широ-
ко выращивается в культуре
15. С. Д/*ан4д - поыело. или и_«лдоя Юг о Всктичиая Азия широки
• и -Ill-н: ИП-Н В I у.- Й0М
16 О parodist —грейпфрут Острова Кзрибского моря; шнрсяо вы-
расяаастся а культуре
Вероятная первоначальная родина каждого из вырашимемых
видов указана в табл. 32J2.
Важно понять, что таксономия рода Citrus, и в особенности
его культурных форм, отнюдь не так проста, как это может
показаться нз табл. 32.2. Признание культурных форм отдельны-
ми видами (вилы 9—16) неизбежно делает эту классификацию
типологической. Тем нс менее такая система полезна в качестве
предварительной схемы, и поэтому мы ее здесь приводим, одна-
ко она не дает точной картины характера изменчивости в этой
группе.
Для изменчивости, с которой столкнулись исследователи
культурных древесных форм Citrus с начала XIX в. и которая
озадачила нх, характерно отсутствие четко выраженных видов.
Вместо этого в данной группе можко видеть ряд различных раз
новидиостей, многие нз которых — гибриды, происходящие от
той или иной комбинации родительских форм и соединяющие
именованные виды в один обширный сингамеон. Именованные
виды 9—16 нз табл. 32.2 служат удобными узловыми точками в
этом переплетении форм.
f л. 32. Агамчый кояыелс
433
Ряд имеиоиаиных видов, перечисленных в табл. 32.2, — не
единственная система, которую можно создать для Citrus
В сущности, уже на протяжении полутора веков в равной мере
компетентпые исследователи плодовых деревьев рода Citrus
делятся на крайних «видсюбъеднннтслсн» н крайних «яидодробн-
телем»
Эти две противоположные тенденции иллюстрируются систе-
мами Танаки [822, 823] и Свингля [818] Танака давал видовые
названия каждой морфологической форме и каждому гибрид-
ному типу. В результате в роде Citrus в целом Танака 1823] раз-
личает 157 видов, тогда как Свингль— только 16 [818]. В свою
очередь Свннгль различает только наиболее ярко выраженные
разновндасстн и гибриды и рассматривает нх как ннфравиловые
единицы, относящиеся к различным таксономическим видам
|8!8Д
При более летальном соиоставлеикм альтернативных систем
Свингля и Танаки можно обнаружить, что эти две системы дос
таточно хорошо совпадают в отношении дикорастущих видов
подрода Papeda, Расхождения между системами наиболее вели-
ки и том, что кесагтся культивируемых древесных форм Citrus
Знаменательно что именно в этой части рода таксономическая
путаница достигает максимума
Основное число хромосом у Citrinae. так же как н у других
Aurantiotdae. обычно равно к-9 Почти все виды Citrus и род-
ствеивых родов — дипломы (2п=18). lei ран ленды И трнпло-
иды встречаются как отклоняющиеся типы у ряда культурных
древеслых фирм Citrus [151, 677].
Виды рода Citrus можно скрещивать искусственно, и они
спонтанно гибридизируют во многих комбинациях. Виды Citrus
скрещиваются также с представителями Poncirus, Fortunclla,
Microcitrus и Eremocitrus. Многие межвидовые и даже некото-
рые межродовые гибриды плодовиты прн половых скрещива-
ниях. Другие гибриды Citrus стермльиы при половых скрепит
ПЯХ, во продуцируют семема бесполым путем [677, 818].
В смысле взаимоотношений по плодовитости группа и/где при-
надлежит к модели Ccanothus.
Большинство культивируемых разновидностей Citrus пред-
сгавляет собой факультативно агамоспермные или псевдогам-
ные формы. Наблюдаемый у них тип агамоспермии — это адвен-
тивная эмбрновия с развитием зародышей из клеток ну цел л уса.
После опыления наряду с зародышами, рззвиваюшнмнел из эм-
готы, мигут раззиватъея и нуделлярные зародыши. В результа-
те семя содержит два нли несколько зародышей - явление. из-
вестное под иазпанисм полиэмбрионии; зародыши могут быть
половыми и (или) бесполыми в различных соотношениях. Опы-
ление может приводить к расщеплению средн потомков нли к
434 Часть «ось«м. Сл/цшиштюмимж сюатжсши акпм»
образованию потом нов толью материнского типа в зависимости
от родительских форм (317, 677, 880].
В одной серии испытания от лимона сорта Эврика после опы-
ления было получено 233 растения, из которых 33% были ага-
моспермными и 67% — половыми. При исныганиих, проведен-
ных иа других цитрусовых, процент всех растений, возникших
путем нупеллярпой змбрионии, составил: у апельсинов сорта
Навель Вашингтонский 97%; у апельсинов сорта Валенсия 85%;
у грейпфрута сорта Марш 96%; у американских лаймов 78% и
у мандаринов сорта Дейся 100% (677].
Культурный Poncirus trifoliata также яЫляетея факультатив
во агамоспермным видом. Нуцелляриая эмбрноеия обнаружена
и у выращиваемых в культуре Forttuulla. Наряду с этим при
Рае. 32.7 Сжнгтаикме гибриды тиоловых деревьев Curia, еаминшне а са-
дах на Делисы Востоке (по [313]).
Родственные е*«в уеТВВОМВНЫ на основе морфологических данных я поле-
вых «следования.
Г л. 32. Л/амммй ка*л.«екс
435
испытаниях по потомству одной линии дикорастущего вида Cit-
rus ictiagensis и нескольких культиваров С. grandis до сих пор
не выявлено никаких признаков агамоспермии [677|.
Для рассматринаемых нами проблем весьма важно, что не-
которые искусственные межкмдояые гибриды Citru* ралмножа*
ются исключительно при помощи нуцеллярной эмбрнонни. Так
обстоит дело у гибридов F| от скрещивания С. parodist и С reti-
culata (танжело), а также С. sinensis и Poncirus trifoliate (пит-
раиж) Эти гибриды размножаются в чистите при помощи семян,
и иногда для получения больших насаждений их разводят семе-
нами [М18].
Гибридные древесные формы Citrus обычны я приусадебных
садах и в деревенских ротах по пссму Дальнему Востоку. Сре-
ди них есть самые разнообразные гибриды (рис 32 7). Некото-
рые нз них встречаются лишь спорадически в локально, а дру-
гие широко распространились благодаря культивированию. Эти
последние, повиднмоыу, возникли спонтанно в смешанных на-
саждениях старых разновидностей Citrus, а затем их сохра-
няли, а иногда и разводили, если они обладали желаемыми при-
знаками [818].
На рис. 32.7. созданном по данным Свин гл я [818]. показана
многочисленность спонтанных гибридных комбинаций, обнару-
женных в восточных районах Азии и Тихоокеанском регионе.
Заключения о родительских формах этих гибридов, возникших
в садах, делались в бслыьинстве случаев па основе морфологи-
ческих данных и полевых наб...1..iк» . _ ПССЮТЬКВХ I
пглтнепл.-.сны результатами искусственных скрещиваний, про-
водикшихся п контролируемых условиях Спонтанное возникнове-
ние некоторых из этих гибридов, а также нескольких новых, как.
например. Eremoctlrus glaucaX Citrus sinensis, наблюдалось так-
же в uHipycuMMx насаждениях в Северной Америке [818]
Юзу — гибрид, возникший, вероятно, в результате скрещива-
нии между Citrus retieulata и С. ictiangensis, ои выращивашея
повсеместно и Китае и Японии. Как свидетельствуют китайские
травники. юзу выращивали еще в древнем Китае начиная по
меньшей мере с 1108 г. я. э.. а возможно, и с 237 г. до и. э [818]
Юзу вреде, а ил нет собой высокоагамоснермное растение, до
95% потомхеп которого возникает из семян, развивающихся в
результате нупеллярион змбрионии [677]. Таким образом, ом мо-
жет воспроизводить свою гибридную конституцию бесполым пу-
тем в ряде последовательных поколений.
Грейпфрут (Citrus paradisi) неизвестен в естественном или
полуестествгпгом состоянии на Дальнем Востоке, где находится
центр распрос-ранения рода Citrus, он возник, по-видимому, в
Вест-Индии. Суля по морфологическим признакам, грейпфрут,
возможно, представляет собой гибрид от скрещивания С. gran-
4Эв Часть ьосьжая, ^-мгп,^глмг систем*
dis и С. sinensis [818]. Если это правдоподобное, но недоказан-
ное предположение справедливо, то грейпфрут мог возникнуть
в результате спонтанной гибридизации между помело н сладким
апельсином, которые выращивались на этих островах в ранний
период заселении их европейцами. Как уже отметалось выше,
грейпфрут — высокоагамоспермиая форма н поэтому, очевидно,
воспроизводит свой гибридный генотип, размножаясь семенами.
Источника изменчивости групп Carns многообразны. О гиб-
ридизации между ранее существовавшими видами, полувидами
или разновидностями уже говорилось. Гибриды при полоном
размножении могут давать расщепляющихся потомков
(рис. 31.4). Часто возникают почковые мутации; у апельсинов
Невель Вашингтонский и других форм они привели к появлению
новых разновидностей. Большая часть культивируемых разно-
видностей Citrus дает некоторую определенную долю автотетра-
плоидиых растений [677]
Новые варианты, независимо от их природы и происхожде-
ния, могут затем размножаться в частоте при помощи семян,
образующихся в результате нуцеллярной эибрионии, и тем са-
мым увеличивать общее число агамоспермных микровидов в
группе Citrus. Бессознательный отбор, вэдавна производившийся
человеком. а в более недавнее время — целенаправленный искус-
ственный отбор превратили некоторые из мелких микровндов в
широко распространенные разновидности и агамоспермные ви-
ды. В результате втих процессов дикорастущие вады Citrus с
несъедобными плодами дали начало разнообразным культурным
плодовым цитрусовым деревьям, которые теперь выращивают
во всех областях земного шара с субтропическим и теплым уме-
ренным климатом.
Citrus служит примером агамиых комплексов третьего типа
У этого типа в отличие от типов Rubus Moriferus и Bouteloua
агамоспермные формы размножаются при помощи адвективной
эмбриоини и остаются преимущественно ва диплоидном уровне.
Pteris
Апогаметня широко распространена среди папоротников, она
встречается у Adianlum. Asplentum. Dryopteris. Poiystichum.
Pteris и у других родов. Особенно часто она наблюдается у об-
ширного, распространенного по всему земному шару рода Pteris,
у которого 34% таксономических видов апогамкые [871. 872].
У рода Pteris имеется несколько различных агамных комп-
лексов Некоторые из иих состоят нз смеси апогамных и пато-
вых форм, к другим относятся преимущественно нли исключи-
тельно облигатно агамоспермные папоротники. Наличие или
отсутствие гибридизирующих половых форм коррелирует, как и
Гл 32 Лгам»шй кодл.исс
4ЭТ
следовало ожидать, с величиной и разнообразием а га много
комплекса (871]
Группа Ptcris Quadriaurita — очень большая и сложная груп-
па. содержащая как половые, так и апогамньм* формы. Две по-
ловые формы. Pteris quad rinur-it a .sensu stricto (2л—58) и
Р. multiaurita (2п —58). гибридизируют на Шри Ланке: кроме
того, эти виды или их гибриды гибридизирую? также с третьим
членом этой группы — а пота мп ын Р. confusa (2п—58). Все это
переплетение гибридизация выходит за пределы нейтральной
группы Р. quadriaurita, так что в него вовлекаются члены дру-
гих групп видов, относящихся к роду Pteris (870. 87IJ.
Группа Pltris quadnaurila. распространенная в восточной
Азии, состоит. насколько это известил, целиком из облигатных
амгамкых ферм. Большинство из них — «дипленды> (2л» 58),
а некоторые — триплонди (2л = 87) Они образуют ряд размно
жагошихся а чистоте микровидов, отличающихся один от друго-
го по второстепенным признакам строения спор и молодых
листьев. Эти морфологические вариации сгруппированы в один
обособленный таксономический вид. Простота таксономической
структуры вгамиото комплекса Р. biaurila связана с отсутствием
в этой группе полового размножения и естественной гибридиза-
ции или их редкойыо (8/1]
Pteris служит примером агамных комплексов четвертого ти-
па. довольно широко распространенных среди Polypoaiaceae.
Для этого типа характерно бесполое размножение при помо
ши апогаметмн. обычно облигатной. Апогамные папоротники мо-
гут быть ДИПЛОИДНЫМИ ИЛИ полиплоидными, но число хромосом
у них всегда высокое. В их ягами ые комплексы часто о ходят
как половые, так и апогамные члены, а иногда они состоят иск-
лючительно из апогамвых форм.
Стадии развития
Относительную стадию развития данного агамиого комплекса
можно определить по степени его таксономической сложности,
широте географического распространения я наличию или отсутст-
вию диплоидных половых предковых форм. На основе этих четы-
рех критериев различают четыре главные стадии развития комп-
лекса. юность. раннюю зрелость, позднюю зрелость и старость.
Parthtnium служит примером агаммого комплекса, находя-
щегося на отиоскттльио юной стадии развития Наблюдаемые
у него морфологические вариации можно сгруппировать всего в
два таксономических вша — Р. argcntalum и Р incanum Этот
комплекс встречается только в одной географической провин-
ции. занимающей область от Техаса до Аризоны и заходящей в
<м
Част» еосямал. Сягцааммроеатые систем»»
прялежащне части Мексика. Половые предковые формы этого
комплекса — тетрайлоидвые формы Р argenlatum Р. inca-
пит — существуют а настоящее время, хотя и имеют несколько
ограниченное распространение [689—691].
Агамкый комплекс, содержащий сотни или тысячи агамо-
спермных микровидов, распространенных но обширной геогра-
фической области, находится на стадии развития, которую мож-
но назвать зрелостью. Зрелые агамиые комплексы можно под-
разделить на комплексы, у которых сохранились многие нли
большая часть диплоидных предковых видов, и комплексы, у
которых многие или большинство диплоидных половых предко-
вых видов вымерло. Считаетси, что первые находятся на стадии
ранней зрелости, а вторые —на стадии поздней зрелости.
В предыдущих разделах этой главы мы рассмотрели несколь-
ко примеров агамных комплексов, находящихся иа стадии ран-
ней зрелости. У родов Crepis. Rubus. Taraxacum. Hieracium и
Citrus достаточно хорошо представлены диплоидные половые
виды вместе с очень разнообразным рядом агамоспермиых про-
изводных.
У агамного комплекса, находящегося на стадии поздней зре-
лости, число диплоидов и половых полиплоидов сокращается, и
они исчезают, ио агамосперииая суперструктура достигает пол-
ного развития. Примеры этой стадии развития можно видеть у
Alchemilla, Potentilta н Роа.
Агамиый комплекс подрода A phones рода Alchemilla содер-
жит одни диплоидный половой вид A. microcarра (2п= 16) и
много агамоспермных полиплоидов. У подрода Alchemilla ро-
ла Alchemilla половое размножение встречается очень редко, а
агамоспермные полиплоиды многочисленны (317J. В группах
Роа pratensis и Р. polustris половые диплонды неизвестны; эти
группы состоят главным образом из агамоспермных и живоро-
дящих форм с полиплоидным и вторично аиеуплондиыы числом
хромосом [317]. Точно так же Poteniilla gracilis (х—7) в запад-
ной частя Северной Америки содержит по крайней мере одни
половой полиплоид, Р. pectinisecta (6л), а возможно, и другие;
однако в этой группе не известно ни одного полового дяплоида,
но встречается много агамоспермных полиплоидных форм [101].
Старый агамный комплекс обеднен не только половыми дн-
плоядвмя и половыми полиплоидами, но также н числом агамо-
спермных мнкровндов. В таксономическом аспекте он представ-
ляется нам изолированным агамоспермным видом, не имеющим
известных половых родичей.
Houltuynla cordata (Saurnraceae) в Восточной Авин —един-
ственный вид данного рода. Он является высоким полиплоидом
(2л—94—98), размножающимся при помощи агамоспермии
и не имеющим известных половых родичей. Вполне возможно,
Гл. 32. Аламннй кодыгаг
4Э9
что он представляет собой остатки какого-то агамного комплек-
са. который в прошлом был более многочисленным и разнооб-
разным. а б настоящее время оказался крайне обедненным (36].
Эволюционный потенциал
В число агамиых комплексов входят некоторые из наиболее про-
цветающих в настоящее время групп растений. Комплексы Ru-
bus, Hieraeium, Роа и подобные им роди заселили обширные и
разнообразные континентальные области с неблагоприятными
климатическими условиями; они сумели осуществить wo благо*
дари необычайно широкой изменчивости, воплощевиой во мно*
ГВх десятках видов н сотнях иди тысячах мнкровндов. Следует,
однако, отчетливо представлять себе, что агамные комплексы,
находящиеся на поздних стадиях развития, утратили большую
часть своего прежнего набора видов н ми крови доя. в значитель-
ной степени утратили способность порождать новые вариации,
а следовательно, почти полностью или полностью лишились спо«
собностя успешно справляться с разного рода изменениями, ко
торые могут возникнуть в среде в будущем.
Из этих противоположных аспектов агамиых комплексов вы-
текают противоположные суждения об их эволюционном потен-
циале Дарлингтон и Стеббинс считают, что эволюционный по-
тенциал aiаыпых ШМПМЖМ сильно ограничен» тогда как Гус-
тафсон подчеркивает, сколь высокого эволюционного успеха они
могут достигнуть и достигают в благоприятных случаях.
В «Эволюции генетических систем» (141] Дарлингтон пишет;
«Такая обраэс.ч. мы виляя. что апоинксвс спасает то. что можно сластя
• тся случаях, когда при пслоаом ра*мп<лж»хмн осаживается
утраченной Пэлсэсч рэгпювгпас Олагодаря сапрг.ножхаюже’ его
рексчОмпэцна оОссоечинае; асам последующем поколениям :>сиэиу для
ал&птаимм. Лночгксж оСсспечнаает этом смысле лишь псколенна мв1юсрад-
ственнъх логомкэо. Ом шчрааяет у некоторых видов кое канле остатки оо
ло»с< систем к, что дает им аозуожиэггк отсрочить вымирание и даже при-
noxHi к кра1л.жр«м«н»<-му рэошмгу игных форм Озязхо с утратой -олонга
рекомСичаоиц для аппмвкта так же как н яля перманентного ггбрила, пути
к длительном, существоьа.-.И1э игреьи Лиочмаак дает возможность кягм-
жоть стерильности, нс ври этом он «анолит а *аслежкой сы* тупик».
Итак. по мнению Дарлинг гона, агамоспермия — это «эволю-
ционный тупик», Стеббинс же считает f 7SOJ, что это «эволюцион-
ный оппортунизм, доведенный до предела». Кроме того (Бэбкок
и Стеббинс [36]):
«Комичную судьбу агач-ого кэмг.х-гя половые яреткп которо-а ожиа
лжь ограничена мми о соосч распрсст.-аиепна или вымерля. можно предска-
зать яс тех гор йока преобладает ило-ця. сушктвоы»дие и период его
формирования, ои 6у^«> процветать, но он оимже'сн неспособным справ-
<10 Чат аопяал. Сяецаалияеротммаи гелепчасаал системы
мяться с ,*ки-«ли6о нсьым» а»ме»е»иачи трехм<и поттоиу с течением ере*
ненн будет •« белен» II бздьше соьраьхатьеа. печ к пнхе конное не к-
*етиет>
Но, как стирает Густафсон {317], мнение о том, что агамо-
спермия кладет конец изменчивости, справедливо лишь отчасти.
А если агамный комплекс обречеи на истощение изменчивости
и вымирание, то «не типично ли это для всех видов и родов?
Подобно отдельным индивидуумам, виды к роды рождаются,
достигают расцвета и умирают» [317].
Рассмотрим эта противоположные точки зрении, связав нх с
основной биологической функцией полового размножения. Пол-
ито генетическая система, обеспечивающая рекомбинацию генов,
которая в свою очередь служит главным источником наследст-
венной изменчивости в популяции (см. гл. 1). Полный отказ от
Полового процесса и переход к облигатному апомиксису лишает
данную популяционную систему ее главного источника новых
вариаций. А промежуточные формы бесполого размножения,
как при факультативном апомиксисе, ограничивают возникнове-
ние изменчивости до некоторого промежуточного объема.
В сущности, механизм полового размножения представляет
собой главны* источник новых вариаций как в группе, размно-
жающейся исключительно половым путем, так и в агамиом
комплексе Агамный комплекс, состоящий, подобно Rubus Morl-
ferus. из факультативных агамоспермных форм и половых дн-
плоидов, может обладать высоким уровнем изменчивости к
большим эволюционным потенциалом. Однако не вес агамные
комплексы обладают этими чертами. У такого изолированного
комплекса облигатно апогампых папоротников, как Pteris Ыаи-
rtta, наблюдается относительно стереотипная картина изменчи-
вости [871]. У комплекса Crepis occidentalis. который в областях,
занимаемых половыми популяциями, и вблизи от этих областей
содержит, помимо факультативно агамоспермных форы н поло-
вых диплоидов, облигатных агамоспермов, изменчивость очень
велика; однако в тех областях, где обитают исключительно об-
лигатные агамоспермы, она сводится к одному или нескольким
типам [36].
Судя но данным фитогеографии, некоторые агаыоспермные
типы остаются неизменными иа протяжении длительных перио-
дов временя. Taraxacum retchenbachU и Т. door еще — два почти
идентичных агамоспермных мнкровнда, встречающихся как ре-
ликты ледникового времени в Альпах и Норвегии соответствен-
но. Со времени последнего межледниковья они днвергпровали
очень незначительно. Древние агаиоспермиые мнкровиды обна-
ружены также у Hieracium и Alchemilta Возраст некоторых
агамоспермных форм, судя по данным фитогеографии, должен
достигать по крайней мере 7S или 100 тыс. лет [37].
Гл. 92. АламтлЛ калллекл
Ш
Как уже говорилось, многие.агамоспермные растения явля-
ются полиплоидами. В такит случаях к ограниченности реком-
бинаций, обусловленной агамоспермией как таковой, следует
добавить ограничения, налагаемые поляплоилмостью.
Сохранение данной группой растений способности к полово-
му размножению не гарантирует ей длительного эволюционного
успеха. Как верно отмечает Густафсон, группы видов, размно-
жающиеся половым путем, приходят в упадок и гибнут подобно
апомиктическим группам, тогда как частично бесполые группы
могут успешно развиваться в течение долгого времени. Кроме
того, полная утрата данной группой способности к шмовому
размножению ое обязательно означает, что она погибнет в бли-
жайшем будущем. Между тем переход к облигатному апомикси-
су. опешило. влечет за собой в конечном счете вымирание.
Приведем в заключение точку зрения Стеббинса [780] по
этому вопросу:
«Нет нх«ак1гх лаенмз о том. что апочкктм когда-либо были способны
создать в процессе мкмажви копий рем ала хотя бы г.терод, В згой смысле
вес аэаекме иоипзезеи — ьакрмтые стсетекы н зоолюллостеие «тупнхз*—
И если половые вялы могут о перки своего суихеетвозаиаз хавать зачало
солеетемно новым тлеем при гхпеопха мутасисииого процесса и рекомбава-
цха гелоз. то агвнмнм кои a.I asа и сужлаяо соадаиать только мам* варка
паи па старую тему»
29-45
33 Вегетативное размножение
гибридов
Среди многолетних растений широко распространены разнооб-
разные способы вегетативного размножения. Многим или. воз-
можно, большинству групп многолетних растений, способных к
естественной гибридизации, присущ тот или иной способ веге-
тативного размножен иа.
Вегетативное размножение — свойство, выгодное для гибри-
дизирующей группы растений. Оно обеспечивав^ сохранение н
размножение гибридного генотипа, обладающего адаптивной
ценностью. Данный гибрид может быть плодовитым, и в этом
случае его пенная в адаптивном отношении комбинация генов
будет утрачена его половыми потомками, или же он может быть
стерильным, и в таком случае вегетативное размножение —
главная надежда на выживание.
Гибрид, размножающийся вегетативным путем, если ои хо-
рошо адаптирован, может увеличивать свою численность и рас-
ширять занимаемую нм область, проходя через последователь-
ные стадии гибридного клона и клонального иикровнда. Ника-
ких объективных различий между этими стадиями не сушест
вует, они различаются только по относительным размерам попу-
ляций. В згой главе будут приведены примеры каждой нз этих
стадий.
В тех случаях, когда в разных линиях дайной группы расте-
ний независимо проходит образование гибридов, так что от трех
нли большего числа исходных родительских видов возникают
два или большее число вегетативно размножающихся гибрид-
ных видов, эта группа развивается как клональный комплекс.
Гибридные комплексы такого типа менее известны, чем агамные
и полиплоидные комплексы. Тем ие менее а этой главе в даль-
нейшем можно будет привести несколько примеров таких комп-
лексов.
Гибридные клоны
Многие культурные разновидности цветковых растений про-
изводят стерильные семена, и поэтому нх размножают вегета-
тивным путем. Хорошо известными примерами служат десерт-
ный банан, картофель и батат —средн плодов и овощей, а сре-
ди декоративных растевнй — некоторые розы и тигровые лилии.
Так. большая часть выращиваемых съедобных бананов —бессе-
мянные трнплоидиые (2п = 3иг=33) разновидности Musa acumi-
nata, а также гибряды М acuminata я М. balbislana, которые
разводят корневищами и луковицами [727, 728].
Гл. Si. Бесе гитиамюг /кдодогсгде гмбдкйов
443
Гибриды, размножающиеся вегетативным способом, обнару-
жены во многих группах дикорастущих растений, хотя у них это
явление петргчастся несколько реже, чем у культурных расте-
ний.
Elytnus trilictndcs и £ rondentaius (Grammcne) — хорошо
дифференцированные виды дикорастущего многолетнего злака»
распространенно!о в Калифорнии. Искусственный гибрид меж*
ту ттиип видами жизнеспособен, по высокое терилен. В некото-
рых местах встречаются также естественные гибриды между ни-
ми; гибриды эти также высокэстернльии, но благодаря образо-
ванию корневищ они смогли распространиться локально. Неко-
торые нз таких клонов морфологически сходны с искусственно
полученным« Г, и представляют собой естественные Г|, однако
Другие, судя по их морфологическим признакам, вищепеицы
из более поздних поколений, возникших нз тех редких семян,
которые образуются у стерильных индивидуумов Fi |171, 786]
Hol с us mollis в Польше — другое высокостсрнльное травяни-
стое растение (возможно, гибрид), размножающееся корневи-
щами [877]. Естественные гибриды Iris fulvaXf, hexagona gigan-
tlcaeculeu в дельте Миссисипи размножаются вегетативно прн
помощи корневищ н образуют локальные клоны [656. 561]
Интересный пример гибридных клонов, часть которых, веро-
ятно, вознцх.щ очень давно, обнаружен у Quercus в Юте и
Аризоне Гибриды между Q gambehi н Q. turbine На встречают-
ся в местах, разбросанных по северной части Аризоны, южной
части Юты и северной ее части, вблизи Соленого озера. Эти рас*
тения не образуют желудей. Они образуют большие гибридные
колонии шириной от 25 до 60 м. Кроме того, эти колонии мор-
фологически однородны, что даст основание считать нх клопа*
ми, образованными прн помети подземных побегов. Такое пред-
ставление подтверждается результатами эксперимента с рас
аростраисянсм радиоактивного индикатора я одной из эп«х ко-
лоний. Клок / эти достаточно обширны, кроме того, они золе :к-
лн давно, по крайней мерс некоторые из них [125]
Quervus gam belli — дерево, распространенное в южной часш
Скалистых гор. тогда как Q turbinclkt — кустарник, растущий d
юго-западных районах США. Гибридные к .топы на севере Ари-
зоны и на юге Юты находятся в пределах тоны симпатричсскою
перекрывай ин ареалов этих двух родительских видов, однако
местность па сеттере Юты, где они также встречаются, лежит
бадсе чем па 400 км севернее современной северной границы
Q f иг b India- следовательно, в прошлом юго-западный вид
Q turb India заходил дальше на север. По всей вероятности.
Q. lurbindia доходил до северных районов Юты в период между
7500 и 4000 гт. до и. э., когда климат был теплее и <\ше, чех
впоследствии Если эта хронология верна, то возраст северных
29*
444 Часть сосьлал. Сплциалилирсюачкыс мпетнчгсяш стйстглм
гибридных клопов равен 4000 лет, а может быть, и больше
[12Л
Осины группы Populus tremuloides размножаются в природе
вегетативно —корневыми побегами, отходящими от корневой
шейки, и образуют хорошо известные клоны; эту свою способ-
ность они передали и естественным гибридам. Гибрид между
Populus tremuloides и Р. grandidenlata ( = P.Xbarnesii) зани-
мает обширную территорию от Миннесоты до Новой Англии и
часто образует большие клоны (45. 866]. Гибридные клоны с та-
кими же признаками встречаются также в северной части Айда-
хо. Р. grandidenlata в настоящее время в данной области не
встречается, ио этот вид или его близкий родич был, вероятно,
распространен здесь прежде [44].
Клональные мнкровнды
Гибридизация между Iris f ultra и 7. hexagona giganlicaerulea'
в районе дельты Миссисипи дает не только гибридов Fi, но так-
же ряд выщепеяиев из последующих поколений к продукты воз-
вратных скрещиваний. Все они часто образуют идонм. Некото-
рые из иих достигли уровня клональных иикровкдов, занимаю-
щих различные микроместообнтанвя в районе дельты (15, 861|
Вариации среди популяций Iris настолько резко выделяются,
что многие из иих были описаны прежде в качестве новых так-
сономических аилов. Смолл и Александер [738] различали в
группе Iris lulva—hexagona в этой области 84 вида. Как Пока-
зал Вноска [861]. все они. за исключением трех представляют
собой различные продукты гибридизации между 7. fulva и 7. he-
хagona gigantUaerulea. Число получивших названия таксономи-
ческих видов — хороший показатель разнообразия клональных
микровидов в этой группе.
Межклональиая изменчивость хорошо выражена также в
группе Populus tremuloides—grandidenlata. В этом случае она
привела к выделению иссколышх новых именованных аилов
(например. Р. aurea), а также многочисленных разновидностей
и форм Р. tremuloides и Р. grandidenlata [45].
Solarium chacoense подвид chacoense— дикорастущий клуб-
необразующий представитель секции Tuberanutn рода Solanum.
распространенный на юге Южной Америки. Эта группа очень
изменчива, и многие ее члены были описаны в литературе в ран-
ге видов, подвидов иля форм с соответствующими латинскими
названиями. Хоне в Яертииг [354] свели 32 таких названия в
1 Iris fulva я 1. jriganlizaeralea обыхпо рассматриваются км два само-
стиятмьимх вил» — Ппим р.п?.
Гл. и. BttrtUJMWt разллохвчил rutfuioe
«45
синонимику подвида 3. cbacoense chacomse, в этой группе они
различают также 13 микровидов.
Solatium chacounsa часто гибридизирует с S. microdonlum и
реже-с некоторыми другими пилами В результате згой гиб
ридизацин у S. chaooensc создается высокий уровень изменчиво-
сти, причем некоторые вариации становятся клональными мик-
ровидами [354].
Известно, что кустарниковые кактусы Opuntia fulgida к
О. spinosior гибридизируют в двух местностях в Аризоне с об-
разованием высоко, но не полностью стерильных гибридов.
У ьтих растений имеет место вегетативное размножение при по-
мощи отчленяющихся частей стебля, которые укореняются и
дают побеги. В начальной стадии гибридизации образуются гиб-
ридные клопы. Затем в результате ограниченного подового раз-
множения гибридов в более поздних поколениях появляются
различные выщепеисы, которые также размножаются вегета-
тивно и образуют «тональные микровиды. Индивидуумы, состав-
ляющие зги мнкроэады, морфологически очень однородны [302].
Млкроаидм, как правило, характеризуются небольшими раз-
мерами и локальным распространением, но один обширный мнк-
ровид гибридного происхождения (мякровнд W) занимает уча-
сток пустыни длиной 32 н шириной около 42 км. Гибриды от
скрещивания между Opuntia fulgida и О. spinosior образуют от-
личительный элемент флоры Аризоны; ПОЗТОМу они были выде-
лены в таксономический вид О. belvintnsis [302].
В Техасе Opuntia lindhetmeri гибридизирует с О. phaeacaniha
та/or и О. eduardsii. Гибриды Fi и вышеоенцы из последующих
поколений размножаются вегетативно при помощи отчленяю-
щихся частей стебля, как это свойственно опунциям вообще.
Гибридные клоны весьма обычны. Для изменчивости в этой
группе характерно также наличие клональных ыккроэндов со
своими отличительными комбинациями признаков и микрогеог-
рафнческими областями распространения. У О. lindheimeri име-
ются десятки или сотни таких мнкроввдов. Как и в других рас-
смотренных выше примерах, систематики прежних лет описыва-
ли многие из этих микровидов как новые таксономические виды
[304, 306).
Клональные комплексы, размножающиеся
ответвлениями стебля
Очень немногие клональные комплексы проанализированы фи-
логенетически, В главе по автополнплоидии был описан кло-
нальный комплекс в группе Solarium tuberosum. Это. однако, нс
просто клональный комплекс, даиная группа характеризуется
также сочетанием некоторой степени межрасовой автополнпло-
44S Часп ахкьлал. Смциамиироейнкъе гспетичегкзи системы
ндяи и вегетативного размножения. Кроме того, «сходные роди-
тельские формы связаны очень близким родством.
Клояапьный комплекс существует также у кустарниковых
кактусов подрода Cylindropuntia рода Opuniia. распространен-
ных иа юго-западе США. У этих растений происходит половое
размножение семенами и вегетативное размножение отменяю-
щимися частями стебля. Соотношение полового и вегетативного
размножения различно у разных видов. Половое размножение
преобладает у О. spinosior, О. versicolor, О. acanthocarpa и
О. echinocarpa, я вегетативное размножение — у О. futgida и
О. bigelovu ([302. 303], а также неопубликованные данные).
Пинкава и его сотрудники [641—643] показали, что все эти виды
являются диплом да мн с числом хромосом 2п— 22.
В пустынях Аризоны и Калифорнии >ти виды гибридизируют
в природе в различных комбинациях я образующиеся гибриды
размножаются вегетативным путем. Структура возникающего
прн этом клонального комплекса изображена на рнс. 33.1. Гиб-
рид Opuntia spinosior и О. futgida дал начало О helvinensls со
стерильными семенами и вегетативным размножением, о чем
уже говорилось в предыдущем разделе. Однако в результате
гибридизации между О. spinosior и О. versicolor образуются
скопления гибридов, размножающихся половым путем [302. 303]
Opuniia bigelovu — очень близки* родич О. fulgida, — воз-
можно, гибридизирует с О. acanthocarpa я О echinocarpa на юге
Калифорнии. Бенсон [58] сообщает о наличии в «той области
двух различных таксонов, яромежуточных по морфологическим
признакам между О. bigtlovii и либо О. acanthocarpa, либо
О. echinocarpa; эти таксоны продуцируют стерильные семена и
размножаются вегетативно. Вполне возможно, что они представ-
ляют собой гибриды, происходящие от родительских форм, ука-
занных на рис. 33.1- Такие предполагаемые гибридные формы
требуют, по мнению Бенсона [58], дальнейшего изучения.
Группа PotentiUa anserine (х-7) содержит таксоны тетра-
илоядного и гексаплоидного уровней. Тетраплоидные популяция
этой группы размножаются половым путей и распадаются по
крайней мере на два интерстернльиых вида. Но у гексаплокд-
кых форм мейоз нарушен, к они’образуют стерильную пыльцу и
стерильные семена. Эти гексапловды усиленно размножаются
ползучими побегами. При помощи такого способа размножения
они образовали ряд клональных микровидов, происходящих от
различных тетраплоидных популяций к соответственно родствен-
ных последним [699).
Группа Stellarta longipes (Caryophyllaceae). к которой отно-
сится несколько именованных видов, широко распространена по
всему Северному полушарию. Собственно Stellaria longipes
(2n—52) образует фертильные семевя. Но S. crassipes— пред-
447
Г A. A3 BWTQTUtHQt poiMMOXTMUf
Рнс 33.1. Фглогсаяя ьасмалыюго помтитегеа у кугт»рнмжовы« яякгумя
ОршлЛ* (подрод CflinJrofuntla).
ставитсль этой группы, произрастающий в Арктике и содержа-
щий большее число хромосом (2 л —104), —образует высоко*
стерильные семена и размножается при помощи ползучих под*
земных побегов. Высокая плотность представителей вяла
S. cfassipts позволяет предполагать, что ом произошел от како-
го-то кизкопломдвого вида дайной группы и, возможно, пред-
ставляет собой амфяплонд. Современное широкое его распрост-
ранение в арктической области наводит на мысль, что этот вид
возник довольно давни и распространился пря помощи вегета-
тивного размножения (317, 388].
<« Уест» nxi-лал- Специимяироеипплн; ммгичкпм аснмы
Клональные комплексы, размножающиеся
при помощи ложного живорождения
Большинство видов Allium размножаются половым путем и
производят фертильные семена. Однако у этого большого рода
широко распространено живорождение или. вернее, ложное жи-
ворождение [639]. Разные виды Allium размножаются при по-
мощи луковичек, образующихся в соцветиях вместо семян. Сте-
пень ложного живорождения варьирует в разных случаях от
частичной до полной, и соответственно варьирует редукция об-
разования семян, вплоть до полного их отсутствия у таких ви-
дов. как A. sahvum и A. paradoxum. Мы рассмотрим здесь вкрат-
це ситуацию в двух группах видов — в группах A. paniculatum
и A. canadense.
Группа Allium paniculatum (*-8) в Евразии содержит виды
A. paniculatum, A. pulchellum, A flavum, A oleraceum и A. сап-
natum. Из них A. paniculatum, A. pulchcUum н Л. flavum— ди-
плоидные половые виды (2л=16). Они распространены на юге
Европы [466]. Ложноживородяшие виды — A. carlnalum и
A oleraceum — встречаются в северной части Европы, и их
плоидиость варьирует от 2х до 5х Ложножнвородяший вид
A. carlnatum. произрастающий в Альпах, диплоиден, а произ-
растающий иа севере Европы — тркплоидеи. Он морфологиче-
ски близок к половым видам A. pule helium и A. flavum. Ложно-
живородящи* вид A. oleraceum с тетраплоидным или пеитапло-
идиым числом хромосом (2п-32, 40) морфологически близок к
половому виду A. paniculatum (151, 317, 466].
В восточных областях Северной Америки распространен дру-
гой клональный комплекс — группа Allium canadenee (х—7),—
содержащий несколько половых форм (2х и 4х) и несколько
ложиоживородящих производных (Зх и 4х). Родственны* комп-
лекс A. geueri (х=7). распространенный в западных областях
Северной Америки, также содержит несколько половых дипло-
идов и половых тетраплоидов, а кроме того, — ряд живородящих
полиплоидных производных (4х, 5х, 6х) |632].
Группа ЗахЦгава stellarls —другой большой комплекс, рас-
пространенный в Северном полушарии. S. stellarts sensu stricto
(2п=28) и S. ferruginea— половые виды, произрастающие в эо-
не с умеренно холодным климатом. S. foliolosa (2л—56, »64)—
ложноживородящий вид. Это бесполое полиплоидное произвол-
ное имеет циркумполярный ареал, который расположен север-
нее, чем ареалы его половых родичей, я заходит в арктическую
область [317].
Густафсон [317] приводит несколько примеров ложиоживоро-
дящих видов, которые обладают высокой плотностью и рас-
пространены в северных или арктических областях, и близкие по-
Гл 33 Вамтатьлмо? ралм*огюши гибриды
4*9
лояыс родичи которых нгмзвсстпы. К числу таких таксоном пчг
ски изолированных ложноживородящих видов принадлежат
Ро/у^олит viviparum (2ч = 83—ПО). Cardamine bulbijera
(2л «96) и Saxifraga cornua (2л «66)
Ложное живорождение обнаружено у нескольких родов ала*
ков, в частности. у Agroftit, Deschampsiu. Eleucine, festuca и
Роа В одной из предыдущих глав уже говорилось о том. что
агамоспермия часто встречается также у Огапмпсас. У Роа в
некоторых грувлах видов одновременно наблюдается па i вно-
ся ср ми я. и ложное живорождение.
Циркумбореальная группа Роа alpina (х-7) содержит поло-
вые. агамоспермвые н ложножнвородяшпе формы. Формы всех
этих трех типов существуют у самого таксономического вида
Роа alpina, у которого имею тем днвлондные половые популяции
(2л—Н), полиплоиды вплоть до гексаiMciUHoro уровня и раз-
личные происходящие от них ложноживородяшне и эгамосиерм-
лыс популяции. Р. bulbosa, принадлежащий к этой группе, пред-
ставлен на юге Европы половыми формами с низкой плонд
костью. а на севере Европы — производными ложможивородяшн*
мн фоэмэми с тетрадлоидным или гсксаплоидным числом хро-
мосом [317].
Группа Deschamps ia caespitosa (х-7) представляет собой еше
одни обширный клональный комплекс, распространенный ии все-
му Северному полушарию и доходящий в некоторых местах до
Южного полушария Половые тегрэплондныс виды или полуви-
ды - £> CGCspitosa. D. arciica, D. bottnica и другие — широко
распространены я связаны между собой гибридизацией. Места-
ми среди половых популяций встречаются лижт.оживородягцие.
Близкий к ним вид D alpina. обитающий в арктической зоне,
содержит несколько высоких полиплоидов н вторичных апеуп-
лоилов (2н — 39. 41. 49 56). н все его популяции почти исключи-
тельно ложноживородяшис (317]
Эволюционный потенциал
Комбинация полового к вегетативного размножения выгодна
естественным гибридам и их потомкам. Ока создает bow «яс-
ность дпн возникновения целого ряда новых типов н сохранении
тех из них. которые обладают ценными адаптивными признака-
ми. Кпмбкпаикя этих двух способов размножении характерна
в числе прочих для гибридов tris fulvaxl hexagona gigant ira ff-
rutea Popufux IremuMdcsxP grandidentala. Solanum chacoen-
scxS microdontum и Opunlia fulgida xO. spinosinr
Гибриды или потемки гибридов, обладающие адаптивной
ценностью, развиваясь, превращаются в тональные мнкрсвиды
и распространяются по нсеыу местообитанию, к которому имя
«80 V«»» «кыш*. mnmectur летмм.
адаптированы. Ряд стадий — от мелких локальных до больших
широко распространенных иикровндов — обнаружен в му*
группах видов опунция, изученных в этом плане [302. 305].
Если подобный процесс повторяется в разливных линиях од-
ной к той же группы видов, то возникает клональный комплекс,
и нам известно несколько примеров таких комплексов. Но это
не «чистые» клональные комплексы. В большинстве случаев ве-
гетативное размножение сочетается у них с другими генетиче-
скими системами; с межрасовой автополколондней - в группе
Sola num tuberosum, с половым размножением — в группе Opun-
iia fulgida — spinosior, с амфиплондией —в группе Potentilla
anserine и с агамоспермией — в группе Роа atplna.
У агамоспермных групп растений и у групп с вегетативным
размножением существует некоторое равновесие между половым
и бесполым размножением Уместно сравнить эти две системы.
Преимущество вегетативного размножения состоит в том. что
оно обычно легко доступно гибридным растениям во многих
группах. В отличие от итого агамоспермия — высокоспецнализи-
рованиая генетическая система, которая обычно должна возник-
нуть de novo у гибридов или гибридных производных.
Вместе с тем семена в общем обеспечивают более эффектив-
ное распространение, чем вегетативные органы. Бесполые фор-
мы данного агамного комплекса используют для распростране-
ния адаптация, присущие семенам н плоидам. и благодаря этому
их микровидам удается ио многих случаях заселить области
средних или больших размеров. Потенциальные возможности
сильно варьируют; так, воздушные луковички, или детки, могут
обеспечить распространение почти на такие же далекие расстоя-
ния. как желуди, а ползучие побеги пригодны для расселения
лишь на небольшие расстояния. Таким образом, следует ожи-
дать. что некоторые клональные мнкровнды, обладающие эффек-
тивкыми способами расселения, достигнут широкого распростра-
нения; однако существуют и многие другие клональные комп-
лексы, состоящие нэ растений, лишенных возможности распро-
страняться иа далекие расстояния при помощи вегетативных
органов расселения. У таких комплексов варианты, возникшие
в результате гибридизации, могут никогда не выйти за преде-
лы той локальной местности, в которой они возникли.
Другим фактором, способным ограничить развитие многих
клональных комплексов, служит старения клонов. Старение кло-
пов обсуждалось с разных точек зрения Молишем (582]. Брин-
ком [70] и Камероном к Фростом [80]. Это явление все еще оста-
ется ие вполне понятным. Несомненно, многие виды растений
могут на протяжении сотен и тысяч лет размножаться вегета-
тивно без заметного понижения жизнеспособности. Несколько
примеров этого было приведено в гл. I, и к инм можно доба-
f'.i 33 Вгггтсгизчое -ибрыаов 4!»
вить много других. Но т>к бывает не всегда [582]. Некоторые
группы растений (например, Citrus, картофель, сахарный трест*
ник) после длительного вегетативного размножения проявляют
симптомы старения или дегенерации [80. 562]. Наблюдаемое
старение клоков, возможно, обусловлено главным образом на-
коплением в цитоплазме клеток растений патогенных вирусов
[80, 562. 625] Возможно также, что одпнм из фактпроа такого
старения служат регулярные направленные изменения генов в
соматических клетках — некая форма парамутировапкя.
У Citrus агамоспермные растения, развившиеся из семян,
бывают более мощными, чем растения, выращенные кз отвод-
ков. Образовавшиеся бесполым путем семена, по-видимому. сво-
бодны от вирусов, которыми заражены ткани стебля того же
самого растения [90. 625]. Это наблюдение представляет боль-
шой интерес для вопроса, который мы сейчас обсуждаем. Оно
позволяет предполагать, что агамные комплексы, избегая старе-
ния Молов благодаря размножению семенами, часто способны
развеваться дольше во времени и шире в пространстве, чем
клональные комплексы.
Клональные комплексы имеют, в общем, более простую так-
сономическую структуру, чем агамные комплексы. Они обычно
содержат меньше микрокодов Число микрооидод ь данном гиб-
ридном комплексе, находящемся в стадии зрелости, может слу-
жить мерой его эволюционного потенциала. Более низкий эво
л юн ионный потенциал клональных комплексов по сравнению с
эволюционным потенциалом агамных комплексов может быть
частично обусловлен такими факторами, как старение хлопов,
а нередко также пониженной способностью к распространению
ортанов веютагнвкиго размножения
В предшествующих плавах были показано, что эволюционный
потен спал агамных комплексов, хотя он, несомненно, велик,
ограничен по сравнению с эволюционным потенциалом групп,
размножающихся патовым путем. Теперь мы видим, что эаолю-
цпоиныЯ потенциал клональных комплексов еще более ограни-
чен.
34 Естественные триплоиды
В очень многих группах растений встречаются спонтанные три-
плоидные индивидуумы и спонтанные трнплоилпые клоны, при-
чем как средн дикорастущих (например. Зх-формы Populus tre-
mula н Clautonia vlrglnica), так и среди культурных видов
(Зх-форыы Руг us communis и Juniperus virginiana). Как и сле-
довало ожидать, спонтанные триплоиды редки среди дикорас-
тущих, но нс Слишком редки среди культурных растений. Неко-
торые примеры приведены в табл. 34.1.
Нас здесь интересуют два вопроса относительно спонтанных
трнплондных растений: хак они возникают и км они эосяроиз
водятся? В этой главе мы попытаемся рассмотреть эти вопросы
на основе данных, имеющихся как по дикорастущим, так и по
культурным видам.
В природе пентаплокдные н гепта плоили не растения встреча-
ются редко. Естественные пентаплондные формы обнаружены у
Lemna minor и L. gibba (2n=50) (851] и у Hotcus mollis
(2л—35) [877], а также а группах Rosa canina н Solanum tube-
rosum, рассматривавшихся выше. Естественные гептаплоиды об-
наружены у Lenina gibba (2n=7O) [851] и у гибридов Opuniia
flcus-indica (536).
Проблемы образования и сохранения этих форм с более вы-
соким нечетным уровнем плоидности мало чем отличаются от
тех же проблем у трнплондов, поэтому эти формы рассматрива-
ются здесь вместе.
Проблема сохранения решается у трнплондов и у других
иечетяых полиплоидов несколькими различными способами
В этом участвуют несколько генетических систем, каждая из
которых рассматривалась в одной из глав части Сельмой.
Возникновение
Существует два основных способа возникновения трнплондных
растений. Один из них — гибридизация между тстраплоидом и
днплондом. Другой — отсутствие редукции у одной из диплоид-
ных родительских форм: при этом обычно происходит образо-
вание нередуцироваяиых яйцеклеток и последующее их слияние
с нормальными редуцированными пыльцевыми зернами.
Как показали эксперименты, проведенные с 121 диплоидной
линией Zea mays, в 18 из этих линий иередуцированные (2л)
яйцеклетки возникают спонтанно с определенной низкой часто-
той. Средняя частота иередуцировакных яйцеклеток в mix
18 линиях составляла 59 на 10000 яйцеклеток (0.59К>). а общая
Таблица 34.1. Некоторые неагамоспермные виды11, у которых
имеются сповтаниые триплоидные индивидуумы и триплоидные
клоны («о данным Дарлингтона и Уайли [151], Готтшока [258]
и другим источникам)
СямяЬстм вял Qaeau х|м»!еви Т^МЛХиистсЛ фир Сосгмим
1 Хвойяые
CupresMc-M* ‘/smiperus cirgtmana д-ll. 2n-33 Кгльт.
11. Даудолыше Thula plicata txcclsa »-H. Sa-33 »
Rarwnculicrj# DrljAtnium motrhtimii x-B, 2л-24 *
Crucifer *c A'e«rw/ivxn oj'idmdt x-K 2Я-24
Cardamtm ir.ssieJa x-6. 2л-24 Дмкор
Portuh<dce«e CtenroflMJ Virginia ж-бу 2л-2« >
Глсчсгйе Opuntlu Nuilarts {release. ж-II ?л-35 *
Ргияил iarrtitaia ж-& 2п-24 Культ.
Malin sgfoextris и дру ГИС ЗИЛЫ ж-IT, SM-S1 »
Руг** ГОЛПНШЛ& х-П. 2л-51
SihtMMe Popalus inmda 1 = 19, 2п-5? Дико?
ЕрвепАавмд Lra сиро gon lunipermus х=4. 2л-12 х — 17. 2я-51 Дл«.ср.
Campost lac Hritnih^s mtsltiftoriM Культ.
Inula х-6. 2Л-21 *
PrtoieJacMe Primula siehatdli х- 12. 2л = 36 >
Solanacea* Solanum /гютдгхиДй и другие аплы х-12. 2п-3б Дибор и аулы.
SrrupbuUrii e**e III Однодольные Limtneila sabulalu X X aguatica г-10. 2и-ЗО Дмдор.
Мим)п*яс Musa attuniMaia х-11. 2п-33 Культ.
Савпдсен Саппа indica х-g, 2п-27 >
Ышяй» Hoaanihus oricatalis х-8, 2л—24
Urgmea tndica Atuam сера i loiparum х«3. 2л» 15 х-А. 2д-24 Дикс^ Культ. Амкор.
Album ruPtilum A ta- 'itmtetm х-3, 2д-24
f.ilium •artZi’&Jju.w х-12. 2л-35 Культ «дикпр
Гм.on £«д»’лалй и дру- гие ВЯХЫ х-12. 2я-36 Культ.
Раги icriidllaia ж-5, 2п- |5 Дн.чор.
Amaryll.daceie Lucorij radiate Narcisiai tart'fa ftemtfocaltis fulva и H dl^lifha SSS 1 1 1 МД i । 7 МММ Культ.
>
lri4Mv»e Crocus salivas другие MJU г-д, 2Л-24 Кужт.
Л/<»емг ^^brids у-11. 2л-ЛЗ в
Gloa.'jtm far/nos us и • д-15. 2п-« »
Лпсвае Агг,<к* eW<z.~>tu ж-IS, 2а —36
CtA’xasia antiguorum х-12, 2л- Ж
LemnacBM I tmna чияс< x-lfl, 2ч-30 Дммор
Gramineae Phlcum nodosum л-7. 2п—21 »
’J у мгех пкретжхемсп шдм <»• •сжлюче* кс* imom: «мгютсх днп-
ЛСМДМЫ» V бониииггц иа Нх «note, 6т< И; тгтрвплойка ЫЯ
фо;мм а у агвкотсрыь яедтн или »г>1М май МХЯ (UCOKM 1О*Я(1»аД1>
4М Часть ласьмая. Спи^иализыроваммыл гсмтклслии системы
средняя частота зля всех 121 линий составила 8,78 на 10 000
яйцеклеток, т. е. 0,088% [3].
Саидфер (706] получил оценки частоты спонтанных триплонд-
ных проростков среди потомков диплоидных растений дли двух
культиваров ячменя; ока составила 0,04% для одного сорта и
2.6% —для другого.
Частоты, подобные обнаруженным у кукурузы н ячменя, дос-
таточны для того, чтобы обеспечить спонтанное возникновение
триплоидных растений во многих больших природных популя-
циях.
Естественным трнпловдом, возникшим, вероятно, таким об-
разом. а именно в результате одностороннего отсутствия редук
цни. является форма gigas вида Populus iremula в Швеции [575,
600]. Если говорить о культурных растениях, то некоторые три-
плоидиые формы Musa acuminata также представляют собой
автотриплонлы. непосредственно происходящие от днплоидов
[726, 727].
Во многих других случаях как среди дикорастущих, так и
среди культурных видов трнплоидные формы возникни, по вили-
моиу, в результате гибридизации между диплоидами и тетра-
плонлами. Большинство триплоидных растений встречается у
видов или у групп видов, содержащих и диплоидные, и тетра-
плоидиые формы (см. табл. 34.1). Во многих реальных случаях,
как мы увидим далее, на основе морфологических, цитологиче-
ских и другого рода данных установлено, что триплоиды пред-
ставляют собой гибриды.
Сохранение
При половом размножении триплоиды стерильны и нестабиль-
ны, Как же в таком случае они сохраняются?
Первое условие, необходимое триплоиду для достижения ус-
пеха, — наличие какого-либо способа вегетативного размноже
имя. Все растения, перечисленные в табл. 34.1, — многолетники,
в большинстве своем многолетние травы, но среди них есть и
несколько древесных форм; многие из этих растений усиленно
размножаются вегетативным способом. Надежные способы ве-
гетативного размножения лежат в основе комбинированного
Хлоиально-полиплондиого комплекса в группе Solatium tubero-
sum. которая содержит несколько триплоидных и пентаплондиых
микровндов, о чем уже говорилось в одной из предыдущих глав.
Второй хороший способ поддержания триплокдиого состоя-
кня нлн нечетной полиплоидии более высокого уровня—это
агамоспермия. Среди агамоспермных членов группы Crepis occi-
dentalis имеются микровнды с числом хромосом Зх. 5х и 7х (см.
Г А. it. гршмошы
465
гл 32). Триплоиды встречаются также у агамоспермнего вида
Taraxacum officinale
Генетическая система Rosa canina преастааляет собой спе-
циализированный способ воспроизведения состояния мечетной
полиплоидии. В группе Roza canina ага система связана с пен-
тзплондией, у Leucopogon jumpennus — с триплопдней. а у
Cardamine insuela — также с триплоидией, но в модифицирован-
ной форме (гл. 30).
Избранные примеры
Обычная форма Populus tremula — диплонд (2п = 38). В отдель-
ных местностях как в южной, так н в северной частях Швеции
встречается форма gigax с 2п = *57, представляющая собой ав-
ютриплоил. у которого при мейозе образуются многочисленные
НТК Он продуцирует полустерильную пыльцу а в своих
природных местообитаниях распространяется прн помощи веге-
тативного размножения. Эти трипловды. вероятно, возникли и
результате соединения исрсдуцнрованной и редуцированной га-
мет Г575, <UM)]
(jpunlia basilaris в пустынях на юго-западе Северной Амери-
ки представлена днплондной формой (л—II). В одной из мест-
ностей в Калифорнии недавно было обнаружено трмплолдное
растение. Этот трпплонд возник, по-видимому, в результате от-
сутствия редукции у одной из родительских форы, поскольку в
остальном—это диплокдвый вид и ни одной тетраплондной фор-
мы у него не i пи [643]
Культурные съедобные бананы в большинстве случаев явля-
ются трмплоидаыи (х=11). Они происходят от дикорастущих
диплоидных видов Musa acuminata и М. balbisiana, плоды кото-
рых содержат схжсиа. Бессемянные трнплондпые сорта размно-
жаются вегетативно клубнелуковицами и корневищами. Неко-
торые триплондные культивары, такие, как Грос Михель,—авто-
трнплоихы, происходящие от М acumiaafa, тогда как другие—
ВЛДОТраплокды, возникшие в результате скрещивания между
Af. acuminata и Л), balhisiana [726, 727]
Lilian lancifotium (х^12) встречается в Китае и Японии*.
Диплоидная форма, по-вилимому, широко распространена Не-
которые культурные клоны— триплокды: в отдельных местах в
Японии триплонды встречаются также в природе. Трнпломлные
растения продуцируют стерильную пыльцу и размножаются ве-
гетативно воздушными луковичками и луковицами. Прежде по-
лагали, что они представляют собой автогрнплоиды. но теперь
' Этет вид лилии встречаете! также у нас на Дмммм Востоке — f7jw«t
456 Часть восьмая. Слецяализированпые гсчетичкюм системы
их считают сегментными аллотрнплондами. Эти триплоиды. ве-
роятно. происходят от гибридов между L. laneifolium llaviflorum
(2х) и L matimotticiil (2х). Один из родительских видов, по-
видимому, внес иередуинроваиную гамету, что и привело к воз-
никновению трнплоидпос-ги. Триплоиды, очевидно, независимо
возникали в Японии несколько раз н распространились локально
в разных местах вегетативными способами [93, 617].
Melianthus mulliflorus — декоративное многолетнее травяни-
стое растение, выращиваемое в Европе. Это триплоид (2л—51)
со стерильными семенами, и его размножают вегетативными
органами. Он был получен в результате гибридизации между
тетраплоидом Н decapetalus (2л—34 II) и диплоидом Н. annum
(2л —17 II). Н. multi/lorus является аллотетра ил оидом, образую-
щим при мейозе 17 II-f-17 1 [573, 5751
Представители Juniper us — преимущественно диплонды
(л—11). У видов 1. oirginuma и /. chinsnsis имеются трнплонл-
иые культивары [333, Ю5|. У /. chlnensis и других видов встре-
чаются также тетраплоидные культивары [333]. Установлено,
что трнплоидпые культивары —это гибриды между 1. chlnensis
(4х) и /. virginlana (2х) [333].
Lemna minor. L. gib Ра и L. perpusllla (x—10) содержат фор-
мы, находящиеся иа различных уровнях плоидвости, в том чис-
ле и нечетные полиплоиды. У L. minor известны полиплоидные
формы 2х, Зх, 4х, 5х, 8х; у L. gibba— 4х, 5х, 7х, 8х, у L. регри-
silla— 2х. 4х. 5х, бх. 8х [851]. Lemna размножается пегетатяв-
ным путем и образует клоны, так что нечетные полиплоиды мо-
гут поддерживаться именно таким способом.
Триплонлпые формы папоротннкоз отличаются, кая и следо-
вало ожидать, высокими хромосомными числами. Например,
группа Pityrogramma triangularis (Pteridacaaa) (х—30) в Ка-
лифорнии состоит главным образом нз диплоидов и тетрапло-
идов. В нескольких местностях обнаружены естественные три-
плонды с 2л«90. Триолоилные растения стерильны, и мейоз у
них нарушен. Установлено, что они являются гибридами, воз-
никшими в результате различных скрещиваний между тетра
плоидами и дкплондами [12, 739]. Сохранение и распростране-
ние этих стерильных гибридных раетеинй обеспечивается пол-
зучими подземными побегами
Часть девятая
Заключение
35 Вид и макроэволюция
Сетчатая эволюция
Филогения многих иля большинства групп высших растений,
которые были тщательно исследованы в природе и в лаборато-
рии. представляется сетчатой, а не исключительно дихотомиче-
ской я ветвящейся. Модель филогенетического древа нельзя пе-
реносить из зоологии в ботанику, так как по многих случаях это
приведет к искажению истинных взаимоотношений между груп-
пами. Во всех крупных группах высших растений нам приходит-
ся HMeib дело не с филогенетическими деревьями, а с филогене-
тическими сетями.
Формулировкой и разработкой концепции сетчатой эволюции
в царстве растений занималось несколько поколений ботаников.
Первыми авторами, высказавшими мнение о важной роли гиб-
ридизации в эволюции растений и сформулировавшими этот те-
зис в виде научно обоснованного обобщения, были Кернер [131]
и Лотси |519] Их ранние формулировки страдали недостатками,
присущими всем первым шагам в любой обиасти. Так. напри-
мер. Лотси вновь определил виды как чистые линии и поэтому
был склотн приравнивать гибридизацию главным образам, хо-
ти по исключительно, к обычному скрещиванию Работы Керне
ра относятся, разумеется, к домеиделевской эпохе, а кроме то-
то. он воздерживался от слишком катс1оричных или слишком
определенных высказываний относительно эволюционных изме-
нений более крупных масштабов, чем видообразование Тем не
менее Кернер и Лотси поставили эту проблему достаточно
четко.
В первой почовиме нашего века ряд специалистов по гене-
тике растений — Вииге, Тэкхолм, Реймер. .Мюнтиинг. Густаф-
сон, Ментон и другие, — подойдя к изучению проблемы гибриди-
зации и эволюции с применением экспериментальных и индук-
тивных методов, стали получать для вес большего числа групп
растений даииыс в пользу сетчатой филогении Средн система-
тиком этого периода, также разделявших мнение о сетчатом ха
рэктере филогении у растений, были Дильс, Хайата и Дю Рис.
45® Часть дгьлтал. За/ииочс/ам
Несколько позднее в результате слияния генетических и таксо
комических подходов возникла современная школа биосястема-
тяки. К числу представителей этой школы, внесших важный
вклад в понимание сетчатой эволюции, принадлежат Андерсон.
Стеббинс, Хейзер. X. Люкс, Эрендорфер, Л. Бенсон, Рейвн и
многие другие.
Для описания одного нз аспектов сетчатой эволюции Зрел-
дорфер предложи.! полезную концепцию циклов дифференциа-
ции—гибридизации. Он подчеркивает, что во многих группах
растений процесс эволюции представляет собой циклическое
чередование двух фаз —дивергенции и гибридизации (187, 188].
Эволюционный потенциал гибридных комплексов
В предшествующих главах было показано, что для гибридных
комплексов большинства типов —полиплоидных, вгамных. кло-
«альиых и гетерогамных —характерен период расцвета, за ко-
торым следует период упадка. Это последовательные стадии
эволюционного продвижения и эволюционного отступления кор-
релируют с генетическими системами, в которых гены замкнуты
» определенных гетерозиготных комбинациях.
Амфиплоидня, агамоспермия, вегетативное размножение,
перманентная гетерозиготность по транслокациям и перманент-
ная нечетная полиплоидия — способы воспроизведения опреде-
ленных благоприятных гетерозиготных комбинаций генов, воз-
никших в результате гибридизации. Такое воспроизведение дос-
тигается путем строгого ограничения рекомбинации генов, а его
результаты выгодны перманентным гибридам в период расцвета
гибридного комплекса Но в конечном счете перманентным гиб-
ридам приходится расплачиваться за свой оппортунизм, ибо,
когда в среде наступают, наконец, глубокие изменения, гибриды
порой оказываются пе способными создавать новые генные ком-
бинации, адаптированные к новым условиям среды. Гибридный
комплекс вступает в период упадка.
Гомогамные комплексы отличаются от гибридных комплексов
всех других типов тем, что входящие в них гибридные произ-
водные обладают такой же неограниченной способностью к ре-
комбинации, как и исходные родительские виды. Гомогамный
комплекс может пользоваться всеми преимуществами, предостав-
ляемыми гибридизацией, ие жертвуя при этом не менее выгод-
ной способностью к свободной рекомбинации генов. Отсюда
следует, что эволюционный потенциал гомогамного комплекса
ис имеет никаких особых ограничений, а может, напротив, про-
грессивно развиваться на протяжении очень длительного време-
ни, давая начало новым крупным ветвям (272]
Га Л5 Bvd и •вкрсмшоллг***
450
Когда я агамном, клональном нлн гетерогамном и — в мень-
шей степени - - пещи плои дном комплексе предковые диплоидные
половые виды вымирают, их гибридные производные лишаются
важного источника новых вариаций. Весьма вероятно, что такая
утрата нанесет урон последующей эволюции данного гибридного
комплекса. В отличие от этого и гомогамном комплексе исход-
ные пилы могут вымереть, нс подвергая опасности эволюцион-
ный питениимл производных видов. В сущности, вполне позмож
но. что предковые виды одного гомогамного комплекса пред-
ставляют собой конечные продукты какого либо предыдущего
гомогамною комплекса [272]
На рис. 21.] показаны различные «слом» (slra(a) гибридного
видообразования в гомогамном комплексе- Нет никаких осно-
ва ним полагать, что о фклетнчсской истории данной группы су
шествовали юлько эти нэзесгиые нам «слон» гибридизации На-
против, по всей вероятности, эволюция этой и многих другие
групп растений представляла собой последовательную смогу го-
могичпых комплексов [2721
Кпроче говоря, если для агамных, клональных и гетерогам-
ных комплексов типична закрытая система эволюции, а для
полиплоидных комплексов — ограниченная система, то можно
сказать. что гомогамные комплексы обладают открытой эволю-
цнолней системой, т. е. системой, ко тиран может существовать
в течение неопределенно долгого времени
Эволюция покрытосеменных — это макроэватюпня: кроме
того, это эволюции сетчатая. Проммдаммм иа пре.
длительного времени прогрессивные эволюционные изменения в
различных основных фнлетнческих линиях покрытосеменных
могли сочетаться с сетпатнм характером филогенетических свя-
зей в том и только в том случае, если эти фнлетичеекис линки
развивались кин последовательные гомогпмлыс комплексы |272].
потому чтп гомогамным комплекс — единственный гни гибрид-
ного комплекса с открытой эволюционной системой
Значение сетчатого характера эволюции
для филогении покрытосеменных
В этой книге рассматривается эволюция растений на уровне
видов и групп видов Эволюция растений на эпю уровне в зпа-
ЧПТеЛЬМОй степени определяется относительно простой органи-
зацией растения, открытой системой его роста, относительной
простотой генных систем, контролирующих многие признаки, лег-
костью естественной гибридизации и тенденцией к унипяреитяль-
ному размножению. Факторы эволюции и се различны* тип.
выявляемые в результате генетического и таксономическою изу-
чения групп видов, позволят, быть может, пролить свет на про-
4Й0 Van» ittxru. Зшикжнвг
блемы, связанные с филогенетической классификацией высших
категорий.
Филогенетическая система классификация многих порядков
н иадпоридков (или подклассов) покрытосеменных после таксо-
номического изучения иа протяжении нескольких столетий все
еще остается неясной. В настоящее время в этой области кает-
ся много успешных исследований представителями нескольких
научных школ, которые понимают трудности, связанные с соз-
данием такой системы, и не скрывают этого 1130, 135, 788. 821,
8281
Трудность и неопределенность классификации покрытосемен-
ных на уровне порядков проистекает из сетчатого характера фе-
иетического сходства между разными группами. Рассмотрим че-
тыре родственные группы, обозначив нх Л, В, С и D. Группы Л
и В имеют несколько общих морфологических признаков; другой
набор признаков является общим для групп В и С: н еще один
набор —для групп С и D. Для всей этой совокупности групп раз-
ные ученые могут н будут предлагать различные системы клас-
сификации— в зависимости от того, каким признакам каждый
Из них придает первостепенное значение. Так, в соответствии с
одной системой, таксонами одного ранга могут считаться группы
Л, В—С и D, в соответствии с другой — группы Л—В и С— D.
Такого рода трудност и частично удовлетворительное их
разрешение характерны для поисков естественной системы по-
рядков в различных крупных группах покрытосеменных. Труд-
ности, возникающие в реальных случаях, больше, чем в нашей
простой гипотетической модели, так как переплетение фенети-
ческих взаимосвязей обычно захватывает дополнительные груп-
пы Е. F, О и т. д. Эта картина хорошо описана Кронквистом
для ряда реально существующих ивдпорялков и порядков в его
сЭволюцни и классификации цветковых растений» [130].
Весьма важно, что описанная выше переплетающаяся таксо-
номическая структура, несмотря на ее широкое распростране-
ние у покрытосеменных, не универсальна. Некоторые крупные
группы покрытосеменных представляют собой ясно выраженные
филетические единицы и уже давно признаны таковыми. Хоро-
шим примером служат очень обширное семейство Compositae
и порядок Aster a les, к которому оно относится, Деление покры-
тосеменных иа две основные группы —иа однодольных и дву-
дольных— признается всеми со времен Джона Рэя. Дихотоми-
ческая филогения высших животных — другой пример системы,
резко контрастирующей с сетчатой системой, характерной для
многих групп покрытосеменных.
Итак, во многих обширных группах покрытосеменных мы
наблюдаем сетчатую модель сходства по морфологическим при-
знакам: это подлинный факт, а ие артефакт, обусловленный
Гл 35. Ялт) в
♦61
недостатками наших методов исследования, поскольку эти же
самые методы позволяют выявить дихотомическую в своей ос-
нове филогению там. где опа действительно существует, даже
в больших и разнообразных группах
Сетчатый характер морфологических взаимосвязей, наблю-
даемый из уровне порядков у покрытосеменных, можно также
наблюдать во многих семействах тех же покрытосеменных на
промежуточном уровне ролов к трмб. Эта сетчатая модель фено-
типического сходства иа всех уровнях заставляет вспомнить сет-
чатую модель генетических взаимосвязей. обнаруженную комп
лексах видов у растений. Такая общая модель побуждает нас
начать поиски лежащих в ее основе общих причин, присущих
как самим растениям, так н формам их эволюции
ОДНО* из причин того, что как макро-, так и микроэеолюиня
растений носит сетчатый хярактер} является конвергенция, т. с.
приближение одной филетической линии к другой по иесколь
Ким или многим признакам
Вернемся к нашс'<у гипотетическому примеру. Вспомним,
что у групп А, В, С. D обнаружено сходство по морфологическим
признакам в комбинациях Л к В, В и С. С и D. Мы часто обна-
руживаем то клм иное приближение к следующей обшей ентуа
□п.। Родственные группы А и В различаются по признакам, ко-
грушш А находятся на мам яркиятамм I
В — на более продвину той стадиях эвотюцпн Сходным образом
у группы С наблюдается прогресс по ряду* признаков по срав
ненпю с родственной, но более примитивной группой D В отли-
чие пт этого сходство между группами В к С касается продви-
нутых или специализированных признаков Иными словами, эти
гКЦПйе признаки — результат КЯИЯСМЯЛТПОЯ ВВОДИМ.
Конвергентная эволюция может быть обусловлена отбором
без гибридизации или же совместным действием гибридизации
и отбора В любом случае полагают, что отдельные линии под
вергаютсм отбору, благоприятствующему сходным адаптивным
реакциям па одни и те же клм сходные условия среды. Оба ти-
па конвергентной эволюции часто встречаются у растений.
Легкость, с которой разные группы растений переходят из
одной экологической ниши и другую, что приводят к Кохвср-
генциям по целым наборам признаков, можно продемонстриро-
вать на сотнях примеров. Многие отдельные роды многолетних
трав в Средиземно мере кой области дали начало однолетникам;
различные группы мезофитов в пустыне коивергентно дали на
чало ксерофитам; там, где живут колибри, у родов, опыляющих-
ся пчелами, неоднократно возникали виды, двсткн которых при
слособлены к опылению колибри, и т д
Тот факт, что линии растений сравнительно свободно передо
лят из одной адаптивной зоны в другую, коррелирует с о тио-
442
Чат Девятая. Заключение
сятельной простотой участвующих * «том генных систем На ра-
совом уровне димргеицнн в некоторых случаях можно обнару-
жить комбинации контрастирующих признаков, адаптированные
к различным экологическим нишам. Поэтому переключения, или
квантовые сдвиги, которые велики на экологическом уровне, но
относительна малы в генетическом плане, могут возникать не-
однократно в разных филетических линиях пол направляющим
влиянием одного лишь параллельного отбора.
В Пределах скрещиваемости, которые во многих группах
растений достаточно широки, дальнейшей конвергенции способ-
ствует гибридизация; в этом заключается вторая важная при-
чина сетчатого характера фенетяческих взаимосвязей у покры-
тосеменных.
Вид как единица макроэволюции
Некоторые молекулярные биологи, пытавшиеся изучать эволю-
цию органического мира, по сути дела, сводил» это явление к
равнодействующей между всего лишь двумя силами — мутаци-
онным процессом и отбором (см., например, [22, 51] и в мень-
шей степени [23]). К тому же представители этой школы нс при-
нимали во внимание существование видов как неких единиц и
существование видообразования кии эволюционного процесса.
Между тем, поскольку множество видов действительно возник-
ло в процессе эволюции, любое рассмотрение этого предмета,
не учитывающее формирование видов, не может внушать до-
верия.
Комбинацией мутационного процесса и отбора можно объяс-
нить биохимическую эволюцию, самые первые этапы биологиче-
ской эволюции и некоторые очень простые эволюционные изме-
нения у высших организмов. Но мутационво-селехтявная теория
не лает удовлетворительного объяснения эволюции более слож-
ных органических структур и функций. В результате одних лишь
мутаций и отбора на нашей планете вряд ли могло произойти
развитие растений и животных в течение того времени, которое
для этого имелось.
Признаки, при помощи которых организмы адаптируются к
условиям окружающей их среды, во всех случаях, за исключе-
нием самых простых, детерминируются не отдельными генами,
а их комбинациями, обычно довольно сложными. На сравни-
тельно ранних этапах эволюционной истории, еще у однокле-
точных форм, возник половой процесс, представляющий собой
механизм, который создает комбинации и рекомбинации раз-
личных генов. В то же самое время в ответ на различные комп-
лексы условий среды в пределах одной общей области обитания
стали возникать дивергентные специализация Воспользоваться
Гл. 35. Вид и дакрсмеолчлт»»
463
преимуществами, которые предоставляет половое размножение,
сохраняя при этом дивергентные специализация, можно было
только путем создания барьеров, препятствующих гибридизации,
как было показано ранее в этой книге.
Короче говори, в поисках адекватного объяснения органи-
ческой эволюции мы никак не можем ограничиваться одними
лишь мутациями н отбором. Для развитии сложных адаптаций
необходимо было половое размножение, а это в свою очередь
потребовало формирования репродуктивно изолированных пи
лов
С тех пор как Даркин дал своей книге название «О проис-
хождении видов». профессиональные эволюционисты признают,
что вид занимает центральное место в процессе эволюции Боль-
шинство биологов-эволюииоиистов разделяет представление о
том, что разнообразие видов на земном шаре —это система,
обеспечивающая непрерывное сохранение различных специали-
зированных образов жизни. Чго же касается соотношения меж-
ду видами и филетической эволюцией, то по этому вопросу бы
ло высказано несколько точек зрения
Хаксли (392] рассматрмвя’ видообразование как процесс,
порождающий разнообразие, ио не имеющий никакого отноше-
ния к прогрессивным эволкипюнным направлениям. Позднее
Майр [56У] пришел к выводу, что если большилсыи? видов — это
в сущности эволюционные тупики, то небольшому, но все же
достаточному их числу досталась такай комбинация благоприят-
ных признаков и экологических возможностей, которая позво-
> пл нм стать основателями новых доминантных групп. Пра-
вильность такой точки зрения отрицать невозможно. В дополне-
ние к ней я высказал предположение [282. 296]. что видообра-
зование неразрывно связано с некоторыми случаями филетнче-
ской эволюции, в которых прогрессивное развитие происходит
путем ряда последовательных этапов видообразования.
За последние годы получает все большее признание так на
зываемос прерывистое представление о макрозвапюиии. Соглас-
но этому представлению, виды и видообразование играют глав-
ную роль в эволюционных взрывах, характеризующих модель
прерывистой мэкроэволюцнн. Недавно Стенли [7*1. 772] разра-
ботал и очень хорошо сформулировал взгляд на видообразова-
ние как и а основной яроцесс в макроэволюцни.
Литература
I Adams R. P., Turner В. L. Chemcoptematie «nd numerical studies of natu-
/al populations of Juiuperw ashei ouch. Така» 19. 729—751 (1970b
2 Addison G., Tavares R. Hybridization and grafting in spacles of Theobroma
which occur in Amazonia, Evolution, 4, 380—386 (1952).
3 . Alexander D. E., Beckett J B. Spontaneous tnploidy and tefraploidy in
rnaae, J. Hered., 34. 103—106 (1963)
4 A dan H. H. Wild species-hybrids tn the phanerogams, Bot. Rev. IS 77—
iOS (1969).
5 . Allard R. IF, Jain 5 K. Population studies in predominantly sell-pollinated
species. П. Analysis of quantitative genetic changes in a bulk hybrid popu-
lation of barley. Evolution, IS, 90-101 (1962)
6 . Allard R. V.. Ashler A. L Alloeyme polymorphisms in plant populations,
Stadler Sympcnla (Univ. Missouri), I, 9—2A (1971).
7 Allard R. T.. Vor Iman P. L Population studies in predominantly aelf-
pollinated spec.ea. IV. Seasonal fluctuation* in estimated valuta of genetk
parameters m lima bean populations. Evolution. 17, 470—460 (1963).
8 АШил R E. Makra T. J. Turner В L Perspective* in rhema taxonomy.
Science. 142. 545-W2 (19631.
9 . Airtcw R. E.. Turner B. L New technique® in analysis of complex natural
hybridization, Proc. Nat. Acad. Sts, 4a. 130—137 (1962).
10 . Alston R £, Turner B. L Biochemical Systematics, Prentice-Hall. Eagle
wood Cliffy N J, 1963
11 Alston R. £u Turner 5 L Natural hybridisation among four species ol Bap-
tIMa (Lcgumtnoaae), Amer. j. Bot., 50, 159—173 (1963).
12 . Alt A A, Grant V. Cytouxononuc observations oo the goldback fern, Brit-
Ionia, 12, 153—170 (1950)
13 . Anderson E Recombination in epeews cross**, Genetics, 24, 668—698
14 . Anderson E. liybndization of the habitat. Evolution. 2. I—9 (1948).
15 . Anderson E. Introgressive Hybridization. Wiley, Nev York. 1949.
16 . Anderton E. Introgressive hybridization, B»ol Reviews, 28, 280—307
(19631.
17 Anderson E, Anderson В R Introgression of Sotafa apiana and Salvia
meUifera, Ana. Miasowri Bot Gard.. 41. 32^-338 (1964).
18 . Anderson E., Gage A Introgressive hybridization In Phlox bifida, Amer.
J Bet, 39 399—404 (1952).
19 . Anderson E, Hubrichl L Hybridlzatloa in T radetraniia. Ill. The evidence
for introgreeeive hybridization, Amer. J. Bot, 28, 396—402 (1938).
20 . Anderson E^ Sax К A eylological monograph at the. Amertcao specie» of
Tradesemtia, Bot. Gat. 97. 433-476 (1936).
21 . Anderson Schafer B. Species hybrids in Aquilegia, Ann. Bot., 45, 639-
646 (1931).
22 Anfiruen С. B. The Molecular Basis of Evolution. Wiley. New York. 1959.
[Ииеегся перевод: Анфжнсея К. Можауляраые оежты зоожзцна. -• М.:
ИЛ, 1962.]
23 . Antonootcs J. The affects of a Iwtcfceaaeou* enviromnent on the geoellc*
ol natural papulatloas. Amer. Sd., 59. ЙЗ—И9 (1971)
24 . AniOfuxlcs Z. B/odshaar A. D. Evolution tn clcody idjacent plant popula-
te». VI11. Clinal pattern* at a mine boundary. Heredity. 25, 349—362
(1970).
25 Aotfg A. G.. SaJlna S. Rletnema J. Blakeslee: The Genus Datura, Ronald
Press. Nev York. 1959.
Лит грату ра
4G5
26 Лоегу Р Cytogenetic evidences of Nicriiaaa phylesis in the alatagroup.
Univ Calif Publ. Bot. I». 153—194 (I9&8)
27 AorvA&v M. ZoAozu [) Genette at’InltleB алтее Ibc Orwocyi/us [rises, BoL
Gat. HI. 107-115 (l^aC)
28 Ayala F / ed Molecular Evolution, Sinauer As«oc*atm, Sunder'.and, Mass.
1976
29 Ayala P /, Valentine 1 IT Evolving The Theory and Processes of Orga-
nic Evohirien. Benjamin Cummings, Menlo Pari;. Calif, 1979
30 Axelrod D t Evolution of the California closed cone pine forest. Proc
S^’P R'°* Calll Islands (Sxn'.a Barbara. Botanical Garden). 93—149
31 Axelrod D i Hisiorv of the coniferous fores л. California and Nevada, Univ
Calif Pub! Sot 7t, 1-62 (1976)
32 . Barbel 6 R. Fain P P Genetic structure of Hbtdftun jatatam 1 Ootcros-
* ng rates and heterozygosity levels, Canad J Genet Cytol, 19, 143—
152 <1977)
33 Bebcock E В Ths genus Creptt. Univ. Calif P, til Bot Vols, 21. 22 (1947)
34 Babcock E B. Jenkins J. A Chromosomes and phylogeny In Crepis III
The relationships ci nw hundred and thirteen species, Univ. Calif. Pubi
Bot. 18. 241—292 (19431
35 Babzvcb E В, Stebbins G L The genua TouisgM, Carnegie Inst Washing-
ton PuM no. 484, 1937.
M BoArorA E В . Stebbins Q. L Tba American species of Crepis. Their Inter-
reUticnahifi $ and distrAutlcn as a Heeled by pofyptoidv and apomixis. Car
Regie Ins1 Washington Publ no 594, 1938
37 . Babcock E В. Stebbins G L. /еп*?«м J A ChronwTMx^s and phvlogenv
in stent- ganar.s of the Crepdinae. Cylo4ogia. Fuji Jubilee Vol ife—210
38 Baetcke К P, Alston R E Tba composition ol a hybridizing population of
Bu^'izia sphaetactrf pa an J Baptism Uucopbaea. Evolution, 22, 157— IGS
39 . Baker H. C Hybridization and natural gene-Поv between higher plants.
Biot Rev . 2®. 3»r?—337 (1951)
40 Bohfui-t J Г. Kzlandioe. th* genns and its chromosomes, Amer J Bot,
26. 572-379 (19381.
41 . Baldvin l J Galax- the g?nus and its chromoaomrt. J Hered. 32. 249—
254 <194!I
42 fta'lca к 4. Fines D The cykigecgiaphy of the locinthaceous mistletoes.
Tstoo. 20. 291-312 (1971)
43 Вагпл» В. V. ТЫ clonal growth habit of Anaarkan aspvas, Ecology. 47.
439-447 (1967)
44 Barnas В К Indications ol potsibk mJCcnozon hybfidisatioc tn the
aspens of the Columbia plateau. Rhodora, BP, 73—81 (1967)
45 Barms В. V Natural cxtUtLon atsd delineation or ckm«s of Poputafus ire
mulaidts and P graiuiidrafata >n northern Lo*xr Mich gan Silvae Gene-
tics. 18. 130- 142 < 19»)
46 Bateman .4 J Genetical aspects of seed-growing. Nature, 1S7, 752—755
(19461
47 Bo/emar. A. J Cryp'Jc aelf-incocnpatibililv in the nallllower Ckriranthas
cheiri U Heredity. M. 25? 261 (1966)
48 Baifagiia E Apomixis In Recent Advances In the Embryology of Anglo-
sperms, td P Matieshwari, lokmat Sue Plant Morph. Delhi. 1963
49 Boar E Eiaffihrung in die esperimeatelh Ve-erbuBgllehre. 2nd ed. B<xn-
traegw. Berlin. 1914.
59 Baur E Adrmgrenzung «nd ArlMIdung W 6*- Gattnng АяИгМНлшя. Sck-
tion Antiefhxncnlram. Zeitachr Ind Atetanwiungi- u Vcrarbuogslebre, 63,
236—302 (1932)
30-45
♦66 Литература
51. Beadle G. W Genetic* in4 Modern Biology, American Philosophical Society.
Philadelphia, Pa, 1963
62. Beaman 1. H The syslanuixs and evolution ol Toviucndlo (CompelПае).
Cootrib. Gray Herbarium (Harvard), Net 133, 1957.
63. Beaudry J. k. The species concept >ts evolution and present status, Revue
Cacadieone d« Bmlogt*, 19. 219—240 (I960).
M. Beehs R. AL Variation and hybridtialion in soalbcm California populations
Of Diplomas (Scrophulariaceaej, ALiso. S, 85-122 <1962).
55. Bell C. R. rhe Sanicula crsasicaulLs complex (Umbel llferae); a study of
variation and potypfoidy, Univ. Calif. Pubt BoL, 27. 1М-2Э0 (1»4).
56 Bennett E. Historical penpedivee in genecology Scottish Plant Breeding
Station Record. 49—115 (1964).
57. Benson L Plant Taxonomy. Ronald Press, New York. 1962.
58. Benson L. The Native Cacti u( California, Stanford University Press. Slan-
tord, 1969.
5fl. Benson L. Phillips E .4 . Vsldrr P A Evolutionary sorting of characters in
a hybrid swarm. I. Direction ut slope. Amer. J. Bat, 54. 1017-10% (1967}
60 . Bergman В Zytoiogische Stvdien uber die Forlpflantang bai den G*tlon-
gen Leontodon und Pieris. Sveask Bot. Ttd&rlft. 2t. IBS—301 (1935).
61 Bernardini J. V.. Uma-de-Faria A. Asynchrony ol DMA replication in th*
chromosomes of Lamia. Chromoooma. 22, 91 100 (1967).
62 Blackhurst H T. Cytogenetic studies on Rasa rubtgtnosa L. and Its hybrid*.
Proc. Amer. Hort Sd . 62. 510—516 (1946)
63 . Bogart J P Evolutionary implications of polyploidy In amphibians and
reptiles In- Polyploidy, Biological Retevaaca, ed. W. H. Lewis, Planum
Press, Nev York and London, I960
64 Bradner AL R., Frakes R. V, Stephen IF P. Effects ot bee species and iso-
lation distance on possible varietal contamination in alfalfa. Agroci J, 57.
247 -248 (1965).
65 . Bradshaw A. D. Population differentiation in Agrostw tenuis Sibth. L Mor-
phological differentiation. Nev Phytol.. 58, 208 -227 (1959).
66 Brainerd E Seane natural violet hybrids of North America. Vermont Agree,
Exp. Sta Bull. No. 239. 1924
67 . Breedlove D E. Th* ayat emetics of Fuchsia section Endiandra (OnagrS-
ceae). Uwtv Calif Puhi Bot, M 1-69 (1969)
68 Bridget С В The origin of venations In sexual and sax-limited character*,
Amer Nat. 66, 51—63 (1922)
69 Rrighi C A. Telocentric chromosomes In Corsican Croau L. (fridaceae).
Plant Sy*t EvoL, 129, 299—314 (1978).
70 . Brink R. A Physe change In higher plants and somatic cell heredity. Quart
Rev. B;ol. 87, 1—22 (1Л2).
71 . Brown S The origin and nature of variability in the Pacific coast black-
berries {Rubus ursinas Cham end Schfoht and R Umar am sp ПОтг ),
Amer. J. Bot, 86, 686—697 (1943)
72 . Brom S Mitosis and meios'e in Landa cam pettris DC, Univ. Calif Publ.
Bot. 87, 231-778 (1954)
73 . Brown V. L. Wilson E. O. Character dteplaceroefit. Systematic Zool. 5,
49—64 (1956).
74 . BttJlon C. de. Histoirc Naturdla. vol. 2, Pare, 1749.
75 BungMkberg de Jong С, M. Polyploidy in animals, Bibitngrephia Genctka,
17. Ill—228 (1967).
76 Burma В H. Reality, existence, and claxaificatfon a discussion ol the «*•
cte* probtem, Madrono, 12, 193-209 (1964).
77 Busk G L JActdsn ot animal socciatlon, Ann. Rev. Ecol. Sys1., 6. 339—364
(I975).
78 Cadman С H Autotet-aplnld Inheritance in thr potato: some new evidence.
J Genetic,. 44. 33 32 (1942).
Литература
467
79 СаГл A I Animal Specie and Thrtr Evolution. Hutchinson. Landon and
Harper and Row, New Yurt, 1934
SO CarsiTon ! Г. />л« W 8 Genetics, breeding, and macellar ernbf jooy
In The Ci Ires Industry. 2d ed. vol 2, eds U Reuther, L D Batcheloe and
H J Webber, University of California Press. Berkeley and Lm An gyle*
fl! G**i^ F H, Gitty C L Ihe structure and origin of species» Br.ttmila, 4.
323-385 (1943).
62 C-~.rlquiil S On the genetic limits of FriophjyJi'i-rn (Composilae) and rela-
ted genet a. Madrono, 13. 225-239 (1956)
83 Co«sa% If L The genetic characteristics ol ma'girul populations of Dro-
C&A Spring Harbor Symp Q-jsaf Biol. 20. 276-287 (1965)
81 Ca'$t>* H L Genetic conditions wlixh prnmol» or ratith* formation
or species, СоИ Spring Harbor Symp Quant Biol. 24. 87—105 (1959)
85 Carso.*! H t CJvnmnsnm» tracers of the origin of spaces. Scfanee. IM.
14U-1418 (19701
86 Carsan H L Spec? I ion and the founder principle. Stadler Genet Sym-
posia (L’uiv Missouri), 3. 51—70(1971).
87 Carson H L The genet rs ol apcciatian at the d.nloid level, Amer Nat,
100. 83-92 (1975)
M Сигчпп H L , Heed F fi Stuclura! hocnotvgo^ty in marginal population!
cl neat cite and neotropical species of Drosophila in Floc. da. Proc. Nat
Acad Sei . 32. 427—430 (19*54)
89 CMpati F Cvlopltsmic inheritance, Advances fn Genetics» 2. 1—66
(1948)
90 Cove M 5. Сапйаясг L Chromosome numbers in the Hvdrophyllaceae
1П. Univ. Calif Publ. Bot. I& 449-466 (1947!
91 Свис A1 5. Convener L Chromosome numbers in 1be HydronhyllactBt
IV, Univ Calif Pnbl Brit 28. 368—382 (19Л0»
92 . Chambers К L. A blosybtanatk study ol the annua! species of kiicraseris
Contnb Dudley Herbarium (Stanford). 4, 207—312 (1955)
93 Chnrwtfer С. Poftc/iield IF .4, Stnvl A В Microsporogenesis in diploid
and Lriplotd types of Ltkum tigfinum with special reference to abortions,
Cyfologia Fu'ii Job. vol 2. 7.^-7M (1937)
94 CAast V C. ffaz-tn P Я L Bl nan snd ecological relationships between
Лфш.'едй) Jormosa and A pubescent (Ranunnilicea*1). two perennial plants.
Evolution. 29. 474 486 (197*1
95 dates*’» / Genet<al and cytclogxal investigations on Viola tricolor L. and
Г тайл Mtfr. HefedHM» 8. 1-155 (1926)
96 CAiusr.i 7 Stages n the Evohlion of Plant Species. Cornell University
Press. Ilheca. X Y. 1951.
97 . Clouarn J Pop sla ic-л studies ol al₽in< and aubalpine races of coeifm and
willows in the Cali'omia high Sierra Nevada, Evolution, 19. 36—68
(19651
98 Ctauvn i. Hincu F Al Erperimental «tiadlcs on the nalare of фс1к
IV Gcnetk structure ol ocOlcgical races. Cornigie Inst Washington Publ.
615, 1958
99 СГал-scn 1, HitSMt F .41 Tbe balance between :o'"eren:e and variation in
evoJulKin. Proc Nat Acad Sa, 44. 494-W6 (1*Л)
100 Оая&еч /. Kerft О О, Hies^u T Л1 The concept of species based an expe-
riment Amer J Bot. 20. 103- 106 (1919)
101 . Clouser. J. Keck D D. Hicsey Г Л1 Eaperimcntal stud « or the nature
rtf species 1 EHec* of varied environments on western American plants.
Carnegie Inst. Washington Publ .529. 194Л
t02 Clwffit J. Ketk D D. Hlwj IF M Experlrenfa: «todies on the nature of
species П Plan muwtiun through araphiolcidy and awtoolo*dy. with exam
plea from 'he Madrin.ie, CariMgie Inat Washington P11SL, 564, 1945
30‘
468 Л м? грогу ре
103 (Ломмп J. Keck D. D.. Hiney Ж. М. Experimental «todies од the nature al
species. II! Environmental responses oi climaiK races o> Achillea, Carne-
gie Inst, Washington PuN., 581. J 948.
104 Clausen К Keek D. D-, iiieseu IF M, Gran P Experimental taxonomy.
Carnegie Inst Washington Yaartxx* 48. 96—106 <1949).
105 . Clausen R E. Goodspeed T. H 1п1егврес-.1к hybrr-dixation in Nicotiana.
11. A letraploid glu&uMo tabacum hybrid, and experimental verification of
Winge’s hypothesis. Genetics. 18. 27»— 284 (1925).
108 Clausen R. T. Sedum ot the Trans Maxican Vokanic Balt, Comatl Univer-
sity Preu, Ithaca, N. Y., 1959
107 , Clayton E. E Male-stenle tobacco, J Heredity, 41, 1T1—173 (I960}
108 . Clegg M. T. Allard R. OF. Viability versus tecundtty selectton tn the slen-
der wild oat. Auerw barbala LM Science. 181. 667-668 (1973).
109 . Cleland R E Some aspects ol the cyto-genetics of Oenothera, Bot Rev.,
2.318—348 (1938).
110 Cleland R E. Phylogenetic relationships tn Oenothera, Heredilas, SuppL
Vol. 173-IM, 1949
111 . Cleland R. E. SlvdSea in Oenothera cytogenetic* and phylogeny. Indiana
Univ. Publ, Science Series, No 16 (I960).
112 . Cleland R E. The cyloganKict ol Oenothera. Advances in Genetics, II.
147—237 (1962).
113 . Cleland R. E. The evolutionary history ot the North American evening pri-
mroses ol the rbiennis group». Proc. Amer. Phil. Soc, 108, M—98
(1964).
114 . Cleland R. E. Oenothera: Cytogenetics and Evolution, Academic Press, Nev
York. 1972.
115 . CUJIord H T. Analysis ol suspected hybrid swarms in the genus £ucoiyp-
tM, Heredity, 8, 259—269 (1964).
116 . CocAruM E. L. A check list and bibliography of hybrid birds in North Ame-
rica north pf Mexico, Wilson Bull. 84, 140-159 (1952)
117 . CoteeU R N. The use o( radioactive Isotopes in determining spore distri-
bution patterns, Amer. J. Bot., M, 511—523 (1961).
118 Conant [) S. Cooper-Ortner 0 Atatogamovx al I о homo p lol dy in Altophila
and Nepheiea (Cyatheaoeae)- a new hypothecs for speciatioa in homoplod
homosporous term, Amer J Bot.. 67. 1ЭУ?—12ЯА (I960).
119 . Constance L. Chromosome number and classification In HydrophyUnceae,
Brittonia, IB. 273-286 (1963).
120 . CocA R E. Patterns of juvenile mortality and recruitment tn plants In:
Topics in Plant Population Biology, ed. O. Solbrig, Columbia University
Press. Naw York. 1979
121 . Cook S. A. Genetic system, variation and adaptation in Ecchsc-Meia Cali-
fornia. Evolution, 18. 278-299 (1962)
122 Cooper rider M Tnlrogree'ive hybridisation tn Quarcsu marilandlca. Amer
J Bot_ 44,804-810 (1957)
123 Cat fell D 5 The Potato and Its Wild Relatives. Section Tuberosum cd the
Genua Sotewu, Texae Research Foundation, Rennar, Texas, 1962
124 Cattam V P Prewnal leafing of aspen tn Utah mountains, J, Arnold
Arboretum. 88. 239-248 (1954).
125 Cation IF P. Tucker J M_ Drab nick R. Same dues Io Great Basin poet-
pluvial climates provided by oak dentribaboos, Ecology, 40, 361—377
(1959).
126 . Cosuens J. E. Variation of some diagnostic characters ot the sessile and
pedunculate oaks and their hybrids in Scotland. WaUonla, 6. 272—286
(1963).
127 . Cfturns J. E. The status ot the pedunculate and sessile oaks in Britain,
Watsonla. 8. 161-176 (1968).
Лиге роторе
4Ю
128 С» on* м в Mather Л The natural cross-polLnation oi crop plant* with
partkuGr relertnce to the radish. Ann Appl Biol. >0. >91—300 (1943)
129 CrttchfMd T В Interspecific hybridiial«on in Finn* a >umm.iry review,
Proc 14 th Meeting Canadian Tree Improvement Asm. 2. 99—1(6 (I9?5|.
130 Cranat^tl A Th» Evolution and Claetiricalioo ol Flowing Plants, Hough-
ton. Boston Мам. I960
131 CraneadsJ Я Once again, what is a species9 DeltMtllc Svmp Aerie Re*
search, 2, 3—2(1 1197Я)
132 Crude* R 8 Pc.ten ovate ratios о conservative indicator of breeding sys-
tems n flowering plants. Evolution, 31. 32—46 (1977)
133 . Cfutnpaiktf D. . Bj.Tur,72j ! S Density, dispersion, and pspuhirloe strut*
tm« m Drmophxla pieudwbszura. EcoL Monogr. 43. 499—538 (1973),
134 Ca/not L L'Evolu’ion btologique, Mrsson. Pans, 1951
135 R. A system of classification of the angiospernn to be used to
demonstrate the distribution ol characters, Bolaniska Notiser. IffJ, 110
147 (1975)
136 Da.ivr ft H Cbf* die Btgr.lle Kompnnuiw. KommiAuum ued Knnvivium
und uber dtp EntMehurigswrisc der Kemvfvicn, Grnetira. II, 399—450
11929)
137 Ds-nuremt P M Etudes шт les hybradea de Cutet VI, Introgression dans
la aerlioa Li if arum, Carud. J. Res, C, 19, 59—67 I'lOt! |.
138 fXi.'/i^gron C D Recent Advances la Cytology, 1st ed, Churchill» London,
1932
139 Dar/.ngrou C. D Rerent Advances in Cytology, 2nd cd Churchill. London.
1937
140 Dari, ng Io* C D The early hybrldtscrx and the origins of genetics, Her her-
til 4 63-69 (13371
Hi DnrtiagKm C D The Evotutlcn of Genetic Syytcnn, 1st ed, Cambridge
University Prers. Cacabr Jge, 1999
142 Dariiagt&i C D Taxonom*: spedes and genetic systems In The New Sys*
tcmataca. ed J Huxier. Carenoon Press. Oxford. 194C
143 Dariinglo* C D Natu'sl populations and ’he breakdown of classical gene-
tics, P oe Rev Sot. В 148. 353-364 (1956)
144 Darlingfv. C D Chromo* о me Dotany. 1st ed. George Alien and L’nw.n.
‘..ondon 1966
145 DarUr.gtfjft C D The Evolution ot Genetic Systems. 2d cd. Bask Books,
New York I96fi
146 Darlrtgioi C D Ch.'omownie Botany and the Origins of Cultivated Plants,
2d ed. Ifafner, New York, I960
147 Darlir.gtcn C D Cytology. Churehtll, London 1945
148 Da'tingtcr. C D Chronwsamc Botany and Ibc Origin of Cultivated Plants»
3d cd. Hafner, Nrr York, 1373
149 Darlington C D. JanaH Animal E К Chromosome Atlas of Cultivated
Plan’s. 1st ed . George Allen and Unwin. London 1943
IbO Dariingl&t C D Mother К The Elemenls of Genetics George Allen and
Unwin. London. 1949
151 . Dartir.gt&n C D. Fp^i'e A P Chromosome Atlas of Flowering Plants. 2nd
ed , George Alien and Urtwin. London. 1955
152 Da/uin C On the Origin ol Spoclcs, tat ed.. Mm Murray, London, 1859
Repclutcil by Watts and Co. London. I960; aud by Harvard University
Pre*a. Cambridge. Mass 1964
153 Detain C The Variation Ы Animals and Plants under Domes lira I ton.
2 vols. John Murray. London. IB®
154 fXiziai>i C On the Ongn of Species, Mh ed, John Murray LonAon, 1862,
various ntodtra rvprnts
155 . Duwdran /FA drptloglstlcated species concept, Madrono. 12. 246—251
(1954).
470 Литература
156 Pot’iM Е Cytology. evolution and origin of the aneuplold aerlea In the
genu* Carex, Heredilas, 42, 349—366 (1956).
157 Daeit P H. Heyetood V H Principle* ol Angfoaperns Taxowomy Van Noet-
rand, Princeton, N. J., 1963,
IM Day A The evolution of a paar ol sibling allotetr apioid sped*» of Cobwebby
Glliae (PutaDOiiiaceae), Aliao, 6. 25—7$ (1965).
169. De/eooeyoa Г. Polyplady in gymnospcrnu In; Polyploidy; Biological Re-
levance, rd W H Lewie, Plenum Prata. New York and London. 1990
160. De Vet /. M. J. Diplold-tetrapkud haploid cycles and the origin ol vartabb
llty in DuJianthium agsmospcrie-i, Evolution. 22, 394—397 (1968)
161. Dempster L T„ Ehs endorses F. Evolution cf the Qalium miutiflorum complex
tn western North America. IL Critical taxonoamc revision. Brtttaiua, 17,
289 334 (1965)
162. Dhillon T. S., Oarber E. D. The genus СсШаяп XIX. <Nrw> genomes
from two fertile interspecific hybrids, Cytoiogla, 27. 189—203 <1962).
163. Diets L. Die Methoden der PtrytograpLie und der Systematic der Pflansen.
In: Abderhaldens Hzndbuch der biologiscben Arbcitsmethoden, Abt. II, Ber-
lin, 1921.
164. Dietrich W. The South American species of Oenothera sect. Oenothera
(Raimannia, Renneria; Onagracaae). Annals Missouri Bot, Gard.. 14. 425—
626 (1977).
165. DnbtfuMitky Th Genetic nature at species difference». Amer. Nat, 71, 404—
420 (1937).
166. Dobzhanshy Th Genetics and the Origin of Specie», 1st. 2d and 3d eds..
Columbia University Press, New York, 1937. 1941. 1961*
161. Dobzhofuhv TA. Speciation at a stage in evolutionary divergence, Amer.
Nat, 74, 212-321 (1940).
165. Dabihansky Th Menddian populations and their evolution, Amer. Nat.,
B4, 401 418 <1960)
169 . Dobzhansky Th Specie» after Darwin. la. A Century of Darwin, ed.
S. A- Barnett. Heinemann. London. 1968
170 . Dobzhansky Th Genetic» of the Evolutionary Proems, Columbia University
Press, New'York. 1970.
171 Dobzhaaskv Th, Ayata F J. Stebbins 0 L. Valentine J, V Evolution,
W H. Freeman. San Francisco.
172 . Dobehaneky Th. Ehr тая L. Patdofsky О Drosophila Insular is, a new
sibling upecbs of the teiliisiotu group. Studies in the geartzs of Drosophila
(Uilv. of Texas). 9. 39-47 (1957),
173 . Dobihanshy Th.. Epllng C Cxxjtribulxjns to the genetics, '.алопату. and
ecvlogv of D’tn.-.-r»hdu oxeudootiscura and its relatives. Carnegie Inst. Wa-
shing- on Publ ,654. 1944
174 Dodds К S Classification of cultivated potatoes. In The Potato and lb
Wild Relatives, by D. S. Correll Texas Research Foundation, Renner, Te-
xas. 1962.
175 . Dodds K. S. Paxman C. J The genetic system ol cultivated diploid pota-
toes, Evolution. 14, 154—167 (196?)
176 Dodson С. H Cytological studies in the genus Oneidiam. Ph D. thwis, Cla-
remont Graduate School Claremont. Calif, 1966
177 Dodson С H. Frymire G. P Prdimdnarv studies Ln the genus S/onAopcu
(Orchbdaceae). Ann. Missouri Bot Gard. 48. 137—172 (1961)
178 . Doit Ц. Revision der sect Expthrosperma Dahtet emend l.lndb f der Gat-
trnig Taraxacum Zinn. Feddee Repertorlum, 83, 673—740, M. 1—180
(1973).
179 Dru/y T H The ecology of the natural origin of a apeeias of Caret by
hybridization, Rhodora, 58. 51—72 (1966).
180 Duffield J T Reb’ionshipv and tMeite hybrtdlfation In the genus Piaui.
Zeltschr. Forst genet Ik a Focatplhnxenxikht, 1, 73—97 (19&2).
Литература
4?f
181 . (J E The lundasnental unit» ol biological Uionociy, Svetuk But
Tidskr. 24. 333—428 (1930)
182 Eaxi £ M Studios on ми inheritance In Nicotiana, GeneLus. |. 164—176
(1916)
183 . Edvardion J R. Cytoplasmic male-sterility, BoL Rew, 22, 666—738
119661
184 . Elueidorfttr F. Zur Pbylogeme dcr Gal lung Gata-m L. 1 Polyploidie und
ceof;raph.sch ukologiact* Linheaten in der Gruppe des Gatium putnilum
Mvr-ay (Scat Lrptogalium Lange sense Roay) in oeterrcichischcfi Alpen-
r»um. Osterr Bot Zatlsdir. M. 109-138 (1949).
185 Ehmdoe'er F Zur Phylogenie der Gattung Gofium II Rtssengli ederung,
Viirialrlinl-rentrcn und gcogcaphiscLc Merkmalaprogrebaioflen a Is Aus-
druck dcr raum-zei'.lkben Eeilaltuflg des horffleture.ses Galium uiconum
S S Oslerr Bot Zeilachr. oe, 427-490 (1951)
ISO £!if»iuicr;tf F Oiiolugibch-geographiache Mikro-DslleteiuiefMg eitne Popu-
lation von ОаИмт рнли/um Mrr s etr Osterr, Boi ZeilMir, 616—
638 (1963)
187 £A'anrfor(r.e F Zur Phvlogeme der Gattung Galium VI Ln VariabiliUs-
zantrum ah «loosien Hybrid Komplex- drv o*t-mad I terran.- GaLum gfat-
CAin L — G lamun Req hxtnenkreti. (Xtarr. Bot Zotsdir. 10Б, 229—279
188 8&U, F Differen ial oo-iybfidkution cycles ir.d polyploidy In AdW*
1м. Cold Spring Harbor Synw Quant Biot, 28, 141—IW (1959)
189. Ehrendorier F EvjIUK'I of the Go£u/n maltiflaruM complex in wrrtrrn
North Arow ca 1. Pipluiih and polvplolds in this dfoccions group. Madrono,
14. 10? 140 (1ЙП
19C £A/«rnif«'i'c/ F Bi.iLigr w Phvlugcrue de.- Gattung Калика (Dipsacaceae).
I Cyi.okigi<chc GruatLagm und allgrmeine Hinwise, Oslerr, Bol Zeitschr,
IM. 278 343 (IМ2)
191 Ehtendoncr f Cytologie, Taxonarrie anil Erolulion bei Samenp (lancets.
Visits in В o’, any. 4. l& (1ЭД)
192 £n/<nrftu|rz F Dispersal medianisma, genetic systems and cotoniting abili-
ties in swiie Towering plant families In The Genetics ol Colonising Spe-
cies. eds H G. Baker and G L Slebbiiix Academic Preset. Near York.
1965
190 Ehrcrtdcf}** F Puhplo.dy and distribution In Polypkady. Biulogiml Re-
levance, td U H. Lewis. Plenum Press. New York and London. 1 МО-
ИН Ekmndarf>rr F. KfanAl F. llabttef F. Sauer T Chrnmncrr-.n numbers and
evolution in onnutivc angKriperms, Taxon. 17, 337 - 353 (1968)
195 Fhtlirh F ft. Haim К IT Patterns and pnpulalions. Science, 137, 652
657 (И62)
195 £ftni(ch F R. Razcu P H Differentiation ol popclat ons, Soence, 166,
1228 -1232 (19691
197 fAzftcA P K. Wmtt R ft Colorado cherberspot butterflies i*oI»Iiqh, neolra-
Шу. arsd .be biusprc.es. Amer. Nat. 115. 328-341 (19№)
198 Ehrmarz f. Hybrid ete*i|ilv as »• lactating mechanism ia the genus Dr<>-
Quart Rev B ol, 37. 279- 302 (1962)
199 EhrmuK I. Direct observation ot sexual Isolation between alfopalric and
bs’wecr sympatric strains of the dlffn’ml DrosapAila pou^slorum races,
Bwtotta. It. 459-461 (1965)
200 £me'sc-.e S H The geaetK natore of De Vries's mutations in Oenothera
lamar-ktana Anw Nat. 60,645-559 (193S).
201 Endler 1 A Geographic rariatxm Specialion, and Clines, Pnncetoe Uni-
versity Press. Prinretom, N. J , 1977
202. Ephng C N'afaral hybridization of Safoia opiona and S mellifera, Evolu-
tion. 1. 69-78 (1947).
472
Литература
KU. Epdng C., Dobehanshy Th- Genetics of natural populations, VI. Microgeo-
graphic races rn Lsnanihus parryae. Genetics. 27. 317 - 332 (1942).
204. Epling C.. Lewis ti. Ball F. M The breeding group and seed storage:
a study in population dynamics, Evolution, 14, 238— 255 (1960).
205. Ernsi A. Bastardierung ale Ursacha der Apogamie dn Pflamenreich. Gustav
Fischer, Jena. 1918
206 Fatgri R. can der Pi)l L. The Principles of Pollination Ecology, 1st and 3d
eds. Pergamon Press Oxford, 1966. 197»
207. FagerUng F. Sind die canina-Rosen agamospermtsche Baatarda? Svenak
Bot. T.dskr.. 34. 334—354 (1940).
206 Fage/hnd F. Die Bastacde der camna Rosen, tire Syndesa und Foitnbil-
dungsverhaltnisse, Acta Hartl Bergum (Uppsala). 14, 7—37 (194b).
209. Fasset: N. C. J им per us ol'ghuana. t. horieordalle and J. scopula rum II.
Hybrid swarms of J. емглмима and J. tcopulorum. Bull. Torrey Bot. Club.
71. 475—483 (1944)
210. Федоры А. А. ред. Хромосомжаае тесл» цжеткояыг расгаяай. Изд-во АН
СССР, Л. 1969
211. Лей/тал М Wheals. In: Evolution of Crop Plants, ed. N. W. Simmonds,
Lnogssan London and New York. 1976
212. Fernandes A Sur la phyioginir des espetes du genre Narcissus L, Both
Soc Broteriana, 25. 113—190 (1961).
213 Fisher R A The Genetical Theory of Natural Selection, Clarendon Press.
Oxford. 1930
214, Fisher R A Retrospect of the crlHdsms of the theory o1 natural selection.
In Evolution as a Process, ed. J. Huxley, George Alien and Unvia, London.
1964.
215. Flake R H.. оол /?ud/off £. Гоглаг В L QuanbUtnra study of clinal va-
riation in J uniper ue virglniana using terpenoid data, Proc. Nat. Acad. Sei.,
64, 487- 494 (1980).
216 Flake R. H., b'rbalscA L. Turner В L. Chemical decumentaUon ol allopat-
ric introgression is J temper as Syst. BoL, 3. 129—164 (1078).
217. Focke w 0 Die PfHnxenmischlHge, Borntraeger, Berlin. 1881
218. Ford £ B. Ecological Genetics, 1st and 3d ed. Methuen and Co, and
Chapman and Hall. Lorad oe. 1984; 1971.
219 Franhel O-. Munday A The evolution of wheat. In: The Evolution of luring
Organisms, Symposium. Royal Society. Victoria. Australia, 1962.
220 Fret er L. E.. Brown F V. A cytotaxooonbc study of Bouleloaa curtipendiela
and В uniflora, Bull. Torrey Bot. Chib. 82. 121-130 (1965)
221. Frisch A Der Farbenelon und Forinenainn d*r B*ene, Zool. Jahrb., 36, I—
182 (1914)
222- Frisch К Uber den Geruchalnn der Bienc und seine blulenbiologiKhe Be-
deutung. Zool. Jahrb. 37, 1-238 (1919)
223 Fry sell P. A. Mode ol reproduction in higher plants, Bot. Rev, 23. I3S—
233 (1957)
224 Ffifnbran? D Cytogmehsctw L'ntrrsuehungen an Гагахассыя In Raumn von
Wien, Otfetr. Bot. Ztttocfar.. 107. 310 3S0 (19ЭД|.
22S FUfnJfa/te D Tarcmazu<n apenninum — nn altes Element medlterraner Ga-
birgv. O»t»r. Bot Zwtschr. HI. 231 -239 (1964).
226 Firnkrant D. Untersucnungea an Papulabonen des Гагажасшв officinale
Komplaxes icn Kontahgebtet der dip lol den und polyploiden Biotypen, Gsterr.
Bot ZaHacbr , 1 IB. 427-447 (1966)
227 Fkrnkfanx D Fine neue meditemne Relilriari der Gathmg Тагаеаеам.
Oatcrr. Bot. Zeitschr. 116. 149—156 (I960).
228 Gajewshi W Cytogenetic investigations on the genus Geum Heredftas,
Suppl Vol,. 578-579 (1949)
229 (ia^ewski V A fertile amphipolyplaid hybrid of Ceu/ч rrtm/r with G macro-
phyUam. Acta Soc. Bot Polon. 22. 411—439 (1953).
Литература
<73
230 Go/ewzAi i* Л cytogenetic study on the genus Gewni L, Monograph nr
BotniiK'we (Warsaw), vol. 4 (1957).
231 (torwiAi » Evuljl un in the gtflua Gturi. Evolution, 11. 378—38b
(1969)
232 Gather E D The genus СоШлша III Th* significance of chiasnuta fre-
quencies as a cytotasoaomic tool, Madrono, 14, 172- 176 (1957).
233 Gofbc* E D The grnuv Cufhn-iui IX hpccintico and chrumusotne r epat ter-
mng. Crefogia. 25. 233 243 (I960).
234. Grtfbt* F. 0 CoUtnbia HandbuoR of Genetics, 2. 323-361 ad R C. King.
Plenum Pre». New York 1«*74
235 Garbtf L D. Garde J The genus CaiLnua 11. Imerspeciiic hybrids invol-
ving C tetefopiuUa. C ihtuMlOf and C tpwstflora, B<jt Una, I IB, 73—77
(1956)
23b Gales R ft The taxonomic un ts in relation to cytogenetics and gene cco-
log?. Amer. NjI. w. 3i BO <19511
237 Gerarutnoz-a II Chrcmosorne alterations as a factor ol divergence of forms.
1 New esperimxmlally produced strains o' C lector urn. which are physiolo-
gically rotated iron» the original tin nis owing Io reciprocal translocation,
Com.pt Rend Acad Set URSS. 25. 148-244 (1939)
238 Guratfei D U Chrcanoeamal translocations in inlcrspecillc hybrids of the
genus Gmvpiwn. Evolution, 7, 234—244 |J9M).
239. D V A new kthal combination In Interspecific eortoo hybrids. Gc-
•Hick 39. 628-43? (1954).
240 Gertie! D U. Pbilhps L L Segregation of synthetic amphiplolds In Gossy-
pittm and Rievli^na. Cold Spring Harbor Symp. Quant Biol., 23. 225—237
(1UM)
241 Gertiri D I1. Sarvtila P A Addibonal cbscrvalions on chrnaiosomal trans-
localions in cotton hybrid?, Evolwtion. 10 Il’S 414 (1958)
242 C-lfijon J н Thcrfoy J M Etiecti of disruptive selection IX Low selection
intrnUv, Heredity, 19. 125-130 (1964).
243 Gi/lrs Л. WandolpO L f Reduction in quadrivalent frequency in autotelra-
plod maize during a period o( 10 years, Anter J Bot. .48. 12—IB (1961).
244 Gu'mcMif f S L, tiesiop Нстчоп I The deme terminology and the units
of micro evolutionary change, Gcnctica, 27. 147 161 (1964).
245 Giais В The germ.nation oi the idea ol t.ologi>al spvc.ts tn Forerunners
of Darwin, 1745 1859. e<ls В Glass. О Trmkin and W L Straus. John
Hcpkins University Press. Baltimore. 1959
246 G&ldLlaP P Polyploidy in 3 igiosperm* menoeotviedofts In Polyploidy:
Biological Relevance, ft! W H Lewis, Plenum Piesa, New Yurfc and Lon-
don, 1980
247. GtMictnid! P В Physiological Genetics. MeGmw-HHI. New York 1933
248 Gvfchctaidf fi В The Material Basis u< Evolution, Так University Press,
New Haven Coiwi. 1M0
249 GofdscftMiidi ft В Thcoreii.al Genet к», tinker m!v of Caliloruia Press
Berkeley. Calil. 1955
250 Gfx&ptwt T H The Genus filcoilana. Chrontca Botanica, Waltham, Mats,
ISM
251 Gctttirb L D Evolut -anary relationships In the outcrossing diploid annual
species oi SttpksA>yrtff:a '(Compasilael. Evolution, 25. 312—3^9 (1971).
252 (jhiiiieb i. I) Generic diiferenliation, svmpatric specirf-.on and the origin
of a diploid species of Supharxcmeria. Amer J Bot. 60. 545 653 (1973)
253 GorfheO L D Erxzvme differentiation and phykrget у in Cixrbia francibcana.
C rabirunda and G a.noena. Evolution. 21. 205 —214 (‘973)
254 Gottlieb I. D Grne4 c confirmation ol the origin о Clariia lingtrlata. Evo-
lul on 28, 944-250 (1974)
255 Gottlieb L D Electraphureic evidence and plant systematics. Annals Mis-
souri Bot Gard . 44. 161—160 (1977)
474
Литература
256 Gonheb L. D. Pbeootypac venation in Siephanomrria txigua sap. eoronaria
(Compositae) and iU recent dsrtvativs kpeau «uiaRrsur antis», Auwr, J. Bot,
64. 873—MO (1977).
257 GottUpb L D. Stephanamxria nuMtiu/f/ий (Compositae), a new speoes
Irons Oregon. Madrone, 35, 44—46 (1976).
258. Gaitichalk F Die Badeutusg der Polyplotdie ftir die Evolution der Pflan-
icn. Gustav ПьсЬес, Stuttgart, 1976,
259. GouM F F Notes on apomixis in sideoats grama, J. Range Management,
12, 25-28 (I9M).
260. Goidd f. w Chromosome numbers ol some .Mexican grasses, Caiud. J. Bot*
44, 16ВД—IW6 (1966).
261. GoiUd F. W-. Aopodia I. J. Biosystemabc studies in the Bouteloua curti-
ptfidaia complex I. The anecplord rtxiuma’.ous B. curh>mdada o( Texas,
Amer. J. BoU 46, 667 ДО (19И).
262. Gould F. F., Kapodin Z. 1. Biovysternat* studies ia the ЯлиГй1оил cartt-
ptndula complex. Il Taxotnaciy, Britlania, It. 182—207 (1964).
263 Grant A.. Grant V. Genetic and taxonomic studies m Giua. Vtll. The Cob-
webby G.lias, Alisa, 2, 203287 (1866).
264 Grant К A.. Grant V .Mechanical isolation ol Satoa apinno and Saloia
/палата (Lab.atae). Erolutxm, 18, 196-212 (1964)
265. Grant V. Pollination anteOW as Isolating mechanisms in towering plants,
Evoljlxjn, 4, 82 - 97 (1949).
266. Grant V Genetic and taxonomic studies in GUin I. Gttia capitate, Alba,
2. 239—316 (195U).
267. Grant V. The flower «nahney ol bees, Bo*. Rev, It. 379 396 (I960).
268. Grant V. Genetic and taxonomic studies in (JiUn. 11. 01йа capital a abro-
tanifolio. Ahso, 2. 361-373 (1ВД.
269. Gran! V. Genetic and taxonomy sluiLea in (Mia. III. The Gih'a tricolor
complex. Ahso. 2. 375-3*3 (l»2).
270. Grant V. Isolator and hybridixatioa between AfWbgia jormota and
A pubticrn*, Alito. 2,341 -Зло (1962).
271, Grant V. Cytog«®ebcs of the hybrid Gtlia mtttirfolialaxacMlicaJalla. I. Va-
riations m metoMa and poiyptaidT rate as affected by n и tri tonal and gene-
tic conditions. Chromoecma, S. 372—390 (1952)
272. Grant К The rob ot hvbridaation ia the evolution ot tba Lsalysteoimed
Giliaa, Evolution. 7. SI-64 (1963).
273. Grant V. Genetic and taxonomic studies in Oilia. IV. GlUa achiliaa folia.
Alito. 3. 1—18 (1954).
274. Grant V Genet* and (axoeoewe studies in GlUa. VI. Interspecific relation-
•hips in the Leafy stemmed Gilba. Aliso, 3, 35—49 (1954).
27S, Grant V. Genetic aad taxeaomic siudus in GiZlo. V. Oilia chtrorum. AHso.
3. 19-34 11954).
27*. Grant V. The genetic structure of races and species in Gilia, Advances in
Genetics, 8, 55-87 (1956).
277. Grant V. Chromosome repatterning and adaptation. Advances ia Genetics,
8. 89—107 (1956).
278. Grant V The influence ol breeding habit on the oulcocne of natural hybri-
disation in plants. Amer. Nat, M, 319 322 (1956).
279. Grant V. The plant species tn theory and practice, la: The Species Problem,
ed E. Mayr. Amer. Assoc. Adv. Set, Washington, D. C, 1957
280. Grant V. Ths regulation of recombination la plants, Cold Spring Harbor
Symposia Quant. BloL 23, 337—363 (1968).
2*1. Grant V. Natural History cf the Phlox Family. M. Nijtoff. Tbs Hauge,
1959.
282. Grant V. Tbe Origin of Adaptation. ColumbU University Presa New York,
1963.
Ъиература
47*
283 Grant V The biological composition ol a taxotiouix spco«s m Gdia. Advan-
ce» ш Genetic*, 12, 281-328 (1964).
284 Огам; I' The Ardai.ecture ot the GeUBRiaam, John Wiley, New York,
1
J8o Слеп/ V Geoc.x and taxonmu^ atndiea 4i Cilia XII. FertIMty relation-
ships of the polyploid Cobwebby G.lias. Alt so, 5, 479—307 (i%4).
2BG Gr*-n; Г Sr-ecies hybrids and spontaneous arr.;.hip|i>ds in the GlUa loci-
niata group. Heredity, 2U, 537— 5э0 |l**5)
287. Gram I* Block inheritance oi vubi-ity genes .n p ant speoea, Amer. Nat.
I<№ »l-60l (1966).
28o Gram V Linkage between viability and fertility in a species cross in Gilja^
M. 8b» *-8OU USKffij
289 Grant k‘ Selecion for vigor arxl fertility in (he progeny ai a highly sterile
species hybrid ui Gliia. Genetic», 83. 767—775 (19o6)
?90 Grant 1 The «elective vng n и iixoanpalability barriers in the plant gen»
GOiu. Amer . 100 49-118 (1966)
291 Grunf V jhe ОП0П ol a new species oi GiLiu in a hybridization experiment.
Genetics, $4. 11Я9—1199 (1966)
292 Grant V. Linkage between morphology 411 Viability in plant species. Amer.
Nat. 101, 125—139 (1У67)
[ч Unt PUnt Sf*aation’ l,t • ^®|un|b»a University Press, New York,
294 Grant V. Geneties of Flowering Plant*. Columba Uni* Press, New York.
1975
295 Grant V Isolation between A^tiu'rgui fcsmoia and A pubaterns. a reply
and recons derackn. Evolution, 30. 625—<»2а |197(ii
296 Gromr V Pcpula xm structure in relation to mar roe volution. Biui. Zefllral-
bial . W. 129-139 (1977)
29/ Grant V Character coherence in natmal hybrid population in plants. Bob
Gas. И9, 443—448 (1979)
298 . Grant V Gene flow and the homogcnnly of species populations. Hiol ZeD-
tratblatt, 99. 157—169 I I98U)
299 , G'lintf V. Gram .4 Genetic and taxonomic зреекя in Glhc VII Th* Wood-
land Шмь Aliao. 3. 59-91 (1954).
300 Grant 1 . Grant A Genet , and taxonomic studies in GiUa, XI. Fertility
reiationshaps oi th* <! ?k>id Cohwwbhy Gilin-. Aliso, 4. 435— 481 (1960)
301 Grant V. Gram К A R<nw Pollination in Им Phlox Family, Columbia
LnivcfS ty Press, New York. 1965
302 Огаш V. Grant К 4 Dynamic» of ckxul nucrospccies tn cbolla cactus.
Evolution. 2D, 144—155 (1971|
333. Gram V. Grant К Л Natural hybridizabon between the cholla cactus
apes tea Opu.irio tpinusliu anc О oerucotur. Proc Nat Acad Set, 88,
19Ю—1МБ H0TIJ
304 Grant F. Grant A A Hybridization and чаш Lion .n the Opuniia ptiacacan-
Uta group in Central Texas, Bo!. Gaz , 140, 233—215 11979|
305 Gran! V . Grant К A Clonal mi»'ro?pceir* of hybrid ong’n in the Opwnlia
harfAsermen group. Bo’ Gat 141. 101—106 (19811)
3116 Grant IT A BttUrn Л1 R. h'cttaiunurt D The cytogenebes of Lnlvr.
I Frnbryo-rut1iL.’6d intecspecliic d pto.d hybnds cxisety related !o L corm-
r.ttlaTt№ l . Canad J Genetics and Cfol ♦, 105—128 (1952)
307 Grau A P. Mammal an Hybr-ds Con.mooweslth Agricultural Bureaux,
Famhirr Royal, England. I MM
33s Grav A P. B.rd Hvb~ids ConuDonweatth Abrkuluaral Bureaux, Farnhim
Royal England. 195ft
309. Grtrnlrt! V. //. Sterile and fertile ampliidiploids their possible relation 1л
the origin of Ntcoftana tabacum. Genetics, 2». 3OI—324 (1941).
476 Литература
310 Grun Р. Cytogenetics studies at Poo. I. Chromosome numbers and mocpho*
logy of inlerspedfic hybrids, Amer. J. Bot4 41. 671—678 (1964).
311. Gfan P Early stages in the formation of internal harriers to gene eiclunge
btfverri diploid aperies b! Sofcnum, Amer. J. Bot , 48. 79—89 (1961)
312. Grun P. A uber tin M. Radio* A. Multiple differentiation of plasmons of
diploid species of ЛЯолаип, Genetics. 47. 1321-1338 (1962).
313. Gulick /, T. Evolution, racial and habitucLnal, Carnegie Inst, Washington
PuM,2fi (1905).
314. Gattakatui A. The origin and properties of the European blackberry flora,
terabits, 78, 249-177 (1942}
315. Geatafnon A. The genesis ul ibe European blackberry flora. Lunds Univer*
sltets Artskrill 30(8), 1—2W (1943)
316. Gaitafxson .4 The constitution of the Rua canina complex. Heredias, 30,
405-428 (1944).
317. Qustajison A Apomixis In higher plants. Lunds Univeraitets Arsskrift,
42 -43. 1—370 (1946, 1947).
318. GuttafMan 4 Polyploidy, life form and vegetative reproduction, Heredilas,
34, 1—22 (1948)
319. Gustakton 4„ НААаяиоя A. Meioeis mi soma Roas-trybrlds, Bottnhka No*
titer. 1942. 331—343 (1942)
320. Gusfofsaon M. Evolutionary trends in the AtnpUx tnaagularls group of
Scandinavia. I. Hybrid stcri.ity and chromosomal differentiation, Bounlska
Notiser, 124 345-392 (1973)
321. Weartnun J Plaatae Hybndae, Uppsala, 1751; AmoenHatea Academics*.
Holm. 1764
322. Hagen C. F. A contribution to the cytogenetics of the genus Oenothera
with special reference to certain forms from South America, Indiana Univ.
Publ. Science Series, 18. 305 348 (1960).
323. Hag er up O. Uber Polyptoidie tn Beziehung zu Klima, Okologie und Phylo-
genle. Heredilas, it, 19—40 (1932).
324. nage/up O. The spontaneous formation of haploid, polyploid and anrupioid
embryos in some orchids, Biologlske Meddelelwr, in, 1—22 (1947).
326 Half 1 В Biosyswnafic* of the New Zealand flora 1945—1964. New Zea-
land J. Bot. 4. 559-545 (1966)
326. Hau 1. В The chromosomes of the Сиргемасеае. I. Tetraciioeae and Acti-
nostrobeae (CaHitroidoa), New Zealand J. Bot., 4. 277-284 (1968).
327. Hair 1. В , Beutenberg E !. Chromosomal «volution fn the Podocarp«etaa.
Nature. 111. 1584-1Ш (1958)
328. Hau ] В. Beuzenfiarg E 1 High polyploidy Ln a New Zealand Poe. Natu-
re, 19». 160 (1951).
329. H&kanston A. Coalributions to a cytologscal analysis of the species diffe-
rences of Godeila amoena and О. Unapi. Heredilas, S3, 235-260 (1947)
330. H&hanston A Meioeis and pollen mitosis in X-rayed and untreated spikelets
of Eleocharij pakutrts. Heredilas, 40, 325—345 (1954).
331 //Mansion A. Hotocentric chromosome in Eltocharis. Hereditas, 44. 531—
$40 (1958)
332 Hall R 4 Genetic analysis of interspecific hybrids in the genus Brontes,
section CeraJochloa. Genetics. 48. 175—192 (1955)
333 Halt M T. Muhhefiee A. Ooa/ltp W. R. Clr-znosome numbers of culti-
vated junipers. Bot G« . 140. 364—370 (1979)
334. Haller i R Variation and hybridization fa ponderoaa and J«f1rty pines,
Univ. Calif Pvbl Bot. 34. 123—166 (1962)
335. Haeuick 1 L. Genetic variation and longevity. In: Topics In Plant Popula-
tion Biology, nd O. Solbrlg, Colixnbia Umvannty Prms, New York, 1979.
336 Hanoi! P Polyploidie Frrquenz uad gnogranhisdie Vrrbreituig bd h&heren
Pflansen. BfciL Rundschau. 4. 183—196 (1966).
Литература
477
337 Hanrii Р., .Merftn D. Cjlosyslcmalische UnlCfsuehungcn in бег Arten-
gninpt uic 1'ic.a sa/icu L II Die KuIturplU/UC (Befkn), 14, 137—161
<i«6)
338. Harbtrd D f Otoervalxm* cn population structure and xmgevlty of Fettu-
ca rubra L . New Phytol, *0, IM—206 (1961)
339 Harbvrd D J. Same ob&enatlons on uatural ctontb in Ftiluca скЯла. New
Phytol, 61, *5—IOC (1962)
340 Hat din G The competitive exc|ws*on principle. Science, 131, 1292— 1297
(196Ю1.
341 . Hardin i DT Studies in *hc НфросахОмссас lit A hybrid swarm in the
buckeye», Rhodora. 59, 45—31 (1957).
342 Ha* din J IT S’.tk-icj in the llippocastaneceae. IV. lly brut nanon in Acslu-
nodora. St. 185—203 (1967)
343 . Hardin J W. Hvbfidiiation and introgressKin in Qoercus aJba, J. Arnold
Arb. M. 336-ЭвЗ (1975).
344 Ha* ding J-, Alla*d R. V . SmcUzer D C. Population studic* in predociican-
lly Mit pal «naled species IX Frequerscy dependent select.on ia Phatedtns
lamu/ws. Proc Nat Acad Sr , И. <*»-)04 (1966)
345 Marian 1 R Natural breeding structure m the Hrrunai caHnatm complex
аь deform ined by population anal ум?» Amer. J, Bot. 32. 142—148 (I'Mb)
346 . Mo'ian I P Apomiiia in side oats grama, Anw. J. Bot. 36. 495 -499
(1949).
347 //urlun / R. de Tet J M J Toward a rational class!fka bon of cultivated
plant». Taxer. 20 509—.517 (1971)
348 Murlun J R, ZuLurv D Ditlnbubon ol wild wheats and barky. Science;
153, IU74—1080 (1966).
349. Ha*iund S C The gcn»'*ca) conception of the species. Biol Rev, 11. 83—
112 (I Mr)
350. Harpe* J L Biological Dora ot the British Hies RnrruncaJos aerts L . Ra-
ftuncuias repeat L., Ranunculdut bullrmaA L, J Erol. 45. 289—342
(19571
351 //с«до / L. Popxilatfon Biology of Plante. Academic Press, New York,
1977
352. H<t*pe* ! L. While 1. The demography of plants. Ann Res* Ecol Sy st, 5.
419-463 (1974).
353 Motaf/on $ С Ep phyllvm Handbook. Abbey Garden Press, Paiadeaa, Ca*
lit. 1931.
354 Hetties / G. Ilfiriing J P Hu* Potato»* of Argentina, Brazil. Paraguay
and Uruguay A Blosystwiatlc Study, Clarendon Press Oxford. 19Й9
3&&. Hayaia В The natural clasutkaban ci ptants according to the dynamic
system, Jcoo, Plant. Fortnoe., 10,97 234 (1921)
356 Hayman fi I. Matter H, The progress ol inbreeding when bnmozygolcs
are al a dtredveniage. Heredity, 7. — 83 (1953)
357. Hecht A Cytogenetic studies ci Oenothera eubgenus Ratmonnla. Indiana
Uriv Publ. Science Series, 16 255—304 (I960)
338 Hrckard L R Taxonotrit MudlPS in the РКлееЛа magHlanira polyploid
complex wrth special reiercnce 1o the California members. Univ. Calif.
Publ Bot. 32. 1-126 < l%0)
Vi9 Hegt G llluslrlerte Flora von Mittel-Furopa. 7 vol* Carl Напьет. Munich,
1909-1931.
350 Heitbarr. О Chromosome numbers and dimensions, spedes formation and
phylogeny In the genus Caret. Hr-edltas, S, 12^—216 (1924)
361. НеЛЬогп О Aneup'.oidv and polyploidy in Carex. Svensk Bat Tldskr. 24
137-146 (1932)
362 Hedbarr. 0 On the OTigis and preservation of polyplcidy, Weredrias, 19.
233-242 (1934)
47В Лыгервгдоа
М3. Heilborn О. Chromosome studjes in Сурегасеае Ш—IV, Нм«!1гая, 25.
224-240 (1939).
364 Htiitr С. В. Hybridization between the sunflower specie* Hetlantluu an
Лиля and H. petwlaris. Evolution. I. 249—262 (1947).
365. H titer С. В Natural hybndixatxm with partxular reference to introgression,
Bot Rev, 15, MS- «7 (1949)
366. Heiser С. B. Study in the avoiutiua of the sunflower species Melianthus
аллй.ис and H boUadeH, Uaiv Calif PuM BoL. 23. 157—20® (1949).
367. Heiser С. B. A cutrparjwn o! the flora aa a whole and the weed flora of
Indiana as to polyploidy and growth habits, Proc. Indiana Acad. Scl., 59.
M-70 (i960).
368. Heiser С. B. Hybridisation In the annual aua f lower v НлИлпНии апликех
XH debdu var eurtunertfolitu. Evolution, 8, 42—51 (1961).
369. Heiser С. B. Hybrldlaaboa in th* annual aunllowcra. ЯдеопМш оплилаХ
XH. argophytius. Amer. Nat, M, 86—72 (1961).
370 /falser С В Variation and subspeciation in the coounon Sun I bo war. Helion-
thtix annuus, Amar. Midland Nat.. 51. 287—306 (1964)
371 Hnur С В The North American sunflowers (H eltanthete), Memobs Torrey
Bot. Club. 22(3). I—218 (1989).
372 /faiser С. В Introgression re examined. Bot. Rev. 38, 347 386 (1973).
373. Hester С. B. The Suolloww, University ol Oklahoma Pre**, Nor man. Okla,
1976.
374, Налег С. B. Martin F C. Smith D M Species crosses in Helianthus.
1. Diploid Special Bfittonla, 14. 137—147 (1962)
375. Heiser C. 6 . Smith D M The origin of Melianthus mofti/farux. Amer. J.
Bot. 47. 860—865 (1980).
3Tb. Herbert W On the production of hybrid vegetables, with the result oi главу
eapemnents made 1л the uivwtigaboo of the subtat, Trank Hurt Soc. Lon-
don. 4,15—50 (iftM).
377. //irsey F. M, A’obs M. A, Bjbrhman O. Experimental studies on the na-
ture ol species V. Biusyalemalics. genetics and physiological ecology of the
Ef^ttuanihr section of Mimeifaa. Carfwgfe Inal. Wash PubL. 628, 1971
378 Hiorlh 0. Eine Serie multi pled Allele fur BlutenxeichR-ongen bei Godetia
amoena Heredias, 26, 441—453 (1940)
379. Hiarth G Zur Geoclik und SystwnaUk de; omoerw Geupp* dcr Gattung
Godetia. Zarischr. Ind Absiammungs- и Vererbeng$UHre. M. 289—349
380. № M C Abrahamson W G Seed and vegetative reproduction In re
latiom to density m Fragana oirginiana (Rosaceaa), Axner. J. Bot., 64,
1003—1007 (1977)
38i. Hoilingihead L. A lethal factor in Crepu efieetive only in an interspecific
hybrid. QenetKx II, 114-140 (19301.
382. Hopper 5. D., Burbidge A. H. Assort alive pollination by red wattlebirds In
a hybrid populatson of Anigozanlhos Labill. (Haemodoraeeac). Austral. J.
BoL, 28. 33S-350 (1978).
383 Hovanitt V. Increased variability in populations (oliowing natural hybri
d'rutkUL In: GeoeticK Paleontology and Evolution, ed. G. L Jepsen, Price-
ton University Press, Princeton. N. J.. 1949.
384. Hoeeard H w*. Potato cytology and genetics, Blbl-ographia GensCxa, 19,
87-216 (1952-1959).
385 Hubbt C. Delco E A Mate preference in cnalae of four species of gambu
stint Itshcs. Evolution. 14. 14S-I52 (1960|.
386. Hvbbt C. L Hybridisation between liah speciea in nature. Systematic Zool,
4. 1-20 (1085)
387. Htibbi C L. Miller R R Mass hybridizatioo hrtwaen two genera of cyprt-
ntd fishes tn the Mohave Dcsen. California, Papers Michigan Acad. 3d.
Arts and Latters, 28. 343—378 (1943).
•Литература
479
м 5 § § 2 § § £ § М X НН
Mullen L HeliAfia longipes Goidie and its «Лам, BoUnHka Kobser, 25’.—
270 ,1943)
Molten E The Crzuifium elpfaiun complex A case of world wide in tro-
gresUvc hybrid ration. Svensk Bot. Tieskr. 50. 411—495 (1994)
/1мгsr С C Embryo sae formation in diploed and plyploid species q| Rq-
*ew. Proc. Roy. Six. 0. 10». 126-14» (1 Ml)
//мгаАхмоя J. В. Stephens S G. The Evolution of Ойодкт Oxford
University Press. London. 1947.
J/uxihy J S Evohilion The Modern Synlhteia. George Allen and Unwin.
London. 1942
/Ашотта У Chromnvomaj evolution in Ц» Mibtribe Asterinat, Evolution,
11, 1НМ-19Э (196W.
/тол A. C, Allard R F Population atudica in predominantly seli-pollina-
tod spedcs. VI. Genetic variability between and within natural populations
of wi.d oata iron» differing habitats in California, Genetics, SI, 49—62
(1965).
/attapn R C New tow chrcnsoxwnc number for planb. Shhkc. 128,
1115-11 lb (I957|.
/тл 5 A' Male sterility in flowering plants. B’.bbograpbi Gen elk a. 18,
101—165 (19591
/ou* S, A*. Cytogenetics ol rye (5c<oJr spy ) BAlwfiiaphu Geoatica, 19,
1-96 (19601.
/•ft S A’ Population structure and the effects of breeding system In Crop
Generic Rnources for Today and Tomorrow rd О. H Hankel and
J G Hawkes. Cambridge University Prate. New York. 1975
/di* S A' Adaptive strategies polymorphism, plasticity and homeostasis.
In Topics m Punt Population Biology, rd О Sol brig. Colunb.a Univar-
lily Press. N«w York, 1979
Jain S A . Allard R. V. Popu:ation studies .n predominantly self-роНлы*
led species. 1 Evidence for hetcroxygo'.e i dv ant age in a closed population
Ol barley, Pnx Nat. Acad Scf 46. 1371—137? (I9o0)
Jain S K. Allard fi F The nature and stability of eqwilibria under opti-
mising selection. Proc. Nat Acad Sci. 64, 1436— 1443 (1963)
Idin S К В'adsnote A D Evolutionary divergence among adjacent plant
population: I The evidence and its thcxuetxal analysis» Heredity, 21,
407—441 (1966).
iaia $ A . /cpi К В L The progress oi inbreeding in a pedigree-bred po-
pulation ol barky, EuMiytica, II. 229—232 (1962)
/out S A. MarvAaH D R Population eludlee in andooiiBanUy self pollina-
ted bpecks. X. Variation in natural population* ol Atwrxi }aJua and A bar-
baa, Amer. Nat. 101. 19-33 (19571.
/ерхоп V L The Trtea ol California. Aaaociated SluJwils Store, Berkeley,
I xli1 M -
/оАл В. Lewis К R Genetie apaciatlon in the gris*Snpper Eypr^pocnanut
plarawt Chramosocm. 16 306—344 (1965)
407 Jrdin B. Lewis K. fi. The MekXlc System. Protoplaunato'ogia, Springer-
Vcrlag. New York, 1965
408 Zahn В. Letois K. R. Chroanosonx variability and geographic dUhibvtion
in insects. Science. 132. 711—721 (1%6|
409. /ohnron A ST. Pociter / G Polyploidy and environment in aretlc Alaska.
Science. 148. 237-239 (1^5).
410. /ttlrtson A F. Packer / G r Reese G. Polyploidy, distribution and environ*
went In The Quaternary of the United States, ed H E. Wright and
D. G Frey. Princeton University Press. PrirKelon. N. J. 1955.
411. Johnson B. L Cytn-laxcnnmSe studies in O/pjopsts. Bat Gar. 107. 1—32
(19461.
412 johruon G. В Ему—ю polymorphism genetic variation in the physiological
480 Литература
phenotype In- Topees in Plant Population Biology. ed. O. Solbrit Coham-
baa University Press, New York, 1909.
413. /«Anson L A S_ Bnggs В 0. Evolution in the Proteaccae. Australian J.
BoL. 11.21—61 (1963).
414. Johnson L. A 5. Or the Proleaeceae — the evolution and dasaification of
a southern lamilr. Bot J. Linnean Soc. (London), 70. 83—182 (1975).
410, Johnston J. S- Herd IF. В DIsmtmJ of desert-adapiad DrascpAdd: the
saguaro-breeding D ntgroapiracvla. Amar Nal, 110. 629—651 (1976).
416. /сама D F Genie and cytoplasm* control ot pollen abortion In maisc.
Brookhaven Symp. B»oi. •, 101—112 (1956).
417. Jonas E £. Pollinator «onstancy aa a pre pollination isolating mechanism
between sympatric spec**» ol Gerodton. Evolution. 32, 1в9—1М (1978).
418. Jones S В Cytogenetics and afftnrtsed of Veraiua (Compcsifae) tram the
Mexican highlands and eastern North America. Evolution. >0, 466-462
(1976).
419. JonsaU В Linnaeus's views on plant classification and evolution. Bataniska
Nntiser, 131. 523—530 (1978).
420. Jordan D. 3. The origin of species through isolation. Science, 22, 545—562
(1905).
421 Jordan К On mechanical selection and other problems. Nwit ZooU 3»
426—5» (1896).
422 Jowrll D Population studies on lead toterant Agrostu tentui. Evolution,
18» 713-81 (IM).
423, Juiai T. H. Molecules and Evolution, Columbia University Press, New York,
l'-Жб
424. Knmemoto H.. Sklndo Jf. Meioszs in interspecific and intergenenc bybn da
of Honda. Bot Cm. 128. 132-138 (1964)
425. Ronnenberg L F.. Allard R IF Population studies in piedominanlly sell-
pollinated speoes. Vlll. Genetic variability jn the feaface Buerostach^
complex. Evolution. 21. 227-240 (19671.
426 Kapodia Z /, Gould F. V. Bioswlematic studies in the Boulrfmta cut Ji-
penduia comptex IV. Dynamics of variation tn B. curtlpendala var. cars pi-
tons. Bull. Torrey Bot. Qub. 91, 466—478 (1964).
427. Карлммекксо Г. Д Помплмцпые гибриды Raphnmsu L хвгахяка adorn*
сеа L Труды бюро зриил. бот, гамет и оелехпмм. 17, 306—410 (1927).
428 Xw* D D Trends in systematic botany, Survey of Вю1 Progress. 3. 47—
107 <19671
429. Kttp E. Interspecific hytridualxm in Яйы, Genetics, 33, 1—23 (1962),
430. Kemper man J. A.. Barnes В V. Clone sisc tn ainenran aspens, Canadian
J. Bot. 84. 2603—2607 (1976).
431 Kerner A The Natural History of Plants. 2 vote Translation, Blackie and
Soo. London. 1894—1896.
(Имеется веэевох. Кернер А. «Форма и жшп. растяпа», т. I, т-ао «Про-
соепепе». Саккт Петербург, 1899. еИсторие растееей». т П. 10021
432, Arrsfrr Н. V.. Levin D. A. Neighborhood site in LiMunpermum caroJiruense.
Genetka, 60. 577-587 (1968)
433. Khashcxj T. N. Polyploidy in gymnoeperniK Evolution. 18. 24—39 (1959)
434. Kkush G. S. Cytogmattea ot Aneueloidy. Acadaeiic Press. New York. 1973.
435. Kiang Y. T. Floral structure, hybridization a ad evolutionary relationships of
two specks of Mimuiu*. Rbodoea, 78. 2»—233 (1973>
436 Kihara H CytologiKhe and gcaetlsche Stedten bd wlchlgcn GetrridesHen
nit besondefer Ruckslcht auf das Verhalten Лет Chrnenoscmcn und dte Ste-
rililat in den Bastarden, Metn Coll SeL Kyoto Univ. BL 1. t—200 (1924).
437 AiЛа/a H Consideratloas an the evolution and distribution of Aegilops
spedtt bssad on the anilyzer-meLhod. Cytologia, 19. 336—357 (1954).
438 KiAara H. Ono T Chromosomcnxahten und systems tfsebe Groppierung dcr
/tones-Arten. Zvitschr. Zellforsch. u mikr. Anatomic, 4. 475—481 (1926).
Литература
481
439 A.mg /. С 'ntersparific relationships within the guarani group ol fkroso-
fMa Evolution, I. 143—153 (1947)
440. King .M. Flfoon A C Evolution al two Inn* in humam and (tiiihpaniees.
Science, 188, 107-116 (1975).
441 Klekou-sbi £ i RcpcodiK'.lve biology of the Pieridop hyla II Theoretical
consider*!ion«, Hot. J L.nrwao S« . 82. 347—359 (1969)
442 Klekauih L J . iMtr it <j Evotutianarr significance oi polyploidy tn
the Puj.odupLyla. Science, 153. 305—307 (1Уб6|.
443 AmigA/ G A , Robertson 4 . Haddington С H Stfectioe ivr sexual isola-
tion within a species. EvoUitiue. It. 14—22 (19У-1
444. KOlreut«T J Q VuiMuliga hiaefoicht von aiwigen da* Getchlei tit dar Pflan-
ren betreffenden Verauchen und Bcobachlungeo Rcpnat. 1893 In Ost-
wald'* Kbs.sker, Leipsif. 1761—1766
445 Kaoptnan, Л. f Natural selection for r<p*odue*.i»t lvoi.it ion between Dro-
sophiia рхеиРаа8леиги and fruc/ifiAtfa Evolution, 4. 135—14»
(I960).
446 KfurMarg A R Intraspecific variability in the response oi certain native
plant specie* to sapratine suit. Amer. J. Bot, 3», 41Ж— 419 (1951)
447. Kracbekerg A. R, (.hromowcnc numbers tn Sdene (Carvophvllaccae) 1.
Madrono, 12. 23*-246 (1Я54).
445 Krucfteberg A. ft. tnlcrspccil* hybridizatMin o4 Sdene, Amer J Bot, 42,
373-378 (1933)
4<9. K'ȣ\tba>g A R Variation ia fertility oi hybrids between populations of
the serpentine speacs, Streptantkus giandulomt Hook. Evolution, 11. 185
211 (1957)
450 Kn^kfberji A. ft Chromosome numbers in Sdene (Caryophylaccac) II Mad-
-ono. 15. 205 - 215 (INC)
451 XrerWurr .1 R Art Firial crosses ol ueatern North American Silenrs, Bril
tonia. 13,306—333 (1961).
452 RtitcMug /4 R Ti e implications of ecology lor plan, syslemata:*, Taxon,
1Я. 97—170 11969)
453 Kugler H Blbtcn£4alc>gte. 2nd ed. Gustiv Fischer. Stuttgart, 1970
454 АигаЬидоЛ. <4 Evolotorr and variation n Jap.ncsc species of Trilham.
Evolution. 12. 296—310 (196*)
455 Kurabu«tt*AJ At Levis II. Raven P fl Л cocnparative «tudy of mitosis in
the Onar-jcear Amrr J Bot 49, 1003 102 H (1962).
456 К®itA A? M P^ll naiicc. hybridization and ctbolcgic»- isolation oi AAinan-
.'Aut Mowor ard R тего!^пат (Rhinandnidcoc ScxopltulariacMe) by bern-
blrbw* (flom^s Latr), Тахоя. V, 145-158 (1978)
457 Ladtgrs P. ¥. Variation in d-oughl resistance tn adieccr.t edophic popula-
tion» of EuMtyptnt rfrtii/tous Labni. AiMtra! J Erol. I. 67—76 (19761
458 La.TtpreiA.' H Ihe Artgrcnze zwiadien FAowofus lutgarH L und mullijlo-
fui I im, 27.51 — 175 (19411
45<O Lanpraik! H Die geniach nlasmafrsdie Gnindlage At' Artborrlare. Agr
I fort Genet. I. 7B-142 (19441
460 latino H A atiady o4 the breeding barrier between Gilia australi* and
Gdia .splcidim Ph D the*)’, СЗагстмг. Graduate School, Claremont,
Cail 1ОУ<
461 /.ms L H A re«14on ol the Padbc coast iria«, AI**Ok 4. 1—72 (1956).
462 /ма L 9 Hybr.dilation and speciation in the Pacific coast Irises. Alisa.
4 237-309 (19691.
463 /.<mu L V !rii trnnii S Wats, a new traiHfer to Ibe subsection Evan*t*.
Mrso.4.311 319(1959)
454. /.м2 L ЧГ. Hfrtibcf D E Hybrid r«I on and inheritance in orchids In-
7Ы Orchid*, ed C W.thne-, Pottild Prets. New York. 1959
455, /тяпу К Alfalfa, lucerne fn Evol-jtiun of Crop Plants» ed. N W Siraunooda,
Longman. London and New York. 1976
31-45
452 Летедодо
466. £«doq A. Cytologies! studies in the AlUum paniadainm group. Heredltaa,
99, 317 -370 (1937).
467. LtfDGA A. Polyploidy in flu, sugar berk and t.moihy. Heredltaa, Suppl
Vod. 46-47 (1949).
468. Lavin D A. Chromatographic evidence of hybridization and evolution in
Phlox mandala. Amer. I Bot, 53. 238-245 (19661.
469. Lavin D A Hybridisation between annual species of PMox; population
structure, Amer J. Bot. 54, 1122—1130 (1967).
470 Lepra D A The genome constitutions of eastern North American Phlox
amphlploid». Evolution. П. 617—632 (19681
471. Lavin D A. Associative pol line lion in Lyihrutn, Amer. J. Bot, 57, I—5
(1970)
472 Lavin D A Рем pressure and recomb nation вуммп In planta, Amer. Nat.
199. 437-431 (1975)
473 Lavin D. A Genic hate* oaygoaity and protein polymorphism among local
populations of Oeno(Aa/u biennis. Genetics, 79, 477—491 (1973)
474 Десил D A. The origin Ы isolating mechanisms in flowering plants, Evol.
ВЫ., II, 155—317 (1975).
475. Levin D A The nature a! plant species, Science. 204. 351-354 (1979).
476 Levin D. A. rd Hybr»dizatxon. An Evolutionary Perspective, Dowden. Hut
•hinson and Poe*. Stroudsburg, Pa, 1979.
477. Levin D A . Howland 0., Steiner E Prolein polymorphism and genic hete-
roxygosity in a population of the permanent translocation bcteroxygole.
Oenothera biennis, Proc Nat Acad. 5ti. 69. 1475—1477 (1972).
478 Levin D. A. Kersiar H W. An analysis of inter specific pollen exchange in
PMoa. Amer. Nat, 101. 387—400 (1967).
479. Levin D A.. Kerslaf H. W Nature: selection for reproductive Holston In
Phlox. Evolution, 21, 579-687 (1967).
40. Levin D A. Kantar H W. Local gene dispersal In Phlox. Evolution, 22,
130-139 (1968).
481 Levin D A. Kantar H V Density dependent gene dispersal in LLatrU.
Amer. Nat, 183, 61—74 (1969).
482. lavin D. A . Karsler ft IF Ciena flow in seed plants, Evol Biol., 7, 139—
220 (1974).
483. Lavin 0 A, Ritter K-, EJlstrnnd N. C. Protein polymorghkan in the narrow
endemic Oenothera огеапеплх, Evolution. 33, 534—542 (1979).
484. Lavin D A.. School B. A. Corolla color as an Inhibitor of Interspecific hyb
rtdixatio® in Phlox, Амет. Nat., 104, 273—283 (1970).
465. Lavin D. A.. Wilson A. C Rates of evolution in seed plants: net increase
in diversity ci chromosome numbers and species cumbers through time.
Proc. Natl. Acad. Sci. 73, 2086-2090 (1976).
486 Levins R Evolution in Changing Environments. Princato® University
Prtas, Princeton. N J. 1968.
487 Lavy M. Levin D A. Genic hricroxygooi'y and variation In permanent
translocation heleforygotcs ol tbc Oatwlkera biennis complex. Genetics, 79,
493—512 (1975)
458 Lewis D Phvuology ol Incompatlblity in plants. 111. Autopolyplolds, J.
Geoefks.46, 171—1« (1943)
489. Lewis D. The Incocnpatibikity sieve for producing polyploid, J. GeodlcK 45.
261—264 П948).
490 Lewis D The g»n*tk Integration of the broading sysiam. Tn: Reproductive
Biology and Taxonomy of Vascular Plants, ed J. Q. Hawkes. Pergamc-n
Prcs.v Oxford, 1988.
491. Lewis ft. The mechanism of evolution in the genus Clarkis, Evolution. 7,
1-20 (1953).
492. Lewis H. ChrotmoKume phylogeny and habitat preference of Clarhia» Evolu
tUm, 7, 102—109 (1963).
483
♦S3 Lnw H Cataslcphic selection as a factor in speciation, Evolution, 18.
257-271 (H№8).
494 irais II Spcdatxxi in Dowering plants, Science. 152. 167—172 (1966).
495. Lews H. Ef/iing C EMptuntHm gypso/fatum, ft diplrk d species oi hybrid
origin. Evolution. 13. 511—525 (1959).
4% Laois H LnMs M. The gFnus Ctarkta. Uilv ClM Puhi Bot, 20. 241 —
392 (1955).
497. Lewis H. Raven P. it. Rapid cvo(ut>on in Clafha, Evpluticai. 12, 319—
336 (1958).
4$в L4CPM H. Raven P if, VrnAaJeiA C 5 Wedbcrg H L Obsci ration» ol
nwioiic diromofumes :n the Onagraceae. Allan. 4. 73—ев (196Й)
4‘X> Lewis H. Roberts AL R. Tbc cr.g.n cm Clarkia Ungulata, Evolution, 10.
126-138 (1ЗД6|.
500 Lraif //. Srweutowsta J The genus ОщорЬр'ат (Onagraceae), Bril Ionia.
16, 343-391 (l<64).
501. Lena X it. John B. Hybridisation in a wild population u( Elevchar is pa-
lustres. ChionwaiXDa. 12, 433—448 11961).
502 Lewis А Я, John R Chf<xnu«omt Market. Little, Brown and Co. Boston.
.Hass, 1963
503 Lewis W fi Chromosomal drift, a mu phenomenon in plants, Science, 168,
1115—1116 (1976).
504 Lewis F. Si Temporal adaptation correlated with plordy in Goyronio tv7-
rioicA. Systematic Rot. I 340—347 (1976)
505 Lew»» F. H. ed Potyplocdj. Biological Relevance. PJenna PrrM. Neo York
and London, I960
505 Lewis F. fi. О licet Я L. Lidkart T 1. Multiple gecatypcs in individuals
of C.'ap/oue airginica. Suenc?. 172, 564—565 (1971)
507, Lewis F. fi demote J, C Geography of Clav! onia virguuca cvtotvpes.
Amer J Bot. 84 1078— 10M2 G9T7).
508 Lfwonrin Л C The Gene-tic Boss of Evolutionary Change. Columbia Uni-
vefbily PreSa, New York, 1971 [Наамтся перевод Леэиятнн P. Гепетиае
сжиг оеноеы «делкмииг — М «Мир». 1978 |
Ismo-da-farta 4 Genetics, origin jnd evolution of kinetochures. Heredilas.
35, 422—444 (15491
510 Idndsa^ D F. Viderjp R К Cumporothe cvoluton in M/tfttJau gatftai'ara
oi the Bonneville basin Evolution. 21, 439-456 (1967)
511 Lmdsr'OM E T GeBeln s oi polyploidy. Bot ftev. 2. I97-JI5 (1936)
512 Lir.tuuus C Cnika Bn taeles, Leiden. 1737
513 Linruuus C Philosnphia B-atanica Stockholm, 1731
314 Linnaeus C Disqulftitio do Sc mj Plantar urn St Petersburg, 1760. Anaoenila-
lei Academkx. Er augen. 1790
515 l.iKnaeus C Genera Plant arum. 6lli ed . Vienna 1767
3)6 Little E L. Pauley S S A natural hybnd bciwccn h.i»ck and whale spruce
ift Minnesota. Amer Midland Nat. 60 202-511 (19581
517. LoLLi К Sflotru Л Polwloidy in insect evolution In Pcbplcidy, Biologi-
cal Relevance, cd W H Lewis. PIm—n Pr**-» N*w York. London. I960
318 LorArxJe Z Some xcuEahtics ol apa.laity amt h>mjsIIt .’«alricted gene
exchange in ;be frebia headerus g*oup. Cold Spring Harbor 5vmp Quant
Bio- . 21 319—323 (I968i
319 Latty J P. Evolut an bv Mc;m< of Habraluation, M Ni-hoii, The Hagur,
1916
529 t.at^y i P Species «• ‘.Jniwon, Geaetica. 6. 48T— 506 (1923|
521 iotty J P On the eper os oi he faxonnmtst in its relation to evolivtion.
Genetics. 13. 1-16 (IfWIi
522 Lne? A Cvtogenetic studies on Pumet. subg Aeetoielia. Hered.tas 30, 1—
136 (1944)
523 Ldte Л Preparatory studies tor breeding k-elaodk Poa irngctn. Hcredilas.
38. 11-32 (IK?)
ЗГ
4А4 Литература
524 Love Л. Subarctic polyploidy, Hereditas, 39, t IS—124 (1953).
525 Love A. The biological speoes concept and its evolutionary structure, Ta-
xon. 13. 33—45 (1964).
326 Live A.. Kapoor в M An allcploid OpAiogfoaaum, Th* Nucleu*. •. 132—
133 (1956)
527 Ldcw A. Kapoor В M The highest plant chromosome number in Europe
Svens* Bot. Tidakrilt, 91, 29-32 < iWf).
•528. Live A, LCoe D The geobotanKal significance of polyploidy. 1. Polyploidy
and latitude. Portugalae Acta Biologic*, Suppl. voL 273 352 (1949).
529. btee A. tdoe D lhe geo-botanical significance ol podypyuidy Proc. 64h In-
ternet. Grasslands Congress, 240—246 (1963)
530 Live A. Live D. KttLc polyploidy. Proc. Genetics Soc. Canada, 2. 23—27
11957).
531. Love A.. Loot D. Polyploidy and altitude: Mt Washington, Biol Zentral-
blatt. Suppl vol. 307 312 (1967).
532. Love A^ Love D Cylotaxooocnical Atlas of the Arctic Flora, J. Cramer.
Vaduz, 1975.
533. Live A.. Live D. Ptchi SermotU R. £. O. Cytotaiooomkal Atlas of the
Pteviodophyta, J. Cramer, Vaduz, 1977.
534 Lice A. Live D, Raymond M Cytotaxcaomy of Cam sec line Capillares.
Caned. J. Bot, M. 715—761 (1957).
535 MacKey J Neutron and X-rav eaptrimaela in wheat and a revision of the
ам!Ш ргоЫяп. Hereditaa, 49. 65-180 (1954)
536 McLeod M G A new hybrid i’.eahy-tru.ted pclckly-jsear in California. Mad'
голо. 23. 96-98 (1975)
537. .МсЛблп H. E. The importance of field hybrids In determining species in
the Genus Ceanot.hia, Proc. Calil. Acad. ScL, », 323 556 (1944).
538 McNaughlcn I. H., Harper J L The comparative biology of closely related
species living m the same area II. Aberrant xiorphology and a virus-likc
syndrome in hybrids between Popover rAoetu L and r. dubium L., New
PhyloL, И. 27—41 (1960).
339. McNaughton / H. Harper J. L The comparative biology of ekuely related
specie* living in the same are* III. The nature of barriers isolating sywi-
Btrte populations of Papaott dtbium and P lecogud. New Phytol.. 59.
9-137 (I960).
540. McNedlu T., AnioaocU» J. Evolution in rioselv adjacent plant populations
IV. Barriers to gene flow. Heredity. 23. 206-218 (1968).
541. M actor L IF rhe pollination ecology ol Pedicidane (Scrophnlariaceae) in
the Sierra Nevada of California. Bull. Torrey Bot. Club, 104, 148—154
(1977).
542 Afogoori iM L, Cooper D. C. Hou gat R W Cytogenetic studies of some
d^plad Solanums section Tuberarium. Amer J Rot. 45 207-921 (1968)
543. Maguire В Apomixis ut the genu* CUteia (Cluaiacaae) — a preliminary
report, Taxon. 23. 241—244 (1978).
544. MaheiKvari P. The male gametophyte of angioeperms, Bot. Rev.. IS. 1—
75 (1949)
545 Mutecka /. Studiea ia tha mod* ot reproduction of the diploid endemic spe-
cies, Taraxacum pienintcum Pawl. Acta Biological Cracovtamla. 4, 25—41
(1961).
546 Maltcka J Proeeaaea of degeneration In the anther»’ tapetum ot two male-
aterile species of Taraxactan. Acta Biol Crecovleaala, But., 14, 1—10
(1971)
547, Atalw-tfl J Cytn-taxonamxa! and emhryoCog.crl investigations on a natu-
ral hybrid between Taraxacum kok eaebz Rebin and 7. oHtdaate Web. and
ther putative parent aperies, Acta Bio! Cracoviensia, Bui. 14. 179—197
(1971).
54 H Malecka i P.'obkms of the mode of reproduction in microspecie» of Tara-
.luiepuij/pa 485
Н П НП i! Н s S 3 5 § з 5 S S
хл.-иля ра^импй Da his led t Acta Biol Cracovicnsia. № Bot, 16, 37—34
(HI73)
649 .Miii.'avm .V. De Castro D, Coxtara A CliroitY)*o‘ies meh lentruirwo Io
cal i*do Q da Lutula pwrpv/ra Link. Agromniua Ги**4ала. 9, 51— 71
&. Gas de N, Garde A Agtnatoploidia no genero Ltftda DC, Gear-
tica Ibc.-fca. 3. 155-176 (1951I
,Мал<><.Глг( P C The fflulageftK* effect <H hytxldtflng maw and tetoiale.
Cold Spring Hat bur Symp Qu и til Biol. 23. 4C&—421 (iWl?)
Ма«и*<ж I lhe probk^t u! tiiMuftUa tafdgata L. Zctlscnr ind Abstinitn,
Ven^oungsicT.rc, 97, 41—57 (19311
.Мишели I Thv proMc.n ul Biecut elia lae-ngMa L II Die rvidvncv It out
ttieiasib, Ann Bot. n *. I, 439—4I»± il937)
.Man/an / Problems M Cytology and E«oiahan in lite Pieridapliyi*. Cam-
bridge University Press. Cambridge, I960
.Ч'яп.'сп / Cylc og} oi Pdyfxxlium in America, Nature. 167. 37 (19511
Mun.'cT I The concept oi the aggregate specie» In Syslema к* of Today,
cd О Hedxrg, Upn'cla, 1958
MurAden /cncjs E Al Th; gcnrtKs of Gca*'i йи'вгявеЛика WiDd baud Ehih
and its he? к-crosses J Gcmrtics. 23. 377—3% (I OSO)
Mason tt L Evide.xc lor the gvnvliv spbnaargonce ol Лла» r.’etv/u'u In
Genetics, paU«wiiol<igy and Evok t im. «I G 1 J»p*r»i, Princeton Uni»er-
Uty Pr*wx PritKclon. N J . 1949
Mason U L Taxonomy. svatr iatic» bataev and biotyMctwka, Madrono.
Id, Itt—M8 I !*• ><»)
МлГЛег A Species crosses .n 4«Wt/rA*nam I Genetic LSfXii-un <il Ute specie»
•Ш/us ifa.'iioxuM and демпмя, Heredity, I, 175— 1Л6 (1947)
MofAer К Palvnccpl, ,щ » ап oo'comc of dh’iipfivc jrketioa. Е»о1и1юп,
9. 52-61 (1965)
МаГАсг * П-ceding systens .-nd rc«ponic to seieelion In Reproductive
Billing» and Tavunomy c Yaacilar Plants, rd J G Hawkes, Pcrgamoci
Press. Oxford 19^
J flbw Inbridogme Sipprn dvr Tanttrn, Bibbidheca Botanica,
moI 25. 19Ы1
-Muyr L Systematic and the Origin o4 Species. Columbia Univarsity
Press Neu- York. 144?
Ua.yr £ Cha.iav oi genetic ciivironnwai und e» al и uon In Evolution as a
Process, «1 J Нихкт, George Allen and Unvin. Loedoe. 1954
Март E The species as a systematic and as a biological problem Itu
Bio-ogy C^foourum «Oregon State College, Corvalbs. Oregon),
Afayr E KJtl Jordan * (on'r?>ution to :tirrrn( concepts n sys1cm»t>.-s and
Т-tins Royal Enbnol. So- (Loud > 1866 4o <r»G (It35)
Muyr F Specie* еолгерЬ a?>d dcliflitiono In Th* Sptcir^ Problem, cd
E Йауг. A:r»e? AflBOC Adv Sei. Washington. 1967
E Animal Spefies jnd Evolution. Harvard University Press. Cmbrid-
g«. Mass. 19ЯЗ
570 M.T'/r £ Populations Spaces and Evolution, Harvard Un v Pres*. Cam-
bridge, Mass, 1970. 'Ииеетса перепад Ma Др Э. Пзяуляимм, виды a w«-
лмшвг — M «Мира 1974 ]
571 Moyr E Ziulclion and '.he Divdsihr of Lite Selected Essays, Harvard
Laiversitv Pres- Cambridge. Мам . 1975
572 AfayiMrJ .S»iiM / Sympalrk speciaUon. Amer Nat. 100. 637—650 (1965).
S73 Atuejr J Past liybridixatiun between Qutfctts triac/ocarpa aad Qiu^cut ganr-
Мй. Brillonla. 20. 321-333 (1968)
574 Mfffetscft P A On the nature of lhe sperm. Systematic Zool. 3. 49^-05
(1954).
4ЭД Лиг rpar у pa
575 Meiunder Y. A new giant Papal as irmuls ia Norrbottesi, Hereditas, 24.
189-194 (1938).
876 Mittin D.. li&nttl P. Uber Spttiaticmvorgange in der Gatling Vida L.
Kultarptlanxe. 21. 25—34 (1973)
577. .MkAariu P. Eat wicklungagr^hirhtlkh-genet ru be Untarauchungta an Epi-
1оЙвт II, Die Bedeutung dee PUams Iflr die PollMi.'ertilrtif des Epilo-
biam haeum kiriolum Bastardes, ZelUchr. bid. Abstaoununga ta. Vercr-
bungslehre, 6Б. 1-71; 353—411 (1933)
578. MUUctd £.. Tkoda9 i. M. El («eta Ы disruptive selection. TV. Gen* flow
and divergence. Heredity. 16. 199—217 (I96Q.
579. Misov N.T. The Genua Pluus. Ronald Press. New York, 1967
580 Maseru P. В Chlauna distribution and the segregation of markers la chro-
mosome 2 of ths tetraplocd tomato, Genetics, 49, 123—133 (19641
581. Motainwd A H., Gould F. V Biosysteaiatic studies ia the Bouteloua curli-
prnduia complex. V. Megasporogenesis and embryo sac development, Amer.
J gj 166 169 119661
u82 Moliich H. The Longevity oi Plants, Translation. E H- Fulling, New York.
1938
583, Moore R. /.. ed. Index to Plan! Chromosome Numbers 1967—1971. Inter-
net Hnraau Plant Teton, Utrecht. 1973
564. Moore V. $ An evaluation of narrow hybrid zooes in vertebrates, Quart
Rev. Biol, ». 263—277 (1977)
585. Mot^tdn T Toward a more uwiul taxonomy for chromosomal races. Brit-
tooia. 18. 2(13 -214 (1966).
586. Моядып T Evidence for autopolyploidy ui Epilobiam angtutifohtfm (Ona-
graceae). Evolution. 21. 713—719 (1967).
587. Afaidstin T, Small E. An example ol parallel evolution in Epdotaum (Ona-
(paceae). Evolution, 25. 678 682 (1971).
5АЯ Muidd S. The chfomojumes cl the earthworms I The evolution of puly-
plc*dy, Heredity. 8. 55—76 (1952)
589. Muller С. H. The oaks of Texas, Con trib Texas Reseat ch Foundation, 1(3),
21—323 (1961)
590 Muller С H The signtikanc* ol vegetative reproduction In Quercia. Mad
reno, |j, 120—137 (1951).
591. Машаг С H- Ecological control ot hybridisation In Qurrcus a factor m
the machaniMTi of evolution. Evolution. 4. 147—161 (1952).
592. Midler С H The role of chenuca) inhibition (allelopathy in v-egelalional
composition. Bull Torrry Bel Club, W. 332—351 (1905).
593. Mullet С. H. Quefctts L In Manual ol the vascular Plants ol Texas.
D. S Correll and M. C. Johnston. Texas Research Foundation, Renner. Te-
xas, 1970.
594. Muller H /. Why polyplocdy is rarer in anunals than in plants, Amer. Nat4
59. 346-353 (1925).
595. Mia/aing A Outlines to a genetic monograpti of the genus Galooptie. He-
r edit as, ft. 185—341 (1930)
596. MUmtring A. Uber Chronaoaomenvermehrung tn Geleopwr-Kreuxungan und
iltre phylogefletiache Bedeutung. Hereditas, 14. 153—172 (1930).
597. М&лЫкр A. Cyto geeetk investigations on synthetic tWeopeir UtraAtt.
Htreditaa, 14, 105—154 (1932).
598. MdnTjtiig A Chrotnosoaie fragmentation in a Crepu hybrid. Heredltas.
It 284-301 (1934).
599. M^Bby^A^The evolutionary significance of autopolyploedy. Heredrtas, 21.
600 MuntMg A The chromosome* of a giant Papula» trrmuia. Hereditas, 21,
383—303 (1936).
601. М(МИлл A Stcnlrty and chromosome pairing In intraspecific QaleopsU
hybrids. Heradltas, 24, 117—188 (1938)
Jiartpaiypa
602. A!«A<a/«vg A Genetics Bass* and Applied. 2nd ed. LTs Pflrlag. Stockholm,
€03 Ai^Mc.ttg A . R Chromosomal fibrrrations in rye pcpn.itons, lie»
redd as, 27, 273—31» (1941J
604. Mum P .4 The Oraofbere hoodren group. Alia©. 2, 1—47 (l*M9)
605. jMuci? PAA Cali.'otBia Flora, Unit a ally ol California PrtM. Berkeley.
Caln, 1959
606 Afunz P 1 Onagr*crx- Koi th Anu'i . Flore. 5, 1—278 (1965)
607 .Ммпг P Д A [ Lara ol Southern California, University ol California Press,
Berkeley, 1974
6C6 AumAwng G Statistical analysis of ink ogress ion. Biometrics, 22, 488--
302 (I966J
60У Auutf.n C Novvelics recftercbes sur I’h) bridle dans 1м* v^Ujx, Ann Sei
Nat, Ser Bonn | Pars). 19 180—203 |) 863)
610 ftiavasJun Л1 5 The d .«location hypothec ol evolution ot «hrmtinsorne num-
bers, Zeitsdi. it.
611 Л’.ч’о Г Adaptive cutisc*fenee and divergence ot subicrranctn пммипаК
Am Ro Ecul S)St. it. Л9-308 < 19T9)
612 АГеаГгж Г C I . PSk* C Pnmttla towntit and its detivattvee, J Ge-
i."
613. .ViAfle C A Stud-rs on the cytology and phylogeny ot the Pteridophyles
VI Observations on lhe Ophiog эдасепе, Cytotogia, 23. 291 316 (1*308)
614 Nobi .4 A Ekperiafetitai studio on species relationships in Ctanothui.
Carnrgk Inst Wash Publ. ti2ii (I963J
615. A’ohs M A. liiocg 0 At Studies on differential selection in Mmiitas
Carnegie in?*. Washington Yearbook, 56, 291—292 (1967)
616 NobiM A. Hiftcy v A< Penciltnanre ol .Mimulwr rater and their hyb
r ds is contrasting cttn.-cin.uentl Cxrncgic (nil. Washington Yearbook, 57.
270-272(1938)
617. AWe S Cyiogctxlir relaliunships between Lilivui '.griruiM and related
snnics ,md the hybrid origin of Lr/rw.w /ignnwm. Lily Yearbook. 27. 98—
111 (1974).
Gt8 Кшйл1М/мьЫ H The soi ^Ik clicuriuvunivs id vya» l.uiaki spaci**, Botaru*
ska Notlse*. 1949 81-92 (1949}
619 S'ofdensk.ini И CvtjI iuMKMi’kal studies in 1he genus Ltauia I. Hercdi*
tas, 37. 325-3M 11 %1)
620 Nord^uki^td H C>io Lavoaomleal studle- In the genus Ьк.2лЛл II Hybrid!*
talioti rtprrinimls in the гггтпссггь-'Ли^гЖагв complex Hered ias, 42, 7—
73 (1956)
621 ftfy'diOik.did ii Tetrad analysis and the course of meiosis In three hybrids
<A Z-urwlu coxnpestrii, HcrediUx, 47, 203 238 (196!)
622 .VwtlenskicMif H Studies <x rseioals in Lwle pvrpurra. Her edit as. 48, 503—
519 (196?)
623 .4 TLe genesis of some Srondinavion species of Culemagroifis.
Hcrtditat. 3?. 131—262 f!946|
fi24 Wgiun A Fo*nt arH Metype fonnMiiM» in CalAMagrMtit Here
dii.is.37. 519-532 <1951)
625 4 AnnmlTls ;fl the агг1г»^с*тп$. II BiM Rev. 20, 577—649
(1Я4).
«25 A Artiienl and wural hybridIxation in European Csiamogrtwu.
Svtnbolae Botinknc UpMlhmee. 17. 1 105
627 A'tigren 4 On Poa Icdensis. Рпв ro«rt«wa. Роа рштйа. Paa xrr&pAlTa and
the passible origin o* Paa islpjna. An no Im Academioe Regt де SdentUrusn
Lpsa liens is, 1902 |в), 1-2? (l&2|.
628 OelHAert F Bastnrdimmgsversurhe in der Gnttung Shepiozazput Lindley.
HI Neve Ergebnisso uber die Gcoetik von Wuchigcatah und Gcrchlechts-
bes^irnmnng. Ber Deuts.h Bot. Ges , 58, 75—91 (1940).
46В * Литература
629, Ottvtari A. M, Jain S К Var.atioa id the HeUanlluis exilis-botanderi
complex a reexammaliun. Madrono, M, 177—189 (1977).
630, Qllgoard H. Nev apeoei of Taraxacum from Denmark, Bot Not. 181,
497—821 (1978).
631. Orndufj fi Concept» ol spee.es (Bock review,). Sytf. Bot, 1, 347 (1976).
632. Ownhey M. Aase H C. Cytotaxoeomic stud»» in Alton. !. The AWum
canodtnse alliance, Research, Studies State Collage ol Washington, Mono-
graph Soppl. I, I—106, 1955
633. Palmer t J. Hybrid oaks of North America. J. Arnold Arboretum. 79. 1—
48 (1948).
634 Pofenuaxf £. Selection far reproductive isolation between two populations
ol maize, Zm mays L Evolution, 28, 534—547 (1969)
635. Pafermani E., Start A. C. Effective maize pollen dispersal in the field. Euphy
tka. 23, 129-134 (1974).
636 Paso B. Zohar у D The process of introgression between Aegilops poly-
plends: natural hybridization between A. variabUis. A. owsia. and A biunaa
bl Evolution, it. 385—394 (1965).
637. Peterson P. A Cytoplaanskally inherited male sterility in Capsicum Amer.
Nil, 92. Ill—119 (1958).
638. PianAa E R On r- and A-wtectlon. Amef Nat. 104. 692—697 (1970)
639 Pi\l L. иал der. Principles ol Dispersal la Higher Plants. 2d cd, Sprlx.gur-
Veriag. Berlin and New York 1972
640. АммГН D. Smith G. L. Soaos 1. A population model of sympatric specia-
tion. Amer Nat, 101. 493-504 (1967).
641. Piakaua D J.. McLeod M. G. Chromosome numbers in some each of wes-
tern North America. Britton»». 23» 171-178 (1971).
642. Pirdtava D К McLeod M. G.. McGill L A, Brcnn R. C. Chromosome num-
bers in some cacti of western North America If Brilionia, 75. 2—9 (1973).
643 Pmltava D. McGill L. A- Reeves T. McLeod M G Chromosome num-
bers In some cacti ol western North America. 111. Bull. Torrey Bot. Club.,
104. 106-110 (1977)
644 Powers L Fertilization without reduction in guayule (Parthenium arg ent Or
turn Gray) and 8 hypothesis as to the evolution of apomiais and polyploidy,
Gtt»Hi«, 80. 323-346 (1945)
645. Prager £. M-. fowler D. P.. Wdsrm A. C. Rates of evolution In conifers
(Ptnaceaei, Evolution, », 637—649 (1976)
646. Proxmo IF Genetic studies on the ger.us Aquilegia L. I. Crouses between
Aquilegia vulgans L. and Aquilegia ecalcarata Maxim. Acta Soc Bot. Po-
Ion., W, 57-П (I960).
647 Praemo V. Genetic studies on the genus AaMfeg.n L II Crosses betwuan
Agaffcffe erakwafa Maxim, and Aquilegia chrysanlha Gray,* Ada Soc.
Bo4. Pokm. 36. 423—442 (1961)
64R Prurmn IF Cytogenetic studies c-n the genus Aquilegia 111 InberiUnce of
the traits distingusshing different rcoiplezes in the gmiM AqmUgia. Acta
Soc Bolan PoJon, 84, 4H3—437 (1965)
649. Prarmo T Cytogenetic studies on the genus Aaudegla. IV.'Fertility rela-
tionahipa among tlx Aquilegia species. Acta Soc Bot Polos»., 84. 667—
6Я6 (1966).
650. Prgor L. D. Species distribution and association in Eucalyptus. In: Blogvo-
Band Ecology In Australia, Monccraphiae Blologicae, 8. 461—47|
teat A. Some observations on male aterilitv in Geranium sylaaiicum
L. var alpvstre Schur. Acta Soc Bot Polon. 31. 723—736 (1962)
652 Raihathy T.. Thanas H. Cytogenetics of oeta (Aeune L). Genetics See.
Canada Мне Publ, 2 (1974).
653. RamaboMom /. Linnaeus and the ароейа conrept, Proc. Llnnean Soc Lon-
don. ISO. 192—219 (1938).
Н з § § s § § i 1 ssSssSbB
Литература
Randolph L. F Cytogenetics oi te<r*ploid maize, J Agr Research. SO. 591 —
60$ (1935)
Лип4сУр6 L F. Л'еЬоч f 5. P/uis'M ₽ L Negative evtionr-e of introgres-
sion affectlr.g toe nubility о' Louisiana Iris sptt_.es, Cornell Univ. Agr. E*p.
Station Memoir. 398 (1967)
Ratter 1 A Cyloaev>e4ac9 studies in Sptff>tdufia IX Summary and conclu-
sions, Notes irom Royal Bo: Gerd (Edinburgh), 34 411—428 (1976)
Raven P ft I nt er specif ic hybc-dilation as an evutuliuaary skmalus in
ОслоГЛгга, Pro: linecan Soc London, 173. 92 98 (19G2).
Raven P H A revision or the genus Садигдепкг (OugYKtac), Contrlb
US Kat Herbtrium, 37, 159—395 (1989)
Raven P H The bases of angiosperm phylogeny cytology- Ann Missouri
Bot Garr. «. 724-764 11975)
Raven P H 5умепы:к:. and plant population biol op. Syst Bot. I, 284 —
316 (1976)
Raven P II Future directions in plant population biology In Topics in
Plant Population Biology, ed О T Solbrig. Columbia L diversity Press.
New York. 1979
Raven P H. Dietrich T. SfnMe W An Catline of the systematics oi
Uenotkffc stA"«-t Luaennthem (Omgrarete). Svst Rot. 4, 242—232
(1979 1988), (Fall I97& Mue. pvtl April. I9tkl
Raven P И . Gcego*y D P Ot>u>'vations of rceiohe chromcMxnes In Gcutra
(Onagraceae), Britton»». 24 71 -86 (1972)
Raven P H., Ryhcs Г) IT Neu evidence concerning the ong.nal baste num-
ber of angiosperms. Evolution, 19 244 248 (1965)
Raven P H . Ratios D IT. Cave M S Chron-osnm? numbers and relation-
ships in Anno*ii florae Taxor 20 479—483 (1971)
Raven P H. Rtn rn T E Th? Genus ГдгГаЬгяле in Autlralaaia. Dept Sci
Indust Research. Chrtstchurc-i. Sew Zealand, 1978
Raven P H Thompson IT / HnpluMly and ang«ospe»«n evolution, Amer
Nal.W. 251 252 (19541
Ray ! H s-oria Plan-arum. London, 15MW
Reddy .M Af . Qa/btr E D Gerelit »ladies ol variant enxymes 111 Compu-
rative cIcet.-eiphn-rK- afadws ol »-Ггта:«'ч and peroxidase* for «pedes. h)b
rids and amphiplouis la the genus Afcufiunu. Bot Gaz. 132. 156 166
Rees H Genc.ypic control oi' clirucncoume foini and behavior, Bot Rev.
27, 288-318 (1961)
Rehfcldf G E Ecolypic diFSeicntiation in pepulaliows uf Aruua monticoia
In north 1 daho-rnyth or reality* Amer Nal. 114. 627—636 (I9?9)
RrmmgJcn C L Suhire-KHM el hybrid interaction balwten rttetilly joined
biuUb. EsObt uOarv bioi. 2. 321-428 ПЮ|
Rittner О Untevsur-iur.gvn uber die (jk orvelle Konstitntion rin ger kompto-
ihHeroeygotischer B>bl»uthr.a Onetiea, 9 1—IG8 (1925)
Renner О Cber die Entstrtung nomr^vgot «rhe- Fornrn aus tamplcxhctcro»
zygociscVn Oenotheren. r ora. 35, 201 238 (1941)
Renner О Europaiscbc Wildarter von Oenothera. Ber Deutsch Bat Ges,
GO 448-466 419421
Rcnsch В Das Pi inzip ct:c>flraphisc*>er Rassenkieise und das Problem der
Arlbikkng. Ro nirargrr, BrrTln. 1929
Reaiher W. Burc^n’cx L D. Vehlxrt It I e«h The C-trus Industry,
vol (1 Anatomy PhysioJop*. Gcnelke and Reproduction 2d ed Univer-
sity of Caliivt nia Picse. Berkeley and Lu- Angeles. Cali 1. 1И8
67Я Reuther Г IFeftb?/- H I. Batchdor L D. Ms The Citrus Industry, vol I.
History. World Dituibulion Bolan у and Varieties 2d cd University of Ca-
litonMa Pros. Berkeley and Los Angelas. CaRf, 1967
490 Лиггрстурв
679 Rhoades И. М. Dempssy Е Induction ot chromosome doubling nt nwionte
by the elongate gene in maize. Genetics. 54, 506—522 <1966)
660 Rhyne C. L Linkage stixli»* in Gaiiypium I. Altered recombination In allo-
telraploid G. /Uranium L. following linkage group bansfercore ircen rela-
ted diploid species.Geoetlo. 4*. 822—834 (1956).
661. Richards A /. The origin of Tartuaium agamoaperiea, Rot J, Linn. Soc.
•8. 189—211 (1973).
682. Richardson M M Structural hybridity in Lilian martagon aibumxL han-
uonli. J. Genetics. 52. 411—450 (1936).
683. Rich С. M. DUferentul zygotic lethality in a tomato species hybrid. Gene-
tic*, 46, 1497—1507 (1963).
684. Piegar R. Die Gcnoamrulatiooefi, Veb Gustav Fischer, Jena, 1963.
688 Right# F I.. Duffield J. T. Intenpecies hybrids tn pine*» J. Heredity. 42.
75—80 (1951).
686. Riley H P. A character analysis ol cotontea of Iris fuloa, Iris hsxag&na vur.
giganiicaerulee end natural hybrids, Amar. J. Bot. 26. 727—738 (1938)
687. Riley R. The diploditialian ol polyploid wheat. Heredity, 15. 407—429
(I960).
688. Roberts H. F. Plant Hybridization before Mendel. Reprint, 1965, Hainer.
New York, 1929. 1965.
689. Radins R C. Evidence for natural hybridity between guayule (Purthemum
argentatum) and marlola (Parthenlum тсапшп), Amer J. Bot. 31, 90—
99(1944)
690. RoUuu R C Interspecific hybridization in PariheiauM I. Crosses between
guayule IP. argentatum) and marioda (P. taconawt), Asoer. J. Bot.. 52. 395—
404 (1945).
691 Rollins R. C. Interspeafic hybridization in ParihetUum IL Crosses invol-
ving P. ar rental urn. P Incuaum. P siranvmium, P. tomenlQMM and
P hysterophorus. Amer. J. Bot, 55. 21—30 (19461
692. Rollins R. C. The evolution and systematica ot Leaoenteorihla (Crwciierae),
Contrlb. Gray Herbarium (Harvard), No 192 (1963).
«S3 Rollins R C, .Shoe £ A The Genus Lesquerella (Cruciferae) in North
America. Harvard Uorvaraity Press, Cambridge, Mass^ 1973.
6£4 Row M L. GoitReb L D Genetic and biochemical conacqucnces of poly-
ploidy in Trasopagan. Evolutice. 30, 818—830 (1976).
696. Roes H H Principles of natural coexistence indicated by leafhopper popu-
lations Evolution. It. 113-129 (1967).
698 Row If ff. Evidence suggesting a hybrid origin lor certain leafhopper spe-
cies, Evolution, 12, 337-346 (1958)
697 Raes 1. G.. Duncan R. E. Cytologies! evidence» ut hybridization between
Junipcrus vagituana and К hceiioaialis. Bell. Torrey Bot. Cl eb. 78. 414—
429 (1949)
698. RotAve# N. V. Aneuploidy in ClagtofUa eirgiiUca, Amer. J. Bot. 44. 353—
360 (1959)
699 Roast A. Bioetystematic studies on the species aggregate Potentllla cwri-
no L., Annake Botamci Fennki, 2, 47—112 (1966).
700 Russell N. H. Three field studies of hybridization In the stemless white vio-
lets, Amer J. BoL. 41, 679—686 (1964)
701. Ruiisltausef A EmbryoXiglc und FartpILanxungsMulogie der Angtcopermen,
Springer Verlag. Vienna, 1969.
702. SocAs 7. History ot Botany, Translation. Oxford University Press, Oxford.
1906.
703. Salisbury E. J. Reproductive Capacity of Plante. О. ВеП and Soea, Lon-
don, 1942
704. Salisbury E J. Weeds and Aliens. Collins, London, 1961.
705. Salisbury E. / A note on shade tolerance and vegetative propagation of
woodland species, Proc. Royal Soc. London. B, IV2.257—258 (1976)
Лнгсмтура
491
706. Sandlast 1 High frequency oi spontaneous Iripfoida in barley. Heredita*.
•4, 131—134 (1970)
707. 5 ало F, Kilo F Genes lor reproductive isolation located on rear fanned
chromoscmcs, Heredity, 41, 377—3S3 (1978)
708 Зало У, СЛ« У fJXa H Genetic studies oi speciation in cultivated rice
(I Character vanatiOfM m backrow denvalie* hetw«a Огула до/й« and
О. elabcfrima M V I.nAagc and hey characters* J. J. Genetics, W. 19-39
(ISM)
700 Salina S A f Cytalogical effects of в gene in fratara whu>
causes dyad furmation in xpcrogencsts Bot Gai. 94. 521—523 11935).
710 Sex H / Polyploidy and apomixis m CoicneosJr’-. J Arnold Arboretum.
35. 331-365 11960. ‘
711 Sox X Chromosoine nog forrr.aLoo m Photo discedor, Cytologia, 3, 36—
33 <1«11)
712 Schaai В A. Lovin D A Morphological diiferмНialion and neighborhood
site in Liatru c^tindxacea. Amer J Bot. 45. 923—928 (1978)
713. SchuJf2 R. J Hjbriluatiuu, un.sexuality and polyploidу in Ute lektxsl
Potahaptix (Poecilitdee) and other vertebrate*, Axnc-t Nat, 103, G06—619
(19691
714 JirWir R I Role □! polyploidy in the evolution ol lishrs In PUyploidy,
Biological Relevance, cd W H Lewis. Plenum Press, Nrw York and Loa
dork l»®0
71Б Sckuanilf F SptiraUldunc i»*i rinrm Bastard twrschcn drel Digitalis .4л-
ЙЯ, Biol Zemraiblatl. 78. 226-231 (l»7).
716. Sears E R Chromosome pairing and terlililv in hybrids and amphiploids in
the Tnlxinae, Lnlv Mlsscarl Agr Exp Station Bull. No 337 (1941)
717. Sears E il Cvtogcneti: studies with polyploid species ct wheat. 11 Add'
Ifonal chrccntwunal abcrr&hons :n TritiCMI vuigorc, GcDtlkb 29, 232
246 (1944)
718 Senn C R The cytology and gene1<* ol the wheats and their relative»,
Advances in GeraHKS. 1. 239 270 (1949)
719 Soars £ R The aystemabes, cytology and genetics of wheat, llandbuch
der Pfltflienxiichtune. 2. 184—187 (IfifiO)
720 Sensplo J C. Bmurllef L Chromowmw numbers and satellite chromosome
morphd ary tn Aslef and LaxaUta. Amer J Boc, 47, 1027—1039 (I960)
721 Shiado R, К&шгтлго ti Chitxnosoene numbe s and genome rehlkmships
of some species »n the S'larnktriatae Section of Dentfrobix’*. Cytologia. 2*.
M 75 (19631
722 Sifr/ry С О Species fo-rnaiKin m the red-eyed towtwes of Mexico, Univ.
Calif Publ Zoo:. И. 109 (I960)
723 SrMey С C Hybridization in the rad-eyed towtsres of Mexico, Evolution,
8. 252-290 (1954)
724 Sifaru/rr / /4. Microevolution and clone structure in Sparttna palcns.
Science. 203. 65A-<r/. (1979)
725. Srmmocds .V F Mclcsls in hopioal Rkom dixnior, Natuic. 1S5. 731
735 .V F Th: Evotatlon oi the Banana Lungmaes Green, London,
1962
727 Snnmcir.fls S' ЗГ Bananas. 2d ed. Longnibtw Green. London, ID6fi
728 Simmonds iV F. cd Evolution ol Crop Planlv, Longman Group Lid. Lon-
don. 1976
T29. Simpson В В ed Mesqui e Its Biolog} in T*o Desert Scrub Ecosystems,
Dowden. Hutchinson and Russ. Strnudshiirg. Pennvytvanta, 1977
730. 5^mpjcv> О О Tempo and Mode in Evolullcn. Columbia Onivenity Press.
New York. I!M4 |И.меека перееэд Симл-оо Дж Темпы а фирмы seofiio-
ими -M ИЛ. 1948 ]
731 Simp» о л G G The species concept. Evolution. 5. 285—298 (1951)
492
732. SjmpsoR О О. Principles ol Animal Taxowomy, Columbia University Preaa,
New York, 1961-
733. SinsAai’o E. N. lhe study of species In their dynamics and Interrelatica
will» different types of vegetation. Ball. Appl. Bo4. Genetics Plant Breeding
(Leningrad). 28, I 97(1931)
734. Skafintka M Seed development alter crosses of AguUegia with Jsopgram.
Studies In Plant Physiol (Prague). 213-221. 1968
736. Skolifuka M Cytological analysis o4 acene Hieraeium species, sdbg. Wo
»etlo from mountains of soulhrru Poland. Ada Bic4ogka Crscoviewsis, Bo
tanica, 10. 127—141 (1967)
736. Склорцол A < И км СССР. ТааоономиаесжнЯ и географячасхжЯ обзор —
М.: «Неука». 1966.
737. SlobodchihoU С h'. ed. ConcasU of Spades. Dowden. Hutchinson and Roes,
Strondanurg, Pennsylvania, 19.6
738. Small /. Aleaxuufar £ J Botanical Intarpratabon of the Iridaceeus plants
of the Gulf states Contrib New York Bot. Card.. No. 327 (1931).
739. Smith D M.. Craig S.. Ssntaroia J Cytotogicil and chemical variation tn
Pltyrvgramma tfioagulufit, Amer. J. Bot, M, 292—299 (1971).
740. SmiM D l-cuin D. A. A chromatographic study of reticulate evolution
in the Appalachian Asplenium complex. Amer J. Bot, SO. 952 958 (1963).
741. Smith F. ff, Clarkson Q D Cytologies' studin of interspecific hy be idea-
tion in !rtt. subsection Califonticae. Amer J. Bot.. 48. 882—588 (1966)
742 Smith H. П. Development of rnocphtxogkallу distinct and genetically iso-
lated populations by interspecific hybridisation end selection, Caryologla,
Suppl Vol. 867-876, 1954.
743. SmM И H Genetic plant tenors in Nieatiana. Aanah New York Acad.
Sei. 71. 1163-1177 (1953)
744. SrruM H. H. Daly K. Discrete populations derived by interspecific bybrl*
dilation and «election In NUotiana. Evolution. is, 476—467 (1969).
745. Smith-White S Polarised segregation In the pollen mother cells of a stable
tnploid. Heredity. 2.119-1» (1948)
746. Smith-White S. The life history and genetic system of Leocopogovs funipe-
rinvs. Heredity, 9, 79-9! (1966).
747. Smt/jh-Ffufa 5 Cytnlogical evolution in the Australian flora, Cold Spring
Harbor Strop Quant Biol.. 24. 273—289 (1959).
748. Smith-White S. Pollen development patterns In the Epaerldaceae. Proc.
Linnean Soc. New South Wales, 84, 8—35 (1959)
749 Snow up S. Studies in the Aegean flora IX Erysimum sect Cheiranlhut.
В Variation and evolution in the sniall-pnpalatlon system. Opera Botanl-
ca. 14. 1-86 (1967)
750. Srvyder L. A Morphological variability and hybrid development in Путли
giaucus. Amer J. Bot., 37, 628-636 (1956)
751. -Snprfar I. A Cytology of inter strain hybrids and the probable origin oi
variability In Flymus glaucus Amer. J. Bot, 38. 196—2C2 11951).
752 Soun$ A. B.. Pimentel D, Sowu J. S Evolution of reproductive isolation
in allopatric and sympatric populations, Amer. Nat, 106. 117—124 (1974)
7t»3. Sofctf R. R Th* species problem reconsidered. 5yst. Zool, 22, 360—374
И973)
754 SoAal R R. Cruvetlo T 7 The biological species concept: a critical eva-
luation, Amer. Nat, IM. 127—?53 (’970).
7ББ. Sciibrig О. T- Cvtotaxooomic and evolutionary studies in the North Ameri-
can species of Ga/herrma (Coenpoaitoe). Coitrlb Gray Herbarium (Har-
vard Univ), 188. 1-63 (I960).
756. Solbrig О 7. Principles and Methods of Pleat Btosystematic*. MacMillan.
New York. 1970.
757. Snibn'f О. T. The population biology of dandelions, Amer. Sd , 59. 686—
694 (1971)
Лкгсратира
493
758 SofAng О Г On the relative advantage-, of cross and soiLfcrtiltralion.
Ana M'rwuriBol Gird , 43, 262— 276 (I976|
759 5offr/i|f О 7 A coat benefit analysis of rcrnnihinalion in plants In Topics
in Plant Pupb.itiari Biology, rd О- T. SoRirg. Columbia University Press
Neu \Ofl. 1070
760 Soib/ig 0 Л. .4r*Jcrron f. C. Kyhns D V. /taoen P H. fiMdatbetg I.
Chromosome nuinbn* m Composita* V Astercae II. Amer J Bot, 31.
513-519 (1964).
761 Solbrig О Г Jain S, JdAnaim Q B, Rauta P /Л» ads Topic* in Plant
Popubtfofl Biology. Co umbia University Press, Neu York. 1979
762 .Snnnebnrrr 7 ?•< breeding systems. reproduclive methods, and species prob-
lems in Protozoa In The Species Problem, ed Г. Маут. Amer Assoc Ad»
Set. ‘infiIon. D C , 1967
7(13 Sar.mJcn 7. Gudir^twn G Spontaneous chromosomcabberrenfs in apo-
mictic Tarataea -wpl’olngical and cytogenetical InvcCigartons. К Dan
skrVidrnd Sd*MI. BiOi .4.1 48 (1946)
704 Soff/nafadwi £ Parttcnogcnes s In inwnaK Advances in Genetics. 3.193—
253 (19101
705 Sprague E Pollinator and evolution in Pedicular и (Scrophulariaetae).
Aina 1 181-264 119*2)
766 S quit lact А Г, Blagha*t К T Localized ecotypac variat'on in western uhile
pine. Fores! Sd . 4. 20 34 (!9бВ)
767 Stare C .4 Hybridization arsd the Flora of the British Is I**, Academic
Prc*»‘. Lindon. 975
7M St a) leu F A Linnaejs and the Linnaeans A OosthoeL Utrecht. 1971
7W9 -StalA/r M D A case of polyploidy in Diptcra Proc Kat Acad Set. 42.
194- 199 (1936).
770 SrulAcr H D The phvtogcnetic relationships ot the species in the Drw
pMa m ria nice gto-ip. Genetics 53 327—342 fl066)
771 StauJe* S -W A t aeors- ol evolution above th» spcr«ca level, Proc Nat-
Arad &cl 72. 646-650 (1975)
772 Stanley 5 Л1 Macroevo1 ution. Patiem and Process. W H Freeman, San
Fra nr i «а I £79
773 Slrbbias G 1 Cytology с* 4е< иядгкт I Nornal snortee. Bot Gaz. 94.
IM—151 (I93?|
774 S/rfrbms 0 L Л nr* sper.es oi Anicnnarta from the Appalachian region.
Rhodora, 37. 229-237 (1935)
775 StabMw G L CytotagicAl characteristics associated with the different
growtli habits in th.»- dicutytedcnv Amer J Bot. 23. 189—196 (1938)
776. Stebbini G L The roJe n* isobtmn In the dFkrentlor of plant species.
Bol Symp. 6 217-233 (1942)
777 SrcbAt<x G i The cvlologkal analysis of specks hybrids, П, Bot. Rev»
11.443-486(1945)
778 S.'ebtaMs G L Types oi polyplosdc their classification о nd Significance.
Advances in GencJicx l, 4СЙ--429 (1947)
779 S/ebMnr G L The origin or the complex of Srcvans cariiatus nnd its phy-
togeograplu: inplicabons. Contrib Gray Hcrbaiium (Harvard), 16Б, 42—
55 <1947)
780 SJt*№>u G L Variation and Evolatioa In Plants. Caiumbia University
Press, hew Yorx. 1Я50
731 SrcMiHs G L Cvtogcnrticn and evolution of the gras* family, A*?»rr J
Bol. 43. 89O-9C6'(19^|
782 Sf^binf G L Self frrtil rat an and population variability m the higher
plants, Amci Nat. »|. 337—354 (1937)
783 G L The hybrid origin ot mirrosner^a in the £/ymm £1амсде
complex, Cyloloeta. 3uppl vol. 336— 340. 1957.
4*4
Литература
784. Stebbins G. L ТЫ in viability, w«akneu> and sterility of interspecific hyb-
rids. Advances in Genetics, 9. 147—2lS (1968)
786. Stebbins G. L Longevity, habitat, and гЫоам of genetк variability In the
higher planti. Cold Spring Harbor Symp Quant. Biol. 23, 385—378 (1958)
786. -Stabbuu G. L. The role of hybridization in evolution, P»oc_ Amer. PtilL
Soc, )03, 231-251 (1969)
787. StebNna G L Chrotnosomal Evolution in Higher Plants. Edvard Arnold.
London, 1971.
788. Stebbins G L Flowering Plants Evolution Above the Sport» Level. Har-
vard University Press. Cambridge, Mess . 1974.
789. Sfrbtau G. L Polyploidy in plants unsolved problems and prospects, In
Potyploldy:Bto4ogical Relevance, ad. W. H. Lewk. Plenum Ргмя. New York
and London, I98tt
790. Stebbins G. L.. Daly K. Changes tn the variation pattern of a hybrid popula-
tion over an etRhi-year period. Evolution. 15. 80—71 (1981).
791. Stebbins G. L. Ferian L Population variability, hybridization, and Intro
gresiian in scene species of OpArpr, Evolution, 10, 32—46 (1956)
792 Stebferis G L . Jenkins I A Apospcrk development in the North American
species ul Crept». Genetka. 21. 191—224 (1939).
793, Stebbins G L. JenJtim / A. Foffara M S Chromosomal and phylogeny
in the Composite, tribe Ctcborteae, Unhr. Calif. Publ. Bot, 2% 401—429
<19M).
794. Stebbins G L. Tobey H. A The cytogenatka of hybrids In flrnnw I Hyb-
rids within the section CeraiDcMoa, Amer J. But, 31. 1—II (1944).
796, Stebbins G. L, Tobgy H A . Harlan I R The cytogenetks of hybrids in
Bromos. IL Bromus carinaius and B'omus arisonicus. Proc. Cahf. Acad.
Sei. 2S. 307-322 (1944)
796. Stebbins 0. L, Vanrama A Artificial and natural hybrids tn the Or a mine a e
tribe Hordeae VH. Hybrids and alloploids between Etyiwus gtaucus and
Sitanion spp. Genetics. W, 378—396 (1964).
797. S/abftwir G L. Lobo's D Cytogcnetk and evolutionary studies In the ge-
nus Dactylis I .Morphology, distribution and interrelationships of the di-
ploid aubspadea, Univ. Сайг. Pubt Bot, 31, I—40 (1959).
796 Steiner E Phylogenetic Matiomhip of certain races of Euoenothera from
Mexico and Guatemala, Evolution. 5. 265- 272 (1951).
790. Steiner E Phylogenetic studies in Eteoenothera, Evolution, 8. 89—80
800. Sfrphrns S G The cytogenetics ol speciation in Gas#vpwn. I. Selective
elimination of the donor parent genotype in interspecific Ьжкспжмк Gene
tics, 34.827—637 (1949)
801 Stepbent 5 G The iruernal mechanism of specuhon In Gassgpium. Bot
Rev. 16. 115—149 (I960).
-НС2 Stephens S. G- Possible significance of duplication in evolution, Advances
in Genetics 4. 247-ЖБ (ItSI).
803. Stephens S 0 «Homologcu» genetic loci in Gosr^puun, Cold Spring
Harbor Symposia Quant. BloL. 14 131—141 (1961).
804. Stephens 3. G. Flnkner M D Natural crossing in coltoa Economic Bot.
7. 257—269 (1953).
806. Stiff M. L A nalurally-cM-rnrring trlplold juniper, Virginia J. Set. N, S, 2,
317 (1951 >
806 Stinson N T. Cytogenetics and phylogeny of Oenothera argil Leola
MarAenz. Genetic*. 38. 389—406 (1953).
807. Strolei/ G B. Systematics «I Omnlhara soft, Kneiffia (Onagrweaae). Annals
Miesouri Bot- Gard.. 64. 381—424 (1977)
808 Sfnr» Я M. Hybndiwttoo. homogamy. and sympatric speciation. Evole-
tion. 9. 441 444 (1955)
8CG. Stra9 R .M. Floral iwlation in Penstemon. Xtzer Nat, 90, 47—S3 (1966)
Литсрлгура
44S
810 St г tarns f. A. PltnrnM D Elfvcta ol ievnugraiioa on the evolution of po-
pulations. Amrr Nat, *». 201— 210 (196! I
811 Send A Studies in the Aegean Пота XVI Bmsyskinatics of the .Vigdfo
artmtrj coriylex v'lti special reference Io the problem of nonadaplive ra
diatiuei. Opera Botanxa. 24. I 169 (1970)
812 Swp-'unj.'i'ijri .4 5 Microsporogenesis in some commercial potato varieties.
J Heredity. 45. 265 272 (1954)
813 .ЧаадлпиогНал .M S. Hougas R V Cytogenetic stud «9 ifl Sotanem verructb
mtn variety apertaMis. Amer J But. 41. 648—651 (1954)
614 5м..чшда/6оп Af 5 лммп( // V The сгойЕУ лг«1 genet <5 ot the po-
tato {S^iantx^ iub^rAfvm) rad related species, Blbbographla (ienetica,
16 1—192 (1953)
815 5ичишла/*ап .M S, Magoon .V L Origin and cvtogenel rs oi the commer-
cial potato, Advsncii in GonUOs It. 217—256 (l»l).
816 5wir/.;ru4a Z Spontaneous polvpIccdtraHon in Pumex hybrids. .Acta Soc.
BiXan Poton . 29. 79—98 (IMO)
817 SKi>>itnsko Z. Zuk 1 Further ubservations on soonianeous polyploid izatloa
in Явглк’х hybrids. Acta Soc Bolan Poton . 34. 439-450 11953)
818 Swingle V T The botany ot G/m and its wild retainm In The Citrue
Industry. 2d ed . vol I. e<b W Reuther. H J Webber and L D Batcbeluf.
Lfkiveraily of Caliiornia Press. Berkeley and Los Angeles. Calif, 1967
819 7m Mota G ZytoiugiNcIte Studiefl uber die Gattung Rosa. Acta Hotli Ber-
gum fUnpMla). 7 97—331 (1922)
820 Jethone .4 Cvlologkal studies co ChsysaatKenuim II &ot Mag Tokyo. 29,
48-50 (1415)
821 ruMltrja.v .4 Flowering Plants Origin and Dispersal Translation, Oliver
and Boyd, Ldinbefgh, 1969
ъ22 Tanaka 1 Specks Problem in Cdrus Japanese Society for the Promotuin
ot Science. Lena, To<va Japan, 1954
823 Tanaka T Ckroiogia Citrologia Supporting Foundation, Osaka. Japan
(Summarized is Sw.ngie. I967|, 1961
824 7>din 0 Ve'trbiing. Variation und Systematic m dor Gattung CoMdiran,
Itacdilth К 273-3*5 flc25)
825 Thnday J 4. gnnnt T В Eifccts of it vuptlve sHeclion If, Polymorphism
and divatgance vithout isolation, Heredity, 11 2tt>-218 (19.Л)
826 ftadii? f a* GiWson J в Isolation by disruptive tclection, Nature. 193.
UM 1168 <19f2)
827 Tkoduu / .4 , Gibxvn J В Tlie probability of isolation by disruptive selec-
tion, Анн-r hat IП4. 210-230 (1970)
828 Thurne R 7 Synapsis oi a putatively phylogenetic classification of the
flowering plants, A Iso 6, 57—66 (1968)
829 Ting Y C Con.mcn inversions in maize and teoiinte. Amer Nat, 101, 87—
89 (19671.
830 Tischler G D e Bederlrng tier PnfyploMH lur if Verbreituftg der Ang»o-
spemien. Bot Jahrb .67. I - 36 110351
831 Tiscb/er G A'lgcmctor PilanzenMcyotogic III Ar.gewandte Pflanrenkarjo*
logй. Htndbucb der Pt’rni^nanatomle, Berlin, 1953
832 УмсАГ/г G Das Problem dt-r Basis Chrcenc^amenzahlen bci den Anglceper*
men -GaH*mgwn trnd -Familien. Cytologia. 19, I 10 (1954)
833 ПмЛГе/ G Det Grad de- Polypfcndfe b*i den Angiuspermen in verschicde*
nr-п GrceiMreilen. Cyto ng.b. 20 ICi —118 (I965t
834 Tobfu H A X cytokigrcal studv of Crepis IttUgfe&sa. C negltcta. and
their Fi hybrid, and its bearing on th* iMcbiniaffi of ohylogenetic redac-
tion in cbroitiiotouie namber, J Genetics. 45. 67—111 (194d)
835. Гл®пег H Г The bioayUemalfcs of Calytopiws (Onagrarcae), Annals
Missouri Bot Gard. 84. 46-120 (1977)
486 Лursparуpa
836. Tucker J. М. Taxonomic int trrelatioeAipa In the Quercus dumosa complex.
Mxdroaa, 11. 234-251 (|%2)
837. Tucker /. M. Two new ask hybrids from California, Madrono, 12. Ilfr—
127 (1963)
836, Tucker 1. M- Identity ot the oak tree Live Oak Tanks Joshua Tree National
Monument. California, Madrono. I«, 256 266 (1968)
639. Tucker I M. Sauer i Aberrant Amenm/kus t> opuUfiuns of the Sacrarneato-
San Joaquin delta, Calilornia. Madrono, 14. 2S2 261 <1958)
840. Turesson 0 The genotypical response of (be plant aperies to the habitat.
Hcrcdlta. 3. 211 -350 (1922).
-841. Taresson G. The plant species in relation to habitat and climate» Hrrditas.
6, 147—236 (1995)
842. Turerxcn G Zur Nat» und Begrtfuung der Artainberien, Haredltaa, 12,
323—334 (1929)
843. Tureseon G Tba selective eltoct od climate аров the plant apectm, Heredl-
tai, 14, 99—152 (1930).
844. Turner В I Summary ol the biology of the Composltae In* The Biology
and Chemistry ot the Composite*, ed. V. H. Heywood, J. B. Harborne and
В L Turner. Academic Pre**. London and New York. 1978.
345 Turner B. L, Alston R. Segregation and recombination ol chemical coostl
tuentb in a hybrid swarm of Bapiuia 1аегчсаиИзхВ. viridis and their taxo-
nomic implications. Amer. J. Bot. 44 678—686 (1959)
846. Turner В L, Horne D. Taxonomy of Machueranthera sect. PrUoctls (Con-
positae-Asterae). Brittortfa. 16. 316—331 (1964)
847 Turner 8. L, PrneeJi A M Deserts, gypsum and endemiwn. Io: Arid Land
Plant Resources, H J R. Goodin end D. К Ncrthington. Texas Tech Uni-
versity, Lubbock. 1979
848 U N Graome analysi* la Brassica with special reference to the experimen-
tal formation nf R парил and peculiar mode o1 fertnixatlon, .Japanese J
Dot., 7, 389—452 (1935).
849 Upcatt M. The nature of tetrapioidy tn Pnmula krwmsts, J. Genetics, 39,
79-100 (1939)
850 Urbans ka-Worutkiemtcz K. L’agamospennie, syitdme de reproduction Im-
5orient dans la spdciation des angiospermes, Bull. Soc Bot France, 121.
29-346 (1974).
851. Urbanska Worytkieuritt К Cytdogicaf variation within Lemna L Aquatic
Botany. 1. 377—394 (1975).
852. Urbanska VoryWfcz К Reproduction h natural Hpjold hybrids (2n«
—24) between Cardamine riuularis Schur and C алии a L. Ser. Geobot.
Inal ETH, Stittimg Rubai (Zurich), 44, 42—36 (1977)
853. UrbanskaWorytkiemicz K.- An auloallohexaplold in Cardamine L, naw to
the Swiss flora, Ber Geobol Inst ETH, Stiftung Rubel (Zilrfch). 44, 86 -
103 (1977)
854. Urbanaka-Worytkiemicz K. Segregation polaris4« cbez 1ч hybrldes naterels
triploides (2n — 24) ettbre Cardamine rival oris Schur (2n—16) et C amara
L (2n-16). Ball Soc. Botan. France. 128.91—93 (1978).
855 Valenhne D. H. LAve A Taionomie and Uosystamatle categories, Britlo-
nia. 10. 153-166 (1958).
856 Умеб F C Outerouing in natural populafierm I The Breckenridge Moun-
tain population of Clarkia exlUs. Evolution. 18. 213—218 (1964).
857. Vzueb F C Outcrossing m natural populations II. Clarkia engaicwJafa,
Evolutma. 19. 152-156 (1965).
858. Uaseb F C. Outerowing m natural populations Ш The Deer Creek pope-
lation ot Clarkia nilu. Evolution. 21. 241-248 (1967)
859 Varr* F. C The relatiomNps of two ecologically marginal sympatric
Clarkia populations. Amer Nat., 102. 25—40 <19M).
Лнгература
497
860 Ftahfry .? К С.яэе studies in the evolution of species complexes in Mila-
lus. Ечо! Biol. 11. 405507 (1978)
86! Hosra P Tli* ir ses ol suulhwsli-.n Louisiana. Bull Amef. Iris Soc, No 57
(19351
86? Waddiagtoa К D Di» argent г in inltorcwoiv bright. an evolutKinary res-
ponse lo pollinator. Otvnfcigia 40. 43—50 (1970]
863 ft wgnr.- Л1 Die Entstehung der Anen (torch rdumlkhc Son dr'll ng. Benno
Schwalbe, Basel. 1889 ,
6€4 B'ugnrv T H Reticulate evolution In Appalachian Aspteniums, Evolution.
8. 103-118 (1954)
WR> IFagne.r IF H The Barnes hybrid aspen, Рорь/олХЕоглезп, hyb nov. A
nomem Isttiral case n point. .Mich Botantst. 9. 53—54 (1970).
856 UTugnee IT H. Зело** D > Shingle oak (Que'cas imbricaria) and its hyb-
rid» in Michigan. Michigan Bnt. 13. 141 — 155 (1976).
867 Il'flAl H A Chromosome numbers and meiosis in the genus Caret Amer
J Bot. 27. 458—473 (1944D
№Л ОГшмгс J G A rode! ter the arsgui ol diploidizing meclwn sms in poly-
ploid spcc-.es. .Amer. Nat. ПО 4 IS—430 (1976)
8У> ITn/4?r 1 IF Chromowir numbers, phylogeny, pkyiogeography of the
АппОПЭСбДе and their beaming пл the (original) bas>r chromosome number
ol angiosperms. Тадпа. 21, Я7 65 (I9T2>
870 Walker T 6 Hybridization in some species o’ Pteris L. Evolution. 12, 82—
92 (19G8)
871 Walker 7 G Cytology and evolution in the fern genus Pteris L, Evolution,
16. 27—43 (l«i)
872 Walker T Ct A .-«omit s and vegetative repfoduvlion in lerns In Repru-
ducliMr Biolugv and Tsxcrimy ol Vascular Plants ed J G Hawkes. Per-
gamon Press (niord. 1966
873 Valtafe 4 R Darwinism An Exposition ol the Theo-у ol Natural 5«4eetioo.
Macmillan. Landon, 1M9
874 Wallace В Genelir divergence o4 isolated popnlul»ons of DtosofMa meta-
nogailef, Caryologia. Supp' vol. 761—764 (1954)
87S WoJl/re В Influence of genets xvstons on geographical diMribiiboa, Cold
Spring Harbor Symr Quant Biol, M. 193-234 (193Л)
876 trailer D M The relative сое!» of s«T- and cross-fertiliied seeds in Лпро-
Hents capnists (BalMtrimeeMi. At^e* J Bot 64 313—320 (1«79>
977 Walter R fJoti ил hmoNm L in Poland II Studies in (he origin o! poyptoid
Крек Acta Biol Crnrar.cnu*. 29. 113—131 (1ЭТ71
878 (rape's Л! S Studies of spontaneous chrociovnme brrkage in Intcrsoccilk
hvbi da of Brontes. Ualv. Caht Publ Bot. 2Я. 33b—447 (1967).
879 FuPf’a Л1 S Raks of rneioeis, spindle •.rrrirularifles and mirrniporocyte di-
vi»»ort in B'Ofr.us coo-afus Chromosoma, 11, 167 204 (19601
MO Webber ll J Rnichcier I. fi The Citrus fndmtry. vol I History. Botany
and Breeding. 1st ed University of Cihlomia Press. Berkeley and Lns An
gclcs Cnllf. !943
881 Writ J, Allard R T The mating avMcir. and genetk va-labili’y In natural
populaton.5 al CbMbuhi ЛеГгорАэЯа- Elution. 18. 515—525 (1964)
882 wicauum A Vorlrig- (Ibo- Э₽:тгПсег71Ъюпр. 3rd cd. Ficher. Jen»,
1913
883 Vi’L’s H \ distance cccMlclent as a hybridization indes an example using
,4i*nu>lux l<mgiilcru\ and M [Icmingil (ScropliuLariaceae) irom Senia Crus
Island California. Taxon, 29. 53—65 (198(1).
HN4 IT.-sf Л’ B. Garber E D Genet c rodier of variant enzymes П The Inhe
ritanre of e*l*r*v>s ?nd leurii: aminopeptidases in МвмЫш valgartsX
XP ccoineus Cana£ J Genet Cvtol.B. 648 655 (19671
885. Weilergaerd M Sludm on cytology and sex dHcrm nation in pc’.ypzjid
form* of Af.*i!amfr:4».vT а/Ълн Damk Bot Arh»v, IP. I —131 (1940)
32—45
Н = I Н Н М I Н ПН sM H U S
Литература
Whalen М. D. Reproductive character dtsplacamMit and floral diversity la
Sotanum section Androcera. Syst. Bot. 3. 77—86 (1978).
White M. J. D. Evolution ot cytogeceix mechanisms in animals. In: Gene-
tics in the Twentieth Centnry, ed. L Q Dunn. Macmillan, New York 1961.
While M. 1. D. Animal Cytology and Evolution, 2d ed. Cambridge Univer-
sity Press. Cambridge, 1954.
While M. J. D. CytogsoelLcs and byMematk entomology, Ann. Rev. Ento-
nd.. 2, 71—00 (1957).
VAir< MJ D Sotn* general ргсЫмпе of chromosomal evolution and spe-
ciation in animate. Survey of Biol. Progress, J, 109—147 (1957).
White M. J. D Soadatmn in animals, Australian J. Set. 22, 32—39 (1967).
While M. J. D. Principles ol karyotype evolution in animals, in: Genetics
Today, Perga mon Press. Oxford, I9M.
White AL Г. D. Animal Cytology and Evolution. 3rd cd.. Cambridge Univer-
sity Press, Cambridge, 19/3.
While M. J. D Modes ol Speciation, W. H. Freeman. San Francisco, 1978.
Wiegand К. M.. 1935. A taxonocnist's experience with hybrids in Ibe wild.
Science, 91. 161—166 (1935)
Wilson A. C Evolutionary impart a txt o! gene regulation. Stadler Sympo-
sia (Univ Missouri). 7. 117—134 1975
OFi/nbrr D. E. The nuclear cytology of bivalent and ring-forming Rhocoe
and their hvbt>ds. Ar>« J Bot, «, 572-674 (1968).
Winge 0. Th»* ehrtxnoeome: their number and general Importance. Comet
Reni Trav. Lab. Carlsberg 18. 131-275 (1917).
Finge 0. Contributions to the knowledge of chrccnosome numbers in plants.
La Cellule. 33. 303-324 (1914).
Winge 0. Inheritance of species characters in Tragopcgon. CompL Read.
Trav. Lab. Carlsberg, Series Physiol.. 22. 155—194 (1938).
Winge 0 Техигкжпк and •voJutioiary studies tn Erophlta based oo cytoge-
netic investigations, Canpt Rend. Trav. Lab. Carlsberg, Series PhyaioL.
M. 41-74 (1940).
IFinAler H Cher die expcrimentelle Erreugwng von Pflanxen nut abwd-
then den Chromoaomeaxahkc. Zedsthr. f Bot, a. 417—531 (1916)
Wolf С. В The Gander oak. a new hybrid oak irnen San Dtego County, Ca-
ll lornla, Proc. Calif. Acad. Sri., 28, 117-188 (1944).
Wolf С. B. Taxoocrmc and distributional studies of the New World cypres*
tee. Altax 1.1250 (1948).
Wright 1. W. Species crossability in spruce In relation to distribution and
laioootny. Forest Sei, 1, 319—349 (1965).
Wright J. W. Genetka ol eastern white pin< Fore»! Service Research Pa-
per US. Rcpf Agncnltura, 1970.
Wright J W., Gabriel V. J. Species hybridization in the hard pines, series
Sylvesttres, SUvae Genetica. 7. 109—lid (1958).
Wright S. Evolutxm in Meodelian peculations. Genetic*. 18, 97—159
(1931).
Wnghl S Isolation by distance. Genetics. 28. 114—138 (1943).
910 IFrigAf S. An analysis of local variability of (lover color in Linanikus pur-
ryot. Genetks. 28, 139—156 (I9U).
911. Wrighl S. IsolaHon by distance under diverse a у st cm a of mating Genetics.
31. 39—59 (1946).
912 Vnght S Adaptation and selection. In: Genetics. Paleontology and Evolu-
tion, ed. G, L. Jepaen. Princeton University and Evolution, cd. G. L. Jep-
M>n, Princeton Vftfvcrrity Prsae. Princeion. N. J., 1949
913. Wright S Evolution and the Genetics oi Population, vol 4 Variability Wi-
tiiiB and Among Natural Populations University of Chicago Press, Chxago.
1974
914. Wutfl H. D. Die Potienent wick lung der Juncaceen nebst elncr Auawertung
Лвтервтура
♦90
der enbfyul^i^rti Bel unde hlcakhtllch der VerwwndUfhaft der Junes,
сад und Cyperscew!. Jahrb. Wim Botanlk, >7. 533—SS6 (1939).
915. Wulff H. D, Ober das apoatatM Auhreten einrr Caninae-MeLnat* be! der
Mlkrospocofcnesc der diploodeo, Яозя ruga Lindt Oslerr. Bot Zdbcfar,
Itl, 539-557 (1954).
915. Yarnell S. H. Cytogenetics of the vegetable crop». IL Crudfers, Bot Rev..
22. 81—IM (1966).
917, Zabel B. The natural hybrid between Coulter and Mfray plnaa» Evdutio®.
5. 405—413 (1951).
918 ZoderjrD.. hid man M HybridiMtlon betwwn amphidiplaida and th* evo-
lution of polyplodds in the wbeal (A/ftfofw-Trtllam) group, Evolution. 18,
♦4-41 (1МП.
919. Zokarg [),. Harloa 1. ILVerdi A. The wild diploid progenitors Ы wtieai
and the* breeding value. Eupkytlra, 1Л, 58—BS (1969).
32*
Указатель авторов
Айала (Ayala) 153
Александер (Alexander) 444
Аллан (Allan) 191
Андерсом (Anderson) 128. 189. 194—
196. 201. 206. 213. 214. 220
birrajue (Battaglia) 393
Бейкер (Baler) Г89
Бейиеябер- (Betuenberg) 284. 347
Беннет (Bennett) 94
Бенсон (Benson) 216. 446. 458
Бишем (Beaman) 424
Бинхем (Biiielwca) 36
Бодри (Beaudry) 86
Браун | Brown) 186, 427
Бредяер (Bradner) 30
Бргкк I Brink) 49u
Dy Шемберг (Bungenberg) 280
Бурма (Burma) 94, 96
Буш (Bush) 148
В»Бжок (Batcock) 415
Бэтке (Baetcke) 200
Бюффе (Bui Ion) 91
Вагнер (Wagner) ISO
Валем (Whalen) 186
ВалейтаАи (Valentine) 153
Ветле (deWei) 89,90
Виганд (Wtegand) 194
Викера (Vickery) 135
Ванге (Winge) 141, 239. 264. 256.
274, 276
Винклер (Winkler) 274
Веские (VK4CI) 444
Гождшмклт (GcJdachoudt) 168, 169
Готлиб (Gottlieb) 180
Готтшалк (Gottschalk) 276, 453
Граи (Grun) 409
Грант (Grant) 24. 106, 109. 114. 161.
153, 167, 170, 243, 250. 363. 366.
367
Грэй (Gray) 197
Гулд |Gould | 426
Густафсон (Gustalsson) 17. 7Э. 81.
301-303. 309. 393. 396-306. 401.
404. 412. 419. 423, 424. 440. 441.
448
Дайрана (Darrin) 84. 153. 184. 185,
Дарлингтон (Darlington) 301. 411.
439.453
Данис (Deris) 360
Джайлс (Gilles) 355
ЛжМм (Jain) 210
Джои (John) 167
Джонсон (Johnson) 17, 305. 306
Джордан (Jordan) ISO
Дилме (DWsl 457
Дитрих (D»etr*ch| 369. 380
Добожамсккй (Do hz hansky) 51, 55»
56. 150. IS3. 160. 167. 170. 184. 247,
276. 278. 286. 289. 292. 296. 301.
302. 310. 349, 362
Доддс (Dodds) 340
Друри (Drury) 331
Дю Pna (Du Rim) 51. 199. 227, 237.
457
Давидсон (Davidson) 94» 96
ДМ (Dey) 65. 66, 325
Гарбер (Garber) 258
ГеАтс (Gates) 94. 95
Ге^денмова (Gerassbnova) 243. 254.
Герберт (Gerber t) 237
Герстел. (Orslel) 294
Гоадблат (Goldblatt) 287
Reprswr (HiertLng) 444
Лордам (Jordan) 160
Камерам (Cameron) 450
К ап адм a (Kapadia) 426
Укаэтм авторов
SOI
Карсон (Cirson) 155
Кек (K^k) 4». 276. 289. 290. 295.
296, ЗЗД
Кернер (Кегмг) 190. 191 193-195.
237. 233. 244. 457
К'срстгр iKentcr) 183
КЬьрейтер (Kai renter) 188. 189.
193. 238
K*n (Kron) 127
Кичлр* (Kitart) 275. 289. 327
Кларвгоя (CUrkson) 134. 135
Клоуэиг (Clausen) 4A, 107, 134, 138,
138. 276. 2Ю. 290. 295. 296, 338.
406
Клеланд (Cleland) 362. 364. 369-374.
376.378
Кокрсч (Cockruml 19?
Кромлло (CroveUo) 94 96
Кроннмст (CriMimtfol) «4. 94
Крукебеэг (Kfucktfwg) 48, 133» 165.
190
Кхошу (Khoehoo) 301317
Kuln (Cain) 55, 8*2
Ла «пре» т (Lanprvchi) 239.254
Латимер (La(iantr) 281
Левен lUria) 27. 94. 115, 183. 189.
Мб, 3U. 379
Леа* (Levy) 276» W. 304. 305» 410
Лею: (Lrnx) 134
Линией (LlnnociK) SO. 81, 91, 92.
188. 189.237,238, 408. 419
Лотси (Lotsу| 227. 237. 457
Лжлг (Lewis) 155 195. 187. №. 276.
280. 30*. 349. 351 4«
Мазер I Maser) 160. 170
Май® (Marr) 51. 60. 151. 152. 154.
155, 160. 151. 189. 197.463
Малсйрос (Malbtiroa) 338
Марсден Джонс (Marsden Jones) 190
Mer.iww (МсцЫэсЬ) 70
Мейсон (Чамп) 94. 91 2.1
Мендель (Mendel) 57. 189. 238
Менмм (Маптел) 2Я(). 233 283. 285.
m W6.301.3M, 4ci. 414
Ml.wp (Muller) 309
Молит (Mn Usch) 450
Моор (Moore I 197
Москуин (Mousain) 04
Myp (Moore) Й1. 287
.Moan (Maha) 369-372
Мжжп.ни: (Munlzing) 242. 243. 275.
286. 789, 296. 30 .. 303
Ново (Nobs) 108. 123. 155. 126
Нодзн (Ntudin) 237, 238
Нордеяшельд (Nordeasktfld) 368,
SO. 360
Нюгрея (Nygf*n) ЗИ. 397
Олглрд (0llgard) 423
Оливьери I Olivieri | 210
Олегов (Alston| 2<Ki
O*o (Ono) 275, 289
Пазя (Раду) 220
Палъиср (Palmer) 229
Патерпиажа (Pakrniani) 177
Пнмхлва (Pinkara) 446
Праэио (Ргмтпо) 199
Прэйор (Pryor) 128
Пит ван дер (Pill) 396
Пэкер (Picker) ЭОо
Радлоф (Radio ff) 39
Райли (Riley) 204. 205
РлЯт (Wright | 33. 148. 154. 164
Рейвн (Raim) 60. 94 95. 369. 458
Реммггон (Refninglon) 189
Рендер (Renner) 361, 364, 367, 368.
376. 377. 380
Репс (Rensdij 31. 150
Риган (Regan) 84
Ригер (Rieger) ?7Ь
Рш (Rrese) ЗМ
Ричардс (Richards) 423
Ричардгэн (Rieturdsnn) 154
Podw-jc (Roberts! 156, 189,349
PynOepr (Rudberg) 237
Ружхаулер (Rutishauser) 393
Рэй (Ray) oO. 91. 4*3
Рэндолф (Randolph) 205. 335
Саде (Sachs) 424
Омдфер (Sandlacr) 454
Сауэр tSawcr) 245
Саням I Swingle) 431. 433. 435
Cnmbcom (Sinrnsnn) 82. Й. 87, 154
Симеиза (Sinskuja) 48
Сире (Se*rs] 293
Скалннсм (SkalitsU) 129. 424
Сауиллас (SquiJace) 36
Смит (SiniLtj) 134. 135, 257
Смолл (SaiaJi 444
Сокал (Sakai) 94. 95
Омбриг (Solbrig) 24, 25.
Солсбери (Salsbury) 2S
Стеббинс (Slebbrns) 167.
189. 190. 191, 194. 220.
250. 258. 276. 278. 286.
301. 306. 392. 335. 342.
27. 157
170.
243.
290.
393.
171.
245.
296.
Ж.
402. 4I&, 43», 441, 458
КЙ Уноатгаь авторов
Стебе (Slice) 189.191
Станли (Stanley) 463
Тикер (Tucker) 245
Таньки (Tanaka) 433
Тахара (Tahaxa) 274
Тернер (Tnrnef) 30
Тогва (Togby) 348
Тодчб (Thoday) 160, 181
Турессон (Tureasoo) 47
Титан» (Tiaehkr) 276. Ж. 334
Тзкхолм (Tacthota) 385—388
Уаклдер (Wild») 216
УаВли (Wylia) 453
УаВт (White) 146. 151
Уклсон (Wltoon) 160. 1^4
Уокер (Water) IW. 401
Уоллас (Wallace) 184-1
167. 280, 367
Xaftati (HayaU) 237. 457
Хиссдя I Huxley 1 184, 463
Ханельт (Haadt) 304. 305
Хардхи (Hardin) 233
Харлан (Harlan) 89. 90, 327, 329
Хегя (Heel) 244. 385
XekJep (Heiser) 189, 191, 1H 210.
303,458
Хейльборм (Wellborn) 368. ЗГЛ
Xd»p (Heir) 284. 347
Хскард (Hsckard) 321, 322
Хёк1*:сои (Накапысо) 387
Хиси (Hiesey) 48. 107, 106. 278, 269.
290. 295. 296, 338
Хоке (Ha«te) 444
Холм (Xohn) 94. 95
ХКИк (Hubbe) 197
♦аесет (FatMtl) 211
Федоров (Fedorov) 291. 287
Федыиан (Feldman) 324, 327
Ферлеи (Гег)аа) 245
Филлипс (Phillipa) 216 294
Фмшк» IFIxch») 184, 18S
Фле*х (Flake) 39
Фокже (Focke) 238. 419
Форд (Ford) 161, 162
Фрстер (Fretar) 427
Фржкелл (Fryxell) 393
Фраз (Frisch) 182
Фрост (Frost) 453
Фюрикранц (FQ.-nkra.nz) 422
Квартили (Haartman) 188. 190
Xarepyx (Hagerup) 304. 404
Цо хари (Zohary) 324, 327, 342
Чамдьашю (Ceaalpino) 50
Шааль (Sdaasl) 183
Шафер (Shafar) 128.
Штуббе (Stubbc) 360
129
Эрлих (Ehrlich) 60. 94, 95
Эриан (Ehrman) 176
Эрнст (Ernst) 402
307. 319,
a
Указатель латинских
названий
я
s
я s'
ЗОв Уллгаггд» лахипашл пахачиб
- bafbadarue 101, 776. 294. 336
— barbsceum 376
Oramineac 22. tn, 301, 330, 349, 380.
399. 400
GrrriDeoideae 349
Gutter rrria 157
Giittlferae 157
Haemodoracw 114
HalpvpapfMU graalit 280
HapiOKglan 127
Harimannia 380
Haynaldia 90, 293
Hf llan'hcae 333
HtUanthus 139. 142. 144. 192. 202.
206. 225. 226, 301
— un-n^f 36. 144. 206. 208, 200. 456
— argОрЛрШк 210
— ftUandari 36. 206. 208- 210
— debiht 210
— dtcaptlalut 456
— giganitus 144
— nuittoffrvut 4Й, 435
— pctiolcra 210
HtiwrocaUis dfsllcko <53
— falta 453
334, 349
Hftarc^lera 358
Htrraclum B, 21, 37. 70. 79, 396. 398
— aurantia^am 397
— ambtllaium 36, 401
HippccasUnactM 333
Hiptagc madablata 400
Hoicus motUt 443. 432
Hc.fdtum jubalum 22
— vulgar* 22
Hott я cosrulra 400
Hosiia 144
Houttuyala vo/daia 399. 438
Hyacinthus ortmtalls 453
HydrophyllacftM 333, 345. 347
HydfOCbatHaceae 20
Hyjttrfcaceae 425
HypaHcum 396, 425
— parfc-ralm 399
ImpaHms сироты 25
Inula briltanica 453
Ipomopsia 79, 216
— aggrtgata 77
Ir-.daceae 279. 347. 353
IrU 115. 130, 134. 14| |G6, 202, 233.
214, 220.228
— tract tala 228
— chrysophyUa 228
— dougtasiaaa 184. 228, 229
— (arnaldii 228
- I»dva 78, 204. 205, 207, 443, 449
— giganticaeruUa 2lM, 2’26
— 228
— tunugona gifAnlicaerulra 78» 204,
207. 441. 449
— 1Л/*с«ш‘лд2а 228
— iHor. oiiprtnri ‘.34. 228. 229
— manzh 228, 229
— purdyi 228
- tmax 115, 134. 135.228
— tonuli 115, 134. 135
— tannaiufma 228
— thompsonii 228
/»rtej 280
laopoda 280
Ifvpurutn 128 129
— thahcirouin 129
!t*ri* denial a 399
Juslandaccic 3M
Juacaccat 356. 368—360
Juncos 360
Junlptrus 192. 203. 213. 220. 278. 279
— aih*i 192
— chincnsts 456
— scopulorum 211, 213
- vlrgumtna 3». 192. 211-213, i52.
KaLaruAoe 280, 285
Knautia 144. 345
Knaiftia 330
LahlatMlH
Laths. 129
Lacheac 129
Lavaaxia 380
Layla 106. I3fc 142
— gaiUardi'ndn 117. 138
— glandal-.'ba 138
— hticsotiicha 138
— IdcratuMdas 117. 138
— paiUachata 138
— fdotygloua 138
faoreneo/rnia 192, 193
La^onunosae 78. 128, 141. 159. 203,
301
Ltrnna gibba 452, 456
— nUn-r 452. 453. 455
— pf/pasilta 456
Lemnacr**» 453
£etmJGdo4 Klspldut 404
Lepidobatamu 231
Leplcoptara 229. 280
Ltpidastris 144
Указатель авгммпсмх нолмлай
509
Lesquereita 345
Lcucvpcgon janiperus М2, 588, 453
£ГаРк aspera 33, 61
— cylid'acta 33. 34
LiUtCM* 331. 333, 345, 347. 433
Ulium 279.310
— hantonii 104
— taadfottam 453. 455
— maria gon 104
— monmoviczu 456
LimcteUa 453
Linaceae 349. 400
LinanJhuj, parryat 34
Linaria 237
Imam 349
Li ster a mate ЗСЮ. 403
Littospermum carotinimse 33, 34
Lossactae 141
Lojamactac 295
Lotas cornlculalia 1И. 193
ГятЬпа'мх 28C
LuraAa 358
— ca^omiris 354. 357
— шефа 359
— paflewns 359
— pdosa 309
— purpurea 356. 359
— Wafa 359
— sadetica 359
Ij/ropowm 109
Lycopodiaks 280
Lpcopcdiaifi 280
Lyco'is radiata 453
Lpttirum ||4
MmU 106» 138. 142, 143» 166. 173.
271. 303, 308
MaanoJiacoae 333, 333
Malphigiaceae 400
Ма.'м1 398
— 399
— bgloegtria 453
Malvaceae 279
Mangijrra indira 400
Maydeae 349
Mcdieago coerulea 343
— softtw 30, 31. 343
.МеГслЛ'£лл 227. 310
Meftfota 109
Meliphagidae ]]{
Menizeha ЗЗД
— dincaul is 141, 142
ЛЬ’Гш/вваи 27$
MUftKiirut 431. 433
Microtis 139, 142. 303
Mimuius 114, 1X5. 142. 159, 192. 216.
228
— car din alls 107
— glob rat us 135
— guitutui 49. 133. 160
— 107
— longiflorat 40-44, 77
— 160
— 4П—43. 77
Moderns 71, 72, 247.419
Musa acumtnaia 442. 433. 454
— bafiлюла 442. 454
Musaceae 356, 453
Masca domestics 161
Myrlaceoe 127, 400
jYutcujhj 347, 348
— lamia 453
hiasJuftia» of I и mat* 453
JittnaucJienti 144
hifuopbUa 333
NepMea portarieenUs 246
fiicoban* 104, 1C6. 116» 116, 139, 142,
167. 239. ж 267, 270, 294, 345
— oJo/a 104
— banariensis 104
— rJuhwaa 276
— TungHiorfiii 104. 11л 257
— 1ы1&1Ьга 99
— saaderae 257
— Jufiocu.4 276
fii&Ua d^anii 38
A'<f rUeXta nigra 400
ActfAajegiu 227
ftoibascordu/rt 35»
— [ragrans 1СЦ. 401
Nupkar 194. 238
Ochnoccac 4410
Ochna serridaia 40C
(khlhiphila 309
— polgetigma 2S>j
Oenothe/a 270. 362, 36^ 381
— vgilliiMla 370, 374
— aasirMBOiWana 370
— bininu 72, 335, 270. 362, 369, 370,
373. 376—377
— Jketipei 113
— dazaeiornis 113
— data 370
— trylhfasrjMLi 370
— fkWi.iin’dfld J70
— grrtndijlora 370. 373, 374
— hooluri 354. 369. 370, 373. 375,
376
— irrig aa 366
я
rnsjtmxw «'ll
619
Улазвнм латинских исвмишЛ
- rob* 233, 234
— rugaia 231
— xtaauita 232
— stdtala 232 233
— turbinctia 211 230-232. 443
— virginiana 12, 18, 232
ftaimaMia 380
Reounmlaceae 128, 399. 453
RanJutfohii 79, 398
- аг/is 102. 193
— ounrorttui 3W. 401
— «оЙома 19?
— reptns 192
Rapbanobrassica 241. 242, 273, 289,
290, 300
Rapbunus sativus 31, 241
Rhtmnarcae 123
RH*anihut 101, 114
Rhadodondran 194. 238. 244
— ffifrugintam 238, 244
— -Wriu/fcvi 238, 244
— LnicrmcaLum 231 244
Rhoeo discolor 381
— «path.».v* 381
Ribot 127. 141, 303
Rom 394
— agrcstis 38t>
— ЛМ1ЛД 7. 236. 249. 270, 277. 382.
384
— ch&wii 386
— coriifdia 386
— dunelorum 385, 386
— Jn nsJUti 386
— лисгщаЬа 385, 386
— mottis liHti
- poniftra Ш
- potato 3M
— ru Ыриюза 387
— rubrifalia 385. 386
— itpiosa 386
— irnmMg 385. 366
— <олмп/«й 388
Rot+crae 131. 279. 382. 399. 453
RbW СогИЦоШ 420, 421
Rubiacrue 279. 319
Rubai 70-72. 79. 194. 300, 399. 402,
418
— acwfuj 407. 408. 410
— aiteghcnu.-^ 420. 421
— alrdcola 420
— Mid 420
— саышо 406 408, 421
— goihicas 408
— idatuo 421
— iAcanw^nr 420
— iruulans 406
— noschas 420
— pfca/u; 4LKi. 407
— 4C6
— poiyC*pai 408, 410
— prumottu 410
— rwdoiuii 406
— tomenioius 420
— uinufcdius 420
— vohibtrgii 421
Rudbtckta aiitt^rnica 399
Rttmex 310
— acrtata 311
— ihgfsiflorut 311
RuUceae 400, 431
Sabcaceae 279
Salix 190, 227. 2791 310
— dapiwid" 191
— Мквда 191
S*lmoDid*e 280
Saloia ill. U4. 194. 238
— apiana 112, 117. 163, 194
- meUUrra 112. 117. 163. 194
Sanicula bipinnoiifida 319
— entxncaulis 319
— hofjnatu* 319
— laciaiata 319
Sarcococai pfunijormit 400
S*ururaceie 399, 438
Saxifraga cernua 449
— (♦T/’Wfi.we 448
— Jolioioxa 448
— «Ыйлй 448
Saadr«fMe«c 127
ScUta 333
Scirpus 35Л
Scroohulori^ccic 40, 77. 114, 131, 135»
160» 349. 453
5 май 90
- i*fait 23, 291, 329
Sedan 192
Sela&Mtlla 288
Smfcaz 2>Ц
— jacobaaa 28
— robtrtifriMU 284
Septnla jumperи«гд« 279. 301—302
SUene 13S. 141, 192, 193
SlmMM 138. 142
SUanim ЛдоГпл 258
- Jitbotum 258. 259
5m Juana гасепюла 400
Soitnarwae 133. 309. 345. 483
Sola^um 111 133. 142. 186. 274. 309
- Afflelr 340 341. 342 .
— rftaewnte 444, 445, 449
X%0MX«ai narwutxux названий 513
- сИгаИЦсЛат IM — u'rmjijwi 342 — g'agi IM — 1тмЫ(х1альт 185 — m^rodcnium 449 — nigrum 271 — rostral urn 186 — rtrnotajuatm 341, 342 — tuberonm 297. 339-341 445, 450 S'ofono&a trigueirrila 280 403 — chamatmespUiu 399 — natrida 3W ScrgAum 301 Sparfina patent 2(1 SpGtkiphgUum paiinii 400 S pergtd aria 142 SblrtMidM 279 57лпЛ<1р*а 114 Static^ alaerrftn»a confuse 399 SlaUaria crauiMs 446 — iangipts 446 Sitpkanomeria 139. 142. ICO — ext ana coronaria 150 — malhrurenjUx 160 SUrculiaceae 112 Stipe 321 Sir epl ant hxi glandulosax 48, 165 Sir rpt Otar сил 116 Styphtlieae 389 Suwtjuj 347 — pral etuis 102 T richer a 144 TridwruidM 144 Triduxen’.rcac 129 TrfcAcwucMS ehzeMnee 2И0 frtfolium pcatense 190 — repent 190 TnUium 333 — kamischaUrtu* 321 TrrjfttfSia auraatiGla 400 Trlttcate 329 Tritium 109, 167, 277, 2ЯЗ, 318, 35Л, 327 - ^iivu.-r. Vi. ЗП-2&. 336 ~ d^um 334 - лмтососсапт 91 327. 329, 334 —- speltoides 328 /ошсЛй 328 — hawpAert’ii 90, 327. 329 — (urgidem 90. 327. 328 — tmlgara 327 Tr.-cftodcMfiMit 333 Tsuga heferaptiyUa 87 — nurttJUiona 87 — /nsricrutoldes К? — tonomenrit 87 ТЫ.ра 279 — gtineriana 4СС, 453 UlnuccK 353 Udnbelh(ee*e 319 Urglnaa litdiea 433
Tongsipfera galatea 60. 155 — hjfdrodwris , Taraxacum 8, 21, 79, 396, 398 402 410, 422 — ttovtense 439, 440 — ко* xaghgt 422 — affefoafe <22. 423. 455 — pttmnieum 422 — retckenbachli 439, 440 Taamannla 333 Taxodiaeeae 279 Tholictrum 279 ГМойготл 142 Thuja piUula cxceixa 433 Thymeluectit 399 Tmesiptcrts 280. 333 — tannensis 281 Tofieldia 333 Tvamsendia 426 Trmlncanlia 225. 226 Trugopogon - ilub'us 102 — parrijoties 102 Urticaceie 399 ItacaftiKm uhguvMam 296 VanJa 130 V*«r банде fAajMua 28 Vermxad I3&. 142 б'агаш’га 349 Vlrte 159 — safk*a 141, 142 Vinr.Q major 14 Viak IOO, 167, 192 — arvensit 100 — Irteotor 130 VcfotteMa 279 Tisandia 333 Vibr/o*m£a 402 — tiridtfofia 399 Winter ке»е 333 132
U-4S
Уяахг* и> .^rvHC-Kut *азмчий
XuiUAoxyium ftunf oi 400
Xtfocopu 130, 131. |fi3
>'мм*<й 343
Zau vfr^rne ЗСО
Zeu 199. I LG,
- гмср» 177. 311. 335
— ДГГНПЯ14
Zfp?!»f6Mtbci 4U3
— tfiane 399
Zevxw ИЮ
21г-й»Ьег»с«яе УэС
Z>nnia 192
Zt'tcpeieJ«« mackayi ICO
Предметный указатель
Аберрантные ярпилки в пернферичс
аик пспулпнкт 184
Автогамия 15, 16. 21—24, 139—141,
159-160. 270 лол в-(1
Аетоаолиплондми 275, 299. 338—343
Аитосегмсаши 410
Ага мн и Й bamiumkc 267. 415—441
----стадия раэыгткя 437—439
----фнл.хсхетмчесжмЯ вналт 414.
418
----«олг'ИНОмвнЛ пстигииэл 439—
441
Агамоспермия 15 16. 20—21, 247,
248, 259, 392- 413. 454, 435. 458
— облигатмя 21
- фамультатявнвя 21
Лгматсслпихии 344. 354—360
Лдаамьжмс геыиые хомбимдцив. рас-
пад 117
Адмитмижая эмбрнояжя 20, 303—
3S6, 405, 435—436
Axjmtxwiccts 102. 103
Алдмирня 118, 119
Лдл&валнялояд гемомныА 290. 291
- ьсгчентиый 290. 29). 295, 296
Аллоплмиия ел Лмфиааонлмя
АмфнСмя 280. 288
Хмфпялонлня 235, 240—242, 267.
275. 298 290
АмеуялОШИ 344—554 См также
Агиятоплоидия
— посюляиыя 347— 349
— птсрячивя 431
— иисходшхвя 344—347
- вояимлоядпыП уровень 347—349,
410. 413. 427—431
Амеуплокдный ряд 287
Хпедьнш 431—436
Лпосвматмв 395. 397. 436. 437
ArxiMHKCHc 247. 248 См также Ага
♦скиермия, Вегетативное раменп*
женне
- геметофитмый 393 - 397, 404—405,
424
— oOjuibiwA 397
— факультативным 397
Апоспории 394—397, 404
Асамаок BII
Ассортятмвние скрещивание 147, Н8.
161, 162
Аутхроссянг оОлягатиы» 308
Б.«ивн 456. 156
Ьарамнси 14
Барьеры несовместимости 177, 179—
— зтоиюгмчесхме 177
Биотипы малые 414
Бвоеистематияесше чатегопяи 82. 88.
89
Блоки тежмв 262. 367, 168
Буж 26
В хромосомы 344
Вегетативные сворты 410
— химеры 274
Взаимоотношения яо плодовитости
123. 141—145
--------модели 141—143
Вадим* лоп>л»иня. гомогенность
39-63, 73—74
Видообоамамиие 146—140, 150—
166. 268-209
• - вллопатрячесмог 157. 160
— апоммктичсмио* 247—248
— бнотитесх» cuiuurnwiH-Moe 160
153
— гкграфи'«еское 147, ISO—153
- гибржджж 235-240. 244-246. 248.
249
— пантам 147-149, 156—157
— яарапатрмческое 148
— перамтгле 151 — 155
— гкчичбннаяис. 1:ос 242—244. 24в.
250-265
— сммпатрячесжое 147, 148, 169-163
- способы 146—149
— сталвпвтрнмесэсое 148—149
1Г
516 ИрлдметлыЛ у лазит ело
Виды аллопатрические 85. 86
— CwCi.iorww<-KHe 45. 50—68, 82. 85.
86. 91-97
— генофонд 59
— двоЛмими 64—68, 92-94, 3IS, 316
— мсрфологмчвскве 83
— послах паателыше 82. 86-87
— симватричсоге 65, 96, 119, 121,
122
— тахсономическяе 82—86, 91—94
— <нолжхь*ониые 83, 87. 88
Ввквивам 15
Водоросли 2KI
Вшцеммцы гэмологгаые 246
Гамггиый фонд 200
ГаиетсфнтныД апомиксис 393, 397,
404, 405. 424
Генетическая идентичность, оосаяя
т*дв 102
Г»*гтичгчжие тотемы. гсеилалв^ярс!
ааииые *>1—384
— — Ottufihfra, 362—367
Генетический д;«*ф 38
-----сочетание с отОэрсм 38
Геиегаческов равковкм* НО, 111
Геннад регуяацхк 169
Г*ыные системы 98—102. 105. ПО—
111
— — сериальные 98—102
Гекотнп нитегрнроалняый 1101 111
Генофонд 59. 89—ад
— вторичный 90
— первичный
— третичный 90
Г«Ы. блоки 367. 368
— жж»п«ж>собностм 17—110
— виожек-тмениые 98. 99
— модификаторы 98—101
— мг.рфшосичкхие 107—НО
— мутатсры 189
— оспкиицмоикые 98-102, 133
— пг.ток 28-30. 33- 36. 59 63. 74.
156—157
— регуляторы 103, 104. 169
Гетерогамны! комплекс 268. 362.
ЗЮ- 575. 3*0-382. 384-388. 391.
392
-----аволюшюннмй потен паз л 391,
392
Гетеражтотмосп структурная 253,
254. 267. 268
I ггерозис 298-300. 411. 412
Гиб^идиадши 72-79. 188-198. 225-
— естественная 1W-190. 227
— и агамоспермия 402—404
— я микромам 72. 73
— интрогрессивная 225, 226
— и полувады 77—79
— мсстообмтакки 195. 196
Гибрид пак мовтоств 298- 300
— hcvodmkth моста 179—181
Гибрид кость кумяпнчсская 237, 249
— структурная 237. 249
Гибридные полумглы 79-80
Гибридный индекс 201-203
— комплекс 267—273
Гифип гяпгдиеоаы 227
Гибриды. мелоинеаюсоби«хть 116
— разрушение 263— 265
— скопления 7В, 190—192
— Сжпгтаикые I9O—192
— стабмдкмши аосгшомлаедсмия
235—237
— стерильность 118, 117
Гинкго 279
Гимеоаогичыые Хроааосоым 240. 292.
336, 349
Гомеостаз фвэнодигичссхжй 298
1омогям«ый ксыхлехс 267. 268. 270—
273. 46Я-459
Гомшиготпость структурчия 2S3, 254
Гомоллоидкма имааепенцм 246
— шбркды 262
— лкпии 250, 262
Ге^хфоут «7—«Ов. «2—<36
Грабы 280
Делец,. 1М
— н1гтер-стнБналм<яч 105
Дивергенция 88, 86, 163—165, 458,
— эвхиоалюннда 117
— ^логическая 122
Диллсжлмлацмя 335—337
Диожкпсфмм 394—397, 404
Дисгармония гевнад 138
Ддсаирслсста (*ф.итэсть) среды .15
Дифференциальное сродство 240. 241
Дифреренциацт расовая 33, 34
— ывирогвографиымжая 36—38
— эколсгмчасхяя 119. 120
Дэждваме «ерзк 260
Дрейф геветжчесий 38
Лронго 155
Дуг« белые 220- 234
— черные 229
Дутилпсациа 106. 240. 299
Ежевика 247. 418—422
— микро яйлы 418, 419
Лрсдмгтнъй указатель
517
— урежим пломдвости 119. 420
Естественная rvtyNUmilNfl 77—79
— система классифвкйивн 93
Живорождение лож л ос 16, 396, 446,
449
Жимееаная форма 143—145. 301. 302
Жорденов 7|
Ж* ж*м* 49
Закрытое сообсжество 26, 27
Замешена;е видов 12]
Зимородок ракетоаьостый 155
Зона вторично* игтерградаиии 190
Идентичность генетическая 102
Иерархия тачсономымаекня катхо-
рМ ЙЗ—8b
Измемчивсстъ ажеупхогдиая 3SI, 352
— гсографхнеская 39—44
— клгсввльная 39
— полйзленяе 334, 335
— кьгктрофоретичигмаи 102. 103.
203. 379
Иэатнркинцм? меиннтиы 51. 89.
112 117. 163 166, 2О4-21С
— — классификация 113
-----врсстраиствежлые 112—113
-----рещхлаухтмвные 112—117
— — редовые 112—113
Издяжин престравегаснлап 151
— рсародуктнммя 69. 151, 163—166
— стенная 163
И мирней ты 20. 102. Юк 233
Инверсии 104—106, 170. 347, 349
Индик гибридный 2О1--2ОЗ, 206
Интерграхапая 43
Hinp^rpeixuMMR гнбрждилаомя 199
Интрогрессия 199-226
Ириты 134-133, ИМ. 204-206.
229-229
Исмусстняпа» система кдассв^/ха
инх УЗ
Искусственный отбор ПУ—178
Кактусы 247, 443
— устарниаж.вне 446—447
Картофель 340—342. 451 См ме-
хе Solojwm
Классификация полиплоидов 289—
291
KotuH >56
Климы 39—44
К.хэиааакые комхэеясы 267. 442,
445—449
Клоны 17—20. 442—444
Клопы 356
Колибри 129. 131
Кольца жроаютомшые 361—369
Комары J22
Кпмпарнум 89
Комплексы агамные 393
- гстгрогтмные 25в, 362, 3W-375.
JAO— ЗЬй. 391-392
— гибридные 267—273, 442-444.
456—459
— гомогамные 267—268, 270—273.
458 459
— клональные 267. 442. 445—449
— паднплсстлиые 315—344
Коивергесция 401—462
Конжурсмтное исключежие 121
Конкуренция межаядоеля 119 121
Клнцекич вида, бволотичеекая 94—
97
— — дуалжам 93
-----вомипалистическая 95
— ггягтитсехмх pBHC^MiHA 154
—^ставилизлили гибридности 411—
Конъюгация гетрогеметичеоля 292
— юаптегетнчвекая 292
— жеяиеномжам 392
— лвруштия 311
Критерий BUDMKtuH4t признаки* 201
К рос . н» мл яр 22V-224
Кукуруза 31. ЭН. 454
Культивары 90. 327. 342. 435. 454-
450
Культурные растения, едмимиы клас-
афикажпп 99—90
К отбор 27
Лайм 432- 434
Лета.» таметофхтяие 366
— л^гопые 366
— сОалазяропднжыс 362. 365—366.
370
Лимони 407—408. 431—436
Локжзьаые расы 28 ЗЯ, ISO
Люаерма 30—31
Макрорекомбнмаиия 101—102
Чакронюхазжия 457-463
.Чалима 421
Чандарин 432—430
.Межмдоме аамкуреядня 119-121
Мсжсмщиомиые гиСриды 125
5ia
Прлдшпшй указатель
Мейоэ пострел ужжиоешы ft 360
Местообитамае открытое 25—17, 196,
200
— гябрнднмши 195—196
Ммкроаиды 71—73, 416- 419
— и нт оса мама 71—73
— агаыосзмржиыа 21, 71 79, 237,
411-412, 421. 424
— гетерогвмнме 71—72. 362. 3Тб-
382
— । «брихные 72—73
— клсшалям* 71—72. 237. 442-444
— л мил мыс 420
Моносомия 345
Мттащпг почковые 410
Мхи 260
Нагледаванже блочное 261—263
— тетрвсомноа 293—2Я4
Н«жмчкклпсоЛя<х~п> гнбрадоа ПО
Номтвм 240
Ниша пищевая 122
— акологннеская 118, 461—462
ООмеж геи а мл. блокирование 112
Оплодотворение верафктмое 15—16
Опознавательные анаки цасгиоа 182—
184
Опущен 445, 450
Опылсжие » 31, 49. 113-115. 128-
121. 185-187
Орхждом 129-130, 166. 4С4
Отбор дихруптивмый 37. 148, 161.
— катастрофический 156
— межвидовой 119—123
— стабмлвмруюшнй 69, 196
— тжпы Г- И Л- 27
Открытая система роста 80 81
Открытое шестообвтанма 25—27
— сообщество 26 -27
Осина 17-19. 444
Осы-поевломааориды 131
Палекжтолос-внеси я лети пись 86—87
Наим висит 15—16
Паимнжтвчакмая единима 31—33
Лапоеетжнкм 192, 246. 269. 285. 298.
307. 319. 397—401, «И. 436 -437,
440
— апосампые 401. 437, 440
Парапатркя 118—119
Партеногенез 20. 394—400. 404-405
Перекрестное оалодотвореам 15—16
Лсрсоончатхмрылые 30
Перестройки млавигнмые 218
Перетяжка центромерная 356
Пермамежткан гетероамготнэсть по
гране.кжаиням 249, 267—270, 361 —
Ml
— гнбфндвость 249, 267
— яеитная аолниоидия 249, 267,
ЗЯЗ—392, 456
Пгчгмокжнкн 280
Подсопевим 206—210
Лолнгаллмди 334, 342
Полиморфизм 39-44, 56, 180, 160—
163
— ш> фа?м«™ 102-103
Полнпломдизааяя непрерывная 315
Пслшиюмджя 271 278. 29$. 301, 396
— днбатнеесжая 351
— и жизненная форма 301—302
— моио6аая««смии 361
- негмбрмдкая 275
— пеметиая <58
— распространение 279, 280
— частота 286 -288
Полипжшдмый ряд 340
— — НвШИЦЫ 328
- г/фое 278. 284 285. 334, 344
Полмплждяые комплексы 267. 315»
430
-----генегкко таксономический а на
яп 317—318
-----адекгмфмкацжя 313—316
-----методы ажалнаа 315—31*
-----остатки д.рсаямх 332—334
-----старые 330-332
-----структур* 316
— — филогенетический амалма 317
-----дитотаксономичеояй аиали
316-321
Полиплоиды янАааич'скме 278
— вплаваемш намегавоста 334—336
— способ» обп*>пвая»я 299—ЗОС
- тяпы 289- $97
Пгалмсошмя 344
П<Х1к*термая 34Ю
Псишла^жчсаше хромосомы 356—
Полхлыбрновжя 433
Лиловое размножение 13—15, 55—
Полуашш 75—78, 151-152, 227—228
Помершсц 432—434
Поттулякии дядькжктивные 76
— П1%1иффм«»сгкме 153—155
- прмродяыс 178—179
- Пром водные 736—737
— степами интеграции 70—71
Лреп'жгтный указатель
619
Поток гено» 28—31. 33—36, 59—63,
74. 156—157
Ппчнлкие ирякмн *10. 436
Пред»пятктс.«.ивя ион ысс^иия 24!
Прерывистая эволюсня
Пржвняки 274—275
Приложи. СММ0СТ6 201—203
СИММИ 120. 186-187. 225 -2Й
Прожито основателя 154
Природные яатоладиоллцды 338—343
Пссахогзмня 20. 396
Пчелиные 130
Пчелы ИЗ. II7. 163. 182-183
— мне» ши оы 131. 163
Пшенньы 326—330
— гечожнаи иоостктуцмя 327
— гаЮЧММ анализ 327—328
— система асжв:сиф«кацим 327
— филогения 328—32!»
Рлвикюеок генетическое 110—111
Рвзчножеяие С*нпаре:гтв» ое 16, 69-
72. 75
— v-rtTITHMOe 14—70. 747. 267—
268. 442—443
— половое 13—16, 55—58
— у ннкареитальи«ге 16. 60—72
Разрушите гхОридое 112—113
Расиобраэовамне 225—226
— г и брили се 235
Рксслшие 28-31. 61—53
Расы биотнческни 48
— п-уерафинеские 38—49. 76
— дмдыаиктимыс 44—45. 189—190
— пнтротрссеквные 225
— лсжалъные 28 38, 86. ;50
— пгрегрыоающкеся 77
— .мафмч&дше 37
— жогеОграфжжческне 76
— зкол о г ические 45—47
Регул sum* 103—104
Редукция чакла хромосом 34 ti - 317
-----— отсутствие 44'
Редька 21
Ртипверсмн 252- 253
Рсчимбанациониая система 24—27
— - закрытая 25—26
-----открытая 26
-----экология 25—27
Ринкера комплекс .361—369
Репродуктивная квамцив М9
— — внешняя 11>—115
— — мутренкяя 11^-117
Реснитсз волпплемхов 317, 325
Реципрокные транслсжацм 361
Рожь 291
Роте собачья 382—368
Рыбы 280
Рид вам) Я.НЖДНЫЫ 350- 353
— аол1Н1.Х1Н.1-ый 276- 279. 159
т-Отбор 27
Саговиими 279
Симоин.сммистямссть 308 — 309
Самооньммгнв 158
Сахарных трглгтмик 451
ССалансмсованные летали 362. 365—
3GU 370
Связность (анагятссмя 106—107,
213—215
— признаков 201—2СЗ. 213—215
Сетчатая филателии 457—458. 459—
461
— эяасаасия 457—139
С и и натрия 118—119. 121—122
— байт ичсема 119, 122
— нежная 119
Скитком 79. 85. 153. 227-234. 432
Слмтстнчсские ачфипюиды 294
Система (ы)гсяядя 96—102, 106—107
— (иыссмфииаими 93
-----ыекусстя«ш|вв 93
— — филсггиетичесмая 160
— размножения 13—27. 308—310
рекамбжиаамонкан 24—27
роста открытая ЙО—86, 459. 460
— схршнвяаия 15—16, 21. 143—147.
• J4. 235-236. 266—2701 401 —
402
тволюцим закрытая 459
— — открытая 459
Системные мутации 168—160
Сксалеиия габридйв 76. 190—192.
402
Скосарв турецкий 339
Скрснниааяяй ассортативное 147—148,
161-162
— cayreftBoc 15—16
Смшцамиы яркая яков 120. 186—187.
225-226
С ома тесное улвоегше 299 300
Сообтество закрытое 26 27
— открытое 26—27
(^осеастеа 31—34
Окна 29. 211 См тяжж? Ришт
Сосуществаяанне 118—119. 120—121
С лари камне МЯЦММШМ 354
СоенмфжчпыА кариотип 167
Среда, ди1”*^сн1>:ть 35
Стабилизация хоспроктпелеияя гиАрм
Лов 235-237
820
Пр*дл*ТМЫЛ рГвЗИГежь
— ПгОрНЕКОСТЯ 411—412
Старение клонов 450, 451
Стерктьнот. гибрид** 115—117
Структурные гошомготы 171—172
(Хвалимы* 106—110. 170—171. 201—
203. 213—215. 217—220, 262 263
— и нгэс атторнее 106—107
- M-V 107-110, 217-220. 260-
262
Тетра сомин 341
Тетра соыныв хромосомы 346
Тимофеевна 285—285
Трансгрессия барьеров стерильности
220-224
Транслокация 104—166, 170. 250—
253. 258. 347-349
— плеча хромосомы 105
— пемлехонатвльвые 362
— решип режиме
Траясппаицш 106. 252—253
Триплоиды 389—391, 452—456
— аглиикловение 452-454
Три со мини 368
Усвоение вторвшие 334
— соматическое 299—300
Уолмов *ф4**т 188—187
Уровня плсвимосги 281—286
♦акторы отбора 47—49
♦клада 143
Фжлгтическая молюция 461—462
Филогенетическая сеть 374—375
— схема 273
Флоксы 183
Фрагменты яиемтр вчинив 350—367
ХаоАнм 279
Хемотаксаиоммчсскне методы 331
Химеры 274
Хлопчатяин 336
Хрсмятографян иа бумаге 321
Хромосомные кольиа 361—369
— мреггроАки 104—106, 116—117,
167—173, 220—224. 240- 242. 250-
254
— числа. диапйю® 280-281
----направления 170, 360
----ОСЯОТШЫЙ 276-279, 280-281,
333 345-349
----— у агам<х2ер|шыл распаяй
398-401. 415—418
Хромосомы, величина 310—311
— гомолегячг.ые 240, 292, 336, 349
— добавочные 344
— <к>*о в ное число высокое 353—354
--------ии-мов 357—358
--------яервомачальиов 364
—-------редукция 345—347
— полидвггричвекив 356—338, 366
-----фра.'ыегтасия 356—ЗбО
— структурные кдмеиення 167—174
— птрасомные 348
— терминальные участия 364. 373
Ценсаиды «2, МН—89
Центромера диффузная 366—358,
Цикадки 122
Инда лнрфеоемцна дкн — гмбрклгта-
ига 468
Цмтэллазматячссхжв факторы 98
Цятрпи 432
Чалломоски 309
Чешуекрылые 122
Числа хромосом оси сем ое 276 261.
333. 345-349
— — удвоение 274—275
— — — соматичееков 275
—------СПСЯТЯНИч'И:
Шмеля 129. 340
Эвазюлноечая хквергемджя 117
Заслоняя конаерггнтная 461—462
— филвтическля 461—162
jrxjjfit 326
Экоаиды 82. 8Й-Я9
Экалзтиж рплиС.«иаияокны1 гнетем
25-27
Экологичен а к ниша 118, 461—462
Экологические ахавгахшм 328 329
Экотип 5Н, 32Н—329
Электрофора 203
Элвмтр-.'.форетнчосмая иэменчнвсстш
102-1Ш, 2Ш, 379
Эмбри амия адвсктявиая 433—436
— куоеллярная 435—436
Эддмакгов 310
Эффект положения 167—170
— иосладователмостн участков 168—
174
— Уодлсся J88^187
Ячмень 454
Яшерижы 280
Оглавление
Предисловие редактора перевода................5
Предисловие к русскому изданию...............10
Преднслонш................................ .11
Часть первая. Популяция в расы
Глава 1. Размножение растений................
Отклонения от случайного перекрестного оплодотво-
Е:»ня........................................
егетатявиое размножение.....................
Агаиоеперм ня ...............................
Автогамия....................................
Рекомбинационные системы.....................
Экология рекомбинационных систем
Глава 2. Локальные расы.......................
Расселение я поток генов......................
Панмиктические единицы и соседства . . . .
Поток генов и расовая дифференциация . . .
Взаимодействие между Потоком гаю* в отбором
Микрогеографическая расовая дифференциация
Сочетание отбора с генетическим дрейфом
Глава 3. Географические расы..................
Непрерывная географическая изменчивость
Дизъюнктивные географические расы
Экологические расы ...........................
Факторы отбора, участвующие в формировании рас
13
15
16
47
Часть вторая. Природа видов
Глава 4. Биологический вид.....................50
Пример.........................................53
Биологический вид как естественное следствие по-
лового размножения в биотическом сообществе . 55
522 Омоалслае
Концепция генофонда вида............................
Погож генов н гомогенность видовых популяций
Виды-двойники . ..............................
Глава 5. Видовые единицы у уиипаренталькых
растений......................................
Системы уииларентальных популяций . . . .
Степени интеграции............................
Микровиды.....................................
Гнбркдиость и микровиды....................
Пересмотр вопроса о гомогенности вида .
Глава 6. Вопрос о ваде у растений.............
Полувиды как результат постепеииого видообразо-
вания ........................................
Полувиды как результат естественной гибридизации
Масштабы проблемы вида........................
Причины и следствия, вытекающие w концепции
вида у растений...............................
Глава 7. Разные типы видов ......
Типы видов............................
Таксоном ичеекиА вид..........................
Биологический вид как одна из стадий процесса ди-
вергенции ....................................
Последовательные виды......................
Эволюционный вид......................
Биосистематическне категории..................
Единицы классификации культурных растений
Биологический вид или таксономический вид?
Критика биологической концепции вида
Часть третья. Генетические и экологические аспекты вида
Глава 8. Генетическая основа различий между
видами .......................................
Генные системы................................
Гены, кодирующие ферменты.....................
Замечания относительно регуляции ....
Хромосомные перестройки.......................
Системы сцепления.............................
Многофакторное сцеилевие................... .
Сцепление М—V............................. .
Интегрированный генотип .......
Омымчиг
523
Глава 9. Изолирующие механизмы................112
Основные формы изоляции........................42
Внешняя репродуктивная изоляция . . . .113
Внутренняя репродуктивная изоляция .115
Выводы....................... .... 117
Глава 10. Экологические взаимодействия .118
Сосуществование . . . ....................4®
Эффекты межвнлоэото отбора .... 119
Сосуществование без межвидового отбора или за
мешение индии . . . . ...................120
Эффекты экологических требований ... 121
Глава II. Формы взаимоотношений по плодовитости 123
Древесные растения............................123
Многолетние травянистые растения е резко выра-
женными различиями в механизме цветка 128
Многолетние травянистые растения с ярко выражен-
ными межвидовыми различиями в механизме
цветка........................................131
Однолетние травянистые растения...............138
Автогамные однолетники........................139
Общие модели..................................141
Общие корреляции..............................143
Выводы...................................... 145
Часть четвертая. Эволюционная дивергенция
Глава 12. Способы видообразования 146
Силы и поля.................................. 146
Способы видообразования .... 147
Обсуждение ....................................148
Глава 13. Первичное видообразование 150
Географическое видообразование .... 150
Квантовое видообразование 153
Квантовое вкдообразование у Clarkia .... 155
Аллопатрическое видообразование, связанное с са
мооплодотвореиием............................. 157
Внотн'зески-симлатрическое видообразование . . 160
Репродуктивная изоляция как побочный продукт
дивергенции....................................163
Выводы................................... 165
5М
01 ЛЛ6 ЛС .4 Uf
Глава 14. Структурные изменения хромосом при
ввдообрпэозанин ............................... 167
Теория эффект» последовательности участков в хро-
мосомкх .......................................168
Теория, основанная на сиеплеиин генов . . . 170
Закрепление новых структурных гомозигот 171
Выводы......................................173
Глава 15. Отбор на репродуктивную изоляцию
Особые условия..................................175
Эксперименты по искусственному отбору . .177
Данные по природным популяция и .... 178
Барьеры несовместимости у Gllla.............179
Опознавательные знаки цветков...............182
Исторические аспекты........................184
Эффект Уоллеев и смещение признаков . . .186
Часть пятая. Естественная гибридизация и ее результаты
Глава 16. Естественная гибридизация . . . .188
Гибридизация как процесс, обратный дивергенции 189
Гибриды и скопления гибридов . . 190
Внутренний контроль иад гибридизацией .192
Внешний контроль над гибридизацией . . .193
Гибридизация местообитания ... 196
Сопоставление высших растений с высшими живот
ными............................................197
Глава 17. Интрогрессия......................199
Факторы, способствующие интрогрессии . . . 199
Методы .... ....... 201
Примеры...................................... 204
Связность признаков как критерий гибридиостн 213
Экологическая сегрегация морфологических типов 215
Сцепление M—V в связи с интрогрессией . .217
Трансгрессия барьеров хромосомной стерильности 220
Интрогрессия и расообрадованис . . . . . 225
Выводы.................'........................226
Глава 18. Сиитамеои . ..........................227
Тихоокеанские ирисы.........................228
Белые дубы..................................229
Глава 19. Гибридное видообразование . . . 235
Стабилизация воспроизведения гибридов . . . 235
История вопроса ............................... 237
Оглавление
г»
Введение и амфнплоидию.......................240
Введение в рекомбинационное видообразпванне 242
Гибридное видообразование при наличии внешних
барьеров.....................................244
Введение в апомиктические видообразование . 247
Выводы......................................248
Глава 20. Рекомбинационное видообразование 250
Гипотеза................................ . . 260
Экспериментальный подход......................254
Раикие эксперименты 255
Последующие эксперименты при существовании
слабых барьеров стерильности..................256
Более поздние эксперименты при существовании
мощных барьеров стерильности..................258
Роль блочного паследовакия в рекомбинационном
видообразовании ............................ .261
О роли рекомбинационного видообразования в при-
роде .........................................263
Глава 21. Гибридные комплексы.................266
Классификация ................................267
Роль системы размножения......................268
Гомогамный комплекс.......................... 270
Гомогамный комплекс облнствениостебельных
гнлий.........................................271
Часть шестая. Полиплоидия
Глава 22. Полиплоидия: распространение и частота 274
Исторический очерк............................274
Масштабы проблемы и ее суть...................275
Полиплоидные ряды.............................276
Распространен не полиплоидии в разных таксономи-
ческих группах................................279
Диапазон хромосомных чисел....................280
Диапазон уровней илоидиостн ... 281
Частота.......................................286
Глава 23. Типы полиплоидов....................289
Разработка классификации......................289
Цитологические критерии ...................... 291
Генетические критерия..........................292
Морфологические критерии.......................294
WK Oitaamst
Относительная частота автополиплондов и амфи-
плоидов в природе ............................
Глава 24 Факторы, способствующие возникновению
полиплоидии ..................................
Преимущества, внутренне присущие амфиплоидио-
му состоянию..................... , . .
Способы образовании ..........................
Полиплоидия и жизненная форма.................
Факторы, имеющие псрвосгепепмое значение .
Широта и высота местности.....................
Экологические факторы.........................
Полиплоидия и система размножения ....
Влияние величины хромосом ....................
Влияние генотипа..............................
Общие выводи..................................
Глава 25. Полиплоидный комплекс ....
Методы анализа................................
Цитотаксономичсские примеры...................
Биосистсматические примеры....................
Комплекс Gllia Inconspicua....................
Trtttcum и Aegilops . .......................
Старые полиплоидные комплексы.................
Остатки древних полиплоидных комплексов
Необратимость.................................
Подавление изменчивости.......................
Дяплоидизация.................................
Глава 26. Природные автополиплоиды
Группа В (scutella laevigata.............. .
Группа Solarium tuberosum.....................
Группа Dactylls glomerata.....................
Другие примеры ................................
Часть седьмая. Аисуп.тондия
Глава 27. Лиеупловдия .......................
Нисходящая анеуплондня на диплоидном уровне
Восходящая анеуплондня на диплоидном уровне
Полиплоидный сброс...........................
Анеуплоидиое увеличение у полиплоидных гибридов
Carex........................................
Claytonla oirginica .........
Высокие основные числа у покрытосеменных
Оглаг.ггпШ Mt
Глава 28. Агматоплоидия................
Полицентрические хромосомы . . ...
Carex..................................
Luzula ............
Сурегасеае и Juncaceae ................
Часть восьмая. Специализированны» генетические системы
Глава 29 Перманентная гетеротитотяость по
трпвсдокацяям .........................
Генетическая система у Oenothera.......
Источники изменчивости ................
Гетерогамный комплекс в группе Oenothera hookeri-
biennis ............
Гетерогамные микровнды в группе Oenothera bien-
nis .............
Гетерогамные микровнды и комплексы в других
группах растений .... ............
Глава 30. Перманентная нечетная полиплоидия
Генетическая система Rota canina.......
Гетерогамный комплекс в группе Rota canine
Leucopogon juniperiaus ........
Cardamtne insueta......................
Эволюционный потенциал гетерогамных комплексов
Глава 31. Агамоспермия.................
Эмбриологические пути..................
Таксономическое распределение .........
Система размножения....................
Гкбрндность и агамоспермия ...
Возникновение гаметофитного апомиксиса
Поведение при скрещиваниях ............
Расщепление........................
Характер изменчивости..................
Преимущества агамоспермии . . . . .411
Ш SgfeoSSSfeg
Глава 32. Агамиый комплекс .... .414
Комплекс С repie occidentalis ...... 415
Европейские ежевики.....................418
Taraxacum...............................422
Аглмиые комплексы типа Rub us Moriferus . . 424
Комплекс Bouteloua curtipendula.........426
Citrus .................................431
SW Оелаалапиа
Pttrls ............
Стадии развития..........................
Эволюционный потенциал...................
Глава 33. Вегетативное размножение гибридов
Гибридные Клоны .........................
Клональные микровнды ....................
Клональные комплексы, размножающиеся ответ-
влениями стебля . . . . .................
Клональные комплексы, размножающиеся при по-
мощи ложного живорождения................
Эволюционный потенциал...................
Глава 34. Естественные триплоиды . . . .
Возникновение ...........................
Сохранение ..............................
Избранные примеры .......................
Часть девятая Заключение
Глава 35. Вид и макроэволюция . . . . .
Сетчатая эволюция........................
Эволюционный потенциал гибридных комплексов
Значение сетчатого характера эволюции для фило-
гении покрытосеменных....................
Вид как единица макроэволюцнн............
Литература...............................
Указатель авторов .......................
Указатель латинских названий.............
Предметный указатель.....................
~ 8 О & ст> * * сп сп ся сл сп ся £ £ * * * * w w w
WWOAWID &АЮЮ <0 00 Ct AWK5 5 М К