






















































































































































































































































Автор: Авцынезы А.П. Жаворонков А.А. Риш М.А. Строчкова Л.С.
Теги: патологическая физиология формы развития заболеваний патогенез учение о происхождении заболеваний общая патология биология микробиология инфекционные болезни издательство медицина микроэлементозы человека
ISBN: 5—225—02128—X
Год: 1991




АКАДЕМИЯ НАУК
СССР
АКАДЕМИЯ '
МЕДИЦИНСКИХ НАУК
СССР
М5д
А.П.Авцын езы
А. А. Жаворонков
М.А.Риш
Л.С.Строчкова
МИКРОЭЛЕМЕНТОЗЫ
ЧЕЛОВЕКА
(этиология, классификация, органопатология)
I юта
^чает
нк
щите
Москва „Медицина" 1991
ггеро-
лазме
3
ББК 52.5
М59
УДК 616-092: 612.126
Рецензент Д. С. САРКИСОВ, акад. АМН СССР
ИЗДАНИЕ ОДОБРЕНО И РЕКОМЕНДОВАНО К ПЕЧАТИ
РЕДАКЦИОННО-ИЗДАТЕЛЬСКИМ СОВЕТОМ
ПРИ ПРЕЗИДИУМЕ АМН СССР
Авцын А. П, и др.
М59 Микроэлементозы человека: этиология, классификация,
органопатология/А. П. Авцын, А. А. Жаворонков, М. А. Риш,
Л. С. Строчкова; АМН СССР. — М.: Медицина, 1991, 496 с.:
ил. ISBN 5—225—02128—X
В монографии изложены современное состояние учения о микроэле-
ментозах человека и структурные основы всасывания микроэлементов.
Предложена оригинальная классификация микроэлементозов человека.
В ней выделены природные, техногенные и ятрогенные формы этих рас-
пространенных заболеваний. Представлена характеристика эссенциаль-
ных, условно эссенциальных, условно токсичных и токсичных микро-
элементов, а также связанных с ними заболеваний. Описаны наследст-
венные и врожденные формы микроэлементозов человека и животных.
Теоретически обосновано значение микроэлементозов для медицины.
Для патологов, физиологов, биохимиков, профпатологов.
4107010000—149
М039(01)—91 56 91
ББК 52.5
ISBN 5—225—02128—X
© Коллектив авторов, 1991
СПИСОК СОКРАЩЕНИИ
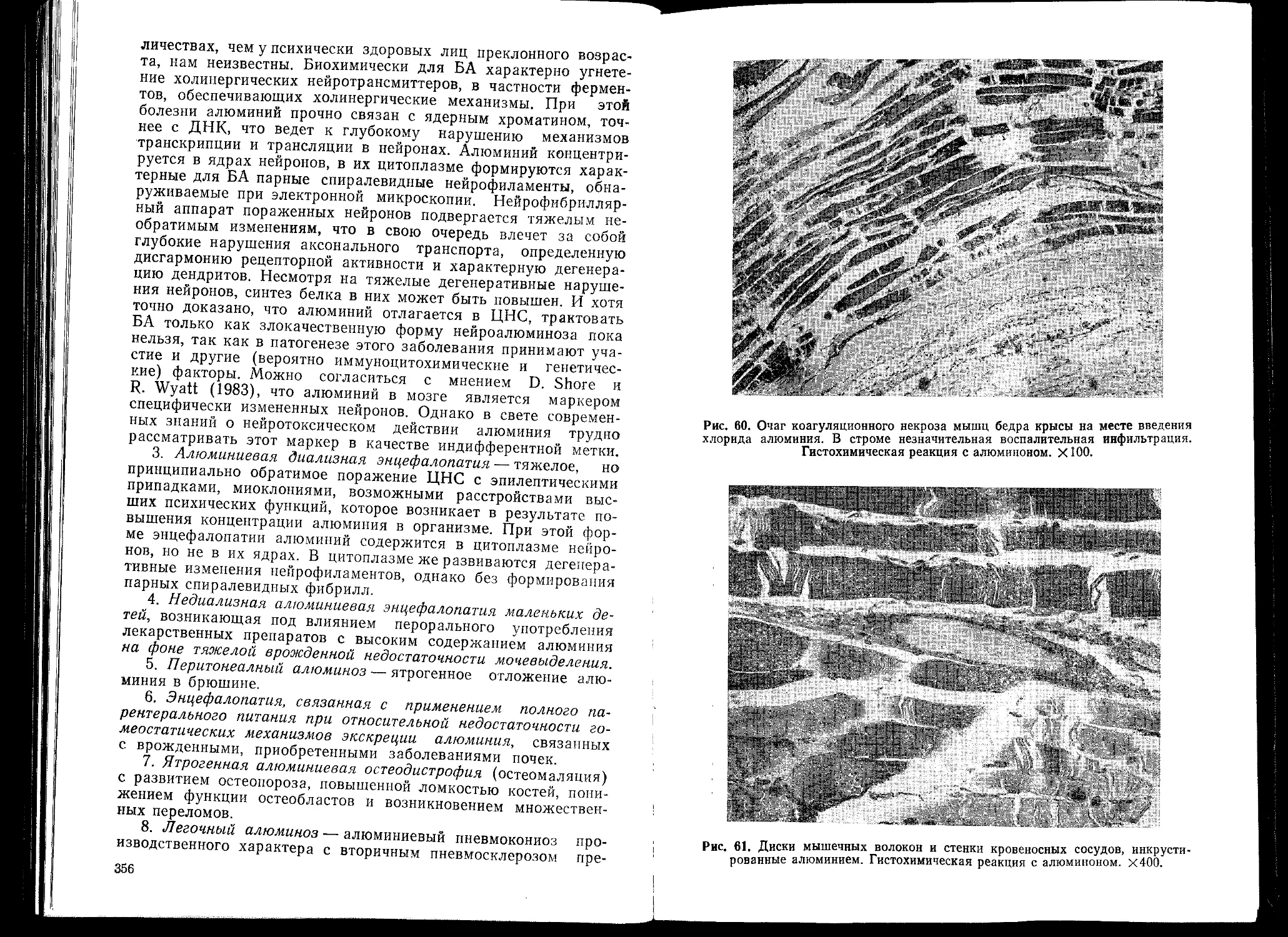
ДАТ АДП АКТГ АМД БА БАС БГГ ВОЗ ГАМК г-цд — а-антитрипсин — алиментарная дистрофия поджелудочной железы — адренокортикотропный гормон — алиментарная мышечная дистрофия — болезнь Альцгеймера — боковой амиотрофический склероз — богатый гистидином гликопротеид — Всемирная организация здравоохранения — гамма-аминомасляная кислота — гепатоцеребральная дистрофия (болезнь Вильсо- на—Коновалова)
ддк ДМБА ДОФА дп ДТПА здт ИЛ-1 и КБП кДНК — диэтилдитиокарбомат натрия — 7,2-диметилбензатрацен — 3,4-дигидроксифенилаланин — синдром деменции-паркинсонизма — 1,1,4,7,7-диэтилентриаминопентауксусная кислота — зоб диффузный токсический ИЛ-2 — интерлейкин 1 и интерлейкин 2 — коэффициент биологического поглощения — ДНК-комплементарная мРНК, которая получает-
кмпс КС лип ЛОНИ лэм МКРЗ мт МТОЗы мэ НДФаза окп ПААГ ПБ пдк пмик ся действием обратной транскриптазы на мРНК — кислые мукополисахариды — коэффициент концентраций — липопротеиды низкой плотности — липопротеиды очень низкой плотности — лейкоцитарный эндогенный медиатор — Международный комитет по радиационной защите — металлотионеины — микроэлементозы — микроэлемент — нуклеозиддифосфатаза — оральные контрацептивные препараты — (электрофорез) в полиакриламидном геле — профессиональные болезни — предельно допустимая концентрация — пузырьки мембраны исчерченной каемки энтеро-
пен цитов — парные спиральные нейрофиламенты (в цитоплазме
РТМЛ СМФ нейронов при болезни Альцгеймера) — реакция торможения миграции лейкоцитов — система мононуклеарных фагоцитов
3
СОД — супероксиддисмутаза
СЭД — синдром Элерса—Данло
Тб — биологический период полувыведения микроэлемен-
та из организма
ТГГ — тиреотропный гормон гипофиза
ТПД — трихополидистрофия (болезнь Менкеса)
ФАД — флавинадениндинуклеотид (кофермент)
ФЕП-карбоксикиназа — фосфоенолпируват-карбоксикиназа
цАМФ — циклический аденозинмонофосфат
цГМФ — циклический гуанозинмонофосфат
ЦНС — центральная нервная система
ЭДТА — этилендиаминтетрауксусная кислота
ЭМГ — электромиограмма
ЭПР — электронный парамагнитный резонанс
APUD-система (amine precursors uptake and decarboxylati-
on)— эндокринные элементы в эпителиях, способ-
ные воспринимать предшественники аминов и де-
карбоксилировать их
BEI (butanol extracted I) — йод, экстрагируемый бутанолом
GSH-Px (glutation-SH peroxidase) — глутатионпероксидаза
GTF (glucose tolerance factor) — фактор толерантности к
глюкозе
DIT — дийодтирозин
DLM25 (dose lethal minimum)—V4 минимальной летальной
дозы
EF-1 и EF-2 (elongation factor)—факторы элонгации белко-
вой цепи
MIT — монойодтирозин
NAD — никотинамидадениннуклеотид (кофермент)
NADP — никотинамидадениндинуклеотидфосфат (кофер-
мент)
PBI (protein bound I) — белок, связывающий йод
SALT (skin-associated lymphoid tissue) —лимфоидная ткань,
связанная с кожей
Se-Met (Se-methionine) — селен-метионин
SeCN — селеноцианид
SMON (subacute myelo-optic neuropathy) —подострая миело-
оптико-нейропатия
SCN — тиоцианат
Тз — трийодтиронин
гТз — неактивные метаболиты, образующиеся из тирокси-
на (Т4)
TBG (thyroxine bound globulin)—глобулин, связывающий
тироксин
ТВРА (thyroxine bound prealbumin) — Т4-связывающий пре-
альбумин
TM-Se — триметилселенид
TRH (thyrotropin-releasing hormone) — тиротропин-рили-
зинг гормон гипоталамуса
4
Посвящается памяти
основоположника биогеохимии
Владимира Ивановича Вернадского
и его выдающихся последователей
Александра Павловича Виноградова
и Виктора Владиславовича Ковальского,
рано оценивших значение
микроэлементов для жизнедеятельности
здорового и больного организма
и много сделавших для развития
этого направления науки
ПРЕДИСЛОВИЕ
Путь к созданию нашего труда был во многих отношениях
нетрадиционным. В основу первоначального замысла данной мо-
нографии легли результаты многолетних наблюдений по геогра-
фической патологии эндемических заболеваний биогеохимичес-
кой природы, а также экспериментально-морфологических ис-
следований токсического действия некоторых микроэлементов
(МЭ) на органы, ткани и клетки человека и лабораторных жи-
вотных. Указанные наблюдения были начаты в 60-х годах и про-
должаются до настоящего времени в лаборатории географичес-
кой патологии Института морфологии человека АМН СССР.
Здесь же были сформулированы первые теоретические положе-
ния об основных закономерностях функционирования системы
МЭ как особого аппарата обеспечения микроэлементного гомео-
стаза и поддержания элементного состава живых организмов.
В краткой форме эти положения были изложены ранее [Ав-
цын А. П., 1972] и неоднократно дополнялись и развивались в
последующих публикациях. Примером того, как в нашем не-
большом коллективе осуществлялось изучение географической
патологии и экспериментальной патоморфологии токсического
действия одного из микроэлементов, а именно фтора, является
книга Патология флюороза [Авцын А. П., Жаворонков А. А.,
1981].
В 1982 г. впервые было сформулировано понятие о микро-
элементозах (МТОЗах), которое обсуждалось на научной кон-
ференции в марте 1983 г. в Институте аналитической химии и
геохимии им. В. И. Вернадского. В 1983 г. была опубликована
первая классификация этих своеобразных и, как выяснилось,
широко распространенных болезней. В коллективной моногра-
фии Патология человека на Севере (1985) А. П. Авцын и
А. А. Жаворонков описали проявления недостаточности жиз-
ненно важных МЭ и влияние некоторых токсичных МЭ в экст-
ремальных зонах техногенного загрязнения на население Севе-
ра СССР. В 1986 г. были охарактеризованы важнейшие алю-
минозы человека. В 1987 г. специально для врачей был опуб-
ликован уточненный вариант классификации МТОЗов человека.
В ряде публикаций и докладов совместно с Л. С. Строчковой
рассматривались вопросы о роли МЭ на клеточном уровне в
5
условиях нормы и патологии и выполнены соответствующие
экспериментальные работы. Из сказанного достаточно ясно, что
проблема МТОЗов возникла в рамках географической патоло-
гии и получила дополнительные стимулы для своего развития
в результате развернувшихся затем исследований по общей и
частной патологии клетки.
Вначале мы предполагали ограничиться публикацией до-
вольно короткого обобщения результатов конкретных наблю-
дений. Однако углубленное изучение данных обширной лите-
ратуры, посвященной всем МЭ, сопоставление этих в чем-то
сходных или резко контрастных материалов, многолетнее обду-
мывание закономерностей полезного или, напротив, токсическо-
го действия МЭ, присутствующих в клетках и жидкостях орга-
низмов в удивительно малых количествах, побудили нас резко
изменить первоначальный план. Мы пришли к выводу, что на
настоящем этапе изучения МЭ необходимо более подробно рас-
сказать читателю об их основных химических и биологических
свойствах. Это в большей степени соответствует потребностям
текущего момента, так как знакомство врачей с данной пробле-
мой является крайне недостаточным.
Все авторы предлагаемого читателю труда являются чле-
нами Научного совета АН СССР по проблемам микроэлементов
в биологии. Двое из нас имеют медицинское, двое — биологичес-
кое образование. В связи с этим вопросы общей и частной па-
тологии, включая проблемы клинических проявлений и класси-
фикации микроэлементозов, изложены автором этих строк и
зав. лабораторией географической патологии НИИ морфологии
человека АМН СССР, проф. А. А. Жаворонковым. Системати-
зированные сведения о биохимии МЭ, а также данные о роли ге-
нетических факторов в происхождении микроэлементозов изло-
жены зав. кафедрой физиологии и биохимии человека и жи-
вотных Самаркандского университета, засл. деят. науки УзССР,
проф. М. А. Ришем — известным специалистом в этой области
знания. Характеристику влияния различных концентраций МЭ
на культуры клеток человека осуществила ст. науч, сотр., докт.
биол. наук Л. С. Строчкова. К счастью, в число изучаемых ею
элементов вошел селен, благодаря чему удалось получить новые
факты об антибластическом влиянии этого МЭ на опухолевые
клетки в условиях прямого контакта.
Пересмотр первоначального плана нашей книги существен-
но отразился на ее структуре. В главах 1 и 2 приведены общие
сведения о МЭ, краткая история их изучения и современные
данные об их содержании в органах, тканях и жидкостях орга-
низма человека. Значительной новизной отличается глава 3,
в которой дана характеристика структурных основ всасывания
МЭ, а также изложены взгляды биохимиков, изучающих этот
вопрос.
В главе 4 обосновано подразделение МЭ на эссенциальные
и токсичные в соответствии с данными современной литературы.
В главе 5 изложены взгляды авторов на специальную термино-
6
логию, содержится предложенная нами классификация
МТОЗов. Таким образом, после ознакомления с важным с прак-
тической и теоретической точки зрения подразделением МЭ чи-
татель встречается с детальной характеристикой их свойств и с
главными особенностями обусловленных ими заболеваний (гла-
вы 6, 7 и 8). В главе 9 рассматривается новый и важный в
практическом отношении вопрос о МТОЗах ятрогенного проис-
хождения. Глава 10 знакомит читателя с наследственными фор-
мами МТОЗов. В главе 11 в краткой форме излагаются совре-
менные данные о врожденных пороках развития, а также о
преждевременной гибели эмбрионов и плодов при дефиците и
избытке МЭ. В общем заключении содержится суммарная оцен-
ка значения МТОЗов для патологии и современной нозологиче-
ской номенклатуры.
Авторы хорошо понимают, что в связи с обширностью и мно-
гокомпонентностью учения о микроэлементах и микроэлементо-
зах далеко не все стороны этой проблемы удалось осветить с
одинаковой полнотой. В современной зарубежной литературе
описанию свойств каждого из важнейших МЭ обычно посвяща-
ется отдельный том. Такова, например, монография R. J. Sham-
berger (1984) о свойствах селена, в которой дана исчерпываю-
щая характеристика этого важнейшего эссенциального, а при
некоторых условиях токсичного элемента.
Мы ставили перед собой совершенно другую задачу. Кратко
характеризуя каждый МЭ, мы в первую очередь стремились
привлечь внимание врачей и биологов к его главным свойствам,
которые в значительной степени обусловливают клиническую
картину и соответствующие патоморфологические изменения.
Цитопатологические данные в этой монографии имеют главным
образом вспомогательный характер. Они в определенной степе-
ни показывают нам многообещающие возможности анализа
механизмов действия различных химических элементов, кото-
рый мы предполагаем осуществить в будущем. Несколько забе-
гая вперед, можно сказать, что уже сейчас экспериментальная
цитопатология МЭ содержит в себе большой познавательный
материал для выделения специфических и неспецифических ре-
акций клеток и их органелл. В еще большей степени он может
быть использован в практическом здравоохранении для созда-
ния клеточных диагностических тестов для выяснения обратимо-
сти повреждающего действия и разработки клеточно-молеку-
лярных основ экспериментальной терапии недостаточности эс-
сенциальных МЭ и микроэлементных токсикопатий.
В разделе «Приложения» читатель найдет сводные таблицы
о важнейших заболеваниях человека при дефиците и при из-
бытке эссенциальных, условно эссенциальных и токсичных
микроэлементов.
Представленный вниманию читателя труд является одной из
первых попыток медико-биологического освещения проблемы
МТОЗов человека. Учитывая значительную сложность всей про-
блемы, авторы вполне отдают себе отчет в том, что упущения
7
и возможные неточности в этой большой работе неизбежны.
Мультидисциплинарное изучение микроэлементов имело и
имеет преимущественно аналитический характер, причем мно-
гие специалисты пока не преодолели периода научной автар-
кии, когда все добытые материалы предназначаются главным
образом для внутреннего пользования. Это в значительной сте-
пени тормозит познавательный процесс. Достаточно сказать,
что все это огромное научное направление еще не имеет офи-
циально признанного названия. Между тем необходимость в
нем ощущают многие исследователи и практики. Таким назва-
нием, по-видимому, является микроэлементология, которая в
свою очередь должна подразделяться на общую и частную в
зависимости от основного предназначения каждой ветви этой
науки. Таким образом, кроме общей микроэлементологии, дол-
жны развиваться биологическая, агрономическая, химическая
и, разумеется, медицинская микроэлементология.
За все критические замечания мы заранее благодарны.
В заключение мы считаем своим приятным долгом выразить
благодарность всем учреждениям и отдельным лицам, способ-
ствовавшим завершению данного многолетнего труда. Очень
большую помощь по уточнению библиографии, подготовке ру-
кописи к печати нам оказали наши дорогие сотрудники
Н. И. Краевская и М. П. Баланова, за что мы им также при-
знательны.
По независящим от нас условиям список литературы приво-
дится в сокращенном виде.
Акад. АМН СССР
А. П. Авцын
ВВЕДЕНИЕ
Учение о микроэлементозах как о заболеваниях, синдромах
и патологических состояниях, вызванных избытком, дефицитом
или дисбалансом микроэлементов в организме человека, пере-
живает принципиально новый этап своего развития. После дли-
тельного периода накопления множества ценных, но слабо си-
стематизированных фактов оно встало перед необходимостью
своего осознания в качестве особого раздела патологии, пред-
метом которого является новый класс болезней человека с уже
установленной этиологией, но с еще неясным патогенезом. Не-
смотря на то что болезни этого происхождения были давно
известны клинической медицине (эндемический зоб, железоде-
фицитные анемии, отравления некоторыми металлами и др.),
под объединяющим названием они никогда не выделялись и
фигурировали в современных классификациях в разных рубри-
ках. Самое важное, однако, что подавляющее большинство бо-
лезней и синдромов этого класса почти не регистрировалось,
так как на них не обращали достаточного внимания.
В настоящее время хорошо известно, что МТОЗы широко
распространены в патологии животных и растений. Более того,
в этих областях знания сделаны ценные наблюдения и одержа-
ны успехи безусловного практического значения. Экономичес-
кий эффект профилактики и терапии МТОЗов в животноводстве
весьма велик, так как в этом случае дело может идти об ис-
коренении определенных болезней и о сохранении большого
числа ценных сельскохозяйственных объектов.
На первый взгляд может показаться, что МТОЗы животных
и растений, включая и соответствующую патологию таких форм
жизни, как вирусы, бактерии и простейшие, — это совершенно
независимые и малоизученные разделы общей биологии, микро-
биологии, вирусологии и всего комплекса дцсциплин сельско-
хозяйственного профиля. На самом деле это, очевидно, не так,
так как принципиальные положения учения о МТОЗах челове-
ка почти безоговорочно приложимы к соответствующим главам
патологии любых представителей живого мира. Это, в частно-
сти, относится и к предложенной нами классификации МТО-
Зов, в которую входят не только патологические формы при-
родного характера, но и многие феномены антропоургического
происхождения. В связи с бурным развитием промышленности
9
В. И. Вернадский (1863—1945 гг.) —
создатель современного учения о био-
сфере, основоположник геохимии и
биогеохимии. В 1891 г. выдвинул
идею об особом значении микроэле-
ментов и их биогенной миграции, ор-
ганизовал в АН СССР Институт гео-
химии и лабораторию по изучению
биогеохимии микроэлементов.
глобальная проблема техно-
генного загрязнения внешней
среды теснит чисто природные
формы патологии всех живых
существ и неизбежно наклады-
вает на них свой искажаю-
щий отпечаток.
Мировая литература о МЭ
по существу необозрима и в
последние годы лавинообраз-
но увеличивается. Медицинс-
кие аспекты учения о МЭ еще
недостаточно разработаны. Относящиеся к нему факты и вы-
сказывания разбросаны в публикациях по различным дисцип-
линам. Это создает большие трудности при попытках их систе-
матизации. Некоторые важные разделы патологии МТОЗов че-
ловека до сих пор остаются белыми пятнами. Для того чтобы
их заполнить, нужна длительная и многосторонняя исследова-
тельская работа. Специальный аспект проблемы — воздействие
радиоактивных МЭ — в этой книге не рассматривается.
В нашей стране учение о МЭ имеет давние и славные тра-
диции, что в первую очередь связано с именем В. И. Вернад-
ского, который пророчески предсказал большое будущее этой
области знания и заложил научные основы ее последующего
развития. Именно он в 1926 г. открыл «Отдел живого вещест-
ва» при Комиссии по изучению естественных производительных
сил. В 1928 г. по его же инициативе организована «Биогеохими-
ческая лаборатория» АН СССР, на долгие годы ставшая основ-
ным центром координации исследований этого профиля. В тече-
ние последних 25 лет ее возглавлял замечательный последова-
тель В. И. Вернадского лауреат Ленинской премии и премии
им. В. И. Вернадского, чл.-корр. ВАСХНИЛ, проф. В. В. Ко-
вальский. В этот период развернулись широкие исследования
самого Виктора Владиславовича и его многочисленных учени-
ков, результаты работ которых были опубликованы в послед-
них 10 томах «Трудов Биогеохимической лаборатории». Науч-
ная и организационная деятельность В. В. Ковальского спо-
собствовала тому, что в нашей стране стали проводиться много-
численные исследования по единому плану, выработанному На-
учным советом АН СССР «По проблемам микроэлементов в
10
А. П. Виноградов (1895—
1975 гг.)—ученик и ближайший
сотрудник В. И. Вернадского, ос-
новоположник учения о биогеохи-
мических провинциях и их роли в
возникновении эндемических забо-
леваний человека и животных.
биологии». В результате
этой работы возникли мно-
гочисленные центры, в кото-
рых велась самостоятель-
ная исследовательская ра-
бота по МЭ. Наиболее
крупные из них находились
в Прибалтийских республи-
ках, главным образом в
Латвии, а также в Туркме-
нии, Иркутске, Улан-Удэ,
Чите, Казани, Самарканде,
Таджикистане, Казахстане,
Чебоксарах, Кишиневе и
др. Медицинские аспекты
учения о МЭ имеют наибо-
лее давние традиции на Ук-
раине и успешно развиваются на кафедре биохимии Ивано-
Франковского мединститута, возглавляемой проф. Г. А. Бабенко.
Кроме биогеохимической лаборатории АН СССР, в Москве
работали коллективы исследователей в Институте общей и ком-
мунальной гигиены им. А. Н. Сысина, при Московском государ-
ственном университете им. М. В. Ломоносова, в Педагогичес-
ком институте им. В. И. Ленина, в Институте минералогии, гео-
химии и кристаллохимии редких элементов АН СССР, в НИИ
морфологии человека АМН СССР и др. Каждое из этих учреж-
дений, имея свои специфические исследовательские задачи, на-
ходилось в контакте с названным выше Научным советом АН
СССР, участвуя в организованных им мероприятиях и внося в
общее дело свой посильный вклад в соответствии со своей спе-
циальностью.
Следует подчеркнуть, что в некоторых городах исследования
МЭ не ограничивались определением их количества в пищевых
продуктах, растительных и животных тканях, а сопровождались
также клинической и биогеохимической характеристикой энде-
мических болезней, являвшихся реакцией на аномальный хими-
ческий состав природной или измененной техногенными вли-
яниями среды. В течение длительного времени в указанном
Научном совете превалировал интерес к биогеохимическим эн-
демиям природного происхождения. Однако в последующие го-
ды наибольшее внимание стали привлекать микроэлементные
аномалии индустриального происхождения. Сравнительно не-
давно объектами изучения уровня микроэлементного загрязне-
ния внешней среды стали промышленные города и окружающие
11
В. В. Ковальский (1899—1984 гг.)—
основоположник геохимической эко-
логии и биогеографии химических
элементов. В течение 25 лет руково-
дил лабораторией биогеохимии им.
В. И. Вернадского и был председате-
лем Научного совета АН СССР по
проблемам микроэлементов. Впервые
осуществил биогеохимическое рай-
онирование СССР.
их регионы. Многочисленные
публикации, в том числе до-
клады экспертных комитетов
ВОЗ, позволяют сделать за-
ключение, что в настоящее
время есть все основания го-
ворить о возрастающей агрес-
сивности внешней среды. Од-
ной из важных сторон этого,
еще в недостаточной степени
контролируемого, процесса яв-
ляются загрязнения микроэле-
ментной природы. Современная нозологическая панорама не
может игнорировать указанный фактор, имеющий фундамен-
тальное значение. В связи с этим наши представления о проис-
хождении и патогенезе многих болезней человека подлежат су-
щественной переработке.
Необходимо указать, что в других учреждениях параллель-
но с активностью врачей, биогеохимиков, биологов и биогеогра-
фов развивалась не меньшая по своему объему и значению ис-
следовательская и организаторская деятельность гигиенистов.
Они внесли в изучение этой проблемы самостоятельные подхо-
ды, в значительной степени продиктованные потребностями
коммунальной гигиены и особенно гигиены труда. Профпатоло-
гам принадлежат крупные труды о повреждающем действии не-
которых токсичных элементов и о рациональных путях профи-
лактики. Особое внимание уделялось определению предельно-
допустимых концентраций (ПДК), а также установлению раз-
личных индикаторов, характеризующих ту или иную степень
контаминации внешней среды. Гигиенисты интересовались, кро-
ме того, характеристикой различных синдромов, возникающих в
результате контакта организма с токсичными МЭ, главным об-
разом металлами в процессе их производства. Это способство-
вало возникновению оригинального аспекта проблемы, который
получил название «учения о ксенобиотиках». Полезные сведе-
ния о загрязнении пищевых продуктов металлами приведены в
книге К. Рейли (1985).
В заключение мы должны вкратце объяснить, почему у нас
возникла потребность осветить эту проблему в виде обобщаю-
щего труда. Поводов к этому было несколько. .Одним из них
было наше ознакомление с замечательными произведениями
12
Эрик Андервуд (1905—
1980 гг.)—один из основополож-
ников учения о микроэлементах.
Автор и редактор многократно из-
дававшегося руководства Микро-
элементы в питании человека и
животных. Во время многолет-
ней работы в Австралии установил
микроэлементную этиологию ряда
заболеваний сельскохозяйственных
животных.
В. И. Вернадского и с кни-
гой А. И. Войнара «Биоло-
гическая роль микроэле-
ментов в организме живот-
ных и человека» (1960), ко-
торые побудили нас занять-
ся прицельным изучением
этой проблемы. Эти труды
привели нас к мысли, что
микроэлементы — это ско-
рее всего не случайные ин-
гредиенты тканей и жидко-
стей живых организмов, а
компоненты закономерно
существующей очень древ-
ней и сложной физиологической системы, участвующей
в регулировании жизненных функций организмов на всех
стадиях развития. Наше понимание некоторых сторон ак-
тивности этой еще недооцененной, но универсальной физио-
логической системы мы выразили в том, что провизорно выде-
лили три основополагающих принципа ее функционирования,
которыми являются: 1) избирательное поглощение определен-
ных МЭ; 2) избирательная концентрация их в определенных ор-
ганизмах, органах, тканях и некоторых органеллах клетки и
3) их селективная элиминация. Вероятно, именно эти механизмы
поддерживают микроэлементный гомеостаз [Авцын А. П., 1972].
Другие стимулы к продолжению этой работы пришли со-
всем с другой стороны. Во время экспедиции в Целинный край
в 1963—1965 гг. А. А. Жаворонков собрал большой материал по
эндемическому флюорозу в Северном Казахстане. Не ограничи-
ваясь природными материалами, он провел многолетнее экспе-
риментально-морфологическое исследование всех органов бе-
лых крыс при хронической интоксикации фтором. Результаты
этого исследования вышли за пределы ожидавшихся. Другими
словами, они не могли быть сведены только к более детально-
му описанию патологической анатомии флюорозного поражения
зубов и костей. Выяснилось, что один и тот же этиологический
агент — избыток фтора — вызывает поражение всего организ-
ма и как следствие этого заболевания различных систем и ор-
ганов.
При изучении мировой литературы о действии алюминия на
13
Вальтер Мертц — выдающийся уче-
ный США, специалист по микро-
элементам, редактор 5-го издания
руководства Микроэлементы в пи-
тании человека и животных (1987).
организм эксперименталь-
ных животных и человека
выяснилось, что алюминоз
человека также выражается
не в одном, а во многих син-
дромах и болезнях, часть из
которых оказалась крайне
неблагоприятной. Сопостав-
ление алюминоза и флюоро-
за позволили сделать пред-
варительное теоретическое
заключение, согласно кото-
рому МТОЗы, как правило,,
выражаются в многообраз-
ных заболеваниях человека
и животных. Это полностью
подтверждается данными
эксперимента. Органотропное влияние МЭ имеет сложную при-
роду и реализуется на уровне глубинных метаболических про-
цессов, затрагивая биологические структуры и содержащиеся в.
них ферменты многих клеток и тканей. Вместе с тем нельзя от-
рицать действие МЭ на регулирующие и интегрирующие систе-
мы, т. е. на весь организм в целом.
Отечественная медицинская наука рано оценила роль МЭ в
патологии. На протяжении последних десятилетий были опуб-
ликованы ценные монографии Г. А. Бабенко (1965, 1972),
В. Я. Шустова (1967), М. Г. Коломийцевой и Р. Д. Габовича
(1970), Л. Р. Ноздрюхиной (1977), В. И. Венчикова (1978).
Количество клинических и экспериментальных работ, посвя-
щенных тем или иным аспектам изучения МЭ, огромно. Есть
основания говорить о взрыве информации, посвященной откло-
нениям от нормы, обусловленным дефицитом или избытком МЭ
в организме человека и экспериментальных животных. В этой
информации непросто разобраться, не имея руководств меди-
ко-биологического профиля.
В 1983 г. вышла в свет коллективная монография Н. Zumk-
ley и соавт., в которой дана раздельная характеристика патоло-
гических процессов, развивающихся при недостаточности и из-
бытке ряда МЭ в организме. В течение многих лет основным
руководством по проблеме МЭ в питании была прекрасная кни-
га Е. Underwood «Trace Elements in Human and Animal Nutri-
tion», выдержавшая 4 издания. В 1987 г. она вышла 5-м изда-
нием под редакцией W. Mertz.
Работая в области изучения патологии, связанной с дейст-
вием МЭ, более 20 лет, мы вместе с сотрудниками только в
1983 г. решились сделать первое обобщение по всей проблеме,
опубликовав работу «Принципы классификации заболеваний
биогеохимической природы». В этой работе был впервые пред-
ложен термин «микроэлементозы» и дана рациональная клас-
сификация этих многообразных заболеваний, которая должна
быть известна работникам здравоохранения. Имеющая прямое
отношение к проблемам эпидемиологии, клинической и профи-
лактической медицины предложенная классификация нацелива-
ет врачей на выработку рациональных методов борьбы с этими
широко распространенными заболеваниями.
Книга имеет подзаголовок: этиология, классификация, орга-
нопатология. Изложение именно этих сторон проблемы пред-
ставляется нам первоочередным и наиболее важным в практи-
ческом отношении. Освещение методов лечения МТОЗов не вхо-
дит в число задач этой книги. Это по силам только клиницис-
там, располагающим многолетним личным опытом. Однако
книга все же имеет прямое отношение к больному человеку,
в частности к диагностике МТОЗов. Можно выразить надежду,
что лечащие врачи, гигиенисты и патологи после ознакомления
с нашим трудом будут гораздо чаще и обоснованней ставить
диагноз определенной микроэлементной недостаточности, а так-
же микроэлементных токсикопатий. Это, очевидно, будет ре-
зультатом более пристального внимания к профессиональному
анамнезу и условиям жизни больных, так как очень многие
МТОЗы человека, за исключением довольно редких наследст-
венных форм, имеют экзогенную природу и связаны с режи-
мом питания и химическими факторами среды обитания и тру-
довой деятельности. Впрочем даже в условиях очевидной экзо-
гении интенсивность и особенности реагирования организмов
неразрывно связаны с генетическими механизмами их гомеоста-
за. Именно это в значительной степени объясняет не всегда
одинаковый ответ организма на дефицит, избыток и дисбаланс
МЭ.
Глава 1
ОСНОВНЫЕ СВЕДЕНИЯ О МИКРОЭЛЕМЕНТАХ
1.1. Определение понятия «микроэлементы».
Краткая история их изучения
Микроэлементы — это группа химических элементов, которые
содержатся в организме человека и животных в очень малых
количествах, в пределах 10-3—10-12%. Именно это определяет
их названия: «следовые элементы» в немецком и английском
языках, «олигоэлементы» — у французских авторов, «рассеян-
ные элементы» — в трудах В. И. Вернадского. По мнению Е. Un-
derwood (1976), единственной характерной чертой МЭ является
их низкая концентрация в живых тканях. «Резко отличаясь
друг от друга по своим физико-химическим свойствам и био-
логическому действию, они вместе с тем не имеют какой-либо
общей химической характеристики, которая давала бы возмож-
ность отличить их от макроэлементов».
Изложенное не вызывает сомнений с позиций аналитичес-
кой химии. Однако в определении МЭ должны отражаться и их
биологические свойства, имеющие, как установлено, исключи-
тельную теоретическую и практическую важность. По мнению
М. Апке и соавт. (1987), в процессе абсорбции неорганических
частей почвы, воды и пищи флорой и фауной, который продол-
жался миллионы лет, следовые элементы, вероятно, приобрели
биологическую функцию компонентов или активаторов фермен-
тов, синтеза белков и других органических соединений. Более
подробно наши представления о биологическом значении МЭ
приведены в общем заключении этой книги.
Необходимо рассмотреть некоторые терминологические во-
просы, так как они прямо связаны с формированием научных
понятий. Принятый в англоязычной литературе термин «следо-
вые элементы» (trace elements) может быть предметом обсуж-
дения. Нам он кажется не самым удачным, так как содержание
некоторых МЭ в человеческом организме исчисляется в грам-
мах и миллиграммах, что противоречит представлению о сле-
довых количествах. Мы считаем, что требованиям рациональной
терминологии в большей степени отвечает термин «микроэле-
менты», так как он образован от корней классических языков
всех научных терминологий. Дискуссия об основном термине
имеет далеко не праздный интерес, так как изучение и приме-
нение МЭ в настоящее время приобрели интернациональный и
междисциплинарный характер. Поднимая этот вопрос, необхо-
димо принять во внимание и задачи здравоохранения, так как
16
5 6 7
BON
28
М
26
78
Pt
46
Pd
47
Ag
А
48
Cd
33
As
51
Sb
83
Bi
Рис. 1. Место микроэлементов (обведены жирной чертой) в периодической си-
стеме Д. И. Менделеева. Буквой М обозначены микроэлементы, избыток или
дефицит которых имеет практическое значение для здоровья человека
[Mertz W., 1987].
предложенный нами термин «микроэлементозы», за которым
стоит очень многообразное и практически важное содержание,
является производным от слова «микроэлементы». Предложить
равнозначный термин, приняв за основу обозначение «trace ele-
ments», не представляется возможным. Тем более нет медицин-
ских терминологических перспектив у нового очень громоздкого
обозначения «ультраследовые микроэлементы» [Frieden Е.,
1984J.
Все живые существа на 99% состоят из 12 наиболее рас-
пространенных элементов, входящих в число первых 20 элемен-
тов периодической системы Д. И. Менделеева. Это основные,
или «структурные», элементы, присутствие которых в живой ма-
терии связано в первую очередь с их огромным содержанием в
биосфере. Кроме того, во всех организмах находится неболь-
шое количество более тяжелых элементов, которые до некото-
рой степени произвольно подразделяются на микро- и ультра-
микроэлементы. Из 92 встречающихся в природе элементов
81 обнаружен в организме человека. При этом 15 из них (же-
лезо, йод, медь, цинк, кобальт, хром, молибден, никель, вана-
дий, селен, марганец, мышьяк, фтор, кремний, литий) признаны
эссенциальными, т. е. жизненно необходимыми. Четыре других
(кадмий, свинец, олово, рубидий) являются «серьезными кан-
дидатами на эссенциальность». Это подразделение МЭ в основ-
ном признается большинством специалистов, хотя и существу-
ют небольшие разночтения (рис. 1).
G. N. Schrauzer (1985), которому принадлежит краткий
очерк истории учения о МЭ, в XX веке выделяет только два
основных периода: первый, или классический, с 1925 по 1956 г.
и второй, современный, с 1957 г. по настоящее время. В клас-
сическом периоде почти все открытия МЭ были сделаны слу-
2—568
17
чайно или при локальных вспышках необъяснимых болезней у
сельскохозяйственных животных. Во втором периоде, в котором
доминировали блестящие исследования К. Schwarz, работа про-
водилась более систематизированно. Были разработаны приемы
эксперимента, вызывавшие специфические дефицитные состоя-
ния в отношении ряда МЭ у лабораторных животных при содер-
жании их на специально созданных синтетических диетах. Успех
этой работы был связан также с появлением в продаже очищен-
ных аминокислот, разработкой высокочувствительных методов
анализа МЭ и применением пластиковых клеток-изоляторов.
По-видимому, история открытия МЭ, в частности установле-
ние их эссенциальности, заслуживает специального освещения.
Хочется подчеркнуть, что в мировой литературе по этому воп-
росу недостаточно оцениваются работы отечественных ученых.
В истории учения о МЭ сложилось своеобразное положение:
вызываемые ими патологические процессы —МТОЗы — были
известны человечеству за несколько тысяч лет до открытия МЭ.
Один из первых МТОЗов — эндемический зоб—упоминается в
индийской и китайской литературе 4000-летней давности. Бо-
лее того, даже в те далекие времена лечебные приемы включа-
ли применение морских водорослей и препаратов из щитовид-
ных желез свиньи и оленя [Langer Р., 1960].
В XIX веке ряд исследователей применяли йод для лечения
зоба. В 1852 г. A. Chatin впервые опубликовал свои новаторс-
кие наблюдения о содержании йода в почвах, водах и продук-
тах питания в Европе. На основании этих данных он пришел к
выводу о связи эндемического зоба у человека с недостаточно-
стью йода в окружающей среде. На эту же возможность еще в
начале XIX века указывали другие исследователи, хотя они и
не располагали убедительными аргументами. Через 44 года пос-
ле публикации работ A. Chatin немецкий химик Е. J. Baumann
(1896) установил, что йод концентрируется в щитовидной желе-
зе, причем его содержание в этом органе при эндемическом зо-
бе оказалось сниженным.
Только во второй половине XIX века было показано, что хи-
мические элементы, содержащиеся в тканях организма в очень
низком (следовых) количестве, способны оказывать опреде-
ленное действие на физиологические процессы. К числу таких
элементов относится цинк [Тимирязев К. А., 1871; Raulin J.,
1869], который, как выяснилось в дальнейшем, обладает уди-
вительно многообразной и, более того, жизненно необходимой
активностью.
Краткий исторический очерк, написанный Г. А. Бабенко
(1965), не устарел до настоящего времени и дает объективную
характеристику важнейших отечественных работ, посвященных
роли МЭ в экспериментальной и клинической медицине. Имен-
но в этой небольшой книге справедливо оцениваются основопо-
лагающие идеи В. И. Вернадского о способностях живого веще-
ства к захвату, аккумулированию и использованию солнечной
энергии, которые лежат в основе его химической активности.
18
Последняя проявляется в непрерывном выделении и захвате
атомов элементов из мертвой материи, принимая таким обра-
зом участие в процессах миграции, концентрирования и рас-
пределения химических элементов в земной коре. Именно
В. И. Вернадский, впервые показавший планетарную роль жи-
вого вещества в геохимических процессах, теснейшим образом
связал их с эволюцией всего органического мира. С 1922 г. на-
учные интересы В. И. Вернадского сконцентрировались на про-
блеме связей живых организмов с различными химическими
элементами и особенно элементами, содержащимися в живых,
организмах в виде «следов». Именно этим элементам он при-
давал огромное значение в жизненных процессах.
В образной форме В. И. Вернадский говорил, что, посколь-
ку все организмы, составляющие в своей совокупности «живое
вещество», выполняют большую работу, пропуская через себя
«атомные вихри», элементный состав их находится в тесной за-
висимости от состава земной коры. Организм нераздельно свя-
зан с механизмом земной коры и должен изучаться в тесной,
связи с последним. «Автономного организма вне связи с зем-
ной корой в природе не существует» [Вернадский В. И., 1940].
Крайне важную мысль о единстве организма и окружаю-
щей среды в середине XIX века высказал И. М. Сеченов (1864).
Современные многосторонние исследования о влиянии окружаю-
щей среды на здоровье человека, выполненные под руководст-
вом ВОЗ, развивают и конкретизируют эти идеи. Особенно де-
тальной разработке подвергается вопрос об опасности химиче-
ских веществ как реальных и потенциальных загрязнителей ат-
мосферного воздуха. Среди последних в первую очередь назы-
вают токсичные МЭ—металлы, переносимые потоками воздуха?
в виде взвешенных пылевых частиц, паров и аэрозолей. Боль-
шое внимание уделяется свинцу, ртути, кадмию, бериллию,,
марганцу, мышьяку, а также фторидам.
Таким образом, мысли В. И. Вернадского оказались проро-
ческими во многих отношениях. Как современно звучат слова
об «атомных вихрях», проникающих в живые организмы в све-
те наших знаний о мощном техногенном прессинге на живую-
природу в эпоху НТР, о различных химических выбросах, воз-
никающих в результате промышленных неожиданных аварий!
МЭ — это важнейшая составная часть многих промышленных:
загрязнителей, которые оказывают на человеческий организм
разнообразное действие, причем не только во время катастроф,,
но и в условиях повседневной и, казалось бы, вполне благопо-
лучной жизни.
«Следовые вещества», или МЭ, играют большую роль не
только в жизненных процессах нормального организма, как
утверждал В. И. Вернадский, но они сохраняют свое важное
значение и в больном организме. Впрочем и эту сторону про-
блемы хорошо понимал В. И. Вернадский, так как именно под
его руководством начались исследования, направленные на вы-
яснение аномального состава химических элементов в регионах
2* 1Э
с зарегистрированными в то время заболеваниями химической
природы. По персональному сообщению акад. АМН СССР
М. А. Скворцова в далекие довоенные годы В. И. Вернадский
по собственной инициативе прочитал доклад патологам Москвы
о связи химического строения земной коры и состояния здо-
ровья человека. Весьма вероятно, что дело идет о том же до-
кладе, который В. И. Вернадский сделал в декабре 1909 г. на
XII съезде русских естествоиспытателей и врачей в Москве
[Добровольский В. В., 1983]. М. А. Скворцов присутствовал на
этом съезде.
Следует подчеркнуть, что патологическая анатомия, гисто-
логия и цитопатология МТОЗов создаются только в настоящее
время. Имеются лишь фрагментарные данные об изменении
структуры тех тканей, которые содержат либо избыточные, либо
крайне недостаточные количества МЭ. Однако единичные све-
дения о повреждающем действии некоторых МЭ на ткань опуб-
ликованы.
Наряду с этим в судебно-медицинской литературе, посвящен-
ной отравлениям и местному токсическому действию различных
химических веществ, приводятся отдельные полезные данные.
Значительно больший интерес представляют руководства по про-
фессиональной патологии и токсикологии, где суммированы не
только патоморфологические описания измененных под воздей-
ствием тех или иных МЭ органов и тканей, но и весьма важные
наблюдения клинического, патофизиологического и патохимиче-
ского характера. Мы обратимся к ним при характеристике каж-
дого из МЭ.
Ценные наблюдения были накоплены при изучении МТОЗов
сельскохозяйственных животных. К сожалению, значительная
часть их сопровождается очень краткими патогистологическими
•сведениями. В лечении и профилактике МТОЗов у сельскохо-
зяйственных животных достигнуты определенные успехи. Они
имеют большой теоретический и практический интерес, так как
материалы сравнительной патологии МТОЗов обладают значи-
тельной познавательной силой.
Один из ближайших учеников В. И. Вернадского — академик
А. П. Виноградов — в течение многих лет исследовал элемент-
ный состав различных видов животных, а также среды их оби-
тания. Он считал, что в живом веществе находятся не только
все известные элементы, но также и их радиоактивные и нера-
диоактивные изотопы. В связи с этим расширение наших зна-
ний о химическом составе организмов зависит от прогресса ме-
тодических возможностей. По его мнению, количественное со-
держание того или иного элемента в организме определяется
его содержанием во внешней среде, а также свойствами самого
элемента. А. П. Виноградов отметил большие различия между
огромными концентрациями алюминия, кремния и титана в зем-
ной коре и их ничтожным количеством в живом веществе. Это,
по его мнению, связано с тем, что указанные элементы входят в
состав труднорастворимых соединений. Вместе с тем элементы,
20
легко образующие газы и водорастворимые соединения (угле-
род, йод, азот, фосфор, калий, кальций, натрий и др.), несмотря
на их сравнительно низкое содержание во внешней среде, кон-
центрируются в организмах вследствие высокой подвижности
их соединений в биосфере. Таким образом, А. П. Виноградов
приходит к заключению, что концентрация элементов в живом
веществе прямо пропорциональна содержанию их в среде оби-
тания с учетом растворимости их соединений. Это важное пра-
вило А. П. Виноградова полностью основано на химических за-
кономерностях. Мы полагаем, что с течением времени в свете
последних достижений это правило будет заново проанализи-
ровано и дополнено данными молекулярной биологии клетки и
тканей с учетом новых фактов о строении и функционировании
специфических клеточных рецепторов и ионных каналов.
А. П. Виноградову (1933) принадлежат еще два крупных
теоретических обобщения. Во-первых, сопоставляя огромный
фактический материал о химическом элементном составе орга-
низмов различных типов, классов, родов и видов, он пришел к
выводу, что «химический состав организма есть его признак —
видовой, родовой и др.». К этому следует добавить, что своеоб-
разие химического состава организмов особенно проявляется в
содержании и локализации МЭ. Распространяется ли это пред-
положение на микроэлементный состав клеток различных жи-
вых существ, покажет будущее. Однако в настоящее время этот
подход уже используется в таксономии растений.
Во-вторых, А. П. Виноградов создал учение о биогеохимиче-
ских провинциях, которое логически завершило его исследова-
ния о значении химического состава среды обитания организ-
мов для их размножения, роста, развития, а также для их мор-
фологической и физиологической изменчивости. Согласно его
концепции, различные области земной поверхности характери-
зуются неодинаковым типом геохимических процессов и поэто-
му отличаются и по количественному элементному составу.
Именно эти области получили название «биогеохимические про-
винции». «Поскольку поступление химических элементов в ор-
ганизм определяется наряду с другими факторами и содержани-
ем элементов во внешней среде, то растения и животные в пре-
делах той пли иной геохимической провинции будут находиться
в различных условиях питания». В связи с этим животные и
растения проявляют своеобразные биологические реакции в от-
вет на элементный химический состав атомов, которые про-
ходят через организм в ходе питания. «В результате длитель-
ного непрерывного воздействия на организм определенного по
химическому составу потока атомов происходит подбор и рас-
пределение организмов по разным зонам земли, а наряду с этим
наступает и изменчивость организмов». «Своеобразие и отличие
этой изменчивости от всех других видов ее состоит в том, что
она обусловливается первичным изменением содержания хими-
ческих элементов в тканях организма». Эти взгляды получили
широкое распространение и легли в основу многолетней работы
21
по биогеохимическому районированию СССР, которая под руко-
водством В, В. Ковальского позволила установить «аномальные
биогеохимические провинции», характеризующиеся понижением
или повышением содержания таких элементов, как барий, бор,
кобальт, кальций, медь, молибден, никель и др. Совершенно;
естественно, что эти важные материалы были использованы В;
эпизоотологии, ветеринарной медицине и в других сельскохо-
зяйственных науках. Были выделены новые формы патологии
животных, растений и, наконец, человека. Более того, на основе
обнаружения почв с низким содержанием кобальта [Under-
wood Е., 1937, 1971], а также меди удалось осуществить целе-
направленную коррекцию пищевых рационов у сельскохозяйст-
венных животных и добиться их выздоровления и оптимального,
развития [Ковальский В. В., 1945, 1955; Берзинь Я. М., 1952;
Underwood Е„ 1977].
Недостаточность йода во многих регионах СССР, в частно-
сти в гористых местностях и по долинам рек, вызывает эндеми-
ческое увеличение щитовидной железы и зоб у людей и живот-
ных. Профилактическое йодирование способствовало предот-
вращению интенсивности зобных эндемий и эпизоотий.
Огромные по охвату обследованных территорий нашей
страны исследования были выполнены В. В. Ковальским, кото-
рый не только открыл конкретные формы биогеохимической па-
тологии людей и животных, но и стал основоположником новой
науки — геохимической экологии.
Среди безбрежной литературы о МЭ особого внимания за-
служивают довольно редкие работы методологического характе-
ра. К ним относится совсем небольшая статья Е. Underwood
(1976), имеющего не только большие заслуги в установлении
микроэлементной этиологии ряда эндемических заболеваний
сельскохозяйственных животных, но и разработавшего общие
принципы исследования МЭ. Он подчеркивает исключительную
сложность работы с МЭ, которая связана с низкой концентра-
цией исследуемого элемента, с загрязнениями, попадающими в
объект изучения из внешней среды, а также из реактивов и
инструментов, с возможностью связывания МЭ с другими ве-
ществами, содержащимися в продуктах или тканях. Более то-
го, многие МЭ соединены координационными или ковалентными
связями с различными органическими веществами, обладающи-
ми неодинаковой устойчивостью и биологической активностью.
Очень большие осложнения обусловлены взаимодействиями
МЭ с макроэлементами, происходящими в процессе всасывания
в кишечнике и на уровне клеток. Кроме того, МЭ могут взаимо-
действовать и с другими веществами, в частности с продукта-
ми питания. По мнению Е. Underwood (1976), при взаимодейст-
вии МЭ граница между их благотворным и токсическим влия-
нием на организм становится трудно уловимой или даже исче-
зает. Это установлено на примере меди и молибдена, а также
цинка и кадмия, селена и ртути.
Разнообразие 19 МЭ, считающихся в настоящее время не-
22
обходимыми для жизнедеятельности животных, простирается
ют галогенов йода и фтора через переходные металлы до крем-
ния 'и ванадия, которые лишь в последнее время включены в
список условно-эссенциальных МЭ. Методические сложности
усугубляются тем, что такие потенциально токсичные элементы,
как свинец, ртуть и кадмий, также подлежат изучению, так как
они способны взаимодействовать с эссенциальными МЭ.
Е. Underwood (1976) выделяет два главных метода, с по-
мощью которых были получены основные знания по физиоло-
гии МЭ. Этими методами являются: 1) изучение «локальных
проблем МЭ» в природных условиях (т. е. эндемических забо-
леваний, связанных с МЭ); 2) использование очищенных диет и
строгого контроля среды обитания. Он специально останавлива-
ется на проблеме локальных заболеваний, вызванных дефици-
том, токсичностью и нарушением баланса МЭ, подчеркивая, что
именно их разгадка оказалась наибольшим научным и эконо-
мическим вкладом в области изучения МЭ. Справедливости ра-
ди следует отметить, что начало этим исследованиям положил
A. Chatin (1852), но поднял их до уровня теории А. П. Вино-
градов (1949).
В 20-х годах XX века был предложен метод эмиссионной
спектрографии, который давал возможность определять малые
количества многих элементов и позволил осуществить сравнение
минерального состава почв, растений и животных тканей в пора-
женных и непораженных эндемическими заболеваниями райо-
нах. После этого были сделаны крупные открытия в области па-
тологии сельскохозяйственных животных, в частности описание
молибденозиса в некоторых районах Англии [Ferguson W. S et
al., 1938]. В 1937 г. Н. W. Bennets, F. Е. Chapman показали, что
энзоотическая атаксия новорожденных ягнят в некоторых
районах Западной Австралии является следствием недостаточ-
ности меди. В 1945 г. А. Т. Dick и L. В. Bull сделали важное
наблюдение о том, что потребление молибдена может оказы-
вать выраженное влияние на обмен меди у травоядных живот-
ных. В 1937 г. Е. Underwood описал истощающее заболевание
крупного рогатого скота в Новой Зеландии и в начале предпо-
ложил, что оно зависит от дефицита железа. В дальнейшем он
же сам выяснил, что истинной причиной этой болезни является
дефицит кобальта (вернее, витамина В12), хотя и терапия же-
лезом оказала высокое профилактическое и лечебное действие.
Таким образом, изучение эндемических заболеваний, сыграв-
ших очень большую роль в развитии учения о МЭ, показало,
что установление их истинной причины сопряжено со значи-
тельными сложностями, и поэтому поспешные суждения об
этиологии вновь обнаруживаемых природных МТОЗов по
меньшей мере неуместны. Это полностью относится и к
МТОЗам техногенного происхождения, которые не всегда яв-
ляются монокаузальными заболеваниями.
Наряду с большими успехами в изучении этиологии микро-
элементных зоонозов неменьшими достижениями явились ра-
23
боты отечественных ученых, заложивших основы новой теоре-
тической науки синтетического характера. Эта наука называ-
ется геохимической экологией. Она является разделом биогео-
химии и экологии, исследует взаимодействие организмов и их
сообществ с естественной и техногенной геохимической средой,
самих организмов между собой в условиях популяций, биоцено-
зов, а также регионов и субрегионов биосферы, биогеохимичес-
ких провинций как структурных частей единой целой экосисте-
мы— биосферы. Основной задачей этой науки является «выяс-
нение процессов приспособления организмов, популяций, сооб-
ществ к условиям окружающей геохимической среды и измене-
ния ими геохимических свойств окружающей среды» [Коваль-
ский В. В., 1983]. Данные геохимической экологии в дальней-
шем послужили основой для разработки системы биогеохими-
ческого районирования гетерогенной биосферы. В свою очередь
биогеохимическое районирование стало необходимым методом
изучения строения биосферы, основанным на признании единст-
ва жизни и геохимической среды в биосфере.
Взаимодействие геохимических факторов среды и организ-
мов осуществляется в последовательных звеньях биогеохимиче-
ской пищевой цепи и зависит от природных концентраций хими-
ческих элементов и пороговой чувствительности организмов.
Понятие о биогеохимических пищевых цепях глубоко разраба-
тывалось В. В. Ковальским, основным объектом исследовательс-
кой деятельности которого была обширная сфера биогеохимии
сельскохозяйственных животных. Эти теоретические положения
привели В. В. Ковальского к учению о регионах биосферы, ко-
торые характеризуются биогеохимической мозаичностью и по
принципу географической непрерывности разделяются на субре-
гионы.
Значительную практическую важность имеют мысли
В. В. Ковальского об экстремальных условиях субрегионов
биосферы, в которых биологические реакции организмов, вызы-
ваемые недостатком определенных естественных или избытком
техногенных элементов, могут быть особенно ярко выражены.
Это совпадает с нашей концепцией о природных и техноген-
ных экстремальных зонах [Авцын А. П., 1966].
«Система биогеохимического районирования биосферы
должна охватить континенты Земли. Живое вещество биосферы
в пределах между верхними и нижними пороговыми концент-
рациями должно находиться в отношениях определенной сла-
женности со средой, в состоянии относительного равновесия,
при котором сохраняется живое вещество как жизнеспособная
система...» [Ковальский В. В., 1983]. Исходя из этих теоретиче-
ских положений, В. В. Ковальский придает исключительное
значение районированию биосферы, в котором биогеохимичес-
кое картографирование, основанное на количественной харак-
теристике звеньев биогеохимической цепи, имеет важное прак-
тическое (прикладное) значение. Со всей определенностью он
указывает на значение этих методов не только для практичес-
24
ких задач сельского хозяйства, но и для медицинской и ветери-
нарной гигиены. С полным основанием он утверждает, что ис-
следование роли геохимических факторов выдвигает новую
большую программу в области изучения условий сохранения
здоровья и причин болезней растений, животных и человека.
Вернемся, однако, к некоторым методическим вопросам. Ис-
следования Е. Underwood (1977) на животных с дефицитом ко-
бальта потребовало разработки новых методов определения его
концентраций в тканях. Перед ним встала трудная задача со-
здания такого метода анализа, который давал бы возможность
определить концентрацию кобальта, равную 1,7 мкмоль/кг.
Простая оптическая колориметрия оказалась для этой цели не-
пригодной. Только через много лет с использованием усовер-
шенствованных методов удалось выяснить, что «кобальтовая не-
достаточность» у жвачных животных является результатом де-
фицита витамина В12, вызванным неспособностью микроорганиз-
мов кишечника (при недостаточности кобальта) синтезировать
необходимые для организма количества этого витамина [Under-
wood Е., 1977J.
Крупные успехи по выяснению эссенциальной роли некото-
рых микроэлементов были связаны с использованием высоко-
очищенных диет. Таким образом удалось получить доказатель-
ства эссенциальности селена, фтора, мышьяка и других МЭ для
роста животных. В частности, для селена пришлось разрабо-
тать адекватную во всех отношениях диету, содержащую менее
0,06 мкмоль/кг.
В настоящее время проблема МЭ освещается на страницах
специальных журналов и в периодически издаваемых моногра-
фиях. Большую информацию содержат труды ежегодно созы-
ваемых международных симпозиумов. Существуют междуна-
родные научные общества, глубоко изучающие отдельные МЭ,
например фтор. Они же созывают регулярные конференции.
С 1968 г. выходит международный ежеквартальный журнал
«Fluoride», посвященный медицинским и биологическим аспек-
там проблемы фторидов.
Отечественные публикации по МЭ сравнительно немногочис-
ленны, специального периодического издания (журнала) не су-
ществует, за исключением трудов биогеохимической лаборато-
рии им. В. И. Вернадского. Однако в настоящее время и они
по необъяснимой причине не издаются. К сожалению, еще нет
специальных обобщающих изданий, посвященных МТОЗам.
Те сведения, в которых затрагивается данный вопрос, фрагмен-
тарны, причем это относится не только к отечественной, но и
зарубежной литературе. Однако есть много признаков того, что
это положение в самом ближайшем будущем изменится. В ча-
стности, число публикаций о нейротоксическом действии алю-
миния и особенно о многообразных свойствах селена стреми-
тельно увеличивается, освещаются данные о ряде других токсич-
ных, а также эссенциальных МЭ. Изобилие разнообразных на-
учных фактов по многим вопросам этой сложной проблемы на-
25
стоятельно диктует необходимость публикации систематичес-
ких и особенно концептуальных обзоров. С 1984 г. выходит еже-
квартальный международный журнал Trace Elements in Medici-
ne, с 1987 г. еще один журнал — J. Trace Elements and Electro-
lytes in Health and Disease и c 1988 r. — J. of Trace Elements
in Experimental Medicine, посвященные изучению роли МЭ в ме-
дицине.
1.2. Содержание микроэлементов
в окружающей природной среде
Общие сведения о распределении МЭ в окружающей среде
в краткой форме изложены в книге А. И. Войнара (1960), где
он довольно подробно останавливается на химическом составе
почв и литосферы. Более новые важные данные, сгруппирован-
ные по несколько иному принципу, приведены в переводной мо-
нографии Дж. Фортескью Геохимия окружающей среды (1985).
Из нее мы приводим данные, отражающие распределение МЭ в
главных геосферах (табл. 1). Это дает общее представление о>
крайнем разнообразии распространенности химических элемен-
тов. Обращает на себя внимание огромное содержание таких
МЭ, как кремний, алюминий и железо в земной коре и их отно-
сительно небольшие концентрации в пресной и морской воде.
Однако общее содержание кремния в биосфере очень велико,
что свидетельствует об активности протекающих с его участи-
ем химических процессов. Это было предметом многолетних ис-
следований М. Г. Воронкова (1968, 1972), который посвятил
данной проблеме, в частности вопросу о роли соединений крем-
ния в биогенезе, интересные и пока еще недостаточно оценен-
ные труды.
В табл. 2 представлено содержание химических элементов в
морских и наземных растениях и животных. Данные этой таб-
лицы также имеют общее ориентирующее значение для сужде-
ний о содержании МЭ в пище растительного и животного про-
исхождения. Содержание МЭ в почве и во внутренних водах
США охарактеризованы в книге К. Рейли (1985). Геохимичес-
кие и биохимические процессы, прямо или косвенно влияющие
на характер распределения МЭ в почвах и растениях, на со-
временном уровне представлены в монографии А. Кабата-Пен-
диас и X. Пендиас (1989), авторы которой не останавливаются
на медицинских аспектах этого вопроса.
Существуют многочисленные данные о загрязнении МЭ био-
сферы, которые систематически освещаются в соответствующей
литературе [Жигаловская Т. М. и др., 1974]. Содержание МЭв
аэрозолях воздушных бассейнов, в частности в Средней Азии,
меняется в значительных пределах. Кроме того, эти изменения
вариабельны и во времени. Расчеты показали, что содержание
бора в этом регионе изменилось в 20 раз, никеля — в 60, тита-
на— в 400, ванадия — в 20, меди — в 40, цинка — в 100, сереб-
ра— в 20, олова — в 200, стронция —в 20, бария — в 20 раз.
26
Таблица 1
Распространенность химических элементов
в четырех главных геосферах [Фортескью Дж., 1985]
Верхняя ли- Гидросфера?
Элемент Тосфера (земная пресные морская Атмосфера3 Биосфера*
кора)1 воды вода
Кислород 456 000 889 000 857 000 755’100 780 000
Кремний 273 000 6,5 3 - (4,0) 21 000
Алюминий 83 600 0,24 0,01 - (3,0) 510
Железо 62 200 0,67 0,01 - (3,0) 1100
Кальций 46 600 15,00 400 - (2,0) 51 000
Магний 27 640 4,1 1350 - (1,0) 4100
Натрий 22700 6,3 10 500 - (1,1) 2100
Калий 18 400 2,3 380 31 000
Титан 6320 0,009 0,001 - (0,01) 81
Водород 1520 111000 108 000 0,35 (300,0) 105 000
Фосфор 1120 0,005 0,07 — • 7100
Марганец 1060 0,012 0,002 - (0,01) ПО
Фтор 544 0,090 1,30 - (0,01) 51
Барий 390 0,054 0,03 310
Стронций 384 0,080 8,10 — — 210
Сера 340 3,7 885,0 — (3,50) 5100
Углерод 180 162 11,0 28,0 460 (164 000) в форме СОг 180 000
Цирконий 0,003 0,00002 —. —. —
Ванадий 136 0,001 0,002 — (0,001) —
Хлор 126 7,8 19 000 - (1,2) 2100
Хром 122 0,0002 0,00005 — (0,002) —
Никель 99 0,01 0,005 — (0,002) 5
Рубидий 78 0,002 0,12 - . — ' » 51
Цинк 76 0,01 0,01 - (0,07) 51
Медь 68 0,01 0,003 — (0,02) 21
Церий 66 —. 0,0004 —
Неодим 40 —- — —
Лантан 35 —, 0,00001 — —
Иттрий 31 — 0,0003 —. —. 1,1
Кобальт 29 0,0009 0,0003 — (0,0007) 2,1
Скандий 25 — 0,000004 —. —
Ниобий 20 — 0,00001 . . .
Азот 19 0,23 0,50 750 100 (9,73X10) 31 000
Галлий 19 0,01 0,00003 -
Литий 18 0,0011 0,18 1,1
Свинец 13 0,005 0,00003 — (0,2) 5,1
Празеодим 9,1 — —
Бор 9,0 0,013 4,6 110
Торий 8,1 0,00002 0,00005 __
Самарий 7,0 .— — — .
Олово 2,1 0,0004 0,003 — (0,01) 5,1
Бериллий 2,0 0,001 0,0000006 — (0,0001)
Мышьяк 1,8 0,0004 0,0003 - (0,01) 3,1
Гольмий 1,3 . 1,1
Ртуть 0,86 0,00008 0,00003
Золото 0,004 0,00006 0,000011 —- — —
‘ А. Б. Роков, А. А. Ярошевский (1972) процентов). (данные рассчитаны на основании весовых
« Л Bowen, 1966 (данные рассчитаны иа основании весовых пооцентов).
. • i хаьоп (1Уоо) (данные рассчитаны на основании весовых пооиентов): в скобках
данные по ri. J. Bowen (1966) (рассчитаны иа осиоваи-ии объемных" пЬопентов).
’ a. в. иерельман (1966) солютно сухого вещества). (данные рассчитаны на основании весовых процентов аб-
27
Таблица 2
Содержание химических элементов [Bowen Н. J., 1966]1
Элемент Морские расте- ния Наземные растения Морские жи- вотные Наземные жи- вотные
Кислород 470 000 410 000 400 000 186 000
Кремний 1500—20 000 200—5000 70—1000 120—6000
Алюминий 60 500 10—50 4—100
Железо 700 140 400 160
Кальций 10 000—300 000 18 000 1500—20 000 200—8500-
Магний 5200 3200 5000 1000
Натрий 33 000 1200 4000—48 000 4000
Калий 52000 14 000 5000—30 000 7400
Титан 12—82 1,0 0,2—20 0,2
Водород 41 100 55 000 52 000 70 000
Фосфор 3500 2300 4000—18 000 17 000—44 000»
Марганец 53 630 1-60 0,2
Фтор 4,5 0,5—40 2 150—500
Барий 20 14 0,2—3 0,75
Стронций 160—1400 26 20—500 14
Сера 12 000 3400 5000—19 000 5000
Углерод 345 000 454 000 400 000 465 000
Цирконий 20 0,64 0,1—1 0,3
Ванадий 2 1,6 0,14—2 0,15
Хлор 4700 2000 5000—90 000 2800
Хром 1 0,23 0,2—1 0,075
Никель 3 3 0,4—25 0,8
Рубидий 7,4 20 20 17
Цинк 150 100 6—1500 160
Медь 11 14 4—50 2,4
Церий — (320) — 0,03
Неодим 5 (064) 0,5 —-
Лантан 10 0,085 0,1 0,0001
Иттрий — 0,6 0,1—0,2 0,04
Кобальт 0,7 0,5 0,5—5 0,03
Скандий — 0,008 — 0,00006
Ниобий — 0,3 0,001 —
Азот 15 000 30 000 75 000 100 000
Галлий 0,5 0,06 0,5 0,006
Литий 5,0 0,1 1,0 0,02
Свинец 8,4 2,7 0,5 2,0
Бор 120 50 20—50 0,5
Торий — — 0,003—0,03 0,003—0,1
Самарий — 0,0055 0,04—0,08 0,01
Данные приведены в условных единицах (часть на 1 млн сухого вещества).
Хотя и неизвестно, насколько отмеченные изменения характер-
ны и постоянны, однако они, несомненно, весьма существенны
[Жигаловская Т. М. и др., 1976]. Недостаточно учитывается
роль внезапных «микроэлементных кризов» в происхождении
неожиданных ухудшений состояния здоровья людей и живот-
ных, связанных с промышленными выбросами и отчасти с не-
благоприятными погодными условиями.
Для оценки миграции элементов в биосфере исследователи
широко используют ряд эмпирических показателей, в частности
так называемый коэффициент биологического поглощения
28
(КБП). А. П. Перельман (1966) определяет его как отношение
содержания определенного элемента в золе организмов (обычно
растений) данного ландшафта к содержанию его в горной поро-
де или почве, на которой произрастает это растение, или же в
земной коре в целом. Элементы с более высоким КБП актив-
нее поглощаются растениями из окружающей среды.
В краткой форме техногенное рассеивание элементов и со-
ответствующие изменения геохимии окружающей среды оха-
рактеризованы В. В. Добровольским (1983). Он специально вы-
деляет вопрос о загрязнении тяжелыми металлами. «...Нараста-
ющие масштабы промышленного производства влекут за собой
далеко идущие последствия. Так, если в первой половине теку-
щего столетия беспокойство вызывали перспективы нехватки
сырья, то во второй половине обнаружилась более серьезная
опасность, а именно: изменение состава среды, окружающей
человека». Действительно, в современной гигиенической и об-
щемедицинской литературе имеются многочисленные сообще-
ния от отравлениях определенных групп населения ртутью, кад-
мием, свинцом. С полным основанием тяжелые металлы рас-
сматриваются в качестве основных промышленных загрязните-
лей.
Наиболее важные для физиологии и патологии сведения о
суточном балансе химических элементов для «условного» чело-
века приведены в книге «Человек. Медико-биологические дан-
ные» (1977). В табл. 3 представлены показатели, характеризую-
щие баланс макроэлементов и МЭ в течение суток. Эти пока-
затели относятся к пищеварительному и аэрогенному путям по-
ступления их в организм. Приведены также данные о выделе-
нии их с мочой и калом, а также с помощью других биосубст-
ратов (пот, молоко, волосы и др.).
При острых и особенно хронических МТОЗах эти показате-
ли могут значительно изменяться, что позволяет сделать соот-
ветствующие медицинские выводы, например об избыточном
поступлении тех или иных МЭ или о задержке их элиминации.
Сравнительные данные о содержании химических элементов
в усредненной диете человека в различных регионах земного
шара (табл. 4) представляют несомненный интерес, так как
позволяют с определенной долей вероятности использовать их
для ориентировочной оценки микроэлементного гомеостаза кон-
кретного человека.
Как видно из табл. 4, поступление химических элементов не-
одинаково в различных странах, причем в большей степени это
относится к МЭ. Так, в Великобритании потребление с пищей
алюминия составляет всего 2,3 мг, в то время как в США оно
достигает 45 мг, т. е. почти в 20 раз больше. Различия по дру-
гим элементам обычно значительно меньше. Неожиданно ве-
лико содержание лития и тория в пище в США. Отражение этих
существенных различий в патологической панораме человека
еще не улавливается либо оценивается в недостаточной степени.
Среди множества МЭ существуют особенно важные для пол-
29
Таблица 3
Суточный баланс микроэлементов для условного человека
(Из книги: Человек. Медико-биологические данные. М.: Медицина, 1977)
Элемент Единица измерения Поступление с Выделение с
пищей и жид- костя - мн воздухом мочой калом потом волосами и др.
Алюминий МКМОЛЬ 1668 3,7 3,7 1594 37 0,022
Барий МКМОЛЬ 5,46 0,0006—0,19 0,36 0,50 0,007 0,054
Бериллий мкмоль 1,33 0,001 о,н 1,11
Бор мкмоль 120 — 90 20 0,1
Бром мкмоль 94 — 88 0,88 2.4 0.025
Висмут нмоль 96 0,048 7,66 86 Неизвестно
Ванадий мкмоль 39 0,004 0,29 38
Германий мкмоль 21 — 19 1,38 __
Железо мкмоль 287 0,54 4,48 269 8,9 0,23
(муж.)
215 0,54 3,58 197 10,8
(жен.) Менстру-
альные по-
-Иод мкмоль 1,58 0,004—0,276 1,34 0,39 0,047 терн 0,176
Кадмий мкмоль 1,33 0,009 0,89 0,44 — -
Кобальт мкмоль 5,09 0,002 3,40 1,53 0,068 0,041
Кремний мкмоль 0,12 0,53 0,36 0,36 — 0,011
Литий ммоль 0,29 — — 0,12 0,17 Следы
Марганец мкмоль 67 0,036 0,55 65 0,71 0,036
Медь мкмоль 55 0,31 0,79 53 0,63—6,30 0,047
Молибден мкмоль 3,13 0,001 1,56 1,25 0,21 0,0001
Мышьяк мкмоль 13,3 0,019 0,67 1,07 —- 0,007
Никель мкмоль 6,81 0,010 0,19 6,30 0,34 0,017
Ниобий мкмоль 6,67 — 3,87 2,80 Следы 0,003
Олово нмоль 33,7 0,003 0,17 29,5 4,21 —
Ртуть нмоль 74,8 4,99 1,74 49,8 Следы 4,48
Рубидий мкмоль 23,4 — - 22,2 3,51 0,58 —
Свинец мкмоль 2,12 0,048 0,22 1,45 3,14 0,145
Селен мкмоль 1,90 Неизвестно 0,63 0,25 1,01 0,013
Серебро мкмоль 0,65 ——• 0,08 0,56 0,004 0,006
Стронций , мкмоль 21,7 —- 3,88 17,1 0,23 0,002
Сурьма мкмоль 410 0,41 329 74 — 8,21
Таллий нмоль 7,34 0,24 2,45 4,89 —— 0,0005
Теллур мкмоль 4,70 0 4,15 0,78 Виды- 0,078
хается
Титан мкмоль 17,7 0,021 6,89 0,23 — 0,0017
Фтор нмоль 94,7 52,6 7,89 34,2 ' 1
Хром мкмоль 2,88 0,002 1,35 1,54 0,02 0,012
Цезий нмоль 75,2 0,19 67,7 7,52 Следы
Цинк ммоль 0,20 0,0015 0,0076 0,17 0,012 0,0005
Цирконий мкмоль 46,0 —. 1,64 43,8
неценного питания человека. Питьевая вода является главным
источником поступления в организм лишь фтора и стронция.
«Для населения развитых стран характерно включение в ра-
цион разнообразных продуктов питания, часть из которых про-
изводится в других биогеохимических районах, в виду чего лик-
видируются условия, способствующие воздействию на человека
.30
Таблица 4
Суточное содержание некоторых химических элементов в пище человека-
в различных регионах земного шара [Hamilton Е. J., 1981]
Химический элемент Единица из- мерения Великобри- тания США Индия ФРГ
Алюминий ММОЛЬ 0,085±0,041 1,67
Серебро МКМОЛЬ 0,25±0,16 0,65
Барий МКМОЛЬ 4,39±1,63 5,46 3,20
Бериллий мкмоль 1,66 1,33 —
Бром ммоль 0,11±0,01 0,09 0,03
Кадмий мкмоль 0,б7±0,27 1,33 __ 0,44
Кальций ммоль 34,9±0,5 27,4 17,7 9,48
Цезий мкмоль 0,098±0,053 0,75 — 0,098
Хлор моль 0,150±0,002 0,148 — 0,126
Хром мкмоль 6,15±3,11 2,88 2,88 1,00
Медь ммоль 0,049±0,012 0,055 0,091 0,042
Железо ммоль °,417±0,002 0,21—0,28 0,71 0,25
Калий ммоль 71,6±0,8 76,7 105 61,4
Литий мкмоль 24,5±7,6 288 14,4 —
Магний ммоль 10,3±0,8 11,0—14,0 30,4
Марганец мкмоль 49,1±14,6 67,3 151 49,1
Молибден мкмоль 1,30±0,35 3,13 — 0,73
Фосфор ммоль 61,3±1,0 58—84 48 —
Свинец мкмоль 1,54±0,73 2,12 — —
Рубидий мкмоль 51,5±17,5 25,7 31,6 22,2
Сера ммоль 29,3±2,1 26,5 37,4 ——
Сурьма мкмоль 0,28±0,22 0,41 — 0,189
Стронций мкмоль 9,79±1,64 21,7 45,6 —•
Таллий нмоль 9,78 7,34 —
Торий нмоль 0,215 12,9 —• —
Титан мкмоль 16,7 17,7 —— —
Уран нмоль 4,16 7,98 —• —
Цинк ммоль 0,218±0,018 0,199 0,246 0,180
биогеохимических особенностей данной местности» [Книжни-
ков В. А. и др., 1981]. Таким образом, разнообразная пища со-
значительной долей привозных продуктов не только предупреж-
дает возникновение эндемических дефицитов МЭ в регионах с
дискомфортными природными условиями, но и является одним
из мощных средств ликвидации эндемических заболеваний био-
геохимического происхождения.
Подробные сведения о содержании МЭ в основных продук-
тах питания изложены в справочнике Химический состав пище-
вых продуктов (1987).
1.3. Перенасыщение организма микроэлементами
в условиях природных и техногенных локусов
Для территории СССР, отличающейся исключительным раз-
нообразием биогеохимической ситуации, важное практическое*
значение имеют исследования регионов с экстремальными усло-
виями обитания. Эти аномальные в биогеохимическом отноше-
нии регионы и локусы природного и антропоургического проис-
311
хождения предъявляют значительные требования к адаптаци-
онным механизмам организма, в том числе и к обеспечивающим
микроэлементный гомеостаз. Важное значение в эпидемиоло-
гической оценке очагов природной и техногенной эндемии
МТОЗов имеют исследования распространенности биогеохими-
ческих эндемий у сельскохозяйственных животных. Они явля-
ются своеобразным индикатором неблагополучия биогеохимиче-
ской ситуации определенного региона и для человека.
При этом установлены видовые, породные, популяционные и
индивидуальные различия чувствительности животных к био-
геохимическим факторам региона, в том числе и к МЭ. В энде-
мичных регионах манифестные признаки заболевания встреча-
ются обычно не более чем у 20% животных, в то время как ос-
новная масса поголовья адаптируется к избытку или дефициту
определенных МЭ в данной геохимической среде [Раец-
кая Ю. И., 1987].
Систематические исследования в этом направлении, прово-
дившиеся в течение нескольких десятилетий В. В. Ковальским,
позволили создать карту биогеохимического районирования
СССР. Из этой карты видно, что районы Крайнего Севера
практически не изучены и представляют собой «белые пятна».
Недостаточная изученность биогеохимической панорамы Севе-
ра нашей страны связана в основном с его слабой заселенно-
стью, что сказывается на отсутствии достоверных сведений о
распространенности и напряженности многих эндемий биогео-
химического происхождения.
Хорошо известно, что рост населения, интенсивное исполь-
зование природных ресурсов и прогрессирующее загрязнение
окружающей среды представляют собой проблемы планетарно-
го характера. На Западе распространено определение загрязне-
ния как «неблагоприятного изменения окружающей среды, ко-
торое целиком или частично является результатом человеческой
деятельности, прямо или косвенно меняет распределение ...энер-
гии, уровня радиации, физико-химические свойства окружаю-
щей среды и условия существования живых существ...» [Ра-
мад Ф., 1981].
В соответствии с современной классификацией загрязнений
биосферы выделяют несколько типов последних, уделяя особое
внимание контаминациям физической и химической природы.
Хотя химические загрязнения давно известны и наиболее много-
образны, систематика их, приводимая в литературе, не може’
считаться полной, так как в ней не выделяются загрязнения
микроэлементной природы. Между тем в последние десятилетия
такие загрязнения становятся все более значимыми, их пате
генный эффект начинает привлекать внимание не только эко-
логов, но и специалистов в области человеческой и ветеринар-
кой патологии. Существенные наблюдения накоплены фитопа-
тологами и специалистами по заболеваниям наземных и океани-
ческих животных. Многие из этих загрязнений по своему значе-
нию приобретают характер «лимитирующих факторов», кото-
32
рые угрожают не только нормальному развитию, но и самому
существованию различных видов [Одум Юд., 1975].
Микроэлементное загрязнение окружающей среды пред-
ставляет наибольшую опасность для индустриально развитых
стран. Известно, что по соседству со многими промышленными
предприятиями образуются постоянно расширяющиеся техно-
генные биогеохимические провинции с повышенным содержани-
ем в биосфере свинца, мышьяка, фтора, ртути, кадмия, мар-
ганца, никеля и других элементов. Однако не только вблизи с
промышленными предприятиями определяются микроэлемент-
ные загрязнения, но и на значительном отдалении, возникаю-
щие в результате трансгрессии загрязнителей воздушными и
водными потоками. А. П. Шицкова (1989) подчеркнула особое
значение ветрового распространения загрязнений в условиях
Севера СССР.
Микроэлементный перенос может быть эпизодическим, на-
пример в результате природных или техногенных катастроф, ли-
бо явиться следствием систематического загрязнения атмо- и
гидросферы отходами производства. Типичным примером транс-
грессии являются кислотные дожди, наблюдающиеся во многих
странах Европы и Северной Америки. Одни из первых сообще-
ний по этому вопросу поступили из Скандинавии, затем из Ка-
нады и ряда других стран. Кислотность осадков в Северо-Вос-
точной Европе за последние 20 лет резко возросла; концентра-
ция ионов водорода в дождях в некоторых частях Скандинавии
с 1956 г. возросла более чем в 200 раз [Likens G. Е. et al.,
1974].
Данные, полученные в зоне озер Канады к западу от Сэд-
бурийских плавильных заводов, говорят о 100-кратном повыше-
нии кислотности их воды. Так, в 1970 г. вода 33 из этих озер
резко повысила свою кислотность,которая достигала величины
pH, равной 4,5. Эти изменения сопровождались массовой ги-
белью рыб, особенно лососевых. Кроме того, окисление водных
экосистем приносит значительный ущерб другим живым объ-
ектам, а также промышленным и архитектурным сооружениям.
По данным G. Е. Likens и соавт. (1974), в США из антропоген-
ных источников попадает в атмосферу около 32Х106 т SO2.
К 2000 г. это количество должно возрасти в 2—5 раз. Возра-
стающее поступление SO2 в атмосферу является предметом
серьезной озабоченности не только органов здравоохранения,
но и руководителей многих предприятий.
Основными загрязнителями атмосферы в Швеции являются
такие отдаленные индустриальные страны и их отдельные ре-
гионы, как Англия и Рур (ФРГ). Более 70% серы в воздухе
Швеции имеет антропогенное происхождение. SO2 до своего
выпадения в виде осадков может проделать путь более 5000 км.
Наряду с SO2 воздух загрязняется также NO3_, NH4+ и реже
НСОз-". Исследование этих процессов в настоящее время ак-
тивно продолжается, они зарегистрированы в других регионах
Земного шара.
3—568
33
Кроме антропогенных кислотных дождей, существуют и не-
антропогенные. В частности, на Гавайских островах на высотах
от 0 до 3400 м собрано большое число проб дождей, причем
большинство из них оказалось кислым. Кислотные дожди об-
наружены также в центре Индийского океана. Часть непро-
мышленных кислотных дождей, очевидно, имеет природное
происхождение и может быть связано с вулканами или ат-
мосферными загрязнениями, источниками которых являются
определенные регионы на дне океанов.
Крупные индустриальные города представляют собой экстре-
мальные зоны обитания, их следует рассматривать как регионы
интенсивного загрязнения. Так, по данным Ю. Е. Саета и соавт.
(1981), средний уровень загрязнений для таких МЭ, как вольф-
рам, ртуть, кадмий, свинец, сурьма, молибден, цинк, медь, ни-
кель и кобальт, во много раз выше в городах по сравнению с
природными ландшафтами. Среди МЭ-загрязнителей преобла-
дают элементы с повышенной «экотоксичностью». Распределе-
ние микроэлементных аномалий, по данным этих авторов, опре-
деляется не столько метеорологическими параметрами, сколько
условиями застройки городов. В настоящее время вся биосфера
загрязнена свинцом индустриального происхождения, основными
источниками которого являются литейное производство, авто-
мобили, а также сжигание промышленных отходов. Высо-
кая концентрация свинца в воздухе обусловлена плотностью
автомобильного движения, а также плотностью застройки. Из-
вестно, что каждый автомобиль ежегодно выбрасывает в воз-
дух 1 кг этого МЭ [Рамад Ф., 1981].
По данным Б. А. Ревича и соавт. (1982), вблизи промыш-
ленных предприятий черной, цветной металлургии и машино-
строения, расположенных в городах среди жилых массивов,
максимальные концентрации свинца превышают фоновые зна-
чения в 14—50 раз, цинка — в 30—400 раз, хрома — в 11—46 и
никеля — в 8—63 раза.
С целью оптимизации мер по контролю и борьбе с загрязне-
нием окружающей среды как в региональных, так и глобальных
масштабах принята «Конвенция о трансграничном загрязнении
воздуха на большие расстояния». Известно, что воздействие за-
грязняющих веществ на биосферу зависит от их концентрации,
физических и химических свойств, а также от продуктов их по-
следующих превращений. В связи с этим выделены три основ-
ных типа выбросов загрязняющих веществ в атмосферу: одни
из них приводят к загрязнению в глобальном масштабе, дру-
гие— к загрязнению в региональном масштабе (регион нередко
охватывает территории нескольких стран), третьи загрязняют в
локальном масштабе, т. е. на относительно небольшой терри-
тории, например в пограничных районах двух соседних стран
[Израэль Ю. А. и др., 1983].
Содержание МЭ в атмосфере зависит от многих причин,
в частности от метеорологических и технологических, и сущест-
венно колеблется во времени. Эти колебания являются опреде-
34
ляющими для соответствующих расчетов кумулятивных доз за-
грязнителей, поступающих в организм [Буштуева К. А., 1976;
Ревич Б. А и др., 1982]. Так, Т. М. Roberts и соавт. (1974), изу-
чая уровни свинца в воздухе, пыли, почве и в крови детей, жи-
вущих вблизи плавильного завода в Торонто, установили, что
среднемесячный уровень этого МЭ (1,0—5,3 мкг/м3) здесь был
вдвое больше, чем в других частях города. Месячные уровни
свинца в выпадающей пыли (200—1500 мг/м2), в почве (16 000—
40 000 мг/кг) вблизи завода были выше, чем на расстоянии
300—400 м от предприятия. При этом концентрация свинца в
крови детей, живущих в радиусе 300 м от заводской трубы, со-
ставила более 400 мкг/л у 13—30% обследованных. Таким об-
разом, эти дети могут составлять группу риска.
Специалистами, изучающими геохимию окружающей среды,
установлено, что в условиях умеренных широт в регионах, зна-
чительно удаленных от урбанизированных зон, суточное фоно-
вое выпадение аэрозольных загрязнений составляет 9,75 кг на
1 км2. В то же время в урбанизированных зонах эти показате-
ли в 20—50 раз возрастают и в среднем достигают 200—500 кг.
Максимальное загрязнение составило 5739 кг/км2, в 600 раз
превышая суточную фоновую величину [Ревич Б. А. и др.,
1982].
Эти же авторы представили суммарные данные по распре-
делению микроэлементных загрязнений в аэрозольных выпаде-
ниях, уловленных снеговым покровом на территории крупной
урбанизированной зоны с разнообразной промышленностью.
Их анализ проводился на основании соответствующего коэф-
фициента концентрации (Кс), который равен отношению вели-
чины среднего выпадения элемента на территории города к ве-
личине среднего выпадения его на площади, удаленной от го-
рода на 100 км и более. Все МЭ в соответствии с этим коэффи-
циентом распределились на три группы. В 1-ю группу вошли
МЭ, накапливающиеся выше фоновых концентраций (Кс в пре-
делах 23—2); 2-ю группу составили МЭ, накапливающиеся на
уровне фоновых концентраций (Кс около 1); МЭ 3-й группы на-
капливаются ниже фоновых концентраций (Кс = 0,6—0,4). Ха-
рактерно, что все МЭ 1-й группы широко используются в про-
мышленности, в то время как МЭ 2-й и 3-й групп мало приме-
няются в промышленности, поэтому их в значительно меньшем
количестве обнаруживают в аэрозольных загрязнениях. Эти
же авторы дали оценочную характеристику концентрации ряда
МЭ в волосах детей в зависимости от их содержания в аэро-
зольных загрязнениях, осажденных снегом. Различия между
средними концентрациями всех исследованных МЭ в волосах
детей по изученным выборкам оказались статистически досто-
верными, что является еще одним доказательством того, что
волосы человека — это накопитель МЭ, причем их концентра-
ция в волосах может служить объективным показателем мик-
роэлементной ситуации в целом организме.
В настоящее время агрессивность внешней среды, обуслов-
3*
35
ленная антропогенным загрязнением, в том числе и микроэле-
ментным, настолько серьезна, что ее нельзя игнорировать. Об
этом, в частности, свидетельствуют обширные исследования
группы авторов [Козлюк А. С. и др., 1987], изучавших иммун-
ный статус детей в местностях, где применяют пестициды с раз-
личной интенсивностью, а также в населенных пунктах Молда-
вии с вредными геохимическими факторами. При этом уста-
новлено, что загрязнения окружающей среды химическими ве-
ществами малой интенсивности обусловливают ряд сопряжен-
ных сдвигов в иммунной системе детей. Отмечены угнетение
преимущественно клеточного звена иммунитета, наличие IgE-
гиперглобулинемии и высокого уровня спонтанной бласттраис-
формации лимфоцитов. Выраженность дисбаланса иммуноло-
гических показателей и степень функциональных нарушений
иммунитета коррелируют с уровнем загрязнения окружающей
среды.
Важно подчеркнуть, что в настоящее время существуют ме-
тоды клинической иммунологии, позволяющие провести имму-
нологический скрининг и выявить в «группе риска» детей, у ко-
торых в результате патогенного действия токсичных факторов
малой интенсивности создаются предпосылки для возможного
развития заболеваний, в той или иной степени связанных с им-
мунопатологией. Вспышки необъяснимых заболеваний детей с
синдромом тотальной алопеции могут представлять собой пря-
мое следствие загрязнения внешней среды М.Э (подробнее см.
главу 8).
Эти своеобразные индикаторы загрязнений не столь без-
обидны, как это может показаться на первый взгляд, так как
при этом отмечаются не только выпадение волос, но и невро-
логическая симптоматика, свидетельствующая о патологии
гипоталамуса.
Проблема развития и здоровья детей в связи с влиянием
МЭ заслуживает самостоятельного рассмотрения. Уже есть
сведения о том, что в условиях микроэлементных загрязнений
происходит интенсивное накопление различных токсичных эле-
ментов в плаценте. Некоторые промышленные регионы с особо
интенсивным микроэлементный загрязнением могут стать зона-
ми экологического бедствия в результате избыточного накопле-
ния в них ртути, кадмия, мышьяка, бериллия и др. Организм
может реагировать не только развитием болезней. Могут по-
явиться признаки дизадаптации, а также нарушения физичес-
кого и психического развития. В. Н. Кувина и соавт. (1989) на
обширном материале доказали, что в промышленных регионах
Восточной Сибири и Севера у детей и подростков в высоком
проценте регистрируются диспластические изменения скелета,
которые также следует считать индикаторами микроэлемент-
ных загрязнений.
Глава 2
СОДЕРЖАНИЕ МИКРОЭЛЕМЕНТОВ В ОРГАНАХ,
ТКАНЯХ И ЖИДКОСТЯХ ОРГАНИЗМА ЧЕЛОВЕКА
2.1. Содержание микроэлементов в организме человека
в норме и в различные периоды его развития
Несмотря на общий интерес к проблеме МЭ, вопрос об их
содержании в организме человека освещен недостаточно. Ста»
рые литературные данные изложены в монографии А. И. Вой-
нара (1960). Многочисленные данные о некоторых МЭ приве-
дены в трудах М. Г. Коломийцевой, Р. Д. Габовича (1970)„
В. В. Ковальского (1974) и их сотрудников. Ценные материалы
о содержании МЭ в различных регионах Сибири и Дальнего
Востока суммировал В. М. Мещенко (1968).
Более новые данные представил D. К- Kalani (1980), кото-
рый показал, что содержание МЭ в органах и жидкостях чело-
века очень неравномерно. Обращает на себя внимание, чтоМЭ,
имеющие заслуженную репутацию токсичных (мышьяк, сурь-
ма, барий, бериллий, висмут, бор, свинец, таллий), содержатся
в норме в органах и жидкостях организма человека в значи-
тельно меньших количествах, нередко на несколько порядков
ниже, чем эссенциальные МЭ. В этот перечень не входят дру-
гие, несомненно, токсичные МЭ, которые по неизвестной пока
причине в единичных органах содержатся в относительно боль-
ших количествах. В частности, пока необъяснимо относительно
высокое содержание золота в головном мозге и в почках
(2,54 мкмоль/кг сухой массы). Также непонятно относительно
высокое содержание бора в почках (46,2 мкмоль/кг) и в пече-
ни (44,4 мкмоль/кг).
Относящийся к токсичным элементам кадмий обнаружива-
ется во всех исследованных органах, причем его концентрация
в почках необычно высока (0,59 ммоль/кг). Широко известная
из-за своей токсичности ртуть содержится во всех изученных
органах, причем в головном мозге ее концентрация достигает
0,014 ммоль/кг; еще выше концентрация этого МЭ в печени
(0,018 ммоль/кг). Содержание таллия, обладающего токсичес-
ким свойством, во всех органах находится почти на одном уров-
не (1,96 мкмоль/кг) и только в головном мозге увеличивается
до 2,44 мкмоль/кг. Содержание олова также необычно высоко
в головном мозге (16,8 мкмоль/кг) и на порядок превышает со-
ответствующие показатели в сердце и почках. Эти различия по-
ка необъяснимы.
Содержание МЭ в крови, по данным D. К. Kalani (1980),
как правило, очень невелико. Эти данные, характеризующие
37
норму, заслуживают пристального внимания при их сопостав-
лении с соответствующими показателями при МТОЗах. Попутно
заметим, что высокие или необычно низкие показатели содер-
жания МЭ в крови могут встречаться и при других заболе-
ваниях.
Важным показателем отравления ртутью может служить
содержание этого элемента в волосах. То же самое относится и
к никелю.
В исследовании В. А. Бацевича (1988) с помощью нейтрон-
но-активационного анализа проведено определение содержания
11 МЭ в волосах у представителей 17 этнотерриториальных
групп населения СССР в зонах, не подверженных техногенно-
му загрязнению. При этом установлено, что в биогеохимических
провинциях с избытком или недостатком определенных МЭ на-
блюдается соответственное изменение их содержания в воло-
сах. Отмечено повышенное поступление в организм человека
селена (Тува, Якутия, Чукотка), ртути (Чукотка), марганца
(Якутия). Кроме того, закономерные возрастные изменения со-
держания МЭ установлены для цинка и марганца в пубертат-
ном периоде.
D. К. Kalani (1980), как и многие другие авторы, различа-
ет эссенциальные, предположительно эссенциальные и физио-
логически инертные МЭ. Критериями необходимости исследуе-
мого МЭ для организма являются: закономерная реакция на
его добавку в состав пищи, возникновение дефицита МЭ при
устранении его из диеты, корреляция состояния дефицита МЭ с
субнормальным уровнем его концентрации в крови или в тка-
нях лабораторных животных. Эти данные в отношении ряда МЭ
отчасти устарели, так как в последние годы показано, что та-
кие МЭ, как фтор, бром, селен, молибден, в действительности
являются и эссенциальными для организма. Патогенное дей-
ствие фтора, селена и молибдена, содержащихся в высоких кон-
центрациях в воде или пище жителей определенных географи-
ческих зон, не противоречит их возможному положительному
эффекту в адекватных, как правило, очень низких дозах. Такие
потенциально токсичные МЭ, как медь, мышьяк и цинк, имею-
щие репутацию важных промышленных контаминантов, наря-
ду с этим являются в определенных дозах жизненно необходи-
мыми.
Действие МЭ на организм в большой степени зависит от на-
личия (обилия) внеклеточной жидкости в тканях, степени раз-
вития подкожной клетчатки, а также от содержания кальция в
костной ткани и от степени кальцификации других тканей.
В настоящее время еще отсутствуют или неполны обобщаю-
щие данные о содержании МЭ в различных органах и тканях
эмбрионов и плодов человека в пренатальном периоде. В про-
цессе онтогенеза определенные органы и ткани способны изби-
рательно концентрировать в своем составе те или иные МЭ.
Большинство исследователей объясняют это физиологической
ролью МЭ и специфической деятельностью органа. Так, в част-
38
ности, показано, что содержание меди, цинка, кремния, марган-
ца, алюминия в сером и белом веществе головного мозга по-
вышается к моменту рождения [Баженко Г. А., 1953]. У ново-
рожденных наибольшее количество таких МЭ, как медь, титан,
алюминий, кремний, содержится в зрительных буграх и продол-
говатом мозге. В зрелом возрасте некоторые МЭ, особенно медь
и титан, концентрируются в коре полушарий большого мозга.
В старческом возрасте отмечены обратные соотношения [Дель-
ва В. А„ 1973; Curzon G„ 1975; Greiner А., 1975].
Возрастная динамика концентрации некоторых МЭ в ЦНС
связана с особенностями обменных процессов, свойственных оп-
ределенному возрастному периоду и в первую очередь с изме-
нением интенсивности окислительных процессов [Райцес В. С.,
1981]. Особенно много МЭ накапливается в костях и зубах,
некоторые из них (фтор, стронций и др.) входят в структуру
апатита высокоминерализованных тканей организма и тем са-
мым изменяют их физико-химические свойства. Накопленные
данные свидетельствуют о том, что обмен МЭ в раннем воз-
расте менее совершенен, чем у взрослых. В частности, в экспе-
рименте на телятах показано, что повышение концентрации
цинка в рационе приводило к резкому увеличению его содержа-
ния в печени и поджелудочной железе, а также к перераспре-
делению его внутриклеточной локализации. В то же время у
взрослых животных отмеченные изменения не наблюдались
[Miller W. J., 1981].
Как у человека, так и у животных в период старения гомео-
статический контроль нарушается. При этом снижается и уро-
вень некоторых МЭ, что находится в соответствии с современ-
ными представлениями об адаптационно-регуляторной теории
старения. Согласно этой теории, старение приводит к затуха-
нию одних и появлению новых механизмов, обеспечивающих
жизнедеятельность организма в измененных условиях [Раец-
кая ГО. И., 1987].
Исследование содержания свинца в крови в онтогенезе по-
казало, что у детей дошкольного возраста оно примерно равно
содержанию этого МЭ у взрослых, в то время как у детей
школьного возраста его концентрация в крови значительно
меньше. При изучении пуповинной крови новорожденных Нью-
Йорка обнаружено, что содержание свинца составляет 0,39±
±0,02 мкмоль/л [Kaul В. et al., 1983]. Установлено, что для
детей европейских стран содержание свинца в крови считается
угрожающим здоровью, если оно превышает 1,2 мкмоль/л, для
взрослых мужчин эта концентрация составляет 1,93 мкмоль/л
и для женщин—‘ 1,45 мкмоль/л [Page R. А., 1984].
Нарушения в содержании МЭ в организме человека могут
прослеживаться на протяжении всей жизни. При этом как их
избыток, так и недостаток в ряде случаев сопровождаются оп-
ределенными морфофункциональными изменениями отдельных
систем и органов. В детском возрасте могут возникать и про-
являться врожденные пороки развития (при дефиците цинка,
39
меди, селена и др.), генетически обусловленные заболевания
(болезнь Вильсона — Коновалова, болезнь Менкеса). В пост-
натальном периоде встречаются алиментарный дефицит МЭ,
недостаточность их при ряде заболеваний, нарушающих усвое-
ние МЭ, а также в результате лечебных мероприятий. Конкрет-
ные формы указанных заболеваний описаны в соответствую-
щих разделах.
2.2. Участие микроэлементов
в гомеостатических функциях организма
Организм здорового человека обладает достаточно четкой
саморегулирующей системой гомеостаза, в которой определен-
ную роль играют и МЭ. Их уровень в крови и тканевой жидко-
сти подчиняется определенным физиологическим закономерно-
стям. Для большинства МЭ основными регуляторными меха-
низмами гомеостаза являются процессы всасывания, преиму-
щественно из желудочно-кишечного тракта, а также экскреция
их с мочой и калом.
О суточном балансе МЭ условного человека можно судить
по данным табл. 3, составленной по материалам Междуна-
родной комиссии по радиологической защите (МКРЗ, Публика-
ция 23, 1977]. Как видно из этой таблицы, МЭ поступают в ор-
ганизм из пищи и в значительно меньшем количестве — из воз-
духа. Выделяются МЭ из организма с мочой, калом и в не-
большом количестве с потом, волосами, ногтями и менструаль-
ной кровью.
Не останавливаясь в деталях на этих процессах, необходи-
мо, однако, отметить, что для отдельных МЭ существуют опре-
деленные особенности, которые оказывают заметное влияние
на гомеостаз. Так, в частности, известно, что для фтора не име-
ют значения пути его введения в организм (ингаляционный, али-
ментарный, парентеральный), так как этот МЭ свободно (без
носителя) проникает в кровь и быстро элиминируется с мочой,
а также селективно абсорбируется в высокоминерализованных
тканях (кости, зубы, петрификаты). Для других же М.Э (цинк,
медь, кадмий, ртуть и др.) процесс всасывания в желудочно-ки-
шечном тракте зависит от наличия специальных носителей-—
металлотионеинов.
Включение металлопротеинов тканями осуществляется по-
средством фиксации их на специфических рецепторах мембран,
в комплексе с которыми они как бы «стекают» в окаймленные
везикулы и поступают в клетки, где и разрушаются лизосомаль-
ными ферментами, а металл восстанавливается и используется
в обмене. Такой механизм, в частности, доказан для трансфер-
рина [Gachelin G., 1980] и церулоплазмина [Kostsak S. A., Be-
arn А. С., 1978]. Есть достаточно оснований полагать, что
именно с помощью этого механизма происходит поступление в
клетку большинства жизненно необходимых МЭ.
Гомеостаз МЭ тесно связан с селективной реабсорбцией или
40
секрецией в тканевую жидкость необходимых количеств воды,
электролитов и неэлектролитов. Известно, что плазматическая
мембрана всех эпителиоцитов по характеру ее проницаемости и
содержанию транспортных белков четко дифференцируется на
апикальную и базолатеральную части. В связи с этим ее про-
ницаемость обусловлена изменением числа ионных каналов и
содержанием транспортных протеинов. Это происходит благо-
даря включению в нее цитоплазматических везикул, мембраны
которых содержат ионные каналы или транспортные белки. По-
добная адекватная реакция на изменение тканевой (окружаю-
щей) среды является одним из многочисленных процессов, уча-
ствующих в поддержании гомеостаза [Stanton В. А., 1984].
Имеющиеся различия в концентрации макроэлементов и МЭ
в межклеточной жидкости и в самой клетке являются объектив-
ным доказательством определенных закономерностей их рас-
пределения. Не подлежат сомнению, что наряду с этим сущест-
вует и случайное загрязнение клеток МЭ.
О неравномерности распределения жизненно необходимых
МЭ в различных органах, а также в структурных компонентах
клетки, следовательно, и об их биологической роли можно су-
дить на примере марганца [Hiromu Sakudai, 1986]. Так, уста-
новлено, что в норме максимальное содержание этого МЭ оп-
ределяется в щитовидной железе, печени, гипофизе и надпо-
чечниках. Максимальная концентрация марганца в указанных
органах может свидетельствовать о том, что они являются в оп-
ределенной степени функциональными тканевыми депо этого-'
МЭ. При нагрузке организма животных марганцем (например,,
МпСЬ) еще больше увеличивается его концентрация в подже-
лудочной железе и печени, принимающих участие в процессах
элиминации этого МЭ.
Фундаментальное исследование экскреции 18 металлов из
организма крыс было проведено A. Gregus и соавт. (1986). Оно
дает достаточно четкое представление о естественных путях и
скорости элиминации внутривенно введенных МЭ. Этими авто-
рами показано, что общая экскреция (с калом и мочой) была
относительно быстрой. Так, через 4 дня для кобальта, серебра
и марганца она составила более 50% введенной дозы, для ме-
ди, таллия, висмута, свинца, цезия, золота, цинка, ртути, селе.-
на и хрома была в пределах 50—20%, а для мышьяка, кадмия;,
железа, метилртути и олова она была менее 20%. Наряду с
этим показано, что серебро, марганец, медь, таллий, свинец,
цинк, кадмий, железо и метилртуть в основном выделялись с
калом, тогда как кобальт, цезий, золото, селен и хром выде-
лялись преимущественно с мочой; такие МЭ, как висмут, ртуть,
мышьяк и олово, в равной степени экскретировались обоими
путями.
Поступление в желчь таких МЭ, как серебро, мышьяк, мар-
ганец, медь, селен, кадмий, свинец, висмут, кобальт, а также
метилртуть, является определяющим фактором для их экскре-
ции с калом. В частности, установлено, что за 2 ч после введе-
41
ния этих МЭ в кровь 45% серебра поступали в желчь. Кон-
центрация МЭ в желчи не зависела от дозы внутривенно вве-
денного серебра, мышьяка, марганца, висмута, метилртути, зо-
лота, цезия, таллия и олова. Однако она заметно возрастала
при увеличении дозы кадмия, цинка и железа. Это, по-видимо-
му, является результатом избыточного накопления ряда МЭ в
печени (кадмий, цинк) вследствие связывания их определенны-
ми компартментами гепатоцитов или же вне печени (железо).
Сравнительный анализ величин экскреции МЭ с желчью и ка-
лом свидетельствует, что мышьяк и селен повторно абсорбиру-
ются кишечником, в то время как таллий и цинк попадают в
кал не через желчь. Наибольшая концентрация большинства
изученных МЭ наблюдалась в печени и почках. Однако.не
установлено прямой корреляции между распределением МЭ в
этих органах и основным путем их выведения из организма.
Важную роль в экскреторных системах играет различие в
скорости выделения молекул — носителей МЭ. В частности, из-
вестно, что крыса ежечасно выделяет в желчь 5—10 мг/кг бел-
ка [La Russo N. F., 1984; Karlowitz N. et al., 1985]. В то же вре-
мя в моче эти вещества практически не определялись. Кроме
того, F. М. Zuyderhoudt (1985) показал, что предпочтительная
экскреция железа в виде ферритина и гемосидерина в желчь
происходит за счет лизосомального экзоцитоза.
Хорошо известно, что в пищеварительном тракте имеются
сложные взаимоотношения между всеми компонентами пищи
(органической и неорганической природы) и поэтому необходи-
мо рассматривать потребность организма в определенном МЭ с
учетом содержания и других элементов в диете. Для полноцен-
ной работы гомеостатической системы организма важное зна-
чение имеет так называемая константа устойчивости хелатов,
образующихся в пищеварительном канале. Она должна быть ни-
же, чем у комплексов МЭ с биологически активными вещества-
ми, которые вовлекают МЭ в промежуточный обмен [Раец-
кая Ю. И., 1979; Miller W., 1975, 1981].
В табл. 5 приведены данные о дифференциальной возмож-
ности всасывания химических элементов периодической системы
Менделеева в желудочно-кишечном тракте. При этом обраща-
ет на себя внимание, что все щелочные элементы первой, седь-
мой и нулевой групп, а также все элементы первого и второго
периодов, за исключением бериллия, обладают максимальной
всасываемостью (1) в желудочно-кишечном тракте. Всасывае-
мость коррелирует с возрастом и обычно приближается к 1 у
молодняка животных и детей. Кроме того, на ее величину ока-
зывают влияние физико-химические характеристики самого эле-
мента, она снижается с увеличением атомной массы элемента и
его металлических свойств. Всасывание в желудочно-кишечном
тракте можно увеличить на 1-—2 порядка, вводя комплексообра-
зователи, в частности, такие как ЭДТА и ДТПА.
Плохая всасываемость тяжелых металлов объясняется их
способностью образовывать коллоиды и нерастворимые комп-
42
Таблица 5 сазатели всасывания химических элементов в желудочно-кишечном тракте в зависимости от их места в периодической системе [Москалев Ю. И., 1985] Группа элементов VIII 1 0 Не 1,0 Ne 1,0 Аг 1,0 Кг 4,0 Xe 1,0 с о JI Се 1 Pr 1 Nd | Pm | Sm | Eu | Gd | Tb 1 Dy | Ho | Er | Tm | Yb I Lu ’ -« — 0,0002—0,005 „ T h„ I „ I a ,1 U Np 1 Pu 1 Am | Cm | Bk | C£ | Es I Fm | Md | No | Lr | 0,0002 1 0,0003 | 0,2 0,02 И 0,0005 . — — “*•
Ni 0,05 £ 1 £
Со 0,3 Rh (0,05) llr 0,01
Ru 0,05 C/1 о
VII | [J-, о_ С1 1,0, 1 Мп 0,1 Вг 1,0 CJ 00 H o' Re (0,8) At (1,0)
> О О GO оо Сг 0,1 Se 0,9 1 Mo 0,85 Те 0,25 W 0,1 Po 0,1
> CU<4 V 0,02 As 1,0 Nb 0,01 Sb 0,1 Ta 0,0001 Bi 0,05
> Si 0,33, Ti 0,03 Ge 0,01 Zr 0,002 Sn 0,2 Hf 0,0001 Pb 0,2 3 I 1
soo'o IV 1 Sc 0,001 Ga 0,001 Y 0,0005 In 0,02 La* 0,0005 TI 0,45 j Ac** 0,0001
Be 0,002 Mg 0,3 Ca 0,3 Zn 0,5 Sr 0,3 Cd 0,1 Ba 0,1 Hg 0,4 Ra 0,2.
- н 1 (1,0) Li 1,0 .Na l,o ! К 1,0 Си 0,5 >Rb 1,0 Ag 0,05 Cs 1,0 Au 0,1 £ °
О С Период 1 т—< сч со to CD b-
4&
лексные соединения. Важно иметь в виду, что большинство жиз-
ненно необходимых макроэлементов и МЭ практически полно-
стью (Н, С, N, О, F, Cl, I, К, Na, Р, Мо) или достаточно хоро-
шо (Mg, Са, Р, S, Со, Fe, Си, Zn, Se) абсорбируются в желу-
дочно-кишечном тракте [Москалев Ю. И., 1985].
Нередко встречающиеся патологические изменения процес-
сов пищеварения и всасывания могут явиться основой наруше-
ний обмена МЭ [Bunch J. et al., 1973; Zalewski H., 1973]. Об
этом, в частности, могут свидетельствовать эксперименты на
крысах по изучению влияния ваготомии на экскрецию цинка
клетками Панета1 тонкой кишки в условиях функциональной
нагрузки [Куликов О. В., 1982]. Отмечено увеличение числа
клеток Панета с большим и максимальным, а также с низким и
умеренным содержанием цинка. Динамика выведения цинка
тонкой кишкой у ваготомированных животных характеризова-
лась одновершинной кривой с минимумом через 1,5 ч после
функциональной нагрузки. Через 7 ч этот показатель прибли-
жался к контрольному, а через 12 ч превышал его. При этом в
норме у крыс при парентеральной нагрузке цинком наблюдает-
ся через 3,5 ч накопление его в клетках Панета, затем проис-
ходит частичное выделение (7 ч), которое сменяется дальней-
шим накоплением этого МЭ.
Как уже отмечалось ранее, уровень абсорбции цинка в же-
лудочно-кишешном тракте связан с содержанием цинксвязыва-
ющего лиганда. Однако, по данным С. В. Ammorinan и соавт.
(1983), при избытке этого МЭ в рационе он хорошо усваивает-
ся и в отсутствие лиганда. Исходя из этого, повышенное со-
держание МЭ в рационе сопровождается увеличением их кон-
центрации в тканях. Данный факт, по мнению М. Kircghesser
и соавт. (1981), является более объективным показателем обес-
печенности организма МЭ, чем данные так называемых балан-
совых опытов.
При избыточном поступлении МЭ вступает в действие си-
стема элиминации. В частности, может происходить блокиро-
вание процессов их всасывания в желудочно-кишечном тракте
и последующее выведение с калом. Всосавшийся в кровь избы-
ток МЭ выводится с мочой, желчью, потом, молоком, а также
откладывается в депо. В связи с этим становится очевидно, что
дефект определенного звена системы, обеспечивающей микро-
элементный гомеостаз, приводит к тому, что в организме может
возникнуть недостаток или избыток МЭ, сопровождающийся
соответствующими клиническими симптомами и развитием за-
болевания.
Таким образом, существующее многообразие путей регуля-
ции микроэлементного гомеостаза должно быть в поле зрения
исследователя. Однако значение каждого из них неодинаково
1 В соответствии с международной гистологической номенклатурой клетки
Панета следует называть энтероцитами с ацидофильными гранулами. — При-
ме ч. р е д.
44
для различных МЭ. Так, для меди, кобальта, железа, цинка и
молибдена основным путем регуляции является изменение уров-
ня абсорбции, в то время как для кадмия и йода наиболее
важный путь их регуляции в организме — экскреция с мочой.
Последний путь для большинства МЭ составляет не более 1—
4% от их количества, выделяемого с калом [Раецкая Ю. И.,
1987].
Отдельные авторы [Adams Е., 1975] придают существенное
значение регуляции микроэлементного гомеостаза путем сек-
реции МЭ с молоком. Показано, что она вариабельна и изме-
няется не только в зависимости от стадии лактации, но и от
уровня поступления МЭ в организм. В то же время содержание
йода и цинка в молоке, по мнению М. Е. Kirchgessner и соавт.
(1978), является важным и надежным тестом обеспеченности
организма этими МЭ.
Пусковым механизмом абсорбции ряда МЭ (железо, цинк
и др.) в желудочно-кишечном тракте является снижение их
концентрации в тканевых депо. Однако путь передачи инфор-
мации от тканей к эпителиоцитам слизистой оболочки желудоч-
но-кишечного тракта еще неизвестен. Логично, что на роль та-
ких передатчиков информации могут претендовать уровень кон-
центрации белков-носителей (лигандов) МЭ в крови и ткане-
вой жидкости, опосредуемый через клеточный компонент им-
мунной системы, а также увеличенное число свободных рецеп-
торов к этим лигандам.
Интересную мысль высказывает Ю. И. Раецкая (1987).
В связи с тем что пики анаболизма МЭ находятся в соответст-
вии с высотой активности определенного звена эндокринной си-
стемы, в частности с высокими уровнями в организме сомато-
тропных, тиреоидных и половых гормонов, можно думать о ве-
дущей роли этой системы в поддержании микроэлементного го-
меостаза. Наиболее изучена взаимосвязь между уровнем со-
держания в организме йода и активностью щитовидной железы,
т. е. показана четкая система обратной связи. При этом нельзя
забывать о центральных механизмах регуляции гомеостаза.
Следовательно, микроэлементный гомеостаз — это частная фор-
ма общей гомеостатической системы организма.
2.3. Адаптация организма в экстремальных условиях
и влияние ее на содержание микроэлементов.
Акклиматизационный дефицит микроэлементов
Адаптация — это процесс приспособления живых организмов
к тем или иным условиям существования. Молекулярные меха-
низмы этого процесса интенсивно изучаются. Так, Ф. 3. Меер-
сон (1973, 1981) отмечает, что повышение потенциала фосфо-
рилирования в клетке сопровождается увеличением синтеза
ДНК и белка. В свою очередь Л. Е. Панин (1978, 1983) боль-
шую роль придает активации нейроэндокринных механизмов,
в частности повышенному синтезу и выбросу в кровь кортико-
45
стероидов и катехоламинов с одновременным ингибированием
синтеза инсулина, что обеспечивает на уровне клеточных мемб-
ран переключение с углеводного типа обмена на жировой как
наиболее эффективный не только в энергетическом, но и в син-
тетическом плане.
Наши исследования [Авцын А. П. и др., 1985] показывают,
что одним из необходимых условий долговременной адаптации
организма к экстремальному воздействию внешней среды явля-
ется модификация липидного состава биомембран, направлен-
ная на изменение информационных и энергетических потоков,
а также функциональной активности клетки. Следовательно,
в основе этой адаптации к различного рода чрезвычайным разд-
ражителям, в том числе и избыточному поступлению в организм
МЭ, лежат не только прирост массы функционирующих струк-
тур, но и их адекватные качественные изменения. Сохранение
достаточного морфофункционального резерва снижает воз-
можность развития дизадаптационных повреждений. В нераз-
рывной связи со сказанным находится выдвинутое Д. С. Сар-
кисовым (1977) положение о временной адекватности реакций
организма на действие повреждающего химического агента.
Установлены закономерные корреляции между климатиче-
скими и ландшафтно-биогеохимическими особенностями от-
дельных регионов страны, с одной стороны, и распространени-
ем ряда заболеваний и синдромов — с другой. Так, природно-
биогеохимические условия Севера, являясь экстремальными,
предъявляют к организму повышенные требования в смысле
его «биосоциальной платы» за достижение адаптированности.
Последняя сопровождается значительной перестройкой обмен-
ных процессов. В частности, в условиях Севера происходят су-
щественные изменения микроэлементного гомеостаза в орга-
низме человека и животных.
Медико-географические исследования, проведенные нами и
другими авторами в различных регионах Севера, показали, что
здесь четко выявляются заболевания (кариес зубов) и синдро-
мы (железодефицитные состояния), обусловленные дефицитом
определенных жизненно важных МЭ, например фтора и железа.
До последнего времени установленный дефицит этих МЭ рас-
сматривался односторонне как недостаточное поступление их с
водой и пищевыми продуктами. Однако исследованиями ряда
авторов [Алексеев В. П. и др., 1976; Панин Л. Е., 1980, и др.]
убедительно показано, что в условиях холодового воздействия
резко нарушается микроэлементный и электролитный гомео-
стаз. Метаболическая перестройка в организме при холодовой
адаптации настолько существенна, что это позволяет нам гово-
рить о развитии акклиматизационного дефицита МЭ. Так,
у крыс, получавших воду с концентрацией фтора 15 мг/л и под-
вергавшихся 5-месячному охлаждению, и у животных, нахо-
дившихся в комфортных температурных условиях и потребляв-
ших воду с концентрацией фтора 0,3 мг/л, содержание этого<
МЭ в зубах и костях — органа-накопителях — существенно не-
46
различалось [Марачев А. Г., Жаворонков А. А., 1987; Тед-
дер Ю. Р., Жаворонков А. А., 1988].
В неразрывной связи с выявленными нарушениями фторово-
го статуса организма в условиях холодового воздействия нахо-
дится и высокая встречаемость железодефицитных состояний
человека на Севере, так как известно положительное влияние
фтора на кроветворение за счет улучшения усвоения железа.
В частности, у животных, получавших питьевую воду с опти-
мальной концентрацией фтора (1 мл/л), отмечено лучшее
усвоение железа и повышение его уровня, а также меди в кро-
ви [Абросимов А. И., 1975; Книжников В. А., 1975; Messer Н.Н.,
1984]. Наряду с этим нашими исследованиями показано, что вы-
сокая встречаемость железодефицитных состояний у жителей
Севера обусловлена повышенной потребностью организма в же-
лезе вследствие усиления метаболических (анаболических и ка-
таболических) процессов под воздействием экстремальных фак-
торов внешней среды, особенно холода [Авцын А. П. и др.,
1985].
Аналогичный эффект был отмечен Л. Е. Паниным (1980),
изучавшим содержание некоторых микроэлементов и МЭ (цин-
ка, меди, железа, марганца, магния, натрия, калия и кальция)
в крови рабочих Норильского медно-никелевого комбината. Так,
содержание в крови цинка, магния, натрия и кальция оказалось
сниженным. В то же время концентрация меди в крови была
повышенной, что объясняется спецификой производственных
условий, избыточным экзогенным поступлением указанного МЭ.
При анализе этих данных следует принять во внимание физио-
логический антагонизм меди и цинка.
Таким образом, физиологические механизмы адаптивной пе-
рестройки организма в экстремальных условиях Севера могут
закономерно приводить к сдвигу микроэлементного гомеостаза
и возникновению акклиматизационного дефицита МЭ. Это в
свою очередь влечет за собой повышенную потребность орга-
низма в жизненно важных МЭ. Акклиматизационный дефицит
МЭ, несомненно, играет важную роль в развитии и широком
распространении па Севере как скрытых, так и выраженных
форм МТОЗов, обусловленных недостатком или дисбалансом в
первую очередь жизненно необходимых МЭ.
Вводя понятие об акклиматизационном дефиците МЭ в экст-
ремальных условиях обитания, мы относим его только к физио-
логическим уровням поступления их в организм. В случаях их
избыточного поступления, очевидно, правильнее говорить об ак-
климатизационной «блокаде» процессов накопления МЭ в ор-
ганах-мишенях или об акклиматизационном сдвиге их метабо-
лизма [Теддер Ю. Р., Жаворонков А. А., 1988]. К проявлениям
акклиматизационного дефицита МЭ, очевидно, относятся и
большие потери цинка с потом в условиях высокой температу-
ры внешней среды, что, вероятно, является одним из факторов
патогенеза болезни Прасада.
Глава 3
СТРУКТУРНЫЕ ОСНОВЫ ВСАСЫВАНИЯ
МИКРОЭЛЕМЕНТОВ
3.1. Абсорбция микроэлементов в желудочно-кишечном тракте.
Мембранное пищеварение. Особенности
метаболизма микроэлементов. Ультраструктурные основы
кишечного всасывания
Общая характеристика абсорбции металлов, МЭ и радио-
нуклидов приведена в книге S. С. Skoryna и D. Waldron-Ed-
ward (1971). М. Freeman и соавт. (1971) применили для иссле-
дования тонких механизмов всасывания карбоната железа эн-
тероцитами метод электронной микроскопии и показали, что
железо расщепляется до мицелл на микроворсинках, затем
транспортируется в цитоплазму и концентрируется в зоне тер-
минальной сети, а также в эндоплазматической сети в виде фер-
ритина. Разработанные данные о всасывании МЭ принадлежат
ряду авторов. Цинк изучали М. М. Меркулова и соавт. (1970),
железо — М. М. Щерба (1977), медь — М. Kirhgessner и
соавт. (1975). Стронций проходит пищевод и желудок, почти
не всасываясь. Быстрая и интенсивная абсорбция этого МЭ от-
мечается в двенадцатиперстной кишке. Однако основным мес-
том его поглощения, по-видимому, являются тощая и подвздош-
ная части тонкой кишки [Cramer С. F. et al., 1959]. У цыплят
наиболее эффективное всасывание стронция наблюдается в две-
надцатиперстной кишке. Следует отметить, что места всасыва-
ния в кишечнике кальция и стронция совпадают.
Сравнительно немногочисленные данные о резорбции МЭ в
кишечнике, полученные в первой половине XX века, основыва-
лись на устаревшей ныне двухзвенной схеме ассимиляции пище-
вых веществ, которая включала в себя этап полостного (или
дистантного) пищеварения и сам процесс всасывания. В 60-х
годах XX века общепринятой стала трехзвенная схема пищева-
рения, которая включает в себя: 1) полостное пищеварение,
2) мембранное пищеварение и 3) всасывание. Внеклеточное пи-
щеварение обеспечивает начальный этап переваривания пище-
вых частиц.
Внутриклеточное пищеварение делится на два этапа:
1) транспорт молекул через клеточные мембраны с последую-
щим расщеплением их ферментами цитоплазмы и 2) лизосо-
мальное переваривание различными гидролазами —фосфата-
зами, глюкозидазами, протеазами, липазами и др. Оно являет-
ся широко распространенным биологическим актом, который
входит в общее понятие «эндоцитоз» и играет большую роль-
уже на самых ранних этапах постнатального развития. После-
дующие работы выяснили, что, кроме названных выше этапов
48
а б в
Рис. 2. Основные типы пищеварения [Уголев А. М., 1985].
а — внеклеточное пищеварение; б — внутриклеточное цитоплазматическое; в — внутрикле
точное вакуоляриое, или экстраплазматическое, связанное с фагоцитозом и пиноцитозом;
г — мембранное пищеварение. 1внеклеточная среда; 2 — внутриклеточная среда; 3 —•
пищеварительная вакуоль; 4—лизосома; 5 — ядро клетки; 6 — клеточная мембрана; 7 —
внутриклеточные ферменты; 8 — частицы пищи.
г
ассимиляции, крайне важное значение имеет мембранное пище-
варение (рис. 2), в раскрытии механизмов которого основную-
роль сыграли многолетние исследования А. М. Уголева (1967,
1970, 1972, 1977, 1985, 1986).
Мембранное пищеварение обеспечивается определенными
ферментами. Они имеют двоякое происхождение: 1) часть из
них, главным образом панкреатические ферменты, адсорбиро-
вана на химусе и 2) собственно кишечные ферменты, которые
синтезируются энтероцитами и инкорпорированы в состав их
липопротеиновой мембраны. За счет мембранного пищеварения
осуществляются промежуточные и заключительные стадии гид-
ролиза пищевых веществ, а также сопряжение конечных этапов
переваривания и начальных этапов всасывания. По А. М. Уго-
леву (1982), это обеспечивается своеобразным пищеварительно-
транспортным конвейером.
Всасывание начинается в ротовой полости, имеет значитель-
ную выраженность в желудке, но достигает наибольшей интен-
сивности в тонкой кишке. Поступления пищевой массы из же-
лудка в кишечник сопровождается изменением величины pH в
результате действия сока поджелудочной железы и желчи. По-
видимому, именно такая среда способствует более активному
всасыванию многих компонентов пищи, в том числе и МЭ.
Для обеспечения тонко отрегулированного процесса всасы-
вания существует сложный анатомо-физиологический аппарат,
схематическое строение которого авторы представляют различ-
ным образом. Наиболее известна и проста схема С. С. Booth
(1967), приведенная А. М. Уголевым (1982). К сожалению, она
далека от реальных анатомических отношений и содержит дан-
ные о всасывании только одного МЭ (железа) и двух макро-
элементов (магния и кальция), которые, согласно этой схемы,
всасываются в верхнем отделе желудочно-кишечного тракта.
4—568 49>
I
Рис. 3. Схематическое изо-
бражение некоторых отделов
пищеварительной системы
человека [Михайлов С. С.,
1982].
Сплошным черным цветом
изображена главная зона
всасывания МЭ в двенадца-
типерстной кишке, серым
цветом обозначены зоны ме-
чее активного всасывания
МЭ в желудке и тонкой
кишке. Сфинктеры пищева-
рительного тракта:
1 — пищеводно-желудочный; 2 —
привратник; 3 — сфинктер пу-
зырного протока (Люткеиса);
4 — сфинктер общего печеночно-
го протока (Морисси); 5 — об-
щего желчного протока; 6 — пе-
ченочно-поджелудочной ампулы;
7 — протока поджелудочной же-
лезы; 8 — терминальной ч-асти
подвздошной кишки (сфинктер
Варолия): 9 — сфинктер Бузи;
10 —сфинктер Гирша; 11 —
сфинктер Кеннона — Бема; 12 —
сфинктер Херста; 13 — сфинктер
Кеннона; 14 — сфинктер Пай-
на — Штраусса; 15 — ссЬннктер
Балли; 16 — сфинктер Росси —
Мютье; 17 — сфинктер Пирого-
ва — Берна; 18 — третий сфинк-
тер прямой кишки (сфинктер
Хепфнера); 19 — внутренний
сфинктер заднего прохода; 20 —
наружный сфинктер заднего
прохода.
.Для большего приближения к истинным анатомическим отно-
шениям мы приводим рисунок С. С. Михайлова (1982). На нем
двенадцатиперстная кишка и верхние отделы тощей кишки не-
сколько распрепарованы, что дает возможность более нагляд-
но нанести на представленные контуры условные обозначения
главных зон всасывания МЭ. Выбор этого схематического ри-
сунка объясняется его обобщающим характером. Как видно из
рис. 3, весь пищеварительный тракт разделен с помощью точно
установленных сфинктеров на определенные анатомо-физиоло-
гические сегменты. В этих сегментах у здорового человека име-
ет место физиологическая ретенция желудочно-кишечного со-
держимого, которая происходит через определенные сроки.
Каждый сегмент пищеварительного тракта обладает присущей
ему системой ангио- и лимфоциркуляции, иннервации, а также
ферментного и нейрогуморального обеспечения [Морозов И. Н.
и др., 1988]. В последние годы выяснено, что в стенке кишечни-
ка, в частности в двенадцатиперстной кишке, находится мощ-
ный аппарат иммунокомпетентных структур [Сапин М. Р.,
:50
1987], принимающих участие в реакциях гиперчувствительностк
[Гайдар Ю. А. и др., 1984].
В тонкой кишке не только завершается переваривание пи-
щи, поступающей из желудка, но и избирательно всасываются
в кровь и лимфу продукты переваривания. Эффективное выпол-
нение функции всасывания требует обширной поверхности, по-
крытой энтероцитами, осуществляющими всасывание веществ.
Существуют специальные анатомические аппараты, способст-
вующие увеличению всасывающей поверхности. Это циркуляр-
ные, или спиральные, складки, называемые клапанами Керкрин-
га. Они наиболее развиты в проксимальном отделе двенадцати-
перстной кишки. Всасывающую поверхность слизистой оболочки
увеличивает огромное количество мелких листовидных выпячи-
ваний, называемых кишечными ворсинками. Высота этих ворси-
нок по направлению от привратника к илеоцекальной заклонке
постепенно снижается.
В еще большей степени увеличивает всасывающую поверх-
ность тонкой кишки множество микроворсинок, расположенных
на апикальной поверхности эпителиоцитов. Большинство эпи-
телиоцитов (энтероцитов) тонкой кишки представляют собой
высокоспециализированные цилиндрические клетки с широкой
исчерченной каемкой. Именно эти клетки называются всасыва-
ющими (рис. 4—6). Кроме этих клеток, в эпителии ворсинок
имеются бокаловидные клетки, продуцирующие слизь, и нейро-
эндокринные клетки, на которые приходится менее 0,5% от об-
щего числа элементов эпителиального пласта. Особый вид эпи-
телиоцитов— это клетки Панета, содержащие крупные грану-
лы зимогена и обладающие специфическим свойством накопле-
ния и выделения цинка (рис. 7). Именно эти клетки выделяют
лизоцим. Особый вид энтероцитов — это олигомукозные клет-
ки, содержащие слоистые глобулы. Относительно немногочис-
ленные «пещеристые клетки» содержат цитоплазматические
«кавеолы», образованные инвагинациями апикальной плазма-
леммы, а также длинные пучки прямых филаментов, которые тя-
нутся от стержня микроворсинки и глубоко проникают в цито-
плазму. Эти элементы иногда называют «щеточными», или «хо-
холковыми», клетками. Между клетками эпителиального плас-
та мигрируют лимфоциты и другие типы лейкоцитов, в том чис-
ле особый клеточный тип, получивший название «глобулярный
лейкоцит». Такие клетки могут быть обнаружены в норме, но
чаще они встречаются у больных, зараженных паразитами.
Глобулы этих лейкоцитов представляют собой крупные включе-
ния. Ядро их несколько уменьшено. Приведенный краткий пе-
речень клеточных вариантов эпителиоцитов и лейкоцитов тон-
кой кишки имеет существенное значение для понимания функ-
ции всасывания и переваривания. Сложный процесс резорбции
МЭ обеспечивается именно этим клеточным ансамблем. Точные
механизмы, при помощи которых МЭ всасываются в энтероцит,
не выяснены. Т. М. Сох и М. W. O’Donnell (1981) в экспери-
менте на животных показали, что мембрана с исчерченной ка-
4* 5t
Рис. 4. Схематическое изображение ультраструктуры энтероцита при голода-
нии (А) и интенсивном всасывании липидов (Б) [Cardcll R. et al., 1967].
емкой имеет по крайней мере два типа участков, связывающих
железо и цинк. Можно полагать, что в недалеком будущем бу-
дут открыты ультраструктурные механизмы селективного вса-
сывания и элиминации МЭ энтероцитами. Большие перспекти-
вы в этом отношении имеет прицельное изучение строения, хи-
мизма и функции исчерченной каемки.
Характеризуя преимущественные места всасывания МЭ в
пищеварительном тракте, необходимо подчеркнуть неполноту и
некоторую противоречивость данных литературы. Многие МЭ
всасываются в нескольких отделах пищеварительной трубки.
Однако в большинстве случаев авторы все же называют наряду
с добавочным, или второстепенным, главное место всасывания.
Наиболее точные данные получены на животных, которым вво-
дили меченые изотопы МЭ. Очень полезным оказался метод
52
Рис. 5. Абсорбция железа в энтероцитах двенадцатиперстной кишки человека.
Сидерофильные энтероциты. Сидеросомы показаны стрелками. X13 900. При
большем увеличении они изображены в правом верхнем углу. Х34 300 [Parm-
ley Р. Т. et al., 1984].
изолированных кишечных петель. Однако эксперименты с пе-
роральным введением меченых МЭ имеют еще большую дока-
зательность. Эти опыты позволили выяснить важный феномен
повторного всасывания (реабсорбции) МЭ в одном и том же
месте или в нижележащих отделах кишечника.
Всасывание стронция происходит в тощей и подвздошной
кишке. Здесь же наблюдается абсорбция кальция, которая име-
ет проксимально-дистальный градиент (в желудке — 2%, в две-
53
Рис. 6. Микроворсинки и апикальная часть сидерофильного энтероцита две-
надцатиперстной кишки человека с всосавшимися депозитами железа. X37 500
[Parmley Р. Т. et al., 1984].
надцатиперстной кишке—15%, в тощей — 23% и в подвздош-
ной— 60%). Магний всасывается по всей длине тонкой кишки,
а у крыс преимущественно в подвздошной кишке. У человека с
калом выделяется 15—25% парентерально введенной дозы маг-
ния [Lengemann F. W., 1959].
Всасывание цинка происходит в верхнем отделе кишечника,
причем в двенадцатиперстной кишке поглощается его основная
масса (40—45%), а в тощей и подвздошной — от 15 до 21%.
В желудке и прямой кишке всасывается только 1—2% этого
важнейшего МЭ. Экскреция цинка происходит на всем протя-
жении желудочно-кишечного тракта. Однако особенно интенси-
вен этот процесс (80%) в тонкой кишке [Methfessel А. Н.,
Spencer Н., 1973]. При пероральном введении радиоактивного
цинка (65Zn) показано, что около 70% поступившей в организм
дозы обнаруживается в кале и только 0,3% —в моче [Feas-
ter J. Р. et al., 1954]. Есть основания полагать, что в тонкой
кишке и особенно в клетках Панета сосредоточен и наиболее
развит регуляторный аппарат поглощения и выведения цинка,
активность которого обеспечивается вегетативной нервной си-
стемой. В пользу этого говорят данные Ю. К. Елецкого и
А. Ю. Цибулевского (1989), обнаружившие изменение выделе-
ния цинка после перерезки блуждающего нерва.
Своеобразно всасывается медь. Основная масса ее абсор-
бируется в желудке (52%), в двенадцатиперстной (16—21%) и
в тощей 10%) кишке. По данным G. Cartwright и соавт.
(1960), из 2—5 мг суточной дозы меди, получаемой взрослым
54
Рис. 7. Цинкпоглощающие и цинквыделяющие клетки Панета слизистой обо-
лочки кишечника (препарат С. П. Луговского).
а — ШИК-реакция и альциановой синий. Х400; б — окраска гематоксилином и эозином,
ХЮОО.
человеком с пищей, 0,6—1,6 мг, или 32%, всасывается в желу-
дочно-кишечном тракте. По данным этих же авторов, с желчью
выделяется 0,5—1,3 мг меди в сутки, 0,1—0,3 мг — слизистой
оболочкой кишечника (из крови) и всего 0,01—0,06 мг экскре-
тируется с мочой.
Железо в отличие от меди всасывается преимущественно в
двенадцатиперстной кишке, меньше в начальных отделах то-
щей кишки. Это впервые показал S. Granick (1946) путем опре-
деления ферритина, который образуется в эпителиоцитах тон-
кой кишки из всосавшегося здесь железа. Скорость абсорбции
железа в проксимальных отделах тонкой кишки в 7 раз выше,
чем в дистальном. По мнению отдельных авторов, всасывание-
железа является единственным регуляторным механизмом,,
обеспечивающим постоянство содержания этого МЭ в организ-
ме в определенные этапы его индивидуального развития ГЩер-
ба М. М„ 1977].
Основной путь поступления фтора в организм — это желу-
дочно-кишечный тракт. Хотя всасывание фтора начинается в же-
лудке, однако его наибольшая абсорбция происходит в тонкой
кишке. Максимальная концентрация фтора в крови после перо-
рального введения наблюдается через 60 мин [Cormis L. D.„
1975] и составляет 80% от введенной дозы. Считается, что фтор
быстро абсорбируется в результате пассивной диффузии по гра-
диенту концентрации [Messer Н. Н., 1984]. На скорость и сте-
пень абсорбции оказывают существенное влияние количество
поступивших фторидов, их растворимость, особенности питания
и физиологическое состояние организма. Так, показано, что 90—
97% фтора питьевой воды всасываются, а фтор пищи абсорби-
руется медленнее и меньше на 20% [Габович Р. Д., МинхА. А.,.
1979]. Фтор может абсорбироваться также в дыхательном трак-
те. Этот путь поступления бывает значительным у людей, нахо-
дящихся в контакте с фтором в производственных условиях.
Р. I. Aggett (1985), которому принадлежит один из наи-
более полных обзоров по физиологии и метаболизму эссенци-
альных МЭ, подчеркивает, что распределение элементов в тка-
нях организма и даже внутри клеток ни в коем случае не явля-
ется случайным и единообразным. Он справедливо считает, что-
это отражает различную функциональную роль МЭ. Как изве-
стно, йод концентрируется в щитовидной железе. Железо нахо-
дится в наибольшем количестве, в печени, селезенке и в костном
мозге. Концентрация цинка в сосудистой оболочке глаза и сет-
чатке на 3 порядка выше, чем в лейкоцитах, что в свою очередь,
в 10 раз меньше, чем в эритроцитах. Если охарактеризовать
эти отношения применительно к содержанию МЭ внутри клет-
ки, то железо преимущественно обнаруживается в митохондри-
ях и пероксисомах, медь и марганец преобладают в митохонд-
риях, в то время как цинк в основном присутствует в цитозоле
и в «предсекреторных продуктах» или в секреторных гранулах.
Ставя вопрос о причинах поразительной селективности мета,-
болизма МЭ, этот автор по существу не находит на него удов-
56
летворителыюго ответа и дает только частичные объяснения.
С одной стороны, он обращает внимание на возможность мно-
жественных взаимодействий между металлами, например, во
время их поглощения слизистой оболочкой кишечника, при пе-
ремещении в транспортных белках крови, в процессе поглоще-
ния металлов гепатоцитами, во внутриклеточных участках свя-
зывания и даже при включении металлов в апопротеины металло-
ферментов. С другой стороны, вслед за R. J. Williams (1981,
1984) он подчеркивает, что изоморфные замены металлов in vi-
vo, по-видимому, не происходят. Это говорит в пользу того, что
живые организмы с удивительной тонкостью используют инди-
видуальные свойства элементов.
Р. I. Aggett условно разделяют МЭ на три группы: 1) ка-
тионные элементы (цинк, железо, марганец и медь), которые
всасываются с различной интенсивностью; гомеостатический
контроль за этой группой элементов осуществляется печенью
и желудочно-кишечным трактом; 2) анионные элементы (хром,
селен, молибден, йод), эффективно абсорбируемые желудком и
выделяемые из организма в основном почками; 3) элементы,
существующие в виде органических комплексов, метаболизм их
затруднен.
Выдвигая такую классификацию МЭ, автор признает также
общий метаболический контроль за этими группами, который
осуществляется путем изменения валентности (состояние окис-
ления) данных элементов и их взаимодействия с возможными
лигандами. Другими словами, создается система, в которой
каждый из элементов попадает в ловушку в цепи дискретных
метаболических и физических «ячеек», в конечном счете обеспе-
чивающих его поступление к функциональным участкам в соот-
ветствующей форме и концентрации [Aggett Р. I., 1985].
Предварительные этапы абсорбции эссенциальных катион-
ных МЭ связаны с их высвобождением из пищевых комплексов
и последующим попаданием в слизистую оболочку кишечника в
той химической форме, которая является приемлемой для по-
глощения и последующей транслокации в организме. По мне-
нию Р. I. Aggett, количество биометаллов в виде свободных ио-
нов при нейтральной величине pH чрезвычайно мало. Железо,
цинк, медь и марганец образуют в просвете кишечника мало-
растворимые гидроокиси и фосфатные соединения. В связи с
этим необходимо, чтобы эти элементы достигали слизистой
оболочки в комбинации с эндогенными или пищевыми лиганда-
ми. W. Forth и W. Rummel (1976) обобщили данные о факто-
рах, влияющих на абсорбцию МЭ. Они выделяют системные
факторы, в частности анаболические потребности, связанные с
ростом организма в грудном и детском возрасте, с состоянием
организма в период беременности и лактации и с посткатабо-
лическими состояниями организма. На поглощение МЭ кишеч-
ником и на их транспорт оказывают определенное действие эн-
докринные влияния, инфекции и стресс, специфические систем-
57
ные резервы металла, генетические влияния, врожденные нару-
шения метаболизма, особенности пищевого режима.
Другая большая группа факторов, влияющих на поглощение
МЭ слизистой оболочкой кишечника, относится к «люменаль-
ным», т. е. действующим в его просвете. Среди них авторы на-
зывают: 1) химическую форму и состояние окисленности
этих элементов в пище; 2) наличие антагонистических лигандов
(фосфаты, карбонаты, таннаты, полифенолы, оксалаты); нали-
чие специализированных лигандов (например, аскорбатов, саха-
ра для железа), карбоновых кислот, некоторых сахаров, амино-
кислот и жирных кислот, а также конкурирующих металлов.
Предварительный гидролиз жиров и белков в кислой среде
желудка способствует отделению металлов от пищевых компо-
нентов. При обычной величине pH содержимого желудка метал-
лы, по-видимому, должны иметь ионную форму. Весьма важно,
что такие металлы, как марганец, цинк, железо, хром и медь,
представлены их хлорными солями.
Кислая среда способствует образованию двухвалентных ио-
нов железа и марганца, что является наиболее благоприятным
состоянием этих металлов для их всасывания кишечником. Вы-
свобождающиеся в процессе желудочного пищеварения глута-
тион и цистеин содействуют восстановлению этих металлов.
В дальнейшем происходит нейтрализация химуса секретом под-
желудочной железы в просвете двенадцатиперстной кишки, что
способствует осаждению нерастворимых гидроокисей металлов,
карбонатов и фосфатов. В процессе пищеварения высвобожда-
ются аминокислоты и другие макромолекулы, образующие рас-
творимые комплексы с металлами, способные к всасыванию
слизистой оболочкой.
При исследовании пищеварения in vitro широко использует-
ся метод изолированных петель кишечника. Процесс абсорбции
меди, марганца, цинка и железа ухудшается в результате обра-
зования комплексных соединений этих металлов с компонента-
ми секрета поджелудочной железы. Желчь улучшает раствори-
мость и абсорбцию железа и меди, поглощаемых из пищи. Жир-
ные и фосфатидные кислоты легко образуют комплексы с пере-
ходными металлами. Данные комплексы обнаруживаются в
мезентериальной лимфе. В связи с этим предполагается, что
они также участвуют в абсорбции эссенциальных МЭ. Влияние
пищевых жиров на абсорбцию МЭ изучено мало. Отмечено
только, что диета с низким содержанием жира у больных с
синдромом «короткого желудка» улучшает абсорбцию цинка.
Аминокислоты и олигопептиды облегчают абсорбцию МЭ,
причем L-аминокислоты в большей степени улучшают абсорб-
цию меди, чем их D-изомеры [Kirchgessner М., Grassman Е.„
1970]. Влияние сахаров на абсорбцию МЭ изучено мало.
Важную роль в осуществлении биологической доставки и
абсорбции биометаллов играют антагонистические лиганды,
в частности органические и неорганические фосфаты. Р. I. Ag-
gett (1985) указывает, что орто- и полифосфаты оказывают си-
58
нергическое действие на всасывание магния и кальция и об-
разуют нерастворимые соли с цинком, медью и железом при pH
содержимого кишечника, ухудшая их абсорбцию. Казеин ко-
ровьего молока связывает 24% железа, 44% меди, 84% цинка
и 67% марганца. В отличие от этого в человеческом молоке
лишь 9% железа, 7% меди, 8% цинка и 11% марганца свя-
зываются с казеином.
Очень большую роль в поглощении биометаллов играет вез-
десущее фосфорсодержащее соединение — фитат, которое обна-
ружив ается’во всех злаках, многих бобовых растениях, орехах,
в некоторых фруктах и корнеплодах [Maga J. А., 1982]. При
почти нейтральном значении pH фитат может образовывать
разнообразные комплексы не только с белками, но и с кальци-
ем, магнием, медью, цинком, никелем, железом, уменьшая
усвоение их организмом [Davies N. Т., Nightingale R., 1975].
Высказывается предположение, что поступление МЭ из белка
овощей может быть снижено не только действием фитата, но и
некрахмальными полисахаридами, в частности целлюлозой, ге-
мицеллюлозой и пектином, которые также могут связывать
медь, цинк и железо и снижать их абсорбцию.
В организм живых существ МЭ поступают различными пу-
тями. В. И. Вернадский указывал, что у низкоорганизованных
форм жизни поступление МЭ осуществляется через наружные
покровы тела. В свете более новых данных можно утверждать,
что у высших животных и, в частности, у человека в ходе эво-
люции сформировался основной путь поступления МЭ — желу-
дочно-кишечный тракт, где в верхнем отделе тонкой кишки,
а именно в двенадцатиперстной кишке, возник особый высоко-
специализированный резорбционный аппарат, широко связан-
ный с кровеносными и лимфатическими сосудами, а также с
центральной и вегетативной нервной и эндокринной системой.
Отличительной особенностью абсорбции МЭ в данной зоне яв-
ляется многократность этого процесса, который начинается в
проксимальных отделах тонкой кишки и неоднократно повторя-
ется в более дистальных отделах [Авцын А. П., Жаворон-
ков А. А., 1989]. Специальные исследования с радиоактивными
изотопами показали, что способностью к абсорбции МЭ обла-
дают и другие отделы кишечника. Какая-то часть абсорбции
МЭ происходит и в ободочной кишке.
J. W. Matseshe и соавт. (1980) в опытах с перфузией кишеч-
ника, проводившихся на добровольцах, показали, что железо
и цинк эффективно поглощаются проксимальной частью тонкой
кишки. Одновременно в просвет двенадцатиперстной кишки по-
ступает значительное количество цинка с панкреатическим со-
ком. Следует обратить внимание, что от 0,12 до 0,23 ммоль
цинка (7,5—14,5 мг) в сутки участвует в энтеропанкреатической
циркуляции, осуществляемой через кровеносные сосуды. С из-
вестным основанием можно говорить о рециркуляционном аппа-
рате дуоденально-панкреатического кровообращения. Именно
этот аппарат способствует насыщению организма цинком. По
59
такому же принципу резобции и реперфузии происходит насы-
щение организма, в частности печени, поджелудочной железы,
другими МЭ, например марганцем.
Многосторонние исследования процессов связывания метал-
лов различными биосубстратами выдвинули на передний план
изучение металлотионеина (МТ). Этот белок, обладающий низ-
кой молекулярной массой, равной 6600, состоит из 61 аминокис-
лоты, из которых почти половина (30) — это цистеин, осталь-.
ное глицин, лизин и серин, причем ароматические аминокисло-
ты в состав этого белка не входят. Его уникальные свойства
выражаются в том, что 1 моль этого апопротеина способен свя-
зать 7—10 моль металла (!). Впервые он был открыт в почке
лошади в 1957 г., когда М. Margoshes и В. Z. Vallee обнаружи-
ли макромолекулу, способную аккумулировать кадмий, в кор-
ковом веществе этого органа [Falck F. J. et. al., 1983]. Данные
изучения биоптатов почек людей, подвергавшихся интоксика-
ции кадмием, показали, что кадмиевая нефропатия развивается
в тех случаях, если концентрация кадмия в корковом веществе
почки превысит 1,78 ммоль/кг ткани. В дальнейшем выяснено,
что МТ широко распространен в природе. Он содержится в пе-
чени плодов и новорожденных, причем в более высоких кон-
центрациях, чем у взрослых животных. После рождения кон-
центрация МТ понижается, причем если он первоначально со-
держится в ядре и цитоплазме клеток, то после рождения этот
белок перераспределяется и локализуется преимущественно в
цитоплазме. Его поразительная металлофильность сразу при-
влекла внимание исследователей, занимающихся изучением ме-
таллоферментов. Так, перераспределение МТ и связанного с
ним цинка, по-видимому, отражает изменение потребности ор-
ганизма в цинке для обмена нуклеиновых кислот и белков.
В дальнейшем МТ были обнаружены не только в тканях и клет-
ках животных, но и в тканях высших растений. Они образуют
комплексы с цинком, кадмием, медью, ртутью, золотом и сереб-
ром. Первоначальная индукция транскрипции специфической
иРНК названными металлами вызывает последующий синтез
МТ. Наибольшей индуцирующей активностью обладает кад-
мий. Другими индуцирующими факторами, вызывающими син-
тез МТ, являются, в частности, инфекция, голодание, введение
эндотоксинов, а также глюкокортикоидов, эстрогенов, глюкаго-
на, удаление надпочечников, гипотермия, физическая нагрузка
и лапаротомия [Webb М. et al., 1982; Bremner J. et al., 1983].
Выяснилось, что индуцированный этими крайне разнообраз-
ными факторами синтез МТ в печени сопровождается накопле-
нием в ней цинка и понижением его содержания в плазме кро-
ви, в частности, в той ее фракции, которая связана с альбуми-
ном. По данным J. R. Riordan и соавт. (1982), большая часть
МТ в растворимых фракциях связана с цинком, в то время как
большая часть нерастворимых фракций представляет собой ли-
зосомальный МТ, образующий комплексы с медью. Окончатель-
но функция МТ не выяснена. Большая часть исследователей ви-
60
дит в них белки, обладающие обезвреживающим свойствомг
предохраняющие организм от чрезмерной интоксикации ме-
таллами.
Другие исследователи считают МТ внутриклеточным дело
цинка для активации апоферментов. R. J. Р. Williams (1984)
рассматривает их как регуляторы метаболизма цинка и меди.
С помощью радиоиммунного анализа удалось показать тесную
корреляцию между концентрацией МТ в печени и его содержа-
нием в плазме. У крыс был обнаружен активный поток цинка
от слизистой оболочки к серозной [Kowarsky S. et al., 1974].
Парентерально введенный цинк быстро появляется в просвете
кишечника [Matseshe J. W. et al., 1980].
На основании систематических исследований гомеостаза-
цинка у животных Е. Weigand и соавт. (1980) показали, что
ресекреция цинка в просвет кишечника, по-видимому, является
преобладающим механизмом регуляции гомеостаза. Однако это
имеет место только при нормальном потреблении цинка. Напро-
тив, при его высоком содержании в пище возникает блокада
переноса данного металла в слизистой оболочке. Этот меха-
низм может быть связан с индукцией внутри клеток слизистой-
оболочки кишечника синтеза МТ.
У больных, которые в течение длительного времени получа-
ли терапевтические дозы цинка, возникали гипокупремия и при-
знаки недостаточности меди. Эти приемы использовались для
снижения абсорбции меди при болезни Вильсона — Коновалова
[Hoodgenraad Т. U. et al., 1984].
По мнению Р. I. Aggett (1985), для оптимального течения*
обменных процессов у человека необходимы как минимум де-
вять МЭ (железо, медь, марганец, йод, цинк, хром, селен, мо-
либден, кобальт). Именно перечисленные элементы выполняют
разнообразные функции, в том числе каталитическую, струк-
турную и регуляторную. В процессе осуществления этих функ-
ций они взаимодействуют с макромолекулами, такими как фер-
менты, прогормоны, а также с предсекреторными гранулами »
биологическими мембранами, участвуя во всех видах обмена
веществ. Уровни обмена, на которых это происходит, столь
фундаментальны, что признаки недостаточности многих МЭ'
оказываются крайне изменчивыми и на первый взгляд кажутся
неспецифическими.
Эти МЭ, как уже говорилось, распадаются на катионы и
анионы. Последние абсорбируются относительно легко и их го-
меостаз регулируется в основном за счет выделения с мочой.
Катионы нуждаются в специфических путях абсорбции и в
их гомеостазе принимает участие желудочно-кишечная и желч-
ная секреция. Некоторые МЭ абсорбируются более эффективно
в виде органических комплексов. Характерной особенностью
метаболических путей для каждого элемента является достав-
ка его к «функциональному сайту» с помощью использования
физико-химических особенностей МЭ.
61!
Концепция Р. I. Aggett вполне логична с позиций физичес-
кой химии. Однако она в неполной степени объясняет селектив-
ную локализацию МЭ, на которую влияют многие другие фак-
торы, о которых автор, разумеется, знает, но, по-видимому, не
придает им существенного значения. С нашей точки зрения, яв-
но недооценивается генетическая предопределенность селек-
тивности путей миграции и конечной локализации МЭ при за-
вершении их сложного транспортного цикла. Принимая во вни-
мание физико-химические взаимовлияния биометаллов, труд-
но избежать биологических гипотез о рецепции МЭ именно в
нужном, т. е. соответствующем физиологическим потребностям,
«месте». Эта рецепция обязательно должна быть связана со
специфическими свойствами клеточных мембран, ионными ка-
налами и другими механизмами, которые определяют селектив-
ное распределение МЭ в строго определенных частях орга-
низма.
Приведенные выше данные показывают, что стройная сис-
тема всасывания и выделения МЭ характеризуется определен-
ными балансовыми параметрами. Другими словами, адекватное
физиологическим условиям всасывание у здорового человека
уравновешивается адекватной элиминацией, в которую включа-
ются не только удаление МЭ из организма, но и отложение их
в определенные тканевые и клеточные депо. Последнее особен-
но относится к эссенциальным МЭ, часть из которых представ-
ляет собой нутритивный и пластический материал. Следует не
упускать из виду, что ряд МЭ элиминируется не только с ки-
шечным содержимым, но и с мочой.
Каждого медика в первую очередь должны интересовать
факторы, способные повредить или даже разрушить механизмы
абсорбции и элиминации МЭ. Практические врачи отдают себе
отчет в том, что чем сложнее устроена система какой-то жиз-
ненно важной функции, тем большее число факторов должно
участвовать в обеспечении ее сохранности. Более глубокое
изучение механизмов абсорбции и элиминации приносит все
новые доказательства участия в них регулирующих систем ор-
ганизма— нервной, эндокринной и иммунной.
Несомненно, даже после беглого ознакомления с механизма-
ми абсорбции и элиминации МЭ в верхнем отделе желудочно-
кишечного тракта крайне распространенный диагноз «дуоде-
нит», который встречается так же часто, как диагноз «гастрит»,
с позиций учения о МТОЗах должен вызывать по меньшей мере
настороженное отношение. Таким образом, воспалительные, ди-
строфические и особенно атрофические изменения слизистой
оболочки двенадцатиперстной кишки неизбежно сопровожда-
ются нарушениями всасывания МЭ. Это прежде всего относит-
ся к хроническим дуоденитам с глубокой перестройкой слизис-
той оболочки.
Причины упомянутых болезней очень разнообразны. К ним
относятся паразитарные, бактериальные, вирусные, а также
послеоперационные осложнения. Классическим примером этой
62
З-Келудочно'
кишечный
Рис. 8. Использование микроэлементов в желудочно-кишечном тракте, в крови
и клетке [Kirchgessner М. et al., 1974].
Клетка
Синтезы'
металлофер -
ментов и дру-
гих субстан-
ций, например
гемоглобин
и др.
патологии является синдром мальабсорбции, обусловленный
нарушением всасывания через слизистую оболочку тонкой киш-
ки одного или нескольких питательных веществ. Он может
быть первичным, т. е. наследственно обусловленным, или вто-
ричным, т. е. приобретенным.
Крайне важно, что мальабсорбция выражается в атрофии
слизистой оболочки тонкой кишки с трансформацией всасыва-
ющего эпителия. Очень характерны уменьшение диаметра вса-
сывающих пор и глубокие нарушения кишечного лимфотока и
микроциркуляции. Ультраструктурно обнаруживают: 1) гипо-
плазию микроворсинок, 2) угнетение микропиноцитоза, 3) ат-
рофию всей клетки и парциальную атрофию ее органелл. Маль-
абсорбция вызывает большие изменения в жизнедеятельности
всего организма, часть из которых связана с гипонатриемией,
гипокалиемией и гипокальциемией. При мальабсорбции прок-
симальных отделов тонкой кишки нарушено всасывание каль-
ция и железа, витаминов группы В. Поражение дистальных от-
делов тонкой кишки сопровождается недостаточной абсорбци-
ей витамина В12 и желчных кислот.
Представленная схема М. Kirchgessner и соавт. (1974) дает
основную ориентировку в процессах утилизации МЭ в желу-
дочно-кишечном тракте, крови и клетке (рис. 8). По мере раз-
работки вопроса данная схема, по-видимому, будет дополнена
данными о роли распределительного пула МЭ и в лимфатиче-
ской системе.
В последние годы наши представления о структурных осно-
вах всасывания в тонкой кишке существенным образом измени-
лись под влиянием систематических исследований И. А. Моро-
зова и соавт. (1988). Еще в 1967 г. А. М. Уголев, характеризуя
ба
поверхность кишечной стенки, указал, что ворсинки увеличива-
ют эпителиальную поверхность в 8 раз, а микроворсинки эпи-
телиоцитов — в 30—60 раз. По определению И, А. Морозова,
кишечная ворсинка является микроорганной структурой, обла-
дающей своим сосудистым, мышечным и нервным аппаратом.
Микроциркуляция в слизистой оболочке тонкой кишки осущест-
вляется в богатой капиллярной сети, находящейся в тесном
контакте с эпителиоцитами. Сами капилляры ворсинок относят-
ся к фенестрированным и располагаются на расстоянии 0,5—
1 мкм от базальной мембраны. Вокруг центрального лимфати-
ческого сосуда ворсинок находятся гладкомышечные клетки.
Их сокращения лежат в основе ритмических движений ворси-
нок и облегчают микроциркуляцию. Энтероциты отличаются
очень высокой скоростью обновления. В тонкой кишке человека
в течение 1 мин образуется и одновременно гибнет, отторгаясь
в просвет, от 10 до 60 млн эпителиальных клеток [Da Costa
L. R., 1972]. Полное обновление эпителиальных клеток у чело-
века происходит в кишечнике каждые 5—6 дней [Eastwood
G. L., 1977]. Общая площадь «пищеварительно-транспортной
поверхности» эпителия достигает 200 м2. Каждый энтероцит
имеет приблизительно 1500—2000 микроворсинок [Eggermont
Е., 1979]. Каждая микроворсинка обладает осевым пучком ак-
тиновых филаментов [Mooseker М. S., Tilney L. G., 1975].
И. А. Морозов и соавт. (1988) пришли к заключению, что
так называемый надэпителиальный слизистый слой обладает
пищеварительными и транспортными функциями, так как в его
слизистых наложениях содержатся панкреатические и собст-
венно кишечные ферменты. Эти авторы подчеркивают особое
значение микровезикулярных механизмов всасывания и секре-
ции в тонкой кишке, которые недооценивались в литературе.
Неменьшее значение имеют новые доказательства участия апи-
кального сократительного комплекса в инициации процессов
всасывания и секреции. Новыми фактами, важными для пони-
мания сложной функции энтероцитов, являются установление
динамики транспорта нутриентов через цитоплазму энтероци-
тов, а также реакция иммунокомпетентных клеток слизистой
оболочки тонкой кишки на пищевые вещества. Так, число инт-
раэпителиальных лимфоцитов в межклеточном пространстве
эпителиального пласта очень велико и составляет от 10 до 30%
всех клеточных элементов [Ferguson А., 1977]. С полным осно-
ванием в современной литературе говорится о «лимфоидной
системе кишечника» [Arnaud-Battandier F., 1984]. Обобщая
эти данные, И. А. Морозов и соавт. (1988) убедительно пока-
зывают, что на протяжении всего процесса всасывания нутри-
ентов имеет место тесный контакт введенных в организм ве-
ществ с лимфоцитами эпителиального пласта, а также с дру-
гими иммунокомпетентными клетками. Особенно активны в
в этом отношении средние лимфоциты.
Авторы анализируемой монографии лишь попутно затрону-
ли вопрос о всасывании МЭ в кишечнике, ограничившись дан-
64
.Сокращение
Расслабление
Рис, 9. Схематическое изображение возможных механизмов регуляции деятель-
ности апикального сократительного комплекса эпителиальных клеток тонкой
кишки [Морозов И. А. и др., 1988].
ными о сорбции изотопов железа и цинка структурными компо-
нентами слизи тонкой кишки. Специально исследуя механизмы
везикулярного транспорта в энтероците, они указывают, что
эндоцитоз и транспорт веществ по существу являются логичес-
ким продолжением сорбционных процессов в гликокаликсе и
мембранного пищеварения. В образующиеся везикулы в про-
цессе всасывания попадают не только крупномолекулярные, но
и низкомолекулярные соединения. Результаты прицельного ис-
следования процесса всасывания микроэлементов в кишечнике
не приводятся.
Образование эндоцитозных везикул связано с активацией
сократительного актомиозинового комплекса апикальной части
энтероцитов. Пиноцитоз является Са2+-зависимым процессом,
5—568
65
>I Полостное
. пищеварение
Мембранное пищеварение|Ц
IV
Всасывание
Внутриклеточное
интравезикулярное
мембранное
пищеварение
Пищеварение
в собственной
пластинке,
органах и
тканях
Рис. 10. Основные этапы пищеварительного конвейера в тонкой кишке [Моро-
зов И. А. и др., 1988].
который в терминальной сети регулируется кальмодулином.
Активность цАМФ находится под регулирующим влиянием ад-
реналина, а цГМФ — ацетилхолина. В конечном счете эти два
медиатора вегетативной нервной системы оказывают влияние
на циклические сокращения и расслабления микроворсинок.
Основные представления о возможных механизмах деятельно-
сти апикального комплекса энтероцитов отражены на рис. 9.
Ссылаясь на данные Г. К. Шлыгина (1974), И. А. Морозов
и соавт. (1988) подчеркивают, что холиномиметические вещест-
ва (ацетилхолин и др.), а также средства, угнетающие ацетил-
холинэстеразу (например, физостигмин), усиливают всасыва-
ние и секрецию, тогда как холинолитики (атропин) и симпато-
миметики (адреналин, норадреналин и др.) тормозят эти про-
цессы. Представляется очевидным, что эти данные должны
быть использованы при изучении всасывания МЭ в условиях
клиники и эксперимента.
Оценивая в целом вклад группы исследователей под руко-
водством И. М. Морозова (1988) в изучение проблемы микро-
физиологии процессов всасывания нутриентов в тонкой кишке,
надо признать его весьма значительным. Особую ценность это-
му труду придает систематическое применение электронно-мик-
роскопического метода, с помощью которого получено много
новых данных.
И. А. Морозов и соавт. (1988) с полным основанием гово-
рят о «пищеварительном конвейере», под которым они понима-
66
ют непрерывно движущуюся транспортную систему для после-
довательной обработки поступающих в организм пищевых ве-
ществ. Этот конвейер включает шесть этапов, что хорошо пред-
ставлено на рис. 10. Принимая это схематическое подразделе-
ние, мы должны вместе с тем указать, что систематическое ис-
следование прохождения МЭ по этим этапам еще не изучено,
недостаточно исследованы также регулирующие влияния на
механизмы этого крайне сложного процесса.
3.2. Микроэлементы и органы дыхания
Воздухоносная система является вторым по значению пу-
тем поступления МЭ в организм, особенно в профессиональной
патологии. Судя по литературным данным, аэрогенный путь
существенно важен для следующих МЭ, перечисленных в по-
рядке увеличения атомной массы: литий, бериллий, фтор, алю-
миний, кремний, титан, ванадий, хром, марганец, железо, ко-
бальт, никель, кадмий, медь, цинк, мышьяк, стронций, молиб-
ден, серебро, олово, йод, вольфрам, осмий, платина, золото,
ртуть, свинец, висмут.
Патогенез ринитов, фарингитов, ларинготрахеитов и брон-
хитов, которые часто бывают начальными проявлениями раз-
личных МТОЗов, связан как с токсическим действием некото-
рых МЭ, так и с дефицитом эссенциальных МЭ, участвующих
в поддержании трофики дыхательных путей. Значение МЭ в
этой патологии, как правило, недооценивается, так как наблю-
даемые клинические симптомы обычно объясняют влиянием
других патогенных факторов (колебания температуры воздуха,
избыточная или недостаточная влажность, запыленность). Тер-
мин «пылевой бронхит» широко используется в профессиональ-
ной патологии. В действительности он представляет собой
сборное понятие, под которым объединяется результат влияния
патогенных факторов с различными физико-химическими и
биологическими свойствами.
Профессиональные заболевания дыхательных путей, вызван-
ные МЭ, крайне многообразны. В основном они входят в пер-
вые две группы общепринятой классификации профессиональ-
ных болезней (ПБ): 1) ПБ, вызываемые действием химических
факторов (острые и хронические интоксикации и их последст-
вия, заболевания кожи, профессиональные новообразования,
профессиональные аллергозы); 2) ПБ, вызываемые действием
пылевого фактора (силикоз, силикатозы, антракоз, металлоко-
ниозы от смешанной пыли, бронхит и др.) [Монаенкова А. М.,
Архипова О. Г., 1983].
Эти заболевания в зависимости от длительности воздействия
и характера профессиональной вредности могут протекать
остро, подостро и хронически. Особенно тревожат клиницистов
сверхострые заболевания, развивающиеся после однократного
воздействия относительно высоких концентраций веществ, в
том числе и МЭ, содержащихся в воздухе рабочей зоны.
5* 67
Подострые и хронические ПБ возникают в результате неод-
нократного воздействия на организм профессиональных вредно-
стей. Особенностью хронических, наиболее часто встречающих-
ся в настоящее время, профессиональных заболеваний является
постепенное нарастание симптомов, причем в течение длитель-
ного периода заболевание может протекать с минимальной
симптоматикой.
Некоторые МЭ вызывают острое аллергическое поражение
верхних и нижних дыхательных путей. Заслуживают внимания
острые бронхоспастические реакции, а также различные формы
раздражения н повреждения эпителиального покрова. При хро-
нических поражениях дыхательных путей МЭ, преимуществен-
но металлами, возникают разнообразные изменения, в частнос-
ти атрофия эпителиального покрова, которая может иметь как
локальный, так и распространенный характер. В этом случае
возникают риниты с затруднением носового дыхания, хроничес-
кие катаральные, гипертрофические и атрофические процессы в
носу, глотке и гортани. Особое место занимают пылевые по-
ражения легких, относящиеся к числу наиболее распространен-
ных форм профессиональной патологии.
Хронические поражения дыхательных путей МЭ (никель,
хром, кобальт, мышьяк) могут вызывать местнодеструирующие
и неопластические процессы. Нередко возникают изъязвления
носовых раковин при воздействии никеля и перфорация носовой
перегородки у рабочих хромовой промышленности.
В течение длительного времени в общей и частной токсико-
логии повреждающее действие многих патогенных факторов
рассматривалось в качестве результата нарушения активности
ферментных систем и сложной ферментной сопряженности
[Правдин Н. С., 1960; Покровский А. А., 1971; Тиунов Л. А.,
1976]. В свете новейших данных становится все более понят-
ным представление о том, что действие ряда экзогенных фак-
торов на организм в целом осуществляется через рецепторный
аппарат некоторых клеток и тканей [Сергеев П. В., 1976]. Это
показано, например, для фтора, свинца, ртути, марганца, кото-
рые изменяют активность мембраносвязанного фермента аде-
нилатциклазы, регулирующей многие звенья внутриклеточного
метаболизма [Бархина Т. Г., Ляпин М. Н. 1978; Слинчен-
ко Н. Н. и др., 1979; Строчкова Л.С. и др., 1987].
Среди профессиональных заболеваний органов дыхания,
вызванных воздействием некоторых металлов, широко извест-
ны острые и хронические кадмиозы, ванадозы, алюминозы, ко-
бальтозы, вольфрамозы, никелезы, молибдеиозы, стибиозы,
цинкозы и другие МТОЗы.
Патогенез МТОЗов, возникающих в результате аэрогенного
проникновения токсичных металлов в организм, чрезвычайно
сложен и относительно некоторых элементов еще не изучен.
Важно подчеркнуть, что наряду с общими чертами в механиз-
ме их развития существуют ясные признаки клинической и
патоморфологической индивидуальности действия многих МЭ.
68
Эта индивидуальность становится более рельефной при глубо-
ком изучении каждой нозологической формы. В качестве при-
мера может служить бериллиоз. Бериллий обладает уникальной
способностью к аллергизации организма. Бериллиоз характе-
ризуется неблагоприятным течением и специфическими морфо-
логическими изменениями в легких. Большие перспективы
для более углубленного изучения патогенеза и терапии каж-
дого из МТОЗов имеют моделирование на экспериментальных
животных и исследование непосредственного воздействия МЭ
на клетки в культуре.
Если острые формы заболевания при ингаляционном поступ-
лении МЭ в токсичных дозах относительно хорошо изучены,
то подострые и хронические формы ингаляционных интоксика-
ций исследованы недостаточно полно. Как известно, основными
промышленными токсикантами являются хлор и его соедине-
ния, соединения серы (сернистый газ, сероводород, диметил-
сульфат, серная кислота), соединения азота (окиелы азота,
азотная кислота, аммиак, гидразин), соединения фтора (фто-
рид водорода, плавиковая кислота и ее соли, перфторизобути-
лен), соединения хрома (хромовый ангидрид, окись хрома, би-
хроматы калия и натрия, хромовые квасцы), карбонильные
соединения металлов (карбонил никеля, пентакарбонил желе-
за), растворимые соединения бериллия. Из перечисленных семи
групп соединений пять относятся к МЭ (фтор, хром, никель,
железо, бериллий). Многие из них вызывают следующие основ-
ные клинические синдромы: 1) острый токсический ларингофа-
ринготрахеит, 2) острый токсический бронхит, 3) острый токси-
ческий бронхиолит, 4) острый токсический отек легких, 5) ост-
рую токсическую пневмонию.
Всегда надо помнить, что при острых интоксикациях может
возникнуть рефлекторный спазм гортани и голосовых связок
с развитием асфиксии и наступлением смертельного исхода
[Иванова И. С. и др., 1983].
Интенсивность интоксикации и ее исход зависят от продол-
жительности экспозиции и дозы поступившего токсичного веще-
ства. Дифференциальная клиническая диагностика, терапия и
прогноз изложены в руководствах по профессиональным забо-
леваниям.
3.3. Микроэлементы и кожа
Поражения кожи, вызванные той или иной формой контак-
та с различными МЭ, обычно описываются дерматологами.
Составляя 33,7% всей профессиональной заболеваемости [Ан-
тоньев А. А., 1965], поражения кожи чаще всего вызываются
различными химическими соединениями и могут иметь как
острое, так и хроническое течение [Алексеева О. Г. и др.,
1978; Braun-Falco О., 1980]. В настоящее время благодаря про-
филактике при воздействии низких доз металлов чаще встре-
чаются хронические поражения кожи химическими веществами.
69
Следует подчеркнуть, что очень часто пострадавшие не замеча-
ют и не обращают внимания на контакт их кожи с МЭ. Только
в случае повреждения кожи с появлением выраженной воспа-
лительной реакции они обращаются к врачу. Все химические
вещества в соответствии с их действием на кожу подразделя-
ются на три группы: вещества, оказывающие раздражающее
действие (1-я группа); фотостимулирующее и фотосенсибили-
зирующее действие (2-я группа); вещества-сенсибилизаторы
(3-я группа) [Антоньев А. А. и др., 1965]. В общем виде это
относится и к МЭ, которые могут оказывать влияние на кожу
как сами по себе, так и в качестве компонентов, входящих в
состав сложных химических соединений. Так, раздражающее
действие на кожу оказывают МЭ в форме солей тяжелых ме-
таллов (например, соли хрома и кобальта). Эти же вещества
обладают мощным сенсибилизирующим свойством, что прояв-
ляется при постановке кожных аллергологических тестов.
Разумеется, что реакция кожи на тот или иной МЭ в сущест-
венной степени зависит от места контакта на кожном покрове,
от концентрации и продолжительности экспозиций действующе-
го начала. Именно это побуждает более подробно остановиться
на современных представлениях о строении и функции кожи.
Известно, что поверхность кожи покрыта роговым слоем
погибающих кератиноцитов, живые функционирующие керати-
поциты преобладают в эпидермисе и, размножаясь, перемеща-
ются в его наружные слои по мере слущивания омертвевших
клеток (рис. 11). Внутриэпидермальные макрофаги (клетки
Лангерганса, дендритные клетки), принимающие непосредст-
венное участие в процессинге антигена, попавшего на кожу,
располагаются над базальным слоем кератиноцитов. Они име-
ют характерное ядро лопастной формы с четко выраженным
гетерохроматином. В их цитоплазме видны многочисленные ор-
ганеллы, в частности специфические гранулы в форме «теннис-
ной ракетки» с ампулярным расширением, продольными и по-
перечными пластинками в области «рукоятки». Другим видом
специализированных клеток в эпидермисе являются тактильные
эпителиоциты (клетки Меркеля), представляющие собой осо-
бый тип чувствительных клеток, структурно связанных с нерв-
ными волокнами. На отдельных участках плазмалеммы они
имеют синапсоподобные щелевидные контакты шириной 30 —
60 нм [Chen S. J. et al., 1973]. Происхождение и функция
этих клеток точно не установлены. Так, одни авторы рассмат-
ривают тактильные эпителиоциты в качестве нервных клеток,
другие связывают их происхождение с дифференцировкой ке-
ратиноцитов [Михайлов И. Н., Виноградова Е. В., 1982]. Дер-
ма представляет собой соединительнотканную сеть, под ней
находится подкожная клетчатка. Хотя Т-лимфоциты рассеяны
по всей толще кожи, но особенно их много в эпидермисе и
поверхностных слоях дермы. Специализированные кератиноци-
ты волосяных луковиц синтезируют и секретируют вещество
волос. Дерма богата нервными волокнами, часть из которых
70
Рис. 11. Гистологическая картина эпидермиса в норме.
а —выраженный роговой слой, чешуйки которого осуществляют элиминацию МЭ. Окраска
гематоксилином и эозином. Х400; б — иммунокомпетентная клетка Лангерганса (стрелка).
Полутонкнй срез эпидермиса. Окраска толуидиновым синим н пироннном. ХЮОО; в —кон-
такты соседних эпителиоцитов шиповатого слоя. При микроэлементпых аллергодермато-
зах происходит резкое расширение межклеточных пространств. ХЮ ООО.
являются чувствительными, а также кровеносными и лимфати-
ческими сосудами.
Важное значение в процессе всасывания МЭ через кожу
имеют микрососуды (рис. 12). Как видно из этой схемы, основ-
ной путь проникновения в просвет жидкости и микрочастиц из
интерстиция пролегает через расширенные межклеточные щели,
или межэндотелиальные люки. Кроме того, здесь происходит
и микровезикулярный транспорт веществ через эндотелий. Не
останавливаясь на морфологической характеристике лимфати-
ческих и кровеносных капилляров, которая детально представ-
71
Рис. 12. Тонкое строение стенки лим-
фатического капилляра кожи (рекон-
струкция по серийным срезам, изу-
ченным в электронном микроскопе).
Большие -стрелки показывают основ-
ной путь проникновения в просвет
капилляра жидкости и микрочастиц
из интерстиция через расширенные
межклеточные щели (люки); малень-
кие — микровезикулярный транспорт
веществ через эндотелий. ЯН — якор-
ные нити [Leak L. V., 1971].
Рис. 13. Пути проникновения веществ
через эндотелий капилляра сосочко-
вого слоя собственно кожи.
1 — диффузия через участки, истонченные
мнкровезикуляцией; 2 — диффузия н уль-
трафильтрация через фенестры; 3 — диф-
фузия и фильтр-ария через сквозные поры;
4 — комплексный механизм (микровезику-
лярный транспорт в обход плотного меж-
клеточного соединения и диффузия по меж-
клеточной щелн); 5 — микровезикулярный
транспорт; 6-—диффузия через всю толщу
клетки (траисцеллюлярная диффузия); 7 —
диффузия и ультрафильтрация через мик-
роканальцы, образующиеся при временном
слиянии цепочки мнкровезикул. БМ — ба-
зальная мембрана; Пц — перицит; ЭН —
эндотелий [Алексеев О. В., 1982].
лена в монографиях В. А. Шахламова (1971, 1982), необходимо
все же показать основные пути проникновения веществ через
эндотелий кровеносного капилляра в сосочковом слое кожи
(рис. 13). Из представленной схемы становится понятным мно-
гообразие путей проникновения веществ как в просвет капил-
ляра, так и из крови в окружающие ткани. Принимая во вни-
мание, что в развитии профессиональных аллергодерматозов
важное значение имеет состояние иммунной системы человека,
следует кратко остановиться на относительно новой трактовке
кожи как составной части иммунной системы [Эдельсон Р. Л.,
Финк Д. М., 1985]. В работе этих авторов показано, что кера-
тиноциты базальной части эпидермиса человека содержат в
высокой концентрации вещество, которое идентично тимопоэти-
72
ну — гормону тимуса. Известно, что именно этот гормон регу-
лирует созревание Т-клеток. Кроме того, внутриэпидермальные
макрофаги несут на своей поверхности вещество Т6, характер-
ное для Т-клеток, созревающих в тимусе. Таким образом, внут-
риэпидермальные макрофаги, являясь по своему происхожде-
нию моноцитами костного мозга, ответственны за развитие им-
мунного ответа на локально нанесенный антиген. В настоящее
время установлено, что интерлейкин (ИЛ-1), вырабатываемый
макрофагами, синтезируется также и кератиноцитами. Следо-
вательно, и кератиноциты являются важным элементом иммун-
ной системы. Они не только обеспечивают создание на поверх-
ности тела защитного кератинового слоя и волосяного покрова,
но и, продуцируя гормоноподобные вещества, способны глубоко
воздействовать на функционирование Т-лимфоцитов кожи. Так,
их реакция проявляется в широком диапазоне: от регуляции
созревания Т-лимфоцитов до усиления специфической реакций
на антигены, которыми могут быть как сами МЭ, так и их
сложные соединения.
В 1984 г. был открыт еще один тип иммунологически актив-
ных эпидермальных клеток. Это также дендритные так назы-
ваемые антигенпредставляющие клетки Гренстейна. Они в отли-
чие от эпидермальных макрофагов более устойчивы к ультра-
фиолетовому облучению и способны взаимодействовать с Т-су-
прессорами, а не с Т-хелперами. Хелперный и супрессорный
ответы находятся в относительном динамическом равновесии,
хотя в норме преобладает хелперный сигнал, который обеспечи-
вает адекватный ответ на антиген, проникающий в кожу.
Все локализованные в эпидермисе компоненты иммунной
системы принято называть единым термином SALT (от англ,
skin-associated lymphoid tissue). Авторами показан координи-
рованный ответ иммунокомпетентных клеток кожи на воздей-
ствие антигена, проникшего через роговой слой. Антиген встре-
чается с внутриэпидермальными макрофагами, которые пред-
ставляют его (в комплексе с антигеном гистосовместимости)
Т-клеткам, обладающим запрограммированным ответом на
данный антиген. Эти Т-клетки экспрессируют рецептор к ИЛ-1.
ИЛ-1, продуцируемый кератиноцитами, а также, возможно, и
внутриэпидермальными макрофагами, побуждает активирован-
ные Т-клетки секретировать интерлейкин 2 (ИЛ-2). Последний
связывается с рецепторами на поверхности других Т-клеток,
отвечающих на данный антиген и находящихся в дерме, в лим-
фатических узлах, селезенке, и инициирует их пролиферацию.
В развитии профессиональных аллергодерматозов, в том
числе вызванных МЭ, необходимо учитывать биологические
свойства продуктов активированных лимфоцитов.' Возникнове-
ние аллергической реакции замедленного или немедленного ти-
па зависит от свойств антигена, его всасываемости, путей про-
никновения в кожу и некоторых других факторов (состояние
реактивности, иммунологического статуса, сенсибилизации и
др.). В частности, при трансдермальном пути попадания аллер-
73-
генов в организм в производственных условиях возникает ал-
лергическая реакция замедленного типа. И, напротив, при про-
никновении аллергенов через дыхательные и пищеварительные
пути характерно развитие аллергической реакции немедленно-
го типа. Среди химических веществ, обладающих выраженны-
ми сенсибилизирующими свойствами и играющих этиологичес-
кую роль в развитии профессиональных аллергодерматозов с
реакцией замедленного типа, определенное значение имеют со-
ли хрома, кобальта, никеля, а также содержащие их материа-
лы — цемент, бетон, легированные стали [Сомов Б. А., Ко-
ган В. Ю., 1983]. Так, показано, что лимфоциты больных, стра-
дающих никелевым дерматитом, при воздействии in vitro ионов
никеля претерпевают бласттрансформацию.
Известно, что соли МЭ, обладающие сенсибилизирующими
свойствами, являются неполными антигенами — гаптенами.
Проникая в кожу, они связываются с белками в виде комплекс-
ных соединений, приобретают свойства полных антигенов. Ан-
тигенная стимуляция вызывает активное кооперативное взаимо-
действие всех заинтересованных компонентов кожи. В частнос-
ти, сенсибилизированные лимфоциты, помимо прямого цитоток-
сического действия на клетки, содержащие антиген, выделяют
фактор, тормозящий миграцию макрофагов. В клинических
условиях широко используется диагностический тест — реакция
торможения миграции лейкоцитов (РТМЛ) [Адо В. А. и др.,
1978].
В патогенезе профессиональных дерматозов важная роль
принадлежит нейроэндокринным и функциональным нарушени-
ям, особенно в ЦНС, которые и проявляются в развитии за-
стойных очагов возбуждения. В клинической картине заболева-
ния у таких больных преобладают эмоциональная лабиль-
ность, нарушения ночного сна, зуд кожи. Факторы организма,
способствующие развитию аллергического поражения кожи при
экземе и других аллергодерматозах, представлены на рис. 14.
При начальных формах профессионального аллергического
дерматита гистологические изменения развиваются преимуще-
ственно в глубоких слоях эпидермиса и характеризуются вы-
раженным межклеточным отеком с формированием везикул.
В дерме отмечается мопонуклеарная, обычно периваскулярная,
инфильтрация. При прекращении действия этиологического
фактора воспалительные изменения довольно быстро стихают,
оставляя щелушение и незначительную пигментацию.
Профессиональные аллергические дерматиты, обусловлен-
ные МЭ и их более сложными соединениями, развиваются у
рабочих следующих специальностей: металлургов, химиков,
никелировщиков, хромировщиков, строителей и др. При этом
профессиональными аллергенами могут быть соли хрома, ко-
бальта, никеля, соединения ртути, соли платины, бериллия,
золота и некоторые другие.
Токсикодермия возникает как результат сенсибилизирую-
74
дни
Слабый иммунный ответ
Слабый иммунитет Частичная толерантность
Сенсибилизация
Рис. 14. Факторы организма, способствующие развитию аллергического воспа-
ления кожи при экземе и других аллергодерматозах [Фролов Е. П., 1982].
щего и токсического воздействия химических веществ, попада-
ющих в организм через дыхательные пути и кожу. При этом
важна повышенная чувствительность организма немедленного
или замедленного типа. Причинами микроэлементной токсико-
дермии нередко являются соли йода и брома. Развивается она
в короткие сроки после поступления в организм аллергена,
75
однако вслед за его элиминацией воспалительные изменения
довольно быстро исчезают.
Профессиональная экзема — аллергическое заболевание ко-
жи, в этиологии и патогенезе которого важную роль играют
воздействие определенного производственного фактора и функ-
циональные нарушения в нервной, сосудистой и других систе-
мах, определяющие динамику аллергических реакций, особен-
ности клинической картины и хроническое рецидивирующее те-
чение. Характерно, что при профессиональной экземе повышен-
ная чувствительность вначале, как правило, бывает монова-
лентной, а в дальнейшем становится би- и поливалентной.
Этиологическими факторами при ней являются хром, никель
(«никелевая чесотка»), платина и другие металлы и их соли.
Необходимо отметить, что кожные поражения при так называе-
мой платиновой экземе нередко сочетаются с бронхиальной
астмой.
При контактном дерматите облигатными раздражителями
являются соли тяжелых металлов (действуют как кислоты) и
щелочных металлов (действуют как щелочи), а также бром,
растворимые соединения бериллия, фториды, хлориды, суль-
фаты, ацетаты и др. Для этой формы дерматита характерны
отеки или изъязвления. Первичная экзема при действии берил-
лия встречается редко.
Фотодерматит возникает при производственном контакте с
веществами, повышающими чувствительность организма к свету.
К ним относятся соли марганца, железа, урана, препараты
ртути и др. Характерной чертой этих заболеваний является
поражение открытых частей тела, подвергающихся загрязнени-
ям профессиональными вредностями и инсоляции. Выражена
сезонность рецидивов фотодерматитов в весенне-летние перио-
ды. Близко к этой группе болезней микроэлементной природы
относятся МТОЗы ятрогенного происхождения.
Глава 4
ЭССЕНЦИАЛЬНЫЕ И ТОКСИЧНЫЕ МИКРОЭЛЕМЕНТЫ
В 1912 г. в классической работе G. Bertrand впервые был
поставлен вопрос о различных ответах организма на эссенци-
альные МЭ. По его данным, организм проходит через несколь-
ко стадий по мере того, как концентрация «эссенциального пи-
тательного вещества» увеличивается от состояния недостаточ-
ности до состояния, характеризующегося избыточным содер-
жанием. При «абсолютном дефиците» наступает смерть. При
ограниченном поступлении «эссенциального вещества» орга-
низм выживает, но при этом появляются признаки «погранич-
ного дефицитного состояния». По схеме, представленной
G. Bertrand, с увеличением приема питательного вещества его
содержание в организме достигает плато. Именно это сопро-
вождается оптимальным функционированием организма. По
мере того как изучаемое питательное вещество начинает по-
ступать в организм в избытке, сперва возникают состояние
«маргинальной токсичности», а затем и проявления «летальной
токсичности». Е. Frieden (1984) справедливо пишет, что ука-
занная тенденция в количественном отношении может сущест-
венно варьировать для каждого эссенциального элемента. Од-
нако ее основные слагаемые до сих пор остаются практически
неизменными для всех элементов. Впоследствии этот же под-
ход был использован при изучении некоторых потенциально
токсичных МЭ. В частности, исследование селена и фтора по-
казало, что существует почти 10-кратный разрыв между необ-
ходимым суточным количеством для выживания подопытных
животных и тем количеством, при котором появляются приз-
наки токсического действия этих МЭ. W. Mertz (1982) подчер-
кивает, что из схемы G. Bertrand можно сделать по крайней
мере два вывода: 1) каждый элемент имеет присущий ему ди-
апазон безопасной экспозиции, который поддерживает опти-
мальные тканевые концентрации и функции-, 2) у каждого МЭ
имеется свой токсический диапазон, когда безопасная степень
его экспозиции превышена.
Как показали последующие исследования, весь этот вопрос
оказался значительно сложнее, чем это представлялось на пер-
вых этапах изучения МЭ. В частности, Klaus Schwarz (1977)
отметил, что высокая токсичность МЭ не исключает его биоло-
гической эссенциальности. Например, фтор и селен первона-
77
чально рассматривались только в свете их относительной ток-
сичности. Однако в дальнейшем было установлено, что они же
являются, несмотря на токсичность, эссенциальными ультра-
микроэлементами. По-видимому, это в известной мере относит-
ся также и к высокотоксичному кадмию. В свете новых данных
нельзя исключить его эссенциальность по крайней мере для
животных. При разграничении эссенциальных и токсичных МЭ
нужно учитывать тот факт, что некоторые металлы, не буду-
чи, строго говоря, эссенциальными, могут обнаруживать стиму-
лирующее влияние при проведении исследований по принципу
доза — ответ. В настоящее время некоторые «стимулирующие»
металлы получили название «горметины» [Luckey Т. D., Venu-
gopal В., 1977]. Во всех группах периодической системы Мен-
делеева обнаружены элементы, стимулирующие рост и выжи-
ваемость животных. К ним относятся литий, титан, галлий, гер-
маний, рубидий, цирконий, сурьма, барий, золото, ртуть и мно-
гие элементы из семейства лантанидов [Frieden Е., 1984].
Все же не подлежит сомнению, что медицинская практика
нуждается в обоснованном разграничении токсичных и эссен-
циальных МЭ. Такие данные, полученные в результате дли-
тельного изучения многочисленных биологических моделей,
имеются в современной литературе.
В большом обзоре Р. I. Aggett (1985) к эссенциальным МЭ
отнесено только 9 (железо, цинк, медь, марганец, селен, молиб-
ден, хром, йод, кобальт). Он приводит основные данные о со-
держании и биохимической функции этих МЭ (табл. 6). Несом-
ненно, автору известно, что число этих МЭ несколько больше,
но он ограничивается рассмотрением только тех, которые при-
знаны необходимыми «для оптимального здоровья человека».
Быстрый прогресс науки позволяет уже в настоящее время
говорить о десяти МЭ, которые признаны эссенциальными для
животных организмов, так как к приведенному перечню добав-
ляется фтор. Однако и это число не является окончательным,
так как существуют данные о необходимости для животного
организма ряда других элементов. В частности, В. В. Ковальс-
кий (1982) указывает, что процессы метаболизма в организмах
происходят при участии многих металлоферментов, которые
обладают микроэлементными координационными центрами, ме-
таллоферментных комплексов, содержащих также разнообраз-
ные металлы (марганец, кобальт, цинк, никель, железо, барий,
кадмий, ртуть и др.). Металлы, поглощаемые организмами и
содержащиеся в тканях и тканевых жидкостях, могут являться
активаторами действия ферментов (цинк, марганец, железо,
кадмий, кобальт, никель, ртуть, рений, цезий, литий, лантан,
алюминий) или их ингибиторами (бериллий, стронций, барий,
кадмий, ртуть, никель, железо, рубидий). Как видно из приве-
денных данных, подход В. В. Ковальского к причислению к
необходимым для жизни МЭ является наиболее широким.
Определенный смысл имеет следующее высказывание
В. В. Ковальского (1982): «Все эти химические элементы ока-
78
Таблица 6
Содержание в организме эссенциальных микроэлементов
и их биологическая роль [Aggett Р. I., 1985; с дополнениями Авцына А. П. и соавт. (1988)]
Элемент и его количе- ство в орга- низме взрос- лого чело- века Биологическая роль
Железо, 4—5 г Транспорт и депонирование кислорода, цитохромы, окислитель- ное фосфорилирование, редокс-ферменты, антиоксидантное действие. (Специфически предупреждает, устраняет или ос- лабляет признаки дефицита железа у человека и животных)1
Медь, ВО—120 мг Оксидазы, окислительное фосфорилирование, антиоксидантное действие, метаболизм жирных кислот, участие в синтезе сое- динительнотканных структур (коллагена, эластина). (Специ- фически предупреждает, устраняет или ослабляет признаки дефицита меди у лабораторных и сельскохозяйственных жи- вотных)
Цинк, 1—2 г Каталитическая, структурная и регуляторная функция в мно- гочисленных ферментах, в экспрессии геиа, в макромолекуляр- ных и полимерных структурах. (Специфически предупрежда- ет, устраняет или ослабляет признаки дефицита циика. Мощ-
Марганец, 12—20 мг ный активатор Т-клеточного иммунитета) Антиоксидантное действие, синтез стерола, гликозилирование, метаболизм углеводов. (Специфически предупреждает, устра- няет или ослабляет признаки дефицита марганца у сельско- хозяйственных животных)
Хром, 6 мг Оптимизация толерантности к глюкозе. (Оказывает положи- тельное влияние на признаки дефицита хрома)
Селей, 14 мг Антиоксидантное действие, оксидазы, обладающие смешанной микросомальной функцией, поддержание сохранности спермы. (Специфически и в комбинации с витамином Е устраняет и предупреждает признаки дефицита селена. Антибластический фактор)
Молибден, 10 мг- Метаболизм пуринов и серы. (Специфически предупреждает, устраняет или ослабляет признаки дефицита молибдена у жи- вотных)
Иод, 19—20 мг Тироидные гормоны. (Специфически предупреждает, устраняет или ослабляет признаки дефицита йода у человека, лабора- торных и сельскохозяйственных животных)
Кобальт, 1,5 мг Количественные данные не приводятся. (Одно из действую- щих начал витамина Bi2 специфически предупреждает, устра- няет или ослабляет признаки дефицита кобальта у человека и животных)
1 В скобках приведены данные литературы последних лет, обобщенные авторами
КГЛ1ГИ.
зывают большое влияние на жизнь организмов, вступая в связь
с органическими веществами, синтезируемыми в живых, клет-
ках. Они влияют на оплодотворение, развитие, рост, жизнеспо-
собность организма, его иммунобиологические свойства, дыха-
тельную функцию гемоглобина, фотосинтетическую деятель-
ность хлорофилла, на фиксацию микроорганизмами атмосфер-
ного азота и прочие важнейшие функции». Он подчеркивает,
что в отличие от всех веществ, синтезируемых организмами,
79
МЭ поступают в организм из геохимической среды, почвообра-
зующих пород, почв, природных вод, атмосферного воздуха
(аэрозоли химических элементов) благодаря первичной орга-
низованности биогенных циклов, которые представляют собой
биогеохимические явления. Именно эти мысли позволили
В. В. Ковальскому с позиций биогеохимика рассматривать
единство организма и геохимической среды, которые представ-
ляют собой настолько зависимые явления в биосфере, что рас-
сматривать отдельно эволюцию жизни и эволюцию среды не-
возможно.
Указав, что количество МЭ, обладающих потенциальной
эссенциальностью, может быть значительно увеличено, необхо-
димо одновременно подчеркнуть, что проявления и синдромы
истинного дефицита МЭ у человека доказаны только для желе-
за, меди, цинка, марганца, хрома, селена, молибдена, йода, ко-
бальта и фтора. Окончательное суждение об эссенциальности
новых МЭ должно выноситься только на основании убедитель-
ных аргументов.
Значительный биологический интерес представляют данные
Р. I. Aggett (1985) (табл. 7), которые привлекают внимание к
синдрому истинного дефицита МЭ главным образом у некото-
рых животных. Изучение этих данных будет иметь значение
Таблица 7
Другие возможно эссенциальные микроэлементы
[Aggett Р. I., 1985; с некоторыми изменениями]
Элемент Модель Признаки недостаточности
Мышьяк Цыплята, мини-свиньи Задержка роста, изменение синтеза белков, повышенное содержание мо- чевой кислоты в плазме
Бор Цыплята Задержка роста, аномалии костей, соединение с холекальциферолом в обмене магния
Бром Цыплята, мыши, человек Может замещать хлорид и йодид. Бессонница у больных во время по- чечного диализа
Фтор Грызуны, человек Субоптимальная утилизация железа, анемия Возникновение и усиление имеющего- ся кариеса зубов
Литий Козы, крысы Замедление роста, пониженная фер- тильность
Никель Цыплята, коровы, козы, мини-свиньи, крысы, ов- цы Задержка роста, нарушение гемопоэ- за, измененный обмен железа, цннка, меди
Кремний Цыплята, крысы Неполноценное формирование колла- гена и энхондрального окостенения
Ванадий Крысы, цыплята Нарушение регуляции ферментов, пе- реносчиков фосфорила, Na, К-АТФ- азы
83
для выяснения механизма различных форм микроэлементного
дефицита, особенно если учесть, что этот вопрос в медицине и
биологии недостаточно исследован.
Огромный интерес представляют МЭ, обладающие способ-
ностью вызывать токсический эффект в определенных концент-
рациях. Кроме того, необходимо обратить внимание на продол-
жительность экспозиции, характер контакта МЭ с организмами
и чувствительность последних в различные периоды их жизнен-
ных циклов.
К общеизвестным токсичным МЭ относятся ртуть, свинец,
фтор, кадмий, бериллий, мышьяк, хром и, как выяснилось в
последние годы, алюминий. Специалисты по геохимическим за-
грязнениям техногенного происхождения [Ревич Б. А. и др.,
1982] выделяют четыре основные группы химических элементов,
которые различаются по типам создаваемой ими нагрузки на
окружающую среду за счет выпадения из атмосферы и харак-
теру формируемых аномалий.
1. Элементы с невысокими содержаниями в выпадениях,
создающие повышенную нагрузку на воздушный бассейн. Хи-
мических аномалий в почвах они не образуют. Сюда входят
преимущественно так называемые литофильные элементы (ти-
тан, барий, цирконий, ниобий). Экологическая токсичность этой
группы элементов невелика.
2. Элементы с невысокими средними содержаниями в выпа-
дениях, но с отдельными (локальными) участками высоких и
очень высоких (экстремальных) концентраций, которые создают
очень большие нагрузки на воздушный бассейн и сравнитель-
но небольшие по размеру аномалии в почвах. Сюда входят
стронций, серебро, кобальт, висмут, хром, ванадий, бериллий,
литий, олово. Для некоторых из этих элементов характерны
токсические эффекты даже при невысоких концентрациях .
3. Элементы с высокими средними концентрациями в выпа-
дениях, сравнительно равномерно распределенными по терри-
тории урбанизированных зон. Участки с экстремальными со-
держаниями МЭ нехарактерны. Эти элементы формируют по-
вышенную общую нагрузку на воздушный бассейн и обширные
аномалии в почвах. Сюда входят свинец, сурьма, частично
медь. Из них особо следует отметить свинец — элемент транс-
портных выбросов и один из опаснейших загрязнителей окру-
жающей среды городов (входит в первый класс опасности по
гигиеническим оценкам).
4. Элементы с высокими средними концентрациями в выпа-
дениях, на фоне которых распространены участки с очень вы-
сокими и даже экстремальными значениями. Эти элементы
создают обширные аномальные поля и локальные экстремаль-
но-аномальные центры. К ним могут быть отнесены ртуть, мо-
либден, цинк, никель, вероятно, кадмий. Из этих элементов
наиболее опасны ртуть и кадмий.
В 1978 г. Г. И. Сидоренко выдвинул концепцию о макси-
мально допустимой нагрузке на население с учетом всех факто-
6—568
81
ров окружающей среды. В этих исследованиях широко изуча-
ется содержание химических элементов в диагностических био-
субстратах человека (кровь, моча, слюна, желчь, волосы, ног-
ти, зубы), а также в «критических» органах (печень, почки,
костная ткань и др.). Эти показатели используются в качестве
необходимых сведений о рекомендуемых и допустимых уровнях
некоторых химических элементов.
* * $
Подразделение МЭ на эссенциальные и токсичные в значи-
тельной степени условно. Так, некоторые в основном токсичные
элементы (мышьяк, свинец и даже кадмий) некоторыми авто-
рами относятся к эссенциальным по крайней мере для лабора-
торных животных. Вместе с тем некоторые, казалось бы, сугубо
эссенциальные элементы при определенных условиях могут вы-
зывать симптомы интоксикации (медь, марганец, селен, молиб-
ден, йод, кобальт). Следует особо подчеркнуть, что результат
действия многих МЭ в большой степени зависит от возраста
человека, а также от функциональной активности реципииру-
ющих, детоксицирующих и элиминирующих систем. В 1975 г.
А. И. Арчаков обобщил свои обширные исследования о деток-
сикации многих ксенобиотиков, в том числе и некоторых МЭ,
с помощью микросомального окисления.
Ряд авторов склонны относить многие МЭ к ксенобиотикам.
В соответствии с содержанием этого термина они должны быть
действительно чужеродными веществами. Так ли это? По-види-
мому, в настоящее время большую часть МЭ, а может быть и
все (?), не следует называть ксенобиотиками, так как вопрос
об их эссенциальности интенсивно изучается.
Глава 5
МИКРОЭЛЕМЕНТОЗЫ ЧЕЛОВЕКА.
ОПРЕДЕЛЕНИЕ ТЕРМИНОВ. КЛАССИФИКАЦИЯ
Заболевания, вызываемые токсическим действием веществ,
находящихся в организме в очень малых количествах, известны
с античных времен, и каждое из них называлось по имени того
химического элемента, с которым было связано их происхож-
дение. Например, отравление ртутью и свинцом именовали со-
ответственно меркуриализмом и сатурнизмом.
Современная научная медицина, начиная с XX века, обра-
тила внимание на те болезни, происхождение которых было
связано с недостаточностью поступления и содержания в орга-
низме определенных веществ, в частности металлов. Одной из
первых болезней этой природы стали железодефицитные состоя-
ния, главным образом анемии. Применение железа при анемии
в XIX веке имело чисто эмпирический характер, так как сведе-
ния о его исключительной биологической роли стали известны
значительно позже. В медицине прошлых веков в виде полуми-
стических представлений высказывались идеи о нарушениях
баланса различных элементов в организме. Истоки этих пред-
ставлений восходят к древнему учению о жидкостях живого
тела.
Благодаря наблюдательности врачей выделены многие болез-
ни, развитие которых сперва связывали с гипотетической, а
впоследствии с вполне доказанной недостаточностью опреде-
ленных веществ. Первым научно доказанным заболеванием,
вызванным элементной недостаточностью, стал эндемический
зоб, после того как в 1895 г. Е. Baumann подтвердил гениаль-
ную догадку J. L. Prevost и А. С. Maffoni (1846) и показал,
что при этом патологическом процессе количество йода в щито-
видной железе оказывается уменьшенным.
В течение многих десятилетий XX века не существовало
никаких попыток объединить между собой такие разнообразные
заболевания, как железодефицитная анемия, эндемический зоб
и отравления тяжелыми металлами. Это происходило потому,
что еще не существовало самого учения о МЭ, получившего на«
учную базу только после новаторских работ В. И. Вернадско-
го. Краткий очерк становления этого крупного научного обоб-
щения в химии, биологии и медицине изложен в первой главе
нашей книги. В настоящее время учение о МЭ — это один из
наиболее активно развивающихся разделов медицины и биоло-
6*
83
гии, многие стороны которого глубоко разработаны и приносят
реальную пользу практическому здравоохранению и всему
комплексу наук сельскохозяйственного профиля. Как уже ука-
зывалось, в истории учения о МЭ сложилось парадоксальное
положение: вызываемые ими патологические процессы стали
известны человечеству за несколько тысяч лет до открытия са-
мих МЭ.
С учетом достижений современной науки настало время
предложить объединяющее название для всех патологических
процессов, вызванных дефицитом, избытком и дисбалансом МЭ
в организме. Таким названием, по нашему мнению, является
термин «микроэлементозы» (МТОЗы) [Авцын А. П. и др.,
1983]. Что же выигрывает практический врач от этого предло-
жения и выигрывает ли он вообще? По нашему мнению, ответ
на этот вопрос должен быть утвердительным. Врач приобрета-
ет одно короткое и вытекающее из современного уровня разви-
тия науки обозначение для логичного объединения обширного
круга внешне очень разнообразных патологических процессов,
имеющих в действительности принципиально единую химичес-
кую этиологию. Применение этого термина центрирует внима-
ние исследователей на причине явлений и таким образом по-
зволяет глубже понять их закономерные следствия, которыми
в данном случае являются многие болезни, синдромы и пато-
логические признаки.
Несмотря на то что термин «микроэлементозы» введен в
литературу недавно, можно констатировать, что он легко при-
нимается представителями многих специальностей. Это объяс-
няется не только его конкретностью и простотой, но и назрев-
шей необходимостью предложить обозначение, позволяющее
объединить многочисленные факты и, более того, поставить
всю эту проблему на рельсы клинической медицины и практи-
ческой эпидемиологии. Термин «микроэлементозы» приняли не
только врачи, но и биологи, а также научные работники сель-
скохозяйственного профиля.
Предложенная нами рабочая классификация микроэлемен-
тозов человека (табл. 8) потребовала разработки определенной
терминологии, которая в основном является общемедицинской.
Прежде всего необходимо остановиться на принципе выде-
ления приемлемого для всех обозначения любых МТОЗов.
С нашей точки зрения, таким принципом должно быть перво-
очередное выделение этиологического фактора, имеющего в
данном случае химическую природу. Каждый МТОЗ следует
именовать в соответствии с латинским (в отдельных случаях
с греческим) названием МЭ, дефицит или токсическое дейст-
вие которого вызвало заболевание. Таким образом выделяются
купродефициты (медьдефицитные болезни, синдромы, состояния
и признаки), цинкдефициты (цинкдефицитные болезни, синдро-
мы, состояния и признаки), манганодефициты (марганецдефи-
цитные болезни, синдромы, состояния и признаки), хромдефи-
цитные заболевания, синдромы, состояния, признаки), селенде-
84
Таблица 8
Микроэлементозы человека
МТОЗы F Основные формы заболеваний Краткая характеристика
Природные 1. Врожденные При врожденных микроэлементозах в осно-
эндогенные 2. Наследствен- ные ве заболевания может лежать микроэле- ментоз матери При наследственных микроэлементозах не- достаточность, избыток или дисбаланс МЭ вызываются патологией хромосом или ге- нов
Природные 1. Вызванные де- Природные, т. е. ие связанные с деятель-
экзогенные фицитом МЭ 2. Вызванные из- бытком МЭ 3. Вызванные дис- балансом МЭ ностью человека и приуроченные к опреде- ленным географическим локусам эндемиче- ские заболевания людей, нередко сопро- вождающиеся теми или иными патологи- ческими признаками у животных и расте- ний
Техноген- 1. Промышлен- Связанные с производственной деятель-
ные ные (профессио- нальные) 2. Соседские 3. Трансгрессив- ные ностью человека болезни и синдромы, вызванные избытком определенных микро- элементов и их соединений непосредствен- но в зоне самого производства; по соседству с производством; в значительном отдалении от производст- ва за счет воздушного или водного пе- реноса микроэлементов
Ятрогенные 1. Вызванные де- фицитом МЭ 2. Вызванные из- бытком МЭ 3. Вызванные дис- балансом МЭ Быстро увеличивающееся число заболева- ний и синдромов, связанных с интенсивным лечением (пероральным, парентеральным, чрескожным, ингаляционным) разных бо- лезней препаратами, содержащими МЭ, а также с поддерживающей терапией (на- пример, с полным парентеральным пита- нием) и с некоторыми лечебными процеду- рами— диализом, не обеспечивающим ор- ганизм необходимым уровнем жизненно важных микроэлементов
фициты (селендефицитные заболевания, синдромы, состояния,
признаки), молибденодефициты (молибдендефицитные заболе-
вания, синдромы, состояния, признаки), йоддефициты (йоддефи-
цитные заболевания, синдромы, состояния, признаки). Следует
учесть, что изредка употребляются термины «гипойодоз», «ги-
пойодное состояние», против чего нет оснований возражать.
Логично выделение железодефицитов, или ферродефицитов.
Однако приходиться считаться с тем, что эти болезни уже име-
ют традиционное название «гипосидерозы», или «сидеропени-
ческие заболевания», синдромы, состояния и признаки. В на-
стоящее время не надо настаивать на их отмене. Дальнейшая
практическая деятельность покажет, какие термины окажутся
более «жизнеспособными» и будут в конце концов приняты
ВОЗ по рекомендации соответствующих экспертов.
i 85
Необходимо оговорить также то, какое содержание следует
вкладывать в термин «микроэлементный дефицит». Разумеется,
речь идет не о полном отсутствии данного МЭ, а об его недо-
статочном для оптимального здоровья содержании в организ-
ме. Микроэлементные дефициты могут быть манифестирующи-
ми, явными, т. е. клинически выраженными, а также латентны-
ми и потенциальными. В последнем случае микроэлементный
дефицит выявляется с помощью лабораторных исследований
или становится манифестирующим под влиянием различных
стрессов.
Аналогичным образом формируются названия заболеваний,
синдромов, состояний и признаков, вызванных дефицитом ус-
ловно-эссенциальных МЭ. Разумеется, что среди исследовате-
лей и практических врачей существует различное отношение к
доказанности эссенциального характера некоторых элементов.
Очень непривычно считать, что один из классических ядов —
мышьяк — в то же время (но совсем в иных дозах и продол-
жительности экспозиции) может быть жизненно необходимым
МЭ. Требуется'время для того, чтобы убедиться в правильности
новых данных об эссенциальности ряда МЭ, полученных только
в эксперименте на животных и о правомерности распростране-
ния их на человеческие популяции.
В настоящее время в эссенциальности фтора и кремния,
следовательно, и в необходимости выделения соответствующих
дефицитных состояний трудно сомневаться. Провизорно выде-
ляемые нами заболевания фтордефицитной природы с рацио-
нальным названием «гипофторозы», приуроченные к различным
периодам жизни человека, отображают вполне реальные пато-
логические процессы. При этом они имеют прямое отношение
к таким часто встречающимся заболеваниям, как кариес зубов
и возрастной остеопороз. Медицинская практика показала, что
проблема кобальтового дефицита (акобальтоза, гипокобальто-
за) полностью покрывается проблемой недостаточности вита-
мина В12. Хорошо известно, что купирующее действие кобальта
при пернициозной анемии достигается только при введении в
организм не самого кобальта, а именно цианкобаламина.
Разработка терминологии микроэлементных токсикопатий
(микроэлементных токсикозов) должна руководствоваться тем
же этиологическим принципом. Следовательно, необходимо вы-
делять алюминиевую, кадмиевую, свинцовую, ртутную, берил-
лиевую, бариевую, висмутовую и другие токсикопатии, т. е. ин-
токсикации, вызванные токсичными МЭ. Наиболее широкое
употребление в профессиональной патологии имеет термин
«интоксикация». Определенного внимания заслуживают также
«микроэлементный токсикоз» и малоупотребляемый термин
«МЭ-токсикопатия», где в каждом конкретном случае вместо
общего обозначения «МЭ» ставится латинское (иногда гречес-
кое) название токсичного МЭ (например, А1-токсикоз, Cd-ток-
сикоз, Hg-токсикоз, Pb-токсикоз и др.). Возможно, целесооб-
разно сохранить термины «меркуриализм» и «сатурнизм»
86
(«плюмбизм») для обозначения именно профессиональных ин-
токсикаций.
Определенные терминологические сложности возникают при
появлении новых публикаций, авторы которых отходят от об-
щепринятого выделения девяти эссенциальных МЭ и добавляют
к ним новые. Так, в частности, поступили A. Shenkin и
G. S. Fell (1985). Вслед за ними М. Апке и В. Groppel (1987)
отнесли к числу классических эссенциальных МЭ фтор, а к
вновь открытым эссенциальным МЭ — кремний, олово, вана-
дий, никель, мышьяк, кадмий, литий и свинец. Важно замеча-
ние этих авторов о том, что симптомы дефицита некоторых
элементов, в частности молибдена и марганца, обнаружива-
ются только в случае парентерального питания или при гене-
тической неполноценности. Большое значение приобретают дан-
ные этих исследователей о том, что при определенных услови-
ях у таких эссенциальных элементов, как молибден, медь,
цинк, железо и марганец, может проявиться токсическое дейст-
вие, которое у животных является крайне различным, а иногда
видоспецифическим. Так, в практике животноводства обнару-
жено, что добавка молибдена к рациону может не только вы-
зывать молибденовую интоксикацию, но и индуцировать вто-
ричную медную недостаточность.
Большие, но все же преодолимые трудности возникают при
разработке номенклатуры заболеваний человека, связанных с
дисбалансом МЭ, а также с нарушением их обмена, завися-
щих от различных макроэлементов. Этот вопрос нами не рас-
сматривается.
Последовательно отстаивая приоритет этиологического
фактора при разработке номенклатуры МТОЗов, мы сознатель-
но стремимся к выделению определенных нозологических еди-
ниц и хорошо охарактеризованных синдромов. Особое внима-
ние должно быть уделено микроэлементный дефицитным со-
стояниям организма, многие из которых не обнаруживают зна-
чительной динамики. Часть их, как уже указывалось, имеет
только потенциальную опасность для здоровья.
В свое время И. В. Давыдовский, работая над курсом част-
ной патологической анатомии, заметил, что «нозологический
синтез требует органопатологического анализа». Это замеча-
ние, несомненно, справедливо и полностью приложимо к нозо-
логическому синтезу МТОЗов, который является одной из ос-
новных задач данной монографии. Действительно, железодефи-
цитные заболевания обладают крайне разнообразной органо-,
гисто- и цитопатологией. В нее входят не только аномалии
кроветворения, но и умеренные и тяжелые поражения эпителия
слизистой оболочки носа, верхних отделов пищеварительного
тракта, включая предраковые изменения пищевода при синдро-
ме Пламмера — Винсона, нарушения кератиногенеза, обуслов-
ливающие койлонихии и т. д. Характерная органопатология
при эндемическом зобе настолько привлекает внимание клини-
цистов, что она в определенной степени даже заслоняет веду-
87
щий этиологический фактор этого массового заболевания, ка-
ким является йодная недостаточность. Настаивая на приорите-
те этиологии при выделении нозологических единиц при раз-
работке номенклатуры патологических процессов, вызванных
дефицитом или избытком МЭ, мы отчетливо представляем себе
огромную практическую полезность органопатологического ана-
лиза и значительную сложность патогенеза каждой нозологи-
ческой формы, каждого синдрома. Крайняя и тем более при-
митивная трактовка каждого заболевания микроэлементной
природы как простейшей нехватки полезного вещества не мо-
жет встретить поддержки. Очень поучительные примеры в этом
отношении дает нам изучение генетики МТОЗов. Достаточно
вспомнить, что существует не менее 5 типов наследственно
обусловленных нарушений поглощения йода и синтеза тиреоид-
ных гормонов, которые могут встречаться при семейных фор-
мах эндемического зоба.
Дефицит многих МЭ связан не только с их недостаточным
поступлением в организм, но и с интенсивностью их всасыва-
ния, с неполноценностью транспорта и метаболизма, с наруше-
ниями активности специфических лигандов и клеточных рецеп-
торов, словом, с деятельностью очень многих систем организ-
ма, которые вовлекаются в патологический процесс на разных
стадиях его развития. Также сложны этапы патологических
процессов при действии токсичных МЭ. Они подлежат конкрет-
ному преимущественно экспериментальному изучению, причем
ведущая роль должна принадлежать биохимическим исследо-
ваниям. С нашей точки зрения важное значение имеет не толь-
ко органопатологический, но и клеточно-молекулярный анализ
нарушений, вызванных избытком (токсическим действием) и
дефицитом МЭ. Именно он должен выяснить глубинные меха-
низмы этих процессов.
К сказанному следует добавить, что многообразие патогене-
тических механизмов при различных вариантах микроэлемент-
ных дефицитов и МЭ-токсикозов не колеблет их нозологическо-
го единства, которое является отправной точкой для их разгра-
ничения, более глубокого понимания и разработки методов их
рациональной профилактики и терапии.
Эндогенные генетические и врожденные МТОЗы (болезнь
Вильсона — Коновалова, болезнь Менкеса, синдром Марфана,
синдром Элерса — Данло) представляют собой сложные ано-
малии обмена меди, давно известные клиницистам. Наряду с
ними существуют менее изученные заболевания, вероятно, той
же природы, связь которых с эндогенным дефицитом меди
окончательно не установлена.
Среди экзогенных МТОЗов биогеохимического происхожде-
ния мы выделяем: 1) природные, 2) техногенные и 3) ятроген-
ные заболевания. Первые из них связаны с характерными осо-
бенностями местной биогеохимической среды. Таковы, в част-
ности, природные эндемии флюороза, селенотоксикоза и селе-
нодефицита, бороза и др., имеющие свою в настоящее время
88
уже довольно определенную географию [Ковальский В. В.,
1980; Ковальский В. В., Сусликов В. А., 1980; Авцын А. П.,
Жаворонков А. А., 1981, 1987].
Среди техногенных эндемий предлагается различать про-
мышленные, соседские и трансгрессивные МТОЗы. Отдельные
формы промышленных МТОЗов давно известны профпатоло-
гам. Заслуживает особого внимания тот факт, что с годами
промышленные МТОЗы становятся все более разнообразными.
Как следует из названия, соседские МТОЗы поражают лиц,
не принимающих прямого участия в производстве, а прожива-
ющих по соседству. Очень часто эти МТОЗы развиваются у
членов семей рабочих вредных производств. Как правило, они
нерезко выражены и могут принимать скрытую и непрогреди-
ентную форму тех или иных патологических состояний, иногда
это только микросимптомы или небольшие аномалии развития
у детей.
Термин «трансгрессивные МТОЗы» введен нами впервые в
1983 г. и заимствован из геологии, в которой им обозначается
наступание моря на сушу (от лат. transgressio — переход, пере-
движение)1. Трансгрессивные техногенные заболевания хими-
ческой природы стали известны сравнительно недавно в связи
с резким возрастанием промышленного загрязнения внешней
среды в индустриально развитых странах. Вспышки их регист-
рируются не по соседству с предприятиями, а на значительном
отдалении от них, причем перенос специфической химической
вредности (в том числе и аэрозолей МЭ) осуществляется за
счет атмосферной или водной циркуляции. Примерами этих
заболеваний людей, животных и растений являются разнооб-
разные патологические процессы, возникающие в результате
массивного выброса в атмосферу в основном сернистых и азо-
тистых окислов, а также других токсичных веществ. Огромное
загрязнение водной среды ртутью имело место в Японии с по-
следующим отравлением морских животных и произраставших
в воде растений. Трансгрессивные МТОЗы непосредственно
связаны с проблемой кислотных дождей, которая имеет по су-
ществу глобальное значение.
Выделяя МТОЗы для их прицельного изучения, мы, разуме-
ется, понимаем, что многие заболевания этой природы имеют
сложное происхождение и зависят не только от МЭ, но и от
очень многих других эндо- и экзогенных факторов. В частнос-
ти, недавно выяснилось, что динамика поглощения и распреде-
ления МЭ находится под влиянием ряда гормонов.
В последние годы возросло значение ятрогенных МТОЗов.
Современная терапия использует МЭ в качестве лечебных пре-
паратов или добавок к различным медикаментам. МЭ могут
загрязнять смеси, употребляющиеся при полном парентеральном
1 Существующий в гидрометеорологии термин «трансграничный» употреб-
ляется для обозначения переноса химических загрязнителей атмосферой через
границы соседних стран.
89
питании. Многие МЭ в течение десятилетий считались без-
вредными. Однако более прицельное и длительное изучение вы-
явило их потенциальную токсичность. Последняя не всегда
объясняется большой дозой МЭ, а может быть связана с инди-
видуальной чувствительностью организма. Эта чувствительность
может иметь и аллергическую природу. Она также может быть
связана с врожденной или приобретенной неполноценностью
физиологических аппаратов обезвреживания или специфичес-
кой элиминации токсичного начала. Существенную роль в воз-
растании числа ятрогенных МТОЗов сыграло стремление вра-
чей к более активному и часто парентеральному применению
лекарственных веществ.
Выделение микроэлементных токсикопатий и микроэлемент-
ных дефицитов позволяет организовать целенаправленные про-
филактические мероприятия против первых и осуществлять
специфическую терапию вторых. Из приведенных данных видны
сложность разработки систематики каждого из МТОЗов и вы-
сокий уровень компетенции врачей, необходимый для их выяв-
ления. Обобщающие теоретические концепции о значении МЭ
в жизненных циклах организмов нуждаются в новых фактах и
организации специальных исследований с использованием всех
возможностей мультидисциплинарного подхода. В связи с этим
суммация и анализ соответствующей литературы для точной
индивидуализации каждого МТОЗа и разработки методов их
профилактики и лечения крайне необходимы.
Рассматривая болезни и синдромы биогеохимической при-
роды, мы выделяем мономикроэлементозы, т. е. заболевания,
обусловленные в основном избытком или дефицитом одного
МЭ (например, алюминозы, селенозы, флюороз и др.). В то
же время необходимо выделить полимикроэлементозы— забо-
левания, в этиологии которых существенную роль играют не-
сколько МЭ или их дисбаланс с некоторыми макроэлементами.
К ним относятся такая массовая болезнь человечества, как ка-
риес зубов, широко распространенная группа мочекаменных
болезней, весьма гетерогенная по этиологии и патогенезу группа
эндемического зоба, группа анемий сложной биогеохимической
природы (включая железодефицитные), а также давно извест-
ная, но до сих пор мало понятная болезнь Кашина — Бека.
Следует обратить внимание на определенную условность
термина «мономикроэлементоз». В случаях тяжелых и хрони-
ческих отравлений ртутью, свинцом, мышьяком и другими ток-
сичными МЭ, которые сохраняют значение ведущего фактора
в течение всего патологического процесса, целесообразность
применения термина «мономикроэлементоз» достаточно ясна.
Однако при других МТОЗах дело обстоит сложнее. Например,
при выраженных кадмиозах именно интоксикация кадмием яв-
ляется главным, поддерживающим патологический процесс,
фактором; вместе с тем при кадмиевой токсикопатии обязатель-
но наступают последовательные изменения обмена других МЭ,
Хорошо известно перераспределение цинка и даже истощение
90
его запасов в организме при нагрузке кадмием. Таким обра-
зом, кадмиевая токсикопатия одновременно является примером
и цинкдефицитного состояния, так как метаболизм цинка при
этом резко нарушается.
Понятие о полимикроэлементозах недостаточно разработано
и должно использоваться только в доказанных случаях, когда
заболевание действительно является результатом сочетанного
воздействия нескольких МЭ. Не подлежит сомнению, что и за
пределами заболеваний, вызванных МЭ, любой патологический
процесс сопровождается теми или иными изменениями обмена
и содержания многих элементов, в том числе довольно значи-
тельными. Однако, если эти изменения не имеют значения ве-
дущих, а представляют собой лишь фоновые явления, нет до-
статочных оснований относить их к полимикроэлементозам.
Наиболее доказательными в качестве болезней микроэле-
эденг.юй недостаточности являются цинкдефицитные состояния.
Следует особо подчеркнуть их крайнее разнообразие и приуро-
ченность к определенным периодам индивидуального развития.
Если цинкдефицитные состояния способны вызывать разнооб-
разную патологию, преимущественно у детей и лиц молодого
возраста, то нейроалюминозы чаще встречаются в пожилом и
старческом возрасте, а также при ятрогенных воздействиях.
Если цинкдефицитные состояния в большинстве случаев не
имеют прямой связи с профессиональными факторами, то кад-
миозы, напротив, являются МТОЗами, которые в значительной
степени зависят от производства, а также от интенсивного за-
грязнения внешней среды.
Не подлежит сомнению, что значительная часть МТОЗов
еще не выделена, и мы должны ожидать новых открытий в
этом направлении. Еще в меньшей степени определен удель-
ный вес каждого из МТОЗов в качестве факторов, отягощаю-
щих течение болезней другого происхождения. В связи с этим
очень перспективным и практически важным является понятие
«вторичный микроэлементов», который может присоединяться
к основному заболеванию на различных стадиях его течения
или иметь ятрогенное происхождение. В частности, это забо-
левание может быть следствием хирургических операций на
желудочно-кишечном тракте с повреждением или даже выклю-
чением основных зон всасывания МЭ. Вторичные МТОЗы мо-
гут также развиться при таких тяжелых гельминтозах, как опи-
сторхоз и особенно дифиллоботриоз, при котором у Уз больных
возникает перциниозная анемия, поддающаяся лечению циапо-
кобаламином.
Известно, что перенасыщение окружающей среды опреде-
ленными МЭ может привести к их более или менее выражен-
ному накоплению в организме. Это не всегда вызывает клини-
чески выраженные ответные реакции. Наиболее интенсивные
загрязнения воды, воздуха, пищи способны вызывать вторичный
микроэлементный гисто- и цитотесаурисмоз.
Глава 6
ВАЖНЕЙШИЕ ЭССЕНЦИАЛЬНЫЕ МИКРОЭЛЕМЕНТЫ
И СВЯЗАННЫЕ С НИМИ ЗАБОЛЕВАНИЯ
В эту группу Р. I. Aggett (1985) относит следующие МЭ:
железо, медь, цинк, марганец, хром, селен, молибден, йод, ко-
бальт. Другие авторы главным образом на основании экспери-
ментальных данных указывают на значительно большее число
эссенциальных МЭ. Особенно важны в этом отношении опыты
К. Schwarz (1972). Он доказал эссенциальность многих МЭ
с помощью сложных экспериментов, в основе которых было
полное предотвращение любых микроэлементных загрязнений
внешней среды, в том числе пищи и воздуха. Воспроизведение
«феномена эссенциальности», в частности поддержания жизни,
нормального роста и развития, репродуктивной способности,
предотвращения заболеваний и преждевременного летального
исхода, было получено и у потомства некоторых лабораторных
и сельскохозяйственных животных [Anke М. et al., 1987]. Этот
автор различает «классические эссенциальные» МЭ, перечень
которых совпадает с приведенным выше (с добавлением фто-
ра), и так называемые новые эссенциальные МЭ (кремний,
олово, ванадий, никель, мышьяк, кадмий, литий, свинец). Эта
точка зрения пока еще не является общепринятой.
Изложение материала этой главы построено по единому
плану — доказательства эссенциальности МЭ, распространен-
ность их в природе, всасывание, транспорт и выделение из ор-
ганизма, физиологическая роль и патологические процессы,
обусловленные дефицитом и избытком МЭ в организме чело-
века и животных.
6.1. Железо
Железо — широко распространенный в природе элемент, ра-
нее его относили к макроэлементам в связи с относительно вы-
соким содержанием в организме (4,2 г в теле «стандартного»
человека). Если, однако, исключить гемоглобиновое железо,
составляющее 75—80% от его общего количества, то концент-
рация этого элемента в тканях окажется равной и даже мень-
шей, чем концентрация такого типичного МЭ, как цинк. Уста-
новлено, что 20—25% железа являются резервными, 5—10%
входят в состав миоглобина, около 1 % содержится в дыхатель-
92
ных ферментах, катализирующих процессы дыхания в клетках
и тканях. Поскольку МЭ осуществляют свою биологическую
функцию главным образом в составе других биологически ак-
тивных соединений, преимущественно ферментов, то железо,
входящее в состав более 70 различных по своей функции фер-
ментов, следует также признать истинным МЭ. Это принято в
большинстве современных специальных руководств [Mertz W.,
1987J.
Железосодержащие биомолекулы выполняют четыре основ-
ные функции: 1) транспорт электронов (цитохромы, железо-
серопротеиды); 2) транспорт и депонирование кислорода (мио-
глобин, гемоглобин, эритрокруорин, гемэритрин, хлорокруорин);
3) участие в формировании активных центров окислительно-
восстановительных ферментов (оксидазы, гидроксилазы, су-
пероксиддисмутаза и Др.); 4) транспорт и депонирование железа
(сидерофилины и среди них наиболее изученный трансферрин,
гемосидерин, ферритин, сидерохромы).
Цитохромы — это гемопротеиды, выполняющие функцию
электронного переноса в процессах дыхания и фотосинтеза,
основанную на обратимом окислении атома железа. В зависи-
мости от типа гема и его спектральной характеристики выделя-
ют четыре (а, Ь, с и d) группы цитохромов. Типы гемов разли-
чаются боковыми цепями порфириновых структур. Цитохромы
а содержат формильную боковую группу, цитохромы b — про-
тогем, нековалентно связанный с белком, цитохром с — гем,
ковалентно связанный двумя цистеиновыми остатками одной
полипептидной цепи, цитохром d содержит дигидропорфирин
(хлорин) [Диксон М., Уэбб Э., 1982J. Различия в пределах
группы, вызванные особенностями структуры апопротеида, обо-
значаются порядковым индексом или величиной длины волны
поглощения света в видимой области. Цитохромы с и аа3 при-
сутствуют в митохондриальной дыхательной цепи, и Р-450—
в микросомах клеток животных, остальные индексиро-
ванные цитохромы и цитохром d(a2) —в растительных и бакте-
риальных клетках. Цитохром Ь3 является переносчиком элект-
ронов для десатуразной системы эндоплазматической сети.
В ней и в митохондриях коры надпочечников обнаруживается
также цитохром Р-450, являющийся типичной монооксигеназой,
участвующей в процессах гидроксилирования стероидов, отщеп-
пления боковой группы холестерина, гидроксилирования широ-
кого круга ксенобиотиков.
В печени млекопитающих присутствует несколько форм ци-
тохрома Р-450, прочно связанных с мембраной агранулярной
эндоплазматической сети и непосредственно реагирующих с
флавопротеидами. Под влиянием отдельных лекарственных пре-
паратов их синтез резко возрастает, например фенобарбитал и
бензофлавон индуцируют 20-кратное повышение активности ци-
тохрома Р-450. Еще большая активность индуцируется диокси-
ном, чем, по-видимому, объясняется хроническое токсическое
действие малых доз этого соединения.
93
Роль цитохрома Р-450 заключается, видимо, в превраще-
нии путем гидроксилирования ксенобиотика в легко экскрети-
руемую форму. Однако гидроксилирование может сообщать
веществу канцерогенные свойства, как, например, в случае ме-
тилхолантрена.
Железосеропротеиды — новая важнейшая группа электрон-
переносящих белков. Они содержат негемовое железо, связан-
ное четырьмя лигандами с серой, образующими кластер с тет-
раэдрической или искаженной тетраэдрической геометрией. Се-
ра кластера принадлежит как цистеиновым остаткам полипеп-
тидной цепи, так и неорганической «лабильной» сере (исключая
рубредоксин). Железосеропротеиды подразделяют в зависи-
мости от числа атомов железа в кластере. В эту группу входят
сукцинатдегидрогеназа, дегидрогеназа электронпереносящего
флавопротеида, NADH-дегидрогеназа и другие ферменты.
Важнейшим кислородпереносящим белком позвоночных яв-
ляется гемоглобин, характеристику которого можно найти в
учебниках физиологии и биохимии. Столь же хорошо изучен
и миоглобин, обеспечивающий депонирование кислорода и его
перенос на дыхательные ферменты.
К оксидоредуктазам, содержащим гемовое железо, относятся
обширные семейства пероксидаз и каталаз. Пероксидазы ката-
лизируют окислительные реакции, в которых в качестве окис-
лителя участвуют перекись водорода или замещенные перекиси.
Из многочисленных представителей этой группы ферментов у
млекопитающих наиболее хорошо изучены лактопероксидаза,
тиреопероксидаза и миелопероксидаза [Hewson W. D., Hager
L. Р., 1979]. Лактопероксидаза, выделенная первоначально из
коровьего молока, была обнаружена затем в слюнных, слезных
и гарднеровых железах, обладающих, кроме того, способностью
концентрировать йод. Биологическая роль фермента недоста-
точна ясна. Он с особой легкостью катализирует окисление
йодидов, в связи с чем используется в лабораторной практике
для метки белков радиоактивным йодом. Допускается также его
участие в образовании меланина.
.Тиреопероксидаза прочно связана с мембранами эндоплаз-
матической сети фолликулярных эндокриноцитов щитовидной
железы и принимает участие в биосинтезе йодтиронинов. Мие-
лопероксидаза — гемопротеид, выделенный из лейкоцитов, гноя,
хлоромы крыс, костного мозга морских свинок. Основной функ-
цией фермента является его участие в процессах фагоцитоза и
лизиса микроорганизмов. Миелопероксидаза находится в азур-
офильных гранулах нейтрофилов и выделяется в фагоцитирую-
щие вакуоли в процессе лизиса гранул [Пигаревский В. Е.,
1982]. Реакция окисления субстратов пероксидазой протекает
в три этапа: нативный фермент + Н2О2-> соединение I. Соеди-
нение 1 + АН2->-соединение П + АН. Соединение II4-АН2-> на-
тивный фермент+АН. Соединение I содержит на два, а соеди-
нение II — на один окислительный эквивалент больше, чем
нативный фермент.
94
Таблица 9
Железопротеиды и ферменты человека и животных,
не содержащие железосерных центров
Наименование Источник полу- чения Молекулярная масса Число атомов железа Простегнческая группа
Цитохром а Сердце крупного 93 000 1 Си, гем А
Цитохром b рогатого скота 93 000 1 Протопорфи-
Цитохром с 13 000 1 рин IX Гем С
Фенила ланингид- Печень крысы 100 000 (димер) 2 Fe (II), птерин
Лактопероксида- Коровье молоко 78 000 (димер, 2 Мезопорфи-
за Тиреоидперокси- Щитовидная желе- гликопротеид) 200 000 (глико- 2—3 рин IX Протопорфи-
даза за протеид) рин IX
Миелопероксида- Лейкоциты 104 000 (димер) 2 Гем А
за Каталаза Эритроциты круп- 250 000 4 Протопорфи-
Цитохром Р-450 кого рогатого ско- та Микросомы печени 46 000 1 рин IX Протопорфи-
Гемоглобин Эритроциты мле- 67 000 4 рин IX Протопорфи-
Миоглобин копитающих Мышцы млекопи- 16 000 1 рин IX Протопорфи-
Эритрокроурин тающих Arenicola 3 000 000 192 рин IX Гем
Гемэритрин Sipunculidae 108 000 (окта- 16 —
Трансферрин Кровь млекопита- мер) 79 000 2
Лактоферрин Кональбумин ющих Молоко Яичный белок 77 000 2 2 ... _
(овотрансферрин) Фосвитин плазма крови птиц Яичный желток 35 000 47 —
Ферритин Печень млекопи- 440 000 4500 —
Гастроферрин тающих Желудочный сок 260 000 —
Каталаза, выделенная из эритроцитов крупного рогатого
скота, катализирует реакцию 2Н2О2->-2Н2О + О2, также прохо-
дящую с образованием промежуточных соединений. При высо-
кой концентрации Н2О2 она катализирует разложение этого со-
единения, а при низкой концентрации в присутствии донора
водорода (метанол, этанол и др.) у нее преобладает перокси-
дазная активность. Рассмотренные соединения железа пред-
ставлены в табл. 9.
Транспорт и депонирование железа осуществляется группой
белков, получивших название «сидерофилины». Это негемовые
гликопротеиды с молекулярной массой около 77 000 и содержа-
нием углеводов около 6%. К ним относится трансферрин из
плазмы крови позвоночных, лактоферрин молока и кональбу-
мин (овотрансферрин) из крови и яиц птиц. Сидерофилины
95
различаются по физическим, химическим и иммунологическим
свойствам, но все имеют по два участка связывания трехва-
лентного железа. Связь железа с белком в сидерофилинах сла-
бее, чем в ферритине. Наиболее изученным представителем
этой группы является трансферрин, относящийся к |3-глобули-
нам. У человека известно 15 генетических вариантов трансфер-
рина, из которых чаще других встречаются варианты А, В и С.
Главная функция трансферрина — это транспорт всосавшегося
железа в его депо (печень, селезенка), в ретикулоциты и их
предшественники в костном мозге. Трансферрин способен так-
же связывать ионы других металлов (Zn, Со, Ga и др.). Транс-
феррин связывается специфическими рецепторами на мембра-
нах ретикулоцитов, отдает клетке железо и возвращается в
циркуляцию в виде лишенного железа апотрансферрина. Из
общего количества трансферрина в организме человека только
25—40% содержит железо. Общая железосвязывающая способ-
ность плазмы крови, определяемая содержанием в ней транс-
феррина, составляет у здорового человека 44,7—71,6 мкмоль/л
[Петров В. Н., 1982]. Молекула трансферрина состоит из од-
ной полипептидной цепи с двумя участками связывания желе-
за. Она содержит 678 аминокислотных остатков и две углевод-
ные цепи с молекулярной массой 79 550; 40% N- и С-концевых
остатков идентичны, что свидетельствует о происхождении
трансферрина в результате дупликации гена более простого
железопереносящего белка. N-концевой домен беднее дисуль-
фидными мостиками, а С-домен содержит обе углеводные цепи,
связанные с остатками аспарагина и оканчивающимися остат-
ками сиаловой кислоты. Участок связывания, находящийся
ближе к N-концу, легко отщепляет железо в кислой среде, а
второй, расположенный ближе к С-концу, устойчив к действию
кислот [Huebers Н. et al., 1981].
В плазме крови человека трансферрин присутствует в че-
тырех формах — апотрансферрина, лишенного железа, двух
моноферриформ, содержащих железо в одном из обоих участ-
ков связывания, и диферритрансферрина. Оба вида монофер-
ритрансферрина функционально эквивалентны, но диферри-
трансферрин способен передавать железо ретикулоцитам крыс
вдвое эффективнее, чем мононасыщенные формы. In vivo кисло-
толабильный участок нагружен железом в большей степени,
чем кислотоустойчивый.
Другая важная роль сидерофилинов — ограничение доступ-
ности железа для бактерий и опухолевых тканей, в связи с чем
они представляют собой самостоятельную систему естественно-
го иммунитета. В курином яйце все железо находится в желтке
в составе фосвитина. Но это железо практически недоступно
для бактерий, так как в яичном белке- они подвергаются воз-
действию кональбумина, прочно связывающего железо и ли-
шающего бактерии этого необходимого для их роста элемента.
На данной особенности в значительной степени основаны бак-
териостатические свойства яичного белка.
66
Лактоферрин присутствует в молоке и большинстве других
секретов, является главным белковым компонентом специфиче-
ских гранул полиморфно-ядерных нейтрофилов. Он высвобож-
дается в процессе дегрануляции этих клеток и затем поглоща-
ется макрофагами после связывания с железом, присутствую-
щим в инфицированных участках. В отличие от других сидеро-
филинов лактоферрин способен удерживать железо при pH 4,0
и обеспечивать таким образом его связывание в условиях по-
вышенного содержания в инфицированных участках молочной
кислоты. Лактоферрин способен связывать избыток железа в
пищеварительном тракте. Исходя из этого у животных, обла-
дающих медленным ростом, как, например, у морской свинки,
а также у человека, лактоферрина в молоке значительно боль-
ше (в 20 раз), чем у быстрорастущих животных — крупного
рогатого скота, свиней, крыс и кроликов, нуждающихся для
своего роста в повышенных количествах железа. Высоким со-
держанием лактоферрина в грудном молоке объясняется зна-
чительно большая устойчивость детей, вскармливаемых грудью,
к инфекциям, чем получающих коровье молоко [Weinberg Е.,
1984]. Аполактоферрин оказывает бактериостатическое дейст-
вие, которое утрачивается при его насыщении железом. Име-
ются также данные, что лактоферрин нейтрофильных лейкоци-
тов играет роль в регуляции миелопоэза [Петров В. Н., 1982].
Основным белком млекопитающих, депонирующих железо,
является ферритин. Вместе с гемосидерином он содержит око-
ло 25% железа, присутствующего в организме. Он находится
в клетках печени, селезенки, костного мозга и ретикулоцитах.
Молекула ферритина — это белковая оболочка сферической
формы диаметром 13 нм, состоящая из 24 субъединиц апофер-
ритина двух видов (Н и L) примерно одинакового размера.
Субъединицы различаются по молекулярной массе, раствори-
мости и иммунологически. При значительной общности амино-
кислотного состава L-субъединица богаче лейцином, фенилала-
нином и аргинином. Изоферритины человека, видимо, возника-
ют в результате посттрансляционной модификации. Субъединица
ферритина, выделенного из селезенки лошадей, состоит из 174
аминокислотных остатков с молекулярной массой 19 824. Внут-
ри сферы может находиться до 4300 атомов трехвалентного
железа. В отсутствие фосфата железо находится в форме фер-
ригидрата (5Fe2Os-9H2O) в виде гексагонального кристалла.
Железо поступает внутрь апоферритина через 6 каналов, имею-
щихся в сфере, преимущественно в двухвалентном виде, после
чего окисляется до трехвалентного состояния. Степень насыще-
ния ферритина железом составляет в среднем 2000 атомов на
молекулу.
Создание радиоиммунологических методов определения
ферритина позволило обнаружить этот белок и в сыворотке
крови, где он, по-видимому, выполняет функцию транспорта
железа от ретикулоэндотелиальных к паренхиматозным клет-
кам печени [Петров В. Н., 1982]. Апоферритин способен в при-
7—568
97
сутствии кислорода катализировать окисление Fe2+ в Fe3+. Об-
ратная реакция — восстановительная мобилизация железа фер-
ритина — катализируется NADFH-зависимой оксидоредукта-
зой, в результате чего повышается всасывание железа в кишеч-
нике. Радиоиммунологическое определение ферритина сыворот-
ки крови является удобным тестом для выявления дефицита
железа в организме и хорошо коррелирует с печеночными запа-
сами этого белка.
Гемосидерин — бурое аморфное вещество, содержащее 26—
37% железа от его массы. Полагают, что гранулы гемосидери-
на являются крупными агрегатами молекул ферритина, частич-
но подвергнутого протеолизу. Он также нерастворим в воде.
Относительное содержание обоих соединений в тканях зависит
от общего количества резервов железа. При содержании в пе-
чени железа более 2000 мг/кг в ней начинает преобладать ге-
мосидерин.
В желудочном соке обнаружен железосвязывающий белок,
получивший название «гастроферрин», содержание которого
снижается при дефиците железа и увеличивается при его нор-
мальном и повышенном поступлении с пищей. Существует ги-
потеза, что этот белок принимает участие в регуляции всасыва-
ния железа, однако она недостаточно подкрепляется фактами
[Underwood Е„ 1977].
Сидерохромы — железосодержащие красно-бурые водораст-
воримые метаболиты бактерий. Они включают серию антибио-
тиков-сидеромицинов (альбомицин, ферримицин, даномицин и
др.) и класс соединений, обладающих свойствами факторов
роста для ряда микроорганизмов, сидераминов (феррихром,
копроген, ферриоксамин, феррихризин, феррирубин и др.). Си-
дерамины конкурентно ингибируют сидерохромы катехолатного
типа, синтезируемые анаэробами, и гидроксаматного типа, син-
тезируемые аэробными микроорганизмами. Основная функция
сидерохромов — обеспечение микробных клеток железом. В ус-
ловиях дефицита железа микроорганизмы усиленно продуциру-
ют сидерохромы. Вирулентность микроорганизмов, нуждающих-
ся в железе главным образом для синтеза железосеропротеи-
дбв и цитохромов, в значительной степени зависит от
способности их сидерохромов успешно конкурировать с сиде-
рофилинами организма хозяина за присутствующее в тканях
железо.
Обмен железа. Гомеостаз железа обеспечивается в пер-
вую очередь регуляцией его всасывания в связи с ограниченной
способностью организма к выделению этого элемента. Сущест-
вует выраженная обратнаяз ависимость между обеспеченностью
железом и его всасыванием в пищеварительном тракте,; меха-
низм которого недостаточно изучен. Гипотеза «мукозного бло-
ка», согласно которой усвоение железа регулировалось уровнем
ферритина в клетках слизистой оболочки, не нашла экспери-
ментального подтверждения.
Всасывание железа зависит от: 1) возраста, обеспеченности
98
организма железом, состояния здоровья; 2) состояния желу-
дочно-кишечного тракта; 3) количества и химических форм
поступающего железа и 4) количества и форм прочих компо-
нентов рациона. У животных с однокамерным желудком вса-
сывание железа происходит главным образом в двенадцатипер-
стной кишке преимущественно в двухвалентной форме, хотя
известны и соединения трехвалентного железа, обладающие бо-
лее высокой биологической доступностью, чем некоторые фер-
роформы. Всасыванию негемового железа предшествует, по-
видимому, распад его координационных соединений, тогда как
гемовое железо всасывается в составе порфиринового макро-
цикла. Для оптимального всасывания железа необходима нор-
мальная секреция желудочного сока. Прием соляной кислоты
способствует усвоению железа при ахлоргидрии.
Пищевые компоненты. Аскорбиновая кислота, вос-
станавливающая железо и образующая с ним хелатные комп-
лексы, повышает доступность этого элемента, так же как и
другие органические кислоты. Другим компонентом рациона,
улучшающим всасывание железа, является «фактор животно-
го белка». Опыты с радиоактивным железом свидетельствуют о
его лучшем усвоении в составе рационов, богатых белком жи-
вотного происхождения, по-видимому, в результате образования
легко усвояемых комплексов железа с аминокислотами и в
первую очередь со свободным или пептидно-связанным цистеи-
ном [Morris Е. R., 1987]. Улучшают всасывание железа про-
стые углеводы — лактоза, фруктоза, сорбит, а также такие
аминокислоты, как гистидин, лизин и упомянутый выше цисте-
ин, образующие с железом тридентатные хелаты.
Десферриоксамин — бактериальный сидерофор — является
мощным ингибитором всасывания железа и может быть ис-
пользован при лечении сидерозов. Всасывание железа угнета-
ется фитином при условии, если образуются 2—4-замещенные
фитаты железа. Однозамещенный фитат, образующийся при
избытке в пище фитина, обладает хорошей усвояемостью. Уг-
нетение всасывания негемового железа пшеничными отрубями,
по мнению ряда авторов, объясняется его связыванием фити-
ном, хотя есть данные, что в отрубях присутствуют и другие
компоненты, затрудняющие всасывание этого элемента, как,
например, клетчатка [Simpson К. М. et al., 1981]. Всасывание
железа угнетается соевым белком и фосфатами, а также при
поедании глины, чем, по-видимому, объясняются анемии, на-
блюдаемые при геофагии. Всасывание железа снижают такие
напитки, как чай и кофе, полифенольные соединения которых
прочно связывают этот элемент. В этой связи было предложе-
но использовать чай для профилактики повышенного усвоения
железа при талассемии [De Alcorn Р. A. et al., 1979].
Заболевания. Большое влияние на усвоение железа
оказывают различные заболевания. Оно усиливается при недо-
статочности железа, при анемиях (гемолитической, апластичес-
кой, пернициозной), гиповитаминозе Вй и гемохроматозе, что
7
99
объясняется усилением эритропоэза, истощением запасов железа
и гипоксией [Morris Е. R., 1987]. Снижение усвоения этого
элемента отмечено при трансфузионной полицитемии в резуль-
тате перегрузки организма железом, а также при дефиците ме-
ди и никеля у крыс [Nielsen F. Н., 1987].
Поступление в энтероцит. Большинство современ-
ных представлений о процессах всасывания железа в кишечни-
ке отводит центральную роль двум видам трансферрина — му-
козному и плазменному, между которыми имеются различия в
иммуннохимических свойствах и устойчивости к протеолитиче-
скому действию трипсина [Ward J. Н. et al., 1984]. Из всех
предложенных вариантов экспериментально наиболее 'обосно-
ван взгляд, что мукозный апотрансферрин секретируется энте-
роцитами в просвет кишечника, где он нагружается железом,
после чего проникает в энтероцит. В нем он освобож-
дается от железа, после чего возвращается к исчерченной ка-
емке и вступает в новый цикл [Huebers Н. et al., 1983]. Источ-
ником мукозного трансферрина является, однако, не сам энте-
роцит, в котором не удалось обнаружить трансферриновую
мРНК, а, по-видимому, печень, из которой этот белок посту-
пает в кишечник с желчью [Jdzerda R. L. et al., 1986]. На ба-
зальной стороне энтероцита мукозный трансферрин отдает же-
лезо своему плазменному аналогу.
Путем применения таких современных методов, как изоли-
рованные энтероциты и пузырьки мембраны исчерченной каем-
ки (ПМИК), было установлено, что первоначальное поступле-
ние железа в эпителий слизистой оболочки является пассивным
процессом, не требующим затраты энергии, но зависит от ве-
личины pH, температуры и ингибируется реагентами, образую-
щими хелаты с этим элементом пропорционально их концент-
рации. Немногим более половины железа, всосавшегося в изо-
лированные энтероциты, оказывается связанным мембраной
исчерченной каемки [Savin М. A., Cook J. D., 1980]. Поступле-
ние железа в ПМИК характеризуется кинетикой насыщения,
свидетельствующей о существовании специфического перенос-
чика для этого элемента, находящегося под биологическим
контролем. После поступления в энтероцит железо включается
в систему внутриклеточного транспорта, переносящую его в
базальный отдел клетки и оттуда в плазму крови. В цитозоле
энтероцита некоторое количество железа включается в ферри-
тин, большая часть его теряется при слущивании клеток сли-
зистой оболочки, происходящем каждые 3—4 дня, и лишь не-
большая часть переходит в плазму крови. Железо может посту-
пать в энтероцит и из крови, о чем свидетельствует наличие
ферритина в бокаловидных клетках и макрофагах слизистой
оболочки, которая таким образом может служить и местом его
выделения. Гемовое железо, поступившее в энтероцит, отщеп-
ляется от гема гемоксигеназой слизистой оболочки. Перед
включением в ферритин или трансферрин двухвалентное желе-
зо превращается в трехвалентное. Этот процесс осуществляет-
100
ся, по-видимому, ксантиноксидазой слизистой оболочки, кото-
рая в этом отношении активнее церулоплазмина.
Наиболее интенсивное всасывание железа происходит в про-
ксимальных отделах тонкой кишки, т. е. в двенадцатиперстной,
тощей, и отсутствует в подвздошной кишке. Регуляция всасы-
вания железа достигается, видимо, за счет синтеза мукозного
трансферрина, концентрация которого в энтероцитах при де-
фиците железа возрастает, и прямо коррелирует с всасыванием
59Fe из изолированной петли двенадцатиперстной кишки. Еще
лучше коррелирует с ним отношение мукозного трансферрина
к ферритину. Гуморальные факторы, по-видимому, не оказыва-
ют влияния на процессы усвоения железа [Morris Е. R., 1987].
Поступление в ткани. У здорового человека в сутки
синтезируется около 10,6 мг трансферрина на 1 кг массы тела.
Распад трансферрина происходит в печени после его десиали-
рования и связывания галактозоконцевыми рецепторами гепа-
тоцитов. Константа связывания железа трансферрином очень
высока, составляет 1024 М-1, в связи с чем в 1 л крови присут-
ствует менее одного свободного иона этого элемента. Трансфер-
рин доставляет железо тканям, имеющим специфические ре-
цепторы. Он необходим для культивирования клеток в средах
в отсутствие сыворотки крови. Применяемый для диагностики
опухолей 67Ga активно связывается трансферрином и перено-
сится им на мембранные рецепторы опухолевых клеток [Seph-
ton R. G., Harris A. W„ 1981].
Включению железа в клетку предшествует связывание
трансферрина специфическими мембранными рецепторами, при
утрате которых, как, например, у зрелых эритроцитов, клетка
теряет способность поглощать этот элемент. Количество желе-
за, поступающего в клетку, прямо пропорционально числу
мембранных рецепторов. Рецепторы трансферрина, выделенные
из ретикулоцитов, плаценты человека, Т-лимфоцитов, культуры
фибробластов, опухолевых клеток являются гликопротеидами,
образованными двумя субъединицами с молекулярной массой
около 90 000 каждая. Ретикулоциты человека могут содержать
до 300 000 рецепторов на одну клетку, а клетки хронического
миелолейкоза человека К562 — до 1 млн [Schulman Н. М. et al.,
1981]. Используя флюоресцентную метку, удалось показать,
что трансферрин, связанный рецептором, поступает внутрь
клетки путем эндоцитоза, где он обнаруживается в лизосомной
фракции. В клетке происходит высвобождение железа из транс-
феррина, чему способствует, по-видимому, кислая среда. Затем
апотрансферрин возвращается в циркуляцию. Повышение по-
требности клеток в железе при их быстром росте или синтезе
гемоглобина ведет к индукции биосинтеза рецепторов трансфер-
рина и, напротив, при повышении запасов железа в клетке чис-
ло рецепторов на ее поверхности снижается.
Внутриклеточный транспорт. Железо, высвобо-
дившееся из трансферрина, связывается специфическим бел-
ком с молекулярной массой около 60 000 или ферритином, ко-
101
торне доставляют железо в митохондрии, где оно включается
в состав гема с участием феррохелатазы. Роль трансферрина
как переносчика железа на митохондрии экспериментально не
доказана [Romslo I., 1980]. Помимо синтеза гема, двухва-
лентное железо используется в митохондриях для синтеза же-
лезосерных центров. Имеются также данные, что перенос же-
леза от трансферрина к ферритину и митохондриям может осу-
ществляться полифосфатами [Konopka К. el al., 1981].
Кинетикд обмена железа. Как показывают ферро-
кинетические исследования, в организме происходит системати-
ческое перераспределение железа. В количественном отноше-
нии наибольшее значение имеет метаболический цикл: плаз-
ма-*-красный костный мозг-*-эритроциты-> плазма. Кроме то-
го, функционируют циклы: плазма->• ферритин, гемосидерин-*•
плазма и плазма-*-миоглобин, железосодержащие фермен-.
ты->• плазма. Все эти три цикла связаны между собой через
железо плазмы (трансферрин), которое регулирует распреде-
ление этого элемента в организме.
Обычно более 70% плазменного железа поступает в кост-
ный мозг [Bothwell Т. Н. et al., 1979], хотя у беременных зна-
чительная часть его направляется в плаценту. За счет распада
гемоглобина в сутки освобождается около 21—24 мг железа, что
во много раз превышает поступление железа из пищеваритель-
ного тракта (1—2 мг/сутки). Более 95% железа поступает в
плазму из системы мононуклеарных фагоцитов (СМФ), кото-
рые поглощают путем фагоцитоза более 10й старых эритроци-
тов в сутки. В опытах с эритроцитами, меченными s9Fe, прове-
денными в культуре перитонеальных макрофагов мыши, пока-
зано, что железо эритроцитов или комплекса трансферрин —•
антитело поступает в плазму в виде ферритина, новообразован-
ного трансферрина или комплекса гем — гемопексин, причем
последний образуется в условиях, когда гемоксигеназа не ус-
певает высвободить железо из порфиринового макроцикла
[Kleber Е. Е. et al., 1978]. Железо, поступившее в клетки
СМФ, либо быстро возвращается в циркуляцию (лабильный
пул) (Т1/2—34 мин), либо откладывается про запас (стабиль-
ный пул). Его Ti/а составляет 7 дней.
У взрослого человека плазма крови транспортирует в сутки
25—40 мг железа, хотя одновременно его общее количество в
полном объеме плазмы не превышает 3—4 мг. Это железо име-
ет довольно короткий период полувыведения (Ti/2 составляет
90—100 мин) [Morris Е. R., 1987]. Меченый гемоглобин обна-
руживается в периферической крови через 4—8 ч после введе-
ния 59Fe, а спустя 1—2 нед в циркуляции присутствует 70—
100% введенного изотопа.
Промежуточный обмен железа связан в первую очередь с
процессами синтеза и распада гемоглобина, в которых цент-
ральную роль играет СМФ. У взрослого человека в костном
мозге железо трансферрина с помощью специфических рецеп-
торов включается в нормобласты и ретикулоциты, использую-
102
ющие его для синтеза гемоглобина. Вместе со зрелыми эрит-
роцитами они составляют эритрон, который содержит около
3000 мг железа всего организма. Гемоглобин, поступающий в
плазму крови при распаде эритроцитов, специфически связы-
вается гаптоглобином, что предупреждает его фильтрацию че-
рез почки. Гем из этого комплекса отщепляется в печени, пос-
ле чего 0-цепь гаптоглобина осуществляет протеолиз глобино-
вой части молекулы. Свободный гем транспортируется в плаз-
ме другим белком — гемопексином. После распада порфирина
железо снова связывается трансферрином и вступает в новый
цикл. В прочие ткани трансферрин доставляет в 4 раза мень-
шее количество железа, что составляет около 6 мг/сутки, и
столько же примерно поступает из них в плазму крови. Общее
количество тканевого железа составляет около 125 мг, из ко-
торых основная часть приходится на долю миоглобина. Печень
содержит около 700 мг железа, представленного преимущест-
венно ферритином, а другие органы СМФ — около 300 мг
[Петров В. Н„ 1982J.
Железо выделяется из организма в основном путем слущи-
вания эпителия слизистой оболочки и с желчью. Оно теряется
также с волосами, ногтями, мочой и потом. Общее количество
выделяемого таким образом' железа составляет у здорового
мужчины 0,6—1 мг в сутки, а у женщин репродуктивного воз-
раста немногим более 1,5 мг. Такое же количество железа ус-
ваивается из съедаемой пищи, что составляет 5—10% от его
общего содержания в рационе. Железо из животной пищи ус-
ваивается в несколько раз лучше, чем из растительной.
Железодефицит (гипосидероз) — один из наиболее распро-
страненных МТОЗов человека. Формы его клинического выра-
жения крайне разнообразны и варьируют от латентных состоя-
ний, проявляющихся только под влиянием дополнительных
неблагоприятных факторов (неадекватное питание, инфекции и
др.), до тяжелых прогрессирующих заболеваний, способных
привести к типичным органным и тканевым повреждениям,
значительному ослаблению жизненных сил организма, стойкой
потере трудоспособности и, наконец, к летальному исходу. От-
сутствие своевременного диагноза и специфического лечения
ухудшает прогноз.
Становление понятия о гипосидерозе проходило с преодоле-
нием многих ошибочных взглядов, в частности, на природу хло-
роза, ранее довольно распространенного, главным образом у
девушек, анемического состояния, сочетающегося с зеленова-
тым цветом лица. Причиной хлороза без достаточных основа-
ний считали эндокринные расстройства. В настоящее время
благодаря установлению этиологии железодефицитных состоя-
ний клинический диагноз хлороза не регистрируется.
Большое значение для углубления понимания алиментар-
ных железодефицитных состояний имели исследования
М. В. Schmidt (1928), которому удалось воспроизвести этот
патологический процесс у мышей. Только осторожность удер-
103
жала его от распространения этого понятия на патологию че-
ловека. Еще большее значение имело открытие L. Heilmeyer и
соавт. (1936) низкого уровня железа в крови у всех больных
анемиями, которые давали положительную реакцию в ответ на
лечение железосодержащими препаратами. У больных анеми-
ей, рефрактерных к препаратам железа, напротив, уровень
содержания этого МЭ в крови был нормальным или даже вы-
соким. В дальнейшем был установлен и другой важный факт,
что при железодефицитных анемиях парентерально введенное
железо очень быстро исчезало из крови, а при других формах
малокровия поглощение железа крови тканями происходило
значительно медленнее. Эти наблюдения нашли полное под-
тверждение и до сих пор используются в клинической диагнос-
тике железодефицитных состояний.
Сведения об их распространенности многочисленны, но не
всегда сопоставимы, так как часть авторов учитывают только
показатели крови, в то время как действительно широко рас-
пространенные и практически важные железодефицитные ане-
мии являются далеко не единственным проявлением гипосиде-
роза. Наиболее надежны данные ВОЗ (1959), которые свиде-
тельствуют о почти повсеместном распространении гипосидеро-
за, причем частота его в различных популяциях колеблется от
20 до 95%. В Индии и в некоторых частях Африки более 50%
населения страдают анемиями и от 10 до 40% материнской
смертности во время беременности и родов связано с анемия-
ми [Menon К., 1958]. По более новым данным ВОЗ (1977),
более половины детей на планете страдает анемиями, большая
часть которых относится к железодефицитным.
Не ставя перед собой задачи полной характеристики рас-
пространения анемических и неанемических форм гипосидеро-
за на Земном шаре, подчеркнем только, что эти патологичес-
кие состояния и заболевания встречаются не только в разви-
вающихся странах. В частности, по данным V. F. Fairbanks и
Е. Beutler (1977), не менее 18 млн жителей США страдают ги-
посидерозом. В это число вошли только больные, обратившие-
ся за медицинской помощью. Он распространен среди лиц мо-
лодого возраста, причем в группе 14—16-летних он чаще реги-
стрируется у девочек. Более высокая частота гипосидероза у
девушек отчетливо заметна уже в возрасте от 17 до 20 лет и
резко преобладает у женщин от 30 до 39 лет. Эти данные пра-
вильно характеризуют частоту анемических состояний в раз-
личных возрастных группах у лиц женского пола, но не отра-
жают всех форм гипосидероза, при котором, как показали бо-
лее новые наблюдения, часто встречаются другие, типичные
органные и тканевые поражения.
По мнению I. Bernat (1983), при железодефиците страдает
весь организм, а гипохромная анемия — это поздняя стадия
болезни (рис. 15—18). Отдельные признаки гипосидероза не-
специфичны и выражаются в легкой утомляемости, головных
болях, повышенной возбудимости или, напротив, депрессии.
104
Рис. 15. Гипохромные эритроциты при железодефицитной анемии. Резкое
уменьшение толщины эритроцитов до 1—2 мкм, анизо- и пойкилоцитоз [Win-
trobe М. М., 1967].
Рис. 16. Массивные отложения железа в митохондриях. Приобретенная сиде-
робластная анемия [Bernat I., 1971].
а в
б
Рис. 17. Различные формы отложения железа в ретикулярных клетках костно-
го мозга.
а — мелкие гранулы железа в макрофаге здорового человека; б — небольшие глыбки же-
леза при гемолитической анемии; в — крупные глыбки железа при хронических инфек-
ционных болезнях и злокачественных опухолях [Heilmeyer L., 19681.
Рис. 18. Эритробластный
островок в мазке костно-
го мозга. Центрально рас-
полагается ретикулярная
клетка с железосодержа-
щими гранулами, окру-
женная эритробластами
[Bernat I., 1971].
Вслед за этим развиваются тахикардия, поверхностное дыха-
ние, боли в области сердца, головокружения и склонность к
обморокам. У многих больных наблюдаются гастроинтести-
нальный дискомфорт, отсутствие или извращение аппетита.
В том числе имеют место и такие признаки, как геофагия,
пагофагия (поедания льда), амилофагия (поедания крахмала)
106
и др. Они получили общее название «pica sideropenica». Более
'новые исследования установили, что поедаемые вещества или
объекты не содержат высоких количеств железа и таким обра-
зом поглощение их нельзя трактовать как попытку аутокоррек-
ции дефицита железа. Ответ на природу этих необычных вле-
чений надо искать в нарушениях невропсихической сферы и в
выяснении роли дефицита железа, а также цинка в патофизио-
логии вкусовых потребностей. J. Н. Ward и соавт. (1984) опи-
сали 2 детей, глотавших волосы. У них был обнаружен дефи-
цит железа. После лечения препаратами железа трихофагия
прошла. Сравнительно редким симптомом гипосидероза явля-
ется кожный зуд, который также поддается терапии железосо-
держащими препаратами.
В настоящее время общепринято, что диагноз железодефи-
цитных состояний надо ставить до развития полной картины
заболевания, т. е. до возникновения гипохромной анемии. К со-
временным методам относится определение концентрации же-
леза и трансферрина в сыворотке крови, а также процента
насыщения трансферрина. К этой триаде надо добавить изме-
рение концентрации ферритина в сыворотке крови, которое
является ранним и надежным диагностическим тестом, в доста-
точной степени отражающим запасы железа в организме
(табл. 10). Дополнительным диагностическим тестом для рас-
познавания гипосидероза считается определение концентра-
ции свободного эритроцитарного протопорфирина. Путем изу-
чения магнитной восприимчивости можно установить содержа-
ние железа в печени [Brittenham G. М. et al., 1984J.
Традиционные гематологические анализы имеют важное
ориентирующее значение при сопоставлении с опубликованны-
ми нормативными данными. Так, средняя концентрация гемо-
глобина у мужчин в норме составляет 165 г/л, у женщин —
145 г/л, у женщин на 8-м месяце беременности—130 г/л, у де-
тей от 6 до 12мес— 120 г/л (сводные данные литературы, цит.
по Bernat I., 1983). Если данные анализа крови указывают на
гипохромную анемию, то диагноз железодефицитного состояния
наиболее вероятен, но он должен быть уточнен определением
степени его выраженности и наиболее вероятных причин. Сре-
ди последних важное место занимают кровопотери при менор-
рагиях, кровоточащих язвах желудка и двенадцатиперстной
кишки, варикозе вен пищевода, опухолях пищеварительного
тракта, изъязвленных полипах, регионарном илеите, при теле-
ангиэктазиях, ущеплении части желудка в диафрагмальной
грыже, тяжелой глистной инвазии анкилостомами, непарным
власоглавом и другими гельминтами. Существуют и другие
причины, вызывающие постгеморрагическое железодефицитное
состояние, поиск их должен быть настойчивым и целенаправ-
ленным. Если повторные носовые кровотечения и гематурия
широко известны, то этого нельзя сказать о кровопотерях при
легочном гемосидерозе у детей и подростков, а также о ла-
тентном или манифестном гипосидерозе постоянных доноров,
107
Таблица 10
Некоторые показатели метаболизма железа у здоровых лиц
и при нарушенном эритропоэзе (синтезе гемоглобина)
[Bernat I., 1983]
Показатель метаболизма железа НсУрма Гипосидероз Инфекцион- ная, опухо- левая ане- мия Нарушение синтеза гема и глобииа
Железо плазмы (PIFe), 115 <70 <70 >180
мкг/дл (80—150) (20—50) (20—65) (180—300)
Общая железосвязываю- 320 >360 200 250
щая способность плаз- (290—350) (360—550) (80—280) (200—360)
мы, мкг/дл Коэффициент насыще- 32 <25 <25 ~100
НИЯ, % Содержание железа в (30—40) (5—25) (10—35) (70—100)
+ 0 ++ +++
ретикулярных клетках костного мозга Сидеробласты, % 30 3 10 80
Кривая нагрузки перо- (20—50) 80—150% (0-5) Крутая (5-15) Ровная (50—90) Ровная
ральным железом (повыше- (>250% по- (низкая (высокая
Абсорбции радиоактив- ного железа из желудоч- но-кишечного тракта, % Содержание железа в моче после внутривен- ной нагрузки 100 мг же- леза, мг/24 ч Содержание железа в ние) 25—35 4,5—5,5 0,5—1 вышение) 30—60 0,9—4,5 0,1—0,5 PIFe нато- щак; <50% повышение) 10—25 5—15 1—8 PIFe нато- щак; <50% повышение) 1—20
моче после внутримы- шечной инъекции 500 мг десферриоксиамина, мг/24 ч Сывороточный ферритин, мкг/л: мужчины женщины Средние значения Крайние значения Экскреция S7Co, % от дозы 100—120 35 12—300 4—11 <10—12 >12 Более 100 <П Более 100— 1000
которые сами недооценивают важность многократных крово-
потерь для их здоровья. По мнению I. Bernat (1983), нельзя
сдавать более 1 л крови в год.
Слизистая оболочка полости рта и языка при сидерозе
обычно отличается сухостью. Красная кайма губ имеет неров-
ную поверхность, с трещинами. Хейлоз в сочетании с болез-
ненными влажными трещинами в углах рта (с гиперкератозом
или без него встречается у 15% больных с дефицитом железа)
[Chisholm М., 1973]. Изменения и болезненность ящчка регист-
108
рируются в 50% всех случаев тяжелого, хронического гипоси-
дероза. Путем изучения биоптатов слизистой оболочки языка
устанавливают атрофию нитевидных сосочков. Она может рас-
.спространиться на всю поверхность языка, делая ее гладкой,
й иногда и блестящей. После лечения препаратами железа
язык приобретает нормальный вид. Эпителий слизистой обо-
лочки щек отличается патологической тонкостью, количество
митозов в нем увеличено, а диаметр клеток по сравнению с
нормой уменьшен. Эти изменения могут быть одной из причин
возникновения повреждений и небольших кровотечений. По-
скольку атрофический процесс может распространяться на
глотку и всю слизистую оболочку гортани и голосовые связки,
могут наступить выраженные нарушения в виде охриплости и
даже полной утраты голоса. Два таких наблюдения при хро-
ническом гипосидерозе, начавшемся еще в детском возрасте,
приводит I. Bernat (1983). Морфологический субстрат сидеро-
пенической афонии пока охарактеризован только ларингоско-
пически и ограничивается атрофией и недостаточной подвиж-
ностью средней трети голосовых связок.
Одним из важнейших проявлений гипосидероза является
сидеропеническая дисфагия, или синдром Пламмера — Винсо-
на, частота которого колеблется от 5 до 20%. Он встречается
как при латентной форме гипосидероза, так и при железоде-
фицитной анемии и выражается клинически в чувстве жжения
в regio postcricoidea или в ощущении застревания пищи. Начи-
ная с 30-х годов, установлено, что причиной этого синдрома
является истинное сужение пищевода за счет очагового мем-
бранозного воспаления, которое может распространяться и на
мышечный слой пищевода. Слизистая оболочка пищевода тон-
кая, бледная, сухая, может кровоточить. Рентгенологические
признаки этого процесса изучены рядом скандинавских авто-
ров, а также С. С. Даниленко (1950) и особенно тщательно
I. Bernat (1966). Постоянным местом сужения является крико-
фарингиальная зона пищевода. Под влиянием специфической
терапии препаратами железа сидеропеническая дисфагия исче-
зает, но это не всегда сопровождается рассасыванием очагов
воспаления. Важно подчеркнуть, что сидеропеническая дисфа-
гия является предраковым состоянием и заканчивается возник-
новением рака в 4—16%' [Chisholm М., 1973J. Со времени
опубликования этих наблюдений они были многократно под-
тверждены, причем показано, что локальные симптомы со сто-
роны пищевода должны вызвать определенную онкологическую
настороженность, особенно если дефицит железа был поздно
диагностирован и больные не подвергались лечению.
Важным проявлением гипосидероза является сидеропениче-
ский ринит, называемый в широких медицинских кругах атро-
фическим. Основополагающие работы по этому вопросу при-
надлежат венгерскому гематологу I. Bernat (1965), который
не только показал, что подавляющее большинство больных си-
деропеническим ринитом излечиваются или получают облегче-
109
ние (в 30% случаев) после терапии препаратами железа, но и
воспроизвел этот патологический процесс у мышей, тщатель-/
но изучил его патоморфологию и пришел к заключению, что
возникновение тяжелого дефицита железа во внутриутробном
периоде и в раннем детском возрасте ведет к определенным
конституционным изменениям черепа. Иногда на фоне этих
изменений возникает зловонный насморк (озена). Сидеропени-
ческая атрофия слизистой оболочки носа распространяется на
придаточные полости и ведет к истончению соответствующих
костных структур. Эти наблюдения и эксперименты, под-
тверждающие более ранние наблюдения F. Reimann (1955),
позволили утверждать, что некоторые конституционные особен-
ности лица и черепа больных озеной являются не наследствен-
ными, а приобретенными и вызваны внутриутробным дефици-
том железа. Особенностью многих больных озеной является их
внешнее сходство, обусловленное брахицефалическим и «мно-
гоугольным» черепом, выступающими лбом и скуловыми костя-
ми, а также характерными чертами лица. На секции установ-
лено уменьшение спонгиозной части костей черепа, а также
истончение его кортикального слоя.
I. Bernat (1983) предлагает называть характерные измене-
ния лица и черепа, возникшие вследствие внутриутробного де-
фицита железа, следующими медицинскими терминами: «гипо-
сидерозный череп» и «гипосидерозное лицо», которыми целе-
сообразно пользоваться в клинической диагностике железоде-
фицита.
В последние годы увеличился интерес к клинической харак-
теристике железодефицитных анемий. В 1983 г. П. М. Альпе-
рин и Ю. Г. Митерев предложили новую классификацию их.
Они выделяют: 1) постгеморрагические, 2) нутритивные (али-
ментарные) анемии, 3) анемии при повышенном расходе желе-
за в организме (например, при беременности, лактации, росте
и созревании), 4) железодефицитные анемии при исходно не-
достаточном уровне железа, 5) железодефицитные анемии при
его недостаточной резорбции (например, постгастрорезекцион-
ные, агастральные, анентеральные), 6) при перераспределении
железа в результате инфекции, при воспалительных и опухо-
левых процессах, 7) при нарушениях транспорта железа (на-
пример, гипотрансферринемические и атрансферринемические).
В основу предложенной классификации положены механиз-
мы, которые могут вызвать дефицит железа. Однако не все ее
пункты одинакового удачны. В частности, в пункте 6 основной
акцент делается на перераспределении железа в результате
инфекций, воспалительных и опухолевых процессов. Следует
напомнить, что эти процессы очень разнообразны и в них
участвуют различные патогенетические механизмы. В частно-
сти, при опухолях в организме появляется особый белок, спо-
собный поглощать железо. Недавно открыты и особые штаммы
кишечных палочек, обладающие повышенной сидерофиль-
ностью.
ПО
В пункте 7 классификации, где речь идет о нарушениях
транспорта железа, имеется в виду только дефицит трансфер-
рина. По-видимому, следовало бы упомянуть и о возможности
блокады или другой формы неполноценности клеточных рецеп-
торов железа. Не исключается, что этот механизм возникнове-
ния железодефицитной анемии заслуживает отдельной руб-
рики.
При длительном течении железодефицитной анемии процесс
не ограничивается гиперплазией эритроидного ростка, но соче-
тается также с нарушением эритропоэза. Большое значение в
патогенезе ферментативных нарушений во внутренних органах
и в ЦНС имеет анемическая гипоксия.
В условиях Севера железодефицитные состояния, в том
числе и анемии, встречаются особенно часто. А. Г. Марачев и
А. Л. Жаворонков (1987) выделяют акклиматизационный де-
фицит железа у детей и взрослых. Анемии, связанные с дефи-
цитом железа в пище, регистрируются у детей, находящихся
на однообразном молочном вскармливании. Железодефицит-
ные состояния новорожденных могут быть связаны с соответ-
ствующим состоянием матери. Этот дефицит может носить
относительный характер, что связано с перераспределением
данного элемента в организме, пораженном инфекциями и
гельминтами. Железодефицитные анемии (главным образом
у детей) распространены также среди населения высокогорных
и среднегорных районов и имеют свои клинические особенности
[Миракплова А. М., 1986].
Избыточное содержание железа в организме, как указыва-
лось, носит название «сидероз» или «гиперсидероз». Он может
иметь местный и генерализованный характер.
Экзогенный сидероз нередко наблюдается у шахтеров, уча-
ствующих в разработке красных железных руд, и у электро-
сварщиков. Сидероз шахтеров может выражаться в массивных
отложениях железа в ткани легких. Местный сидероз встреча-
ется при внедрении в ткани железных осколков. В частности,
выделен сидероз глазного яблока с отложением гидрата окиси
железа в цилиарном теле, эпителии передней камеры, хруста-
лике, сетчатке и в зрительном нерве.
Эндогенный сидероз чаще всего имеет гемоглобиновое про-
исхождение и возникает в результате разрушения этого пиг-
мента крови в организме. Гемосидерин представляет собой
агрегат гидроокиси железа, соединенного с белками, гликоза-
миногликанами и липидами. Гемосидерин образуется внутри
клеток мезенхимной и эпителиальной природы. В. В. Серов и
соавт. (1986) называют эти клетки сидеробластами. Очаговое
отложение гемосидерина, как правило, наблюдается на месте
кровоизлияний. От гемосидероза надо отличать тканевое «оже-
лезнение», которое наблюдается при пропитывании некоторых
структур (например, эластических волокон) и даже нейронов
головного мозга коллоидным железом. Это имеет место при
некоторых психических заболеваниях, в частности при болезни
111
Пика, некоторых гиперкинезах, а также при бурой индураций
легких. '
Особой формой наследственных отложений гемосидерин^,
возникающего из ферритина в результате изменения клеточно-
го обмена, является гемохроматоз. При этом заболевании осо-
бенно большие отложения железа наблюдаются в печени, под-
желудочной железе, почках, в клетках СМФ, а также в сли-
зистых железах трахеи, в щитовидной железе, эпителии языка
и мышцах. Наиболее известен первичный, или идиопатический,
гемохроматоз — наследственное заболевание, для которого ха-
рактерны нарушение обмена железосодержащих пигментов,
повышенное всасывание в кишечнике железа и накопление его
в тканях и органах с развитием в них выраженных изменений.
Важно подчеркнуть, что при гемохроматозе имеет место от-
ложение большого количества гемосидерина, а также пигмен-
тов, не содержащих железа (гемофусцина и меланина), внутри
клеток паренхиматозных органов. Заболевают преимуществен-
но мужчины с явлениями гипогенитализма и симптомах феми-
низации. К ранним симптомам относится увеличение печени,
к которому присоединяются сахарный диабет (за счет фиб-
роза поджелудочной железы) и прогрессирующее потемнение
кожи (бронзовый диабет). Нередко встречается тяжелое и
быстро прогрессирующее поражение миокарда с увеличением
размеров сердца и симптомами декомпенсации, обусловленной
накоплением железа в миокарде и субэндокардиальным фиб-
розом. Характерны поражения периферических нервов, а в
терминальной стадии асцит, тромбоз сосудов печени и селе-
зенки, печеночная кома, кровотечения из расширенных вен
пищевода. У детей гемохроматоз может протекать в латентной
форме. Характерны для гемохроматоза поражения эндокринной
системы, с которым связаны признаки инфантилизма и гипо-
плазии половой системы.
В течение длительного времени наследственный гемохрома-
тоз и другие формы гиперсидерозов рассматривались в качест-
ве очень редких заболеваний. В последние 20 лет показано, что
это не соответствует действительности, так как выяснилось зна-
чительное разнообразие их клинических проявлений. Очень по-
лезной оказалась клиническая классификация гиперсидерозов,
предложенная J. Н. Dagg и соавт. (1971). Различают парен-
химатозные формы гиперсидероза (с преимущественным отло-
жением железа в клетках паренхимы), к которым относятся
первичный наследственный гемохроматоз, сидероз при некото-
рых видах цирроза печени, вторичный сидероз при портока-
вальном анастомозе, сидероз при врожденной атрансферрине-
мии, врожденный гиперсидероз, описанный L. Vitale и соавт.
(1969). К преимущественно «ретикулоэндотелиальным» формам
гиперсидероза относятся генерализованные отложения железа
при хронических рефрактерных (к специфическому лечению)
анемиях, гемолитических анемиях, многократных гемотрансфу-
зиях, при избыточном парентеральном введении железа. При
112
широко распространенном в Южной Африке так называемом
сидерозе банту отложение железа имеет более разнообразный
Характер. К локальным формам гиперсидероза относятся:
идиопатический гемосидероз легких, легочно-почечный синдром
Гудпасчера и гемосидероз почечного происхождения, который
выражается в ночной пароксизмальной гемоглобинурии.
Наибольшие изменения претерпело понятие о наследствен-
ном (идиопатическом) гемохроматозе. До 1955 г. в литературе
было зарегистрировано только 1200 случаев этого заболевания.
В настоящее время оно встречается значительно чаще. Обще-
принято. что гемохроматоз проявляется циррозом печени, пиг-
ментацией кожи, поражением суставов, кардиомиопатией и
эндокринными нарушениями, включая сахарный диабет.'Боль-
шая часть этих симптомов зависит от отложения значительных
количеств железа в органах, однако они характерны только
для конечной стадии заболевания, которая развивается через
много лет. Наследственная природа гемохроматоза не вызыва-
ет сомнения. Частота встречаемости генов гемохроматоза со-
ставляет 0,05—0,07% при частоте гетерозиготности в популя-
циях европейцев около 10% (состояние носительства), а забо-
леваемости (гомозиготность)—от 0,3 до 0,5% [Ward J. Н. et
al., 1984].
Очень важным методом лечения гемохроматоза является
кровопускание. Оно предупреждает большинство осложнений.
Однако под влиянием кровопусканий у больных гемохромато-
зом не исчезают симптомы хондрокальциноза и гипогонадизма,,
а также риск развития злокачественной гепатомы, которая
встречается у 7—33% этих больных при наличии цирроза пе-
чени.
Вторичный гемохроматоз при циррозе печени встречается
главным образом у мужчин, страдающих хроническим алкого-
лизмом, часто употребляющих неполноценную по содержанию
белков пищу. Это способствует развитию хронического панкре-
атита, который в свою очередь ведет к более высокой абсорб-
ции железа.
Среди перечисленных выше форм гиперсидероза особое
внимание привлекает сидероз банту. Он развивается у 40—88%
аборигенов Южной Африки, преимущественно у мужчин. Это
объясняется очень высоким содержанием железа в их пище.
Значительное количество этого МЭ попадает из железных кот-
лов для приготовления пищи. Особенно большое количество
этого металла содержится в различных видах алкогольных на-
питков, которые варятся в такой же посуде. Соответствующие
подсчеты показали, что банту потребляют от 50 до 100 мг
железа ежедневно только за счет пива домашнего приготовле-
ния. Около 2—3% железа всасывается [Bothwell Т. Н. et al.,
1979].
Пищевая перегрузка организма железом встречается и в
других частях света. Например, в некоторых районах Италии
вина очень богаты железом. Семейная перегрузка организма
8—568 ИЗ
Рис. 19. Массивные отложения железа в звездчатых макрофа'гоцитах печени
яри синдроме Гудпасчера у мальчика 5 лет. Реакция Перлса с докраской кар-
мином. Х400.
Рис. 20. Резко выраженный гемосидероз селезенки при синдроме Гудпасчера
у мальчика 5 лет. Реакция Перлса. Х640.
Рис. 21. Сочетание склероза клубочков (слева), облитерирующего гломерулита
и интерстициальных лимфоидных инфильтратов при синдроме Гудпасчера
у мальчика 5 лет. Окраска гематоксилином и эозином. Х400.
железом в данных случаях не имеет наследственной обуслов-
ленности и зависит от местных обычаев.
Клиническая картина гемохроматоза в виде редкого исклю-
чения развивается после перорального применения железосо-
держащих препаратов.
Редкая форма гиперсидероза — это синдром Гудпасчера,
который представляет собой заболевание аутоиммунной при-
роды. Антигеном, очевидно, являются белки базальных мем-
бран легочных и гломерулярных капилляров. В клинической
картине отмечаются кровохарканье, гематурия и анемия.
В легких наблюдаются отложения гемосидерина. Болеют пре-
имущественно лица старше 16 лет, но возможно развитие за-
болевания у подростков и детей [Геллер А. И., 19691 (рис.
19—21).
В 1947 г. А. И. Абрикосов впервые в СССР описал клини-
ко-анатомическое наблюдение, относившееся к идиопатическо-
му гемосидерозу легких (синдром Целена — Геллерстедта).
Это преимущественно детское заболевание, характеризующие-
ся повторными геморрагиями в легочную ткань и железодефи-
цитной гипохромной анемией. Этиология этого заболевания не-
известна, предполагается иммунный или аутоиммунный пато-
генез.
Ряд авторов [Strohmeyer G. et al., 1983] наряду с первичным
гемохроматозом выделяют несколько вторичных форм, при ко-
торых пигментный цирроз встречается в качестве осложнения
8*
115
талассемии, сидеробластической анемии, наследственного сфе-
роцитоза, пиридоксинчувствительной анемии и др.
Ятрогенные формы сидероза описаны в главе 9.
6.2. Медь
Медь — один из важнейших незаменимых МЭ, необходи-
мых для жизнедеятельности человека, животных и растений.
В организме взрослого человека содержится 1,57—3,14 ммоль
меди, причем половина этого количества приходится на мыш-
цы и кости, a 10% на ткани печени. В частности, печень, мозг,
сердце и почки содержат 0,230±0,014; 0,088 + 0,003; 0,052 +
±0,005; 0,33 + 0,006 ммоль/кг свежего вещества меди соответ-
ственно. Небольшие количества этого элемента находятся в лег-
ких, кишечнике, селезенке, коже и волосах. Все эндокринные
органы содержат небольшие количества меди. В крови в сред-
нем имеется около 100 мкг меди, причем в эритроцитах и лей-
коцитах— 60 мкг. Значительная часть меди плазмы крови
находится в церулоплазмине — важнейшем медьсодержащем
белке. Медь найдена в супероксиддисмутазе эритроцитов и
лейкоцитов [Шапошников А. М. и др., 1980].
Важно подчеркнуть, что концентрация меди в печени, мыш-
цах и других тканях новорожденных в несколько раз выше,
чем в соответствующих тканях взрослых. Медь плода имеет
материнское происхождение, причем во время беременности ее
всасывание у матери увеличивается, а выведение снижается.
Только к 6—12 мес уровень тканевой меди у детей достигает
уровня взрослого.
Поступление меди с пищей должно составлять 2—5 мг
(0,031—0,079 мкмоль) в сутки, причем суточное потребление
менее 2 мг (0,031 мкмоль) опасно в связи с возможностью
развития медьдефицитных состояний. Около 30% суточного по-
ступления меди усиливается, а остальная медь в желудочно-
кишечном тракте превращается в нерастворимые соединения,
выводящиеся с калом. Из общего количества резорбируемой
меди около 80% выводится с желчью и около 16% стенками
желудочно-кишечного тракта. С мочой выделяется около 4%
всосавшейся меди. Незначительное количество этого элемента
выделяется с потом [Шапошников А. М. и др., 1980].
Основные процессы всасывания меди происходят в желудке
,и тонкой кишке, слизистая оболочка которой содержит метал-
лотионеин, образующий комплексные соединения с медью.
Пройдя через эпителий слизистой оболочки желудка и тонкой
кишки или введенная внутривенно, медь связывается недавно
открытым белком транскупреином [Weib К. С., Linder М. С.,
1985] и альбумином в отношении 2: 1 соответственно и в
меньшей степени аминокислотами, преимущественно гистиди-
ном. Из плазмы крови человека в количестве около 200 нг/мл
выделен также связанный с медью (или железом) трипептид —
глицилгистидиллизин, ассоциированный с альбуминовой и а-
116
Глобулиновой фракцией. Этот трипептид образует тройной
комплекс с медью и альбумином и обеспечивает поступление
меди( и железа) внутрь клеток в нетоксичной и доступной для
них форме. У плода функцию транспорта меди вместо альбу-
мина выполняет а-фетоглобулин, имеющий близкую к альбу-
мину первичную структуру. Ген а-фетоглобулина возник, по-ви-
димому, в результате дупликации альбуминового гена [Frie-
den Е., 1986].
Ключевую роль в обмене меди играют печень и ее основные
структурные элементы — гепатоциты. Поступающая в них че-
рез систему воротной вены медь связывается первоначально
металлотионеином, найденным в печени человека и большинст-
ва исследованных животных. Противоречивость данных о при-
роде белка, связывающего медь в печени, объясняется его
окислением в процессе выделения. Синтез металлотионеина
регулируется содержанием цинка и меди в печени на уровне
транскрипции мРНК. Тионеин выполняет функции детоксика-
ции меди и ее внутриклеточного транспорта. Повышение со-
держания меди может привести к амплификации тионеиновых
генов и резкому повышению синтеза данного белка [Evans
G. W., 1978].
Медь, первоначально связанная металлотионеином, в даль-
нейшем включается в церулоплазмин, другие медьсодержащие
ферменты и компоненты желчи. Церулоплазмин наряду со
своими функциями оксидазы выполняет также роль транспорт-
ного белка, переносящего медь на тканевые ферменты, в пер-
вую очередь на цитохромоксидазу. В пользу такого допущения
свидетельствуют эксперименты, показавшие, что единственным
соединением меди, эффективно восстанавливающим активность
цитохромоксидазы у крыс, испытывающих недостаточность ме-
ди, является именно церулоплазмин. Важную роль в гомеоста-
зе меди играют лизосомы гепатоцитов, удаляющие комплекс
медь — тионеин из цитоплазмы. Последний превращается в них
в водонерастворимую полимерную форму, ошибочно описанную
ранее как митохондрокупреин. Лизосомы захватывают частич-
но десиалированный церулоплазмин, который подвергается
глубокому протеолизу и затем удаляется из гепатоцитов в
желчные протоки. Большая часть асиалоцерулоплазмина
( — 70%), однако, поступает в желчь, минуя лизосомы. Кроме
того, часть интактного церулоплазмина может поступать в
желчь непосредственно из печеночных кровеносных сосудов
через синусоиды между печеночными балками [Kressner М. S.
et al., 1984].
Церулоплазмин, утративший сиаловую кислоту, немедленно
узнается галактозоспецифическим рецептором гепатоцита и по-
ступает внутрь клетки в результате эндоцитоза. Интересно,
что рецепторы для церулоплазмина присутствуют и в клетках
других тканей, например в миоцитах и эритроцитах. Из послед-
них выделен специфический рецепторный белок с молекуляр-
ной массой 60 000. Церулоплазмин, связанный этим рецепто-
117
Рис. 22. Метаболические изменения меди в гепатоците.
ЖК — желчный капилляр; ЦП — церулоплазмин; TH — тионеин; Си — ак — Си — амино-
кислота; ЭС— эндоплазматическая сеть.
ром, обеспечивает эритроцитам заметную устойчивость к гемо-
лизу [Frieden Е., 1984].
Содержимое лизосом после частичного протеолиза высво-
бождается в желчные протоки, прилегающие к гепатоциту, и
выводится наружу. Считается, что медь-тионеин с трудом под-
дается действию протеолитических ферментов. Первоначально
медь в желчных протоках находится в составе низкомолекуляр-
ных соединений — комплексов с аминокислотами и желчными
кислотами [Lewis A. J., 1973]; по мере продвижения в желчных
ходах образует соединение с высокомолекулярными белками
[Samuel A. R. et al., 1983] или конъюгатами билирубина [Мс
Cullars G. М. et al., 1977], недоступными для обратного всасы-
вания.
Для гомеостаза меди существенное значение должна иметь
скорость биосинтеза металлотионеина (рис. 22). Чем больше
синтезируется этого белка, тем больше меди накапливается
в клетке. Нарушение регуляции синтеза металлотионеина, ве-
дущее к накоплению избытка меди в печени, происходит в
первую очередь при некоторых генетических заболеваниях (см.
ниже), а также под воздействием токсичных веществ, вызыва-
ющих дерепрессию тионеиновых генов. К появлению избытка
меди приводят также нарушения выделительной функции ли-
зосом в результате дефектов мембран или цитоскелета. Следу-
ет отметить, что любая задержка выделения меди из клетки
приводит к индукции биосинтеза металлотионеина, создавая
механизм порочного круга. Медь повреждает мембраны и ци-
118
Рис. 23. Обмен меди и церулоплазмина в гепатоците [Cousins R. J., 1985].
•ЛЭМ — лейкоцитарный эндогенный медиатор; ак — Си—-аминокислота — Си, ААА — по-
ли-А-конец мРНК.
тоскелет, что в свою очередь сопровождается дальнейшим на-
коплением меди в клетке.
Радиоактивная медь, обнаруживаемая в плазме в первые
часы после ее перорального введения, рыхло связана с фрак-
цией альбуминов, транскупреином и аминокислотами («пря-
мая» медь). Через 2—3 ч после введения 65Си радиоактивность
плазмы начинает понижаться, а затем снова возрастает, дости-
гая второго максимума через 24 ч. Эта радиоактивность свя-
зана уже главным образом с церулоплазмином.
Синтез церулоплазмина происходит на рибосомах печени
[Scheinberg I. Н., 1966]. В ней осуществляются по меньшей ме-
ре три процесса: синтез самого церулоплазмина, депонирова-
ние меди в составе металлотионеина и превращение меди в
недиализируемую форму, выделяемую через желчь. Эти про-
цессы происходят в пространственно разобщенных компартмен-
тах (отсеках) ткани печени, причем между первым и вторым
компартментом происходит непрерывный обмен «рыхло» свя-
занной меди [Owen Е. С., 1980; Owen Е. С., Ludwig J., 19821
(рис. 23).
В норме в организме человека ежедневно около 0,5 мг ме-
ди включается в состав церулоплазмина и выделяется с жел-
чью в виде труднодиализируемого соединения, почти не под-
вергающегося обратному всасыванию [Farrer Р. A., Mistilis
S. D., 1967]. У человеческого плода обратное всасывание меди,
выделенной с желчью, по-видимому, имеет место, так как у
недоношенных детей меконий в 2—3 раза и более богаче
медью, чем у доношенных [Kleinbaum Н., 1962].
119
Содержание меди в плазме регулируется нейрогуморальны-
ми механизмами, причем неодинаково у человека и различных
животных. У крыс, например, адреналэктомия ведет к повы-
шению уровня меди в плазме крови, которое сохраняется даже
через 10 мес после операции. Кортикостерон и тироксин вызы-
вают снижение содержания меди в крови. У человека, напро-
тив, отмечены повышение концентрации этого МЭ в крови при
гипертиреозе и ее снижение при гипофункции щитовидной же-
лезы.
При даче эстрогенов или при ^физиологическом повышении
их содержания в крови (например, при беременности) и ис-
пользовании противозачаточных средств с эстрогенным дейст-
вием повышается содержание меди в крови, связанное с ин-
дукцией синтеза церулоплазмина. Повышение концентрации це-
рулоплазмина при удалении гипофиза, надпочечников и
щитовидной железы объясняется задержкой выделения меди
с желчью, ведущей к повышению запасов меди в печени, что
в свою очередь вызывает индукцию синтеза церулоплазмина
[Henkin R. J., 1976].
Болевое раздражение, стрессовые ситуации и инфекционные
заболевания вызывают повышение содержания в крови меди
и церулоплазмина, действуя на обмен этого металла отчасти
через нейрогуморальную систему.
У человека и всех изученных видов животных преобладаю-
щая часть поступающей в организм меди выделяется с калом.
Большая часть этого МЭ, присутствующая в кале, является
невсосавшейся медью пищи и меньшая часть — эндогенной
медью, выделенной с желчью и стенками желудочно-кишечного
тракта.
Выделение меди с мочой, как уже отмечалось, весьма ог-
раничено. У человека оно составляет в норме 20—60 мкг в сут-
ки [Giorgio А. Р. et al., 1964; Schroeder Н. A. et al., 1966].
При болезни Вильсона — Коновалова, однако, количество меди,
выделяемой с мочой, может достигать 1500 мкг в сутки. АКТГ
и гормоны коры надпочечников повышают выделение меди с
мочой, а адреналэктомия, напротив, ведет к снижению ее выде-
ления почками.
Медь участвует в биохимических процессах как составная
часть электронпереносящих белков, осуществляющих реакции
окисления органических субстратов молекулярным кислородом.
Этой ролью она обязана своим особым свойством как переход-
ного металла. Имея два обычных валентных состояния, она в
зависимости от природы и расположения лигандов позволяет
медьсодержащим белкам охватывать широкий интервал окис-
лительно-восстановительных потенциалов, а также обратимо
связывать кислород и окись углерода. Каталитические центры
медьпротеидов могут быть одно- и двуядерными. Многие белки
содержат по одному такому центру. Например, в галактозо-
оксидазе присутствует одиночный одноядерный медный центр,
а в тирозиназе — двуядерный. Ряд белков содержит четыре и
120
Таблица 11
Медьсодержащие ферменты животных и человека
Ферменты Молекуляр- ная масса Функция Источник
Церулоплазмин (КФ 1.16.3.1) 132 000 Окисление железа, биогенных аминов, транспорт меди Плазма крови жи- вотных и человека
Супероксиддисмутаза (КФ 1.15.1.1) 32 000 Дисмутация супе- роксида Эритроциты, ткани животных
Тирозиназа (КФ 1.1.18.1) 46 000 Гидроксилирова- ние тирозина Кожа, меланома, ткани животных
Дофаминф-гидроксн- лаза (КФ 1.14.17.1) 290 000 Гидроксилирова- ние дофамина Кора надпочечни- ков крупного ро- гатого скота, плаз- ма крови человека
Аминоксидаза, бензил- 180 000— Окисление первич- Плазма крови,
аминоксидаза, спермино- ксндаза, гистаминаза и др. (КФ 1.4.3.6) —195 000 ных аминов почки
Аминоксидаза (лизилок- 30 000— Окисление Плацента челове-
сидаза) (КФ 1.4.3.6) (КФ 1.4.3.4) —60 000 6-аминогруппы лизина ка, аорта, хрящ человека и живот- ных
Цитохромоксидаза (КФ 1.9.3.1) 140000 Терминальное окисление Митохондрии жи- вотных и человека
Уратоксидаза 112 000— Окисление моче- Печень, почки мле-
(КФ 1.7.3.3) —125 000 вой кислоты в ал- лантоин копитающих (у че- ловека отсутству- ет)
более ионов меди, входящих в состав центров обоих типов, их
характеризует необычно интенсивная голубая окраска, благо-
даря которой они получили название голубых или мультимед-
ных оксидаз. Медь в этих оксидазах с учетом лигандного ок-
ружения бывает трех типов: 1) интенсивно голубая, обнаружи-
ваемая ЭПР, 2) неголубая, обнаруживаемая ЭПР, и 3) него-
лубая, не обнаруживаемая ЭПР (табл. 11).
Медьсодержащие аминоксидазы с одноядерным
медным центром катализируют общую реакцию: R'CH2NR2 +
+ O2 + H2O->R!CHO + HNR2 + H2O2. Они широко распростране-
ны в природе. В связи с тем что аминоксидазы животных при-
нимают участие в катаболизме и инактивации многих физиоло-
гически активных аминов, таких как гистамин, адреналин, ти-
рании, путресцин, спермин и др., они подвергались разносто-
роннему интенсивному изучению [Горкин В. 3., 1977].
Реагенты, образующие комплексные соединения с металла-
ми (О-фенантролин, 2,2-дипиридил, 8-гидроксихинолин, цианид
и др.), и реагенты на карбонильные группы (фенилгидразин,
семикарбазид, гидроксиламин, аминогуанидин) являются мощ-
ными ингибиторами медьсодержащих аминоксидаз. Молекула
образована двумя субъединицами, содержащими каждая один
атом меди, которые в процессе катализа не меняют своей ва-
121
лентности. Аминоксидазы имеют розовую или желтоватую
окраску, обусловленную как самой медью, так и органическим
кофактором, природа которого окончательно не установлена.
Предположение, что этим кофактором является пиридоксаль,
не нашло подтверждения [Malmstrom В. G., 1981].
Г емоцианины — ярко-голубые белки с двуядерным медным центром,
присутствующие у членистоногих (краб, лангуст, омар и др.) и моллюсков
(улитка)—хорошо изученная группа медьпротеидов, способных обратимо
связывать кислород. При физиологических условиях оии представляют собой
высокомолекулярные агрегаты (их мол. масса достигает 6 000 000), имеющие
кооперативный характер связывания кислорода. В то же время они значитель-
но различаются по структуре молекул и взаимодействию с кислородом и дру-
гими низкомолекулярными соединениями. В частности, гемоцианин моллюсков
в отличие от аналогичного белка членистоногих обладает выраженной ката-
лазной активностью.
Ц и т о х р о м-с- оксидаза — фермент внутренней мембра-
ны митохондрий — имеет молекулярную массу, близкую к
140 000, и состоит из 7 субъединиц, из которых предположи-
тельно субъединица I содержит гем и кардиолипин, субъеди-
ница II — второй гем, два остатка меди и, по-видимому, взаи-
модействует с цитохромом с. Субъединица III, которую можно
удалить без потери ферментом каталитической активности
участвует в трансмембранном переносе протонов. Функции
остальных четырех субъединиц неизвестны. Крупные субъеди-
ницы I—III кодируются митохондриальной ДНК и синтезиру-
ются в митохондриях, а единицы IV—VII — в цитоплазме.
Молекула цитохромоксидазы имеет Y-образную форму и про-
шивает митохондриальную мембрану, в которой присутствует
в виде димера. Она является терминальным ферментом дыха-
тельной цепи и осуществляет четырехвалентное восстановле-
ние кислорода с образованием воды. Перенос электронов ци-
тохромоксидазой — экзергонический процесс и выделяемая при
этом энергия накапливается в виде трансмембранного протон-
ного градиента, используемого для синтеза АТФ или транспор-
та ионов [Capaldi A. et al., 1983].
Церулоплазмин человека — <х2-гликопротеид плазмы
крови — единственная оксидаза млекопитающих, имеющая го-
лубой цвет, этот фермент образован одной полипептидной
цепью с установленной первичной структурой (1046 аминокис-
лотных остатков), имеет молекулярную массу 132 000 и содер-
жит 6 атомов меди трех .различных типов и четыре цепочки
олигосахаридов, построенных из остатков гексозамина и сиа-
ловой кислоты. Церулоплазмин — мультифункциональный бе-
лок, обладающий активностью ферроксидазы, аминоксидазы
и частично супероксиддисмутазы, участвующий в гомеостазе
меди и играющий роль реактанта острой фазы в воспалитель-
ных, процессах, защищает липидные мембраны от перекисного
окисления [Lovstad R. А., 1984; Nakano Н., 1984]. Полипеп-
тидная цепь церулоплазмина образована тремя гомологичными
участками по 350 аминокислотных остатков, возникшими в ре-
зультате тандемной трипликации исходного гена. Каждый
122
участок включает три домена, различающихся по своей струк-
туре и функции [Ortel Т. L. et al., 1984].
Дефицит меди (гипокупреоз) был впервые обнаружен у ла-
бораторных животных в 1927 г. Вскоре появились сообщения
об эндемичных заболеваниях овец и крупного рогатого скота
в ряде стран, связанных с недостатком меди в пастбищных
растениях [Bennet J. С., Chapman F. Е., 1937, и др.] и преду-
преждаемых включением в рацион животных небольших ко-
личеств соединений данного элемента. Эти наблюдения показа-
ли, что медь, кроме участия в кроветворении, необходима так-
же для нормального течения многих биохимических процес-
сов — пигментации и кератинизации шерсти, формирования
миелина, синтеза различных производных соединительной тка-
ни и др.
Было установлено, что медь — это незаменимый компонент
многих ферментов и белков, играющих важную роль в окисли-
тельно-восстановительных процессах. Медная недостаточность
у человека возможна при полном парентеральном питании и
физиологической анемии беременных [Приев И. Г., 1985].
У большинства млекопитающих этот элемент может депониро-
ваться на случай его дефицита [Evans G. W1978].
Экзогенные факторы, влияющие на усвоение
меди. Усвоение и обмен меди тесно связаны с содержанием
в пище других макроэлементов и МЭ и органических соедине-
ний. Установлено, что существует физиологический антагонизм
между медью, с одной стороны, и молибденом и сульфатной се-
рой— с другой [Dick А. Т., 1953]. Нормальным соотношением
меди и молибдена следует считать 10: 1 при 0,1—0,2% сульфа-
тов в сухом корме. Другими элементами, влияющими на усвое-
ние и обмен меди у животных, являются марганец, цинк, сви-
нец, стронций, кадмий, серебро и др. В свете сложного физио-
логического антагонизма между медью, марганцем, цинком и
кальцием следует рассматривать так называемую мясную ане-
мию, появляющуюся у мышей и крыс при скармливании им
сырого рубленого мяса. При этом у молодых крыс возникает
синдром кальциевой и медной недостаточности, а у взрослых —
медной недостаточности (ахромотрихия, анемия, пигментация
зубов). Варка мяса или добавка к сырому мясу 10 мг/кг меди
предупреждает возникновение данного заболевания [Moore F. D.
et al., 1964].
Усиление явлений медной недостаточности у цыплят наблюдалось при да-
че им небольших количеств цинка, кадмия и серебра [Hill С. Н. et al., 1964]
и выражалось уменьшением количества эластина в аорте, задержкой роста
и понижением концентрации гемоглобина. Эти явления отсутствовали, если
одновременно с указанными элементами в рацион добавляли медь (10—
25 мг/кг). Отмечено взаимодействие между медью и ртутью: соли ртути
оказывались ядовитыми для цыплят только при одновременном скармливании
солей меди. Показано, что явления медной недостаточности могут усиливать-
ся при скармливании цыплятам восстанавливающих веществ — аскорбиновой
кислоты и димеркаптопропанола (британский антилюзит).
Существует еще ряд соединений, вызывающих дефицит меди у животных.
К их числу относятся сульфиды, карбонаты, соединения железа [Cousins R. J.,
123
1985]. Карбонат кальция « организме животных понижает кислотность средь»
в верхних отделах пищеварительного тракта, ограничивая всасывание меди,
происходящее главным образом при величине pH ниже 7,0. Усвоение меди
зависит от типа ее соединений в пище.
Доступность меди из пищеварительного тракта определяется в первую
очередь характером лигандов, связывающих этот элемент [Kirchgessner М.
et al., 1970]. В качестве последних могут фигурировать щавелевая и фумаро-
вая (но не лимонная) кислоты, комплексы которых с медью всасываются на
20% быстрее, чем сульфат меди, а также комплексы этого элемента с амино-
кислотами, особенно с лейцином. Комплексы с синтетическими пептидами
усваиваются в обратной зависимости от степени полимеризации. Фитиновая
кислота и ее соли образуют очень прочные комплексы с медью и понижают
всасывание этого элемента. Другими природными соединениями, снижающи-
ми усвоение медн, являются растительные пигменты, особенно хлорофилл,
комплексы которых с медью малодоступны для животных при величине pH
выше 3,2 [Cousins R. J., 1985].
Из органических компонентов важное значение для отложения меди в ор-
ганизме имеет содержание в корме белка. Высокий уровень белка способству-
ет более быстрому наступлению медной недостаточности у овец н тормозит
накопление этого МЭ в печени крыс н свиней. Отмечено также, что ухудшение
белкового питания является фактором, провоцирующим наступление гемоли-
тического криза при хроническом отравлении медью овец [McPherson Е. A. et
al., 1964] и свиней [Lukas F. V. et al., 1965].
На усвоение меди оказывает сильное влияние кальций, снижающий вса-
сывание этого элемента у жвачных и практически не влияющий на этот про-
цесс у свиней и кур, что объясняется высоким значением pH желудочного сока
последних [Grabman Е., 1973]. Используя физиологический антагонизм между
медью, цинком и железом, удается в ряде случаев предупредить токсическое
действие меди при включении повышенных доз этих элементов в рацион
свиней. Как отмечалось выше, вопрос о существовании специфических эндоген-
ных лигандов для меди в желудочно-кишечном тракте остается открытым. Дан-
ные о том, что аминокислоты способствуют усвоению меди из пищи [Kirchges-
sner М., Grassman Е., 1970], не подтверждаются другими авторами [Mar-
ceau N. et al, 1970].
Медь участвует в обмене веществ животного организма как
компонент семи оксидоредуктаз, а у моллюсков и некоторых
членистоногих выполняет роль переносчика кислорода, которым
является гемоцианин. Из них для человека важное значение
имеют только 6 ферментов, поскольку седьмой — уратоксида-
за — у приматов не найден [Hamilton J. W., 1981]. Не нашло
подтверждения присутствие каталитически активной меди и
в ряде других ферментов — каталазе, 6-аминолевулинат-синта-
зе, триптофан-2,3-диоксигеназе, бутирил-СоА-дегидрогеназе, мо-
ноаминоксидазе, допускавшееся ранее отдельными авторами.
Остается неясной роль некоторых медьпротеидов, найденных
в мозге, таких как альбокупреины I и II и нейрокупреин [Sha-
roian S. G. et al., 1977]. В то же время есть основание пола-
гать, что в животном организме имеются еще и некоторые пока
неидентифицированные медьсодержащие ферменты. Физиологи-
ческое значение медьсодержащих ферментов становится более
понятным при рассмотрении метаболических нарушений, выз-
ванных их недостаточностью (табл. 12).
Если физиологическое значение четырех медьсодержащих
ферментов удовлетворительно объясняется их специфической
биологической функцией, то роль супероксиддисмутазы пока
трудно связать с конкретной патологией. Вызывает сомнение
124
Таблица 12
Нарушения метаболизма при недостаточности меди
Патология Метаболический дефект Предполагаемая фермент- ная недостаточность
Ахромотрихия Нарушения формирова- ния сердечно-сосудистой системы, скелета, колла- гена и эластина Поражение ЦНС Поражение ЦНС Нарушение синтеза ме- ланина Нарушение образования поперечных сшивок кол- лагеновых и эластичес- ких волокон Гипоплазия миелина Нарушение синтеза ка- техоламинов Тирозиназа Аминоксндаза соедини- тельной ткани (лизнлок- сндаза) Цитохром-с-оксндаза Дофамин-Р-гндрокси- лаза
представление о роли супероксиддисмутазы в устранении токси-
ческого действия супероксидного ион-радикала, поскольку он
является короткоживущим и малоактивным в водной среде при
физиологическом значении pH. Кроме того, его дисмутация
легко осуществляется природными комплексами переходных ме-
таллов [Valentine W. N., 1981]. Физиологическое действие це-
рулоплазмина как «ферроксидазы» также вызывает сомнение,
так как апотрансферрин легко присоединяет Fe3+, образующий-
ся из Fe2+ в аэробных условиях без участия церулоплазмина.
Известны многочисленные случаи, когда при недостаточности
церулоплазмина нарушения обмена железа и сопутствующие
анемии отсутствуют, например в младенческом возрасте, при
болезнях Вильсона — Коновалова и Менкеса [Reinhammer В.,
Malmstrom В. G., 1981]. В то же время оба белка обладают
важным свойством депонировать и переносить медь. В связи
с этим не исключено что именно данная функция у них являет-
ся главной, а оксидазная функция — побочной.
При медной недостаточности у овец появляются светлые по-
лосы на пигментированной шерсти, у индюков — депигментация
оперения, у грызунов — ахромотрихия [Underwood Е., 1977],
Эти явления связаны с нарушением синтеза фермента тирозина-
зы, катализирующего две реакции биосинтеза меланина: гидр-
оксилирование тирозина до 3,4-дигидроксифенилаланина
(ДОФА) и окисление ДОФА в ДОФА-хинон.
Другое хорошо известное проявление дефицита меди — это
дефектный синтез коллагена, сопровождающийся ломкостью
костей и деформацией скелета у овец, крупного рогатого скота,
собак, домашней птицы, лабораторных животных и при некото-
рых заболеваниях человека. В костной ткани животных при де-
фиците меди повышено содержание растворимого коллагена
(тропоколлагена) и снижено число альдегидных групп, что
свидетельствует о нарушении процессов образования костного
коллагена ([а! (1)]2а2(1), требующего образования прочных
поперечных связей между отдельными молекулами тропокол-
лагена для формирования коллагеновых фибрилл.
125
Процесс превращения тропоколлагена в зрелый коллаген
сопровождается появлением внутри- и межмолекулярных свя-
зей, образующихся за счет боковых цепей остатков лизина в оп-
ределенных участках молекул тропоколлагена. Этому процессу
предшествует окисление лизилоксидазой s-аминогрупп лизина
и оксилизина с образованием полуальдегида аминоадипиновой
кислоты — аллизина и оксиаллизина. Альдегидные группы всту-
пают в реакции альдиминной и альдольной конденсации с обра-
зованием лизин- и оксилизиннорлейцина, производных двух мо-
лекул аллизина (синдесмина), двух молекул аллизина и моле-
кулы лизина (меродесмозин), а также специфического для кол-
лагена гистидиноксимеродесмозина.
Аналогичные изменения в соединительной ткани описаны
при латиризме — отравлении, вызванном употреблением в пищу
душистого горошка (Lathyrus odoratus), содержащего пред-
шественник мощного ингибитора лизилоксидазы — ^-аминопро-
пионитрил. Латирогенные факторы широко используются в экс-
периментах на животных. С их помощью удается вызывать
различные повреждения соединительной ткани, главным обра-
зом у молодых животных. Эти повреждения можно относить
к экспериментальному 'десмолатиризму.
В патологии человека латиризм представлен довольно редки-
ми заболеваниями нервной системы, которые целесообразно от-
носить к нейролитиризму. Наиболее известны случаи отравления
при потреблении особого вида кормового гороха, так называе-
мой чины (Lathyrus sativa L.). Небольшие вспышки латиризма
с симптомами тяжелого органического поражения ЦНС (пара-
парезы и параплегии нижних конечностей) зарегистрированы
в Татарской АССР и в среднеазиатских республиках.
Это заболевание встречается также в Индии, Сирии, Алжи-
ре и крайне редко в Испании, Италии и Франции. Патогенез
нейролатиризма до сих пор мало исследован. Обращает на себя
внимание избирательная заболеваемость отдельных индиви-
дуумов. Очевидно, это объясняется избирательной чувствитель-
ностью некоторых людей к потенциальному токсическому дей-
ствию латирогенов, которая, вероятно, имеет генетическую при-
роду [Авцын А. П., 1959, 1972]. Существует известная аналогия
между латиризмом и фавизмом-—пищевым отравлением кон-
скими бобами (Vicia fava). К фавизму особенно чувствительны
субъекты с наследственным дефицитом глюкозо-6-фосфат —
дегидрогеназы, у которых процесс интоксикации протекает
значительно тяжелей, чем у других практически здоровых
людей.
Нарушение синтеза коллагена III типа [а1(Ш)]3 и особен-
но эластина проявляется обширными внутренними кровоизлия-
ниями в результате образования аневризм крупных артерий.
Как и в случае скелетных нарушений, основным биохимическим
дефектом при данной патологии является неспособность к син-
тезу соединений, обеспечивающих сшивку фибрилл этих белков.
В аорте свиней и кур, испытывающих медную недостаточность,
Л 26
найдена растворимая форма эластина (тропоэластин), не обна-
руживаемая у здоровых животных и отличающаяся от нормаль-
ного белка отсутствием продуктов конденсации четырех боковых
цепей лизина — циклических аминокислот десмозина и изодес-
мозина, образующих межмолекулярные связи в эластине..
В этом белке у животных, испытывающих дефицит меди, обна-
ружена в 6—10 раз большая, чем в норме, концентрация лизи-
на (37—47 остатков против 3—9).
Нарушения формирования и повышенная повреждаемость-
соединительнотканных структур, особенно коллагеновых и эла-
стических волокон, а также кости и хряща при различных фор-
мах медной недостаточности довольно хорошо изучены у раз-
личных видов животных и воспроизведены в эксперименте. Это-
го нельзя сказать о соответствующей патологии человека, кото-
рая пока еще не находится в центре внимания клиницистов и
патологоанатомов.
В 1891 г. А. Н. Черногубов описал 2 случая «слабости со-
единительной ткани», которая имела наследственный характер.
Она выражалась в необычной растяжимости и повышенной ра-
нимости кожи, гиперподвижности и вывихах суставов, зиянии
ран, прорезывании швов, в аномалиях строения кожных рубцов,
в мягких опухолевидных образованиях на коленях, на нижних.
конечностях, в резкой атрофии подкожной жировой клетчатки.
Через 10 лет Е. Ehlers обратил внимание на склонность таких
больных к кровоизлияниям. В 1908 г. Н. A. Danlos дополнил
описание этого синдрома указанием на наличие «псевдоопухо-
лей» в области локтей и коленей [Блинникова О. Е., 1985].
В дальнейшем это заболевание получило название «синдром-
Элерса— Данло» (СЭД) и было неоднократно охарактеризова-
но дерматологами, хирургами, терапевтами, педиатрами, артро-
логами, офтальмологами, медицинскими генетиками и др. В свя-
зи с тем, что в различных случаях заболевания на передний
план выходят то один, то другие признаки, в настоящее время
выделяют не менее одиннадцати клинических типов СЭД. Ис-
следования по выяснению его морфологических, биохимических
и генетических основ продолжаются. В настоящее время уста-
новлено, что распространенность СЭД составляет не менее 1 на
100 000 новорожденных, но стертые формы, очевидно, не рас-
познаются [McKusick V., 1983]. Важное значение имеет патоло-
гия внутренних органов, в частности аномальное формирование-
кровеносных сосудов (аневризмы, артериовенозные фистулы,
варикозные расширения), возможность их разрыва, эмфизема-
легких со спонтанным пневмотораксом, дивертикулы пищевари-
тельного тракта, спонтанные перфорации кишечника, пороки
сердца и многие другие аномалии развития. При патологоана-
томических исследованиях установлена аномалия развития кол-
лагеновых волокон, сочетающаяся или не сочетающаяся с пато-
логией эластических структур. Несмотря на то что для изуче-
ния соединительнотканных структур были применены современ-
ные методы, вопрос о патоморфологическом субстрате СЭД
127
нуждается в дополнительном исследовании. Важно отметить,
что у некоторых животных (собаки породы спаниель, норки)
встречается аналогичное заболевание, имеющее наследственную
породу. Сходное заболевание у коров, характеризующееся рас-
тяжимостью и хрупкостью кожи, носит название «дерматопара-
ксис» [Блинникова О. Е., 1985]. Среди клинических типов СЭД
у человека точно установлена неполноценность лизилоксидазы
при IX типе, сниженная активность которой препятствует пре-
вращению вновь синтезированного коллагена в его растворимую
форму. Роль дефицита меди при других формах и клинических
типах СЭД изучается.
В 1896 г. французский педиатр А. В. Marfan описал редкий
порок развития соединительной ткани, названный им «врожден-
ной гипопластической мезодермальной дистрофией», для кото-
рого были типичны резко выраженная астеническая конститу-
ция, «птичье лицо», узкий череп, срезанный подбородок, истон-
чение ушных раковин, синеватая окраска склер, вывих или
подвывих хрусталика, слабое развитие мышц и подкожной клет-
чатки, аномалии формирования грудной клетки, диспропорция
роста туловища и конечностей, арахнодактилия, тяжелая пато-
логия аорты и (или) ствола легочной артерии с формированием
аневризм. Только позднее было установлено, что заболевание
наследуется по аутосомно-доминантному типу. Еще позднее
были обнаружены значительные нарушения обмена, состоящие
в накоплении в организме свободных или связанных с белком
кислых гликозаминогликанов, которые вместе с оксипролином
в повышенных количествах экскретировались с мочой.
Очень поучительны данные патологоанатомического исследо-
вания, при котором были обнаружены изменения эластических
волокон, в том числе в средней оболочке аорты (рис. 24), ди-
строфические процессы в миокарде, аномалии структуры хряща,
разрежения костных балок с неравномерным отложением в них
солей кальция, а также недоразвитие гонад (рис. 25).
Только через несколько десятилетий после первого клиниче-
ского описания было установлено, что синдром Марфана отно-
сится к медьдефицитным заболеваниям, так как у этих боль-
ных имеется конституциональная недостаточность лизилоксида-
зы. При электронно-микроскопическом исследовании в участках
аорты, подвергшихся аневризматическому расширению, были об-
наружены деструкция эластических мембран и их глыбчатый
распад с образованием микрофибриллярного компонента, ли-
шенного эластина [Saruk М., Eisenstein R., 1977]. В. Oakes
и соавт. (1976) описали патологические изменения эластических
волокон при болезни Менкеса, которая также относится к медь-
дефицитным заболеваниям, но имеет своеобразную патолого-
анатомическую картину, в которую входят аномалии развития
ЦНС, в частности своеобразные изменения грушевидных нейро-
нов (клеток Пуркинье) в мозжечке. Патогенез синдрома Мен-
кеса описан в главе о генетике МТОЗов.
Многие авторы установили патологические изменения эла-
128
стических волокон при экспериментально вызванных медьдефи-
цитных состояниях у свиней и крыс. В. В. Серов и А. Б. Шсх-
тер (1981) специально останавливаются на проблеме эластоза
и катаболизма эластина, подчеркивая, что «эластическая ткань
и во взрослом организме не является инертной, а подвергается
процессу разрушения и восстановления, особенно интенсивному
в условиях патологии». А. Б. Шехтер (1981) полагает, что глав-
ную роль в эластолизе играют локальные ферментные процес-
сы, хотя нельзя исключить и диффузию других эластолитиче-
ских ферментов. Особый интерес к ферментному эластолизу,
который появился после серии открытий панкреатической, лей-
коцитарной, макрофагальной и тромбоцитарной эластазы, впол-
не закономерен, однако это лишь одна сторона очень большой
проблемы гомеостаза эластических структур организма в раз-
личные возрастные периоды в норме и в патологии.
Вполне понятно, что в поле зрения исследователей попала
и прогрессирующая биодеградация эластического каркаса лег-
ких при одном из частных и пока еще недостаточно изученном
заболевании человека — идиопатической эмфиземе. R. Senior и
соавт. (1977) считают, что патология эластолиза при эмфиземе
легких обусловлена эластазой лейкоцитов и макрофагов, а не
панкреатическим ферментом. Существуют и другие точки зре-
ния на патогенез эмфиземы легких.
При дефиците меди в организме отмечается снижение актив-
ности такого медьзависимого фермента, как лизилоксидаза. При
экспериментальном дефиците меди N. Soskel и соавт. (1982)
установили достоверное снижение содержания легочного эла-
стина у свиней (11,2 мкг/мг сырой ткани, при норме 16,7 мкг/мг).
В то же время выявлено почти двукратное увеличение концент-
рации коллагена. Авторы не без основания полагают, что при
этом может ингибироваться активность и другого медьсодержа-
щего фермента — супероксиддисмутазы, ответственного за инги-
бирование процессов перекисного окисления липидов мембран
клеток [Soskel N. Т. et al., 1984; Soskel N. T., Sandberg L. В.,
1985].
Полностью оценивая значение возможности ферментной ауто-
агрессии по отношению к таким высокоспециализированным
структурам, как эластические мембраны, мы должны, однако,
задуматься и над ролью тех эндогенных факторов, которые
обеспечивают их трофику, функционирование и сохранность в
организме. Очень многие точно доказанные факты эксперимен-
тальной и клинической патологии показывают, что одним из
этих факторов является медь, входящая в состав упомянутых
выше ферментов, в частности медь лизилоксидазы. Легко допус-
тить, что ферментная атака оказывается наиболее разрушитель-
ной именно для конституционально-неполноценных, в частности
не обеспеченных нормальным микроэлементный составом,
структур.
Не подлежит сомнению, что в недалеком будущем список
медьдефицитных синдромов и болезней, который уже сейчас
9—5G8
129
Рис. 24. Субтотальная биодеградация эластического каркаса аорты вблизи
разрыва аневризмы при синдроме Марфана у мальчика 14 лет. В единичных
сохранившихся волокнах — фрагментация, зернистый распад, утрата тинкто-
риальных свойств. Окраска фукселином. Х640.
достаточно внушителен, будет увеличиваться и главное уточ-
няться. Следует упомянуть о том, что изменения эластических
волокон часто сочетаются с той или иной патологией коллагена.
Об этом, например, свидетельствуют данные С. Lapiere и соавт.
(1976), систематизировавшие факты о нарушениях коллагена
у человека и лабораторных животных. В этих материалах при-
влекают внимание такие экзогенные влияния, как интоксикация
латирогенами и недостаточность содержания меди в диете.
Первостепенное значение будут иметь поиски объяснения
причин биодеградации эластических волокон артериальной стен-
ки, являющейся интегральным компонентом и, возможно, одним
из наиболее важных факторов в патогенезе и инициации атеро-
склероза. Взгляд на эту проблему с точки зрения учения о
МТОЗах совершенно закономерен и имеет много доказательств
эпидемиологического и медикогеографического характера. Одна-
ко он почти не разработан патологами и еще мало известен
клиницистам. Дефицит МЭ (в частности, недостаточность меди
и селена) необязательно должен выражаться в генерализован-
ных проявлениях. Нетрудно представить себе и локализованные
формы микроэлементной (энзимопатической) недостаточности,
связанные с неполноценностью определенных отделов сосуди-
стой системы (и других систем). Именно этим легче всего объ-
яснить удивительную разницу между закономерным пораже-
130
Рис. 25. Гипоплазия сперматогенного эпителия, отек межуточной ткани яичка
яри синдроме Марфана; тот же случай. Окраска гематоксилином и эозином.
Х400.
нием при атеросклерозе крупных артерий эластического типа
при практической интактности венозной системы, хотя атероген-
ные липопротеиды циркулируют во всем кровяном русле. Не
в этом ли разгадка резкого контраста между тяжелым пораже-
нием селезеночной артерии при распространенном атеросклеро-
зе и полным отсутствием атеросклеротических изменений в си-
стеме печеночной артерии? Взгляд на значение местных энзимо-
патий неоднократно обсуждался в литературе. Так, его развивала
и оригинально аргументировала наша сотрудница Ф. Л. Лей-
тес (1967), которая обнаружила местный дефицит липопротеид-
липазы и связывала с этим избирательное поражение опреде-
ленных отделов сосудистой системы.
Новое рассмотрение этой проблемы с целью выяснения воз-
можной роли дефицита меди в патогенезе атеросклероза необ-
ходимо в связи с обнаружением того, что в активный центр ли-
попротеидлипазы входит медь. Вместе с тем содержащая медь
лецитинхолестеринацилтрансфераза — один из важных фермен-
тов липидного обмена — характеризуется пониженной активно-
стью при медьдефицитных состояниях.
Одна из наиболее известных форм патологии медного обме-
на— гепатоцеребральная дистрофия, ныне называемая болезнью
Вильсона — Коновалова, представляет собой прогрессирующее
дегенеративное наследственное заболевание ЦНС, сочетающее-
ся с крупноузловым циррозом печени (его генетика и биохими-
9*
131
ческие нарушения характеризуются в главе 12). При этой бо-
лезни особенно типичны экстрапирамидные симптомы в виде
мышечной ригидности, разнообразных гиперкинезов, расстрой-
ства психических функций, отложение по периферии роговицы
медьсодержащего зеленоватого пигмента (роговичное кольцо
Кайзера — Флейшера), частые проявления геморрагического
диатеза, атония капилляров и застой кровотока, остеопороз,
ломкость костей. Патология печени выявляется примерно
у Уз больных, главным образом с помощью функциональных
проб.
Путем диагностической пункции печени обнаружены гисто-
логические признаки цирроза и высокое содержание меди.
Очень характерны гиперкупрурия и гиперацидоаминоурия.
В крови, как правило, понижена концентрация белка и резко
уменьшено содержание церулоплазмина.
При патологоанатомическом исследовании обнаруживают
размягчение и мелкие кисты в чечевичных ядрах головного моз-
га, в хвостатом ядре, глубоких слоях коры, в зубчатых ядрах
мозжечка, в подбугорных ядрах. Все изменения в ЦНС
Н. В. Коновалов (1948, 1960) подразделяет на ангио- и цито-
токсические. Они выражаются в распространенных стазах, со-
провождающихся периваскулярным отеком, вызывающим анок-
сию и гибель нервной ткани. Цитотоксический компонент вы-
ражается в дистрофических изменениях астроцитов и нервных
клеток, вплоть до их некробиоза; характерным морфологическим
признаком болезни Вильсона — Коновалова является патологи-
чески измененная глия Альцгеймера, возникающая из обычных
астроцитов. Последние в процессе прогрессирующей дегенера-
ции постепенно утрачивают цитоплазму, причем объем их ядер
резко увеличивается, содержание хроматина уменьшается.
Н. В. Коновалов связывал эти изменения с нарушениями ну-
клеинового обмена под влиянием токсичного фактора, которым,
наиболее вероятно, является сама медь (рис. 26, 27).
Избыток свободной меди угнетает активность окислительных
ферментов, что приводит к гибели клеток. Тяжелые нарушения
кровообращения усиливают имеющуюся тканевую гипоксию.
Погибают именно те элементы ЦНС, потребность которых в
кислороде особенно велика.
В течение многих лет, помогая Н. В. Коновалову в патолого-
анатомическом исследовании этого своеобразного заболевания,
А. П. Авцын обратил внимание на характерную особенность па-
тогистологической картины, которая в те далекие времена
(в первые послевоенные годы) не казалась особенно существен-
ной. Она проявляется закономерным отсутствием заместитель-
ного клеточного и волокнистого глиоза, несмотря на явную вы-
раженность довольно грубых деструктивных изменений в ЦНС.
Логично связать обнаруженную глиальную афиброплазию с глу-
боким поражением самих астроцитов. Способность олигодендро-
цитов к пролиферации при болезни Вильсона — Коновалова
даже в очагах размягчения сохраняется.
132
Можно предполагать, что эти изменения мозга являются
истинными маркерами дефицита меди, а также токсического
действия меди, которая при болезни Вильсона — Коновалова
аномальным образом пропитывает многие ткани, в частности
край роговицы, ЦНС, печень и органы ее основного выделе-
ния — почки. Известная роль в происхождении этих поврежде-
ний принадлежит характерным для данного заболевания нару-
шениям аминокислотного обмена. Весьма сходные изменения
ядер астроцитов наблюдаются в ЦНС умерших от тяжелых
поражений печени [Коновалов Н. В., 1960].
Крупный успех в лечении одного из тяжелейших наследств
венных неуклонно прогрессирующих МТОЗов, которым по суще-
ству является болезнь Вильсона — Коновалова, имеет принци-
пиальное значение. Наилучшие результаты дает длительное при-
менение L-пеницилламина. Оно может вести к полному исчезно-
вению клинических симптомов. Значительный эффект дают
тиоловые препараты, связывающие медь и выводящие ее из орга-
низма. Это показывает, что именно медь играет главную пато-
генетическую роль в возникновении основных симптомов заболе-
вания. В настоящее время мысль о первичном поражении пече-
ни, которая казалась не очень убедительной 40 лет назад, все
же нашла по крайней мере частичное подтверждение, так как
признаки патологии печени обнаруживаются значительно рань-
ше, чем появляются неврологические симптомы. Более вероят-
но, однако, что начальные стадии поражения мозга и печени
возникают одновременно и связаны с тяжелыми наследственно
обусловленными нарушениями обмена меди. Поскольку имеет
место не первичная недостаточность меди и не экзогенный из-
быток этого МЭ, а главным образом его патологическое пере-
распределение и нарушение выделения, термин эндогенный
дискупреоз кажется нам заслуживающим внимания для краткой
характеристики этой патологии. Е. В. Шмидт (1977) справедли-
во указывает, что до появления тиоловых препаратов 90% боль-
ных умирали в возрасте до 30 лет. Лечение тиоловыми препа-
ратами значительно улучшило прогноз. Успех этого способа ле-
чения вселяет надежду, что и при других эндогенных МТОЗах
этиотропная и патогенетическая терапия окажется эффективной.
Недостаточность меди вызывает у сельскохозяйственных и
лабораторных животных поражение ЦНС, примером которого
может служить эндемическая атаксия ягнят, описанная в раз-
ных странах мира, включая СССР [Риш М. А. и др„ 1980].
Причиной ее возникновения являются как первичный дефицит
меди на пастбищах, так и ее вторичный недостаток, вызванный
избытком физиологических антагонистов этого металла — мо-
либдена, сульфатов, свинца, кадмия и др. Характерный признак
атаксии — частичный паралич задних конечностей, которому это
заболевание обязано своим названием (swayback). При эндеми-
ческой атаксии отмечается целый ряд конституциональных де-
фектов, среди которых ведущими являются аплазия миелина в
стволе головного мозга и спинном мозге, хроматолиз и гибель
133
Рис. 26. Изменения в ЦНС при болезни Вильсона — Коновалова.
а — пролиферация капилляров в зоне начинающегося размягчения в чечевичном ядре
Импрегнация по методу Тибор-Паппа. Х60; б — разрастание атипических капилляров и
ретикулярных волокон в чечевичном ядре. Импрегнация по методу Тибор-Йаппа. Х250;
в—пролиферация олигодендроглии в зубчатом ядре мозжечка. Окраска по методу Нис-
сля. Х400 (Препараты Н. В. Коновалова).
Рис. 27. Изменения астроцитов при болезни Вильсона — Коновалова.
а —динамика превращения астроцитов в глию Альцгеймера: (1—5)—препараты, окра-
шенные по методу Ниссля; (6—9) — препараты, импрегнированные карбонатом серебра;
(10—13) — гибель астроцитов — препараты, импрегнированные карбонатом серебра [Коно-
валов Н. В., I960]; б — превращение астроцитов в глию Альцгеймера, резкое увеличение
ядра, зернистый распад отростков. Импрегнация карбонатом серебра. XI000 (Йрепарат
Н. В. Коновалов-а); в — набухание и слабовыраженная окраска ядер астроцитов. Метод
Ниссля. Х400 (Препарат Н. В. Коновалова).
нейронов. Характерны также изменения в красных и вестибу-
лярных ядрах. В случаях острой формы эндемической атаксии
в головном и спинном мозге наблюдается образование полостей,
вызванное отеком и венозным застоем с последующими некро-
тическими изменениями. Последние связаны, по-видимому, с по-
вышенной проницаемостью стенок сосудов и замедлением про-
цессов образования дисульфидных мостиков в нейрокератине
[Риш М. А., 1973]. Аналогичные изменения в ЦНС описаны
у морских свинок и крыс. Аплазия миелина рассматривается
как прямой результат угнетения фермента цитохромоксидазы,
обеспечивающего генерирование митохондриями АТФ, необхо-
димого для синтеза фосфолипидов [Underwood Е. 1977].
Сходство симптомов эндемической атаксии овец с паркинсо-
низмом у человека и другими экстрапирамидными заболевания-
ми дало основание предположить, что при атаксии нарушен об-
мен катехоламинов [O’Dell В., 1982]. Действительно, у больных
ягнят и крыс при дефиците меди содержание дофамина и но-
радреналина в головном мозге существенно понижено, что, види-
мо, связано с угнетением ферментов тирозиназы и дофамин-^-
монооксигеназы, участвующих в биосинтезе адреналина. Воз-
можно также, что недостаточность супероксиддисмутазы приво-
дит к накоплению супероксидного ион-радикала, ускоряющего
процесс превращения адреналина в адренохром.
Медь играет определенную роль в процессах передачи нерв-
ного импульса. При ее дефиците в препаратах синаптосом моз-
га крысы существенно повышается связывание ГАМК мускари-
новыми рецепторами и снижается связывание бензодиазепина,
что может служить еще одной из причин возникновения рас-
стройств ЦНС при недостаточности меди. Угнетается при этом
и функция опиоидных рецепторов мозга [Geiger G. D. et al.,
1984]. Прием препаратов меди ведет к нормализации описан-
ных нарушений, но не устраняет изменений, связанных с гипо-
миелинизацией. В последнее время [Bhathena S. J. et al., 1986]
установлено, что содержание меди в организме человека и жи-
вотных оказывает заметное воздействие на уровень нейропепти-
дов, влияя на процессы их биосинтеза или высвобождения. Уме-
ренному гипокупреозу у человека сопутствуют заметное сниже-
ние концентрации плазменного энкефалина, прогрессирующее
повышение в поджелудочной железе количества пептидов, со-
держащих лей-энкефалин наряду со снижением концентрации
свободных лей- и мет-энкефалинов.
Недостаточность меди вызывает анемию у всех исследован-
ных видов животных, однако конкретная роль меди в процессах
гемопоэза остается неясной. Принято считать, что медь необхо-
дима для образования ретикулоцитов и мобилизации железа из
его тканевых депо. Медьдефицитная анемия у человека отмечена
при продолжительном полном парентеральном питании и
вскармливании детей молочными смесями, бедными медью.
Прием препаратов меди приводит к полной нормализации всех
указанных нарушений [Cordano А., 1978]. Предполагалось, что
136
действие меди на обмен железа реализуется через феррохела-
тазу, включающую железо в состав гема, или через 6-аминоле-
вулинатсинтетазу, участвующую на первых этапах его синтеза,
но этот механизм не нашел экспериментального подтверждения.
Нет убедительных данных и в пользу функции церулоплазмина
в качестве ферроксидазы, как это предполагали S. Osaki
и соавт. (1966), даже если допустить присутствие в плазме кро-
ви еще одного медьсодержащего фермента — ферроксидазы II,
осуществляющей окисление железа в отсутствие церулоплазми-
на [Frieden Е., 1979].
Характерным признаком дефицита меди у овец и при болез-
ни Менкеса у человека и мышей является нарушение процессов
кератинизации. Шерсть овец теряет извитость, эластичность.
В ней находят больше N-концевых групп серина и глутамино-
вой кислоты, значительное количество неокисленных сульфгид-
рильных групп [Риш М. А., 1980; Underwood Е., 1977]. Нару-
шение образования дисульфидных мостиков в кератине объяс-
няли ранее недостаточностью особого гипотетического медьсо-
держащего фермента, условно названного сульфидоксидазой,
который, однако, так и не был обнаружен. Таким образом, хотя
медь сама по себе и является эффективным катализатором-
окисления сульфгидрильных групп, конкретный механизм ее
участия в процессах кератинизации продолжает оставаться не-
ясным.
Одним из первых конституциональных дефектов, обнаружен-
ных при недостаточности меди, была симметричная гипо миели-
низация головного и спинного мозга. Фосфолипиды являются
главным компонентом миелина. Их содержание в митохондриях
печени значительно снижается, хотя общее число и распределе-
ние фосфолипидов в мозге существенно не меняются. Выражен-
ная недостаточность меди ускоряет процесс «старения» мито-
хондрий, потерю ими пиридиновых кофакторов, приводит к по-
вышенной проницаемости митохондриальной мембраны и утра-
те ею способности связывать адениновые нуклеотиды, что нару-
шает транспорт АТФ через внутреннюю мембрану. Митохонд-
рии значительно увеличиваются, занимая большую часть цито-
плазмы гепатоцита, и деформируются. Основным биохимиче-
ским дефектом считают нарушение образования фосфатидных
кислот из ацил-СоА и L-глицерол-З-фоофата, вызванное нехват-
кой АТФ при нормальном содержании и активности глицерофос-
фатацилтрансферазы (КФ 2.3.1.15) [Gallagher С. Н., 1979].
Другими проявлениями недостаточности меди у животных
являются особая болезнь крупного рогатого скота в Западной
Австралии, вызванная дегенерацией и фиброзом миокарда,
а также внезапная остановка сердца, связанная с его гипертро-
фией у свиней и крыс, нарушениями структуры эластических
волокон аорты и разрывом крупных кровеносных сосудов у цып-
лят.
Медь обладает выраженным противовоспалительным свойст-
вом, смягчает проявление аутоиммунных заболеваний, таких,
137
например, как ревматоидный артрит [Sorenson J. R„ Kishore V.,
1984]. Связывание меди с недоступными для лизилоксидазы хе-
латными комплексами, различными лекарственными препарата-
ми, такими как тубазид, апрессин, изониазид, новокаинамид, ди-
фенин, лежит, по-видимому, в основе заболевания человека не-
которыми формами «лекарственной волчанки», которую исходя
из этого, можно рассматривать как лигандную патологию. Эндо-
генными лигандами, связывающими медь и блокирующими ли-
зилоксидазу при «спонтанной» волчанке, являются, возможно,
фрагменты гистонов, другие полипептиды, нуклеиновые кисло-
ты, поступающие в кровь из очагов кожных поражений [Поды-
мов В. К., 1982]. Аналогичный процесс лежит, вероятно, в ос-
нове других коллагенозов. Следует иметь в виду, что многие
лекарственные вещества, например ацетилсалициловая кислота,
проявляют свой лечебный эффект в виде комплексных соедине-
ний с МЭ-металлами, но этот вопрос относится к фармакологии
и рассматривается в специальных руководствах [Sigel Н., 1982].
Дефицит меди отражается и на липидном составе плазмы
крови. Он выражается повышенным содержанием холестерина,
триглицеридов и фосфолипидов, не связанным с усилением био-
синтеза холестерина или задержкой выделения стероидов
желчью [Allan S. G., Klevay L. М., 1980]. Повышение концент-
рации холестерина в крови объясняется ускорением его поступ-
ления из печени и задержкой в кровяном русле [Shao М. S.,
Lei К, 1980]. Изучение механизма возникновения гиперхолесте-
ринемии показало, что при недостаточности меди снижена ак-
тивность фермента лецитинхолестеринацилтрансферазы
(КФ 2.3.1.43), катализирующего перенос остатков жирных кис-
лот лецитина в ^-положении на гидроксильную группу холесте-
рина. В результате этой реакции образуется основное количе-
ство холестеридов, которые транспортируются липопротеидами
низкой плотности (ЛНП) и липопротеидами очень низкой плот-
ности (ЛОНП) в печень, где они вовлекаются в процессы ката-
болизма [Lau W. М., Klevay L. М., 1981]. Механизм участия ме-
ди в функционировании трансацетилазы неизвестен. Авторы
полагают, что она необходима для синтеза апофермента или вхо-
дит в состав его простетической группы, что, однако, малове-
роятно, поскольку медь присутствует только в активном центре
оксидоредуктаз, а в трансферазах не обнаружена.
При дефиците меди у крыс имеет место угнетение еще одно-
го фермента липидного обмена — диацилглицерол-липазы (ли-
попротеинлипазы) (КФ 3.1.1.34). В ее активный центр, возмож-
но, входит медь [Lau W. М., Klevav L. М., 1982]. Существует
гипотеза, согласно которой всасывание и внутриклеточный син-
тез холестерина контролируются специфическим рецептором
для ЛНП. Связывание ЛНП, богатых холестерином, на наруж-
ной мембране гепатоцита запускает серию реакций, приводя-
щих к ингибированию р-гидроксиф-метилглутарил-СоА — ре-
дуктазы (КФ-L1.1.88) и подавлению биосинтеза холестерина.
Липопротеинлипаза действует на ЛОНП, превращая их в струк-
138
туры, близкие ЛНП. Важно подчеркнуть, что при недостаточно-
сти фермента этот процесс нарушается, что приводит к возник-
новению гиперхолестеринемии. Имеются также данные, что медь
входит в состав аполипопротеида ЛНП (апо-В) и необходима
для его перевода в растворимую форму. Дефицит меди может
вызывать структурные изменения апо-В, затрудняющие его свя-
зывание рецепторным белком.
Липопротеинлипаза, локализованная на эндотелии сосудов,
катализирует гидролиз белково-связанных триглицеридов с об-
разованием свободных жирных кислот, моно- и диацилглицери-
нов. Снижение ее активности при дефиците меди может быть
также причиной повышенной концентрации в крови триглицери-
дов [Lau W. М., Klevay L. М., 1982]. Отмеченные изменения
активности ферментов в липидном обмене наступают не только
при первичном дефиците меди, но и при неблагоприятном соот-
ношении этого элемента с цинком. Изменения, наблюдаемые
в опытах на крысах, имеют близкое сходство с биохимическими
сдвигами у человека, рассматриваемыми в качестве факторов
риска возникновения ишемической болезни сердца. К их числу
относятся гиперхолестеринемия, изменения на ЭКГ, гиперурике-
мия, отсутствие толерантности к глюкозе. В этой связи вполне
возможно, что дефицит меди в рационе может служить одним
из этиологических факторов этого массового заболевания. Это
тем более вероятно, что, например, в США не более 25% на-
селения потребляют с пищей 2—3 мг меди в сутки, т. е. коли-
чество, считающееся нижним порогом содержания этого элемен-
та в рационе человека [Klevay L. М., 1982]. Приведенные на-
блюдения дают основание обратить пристальное внимание на
необходимость нормирования питания человека по меди и цин-
ку, а также селену, обеспеченность которыми его рационов до
недавнего времени не вызывала никаких сомнений. Не исключа-
ется, что систематическое адекватное потребление этих МЭ с пи-
щей и лекарственными препаратами в будущем станет одним
из путей профилактики ишемической болезни сердца и атеро-
склероза.
Кроме того, медь необходима для образования ненасыщен-
ных жирных кислот. Первые наблюдения о роли меди в этом
процессе были сделаны в ходе опытов по подкормке свиней по-
вышенными дозами этого элемента, вызвавшие у них увеличе-
ние мягкости спинного сала и показавшие, что в нем повыси-
лось содержание мононенасыщенных и снизилось содержание
насыщенных жирных кислот. Аналогичные наблюдения были
сделаны и на крысах [Balevska Р. S. et al., 1985; Cunnane S. С.
et al., 1985]. Однако имеются и прямо противоположные данные
[Singh N. Р„ Medeiros D. М„ 1984].
Дальнейшие исследования показали, что при дефиците меди
понижена активность десатуразы стеариновой кислоты (ацил-
СоА-десатураза, А-9-десатураза, КФ 1.14.99.5). Важно, что для
нормального функционирования Д-6-десатуразы, превращающей
линолевую кислоту в ^-линоленовую, необходим цинк. В этой
139
связи медь и цинк оказывают разное влияние на метаболизм
жирных кислот. Цинк стимулирует превращение незаменимых
жирных кислот пищи в простагландины, тогда как медь способ-
ствует превращению стеариновой кислоты в полиненасыщенные
жирные кислоты семейства олеиновой кислоты. Следует также
отметить, что жирные кислоты обоих метаболических путей име-
ют общие ферментативные стадии, на которых они выступают
как антагонисты.
Антагонизм меди и цинка проявляется не только при биосин-
тезе простагландинов, но и на уровне их функционирования.
Так, медь усиливает действие простагландина PGI2 (простацик-
лина), способствуя его связыванию специфическим рецептором,
тогда как цинк является синергистом простагландина PGIi,
ослабляющим эту связь [Curmane S. С., 1982]. Антагонизм
между медью и цинком может играть важную роль в патологи-
ческих процессах. Известно большое число заболеваний, сопро-
вождающихся нарушением баланса этих элементов в организме,
что влечет за собой сдвиги в обмене незаменимых жирных кис-
лот и синтезе простагландинов. В этой связи представляют не-
сомненный интерес исследования возможности применения дан-
ных элементов в клинической практике.
При дефиците меди отмечено двукратное усиление перекис-
ного окисления липидов в митохондриях и микросомах печени
крыс при одновременном снижении активности СОД, каталазы
и глутатионпероксидазы [Balevska Р. S. et al., 1981; Fields М.
et al., 1984]. Природа взаимодействия меди с последними дву-
мя ферментами остается неясной. Возможно, что в восстановле-
нии глутатионпероксидазы определенную роль играют продукты
супероксиддисмутазной реакции.
Влияние избыточных количеств меди (гиперкупреоз) мало
исследовано в патогенетическом отношении. Известно, что из-
быток меди может вызывать признаки отравления, сопровож-
дающиеся снижением активности и биосинтеза некоторых фер-
ментов.
Ммлъ альбуминовой фракции легко доступна для тканей и
при избыточном ее содержании постепенно в них накапливает-
ся, вызывая ряд патохимических процессов, из которых основное
значение имеют, по-видимому, угнетение мембранной АТФазы,
а также ингибирование некоторых ферментов и кофакторов, со-
держащих сульфгидрильные группы (глутатион, липоевая кис-
лота), что в свою очередь ведет к задержке окисления в тканях
пировиноградной кислоты и других метаболитов углеводного об-
мена [Peters Т. J., Blumenstock F. А„ 1967]. Следует также
иметь в виду, что избыток меди угнетает активность цитохро-
моксидазы и аминоксидазы (КФ.1.4.3.6) [Mondovi В. et al.,
1967]. В хрусталике глаза человека и крупного рогатого скота
избыток меди ведет к полному или частичному угнетению 9 фер-
ментов гликолитического и пентозного путей окисления глюкозы
[Awasthi Y. С. et al., 1975].
Интересно, что у кроликов молибден и сульфаты снижают
140
Синтез церулоплазмина и повышают количество «прямой» меди
в плазме, в результате чего происходит обогащение этим эле-
ментом мозга и почек [Gabbalah S. S. et al., 1965]. Повышение
содержания меди в почках при длительном скармливании мо-
либдена и сульфатов наблюдается также у овец [Даминов Р. А.,
1967].
По данным А. М. Шапошникова (1980), «...у здоровых лю-
дей концентрация меди в крови относительно постоянна и уве-
личивается только при беременности и при стрессе... Концентра-
ция меди в крови увеличивается при острых и хронических
воспалительных заболеваниях, шизофрении, алкоголизме, ревма-
тизме, бронхиальной астме, заболеваниях почек, печени, ин-
фаркте миокарда, при некоторых типах анемий, лимфогрануле-
матозе, лейкозах, злокачественных новообразованиях, при хро-
нических инфекционных заболеваниях и после обширных опера-
тивных вмешательств». Весьма вероятно, что столь обширное
число гиперкупремических состояний имеет как общие, так и
специфические факторы патогенеза. Одним из таких общих фак-
торов является стресс.
Поступление меди в организм человека и животных с пищей
подвержено непрерывным изменениям. Описаны случаи как не-
хватки меди в рационе взрослых [Klevay L. М., 1975; Wolf Р.,
1982] и детей [Приев И. Г., 1985], так и ее избытка, например
при потреблении кислых продуктов, хранившихся в металличе-
ской таре. Избыток меди может возникнуть и в результате при-
сутствия этого элемента в разного рода протезах, пестицидах,
а также при гемодиализе и применении противозачаточных
средств. При контакте с медьсодержащими рудами и использо-
вании соединений меди в промышленности и сельском хозяйстве
возможно избыточное поступление ее в организм. При соприкос-
новении с медьсодержащими фунгицидами возможно отравле-
ние, сопровождающееся ознобом и высокой температурой. Кли-
нически картина «медной лихорадки» напоминает «литейную»
лихорадку. При длительном соприкосновении с рудной пылью
у рабочих может возникать пневмокониоз. Окислы меди могут
ускорять развитие силикоза легких, специфически поражая со-
суды и способствуя развитию воспалительной реакции.
Сульфат меди широко используется в медицине, в частности
для лечения гипохромных анемий. Привлекает внимание его
пока еще не вполне объяснимое лечебное действие (в комплексе
с витаминами) при экземе. Микродозы меди успешно исполь-
зуются в комплексном лечении больных склеромой [Баби-
чев В. Н., 1980]. Это говорит в пользу того, что медь, по-види-
мому, способна усиливать ферментативный процесс внутрикле-
точного переваривания возбудителя склеромы. Следует прове-
рить эту гипотезу экспериментально, испытав действие препара-
тов меди при тех инфекциях, для которых характерен внутри-
клеточный паразитизм возбудителей (риносклерома, лепра, не-
которые формы туберкулеза, риккетсиозов и хламидиазов). Сле-
дует также отметить, что в отечественной литературе примене-
141
нию меди и ее солей в лечебной практике уделяется определен?
ное внимание [Вельховер Е. С. и др., 1982].
Изложенные выше факты подтверждаются данными экспе-
риментальной и клинической патологии. Медь имеет важное
'значение как жизненно необходимый МЭ. В связи с этим проб-
лема дефицита меди выдвигается в клинической и профилакти-
ческой медицине на передний план.
Токсические повреждения организма медью и ее соединения-
ми представляют собой существенный вопрос профессиональной
патологии. Дисбаланс меди и других МЭ и макроэлементов ши-
роко изучается в физиологии и патологии сельскохозяйственных
животных, имеет большое практическое и познавательное значе-
ние для проблемы рационального питания. Разные формы дис-
баланса меди и других МЭ имеют свои аналоги в клинической
медицине.
6.3. Цинк
Биологическая роль цинка была установлена около 120 лет
назад, когда J. Raulin (1869) показал, что данный элемент не-
обходим для роста Aspergillus niger. В дальнейшем это поло-
жение было подтверждено для растений и животных. Принци-
пиальное значение для понимания биологической роли цинка
в животном организме имело открытие D. Keilin и соавт. (1940).
Они установили, что цинк является составной частью карбонат-
дегидратазы (4.2.1.1). Изучение на животных недостатка цинка
позволило Е. Underwood (1971) выделить следующие симптомы:
изменение аппетита, нарушение поведения животных и умень-
шение способности к дрессировке, задержку роста и полового
созревания с прекращением развития яичек и сперматогенеза
у самцов, блокирование эструса, бесплодие, патологические ро-
ды у самок, паракератоз с облысением, лимфопения и увеличе-
ние показателя гематокрита. Цинкдефицитные состояния у чело-
века были впервые идентифицированы как синдром гипогона-
дизма и карликовости у молодых иранцев [Prasad A. S. et al.,
1961].
Запасы цинка в организме невелики, у взрослого человека
содержится всего 22,9—30,6 ммоль, т. е. 1,5—2 г [Prasad A. S.,
1979]. Он обнаружен во всех органах и тканях, однако его ко-
личество колеблется в широких пределах: от 0,15 до 3,3 ммоль
на 1 кг сырой ткани [Henkin R. I. et al., 1971]. Скелетные мыш-
цы наиболее богаты цинком, на их долю приходится 62,6% от
всего количества этого МЭ [Schroeder Н. A. et al., 1967]. Важ-
но отметить, что выходу цинка из его тканевых депо способст-
вуют глюкокортикоиды [Dorn F. et al., 1970]. По данным МКРЗ
(1977), баланс цинка для условного человека составляет: по-
ступление с пищей 13 мг, с вдыхаемым воздухом менее 0,1 мг,
выделение с калом 11 мг, с мочой 0,5 мг, с потом 0,78 мг в сут-
ки. Общее содержание в организме составляет — 2300 мг и из
них в мягких тканях— 1800 мг.
142
На клеточном уровне цинк стимулирует образование поли-
сом, тормозит катализируемое железом свободнорадикальное
окисление. Показано, что для перехода из одной фазы клеточ-
ного цикла в другую необходимо наличие цинка, его недоста-
ток блокирует этот процесс [Riordan J. R. et al., 1976].
В непосредственной связи с указанным физиологическим
фактом находится также высокое (1900 мкг/г) содержание цин-
ка в сперматозоидах [Underwood Е., 1977]. Несомненно, что
именно они являются носителями запаса этого жизненно важно-
го МЭ, необходимого для нормального прохождения всех фаз
дробления оплодотворенной яйцеклетки до ее фиксации в поло-
сти матки, когда развивающийся эмбрион начинает получать
все необходимое с кровью матери. Так как цинк и некоторые
другие химические элементы оказывают свое действие как кри-
тические кофакторы на рост клеток, особенно в фазах репродук-
ции и дифференцировки, высказывается предположение, что
с этим связан и опухолевый рост. В частности, в условиях экс-
перимента показано, что содержание животных на диете, бедной
цинком по сравнению с нормой в 30 раз, статистически досто-
верно угнетает рост некоторых опухолей [Бабенко Г. А., 1981].
Особый интерес к цинку в последние 15 лет связан с открытием
его роли в нуклеиновом обмене, процессах транскрипции, ста-
билизации нуклеиновых кислот, белков и особенно компонен-
тов биологических мембран [Пейве Я. В., Жизневская Г. Я.,
1975], а также в обмене витамина А.
Всасывание цинка. Основным регуляторным механиз-
мом гомеостаза цинка является его всасывание, которое проис-
ходит в тонкой кишке. Цинк усваивается отчасти с помощью
металлотионеина, вырабатываемого, очевидно, в слизистой обо-
лочке кишечника, почках и печени. По мнению G. W. Evans и
Е. С. Johnson (1980), это белок имеет панкреатическое проис-
хождение; у грудных детей в абсорбции цинка принимает уча-
стие простагландин Е2, содержащийся в материнском молоке,
но отсутствующий в коровьем. На абсорбцию цинка из кишеч-
ника большое влияние оказывают обилие пищевых фитатов,
а также содержание других элементов, в первую очередь кад-
мия, меди и кальция, с которыми он вступает в конкурентные
взаимоотношения. Как свидетельствуют эксперименты на жи-
вотных и клинические наблюдения, обмен цинка и железа нахо-
дится под эффективным гомеостатическим контролем.
У человека и крыс цинк, как и кальций, всасывается в ос-
новном в тонкой кишке. У крупного рогатого скота около '/з ра-
зовой дозы цинка, поступившей per os, всасывается в сычуге.
У кур всасывание цинка происходит как в железистом желудке,
так и в тонкой кишке [Hambidge К. М., 1986]. У крыс при го-
лодании 60% дозы 6SZn всасываются в двенадцатиперстной,
30% — в подвздошной и 10% — в тощей кишках, тогда как у не-
голодающих животных основным участком всасывания цинка
является подвздошная кишка (60%) и только 20% приходятся
на двенадцатиперстную и тощую кишки. Часть всосавшегося
143
цинка задерживается в самой слизистой оболочке. В двенадца-
типерстной и подвздошной кишках она составляет 10% и в то-
щей кишке— 30%. /
Всасывание цинка через исчерченную каемку происходит
частично по механизму регулируемой облегченной диффузии.
У крыс, испытывающих дефицит цинка, скорость поступления
этого элемента в изолированные ПМИК вдвое выше, чем
у контрольных животных, обеспеченных цинком. Это свидетель-
ствует о том, что гомеостатический контроль всасывания цинка
существует даже на уровне исчерченной каемки. Данные изуче-
ния кинетики поступления в нее цинка позволяют также пред-
положить, что наряду с насыщаемым (опосредованным) меха-
низмом существует и пассивный (неопосредованный) механизм
транспорта этого элемента. В ПМИК, полученных от крыс, ис-
пытывающих дефицит цинка, преобладает всасывание по перво-
му из указанных механизмов. Не исключено, однако, что так
называемая пассивная диффузия является артефактом, объяс-
няемым поступлением цинка в пузырьки через поврежденную
мембрану [Cousins R. J., 1985].
При дефиците цинка в исчерченной каемке отмечено резкое
усиление синтеза особого мембранного белка с молекулярной
массой около 45 000, который, по-видимому, участвует в гомео-
статической регуляции всасывания этого элемента. Данный бе-
лок синтезируется также при ограниченном кормлении живот-
ных, что отчасти объясняет повышенное усвоение цинка, а так-
же ряда других элементов при голодании. Возможно, что ана-
логичным образом объясняется и усиление всасывания цинка
беременными и лактирующими животными. Аналогичный мемб-
ранный белок, найденный в вилочковой железе крыс, получил
название «цинкин» [Ribas В. et al., 1987].
Цинк всасывается в две фазы, одна из которых, возможно,
сопряжена с затратой энергии, что, однако, признается не все-
ми авторами [Solomons N. W., Cousins R. J., 1984]. Первая фа-
за протекает быстро и отражает поступление цинка через исчер-
ченную каемку, тогда как вторая, более медленная, фаза харак-
теризует, очевидно, транспорт этого элемента через базолате-
ральную мембрану. Всасывание цинка в известных пределах
обратно пропорционально его концентрации в содержимом кишеч-
ника, что объясняется постепенным насыщением участков свя-
зывания этого элемента на мембране исчерченной каемки. При
высоких концентрациях цинка возможно, однако, нарушение
барьерной функции мембраны и его неспецифическое связыва-
ние клеточными белками и другими лигандами. Существует,
по-видимому, два различных механизма всасывания цинка, один
из которых функционирует при низких концентрациях этого эле-
мента. В этой связи количественное поступление цинка в эпи-
телий слизистой оболочки может быть сходным как при низком,
так и при высоком содержании этого элемента в содержимом
кишечника.
Интенсивные исследования форм цинка, поступающих в эпи-
144
Таблица 13
\ Влияние эндогенных и экзогенных лигандов на всасывание цинка
\ [Cousins R. J., 1985]
; Лиганд Эффект Лигаид Эффект
ЭДТА, о-оксихинолин t f Аскорбиновая кислота f О
Высокий уровень белка f Лимонная кислота К О 1
в рационе
Лнзин, цистеин, глицин, t Пиколиновая кислота 1. О 1
глутаминовая кислота
Низкий уровень железа Глутатион 0
в рационе
Лактоза t Высокое содержание |, о
кальция в рационе
Г истидин f, 0 Клетчатка |, О
Сок поджелудочной же- f, 0 Низкое содержание бел- 4
лезы ка в рационе
Простагландин Е2 f, 0 Фитаты
Повышенный уровень 1 Медь 1, О
железа в рационе
Условные обозначения: ф — повышение; f — понижение; о — отсутствие
эффекта.
телий слизистой, не дали определенных результатов. Показано»
что этот элемент может всасываться как в ионной форме из
изотонического раствора NaCl, так и в виде комплексов с раз-
личными низкомолекулярными лигандами, причем разные авто-
ры дают нередко прямо противоположную оценку роли этих ли-
гандов в данном процессе (табл. 13).
ЭДТА значительно повышает всасывание цинка, в связи
с чем используется для предупреждения паракератоза у свиней.
Аналогичным свойством обладает о-оксихинолин, применяемый
с успехом при лечении энтеропатического акродерматита. Комп-
лекс цинк — ЭДТА, не разрушаясь, проходит через слизистую
оболочку в систему воротной вены. Повышенное содержание
белка в рационе улучшает всасывание цинка, по-видимому, за
счет образующихся аминокислот. При низкобелковом рационе
усвоение цинка, напротив, снижается. Лизин, цистеин, глицин и
глутамат стимулируют этот процесс, тогда как положительное
действие гистидина признается не всеми авторами. Повышенный
уровень кальция в рационе затрудняет всасывание цинка и не-
редко служит причиной заболевания свиней паракератозом.
У человека, однако, подобное действие кальция не наблюдается
[Snedeker S. М. et al., 1982].
В связи с лечебным действием грудного молока при энтеро-
патическом акродерматите делались попытки выделить из него
специфические лиганды для цинка, в качестве которых указыва-
лись метаболит триптофана — пиколиновая кислота — и цитра-
ты [Evans G. W., Johnson Р. Е., 1980]. Однако эти данные не
получили подтверждения в опытах с перфузией изолированного
10—568
145,
кишечника крыс. Оба эти соединения вызвали также существен-
ное угнетение всасывания цинка в ПМИК. Пиколевая кислота
оказалась неэффективной в опытах на крысах и в предупрежде-
нии болезни Адема (наследственной недостаточности цинка у те-
лят) [Flagstad Т., 1981]. В то же время имеются данные о том,
что пиколиновая кислота способствует усвоению цинка из ра-
ционов с ограниченной доступностью этого элемента, а в опы-
тах с вывернутыми сегментами кишечника повышает поступле-
ние в них радиоактивного цинка. Лимонная кислота в этих
опытах оказала прямо противоположное действие. Лечебные свой-
ства грудного молока связаны, по-видимому, не столько с низ-
комолекулярными лигандами, сколько с типом и количеством
белка. Оно содержит значительно меньше белка и некоторых
других компонентов, чем коровье. Кроме того, основными бел-
ками грудного молока, связывающими цинк, являются лакто-
феррин и альбумин, тогда как в коровьем молоке этот элемент
преимущественно связан с казеином. Исходя из этого большая
доступность цинка из грудного молока объясняется лучшей пе-
ревариваемостью данных белков и меньшей прочностью связы-
ваниями ими указанного МЭ.
В ряде работ допускалось, что для всасывания цинка, осо-
бенно у новорожденных, имеет важное значение липидная фрак-
ция молока. При этом главная роль отводилась незаменимым
жирным кислотам и в первую очередь метаболитам линолено-
вой кислоты (у-линоленовая кислота, дигомо-у-линоленовая кис-
лота и простагландин Ej) [Cunnane S. С., 1982]. Утверждалось
также, что в качестве эндогенного лиганда для цинка может
служить простагландин Е2 [Song М. К., Adham N. F., 1978].
Однако эти данные также не нашли подтверждения.
Из пищи, бедной цинком, усваивается до 85% от общего ко-
личества этого элемента, а из обычной пищи — всего 10—30%.
Усвоение цинка из различных кормов неодинаково. В составе
кукурузы он доступен на 52%, пшеницы — на 60%, гороха, яч-
меня, бобов — на 66—68% и люпина — на 80%. Из полусинте-
тического казеинового рациона, содержавшего 18 мг цинка, его
всасываемость составила 83% [Weigand Е., Kirchgessner М.,
1980]. На всасывание цинка влияют объем съедаемой пищи
и содержание в ней клетчатки, которая, ускоряя прохождение
пищевых масс по желудочно-кишечному тракту, уменьшает кон-
такт цинка с участками его абсорбции [Ковальский В. В.,
Риш М. А., 1979]. Всасывание цинка значительно снижается
при воспалительных процессах под влиянием лейкоцитарного
эндогенного медиатора ИЛ-1, вызывающего одновременно сни-
жение уровня цинка в плазме крови и накопление его в печени
[Pekarek R. S., Evans G. W., 1976].
Наряду co специфическим транспортом возможны и другие
пути всасывания цинка через стенку кишечника. У крыс, испы-
тывающих дефицит железа, всасывание цинка повышено, по-ви-
димому, за счет связывания его внутриклеточным трансферри-
ном. Транспорт некоторого количества цинка через эпителий
146
слизистой оболочки возможен также кальцийсвязывающим бел-
ком.
К числу интересных результатов о влиянии витаминов на
обмен МЭ следует отнести данные о взаимосвязи между обме-
ном цинка и железа, с одной стороны, и витамина А— с другой.
Первыми на эту взаимосвязь указали В. К. Бауман и соавт.
(1975), обнаружившие нарушение всасывания цинка при гипо-
витаминозе А у цыплят, а в 1981 г. В. К. Бауман выделила
специфический цинксвязывающий белок из кишечной стенки
цыплят, синтез которого индуцируется витамином А. Этот ви-
тамин выполняет, таким образом, в отношении цинка ту же
функцию, что витамин D в отношении кальция, и не исключено,
что механизм его действия близок к таковому стероидных гор-
монов. Вместе с тем G. W. Evans и соавт. (1976) показали, что
при дефиците цинка в печени нарушен синтез ретинолсвязываю-
щего белка, необходимого для транспорта витамина А в кровя-
ном русле. Эти явления запускают каскадный механизм пороч-
ного круга, при котором дефицит цинка или витамина А уве-
личивает их недостаток. Особенно сильное влияние оказывает
одновременный дефицит обоих пищевых факторов, приводящий
к срыву гомеостатической регуляции организма. Этим, по-види-
мому, объясняется сезонный характер некоторых гипомикроэле-
ментозов, таких как, например, эндемический паракератоз круп-
ного рогатого скота в Узбекистане, наступающий в поздний зим-
ний и ранний весенний период [Риш М. А. и др., 1979], и энде-
мическая иктерогемоглобинурия у каракульских овец, прекра-
щающаяся в период их перехода на зеленый подножный корм.
Следующей стадией усвоения цинка является его взаимодей-
ствие с внутриклеточными лигандами энтероцита. На этой ста-
дии существенное значение принадлежит металлотионеину.
Установлено, что цинк, поступивший в клетку слизистой обо-
лочки, связывается металлотионеином, причем повышение со-
держания цинка в рационе индуцирует транскрипцию металло-
тионеиновой мРНК и синтез новых количеств этого белка, кото-
рый связывает избыток цинка. При слущивании эпителия боль-
шая часть связанного цинка теряется. При низком содержании
цинка в рационе уровень металлотионеина невысок, и транс-
порт к базальной мембране этого элемента осуществляют его
комплексы с низкомолекулярными лигандами. Таким образом,
имеет место второй гомеостатический механизм, связанный с ос-
новным внутриклеточным лигандом цинка металлотионеином.
При введении антител против металлотионеина они обнаружи-
ваются в эпителии, выстилающем ворсинки, и в клетках Панета.
Дополнительный синтез металлотионеина происходит, одна-
ко, только при резком повышении количества цинка в рационе
и является кратковременным процессом. В отсутствие столь рез-
ких изменений уровня этого элемента экспрессия металлотио-
иеиновых генов снижается и транскрипция металлотионеиновой
мРНК возвращается к стационарному уровню, В этой связи
в настоящее время полагают, что продолжительные изменения
10* 147
процесса всасывания цинка (и меди) в меньшей степени зави-
сят от синтеза металлотионеина, а происходят за счет эндоген-
ной секреции этого элемента, который, поступая из крови в эпи-
телий слизистой оболочки, блокирует всасывание цинка из со-
держимого кишечника. Вполне возможно, что металлотионеин,
обнаруживаемый в эпителиоцитах слизистой оболочки и в клет-
ках Панета, регулирует не столько всасывание цинка, сколько
его выделение.
Не исключено, что физиологический антагонизм меди и цин-
ка разыгрывается отчасти на уровне металлотионеина. Синтез
данного белка индуцируют оба эти элемента, однако цинк более
активен, а медь образует более прочные комплексы с указан-
ным белком. В связи с этим при даче умеренных количеств пре-
паратов цинка реально ожидать в первую очередь образование
медь-тионеина и снижение поступления в кровь этого элемента.
Взаимодействие обоих металлов может иметь клиническое зна-
чение, например, при приеме массивных доз препаратов цин-
ка при . болезни Вильсона — Коновалова [Prasad A. S. et al.,
1978].
Перенос цинка через базолатеральную мембрану представ-
ляет собой, по-видимому, активный процесс, требующий присут-
ствия кислорода и затраты энергии. Он ингибируется метаболи-
ческими ядами [Kowarsky S. et al., 1974] и может зависеть так-
же от доступности металлосвязывающих участков на плазменном
альбумине. Цинк содержится в плазме и эритроцитах в от-
ношении 1 : 8 или 1 : 9. В эритроцитах он представлен главным
образом как компонент карбоангидразы и в меньшей степени
супероксиддисмутазы. Некоторое количество цинка связано,
по-видимому, с мембраной, участвуя в стабилизации ее струк-
туры. Лейкоциты содержат до 0,3% цинка, который в отличие
от эритроцитарного цинка не обменивается с его плазменными
запасами и не реагирует на дефицит этого элемента. Основным
транспортным белком плазмы крови, переносящим 2/з метабо-
лически активного цинка, является альбумин. Содержание цин-
ка в плазме крови тесно коррелирует с количеством цинка, свя-
занного альбумином. Основное количество цинка, присутствую-
щего в амниотической жидкости, также связано с альбумином.
Цинк присутствует также в «2-макроглобулине, метаболическая
функция которого недостаточно изучена. Одно время допускали,
что транспорт цинка в системе воротной вены осуществляется
трансферрином [Evans G. W., Winter Т. W., 1975], но оказа-
лось, что связь цинка с альбумином прочнее, чем с трансферри-
ном [Charlwood Р. А., 1979], причем при перфузии изолирован-
ных сегментов кишечника всосавшийся цинк связывается не
трансферрином, а альбумином [Smith С. J., Cousins R. J., 1980].
Степень всасывания цинка коррелирует с содержанием альбу-
мина, а не трансферрина. Таким образом, уровень циркулирую-
щего альбумина может иметь существенное значение в процес-
сах всасывания цинка. Более того, оказалось, что культура
гепатоцитов усваивает цинк только из сред, содержащих ами-
148
нокислоты И альбумин, но не трансферрин [Failla A. L., Сои-
sinsvR. J., 1978].
Эти наблюдения свидетельствуют, что именно альбумин яв-
ляется основным физиологическим лигандом, транспортирующим
цинк из кишечника к печени. Выше отмечалось, что альбумину
принадлежит важная роль и в транспорте меди в системе ворот-
ной вены, но оказалось, что участки связывания обоих элемен-
тов на альбумине не совпадают. Относительно небольшая часть
цинка в плазме крови связана аминокислотами, преимуществен-
но гистидином и цистеином. Физиологическое значение этих ли-
гандов неизвестно, но вполне возможно, что в некоторые органы
(например, в мозг) цинк поступает в составе низкомолеку-
лярных комплексов. Эти же комплексы могут играть определен-
ную роль и в процессах выделения цинка почками, причем боль-
шая часть упомянутых комплексов должна подвергаться реаб-
сорбции во избежание повышенных потерь цинка [Victeri W.
et al., 1984].
Поступление цинка в гепатоциты было изучено на первичных
однослойных культурах этих клеток [Failla A. L., Cousins R. J.,
1978]. Оно является двухстадийным энергозависимым процес-
сом, требует наличия свободных сульфгидрильных групп и до-
стигает насыщения при концентрации цинка в культуральной
среде, равной 12 мкмоль. Первая стадия совершается быстро и
заключается, очевидно, в связывании цинка специфическими
лигандами на поверхности плазматической мембраны, куда он
доставляется альбумином. Вторая стадия протекает значительно
медленнее и носит линейный характер. Первая стадия проис-
ходит по принципу облегченной диффузии с насыщением и
является, ,по-видимому, общей для цинка и кадмия.
В последние 15 лет выполнен большой объем исследований
по внутриклеточному обмену цинка. Установлено, что глюкокор-
тикоиды повышают содержание цинка в гепатоцитах, причем
это накопление связано с синтезом металлотионеина, в селек-
тивной стимуляции синтеза которого участвуют также инсулин
и глюкагон [Di Silvestro R. A., Cousins R. J., 1983]. Высвобож-
дение цинка из клетки обусловлено распадом его внутриклеточ-
ных лигандов, в частности металлотионеина, соединение которо-
го с цинком распадается в 5 раз быстрее, чем с кадмием. Отток
цинка из клеток зависит также от содержания в плазме альбу-
мина и аминокислот (рис. 28).
В основном цинк из организма выводится с калом. У взрос-
лого человека при суточном поступлении цинка в пределах 10—
15 мг около 90% этого количества выводится с калом и 2—10%
(300—500 мкг) — с мочой. С калом удаляется также большая
часть 65Zn, введенного внутривенно. У овец и крупного рогатого
скота концентрация цинка в моче мало зависит от его поступле-
ния с кормом. Цинк, выделяемый с калом, состоит из неусвоен-
ного и эндогенного цинка, причем количество последнего отра-
жает уровень поступления этого элемента с пищей и играет
важную роль в его гомеостазе.
149
Рис. 28. Регуляция обмена циика и синтеза металлотионеииа в гепатоците
[Cousins R., 1985].
Эндогенный цинк в отличие от меди подвергается частичной
реабсорбции. Так, было показано, что верхние отделы пищева-
рительного тракта человека содержат на 144—300% больше
цинка, чем его поступает с пищей [Matsesche J. W. et al., 1980],
что значительно превышает его выделение с калом. Источни-
ками эндогенного цинка являются поджелудочная железа,
желчь и стенка кишечника. У свиней и собак желчь бедна цин-
ком. У крыс он выделяется поджелудочной железой преимуще-
ственно в составе белков, тогда как в желчи он связан с низко-
молекулярными компонентами, главным образом с глутатионом.
После оперативных вмешательств содержание цинка в желчи
существенно возрастает [Schneeman В. О. et al., 1983].
При распаде тканей, например при ожогах, хирургических
операциях и других травмах, голодании, содержание цинка
в моче существенно возрастает. Наблюдается гиперцинкурия и
при печеночной порфирии, посталкогольном циррозе и хелатоте-
рапии [Prasad A. S. et al., 1965; Spencer Н. et al., Kostoff, 1966].
Цинк поступает в мочу в основном из ультрафильтрата
плазмы крови. В дистальных почечных канальцах он обычно на
95% подвергается реабсорбции и его количество в моче хорошо
коррелирует с ее объемом и содержанием в ней креатинина.
При увеличении объема мочи и при беременности происходит
некоторое повышение выделения цинка, связанное, по-видимому,
с усилением тубулярного тока. Цинкурия повышается также при
даче некоторых диуретиков и натрийуретиков (например, хлоро-
тиазида). Она наступает нередко и при полном парентеральном
питании, при котором может выводиться в сутки до 23 мг этого
элемента [Kay R. et al., 1983]. Значительное выделение цинка
происходит при инфузиях аминокислот, особенно цистеина и
150
гистидина [Junice A. A. et al., 1978; Van Roj A. M. et al., 1979;
Abu-Hamdan et al., 1981].
У здоровых лиц, живущих в умеренном климате, в сутки
с потом теряется 0,4—2,8 мг цинка [Schraer К. К., Callo-
way D. Н., 1974; Hess F. М. et al., 1977; Bauer М. Т., King J. С.,
1984; Milne D. В. et al., 1984], причем его количество отражает
содержание этого элемента в рационе. В жарких странах поте-
ри цинка с потом могут служить одной из причин наступления
дефицита этого элемента [Prasad A. S., 1963]. Ежемесячные по-
тери цинка с менструальной кровью невелики и составляют
0,1—0,5 мг на период. Небольшое количество этого элемента
удаляется также с волосом и ногтями [Milne D. В. et al., 1984;
Umoren J., Kies C., 1982]. По мнению О. Guillard (1980), опре-
деление цинка в волосах является важным диагностическим
тестом стертых (скрытых) форм его недостаточности. Довольно
значительные количества цинка выводятся с молоком. В нем
содержится у человека от 7 (на 1-е сутки) до 2,5—3 мг/л в кон-
це 1-го месяца лактации. Далее следует постепенное снижение
концентрации этого элемента в молоке, составляющее в конце
1-го года 0,65 мг/л.
Ферменты, содержащие цин к. Первые данные о ме-
ханизмах, лежащих в основе биологической активности цинка,
были получены D. Keilin и Т. Mann (1940), показавшими, что
цинк входит в состав карбоангидразы эритроцитов. Через
15 лет В. L. Vallee и Н. Neurath (1955) обнаружили цинк в со-
ставе карбоксипептидазы А поджелудочной железы крупного
рогатого скота. Вскоре был открыт еще целый ряд цинксодер-
жащих ферментов [Vallee В. L., Wacker W. Е. С., 1970].
В настоящее время цинк найден более чем в 200 металло-
ферментах, участвующих в самых различных метаболических
процессах, включая синтез и распад углеводов, жиров, белков
и нуклеиновых кислот. Цинксодержащие ферменты относятся ко
всем шести известным классам [Vallee В. L., Galdes А., 1984],
но в наибольшем количестве представлены в классе гидролаз.
Они катализируют 27 различных реакций. Гомологичные фер-
менты получены из самых разных источников. Так, щелочная
фосфатаза изучена у 11, аминопептидаза — у 17, а нейтральная
протеаза — у 25 различных видов живых организмов
[O’Dell В. L. 1984].
Следующая по численности группа, содержащая более
20 ферментов, относится к лиазам. Среди ее представителей на-
ходятся альдолаза, карбоангидраза и аминолевулинат-дегидра-
таза (КФ 4.2.1.24).
Более 10 цинксодержащих ферментов относятся к подклассу
фосфотрансфераз. Это тимидинкиназа, ряд нуклеотидилтрансфе-
раз: РНК- и ДНК-полимераза, полученные из И различных ис-
точников, обратная транскриптаза миелобластоза птиц, поли(А) -
полимераза из 8 источников и др. (табл. 14).
В связи с тем что цинк имеет заполненную d-подоболочку
(О10-электронная конфигурация), он не обладает переменной
151
Фермент
Источник
Таблица 14
Металлоферменты, содержащие цинк [Vallee В. L., 1985]
Оксидоредуктазы
Алкогольдегидрогеназа
Алкогольдегидрогеназа
D-лактатдегидрогеназа
D-лактатцитохромредуктаза
Супероксиддисмутаза
Дрожжи
Позвоночные, растения
Усоногий рак, бактерии
Дрожжи
Позвоночные, растения, грибы, бак-
терии
Трансферазы
Транскарбоксилаза
Аспартаттранскарбамилаза
Фосфоглюкомутаза
РНД-полимераза
ДНД-полимер аза
Обратная транскриптаза
Ядериая поли (А)-полимераза
Концевая дезоксинуклеотидилтранс-
фераза
Меркаптопируватсульфидтрансфера-
за
Фосфодиэстераза (экзонуклеаза)
Фосфодиэстераза циклических нукле-
отидов
Нуклеаза Pi
а-амилаза
a-D-маннозидаза
Аминопептидаза
Аминотрипептидаза
Дипептидаза
Ангиотензиназа
Прокарбоксипептидаза А
Прокарбоксипептидаза В
Карбоксипептидаза А
Карбоксипептидаза В
Прочие карбоксипептидазы
DD-карбоксипептидаза
Эластаза
Penicillum shermanii
Е. coli
Дрожжи
Зародыши пшеницы, бактерии, ви-
русы
Морской еж, фаг Та
Ретровирусы
Печень крысы, вирусы
Тимус теленка
Е. coli . .
Яд змеи
Дрожжи
Микроорганизмы
Bacillus subtilis
Млекопитающие, растения
Млекопитающие, грибы, бактерии
Кишечник кроликов
Млекопитающие, бактерии
Млекопитающие
Поджелудочная железа
» »
Позвоночные, ракообразные
» »
Позвоночные, ракообразные, расте-
ния, бактерии
S. albus
Pseudomonas aeruginosa
Гидролазы
Щелочная фосфатаза
б'-нуклеотидаза
Фруктозо-1,6-дифосфатаза
Фосфолипаза С
Р-Лактамаза
Креатининаза
АМФ-деаминаза
Неорганическая пирофосфатаза
Нуклеотидпирофосфатаза
Млекопитающие, бактерии
Бактерии, лимфобласты, плазма
Млекопитающие
Bacillus cereus
Bacillus cereus
P. putida
Мышцы кролика
Дрожжи
Дрожжи
152
Продолжение
Фермент
Источник
Лиазы
Альдолаза фруктозо-1,6-дифосфата
Альдолаза Ь-рамиулозо-1-фосфата
Карбоангидраза
Дегидратаза а-аминолевулиновой
кислоты
Глиоксалаза
Нейтральная протеаза
Коллагеназа
Аминоацилаза
Дигидропиримидин амииогидролаза
Дигидрооратаза
Дрожжи, бактерии
Е. coli
Животные, растения, человек
Печень, эритроциты млекопитающих
Млекопитающие, дрожжи
Позвоночные, грибы, бактерии
Млекопитающие, бактерии
Почки свиньи, микроорганизмы
Печень крупного рогатого скота
Clostridium oroticum
Изомеразы фосфоманнозы
мРНК -синтетаза
Пируваткарбоксялаза
Изомеразы
| Дрожжи
Лигазы
Е. coli, Bacillus stearothermophilus
Дрожжи, бактерии
валентностью и в окисленном виде всегда двухвалентен. Неко-
торые молекулярные параметры ферментов, содержащих цинк,
приведены в табл. 15.
Координационное число цинка обычно равняется четырем,
что позволяет ему образовывать связи с четырьмя лигандами.
Подобные комплексы цинка имеют тетраэдрическое строение и
отличаются стерически от квадратных планарных комплексов
двухвалентной меди. Известны и октаэдрические комплексы
цинка, в которых участвует 6 лигандов. Ионный радиус цинка
меньше, чем у двухвалентной меди (0,074 и 0,092 нм соответ-
ственно). По этой причине ион цинка несет более концентри-
рованный заряд, чем ион двухвалентной меди, и обладает
большим сродством к электронам. Химическая стабильность
цинка лежит, видимо, в основе его широкого участия в самых
различных биологических процессах, таких как гидролиз, ре-
акции переноса, присоединения к двойным связям и даже окис-
ления — восстановления. Однако цинк в редокссистемах, напри-
мер в алкогольдегидрогеназе, выступает не как переносчик
электронов, а как сильная льюисовская кислота, оттягивающая
на себя электронные пары.
В связи с заполненностью d-оболочки у двухвалентного цинка отсутству-
ют хромоформные свойства в видимой области и другие спектральные харак-
теристики таких переходных металлов, как железо, медь, марганец и кобальт.
Этой особенностью объясняется отчасти относительно позднее обнаружение
его ^значения для биологических процессов. Отсутствие у цинка парамагнитных
свойств не позволяет использовать для изучения его соединений ЭПР-спект-
роскопию. В этой связи впервые на цинксодержащих ферментах был разрабо-
153
Таблица 15
Молекулярные характеристики некоторых цинксодержащих белков
и ферментов [Vallee В. L., Galder А., 1984]
Наименование Источник Содержа- ние цинка, число ато- мов на мо- лекулу Моле- куляр- ная масса, хюз Кофактор
Алкогольдегидрогенеза Печень человека, лоша- 4 £801 2NAD
D-лактатцитохромредук- ДИ Дрожжи 4—6 50J —
таза D-глицеральдегидрофос- Мышцы свиньи 3 137 3NAD
фат дегидрогеназа Г устин Слюна человека 2 37 —
РНК-полимераза Е. coll, дрожжи 2—2,4 700 —•
ДНК.-полимераза Euglena gracilis, мор- 1—4 109 —
Обратная транскриптаза ской еж Вирус миелобластоза 2 180 —
Меркаптопируват-S- птиц, млекопитающих Е. coif 1 24 —
трансфераза Щелочная фосфатаза Почки, печень, плацента 2 80 —
Аминопептидаза млекопитающих Почки свиньи 6 300
Карбоксипептидаза А Поджелудочная железа 1 35 —
Карбоксипептидаза В человека, быка, свиньи Поджелудочная железа 1 35 —..
Дипептидаза человека, быка, свиньи Почки свиньи, опухоле- 1 47 —-
Нейтральная протеаза вые клетки мыши Яд змеи 1 22,5 2СА
5'-АМФ-аминогидрола- Мышца крысы, кролика 2 290 —
за Альдолаза Дрожжи, 2 80
Кар бон ат-дегидр ат аз а Aspergillus niger Эритроциты млекопитаю- 1 29
(карбоангидраза) Супероксиддисмутаза Фосфоманнозонзомера- 1ЦИХ Дрожжи 2 1 33,6 450 2 Си
за Пируваткарбоксилаза Печень свиньи, дрожжи 4 600 Биотин
Термолизин Bacillus thermoproteoly- 1 35
5'-Нуклеотидаза ticus Е. coli 1 52
а2-Макроглобулин Сыворотка крови челове- ка 2 725 —
тан широко используемый в настоящее время метод замещения цинка пара-
магнитными металлами. Играя роль спиновой метки, эти металлы позволили
выяснить строение активного центра ферментов, включающих цинк. Этот эле-
мент был замещен кобальтом в карбоксипептидазе, карбоангидразе, термоли-
зине, щелочной фосфатазе без потери ими ферментативной активности. Таким
путем было показано, что вследствие гетерогенности лигандов в активном
центре, напряжений, вызванных трехмерной структурой белка, различной сте-
пени гидрофобности в непосредственном окружении металлосвязывающего
участка его геометрия существенно отличается от картины, наблюдаемой на
модельных комплексах металла с более простыми лигандами. В результате
активный центр фермента приобретает термодинамически менее выгодную
154
корформацию по сравнению с более простыми комплексными соединениями
данного металла в растворе, но более приспособленную для реализации метал-
лом его метаболической функции — осуществления им определенной катали-
тической реакции. Это предопределяемое генетически напряженное состояние
иона цинка в цинксодержащих ферментах было обозначено как энтатическое
{Vallee В. L., Williams R. J., 1968]. Термин «энтазис» в данном случае служит
для обозначения состояния напряжения в структуре металлофермента, возни-
кающее еще до его соединения с субстратом.
Цинк в металлоферментах присутствует в активном центре и непосредст-
венно участвует в каталитическом [процессе. Но он может выполнять в фер-
ментах и иную функцию, например участвовать в стабилизации их третичной
структуры, как в а-амилазе Bacillus subtilis, или выполнять регуляторную
роль, как в аспартат — карбамилтрансферазе. Цинк в одном ферменте может
функционировать одновременно и в двух процессах — катализе и поддержа-
нии структуры, как, например, в алкогольдегидрогеназе из лошадиной печени,
или катализе и регуляции активности, как в аминопептидазе из бычьего хру-
сталика.
Алкогольдегидрогеназа, присутствующая в печени и других
органах, окисляет этанол, другие первичные и вторичные спир-
ты, стероиды, а также витамин А. Ретинолдегидрогеназа сетчат-
ки глаза, по-видимому, идентична или очень близка по своей
•структуре к этому ферменту. Алкогольдегидрогеназы из печени
лошади и человека содержат по четыре атома цинка на молеку-
лу, причем цинк выполняет в их составе как каталитическую,
так и структурную функции. Алкоголыдегидрогеназа печени че-
ловека участвует в первичном механизме детоксикации спир-
тов.
Эритроциты содержат высокую концентрацию карбонат-
дегидратазы (КФ 4.2.1.1), катализирующую реакцию
СО2+Н2О Н2СОз. Этот фермент обнаружен и во многих
других тканях. В его отсутствие скорость удаления СОг из орга-
низма недостаточна для поддержания жизни. Все карбонат-де-
гидратазы из эритроцитов млекопитающих, включая человека,
содержат по одному атому цинка на молекулу фермента. Столь-
ко же металла присутствует и в молекулах карбоксипептидаз А
и В, выделяемых с соком поджелудочной железы в двенадцати-
перстную кишку и осуществляющих деградацию полипептидных
цепей с карбоксильного конца. Содержание карбоангидразы ис-
следовано при многих болезнях человека.
Щелочная фосфатаза Е. coli содержит четыре атома цинка.
Она состоит из двух субъединиц, в которых один атом цинка
участвует в активном центре, а второй — в поддержании струк-
туры. Кроме того, для поддержания активности голофермента
необходимы еще два атома магния. При дефиците цинка у жи-
вотных активность щелочной фосфатазы существенно понижена.
По-видимому, цинк, составляющий 0,3% сухой массы лейкоци-
тов человека, преимущественно присутствует в них именно в со-
ставе щелочной фосфатазы [Vallee В. L., Galder А., 1984].
Цинку принадлежит важная роль в синтезе белка и нуклеи-
новых кислот. Он присутствует во всех 20 изученных в настоя-
щее время нуклеотидилтрансферазах, а его открытие в обрат-
ных транскриптазах впервые позволило установить тесную
взаимосвязь с процессами канцерогенеза. Цинк необходим для
155
стабилизации структуры ДНК, РНК и рибосом и, входя в со-
став аминоацил-тРНК-синтетаз и фактора элонгации белковой
цепи у млекопитающих (EF-1), играет важную роль в процессе
трансляции и незаменим, таким образом, на многих ключевых
этапах экспрессии гена. В этой связи становится понятной за-
держка роста и развития у человека и лабораторных животных
при алиментарной или наследственной недостаточности цинка.
Необходимость цинка для нормального протекания всех этапов
клеточного цикла и для функционирования генетического аппа-
рата^ продемонстрирована на примере эвглены [Vallee В. L.,
Следует иметь в виду, что целый ряд ферментов, аналогич-
ных по функции указанным в табл. 15, не содержит цинка. Так,
альдолаза из дрожжей содержит этот элемент, а из мышц — не
содержит его. Отсутствует он и в карбоксилазе пировиноград-
ной кислоты печени, карбоангидразе из шпината, супероксид-
дисмутазе митохондрий и бактерий. Предполагалось, что боль-
шинство дегидрогеназ, коферментами которых являются пириди-
новые нуклеотиды, представляют собой цинксодержащие
ферменты. Однако ранние сообщения о присутствии цинка во мно-
гих ферментах этой группы, как, например, в лактатдегидроге-
назе из печени крысы и сердца свиньи, глицеральдегид-3-фос-
фатдегидрогеназе из мышц кролика и дрожжей, глюкозо-6-фос-
фатдегидрогеназе из дрожжей, глутаматдегидрогеназе из печени
быка [Джонс М. М., Хикс Дж. Э., 1978], не нашли подтверж-
дения при изучении очищенных препаратов.
Токсичность цинка невелика, при введении его в избытке он
не кумулируется, а выводится. Однако в литературе имеются
отдельные сообщения о токсическом влиянии цинка. При прие-
ме внутрь 12 г ZnSO4 у юноши 16 лет были отмечены сонли-
вость, рвота, повышение активности липазы в сыворотке крови
и амилазы слюны. Зарегистрировано отравление со смертель-
ным исходом при приеме внутрь 45 г ZnSO4 [Prasad A. S.,
1979].
В противоположность этому большое значение для клиниче-
ской медицины имеют цинкдефицитные состояния организма.
У человека классификация цинкдефицитных состояний разрабо-
тана А. А. Жаворонковым (1983). Наиболее удобно подразде-
лять цинкдефицитные состояния человека в соответствие с раз-
личными возрастными периодами. Так, по данным ряда авторов,
цинкдефицитная патология разных периодов индивидуального
развития представлена следующим образом.
Антенатальный период. У 13—18% беременных с дефицитом
цинка отмечается наличие пороков у плода и новорожденных:
гидроцефалия, микро- и анофтальмия, расщепление неба, ис-
кривление позвоночника, образование грыж, пороки сердца и др.
(подтверждено экспериментально).
Постнатальный период. Эндогенный дефицит цинка при
врожденных и генетических заболеваниях: 1) энтеропатический
акродерматит — аутосомно-рецессивное заболевание, сопровож-
156
дающееся нарушением синтеза белка — лиганда для связыва-
ния, всасывания и транспорта металла; 2) клинические формы
серповидно-клеточной анемии, сопровождающиеся хроническим
гемолизом и цинкурией вследствие нарушения клубочковой
фильтрации и канальцевой реабсорбции.
Экзогенный дефицит цинка. К нему относятся: 1) али-
ментарная недостаточность: а) симптомокомплекс тяжелой же-
лезодефицитной анемии с гепатоспленомегалией, карликовостью,
половым недоразвитием, нарушением нормального оволосения,,
атрофией яичек и предстательной железы — болезнь Прасада;
б) синдром идиопатической гипогевзии и гипосмии с дизосмией;.
в) у беременных — прямая корреляция между снижением кон-
центрации цинка в сыворотке крови и частотой слабости родо-
вой деятельности, атонических кровотечений, преждевременных
родов и врожденными уродствами новорожденных; 2) при за-
болеваниях: а) желудочно-кишечного тракта (постгастрэктоми-
ческом синдроме, хронических энтеритах, колитах, болезни Кро-
на и др.) —дерматит, гипогевзия с гипосмией; б) печени (алко-
гольный цирроз и др.)—дерматит, гипогевзия с дизгевзией и
гипосмия с дизосмией.
Ятрогенный дефицит цинка описан в главе 9. Имеющие-
ся клинические и экспериментальные материалы свидетельству-
ют о том, что уже эмбрион и плод в критические периоды своего
развития чувствительны к недостатку цинка в организме мате-
ри. Возникающие при этом цинкдефицитные состояния сопро-
вождаются рождением незрелого плода, а также формирова-
нием разного рода аномалий. Концентрация же этого МЭ в тка-
нях зрелого новорожденного идентичны таковым у взрослого
организма.
В постнатальном периоде цинкдефицитные состояния могут
быть обусловлены эндогенными, экзогенными и ятрогенными
причинами. В первом случае они сопровождают некоторые
врожденные и генетические заболевания. Во втором они возни-
кают при первичном алиментарном недостатке этого МЭ, а так-
же при многих хронических заболеваниях органов пищеварения.
Ятрогенный дефицит цинка развивается при несбалансирован-
ном в отношении МЭ парентеральном питании, а также при
длительном лечении цитостатиками, D-пеницилламином, L-гис-
тидином.
Алиментарная недостаточность данного МЭ у детей приво-
дит к симптомокомплексу, впервые описанному в 1961 г.
A. S. Prasad и соавт. При этом у И молодых мужчин-иранцев
отмечены железодефицитная анемия, гепатоспленомегалия, кар-
ликовость, гипогонадизм (атрофия яичек и предстательной же-
лезы) с почти полным отсутствием лобкового, аксиллярного и
лицевого оволосения. Анемия у этих больных была гипохромной
микроцитарной и легко корригировалась патогенетической те-
рапией, в то время как другие симптомы, напоминавшие призна-
ки цинкдефицитного состояния у экспериментальных животных,
не поддавались коррекции. Через 2 года была опубликована
157
работа, в которой описывалась аналогичная клиническая кар-
тина у 23 мужчин-египтян 17—19 лет [Prasad A. S. et al., 1963].
Питание больных как в Иране, так и в Египте характеризова-
лось недостаточностью белков животного происхождения и пре-
обладанием углеводов. При клиническом обследовании у этих
больных обнаружены низкие концентрации цинка в плазме,
эритроцитах и волосах. Успешное лечение этих больных сульфа-
том цинка подтвердило ведущую роль недостаточности цинка
в этиологии данного синдрома. В связи с тем что он является
не единственным показателем алиментарной недостаточности
.цинка, мы предложили этот синдром назвать болезнью Прасада
ло имени автора, впервые его описавшего [Жаворонков А. А.,
1983].
Дефицит цинка у взрослых сопровождается развитием синд-
ромов, связанных с поражением кожных покровов (типа так
называемого энтеропатического акродерматита) и с нарушением
вкуса и обоняния.
Гистологическая картина энтеропатического акродерматита
•напоминает псориаз, однако при последнем отсутствуют акан-
толические клетки [Lever W. F., 1975]. В то же время она ана-
логична дерматиту, развивающемуся при приобретенных цинк-
дефицитных состояниях. При этом выделяют два типа (эрите-
матозный и буллезный) кожных изменений [Katsumi Hanada,
1981]. При первом типе отмечаются гипер- и паракератоз, акан-
тоз и незначительная периваскулярная инфильтрация лимфоци-
тами в верхних слоях дермы. Иногда встречаются псевдорого-
вые кисты и базофильная дегенерация в эпидермисе. В роговом
и чешуйчатом слоях выявляются липидные включения. При бул-
лезном типе имеют место интраэпидермальные пузырьки с не-
большим количеством акантолитических клеток, паракератоз и
губчатые пустулы Когои вокруг пузырьков. В сосочковом слое
дермы отмечаются отек и инфильтрация лейкоцитами. Липид-
ные включения выявляются не только в верхушке пузыря, но и
в нижних слоях эпидермиса. Сходные изменения обнаруживают-
ся при паракератозном дерматозе у больных с цинкдефицитным
состоянием, возникающим после длительного применения пени-
цилламина (рис. 29).
Электронно-микроскопическое исследование кожи при приоб-
ретенном цинкдефицитном состоянии впервые провел Katsumi
Hanada (1981), который в частности, показал, что в очаге эри-
тематозных изменений роговой слой содержит остатки ядер, ор-
ганелл и тонофиламентов. В поверхностных чешуйчатых клет-
ках увеличено число свободных рибосом и отсутствуют керато-
гиалиновые гранулы. Многочисленные овальные тельца низкой
электронной плотности диаметром 0,2—0,8 мкм, связанные
с плазмалеммой, были обнаружены в клетках рогового слоя,
в поверхностных клетках чешуйчатого слоя и даже в клетках
Лангерганса. Десмосомальные соединения в поверхностных эпи-
дермальных клетках ослаблены, межклеточные пространства
расширены. В базальном слое эпидермиса патологии не выявле-
158
Рис. 29. Гистологическая картина выраженного паракератозного дерматоза
при циикдефицитном состоянии у больного, длительно лечившегося пеницилла-
мином по поводу болезни Вильсона — Коновалова [Klingberg W. et al., 1976].
но. В клетках верхушки интраэпидермального пузырька видны
вакуолизированные митохондрии, много свободных рибосом и
единичные тонофиламенты.
Ультраструктурные изменения эпителия слизистой оболочки
щек крыс при экспериментальном циикдефицитном состоянии
характеризовались избыточной пролиферацией эпителиоцитов и
патологической кератинизацией [Ashraf S. Н. et al., 1980] и-
свидетельствовали об аналогичных изменениях, развивающихся
в ороговевающем эпителии при дефиците цинка как врожденно-
го, так и приобретенного характера.
При исследовании слизистой оболочки двенадцатиперстной
кишки больных энтеропатическим акродерматитом [Loembeck I.
et al., 1974], а также при приобретенном дефиците цинка [Su-
zuki Н. et al., 1979] отмечено наличие так называемых апопто-
тических включений в клетках Панета диаметром до 7 мкм.
Они представляли собой конденсированный миелиноподобный
материал в филаментозном матриксе. Явление апоптоза, т. е.
гибели клеток с конденсацией ядра и цитоплазмы и последую-
щей их десквамацией или фагоцитозом соседними клетками,
было описано J. F. Kerr и соавт. (1972).
При исследовании слизистой оболочки тонкой кишки подо-
пытных крыс с недостатком цинка установлены дистрофические-
изменения энтероцитов на всем протяжении ворсинок и крипт.
Отмечались просветление гиалоплазмы, деструкция микроворси-
15»
«ок и редукция большей части органелл. Апоптотические вклю-
чения отмечены в цитоплазме энтероцитов и макрофагов, ин-
фильтрирующих слизистую оболочку [Elmes М. Е. et al., 1980].
Эти авторы полагают, что изменения эпителия слизистой обо-
лочки тонкой кишки при дефиците цинка у животных сходны
с таковыми у человека при энтеропатическом акродерматите.
Апоптоз встречается и при других патологических состояниях,
не связанных с дефицитом цинка.
Электронно-микроскопически в области пор вкусового сосоч-
ка при идиопатической гипогевзии обнаружены резкое уменьше-
ние числа и диаметра секреторных гранул с электронно-плот-
ным гомогенным содержимым, наличие крупных вакуолей и
большого количества везикул, а также отсутствие пальцевидных
отростков и плотного экстрацеллюлярного вещества. При обыч-
ном гистологическом исследовании вкусового сосочка методом
световой микроскопии никакие изменения в нем не обнаружены
[Henkin R. I. et al., 1971].
Диагноз «цинк-дефицитное состояние» ставится в том случае,
если содержание цинка в крови менее 13 мкмоль/л, еще более
низкая концентрация этого элемента в сыворотке крови
(8,2+0,9 мкмоль/л') является неблагоприятным прогностическим
признаком [Карлинский В. М., 1979]. Надо иметь в виду, что
существует прямая коррелятивная связь между понижением
активности цинкзависимых ферментов и сниженным содержа-
нием этого МЭ в тканях с симптомами недостаточности цинка.
Представляется целесообразным в группу риска в отноше-
нии цинкдефицитных состояний включить подростков с задерж-
кой роста и полового созревания, беременных и кормящих жен-
щин с микросимптоматикой энтеропатического акродерматита
и нарушениями вкусовой чувствительности и обоняния, а так-
же больных хроническими заболеваниями печени, желудочно-
кишечного тракта, талассемией (встречается на юге нашей
страны), больных с длительным парентеральным питанием и
подвергавшихся продолжительному лечению цитостатиками.
Весьма вероятно, что у очень многих больных хирургического и
терапевтического профиля с синдромом истощения, сопровож-
дающимся анорексией и стойкими нарушениями вкусовой функ-
ции в форме агевзий и парагевзий с соответствующими атрофи-
ческими изменениями вкусовых сосочков языка, имеется недиаг-
ностированная недостаточность цинка. Это, в частности, надо
отнести к синдрому дистрофии раненых, очень часто осложняв-
шему нагноительные процессы различной интенсивности и дли-
тельности течения. Нарушения вкусовой чувствительности были
объективно зарегистрированы у них с помощью клинических
тестов [Авцын А. П., 1944, 1946]. Ликвидация лихорадки, обус-
ловленной нагноением, сопровождалась быстрым улучшением
вкусовой чувствительности и постепенной регенерацией сосочков
языка. Эти имеющие практическое значение наблюдения отно-
сятся и к кахексии, иногда сопровождающей обширные длитель-
но не заживающие ожоги.
160
Естественно полагать, что явления недостаточности цинка
объясняются нарушением конкретных биохимических функций,
которые обеспечиваются данным элементом. Однако установле-
ние такой связи не всегда возможно. Наблюдения над явления-
ми недостаточности цинка позволяют говорить о его участии
в следующих биологических процессах.
Рост и деление клеток. Наиболее четко прослежива-
ется связь между задержкой роста и деления клеток с угнетени-
ем активности ферментов нуклеинового и белкового обмена [Val-
lee В. L., 1983]. Содержание цинка в нуклеиновых кислотах
превышает концентрацию марганца и меди и достигает в ДНК
18,4 ммоль/кг и в РНК печени крысы—19,9 мкмоль/кг. Уста-
новлено, что марганец и цинк образуют наиболее прочные связи
с фосфатными группами. Цинк обеспечивает обратимость про-
цессов денатурации ДНК. Это действие цинка объясняется тем
обстоятельством, что в нативной ДНК он связан с фосфатными
группами, повышая тем самым ее стабильность, а при расхож-
дении нитей ДНК этот МЭ образует, подобно меди, координа-
ционные комплексы с азотистыми основаниями, удерживая их
на близком расстоянии и способствуя их быстрому объединению
в двойную спираль при охлаждении. Это может иметь важное
значение для ренатурации молекул ДНК [Zimmer С., 1973].
Цинк оказывает влияние на деление клеток и через воздейст-
вие на цитоскелет. Показано in vitro, что он может стабилизи-
ровать микротрубочки в результате образования меркаптидных
мостиков между субъединицами димеров тубулина. Повышение
митотического индекса отмечено при дефиците цинка в эпите-
лиоцитах слизистой оболочки пищевода и поджелудочной желе-
зы крыс. В веретене и хромосомах во время митоза обнаружено
повышенное содержание цинка. Влияние этого МЭ на созрева-
ние сперматид также связывают с его способностью стабилизи-
ровать микротрубочки их манжетки [Chesters J. К., 1978]. Уча-
стием цинка в функционировании генетического аппарата и про-
цессах деления клетки определяется его важное значение для
роста и развития животного организма, показанное впервые на
крысах еще в 1934 г. [Todd W. R. et al., 1934] и многократно
подтвержденное в опытах на сельскохозяйственных животных
[Mertz W., 1986].
Кератогенез. Уже при первых исследованиях недостаточ-
ности цинка у крыс и мышей наблюдалось облысение, сопро-
вождающееся обширными повреждениями эпителия кожи и пи-
щевода. Гистологическими методами при недостаточности цинка
обнаруживают состояние паракератоза с характерной задерж-
кой гибели ядер и слущиванием эпителия. Наиболее полно
паракератоз при недостаточности цинка изучен у свиней, у кото-
рых гистологические изменения кожи имеют большое сходство
с таковыми при псориазе у человека [Anderson Е. et al., 1967].
Чаще всего поражения кожи наблюдаются вокруг глаз и рта,
мошонки и с внутренней стороны задних конечностей, откуда
они распространяются на остальные участки тела. Кожа утол-
11—563
161
щается, собирается в складки и покрывается струпьями. Все
описанные явления исчезают в короткие сроки после приема
препаратов цинка и их парентерального введения.
Установлено, что целый ряд кожных заболеваний у челове-
ка и животных связан с недостаточностью цинка и поддаются
лечению соединениями этого МЭ. Так, отмечена связь между
хроническим язвенным дерматитом различной этиологии и низ-
ким уровнем цинка в плазме крови у человека. Препараты цин-
ка дают выраженный лечебный эффект и при энтеропатическом
акродерматите.
Остеогенез. Цинку принадлежит важная роль в разви-
тии скелета. Нарушения скелета при дефиците цинка описаны
у плодов крыс и у домашней птицы [Blumberg А. В. et al.,
1960]. У телят при экспериментальной недостаточности цинка
наблюдаются ригидность, опухание суставов и хромота, которые
легко устраняются дополнительной дачей препаратов цинка.
У поросят W. J. Miller и соавт. (1968) описали уменьшение
размеров и прочности бедренной кости.
При дефиците цинка отмечено угнетение щелочной фосфата-
зы в хондроцитах эпифизарного хряща, что, по мнению S.J. Hen-
ning (1976), и является основным биохимическим дефектом
развития кости. Зольность кости при недостаточности цинка сни-
жается или остается нормальной [Rahmat A. et al., 1961; Star-
cher В. С. et al., 1963]. Нет сомнения, что цинк принимает уча-
стие в процессах кальцификации, однако, раскрытие конкретно-
го механизма его действия является делом будущего.
Заживление ран. Роль цинка в заживлении ран впер-
вые была показана W. Н. Strain и соавт. (1960). Было также
отмечено, что в волосах больных с обширными ожогами содер-
жится пониженное количество цинка. Разработав количествен-
ный метод оценки скорости лечения ран, авторы показали, что
она прямо коррелировала с уровнем цинка в волосах. Прием
50 мг сульфата цинка с пищей 3 раза в сутки повышал ско-
рость заживления ран у молодых людей после операции удале-
ния пилоноидальных кист [Pories W. J. et al., 1967]. Особенно
ускорялось заживление на стадии эпителизации через 15 дней
и более после операции.
Прием сульфата цинка способствовал ускорению заживления
язв на нижних конечностях [Hussain S. L., 1969]. При этом
отмечено повышение содержания цинка в сыворотке крови, что
дает основание предположить наличие в организме пациентов
дефицита цинка и в начале лечения. Аналогичные данные при-
водят G. R. Sergeant и соавт. (1970), обследовавшие больных
серповидно-клеточной анемией с язвами на нижних конечностях.
Цинк при заживлении ран оказывает, по-видимому, стаби-
лизирующее действие на цитоплазматические мембраны, препят-
ствуя высвобождению гидролитических ферментов, таких как
катепсин D и коллагеназа, контролирующих скорость распада
поврежденных тканей. Имеются также данные, что цинк видо-
изменяет течение воспалительного процесса и ускоряет синтез
162
коллагена в заживающей ране [Tengrup В., 1977]. Не следует
сбрасывать со счетов и роль цинка в синтезе ДНК и РНК, осо-
бенно интенсивно протекающем в регенерирующей ткани.
Воспроизводительная функция. Известно боль-
шое количество работ о роли цинка в процессах воспроизведе-
ния потомства. При дефиците цинка наступает угнетение спер-
матогенеза и развития первичных и вторичных половых призна-
ков у самцов, всех этапов полового цикла у самок — от течки
до родов, а также лактации. При этом чувствительность к де-
фициту цинка у самцов выше, чем у самок, а у молодых живот-
ных значительно выше, чем у взрослых. Недостаточность цинка
влечет за собой замедленное развитие семенников, их придат-
ков, предстательной железы и гипофиза, а также атрофию спер-
матогенного эпителия семенников. Задержка развития семенни-
ков является характерным признаком недостаточности цинка
у коз, телят, овец [Underwood Е. J., 1987] и человека.
Данные A. S. Prasad и соавт. (1969), а также Н. Sandstead
(1982) о недостаточности цинка у подростков в Иране и Египте,
вошедшие во все обзоры литературы по биологической роли
цинка, первоначально подтверждались не всеми авторами. Так,
анализируя результаты ряда авторов, J. Е. Caughey (1973) под-
верг сомнению данные A. S. Prasad и пришел к выводу, что
в основе наблюдавшегося им явления лежит не недостаточность
цинка, а неполноценность питания по белку и калорийности,
которая, по данным ВОЗ, отмечена у более 70% семей, где про-
водились наблюдения. В настоящее время, однако, данные
A. S. Prasad являются общепризнанными.
Взаимодействие с сульфгидрильными груп-
пами. Ряд проявлений биологической активности цинка объяс-
няется его высоким сродством к сульфгидрильным группам,
являющимся важными детерминантами структуры и функции бел-
ков. Поскольку ионы цинка не участвуют в окислительно-восста-
новительных реакциях, они способствуют стабилизации сульф-
гидрильных групп, предупреждая их окисление ионами меди и
железа. -1
Содержание цинка в органах и тканях подвергается обычно
незначительным изменениям даже при выраженной недостаточ-
ности этого МЭ. Существенные колебания концентрации цинка
наблюдаются только в плазме крови, что может иметь опреде-
ленное значение в процессах регуляции мобильности макрофа-
гов. Так, установлено, что при повышенном уровне цинка в
плазме крови отмечается замедление миграции макрофагов,
а при дефиците.— ее значительное повышение [Chvapil М.,
1976]. Способность 1 моль цинка стабилизировать клеточные
мембраны используется при выделении форменных элементов
крови — эритроцитов и лимфоцитов с параллельным повыше-
нием содержания в них цинка примерно в 10 раз.
Иммунный ответ. Цинку принадлежит важная роль в
иммунном ответе. Недостаточность этого МЭ у лабораторных и
сельскохозяйственных животных сопровождается угнетением об-
Hl
163
разования антител, снижением числа лимфоцитов, циркулирую-
щих в крови, и существенным уменьшением массы тимуса.
У больных энтеропатическим акродерматитом отмечено почти
полное отсутствие лимфоидной ткани, включая тимус, минда-
лины, лимфатические узлы, селезенку. У крыс при дефиците
цинка В-лимфоциты утрачивают способность к трансформации
под влиянием фитогемагглютинина и других митогенов. Сход-
ным образом у человека при энтеропатическом акродерматите
нарушение синтеза антител связано не столько с недостатком
В-лимфоцитов, сколько с их неспособностью к трансформации
в плазматические клетки при воздействии антигена, объясняе-
мой нарушением механизма дерепрессии генов иммуноглобули-
нов [Chesters J. К., 1978].
Для функции Т- и В-лимфоцитов важное значение имеет
цинксодержащий фермент нуклеозидфосфорилаза (5'-нуклеоти-
даза, КФ.3.1.3.5), участвующий в катаболизме пуринов. Недо-
статочность этого фермента отмечена при врожденном дефекте
Т-лимфоцитов и в лимфоцитах лиц пожилого возраста. По ана-
логии с бактериальной 5'-нуклеотидазой нуклеозидфосфорила-
за, присутствующая в эритроцитарной мембране, снижает свою
активность при дефиците цинка, и она может служить в каче-
стве диагностического теста для оценки цинкового статуса
[Prasad A. S., Rabbani Р., 1981]. Не исключено, что усиление
иммунного ответа лимфоцитов в пожилом возрасте при приеме
препаратов цинка связано с активацией данного фермента.
В 1974 г. было показано, что при дефиците цинка резко сни-
жается активность тимидинкиназы — фермента, катализирующе-
го реакцию образования ТМФ из тимидина и АТФ [Prasad A. S.,
Oberleas D., 1974]. Это наблюдение было подтверждено и дру-
гими авторами [Hurley L. S., 1980]. Активность тимидинкиназы
резко возрастает при вирусной инфекции, клеточном делении
и снижается до минимума в дифференцированных клетках.
В связи с этим допускалось, что действие цинка на пролифери-
рующие и эмбриональные ткани реализуется отчасти через дан-
ный фермент. Однако в этом вопросе нет ясности, поскольку
в синтезе ТМФ принимает также участие тимидинфосфорилаза,
катализирующая синтез тимидина из тимина и рибозы и близ-
кая по своему действию к описанной выше нуклеозидфосфори-
лазе.
Клеточное дыхание. Цинк угнетает транспорт электро-
нов в дыхательной цепи митохондрий, что свидетельствует о спе-
цифической роли этого металла в процессах дыхания [Чистя-
ков В. В., 1969]. Это свойство цинка, возможно, реализуется
in vivo на сперматозоидах человека, содержащих высокую кон-
центрацию этого МЭ и отличающихся весьма низкой дыхатель-
ной активностью. При удалении из них цинка наблюдается уси-
ление дыхания [Eliassen A. et al., 1971]. Как отмечают
В. П. Скулачев и соавт. (1967), цинк блокирует дыхательную
цепь между цитохромом b и с, воздействуя на белки, содержа-
щие негемовое железо. В. В. Чистяков (1969) на основании
164
изучения ЭПР-спектров считает, что ионы цинка, будучи более
сильными акцепторами электронов по сравнению с ионами же-
леза, вытесняют его из соединения с белком, нарушая тем са-
мым функции негемового железа в транспорте электронов.
Развитие мозга и поведение. Важной особенностью
цинка является его влияние на выработку поведенческих реф-
лексов. Как показали D. Caldwell и D. Oberleas (1970), у по-
томства самок крыс, испытывавших умеренную недостаточность
цинка, значительно медленнее вырабатываются условные реф-
лексы, чем у потомства матерей, получавших с пищей нормаль-
ное количество этого МЭ. Помимо того, у крыс, получавших
подкормку препаратами цинка, по сравнению с животными, ис-
пытывавшими недостаточность цинка, вырабатывалось значи-
тельно больше условных рефлексов и проявлялось меньше без-
условных рефлексов. Повышенной способности к обучению соот-
ветствовал и более высокий «индекс активности», который мож-
но интерпретировать как пониженную эмоциональность. Причи-
ной такого действия цинка, по-видимому, является его виталь-
ная необходимость для развития тканей головного мозга. Недо-
статочность цинка в критический период развития мозга неиз-
менно приводит к угнетению его функции. При дефиците этого
МЭ в последней трети беременности уменьшаются объем мозга
и общее число нервных клеток, изменяется ядерно-цитоплазма-
тическое соотношение, что указывает на угнетение клеточного
деления в период формирования крупных нейронов. Также по-
казано, что недостаточность цинка у крыс с момента рождения
и до 12-го дня жизни задерживает развитие мозга, судя по
активности включения тимидина в ДНК и серы в белки мозга
и учитывая общее содержание в нем липидов. Авторами были
отмечены также изменения характера полирибосом, что указы-
вает на сдвиги в синтезе белка. У взрослых самцов из потом-
ства крыс, испытывавших дефицит цинка, было отмечено нару-
шение способности активно избегать столкновений, а у самок—•
повышение агрессивности.
У макак резусов при лишении их цинка на НО—150-й день
беременности изменялись взаимоотношения между матерью и
потомством. У детенышей снижались уровень активности, игри-
вость и наклонность к поиску. Мать проводила больше времени
с детенышами, чаще их вскармливала. Если при кормлении
крысят молоком не хватало цинка, то у них задерживалось раз-
витие мозга, особенно мозжечка, снижался синтез ДНК и ги-
стонов. У взрослых самцов ослабевала способность к ориента-
ции. Эти опыты свидетельствуют о том, что дефицит цинка
в критические периоды развития мозга задерживает его созре-
вание и вызывает стойкие изменения поведения потомства крыс
и обезьян [Sandstead Н., 1985].
Вкус, обоняние. Недостаточность цинка у человека мо-
жет быть вызвана приемом гистидина, используемым при
лечении ожирения. Гистидин высвобождает цинк из его соеди-
нения с альбумином, и он выделяется с мочой в виде цинк-гис-
165
тидинового комплекса. На фоне дефицита цинка вначале ухуд-
шается и теряется аппетит, а затем снижаются и извращаются
обоняние и вкус. Механизм действия цинка на вкусовой анали-
затор объясняется наличием его ,в специальном белке — густине,
вырабатываемом в околоушных слюнных железах. Молекуляр-
ная масса его равняется 37 000. В одной молекуле имеются два
атома цинка и 8% остатков гистидина. Показано, что этот бе-
лок специфически связывается с мембранами вкусовых сосоч-
ков, регулируя скорость их роста и поступление в них питатель-
ных веществ [Henkin R. I., 1977].
Назначение препаратов цинка способствует восстановлению
вкусового восприятия у детей с его врожденным нарушением.
Снижение содержания цинка в организме детей влечет за собой
ухудшение функции вкусового анализатора, которая восстанав-
ливается при введении в рацион цинксодержащих продуктов.
У лиц с гипогевзией в слюне околоушных' желез существенно
понижено содержание этого МЭ, что объясняется нарушением
синтеза густина. Слюна необходима для поддержания остроты
вкусового восприятия и сохранности структуры вкусовых луко-
виц. Влияние ионов цинка на вкусовой анализатор также осно-
вывается, по-видимому, на его взаимодействии с сульфгидриль-
ными группами.
Цинк и стресс. При стрессах различного рода, как, на-
пример, при инфекциях, травмах, низком содержании белка в
пище, беременности, отмечается снижение концентрации цинка
в плазме крови и его перераспределение между органами и тка-
нями. Эти изменения регулируются особым термолабильным
фактором, получившим название «лейкоцитарный эндогенный
медиатор» (ЛЭМ). Он вырабатывается фагоцитирующими клет-
ками (нейтрофилы, макрофаги, моноциты). При введении здо-
ровым животным ЛЭМ вызывает быстрое поступление железа,
цинка и большей части свободных аминокислот в печень, сни-
жение содержания железа и цинка в плазме крови и повыше-
ние в ней уровня меди. В печени вскоре отмечаются усиление
синтеза ядрышковой РНК, ее включение в состав рибосом и
ускорение синтеза ряда гликопротеидов плазмы, в частности це-
рулоплазмина. ЛЭМ — низкомолекулярный белок, близкий по
своим свойствам к эндогенным пирогенам [Beisel W. R., Peka-
rek R. S., 1974]. Недавно было показано, что этот белок иден-
тичен с ИЛ-1.
Кроме того, уровень цинка в организме могут снижать про-
дукты распада тканей, образующиеся при упомянутых выше
стрессовых состояниях. Связывая цинк, эти продукты, прежде
всего аминокислоты, могут повысить содержание соединений
цинка в крови и усилить выделение их с мочой. Стресс приво-
дит также к увеличению выделения в плазму кортикостероидов,
стимулирующих поступление цинка в ткани.
Наибольшее внимание ученых привлекала взаимосвязь
цинка с инсулином. Распространенное в прошлом мнение
о наличии цинка в структуре молекулы инсулина не нашло под-
566
тверждения. По-видимому, этот элемент необходим для образо-
вания гексамера и других кристаллических форм инсулина,
в виде которых происходит депонирование инсулина ^клетками
поджелудочной железы [Maske Р., 1964]. Изучение влияния
цинка на активность инсулярного аппарата показало связь меж-
ду содержанием в нем цинка и его функциональным состоянием.
Особый интерес в этом отношении представляют гистохимиче-
ские исследования И. А. Шевчука (1970). Включение цинка в
инсулин пролонгирует его физиологическое действие и гипогли-
кемическое состояние. С учетом этого явления был приготовлен
препарат протаминцинкинсулин, оказывающий пролонгирован-
ное гипогликемическое действие. Нарушение гомеостаза глюко-
зы в плазме крови при недостаточности цинка у крыс, описан-
ное ранее, в более поздних опытах не подтвердилось [Che-
sters J. К-, 1982].
Извращение чувства материнства у крыс при цинковой не-
достаточности является, по-видимому, также результатом нару-
шения выработки половых гормонов. В период, предшествую-
щий беременности, у кроликов наблюдается заметное повыше-
ние концентрации цинка в матке, особенно в эндометрии. Цинк
мобилизуется в тканевых депо и транспортируется в матку для
обеспечения развития эмбриона. Поскольку матка на ранних
стадиях беременности у кролика изменяется главным образом
под влиянием прогестерона, то вполне вероятно, что изменение
уровня цинка и активности карбоангидразы в матке происходит
под влиянием этого гормона. В соответствии с этим предполо-
жением J. Apgar (1970) нашел, что введение крысам, испытыва-
ющим недостаточность цинка,прогестерона и эстрогенов способ-
ствует нормальному исходу беременности. Имеются также дан-
ные о влиянии хорионального гонадотропина и тестостерона на
поступление цинка в дорсолатеральном отделе предстательной
железы и дополнительные половые железы у самцов крыс.
Таким образом, цинкдефицитные состояния представляют со-
бой обширную группу эндогенных и экзогенных МТОЗов. В ка-
честве самостоятельных нозологических форм они еще мало
известны, однако широкий спектр биологической активности
цинка, его большое значение как в антенатальном, так и в пост-
натальном развитии организма позволяют думать, что в неда-
леком будущем обнаружат новые синдромы этой природы.
Акцессорная роль других МЭ, в первую очередь сопряженно
связанных с обменом цинка в организме, вероятно, проявляется
в полиморфизме клинических симптомов. Это свидетельствует
о чрезвычайно сложном патогенезе цинкдефицитных состояний
в каждом конкретном случае.
6.4. Марганец
Марганец был обнаружен в тканях животных более 70 лет
назад [Bertrand G„ Medigreceanu М. F., 1913], однако его жиз-
ненно важная роль для животного организма была установлена
лишь 18 лет спустя, когда было показано, что он необходим для
роста и развития крыс и мышей [Kemmerer A. R. et al., 1931;
Orent Е. R., McCollum E. V., 1931]. Вскоре было найдено, что
этот МЭ предупреждает развитие аномалии скелета у кур, из-
вестных под названием «перозис» [Wilgus Н. R. et al., 1939].
Явления недостаточности марганца затем были обнаружены
у коз [Anke М., Groppel В., 1970] и практически у всех изу-
ченных лабораторных и сельскохозяйственных животных [Un-
derwood Е., 1977]. Впервые дефицит марганца у человека опи-
сан как марганецзависимый диабет в 1972 г. Е. A. Doisey.
В норме в организме мужчины массой 70 кг имеется 10—
20 мг (0,18—0,36 ммоль) марганца. В норме, по данным
L. S. Hurley и соавт. (1987), в костях, особенно трубчатых
а также в печени и почках находятся более высокие концентра-
ции марганца, чем в других органах. Это справедливо для че-
ловека, крупного рогатого скота и крысы. Содержание марган-
ца в мышцах человека очень низкое, еще ниже оно в цельной
крови и особенно в плазме. Концентрация марганца в больше-
берцовой кости в большей степени зависит от его содержания
в пище, чем в тканях других органов. В печени здоровых лю-
дей всех возрастов находится 6—8 мг/кг (расчет на сухую мас-
су). Концентрация марганца в реберных хрящах человека с воз-
растом уменьшается. Данные о количестве марганца в цельной
крови очень вариабельны. Уровень его в сыворотке крови повы-
шен при заболеваниях сердца, инфекциях и некоторых психозах.
Повышение уровня марганца в цельной крови зарегистрировано
при избыточном потреблении этого МЭ, а также при ревма-
тоидном артрите и железодефицитных состояниях.
Марганец, по-видимому, имеет жизненно важное значение
для функции мозга. Наиболее высока его концентрация в эпи-
физе, обонятельных луковицах, срединном возвышении гипота-
ламуса и в базальных ганглиях [Barbeau A. et al., 1975]. Он
аккумулируется преимущественно в меланинсодержащих струк-
турах ЦНС, как, например, в черном веществе.
В отличие от других МЭ марганец не накапливается в нор-
ме в печени плода. Характерно его более высокое содержание
в клетках органов, богатых митохондриями. Дефицит марганца
ведет к селективному поражению этих органелл, они удлиняют-
ся, нарушается правильная ориентация их крист [Hurley L. S.
et al., 1987]. Это, очевидно, специфическое повреждение мито-
хондрий может быть предотвращено путем включения солей
марганца в пищу.
Всасывание. Обмен марганца был изучен преимущест-
венно с помощью его изотопа 54Мп с периодом полураспада
312 дней. Марганец в виде двухвалентного катиона всасывается
одинаково на протяжении всей тонкой кишки [Thomson А. В. R.
et al., 1971]. Механизм этого процесса изучен мало. Известно,
что в норме у человека, крупного рогатого скота, мышей и кур
всасывается лишь небольшая (3—4%) часть этого МЭ незави-
симо от его содержания в рационе; у молодых крыс всасывание
S68
марганца достигает 20%. Этот процесс играет, по-видимому, не
основную роль в поддержании гомеостаза марганца, который
в отличие от железа контролируется механизмами выделения
этого элемента.
На всасывание марганца оказывает влияние целый ряд компонентов кор-
ма. Избыток кальция и фосфора повышает потребность в марганце у свиней
[Miller R. С. et al., 1940], домашних птиц [Schaible Р., Bandemer S., 1942]
и телят [Hawkins G. Е. et al., 1955], а цитрат железа усиливает перозис
у цыплят. Марганец в свою очередь угнетает всасывание железа и может
в повышенных дозах привести к снижению синтеза гемоглобина [Matrone G..
et al., 1959]. Имеет место также взаимное угнетение всасывания марганца
и селена [Burch R. et al., 1975]. У крыс при дефиците железа с наибольшей
скоростью всасывается кобальт, затем следуют железо и марганец, причем
участки связывания этих металлов на апикальной и базальных частях эпи-
телиоцитов не идентичны. Этанол усиливает всасывание марганца в кишечнике
[Schafer D. F. et al., 1974]. В опытах с этанолом и различными ингибиторами
было также показано, что всасывание марганца, по-видимому, происходит
не в результате активного транспорта, а путем облегченной диффузии с уча-
стием переносчика [Leach R. М. et al., 1978]. .
Транспорт и распределение в тканях. В крови
воротной вены большая часть марганца связана с белком, отно-
сящимся к а2-макроглобулинам. Свободный и белково-связан-
ный марганец эффективно улавливается печенью. Небольшая
часть марганца окисляется до Мп3+, связывается трансферри-
ном и поступает из кровяного русла в различные ткани орга-
низма. Марганец окисляется, видимо, церулоплазмином, так
как его ионы in vitro в отсутствие окисляющих агентов с транс-
феррином не связываются [Gibbons R. A. et al., 1976].
Марганец очень быстро покидает кровеносное русло. Через
10 мин после внутривенного введения S4Mn в крови удается об-
наружить только около 1% этого изотопа. С учетом профессио-
нальной вредности допускается содержание марганца в крови
не более 18 мкмоль/л.
Попав в клетку, этот МЭ включается главным образом в ми-
тохондрии [Maynard R. S., Catzias G. С., 1955], в связи с чем
клетки печени, почек, поджелудочной железы, богатые этими,
органеллами, содержат повышенное количество марганца. Глю-
кокортикоиды и АКТГ вызывают переход марганца из печени
в другие органы и ткани.
В гепатоцитах содержится как белково-связанный, так и свободный мар-
ганец, выявляемый методом ЭПР-спектроскопии, причем соотношение между
этими двумя формами зависит от характера питания животных. У голодающих
крыс содержание свободного марганца во внутриклеточной жидкости в 3 раза
ниже, чем у сытых животных, поскольку при голодании марганец включается
в состав ферментов глюконеогенеза, таких как фосфоенолпируват (ФЕП)-карб-
оксикиназа, нуждающихся в этом элементе. Активность последней под влия-
нием марганца способна возрасти в 10 раз [Ash D., Schramm V., 1982]. Пред-
полагают, что .изменение концентрации свободного марганца, возможно, пред-
ставляет собой важный механизм регуляции внутриклеточного метаболизма,
?0Я9?ГИгтНЫИ деиствию свободных иоиов кальция и магния [Williams R. J.,
1Уо2]. Поступление марганца из митохондрий в цитозоль обеспечивается,
по-видимому, той же транспортной системой, что и поступление кальция, ко-
торый активирует фосфорилазу, также участвующую в процессах глюконео-
генеза [Jeng A. Y., Shamoo А. Е., 1980].
16»
Марганец в незначительном количестве выделяется с мочой.
В норме этот МЭ, поступая в желчь, экскретируется из организ-
ма преимущественно с калом. Однако при поступлении в повы-
шенных количествах он элиминируется также через стенку ки-
шечника, причем в первую очередь через проксимальный отдел
подвздошной кишки.
Некоторое количество марганца экскретируется также под-
желудочной железой, которая в случае холестаза может стать
даже основным органом выделения этого МЭ. У мышей способ-
ность поддерживать гомеостаз марганца появляется только
через 17—18 дней после рождения [Miller S. Т. et al., 1975].
Скорость выделения марганца определяется его содержанием
в пище и не зависит от ионов других металлов. Выделение 54Мп
из организма животных повышается только при введении его
стабильного изотопа. Марганец, выделяемый с желчью, подвер-
гается частичной реабсорбции, в связи с чем устанавливается
энтерогепатическая циркуляция этого элемента [Keen С. L.
et al., 1984].
Некоторое количество марганца выделяется с молоком. Со-
держание марганца в грудном молоке в 10 раз ниже, чем в мо-
локе других млекопитающих. В начале периода лактации оно
составляет 6 мкг/л, затем снижается до 4, а к концу лактации
снова возрастает до 6—8 мкг/л. В коровьем молоке содержится
20—50 мкг/л этого МЭ. В грудном молоке 75% марганца свя-
заны с лактоферрином, а в коровьем молоке 67% этого МЭ об-
разуют комплекс с казеином [Lonnerdal В. et al., 1984].
У взрослых лиц период полувыведения одноразовой дозы
марганца, данной через рот, составляет около 10 дней, тогда
как при внутривенном введении он достигает 40 дней. Это раз-
личие объясняется тем, что при приеме per os большая часть
марганца поступает в печень, где он быстро включается в про-
цессы обмена, тогда как при внутривенном введении марганец
поступает и в те органы, где его обмен протекает значительно
медленнее.
Ферменты, содержащие марганец. В растворах и
биохимических структурах находится преимущественно двухва-
лентный марганец [Ash D., Schramm V., 1982]. Он, подобно
трехвалентному железу, обладает значительным сродством
к имидазолу в отличие от двухвалентных катионов меди, цинка
и кадмия, предпочитающих SH-группы, и не замещает эти ионы
в их комплексах с белками. Марганец и другие жизненно необ-
ходимые переходные металлы выступают либо как прочносвя-
занные компоненты молекул ферментов, которые в этом случае
относятся к истинным металлоферментам, либо служат в каче-
стве активаторов ферментов, образуя с ними легко распадаю-
щиеся комплексы.
Металлоферменты. В отличие от железа, меди и цинка
для марганца существует лишь три фермента первого типа,
а именно аргиназа, пируваткарбоксилаза и супероксиддисмута-
за. Аргиназа (КФ 3.5.3.1)—фермент с молекулярной массой
170
120 000, присутствующий в цитозоле и катализирующий расщеп-
ление аргинина с образованием мочевины. В его молекуле со-
держится четыре атома Мп2+, из которых два могут быть уда-
лены диализом с 50% потерей активности, полностью восста-
навливающейся при добавлении в систему марганца. При уда-
лении всех четырех атомов металла фермент необратимо инак-
тивируется. При дефиците марганца активность фермента в пе-
чени снижается в 3 раза [Ковальский В. В., 1974; Kirchgess-
ner М., Heiske D., 1978].
Функциональная значимость низкого уровня аргиназы у жи-
вотных, испытывающих дефицит марганца, неизвестна. Очевид-
но, что по крайней мере у птиц патологические изменения, на-
блюдаемые при недостаточности марганца, не могут быть выз-
ваны нарушением функции данного фермента, поскольку у них
отсутствует синтез мочевины. При повышении содержания мар-
ганца в организме активность аргиназы может существенно воз-
растать. Так, активность печеночной аргиназы у крыс с экспе-
риментальным диабетом, вызванным инъекцией стрептозотоци-
на, повышена в 2—3 раза по сравнению с нормой параллельно1
с аналогичным увеличением концентрации в печени марганца
[Bond G. Н. et al., 1983].
В одной молекуле пируваткарбоксилазы, выделенной из ми-
тохондрий печени птиц, содержится четыре атома марганца,,
а также четыре молекулы биотина. Пируваткарбоксилаза необ-
ходима для синтеза углеводов из пировиноградной кислоты.
Было показано, что марганец находится на внешней поверхно-
сти фермента и, по-видимому, активирует метильную группу пи-
рувата, «оттягивая» на себя электроны. В результате метиль-
ная группа приобретает способность акцептировать двуокись,
углерода с образованием оксалацетата [Fung С. Н. et al., 1973].
У цыплят при дефиците марганца вместо него в состав пи-
руваткарбоксилазы включается Mg2+, что не оказывает сущест-
венного влияния на активность фермента [Scrutton М. С. et al.,.
1972]. Пируваткарбоксилаза из печени млекопитающих содер-
жит ионы обоих металлов. Ионы марганца участвуют не только
в стадии транскарбоксилирования пирувата, но и в начальной
стадии карбоксилирования биотина. Дефицит марганца вызы-
вает снижение активности фермента у голодающих взрослых
крыс, чего не отмечается у сытых животных [Baly D. L. et al...
1984].
У человека описан наследственный дефект синтеза пируват-
карбоксилазы, проявляющийся клинически в виде лактат- и пи-
ру ват ацидоза, неврологических расстройств и прогрессирующей’
умственной отсталости, известный под неточным названием
п°д°!;тРЬ1й энЦеФаломиелит», или болезнь Ли [Lestradet М.,
1978].
Третьим металлоферментом, содержащим марганец, являет-
ся Мп-супероксиддисмутаза (Mn-СОД), катализирующая дис-
мутацию супероксидного анион-радикала с образованием пер-
оксида (КФ 1.15.1.1). У млекопитающих и птиц найдены две-
171
формы супероксиддисмутазы, из которых одна содержит марга-
нец, а вторая —медь и цинк (Си, Zn-СОД). Локализация этих
ферментов в клетке различна. Си, Zn-СОД присутствует в цито-
золе, а Mn-СОД преимущественно в митохондриях. Мп-СОД
(мол. масса 80 000), выделенная из печени кур, состоит из че-
тырех идентичных субъединиц, каждая из которых имеет по
одному атому металла. В отличие от описанных выше двух фер-
ментов неактивная Mn-СОД содержит трехвалентный марганец.
Каталитический цикл этого фермента включает поочередное вос-
становление и окисление марганца в ходе его взаимодействия
с радикалами супероксида. Найдены изоферменты Мп-СОД,
однако неизвестно, различаются ли они по своей биохимической
функции [Lonnerdal В. et al., 1979].
При недостаточности Mn-СОД супероксид вступает в реак-
цию с пероксидом с образованием гидроксильного радикала,
который способен вызвать перекисное окисление липидов и по-
вреждение мембран митохондрий. Это согласуется с наблюде-
нием, что в период постнатального развития в печени крыс
активность Mn-СОД постепенно возрастает, а степень перекис-
ного окисления липидов снижается [Yoshioka Т., 1977]. Актив-
ность Mn-СОД при голодании возрастает, а при дефиците мар-
ганца снижается [Zidenberg-Cherr S. et al., 1983].
При гипербарической оксигенации, воздействии озона, про-
должительном потреблении этанола и других условиях, способ-
ствующих образованию супероксидного радикала, активность
Mn-СОД существенно возрастает [Dreosti Т. Е. et al., 1982;
Keen С. L. et al., 1983].
Известен ряд белков, содержащих в своем составе марганец.
К ним относятся авиманганин, манганин и конканавалин. Неко-
торые их характеристики, а также указанных выше металло-
ферментов приведены в табл. 16.
Таблица 16
Белки и ферменты, содержащие марганец
Наименование Молекуляр- ная масса, ХЮ3 Число ато- мов марган- ца н/а моле- кулу Источник
Супероксиддисмутаза КФ 1.15.1.1 Ачгиназа КФ 3.5.3.1 Пируваткарбоксилаза КФ 6.4.1.1 Авиманганин Манганин Конканавалин 80 39,5 120 500 89 56 190 4 2 4 4* 1 1 1 Митохондрии птиц, дрожжи Escherichia coli Печень животных Митохондрии печени жи- вотных Печень птиц Арахис Бобы канавалии мече- видной
* В состав входят четыре молекулы биотина.
172
В молекуле авиманганина содержится один атом марганца,
причем количество этого МЭ в белке снижается при дефиците
марганца, так же как и содержание в печени самого белка.
Ферменты, активируемые марганцем. Марганец
выполняет функцию активатора целого ряда ферментов, относи-
мых в первую очередь к киназам, кабоксилазам и трансфера-
зам. В случае реакций, активируемых марганцем, ион металла
вступает во взаимодействие с субстратом, содержащим фосфат-
ный остаток (особенно АТФ), образуя хелат, или реагирует не-
посредственно с белком. Марганец химически близок к магнию.
Исходя из этого, активация большинства ферментативных реак-
ций этим элементом носит неспецифический характер и он мо-
жет быть заменен магнием [Keen С. L. et al., 1984]. В то же
время действие Мп2+ и Mg2+ может быть совершенно различ-
ным. Так, оба иона образуют комплексы с АТФ. Однако марга-
нец взаимодействует со всеми тремя фосфатными группами и
с пиридиновым циклом или молекулой воды, тогда как магний
связывается только с и у-фосфатными группами.
Для активируемых обоими металлами ферментов была пред-
ложена классификация, согласно которой их можно подразде-
лить на три группы. В 1-ю группу вошли ферменты, активируе-
мые марганцем в большей степени, чем магнием; во 2-ю—ак-
тивируемые обоими металлами в одинаковой степени; в 3-ю груп-
пу — активируемые магнием в большей степени, чем марганцем
[McEuen A. R., 1981]. Важное исключение составляет фосфо-
енолпируват-карбоксикиназа (КФ 4.1.1.32). В ней марганец кон-
курирует не с магнием, а с двухвалентным железом, которое
в условиях интенсивного глюконеогенеза, например при голо-
дании или диабете, может функционировать в качестве регуля-
тора активности этого фермента.
Несмотря на то что активирование ферментов металлами
носит обычно неспецифический характер, все же целый ряд
явлений, наблюдаемых при недостаточности марганца, можно
связать с нарушением функции ферментов, активируемых мар-
ганцем. В первую очередь это относится к гликозилтрансфера-
зам.
Глик оз и л трансферазы играют важную роль в син-
тезе гликозаминогликанов и гликопротеидов. При дефиците
марганца отмечено снижение активности этих ферментов, обус-
ловливающее аномалии развития скелета, строения отолитов
у грызунов и яичной скорлупы у птиц [Leach R. М. et al., 1978].
Перечень гликозилтрансфераз, нуждающихся в марганце в
качестве кофактора, весьма обширен. К числу этих ферментов
относятся галактозил- и N-ацетилгалактозаминилтрансферазы и
ксилозилтрансфераза, осуществляющая перенос ксилозы на бел-
ковую часть гликопротеидов. При дефиците марганца нарушено
также включение сульфата в хрящевую ткань. Некоторые
ферменты, активируемые марганцем, приведены в табл. 17.
Недостаточность и функции марганца. Марга-
нец оказывает заметное влияние на процессы глюконеогенеза и
173
хондриальной мембраны, а также нарушения в системе моно-
нуклеарных фагоцитов. Ультраструктурные нарушения, наблю-
даемые в клетках, вызваны, по-видимому, угнетением синтеза
митохондриальной СОД (Mn-СОД), активность которой в серд-
це мышей и печени птицы при дефиците марганца резко пони-
жена [De Rosa G. et al., 1980; Paynter D. I., 1980].
При окислительном стрессе, вызванном этанолом, в печени
крыс, получавших адекватное количество марганца, индуцирует-
ся дополнительный синтез Mn-СОД, тогда как при дефиците
этого элемента уровень СОД повышается лишь незначительно,
и у животных развиваются признаки тяжелого алкогольного от-
равления [Ziderberg-Cherr S. et al., 1984]. Другой причиной
«дополнительного окислительного стресса» при даче марганец-
дефицитным животным этанола может являться накопление
в их печени повышенных количеств железа, являющегося доста-
точно сильным окислителем [Hurley L. S., 1984].
СРоль супероксиддисмутазы заключается в устранении супер-
оксидного ион-радикала, вызывающего повреждение тканей,
в первую очередь тех, которые содержат повышенное количе-
ство липидов. Сходную защитную функцию выполняет глута-
тионпероксидаза (GSH-Px) — селенсодержащий фермент, раз-
рушающий перекись водорода и органические гидроперекиси.
Удалось показать, что у свиней при дефиците марганца сущест-
венно понижено содержание в тканях селена, так что усиление
перекисного окисления липидов при дефиците марганца может
быть результатом недостаточности обоих указанных ферментов
(СОД и GSH-Px) [Burch R. Е. et al., 1975].
Остеогенез. Дефицит марганца у животных отражается
в первую очередь на формировании скелета как во внутриутроб-
ный, так и постнатальный периоды жизни. У цыплят поражение
скелета, известное под названием «перозис», или «соскальзываю-
щее сухожилие», представляло для птицеводства важную эконо-
мическую проблему, пока не было показано, что заболевание
полностью предупреждается введением в рацион адекватного ко-
личества марганца. Сходное влияние дефицита марганца на
скелет отмечено и у крыс, у которых он вызывает явления хонд-
родистрофии, непропорциональный рост, деформацию трубчатых
костей и позвоночника, дисплазию коленных суставов. Эти явле-
ния вызваны нарушением хондрогенеза, обусловленным угнете-
нием синтеза гликозаминогликанов. При дефиците марганца су-
щественно снижается включение 35S в организм плода, что сви-
детельствует о важной роли этого элемента и в синтезе сульфо-
мукополисахаридов.
Привлекает внимание, что клиническая патология в настоя-
щее время еще не знает каких-либо аналогов перозиса и других
марганецдефицитных аномалий скелета у человека. Это тем бо-
лее странно, что педиатры сталкиваются с большим числом раз-
нообразных деформаций хрящевых и костных структур у детей
раннего возраста. Природа их обычно остается нерасшифрован-
ной. По-видимому, часть из них относят к витамин-О-резистент-
176
ному рахиту. Выявлению дефицитов МЭ, проявляющихся в той
или иной патологии костно-хрящевой ткани, могло бы способ-
ствовать клиническое исследование содержания и синтеза гли-
козаминогликанов и уровня марганца в тканях и биологических,
жидкостях.
При дефиците марганца в природно-хозяйственных условиях
у крупного рогатого скота наблюдается целый ряд скелетных,
аномалий и нарушений воспроизводительной функции, большая
часть которых также связана с дефектным синтезом гликозами-
ногликанов. к их числу относятся: отсутствие либидо при нор-
мальной овуляции у коров, снижение оплодотворяемости, повы-
шенная частота выкидышей и мертворождений, преобладание
потомков мужского пола, удлинение сроков стельности, пони-
женная живая масса и повышенная смертность новорожденных,
повреждения опорно-двигательного аппарата, особенно хряще-
вой ткани коленных суставов, неврологические симптомы (щел-
канье языком, параличи), сокращение продолжительности
жизни.
Меньшие по размерам спермин, несущие Y-хромосому, об-
ладают в половых путях самки, бедных слизью, большей по-
движностью, чем более крупные спермин, несущие Х-хромосому,
и быстрее достигают яйцеклетки [Anke М., 1977]. Остеохондро-
дистрофия крупного рогатого скота, связанная с дефицитом'
марганца, описана в СССР на Южном Урале [Кабыш Г. М.,
1966] и в промышленных комплексах в Узбекистане [Наза-
ров Ш. Н. и др., 1985].
Соответствующие материалы о нарушении репродуктивной
функции у человека в условиях дефицита марганца пока не ре-
гистрируются в связи с отсутствием целенаправленных иссле-
дований.
Атаксия новорожденных. При дефиците марганца
во внутриутробном периоде рождается потомство с признаками:
атаксии, характеризующейся потерей равновесия, нарушением
координационных рефлексов, запрокидыванием головы^ Эти на-
рушения проявляются наиболее четко в неспособности мышей
держаться на поверхности воды, в результате чего они тонут.
Причиной указанных дефектов является аномальное развитие-
отолитов во внутреннем ухе, необходимых для нормального-
функционирования вестибулярного аппарата. Аналогичное нару-
шение наблюдается при генетическом дефекте у линии мышей,
носящей название «pallid» (см. главу 10). Важно, что потреб-
ность мышей и крыс в марганце тоже находится под генетиче-
ским контролем [Dungan D. D. et al., 1984].
Центральная нервная система. Недостаточность
марганца отражается и на функции мозга. Крысы, испытываю-
щие недостаток в этом МЭ, в большей степени подвержены су-
дорогам, чем нормальные животные, имеют пониженный порог
чувствительности к электрошоку и такие же энцефалограммы,,
как при эпилепсии.
12—568 177'
У 30% детей со склонностью к судорогам обнаружено пони-
женное содержание марганца в крови [Tanaka К., 1977],
составившее около !/3 от нормы. Аналогичные наблюдения были
сделаны и на взрослых, страдающих эпилепсией [Papavasiliou
Р. S. et al., 1979], причем показано, что снижение уровня
марганца в крови не было вызвано применением антиконвуль-
сивных препаратов. У лиц с частыми приступами эпилепсии со-
держание марганца в крови оказалось существенно ниже, чем
у больных с редкими приступами. Поскольку между уровнем
марганца в крови и печени .существует тесная корреляция, то по
данным анализа крови можно судить об обеспеченности орга-
низма этим МЭ.
Биологическая роль марганца в ЦНС может быть связана
с обеспечением нормальной структуры и стабильности мембран.
Известно также, что марганец необходим для поддержания мак-
симальной активности галактозилтрансферазы, участвующей
в нормальном синтезе ганглиозидов [Yip G. В., Dain J. А.,
1970]. Этот МЭ необходим также для нормального синтеза био-
генных аминов. Он является активатором катехол-метилтранс-
феразы, фермента, переносящего метильную группу 5'-аденозил-
метионина на З'-гидроксильную группу катехола. Высказыва-
лось предположение, что некоторые лекарственные препараты,
угнетающие этот фермент, возможно, образуют хелаты с мар-
ганцем [Tagliamonte Е. et al., 1970].
Имеются убедительные доказательства, что воздействие мар-
ганца на обмен катехоламинов частично опосредуется через си-
стему цАМФ. В бесклеточных системах марганец может выз-
вать повышенное образование цАМФ отчасти благодаря замед-
лению обмена АТФ и АДФ. Марганец способен in vitro оказы-
вать двоякое воздействие на аденилатциклазу, непосредственно
активируя фермент либо снижая стимулирующее действие на
него дофамина. Эти наблюдения позволяют считать, что путем
изменения соотношения в мозге марганца и дофамина можно
оказывать регуляторное воздействие на аденилатциклазу и
через нее на уровень цАМФ [Walton К- G., Baldessarini R. J.,
1976]. Биогенные амины вызывают повышение концентрации
марганца в крови, что связано, по-видимому, с образованием
комплексов с этим металлом, участвующих в их транспорте, свя-
зывании и депонировании [Papavasiliou Р. S. et al., 1981].
Из ферментов, активируемых марганцем, следует отметить
глутаминсинтетазу, играющую важную роль в процессах деток-
сикации аммиака. Фермент состоит из 12 идентичных субъеди-
ниц, образующих два шестигранника по шесть субъединиц.
Активная глутаминсинтетаза нуждается для проявления своего
каталитического действия в Mg2+. При низком содержании
а-глутарата в ферменте аденилируется один из тирозиновых
остатков, после чего он для проявления активности нуждается
уже не в магнии, а именно в Мп2+. Модифицированная форма
фермента в значительно большей степени подвержена регуля-
торному воздействию '(ретроингибированию) целым рядом про-
178
дуктов метаболизма глутамина, чем исходная форма. Фермент
имеет 12 участков прочного связывания марганца, 12 — проме-
жуточного и 48 участков слабого связывания этого элемента.
Возможно, что марганец выполняет регуляторную функцию и
для целого ряда других ферментов [Мецлер Д. Е., 1980]. К чис-
лу ферментов, активируемых марганцем, относится также лак-
тозосинтетаза, переносящая галактозильный остаток с УДФ-га-
лактозы на глюкозу.
Известно, что полимеризация ДНК, осуществляемая ДНК-
полимер азами, требует присутствия магния. Ионы марганца
способны заменить ионы магния, существенно снижая при этом
избирательность реакции. Так, если ионы магния допускают
включение в ДНК только дезоксирибонуклеотидов, то ионы
марганца вызывают ошибочное встраивание и рибонуклеотидов.
Показано, что один ион марганца прочно связывается с ДНК-
полимеразой, три другие — менее прочно и еще шесть — совсем
слабо.
Марганец способен ускорять процесс транскрипции, активи-
руя РНК-полимеразу. Однако, как и в случае репликации ДНК,
он может вызвать ошибочное включение в РНК наряду с рибо-
нуклеотидами и дезоксирибонуклеотидов. Замена Mg2+ на Мп2+
снижает также избирательность тРНК-нуклеотидилтрансфера-
зы, катализирующей синтез З'-ЦЦА-концов тРНК, что ведет
к образованию нетипичных концевых последовательностей, со-
держащих урацил вместо цитозина. В настоящее время, однако,
нельзя судить, оказывают ли ионы марганца аналогичное дейст-
вие в интактном организме или же оно проявляется только
in vitro. В печени крыс имеются несколько различных ДНК-ио-
лимераз, из которых одни специфически активируются ионами
магния, а другие — марганцем [Эйхгорн Г. Б., 1978]. Активи-
руемая марганцем ДНК-полимераза находится также в плазме
крови.
Содержание марганца в продуктах питания, как правило,
достаточно для обеспечения потребности организма. В связи
с этим у человека почти не встречается заболеваний, связанных
с алиментарным недостатком марганца. Его содержание в рас-
тительных продуктах колеблется от 8 до 50 мг/кг, в продуктах
животного происхождения оно значительно ниже — 0,1—2 мг/кг.
Только печень и почки так же богаты марганцем, как и расте-
ния.
Манганозы — промышленные МТОЗы, они обусловлены
избыточным поступлением марганца в условиях производства
(при добыче, переработке и применении марганецсодержащей
руды в металлургии, в производстве сухих элементов для элек-
трических батарей и перманганата калия).
Основным путем поступления марганца в производственных
условиях являются дыхательные пути. Определенное значение
могут иметь энтеральный путь и в незначительной степени по-
ступление через кожу. Марганец обладает выраженными куму-
лятивными свойствами, накапливаясь в печени, почках, железах
179
внутренней секреции, в меньшем количестве он накапливается
•в костях, в головном и спинном мозге. Необходимо подчеркнуть,
что этот МЭ свободно проникает через гематоэнцефалический
•барьер и обладает тропизмом к подкорковым структурам голов-
ного мозга. Именно с этим связано его патогенное действие.
Крайним выражением профессионального манганоза — хро-
нической интоксикации марганцем — является синдром паркин-
сонизма. Он характеризуется тяжелыми расстройствами двига-
тельной активности, маскообразностью лица, дизартрией, нару-
шением письма. Специфична для этого синдрома «петушиная»,
или спастико-паретичеокая, походка, нередко в мышцах ног от-
мечаются повышение тонуса, реже гипотония и дистония. Сухо-
жильные рефлексы, как правило, повышены.
Характерны также психические нарушения, проявляющиеся
эйфорией, благодушием, снижением критики к своему состоя-
нию. Иногда больные апатичны, периодически беспокойные. Тя-
желым проявлениям манганоза ЦНС предшествует стадия функ-
циональных нарушений нервной системы (астеновегетативный
.синдром) с характерными психопатологическими симптомами
в форме ослабления ассоциативных процессов. За этой стадией
следует вторая, для которой типично нарастание признаков ток-
сической энцефалопатии, проявляющейся симптомами экстра-
пирамидной недостаточности, мнестико-интеллектуальными де-
фектами психики, полиневритами и др. Следует подчеркнуть,
что поражение ЦНС при хроническом манганозе сопровождает-
ся признаками угнетения функций гонад, а также функциональ-
ными нарушениями щитовидной железы, печени и желудочно-
кишечного тракта. Установлена связь экстрапирамидных и, воз-
можно, психических расстройств с нарушением синтеза и депо-
нирования дофамина [Рыжкова М. Н., Тарасова Л. А., 1975].
Патологоанатомические изменения при интоксикации мар-
ганцем изучены главным образом на экспериментальных жи-
вотных. Секционные случаи единичны. Установлено, что при
этом наиболее тяжело поражаются стриопаллидарная система
и лимбико-ретикулярный комплекс, кора большого мозга и
спинной мозг изменяются меньше. Нерезко выраженная дистро-
фия отмечается в печени, почках и железах внутренней секре-
ции. М. Yamada и соавт. (1986) описали типичный случай хро-
нического поражения ЦНС марганцем у мужчины 52 лет, сопро-
вождающегося тяжелым изменением базальных ганглиев, осо-
бенно бледных шаров при интактном черном веществе. Содер-
жание и задержка марганца в крови и веществе мозга зависят
от формы, в которой этот металл поступает в организм. Так,
G. Gainutsos и соавт. (1985) в эксперименте на мышах показа-
ли, что дихлорид маргайца вызывает наиболее длительную за-
держку этого металла в тканях мозга.
Если конкретные формы интоксикации марганцем в основном
изучены методами патологической анатомии, то этого нельзя
сказать об общей гистопатологии отравлений марганцем у че-
ловека. Ценные материалы для характеристики тканевых реак-
<80
Рис. 30. Очаговый некротически-язвенный ларингит. Выраженная воспалитель-
ная реакция у 11-месячной девочки при отравлении марганцем. Окраска ге-
матоксилином и эозином. Х250.
Рис. 31. Бронхопневмония, резкая гиперемия капилляров, макрофагально-лей-
коцитарная реакция у 11-месячной девочки при отравлении марганцем. Окра-
ска гематоксилином и эозином. Х400.
в печени при внутрибрюшинной инток-
мг/100 г массы тела крыс в течение-
Рис. 32. Патогистологические изменения
сикации хлоридом марганца в дозе 5
20 сут.
'’S’ "1 —‘-V—• ->—<«>-- — . - . ____—— _
а — мостовидные некрозы в печени с умеренной воспалительной"реакцией ’ Окп?скя™гей’я-
токсилином и эозином. Х120; б — резкое утолщение капсулы печени. Окр'аска гсматокси-
лином и эозином. Х250; в — то же самое при окраске по методу М-аллоои .Х2ЭД <пре-
параты С. П. Луговского). ’ ' р
Рис. 33 (подпись см. на с. 184).
Рис. 34 (подпись см. на с. 184),
Рис. 35. фагоцитоз двух апоптозных телец звездчатым макрофагоцитом (стрел-
ка) при подострой внутрибрюшинной интоксикации хлоридом марганца в дозе-
5 мг/100 г массы тела крыс в течение 20 сут. Х5000 (препарат С. П. Лугов.-
ского).
ций на поступление этого МЭ из внешней среды дают редкие:
случаи отравлений человека. Установлено, что KMnCU оказыва-
ет быстрое местное некротизирующее действие на слизистую»
оболочку верхних пищеварительных и дыхательных путей в фор-
ме острого фарингита, эзофагита, трахеобронхита. Развиваю-
щаяся при этом марганцевая бронхопневмония сопровождается
образованием экссудата, содержащего большое число альвео-
лярных макрофагов (рис. 30, 31).
Понимание тканевых и клеточных процессов существенно
углубляется благодаря воспроизведению острого и хронического»
Рис. 33. Набухание и мелкокапельное ожирение цитоплазмы гепатоцитов.
Анизоморфизм ядер и крупноглыбчатый метаморфоз хроматина. Набухание-
цитоплазмы звездчатых макрофагоцитов, сужение просвета синусоидов и рас-
ширение перисинусоидальных пространств (Диссе). Подострая интоксикация!
хлоридом марганца в дозе 5 мг/100 г массы тела крыс в течение 20 сут. Полу-
тоикий срез. Окраска метиленовым синим, азуром II и основным фуксином»
Х640 (препарат С. П. Луговского).
Рис. 34. Макрофаг в просвете вены, содержащий крупную фагосому. Подост-
рая интоксикация хлоридом марганца в дозе 5 мг/100 г массы тела крыс в те-
чение 20 сут. Х5000 (препарат С. П. Луговского).
184
Рис. 36. Апоптоз эпителиоцита крипты тощей кишки. Резко выраженное смор-
щивание клетки и осмиофилия цитоплазмы. Относительная сохранность внут-
риклеточных органелл при подострой интоксикации хлоридом марганца в дозе
5 мг/100 г массы тела крыс в течение 20 сут. Х6600.
манганоза в эксперименте на животных. Так, С. П. Луговской
(1989) в эксперименте на крысах при внутрибрюшинной инток-
сикации хлоридом марганца в течение 20 сут в ряде случаев
выявил тяжелое повреждение печени в виде мостовидных не-
крозов, сопровождавшихся выраженной воспалительной инфиль-
трацией (рис. 32). Наряду с этим наблюдались мелкокапельное
ожирение гепатоцитов, резкое набухание звездчатых ретикуло-
эндотелиоцитов с сужением просвета синусоидов и выраженная
макрофагальная реакция (рис. 33). Цитоплазма макрофагов
часто содержала крупные фагосомы (рис. 34). В отдельных
звездчатых ретикулоэндотелиоцитах выявлялись апоптозные
тельца (рис. 35). Апоптоз выражен также в криптах тонкой
кишки (рис. 36).
Явления апоптоза в эпителии крипт тонкой кишки обнару-
жены и при пероральной интоксикации марганцем, однако вы-
раженность его была значительно ниже. Хроническая перораль-
ная интоксикация хлоридом марганца в течение 40 и 70 сут
сопровождалась повышенной секреторной активностью эпите-
лиоцитов слизистой оболочки тонкой кишки, особенно бокало-
видных клеток и клеток Панета (рис. 37). В поджелудочной
железе отмечены неравномерное содержание зимогенных гра-
нул в ацинозных клетках, а также некробиотические изменения
185
Рис. 37. Клетка Панета в крипте двенадцатиперстной кишки крысы при хро-
нической пероральной интоксикации хлоридом марганца в дозе 20 мг/100 г
массы тела крыс в течение 40 сут. В цитоплазме большое количество гранул
различной плотности. Х5000 (препарат С. П. Луговского).
отдельных клеток или их групп. Наблюдалось резко выраженное
накопление фуксинпозитивных гранул в В-клетках панкреатиче-
ских островков (рис. 38). Таким образом, автору удалось пока-
зать, что гистопатология экспериментального манганоза суще-
ственным образом зависит от способа введения хлорида мар-
ганца.
В настоящее время благодаря герметизации производствен-
ных процессов, обеспыливанию и применению влажного бурения
выраженные формы отравления марганцем встречаются очень
редко. В начальных стадиях и при умеренных формах этого-
процесса необходимы своевременный диагноз и полное прекра-
щение дальнейшего контакта с марганцем. Это особенно важ-
но, так как для диагностики начальных стадий марганцевой эн-
цефалопатии требуется хорошее знание ее инициальной симпто-
матики. Значительную помощь в этих случаях могут оказать
электроэнцефалография и особенно электромиография мышц
нижних конечностей, позволяющие на ранних стадиях отравле-
ния обнаружить характерные изменения биоэлектрической ак-
тивности.
По сравнению с богатой и довольно тяжелой патологией,
обусловленной дефицитом марганца у сельскохозяйственных и
лабораторных животных, соответствующая симптоматика пато-
186
Рис. 38. Патогистологические изменения поджелудочной железы при хрониче-
ской пероральной интоксикации хлоридом марганца в дозе 20 мг/100 г массы
тела крыс в течение 70 сут.
^ДераВНОМерДое с°ДеРжание зимогенных гранул в ацинозных клетках поджелудочной
тм л КРЫС- Пикноз отдельных ядер с перинуклеарным отеком (стрелка). Полутонкий
но?' икраска метиленовым синим, азуром II и основным фуксином. Х400; б — умерен-
ное содержание альдегид-фуксинпозитивных гранул в p-клетках панкреатических остров-
ков контрольных крыс; в — большое число альдегид-фуксинпозитивных гранул в [3-клет-
ках панкреатических островков подопытных крыс. Окраска альдегид-фуксином по методу
Гомори. Х320 (препараты С. П. Луговского).
логии человека выглядит либо бедной, либо несущественной.
Вряд ли это отражает действительное положение вещей. Надо
полагать, что очень скоро будет раскрыта роль марганца в фи-
зиологии и патологии репродукции человека, в этиологии и па-
тогенезе болезней костного и хрящевого скелета, в развитии и
функционировании ЦНС. В связи с необходимостью оптималь-
ного содержания марганца для формирования отолитов напра-
шивается мысль о связи генетических, ввожденных и некоторых
приобретенных вестибулопатий человека с нарушениями марган-
цевого гомеостаза. Заслуживает внимания гипотеза о целесооб-
разности поддержания оптимального состояния отолитов с по-
мощью профилактического применения марганецсодержащих
препаратов в экстремальных условиях обитания и трудовой дея-
тельности, в частности у членов экипажей космических кораб-
лей.
6.5. Хром
Хром относится к числу «классических» МЭ. Его токсичес-
кое действие было установлено более 160 лет назад [Grtie-
lin С. G., 1826], вскоре после открытия этого металла, а жиз-
ненно важная необходимость для животного организма доказа-
на только в 1957—1959 гг. в опытах К. Schwarz и W. Mertz
(1959). В то же время механизм физиологического действия
хрома продолжает оставаться нераскрытым, несмотря на то что
за последние 30 лет ему посвящено более тысячи научных пуб-
ликаций.
Длительное время полагали, что хром осуществляет свою
биологическую функцию в составе особого витаминоподобного
соединения, названного фактором толерантности к глюкозе
(GTF). Утверждалось, что этот фактор присутствует в повы-
шенном количестве в пивных дрожжах и что в его составе на
один атом хрома приходятся две молекулы никотиновой
кислоты и остатки цистеина, глицина и глутамата [Mertz W.,
1974]. Соединение с такой же структурой найдено в печени и
в грудном молоке [Jamamoto М. et al., 1988], но синтетические
препараты аналогичного состава оказались в сотни раз менее
активными, чем препараты из дрожжей. В связи с этим вопрос
об истинной структуре указанного фактора остается открытым.
Есть также работы, где под сомнение ставится само его су-
ществование [Haylock S. I., 1983]. Не обнаружены также фер-
менты, содержащие в своей молекуле хром.
В связи с совершенствованием аналитических методов сов-
ременные данные по содержанию хрома в биологическом мате-
риале значительно снизились. Так, именно благодаря улуч-
шению методов анализа показатели содержания хрома в крови
уменьшились по сравнению с 1962 г. более чем в 3000 раз, а
за последние 10—12 лет—более чем в 10 раз. Наиболее досто-
верные данные, полученные методом нейтронно-активационного
анализа и беспламенной атомно-абсорбционной спектроскопии
188
в 1978—1983 гг., свидетельствуют о содержании хрома в крови,,
равном 1,44—3,08 нмоль/л против 2,88—10 мкмоль/л в 60-х го-
дах и 0,03—0,27 мкмоль/л в начале 70-х годов.
Аналогичные изменения произошли также с аналитическими
данными о содержании хрома в органах, тканях и моче. Это
обстоятельство заставляет относиться с большой осторожно-
стью к работам, в которых сообщается об изменениях концент-
рации хрома в организме, а также в моче при диабете, сердеч-
но-сосудистой патологии и других заболеваниях. В то же врем®,
нет оснований сомневаться в том, что хрому принадлежит важ-
ная биологическая роль в животном организме, основными-
проявлениями которой являются его взаимодействие с инсули-
ном в процессах углеводного обмена, участие в структуре и
функции нуклеиновых кислот и, очевидно, щитовидной железы.
Судя по опытам с перфузией кишечных сегментов 51СгС13
[Chen N. S. et al., 1973], хром всасывается в основном в тощей
кишке. В норме усвоение этого элемента не превышает 0,4—
0,7% от введенной дозы, тогда как у больных диабетом оно
выше в 3 раза. Всасывание хрома зависит от присутствия хе*
латирующих агентов. Так, у крыс щавелевая кислота повышает,,,
а фитиновая — заметно снижает усвоение этого МЭ [ChenN. S.
et al., 1973]. Лиганды, повышающие всасывание хрома, преду-
преждают его выпадение в осадок в виде основных солей в
близкой к нейтральной среде кишечного тракта. На всасывание
хрома оказывают влияние также цинк и железо. При недоста-
точности этих МЭ усвоение хрома возрастает, а при их одно-
временном поступлении снижается, что свидетельствует об
общих путях усвоения хрома, цинка и железа. Взаимный
антагонизм на уровне всасывания существует и между хромом
и ванадием [Hill С. Н., 1975]. ?
В кровяном русле хром специфически связывается с транс-
феррином, который служит переносчиком не только железа,,
но и хрома. Из двух участков связывания металла на транс-
феррине с различным сродством к железу последнее связыва-
ется преимущественно участком А, а хром — участком В. При
высоком насыщении трансферрина железом оно конкурирует
с хромом за этот участок, в связи с чем у больных гемохрома-
тозом при насыщении трансферрина более 50% в крови и-
организме в целом понижено содержание хрома. Поскольку
диабет нередко сопутствует гемохроматозу, то нарушение тран-
спорта хрома трансферрином может вызывать дальнейшее
ухудшение течения этого заболевания.
Содержание хрома в органах и тканях в 10—100 раз выше,
чем в крови. Однако, как уже отмечалось, приводимые цифры
требуют критического подхода и не только в связи со сложно-
стью аналитических методов, но и в связи с тем, что при взя-
тии проб тканей нередко используются инструменты из нержа-
веющей стали, содержащей около 18% хрома. Наибольшего
доверия заслуживают результаты, полученные с использовани-
ем нескольких различных эталонных образцов.
18Э'
Концентрация хрома в тканях новорожденных, как правило,
значительно выше, чем у взрослых, и быстро снижается в тече-
ние первых месяцев жизни. Исключение составляют печень и
почки, в которых концентрация этого МЭ сохраняется на одном
уровне от рождения до 10-летнего возраста ([Schroeder Н. А.,
1962]. При сахарном диабете содержание хрома в тканях, как
правило, понижено.
Распределение 5!Сг в органах и тканях зависит от возраста, вида живот-
ных, химической формы хрома и наличия заболеваний, в частности диабета.
Так, при внутривенном введении Na25!CrO2 почти 100% радиоактивной метки
включается в клетки СМФ, тогда как прн введении 5!СгС13 55% 51Сг поступает
в печень. При введении СгС13 в цитратном или ацетатном буферах в печени
удается обнаружить менее 5% 5!Сг, а преобладающая часть дозы выделяется
с мочой. Распределение радиоактивного хрома в тканях крыс в норме и при
диабете не обнаруживает существенных различий, но при экспериментальном
диабете у крыс в гепатоцитах ядра и супернатант содержат больше 5!Сг, а ми-
тохондриальная и микросомные фракции меньше, чем у здоровых животных
[Mathur R. К., Doisy R. J., 1972]. Аналогичная картина наблюдается и у здо-
ровых крыс, получающих пищу, богатую жирами. При нормальном или уси-
ленном липогенезе отмечена тенденция к переходу хрома из ядер в микросо-
мальную фракцию гепатоцитов.
Изучение кинетики хрома и его компартментации в организме свидетель-
ствует, что после внутривенного введения 51СгС13 независимо от дозы и пред-
шествующей нагрузки организма этим элементом его выделение происходит
в три последовательных этапа с полупериодом выведения 0,5; 5,9 и 83,4 дней
[Mertz W. et al., 1965]. Существование метаболических компартментов для
хрома показано и при исследовании распределения в организме его стабильно-
го и радиоактивного изотопов, позволивших обнаружить наличие его внекле-
точной фазы, находящейся в равновесии с быстро- и медленнообменнвающи-
мися клеточными фазами и очень медленнообменивающимся внутриклеточным
пулом хрома [Jain R. et al., 1981].
Содержание хрома в волосах хорошо отражает концентра-
цию этого МЭ в организме. В волосах доношенных детей кон-
центрация хрома в 2’/2 раза выше, чем в волосах у их матерей,
и составляет в среднем 0,9 мг/кг против 0,44 мг/кг у детей
2—3-летнего возраста [Hambidge К. М., 1977]. Роды, диабет и
атеросклероз приводят к снижению концентрации хрома в
волосах [Cote М. et al., 1979].
Выделение. Всосавшийся хром выводится из организма в
основном с мочой и лишь небольшая его часть удаляется с
желчью, потом и волосами. Невсосавшийся хром, составляю-
щий, как правило, 99% его содержания в принимаемой пище,
выделяется с калом. Поскольку содержание хрома в рационе
не превышает 100 мкг и составляет 62+28 мкг/сутки в рацио-
нах, богатых жирами, и 89+56 мкг в рационах, бедных жирами
[Kumpulainen J. Т. et al., 1979], то суточная моча содержит
обычно менее 1 мкг этого элемента. Выделение хрома с мочой
удобно представить как отношение его к содержанию креатини-
на, количество которого в норме характеризуется значительным
постоянством, мало зависит от питания, объема мочи и опре-
деляется только мышечной массой.
Комплекс хрома с ЭДТА не подвергается реабсорбции в
почках и может служить хорошим индикатором клубочковой
:190
фильтрации. Ультрафильтруемая часть хрома составляет у че-
ловека 6—28%, а у собак — 41% от общего количества этого-
элемента, находящегося в плазме крови. В моче человека и
крыс обнаружено низкомолекулярное соединение с молекуляр-
ной массой 1500, ковалентно связывающее трехвалентный
хром. Аналогичные соединения найдены также в печени и дру-
гих органах и тканях кроликов и собак.
Существует определенная связь между обменом хрома и
глюкозой, хотя мнения о характере этой связи расходятся.
В ряде работ сообщается, что нагрузка организма глюкозой ве-
дет к мобилизации хрома из органов-депо, его поступлению в-
плазму крови и последующему выведению с мочой [Mertz W.,
1969]. У больных диабетом с пониженной толерантностью к
глюкозе такое повышение наблюдалось только после обогаще-
ния рациона хромом. Другие авторы [Lin V. J. К., Morris J. S.,
1978] отмечают, что концентрация хрома в плазме крови после
нагрузки глюкозой зависит от запасов этого МЭ в организме.
Имеются также наблюдения о снижении концентрации хрома
в плазме крови здоровых лиц после введения глюкозы [David-
son I. W. F., Burt R. L., 1973]. У беременных уровень хрома
в плазме крови натощак ниже, чем у небеременных жен-
щин.
Аналогичным образом расходятся данные и о влиянии на-
грузки глюкозой на уровень хрома в моче. Эти расхождения,
как отмечалось выше, объясняются, по-видимому, в первую
очередь несовершенством применявшихся ранее аналитических
методов. Все же в свете современных данных можно считать,
что концентрация хрома в моче после нагрузки глюкозой, как
правило, повышается. Так, в одном из опытов было показано,
что концентрация хрома в моче здоровых лиц натощак состави-
ла в среднем 30 нмоль на 1 мкмоль креатинина, а через 90 мин
после нагрузки глюкозой — 0,4 нмоль на 1 мкмоль креатинина.
Однако повышение концентрации хрома в моче было отмечено
только у 61 из 76 обследованных лиц, что свидетельствует
о ненадежности данной пробы для оценки состояния обмена
хрома [Anderson R. А., 1984].
На обмен хрома оказывают заметное влияние и различные
виды стресса, так же как белковое голодание, инфекция, фи-
зическая нагрузка и др., приводящие к снижению концентрации
. этого элемента в плазме крови и его повышенному выделению
: с мочой {Anderson R. А., 1984].
Недостаточность хрома и его ф у н к ц и и. Наи-
более хорошо изучено, что хром способен усиливать действие
инсулина во всех метаболических процессах, регулируемых
этим гормоном. Хром в присутствии инсулина ускоряет окисле-
ние глюкозы в жировой ткани придатков яичников хромдефи-
цитных крыс, повышает скорость проникновения глюкозы в.
клетки и ее превращение в жир, стимулирует поступление глю-
козы в изолированный хрусталик глаза и синтез гликогена, но
не оказывает влияния на процессы, не зависящие от инсулина,
191
как, например, на включение в жир ацетата. Хром усиливает
также поступление в клетки жировой ткани яичников галакто-
зы, но поскольку в этой ткани не происходит фосфорилирование
данного сахара, то галактоза позволяет более четко разграни-
чить действие хрома и инсулина на транспортные системы от
их влияния на внутриклеточный обмен углеводов. Действие
хрома на транспорт сахаров объясняется, по-видимому, его
участием в образовании комплекса между инсулином и его ре-
цептором на клеточной мембране.
Описаны признаки выраженной недостаточности хрома у
2 больных, находившихся на полном парентеральном питании,
у которых наблюдались периферические невропатии и потеря
массы тела, несмотря на адекватное питание, утрата толерант-
ности к глюкозе. При этом толерантность к глюкозе повыша-
лась при включении в питательный раствор 150—250 мкг хро-
ма в сутки. В человеческих популяциях значительно чаще
встречаются случаи умеренной недостаточности хрома, прояв-
ляющиеся понижением толерантности к глюкозе и повышением
уровня липидов в крови, поддающиеся коррекции препаратами
хрома.
Опыты на крысах, испытывающих недостаточность хрома,
показывают, что этот МЭ повышает эффективность действия
инсулина на белковый обмен, способствуя поступлению в ткани
меченых глицина, серина, метионина и их включению в белки
сердечной мышцы [Mertz W., Roginski Е. Е., 1969].
Исследования на животных и клинические наблюдения сви-
детельствуют, что хром играет также определенную роль в ли-
пидном обмене и что дефицит этого элемента может привести
к развитию атеросклероза. В середине 60-х годов было показа-
но, что в плазме крови крыс, содержащихся на дефицитной по
хрому диете, с возрастом повышаются концентрация глюкозы
в крови, взятой натощак, и уровень холестерина, снижается
толерантность к глюкозе и увеличивается количество липидов
и бляшек в стенках аорты [Schroeder Н. А., 1965].
При экспериментальном холестериновом атеросклерозе кро-
ликов применение препаратов хрома способствует значительно-
му уменьшению размеров бляшек и концентрации холестерина
в стенке аорты [Abraham A. S. et al., 1982]. У лиц с расстройст-
вами коронарного кровообращения концентрация хрома в плаз-
ме крови значительно ниже, чем в условиях нормы [New-
man Н. A. I. et al., 1978], достоверно коррелирует с наступле-
нием заболевания (р<0,01) и содержанием триглицеридов в
плазме крови (р<0,05). Корреляция уровня этого элемента с
.другими факторами риска, такими как концентрация холестери-
на, повышенное артериальное давление и увеличение массы
тела, отсутствует.
Убедительные доказательства влияния хрома на липидный
обмен зарегистрированы у 12 взрослых мужчин, получавших по
.200 мкг этого элемента 5 раз в неделю в течение 4 мес [Ria-
les R., Albrink M. J., 1984]. У них были отмечены существенное
<92
понижение концентрации триглицеридов в плазме крови и уве-
личение содержания холестерина в составе липопротеидов вы-
сокой плотности. При атеросклерозе и в период, предшествую-
щий этому заболеванию, в плазме крови нередко наблюдается
повышенная концентрация инсулина, которая на чувствитель-
ных к этому гормону участках артерий может привести к воз-
никновению липидных отложений, аналогичных структурам,
наблюдаемым на ранних стадиях атеросклероза [Stout R. W.,
1977]. Поскольку хром, усиливая действие инсулина, участвует
в регуляции содержания этого гормона в крови, то недостаток
этого элемента вполне может служить одной из причин повы-
шения в ней содержания инсулина [Borel J. S., Anderson R. А.,
1984].
Роль хрома в обмене нуклеиновых кислот начала вырисовы-
ваться более 25 лет назад, когда было обнаружено его высокое
содержание в некоторых «нуклеопротеиновых» фракциях печени
крупного рогатого скота, превосходившее концентрацию всех
остальных металлов и достигавшее 19 ммоль/кг [Wac-
ker W. Е. С., Vallee В. L., 1959]. Было также показано, что
хром прочно связан с нуклеиновыми кислотами и эффективно
защищает их от денатурации. Обработка нуклеиновой кислоты
ЭДТА, удаляющей большинство металлов, не вызывает замет-
ного снижения концентрации в ней хрома [Mertz W., 1969].
Инкубация ДНК или хроматина гепатоцитов мышей с мик-
ромолярными концентрациями СгС13 до добавления РНК-поли-
меразы существенно повышает скорость синтеза РНК, но ин-
гибирует его при предварительной обработке фермента этим
соединением. Это свидетельствует об отсутствии данного метал-
ла в активном центре РНК-полимеразы. Указанное действие
хрома имеет, по-видимому, функциональное значение, так как
во вновь синтезированной РНК обнаруживают большее число
нуклеотидных замен, чем в контроле. При этом изменяется
отношение РНК-синтезирующей системы к отдельным ингиби-
торам {Okada S. et al., 1983]. Показано также, что в регене-
рирующей после частичной гепатэктомии печеночной ткани
синтезируется белок с молекулярной массой около 70 000, в
молекуле которого содержится 5—6 атомов хрома. Он оказы-
вает стимулирующее действие на транскрипцию РНК и после-
дующий биосинтез белка. При полном насыщении хромом
данный белок в несколько раз богаче этим элементом, чем
«нуклеопротеид» Вэлли (77 ммоль/кг против 21 ммоль/кг бел-
ка).
Соединения трехвалентного хрома оказывают выраженное
мутагенное и канцерогенное действие на незащищенный гене-
тический аппарат клетки. Трехвалентный хром всасывается с
большим трудом, но его соединения могут появиться в ядре при
длительном воздействии на организм низких концентраций не-
которых шестивалентных форм этого металла, легко проника-
ющих через клеточные мембраны и восстанавливающихся затем
до трехвалентного состояния. Хромсодержащий белок Окада,
13—568
193
продуцируемый при необходимости интенсивного синтеза РНК,
защищает клетку от повреждения трехвалентным хромом и
доставляет этот МЭ в незначительном количестве только в
определенные участки ядерного аппарата в специфической
химической форме, выполняя, по-видимому, функции физиоло-
гического переносчика хрома в клетке и регулятора синтеза
нуклеиновых кислот.
Еще одним проявлением биологической активности хрома
считается его взаимодействие со щитовидной железой. При
определенных условиях хром способен замещать йод в тиреоид-
ных гормонах. Дача физиологических доз хрома крысам, испы-
тывающим недостаточность йода, судя по гистологической
картине щитовидной железы, повышает ее функциональную
активность [Гончаров А. Т., 1968], тогда как повышенные дозы
хрома угнетают функцию этой железы у животных, получаю-
щих нормальное количество йода. Установлено, что в щитовид-
ной железе человека и животных, обитающих в йоддефицитных
биогеохимических провинциях СССР, накапливается значитель-
но большее количество хрома, чем в норме за счет снижения
концентрации хрома в прочих органах и тканях [Соркина А. И.
и др., 1963]. Имеются также наблюдения, что при тиреоидэкто-
мии содержание хрома в организме животных снижается, а
при даче тиреоидных гормонов снова возрастает [LifschitzM. L.
et al., 1980]. Таким образом, при недостаточности хрома у
животных и человека могут быть обнаружены следующие приз-
наки: снижение толерантности к глюкозе, повышение концент-
рации инсулина в крови, глюкозурия, гипергликемия натощак,
задержка роста, уменьшение продолжительности жизни, повы-
шение концентрации триглицеридов и холестерина в сыворотке
крови, увеличение числа атеросклеротических бляшек в аорте,
периферические нейропатии, нарушение высшей нервной дея-
тельности, снижение оплодотворяющей способности и числа
спермиев.
Потребность человека в хроме колеблется, по имеющимся
данным, в пределах 50—200 мкг в сутки. В то же время в
общепринятой диете содержится 33—125 мкг хрома, а для лиц
пожилого возраста даже 5—115 мкг. Особенно бедны хромом
высокоочищенные продукты, такие как сахар-рафинад
(3,85 мкмоль/кг), пшеничная мука тонкого помола и выпечен-
ный из нее хлеб (2,7 мкмоль/кг). Если учесть, что сахар, кро-
ме того, усиливает потери хрома из организма, то вполне можно
допустить, что в человеческой популяции имеются значитель-
ные группы лиц, испытывающих недостаточность этого элемен-
та. К ним в первую очередь относятся лица пожилого возраста.
Соединения хрома широко используются в металлургии, в
химической, текстильной, кожевенной промышленности и других
отраслях народного хозяйства. Имеются многочисленные сооб-
щения о хроме как о профессиональной вредности. Многие
авторы показывают, что этот МЭ в экспериментальных услови-
ях не только вызывает патологические процессы во внутренних
194
органах, но и способствует возникновению изменений иммуно-
логической реактивности организма. Случаи острого отравления
•соединениями хрома с высокой летальностью описали Н. К. Пер-
мяков и Л. Н. Зимина (1976). Избыточные количества хрома,
поступающие в организм в производственных условиях, нару-
шают процессы биологического окисления, в частности цикл
трикарбоновых кислот.
По данным А. В. Рощина и соавт. (1982), хром способен
аккумулироваться в организме человека, легко проникает в
ткань легких и накапливается в них. У работающих в контакте
•с хромом этот МЭ можно обнаружить в легких через много лет
после прекращения контакта. Токсическое действие хрома за-
висит от его валентности: чем выше валентность, тем сильнее
токсическое действие. Окислы шестивалентного хрома значи-
тельно токсичнее окислов трехвалентного хрома при их воздей-
•ствии на слизистую оболочку дыхательных путей и другие
ткани. Именно шестивалентный хром оказывает общетоксичес-
кое, нефротоксическое и гепатотоксическое действие. Этот МЭ
вызывает специфические поражения кожи (дерматит, язвы),
изъязвления слизистой оболочки носа с характерной перфора-
цией хрящей носовой перегородки, пневмосклероз, гастриты
(фундального и антрального отделов), неблагоприятно проте-
кающую, склонную к перфорации и пенетрации язву желудка
и двенадцатиперстной кишки, хромовый гепатоз, нарушения
регуляции сосудистого тонуса и сердечной деятельности. Соеди-
нения шестивалентного хрома по сравнению с трехвалентным
обладают большей мутагенной способностью [Рудных А. А.,
1986]. Они наряду с общетоксическим действием способны вы-
зывать мутагенный [Виталиев А. Б., 1982] и канцерогенный
[Ashby J. et al., 1985] эффекты. Высказывается предположе-
ние, что при этом хром выступает в роли комутагена ,[Ross-
ma Т. С., 1981], снижая репаративные процессы в клетках
человека и животных. Аллергизирующее действие хрома связы-
вается с его способностью образовывать комплексные соедине-
ния.
Многостороннее патогистологическое и гистоэнзиматическое
изучение хромовой интоксикации в эксперименте на собаках
и крысах осуществил А. М. Шахназаров (1980). Из выводов
его работы наибольшего внимания заслуживают установление
при хронической интоксикации хромом ингибирования тиоло-
вых ферментов мембран, а также ферментов аэробного глико-
лиза, цикла Кребса и терминального окисления, угнетение
синтеза аминокислот и белков, возникновение гипоксии в ре-
зультате склерозирования биологических мембран и микроцир-
куляторного русла, активации ферментов анаэробного гликоли-
за, пентозофосфатного шунта и неспецифических фосфатаз.
В 1986 г. R. Laborda и соавт. прицельно изучали патомор-
фологию ° нефротоксического и гепатотоксического действия
соединений хрома на крысах Вистар. Они вновь подтвердили
большую токсичность шестивалентного хрома по сравнению с
'53* 195
трехвалентным, но вместе с тем отметили, что некоторые соли
последнего, как, например, ацетат хрома и др., при их вдыха-
нии, поглощении или в случае контакта с тканями обладают
канцерогенным свойством [Hneper W. С. et al„ 1962; Steffes Р.
et al., 1965]. Тяжелые поражения печени и почечных канальцев
наблюдались при внутрибрюшинном введении шестивалентного
хрома.
Установлено, что поступающий в организм человека хром,
помимо легочной ткани, накапливается в органах системы мо-
нонуклеарных фагоцитов, т. е. в печени, почках, селезенке, кос-
тях и костном мозге. Шестивалентный хром проникает через
плацентарный барьер, может откладываться в органах и тка-
нях плода, а также поступать в организм ребенка с грудным
молоком [Рощин А. В. и др., 1986].
6.6. Селен
Одним из первых сообщений о синдроме, связанном с ис-
пользованием в пищу растений с избыточным содержанием се-
лена, являются записки Марко Поло в 1295 г. [Komroff М.,
1926]. В журнале путешественника указано, что вьючные жи-
вотные страдали от поражений ног и копыт при поедании
некоторых трав, произрастающих на границе Туркестана и
Тибета. В 1860 г. военный хирург Т. С. Madison описал харак-
терное заболевание лошадей, пасущихся в некоторых степях
Южной Дакоты (США). Болезнь начиналась примерно через
10 дней после начала выпаса скота и прекращалась при замене
фуража. Для нее была характерна, помимо «воспалительных
процессов в ногах», потеря волос гривы и хвоста.
Внимание к селену вновь было привлечено в 1931 г., когда
стало известно, что в основе ряда эндемических заболеваний
крупного рогатого скота, свиней и домашних птиц, регистрируе-
мых на отдельных пастбищных массивах Великих Равнин се-
веро-запада США («алкалоз», «слепая хромота»), лежит отрав-
ление селеном, присутствующим в повышенных количествах в
пастбищных растениях (особенно в некоторых видах астрага-
лов) и в зерне, выращенном на богатых селеном почвах [By-
ers Н. G., 1934].
Жизненная необходимость селена была установлена лишь
26 лет спустя [Schwarz К., Foltz С. М., 1957] в ходе класси-
ческих экспериментов, продемонстрировавших, что именно
этот МЭ предупреждает возникновение некрозов в печени у
крыс. Вскоре был открыт еще ряд заболеваний, излечиваемых
селеном, к числу которых относятся беломышечная болезнь
овец и крупного рогатого скота, алиментарный гепатит свиней,
экссудативный диатез домашней птицы, причинявших значи-
тельный ущерб животноводству на обширных сельскохозяйст-
венных угодьях, где почвы и растения содержали пониженное
количество селена (штат Орегон, США; Новая Зеландия, Фин-
ляндия; Эстонская ССР, Бурятская АССР и др.).
1 96
В связи с близостью биологического действия селена и ви-
тамина Е одно время допускалась их взаимозаменяемость.
Однако в ряде работ было установлено, что животные испыты-
вают потребность в селене даже при их обеспеченности витами-
ном Е. Явления специфической недостаточности селена вклю-
чают некроз и фиброз поджелудочной железы у цыплят, облы-
сение, повреждение стенок кровеносных сосудов, катаракту,
задержку роста и нарушение репродуктивной функции у крыс
[Wu A. S. Н. et al., 1979].
Роль селена в патологии человека была установлена после
описания селенодефицитной кардиомиопатии в Китае (болезнь
Кеилана), а также явлений недостаточности селена при полном
парентеральном питании [Johnson R. A et al., 1981]. Характе-
ристика селенодефицитных заболеваний устанавливается толь-
ко в последние годы. Представляют интерес также антибласти-
ческое действие селена и его способность противодействовать
токсическому влиянию тяжелых металлов.
Всасывание селена может осуществляться через пищевари-
тельный тракт, кожу и легкие. Усвоение растворимых форм
селена в пищеварительном тракте происходит на 80—100%.
В частности, С. D. Thomson и соавт. (1972, 1973) в эксперимен-
тах на крысах с введением изотопа 75Se показали, что 91—93%
этого МЭ всасываются в желудочно-кишечном тракте. В то же
время у 3 мужчин-добровольцев всасывание составляло только
70, 64 и 44% соответственно. Особого внимания заслуживают
данные, приведенные в обзоре О. Levander (1984). Так, оказа-
лось, что у новозеландских женщин, для которых характерна
пища с содержанием селена (24 мкг/день), абсорбируются
79% пищевого селена. У взрослых людей всасывается от 76
до 100% введенного per os селена в виде 755е-метионина. Ря-
дом исследователей [Сучков Б. П., 1981; Jaakkola et al., 1983]
установлено, что органические соединения селена усваиваются
лучше, чем неорганические.
Наиболее интенсивно всасывание селена происходит в две-
надцатиперстной и в несколько меньшей степени--—в тощей и
подвздошной кишках. В желудке этот МЭ практически не ус-
ваивается. Из почек селен усваивается на 87%, а из мышц
рыбы — всего на 64% [Richold М. et al., 1977]. Относительно
низкая усвояемость «рыбьего» селена подтверждается и рядом
других авторов.
Транспорт и депонирование селена осуществляются, по-ви-
димому, особыми белками, содержащими селеноцистеин, и
названными соответственно селенопротеином Р из плазмы
крови и селенопротеином Рх из почек и печени [Motsenboc-
ker М. A., Tappel A. L., 1984]. Селенопротеин Р найден у крыс
и макак резусов. Он имеет молекулярную массу 85 000, синте-
зируется в печени и переносит селен в семенники, селезенку и
прочие ткани, в которых найден еще ряд белков, содержащих
селен. Взаимоотношение между указанными соединениями селе-
на представлены на схеме 1. Содержание селенонопротеина Р
197
С х е м a 1
СИНТЕЗ И ФУНКЦИИ СЕЛЕНОЦИСТЕИНСОДЕРЖАЩЕГО БЕЛКА Р
[Motsenbocker М.А., Tappei A.L.,
Печень
в плазме зависит от уровня селена, поступающего в организм
животного. При высоких дозах селена в тканях откладывается
неидентифицированное соединение этого МЭ, не извлекаемое
вместе с белками.
Селен обладает большим сходством с серой. Их атомы име-
ют близкую конфигурацию наружных электронных оболочек,
почти одинаковые размеры и сходные энергию связи, ионизаци-
онные потенциалы и сродство к электронам. В то же время в
живых системах эти два элемента ведут себя зачастую совер-
шенно по-разному. В частности, в организме животных соеди-
нения селена подвергаются обычно восстановлению, а серы —
окислению. Селеноводород является более сильной кислотой,
чем сероводород, и диссоциирован в большей степени, чем пос-
ледний. Это относится и к степени диссоциации селено- и
сульфгидрильных групп.
В живой природе найдены различные соединения селена,
являющиеся главным образом производными селеносодержа-
щих аминокислот и продуктами метилирования селена. В бакте-
риях, грибах, ряске, клевере, плевеле, пшенице найдены
селенометионин и селеноцистеин. Последний присутствует так-
же в глутатионпероксидазе млекопитающих и в ряде белков
плазмы крови, семенников, мышц. В орехах и растениях-кон-
центраторах селена (виды Astragalus, Morinda) присутствует
селеноцистатионин. В астрагалах встречаются также Se-метил-
селеноцистеин, Se-метилселенометионинселеноний, диметилселе-
нид, селеногомоцистин, в чесноке — Se-пропенилселеноцистеин-
селеноксид (табл. 18). Открыты также азотистые основания у
некоторых анаэробных бактерий, содержащие в своем составе
этот МЭ [Tsai L. et al., 1984]. В тРНКглу из Cl. sticklandii
селеноуридин встречается наряду с 4-тиоуридином.
В качестве кофактора в ряде окислительно-восстановитель-
198
а б л и ц a
Селеносодержащие соединения, найденнные в живой природе
Наименование Формула Источник
Диметилдиселенид Бе-метилселеноцистеин Селеноцистеинселенино- вая кислота Селенометионин Селеноцистин Селеноцистатионин Селеногомоцистин Селенометионинселе- ноксид Бе-пропенилселеноцн- стеинселеноксид Селенотрисульфид Г лутатионселенотри- сульфид Г лутатионселенопер- сульфид Диметилселенид Триметилселеноний Селен-РНКглу MeSeSeMe CysSeMe CysSeOH CysCHzSeMe CysSeSeCys CysCH2SeCys CysCH2SeSeCH2Cys CysCH2Se(Me) =0 CysSeOCHz— CHMe R—S—Se—S—R G—S—Se—S—G G— S— SeH (Me)2Se (Me)3Se+ 5- [ (метиламино) ме- тил] -2-селеноуридин Грибы, астрагалы Астрагалы, Stranleya bipinnata Плевел, клевер Бактерии, грибы Ряска, плевел, клевер, пшеница Астрагалы, Моринда Astragalus crotolaria Плевел, клевер Лук Млекопитающие » Выдыхаемый воздух Моча Е. coli, Cl. stricklandii, Methanococus vanniellii
ных ферментов селен присутствует один или вместе с железом
и молибденом (табл. 19).
Кроме указанных, найден ряд недостаточно идентифициро-
ванных селеносодержащих белков в семенниках, селезенке,
других органах и тканях. Селен повышает активность еще двух
ферментов, выделенных из Cl. barkeri, — гидролазы никотиновой
кислоты и ксантиндегидрогеназы [Shamberger R., 1984].
Глутатионпероксидаза — первый селеносодержа-
щий фермент, найденный в организме млекопитающих. Он пре-
дохраняет клетки от токсического действия перекисных радика-
лов. Фермент не обладает строгой специфичностью по отноше-
нию к перекисям и нуждается в качестве кофактора в глута-
тионе, который в ходе ферментативной реакции подвергается
окислению:
H2O2 + 2GSH —> H2O + GSSG
ROOH+2GSH —> ROH + Н2О + GSSG
В этой реакции электрон, однако, переносится на перекись
не с восстановленного глутатиона, а с селенола, который при
этом переходит в селенистую кислоту, а затем последняя реге-
нерируется в селенол восстановленным глутатионом.
Взаимосвязь между селеном и витамином Е объясняется их
воздействием на разные этапы образования органических пере-
199
Таблица 19
Селеносодержащие ферменты и белки [Favier, A., Maljournal В.,
1980; с дополнениями]
Наименование Молекуляр- ная масса, 103 Кофакторы Источник
Формиатдегидрогеназа Глицинредуктаза Г лицинредуктаза Селенопротеид из мы- шечной ткани Селенсодержащий фер- редоксин Селеносодержащий ци- тохром с Г лутатионпероксидаза Белок Р Белок Pi Белок X, участвующий в цепи переноса электро- нов между флавопротеи- дами и системой цито- хромов Р-450 590 (3—4 субъе- диницы) 12 12 10 83 (4 субъ- единицы) 85 Неизвест- на » Se, Mo, Fe Se, Mo Se Se Se, Fe Se Se Se Se Se Se, Fe Cl. thermoaceticum, Methanococcus vaniellii E. coli Cl. thermoaceticum Cl. sticklandii Овцы Крысы Овцы Человек, крупный рога- тый скот (эритроциты) Крысы, обезъяны (плаз- ма крови) Крысы (почки и печень) Крысы
кисеи. Токоферолы служат антиоксидантами по отношению к
ненасыщенным липидам плазматической мембраны, предохра-
няя их от разрушения свободными радикалами, образующимися
под действием ферментов и различных окислительных агентов
и индуцирующими автокаталитическую цепную реакцию окис-
ления ненасыщенных жирных кислот. Токоферолы ингибируют
эти процессы, перехватывая, по-видимому, образующиеся ради-
калы. Селеносодержащая глутатионпероксидаза разрушает как
перекись водорода, так и перекиси липидов, тогда как субстра-
том не содержащей селен глутатионпероксидазы, (синоним:
глутатион-Б-трансфераза В) и каталазы является только пере-
кись водорода. При перекисном окислении липидов возникает
малоновый диальдегид, который выступает в качестве сшиваю-
щего агента, образуя шиффовы основания с аминогруппами
белка. В результате появляются белково-липидные комплексы,
относимые к липофусцинам.
Применение витамина Е позволяет значительно снизить
уровень селена, необходимый для предупреждения экссудатив-
ного диатеза домашних птиц, всего до 10 нг/г МЭ в рационе,
тогда как без добавления витамина Е аналогичный эффект
оказывали только 50 нг/г селена. Беломышечная болезнь ягнят,
получавших витамин Е и 100 нг/г селена, носила лишь умерен-
ный характер, тогда как без добавки этого витамина при дан-
ной концентрации селена в норме наступала гибель животных.
200
Аналогичные наблюдения имеются и в отношении миопатии и
зоба у индюков и недостаточности селена у свиней [Hakka-
rainen J. et al., 1978].
Селен в железосеропротеидах. В опытах in vit-
ro установлено, что селен может замещать кислотно-лабильную
серу в путидаредоксине, адренодоксине и ферредоксине, сохра-
няющих при этом свою биологическую активность. В этой свя-
зи предложена гипотеза [Diplock А. Т., Lucy J. А., 1973], сог-
ласно которой одной из биохимически активных форм селена
может являться селенид, находящийся в активном центре еще
неидентифицированных негемовых железопротеидов. Было
установлено, что кислотно-лабильный селен связан главным
образом с мембранами митохондрий и агранулярной эндоплаз-
матической сети. Селену принадлежит специфическая, завися-
щая от витамина Е, роль в эндоплазматическом ретикулуме,
где этот элемент входит в состав негемового железосеропротеи-
да X. Он функционирует в качестве переносчика электронов
между флавопротеидом и цитохромом Р-450, NADFH-зависимой
цепи переноса электронов микросомной фракции печени крыс
[Levander О., 1976]. В соответствии с этой гипотезой устанав-
ливается еще один аспект взаимодействия между селеном и
витамином Е, а именно защита этим витамином кислотно-
лабильного селена в составе железосеропротеидов от окисления
кислородом. Следует, однако, отметить, что попытки выделения
из микросомной фракциив итамина Е и селензависимого желе-
зосеропротеида, неустойчивого к окислителям, пока не увенча-
лись успехом.
Индукция цитохрома Р-450 в печени повтор-
ным введением крысам фенобарбитала заметно угнетается при
их длительном содержании на дефицитном по селену рационе.
Понижена также индукция фенобарбиталом активности деме-
тилазы этилморфина, в то время как активность цитохрома Ь-,
NADFH-цтихром с редуктазы и бифенилгидроксилазы сохра-
няется без изменения. У селендефицитных крыс при введении
фенобарбитала в 6—8 раз возрастает активность гем — оксиге-
назы (КФ 1.14.99.3) по сравнению с нормальными животными.
Конкретный механизм влияния селена на активность гем —
оксигеназы остается, однако, неясным.
Митохондриальный транспорт электронов.
Одним из наиболее ранних биохимических сдвигов, описанных
при дефиците селена и витамина Е у крыс, была неспособность
гомогенатов и срезов печени поддерживать нормальный уровень
дыхания после длительной инкубации in vitro. Селен оказался
прекрасным катализатором восстановления цитохрома с тиола-
ми, и в мышцах овец был описан селенопротеид, содержащий
гем, близкий по своей структуре к цитохрому с. Все же для
выяснения роли селена в дыхательной цепи митохондрий не-
обходимо проведение дополнительных исследований.
По нашим данным [Авцын А. П. и др., 1986], воздействие
селенита натрия на культуру клеток HeLa вызывало деструк-
201
тивные изменения митохондрий: при низких концентрациях
МЭ — локальное' просветление их матрикса, расширение меж-
кристных промежутков, усиление полиморфизма, а при больших
концентрациях (10 мкмоль/л)—набухание митохондрий, лизис
крист и нередко полную утрату структурированности этих ор-
ганелл. Часть митохондрий переходила в так называемое кон-
денсированное состояние.
Биотрансформация соединений селена. Селе-
нат образуется в организме животных лишь в незначительных
количествах, а при введении через рот или парентерально, он
быстро выводится с мочой, причем его выделение усиливается
сульфатами. Селенит в отличие от сульфита — довольно силь-
ный окислитель и легко восстанавливается органическими
соединениями до серого или красного элементного селена.
Окисление селенита сульфитоксидазой, катализирующей
превращение сульфита в сульфат, в организме животных, по-
видимому, не происходит. Отсутствуют данные и об образова-
нии селенита из селеноаминокислот, тогда как сульфит образу-
ется из цистеина через цистеинсульфин ат и [3-сульфинилпируват.
Восстановление селенита в клетках катализируется ферментом
глутатионредуктазой после неферментативной реакции селенита
с глутатионом [Ganther Н., 1971].
HzSe способен вступать в реакцию с металлами, образуя
нерастворимые комплексы, понижающие биологическую доступ-
ность селена и металла. Это взаимодействие лежит в основе
снижения токсичности металлов повышенными дозами селена.
Образованием биологически недоступных соединений селена с
металлами объясняется способность серебра, кадмия и др.
вызывать у животных вторичную недостаточность селена и
блокировать синтез глутатионпероксидазы даже при рационах,
содержащих адекватное количество селена [Ganther Н., 1980].
Детоксикационное действие селена по отношению к металлам
при их большом избытке может быть связано со способностью
этого МЭ восстанавливать дисульфидные связи в белках в
SH-группы, которые затем «улавливают» тяжелые металлы
[Nuttall К. L„ Allen F., 1984].
Предполагается, что селен вовлекается в целый ряд анабо-
лических процессов в форме H2Se, например при синтезе глута-
тионпероксидазы, при перфузии печени селенитом [Sunde R. А.,
Hoekstra W. G., 1980]. Аналогичным путем происходит, по-ви-
димому, синтез и других функциональных и нефункциональных
селенопротеидов. Существуют две гипотезы о включении
селена в состав глутатионпероксидазы — трансляционная и
посттрансляционная. Согласно первой, синтез селеноцистеина
происходит путем переноса селеноводорода на соответствую-
щий акцептор, например на О-ацетилсерин. Затем образовав-
шийся селеноцистеин с помощью специфической транспортной
РНК может быть включен в полипептидную цепь.
Посттрансляционная гипотеза допускает включение неорга-
нического селена в предобразованную полипептидую цепь в
202
процессе ее посттрансляционной модификации. Имеются экспе-
риментальные данные, свидетельствующие, что при перфузии
печени селен из его неорганических соединений включается в
состав глутатионпероксидазы с большей скоростью, чем из
селеноцистеина [Hoekstra W. G., 1980]. Это предлагает сущест-
вование особой формы ферментапреглутатионпероксидазы,
содержащей вместо селеноцистеина цистеин или серин, который
подвергается превращению в селеноцистеин под влиянием спе-
цифической селеноцистеинилсинтетазы.
Важным аспектом метаболизма селена является его метили-
рование [Ganther Н., 1979]. Так, селен выделяется из организ-
ма животных в виде диметилселенида и триметилселенония.
Имеются также данные, что диметилселеноксид является глав-
ным компонентом соединений селена, выделяемых с мочой у
крыс [Hasegawa A. et al., 1977]. Метилированные соединения
селена могут образовываться не только из селеноводорода, но
и в процессе катаболизма селеноаминокислот. В частности,
метилселенол может образовываться из селенометионина и
селеноцистеина.
Из организма селен элиминируется тремя основными путя-
ми — через почки, кишечник и выдыхаемый воздух. Однако в
большинстве экспериментов и клинических наблюдений уста-
новлено, что в физиологических условиях гомеостаз селена ре-
гулируется в основном экскрецией этого МЭ с мочой [Burk R.
et al., 1972; Downes С. P. et al., 1979; Robberecht H. J., Deelst-
la H. A., 1984] в течение 24 ч после его введения. При этом
уточнены вариации концентраций селена в моче в пределах
0,9—3900 мкг/л. Последняя самая высокая индивидуальная
концентрация иона селена в моче зарегистрирована в Венесуэле
в селеновой биогеохимической провинции, где часто встречают-
ся случаи селенового токсикоза.
Выделение селена с мочой в основном в виде триметилселе-
нида (TM-Se) отмечено при поступлении его в организм в виде
селенита. Элиминация газообразного селена (диметилселенида)
через легкие становится значительной только при высоких до-
зах этого МЭ [Jiang S. et al., 1983].
Единственной точно установленной формой селена в моче
является TM-Se, найденный у крыс в 1969 г. [Byard J. L.,
1969; Palmer I. S. et al., 1969]. Он долгое время рассматривался
как основной метаболит селена у всех видов животных. Однако
прямые обследования человека показали, что если использовать
в качестве источника селена мясную пищу, а не его неоргани-
ческие соединения, как это практиковалось в опытах на лабо-
раторных животных, то из общего количества селена, выде-
ляемого с мочой, на долю TM-Se приходится всего около 12%
[Ganther Н., 1983], тогда как у крыс, получавших физиологи-
ческие дозы селенита, оно достигало 40% [Palmer I. S. et al.,
1969]. Имеются более поздние данные [Nahapetian А. Т. et al.,
1983], что при низких дозах селенита (16 мкг/кг живой массы)
в моче крыс также присутствует лишь незначительное количе-
203
ство TM-Se, которое, однако, существенно возрастает при повы-
шении дозы этого соединения до 1500 мкг/кг. В моче обнару-
жены и другие соединения селена, в частности метаболит
U-2, природа которого не установлена [Burk R., 1976; Pal-
mer I. S. et al., 1977]. Нет сомнения, что будут найдены и дру-
гие метаболиты селена, аналогичные конечным продуктам
обмена серы, причем наиболее вероятно нахождение соедине-
ний селена со степенью окисления (4-4).
Элементный селен труднорастворим и в пищеварительном
тракте не усваивается. При парентеральном введении, однако,
его частички могут подвергаться фагоцитозу и под воздействи-
ем пероксидгенерирующей системы фагоцитов этот МЭ стано-
вится растворимым, доступным для организма. Это может
иметь определенное значение при полном парентеральном пи-
тании, если селенит, добавляемый в питательный раствор,
восстанавливается до элементного селена. Точно так же селен,
добавляемый в виде SeSs в некоторые виды шампуней против
перхоти, может частично всасываться через кожу и подвергать-
ся аналогичному превращению.
Имеются немногочисленные данные о распределении селена
у человека. Так, после инъекции 758е-метионина (или селенита)
содержание этого МЭ в печени и поджелудочной железе уве-
личивалось в 1,7 раза по сравнению с исходным уровнем в
течение 2 дней и затем постепенно уменьшалось [Lathrop К. А.
et al., 1972]. При этом М. Wenzel и соавт. (1971) определили
биологические полупериоды существования селена в тканях.
В частности, для мышц этот срок составил 100 сут, для пече-
ни — 50 сут, почек — 32 сут и для сыворотки крови — 28 сут.
После введения радиоактивного селена значительная его
часть связывается с белками плазмы. При этом оказалось, что
эритроцитам в данном процессе принадлежит важнейшая роль,
так как 75Se в виде селенита чрезвычайно быстро (в пределах
нескольких секунд) проникает через их мембраны [McMur-
ray С. Н., Davidson W., 1974]. Уже через 1—2 мин в эритроци-
тах концентрируются 50—70% всего селена крови. Затем в
течение 15—20 мин почти весь селен выходит из эритроцитов,
связываясь сначала с альбуминами, а затем и с глобулинами
плазмы крови. Важно подчеркнуть, что в отсутствие эритроци-
тов белки плазмы крови человека мало связывают этот МЭ
и 97—99,5% 75Se диализуются [Петрович Ю. А., Подорож-
ная Р. П„ 1981; Ворр В. A. et al., 1982].
Эритроцитарный «насос» для селенита обнаружен у челове-
ка и ряда животных [Wright Р. L., Bell М. С., 1963; Jenkins К. J-,
Hidiroglou М., 1972; Sandholm М., 1975]. В эритроцитах под
влиянием системы глутатион — глутатионпероксидаза селенит
подвергается превращению с образованием комплекса селена
с глутатионом. При последующем восстановлении селен катали-
зирует транспорт электронов к кислороду. Выйдя из эритроци-
та, возможно, в составе селеноглутатионового комплекса, этот
МЭ фиксируется в белках плазмы.
204
Липопротеиды плазмы крови человека содержат примерно
€% селена от общего его количества в плазме {Moriss S. G.
et al., 1972]. Селен является естественной составной частью
ЛИП '[Burk R. F., 1976]. Кроме того, установлено, что у мла-
денцев в первые месяцы жизни уровень селена довольно низок
и с возрастом постепенно увеличивается [Verlinder М. et al.,
1983]. По данным О. Levander (1975), средняя концентрация
селена в цельной крови составила 2,6; 2,3 и 4,05 мкмоль/л
соответственно для американцев, канадцев и англичан. Наряду
с этим у детей, проживающих в селеновых биогеохимических
провинциях Венесуэлы, его концентрации в крови повышались
до 10,3 мкмоль/л. Эти данные нашли подтверждение в более
поздних исследованиях К. Howe (1979) в различных регионах
США, а также М. Verlinder и соавт. (1983) в различных воз-
растных группах населения Бельгии.
С. Е. Casey и соавт. (1982) представили концентрации селе-
на в различных тканях и органах в зависимости от возраста.
Так, у эмбрионов уровень селена в печени равен 1,26 мг/г
•сухой массы, у детей в возрасте 4—54 мес в печени и почках
содержится 0,58 и 2,07 соответственно. У лиц в возрасте от 15
до 74 лет концентрация селена составляет в печени, почках,
мышцах, сердце и легких 0,37—0,72; 0,64—6,17; 0,11—0,43 и
0,57—0,8 мкг/г массы соответственно.
Наиболее известным проявлением недостаточности селена
у животных является алиментарная мышечная дистрофия (бе-
ломышечная болезнь), причиняющая значительный экономи-
ческий ущерб животноводству в различных странах. Она была
описана в ряде селендефицитных регионов Новой Зеландии,
Скандинавии, США, Турции, Эстонской ССР, Бурятской АССР,
поражает крупный рогатый скот, лошадей, свиней, ягнят и до-
машних птиц. Заболевают в основном новорожденные живот-
ные и молодняк до 12-месячного возраста, болезнь может иметь
длительное течение, часто заканчивается летально, характери-
зуется очаговой дегенерацией различной тяжести и некрозом
скелетной и сердечной мышц невоспалительного характера (рис.
39); она предупреждается и излечивается включением в рацион
селена.
Патоморфологические изменения при этом хроническом за-
болевании характеризуются глубокими нарушениями скелетных
мышц и миокарда. В частности, наблюдается пестрая патоги-
стологическая картина за счет неравномерного полнокровия,
дистрофических и некробиотических изменений кардиомиоцитов,
нередко с явлениями дистрофического обызвествления (рис.
40). По мнению А. П. Авцына (1972), белая окраска мышц
обусловлена исчезновением из них миоглобина и вторичным
коагуляционным некрозом миоцитов. В печени и почках посто-
янно отмечаются дистрофические процессы. Изменения в легких,
кишечнике и других органах неспецифичны и непостоянны.
Изменения миокарда и скелетных мышц имеют дегенеративно-
некробиотический характер. Часто встречается гиалиновая деге-
205
Рис. 39. Очаговые некрозы миокарда правого желудочка при селенодефицит-
ной беломышечной болезни у теленка. Окраска гематоксилином и эозином.
Х400.
нерация мышечных волокон. Ткань печени и почек в большин-
стве случаев находится в состоянии зернистой, реже жировой,,
дистрофии {Ермаков В. В., Ковальский В. В., 1974]. Другими
исследованиями было показано, что недостаток селена играет
специфическую роль также в развитии токсической дистрофии
печени животных и птиц [Кудрявцев А. А. и др., 1966], экссу-
дативного диатеза птиц [Георгиевский В. И., 1972] и ряда дру-
гих заболеваний. Содержание мышей и крыс на полусинтети-
ческом аминокислотном рационе с низким содержанием селена
сопровождалось резким понижением уровня других МЭ, напри-
мер цинка в клетках панкреатических островков, в паренхиме
почек и в клетках сперматогенного эпителия [Сучков Б. П.,
1981].
Биохимические исследования при беломышечной болезни
продемонстрировали резкое снижение количества миозина в
мышцах и фракции водорастворимых белков одновременно с
увеличением количества коллагена, что согласуется с гистоло-
гическими наблюдениями, показавшими замещение мышечных
волокон соединительной тканью. Одновременно отмечено сни-
жение уровня креатинфосфорной кислоты, АТФ, гликогена с
параллельным возрастанием неорганических фосфатов, увели-
чением активности аминофераз, в некоторых случаях АТФазы
и пирофосфатазы мышц [Ермаков В. В., Ковальский В. В.,
1974].
При беломышечной болезни ягнят резко активируются кол-
206
Рис. 40. Некроз и обызвествление мышечных волокон левого желудочка серд-
ца в зоне расположения проводящей системы при селенодефицитной беломы-
шечной болезни у теленка. Окраска гематоксилином и эозином. Х250.
лагеназа мышц [Broderius М. A. et al., 1973], липопротеин-
липаза миокарда [Kelley Т. F., 1968] и ряд других ферментов.
R. Poukka (1968) отметил, что в сердечной мышце телят с этим
заболеванием изменяется состав фосфолипидов (например, от-
мечено увеличение фосфатидилхолина). По мнению Р. D. Whan-
ger и соавт. (1973), именно исчезновение одного из селенопро-
теидов— белка с молекулярной массой 10 000 в скелетных
мышцах овец при недостаточности селена в известной мере
обусловливает возникновение этого заболевания скота.
В одной из работ [Reiter Р., Wendel А., 1983] показано, что
содержание мышей-альбиносов на диете с очень низким содер-
жанием селена (менее 10 частей на биллион) сопровождалось
существенными изменениями ксенобиотического обмена печени.
Так, содержание селена в этом органе было менее 10% от
контроля, глутатионпероксидазы — менее 2%. В микросомаль-
ной фракции активность NADPH-цитохром Р-450 — редуктазы
уменьшилось на 50 % • Темп N-окисления флавинсодержащих
монооксигеназ снизился на 35%, скорость N-деметилирования —
на 50%. Хотя следует отметить, что активность ряда других
ферментов при дефиците селена не только не изменилась, но
даже повысилась. Эти данные еще раз подчеркивают, что про-
должительное селендефицитное состояние сопровождается не
только уменьшением активности глутатионпероксидазы, но и
влечет за собой целый спектр ферментных нарушений. По-види-
мому, эта болезнь сопровождается изменением обмена многих
веществ. В частности, нарушаются метаболизм аминокислот и
207
Рис. 41. Ребенок 3 лет, стра-
дающий селенодефицитной
кардиомиопатией (болезнь
Кешана). Асцит, резкое уве-
личение печени, выбухание
прекордиальной области
[Guangqi Yang et al., 1984].
Рис. 42. Резкое расширение полостей сердца
и гипертрофия миокарда. Застойное полно-
кровие легких. Рентгенограмма грудной
клетки девочки 12 лет, страдающей бо-
лезнью Кешана с 5-летнего возраста [Guan-
gqi Yang et al., 1984].
кетокислот, фосфорный и углеводно-фосфорный обмен в мыш-
цах, снижается гликолиз.
Биологическая роль селена может быть также связана с его
влиянием на метаболизм серы. При беломышечной болезни в
мышечной ткани ягнят отмечены значительное снижение общего
количества SH-групп и их содержания в белках и существенное
увеличение числа небелковых SH-групп и концентрации восста-
новительного глутатиона [Broderius М. A. et al., 1973].
Повышенное количество небелковых SH-групп и снижение их
числа в белках печени отмечены также у крыс при дефиците
селена. Из этих опытов был сделан вывод, что при дефиците
этого МЭ нарушен процесс образования дисульфидных связей
[Underwood Е., 1977]. Участие селена в процессах обмена серы
подтверждается и более поздними наблюдениями.
Известно, что цистеин предупреждает алиментарную мы-
шечную дистрофию (АМД) при Е-авитаминозе цыплят. Ана-
логичным, но менее выраженным свойством обладает метионин.
Факторы, замедляющие превращение метионина в цистеин
(креатин, холин), снижают способность метионина предупреж-
дать АМД, тогда как соединения, ускоряющие его конверсию
(гуанидинуксусная кислота, NAD), напротив, повышают его
эффективность. Соединения, замедляющие катаболизм цистеи-
на (например, меркаптоэтаноламин), облегчают течение АМД,
тогда как соединения, способствующие усилению его катаболиз-
ма, усиливают тяжесть этого заболевания. В опытах на цыпля-
тах было показано, что способностью повышать эффективность
208
Рис. 43. Типичная картина поражений миокарда при подострой форме болезн»
Кешана. Вверху слева — сохранившиеся мышечные волокна, справа—резка
выраженный лизис кардиомиоцитов [Guangqi Yang et al, 1984].
конверсии метионина в цистеин обладает селен. В плазме крови
цыплят при дефиците селена практически отсутствовали в
свободном виде цистеин, цистатионин и гомоцистеин, которые
появлялись вновь только после включения в пищу селена [Hal-
pin К. М. et al., 1986]. Из этих опытов авторы справедливо
заключают, что селен, по-видимому, играет определенную роль,
в процессах транссульфурирования.
В последние годы дефицит селена рассматривают как воз-
можный этиологический фактор при некоторых сердечно-сосу-
дистых заболеваниях. Еще в 1935 г. у людей была впервые
выявлена болезнь Кешана. Однако систематическое изучение
этого заболевания было проведено в 60-х годах только в Китае,
где оно носит эндемический характер и предотвращается введе-
нием селенита натрия [Chen X., 1980; Diplock А., 1981]. Болезнь
Кешана представляет собой эндемическую фатальную миокар-
диопатию, для которой характерны аритмии, увеличение раз-
меров сердца, фокальные некрозы миокарда, за которыми сле-
дует сердечная недостаточность (рис. 41, 42, 43). Электронно-
микроскопические исследования дополнили картину поражения
миокардиоцитов [Ge К- et al., 1983]. Так, наряду с увеличением
числа митохондрий выявлены их набухание с резким просвет-
лением матрикса и разрушением крист, а также появление так
называемых конденсированных форм этих органелл. Авторы
показали дезорганизацию саркомеров, разрывы сарколеммы,
изменение морфологии вставочных дисков, увеличение числа
14—568
209
лизосом и появление «специфических гранул», которые, воз-
можно, имеют вирусную природу.
Наиболее типично это заболевание для детей обоих полов
и беременных женщин. В отдельных поселках болезнь поража-
ла до 10—12% детей и женщин репродуктивного возраста
[Chen X. et al., 1980]. Как установлено, это заболевание имеет
биогеохимическую природу и встречается только в определен-
ных ареалах 40 провинций, простирающихся от северо-востока
до юго-запада Китая, который можно определить как селено-
дефицитный пояс. При этом были выявлены многочисленные
«спасательные острова», окруженные территорией, где была
отмечена высокая частота этой болезни. У больных также об-
наруживают дегенеративные изменения мышц нижних конеч-
ностей, по гистологической картине напоминающие беломышеч-
ную болезнь. По мнению A. Diplock (1984), молекулярный
механизм этих изменений обусловлен поражением клеточных
мембран свободными радикалами при отсутствии селена. Дей-
ствительно, при болезни Кешана в крови выявляется необычно
низкий уровень (0,063—0,127 мкмоль/л) этого МЭ, что примерно
в 10 раз меньше, чем у адекватно питающегося населения
(1,14—1,9 мкмоль/л) [Oster О., Prellwitz W., 1982; Tan Yian
Au, 1982; Diplock A., 1984; Levander O., 1984]. Дополнительные
исследования подтвердили, что в очагах распространения бо-
лезни Кешана низкое содержание селена в почве коррелирует
с низким уровнем его в волосах людей, проживающих в данных
регионах. Так, среднее содержание селена в волосах людей
эндемичных зон равно 0,085+0,032, а в неэндемичных—0,187+
+0,086 мкг/г.
Крупные биогеохимические регионы селенодефицита на тер-
ритории СССР установлены в Забайкалье [Вощенко А. В. и др.,
1988]. Отдельные очаги природного селенодефицита выявлены
в Ярославской области, Удмуртской АССР. Потенциально такой
же биогеохимической провинцией, по-видимому, является юго-
запад Карельской АССР. В Читинской области в 1986—1987 гг.
установлено наличие болезни Кешана — эндемической селеноде-
фицитной кардиопатии, имеющей определенные особенности
клинического течения. В Забайкалье она имела стертые формы
с латентным и хроническим течением. Острые и подострые фор-
мы болезни Кешана в этом регионе еще не выявлены. При
латентной форме доминировал астеновегетативный синдром.
На ЭКГ чаще всего выявлялись блокада правой ветви пред-
сердно-желудочкового пучка (Гиса), экстрасистолия, замедле-
ние атриовентрикулярной проводимости, а также метаболичес-
кие и диффузно-дистрофические изменения в миокарде и гипер-
трофия миокарда. Проба с физической нагрузкой выявила
скрытую сердечную недостаточность.
Хроническая форма заболевания характеризовалась в основ-
ном болевым синдромом и явлениями хронической сердечной
недостаточности. На ЭКГ наиболее часто регистрировались
изменения, свидетельствующие о метаболических и диффузно-
210
дистрофических нарушениях. Рентгенографически увеличение
размеров сердца отмечено у 7 из 12 больных хронической фор-
мой и у 32 из 56 больных латентной формой болезни Кешана.
У 72,5% больных селенодефицитной кардиопатией выявлено
снижение кислотности желудочного сока, что может способст-
вовать снижению абсорбции селена в желудочно-кишечном
тракте. Содержание селена в крови больных составило 0,53—
0,82 мкмоль/л (латентная форма) и 0,18—0,42 мкмоль/л (хро-
ническая форма), т. е. в несколько раз ниже нормы (1,77—
2,79 мкмоль/л). Лечение больных с этими формами болезни
Кешана селенитом натрия оказалось достаточно эффективным
[Обухова Т. И., 1988].
В литературе обобщены данные эпидемиологических наб-
людений об обратных взаимоотношениях между содержанием-
селена в крови и летальностью при других болезнях сердца,,
проведенные в 19 штатах США [Shamberger R. et al., 1979].
Эти результаты подтверждаются работами J. Т. Salonen и со-
авт. (1982), О. Oster и соавт. (1983). Так,, J. Т. Salonen и соавт.
(1982) продемонстрировали, что при концентрации селена в сы-
воротке ниже 0,4 мкмоль/л вероятность возникновения инфарк-
та миокарда увеличивается в 7 раз, а при содержании 0,4—
0,6 мкмоль/л — в 3 раза.
Эти наблюдения обусловили интерес к определению уровня
селена как у больных, так и в группах повышенного риска сер-
дечно-сосудистых заболеваний. Так, снижение уровня селена
(почти в 2 раза) в сыворотке людей при застойной миокардио-
патии отметили О. Oster и соавт. (1983). По мере прогрессиро-
вания миокардиодистрофии у 22 больных концентрация селена
в сыворотке также снижалась [Orndahl G. et al., 1982].
Однако следует заметить, что существуют работы, в которых
не отмечено корреляции между концентрацией селена в крови
у человека и факторами риска серечно-сосудистых заболеваний
[Miettinen Т. et al., 1983]. В этом случае зафиксирована поло-
жительная корреляция между полиненасыщенными жирными
кислотами, холестерином, фосфолипидами и концентрацией это-
го МЭ.
Низкий уровень селена в волосах и в крови и активности
глутатионпероксидазы у детей встречается не только при болез-
ни Кешана, но и при полном парентеральном питании, кваши-
оркоре, при содержании детей с фенолкетонурией и «болезнью-
запаха кленового сиропа» на полусинтетической диете. Во всех
этих случаях дача селена ведет к нормализации биохимических
параметров, а иногда сопровождается и положительным тера-
певтическим эффектом [Loembeck I. et al., 1984].
Изучение терапевтического влияния селена на течение эк-
спериментального инфаркта миокарда у крыс позволило четко
установить его благоприятное действие [Кактурский Л. В.,
1986]. Заживление инфаркта миокарда продемонстрировано в
эксперименте на крысах, у которых инфаркт воспроизводили
лигированием венечной артерии (120 животных). В течение
14*
211
'Рис. 44. Морфологические проявления влияния селена на экспериментальный
инфаркт миокарда у крыс.
а — миокард внеинфарктной зоны через 2 ч после лигирования венечной артерии у не-
леченой крысы (контроль). Реакция на сукцинатдегидрогеназу. Наряду с линейным рас-
положением диформазана отмечаются мелко- и крупнозернистый его осадок, а также
диффузное прокрашивание саркоплазмы в некоторых участках. Разобщение мышечных
'волокон вследствие интерстициального отека. Х400; б — миокард внеинфарктной зоны
через 2 ч после лигирования венечной артерии у крысы, леченной селенитом натрия в
сочетании с а-токоферолом. Реакция на сукцинатдегидрогеназу. Линейное расположение
продукта реакции по ходу миофибрилл Х400; в — ультраструктура миокарда внеинфаркт-
ной зоны нелеченой крысы (контроль) через 2 ч после лигирования венечной артерии.
Выражено набухание митохондрий с деструкцией крист и просветлением матрикса.
Х15 000; г — ультраструктура миокарда внеинфарктной зоны у крысы, леченной селени-
том н-атрия в сочетании с а-токоферолом, через 2 ч после лигирования венечной артерии.
Сохранность ультраструктурной организации кардиомиоцита. X15 000: Д— гистоструктура
постинфарктной зоны на 30-е сутки после лигирования венечной артерии у нелеченой
•крысы (контроль). Преобладание рыхлой волокнистой соединительной ткани с умерен-
ной клеточной инфильтрацией. Окраска гематоксилином и эозином. Х70; е — гистострук-
тура постинфарктной зоны на 30-е сутки после лигирования венечной артерии у крысы,
леченной селенитом натрия и а-токоферолом. Компактное строение постинфарктного
участка, преобладание плотной соединительной ткани, большое число кардиомиоцитов.
Окраска гематоксилином и эозином. Х70 (Препараты Л. В. Кактурского).
недели после начала инфаркта крысам подкожно вводили вод-
ный раствор селенита натрия из расчета 30 мкг/кг в сутки. Для
потенцирования эффекта селена одновременно внутримышечно
вводили а-токоферол в дозе 50 мг/кг в сутки. Крыс забивали
через 2 ч, 1, 3, 7, 14 и 30 сут.
У леченых крыс отмечено уменьшение зоны инфаркта мио-
карда, особенно выраженное в ишемической стадии (4,52%
против 31,7% в контроле, р<0,001). Величина зоны инфаркта
коррелировала с уровнем перекисного окисления липидов в
этой же зоне, определявшимися хемилюминесцентным методом.
Гистостереометрически показано ускорение созревания гра-
нуляционной ткани у леченых животных. Это выражалось уве-
личением числа клеток фибробластического ряда (7,84% про-
тив 2,08% в контроле на 3-и сутки и 17,64% против 11,96% в
контроле на 7-е сутки, р<0,001), уменьшением доли сегменто-
ядерных лейкоцитов (0,28% против 1,92% в контроле на 3-и
сутки и 0,20% против 0,96% в контроле на 7-е сутки, р<0,001),
увеличением доли сосудистого компонента на 3-и сутки (24,8%
против 12,92%, р<0,001) и уменьшением этой доли на 7-е
сутки (5,76% против 9,88%, р<0,001).
Во внеинфарктной зоне миокарда в отличие от контроля
под влиянием селена отмечены нормальная активность окси-
доредуктаз (рис. 44 а, б), а также сохранность ультраструктур-
ной организации кардиомиоцитов (рис. 44 в, г), т. е. отмечен
протективный эффект селена на кардиомиоциты на гисто- и
ультраструктурном уровне.
Сформированная на 30-е сутки постинфарктная зона у неле-
ченых животных имела рыхлое строение (рис. 44, д), а у крыс,
получавших селен,— компактное строение со значительным
содержанием мышечных элементов (рис. 44, е).
Фиброз экзокринной части поджелудочной
железы, вызванный дефицитом с е л е н а, был впер-
вые описан в 1970 г. [Thompson J. N., Scott М. L., 1970] у цып-
лят, получавших синтетический корм на основе кристалличес-
ких аминокислот, содержавший менее 0,015 мг/кг селена и
повышенные дозы витамина Е. Алиментарная дистро-
фия поджелудочной железы (АДП) характеризуется
выраженной стеатореей (главным образом за счет повышенного
выделения с экскрементами нейтральных липидов), обусловлен-
ной снижением активности панкреатической липазы и пониже-
нием уровня токоферола в плазме крови, который возвращается
к норме при включении в пищу свободной линолевой кислоты,
моноолеилглицерина и таурохолата натрия.
При электронно-микроскопических исследованиях ацинар-
ных клеток поджелудочной железы цыплят с недостаточностью
селена отмечены разрывы внешней и внутренней мембран
митохондрий, а также снижение на 27% количества митохонд-
риальных белков. Эти данные позволяют считать, что причиной
АДП является уменьшение числа функционально активных
митохондрий в ацинарных клетках [Whiteacre М. Е.,
213
Рис. 45. Кистозный фиброз поджелудочной железы при врожденном гипоселе-
нозе (?). Резко выраженное расширение протоков с очаговой гибелью эпи-
телия. Сверху справа — плотный белковый конкремент в просвете протока.
Фиброз стромы. Окраска гематоксилином и эозином. Х120.
Combs G. F. 1984]. Однако неясно, связано ли повреждение
митохондрий с дефицитом глутатионпероксидазы или другого
селеносодержащего фактора. Против исключительной роли глу-
татионпероксидазы в этом процессе свидетельствуют, в частно-
сти, отсутствие корреляции между активностью данного фер-
мента в плазме крови и в ткани поджелудочной железы и тече-
нием болезни, а также больший лечебный эффект при АДП
селена, находящегося в составе селенометионина и пшеницы,
чем селенита и селеноцистеина, наиболее эффективных при
беломышечной болезни [Underwood Е., 1977].
Кистозный фиброз поджелудочной железы
(муковисцидоз) является довольно распространенным
заболеванием детей младшего возраста и крайне редко регист-
рируется у подростков и молодых людей. Патологическая ана-
томия этого заболевания хорошо изучена. Ее основные черты:
1) аномалии развития выводных протоков поджелудочной же-
лезы, подвергавшихся резкому расширению с образованием
кист, обычно заполненных густым и вязким секретом; 2) актив-
ная пролиферация фибробластов перидуктальной соединитель-
ной ткани (рис. 45, 46). В течение длительного времени кистоз-
ный фиброз поджелудочной железы относили к аутосомно-
рецессивным заболеваниям. В настоящее время эта точка зре-
ния подвергается сомнению, так как получены достоверные
данные о врожденном генезе этого заболевания.
214
Рис. 46. Кистозный фиброз поджелудочной железы при врожденном гипосе-
ленозе (?). Протоки заполнены густым секретом, имеющим слоистое строение.
Умеренная гипертрофия панкреатических островков. Окраска гематоксилином
и эозином. X120.
S. Wallach и соавт. (1979) провели обследование ряда се-
мей в 33 штатах США и 4 провинциях Канады, в которых ро-
дились дети с кистозным фиброзом поджелудочной железы.
При этом клинические и экспериментальные исследования по-
казали, что этиология указанного заболевания на самом деле
обусловлена дефицитом ряда элементов, особенно селена, в
перинатальном периоде. Кроме того, установлено, что диетоте-
рапия с добавлением МЭ значительно улучшала состояние
больных.
В 1976 г. было установлено [Prohaska J. R., Ganther Н. Е.,
1976], что наряду с селеносодержащей глутатионпероксидазой
существует еще один фермент с аналогичным действием —
глутатион-Э-трансфераза В (КФ 2.5.1.18), не содержащий селе-
на, причем органы и ткани с высокой активностью одного фер-
мента, как правило, имеют пониженную активность другого.
Так, у человека наиболее богаты селеносодержащим фермен-
том сердце и мозг, тогда как печень, скелетные мышцы и почки
содержат преимущественно глутатион-5-трансферазу. В осталь-
ных органах и тканях присутствуют различные количества
обоих ферментов [Carmagnol F. et al., 1983].
Алиментарный гепатоз (Hepatosis dietetica) полу-
чен в экспериментах на крысах, а в природно-хозяйственных
условиях зарегистрирован в Новой Зеландии и скандинавских
странах при скармливании свиньям зерна местного производст-
215
ва, бедного селеном. Заболевание встречается наиболее часто
у поросят в возрасте 3—15 нед и характеризуется высокой
смертностью. При вскрытии отмечены тяжелые некротические
изменения печени, обширный отек и отложение цероидного
пигмента в жировой ткани, придающего ей желто-коричневую
окраску. Селен полностью предохраняет от этого заболевания,
но витамин Е более эффективно предупреждает сопутствующие
мышечную дистрофию и отложения пигмента [Underwood Е.,
1977].
Экспериментальные исследования С. В. Hong и С. К- Chow
(1988) привлекли внимание к еще одной форме селенодефици-
та, протекающего на фоне недостаточности витамина -Е —
эозинофильному энтериту (ЭЭ) человека, домашних и лабора-
торных животных. Его впервые описали A. L. Ureles и соавт.
в 1961 г. под названием «идиопатическая эозинофильная ин-
фильтрация». Эти же авторы подвергли экспериментальному
анализу данную патологию на крысах породы Sprague-
Dawley, что позволило выяснить новые патогенетические фак-
торы. ЭЭ с эозинофилией периферической крови возникал в
условиях пищевого дефицита селена и витамина Е и законо-
мерно излечивался с помощью добавки этих веществ в корм.
Очаговая или диффузная инфильтрация наблюдалась в кишеч-
нике, желудке, брыжейке и регионарных лимфатических узлах,
но она достигала наибольшей выраженности вокруг очагов
некроза гладких мышц в стенке кишки. Первоначальные пред-
положения о связи этой своеобразной инфильтрации с пищевой
аллергией и глистной инвазией не подтвердились. Более веро-
ятна ее зависимость от очагов некроза. В 1975 г. были открыты
мощные хемотаксические для эозинофилов вещества в виде
продуктов окисления арахидоновой кислоты, в метаболизме
которой играют роль селен и витамин Е [Reddy С. С., Massa-
ro Е. J., 1983]1. Таким образом, ЭЭ и эозинофилия, вероятно,
возникают в результате избыточного образования продуктов
окисления. Селено дефицит может вызвать у крыс также некроз
мышц, тубулярный нефроз, очаговые некрозы печени и гемолиз
эритроцитов, которые излечиваются препаратами селена и
витамина Е. Последний проявляет антиокислительное действие
с помощью механизмов, инактивирующих свободные радикалы,
в то время как селен, входящий в состав глутатионпероксидазы,
катализирует восстановление токсичных перекисей водорода.
У жвачных животных описан еще ряд заболеваний,
излечиваемых селеном. К их числу относятся задержка роста
и развития овец и крупного рогатого скота в Новой Зеландии,
западных штатах США и в Шотландии, паралитическая
миоглобинурия крупного рогатого скота, задержка отделения
плаценты у коров, периодонтоз у овец, причем во многих случаях
получен лечебный эффект и от применения витамина Е. Селен
обладает также способностью в дозах, превышающих физиоло-
гическую потребность (0,1 мг/кг), стимулировать иммуногенез.
Так, при даче селена в дозах 0,7—2,8 мг на 1 кг корма у мышей
216
повысился титр антител против овечьих эритроцитов в 7 и
30 раз соответственно. Сходное действие оказало и внутрибрю-
шинное введение 3—5 мкг селенита натрия, причем наиболее
высокий эффект был получен при введении его одновременно
с антигеном или до него ([Spa'llholz J. Е. et al., 1975].
Воспроизводительная функция. У всех изучен-
ных видов животных дефицит селена вызывает нарушение вос-
производительной функции. Крысы при дефиците селена не
утрачивают способность приносить потомство, но уже первое
поколение лишено этой способности. У одной части самцов
сперматозоиды теряют подвижность, хвост отрывается от голов-
ки. У другой части животных сперматозоиды полностью отсут-
ствуют. Самки приносят всего 1—2 нежизнеспособных детены-
ша. Разрыв сперматозоида происходит обычно посередине хво-
ста, где обнаружен селенопептид с молекулярной массой 17 000,
который, по-видимому, играет важную роль в формировании
хвоста [Calvin Н. J., 1978]. У кур и индюков дефицит селена
приводит к снижению яйценоскости и выводимости цыплят.
В Новой Зеландии установлена тесная связь между распро-
странением беломышечной болезни и яловостью овец, которая
предупреждается дачей 4 мг селенита натрия на животное с
месячными интервалами. При дефиците селена течка, овуляция,
оплодотворение и раннее развитие плода у овец протекают
нормально, но на 3—4-й неделе суягности (примерно ко време-
ни имплантации) эмбрионы погибают по неизвестной причине.
Во всех случаях нарушений воспроизводительной функции вве-
дение животным одного витамина Е не дает положительного
эффекта. Улучшение воспроизводительной функции у овец и
крупного рогатого скота при совместной даче селена и витамина
Е отмечено в ряде селенодефицитных биогеохимических про-
винций Шотландии и Калифорнии ;[Underwood Е., 1977].
К числу селенодефицитных заболеваний относятся также экс-
судативный диатез цыплят и индюков, характеризующийся
отеком тканей, особенно на груди и животе, и кровоизлияниями
в наружные покровы, а также приводящая к внезапной смерти
«болезнь тутового сердца» у свиней, получившая свое название
от обширных кровоизлияний в миокарде, придающих сердцу
красновато-пурпурную окраску, напоминающую ягоду шелко-
вицы.
Следует отметить, что токсический эффект селена к настоя-
щему времени хорошо изучен на животных в ареалах с избыт-
ком селена, в то время как в литературе имеются лишь немно-
гочисленные данные об отравлении селеном человека. Так,
можно упомянуть сообщения М. I. Smith и соавт. (1936),
М. I. Smith, В. В. Westfall (1937); R. Е. Lemley (1943) о селе-
новом токсикозе у жителей штатов Южная Дакота и Небраска.
W. G. Jaffe и соавт. (1972) выявили ряд клинических отличий
у детей контрольной группы и у проживающих в местности с
высоким содержанием селена в Венесуэле. I. Rosenfeld и
О. Beath (1964) описали отравление селеном людей, употреб-
217
лявших питьевую воду с концентрацией 0,11 ммоль/л этого
МЭ. Однако наиболее существенные данные о токсичности
селена для человека поступили из Китая [Jang G. et al., 1983].
Они были собраны в период с 1960 по 1964 г.
Так, средняя концентрация селена в моче (34 мкмоль)л)
при хроническом селенозе у людей, проживающих в провинци-
ях с высоким уровнем селена в почве, примерно в 20 раз боль-
ше, чем у людей без селеноза, но также проживающих в бога-
тых селеном ареалах, и в 100 раз больше, чем у людей в
местности с оптимальным содержанием этого МЭ, и, наконец,
в 400 раз больше, чем у людей с болезнью Кешана.
Наиболее типичными симптомами селенового токсикоза яв-
ляются поражения ногтей и волос. Кроме того, наблюдаются
желтушность, шелушение эпидермиса, повреждения эмали зу-
бов, артриты, анемия, нервные расстройства. В биогеохимичес-
ких селеновых провинциях у людей встречаются хронические
дерматиты, постоянная усталость и потеря аппетита [Суч-
ков Б. П., 1981; Rosenfeld I., Beath О., 1964; Schroeder Н. А.
et al., 1970; Harr J., Muth О., 1972], депрессия, гастроэнтериты,
дегенерация печени и увеличение размеров селезенки [Buell D.,
1983]. Избыток селена в среде неблагоприятно влияет на про-
цессы оссификации и на состояние зубов. При высоком содер-
жании этого МЭ в питьевой воде у людей нарушается форми-
рование эмали, снижается поступление кальция без изменения
усвоения фтора [Hajimarkos D. М., 1973].
Наконец, существуют немногочисленные данные о тератоген-
ном действии селена. Этот МЭ в органической или неоргани-
ческой форме относительно легко проникает через плаценту
в плод [Underwood Е., 1977]. Селен в концентрации 3 мкг/г
вызывает уменьшение числа особей в помете третьего поколе-
ния у мышей и крыс [Schroeder Н. A., Mitchener М., 1971].
При экспериментальном повышении уровня этого МЭ в орга-
низме матери отмечаются дефекты развития эмбрионов у кур,
крыс, свиней, овец и кошек [Rosenfeld I., Beath О., 1964; Pu-
zanova L. et al., 1979]. В селеновых ареалах пороки развития
отмечены также у цыплят и ягнят.
Таким образом, анализ данных литературы позволяет счи-
тать твердо установленным существование в организме сба-
лансированного механизма, поддерживающего гомеостаз селена.
Этот МЭ интенсивно включается в обменные процессы клетки.
По-видимому, частично его метаболизм сходен с обменом серы,
но по ряду признаков между ними есть и коренные отличия.
Для селена характерны активное участие в окислительно-вос-
становительных и антиоксидантных процессах, дыхании клетки,
а также в синтезе специфических функциональных белков,
содержащих селениды.
Селен как антиканцерогенный фактор. Исто-
рия изучения связи селена с онкогенезом удивительна и полна
неожиданностей. Так, пионером терапевтического использования
селена при злокачественных новообразованиях является A. Was-
218
serman [Ермаков В. В., Ковальский В. В., 1974]. Еще в 1911г.
ему удалось с помощью внутривенного введения селен-эозина
полностью излечить спонтанные и перевиваемые опухоли мы-
шей. Этот эксперимент послужил толчком к целой серии иссле-
дований по противоопухолевому действию селена, продолжав-
шихся до середины 30-х годов XX века.
В начале 40-х годов в ходе экспериментов с высокими до-
зами селена были получены данные, что этот МЭ обладает
бластомогенными свойствами. Все началось с опытов R. D. Lil-
lie и М. Т. Smith (1940), которые показали, что при скармлива-
нии крысам зерна с 5 мг/кг иона селена развивается узелко-
вый цирроз печени. В 1943 г. A. Nilson и соавт. содержали крыс
на диете с очень низкой концентрацией белка, содержащей 5,7
и 10 мг/кг селена в виде селенида или органического селена.
Они обнаружили, что возникновение цирроза печени сопровож-
дается у крыс, проживших более 18 мес, последующим разви-
тием аденом и «маловыраженного рака» (аденом и карцином
печени, аденоматозной гиперплазии). А. Л. Черкес и соавт.
(1962) представили данные, что 4,3 мг/кг селена в виде селени-
та натрия в пище с низким содержанием белка вызывали уве-
личение возникновения опухолей у крыс, живших более 18 мес.
К сожалению, как в этом эксперименте, так и в ему подобных
не оценивалось бластомогенное действие самой диеты без до-
бавок селена. Из этих данных видно, что патогистологический
контроль осуществлялся неточно.
Следующий этап познания роли селена в патогенезе рака
связан с серией работ, проведенных в конце 60-х годов. Из
результатов, полученных в ходе этих исследований, вытекало,
что селен вообще не обладает канцерогенными свойствами.
Так, J. R. Harr и соавт. (1967) изучали хроническую селеновую
интоксикацию у крыс линии Вистар, которым скармливали се-
ленит или селенат натрия в различных концентрациях от 2,9
до 203 мкмоль/кг. Около 175 крыс прожили более 2 лет. В эту
группу входили 76 контрольных и 99 подопытных крыс,
которые получали от 6,3 до 101 мкмоль/кг селена. У животных,
которым давали более 200 мкмоль/кг иона селена, была сни-
жена выживаемость. При концентрации этого МЭ, равной
50 мкмоль/кг, у животных возникали хронические гепатиты и
нефриты. Никаких признаков неоплазии эти исследователи не
отметили. Более того, среди 1437 экспериментальных животных
не выявлено разницы между контрольной и обработанной селе-
ном группами по частоте возникновения лимфом и аденокарци-
ном молочной железы, т. е. тех опухолей, которые часто
спонтанно возникают у крыс линии Вистар. К сходным выводам
пришла группа исследователей во главе с J. Tinsley [Tinsley J.
et al., 1968]. Они провели эксиерцшенты на четырех линиях
крыс, содержавшихся на рационе с'0,5—16 мг/кг селена. Хотя
в таких дозах этот МЭ и задерживал рост животных, снижал
их выживаемость, однако канцерогенного действия он не про-
являл. J. R. Glover (1970), обследовав около 5000 рабочих в
219
условиях хронического воздействия селена на производстве, не
отметил увеличения смертности от злокачественных новообра-
зований по сравнению с контрольным контингентом.
Критический анализ этих работ позволяет прийти к выводу,
что мнение о бластомогенном действии селена является скорее
всего преувеличенным. Противоречивость данных по канцеро-
генности этого МЭ может быть связана с некорректностью
экспериментов, в которых игнорировалось содержание селена
и других токсичных соединений в контрольном рационе и его
ингредиентах. На полученные результаты могли повлиять вид
и возраст животных, пол, продолжительность воздействия и ряд
других факторов. Возможно, имела место неточная интерпрета-
ция результатов, выраженные регенераторные гиперплазии при
хронических токсических гепатитах трактовали как появление
злокачественных новообразований. Более того, А. Н. Кудрин
(1975) считает, что, как правило, токсические дозы селена в
хроническом эксперименте вызывают некроз и цирроз печени,
которые являются условием для канцерогенеза, в чем нет ниче-
го удивительного. В 1958 г. И. М. Эммануэль и Л. П. Липчина
выдвинули тезис, согласно которому механизм канцерогенеза
может быть связан с появлением в биосубстрате свободных
радикалов и с возникновением цепных радикальных реакций.
Исходя из этого, по мнению указанных авторов, например, при
лечении лейкозов целесообразно проводить терапию с помощью
веществ, легко взаимодействующих со свободными радикалами
и тем самым обрывающих цепные реакции. Так, в 1966 г. по-
явилось сообщение R. Shamberger, в котором не только разви-
валось теоретическое положение предыдущих авторов, но и
была предпринята попытка лечения опухолей у мышей Swiss 30,
индуцированных 1,2-диметилбензатраценом (ДМБА). Этот
американский исследователь полагал, что в ходе взаимодейст-
вия канцерогенов с клеткой возникающие свободные радикалы
и продукты окисления липидов мембран могут вызвать целый
спектр клеточных изменений, в частности деструкцию фермен-
тов и цитохромов, участвующих в переносе электронов, распад
лизосомальных и микросомальных мембран с высвобождением
гидролитических ферментов.
В дальнейшем было показано, что введение животным тако-
го антиоксиданта и стабилизатора мембран, как селенит нат-
рия (0,0005%), вместе с кротоновым маслом сопровождалось
заметным снижением числа опухолей. При этом по сравнению
с другим антиоксидантом — а-токоферолом — соединение се-
лена было в 735 раз эффективнее [Shamberger R., 1972].
Упомянутые экспериментальные данные нашли подтвержде-
ние в серии эпидемиологических наблюдений. Так, R. Shamber-
ger и D. Frost (1969) указали, что в 31 штате США, где содер-
жание селена в фуражном зерне составляет 0,76 мкмоль/кг или
выше, отмечен более низкий уровень смертности людей от рака.
Влияние вариаций содержания селена в различных географи-
ческих регионах на частоту возникновения раковых опухолей
220
убедительно продемонстрировано в работе R. Shamberger и
С. Willis (1971). Оказалось, что смертность от лимфом, рака
пищеварительных органов, рака легких и молочных желез
уменьшается в регионах с высоким и средним содержанием
селена в почве (0,76—1,27 мкмоль/кг и более). Между тем в
зонах с низкой концентрацией (0,25—0,63 мкмоль/кг) этого
МЭ смертность людей от злокачественных новообразований по-
вышена. Обнаруженная этими авторами тенденция была под-
тверждена позднее другими исследователями в США [Jans-
son, 1976; Cowgill U. М., 1983], в Бельгии [Deelstra Н., 1982],
в Новой Зеландии [Schrauzer G., 1980].
Аналогичные исследования велись и в СССР [Сучков Б. П.,.
1981]. Так, в Черновицкой области УССР заболеваемость сель-
ского населения злокачественными новообразованиями превы-
шает среднереспубликанскую, причем в горной зоне она ниже,,
чем в лесостепной. Статистический анализ показал, что между'
содержанием селена в почве естественных угодий и заболевае-
мостью сельского населения этого региона существует тесная
отрицательная корреляция (г = —0,85). G. Schrauzer и соавт,
(1977) провели статистический анализ связей между содержа-
нием селена в пище, его ежесуточным потреблением и частотой
возникновения рака в 27 странах мира. Им была также отмече-
на значительная отрицательная корреляция между потреблени-
ем селена и смертностью от таких злокачественных новообразо-
ваний, как рак толстой и тонкой кишок, молочной железы,,
яичников и легких.
Впоследствии было установлено, что у больных с некоторы-
ми формами неоплазии содержание селена в цельной крови и-
ее сыворотке снижено по сравнению со здоровым континген-
том. Так, у 48 здоровых обследованных лиц уровень селена в
крови составлял в среднем 2,9 мкмоль/л. Значения были почти-
одинаковыми в двух возрастных группах (18—49 и 50—80 лет)
как у мужчин, так и у женщин [Shamberger R. et al., 1973].
У больных раком толстой кишки, желудка, поджелудочной'
железы был отмечен значительно более низкий уровень селена
в крови по сравнению с контролем (2; 1,94; 1,67 мкмоль/л>
соответственно), впрочем как и у больных циррозом и гепати-
том (1,72 и 1,84 мкмоль/л соответственно).
W. Broghamer и соавт. (1976) с помощью нейтронно-актива-
ционного анализа выявили снижение концентрации селена
(1,61 мкмоль/л) в сыворотке ПО больных карциномами (в кон-
троле 1,89 мкмоль/л). При этом было отмечено, что при гисто-
логически высокодифференцированных опухолях наблюдалась
низкая концентрация селена, а низкодифференцированные опу-
холи иногда выявлялись и при высоких концентрациях этого
МЭ. Если же уровень селена в организме оставался достаточно
высоким, опухоли были менее злокачественными, реже давали
метастазы, с ослаблением рецидивирования опухолей увеличи-
валась продолжительность жизни больных.
Хотя, по мнению R. Shamberger и соавт. (1973), в органах,.
22 Ь
контактирующих с пищевым селеном, рак возникает реже, чем
-в органах, не соприкасающихся с этим МЭ, данное положение
не всегда справедливо. Так, К. Р. Me Connell и соавт. (1980)
сравнили концентрацию селена в сыворотке 35 больных раком
молочной железы и здоровых. Понижение уровня этого МЭ при
.данном заболевании было статистически достоверным (р<
<0,001). Б. П. Сучков (1981) с помощью корреляционного ана-
лиза показал, что между средним содержанием селена в крови
практически здоровых лиц и заболеваемостью сельского населе-
ния Черновицкой области злокачественными новообразованиями
существует тесная отрицательная корреляция (г=—0,93). Как
при предраковых заболеваниях (за исключением заболеваний
желудочно-кишечного тракта), так и при злокачественных опу-
холях различной локализации среднее содержание селена в
цельной крови больных было ниже, нежели в цельной крови
контрольной группы практически здоровых лиц. Однако разли-
чия были статистически достоверными лишь между уровнем
селена в крови больных, страдающих раком желудка, и между
содержанием селена в крови практически здоровых лиц.
В 1983 г. К. О. Goodwin и соавт. привели неожиданные
данные об изменении концентрации селена и активности селе-
носодержащего фермента — глутатионпероксидазы — у 50 боль-
ных раком слизистой оболочки ротовой полости и глотки. Так,
в этой группе отмечено снижение содержания селена и актив-
ности глутатионпероксидазы в эритроцитах, а в плазме уровень
этого МЭ даже повысился. Авторы рассмотрели влияние такого
фактора, как курение, на исследуемые показатели, поскольку,
например, у курящих больных уровень селена в эритроцитах
был повышен по сравнению с некурящей группой. Было выска-
зано предположение, что некоторые сигареты могут быть
экзогенным источником этого МЭ, накапливающегося в клетках
крови. Вместе с тем у этих больных рак все же развивается из-
за сильного воздействия канцерогенов даже при высоком уров-
не селена в организме.
В другой работе американские исследователи [Willett W. С.
et al., 1983], обобщили данные о содержании селена в сыворот-
ке крови, собранной в 1973 г. у 111 пациентов, у которых в
последующие 5 лет появились злокачественные новообразова-
ния. В качестве контроля были обследованы 210 практически
здоровых лиц, причем во внимание принимались такие факторы,
как возраст, раса, пол и курение. Оказалось, что уровень селе-
на у больных значительно ниже 1,63 мкмоль/л по сравнению с
нормой (1,72 мкмоль/л). По расчетам авторов, контингент
людей с низким содержанием этого МЭ имеет в 2 раза
больший риск заболеть раком, чем люди с высоким уров-
нем селена в организме. Особенно сильные корреляционные
связи выявлены между низким уровнем селена и раком желу-
дочно-кишечного тракта и простаты. Содержание липидов,
витаминов А и Е в сыворотке незначительно влияло на установ-
ленные отношения.
222
В 1984 г. появилось сообщение финских исследователей
J. Т. Salonen и соавт., в котором также исследовалась патоге-
нетическая роль селена в возникновении неоплазий. Для этого-
в 1972 г. брали пробы крови у 8113 человек (в возрасте от 31
до 59 лет), проживающих в двух регионах Финляндии. Только
в 1983 г. эти пробы сыворотки, которые хранились при —20°C,
подвергли аналитическому исследованию. При этом принима-
лось во внимание, что за прошедший период из обследуемого
контингента 43 человека умерли от рака и 85 были госпита-
лизированы по поводу злокачественных новообразований. Сред-
нее содержание селена в этих 128 случаях составляло 0,64 мк-
моль/л (в контроле 0,69 мкмоль/л). Сложный мультифакторный
статистический анализ показал, что при содержании селена в
сыворотке менее 0,57 мкмоль/л риск заболеть раком в среднем
возрасте увеличивается в 3,1 раза. Одновременно появилась
другая работа [Sundstrom Н. et al., 1984], в которой определя-
ли уровень селена и активность глутатионпероксидазы в сыво-
ротке крови больных некоторыми формами гинекологического-
рака. В этой работе также зафиксировано снижение концент-
рации этого микроэлемента (1,15 мкмоль/л) и активности се-
леносодержащего фермента (404 ед/л) по сравнению с кон-
трольной группой (1,25 мкмоль/л, 444 ед/л) соответственно
Введение больным в сутки 96 мкг одного селена, а также в
комбинации с витамином Е в течение 1 нед снижало концент-
рацию перекисей липидов и повышало активность глутатион-
пероксидазы в сыворотке крови.
Однако до сих пор остается не ясным, является ли снижен-
ное содержание селена в сыворотке фактором, способствующим
канцерогенезу, или уже результатом развития неоплазм [Wil-
lett W. С., Mac Mahon В., 1984]. Поскольку, например, по дан-
ным Б. П. Сучкова (1981), у больных раком желудка и кишеч-
ника наиболее часто отмечают отвращение к мясным и рыбным
блюдам и из их рациона практически исключают черный хлеб,
т. е. в организм не поступают основные продукты, содержащие
наибольшее количество селена. Кроме того, уменьшение селена
в организме может быть связано и с его повышенной концентра-
цией в опухолевой ткани. Так, A. Baumgartner и соавт. (1978)
показали, что развитие асцитной опухоли Эрлиха сопровожда-
ется резким уменьшением содержания и скорости обмена
селена в печени, почках и тимусе крыс.
Целый цикл фундаментальных работ посвящен изучению
влияния селена на опухолевую ткань при индуцированном хи-
мическом канцерогенезе. Так, число папиллом на коже мышей-
альбиносов ICR Swiss, индуцированных ДМБА, значительно
уменьшалось после смазывания пораженной кожи селенитом
натрия [Shamberger R., 1970]. Если селенит натрия
(5,78 мкмоль/кг) добавляли в корм мышей, обработанных
ДМБА, то уменьшалось и число мышей-опухоленосителей, и
число папиллом у каждой особи. Аналогичные опыты с бенз(а)-
пиреном подтвердили выявленную тенденцию. После обработки
22Э
этим канцерогеном опухоли кожи возникали у 86% мышей.
Если к пище добавляли 5,78 мкмоль/кг селенита натрия, опухо-
ли развивались только у 44% подопытных животных. Апроби-
рование других антиоксидантов (аскорбиновая кислота) и
стабилизаторов мембран лизосом (гидрокортизон) не дало
положительного эффекта. На основании этих данных R. Sham-
berger (1970) сделал вывод, что противоопухолевый эффект
селена является высокоспецифическим и, по-видимому, не свя-
зан с антиоксидантной функцией этого МЭ.
Интересные опыты были поставлены С. 1р и D. К. Sinha
(1981). Скармливание самкам крыс Спрейг-Доули корма с раз-
личным содержанием и типом жиров на фоне относительно
низкого содержания селена (1,27 мкмоль/кг) показало, что
такой характер питания способствует повышению частоты воз-
никновения аденокарцином у животных после введения ДМБА.
При этом крысы, получавшие высоконенасыщенные жиры, были
более чувствительным к дефициту селена, что проявилось в
усилении у них химического канцерогенеза. Авторы полагали,
что в ходе повышенного перекисного окисления ненасыщенных
липидов при низком уровне селена образуется малондиальде-
гид, обладающий канцерогенными и мутагенными свойствами,
обусловливающими увеличение частоты развития неоплазм у
животных.
Антиканцерогенные свойства селена продемонстрированы в
работе L. A. Witting и соавт. (1982), которые давали под-
опытным мышам BALB/c питьевую воду с 23 мкмоль/л селенита
натрия на протяжении 12 мес. Затем животным однократно под-
кожно вводили бенз(а)пирен. Через несколько недель установ-
лено, что у подопытных животных саркомы развиваются реже
и они меньше по размерам, чем у контрольных животных.
D. Birt и соавт. (1982) крыс сублинии Вистар содержали на ра-
ционе с 0,58 или 11,5 мкмоль/кг селенита натрия. После 50
инъекций бис (2-оксопропил) нитрозамина оказалось, что данный
канцероген способствовал уменьшению содержания селена в
печени при низкой пищевой дозе этого МЭ и увеличению его
концентрации в поджелудочной железе при высокой пищевой
нагрузке этого МЭ. Повышенный фоновый уровень селена в
организме способствовал уменьшению частоты возникновения
аденокарцином в легких и прямой кишке крыс.
В 1982 г. появилось сообщение американских исследователей
J. N. Thompson и соавт., которые содержали самок крыс
Спрейг-Доули в течение 20 дней на специальной диете с раз-
личным количеством селенита натрия (0,29; 0,87; 6,07 и 11,9
мкмоль/кг пищи). Затем животным вводили per os одну из
двух навесок ДМБА (7,5 и 15 мг) и через несколько дней крыс
переводили на постоянный рацион с 2,66 мкмоль/кг селенита
натрия. Наблюдения, продолжавшиеся несколько месяцев, по-
казали, что, помимо уменьшения числа опухолей молочной же-
лезы, у самок удлиняется латентный период возникновения
•опухоли именно при повышенной дозе селена. Эти авторы выд-
224
винули гипотезу, согласно которой селен подавляет инициацию
и/или промоцию рака молочных желез, индуцированного
ДМБА. Эту гипотезу подтвердили М. Chatterjee и
М. R. Banerjee (1982), которые исследовали трансформацию
клеток молочных желез самок мышей Balb/c в органной культу-
ре после введения в инкубационную среду ДМБА. О трансфор-
мации клеток канцерогеном судили по появлению in vitro
альвеолярной дисплазии, аналогичной так называемым альвео-
лярным узелкам, возникающим in vivo. Так, оказалось, что
добавление селенита натрия (10_5 М) в инкубационную среду
умеренно снижает частоту трансформации (на ~ 18%) в ста-
дии инициации (0—4 дня). Наряду с этим та же концентрация
МЭ вызывает выраженное подавление этого процесса (на
~84°/о) в стадии промоции (4—10 дней культивирования с
канцерогеном). В противоположность этому феномену в низком
диапазоне концентраций (0,01—0,1 мкмоль/л) селен усиливает
процессы опухолевой трансформации.
Механизм антибластического действия селена в ходе хими-
ческого канцерогенеза до сих пор не совсем понятен. Так, одной
из причин считают влияние селена на ксенобиотический обмен
в печени. Действительно, в ряде работ, проведенных in vitro,
установлено, что этот элемент уменьшает N-гидроксилирование
2-ацетиламинофлуорена и других канцерогенов в микросомаль-
ной фракции печени крыс, тем самым направляя их обмен по
пути детоксикации [Marshall М. V. et al., 1979; Jacobs М. М.,
1980; Sinha D. К., Ip С., 1981]. Не исключено, что селен подав-
ляет метаболическую активацию проканцерогенов в клетке
[Daoud А. Н., Griffin А. С., 1978]. Так, модификации процесса
активации проканцерогенов с помощью селена выявлены в
тесте Эймса {Martin J. L. et al., 1981]. Другой причиной анти-
канцерогенного действия селена в стадии инициации, по мне-
нию J. N. Thompson и соавт. (1982), может быть уменьшение
числа клеток, взаимодействующих с канцерогеном, или же
уменьшение скорости пролиферации в ткани-мишени. Наконец,
полагают, что этот МЭ может стимулировать репаративный
синтез ДНК, поврежденной канцерогенами, хотя этот феномен
наблюдали пока только на дрожжевых клетках [Rosin М. Р.,
1981]. При перевивании опухолей дача селена оказалась менее
эффективной [Medina D., Sheperd F., 1979]. Авторы заключают,
что этот МЭ проявляет свое действие только на этапе химичес-
кой или вирусной опухолевой трансформации нормальных
клеток. Биохимический механизм, лежащий в основе данного
действия селена, неизвестен, но он может быть связан со сти-
муляцией иммунного ответа организма, защитным влиянием
Se на цитоплазматические мембраны.
Наряду с антиканцерогенным свойством селен обладает
также и антимутагенным эффектом. Так, он снижает частоту
обмена сестринскими хроматидами, индуцированного метилме-
тансульфонатом, М-гидрокси-2-ацетиламинофлуореном [Ray
W. J. et al., 1978] и ртутью {Morimoto К. et al., 1982], уменьшая
15—568
225
тем самым их цитотоксичность. Возможно, что аналогичное
действие лежит в основе его защитного эффекта и при хими-
ческом мутагенезе.
Важным свойством селена является его особое сродство
к опухолевым клеткам организма. Этот факт был установлен
в 60-х годах и с тех пор неоднократно подтверждался [Забро-
дин Н. И., Ермаков В. В., 1963; CavaRery R. R. et al., 1966;
Hara Т. et al., 1973]. Так, установлено, что через 24 ч после
инъекции изотопа 75Se крысам-опухоленосителям этот МЭ эли-
минируется из скелетных мышц, ткани мозга и концентрирует-
ся в опухоли [Cavaliery R. R. et al., 1966]. Это свойство селена
положено в основу метода обнаружения некоторых типов опу-
холей у человека [Cavaliery R. R. et al., 1966]. Показано, что
концентрация селена в 5—10 раз больше в жизнеспособной
опухолевой ткани, чем в некротизированной [Broghamer W.
et al., 1978].
Антибластомогенное действие различных (0,01; 0,1 и
0,25 мкг/мл) концентраций селена изучено нами {Авцын А. И.
и др., 1986] в культуре клеток HeLa и Нер. Исследование ми-
тотического режима клеток HeLa показало, что инкубация их
со всеми указанными концентрациями селена приводит к по-
давлению митотической активности, особенно к концу 2-часово-
го воздействия. При этом выявился парадоксальный эффект
малых концентраций селена, действие которых в короткие (2—
4 ч) сроки инкубации характеризовалось более низким мито-
тическим индексом, чем при введении более высоких концен-
траций. Отмечено также изменение продолжительности от-
дельных фаз митоза. При дозе 0,01 мкг/мл иона селена пер-
воначальная телофазная задержка впоследствии сменяется
удлинением времени профазы. Повышение дозы в 10 раз при-
водит к непродолжительной метафазной задержке, а к концу
1-х суток опыта при концентрации 0,25 мкг/мл происходит
резко выраженная телофазная задержка. При токсической дозе
МЭ определяется значительное увеличение продолжительности
метафазы уже при малых сроках инкубации. Выявлено возра-
стание числа некоторых форм патологических митозов —
К-митозов, трехгрупповых метафаз, хромосомных и хроматид-
ных мостов, многополюсных митозов, а также отставаний хро-
мосом в метакинезе. Особенно характерно появление профаз
с хромосомами типа барабанных палочек и телофаз в форме
песочных часов. Эти результаты могут по-новому объяснить
данные L. Но и соавт. (1978) об индукции селенитом натрия
фрагментации хроматина, увеличении числа хромосомных абер-
раций и репаративного синтеза ДНК в фибробластах человека.
Исследование динамики индекса метки 3Н-тимидина, а так-
же синтеза РНК и белка в культуре клеток HeLa при введении
0,1 и 0,25 мкг/мл селена показало, что при этом наблюдается
определенная мозаичность в проявлении метаболических про-
цессов.
Изучение клеточного монослоя на светооптическом уровне
226
выявило локальный характер поражения клеток HeLa и Нер.
Так, если в контроле монослой формирует клетки с четко выра-
женными границами, гомогенной цитоплазмой с единичными
вакуолями, резко очерченным структурированным ядром, то
воздействие малых концентраций селена уже на ранних сроках
приводит к появлению скоплений клеток с крупными множест-
венными вакуолями, расположенными преимущественно в
парануклеарной зоне и с достаточно высоким митотическим
индексом. Более высокие концентрации иона селена через 2 ч
приводят к появлению зернистости в цитоплазме, границы
клеток становятся нечеткими. Удлинение срока воздействия
-сопровождается очаговой деструкцией клеток. В частности,
через 24 ч инкубации в зонах некроза происходили «расплавле-
ние» цитоплазмы с образованием очагов детрита, а также фор-
мирование поврежденных цитоагломератов (рис. 47).
Следует подчеркнуть, что клетки монослоя, тесно контакти-
рующие между собой, в меньшей степени подвергаются цито-
лизу. Это в определенной степени свидетельствует о том, что
«клеточная кооперация» способна в большей степени противо-
стоять токсичному агенту. Важно, что в сильно вакуолизирован-
ных и дистрофически измененных клетках все же встречаются
различные картины митоза, что подтверждает тезис об относи-
тельной независимости протекания митоза от состояния
цитоплазмы.
Исследование монослоя в сканирующем электронном микро-
скопе позволило показать, что стереоультраструктура поверх-
ности клеток HeLa при инкубации с малой концентрацией селе-
на изменяется за счет уменьшения числа микроворсинок и
филоподий. При повышении концентрации селена происходит
увеличение числа полиморфных бульбарных образований и
погибающих клеток, лишенных микроворсинчатого покрова
(рис. 48).
При инкубации с малыми концентрациями селена в клетках
HeLa отмечены полиморфизм митохондрий с очаговым просвет-
лением их матрикса, уменьшение числа полисом. Наиболее
характерны появление очагов парциального некроза и скопле-
ние около них миелиноподобных структур, аутофагосом, липид-
ных гранул.
Известно, что в условиях повышения уровня антиоксидантов
имеют место стимуляция клеточной пролиферации [Воскресен-
ский О. Н. и др., 1982] и стабилизация мембран клеток. Однако
этого не происходит при действии селена на опухолевые клетки
in vitro, напротив, в данном случае ярко проявляется его ток-
сическое действие. Необходимо отметить, что механизм этого
действия, по-видимому, не связан с клеточной антиоксидантной
системой и пока не совсем понятен. Однако в литературе име-
ются данные о способности селена извращать нормальный
белковый синтез, замещая серу в серосодержащих аминокисло-
тах [Ермаков В. В., Ковальский В. В., 1974], инактивировать
некоторые ферменты окислительно-восстановительных процес-
15*
227
Рис. 47. Изменение морфологии культуры клеток HeLa после инкубации с раз-
личными концентрациями селена.
И _ кпятпольвая культура; б —через 4 ч после добавления 1,27 мкмоль/л селена; в, г—
через 2 и 24 ч после добавления 3,17 мкмоль/л селена. Х400 (а); Х700 (б, в, г).
Рис. 48. Стереоультраструктура поверхности клеток HeLa в контроле (а)’
и после воздействия (б—г) различных концентраций селена.
а — контрольная культура. Х2000; б — через 2 ч при концентрации 1,27 мкмоль/л селена.
Х1000; в, г — через 24 и 4 ч соответственно при концентрации 3,17 мкмоль/л селена.
Х3000.
сов, блокируя их активные центры путем простой адсорбции или
же связывая их сульфгидрильные группы [Hoffmann G. R.,
1982]. Можно предположить, что в свою очередь это приводит,
во-первых, к ингибированию перекисного окисления и липолиза,
в частности распада нейтральных липидов, в результате чего
мы наблюдали скопления липидных гранул вблизи очагов
парциального некроза. Во-вторых, происходит усиление протео-
лиза за счет активирования протеаз в цитозоле и фосфолипаз
митохондрий, в связи с чем мы отмечали появление очагов
229
парциального некроза, полиморфизма митохондрий и образова-
ние пластинчатых миелиноподобных структур как результата
их деградации.
При этом установлено, что клетки HeLa более чувствитель-
ны к токсическому действию селена, чем культура опухолевых
клеток Нер. Добавление же селена в токсической дозе в первич-
ную культуру нормальных фибробластов человека приводило
только к усилению очаговой эозинофильной дегенерации клеток
при максимальном сроке воздействия.
Таким образом, полученные нами данные однозначно свиде-
тельствуют о непосредственном повреждающем влиянии этого
МЭ на опухолевые клетки человека, причем не только на про-
лиферирующие клетки, но и на интерфазные опухолевые
клетки.
В заключение необходимо отметить, что уменьшение частоты
возникновения спонтанных и химически индуцированных опухо-
лей у лабораторных животных при введении селена, а также
закономерное снижение его уровня в организме онкологических
больных еще раз свидетельствуют о его потенциальной значи-
мости в качестве важного компонента сбалансированного пи-
тания.
Более того, есть достаточно веские основания считать, что
дефицит селена в окружающей среде, обусловливающий его
низкое содержание в организме, способен вызвать тяжелые
прогрессирующие поражения миокарда, которые могут быть
предотвращены назначением селеносодержащих препаратов.
Это с наибольшей отчетливостью показано на примере эндеми-
ческой селенодефицитной кардиомиопатии — болезни Кешана.
Важная роль селена в патогенезе сердечно-сосудистых заболе-
ваний не ограничивается только этой патологией, впервые опи-
санной в Китае. Финские, затем и западногерманские исследо-
ватели показали, что такие широко распространенные заболе-
вания человека, как конгестивная кардиомиопатия и
коронарогенная кардиопатия, включая инфаркт миокарда, в
некоторых регионах мира (восточная Финляндия, часть терри-
тории ФРГ) чаще возникают на фоне селенодефицитных
состояний, которые способствуют более высокому уровню
летальных исходов. С этих позиций многократно доказанное в
ветеринарной практике положительное влияние препаратов се-
лена при беломышечной болезни, а также экспериментальные
материалы о селенозависимом ускорении процесса рассасыва-
ния и заживления некротической зоны инфаркта миокарда
привлекают особое внимание.
В последние годы китайский ученый F.-Y. Yang (1989)
обнаружил, что у больных, страдающих болезнью Кашина —
Бека, которая встречается не только в Забайкалье, но и на
обширных территориях КНР, выявляются аномалии мембран
эритроцитов. В эритроцитах больных детей уровень селена,
активность Na+, К+-АТФазы, текучесть липидов их мембран
отличаются от соответствующих показателей детей контрольной
230
группы, проживавших в том же регионе. На основании этих
данных предполагается, что недостаточность селена является
одним из патогенетических факторов болезни Кашина — Бека.
Автор обращает внимание, что эта болезнь в основном распро-
странена в местностях с низким содержанием селена в при-
родной среде и у населения эндемичных районов.
6.7. Молибден
Молибден — единственный элемент второй серии переходных
металлов, биологическая функция которого в настоящее время
известна. Впервые внимание на этот МЭ, как и на селен, было
обращено в связи с его токсическим действием на организм
сельскохозяйственных животных: было установлено, что извест-
ный более 100 лет в Англии «пастбищный понос» крупного ро-
гатого скота (teart) представляет собой не что иное, как молиб-
деноз [Ferguson W. et al., 1938]. Вскоре молибденоз был опи-
сан как «болотный понос» в Новой Зеландии, Нидерландах,
США, ГДР и других странах мира. Во всех случаях он встре-
чался в определенных ограниченных пастбищных массивах,
отличающихся повышенным уровнем этого МЭ в почвах и
растительности. Первые исследования молибденоза позволили
предложить и эффективное средство его предупреждения —
дачу животным сульфата меди. Таким образом, с самого начала
был обнаружен физиологический антагонизм этих двух метал-
лов, который, как вскоре оказалось, имеет различные формы
проявления в зависимости от содержания в корме сульфатов.
Явления недостаточности молибдена у сельскохозяйственных
животных в естественных условиях не обнаружены, если не
считать повышение содержания меди в организме животных
при низком уровне молибдена в пастбищных растениях [Un-
derwood Е., 1977]. Дефицит молибдена удалось вызвать экспе-
риментальным путем у жвачных, которым он необходим для
нормального функционирования микрофлоры преджелудков
[Anke М. et al., 1977], и у домашних птиц, нуждающихся в по-
вышенном количестве этого МЭ в связи с интенсивным течением
у них пуринового обмена [Payne С. G., 1977].
Выяснение роли молибдена в обмене веществ животного
организма связано с открытием трех молибденсодержащих фер-
ментов— ксантиноксидазы [Richert D. A., Westerfeld W. W.,
1953], альдегидоксидазы [Mahler Н. R. et al., 1954] и сульфи-
токсидазы |[Cohen Н. J. et al., 1971].
Молибден хорошо всасывается как из продуктов питания,
так и из большинства своих неорганических соединений. Осо-
бенно хорошо усваиваются жвачными водорастворимые соеди-
нения шестивалентного молибдена и его соединения, присутст-
вующие в зеленых растениях. Кролики и морские свинки
хорошо усваивают даже такие слаборастворимые соединения,
как МоО3 и СаМоО4 (но не M0S2), если их добавлять в корм
большими дозами. Молодняк крупного рогатого скота усваи-
231
Бает поступившие per os дозы 49Мо значительно хуже, чем
растущие свиньи. Основным местом всасывания молибдена
является тонкая кишка.
У свиней, крыс и человека молибден выделяется преимуще-
ственно с мочой, тогда как у крупного рогатого скота и овец,
получающих корм, бедный сульфатами, в моче появляются
всего 3—5% молибдена от поступившей дозы. При повышении
суточной дозы сульфатов до 0,3% в сухом веществе рациона
количество молибдена, выделяемое с мочой, у жвачных воз-
растает до 40—50%. Действие сульфатов носит высокоспецифи-
ческий характер, не связано с усилением диуреза, и их замена
целым рядом других солей, таких как вольфраматы, селенаты,
фосфаты, цитрат и перманганат, не оказывает подобного влия-
ния на выделение молибдена. Эндогенный сульфат столь же
эффективен, как и сульфат экзогенного происхождения, о чем
свидетельствует влияние на обмен молибдена пищи с высоким
содержанием белка. Дача овцам тиосульфата, цистеина и
метионина оказывает защитное действие при молибденозе. Оп-
ределенная часть молибдена выделяется с молоком и желчью.
Обмен молибдена в значительной мере зависит от деятель-
ности микрофлоры желудочно-кишечного тракта. Молибден
длительное время задерживается в пищеварительном канале
жвачных, так как включается в состав его микрофлоры. Будучи
ростовым фактором для бактерий, этот МЭ в повышенных
количествах вызывает их усиленное размножение в кишечнике,
что приводит к поносам, наблюдаемым у жвачных при молиб-
денозе.
В физиологическом антагонизме меди и молибдена определенное значение
имеет, по-видимому, их действие на активность сульфидоксидазы. Сульфиды,
всосавшиеся в кровь из содержимого рубца, окисляются в печени до сульфа-
тов и в этом виде выделяются с мочой [Huisingh J., Matrons G., 1976]. Медь
активирует этот фермент, тогда как молибден вызывает его угнетение, что
ведет к накоплению, сульфидов в печени и образованию в ней нерастворимого
CuS. Сульфаты (препятствуют всасыванию молибдена через клеточные мем-
браны [Huisingh J., Matrone G., 1976]. Как правило, у жвачных молибден
всасывается лучше, чем у животных с однокамерным желудком, так как у пер-
вых большее количество сульфатов превращается в сульфиды рубцовой мик-
рофлорой.
Еще одним аспектом физиологического взаимодействия меди, молибдена
и сульфатов является образование медно-молибденовых комплексов, проис-
ходящее в рубце жвачных. Первоначально допускалось, что эти элементы об-
разуют между собой труднорастворимый молибдат меди, близкий по своему
составу к редкому минералу линдгрениту — Си2МоО4 (ОН) 2. В более поздних
работах, однако, было установлено, что наряду с этим соединением или вме-
сто него образуются комплексы меди с тиомолибденовой кислотой, имеющие
общую формулу лиганда — MoOxS4_x [Dick А. Т. et al., 1975]. Было показано,
что тиомолибдаты образуются in vitro содержимым рубца при взаимодействии
в анаэробных условиях молибдата аммония и соединений серы. Тиомолибдаты
существенно понижают всасывание меди, тогда как молибдаты и сульфаты как
совместно, так и в отдельности не влияют на ее усвоение [Mills С. F. et al.,
1978].
Всосавшись в кровь, тиомолибдаты прочно связываются с альбумином
и другими плазменными белками и делают медь недоступной для метаболиче-
ских процессов. Продолжительное введение крысам тиомолибдата натрия
позволило воспроизвести у них в эксперименте признаки, имевшие близкое
232
сходство с молибденозом у жвачных и характеризовавшиеся повышением
уровня меди в плазме крови, снижением активности церулоплазмина и общего
содержания меди в организме животных [Kirchgessner М. et al., 1982]. Тиомо-
либдат в концентрациях 1—5 мкмоль/л in vitro вызывает необратимое угне-
тение пяти медьсодержащих ферментов — церулоплазмина, аскорбиноксидазы,
цитохромоксидазы, супероксиддисмутазы и тирозиназы, но не действует на
карбоангидразу, содержащую цинк [Chidambaram М. V. et al., 1984]. Внутри-
венное введение овцам тиомолибдата оказалось эффективным средством
предупреждения и лечения хронического отравления медью, причем в лизо-
сомах гепатоцитов животных удалось обнаружить скопление молибденовобел-
ковых комплексов меди [Jones Н. В. et al., 1984].
В настоящее время известно 15 молибденсодержащих фер-
ментов, три из которых встречаются в животном организме. Это
альдегидоксидаза, ксантиноксидаза и сульфитоксидаза. Молиб-
денсодержащие ферменты образованы обычно несколькими
субъединицами, содержат два атома молибдена и дополнитель-
ные простетические группы (молибдоптерин, ФАД, Fe, гем,
Se). В биологических системах молибден присутствует в четы-
рех состояниях окисления (IV—VI), из которых соединения
Mo(V) имеют отчетливый ЭПР-сигнал, легко распознаваемый
по его характерной сверхтонкой структуре, содержащей шесть
сателлитных пиков.
Дача крысам вольфрама приводит к инактивации молибденсодержащих
ферментов. Вольфрам частично замещает молибден в сульфитоксидазе, но
значительная часть фермента остается полностью лишенной металла. Воль-
фрам в биологических системах не способен восстанавливаться, подобно мо-
либдену, в четырехвалентную форму из пятивалентной, что, по-видимому,
и является причиной инактивации им молибденсодержащих ферментов. Что же
касается ксантиноксидазы, то в присутствии вольфрама образуется ее не со-
держащий металла апопротеид, лишенный ферментативной активности. Мо-
либденовый кофактор восстанавливает активность обоих апопротеидов.
Из перечисленных в табл. 20 ферментов остановимся на
ксантиноксидазе и сульфитоксидазе, которые имеют существен-
ное значение в патологии человека, а также упомянем молиб-
деновый кофактор, содержащий молибден, железо и серу.
Ксантиноксидаза коровьего молока (фермент Шар-
дингера) представляет собой димер с молекулярной массой
283 000, содержащий на каждую субъединицу молекулу ФАД,
кластер Fe4S4 (или два кластера Fe4S4), персульфидную груп-
пу и один атом молибдена. Фермент катализирует окисление
гипоксантина в ксантин и ксантина в мочевую кислоту.
Ксантиноксидаза — фермент, катализирующий окисление
ксантина, гипоксантина и альдегидов с поглощением кислорода
и образованием соответственно мочевой кислоты, ксантина или
карбоновых кислот и супероксидных радикалов [Горкин В. 3.,
1980]. Она является важным ферментом обмена пуринов, ката-
лизирующим реакцию, завершающую образование мочевой
кислоты в организме человека и животных. При генетическом
дефекте ксантиноксидазы и нарушении реабсорбции ксантина в
почечных канальцах возникает ксантинурия, которая характе-
ризуется выделением с мочой очень большого количества
ксантина и тенденцией к образованию ксантиновых камней; при
233
Молибдеисодержащие ферменты [Schrauzer G. N., 1984]
сч
s
ч
ю
w
Н
234
этом содержание мочевой кислоты в сыворотке крови и в моче
в суточном количестве резко снижено (менее 1,19—2,98 мк-
моль/л и 2,38—5,95 ммоль/л).
Ксантиноксидаза окисляет большое число соединений, вклю-
чающих пурины, птеридины, пиримидины и альдегиды [Raja-
gopalan К- V., 1980]. Однако физиологическое значение некото-
рых из этих реакций неизвестно. Аллопуринол — пиразолопири-
мидин, используемый в клинике при лечении подагры, окисля-
ется ферментом до оксипуринола, блокирующего молибденовый
центр и реакцию образования уратов. Молибден фиксируется
при этом в состоянии окисления +4 [Coughlan М. Р., 1980].
Оксидазная активность свойственна ферментам млекопи-
тающих, тогда как аналогичные ферменты из тканей пищи,
грибов и микроорганизмов являются дегидрогеназами, исполь-
зующими в качестве физиологических акцепторов электронов
NAD+ или другие переносчики электронов. Ферменты млекопи-
тающих претерпевают превращение в оксидазу из дегидрогена-
зы в процессе выделения [Rajagopalan К- V., 1987]. В этой
связи следует иметь в виду, что in vivo образование супероксид-
ного иона-радикала, по-видимому, весьма ограничено.
Сульфитоксидаза превращает сульфит в сульфат и
строго специфична к своему субстрату. Фермент присутствует
преимущественно в печени, где он локализуется в межмембран-
ном пространстве митохондрий. Его физиологическим акцепто-
ром электронов является митохондриальный цитохром с. Суль-
фитоксидаза выделена из печени человека, крупного рогатого
скота, кроликов и крыс. Она представляет собой димер, образо-
ванный двумя идентичными субъединицами с молекулярной
массой 55 000—60000, из которых каждая содержит по одному
атому молибдена и одной молекуле цитохрома Ь5-типа.
Окисление сульфита осуществляется молибденовым цент-
ром, что приводит к восстановлению Мо6+ до Мо4+, реокисление
которого совершается одноэлектронными этапами и включает
образование Мо5+. Две простетические группы сульфитоксида-
зы расположены в разных доменах. С помощью мягкого проте-
олиза сульфитоксидазы из печени крысы удалось выделить
молибденовый домен с сохранением его каталитической актив-
ности [Johnson J. L., Rajagopalan К- V., 1977]. Этот домен
является единственным известным молибдопротеидом, не имею-
щим, кроме Мо, других простетических групп.
Генетический дефект сульфитоксидазы у человека характе-
ризуется выраженными аномалиями мозга, умственной отста-
лостью, эктопией хрусталика и повышенным выделением с
мочой сульфитов, S-сульфоцистеина и тиосульфата при замет-
ном снижении количества сульфатов. Молекулярная основа этой
патологии неизвестна. Можно предположить, что она наступает
либо в связи с накоплением токсических количеств сульфитов
в одном из критических органов, либо из-за отсутствия сульфа-
та, необходимого для образования сульфолипидов, белков и
мелких молекул. Тяжелые патофизиологические нарушения при
235
этом дефекте свидетельствуют о незаменимости молибдена для
организма человека.
Ксантиноксидаза, альдегидоксидаза, пурингидроксилаза и
1пир|идоксальоксидаэа объединяются обычно под названием
«молибденовые гидроксилазы» [Coughlan М. Р_, 1980], посколь-
ку они катализируют гидроксилирование субстратов за счет
молекулы воды.
Молибденовый кофактор. Одним из наиболее впе-
чатляющих событий в исследованиях биохимии молибдена
последних лет является расшифровка природы молибденового
кофактора. Этот кофактор, рассматривавшийся вначале как
чисто гипотетическое соединительное звено двух молибденовых
ферментов у Aspergillus nidulans, вскоре оказался незамени-
мым компонентом всех (кроме одного) известных в настоящее
время молибденсодержащих ферментов.
Существование молибденового кофактора было открыто в ходе биохими-
ческих и генетических исследований Aspergillus nidulans [Pateman J. A. et al.,
1964], у которого был обнаружен класс плейотропных мутантов с дефектом
синтеза нитратредуктазы и ксантиндегидрргеназы. Эти CnX-мутаиты принад-
лежали к пяти различным генетическим локусам, одновременная экспрессия
которых была необходима для синтеза постулированного общего кофактора.
Химическая структура молибденового кофактора была выяснена в ходе иссле-
дований аналогичного мутанта у Neurospora crassa, обозначенного как nit-1
[Nason A. et al., 1970]. Было показано, что неактивная нитратредуктаза
в экстрактах индуцированных культур мутанта активировалась экстрактами
из дикого штамма, а также из ряда мутантов N. crassa, у которых отсутство-
вала только нитратредуктаза. Еще более важным оказалось наблюдение, что
активация фермента удавалась препаратами молибденсодержащих ферментов,
обработанных кислотой из самых различных источников [Nason A. et al., 1971].
При этом сам молибден и целый ряд его серосодержащих комплексов оказа-
лись неэффективными. Кофактор оказался исключительно лабильным. Совре-
менное представление о его химической природе было получено в ходе изуче-
ния его неактивных форм.
Было показано, что в моче больных отсутствуют молибдено-
вый кофактор и уротион [Johnson J. L., Rajagopalan К. V.,
1982]. Это свидетельствует, что у обоих соединений имеются
общие пути биосинтеза. Молибденовый кофактор получил наз-
вание «молибдоптерин».
В литературе уже давно обсуждается вопрос о возможной
связи избытка молибдена в пище с возникновением подагры.
Предполагается, что повышенный синтез ксантиноксидазы и
интенсификация пуринового обмена ведут к накоплению избы-
точных количеств мочевой кислоты, с выделением которых не
справляются почки. В результате этого мочевая кислота и ее
соли откладываются в сухожилиях и суставах. Интерес к это-
му вопросу был активирован исследованиями В. В. Ковальского
и Г. А. Яровой (1979), впервые описавших молибденовую по-
дагру в Горно-Анкаванском районе Армянской ССР. Это
заболевание характеризуется соответствующими биохимически-
ми изменениями в крови. Однако в настоящее время еще нет
определенного мнения о его истинной природе.
236
В частности, в4некоторых интенсивно обогащенных молибде-
ном техногенных провинциях такая патология еще не регист-
рируется. Вместе с тем клиническая и патологоанатомическая
картина пастбищного молибденов а сельскохозяйственных
животных не имеет сходства с подагрой и в первую очередь
характеризуется поносом, анемией и депигментацией волосяного
покрова.
При экспериментальном молибденозе у подопытных живот-
ных наблюдаются задержка роста, деформация костей, анемия,
анорексия и утрата оплодотворяющей способности у самцов.
Интратрехеальное введение молибдена вызывает небольшой
фиброз легочной ткани. Цинк и сероводород потенцируют ток-
сичность молибдена, а медь и неорганический сульфат ее
уменьшают.
По данным О. Д. Лопиной и Р. С. Воробьевой (1981), хро-
ническая профессиональная интоксикация молибденом вызыва-
ет функциональные изменения печени, повышение содержания
мочевой кислоты и самого молибдена в сыворотке крови.
Затем появляются полиартралгии, артрозы, гипотония, сниже-
ние концентрации гемоглобина, числа эритроцитов и лейкоци-
тов в периферической крови. Таким образом, данные профпа-
тологов все же говорят о том, что некоторые формы патологии
суставов могут быть связаны с хроническим поступлением в
организм молибдена. Аэрозоли молибдена могут вызывать
поражение бронхов и приводить к пневмокониозу. Патологичес-
кая анатомия молибденоза и молибдендефицита не изучена.
6.8. Йод
Первые данные о том, что йод является необходимым ком-
понентом тканей щитовидной железы, были получены в 1896 г.
Е. Baumann, а в начале XX века A. Oswald установил, что
основным йодсодержащим белком щитовидной железы является
тироглобулин. В 1918 г. Е. С. Kendall выделил тироксин, в
1926 г. С. R. Harington установил его истинное строение. Дру-
гой важной вехой в химии гормонов щитовидной железы
явилось открытие в 1952 г. J. Gross и R. Pitt-Rivers трийодтиро-
нина, который оказался наиболее активным гормональным
препаратом щитовидной железы [Frieden Е., 1981].
Основным источником йода на планете является мировой
океан, куда этот элемент приносится из атмосферы, водными
потоками и ледниками. Иодид-ионы окисляются под влиянием
света (с длиной волны до 560 нм) до элементного йода [Gol-
dschmidt V. М„ 1954]. Ежегодно с поверхности мирового океа-
на испаряется около 400000 тонн йода [Koutras D. A. et al.,
1980], концентрация которого в морской воде составляет 0,39—
0,47 мкмоль/л — почти столько же, сколько в плазме крови
человека. Атмосферный йод, как правило, поступает в почву с
осадками (5,5 нмоль/л). Соединения йода легко вымываются
дождем, снегом из почвы, в связи с чем в местностях с обиль-
237
ними осадками и хорошо дренируемыми почвами возможна
возникновение дефицита йода у человека и животных. Количе-
ство йода в воде отражает его содержание в почве, причем'
концентрация йода <1,58 нмоль/л свидетельствует о дефиците
йода.
Йод выполняет свою биологическую функцию как составная
часть тиреоидных гормонов, что отчасти объясняется его высо-
ким сродством к тирозину. В богатой йодом морской среде'
образование монойодтирозина (М1Т) и дийодтирозина (DIT)
происходит во многих тканях и у многих организмов.
Щитовидная железа имеется у всех позвоночных животных,.
Это во многих отношениях уникальный специфический орган,
концентрирующий йод и синтезирующий тиреоидные гормоны.
Однако ее роль у различных видов животных неодинакова,
кроме общей трофической функции. У рыб она участвует в се-
зонных изменениях, у амфибий связана с процессами метамор-
фоза, а у млекопитающих необходима для роста, развития и
дифференцировки клеток. Дефицит йода у человека, по-прежне-
му, представляет собой серьезную проблему, особенно в раз-
вивающихся странах Азии, несмотря на широкое распростране-
ние профилактики гипойодоза.
Йод—истинный биомикроэлемент. По данным Л. К- Старо-
сельцевой (1978), в организме взрослого человека он содержит-
ся в количестве 20—30 мг, в том числе и в щитовидной желе-
зе— около 10 мг. Орган — концентратор йода — щитовидная
железа, микроскопически ее паренхима состоит из трех видов;
клеток (А, В и С), из которых только А- и В-тироциты обла-
дают выраженным свойством специфически захватывать неор-
ганические соединения йода из протекающей через железу
крови и синтезировать органические соединения йода — тирок-
син (Т4) и трийодтиронин (Т3), обладающие гормональной
активностью. С-клетки щитовидной железы, выделяющие гор-
мон кальцитонин, неорганического йода не захватывают. По-
ступление йода в щитовидную железу регистрируется уже в
антенатальном периоде, в частности у человека на 10-й неделе-
эмбрионального развития.
В крови здорового человека содержится 0,67+0,28 мкмоль/л
йода; из этого количества 35% находятся в плазме крови, глав-
ным образом в виде органических соединений йода. При гипер-
тиреозе содержание йода в крови может возрастать до
7,88 мкмоль/л.
Суточная потребность человека в йоде составляет 50—-
60 мкг, что многими исследователями признается недостаточ-
ным, так как оптимальная деятельность щитовидной железы и
жизнедеятельность всего организма требуют значительно боль-
ших количеств йода, не менее 300 мкг.
Из крови йод проникает в различные органы и ткани, а так-
же частично депонируется в липидах. Он в значительных коли-
чествах накапливается в почках и слюнных железах, которые
осуществляют его выделение из организма. Главными путями
238
выделения йода являются почки. Повышенные концентрации
этого МЭ найдены также в яичниках, гипофизе, желчи и слюн-
ных железах. В мышцах концентрация йода в 1000 раз ниже,
чем в щитовидной желёзе [Brown-Grant К., 1961].
Щитовидная железа взрослых млекопитающих, в
том числе и человека, содержит в норме 15,8—39,4 ммоль/кг
йода (на сухое вещество).
При эндемическом зобе содержание йода в щитовидной же-
лезе может снижаться до 7,9 нмоль и менее. Гиперплазия же-
лезы наблюдается обычно при концентрации в ней йода менее
7,9 ммоль/кг. У овец выраженная гиперплазия фолликулов от-
мечена при концентрации йода, равной 0,79 ммоль/кг, умерен-
ная— при 3,16 ммоль/кг и очень слабая — при 8,7 ммоль/кг
(Arrington L. R. et al., 1967]. Сходные данные получены также
на телятах [Setchell В. Р. et al., I960; Sinclair D. P., And-
rews E. D., 1961]. Йод присутствует в щитовидной железе в виде
MIT, DIT, Т4, Т3, полипептидов, содержащих Т4, тироглобулина
и, вероятно, еще ряда других йодсодержащих соединений, в том
числе гистидина [Roche J. et al., 1955].
Тироглобулин — основной компонент коллоида фолликулов —
является гликопротеидом с молекулярной массой 660 000, со-
стоящим из четырех идентичных субъединиц, содержит 0,5—
1% йода и 8—10% углеводов. Он представляет собой запасную
форму тиреоидных гормонов и содержит обычно 90% от общего
количества йода, присутствующего в щитовидной железе.
Количество и соотношение различных форм йода в щитовидной
железе зависят от множества факторов — скорости поступления
в нее йода, присутствия струмогенных веществ — гойтерогенов,
которые могут нарушить механизм улавливания йода, от неко-
торых патологических состояний, включая влияние генетических
факторов.
В крови йод также присутствует в неорганической и орга-
нической формах. Содержание йодидов в плазме крови в норме
колеблется в пределах 6,3—47,3 нмоль/л ;[Wayne F. J. et al.,
1964]. Органические соединения представлены преимуществен-
но Т4, отсутствующим в эритроцитах. Т4 обычно связан с белка-
ми плазмы крови и только очень незначительная его часть
(менее 0,5%) находится в свободном виде [Sterling К-, Bren-
ner М. А., 1966]. Около 10% органического йода плазмы пред-
ставлено Т3 и DIT [Gross J., Pitt-Rivers R., 1952; Roche J. et
al., 1955]. Тироглобулин встречается в плазме только при
патологических состояниях, тогда как йодтирозины появляются
в периферической крови при стимуляции щитовидной железы
тиреотропным гормоном (ТТГ) или при гипертиреозе [Wil-
Iby М. L., O’Halloran М. W., 1966]. Йод, связанный с белками
плазмы крови, обозначается как PBI (от англ, protein bound I)
или экстрагируемый бутанолом йод (BEI), и его содержание
хорошо коррелирует с функциональной активностью щитовид-
ной железы, в связи с чем его определение используется в
диагностических целях. У взрослых лиц содержание PBI
239
колеблется в пределах 0,32—0,64 мкмоль/л [Wellby М. L. et al.,
1981].
Тироксин связывается в крови тремя различными белками.
Его специфическим переносчиком является гликопротеид с
молекулярной массой 55 000, состоящий из четырех идентичных
субъединиц и имеющий один участок связывания гормона. Он
мигрирует при электрофорезе между аг и «г-гл обул инами и.
носит название <интер-а-глобулин» или «тироксинсвязывающий
глобулин» (TBG). При повышенных концентрациях тироксина
и pH 8,6 он частично связывается преальбумином, который,
способен переносить в 4 раза больше йода, чем TBG. Неболь-
шие количества тироксина могут связываться также альбуми-
ном. Сродство Т3 к TBG почти в 10 раз ниже, чем у Т4, что
благоприятствует поступлению Т3 в ткани и определяет период
его полувыведения, составляющий около 2 сут против 6—7 сут
у Т4. Концентрация свободных Т3 и Т4 в плазме крови составля-
ет 6 и 20 нг/л соответственно. В норме 85—90% йода плазмы
крови приходятся на долю Т4 и только 4—5%—на Т3.
Определение общего содержания Т4 в сыворотке крови, ос-
нованное на конкурентном связывании гормона белком, позво-
ляет дать достоверную оценку функциональной активности
щитовидной железы [Ekins R. Р., 1960], но при изменении соот-
ношения Т4-связывающих белков может привести и к непра-
вильным выводам. Например, при повышении содержания TBG
в плазме крови эутиреоидных женщин, принимающих противо-
зачаточные средства или в период беременности, отмечается
ложный подъем Т4 в сыворотке крови [Goolden A. W. et al.,
1967]. В этой связи предлагается использовать для оценки
функциональной активности щитовидной железы определение
концентрации Т4, позволяющее четко различать гипотиреоз и
тиреотоксикоз.
Содержание йода в коровьем молоке колеблется в пределах
72—136 и в женском — в пределах 40—80 мкг/л. В молозиве
концентрация йода выше в 3—4 раза. У жвачных в молоке
присутствуют только неорганические соединения йода {Bernal J.,
Refetoff S., 1977], так как в молочную железу жвачных не
проникает Т4. В молоке крыс, кроликов и собак после введения
1S'I удается обнаружить белок, содержащий йод [Middles-
worth I., 1956]. Содержание йода в молоке в сильной степени
зависит от поступления этого элемента с пищей [Henkin R. W.
et al., 1972].
Всасывание. В пище и воде йод присутствует в виде
йодидов, которые всасываются на протяжении всего пищева-
рительного тракта. Йодиды, поступившие через рот, быстро и
почти полностью всасываются и лишь незначительная их часть
выделяется с калом [Vought R. L. et al., 1963]. Йодированные
аминокислоты, включая йодтиронины, также хорошо усваива-
ются, хотя и с меньшей скоростью и полнотой, чем йодиды.
Некоторая часть связанного ими йода может быть обнаружена
в кале, в составе органических метаболитов, а остальное коли-
240
чество распадается\в пищеварительном тракте с образованием
йодидов. Прочие соединения йода, прежде чем всосаться, также
восстанавливаются доХйодидов. У жвачных животных участком
всасывания йодидов является рубец, а сычуг — основное место
его эндогенной секреции' [Barua J. et al., 1964; Miller J. К- et
al., 1974].
Выделение. Иод выделяется из организма главным об-
разом с мочой. Содержание этого МЭ в моче хорошо коррели-
рует с его концентрацией в крови и поступлением 1311 в щито-
видную железу. Выделение йода происходит также с молоком,
потом и калом. В тропических странах при низком уровне пода
в среде его потери с потом могут привести к существенному
снижению содержания этого МЭ в организме. В норме уровень-
йода в моче подростков составляет более 0,051 мкмоль на
1 ммоль креатинина, детей 5—10-летнего возраста — 0,034 мк-
моль и взрослых — 0,077 мкмоль [Follis R. Н., 1963]. Содержа-
ние йода в суточном количестве мочи менее 0,31 мкмоль можно
рассматривать как показатель недостаточности этого МЭ при
условии нормальной функции почек [Koutras D. А., 1968].
Йодид-ионы, подобно хлорид-ионам, легко проникают через-
клеточные мембраны, в связи с чем общий неорганический пул
йода в организме включает как йодиды, присутствующие во
внеклеточном пространстве и эритроцитах, так и в концентри-
рующих йод железах — щитовидной, слюнных и слизистой обо-
лочке желудка. Как показывают опыты на крысах, равновесие
в пуле неорганического йода достигается в период полувыведе-
ния, равный 6—7 ч, в течение которого происходит выделение
с мочой около 52% 1311, введенного через рот или внутрибрю-
шинно. Метаболическое равновесие между удержанной частью
1311 и общим йодным пулом достигается в течение 14 сут, а
период полувыведения этой части 1311 составляет 9,5 дней, кото-
рые можно рассматривать как время обмена органического
йода [Heinrich Н. С. et al., 1964].
Восполнение метаболического пула йодидов в организме происходит
непрерывно как за счет экзогенных источников — пищи и воды, так и эндо-
генных— секреции щитовидной железы, слюны, желудочного сока и, наконец,
всасывания продуктов распада тиреоидных гормонов. У собак клиренс йоди-
дов в желудке превышает клиренс хлоридов в 10—50 раз [Underwood Е„.
1977], а у подсосных телят суммарное выделение йодидов в сычуге превышает
таковое хлоридов в 15 раз [Miller J. К., 1966]. Свойство сычуга концентриро-
вать йод способствует удержанию этого МЭ в организме и созданию допол-
нительного резерва йодидов, не удаляемых с мочой [Miller J. К- et al., 1974].
Йодиды непрерывно покидают свой метаболический пул в ре-
зультате деятельности щитовидной и слюнных желез, почек и-
желез слизистой желудка, конкурирующих между ’ собой за-
доступные количества этих ионов. В обмене йода принимают
участие,, три метаболических пула: неорганический йод плазмы
крови, йод щитовидной железы и пул гормонального йода, при-
сутствующего в плазме крови и в клетках других тканей [Kout-
ras D. А., 1968]. Скорость поступления йодидов из первого-
16—568
241?
Рис. 49. Пути синтеза тиреоидных гормонов в щитовидной железе [Hetzel В. S.,
Maberly G. F., 1986].
пула в почки и щитовидную железу соответственно обозначает-
ся как ренальный и тиреоидный клиренс, вычисляемый как
отношение количества йодидов, накопленных в органе за едини-
цу времени, к их концентрации в плазме. В норме у человека
суммарный клиренс йодидов из неорганического пула имеет
значение 50 мл/мин, а ренальный клиренс — около 35 мл/мин
при самых различных концентрациях йодидов в плазме [Under-
wood Е., 1977]. Тиреоидный клиренс, напротив, зависит от
колебаний уровня йодидов в плазме и функциональной актив-
ности щитовидной железы. У здоровых лиц он составляет 10—
20 мл/мин, тогда как при токсическом зобе (болезнь Грейвза)
он обычно составляет 100 мл/мин, а может превысить и
1000 мл/мин.
Тиреоидные гормоны и обмен йода в щи-
товидной железе. Щитовидная железа человека должна
улавливать около 60 мкг йода в сутки, чтобы обеспечить адек-
ватное снабжение организма тиреоидными гормонами. Эффек-
тивность работы железы, масса которой составляет 15—25 г
(0,28% от общей массы тела), обеспечивается густой сетью
кровеносных сосудов и совершенством механизма улавливания
йода («йодного насоса»). Последний в условиях физиологичес-
кой нормы функционирует при разности концентраций йода
между внеклеточной жидкостью и цитоплазмой, равной 1:100,
а при токсическом зобе или при йодной недостаточности — даже
более чем 1 : 400 [Wolff W., 1982].
Этапы биосинтеза тиреоидных гормонов и их секреции щи-
товидной железой представлены схематически на рис. 49.
Иодный насос является активным транспортным механизмом,
242
сопряженным с Nat, К+-АТФазой, подавляется оубаином
[Kendall — Taylor Р.,' 1972] и находится под регуляторным воз-
действием ТТГ. Крупные неорганические ионы могут играть
роль конкурентных ингйбиторов транспорта йода. К их числу
относятся ионы галоидов, такие как астат (At-) или Вг-, «псев-
догалоидные» ионы — тиоцианат (SCN) и селеноцианид
(SeCN), а также ионы негалоидной природы — нитрат, перхло-
рат, перренат (ReO4) и пертехнетат (ТсО4). В клиническом от-
ношении наиболее важен тиоцианат — зобогенное вещество
(гойтероген), образующийся из цианидов, высвобождающихся
при гидролизе цианосодержащих гликозидов пищи [Bour-
doux F. et al., 1980]. Для диагностических целей (сканирование
щитовидной железы) имеет определенное значение также пер-
технетат в виде "тТсО4, чему благоприятствует его короткий
период полураспада (6 ч), отсутствие ^-излучения и 7-радиации
с энергией 140 KeV.
Йодиды после выхода из эпителиальной клетки фолликула
в его внутреннее пространство, занятое коллоидом, окисляются
тироидпероксидазой до «активного» йода [Alexander N. М.,
1984], приобретая способность реагировать с остатками тиро-
зина полипептидной цепи тироглобулина с образованием MIT и
DIT. Тироидпероксидаза — гликопротеид — фермент, выделен-
ный из щитовидной железы свиней, содержит феррипротопорфи-
рин IX и имеет молекулярную массу, близкую к 200 000. Он
прочно связан с гранулярной эндоплазматической сетью эпите-
лиальных клеток фолликулов и состоит из субъединиц с моле-
кулярной массой около 70 000.
«Активный» йод представляет собой либо молекулярный
йод (I), либо связанный с пероксидазой ион йодиния (Е—FL
возможно, в виде сульфофенилйодида (Е—SI), либо связанный
с ферментом свободный радикал. Наиболее вероятной формой
представляется ион йодиния, поскольку остальные две формы
малоспецифичны. Согласно последним данным, йодирование
тирозина пероксидазой в присутствии Н2О2 и йодида протекает
с образованием в качестве промежуточного продукта йоднова-
тистой кислоты (HOI) [Alexander N. М., 1984[.
В ходе дальнейшего окисления пероксидазой происходит
сопряжение DIT и MIT с образованием различных йодтирони-
нов [Taurog А., 1978], из которых в количественном отношении
преобладает тетрайодтиронин (Т4). Процесс сопряжения, ве-
роятно, происходит следующим образом: из йодированного
тирозина образуется радикал с дефицитом электронов, который
подвергается ^-элиминированию с образованием дегидроалани-
на и ароматического радикала. Последний реагирует с остатком1
йодтирозина, связанным с другим участком фермента, образуя
йодтиронин i[Gavaret J. М. et al., 1980]. На 5000 аминокислот-
ных остатков тироглобулина приходится всего 2—5 остатков
тироксина, что указывает на специфический характер йодирова-
ния в участках молекулы белка с достаточно близко располо-
женными остатками тирозина. Лактопероксидаза и пероксидаза
16'
243
Высшие центры
глобулин и тиреоидные гормоны
Рис. 50. Гормональные взаимо-
действия высших центров в моз-
ге, гипофизе, щитовидной же-
лезе, кровеносной системе и
клетках периферических орга-
нов.
хрена также способны
йодировать белки, что
используется для их ра-
дио актвной метки 1251 и
131 j
Процесс включения
йода в состав молекулы
тирозина значительно ме-
нее Эффективен, чем
улавливание йода эпите-
лиальной клеткой, и бло-
кируется большим чис-
лом редуцирующих аген-
тов, включая лекарствен-
ные препараты, например
пропилтиоурацил и кар-
бимазол, а также гойте-
рогенными веществами
типа винилоксазолидина
[Taurog А., 1978].
Иодированный тиро-
подвергаются реабсорб-
ции эпителиальными клетками фолликулов, где происхо-
-дят их протеолиз и частичная деградация. Большая часть
тиреоидных гормонов при этом сохраняется и секретиру-
ется в кровеносное русло [Wullman S. Н., 1969; Dumont J.,
1971; Burke G., 1971]. При гидролизе тироглобулина образуется
также йодированное производное L-гистидина. Щитовидная
железа способна к реутилизации йода. Она содержит микросо-
мальную дегалогеназу, зависящую от NADP, катализирующую
дейодирование M.IT и DIT, не действует на йодтиронины.
Регуляция синтеза и гормональной актив-
ности йодтиронинов осуществляется в процессе сложного
взаимодействия нейромедиаторов, гормонов и ферментов на
уровне ЦНС, гипофиза, щитовидной железы, крови и перифери-
ческих тканей, как схематически представлено на рис. 50. ЦНС
воздействует на гормональную активность через нейронные
пути, идущие от таламуса и прилегающих к нему ядер, к гипо-
таламусу. Характер взаимодействия между ними пока не ясен.
По современным представлениям, адренергические и неадренер-
гические медиаторные вещества (катехоламины), серотонин
(моноамины) и опиоидные вейромедиаторы стимулируют сек-
рецию тиротропин-рилизинг гормона (TRH) гипоталамусом,
244
тогда как дофамин ее угнетает [Cooper D. S. et al., 1983].
Наряду с обычной нормальной регуляцией существует меха-
низм реагирования тиреоидной оси на стрессовые ситуации,
такие как патологические состояния, шок, голодание, понижен-
ная температура [Hughes J. N. et al., 1982; Mendewics J., 1982;
Wecke J., Grunderson B. J., 1983]. TRH, попадая в гипоталамо-
гипофизарную циркуляцию, стимулирует синтез и секрецию
ТТГ, вырабатываемого тиреотропными клетками аденогипофи-
за. Допускается существование механизма отрицательной об-
ратной связи между ТТГ, гормонами щитовидной железы и
секреции TRH, однако этот вопрос окончательно не решен.
TRH представляет собой трипептид (пироглутаминилгисти-
дилпролинамид), функция которого не ограничивается только
стимуляцией секреции ТТГ. Он обнаружен в повышенных ко-
личествах и в других отделах ЦНС, особенно в эпифизе, а
также в поджелудочной железе [Chopra I. J., 1981; LechanR. М.
et al., 1983]. Это дает основание полагать, что TRH может
функционировать как нейрогормональный регулятор и в других
системах, помимо тиреоидной оси. Однако без постоянной сти-
муляции со стороны TRH наступает снижение секреции ТТГ
тиреотропными клетками, ведущее к снижению активности щи-
товидной железы (гипотиреоз). TRH подвергается быстрой
инактивации протеиназами плазмы крови, активность которых
пропорциональна содержанию в крови тиреоидных гормонов.
Циркулирующие в крови тиреоидные гормоны тормозят сек-
рецию ТТГ по принципу отрицательной обратной связи, про-
тиводействуя тем самым постоянному стимулирующему эффек-
ту TRH, который в противном случае мог бы повлечь за собой
гиперфункцию железы. Тиреотропные клетки, регулирующие
синтез и секрецию ТТГ, в ответ на уровень циркулирующих в
крови тиреоидных гормонов определяют соответствующий уро-
вень функциональной активности щитовидной железы. ЦНС в
ответ на стресс и ряд других факторов способна видоизменять
активность железы, не оказывая на нее, однако, прямого воз-
действия.
Тиреотропный гормон гипофиза представляет собой глико-
протеид с молекулярной массой около 28 000 [Condliffe Р. G.,
Weintraub В. D., 1979]. Он образован двумя полипептидными
цепями (а и р), присутствие которых необходимо для полного
проявления гормональной активности. Принципиально : его
структура аналогична таковой других гормонов аденогипофи-
за— лютеинизирующего и фолликулостимулирующего, а также
хорионического гонадотропина человека. У животных одного
вида а-субъединицы этих гормонов почти идентичны, но все же
обнаруживают видовую специфичность. ^-Субъединицы, напро-
тив, у каждого гормона уникальны и определяют его специфи-
ческие свойства. ^-Субъединицы ТТГ у разных животных близки
по своей первичной структуре [Condliffe Р. G., Weintraub В. D.,
Проявление гормональной активности ТТГ начинается со
245
связывания его ^-субъединицы со специфическими рецепторами
мембраны эпителия щитовидной железы. Дальнейшая цепь
событий не вполне ясна, но считается, что они протекают по
механизму с участием цАМФ [Burke G., 1971] и запускаются в
результате повышения проницаемости клеточной мембраны для
ионов кальция и магния. Далее следует активация аденилат-
циклазы, что в свою очередь влечет за собой активирование и
фосфорилирование множества внутриклеточных белков и фер-
ментов. Помимо прямого воздействия на метаболические про-
цессы, активация этих ферментов приводит к фосфорилирова-
нию ядерных негистоновых белков, делая доступными для
транскрипции определенные генные структуры с последующим
синтезом специфических белков [Cooper Е., Spaulding S. W.,
1983; Kawano Н. et al., 1983].
ТТГ воздействует на все этапы йодного обмена в щитовид-
ной железе, начиная от поступления йода до выделения Т3 и
Т4. Транспорт йода непосредственно связан с окислительным
фосфорилированием '[Tong W., 1974; Eastman С. J., 1977].
ТТГ не только способствует образованию органических соеди-
нений йода, но и повышает скорость сопряжения DIT и MIT с
образованием Т3 и Т4. ТТГ влияет также на скорость пиноцито-
за коллоида и его слияния с лизосомами [Burke G., 1971].
Регуляция всех этих процессов достигается, видимо, через воз-
действие на механизм кальмодулина, который контролирует
функцию микротрубочек и микрофиламентов в эпителиальной
клетке щитовидной железы [Wolff J., Williams J. А., 1973].
Тиреоидные гормоны в кровообращении.
В условиях физиологической нормы большая часть тиреоидных
гормонов, секретируемых щитовидной железой, представлена
Т4 (80 мкг/сутки) и лишь небольшое количество (~3 мкг/сут-
ки) — Т3 [Chopra I. J., 1981]. Эти гормоны связываются преи-
мущественно с тремя белками плазмы крови, из которых наи-
большим сродством к тиреоидным гормонам обладает, как
отмечалось выше, TBG, за ним следуют Т4, связывающий пре-
альбумин (ТВРА) и альбумин. Сывороточные белки, связываю-
щие тиреоидные гормоны у человекообразных обезьян и лоша-
дей, аналогичны таковым человека. У крупного рогатого скота,
овец, коз, свиней и собак имеется белок, близкий по своей
структуре к TBG, но ТВРА отсутствует или найден в очень
малых количествах. У голубей, кур и кенгуру имеются ТВРА-
подобный белок и альбумин, но отсутствует TBG. У морских
свинок и крыс отсутствуют как TBG, так и ТВРА и тиреоидные
гормоны связываются только альбумином [Farer L. S. et al.,
1962; Refetoff S. et al., 1970]. У человека названные три белка
связывают 99,97% Т4 и 99,7% Т3, причем сродство этих белков
к Т4 значительно выше, чем к Т3, что объясняет более длитель-
ный период полужизни Т4 (8 сут) против 8 ч у Т3. Свободные
гормоны Т4 и Т3 непосредственно доступны тканям.
При прохождении тиреоидных гормонов через ткани, особен-
но почки, печень, легкие и мышцы, происходит дейодирование
246
Т4, в результате которого в кровяном русле повышается кон-
центрация Т3 {Braverman L. Е. et al., 1970; Pittman J. A. et al.,
1971]. Благодаря этой периферической конверсии Т4 образуется
около 80% Т3> присутствующего в кровеносном русле. В итоге
у взрослых содержание Т4 в сыворотке крови составляет 70—
160, а Тз— 1,2—2,8 нмоль/л [Sterling К-, 1970; Pittman J. А.
et al., 1971].
Печень является главным органом, регулирующим обмен
Тз и Т4. В ней происходят дезаминирование, дейодирование Т3
и Т4, а также образование парных глюкуроновых и серных кис-
лот гормонов, которые удаляются с желчью. Этим путем поки-
дает печень и небольшое количество Т4 и Т3. Конъюгаты гор-
монов, попавшие в кишечник, частично гидролизуются фермен-
тами бактерий, и свободные гормоны снова всасываются
Печень также способна осуществлять ферментативное метили-
рование гидроксильной группы тетрайодтироуксусной кислоты,
образовывать из тиронинов тирозин, З'-окситиронин, 3,4-диокси-
фенилаланин, тироуксусную и п-оксифенилпировиноградную
кислоты. Продукты деградации йодтиронинов образуют конъ-
югаты, выделяемые с желчью.
Тиреоидные гормоны проявляют свое действие в первую оче-
редь на уровне генетического аппарата клеток-мишеней. Здесь
происходит связывание Т3 специфическим ядерным рецептором
с молекулярной массой 48 000—70 000, относящимся к кислым
негистоновым белкам. Рецептор обладает высоким сродством
к Т3, но ограниченной емкостью. Комплекс Т3 — рецептор всту-
пает во взаимодействие с компонентами хроматина, инициируя
процесс транскрипции определенных генов с последующим
синтезом белков и ферментов. Существует хорошее соответствие
между связыванием Т3 в ядре и синтезом целого ряда фермен-
тов, таких, например, как а-глицерол-3-фосфат — дегидрогена-
за, малатдегидрогеназа, глюкокиназа, ферменты орнитинового
цикла и др. [Oppenheimer J. Н. et al., 1976].
Действие тиреоидных гормонов не ограничивается, однако,
только ядерным аппаратом. В клетке имеются другие специфи-
ческие рецепторы Т3, отличающиеся от ядерных. Они найдены
в цитоплазматической мембране, мембране митохондрий и цито-
плазме. Митохондриальный рецептор участвует в регуляции
процессов энергетического обмена, калоригенного действия гор-
монов. Показано, что Т3 активирует натриевый насос, отдель-
ные этапы процесса трансляции (связывание аминоацил-тРНК
с рибосомами, образование пептидной связи, транслокацию).
Еще одним интересным биологическим действием тиреоидных
гормонов является их способность индуцировать синтез специ-
фического белка, вызывающего неопластическую трансформа-
цию клеток, подвергнутых воздействию рентгеновского излуче-
ния [Guernsey D. L. et al., 1981]. Рецепторы на плазматической
мембране, по-видимому, участвуют в транспорте гормонов
внутрь клетки или в событиях, связанных с мембранными бел-
ками.
247
Важным механизмом локальной регуляции действия тире-
оидных гормонов является дейодирование Т4 с образованием
его различных производных. В тканях найдены две различные
монодейодазы.
Как было показано на крысах, метаболизм тиреоидных гор-
монов в различных тканях протекает неодинаково [Braver-
man L. Е. et al., 1970; Silva J. E., 1983]. В коре большого мозга,
мозжечке, гипофизе преобладающее количество Т3, связываю-
щегося с ядерным рецептором, образуется на месте внутрикле-
точной Т4-дейодазой, действующей на внешее кольцо гормона.
В этих тканях лишь незначительное количество Т4 теряется на
образование неактивных метаболитов типа гТ3, так что насы-
щенность ядерных рецепторов достаточно высока. Напротив, в
таких органах, как почки и печень, преобладает Т4-монодейода-
за, действующая на внутреннее кольцо. В этой связи значитель-
ная часть внутриклеточного Т4, поступающего извне, превраща-
ется в гТз и большая часть Тз, связываемая ядерными рецепто-
рами, происходит из гормона, присутствующего в плазме крови.
По этой причине занятость ядерных рецепторов периферических
тканей Т3 обычно ниже, чем в мозге и гипофизе ([Braver-
man L. Е. et al., 1970].
В эмбриональный период, начиная с 12-й недели, преобла-
дает активность дейодазы внутреннего кольца, что приводит
к образованию гТ3 из Т4. В связи с этим в крови эмбриона
отмечается пониженный уровень Т3 [Chopra I. J. et al., 1975].
В конце пренатального периода активность данного фермента
снижается и возрастает активность дейодазы внешнего кольца
(Wu S. J. et al., 1977; Hollingsworth D. R. et al., 1980]. Это вле-
чет за собой быстрое повышение в плазме крови концентрации
Т3 и снижение концентрации гТ3, в связи с чем уже на 2-й не-
деле постнатального развития концентрация Т3выше, чем гТ3.
Начиная со второй половины беременности, тиреоидная си-
стема плода не зависит от материнской тиреоидной системы,
поскольку ТТГ, Т4 и Т3 не способны проникать через плацен-
тарный барьер [Fisher D. A. et al., 1977]. В то же время гой-
терогенные вещества, а также лекарственные препараты, бло-
кирующие или, напротив, стимулирующие функцию щитовидной
железы, преодолевают его и действуют в одинаковой степени на
организм матери и плода. Это относится и к избытку или дефи-
циту йода в среде (Hollingworth D. R. et al., 1980]. Тиреоидный
эпителий зародыша реагирует на дефицит йода в организме
матери, о чем свидетельствует возникновение врожденной мик-
седемы.
Повышение концентрации эстрогенов в крови, связанное с
беременностью или приемом противозачаточных средств, влечет
за собой усиление синтеза TBG, что в свою очередь вызывает
повышение содержания в плазме крови Т4 и увеличение про-
дукции тиреоидных гормонов, особенно Т4, для поддержания на
должном уровне концентрации свободных гормонов (EggoM. С.а
Burrow G. N., 1982].
248
Повышенный уровень в крови новорожденных Т4 и Т3 сохра-
няется на протяжении всего детского возраста, а в период
полового созревания постепенно снижается до величин, свой-
ственных взрослому организму [Corcoran J. М. et al., 1977;
Penny R. et al., 1983]. В пожилом возрасте наблюдается посте-
пенное снижение уровня Т3 в плазме крови без каких-либо
сдвигов в концентрации Т4 и TBG, что, по-видимому, связа-
но со снижением активности дейодазы внешнего кольца в
тканях.
Иодная недостаточность (гипойодоз). Наибо-
лее частым МТОЗом человека и млекопитающих является ги-
пойодоз, который, как правило, проявляется в эндемическом
увеличении щитовидной железы и эндемическом зобе. Величина
этих изменений варьирует в значительных пределах. Недоста-
точное поступление йода в организм влечет за собой гипофунк-
цию щитовидной железы, проявляющуюся замедлением обмен-
ных и в первую очередь окислительных процессов, снижением
основного обмена, температуры тела. Длительный дефицит
йода в детском возрасте ведет к кретинизму, дети резко отстают
в умственном и физическом развитии, плохо развиваются мозг
и костная система. У взрослых гипотиреоз приводит к слизисто-
му отеку (микседеме), компенсаторному разрастанию эпителия
щитовидной железы (эндемическому зобу), связанному с сек-
рецией гипофизом повышенных количеств ТТГ. При эффектив-
ности компенсаторного механизма наличие зоба не сопровож-
дается нарушением метаболических процессов. Оно наступает
только при неспособности железы продуцировать необходимое
организму количество гормонов.
Гипофункция щитовидной железы может возникнуть и в
результате действия зобогенных веществ, которые вызывают
нарушение синтеза тиреоидных гормонов и соответственно по-
вышение продукции ТТГ, приводящее к компенсаторной гипер-
трофии щитовидной железы, зависящей от дозы гойтерогена и
уровня поступления в организм йода.
Зобогенные вещества. Первые сведения о существо-
вании в пище зобогенных веществ были получены еще 60 лет
назад, в 1928 г., при скармливании кроликам свежей капусты.
Возникновение зоба предупреждалось дачей 59 мкмоль йода
на одно животное в неделю [Chesney А. М. et al., 1928].
Впоследствии «капустный» зоб удалось вызвать и у других
видов животных [Blum F., 1971], а гойтерогенная активность
была обнаружена в большом числе растительных продуктов и
кормов, включая практически все виды крестоцветных. Было
также показано, что зоб, вызываемый семенами рапса, в отли-
чие от «капустного» зоба только частично предупреждается
добавками йода и возникает в результате нарушения процесса
гормоногенеза [Griesbach W. Е., Purves Н. J., 1943]. В дальней-
шем„из брюквы было выделено и идентифицировано соединение
с гойтерогенным действием, названное гойтрином [Greer М. А.
et al., 1967].
249
Кроме того, крестоцветные содержат еще ряд тиогликозидов
с зобогенным действием — производных тиоцианата, к числу
которых относятся синальбин из белой и синигрин из черной
горчицы. Под действием фермента мирозиназы (тиогликозида-
за, КФ 3.2.3.1) происходит гидролиз этих соединений с образо-
ванием свободных горчичных масел, являющихся собственно
гойтерогенами. Рассмотренная группа тиогликозидов объединя-
ется в настоящее время под названием «гликозинолаты».
Известен еще ряд гойтерогенов растительного происхожде-
ния [Bourdoux L. et al., 1980; Delangs F. et al., 1982], которые
присутствуют в таких распространенных продуктах питания на-
селения развивающихся стран, как маниок, кукуруза, побеги
бамбука, батат, просо, некоторые сорта бобов. Они содержат
цианогенные гликозиды, освобождающие при гидролизе повы-
шенное количество цианидов. При этом зобогенное действие
оказывают не столько последние, сколько образующийся из них
тиоцианат (—SCN, роданид). Роль маниока в возникновении
эндемического зоба показана рядом исследователей в Заире
и Сицилии [Delange F., 1974] и Малайзии {Maberly G. F. et al.,
1983]. Цианогенные гликозиды присутствуют также в белом
клевере (Trifolium repens), поедание которого вызывает нару-
шение функции щитовидной железы у сельскохозяйственных
животных. Другим источником тиоцианатов могут быть горчич-
ные масла (изотиоцианаты) и органические нитрилы, способные
превращаться в роданид в процессе метаболизма.
Действие гойтерогенов неодинаково. Тиоцианаты затрудня-
ют поступление йода в щитовидную железу, но их зобогенный
эффект предупреждается дачей йода, тогда как винилоксазоли-
дин ингибирует тирозинйодиназу, и йодные добавки неспособны
полностью нормализовать его действие на щитовидную железу.
Гойтерогенные вещества переходят в молоко и могут оказывать
отрицательное влияние на щитовидную железу детей [Under-
wood Е. J., 1977].
Гойтерогенным свойством парадоксальным образом облада-
ет также избыток йодидов, вызывающий зоб у лиц, питающихся
морской пищей, богатой йодом, как, например, в некоторых
прибрежных районах Японии [Suzuki Н., 1980]. Это неожидан-
ное действие йодидов, известное под названием «эффект Воль-
фа— Чайкова», объясняется, по-видимому, превращением ак-
тивной формы йода в сложный анион 13_. На свойстве йодидов
блокировать поступление йода в щитовидную железу основыва-
ется применение стабильного йода (К127Ц для предупреждения
поступления в щитовидную железу радиоактивного йода, выб-
расываемого в атмосферу при авариях на атомных электростан-
циях и при испытаниях атомного оружия [Hippel F. et al.,
1982].
В возникновении эндемического зоба определенное значение
могут иметь геохимические факторы. Как показывают исследо-
вания В. В. Ковальского, распространение эндемического зоба
совпадает не только с недостатком йода в среде, но и с избыт-
250
ком или дефицитом таких элементов, как кобальт, марганец,
кальций и стронций [Ковалевский В. В., 1974]. Автор считает,
что кобальт усиливает один из механизмов, компенсирующих
недостаточность йода, а именно синтетическую активность щи-
товидной железы, не влияя на ее массу.
Щитовидная железа обладает способностью концентриро-
вать не только йод, но и ряд других элементов, таких как ртуть,
мышьяк, сурьма (Фузайлов Ю. М, 1985; Underwood Е, 1977].
В этой связи важно отметить, что зобная эндемия в Горно-
Алтайской автономной области наиболее выражена в пределах
ртутной биогеохимической провинции, несмотря на достаточный
уровень йода в среде и проводимую противозобную профилак-
тику. Выраженная зобная эндемия отмечена также в Южной
Фергане, где в среде также содержатся повышенные количества
ртути и сурьмы.
Другим примером антитиреоидной роли геохимических фак-
торов могут служить данные, полученные в Западной Колумбии
(США), в районах с высоким уровнем эндемического зоба
(42%). Здесь гойтерогенными веществами оказались продукты,
поступавшие в питьевую воду из осадочных пород (сланцы,
уголь, лигнит) и представляющие собой фтор- и серосодержа-
щие органические соединения гуминовой природы [Gaitan Е.
et al, 1981].
Нарушение функции щитовидной железы может иметь также
генетическую природу. Известно пять основных типов генети-
ческих дефектов тиреоидных гормонов и обмена йода, сопро-
вождающихся зобом (семейный зоб, врожденный гипотиреоз):
1) неспособность железы концентрировать йод; 2) нарушение
синтеза тиреоидных гормонов, вызванное дефектом синтеза
тироидпероксидазы, сочетающееся с тугоухостью (синдром
Пендреда); 3) неспособность к дейодированию MIT и DIT,
вызванная дефектом синтеза специфической дегалогеназы (дей-
одазы), приводящим к нарушению реутилизации йода щитовид-
ной железой и его повышенным потерям из организма; 4) не-
способность синтеза Т4 из MIT и DIT; 5) циркуляция в крови
атипичного белка, прочно связывающего йод, который теряет
способность экстрагироваться бутанолом. Предупреждение ги-
потиреоза при этих дефектах достигается назначением тирокси-
на и препаратов йода (Бадалян Л. О, 1980] при условии
ранней диагностики (на 1-м месяце жизни или до рождения),
успешно осуществляемой в СССР.
Кроме первичного гипотиреоза, вызванного перечисленными
выше дефектами синтеза ферментов щитовидной железы, из-
вестны и редкие случаи недостаточности синтеза ТТГ (вторич-
ный гипотиреоз) и TRH (третичный гипотиреоз). Имеются
также заболевания щитовидной железы, связанные с наруше
нием иммунной системы. В США, например, одной из наиболее
распространенных причин гипотиреоза и зоба у детей старше
6 лет является аутоиммунное поражение щитовидной железы
(болезнь Хашимото), при которой в крови больных обнаружи-
251
ваются антитела к микросомам щитовидной железы и тирогло-
булину [Alexander N. М., 1984].
В горных районах с резко выраженной недостаточностью
йода в природной среде как эндемическое увеличение щитовид-
ной железы, так и зоб различной степени выраженности встре-
чаются с большой частотой. Картосхемы распространения эн-
демического зоба в Европе и Азии демонстрируют совпадение
регионов с наибольшей выраженностью зобной эндемии с вы-
соко- и среднегорными, главным образом континентальными,
регионами [Kelly F. С., Snedden W. W., 1963]. В СССР имеется
несколько очагов зобной эндемии, причем некоторые из них
расположены и в равнинных регионах.
По данным ВОЗ, на земном шаре насчитывается около
200—400 млн больных эндемическим зобом, причем наиболее
выраженные эндемии зарегистрированы в Альпах, на Алтае,-
в Андах, Гималаях, на Кавказе, в Карпатах, Кордильерах ина
Тянь-Шане. В равнинных районах эндемический зоб встреча-
ется в тропических районах Африки, Южной Америки, а также
в странах Восточной Европы, В СССР районами эндемии яв-
ляются Западная Украина, Полесье, верховья Волги, Урал,
Центральный и Северный Кавказ, некоторые районы Средней
Азии, Алтай, ряд районов Забайкалья и Дальнего Востока, до-
лины больших сибирских рек [Мазовецкий А. Г., 1978].
В районах эндемического зоба, как правило, наблюдаются
недостаточность йода в почве и связанное с этим очень низкое
содержание его в питьевой воде. «...Зоб может рассматриваться
как способ приспособления железы к дефициту йода. Эта адап-
тация может развиваться и вследствие действия факторов, на-
рушающих утилизацию или абсорбцию йода пищи» [De Smet
М. Р., 1963]. Таким образом, есть все основания рассматривать
эндемический зоб как заболевание адаптационной природы
[Авцын А. П., 1959].
Распространенность и тяжесть зобной эндемии зависят не
только от дефицита йода, но также от поступления с пищей не-
которых зобогенных веществ.
Патологоанатомически в основе эндемического зоба лежат
гипертрофия и гиперплазия тиреоидного эпителия в сочетании с
довольно выраженными разрастаниями тиреоидной стромы,
а также перестройка микроциркуляторного русла. Значитель-
ную роль в увеличении щитовидной железы играет также ретен-
ция ее коллоида, нередко выходящая на передний план (кол-
лоидный зоб). Будучи хроническим патологическим процессом,
эндемический зоб, как правило, имеет сложную и мозаичную
гистоархитектонику, в которой наблюдаются различные прояв-
ления нарушений специфической секреции тироцитов, а так-
же дистрофические изменения, иногда доходящие до гибели
участков паренхимы, впоследствии замещаемых соединительно-
тканными разрастаниями. Изложение данных о деталях строе-
ния и цитопатологии тиреоидного эпителия не входит в наши
задачи. Упомянем только, что в ткани щитовидной железы мо-
252
гут наблюдаться признаки нарушения минерального обмена,,
в частности кальцинозы, которые могут быть и внутриклеточны-
ми с выпадением солей кальция в дистрофически измененных
тироцитах. Этот процесс начинается с обызвествления митохонд-
рий и лизосом тироцитов. Причиной этих изменений может быть
гиперкальциемия при дефиците кальцитонина или при избыточ-
ной продукции паратгормона.
Эндемический зоб — это основное местное выражение слож-
ного патологического процесса, вызванного эндемическим гипо-
йодозом, который во многих отношениях еще не изучен. Хрони-
ческое течение заболевания с периодами обострения и возмож-
ным усилением гормональной активности зобно-измененной щи-
товидной железы сменяется периодами выраженного гипотирео-
за, что имеет соответствующее отражение в строении и функции
других систем организма.
Очень большое клиническое и общепатологическое значение
имеет диффузный токсический зоб (ЗДТ) (базедова болезнь,,
болезнь Грейвса). Это одно из самых распространенных эндо-
кринных заболеваний (встречается у 0,19% женщин и 0,03%
мужчин). Первичный дефицит йода при ЗДТ не регистрируется.
Этиология заболеваний не установлена. Неблагоприятными фак-
торами являются психические травмы (у 80% больных), ин-
фекционные заболевания (у 10%) [Шерешевский Н. А., 1957]..
По мнению В. Г. Баранова (1966), ЗДТ возникает на фоне ней-
роциркуляторной дистонии, которую автор считает предстадией.
Имеют значение наследственные факторы. В 1958 г. в крови
больных ЗДТ был выявлен «длительно действующий тиреоид-
ный стимулятор», который отличается от ТТГ продолжитель-
ностью своего эффекта. Этот стимулятор является иммуноглобу-
лином, взаимодействует с тканью щитовидной железы, выраба-
тывается лимфоцитами, что послужило основой для аутоиммун-
ной теории патогенеза ЗДТ. В патогенезе этого заболевания зна-
чительную роль играют нарушения симпатико-адреналовой си-
стемы. Иодный обмен значительно нарушен, что выражается в
избыточной продукции тироксина, трийодтиронина, а также
повышением чувствительности периферических тканей к дейст-
вию этих гормонов. Обмен веществ характеризуется ускорением
катаболизма белков, липолиза и окисления жирных кислот, от-
рицательным азотистым балансом, усилением гликогенолиза в
печени и мышцах, значительным обезвоживанием организма
[Кличко В. Р., 1978].
Проницаемость сосудистой стенки при ЗДТ возрастает, что
сопровождается выходом грубодисперсных белков, затрудняю-
щих обмен в тканях и приводящих к тканевой гипоксии. Глав-
ной причиной стойкого экзофтальма, по-видимому, является на-
копление кислых мукополисахаридов (КМПС) в ретробульбар-
ной клетчатке, которая активно связывает воду, увеличивается;
в объеме и вызывает протрузию глазного яблока. Накопление1
КМПС наблюдается и при так называемой претибиальной мик-
седеме.
253
Патологическая анатомия щитовидной железы при ЗДТ ха-
рактеризуется полиморфизмом фолликулов, изменением их про-
света, гипертрофией и гиперплазией клеток фолликулярного
эпителия, приобретающих цилиндрическую форму и образую-
щих в просвете фолликулов папилломатозные разрастания. Кол-
лоид жидкий, слабоэозинофильный. Кровеносные сосуды пере-
полнены кровью. В соединительной ткани обнаруживаются
скопления лимфоцитов с образованием лимфатических фолли-
кулов. Гистопатология ЗДТ при лечении препаратами йода,
а также тиреостатическими препаратами резко изменяется.
За пределами щитовидной железы при ЗДТ обнаружены
атрофические и дистрофические изменения в скелетных мыш-
цах, выраженный отек и мелкоклеточная инфильтрация стромы
глазодвигательных мышц при характерной для ЗДТ офтальмо-
патии [Одинокова В. А., 1978].
Спорадический зоб — особая клиническая форма, ко-
торая возникает у людей, проживающих вне эндемичных по
зобу районах. С позиции учения о МТОЗах она относится к
йоддефицитным состояниям. Именно при этой форме регистри-
руется: 1) поступление в организм зобогенных веществ, блоки-
рующих утилизацию йода щитовидной железой; 2) поражения
печени и желудочно-кишечного тракта, нарушающие всасыва-
ние и метаболизм йода; 3) поступление в организм йода в не-
доступной для всасывания форме (высокое содержание в воде
и почве зобогенных веществ); 4) нарушения интратиреоидного
обмена йода и биосинтеза гормонов в результате врожденного
понижения способности тироцитов к поглощению и депонирова-
нию йода, недостаточность ферментных систем при окислении
йодида в атомарный йод, нарушение комплексации йодтирози-
нов в йодтиронины [Мазовецкий А. Г., 1978].
По данным В. А. Одиноковой (1978), патологическая анато-
мия характеризуется наличием трех морфологических форм:
диффузной, узловой и смешанной. Микроскопически эти формы
характеризуются значительной гетерогенностью.
В. А. Одинокова (1965), прицельно изучавшая варианты
смертельных исходов при эндемическом и диффузном токсиче-
ском зобе, выделяет наиболее частые смертельные исходы в ре-
зультате сердечно-сосудистых нарушений, поражений паренхи-
мы печени и, более редко, поражений ЦНС. Нервная форма
встречается главным образом при тиреопатиях, сочетающихся с
резко выраженным гипертиреозом. Она была специально изу-
чена Д. И. Фридбергом, который обратил особое внимание на
патологию мотонейронов спинного мозга, а также некоторых от-
делов периферической нервной системы. Патологическая анато-
мия нервных осложнений при гипертиреозе была глубоко изуче-
на в экспериментах Ф. А. Айзенштейна (1964). Автор во многих
отношениях подтвердил данные Д. И. Фридберга. В настоящее
время не вызывает сомнений, что хронические тиреопатии,
в том числе эндемический зоб, вызывают многие патологические
процессы в различных органах и системах организма.
254
6.9. Кобальт
Жизненная необходимость кобальта была установлена в.
ходе изучения эндемической болезни овец и крупного рогатого
скота, встречающейся в определенных пастбищных массивах
Новой Зеландии и Австралии и известной в первой под назва-
нием «кустарниковая болезнь», «болезнь Мортона Майнса» и во
второй — как «береговая» или «изнуряющая болезнь», «энзооти-
ческий маразм». Первоначально предполагалось, что эта бо-
лезнь вызвана дефицитом железа, поскольку она предупрежда-
лась и успешно лечилась его неочищенными препаратами пру-
дами (лимонит). В дальнейшем, однако, оказалось, что лечеб-
ным эффектом обладает не само железо, а содержащийся в его
соединениях кобальт [Underwood Е., Filmer J., 1935], дефицит
которого был вскоре установлен в почвах и растениях «боль-
ных» пастбищ. Было также показано, что существующая в те-
чение столетий в Шотландии болезнь жвачных, известная как
«сухотка» (pining), тоже представляет собой недостаточность
кобальта. В СССР биогеохимические провинции с дефицитом
этого МЭ выявлены в Латвийской ССР [Берзинь Я. М., 1950;
Пейве Я- В., 1950] и Ярославской области [Ковальский В. В.,
Чебаевская В. С., 1949], где это заболевание также было из-
вестно под названием «сухотка» (лизуха). В Эстонии [Каар-
де И., 1950] ее называли болотной болезнью. В настоящее вре-
мя недостаточность кобальта описана в 25 странах мира, вклю-
чая СССР, США, Канаду, Англию, Африку.
Изучая кобальтовую недостаточность у жвачных, J. Filmer*
Е. Underwood (1937) обратили внимание на лечебный эффект
печени и высказали предположение, что кобальт способствует
синтезу особого эндогенного фактора, необходимого для жизне-
деятельности животного организма. В 1926 г. был установлен
лечебный эффект печени при злокачественной (пернициозной)
анемии у человека (болезнь Аддисона — Бирмера) (Minot G.R.,»
Murphy W. Р., 1926], а 22 года спустя из этого органа был вы-
делен в чистом виде витамн Bi2, содержащий 4% кобальта
[Smith Е. L., 1948]. В том же году витамин Bj2 был выделен из
ферментационной среды Streptomyces griseus, наиболее эффек-
тивного продуцента этого витамина [Riekes Е. L. et al., 1948].
Во многих исследованиях было показано, что животные, содер-
жащиеся на рационе, бедном животным белком, и лишенные до-
ступа к своим экскрементам, испытывают потребность в особом
факторе, получившем название «фактор животного белка».
Он оказался идентичным витамину В12. Важную роль в изучении-
витамина В12 сыграло установление его необходимости в качест-
ве ростового фактора для молочнокислых бактерий (Lactoba-
cillus lactis). Это позволило разработать высокочувствительный;
микробиологический метод его определения. В 1955 г. методом
рентгеноструктурного анализа была установлена структура ви-
тамина В!2 [Hodgkin D. С. et al., 1955], а в 1972 г. был осу-
ществлен его полный лабораторный синтез [Babior В. М., 1975].
255»
Все имеющиеся в настоящее время данные свидетельствуют,
что основной биологической функцией кобальта в организме
животных является его присутствие в молекуле витамина В12
и что, по мнению многих авторов, недостаточность кобальта яв-
ляется не чем иным, как недостаточностью витамина В12-
Всасывание. Кобальт усваивается организмом человека
и животных лучше других МЭ-металлов. Так, мыши усваивают
26,2% разовой дозы, а человек, по данным балансовых исследо-
ваний, от 20 до 95% кобальта, содержащегося в рационе. Кры-
сы в течение 3 дней усваивают около 30% кобальта, получен-
ного с кормом, а из молока — до 40%. При даче повышенных
доз кобальта (до 100 мкг на одно животное) его усвоение сни-
жается до 16%, но при дальнейшем повышении дозы относи-
тельное снижение всасывания этого МЭ не происходит, что сви-
детельствует об ограниченной способности организма животных
регулировать поступление кобальта из пищеварительного
тракта.
Исследованиями G. К. Murthy и соавт. (1971) показано, что
в различных регионах США концентрация кобальта в пище
варьирует от 0,25 до 0,69 мг/кг, а в целом в организм человека
поступает 0,30—1,77 мг кобальта в день. Потребление его с во-
дой может доходить до 10 мкг в сутки; 90% всего количества
потребляемого кобальта приходится на продукты растительного
происхождения. Уровень этого МЭ в коровьем молоке низок и
варьирует в пределах 0,4—1,1 мкг/л. Печень и почки животных
обычно содержат 0,15—0,25 мкг/г сухого вещества, а мышечная
ткань—-только половину этого количества (Underwood Е.,
1975].
Всасывание кобальта и железа существенно повышается при
дефиците последнего, а также при портальном циррозе и гемо-
хроматозе, что указывает на общность механизма всасывания
обоих МЭ. Об этом же свидетельствует их взаимный антаго-
низм на уровне всасывания из сегментов тощей кишки
[Forth W., Rummel R., 1971]. Кобальт концентрируется в пече-
ни и в меньшей степени — в щитовидной железе, надпочечни-
ках, почках, лимфатических узлах и поджелудочной железе.
Выведение кобальта у человека и крыс происходит главным
образом с мочой, причем существует прямая связь между до-
зой и содержанием его в моче. В частности, у рабочих, занятых
на кобальтовом производстве, содержание этого металла в моче
значительно превышало его уровень у рабочих контрольной
группы [Morgan L. G., 1983]. При парентеральном введении до
90% кобальта выделяется с мочой и остальная часть — с
желчью, молоком, секретом поджелудочной железы и кишеч-
ника. С потом и волосами удаляются лишь незначительные ко-
личества кобальта. С калом выделяется все невсосавшееся ко-
личество этого элемента, составляющее 10—80%.
Из желудочно-кишечного тракта кобальт поступает в кровь,
где его содержание колеблется от 0,07 до 0,6 мкмоль/л и зави-
сит от сезона и времени суток. Оно несколько выше летом, что
255
связано с употреблением в пищу свежих овощей и фруктов, бо-
гатых этим МЭ [Ноздрюхина Л. Р., 1977]. Концентрация ко-
бальта значительно выше в эритроцитах, чем в плазме. Подан-
ным Т. Gunther и соавт. (1974), в сыворотке крови кобальт свя-
зывается с альбуминовой фракцией.
Всасывание витамина Bi2 существенно отличается от всасы-
вания ионов кобальта. Оно зависит от так называемого внут-
реннего фактора Касла — одного из нескольких мукопротеидов,
вырабатываемых слизистой оболочкой желудка. Очищенный
фактор Касла имеет молекулярную массу —50 000 и содержит
13% углеводов. Он способен связывать одну молекулу витами-
на В12, образуя устойчивый к пепсину комплекс. Внутренний
фактор выделяется добавочными клетками желудка, секрети-
рующими также соляную кислоту. Комплекс внутренний фак-
тор— витамин В12 поступает в клетки слизистой оболочки, где
витамин высвобождается особым ферментом, а фактор либо
подвергается протеолизу, либо поступает в просвет кишечника.
При пернициозной анемии имеет место генетический дефект
синтеза этого фактора. Нарушение всасывания витамина В12
наблюдается также при резекции желудка и заражении гель-
минтами, конкурирующими с организмом хозяина за доступный
витамин В12.
В крови витамин В12 переносится особыми белками — транс-
кобаламинами, из которых транскобаламин II транспортирует
его в крови портальной системы, а транскобаламин I — в плаз-
ме большого круга кровообращения (транскобаламин I —
а-глобулин с молекулярной массой 121 000; транскобаламин II —
глобулин с молекулярной массой —35 000). Считается, что
транскобаламин I, более прочно связывающий витамин В12, вы-
полняет функцию его депонирования [Уайт А. и др., 1981].
Известен также транскобаламин III, функция которого изуча-
ется.
Витамин В12 превращается в печени в гидроксокобаламин
(Со3+), затем в митохондриях кобальт восстанавливается фер-
ментативно до Со+, после чего витамин превращается дезокси-
аденозилтрансферазой в кофермент 57-дезоксиаденозил-В12 и
далее 5-аденозилметионин-В12-метилтрансферазой — в метилко-
баламин. Последние два кофермента входят в состав двух об-
наруженных в организме животных ферментов — метилмало-
нил-СоА-мутазы и тетрагидроптероилглутамат-метилтрансфера-
зы (КФ 2.1.1.13).
Витамин В12, как и ионы кобальта, выводятся из организ-
ма главным образом с мочой (схема 2).
Уникальной особенностью структуры витамина Bi2 является
a-связь между кобальтом и атомом углерода длиной 0,205 нм,
не найденная ни в одном другом природном металлоорганиче-
ском соединении. 5'-Дезоксиаденозилкобаламин преобладает
среди других форм витамина В12, найденных в печени и бак-
териях, но наряду с ним присутствует также псевдовитамин Bi2,
содержащий вместо диметилбензимидазола аденин и фактор А,
17—568
257
Схема 2
ОБМЕН ВИТАМИНА В]2 В ОРГАНИЗМЕ ЖИВОТНЫХ
[Favier A., Maljour В., 1980]
Просвет кишечника
Мембрана гепатоцита
Метил-В]? (Со 3+ )
Метил--
тетра-
ГИДрО —
фолат
Тетра-
гидро—
фолат
СТ етрагидр оптор ои л глутамат—
метилтрансфераза
представляющий собой форму витамина В12, содержащую 2-ме-
тиладенин.
Циклическая структура витамина Bi2 близка к порфиринам и имеет с ни-
ми общих предшественников, в частности уропорфириноген III и сирогидро-
хлорин. Она образована четырьмя замещенными пиррольными кольцами, об-
разующими сопряженную, стерически затрудненную корриновую систему, ли-
шенную по сравнению с порфиринами одного метионииового мостика между
кольцами А и D. В состав молекулы витамина В]2 входит остаток диметил-
бензимидазола. Источником диметилбензимидазола является 7,8-диметил-10-
рибитиллюмазин — предшественник рибофлавина.
Метаболические функции кобамидных коферментов и реак-
ции, протекающие с участием коферментов В12) а также соот-
ветствующие ферменты приведены в табл. 21.
Известны ферменты, содержащие в своем активном центре кобальт, не
входящий в состав корринового цикла. Это, во-первых, 2 биотииилпротеида —
метилмалонил-СоА — карбоксилтрансфераза (КФ 2.1.3.1), превращающая ме-
тилмалонил-СоА и пируват в пропионил-СоА и щавелевоуксусную кислоту,
и пропионил-СоА-карбоксилаза (КФ 6.4.1.3), катализирующая реакцию обра-
зования метилмалонил-СоА из пропионил-СоА в присутствии АТФ, Н2О и СО2.
Этот фермент, получаемый из пропионовокислых бактерий, как и предыдущий,
является крупным белком с молекулярной массой —800 000 и состоит из
258
Таблица 21
Ферменты, содержащие кофермент В12 и катализируемые ими реакции
Фермент Катализируемая реакция Источник
Этаноламиноксидаза (КФ 1.4.3.8) Этаноламин+Н2О+О2—>гли- кольа льдегид+NH3+Н2О2 Бактерии
Рибонуклеозидтрифосфат- 2/-дезоксирибонуклеозидтри- Животные,
редуктаза (КФ 1.1/.4.2) фосфат—>2/-рибонуклеозид- трифосфат бактерии
Тетрагидроптероилглута- 5-метилтетрагидроптероил-1 - Человек, мле-
мат — метилтрансфераза глутамат+L-гомоцистеин—► копитающие,
(КФ 2.1.1.13) тетрагидроптероилглутамат+ -f-L-метионин бактерии
Пропандиолдегидратаза (диолдегидратаза) (КФ 4.2.1.28) 1,2-пропандиоЛ'—>пропион- альдегид+Н2О; этиленгли- коль—>а цет а льдегид Бактерии
Г лицеролдегидратаза (КФ 4.2.1.30) Глицерол'—>3-гидроксипропион- альдегид-|-Н2О »
Метиласпартат — аммиак- лиаза (КФ 4.3.1.2) L-Tpeo-3-метиласпартат—► —>мезаконат+КН3
Этаноламин — аммиак- лиаза (КФ 4.3.1.7) Этаноламин—>ацетальдегид + + NH3
₽- Лизин-5,6-аминомутаза (КФ 5.4.3.3) Ь-3,6-диаминогексаноат—> —>3,5-диаминогексаноат
1)-Лизин-5,6-аминомутаза (КФ 5.4.3.4) D-Лизин—>2,5-диаминогекса- ноат
Метиласпартатмутаза (глу- таматмутаза) (КФ 5.4.99.1) L-Tpeo-3-метиласпартат—> —>Ь-глутамат »
Метилмалонил-СоА — му- Метилмалонил-СоА—>сукци- Человек, мле-
таза (КФ 5.4.99.2) нил-СоА копитающие, бактерии
2-Метиленглутаратмутаза (КФ 5.4.99.4) 2-Метиленглутарат—>2-мети- лен-3-метилсукцинат Бактерии
18 субъединиц, содержит шесть молекул биотина и шесть атомов металла,
в число которых входит кобальт и цинк. Кобальт способен выступать в каче-
стве кофермента также в составе некоторых пирофосфатаз, пептидаз и арги-
назы [De Gruyter W., 1983].
Метилкорриноиды участвуют в реакциях метилирования
ртути, мышьяка, селена и теллура с образованием высокоток-
сичных метильных производных (метил- и диметилртуть, диме-
тиларсин, триметилселеноний и др.), что имеет важное значение
в круговороте токсичных элементов, поступающих в окружаю-
щую среду в результате ее техногенного загрязнения. Так, на-
пример, превращение ртути в метилртуть явилось причиной мас-
совой гибели рыб в заливе Минамата и заболевания употреб-
лявших их в пищу животных и местных жителей.
Недостаточность витамина В12 у жвачных животных насту-
пает при снижении его содержания в рубцовой жидкости ниже
5 нг/л и в крови — ниже 0,2 нг/л.
Витамин В12, синтезируемый микрофлорой слепой и толстой
кишок, у жвачных, по-видимому, не усваивается. Микрофлора
17* 259
рубца продуцирует множество витамин В12-подобных соедине-
ний, причем синтез его биологически активной формы происхо-
дит с относительно низкой эффективностью, составляющей при
дефиците кобальта около 15% от общего количества соедине-
ний, содержащих этот элемент, а при адекватном содержании
в рационе — всего 3%.
Другой особенностью жвачных является их ограниченная
способность усваивать витамин В12, составляющая всего около
3—5% от заданной дозы. Таким образом, жвачные очень неэф-
фективно используют кобальт как при синтезе витамина В12,
так и в процессе его усвоения организмом. В то же время они
испытывают повышенную потребность в витамине В12, что
обусловлено особенностями их энергетического обмена
[Mertz W., 1987].
У жвачных основным источником энергии является не глю-
коза, а низшие жирные кислоты — уксусная, пропионовая,
в меньшей степени масляная и др., образуемые микрофлорой
рубца в процессе сбраживания корма. В этой связи у жвачных
особое значение приобретает фермент метилмалонил-СоА-мута-
за, превращающий метилмалонил-СоА в сукцинил-СоА. По-
скольку путь метаболизма пропионовой кислоты лежит через
образование метилмалонил-СоА, то в отсутствие витамина В12
она вместе с метилмалоновой кислотой накапливается в крови
и вызывает у жвачных потерю аппетита и истощение, столь ха-
рактерные для дефицита кобальта. Содержание этих кислот в
моче жвачных может служить надежным признаком недоста-
точности кобальта еще задолго до наступления клинических
признаков акобальтоза. Таким образом, у жвачных высокая по-
требность в витамине В12 сочетается с низкой эффективностью
его синтеза и усвоения, делающими их особенно восприимчивы-
ми к дефициту кобальта по сравнению с другими видами жи-
вотных.
Отдельные признаки недостаточности витамина В12 у овец
объясняются, по-видимому, угнетением второго витамин В12-за-
висимого фермента тетрагидроптероилглутамат—метилтранс-
феразы (КФ 2.1.1.13), обеспечивающего восстановление метио-
нина после утраты им метильной группы. Активность этого фер-
мента в печени В12-дефицитных овец понижена и его дефицит
сочетается с пониженным уровнем S-аденозилметионина, жиро-
вой печенью и истощением запасов холина. Эти дефекты устра-
няются инъекциями метионина. Дефицит метионина может быть
также причиной замедления усвоения азота у овец, наблюдае-
мого при дефиците витамина Bi2, и выступать в качестве факто-
ра, лимитирующего рост овец и их волосяного покрова.
Дальнейшим следствием недостаточности витамина В12 яв-
ляется заметное снижение в печени уровня фолиевой кислоты,
наблюдаемое у крыс и овец, который восстанавливается при
введении метионина. Эти наблюдения свидетельствуют о том,
что влияние витамина Bi2 на обмен липидов и фолиевой кисло-
ты опосредуется через метионин, способствующий повышению
260
запасов фолиевой кислоты, улучшая ее поступление в гепато-
цит [Underwood Е., 1977].
На некоторых пастбищах в Австралии, поросших многолет-
ним злаком канареечником клубненосным (Phalaris tuberosa),
у овец и в меньшей степени у крупного рогатого скота возни-
кает заболевание, приводящее к быстрой гибели или принимаю-
щее хроническое течение с признаками нервного расстройства,
некоординированностью движений, шаткой походкой, мышеч-
ной дрожью, учащенным дыханием и сильным сердцебиением.
Это заболевание, известное как Phalaris staggers (канареечни-
ковая вертячка), встречается также в Южной Африке и Новой
Зеландии на пастбищах с канареечником малым (Phalaris mi-
nor) и во Флориде (США), где распространен гибрид Ph. tu-
berosa X Ph. minor. Было показано, что отравление вызывается
триптаминовыми алкалоидами, близкими по строению к серо-
тонину, обнаруженными в составе этих злаков. Токсическое
действие этих алкалоидов проявляется, однако, только при де-
фиците кобальта. Витамин В12 не эффективен против отравле-
ния Phalaris staggers и потребность в кобальте возрастает с
повышением концентрации и токсичности алкалоидов. Там, где
уровень кобальта в растениях удовлетворяет потребность жвач-
ных в этом МЭ, их выпас на канареечнике не имеет отрицатель-
ных последствий. Эти наблюдения показывают, что кобальт
обладает специфической способностью предупреждать дегенера-
тивные изменения нервной системы, вызываемые действием ал-
калоидов.
Другим проявлением дефицита кобальта является «болезнь
белой печени» овец, описанная в 1974 г. в Новой Зеландии и
найденная затем в Австралии, Ирландии и в других странах
Европы [Mertz W., 1987]. Поражаются, как правило, 2—6-ме-
сячные ягнята, большая часть которых обычно погибает. Бо-
лезнь излечивается введением витамина В12, но не исключено,
что в ее возникновении играют определенную роль растительные
токсины. В числе симптомов заболевания, кроме обычных при
дефиците кобальта исхудании, потери аппетита, бледной окрас-
ки печени, у части овец отмечены фотосенсибилизация организ-
ма, реже неврологические расстройства. Жировая инфильтрация
печени возникает, видимо, в результате снижения уровня холи-
на и метионина. Наблюдаемые изменения имеют близкое сход-
ство с заболеванием печени у человека при алиментарной недо-
статочности, вызванной дефицитом в диете липотропных
веществ.
Животные с однокамерным желудком нуждаются только в физиологиче-
ски активной форме кобальта, т. е. в витамине Bi2. Их ткани ие способны
к синтезу этого витамина, так же как и микрофлора тех отрезков пищевари-
тельного тракта, где происходит всасывание э,того витамина. Все же у моно-
гастричных животных недостаточность кобальта до сих пор ие обнаружена,
если не считать отставание в росте свиней и домашних птиц, содержащихся
исключительно на растительных кормах и лишенных доступа к поеданию соб-
ственных экскрементов. Лошади чувствуют себя прекрасно иа пастбищах
с низким содержанием кобальта, на которых крупный рогатый скот и овцы
гибнут от дефицита этого МЭ.
261
Ранее соли кобальта использовались эмпирически как не-
специфический стимулятор эритропоэза у человека в дозах 20—
30 мг/сутки. Введение этого МЭ в указанных дозах вызывает
полицитемию и ряд токсических явлений, в связи с чем его ши-
рокое применение для лечения анемий было прекращено. Осо-
бое внимание к токсическому действию кобальта было привле-
чено после установления того факта, что добавление хлорида
кобальта к пиву в количестве 1,2—1,5 мг/л, практиковавшееся
в Квебеке (Канада), США и Бельгии, для улучшения пенообра-
зования, приводило к кобальтовой кардиопатии с тяжелой сер-
дечной недостаточностью и смертельным исходом, а также поли-
цитемии, гиперплазии щитовидной железы и истощению кол-
лоида. Явления гипотиреоза наступают в результате ингибиро-
вания кобальтом тироидпероксидазы, осуществляющей йодиро-
вание тирозиновых остатков тиреоглобулина. На щитовидную
железу оказывает заметное влияние не только избыток, но и
дефицит этого МЭ. Установлено, что в физиологических кон-
центрациях кобальт необходим для синтеза тиреоидных гормо-
нов у крыс. У животных, испытывающих дефицит кобальта,
дача этого МЭ приводит к уменьшению размеров фолликулов
щитовидной железы и увеличению высоты выстилающего их
эпителия. У сельскохозяйственных животных и человека из
биогеохимических провинций с пониженным уровнем кобальта
в среде или неблагоприятным его соотношением с йодом наб-
людаются эндемические нарушения функции щитовидной желе-
зы [Ковальский В. В., 1974].
При изучении факторной связи 12 классов болезней чело-
века с содержанием некоторых МЭ, в том числе и кобальта в
почве различных географических зон Азербайджана, К. Ахундов
и соавт. (1979) обнаружили важные закономерности. Так, вы-
явлена очень сильная обратная корреляционная связь частоты
заболеваний эндокринной системы с дефицитом содержания ко-
бальта в почве. Подобная же закономерность установлена меж-
ду заболеваниями системы кровообращения и низким уровнем
этого МЭ.
Со способностью кобальта угнетать тканевое дыхание, в том
числе и клеток костного мозга, связывается развитие компенса-
торной полицитемии [Ноздрюхина Л. Р., 1977] с выраженной
гиперплазией костного мозга и появлением очагов экстрамедул-
лярного кроветворения. Одним из возможных механизмов сти-
муляции эритропоэза является влияние кобальта на образова-
ние эритропоэтинов [Федоров Н. А. и др., 1970]. Это действие
кобальта объясняется, по-видимому, блокированием SH-rpynn
некоторых оксидоредуктаз, приводящим к кислородному голо-
данию костного мозга, что побуждает его к усиленной деятель-
ности либо непосредственно, либо через усиление синтеза эрит-
ропоэтина. Последний вырабатывается из находящегося в кро-
ви неактивного предшественника в ответ на гипоксию под влия-
нием образующегося в почках эритрогенина. В этой связи при-
менение кобальта как стимулятора кроветворения может быть
262
оправдано только в случаях пониженного содержания в крови
эритропоэтина [Waldron Н. А., 1980]. Однако до настоящего
времени не выяснен вопрос, связано ли действие кобальта толь-
ко с его влиянием на выработку эритропоэтина, или же наблю-
даемый в клинике эффект терапиии кобальтом обусловлен дефи-
цитом этого МЭ в организме.
Кобальт обладает низкой токсичностью для исследованных
экспериментальных животных и человека. Необходимо подчерк-
нуть, что 1000-кратный избыток кобальта почти полностью тор-
мозит поглощение железа. На модели железодефицитных крыс
было показано, что избыток кобальта подавляет выход железа
из клеток слизистой оболочки кишечника в кровь, в то время как
проникновение железа из просвета кишечного тракта не отли-
чалось от контроля при отсутствии кобальта. Субклеточное
распределение этих металлов в энтероцитах двенадцатиперст-
ной кишки было сходным [Becker G. et al., 1979]. На основе
этих данных авторы заключают, что кобальт тормозит абсорб-
цию железа не путем влияния на связывающие сайты на апи-
кальной поверхности энтероцитов, а воздействуя на процессы
высвобождения, протекающие в базальной части клеток.
Органические соединения кобальта оказывают гипотензив-
ное и коронарорасширяющее действие. Н. Г. Леонтьева (1971)
обнаружила, что длительная ингаляция паров металлического
кобальта увеличивает концентрацию в сыворотке крови общего
холестерина и липидного фосфора, а также уменьшает величи-
ну лецитин/холестеринового коэффициента. Лекарственные пре-
параты, содержащие этот МЭ, способствуют усвоению железа
и оказывают благоприятное влияние на иммунологическую ре-
активность.
Кобальт является промышленным ядом. В организм он мо-
жет поступать через органы дыхания, желудочно-кишечный
тракт, а также через кожу. Токсична пыль, содержащая ко-
бальт, причем смесь этого МЭ с вольфрамом и титаном более
токсична, чем пыль каждого из этих металлов. Наиболее ток-
сичны хорошо растворимые в воде соли кобальта, а также ме-
таллический кобальт. Токсическое действие проявляется пора-
жением органов дыхания, кроветворения, нервной системы и
органов пищеварения. Повышенная температура воздуха усили-
вает токсическое действие кобальта. У рабочих, контактирую-
щих с этим МЭ, наблюдаются кашель, диспепсические расстрой-
ства, нарушения обоняния, хронические риниты, ларингиты,
фарингиты. Длительный контакт с кобальтом может вызвать
хронический бронхит, пневмонии и пневмосклероз, а также мио-
кардиопатию.
Морфологические изменения у человека проявляются в виде
резкой гипертрофии кардиомиоцитов с явлениями вакуолизации
и диффузных разрастаний волокнистой соединительной ткани.
Кроме того, выявляются мелкокапельная жировая дистрофия и
мелкие очаговые скопления лимфоцитов в эпикарде [Агеев А. К.
и др., 1981; Kennedy A. et al., 1981]. М. П. Чекунова и соавт.
263
(1978) при кобальтовой кардиомиопатии у крыс отметили пол-
нокровие микроциркуляторного русла и периваскулярный отек.
Высокие дозы этого МЭ приводят к формированию очаговых
некрозов в миокарде. Подострое действие металла характеризу-
ется менее выраженной степенью повреждения, развивающего-
ся по типу белковой дистрофии.
В реализации повреждений миокарда на субклеточном уров-
не особую роль могут играть нарушения проницаемости мем-
бран лизосом [Чекунова М. П. и др., 1978]. По мнению ряда
авторов [Чекунова М. П. и др., 1978; Sundusky A. et al., 1981],
при кардиотоксическом действии кобальта наиболее уязвимы
ферментные системы, осуществляющие декарбоксилирование и
дегидрирование кетокислот, следствием чего является блокиро-
вание их окислительного превращения. Отмечены также нару-
шения обмена катехоламинов, особенно норадреналина. Элект-
ронно-микроскопически выявлены набухание и дезорганизация
митохондрий с исчезновением крист и разрывами внутренней
мембраны. Эндоплазматическая сеть содержала большое коли-
чество расширенных цистерн, особенно в тех кардиомиоцитах,
где повреждены митохондрии. Отмечен целый диапазон повреж-
дений миофибрилл —от локальных утолщений Z-полос до по-
тери исчерченности и гранулярного распада миофиламентов.
Повреждения миокарда больше выражены в предсердиях
[Sundusky A. et al., 1981].
Установлено, что соединения кобальта могут вызывать сен-
сибилизацию организма, которая может быть причиной возник-
новения дерматитов с характерным гиперкератозом, а также
интерстициального фиброза легких [Reinl W. et al., 1979].
Электронно-микроскопическое исследование биоптатов легких
рабочих, связанных на производстве с кобальтом, позволило
выявить также наличие макрофагов, содержащих кристаллы
этого металла [Coates Е. О. et al., 1973].
Интересные данные о глубоком нарушении баланса кобаль-
та у детей с хроническими заболеваниями органов пищеварения
представили А. П. Решеткина и соавт. (1978). Так, установлено,
что у детей с хроническим гастритом в результате
увеличения ренальной экскреции МЭ в Р/г—2 раза возникает
эндогенный дефицит его. Подобная тенденция отмечена у боль-
ных язвенной болезнью двенадцатиперстной кишки и хрониче-
ским холангиохолециститом. Только в этих случаях имеет ме-
сто снижение алиментарной обеспеченности МЭ, вследствие
чего возникает их сочетанный экзо- и эндогенный дефицит.
Кобальт может оказывать токсическое действие на организм
беременной и развивающийся плод. До сих пор спорной и дис-
куссионной остается связь этого металла с канцерогенезом. Так,
согласно тестам in vitro, канцерогенный эффект этого МЭ бли-
зок к действию такого хорошо известного канцерогена, как ни-
кель [Sunderman W., 1984]. Действительно, в опухолевой ткани
людей и животных [Хусейнов Ф. и др., 1978], в крови онколо-
гических больных [Нурягдыев С. К-, 1972] содержание кобаль-
264
та повышено в Р/2—2!/2 раза по сравнению с нормой. Наряду
с этим у больных лейкозами концентрация этого МЭ в цельной
крови снижена [Шевцова Н. Г. и др., 1978; Mangal Р. С. et al.,
1981]. Наиболее выраженное понижение его уровня отмечено
при недифференцируемой и лимфобластной формах лейкоза.
В период ремиссии уровень кобальта приближается к контроль-
ным цифрам, а при ухудшении состояния больных — значитель-
но снижается в сыворотке, эритроцитах и в цельной крови. Вме-
сте с тем в экспериментах на животных введение кобальта в
концентрации 0,01 мг/кг тормозило рост лимфосаркомы Плисса
у крыс и асцитной карциномы Эрлиха у мышей [Хусейнов Ф.
и др., 1978]. Таким образом, возможная бластомогенная роль
кобальта требует дальнейших исследований.
Болезнь Аддисона — Бирмера в связи с эффективной тера-
пией в настоящее время реже встречается в практике патолого-
анатома. Патогенез ее до сих пор во многих отношениях неясен,
хотя значение генетических факторов не подлежит сомнению.
Это выражается не только в конституциональной неполноцен-
ности секреции мукопротеидов слизистой оболочкой желудка,
но также в отягощенности наследственности больных некото-
рыми нервными и психическими заболеваниями.
Как известно, диагноз «пернициозная анемия» необходимо
дифференцировать от фолиеводефицитных анемий, а также от
симптоматических мегалобластических пернициозноподобных
анемий при опухолях желудка, тонкой и толстой кишок, при
обширных резекциях желудка и тонкой кишки. Еще большее
практическое значение имеет дифференциальный диагноз от
глистной В12-дефицитной анемии. Именно глистная мегалоблас-
тическая анемия в настоящее время привлекает особое внима-
ние в связи со значительным увеличением географического рас-
пространения дифиллоботриоза в СССР. Под энным Н. Н. Дар-
ченковой (1976) и А. А. Жаворонкова (1985), почти вся терри-
тория Севера нашей страны представляет собой зону высокого
распространения дифиллоботриоза и закономерно связанного
с ним гипокобальтоза. Новые очаги дифиллоботриоза обнару-
жены, кроме того, в зоне искусственных водохранилищ волж-
ского каскада.
Глава 7
УСЛОВНО ЭССЕНЦИАЛЬНЫЕ МИКРОЭЛЕМЕНТЫ
И СВЯЗАННЫЕ С НИМИ ЗАБОЛЕВАНИЯ
В эту группу в соответствии с классификацией Р. I. Aggett
(1985) вошли следующие МЭ: мышьяк, бор, бром, фтор, литий,
никель, кремний, ванадий. Изложение материала, как и в
предыдущей главе, построено по единому плану — доказатель-
ства эссенциальности МЭ на животных, распространенность его
в природе, всасывание, распределение по тканям, органам и
жидкостям организма, а также транспорт и выделение, физио-
логическая роль в животном организме, патологические состоя-
ния, обусловленные дефицитом и избытком МЭ в организме че-
ловека и животных.
7.1. Мышьяк
К соединениям мышьяка с древнейших времен существовал
двоякий подход. Их рассматривали как сильнейшие яды, но и
как вещества, обладавшие целебными тонизирующими свойст-
вами.
В 1786 г. Th. Fowler вел в медицинскую практику 1% рас-
твор арсенита натрия как общеукрепляющее и тонизирующее
средство, а в 1905 г. Р. Ehrlich впервые осуществил синтез ор-
ганических соединений мышьяка и продемонстрировал их тера-
певтический эффект при трипаносомозе. Вскоре им были синте-
зированы сальварсан и неосальварсан, которые произвели на-
стоящую революцию в лечении сифилиса, фрамбезии, куриного
спирохетоза, возвратного тифа. В настоящее время известны
более 8000 лекарственных препаратов, содержащих мышьяк
[Nielsen F. Н., Uthus Е. О., 1984] Л)В середине 40-х годов в жи-
вотноводстве нашли применение мышьяковые препараты как
стимуляторы роста. Соединения мышьяка получили также ши-
рокое распространение в качестве гербицидов.
Первые данные о жизненной необходимости мышьяка для
животного организма были получены в 1975—1976 гг. в опытах
на крысах [Nielsen F. Н. et al., 1975], козах и карликовых
свиньях [Anke М. et al., 1976]. Продолженные в последние годы
подобные эксперименты способствовали созданию современных
представлений о патологии арсенодефицитных состояний, кото-
рая у животных в основном выражается в тех или иных нари-
266
шениях репродуктивной сферы и в преждевременной гибели по-
томства. Арсенодефицит у человека не доказан.
Широко распространенным путем биотрансформации мышья-
ка является его метилирование. Первым идентифицированным
метильным производным этого элемента оказался триметилар-
син — ядовитый газ, образуемый плесневыми грибами Scophu-
lariopsis brevicaulis из мышьяковистого ангидрида [Challen-
ger F. et al., 1933]. Это соединение, имеющее характерный чес-
ночный запах, было обнаружено в помещениях с отсыревшими,
покрытыми плесенью обоями, окрашенными мышьяк-содержа-
щими зелеными пигментами (зелень Шееле, Швайнфуртская
зелень). Обнаружение в волосах Наполеона повышенного со-
держания мышьяка (3—10 мг/кг) связывают с наличием таких
обоев в его комнате на острове Святой Елены. Плесневые грибы
из родов Penicillium, Fusarium, Aspergillus способны в аэроб-
ных условиях превращать мышьяк в триметиларсин и ряд дру-
гих соединений. Они способны метилировать также моно- и ди-
метиларсоновые кислоты с образованием различных алкилар-
синов (этилдиметиларсин, пропилдиметиларсин, аллилдиметил-
арсин и др.). Арсенаты восстанавливаются и подвергаются ме-
тилированию и в анаэробных условиях метаногенными бакте-
риями (Methanobacterium) в присутствии метилкобаламина в
качестве донора метильных групп [McBride В. С., Wolfe R.S.,
1971]. Они способны образовывать как диметиларсин, так и
метиларсоновую кислоту.
Сравнительно недавно метилированные производные мышьяка были най-
дены и у целого ряда других организмов. Среди обнаруженных соединений
описаны и жиро-, и водорастворимые формы. К числу первых относится груп-
па необычных арсенолипидов, найденных в фитопланктоне бедных фосфором
тропических морей. Ими оказались 5-диметиларсенозорибозиды глицерина
и глицерннсульфата [Edmonds J. S., Francesconi К. А., 1984]. В морских во-
дорослях был найден также арсенофосфолипид, описанный первоначально как
лактат о-фосфатидилтриметиларсония [Benson A. A. et al., 1978], но иденти-
фицированный впоследствии как 5-диметиларсенозорибозид фосфатидилглице-
рина [Edmonds J. S., Francesconi К. А., 1981]. Другой арсенофосфолипид —
3-0-5' -диметиларсенозо-5'-деоксирибозилфосфатидилглицеринкакодилат — опи-
сан среди компонентов морской воды и продуктов метаболизма мышьяка, вы-
деляемых водорослями. К ним относится также и сама какодиловая кислота,
отщепляемая, по-видимому, бактериальными оксидазами от арсенофосфолипи-
дов наружных клеточных мембран. У диатомовой водоросли Chaetoceros gra-
cilis найдено еще одно водорастворимое соединение мышьяка (продукт D),
являющееся, по-видимому, предшественником арсенобетаииа [Benson А. А.,
1984]. Органические соединения мышьяка, найденные у моллюсков н асцидий,
синтезируются не ими самими, а симбиотическими микроскопическими водо-
рослями. Так, у крупных моллюсков Tridacna и Hippopus присутствует водо-
росль зооксантелла, а у Didemnumo ternatanum — необычная прокариотиче-
ская зеленая водоросль Prochloron, которая аккумулирует мышьяк из мор-
ской воды и превращает его в органические соединения, поступающие
в организм хозяина. Выделение соединений мышьяка моллюсками происходит
под влиянием окислительных бактериальных ферментов.
Присутствие органических форм мышьяка у некоторых морских ракооб-
разных объясняется также не только самостоятельным синтезом, но и поступ-
лением этих соединений с водой или пищей. В экстрактах из фитопланктона
водоемов, богатых фосфором, мышьякосодержащие соединения не обнаруже-
ны, а введенный радиоактивный мышьяк (74As) прочно удерживается клеточ-
267
ними структурами, связанный, по-видимому, с белками. Представляется веро-
ятным, что у водорослей, обитающих в среде, богатой фосфором, отсутствует
способность к детоксикации мышьяка, и они погибают при концентрациях
этого элемента, безвредных для водорослей тропических морей. Наземные рас-
тения способны метилировать мышьяк с образованием метиларсоновой, метил-
и диметиларсиновой кислот, но не синтезируют арсенолипидов, тогда как рас-
тения пресных водоемов обладают этой способностью.
У целого ряда обитателей морей — ракообразных (лангус-
тов, омаров, креветок), а также в печени акул мышьяк присут-
ствует в виде арсенобетаина [Shiomi К- et al., 1984]. Употреб-
ление в пищу морских животных не представляет, однако, опас-
ности для человека, поскольку арсенобетаин быстро выделяется
из организма почками. Биотрансформация мышьяка обеспечи-
вает таким образом гидробионтам возможность существования
в водной среде, а наземным животным — безопасное употребле-
ние в пищу водных организмов.
'Обмен мышьяка у млекопитающих зависит от
формы его соединений, их количества и путей поступления это-
го МЭ в организм. Органические соединения мышьяка как ес-
тественного, так и синтетического происхождения не претерпе-
вают в организме животных каких-либо существенных превра-
щений и выводятся почками в основном в неизмененном виде.
Это относится и к арсенобетаину, присутствующему в органах и
тканях морских животных. Так, у человека из 25 мг мышьяка,
поступивших с мясом лангустов, уже через 48 ч 74% выделяют-
ся с мочой, а мышьяк из креветок полностью выводится за
4 сут. При употреблении в пищу камбалы, содержащей мышьяк
в основном в виде арсенобетаина, показано, что через 5 дней
после ее приема выделение этого МЭ с мочой составило 69—
85% от поступившего количества [Luten J. В. et al., 1982].
Неорганические соединения мышьяка, поступившие через
рот, появляются в моче как в неизмененном, так и в метилирован-
ном виде. Так, в моче человека после приема разовой дозы ар-
сенита натрия найдены 51% мышьяка в виде диметиларсино-
вой кислоты, 21% в виде монометиларсиновой и 27% в виде
неорганической формы этого элемента. Кроме того, показано,
что через 4 дня после приема 500 мкг мышьяка в виде арсени-
та натрия, монометиларсоната или диметиларсиновой кислоты
у обследованных лиц в моче были найдены соответственно 46,
78 и 75% от введенной дозы, причем какодиловая кислота вы-
делилась в неизмененном виде, моиометиларсонат оказался ме-
тилированным на 13%, а арсенит —на 75% [Tam G. К- Н. et al.,
1979].
При изучении биотрансформации мышьяка у млекопитаю-
щих следует различать превращение этого МЭ микрофлорой
пищеварительного тракта и метаболизм мышьяка в клетках
организма-хозяина. Влияние кишечной микрофлоры на процес-
сы метилирования мышьяка не следует, однако, переоценивать.
Так, нМтример, степень метилирования арсената, поступившего
через рбт, у безмикробных и обычных мышей, судя по количест-
ву диметиларсиновой кислоты, выделенной с мочой, оказалась
268
практически одинаковой^ (84,5 и 82,3% соответственно) [Vah-
ter М., Nerin Н., 1980] W собак выделение мышьяка происхо-
дит также в основном (>95%) с мочой. Через 50 мин после
дачи мышьяка в плазме и эритроцитах обнаруживается диме-
тиларсиновая кислота, а спустя 6 ч в виде этого соединения на-
ходилось более 90% всего мышьяка, присутствовавшего в плаз
ме, эритроцитах и моче [Charbanneau S. М. et al., 1979]^Меха-
низм всасывания мышьяка в желудочно-кишечном тракте изу-
чен недостаточно.
^Всасывание мышьяка обладает видовыми особенно-
стями. У кур всасывание 76As2O3 происходит быстрее, чем у
жвачных, у которых микрофлора рубца задерживает процессы
усвоения, выделения и отложения мышьяка в организме. Козы,
испытывающие недостаточность этого МЭ, усваивают большие
его количества, чем животные, обеспеченные мышьяком
[Anke М., 1986].
b Распределение мышьяка в тканях зависит от его уровня в
окружающей среде. В норме в легких человека содержится
0,08—0,17 мг/кг мышьяка (на свежую ткань), а при нагрузке
этим МЭ 2,3—2,6 мг/кг. Соответствующие величины для печени
составляют 0,09—0,30 и 4,4—6,9 мг/кг. Волосы и ногти относи-
тельно богаты мышьяком. Фоновое содержание этого элемента
в волосах человека (по данным анализа 1000 проб) составляет
0,51 мг/кг, причем концентрация его в волосах мужчин почти
вдвое выше, чем у женщин (0,62 и 0,37 мг/кг соответственно).
Содержание мышьяка в волосах, превышающее 3 мг/кг, указы-
вает на возможное отравление этим элементом.
А Выделение 74As2O3 из организма человека происходит в
соответствии с трехкомпонентной показательной функцией. Око-
ло 66% мышьяка имеет полупериод выделения, равный 2,1 сут,
30% — 9,5 сут и 4%—38 сут [Pomroy С. et al., 1980]. Выделе-
ние мышьяка возрастает с увеличением его поступления, и об-
щее содержание этого МЭ в моче может служить удобным по-
казателем нагрузки организма мышьяком [Martinez G. et al.,
1983]. У собак 85% дозы мышьяка имели полупериод выведе-
ния, равный 5,9 ч, и 14% —2,4 сут.
Выделение мышьяка с желчью зависит от пути его поступле-
ния в организм и химической формы. Оно весьма незначитель-
но при приеме этого МЭ через рот и превышает 10% при па-
рентеральном введении. Для некоторых мышьяксодержащих
препаратов, вводимых внутривенно, как, например, для меларсо-
прола, желчь представляет основной путь выделения из орга-
низма, тогда как мышьяк из других соединений (ацетарсола,
трипарсамида) выводится главным образом с мочой.
С середины 40-х годов XX века производные фениларсоновой
кислоты — арсаниловая, З-нитро-4-гидроксиарсоновая, 4-нитро-
фениларсоновая и 4-уреидофениларсоновая кислоты и их соли —
нашли применение в птицеводстве и свиноводстве в качестве
кормовых добавок для повышения привесов и профилактики
заболеваний.
269
Эти соединения легко всасываются, но не задерживаются
тканями и удаляются из организма в основном с калом. Через
5—7 дней после прекращения скармливания их содержание в
мышцах и жировой ткани снижается до 0,5 мг/кг, в печени и
почках — до 2 мг/кг, что допустимо по гигиеническим нормам.
В этой связи дачу животным стимуляторов роста, содержащих
мышьяк, прекращают за 7 дней до забоя [Ferslew К. Е.,
Edds G. Т„ 1979].
У крыс процессы метилирования мышьяка выражены значи-
тельно слабее, чем у других изученных млекопитающих, что ока-
зывает существенное влияние на характер обмена этого МЭ.
Неорганические соединения мышьяка концентрируются в эрит-
роцитах, связываясь на 99% с гемоглобином, дольше задержи-
ваются тканями и прочно связываются с белками плазмы кро-
ви. Эти особенности обмена мышьяка делают крыс непригодны-
ми в качестве модели для изучения метаболизма мышьяка у
человека [Nielsen F. Н., Uthus Е. О., 1984].
Изучая жизненную необходимость мышьяка, следует иметь
в виду, что дефицит этого МЭ наступает только при его содер-
жании в рационе животных менее 50 мкг/кг, что в 7—10 раз-
ниже его обычного содержания в кормах, а концентрации, рав-
ные 3,5—5 мг/кг, оказывают на организм уже не физиологиче-
ское, а фармакодинамическое действие. В этой связи целый ряд
опытов, в которых контрольные животные получали повышен-
ные дозы машьяка [Nielsen F. Н., 1980; Uthus Е. О. et ai.,.
1983], нельзя признать корректными.
Жизненная необходимость мышьяка была продемонстриро-
вана на животных, полусинтетический корм которых содержал
менее 10 мкг/кг этого МЭ при 350 мкг/кг в контроле [Anke М.
et al., 1976]. Особенно убедительные результаты дали экспери-
менты, продолжавшиеся в течение нескольких поколений под-
опытных животных. Приплод коз, свиней и крыс, испытывав-
ший дефицит мышьяка еще во внутриутробный период, рождает-
ся с пониженной против контроля живой массой. В подсосный
период эти различия, однако, постепенно сглаживаются, по-
скольку содержание мышьяка в молоке существенно выше, чем
в полусинтетических рационах, даже если матери испытывают
дефицит этого МЭ. У растущих, суягных и лактирующих коз де-
фицит мышьяка ведет к существенному снижению потребления
корма, задержке роста, образования молока, увеличению числа
выкидышей, повышенной смертности матерей и потомства, сни-
жению воспроизводительной функции [Anke М., 1986].
Содержание белка и жира в молоке снижается на 25—30%..
В крови уменьшается концентрация триглицеридов. Наблюдает-
ся пока еще необъяснимая внезапная смерть лактирующих коз,,
которая наступает обычно на 17—35-й день второй лактации..
Гибель козлят наступает на 8—91-е сутки после рождения. Ана-
логичные результаты ; получены в опытах на крысах [Niel-
sen F. Н. et al., 1977]'РУ цыплят при дефиците мышьяка повы-
шенная смертность не отмечена [Uthus Е. О., Nielsen F. Н.„
270
1983]. Одним из ранних проявлений недостаточности мышьяка
у коз являются атрофические изменения гистоструктур миокар-
да и скелетных мышц, характеризующиеся повреждением мито-
хондриальных мембран, их разрывом и поступлением в цито-
плазму содержимого митохондрий, в которых наблюдаются
скопления электронно-плотного материала, представляющего со-
бой, по-видимому, фосфат калия [Schmidt A. et al., 1984]. Дан-
ные о влиянии мышьяка на гемопоэз противоречивы.
А Дефицит мышьяка влечет за собой повышение содержания
меди и особенно марганца в органах и тканях коз, свиней и
снижение зольности костной ткани [Anke М., 1986]. Повышение
уровня марганца отмечено и в печени мышьякдефицитных крыс
[Uthus Е. О. et al., 1983]. У взрослых крыс дефицит мышьяка
приводит к существенному снижению активности метилтранс-
фераз фосфатидилэтаноламина и фосфатидилметилэтаноламина
и фосфотрансферазы холина [Cornatzer W. Е. et al., 1983].
Однако это действие не носит, по-видимому, специфического
характера, а объясняется угнетением синтеза белка или обмена
аминокислот, о чем свидетельствует и общее снижение содер-
жания белка в микросомах печени подопытных животных.
Содержание мышьяка в природных средах.
Почвы содержат в среднем 5—6 мг/кг мышьяка с колебаниями
1—40 мг/кг. Повышенное фоновое содержание этого элемента
найдено в почвах Кавказа и Среднеазиатских республик, где
оно составляет 10—30 мг/кг [Атаджанов С. и др., 1982]. В ре-
зультате применения мышьяксодержащих пестицидов и загряз-
нения среды предприятиями цветной металлургии и тепловыми
электростанциями, сжигающими уголь, богатый мышьяком, со-
держание этого элемента в почвах может существенно воз-
растать.
Концентрация мышьяка в морской воде составляет 2—
5 мкг/л. В пресных водоемах содержание этого МЭ может из-
меняться в тысячи раз в зависимости от геологической ситуа-
ции. При ПДК мышьяка в воде, равной 50 мкг/л, отдельные во-
доисточники, например в Чили, Аргентине, Орегоне (США), на
Тайване, содержат 800—2000 мкг/л этого элемента. Вода из них
при приеме внутрь может вызвать отравление у человека и жи-
вотных.
Бактерии способны превращать органические соединения
мышьяка в неорганические, деметилируя, в частности, метилар-
соновую кислоту, используемую в качестве пестицида. Рубцо-
вая микрофлора обладает повышенной чувствительностью к
мышьяку, который ингибирует ее рост и ферментативную актив-
ность при концентрациях, безвредных для организма хозяина
[Forsberg С. V., 1978]. Известны штаммы бактерий из сточных
и загрязненных мышьяком вод, обладающие повышенной резис-
тентностью к арсенатам [Silver S„ Nakahara Н., 1983].
Фоновые содержания мышьяка в растениях составляют
0,13—66,7 мкмоль/кг, однако на почвах, загрязненных этимМЭ,
271
они могут достигать и в 10 раз больших величин. Особенно вы-
сокие концентрации мышьяка найдены в водорослях (0,25—
2,3 ммоль/кг) [Watanabe Т. et al., 1979]. Богаты этим МЭ и
другие морепродукты. В пище человека содержание мышьяка
составляет 0,13—1,73 мкмоль/кг при минимальной потребности,
вычисленной по данным опытов на козах и карликовых свиньях
(из расчета 0,08 мкмоль на 1000 ккал)—0,16—0,33 мкмоль в
сутки [Nielsen F. Н., Mertz W., 1984].
Токсичность соединений мышьяка зависит от скорости их
выделения из организма и накопления в органах и тканях. Наи-
более токсичны соединения, задерживающиеся в организме
дольше других. Соединения мышьяка можно по их токсичности
расположить в следующий ряд: AsH3>As3+>As5+>R-As-X>
>As°; неорганический арсенит значительно токсичнее для жи-
вотных, чем арсенат. Механизм токсического действия соедине-
ний трехвалентного мышьяка заключается в связывании им
сульфгидрильных групп тканевых белков и ферментов, что при-
водит к их ингибированию. Добавление тиолов снимает этот эф-
фект. Токсичность арсената объясняется, видимо, конкурентным
замещением неорганического фосфата в реакциях фосфорилиро-
вания с образованием нестабильных эфиров мышьяковой кисло-
ты, способных к спонтанному распаду.
Мышьяк широко известен как профессиональная вредность
в цветной металлургии, в промышленности, перерабатывающей
различную руду, содержащую примеси этого элемента, в про-
изводстве инсектофунгицидов. Клиника острых и хронических
отравлений мышьяком хорошо изучена профпатологами и ги-
гиенистами. По распространенному мнению общий характер от-
равления соединениями мышьяка человека и животных состоит
в первоочередном действии их на нервную систему и стенки со-
судов, что ведет к увеличению проницаемости и параличу ка-
пилляров. Считается, что «в результате нарушения трофики»
развиваются некробиотические очаги в печени, сердце, кишечни-
ке, почках, ногтях, на коже отмечаются экзема, гиперкератоз,
бородавки. По-видимому, патогенез возникающих в результате
отравления мышьяком очень многообразных изменений сложен
и нуждается в новом и более детальном исследовании. В произ-
водственных условиях этот МЭ проникает в организм главным
образом через органы дыхания и в меньшей степени — через
неповрежденную кожу и пищеварительный тракт.
£)При хроническом отравлении соединениями мышьяка отме-
чаются диспепсические явления, гастрит, гепатит, раздражение
конъюнктивы, слезотечение и светобоязнь, отек век, помутнение
стекловидного тела и роговицы, сухость в носоглотке, насморк,
иногда изъязвление (вплоть до прободения) носовой перегород-
ки, стоматит, ларингит, трахеит, бронхит. Характерны папулез-
ная и пустулезная сыпь, чаще между пальцами, изъязвления на
мошонке, а также фурункулез, рецидивирующая экзема, атро-
фический акродерматит, гиперкератоз ладоней и подошв, пиг-
ментация кожи, атрофия и ломкость ногтей, преждевременное
272
поседение и выпадение волос [Книжников В. А. и др., 1981]..
В специальном выпуске экспертов ВОЗ Мышьяк (1985) обра-
щено внимание на симметричный бородавчатый кератоз ладо-
ней и подошв в качестве характерных симптомов после дли-
тельного поступления в организм неорганического мышьяка с
питьевой водой или лекарственными препаратами. Характерен-
также меланоз, сочетающийся с участками депигментации кожи.
В производственных условиях эти кожные изменения встреча-
ются реже. Японские авторы [Hamada Т., Horiguchi S., 1976]
описали симметричный точечный ладонно-подошвенный гипер-
кератоз и бронзовую гиперпигментацию у рабочих, занятых в
производстве мышьяксодержащих инсектицидов.
Большое практическое значение имеют неврологические
симптомы, в частности вызванные хронической интоксикацией-
мышьяком интеллектуально-мнестические и речевые расстрой-
ства, депрессии, полиневриты (чаще симметричные, особенно
малоберцового и лучевого нервов), заканчивающиеся парезами
с атрофическими изменениями мышц, а также ретробульбарный
неврит, нарушение вкуса и обоняния. Природа этих расстройств
при недостаточном учете анамнестических данных не всегда-
своевременно распознается.
Менее известны непрофессиональные интоксикации мышь-
яком, связанные с употреблением загрязненной питьевой воды,
а также мышьяксодержащих пестицидов. Еще в конце XIX века
были описаны мышьякзависимые поражения периферических
сосудов у жителей Силезии, которые приводили к возникнове-
нию облитерирующего эндартериита. Эти наблюдения были под-
тверждены в 1976 г. J. W. Gro-le, указавшим на их высокую час-
тоту у виноградарей. Однако наибольшее внимание привлекла
эндемическая болезнь «черных ног» (черных стоп) на Тайване,
где концентрация мышьяка в питьевой воде достигала 0,4—
0,6 мг/л. Показатель распространенности заболевания (8,9 слу-
чаев на 1000 населения) увеличивался с возрастом и зависел
от уровня содержания мышьяка в воде. Общая поглощенная
доза этого МЭ достигала примерно 20 г. Тяжесть заболевания
зависела от продолжительности потребления загрязненной
питьевой воды.
Аналогичное заболевание периферических сосудов описано
на севере Чили, где жители Антофагасты потребляли питьевую
воду, содержащую до 0,6 мг/л мышьяка, в течение 15 лет.
J. М. Borgono и соавт. (1977) обнаружили высокую распростра-
ненность сердечно-сосудистых заболеваний, гиперкератоз, синд-
ром Рейно и акроцианоз. У детей эти симптомы были более
выражены, чем у взрослых. У 2 детей были описаны системные
поражения артерий и инфаркт миокарда. В мелких и средних
артериях большинства органов наблюдалось выраженное утол-
щение интимы.
Есть определенное сходство этих изменений с облитерирую-
щей ангиопатией Кавасаки, распространенной среди детей в
Японии,
18—568
273.
Большую практическую ценность имеют данные о массовых
'отравлениях неорганическим мышьяком в Японии при употреб-
лении молочного порошка, содержащего мышьяк в концентра-
ции 15—24 мг/кг. Отравились 12 000 грудных детей, из которых
погибли 130. В клинической симптоматике отравления отмеча-
лись набухание печени, меланоз кожи, анемия с относительным
лимфоцитозом, микрогематурия, расстройства сердечной деятель-
ности с патологическими изменениями на ЭКГ. В других слу-
чаях пищевых отравлений мышьяком отмечались аналогичные
симптомы, а также отек лица, в поздних стадиях — перифери-
ческие невропатии. Диагностическое значение имеют «линии
Мее» — поперечные белые полосы на ногтях, а также повышение
содержания мышьяка в волосах. Этот МЭ связывают SH-rpyn-
пы, содержащиеся в кератине волос. Указанные данные много-
кратно подтверждены на примере соседского арсеноза детей,
проживающих вблизи от производств с высоким выбросом
мышьяка в окружающую среду (медеплавильные заводы, золо-
тодобывающие рудники, тепловые электростанции и т. д.). Важ-
ным признаком профессионального контакта с неорганическим
мышьяком могут быть периферические невропатии, а также
снижение слуха и изменение на ЭКГ и ЭМГ.
% Канцерогенное действие мышьяка изучается в
течение 100 лет. Группа экспертов МАИР признала, что сущест-
вуют достаточные доказательства канцерогенности неорганиче-
ских соединений мышьяка для кожи и легких человека. Высо-
кий уровень смертности от рака легких зарегистрирован среди
рабочих, подвергавшихся воздействию неорганического мышьяка
при производстве и применении пестицидов, добыче золота и вы-
плавке цветных металлов, особенно меди. Эти данные совпада-
ют с основными результатами эпидемиологических исследова-
ний. У рабочих плавильных заводов с большой частотой наблю-
дался низко дифференцированный рак эпидермоидного типа.
Среди неоплазий кожи принято связывать с возможным воз-
действием мышьяка болезнь Боуэна, базально-клеточный и
плоскоклеточный рак. Плоскоклеточному раку, расположенно-
му на любой части тела, предшествует арсеногенный кератоз
кожи конечностей. В настоящее время раку кожи, связанному
с воздействием неорганического мышьяка, посвящено более.
1000 работ из различных районов мира. Он возникал, как пра-
вило, после очень длительного перорального употребления за-
грязненной воды или лекарственных препаратов. Повышенная
заболеваемость раком зависела от общей дозы принятого фау-
лерова раствора, которая при потреблении 1000 мл этого лекар-
ственного препарата была эквивалентна 7,6 г мышьяка (ВОЗ,
А1ышьяк, 1985).
На Тайване при обследовании 40 000 жителей была установ-
лена положительная зависимость между содержанием мышьяка
в колодезной воде и распространенностью рака кожи. Посколь-
ку ладонно-подошвенный гиперкератоз является характерным
признаком хронического арсеноза, именно этот элемент законо-
274
мерно накапливается в кератине волос, необходимо поставить
вопрос о повышенном содержании мышьяка в кератоцитах брон-
хиальных карцином эпидермоидного типа и в клетках различных
типов рака кожи. Прямых доказательств внутриклеточного на-
копления мышьяка пока не имеется. Трудно сомневаться в том,
что возникновение рака представляет собой следствие местных
изменений генетического аппарата клеток под влиянием соеди-
нений мышьяка. Однако есть основания полагать, что этому эта-
пу канцерогенеза в данном случае предшествует длительный
период депонирования мышьяка в кератине, что должно выра-
жаться в его биохимических и, возможно, цитопатологических
изменениях.
Разделяя осторожность экспертов ВОЗ в отношении припи-
сывания мышьяку важной роли в происхождении опухолей дру-
гой локализации (т. е. нелегочной и некожной), мы, однако, по-
лагаем, что такая редкая опухоль, как гемангиоэндотелиома
печени, в действительности является арсенозависимой. Большая
часть новообразований этого типа описана именно у больных,,
которые в течение 20—25 лет лечились фаулеровым раствором.
Это, разумеется, не снимает проблемы индивидуальной чувстви-
тельности к бластомогенному действию мышьяка. Справедливым
является то, что большинство патологов, которые ставят диаг-
ноз гемангиоэндотелиомы печени, в недостаточной степени ин-
тересуются терапевтическим и профессиональным анамнезом
больных в каждом конкретном случае. Мышьяк может высту-
пать и в качестве коканцерогена, угнетая иммунную систему
или воздействуя на обмен цинка или других компонентов пищи.
7.2. Бор
Бор жизненно необходим для некоторых растений и предста-
вителей морского и пресноводного фитопланктона (Chlorella
vulgaris), видов диатомовых водорослей (Dinophycae, Prasi-
nophycae). Его незаменимость для бобовых растений была про-
демонстрирована более 65 лет назад [Warrington К., 1923]. Де-
фицит этого МЭ вызывает у свеклы и других корнеплодов забо-
левание, известное как гниль сердечка, в связи с чем этот эле-
мент нашел применение в качестве микроудобрения для данных
культур. Механизм действия бора не раскрыт, несмотря на мно-
гочисленные данные о его влиянии на обмен углеводов, фено-
лов, лигнина, содержание ауксинов, фотосинтез, функцию кле-
точных мембран и синтез нуклеиновых кислот [Lovatt С. J.,
Dugger W. М„ 1984].
Установлено, что при дефиците бора в растениях накапли-
вается избыток фенольных соединений, вызывающий значитель-
ные тератологические изменения [Школьник М. Я., 1983].
Единственным известным соединением бора, встречающимся в
живой природе, является антибиотик боромицин, отвечающий
формуле C45H74O15NB, выделенный из культур стрептомицетов
[Dunitz A. et al., 1971]. Эссенциальность бора для животного
18*
275
организма долгое время отрицалась и была установлена только
в начале 80-х годов [Hunt С. D., Nielsen F. Н., 1981].
Как и в случае других «новых» МЭ, данные о содержании
бора в суточном рационе человека неоднозначны и различаются
почти на 2 порядка. Так, по мнению одних авторов, они состав-
ляют 7—20 мг [Kent N. L., McCance R. А., 1941; Ploquin J.,
1967], а по другим — всего 0,35—0,42 мг [Tipton I. J. et al.,
1966]. Промежуточное положение занимают финские рационы —
1,7 мг (Varo Р., KRivistoinen Р., 1980], 2,1—43 мг {Zook Е. J.,
Lehmann J., 1965] и английские — 2,8± 1,5 мг [Hamilton Е. J.,
Minski М. J., 1972—1973].
Бор из пищи, борной кислоты и ее натриевой соли быстро
всасывается и выводится в основном с мочой, у человека этим
путем удаляется более 90% от поступающего количества эле-
мента. Однако при меньших дозах выделение с мочой может
составить и 30%. В опытах на животных получены меньшие
величины. Так, в моче собак за 48 ч обнаружены 40% от вве-
денной дозы, у коров — 57 и 71%, у морских свинок и кроликов
за 48 ч — более 70% и за 120 ч—более 80%, у телок за сут-
ки— от 30 до 67—69%. Более высокие цифры соответствуют
большим введенным дозам и более длительным периодам наб-
людений. По данным Ю. И. Москалева (1985), условный чело-
век экскретирует с мочой около 77% бора, поступившего с пи-
щей и водой, а с калом — около 21%. Предполагают, что пери-
од полувыведения бора составляет около И сут. Эта оценка ис-
ходит из суточного поступления бора, равного 0,12 ммоль, и,
возможно, нуждается в уточнении.
Распределение в организме. В организме челове-
ка имеется около 20 мг бора [Schrauzer G. N., 1984]. Свежая
ткань различных органов содержит следующие концентрации
этого МЭ: печень—18,5 мкмоль/кг, почки, легкие, лимфатиче-
ские узлы — 55,5 мкмоль/кг; мышцы — 9,2 мкмоль/кг; мозг —
5,5 мкмоль/кг; семенники — 9,2 мкмоль/кг. Наиболее богата
этим МЭ костная ткань, содержащая 0,94±0,46 ммоль/кг бора
в районах с жесткой и 0,57±0,18 ммоль/кг в районах с мягкой
питьевой водой [Hamilton Е. J. et al., 1972, 1973]. По другим
данным, содержание бора в костной золе человека значительно
выше и колеблется в пределах 1,47—12,8 ммоль/кг при среднем
значении 5,64±0,16 ммоль/кг [Alexander G. V. et al., 1951]. Со-
держание бора высокое также в зубной эмали, составляющее
в среднем 1,68±0,24 мкмоль/кг [Losee F. et al., 1974] и
0,78 мкмоль/кг [Curzon М. Е., Crocker D. С., 1978], однако оно
очень сильно колеблется. Концентрация бора в волосах состав-
ляет 64,7—92,5 мкмоль/кг [Creason J. Р. et al., 1977]. У взрослых
в цельной крови содержится 9,2—18,4 мкмоль/л, в плазме —
7,4—19,4 мкмоль/л бора, а у новорожденных — 46,2—
55,5 мкмоль/л. Коровье молоко содержит 46,2—92,5 мкмоль/л
бора и в 100 раз богаче этим МЭ, чем женское молоко (5,55—
7,4" мкмоль/л). Более высокая концентрация бора свойственна
щитовидной железе овец — 2,31—2,77 ммоль/кг. Концентрация
276
бора в органах и тканях в значительной степени зависит от его
содержания в пище [Nielsen F. Н., 1986].
Жизненная необходимость бора. Первые опыты
на крысах, преследовавшие цель доказать жизненную необходи-
мость бора для организма животных [Orent-Keiles Е., 1941; Те-
resi J. D. et al., 1944; Skinner J. T., McHarque J. S., 1945], не
увенчались успехом, несмотря на то что в них использовались
рационы с очень низким содержанием этого элемента — 0,155—
0,163 мг/кг. Только почти 40 лет спустя это удалось сделать в
опытах на цыплятах, продемонстрировавших, что при гиповита-
минозе D3 именно дефицит бора снижает прирост живой массы
и повышает активность щелочной фосфатазы в плазме крови
[Hunt С. D., Nielsen F. Н., 1981]. В дальнейших опытах было
показано, что при гиповитаминозе D3 повышается потребность
животных в боре и что этот МЭ независимо от витамина D3
оказывает заметное влияние на обмен кальция, фосфора и осо-
бенно магния. Недостаточность магния излечивается бором.
Действие этого МЭ на обмен магния опосредуется по-ви-
димому, околощитовидными железами. Известно, например, что
дефицит магния вызывает у крыс гипопаратиреоидное состоя-
ние, которое при включении в рацион бора (3 мг/кг) наступает
значительно позже, чем при дефиците (0,3—0,4 мг/кг) этого МЭ.
Приведенные данные и ряд других дают основание полагать,
что физиологическая функция бора заключается в регуляции ак-
тивности паратгормона и через него — обмена кальция, магния,
фосфора и холекальциферола. Эта гипотеза подтверждается
также результатами исследований, показавших, что повышенное
содержание бора в рационе ослабляет признаки гиперпарати-
реоидизма у кроликов, вызванные дачей фтора и проявляющие-
ся гиперкальциемией, гипофосфатемией и угнетением почками
реабсорбции фосфора [Elsair J. et al., 1980].
У свиней борат натрия и в меньшей степени силикат магния
предупреждали утолщение компактной части трубчатых костей,
вызываемое фтором; повышенное содержание бора в рационе
вызывало изменения гистологической структуры околощитовид-
ной железы [Seffner W. et al., 1983]. Было также показано, что
этот МЭ нормализует гистологическую картину костной ткани
при отравлении фтором [Baer Н. Р. et al., 1977]. Высказыва-
лось предположение, что регуляция активности паратгормона
осуществляется бором на уровне клеточной мембраны путем
воздействия на синтез вторичного посредника, подобного цАМФ
[Jackson J. F., Chapman К. S. R., 1975], как это наблюдается
при регуляции бором активности растительных гормонов — аук-
сина, гибберелловой кислоты и цитокинина.
Влияние этого МЭ на обмен кальция подтверждается и це-
лым рядом других исследователей, которые также допускают,
что оно реализуется на уровне клеточной мембраны. Так, пока-
зано, что бор, как и кальций, способствует связыванию палит-
оксина (высокотоксичного яда кораллов) клеточными мембра-
нами [Habermann Е., 1983]. Аплазмомицин—-новый макро-
277
циклический антибиотик-ионофор, выделенный из Streptomyces
griseus, представляет собой симметричный димер, в котором в
качестве центрального атома присутствует бор [Chen Т. S. et
al., 1980]. Имеется также целый ряд наблюдений, показываю-
щих, что бор влияет на конформацию, активность компонентов
мембраны и ионный транспорт [Nielsen F. Н., 1986].
О роли бора в обмене кальция свидетельствуют данные, что
бороглюконат кальция эффективен при родовом парезе у ко-
ров, характеризующемся низким уровнем кальция и фосфора и
повышенной концентрацией магния в плазме крови животных,
тогда как лечение одними солями кальция значительно менее
эффективно. Аминоцианобораны и аминокарбоксибораны (бор-
ные аналоги а-аминокислот) эффективно предупреждают экспе-
риментальный артрит [Hall I. J. et al., 1980], а таблетки, содер-
жащие карбонат магния и борат натрия, рекомендуются при ле-
чении артрита (Newnham R. Е„ 1984].
В опытах in vitro показано, что ион бора может принимать
участие в ферментативных процессах. Он вызывает конкурент-
ное ингибирование ферментов, относящихся к двум различным
классам. Один из них включает флавин и пиридинзависимые
оксидоредуктазы—алкогольдегидрогеназу дрожжей, альдегид-
дегидрогеназу, ксантиндегидрогеназу, редуктазу цитохрома bs,
в которых борат-ион, видимо, конкурентно замещает NAD и
флавин, так как имеет выраженное сродство к цис-гидроксиль-
ным группам. Другой класс — это ферменты, у которых борат
или производные борониевой кислоты связываются с активным
центром фермента. Эта группа включает химотрипсин, субти-
лизат, глицеральдегидфосфатдегидрогеназу (КФ 1.2.1.12) [Wol-
ny М„ 1977].
Борозы — МТОЗы человека и животных, возникающие при
употреблении пищи с избыточным содержанием бора.
Распространенные представления о химической инертности
этого МЭ нуждаются в сопоставлении с данными медицинских
наблюдений. В частности, широко используемая в медицине
борная кислота, ошибочно считавшаяся безвредной, легко всасы-
вается и концентрируется в мозге, печени, жировой ткани.
Острая интоксикация соединениями бора, в частности дибора-
ном, вызывает острый бороз с симптомами литейной лихорад-
ки— чувством сдавления грудной клетки, кашлем, тошнотой,
ознобом. Еще более токсичен пентаборан, вызывающий пора-
жение ЦНС (возбуждение, тремор, судороги, миоз), а также
снижение артериального давления, аритмию, сердечную недо-
статочность, нарушение дыхания, функции печени, почек. Ост-
рая интоксикация декабораном вызывает беспокойство, угне-
тение дыхания, нарушение координации, судороги, брадикар-
дию, гипотонию, помутнение роговицы. При хронической инток-
сикации— выраженное нейротоксическое действие, некроз и
ожирение печени, гематурия, изменение почечных канальцев.
Лечение симптоматическое [Каспаров А. А., 1976].
Хронический бороз сопровождается патологическими изме-
278
нениями верхних дыхательных путей и легких, в том числе пнев-
мокониозом, которые возникают в результате длительного вды-
хания борсодержащих соединений. Профилактика отравлений
требует герметизации оборудования, механизации ручных опе-
раций, а при работе с гидридами бора применения средств ин-
дивидуальной защиты (противогазов) с использованием специ-
альных поглотителей, а также спецодежды, перчаток.
Борный энтерит — эндемическое заболевание желудочно-ки-
шечного тракта людей и животных в регионах с повышенным
содержанием бора в окружающей среде, особенно в растениях.
Впервые описан В. В. Ковальским и соавт. (1978). Клиническая
картина и патологическая анатомия борного энтерита у людей
недостаточно изучены.
В Западной Сибири типичная борная биогеохимическая про-
винция занимает степные районы Омской, Новосибирской, Пав-
лодарской областей, а также Алтайского края. В воде озер этой
провинции много бора. Органы и ткани, а также кровь живот-
ных, вскармливающихся растительностью этого региона, содер-
жат повышенные количества бора. Кроме энтеритов, у живот-
ных регистрируются анемии, легочные заболевания, наблюдает-
ся падеж молодняка, особенно в засушливые годы. Этиологи-
ческим фактором этих заболеваний являются избыток бора и
дефицит меди в биосфере данной провинции [Скуковский Б. А.,
Ковальский В. В., 1985].
К- И. Плотников и соавт. (1985) изучили микроэлементный
состав крови и печени животных Кулунды и показали, что со-
держание меди и кобальта у них значительно понижено, а кон-
центрация бора и молибдена существенно повышена. Различа-
ют несколько клинических форм массовых заболеваний жи-
вотных.
Из данных этих исследований очевидно, что заболевание,
описанное под названием «борный энтерит», вероятно, пред-
ставляет собой полимикроэлементоз, причем органами типично-
го поражения являются не только желудок и кишечник, но так-
же легкие и, по-видимому, в не меньшей степени печень.
7.3. Бром
Бром — представитель группы галогенов, содержание его в
земной коре достигает 1,6-10—4%. Соединения брома встреча-
ются в воде некоторых соляных озер, в морской воде его кон-
центрация составляет 0,005%. Кроме того, он входит в состав
ряда хлорсодержащих минералов. Наиболее богаты бромом бо-
бовые растения, такие как фасоль, горох и чечевица.
В промышленных условиях бром получают из морской воды
и концентрированных рассолов соляных озер. Бром и его со-
единения используются в производстве антидетонаторов горюче-
го, в органическом синтезе, а также в фармацевтической, фото-
и кинопромышленности. Некоторые броморганические соедине-
ния применяются в качестве инсектицидов. Неорганические со-
единения брома в виде бромида калия, натрия, кальция и аммо-
279
ния, а также броморганические препараты (бромкамфора,
бромизовал и др.) широко используются в медицинской прак-
тике.
Бром, как и другие галогены, хорошо всасывается в кишеч-
нике и сравнительно равномерно распределяется в организме.
Однако наибольшие его концентрации определяются в щитовид-
ной железе и почках. Необходимо подчеркнуть, что концентра-
ция ионов брома выше в тканевой жидкости, чем в клетках.
Выделение брома из организма происходит с мочой сравни-
тельно долго, в течение нескольких недель. Скорость экскреции
этого МЭ находится в значительной степени в зависимости от
содержания хлоридов в организме. Так, при низком содержа-
нии последних наблюдаются аккумуляция брома, а также пони-
женное его выделение с мочой. Повышенное поступление хло-
ридов с пищей сопровождается ускоренным выделением брома.
Физиологическая роль брома в организме связана с его из-
бирательным усиливающим влиянием на тормозные процессы
в нейронах коры большого мозга.
В условиях воздействия профессиональных вредностей в
случае вдыхания паров брома наблюдаются кашель, нередко
носовые кровотечения, головокружение, головные боли, иногда
рвота, понос, миалгии. При хроническом поступлении брома по-
является аллергическая или кореподобная кожная сыпь, слизи-
стая оболочка рта принимает коричневую окраску, наблюдают-
ся конъюнктивит, явления бронхоспазма с осиплостью голоса.
Поступление с воздухом больших концентраций брома может
привести к химическому ожогу легких и смертельному исходу.
Попадание жидкого брома на кожу сопровождается ожогом, ее
пигментацией и образованием плохо заживающих язв [Ленке-
вич М. и др., 1976].
Бром изм — хроническое отравление бромом и его соедине-
ниями (катаральный ринит, бронхит, конъюнктивит, энтерит).
Неврологические симптомы бромизма — сонливость, атаксия,
снижение болевой чувствительности, слуха, зрения, ослабление
памяти и др.; психотические нарушения в форме делирия со
зрительными, слуховыми, тактильными и вкусовыми галлюци-
нациями [Муратов В. К и др., 1976].
Бромодерма — специфическое поражение кожи при дли-
тельном приеме препаратов брома, особенно бромида калия,
у лиц с повышенной чувствительностью к этому МЭ.
Врожденная бромодерма встречается у грудных
детей, матери которых принимали бромиды во время беремен-
ности.
Ятрогенные формы бромизма более подробно изложены в
главе 9.
7.4. Фтор
В основе биологического действия иона фтора лежит его
способность эффективно замещать ион гидроксила не только
в апатите костной ткани, но и в неминерализованных тканях,
280
а также, по-видимому, в активном центре ферментов. Разные
ученые придерживаются неодинаковых взглядов на значение
фтора как биоэлемента. Эссенциальность фтора для животного
организма нельзя считать окончательно доказанной, хотя в ряде
случаев при дефиците этого МЭ у животных отмечены задерж-
ка роста, снижение плодовитости и продолжительности жизни
[Schroeder Н. А., 1968; Schwartz К-, Milne D. В., 1972]. В.В.Ко-
вальский твердо считал, что фтор является незаменимым био-
элементом.
Почти 99% всего организменного фтора находится в твер-
дых тканях в составе апатита — основного фосфата кальция,
отвечающего формуле Саю(РО4)6(ОН)2. В минеральной фазе
преобладают мелкие кристаллы, погруженные в органическую
матрицу. Фторид-ион превосходит все прочие ионы по своей спо-
собности замещать ОН" благодаря близости их ионных радиу-
сов, одинаковым заряду и степени гидратации, равной двум.
Фтор включается в апатит либо в период формирования первич-
ного кристалла, либо путем замещения ОГК в преобразован-
ном минерале. In vivo преобладает второй из указанных про-
цессов, особенно у взрослых особей. В результате реакции за-
мещения образуется смешанная форма апатита, отвечающая
формуле Саю(РО4)6(ОН)2-хЕх. Содержание фтора в чистом
фторапатите составляет приблизительно 2 моль/кг — величина,
почти не встречающаяся в твердых тканях. Содержание фтора
в кости и зубной эмали составляет обычно 0,05 моль/кг и сви-
детельствует об отношении ОН- к фтору в молекуле апатита
как 40:1. Большинство минерализованных тканей содержит
менее 0,5 моль/кг фтора, исключая эмалоид рыб, очень богатый
этим МЭ. Среди твердых тканей первое место по содержанию
фтора занимает цемент зуба, за ним следуют кость, дентин и
эмаль.
Содержание фтора в кости колеблется в широких пределах:
от 50 мг/кг в ребре новорожденного до 1500 мг/кг в ребре боль-
ного флюорозом [Singer L. et al., 1974]. Еще большие концен-
трации этого МЭ найдены в костях экспериментальных живот-
ных. При умеренной нагрузке фтором в костях взрослых содер-
жится 1000—5000 мг/кг этого МЭ, причем губчатые кости бога-
че фтором, чем трубчатые. Основными факторами, влияющими
па содержание фтора в костях, являются возраст и нагрузка
фтором. Кость играет важную роль в регуляции концентрации
фтора во внеклеточной жидкости благодаря способности быст-
ро связывать избыток этого МЭ. В этой связи в костной ткани
происходит постепенное накопление фтора на протяжении всей
жизни человека, хотя оно и ослабевает в пожилом возрасте.
При дефиците фтора происходят его мобилизация из костной
ткани и поступление во внеклеточную жидкость.
Мягкие ткани бедны фтором, если они не находятся с ним
в непосредственном контакте, как, например, легкие, или не за-
тронуты патологической кальцификацией, как, например, стен-
ка аорты при атеросклерозе.
281
Содержание фтора в клетках зависит от градиента pH.
Внутри клеток млекопитающих значение pH обычно ниже, чем
во внеклеточной жидкости (7,2—7,4 — в гепатоцитах; 6,9—
7,0 — в мышце против 7,4 в плазме крови). В связи с этим про-
исходит изменение направления градиента концентрации фто-
рид-иона [Armstrong W., Singer L., 1980]. По этой же причине
мышца, имеющая по сравнению с печенью менее высокое значе-
ние pH, беднее фтором, чем гепатоциты.
Плазма содержит около 7,5% фтора от его общего количест-
ва в крови в его физиологически наиболее активной форме.
В плазме присутствуют как ионная, так и органическая формы
фтора, из которых только уровень первой подлежит физиологи-
ческой регуляции.
Концентрация фторид-иона в плазме или сыворотке крови
колеблется на уровне нескольких микромолей и в норме зависит
от длительности поступления в организм этого МЭ и ряда дру-
гих факторов. С возрастом концентрация фтора в плазме кро-
ви умеренно повышается [Hanhijahrvi Н., 1975; Cowell D. С.,
Taylor W. Н., 1984] одновременно с накоплением фтора в ске-
лете.
Фтор обнаруживает выраженную избирательность к твердым
тканям и принимает участие в начальных этапах их минерали-
зации. В то же время никаких нарушений этого процесса у под-
опытных животных, содержавшихся на дефицитном по фтору
рационе в течение нескольких поколений подряд, обнаружить
не удалось, так же как и добиться снижения его концентрации
в костях ниже уровня, необходимого, согласно теоретическим
расчетам, для наступления процесса минерализации (0,01—
0,1 мкмоль/л). Фтор обладает высоким сродством к белку мат-
рикса эмали и, включаясь в эмаль зубного зачатка еще до на-
чала его минерализации, может способствовать формированию
центров кристаллизации (нуклеации) апатита [CrenshawМ. А.,
Bawden J. W., 1981].
К появлениям недостаточности фтора большинство исследо-
вателей относят остеопороз и кариес зубов, имеющих весьма
сложную природу. Предупреждение их возможно как при недо-
статочности, так и избытке фтора. Таким образом, имеющиеся
в настоящее время данные дают основание рассматривать явно
выраженный защитный эффект фтора при этих заболеваниях
скорее как результат фармакологического, а не физиологическо-
го действия этого МЭ. Несмотря на эти сомнения, в медицине
распространена точка зрения, что именно кариес зубов являет-
ся наиболее известным маркером гипофтороза у человека.
У животных, испытывающих недостаточность фтора, в боль-
шинстве случаев отмечается снижение активности ряда фермен-
тов, таких как щелочная и кислая фосфатаза костной ткани,
изоцитратдегидрогеназа печени и др., на 10—20%. Однако эти
заметные различия в активности ферментов пока не удается
связать с клиническими проявлениями дефицита фтора, что яви-
лось бы решающим критерием его жизненной необходимости.
282
Фтор может играть существенную роль не только в началь-
ных стадиях минерализации твердых тканей, но и предупреж-
дать их деминерализацию. При кариесе зубов и остеопорозе ми-
неральная часть костной ткани растворяется под воздействием
кислот. В первом случае кислая среда создается бактериями,
населяющими поверхностные слои эмали, а во втором — остео-
кластами и другими костными клетками, резорбирующими ми-
неральные компоненты кости. Твердо доказано, что фтор при-
дает кристаллам фторапатита большую упорядоченность, сни-
жая тем самым их растворимость при физиологическом значе-
нии pH. Вместе с тем поскольку экзогенный фтор замещает
гидроксил-ион в предобразованных кристаллах гидроксилапа-
тита, то это замещение происходит в первую очередь в наруж-
ных слоях эмали толщиной 1—5 мкм, что снижает их раство-
римость даже при незначительном общем повышении концен-
трации фтора в зубах. Рассматриваются и другие более слож-
ные причины защитного действия фтора, такие как изменение
термодинамических характеристик эмали, а также облегчение
процессов реминерализации эмали под влиянием этого МЭ, не
говоря уже о его антибактериальном действии [Messer Н. Н.,
1984].
Как свидетельствуют эпидемиологические данные, среди на-
селения, использующего воду с повышенным содержанием фто-
ра (4—6 мг/л), отмечаются снижение тяжести атеросклероза и
уменьшение летальности от сердечно-сосудистых заболеваний
[Taves D. R., 1978].
Механизма активного транспорта фтора через плазматиче-
скую мембрану, подобного таковому для хлора, по-видимому,
не существует, а различие между обоими ионами по степени
гидратации и ионным радиусам не позволяет фтору использо-
вать транспортную систему хлора. В этой связи всасывание
фтора происходит по принципу простой диффузии.
Всасывание. Фторид-ион, поступающий с пищей, всасы-
вается в желудке и тонкой кишке, причем скорость и степень
этого процесса определяются величиной дозы, типом соедине-
ний фтора, качественным и количественным составом пищи.
Фтор быстро всасывается, хотя и со значительно меньшей ско-
ростью, чем хлор и вода. Умеренные дозы растворимых соедине-
ний фтора всасываются в пищеварительном тракте на 50% в
среднем за 30 мин и полностью за 90 мин. В норме человек
усваивает около 80% фтора, содержащегося в пище. Раствори-
мые соединения этого МЭ (NaF и Na2SiF6), используемые для
фторирования воды, усваиваются почти полностью. Фтор из
костной муки доступен в меньшей степени и усваивается менее
чем на 50%. У человека в отличие от животных кальций, маг-
ний и фосфаты не оказывают существенного влияния на всасы-
вание фтора при его обычном содержании в пище [Spencer Н.
et al., 1980]. Гидрат окиси алюминия, широко используемый в
медицинской практике для понижения кислотности желудочно-
го сока, снижает усвоение фтора более чем на 50%. Пища, бо-
283
гатая липидами, задерживая опорожнение кишечника, повыша-
ет усвоение этого МЭ, что имеет особое значение при его ток-
сических дозах. Фтор, кроме того, весьма эффективно усваива-
ется легкими, что может существенно повысить его поступление
в организм в случаях загрязнения воздуха этим МЭ.
Транспорт и регуляция обмена. Поскольку ион
фтора способен связываться с амидами, допускалось, что он мо-
жет транспортироваться белками плазмы крови, однако это не
нашло экспериментального подтверждения [Armstrong W. D.,
1982]. После всасывания фтора из пищеварительного тракта
наблюдается кратковременное возрастание его концентрации в
кровеносном русле, которая, однако, вскоре снова возвращается
к норме. Концентрация фтора в мягких тканях стабильна, ко-
леблется в узких пределах и, по-видимому, мало зависит как от
поступления этого МЭ в организм, так и от изменений его со-
держания в плазме крови. В то же время фториды тканей легко
обмениваются с фторидами внеклеточной жидкости, о чем сви-
детельствует быстрое перераспределение между ними 18F~ пос-
ле его внутривенного введения. Однократный прием 1,5 мг фтора
(в виде NaF), соответствующий суточному количеству этого МЭ
во фторированной питьевой воде, вызывает у человека 5-крат-
ное повышение содержания фтора в плазме крови с максималь-
ным подъемом через 1 ч и возвращение к исходному уровню
через 3—5 ч [Ekstrand J., 1977]. Более высокие дозы (до 10 мг
фтора) вызывают более значительный подъем его концентра-
ции и большую продолжительность его нахождения в крови.
Имеющиеся данные свидетельствуют, что уровень фтора в плаз-
ме крови, видимо, не находится под строгим гомеостатическим
контролем в отличие от кальция, натрия и хлора. Однако меха-
низмы, регулирующие содержание фтора в плазме крови, все
же существуют. Эта регуляция осуществляется в основном дву-
мя системами — скелетом и почками [Smith F. А., 1966]. Огра-
ниченная реабсорбция фтора в почечных канальцах ведет к
его быстрому выделению с мочой, однако основным механизмом
удаления этого МЭ из кровотока является его фиксация скеле-
том. У человека при умеренном поступлении в организм фтора
около 50% этого МЭ депонируется в скелете и столько же уда-
ляется с мочой [Largent Е. J., 1961]. Величина дозы, депонируе-
мая скелетом, снижается с возрастом и по мере насыщения ор-
ганизма фтором. При высокой степени насыщения с мочой вы-
деляется почти 90% от введенной однократной дозы этого МЭ.
При пониженном поступлении фтора происходит его мобилиза-
ция из костной ткани, способствующая сохранению нормаль-
ного уровня фтора во внеклеточной жидкости.
Выделение. Основным путем выделения фтора из орга-
низма является моча, с которой удаляется около 90% этого эле-
мента. С калом теряется 7—15%, включающие как неусвоен-
ный, так и эндогенный фтор. С потом выделяется лишь не-
большое количество этого МЭ, хотя в отдельных случаях оно
может быть и весьма значительным. Фтор очень быстро выво-
284
дится с мочой. У человека I8F появляется в моче через несколь-
ко минут после его приема per os, а в течение 3—4 ч с мочой
выводятся 20—30% полученной дозы [Carlson С. Н. et al.,
1960; Hodge Н. С. et al., 1970]. Фтор покидает кровоток в ре-
зультате клубочковой фильтрации и ограниченной реабсорб-
ции его в почечных канальцах. Так, в опытах на 2 доброволь-
цах было показано, что у человека реабсорбция фтора в почеч-
ных канальцах составляет 51—63%, тогда как для хлора она
равна 99,5%. У экспериментальных животных разброс данных
значительно выше и колеблется в пределах 20—70% [Whit-
ford G. М., Pashley D. Н., 1979]. Клиренс фтора в 26—230 раз.
превышает клиренс хлора [Габович Р. Д., Минх А. А., 1979],
и у здоровых взрослых лиц существует прямая связь между со-
держанием фтора в питьевой воде и моче [Smith F. А., 1970].
Механизм реабсорбции этого МЭ в почечных канальцах
включает, по-видимому, диффузию недиссоциированного HF.
Изменяя кислотно-щелочное равновесие у крыс, собак и чело-
века, было показано, что реабсорбция фтора в почечных каналь-
цах обратно пропорциональна величине pH мочи [Whitford G. М.,.
Pashley D. Н., 1979]. Алкалоз благоприятствует выведению фто-
ридов, тогда как ацидоз приводит к их задержке, повышению-
концентрации в крови и отложению в скелете.
Кислотно-щелочной статус является, таким образом, основ-
ным фактором, определяющим баланс фтора в организме, од-
нако он в значительной степени зависит и от объема выделяе-
мой мочи, который в нормальных физиологических условиях
имеет решающее значение в выведении этого МЭ [Schiffle Н. Н„
et al., 1981]. При обследовании 400 взрослых лиц, употребляю-
щих пищу с пониженным уровнем фтора, не обнаружено сущест-
венной корреляции между величиной pH мочи и количеством
выделяемого фтора [Massmann W., 1981], откуда следует, что
кислотно-щелочной баланс играет главную роль в выделении;
фтора только в экстремальных ситуациях.
Выделение фтора через желудочно-кишечный тракт весьма*
вариабельно, составляет 5—30% и зависит как от содержания-
неабсорбированных фторидов пищи, так и от его содержания
в организме. Это показано исследованием радиоактивного изо-
топа I8F после внутривенного его введения [Messer Н. Н., 1984]..
Другие пути выведения фтора (пот, слюна, слущивание клеток
эпидермиса, волосы), играют, как правило, второстепенную роль.
Фтор легко проникает через плаценту человека. 18F, введен-
ный матери внутривенно, обнаруживается в первой же пробе-
крови плода, взятой через 4 мин после инъекции [Ericsson Y.,
et al., 1962]. Несмотря на легкость проникновения фтора через!
плаценту, неоднократно высказывалось мнение о существова-
нии плацентарного барьера по отношению к этому элементу
[Gedalia I., 1970]. Так, приводились данные, что у человека кон-
центрация фтора в пупочной вене составляет лишь 75% от его
концентрации в крови матери [Chen N. S., Taves D., 1974], не
285
Рис. 51. Картосхема географического распространения различных концентраций фтора в водоисточниках СССР [Жаворон-
ков А. А., 1977].
согласующиеся, однако, с данными другой работы, в которойне
были обнаружены достоверные различи по содержанию фтора
между материнской кровью и кровью пупочной вены, взятой при
кесаревом сечении [Armstrong W. D. et al., 1970].
Содержание фтора в скелете плода обычно невелико, хотя
и возрастает в процессе внутриутробного развития. Отложение
I8F в скелете плода после введения этого изотопа матери про-
исходит в значительно меньшей степени, чем в материнском ске-
лете, что также рассматривалось как свидетельство существо-
вания плацентарного барьера. Однако этот вопрос осложняется
еще и тем обстоятельством, что в плаценте ко времени родов
образуются петрификаты, способные связывать фтор. Причиной
ограниченного поступления фтора в организм плода может слу-
жить не столько существование активного барьера, сколько
способность скелета и почек матери исключать фтор из крове-
носного русла.
Контакт человека с фтором и его соединениями может быть
чрезвычайно разнообразным. Это обусловливает также разно-
образие клинических форм заболеваний, вызванных избытком-
данного МЭ. Если флюороз в настоящее время достаточно изу-
чен, то этого нельзя сказать о фтордефицитных состояниях че-
ловека, которые мы предлагаем выделять под названием «гипо-
фтороз». Медицинская практика накопила достаточно наблюде-
ний, согласно которым кариес зубов следует рассматривать в
качестве маркера гипофтороза внешней среды, в первую оче-
редь питьевой воды (рис. 51). Кариес зубов является также
частным выражением гипофтороза организма, тем более что
уже в течение многих лет он поддается успешной профилактике
путем искусственного фторирования воды. Изучение клиниче-
ского разнообразия кариеса зубов у различных контингентов
людей позволяет по крайней мере провизорно говорить о раз-
личных формах гипофтороза. Действительно, клиническая прак-
тика знает примеры как чрезвычайно медленно текущего, так
и крайне быстрого и разрушительного развития кариеса зубов.
Это совпадает с определенными физиологическими состояния-
ми, а также с особо неблагоприятными условиями существова-
ния, в частности с голоданием.
Течение кариеса в различные возрастные периоды имеет свои
характерные особенности. С нашей точки зрения, есть основа-
ния различать следующие формы гипофтороза: внутриутробный
гипофтороз, гипофтороз детей грудного и раннего дошкольного
возраста, гипофтороз детей школьного возраста, гипофтороз
подростков и юношей, гипофтороз взрослых, гипофтороз в стар-
ческом возрасте, гипофтороз беременных. Возникают законные
сомнения, в какой степени оправдано подразделение гипофторо-
зов по определенным возрастным периодам и физиологическому
состоянию. Несмотря на то что проблема гипофтороза еще не-
достаточно изучена в патогенетическом и нозологическом отно-
шении, нельзя упускать из виду, что одно из основных свойств
фтора как биоэлемента выражается в его свойстве поддержи-
287"
зать и стимулировать рост и развитие организма. Это установ-
лено на Животных.
В настоящее время мало изучены клиническая картина и
патоморфология врожденного гипофтороза. Они должны выра-
жаться в признаках известного недоразвития организма, в част-
ности его скелета. Более детальные исследования этой формы
пока отсутствуют. Очень большой интерес вызывают функцио-
нальное состояние ферментов слюны и морфология (созрева-
ние) тканей слюнных желез и эпителия полости рта. Врожден-
ный гипофтороз, очевидно, наблюдается в эндемичных зонах,
которые топографически совпадают с зонами широкого распро-
странения кариеса зубов у детей и взрослых.
Гипофтороз детей грудного и раннего дошкольного возраста
характеризуется теми же признаками, рост детей в той или в
иной степени замедлен. В этом периоде регистрируются такие
признаки гипофтороза, как запаздывание прорезывания зубов
и специфическое поражение зубов молочного прикуса карие-
сом. Причинная связь замедленного прорезывания зубов и ги-
пофтороза требует дальнейших исследований, тем более что в
этом периоде детства на передний план выступает такая рас-
пространенная патология, как рахит. Вопрос о гипофторозном
компоненте в патогенезе гиповитаминоза D требует специаль-
ных исследований. Заслуживает внимания также влияние гипо-
фтороза на эндокринный и метаболический статус грудных де-
тей, в особенности на системы, регулирующие кальциевый го-
меостаз. Пристального внимания заслуживает исследование
ферментов слюны, а также всего ферментного комплекса, ко-
торый участвует в процессах минерализации тканей зубоче-
люстного аппарата и всего скелета в целом.
Гипофтороз детей школьного возраста, выражающийся в ка-
риесе зубов, по-видимому, представляет собой одно из наиболее
массовых заболеваний молодого поколения планеты. Не подле-
жит сомнению, что кариес зубов постоянного прикуса является
не единственным выражением гипофтороза. Общеизвестно также,
что дефицит фтора — это одна из причин кариеса, но роль его
в происхождении этой патологии очень велика. К сожалению,
среди педиатров еще не существует прицельного интереса к не-
стоматологическим проявлениям гипофтороза, а они, безуслов-
но, имеются, хотя и мало изучены. Существуют косвенные ука-
зания на связь гипофтороза с тонзиллопатиями, рахитом, не-
полноценностью иммунного статуса и нарушениями обмена
кальция. Точно установлено, что небольшие дозы фтора стиму-
лируют иммунные реакции. Учитывая, что гипофтороз и гипо-
витаминоз D, а также неадекватный режим питания часто соче-
таются в одних и тех же группах детского населения, анализ
причинно-следственных отношений в этих случаях требует осо-
бой осторожности. Специального внимания, по-видимому, за-
служивает связь аденоидов и гипофторозных состояний. При-
цельные исследования этого вопроса нам неизвестны.
Внедрение в практику здравоохранения фторирования воды
288
показало, что среди детей младшего школьного возраста явно
снижается заболеваемость кариесом зубов спустя 2—3 года
после начала этого мероприятия. К сожалению, другие поло-
жительные результаты влияния фторирования на выраженность
и течение гипофтороза в детском и подростковом возрасте пока
мало изучены. С этих позиций необходимо организовать глубо-
кое изучение влияния различных доз фтора на организм. Пос-
ле многолетних исследований А. А. Жаворонков (1977) пришел
к выводу, что оптимальные дозы фтора оказывают в широком
смысле слова трофическое, т. е. стимулирующее, действие на
все производные соединительной ткани. Это, в частности, отно-
сится к различным видам коллагена, костной и хрящевой
ткани,
Гипофтороз взрослых представляет собой реальную пробле-
му во всех обширных природных зонах эндемического кариеса
зубов. Однако он еще прицельно не исследован. Вероятно, его
проявления будут обнаружены при заживлении ран и костных
переломов.
Гипофтороз в старческом возрасте представляет собой ре-
альное явление для эндемичных зон кариеса зубов, а также
для всех обширных контингентов населения, находящегося в
условиях несбалансированного по МЭ пищевого режима. Осо-
бую остроту проблема старческого гипофтороза имеет для
столь частых в преклонном возрасте костных переломов. Это,
по-видимому, в наибольшей степени касается женщин старче-
ского возраста, значительное число которых погибает после
переломов шейки бедра. Патологам, знакомым с особенностями
строения костной ткани в старческом возрасте, хорошо извест-
ны резкие атрофические изменения костных балок в сочетании
с хрупкостью не только спонгиозы, но и компакты длинных труб-
чатых костей. Хрупкость зависит не только от деминерализации
коллагена кости, но и от ослабления его механической прочно-
сти. На этом основано лечебное применение оптимальных доз
фтора при старческом остеопорозе. Экспериментально показа-
но, что большие дозы фтора способствуют интенсивной про-
дукции остеоидной ткани, которая, однако, не всегда оссифици-
руется [Жаворонков А. А., 1977].
Известно, что ранняя диагностика остеопороза сложна, так
как он протекает бессимптомно до тех пор, пока не вызовет
тяжелых осложнений, часто переломов. У большинства взрос-
лых людей потеря минеральных веществ из скелета происходит
постоянно на протяжении всей жизни. Однако у женщин в те-
чение 1—2 лет постклимактерического периода потеря мине-
ральных веществ особенно велика. У мужчин соответствующая
ускоренная потеря минеральных веществ может наблюдаться
в возрасте 60—65 лет.
Впервые фторид натрия был использован для лечения остео-
пороза С. Rich и J. Ensinck (1961), он улучшал минерализацию
костей, однако не снижал числа переломов. Последующие рабо-
ты в этом направлении дали неоднозначные результаты. И толь-
19—568
289
ко после работ J. Inkovaara и соавт. (1975), которые показали
необходимость комбинации фтортерапии и введения кальция,
клиницисты стали получать определенный положительный эф-
фект. В ряде случаев в комплексную терапию стали включать
витамин D [Jowsey L. I. et al., 1972; Riggs В. L. et al., 1982].
Наиболее эффективной в лечении постклимактерического остео-
пороза оказалась комбинация кальция, фтора, витамина D в
эстрогенов. При этом важно подчеркнуть, что наибольший эф-
фект проявляется на 2-м году лечения. Это нашло свое отраже-
ние в рентгенологической картине повышенной плотности кос-
тей [El-Khoury G. Y. et al., 1982].
В лечении остеопороза использовались, как правило, высо-
кие дозы (40—100 мг) фторида натрия в день, в результате
чего у некоторых больных отмечались побочные явления, в пер-
вую очередь со стороны желудка и кишечника [Riggs В. L. et
al., 1982]. В связи с этим в настоящее время выпускают таблет-
ки с фтором, покрытые оболочкой. Исследованиями A. Dixon
(1983) установлено, что минимальной эффективной дозой для
взрослого человека является 30 мг фторида натрия в сочета-
нии с 1 г кальция в день. Высокая эффективность терапии фто-
ром выявлена при отоспонгиозе (доза 40—60 мг в день), одна-
ко необходимо многолетнее применение (до 8 лет) [Caus-
se J. R„ 1980].
Эффективность использования фторированной питьевой воды
при остеопорозе изучена в Финляндии на группе женщин с
длительной неподвижностью [Alhava Е. М., 1980]. Установле-
но, что 20-летнее потребление фторированной воды приводило-
к улучшению структуры губчатых костей у этих больных по
сравнению с соответствующей группой женщин из местности с
низким содержанием фтора в воде.
Таким образом, представленные данные свидетельствуют о
четком положительном действии комплексной терапии фтором
старческого и постклимактерического остеопороза и отосклеро-
за. Однако необходимы дальнейшие исследования этого вопро-
са с целью выработки оптимальных сроков и наиболее безопас-
ных и эффективных остеотропных компонентов.
Гипофтороз беременных — предположительно выделяемая
форма, так как прямых исследований влияния дефицита фто-
ра на организм беременной в разные периоды беременности
пока проведено мало. Вместе с тем общеизвестно, что беремен-
ность протекает со значительным напряжением минерального
обмена и в редких случаях на ее фоне возникает тяжелая па-
тология костной системы в виде остеомаляции. Важно, что и
при нормальной, и при патологической беременности наблюда-
ются явные клинические признаки ухудшения течения кариеса
зубов. Нередко у практически здоровых беременных женщин на
вестибулярной поверхности зубов появляются множественные
меловидные пятна, на месте которых образуются дефекты эма-
ли, сопровождающиеся значительной болезненностью.
Степень развивающейся в этих участках деминерализации
290
тканей зуба варьирует. Специальные исследования кариеса бе-
ременных немногочисленны. Известно, что так называемый ост-
рый кариес, а также его хронические формы, по предложению
И. Г. Лукомского (1936), лечатся с применением соединений
фтора. Определенное значение имеет предложенное И. Г. Лу-
комским профилактическое фторирование зубов. По-видимому,
эти мероприятия целесообразны в тех случаях, когда в резуль-
тате гормональных сдвигов возникает угроза эндогенно обус-
ловленной деминерализации тканей зубов.
Проблема эндемического гипофтороза в настоящее время
недооценивается главным образом из-за ее недостаточной изу-
ченности. Несмотря на огромную распространенность кариеса
.зубов, фторирование питьевой воды осуществляется в недоста-
точных объемах. По-видимому, неправильно возлагать особые
надежды при борьбе с гипофторозом только на фторирование
питьевой воды. Это мероприятие нужно сочетать с какими-то
другими мероприятиями, направленными на коррекцию содер-
жания фтора в организме. В этом отношении большого внимания
заслуживает предложение А. И. Рыбакова, разработавшего дие-
ты с разным содержанием фтора и других МЭ для населения,
проживающего в зонах с низким содержанием фтора, а также
для жителей регионов с оптимальным содержанием этого МЭ.
В этом разделе уместно затронуть вопрос о возможных пу-
тях профилактики переломов шейки бедра у пожилых и старых
людей. Общеизвестно, что из общего числа больных с этой тя-
желой патологией большинство из них являются лицами жен-
ского пола. Весьма вероятно, что эффективная профилактика
переломов шейки бедра должна быть комплексной. Кроме про-
филактического фторирования организма (в разумных дозах и
при допустимых интервалах времени), свое место в ней долж-
но найти и применение анаболических стероидов в сочетании
с приемом витаминов, введением солей кальция и других остео-
тропных МЭ.
Флюороз — МТОЗ, вызываемый избыточным
поступлением фтора в организм, В течение дли-
тельного времени считалось, что флюороз выражается только
элективным поражением зубов и скелета. В настоящее время
в значительной степени благодаря многолетним исследованиям
отечественных ученых выяснено, что, кроме флюороза скелета
и зубов, существуют многие другие формы этого распростра-
ненного заболевания [Авцын А. П., Жаворонков А. А., 1981].
В многочисленных исследованиях Е. Я. Гирской (1971),
Д. М. Зислина и В. И. Токаря (1977), Н. А. Богданова и
Е. В. Гембицкого (1975) существуют указания, что при профес-
сиональном флюорозе могут регистрироваться признаки пора-
жения многих органов и систем.
В большом числе исследований, посвященных токсикологии
фтора, доказано его воздействие на ферментативные процессы.
Следует, однако, иметь в виду, что концентрации фтора, ис-
пользуемые в биохимических исследованях по ингибированию
19* 291
ферментов, находятся в пределах 1—10 ммоль/л, что значитель-
но превышает физиологические концентрации этого МЭ, не вы-
ходящие за пределы микромолекулярного уровня. Внутрикле-
точные концентрации фтора, способные воздействовать на фер-
ментативные процессы, могут быть достигнуты только при ост-
ром отравлении фтором. Фтор может оказывать на ферменты
ингибирующее, в более редких случаях и активирующее дей-
ствие.
Ингибирование ферментов. Неорганические' со-
единения фтора как неспецифические ингибиторы ферментов
были предложены D. Keilin и Е. Hartree в 1939 г. и с тех пор
широко используются в энзимологии [Строчкова Л. С., Соро-
ковой В. И., 1983]. Наиболее высокой чувствительностью к фто-
ру отличаются эстеразы микросом печени, угнетение которых
наступает при низких концентрациях фтора, равных 0,05—
0,2 ммоль/л [Haugen D. A., Suttie J. М., 1974]. В то же время
панкреатическая и кишечная эстеразы резистентны к действию
этого МЭ, что объясняется, видимо, различной потребностью
ферментов в металлах-активаторах и сродством последних к
фтору. Фтор угнетает также активность липаз, воздействуя на
их лабильный кофермент [Строчкова Л. С., Сороковой В. И.,
1983]. Ферменты считаются чувствительными к фтору, если он
ингибирует их в концентрациях 0,01—0,1 мкмоль/л. При более
высоких концентрациях ингибирование может являться арте-
фактом и возникать в результате образования нерастворимых
комплексов фтора с металлами, как, например, в случае обрат-
ной транскриптазы (Srivastava S. К- et al., 1981].
Ингибирование фтором может носить конкурентный, некон-
курентный и смешанный характер [Cimasoni G., 1976]. Этот
МЭ ингибирует металлоферменты, содержащие медь, марганец,
цинк, никель, гемовое и негемовое железо, связываясь с этими
металлами в активном центре [Novak Т., Maurer Р. J., 1981].
Часто происходит также ингибирование ферментов, активируе-
мых металлами, особенно Mg2+, причем оно может усиливаться
фосфатами, как, например, в случае энолазы и сункцинатдегид-
рогеназы. Известны, однако, и металлактивируемые ферменты,
не ингибируемые фтором. Лактатдегидрогеназа — металлофер-
мент, содержащий цинк, ингибируется фтором, но не за счет
его соединения с металлом, а в результате реакции фтора с гис-
тидином или аргинином в центре связывания пирувата [Ander-
son S. R., 1981]. Подобным же образом фтор способен ингиби-
ровать и другие ферменты, особенно если в их активном центре
содержатся положительно заряженные остатки лизина и арги-
нина.
Среди всех ферментов, ингибируемых фтором, энолаза явля-
ется наиболее изученным. Первое сообщение о действии на нее
фтора появилось более 45 лет назад [Варбург О., Христиан В.,
1942]. Чувствительность фермента к фтору относительно невы-
сока и для его ингибирования требуется концентрация этого МЭ
порядка 0,1 — 10 ммоль/л [Wiseman А., 1970]. Энолазакатализи-
292
рует превращение 2-фосфоглицерата в фосфоенолпируват с вы-
делением молекулы воды. Активный центр фермента содержит
участки связывания двухвалентного катиона (Mg2+ или Мп2+)
и фосфорильной группы. Ингибирование фтором носит конку-
рентный характер и усиливается неорганическим фосфатом. Оно
наступает в результате образования нераспадающегося комп-
лекса между ферментом, ионом металла, фтором и фосфориль-
ной группой, в котором фтор непосредственно взаимодействует
с металлом в активном центре фермента и косвенно —с фосфа-
том в центре связывания фосфорильной группы [Novak Т.,
Maurer Р. J., 1981]. Фтор, по-видимому, выступает как аналог
ОН-иона, участвующего в ферментативной реакции присоедине-
ния или отнятия молекулы воды. Подобный же механизм заме-
щения ОН- на F" был предложен для ингибирования неоргани-
ческой пирофосфатазы (КФ 3.6.1.1), при котором образуется не
распадающийся комплекс фермент — ион металла — субстрат —
фтор. Этот механизм аналогичен замещению фтором ОН~-груп-
пы в минерализованных тканях, также способствующему повы-
шению стабильности образующегося соединения.
В качестве ингибиторов ферментов могут выступать и неко-
торые фторорганические соединения, из которых наиболее хо-
рошо известен фторацетат. Не будучи токсичным, он в процессе
метаболизма превращается во фторцитрат, являющийся высо-
коспецифическим ингибитором аконитат-гидратазы (аконита-
зы). Изомерный фторцитрат, образующийся из фтороксалоаце-
тата и ацетил-СоА, оказывает лишь слабое ингибирующее дей-
ствие на этот фермент, а дифторацетат HCF2—COO- вообще
нетоксичен. Образование в процессе метаболизма смертельного
яда из нетоксичного предшественника известно как «летальный
синтез». Выступая как ингибитор аконитат-гидратазы, фторцит-
рат образует прочный комплекс с ионом железа, присутствую-
щим в активном центре фермента.
Активирующее воздействие фтора установлено в отношении
ряда ферментов, из которых наиболее изучена аденилатцикла-
за. Другими являются фосфодиэстераза циклического ГМФ из
фоторецепторов позвоночных и бактериальная ферментная си-
стема, катализирующая образование фитоена из мевалоновой
кислоты. Фтор активирует аденилатциклазу значительно эффек-
тивнее гормонов, в связи с чем его используют в концентрации
10—20 ммоль/л для выявления максимальной активности пре-
паратов этого фермента. Аденилатциклазы образованы двумя
субъединицами: каталитической, связывающей АТФ, и регуля-
торной (N-белок), связывающей гуаниновые нуклеотиды и от-
ветственной за активацию фермента. Именно с последней про-
исходит связывание фтора, а не с каталитической субъедини-
цей, как считалось ранее (Downs R. W. et al., 1980]. Фтор, по-
видимому, взаимодействует с ионом магния и гуаниновыми
нуклеотидами, связанными с регуляторным белком, однако ме-
ханизм активации каталитической субъединицы остается нерас-
крытым. ,
293
Не вполне ясно, происходит ли активация аденилатциклазы
фтором в интактном организме или только in vitro. Первона-
чально считалось, что этот эффект возможен только при повреж-
дении цитоплазматической мембраны {Perkins J. Р., 1973] и
высоких концентрациях фтора, не встречающихся in vivo. Одна-
ко в более поздних работах было показано, что этот эффект мо-
жет быть достигнут и при культивировании неповрежденных
клеток в среде с повышенным содержанием фтора [А1-
Imann D. W., Sahed A. R, 1977]. Активация наблюдается толь-
ко в культуре клеток определенного типа, например гепатоци-
тов, и отсутствует в таких клетках, как фибробласты [Hol-
land R. I., 1980]. Повышение активности аденилатциклазы и
уровня цАМФ в тканях человека и экспериментальных живот-
ных при концентрациях фтора в питьевой воде от 52,6 мкмоль/л
до токсического уровня, отмечавшееся одними авторами
{Susheela А. К-, Singh М, 1982; Kleiner Н. S., Allmann D. W.,
1982], не подтверждается другими исследователями (Ophaug
R. Н. et al, 1979].
Повышенная активность аденилатциклазы в тканях кроликов,
получавших токсические дозы фтора в течение 6 мес [Sushee-
1а А. К, Singh М, 1982], отражает, по-видимому, не столько ак-
тивацию самого фермента, сколько усиление его биосинтеза.
Отсюда напрашивается вывод, что повышение активности фер-
мента отражает какие-то вторичные проявления токсического
действия фтора. Учитывая фундаментальное значение цАМФ в
регуляции ферментативных процессов, можно было полагать,
что сдвиги в его концентрации вызовут заметные изменения в
клеточном метаболизме. Однако до настоящего времени дока-
зательства подобных изменений не получены, хотя нельзя ис-
ключить и вероятность участия аденилатциклазы в патогенезе
флюороза.
Целесообразно выделять следующие виды флюороза: эндеми-
ческий, профессиональный, соседский и ятрогенный. Наиболее
изучены эндемический и профессиональный флюороз.
Эндемический флюороз встречается в природных зо-
нах с высоким содержанием фтора в окружающей среде. До-
статочным указанием на пораженность местности является вы-
сокое содержание фтора в водоисточниках. Концентрация фто-
ра выше 1,5 мг/л является потенциально флюорозогенной, осо-
бенно в условиях жаркого климата. Существуют природные
зоны с исключительно высокой концентрацией гидрофтора.
В СССР это юго-восточные районы Украины, некоторые регио-
ны Молдавии, Казахстана и др. Географическое распростране-
ние гидрофтора и эндемического флюороза на земном шаре до-
статочно подробно изложено в книге «Патология флюороза»
[Авцын А. П, Жаворонков А. А, 1981].
Происхождение природного флюороза, как правило, связа-
но с районами старой или современной вулканической деятель-
ности, а также с так называемыми фосфатными зонами. Этим
объясняется топографическое совпадение зон залегания грани-
294
Рис. 52. Типичная картина эрозивно-пигментированного флюороза зубов у ре-
бенка (а) и взрослого (б).
тов и других пород вулканического происхождения с зонами
эндемического флюороза.
Необходимо различать эндемический флюороз у детей и у
взрослых (рис. 52). Первый характеризуется специфическим по-
ражением зубов молочного и постоянного прикуса, а также уме-
ренными изменениями скелета. В странах с жарким климатом
(Индия, Китай) клинические симптомы флюорозного остеоскле-
роза скелета встречаются чапХе и они более выражены.
Оценка тяжести флюорозного поражения зубов в нашей
стране обычно проводится по классификации, предложенной
Р. Д. Габовичем (1957). Отечественные и зарубежные исследо-
ватели единодушны в том, что у зубов с легкими (I и II) степе-
нями флюороза не изменены их функциональные свойства—•
прочность, стираемость, форма; они в меньшей степени подвер-
295
жены кариесу. В то же время для зубов с тяжелыми (III и IV)
степенями флюороза характерны выраженные расстройства ми-
нерализаци, в результате чего они становятся более хрупкими,
ломкими и быстро стираются. Все это приводит к разрушению
зубов, которое нередко ошибочно расценивается как кариозное
поражение.
Установлено разнообразие макроскопической картины флюо-
розных зубов, особенно у детей старших возрастных групп.
Наиболее часто зубы поражаются в такой последовательности:
резцы, малые коренные, большие коренные, клыки.
В зонах выраженной эндемии обычно выявляются все сте-
пени флюороза. Характерно, что нижние резцы реже поража-
ются эрозивно-крапчатой формой флюороза. Нормальный блеск
эмали в пришеечной области теряется и она становится молоч-
но-матовой. С нарастанием тяжести флюороза площадь пора-
жения эмали увеличивается. Иногда встречается зубной ряд
полностью молочно-белого цвета, фарфоровидный. Под лупой
поверхность эмали в них имеет мелко волнистый вид.
В очаге эндемии (Северный Казахстан), где дети с рожде-
ния получали фтор с питьевой водой в концентрации 4 мг/л, ус-
тановлено, что по мере прорезывания постоянных зубов интен-
сивность флюороза нарастала, достигая максимума к 11 годам,
и составила 98,2+1 %. Необходимо также отметить исключи-
тельную симметричность поражения зубного ряда флюорозом
у близнецов. В то же время дети одной семьи, а тем более
разных семей, хотя и одного возраста, имели различную кар-
тину поражения. Кроме того, у отдельных детей флюорозные
изменения зубов совсем не обнаруживались. Следовательно, ин-
дивидуальная чувствительность организма к фтору является
определяющей для оценки выраженности флюорозного пораже-
ния. Рентгенография костей предплечья, кис; и, позвоночника и
таза у детей 6—14 лет в зоне эндемического флюороза позво-
лила определить рентгенологические признаки более ранней
дифференцировки костного скелета. Необходимо указать на от-
сутствие прямой зависимости между интенсивностью изменений
в скелете и зубах.
Выраженные формы эндемического флюороза скелета у де-
тей выявлены в некоторых регионах субтропиков (Китай, Индия,
Танзания), где питьевая вода содержала более 5 мг/л фтора.
Заболевание начинается с 10—13 лет с постепенного нараста-
ния тугоподвижности в суставах, особенно в позвоночнике,
Рентгенологически кости у таких больных были очень плотные,
трабекулы утолщенные, костномозговой канал сужен. На кон-
цах некоторых длинных трубчатых костей отмечен остеопороз.
Частота поражения зубов эндемическим флюорозом с воз-
растом изменяется, число больных кариесом зубов увеличива-
ется. В частности, в Северном Казахстане в возрастной группе
старше 50 лет эти показатели соответственно составили 36,5 и
94,1%. В среднем же среди взрослого населения распространен-
ность флюороза зубов достигает 46,8 + 6,5%, а кариеса 72,4±
296
+ 10,5%. Сочетанное поражение зубов флюорозом и кариесом
в различных возрастных группах колеблется от 27,2 до 35%,
в среднем составляя 31,4%- Необходимо также отметить, что
интенсивность поражения флюорозом зубов с возрастом снижа-
ется как у местных жителей, так и особенно у приезжих. В сред-
нем в группе местных жителей этот показатель почти в 4 раза
был выше, чем среди приезжих (13,4 + 0,9 и 3,5+1% соответ-
ственно). При рентгенологическом исследовании костного ске-
лета в возрастной группе 20—50 лет отмечено небольшое
огрубление трабекулярного рисунка строения костей по сравне-
нию с контролем. Иногда встречалось умеренное сужение кост-
номозгового канала, которое было выраженным в локтевой и
пястных костях. В возрастной группе старше 50 лет выявлены
выраженные дистрофические изменения в позвоночнике в виде
обызвествления межпозвоночных связок, остеофитоза тел пояс-
ничных позвонков (Lin-v). В ряде случае выявлены периосталь-
ные разрастания в области межкостных гребешков костей пред-
плечья. Клинически это сопровождалось жалобами на постоян-
ные боли в поясничной области, по поводу которых больные
лечились в течение нескольких лет [Авцын А. П., Жаворон-
ков А. А., 1981].
Наиболее высокая частота эндемического флюороза скеле-
та установлена в Индии [Reddi D. R. et al., 1970; Siddiqui A. H.,
1970; Teotia S. P. S. et al., 1978]. Клинически он характеризу-
ется повторяющимся чувством покалывания в конечностях и во
всем теле. Затем появляются боли и скованность движений, осо-
бенно в пояснице, а также в грудном и шейном отделах. В да-
леко зашедших случаях сопровождается резко выраженной де-
формацией скелета с развитием кифоза, сколиоза. Скованность
может увеличиваться до тех пор, пока позвоночник, включая
шейный отдел, не превратится в неподвижный костный столб.
Отведение конечностей более болезненно, чем приведение. На-
растающая тугоподвижность обусловлена обызвествлением свя-
зочного аппарата, мест прикрепления мышц и межкостных фас-
ций. В связи с неподвижностью грудной клетки появляется
брюшной тип дыхания. В результате хронической гипоксии у
5% больных флюорозом в Северной Танзании обнаружен синд-
ром «барабанных палочек» [Latham М. С., 1966].
Описанная картина прогрессирующего эндемического флюо-
роза костей скелета наблюдается уже в возрасте 30—40 лет.
Критическим уровнем для развития патологических изменений
в костном скелете является 4000 мг фтора на 1 кг массы кости.
Фтор обладает сродством не только к высокоминерализованным
тканям, но и к производным эктодермы, в частности к волосам,
в которых он накапливается в повышенных количествах. Так,
в очаге эндемии его содержание достигает 480—830 мг/кг,
а вне очага — 53—72 мг/кг.
Распространенность эндемического флюороза скелета нерав-
номерная— от 2,8 до 83,7 % в индийском штате Пенджаб
[Jolly S. S. et al., 1973]. Деформирующий флюороз с выражен-
297
ной неврологической симптоматикой чаше наблюдается у муж-
чин. В неврологической симптоматике доминируют признаки
радикуломиелопатии и шейного спондилоза. В противополож-
ность этому в условиях Северной Африки эндемический флюо-
роз почти не сопровождается неврологическими осложнениями
(Vittori F. et al., 1961]. Главными причинами, способствующи-
ми развитию флюороза скелета, являются повышенные концен-
трации фтора в питьевой воде (0,12—0,49 ммоль/л), продолжи-
тельная экспозиция в сочетании с усиленной физической на-
грузкой— «физический стресс».
Особую форму флюороза скелета описали М. Soriano и
F Manchon (1966), назвав ее винным флюорозом (la fluorosis
vinica). Она возникает при употреблении вина домашнего при-
готовления, в которое в некоторых странах (Испания) в качест-
ве консерванта добавляются соединения фтора. Для винного
флюороза характерны симметричное поражение костей и соче-
тание выраженных участков остеосклероза, остеопороза и ос-
теомаляции. При остеобластической стадии наблюдается фор-
мирование периостальных пролифератов. В остеопластической
стадии происходит уменьшение их размеров, а иногда и полное
исчезновение (атрофическая стадия).
Профессиональный флюороз. Клиническая карти-
на и патологическая анатомия профессионального флюороза
изучаются более 50 лет профпатологами и детально представ-
лены в работах отечественных и зарубежных авторов
[Roholm К-, 1937; Гринберг А. В., 1955; Молоканов К- Н., 1961;
Гирская Е. Я., 1967; Богданов Н. А., Гембицкий Е. В., 1975].
Профессиональный флюороз скелета развивается быстрее в мо-
лодом возрасте даже при небольшом стаже работы с фтором
иа производстве [Григорьев Э. Э., 1967]. При этом в скелете
наряду с фторовым остеосклерозом находят признаки выражен-
ного остеопороза.
У лиц, длительно подвергавшихся интоксикации фтором,
наблюдаются признаки раннего старения, несоответствия воз-
раста, определяемого по внешнему виду, с фактическим. При
крайнем выражении этого явления можно говорить о «фторной
прогерии».
До недавнего времени в клинике фторной интоксикации на-
рушения мышечной системы расценивались как вторичный эф-
фект флюороза скелета (миелорадикулопатия). Однако иссле-
дование биоптатов мышц больных флюорозом выявило сниже-
ние активности сукцинатдегидрогеназы, уменьшение числа
митохондрий и диаметра мышечных волокон. Кроме того, были
обнаружены очаговые некрозы мышечных волокон с фиброзом
этих участков, пролиферацией фибро- и саркобластов [Fran-
ke J., 1972; Bogin Е., 1978]. Все это может свидетельствовать
и о прямом токсическом эффекте фтора на мышечную ткань.
При работе с соединениями фтора отмечаются носовые кро-
вотечения у 77% обследованных рабочих. Следует отметить,
что в проведенных нами экспериментах у крыс, получавших
298
фторид натрия в виде подкожных инъекций, также отмечался
геморрагический ринит. Таким образом, не только местное, ио
и общее действие фтора вызывает повышение проницаемости
капилляров верхних дыхательных путей. Наряду с этим у рабо-
чих вначале наблюдали гипертрофические, затем атрофические
риниты, ринофарингиты и фаринголарингиты с язвенно-иекро-
тическими изменениями слизистой оболочки. Изменения трахеи
и бронхов дают картину хронического неспецифического воспа-
ления. Профессиональный флюорозный пневмокониоз характе-
ризуется катарально-десквамативным бронхитом с утолщением
базальной мембраны. Особенностью гистологической картины
флюорозного пневмокониоза по сравнению с силикозом явля-
ется выраженная склонность коллагеновых волокон к дистро-
фии и распаду за счет более тяжелого поражения фтором сте-
нок сосудов.
Наличие у больных профессиональным флюорозом выра-
женных симптомов поражения органов пищеварения дает ос-
нование в первую очередь думать о развитии хронического
гастрита. Действительно, данные аспирационных гастробиопсий
свидетельствуют об субатрофическом и атрофическом гастрите
с перестройкой слизистой оболочки желудка по кишечному типу
у лиц, имеющих профессиональный контакт с фтором на произ-
водстве более 10 лет.
В специальной литературе нет сведений о морфологическом
состоянии поджелудочной железы при флюорозе у человека.
Однако экспериментально-морфологические исследования при
хронической интоксикации фтором выявили наличие микроцир-
куляторных, дистрофических и склеротических процессов, кото-
рые позволяют нам говорить о фторной панкреатопатии [Жа-
воронков А. А., 1975].
Патология печени наиболее часто отмечается при профес-
сиональном флюорозе и трактуется в клинике или как токсиче-
ский гепатит, или как токсическая дистрофия печени, или ток-
сическая гепатопатия. Исходя из того, что морфологический
субстрат этих поражений—невоспалительные изменения пече-
ни, более адекватны термины «фторный гепатоз» или «фторная
гепатопатия», в которых представлены этиологический фактор
и характер изменений. Необходимо также иметь в виду, что
у V3 больных профессиональным флюорозом отмечается гепато-
мегалия. При этом гистологически выявляется зернистая и жи-
ровая дистрофия печени.
При хронической производственной интоксикации фтором
изменения сердечно-сосудистой системы обнаруживают непо-
стоянно и оценивают их нередко противоречиво. В генезе фтор-
ной миокардиодистрофии (Богданов Н. А., Гембицкий Е. В.,
1975] выделяют два механизма — опосредованный и кардиоток-
сический. Последний связывают с прямым токсическим влия-
нием фтора на кардиомиоциты. Гистологические изменения со-
судистой системы касаются в основном фиброза и гиалииоза
стенок внутриорганиых разветвлений артерий.
299
Биохимические исследования гормонального статуса рабочих
в условиях профессиональной интоксикации фтором [То-
карь В. И., 1983], а также морфофункциональные данные по
экспериментальному флюорозу [Жаворонков А. А., 1977] сви-
детельствуют о разнонаправленных изменениях в эндокринной
системе. Так, отмечено снижение эндокринной функции гонад
и поджелудочной железы и, напротив, активация паращитовид-
ных желез и С-клеток щитовидной железы. В то же время из-
менения тиреоцитов и клеток коры надпочечников характери-
зовались определенной фазностью и зависели от тяжести ин-
токсикации фтором. Эти морфофункциональные изменения ор-
ганов нейроэндокринной системы связаны не только с прямым
токсическим эффектом фтора, но обусловлены также эндокрин-
ным дисбалансом организма.
Для обозначения флюорозных поражений, выявляемых
вблизи промышленных предприятий, выбрасывающих в окру-
жающую среду фтор и его соединения, был предложен термин
«соседский флюороз» [Murray М. М., 1950]. Эта форма явля-
ется как бы промежуточной между профессиональным и энде-
мическм флюорозом. При соседском флюорозе наиболее тяже-
ло страдают сельскохозяйственные животные и растения. У лю-
дей, проживающих вблизи алюминиевых, сталелитейных и су-
перфосфатных заводов, в выбросах которых имеются примеси
фтора и его соединений, выявлены поражения кожи. Впервые
они обнаружены в Италии, поэтому их в соответствии с назва-
нием местности первого обнаружения стали называть «пятна
Чиззола». Они представляют собой поверхностные пятна серо-
коричневого, голубого или даже синего цвета, округлой или
овальной формы, диаметром 1—2 см, не меняющие своего цве-
та при надавливании. Они спонтанно исчезают через 3—
10 дней. Гистологически в них определяется повышенная пороз-
ность капилляров в сочетании со скоплениями лимфоцитов и
тканевых базофилов (тучных клеток), а также меланофоров
около сосудов [Waldbott G. L., Cecilioni V. А., 1969, 1970]. По-
пытка воспроизвести это поражение в эксперименте не увенча-
лась успехом [Cavagna G. et al., 1969].
Кроме указанных выше форм гиперфтороза, необходимо
упомянуть о тех его формах, которые сочетаются с другими за-
болеваниями различных органов и систем. Здесь уместно вспом-
нить об исключительном сродстве фтора ко всем кальциниро-
ванным тканям и внетканевым отложениям кальция. По-види-
мому, есть определенные основания говорить об атерокальци-
нозе аорты в сочетании с местными отложениями фтора. Таким
же вторичным гиперфторозом может сопровождаться желчно-
каменная и мочекаменная болезнь.
Обширные эпидемиологические обследования [Steven-
son С. A., Watson R., 1960; Bernstein D., 1966] населения США,
употреблявшего в течение продолжительного времени (в сред-
нем 37 лет) питьевую воду с повышенным содержанием фтора,
не выявили ухудшения общего состояния здоровья. Напротив,
300
рентгенологически установлено, что фтор благоприятно влияет
на костную ткань взрослого человека, так как предохраняет ее
от возрастного остеопороза.
Ятрогенные формы флюороза возникают при длительном
лечебном применении фторсодержащих препаратов (см. гла-
ву 9).
7.5. Литий
Содержание лития в почвах СССР колеблется от 1,4 до
9,9 ммоль/кг. По данным А. П. Виноградова (1949), содержание
лития в морской воде составляет 14,4 мкмоль/л. Литий широко
распространен в природе; растения, концентрирующие этотМЭ,
относятся к различным семействам. Известна так называемая
литиевая флора, содержащая в десятки раз больше лития, чем
другие совместно произрастающие растения. К ней относятся
представители пасленовых —табак (Nicotiana tabacum), дере-
за (Lycium ruthenicum), лютиковых—василистник (Thalictrum
minus) и другие растения, накапливающие особенно много ли-
тия при произрастании на почвах, обогащенных этим МЭ. Мор-
ские животные, в том числе рыбы, концентрируют литий в своих
органах и тканях. Ранние литературные данные о содержании
лития в организме человека обобщены А. И. Войнаром (1960).
Литий относится к числу «новейших» МЭ, хотя история
его терапевтического применения насчитывает более 120 лет.
Во второй половине XIX столетия литий начали применять при
лечении подагры, используя его свойства образовывать водо-
растворимые соли мочевой кислоты. Бромид лития назначали
как седативное средство. Хлорид лития в США применяли как
заменитель хлорида натрия, что привело к случаям острого от-
равления лиц с сердечными и почечными заболеваниями со
смертельным исходом и вскоре было прекращено [Samuel D.,
Gottesfeld Z„ 1973].
В 1949 г. J. F. J. Cade было открыто свойство лития вызы-
вать у морских свинок сонливость и потерю возбудимости при
полном сохранении ориентировки. Это свойство затем было с
успехом использовано для лечения больных маниакально-де-
прессивным психозом. В настоящее время карбонат лития ши-
роко применяется для снятия острого психического возбужде-
ния при условии систематического контроля за содержанием
этого МЭ в крови. Жизненная необходимость лития была впер-
вые продемонстрирована в 1976—1980 гг. [Anke М. et al., 1980].
Биологическое действие лития может проявляться физиоло-
гическими, фармакодинамическими и токсическими признака-
ми. Первые из них обнаруживаются при концентрациях этого
элемента в плазме крови от 0,14 мкмоль/л до 1,4 мкмоль/л,
вторые—при 1 ммоль/л, а третьи — при удвоении этой концен-
трации.
Сведения об обмене лития весьма ограничены. Он от-
носится к числу элементов, изотопы которых в живых организ-
301
мах метаболически не идентичны. Количество лития, содержа-
щееся в суточном рационе человека, точно не установлено.
По одним данным, оно близко к 2 мг [МКРЗ, 1975], а по дру-
гим,— в 20 раз меньше—107 мкг [Hamilton Е. I., 1979] и
100 мкг [Soman S. D. et al., 1969]. Нет единого мнения и о сте-
пени усвоения лития из рациона, которое оценивается как
20 мкг/сутки {Wittrig J. et al., 1970] и 2—5 мкг/сутки [Pic-
kett Е. Е., Hawkins J. L., 1981].
Ионы лития всасываются в желудочно-кишечном тракте.
Он выделяется с мочой (95% от введенной дозы), около 1%
выводится с калом и до 5%—с потом [Ljiumberg S., Paal-
zov L., 1969; Schou M., 1976].
Концентрация лития в слюне значительно выше его концен-
трации в плазме. Гематоэнцефалический барьер проницаем
для лития, причем концентрация его в спинномозговой жидко-
сти составляет около 40% от содержания в плазме.
Существуют многочисленные наблюдения о тяжелых отрав-
лениях кошек и собак солями лития. Токсическая концентрация
лития в плазме крови человека равна 3 ммоль/л, летальная —
4—5 ммоль/л. Содержание лития в органах и тканях крыс при
различных его уровнях в рационе представлено в табл. 22.
Из таблицы следует, что одни органы и ткани реагируют на
содержание лития в рационе увеличением концентрации этого
элемента, например почки. Другие не реагируют на его содер-
жание в рационе. К ним относятся мозг и железы внутренней
секреции, через которые литий, по-видимому, и осуществляет
свою физиологическую функцию [Mertz W., 1986]. В организме
функцию депо лития выполняет скелет, содержащий 14,4—
28,8 мкмоль/кг лития при рационах, богатых этим элементом,
и 0,72—2,88 мкмоль/кг при его дефиците. Данные, приведенные
в табл. 22, получены методом эмиссионного спектрального ана-
лиза с использованием пламени закись азота — ацетилен при
тщательном исключении возможных источников загрязнения.
Таблица 22
Зависимость между дозой лития и его содержанием в органах
и тканях крыс [Patt Е. L. et al., 1978]
Орган, ткань1 Доза, мкмоль/кг
менее 1,44 более 72
Кровь (плазма) 0,52 0,84
Цельная кровь 0,85 1,58
Печень 0,39 2,02
Почки 0,66 0,91
Вилочковая железа 2,31 2,59
Мозг 2,88 3,60
Надпочечники 8,21 8,79
Гипофиз 20,20 18,70
Содержание литня определяли свежей тканн.
302
Таблица 23
Содержание лития в органах и ткаиях человека [Bauman W. et al., 1983],
коз [Kronemann A. et al., 1983], обезьян макак резусов
[Bauman W. et al., 1983] и свиней
[Latvietis J. et al., 1983] в ммоль/кг сухой массы
Орган; *гкань Человек Коза Обезьяна Свинья
женщина мужчина обеспечен- ная литием с дефици- том лития
Легкие 2,88±1,44 2,74+1,58 2,59+2,16 0,79+0,55 5,19+0,76
Ребро 3,31+2,16 2,88+1,87 3,46+3,17 1,44+0,66 4,47+3,17 0,16±0,03
Печень 3,17±2,31 3,02+1,87 3,31 + 1,87 1,58+1,58 4,32±2,88 3,85±1,02
Почки 4,32±2,59 3,75±3,03 5,04+3,60 2,45+2,45 7,92+8,21 1,07+0,12
Мозг 4,61+2,45 3,60±1,73 3,75+3,46 2,59±0,99 4,03+2,02 0,76+0,14
Поджелу- дочная же- леза 2,02+1,73 2,59+1,58 5,19±2,74 4,47+3,03 8,36+7,78
Сердечная мышца 3,31+3,75 2,74+1,87 3,17+1,87 3,31+3,03 —- 1,77+0,86
Эти данные на несколько порядков отличаются от значений, по-
лученных для других видов животных и человека методом
атомно-абсорбционной спектроскопии (табл. 23).
Следует сказать, что при низких концентрациях лития в тка-
нях животных значительно возрастает вероятность аналитиче-
ских ошибок. Этим, по-видимому, и объясняются значительные
расхождения между данными табл. 22 и 23. Предпочтение, на
наш взгляд, следует отдать методу, использованному Е. L. Patt
и соавт. (1983), применявшими коррекцию фона и платиновую
посуду, исключающую возможность загрязнения пробы.
Жизненная необходимость лития была впер-
вые продемонстрирована М. Апке и соавт. (1980). Опыты про-
водились на козах и охватывали весь их жизненный цикл,
включая период внутриутробного развития. Животные получа-
ли в сутки 1—2 мг лития в составе полусинтетического рациона
против 20 мг этого МЭ в контроле. По данным 5 опытов при-
рост живой массы коз, получавших бедный литием рацион в
течение первых 6 мес жизни, оказался в среднем на 28% ниже,
чем в контроле (77 г против 107 г в сутки). Если же живот-
ные испытывали дефицит лития и в течение внутриутробного
периода (84 дня), то различия в приросте живой массы были
бы еще более значительными. Козлята, рожденные у самок,
получавших с пищей недостаточное количество лития, сущест-
венно уступали по живой массе и выживаемости потомству от
контрольных животных. У коз, содержавшихся на дефицитном
по литию рационе, отмечены снижение удоя, повышенные
смертность и частота выкидышей, меньшая эффективность пер-
вого осеменения.
Доказательства жизненной необходимости лития, получен-
ные на крысах, оказались менее убедительными, чем в преды-
303
дущем случае. Крысы опытной группы получали рацион, со-
державший всего 5—15 нг/г лития в сутки против 500 нг/г в
контроле, но у них были отмечены только нарушения воспро-
изводительной функции, тогда как различия по живой массе
были статистически не достоверны [Patt Е. L. et al., 1983].
Из биохимических сдвигов у литийдефицитных животных ока-
зались существенно повышенная концентрация холестерина и
сниженный уровень триглицеридов в сыворотке крови. Клиниче-
ских признаков недостаточности лития установить не удалось,
не считая воспаления кожных покровов у отдельных животных.
Таким образом, приведенные данные позволяют отнести литий
к числу элементов с весьма вероятной жизненно необходимой
функцией.
Литий специфически накапливается в тиреоцитах и вызыва-
ет у человека увеличение щитовидной железы, а у крыс пока-
зано формирование коллоидного зоба [Sheard М. Н., 1980;
Wenzel R., 1984]. Экспериментальными исследованиями
Л. В. Гербильского (1987) установлено, что под действием ли-
тия происходят ретенция коллоида в фолликулах щитовидной
железы крыс, а также достоверное уменьшение числа резорб-
ционных вакуолей, высоты тиреоцитов и уменьшение диаметра
их ядер. Кроме того, резко понижается включение радиоактив-
ного йода в щитовидную железу, снижается уровень тироксина
и трийодтиронина в сыворотке крови, что свидетельствует о за-
держке выведения гормонов из щитовидной железы, характер-
ной для коллоидного зоба.
В профессиональной патологии известны случаи отравления
аэрозолями лития, которые могут вызвать трахеит, бронхит, ин-
терстициальную (межуточную) пневмонию и диффузный пнев-
москлероз. Попадание лития на кожу и слизистые оболочки
способно вызвать ожоги. Симптомы хронической интоксикации
литием выражаются в общей слабости, сонливости, головокру-
жении, утрате аппетита, боли при глотании, треморе. Число сер-
дечных сокращений уменьшено, мышечная возбудимость, боле-
вая и осязательная чувствительность кожи повышены. Эти симп-
томы указывают на нейротоксическое действие лития.
Ион лития угнетает подвижность и метаболизм сперматозои-
дов человека. Эритроциты человека наименее устойчивы к изо-
тоническим растворам хлорида лития. Токсичность солей лития
усиливается при исключении из пищи солей натрия.
В настоящее время карбонат лития широко применяется при
лечении маниакально-депрессивного психоза в дозах порядка
2,5 г упомянутой соли, что составляет около 72 ммоль этого МЭ
в сутки и повышает его концентрацию в плазме крови до 0,5—
1,5 ммоль/л. При этом следует, однако, иметь в виду, что в
ряде случаев при концентрациях, равных 1,6 ммоль/л, уже мо-
гут наступать токсические явления.
Важно, что терапевтические дозы лития на психически здо-
ровых лиц психотропного действия не оказывают. Лечебный эф-
фект, по-видимому, связан с изменением обмена биогенных ами-
304
нов в ЦНС. Под влиянием лития высвобождение норадренали-
на и серотонина уменьшается. Усиливаются захват норадрена-
лина нейронами и его внутриклеточное дезаминироване. Кроме'
того, в больших дозах литий способен угнетать активность аде-
нилатциклазы, снижать концентрацию глютамата и ГАМК в го-
ловном мозге. Таким образом, терапия литием направлена на
нормализацию обмена медиаторов в ЦНС. Кроме того, ионы
лития оказывают влияние на некоторые звенья эндокринной
системы, в частности на кору надпочечников, а также на секре-
цию антидиуретического гормона. В психатрической практике
наибольший эффект достигается при профилактике аффектив-
ных расстройств. Влияние терапии литием при депрессивных
состояниях менее выражено. Противопоказанием к терапии ли-
тием являются тяжелые нарушения сердечного ритма.
Ятрогенный токсический эффект препаратов лития проявля-
ется у отдельных лиц, особенно у женщин, угнетением функции
щитовидной железы [Mertz W., 1986], развитием нефрогенного
несахарного диабета [Sox М., Singer J., 1975], поражением сер-
дечно-сосудистой системы [Albrecht J., Muller-Oerlinghausen В.,
1977]. Более подробно синдромология ятрогенного литиевого-
МТОЗа представлена в главе 9.
7.6. Никель
Никель относится к числу «новых» МЭ, жизненная необхо-
димость которых была установлена только после 1970 г. Первые-
данные о его эссенциальности для животного организма, полу-
ченные в опытах на цыплятах [Nielsen F. Н., 1970], вскоре
были убедительно подтверждены на козах, карликовых свиньях
[Anke М„ 1973] и крысах [Kirchgessner М., Schnegg А., 1975].
Первым ферментом, в составе которого был обнаружен никель,
оказалась уреаза из бобов канавалии мечевидной [Dixon N. Е.
et al., 1976]. Затем он был найден у архебактерий в составе фер-
ментов метанового брожения [Shonheit Р., Rhauer Р. К-, 1979],,
что позволяет отнести его к числу важнейших биоэлементов.
По механизму своего биологического действия никель обнару-
живает заметное сходство с другими Зй-элементами— железом
и кобальтом.
В последнее время биологический эффект МЭ интенсивно
исследуется в связи с глобальным загрязнением окружающей
среды. В частности, по данным Я. М. Грушко (1972), ежегодно
в атмосферу и на поверхность почвы поступало в среднем
15 000 т никеля. Более поздние данные [Hardy J. et al., 1982]
свидетельствуют о возрастании выбросов никеля в атмосферу
(47x103 т никеля в год), причем это количество будет увели-
чиваться в течение последующих 20 лет. В программе глобаль-
ного мониторинга, принятой ООН в 1980 г. [Доброволь-
ский В. В., 1983], никель упомянут как один из наиболее опас-
ных загрязнителей окружающей среды. Действительно,
к 2000-му году ожидается десятикратное увеличение поступ-
20—568
305-
ления никеля в атмосферу только в результате сжигания камен-
ного угля. Значительная часть растворимых соединений никеля
поглощается как фотосинтезирующими организмами мирового
океана, так и морскими животными. Ориентировочно можно по-
лагать, что на протяжении года в биологическую миграцию во-
влекаются сотни тысяч тонн никеля. Потребление в пищу про-
дуктов животного и растительного происхождения приводит к
повышенному поступлению в организм человека этого микроэле-
мента. Другой источник проникновения никеля в организм —
локальные техногенные геохимические аномалии, формирующи-
еся вблизи предприятий по переработке никеля.
Концентрация никеля в органах и тканях животных невы-
сока. Наибольшее содержание этого элемента найдено в кости,
легких, причем в последних с возрастом его содержание воз-
растает. По данным F. М. Sunderman (1980), в легких, почках,
печени, сердце и костях человека обнаружено соответственно
1,45 ±1,11; 0,18±0,02; 0,14±0,04; 0,11 ±0,03 и 5,68±2,5 мкмоль/кг
свежей ткани.
Обмен никеля. Всасывание никеля даже при его повы-
шенном содержании в рационе колеблется в пределах 1—10%,
требует затраты энергии и осуществляется, по-видимому, общей
с железом транспортной системой, функционирующей в прокси-
мальной части тонкой кишки. При дефиците железа у крыс ско-
рость всасывания никеля существенно возрастает. Большая
часть никеля выделяется с калом и только ограниченное его
количество — с мочой. С потом удаляется в сутки около 8 мкг
никеля, но при обильном потоотделении это количество сущест-
венно возрастает. У собак, как и у крыс, 90% никеля, посту-
пающего с кормом, выделяются с калом и около 10%—с мо-
чой без какой-либо длительной задержки в организме [Tede-
schi R. Е., Sunderman F. W., 1980]. У человека с калом выде-
ляется в сутки около 260 мкг никеля, что в 20 раз превышает
количество этого МЭ, выделяемого с мочой. В балансовых опы-
тах на 10 добровольцах, проведенных Р. I. Nodya (1972), из
289±23 мкг никеля, содержавшихся в суточном рационе, 258±
±23 мкг (90%) было найдено в кале, причем это количество
включает, по-видимому, также и никель, ранее усвоенный орга-
низмом и выделенный с желчью, в которой, как было показано,
содержится этот металл.
Иная картина наблюдается при парентеральном введении
радиоактивного никеля (63Ni) мышам, крысам и кроликам. Изо-
топ распределяется между органами и тканями прямо пропор-
ционально содержанию в крови, после чего удаляется с мочой и
в значительно меньшем количестве с желчью. 63Ni находится в
плазме крови не более 48 ч после инъекции, причем уже в те-
чение 1-х суток из организма удаляется 68—78% дозы. Осталь-
ное количество выводится на 3—8-е сутки [Underwood Е., 1977].
Никель может поступать в организм и через легкие, особен-
но у рабочих, находящихся в профессиональном контакте с этим
металлом. Если вдыхаемые соединения никеля достаточно хо-
306
рошо растворимы в воде, то они вскоре выделяются с мочой,
тогда как нерастворимые соединения могут задерживаться в
легких в течение длительного времени. По некоторым данным,
период полувыведения никеля из легких человека может дости-
гать 330 дней [Levin I. F., 1981].
J Транспорт этого МЭ в крови осуществляется в основном
альбумином, который образует с ним пентакоординированный
комплекс, включающий в качестве лигандов азот «-аминогруп-
пы, два депротонированных азота пептидных связей, имидазоль-
ный азот и карбоксильную группу боковой цепи остатка аспа-
рагиновой кислоты. Это те же лиганды, которые участвуют и в
транспорте иона меди. В транспорте никеля принимает участие
и тройной комплекс этого МЭ с альбумином и гистидином. По-
скольку связь никеля с последним прочнее, чем с альбумином,
то эта аминокислота способна через образование тройного комп-
лекса высвободить никель из его соединения с транспортным
белком и осуществить его перенос через клеточную мембрану
в виде низкомолекулярного комплекса с гистидином. Различие
между медью и никелем в данном случае заключается в том,
что последний имеет меньшее сродство к альбумину, чем медь,
и большее сродство к гистидину, физиологическая концентрация
которого в плазме крови вполне достаточна для удаления его
из комплекса с альбумином. Возможно, что именно этим объяс-
няется быстрое выделение с мочой никеля, введенного паренте-
рально, в то время как выделение меди происходит в основном
с желчью [Clennon J. D., Sarkar В., 1982].
В ультрафильтрате сыворотки крови кроликов обнаружено
5 различных комплексов никеля с цистеином, гистидином и ас-
парагиновой кислотой, которые также участвуют во внеклеточ-
ном транспорте этого МЭ [Sunderman F. W., 1981].
В транспорте и обмене никеля принимают участие, по-види-
мому, еще два белка. Это богатый гистидином гликопротеид.
(БГГ) и никелеплазмин. БГГ способен в физиологических усло-
виях, т. е. в присутствии гистидина и альбумина, связывать зна-
чительные количества никеля [Guthaus S. L., Morgan W. Т.,
1982]. Никелеплазмин — это аг-макроглобулин, выделенный из.
сыворотки крови человека и кролика. У человека этот белок
содержит 43% от общего количества сывороточного никеля
[Nomoto S., 1980], имеет молекулярную массу около 700 000 и
0,9 моль никеля на 1 моль белка. Он обладает эстеразной ак-
тивностью. Никель, присутствующий в этом белке, не замещает-
ся S3Ni2+ ни в животном организме, ни in vitro и его физиологи-
ческая функция неизвестна.
Еще в 20-х годах высказывалось предположение, что никель
играет определенную физиологическую роль у животных и чело-
века. Впоследствии оказалось, что уже в период эмбриогенеза
он концентрируется в тех органах и тканях, где происходят ин-
тенсивные обменные процессы и где сосредоточен биосинтез
гормонов, витаминов и других биологически активных соедине-
ний. Так, в 1948 г. Е. Lederer и соавт. представили косвенные
20*
307-
доказательства участия никеля в гемопоэзе. Было высказано
предположение, что он при этом активирует некоторые фермен-
ты. В 70-х годах были получены прямые доказательства необ-
ходимости никеля для нормального развития и существования
организма [Nielsen F. Н. et al., 1970]. Кроме того, было пока-
зано, что внутрибрюшинное введение соединений никеля уси-
ливает эритропоэз у кроликов с гемолитической анемией. С тех
пор появилось много сообщений о том, что никель влияет на
гемопоэз через обмен железа.
Из крови по градиенту концентрации никель поступает в
межклеточную жидкость, а затем с помощью специальных бел-
ков проникает в клетки. Этот МЭ способен вызвать индукцию
синтеза металлотионеинов, механизм которой, однако, неизвес-
тен. F. W. Sunderman и соавт. (1983) предложили несколько ги-
потез для объяснения данного явления. Во-первых, никель мо-
жет усиливать синтез этого белка опосредованно, путем увели-
чения включения цинка, меди и других эндогенных металлов
в клетки организма. Во-вторых, усиление синтеза металлотио-
неинов в присутствии никеля могло быть детерминировано гор-
мональной регуляцией. Это предположение внушает доверие,
так как данный МЭ вызывает гиперглюкагонемию у крыс, а
•глюкагон, как известно, индуцирует синтез металлотионеинов
в печени этих животных. И наконец, никель может непосредст-
венно влиять на синтез этой группы белков путем стимуляции
транскрипции их мРНК в клетках печени и почек.
Как отмечалось выше, никель найден в уреазе — первом
•ферменте, полученном в кристаллическом виде из бобов кана-
валии мечевидной [Samner J. В., 1926], однако присутствие ни-
келя в его активном центре было установлено лишь 50 лет
спустя, хотя уже первые спектры поглощения указанного фер-
мента позволяли предполагать в его составе наличие этого ме-
талла [Dixon N. Е. et al., 1976]. Уреаза найдена также в сое,
табаке, рисе, ряде микроорганизмов. Никель прочно связан ок-
таедрически в активном центре фермента и принимает непо-
средственное участие в фиксации мочевины.
К числу микроорганизмов, нуждающихся в никеле, относит-
ся группа бактерий, использующих в качестве источника энер-
гии реакцию гремучего газа: 2Н2 + О2-^-2Н2О, и в качестве
единственного источника углерода — СО2. Катализирующая эту
реакцию гидрогеназа содержит никель или нуждается в нем для
своего синтеза [Thauer R. К- et al., 1980]. Никель необходим
для дерепрессии гидрогеназной активности, причем для участия
никеля в экспрессии фермента у свободноживущего азотфикса-
тора Rhizobium japonica требуется, по-видимому, синтез белка
[Nielsen F. Н., 1987].
Этот МЭ входит также в состав гидрогеназ метанобразую-
щих бактерий Methanobacterium bryantii, М. thermoautotrophi-
cum, С. vinosum и восстанавливающей сульфат бактерии Desul-
fovibrio gigas, в которых преобладает трехвалентный никель
[Le Gall J. et al., 1982]. У метанобразующих архебактерий бо-
308
Таблица 24
Ферменты, содержащие никель [Nielsen F., 1987]
Фермент (кофактор) Источник Молекулярная масса фермента и содержание никеля
Уреаза (КФ 3.5.1.5) Канавалия, соя, рис, та- бак, ряска, микрофлора рубца 483 000, 2 моль NI на субъединицу фермента
Метил-кофермент-М- метилредуктаза (фактор F43o) Метаногенные бактерии 2 моль Ni на 1 моль фермента. F430 — тетра- пиррольное соединение
Гидрогеназы: Alcaligenes eutrophus 1—2 моль NF+ на 1 моль фермента массой 89 500
растворимая
мембраносвязанная Alcaligenes eutrophus 1 моль NF+ на 1 моль
периплазматическая Desulfovibrio gigas фермента массой 89 500
восстанавливающая Methanobacterium 1 моль NP+ на 1 моль
одноэлектронная (вио- thermoautotrophicum фермента
логен) восстанавливающая двухэлектронная (фак- тор 420)
мембранная Vibrio succinogenes 1 моль Ni на 1 моль фермента массой 100 000
Гидрогеназа I Desulfovibrio desulfuricans 0,6 моль Ni на 1 моль фермента массой 77 600
Гидрогеназа II Тот же 0,6 моль Ni на 1 моль фермента массой 75 500
.Дегидрогеназа окиси уг- Cl. thermoaceticum, Фермент массой 440 000
лерода Acetobacterium woodii 2 моль Ni на 1 моль ди- мера в составе порфири- ноподобного фактора
лее 50% никеля находится в составе низкомолекулярного тет-
рапиррольного макроцикла, получившего название фактора
F43o- Этот фактор представляет собой составную часть метил-
кофермент-М-метилредуктазы, восстанавливающей СН3—S—
—СоМ до метана и HS—СоМ [Elefson W. L. et al., 1982]
{табл. 24).
Представители бактерий другой группы — Cl. thermoaceti-
cum, Cl. formiaceticum, CI. aceticum — являются анаэробами,
использующими CO2 в качестве акцептора электронов, восста-
навливая его до уксусной кислоты по уравнению: 4Н++2СО2 +
+ 4ё^СН3СООН.
Восстановление СО2 до ацетата катализируется мультифер-
ментной системой, один из компонентов которой обладает актив-
ностью дегидрогеназы окиси углерода [Dickert G., Ritter М.,
1983] и содержит в составе своей простетической группы тет-
рапиррольное соединение с никелем в качестве центрального
атома. Это соединение по своей структуре приближается к ви-
тамину В12.
Рассматривая жизненную необходимость никеля для живот-
ных и человека, следует, как и в случае кобальта, различать
309
два главных аспекта: его косвенное воздействие на организм
через симбиотические микроорганизмы и экзогенные ферменты
пищеварительного тракта и его прямое участие в метаболизме.
Населяющая желудочно-кишечный тракт микрофлора содер-
жит целый ряд ферментов, в составе которых обнаружен ни-
кель. Эти ферменты, принимая участие в пищеварительных про-
цессах животных, могут оказывать заметное влияние на физио-
логическое состояние их организма. Особую роль они играют у
жвачных и животных, у которых интенсивные микробиологиче-
ские процессы совершаются в слепой кишке [Anke М- et al.,.
1983].
У животных не обнаружены ферменты, содержащие никель, хотя извест-
но, что он способен активировать целый ряд ферментов животного происхож-
дения in vitro ([Nielsen F. H., 1980]. Наибольшее значение он может иметь по
меньшей мере для ферментов—кальцийнейрина и кальмодулинзависимо»
фосфопротеинфосфатазы, а также для образования двух СЗ-конвертаз систе-
мы комплемента: СЗЬ, ВЬ альтернативного пути и С4Ь, 2а — классического пу-
ти, в которых он оказывается эффективнее замещаемого им Mg2+. Более того,
в присутствии никеля с эритроцитами овец, несущими СЗЬ(ЕСЗЬ), специфиче-
ски связывается в 9 раз большее количество фактора В, чем в присутствии-
магния. Для образования эффективного литического участка на одну клетку
ЕСЗЬ в присутствии никеля требуется в 3 раза меньше фактора В, в 12 раз
меньше фактора D и в 66 раз меньше ионов металла, чем при добавлении Mg2+
[Fishelson Z. et al., 1983]. Эти данные позволяют считать, что никель играет
определенную роль в системе комплемента. Показано также, что АТФ и Ni2+
участвуют в реакции образования пирофосфатидной кислоты из фосфатидной
в процессе биосинтеза фосфатидилсерина в микросомах мозга крысы [Pullar-
kat Р. К- et al., 1981].
Никель способствует всасыванию железа в пищеваритель-
ном тракте, будучи кофактором неидентифицированного био-
лиганда, связывающего железо, или участвуя в ферментном
механизме, превращающем Fe3+ в легкоусвояемое Fe2+ [Niel-
sen F. Н., 1987]. Имеются наблюдения, что сразу после родов,
но до отделения плаценты содержание никеля в сыворотке кро-
ви рожениц повышается в 20 раз. Однако уже через 60 мин
возвращается к норме [Rubanyi G. et al., 1982]. Никель, по-
добно окситоцину, вызывает сокращение матки крыс. Эти на-
блюдения дают основание полагать, что этот МЭ необходим
для отделения плаценты или предупреждения атонических кро-
вотечений в послеродовом периоде [Rubanyi G„ Balogh I.,
1982].
По другим данным никель необходим для восстановления
реакции лимфоцитов свиней на действие митогенов, утрачен-
ную после их обработки ЭДТА [Yang W. С., Schultz R. D.,
1982]; низкие концентрации никеля способствуют полимериза-
ции in vitro очищенного тубулина из мозга коз и ингибируют
ее при повышенной концентрации (~ 0,1 ммоль/л) [Roychaud-
hury S. et al., 1982]; сообщается также о роли никеля в ста-
билизации цитоплазматических мембран [Nielsen F. Н., 1987],
в которых он играет роль структурного компонента. Представ-
ляет интерес сообщение [Reeves Р. G. et al., 1981] о том, что
310
этот МЭ необходим для роста Legionella pneumophilia — бак-
терии, вызывающей у людей «болезнь легионеров».
Биологическое действие никеля на животный организм объ-
ясняется, по-видимому, также и тем, что он принимает участие
в структурной организации и функционировании основных
клеточных компонентов — ДНК, РНК и белка. Наряду с этим
высказано предположение о том, что никель участвует в гормо-
нальной регуляции организма В частности, имеются данные,
что он вовлекается в обмен пролактина [Skreb J. et al., 1984;
Smialowicz R. et al., 1984]. Некоторые изменения организма
при дефиците никеля могут быть объяснены именно дисбалан-
сом гормонов. Наиболее полные данные о влиянии никеля на
организм животных и человека представлены в обзоре
[Строчкова Л. С. и др., 1987]. Суточная потребность в никеле
составляет у жвачных 5,11—5,96 мкмоль/кг, у животных с од-
нокамерным желудком — менее 3,41 мкмоль/кг и человека —
около 60 мкмоль в сутки.
Никельдефицитные состояния у человека не
описаны, хотя принципиально они возможны. В частности,
группы риска могут составлять больные с явлениями наруше-
ния всасывания МЭ слизистой оболочкой желудочно-кишечно-
го тракта при хронических гастроэнтероколитах, с симптома-
ми мальабсорбции и при высоком содержании фитатов в ра-
ционе.
Признаки недостаточности никеля у жвачных сельскохо-
зяйственных животных во всех случаях были сходными: за-
держка роста, повышенная смертность матерей и их потомства,
снижение уреазной активности рубцовой микрофлоры, уровня
гемоглобина, гематокритного числа и сходные с паракератозом
изменения кожных покровов, особенно на вымени лактирую-
щих животных.
У коз при внутриутробном дефиците никеля масса потомст-
ва оказалась сниженной на 10% по сравнению с этим показа-
телем у контрольных животных. Явления недостаточности у
них наступают при содержании никеля в сухом веществе раци-
она менее 0,1 мг/кг без снижения, однако, концентрации его
в молоке. В этой связи козлята, родившиеся с пониженной
живой массой от никельдефицитных матерей, догоняют по жи-
вой массе контрольных животных в течение 4 нед постнаталь-
ного развития.
Воспроизводительная функция животных при дефиците ни-
келя изменяется в меньшей степени, чем при дефиците других
жизненно необходимых МЭ. Все же недостаточность никеля
приводит к существенному снижению эффективности первого
осеменения, оплодотворяемости, повышению числа яловых жи-
вотных, выкидышей.
У козлов дефицит никеля вызывает атрофию семенников,
снижение продукции спермы и ослабление полового инстинкта.
При дефиците никеля повышается (приблизительно в Р/г
раза) выделение из организма железа и кальция. У карлико-
311
вых свиней при этом снижается содержание в скелете кальция1,,
который частично замещается магнием. Нарушение обмена
кальция у коз и крыс приводит к обеднению организма цин-
ком, паракератозу, а нарушение обмена железа — к анемии.
В данном случае имеет место довольно редкий пример физио-
логического синергизма между МЭ, когда дефицит одного из
них влечет за собой дефицит другого и наоборот. Поскольку
никель входит в состав уреазы, он необходим для обмена мо-
чевины у жвачных и других травоядных. Показано (Spears J. W.
et al., 1978], что при дефиците никеля в сыворотке крови ягнят
повышается содержание мочевины, а при включении в рацион
этого элемента ее концентрация возвращается к норме. Одно-
временно повышается и активность уреазы в содержимом руб-
ца. При дефиците этого МЭ она снижается до 2% от нормаль-
ной величины. Повышение активности уреазы и прироста мас-
сы при добавке никеля наблюдалось также у бычков, содер-
жавшихся на малобелковом рационе. При скармливании ра-
циона с нормальным уровнем белка эффект от никеля был ме-
нее выражен. Приведенные данные можно рассматривать как
свидетельство способности этого МЭ усиливать рециклизацию»
азота в рубце путем повышения активности уреазы в его со-
держимом [Anke М., 1985].
В тканях животных не обнаружены ферменты, содержащие
никель. Тем не менее он оказывает заметное влияние на обмен-
ные процессы в их организме. При дефиците этого металла у
крыс снижается на 40—75% активность семи дегидрогеназ,
двух аминотрансфераз и а-амилазы [Kirchgessner М., Sch-
negg А., 1980]. Дефицит никеля мало отражается на обмене
белков, но вызывает заметные сдвиги в углеводном и липидном
обмене. Так, в печени крыс, испытывающих дефицит никеля,
содержится в 10 раз меньше гликогена и глюкозы, чем в нор-
ме. Понижен был также уровень триглицеридов и повышена
концентрация холестерина. У коз при остром дефиците никеля
отмечено только существенное снижение активности D-сорби-
тол-6-фосфат—дегидрогеназы (КФ 1.1.1.14) при сохранении
нормального уровня прочих изученных дегидрогеназ, амино-
трансфераз и а-амилазы.
F. Н. Nielsen и соавт. (1983) полагают, что признаки нике-
левого токсикоза и дефицита его у человека могут быть ана-
логичны. Токсический эффект никеля сопровождается сниже-
нием активности ряда металлоферментов, нарушением синтеза
белка, РНК и ДНК- Выявлены разрывы одиночных нитей ДНК
(в легких, почках). Внутриклеточная концентрация никеля
тесно сопряжена с уровнем повреждения ДНК и с интенсив-
ностью ее репарации. Степень нарушения структуры ДНК (на-
пример, в почках крыс) равна примерно одному повреждению
на 107—108 нуклеотидов. Повреждающее действие иона никеля
в нуклеиновых кислотах проявляется в месте расположения
аденина и гуанина, с которыми, как показано in vitro, взаимо-
действует этот МЭ. Вначале никель в основном концентриру-
312
ется во фракциях негистоновых белков или РНК, затем отме-
чено его перемещение в ДНК.
Никель уменьшает жизнеспособность альвеолярных макро-
фагов, вызывает снижение содержания лизоцима, вырабатыва-
емого альвеолярными макрофагами и трахеобронхиальными
слизистыми железами, а также приводит к замедлению колеба-
тельных движений ресничек мерцательных клеток респиратор-
ного эпителия. Накопление этого МЭ в легких и почках связы-
вается с его канцерогенным потенциалом, так как известны
случаи никелевого рака почек и легких, но неизвестен никеле-
вый рак печени. В рабдомиосаркомах, индуцированных нике-
лем, последний в 70—90% выявляется в ядрах опухолевых
клеток. Кроме того, он концентрируется в ядрах фибробластов
кожи. Зарегистрировано также накопление никеля в ядрыш-
ках.
Экспериментально показано, что экзогенно введенный ни-
кель в низких дозах (0,02 мг/кг) вызывает четко выраженный
коронарный вазоконстрикторный эффект [Belloni F., 1979].
Наряду с этим установлено, что из ишемизированного миокар-
да высвобождается эндогенный никель [Rubonyi G. et al., 1981;
1984], который способствует формированию порочного круга
ишемии миокарда. Таким образом, становится очевидным, что
никель является новым эндогенным вазоактивным элементом и
играет, вероятно, важную роль в патогенезе некоторых форм
хронической ишемической болезни сердца.
Субмикроскопическими исследованиями [Ong S. et al., 1973]
установлено, что электронно-плотные отложения в тканях, со-
ответствующие накоплению никеля (метод с диметилглиокси-
мом и рентгеноопектральный анализ), имели вид миелинопо-
добных телец и локализовались в просвете и эндотелии капил-
ляров, а также в цитозоле и митохондриях кардиомиоцитов.
При этом характерно скопление гликогена в расширенных цис-
тернах эндоплазматической сети, Т-трубочках и во внешнем
компартменте поврежденных митохондрий.
Таким образом, действие никеля реализуется в основном
на клеточном и субклеточном уровне. Определенные аспекты
этой проблемы были исследованы А. В. Юровой (1989). Так,
с помощью включения меченых предшественников PHR и бел-
ка ей удалось показать, что клетки печени и двенадцатиперст-
ной кишки белых крыс различаются по чувствительности к
действию перорально введенного никеля. В частности, в эпите-
лиоцитах ворсинок двенадцатиперстной кишки были обна-
ружены очаговые поверхностные повреждения исчерченной ка-
емки в сочетании с отеком и другими неспецифическими изме-
нениями (рис. 53). Более своеобразные процессы, вызванные
интоксикацией никелем, удалось установить на ультраструк-
турном уровне. Среди них особое внимание привлекают резко
выраженная гипертрофия ядрышка гепатоцитов, увеличение
доли агранулярной эндоплазматической сети, появление юных
митохондрий (рис. 54). Очень чувствительным тест-объектом к
313
Рис, 53. Участки разрушения исчерченной каемки (стрелки). Отек, вакуолиза-
ция, зернистость цитоплазмы эпителия, расширенные межклеточные простран-
ства. Отек собственной пластинки слизистой оболочки двенадцатиперстной
кишки крысы через 3 нед после введения хлорида никеля в концентрации
5 мг/100 г массы. Полутонкий срез. Окраска метиленовым синим, азуром II
и основным фуксином. Х400 (препарат А. В. Юровой).
повреждающему действию предельно допустимых и более низ-
ких концентраций никеля оказалась культура клеток HeLa,.
в которых найдены, по-видимому, специфичные для интоксика-
ции этим МЭ изменения; появление атипичных полисом (в ви-
де спиралей и цепочек) и кристаллоподобных включений, окру-
женных мембраной, а также деструкция митохондрий в соче-
тании с гипертрофией ядрышек, комплекса Гольджи и расши-
рением цистерн эндоплазматической сети с накоплением в них
электронно-плотного содержимого (рис. 55—58). Кроме харак-
терного цитопатического эффекта никеля, А. В. Юровой уда-
лось подтвердить повреждающее влияние этого элемента на
сперматогенный эпителий крыс и установить наличие наруше-
ний физиологической проницаемости гематотестикулярного
барьера, сочетавшихся с повышением концентрации тестостеро-
на и снижением концентрации пролактина в сыворотке крови
подопытных животных. Изложенные данные показывают, что
цитопатическое действие никеля даже в ПДК/2 может быть
установлено с помощью современных морфологических мето-
дов.
Более существенное значение для практической медицины
314
Рис. 54. Отек гиалоплазмы с дискомплексацией гранулярной эндоплазматиче-
ской сети, частичной дегрануляцией ее мембран; очаговые просветления мат-
рикса митохондрий с дискомплексацией и лизисом крист, юные митохондрии
(стрелки) гепатоцита крыс через 3 нед после введения хлорида никеля в кон-
центрации 5 мг/100 г массы. Х7000 (препарат А. В. Юровой).
Рис. 55. Большое количество ограниченных мембраной электронно-плотных
кристаллоподобных включений (КВ), цистерны эндоплазматической сети рас-
ширены, иногда с мелкозернистым электронно-плотным содержимым и частич-
ной дегрануляцией мембран (стрелка), атипичные формы полисом (стрелки)
в клетках HeLa при воздействии никеля в токсической концентрации (4 мкг/мл)1
в течение 24 ч. X16 000 (препарат А. В. Юровой).
Рис. 56. Гипертрофия ядрышка с фибриллярным (Ф) и гранулярным (ГР>
компонентом в ядре клетки HeLa при воздействии никеля в концентрации
ПДК/2 (0,05 мкг/мл) в течение 24 ч. X13 000 (препарат А. В. Юровой).
Рис. 57. Кристаллоподобные включения (КВ), ограниченные мембраной; пар-
циальный отек митохондрий с избирательным лизисом крист и выпадением
электронно-плотного детрита в их матриксе в цитоплазме клеток HeLa при
воздействии никеля в концентрации ПДК/2 (0,05 мкг/мл) в течение 24 ч.
X10 000 (препарат А. В. Юровой).
Рис. 58. Слияние митохондрий (М); парциальный и субтотальный лизис крист
в клетках HeLa при воздействии никеля в токсической концентрации (4 мкг/мл^
в течение 2 ч. X 15 000 (препарат А. В. Юровой).
-представляют наблюдения, согласно которым повышенное по-
ступление никеля и его соединений в организм приводит к ток-
сикопатическому эффекту. Однако сведений о токсическом
действии никеля на организм человека в литературе немного,
касаются они в основном профессиональной патологии.
Никелевая пыль при длительном вдыхании вызывает раз-
дражение слизистых оболочек дыхательных путей, носовые
кровотечения, гиперемию зева, развитие пневмокониоза. Вды-
хание паров никеля и его окислов может привести к острым
приступам литейной лихорадки [Рощин А. В., 1981].
В обзорной работе F. W. Sunderman (1984) приводятся дан-
ные о том, что ряд соединений никеля в субмиллимолярных
концентрациях индуцируют переход ДНК из В-формы в Z-фор-
му. Стабилизация Z-формы ДНК также может явиться одним
из факторов в механизме никелевого канцерогенеза. Другим
важным инициальным этапом этого механизма служит инги-
бирование никелем и его соединениями в легких и печени ак-
тивности бенз(а)пирен-гидроксилазы — микросомального фер-
мента, превращающего канцерогенный бенз (а) пирен в гидрок-
силированные неканцерогенные метаболиты [Сидоренко Г. И.,
Ицкова А. И., 1980].
Из неорганических соединений NijSs обладает наиболее вы-
раженным канцерогенным свойством, особенно при внутримы-
шечной имплантации. Вместе с тем карбонил никеля при ин-
галяционном поступлении вызывает развитие рака легких в
течение 2 лет у крыс линии Вистар, обладающих высокой про-
тивоопухолевой резистентностью. В противоположность этому
дИэтилдитиокарбамат натрия (ДДК) увеличивает способность
.металлотионеина связывать никель в организме, предотвращая
у значительного числа крыс развитие опухолей. Этот эффект
ДДК больше выражен у самок, чем у самцов [Sunderman
F. W., 1984].
Исследования последних лет показали, что нормальные
чклетки млекопитающих содержат так называемые группы он-
когенов, которые при определенных условиях могут быть вовле-
чены в неопластическую трансформацию. К настоящему вре-
мени практически ничего неизвестно о взаимоотношениях кан-
щерогенных металлов и онкогенов. Однако допустимо предпо-
ложение: связывание металла со специальным участком гена
может вызвать последующие мутации или активацию онкоге-
нов, что рассматривается как начальный этап канцерогенеза,
индуцированного металлом. В этом смысле удобно использо-
вать соединения никеля и в первую очередь его сульфид как
индикатор для подобных исследований в связи с его необычай-
ной канцерогенностью и пригодностью его радиоизотопа.
Наиболее тяжелой формой профессиональной патологии,
обусловленной токсикопатическим действием никеля и его со-
единений, является рак легкого. Первыми отечественными ра-
ботами по данной проблеме были сообщения С. В. Знаменско-
"320
го (1967) и С. В. Знаменского и соавт. (1967), в которых про*
анализированы 179 наблюдений рака легких в Норильске за
период с 1955 по 1966 г. При этом в 79 случаях рак легкого
выявлен у рабочих, непосредственна контактирующих с нике-
лем. В 58 наблюдениях рак легкого установлен у рабочих дру-
гих предприятий, не имевших ирямого контакта с никелем, и
в 42 случаях он имел место у других жителей Норильска.
Показано также, что заболеваемость никелевым раком лег-
ких повышена среди рабочих, занятых на всех трех этапах
технологического процесса, и составила 29,0; 73,1; 81,5 на
100 000 мужчин, занятых добычей никелевой руды, ее обогаще-
нием и переработкой соответственно. В то же время в конт-
рольной группе этот показатель был равен всего 8,6. Особенно
высокая заболеваемость установлена на никелевом заводе (115
случаев заболеваний в год на 100 000 мужчин), особенно в
электролизном цехе, где она достигала 210,7 случая. Гермети-
зация производства в электролизных цехах резко снижает
уровень заболеваемости этой формой профессионального рака.
Никелевый рак легких в 65% случаев выявлялся у лиц,
считавших себя практически здоровыми, без выраженных прод-
ромальных явлений. В 98% наблюдений это был центральный
рак с преимущественной локализацией в главном (в 20% слу-
чаев) или долевых (в 70%), значительно реже в сегментарных
(в 8%) и дистальных (в 2%) бронхах. Правое легкое поража-
лось чаще (в 61%), чем левое. По гистологическому строению
никелевый рак легких относится в подавляющем большинстве
случаев к различным формам плоскоклеточного с преоблада-
нием малодифференцированного рака без склонности к орого-
вению. А. П. Авцын в 1965 г. наблюдал случаи никелевого ра-
ка трахеи и бронхов и обратил внимание на крупные ядра и
крупные ядрышки опухолевых клеток, которые имели амфо-
фильную окраску.
Рабочие никелевых предприятий заболевают раком легких
не только чаще, но и в более молодом возрасте, чем рабочие
других предприятий. Так, средний возраст больных раком лег-
ких на никелевых предприятиях составил 44,9 года, на других
предприятиях — 48,4 года. Отмечено, что канцерогенный эф-
фект никелевой руды обусловлен именно соединениями никеля,
а не другими ее составными частями, такими, например, как
медь и кобальт. Исследователи приходят к обоснованному вы-
воду о том, что рак легких возникает при наличии в легочной
ткани очагов отложения никеля, образующих депо канцероген-
ного металла. Средний стаж работы у больных никелевым ра-
ком составил 10,1 года, а средняя продолжительность латент-
ного периода равнялась 12,7 года. Загрязнение окружающей
среды пылевыми выбросами никелевого завода существенно
влияет также на заболеваемость раком легких рабочих других
предприятий, расположенных по соседству с никелевым заво-
дом. Таким образом, тяжелой форме промышленного никеле-
вого МТОЗа в виде профессионального рака легких всегда со-
21—568
321
С х е м a 3
КАНЦЕРОГЕННОЕ (I) И ТОКСИЧЕСКОЕ (II) ДЕЙСТВИЕ НИКЕЛЯ НА ОРГАНИЗМ
[Юрова А.В., 1989]
путствует соседская его форма, отличающаяся более продол-
жительным латентным периодом.
Как экспериментальные, так и клинические наблюдения
свидетельствуют о том, что в химическом канцерогенезе ини-
циальным моментом является наличие в тканях депо экзоген-
ного канцерогенного материала, т. е. накопление его до опре-
деленного критического уровня. Этот критический уровень кан-
церогена индивидуален и зависит от многих приводящих мо-
ментов, в первую очередь от резистентности, иммунологическо-
го и нейроэндокринного статуса организма. Этим во многом
322
определяется продолжительность латентного периода бласто-
могенеза, который, в частности, для никелевого рака, по дан-
ным разных авторов, колеблется от 13 до 20 лет и более
[Знаменский С. В., 1967; Сидоренко Г. И., Ицкова А. И., 1980].
Приведенные выше данные позволяют схематически выде-
лить основные моменты повреждающего действия никеля и его
•соединений на ткани животного организма и человека (схема
3). Токсический эффект избытка никеля в организме сопровож-
дается развитием выраженных повреждений во многих орга-
нах и тканях. Особенно важными исходами хронической нике-
левой интоксикации являются метапластические, диспластичес-
кие и неопластические процессы. Морфологическим выражени-
ем последних считается никелевый рак носовой полости, ни-
келевый рак легкого, рак почек и саркомы.
Экспериментально установлена эмбриотоксичность никеля,
выражающаяся патогистологическими изменениями в печени,
почках, семенниках, миокарде [Mathur A. et aL, 1977], а так-
же развитием сарком [Sunderman F. W., 1978]. Ятрогенные
формы никелевых токсикопатий представлены в главе 9.
7.7. Кремний
Долгое время интерес к действию кремния на животный
«организм объяснялся только способностью этого МЭ вызывать
силикоз и лишь в 1972 г. одновременно и независимо друг от
друга в разных лабораториях Калифорнийского университета
К. Schwarz и Е. Carlisle показали, что кремний необходим для
нормального роста и развития крыс и цыплят, формирования
хряща, кости и соединительной ткани в целом и что он участ-
вует в ряде важных метаболических процессов.
Содержание в тканях. Данные о содержании крем-
ния в тканях млекопитающих, приводимые в ранних работах,
как правило, завышены, поскольку большинство анализов вы-
полнялось в стеклянной посуде, без исключения попадания
пыли и с использованием устаревших колориметрических мето-
дов. Сравнивая цифры, полученные в лаборатории Е. Carlisle
и характеризующие содержание кремния в свежей ткани по-
чек, мышц и печени крыс в 1974 г. (0,14; 0,11; 0,07 ммоль/кг
соответственно) с данными, приводимыми ею же в 1986 г.
(0,02±0,02; 0,03±0,02; 0,06 + 0,02 ммоль/кг), можно заметить,
что для первых двух органов они оказались ниже в 7 и 4 раза
и подтвердились только для печени. Содержание этого МЭ в
органах и тканях человека в десятки, а в отдельных случаях
в сотни раз выше, чем у крыс, и особенно высоко в лимфати-
узлах (21±7,6 ммоль/кг свежей ткани) [Carlisle Е. М.,
1Уоо], где кремний может присутствовать в виде отдельных
скоплении мелких зернышек кварца. Концентрация кремния в
большинстве мягких тканей с возрастом существенно не меня-
ется, если не считать легкие, в которых содержание этого МЭ
возрастает.
21*
323
Умеренное повышение концентрации кремния в тканях крыс
отмечено при скармливании им метасиликата натрия, а су-
щественное повышение при даче органических соединений
кремния. Среднее содержание кремния в плазме крови крыс
и обезьян близко к 1 мг/л. У овец оно значительно выше —
5 мг/л, а у человека составляет всего 0,5 мг/л. Плазма крови
и другие жидкие среды организма содержат почти весь крем-
ний в виде растворимой свободной кремниевой кислоты и мало
различаются между собой по ее концентрации, что свидетель-
ствует о легкости ее диффузии через мембраны. Как показы-
вают наблюдения на нескольких сотнях людей, концентрация
кремния в крови не меняется в зависимости от возраста, пола,
профессии, состояния дыхательного тракта, но повышается при
введении соединений этого элемента [Worth G., 1952].
Наиболее богаты кремнием соединительная ткань аорты,
трахеи, сухожилия, кость, кожа и эпидермальные образования
[Carlisle Е. М., 1974]. У крыс, например, аорта, трахея и сухо-
жилия содержат в 4—5 раз больше кремния, чем печень мыш-
цы и сердце. С возрастом концентрация этого МЭ в соедини-
тельной ткани снижается, что может иметь определенное от-
ношение к развитию атеросклероза [Schwarz К. et al., 1977;
Davison Е. В. et al., 1978]. Так, у кроликов в возрасте 3 мес,
1,5 и 2 лет содержание кремния в сердце, печени и мышцах
колебалось в пределах 0,18—0,53 ммоль/кг сухого вещества
(0,05—0,15 ммоль/кг свежей ткани), тогда как в аорте оно
снизилось с 2,85 до 0,53 ммоль/кг и в коже — с 1,64 до
0,32 ммоль/кг. Аналогичное уменьшение концентрации отмече-
но в коже свиней (от 3,38 ммоль/кг у новорожденных поросят
до 0,36 ммоль/кг у взрослых животных), а также в коже крыс.
У человека происходит снижение концентрации кремния в
аорте не только с возрастом, но и в процессе развития атеро-
склероза. Высокие концентрации этого МЭ найдены в эпидер-
мисе и волосах. Кремний концентрируется в роговом слое ко-
жи и кутикуле волоса. Повышенные концентрации этого МЭ
найдены также в волосяном покрове животных и перьях птиц
в составе не растворимого в щелочи компонента. На его долю
приходится около 0,4—1,7% их массы, придающего им высо-
кую химическую стойкость и снижающего их адсорбционные
свойства [Fregert S., 1959]. Высокое содержание кремния
(8,65 ±0,64 ммоль/кг сухой массы) свойственно также зубной
эмали человека (Losee F. et al., 1974] и эпифизам бедренной
кости обезьян резусов (16,2±2,5 ммоль/кг сухой массы) [Le
Vier R. R., 1975].
Высокое содержание кремния в соединительной ткани сея*
зано, по-видимому, с его присутствием в качестве структурного-
компонента в составе гликозаминогликанов и их белковых
комплексов, образующих остов данных тканей и придающих
им прочность и упругость. При фракционировании экстрактов
различных видов соединительной ткани, таких как хрящ, кость,
и кожа, были получены белково-гликозаминогликановые комп-
324
лексы, а также гликозаминогликаны с высоким содержанием
кремния (Carlis Е. М., 1986]. Гидролиз сильными кислотами
и щелочами ведет к разрыву связей кремний — полисахарид,
высвобождая способный к диализу силикат, в то время как
ферментативный гидролиз гиалуроновой кислоты или пектина
не отщепляет кремниевую кислоту, но приводит к образова-
нию низкомолекулярных продуктов, содержащих связанный
кремний.
Дисахариды, полученные ферментативным гидролизом из
хондроитинсульфата А хряща носовой перегородки, оказались
богаче кремниевой кислотой, чем дисахариды из хондроитин-
сульфата С и кератинсульфата [Carlisle Е. М., 1986]. Некото-
рые другие гликозаминогликаны, такие как гиалуроновая кж>
лота и дерматансульфат, выделенные из кожи свиней и крыс, и
гиалуроновая кислота из петушиного гребня и пупочного ка-
натика богаты кремнием и сходны по его содержанию с хонд-
роитинсульфатом А, тогда как гиалуроновая кислота из стекло-
видного тела и роговицы глаза бедны этим МЭ. Ранее сооб-
щалось, что гиалуроновая кислота пуповины содержит
54,5 ммоль/кг свободного и 12,8 ммоль/кг связанного кремния
[Schwarz К., 1973]. Однако впоследствии эти данные не под-
твердились, поскольку материал оказался загрязненным крем-
нием [Schwarz К. et а]., 1977].
Ежедневная потребность организма в кремнии составляет
20—30 мг кремнезема. С пищей и водой в сутки поступает
3,5 мг, с воздухом —15 мг. Организм человека усваивает в
сутки 9 —14 мг кремния, что хорошо соответствует количест
ву его суточного выделения с мочой — 9 мг. Усвоение кремния
из пищи, богатой клетчаткой, почти в 2 раза выше, чем бедной
клетчаткой. Как свидетельствуют балансовые опыты на жи-
вотных, почти весь кремний, поступающий с пищей, проходит
транзитом через пищеварительный тракт и выводится с ка-
лом, а всосавшееся небольшое количество этого МЭ выводит-
ся с мочой. У шахтеров, имеющих длительный контакт с крем-
ниевой пылью, концентрация его в моче в десятки раз превы-
шает нормы, составляя от 3,9 до 76 мг/сутки. Отсутствуют дан-
ные о механизме всасывания кремния из таких соединений, как
кремнезем, ортокремниевая кислота, пектин, мукополисахари-
ды, органический кремний. Величина всасывания кремния из
желудочно-кишечного тракта составляет для всех соединений
этого элемента около 1% [МКРЗ, публикация 30, 1984].
У человека кремниевые кислоты, присутствующие в пище и
напитках, легко всасываются и быстро выводятся с мочой
[Baumann Н., 1960]. У морских свинок этот элемент усваивает-
ся преимущественно в виде ортокремниевой кислоты [Sauer F.
et al., 1959], часть которой образуется из кремнезема растений
под действием пищеварительных соков. Значительное влияние
на усвоение кремния оказывает форма его соединений, от ко-
торой, по-видимому, зависит содержание растворимых соедине-
ний этого МЭ в пищеварительном тракте [Benke G. М., Osborn
325
T. W., 1978]. Кроме того, на усвоение кремния у крыс влияют
возраст, пол и активность пищеварительных желез [Charnot
J. et al., 1979].
Как и в случае большинства других МЭ, всасывание крем-
ния в сильной степени зависит от различных минеральных
компонентов рациона, которые способны снизить раствори-
вость этого МЭ. К числу таких компонентов относятся Ре20з
Са(ОН)г, MgO и особенно алюминий, осаждающие, по-видимо-
му, кремниевую кислоту в виде нерастворимых силикатов.
Кремний в свою очередь затрудняет всасывание марганца, так
как он образует с ним неусваивающийся силикат [Under-
wood Е., 1977].
Установлено также существование физиологического анта-
гонизма на уровне всасывания между кремнием и молибденом,
проявляющегося как на полусинтетических, так и полностью
синтетических рационах, составленных из аминокислот. Так,
при включении в рацион крыс печени, содержащей 3 мг/кг мо-
либдена, концентрация кремния в плазме крови снизилась на
348% по сравнению с контрольными животными, получавшими
казеин с 1 мг/кг молибдена. Прием молибдена вызывает также
снижение содержания кремния в органах и тканях. Вместе с
тем включение в рацион кремния приводит к существенному
снижению содержания в крови молибдена.
В лимфатических узлах и мочевых камнях овец обнаруже-
ны микроскопические частицы аморфного кремнезема расти-
тельного происхождения (фитолиты). Мелкие фитолиты спо-
собны проникать через стенки пищеварительного тракта чело-
века и животных, поступать в кровь и лимфу и мигрировать в
организме. Присутствие фитолитов в органах и тканях, особен-
но таких как легкие, печень, лимфатические узлы, селезенка,
затрудняет оценку физиологических концентраций в них крем-
ния. Интактная кишечная стенка человека также проницаема
для таких мелких частичек кремнезема, как панцири диатомо-
вых водорослей. Они разносятся с кровью и проникают через
мембраны, а также плаценту, о чем свидетельствует их обна-
ружение в органах и тканях недоношенных и мертворожден-
ных детей [Volkenheimer G. Z., 1964]. Проникновение этих час-
тичек в организм возможно и через легочные альвеолы.
Транспорт. Жидкие среды организма свободно обмени-
ваются кремнием. Ортокремниезая кислота, попав в организм,
поступает во все биологические жидкости и ткани в концент-
рациях, ограниченных только растворимостью, и быстро выво-
дится из организма [Baumann Н., 1960]. Концентрация крем-
ния в жидких средах организма, исключая мочу, близка к его
концентрации в плазме крови. Основной транспортной формой
этого А4Э в крови являются, по-видимому, поликремниевые
кислоты, состоящие из 4—5 остатков кремниевой кислоты
[Policard A. et al., 1961].
Выделение. У человека, крыс, морских свинок, коров и
овец количество кремния, выводимое из организма, зависит от
326
его содержания в пище, но имеет предел, определяемый ско-
ростью и степенью усвоения этого МЭ в пищеварительном
тракте. Количество кремния в моче не лимитируется скоростью
его выведения почками, так как при внутрибрюшинном вве-
дении кремния его выделение существенно возрастает [Sauer F.
et al., 1959]. У овец выделение этого МЭ с мочой пропорцио-
нально его содержанию в пище до 83 мг/сутки и достигает
предела при поступлении 200—250 мг/сутки [Nottle М. С.,
Armstrong J. М., 1966].
Поступив в кровь, кремний не задерживается в организме
и быстро выводится с мочой, благодаря чему его концентрация
в крови даже при самых широких колебаниях содержания это-
го МЭ в пище меняется незначительно. Как свидетельствуют
данные балансовых опытов на добровольцах, у человека усвое-
ние кремния выше, чем у исследованных животных, и может
наблюдаться отрицательный баланс этого МЭ [Kelsay J. L. et
al., 1979].
О физиологической роли кремния в организме млекопитаю-
щих, в том числе и у человека, известно очень мало (Ворон-
ков М. Г. и др., 1978]. Первые данные, свидетельствующие об
эссенциальности кремния, были получены in vitro с помощью
электронного микрозондирования кости, показавшего, что крем-
ний специфически концентрируется в участках активного роста
молодой костной ткани крыс и мышей [Carlisle Е. М., 1969].
На самых ранних стадиях кальцификации содержание крем-
ния и кальция в этих участках остеоидной ткани крайне неве-
лико. Затем в процессе минерализации кости оно сперва воз-
растает, а по мере достижения кальцием концентраций, соот-
ветствующих зрелому костному апатиту, снова снижается до
минимума. Отношение кальция к фосфору в участках, богатых
кремнием, не достигает 1, тогда как в зрелом костном апатите
оно составляет 1,67. Эти данные позволяют считать, что крем-
ний наряду с фосфором принимает участие в формировании
органической матрицы кости на начальных этапах ее кальци-
фикации.
Необходимость кремния для процессов оссификации была
показана также в опытах in vivo, продемонстрировавших, что
включение кремния в рацион ускоряет процесс минерализации
кости у крыс и повышает ее зольность. У цыплят, испытываю-
щих дефицит кремния, утончаются трубчатые кости конечности
и компактное вещество диафизов, снижается их эластичность.
Череп уменьшается в размерах и деформируется, 1 его кости
уплощаются [Carlisle Е. М.., 1972]. Деформация черепа при
дефиците кремния отмечена также в опытах на крысах
(Schwarz К., Milne D., 1972]. В дальнейших опытах установлено,
что в процессе формирования кости кремний взаимодействует
с магнием и фтором. Выявлена также зависимость действия
кремния от гормонального баланса организма и состояния его
минерального обмена [Charnot J., Peres G., 1978].
Деформация черепа наступает в результате уменьшения
327
числа остеобластов и содержания коллагена в его костях. При
дефиците кремния происходят полная утрата основным веще-
ством кости регулярной трабекулярной структуры, ее замена
примитивной узловатой костной тканью, в которой трабекулы
появляются только после слияния узелков. Действие этого МЭ
на процессы остеогенеза осуществляется независимо от витами-
на D и направлено в первую очередь на синтез коллагена в
основном веществе, на который витамин D не оказывает влия-
ния.
При дефиците кремния отмечены также патологические из-
менения хрящевой ткани, обусловленные нарушением синтеза
гликозаминогликанов, основного вещества хряща и соедини-
тельнотканных волокон. Коленные и тибиометатарзальные сус-
тавы заметно уменьшены в размерах, эпифизы берцовой кости
и цевки на !/3 беднее водой, хрящи развиты слабо и истончены.
Снижение содержания соединительной ткани и гликозаминогли-
канов, а также кремния отмечено в гребешках петушков при
дефиците кремния [Carlisle Е. М., 1984].
Приведенные выше данные свидетельствуют, что кремний в
первую очередь необходим для формирования основного веще-
ства кости и хряща, хотя может принимать и непосредственное
участие в процессе минерализации костной ткани. Физиологи-
ческая роль кремния при этом связана в основном с синтезом
гликозаминогликанов и коллагена. При репаративных процес-
сах в костной ткани отмечено увеличение (иногда в 50 раз)
содержания этого МЭ. И напротив, иммобилизация конечности
сопровождается снижением содержания кремния в мышцах,
сухожилиях и в костной ткани [Иваненко А. Н., 1971]. Эти на-
блюдения, сделанные на живых организмах, получили дальней-
шее подтверждение в опытах с культурами тканей, показав-
ших, что включение кремния в культуральную среду вызывает
за 12 сут двукратное повышение содержания коллагена в кост-
ной ткани по сравнению с контролем [культуральная среда без
кремния). Кремний также существенно ускоряет синтез гли-
козаминогликанов, количество которых па 8-й день опыта в
3 раза превосходило контроль и сравнялось с ним только на
12-е сутки.
Аналогичные данные получены на хрящевой ткани куриных
эмбрионов, причем действие кремния на синтез коллагена бы-
ло наиболее заметно в опытах с хрящом 14-дневных, а на син-
тез гликозаминогликанов — с хрящом 12-дневных эмбрионов.
Стимуляция кремнием прироста массы кости, синтеза гексоз-
аминов и образования пролина усиливается в присутствии ас-
корбиновой кислоты, тогда как его действие на синтез окси-
пролина, общего и неколлагенового белка не требует участия
данного витамина [Carlisle Е. М., Suchil С., 1983]. Наиболее
высокая продукция гексозаминов достигается при взаимодей-
ствии кремния и аскорбиновой кислоты.
Влияние кремния на образование межклеточного вещества
хрящевой ткани было также показано на эпифизарных хондро-
328
цитах, культивировавшихся в средах с низкой и высокой кон-
центрацией этого МЭ. Через 18 дней после начала опыта изу-
чали содержание в клетках ДНК, гексозаминов, оксипролина,
пролина и неколлагеновых белков. Установлено, что наличие
кремния в первую очередь отражается на содержании оксипро-
лина, входящего в состав проколлагена, количество которого
возросло в кремнийсодержащей культуре по сравнению с бес-
кремниевой на 243% [Carlisle Е. М., Garvet D. L., 1982].
В меньшей степени изменяется содержание межклеточных по-
лисахаридов, оно повышается на 152%. Поскольку культуры
не отличались по числу клеток, то наблюдаемый эффект следу-
ет полностью отнести за счет действия кремния на клеточную
систему, продуцирующую эти вещества.
Роль кремния в метаболизме соединительной ткани на
клеточном уровне подтверждается данными рентгеноспектраль-
ного микроанализа [Carlisle Е. М., 1975], свидетельствующи-
ми, что этот МЭ присутствует в участках активного роста мо-
лодой кости и в составе изолированных остеобластов в коли-
чествах, близких к содержанию кальция, фосфора и магния.
Особенно высокое содержание кремния в остеобластах обна-
ружено в период их повышенной метаболической активности.
Внутри остеобластов этот МЭ локализуется преимущественно
в митохондриях.
Функция кремния в синтезе коллагена связана, по-видимо-
му, со способностью данного МЭ активировать пролин, 2-оксо-
глутарат — диоксогеназу (пролингидроксилазу) (КФ 1.14.11.2).
Удалось показать, что активность этого фермента, выделенного
из лобных костей 14-дневных куриных эмбрионов, возрастает
при инкубации в среде, содержащей 0,2 ммоль/л кремния, по
сравнению с контролем в 5 раз, при концентрации 0,5 ммоль/л—
в 7 — 8 раз и при 2 ммоль/л — в 10 раз [Carlisle Е. М., 1984],
Коллоидная двуокись кремния ингибирует ряд ферментов,
к числу которых относятся ацетилхолинэстераза, сукцинатде*,
гидрогеназа, эстераза А и В печени, гиалуронидаза, что при-
влекалось для объяснения патохимического механизма разви-
тия силикоза [King Е. J., 1956]. В то же время, по другим
данным [Rowsele Е. V., Leonard R. Е., 1957], она способна
усиливать поглощение кислорода гомогенатом печени крыс.
В поджелудочной железе, желудке и почках обнаружен фер-
мент силиказа, способный отщеплять кремниевую кислоты от
синтетического субстрата — триметоксидодеканоксисилана.
Фермент локализован в мембранах митохондрий и микросом,
из которых может быть выделен обработкой некоторыми не-
ионными детергентами [Schwarz К., 1978].
В связи с тем что кремний является одним из жизненно
важных МЭ, патологические изменения у человека могут быть
обусловлены как его недостаточностью, так и избытком в ор-
ганизме. С медицинской точки зрения, убедительных данных о
наличии у человека патологических процессов, обусловленных
дефицитом кремния, до настоящего времени нет, хотя в лите'
329
ратуре имеются отдельные сообщения, свидетельствующие о
пониженном содержании этого МЭ при целом ряде патологи-
ческих состояний. Более того, добавление к пище соединений
кремния нередко оказывает благотворное влияние на течение
этих патологических процессов. Вместе с тем достоверные све-
дения об инициальных механизмах, связанных с дефицитом
кремния, в организме отсутствуют. Можно с достаточной долей
вероятности ожидать развития кремнийдефицитных состояний
при длительном несбалансированном в отношении этого жиз-
ненно важного МЭ парентеральном питании больных. Счита-
ется, что одним из простых диагностических признаков недо-
статочности кремния в организме может служить повышенная
ломкость ногтей. При этом ногти утрачивают нормальную
флюоресценцию в ультрафиолетовом свете.
Наиболее известной и изученной патологией, связанной с
избыточным поступлением в организм кремния, является сили-
коз. Этот термин впервые был предложен в 1870 г. Visconti.
О сложности патогенеза силикоза свидетельствует большое
число предложенных теорий. J. W. Schepers (1960) насчитал
около 50 концепций, подразделив их на физические, химичес-
кие и биологические. Широкое распространение имеет токсико-
химическая теория, согласно которой развитие силикоза объяс-
няется действием свободной или связанной двуокиси кремния.
При этом считалось, что другие пыли, не содержащие кремне-
зема, не оказывают выраженного фиброгенного действия.
По данным Б. Т. Величковского (1980), самая крупная
(5—7 мкм) кварцсодержащая пыль обычно приводит к разви-
тию пылевого бронхита. Частицы кварца диаметром 0,5—2 мкм
обладают наиболее выраженным цитотоксическим эффектом и
приводят к формированию узелковых форм силикоза. Введение
в легкие высокодисперсных (0,2—0,3 мкм) кварцевых частиц
сопровождается развитием диффузных склеротических измене-
ний.
Кремнезем представляет собой решетку из кремнекислород-
ных тетраэдров (S1O2)4, где каждый атом кремния окружен че-
тырьмя атомами кислорода, а каждый атом кислорода при-
надлежит одновременно двум атомам кремния. В биологиче-
ских средах каждая кварцевая пылевая частица несет на своей
поверхности ОН-группы, в результате чего она обладает опре-
деленными физико-химическими свойствами, позволяющими
отнести кремнезем к полиэлектролитам. С наличием ОН-групп
на поверхности кремнезема связывается фиброгенность кварце-
вой пыли [Величковский Б. Т., 1978]. Следует признать спра-
ведливым его высказывание о том, что «выяснение природы
активных центров поверхности кремнезема, хотя и приближает
нас к пониманию механизма первичного взаимодействия пыли
с биосубстратом, само по себе еще не дает однозначного отве-
та на эту проблему.»
Пусковым механизмом развития силикотических изменений
как в легких, так и в других органах и тканях являются фаго-
330
цитоз фастиц кремнезема макрофагами, гибель последних и
активация фибробластов. Без установленной многочисленными
исследованиями [О. Г. Алексеева, 1970; Б. Т. Величковский,
1974; Б. А. Кацнельсон, 1976; И. М. Шнайдман, 1978; Viglia-
ni Е. С., Pernis В., 1962, и др.] последовательной смены про-
цесса фагоцитоза, гибели и распада макрофагов пыль, даже
кварцевая, фиброгенным действием не обладает.
На основании фактических данных [Кацнельсон Б. А.,
1976; Шнайдман И. М., 1978; Величковский Б. Т., 1980; Соро-
ковой В. И., 1984, и др.], касающихся тонких механизмов ги-
бели клеток, в том числе и макрофагов, при воздействии фиб-
рогенных пылей и гипоксии, нами представлена схема патоге-
неза силикоза (схема 4). Как видно из этой схемы, в нее вклю-
чены наиболее узловые моменты патогенеза, не вызывающие
принципиальных возражений и получившие эксперименталь-
ную проверку и подтверждение. Необходимым условием воз-
никновения пневмокониоза, в том числе и силикоза, является
образование так называемого пылевого депо. Установлено, что
«критическая» его величина, за которой следует развитие си-
ликоза, равняется 150 мг/100 г сухой ткани легких [Einbrodt
Н. J., 1965].
Основное значение в самоочищении легких от пыли прида-
ется активности альвеолярных макрофагов. Чем большей ци-
тотоксичностью обладает пыль, тем больше ее скапливается в
регионарных лимфатических узлах. Накопление пылевых час-
тиц кремния в интерстиции осуществляется как в результате
их пенетрации, так и с помощью макрофагов. Установлено, что
вирусная инфекция, способная подавлять функциональную ак-
тивность макрофагов, приводит к значительному снижению
самоочищения легких от пыли.
Для силикоза легких типичной является узелковая форма.
Силикотические узелки большей частью располагаются пери-
бронхиально и периваскулярно, однако при высоком содержа-
нии в пылевых частицах свободной кристаллической двуокиси
кремния они могут формироваться и в просвете альвеол. Гис-
тологически на первых этапах узелок представлен пылевыми
частицами и клеточными элементами, затем он становится
клеточно-фиброзным и в конечной стадии склеротическим. Ха-
рактерной особенностью развитых склеротических силикотиче-
ских узлов являются концентрическое расположение волокон
фиброзной ткани и клеточных элементов, а также наличие на
его периферии зоны эксцентрического роста, где концентриру-
ются пылевые частицы и клеточные элементы. Именно за счет
этой зоны происходит рост узелков, нередко они сливаются
между собой, образуя конгломераты. Экспериментально [Нер-
pleston A. G., 1962] показано, что кремнезем в узелках нахо-
дится постоянно, он может подвергаться захвату мигрирующи-
ми макрофагами и служить основой для формирования новых
свежих силикотических узелков.
Диффузно-склеротическая форма силикоза связана с накоп<
331
ПАТОГЕНЕЗ СИЛИКОЗА
Схема 4
лением в легких пыли, содержащей малые концентрации сво-
бодного кремнезема.
Коллагенообразование при силикозе осуществляется за
счет активной синтетической деятельности фибробластов. Элек-
тронно-микроскопическими исследованиями установлена фаз-
ность их морфофункциональной активности [Шнайдман И. М.,
332
\
х
'\
1971Д1984]. Превращение формирующегося фибробласта в зре-
лый характеризуется увеличением объемной плотности ультра-
структур, ответственных за синтез специфического белка. В ча-
стности) наблюдаются развитие цистерн гранулярной эндоплаз-
матической сети, увеличение числа везикулярных образований,
содержащих материал, идентичный по структуре таковому в
просвете цистерн. Везикулы осуществляют транспорт синтези-
рованного тропоколлагена в межклеточные пространства, где
происходит формирование коллагеновых фибрилл. Отмечено
также, что в зрелом фибробласте синтез коллагена превышает
скорость его выведения. В результате этого цистерны эндо-
плазматической сети представляются резко расширенными и
переполненными вновь синтезированным тропоколлагеном. Та-
кой фибробласт, приобретая ячеистость цитоплазмы, превра-
щается в клетку — депо коллагена [Шнайдман И. М, 1984].
В гиалине силикотических узелков содержится только 30%
коллагена, а 50% его составляют ^-глобулины, а именно гло-
булины G- и М-классов [Pernis В., 1966].
При выраженном силикозе легких рентгенологически не-
редко наблюдается своеобразная картина поражения прикорне-
вых лимфатических узлов в виде так называемого скорлупко-
образного обызвествления их [Молоканов К. П., 1961]. При
этом лимфатические узлы фиброзно изменены и петрифициро-
ваны в краевой зоне. Рентгенологически в зависимости от мас-
сивности обызвествления в области корня легких определяют-
ся множественные кольцевидные или скорлупкообразные тени,
контуры их ровные или пунктирные (при неполном обызвеств-
лении). Высокая частота встречаемости этого рентгенологичес-
кого признака является патогномоничной для тяжелого сили-
коза легких. Так, у 37 из 39 обследованных больных силикозом
и антракосиликозом он был положительным [Фастыковс-
цая Е. Д. и др, 1967].
Патогистологически в органах и тканях при силикозе обна-
ружены преимущественные изменения соединительной ткани и
сосудов. В. частности, глаза человека, не защищенные от вред-
ного воздействия частичек кремнезема, находящихся в воздухе,
нередко подвергаются поражению [Пономарева Т. Б, 1982].
При этом отмечены мукоидное набухание, фибриноидный не-
кроз соединительной ткани, примыкающей к лимбу, с макро-
фагальной или лимфогистиоцитарной реакцией по периферии
очагов. Выявлено также статистически достоверное увеличение
толщины роговицы, склеры, сосудистой и сетчатой оболочек у
больных силикозом по сравнению с контрольной группой. При
кристаллооптическом исследовании глазного яблока обнаруже-
ны частицы свободной двуокиси кремния диаметром 1—2 мкм
в роговице, радужной, сосудистой и сетчатой оболочках, что
свидетельствует как об аэрогенном, так и гематогенном путях
их распространения. Экспериментальными исследованиями
A. Poole (1985) удалось показать, что динамика в организме
таких ферментов, как пролингидроксилаза и лизилоксидаза,
333
может служить ранним прогностическим биохимическим тес-
том, дающим возможность судить о выраженности колларено-
образования на ранних стадиях силикоза.
Эпидемиологические исследования и биогеохимическая,
оценка окружающей среды [Schroeder Н. A., Kraemer L. А.,.
1974; Schwarz К. et al., 1977] свидетельствуют о том, что крем-
ний играет определенную роль в процессе атеросклероза.
В частности, J. Loeper и соавт. (1979) показали, что в аорте
содержится наиболее высокая концентрация этого МЭ. Крем-
ний, введенный per os или внутривенно, тормозит развитие у
кроликов экспериментального атеросклероза. Yasuhide Nakas-
hima и соавт. (1985), исследовавшие в секционном материале
содержание кремния, различных липидов и гликозаминоглика-
нов в нормальной аорте, липидных и атероматозных бляшках
человека, показали, что содержание кремния в интиме аорты
возрастает со степенью как первичного, так и вторичного ате-
росклероза. При этом отмечена положительная корреляция
между содержанием кремния и дерматансульфатом — одним
из компонентов гликозаминогликанов. Предполагается, что
кремний может защищать интиму аорты человека от ее ин-
фильтрации липидами за счет уменьшения проницаемости со-
единительной ткани и межклеточного цемента.
На этиологическую связь между высоким содержанием
кремния в растительной пище (до 50 мг/кг) и эндемическим
уролитиазом в Грузии указывает В. С. Цинанцадзе (1973).
Обобщенные данные литературы и материалы собственных ис-
следований о распространенности мочекаменной болезни на
территории СССР с биогеохимических позиций представлены
в работе В. В. Ковальского и В. Л. Сусликова (1980). Они
указывают на совпадение очагов эндемического уролитиаза с
месторождениями кремниевых пород, с кремниевыми биогео-
химическими провинциями и субрегионами биосферы. По мне-
нию этих авторов, эндемический уролитиаз следует рассматри-
вать как один из синдромов дизадаптации человека к условиям
повышенного поступления в организм кремния на фоне избыт-
ка других макроэлементов и МЭ или их соединений (кальция,
фтора, марганца, сульфатов, нитратов, хлоридов). Следует
признать справедливым выделение подобной формы моче-
каменной болезни в виде эндемического кремниевого уроли-
тиаза.
Имеются отдельные сообщения, свидетельствующие о нару-
шении обмена кремния при развитии злокачественных опухо-
лей [Воронков М. Г. и др„ 1978]. В частности, установлено
высокое содержание (37 мг/кг) этого МЭ в опухолевой ткани.
В некоторых географических регионах с высокой заболевае-
мостью раком пищевода (отдельные районы Туркменской и
Казахской ССР, Каракалпакской АССР и др.) почвы богаты
кремнием. В наших исследованиях (А. А. Жаворонков и Н. Са-
хипов) эзофагобиопсий при изучении патологии пищевода у
жителей Гурьевской области Казахстана выявлено в его сли-
334
ристЗй оболочке наличие диатомовых водорослей. Последние
попадают в пищевод с питьевой водой.
Диатомовые водоросли, или диатомеи, представляют собой
одноклеточные микроскопические организмы, их размеры ко-
леблются в широких пределах (от 4 до 1000 мкм и более)
[Макарову И. В., 1977]. В наших наблюдениях они имели
диаметр 10 мкм. Клетка диатомеи состоит из протопласта, ок-
руженного панцирем, или кремнеземной оболочкой. Панцирь
состоит из аморфного кремнезема. Стенки панциря пронизаны
мельчайшими отверстиями, которые обеспечивают обмен ве-
ществ между протопластом клетки и окружающей средой.
Большое количество диатомей, попадающих с водой в про-
свет пищевода и оседающих в складках его слизистой оболочки,
могут вызывать микротравмы эпителиальных клеток с после-
дующим развитием реактивных воспалительных изменений. По-
мимо травматического действия плотным, нередко окаймленным
крючками, панцирем, возможны и другие вредные влияния ди-
атомей, в частности, обусловленные избытком кремнезема
(рис. 59). Таким образом, связь предкарциноматозного эзофа-
гита с инвазией диатомовых водорослей представляется веро-
ятной. Необходимо отметить, что частота встречаемости диато-
мей в эзофагобиоптатах у жителей г. Гурьева оказалась до-
вольно высокой. Диатомеи при исследовании в сканирующем
микроскопе обнаружены в 6 (17,6%) из 34 изученных биопта-
тов. В литературе нам не встретилось указаний на этиологи-
ческую связь между наличием диатомовых водорослей и па-
тологией пищеварительного тракта, однако приведенные дан-
ные с несомненностью могут свидетельствовать о такой воз-
можности. Этот вопрос требует дальнейшего прицельного изу-
чения.
На основании изложенного могут быть выделены следую-
щие основные формы патологии человека, обусловленные из-
быточным или недостаточным поступлением кремния в орга-
низм.
1. Общий гиперсиликоз организма (избыточное поступле-
ние или замедленное выделение кремния).
2. Локальные формы силикозов:
а) пневмокониоз;
б) нефросиликоз — эндемический кремниевый нефролитиаз;
в) силикокальциноз лимфатических узлов корня легкого;
г) окулосиликоз.
3. Силикопривные состояния (гипотетически):
а) силикопривная дерматопатия с недоразвитием придатков
кожи;
б) силикопривная онихопатия;
в) силикопривная коллагенопатия;
г) силикопривные ангиопатии (типа синдрома Марфана?);
д) силикопривная аортопатия (участие в патогенезе атеро-
склероза?) ;
335
Рие. 59. Патогистологическая и стереоультраструктурная характеристика сли-
зистой оболочки пищевода с инвазией диатомей:
а — хронический эзофагит, отек и дистрофия эпителиального пласта, отек и воспалитель-
ные инфильтраты в субэпителиальиой ткани. Окраска гематоксилином и эозином X2S0;
б — эндоскопически-атрофический эзофагит (биопсия); на поверхности эпителиального
пласта расположены бактерии и 4 диатомеи (стрелки), Х1400; в — инвазия диатомеи в
разрыхленную дистрофически измененную поверхность слизистой оболочки у больного-
раком пищевода (биопсия). ХЮ ООО: г — эндоскопическая биопсия у больного атрофи-
ческим эзофагитом с явлениями паракератоза н лейкоплакии. ХЗООО.
е) силикопривные хондропатии;
ж) локальная и генерализованная силикопривная остеопа-
тия {недоразвитие скелета).
7.8. Ванадий
Ванадий концентрируется многими живыми организмами.
Он накапливается красным мухомором (Amanita muscapa),
из которого выделен ванадиевый комплекс амаванадин. /Ва-
надий необходим для роста и развития черной плесени (Asper-
gillus niger), зеленой водоросли (Scenedesmus obliqus). (^Qh
присутствует в повышенных количествах в нефти, битуме, ас-
фальте и горючих сланцах, зола которых обогащена этим МЭ.
В нефти и асфальте ванадий встречается в составе порфирино-
вого макроцикла. !
Особенно высокие концентрации ванадия обнаружены в не-
которых морских организмах. Среди них выделяются предста-
вители оболочечников — асцидии (порядки Phlebobranchia и
Aplousbranchia), у которых этот МЭ входит в состав зеленых
кровяных телец — ванадоцитов, содержащих до 4% ванадия и
специфический восстанавливающий агент — тунихром. Ранее
полагали, что внутри этих клеток ванадий присутствует в со-
ставе белкового комплекса гемованадина (ванадохрома)
[Califano L., Caselli Р., 1948J, однако при исследовании мето-
дами ЭПР-спектроскопии обнаружены связи этого МЭ с бел-
ком или другими макромолекулами [Dingley A. L. et al., 1981].
Ванадий найден в ванадоцитах в составе низкомолекулярного
комплекса. Не подтвердилось также предположение о роли ге-
мованадина в транспорте кислорода [Califano L., Boeri Е.,
1950], а сам гемованадин оказался, по-видимому, артефактом,
В ванадоцитах одного вида асцидий (Ascidia aspersa) наблю-
дается динамическое равновесие между V3+ и V5+, что указы-
вает на возможное участие этого МЭ в окислительно-восстано-
вительных процессах [Uderwood Е., 1977]. Ванадоциты оболо-
чечников способны в 106—107 раз концентрировать ванадий по
сравнению с его содержанием в морской воде с помощью ме-
ханизмов облегченной диффузии этого МЭ через плазматиче-
скую мембрану и восстановления ванадия внутри клетки до
малоподвижной катионной формы.
Высокие концентрации ванадия имеются у голотурий и не-
которых моллюсков (Pleurobranchus plumula). Не исключено,
что в отдельных случаях высокое содержание ванадия в морс-
ких организмах объясняется загрязнением моря нефтепродук-
тами(25изненная необходимость ванадия для животного орга-
доказ.-ша в 1971 г. в опытах на крысах (Strasia С А.,
1971] и курах [Hopkins L. L„ Mohr Н. Е„ 1971] и подтверж-
дена ^Дальнейших исследованиях [Schwarz К-, Milne D. В,
19?1; Nielsen F. H„ Ollerich D. A., 1972].
В £ a с ы в а н и e ванадия в желудочно-кишечном тракте за-
висят от растворимости соединений этого МЭ и его химической
22—568
337
формы. Водорастворимые анионы ванадия усваиваются в боль-
шей степени, чем катионные и ванадий, присутствующий в пи-
ще. У крыс всасывание 48УгО5 в кишечнике составляет 2,6±
±1,6%. У человека из 100 мг ванадия (в виде растворимого
ортованадата аммония) в кишечнике всасывается всего 0,1 —
1% принятой дозы [Curran G. L. et al., 1959]. По данным Меж-
дународного Комитета радиационной защиты (МКРЗ, публи-
кация 30, 1984], всасывание ванадия в желудочно-кишечном
тракте человека составляет в среднем 1%. В 1-е сутки 60%
всосавшегося ванадия выделяются с мочой, а остальная часть
откладывается в печени, костях и частично выводится с калом
[Talvitie N. A., Wagner W. D., 1954].
При парентеральном введении соединения ванадия быстро
всасываются и переходят в кровь. После подкожного введения
крысам VCh он выводится преимущественно с мочой. За 1 сут
удаляются 18%, а за 2 сут — 25% этого соединения. При этом
с мочой выделяется в 5 раз больше ванадия, чем с калом [Ро-
щин А. В., 1968]. У мышей после внутривенного введения
4SV2O5 наибольшее накопление этого изотопа происходит в
костях и зубах, особенно в дентине [Soremark R., Ulberg S.,
1962], а у цыплят — в костях и почках [Hathcock J. N. et al.,
1964]. По другим данным, у крыс основными органами — нако-
пителями ванадия являются почки, печень и легкие [Ро-
щин А. В., 1968].
В течение 4 ч после внутривенного введения 4SV у крыс
происходит повышение концентрации этого изотопа в печени,
почках, селезенке и семенниках. Большая часть радиоактивно-
сти сохраняется в них на 4-е сутки эксперимента, тогда как
в остальных органах и тканях происходит ее существенное
уменьшение. К этому времени 46% 48V выводятся с мочой и
8,6%—с калом. За этот период концентрация 48V в цитозоле
печени снижается с 57 до 11 %, тогда как в митохондриальной
и ядерной фракциях она повышается с 14 до 40%. Этим, по-ви-
димому, и объясняется задержка радиоактивного ванадия в
печени [Hopkins L. L., Tilton В. Е., 1966]. Степень окисления
ванадия не оказывает существенного влияния на скорость и
количество поступления этого МЭ в органы и ткани.фНаиболее
быстрое усвоение ванадия происходит при вдыхании его соеди-
нений или их введении в трахею. В последнем случае около
40% V2O5 удаляется из легких в течение первых 15 мин после
введения, а через 7 сут в них обнаруживается только около
9% введенной дозы. С мочой за 7 сут .выводится около 50%
48V. Оставшееся количество изотопа обнаруживается в основ-
ном в скелете (27% через 6 ч после введения), печени (10%
через 2 ч) и почках (5% через 6 ч). Всасывание ванадия, как
и фосфора, в желудочно-кишечном тракте снижается при од-
новременном поступлении гидроксида алюминия в связи со
сходством анионов этих элементов.
Выделение. В ранних работах указывалось, что у здо-
рового человека в сутки выделяется с мочой 8—15 мкг ванадия
338
[Lewis С. E., 1959; Schroeder H. A. et al., 1963]. Если допус-
тить, что этим путем выводятся 0,75% ванадия, содержащегося
в суточном рационе [Москалев Ю. И., 1985], то в нем должно-
содержаться 1—2 мг этого МЭ, что в 50—100 раз превышает
величины, обнаруживаемые современными аналитическими ме-
тодами. В этой связи цифры, характеризующие содержание ва-
надия в моче, приводимые в ранних работах, приходится счи-
тать сильно завышенными.(В суточное количество мочи пере-
ходят 17% ванадия, содержащегося в цельной крови, и около
50% ванадия, присутствующего в плазме [Москалев Ю. И.,
1985]. Аскорбиновая кислота ускоряет выведение ванадия
[Архипова О. Г. и др., 1985]. Биологический период полувыве-
дения ванадия из организма в целом, а также из почек, селе-
зенки, печени и костей человека составляет [МКРЗ, публ. 2,
1961] соответственно 42, 74, 90, 70 и 135 сут, однако эти вели-
чины нуждаются в свете последних данных в существенной
корректировке. При повышенном содержании ванадия в плаз-
ме крови небольшое количество его выводится с желчью.
Оно, по данным опытов с WsOs, составляет 7ю—Vs часть
этого элемента, выделяемого с мочой [Sabbioni Е. et al.,.
1981].
Распределение радиоактивного ванадия в органах и тканях
происходит в следующем порядке: кость>почки>печень>селе-
зенка > кишечник > желудок > семенники > легкие >мозг.
Концентрация этого МЭ в органах и тканях человека невысо-
ка. В мозге, мышцах, печени, семенниках, легких и лимфати-
ческих узлах она составляет 0,59±0,16; 1,18±0,06; 0,78±0,2;
3,92±1,58; 1,96±0,39 и 7,85±3,92 мкмоль/кг соответственно.
В зубной эмали имеется менее 1,93 мкмоль/кг [Underwood Е.,
1977]. Некоторое повышение содержания ванадия в легких мо-
жет быть связано с вдыханием атмосферной пыли. Оценка со-
держания ванадия в плазме крови за 20 лет снизилась пример-
но на 2 порядка — с 6,87—9,42 ммоль/л [Schroeder Н. A. et al.,
1982] и 0,9 мкмоль/л [Ноздрюхина Л. Р. и др., 1973] до 0,066 ±
±0,012 — 0,13±0,006 мкмоль/л [Cornelis R., Versieck J., 1963].
Это объясняется в первую очередь появлением новых, более
совершенных аналитических методов. Снизилась также оценка
количества ванадия, содержащегося в суточном рационе чело-
века. Если 25 лет назад оно принималось равным в среднем
2 мг [Schroeder Н. A. et al., 1963], величине принятой также
Международным Комитетом радиационной защиты (МКРЗ,
публикация 23, 1977), то вскоре были получены в 15—18 раз
меньшие величины— 108—137 мкг, а спустя еще 10 лет 12,4—
30,1 мкг [Myron D. R. et al., 1978], что почти в 100 раз ниже
первоначальных результатов. Концентрация ванадия в эритро-
цитах выше, чем в плазме крови (МКРЗ, публикация 23, 1977).
Половые различия по содержанию ванадия в цельной крови
(0,63—1,16 мкмоль/л у мужчин и 0,47—1,12 мкмоль/л у жен-
щин) [Ноздрюхина Л. Р., 1977] невелики и нуждаются в под-
тверждении.
22* оо э
Недостаточность ванадия и его роль в орга-
низме. Как впервые установили L. L. Hopkins и Н. Е. Mohr
(1971), при дефиците ванадия (10 мкг на 1 кг корма) у цыплят
замедляется оперение крыльев и хвоста. Явления недостаточ-
ности ванадия, выражавшиеся в задержке роста и аномальном
развитии скелета, были отмечены у цыплят при содержании в
рационе 30—35 мкг/кг этого МЭ [Nielsen F. Н., Ollerich D. А.,
1973]. Задержка роста была отмечена также у крыс, получав-
ших менее 100 мкг ванадия на 1 кг корма, по сравнению с
животными, в рацион которых было дополнительно включено
0,5 мг/кг этого МЭ [Strasia С. А., 1971]. Стимулирование рос-
та крыс при включении 0,25—0,5 мг ванадия на 1 кг высоко-
очищенного рациона наблюдалась также в лаборатории
К. Schwarz [Schwarz К-, Milne D. В., 1971]. В свете приведен-
ных данных может оказаться, что содержание ванадия в раци-
оне человека не всегда удовлетворяет его потребность в этом
элементе.
При дефиците ванадия (менее 10 мкг/кг рациона), поддер-
живавшемся на протяжении 3—4 поколений крыс, отмечены
снижение плодовитости самок, уменьшение числа беременнос-
тей за период спаривания, повышенная смертность потомства
[Hopkins L. L., Mohr Н. Е., 1976]. Дефицит ванадия вызывает
так называемый алиментарный отек, который объясняется
влиянием этого МЭ на почечный натриевый насос [Gol-
den М. Н., Colden В. Е., 1981]. Инфузии ванадия повышают
диурез и натрийурию [Balfour W. Е. et al., 1978; Grantham J.,
1980]. Вполне вероятно, чтоУлюбое заболевание, ведущее к по-
тере ванадия из организма, способно вызвать задержку в нем
натрия и воды. Снижение концентрации ванадия в крови отме-
чено у детей при квашиоркоре [Golden М. Н., Golden В. Е.,
1981].
Имеются данные, что ванадий может играть роль этиологи-
ческого фактора при маниакально-депрессивном психозе [Nay-
lor G. Н., Smith А. Н., 1981]. При этом заболевании имеет мес-
то нарушение обмена натрия в эритроцитах, вызванное, по-ви-
димому, угнетением Na+, К+-АТФазы повышенной внутрикле-
точной концентрацией ванадата. В этой связи больным были
назначены массивные дозы витамина С (3 г/сутки) или парен-
теральное введение ЭДТА, направленные на снижение концент-
рации эритроцитарного ванадия, что дало положительный
лечебный эффект./Витамин С при этом изменял окислительно-
восстановительный потенциал, происходило восстановление ва-
надата до ванадила, а ЭДТА ограничивал поступление этого
МЭ в эритроциты. Было также высказано предположение, что
литий, используемый при лечении маниакально-депрессивного
психоза, снимает ингибирующее действие ванадия на Na+, К+-
АТФазу [Naylor G. Н., Smith А. Н., 1981]. Однако оно не по-
лучило экспериментального подтверждения [Mac Donald Е. et
al., 1982]. Имеются также данные, что роль Na+, К+-АТФазы
в этиологии аффективных состояний сильно преувеличена, в
540
связи с чем «ванадиевая» гипотеза их возникновения нуждает-
ся в пересмотре, как и попытки снижения уровня ванадия в
тканях при указанных заболеваниях.
При подкожном введении радиоактивный ванадий концент-
рируется в участках активной минерализации костной ткани
и дентина [Soremark R„ Ullberg S., 1962]. Имеются даже дан-
ные, что включение ванадия (и стронция) в синтетические ра-
ционы крыс и морских свинок ускоряет процесс минерализации
костей и зубов и уменьшает частоту возникновения кариеса
[Kruger В. J., 1958]. В опытах на хомячках было показано,
что парентеральное или пероральное введение V2O5 существен-
но снижает появление кариеса при скармливании кариогенно-
го рациона [Geyer С. F., 1953]. Имеются, однако, и прямо про-
тивоположные данные об увеличении под влиянием ванадия
частоты кариеса у обезьян [Bowen Н. J., 1972], а также о его
неспособности снизить частоту кариеса [Mahler J. С., 1957;
Buttner W., 1963]. Исходя из этого, данный вопрос заслужива-
ет дальнейшего изучения.
Повышенные дозы ванадия усиливают сокращение сердеч-
ной мышцы. Так, ванадат аммония (20—500 мкмоль/л) при
электростимуляции усиливал сокращение папиллярных мышц
правого предсердия кошки, причем этот эффект не был вызван
ионом аммония или стимуляцией p-рецепторов [Hackborth I.
et al., 1978]. В то же время инотропное действие ванадия за-
висит от вида животных, ткани, концентрации и химической
формы ванадия, присутствия других ионов и активности аде-
нилатциклазы. Описаны также случаи отрицательного инотроп-
ного действия этого МЭ [Ramasarma Т„ Crane F., 1981]. Из-
вестен фермент NADH-ванадатоксидоредуктаза [Erdman Е.
et al., 1984], катализирующий превращение ванадата в вана-
дил, ингибирующее действие которого на Na+, К+-АТФазу зна-
чительно слабее, чем у ванадата. Этот фермент может таким
образом выступать в качестве регулятора активности ванадий-
зависимых ферментов, например Na+, К+-АТФазы.
Ванадат концентрируется клетками почек, что влечет за
собой угнетение в них Na+, К+-АТФазы [Grantham J. J., 1980].
Так, у крыс, находящихся под эфирным наркозом, внутривен-
ное введение 1,5 мкмоль ортованадата вызывало 9—25-кратное
повышение выделения мочи, что свидетельствует о воздействии
ванадия на клетки проксимальных почечных канальцев. Вы-
ведение натрия при этом соответствовало количеству отделяе-
мой мочи [Balfourt W. Е. et al., 1978]. У крыс диурез и нат-
рийурез особенно выражены при повышении концентрации в
сыворотке крови ионов калия. У собак и кошек введение вана-
дия не оказывает столь выраженного диуретического эффекта,
как у крыс, поскольку проявляется в первую очередь сосудо-
суживающее действие [Grantham J. J„ 1980].
Как уже отмечалось, ванадат может участвовать в клеточ-
ном механизме регуляции натриевого насоса. Увеличение кон-
центрации внеклеточного К+, так же как и уменьшение содер-
341
жания Na+ ниже физиологического уровня, усиливает ингиби-
рующее действие ванадия. Для ингибирования насоса необхо-
димо наличие внутри клеток магния. Его ингибирование ва-
надием происходит с цитоплазматической стороны интактных
клеток. Отношение ванадата к ванадилу может представлять
собой главный механизм регуляции натриевого насоса [Grant-
ham J. J., 1980].
Влияние на обмен глюкозы. В опытах in vitro ус-
тановлено, что ванадий способен оказывать влияние на обмен
глюкозы. В культурах адипоцитов оптимальные концентрации
этого МЭ в среде стимулируют окисление глюкозы столь же
эффективно, как и оптимальные концентрации инсулина [Du-
byak G. R., Kleinzeller А., 1980; Schechter J., Karlisch S. J.,
1980]. При предварительном воздействии на адипоциты инсу-
лином в концентрации, отвечающей максимальной активации,
дополнительной стимуляции окисления глюкозы под влиянием
ванадия не происходит [Dubyak G. R., Kleinzeller А., 1980].
При повышении концентрации глюкозы в культуральной среде
степень стимуляции как инсулином, так и ванадатом снижа-
ется. Это свидетельствует, что они активируют транспортную
систему глюкозы, которая достигает насыщения [Schechter J.,
Karlisch S. J., 1980]. Было обнаружено, что инсулинмиметиче-
ский эффект ванадия не связан с ингибированием Na+, К+-
АТФазы, поскольку ванадат оказался эффективным и в среде,
не содержащей К+ [Schechter J., Karlisch S. J., 1980], не на-
блюдалось потери клеточного К+ и, как свидетельствовало по-
ступление в клетку 86Rb, не происходило угнетения функции
Na+, К+-АТФазы [Dubyak G. R., Kleinzeller А., 1980]. Стимуля-
ция окисления глюкозы осуществлялась, по-видимому, главным
образом ионом ванадила. Проведенные опыты дают основание
полагать, что, поступив в клетку, ванадат фиксируется или ви-
доизменяется, в связи с чем его транспорт сохраняет продол-
жительное время линейный характер. Вероятно, внутри клетки
ванадат превращается в ванадил [Dubyak G. R., Kleinzeller А.,
1980]. Показано также, что при включении в среду глутатиона
концентрация ванадата, требуемая для достижения полумак-
симальной стимуляции, снижается почти на порядок. Глутатион,
поступая в адипоциты, способствует, по-видимому, эндогенному
восстановлению ванадата [Schechter J., Karlisch S. J., 1980].
Обмен холестерина. Заметное место занимают иссле-
дования влияния ванадия на обмен холестерина в связи с его
значением в патогенезе атеросклероза, однако единое представ-
ление о характере действия этого МЭ отсутствует [Ramasarma
Т., Crane F. L., 1981]. Так, при дефиците ванадия наблюдали
снижение уровня холестерина в плазме крови 28-дневных цып-
лят и его повышение у 49-дневных животных [Hopkins L. L.,
Mohr Н. Е., 1974]. По другим данным, у 4-недельных цыплят
при дефиците этого МЭ концентрация холестерина в сыворотке
крови, напротив, возрастает. Увеличенный уровень холестерина
в печени и плазме крови отмечен и при высоком содержании
342
в рационе ванадия (1,96 ммоль/кг), близком к. токсическому
[Zenz С., 1980]. Сходные противоречия наблюдались и в более
ранних исследованиях. Так, на кроликах было показано, что
дефицит ванадия ускоряет мобилизацию холестерина стенки
аорты [Curran G. L„ Costello R. L„ 1956]. Аналогичное наблю-
дение было сделано и на людях [Curran G. L. et al., 1959;
Lewwis С. E„ 1959; Blankenhord D. H„ Chin H. P., 1964]. Одна-
ко, по другим данным, ванадотартрат аммония, вводившийся в
течение 6 мес больным с гиперхолестеринемией и ишемией
миокарда, не оказал никакого влияния на содержание в сыво-
ротке крови холестерина и липопротеидов [Sommerville J.,
Davis В., 1962]. Не было обнаружено влияния ванадия и на
уровень холестерина в сыворотке крови у лиц старше 60 лет
[Schroeder Н. A. et al., 1963].
У крыс массой 400—600 г дача ванадия вызывала угнете-
ние синтеза холестерина [Curran G. L., Burch R. Е., 1967].
Существует несколько гипотез, объясняющих наблюдаемые
расхождения. Так, в ряде работ было показано, что ванадий
ингибирует скваленсинтетазу, катализирующую превращение
фарнезилпирофосфата в предшественник холестерина — сква-
лен [Azarnoff D. L. et al., 1961 j. Вместе с тем оказалось, что
ванадий активирует митохондриальную ацетоацетил-СоА —
деацилазу (гидролазу) — фермент, ограничивающий использо-
вание ацетата для синтеза холестерина. Эта активация наблю-
далась только у молодых животных, тогда как с возрастом
происходит ингибирование этого фермента ванадием [Curran
G. L., Burch R. Е., 1967]. Этим, по мнению авторов, можно
объяснить возрастные различия в действии ванадия на синтез
холестерина.
Другая гипотеза исходит из того, что действие ванадия за-
висит от конентрации других клеточных компонентов, в част-
ности железа и цистеина, различные комбинации которых по-
разному влияют на уровень липидов в плазме крови крыс,
концентрацию в печени железа, ванадия и меди, отношение
массы сердца к массе тела, гематокритное число [Nielsen F.
et al., 1983].
Наблюдаемые различия могут быть связаны и с воздейст-
вием ванадия на ряд других ферментов липидного обмена. Так,
в 1980 г. было показано, что этот МЭ ингибирует микросомаль-
ную гидроксиметилглютарил-СоА — редуктазу (КФ 1.1.1.34) и
инактивирующий ее фермент [Menon D. R. et al., 1980]. По-
следний оказался более чувствительным к ванадату и ингиби-
ровался в значительной степени уже при его концентрации,
равной 1 ммоль/л, тогда как для ингибирования гидроксиметил-
глютарил-СоА — редуктазы требовалось 10 ммоль/л ванадата.
Поскольку эта редуктаза является ферментом, лимитирующим
скорость биосинтеза холестерина в печени, то вполне вероятно,
что его синтез может повышаться при низких и снижаться при
более высоких концентрациях ванадия [Menon D. R. et al.,
1980]. Показано также, что сульфит и ванадат способны вызы-
вать in vitro перекисное окисление липидов, которое предуп-
реждается гидрохиноном, а-токоферолом и супероксиддисмута-
зой, устраняющих супероксидный ион-радикал.
Действие на АТФазы. В 1977 г. было показано, что
ванадат является ингибитором Na+, К+-АТФазы в наномоляр-
ных концентрациях, встречающихся в естественных условиях в
тканях млекопитающих [Cantley L. С. et al., 1977]. В опытах
на тенях эритроцитов было установлено, что ванадат поступает
в клетку с помощью той же системы обмена анионов, что и
фосфат, и ингибирует Na+, К+-АТФазу с цитоплазматической
стороны мембраны [Cantley L. С. et al., 1978]. В дальнейших
опытах было установлено, что большая часть ванадия цито-
плазмы присутствует в виде ионов ванадила, который менее
эффективен как ингибитор К+, Ыа+-АТФазы, чем ванадат. Вос-
становление последнего снижает его ингибирующее действие на
данный фермент [Cantley L. С., Aisen Р., 1979]. Ванадил в ци-
топлазме эритроцитов связан с крупными молекулами, по-ви-
димому, с гемоглобином. Известно, что в процессе функциони-
рования натриевый насос принимает различные конформации.
Удалось выяснить, что ион ванадата фиксирует фермент в пе-
реходном состоянии, образующемся при отщеплении фосфо-
рильной группы, так как он способен принять ту же бипирами-
дальную структуру, что и фосфат.
Ванадат ингибирует также Са2+, Мд2+-АТФазу мозга, при-
чем это ингибирование усиливается КС1 и диметилсульфокси-
дом. Усиление ингибирования ванадием этого фермента калием
сближает его с Na+, К+-АТФазой плазматической мембраны и
Са2+, М§2+-АТФазой саркоплазматического ретикулума [Ro-
binson J. D., 1981]. Ванадий ингибирует также ряд других
АТФаз [Ramasarma Т., Crane F. L., 1981] с неодинаковой ско-
ростью в разных тканях, причем этот эффект зависит от кон-
центрации ванадата и других ионов. Ванадий активирует аде-
нилатциклазу. Активация ферментного комплекса происходит,
по-видимому, через регуляторную субъединицу G в результате
образования стабильного комплекса фермент — ГДФ — вана-
дат. Неясно, однако, возможна ли подобная активация в фи-
зиологических условиях, поскольку для нее требуется относи-
тельно высокая концентрация ванадия, значительно большая,
чем для ингибирования АТФазы [Nielsen F., 1987]. Этот МЭ
усиливает синтез ДНК, эффективность инсулина и эпидермаль-
ного ростового фактора. Действие на ростовой фактор объясня-
ется, по-видимому, способностью этого фермента активировать
протеинкиназу, специфически фосфорилирующую тирозиновые
остатки [Brown D. J., Gordon J. А., 1984] и ингибированием
фосфатазы фосфотирозина [Gentleman S. et al., 1985].
Ванадий усиливает in vitro связывание кислорода гемогло-
бином и миоглобином, видимо, в результате восстановления
Fe3+Hb в Fe2+Hb, более прочно удерживающего кислород [Sa-
kurai Н. et al., 1982]. Усиление связывания кислорода гемопро-
теидами, возможно, является физиологической функцией ва~
344
надия. Ванадат вызывает 10—15-кратное повышение проницае-
мости эритроцитов, лишенных АТФ, для ионов калия. Этот
эффект наблюдается после некоторого лаг-периода, в течение
которого, по-видимому, ванадат превращается в ванадил, вызы-
вающий раскрытие мембранных калиевых каналов [Siemon Н.
et al., 1982].
В ранних работах, посвященных биохимии ванадия, отме-
чается, что при его поступлении в организм нарушается синтез
цистеина и цистина. Было показано, что у крыс, получавших
корм, богатый ванадием (0,49—19,6 ммоль/кг), понижена кон-
центрация этих аминокислот в когтях и шерсти [Mountain J.T.
et al., 1955]. Оказалось, что в присутствии ванадия происходит
также их ускоренный распад [Bergel F. et al., 1958]. В этой
связи можно было ожидать, что при избыточном поступлении
в организм ванадия произойдет угнетение целого ряда метабо-
лических процессов, прямо или косвенно связанных с обменом
цистеина. Действительно, при поступлении метаванадата нат-
рия с кормом или в виде внутрибрюшинных инъекций в печени
крыс наблюдалось существенное снижение содержания СоА,
для биосинтеза которого необходим тиоэтаноламин, образую-
щийся путем декарбоксилирования цистеина [Mascitelli-Corian-
deli Е. et al., 1959]. Не исключено также, что угнетение синтеза
СоА связано и с нарушением обмена пантотеновой кислоты, вы-
зываемым ванадием [Aiyar A. S., Sreenivasan А., 1961].
Снижение синтеза СоА в свою очередь ведет к угнетению
образования кофермента Q, количество которого в митохонд-
риях крыс при даче ванадия также снижается. При введении
одновременно с ванадием цистеина этот эффект ослабляется,
а при совместной даче ванадия, СоА, АТФ и цистеина исчезает
совсем. Другим коферментом, синтез которого угнетается вана-
дием, является липоевая кислота в печени крыс.
Соли ванадия активируют тканевую моноаминоксидазу.
Они способны в концентрации 10 мкмоль/л ускорять на 125%
окисление триптамина в присутствии неочищенных препаратов
моноаминоксидазы из печени морских свинок, в то время как
ионы железа и кобальта повышают активность этого фермента
всего на 25 и 33% соответственно. Ванадий ускоряет также
окисление серотонина—другого субстрата данного фермента
и может играть определенную роль в гипертонической болезни
[Schroeder Н. А., 1960]. In vitro ванадий способен катализиро-
вать окисление серотонина и близких к нему аминов — гидрок-
сииндола, тирамина, адреналина и др. [Martin G. М. et al.,
Ванадий оказывает влияние на целый ряд других фермен-
тов (РНКаза, щелочная фосфатаза, аденилатциклаза, фосфо-
рофруктокиназа, сукцинатдегидрогеназа), снижая их актив-
ность [Ramasarma Т., Crane F. L., 1981].
Этот МЭ широко используется в металлургии, резиновой,
стекольной и химической промышленности. В связи с этим
встречаются профессиональные отравления ванадием. Наибо-
345
лее токсичны V2O5, ванадаты аммония и натрия, хлориды ва-
надия, менее токсична V2O3, в то время как металлический ва-
надий, феррованадий и карбид ванадия малотоксичны.
' 'Острые ванадозы, как правило, обусловлены вдыханием
соединений “ванадия и характеризуются выраженным насмор-
ком, чиханьем, слезотечением, сухостью в горле, загрудинными
болями, общей слабостью, головной болью, иногда повышением
температуры. При нарастании тяжести отравления развивают-
ся бронхоспазм, бронхит, бронхопневмония, кашель, крово-
харканье, иногда кровотечение. Имеют место аллергические
реакции в виде бронхиальной астмы и экземы.
•' ~ Хронические ванадозы связаны с длительным токсическим
воздействием этого МЭ и сопровождаются синдромами рино-
фарингита, бронхита, нерезко выраженного пневмосклероза,
эмфиземы легких, а также изменениями нервной и сердечно-
' сосудистой системы, в частности отмечается повышенная за-
j болеваемость гипертонической болезнью. Часто наблюдаются
, головная боль, головокружения, истерия, умеренная анемия и
j лейкопения. Редко встречаются расстройства зрения (вплоть
; до слепоты) в результате воспалительных изменений зритель-
ного нерва и сетчатки. Пониженная резистентность организма
сопровождается повышенной частотой инфекционных заболева-
ний [Рощин А. В. и др., 1976].
В обзоре новейших данных о действии ванадия [Nechay
В. R. et al., 1989] этот вопрос оценивается очень осторожно.
Авторы приходят к заключению, что ванадий является ультра-
следовым элементом с последней определяемой концентрацией
в сыворотке крови человека, равной 5-10~9 моль, которая яв-
ляется оптимальной для роста фибробластов в тканевых куль-
турах. Этот МЭ может быть эссенциальным нутриентом, но его
роль для высших животных еще не установлена. Предполага-
ется, что он может иметь отношение к процессам клеточного
роста, дифференцировки и восстановления тканей. У растений
ванадий играет роль в фиксации азота. Хотя этот элемент по-
всеместно используется в промышленности, однако не наблюда-
ется широко распространенных вредных эффектов. Такие эф-
фекты могут быть получены в эксперименте, но при более вы-
соких концентрациях этого МЭ. В противовес более ранним
наблюдениям считается, что ванадий, очевидно, не регулирует
Na+, К+-АТФазную активность.
Ятрогенные формы ванадоза связаны с применением лекар-
ственных препаратов, содержащих в своем составе ванадий
(см. главу 9).
Глава 8
ТОКСИЧНЫЕ МИКРОЭЛЕМЕНТЫ
И ВЫЗЫВАЕМЫЕ ИМИ ЗАБОЛЕВАНИЯ
В эту группу в соответствии с классификацией Р. I. Aggett
(1985) входят следующие МЭ: алюминий, кадмий, свинец,
ртуть, бериллий, барий, висмут. Изложение материала, как и
в предыдущих двух главах, построено по единому плану —
распространенность МЭ в природе, всасывание их в организме,
распределение по тканям, органам и жидкостям организма,
выделение, патологические состояния, обусловленные избыточ-
ным поступлением их в организм человека и животных.
8.1. Алюминий
Алюминий — один из самых распространенных металлов, на
долю которого приходится более 8% массы земной коры. В ви-
де МЭ он входит в состав живых организмов. Этот МЭ широко
используется в промышленности и медицине. Источниками
алюминия для человека являются запыленный воздух, дезодо-
ранты, питьевая вода, пища, лекарственные вещества (главным
образом антациды), бумажные полотенца.
Данных о том, что алюминий выполняет какие-либо полез-
ные функции в организме человека, очень мало. Дефицит его
в пище, по-видимому, не ведет к возникновению серьезных па-
тологических изменений. В течение длительного времени ток-
сическое действие алюминия недооценивалось или даже игно-
рировалось, хотя первые публикации по этому вопросу появи-
лись еще в XIX веке. Алюминозом называли только профессио-
нальный пневмокониоз.
За последние 30 лет взгляды на патогенность алюминия для
человека коренным образом изменились. В 1965—1968 гг.
I. Klatzo и соавт. установили, что при введении растворов со-
лей алюминия в мозг кроликов и кошек в их нейронах образу-
ются «узелки», или клубки, нейрофибрилл (tangle), подобные
тем, которые наблюдаются у человека при тяжелой форме
старческого слабоумия — болезни Альцгеймера (БА). Эти дан-
ные были подтверждены и значительно дополнены другими
исследователями. В частности, такие структуры возникали в
цитоплазме нейронов при культивировании клеток мышиной
нейробластомы в питательной среде, содержавшей повышенное
количество алюминия. Несмотря на очевидное сходство свето-
347
оптических изменений нейрофибриллярного аппарата, вызван»
них алюминием, с морфологическими признаками, характер-
ными для БА, об их полной идентичности говорить было пре-
ждевременно. Н. Wisniewski и соавт. (1980) показали, что оп-
ределенные различия этих изменений выявляются на ультраст-
руктурном уровне. В частности, для БА характерно образова-
ние парных спиральных нейрофиламентов (ПСН), которых
долго не находили при экспериментальном алюминиевом ней-
ротоксикозе у животных. Однако недавно Р. Bosh de Aquilar и
J. Gocmacre-Vanneste (1984) обнаружили ПСН в цитоплазме
нейронов спинальных ганглиев у старых крыс.
Блестящие экспериментальные работы, направленные на
изучение патогенеза нейротоксичности алюминия, выполнены
морфологическими, патофизиологическими, цитохимическими и
молекулярно-биологическими методами группой канадских
ученых под руководством D. Сгаррег (1973) (он же известен
как D. Crapper-McLachlan). Они показали, что после однократ-
ного введения небольшого количества хлорида алюминия в
мозг кошек у животных развивается тяжелое и неуклонно про-
грессирующее заболевание ЦНС, симптомы которого, в част-
ности утрата способности к обучению и памяти на недавние
события (животные не могут найти спрятанную ими пищу),
имеют значительное сходство с альцгеймеровской формой се-
нильной деменции у человека. В нейронах этих животных об-
разуются такие же «узелки» нейрофиламентов, как при БА.
Однако экспериментальный нейроалюминоз у животных про-
текает быстрее и тяжелее, чем БА у людей. Через 2—3 нед
кошки погибают при явлениях отказа от еды и наличии эпи-
лептических припадков. Продлить жизнь животных удается
только с помощью особого ухода. D. Сгаррег и соавт. (1978)
считают, что и у человека в патогенезе БА важную роль игра-
ет алюминиевый нейротоксикоз. Они показали, что алюминий,
введенный в спинномозговую жидкость некоторых видов жи-
вотных, быстро накапливается в ядерном хроматине, а изме-
нения нейронов с указанной патологией нейрофибриллярного
аппарата возникают уже через 1 нед после введения хлорида
алюминия в ЦНС. В ядрах нейронов алюминий прочно связы-
вается с ДНК.
Данные этих экспериментов были неоднократно подтверж-
дены. В 1984 г. S. Crusa и Н. Wisniewski в опытах с интрацис-
териальным введением кроликам 1% раствора хлорида алю-
миния в физиологическом растворе исследовали состояние
органелл нейронов гиппокампа и мотонейронов спинного мозга.
Проведенные эксперименты показали, что через 8—12 дней
после введения алюминия, когда у животных развивались на-
рушения походки и эпилептические припадки, в нейронах по-
являлись вторичные лизосомы, а общее количество лизосом
увеличивалось. Основной вывод этих исследований состоит в
том, что при экспериментальном алюминиевом нейротоксикозе
активируется система внутриклеточного переваривания, но по-
348
давляется активность тиаминпирофосфатазы и нуклеозидди-
фосфатазы в комплексе Гольджи. В эндоплазматической сети
не обнаружено изменений реакции на НДФазу. Попытка трак-
товать патогенез БА как прямое следствие интоксикации алю-
минием вызвала критические замечания, суть которых сводится
к тому, что экстраполяция данных эксперимента на патогенез
БА у человека является слишком прямолинейной. В частности,.
D. Shore и R. Wyatt (1983), не обнаружив повышенного содер-
жания алюминия в спинномозговой жидкости, сыворотке и во-
лосах больных БА, воздерживаются от такой трактовки.
Н Wisniewski и соавт. (1980) отрицают связь слабоумия при
БА с токсическим воздействием этого металла. Несмотря на
скептическое отношение многих исследователей к изложенным-
выше экспериментальным данным, учение о нейротоксичности
алюминия для человека получило неожиданное и крайне важ-
ное подтверждение. В 1976 г. А. С. Alfrey и соавт. обнаружи-
ли, что у ряда больных, подвергавшихся гемодиализу в связи
с недостаточностью функции почек, развивается своеобразная
тяжелая энцефалопатия (не БА!), связанная с высоким содер-
жанием алюминия в водопроводной воде, которую использова-
ли для приготовления диализата. Эти наблюдения вскоре бы-
ли подтверждены во многих странах Европы. Практические
рекомендации, направленные на уменьшение содержания алю-
миния в диализате, спасли жизнь многим больным, предупре-
дили диализную энцефалопатию и привели к ее обратному раз-
витию. В отличие от БА при алюминиевой деменции, осложня-
ющей гемодиализ, как показали D. Сгаррег и соавт. (1978,
1981), содержание алюминия повышено в цитоплазме нейронов,
но не в их ядрах. Установление нейротоксичности этого МЭ
вызвало необходимость пересмотра старых данных о содержа-
нии и распределении его в организме.
Общее количество алюминия в организме практически здо-
рового человека составляет менее 50 мг. При этом большая
часть тканей содержит менее 4 мг алюминия на 1 кг их сухой
массы. Так, в печени алюминий содержится в количестве
4,1 ±1,7 мг/кг, в селезенке —2,6±2,1 мг/кг, в костях — 3,3±
±2,9 мг/кг, в сердце— 1,0±0,8 мг/кг, в мышцах—1,2±
±1,2 мг/кг, в сухой обезжиренной ткани головного мозга—;
2,4± 1,3 мг/кг. Только в легких алюминий содержится в коли-
честве 43 мг/кг. Содержание его в мозге и легких увеличива-
ется с возрастом.
Несмотря на высокое содержание алюминия в природной
среде, в организм поступает ежедневно лишь 2—3 мг этого
МЭ. Только небольшая часть этого количества всасывается в
желудке и в проксимальном отделе двенадцатиперстной кишки,
откладывается в печени, костях и сером веществе головного
мозга. Избыточному поступлению алюминия в организм пре-
пятствует гомеостатический механизм, который способствует
выделению этого МЭ. Так, с мочой выводится в среднем от 10-
до 15 мг/сутки. Если поступление экзогенного алюминия значи-
349»
тельно возрастает, то в 20—40 раз увеличивается его выделе-
ние с мочой, достигающее 200—500 мг/сутки. Определение су-
точного выделения алюминия с мочой является одним из важ-
ных показателей функции почек, так как при их недостаточ-
ности, в частности у 80% больных с уремией, концентрация
алюминия в моче повышена. В 1980 г. также показано, что
парентеральное ятрогенное избыточное поступление алюминия
в организм наблюдается не только при гемодиализе. Другой
причиной может быть полное парентеральное искусственное
питание, в том числе у больных с нормальной функцией почек.
Внутривенное введение больным алюминия, являющегося за-
грязнителем растворов казеина, может быть очень значитель-
ным. В этих случаях организм больных не способен освобо-
диться от избыточного количества алюминия, который накапли-
вается в печени и костях. Если в норме среднее содержание
алюминия в печени равно 6 мг/кг, то при задержке его в орга-
низме больных с полным парентеральным питанием содержа-
ние алюминия достигает 84±21 мг/кг. Кроме того, увеличива-
ется его концентрация в костях. Так, вместо 10 мг/кг в норме
в некоторых случаях при парентеральном гипералюминозе его
содержание повышается до 265 мг/кг.
Уже в 1978 г. было выяснено, что диализная энцефалопатия
может сопровождаться остеомаляцией и связанными с ней
неожиданными переломами костей. Эта тяжелая патология
также является прямым следствием системного токсического
воздействия алюминия, обусловленного его накоплением и
повышенной концентрацией в связи с этим в костной ткани.
Литература, посвященная этому вопросу, содержит многочис-
ленные экспериментальные факты и относительно небольшое
число патологоанатомических исследований, которые в основ-
ном аргументируются морфометрическими данными. Авторы
установили в эксперименте на крысах, что на ранних стадиях
интоксикации алюминий вызывает не остеомаляцию, а задерж-
ку костеобразования. Она зависит от токсического воздействия
этого МЭ на клеточную активность остеобластов. Методом
спектрофотометрии при алюминиевой интоксикации у живот-
ных в костной ткани обнаружено, что алюминий накапливается
на границе между остеоидной и зрелой костной тканью, в ча-
стности во фронтальных зонах обызвествления. С целью выяв-
ления начальных деструктивных изменений A. Hodsman и со-
авт. (1982) предложили исследовать у больных костную ткань
передневерхней ости подвздошной кости.
В 1983 г. М. Randall показал, что при почечной недостаточ-
ности у маленьких детей тяжелая энцефалопатия может воз-
никать после перорального применения карбоната алюминия и
без диализа. В 1979 г. R. Lahl и U. Lange описали так назы-
ваемый перитонеальный алюминоз, вызванный алюграмином
(смесью порошкообразного алюминия и растительного масла),
применявшимся для лечения пролежней в течение 2,5 мес. На
вскрытии обнаружены распространенный алюминоз висцераль-
350
ной брюшины, брыжейки, париетальной плевры, перимизпя
брюшных мышц с появлением игольчатых и копьевидных обра-
зований. Макрофагальная реакция отсутствовала. Предполага-
ется лимфогенный путь распространения алюминия.
Важной проблемой современной психиатрии являются стар-
ческое слабоумие и вероятная связь этой распространенной па-
тологии с патогенным воздействием алюминия. В настоящее
время в результате увеличения продолжительности жизни на-
блюдается общее постарение населения. Психическая неполно-
ценность лиц пожилого возраста привлекает значительное вни-
мание не только невропатологов и психиатров, но и организа-
торов здравоохранения. Американская психиатрическая ассо-
циация считает, что первичное дегенеративное слабоумие с
началом его в пресенильном и старческом возрасте является
именно тем заболеванием, которое в свое время описал
А. Альцгеймер. Советские психиатры все же склонны разграни-
чивать старческое слабоумие и БА на основании некоторых
клинических признаков. В США это тяжелое заболевание ЦНС
поражает примерно 3% всего населения старше 65 лет. В свя-
зи с этим R. Katzman (1976) и F. Plum (1979) указывают на
возможность возникновения в ближайшие годы «эпидемии»-
БА. В предположении нет преувеличения, так как БА действи-
тельно представляет собой массовый нейрогеронтоз, превраща-
ющий известную часть общества в беспомощных и слабоумных
стариков. Данные морфологического исследования показали,
что изменения при этой форме слабоумия четко отличаются от
тех изменений, которые свойственны атеросклерозу или гипер-
тонической болезни. Основными при БА является распростра-
ненная мелкоочаговая дегенерация нейронов головного мозга
с формированием так называемых сенильных бляшек Редлиха,
спутанных нейрофибрилл и микротрубочек в нейронах с обра-
зованием парных спиралей. Первоначально считалось, что узел-
ки из нейрофибрилл в цитоплазме центральных нейронов —
специфический признак БА. Однако в дальнейшем выяснилось,
что эти изменения неспецифичны и могут обнаруживаться у
больных, погибших от других заболеваний, более того, у пси-
хически здоровых лиц пожилого и старческого возраста и
даже у сравнительно молодых людей. Однако в таких случаях
число сенильных бляшек и пораженных нейронов невелико.
При БА указанные изменения имеют массовый характер, а
степень слабоумия в высокой степени коррелирует с выражен-
ностью этих морфологических признаков. В 1980 г. D. Perl и
A.„Brody методом рентгеноспектрофотометрии показали, что в
нейронах, содержащих нейрофибриллярные узелки, накаплива-
ется алюминий.
В 1982 г. D. Perl и соавт. установили, что при боковом
амиотрофическом склерозе (БАС) и при синдроме деменции-
паркинсонизма (ДП), часто встречающихся на острове Гуам,
имеет место внутриядерное накопление алюминия в нейронах.
Именно на этом острове, а также на японском полуострове
351
Кии вода содержит повышенные концентрации данного МЭ.
Попутно заметим, что это может вызвать определенные сомне-
ния в природе БАС и синдрома ДП, которую некоторые иссле-
дователи считали вирусной. Наиболее вероятно, что на острове
Гуам эти заболевания имеют комбинированное происхождение,
представляя собой результат сочетания элективного отложения
алюминия в нейронах и повреждающего влияния какого-то
другого патогенного (вирусного? генетического?) фактора.
Может быть, что еще не установленные вирусы способствуют
повышенной проницаемости гематоэнцефалического барьера
и самих ядер нейронов для алюминия. Поскольку эти тяжелые
эндемические заболевания поражают только туземное населе-
ние, нельзя исключить, что механизм алюминиевого цитотеза-
урисмоза имеет и генетическое происхождение. В пользу воз-
можности генетических влияний на интенсивность отложения
алюминия свидетельствует обнаружение этого металла в ядрах
нейронов при трисомии 21 — синдроме Дауна. Вместе с тем
возможное влияние экзогенных факторов на отложение этого
МЭ подтверждается обнаружением алюминия в нейронах при
постэнцефалитическом паркинсонизме, а также при подостром
склерозирующем панэнцефалите.
Вопросу о характере и локализации дегенеративно-атрофи-
ческих изменений при БА уделялось немало внимания. Обще-
признанными фактами являются уменьшение числа нейронов в
коре большого мозга, их гранулярно-вакуолярная дистрофия,
альцгеймеровские изменения нейрофиламентов и микротрубо-
чек, сенильные бляшки, отложение липофусцина и меланинопо-
добного пигмента в цитоплазме нейроцитов, амилоидоз сосудов,
мозговых оболочек, сенильных бляшек, внутренняя гидроцефа-
лия [Ойфа А. И., 1987].
Наряду с дегенеративными изменениями нейронов и бляш-
ками в сером веществе коры большого мозга, а также в ба-
зальных ядрах уже давно замечено, что при старческом слабо-
умии зонами предпочтительного поражения является гиппо-
камп. Однако клиническое значение этих находок в течение
долгого времени не находило объяснения и только после уста-
новления роли гиппокампа в формировании и поддержании
механизмов памяти данный вопрос стал привлекать внимание
исследователей. В настоящее время установлено, что при БА
именно в гиппокампе концентрируется наибольшее количество
алюминия. В белом веществе головного мозга, в частности в
мозолистом теле, его значительно меньше. Способность различ-
ных нейронов формировать при контакте с алюминием харак-
терные изменения нейрофибрилл неодинакова. Так, если ней-
роны гиппокампа и коры большого мозга легко поражаются
алюминием, то существуют и более стойкие к нему нервные
элементы. Чувствительность к токсическому воздействию алю-
миния у различных животных неодинакова. Так, обезьяны, по-
видимому, к нему полностью резистентны.
Сравнительно недавно было показано, что ацетилхолин
352
участвует в механизмах памяти и поддержания сознания. В
этой связи заслуживает особого внимания тот факт, что при
БА резко уменьшается активность ацетилхолинтрансферазы не
только в коре большого мозга, но и в гиппокампе. Уже в 1979 г.
р, Davies показал, что старческое слабоумие типа БА сопро-
вождается изменениями уровня ферментов, участвующих в
функции нейротрансмиттеров. В 1981 и 1982 гг. Р. Whitehouse
и соавт. обнаружили, что при БА имеет место селективная
дегенерация тех базальных холинергических нейронов передне-
го мозга, которые иннервируют кору большого мозга и гиппо-
камп. В частности, в базальном ядре Мейнерта при БА число
нейронов уменьшается на 80% (по сравнению с мозгом конт-
рольных лиц того же возраста), причем часть сохранившихся
нейронов содержит нейрофибриллярные узелки. Как уже упо-
миналось, в ядрах таких нейронов обнаруживали алюминий.
Эти находки имеют принципиальное значение для понимания
природы нейропсихической неполноценности при БА. Наблюда-
ющиеся при БА симптомы, вероятно, не только связаны с про-
стым выпадением функции нейронов коры в результате их
прогрессирующей дегенерации, но по крайней мере частично
могут быть объяснены ослаблением межнейрональных тонизи-
рующих влияний холинергической иннервации самого головно-
го мозга.
Экспериментальная проверка действия алюминия на актив-
ность ацетилхолинтрансферазы в опытах на животных, а так-
же in vitro дала противоречивые данные. В то же время очень
убедительны приведенные выше результаты опытов С. Miller и
Е. Levine (1974). Показано образование нейрофибриллярных
узелков в нейронах культуры тканей мышиной нейробластомы
вследствие введения алюминия в питательную среду. Уже че-
рез 6 дней после контакта с алюминием, несмотря на общее
увеличение синтеза белка, активность ацетилхолинэстеразы,
синтезируемой холинергическими нейронами, понизилась на
36% по сравнению с таковой в контрольных культурах нейро-
нов, не соприкасавшихся с алюминием и не формировавших
нейрофибриллярных узелков. Комплекс реактивных изменений
холинергических нейронов в ответ на контакт с этим МЭ был
в дальнейшем подтвержден рядом исследователей и в настоя-
щее время рассматривается как один из основных механизмов
алюминиевого нейротоксикоза. В культурах клеток нейроблас-
томы, находившихся в контакте с алюминием, включение ти-
мидина не изменяется, тогда как интенсивность включения ури-
дина и лейцина значительно повышается.
Следует отличать простое отложение алюминия в мозге
старых людей, не сопровождающееся признаками нейропсихи-
ческой неполноценности, от тяжелого дегенеративно-атрофиче-
ского процесса, сопровождающегося слабоумием, характерным
для БА. Само накопление алюминия в головном мозге не-
специфично для БА, так как оно встречается и при других за-
болеваниях, например при недостаточности печени и почек,
23—3G8
353
метастазах рака. В то же время нет оснований считать, что
накопление этого металла в ЦНС вообще сопровождает любые
дегенеративные изменения в нервной системе. Так, накопления
алюминия не наблюдаются при таком хроническом заболева-
нии мозга, как болезнь Крейтцфельда — Якоба. Группа авто-
ров показала, что деструкция нервной ткани сама по себе не
вызывает повышения содержания алюминия в головном мозге
[Traub R. D. et al., 1981].
Разумеется, что открытие нейротоксичности алюминия не
могло не вызвать живого интереса медицинской обществен-
ности к возможным источникам его поступления в организм.
В обзорной работе R. Boegman и L. Bates (1984) уже указы-
валось на обычные источники поступления алюминия в орга-
низм. В частности, среди них называлась и питьевая вода. Как
известно, сульфат алюминия используется для очистки питье-
вой воды. Содержание алюминия в растительной и животной
пище зависит от количества этого МЭ в окружающей среде.
По данным ряда авторов, нейротоксичность алюминия должна
привлечь особое внимание в тех географических районах, в ко-
торых он содержится в большом количестве в почве. Важно
подчеркнуть, что под влиянием кислотных дождей, в результате
атмосферного загрязнения алюминий может выщелачиваться.
Биогеохимические изменения, связанные с загрязнением почвы
алюминием, вызывают цепь последовательных явлений. Напри-
мер, растения, произрастающие на такой почве, накапливают
меньше селена, в результате чего активность глутатионперок-
сидазы снижается.
Некоторые авторы защищали точку зрения, согласно кото-
рой необходимо снижать содержание алюминия в пище, другие
рекомендовали ограничить использование алюминиевой кухон-
ной посуды. Исследователи, специально изучавшие этот воп-
рос, считают, что для излишней тревоги нет никаких основа-
ний, так как содержание алюминия в суточном рационе в
среднем равно 22 мг, из которых только 1 мг усваивается. Нет
никаких сведений, что больные БА ранее употребляли боль-
шее количество алюминийсодержащих антацидов, чем здоро-
вые лица. Среди специалистов все же существует известная
озабоченность потенциальной вредностью длительного и мно-
гократного применения алюминиевых антацидов [Ulmer D. D.,
1976].
В литературе имеются работы, авторы которых рекоменду-
ют использовать различные химические вещества для пониже-
ния содержания алюминия в мозге. Этими веществами явля-
ются дезфероксамин и соединения фтора. Еще в 1968 г. J. Kor-
tus и J. Mayer показали, что распределение алюминия в орга-
низме изменяется под влиянием повышенного поступления ио-
на фтора. В частности, если крысам давать фтор в течение
540 дней, то у них уменьшается содержание алюминия в кос-
тях. С позиции географической патологии весьма важно, что
в тех регионах США, где содержание фтора в воде высокое,
354
число больных старческим слабоумием значительно меньше,
чем в регионах с низким содержанием фтора. В противополож-
ность этому не обнаружено подобных различий в распростра-
ненности слабоумия сосудистой природы. Разумеется, эти эпи-
демиологические данные, а также попытки лечения БА с по-
мощью фторидов нуждаются в дальнейшем развитии и про-
верке. С нашей точки зрения, профилактическое применение
фторированной воды в регионах с повышенным содержанием
алюминия в природной или производственной сфере заслужи-
вает внимания не только гигиенистов, но и невропатологов и
психиатров. Важно организовать исследования нейропсихиче-
ского статуса контингентов, находившихся в условиях длитель-
ного и тесного контакта с высокими концентрациями алюми-
ния. Особое внимание необходимо уделить лицам пенсионного
и предпенсионного возраста с целью выявления у них ранних
неврологических и психопатологических признаков патологиче-
ского старения. Следует также проверить предположение о
возможной эффективности лечебно-профилактического фтори-
рования организма в группах риска, в частности с использова-
нием местных фторсодержащих минеральных вод.
Болезни и синдромы, связанные с токсическим действием
алюминия, представляют собой широко распространенные фор-
мы МТОЗов различной степени тяжести. Так как организм
человека защищен от возможного патогенного воздействия
алюминия рядом физиологических механизмов, среди которых
основное место занимает его регулярная 'экскреция, то комп-
лекс патологических изменений в результате избыточного по-
ступления данного МЭ в организм чаще всего возникает под
влиянием серьезных нарушений гомеостаза или связан с ле-
чебными мероприятиями. Патология, вызванная аномальным
накоплением алюминия, многообразна. Провизорно можно
выделить следующие формы этого МТОЗа.
1. Простое накопление алюминия в ЦНС— хронический доб-
рокачественный старческий алюминоз с минимальными или
умеренными признаками снижения нейропсихических функций
у лиц старше 65 лет. Такой нейроалюминоз, как правило, не
диагностируется и относится к проявлениям нормальной ста-
рости. Патологоанатомически при доброкачественном старчес-
ком нейроалюминозе в ЦНС обнаруживают относительно не-
большое повышение содержания алюминия в нейронах, уме-
ренное количество сенильных бляшек и нервных клеток с ха-
рактерной патологией нейрофибриллярного аппарата.
2. Отложение алюминия при болезни Альцгеймера, т. е. при
-сенильном и пресенильном слабоумии с тяжелыми поражения-
ми нейропсихической сферы, более ранними и быстро протека-
ющими дегенеративными изменениями в коре большого мозга,
подкорковых ганглиях с вторичной гидроцефалией, элективной
деструкцией гиппнокампа, ядер переднего мозга, в частности
базального ядра Мейнерта. Достоверные данные о том, что при
БА алюминий накапливается в головном мозге в больших ко-
23*
личествах, чем у психически здоровых лиц преклонного возрас-
та, нам неизвестны. Биохимически для БА характерно угнете-
ние холинергических нейротрансмиттеров, в частности фермен-
тов, обеспечивающих холинергические механизмы. При этой
болезни алюминий прочно связан с ядерным хроматином, точ-
нее с ДНК, что ведет к глубокому нарушению механизмов
транскрипции и трансляции в нейронах. Алюминий концентри-
руется в ядрах нейронов, в их цитоплазме формируются харак-
терные для БА парные спиралевидные нейрофиламенты, обна-
руживаемые при электронной микроскопии. Нейрофибрилляр-
ный аппарат пораженных нейронов подвергается тяжелым не-
обратимым изменениям, что в свою очередь влечет за собой
глубокие нарушения аксонального транспорта, определенную
дисгармонию рецепторной активности и характерную дегенера-
цию дендритов. Несмотря на тяжелые дегенеративные наруше-
ния нейронов, синтез белка в них может быть повышен. И хотя
точно доказано, что алюминий отлагается в ЦНС, трактовать
БА только как злокачественную форму нейроалюминоза пока
нельзя, так как в патогенезе этого заболевания принимают уча-
стие и другие (вероятно иммуноцитохимические и генетичес-
кие) факторы. Можно согласиться с мнением D. Shore и
R. Wyatt (1983), что алюминий в мозге является маркером
специфически измененных нейронов. Однако в свете современ-
ных знаний о нейротоксическом действии алюминия трудно
рассматривать этот маркер в качестве индифферентной метки.
3. Алюминиевая диализная энцефалопатия — тяжелое, но
принципиально обратимое поражение ЦНС с эпилептическими
припадками, миоклониями, возможными расстройствами выс-
ших психических функций, которое возникает в результате по-
вышения концентрации алюминия в организме. При этой фор-
ме энцефалопатии алюминий содержится в цитоплазме нейро-
нов, но не в их ядрах. В цитоплазме же развиваются дегенера-
тивные изменения нейрофиламентов, однако без формирования
парных спиралевидных фибрилл.
4. Недиализная алюминиевая энцефалопатия маленьких де-
тей, возникающая под влиянием перорального употребления
лекарственных препаратов с высоким содержанием алюминия
на фоне тяжелой врожденной недостаточности мочевыделения.
5. 77еритонеалный алюминоз — ятрогенное отложение алю-
миния в брюшине.
6. Энцефалопатия, связанная с применением полного па-
рентерального питания при относительной недостаточности го-
меостатических механизмов экскреции алюминия, связанных
с врожденными, приобретенными заболеваниями почек.
7. Ятрогенная алюминиевая остеодистрофия (остеомаляция)
с развитием остеопороза, повышенной ломкостью костей, пони-
жением функции остеобластов и возникновением множествен-
ных переломов.
8. Легочный алюминоз — алюминиевый пневмокониоз про-
изводственного характера с вторичным пневмосклерозом пре-
356
Рис. 60. Очаг коагуляционного некроза мышц бедра крысы на месте введения
хлорида алюминия. В строме незначительная воспалительная инфильтрация.
Гистохимическая реакция с алюминоном. X100.
Рис. 61. Диски мышечных волокон и стенки кровеносных сосудов, инкрусти-
рованные алюминием. Гистохимическая реакция с алюминоном. х400.
имущественно верхних долей легких. Алюминиевые бронхиты
и пневмонии.
9. Астмоидный алюминоз — бронхоспастический синдром у
плавильщиков алюминия.
10. Тяжелая, но обратимая алюминий зависимая микроци-
тарная анемия, осложняющая гемодиализ.
11. Накопление алюминия в сердце с нарушением его рит-
мической деятельности. Учащающиеся в последние годы край-
не тяжелые токсические поражения миокарда, вызванные от-
равлением фосфидом алюминия (А1РО4), широко используе-
мым для защиты пищевого зерна от грызунов и других вреди-
телей [Khosla N. et al., 1988].
12. Вторичный алюминоз ЦНС при боковом амиотрофичес-
ком склерозе и синдроме деменции-паркинсонизма коренных
обитателей острова Гуам.
Кроме развития перечисленных алюминийзависимых заболе-
ваний, есть основания полагать, что накопление алюминия име-
ет патогенное значение при некоторых формах рака мочевого
пузыря, а также при пигментной дистрофии сетчатки глаз, со-
четающейся с высоким содержанием этого МЭ в крови.
Уже в 1983 г. мы отмечали, что алюминий может перено-
ситься с воздухом и водой на большие расстояния и таким
образом участвовать в появлении трансгрессивных МТОЗов,
патология которых у человека и животных заслуживает спе-
циального рассмотрения в качестве проблемы большого эколо-
гического значения [Авцын А. П. и др., 1983].
При экспериментальном исследовании местной реакции тка-
ней на внутримышечное введение хлорида алюминия крысам
обнаружено его повреждающее действие на структуру скелет-
ных мышц и соединительнотканных производных, вплоть до
возникновения очагов коагуляционного некроза мышц с мини-
мальной воспалительной реакцией (рис. 60, 61). Эти данные в
основном подтвердились и при введении хлорида алюминия в
полость желудка, при котором наблюдалась локальная инкрус-
тация алюминием дистрофически измененной слизистой оболочки
(рис. 62). Алюминий, введенный внутримышечно крысам, на-
капливался в нейронах гиппокампа, а также в пирамидных
клетках коры большого мозга (рис. 63). Электронно-микроско-
пическое исследование показало, что в различных тканях алю-
миний откладывается в виде электронно-плотного материала
Рис. 62. Патогистологическая картина слизистой оболочки желудка и двенад-
цатиперстной кишки при внутрижелудочном введении хлорида алюминия в до-
зе 10 мг алюминия на 100 г массы крысы. Гистохимическая реакция с алю-
миноном:
а специфическая инкрустация алюминием дистрофически измененной слизистой обо-
лочки желудка через 2 ч после введения. Х400; б — двенадцатиперстная кишка через
.. ч после введения алюминия. Продукт реакции на алюминий определяется в эпители-
альном пласте и в собственной пластинке слизистой оболочки. Х640; в — гистохимическая
реакция на алюминий в двенадцатиперстной кишке через 3 ч после введения. Х250; г —
то же самое при большем увеличении. Х640.
359
Рис. 63. Накопление алюминия в нейро-
нах гиппокампа и пирамидных клетках
коры большого мозга крысы при внут-
римышечном введении алюминия в дозе
4 мг на 100 г массы животного. Флюорес-
центный метод выявления алюминия с
помощью морина.
а — общая картина отложения алюминия в
гиппокампе. Х80; б — то же самое при боль-
шем увеличении. 1X400; в — свечение алюминия
в пирамидных клетках коры большого мозга.
Х800.
Рис, 64. Скопление электронно-плотного материала на поверхности ядерной
мембраны эпителиоцита тонкой кишки крысы через 20 дней после внутримы-,
шечного введения 0,2 мл 0,01% раствора хлорида алюминия. X 40 000 (пре-
парат Г. П. Поляковой).
различной формы. Характерны коагулирующее действие алю-
миния на тканевые структуры, а также инкрустация им цито-.
плазматических мембран, в частности ядерной оболочки раз-
личных клеток (рис. 64) [Полякова Г. П., 1988].
8.2. Кадмий '
В п@следние 25 лет значительно возрос интерес к кадмию
как к одному из продуктов радиоактивного распада, накапли-
вающемуся в организме человека и животных, токсичному эле-
менту и антиметаболиту ряда химических элементов, а с не-
S61
давних пор и как, по-видимому, к жизненно необходимому
компоненту животного организма. В 1952 г. были впервые об-
наружены высокие концентрации кадмия в почках человека и
животных, а в конце 60-х годов было показано, что загрязне-
ние окружающей среды кадмием является причиной эндемичес-
кого заболевания итаи-итаи в Японии, гипертонии у лабора-
торных животных и, возможно, у человека. Интерес к кадмию
объясняется также открытием низкомолекулярного белка ме-
таллотионеина, концентрирующего повышенное количество это-
го металла [Webb М., Cain К.., 1979]. Кадмий принадле-
жит к числу МЭ, которые широко используются
в технике, особенно в металлургии. Наибольшие
поступления Cd в атмосферу в Европе связаны с работой ста-
лелитейных заводов (34 т в год) и сжиганием разнообразных
отходов (31 т в год). Таким образом, в глобальном обогащении
окружающей среды кадмием антропогенный вклад в 3 раза
превышает вклад естественных источников.
Анализ диет в нескольких странах Европы показал, что
взрослый человек потребляет в среднем с пищей
от4до84мкг кадмиявдень. Высокий уровень
его в пищевых продуктах зависит от значитель-
ного загрязнения природной среды этим МЭ.
Так, в Японии, где рис является одним из основных пищевых
продуктов, концентрация кадмия в загрязненном рисе может
превышать 1 мкг на 1 г (!). В результате уровень ежедневного
поглощения кадмия может превышать 500—800 мкг. В других
странах этот показатель значительно ниже и для человека
массой 70 кг в среднем составляет 25—60 мкг.
Табачный дым содержит довольно высокие концентрации
кадмия, причем у курильщиков до 50% поступившего в орга-
низм металла абсорбируется в легких. По подсчетам Е. A. Doi-
sy (1972), суммарное ежедневное поступление кадмия в орга-
низм может колебаться от 6 мкг (для некурящих людей, живу-
щих в сельской местности и потребляющих незагрязненную
кадмием пищу) до 115 мкг у курильщиков, живущих в загряз-
ненной кадмием местности и потребляющих контаминирован-
ные продукты.
Основные пути поступления кадмия в орга-
низм— это желудочно-кишечный тракт и органы
дыхания. Только 4—5% кадмия, поступившего в организм
per os, усваиваются, а остальная часть выводится с калом.
Кадмий, поступивший этим путем, лучше усваивается в орга-
низме женщин. На поглощение кадмия из пищеварительного
канала оказывают влияние различные факторы. При беремен-
ности и лактации усиливается его отложение в почках, в две-
надцатиперстной кишке и в молочных железах.
Обмен кадмия характеризуется следующими основны-
ми особенностями: 1) отсутствием эффективного механизма
гомеостатического контроля; 2) длительным удержанием в ор-
ганизме с необычно долгим периодом полувыведения, состав-
362
ляющим у человека в среднем 25 лет [Москалев Ю. И.,
1985]; 3) преимущественным накоплением в печени и почках
и 4) интенсивным взаимодействием с другими двухвалентны-
ми металлами как в процессе всасывания, так и на тканевом
уровне [Kostial К., 1986].
Всасывание кадмия в респираторном тракте зависит от хи-
мического соединения, размера частиц и их растворимости.
Кадмий, присутствующий в папиросном дыме, усваивается на
50%. При выкуривании одной пачки сигарет в организм ку-
рильщика поступает около 1 мкг кадмия. Другие соединения
кадмия, поступающего с вдыхаемым воздухом, усваиваются:
на 10—40%. Однако в связи с тем что содержание кадмия в;
воздухе, не загрязненном этим МЭ, как правило, весьма не-
значительно, то этим путем поступает в организм человека
всего около 0,26 мкг кадмия в сутки, что в 10—12 раз ниже
количества, поступающего из пищеварительного тракта [Smith
Н. А., 1984].
С у т о ч н ы й р а ц и о н ч е л о в е к а содержит около
0,44 ммоль кадмия, из которых усваивается 17,8—•
26,7 нмоль (4—6%). Всасывание кадмия из кишечника
включает два этапа: относительно быстрое поступление через
исчерченную каемку и более медленный переход из слизистой
оболочки в кровь. Всасывание зависит от множества факторов:
оно угнетается цинком, конкурирующим с кадмием за участки
связывания, а также молоком и солями желчных кислот.
Действие молока объясняется повышенным содержанием в нем
кальция, а солей желчных кислот — мицеллообразованием.
В целом ряде работ рассматривается роль металлотионеина,
присутствующего в слизистой оболочке, как отрицательного ре-
гулятора всасывания кадмия и белка, с помощью которого ре-
ализуется физиологический антагонизм между этим МЭ и цин-
ком, а также с другими металлами. Однако имеющиеся наблю-
дения относятся к повышенным концентрациям кадмия,
способным индуцировать транскрипцию тионеиновой мРНК, и
не могут быть перенесены на физиологические концентрации
этого МЭ, связывание которого, по-видимому, не зависит от
металлотионеина [Cousins R., 1985]. Всасывание кадмия про-
исходит в тонкой кишке, причем уровень резорбции этого ме-
талла в подвздошной кишке значительно выше, чем в тощей.
Величина всасывания для кадмия существенно ниже, чем для
меди и цинка, с которыми он, по-видимому, имеет общий
транспортный механизм. У новорожденных уровень всасывания
кадмия в кишечнике значительно выше, чем у взрослых. Кад-
мий может всасываться через кожу, но этот путь играет по
сравнению с предыдущим второстепенную роль. Кадмий может
проникать также через плаценту [Цветкова Р. П., 1970, Waal-
kes М. et al., 1982], хотя на это свойство оказывают сильное
влияние видовые различия. Так, у коз существует, по-видимо-
му, исключительно эффективный плацентарный барьер по от-
ношению к этому МЭ [Anke М., 1970].
363
Способность кадмия проникать через плаценту имеет важ-
ное значение, поскольку этот МЭ в известных дозах обладает
выраженным тератогенным свойством, нарушая поступление в
плод других эссенциальных элементов, не затрагивая, однако,
при этом транспорт сахаров, аминокислот и предшественников
нуклеиновых кислот [Webb М., Samarawickrama G. Р., 1984].
При тератогенных дозах (11,1 мкмоль/кг массы), введенных
парентерально, кадмий в течение 48 ч ингибирует синтез ДНК
в эмбриональных тканях крыс параллельно со снижением по-
ступления в них цинка, что в первую очередь объясняется, по-
видимому, угнетением активности тимидинкиназы, которая, как
показали опыты in vitro, возвращается к норме при введении
цинка. Нарушение обмена цинка лежит в основе угнетения син-
теза ДНК в тканях плода, наступающего при нагрузке орга-
низма матери кадмием. И в данном случае парентеральное
введение цинка в отношении 2: 1 к кадмию предупреждает
тератогенный эффект последнего.
Нелетальные дозы кадмия вызывают уменьшение массы и
длины тела, почек и печени плода без видимого поражения
плаценты. Содержание металлотионеина в эмбриональных
тканях крыс снижается, а у кроликов, напротив, возрастает.
При этом у них повышается также концентрация цинка в ци-
тозоле эмбриональных гепатоцитов за счет его поступления из
других тканей без повышения общего содержания этого метал-
ла в организме плода [Waalkes М. Р., Bell J. V., 1980].
После резорбции из легких и кишечника кадмий транспор-
тируется кровью в печень, почки, слизистую оболочку кишеч-
ника и другие мягкие ткани. Вскоре после парентерального
введения около 95% кадмия крови обнаруживается в плазме в
составе а-глобулинов, альбумина и других белков с молекуляр-
ной массой 48000—77 000. Плазма быстро теряет большую
часть кадмия, а оставшееся количество включается в состав
низкомолекулярного белка, близкого по своим физико-химиче-
ским свойствам к металлотионеину. Подобный же белок начи-
нает накапливаться и в эритроцитах. При хроническом воздей-
ствии кадмия на организм наблюдается вторичное, менее быст-
рое, снижение концентрации кадмия в крови, за которым сле-
дует подъем его уровня в плазме и эритроцитах, наступающий
после усиления синтеза металлотионеина в печени и других
тканях. На основании этих наблюдений допускалось, что метал-
лотионеин участвует в транспорте кадмия и доставке его из
печени в почки. Однако эта точка зрения окончательно не до-
казана. Металлотионеин легко фильтруется через мембрану
клубочков и подвергается реабсорбции в почечных канальцах
[Webb М„ 1979].
К. G. Danielson и соавт. (1982) иммуноцитохимически про-
демонстрировали присутствие металлотионеинов в различных
органах крыс при кадмиевой интоксикации. Так, внутриклеточ-
ная метка обнаружена в эпителиальных паренхимных клетках
печени и почек. Почти во всех гепатоцитах и большинстве
364
клеток проксимальных канальцев выявлен этот тип белка.
В соединительной ткани, окружающей кровеносные сосуды или
почечные канальцы, металлотионеин не обнаруживался. Вне-
клеточная локализация этого белка выявлялась в синусоидах
печени и в просвете почечных канальцев, что подтверждает
возможность секреции и транспорта металлотионеинов. В ки-
шечнике иммунная метка локализовалась в эпителиальных
клетках ворсинок, в клетках Панета и отсутствовала в соеди-
нительной ткани собственной пластинки слизистой оболочки.
В легких белок выявлялся в респираторном эпителии. В се-
менниках металлотионеин локализовался в клетках Сертоли и
в интерстициальных клетках, но не в сперматогониях. В. Ro-
berts и соавт. (1982) обнаружили увеличение содержания ме-
таллотионеинов после однократной внутрибрюшинной инъекции
кадмия крысам в дозе 1,87 нмоль/кг. Так, оказалось, что мак-
симальный уровень синтеза этого типа белков имеет место че-
рез 2 ч после введения МЭ. Тотальная концентрация металло-
тионеинов нарастает в период между 8 и 67 ч после начала
эксперимента и снижается к 336 ч до контрольного уровня.
Металлотионеин, присутствующий в эритроцитах, поступа-
ет в них либо из печени, либо индуцируется кадмием в их
предшественниках. Менее вероятно присутствие в эритроцитах
преобразованной тионеиновой матричной РНК, регулирующей
синтез Cd-тионеина на уровне трансляции [Smith Н. А., 1984].
Распределение кадмия в тканях. При фоновых
концентрациях кадмия в окружающей среде он накапливается
в первую очередь в почках и печени. Если кадмий поступает в
организм в виде солей, то его накопление отмечается прежде
всего в печени, если же его вводят в составе металлотионеина,
то наблюдаются его концентрирование почками и повышенное
выделение с мочой. Содержание кадмия в почках составляет
п-10~2% в расчете на свежую ткань, причем кадмий концент-
рируется в основном в корковом веществе, а мозговое вещест-
во беднее этим МЭ в 2—5 раз. Наиболее богаты кадмием поч-
ки лошади, а почки и печень крупного рогатого скота, овец и
свиней содержат почти на два порядка меньшее количество
этого МЭ. При повышенном содержании кадмия в среде его
концентрация в почках клинически здоровых животных может
возрастать в десятки и сотни раз и достигать у лошадей и
крупного рогатого скота 1603 и 116—74 мг/кг сухой массы со-
ответственно [Anke М. et al., 1976]. В почках человека верхним
безопасным порогом кадмия считается 200 мг/кг.
Колебания уровня кадмия в почках человека зависят от
пола, возраста и географических условий. Повышенные кон-
центрации кадмия отмечены у жителей Дальнего Востока, в
пище которых значительное место занимают продукты морско-
го происхождения, богатые кадмием. Относительно низкие
концентрации этого МЭ найдены у жителей Африки и Узбекис-
тана [Мусаева Г. М„ 1979].
Возрастная динамика содержания кадмия в почках и пече-
365
ни характеризуется минимальным количеством этого МЭ у по-
врежденных (0,01—0,03 мкг/кг), постепенным его повышением
с момента рождения до 40—50 лет (49 + 29 мг/кг у женщин и
92+32 мг/кг у мужчин) и снижением в пожилом и старческом
возрасте [Гулько В. В., 1974; Anke М. et al., 1976, и др.].
Содержание кадмия в организме обнаруживает половой ди-
морфизм. Организм самцов богаче кадмием, чем организм са-
мок. У человека половые различия по содержанию кадмия в
почках и печени обнаружены в возрасте между 30 и 80 года-
ми, причем почки мужчин содержат на 40—50% больше кад-
мия, чем почки женщин. Эти различия объясняются специфиче-
ским повышением выделения кадмия из их организма под
влиянием эстрогенных гормонов.
Парентеральное введение Cd-тионеина вызывает поврежде-
ние плазматических мембран эпителиоцитов почечных каналь-
цев и их митохондрий, тогда как соли кадмия и Zn-тионеин в
эквивалентных количествах не обладают нефротоксическим
свойством. Однако не вполне ясно, вызван ли этот эффект не-
посредственно самим Cd-тионеином или ионами кадмия, высво-
бождаемыми при его распаде. Кадмий легко поступает в изо-
лированные гепатоциты в результате процесса, сходного с про-
стой диффузией, линейно зависящего от дозы этого металла.
Внутри клетки кадмий первоначально включается в состав вы-
сокомолекулярных белков, а спустя 24 ч 90% его обнаружива-
ются в составе металлотионеина. Поступив в гепатоцит, кадмий
проникает в ядро, из которого затем удаляется вместе с ме-
таллотионеином с периодом полувыведения, равным 0,11 в
1 ч [Bryan S. Е. et al., 1979].
С помощью иммуногистохимического метода показано, что
в отсутствие нагрузки организма кадмием металлотионеин об-
наруживается в цитоплазме гепатоцитов, эпителии собиратель-
ных трубочек и дистальных извитых канальцев почек. При
ежедневной даче кадмия в течение 2 нед металлотионеин вы-
является главным образом в ядрах, а в течение 4—8 нед — в
ядрах и цитозоле. При продолжительном введении кадмия
металлотионеин появляется и в ряде других тканей, включая
щитовидную железу. Ядерный металлотионеин совместно с тио-
неином, не содержащим металла, играет, по-видимому, важную
роль в регуляции транскрипции и трансляции, обеспечивающих
синтез данного белка в ответ на поступление в клетку тяже-
лых металлов [Cherian G. М., Nordberg G. F., 1983].
Выделение. Экскреция с калом представляет собой ос-
новной путь выделения кадмия из организма. Так, показано,
что после однократной дачи металла козам через рот или
после однократной или повторных доз коровам около 80—90%
введенного МЭ выделяется с калом на протяжении 14 дней
после окончания воздействия [Van Bruwaene R. et al., 1984].
После внутривенного введения кадмия тем же животным око-
ло 5—6% дозы выводится через кишечный тракт в течение
5—10 дней [Miller et al., 1968; Van Bruwaene R. et al., 1982].
366
Исследования, проведенные в Японии, показали, что в норме
человек в среднем выделяет с калом 25—80 мкг кадмия в день,
что согласуется с ежесуточным потреблением этого МЭ с пи-
щей [Tsuchiya К., 1978]. Однако у людей, проживающих в за-
грязненных кадмием провинциях, через кишечник экскретиру-
ется значительно больше этого металла, чем в контроле. Так,
в этой группе у 16,1% населения выделяется более 300 мкг
кадмия в день [Kojima S. et al., 1977J. К настоящему времени
неизвестно, какое количество эндогенного кадмия выделяется
эпителием кишечника и желчью. Н. D. Stowe (1976) исследо-
вал желчную экскрецию и показал, что у крыс в желчи может
содержаться до 22 ±3 мкг металла на 1 кг массы. При кадмие-
вой интоксикации (100 мкг/мл) с желчью выделяется до 50±
±6 мкг/кг [Manaffey К-, 1984]. Слюна может содержать до
0,1 мкг/г кадмия [Dreizen A. et al., 1970], что может сущест-
венно влиять на уровень кишечного выведения, поскольку у
нормального человека выделяется 1000—1500 мл слюны в день.
Но следует учитывать, что уровень желудочно-кишечной реаб-
сорбции кадмия неизвестен.
В ряде исследований показано, что экскреция кадмия с
мочой в норме составляет примерно 1—2 мкг в день [Hallen-
beck W., 1984; Sangster В. et al., 1984]. С возрастом (до 50—
60 лет) уровень кадмия в моче увеличивается [Kowal N. et al.,
1979]. У рабочих и жителей, подвергающихся воздействию
кадмия, уровень почечной экскреции повышается без пораже-
ния этого органа и может достигать нескольких сотен микро-
граммов в день (10—15 мкг/г креатинина) [Friberg L. et al.,
1974; Lauwerys R. et al., 1979; Hassler E. et al., 1983]. По-
скольку кадмий реабсорбируется преимущественно в прокси-
мальных канальцах почек, то при их поражении выделение это-
го МЭ усиливается. До сих пор дискутируется вопрос, отража-
ет ли уровень кадмия в моче повышение его концентрации в
организме. По мнению W. Hallenbeck (1984), выделение кад-
мия с мочой является ненадежным показателем его накопле-
ния в целом организме или в почках. Однако, как в опытах на
животных (кролики, получавшие в день 1,5 мг кадмия на 1 кг
массы в течение 5 нед) [Nomiyama К., Nomiyama Н„ 1976],
так и в эпидемиологических обследованиях популяций людей
показана значительная корреляция между уровнем металла в
коре почек и его содержанием в моче [Roels Н. et al., 1981].
Благодаря детальным работам R. Lauwerys и соавт. (1976,
1979) противоречие между изложенными выше крайними точ-
ками зрения в определенной степени нивелируется. Так, ими
показано, что концентрация кадмия в моче действительно отра-
жает его накопление в организме при длительном воздействии
малых доз, а при острой интоксикации (поступление высоких
доз) экскреция кадмия через почки служит показателем имен-
но его кратковременного непосредственного эффекта. В случае
наступления канальцевой дисфункции экскреция кадмия с мо-
чой усиливается и по его содержанию в выделениях уже нельзя
367
судить об обмене МЭ в организме. По мнению Z. A. Shikh и
L. М. Smith (1984), в этом случае почечное выделение металла
обусловлено двумя причинами: поражением эпителия каналь-
цев и уменьшением реабсорбции.
Болосы представляют собой дополнительный экскреторный
путь кадмия и могут содержать его от 0,5 до 3,5 мкг/г [Fri-
berg L. et al., 1974]. У детей волосы, как было показано, мо-
гут содержать относительно высокую концентрацию этого МЭ,
которая постепенно снижается в течение жизни [Gross R. Т.
et al., 1976]. Как полагают, содержание кадмия в волосах мо-
жет служить биологическим индикатором задержки его в ор-
ганизме. Так, показано, что у мышей и крыс содержание кад-
мия в шерсти качественно отражает его запасы в печени и поч-
ках [Nordberg G., 1972]. К- J. Ellis и соавт. (1981) выявили
подобную тенденцию у рабочих при контакте с соединениями
кадмия. Однако следует учитывать, что установленные взаи-
моотношения носят больше качественный, чем количественный,
характер. Следует упомянуть данные N. Kowal и соавт. (1979),
которые сообщили о негативной корреляции между уровнем
кадмия в волосах, в крови и моче у людей, проживающих в
США.
С молоком выделяется очень незначительное количество
кадмия. Исследования на животных показали, что этот МЭ
обладает сродством к белковой фракции молока, связываясь
примерно в равных соотношениях с казеином и альбумином
[Van Bruwaene R. et al., 1984]. В женском молоке в среднем
содержится 0,1 мкмоль/л (колебания от 0,08 до 1,19 мкмоль/л),
что установлено в 23 образцах, собранных у 14 доноров
[Pinkerton С. et al., 1972]. Более поздние исследования, прове-
денные в ФРГ, показали, что у матерей, проживающих в сель-
ской местности, содержание кадмия в молоке равно 0,15±
±0,04 мкмоль/л, а у жительниц городов — 0,22±0,06 мкмоль/л
[Sternowsky Н., Wessolowski R., 1985]. Поскольку лактация в
среднем продолжается 7 мес, то за этот период секретируется
примерно 1699 мкг кадмия. У курильщиц не отмечено увеличе-
ния этого показателя [Ryan J. et al., 1982]. Кадмий может вы-
водиться с желчью, в которую он поступает из плазмы крови
против градиента концентрации [Klaasen С. D., 1976] в составе
низкомолекулярного соединения, представляющего собой, по-
видимому, комплекс кадмия с глутатионом [Cherian М. G.,
Vostal J. J., 1977]. Анализ кинетики выведения радиоактивного
кадмия (109Cd, 115Cd) в организме человека свидетельствует о
существовании трех компартментов для этого МЭ с периодами
полувыведения, равными 1,58, 33,7 и 9605 сут.
Для человеческого организма в целом период полувыведе-
ния кадмия тоже очень велик и составляет по разным оценкам
15—30; 18—38 и 10—30 лет [Kostial К-, 1986]. На основании
2-летних наблюдений за добровольцем, получившим однократ-
ную дозу радиоактивного кадмия, установлен период полувы-
ведения, равный 26 годам. По другим расчетам он равен
368
12,6 лет [Москалев Ю. И., 1985]. Содержание кадмия в орга-
низме новорожденного ничтожно мало и не превышает 1 мкг.
С возрастом концентрация этого МЭ постепенно повышается и
достигает к 50 годам 15—30 мг или 0,2—0,4 мг/кг свежей тка-
ни. Наивысшие концентрации найдены в почках, содержащих
15—20 мг/кг, причем преимущественно в корковом веществе.
Кадмий оказывает заметное влияние на обмен нуклеиновых
кислот и белка. Он ингибирует в опытах in vivo включение
тимидина в ДНК регенерирующей печени, повышает in vitro
поглощение ультрафиолетовых лучей при 260 нм в процессе
денатурации ДНК, не изменяя ее точки плавления. Низкие
дозы кадмия (0,34 мг/кг) стимулируют in vivo включение оро-
товой кислоты в РНК в течение 1—72 ч после введения ме-
талла. Более высокие дозы (34 мг/кг) оказывают аналогичное
действие только в течение 1 ч, после чего наступает угнетение
этого процесса. Снижение синтеза РНК кадмием in vitro на-
ступает при значительно меньших его концентрациях, чем in
vivo [Smith Н. А., 1984].
Кадмий угнетает синтез белка в печени крыс на стадии ини-
циации трансляции, нарушая образование полирибосом, тогда
как процесс элонгации, напротив, ускоряется в результате ак-
тивирования факторов EF-1 и EF-2 [Smith Н. A. et al., 1982].
При даче радиоактивного кадмия он обнаруживается в рибо-
сомах, из которых затем высвобождается по экспоненциаль-
ной кинетике, как описано выше для клеточных ядер. Обработ-
ка полирибосом протеазой ведет к высвобождению из них 70%
связанного с ними кадмия, что свидетельствует о его преиму-
щественной ассоциации с белками. Действие РНКазы, напро-
тив, не ведет к удалению кадмия из рибосом, что, однако, не
исключает возможности его связывания участками РНК внут-
ри рибосом, недоступными для этого фермента.
Кадмий обладает способностью индуцировать синтез метал-
лотионеина на уровне транскрипции. Это доказано как в опы-
тах с использованием ингибиторов данного процесса, так и
прямым выделением новообразованных молекул тионеиновых
мРНК. Поскольку синтез металлотионеинов может быть выз-
ван не только введением кадмия, но и целым рядом других
причин, включая стресс, бактериальную инфекцию, алкилиру-
ющие агенты, дексаметазон, эндотоксины, гепат- и адреналэк-
томию, воспалительные процессы, изменение концентрации
цинка и др. [Maitani Т., Suzuki К- Т., 1984], то долгое время
оставалось неясным, является ли действие кадмия на транс-
крипцию мРНК металлотионеина in vivo прямым или оно
опосредованно через органные или системные эффекты. Для
исключения системного действия гормонов ряд исследователей
использовали в своих опытах суспензии и культуры клеток
[Bryan S. Е. et al., 1979]. Было' показано, что при воздействии
кадмия изолированные гепатоциты и клетки гепатомы сохраня-
ют способность синтезировать тионеин, однако, без лаг-перио-
да, наблюдаемого in vivo.
24—568
369
Доказательством того, что синтезу Cd-тионеина предшеству-
ет транскрипция тионеиновой мРНК, а не замещение этим
металлом цинка в предобразованном Zn-тионеине, может слу-
жить ингибирование синтеза этого белка циклогексимидом и
актиномицином D, которые проявляют свое действие на уровне
трансляции и транскрипции соответственно. Новые успехи в
выявлении действия кадмия на транскрипцию генов тионеина
были достигнуты в результате использования для этой цели
современных методов молекулярной биологии. Были получены
кДНК и геномные клоны гена металлотионеина-1, позволившие
непосредственно наблюдать изменения в состоянии этих генов,
а также накопление тионеиновой мРНК под влиянием кадмия
и глюкокортикоидов [Durnam D. М. et al., 1980]. С помощью
данного метода было показано, что добавка кадмия повышает
транскрипцию этого гена в печени в 25 раз, в почках — в 17
раз, причем повышение наблюдается в течение 9 и 6 ч после
приема разовой дозы, по прошествии которых транскрипция
снова возвращается к норме. Синтез мРНК усиливается после
30-минутного лаг-периода и достигает максимума через 4—5 ч
после введения МЭ.
При более высоких дозах (0,044 и 0,222 ммоль/кг массы)
наблюдается индукция синтеза металлотионеина и в ряде дру-
гих органов, включая кишечник, сердце, мозг и селезенку, ио не
в семенниках. По своей способности индуцировать синтез ме-
таллотионеина в печени тяжелые металлы можно расположить
в следующий ряд: Cd>Zn>Cu>Hg; в почках он выглядит так:
(Cd, Hg)>Zn>Cu. Как показывают описанные исследования, в
печени и почках имеют место транскрипция тионеинового ге-
на-1 и ее регуляция тяжелыми металлами, тогда как ранее
считалось, что кадмий влияет только на трансляцию этого бел-
ка в почках [Shikh Z. A., Smith L. М., 1977].
Далее было установлено, что механизм индукции транс-
крипции тионеиновой мРНК кадмием и глюкокортикоидами
неодинаков. Методами генной инженерии была получена ре-
комбинантная плазмида, в которой промоторно-регуляторная
область металлотионеиновых генов была встроена перед струк-
турным геном вирусной тимидинкиназы. При введении плазми-
ды в яйцеклетку мыши металлотионеиновый промотор функци-
онировал с такой же эффективностью, как и собственный про-
мотор тимидинкиназы. При этом добавление кадмия к яйце-
клеткам оказывало стимулирующее действие на синтез данного
фермента. Внутриматочная трансплантация оплодотворенных
яйцеклеток самкам мышей дала начало трансгенной линии
мышей, которые реагировали на введение металлов повышен-
ной активностью тимидинкиназы [Palmiter R. D., 1982], тогда
как глюкокортикоиды не оказывали на этот фермент никакого
влияния. Это свидетельствует, что регуляция транскрипции тио-
неиновой мРНК металлами и глюкокортикоидами может осу-
ществляться независимыми генетическими элементами, один из
которых, по-видимому, теряется при конструировании плазми-
370
ды. В перспективе могут представить большой интерес иссле-
дования характера метилирования тионеиновых генов в эмбри-
ональном периоде и во взрослом состоянии, а также возможно-
сти их независимой регуляции различными металлами.
Данные о биохимических сдвигах при кадмиевом токсикозе
получены, как правило, на лабораторных животных и не всег-
да могут быть перенесены на организм человека. Этой задаче
наиболее близко соответствуют результаты опытов, в которых
животных подвергают хроническому воздействию малых доз
кадмия и в меньшей степени кратковременных опытов in vitro.
Воздействие кадмия на белковый и нуклеиновый обмен рас-
смотрено выше. Установлены также: Q стимулирующее дейст-
вие этого МЭ на синтез цАМФ и активность анаболических
ферментов in vivo и обратный эффект в опытах in vitro; 2) раз-
общение окислительного фосфорилирования и дыхания с уско-
рением его IV фазы; 3) ингибирующее действие на активность
каталазы и карбоангидразы и в меньшей степени щелочной и
кислой фосфатазы; 4) снижение в малых дозах активности
1,25-дигидрохолекальциферола (витамин Эз) и 5) уменьшение
фагоцитирующей способности макрофагов.
Обнаружено также влияние кадмия на ферменты углевод-
ного обмена, выражающееся в стимуляции активности альдо-
лазы, фосфорилазы и пируваткарбоксилазы, фосфоенолпиру-
ваткарбоксикиназы и фруктозо-бисфосфатазы (КФ. 3.1.3.11),
наряду с угнетением секреции инсулина. В итоге при действии
кадмия намечается тенденция к повышению в организме синте-
за глюкозы и худшему ее использованию, что способствует
возникновению гипергликемии. Стимулирующим действием
кадмия на синтез цАМФ можно объяснить, по-видимому, об-
щее усиление этим металлом процессов гликонеогенеза, глико-
генолиза и его диабетогенный эффект.
Кадмий ингибирует активность целого ряда ферментов, к
числу которых относятся алкоголь- и глутаматдегидрогеназы,
аденилаткиназа, глутатионредуктаза и др., в результате обра-
зования меркаптидов с цистеиновыми остатками их активных
центров. Большинство отмеченных нарушений метаболизма,
вызываемых кадмием, можно предупредить или нормализовать
введением селена или цинка.
Подавляющее большинство исследований биологической ро-
ли кадмия посвящено его токсическому действию на организм
человека и животных [Friberg L., 1974; Venugopal В., Luckey
Т„ 1978; Nriagu J., 1980]. В последние 10 лет, однако, также
появились данные о жизненной необходимости этого элемента
для животного организма. Стимулирующее действие низких
доз кадмия (0,01—10 мкмоль/л) на рост аспергиллов и неко-
торых водорослей было известно более 50 лет назад [Scharrer
К., Schropp W., 1934; Pirschle К-, 1934]. Эти дозы значительно
ниже стимулирующих доз цинка, составляющих 1 ммоль/л.
Эссенциальность кадмия для организма млекопитающих
была впервые продемонстрирована с помощью пластикового
24*
371
«изолятора» [Schwarz К., Spallholz Т., 1976]. Было установ-
лено, что этот МЭ в нетоксических концентрациях, свойствен-
ных нормальным рационам, стимулирует рост крыс. Еще более
убедительные доказательства жизненной необходимости этого
элемента были получены в ГДР [Anke М. et al., 1978] в опы-
тах на козах, у которых дефицит кадмия в рационе (20 мкг/кг)
вызвал задержку роста и полового созревания.
Уже более 100 лет известно, что кадмий может вызывать
острое отравление у животных и человека. Однако первое
подробное описание острой кадмиевой интоксикации среди
промышленных рабочих появилось только в 1938 г. [Bulmer
F. М., Rothwell Н. Е., 1938]. Обычно одним из первых симпто-
мов было раздражение глотки. Через несколько часов или дней
появлялись другие симптомы: боль при дыхании, одышка, силь-
ный кашель, тошнота, рвота, цианоз, отек легких, сопровожда-
ющийся часто бронхопневмонией, повышение температуры (до
38,9°C) и учащение пульса. В описанном наблюдении интен-
сивное поступление высоких доз МЭ в организм сопровождалось
несколькими смертельными исходами. Наиболее выраженным
симптомом была сильная одышка, которая проявлялась через
несколько часов или даже дней после контакта с металлом.
Вследствие замедленного проявления серьезных симптомов ре-
альна возможность спутать ингаляционную кадмиевую инток-
сикацию с некоторыми другими заболеваниями, например с
гриппом. Авторами была определена летальная концентрация
(2900 мг/м3 в 1 мин) атмосферной окиси кадмия для людей,
выполняющих легкую работу. В более позднем исследовании в
1966 г. A. Beton и соавт. представили сходную концентрацию—
2589 мг/м3 в минуту (8,6 мг/м3 в течение 5 ч), которая вызы-
вала смертельный исход у рабочих при вдыхании аэрозоля
кадмия. Только с 1958 по 1976 г. в литературе появились сооб-
щения о 24 случаях острого отравления окисью кадмия. Более
100 рабочих подверглись вредному воздействию, из которых
17 погибли [Bernard A., Lauwerys R., 1984], т. е. летальность
при острой кадмиевой интоксикации достигает 15—20%. Отда-
ленные последствия короткого контакта организма с высокой
концентрацией атмосферного кадмия приводят к легочному
фиброзу, перманентному нарушению легочных и печеночных
функций [Ryan J. et al., 1982]. Ряд авторов описали случаи
острого отравления кадмием при его оральном поступлении в
организм [Bernard A., Lauwerys R., 1984]. При этом в основ-
ном поражается желудочно-кишечный тракт. Благодаря быст-
рому обновлению клеток слизистой оболочки кишечника основ-
ная масса кадмия теряется вместе со слущивающимися клет-
ками верхушек ворсинок. Полагают, что при определенном
критическом уровне кадмия в клетках слизистой оболочки про-
исходят структурные изменения в них и нарушение всасывания
кадмия и других веществ [Fox М. R., 1976]. Симптомы кадмие-
вого токсикоза сводятся к сильной тошноте, рвоте, слюнотече-
нию, спазмам кишечника и диарее. Фатальный исход для че-
372
ловека обусловлен шоком из-за потери жидкости или острого
поражения почек и сердечно-легочной недостаточности и на-
блюдается при дозе кадмия от 350 до 8900 мг [Lauwerys R.,
1978].
Хотя острое отравление кадмием иногда со смертельным
исходом имеет место и в настоящее время, однако все же оно
встречается реже, чем синдром хронической интоксикации этим
МЭ, который начали исследовать лишь в 50-х годах [Friberg
L., 1949, 1950]. В последующем в многочисленных опытах на
животных было показано, что подострый и хронический эффект
этого МЭ проявляется в снижении массы тела, уменьшении
потребления пищи, анемии, артериальной гипертензии, проте-
инурии, ухудшении минерализации костей, некрозе яичек, уве-
личении неонатальной смертности и появлении молодняка с
врожденными уродствами [Perry Н. М., 1976; Piscator М.,
1976; Doylei J. J., 1977]. R. Van Bruwaene и соавт. (1984) обоб-
щили данные по кадмиевому отравлению домашних животных.
Л. М. Михалева и соавт. (1988, 1989) изучали гистопатоло-
гию хронической кадмиевой интоксикации крыс, которым еже-
дневно внутрижелудочно вводили раствор хлорида кадмия в
дозе 2,5 мг на 100 г массы тела в течение 12 нед.
Масса тела крыс по мере увеличения срока интоксикации
достоверно уменьшалась по отношению к соответствующему
контролю в среднем на 84 г с заметным уменьшением около-
почечного жира. Отражением стрессорного воздействия вводи-
мого раствора кадмия явилось достоверное увеличение массы
надпочечников на 33%, а показателя относительной массы их—
на 80%. Гистологически в печени отмечены выраженная жиро-
вая дистрофия гепатоцитов и неравномерное полнокровие си-
нусоидов. В почках обнаружены единичные коллабированные
и «лапчатые» клубочки с утолщением наружного листка капсу-
лы почечного клубочка (Боумена — Шумлянского), зернистая
дистрофия и десквамация эпителиоцитов извитых канальцев,
единичные гиалиновые цилиндры; на ультраструктурном уров-
не в эпителиоцитах выявлены просветление ядер, маргинация
хроматина, выраженная центральная деструкция митохондрий
(рис. 65, 66).
Через 2 нед у крыс было обнаружено достоверное повыше-
ние систолического артериального давления до 133± 1 мм рт.
ст. (в контроле 116± 1 мм рт. ст.) с дальнейшим его повыше-
нием до 149±2 мм рт. ст. (контроль 115±2 мм рт. ст.) на 6-й
неделе интоксикации. В последующие 4 нед уровень артери-
ального давления стабилизировался. После прекращения вве-
дения хлорида кадмия выявлено достоверное падение артери-
ального давления на 7-е сутки до 129±2 мм рт. ст., а к 3-й
неделе реабилитации — до 115±2 мм рт. ст.
Впервые морфологически гипертрофия левого желудочка
выявлена на 30-е сутки интоксикации; при раздельном взвеши-
вании сердца на 84-е сутки опыта желудочковыц индекс сни-
зился до 0,346±0,007 (в контроле 0,400±0,010). При гистоло-
373
Рис. 65. Вакуольно-зернистая дистрофия, десквамация эпителиоцитов каналь-
цев, наличие ШИК-позитивного материала в просвете расширенных канальцев
почки при пероральном введении хлорида кадмия в концентрации 2,5 мг/100 г
массы через 20 сут. ШИК-реакЦия. Х400 (препарат Л. М. Михалевой).
гическом исследовании сердца у подопытных крыс отмечены
умеренное расширение и полнокровие капилляров, межклеточ-
ный отек, очаговые повреждения кардиомиоцитов (рис. 67).
Электронно-микроскопически выявлены просветление сарко-
плазмы, деструкция митохондрий и фрагментация миофибрилл
(рис. 68). К 20-м суткам интоксикации четко регистрируется
повышение синтетической активности миоэндокринных клеток
предсердий, выражавшееся в увеличении числа секреторных
гранул, в уменьшении их диаметра и площади (рис. 69).
Таким образом, проведенное исследование показало, что в
условиях данного эксперимента имеются хорошо выраженные
признаки хронической кадмиевой интоксикации с последующим
развитием обратимой артериальной гипертензии.
Хроническое воздействие кадмия на человека привлекает
особо пристальное внимание в связи с нарушениями почечной
функции, легочной недостаточностью, остеомаляцией, анемией
и потерей обоняния. Существует ряд данных о возможном кан-
церогенном эффекте этого МЭ, о его вероятном участии в раз-
витии сердечно-сосудистых заболеваний, в частности гиперто-
нии.
Еще в 1940 г. установлено, что среди рабочих, контактирую-
щих с кадмием при электролизе, наблюдались частые хрони-
374
Рис. 66. Резкий отек цитоплазмы, просветление матрикса митохондрий с пар-
циальным и субтотальиым лизисом крист в эпителиоците проксимального
канальца почки после 40-суточиой интоксикации кадмием. X 18 000 (препарат
Л. М. Михалевой).
ческие риниты и фарингиты, сухость и раздражение глотки,
ощущение жжения в носу, носовые кровотечения, изъязвления
в хрящевой части носа и носоглотке. Этиологическая роль кад-
мия в возникновении анемии у людей, контактирующих с кад-
мием на производстве, до сих пор изучена недостаточно [Ка-
zantzis G. et al., 1963; Tsuchiya К., 1967; Friberg L. et al.,
1974]. Судя по опытам на животных, анемия обусловлена по-
давлением кадмием всасывания железа в кишечнике и неболь-
шим гемолизом [Friberg L. et al., 1974].
Наибольшее внимание при хроническом воздействии кадмия
привлекает повреждение почек. Так, L. Friberg (1948, 1950)
впервые описал кадмиевую нефропатию у промышленных ра-
бочих. Одним из первых лабораторных признаков этого заболе-
вания является наличие в моче низкомолекулярных сывороточ-
ных белков (мол. масса 20 000—25 000, в целом ниже 40 000),
лизоцима, ретинолсвязывающего белка, особенно Рг-микрогло-
булина [Squibb К. et al., 1984]. Их экскреция увеличивается в
1000 раз. Минимальный латентный период до появления проте-
инурии равен примерно 1 году после начала воздействия
[Tsuchiya К., 1967], но может длиться до 10 лет. Однако пер-
вые признаки болезни могут проявляться спустя много лет
375
парат Л. М. Михалевой). (пре
р,“ .га «>.сут„,„,ов *^2г6ийТ(Х«ар“’л.Т%
J
Рис. 69. Очаговое просветление матрикса митохондрий, в некоторых из них
центральный лизис крист, выраженная гиперплазия комплекса Гольджи, боль-
шое количество секреторных гранул, различных по электронной плотности
и размеру, в миоэндокринных клетках правого предсердия после 30-суточной
интоксикации кадмием. Х37 ООО (препарат Л. М. Михалевой).
после окончания контакта с кадмием. Так, если в норме экс-
креция белка с мочой составляет в среднем 50 мг в день, то
при кадмиевой интоксикации она возрастает до 70—2600 мг в
день [Piscator М., 1962]. Концентрация р2-микроглобулина мо-
жет превышать 290 мкг/л (Kjellstrom Т. et al., 1977]. Как уже
упоминалось, концентрация кадмия в проксимальных отделах
почечных канальцев подавляет реабсорбцию этих белков в них,
но белки свободно фильтруются через клубочки. По мнению
377
H. Gonick и соавт. (1980), механизм торможения связан с по-
давлением Na+, К+-АТФазной транспортной системы. Сущест-
вует значительная корреляция между повышенным уровнем
кадмия в крови и увеличением ^г-микроглобулина. На после-
дующих этапах развития заболевания в результате торможения
канальцевой реабсорбции развиваются глюкозурия, генерализо-
ванная аминоацидурия и энзимурия [Linder G., Foulkes Е.,
1985]. Этот комплекс признаков известен как синдром Фанко-
ни [Ryan J. et al., 1982]. По поводу их диагностического зна-
чения при кадмиевой интоксикации до сих пор ведутся дискус-
сии. Так, К. Nomiyama и соавт. (1975) считают, что аминаци-
дурия является одним из самых ранных признаков отравле-
ния кадмием у кроликов, которая появлялась через 16 нед
после начала контакта с этим МЭ, в то время как протеинурия
и глюкозурия имели место только через 27 нед. Однако, по
мнению G. Axelsson и М. Piscator (1966), у человека амина-
цидурия появляется позже. То же отмечено среди японских
рабочих на свинцово-кадмиевом производстве [Goyer Р. A et
al., 1972].
S. Tanaguchi и соавт. (1979) представили данные, что со-
держание карбонат-дегидротазы С увеличивается в моче в
10—250 раз у контактирующих с кадмием людей по сравнению
с нормальной популяцией. Это наблюдение чрезвычайно важ-
но, поскольку указанный фермент выделяется из поврежденных
клеток канальцев. На основе анализа ряда публикаций
Z. A. Shaikh и L. М. Smith (1984) считают, что энзимурия яв-
ляется реальным показателем неспецифического повреждения
почек, причем очень ранним диагностическим тестом ренальной
дисфункции, предшествующим выраженным изменениям в
проксимальных канальцах и клубочках. Глюкозурия, развиваю-
щаяся у ряда животных при нагрузке кадмием, у рабочих и
жителей, подвергающихся воздействию этого МЭ [Shigematsu
J. et al., 1980], является одним из поздних проявлений нефро-
токсичности. На определенных стадиях почечных повреждений
к экскреции низкомолекулярного белка присоединяется выде-
ление с мочой высокомолекулярного белка, что отражает по-
явление нарушений в клубочковой фильтрации. Так, A. Bernard
и соавт. (1979) обнаружили у рабочих, занятых на производст-
ве кадмия, повышение уровня высокомолекулярных белков
(орозомукоида, иммуноглобулина G, альбумина, трансферри-
на) — смешанную протеинурию и снижение клиренса креатини-
на, что является признаком дезорганизации почечных клубоч-
ков. Существуют даже данные, что клубочковая дисфункция
может предшествовать классической дисфункции проксималь-
ных канальцев. Так, по данным Н. Roels и соавт. (1974), у ра-
бочих со стажем работы на кадмиевом производстве менее
20 лет наблюдаются выделение с мочой только высокомолеку-
лярных белков, а при стаже работы более 20 лет — смешан-
ная протеинурия. Это наблюдение было подтверждено в экспе-
риментах на крысах [Bernard A. et al., 1978]. Однако боль-
378
щинство исследователей считают, что выделение с мочой низко-
молекулярных белков, особенно р2-микрогл°булинов, более
показательно как индикатор кадмиевой интоксикации. Опреде-
ленный интерес вызывают другие низкомолекулярные белки,
выделяющиеся из организма с мочой при кадмиевой интокси-
кации — ретинолсвязывающий белок и лизоцим, но все же пре-
обладает экскреция р2-микроглобулина. По данным F. Falck и
соавт. (1984), протеинурия при кадмиевой интоксикации отли-
чается от таковой при неспецифических нефропатиях отсутст-
вием ^-глобулинов, к другим признакам нарушения кадмием
почечной функции относят гипокальциемию, гипофосфатемию,
гиперфосфатурию, нефрокальциноз (почечные камни) и гипер-
кальииурию [Kazantzis G. et al., 1963; Kazantzis G., 1979].
Анализ результатов исследований последних лет косвенно под-
тверждает возможную этиологическую роль кадмия в возник-
новении гипертонии у людей. Так, имеется наблюдение, что у
населения, проживающего в Англии в зонах с высоким содер-
жанием кадмия в почве, отмечена повышенная частота гипер-
тонии [Carruthers М., Smith В., 1979]. У белых и темнокожих
жителей США, страдающих этим заболеванием, отмечено по-
вышение концентрации кадмия в коре почек (секционный ма-
териал) [Horvath D. et al., 1984], волосах [Medeires D., Pellum
L., 1984, 1985], в цельной крови [Tulley R., Lehmann H., 1982].
Таким образом, этиологическая роль кадмия в возникновении
гипертонии у человека остается к настоящему времени до кон-
ца не выясненной и требует дальнейших исследований.
Изучение состояния легких после ингаляционного или инт-
ратрэхеалыюго введения кадмия крысам показало, что металл
довольно быстро выводится, но некоторая его часть (от 1 до
20%) задерживается в этом органе. По сравнению с другими
МЭ (селен, ванадий, мышьяк) кадмий обладает наибольшей
токсичностью [Rhoads К., Sanders С. L., 1983]. У эксперимен-
тальных животных после ингаляции соединений кадмия воз-
никновение острой пневмонии сопровождалось развитием пери-
васкулярного и перибронхиального фиброза. У человека это
заболевание, этиологически связанное с кадмием, было описа-
но Н. A. Smith и соавт. только в 1976 г. при рентгенологичес-
ком обследовании рабочих, контактирующих на производстве
с кадмием. К сожалению, авторы не описали анамнез этих
больных, поскольку фиброз легких у человека может быть свя-
зан с рядом сопутствующих заболеваний (туберкулез, грипп,
пневмония, хронический бронхит и др.). Хроническая ингаля-
ция кадмием у кроликов сопровождалась увеличением легких
и интерстициальной инфильтрацией нейтрофилами и лимфо-
цитами, а также накоплением в альвеолах больших вакуолизи-
рованных макрофагов. Морфометрическое измерение объемной
плотности альвеолоцитов II типа показало более чем двукрат-
ное увеличение за счет гипертрофии и гиперплазии клеток.
Содержание фосфолипидов возросло в легочной ткани пример-
но на 40%, в основном за счет фосфатидилхолинов [Johans-
379
son A. et al., 1984]. После инъекции хлорида кадмия мышам
(0,12 ммоль/кг) через несколько часов были обнаружены эм-
физемоподобные изменения легких с растяжением альвеол.
L. Friberg (1948, 1950) принадлежит заслуга в выявлении эм-
физемы у рабочих, подвергающихся воздействию кадмия.
Впоследствии это наблюдение было подтверждено рядом ис-
следователей при эпидемиологическом обследовании рабочих и
изучении секционного материала [Smith Н. A. et al., 1960; Ка-
zantzis G. et al., 1963].
На основе анализа работ создается впечатление, что этот
вопрос требует дальнейших детальных экспериментальных,
эпидемиологических и ретроспективных исследований. В целом
же можно отметить, что легочные изменения при кадмиевой
интоксикации незначительны, встречаются нечасто и, вероятно,
проявляются позже, чем почечная дисфункция [Piscator М.,
1976; Mahaffey К., 1984].
Наиболее тяжелой формой хронического
отравления кадмием является болезнь итаи-
итаи. Впервые она была обнаружена в 1946 г. у женщин
старше 45 лет в г. Фуху, префектура Тояма (Япония). Кадмий
в организм попадал с рисом, выращенным на полях, орошав-
шихся водой из реки, в которую кадмий поступал из рудника,
расположенного значительно выше по течению. Концентрация
кадмия в рисе в течение многих лет была около 1 мкг/г, в свя-
зи с чем в сутки поступало 300 мкг и более [Piscator М„
1976]. В 1948 г. было установлено, что основной причиной за-
болевания является остеомаляция. В свою очередь возникнове-
ние остеомаляции связывают с дисфункцией проксимальных
отделов почечных канальцев из-за отложения кадмия, в ре-
зультате чего имеют место потеря минеральных элементов кос-
ти через почки и гиповитаминоз D [Blainey J. D. et al., 1980;
Kwast M., 1981]. Поскольку в основном итаи-итаи заболевали
женщины, ранее имевшие многочисленные беременности и лак-
тацию, то полагают, что недостаточное поступление этого ви-
тамина и кальция в организм, их истощение во время беремен-
ностей и кормления детей грудью, недостаточное воздействие
ультрафиолетовых лучей также являются предрасполагающи-
ми патогенетическими факторами. Болезнь характеризуется
деформацией скелета с заметным уменьшением роста, пояс-
ничными болями, болезненными явлениями в мышцах ног, ути-
ной походкой. Кроме того, отмечаются частые переломы раз-
мягченных костей даже при кашле, а также нарушение функ-
ции поджелудочной железы, изменения в желудочно-кишечном
тракте, гипохромная анемия, дисфункция почек (выделение
низкомолекулярных белков), пониженный уровень железа, каль-
ция и неорганического фосфора в крови, а также высокий
уровень щелочной фосфатазы [Hallenbeck W., 1984]. Пораже-
ния почек при итаи-итаи сходны с теми симптомами, которые
отмечаются в условиях хронического профессионального от-
равления кадмием. У мужчин в этом районе также отмеча-
380
ются протеинурия и глюкозурия, но остеомаляция встречается
значительно реже, что может быть объяснено лучшим кальцие-
вым балансом [Piscator М., 1975]. В случае итаи-итаи измене-
ния костей (остеомаляция) являются вторичными из-за нару-
шений функции почек [Korte F., 1983]. Однако в эксперимен-
тах было показано, что кадмий непосредственно влияет на
остеопорозные изменения костей [Takashima М. et al., 1980].
Действительно, в эксперименте было показано, что в печени
цыплят гидроксилирование витамина D подавляется кадмием.
Это приводит к дефициту активной формы данного витамина
[Feldman F., Cousins R., 1973]. Кроме этого, на крысах было
обнаружено, что кадмий ускорял развитие остеопороза, вызван-
ного дефицитом кальция. Это могло объясняться влиянием
кадмия на всасывание кальция в кишечнике и на функцию
эндокринных желез.
К настоящему времени стало ясно, что Япо-
ния является не единственной страной, где
кадмий окружающей среды влияет на здоровье
большой группы населения. Так, исследования, про-
веденные в Бельгии в провинции Льеж, показали, что в ре-
зультате контакта человека с природным кадмием в индустри-
ально загрязненных этим металлом районах может изменяться
функция почек [Lauwerys R. R. et al., 1980; Roels H. et al.,
1981]. Более того, ретроспективный анализ летальности, прове-
денный вскоре после этих наблюдений, показал увеличение
частоты смертельных исходов от «нефритов» и «нефрозов» в
этом районе [Lauwerys R. R., De Wais Р., 1981], что коррели-
рует с удвоением концентрации кадмия в коре почек у жителей
Льежа по сравнению с остальной популяцией этой страны
[Bernard A., Lauwerys R.. 1984].
В ряде работ проанализировано действие этого МЭ на
нервную систему. Так, Р. С. Воробьева (1957) при обследова-
нии рабочих, контактирующих с кадмием на производстве, от-
метила увеличение частоты приступов головных болей, голово-
кружений, усиление коленного рефлекса, тремор, дермогра-
физм, нарушение сенсорной и моторной хронаксии. Кроме того,
при воздействии высокой концентрации кадмия у рабочих вы-
явлено нарушение обонятельной функции, в частности потеря
обоняния [Friberg L., 1950]. G. Gabbiani и соавт. (1967) сооб-
щили о кровоизлияниях в подкорковые ганглии, патологичес-
ких изменениях в головном мозге и мозжечке у новорожденных
животных при введении этого МЭ. Таким образом, было обна-
ружено, что новорожденные животные по сравнению со взрос-
лыми особями более чувствительны к нейротоксическому дейст-
вию кадмия. A. Rohrer и соавт. (1978) сообщили об изменени-
ях сосудистой системы головного мозга у плодов животных под
воздействием кадмия. При внутривенном введении кадмия но-
ворожденным крысятам в их мозге была зафиксирована отно-
сительно высокая его концентрация. Это указывает на то, что
гематоэнцефалический барьер по отношению к кадмию у ново-
381
рожденных полностью не сформировался [Wong К., Klaassen
С., 1981]. Кроме того, было обнаружено, что этот металл вы-
ступает в роли агента, блокирующего как адренергические,
так и холинергические синапсы [Cooper G. et al., 1978].
В последние годы дискутируется вопрос о канцерогенном
потенциале кадмия. Ведутся работы по выявлению мутагенного
эффекта кадмия in vivo и in vitro, канцерогенного действия в
эксперименте на животных, а также эпидемиологические об-
следования по выявлению частоты рака среди населения, кон-
тактирующего с этим МЭ в естественных условиях и на про-
изводстве. Так, J. Shiraishi (1972) обнаружил некоторые хро-
мосомные аберрации у больных итаи-итаи, но это наблюдение
не было подтверждено [Bui Т. et al., 1975]. Т. Bui и соавт. не
обнаружили хромосомных нарушений и у рабочих, контактиру-
ющих с кадмием. Однако, как G. Deknudt и A. Leonard
(1975), так и М. Bauchinger и соавт. (1976) выявили выражен-
ное увеличение числа хромосомных аберраций у человека при
воздействии этого металла. Позднее М. O’Riordan и соавт.
(1978) не выявили хромосомных отклонений в лимфоцитах пе-
риферической крови у рабочих кадмиевого производства. Они
предположили, что изменения хромосом, отмеченные в более
ранних работах, возникли в результате синергического дейст-
вия тяжелых металлов и мутагенов окружающей среды. Одна-
ко в 1981 г. К. Gasiorek и М. Bauchinger отметили значитель-
ное увеличение хромосомных аберраций в лимфоцитах челове-
ка именно при воздействии кадмия. Таким образом, противоре-
чивость полученных результатов затрудняет окончательный
вывод о мутагенных свойствах этого МЭ.
К настоящему времени не существует и окончательного за-
ключения о канцерогенности кадмия для человека. С. Potts
(1965) одним из первых обратил внимание на возможность
возникновения рака у человека в результате воздействия этого
металла. Так, при обследовании им 70 рабочих кадмиевого
производства, имеющих стаж работы более 10 лет, было уста-
новлено, что 3 умерли от рака предстательной железы, 1 — от
рака бронхов и 1 — от распространенного карциноматоза.
Позднее М. Kipling и J. Waterhouse (1967) обследовали то же
производство, что и С. Plotts. Из 248 рабочих, по его данным,
12 умерли от рака, в том числе — 4 от рака предстательной
железы, что несколько превышало региональный показатель.
Однако поскольку выборка была невелика, то окончательное
заключение сделать трудно. К тому же данные не были под-
креплены ни изучением секционного материала, ни определени-
ем числа курящих, ни установлением возможного воздействия
других канцерогенных МЭ (например, никеля).
Заслуживает внимания ретроспективное обследование попу-
ляции людей (292 человека), контактировавших с кадмием на
производстве с 1940 по 1969 г., со стажем работы не менее
2 лет, проведенное R. Lemen с соавт. (1976). Эти авторы обна-
ружили значительное увеличение летальности от «общей
382
малигнизации», от рака легких и предстательной железы в
обследованной группе по сравнению с контрольной группой.
R. Lemen и соавт. (1976) провели подробный анализ типов
злокачественных опухолей. Однако и в этом случае число изу-
ченных опухолей предстательной железы было невелико, для
того чтобы сделать окончательное заключение. G. Вако и соавт.
(1982) сообщили об увеличении частоты рака предстательной
железы среди мужчин, проживающих в ареалах с высоким
уровнем этого металла в почвах некоторых районов Канады.
Эту же особенность отметил D. W. Flanders (1984) в США.
В. Kolonel (1976) предположил существование связи между
кадмием и малигнизацией почек, причем ему удалось устано-
вить 4-кратное увеличение частоты рака почек у курильщиков
по сравнению с контрольной популяцией. В 1979 г. Т. Kjellstrom
и соавт. обследовали людей, контактировавших с кадмием с
1959 по 1975 г. и имеющих стаж работы на этом производстве
не менее 5 лет. Статистически достоверного повышения часто-
ты онкологических заболеваний не обнаружено. Отмеченное
авторами повышение частоты рака носоглотки может объяс-
няться влиянием примесей никеля, поскольку именно этот тип
опухолей наиболее часто встречается на никелевом производ-
стве [Hailenbeck W., 1984]. Тем не менее, несмотря на проти-
воречивые данные, в 1976 г. кадмий включен в список агентов,
которые могут быть канцерогенными для человека.
В последние годы внимание исследователей привлекает воз-
действие кадмия на иммунную систему. Так, хроническое
оральное введение кадмия мышам (диапазоны концентраций —
0,27—5,34 ммоль/л в течение 10 дней) подавляло клеточно-за-
висимый иммунитет в виде гиперчувствительности замедленно-
го типа, индуцированный эритроцитами барана [Muller I. et al.,
1979]. Наряду с этим гуморальный ответ на этот антиген не
изменялся. A. Nelson и соавт. (1982) обнаружили, что кадмий
в концентрации 0,1 — 1 мкмоль/л в отличие от свинца и цинка
значительно подавляет цитолитическую активность иммунных
макрофагов мышей C57B1/6J по отношению к клеткам сарко-
мы. Анализ приведенных данных свидетельствует о влиянии
этого МЭ на функцию защитных систем организма. Более того,,
по данным A. Gualde и С. Delage (1979), существует индивиду-
альная чувствительность к кадмию. Так, у людей с фенотипом
HLA-A3 уровень кадмия в крови повышен по сравнению с кон-
трольной выборкой. Используя тест бласттрансформации с
ФГА, авторы показали, что клетки АЗ более чувствительны к
кадмию. Для объяснения этого феномена высказано предполо-
жение, что, во-первых, молекулы HLA могут играть роль не-
иммунных лигандов, во-вторых, лимфоциты АЗ могут обладать
выраженной способностью к синтезу металлотионеинов.
Таким образом, важнейшими кадмиозами человека являют-
ся острое и хроническое отравление этим МЭ. Четко выделены:
1) кадмиевый ринит; 2) кадмиевая нефропатия с типичной про-
теинурией; 3) кадмиевая остеомаляция (болезнь итаи-итаи);
383
4) нейротоксический синдром. Кадмиевые кардиомиопатия, ги-
пертония, эмфизема легких и поражение печени, а также кад-
миевый рак предстательной железы требуют дополнительных
доказательств не только на экспериментальном, но и на сек-
ционном материале их прямой зависимости от токсического
действия этого МЭ.
Кадмий оказывает выраженное действие на обмен ряда
МЭ, в первую очередь цинка, меди, железа и селена. Это вли-
яние проявляется на уровне ферментативных процессов, вса-
сывания, отложения и выделения элементов, а также функции
целого организма. Кадмиевое отравление в значительной степе-
ни зависит от его взаимоотношения с цинком. В связи с бли-
зостью строения атомов обоих элементов и сходством образуе-
мых ими тетраэдрических комплексов кадмий способен заме-
щать цинк в хелатах этого металла и превосходит в этом от-
ношении все остальные металлы, в том числе близкие к нему
по химическим свойствам ртуть, медь и галлий. Исходя из это-
го кадмий можно рассматривать как специфический антимета-
болит цинка. Целый ряд токсических явлений, вызываемых
действием кадмия, такие как гипертония, поражение кожных
покровов, семенников, яичников, нервных ганглиев, удается в
эксперименте предупредить одновременной дачей цинка. Хоро-
шо известен антагонизм кадмия и по отношению к ряду других
элементов. Удалось показать, например, что при избытке кад-
мия происходит обеднение организма животных медью и они
погибают с выраженными признаками недостаточности меди
[Mills С. F., Delgarno А. С., 1972; Anke М. et al., 1976].
Поражение семенников, вызываемое кадмием, удается пре-
дупредить дачей селена. Защитное действие последнего сопро-
вождается повышением концентрации кадмия в тканях, причем
доля кадмия, присутствующего в составе низкомолекулярной
белковой фракции (молекулярная масса около 10000), снижа-
ется в 2—3 раза и соответственно увеличивается его количест-
во в составе недавно открытого весьма нестабильного белка с
молекулярной массой около 30 000, в котором присутствует
также селен. Аналогичное явление отмечено также в отноше-
нии почек и печени [Chen К. W. et al., 1975]. Селен эффектив-
но предупреждает токсемию беременности, вызываемую кадми-
ем [Parizek J. et al., 1968].
Физиологический антагонизм кадмия и ряда других МЭ про-
является уже на уровне всасывания. Важно, что низкие кон-
центрации цинка (0,01—1 ммоль/л) повышают скорость всасы-
вания кадмия в кишечнике крыс, что связано, по-видимому,
с индукцией цинком биосинтеза металлотионеина. При более
высоких концентрациях цинк, напротив, тормозит всасывание
кадмия [Sahagian В. М.. et al„ 1966]. Кроме того, четко выра-
женный антагонизм в процессе всасывания установлен на кры-
сах между кадмием и ртутью, кадмием и медью [Van Cam-
pen D. R., 1966]. На цыплятах и мышах продемонстрировано
угнетение кадмием всасывания S9Fe и кобальта. Перечисленные
384
выше двухвалентные металлы в свою очередь снижают всасы-
вание кадмия, чем отчасти можно объяснить их защитный
эффект яри кадмиевом отравлении. Этот физиологический ан-
тагонизм может иметь, таким образом, определенное практиче-
ское значение для предупреждения кадмиевого токсикоза в ре-
гионах, загрязненных кадмием.
8.3. Свинец
Интерес к свинцу в медицине и биологии определяется почти
исключительно(«го свойствами как кумулятивного яда, извест-
ного человечеству свыше 4000 лет. Отравление этим элементом
было знакомо людям еще в античном мире как сатурнизм или
плюмбизм, отдельные клинические признаки которого были
описаны Гиппократом в 370 г. до новой эры. Использование
свинца в древнем Риме достигало 4 кг в год на душу населения,
что сопоставимо с современным потреблением этого элемента
в США — 6 кг в год. Свинец использовали главным образом
при строительстве акведуков и изготовлении водопроводных
труб, лужений бронзовых котлов для варки пищи и придания
ей сладости. Вина и соки также изготовляли в посуде со свин-
цовым покрытием, что улучшало их вкус. Широкое применение
свинца в быту древних римлян, несомненно, влекло за собой
загрязнение их пищи и напитков этим элементом. Существует
даже гипотеза [Gilfillan S. С., 1965], согласно которой упадок
Римской империи в значительной мере обусловлен отравлением
ее граждан свинцом. Этот взгляд подтверждается данными о
высоком содержании свинца в костных останках римлян и осо-
бенно римских патрициев.
Первые документированные случаи массового свинцового
токсикоза относятся к XVI веку. Наиболее значительные из них
имели место в Амстердаме, где население пило водопроводную
воду, загрязненную свинцом, и в графстве Девоншир (Англия),
где при изготовлении сидра использовали прессы и посуду, по-
крытые свинцом.
В настоящее время свинец вызывает повышенный интерес
как приоритетный загрязнитель окружающей среды, ежегодные
промышленные и транспортные выбросы которого превышают
400 000 т, угрожая здоровью миллионов людей, особенно детей
[Nriagu J. О., 1978; Chisholm J. J. et al., 1985]. Всасывание это-
го МЭ из пищеварительного тракта у детей происходит в 3 раза
интенсивнее, чем у взрослых.
Жизненная необходимость свинца для животного организма
была впервые показана на крысах [Schwarz К., 1974] и под-
тверждена в серии исследований, выполненных 10 лет спустя
[Reichlmayer-Lais А. М., Kirchgessner М., 1984]. В то же время
механизмы как токсического, так и физиологического действия
свинца до настоящего времени изучены весьма слабо и рас-
крыты значительно хуже, чем для других жизненно необходи-
мых МЭ.
25—568
385
Обмен свинца. В норме содержание свинца в пищевых
продуктах колеблется в пределах 0,02—3 мг/кг свежего веще-
ства, в питьевой воде 0,01—0,03 мг/л и атмосфере 0,03—
0,1 мкг/м3. Выбросы свинца могут многократно повысить его
содержание в окружающей среде. Источниками свинца могут
служить также антидетонационные присадки (тетраэтил- и
тетраметилсвинец), краски, содержащие этот МЭ, посуда. Осо-
бенно много свинца поступает в окружающую среду при вы-
плавке и переработке цветных металлов, сжигании угля, нефти,
сланцев, изготовлении аккумуляторов и др.
^Всасывание. Основным путем поступления свинца в ор-
ганизм человека и животных является пищеварительный тракт.
Всасывание соединений этого металла зависит в первую оче-
редь от их растворимости и составляет 5—15% от его содержа-
ния в рационе. Хорошо усваивается свинец в виде ацетата, хло-
рида, окиси и тетраэтила. Менее растворимы хромат, сульфид,
сульфат и карбонат свинца. Часть свинца, поступающего с пи-
щей, превращается в хлорид и комплексы с желчными кислота-
ми, которые всасываются как таковые. Желчь стимулирует
транспорт свинца через эпителий слизистой оболочки [Con-
rad М. Е., Barton J. С., 1978]. Аскорбиновая кислота и цистеин
повышают растворимость и всасывание этого МЭ. Усвоение
свинца усиливается при полном или частичном голодании [Ra-
binowitz М. В. et al., 1980]. В опытах с радиоактивным свинцом
(210РЬ), проведенных на добровольцах, коэффициенты всасыва-
ния его из пищеварительного тракта составили 0,36±0,065,
а у крыс — 0,23±0,02 [Ермолаева-Маковская А. П., Лит-
вер Б. Я., 1978]. Кальций, железо, магний, фосфаты, этанол и
жиры уменьшают всасывание свинца [Ragan Н., 1977]. Это сни-
жение объясняется главным образом конкуренцией металлов
на участках связывания и переносчиках в кишечном эпителии.
У детей и молодых животных всасывание свинца из желудочно-
кишечного тракта повышено [Kostial К. et al., 1971], чем отча-
сти объясняется их большая чувствительность к токсическому
действию свинца.?/Чеханизм всасывания свинца изучен недоста-
точно. Ряд экспериментальных данных свидетельствует о суще-
ствовании активного транспортного механизма для этого эле-
мента, а также в наличии акцептора, объясняющего обратную
корреляцию между величиной дозы и степенью усвоения свин-
ца. При высоких дозах этого МЭ могут преобладать процессы
пассивной диффузии.
[) Свинец может поступать в организм и через легкие. Это от-
носится в первую очередь к мелким аэрозольным частицам раз-
мером <0,5 мкм. Более крупные частицы удаляются реснитча-
тым эпителием респираторного тракта. Некоторая их часть
может поглощаться легочными макрофагами.
^Выделение. Свинец выделяется с калом, мочой, потом,
молоком и слюной. Основное его количество (в среднем 0,2 мг
в сутки) выводится с калом, который содержит весь невсосав-
шийся свинец. Через почки удаляется в среднем 30 мкг в сутки
386
цутем клубочковой фильтрации. При высоких концентрациях
свинца в крови имеет место его канальцевая экскреция [Vos-
tal J. J-> Heller J., 1968]. С потом выделяется довольно значи-
тельное количество свинца, достигающее в среднем 60 мкг в
сутки.
Транспорт и распределение в организме. Содер-
жание свинца в организме взрослого человека в норме близко
к 130 мг. Кинетические исследования удаления радиоактивного
свинца из органов и тканей свидетельствуют о существовании
в организме трех основных метаболических компартментов это-
го элемента. Самый короткий период полувыведения свинца
установлен для крови. Мягкие ткани, включая скелетные мыш-
цы, представляют собой компартмент (пул) со средней продол-
жительностью полувыведения свинца, равной нескольким неде-
лям, а скелет — пул с очень продолжительным периодом полу-
выведения, продолжающимся месяцы и годы [Castellino N,
Aloj S., 1964]. Различают мобильную, или способную к диффу-
зии, и фиксированную, или не способную к диффузии, формы
свинца в организме. Часть свинца, присутствующего в крови и
внутри клеток, способна к мобилизации и транспорту через
клеточные мембраны [Goyer R. A., Mahaffey К-, 1972]. Свинец
крови в значительной степени обменивается с тканевым свин-
цом. В норме наиболее высокие концентрации свинца наблю-
даются в костной ткани, почках и печени [Kehoe R. Е., 1964].
Аналогичным образом происходит и распределение в организме
210РЬ. Через 1 ч после внутривенного введения крысам радио-
активного свинца в почках обнаруживаются 8,29%, печени —
2,2%, крови—1,45% и диафизе бедренной кости— 1,35% вве-
денной дозы, а через сутки 6,34; 0,71; 0,4 и 3,22% соответствен-
но. Наименьшие концентрации найдены в головном мозге
(0,02%) и слизистой оболочке тонкой кишки [Москалев Ю. И.,
1985].
Более 90% свинца, присутствующего в крови, связано с эри-
троцитами [Castellino N., Aloj S., 1964] и отчасти представляет
собой транспортную форму этого элемента, так как удаляется
из них со скоростью, не соответствущей продолжительности
жизни эритроцитов [Conrad М. Е., Barton J. С., 1978]. Свинец
находится преимущественно в содержимом эритроцитов и в
меньшей мере в их строме. Основным эритроцитарным белком,
связывающим свинец, является гемоглобин, однако в них най-
ден и белок меньшей молекулярной массы (10 000), содержа-
щий этот элемент. Свинец, присутствующий в плазме, комплек-
сируется преимущественно с трансферрином в тех же участках,
которые связывают железо. Насыщение этого белка железом
снижает поступление свинца в плазму крови и повышает его
доставку в печень. Колебания уровня свинца в цельной крови
мало влияют на его концентрацию в плазме [Rosen J. F. et al„
1974]. Содержание свинца в крови отражает нагрузку им орга-
низма.
Концентрация свинца в цельной крови человека в норме
25*
387
колеблется в пределах 1,45—1,93 мкмоль/л. Концентрации
2,9—3,86 мкмоль/л отражают нагрузки этим элементом, способ-
ные вызвать определенные биохимические сдвиги без проявле-
ния, однако, клинических симптомов свинцового токсикоза.
Критической концентрацией свинца для взрослых считается
3,86 мкмоль/л [Goyer R. A., Rhyme В. С., 1973]. Для детей этот
порог существенно ниже.
Этот МЭ обнаружен во всех субклеточных фракциях. Боль-
шая часть поступившей дозы свинца обнаруживается в ядре и
цитозоле, причем в первом он локализуется преимущественно
в ядерной мембране и в меньшей степени в хроматине, главным
образом во фракции гистонов [Sabbioni Е„ Marafante Е., 1976].
Повреждение органелл, вызванное свинцом, привлекает вни-
мание ученых. В митохондриях свинец связывается с внут-
ренней мембраной и частично с матриксом, обнаруживая
кинетику насыщения. Высокие концентрации этого металла
вызывают функциональные и морфологические изменения этих
органелл, угнетают дыхание и фосфорилирование, процессы
активного транспорта, чем, видимо, объясняется ряд патологи-
ческих эффектов этого элемента. В эндоплазматической сети
свинец входит в состав отдельных компонентов мембран и ри-
босом. В цитозоле он связан, по-видимому, с высокомолекуляр-
ными белками.
При токсических дозах свинец в печени, почках и селезенке
частично связывается с ферритином и нерастворимым гидрок-
сидом железа. Происходит ли аналогичное связывание в норме,
неизвестно [Russel Н. А., 1970]. В мозге свинец концентрируется
в сером веществе и базальных ганглиях [Klein М. et al., 1970].
Внутри нервных клеток этот МЭ локализуется преимуществен-
но в митохондриях, занимая, по-видимому, участки связывания
кальция. В норме у человека более 20% свинца находится в
скелете. Из них около 70% приходится на трубчатые кости.
В их компактной части концентрация свинца выше, чем в губ-
чатой, т. е. кроветворной, части [Barry Р. S., 1978]. Свинец от-
кладывается в кости сперва в легкорастворимой форме, затем
превращается в трифосфат. Обмен этого элемента во многом
аналогичен обмену кальция. В ситуациях, ведущих к деминера-
лизации кости, происходит мобилизация свинца из его костных
депо, способная привести к свинцовому токсикозу [Six К. Н.,
Goyer R. А., 1970]. Концентрация этого МЭ в зубах выше, чем
в костной ткани, причем находится он в них в прочно связанной
форме.
Соединение свинца с белками происходит в первую очередь
за счет свободных SH-групп. По этой причине свинец прочно
связывается металлотионеином, содержащим много цистеина
[Ulmer D. D., Vallee В. L., 1969]. Однако остается неясным,
участвует ли этот белок в детоксикации свинца и способен ли
свинец индуцировать синтез металлотионеина, как это установ-
лено для цинка, меди и кадмия. Свинец вступает в реакцию не
только с SH-группами белка, но способен к образованию менее
388
стабильных комплексов с другими боковыми группами амино-
кислот, например с е-аминогруппой лизина, карбоксильной груп-
пой глутаминовой и аспарагиновой кислот, феноксигруппой ти-
розина и имидазолом гистидина [Wong Р. R. et al., 1979].
На обмен свинца оказывает влияние множество факторов и
прежде всего элементы, близкие к нему по своим физико-хими-
ческим свойствам. К их числу относятся в первую очередь каль-
ций и железо и в меньшей степени цинк, медь, магний и кадмий.
Всасывание свинца снижается при высоком содержании каль-
ция в пище и наоборот [Shields J. В., Morrison J. N., 1975], что
объясняется взаимодействием между этими элементами на уров-
не всасывания в пищеварительном тракте [Quarterman J., 1986].
Высокое содержание свинца в пище снижает концентрацию
кальция в плазме крови после ее кратковременного подъема
[Six К- Н., Goyer R. А., 1970]. Свинец конкурирует с кальцием
за участки связывания на эритроцитах и инсулине [Blun-
dell В. Т., Jenkins J. А., 1977]. На культуре гепатоцитов показа-
но, что ацетат свинца способен нарушить функцию кальция как
вторичного посредника [Pounds G. G. et al., 1982]. Усвоение
свинца повышается при низком содержании фосфора в пище
[Quarterman J., 1986].
Витамин D, индуцируя синтез кальцийсвязывающих белков,
стимулирует также всасывание свинца [Smith С. F. et al., 1978].
Свинцовый токсикоз ведет к угнетению синтеза метаболитов
витамина D (например, 1,25-дигидроксихолекальциферола) и
индукции кальцийсвязывающих белков в слизистой оболочке
[Smith С. F., 1981]. Хорошо известен также физиологический
антагонизм между свинцом и железом. При дефиците последне-
го у детей существенно возрастает риск отравления свинцом.
Повышенное содержание железа в пищеварительном тракте
ограничивает всасывание свинца и связи с конкуренцией за
общие акцепторные участки на слизистой оболочке [Barton J. С.
et al., 1978]. Троекратное увеличение содержания железа в пи-
ще вызывает существенное снижение количества свинца в поч-
ках, бедренной кости и крови [Mahaffey К. R. et al., 1978].
Другим физиологическим антагонистом свинца является
цинк, который ослабляет токсическое действие свинца и снижа-
ет содержание его в тканях животных [Cerklewsky F. L., For-
bes R. М., 1976], а также снимает ингибирование свинцом ак-
тивности порфобилиногенсинтазы [Meredith Р. A. et al., 1974].
Цинк видоизменяет, кроме того, характер распределения свин-
ца между органами и тканями, снижая его содержание в скеле-
те и повышая в почках и печени. Уменьшение токсического
действия свинца цинком объясняется, по-видимому, его способ-
ностью индуцировать синтез металлотионеина, который связы-
вает избыток свинца, чем способствует его детоксикации
[Bremner I., 1974]. Повышение содержания цинка в рационе
крыс (1,84 ммоль/кг) ведет к снижению содержания свинца в
крови и печени плодов и их матерей, получивших высокие дозы
свинца [Quarterman J., 1986].
389
Ь| Недостаточность свинца. Данные о жизненной не-
обходимости свинца весьма ограничены. Сообщалось о стиму-
лирующем действии низких концентраций этого элемента на
гемопоэз [Pecora L. et al., 1968] и рост микроорганизма Sarci-
na flava [Devigne J. Р., 1968]. Задержку роста животных при
дефиците свинца в рационе впервые наблюдал К- Schwarz
(1974) в опытах на крысах с использованием пластикового
«изолятора», в котором контролировался не только состав ра-
циона и воды, но и вдыхаемого воздуха. С помощью этой тех-
ники удалось установить жизненную необходимость еще пяти
«новых» микроэлементов (ванадия, кадмия, кремния, олова и
фтора). Следует сказать, что ростовой эффект свинца, достиг-
нутый в этих опытах, составил всего 10—12% и был получен
при относительно высоком содержании в рационе этого элемен-
та (0,5 мг/кг).
Убедительное подтверждение жизненной необходимости свин-
ца было получено А. М. Raichlmayer-Lais и М. Kirchgessner
(1984) в опытах с использованием рациона, содержащего в
10 раз меньшее количество свинца, чем в опытах К- Schwarz
(0,045 мг/кг). Авторы проследили влияние дефицита этого эле-
мента на трех последовательных поколениях крыс. В первом
поколении, полученном от матерей, содержавшихся на дефицит-
ной по свинцу пище, были отмечены снижение на 10—15% ге-
матокритного числа и концентрации гемоглобина. При дальней-
шем снижении концентрации свинца в рационе (0,02 мг/кг) эти
изменения достигли соответственно 22, 23 и 28% по сравнению
с контролем при снижении массы тела на 20%.
Интересным результатом этих исследований явилось уста-
новление неожиданного физиологического синергизма между
железом и свинцом при низких концентрациях последнего.
У всех животных при дефиците свинца оказались существенно
пониженными уровень железа в крови и степень насыщения им
трансферрина. Снижение содержания железа было отмечено
также в печени, селезенке и тушке подопытных крыс наряду с
угнетением некоторых железосодержащих ферментов и возник-
новением ряда признаков недостаточности этого элемента, ко-
торые исчезли при включении свинца в рацион животных.
Токсическое действие. При свинцовом токсикозе по-
ражаются в первую очередь органы кроветворения (анемия),
нервная система (энцефалопатия и нейропатия) и почки (не-
фропатия). Наиболее восприимчива к свинцу гематопоэтическая
система, особенно у детей. Анемия, вызванная свинцом, явля-
ется микроцитарной и морфологически не отличима от железо-
дефицитной анемии. Ее возникновение обусловлено тремя при-
чинами: сокращением продолжительности жизни эритроцитов,
подавлением синтеза глобина (особенно a-цепи) и гема. Наи-
более чувствительна к свинцу порфобилиногенсинтаза, умень-
шение активности которой ведет к снижению синтеза порфоби-
линогена и протопорфирина. Это угнетение наблюдается, по
некоторым данным, при концентрациях свинца в крови менее
390
0 483 мкмоль/л [Waldron Н. А., 1980] задолго до возникновения
клинических проявлений свинцового токсикоза, причем начиная
с концентрации 1,69 мкмоль/л и устанавливается линейная кор-
реляционная зависимость между содержанием свинца в крови
и снижением активности фермента. Она является наиболее
надежным критерием при оценке нагрузки организма свинцом
[Chisholm J. J. et al., 1985].
Другим ферментом, реагирующим на свинцовую интоксика-
цию, считается 6-аминолевулннат-синтаза (К,Ф 2.3.1.37), актив-
ность которой существенно возрастает. Этот фермент лимити-
рует скорость биосинтеза гема и его регуляция осуществляется
по принципу отрицательной обратной связи конечным продук-
том, которым является сам гем. Снижение его концентрации
влечет за собой повышение активности фермента.
Токсические дозы свинца угнетают также активность фер-
мента феррохелатазы, катализирующей включение железа в
молекулу протопорфирина [Otrzonsek N., 1967]. В результате
в митохондриях ретикулоцитов костного мозга происходит на-
копление железа в форме ферритина и железосодержащих ми-
целл [Bessis М. С., Jenson М. М., 1965]. Анемия при свинцовом
токсикозе постоянно сопровождается увеличением числа сиде-
робластов в костном мозге. Образуется также повышенное ко-
личество так называемого свободного эритроцитарного прото-
порфирина, представляющего собой фактически цинкпорфирин.
Последний вступает в соединение с глобином вместо гема [La-
mola A. A. et al., 1975] и может быть обнаружен по интенсивной
флюоресценции, служащей чувствительным тестом на свинцовое
отравление.
Нарушение путей биосинтеза гема токсическими дозами
свинца приводит к накоплению в крови и костном мозге проме-
жуточных продуктов его метаболизма и в первую очередь
6-аминолевулиновой кислоты и копропорфирина и к их выде-
лению с мочой [Nakao К. et al., 1968] даже в отсутствие явных
проявлений токсикоза. В этой связи определение данных соеди-
нений в моче может быть использовано как чувствительный
диагностический тест при отравлении свинцом.
При свинцовом токсикозе наблюдается также угнетение ряда
гемсодержащих ферментов, например цитохрома Р-450. Дейст-
вие свинца на процессы биосинтеза гема удобно представить в
виде схемы (схема 5). Накапливаясь в эритроцитарной мембра-
не, свинец увеличивает ее хрупкость и осмотическую резистент-
ность, снижает активность Na+, К+-АТФазы и вызывает переход
калия из эритроцитов в плазму, что в конечном счете приводит
к их лизису [Raghavan S. R. et al., 1981]. Ранним признаком
свинцового отравления считается появление в цитоплазме по-
лихроматофильных эритробластов и ретикулоцитов базофиль-
ной зернистости, представляющей собой агрегаты целых и час-
тично разрушенных рибосом, возникающие в результате инги-
бирования фермента пиридин-5'-нуклеотид — нуклеозидазы,
подобно тому, что наблюдается при генетическом дефекте этого
391
С х е м а 5
Действие свинца на ферменты, участвующие в синтезе гема
[Chisholm J. J.,O'Hara О,М.,1982]
фермента. Отмечается также повреждение плазматической мем-
браны и увеличение комплекса Гольджи.
Биохимические механизмы нейротоксичности свинца изуче-
ны в значительно меньшей степени, чем в случае нарушений
кроветворения. Имеющиеся данные свидетельствуют, что свинец
видоизменяет скорость проведения нервного импульса через
синапсы, функционирующие с помощью химических нейроме-
диаторов— ацетилхолина, катехоламинов, ГАМК. Он замедляет
холинергическую нейротрансмиссию в результате изменения
скорости поступления предшественников медиатора через пре-
синаптическую мембрану или его синтеза и выделения. Катехол-
аминергическая трансмиссия, напротив, ускоряется, причем
ускорение сопровождается усилением обмена норадреналина,
снижением концентрации этого гормона и дофамина, повыше-
нием активности тирозин-3-монооксигеназы, задержкой обрат-
ного поступления в синаптосому дофамина и повышенным ко-
личеством метаболитов катехоламинов. В опытах in vitro пока-
зано, что свинец способствует высвобождению дофамина из
синаптосом в состоянии покоя и ингибирует как его обратное
поступление, так и активность чувствительной к дофамину аде-
нилатциклазы. Известно, что свинец in vivo вызывает задержку
поступления ГАМК в синаптосомы, а также ее выделения из
них и повышенную сенсибилизацию к этому медиатору.
Патохимические сдвиги при свинцовом токсикозе в целом
ряде случаев объясняются взаимодействием этого элемента с
другими металлами и в первую очередь с кальцием, магнием и
натрием. Кальций стимулирует депонирование и высвобождение
медиаторов под воздействием нервного импульса, регулирует
активность ряда ферментов, участвующих в их синтезе, а также
циклаз, опосредующих действие гормонов. Повышенная кон-
центрация свинца, нарушая трансмембранный перенос кальция,
может помешать выполнению этим элементом его физиологиче-
ских функций. Взаимодействием свинца с натрием можно объ-
392
яснить накопление кальция в синаптосомах ЦНС и повышенное
выделение из них дофамина [Silbergeld Е. A., Adler Н. S., 1978],
а взаимодействием с магнием — влияние свинца на процессы
фосфорилирования. Свинец может также непосредственно ин-
гибировать активность ферментов биосинтеза миелина.
В условиях производства свинец попадает в организм через
органы дыхания в виде его паров, пыли и аэрозоля. Поступле-
ние этого элемента через рот возможно только при грубых на-
рушениях санитарно-гигиенических требований или через за-
грязненную свинцом воду и пищу. В настоящее время острые
отравления свинцом не регистрируются, а хроническая свинцо-
вая интоксикация встречается реже и в менее тяжелой форме.
Есть основания говорить о патоморфозе этого МТОЗа, вызван-
ном успехами гигиены и профессиональной патологии. Совре-
менный сатурнизм характеризуется главным образом наруше-
ниями функции нервной системы, кроветворения и пищевари-
тельного тракта. Клиническая симптоматика сатурнизма
классифицирована Институтом гигиены труда и профзаболева-
ний АМН СССР и подробно изложена в соответствующих руко-
водствах и в специальном издании ВОЗ «Свинец» (Женева,
1980).
8.4. Ртуть
Ртуть с древнейших времен используется в хозяйственной
деятельности человека, а также в медицинских целях. Мерку-
риализм был одним из первых профессиональных заболеваний
среди ювелиров, зеркальщиков, скорняков, а также врачей, ши-
роко применявших ртутную мазь при лечении сифилиса. Таким
образом, отравление ртутью является серьезным профессиональ-
ным и ятрогенным заболеванием с античных времен и до на-
стоящего дня. В данное время оно приобрело иную, более опас-
ную форму в результате антропогенного загрязнения окружаю-
щей среды в глобальном масштабе. Крайним выражением
современного хронического ртутного отравления, связанного с
экологической проблемой, является болезнь Минамата. Впервые
она описана в 50-х годах в Японии, встречаясь в основном у
рыбаков и членов их семей, живших вблизи залива Минамата,
в который сбрасывались отходы промышленных предприятий,
содержащие высокие концентрации ртути. Последняя ассимили-
ровалась планктоном-> мелкими рыбами-> крупными хищны-
ми рыбами. Эпидемия отравлений алкилртутью в Японии де-
тально рассматривалась в отчете группы экспертов по изучению
болезни Минаматы [Katsuna М., 1968].
В окружающую среду ртуть попадает как в результате ес-
тественного испарения ее из земной коры (среднее содержание
составляет 50 мкг/кг) от 25 до 12 000 т ежегодно, так и в ре-
зультате промышленного загрязнения. Известно, что в 1973 г.
мировая добыча ртути достигала 10 000 т и ежегодно увеличи-
валась примерно на 2%. Металлическая ртуть и ее соединения
393
используются в производстве хлора (25%), красителей (15%),
в электротехнической промышленности (20%), а также в сель-
ском хозяйстве (15%), оборонной промышленности (20%) и в
медицине (10%), в том числе в стоматологии (3%).
Ртуть широко распространена в земной коре.
Она встречается в виде киновари (сульфида ртути), в которой
среднее содержание ртути составляет 0,1—0,4%, иногда до 20%.
Основные ее месторождения находятся в Испании, Югославии,
Италии, США (Калифорния), Мексике и Перу. В нефтяных
месторождениях Калифорнии содержание ртути в незагрязнен-
ной пресной воде ниже 200 нг/л, а питьевая вода в ФРГ содер-
жит менее 30 нг/л [Bouquiaux J., 1974]. Максимальное содер-
жание ртути отмечено в донных отложениях залива Минамата
в Японии, где оно составило 20 000 мкг/кг. При этом показано,
что средний уровень ртути в рыбе, выловленной в этом заливе,
равнялся 11 мг/кг, а иногда достигал 36 мг/кг, в то время как
этот показатель в аналогичной морской рыбе, пойманной в не-
загрязненных местах Японии, — менее 1 мг/кг [Takeuchi Т.,
1972]. В рыбе, пойманной в водах Северного моря, удаленных
от материка, общие уровни ртути находились в пределах 0,05—
0,11 мг/кг [Kazantzis G., 1978].
Наибольшее практическое значение в экологической патоло-
гии представляет не естественное поступление ртути в окружа-
ющую среду, а локальное загрязнение, связанное с производст-
венной деятельностью человека. Процесс метилирования неор-
ганической ртути в донных отложениях озер, рек и других во-
доемов является ключевым звеном в пищевой цепи водных
экосистем, обусловливающих ее поступление в организм чело-
века. Другим важным источником поступления алкилированной
ртути являются фунгициды, применяющиеся для протравлива-
ния семян.
Средние концентрации ртути в атмосфере составляют
20 нг/м3. При содержании в воздухе 50 нг/м3 человек в сутки
получает около 1 мкг ртути. Однако в условиях профессиональ-
ной вредности оно может достигать 480 мкг/сут. С питьевой
водой в организм человека поступает менее 0,4 мкг ртути. Ос-
новная масса населения потребляет с пищей от 1 до 20 мкг
ртути в сутки, при этом на другие продукты, кроме рыбы, при-
ходится около 5 мкг/сут.
Около 80% неорганической ртути, поступающей с воздухом,
задерживается в организме [Nielsen-Kudsk F. N., 1965], а из
желудочно-кишечного тракта — всего 7%. Необходимо, однако,
подчеркнуть, что метилртуть практически вся всасывается в
пищеварительном тракте. Она также хорошо всасывается и че-
рез кожу. В результате трансплацентарного переноса ее содер-
жание в эритроцитах плода на 30% выше, чем матери. Период
полувыведения ртути из крови составляет около 70 дней.
Известно два биохимических пути метилирования ртути —
анаэробный и аэробный. Первый, протекает без участия фер-
ментов и обусловлен метилкобаламином [Wood К. et al., 1968].
394
Аэробный путь описан L. Landner (19П). Метилртуть обладает
высокой способностью к кумуляции в биологических субстра-
тах благодаря высокому сродству к SH-группам. Поступившая
в клетку метилртуть связывается с белком и не подвергается
дальнейшей диффузии, однако незначительная ее часть оста-
ется в диффузионно-способной форме.
Пары металлической ртути, а также метилртуть растворя-
ются в липидах и поэтому легко проникают через альвеолярную
мембрану. В эксперименте J. К- Miettinen (1973) на доброволь-
цах показано, что 95% радиоактивной метилртути, скормленной
рыбе, абсорбируются в желудочно-кишечном тракте.
Метилртуть выделяется с молоком у лактирующих женщин,
при этом новорожденный, уже имеющий повышенное содержа-
ние ртути в организме, продолжает получать ртуть с молоком
матери. У некоторых младенцев уровни ртути в крови превы-
шали 0,5 мкмоль/л [Amin-Zaki L. et al., 1974].
Независимо от пути поступления ртути и ее формы этот эле-
мент накапливается в почках (до 90% общего ее содержания
в организме) [Hayes A. L. et al., 1962]. В основном ртуть куму-
лируется в эпителии почечных канальцев. Наиболее высокие ее
концентрации в моче (10,8 мкмоль/л) отмечены у горнорабо-
чих, занятых на добыче ртутных руд. Не выявлено корреляции
между общим содержанием ртути в моче и ее концентрацией
в крови [Bakir F. et al., 1973]. В условиях профессиональной
вредности высокое содержание ртути отмечено также в вещест-
ве головного мозга, печени, щитовидной железе и гипофизе
[Kosta L. et al., 1975]. Приблизительно 90% общего количества
абсорбированной метилртути выделяется из организма с фека-
лиями (желчью) и меньше с мочой [Miettinen J. К-, 1973].
организма человека ртуть выделяется с мочой, калом,
'бьгдыхаемым воздухом, потом, молоком (в период лактации),
волосами. Она теряется также в результате попадания в пла-
центу и через последнюю — в плод.
Ртутная интоксикация обусловлена высоким сродством это-
го элемента к SH-группам, в связи с чем снижается активность
многих ферментов, а также уменьшается синтез белка. Кроме
того, наблюдается нарушение функции плазмалеммы клеток
(ироницаемость и мембранный транспорт). Высказывается
предположение, что токсичность ртути обусловлена повышенной
проницаемостью мембран лизосом и вследствие этого выходом
в цитозоль гидролитических ферментов [Fowler В. A. et al.,
1974]. Отмечен повышенный синтез металлотионеиноподобного
белка в почках, но не в печени, как это бывает при воздействии
кадмием [Piotrowski J. et al., 1973]. Повышенный синтез этого
белка является компенсаторно-приспособительной реакцией,
направленной на связывание ртути и защиту организма от ин-
токсикации.
Ингаляционное поступление паров ртути (концентрации вы-
ше 0,1 мг/м3) сопровождается симптомами острого, нередко
язвенного, бронхита, бронхиолита и пневмонии. Хроническое
395
отравление характеризуется наличием астеновегетативного
синдрома с отчетливым тремором, лабильным пульсом, тахи-
кардией, возбуждением, психическими нарушениями, гингиви-
том, а также изменениями крови и экскрецией ртути с мочой
выше нормы [Трахтенберг И. М., 1981; Smith R. G. et al., 1970].
При пероральном поступлении ртути (например, сулемы)
наблюдается язвенно-некротический гастроэнтерит. В дальней-
шем развивается некротический нефроз с гибелью эпителия
проксимальных отделов почечных канальцев. Если больной не
погибает в фазу анурии, то на 2-й неделе от начала отравления
происходит регенерация эпителия почечных канальцев.
Клинические признаки отравления органическими соедине-
ниями ртути (метилртуть) были подробно освещены после эпи-
демий в Японии (Минамата) и в Ираке. В клинической картине
доминировали поражение ЦНС, мозжечковая атаксия и нару-
шение зрения. Наиболее ранними и распространенными симпто-
мами были парестезии (в 38% случаев) с покалыванием и оне-
мением пальцев рук и стоп, а также вокруг рта и в передней
части грудной клетки. Изменения походки отмечены у 31% из
415 обследованных [Al-Mufti A. W. et al., 1976]. Кроме того,
имели место постоянные боли в конечностях, нарушение зрения,
затрудненная речь, сыпь и снижение слуха.
Клиническая картина отравления метилртутью в Ираке в
целом соответствовала таковой в Японии, за одним важным
исключением, связанным с прогрессирующим течением заболе-
вания в Японии, и, напротив, с более быстрым улучшением
общего состояния отравившихся в Ираке. Психические симпто-
мы нивелировались значительно, потеря зрения восстанавлива-
лась умеренно, атаксия, расстросйтво артикуляции и парестезии
восстанавливались в меньшей степени [Damluji S. et al., 1976].
Различие в прогрессировании клинической картины можно в
определенной степени связать с разными условиями абсорбции
метилртути во время эпидемии болезни в Минамата и вспышек
отравлений в Ираке. Так, если в Японии наблюдалось длитель-
ное, многолетнее поступление в организм малых доз ртути, то
в Ираке оно было связано с более высокими дозами метилртути
из протравленного зерна, но продолжалось максимально в те-
чение 3 мес.
Ранее было отмечено, что органические соединения ртути
легко проходят плацентарный барьер и накапливаются в тканях
плода. В связи с этим особый интерес представляют обследова-
ния Y. Harada (1968) 22 новорожденных в Минамата, у кото-
рых выявлены клинические признаки церебрального паралича.
Необходимо при этом подчеркнуть, что только у небольшой ча-
сти их матерей имелся единственный признак ртутного отрав-
ления — парестезии. Кроме основной патологии, у 1 новорож-
денного отмечено изменение сетчатки и еще у 1 — неправильно
сформированное наружное ухо. Позднее в Ираке L. Amin-Zaki
и соавт. (1974) провели обследование 15 матерей и их новорож-
денных детей. При этом было установлено, что у 5 матерей
396
имели место парестезии и у 4 — сужение поля зрения. У 4 мла-
денцев были тяжелые поражения ЦНС с генерализованными
судорогами, повышенными рефлексами, слепотой и глухотой,
у 1 ребенка из 5 отмечены ослабление хватательного рефлекса
и неспособность держать головку.
Т. Takeuchi (1972) при патоморфологическом исследовании
умерших новорожденных, матери которых потребляли пищу,
содержащую метилртуть, отметил микроцефалию с уменьшени-
ем массы головного мозга на ]/г или 2/з- В этих случаях пора-
жение нейронов коры большого мозга было тотальное, а не ло-
кальное, как у взрослых при болезни Минамата. Оно характе-
ризовалось неправильностями цитоархитектоники, гипоплазией
коры мозжечка и мозолистого тела.
Патологические изменения у взрослых, возникающие в ре-
зультате интоксикации метил- и этилртутными соединениями,
свидетельствуют об их выраженной нейротоксичности и схо-
жести.
В случаях хронического отравления метилртутью отмечена
атрофия извилин головного мозга, наиболее выраженная в за-
тылочных долях, а также в мозжечке. Боковые желудочки мозга
расширены, особенно их задние рога. Патогистологические из-
менения характеризуются деструкцией нейронов коры, особенно
в зрительных центрах затылочных долей. При этом в тяжелых
случаях отмечается полное опустошение нейронов коры с оча-
гами спонгиоза. В менее тяжелых случаях дезинтеграция и вы-
падение нейронов иногда сопровождаются диффузной проли-
ферацией нейроглии. Характерным в гистологической картине
мозжечка является гибель нейронов зернистого слоя. Кроме
того, в тяжелых случаях наблюдается также гибель грушевид-
ных нейронов (Пуркинье), иногда с очаговой пролиферацией
нейроглии. Нередки выраженные гидропические изменения ней-
ронов и явления нейронофагии, встречаются мелкие очаги кро-
воизлияний и периваскулярный отек. В мозговом стволе, спин-
ном мозге и черепных нервах гистологических изменений не
выявлено [Hunter D. et al., 1954; Takeuchi T., 1968, 1972].
Клинические признаки поражения периферических нервов
нашли соответствующее морфологическое подтверждение на
секционном материале. Так, в сообщении К. Eto и Т. Takeuchi
(1971) говорится об изменении диаметра периферических (ик-
роножных) нервов у умерших с болезнью Минамата, гистоло-
гически обнаружено исчезновение осевых цилиндров с проли-
ферацией нейролеммоцитов (шванновских клеток) и разраста-
нием коллагеновых волокон.
Ятрогенные формы меркуриализма представлены в главе 9.
8.5. Бериллий
Этот МЭ широко используется в промышленности, приборо-
строении, авиационной и космической технике, производстве
люминесцентных ламп. Очень токсичен. Соли бериллия в кон-
897
центрации 1 мкмоль/л специфически ингибируют активность
щелочной фосфатазы; оказывают угнетающее действие на дру-
гие ферменты, в частности на аденозинтрифосфатазу. В резуль-
тате изменений в белковом обмене под влиянием бериллия раз-
вивается аутоиммунный процесс, который играет очень большую
роль в патогенезе вызванных данным МЭ поражений.
Установлено, что в естественных условиях бериллий в чело-
веческом организме содержится в очень малых количествах
(в среднем около 11 мкг, в легких —до 2,5 мкг). Концентрация
бериллия в суточной моче в среднем равняется 0,024 мкг, в ка-
ле— 0,078, а суточное выделение составляет 0,162 мкг. Безус-
ловно, что даже максимальные естественные концентрации явля-
ются безвредными для человека [Бурназян А. И., 1983]. Таким
образом, высокотоксичные свойства бериллия и его соединений
проявляются только при превышении определенного уровня.
Именно это лежит в основе принципов профилактики пораже-
ний бериллием.
Достоверных сведений о клинических проявлениях, связан-
ных с дефицитом бериллия в организме человека, в доступной
нам литературе обнаружить не удалось. В производственных
условиях бериллий проникает в организм в основном через ор-
ганы дыхания. Легкие, кости, лимфатические узлы, печень и
миокард могут быть органами, где депонируется этот МЭ. Осво-
бождение организма от бериллия происходит главным образом
через кишечник и отчасти через почки. В профессиональной
патологии принято различать острые отравления бериллием
(главным образом при вдыхании ВеС12, BeF2, ВеО4, Ве(О3)2
и др.), в результате чего развивается так называемая литейная
лихорадка, сопровождающаяся раздражением дыхательных
путей, конъюнктивитами. При остром бронхиолите может на-
ступить бронхоспазм. К хроническим формам относятся хрони-
ческий бронхит, бронхоэктазы, так называемые химические
пневмонии, пневмосклероз, а также бериллиоз — особая форма
профессиональной патологии, вызванная преимущественно ма-
лорастворимыми соединениями бериллия. Характерно появле-
ние клинических симптомов через длительный период после
прекращения контакта с этим МЭ. Клиника и рентгенологиче-
ская симптоматика бериллиоза изучены К- П. Молокановым и
соавт. (1972), патологическая анатомия — П. П. Движковым
и соавт. (1965).
М. С. Толгская и А. А. Чумаков (1976) следующим образом
описывают патологическую анатомию бериллиоза. Характерно
двустороннее диффузное поражение легких и плевры с «...рас-
пространенным гранулематозом и хроническим межуточным
пролиферативным процессом, локализующимся в перегородках
альвеол, под плеврой, перибронхиально, периваскулярно». Гра-
нулемы имеют различные размеры (от 0,2 до 1,5 см). Клеточ-
ное строение их зависит от стадии процесса. Более свежие гра-
нулемы состоят из гистиоцитов, эпителиоидных клеток, лимфо-
цитов, плазматических клеток, клеток Пирогова — Лангханса,
398
а также многоядерных элементов типа гигантских клеток ино-
родных тел, в цитоплазме которых содержатся кристаллические
Структуры. В старых гранулемах преобладает процесс их кол-
дагенизации, который может заканчиваться гиалинозом. Харак-
терным признаком бериллиоза являются так называемые кон-
хоидальные тельца, окрашивающиеся гематоксилином, иногда
дающие положительную реакцию на железо. Диффузный про-
лиферативный процесс в межальвеолярных перегородках также
приводит к пневмосклерозу. В процессе хронического бронхита
возможно формирование бронхоэктазов, появление эмфизема-
тозных и ателектатических очагов, тромбозов, флебитов, эндар-
териитов. Эти тяжелые изменения в легочной ткани приводят к
образованию эмфизематозных булл, разрыв которых вызывает
спонтанный пневмоторакс (в 5—15% всех наблюдений), и к
раннему формированию легочного сердца. Поскольку бериллий
и продукты его метаболизма широко распространяются за пре-
делы легочной ткани, характерные для бериллиоза гранулемы
могут быть обнаружены и в печени, почках, миокарде, лимфа-
тических узлах, скелетных мышцах и костном мозге. Указанные
черты строения бериллиозных гранулем позволяют дифферен-
цировать их от туберкулов, от гранулем при саркоидозе и дру-
гих заболеваниях, что не всегда представляет собой простую
задачу [Калитиевский П. Ф., 1987].
Конхоидальные тельца или так называемые тельца Шауман-
на при хроническом бериллиозе обнаруживаются в 80% случа-
ев. Однако они изредка встречаются при туберкулезе и саркои-
дозе. Возникновение конхоидальных телец связано с гигантски-
ми клетками типа клеток инородных тел и клетками типа
Пирогова — Лангханса. После разрушения этих элементов кон-
хоидальные тельца находятся внеклеточно. Гранулемы при бе-
риллиозе могут сливаться и образовывать конгломераты
(рис. 70—72). В цитоплазме гигантских клеток обнаруживаются
холестериновые и игольчатые двоякопреломляющие кристаллы.
На гистологических срезах вокруг кристаллов имеются скопле-
ния солей кальция и отложения железа. Эти кристаллы в гигант-
ских клетках неспецифичны, так как встречаются и при других
патологических процессах, в частности при саркоидозе.
По литературным данным наиболее характерным патомор-
фологическим признаком бериллиоза, отличающим его от сар-
коидоза, является диффузный интерстициальный
процесс, сопровождающийся утолщением альвеолярных пе-
регородок за счет скопления в них лимфоидных и плазматиче-
ских клеток, ведущий к нарушению диффузионных явлений в
легких и развитию альвеолярно-капиллярного блока [Бурна-
зян А. И., 1983].
Кроме указанных изменений в легких, необходимо обратить
внимание на следующие патологоанатомические признаки: ле-
гочное сердце, тромбоз расширенного ушка правого предсердия,
застойное полнокровие печени с формированием центролобуляр-
ных геморрагических некрозов, увеличение селезенки с гипер-
399
Рис. 70. Интерстициальная пневмония с резко выраженной лимфоцитарной
инфильтрацией межальвеолярных перегородок. Рассеянные бериллиозные гра-
нулемы в стадии склерозирования. В отдельных гранулемах — конхоидальные
тельца. Окраска гематоксилином и эозином. Х250.
Рис. 71. Гигантская клетка инородных тел при бериллиозе. Окраска гематокси-
лином и эозином. Х400.
Рис. 72. Субтотальный лизис и зернистый распад эластических волокон в лег-
ких при бериллиозе. Окраска орсеином. Х400.
плазией ее пульпы, утолщение надкостницы ребер и длинных
трубчатых костей. Существуют указания на возможность фор-
мирования бериллиозных гранулем не только в упомянутых
органах, но и в головном мозге. Кроме того, их часто можно
обнаружить в коже и подкожной клетчатке.
Бериллиоз кожи представляет собой особую форму профес-
сиональной патологии, обусловленной контактом человека с бе-
риллием. По данным литературы, на ее долю приходится от
24,7 до 84,4%. Клинически эти заболевания неоднородны,
к ним относятся контактные дерматиты, изъязвления кожи,
конъюнктивиты, аллергические дерматиты (включая экзему),
бериллиевая гранулема кожи. По заключениям специалистов
диагностика кожной бериллиевой гранулемы довольно трудна,
так как она имеет сходство с саркоидозом и туберкулезом ко-
жи, с кольцевидной и ревматоидной гранулемами, с грануле-
мами инородных тел и, наконец, с лепрой. Несомненную помощь
в правильной диагностике бериллиоза могут оказать установ-
ление в анамнезе контакта с бериллием, наличие положитель-
ной кожной пробы с растворимыми соединениями этого МЭ,
а также обнаружение его в биопсийном материале.
Если профессиональные формы бериллиоза кожи известны
профпатологам и отчасти дерматологам, то этого нельзя сказать
о врачах общего профиля, а также о педиатрах. С нашей точки
зрения, население должно знать, что перегоревшие люминес-
центные лампы не должны попадать в руки детей. В литературе
существует лишь несколько описаний бериллиевой патологии у
26—568
401
детей, поранившихся во время игры с люминесцентными лам-
пами. Нам думается, что этот вопрос пока не привлек доста-
точного внимания гигиенистов и педиатров, тем более что бе-
риллиевая патология проявляется не сразу, а через определен-
ный латентный период, в связи с чем об имевшем место в
анализе контакте ребенка с содержащим бериллий материалом
можно забыть. К счастью, бериллиевые поражения кожи быто-
вого характера, в том числе и кожные гранулемы, редки.
Клиника бериллиозных поражений очень многообразна. Ра-
бочая классификация интоксикаций бериллием в нашей стране
была разработана А. М. Рашевской и соавт. (1983). Эта клас-
сификация разработана с учетом этиологии и клинико-рентге-
нологических форм патологического процесса. Из нее видно, что
растворимые соединения бериллия вызывают острые и подост-
рые заболевания с различным исходом, зависящим от продол-
жительности и интенсивности воздействия. Малорастворимые и
нерастворимые соединения бериллия при их хроническом воз-
действии вызывают преимущественно гранулематозные и ин-
терстициальные изменения с быстрым или медленным прогрес-
сированием. Растворимые, малорастворимые и нерастворимые
соединения при их хроническом воздействии обусловливают
хронический токсический бронхит в сочетании с генерализован-
ным бериллиозом, а также дерматитом. Наконец, одновремен-
ное хроническое воздействие соединений бериллия с кварцем,
никелем, медью и др. вызывает хронический силикобериллиоз,
хронический бронхит, токсический пневмосклероз.
Действие бериллия на организм многообразно. В настоящее
время доказаны его токсический, в том числе и цитотоксиче-
ский, сенсибилизирующий, эмбриотоксический и кацерогенный
эффекты. Последний установлен в эксперименте на животных
определенных видов и обсуждается в отношении человека. Для
патогенеза бериллиоза у человека наибольшее значение имеет
его токсическое и мощное сенсибилизирующее действие, что
подвергалось многостороннему изучению. Бериллий и его сое-
динения обладают способностью проникать во все органы, клет-
ки и их ядра, в клеточные органеллы, в частности в митохонд-
рии. Этот МЭ повреждает клеточные мембраны, в том числе и
их липидные компоненты, нарушая микровязкость. Бериллий
тормозит активность АТФазы саркоплазматической сети путем
ингибирования транспорта магния и кальция.
Проникая в ядра клеток, бериллий снижает активность фер-
ментов синтеза ДНК, в частности ДНК-полимеразы [Gold-
blatt Р. J. et al., 1975; Luke М. Z. et al., 1975]. С. Ф. Харламова
и И. В. Павлова (1975) указывают на возможность молекуляр-
ных ошибок при полимеризации белков в процессе деления кле-
ток и репликации ДНК- О. Г. Алексеева (1979) обратила вни-
мание на значение нарушений синтеза ДНК для появления
аномальных белков, играющих роль аутоантигенов.
Цитотоксический эффект соединений бериллия изучали на
фагоцитах. В частности, по данным J. Vacher и соавт. (1974),
402
внутривенное введение сульфата и цитрата бериллия вызыва-
ет блокаду клеток СМФ и снижает индекс фагоцитоза на 65—
75%. Введение фосфата бериллия подавляет воспалительную
реакцию.
При внутритрахеальном введении соединений бериллия про-
исходит повышенный выход макрофагов и полинуклеаров в про-
свет альвеол. Однако подвижность макрофагов при этом сни-
жается, их органеллы повреждаются и снижается синтез ДНК
[Fritsch Р. et al., 1975].
Н. Н. Литвинов (1975) показал, что при ингаляции раство-
римых солей бериллия разрастается соединительная ткань пре-
имущественно в периваскулярных и перибронхиальных зонах.
По данным А. С. Ивановой (1971), фиброз развивается в ответ
на проникновение бериллия в легкие, причем этот процесс име-
ет максимальную скорость в течение 1-го месяца после внутри-
трахеального введения гидроокиси бериллия. Склероз легочной
ткани, как правило, сочетается с возникновением своеобразных
гранулем. Электронно-микроскопическое и гистохимическое ис-
следование последних показали сходство их с гранулемами
аллергического характера [Williams W. J., 1971]. Доказано, что
в лимфоцитах гранулем число органелл увеличено. Этот факт
и наличие большого числа свободных рибосом свидетельствуют
об их активном состоянии. Эпителиоидные клетки гранулем
возникают из мононуклеаров и лимфоцитов. Н. Н. Литвинов
(1975) показал, что уже в первые месяцы после ингаляции рас-
творимых соединений бериллия развиваются гранулемоподоб-
ные узелки, состоящие из лимфоидно-гистиоцитарных элемен-
тов. В центре таких узелков обнаруживают распадающиеся
макрофаги и клеточный детрит. Это трактуется как результат
выхода бериллия при гибели поглотивших его макрофагов.
Важно, что аллергическая реакция легких на бериллий воз-
никает и при энтеральном, и внутрикожном путях его введения
[Kang К. J- et al., 1977]. Комментируя эти данные, С. А. Кайзер
и соавт. (1983) считают, что, попадая в ткань легких и вступая
в контакт с легочными фагоцитами, бериллий обязательно вы-
зывает в них ответную реакцию. Этот факт важен, так как
в последние годы легкие рассматриваются в качестве своеобраз-
ного иммунокомпетентного органа. Патогенное действие берил-
лия и его соединений наиболее выражено в легких. Именно в
них он накапливается в количествах, которые значительно пре-
вышают его содержание в других органах. При ингаляции бе-
риллия в концентрациях, которые превышают ПДК в воздухе
в 2—40 раз, наступает его патогенное действие. Тонкие меха-
низмы влияния этого МЭ на клеточные элементы аэрогемати-
ческого барьера требуют дальнейшего изучения.
Исследования иммунологической реакции здоровых людей
на контакт с бериллием и его соединениями были впервые осу-
ществлены в СССР сотрудниками НИИ гигиены труда и проф-
заболеваний АМН СССР. Первичная иммунная реакция на
бериллий, направленная на адаптацию организма к воздейст-
26*
403
вию этого вещества, обнаружилась у 45—75% обследованных
уже в течение 1-го года профессионального контакта с этим МЭ.
Появлялись не только признаки сенсибилизации к бериллию,
но и чаще обнаруживались антитела против тканей тимуса и
надпочечников, увеличивалась концентрация IgM в сыворотке
и активировался антимикробный естественный иммунитет.
8.6. Барий
Этот МЭ в природе встречается только в виде соединений,
наиболее важные из них минералы барит (BaSOi) и витерит
(ВаСОз). Барий широко применяется в производстве стекла,
бумаги, резины, керамики, а также в металлургии, сельском хо-
зяйстве и медицине. В организм он поступает ингаляционно в
производственных условиях и через желудочно-кишечный тракт.
Выделение бария происходит в основном с калом и в меньшей
степени с мочой. Водорастворимые соединения этого МЭ доста-
точно хорошо абсорбируются организмом и определяют его ток-
сичность. Ткани, содержащие большие количества кальция,
обычно содержат больше бария, напротив, ткани, богатые маг-
нием, содержат мало кальция и бария.
Всосавшийся в желудочно-кишечном тракте и легких барий
откладывается в костях (65%) и оказывает выраженное дейст-
вие на гладкую мышечную ткань и миокард, по своему дейст-
вию напоминающий эффект ацетилхолина. Барий является
синергистом последнего. Своеобразным органом — накопителем
этого МЭ в организме является пигментная оболочка глаза,
однако физиологическое значение этого необычного факта
остается невыясненным [Войнар А. И., 1960].
Баритозы — МТОЗы, обусловленные интоксикацией ба-
рием и его солями.
Острый б ар ито з — синдром острого отравления водо-
растворимыми солями бария. При поступлении в организм через
органы дыхания и желудочно-кишечный тракт могут возникнуть
следующие симптомы: обильное слюноотделение, жжение во рту
и пищеводе, боли в животе, тошнота, рвота, понос, повышение
артериального давления, судороги, возможны параличи, резкий
цианоз лица и конечностей, обильный холодный пот, общая мы-
шечная слабость, расстройства походки и речи вследствие па-
ралича мышц глотки и языка, одышка, головокружение, нару-
шение зрения. Смерть может наступить в течение 1-х суток.
Несмотря на экскрецию основной массы солей бария, воз-
можно отложение его незначительных количеств в печени, мозге,
железах внутренней секреции. Откладывающийся в костях ба-
рий превращается в нерастворимые соединения сульфата бария.
При использовании в рентгенологической практике сульфата
бария возможны отравления за счет примеси его растворимых
солей.
Хронический баритоз характеризуется резкой слабо-
стью, одышкой, воспалением слизистой оболочки полости рта,
404
насморком, конъюнктивитом, диареей, кровоизлияниями в же-
лудке, повышением артериального давления, тахикардией, не-
правильным пульсом, нарушениями мочевыделения, а также
выпадением волос на голове и бровях.
Баритовый пневмокониоз осложняется воспалитель-
ными процессами в легких и бронхах. При отравлении карбо-
натом бария, кроме пневмокониоза, имеет место общетоксиче-
ское действие, проявляющееся нарушением кроветворения,
функций сердечно-сосудистой системы ц обменных процессов.
Растворимые сол-и бария вызывают симптомы менингоэнце-
фалита.
8.7. Висмут
Висмут содержится (2-10~5 мае. %) в земной коре в виде
соединений и в самородном состоянии. Наиболее часто встре-
чаются минералы — висмутовый блеск (BiaSs) и висмутовая
охра (В120з).
Висмут используется в металлургии, а также в медицине,
косметике, фотографии. Профессиональные висмутозы встреча-
ются редко.
Наибольшее практическое значение в медицине имеют пре-
параты висмута — дерматол, ксероформ, бийохинол, бисмоверол
и пентабисмол.
Всосавшийся в кровь висмут кумулируется в почках, печени,
селезенке и костях. Экскреция его из организма происходит
длительное время, в основном с мочой (до 75% введенной до-
зы), в меньшей степени через желудочно-кишечный тракт (до
25%), а также с потом. При длительном применении висмут-
содержащих лекарственных форм характерны висмутовый гин-
гивит и стоматит.
Следует подчеркнуть, что клинические признаки отравления
висмутом почти не имеют специфического характера, и диагноз
основывается главным образом на данных анамнеза и химико-
токсикологического анализа.
Ятрогенные формы висмутозов представлены в главе 9.
8.8. Таллий и другие
потенциально-токсичные микроэлементы
Таллий в природе встречается в полиметаллических ру-
дах, среднее его содержание в земной коре составляет
4,5-Ю-5% по массе. Известны два стабильных природных изо-
топа— 203Т1 (29,5%) и 205Т1 (70,5%). Таллий и его соединения
применяются в приборостроении, в производстве оптики, полу-
проводников, химической и других отраслях промышленности.
Он входит в состав некоторых сплавов с магнием., селеном и
цинком. Наиболее широко используются галлоидные соли тал-
лия, а также еульфаты, су.’”’ фиды, карбонаты, селениды и му-
405
равьино-малоновокислый таллий (жидкость Клеричи). В част-
ности, сульфат, карбонат и ацетат таллия входят в состав ядов
(родентицидов) для истребления грызунов. Ацетат таллия яв-
ляется компонентом мази для депилляции, которая широко
применяется в животноводстве и медицине. Соли таллия ис-
пользуются как антидетонаторы топлива в двигателях внутрен-
него сгорания, а также в качестве катализаторов в различных
химических реакциях и процессах [Эйтингон А. И., 1983].
Таллий часто обнаруживают в свинцовых и полиметалличе-
ских месторождениях Урала, Алтая, Казахстана, Таджикиста-
на, Забайкалья, Дальнего Востока и др. В подземных водах,
дренирующих эти месторождения таллия, его содержание может
достигать 13,2 мкмоль/л [Шварцев С. Л., Гомонова Л. Н., 1975].
При этом необходимо иметь в виду, что пороговая его, концент-
рация в водоемах составляет 1,47 мкмоль/л, а ПДК в питьевой
воде — 0,49 нмоль/л. В воздухе рабочей зоны ПДК йодида и
бромида таллия равна 0,01 мг/м3. Таким образом, соединения
таллия относятся к I классу опасности. Отравления им назы-
ваются таллотоксикозом [Kamerbeek Н. Н. et al., 1971].
В эксперименте с радиоактивным таллием установлена сле-
дующая тропность органов и тканей к этому МЭ: почки > се-
менники > печень > селезенка > предстательная железа > го-
ловной мозг > шерсть. Наряду с этим отмечается прямая
корреляция между тропностью таллия и выраженностью мор-
фологических изменений в этих органах на основании гистоло-
гических, гистоферментативных и гистохимических исследова-
ний [Кенесариев У. И., Чарыев О. Г., 1979]. Более новые данные
показывают, что таллий является элективным ядом для спер-
матогенных клеток и клеток Сертоли у крыс, а также для кле-
ток волосяных фолликулов [Lehmann Р. A. et al., 1985],
В норме концентрация таллия в волосах человека составляв
ет 0,049 мкмоль/кг, в моче — 0,049 нмоль/л. Из организма по-
ступивший таллий выводится медленно (до 1 года), в основном
с калом и мочой. Среднесмертельная доза таллия для человека
составляет 0,029—0,196 ммоль/кг, а смертельная — 3,62 ммоль
[Эйтингон А. И., 1983].
Таллотоксикоз — М.ТОЗ человека, обусловленный из-
быточным поступлением солей таллия в организм. Токсичность
таллия для человека была доказана A. Lamy (1863). В течение
многих лет появлялись в литературе сообщения лишь об отдель-
ных отравлениях солями таллия со смертельным исходом [Ca-
vanagh J. В. et al., 1974]. Однако, по данным J. С. Munch,
к 1934 г. собраны 778 случаев отравления таллием, а 6 % из
них закончились летально. В большинстве этих наблюдений
таллий использовался для лечебной эпиляции. Доза 8 мг/кг
достаточна для выпадения волос через 2—3 нед, но при этом
регулярно регистрировались параличи. Подобно ртути и мышья-
ку, таллий накапливается в тканях, которые медленно освобож-
даются от этого токсиканта. Повторные введения малых доз
могут вызывать кумулятивный токсический эффект. В сравни-
406
тельно редких наблюдениях, как это, например, было в малень-
кой вспышке в Объединенных Арабских Эмиратах, таллий при-
менялся в криминальных целях [McCormack J. et al., 1983].
Гораздо большее значение имеют неврологические осложнения,
развивающиеся после медикаментозного применения таллия,
т. е. ятрогенные формы таллотоксикоза. Как правило, невроло-
гическим симптомам при этом предшествовали преходящие
рвота и диарея, затем развивались болезненные парестезии
пальцев рук и ног, а позднее слабость конечностей. Скорость
прогрессирования неврологических симптомов различна. В ле-
тальных случаях может наступать поражение всех 4 конечно-
стей, а также черепных нервов и дыхательных мышц [Эйтингон
А. И., 1983].
Первые симптомы отравления таллием почти во всех случа-
ях— это болезненные парестезии, которые возникают через 1 —
5 дней после отравления. В отдельных наблюдениях эти при-
знаки обнаруживались уже через 12 ч. Нижние конечности,
особенно стопы, обычно поражаются сильнее, чем верхние ко-
нечности. Характерны болезненные парестезии подошвенной
поверхности стоп, что обычно не встречается при других невро-
патиях. Онемение пальцев рук и ног может сочетаться с потерей
болевой и тактильной чувствительности. Характерно также со-
хранение рефлексов на ранних стадиях болезни, что может быть
дифференциальным признаком от синдрома Гийена — Барре.
Описывается невропатия глазных нервов преимущественно на
более поздних этапах заболевания. Вслед за этим может на-
ступить атрофия зрительного нерва. Наиболее характерен птоз,
особенно у детей. Нарушение функций IV и VI пар черепных
нервов встречается часто в сочетании с нистагмом. Редко по-
ражается V пара черепных нервов, а IX пара, как правило, не
поражается. Иногда встречается поражение XI и XII пар череп-
ных нервов, отсутствуют гипертензии и тахикардия, которые
более характерны для полиневритов, сочетающихся с острой
перемежающейся порфирией. В отдельных наблюдениях отме-
чалось тяжелое повреждение блуждающего нерва.
Многие неврологические симптомы при таллотоксикозе свя-
заны с поражением центральной, а не периферической нервной
системы. Это относится к часто встречающейся атаксии, при
которой потеря равновесия более выражена, чем нарушение
координации движений конечностей. Часто встречаются тремор,
при тяжелых отравлениях и судороги. Характерны потеря со-
знания (до глубокой комы), чувство беспокойства, бессонница.
Редко развивается психоз с паранойяльными идеями, депресси-
ей, агрессивностью и галлюцинациями.
При хроническом отравлении таллием наблюдается анемия.
Поражение почек сопровождается снижением клиренса креати-
нина, повышением содержания мочевины в крови, а также про-
теинурией. В ряде случаев отмечена диабетическая кривая то-
лерантности к глюкозе, иногда — глюкозурия. Типичным для
тяжелых форм таллотоксикоза считается повышение уровня
407
трансаминаз в крови. При этом отмечена корреляция с выра-
женностью дистрофии, особенно жировой, в печени. Уровень
щелочной фосфатазы в сыворотке крови повышается.
Данные электрофизиологических исследований свидетельст-
вуют о развитии периферической невропатии «затухающего»
типа. Особенно показательны электроэнцефалографические из-
менения при тяжелой таллиевой интоксикации у детей. При
этом характерна неспецифическая медленная волновая актив-
ность ЦНС. На ЭКГ определяются синусовая тахикардия, упло-
щение или инверсия зубца Т. Последнее связывается с повы-
шенным содержанием ионов таллия в миокарде и его конкурен-
цией с калием.
В литературе, как правило, указывается на отсутствие спе-
цифических морфологических изменений при таллотоксикозе.
Однако постоянно выявляется резко выраженная жировая дис-
трофия центральной части печеночных долек [Cavanagh J. В.
et al., 1974]. При подостром и хроническом отравлении крыс
таллием отмечаются ацидофилия цитоплазмы гепатоцитов, ва-
куольная и жировая дистрофия печени. Электронно-микроско-
пически выявлены расширенные цистерны гранулярной эндо-
плазматической сети, жировые вакуоли в цитоплазме. Актив-
ность аминотрансфераз в сыворотке крови увеличена [Danile-
wicz М. etal., 1981]. Наряду с этим в эксперименте обнаружены
дистрофические и атрофические изменения семенников у крыс
[Jormigli L. et al., 1986]. При хроническом (6 мес) эксперимен-
тальном таллотоксикозе светооптически в мышцах обнаружено
исчезновение поперечной исчерченности, а электронно-микро-
скопически выявлены разрушение миофиламентов, дегенерация
эндоплазматической сети, разрывы мембран митохондрий, ва-
куолизация их, а также деструкция крист. Вакуолизация мито-
хондрий отмечена также в нейронах подкорковых узлов голов-
ного мозга [Motoroni D. et al., 1977].
Поражения нервной системы, характеризующиеся развитием
периферических и центральных невропатий, выявлены как у
человека, так и у экспериментальных животных. Отмечаются
дегенерация нервных волокон и хроматолиз в двигательных
нейронах. Наиболее выраженные изменения при таллотоксико-
зе у человека обнаружены в дистальных отделах нервных ство-
лов по сравнению с проксимальными. В частности, такие изме-
нения имеют место в дистальных отделах блуждающего нерва,
икроножного нерва и в его крупных разветвлениях внутри
мышц. Они проявлялись разрушением аксонов и миелиновой
оболочки, реакцией нейролеммоцитов (шванновских клеток)
и макрофагов. Необходимо подчеркнуть, что изменения в череп-
ных нервах, в частности в глазодвигательном, развивались в
более поздние сроки токсикоза и носили менее выраженны-й
характер, чем в нервных стволах конечностей. Набухание аксо-
нов является наиболее ранним манифестирующим признаком
таллотоксикоза [Spencer Р. S., 1973].
Кожные нарушения при таллотоксикозе обычно наступают
408
через 2—3 нед и характеризуются атрофическими изменениями
кожи и подкожной клетчатки, а также расстройствами орого-
вения с шелушением и тотальным выпадением волос на голове,
лице, под мышками и на лобке. Гистологически в выпавших во-
лосах выявляют веретенообразное утолщение корневой части с
массивным отложением черного пигмента. Следует иметь в ви-
ду, что отложения черного пигмента наблюдаются также и в
собственно коже вблизи корня волос и, по мнению ряда иссле-
дователей, являются патогномоничным признаком таллиевого
отравления [Будрин Ю. П., Мещерская К. А., 1975; Пфайфер Г.,
1981]. Исследования N. Thyresson (1951) показали, что таллий
концентрируется в волосяных фолликулах, и с этим он связыва-
ет его прямое токсическое действие на кератосинтезирующий
аппарат, а не на митотическое деление фолликулярных клеток,
как это предполагают Р. Gross и соавт. (1948) и Р. Flesch
(1963). На ногтях через 1—2 мес появляются поперечные по-
лосы.
В понимании патогенеза таллотоксикоза определенное зна-
чение представляют данные, свидетельствующие о химической
и биологической взаимосвязи ионов таллия и калия, так как их
ионные радиусы очень близки. В частности, хлориды обоих
элементов с химической точки зрения должны взаимодейство-
вать, образуя смешанные кристаллы. При этом таллий может
замещать калий в ряде минералов. Показано, что ионы таллия
входят в мышцы и деполяризуют мембраны, они также способ-
ны замещать ионы калия в активированной Na+, К+-АТФазе,
так как имеют в 10' раз большее сродство к этому ферменту,
чем ионы калия [Inturrisi С. Е., 1969]. Изучение влияния тал-
лия на ионный состав и транспортные свойства эпителиальных
клеток кожи лягушки с помощью рентгеноспектрального микро-
анализа показало, что распределение ионов таллия в эпителии
было аналогичным распределению ионов калия. Кроме того,
отмечено, что эти ионы стехиометрически заменяются на ионы
таллия. Изучение электрического сопротивления кожи не вы-
явило блокады натриевых каналов ионами таллия [Скуль-
ский И. А., 1980]. Эти авторы приходят к заключению, что тал-
лий ингибирует внутриклеточный механизм, сопряженный с од-
нонаправленным транспортом натрия, но не влияет на систему,
ответственную за поддержание электролитного состава эпите-
лиальных клеток.
В нато- и морфогенезе центральных и периферических тал-
лиевых невропатий определенное значение придается выражен-
ным ультраструктурным изменениям митохондрий соответству-
ющих нейронов и аксонов [Spencer Р. S. et al., 1973; Cava-
nagh J. В. et al., 1974].
В заключение следует отметить, что исход таллотоксикоза в
значительной степени зависит от дозы таллия, поступившего
в организм, а также от своевременности лечения. При этом не-
обходимо, однако, подчеркнуть, что специфического антидота
при отравлении таллием не существует, хотя имеется опреде-
409
ленный набор средств, обеспечивающих его эффективную тера-
пию [Эйтингон А. И., 1983].
В 1988 г. в Черновцах (СССР) наблюдалась вспышка забо-
леваний среди детей, сопровождавшаяся алопецией. Исследова-
ние почвы, растений, а также крови, мочи и волос заболевших
выявило повышенное содержание ряда МЭ, в том числе и тал-
лия [Гуцуляк В. Н., Прысакарь В. Б., 1989; Зербино Д. Д. и др.,
1989]. В судебно-медицинской литературе эпизодические слу-
чаи таллотоксикоза были описаны ранее.
Сведения о возможном токсическом влиянии других менее
изученных МЭ встречаются в специальной литературе по про-
фессиональной патологии, а также в клинической и судебно-
медицинской литературе. Попытка систематизации этих мате-
риалов осуществлена Н. Zumkley (1983). Этот автор относит к
«другим следовым элементам» сурьму, барий, свинец, бор, гер-
маний, золото, индий, литий, платину, палладий, ртуть, рубидий,
серебро, титан, теллур, кремний, таллий, уран, висмут, воль-
фрам, олово и цирконий. Как видно из перечисления, в эту
группу попали весьма различные вещества, в частности такие
МЭ, как бор, литий и кремний, которые по классификации
Р. I. Aggett (1985), могут быть эссенциальными. Здесь же на-
ходятся такие токсичные МЭ, как свинец, ртуть и олово.
Поскольку характеристика бария, свинца, бора, лития, рту-
ти, кремния, висмута приведена ранее (см. главы 7,8), кратко
остановимся на некоторых сведениях, приведенных в книге
Н. Zumkley (1983).
Определенный интерес представляет сурьма, которая ис-
пользовалась в медицине в глубокой древности и в средневе-
ковье. В начале XX века ее назначали как отхаркивающее и
антипаразитарное средство. Важно подчеркнуть, что в промыш-
ленности сурьма широко используется главным образом для
производства сплавов металлов, изготовления стекла, батарей
и печатного шрифта. Всасывание сурьмы ничтожно, она связы-
вается с эритроцитами и медленно выводится из организма,
преимущественно почками. Симптомы острого отравления сурь-
мой обусловлены ее избыточным поступлением. Они выража-
ются наличием металлического привкуса во рту, появлением
кишечных колик, рвоты, экземы, утратой аппетита, а также
воспалительными изменениями слизистой оболочки рта, носа и
гортани. Вдыхание аэрозолей сурьмы может вызвать опреде-
ленные изменения в легких, в том числе профессиональный
пневмокониоз. Подробные эпидемиологические исследования
содержания сурьмы в пищевых продуктах, воде, а также в ор-
ганах и тканях людей, погибших от случайных причин в Южной
Фергане, произвели Ю. М. Фузайлов и В. В. Ковальский (1981),
при этом специфическая сурьмяная патология не была уста-
новлена.
О поражении организма человека германием сведений
нет, у животных он может вызывать некрозы печени и почек.
410
Золото. Этот МЭ довольно широко используется в меди-
цине, в частности при ревматических и онкологических заболе-
ваниях, а также в стоматологии. Наиболее высокая концентра-
ция этого МЭ в организме обнаруживается в почках. Призна-
ками интоксикации являются поражения кожи, органов крове-
творения, органов дыхания и почек (см. главу 9).
Индий. Действие этого МЭ до сих пор не известно, однако
при использовании коллоидного индия для рентгеноконтрастной
диагностики заболеваний печени наблюдались тяжелые реакции
в виде шока. В эксперименте на животных внутривенное вве-
дение индия вызывало некрозы в печени, селезенке, а также в
проксимальном отделе почечных канальцев.
Физиологические и биологические функции палладия до
сих пор не изучены, однако предполагается, что палладий,
а также платина в будущем приобретут большее значение в
качестве факторов внешней среды в связи с возрастающим ис-
пользованием этих МЭ в промышленности.
Рубидий. Открыт в середине XIX века, до сих пор мало
изучен. Установлено его значительное сходство с калием. Оба
элемента частично замещают друг друга в процессе обмена ве-
ществ. До сих пор не выяснена причина гиперактивности и по-
вышенной агрессивности животных, получавших этот МЭ.
В 1877 г. С. П. Боткин и сотр. отметили замедление пульса и
повышение артериального давления при введении рубидия и
связали эти явления с его влиянием на ядра блуждающего
нерва. Поскольку самочувствие этих лиц улучшалось, возникла
мысль об использовании данного МЭ в качестве снотворного,
а также при эпилепсии.
Серебро. Этот МЭ применялся уже в средние века «для
очищения крови», усиления деятельности сердца, а также для
лечения душевнобольных. В настоящее время препараты сереб-
ра используются главным образом для местного применения.
Интересной особенностью этого МЭ является его накопление
в субэпителиальных слоях кожи (аргироз). Отложение серебра
происходит также в клубочках почек.
Титан. Этот МЭ считается нетоксичным, однако известно,
что у рабочих соответствующих отраслей производства может
наблюдаться отложение титана в легких и в лимфатических
узлах.
Теллур. Является одним из побочных продуктов при «об-
лагораживании» меди, а также при работе со свинцом, сереб-
ром и золотом. Он употребляется в стекольном и керамическом
производстве, а также при приготовлении клея. Описаны случаи
острых и хронических профессиональных интоксикаций.
Уран. Этот МЭ оказывает токсическое действие на почки
человека.
Вольфрам. У рабочих, имеющих дело с карбидом воль-
фрама, наблюдается фиброз легких, который, вероятно, обус-
ловлен наличием примесей кобальта.
411
Олово. Известно с глубокой древности. К. Schwarz (1975)
впервые установил эссенциальность олова, оно обнаруживается
в тканях человека и животных. Его концентрации зависят от
географических факторов, а также от особенностей диеты. Оно,
по-видимому, является катализатором окислительно-восстанови-
тельных процессов. Несмотря на то что ежедневное потребление
олова у взрослых колеблется от 4 до 20 мг в день, избыточно-
го обогащения организма оловом не происходит. Специфический
дефицит олова пока еще не описан. К. Schwarz предполагает,
что этот МЭ имеет отношение к регуляции роста и нормального
развития зубов. Олово, по-видимому, не играет роли в развитии
эмбриона, так как плацента является барьером для его поступ-
ления. Однако после рождения обнаруживается определенное
количество олова в тканях новорожденных. Вероятно, этот МЭ
участвует в специфических ферментативных реакциях белково-
го синтеза, но только после рождения. Описаны профессиональ-
ные отравления. Употреблявшиеся во Франции оловосодержа-
щие соединения для лечения фурункулеза иногда жриводили к
трагическим исходам. Для тяжелой интоксикации оловом ха-
рактерны стойкие головные боли, рвота, боли в животе, зри-
тельные и психические нарушения, парезы.
Цирконий. Этот МЭ является одним из распространенных
металлов земной коры. В связи с жаростойкостью цирконий
часто употребляется в качестве защитного материала в ядерной
физике, а также при изготовлении ядерного оружия. Кроме то-
го, он используется в стекольной, керамической и космической
промышленности. Физиологическое действие циркония незивест-
ыо, в крови он связывается с эритроцитами, токсичность его не-
значительна. Однако у людей при использовании содержащих
цирконий дезодорантов могут появиться гранулемы в подмы-
шечных областях как аллергическая реакция на этот МЭ.
Приведенные краткие сведения о потенциально токсичных
МЭ, которые в настоящее время еще мало изучены, должны
иметь ориентирующее значение. Вероятно, что в скором време-
ни они станут более полными и в корне изменят наши пред-
ставления о возможных токсических свойствах этих МЭ.
Глава 9
ЯТРОГЕННЫЕ МИКРОЭЛЕМЕНТОЗЫ
К ятрогенной патологии в настоящее время относят «все те
заболевания и патологические процессы, которые возникают под
влиянием медицинских воздействий, произведенных с профилак-
тическими, диагностическими и лечебными целями» [Смольян-
ников А. В. и др., 1988]. В соответствии с классификацией
П. Ф. Калитиевского и соавт. (1979), ятрогенные МТОЗы [Ав-
цын А. П. и др., 1983] должны относиться к так называемой
первой группе ятрогений, связанных с лечением, большая часть
из них является токсикопатиями.
Следует подчеркнуть, что относить те или иные патологиче-
ские реакции к ятрогенным МТОЗам необходимо с определен-
ной осторожностью, так как состав лекарственных средств, как
правило, сложен и способы их введения в организм очень раз-
нообразны. Среди ятрогенных МТОЗов большое значение имеют
ятрогенные МЭ-дефицитные состояния, возникающие в процессе
кратковременного и особенно длительного парентерального пи-
тания. Среди них различают как моноэлементные, так и поли-
элементные дефициты. Углубленный анализ патогенеза этих
состояний должен производиться в каждом конкретном случае
клиницистом и биохимиком. Разумеется, что биохимические
исследования при этих часто тяжелых состояниях не могут быть
направлены только на констатацию микроэлементного и элек-
тролитного дисбаланса.
Многие МЭ в течение десятилетий считались химически ин-
дифферентными или безвредными. Однако при более прицель-
ных и длительных клинических наблюдениях обнаружена их
потенциальная токсичность. Она не всегда обусловлена дозой
МЭ, так как зависит от индивидуальной чувствительности орга-
низма, которая может иметь и аллергическую природу. Иногда
токсичность является выражением врожденной или приобретен-
ной неполноценности физиологических аппаратов обезврежива-
ния или специфической элиминации токсичного МЭ. Существен-
ную роль в возрастании числа ятрогенных МТОЗов сыграло
стремление врачей к более частому парентеральному введению
лекарственных препаратов, содержащих МЭ. Оно, кроме того,
зависит от вида поддерживающей терапии. Следует также учи-
тывать, что при полном парентеральном питании организм не
всегда обеспечивается необходимым количеством эссенциальных
413-
МЭ. Избыток МЭ в организме может иметь место при перораль-
ном, парентеральном, чрескожном и ингаляционном поступле-
нии их с лекарственными препаратами.
9.1. Патологические реакции, связанные
с лечебным использованием эссенциальных микроэлементов
В этом разделе освещаются отдельные клинические наблю-
дения, относящиеся к таким эссенциальным МЭ, как железо,
медь, цинк, марганец, хром, селен и йод, дефицит или избыток
которых в организме, обусловленный теми или иными лечебны-
ми мероприятиями, приводит к развитию ряда симптомов и
синдромов.
Железо. Ятрогенные сидерозы у человека могут разви-
.ваться при повышенном содержании железа в пище, при не-
адекватном парентеральном введении препаратов железа в
случаях рефрактерных форм анемий или при многочисленных
трансфузиях крови (трансфузионный сидероз). Избыток железа
может возникнуть всегда, если имеется его положительный ба-
ланс в организме [Bernat I., 1983]. Реже встречается диализный
железодефицит [Blumbegr А. В. et al., 1983].
Инъекции препаратов железа, в частности декстрана железа
(Impheron), являлись причиной развития системных поражений
в виде лекарственной пурпуры [Berliner S. et al., 1983], а также
местных воспалительных и дистрофических изменений [Amitai I.,
Acker М., 1982]. У 1 больного с лекарственной пурпурой число
тромбоцитов и время коагуляции крови были в норме. Гемор-
рагическая сыпь исчезла через 1 нед. Кроме того, авторы опи-
сали 5-месячного ребенка, у которого через 1 мес после внутри-
мышечных инъекций препарата железа обнаружено исчезнове-
ние жировой и мышечной ткани в месте инъекций с формиро-
ванием углубления на ягодице, что, очевидно, было результатом
локальной липомиодистрофии. Постепенно эта ямка исчезла и
ягодица восстановила свою нормальную форму.
D. Ben-Shachar и соавт. (1985) показали, что Па-рецептор
дофамина содержит железо, которое, по-видимому, необходимо
для нормальной его функции, а хелатирующие железо препара-
ты, такие как десферриоксамин и 1,10-фенантролин, инактиви-
руют рецептор Ds. Предполагается, что рецептор лишается
железа.
Известно, что в состав некоторых комплексных витаминов
входят соединения железа. В частности, глутамевит содержит
0,01 г железа сульфата закисного. G. R. Henniger и соавт.
(1979) описали больного 58 лет с гемохроматозом, вызванным
чрезмерным употреблением в течение нескольких лет препарата,
содержавшего железо (500 таблеток за 1—2 нед). У него в
биоптате печени (по поводу гепатоспленомегалии) выявлены
портальный цирроз, не зависевший от гемохроматоза, а также
отложения железа в гепатоцитах, звездчатых макрофагоцитах
(клетки Купфера) и в меньшей степени в эпителии желчных
.414
протоков. При электронно-микроскопическом исследовании об-
наружены множество плеоморфных сидеросом, железо в раз-
личной стадии кристаллизации и ферритин на мембранах рези-
дуальных телец. Впоследствии больной умер от сердечно-сосу-
дистой недостаточности. Очень редко в месте внутримышечно-
го введения железосодержащих препаратов возникают опухоли..
Медь. Известно, что медь является незаменимым компонен-
том многих ферментов и белков. При ее дефиците в организме
нарушаются окислительно-восстановительные процессы, а так-
же развивается выраженная анемия. Описаны случаи дефицита
меди при полном парентеральном питании. Ятрогенные формы
гиперкупреоза могут возникать в результате присутствия этого
элемента в различных протезах, при применении противозача-
точных средств, кроме того, при гемодиализе в результате кон-
таминации соединениями меди [Приев И. Г., 1985].
Цинк. Для клинической медицины большее значение имеют
цинкдефицитные состояния, а не интоксикации этим-
элементом. Имеющиеся клинические и экспериментальные ма-
териалы свидетельствуют о том, что уже эмбрион и плод в
«критические периоды» своего развития чувствительны к дефи-
циту цинка в организме матери. Возникающие при этом цинк-
дефицитные состояния сопровождаются рождением незрелого
плода, а также формированием разного рода аномалий.
Ятрогенные формы дефицита цинка могут возникать при
длительном парентеральном питании и поступлении в организм
менее 19,2 мкмоль цинка в сутки (развивается энтеропатический
акродерматит); при постгастроэктомическом синдроме появля-
ются дерматит, гипогевзия с гипосмией; при лечении склеродер-
мии L-гистидином — гипогевзия и гипосмия; при длительной
терапии D-пеницилламином — паракератоз, кератит, централь-
ная скотома, алопеция; при лечении цитостатиками — анорек-
сия, гипогевзия, гипосмия, язвенный дерматит [Prasad A. S.,.
1979; Wolman S. L. et al., 1979; Dreosti I. E., 1980; Guillet G.
et al., 1980].
Диагноз цинкдефицитного состояния подтверждается при.
содержании цинка в крови менее 12,3 мкмоль/л, а концентра-
ция его в сыворотке крови менее 8,46 мкмоль/л рассматрива-
ется как неблагоприятный прогностический признак [Карлин-
ский В. М., 1979]. Необходимо иметь в виду, что существует
прямая корреляционная связь между понижением активности
цинкзависимых ферментов и сниженным содержанием этого МЭ-
в тканях, а также симптомами дефицита цинка. Представляет-
ся целесообразным включать в группу риска в отношении цинк-
дефицитных состояний всех больных, подвергавшихся резекции
проксимальных отделов тонкой кишки и желудка, а также
больных, находившихся на длительном парентеральном пита-
нии и подвергавшихся лечению цитостатиками.
Известно, что 75% цинка плазмы крови связаны с альбуми-
ном. В результате того что оральные контрацептивные препа-
раты (ОДП) снижают содержание альбумина в сыворотке,
415-
«стественно при этом происходит и снижение концентрации
цинка в плазме. В связи с этим возникает вопрос, нуждаются
.ли женщины, употребляющие ОК.П, в экзогенном поступлении
цинка? Исследования последнего времени [King J. С., 1987]
лозволили прийти к заключению, что потребление современных
ОКП не сопровождается в организме женщин изменениями
цинкового гомеостаза и не требует дополнительного поступле-
ния этого МЭ. Однако вопрос о потенциальных вредностях
ОКП нуждается в дополнительных исследованиях.
Активность такого мембраносвязанного цинксодержащего
•фермента, как нуклеозидфосфорилаза (5'-нуклеотидаза), быва-
ет сниженной при врожденной недостаточности Т-клеточного
иммунитета [Giblett Е. R. et al., 1975], а также у пожилых лю-
дей при нормальном старении [Boss G. R. et al., 1980]. В экс-
перименте на животных [Prasad A. S. et al., 1984] с алиментар-
но вызванным дефицитом цинка показано снижение акти-вности
нуклеозидфосфорилазы в эритроцитах. Таким образом, в настоя-
щее время имеются достаточно веские основания для исполь-
зования в клинической практике определения активности этого
фермента для оценки тяжести цинкдефицитных состояний.
Марганец. В группу риска развития манганодефицита,
несомненно, следует отнести больных, длительное время нахо-
дящихся на полном парентеральном питании. Это особенно
важно для грудных детей.
Хром. Развитие хромовой недостаточности у человека мож-
но наблюдать при длительном парентеральном питании. Она
сопровождается гипергликемией и глюкозурией без предшест-
вовавшего диабета. Так, у больной, находившейся в течение
нескольких месяцев на парентеральном питании вследствие не-
адекватного содержания (всего 6 мкг/сут) хрома, его уровень
в плазме крови составил 0,1 мкг/100 мл (при норме 1,8—
3,8 мкг/100 мл). Введение больной хлорида хрома в дозе
200 мкг/сут в течение 2 нед привело к нормализации уровня
этого МЭ и обмена глюкозы, что, несомненно, свидетельствова-
ло о наличии у больной хромдефицита [Brown R. О. et al.,
1986]. Следует, однако, подчеркнуть, что у данной больной не
отмечались ни периферическая нейропатия, ни энцефалопатия,
которые обычно характерны для хромдефицитных состояний,
а имел место только симптом непереносимости глюкозы.
Селен. Гипоселеноз установлен [Johnson Н. et al., 1981]
у больного, которому на протяжении 2 лет внутривенно вводи-
ли питательные вещества, не сбалансированные по содержанию
жизненно важных МЭ. У него были выявлены миокардиопатия,
напоминающая по своим патогистологическим изменениям бо-
лезнь Кешана, а также резко уменьшенный уровень селена и
активности глутатионпероксидазы в эритроцитах и в тканях
сердца.
В другом сообщении A. Collip и X. Chen (1981) описали
клиническую картину с типичными чертами болезни Кешана
416
у двухлетней девочки. Кроме дефицита селена в крови, у нее
был выявлен дефицит цинка за счет повышенной экскреции его
с мочой. После введения селена в течение 4 нед девочка попра-
вилась. Ряд аналогичных случаев описан в обзоре О. Levander
(1982).
В эксперименте показано, что в условиях дефицита селена
в организме применение таких препаратов, как адриамицин,
сопровождается усилением кардиомиотропности (токсичности)
этого антибиотика. Кроме того, у 70% селенодефицитных жи-
вотных отмечено развитие нефропатии, в то время как у конт-
рольных животных она выявлена только в 10% [Chen et al.,
1986]. Таким образом, в случаях использования адриамицина
в качестве терапевтического средства необходимо учитывать
селеновый статус организма.
Длительное парентеральное питание, как отмечено выше,
может сопровождаться дефицитом селена с развитием типичных
симптомов болезни Кешана. Наряду с этим иногда наблюдается
синдром селеновой миопатии, проявляющейся слабостью круп-
ных мышц конечностей [Brown М. R. et al., 1986]. Клинически
при этом выявлено резкое снижение активности глутатионпер-
оксидазы плазмы, эритроцитов, лейкоцитов и тромбоцитов. По-
сле внутривенных инъекций селеновой кислоты (400 мкг/сут)
указанные симптомы исчезали и уровень селена в крови в тече-
ние 6 нед нормализовался, однако активность глутатионперок-
сидазы эритроцитов достигала нормальных показателей позже
(через 3 мес).
У больных, находящихся на длительном парентеральном пи-
тании, риск развития селеновой недостаточности особенно велик.
Это диктует необходимость тщательного контроля за содержа-
нием селена в организме. Однако до настоящего времени точно
разработанных тестов, а также адекватных доз, путей введения
и лекарственных форм селеновых препаратов с целью профи-
лактики и лечения селенодефицита у человека еще нет. Очевид-
но, что работа I. Neve и соавт. (1986) в этом плане является
основополагающей. В частности, авторы показали, что уровни
селена в плазме и эритроцитах все же являются достаточно
чувствительными тестами для оценки селенового статута орга-
низма. В то же время показатели активности селеносодержа-
щего фермента глутатионпероксидазы не в достаточной степе-
ни информативны. Оптимальные уровни селена в плазме крови
у селенодефицитных больных гораздо быстрее достигались с
помощью Se-метионина (Se-M.et) в дозе 50 или 100 мкг Se в
сутки, чем с помощью селенита натрия в дозе 130—150 мкг Se
в сутки (максимально через 20 или 30 дней соответственно).
Обычные уровни селена в эритроцитах достигались максималь-
но через 40 дней при применении Se-Met (100 мкг Se) и в более
поздние сроки при использовании других препаратов селена.
После 100-дневной терапии селеном отмечено следующее:
1) у больных, получавших селенит натрия, оба показателя уров-
ня селена оставались в пределах нормы; 2) у больных, получав-
27—568
417
1пих Se-Met в дозе 50 мкг/сут, концентрация селена в плазме
достигала и оставалась на уровне верхней границы нормы,
а содержание селена в эритроцитах колебалось в обычных пре-
делах; 3) у больных, получавших Se-Met (доза 100 мкг/сут),
его концентрация как в плазме, так и в эритроцитах повыша-
лась выше нормы.
Как видно из этих данных, у селенодефицитных больных
наиболее быстрое и стабильное восстановление селенового ста-
туса происходит при лечении Se-Met.
Йод. Ятрогенные й о доз ы (й о д из м) у человека воз-
никают при лечебных мероприятиях и нередко обусловлены
измененной реактивностью организма. Лекарственные препара-
ты йода обладают неодинаковой токсичностью. При повышен-
ной чувствительности к йоду введение его в организм сопро-
вождается аллергическими реакциями разной тяжести (крапив-
ница, отек Квинке, йододерма и др.). Радиофармакологические
препараты (их более 30) с изотопами йода выпускаются в раз-
нообразных формах. Они позволяют проводить радиодиагности-
ческие исследования сердечно-сосудистой и гепатобилиарной
систем, почек, легких, желудочно-кишечного тракта, крови,
костного и головного мозга и др.
Препараты элементного йода оказывают выраженное мест-
нораздражающее действие на ткани. Резорбтивный эффект
связан с его влиянием на функцию щитовидной железы. В ма-
лых дозах (микройод) йод тормозит функцию щитовидной же-
лезы, а в больших стимулирует, принимая участие в синтезе
ее гормонов.
Йододерма — поражение кожи, развивающееся при длитель-
ном применении больших доз йода, а также после однократного
приема малой дозы при наличии идиосинкразии или сенсибили-
зации к нему. Патогенез заболевания сложен, наиболее вероятен
аллергический механизм. По мнению некоторых авторов, он
связан с раздражением вазомоторного аппарата и воспалитель-
ными изменениями сосудов, йододерма, как правило, локали-
зуется на лице, шее, конечностях, реже на туловище и волосис-
той части головы. Наиболее частой формой являются йодистые
угри, которые представляют собой папулы и пустулы, сопровож-
дающиеся зудом и жжением. Сливаясь между собой они обра-
зуют мягкие болезненные опухолеподобные инфильтраты сине-
багрового цвета диаметром 0,5—3 см. Редко встречается тяже-
лая, узловатая форма йододермы, впервые описанная П. Унной
в 1894 г. Наблюдаются также распространенные разнообразные
сыпи типа крапивницы, краснухи, пятнистой эритемы, коре- и
скарлатиноподобной экзантемы, рожистого воспаления, вегети-
рующей пузырчатки, вакцинной сыпи, полиморфной экссудатив-
ной эритемы, узловатой эритемы, пурпуры, экземы и даже
ограниченной гангрены. Они могут сочетаться между собой.
При сочетанном поражении кожи и слизистых оболочек полости
носа возможны носовые кровотечения, а также отек гортани,
иногда требующий трахеотомии.
418
При гистологическом исследовании йододермы обращают на
себя внимание выраженный акантоз и межклеточный отек,
иногда создающие впечатление картины псевдоэпителиоматоз-
ной гиперплазии. В эпидермисе видны микроабсцессы, отмеча-
ется обильная инфильтрация сосочкового и сетчатого слоев
дермы полинуклеарами, лимфоцитами, гистиоцитами, эозино-
филами и единичными плазматическими клетками, гигантские
многоядерные клетки встречаются редко. Часто наблюдаются
эндо- и периваскулиты [Пер М. И., 1978].
9.2. Патологические реакции,
связанные с лечебным использованием
условно эссенциальных микроэлементов
В эту группу вошли следующие МЭ: мышьяк, бром, фтор,
литий, никель, ванадий.
Мышьяк. Широкое применение мышьяксодержащих пре-
паратов в терапии сифилиса привело к постановке вопроса о
ятрогенной патологии, связанной с использованием соединений
мышьяка. Статистическим методом были обследованы [Lees R. Е.
et al., 1985] 83 больных раком предстательной железы, для
каждого из которых были подобраны 2 контрольных больных
одного и того же возраста (один с доброкачественной гипер-
трофией предстательной железы, другой с неонкологическим
заболеванием). Установлена достоверно большая частота си-
филиса в анамнезе в основной группе по сравнению с контро-
лем (р<0,05). При этом парный корреляционный анализ также
подтвердил большую достоверность частоты между основной
и первой контрольной группами (р<0,05). Кроме того, было
показано, что средний возраст больных в момент установления
диагноза рака предстательной железы был ниже при наличии
сифилиса в анамнезе.
Органические соединения мышьяка до настоящего времени
используются в лечении паразитарных болезней. При этом от-
мечен ряд осложнений, связанных с токсическим действием
мышьяка. Особенно тяжелыми являются энцефалопатии, кото-
рые обнаружены у 1,5% из 1066 больных африканским трипа-
носомозом, леченных арсобалом [Sina G. et al., 1977]. При этом
необходимо указать на высокую летальность (62,5%) больных
с этим видом энцефалопатии. Арсенозная энцефалопатия на-
блюдалась также у больных, леченных и другими органически-
ми соединениями мышьяка, в частности гликобиарсолом [ColeM.
et al., 1966]. Нередкой формой ятрогенного арсеноза является
атрофия зрительного нерва, которая отмечена у 3—4% больных,
леченных трипарсамидом [Neujean G. et al., 1948]. Из других
осложнений при терапии препаратами мышьяка следует ука-
зать на дерматиты, гиперкератозы, меланоз и поражение пе-
чени. Особый интерес представляет кератодермия, развиваю-
щаяся в результате длительного приема неорганических соеди-
нений мышьяка. Гиперкератоз ладоней и подошв может
27* 419
встретиться у 50% больных, длительно использовавших фауле-
ров раствор (арсенит калия), содержащий 3,8 г/л As [Fierz U.,
1965]. В работе J. Cuzick и соавт. (1982) представлен анализ
группы больных (478 наблюдений) на наличие арсенокератоза,
гиперпигментации и рака кожи. Около половины больных име-
ли один из этих признаков и более. Хотя общая смертность от
рака во всей группе была ожидаемой (среднестатистической),
во всех этих случаях имелись первичные признаки поражения
мышьяком.
Бром. При лечении некоторых неврологических синдромов
(неврозы, истерия, бессонница и др.) широко применяются
бромиды калия, натрия, кальция и аммония, а также ряд бром-
органических соединений (бромкамфора, бромизовал и др.).
Избыточное накопление брома в организме сопровождается
развитием кожных поражений в виде бромодермы, а также
неврологических и психических расстройств. Хроническое отрав-
ление бромом (бромизм) проявляется катаральным ринитом,
бронхитом, конъюнктивитом, энтеритом. Кожные поражения
обычно развиваются на фоне общих нарушений в виде сонли-
вости, атаксии, снижения болевой чувствительности, слуха,
зрения, ослабления памяти и других неврологических рас-
стройств. Отмечаются острые психопатологические нарушения
в виде делирия со зрительными, слуховыми, иногда тактильны-
ми, обонятельными и вкусовыми галлюцинациями [Мура-
тов В. К-, Лукомский И. И., 1976].
Бромодерма возникает при длительном приеме препаратов
брома, особенно бромида калия у лиц с повышенной чувстви-
тельностью к ним. Известны случаи врожденной бромодермы,
а также бромодерма у грудных детей, матери которых прини-
мали бромиды. Принято различать три формы: генерализован-
ные сыпи, угревидно-пустулезную и узловую бромодерму. Мак-
ро- и микроскопическая картина бромодермы во многом сходна
с йододермой. Однако анамнестические данные, а также нали-
чие брома в моче (реакция Белота) позволяют поставить пра-
вильный диагноз [Пер М. И., 1976].
Бромид ипратропия — синтетический антагонист холинерги-
ческих препаратов применяется в виде аэрозоля для лечения
острых и хронических заболеваний дыхательных путей. При по-
падании в глаза он может вызывать острую глаукому с времен-
ным мидриазом [Helprin G. A., Clarke G. К-, 1986].
В последние годы известны более 30 случаев применения
этого препарата в виде ингаляционного раствора, после чего у
больных наступало сужение, а не расширение бронхов.
С. К. Connolly (1982) предположил, что такая бронхоконстрик-
ция является вторичной и связана с вязкостью мокроты,
а О. М. Р. Jolobe (1982) объясняет его идиосинкразией к анти-
холинергическим препаратам [Schoeffel R. Е. et al., 1981]. Од-
нако исследованиями J. S. Mann и соавт. (1984) и Р. Rafferty
и соавт. (1986) было показано, что замена гипотонического
раствора данного препарата на изотонический почти во всех
420
pnvuaax устраняла бронхоконстрикторный эффект бромида
ипратропия. Тем не менее К. R- Patel и W. М. TuIIetf (1983),
представившие описание бронхоконстрикции в ответ на введе-
ние изотонического раствора бромида ипратропия, предполага-
ют что она, вероятно, произошла в результате вдыхания ионов
брома.
фтор. Значительный интерес представляют сообщения о
применении фтора, как правило, в виде фторида натрия с те-
рапевтической целью при различных формах остеопороза: пре-
сенильном, стероидиндуцированном, при отоспонгиозе [Caus-
se J. R. et al., 1981; Chlud K., 1981; Franke J., 1981]. Результаты
оказались вполне удовлетворительными. Однако не отмечено
положительного эффекта терапии фтором у больных с несовер-
шенным остеогенезом и идиопатическим остеопорозом.
В то же время очень длительное (10 лет) лечение фторсо-
держащим препаратом нифурилом в дозе 750 мг в день при-
водило в ряде случаев к развитию гистологически подтвержден-
ного флюороза скелета [Baud С. A. et al., 1981]. Обзор по
терапии остеопороза фторсодержащими препаратами и сопут-
ствующим осложнениям представлен С. К. Whitmore и J. Hag-
gerty (1983).
При использовании в анестезиологической практике фторор-
ганических соединений, например метоксифлурана, иногда на-
блюдается острое отравление фтором. Это возникает в резуль-
тате метаболического разрыва в этом веществе фторуглеродной
связи. Главными симптомами таких отравлений являются по-
лиурия и полидипсия, которые обусловлены токсическим влия-
нием освободившегося фтора на гипофиз, печень и почки [Pan-
ner В. J. et al., 1970; Gottlieb L. S. et al., 1974]. Если появилось
предположение о подобной интоксикации, то необходимо иссле-
довать кровь и мочу на содержание в них фтора, что позволит
аргументировать причину острого отравления. Действие пер-
фторуглеродных кровезаменителей изучается [Маджидов Ф.,
1988].
Литий. Этот МЭ широко используется с лечебной целью в
психиатрической практике. Однако со временем накопилось
значительное число сообщений, свидетельствующих о том, что
его высокие дозы могут сопровождаться развитием различных
форм литиевого МТОЗа. В частности, можно выделить следую-
щие основные синдромы: светобоязнь, нефро- и тиреопатия,
диарея, волчаночноподобный, литиевый гиперпаратиреоз, ней-
ропатия.
Так, лечение больных маниакально-депрессивным психозом
высокими дозами лития привлекло внимание клиницистов раз-
личных специальностей в связи с тем, что литиевая интоксика-
ция сопровождается крайне разнообразными симптомами. В ли-
тературе описывается резко выраженная светобоязнь у больной,
сопровождающаяся тяжелыми болями в глазных яблоках при
освещении их офтальмоскопом. Отмена лития сопровождалась
421
быстрым выздоровлением. Диагноз литиевой интоксикации был
подтвержден исследованием сыворотки крови, в которой кон-
центрация этого МЭ достигла 1,6 ммоль/л (1,1 мг/л). Следует
подчеркнуть, что при этом имел место неблагоприятный эндо-
кринный статус: поликистоз яичников с аменореей, ожирением,
гиперсутизмом и бесплодием [Caplan R., Fry А., 1982].
Нефротоксичность препаратов лития известна давно. Она
установлена как в клинике [Davies В., Kincaid-Smith Р., 1979;
Lippman S., 1982; Waller D., Edwards J., 1984], так и в экспе-
рименте на животных [Sidiropoulou-Skokou S. A., Issidori-
des М. К., 1983; Walker R. G. et al., 1986]. В частности, при
сравнительных исследованиях биоптатов почек, полученных от
16 больных, принимавших литий в течение 5,5 лет, и от 9 боль-
ных, не леченных литие*м, были установлены четкие различия
в патогистологической картине. При светооптических исследо-
ваниях биоптатов почек больных, леченных литием, отмечено
увеличение ШИК-позитивных гранул в цитоплазме эпителия
дистальных канальцев. Электронно-микроскопически найдены
отек цитоплазмы, митохондрий и патологические изменения
внутриклеточных органелл. В то же время в биоптатах почек
больных, не применявших литий, а также в почках в случае
смертельных исходов обнаружены только неспецифические из-
менения [Davies В., Kancaid-Smith Р., 1979]. Эти авторы пола-
гают, что литий может локализоваться на рецепторах вазопрес-
сина и служить причиной развития несахарного диабета.
Фундаментальная экспериментальная работа [Walker R. G.
et al., 1986] о хронической прогрессирующей недостаточности
почек, индуцированной литием, была проведена на новозеланд-
ских белых кроликах, которые получили 50—250 ммоль Li на
1 кг пищи. Исследовались биоптаты почек через 1, 3, 6 и 12 мес.
При этом макроскопически «литиевая почка» была бледной,
поверхность ее зерниста, в ней обнаружены микрокисты. Свето-
оптически выявлены вакуолизация цитоплазмы и накопление
гликогена в эпителии извитых канальцев и собирательных тру-
бочках. Гистологические признаки очаговой нефропатии в ви-
де интерстициального фиброза, атрофии канальцев с расшире-
нием дистальных трубочек и образованием микрокист, а также
гломерулярного склероза обнаружены уже через 1 мес от на-
чала опыта и наблюдались в течение всего срока исследования
(до 1 года). Биохимически к концу срока наблюдения отмечено
повышение содержания мочевины в крови и креатинина в сы-
воротке. Аналогичные данные получены в экспериментах на
крысах [Christensen S. et al., 1986].
Терапия литием сопровождается различными морфофунк-
циональными нарушениями щитовидной железы. В частности,
отмечено развитие эутиреоидного зоба [Schon М. et al., 1968]
и особенно часто гипотиреоза, который наблюдается у 70—30%
больных, длительно лечившихся литием [Emerson С. Н. et al.,
1973; Lindstedt G. et al., 1977; Amidsen A., Anderson C. J., 1982].
Однако индуцированный литием гипотиреоз у половины этих
422
больных имел нестойкий характер [Emerson С. Н. et al., 1973;
Fyro В. et al., 1973]. Показано также, что кратковременное
введение этого МЭ здоровым добровольцам [Lauridsen U. В.
et al., 1974] и, более того, больным с тиреотоксикозом [Temp-
le R.'et al., 1972; Kristensen О. et al., 1976] сопровождается
быстрым снижением уровней тиреоидных гормонов в крови.
Особый интерес в этом плане представляет наблюдение D. Ве-
augeard и соавт. (1983): больной 59 лет проводили лечение де-
прессии препаратом лития, что привело к маскированию зоба
с предшествовавшим гипертиреозом. Однако после оперативного
удаления зоба возобновление лечения депрессии литием при-
вело к выраженному гипотиреозу. Следовательно, если вначале
эффект лития был терапевтическим, то после операции — ятро-
генным, приведшим к гипотиреозу.
Патогистологическое исследование щитовидных желез
5 больных, леченных литием [Fauerholdt L„ Vendsborg Р., 1981],
установило, что в большей части желез имелись узлы макро-
фолликулярного строения с характерным полиморфизмом ти-
реоцитов и их ядер. Последние были крупными, гиперхромными,
отличались необычной формой с хорошо выраженными ядрыш-
ками. Часто встречались пикнотические ядра. Митозы не обна-
ружены. Выраженность описанных изменений варьировала в
значительных пределах, что, очевидно, связано с неодинаковой
продолжительностью (от 3 мес до 15 лет) лечения литием.
Экспериментальные исследования на крысах при сочетанном
введении хлорида лития и тиреотропного гормона показали от-
сутствие развития экспериментального гипертиреоза. Антити-
реоидный эффект лития пропорционален вводимой дозе и за-
висит от исходного состояния щитовидной железы [Семенов В. В.
и др., 1981]. При электронно-микроскопическом исследовании
щитовидной железы мышей, получавших глюконат лития в те-
чение 6 мес, установлены увеличение числа и размеров лизосом,
перераспределение их, коллоидных пузырьков, а также цито-
плазматических включений [Larras-Regard Е. et al., 1983].
Вероятность того, что литий вызывает или усиливает ауто-
иммунное поражение щитовидной железы, изучена недостаточ-
но. Однако в ряде случаев это предположение получило клини-
ческое подтверждение [Reus V. I. et al., 1979; Kontozoglou T.,
Mambo N., 1983]. Так, названные авторы приводят наблюдение,
когда у больной 62 лет после введения карбоната лития с ле-
чебной целью в связи с депрессивным состоянием было отмече-
но диффузное увеличение щитовидной железы. В оперативно
удаленной левой доле ее микроскопически установлено, что па-
ренхима разделена прослойками плотной соединительной ткани
на различной величины островки тиреоидных фолликулов. По
периферии доли железы обнаружено большое число лимфоидных
фолликулов со светлыми центрами. Между тиреоидными фол-
ликулами располагались лимфоциты и гистиоциты. Одни ти-
реоидные фолликулы сохранили нормальную структуру и были
заполнены коллоидом, другие подвергались атрофии. Часть
423
тиреоидных фолликулов гиперплазирована, выстлана кубиче-
ским эпителием. Авторы предполагают, что этот «тиреоидит»
обусловлен не непосредственным токсическим влиянием лития,
а индукцией иммунологических механизмов. Возможно, что ли-
тий играет роль гаптена, конъюгирующегося с тиреоидными
антителами. Аутоиммунный механизм поражения щитовидной
железы при введении лития может быть сходен с таковым при
введении бериллия, роль которого как гаптена установлена.
R. S. Segal и соавт. (1973) показали, что из 100 больных
с эутиреоидным состоянием щитовидной железы, длительно ле-
ченных литием, у 9 отмечен тиреотоксикоз с экзофтальмом.
Н. М. Dosch и соавт. (1980) как in vitro, так и in vivo устано-
вили, что литий тормозит активность Т-супрессоров и может
посредством угнетения активности аденилатциклазной системы
[Shenkman L. et al., 1980] вести себя как иммунологический
адъювант. М. Т. McDermott и соавт. (1986) описали больную
57 лет, в течение 8 лет принимавшую карбонат лития. После
этого у нее появился гипотиреоз, который через 9 мес сменился
гипертиреозом, хотя изменений в курсе лечения литием не было.
Описаны 24 наблюдения гипертиреоза у больных, длительно
получавших литиевую терапию. При этом отмечено, что нет до-
статочно четких объяснений связи между литием и тиреотокси-
козом. Некоторые авторы высказывают предположение, что
тиреотоксикоз у этих больных мог развиться за счет «сверхком-
пенсации» в ответ на отклонения от нормы в кинетике йода,
вызванные литием [Brownlie Е. W. et al., 1976]. Кроме того,
возможно, что литий вызывает тиреотоксикоз не непосредствен-
но, а косвенно путем влияния на интратиреоидный йодид
[Fradkin J., Wolf J., 1983].
Гипертиреоз у ряда больных развился только после завер-
шения приема лития, т. е. у этих больных внезапное прекраще-
ние ингибирующего влияния лития на щитовидную железу мог-
ло привести к эффекту «рикошета». Возможно также, что литий
раньше подавлял слабый скрытый гипертиреоз, который про-
явился в полную силу после прекращения лечения. Учитывая
большое число больных, принимающих литий, следует согла-
ситься с утверждением М. Т. McDermott (1986) о том, что ли-
тиевый тиреотоксикоз представляет собой осложнение, которое
будут диагностировать все чаще. О таких осложнениях должны
знать врачи, в том числе и патологи.
Давно установлено, что литий оказывает существенное влия-
ние на кальциевый обмен [Crammer J., 1975; Christiansen С.
et al., 1978]. При этом наличие гиперкальциемии у ряда боль-
ных, длительно принимавших препараты лития, а также умень-
шение экскреции кальция с мочой связывают с развитием ги-
перпаратиреоза. Действительно, у отдельных больных, длитель-
но лечившихся препаратами лития, выявлены аденомы паращи-
товидных желез [Christensson Т., 1976; Ananth J. et al., 1983].
Показано, что литий стимулирует выработку кальцитони-
на, однако это может и не отражаться на балансе кальция в
424
организме, так как одновременно увеличивается и продукция
паратгормона [Петров Н. М. и др., 1984].
терапия литием депрессивных состояний у женщин детород-
ного возраста в ряде случаев имеет выраженные особенности.
Так, установлено, что в пременструальной фазе менструального
цикла (за 5—10 дней до менструации) наблюдается енижение
концентрации лития в сыворотке крови, которое сопровождает-
ся усилением симптомов психического расстройства. Этот по-
вторяющийся пременструальный психоз характеризуется воз-
буждением, аффективной лабильностью, галлюцинациями, ги-
персексуальностью и паранойей [Conrad С. D. et al., 1986].
Характерные признаки пременструальных рецидивных симпто-
мов могут широко варьировать [Williams Е. Y. et al., 1952;
Endo М. et al., 1978; Glick L. D. et al., 1980; Berlin F. S. et al.,
1982].
Патофизиологические механизмы, ответственные за премен-
струальное снижение концентраций лития в сыворотке крови,
неизвестны. Однако D. С. Ostrov и соавт. (1982) отметили, что
в этот период содержание этого МЭ в эритроцитах существенно
возрастало. Проявление клинических симптомов в пременстру-
альной фазе может быть связано с субфизиологическими кон-
центрациями лития в сыворотке. Исходя из этого, С. D. Conrad
и J. I. Hamilton (1986) рекомендуют контролировать уровни
лития в сыворотке, увеличивая его дозу в пременструальной
периоде и снижая ее во время менструальной, фолликулярной
и овуляторной фазах цикла, чтобы избежать его токсического
действия.
Описано [Virkam R. et al., 1982] всего одно наблюдение,
когда у больной 44 лет с отягощенным аллергологическим
анамнезом (бронхиальная астма, непереносимость сульфанил-
амидов, противостолбнячной сыворотки и цитрусовых) после
назначения карбоната лития в дозе 86,5 ммоль 3 раза в день
по поводу депрессивного состояния развился волчаночыоподоб-
ный синдром. Последний появился через 48 ч после начала
лечения и характеризовался кожной сыпью на лице, животе и
шее. Сыпь на коже лица имела форму «бабочки». Через 1 нед
лечение литием прекратили, еще через 1 нед «бабочка» на лице
стала исчезать. Через 9 нед после отмены лития кожные изме-
нения в виде немногочисленных папул остались только на шее.
Таким образом, существует настоятельная необходимость уста-
новления причин развития волчаночноподобного синдрома у
больных с отягощенным аллергологическим анамнезом.
R. Balon и соавт. (1986), изучая влияние лития на число
тромбоцитов в периферической крови, показали, что лечение
карбонатом лития вызывает недостоверное их увеличение. Од-
нако имеются противоположные результаты — развитие тром-
боците- и нейтрофилопении при лечении литием [Richman С.
et al., 1984]. Индуцированное литием in vitro выделение из гра-
нулоцитов человека белка, связывающего фолат, происходит,
вероятно, в результате их дегрануляции при наличии высоких
425-
концентраций этого МЭ в крови. Длительное лечение литием
/может сопровождаться in vivo серьезными изменениями функ-
ции гранулоцитов [Holm J. et al., 1985]. Таким образом, вопрос
о ятрогенном эффекте лития на организм, в частности на кровь,
требует дальнейшего изучения.
Известно также, что литиевая терапия у 20% больных вы-
зывает диарею [Hollister L., 1982]. В меньшей степени диарея
наблюдается у больных, которым одновременно с литием на-
значают трициклические антиоксиданты, обладающие антихо-
линергическим свойством. Диарею принято связывать с быстрым
увеличением концентрации лития в сыворотке крови (более
0,7 ммоль/л) после его перорального введения. A. J. Mander
(1986) на вопрос, существует ли «синдром отмены лития», от-
вечает утвердительно.
Ятрогенные синдромы, обусловленные токсикопатическим
действием лития, дозозависимы. Так, умеренная интоксикация
литием наблюдается, если уровни его содержания в сыворотке
крови колеблются в пределах 1,5—2,5 ммоль/л. Средняя сте-
пень интоксикации отмечается при концентрации лития 2,5—
3,5 ммоль/л, а высокая угрожающая жизни — при содержании
лития более 3,5 ммоль/л. Большая часть симптомов исчезает
сравнительно быстро после отмены терапии литием.
Внедрение лития в клиническую практику проходило с
чрезвычайной осторожностью в связи с его токсичностью. По-
казано, что литий способен вызывать обратимые изменения в
миокарде, регистрируемые на ЭКГ. При литиевой терапии по-
казано, что смертность по сравнению со средним уровнем была
.в 2,83 раза выше. Высокая летальность объяснялась большим
числом самоубийств и сердечно-сосудистыми заболеваниями.
Смертность от нефропатии, рака, лейкозов не была повышенной
[Norton В., Whalley L. J., 1984]. J. S. Hammond и соавт. (1980),
однако, полагают, что длительная терапия литием может вы-
звать лейкоз. Хотя статистические данные по этому вопросу
недостаточны, немногочисленные наблюдения показали, что во
время лечения литием может возникнуть либо острый миелоид-
ный лейкоз, либо хронический миелолейкоз с наличием фила-
дельфийской хромосомы. У здорового человека литий вызывает
лейкоцитоз. Новые данные показывают, что этот МЭ стимули-
рует пролиферацию миелоидных, эритроидных и лимфоидных
лейкозных клеток in vitro в тех концентрациях, которые име-
ются в крови при лечении литием [Gauwerky С. Е., Golde D. W.,
1982]. Таким образом, опасность активации лейкоза у человека
под влиянием лития все же реальна.
Никель. В непосредственной связи с высоким загрязнени-
ем внешней среды рассеянными металлами, в частности нике-
лем, находится описанное в Японии новое неврологическое за-
болевание подострая миелооптиконейропатия (SMON). Клини-
ческая картина ее характеризуется болями в животе, наруше-
нием чувствительности, моторным параличом и уменьшением
остроты зрения [Kono S., 1975]. Преимущественно поражается
•426
периферическая нервная система. Периферические нейропатии,
сходные со SMON, описаны при контакте и с другими ме-
таллами.
Эпидемия SMON в Японии связывается с широким исполь-
зованием таких фармакологических препаратов, как клиокви-
нол и других производных 8-гидроксиквинолина, применяемых
для лечения инфекционных заболеваний, а также энтеропати-
ческого акродерматита. Исследованиями Н. Tjaler и К- Sthahl
(1984) показано, что клиоквинол увеличивает аб-
сорбцию никеля из кишечника и способствует повышению
уровня его концентрации в тканях. Однако избирательного на-
копления этого МЭ в ЦНС все же не установлено, очевидно,
за счет функции гематоэнцефалического барьера. Клиоквинол,
кроме никеля, образует липофильные хелаты и с другими ме-
таллами (цинк, ртуть), а также обеспечивает прохождение их
через клеточные мембраны и накопление в тканях.
Ванадий. В 1977 г. L. С. Cantley и соавт. выделили из
препарата АТФ («Sigma Grade») ингибитор активности Na+,
К+-АТФазы, который оказался идентичным Na3VO4. Было по-
казано, что ванадий присутствует в коммерческих препаратах
АТФ, приготовленных из мышц, а в препаратах АТФ из дрож-
жей его нет [Erdmann Е. et al., 1984]. В связи с этим многие
исследователи стали рассматривать ванадий как важный агент,
играющий роль эндогенного регулятора натриевого насоса. Про-
ведено большое число экспериментальных исследований по изу-
чению патобиохимии и фармакологического действия солей
ванадия. В частности, эффект ванадата натрия на миоциты
различных сосудов, по мнению Т. Shimada и соавт. (1986), за-
ключается в инициации сокращения гладких мышц различных
артерий при непосредственном его действии на миоциты. Вана-
дат натрия увеличивает концентрацию свободного внутрикле-
точного кальция предположительно путем высвобождения его
из внутриклеточных участков связывания и/или в результате
торможения Са2+—насоса мембранной системы. Вариации от-
ветных реакций можно объяснить в основном за счет различий
проницаемости ванадата через мембраны.
9.3. Патологические реакции,
связанные с лечебным использованием
условно токсичных и токсичных микроэлементов
В эту группу включены следующие микроэлементы: алюми-
ний, ртуть, висмут, платина (цис-платин), золото.
Алюминий. Широкую известность в последнее время
приобрели алюминиевая диализная энцефалопатия, алюминий-
зависимая микроцитарная анемия, ятрогенная алюминиевая
остеодистрофия, алюминиевый псевдогиперпаратиреоз при хро-
нической почечной недостаточности [Авцын А. П., 1986].
В 1972 г. у больных с уремией, подвергавшихся гемодиализу,
был обнаружен новый неврологический синдром, получивший
427
название «диализная энцефалопатия», выражавшаяся в расст-
ройствах речи, судорожных припадках, нередко приводящий к
летальному исходу [Alfrey А. С. et al., 1972]. Дальнейшее изу-
чение этого синдрома позволило установить, что этиологическим
фактором является избыточное накопление в ЦНС алюминия
вследствие его высокого содержания в диализной жидкости
[Alfrey А. С. et al., 1976]. Биохимические и эпидемиологические
исследования подтвердили эти данные [Platts М. et al., 1977;
McDermott J. et al., 1978; Parkinson I. et al., 1979].
Наряду с диализной энцефалопатией у этих же больных не-
редко развивается алюминиевая остеодистрофия (остеомаля-
ция) с повышенной ломкостью костей [Ward М. К. et al., 1978].
При этом алюминий накапливается в костной ткани как при
диализе, так и при его пероральном введении. Это особенно от-
носится к детям дошкольного возраста, больным уремией и
леченным с помощью перорального введения алюминий-фосфат-
ного геля [Andreoli S. Р. et al., 1984]. Гистохимически доказано,
что особенно много алюминия откладывается на границе меж-
ду остеоидом и кальцифицированной костью. Установлено, что
уровень этого МЭ в плазме крови, превышающий 3,70—•
5,56 мкмоль/л, расценивается как фактор риска развития алю-
миниевых энцефалопатии и остеомаляции [Andreoli S. Р. et al.,
1984; Alfrey А. С., 1984].
Весьма показательны сравнительные данные о концентрации
алюминия в некоторых органах у больных, умерших от уремии,
подвергавшихся и не подвергавшихся гемодиализу, а также у
контрольной группы больных без уремии и не подвергавшихся
гемодиализу. Наибольшие концентрации алюминия отмечены
у больных, умерших от диализной энцефалопатии. При этом
содержание алюминия в сердце, головном мозге и в костях со-
ставило 1,59; 0,91; 10,4 ммоль/кг сухой массы ткани соответст-
венно. В то, же время в контрольной группе эти показатели бы-
ли 0,04; 0,08; 0,12 ммоль/кг, т. е. во много раз меньше.
Кроме диализных форм алюминозов, в современной литера-
туре имеются сообщения, свидетельствующие о неблагоприят-
ном токсическом влиянии на ЦНС некоторых распространенных
лекарственных средств, содержащих в своем составе алюминий.
К ним, в частности, относятся антациды. Среди специалистов
имеется настороженность в связи с потенциальной опасностью
их длительного и многократного применения, особенно при на-
рушении функции почек. Каждая доза этих лекарственных
препаратов содержит от 1,3 до 10,4 ммоль алюминия.
Описана больная 61 года с миопатическим синдромом, дли-
тельное время применявшая антациды в связи с повышенной
кислотностью желудочного сока. У нее в биоптатах скелетных
мышц были выявлены атрофические изменения, электронно-
плотные включения, содержащие силикаты алюминия и магния,
а также частицы железа [Dodson A. F. et al., 1981]. Авторы
считают, что миопатический синдром в этом случае мог быть
связан с отложением этих элементов в миоцитах. Существует
428
мнение, что для лечения алюминозов перспективен метод хела-
тотерапии, в частности с экспериментально апробированным
дефероксамином [Alfrey А. С., 1984].
Алюминиевая остеодистрофия у почечных больных, длитель-
ное время принимавших антациды, может подвергаться обрат-
ному развитию после пересадки почки [Nordal К- Р.. et al., 1986].
Так, через 1 год после трансплантации трупной почки выявлено
резкое уменьшение содержания алюминия в биоптате подвздош-
ной кости, а у больных с пересаженной почкой от живого род-
ственного донора избыток этого МЭ в кости вообще не обнару-
жен. До пересадки почки уровень алюминия в плазме крови в
этих группах больных составлял 7,25 и 5,60 мкмоль/л, а через
1 год после пересадки—1,45 и 0,63 мкмоль/л соответственно.
Ртуть. Значение лекарственных препаратов ртути резко
уменьшилось во второй половине XX века. Местное и общере-
зорбтивное действие препаратов ртути в значительной степени
зависит от их растворимости в воде и биологических жидкостях.
Растворимость солей ртути увеличивается в присутствии гало-
генов. Из организма ртуть выводится достаточно быстро, при-
чем ее органические соединения выводятся значительно быст-
рее, чем неорганические. Однако этот МЭ обладает и выражен-
ными кумулятивными свойствами, которые и обусловливают
токсический эффект при длительном его поступлении в ор-
ганизм.
Применение лекарственных препаратов ртути может сопро-
вождаться местнораздражающим действием, а резорбтивное —
симптомами микромеркуриализма. Ртутная серая мазь содер-
жит в своем составе около 30% металлической ртути, она упо-
требляется обычно в качестве противопаразитарного средства.
Концентрации ртути в моче более 0,05—0,25 мкмоль/л имеют
клиническое значение для диагноза микромеркуриализма при
наличии соответствующей симптоматики. Последняя характе-
ризуется выраженными морфологическими изменениями крови
и функциональными нарушениями вегетативной нервной систе-
мы, особенно ее симпатического звена. Нередко наблюдаются
изменения эмоциональной сферы, характерно длительное сохра-
нение фазы повышенной возбудимости коры большого мозга в
сочетании с патологической лабильностью ЦНС [Мишин В. П.
и др., 1984].
Висмут. Висмутозы — МТОЗы, вызванные контактом с
соединениями этого элемента. Входными воротами могут быть
кожа, пищеварительный тракт, дыхательные пути. Однако ос-
новным путем поступления висмута в организм является парен-
теральный, так как висмутсодержащие препараты широко ис-
пользуются в лечебных целях, в частности для внутримышечно-
го введения при терапии сифилиса.
Хронический висмутоз — типичное осложнение при
лечении висмутсодержащими препаратами (бийохинол, бимо-
верол, пентабисмол). Эти препараты применяются парентераль-
но главным образом для лечения сифилиса и неспецифических
429
воспалительных процессов. Характерно депонирование висмута
в почках, печени, селезенке и в костях. Висмутовые препараты
в основном выводятся с мочой, а также через желудочно-ки-
шечный тракт и потовые железы. Процесс выведения висмута
из организма очень длительный. Характерно отложение этого
элемента по краям десен, особенно у кариозных зубов, что яв-
ляется признаком насыщения организма. Висмутовый гингивит
может осложняться стоматитом, При хроническом висмутозе
характерно поражение почек и мочевыводящих путей с появ-
лением в моче эпителиальных клеток, содержащих темные
кристаллы.
Висмутовые токсикодермии и артралгии встречаются редко.
Пероральное применение таких висмутсодержащих препаратов,
как викалин, викаир, при язвенной болезни, энтеритах и коли-
тах может сопровождаться адсорбцией сульфата висмута на
стенках кишок и уменьшением перистальтики.
Описана «висмутовая энцефалопатия» у больной
20 лет со смертельным исходом, развившаяся в результате при-
менения висмута в качестве лекарственного препарата. При
этом на вскрытии выявлены многочисленные гнойные очаги в
легких, селезенке, сердце, почках и в теле позвонка (TIV). Наи-
более выраженные гистологические изменения отмечены в цент-
ральной и периферической нервной системе. Они проявлялись
в головном мозге псевдоламинарным разрежением корковых
нейронов и пролиферацией микроглии. В гиппокампе обнаруже-
ны небольшие некротические очаги, а в белом веществе полу-
шарий отдельные периваскулярные круглоклеточные инфиль-
траты. В подкорковых ядрах наблюдались потеря нейронов,
пролиферация микроглии и гипертрофия астроцитов. В то же
время таламические, гипоталамические отделы и область блед-
ного шара были интактны.
Наиболее тяжело пораженным оказался мозжечок, в кото-
ром почти все грушевидные нейроны оказались разрушенными.
Отмечено также разрежение клеток молекулярного слоя и, на-
против, количество клеток Бергмана увеличено. В спинном
мозге на уровне Суш—Lv имело место выпадение нейронов
передних рогов. В седалищном и малоберцовом нервах обна-
ружены «расширение» аксонов, а также повреждение миелино-
вых оболочек. Необходимо указать, что содержание висмута в
биологических жидкостях, внутренних органах и веществе мозга
было повышено в 3—500 раз по сравнению с нормой [Lies-
sens J. L. et al., 1978].
Висмут обладает выраженным цитотропизмом: при введе-
нии его солей лабораторным животным он образует внутрикле-
точные включения в ряде органов. В эксперименте А. И. Едем-
ского, проведенном по нашему предложению, наибольшие
отложения солей висмута имели место в эпителии почечных ка-
нальцев.
Платина. Другим широко известным МТОЗом ятрогенно-
го происхождения является нефропатия, связанная с примене-
430
нием противоопухолевого препарата цисплатин. После экс-
периментально установленного эффекта этого препарата при
многих опухолях животных были проведены клинические испы-
тания в Национальном институте рака США в 1972 г. Однако
при этом выяснились определенные ограничения использования
цисплатина из-за его токсического действия на костный мозг и.
почки [Roberts J. J., 1981].
Кратко остановимся на тех изменениях, которые возникают
в организме при применении этого препарата. Известно, что-
максимальная противоопухолевая активность цисплатина про-
является в опытах на животных при его внутрибрюшинном и
внутривенном введении. При подкожном и внутримышечном
введении его активность снижается, а при пероральном поступ-
лении он вообще неактивен. Терапевтическая доза цисплатина
для мышей равна 35,9 мкмоль/кг и составляет !Д от минималь-
ной летальной дозы (DLM25). На животных изучен механизм
нефротоксического действия этого препарата. В частности, по-
казано, что цисплатин снижает фильтрационную функцию по-
чек и способствует накоплению в крови азота мочевины и сни-
жению клиренса креатинина, что сопровождается потерей ап-
петита и рвотой.
Цисплатин пассивно транспортируется через плазмалемму
из внеклеточной жидкости. Внутри клетки он подвергается гид-
ратации, в результате чего изменяется его реакционная способ-
ность. Он накапливается в почках, печени, надпочечниках, лег-
ких, костях и коже. Способностью платины взаимодействовать
с молекулой ДНК обусловлена ее мутагенность. В эксперименте
показано, что при внутрибрюшинном введении в; течение 41 нед
она индуцирует у мышей развитие аденом легких. По степени
выраженности канцерогенного действия платина отнесена к
канцерогенам средней активности [Rosenberg В., 1985].
В экспериментальной работе В. Т. Бахтеева и соавт. (1987}
по изучению морфофункциональных корреляций нефротоксиче-
ского действия цисплатина показано, что однократное внутри-
брюшинное введение препарата в дозе 25,6 мкмоль/кг белым
крысам уже через 2 сут вызывает очаговые изменения парен-
химы почек. Это прямо коррелирует со снижением клубочковой
фильтрации, нарастанием концентрации мочевины крови и
уменьшением экскреции при водной нагрузке. На 5-е сутки
после введения цисплатина на фоне дисфункции почек в их па-
ренхиме отчетливо выявляются признаки репарации. Ультра-
структурные изменения почек характеризуются дистрофией
клеток извитых и прямых проксимальных канальцев нефрона,
резким утолщением базальных мембран поврежденных клеток,
расширением межклеточного пространства. Отмеченные изме-
нения являются морфологическим субстратом нарушения секре-
ции органических кислот и увеличения массы органа. Насту-
пающая азотемия обусловлена снижением клубочковой фильт-
рации, однако структурные изменения клубочков сравнительно
малы. Имеется два объяснения снижения клубочковой фильт-
431
рации. Это либо первичное поражение сосудистого аппарата
{Offerman J. J. et al., 1984], либо токсическое повреждение ка-
нальцев, которые приводят к вторичному снижению фильт-
рации.
Детально патологическая анатомия острой и хронической
цисплатиновой нефропатии у экспериментальных животных
представлена в работе D. D. Choie и соавт. (1981). При этом
отмечено, что наибольшие концентрации платины обнаружены
в кортико-медуллярной зоне, в почках содержится около 2%
введенной дозы. По мнению этих авторов, токсический некроз
канальцев, возникший под действием цисплатина, напоминает
изменения после введения ртути и кадмия.
Из осложнений, появляющихся при применении цисплатина,
особого внимания заслуживает ее ототоксичность. Так, S. Schae-
fer и соавт. (1985) при обследовании 24 больных с опухолями
головы и шеи обнаружили дозозависимые необратимые и часто
встречающиеся нарушения слуха. Больные были отобраны из
большой популяции на основе однородности химиотерапии, по-
чечного статуса и отсутствия применения других ототоксиче-
ских агентов в анамнезе. Электронно-микроскопически в 4 слу-
чаях в кусочках височной кости, взятых через 3—5 ч после
смерти, были найдены изменения слухового (кортиевого)
органа.
В экспериментальных исследованиях ототоксичности цис-
платина на морских свинках, проведенных S. Котипе и соавт.
(1981), обнаружено разрушение наружных волосковых клеток
в базальном витке улитки и сохранность их в верхних витках.
Внутренние волосковые клетки были полностью интактны во
всех витках, однако было выявлено тяжелое повреждение пред-
дверной (Рейснера) мембраны в базальном витке улитки.
Цисплатина потенциально нейротоксична и ее применение с
лечебной целью приводит не только к заболеванию внутреннего
уха, но и к ретробульбарным невритам, периферическим нейро-
патиям [Hall Т. С., 1974]. Этиология и патогенез нейропатий
неизвестны, хотя имеются предположения об их связи с изме-
нениями в иммунной системе, с инфекцией и токсическими ве-
ществами, секретируемыми опухолью, например бронхогенным
раком [Black L. F., 1982].
У большинства больных индуцированная цисплатиновая ней-
ропатия развивалась после 3—4 курсов терапии [Hadley D.,
Herr Н. W., 1979]. Обычно неврологические симптомы были об-
ратимы, однако в ряде случаев они держались долго и, как
правило, усиливались после дополнительных курсов цисплати-
новой терапии. Клинически нейропатия проявлялась болезнен-
ными парестезиями по типу «носков» и «перчаток», потерей
вибрационной чувствительности, а также понижением глубоких
сухожильных рефлексов.
Особенности клинических симптомов нейропатии, вызванной
цисплатином, описаны в наблюдении С. Rosenfeld и L. Broder
(1984). Так, у больного бронхогенным раком через 1 нед после
432
второго курса цисплатиновой химиотерапии отмечены атаксия,
парестезии и пойкилотермия, а также снижение рефлексов на
верхних конечностях, следовые рефлексы на уровне колен и от-
сутствие таковых на уровне лодыжек. Тактильная и болевая
чувствительность были понижены по типу «носков». После того
как больной стал получать декстроамфетамин (10 мг ежеднев-
но), а цисплатин был отменен, симптомы нейропатии исчезли
в последующие несколько недель.
Таким образом, цисплатиновая терапия онкологических
больных может сопровождаться развитием следующих синдро-
мов: нефропатии, отопатии, нейропатии периферического и ве-
гетативного типа.
Золото. При клиническом испытании лечебного препарата
ауранофина, содержащего соли золота, в дозе 0,51 —
1,02 мкмоль/кг в день у детей, страдавших ювенильным ревма-
тоидным артритом, обнаружено, что концентрация золота в сы-
воротке крови больше зависела от ежедневной дозы, чем от его
кумуляции [Giannini Е. Н. et al., 1984]. Лечение больных рев-
матоидным артритом ауранофином (3 мг 2 раза в день —
0,43 мкмоль золота) сопровождалось у некоторых из них раз-
витием преходящей диареи средней степени выраженности,
а также снижением активности кислой фосфатазы крови [Lig-
nere G. С., Giavarini Е., 1984]. Кроме того, описано наблюдение
[Wilson А. Р. R. et al., 1984], когда у 63-летней женщины, ус-
пешно лечившейся ауранофином в течение 12 мес по поводу
серопозитивного ревматоидного артрита, при биопсии почки
выявлена мембранозная гломерулонефропатия с отложением
золота в эпителиоцитах канальцев.
Установлено также, что золото, поступающее в организм в
составе лекарственных препаратов, откладывается в фагоцити-
рующих клетках, в печени и лимфатических узлах. В макрофа-
гах оно локализуется в лизосомах или лизосомоподобных струк-
турах [Danscher G., 1981]. В эксперименте показано, что акти-
вированные вирусом herpes simplex II типа макрофаги мышей
активно захватывают золото, которое локализуется в фаголизо-
цомах. Однако этот МЭ не был выявлен в полинуклеарах и
лимфоцитах [Moller-Madsen В. et al., 1984].
В литературе остро ставится вопрос о контроле чистоты и
оптимального содержания МЭ в первую очередь в растворах,
используемых в клинической практике с терапевтической целью,
а также для парентерального питания больных [Adan L. et al.,
1985; Fell G. S. et al., 1986]. Это связано с тем, что при изуче-
нии разных белковых жидкостей, используемых для указанных
целей, выявлено 10—1000-кратное увеличение содержания в них
алюминия, хрома, марганца, никеля, железа. Кроме того, от-
мечено существенное снижение таких жизненно важных элемен-
тов, как магний, цинк, медь, селен. Длительное и многократное
введение этих жидкостей больным является одним из основных
причин ятрогенных МТОЗов. В первом случае они обусловлены
избыточным поступлением токсичных МЭ, а во втором, напро-
28—568
433
Таблица 25
Ятрогенные мнкроэлементозы
Микроэле- мент Лекарственный препарат, лечебное вмешательство, процедура Симптомы, синдромы, патологи- ческие состояния
Железо Эссенциальные микроэ Препараты железа для перо- лементы Сидерозы, в частности транс-
Медь рального применения, комп- лексные витамины, например глютамевит (0,066 ммоль сульфата закисного железа); парентеральное введение Полное парентеральное питание фузионный сидероз печени и селезенки; аллергическая пур- пура. В очень редких случаях на месте внутримышечного введения железосодержащих препаратов возникают злока- чественные опухоли Медьдефицитиая анемия и дру«
Цинк с дефицитом меди. Применение медьсодержащих противозача- точных средств; имплантация медьсодержащих протезов; ге- модиализ Длительное парентеральное пи- гие симптомы гипокупреоза. Г иперкупреоз — снижение ак- тивности и биосинтеза некото- рых ферментов Энтеропатический акродерма-
Марганец тание (цинка менее 0,019 ммоль/сут); гастрэктомия. Ле- чение L-гистидином, D-пени- цилламином Экспериментальная диета, ли- тит. Постгастрэктомический синд- ром с дерматитом, гипогевзией с гипосмией; гипогевзия и гипосмия; пара- кератоз, кератит, центральная скотома, алопеция, анорексия; снижение активности нуклео- зидфосфорилазы — маркера цинкдефицитного состояния Гнпоманганоз: потеря массы
Хром шенная марганца Парентеральное питание с де- тела, снижение концентрации холестерина, триглицеридов н фосфолипидов крови, увеличе- ние протромбинового времени Гипохромоз: гипергликемия,
Селен фицитом хрома Длительное парентеральное пи- глюкозурия; энцефалопатия, периферическая нейропатия Гипоселеноз — симптомы бо-
Иод тание; введение противоопухо- левого препарата адриамицина в условиях селенодефицита ор- ганизма Раствор йода спиртовой, рас- лезнн Кешана, миокардиопа- тия; миопатия крупных мышц конечностей; усиление кардио- тропности и иефротропиости лекарства Иододерма прн длительном
твор Люголя; йодинол, йодо- форм; калия и натрия йодид; йодолипол, билитраст и др.; радиоактивные препараты йода введении йодистых препаратов; отек Квинке, крапивница и другие кожные экзантемы на фоне аллергии
Мышьяк Условно эссенциальные ми Мышьякорганические препара- кръэлементы Энцефалопатия; атрофия зри-
ты (арсобал, трипарсамид и др.); раствор Фаулера (арсе- нит калия) тельного нерва; арсенокератоз, меланоз кожи, бородавки, рак кожи и бронхов, гемангиоэндо- телиома печени (?)
434
Продолжение
— Микроэле- мент Лек-арственный препарат, лечебное вмешательство, процедура Симптомы, синдромы, патологи- ческие состояния
Бром Бромиды калия, натрия, каль- ция, аммония Инъекционные препараты: бромкамфора, бромизовал и др. Бромид ипратропия (Atrovent) Бромизм; бромодерма — при длительном введении бромидов Неврологические расстройст- ва — сонливость, снижение бо- левой чувствительности, слуха, зрения, ослабление памяти и др. Острые психопатологиче- ские нарушения в виде делирия со зрительными, слуховыми, иногда тактильными, обоня- тельными и вкусовыми галлю- цинациями При попадании аэрозоля на конъюнктиву — острая глауко- ма с временным мидриазом
Фтор Фторид натрия, нифлурил Анестетик метоксифлуран При длительном лечении остео- пороза высокими дозами фто- ридов — скелетный флюороз. Острое отравление фтором — полиурия, полидипсия
Никель При Лечении клиоквинолом и другими производными 8-гид- роксиквинолина повышается абсорбция никеля Гиперникелез в виде подострой миелооптиконейропатии (SMON)
Литий Хлорид лития, карбонат лития и др. Умеренная степень интоксика- ции (уровни лития в сыворот- ке 1,5—2,5 ммоль/л). Средняя степень интоксикации (концентрация лития 2,5— 3,5 ммоль/л). Высокая угрожающая жизни степень интоксикации (кон- центрация лития более 3,5 ммоль/л). Неврологические симптомы: светобоязнь, нис- тагм, тремор преходящий, стойкий у 4% больных; мы- шечная слабость преходящая, умеренно выраженная. Желудочно-кишечные симпто- мы: анорексия, тошнота, рвота, диарея, боль и вздутие живо- та, преходящиеу 20% больных. Нефропатия (стойкая у 25% больных) — нефрогенный вя- лый диабет, полиурия и поли- дипсия, олигурическая почеч- ная недостаточность. Кардиомиопатия (редко) — экстрасистолия, дисфункция синусового узла, миокардиты. Тиреопатия: эутиреоидный зоб у 3% больных, гипотиреоз у 7—30% больных, аденомы щитовидной железы, аутоим- мунный тиреоидит, гипертире- оз (редко).
28*
435
Продолжение
Микроэле- мент Лекарственный препарат, лечебное вмешательство, процедура Симптомы, синдромы, патологи- ческие состояния
Ванадий NasVOi — компонент в коммер- ческом препарате АТФ (Sigma Grade) из мышечной ткани Гиперпаратиреоз, в том числе аденомы паращитовидных же- лез. Волчаночноподобный синдром у больных с отягощенным ал- лергологическим анамнезом Вызывает усиленное сокраще- ние сердечной мышцы и вазо- констрикторный эффект, тор- мозит активность Na+, К+- АТФазы
Условно токсичные и токсичные микроэлементы
Алюминий Диализная жидкость; альбуми- новые препараты для паренте- рального введения; антаци- ды — алюграмин, алюминий- фосфатный гель Алюминиевая диализная энце- фалопатия, алюминиевая остео- дистрофия, алюмииийзависимая микроцитарная анемия, алюми- ниевый псевдогиперпаратиреоз, при хронической почечной не- достаточности, миопатический синдром, алюминиевая гасгро- патия; перитонеальный алюми- ноз
Ртуть Меркузал, промеран, иовурит. Дихлорид ртути, оксицианид ртути, амидохлорид ртути. Мазь ртутиая серая Местное — раздражающее действие. Общерезобтивное — симптомы микромеркуриализма. Содер- жание в моче 0,05—0,25 мкмоль/л ртути при длитель- ном поступлении ее микродоэ является доказательством мик- ромеркуриализма при соответ- ствующей симптоматике (мор- фологические изменения кле- ток крови, нарушения вегета- тивной нервной системы)
Висмут Бийохииол, бисмоверол, пеита- бисмол, соли висмута Хронический висмутоз, висму- товый гингивит, стоматит; эи- цефалопатия
Платина Цисплатин Острая и хроническая циспла- тиновая нефропатия; ототок- сикоз; энцефалопатия, перифе- рическая и вегетативная ней- ропатия
Золото Ауранофин Преходящая диарея; мембра- нозная гломерулоиефропатия с отложением частиц золота в эпителии канальцев
436
тиб, это объясняется дефицитом эссенциальных МЭ с последую-
щим развитием гипомикроэлементозов различной степени тя-
жести.
В табл. 25 представлены сводные материалы по некоторым
формам ятрогенных МТОЗов, возникающих у больных, которым
с лечебной целью вводились препараты, содержащие в своем
составе избыточное или недостаточное количество определенных
МЭ. Число описанных в литературе симптомов и синдромов
неполно и с каждым годом оно будет увеличиваться как за счет
более широкого внедрения новых лекарственных препаратов,
в том числе и для инъекций, содержащих в своем составе опре-
деленные МЭ, так и за счет большего знакомства врачей с ука-
занной патологией.
В этой главе суммированы материалы по ятрогенным мик-
роэлементозам, зарегистрированным официальной медициной.
Однако наряду с этим МЭ широко используются в нетрадици-
онной медицине, в частности в гомеопатии, где они в течение
длительного времени считаются одними из основных фармако-
логических средств. Для научной разработки этот материал не
используется. Обобщающие работы, написанные по этому воп-
росу, нам не известны.
Глава 10
НАСЛЕДСТВЕННЫЕ ФОРМЫ МИКРОЭЛЕМЕНТОЗОВ
ЧЕЛОВЕКА И ЖИВОТНЫХ
Исследования генетических дефектов обмена МЭ у человека
и животных развивались в последние годы главным образом
в направлении расшифровки молекулярных механизмов извест-
ных нарушений, открытия новых генетических дефектов и раз-
работки методов их ранней пренатальной и постнатальной диаг-
ностики. Значительное развитие получили также исследования
генетических дефектов микроэлементного обмена на животных
моделях.
Генетические нарушения обмена МЭ можно подразделить
на три категории: первая затрагивает все обменные процессы
организма, вторая — только отдельные этапы метаболизма оп-
ределенных МЭ и третья — вторичные изменения микроэлемент-
ного обмена, вызванные иными генетическими дефектами
[Chan W. J., Rennert О. М., 1985].
10.1. Первичные нарушения обмена микроэлементов
при наследственных болезнях
Микроэлементы и генетический аппарат
клетки. Воздействие МЭ на обменные процессы реализуется
прежде всего через их влияние на функционирование генетиче-
ского аппарата. МЭ принадлежит важнейшая роль в процессах
метаболизма нуклеиновых кислот. Связываясь с нуклеотидами,
ионы металлов вызывают существенные изменения в структуре
нуклеиновых кислот. Присоединение металла может ослабить
электростатическое отталкивание между нуклеотидными цепя-
ми, привести к разрыву водородных связей между парами осно-
ваний и к нарушению третичной структуры полинуклеотидов.
Отсутствие ионов металлов ведет к дестабилизации двойной
спирали и ее частичной денатурации. Переходы между различ-
ными формами ДНК (Z, В и А) также зависят от ионов метал-
лов. Медь, хром и железо могут включаться в состав ДНК, об-
разуя поперечные сшивки между ее нитями и существенно ви-
доизменять ее структуру [Eichhorn G. L., 1984].
Точность передачи генетической информации в процессах
репликации, транскрипции и биосинтеза белка зависит от обра-
зования водородных связей между комплементарными парами
438
азотистых оснований. Установлено [Loeb L. A., Zakour R. А.,
1980], что в этом процессе принимают участие ионы металлов.
При их повышенных концентрациях наблюдаются неправильное
спаривание и ошибки в считывании информации. Структура
хроматина также зависит от концентрации ионов металлов, ко-
торые участвуют в процессах его конденсации и декомпактиза-
ции и влияют тем самым на надежность процессов репликации
ДНК и транскрипции РНК [Eichhorn G. L., 1984]. Ионы метал-
лов стабилизируют ядерные структуры. Показано, например,
что ионы Цинка и некоторых других металлов способны повы-
шать устойчивость ядер гепатоцитов амфибий и млекопитающих
к неблагоприятным воздействиям [Doyle М. J. et al., 1981].
Ряду МЭ присущи мутагенные и канцерогенные свойства,
объясняемые их способностью образовывать координационные
соединения с белками и малыми молекулами, что приводит
к нарушению их естественных функций. Сходный эффект может
возникнуть и в результате их воздействия на структуру нуклеи-
новых кислот и снижения надежности процессов репликации.
Способность металлов связываться с ДНК лежит в основе
успешного лечения некоторых видов рака препаратами циспла-
тина, которая связывает два соседних остатка гуанина. Ионы
металлов играют также важную роль в процессах старения
[Lawton А. Н., 1976]. С возрастом они накапливаются в клет-
ках, что объясняется несовершенством процессов их выделения
и транспорта. Повышенные концентрации ионов металлов влия-
ют на перенос генетической информации и ускоряют мутагенез
и канцерогенез.
Цинк является составной частью большого числа ферментов
нуклеинового обмена, в частности ДНК- и РНК-полимераз,
обратной транскриптазы, терминальной дезоксинуклеотидтранс-
феразы, тРНК-синтетазы, фактора инициации трансляции и не-
которых других [Vallee В. L., 1985]. В состав ряда этих фер-
ментов, кроме цинка, входят также магний и марганец. Це-
лостность рибосом, связывание тРНК и сохранение их структу-
ры также требуют наличия ионов металлов. В опытах in vitro
показано, что избыток ионов металлов вызывает ошибки в про-
цессе трансляции в результате неправильного спаривания азо-
тистых оснований либо ошибочного аминоацилирования тРНК.
Генетические нарушения обмена меди. Наи-
большее число выявленных генетических дефектов у человека
и животных относится к обмену меди. Они затрагивают, види-
мо, не только структурные гены, кодирующие синтез важней-
ших медьпротеидов (металлотионеина, церулоплазмина и др.),
но и регуляцию их экспрессии.
Первым генетическим дефектом обмена меди, обнаруженным
У человека, является болезнь Вильсона — Коновалова. Заболе-
вание носит аутосомно-рецессивный характер и встречается
только у гомозигот, составляющих по последним данным около
29 человек на 1 млн. Гетерозиготные носители этого аллеля
встречаются с частотой примерно 1 на 200 человек.
439
Ряд гипотез, предложенных для объяснения механизма раз-
вития этого заболевания, рассмотрен ранее [Риш М. А., 1983].
Ниже будут приведены только данные последних лет. В свете
этих данных сохраняется представление о центральной роли
церулоплазмина в патогенезе указанной болезни. Однако оно
дополнилось сообщениями о путях его выделения из организма,
остававшихся долгое время неизученными. Так, М. S. Kjressner
и соавт. (1984) показали, что десиализированный церулоплаз-
мин быстро исчезает из кровяного русла и поступает в гепато-
цит в результате эндоцитоза, происходящего с участием специ-
фического рецептора. При этом большая часть данного белка
переходит в желчь, минуя лизосомы, о чем свидетельствует
реакция иммунопреципитации радиоактивной меди в желчи
с антителом против церулоплазмина. Небольшая доля церуло-
плазмина, не утратившего углеводный компонент, поступает в
желчные канальцы через синусоидные сосуды печени. Таким
образом, при дефекте синтеза церулоплазмина неизбежно за-
трудняется выведение меди из организма, так как путь выделе-
ния ее с участием лизосом значительно менее эффективен.
В случае нормального содержания церулоплазмина в сыво-
ротке крови при болезни Вильсона — Коновалова предполага-
ют существование дефекта синтеза рецептора церулоплазмина
в мембране гепатоцита, что ведет к нарушению эндоцитоза это-
го белка и его задержке в организме.
Таким образом, генетический дефект при болезни Вильсо-
на — Коновалова ведет к нарушению выделения меди печенью
при почти нормальном ее поступлении. Неспособность печени
к нормальному выделению меди с желчью достоверно установ-
лена и в опытах С. Т. Walsh (1985), показавших, что при этой
болезни способность гепатоцитов выделять медь снижена по
сравнению со здоровыми лицами в десятки и сотни раз. Осо-
бенно наглядно это заметно при анализе пигментированных
желчных камней, которые у лиц с болезнью Вильсона—Коно-
валова содержат в среднем в 20 раз меньше меди, чем в норме
(0,79 против 16,8 ммоль/кг соответственно). Неспособность пе-
чени к выделению меди четко прослеживается при наружном
сканировании организма больного во времени с радиоактивным
изотопом меди 64Си. Сканирование проводят в положении лежа
на спине от головы к ногам. На сканограмме здоровых лиц
через 24 ч после внутривенного введения радиоизотопа в обла-
сти печени появляется четкий пик, который через 48 и 72 ч до-
полняется вторым пиком в нижней части живота. У больных
второй пик как при поступлении, так и после лечения хелати-
рующими препаратами выражен очень слабо.
Болезнь Менкеса. В 1962 г. J. Menkes впервые описал
у человека редкое (частота 1 на 35 000 новорожденных) наслед-
ственное заболевание, сцепленное с полом, получившее назва-
ние трихополидистрофии (ТПД) [Menkes J. et al., 1962].
Клинические симптомы заболевания легко объяснимы выпа-
дением функций ряда медьсодержащих ферментов — тирозина-
440
зы (депигментация волоса), сульфидоксидазы (нарушение
процессов кератинизации), лизилоксидазы (поражение соедини-
тельной ткани), дофаминф-гидроксилазы (синтез адреналина и
норадреналина), цитохром-с-оксидазы (нейродегенеративные
явления).
Основным метаболическим дефектом при этом заболевании
является неспособность клеток слизистой оболочки обеспечить
поступление всосавшейся меди в кровяное русло. При этом
в энтероцитах накапливается значительное количество данного
МЭ. Указанный дефект встречается и в ряде других тканей.
Повышенное содержание меди обнаружено в почках (при за-
трудненной реабсорбции этого МЭ), поджелудочной железе,
костях, мышцах, амниотических клетках и жидкости. Аномаль-
но высокое содержание меди было найдено также в мозге пло-
да с подозрением на ТПД, в культурах фибробластов и почеч-
ных клеток. Исключение составляет печень, которая бедна этим
элементом.
Наиболее вероятный патогенетический механизм ТПД пред-
ложен канадскими исследователями [Riordan J. R.. Jolicour-
Paquet L., 1982], установившими, что мутантные лимфобласты,
культивируемые в среде с низким содержанием меди (0,015—
15,7 мкмоль), накапливают в 10 раз, а при более высоких кон-
центрациях (157 мкмоль/л) в 2—3 раза большее количество
этого элемента, чем в норме. Мутантные лимфобласты отлича-
ются также повышенным (в 5—6 раз) содержанием металлотио-
неина, причем все добавочное количество металлов, накапли-
вающихся в клетке, обнаруживается в составе данного белка.
Авторы заключают, что основным проявлением метаболиче-
ского дефекта ТПД является неспособность клеток ограничи-
вать биосинтез металлотионеина, причем эта особенность рас-
пространяется на все органы и ткани, исключая печень. Проч-
ное связывание клеточной меди металлотионеином, делающее
ее недоступной для других, нуждающихся в ней белков, может
служить объяснением парадоксального факта — появления не-
достаточности меди при повышенном содержании данного эле-
мента в клетке. Опыты с культурой фибробластов, выполнен-
ные в трех различных лабораториях, подтверждают данную
гипотезу [Chan W. J., Rennert О. М., 1985]. Они свидетель-
ствуют, что в фибробластах детей, страдающих болезнью Мен-
кеса, при повышенном уровне меди в клетке существенно сни-
жена активность диаминоксидазы, лизилоксидазы, цитохромо-
ксидазы, что указывает на дефект внутриклеточного транспорта
меди. Важно, что при болезни Вильсона — Коновалова фибро-
бласты также накапливают высокие концентрации меди, однако
активность медьсодержащих ферментов при этом не снижается.
Поскольку ген МТ-1 у мышей локализован в хромосоме 8,
а «пятнистая» мутация (mottled), аналогичная дефекту Менке-
са, находится в Х-хромосоме, то связь между обоими генами
остается неясной. Нет связи и между геном болезни Менкеса,
находящемся в Х-хромосоме, и генами металлотионеина, лока-
441
лизованными на хромосоме 16 человека [Schmidt Р. et al.,
1984]. В этой связи при болезни Менкеса наиболее вероятно
нарушение регуляции экспрессии генов металлотионеина. Пока-
зано, что кадмий, цинк, медь и ртуть способны индуцировать
транскрипцию гена МТ-1 [Durman D., Palmiter R., 1981], опре-
деляющую скорость биосинтеза металлотионеина. Повышенный
неконтролируемый синтез этого белка может привести к его
накоплению в клетке и прочному связыванию всех присутствую-
щих в ней металлов.
Конкретный механизм запуска и остановки синтеза металло-
тионеина изучен недостаточно. В почках и печени плода метал-
лотионеин обнаружен иммуногистохимическими методами в
ядре, а не в цитоплазме, причем в ядре его находят только
после введения тяжелых металлов [Cherian М. G., 1982]. Эти и
некоторые другие наблюдения позволяют считать, что ядерный
металлотионеин играет важную регуляторную роль, контроли-
руя вместе с апотионеином транскрипцию и трансляцию данно-
го белка в клетке. Тяжелые металлы, поступающие в цитозоль,
образуют комплекс с металлотионеином, дерепрессируя транс-
крипцию его мРНК. Комплекс металла с тионеином также по-
ступает в ядро, где он связывается с негистоновыми белками и
индуцирует синтез дополнительных количеств тионеиновой
мРНК- Когда в норме количество тионеина превышает таковое
несвязанных ионов металлов, его дальнейший синтез прекра-
щается.
Несколькими группами исследователей получены клоны
к ДНК и геномной ДНК тионеина мыши [Cousins R., 1985],
с помощью которых оказалось возможным осуществить гибри-
дизацию ДНК с РНК для ее количественного определения и
проведения пренатальной и ранней постнатальной диагностики
заболеваний, связанных с нарушениями обмена металлотионеи-
на, как это уже практикуется при серповидно-клеточной анемии
[Chang J. С., Kan J. W., 1982]. Методами гибридизационного
анализа в ПААГ обнаружены тионеиновые гены у человека
в хромосоме 16 [Schmidt Р. et al., 1984].
В другом методе пренатального диагноза используется в ка-
честве генетического маркера у гемизигот с дефектом Менкеса
способность культуры клеток, полученных из амниотической
жидкости не позднее 17-й недели беременности, накапливать
троекратное против нормы количество радиоактивной меди. Ана-
логичный метод с использованием культуры фибробластов по-
зволяет выявить женщин — гетерозиготных носителей данного
дефекта. Учитывая, однако, спонтанную мозаичность клеток по
Х-хромосоме, согласно правилу Лайон, картина поглощения ра-
диоактивной меди в данном случае выражена менее четко, чем
у гемизигот [Horn N., 1983].
Недостаточность а-антитрипсина. Накопление
меди в печени отмечено и при наследственной недостаточности
а-антитрипсина (ААТ) в плазме крови человека [Callea F. et al.,
1981]. Это заболевание носит аутосомно-рецессивный характер
442
и наблюдается у лиц, гомозиготных по двум аллелям (Piz и Pis)
23-аллельной системы генного локуса Pi (ингибитор протеаз).
У 10—20% новорожденных детей с генотипом Pis/Z развивается
цирроз печени с летальным исходом, а у взрослых с данным ге-
нетическим дефектом — эмфизема легких.
Связь между недостаточностью ААТ и медным токсикозом
показана и при индийском детском циррозе, при котором в пе-
чени детей накапливается медь в количестве 22,0—75,5 ммоль/кг
(на сухую массу).
Известен и ряд других генетических дефектов обмена меди
у человека. К их числу относятся как умеренные формы болез-
ни Менкеса [Camacaris Т. et al., 1983; Willemse J. et al., 1982],
так и генетические заболевания с признаками неврологических
расстройств, отличающихся по своим клиническим проявлениям
и биохимическим сдвигам от данной болезни [Haas R. et al.,
1981].
Присутствие меди в составе ряда ферментов, принимающих
участие в метаболизме соединительной ткани, таких как лизил-
оксидаза и оксидаза аскорбиновой кислоты, допускает мысль
о роли этого металла в происхождении синдрома Элерса — Дан-
ло IX с дефектом лизилоксидазы, а также в патогенезе таких
расстройств, как идиопатический ювенильный сколиоз, синдром
Марфана, аневризмы аорты и других сосудов.
Как уже отмечалось, многие генетические дефекты обмена
меди у животных обнаруживают близкое сходство с наследст-
венными заболеваниями у человека и могут рассматриваться как
их модели. Это в первую очередь относится к наследственный
дефектам обмена меди у мышей и собак.
Генетические дефекты обмена цинка. Энтеропа*
тический акродерматит, описанный как самостоятельная нозо^
логическая единица в 1942 г., представляет собой редкое на<
следственное заболевание аутосомно-рецессивного типа, хараю
теризующееся типичными признаками цинковой недостаточно-
сти — нарушениями кожного покрова, диареей и облысением,
отставанием в росте и развитии и ослаблением иммунитета
[Hambidge К- М. et al., 1978]. Начиная с 1953 г., для лечения
этого заболевания чисто эмпирически с успехом начали приме-
нять галоидопроизводные ортооксихинолина, образующие с цин-
ком легко усвояемые комплексы. Лечебный эффект оказывает
также женское (но не коровье) молоко. Связь данного заболе-
вания с нарушением обмена цинка и высокий лечебный эффект
соединений этого элемента были открыты около 17 лет назад
[Moynahan Е., Barnes Р. М., 1973]. Тогда же было показано,
что при нем нарушен также обмен триптофана [Robertson А.
et al., 1973].
Рецессивные генетические дефекты обмена меди у мышей
представлены: 1) пятью мутантными аллелями локуса «mottled»
(«крапчатый»), расположенного в Х-хромосоме (Мо, МоЬг,
Modp, Mobl°, Movbr). Они аналогичны болезни Менкеса; 2) ауто-
сомной мутацией «crincled» («сморщенный») (сг), вызывающе
44
недостаточность меди, нарушение пигментации, истончение ко-
жи, аномальный состав липидов мозга; 3) аутосомной мутацией
«quaking» («дрожащий») (qk), проявляющейся медной недоста-
точностью, аномалиями строения миелина, тремором; 4) ауто-
сомной мутацией «токсичное молоко» (tx), характеризующейся
накоплением меди в печени матерей, медной недостаточностью
у потомства.
У бурых норвежских крыс описана аутосомная мутация, ве-
дущая к медному токсикозу, а у бедлингтонтерьеров, хайланд-
терьеров, доберманпинчеров и других пород собак аутосомная
мутация, ведущая к медному токсикозу, аналогичная болезни
Вильсона — Коновалова.
* Клинические симптомы проявляются обычно в первые меся-
цы жизни и наступают при отнятии ребенка от груди и перехо-
де на коровье молоко.
Поскольку предполагалось, что при энтеропатическом акро-
дерматите нарушено всасывание цинка в пищеварительном
тракте [Lombeck I. et al., 1975], несколько групп исследовате-
лей поставили цель выделить из грудного молока и пищевари-
тельных секретов специфический лиганд, связывающий цинк и
способствующий его проникновению в эпителий слизистой обо-
лочки. В качестве такого лиганда в разное время назывались
аминокислоты, пептиды, триметилэтандиамин, простагландин Ег,
низкомолекулярные белки, лимонная и пиколиновая кислоты.
Допускалось также, что этим лигандом является продукт рас-
пада металлотионеина. Однако многие из перечисленных лиган-
дов оказались артефактом, возникшим в результате погрешно-
стей процедуры выделения, не учитывавшей действие протеаз.
В настоящее время считается, что различие действия ко-
ровьего и грудного молока объясняется тем обстоятельством,
что в первом цинк связан преимущественно с казеином, а в груд-
ном — с лактоферрином. Створаживание казеина в желудке ре-
бенка может сделать недоступным связанный с ним цинк, тогда
как лактоферрин распределяется тонким слоем на поверхности
слизистой оболочки кишечника и может выступать не только
в роли донора железа, но и цинка. Исходя из этого авторы за-
ключают, что регуляция всасывания цинка осуществляется не
столько низкомолекулярными небелковыми комплексами цинка,
сколько его соединениями с белками и их различной перевари-
мостью. Существует и ряд других гипотез о патогенетическом
механизме энтеропатичеокого акродерматита. Допускается, что
при этом заболевании преобладают процессы выделения цинка
стенкой кишечника над его всасыванием. Кроме того, возмож-
но, что в эпителии кишечника происходит прочное связывание
цинка внутриклеточным лигандом, подобным металлотионеину
[Cousins R., 1983]. Имеются также данные, что метаболический
дефект при энтеропатическом акродерматите вызван наруше-
нием всасывания полиненасыщенных жирных кислот или неспо-
собностью организма к превращению линолевой кислоты в
7-линоленовую с последующим синтезом простагландина Ei
444
[Cunnane S. C., 1982]. Это предположение основывается на ано-
мальном составе плазменных липидов при данном заболевании,
положительном лечебном действии парентерального введения
хлопкового масла, богатого линолевой кислотой, и отсутствии
подобного эффекта при введении арахидоновой кислоты.
Автор объясняет эти наблюдения необходимостью цинка для
всасывания ненасыщенных жирных кислот или конверсии "(-ли-
ноленовой кислоты в простагландины, что позволило бы также
объяснить лечебное действие хлопкового масла. Однако следует
признать, что окончательной ясности относительно метаболиче-
ского дефекта при данном заболевании пока нет.
Дефицит цинка и его положительный лечебный эффект отме-
чены и при других наследственных и врожденных заболеваниях
человека, таких как болезнь Крона, серповидно-клеточная ане-
мия, кистозный фиброз поджелудочный железы, нефротический
синдром Фанкони и др. Однако в этих случаях дефицит носит
вторичный характер и не является ведущим патогенетическим
фактором.
Заболевание, близкое к энтеропатическому акродерматиту,
обнаружено у крупного рогатого скота. Оно имеет несколько
названий: «летальный синдром А-46», «наследственный параке-
ратоз», «наследственная гипоплазия тимуса», «наследственная
цинковая недостаточность». Заболевание полностью излечивает-
ся ежедневной дачей животным 100 мг цинка.
Было установлено, что все пораженные животные имеют
одного общего предка — быка Адема, в связи с чем это забо-
левание получило также название «болезнь Адема». Заболева-
ние обусловлено рецессивным аутосомным геном и проявляет-
ся только у гомозигот.
Отличительной чертой болезни является выраженная гипо-
плазия вилочковой железы. Гистологически обнаружено резкое
снижение числа малых лимфоцитов, особенно в кортикальной
области железы. Отмечается также гипоплазия селезенки, ре-
гиональных лимфоузлов и лимфоидной ткани, расположенной
вдоль кишечника. Клинические и морфологические данные сви-
детельствуют о значительном угнетении иммунной системы, ко-
торая полностью восстанавливается при даче цинка. Как и при
энтеропатическом акродерматите, ключевым дефектом при дан-
ном синдроме является нарушение всасывания цинка в желу-
дочно-кишечном тракте, что дало основание отдельным авторам
рассматривать болезнь Адема как модель энтеропатического
акродерматита.
Следует, однако, заметить, что большинство схем патогене-
тического механизма энтеропатического акродерматита учиты-
вают различия связывания цинка в грудном и коровьем молоке.
Поскольку в норме подсосные телята не испытывают цинкового
голодания, то либо у них функционирует иной механизм всасы-
вания цинка, чем у человека, либо существует еще другое объ-
яснение этого генетического дефекта, общее для обоих заболе-
ваний. В качестве такого объяснения можно принять гипотезу,
445
согласно которой, как уже отмечалось, при энтеропатическом
акродерматите и болезни Адема имеет место нарушение функ-
ции клеточных мембран, которая восстанавливается рядом ли-
пофильных соединений, таких как дийодохин, полиненасыщен-
ные жирные кислоты и витамин Е, способствующих заметному
ослаблению симптомов цинковой недостаточности, не изменяя
существенно цинковый статус организма [Bettgor W. J., 1979].
Генетические дефекты обмена цинка описаны и у лабораторных
животных. Ниже мы приводим рецессивные аутосомные мута-
ции, затрагивающие обмен цинка у животных. Это: 1) болезнь
Адема у крупного рогатого скота, аналогичная энтеропатиче-
скому акродерматиту; 2) мутация «летальное молоко» у мышей
(im), ведущая к гибели потомства мутантных мышей; 3) синд-
ром тестикулярной феминизации у крыс (Tfm), характеризую-
щийся нечувствительностью к андрогенам, крипторхизмом, ана-
логичный синдрому тестикулярной феминизации у человека;
4) мутация SM. («супермыши»), при которой происходит на-
копление цинка в организме и экономное расходование этого
элемента; 5) устойчивость мышей к токсическому действию кад-
мия (cdm).
Генетические дефекты обмена железа. У чело-
века известны два генетических дефекта, затрагивающие обмен
железа, — гемохроматоз и атрансферринемия. Гемохроматоз
характеризуется накоплением в организме повышенного количе-
ства железа. Экспрессия этого дефектного гена зависит от его
наличия в гомозиготном состоянии, количества железа в пище
и периодического расходования этого МЭ. Несмотря на то что
этот генетический дефект известен уже более 50 лет, его при-
рода остается не изученной. Допускается [Walters G. О. et al.,
1975], что причиной повышенного поступления в организм же-
леза является нарушение синтеза ферритина в энтероцитах при
одновременном повышенном образовании его транспортной фор-
мы, ускоряющей усвоение этого металла. В клетках СМФ,
а также в слизистой оболочке кишечника нарушена способность
депонировать железо, в результате чего оно накапливается
в печени и других органах.
Выяснить характер исследования этого синдрома было труд-
но в связи с его значительной вариабельностью у гомозигот и
преимущественной встречаемостью у мужчин. Окончательно
этот вопрос был решен в результате установления тесного сцеп-
ления между геном гемохроматоза и отдельными антигенами
гистосовместимости системы HLA [Simon М., Botirel М., 1984].
Было показано, что у больных гемохроматозом тканевые анти-
гены HLA-A3 и HLA-B14 встречаются со значительно большей
частотой, чем у здоровых лиц. Это сделало возможным проведе-
ние посемейного анализа, выявившего, что ген первичного гемо-
хроматоза не только расположен в непосредственной близости
от локуса МНС на коротком плече хромосомы 6, но обычно на-
следуется вместе с соседним гаплотипом HLA [Kravitz К. et al.,
1979]. Далее было показано, что у сибсов с симптоматическим
446
гемохроматозом всегда встречаются оба гаплотипа HLA-систе-
мы. Это подтвердило, таким образом, аутосомно-рецессивный
характер наследования данного дефекта [Simon М., Bourel М.,
1984]. Выявление гетерозиготных носителей среди родственни-
ков больных гемохроматозом по типам HLA позволило устано-
вить частоту встречаемости этого гена. Она составляет 1 на
300—400 человек [Garnica A. D. et al., 1985].
Другим генетически обусловленным нарушением обмена же-
леза является атрансферринемия, обнаруженная впервые у трех-
месячной девочки с гипохромной анемией. Механизм возникнове-
ния атрансферринемии не изучен, но имеются данные, что струк-
тура синтезируемого в ограниченном количестве трансферрина
не отличается от нормы. Следует также отметить, что ни один
из множества типов трансферрина, обнаруженных у мыши и че-
ловека, не связан с функциональными нарушениями обмена же-
леза [Bannerman R. М., 1976]. У животных известны следую-
щие генетические дефекты обмена железа: 1) «анемия, сцеп-
ленная с полом» у мышей (sla), тяжелая гипохромная анемия
у гомозиготных самок и гемизиготных самцов, излечивается со-
лями железа («антигемохроматозная модель»). Дефект обмена
железа наблюдается «на выходе» из клетки; 2) микроцитарная
анемия у мышей (mk), обусловленная аутосомной рецессивной
мутацией, вызывающей тяжелую гипохромную анемию. Дефект
обмена железа наблюдается «на входе» в клетку; 3) белград-
ская анемия у крыс (Ь) — аутосомная рецессивная мутация, ве-
дущая к тяжелой гипохромной анемии.
Генетические дефекты обмена марганца у
человека неизвестны. Они наблюдаются у мышей с аутосомной
рецессивной мутацией, носящей название «pallid» (блеклый),
генетический символ — ра. Мыши, гомозиготные по этому гену,
приносят потомство с признаками выраженной марганцевой не-
достаточности: врожденной атаксией, нарушением координации.
Эти нарушения объясняются аномальным развитием отолитов
во внутреннем ухе, необходимых для нормального функциониро-
вания вестибулярного аппарата.
Для предупреждения атаксии и дефекта отолитов у потом-
ства самок, гомозиготных по гену ра, им необходимо скармли-
вать не позднее 14—17-го дня беременности корм, содержащий
27,3 — 36,4 ммоль/кг марганца (в норме 0,82 ммоль/кг). Му-
тантный ген, аналогичный гену ра, найден у пастельных норок
светло-коричневой окраски.
Генетические дефекты обмена молибдена.
В организме человека молибден является необходимым струк-
турным компонентом по меньшей мере трех ферментов: суль-
фитоксидазы (КФ 1.8.3.1), ксантиндегидрогеназы (КФ 1.2.1.37)
и альдегидоксидазы (КФ 1.2.3.1). Нарушение синтеза каждого
из этих ферментов проявляется специфическими клиническими
признаками. Так, недостаточность ксантиндегидрогеназы, опи-
санная у более чем 50 больных, приводит к уролитиазу в связи
с низкой растворимостью ксантина. Недостаточность сульфиток-
447
сидазы приводит к значительно более тяжелым последствиям:
организм утрачивает способность превращать сульфит в суль-
фат, что приводит к активизации побочных путей обмена серы.
В результате в органах и тканях происходит накопление S-суль-
фо-Ь-цистеина, сульфита и гипосульфита при практическом от-
сутствии сульфата. В моче повышается количество названных
соединений, а также таурина при низком уровне сульфатов. От-
мечаются прогрессирующие нарушения нервной системы, под-
вывих хрусталика, умственная отсталость. Дети погибают при
этом дефекте в возрасте 2—3 лет. Недостаточность альдегид-
оксидазы отдельно не описана. Известно только о существова-
нии ее изоферментов у некоторых этнических групп.
Перечисленные ферменты млекопитающих, как и молибден-
содержащие ферменты у других организмов, содержат этот ме-
талл в составе молибденового кофактора — молибдоптерина
[Johnson G. F. et al., 1980]. Нарушение биосинтеза молибдено-
вого кофактора приводит к дисфункции сразу всех трех пере-
численных ферментов.
Генетические дефекты йодного о б м е н а. Извест-
но пять основных типов генетических дефектов синтеза тиреоид-
ных гормонов и обмена йода, сопровождающихся зобом (семей-
ный зоб, врожденный гипотиреоз): 1) неспособность железы
концентрировать йод; 2) нарушение синтеза тиреоидных гормо-
нов, сочетающееся с тугоухостью (синдром Пендреда), вызван-
ное дефектом синтеза тироидпероксидазы; 3) неспособность
к дейодированию M.IT и DIT, вызванная дефектом синтеза спе-
цифической дегалогеназы (дейодазы), приводящим к наруше-
нию реутилизации йода щитовидной железой и его повышенным
потерям из организма; 4) неспособность синтеза Т4 из M.IT и
DIT; 5) циркуляция в крови атипичного белка, прочно связы-
вающего йод, который теряет способность экстрагироваться бу-
танолом. Предупреждение гипотиреоза при этих дефектах до-
стигается назначением тироксина и препаратов йода [Бада-
лян Л. О., 1980] при условии ранней диагностики (на первом
месяце жизни или во внутриутробном периоде), успешно осу-
ществляемой в СССР.
Кроме первичного гипотиреоза, вызванного перечисленными
выше дефектами синтеза ферментов щитовидной железы, извест-
ны и редкие случаи недостаточности синтеза ТТГ (вторичный
гипотиреоз) и TRH (третичный гипотиреоз). Имеются также
заболевания щитовидной железы, связанные с нарушением им-
мунной системы. В США, например, наиболее распространенной
причиной гипотиреоза и зоба у детей старше 6 лет является
аутоиммунное поражение щитовидной железы (болезнь Хаши*
мото), при которой в крови больных обнаруживаются антитела
против микросом щитовидной железы и тироглобулина [Alexan-
der N. М., 1984].
В человеческих популяциях обнаружен генетический поли-
морфизм по способности ощущать вкус фенилтиомочевины и
других близких гойтерогенов типа гойтрина. Не исключена воз-
448
цожность, что лица, ощущающие присутствие в пище гойтероге-
нов, в меньшей степени подвергаются их воздействию.
Наследственные нарушения обмена йода и врожденный зоб
описаны у большинства видов сельскохозяйственных живот-
ных— мериносовых овец в Австралии, у коз в Нидерландах^
у которых обнаружена нефункциональная мРНК тиреоглобули-
на. Широкое распространение врожденный зоб получил среди
крупного рогатого скота породы Африкандер [Ricketts М.. Н.
et al., 1985] в Южной Африке, у которых этот генетический де-
фект носит аутосомно-рецессивный характер.
Генетические дефекты обмена кобальта. У че-
ловека известен ряд генетических дефектов, затрагивающих
синтез кобаламинзависимых ферментов и его транспортных бел-
ков. К кобаламиновым энзимопатиям относятся: дефицит тетра-
гидроптероилглутаматметилтрансферазы, приводящий к повы-
шенному выделению с мочой гомоцистеина, цистатионина и ме-
тилмалоната, сочетающийся с пониженной резистентностью
к инфекциям, отставанием в росте, умственном развитии и ма-
локровием; недостаточность метилмалонил-СоА-мутазы, при-
водящая к повышенному содержанию в крови и моче метилма-
лоновой кислоты, ацидозу и кетонурии. Кроме того, аналогич-
ные биохимические сдвиги наблюдаются при дефекте синтеза
дезоксиаденозилкобаламинтрансферазы. Гомоцистеинурия и по-
вышение содержания в крови метилмалоновой кислоты встреча-
ются также при нарушении синтеза редуктазы кобаламина.
При дефекте синтеза транспортных белков отмечены: 1) пер-
нициозная анемия (болезнь Аддисона — Бирмера) у взрослых »
значительно реже у детей, характеризующаяся недостаточно-
стью внутреннего фактора Касла; 2) специфическая мальаб-
сорбция витамина Bi2 (болезнь Имерслунд—Грасбека); 3) де-
фицит транскобаламина II, сопровождающийся тяжелыми нев-
рологическими расстройствами и 4) дефицит транскобаламина I,
приводящий при адекватном уровне витамина Bi2 лишь к не-
значительным патологическим изменениям [Rajagopalan К. V.,
1984].
Генетические дефекты обмена селена. Описано
10 случаев наследственного дефицита глутатионпероксидазы
эритроцитов у человека, причем допускается, что лица с актив-
ностью фермента, составляющей 47—78% (в среднем 66%) от
нормы, являются гетерозиготными, а с активностью 17—34% —
гомозиготными носителями аллеля низкой активности GSH-Px.
У тех и других наблюдаются хроническая гемолитическая ане-
мия, метгемоглобинемия различной тяжести, тельца Гейнца и
повышенная чувствительность к лекарственным препаратам
(ацетилсалициловой кислоте, сульфисоксазолу, фурадонину
и др.). Наследственный дефицит GSH-Px обнаружен также
у новорожденных, среди которых его частота составляет около
2% [Emerson Р. М. et al., 1972]. У детей с этим дефектом опи-
саны умеренный гемолиз и гипербилирубинемия [Ganther Н.
et al, 1976].
29—568
449'
Наследственный дефицит GSH-Px в тромбоцитах описан
у 3 лиц с геморрагической тромбоцитоастенией Гланцманна —
Риникера [Karpatkin S., Weiss Н., 1972]. Дефицит фермента со-
четается с двукратным повышением содержания кислотораство-
римых тиолов в тромбоцитах, свидетельствующим о их неспо-
собности метаболизировать глутатион, что повышает восприимчи-
вость тромбоцитов к окислительному стрессу. Авторы отмечают,
что в норме тромбоциты относительно богаты GSH-Px и глута-
тионом и что при дефиците фермента могут иметь место пере-
кисное окисление мембраны тромбоцитов или окисление
SH-групп тромбоцитарных белков, в частности тромбоксана,
участвующего в агрегации тромбоцитов. Есть предположение,
что дефицит GSH-Px встречается и при синдроме Германско-
го— Пудлака [Witkop С. et al., 1973].
Генетический контроль активности GSH-Px описан также
у животных. Так, установлены достоверные различия между
породами по активности GSH-Px эритроцитов и уровня селена
крови у овец [Wiener G. et al., 1983] и высокая наследуемость
у них активности этого фермента, составляющая г = 0,5+0,2. На-
следственные различия в реакции на дефицит селена описаны
также у кур.
10.2. Вторичные нарушения обмена микроэлементов
при наследственных болезнях
Разделение генетических нарушений обмена МЭ на первич-
ные и вторичные весьма условно, так как если изменение
лигандного окружения элемента приводит к нарушению его ме-
таболических путей, то любая аномалия элементного состава
в свою очередь отражается на структуре и функции самих ли-
гандов. Не всегда можно руководствоваться также принципом
ведущего патогенетического фактора, так как при ряде наслед-
ственных болезней «вторичный» дефицит или избыток МЭ мо-
жет определять тяжесть или даже исход заболевания. Все же
этим термином удобно пользоваться для описания нарушений
микроэлементного обмена при известных генетических дефектах,
для которых этот аспект болезни ранее оставался в тени.
Среди «вторичных» генетических дефектов микроэлементно-
го обмена важное место занимают явления цинковой недоста-
точности. Они наблюдаются при болезнях, сопровождаемых
мальабсорбцией, как, например, при целиакии, при которой на-
рушено усвоение и ряда других компонентов пищи, и во всех
случаях гиперцинкурии, отмечаемой при целом ряде состояний,
связанных с усилением катаболических процессов. Гиперцинку-
рия наблюдается при хронических заболеваниях печени, нефро-
тическом синдроме, серповидно-клеточной анемии и др. Удалось
показать, что ряд клинических симптомов, наблюдаемых при
этих заболеваниях, вызван дефицитом цинка и поддается лече-
нию его препаратами. В частности, при серповидно-клеточной
анемии задержку физического развития и полового созревания
450
у подростков, недоразвитие волосяного покрова, истощение, по-
терю аппетита можно рассматривать как явления цинковой не-
достаточности. У многих больных серповидно-клеточной анемией
на нижних конечностях образуются стойкие изъязвления, под-
дающиеся лечению препаратами цинка. Цинк способен не толь-
ко снимать вторичные явления при этом недуге, но и преду-
преждать его первичные нарушения — полимеризацию гемогло-
бина в эритроцитах и улучшать их фильтрацию через капилля-
ры. Это действие цинка объясняется его способностью стаби-
лизировать эритроцитарные мембраны, ограничивать поступле-
ние в клетку кальция, участвующего в процессе полимеризации
гемоглобина и образовании дрепаноцитов (серповидных эритро-
цитов) [Prasad A. S., 1983].
Причиной истинно вторичных МТОЗов может быть примене-
ние лекарственных препаратов и полусинтетических рационов,
используемых при лечении отдельных генетических недугов.
Так, при фенилкетонурии и «болезни запаха кленового сиропа»
у больных наблюдается снижение содержания в организме се-
лена. При фенилкетонурии, повышенном уровне в крови метил-
малоновой кислоты, галактоземии отмечается снижение уров-
ня цинка и марганца, обнаруживаемое при анализе содержания
этих элементов в волосах [Harry V. J., Gibson R. S., 1982]. При
длительном лечении болезни Вильсона — Коновалова пени-
цилламином может развиться выраженная недостаточность
цинка.
Другим генетическим дефектом, тесно связанным с наруше-
нием обмена МЭ, по-видимому, являются некоторые формы бо-
лезни Альцгеймера, характеризующиеся прогрессирующей деге-
нерацией клеток головного мозга в пресенильном и старческом
возрасте. Существует шесть разных гипотез о причинах болез-
ни, допускающие, что она обусловлена генетическим дефектом,
отложением аномального белка, инфекцией, токсическим дейст-
вием алюминия, сосудистой недостаточностью, дефицитом аце-
тилхолина.
Гипотеза, рассматривающая в качестве причины болезни
влияние алюминия, основана на наблюдении, что в измененных
нейронах, содержащих клубочки нейрофибрилл, накапливается
повышенное количество этого МЭ. Она подкрепляется и экспе-
риментами на кроликах и кошках, у которых после введения
солей алюминия в нейронах мозга образуются клубки нейро-
фибрилл, аналогичные «альцгеймеровским», причем этот эф-
фект достигается при концентрациях металла, наблюдаемых
при данной болезни у человека. В пользу данной гипотезы го-
ворит также накопление алюминия в мозге людей, подвергав-
шихся гемодиализу растворами с повышенным содержанием
этого МЭ, у которых развивается необратимое слабоумие. Ме-
ханизм действия алюминия связывают с его способностью инги-
бировать ряд ферментов головного мозга и нарушать транспорт
белков, из которых формируются нейрофибриллы. Наиболее
изученными ферментными нарушениями при болезни Альцгей-
29*
451
мера являются недостаточность фосфофруктокиназы и холина-
цетилтрансферазы. Найдено, что алюминий связан с хромати-
ном и образует сшивки между белком и ДНК [Crapper-Mc-La-
chlan D., Eichhorn G. L.( 1984].
Против «алюминиевой гипотезы» говорят различия в строе-
нии нейрофиламентов при болезни Альцгеймера и в модельных
экспериментах. Вполне возможно, что связывание алюминия
клубками нейрофибрилл объясняется их сродством к данному
металлу и таким образом является вторичным процессом. Не ис-
ключено, однако, что алюминий, не будучи ведущим фактором,
способствует усилению последствий первичного дефекта, какой
бы ни была его природа.
А. И. Ойфа (1987) придает основное значение амилоидозу
головного мозга, хотя и указывает, что в ЦНС старых людей
и, в частности, при сенильном слабоумии количество алюминия
повышено.
Дегенеративными изменениями мозга, связанными с накопле-
нием другого МЭ — цинка, характеризуется болезнь Пика.
Накапливаясь в головном мозге, этот МЭ может вызвать по-
вреждение нейронов либо сам, либо через активацию цинксо-
держащей глутаматдегидрогеназы, что влечет за собой нару-
шение обмена глутаминовой кислоты — основного медиатора
возбуждения ЦНС. Это приводит к поражению синапсов, тел
нейронов и появлению в них аргирофильных включений. Пред-
положение о роли цинка в патогенезе болезни Пика, очевидно,
подтверждается лечебным эффектом ЭДТА на ее ранних ста-
диях, способствующим выведению из организма избытка этого
МЭ [Constantinidis J., Tisset R., 1981].
Несомненно, что существует множество других «вторичных»
нарушений микроэлементного обмена, но приведенных выше
примеров вполне достаточно, чтобы привлечь внимание исследо-
вателей и практических врачей к изучению обмена МЭ при
любых генетических аномалиях.
Глава И
ВРОЖДЕННЫЕ ПОРОКИ РАЗВИТИЯ
ПРИ ДЕФИЦИТЕ И ИЗБЫТКЕ
МИКРОЭЛЕМЕНТОВ
В настоящее время пороки развития рассматривают как
«стойкие морфологические изменения органа или организма,
выходящие за пределы вариации их строения и возникающие
внутриутробно в результате нарушений развития зародыша,
плода или иногда после рождения ребенка как следствие нару-
шения дальнейшего формирования органов». Автор приведенно-
го определения Г. И. Лазюк (1983) ссылается на расширенное
толкование этого понятия С. Berry и соавт. (1978), согласно ко-
торым пороки развития — это «отклонение формы, химического
состава или функции». В более узкой трактовке D. W. Smith
(1975) пороки развития — это только «первичные структурные
дефекты, являющиеся результатом локализованного неправиль-
ного морфогенеза» [Лазюк Г. И., 1983].
Все приведенные определения пороков развития по своему
рациональны, но они не содержат отграничения тератогенеза
от наследственных форм патологии возникающего и развиваю-
щегося организма. У многих авторов пороки развития находят-
ся в одной рубрике с наследственными заболеваниями. Вместе
с тем в докладах ВОЗ (1978) специально выделяются «немен-
делевские пороки развития» и учитывается их самостоятельная
роль среди болезней человека.
Синонимом термина «пороки развития» является термин
«врожденные уродства», которые в англоязычной литературе
обозначаются как мальформации. Этот термин перекочевал и
в отечественную медицинскую литературу Р. I. Aggett и соавт.
(1987), основываясь главным образом на результатах модель-
ных опытов и широком изучении этого вопроса на сельскохозяй-
ственных животных, к врожденным мальформациям относят
очень большое число патологических состояний и процессов,
которые возникают в связи с воздействием факторов окружаю-
щей среды на почву, воду, растения и животных. Оно способ-
ствует проникновению в организм многих необходимых для
жизни неорганических веществ, а также поступлению тех МЭ,
которые накапливаются в окружающей среде в результате ан-
тропогенной деятельности, в частности при плавке металлов,
сооружении и эксплуатации шахт, водохранилищ и др. Разу-
меется, химические вещества из биогеохимических провинций
с превышающим норму содержанием различных МЭ могут
453
включаться в пищевую цепь и оказывать влияние на здоровье
человека. Пороки развития, вызванные различными формами
дисбаланса МЭ у сельскохозяйственных животных, в течение
ряда лет изучает А. А. Кабыш (1986).
Согласно Р. I. Aggett и соавт. (1987), следует выделить МЭ,
дефицит которых у матери обусловливает то или иное терато-
генное воздействие на потомство.
Цинк — бесплодие, хромосомные аномалии, нарушения или даже отсутст-
вие имплантации, смерть плода, микро- и анофтальмия, синдакти-
лия, дефекты формирования нервной трубки, диафрагмальные
и брюшные грыжи, аномалии скелета, нарушения иммунных
и психических функций, синдром респираторного дистресса у но-
ворожденных, отставания роста, анемия.
Медь —бесплодие, смерть плода, спастические параличи, отсутствие ко-
ординации движений; тонкие, ломкие кости; сердечная недоста-
точность, аневризмы, дефекты соединительной ткани, аномалии
кожи и волос, слепота, анемия, нарушение иммунных функций,
низкая масса тела при рождении.
Марганец — хондродистрофия, изогнутые (кривые) длинные кости, куполооб-
разный череп, отсутствие отолитов, атаксия, склонность к конвуль-
сиям.
Железо — бесплодие, смерть плода, низкая масса тела при рождении, функ-
циональные нарушения иммунитета и интеллекта. Аномалии раз-
вития скелета, в частности костей черепа.
Иод — бесплодие, задержка психомоторного развития, глухонемота, спа-
стические параличи, связанное с зобом замедление роста, врожден-
ная микседема.
Селен — гибель плода. ;
Приведенные данные представляют определенный интерес
не только с позиции тератологии, но в первую очередь имеют
большое значение для педиатрии. Зная клиническую картину
пороков развития у новорожденных, обусловленных дефицитом
МЭ, их можно диагностировать и организовать разработку про-
филактического применения определенных МЭ. Если предпо-
лагают врожденный дефицит МЭ, то необходимо своевременно
корригировать это состояние у беременных. Такие данные долж-
ны интерпретироваться осторожно, так как низкие показатели
некоторых МЭ, например цинка в сыворотке крови, не обяза-
тельно означают, что у беременной имеется дефицит цинка
[Burch R. Е. et al., 1976]. Не меньшее теоретическое и практи-
ческое значение имеют результаты воздействия избыточного ко-
личества некоторых МЭ на репродуктивную функцию, о чем
сообщают шотландские исследователи R. Е. Burch и соавт.
(1987). Согласно их данным, избыток следующих МЭ может
оказывать эмбрио- или фетотоксическое действие различной сте-
пени тяжести, проявляющееся в те или иные периоды раннего
индивидуального развития.
Медь —эмбриоцидное действие.
Цинк —эмбриоцидное действие и перинатальная смерть.
Марганец —-эмбриоцидное действие.
Свинец — стерильность, некроз плаценты, выкидыши, перии-атальная и нео-
натальная смерть, замедление роста, аномалин поведения, связан-
ные с нейротоксическим действием свинца.
454
Кадмий — хромосомные аномалии, эмбриотоксическое действие, аномалии
строения лица и нервной трубки, изменение скелета.
Ртуть — неорганические формы — относительно низкая токсичность; орга-
нические соединения (например, метилртуть) — нейротоксическое
действие.
Мышьяк —дефекты формирования нервной трубки, микрогнатия, аномалия
скелета и мочеполовой системы, замедление роста.
Селен —эмбриотоксическое действие, аномалии скелета и конечностей, де-
фекты формирования глаз.
Литий — понижение фертильности, расщепление нёба, аномалии глаза
н уха, поражение печени.
Железо — аномалии поведения, связанные с нейротоксическим действием
железа; нарушения иммунных функций.
Приведенные данные с определенностью говорят о том, что
ни на одной стадии своего развития эмбрион и плод полностью
не защищены от потенциально вредного избытка некоторых МЭ.
Р. I- Aggett и соавт. (1987) указывают, что даже трофобласт
на стадии предимплантации обладает способностью метаболи-
зировать некоторые ксенобиотики. На чувствительность к тера-
тогену могут влиять генетический фон, способность тератогена
к прямому контакту с плодом, доза тератогена, стадия развития
зародыша. Различные тератогены вызывают неодинаковые,
в том числе и специфические, эффекты. В других случаях раз-
личные агенты индуцируют сходные или даже идентичные мор-
фологические и функциональные аномалии.
Особо следует подчеркнуть, что смерть эмбрионов может
быть связана как с избытком некоторых элементов, так и с их
дефицитом. Вместе с тем при дефиците цинка, меди, железа и
селена смерть плода наступает позднее.
Все эти формы патологии эмбриона, плода и новорожденно-
го имеют исключительное значение с позиции органо- и гисто-
генеза. Весьма вероятно, что многие морфологи увидят в них
эксперименты, поставленные самой природой. Неменьшее вни-
мание должны привлечь не только макроскопические аномалии,
но и, казалось бы, чисто функциональные изменения иммунного
статуса и невропсихические нарушения при врожденных
МТОЗах, которые, несомненно, имеют свой структурный суб-
страт, требующий тщательного и многостороннего изучения.
С позиции практического здравоохранения проблема эмбрио-
и фетотоксичности МЭ имеет очень большое значение, так как
число работающих в промышленности женщин резко увеличи-
лось. Она особенно актуальна в отношении токсического дейст-
вия тяжелых металлов, с которыми женщины нередко соприка-
саются в условиях производства. Специальный доклад ВОЗ
(1984) подчеркивает высокий риск для плода (и вообще для
потомства) одновременного контакта матерей не с одним,
а с несколькими воздействующими химическими агентами.
Именно этот аспект проблемы был прицельно исследован груп-
пой японских ученых [Tsuchiya К. et al., 1987], которые подчер-
кивают, что однократное комбинированное воздействие кадмия,
марганца, меди, цинка, железа и свинца в виде внутрибрюшин-
455
ных инъекций тяжело поражало плод и практически не вре-
дило организму матери. Кадмий проникал через плаценту и
накапливался во всех изученных тканях.
ОБЩЕЕ ЗАКЛЮЧЕНИЕ
Учение о микроэлементозах вводит в теоретическую и
практическую медицину огромную и крайне разнообразную ин-
формацию, которая на первый взгляд создает впечатление пест-
роты или даже хаоса фактов. В действительности такое впечат-
ление ошибочно, так как эти экспериментальные, клинические
и эпидемиологические наблюдения естественным образом груп-
пируются вокруг трех больших проблем патологии: микроэле-
ментная недостаточность (дефицит МЭ), микроэлементная
интоксикация (избыток МЭ), микроэлементный дисбаланс (на-
рушения нормальных соотношений самих МЭ, а также МЭ и
макроэлементов в организме). Разумеется, что понятия «мик-
роэлементный избыток» и «микроэлементная интоксикация» не-
равнозначны, так как интоксикация, т. е. объективные признаки
отравления, могут появиться и в результате действия неболь-
шого количества потенциально токсичного МЭ при условии по-
вышенной чувствительности организма или нарушении его эли-
минации.
Создание теории микроэлементной недостаточности обычно
относят к достижениям науки XX века. Это в основном пра-
вильно, но не следует забывать и о том, что в 1880 г. Н. И. Лу-
нин предсказал наличие в пище незаменимых алиментарных
факторов, которые отличаются от ранее известных белков, жи-
ров, углеводов и минеральных веществ. С полным основанием
его можно считать одним из тех, кто предсказал открытие ви-
таминов. Однако, по-видимому, в не меньшей степени это долж-
но относиться и к МЭ, указание на роль которых в жизнедея-
тельности организмов было впервые высказано В. И. Вернад-
ским еще в 1891 г. В дальнейшем именно эти представления
вошли как интегральная часть в его учение о биосфере.
В 1912 г. польский ученый С. Funk впервые предложил тер-
мин «болезни недостаточности» и выдвинул свой, получивший
широкую известность термин «витамины», который оказался не
вполне точным в химическом отношении. В этом же году фран-
цузский исследователь G. Bertrand дал одно из первых определе-
ний МЭ как веществ, необходимых для поддержания жизни.
Близко к этой формулировке подошел американский патолог
Е. Y. McCollum, который говорил о диете, способной вызвать
нарушения питания — мальабсорбцию, и о необходимости выяв-
ления «защитных видов пищи».
Выяснением эссенциальных факторов питания занимались
многие ученые, но далеко не все из них интересовались именно
МЭ. Среди тысяч химических соединений «количество эссенци-
альных питательных веществ, вероятно, не превышает 50, если
456
принимать во внимание аминокислоты, минералы, витамины,
воду и несколько микроэлементов» [Klevay L. М., 1987]. Кри-
терии эссенциальности МЭ в трактовке различных авторов не
вполне совпадают. Наиболее широким признанием пользуется
способность обеспечивать нормальный рост и развитие [Heg-
sted D. М., 1967], а также свойства химического вещества, от-
сутствие которого несовместимо с жизнью [Mertz W., 1982].
«Вещество считается эссенциальным и питательным для жи-
вого организма, если этот организм не может ни расти, ни за-
вершать свой жизненный цикл в его отсутствие. Морфологиче-
ские, биохимические и физиологические отклонения от нормы,
вызываемые специальными ограничительными диетами и норма-
лизующиеся при диетах с определенными пищевыми добавками,
считаются достаточными доказательствами эссенциальности»
[Klevay L. М., 1987]. Следует иметь в виду, что дефицит одного
питательного вещества способен вызвать нарушения не одной,
а нескольких функций.
Концепция о возможности существования микроэлементной
физиологической системы гомеостаза, входящей в общую регу-
ляторную систему организма, насколько нам известно, не изла-
галась в литературе. Нет оснований думать, что эта гипотети-
ческая система функционирует автономно. Однако имеется до-
статочное число фактов, позволяющих предположить ее древнее
происхождение в процессе биогенеза, ее универсальное значе-
ние для регуляции жизнедеятельности органов, тканей и клеток
организмов, относящихся к представителям всех царств приро-
ды. Мы считаем, что Е. Frieden (1984) прав, указывая на то,
что учение о микроэлементах дополняет молекулярную биоло-
гию на уровне атомов. Некоторые из них стабилизируют тре-
тичную структуру молекулы ДНК, различных видов РНК и
ряда структурных белков. Более того, некоторые МЭ обеспечи-
вают структурную целостность определенных органелл.
Хорошо известно, что многие ферменты либо содержат
встроенные в них металлы, либо специфически активируются
(или подавляются) с помощью МЭ. W. J. Dann и W. J. Darby
(1945) проанализировали историю изучения физиологических
факторов питания и обратили особое внимание на витамины,
которые в функциональном отношении имеют общие черты
с МЭ. Эти авторы выделяют пять состояний питания: 1—на-
сыщенное состояние (данным питательным веществом); 2 — не-
насыщенное состояние, но без функциональных нарушений;
3 — болезнь потенциальной недостаточности; 4 — латентная бо-
лезнь недостаточности; 5 — клинически выраженная болезнь
недостаточности. Эти состояния наблюдаются при различных
степенях насыщенности и дефицита витаминов в организме.
Однако они могут иметь не только теоретическое, но и точно
доказанное практическое значение при клиническом анализе
предполагаемого дефицита МЭ. Очень близкую и более про-
стую ранжировку этих состояний предложили М. Kirchgessner
и А. М. Reichlmayer-Lais (1985).
457
К сожалению, диагностика этих состояний и вызываемых
ими патологических процессов в настоящее время еще мало
разработана и главное пока недостаточно известна практиче-
ским врачам. Определенным исключением из этого правила яв-
ляются железодефицитные состояния, органопатология и синд-
ромология которых изучаются в течение многих десятилетий и
отражены в общепризнанных монографиях. Некоторые наиболее
выраженные проявления железодефицитных состояний зарегист-
рированы в Международной классификации болезней ВОЗ (на-
пример, анемия, сидеропеническая дисфагия Пламмера — Вин-
сона). По-видимому, все еще недооценивается железодефицит-
ная миокардиопатия, несмотря на существование достаточных
доказательств того, что при дефиците железа миокард изме-
няется не только вследствие тканевой и циркуляторной гипок-
сии, но и в результате снижения в нем концентрации миогло-
бина.
Несмотря на многочисленные прямые экспериментальные и
эпидемиологические доказательства, клинические границы стой-
ких медьдефицитных состояний до настоящего времени еще не
очерчены. Общепризнаны только их генетические формы (бо-
лезни Менкеса, Марфана, Вильсона — Коновалова, отчасти
синдром Элерса — Данло), которые как редкие заболевания
пока мало известны клиницистам.
Познавательное значение этиологии и патогенеза генетиче-
ских МТОЗов чрезвычайно велико. Они представляют собой
природные модели и крайние формы дефицитов МЭ и других
метаболических аномалий, как правило, связанных с неполно-
ценностью тех или иных ферментов. При этой недостаточности
возможны смерть в детском или юношеском возрасте, повреж-
дение функций и структуры определенных органов, тканей, кле-
ток, органелл и специфические биохимические изменения. Леча-
щий врач должен знать, что в основе разрыва аорты со смер-
тельным исходом при болезни Марфана лежит специфическая
аномалия развития эластических структур этой самой крупной
артерии человека, обусловленная отсутствием или недостаточ-
ным количеством медьсодержащего фермента лизилоксидазы.
Разрывы аневризм аорты и легочной артерии при дефиците
меди в организме человека вызывают законный интерес к ана-
логичным процессам в артериях головного мозга и ставят во-
прос о роли возможных метаболических (микроэлементных?)
факторов в формировании церебральных сосудистых пороков.
Вместе с тем пример болезни Вильсона — Коновалова показы-
вает, что исход генетических МТОЗов не обязательно является
фатальным, если своевременно применяется рациональная тера-
пия. Крайне интересный аналог внезапных разрывов наиболее
мощных отделов сердечно-сосудистой системы представляет экс-
периментальная патология медьдефицитных состояний у крыс.
Многие авторы показали, что сердце таких животных увеличе-
но, ткани его отличаются хрупкостью и дряблостью. Под влия-
нием нагрузки оно становится отечным, сокращается аритмично
458
с типичными изменениями на ЭКГ. Очень часто при дефиците
меди у крыс наблюдается разрыв левого желудочка. Биохими-
ческие исследования сердца обнаруживают изменения колла-
гена, уменьшение содержания норадреналина и нуклеотидтри-
фосфатов, а также энергетической способности миокарда. Соче-
тание дефицита меди с дефицитом селена или высокое соотно-
шение Zn/Cu резко увеличивают летальность крыс от разрыва
левого желудочка. Медьдефицитная диета у детей, питающихся
разведенным коровьим молоком и находящихся в неблагоприят-
ных социальных условиях, оказывает влияние на развитие ске-
лета и соединительной ткани, на сохранность сердечно-сосуди-
стой системы и ЦНС, а также на состояние волос [Mills С. F.,
.1987]. У 7s молодых здоровых волонтеров, находившихся на
умеренной медьдефицитной диете в течение 7—15 нед, обнару-
жена явная патология миокарда, выражавшаяся в тахикардии
и изменениях на ЭКГ. Таким образом, в настоящее время есть
достаточно данных, для того чтобы считать, что медь является
одним из важных факторов гомеостаза сердечно-сосудистой си-
стемы.
Другим МЭ с аналогичным, возможно, с более выраженным
кардиотрофическим свойством является селен, снижение кон-
центрации которого в организме является этиологическим фак-
тором болезни Кешана. Эффективную профилактику этой бо-
лезни с помощью специфической терапии селенитами следует
рассматривать как одно из крупных достижений медицинской
практики. Гносеологическое значение этого результата работ
китайских ученых далеко выходит за пределы болезни Кешана.
Оно показывает принципиальное единство этиологии МТОЗов
человека и сельскохозяйственных животных и доказывает воз-
можность их специфического лечения.
Как следует из конкретных материалов данной монографии,
в условиях дефицита МЭ и микроэлементного токсикоза возни-
кают достаточно характерные заболевания и синдромы, знать
которые обязаны клиницисты, патологи, а также специалисты
по клинической биохимии. Часть заболеваний, обусловленных
дефицитом МЭ, протекает латентно, что имеет особенно боль-
шое значение для контингентов с повышенной чувствительно-
стью ко всем колебаниям гомеостаза МЭ. Это относится в пер-
вую очередь к детям, пожилым, ослабленным, в том числе ли-
цам, подвергшимся хирургическим операциям и перенесшим
истощающие заболевания, а также беременным.
В настоящее время есть достаточно оснований утверждать,
что поступление, метаболизм, специфическое накопление и вы-
деление МЭ регулируются специальной биологической системой
микроэлементного гомеостаза (БСМЭГ) [Авцын А. П., 1972,
1989]. Основные элементы этой системы представлены ниже.
I. Входной путь БСМЭГ
1. Наследственное обеспечение возникающего организма
; МЭ.
459
2. Трансплацентарное обеспечение развивающегося орга-
низма МЭ. (
3. Постнатальное обеспечение организма МЭ:
а) через пищеварительный тракт;
б) через органы дыхания;
в) через кожу;
г) через ткани гениталий у женщин;
д) другие пути поступления МЭ.
II. Внутренний отдел БСМЭГ
1. Специфические и неспецифические рецепторы и сайты.
2. Специфические и неспецифические лиганды. Металло-
тионеины.
3. Пути транспорта МЭ в организме (с кровью, лимфой,
желчью, спинномозговой жидкостью, аксональный, внут-
риклеточные и др.).
4. Абсорбция, ее главные зоны и аппараты. Ферменты.
5. Ферменты МЭ-содержащие и МЭ-зависимые. Метабо-
лизм.
6. Тканеспецифическое и неспецифическое накопление.
III. Выводящие пути БСМЭГ
1. Кишечник, включая специфический аппарат концентра-
ции и выведение цинка и других металлов клетками
Панета.
2. Почки.
3. Потовые и сальные железы, секреты и экскреты других
желез (слюна, желчь, панкреатический сок), волосы,
слущивание отмирающего эпителия.
4. Другие пути элиминации.
IV. Высшие регуляторные влияния на БСМЭГ
1. Генетическая программа организма в каждой живой
клетке.
2. Гипоталамус и эпифиз.
3. Стволовые и спинальные регулирующие аппараты.
4. Симпатическая и парасимпатическая нервная система.
5. Эндокринные железы.
6. Диффузная нейроэндокринная система.
7. Система иммунного гомеостаза.
8. Другие регуляторные влияния.
Проблема микроэлементного гомеостаза на ранних стадиях
онтогенеза, несомненно, имеет большое медицинское и биологи-
ческое значение, однако многое в ней до сих пор еще неясно. Это
особенно относится к периодам внутриутробного развития, ново-
рожденности и раннего детства. Вместе с тем накоплены неко-
торые материалы, которые позволяют сделать предварительные
выводы, имеющие по крайней мере ориентирующее значение.
Из опытов на животных известно, что в физиологических усло-
виях плод в достаточных количествах накапливает все необхо-
димые питательные вещества, в том числе МЭ в конце периода
гестации. В. Meinel и соавт. (1979) показали, что в печени ре-
бенка к моменту рождения концентрации многих МЭ во много
460
раз выше, чем в любой другой период его последующей жизни,
В частности, содержание меди в 16 раз, а цинка и железд поч-
ти в 2 раза больше. Это период известного микроэлементного
благополучия, когда новорожденный снабжен даже избытком
этих веществ. Однако уже в течение первых месяцев жизни на-
ступает значительное уменьшение этого резерва МЭ. Пополне-
ние их содержания увеличивается только во второй половине
1-го года жизни.
Среди детей существуют определенные группы риска, в кото-
рых дефицит МЭ встречается более часто. Н. В. Stockhausen
(1987) показывает, что причинами относительного дефицита МЭ
являются гипотрофия и незрелость недоношенного ребенка (мас-
са тела 1000 г и менее), продолжительное парентеральное пи-
тание, рецидивирующая диарея и кишечная мальабсорбция,
длительный катаболизм, синдром короткого кишечника, очень
низкая концентрация МЭ в грудном молоке. Вместе с тем, как
указывает этот автор, дефицит МЭ у детей раннего возраста
за пределами перечисленных групп риска встречается довольно
редко и коррекция его должна производиться с большой осто-
рожностью лишь при условии обязательного мониторинга содер-
жания цинка, меди, железа, селена.
Изучение литературы показывает, что алиментарная и мета-
болическая коррекция микроэлементного статуса представляет
собой отнюдь непростую задачу. Далеко не всегда она может
быть решена только с помощью простого замещения отсутст-
вующего или умеренно дефицитного МЭ. Экспериментальная,
лечебная и ветеринарная практика обнаружила, что действие
МЭ нельзя рассматривать только с позиций их реальной или ги-
потетической активности. Дело в том, что в природе широко
распространены координированные влияния нескольких МЭ.
Есть основания обратить внимание на пары и даже триады МЭ,
которые оказывают синергическое или антагонистическое влия-
ние на различные физиологические и патологические показате-
ли. Так, R. Shamberger (1987) выделяет закономерные взаимо-
действия меди и цинка при сердечно-сосудистых заболеваниях,
неоплазиях, серповидно-клеточной анемии и болезни Вильсона —
Коновалова. Проблема взаимовлияний МЭ стала предметом ис-
следований Н. Н. Sandstead (1981), который выразил их в гра-
фической форме.
Существуют также взаимодействия между железом и мар-
ганцем, железом и цинком, кадмием и медью. Закономерные-
взаимодействия между собой установлены для меди, молибдена
и сульфата, а также для фтора, кальция и фосфата. Таким об-
разом, проблема взаимодействия МЭ выходит за пределы толь-
ко этой группы веществ и естественным образом связывается
с активностью ряда макроэлементов. Примеры такого рода
очень многочисленны.
Современная экспериментальная и лечебная практика умело
использует эти взаимодействия. Так, при серповидно-клеточной
анемии число дрепаноцитов в крови снижалось благодаря при-
461
ему 0,38 ммоль цинка через каждые 4 ч [Brewer G. J. et al.,
1985]. Это, однако, вызвало неожиданный побочный эффект
в виде медьдефицитной гипохромной микроцитарной анемии и
снижения уровня церулоплазмина в крови, что удалось купиро-
вать с помощью добавок препаратов меди. Взаимодействие МЭ
происходит на уровне кишечной абсорбции или на уровне их
тканевой утилизации [Shamberger R., 1987].
Пристальное внимание привлекает экспериментальная ги-
перхолестеринемия у крыс, вызванная алиментарным дефици-
том меди, связанная с повышенным синтезом холестерина и по-
ступлением его в плазму крови, поскольку именно гиперхолесте-
ринемия является одним из важнейших факторов риска ишеми-
ческой болезни сердца. В США существует твердое убеждение,
что снижение уровня холестерина в сыворотке крови на 1%
должно снижать и риск ишемической болезни на 2% [Shamber-
ger R., 1984]. Алиментарный дефицит меди вызывает повышение
уровня холестерина и триглицеридов плазмы крови в экспери-
менте. Существуют достаточно серьезные предпосылки для ис-
следования связи атеросклероза человека и медной недостаточ-
ности. L. М. Klevay (1987), выдвинувший гипотезу о зависимо-
сти частоты сердечно-сосудистых заболеваний от медно-цинко-
вого статуса организма, утверждает: подавляющее большинство
пищевых рационов в США содержит менее 0,02 ммоль меди
в день, что является минимальным показателем. По мнению На-
ционального исследовательского комитета США, безопасное и
адекватное суточное потребление меди должно составлять
0,031—0,047 ммоль.
Возвращаясь к механизму сопряженного антагонизма меди
и цинка, необходимо указать, что он реализуется на уровне
кишечного всасывания. Цинк мешает абсорбции меди из кишеч-
ника. Медь защищает цинк и сохраняется в слизистой оболочке
только до тех пор, пока она (медь) не удалится вместе с энте-
роцитами в просвет кишечника. Это привлекает внимание к фи-
зиологической и патологической десквамации как процессам,
способным вызвать местное уменьшение содержания МЭ. Оче-
видно, это относится не только к десквамации энтероцитов, но
и к слущиванию эпидермиса, роговых чешуек и утрате волос,
которые сопровождают некоторые МТОЗы. В этих случаях по-
стоянная утрата волос может играть некоторую положительную
роль в удалении токсичных МЭ из организма.
Связь микроэлементного статуса с возникновением и про-
грессированием злокачественных опухолей привлекала внима-
ние многих ученых. По обобщающей сводке К. Windstosser
(1982), натрий, калий, марганец, железо и цинк содержатся в
сыворотке крови больных раком в субнормальных количествах,
в то время как уровень кальция обычно повышен, особенно при
костных метастазах. Уровень меди различен в зависимости от
природы опухоли и стадии процесса. Повышенное содержание
меди характерно для быстрорастущих опухолей. Оно зависит
от воспалительной реакции на рост опухоли, что наблюдается
при так называемом раннем раке легких, при меланомах, лим-
фогранулематозе, раке желудка, а также при остеосаркомах.
Уровень цинка при далеко зашедших стадиях рака легких обыч-
но субнормальный. При экспериментальных опухолях у лабора-
торных животных с алиментарным дефицитом цинка рост опухо-
лей задерживается, выживаемость животных увеличивается, что-
показано в экспериментах Г. А. Бабенко (1981).
Уровень сывороточного железа низок при всех формах рака
у человека, а свойственная онкологическим больным анемия
в значительной степени объясняется дефицитом железа. Оно
является необходимым элементом для роста клеток и в норме
поступает в них -с помощью трансферрина. Некоторые опухоле-
вые клетки способны продуцировать белок со свойствами тран-
сферрина. Такие опухоли могут захватывать достаточное коли-
чество железа, вызывая системный дефицит этого МЭ. В соче-
тании с другими факторами неполноценности питания это спо-
собствует возникновению дисфагичеокого синдрома Пламмера —
Винсона. В свою очередь этот синдром в сочетании с варикоз-
ными изменениями вен пищевода увеличивает риск появления-
рака верхних отделов пищеварительного тракта. Вместе с тем
и избыток железа способствует возникновению рака, так как
железо может взаимодействовать с защитными противоопухоле-
выми факторами [Schrauzer G. N., 1987].
Широко известны данные о связи дефицита магния с высо-
ким уровнем заболеваемости злокачественными опухолями. Не-
смотря на критические замечания ряда экспериментаторов, но-
вые эпидемиологические наблюдения подтвердили эти сведения
и позволяют считать, что избыток во внешней среде и в ра-
ционе магния является одним из факторов, уменьшающих риск
заболевания раком. В -частности, низкая заболеваемость раком
зарегистрирована в некоторых богатых магнием регионах Егип-
та. Очень высокая частота рака желудка в отдельных регионах
связывается с низким содержанием магния. Высокая частота
лейкозов у молодняка рогатого скота зарегистрирована в Поль-
ше. У молодых крыс с дефицитом магния наблюдались опухоли
тимуса, классифицированные как злокачественные лимфомы или-
лимфосаркомы. Злокачественные лимфомы наблюдались толь-
ко при тяжелой степени дефицита магния у животных, причем
преимущественно у новорожденных. У магнийдефицитных крыс
развивался миелоидный лейкоз. Многие ферменты, включая
АТФ и щелочную фосфатазу, нуждаются в наличии магния как
кофактора. Он входит в состав нуклеиновых кислот и, вероятно,
поддерживает их структурную стабильность. Гипомагнезия свя-
зана с гормональным дисбалансом, в частности с гиперальдосте-
ронизмом, гипертиреозом и диабетическим ацидозом [Schrau-
zer G. N., 1987].
В настоящее время особое внимание привлечено к антибла-
стическим свойствам селена (см. главу 6). К изложенным дан-
ным следует добавить новые наблюдения -о снижении уровня
селена в цельной крови у японских женщин, страдающих раком
46®
молочной железы. При такой форме предрака, как фиброзно-
кистозная мастопатия, которая в 3 раза увеличивает риск воз-
никновения рака молочной железы, уровень селена в крови
снижен по сравнению со здоровыми.
Эпидемиологические исследования с полной очевидностью
доказали роль мышьяка, хрома, кадмия и никеля в качестве
этиологических факторов рака главным образом профессио-
нального. Это нашло подтверждение в соответствующих экспе-
риментальных работах, где указанные токсичные МЭ изучались
.на различных моделях. Все эти элементы в той или иной степе-
ни обладают мутагенностью и тератогенностью.
В настоящее время внимание ученых привлечено к дефициту
и низкому уровню потребления ряда МЭ, которые могут быть
важным фактором риска возникновения злокачественных опухо-
лей. Мы уже характеризовали эту роль железа и селена.
С. S. Yang (1980) указывает на дефицит молибдена в качестве
одного из этиологических факторов рака пищевода. У крыс,
подвергавшихся действию N-нитрозосаркозина, небольшая до-
бавка молибдена к питьевой воде уменьшала частоту возникно-
вения опухолей пищевода и преджелудка. Ингибирование кан-
церогенеза в молочной железе также было результатом влияния
молибдена, однако при этом увеличивается частота аденом лег-
ких. Дефицит молибдена в окружающей среде, очевидно, явля-
ется одним из факторов высокого риска возникновения опухолей
пищевода.
Так, целенаправленное изучение биогеохимической ситуации
в регионах, эндемичных .по раку пищевода в Казахстане, в част-
ности в Гурьевской области [Молдакулова М. М., 1983], позво-
лило установить прямую корреляционную связь между заболе-
ваемостью населения раком пищевода и содержанием в окру-
жающей среде, прежде всего в воде, почве и растениях, меди,
марганца, ванадия, стронция, сульфатов. При этом выявлена
обратная корреляция между частотой заболеваемости и содер-
жанием .молибдена. Важно также подчеркнуть, что медь и мар-
ганец, являясь антагонистами молибдена, способствуют разви-
тию выраженной формы его дефицита в организме. Дефицит
марганца рассматривается в качестве одного из возможных
факторов предрасположения к раку [Marjanen Н., Soini S.,
1972].
Изложенные материалы показывают, что этиология и пато-
генез некоторых онкологических заболеваний связаны, как с дей-
ствием. ряда токсичных МЭ, так и в неменыией степени с де-
фицитом эссенциальных МЭ. Как избыток, так и дефицит МЭ
играют существенную роль в течении неопластического процес-
са, что должно учитываться при лечении онкологических боль-
ных современными химиотерапевтиескими препаратами. Един-
ственным МЭ, который обладает доказанным антибластическим
действием, очевидно, является селен. Следует иметь в виду,
что низкое содержание селена в сыворотке крови у онкологиче-
ских больных указывает на пониженный уровень антиоксидант-
464
ной защиты организма. Это 'Способствует большим поврежде-
ниям тканей печени и миокарда при цитотоксической химиотера-
пии. Необходимо обратить внимание на свойства тяжелых
металлов в качестве антагонистов селена. Этими свойствами об-
ладают свинец, ртуть, кадмий, а также мышьяк [Schrauzer G. N.,
1987].
Связь рака и дефицита определенных эссенциальных МЭ
уже сейчас имеет практическое значение, так как это позволяет
выделять в популяции достаточно достоверные группы риска.
Сам феномен возникновения рака определенных локализаций
при дефиците МЭ пока недостаточно понятен. Наиболее извест-
на сидеропеническая эзофагопатия при синдроме Пламмера —
Винсона, которая у 4—16% больных приводит к раку пищевода.
Исходя из общих соображений, можно думать, что дефицит
некоторых эссенциальных МЭ способствует снижению противо-
опухолевого иммунитета. Не 1менее вероятно, что трофика и
антибластическая резистентность каждого органа зависят от мест-
ной насыщенности его тканей МЭ. Обе эти гипотезы не противо-
речат друг другу. Однако они требуют соответствующих иссле-
дований, которые еще не завершены.
Клиническая медицина пока недооценивает значения дефи-
цита МЭ. Статистики этих своеобразных патологических про-
цессов у человека не существует. Между тем число больных
с латентными и манифестирующими формами микроэлементного
дефицита очень велико. Так, только больные с эндемическим зо-
бом составляют не менее 400 000 000. Огромно число страдаю-
щих железодефицитными анемиями, которые в отдельных стра-
нах встречаются у 50—90% популяции.
Гипофтороз, если судить по выраженности фторзависимого
кариеса зубов, — это один из наиболее массовых патологических
процессов современного человечества. Селенодефицитные состоя-
ния организма широко распространены и играют важную роль
в этиологии и патогенезе сердечно-сосудистых и онкологических
болезней. Их диагностика и коррекция — это реальные пути
повышения уровня здоровья населения.
Цинкдефицитные состояния — это крупная проблема опти-
мального развития детей и подростков. Цинк — один из важ-
нейших факторов гомеостаза органов иммуногенеза, репродук-
ции и ЦНС. Кобальтсодержащий витамин Bi2 — один из важ-
нейших факторов гомеостаза кроветворения, ЦНС и тканей ске-
лета.
Медьдефицитные состояния вызывают тяжелую патологию
эластических и коллагеновых волокон, что наиболее очевидно
при некоторых наследственных заболеваниях. Роль дефицита
меди в патологии сердечно-сосудистой системы и при наследст-
венной эмфиземе легких требует неотложного изучения. Ряд
МЭ — фтор, кадмий, свинец, кремний и другие — обладает ос-
теотропными свойствами, что имеет большое значение в клини-
ческой патологии.
30—5G8
465
Не меньшее значение имеют многочисленные заболевания,
вызванные избытком и токсическим действием МЭ, что с осо-
бенной отчетливостью видно при изучении МТОЗов техногенного
происхождения, в частности кадмиозов, алюминозов, интоксика-
ций ртутью, свинцом, 'мышьяком, бериллием и др. При опреде-
ленных условиях и эссенциальные МЭ могут проявить токсиче-
ское действие. Вместе с тем некоторые МЭ, например кадмий и
свинец, имеющие давнюю репутацию токсичных, при определен-
ных экспериментальных условиях обладают некоторыми свой-
ствами эссенциальных МЭ.
Проблема коррекции микроэлементного статуса организма
не может быть решена без учета данных о взаимодействии МЭ
и макроэлементов, которое может быть антагонистическим и си-
нергическим. В настоящее время в связи с возрастающим за-
грязнением внешней среды исключительное значение имеет про-
филактика техногенных и соседских МТОЗов. Гигиена труда и
профпатология достигли в этом направлении определенных
успехов.
Острой проблемой клинической медицины являются ятроген-
ные МТОЗы, число которых резко увеличилось в связи с появ-
лением новых лекарственных средств ,и увеличивающейся инва-
зивностыо их применения. Эта проблема требует постоянного
контроля лекарственных средств, обязательной регистрации их
токсических эффектов и разработки профилактических мер. Осве-
домленность врачей в этом вопросе пока еще недостаточна. Под
особый контроль должно быть взято предупреждение возмож-
ной контаминации всех инфузионных препаратов соединениями
алюминия. Пристального внимания заслуживает постоянное на-
блюдение за функцией аппаратов физиологической элиминации
токсичных МЭ. Некоторые МТОЗы возникают именно в резуль-
тате неполноценной работы почек, как это наблюдалось при
перитонеальном диализе.
Концепция МТОЗов человека имеет солидное эксперимен-
тальное подкрепление в результате многочисленных наблюдений
на сельскохозяйственных и лабораторных животных. Именно в
этой области удалось добиться больших практических успехов.
Есть все основания думать, что такие же, а может быть и еще
большие успехи будут достигнуты клинической и особенно про-
филактической медициной. Освещаемая проблема является од-
новременно и очень старой, и очень новой. Многие МТОЗы
человека еще не описаны. Роль некоторых МЭ в физиологии
и патологии человека остается недостаточно ясной. Однако уже
сейчас ясно видны большие контуры этой многообещающей
проблемы.
я
ч
та
я
4)
й
О
3
g
я
4>
3
а
я
та »sa
к
я
S*
2
об
см
к
is
§1
а ю
я 55
S g
к
№
и
Ф
п
&
й-
с
»К
со
см
3
!=Г
S
ч
\о
5 йз
ю 8
CO SJ2
я к
я
та
s §
9 ч
к
о
я
*3
та Р
Sg
& я
я
3
5
S
я
S
$
я
-В
ЭЯ
4>
я
я
та
sa
ж
св
ж
«
ж
А
с
ж
3 о
s
о ®
ч s
и «
ж
!7 /п
ж
ж
ж
ж
ж
2
ч
ж
3
я
S
*
9
tf
ж
ж
св
ж
СО
ж
Л
с
ж
ж
з 2
s S
О О
3 S
Ч <у
ж
»
»
ч
ж
3
*
ч
„ Я
*=< о
3
о
СО '
о 3
3
та
я
ч
к
3
Я
х Я
§ ®
»я Р
я ч
ж
ж
S
3 s
ж
3
fct
Я
Я
Я
3
fct
я
я
к
я
я
g и а
О’ ж
4> 5
ч В ®
tj ’©< о
о
Я и
§
Я к
5 «
СЯ я
о 3-
я
4
к
я
4
и
3
я
я
ч
w
я
3
5
s
и
8
3
3 *я
“ о
я
к
а*
я
я
ч
я
сх
и
§
»я
3
я
3
я
о
ч
я
я я
§s
к g>s
с з
ч Я и
<D t_. 4)
Ч и
К к
я
Я
я
к
я
о
я
и
к
я
я
•Я
3
я
та
я
о
я
и
о
g СА.-
Ч s
я
я
ч
и
ч
я
я я
3
о
и
я
ч
к
®я
я
я
я
О
tA
я
я
я
я
3
3
Я
ч
3
я
Я
я
ч
та
о
о
3
и
я
и
ни !__.
л
»я
3
я
я
я
к
я
я
я
ч
#я
о
а
3
Я
3
Я
ч
я
я
ч
я
я
я
я
та « »я
•&?3
я
я
3
Я
з
со
Я
ч
я
я
о
я
К
X
X 3
со
я
3
QJ
я
я
ф
*
я
я
я
я
3 со
Я О
Я <1>
g ся
3*
вЯ
3
Я ш
к d)
я
ч
я
Я
0J Л Р-Г i^ А. г'*'
ЕН О С £z^O £—I
л «
* я
4*
я
я
Я <L>
о Ь
w та
Й Я
55 h—Г
К
та
я
я
я
Я’Я
я
к
я
ч
я
я
ч
ч*я
а> о
Ч Е-
Я ГУ м
Ю
О Я
ч я
о’в'н
2 о £
ж е-®
< Ч »я
.<С о
я
я
3
<L>
Я
я
я
я
я
я
я
я
я
та ’Я
Я я я
s й и
я х ®
.S 8. .ч
о
§
я
сп
я
к
я
я
я
X
я я
я
я
а
3
3
Й’§
о
я
я
ч
я
я
М« QJ А
о ч §
я
я
я
я
ч
я
я
я
я
я
ч
я
я
ч
я
2
к
я
я
О)
3
«
я
о>
я
я
о>
я
ч
я
к
я
и к
та g
« Й
Я О
3
в
о
я
я
я
я
и
3
о
я
я
о
S3
о ч
ф <1>
ся«
я
о
® о
я к
►П Г5
X
ч
я
я
я
з
3
ч
О)
ч
А
ч
о
я
я
я
я
ч
я
3
3
3
сп
я
И <L>
tH £Д
я
я
я
ч
О)
я
я
я
►-Г '•'*
Я в
я
НН О
2-^
я’Я
3
я
о
ч Я
я
я
ч
я
я я
ч
3
* g а« g
з» .«
я
Б£§§
». °й
3
ч
к 3
я
я
я
ч
я
3
о
я
я
о
ч
3
я
я
ч
к
я
ч
О)
2
я
о
3
Я
ч
Я
к
я
я
та
д
> «у
з о
Я ю
S Я
? *
я
я
3
Я
та
я
я
я
3
я
я
я
Sg
та g
и Я
3
я
я
я
я
я
о.
и
я
я
к
я 2
я g
я 3
£•3 3 э §
30*
467
Продолжение
Микроэле- мент Важнейшие заболевания, синдромы и признаки дефицита микроэлементов Важнейшие заболевания, синдромы и признаки избытка микроэлементов
Цинк Эндогенный дефицит цинка: врожденные пороки развития плодов и новорожденных (гидроцефалия, микро- и анофталь- мия, расщепление неба, искривление позвоночника, пороки сердца и др.). Наследственный энтеропатический акродерматит. Нарушение клубочковой фильтрации и канальцевой реаб- сорбции при серповидно-клеточной анемии с цинкурией. Экзогенный дефицит цинка алиментарного происхождения и при нарушениях всасывания — болезнь Прасада (тяжелая железодефицитиая анемия, карликовость, половое недоразви- тие) . Идиопатическая гипогевзия и дизосмия. Дерматит, гипогевзия и гипосмия при хронических заболе- ваниях желудочно-кишечного тракта, после гастрэктомии и др„ а также при алкогольном циррозе и других заболеваниях пе- чени. Замедленное заживление ран. Цшжодефицитиое атоническое кровотечение у матери, преждевременные роды и врожденные уродства плода. Ятрогенный дефицит цинка при парентеральном питании и длительном лечении цитостатиками, L-пеницилламином и D-гистидином. Снижение активности , нуклеозидфосфорилазы — маркера цинкдефицита в организме. Отравления цинксодержащими соединениями
Марганец
Хром
Селен
Диабет у человека нечувствительный к инсулину (эффек-
тивен хлорид марганца, что подтверждено эксперимен-
тально).
Гипохолестеринемия у человека при марганцевой недоста-
точности.
У крупного рогатого скота в природнохозяйственных усло-
виях манганодефицита наблюдается: отсутствие либидо при
нормальной овуляции у коров, снижение оплодотворяемости,
повышенная частота абортов и мертворождений, преоблада-
ние потомков мужского пола, удлинение сроков стерильности,
пониженная упитанность, повышенная смертность новорож-
денных; выраженные нарушения опорно-двигательного аппа-
рата, особенно хрящевой ткани коленного сустава вследст-
вие угнетения синтеза гликозаминогликанов; сокращение про-
должительности жизни. Атаксия новорожденных (аномальное
развитие отолитов). Перозис (соскальзывающее сухожилие)
у цыплят.
Хромодефицитные состояния у человека характеризуются
снижением толерантности к глюкозе, повышением концент-
рации инсулина в крови, глюкозурией, гипергликемией нато-
щак, задержкой роста, повышением концентрации триглице-
ридов и холестерина в сыворотке крови, увеличением атеро-
склеротических бляшек в аорте, периферическими невропатия-
ми, нарушением высшей нервной деятельности, снижением
оплодотворяющей способности и числа спермиев.
Наследственные селенодефицитные ферментопатии (дефи-
цит глютатионпероксидазы эритроцитов и тромбоцитов).
Наследственный кистозный фиброз поджелудочной железы
(муковисцидоз).
Наследственная миотоничская дистрофия.
Манганозы — техногенные микроэлементозы,
обусловленные избыточным поступлением марган-
ца в условиях производства.
Крайнее выражение профессионального манга-
иоза—синдром паркинсонизма: расстройства
двигательной активности (маскообразность ли-
ца, дизартрия, нарушение гисьма, специфична
«петушиная» походка); психические нарушения
(эйфория, благодушие, снижение критики к свое-
му состоянию); астеновегетативиый синдром
с угнетением функции гонад.
Шестивалентный хром обладает наиболее вы-
раженным общетоксическим, нефротоксическим,
гепатотоксическим, мутагенным и канцерогенным
действием. Профессиональные хромовые дермати-
ты, изъязвление слизистой оболочки носа с пер-
форацией хрящей носовой перегородки. Хромо-
вый гепатоз.
Встречаются в обогащенных селеном регионах
ряда стран: алиментарный селеновый токсикоз
(селеноз) с дерматитом, повреждением эмали
зубов, анемиями и нервными расстройствами; се-
лепотокспческая дегенерация печени и увеличе-
ние селезенки; типичное поражение ногтей и во-
лос. Профессиональный селеноз (?)
ПроволМеНив
470
Микроэле- мент Важнейшие заболевания, синдромы и признаки дефицита микроэлементов Важнейшие заболевания, синдромы я признаки избытка микроэлементов
Алиментарный дефицит селена: эндемическая кардиомиопа- тия (болезнь Кешана) в селеиодефицитном поясе Китая, в Забайкалье (СССР); — фактор патогенеза застойных кардиопатий и фактор риска коронарной болезни и инфаркта миокарда в Восточной Финляндии, в ряде штатов США и некоторых районах ФРГ, — фактор риска возникновения и повышенной смертности при злокачественных новообразованиях желудка, кишеч- ника, молочной железы, яичников, простаты, легких. У животных селенодефицит характеризуется: — алиментарной мышечной дистрофией (беломышечная бо- лезнь сельскохозяйственных животных, — алиментарным гепатозом свиней, — экссудативным диатезом домашней птицы, — некрозом и фиброзом поджелудочной железы у цыплят, — задержкой роста и развития, — нарушением репродуктивной функции со специфической дегенерацией сперматозоидов. Для многих видов животных характерны про- явление селеноза в виде «щелочной болезни», очагового некроза и цирроза печени, а также по- роков развития у эмбрионов и плодов.
Молибден Молибдендефицит характеризуется снижением активности трех молибдеисодержащих ферментов: альдегидоксидазы, ксантииоксидазы и сульфитоксидазы. При генетическом дефекте ксантииоксидазы и нарушении реабсорбции ксантина в почечных канальцах возникает ксан- тинурия, при этом содержание мочевой кислоты в сыворотке крови и моче резко снижено. Эпидемическая молибденовая подагра (болезнь Ковальского), встречается в Армянской ССР. Хронический профессиональный молибденоз ха- рактеризуется повышением содержания мочевой кислоты и молибдена в сыворотке кровн, полиар- тралгиями, артрозами, гипотонией, анемией и лейкопенией.
Иод
Гипойодоз — эндемическое увеличение щитовидной железы
и эндемический зоб.
Профессиональные и ятрогенные интокеикации
йодидами.
Кобальт
Длительно протекающий гипойодоз у детей
— кретинизм, недоразвитие мозга и костной системы.
Гипотиреоз взрослых—эндемический зоб, микседема.
Гипотиреоз, возникающий под влиянием гойтерогенных (зо-
богенных) веществ.
Генетические дефекты синтеза тиреоидных гормонов и об-
мена йода (семейный зоб, врожденный гипотиреоз): 1) не-
способность железы концентрировать йод; 2) нарушение син-
теза тиреоидных гормонов в результате дефекта синтеза
тироидпероксидазы; 3) неспособность к дейодированию МИТ
и ДИТ, вызванная дефектом синтеза специфической дейода-
зы; 4) неспособность синтеза Т4 из МИТ и ДИТ; 5) циркуля-
ция в крови белка, прочно связывающего йод.
Аутоиммунное поражение щитовидной железы — болезнь
Хашим ото.
У многих животных гипойодоз проявляется в виде энде-
мического зоба
Иодизм — аллергическая реакция организма
на контакт и поступление йодидов.
Иододерма.
Недостатоточность витамина В12.
Наследственная пернициозная анемия, атрофия слизистой
оболочки желудочно-кишечного тракта, фуникулярный мие-
лоз.
Пернициозная анемия при инвазии широким лентецом.
Акобальтоз (энзоотический маразм, кустарниковая болезнь,
солевая болезнь, сухотка, болотная болезнь и др.) —эндеми-
ческое заболевание, особенно у овец, коз, крупного рогатого
скота, характеризуется истощением, недостатком витамина
В12, анемией, стеатозом печени, остеодистрофией
Профессиональные отравления с поражением
органов дыхания, кроветворения, сердечно-сосу-
дистой и нервной систем, сопровождающиеся ал-
лергическим дерматитом, хроническим бронхитом,
пневмонией и пневмосклерозом.
Кобальтовая миокарднопатия. Алиментарно-
токсическая кобальтовая миокардиодистрофия
с декомпенсацией, полицитемией и гиперплазией
щитовидной железы («болезнь любителей пива»)
т
Таблица 2?
Важнейшие заболеваний, синдромы и главные признаки дефицита и избытка условно эссенциальных микроэлементов у человека и некоторых животных
Микроэле- мент Важнейшие заболевания, синдромы н признаки дефицита микроэлемента Важнейшие заболевания, синдромы и признаки избытка микроэлемента
Мышьяк Мышьякдефицнтные состояния у человека не известны. Арсенозы характеризуются диспепсическими расстройствами, конъюнктивитом, помутнением стекловидного тела и рогови- цы, изъязвлением носовой перегородки, стоматитом, ларинги- том, трахеитом, бронхитом, папулезной и пустулезной сыпью (часто между пальцами), рецидивирующей экземой, атрофи- ческим акродерматитом, симметричным точечным ладонно- подошвенным гиперкератозом и бронзовой гиперпигментацией, ломкостью ногтей, преждевременным поседением и выпаде- нием волос. Неврологические нарушения в виде интеллекту- ально-мнестических расстройств, депрессии, полиневритов с атрофическим изменением мышц, а также в виде ретро- бульбарного неврита, нарушения вкуса и обоняния. Эндемическое отравление мышьяком, поступающим с питье- вой водой и пищей, — болезнь «черной стопы» (изменения периферических сосудов по типу облитерирующего эндарте- риита), бородавчатый кератоз ладоней и подошв, анемия, расстройства сердечной деятельности, периферические невро- патии. Возможно развитие рака кожи и легких у человека.
Бор Признаки дефицита бора зарегистрированы только в эксперименте. Бор действует через паращитовидные железы на обмен кальция, фтора и особенно магния. Бор регулирует активность паратгормона на уровне клеточной мембраны, отсутствие его рас- страивает эту регуляцию. Бороз — острая интоксикация соединениями бора (синдром литейной лихорадки), нейропатия, некроз и ожирение пече- ни, повреждение почечных канальцев—гематурия. Хронический бороз — заболевание верхних дыхательных путей и легких; пневмокониоз. Борный энтерит — эндемическое заболевание желудочно- кишечного тракта людей и животных в Западной Сибири и Алтайском крае (возможно полимикроэлементное происхож- дение)
Бром У человека признаки дефицита брома не заре- гистрированы. Острое и хроническое отравление бромом (катаральный ринит, бронхит, конъюнктивит, энтерит). Неврологический синдром бромизма (сонливость, атаксия, снижение болевой чувствительности, ослабление слуха, зрения и памяти, делирий с галлюцинациями). Бромодерма — специфическое поражение кожи у лиц с по- вышенной чувствительностью. Врожденная бромодерма у грудных детей, матери которых принимали бромиды во время беременности.
Фтор Врожденный гипофтороз (?). Гипофтороз де- тей грудного и дошкольного возраста — запазды- вание прорезывания зубов и специфическое по- ражение зубов молочного прикуса кариесом. Гипофтороз у детей школьного возраста — типичное поражение зубов кариесом; нестомато- логические проявления почти не изучены. Гипофтороз у взрослых — типичный кариес зу- бов в эндемических зонах с низким содержанием фтора в питьевой воде. Гипофтороз в старческом возрасте — прогрес- сирование типичного кариеса зубов, фторзависи- мый остеопороз скелета, склонность к переломам, особенно у женщин. Гипофтороз беременных (?). Акклиматизационный гипофтороз у приезжих жителей Севера. Эндемический флюороз зубов. Эндемический флюороз костей скелета, в том числе кале- чащие его формы в Индии и Китае. Миелорадикулопатии. Профессиональный флюороз: фторный ринит с носовыми кровотечениями; ринофарингит и фаринголарингит с язвенно- некротическими изменениями слизистой оболочки; остеоскле- роз скелета с петрификацией связочного аппарата; субатро- фический и атрофический гастрит, фторный гепатоз, фторный гиперпаратиреоз, гипогонадизм, миокардиодистрофия, фтор- ная панкреатопатия (?), фторная ретинопатия (?).
Литий Признаки недостаточности лития зарегистриро- ваны только у коз и крыс, находившихся в ус- ловиях дефицита лития. Они выражались в нару- шении репродуктивной функции, повышении смертности и частоты выкидышей. Отравление аэрозолями лития — трахеит, бронхит, межу- точная пневмония, пневмосклероз; ожоги кожи и слизистых оболочек. Неврологический синдром при хронической интоксикации (сонливость, головокружение, боль при глотании, тремор, за- медление пульса, гиперестезия и гипералгезия кожи).
ПроЗблжкии
Микроэле-
мент
Важнейшие заболевания, синдромы и признаки
дефицита микроэлемента
Важнейшие заболевания, синдромы и признаки избытка
микроэлемента
Микель
Кремний
Ванадий
Никельдефицитные состояния у человека не
зарегистрированы.
У животных отмечаются задержка роста, повы-
шенная смертность самок и их потомства, сниже-
ние уровня гемоглобина, гематокритного числа;
паракератозоподобные изменения кожи иа выме-
ни лактирующих животных.
Нарушение репродуктивной функции у коз,
атрофия семенников у козлов.
Убедительных данных о дефиците кремния у
человека не имеется. Повышенная ломкость ног-
тей, которые утрачивают нормальную флуорес-
ценцию в ультрафиолетовом свете.
Гипотетические силикопривиые состояния: си-
ликопривный дерматоз с недоразвитием придат-
ков кожи, силикопривная онихопатия, силико-
привная коллагенопатия, силикопривная ангиопа-
тия (типа синдрома Марфана?), силикопривная
аортопатия (атеросклероз?), силикопривная ос-
теопатия (недоразвитие скелета).
Ванадийдефицитные состояния у человека не
зарегистрированы.
Добавки ванадия при анемиях ускоряли про-
цессы регенерации костного мозга у человека и
животных. Однако этот эффект ванадия иеспеци-
фичен.
У экспериментальных животных при дефиците
ванадия происходит усиление кариеса зубов.
Нарушение подвижности сперматозоидов. Увеличение щито-
видной железы. Угнетение активности аденилатциклазы.
Профессиональные интоксикации никелем сопровождаются
литейной лихорадкой, недомоганием, слабостью, головной
болью, головокружением, стеснением в груди, ринитом, носо-
выми кровотечениями, отеком легких, токсической пневмонией,
пневмосклерозом, гепатитом, постнекротическим циррозом;
никелевым дерматитом (никелевая чесотка, никелевая экзе-
ма) ; изъязвлением слизистой оболочки иоса.
Никелевый рак слизистой оболочки иоса, его придаточных
полостей, бронхогенный рак легких.
У животных (овец) отмечена эндемическая катаракта в
биогеохимических провинциях, обогащенных никелем.
Общий гиперсиликоз организма при избыточном поступле-
нии кремния, а также при нарушении его экскреции.
Локальные формы силикоза: пневмокониоз (силикоз), ие-
фросиликоз (эндемический кремниевый нефролитиаз), силико-
кальцииоз лимфатических узлов корня легкого, окулосиликоз.
В производственных условиях V2O5 оказывает выраженное
токсическое действие иа дыхательную, сердечно-сосудистую
системы, печень и почки.
Таблица 28
Важнейшие заболевания, синдромы, признаки дефицита и избытка токсичных и условно токсичных микроэлементов
у человека и некоторых сельскохозяйственных животных
Микроэле-
мент
Важнейшие заболевания, синдромы н признаки
дефицита микроэлемента
Важнейшие заболевания, синдромы н признаки избытка
микроэлемента
Алюминий
Алюмннийдефицитиые состояния у человека не
зарегистрированы
Кадмий
Кадмийдефицнты у человека не зарегистриро-
ваны.
Дефицит кадмия в эксперименте на козах
вызывает задержку роста и полового созревания
Свинец
Свинецдефицитные состояния (гипосатуриизм)
у человека не известны.
В эксперименте при дефиците свинца у живот-
ных отмечено нарушение обмена железа с раз-
витием признаков железодефицитного состояния.
Это состояние корригировалось введением свинца
в рацион животных.
Алюминиевый пневмокониоз в профессиональной патологии.
Ятрогенные формы алюминоза: алюминиевая диализная эн-
цефалопатия, алюминиевая остеодистрофия, алюмииийзависи-
мая микроцитарная анемия, алюминиевый псевдогиперпара-
тиреоз при хронической почечной недостаточности, миопатиче-
ский синдром, алюминиевая гастропатия, перитонеальный
алюминоз.
Алюминоз сердца, вызванный фосфидами алюминия.
Острое отравление в производственных условиях характе-
ризуется раздражением воздухоносных путей, сильным каш-
лем, одышкой, болезненностью в грудной клетке при дыхании,
тошнотой, рвотой, цианозом, отеком легких, бронхопневмо-
нией.
Хронические формы кадмиоза: кадмиевый ринит, кадмиевая
нефропатия с типичной протеинурией, кадмиевая остеомаля-
ция (болезнь итаи-итаи), нейротоксический синдром.
Кадмиевая кардиомиопатия, гипертония, развитие эмфизе-
мы легких, поражение печени и кадмиевый рак предстатель-
ной железы требуют дополнительных доказательств.
Хроническое свинцовое отравление: микроцитарная анемия,
морфологически не отличимая от железодефицитиой анемии
(ингибирование активности порфобилииогеисиитазы, появле-
ние в крови и моче предшественников гема, характерно повы-
шенное выделение копропорфирииа III с мочой).
Неврологические расстройства — свинцовая энцефалопатия
(заторможенность, беспокойство, раздражительность, голов-
ные боли, тремор, галлюцинации, потеря памяти).
Периферическая свинцовая невропатия связана с наруше-
нием проводимости нерва,
Продолжение
Микроэле-
мент
Важнейшие заболевания, синдромы и признаки
дефицита микроэлемента
Важнейшие заболевания, синдромы и признаки избытка
микроэлемента
Ртуть
Ртутьдефицитные состояния
лизм) у человека не известны.
(гипомеркуриа-
Бериллий
Бериллийдефицитиые состояния (гипоберилли-
оз) у человека не известны.
Свинцовая нефропатия (аминоацидурия, гипофосфатемия
с относительной гиперфосфатурией, а также глюкозурией)
сопровождается медленным сморщиванием почки.
Колики — ранний симптом отравления свинцом. Свинцовая
кардиомиопатия. Канцерогенность свинца для человека не ус-
тановлена.
Острые ингаляционные отравления ртутью характеризуются
признаками острого, нередко язвенно! о бронхита, бронхиоли-
та, пневмонии
Оральные отравления ртутью: язвенно-некротический гаст-
роэнтерит, некротический нефроз. При хроническом отравле-
нии ртутью развиваются астеновеге!агпвный синдром, тре-
мор, психические нарушения, эретизм, лабильный пульс, та-
хикардия, гингивит, изменения клеточного состава крови, про-
теинурия.
При хроническом отравлении органическими соединениями
ртути развиваются болезнь Минамата, энцефалопатия, моз-
жечковая атаксия, нарушение зрения и слуха, парестезии.
Врожденный меркуриализм (микроцефалия с гипопластиче-
скими и диспластическими изменениями ЦНС) в результате
трансплацентарного поступления метилртути от матери, под-
вергавшейся хронической интоксикации.
Острые профессиональные отравления бериллием: симпто-
мы литейной лихорадки, раздражение дыхательных путей,
конъюнктивиты, острый бронхит н бронхиолит, бронхоспазм.
Хроническое отравление характеризуется хроническим
бронхитом, бронхоэктазами, «химическими пневмониями»,
пневмосклерозом.
Бериллиоз — особая форма профессиональной патологии,
вызываемой нерастворимыми (малорастворимыми) соедине-
ниями бериллия. Это хронический гранулематозный пролифе-
ративный воспалительный процесс (бериллиоз легких, берил-
лиоз кожи).
Барий
Барийдефицитное состояние (гипобаритозы)
у человека не известны.
Вйсмут
Стронций
Висмутдефицитные состояния (гиповисмутозы)
у человека не известны.
Стронцийдефицитные состояния у человека не
известны.
Сурьма
Сурьмяиодефицитное состояние у человека не
известно.
Баритозы — МТОЗы, вызываемые интоксикацией барием
и его солями.
Острый баритоз характеризуется обильным слюноотделени-
ем, жжением во рту и пищеводе, болями в животе, тошнотой,
рвотой, поносом, повышением артериального давления, судо-
рогами (возможны параличи), цианозом, «холодным» потом,
расстройствами походки и речи, одышкой, нарушением зре-
ния. Смерть в 1-е сутки.
Хронический баритоз характеризуется резкой слабостью,
одышкой, ринитом, конъюнктивитом, диареей, повышением
артериального давления, неправильным пульсом, нарушением
мочеиспускания, выпадением волос.
Баритовый пневмокониоз с явлениями хронического брон-
хита и пневмонии.
Хронические формы висмутоза, возникающие при длитель-
ном применении препаратов висмута (бийохинол, бисмоверол,
пентабисмол; викалин, викаир и др.),—энцефалопатия, вис-
мутовый гингивит, стоматит.
Высокие концентрации строиппя угнетают процессы оссифи-
кации.
Болезнь Кашина— Бека — страдание биогеохимической
природы с окончательно неустановленной этиологией.
Эндемические остеодистрофии (стронциевый рахит) сель-
скохозяйственных животных. Причины: недостаток каротина,
фосфата, магния, марганца, цинка, меди, йода, кобальта,
а также избыток стронция.
В условиях сурьмяных биогеохимических провинций (Уз-
бекистан) несколько увеличены рост, объем груди и масса
тела детей обоего пола, свидетельствующие об улучшении
физического развития.
Специфическая сурьмяная патология в субрегионе биосфе-
ры не выявлена, однако отмечено повышенное накопление
сурьмы в щитовидной железе в районах распространения эн-
демического зоба.
Список основной литературы
А. Руководства, труды конференций и симпозиумов
Микроэлементозы человека. Материалы Всесоюзного симпоз.— Москва, 15—
17 ноября 1989 года. — М., 1989.—353 с.
Биологическая роль микроэлементов/Ред. В. В. Ковальский, И. Е. Воротниц-
кая. — М.: Наука, 1983.
Биологическая роль микроэлементов и их применение в сельском хозяйстве
и медицине. — VIII Всесоюз. конф. Тез. докл. — Ивано-Франковск, 1978.—
Т. I—II.
IX Всесоюзная конференция по проблемам микроэлементов в биологии. — Тез.
докл. — Кишинев: Штиинца, 1981.
Методы и достижения бионеорганической химии/Ред. Мак-Олиффа К.: Пер.
с англ. — М.: Мир, 1978.
Микроэлементы в биологии и их применение в медицине и сельском хозяйст-
ве.— X Всесоюз. конф. Тез. докл. Т. I—III. — Чебоксары, 1986.
Микроэлементы в медицине/Ред. Г. А. Бабенко.— Киев: Здоров’я, 1973.—
Вып. 4. — 142 с.; 1974. — Вып. 5.— 146 с.; 1975. — Вып. 6.— 146 с.; 1978.—
Вып. 7. — 137 с.
Микроэлементы в питании человека. Докл. Ком. экспертов ВОЗ. — М.: Меди-
цина, 1975. •—74 с.
Биогеохимическая лаборатория. Труды АН СССР/Ред. А. П. Виноградов»
В. В. Ковальский, —М.: Наука, 1960—1968—1974—1976—1978—1980-
1979—1980—1981 — 1985. Т. 11—20.
Неорганическая биохимия/Ред, Г. Эйхгорн. — Пер. с аигл. — М.: Мир, 1978»
Т. 1, —687 с.
Человек. Медико-биологические даииые/Пер. с англ. — М.: Медицина, 1977.
Advances in nutritional research. Vol. 6/Ed. H. H. Draper. — New York — Lon-
don: Plenum Press, 1984.
Biochemistry of the essential ultratrace elements/Ed. E. Frieden. — New York —
London: Plenum Press, 1984.
Spurenelemente. Grundlagen — Atiologie — Diagnose — Therapie/Hrsg. von
H. Zumkley. — Stuttgart: G. Thieme, 1983.
Toxicity of heavy metals in the environment. Pt. I/Ed. F. W. Oehme. — New
York — Basel: Marcel Dekker, Inc., 1978.
Trace substances in environmental health. Vol, VI/Ed. D. D. Hemphill. — Colum-
bia: Univ, of Missouri, 1973.
Trace elements in man and animals. Proc. 5th Int. Symp. Aberdeen, June — July»
1984. — London, 1985. — 977 p.
Trace elements in human and animal nutrition. — Fifth Ed. Vol. I—II/Ed.
W. Mertz. — New York: Acad. Press, 1987.
Trace element analytical chemistry in medicine and biology. Vol. 4/Ed. P. Brat-
ter, P. Schramel. Proc, of the Fourth Intern. Workshop Neunerberg, Fed.
Rep. of Germany, April 1986.— Walter de Gruyter. — Berlin — New York»
1987.
Trace elements in human health and disease. Vol. II. Essential and Toxic Ele-
ments/Ed. A. S. Prasad. — New York: Ac3d. Press, 1976.
Zinc deficiency in human subjects. Proc, of an Intern. Sym. held, in Ankara,
Turkey, April 29—30, 1982/Ed. A. S. Prasad et al.— New York: Alan
R. Liss, 1983,
478
Б. Монографии и периодическая литература
Авцын А. П. Введение в географическую патологию. — М.: Медицина, 1972,
Авцын А. П. Важнейшие алюминозы человека//Арх пат.— 1986. — №5.—•
С. 1—10.
Авцын А. П. Микроэлементозы человека//Клин. мед.— 1987. — № 6. — С. 36.
Авцын А. П. Синтезирующие подходы в изучении микроэлементов//Микро-
элементозы человека. — М„ 1989. — С. 4—10.
Авцын А. П„ Жаворонков А. А. Патология флюороза. — Новосибирск: Наука,
1981 — 335 с.
Авцын А. П., Жаворонков А. А., Строчкова Л. С. Принципы классификации
заболеваний биогеохимической природы//Арх. пат.— 1983.—№9.—
С. 3—14.
Авцын А. Н., Жаворонков А. А., Строчкова Л. С., Корнев А. В. Воздействие
селенита натрия на опухолевые клетки в культуре//Вопр. онкол. — 1986. —•
№ 8. — С. 77—88.
Авцын А. П., Жаворонков А. А., Марачев А. Г., Милованов А. П. Патология
человека на Севере. — М.: Медицина, 1985.—415 с.
Алексеева О. Г. Роль иммунологических механизмов в развитии пневмоко-
ниозов//Патогенез пневмокониозов. — Свердловск, 1970. — С. 185—196.
Алексеева О. Г., Дуева Л. А. Аллергия к промышленным химическим соеди-
нениям.— М.: Медицина, 1978. — 271 с.
Бабенко Г. А. Микроэлементы головного мозга человека и животных: Авто-
реф. дис. канд. — Донецк, 1953. — 16 с.
Бабенко Г. А. Микроэлементы в экспериментальной и клинической медицине. —
Киев: Здоров’я, 1965. — 183 с.
Балыкин Н. С„ Седов К. Р. Изменения ферментов крови при воздействии ток-
сического комплекса ртути и хлора//Сов. мед. — 1982. — № 8. — С. 12—14.
Бацевиц В. А. Антропо-экологическое изучение микроэлементиого состава во-
лос у некоторых групп населения; Автореф. канд. дис. —М., 1988.
Бегалиев А. Б. Генетический эффект солей тяжелых металлов как загрязните-
лей окружающей ореды//Успехи соврем, геиет.— 1982. — № 10. — С. 104.
Беренштейн Ф. Я. Микроэлементы, их биологическая роль и значение для жи-
вотноводства.— Минск: Ураджай, 1955. — 232 с.
Блинникова О. Е. Клинико-генетическая характеристика синдрома Элерса —
Данлоса//Обзорная ииформация/ВНИИМИ. Медицина и здравоохр. Се-
рия: Мед. генетика и иммунология. — Вып. 6. — 1985. — 26 с.
Богданов Н. А., Гембицкий Е. В. Производственный флюороз. — М.: Медици-
на, 1975, —95 с.
Величковский Б. Т. О физико-химических свойствах, определяющих цитоток-
сичность и фиброгеииость кремиеземсодержащих пылей//Патогенез пнев-
мокониозов. Тр. II Всесоюз. симпоз. — Караганда, 1978. — С. 10—17.
Величковский Б. Т. Фнброгенные пыли. Особенности строения и механизма
биологического действия. — Горький, 1980. — 159 с.
Вельховер Е. А., Ромашов Ф. Н., Селюкова В. В. Применение меди и ее солей
в лечебной практике. — М.: УДН им. Лумумбы, 1982. — 44 с.
Венчиков А. И. Биотики (К теории и практике применения микроэлементов).—
Ашхабад: Ылым, 1978. — 279 с.
Вернадский В. И. Химические элементы и механизм земной коры//Приро-
да. — 1922. — № 3/5. — С. 31—40.
Вернадский В. И. Биогеохимические очерки 1922—1932 гг. — М. — Л.: АН
СССР, 1940, —249 с.
Вернадский В. И. Очерки геохимии.—М.: Наука. — 1983. — 422 с.
Виноградов А. П. Биогеохимические провинции//Труды Юбилейной сессии,
посвященной 100-летию со дня рождения В. В. Докучаева. — М. — Л.,
1949.
Войнар А. И. Биологическая роль микроэлементов в организме животных
и человека. — М.: Высшая школа, 1960. — 544 с.
Волъкенштейн М. В. Биоиеорганическая химия и молекулярная биология//
Молекул, биол. — 1982.—Т. 16. —№ 5. — С. 901—929.
Воронков М. ГЗелчан Г. И., Лукевиц Э. Я. Кремний и жизнь, — Рига: Зинат-
не, 1978. — 587 с.
479
Габович Р. Д., Минх А. А. Гигиенические проблемы фторирования питьевой
воды. — М.: Медицина, 1979. — 200 с.
Движков П. П. Пневмокониозы. — М.: Медицина, 1965. — 423 с.
Добровольский В. В. География микроэлементов. Глобальное рассеяние.—
М.: Мысль, 1983. — 160 с.
Дорожко О. В., Ротшильд Е. В. Микроэлементы в жизнедеятельности пато-
генных и некоторых других микроорганизмов//Успехи соврем, биол. —
1985. — Т. 99. — Вып. 2. — С. 313—319.
Елецкий Ю. К., Цибульский А. Ю. Роль парасимпатической нервной системы
в регуляции обмена некоторых макро- и микроэлементов (эксперименталь-
ное исследование)//Микроэлементозы человека. — М„ 1989. — С. 90—91.
Ермаков В. В., Ковальский В. В. Биологическое значение селена. — М.: Наука,
1974. —294 с.
Ещенко В. А. Блокирование циика как возможная причина клеточной альтера-
ции//Арх. пат.— 1977. — № 9. — С. 55—60.
Жаворонков А. А. Патологическая анатомия, географическая патология и не-
которые вопросы патогенеза эндемического флюороза: Автореф. дис.
докт. — М., 1977. — 43 с.
Жаворонков А. А. Циикдефицитиые состояния у человека//Арх. пат.— 1983.—
№ 9. —С. 77—80.
Зербино Д. Д., Шульгин Я. И., Гаврилюк Е. М. Хронические расслаивающие
аневризмы аорты//Дрх. пат.— 1986. — № 11. — С. 15—20.
Зислин Д. М., Гирская Е. Я- К классификации профессионального флюороза//
Гиг. труда и проф. заболевания. — 1971. — № 2. — С. 24—28.
Израэль Ю. А., Назаров И. М„ Пресиан А. Я- и др. Кислотные дождя —Л.:
Гидрометеоиздат, 1983. — 206 с.
Кабата-Пендиас А., Пендиас X. Микроэлементы в почвах и растеииях//М.:
Мир, 1989. — 439 с. Пер. с аигл.
Кактурский Л. В., Бескровнова Н. Н., Кудрин А. Н. и др. Морфологические
показатели влияния селена и витамина Е на течение экспериментального
инфаркта миокарда//Кардиология.— 1976. — № И.-—С. 31—36.
Карлинский В. М. Циикдефицитиые состояния: Автореф. дис. докт. — М.,
1979. —38 с.
Кацнельсон Б. А. Отложение, элиминация и задержание пыли в легких//Ток-
сикология. — М., 1976. — Т. 7. — С. 7—24.
Кейне Р. Д. Ионные каналы в мембране ие.рвиой клетки//Молекулы и клет-
ки/Пер. с аигл. — М., 1982. — С. 176—190.
Клиника и лечение. Острые отравления соедииеииями тяжелых металлов
и мышьяка: Метод, рекомендации. М3 РСФСР. — М., 1980. — 25 с.
Книжников В. А., Муравьев А. А., Тюрин Н. А. Микроэлемеиты//Б. М. Э. —‘
3-е изд.— 1981. — Т. 15. —С. 674—685.
Ковальский В. В. Биогеохимические провинции СССР и методы их изучения//
Труды биогеохимической лаборатории. АН СССР.— 1960. — Т. 11.—
С. 8—33.
Ковальский В. В. Геохимическая экология. Очерки. — М.: Наука, 1974. — 299 с.
Ковальский В. В. Геохимическая среда и жизнь. — М.: Наука, 1982. — 77 с.
Ковальский В. В. Современные направления и задачи биогеохимии//Биологи-
ческая роль микроэлементов. — М., 1983. — С. 3—17.
Ковальский В. В., Сусликов В. Л. Кремниевые субрегионы биосферы СССР//
Биохимическое районирование и геохимическая экология. — М„ 1980.—
С. 3—58.
Коломийцева М. Г., Габович Р. Д. Микроэлементы в медицине. — М.: Медици-
на, 1970. —286 с.
Коновалов Н. В. Гепато-церебральиая дистрофия. — М.: Медгиз, 1960. — 556 с.
Критерии санитарно-гигиенического состояния окружающей среды/ВОЗ. 1.
Ртуть. — М., 1979. — 149 с.
Лейтес Ф. Л. Гистохимия липолитических ферментов в норме и при патологии,
липоидного обмена. — М.: Медицина, 1967. — 248 с.
Лихачев Ю. П. Пневмокониозы (пат. анатомия)//БМЭ.— 3-е изд.— 1982.—
Т. 19. —С. 1450.
Луговской С. П. Морфологические изменения в слизистой оболочке тонкого
480
кишечника крыс при .хронической марганцевой иитоксикации//Микроэле»-
меитозы человека. — 1989.— С. 175—176.
Марачев А. Г., /К.аворонковА. А. Акклиматизационный дефицит железа//’
Физиология человека. — 1987. — Т. 43, № 4. — С. 640—646.
Меерсон Ф. 3. Адаптация, стресс и профилактика. —М.: Наука, 1981.—
276 с.
Миракилова А. М. Клинико-гематологические особенности железодефицитных
анемий у детей в высокогорных районах Памира: Автореф. дис. докт. —-
М„ 1986, —28 с.
Михалева Л. М. Кадмийзависимая патология человека//Арх. пат.— 1988.—
№ 9.— С. 81—85.
Михалева Л. М., Черняев А. Л. Патоморфологическая характеристика экспе-
риментальной кадмиевой иитоксикации//Микроэлемеитозы человека. — М.>
1989, —С. 194—195.
Морозов И. А., Лысиков Ю. А., Питран Б. В., Хвыля С. И. Всасывание и сек-
реция в тонкой кишке (Субмикроскопические аспекты). — М.: Медицина.
1988, —224 с.
Москалев Ю. И. Минеральный обмен. — М.: Медицина, 1985.— 287 с.
Ноздрюхина Л. Р. Биологическая роль микроэлементов в организма животных
и человека. — М.: Наука, 1977. — 183 с.
Ноздрюхина Л. Р., Нейко Е. М., Ванджура И. П. Микроэлементы и атеро-
склероз.— М.: Наука, 1985.— 221 с.
Одум Ю. Основы экологии/Пер. с англ.—М.: Мир, 1975. — 740 с.
Панин Л. Е. Изменения обмена витаминов, солей и микроэлементов//Мехаиизм
адаптации человека в условиях высоких широт/Под ред. В. П. Казна-
чеева. — Л., 1980. — С. 80—108.
Перельман А. И. Геохимия биосферы и иоосферы//Биогеохимические циклы
в биосфере. — М., 1976. — С. 86—97.
Пермяков И. К., Зимина Л. В. Острая почечная недостаточность при экзоген-
ных интоксикациях (вопросы классификации и патогенеза)//Арх. пат.—
1976,—Т. 38, № 11, —С. 3—11.
Раецкая Ю. И. Биогеохимия и проблемы микроэлементов в животноводстве//
Современные задачи и проблемы биогеохимии. — М., 1979. — С. 178—182.
Райцес В. С. Нейрофизиологические основы действия микроэлементов. — Лк
Медицина, 1981.— 152 с.
Рамад Ф. Основы прикладной экологии. Воздействие человека на биосферу. —
Л.: Гидрометеоиздат.— 1981. — 543 с.
Ревич Б. А., Сает Ю. Е., Смирнова Р. С., Сорокина Е. П. Методические реко-
мендации по геохимической оценке загрязнения городов химическими
_ элементами. — М.: ИМГРЭ, 1982. — 110 с.
Рейли К. Металлические загрязнения пищевых продуктов/Пер. с аигл. — М.:
Агропромиздат, 1985. — 183 с.
Риш Л1. А. Геохимическая экология животных и проблемы геиетики//Совре-
меииые задачи и проблемы биогеохимии. — М., 1979. — С. 121—133.
Риш М А. Биогеохимическое районирование Узбекистана. — Ташкент’ Фаи,
1979,— 190 с.
Риш М А. Геохимическая экология животных и проблемы генетики//Биолоси-
ческая роль микроэлементов. — М., 1983.— С. 17—28.
Рощин А. В., Орджоникидзе Э. К-, Припуцкая Л. Л. К вопросу о судьбе хро-
ма в оргаиизме//Гиг. труда и проф. заболевания.— 1982. — №9.—
С. 14—17.
Рудных А. А. Репаративный синтез ДНК в лимфоцитах человека и животных
при действии соединения хрома: Автореф. дис. канд. биол. наук. — М,
1986, —24 с.
Сает Ю. Е., Ревич Б. А., Смирнова Р. И., Сорокина Е. П. Город как техноген-
ная геохимическая провииция//1Х Всесоюз. конф, по проблемам микро-
элементов в биологии. Доклады. — Кишинев, 1981. — С. 42—45.
Сапин^М. Р.^Иммуииые структуры пищеварительной системы. — М.: Медицина,
Саркисов Очерки по структурным основам гомеостаза. — М.: Медицина,.
31—568
481,
'Семенов Н. В. Биохимические компоненты и константы жидких сред и ткаией
челввека. —М.: Медицина, 1971. — 151 с.
Серов В. В., Шехтер А. Б. Соединительная ткань (Функциональная морфоло-
гия и общая патология). — М.: Медицина, 1981. — 312 с.
Сидоренко Г. И., Ицкова А. И. Никель. — М.: Медицина, 1980. — 172 с.
Сороковой В. И. Механизмы развития повреждения ультраструктуры и функ-
ции митохондрий и плазмалеммы при деэнергизации клетки: Автореф.
дис. докт. — М„ 1984. — 31 с.
Строчкова Л. С., Сороковой В. И. Влияние соединений фтора на ферменты
клетки//Успехи соврем, биол.— 1983.— Т. 96, №2(5). — С. 211—223.
Строчкова Л. С., Юрова А. В., Жаворонков А. А. Влияние никеля на организм
животных и человека//Успехи соврем, биол.— 1987. — Т. 103, № 1.—
С. 142—155.
Теддер Ю. Р., Жаворонков А. А. Особенности метаболизма фтора при адап-
тации организма к охлаждению//Физиология человека.— 1988. — Xs 1,—
6. 123—128.
Токарь В. И. Состояние системы гипофиз — гонады у мужчин в условиях хро-
нической фтористой интоксикации: Автореф. дис. докт. — М., 1983.—
31 с.
Трахтенберг И. М., Коршун М. И., Терещенко Л. Г. и др. Новые данные о ток-
сичности неорганических соединений ртути и определяющих ее факторах//
Гиг. труда и проф. заболеваний. — 1981. — № 7. — С. 27—30.
Уголев А. М. Эволюция пищеварения и принципы эволюции функций. — Л.:
Наука, 1985. — 544 с.
Физиология всасывания (Руководство по физиологии) /Отв. ред. А. М. Уго-
лев. — Л.: Наука, 1977. — 667 е.
Фортескью Дж. Геохимия окружающей среды/Пер. с англ. — М.: Прогресс,
1985.— 360 с.
Чернух А. М., Фролов Е. П. (Ред.) Кожа. — М.: Медицина, 1982. — 336 с.
Шапошников А. М. и др. Медь//БМЭ. — 3-е изд. — 1980. — Т. 4. — С. 460—
463.
Шахламов В. А. Капилляры. — М.: Медицина, 1971. — 200 с.
Шахламов В. А., Цамерян А. П. Очерки по ультраструктурной организации
сосудов лимфатической системы. — Новосибирск: Наука, 1982.— 120 с.
Школьник М. Я. Физиологические причины тератологических изменений у рас-
тений//Современные проблемы микроэлементов в биогеохимии, сельском
хозяйстве и медицине. — М., 1980.
Шмидт Е. В. Гепато-церебральная дистрофия//БМЭ. — 3-е изд.— 1977,—
Т. 5, —С. 922—931.
Шнайдман И. М. Морфопатогенез силикоза и изыскание путей его патогене-
тической профилактики и терапии: Автореф. дис. докт.—М„ 1984.—
40 с.
Щерба М. М. Всасывание железа//Физиология всасывания. — Л., 1977.—
С. 223—248.
Щерба М. М., Петров В. И., Рысс Е. С. и др. Железодефицитные состояния. —
Л.: Наука, 1975. — 264 с.
Эдельсон Р. Л., Финк Д. М. Иммунологическая функция кожи//В мире нау-
ки. — 1985. — № 8. — С. 16—24.
Юрова А. В. Структурно-функциональная оценка влияния никеля на организм
животных и культуру клеток: Автореф. дис. канд. — М., 1989.— 19 с.
Ягодин Б. А., Муравин Э. А. Основные направления развития исследований по
агрохимии микроэлементов//Современные проблемы микроэлементов
в биогеохимии, сельском хозяйстве и медицине. — М., 1980. — С. 12—24.
Яцимирский К. О. Введение в бионеорганическую химию. — Киев: Наук. Дум-
ка, 1976, —248 с.
Aggett Р. J. Physiology and metabolism of essential trace elements: An outline//
Clin. Endocrinol. Metab. — 1985. — Vol. 14, N 3, —P. 513—543.
Aggett P. J., Rose S. Soil and congenital malformations//Experientia. —1987.—•
Vol. 43, —P. 104—108.
Alfrey A. C. Aluminum toxicity//Bull, N. Y. Acad. Med. — 1984. — Vol. 60,
N 2. — P. 210—212.
"482
Alfrey A. C„ LeGendre G. R., Raehny W. D. The dialysis encephalopathy syndro-
me . Possible aluminitim intoxication//New Engl. J. Med.— 1976,—-
Vol. 294, —P. 184—188. \ £. , t
Amitai I., Acker M. Adverse Effects of Intramuscular Iron Injection//AcLa
haemat. — Basel. — 1982. — Vol. 68. — P. 341—342.
Ananth J., Dubin S. E. Lithium and-symptomatic hyperparathyroidisim//J. roy,
Soc. Med. — 1983, —Vol. 76. — P. 1026—1029.
Appropriate use of fluorides for human health/Ed. J. J. Murray. — Geneva: WHO,
1986. ±
Arnaud-Battandier F. Le systeme lymphoide intestinal: conceptious actuelles//
Gastroent. clin. biol. — 1984. — Vol. 8. — P. 632—640.
Ash S., Schramm M. Determination of free and bound manganese (II) in hepa-
tocytes from fed and fasted rats//J. biol. Chem. — 1982. — Vol. 257.—
P. 9261—9264. , , ,
Babich H., Schiffenbauer M., Stotzky G. Effects of sterilization method on the
toxicity of Cr3+ and Cr6+ to fungi//Microbios Lett. — 1982b. — Vol. 20,—
P. 55—64.
Babich H„ Devanas M. A., Stotzky G. Review. The mediation of mutagenicity and
clastogenicity of heavy metals by physiochemical factors//Environm. Res.—
1985. — Vol. 37. — P. 253—286.
Baton R., Berchove R., Lucaki A., Pohl R. B. The effect of lithium on platelad
count//Acta psychiat. scand.— 1986. — Vol. 74. — P. 474—478.
Bannerman R. M. Genetics defects of iron transport//Fed. Proc.— 1976.—
Vol. 35, N 11, —P. 66—72.
Barnes G., Frieden E. Ceruloplasmin receptors of erythrocytes//Biochem. bio-
phys. Res. Commun.—‘ 1984. — Vol. 125, N 1. — P. 157—162.
Barton J. C., Conrad M. E., Holland R. Iron, lead and cobalt absorption: simila-
rities and dissimilariteis//Proc. Soc. exp. Biol. Med.— 1981. — Vol. 166.—
P. 64—69.
Batey R. G., Shamir S., Wilms J. Properties and hepatic metabolism of non-
transferrin-bound iron//Dig. Dis. Sci. — 1981. — Vol. 26. — P, 1082—1088;
Baud C. A., Boivin G., Demeurisse C. Drug-induced skeletal fluorosis//Abstr.
XI Ann. Conf. Intern. Soc. Fluoride Res. — Dresden, 1981. —P. 3—4.
Ben-Shachar D., Finberg J. P. M., Youdim M. B. Effect of iron chelators on
dopamine Da receptor//.!. Neurochem. — 1985. — Vol. 45. — P. 999—1005.
Berliner S., Shoenfeld Y., Livni E., Pinkhas J. Allergic purnura following intra-
venous administration of iron dextran//Acta haemat.— 1979. — Vol. 62.—
P. 178—180.
Berndt I. Iron metabolism. — Budapest: Akad. Kiado, 1983. — 415 p.
Bhathena S., Recant L., Voyled R. et al. Decreased plasma enkephalins in cop-
per deficiency in man//Amer. J. clin. Nutr.— 1986. — Vol. 43, N 1. — P. 42.
Bianchi V; Celotti L., Lanfranchi G. et al. Genetic effects of chronium compo-
unds//Mutat. Res. — 1983. — Vol. 117. — P. 279—300.
Blumberg A. B., Marti H. R., Graber C. G. Serum ferritin and bone marrow iron
in patients undergoing continuous ambulatory peritoneal dialysis//
J.A.M.A. — 1983. —Vol. 250, N 24,—P. 3317—3319.
Boegman R. J., Bates L. A. Neurotoxicity of aluminium//Canad. J. Physiol.
Pharmacol. — 1984, —Vol. 62, —P. 1010—1014.
Bonatti S., Lohman P.-H. M., Berends F. Induction of micronuclei in Chinese-
hamster ovary cells treated with Pt. coordination compounds//Mutat. Res. —
1983, —Vol. 116, N 2. — P. 149—154.
Bond G. H., Hudgins P. M. Inhibition of red cell Pa2+ ATPase by vanadate//
Biochim. biophys. Acta. — 1980. — Vol. 600. — P. 781—790.
Bonewitz R. F., Foulkes E. C., O’Flaherty E. J., Hertzberg V. S. Kinetics of zine
absorption by the rat jejunum: effects of adrenalectomy and dexamethasone//
Amer. J. Physiol. — 1983, —Vol. 244.— P. G314—G320,
Bosch de Aquilar P., Gocmacre-Vanneste J. Painre Helical filaments in spinal
ganglion neurons of eldirly rates//Virch. Arch. Abt. В Zellpath. — 1984. —
Vol. 47, —P. 217—222.
Bothwell T. H., Charlton R. W., Cook J. D., Finch C. A. Iron metabolism in
man. — Oxford: Blockwell, 1979.
31*
483
Bragadine M-, Pozzan T.t Azzone G. The nature of the electron spin resonance
signal during aerobic uptake of Mn+2 in mitochondria from a rat liver//
Europ. J. Biochem. — 1983. — Vol. 134. — P. 385—390.
Bremner J., Mehra R. K. Metallothionen some aspects of its structure and func-
tion with special regard to its involvement in copper and zinc metabolism//
Chem. Scripts. — 1983.—Vol. 21, —P. 117—121.
Butzow /. J., Eichhorn G. L. Degradation of polyribonucleotides by zinc and
other divalent uietal ions//Biopolymers.— 1965.'—Vol. 3. — P. 95—107.
-Cairo G., Bardelle L., Schiaffonati L. et al. Multiple mechanisms of iron-induced
ferritin synthesis in HeLa cells//Biochem. biophys. Res. Commun.— 1985.——
Vol. 133, N 1, —P. 314—321.
Cainakaris T., Phillips, Danks D. M. et al. Mutations in humans and aminals
which affect copper metabolism//J. Inner, Metab. Dis.— 1983. — Vol. 6,—
P. 44—50.
Cantoni O., Costa M. Correlations of DNA strand breaks and their repair with
cell survival following acute exposure to mercury (II) and X-rays,//Molec,
Pharmac. — 1983. — Vol. 24. — P. 84—89.
Cantoni O., Christie T., Robinson S„ Costa M. Characterization of DNA lesions
urodused by HgCl2 in cell culture systems//Chem. Biol. Intern.— 1984.—
Vol. 49. — P. 209—224.
Caplan R. P., Fry A. H. Photophobia in lithium intoxication//Brit. med. J. —
1982, —Vol. 285, —P. 1314—1315.
Cariwright G. E„ Markowitz H., Shields G. S., Wintrobe M. M. Studies on copper
metabolism XXIX. Acritical analysis of serum copper and ceruloplasmin
concentrations in normal subjects, patients with Wilson’s disease//Amer. J.
Med. — I960. —Vol. 28. —P. 555—563.
Chan IF. У., Rennert О, M. Genetic trace metal disturbances//.!. Amer. cell.
Nutr. — 1985. — Vol. 4. — P. 39—48.
Chang У. С., Кап У. IF. A sensitive new prenatal test for sicklecells anemia//
New Engl. J. Med. — 1982, —Vol. 307. — P. 30—32.
-Chen X; Xue A., Morris V. C. et al. Effect of selenium deficiency on the chronic
toxicity of Adriamycin in rats//J. Nutr.— 1986. — Vol. 116, N 12.—
P. 2453—2465.
Chesters J. K. Metabolism and biochemistry of zinc//Current topics in nutrition
and disease. — New York, 1982. — P. 221—238.
Chisholm У. У., Thomas D. Y., Hamill T. G. Erythrocyte porphobilinogen synthase
activity as an indicator of lead exposure in children//Clin. Chem.— 1985.—
Vol. 31, N 4, —P. 601—605.
Chlud K. Therapy of osteoporosis with NaF//Abstr. XI Ann. Conf. Intern, Soc.
Fluoride Res. — Dresden, 1981. —P. 6—7.
Choie D. D., Longnecker D. S., Del Campo A. A. Acute and chronic cisplatin
nephropathy in rats//Lab. Invest.— 1981. —Vol. 44, N S.— P. 397—402.
Christiansen C., Baastrup P. C., Lindgreen P., Transboll B. Endocrine effeet of
lithium. II. Primary hyperparathyroidism//Acta endocr. (Kbh.).— 1978.—
Vol. 88, —P. 528.
Chrlstensson T. A. T. Lithium hypercalcemia and hyperparathvroidism//Lancet.—
1*9®. —Vol. 2, —P. 144.
Clark P., Eichhorn G. A predictable modification of enzyme specificity. Selected
alteration of DNA bases by metal ions to promote cleavage specificity by
deoxyribonuclease//Biochemistry. — 1974. — Vol. 13. — P. 5098—5102.
Conrad C. D., Hamilton Y. A. Case report. Recurrent premenstrual decline in
serum lithium concentration: Clinical correlates and treatment implications//
J. Amer. Acad. Child. Psychiat. — 1986. — Vol. 26, N 6. — P. 852—853.
Cousins R. J. Regulatory aspects of zinc metabolism in liver and metabolism in
liver and intestine//Nutr. Rev.— 1979. — P. 97—103.
Cousins R. J. Metallothionein-Aspects related to copper and zinc metabolism//
J. Inher. Metab, Dis, — 1983. Vol. 6. — Suppl. 1. — P. 15—21.
Cousins R. J. Absorption, transport and hepatic metabolism of copper and zinc:
special reference to metallothionein and ceruloplasmin//Physiol. Rev.—
1985.—Vol. 65, —P. 238—310.
'Cox T. M., O’Donnell M. IF. Studies on the binding of iron by rabit intestinal
microvillus membranes//Biochem. J. — 1981, —Vol. 194. — P, 753—759.
484
Crupper D. R., I\arlik S., DpBoni U. Aluminium and other metals in senile
(Alzheimer) dimentia//Alzhelmer's disease: senile dementia and related
disordes/Ed. R. Katrman et ''al. — New York, 1978. — P. 471—485.
Crapper-McLachlan D. R., De Boni U. Aluminium in human brain disease/,'
Neurotoxicology (Park forest, II), 1981. — Vol. 1. — P. 3—16.
Crusa S„ Wisniewski H. M. Cytochemical study on the effect of aluminium on
Neuronal Golgi apparatus and lysosomes//Acta Neuropathol. (Berl.).—
1984.— Vol. 63. —P. 287—295.
Cunnane S. C. Differential regulation of essential fatty acid metabolism to the
prostaglandins: passible basis for the interaction of zinc and copper in
biological systems//.!. Prog. Lipid. Res. — 1982. — Vol, 21. — P. 73 90.
Cuzick J Evans S. Gillman M. et al. Medicinal arsenic and internal malignan-
cies//Brit. J. Cancer. — 1982.—t Vol. 45.-P. 904-911.
Danielsson K. G., Seigo Ohl, Huang P. C. Immunochemical detection of metai-
lothionein in’specific epithelial cells of rat organs//Proc. nat. Acad. Soi. —
1982. — Vol. 79. — P. 2301—2304.
Danscher G. Localization of gold in biological tissue, A photochemical method
for light and electron microscopy//Histochemistry. — 1981. — Vol. 71.—
P. 81—88.
Darwish H. M., Schmitt R., Cheney J., Ettinger M. Copper efflux kinetics from
rat hepatocytes//Amer. J. PhvsioL — 1984. — Vol. 246. — P. G48—G55.
Davis N. T., Nightingale R. The effects of phytate in intestinal absorbtion and
secretion of zinc and whole body retention of zinc copper and manganes
in rats//Brit. J. Nutr. — 1975. — Vol. 34. — P. 243—258.
Dekundt G., Leonard A. Cytogenetic investigations on leucocytes of workers
occupationally exposed to cadmium//Environm. Physiol. Biochem,— 1975.—
Vol. 5, —P. 319—327.
De la. Torre R., Krompotic E. The in vivo and in vitro effects of lithium on
human chromosomes and cell replication//Teretology.— 1976. — Vol. 13,
N 2, —P. 131 — 138.
De Pauw-Glllet M. Cl., Houssier C1'., Hacha R., Fredericq E. Interactions de
1’ADN et de la chromatine avec le cis-PttNHjhClztll) un agent anthnitoti-
que//Biol. Cell. — 1979. — Vol. 35, N 3, —P. 59.
Disilvestro R. A.. Cousins R. J. Physiological ligands for copper and zinc//Ann.
Rev. Nutr. — 1983. — Vol. 3. — P. 261—288.
Dodson R. F., Castitlo-Mozun P., Hieger L. R., Williams M. G. Ultrastructural
and energy dispersive analysis of inorganic inclusions in a muscle biopsv//
Ultrastruct. Path. — 1981, —Vol. 2, N 4, —P. 365—372.
Doyle M. J., Price M. P., Friedem E. Stabilisation of amphibian and mammalian
liver nuclei by zinc and other metal ions//Comp. Biochem. Physiol. — 1981. —
Vol. 680, —P. 115—120.
Durman D., Palmiter R. Transcriptional regulation of the mouse metallethio-
nein-I gene by heavy meta[s//J. biol. Chem.— 1981. — Vol. 256. — P. 5712.
Eastwood G. L. Gastrointestinal epithelial renewal//Gastroenterology.— 1977 —
Vol. 72. — P. 962—975.
Eggermont E. The intestinal brush border membrane//Acta paediat. Belg —
1979-. —Vol. 32, —P. 163—172.
Egle P., Shelton K. Chronic lead intoxication causes a brain-speciiic nuclear
protein to accumulate in the nuclei of cells linning kidnev tubules//J. biol.
Chem. — 1986. — Vol. 261. — P. 2294—2298.
Eichhorn G. L. Intra metal ions and genetic regulation//Metabolism of trace
metal in man/Ed. О. M. Rennert, W. Y Chan. — Boca Raton, 1984 —
P. 1—6.
Elmes M. E., /ones J. G. Ultrastructural changes in the small intestine of zinc
deficient rats//J. Path.— 1980, —Vol. 130, N 1, —P. 37—43.
English L., Macara I., Cantley L. Vanadium stimulates the (Na, K)-ATPase in
Friend erythroleukemia cells and blocks erythropoiesis//,!. Cell. Biol —
1983. — Vol. 97. — P. 1299—1302.
Erdmann E., Werdan K., Krawietz W. et al. Vanadate and its significance in
biochemistry and pharmacology//Biochem. Pharmacol. — 1984. — Vol 33.
N 7. — P. 945—950.
485
Evans G. W. New aspects of the biochemistry and metabolism of copper//Zinc
and copper in clinical medicine. — New York — London, 1978. — Vol. 2.—
P. 113—118.
Evans G. W., Johnson P. E. Characterization and quantitation of a zinc-binding
ligans in human milc//Ped. Res.— 1980. — Vol. 14. —P. 876—880.
Evans G. W., Johnson E. C. Zinc absorption in rats fad a lowprotein diet and
a lowprotein diet supplemented with tryptophan or picolinic acid//J. Nutr.—
1980, —Vol. НО. —P. 1076—1080.
Falck F. J., Fiueh G., Smith R. J. Metallothionein and accupational exposure
to cadmium//Brit, J. Industr. Med. — 1983. — Vol. 40. — P, 305—313.
Fauerholdt L„ Vendsborg P. Thyroid gland morphology after lithium treatment//
Acta path, microbiol. scand.— 1981. — Vol. A89, N 4. — P. 339—341.
Fradkin J., Wolff J. Iodide — indiced thyrotoxicosis//Medicine (Baltimore).—
1983.— Vol. 62, —P. 1—20.
Franke J. Sodium fluoride therapy of osteoporosis-10 years study//In: Abstr. XI
Ann. Conf. Intern. Soc. Fluoride Res. — Dresden, 1981. — P. 16—17.
Frazier J., Caskey J., Yoffe M., Seligman P. Studies of the transferrin receptor
on both human reticulocytes and nucleated human cells in culture//,!, din.
Invest. — 1982. — Vol. 69. — P. 853—865.
Frieden E. A survey of the essential biochemical elements//Biochemistry of the
essential ultratrace elements/Ed. E. Frieden. — New York, London: Plenum
Press. 1984, —P. 1—16.
Farst A. Bioassay of metals for carcinogenesis: Whole animals//Environm.
With. Perspect — 1981. —Vol. 40. — P. 83—91.
Gachelin G. Le cholesterol; un materiau pour le ce!luIes//Recherche.— 1980.—
Vol. 112, —P. 725—731,
Garnica A. D., Chan W. Y., Rennert О. M. Role of metallothioneins in copper
transport in patients with Menkes’ syndrome//Ann. Clin. Lab. Sci. —
1978, —Vol. 8, N 4, —P. 302—310.
Gauwerky С. E., Golde D. W. Lithium enhances growth of human leukaemia-
cells in vitro//Brit. J. Haemat.— 1982. — Vol. 51. — P. 431—438.
Geiger G. D., Seth P. K., Klevay L. M., Parmar S. S. Receptor-binding changes
in copper-deficient rats//Pharmacology.— 1984.—Vol. 28, —P. 196—202.
Gentile J. M., Hyde K., Schubort J. Chromium genothoxicity as influenced by
complexation and rate effects//Toxicol. Lett.— 1981. — Vol. 7. — P. 439.
Ghinea N., Hasu M. Charge effect on binding, uptake and transport af ferritin-
through fenestrated endothelium//J. Submicrosc. Cytol. — 1986. — Vol. 18,
N 4. — P. 647—659.
Giannini E. H. Brewer E., Person D. A. Blood gold concerntrations in children
with juvenile rheumatoid arthritis undergoing long-term oral gold therapy//
Ann. Rheum. Dis.— 1984. — Vol. 43. — P. 228—231.
Goyer R. A., Moore J. F. Protein-metal interactions//Adv. exp. Med. Biol.-—-
New York, 1974. — P. 447-—462.
Gottlieb L. S., Trey Ch. The effects of fluorinated anesthetics on the liver and
kidneys//Ann. Rev. Med. — 1974. — Vol. 25. — P. 411—429.
Hair zinc in normal populations//Nutrition Rev.— 1982. — Vol. 40, N 3.—
P. 74—76.
Halpin К. M., Chanson D. G., Beker D. H. Efficiency of manganese absorption
in chicks fid corn-soy and Casein diets//J. Nutr. — 1986. — Vol. 116-—
P. 1747—1751.
Hambidge К. M. The role of zinc and other trace metals in pediatric nutrition
and health//Pediatr. clin, N. Amer. — 1977.—Vol. 24.—P. 95—107.
Hamilton E. I. An overview:the chemical elements, nutrition, disease and the
health of man. Research meeded on mineral content of human tissues//Fed.
Proc. — 1981, —Vol. 40, N 8. — P. 2126—2130.
Hara TTipbary R., Freed B. et al. Production of Se79 in cyclotron and its
uptake in tumors of mice//Int. J. Appl. Radiat. Isot. — 1973. — Vol. 24,
N 7. — P. 377—384.
Henkin R. I., Foster D. M., Aamodt R. L., Berman M. Zinc metabolism in adrenal
cortical insufficiensy: effects of carbohydrate-active steroids//Metabblism.—
1984. —Vol. 33, N 6. —P. 491—501,
Hennigar G. R., Greene IE. B., Valker E. M., de Saussure C. Hemochromatosis»
486
caused by excessive vitamin iron intake//Amer, J. Path. — 1979.—
Vol. 96. — N 2. — P. 611—625.
Helprin G. A., Clarke G. K. Unilateral fixed dilated pupil associated with nebu-
lised ipratropium bromide//Lancet.— 1986. — Vol. 2. — P. 1469.
Hershko C., Cook J. D., Finch C. A. Storage iron kinetics. II. The uptake ot
hemoglobin iron by hepatic parenchymal cells//J. Lab. clin. Med.— 1972.—
Vol. 80.— P. 624—634.
Higa Y., Oshiro S., Kino K. et al. Catabolism of globin-haptoglobin in liver cells
after intravenous administration of hemoglobin-haptoglobin in rats//J. biol.
Chem. — 1981. — Vol. 256. — P. 12 322—12 328.
Hinks L. J., Inwards К D., Lloyd B., Clayton B. Reduced concentrations of
selenium in mild Crohn’s disease//.!, clin. path. — 1988. — Vol. 4, N 2.-—
P. 198—201.
Hodsman A. B., Sherrard D. J., Alfrey A. C. et al. Bone aluminium and histo-
morphometric features of rend osteodystrophy//.!, clin. Endocr. — 1982.—
Vol. 54. — P. 539—545.
Hoffmann G. R. Mutagenicity testing in environmental toxicology//Environm,
Sci. Teehnol. — 1982. — Vol. 16, —P. 560A—574A.
Hollister L. E. Lithium—induced Diarrhea//J.A.M.A.— 1982. — Vol. 248, N 2.
Holm J., Hansen S. I., Lyngbye J. Li+-induced rease of the folate binding
protein from human granulocvtes//Scand. J. Haematol.— 1985. — Vol. 35,—
P. 109—114.
Hong С. B., Chow С. K. Induction of eosinophilic enteritis and eosinophilia in
rats by vitamin E and selenium deficiency//Exp. Molec. Path.— 1988.—
N2.-P. 182—192.
Hoodgenraad T. U., van den Hamer C. J. A., van Hattum J. Effective treatment
of Wilson’s disease with oral zinc sulphate: two case reports//Brit. Med.
J. _ 1984. — Vol. 289. — P. 273—276.
Hori C., Oka T., Vanadate enhances the stimulatory action of insulin on DNA
synthesis in cultured mouse mammary cells//Biochim. biophys. Acta.—
1980, —Vol. 610. —P. 235—240.
Hu H., Aisen P. Molecular characteristics of the transferrin-receptor complex of
rabbit reticulocytes//.!. Supramelec. Struct.— 1978. — Vol. 8. — P. 349—360.
Johansson A., Curstedt T., Robertson В., Catnner P. Lung morphology and
phospholipids after experimental inhalation of soluble cadmium, copper and
cobalt//Environm. Res.— 1984. — Vol. 34. — P. 295—309.
Kagi J., Nordberg M. (Eds.). Metallothionein — Basel: Birhouser Verlag, 1979.
Karin M., Herschman H. Dexamethasone stimulation of metallothionein synthesis
in HeLa cell cultures//Science.— 1979.—Vol. 204. — P. 176—177.
Kaul B., Davidow B., Eng Y., Gewirtz M. H. Lead, Erythrocyte protoporphyrin
and ferritin levels: in cord blood//Arch. Environm. Hlth.— 1983. — Vol. 38,
N 5, —P. 296—300.
Kay R„ Knight G. Trace meta!s//Surgical nutrition. — Boston etc., 1983.—
P. 283—329.
King J. C. Do women using oral contraceptive agents require extra zinc?//J.
Nutr. — 1987. — Vol. 117. — P. 217—219.
Kipling M., Waterhouse J. Cadmium and prostate carcinoma//Lancet.— 1967.—
Vol. 1, —P. 730—731.
Kino K-, Tsunoo H., Higa Y. et al. Hemoglobin-haptoglobin receptor in rat liver
plasma membranes//.!, biol. Chem. — 1980. — Vol. 255. —P. 9616—9620.
Kirchgessner M., Schwartz F., Schnegg A. Interactions of essential metals in
human phvsiology//Current topics in nutrition a disease. — New York,
1982, —P. 477—512.
Kirchgessner M., Grassman E. The dynamics of copper absorption//Trace element
metabolism in animal. — Edinburgh: Livingstone, 1970.— P. 277—286.
Kiaaser S., Kagi J., Wilson K. Characterization of iso protein petterns in tissue
extracts and isolated samples of metallothioneins by reverse-phase high liquid
ehromatography//Biochem. J. — 1983. — Vol. 209. — P. 71—80.
Котипе S., Asacutna S., Snow J. B. Pathophysiology of the ototoxicity of cis-
diamminedichloroplatinum//Otolaryngol. — Head a Neck Surg—1981 —
Vol. 89, N 2. — P. 275—282. ' к- •
487
Kontozoglou The., Mambo N. The histopathologic features of lithium-associated
thyroiditis//Hum. Path. — 1983. — Vol. 14, N 8, —P. 737—739.
Kortus J., Mayer J. Verteilung des Aluminium in Organismus bei erhohter Zufuhr
von Aluminium und Fluorionen//Nahrung.— 1969. — Bd. 12.— S. 441.
Kostsak S. A., Bearn A. C. Hereditary disoeders of cooper metabolism//The meta-
bolic basis of inherited disease/Ed. I. B. Stanburg. — New York, 1978.—
P. 1098—1125.
Kowarsky S., Blair-Stanek C. S., Schachter D. Active transport of zinc and
identification of zinc-binding protein in rat jejunal mucosa//Amer. J. Phy-
siol. — 1974. — Vol. 226. — P. 401—407.
Kressner M. S., Stocked R. J., Morell A. G., Sterlieb 1. Origin of Biliary copper//
Hepatology.— 1984. — Vol. 4, N 5. — P. 867—870.
Kristensen O., Harrestrup-Andersen H., Pallisgaard G. Lithium carbonate in the-
treatment of thyrotoxicosis: a controlled trial//Lancet.— 1976. — Vol. 1.—
P. 603—605.
Laborda R., Diaz-Mayans G., Nunez A. Nephrotoxic and hepatotoxic effects of
chromium compounds in rat//Bul. Environm. Contum. Tox’/col. — 1986.—
Vol. 36, —P. 332—336.
Larras-Regard E., Niederst Ch., Siami K- Physiologie cellulaire animale. Modi-
fications ultrastructurales de la thyroide de Souris traitee au gluconate de
lithium//C. R. Acad. Sci. (Paris). — 1983. — Ser. 3. — Vol. 294, N 6 —
P. 289—294.
Larrick J., Cressell P. Modulation of cell-surface iron-transferrin reeeptors by
cellular density and state of activation//.). Supermolec. Struct.— 1979.—
Vol. 11, —P. 579—586.
Lau B. IF. C., Klevay L. M. Plasma lecithin: cholesterol acyltransferase in
copper-deficient rats.//J. Nutr. — 1981. — Vol. Ill, N 10. — P. 1698—1703.
Lau B. W. C., Klevay L. M. Postheparin plasma lipoprotein lipase in Copper-
deficient rats//J. Nutr.— 1982. — Vol. 112, N 5. —P. 928—933.
Lauridsen U. B., Kirkegaard C., Nerup J. Lithium and the pituitarythyroid axis
normal subjects//.), clin. Endocr.— 1974. — Vol. 39. — P. 383—385.
Layton IF. M., Layton M. IF. Cadmium induced limb defects in mice strainasso-
ciated differences in sensitivity//Teratology.— 1979. — Vol. 19. — P. 229.
Lawion A. H. Trace elements in aging//The Biomedical role of trace elements in
aging/Ed. J. M. Hsu, R. L. Davis, R. W. Neithmer. — St.Petersburgh,1976. —
P. 1—6.
Leak L. V. Studies on the permeability of lymphatic capillaries//.), cell. Biol —
1971, —Vol. 50, —P. 300—323.
Lees R., Steele R., Wardle D. Arsenic, syphilis, and cancer of the prostate//
J. Epidem. Commut. Hlth.— 1985, —Vol. 39. — P. 227—230.
Lehto V. P., Hovi T., Vartio T. et al. Reorganization of cytoskeletal and contrac-
tile elements during transition of human monocytes into adherent macro-
phages//Lab. Invest.— 1982, —Vol. 47. — P. 391—399.
Levander O. Selenium: biochemical actions interactions, and some human health
implications//Current topics in nutrition a. disease. — New York, 1982 —
P. 345—368.
Levander O. Consideration in the design of selenium bioavailability studies//
Fed. Proc. — 1983. — Vol. 42, N 6. — P. 1721—1725.
Levander O. A. A global view of human selenium nutrition//Ann. Rev Nutr —
1987. — Vol. 7. — P. 227—250.
Liessens I. L., Monstrey /., Vanden E. E. et al. Bismuth encephalopathy. A clinical
and anatomo-pathological report of one case//Acta neurol.— 1978.—
N 5, —P. 301—309.
Lignere G. Ch., Giavarini S. Initial experience with oral gold salts in the treat-
ment of rheumatoid arthritis in patients followed up for one year//J. Int.
Med. Res.— 1984.— Vol. 12.— P. 193—197.
Lim В. C., Morgan E. H. Transferrin endocytosis and iron uptake by developing
erythroid cells in the chicken (Gallus domesticus)//J. comp. Physiol.—
1985, —Vol. B155, N 2, —P. 201—210.
Lin P.-Y., Gruenstein E. Pathways of chloride transport in human fibroblasts//
J. Cell. Biol. — 1985. —Vol. 101, N 5, Pt. 2, —P. 189,
Lindstedt G., Nilsson L. A., Walinder J. et al. On the prevalence, diagnosis and
48
management of lithiuminduced hypothyroidism in psychiatric patients//Brit.
J. Psychiat.— 1977. —Vol. 130. — P. 452—458.
Lippman S. Lithium’s effects on the kidney//Postgrad. Med.— 1982. — Vol. 71,
N 3. — P. 99—116. ki „ x,
Loeb L. A., Zakour R. A. Metals and genetic miscoding//Nucleic acid-metal ion
interactions/Ed. T. G. Spiro. — New York, 1980. — P. 145—178.
Loembeck I., Ebert К. H., Kasperek K- et al. Selenium intake of infants and
young children, healthy children and dietetically treated patients with phenyl-
ketonuria//Europ. J. Pediat. — 1984. — Vol. 143, N 2. — P. 99—102.
Loeper J., Goy-Loeper J-, Rozennsztajn Fragny M. The antiatheromatous action
of silicon//Atherosclerosis.— 1979. — Vol. 33. — P. 397—408.
Lovstad R. A. Catecholamine stimulation of copper dependent haemolysis: pro-
tective action of superoxide dismutase, catalase, hydroxyl radical scavengers
and serum proteins (ceruloplasmin, albumin and apotransferrin)//Acta phar-
macol. toxicol. — 1984. — Vol. 54. — P. 340—-345.
Mahaffey Ph. Toxicity of lead, cadmium and mercury: consideration for total
parenteral support//Bull. N. Y. Acad. Med.— 1984. — Vol. 60, N 2.—
P. 196—209.
Mander A. I. Is there a lithium withdrawal syndrome?//Brit. J. Psychiat.—
1986, —Vol. 149. —P. 498—501.
Matseshe J. W., Philips S. F., Malagelada J. R., McCall J. T. Recover dietary
iron and zinc from the proximal intestinal of healthy man: studies of current
meals and supplements//Amer. J. Clin. Nutr.— 1980. — Vol. 33. — P. 1946—
1954.
McCall D., Zimmer L., Katz A. Kinetics of thallium exchange in cultured rat
myocardial cells//Circulat. Res.— 1985. — Vol. 56, N 3. — P. 370—376.
McDermott J. R., Smith A., Ward M. et al. Brain aluminum concentration in
dialysis encephalopathy//Lancet.— 1978. — Vol. 1. — P. 901—904.
McDermott M. T., Burman K. D., Hofeldt F. D., Kidd C. S, Lithium associated
:hyrotoxicoses//Amer. J. Med.— 1986. —V. 80, June. — P. 1245—1248.
Menkes J. H., Allerx M., Steigleder G. K. et al. A sex-linked recessive disoder
with retardation of growth. Peculiar hair and focal cerebral and cerebellar
degeneration//Pediatrics.— 1962. — Vol. 29. — P. 764—779.
Mertz W. Clinical and public health significance of chronium//Current topics in
nutrition a. disease.—-New York, 1982. — P. 315—323.
Methfessel A. H., Spencer H. Zinc metabolism in the rat. II Secretion of zinc into
intestine//.!, appl. Physiol.— 1973, —Vol. 34. — P. 63—67.
Mikio K-, Tavassoli M. Ceruloplasmin receptors in liver cell suspensions are
limited to the endothelium//Exn. Cell Res.— 1984. — Vol. 155, N 1.—
P. 232—240.
Miller C. A., Levine E. M. Effects of aluminium salts on cultured neuroplastoma
cells//J. Neurochem. — 1974. — Vol. 22. — P. 751—758.
Mingyuan W., Shen Z. Biochemical province and endemia//Sci. sinica.— 1986,—
Vol. 29.— P. 389—396.
Mircevova L. The role of Mg2+-ATPase (actomyosin-like proteins) in maintaining
the biconcave shape of ervtrocvtes//Blut.— 1977.—Vol 35 —P. 323—
327.
Mircevova L., Viktora L., Hermanova E. Inhibition of phagocytosis of poly-
morphonuclear leucocytes bv adenosine and H0CI3 in vitro//Med. Biol. —
1984.— Vol. 62.— P. 326—330.
Moller-Madsen B., Mogensen S. C., Danscher G. Ultrastructural Localization of
Gold in Macrophages and Abast Cells Exposed to Aurothioglucose//Exp.
Molec. Path.— 1984, —Vol. 40,—P. 148—154.
Morgan E. H. Transferrin, biochemistry, physiology and clinical significance//
Molec. Aspects. Med. — 1981. — Vol' 4. — P. 1—123.
Morgan E., Baker E. Iron uptake and metabolism by hepatocytes//Fed. Proc.—
1986.— Vol. 45, N 12, —P. 2810—2816.
Morimoto K-, Lijima S., Koizumi A. Selenite prevents the induction of sister-
chromatid exchanges by methyl mercury and mercuric on evrid in human
whole-blood cultures//Mutat. Res. — 1982. — Vol. 102, —P. 183—192.
Motenbocker M. A., Tappel A. L, Effect of dietary selenium on plasma seleno-
489
protein P, selenoprotein P 1 and glutathion peroxidase in the rat.//J. Nutr.—
1984. — Vol. 144. — P. 279—285.
Moynahan E. Y., Barnes P. M. Zinc deficiency and synthetic diet for lactose
intolerance//Lancet.— 1973. — Vol. 1. — P. 676—677.
Munchausen L., Rahn R. Biological and chemical effects of eisdichlorodiammine-
platinum (II) on DNA//Cancer Chemother. Rep.— 1975. — Vol. 59.—
P. 643—646.
Nielsen F. Possible future implications of nickel, arsenic, silicon, vanadium,
and other ultratrace elements in human nutrition//Current topics in nutrition
a disease. — New York, 1982. — P. 379—404.
Nielsen F. H. The ultratrace elements//Trace Minerals in Foods/Ed.
К. T. Smith. — New York: Marcel Dekker, 1988. — P. 357—428.
Nishimura M., Umeda M. Mutagenic effect of some metal compounds on cultured
mammalian cells//Mutat. Res.— 1978. — Vol. 54. — P. 246—247.
Nordal К. P., Dahl E., Haise J. et al. Kidney transplantation may cure aluminium
osteodystrophy//Acta pharmacol. et toxicol.— 1986. — Vol. 59. — Suppl. 7.—
P. 289—292.
Norton B., Whalley L. I. Mortality of a Lithium-treated population//Brit, J.
Psychiat. — 1984. — Vol. 145. — P. 277—282.
Oakes B., Danks I., Campbell P. Human copper deficiency: ultrastructural studies
of the aorta and skin in a child with Menkes syndrome//Exp. mol. Path.—
1976. — Vol. 25. — P. 82—98.
O’Dell B. Biochemical basis of the clinical effects of copper deficiency//Current
topics in nutrition a disease.— New York, 1982. — P. 301—313.
Okada S., Suzuki M., Ohba H. Enhancement of ribonucleic acid synthesis by
chromium (III) in mouse liver//J. Inorg. Biochem. — 1983. — Vol 19.—
P. 95—103.
Owen C. A., Ludwig 1. Inherited copper toxicosis in Bedlington terriers Wilson’s
disease (hepatolenticular degeneration)//Amer. J. Path. — 1982. — Vol. 106. —
P. 432—434.
Page R. A. Limits for lead//Environm. Hlth.— 1984. — Vol. 92, N 7. — P 181—
182.
Pall H .S., Williams A. C., Blake D. R. Iron, akathisia, and antipsychotic drugs//
Lancet. — 1986. — Vol. 2. — P. 1469.
Panner B. Freeman R. B., Roth-Moyo L. A., Markowitch Jr. W. Toxicity
Following Methoxyfluorene Anesthesia. — I. Clinical and Pathological Obser-
vations in Two Fatal Cases//J.A.M.A.— 1970. — Vol. 21, N 5. — P. 86—90.
Parlzek J., Ostadalova I., Bones J., Baricky A. Pregnancy and trace elements the
protective effect of compounds of an essential trace element-selenium against
the peculias toxic effects of cadmium during pregnancy//,!. Reproduct a. Fer-
tifty. — 1969. — Vol. 16, N 3. — P. 507—509.
Parkinson I. S., Ward M. K., Feest T. G. et al. Fracturing dialysis osteodystrophy
and dialysis encephalopathy. An epidemiological survey//Lancet.— 1979.—
Vol. 1, —P. 406—409.
Parmley R. T., Barton J. C., Conrad M. E., Austin R. L. Ultrastructural cyto-
chemistry of iron absorption//Amer. J, Path. — 1978. — Vol 93 N 3 —
P. 707—727.
Parmley R. T., Barton J. C., Conrad M. E. et al. Ultrastructural cytochemistry
and radioautography of hemoglobin — iron absorption//Exp. Molec Path —
1981.“ —Vol. 34, N 2, —P. 131—144.
Parmley R. T., May M. E., Spicer S. S. et al. Ultrastructural distribution of
inorganic iron in normal and iron-loaded hepatic cells//Lab. Invest.—
1981, —Vol. 44, N 4, —P. 475—485.
Parmley R. T., Barton J. C„ Conrad M. E. Ultrastructural cytochemical identi-
fication of the siderophilic enterocyte//J. Histochem. a. Cytochem — 1984 —
Vol. 32, N 7. — P. 724—730.
Pattison S. E„ Cousins R. J. Kinetics of zinc uptake and exchange by primary
cultures of rat hepatocytes//Amer. J. Physiol. —1986.—Vol 250 —
P. E677—E685.
Perl D. P., Brody A. R. Alzheimer’s disease: X-ray spectrometric evidence of
aluminium accumulation in neurofibrillary tanglebearing neurous//Scien-
ce. — 1980, — Vol. 208, — P. 297—299,
490
Ptletz J. E., Herschman H. R. Hepatic metallothionein synthesis in neonatal mot-
tled-brindled mutant mice//Biochem. Gen. — 1982. —Vol. 21, N 5/6.—
P. 456—475.
Piskator M. Cadmium and hypertension//Lancet. — 1976. — Vol. 2, N 7981,—
P. 370—371.
Planas-Ronhe F. Role of transferrin in metal uptake by human lymphoblasts in
vitro//Cell Biochem. a. Funct.— 1985.—Vol. 3, N 3. — P. 217—222.
Platts M., Good G. C„ Hislop J. S. Composition of domestic water supply and
the incidence of fractures and encephalopathy in patients in home dialysis//
Brit. Med. J. — 1977. — P. 675—660.
Plooy A., van Djk M., Lohman P. Platinum compounds with antitumour activity//
Mutat. Res. — 1982. — Vol. 97. — P. 212—213.
Plum F. Dementia: The approaching epidemic//Nature. — 1979. — Vol. 279,—
P. 372—373. x t .
Pollack S., Weaver J. Iron release from transferrin: synergistic interaction bet-
ween adenosine triphosphate and an ammonium sulfate fraction of hemoly-
sate//J. Lab. clin. Med. — 1986. — Vol. 108, N 5. — P. 411—415.
Poole A. Measurements of enzymes of collagen synthesis in rats with experimen-
tal silicosis//Brit. J. exp. Path. — 1985. — Vol. 66. — P. 89—94.
Pounds G. G., Wright R., Kodell R. L. Cellular metabolism of lead: A kinetic
analysis in the isolated rat hepatocyte//Toxicol. appl. Pharmacol. — 1982.—
Vol. 66. — P. 88—101.
Prasad A S Clinical, biochemical and pharmacological role of zinc//Annu. Rew.
Pharm. Toxicol. — 1979, —Vol. 19, —P. 241—269.
Prasad A. S., Zafrallah T. Cossach. Zinc supplementation and growth in sickel
cell disease//Ann. Int. Med.— 1984. — Vol. 100.— P. 367—371.
Pusanova L., Doskocil M. Teratogenni pusobeni soli lithia na mlada kureci
zarodky//Sb. lek.— 1979, —Vol. 81, N 7,— P. 193—199.
Puzanova L., Doskocil M. Teratogenic effect of lithium salts on young chick
embryos//Folia morphol. (CSSR).— 1979, —Vol. 27, N 1.—P. 26—30.
Rahn R. 0., Johnson N. P., Hsie A. W. et al. The interaction of platinum
compounds with the genome: correlation between DNA binding anf biologi-
cal effects//Sci. basis toxicity asses. Proc. Symp., Galtinburg. Tenn., 1979.—
Amsterdam etc,, 1980. — P. 153—162.
Randall M. S. Aluminium toxicity in an infant not on dialysis//Lancet, —
1983.— Vol. 1, —P. 1327.
Reus V. I., Gold P., Post R. Lithium — induced thyrotoxicosis//Amer. J. Psy-
chiat. — 1979. — Vol. 136. — P. 724—725.
Richman C„ Makii M., Weiser P„ Herbst A. The effect of lithium carbonate on
chemotherapy — induced neutropenia and thrombocytopenia//Amer. J.
Hemat. — 1984. — Vol. 16, N 6. — P. 313—325.
Riordan J. R., Richards V. Human fetal liver contains both zinc- and copper-rich
forms of metallothionein//J. biol. Chem. — 1980. — Vol. 255. — P. 5380—
5383.
Roberts J. J. The mechanism of action of antitumor platinum coordination com-
pounds//Metal ions in genetic information transfer/Ed. G. Eichhorn, L. Mar-
zilli. — New York etc., 1981. — P. 274—326.
Roberts T. M., Hutchinson T., Paciga !. et al. Lead contamination around
secondary smelters: Estimation of dispersal and accumulation by human//
Science, — 1974. —Vol. 186.— P. 1120—1123.
Rosenberg B. Fundamental studies with cisplatin//Cancer (Philad.) — 1985,—
Vol. 55, N 10. — P. 2303—2311.
Rosenfeld C., Broder L. Cisplatin — induced autonomic neuropathy//Cancer
treatm. Rep. — 1984. — Vol. 68, N 4. — P. 659—661.
Samuels A. R., Freedman J. H., Bhargava M. M. Purification and characteriza-
tion of a novel abundant protein in rat bile that binds azo dye metabolites
and copperZ/Bjochim. biophys. Acta. — 1983. — Vol. 759. — P. 23—31.
Sandstead H. Availability of zinc and its requirements in human subjects//
Current topics in nutrition a disease.— New York, 1982, —P. 83—101.
Saruk M„ Eisenstein R. Aortic lesion in Marfan syndrome: The ultrastructurel of
cystic medial degeneration//Arch. path. Lab. Med. — 1977. — Vol 101 —
P. 74—77.
491
Sauve R., Simoneau C., Monette R., Roy G. Single-channel analysis of the
potassium permeability in HeLa cancer cells: Evidence lor a calcium-activi-
ted potassium channel of small unitary conductance//.). Memra a. Biol.—
1986, —Vol. 92, N 3, —P. 269—282.
Sayed A. K, Edwards J. A., Bannerman R. M. Copper metabolism of cultured
fidroblasts from the brindled mouse (gene symbol Mohr) (41038)//Pros.
Soc. exp. Biol. (N. V.). — 1981. — Vol. 166, N 1, —P. 153—156.
Schaefer S., Post У., Close L., Wrigert C. Ototoxicity of low and moderatedose
eisplatin//Cancer.— 1985. — Vol. 56, N 8. — P. 1934—1940.
Schiff L., Graham J. Cytotoxic effect of vanadium and oil-bired fly ash on
hamster tracheal epithelium//Environm. Res.— 1984. — Vol. 34. — P. 390—
402.
Schoa M., Amdisen A., Eskjaer J. S., Olsen T. Occurence of goitre during lithium
treatment//Brit. Med. J. — 1968. — Vol. 3. — P. 710—713.
Schrauzer G. N. The discovery of the essential trace elements: An outline of the
history of biological trace element research//Biochemistry of the essential
ultratrace elements/Ed. E. Frieden. — New York—London: Plenum Press,
1984. —P. 17—32.
Schroeder H. A., Nason A. P., Tipton I. H., Balassa J. J. Essential trace metals
in man: Copper//). Chron. Dis.— 1966. — Vol. 19. — P. 1007—1009.
Schroeder H. A., Kraemer L. A. Cardiovascular mortality, municipal water, and
corrosion//Arch. Environm. Hlth.— 1974. — Vol. 28. — P. 303—311.
Schultz P. N., Warren G., Kosso C., Rogers S. Mutagenicity of a series of hexa-
coordinate cobalt (III) compounds//Mutat. Res. — 1982. — Vol. 102.—
P. 393—400.
Schwarz Miline D. B. Fluorine requirement for growth in the rat//Bioinorg.
Chem.— 1972, —Vol. 1, —P. 331—338.
Schwarz K, Ricci B. A., Punsas S., Karkonen M. J. Inverse relation of silicon in
drinking water and atherosclerosis in Finland//Lancet.— 1977. — Vol. 1.—
P. 538—539.
Segal R. S., Rosenblatt S., Eliasoph I. Endocrine exophthalmos during lithium
therapy of manic-depressive disease//New Engl. J. Med.— 1973. — Vol. 289.—
P. 136—138.
Shamberger R. Trace metals in health and disease//Nutritional elements and
clinical biochemistry. — New York etc., 1980, — P. 241—275.
Shenkman L., Borkowsky W., Shopsin B. Lithium as an animmunologic adju-
vant//Med. Hypotheses. — 1980. — Vol. 6. — P. 1—6.
Shelton K-, Egle P. The proteins of lead-induced intranuclear inclusions bodies//
J. biol. Chem. — 1982, —Vol. 257. — P. 11 802—11 807.
Shimada T„ Shimamura K-, Sunano S. Effects of sodium vanadate on various
types of vascular smooth muscles//Blood Vessels. — 1986. — Vol. 23,—
_ P. 113—124.
Shiraishi Y., Yosida T. Chromosome abnormalities in cultured leucocyte cells
from itai-itai disease patients//Proc. Jap. Acad. Sci. — 1972. — Vol 48.—
P. 248—251.
Shore D., Wyatt R. 1. Aluminium and Alzheimer’s disease//). Nerv. Ment. Dis. —
1983. — Vol. 171, N 9. — P. 553—558.
Sidiropoulou-Skokou S., Yssidorides M. R. Lithium carbonate and problem of
kidney damage//Arch. Path. Lab. Med. — 1983.— Vol. 107, N 5, —P. 270—
273.
Simon M., Bourel M. Clinical and biochemical studies of iodopathic hemochro-
marosis//Metabolism of trace metal in man/Eds. Rennert О. M-, ChanW, Y.
Vol. 11, —Boca Raton, 1984, —P. 82—107.
Skulskii J. A., Manninen J., Jarnefelt J. Factors afftcting the relative magnitudes
of the ouabain-sensitive and oubain-insensitive fluxed of thallium ion in
erythrocytes//Biochim. biophys. Acta. — 1978. — Vol. 506. — P. 233—241.
Smith A., Morgan W. Hemopexin-mediated transport of heme into isolated rat
hepatocytes//), biol. Chem. — 1981. — Vol. 256. — P. 10 902—10 909.
Smith J. Vanadium ions stimulate DNA synthesis in Swiss mouse 3T3 and 3T6//
Proc. nat. Acad. Sci. USA. — 1983. — Vol. 80. — P. 6162—6166.
Song M. K-, Adham N. F. Role of prostaglandin E2 in zinc absorption in the
rat//Amer. j. Physiol. — 1978. — Vol. 234. — P. E99—105.
492
Sorenson J. R., Kishore V. Antirheumatic activity of copper comp!exes//Tracer
elements in medicine. — New York—London, 1984. — Vol. 1. — P. 93—102.
Soskel N. T., 'Watanabe S., Sandberg. L. B. Mechanisms of lung injury in_the-
eopper-deficient hamster model of emphysema//Chest.— 1984. — Vol. 85.—-
P. 70—73.
Soskel N. T., Sandberg L. B. Lysyl Oxidase activity in lung ot copper deficient/
hamsters//Connect. Tis. Res. — 1985. — Vol. 13. — P. 127—133.
Spenser H., Osis D. Excretion of retained fluoride in man//J. appl. physioi.—
1975. — Vol. 38, N 2. — P. 282—287.
Stacey W., Kappus H. Cellular toxicity and lipid peroxidation in response to
mercury//Toxicol. appl. Pharmacol.— 1982. — Vol. 63. — P. 29—35.
Stanton B. A. Regulation of Ion Transport in Epithelia: Role of Membrane-
Recruitment from Cytoplasmic Vesicles//Lab. Invest. — 1984 — Vol. 51,
N 3. — P. 255—257.
Stein B-, Sussman H. Peptide mapping of the human transferrin receptor tn nor-
mal and transformed cells//J. biol. Chem.— 1983. — Vol. 258. — P. 2668—
2673.
Still C. N., Kelley P. On the incidence of primary degenerative dementia versus-
water fluoride content in South Caroline//Neurotoxicology (Parc Forest,
II).— 1980, —Vol. 1, —P. 125—132.
Sunderman F. W. Carcinogenesis effects of metals//Fed. Proc — 1978.—-
Vol. 37. — P. 40—46.
Sunderman W., Fraser G. Effects of nickel chloride and diethyldithiocarbamate
on metallothionein in rat liver and kidney//Ann. clin. Lab. Sci. — 1983. —
Vol. 13, N 6, —P. 489—495.
Sutherland R., Delia D., Schneider C. et al. Ubiquitous cell-surface glycoprotein
on tumour cells in proliferation associated receptor for transfemn//Proc.
nat. Acad. Sci. USA. — 1981, —Vol. 78. — P. 4515—4519.
Suzuki T., Yamomura M. Changes of metal contents and isometallothicmein-
levels in rat tissue after cadmium Ioading//Biochem. Pharmacol. — 1980а,—
Vol. 29. — P. 2407—2412.
Suzuki T., Yamamura M. Induction of zincthionein in rat liver and kidneys by
zinc loading as studied at isometallothionein levels//Toxicol. Lett. — 1980b.—-
Vol. 6. — P. 59—65.
Swarup G., Cohen S., Garbers D. Inhibition of membrane phosphotyrasyl-protein
phosphatase activity by vanadate//Biochem. biophys. Res. Commun.—
1982, —Vol. 107, —P. 1104—1109.
Tanaka K-, Min K-, Onosaka S. et al. The origin of metallothionein in red blood
cel!s//Toxicol. applied Pharmacol. — 1985. — Vol. 78. — P. 63—68.
Taylor R. T., Carwer J. H., Hanna M. L., Wandres D. L. Platinum-induced muta-
tions to 8-azaguanine resistance in Chinese hamster ovary ceils//Mutat..
_ Res. — 1979. — Vol. 67, N 1. — P. 65—80.
Tei T., Makino J., Kadofuku T. et al. Increase of transferrin receptors in rege-
nerating rat liver cells after partial hepatectomyZ/Biochem. biophys. Res.
Commun.— 1984, —Vol. 121, — P. 717—721.
Thorstensen K-, Romslo 1. Albumin prevents nonspecific transferrin binding and
iron Uptake by isolated hepatocytes/ZBiochiin. biophys. Acta — 1984 —
Vol. 804,—P. 393—397.
Tjaler H., Stahl K. Effect of 5-chloro-7-iodo-8-hydroxyquinoline (clioquinol) on
the uptake and distribution of nickel, zinc and mercury in mice/.'Acta Phar-
macol. Toxicol. — 1984, —Vol. 55, —P. 65—72.
Traub R. D., Rains T. C„ Garruto R. M. et al. Brain destruction alone does not
elevate brain aluminium//Neurology. — 1981. — Vol. 31, N 9, —P. 986.
Tsapakos M., Hampton T., Wetterhahn K. Chromium (Vl)-ibduced' DNA lesions-
and chromium distribution in rat kidney, liver and Iung//Cancer Res —
1983. — Vol. 43. — P. 5662—5667.
Underwood E. G. Trace elements in human and animal nutrition//4rd Ed.—
New York: Acad. Press, 1977.— 402 p.
Unger A., Hershko C. Hepatocellular uptake of ferritin in the rai//Brit. J.
Haemat. — 1974. — Vol. 28. — P. 169—179.
Ulmer D. D. Toxicity from aluminium antacids//New Engl J. Med — 1976 —
493»
Van Bruwaene R., Kirchmann R., Impens R. Cadmium contamination in agri-
cultural and zootechnology//Experientia.— 1984. — Vol. 40. — P. 43—52.
Venter P., Montaldi A., Majone F. et al. Cytotoxic, mutagenic and clastogenic
effects of industrial chromium compounds//Carcinogenesis. —1982.—>
Vol, 3, —P. 1331—1338.
Victery W., Miller C. R., Fowler B. A. Lead accumulation by rat renal bruch
border membrane vesicles//Pharmacol. exp. therap. — 1984.— Vol. 231,
N 3. — P. 589—596.
Vigliani E. C., Pernis B. Studien uber die Pathogenese der Silikose//Int. Arch.
Gewerbepath. Gewebchyg. — 1962. — Bd. 19, N 5. — S. 507—528.
Villeneuve A., Gartier Y., Yus A., Perron D. Effect of lithium on thyroid in man//
Lancet. — 1973. — Vol. 2. — P. 502.
Waller D., Edwards Y. Investigation of renal tubular function during treatment
with lithium//Neuropharmacology.— 1984. — Vol. 23, N 2. — P. 277—281.
Walker R. G., Escott M., Birchall I. et al. Chronic progressive renal lesions
induced by lithium//Kidney Int. — 1986. — Vol. 29. — P. 875—881.
Walsh J. M. Hudson memorial lecture: Wilson’s disease: Genetics and Bioche-
mistry— their relevance to therapy//J. Inher. Metab. Dis.— 1983.—।
Vol. 6, Suppl. 1. — P. 51—68.
Walsh С. T., Harnett К. M. Inhibitory effect of lead Acetate on contractility of
longitudinal smooth muscle from rat ileum//ToxicoI. appl. Pharmacol. —
1986. — Vol. 83. — P. 62—68.
Watanabe M., Honda S., Hayashi M., Matsuda T. Mutagenic effects of combi-
nations of chemical carcinogens and environmental pollutants in mice as
shown by micronucleus test//Mutat. Res.— 1982.—Vol. 97.— P. 43—48.
Webb M., Cain K. Functions of metallothionein//Biochem. Pharmacol.— 1982 —
Vol. 31, —P. 137—142.
Weigand E., Kirchgessner M. Total true efficiency of zinc utilisation: determina-
tion and homeostatic dependence upon the zinc supply states in young rats//
J. Nutr.— 1980. —Vol. ПО, —P. 469—480.
Whitehouse P. J., Price D. L., Clark A. IF. et al. Selective loss of cholinaryic
neurons in basel nucleus of Meynert in a patient with familial Alzheimer’s
disease//Amer. Neurol.— 1981. — Vol. 10. — P. 122.
Whiting R. F., Wei L., Stich H. F. Enhancement by transition metals of unsche-
duled DNA synthesis induced by isoniazid and related hydrazines in cultu-
red normal ane xerodermapigmentosum human cells//Mutat. Res.— 1979,—
Vol. 62, —P. 505—515.
"Whitmore С. K., Haggerty J. A. Osteoporosis therapy//Fluoride. — 1983.—
Vol. 16, N 3, —P. 189—190.
Willemse J., Van den Hamer C. J. A., Prins H. W., Yonker P. L. Menkes’kinky
hair disease. Comparison of classical and unusual clinical and biochemical
features in two patients//Brain. Dev.— 1982. — Vol. 4.—P. 105—114.
Williams D. Clinical significance of copper deficiency and toxicity in the world
population//Current topics in nutrition a. disease.— New York 1982.—
P. 277—299.
Williams R. J. P. Physico-chemical aspects of inorganic element transfer through
membranes//Philosophical Transactions of the Royal Society of London —
London, 1981, —P. 57—74.
Williams R. J. P. Zinc: what is its role in biology?//Endeavour. — 1984 —
Vol. 8. — P. 65-70.
Wills M. R., Brown C. S., Bertholt R. L. et al. Serum and lymphocyte aluminium
and nickel in chronic renal failure//Clin. Chim. Acta. — 1985. — Vol. 145. —
P. 193—196.
Wilson A. P. R,, Prouse P. J., Gurnpel J. M. Gold nephropathy due to auranofin
obscured by tolmetin pseudoproteinuria//Ann. Rheum. Dis.— 1984. —
Vol. 43, — P. 511— 512.
Wisniewski H. M., Ygbal K., Dermott J. R. Aluminium induced neurofibrillary
changes — its relationship to senile dementia of the Alzheimer’s type//Neuro-
toxicology (Park Forest, II).— 1980. —Vol. 1.—P. 121—124.
Wolman S. L., Anderson G. H., Marliss E. B., Jetjeebhoy K. N. Zinc in total
parenteral nutrition: requirements and metabolic effects//Gastroenterolo-
gy. — 1979. — Vol. 76. — P. 458—467.
ОГЛАВЛЕНИЕ
Предисловие . . . • ...... . > . .
о.
Введение ... • ...................................
Глава 1. Основные сведения о микроэлементах......................16
1.1, Определение понятия «микроэлементы». Краткая история их
изучения ...............
1.2. Содержание микроэлементов в окружающей природной среде /0
1.3. Перенасыщение организма микроэлементами в условиях при-
родных и техногенных локусов............................ . 31
Глава 2. Содержание микроэлементов в органах, тканях и жидкостях
организма человека.................................................37
2.1. Содержание микроэлементов в организме человека в норме
и в различные периоды его развития ....... 37
2.2. Участие микроэлементов в гомеостатических функциях орга-
низма , ,............................................- 4(У
2.3. Адаптация организма в экстремальных условиях и влияние ее.
на содержание микроэлементов. Акклиматизационный дефицит
микроэлементов ........ ....................................45-
Глав а 3. Структурные основы всасывания микроэлементов ... 48
3.1. Абсорбция микроэлементов в желудочно-кишечном тракте.
Мембранное пищеварение. Особенности метаболизма микроэле-
ментов. Ультраструктурные основы кишечного всасывания 48
3.2. Микроэлементы и органы дыхания ........ 67
3.3. Микроэлементы и кожа . . ........................... 69
Глава 4. Эссенциальные и токсичные микроэлементы .... 77
Глава 5. Микроэлементозы человека. Определение терминов. Клас-
сификация ................................................. .... 83
Глава 6. Важнейшие эссенциальные микроэлементы и связанные с ни-
ми заболевания.........................................92
6.1. Железо...................................... 92
6.2. Медь.........................................116
6.3. Цинк . ,..................................142
6.4. Марганец.....................................167
6.5. Хром . .188.
*6.6. Селен ........................................196 Se
6.7. Молибден . 231
6.8. Иод . 237
6.9. Кобальт . . . ........................... 255-
Глав а 7. Условно эссенциальные микроэлементы- и связанные с ними
заболевания..................................................266
* 7.1. Мышьяк . ................ 266
7.2. Бор ’.....................................275
7.3, Бром ........................................................ 2791
495»
Van
с
Wenit
e
Vide
b
b
Viglu
(
Villet
L
Walle
v
Walki
ii
Wais)
и
V
Walsh
1c
1!
Watar.
n<
si
Webb
V
Weiga
ti<
J.
White.
ni
di'
Whitir.
di
re
V<
Whitm
V<
WiUerr,
ha
fei
Williar
po
P.
Willi ar.
mt
Lo
W Lilian
Vo
Wills 4
ani
P.
Wilson
obs
Vo
Wisnlei
ch a
tox
Wolmat
par
gy.i
'7.4. Фтор . ..............................................2S(
7.5. Литий ..................................................
7.6. Никель....................................................3Q;
_$; • 7.7. Кремний............................................ . 32;
,< 7.8. Ванадий..................................................33;
Глава 8. Токсичные микроэлементы и вызываемые ими заболевания 34
' 8.1. Алюминий . , . ,....................................Ззз
«• 8.2. Кадмий................................................ 36;
‘ • 8.3. Свинец..................................................38;
• •' 8.4. Ртуть .................................................39;
8.5. Бериллий . 391
8.6. Барий................................................... 401
8.7. Висмут....................................................403
8.8. Таллий и другие потенциально-токсичные микроэлементы . . 4о;
Т лава 9. Ятрогенные микроэлементозы.............................45
9.1. Патологические реакции, связанные с лечебным использованием
эссенциальных микроэлементов.................................. 4 Г
9.2. Патологические реакции, связанные с лечебным использовани-
ем условно эссенциальных микроэлементов......................412
9.3. Патологические реакции, связанные с лечебным использованием
условно токсичных и токсичных микроэлементов .... 42’
Глава 10. Наследственные формы микроэлементозов человека и жи-
вотных ...........................................................43й
10.1. Первичные нарушения обмена микроэлементов при наследст-
венных болезнях............................................433
10.2. Вторичные нарушения обмена микроэлементов при наследст-
венных болезнях........................................... 450
Глава 11. Врожденные пороки развития при дефиците и избытке мик-
роэлементов ......................................................453
Общее заключение ................................................ 458
Приложения....................................................... 46”
Список основной литературы ........................................47$
Монография
Александр Павлович Авцын,
Алексей Александрович Жаворонков,
Марк Арнольдович Риш,
Лариса Сергеевна Строчкова
МИКРОЭЛЕМЕНТОЗЫ ЧЕЛОВЕКА э
(этиология, классификация, органопатология)
Зав. редакцией Ю. В. Махотин
Научный редактор С. В. Кузьмина
Мл. редактор Н. И. Нестерова
Художественный редактор С. М. Лылшна
Технический редактор И. М. Клепикова
Корректор Н. П. Проходцева
И Б № 5004
Сдано в набор 19.10.90. Подписано к печати 25.12.90. Формат бумаги бОХЭО'/щ. Бумага
тип, № 1, Гарнитура Литературная. Печать высокая. Усл. печ. л. 31,0. Усл. кр.-отт 313
Уч.-изд. л. 36,52. Тираж 10000 экз. Заказ 568. Цена 3 р. 20 к.
Ордена Трудового Красного Знамени издательство «Медицина»,
10Ю00, Москва, Петроверпгекий пер., 6/8.
Московская типография № 11 Государственного комитета СССР по печати.
113105, Москва, Нагатинская ул., д. I.