



























































Автор: Петров Ю.Е.
Теги: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез биология ботаника водоросли
Год: 1986




XXXV
http://biotex.ibss.org.ua
Ю. Е. ПЕТРОВ
ЭВОЛЮЦИЯ циклов
РАЗВИТИЯ У ВОДОРОСЛЕЙ
АКАДЕМИЯ НАУК СССР
БОТАНИЧЕСКИЙ ИНСТИТУТ им. В. Л. КОМАРОВА
КОМАРОВСКИЕ ЧТЕНИЯ
XXXV
Ю. Е. ПЕТРОВ
ЭВОЛЮЦИЯ циклов
РАЗВИТИЯ У ВОДОРОСЛЕЙ
Доложено на тридцать пятом ежегодном
Комаровском чтении
14 февраля 1983 г.
6
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1986
ACADEMIA SCIENTIARUM URSS
INSTITUTUM BOTANICUM NOMINE V. L. KOMAROVII
J. E. Petrov
EVOLUTIO CYCLI VITALIS ALGARUM
Relatio in sessione Komaroviana annua XXXV
die 14 mensis Februarii anno 1983
УДК 575.8 : 581.16 : 582.26
Петров Ю. E. Эволюция циклов развития у водорослей. (Комаров-
ские чтения, XXXV). - Л. : Наука, 1986. - 61 с.
На основании рассмотрения циклов развития представителей всех
отделов водорослей автор по-новому трактует циклы развития и
смену форм развития (чередования поколений) ряда групп водорос-
лей. Эволюция циклов развития водорослей с появлением полового
размножения прошла следующие основные этапы: отсутствие смены
форм развития, неупорядоченная смена гаметоспорофитов и возобнов-
ляющихся спорофитов, строгое чередование гаметофитов и спорофи-
тов, рост одной формы развития на другой и появление организмов
двойственной природы (циклоспоровые, харовые, некоторые красные
водоросли). Библиогр. 146 назв. Ил. 14.
Ответственный редактор
К. Л. Виноградова
Рецензенты
Т. Ф. Козыр ен ко, Л. П. Перестенко
257-86 - I © Издательство „Наука”, 1986 г.
042 (02) “ои
ВВЕДЕНИЕ
Циклом развития любого вида организмов можно назвать ряд по-
следовательно развивающихся индивидов, различающихся биологиче-
ски или морфологически, или совокупность стадий развития одного ор-
ганизма, начинающихся и заканчивающихся одними и теми же момента-
ми. В литературе распространен еще термин „жизненный цикл”, кото-
рый нередко считают синонимом цикла развития (БСЭ, 3-е изд., т. 9).
По нашему мнению, этот термин лучше применять для обозначения пре-
вращений, претерпеваемых отдельным индивидом с момента зарожде-
ния до отмирания.
В тех случаях, когда цикл развития включает несколько индиви-
дов, различающихся способами размножения, принято говорить о чере-
довании поколений. Чередование поколений — это „закономерная сме-
на у организмов генераций, различающихся типом размножения” (БСЭ,
3-е изд., т. 29, с. 67). В ботанике учение о чередовании поколений соз-
дал Гофмейстер (Hofmeister, 1851), открывший чередование гаплоидно-
го гаметофита и диплоидного спорофита в цикле развития споровых
растений. С тех пор у растений „под чередованием поколений понимают
чередование в цикле развития диплоидной и гаплоидной фаз” (БСЭ,
3-е изд., т. 29, с. 67).
В 50-х годах рядом советских авторов (Голлербах, 1951; Козо-По-
лянский, 1951; Полянский, 1958; Левин, 1959) была предпринята по-
пытка пересмотра учения о чередовании поколений у растений. Боль-
шинство авторов пришли к выводу, что термин „чередование поколе-
ний” не отражает всего многообразия явлений. Так, например, трудно
говорить о чередовании поколений у покрытосемянных, так как у них
произошло слияние гамето- и спорофита в единый организм. Учитывая,
что чередование поколений является частным случаем смены бесполой
и половой форм развития, был предложен термин „смена форм разви-
тия” (Голлербах, 1951).
В отношении организмов, имеющих только бесполое размножение,
но претерпевающих в цикле развития смену морфологически различаю-
щихся индивидов, пользуются термином „цикломорфоз”. В тех случа-
ях, когда такого рода изменения не носят закономерный циклический
характер, а определяются целиком внешними условиями, лучше ввести
термин „смена форм существования”.
При наличии в цикле развития полового размножения принято де-
лить особи на споро- и гаметофиты. Гаметофитом называют фазу разви-
тия (или отдельное поколение), дающую гаметы (независимо от того,
образуются ли наряду с этим споры или нет). „Спорофит — это фаза
1/2 1. Зак. 786
3
развития (или иногда отдельное поколение), дающая только споры”
(Полянский, 1958, с. 625). Подобные определения нельзя назвать одно-
значными, поскольку каждый такой термин, как увидим в дальнейшем,
объединяет разные организмы. Талломы многих водорослей при одних
условиях образуют органы бесполого размножения, т. е. функционируют
как спорофиты, а при других условиях — как гаметофиты. Поскольку
такие растения выполняют две функции — споро- и гаметофитов, то в от-
личие от настоящих гаметофитов их лучше называть гаметоспорофитами.
Спорофиты бывают тоже двух типов. Из спор одних вырастают обяза-
тельно гаметофиты, и за ними следует сохранить название „спорофиты”.
В других случаях из тех же спор или особых происходит возобновление
слоевищ самих спорофитов. В таких случаях лучше говорить о возобнов-
ляющихся спорофитах. Когда речь идет об одноклеточных водорослях,
окончание „фит” можно заменять на „цит” (гаметоцит, гаметоспороцит,
спороцит, возобновляющийся спороцит).
Водоросли демонстрируют большое разнообразие циклов развития.
Они бывают одного типа у всех представителей крупного таксона
(Fucales, Laminariales) или могут различаться у видов одного рода и быть
различными у индивидов одного вида в зависимости от географического
пункта. От особенностей цикла развития конкретных водорослей за-
висит специфика изучения их строения и развития, экологии, у стано вл е-
ние филогенетических связей и т. д. Хансгирг (Hansgirg, 1885) писал,
что без знания циклов развития не может быть настоящей систематики
водорослей. Известно много случаев, когда разные виды или роды со
временем оказывались формами одного вида. Познание эволюции цик-
лов развития водорослей является основой для выяснения путей возник-
новения их у высших растений.
4
ЦИКЛЫ РАЗВИТИЯ АГАМНЫХ ВОДОРОСЛЕЙ
О циклах развития и смене форм развития (чередовании поколений)
обычно говорят в отношении водорослей, имеющих половой процесс.
Однако их циклы возникли как результат эволюции циклов развития их
предков, не имевших полового процесса. В простейших случаях явле-
ния цикличности, наблюдаемые в развитии низкоорганизованных водо-
рослей, проявляются в виде повторяющихся изменений формы строения
тела в зависимости от условий существования. Морская синезеленая во-
доросль Hyella caespitosa в зависимости от условий существования
и возраста может развиваться в виде нитей, стелющихся по грунту, или
нитей,внедряющихся в известковый субстрат, а эпилитические нити могут
совсем отсутствовать. У старых экземпляров нити способны распадаться
на хроококковидные или глеокапсовидные клетки (Надсон, 1900).
Явление адаптивного полиморфизма широко распространено у водо-
рослей независимо от отсутствия или наличия полового процесса. Многие
криптомонады в холодной воде становятся значительно крупнее, чем
в теплой. Полиморфизм динофитовых водорослей выражается в измене-
нии окраски, формы и размера клеток, образовании защитной оболочки,
цистировании. Stylodinium sphaera и Cystodinedria inermis (Dinophyta) —
коккоидного строения, ведущие факультативно то автотрофный, то фа-
готрофный образ жизни, имеют амебо- и хелиозоидную стадии, питаясь
другими одноклеточными организмами. Некоторые их стадии идентичны
видам протозоа из родов VampyreIla, Amoeba, Dinamoeba, Actinophrys,
Raphidiocystis, Trichophrya (Popovsky, Pfiester, 1982). Heterochloris mutabi-
lis (Xanthophyta) может существовать и размножаться в виде трех форм
строения: монадной, амебоидной со жгутиком или без него, ризоподи-
альной. Такой цикл можно вполне назвать триморфным. Chloechloris
smithiana (Xanthophyta) встречается в виде подвижных монад и прикреп-
ленных или плавающих колоний.
Полиморфизм организмов, вегетирующих круглый год, принимает
соответственно смене времен года циклический характер (цикломорфоз).
Colacium libellae (Euglenophyta) в теплое время года представлен свобод-
но плавающими одноклеточными жгутиковыми организмами или коло-
ниями из групп клеток, сидящих на концах ветвей стебелька. Размножа-
ется он, как все эвгленовые, делением клеток на две части. С наступлени-
ем зимы клетки С. libellae проникают в прямую кишку личинок одного
из видов стрекоз, где он существует в пальмеллоидном состоянии как
скопление неподвижных клеток в слизи. С появлением льда личинки
стрекоз уходят в глубокие места. Весной они поднимаются в теплые слои
воды и клетки водоросли выходят наружу (Rosowski, Willey, 1975). Слож-
ная смена форм существования имеется у одноклеточных водорослей и
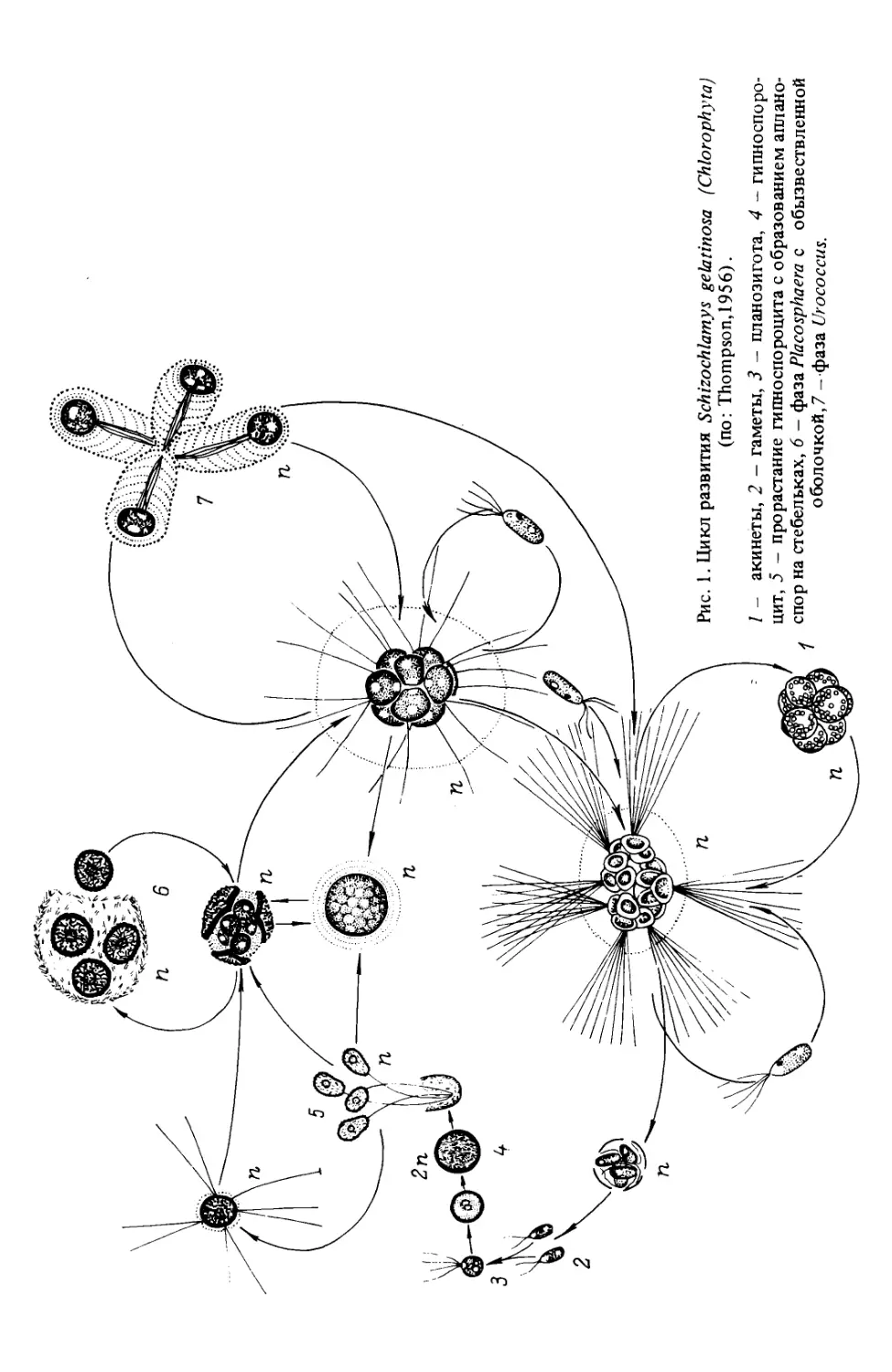
в том случае, когда у них существует половой процесс (рис. 1).
5
Рис. 1. Цикл развития Schizochlamys gelatinosa (Chlorophyta)
(по: Thompson, 1956).
/ - акинеты, 2 - гаметы, 3 - планозигота, 4 - гипноспоро-
цит, 5 - прорастание гипноспороцита с образованием аплано-
спор на стебельках, 6 - фаза Placosphaera с обызвествленной
оболочкой, / - фаза Urococcus.
Примитивные красные водоросли с переходом к многоклеточному
состоянию размножаются бесполым путем за счет выхода наружу одиноч-
ных вегетативных клеток. Asterocytis, имеющий слоевище в виде про-
стых или разветвленных нитей, при определенных экологических услови-
ях, например при низкой солености морской воды, переходит к сущест-
вованию в одноклеточном или пальмеллоидном состояниях. Porphyropsis
coccinea var. dawsonii имеет макроскопическое мешковидное слоевище
из одного слоя клеток. Размножение происходит моноспорами, которые
образуются по одной в вегетативных клетках путем округления их со-
держимого, или самими вегетативными клетками, выскальзывающими
из слоевища, или делением вегетативных клеток косой перегородкой и
отделением моноспоры. Моноспоры, возникшие из целых вегетативных
клеток, и вышедшие вегетативные клетки посредством почкования мо-
гут в свою очередь производить моноспоры. Все эти диаспоры прораста-
ют в микроскопические нитчатые или дисковидные слоевища, которые
способны производить моноспоры, дающие снова нитчатые или диско-
видные слоевища. Макроскопические слоевища образуются на дисковид-
ных в виде выростов (Murray et al.,1972, цит. по: Lee, 1980).
Смена форм существования у некоторых одноклеточных водорослей
может происходить ежесуточно и дополняется изменениями в поведении
клеток. У одноклеточных подвижных водорослей, живущих в приливо-
отливной зоне (литорали) морей, известно явление циркадного ритма.
С наступлением прилива водоросли активно перемещаются в толщу
грунта, а на период отлива поднимаются к его поверхности. Peridinium gre-
garium, живущий в литоральных лужах, ночью собирается в группы, при-
крепленные ко дну при помощи слизи. Двигающиеся клетки вытягивают
слизь в нити, переплетающиеся между собой. Утром с началом фотосинтеза
вся масса нитей под действием пузырьков кислорода поднимается к по-
верхности. С наступлением вечера клетки выскальзывают из этой массы
и плывут ко дну, где образуют новые скопления. В период низких темпе-
ратур и высоких приливов клетки остаются на дне все время. Этим пред-
отвращается вымывание их из литоральных луж (Lombard, Capon, 1971).
В размножении одноклеточных прикрепленных водорослей семейст-
ва Characiaceae (Chlorophyta) можно проследить три этапа эволюции цик-
ла развития: 1) содержимое целиком делится на диаспоры, и они все по-
кидают оболочку материнской клетки (Characiump, 2) содержимое це-
ликом делится на диаспоры, но одна зооспора остается внутри оболочки
материнской клетки и развивается в новый организм (Hydrianump, 3) на
зооспоры делится только верхняя половина клетки, оставшаяся часть
вырастает до прежних размеров и снова производит зооспоры (Acrobasma).
Такое направление эволюции цикла развития ведет к увеличению срока
произрастания одноклеточных водорослей на однажды занятом месте и
увеличивает вероятность воспроизведения вида в подходящих условиях.
Основная масса диаспор прикрепленных организмов гибнет в период по-
иска места для поселения из-за выноса течением за пределы прибрежной
зоны и выедания животными-фильтраторами. Гибель диаспор наступает и
в случае, когда они не находят в течение определенного срока подходя-
щего для прикрепления места.
2 Зак. 768.
7
ЦИКЛЫ РАЗВИТИЯ И СМЕНА ФОРМ
РАЗВИТИЯ ЭУГАМНЫХ ВОДОРОСЛЕЙ
В качестве наиболее примитивных циклов развития у водорослей,
обладающих половым процессом, обычно рассматривают циклы видов
с преобладанием гаплоидной фазы, наличием зиготы, проходящей пери-
од покоя для прорастания, и с зиготической редукцией. Было принято
считать, что слияние гамет и их ядер приводит к таким нарушениям в об-
мене веществ вновь возникшего диплоидного образования, что на пер-
вых этапах эволюции был необходим период покоя и вслед за этим воз-
вращение к гаплоидному состоянию. По мнению Н. П. Дубинина (1970) ,
мейоз на первых порах осуществлялся сразу же после образования зигот.
Прежде подобными наиболее примитивными считали циклы развития
Chlamydomonas и Ulothrix, а также других водорослей, в основном одно-
клеточных и пресноводных. Однако циклы развития пресноводных водо-
рослей нельзя признать первичными, поскольку они должны быть сред-
ством приспособления к менее стабильным условиям, чем в море (вы-
сыхание и промерзание мелких водоемов, колебание уровня воды и про-
зрачности в реках и водохранилищах, большие колебания температуры
и т. д.). Поэтому более примитивные циклы развития должны быть у
морских водорослей.
Рассмотрим по возможности все многообразие циклов развития
эукариотических водорослей на примере представителей разных отделов.
ДИНОФИТОВЫЕ ВОДОРОСЛИ (DINOPHYTA)
На циклах развития динофитовых водорослей мы остановимся более
подробно по сравнению с другими группами водорослей, поскольку о
них почти ничего не говорится в ботанической литературе. Между тем
именно их циклы развития с участием полового процесса можно считать
наиболее примитивными.
Бесполое размножение у подавляющего большинства динофитовых
водорослей осуществляется делением клеток на две части. Клетки пара-
зитических видов делятся на большее число частей, и,кроме этого,суще-
ствует размножение мелкими амебоидами (Pfiester, Lynch, 1980). Поло-
вое размножение наблюдается нечасто. Половой процесс - изогамия или
слабо выраженная анизогамия. В качестве гамет могут функционировать
вегетативные клетки или в результате их деления в каждой клетке обра-
зуется несколько гамет, часто внешне подобных вегетативным клеткам.
У Ceratium horridum мужские гаметы мельче женских. Перед слияни-
ем гаметы соединяются брюшными сторонами. Мужская гамета
8
сбрасывает щитки панциря, и ее цитоплазма вливается в цитоплазму жен-
ской гаметы. Интересно отметить, что при фаготрофном питании динофи-
товые захватывают пищевые частицы тоже брюшной стороной. Образо-
вавшаяся зигота остается подвижной (планозигота). Внешне она подобна
вегетативной клетке, т. е. ее поверхность покрыта панцирем из щитков и
имеет три длинных выроста. Таким образом, возникает диплоидный
организм, который в отличие от гаплоидных индивидов С. horridumHe
размножается бесполым путем. Планозигота растет несколько дней, в ре-
зультате чего образуется большая клетка с необычно длинными выроста-
ми. При первом делении мейоза возникают ядра с половинным набором
хромосом и образуются две жгутиковые клетки: одна с нормальными
антапикальными и ненормально длинным апикальным выростами, вто-
рая с нормальным апикальным и ненормально длинными антапикаль-
ными выростами. Через 2—3 дня происходит второе деление мейоза, про-
изводящее гаплоидные вегетативные клетки (Stosch, 1972) (рис. 2).
Пресноводные виды рода Ceratium проводят зиму в виде толстостен-
ных цист. У С. cornutum циста образуется в результате полового процесса.
Она прорастает с образованием одной жгутиковой клетки, которая в
дальнейшем делится мейотически (Christensen, 1980).
Гаметы пресноводного Gymnodiniumpseudopalustre отличаются от
вегетативных клеток несколько меньшими размерами и более светлой
окраской. Они не способны к длительному существованию и погибают,
если не произойдет слияния. После слияния образуется клетка, подобная
вегетативной, но с различимыми ядрами гамет. Растущая зигота вначале
имеет форму вегетативной клетки, потом происходит удлинение верхней
половины (эпикона). При этом поперечный жгутик теряется, но два про-
дольных остаются. Затем зигота становится неподвижной и округляется,
образуя первичную оболочку. Позднее формируется шиповатая оболоч-
ка гипноспоры, хлоропласты обесцвечиваются, накапливается много
красного масла, происходит слияние ядер. Зигота прорастает в лабора-
торных условиях через 4 нед после пребывания в темноте при 3 °C и по-
сле перемещения ее на свет и повышения температуры. Из зиготы выхо-
дит одножгутиковая клетка — мейоцит. Вначале она овальная и почти
бесцветная, позднее появляются бурые пигменты и ее форма становится
такой же, как у вегетативных клеток, образуются два продольных жгу-
тика, как до образования гипноспоры. После этого происходят два деле-
ния с мейозом, в результате чего образуются четыре гаплоидные жгути-
ковые клетки (Stosch, 1973) (рис. 3).
У пресноводного вида Peridinium willei гаметы имеют вид мелких го-
лых клеток (без теки). Зигота сохраняет подвижность 13—14 сут, затем
проходит период покоя и прорастает одной жгутиковой клеткой, сохра-
няющей капли красного масла, характерного для зиготы. В течение 24 ч
клетка делится на две дочерние, тоже с каплями красного масла. Через
2—3 сут происходит деление на обычные вегетативные клетки (Pfiester,
1976). У видов рода Woloszynskia мейоз совершается в гипнозиготе, и
при ее прорастании выходят четыре жгутиковые клетки (Stosch, 1973).
Гаметами у сапрофитного Crypthecodiniumcohnii служат вегетативные
подвижные клетки. Зигота неподвижная, при созревании растет до удво-
9
Рис. 2. Цикл развития Ceratium horridum (по: Stosch, 1972).
1 - планозигота.
ения своего, первоначального размера. Во время прорастания происходит
мейоз и образуются 4 (реже 8) подвижные клетки (Tuttle, 1980).
При половом размножении клетки фаготрофной Noctilica miliaris де-
лятся на 256-2024 жгутиковые гаметы. Первое деление ядра, по-видимо-
му, мейотическое. После конъюгации гамет возникает зигота, которая
без образования толстой оболочки проходит период покоя и прямо раз-
вивается в вегетативную диплоидную клетку (Zingmark, 1970).
Как видим, уже у динофитовых водорослей, относящихся к мезо-
кариотам, наблюдаются циклы развития с преобладанием гаплоидной
или диплоидной формы развития. В случае преобладания гаплоидной фа-
зы диплоидная форма развития существует относительно короткий
10
3
Рис. 3. Цикл развития Gymnodinium pseudopalustre (по: Stosch, 1973).
7 - планоэигота, 2 - гипноспороцит, 3, 4 - прорастание гипноспороцита.
период. Индивиды, принадлежащие к ней, активно фотосинтезируют и
увеличиваются в размерах. Активное питание существует у планозигот
фаготрофного Gymnodinium fungiforme (Spero, Moree, 1981). Одно из от-
личий их от гаплоидных индивидов заключается в неспособности к бес-
полому размножению с сохранением диплоидности. Переход зиготы в
гипнозиготу (гипноспору) существует не у всех представителей. Гипно-
зиготы возникли у динофитовых водорослей не в связи с переходом от
гаплоидного состояния к диплоидному, а как форма приспособления к
переживанию неблагоприятных условий. Как мы видели выше, гипнози-
гота в ряде случаев составляет один из этапов существования диплоид-
ной формы развития. У динофитовых водорослей с преобладанием гап-
лоидной фазы можно проследить, как мейоз переместился с деления
жгутиковой клетки, появляющейся при прорастании гипнозиготы, на мо-
мент деления самой гипнозиготы.
ЗОЛОТИСТЫЕ ВОДОРОСЛИ (CHRYSOPHYTA)
Одноклеточные золотистые водоросли размножаются делением на
две клетки. Многоклеточные представители (Phaeothamniales) способны
образовывать зооспоры по 4—8 в клетке. Половое размножение наблю-
11
дается редко. В основном оно известно у представителей, тело которых
располагается в домике (Kephyrion, Kephyriopsis, Dinobryon и др.), или
покрытых чешуйками (Mallomonas caudata, Synura petersenii). Половой
процесс сводится к слиянию двух вегетативных клеток (гологамия). У
видов, обладающих домиком, одна из клеток перед слиянием остается
в нем, домик же оказывается прикрепленным к зиготе (Bold, Wynne,
1978; Sandgren, 1981). Считается, что вегетативные клетки гаплоидны и
мейоз происходит в зиготе. Известны случаи аутогамного полового про-
цесса. Во всех случаях зигота превращается в цисту типичного для золо-
тистых водорослей строения. Цисты бывают: 1) одноядерные бесполые,
2) двухъядерные бесполые (потенциально аутогамные), 3) двухъядер-
ные половые (зиготические). Все цисты морфологически идентичны.
Отдельные клоны в пределах видов способны к образованию цист всех
типов или только одного (Sandgren, 1981).
ПРИМНЕЗИЕВЫЕ ВОДОРОСЛИ (PRYMNESIOPHYTA)
Примнезиевые водоросли объединяют в основном одноклеточные
жгутиковые и коккоидные организмы, которые прежде относились к зо-
лотистым водорослям. Этой точки зрения придерживаются некоторые
авторы и сейчас (Bold, Wynne, 1978). Поверхность клеток примнезиевых
водорослей может быть голой или покрытой органическими чешуйками
или известковыми пластинками (кокколитами). Некоторые примне-
зиевые образуют многоклеточные образования в виде пальмеллоидных
(Phaeocystis) и кубических (Sarcinochrysis) колоний, нитей (Apistonema).
У примнезиевых водорослей широко распространено явление поли-
морфизма. В цикле развития Gephyrocapsa huxleyi существуют три типа
различных клеток: 1) безжгутиковые голые, 2) безжгутиковые покры-
тые кокколитами, 3) жгутиковые покрытые органическими чешуйками.
В чистых культурах установлено, что один тип клеток может переходить
в другой. Все они размножаются делением на две клетки (Bold, Wynne,
1978).
В цикле развития Coccolithus pelagicus имеются жгутиковые организ-
мы с кокколитами иного типа, именуемые Crystallolithus hyalinus. Причи-
ны, вызывающие их смену, неизвестны (Bold, Wynne, 1978).
Половой процесс у примнезиевых водорослей, там, где он известен,-
изогамия или слабо выраженная анизогамия. Isochrysis galbata имеет
в своем цикле развития двужгутиковые клетки. В качестве гамет функ-
ционируют эти же вегетативные клетки, сбросившие жгутики. Зигота ра-
стет, значительно увеличиваясь в размерах. Прорастает она с образовани-
ем четырех коккоидных клеток, заключенных в общую слизистую мас-
су. По-видимому, их появлению предшествует мейоз. Каждая коккоид-
ная клетка производит одну жгутиковую (Gayral et al., 1972). Таким об-
разом, в цикле развития указанного вида существует чередование гапло-
идных монадной и пальмеллоидной форм существования. Диплоидная
фаза занимает небольшой отрезок всего цикла развития вида.
Цикл развития с гетероморфной сменой хорошо изучен у Ну тепо-
monas carterae — одноклеточного диплоидного организма с клетками, по-
крытыми кокколитами. Обычно он размножается делением клеток на две
12
или зооспорами. В определенных условиях клетки увеличиваются в раз-
мерах и делятся мейотически с образованием четырех гаплоидных кле-
ток, которые производят по одной зооспоре. Они в свою очередь, увели-
чиваясь в размерах и покрываясь кокколитами, превращаются в жгути-
ковые вегетативные клетки. Эти клетки и клетки, не превратившиеся
после мейотического деления в зооспоры, дают начало нитчатой гаплоид-
гой стадии, принимаемой ранее за самостоятельный организм Apistonema.
Нитчатая стадия размножается зооспорами с органическими чешуйками.
Кроме зооспор образуются гаметы. Зигота превращается в клетку, ти-
пичную для Hymenomonas carterae (Gayral et al., 1972). Таким образом,
цикл развития этого вида заключается в существовании одноклеточной
диплоидной стадии - возобновляющегося спорофита и многоклеточной
гаплоидной стадии — гаметоспорофита. Аналогичный цикл развития уста-
новлен и у других кокколитофорид (Gayral, Fresnel, 1983) .
Неподвижная многоклеточная стадия примнезиевых водорослей от-
личается устойчивостью к неблагоприятным факторам среды. Например,
донная стадия Cricosphaera sp. растет около верхнего уреза воды и под-
вержена значительным и резким колебаниям условий внешней среды.
Она выдерживает замерзание в течение 4 сут, повышение температуры до
35—40 °C более часа, повышение солености воды до кристаллизации со-
лей. Подвижная стадия устойчива к изменениям солености в пределах
4- 5О%о (Lee, 1980).
ДИАТОМОВЫЕ ВОДОРОСЛИ (BACILLARIOPHYTA)
Диатомовые водоросли, будучи одноклеточными организмами, раз-
множаются в основном делением клеток на две. Из-за наличия у них пан-
циря, состоящего из двух половинок, из которых одна краями входит
в другую (подобно чашке Петри), в процессе деления размеры клеток
постепенно уменьшаются. Восстановление прежней величины клеток
диатомовых происходит во время прорастания покоящихся спор, поко-
ящихся клеток, а также посредством ауксоспор, возникающих в резуль-
тате полового процесса. Стош (Stosch, 1965) обнаружил вегетативное
увеличение размеров мелких клеток до максимальных размеров посред-
ством частичного или полного выхода протопласта наружу и регенерации
нового панциря. Галагхер (Gallagher, 1983) установила, что у Sceletonema
costatum увеличение размеров клеток происходит чаще вегетативным
путем, чем в результате образования ауксоспор. Объем клеток при этом
увеличивается в 8 раз. По данным Н. Г. Кустенко (1984), при вегетатив-
ном укрупнении клеток их размеры в 2—2.8 раза превышают размеры
крупных клеток, возникающих из ауксоспор.
Половой процесс у диатомовых водорослей — оогамия (Centrophyсеае
и один род Pennatophyceae) и изогамия (остальные Реппаtophyceae). Ве-
гетативные клетки диатомовых диплоидные. Мейоз происходит при об-
разовании гамет. Из диплоидной зиготы через стадию ауксоспоры возни-
кает одна вегетативная клетка. Как правило, при оогамии в одной жен-
ской клетке формируется одна яйцеклетка, реже две. Оплодотворение
происходит внутри материнской клетки. При длительном отсутствии
мужских гамет яйцеклетки выходят наружу, где тоже возможно оплодо-
13
творение. Мужские гаметы (сперматозоиды) центрических диатомовых
имеют по одному жгутику.
Сперматозоиды у большинства изученных представителей центриче-
ских диатомовых образуются посредством сперматогониев (микроспор,
материнских клеток сперматозоидов), голых или окруженных нежным
панцирем. Содержимое мужской клетки Biddulphia mobiliensis, по наблю-
дениям Стоша (Stosch, 1954), делится без мейоза на две части — сперма-
тангии, которые вырабатывают редуцированную оболочку и расходятся
к противоположным концам материнской клетки. Содержимое каждого
сперматангия в свою очередь делится на 2, 4, 8 или 16 шаровидных спер-
матогониев, хлоропласты при этом уменьшаются в размерах и теряют
пигменты. Сперматогонии остаются внутри, в них происходит мейоз и
образуются по четыре сперматозоида с пластидами. Содержимое женской
клетки после мейоза делится на две части, при этом два ядра дегенериру-
ют, а два становятся ядрами двух яйцеклеток (рис. 4). У других видов
рода Biddulphia (Stosch, 1956) в материнской клетке (в двух сперматан-
гиях) образуются 2—4 (В. rhombus) или (2) -4-8 (В. granulata) дисковид-
ных сперматогониев с сильно редуцированной текой. Одна яйцеклетка
у В. rhombus дегенерирует.
Содержимое мужской клетки Chaetoceros didymum делится на 8 го-
лых сперматогониев, выходящих наружу. После первого мейотического
деления ядра цитоплазма сперматогония делится и возникают два спер-
матоцита, имеющих по два жгутика. Затем происходит второе деление
мейоза и обособляются два однопластидных одножгутиковых спермато-
зоида. Следует отметить, что такая же картина образования спермато-
цитов с двумя жгутиками наблюдается у видов других родов (Biddulphia,
Melosira). Сперматозоид Ch. didymum проникает в женскую клетку, похо-
жую на вегетативную и содержащую одно диплоидное ядро. С проникно-
вением мужской гаметы в ней начинается мейоз. После каждого деления
ядра одно из двух образовавшихся ядер дегенерирует. В результате в яй-
цеклетке оказываются одно функционирующее ядро и два дегенерирую-
щих (Stosch et al., 1973).
У Stephanopyxis palmeriana содержимое мужской клетки делится на
4, 6, 8 шаровидных сперматогониев, окруженных двумя рудиментарны-
ми створками. Во время мейоза содержимое сперматогония разбухает и
раздвигает створки. Четыре гаплоидных ядра с небольшими участками
цитоплазмы, лишенной хлоропластов, отпочковываются от остальной
массы сперматогония, образуют по жгутику и превращаются в бесцвет-
ные сперматозоиды. Оставшаяся цитоплазма сперматогония с хлоро-
пластами погибает. Мейоз в оогонии проходит как у Ch. didymum (Dre-
bes, 1966).
При образовании мужских гамет у Melosira varians (Stosch, 1951) со-
держимое вегетативной клетки распадается на 2 или 4 цилиндрических
сперматангия, в каждом из которых образуются четыре гаплоидных яд-
ра, которые с небольшой частью цитоплазмы дают четыре сперматозоида,
лишенные пластид. Оставшаяся большая часть содержимого материнской
клетки, включающая пластиды и лишенная ядра, погибает. То же самое
наблюдается у М. moniliformis (Stosch, 1958а).
14
Рис. 4.Половое размножение Biddulphia mobiliensis (по: Stosch, 1954).
1 - сперматангии, 2 - сперматогонии, 3 — сперматозоиды, 4 - яйцеклетки, 5 -
ауксоспоры.
3 Зак.786.
15
Если сопоставить образование сперматозоидов у разных центриче-
ских диатомовых — от Biddulphia до Melosira, то можно заметить посте-
пенную редукцию образования сперматогониев и утрату сперматозоида-
ми пластид.
Среди Pennatophyceae только у Rhabdonema adriatica обнаружена
оогамия (Stosch, 1958b). В женских клетках возникает по одной яйце-
клетке. Образованию мужских гамет предшествуют два или более деле-
ния вегетативной клетки. Появляются короткие клетки с небольшим
числом вставочных поясков и небольшим числом бледных хлоропластов.
Такие клетки Стош называет сперматогониями. В них формируются по
два безжгутиковых спермация, способных к амебоидному движению.
Половой процесс у остальных пеннатных диатомовых водорослей
сводится к конъюгации изоморфных безжгутиковых гамет. Две вегета-
тивные клетки приходят в соприкосновение по длинной стороне створок
и окружаются слизью. Ядро каждой клетки делится мейотически на че-
тыре гаплоидных ядра. Содержимое каждой клетки делится на две гаме-
ты, имеющие по одному функционирующему ядру и одному дегенериру-
ющему. От каждой клетки одна гамета перемещается к пассивной гамете
в другой клетке. Так в конъюгирующих клетках образуется по одной зи-
готе (ауксоспоре) . Наблюдаются различные видоизменения такого поло-
вого процесса. Например, в конъюгирующих клетках после мейоза оста-
ется по одному функционирующему ядру и пара клеток дает одну ауксо-
спору. Встречаются случаи аутогамии в форме цито- и ацитогамии. При
цитогамии в одной материнской клетке образуются две гаметы, которые
сливаются между собой с образованием зиготы. В процессе ацитогамии
содержимое не делится на две гаметы. Происходит мейоз; два ядра де-
генерируют, два других сливаются, .образуя диплоидное ядро зиготы.
Таким образом, цикл развития диатомовых водорослей проходит
в диплоидной фазе с гаметической редукцией. При этом необычным для
водорослей путем образуются мужские гаметы у центрических диатомо-
вых. Предварительно возникает большое число мелких клеток (сперма-
тогониев) , остающихся диплоидными. И только в самый последний мо-
мент, непосредственно перед обособлением гамет, происходит мейоз.
У других водорослей при гаметической или спорической редукции сна-
чала совершается мейоз, затем за счет митотических делений увеличива-
ется число ядер и только после этого одновременно появляется большое
число гамет (Fucales) или гаплоидных зооспор („зиготы” зеленых водо-
рослей, спорангии бурых водорослей и т. д.).
ЖЕЛТОЗЕЛЕНЫЕ ВОДОРОСЛИ (XANTHOPHYTA)
Подавляющее большинство желтозеленых водорослей — одноклеточ-
ные микроскопические организмы, и лишь ограниченное число родов
объединяет представителей многоклеточного или сифонного строения.
Половой процесс описан у видов трех родов: Tribonema, Botrydium, Vau-
cheria. У Tribonema известен один случай наблюдения полового процесса
(Scherffel, 1901). Гаметы имели одинаковые размеры, но различались по
поведению. В циклах развития трех перечисленных родов отсутствует
специализация особей на осуществление бесполого или полового
16
размножения. Зато они отличаются многообразием диаспор для беспо-
лого размножения. Нити Tribonema могут размножаться: 1) распадени-
ем на части, 2) двужгутиковыми зооспорами, 3) амебоидными клетка-
ми, 4) апланоспорами, 5) типичными для желтозеленых водорослей ци-
стами с окремненной оболочкой, 6) акинетами, 7) крупными цистами с
неокремненной оболочкой, которые производят большое число зооспор.
Нити Tribonema способны разрушаться с распадением оболочек клеток
на две половинки и освобождением содержимого клеток, переходящих
в пальмеллоидное состояние. Такое же многообразие форм бесполого
размножения наблюдается у видов рода Botrydium, живущих на сырой
почве. У них только отсутствуют крупные цисты и имеется половой про-
цесс в виде изо- или анизогамии. Зигота прямо развивается в новое слое-
вище. По-видимому, при этом происходит мейоз.
Своеобразные способы размножения существуют у видов рода
Vaucheria. Они размножаются крупными многоядерными многожгутико-
выми зооспорами или крупными апланоспорами (до 100Х140 мкм). Эти
же нити производят на своей поверхности оогонии и антеридии. В оого-
ниях образуется по одной яйцеклетке. Оогонии и антеридии располагают-
ся на нити в непосредственной близости. Оплодотворение яйцеклетки
происходит в оогонии на материнской нити. У V hercyniana оболочка
оогония разрушается и яйцеклетка выходит наружу, но остается при-
крепленной слизью к плодущей ветви (Rieth, 1974). Вегетативные нити
Vaucheria диплоидные, мейоз происходит во время гаметогенеза
(Al-Kubaisy et al., 1981).
ЗЕЛЕНЫЕ ВОДОРОСЛИ (CHLOROPHYTA)
Зеленым водорослям свойственно большое разнообразие форм раз-
множения и циклов развития, по-разному протекающих у представите-
лей разных порядков.
Порядок Volvocales
Вольвоксовые бывают одноклеточными и колониальными. Цикл
развития, характерный для одноклеточных вольвоксовых, имеют виды
рода Ch la ту do monas. Бесполое размножение Chlamydomonas осуществ-
ляется делением содержимого клеток на 4—8 зооспор. При половом
размножении наблюдаются гологамия, изогамия, анизогамия и оогамия.
Рассмотрим цикл развития Chlamydomonas на примере хорошо изучен-
ного вида Ch. moewusii. Роль гамет у него выполняют вегетативные клет-
ки. Зигота первое время сохраняет жгутики и плавает, позднее она сбра-
сывает жгутики и образует первичную оболочку. После слияния ядер
появляется вторичная скульптурированная оболочка. Такие зиготы на-
зывают зигоспорами или гипнозиготами. Во время так называемого пе-
риода покоя происходит созревание зиготы. В зависимости от вида она
остается прежних размеров или увеличивается. Во всех случаях в ней на-
капливается большое количество крахмала и масла, из-за чего она приоб-
ретает красную окраску. По данным Люина (Lewin, 1957), для развития
(созревания) зигот абсолютно необходимы свет и двуокись углерода.
17
Рис. 5. Цикл развития Chlamydomonas sp.
1 - планозигота, 2 - зигота, 3 - гипноспороцит, 4 - прорастание гипноспороцита,
5 - размножение зооспорами.
Он нашел, что к моменту созревания объем зигот Ch. moewusii увеличи-
вается в 30—60 раз. Скорость увеличения их объема была сопоставима со
скоростью возрастания объема вегетативных клеток в процессе роста и
размножения. Зиготы обладали повышенной стойкостью к внешним
воздействиям. В течение 60 с они выдерживали 52 °C, вегетативные клет-
ки — 46 °C. Многие зиготы Ch. moewusii, сохраняемые в течение несколь-
ких недель в ацетоне, прорастали при перенесении в свежую среду. При
прорастании зигот Chlamydomonas происходит мейоз и образуется от 4
до 32 зооспор в зависимости от накопления продуктов фотосинтеза за
период покоя (рис. 5).
Таким образом, зрелая зигота сильно отличается от того, что возни-
кает первоначально в результате слияния гамет. Фактически появляется
самостоятельно существующий организм, производящий в конце своей
18
жизни зооспоры. Поэтому его вполне можно называть спорофитом
(Петров, 1971, 1975; Виноградова, 1976). Как увидим ниже, у зеленых
водорослей широко развито образование одноклеточных спорофитов,
особенно у морских представителей. У пресноводных их заменяют гипно-
зиготы. По нашим представлениям, цикл развития Chlamydomonas вклю-
чает гаплоидный гаметоспорофит, способный к бесполому размножению
в течение многих поколений, и диплоидный одноклеточный спорофит.
Поскольку он сформировался путем разрастания зиготы, то его можно
называть зигоспорофитом (по аналогии с зигоспорой). В отличие от од-
ноклеточных спорофитов Ulothrix, Urospora и других у него не формиру-
ются диск для прикрепления и ножка. Считать, как принято до сих пор,
что у Chlamydomonas существует просто зигота, прорастающая после пе-
риода покоя с зиготической редукцией и образованием зооспор, методо-
логически неверно. При этом совершенно выпускают из вида, что проис-
ходит функционирование диплоидного организма, сопровождающееся
его ростом или по крайней мере накоплением в нем питательных ве-
ществ. Зигоспорофиту также необходим свет, как и вегетативным клет-
кам. Без света он не созревает и не производит зооспор. Чем меньше све-
та он получает, тем медленнее идет его развитие. Без учета всех этих об-
стоятельств остается неясным, каким образом продукт слияния двух
гамет — зигота — производит до 32 зооспор, каждая из которых близка
по размерам к средним гаметам.
В качестве примера колониальных вольвоксовых можно рассмо-
треть цикл развития Volvox. Колониальным Volvox считается не совсем
правильно. Этому противоречит следующее. Клетки его соединены плаз-
модесмами и дифференцированы на соматические и репродуктивные.
Расположенные на переднем конце организма клетки имеют более круп-
ный глазок. По существу это многоклеточный организм, состоящий из
двужгутиковых клеток. Колонии вольвоксовых, так же как и некото-
рых хлорококковых, часто называют ценобиями. Это вызвано тем, что
у них при бесполом размножении зооспоры или споры (в случае хлоро-
кокковых) в материнской клетке складываются в крохотную новую
колонию, которая выходит наружу и растет до обычных размеров
(рис. 6).
Ценобии у видов рода Volvox двух типов - бесполые и половые, по-
следние бывают однополыми и обоеполыми. У К carteri, например, цено-
бии трех типов: бесполые, женские и мужские (Lee, 1980) . Бесполые це-
нобии содержат крупные клетки — гонидии 80—100 мкм в диам., в то
время как вегетативные клетки — около 10 мкм в диам. В ценобии из
2—4 тыс. клеток насчитывается 16 гонидий. Зрелые гонидии в отличие от
других клеток не имеют жгутиков. Гонидии путем повторных делений
производят все три типа ценобиев. Начальные этапы роста и развития до-
черних ценобиев протекают внутри материнского ценобия; сформиро-
вавшись, они выходят наружу. При образовании женских и мужских це-
нобиев на стадии 16—32-клеточного проростка в результате неравного
деления одной—двух клеток появляется ограниченное число крупных
женских и мужских инициальных клеток. Мужские ценобии мельче ос-
тальных. Уже перед выходом из материнского ценобия инициальные
19
Рис. 6. Цикл развитая Volvox carteri.
1-4 - спорофит: 2-4 - развитие новых ценобиев; 5 - мужской гаметофит;
6 - женский гаметофит; 7 - группа сперматозоидов; 8 - зигота; 9, 10 - про-
растание зиготы.
клетки делятся и образуют пучки из 64—128 мужских гамет. По выходе
мужских ценобиев наружу пучки гамет тоже покидают свой ценобий.
Оплодотворение яйцеклеток происходит внутри материнского ценобия.
Зигота растет и одевается толстой оболочкой. Ценобий, по-видимому,
продолжает существовать некоторое время, затем разрушается, и зиготы
освобождаются. При прорастании в зиготе образуется одна двужгутико-
вая зооспора. Экзо- и мезоспории сбрасываются, и зооспора остается в
оболочке эндоспория. Зооспора функционирует подобно гонидии, делит-
ся и образует молодую бесполую колонию. Мейоз происходит в зиготе.
Мужские ценобии возникают у многих видов Volvox спонтанно. Женские
20
не возникают, пока родительские ценобии не окажутся среди мужских
или в среде, где они росли (Lee, 1980).
При описании размножения Volvox используются понятия „вегета-
тивные и фертильные ценобии, или колонии” (Schussnig, 1960; Lee, 1980)
или „бесполые и половые ценобии”. Все эти обозначения соответствуют
споро- и гаметофитам многоклеточных водорослей. И последние терми-
ны полностью приложимы к ценобиям Volvox. Поскольку бесполые це-
нобии производят не только половые, но и себе подобные, то их лучше
называть возобновляющимися спорофитами. Итак, в цикле развития
Volvox, по нашим представлениям, существуют возобновляющиеся спо-
ро- и гаметофиты. Подобно эволюционно продвинутым организмам це-
нобии Volvox обеспечивают развитие новых ценобиев, например муж-
ских, до стадии почти полного их сформирования.
Порядок Ulotrichales
Циклы развития улотриксовых лучше всего изучены у видов рода
Ulothrix. Обычно этот род приводят как пример многоклеточных зеле-
ных водорослей, проходящих цикл развития в гаплоидной фазе с дипло-
идной зиготой, в которой при прорастании происходит мейоз (зиготи-
ческая редукция). У ряда пресноводных видов это действительно так.
Например, у U. zonata клетки нитей производят 2—32 зооспоры с че-
тырьмя жгутиками, реже апланоспоры. Из них вырастают новые нити.
Гаметы образуются подобно зооспорам, но имеют по два жгутика. По-
ловой процесс — изогамия. Зигота одевается толстой оболочкой и прохо-
дит период покоя, в течение которого накапливает большое количество
запасных питательных веществ. При прорастании в зиготе происходит
мейоз и образуется от 4 до 16 зооспор или апланоспор.
Зиготы морских видов, например U. subflaccida, часто развиваются в
одноклеточные спорофиты на длинной ножке с диском при основании
для прикрепления к грунту (Kornmann, 1964). Локхорст и Вроман
(Lokhorst, Vroman, 1972) указывают для пресноводных видов Ulothrix
в Голландии, что зигота развивается в одноклеточный спорофит. В поль-
зу такого утверждения свидетельствует разница в размерах гамет и спо-
рофитов. Например, у U. subtilis гаметы 1.7—3.4X5.1—8.5 мкм, спорофи-
ты 15—45X20—70 мкм, у U. albicans гаметы 1.7—5.1X8.5—13.6 мкм, спо-
рофиты 21-45X25-65 мкм. Одноклеточные спорофиты — не такая уж
кратковременная стадия. Авторы указывают, что в Голландии летом
Ulothrix существует в виде спорофитов. Действительно, и в других стра-
нах нити Ulothrix встречаются в определенные сезоны и маловероятно,
чтобы остальное время года он полностью прекращал вегетацию. Иногда
спорофиты Ulothrix из-за их одноклеточное™ называют коккоидными
(Berger-Perrot, 1981).
В циклах развития видов рода Ulothrix наблюдаются различные от-
клонения. U. acrorhiza размножается только зооспорами. Нити U specio-
sa и U. flexuosa производят только гаметы (Kornmann, 1964). У U. albicans
оплодотворение иногда происходит внутри гаметангия и зигота развива-
ется тоже внутри (Lokhorst, Vroman, 1972).
21
Изучение U.flaccida из Роскофа (Франция) в культуре показало, что
существуют две экологические формы, идентичные морфологически и
цитологически, но различающиеся размножением и развитием. Форма из
верхнего горизонта литорали характеризуется однодомностью, изога-
мией и изоморфной сменой форм развития. Гаметы, копулируя, дают
планозиготу, которая без периода покоя прорастает. Одни зиготические
растения продуцируют апланоспоры, другие — зооспоры. Зооспоры дают
несколько идентичных поколений, прежде чем возникающие из них рас-
тения производят апланоспоры, которые проходят стадию покоя и про-
растают в половые растения. Форма из нижней литорали отличается дву-
домностью, анизогамией и гетероморфной сменой форм развития. Зиго-
та проходит длительный период покоя, после чего делится и производит
6—16 апланоспор, прорастающих в половые нитчатые растения (Perrot,
1968).
Так называемые половые растения Ulothrix постоянно образуют зоо-
споры, воспроизводящие материнские растения. Зиготы всегда произво-
дят много зооспор или апланоспор, превышающих по объему две проко-
пулировавшие гаметы. Независимо от строения (коккоидные или на
ножке) зиготы являются спорофитами. Поэтому можно утверждать, что
полный цикл развития Ulothrix включает смену гаплоидных гаметоспо-
рофитов одноклеточным диплоидным спорофитом.
Порядок Ulvales
Ульвовые водоросли дают примеры различных типов изо- и гетеро-
морфной смен форм развития, а также выпадения ее в результате утраты
половой или бесполой форм развития.
Изоморфную смену форм развития имеют виды рода Ulva. Их пла-
стинчатые слоевища бывают дипло- и гаплоидными. Диплоидные слоеви-
ща производят четырехжгутиковые зооспоры двух типов: диплоидные,
из которых снова вырастают диплоидные слоевища спорофитов, и гапло-
идные, образующиеся в родительских клетках после мейоза. Из гаплоид-
ных зооспор вырастают слоевища гаметофитов. В них образуются дву-
жгутиковые зооиды, которые в зависимости от внешних условий и гено-
типических свойств функционируют или как зооспоры, производя снова
слоевища гаметофитов, или как гаметы. Половой процесс — изо- или ани-
зогамия. Зигота без периода покоя прорастает и дает начало слоевищу
спорофита. Широко распространено партеногенетическое развитие гамет
в гаплоидные слоевища.
У видов Ulva наблюдаются различные изменения в цикле развития.
Особенно много в этом отношении изучалась U. mutabilis. Двужгутико-
вые зооиды, образующиеся в слоевищах ее гаметофитов, ведут себя как
гаметы или без оплодотворения дают начало гаплоидным гаметофитам
или гаплоидным партеноспорофитам, в которых каким-то путем восста-
навливается диплоидный набор хромосом. При мейозе они производят
четырехжгутиковые гаплоидные зооспоры, дающие слоевища гаметофи-
тов только одного знака, того, какой имел родительский зооид
(Hoxmark, 1975). Хохмарк нашел, что 1% гамет развивается в новые
22
гаметофиты, чьи гаметы имеют еще больше шансов стать гаметофитами.
Такие линии гаметофитов возвращаются к нормальной смене форм раз-
вития через 2—5 поколений. Зооспоры, образующиеся в результате мейо-
за, никогда не превращались в спорофиты.
Виды рода Monostroma имеют гетероморфную смену форм разви-
тия. Макроскопические мешковидные слоевища, при разрыве превра-
щающиеся в однослойные пластинки, гаплоидны и производят двужгути-
ковые зооиды, функционирующие как гаметы или воспроизводящие ма-
теринские слоевища. Зигота растет и превращается в одноклеточный спо-
рофит. У М. latissimum он достигает 55—80 мкм в диам. (Schussnig, 1960).
Мейоз происходит в спорофите при образовании зооспор или апланоспор.
Спорофиты Monostroma способны после деления содержимого разви-
ваться непосредственно в многоклеточные диски, на которых вырастают
макроскопические слоевища гаметофитов. В цикле развития М. (Protomo-
nostroma) undulatum отсутствует половой процесс. Пластинчатое слое-
вище производит зооиды, которые превращаются в цисты, в свою оче-
редь дающие зооспоры, воспроизводящие слоевища (Tatewaki, 1972).
Kommannia zostericola имеет макроскопические спорофиты и гаметофи-
ты в виде многослойных дисков, прижатых к грунту. Спорофиты могут
созревать на стадии стелющихся нитей или базального многослойного
диска (Виноградова, 1974), и тогда споро- и гаметофиты оказываются
фактически изоморфными.
Довольно обычное явление у ульвовых — выпадение половой формы
развития. При этом существует ряд изо- или гетероморфных поколений.
У некоторых видов разных родов бесполое размножение становится
единственным способом воспроизведения (Виноградова, 1974).
Как видим, у ульвовых нет строго упорядоченной смены форм раз-
вития, поскольку диплоидные слоевища являются возобновляющимися
спорофитами, а гаплоидные — гаметоспорофитами, поведение которых
зависит от условий существования.
Порядок Schizogoniales
К этому порядку относится род Prasiola. Ряд видов его (Р stipitata,
Р. meridionalis) обладает своеобразным циклом развития. Пластинчатые
слоевища у них диплоидные, размножаются бесполым и половым путем.
При бесполом размножении клетки верхней части слоевища делятся и в
них образуются апланоспоры, из которых снова вырастают диплоидные
слоевища, подобные родительским. При половом размножении клетки
верхней части слоевища делятся мейотически. Из одной диплоидной
клетки возникают четыре гаплоидных. В результате последующих деле-
ний клеток верхняя часть слоевища становится многослойной. Часть
клеток оказывается женскими, другая мужскими. Половой процесс —
оогамия. В женских клетках образуется по одной крупной безжгутико-
вой гамете. Мужские гаметы двужгутиковые. Зигота прорастает в дипло-
идное слоевище (Friedmann, 1959; Cole, Akintobi, 1963). Диплоидные
слоевища, появляющиеся из апланоспор и зигот, первоначально явля-
ются гаметоспорофитами. С переходом к размножению одни из них ста-
4 Длк. 78&.
23
Рис. 7. Цикл развития Prasiola stipitata (по: Friedmann, 1959).
1 — спорофит, 2 — гаметофит.
новятся спорофитами, другие гаметофитами. Вероятно, это определяется
условиями, в которых они растут. Prasiola stipitata, например, растет на
берегу моря в зоне, орошаемой брызгами прибоя, и спорофиты распола-
гаются на берегу выше гаметофитов (рис. 7). У пресноводного вида
Р. japonica мейоз происходит в зиготе (Tanner, 1981).
Порядок Cladophorales
Среди представителей этого порядка (в старом понимании) наблю-
дается большое разнообразие в смене форм развития и чередовании
ядерных фаз. Виды родов Cladophora и Chaetomorpha обладают изо-
морфной сменой диплоидных возобновляющихся спорофитов и гаплоид-
ных гаметоспорофитов. Иногда, как у Ch. аегеа, половое размножение
может отсутствовать (Kornmann, 1972). У видов рода Lola весь цикл бы-
вает в диплофазе. Зигота L. implexa прямо прорастает в диплоидное сло-
евище гаметоспорофита, которое производит двужгутиковые диплоид-
ные зооспоры и гаплоидные анизогаметы (Perrot, 1965).
В родах Urospora, Acrosiphonia и Spongomorpha гетероморфный
цикл развития состоит из макроскопических гаплоидных гаметоспоро-
24
фитов и диплоидных одноклеточных спорофитов. Спорофиты, как и
вегетативные клетки макроскопических слоевищ других кладофоровых,
многоядерные. Когда не был полностью известен цикл развития видов
перечисленных родов, их спорофиты были описаны в качестве самосто-
ятельных родов: рода Codiolum, имеющего стебелек и живущего подоб-
но другим донным водорослям, и рода Chlorochytrium, представляющего
собой крупную клетку (до 400 мкм дл.), живущую в красных и бурых
водорослях. Развитие одноклеточных спорофитов имеет явно приспосо-
бительное значение для видов указанных родов. Все они растут в холод-
ных морях в летнее время. Осенью макроскопические слоевища гамето-
спорофитов исчезают, но на камнях и скалах появляется зеленый налет,
образованный Codiolum. Из-за небольших размеров его слоевища уме-
щаются среди неровностей субстрата, и благодаря этому отсутствует
опасность полного уничтожения льдом.
Codiolum у Urospora penicilliformis у берегов Западной Европы мо-
жет появляться партеногенетически из мужских и женских гамет. В та-
ких случаях содержимое Codiolum делится на апланоспоры, которые
способны прорастать внутри оболочки материнской клетки (Lokhorst,
Trask, 1981). Джонсон (Jonsson, 1969) наблюдал у Acrosiphonia sonderi,
как после мейоза в Codiolum на стадии восьми ядер начиналось клеточ-
ное деление, приводящее к появлению гаметоспорофита. У небольшой
части зигот автор не наблюдал слияния ядер гамет. Ранее Джонсон
(Jdnsson, 1964, 1965) установил случаи неслияния пронуклеусов в зиго-
те, из-за чего в южных популяциях A. spinescens наблюдается тенденция
развития нитчатых миктогаплоидов. По мнению автора, во всех этих слу-
чаях за счет подавления спорогенеза в зиготах идет процесс возникнове-
ния моногенетического гаплоидного цикла из дигенетического гетеро-
морфного.
Порядок Oedogoniales
Своеобразие циклов развития многих видов эдогониевых водоро-
слей заключается в существовании карликовых мужских растений.
Бесполое размножение эдогониевых водорослей осуществляется
зооспорами, которые образуются по одной в клетке. Иногда могут фор-
мироваться апланоспоры. Половой процесс — оогамия. Он различается
у разных видов по поведению зооидов, образуемых мужскими клетка-
ми. Различают макрандровые и наннандровые виды. Процесс образова-
ния сперматозоидов у макрандровых видов сходен с другими водорос-
лями. Вегетативная клетка отчленяет короткую клетку, которая делится
повторно, образуя цепочку из 2—40 антеридиев. В антеридиях образуют-
ся по два сперматозоида с венчиком жгутиков, подобно зооспорам эдо-
гониевых. Сперматозоиды направляются к оогониям, ориентируясь на
выделяемые ими вещества (рис. 8, А).
Мужские нити наннандровых видов образуют вместо антеридиев
андроспорангии, возникающие таким же образом, но вместо двух спер-
матозоидов в них формируется по одной андроспоре в клетке. Андро-
споры привлекаются к материнским клеткам оогониев выделяемыми
25
Рис. 8. Цикл развития Oedogonium.
А - макрандровый вид, Б — наннандровый вид. 1 - обоеполый гаметоспорофит,
2 - женский гаметоспорофит, 3 - мужской гаметофит.
26
ими веществами. Прикрепление андроспоры вызывает деление материн-
ской клетки оогония на две: на оогоний и поддерживающую клетку,
к которой оказывается прикрепленной андроспора. После этого прекра-
щается выделение вещества, привлекающего андроспоры. Если материн-
ская клетка оогония покрыта слоем агара и андроспора не может при-
крепиться к ней, то выделение привлекающих веществ продолжается,
деление материнской клетки не происходит и оогоний не образуется.
Таким образом, прикрепление андроспоры служит пусковым механиз-
мом для развития оогония (Lee, 1980).
Андроспора, прорастая, образует двух-трехклеточную нить — кар-
ликовое мужское растение. Под влиянием веществ, выделяемых оогони-
ем, она направлена верхним концом к нему. Апикальные клетки муж-
ской нити образуют антеридии, в которых появляются по два спермато-
зоида (рис. 8, 5).
По мнению Ли (Lee, 1980), наннандровые виды у эдогониевых про-
изошли от макрандровых посредством партеногенетического прораста-
ния сперматозоидов с образованием карликовых мужских растений.
Цикл развития наннандровых видов эдогониевых, по нашему мнению,
иного происхождения. Андроспоры появляются в материнской клетке
по одной, как и обычные зооспоры эдогониевых, в то время как сперма-
тозоиды образуются по два в антеридиях наннандровых и макрандровых
видов. Поэтому андроспоры представляют собой зооспоры, у которых
появился хемотаксис на вещества, выделяемые материнскими клетками
оогониев. Судя по тому, что выделение веществ этими клетками пре-
кращается после прикрепления андроспор, а при выходе сперматозоидов
снова появляются вещества, на этот раз привлекающие сперматозоиды
к отверстию на оогонии, то, вероятно, привлечение андроспор и спер-
матозоидов происходит разными веществами.
Таким образом, если нити эдогониевых можно считать гаметоспоро-
фитами, поскольку они производят зооспоры и гаметы, то карликовые
мужские растения являются самыми настоящими мужскими гаметофи-
тами.
Зигота эдогониевых по мере созревания накапливает масло красного
цвета. Прорастает она после периода покоя. В ней происходит мейоз,
и образуются четыре зооспоры, прорастающие в нити.
Порядок С о 1 е о с h а е t а 1 е s
Из представителей этого порядка наибольший интерес с точки зрения
эволюции цикла развития представляет род Coleochaete. Виды его имеют
наиболее продвинутый цикл развития среди зеленых водорослей. Слое-
вище Coleochaete состоит из разветвленных нитей, которые у ряда видов
образуют псевдопаренхиматический диск. Бесполое размножение осуще-
ствляется двужгутиковыми зооспорами, образующимися по одной в
клетке. Половой процесс — оогамия. Антеридии развиваются на концах
ветвей и содержат по одному сперматозоиду. Оогонии у видов с талло-
мом из разветвленных нитей формируются из одноклеточных ветвей.
Нижняя утолщенная часть такой ветви становится оогонием, а вытянутая
27
Рис. 9. Coleochaete pulvinata (no: Oltmanns).
1 - оогоний после оплодотворения, 2, 3 - развитие спорофита на материнском
талломе.
верхняя часть — трихогиной. Зигота после оплодотворения остается в
оогонии и образует толстую оболочку. В дальнейшем зигота растет, уве-
личиваясь в размерах. Одновременно соседние нити дают выросты в ее
сторону и образуют вокруг нее псевдопаренхиматический слой. Возник-
шее образование иногда называют сперматокарпом. У видов с псевдопа-
ренхиматическим талломом оогоний возникает из периферической ме-
ристематической клетки. В таких случаях оболочка зиготы у некоторых
видов (например, С. orbicularis) образует выросты в примыкающие па-
ренхиматические клетки таллома. Клетки, покрывающие зиготу, по
своей морфологии, локализации и времени развития напоминают плацен-
тарные передающие клетки семенных растений (Graham, Wilcox, 1983).
После периода покоя в зиготе происходит мейотическое деление ядра и
содержимое делится на 8—32 клетки. В каждой из них образуется по од-
ной двужгутиковой зооспоре, дающей начало новому слоевищу. Клетки
таллома Coleochaete всегда гаплоидны, диплоидна только зигота
(Hopkins, McBride, 1976) (рис. 9).
В отношении Coleochaete мы имеем все основания говорить о разви-
тии на гаплоидных гаметоспорофитах диплоидных спорофитов, в кото-
рых происходит соматический мейоз. После развития из зигот спорофи-
ты остаются одноклеточными и диплоидными, затем после мейоза и об-
разования материнских клеток зооспор они становятся гаплоидными
и многоклеточными.
Порядок Dasycladales
Дазикладовые водоросли выделяются среди зеленых и других водо-
рослей морфологией и своеобразным циклом развития. Таллом макро-
скопический, неклеточного строения, в вегетативном состоянии с одним
крупным ядром (Acetabularia, Batophora) или многоядерный (Cymopolia).
С наступлением периода размножения на талломе образуются булавовид-
28
ные вместилища, собранные в виде зонтика на вершине (Acetabularia),
или шаровидные, расположенные среди периферических веточек. В это
время единственное ядро у Acetabularia, цикл развития которой изучен
наиболее полно, делится мейотически с образованием большого количе-
ства мелких ядер. Момент прохождения мейоза был установлен без под-
счета числа хромосом спектрофотометрически по количеству ДНК в яд-
рах (Коор, 1975). Ядра цитоплазмой заносятся в образовавшиеся к пери-
оду размножения вместилища. Вокруг каждого ядра обособляется уча-
сток цитоплазмы, покрывающийся оболочкой. Возникшие таким спосо-
бом образования называют цистами; со временем у большинства родов
их оболочки пропитываются известью. Молодые цисты увеличиваются
в размерах, в них происходит несколько делений ядра, и они становятся
многоядерными. Сформированная циста имеет в оболочке крышку.
Для прорастания цист требуется период покоя продолжительностью от
10 сут до 12—15 нед. У видов, живущих в субтропических морях, он со-
впадает с зимой. Содержимое цист делится с образованием большого
количества жгутиковых изогамет в зависимости от вида перед прораста-
нием (Lee, 1980) или сразу по выходе, или еще до выхода из материн-
ского растения (Tang, Loo, 1983). Гаметы выходят наружу после откры-
тия крышки в оболочке цисты. Зигота развивается в новый таллом
(рис. 10).
Нормальный проросток и таллом образуются только при участии
гамет не менее чем из двух цист. При выходе гамет из одиночной цисты
вместо вертикальных проростков партеногенетически развиваются шары
неправильной формы, со временем только у некоторых из них возникает
стебелек (Green, 1976).
Образование цист не происходит у Dasycladus и Cymopolia, гаметы
формируются прямо во вместилищах. Это рассматривается как вторич-
ное явление — результат неразвития мембраны цист. У некоторых вы-
мерших древних представителей дазикладовых цисты образовывались
внутри по всей центральной оси (Deloffre, Ge'not, 1982) . Коп (Koop,1979),
изучая развитие таллома Acetabularia из участков протоплазмы с оста-
точными вторичными ядрами, обнаружил появление цист в некоторых
талломах не только в зонтике, но и во всех их участках. Шуссниг
(Schussnig, 1960) полагал, что подобные эндогенные споры образовыва-
лись в прошлом в результате сегрегативного деления. Он считал цисты
самостоятельным поколением в обычном смысле слова, поскольку
им присуще индивидуальное развитие, самостоятельное существование
определенный отрезок времени и они имеют значение для экологии и
размножения вида. В то же время, исходя из практических соображений,
Шуссниг допускал возможным называть цисты гаметангиями.
Многие процессы, приводящие к созреванию „гаметангиев”, прохо-
дят во время, когда они находятся вне материнского организма, к тому
же имеются данные о прямом прорастании цист (Valet, 1969). Поэтому
нам кажется более логичной терминология, используемая Тайлором
(Taylor, 1960). Он пишет о размножении дазикладовых апланоспорами
или цистами, производящими гаметы.
По нашему мнению, для лучшего отражения сущности явлений, про-
исходящих в цикле развития дазикладовых водорослей,их таллом можно
ол
Рис. 10. Цикл развития Acetabularia.
1 - зигота, 2-5 - спорофит, 6-9 - апланоспоры (цисты) с развивающимися
внутри одноклеточными гаметофитами.
называть спорофитом, в котором происходит соматический мейоз и об-
разуются апл ано спорангии (лучи зонтика у Acetabularia) с апланоспора-
ми. В апланоспорах происходит развитие гаметофитов, завершающееся
образованием гамет. Возможно, указанием на принадлежность таллома
30
и цист дазикладовых к разным формам развития может служить разли-
чие в химическом составе стенок. У цист Acetabularia они состоят из цел-
люлозы, а единственный структурный полисахарид в других частях тал-
лома — маннан (Herth et al., 1975). Различия в химическом составе обо-
лочек споро- и гаметофитов известны у Porphyra tenera (Mukai et al.,
1981), у Bryopsis и Derbesia, что, вероятно, связано с различиями в числе
хромосом (Bold, Wynne, 1978).
Порядок Siphonales
Представители этого порядка имеют два типа циклов развития:
1) с гетероморфной сменой форм развития в дипло- и гаплофазах и
2) без смены форм развития с образованием диплоидных слоевищ.
Первый тип цикла развития хорошо изучен у видов некогда самосто-
ятельных родов Derbesia, Halicystis, Bryopsis. Слоевище Derbesia нитчатое,
слаборазветвленное. Это спорофит видов двух других перечисленных
родов. В Derbesia после мейоза образуются крупные зооспоры с венчи-
ком жгутиков. Интересно, что у D. marina толы о одно ядро из четырех,
возникающих при мейозе, участвует в образовании зооспор, остальные
три разрушаются (Neuman, 1969). Из зооспор в зависимости от вида
Derbesia вырастают гаметофиты, имеющие пузыревидное слоевище
(Halicystis) или перисторазветвленное (Bryopsis). Гаметофиты произво-
дят двужгутиковые анизогаметы. Зигота развивается в стадию Derbesia.
Часто в морях встречаются районы, где произрастает только одна из
форм развития вида. Такого рода явление подтверждается наблюдения-
ми за указанными сифоновыми в культуре. Например, у В. hypnoid.es
с атлантического побережья Франции существует нормальный цикл,
включающий Derbesia в качестве спорофита, а у Bryopsis с берегов Гол-
ландии зигота развивается в Bryopsis, минуя стадию Derbesia (Rietema,
1969, 1971). Только половым путем размножается В. plumosa у берегов
Исландии (Rietema, 1969,1971). Halicystis может воспроизводитьDerbesia
посредством слияния гамет и образования зигот или в результате парте-
ногенетического развития гамет. При этом часть зооспор из Derbesia вме-
сто Halicystis развивается в слоевища родительского типа, которые на
протяжении многих поколений размножаются зооспорами, производя
каждый раз только слоевища Derbesia (Kornmann, 1966). Было установ-
лено, что это гаплоидные мутанты, производящие гаплоидные зооспоры
(Kornmann, 1970). Прямое развитие Derbesia существует у берегов Но-
вой Англии (Sears, Wilce, 1970).
Цикл развития без смены форм развития с образованием диплоид-
ных слоевищ и гаметической редукцией известен у видов родов Codium,
Udotea, Halimeda, Caulerpa. Половой процесс — анизогамия, гаметы с дву-
мя жгутиками. Зигота без периода покоя прорастает в новое слоевище.
Своеобразное размножение среди сифоновых водорослей наблюдает-
ся у Dichotomosiphon, который по строению таллома и размножению на-
поминает Vaucheria. Бесполое размножение у него происходит крупны-
ми акинетами, образующимися по одной на концах ветвей. Половой про-
цесс — оогамия. Антеридии образуются рядом с оогониями на одной ни-
ти. Мужские гаметы двужгутиковые. Место мейоза неизвестно. У пресно-
31
Рис. W.Nitella (по: Oltmanns).
1-4 - развитие антеридия, 5 - молодой оогоний, а - поворотные клетки.
водного вида Dichotomosiphon tuberosus способ размножения зависит от
условий произрастания. В глубоких местах размножение бесполое, на не-
большой глубине - половое (Lee, 1980).
ХАРОВЫЕ ВОДОРОСЛИ (CHAROPHYTA)
Харовые водоросли имеют вегетативное и половое размножение.
Вегетативное размножение идет за счет нитей, растущих в грунте, и обра-
зующихся на них клубеньков. Бесполое размножение при помощи спор
и зооспор полностью отсутствует. Половой процесс — оогамия. Органы
полового размножения расположены на узлах, откуда отходят мутовки
вегетативных ветвей. Антеридии и оогонии харовых имеют своеобразное
строение и развитие, иное, чем у других водорослей и высших растений.
Антеридий имеет вид шара, сложенного снаружи из восьми или четырех
клеток, называемых щитками. Внутри от каждого щитка отходит
по одной удлиненной клетке (рукоятке). На конце такой клетки нахо-
дится округлая клетка (головка), на которой образуется примерно
шесть клеток меньшего размера (вторичные головки). Вторичные голов-
ки в свою очередь несут по четыре сперматогенные нити, в клетках кото-
рых образуются по одному сперматозоиду.
Развитие антеридия у харовых происходит следующим образом. Ини-
циальная клетка шаровидная, расположена на 1—2 клетках, отходящих
от узла. Инициальная клетка делится двумя продольными перегородка-
ми, расположенными взаимно перпендикулярно (рис. 11, 7). Образуют-
ся четыре клетки (квадранты), которые делятся поперечными перего-
родками на две половины. Возникают четыре нижних и четыре верхних
октанта. Каждый октант делится периклинально на наружную и внутрен-
32
нюю клетки, последняя еще раз делится периклинально. Теперь каждый
октант состоит из трех клеток: наружной, средней и внутренней (рис. 11,
2). Наружные клетки октантов разрастаются, становятся широкими и
плоскими и превращаются в щитки. Средние клетки удлиняются, пре-
вращаясь в рукоятки. Внутренние клетки (головки) тоже увеличивают-
ся в размерах (рис. 11, 3). В это время на головках появляются вторич-
ные головчатые клетки с антеридиальными (сперматогенными) нитями,
которые делятся поперечными перегородками на клетки, производящие
по одному сперматозоиду (рис. 11,4). Шуссниг (Schussnig, 1960) справед-
ливо заметил, что если сравнивать с другими водорослями, то антериди-
ями у харовых являются клетки, непосредственно производящие спер-
матозоиды. Все многоклеточное образование он называет андротеций.
Оогоний харовых состоит из группы генеративных клеток, оброс-
ших видоизмененными однорядными веточками, обвивающими его
плотной спиралью. Развитие оогониев начинается так же, как антеридиев.
Группа генеративных клеток возникает из апикальной клетки генера-
тивного побега, а обвивающие их ветви — из нижележащей узловой клет-
ки. У Nitella верхушечная клетка генеративного побега делится с обра-
зованием одной крупной клетки и трех небольших (дополнительных).
Первая дополнительная клетка возникает на вершине. Позднее ниже и
сбоку образуется вторая, а третья отделяется в основании. Самая круп-
ная клетка превращается в яйцеклетку, при этом три дополнительные
клетки оказываются оттесненными к основанию (рис 11, 5) .У Chara
имеется одна дополнительная клетка, которая появляется на последней
стадии роста яйцеклетки в ее основании. Гебель (Goebel, 1902) устано-
вил, что клеточные перегородки в оогонии гомологичны тем, что делят
инициальную клетку антеридия на квадранты. В оогонии харовых всегда
образуется одна яйцеклетка. После оплодотворения оболочка оогония
утолщается. Зигота прорастает после периода покоя. Ядро делится мейо-
тически на четыре ядра (Bold, Wynne, 1978). На вершине зиготы отделя-
ется небольшая клетка с одним ядром. Другие три ядра остаются в боль-
шой части зиготы, где дегенерируют. При прорастании верхушечная клет-
ка дает два выроста: один вверх, из него образуется вертикальная часть
таллома, другой вниз в качестве ризоида (Fritsch, 1935). Развитие про-
ростка обеспечивается веществами, находящимися в оставшейся боль-
шей части зиготы, имеющей при обособлении триплоидную природу.
Существует несколько точек зрения на происхождение органов по-
лового размножения харовых водорослей. Некоторые сейчас оставлены.
Челаковский (6elakovsky, 1878) полагал, что женский гаметангий харо-
вых представляет собой оогоний, окруженный оболочкой. То же самое
утверждал и Гоетц (Goetz, 1899), рассматривая дополнительные клет-
ки в женских гаметангиях харовых как остатки прежней многоклеточ-
ной стенки вокруг яйцеклетки.
Наиболее распространена в настоящее время теория Гебеля (Goebel,
1930), по которой гаметангии харовых представляют собой редуциро-
ванные ветви различных порядков (Pal et al., 1962). По мнению Гебеля,
в антеридии харовых каждый щиток с рукояткой и головкой соответст-
вует веточке вегетативного побега, состоящей из трех клеток; из них
33
верхушечная клетка превратилась в щиток, средняя — в рукоятку, а
нижняя — в головку.
Шуссниг (Schussnig, 1960) предложил теорию, в соответствии с кото-
рой антеридий харовых построен из четырех матеморфизированных срос-
шихся ветвей. Каждая из них образована двумя клетками. К центру
всего образования от каждой клетки отходит по одной двухклеточной
фертильной веточке, состоящей из междоузлия (рукоятки) и узла (го-
ловки). Шуссниг считал, что оогонии харовых возникли подобно антери-
диям, только подверглись еще большей редукции.
Если в соответствии с теориями Гебеля и Шусснига попытаться пред-
ставить органы полового размножения предков современных харовых
водорослей, то получаются образования, совершенно не свойственные
харовым. По Гебелю, верхушечная клетка вегатативного побега, образо-
вывавшего веточки с гаметангиями, должна была делиться на 4—8 частей,
выраставших в соответствующее число ветвей. Этого у харовых не на-
блюдается. Верхушечная клетка побега у них отчленяет поперечной пере-
городкой только одну клетку вниз, а она уже может делиться продоль-
ными перегородками. Гебель и другие авторы наблюдали случаи ненор-
мального развития антеридиев и оогониев харовых, когда они прораста-
ли в обычные вегетативные ветви, и никогда не бывало, чтобы они при-
нимали вид образований, напоминающих гипотетические структуры
Гебеля или Шусснига. Не согласуется с теорией Гебеля отделение и рас-
хождение головок друг от друга по мере созревания антеридия. По Гебе-
лю, головки должны составлять узел, от которого отходят короткие
веточки.
Поразительным в развитии органов полового размножения харовых
является деление их инициальных клеток на число дочерних клеток,
кратное 4 (за исключением явного сокращения числа делений в оогони-
ях Chara). Между тем в вегетативном узле харовых образуется любое
число ветвей: 1—3, 5—7 и более до 13, но крайне редко 4. Число вторич-
ных головок в антеридии на каждой головке равно шести, что совпадает
с числом ветвей в узле.
Для объяснения всех этих фактов можно предположить, что сфор-
мированные инициальные клетки антеридия и оогония являются микро-
и макроспорангиями. В случае антеридия каждый октант представляет
спору, прорастающую in situ в мужской многоклеточный гаметофит.
В оогонии Nitella дополнительные клетки отчленяются последовательно.
Поэтому приходится считать его содержимое одной спорой, прорастаю-
щей in situ в женский гаметофит, состоящий из четырех клеток у Nitella
и из двух клеток у Chara, из которых только по одной клетке превраща-
ются в женскую гамету.
КРАСНЫЕ ВОДОРОСЛИ (RHODOPHYTA)
Красные водоросли объединяют организмы от одноклеточных и ко-
лониальных до крупных многоклеточных сложного анатомического
строения. Соответственно изменяются форма размножения и цикл раз-
вития.
34
Одноклеточные красные (Porphyridium) размножаются делением
клеток на две части. У колониальных (Goniotrichum, Asterocytis) размно-
жение совершается моноспорами, представляющими собой вегетативные
клетки, выскальзывающие из слизистых трубок наружу. При не вполне
благоприятных условиях такие колониальные организмы могут перехо-
дить к существованию в одноклеточном состоянии. Наиболее примитив-
ные многоклеточные красные водоросли (Erythropeltidales) размножают-
ся моноспорами. Возможна смена форм существования, как например у
Porphyropsis coccinea var. dawsonii, с пластинчатого вертикального слоеви-
ща на разветвленное нитчатое и дисковидное.
Смена бесполого и полового размножений среди просто организо-
ванных красных водорослей имеется у представителей порядков
Rhodochaetales и Bangiales. У Rhodochaete parvula — простейшая для крас-
ных водорослей форма полового размножения, тем не менее это оога-
мия, как и у всех остальных Rhodophyta. Rh. parvula имеет однорядное
нитевидное слоевище с клетками, соединенными плазмодесмами. Спер-
мации образуются при отделении изогнутой перегородкой части содер-
жимого вегетативной клетки. Подобным же способом у ряда примитив-
ных красных водорослей отделяются моноспоры. Спермации попадают
на женские клетки, не отличимые от вегетативных. После оплодотворе-
ния образуется зигота, которая отделяет посредством изогнутой перего-
родки большую карпоспору, как это происходит со спермацием (Magne,
1960). Из карпоспоры развивается спорофит сходного с гаметофитом
строения. Гаметофит кроме карпоспор производит моноспоры, служа-
щие для его воспроизведения. Гаметофиты Rh. parvula гаплоидные, спо-
рофиты диплоидные (Boillot, 1969). У всех остальных красных водорос-
лей развитие зиготы проходит на материнском растении таким образом,
что в результате одного оплодотворения образуется большое количество
диаспор, называемых карпоспорами. Иногда такое развитие зиготы заме-
няется прямым выходом и прорастанием в спорофит (Acrochaetium pecti-
natum) (Hafez, Magne, 1983).
Просто организованные красные водоросли, относящиеся к Bangiales
(Porphyra, Bangia), имеют гетероморфную смену форм развития в виде
вертикального пластинчатого или нитевидного таллома и микроскопиче-
ских нитей, внедряющихся в известковый субстрат. Обе эти формы раз-
вития сильно различаются по требованиям к внешним условиям.
Вертикальные слоевища растут в условиях сильного освещения в при-
ливо-отливной зоне. Нити, сверлящие известковый субстрат, растут зна-
чительно глубже. Макроскопические слоевища Porphyra и Bangia — гаме-
тоспорофиты - производят моноспоры, которыми воспроизводятся сло-
евища материнского типа. Гаметы образуются на них в более узком тем-
пературном диапазоне. У Р. tenera они появляются при температуре
15 °C и ниже, а моноспоры — при температуре 15 °C и выше. Это неболь-
шое совпадение температуры приводит к тому, что в море встречаются
талломы с органами размножения обоих типов. Летом, когда температу-
ра становится выше 20 °C, развиваются карликовые талломы Р. tenera
только с моноспорами (Tseng, Chang, 1955). При половом размножении
содержимое клеток бангиевых дробится на 16—128 бесцветных сперма-
35
циев, другие с небольшим выступом на наружной стороне функциони-
руют как оогонии. После оплодотворения зигота, находясь на материн-
ском талломе, дробится без мейоза на 4—32 карпоспоры. Из карпоспор
развиваются нити, внедряющиеся в известковый субстрат. Эта нитчатая
стадия раньше была известна как самостоятельный род Conchocelis. Сей-
час ее называют стадией конхоцелис. В отличие от эфемерных пластинча-
тых талломов нити стадии конхоцелис способны существовать несколь-
ко лет, но продуцирование ими спор с годами снижается.
Нити Conchocelis в условиях длинного дня производят моноспоры,
прорастающие снова в сверлящие нити. На коротком дне и при понижен-
ной температуре появляются спорангии, где происходит мейоз и образу-
ются четыре конхоспоры, дающие пластинчатые талломы. По наблюдени-
ям Корнмана (Kornmann , 1961) за развитием Р. leucosticta в культуре,
незначительная часть карпоспор сразу развивается в пластинчатое расте-
ние. Карпоспоры Bangia fuscopurpurea на коротком дне (освещение ме-
нее 12 ч в сутки) прорастают в вертикальные талломы; при таком же
освещении стадия конхоцелис производила из своих клеток вертикаль-
ные талломы (Richardson, Dixon, 1968). Таким образом, в цикле разви-
тия Bangiales имеется гаплоидный гаметоспорофит и диплоидный во-
зобновляющийся спорофит.
Смена ядерных фаз у видов Porphyra и Bangia, подвергнутых изуче-
нию, выяснена не во всех случаях. Были сведения о мейозе в оплодотво-
ренном карпогоне (Tseng, Chang 1955). В ряде исследований не было
обнаружено свидетельств оплодотворения и мейоза. Предполагают, что
карпогон партеногенетически производит карпоспоры, из-за чего отсут-
ствует мейоз (Chihara, 1975). Поэтому нередко пишут не о карпоспорах
и спермациях у бангиевых, а об альфа-спорах (крупных) и бета-спорах
(мелких). Одинаковый набор хромосом в клетках пластинчатого талло-
ма и стадии конхоцелис обнаружен у Р. papenfussii (Conway, Cole, 1973).
У этого же вида авторы открыли необычный способ размножения. Веге-
тативные клетки пластинчатого таллома в определенных местах делились
продольно на четыре кар по го но подобные клетки с длинным выростом,
напоминающим трихогину. Эти клетки выходили наружу и округлялись
(25—30 мкм в диам.), у них отсутствовала клеточная оболочка, и они
были способны к амебоидному движению. В течение нескольких суток
после выхода клетки увеличивались до 60 мкм, некоторые имели
тенденцию образовывать почкованием дочерние клетки сходной формы.
Через 25 сут большие клетки дробились на мелкие клетки (карпоспо-
ры). Спустя 5—6 сут мелкие клетки начинали прорастать в стадию кон-
хоцелис, производящую моноспоры.
У других красных водорослей, более эволюционно продвинутых и
объединяемых в класс Florideophyceae, зигота остается на гаметофите
и производит однорядные нити (нити гонимобласта), клетки которых
превращаются в карпоспорангии с одной диплоидной карпоспорой. Из
карпоспор вырастают диплоидные тетраспорофиты. На них образуются
тетраспорангии, где происходит мейоз и появляются четыре тетраспоры,
из которых вырастают гаметофиты.
36
Все, что развивается на гаметофите из карпогона (нити гонимобласта
с карпоспорами), в литературе часто называют карпоспорофитом и счи-
тают третьим поколением у красных водорослей наряду с гамето- и тет-
распорофитами (Fritsch, 1945; Lee, 1980, и др.) . Христенсен (Christensen,
1980) называет карпоспорофит паразитирующим спорофитом. Эти авто-
ры полагают, что карпоспорофит у предков современных красных водо-
рослей был самостоятельно живущим поколением, которое редуцирова-
лось и перешло к паразитическому существованию на гаметофите. Для
признания такого утверждения нет веских оснований. Вероятнее всего
такое явление возникло как приспособление, направленное на образова-
ние множества спор половым путем в условиях отсутствия подвижных
мужских гамет и снижения в связи с этим вероятности оплодотворения
оогониев. Даже если не считать карпоспорофит особым поколением, то
и тогда его название по смыслу остается созвучным со споро- и гамето-
фитами. Вместо него Шуссниг (Schussnig, 196U) предложил термин „кар-
пофит” или, что он считал еще лучше, „гонимофит”. Более подходящим
был бы термин „карпосома”, обозначающий плодовое тело в микологии.
Ван ден Хоек (Van den Hoek et al., 1972) предложил термин „гонимо-
бласт”, который вошел в литературу по красным водорослям.
Эволюция гаметофитов красных водорослей происходила в основ-
ном в направлении лучшего обеспечения развития карпоспор питательны-
ми веществами и образования их в большем количестве от одного карпо-
I она. Для этого в слоевище гаметофита появились ауксилярные (вспо-
могательные) клетки. Среди них различают питающие и генеративные,
последние, как правило, и называют ауксилярными. В случае наличия
питательных клеток нити гонимобласта получают от них дополнительные
вещества (Nemaliales, Gelidiales). При развитии генеративной ауксиляр-
ной клетки (Cryptonemiales, Gigartinales, Rhodymeniales, Ceramiales)
диплоидное ядро, образовавшееся в результате оплодотворения, перехо-
дит в эту клетку и нити гонимобласта развиваются от нее. Ауксилярных
клеток может быть несколько вблизи одного карпогона. Тогда диплоид-
ное ядро зиготы делится, и ядра по специально образовавшимся соедини-
тельным нитям попадают в ауксилярные клетки, причем от каждой раз-
вивается гонимобласт с карпоспорами. Эволюция гаметофитов красных
водорослей пошла дальше — в направлении экономного расходования
питательных веществ на генеративные ауксилярные клетки. У Rhodyme-
niales они образуются, но не развиваются до оплодотворения. Ауксиляр-
ные клетки у Ceramialis возникают только после оплодотворения. Види-
мо, поэтому к данному порядку относятся почти половина родов и боль-
ше половины видов красных водорослей.
У Florideophyceae наблюдается образование диаспор различных ти-
пов. На талломах Nemaliales наряду с карпо- и тетраспорами образуются
моноспоры, служащие для возобновления родительских слоевищ гамето-
спорофитов и возобновляющихся спорофитов. Но основная масса
Florideophyceae размножается карпо- и тетраспорами. На талломах
Corallinaceae часто образуются биспоры в результате деления тетраспо-
рангиев на две части. У некоторых представителей Ceramiales, реже Rho-
dymeniales встречаются пара- и полиспоры. Они значительно мельче тет-
37
распор и образуются в спорангиях в большом количестве. Полиспоры
в спорангии расположены радиально, а параспоры - иногда разветвлен-
ными пучками. Боулд и Вин (Bold, Wynne, 1978) определяют полиспо-
рангии как гомологичные тетраспорангиям, а параспорангии — как него-
мологичные последним. По-видимому, поли- и параспоры образуются на
талломах, в которых произошло нарушение в наборе хромосом (тетра-,
триплоиды и т. д.).
Среди Florideophyceae наблюдается большое разнообразие циклов
развития. В настоящее время, пожалуй, у большинства изученных видов
наблюдается изоморфная смена форм развития. У представителей поряд-
ка Ceramiales, наиболее эволюционно продвинутого среди красных водо-
рослей, известна только эта смена форм развития. В других порядках
наряду с изоморфной встречается гетероморфная смена. В последнее
время ее все чаще обнаруживают у видов (особенно Nemaliales, Cryptone-
miales, Gigartinales), у которых не была известна одна из форм развития.
Споро- и гаметофиты могут иметь макроскопические талломы одинако-
вого размера, но различающиеся морфологически. Иногда различия меж-
ду спорофитом и гаметофитом достигают такой степени, что раньше их
относили к разным порядкам. Например, A sparagopsis из Nemaliales явля-
ется гаметофитом, а его спорофит Falkenbergia относили к Ceramiales.
Гаметофиты ряда представителей Nemaliales, Cryptonemiales и Gigarti-
nales имеют вертикально растущие талломы, а спорофиты образуют не-
большие корочки, наблюдается развитие редуцированных карликовых
тетраспорофитов на гаметофитах. Иногда у видов одного рода одни виды
с изоморфной сменой форм развития, другие — с гетероморфной или
происходит прямое возобновление одной из форм развития. У Gigartina
stellata известны прямое развитие листоватых гаметофитов и гетеро-
морфный цикл развития с корковидным тетраспорофитом. Прямое раз-
витие найдено у растений из Исландии и Дании, гетероморфный цикл
развития — у растений из Испании и Португалии, и оба типа цикла разви-
тия обнаружены для растений из США, Великобритании, севера Франции
(Guiry, West, 1983) . В пределах рода Phyllophora гаметофит и тетраспо-
рофит Ph. pseudoceranoides сходного строения, а у Ph. truncate тетраспо-
рофит развивается на гаметофите в виде нематеция. Карпогон у н?“ по-
сле оплодотворения сливается с ауксилярной клеткой, от которой выра-
стают нити. Одни внедряются между клеток гаметофита, другие растут
вверх,в результате на поверхности появляется подушечка. Несколько
таких подушечек смыкаются, образуя через несколько месяцев большой
шаровидный нематеций, в котором развиваются тетраспорангии. Этот не-
матеций представляет собой сильно редуцированный тетраспорофит, рас-
тущий на гаметофите (Newroth, 1971, 1972), точнее даже несколько тет-
распорофитов, слившихся в один. У Gymnogongrus chiton, образующего
тетраспоры в нематециях на гаметофитах, удалось иммуно химическим
путем установить прохождение мейоза в тетраспорангиях (McCandless,
Vollmer, 1984). Еще большая редукция тетраспорофита произошла
у Liagora tetrasporifera и Helminthocladia hudsoni. Верхушечные клетки их
гонимобласта образуют не карпоспоры, а тетраспорангии, в которых про-
исходит мейоз при образовании тетраспор (Kylin, 1930, Feldmann, 1939).
38
Рис. 12. Цикл развития Palmaria palmata (по: Van der Meer, Todd, 1980).
1 - тетраспоры, 2 - спсрмации, 3 - женский гаметофит с трихогинами, 4 -- жен-
ский гаметофит с развивающимся тетраспорофитом, 5,6- тетраспорофит, 7-8 -
мужской гаметофит.
Один из наиболее простых циклов развития среди Floridophyceae об-
наружен у Rhodophysema elegans — микроскопической корковидной во-
доросли из Cryptonemiales. Оплодотворенный карпогон у нее делится на
стебельковую клетку и тетраспорангий с четырьмя тетраспорами. Сте-
бельковую клетку еще называют материнской клеткой тетраспорангиев,
так как она производит новые тетраспорангии после выхода тетраспор из
старого (De Cew,West, 1982).Поскольку слоевище гаметофита Rh. elegans
имеет крайне упрощенное строение, то трудно решить, каков цикл ее
развития: первично примитивный или это результат сильной редукции.
Своеобразный цикл развития имеют представители порядка Palmaria-
les. У них сильно редуцирован женский гаметофит, а мужской гаметофит
и тетраспорофит с крупными талломами. Например, Palmaria palmata ра-
стет в прибрежье холодных морей в виде крупных пластинчатых талло-
мов диплоидных тетраспорофитов и внешне похожих на них гаплоидных
мужских гаметофитов. Органы размножения появляются на них осенью.
В тетраспорангиях происходит мейоз и образуются гаплоидные тетраспо-
39
ры. Из них развиваются крохотные дисковидные талломы мужские и
женские. Мужские растения весной и летом вырастают в крупные талло-
мы. В женских дисковидных талломах вскоре после их появления обра-
зуются карпогоны и карпогонные ветви, ауксилярные клетки отсутству-
ют. Оплодотворение производится спермациями с крупных мужских
талломов, появившихся из тетраспор прошлого года и созревших за
летний сезон. Из карпогона после оплодотворения на женском гаметофи-
те развивается тетраспорофит. В зимнее время он остается малозамет-
ным, а за весну и лето превращается в крупный вертикально растущий
таллом (Van der Meer, Todd, 1980) (рис. 12). Своеобразие цикла разви-
тия Palmariales заключается в том, что женская гамета оплодотворяется
мужскими гаметами с талломов, зародившихся осенью предшествующе-
го года и просуществовавших год. Таким образом, сливаются гаметы ра-
стений не только разного пола, но и росших в разное время и при разных
погодных условиях. Такой тип смены форм развития оказался, по-види-
мому, особенно удачным для морских водорослей холодных морей.
Представители Palmariales часто доминируют в нижней литорали и верх-
ней сублиторали над многими другими водорослями, образуя нередко
сплошной пояс.
Среди красных водорослей есть представители, смена форм раз-
вития которых заключается в росте гаплоидного макроскопического
гаметофита на диплоидном микроскопическом спорофите в результате
соматического мейоза. У обитателей пресных вод видов родов Lemanea
и Batrachospermum из карпоспор вырастают диплоидные нити (стадия
Chantransia). Апикальные клетки этих нитей подвергаются мейотическо-
му делению. После первого деления мейоза от апикальной клетки отде-
ляется маленькая клетка, содержащая одно из двух ядер. То же самое
происходит при втором делении мейоза. Апикальная гаплоидная клетка
дает начало макроскопическому вертикально растущему таллому гаме-
тофита. В начале своего роста он держится на диплоидной нити, в даль-
нейшем из него вырастают ризоиды, обеспечивающие более прочное при-
крепление к грунту (Hurdelbrink, Schwantes, 1972;Stosch, Theil, 1979; Lee,
1980) (рис. 13).
Циклы развития Florideophyceae не всегда такие четкие, как их изо-
бражают во многих руководствах состоящими из двух поколений, обяза-
тельно воспроизводящих и сменяющих друг друга. Например, тетра-
споры Gloeophycus когеапит одновременно производят проростки двух
типов - гамето- и тетраспорофитов (Notoya, 1983). Иногда наблюдается
развитие нормально функционирующих тетраспорангиев на гаплоидных
гаметофитах, а также мужских и женских гаметангиев на диплоидных
тетраспорофитах. Такие факты известны в литературе по красным водо-
рослям с конца прошлого века, и сведения о них появляются ежегодно
(Van der Meer, 1977). По наблюдениям в культуре за Platythamnion yezo-
ense, 2% тетра- и карпоспор развиваются в ненормальные проростки. Из
карпоспор вырастают растения, производящие тетраспорангии и сперма-
тангии, из тетраспор появляются растения с тетраспорангиями, карпого-
нами и сперматангиями (Notoya, Yabu. 1980).
40
Рис. 13. Цикл развития Batrachospermum (по: Hurdelbrink, Schwantes, 1972; Stosch,
Teil, 1979).
1 - спорофит (Chantransia),2 - гаметофит (Batrachospermum)3 _ фрагмент талло-
ма с карпогоном, оплодотворяемым спермацием, 4 - гонимобласт с карпоспорами.
В результате оплодотворения гапло- и диплоидных карпогонов спер-
мациями с гапло- и диплоидных растений у разных красных водорослей
в природе могут встречаться тетра- и триплоидные талломы. Последние,
по-видимому, способны в основном производить пара- и полиспоры и
этим возобновлять себя. У Spermothamnion turneri в природе довольно
обычны особи с 30, 60, 120, 90 и даже 45—50 хромосомами, встречаются
популяции из триплоидных особей (Drew, 1948). Дру даже предполагала,
что для этого вида определенное число наборов хромосом не имеет суще-
ственного значения.
Самостоятельно растущие гаме то-и спорофиты красных водорослей
представлены в природе не всегда в равной степени. Имеются сведения
о преобладании среди Cryptonemiales и Gigartinales гематофитов. Тетра-
41
спорофиты Gloiosiphonia capillaris встречаются у побережья Швеции, но
их нет у побережий Дании и Франции. У многих Ceramiales гаметофиты
представлены скудно. Имеются данные, что это зависит от температур-
ных условий и, вероятно, от других факторов среды. Для Балтийского
моря упоминаются только тетраспорофиты у Callithamnion furcellarieae,
Polysiphonia nigrescens, Rhodomela subfusca, Spermothamnion roseolum.
Имеются многочисленные данные об одновременном произрастании га-
метофитов и тетраспорофитов в одном месте, хотя они должны были бы
чередоваться (Fritsch, 1945). По нашим наблюдениям, в последнем слу-
чае можно уловить хоть незначительные различия в экологической при-
уроченности. Одна форма развития может преобладать на дне выше или
ниже другой и т. д.
Все многообразие циклов развития Florideophyceae Диксон (Dixon,
1982) свел к пяти типам, обозначенным по названиям типичных родов
или видов. 1. Polysiphonia. Гаметофиты и тетраспорофиты изоморфные.
2. Bonnemaisonia hamifera. Гаметофит крупный, тетраспорофит мелкий.
3. Lemanea. Гаплоидный гаметофит развивается на диплоидном основа-
нии (спорофите) . 4. Liagora tetrasporifera. Тетраспорофиты отсутствуют.
Тетраспоры образуются на гонимобласте. 5.Palmaria. Женский гаметофит
микроскопический. Мужской гаметофит макро-или микроскопический.
Тетраспорофит макроскопический, начинает развиваться на женском
гаметофите.
Под эту классификацию подпадают циклы развития Bangiophyceae
с половым процессом.
Все многообразие циклов развития в сочетании с возможными гено-
типическими и фенотипическими отклонениями создает в целом для
красных водорослей большие адаптивные возможности. Это единствен-
ный отдел почти исключительно прикрепленных водорослей без подвиж-
ной стадии. Тем не менее Rhodophyta более чем в два раза превосходят
Phaeophyta по числу родов и видов.
БУРЫЕ ВОДОРОСЛИ (PHAEOPHYTA)
Все бурые водоросли — многоклеточные организмы. Органы размно-
жения у них много- и одногнездные вместилища, именуемые в литерату-
ре многогнездными гаметангиями или спорангиями и одногнездными
Спорангиями. Известны случаи слияния зооидов из одногнездных вме-
стилищ у Myrionema feldmannii (Loiseaux, 1967). Поэтому все эти органы
размножения, если неизвестна точно выполняемая ими функция, лучше
именовать зоидангиями, поскольку они производят жгутиковые клетки.
Одногнездные зоидангии служат местом мейоза. Половой процесс у
Phaeophyta — изогамия, анизогамия и оогамия. Смена форм развития
бывает изо- или гетероморфной. Наличие той или иной смены форм раз-
вития у представителей порядков использовалось для деления Phaeophyta
на классы.
По системе Кюлина (Kylin, 1933), принимаемой некоторыми автора-
ми и сейчас (Курс низших растений, 1981), Phaeophyta состоят из трех
классов: Isogeneratophyceae (Ectocarpales, Tilopteridales, Sphacelariales,
Dictyotales, Cutleriales), Heterogeneratophyceae (Chordariales, Scytosiphona-
42
les, Dictyosiphonales, Sporochnales, Desmarestiales, Laminariales), Cyclosporo-
phyceae (Ascoseirales, Durvilleales, Fucales). У всех бурых водорослей на-
блюдается спорический мейоз, кроме Cyclosporophyceae, имеющих гаме-
тический мейоз.
Со времени работ Кюлина было выполнено много исследований, по-
казавших относительную принадлежность порядков к классам Isogenera-
tophyceae и Heterogeneratophyceae. Например, Cutleriales включает два
рода: Cutleria с гетероморфной сменой форм развития и Zanardinia сизо-
морфной. В ряде других порядков, относимых к Isogeneratophyceae, фор-
мы развития при сходстве размеров талломов имеют морфологические
различия, позволившие ранее описать их как разные виды одного рода
(Ectocarpua fasciculatus var. refractus — спорофит, E. draparnaldioides — его
гаметофит) или как виды разных родов (Haplospora и Scaphospora из
Tilopteridales).
Прежде было принято считать, что спорофиты бурых водорослей
имеют одногнездные спорангии, а гаметофиты — многогнездные гаметан-
гии. На самом деле у бурых водорослей встречаются одногнездные или
многогнездные зоидангии на одном талломе в зависимости от температу-
ры.На талломах Ectocarpus fasciculatus при 20 °C одногнездные зоидан-
гйи образуются в ограниченном количестве среди многогнездных, при
13 °C возникают только одногнездные зоидангии (Muller, 1972) . У видов
Ectocarpales, Sphacelariales, Chordariales, Scytosiphonales, Dictyosiphonales
и других нередко наблюдается партеногенетическое развитие гамет в га-
метофиты или спорофиты, и в цикле развития у них существует смена
гаметоспорофитов возобновляющимися спорофитами. Среди видов с ге-
тероморфной сменой форм развития часто наблюдается выпадение одной
из них по всему ареалу или в отдельных географических районах (South,
Hooper, 1976; Yotsui, 1982). Половой процесс у представителей перечис-
ленных порядков наблюдается не всегда, о смене ядерных фаз известно
в ограниченном числе случаев. Поэтому обычно говорят о наличии в их
циклах развития макро- и микроталломов или о существовании верти-
кальных и корковидных талломов. Возобновление макроталломов до-
вольно часто происходит посредством выроста на микроталломе. Появ-
ление и рост талломов разного типа зависят от температуры, длиньГдня,
"солености и других факторов среды (Pedersen, 1980).
Своеобразный цикл развития наблюдали Коеман и Кортель-Бреман
(Koeman, Cortel-Breeman, 1976) у Elachista fucicola. В условиях культуры
росли две фазы — макро- и микроталломы, обе диплоидные. Гаплоидный
набор хромосом обнаруживали в очень молодых микроталломах. На
макроталломах образовывались одногнездные спорангии. Зооспоры раз-
вивались в микроталломы с многогнездными зоидангиями, зооиды
которых снова производили микроталломы. Переход к макроталломам
происходил путем роста их на микроталломах. Авторы предполагают,
что в старых микроталломах осуществляется вегетативная диплоиди-
зация.
Спонтанное увеличение числа хромосом в цикле развития Ectocarpus
siliculosus в лабораторных культурах наблюдал Мюллер (Muller, 1967).
Им были обнаружены гапло-, дипло- и тетраплоидные спорофиты. Опре-
43
деленные зооиды гаплоидных растений спонтанно давали гомозиготные
диплоидные и тетраплоидные спорофиты. Из зооспор, образовавшихся
после мейоза в те трапп оидных спорофитах, возникали диплоидные
растения, подобные мужским гаметофитам. Их гаметы копулировали
с гаплоидными женскими гаметами других растений. При партеногенезе
появлялись диплоидные спорофиты.
Самовозобновление форм развития существует у некоторых пред-
ставителей таких эволюционно продвинутых порядков, как Cutleriales и
Dictyotales. Это проявляется в произрастаниии в отдельных районах
только одной формы развития. В Черном море встречается гаметофит
Сиtleria adspersa, а ее спорофит Aglaozonia неизвестен (Зинова, 1967).
Тетр а споро фи ты Dictyotajiichotoma возобновляются при прорастании
I содержимого тетраспорангиев в проростки с ризоидами на материнских
; растениях, после чего происходит их отделение. Такое явление приобре-
тает иногда массовый характер в бухтах залива Петра Великого (Япон-
ское море) . Среди Sporochnales наблюдались случаи образования мик-
роскопических талломов, возобновляющих мужские гаметофиты минуя
макроскопические спорофиты (Wynne, Loiseaux, 1976).
У четырех видов рода Syringoderma (Syringodermatales), имеющих
макроскопические спорофиты, обнаружены четыре типа циклов разви-
тия: 1) с микроскопическими многоклеточными нитчатыми гаметофита-
ми, 2) прямое возобновление спорофитов, минуя гаметофиты, 3) редук-
ция гаметофитов до 2—4 клеток и развитие их вне спорангиев, 4) разви-
тие 2—4-клеточных гаметофитов внутри спорангиев (Henry, 1984).
Обязательное чередование двух форм развития существует в циклах
Desmarestiales и Laminariqles. У них на макроскопических спорофитах
в одногнездных спорангиях после мейоза образуются зооспоры, дающие
микроскопические гаметофитй. Их развитие происходит следующим об-
разом. Зооспора прикрепляется к субстрату, теряет жгутики, округляет-
ся и вырабатывает хорошо заметную клеточную оболочку (стадия эм-
бриоспоры) . Эмбриоспора дает вырост (проростковую трубку), куда
переходит все ее содержимое, отделяющееся поперечной перегородкой
от опустевшей эмбриоспоры. Так появляется первая клетка гаметофита.
Женские гаметофиты Laminariales при оптимальных условиях нередко
остаются одноклеточными. Эта же единственная клетка после роста и
накопления продуктов фотосинтеза превращается в оогоний с одной
яйцеклеткой. Наряду с одноклеточными женскими гаметофитами появ-
ляются многоклеточные из разветвленных нитей. Они растут дольше,
позднее созревают и производят по нескольку оогониев на концах
ветвей. Мужские гаметофиты всегда многоклеточные и мельче женских.
Антеридии образуются на поверхности в виде мелких клеток, содержа-
щих по одному антерозоиду. Яйцеклетка выходит из оогония и остается
прикрепленной к краям отверстия. В таком положении происходит опло-
дотворение и развитие спорофита. По мере роста спорофита на его ниж-
нем конце появляются ризоиды, обеспечивающие прикрепление таллома
к грунту.
Подобным образом на микроскопическом гаметофите развивается
спорофит у Sporochnales. Спорофиты Sporochnales, Desmarestiales, Lamina-
44
riales не получают никаких питательных веществ от гаметофита, так как
яйцеклетка и проросток отделены от его живых клеток пустой оболоч-
кой оогония. Тем не менее такое развитие спорофита дало дополнитель-
ные преимущества видам указанных порядков, так как отпала необходи-
мость в поиске нового места для прикрепления зиготы. При пересе-
вах диаспор происходит наибольшая гибель зачатков бентосных
водорослей из-за ограниченности незанятых площадей в фотической зо-
не, конкуренции других организмов и выедания фильтраторами.
Накахара и Накамура (Nakahara, Nakamura, 1973) в эксперименталь-
ных условиях наблюдали у Alaria crassifolia партеногенетическое и апо-
гамное (из клеток мужских гаметофитов) развитие нормальных гапло-
идных спорофитов. Апоспорические диплоидные гаметофиты были полу-
чены ими из вегетативных клеток диплоидных спорофитов. Однако та-
кого рода явления едва ли распространены в природе. Laminariales сейчас
широко культивируют для научных целей и на плантациях, и всегда цикл
развития у них протекает с половым процессом.
Своеобразный цикл развития и смену фаз развития имеют представи-
тели Cyclosporophyceae (Fucales и др.) . Их макроскопические диплоид-
ные талломы производят в концептакулах (углублениях) антеридии и
оогонии. Мейоз происходит при первом делении ядра гаметангия. В оого-
ниях образуется от 2 до 8 яйцеклеток, в антеридиях 64антерозоида. По-
сле оплодотворения зигота без периода покоя развивается в макроско-
пический таллом. У Sargassaceae и некоторых Cystoseiraceae и Fucaceae
единственная яйцеклетка после созревания выходит из оогония и остает-
ся прикрепленной снаружи тяжем, образованным внутренней оболочкой
оогония. В таком положении происходит оплодотворение и развитие
проростка до стадии появления ризоидов. Таким образом, возобновле-
ние и расселение видов осуществляется здесь не гаметами и зиготами,
а сформировавшимися проростками спорофитов. —
Впервые соображения о природе цикла развития Cyclosporophyceae,
а именно Fucales высказал Страсбургер (Strasburger, 1906). Он предпо-
ложил существование у них смены форм развития. По его мнению, моло-
дой оогоний — это одногнездный спорангий, где в результате мейоза
образуются четыре споры, каждая из которых, делясь, дает эндоспори-
ческий гаметофит. Кюлин (Kylin, 1940) считал, что Fucales всегда были
диплоидными с гаметическим мейозом или гаметофиты исчезли у них
в процессе эволюции и мейоспоры функционируют у них как гаметы.По-
следней точки зрения придерживался Фритч (Fritsch, 1945) . По Гутенбер-
гу (Guttenberg, 1955), у Fucales нет гаметофита, а антеридии и оогонии
развиваются апоспорически на спорофите, который не производит ника-
ких спор бесполого размножения. Напротив, Л. П. Пересхенко (1980)
считает таллом фукусовых гаметофитом.
~~ Развитие концептакулов и ветвей у некоторых Cystoseiraceae и Sar-
gassaceae подробно изучала Йенсен (Jensen, 1974). На основании образо-
вания оболочек в оогониях на стадии четырех ядер у Bifurcariopsis и вось-
ми ядер у Durvillea, Hormosira, Xiphophora, Ascophyllum, Pelvetia Йенсен
пришла к такому же выводу, как и Страсбургер, что_рогрний — фактиче-
ски одногнездный спорангий, который посредством мейоза дает четыре
45
ядра спор. Каждое ядро делится и дает начало двухъядерному гаметофи-
ту, который дифференцируется прямо в оогоний. Поскольку в антери -
диях не обнаружены такие оболочки, как в оогониях, то Йенсен считает
мужской гаметофит полностью подавленным и зооспоры превратились
в антеризоиды или мужской гаметофит представлен 16 ядрами. Йенсен
придает большое значение образованию оболочек внутри оогония в про-
цессе его развития. Однако у всех перечисленных родов они возникают
по-разному. У Durvillea и Xiphophora оболочки плотные, но у Durvillea
на один слой меньше, чем у Xiphophora. Яйцеклетки Hormosira окружены
слизистым футляром, а у Ascophyllum оболочки неизвестной природы
(Jensen, 1974). В пределах рода Pelvetia у Р. canaliculata единственная пере-
городка расположена поперек, а у Р. wrightii — вдоль оси. По-разному
в оогониях различных родов ведут себя лишние ядра, не принимающие
участия в образовании яйцеклеток. Все роды Cyclosporophyceae, в чьих
оогониях есть перегородки, встречаются в приливо-отливной зоне, под-
верженной сильным колебаниям внешних факторов (температура, соле-
ность, уровень воды) и воздействию прибоя. Наиболее примитивным
представителем Fucales Йенсен считает Bifurcariopsis capensis. Он растет
только в приливо-отливной зоне, имеет массивную нижнюю часть и су-
жающиеся к вершине ветви. Этим он подобен Fucus distichus f. anceps,
растущему, например, в Баренцевом море на выступах скал в местах с
сильными ударами волн. Все это позволяет считать, что перегородки
в оогониях Cyclosporophyceae, вполне вероятно, возникли в разных ро-
дах независимо и имеют приспособительное значение.
Решение вопроса о природе цикла развития Cyclosporophyceae зави-
сит больше всего от выяснения происхождения слоя, выстилающего по-
лость концептакула и несущего гаметангии. Давно известно, что высти-
лающий слой концептакулов Fucales развивается от одной клетки, назы-
ваемой инициальной клеткой концептакула. Отчленяется она от апикаль-
ной клетки, расположенной в углублении на конце побега и ответствен-
ной за его рост (рис. 14). Инициальная клетка концептакула делится
поперечной перегородкой на небольшую верхнюю клетку — язычковую и
крупную базальную. Язычковая клетка у некоторых Fucales вырастает
в волосок с базальной зоной роста. Базальная клетка дедится антикли-
нально и образует слой клеток, раздвигающих окружающие ткани талло-
ма с образованием полости. Такой способ развития концептакула от од-
ной инициальной клетки существует у Fucales и других Cyclosporophy-
ceae у которых вместо одной апикальной клетки на вершине находится
несколько клеток (Hormosira из Fucales) или верхушечный рост осущест-
вляется посредством диффузного деления клеток (Durvillea), а также
при базальном росте таллома (Ascoseira).
Выстилающий слой концептакула Hormosira развивается из одной
клетки с образованием в самом начале одного волоска (Fritsch, 1945)
Наличие одного волоска в молодых концептакулах Hormosira наблюда-
лось и нами. У Durvillea antarctica, обладающей пластинчатым талломом
с лопастными выростами, инициальные клетки концептакулов были
встречены из расстоянии 8.5 см от вершины лопасти и на глубине
40—70 мкм от поверхности таллома. Отдельная инициальная клетка
46
4
3
Рис. 14. Cystoseira sp.
1-5 - развитие выстилающего слоя концептакула, а
апикальная клетка таллома.
концептакула образуется из конечной клетки бокового выроста нити
меристодермы. После ее деления появляется однослойная пластинка,
состоящая из четырех клеток. Она разрастается и раздвигает окружаю-
щие ткани, образуя полость концептакула. С появлением и делением
инициальной клетки концептакула нити меристодермы дают боковые
выросты в ее сторону, которые, разрастаясь и переплетаясь, образуют
оболочку вокруг развивающегося концептакула (Петров, 1967) .
Развитие концептакулов Ascoseira mirabilis, имеющей пластинчатый
таллом с интеркалярной зоной роста, происходит из одиночных клеток,
появляющихся в меристодерме выше зоны роста пластины. Первоначаль-
но в нашем распоряжении была часть узкой лопасти таллома. При изуче-
нии ее с поверхности казалось, что сначала образуются „спорангиальные
клетки”, которые, дробясь, производят инициальные клетки концепта-
кулов (Петров, 1963а). Позднее, располагая большим количеством жи-
вых и фиксированных талломов A. mirabilis, автор убедился в отсутствии
„спорангиальных клеток”. Моэ и Хенри (Мое, Henry, 1982) предположи-
ли, что за спорангиальные клетки были приняты крупные клетки эндо-
фитных зеленых водорослей. Инициальная клетка концептакула A. mira-
bilis делится антиклинально перегородками на четыре клетки, которые
в свою очередь делятся периклинальными перегородками и образуют
четыре ряда клеток, врастающих перпендикулярно поверхности в мате-
ринский таллом. С этого момента окружающие клетки таллома отчленя-
ют в сторону полости канала, занятого концептакулом, клетки, высти-
лающие его стенки. Указанные авторы приняли их за клетки выстилаю-
щего слоя концептакула. В основании четырех нитей молодого концеп-
такула, по нашим наблюдениям, располагается по крупной продолгова-
той клетке, отчленяющей клетки вверх, но ни разу не удавалось наблю-
дать, чтобы они отчленяли клетки на образование стенок канала. Это же
подтверждают микрофотографии, опубликованные Моэ и Хенри (Мое,
Henry, 1982). Скотсберг (Skottsberg, 1907) считал, что стенки канала,
47
соединяющего полость концептакула с внешней средой, являются про-
должением меристодермы. Когда молодой концептакул A. mirabilis в ви-
де четырех нитей пройдет плотные слои коры и промежуточного слоя
таллома и достигнет границы рыхлой сердцевины, его основание разрас-
тается в выстилающий слой шаровидной полости концептакула. Одно-
временно вокруг этой полости развивается оболочка из клеток таллома.
Среди альгологов сейчас распространена точка зрения Фритча (Fritsch,
1945), считавшего выстилающий слой концептакула Fucales видоизме-
ненной или задержавшейся в росте ветвью. Йенсен (Jensen,1974) придер-
живается такого же мнения и подобрала новые факты, казалось бы,
подтверждающие это. По ее данным, концептакулы закладываются на
вершине в том же порядке, как и ветви. У Bifurcaria и Bifurcariopsis за-
кладка концептакулов прерывается, когда возникают ветви. Боковые
ветви у Bifurcaria brassicaeformis появляются в тех точках, где должны
были располагаться концептакулы. Образование апикальной клетки про-
ростка Fucales из базальной клетки волоска и появление волоска в нача-
ле развития концептакула, по мнению Йенсен, свидетельствуют о гомо-
логичности этих двух клеток.
Против признания выстилающего слоя концептакула видоизменен-
ной ветвью свидетельствуют следующие факты. 1. Инициальная клетка
концептакула отличается формой и размерами от апикальной клетки
ветви, и после первого периклинального деления она перестает существо-
вать. Таким образом, в выстилающем слое концептакула отсутствует
клетка, аналогичная апикальной клетке побега. При развитии боковой
ветви наблюдается обратная картина. Ее апикальная клетка с началом
формирования ветви постепенно дифференцируется и приобретает
характерную форму. Боковые ответвления проростков и регенератив-
ные побеги на взрослых талломах Fucales образуются без участия специ-
альных одиночных инициальных клеток, апикальные клетки формируют-
ся в них позднее. 2. Непогруженные зоидангии у бурых водорослей
с супротивным ветвлением возникают в таком же положении, как и вет-
ви, т. е. супротивно или напротив ветви (Isthmoplea). Это наводит на
мысль о гомологичности инициальных клеток концептакулов одно-
гнездным спорангиям. 3. Гипотеза Фритча и Йенсен не объясняет возник-
новение концептакулов у Hormosira, Durvillea, Ascoseira.
Нами была высказана гипотеза, по которой инициальная клетка кон-
цептакула Cyclosporophyceae является видоизмененным спорангием
с моноспорой, прорастающей in situ в диплоидный гаметофит в виде
выстилающего слоя концептакула (Петров, 1962, 1963а, 19636, 1964,
1967). В подтверждение этой гипотезы можно привести следующее.
1. У некоторых водорослей наблюдается прорастание спор и развитие
гаметофитов на материнском талломе (Cabioch, 1972; Umezaki, 1972;
West, Hommersand, 1981). 2. Клетки выстилающего слоя концептакула
на разных этапах его развития отличаются от клеток меристодермы и ко-
ры формой, нежной оболочкой, содержимым, восприятием красителей.
3. Развитие концептакулов у Cyclosporophyceae в ряде случаев напоми-
нает внедрение одного организма в другой. Лучше всего это выражено у
Ascoseira mirabilis — наиболее примитивного вида этого класса. В отноше-
48
нии представителей Fucales можно предположить, что у них ткани ре-
цептакула обрастают концептакулы, поскольку размеры тех и других со
временем увеличиваются. На самом деле концептакулы становятся мно-
гоклеточными и внедряются в рецептакул уже на его вершине при выхо-
де из апикальной ямки, где невозможно утолщение рецептакула. Такая
картина наблюдалась, например, у Р. А. Ковалевской
(неопубликованные данные) (рис. 14, 4). 4. Развитие волоска в резуль-
тате первых делений инициальной клетки концептакула Fucales напоми-
нает появление волоска в первые моменты развития проростков некото-
рых бурых водорослей (Schussnig, I960;Pedersen, 1980).УPhaeostroma
pustulosum волосок растет от эмбриоспоры (Pedersen, 1981). 5. Циклы
развития многих Phaeophyta свидетельствуют о том, что уровень плоид-
ности не всегда служит определяющим фактором в развитии макро- и
микроскопического талломов (Wynne, Loiseaux, 1976).
Все это делает вполне допустимым существование диплоидных гаме-
тофитов в талломах Cyclosporophyceae в виде выстилающего слоя кон-
цептакулов.
49
ЭВОЛЮЦИЯ ЦИКЛОВ РАЗВИТИЯ
И СМЕНЫ ФОРМ РАЗВИТИЯ У ВОДОРОСЛЕЙ
До сих пор во всех группах водорослей, обладающих половым раз-
множением, исходным признается цикл развития с фиксированным по-
ложением мейоза и моментов образования спор и гамет. Наиболее при-
митивным циклом считается гаплофазный с зиготической редукцией
(Chapman, 1962; Favre-Duchartre^ 1967; Лодкина, 1969). В качестве пер-
вичной смены форм развития рассматривается изоморфная со спориче-
ской редукцией (Feldmann, 1952). Ли (Lee, 1980), перечисляя основные
типы циклов развития водорослей, без всяких комментариев располо-
жил их в следующем порядке: 1) преобладание диплоидной фазы с гаме-
тической редукцией, 2) преобладание гаплоидной фазы с зиготической
редукцией, 3) изоморфная смена форм развития с чередованием гапло-
идной и диплоидной форм развития, 4) гетероморфная смена форм раз-
вития. По мнению большинства авторов, эволюция ведет к редукции и
полному исчезновению гаплоидной формы развития, представленной
гаметофитом.
Ход решения многих вопросов эволюции циклов развития водорос-
лей зависит от того, можно ли называть зиготой то, что образуется у од-
ноклеточных и нитчатых (в основном пресноводных зеленых) водорос-
лей в результате слияния гамет и после мейоза производит споры, часто
в большом количестве. Как видно из вышеописанных циклов развития
Chlamydomonas и Ulothrix , после слияния гамет у них образуется расту-
щий одноклеточный организм — спорофит. Зиготическая редукция в
чистом виде встречается у водорослей не так часто (Charophyta), Вероят-
но, она существует у Sphaeroplea, Oedogoniales, Conjugatophyceae, если до
момента мейоза зигота, несмотря на отсутствие видимого роста, не на-
капливает в большом количестве продукты фотосинтеза. Истинная зиго-
тическая редукция установлена у видов Spirogyra. У исследованных четы-
рех видов этого рода мейоз происходит в молодой зигоспоре ^заканчи-
вается до ее созревания (Harada, Yamagishi, 1984). На оснований поведе-
ния хромосом неслившихся ядер в зиготе авторы утверждают, что ядро
вегетативной клетки Spirogira диплоидное, а не гаплоидное.
На основании вышеизложенного нет никаких оснований считать
зиготическую редукцию в качестве первичной формы смены ядерных
фаз в циклах развития водорослей. Наличие всех возможных вариантов
положения мейоза (спорического, гаметического, зиготического, сома-
тического) в циклах развития водорослей различных отделов и в преде-
лах их у таксонов, стоящих на разных уровнях эволюционного развития,
дает основание для утверждения, что с возникновением полового процес-
са сначала существовала неупорядоченная смена ядерных фаз.
50
Определенное положение в цикле развития мейоз мог занять только
в результате естественного отбора, который в разных группах водорос-
лей имел различные последствия. Причина этого в том, что гаплоидное
и диплоидное состояния имеют свои положительные и отрицательные
стороны. Диплоидия посредством компенсаторного эффекта генов спо-
собствует более устойчивому развитию организмов. Несмотря на гене-
тические достоинства, диплоидия не стала достоянием всех организмов.
Для многих простоорганизованных существ достаточно гаплоидного на-
бора хромосом. Диплоидия для них означала бы дополнительные затраты
энергии на поддержание ненужной избыточной информации, что имело
бы отрицательные последствия для их существования (нарушение прин-
ципа Данкова).
. У первичных эугамных организмов в результате неупорядоченной
смены ядерных фаз вначале одновременно существовали индивиды
с разным набором хромосом (гапло-, дипло-, тетраплоидным и т. д.).
Такого рода явления наблюдаются сейчас у некоторых красных и бурых
водорослей (Drew, 1948; Muller, 1967). Вероятнее всего, в результате
естественного отбора в разных группах водорослей одновременно могли
закрепиться зиготическая, спорическая и соматическая редукции. Прав-
да, у одноклеточных водорослей, какими, по-видимому, были первичные
эугамные водоросли, различия в положении мейоза были относительны,
как относительны различия между фотосинтезирующими растущими зи-
готами и одноклеточными диплоидными спорофитами, между зооспора-
ми и молодыми одноклеточными жгутиковыми организмами, между
делением одноклеточных диплоидных индивидов с мейозом на четыре
споры (спорическая редукция) или на четыре дочерние одноклеточные
особи (соматическая редукция). Гаметическая редукция явно вторично-
го происхождения, так как она связана с возникновением гамет и зара-
нее подготавливаемым половым процессом. У первичных одноклеточ-
ных эугамных организмов половым процессом была гологамия, которая
носила случайный характер, как следствие их случайных столкновений,
соединений и слияний под влиянием внешних и внутренних факторов.
Положение мейоза могло меняться в процессе эволюции отдельных
групп водорослей. Наиболее широкое распространение среди водорос-
лей, как и среди других растений, получил спорический мейоз, но наи-
более массового развития в природе смогли добиться водоросли с гаме-
тическим мейозом и прохождением цикла в диплофазе (Bacillariophyta,
Fucales, Siphonales, Vaucheria).
У водорослей наблюдается огромное разнообразие циклов развития,
обусловленное независимым сочетанием в ходе эволюции всевозможных
вариантов смены ядерных фаз, форм размножения и невзаимосвязанной
дифференциацией талломов разных форм развития. О циклах развития
можно говорить уже применительно к прокариотам и агамным эукарио-
там, у которых существует цикломорфоз (циклогения). У эукариотных
водорослей цикломорфоз дополнился сменой ядерных фаз и способов
размножения. По мере совершенствования регуляторных механизмов
первичный цикломорфоз исчезает даже у некоторых агамных водорос-
лей (Chlorococcales) или переходит в более упорядоченную смену форм
развития.
s 1
На первых этапах эволюции у эугамных водорослей не существовало
специализации особей на образование спор и гамет. Среди современных
водорослей это проявляется в существовании гаметоспорофитов, когда
одни и те же талломы в зависимости от внешних условий (температура,
интенсивность света, длина дня, соленость) производят споры или гаме-
ты. Со временем произошла специализация части особей на образование
только спор. Так в цикле развития водорослей появились спорофиты.
Другие особи остались гаметоспорофитами. С этого момента возникла
смена форм развития, если не считать возможного сохранения цикломор-
фоза, не связанного с размножением.
Спорофиты примитивных водорослей производят споры, из которых
развиваются гаметофиты, и споры для возобновления талломов споро-
фитов. Это могут быть морфологически одинаковые споры, различаю-
щиеся поведением под влиянием внешних и внутренних факторов, или
разные споры, как например у красных водорослей тетраспоры для раз-
вития гаметофитов и моноспоры для возобновления тетраспорофитов
(Rhodochortori). После утраты спорофитами способности к самовозоб-
новлению появился цикл со строгим чередованием двух форм развития,
который отождествляют с чередованием поколений.
С момента возникновения двух форм развития (половой и беспо-
лой) у водорослей началась дифференциация циклов развития на изо- и
гетероморфные. Приобретение талломами, принадлежащими к различ-
ным формам развития, разных величины и строения имеет большое адап-
тивное значение. По данным Любченко и Кубит (Lubchenko, Cubit, 1980),
вертикальные, корковидные и сверлящие стадии Ulothrix, Scytosiphon,
Petalonia, Bangia, Porphyra обладают разной адаптацией к действию фито-
фагов. Вертикальные стадии адаптированы к высоким скоростям роста
и размножения в условиях низкого выедания их животными. Корковид-
ные и сверлящие стадии приспособлены к переживанию периода сильно-
го выедания или, по нашим наблюдениям, к неблагоприятным условиям
осенне-зимнего периода (усиление штормов, торошение льда у берега
и т. д.). У Scytosiphon lomentaria (Littler, Littler, 1983) морские ежи вые-
дали за 48 ч 81.4—82.7% вертикальных талломов и 16.2% корковых, что
совпадало с их разной калорийностью (вертикальные талломы — 4.46—
4.97 ккал/г сухого вещества, корковые — 3.55 ккал/г сухого вещества)
и твердостью (вертикальные — 12—15 г/мм2, корковые — 26 г/мм2),
но зато вертикальные талломы обладают несравненно более высокой
первичной продукцией — 6.5—8.1 мг С/(г сухого вещества • день), чем
корковые — 0.5 мг С/ (г сухого вещества • день).
Водоросли, имеющие изоморфную смену форм развития, образуют
нередко в неблагоприятных условиях (осенне-зимний период) или во
время усиленного выедания гидробионтами стелющиеся или низко-
рослые талломы. Это наблюдается у красных и бурых водорослей корал-
ловых рифов, где обитает много растительноядных рыб. Талломы из-
вестковых красных водорослей с изоморфной сменой форм развития,
вроде Corallina officinalis, имеют мощную базальную корку, которая дает
им ряд преимуществ по сравнению с негетероталломными водорослями
(Littler, Kanker, 1984). Корка сохраняет первичное пространство при
52
повреждении вертикальной части таллома, за счет выростов ускоряет
его восстановление. Вертикальная часть таллома и его базальная корка
различались, как у вышеупомянутого Scytosiphon, по первичной продук-
ции, калорийности, твердости, сопротивляемости выеданию ежами.
Гетероморфные циклы развития водорослей эволюционировали
в двух направлениях: 1) образования крупного гаметофита и мелкого
спорофита (распространено у Rhodophyta и части Chlorophyta); 2) обра-
зования крупного спорофита и мелкого гаметофита (распространено у
Phaeophyta). В обоих типах гетероморфного цикла водорослей возникло
развитие одной формы на другой. Формирование крупного гаплоидного
гаметофита на небольшом спорофите не получило распространения
и имеется у ограниченного круга пресноводных Rhodophyta (Lemanea,
Batrachospermum). Существование на крупных спорофитах микроскопи-
ческих гаметофитов привело к возникновению и широкому распростра-
нению особой линии эволюции в пределах Phaeophyta — класса Cyclospo-
rophyceae (Fucales), представители которого расселились по всем морям
(кроме Каспийского и Аральского) и часто образуют крупные заросли.
Менее широкие возможности для эволюции оказались у Charophyta с та-
ким же, по нашим представлениям, циклом развития, но и они смогли
заселить илистые грунты в пресных водоемах — биотопы, малопригод-
ные для поселения других водорослей.
В случае роста небольшого спорофита на гаметофите, что присуще
Rhodophyta, виды, видимо, не получили большого преимущества, по-
скольку гаметофиты Rhodophyta и так производят через посредство
гонимобласта большое количество спор, и здесь произошла замена одних
спор (диплоидных карпоспор) на другие (гаплоидные тетраспоры).
Среди Rhodophyta такой цикл возникал многократно у отдельных ви-
дов разных родов, главным образом среди Gigartinales, и не привел к воз-
никновению крупных таксонов, представляющих особые ветви эволю-
ции. В противоположность этому развитие макроскопических спорофи-
тов на микроскопических гаметофитах оказалось более прогрессивным
и привело к появлению крупных таксонов (порядков) бурых водорос-
лей (Sporochnales, Desmarestiales, Laminariales). Представители двух по-
следних порядков заняли наряду с Fucales доминирующее положение
в донной растительности умеренных и холодных морей обоих полушарий
Земли.
Среди водорослей с развитием микроскопических гаметофитов на
крупных спорофитах (Fucales) оказалось возможной дальнейшая эво-
люция, которая привела к сохранению яйцеклеток на материнских
растениях с последующим оплодотворением и развитием из них много-
клеточных проростков с ризоидами, которые только после этого отделя-
ются от таллома (Sargassaceae).
В разных отделах водорослей неоднократно возникали следующие
наиболее важные прогрессивные явления в цикле развития.
1. Оплодотворение внутри или на поверхности материнского орга-
низма (Chlorophyta: Sphaeroplea, Oedogonium, Coleochaete; Charophyta;
все Rhodophyta; Xanthophyta: Vaucheria; Phaeophyta: Sporochnales, Desma-
restiales, Laminariales, Sargassaceae и др.) .
53
2. Образование женских гамет (яйцеклеток) после контакта мате-
ринской клетки с мужской гаметой (Oedogoniales, некоторые центри-
ческие Bacillariophy ta) или соседства мужских организмов (Volvox).
3. Появление питающих клеток для обеспечения развития образова-
ний, возникающих из зигот (Chlorophyta: Coleochaete; Florideophyceae).
4. Сокращение числа пересевов диаспор (Chlorophyta; некоторые
Characeaceae, виды, образующие ценобии, Coleochaete; продвинутые
порядки Phaeophy ta; некоторые Rhodophyta)
5. Прохождение начальных этапов развития особей следующего по-
коления внутри или на поверхности материнского организма (Chlorophy-
ta — виды, образующие ценобии; Sargassaceae).
6. Полное развитие одной формы развития на другой и образование
организма двойственной природы — дибионта (Chlorophyta: мужские
ценобии Volvox, Coleochaete; Charophyta; Phaeophyta: Fucales и др.; Rho-
dophyta — виды с редукцией тетраспорофитов и развитием их на гамето-
фитах) .
В эволюции циклов развития и смены форм развития у водорослей
можно проследить следующие основные этапы. 1. Цикломорфоз с беспо-
лым размножением и делением клеток на неопределенное число частей
(Zakr^s, 1983). 2. Упорядочивание числа делений и образование опреде-
ленного числа дочерних клеток. Появление спор. 3. Возникновение поло-
вого процесса на основе гологамии. 4. Появление гамет и возникновение
изо-, анизо- и оогамии. Специализация особей на образование спор или
гамет отсутствовала. Все особи — потенциальные гамето споро фиты.
5. Утрата частью особей способности образовывать гаметы. Появление
возобновляющихся спорофитов. Неупорядоченная смена форм разви-
тия — гаметоспорофитов и возобновляющихся спорофитов. 6. Возникно-
вение гаметофитов. Спорофиты сохраняют способность к самовозобнов-
лению. 7. Утрата спорофитами способности к самовозобновлению. Возни-
кает чередование споро- и гаметофитов. 8. Сокращение числа пересевов
диаспор. Прохождении начальных этапов формирования одной формы
развития на другой с последующим самостоятельным прикреплением
и ростом на грунте. 9. Полное развитие одной формы развития на дру-
гой. Возникновение организмов двойственной природы (дибионтов).
10. Образование на дибионтах проростков следующего поколения.
Эволюция смены ядерных фаз у водорослей должна была пройти два
этапа: положение мейоза не фиксировано в цикле развития; прохожде-
ние мейоза приурочено к определенному моменту в цикле развития. У
большинства водорослей наблюдается спорический мейоз. Но высшие
представители наряду со спорическим (Laminariales) имеют гаметический
мейоз (Fucales). В обоих случаях происходит чередование крупных
диплоидных спорофитов с микроскопическими гаметофитами, гаплоид-
ными у Laminariales и диплоидными, растущими в спорофитах у Fucales.
54
ЛИТЕРАТУРА
Виноградова К. Л. Ульвовые водоросли (Chlorophyta) морей СССР. Л. 1974.
166 с. - Виноградова К. Л. Циклы развития Chlorophyta и некоторые вопросы их
эволюции. - Ботан журн., 1976, т. 61, № 8, с. 1041-1048. - Голлербах М. М. О не-
которых принципиальных вопросах в понимании циклов развития у водорослей. -
В кн.: Тез. докл. делегат, совещ. Всесоюз. ботан. о-ва (28 января-1 февраля 1951 г.).
М.; Л., 1951, № 4, с. 20-26. - Дубинин Н. П. Общая генетика. М., 1970. 487 с. -
Зинова А. Д. Определитель зеленых, бурых и красных водорослей южных морей
СССР. М.; Л., 1967. 398 с. - Козо-Полянский Б. М. Современное состояние про-
блемы чередования поколений у высших растений. - В кн.: Тез. докл. делегат,
совещ. Всесоюз. ботан. о-ва (28 января-1 февраля 1951 г.). М.; Л., 1951, № 4,
с. 3-17. - Курс низших растений/Под ред. М. В. Горленко. М., 1981. 520 с. -
Кустенко Н. Г. Об укрупнении клеток Ditylum brightwellii и Biddulphia mobiliensis
в культуре. - Ботан. журн., 1984, т. 69, № 8, с. 1074-1076. - Левин Г. Г. Чередо-
вание поколений, цикл развития и онтогенез растений. - Ботан. журн., 1959, т. 44,
№ 7, с. 943—953. — Лодкина М. М. Значение спорогенеза на разных этапах эволюции
растений. - Ботан. журн., 1969, т. 54, № 7, с. 1032-1049. - Надсон Г. А. Сверлящие
водоросли и значение их в природе. - Ботан. зап., 1900, вып. 18, с. 1-40. - Пере-
стенко Л. П. Водоросли залива Петра Великого. Л., 1980. 232 с. - Петров Ю. Е.
О чередовании форм развития у фукусовых водорослей. - Ботан. журн., 1962,
т. 47, № 9, с. 1380-1383. - Петров Ю. Е. Развитие концептакулов у Ascoseira mirabi-
lis Skottsb. и происхождение фукусовых водорослей (Cyclosporophyceae). - Ботан.
журд., 1963а, т. 48, № 9, с. 1298-1309. - Петров Ю. Е. Смена форм развития и про-
исхождение органов полового размножения у циклоспоровых (фукусовых) водо-
рослей: Автореф. дис. ... канд. биол. наук. Л., 19636. 20 с. - (Петров Ю. Е.) Pet-
rov J.E. The life-history and taxonomy of Cyclosporophyceae. - In: Tenth Intern, botan.
congr.: Abstr. of papers. Edinburg, 1964, p. 446. - Петров Ю. E. Развитие концептаку-
лов (скафидиев) у Ascophyllum nodosum (L.) Le Jolis и Durvillea antarctica (Chamis-
so) Hariot. - Ботан. журн., 1967, т. 52, № 3, с. 348-350. - Петров Ю. Е. Основные
этапы эволюции циклов развития у водорослей. - В кн.: Тез. докл. IV Москов. со-
вещ. по филогении растений. М., 1971. кн. 11, с. 77-80. - (Петров Ю. Е.) Petrov J. Е.
Evolution of life-history of algae. - В кн.: Тез. докл. XII Междунар. ботан. конгресса.
Л., 1975, с. 43. - Полянский В. И. Жизненный цикл, смена форм развития и чере-
дования ядерных фаз (в связи с некоторыми особенностями этих процессов у во-
дорослей) . - Ботан. журн., 1958, т. 43, № 5, с. 621-628. - Al-Kubaisy К. И., Schwan-
tes Н.О., Seibold G. Cytophotometrische Untersuchungen zum Generationswechsel auto-
tropher und heterotropher siphonaler Organismen (Vaucheria sessilis und Saprolegnia fe-
rax). - Nova Hedvigia, 1981, Bd 34, H. 1-2, S. 301-316. - Berger-Perrot Y. Mise au
point sur le probleme concernant 1’Ulothrix speciosa (Carm. ex Harvey) Kiitzing et Uro-
spora kormnannii Berger-Perrot. - Phycologia, 1981, vol. 20, N 2, p. 147-164. - Boil-
lot A. Sur le cycle de Rhodochaete parvula Thuret. - C. R. Acad. Sci., 1969, t. 269, N 22,
p. 2205-2207. - Bold H. C., Wynne M. J. Introduction to the algae structure and repro-
duction. New Jersey, 1978. 706 p. - Cabioch J. Un nouveau cas dinomalie du cycle
des Gigartinales (Algues Floridees). - C. R. Acad. Sci., 1972, t. 275, N 18, p. 1979-
1981. - Celakovsky L. Ueber die morphologische Bedeutung der sog. Sporensprosschen
der Characeen. - Flora, 1878, Bd 61, S. 49-57, 73-76. - Chapman V. J. The algae.
London, 1962. 472 p. - Chihara M. Rhodophyta, their life-histories. - In:Advance of
phycology in Japan. Jena, 1975, p. 137-150. - Christensen Y. Algae: A taxonomic
survey. Odense (Denmark), 1980. 228 p. - Cole K., Akintobi S. The life cycle of Prasiola
55
meridionalis Setchell and Gardner. - Can. J. Bot., 1963, vol. 41, N 5, p. 661-668. -
Gonway E., Cole K. Observations on an unusual form of reproduction in Porphyra (Rho-
dophyceae, Bangiales). - Phycologia, 1973, vol. 12, N 3/4,p. 213-225. -De Cew T. C.,
West J. A. A sexual life history in Rhodophysema (Rhodophyceae): a re-interpretation. -
Phycologia, 1982, vol. 21, N 1, p. 67 --74. - Deloffre R., Genot P. Les algues Dasycladales
du Cenozoique. - Bull. Cent.rech. Explor.-Prod. Elf-Aquitaine, 1982, Mem.4,p.5-247. -
Dixon P. S. Life histories in the Florideophyceae with particular reference to the Nema-
Jiales sensu lato. - Bot. Mar., 1982, vol. 25, fasc. 12, p. 611 -621. - Drebes G. On the
life history of the marine plankton diatom Stephanopyxis palmeriana. - Helgoland.
Wiss. Meeresuntersuch., 1966, Bd 13, N 1-2, S. 101-114. - Drew К. M. Genetics and
algal life-histories. - Nature, 1948, vol. 161, N 4085, p. 223. - Favre-Du chartre M.
Theories d’interpretation des cycles biologiques des plantes. - Phyto morphology, 1967,
vol. 17, N 1—4, p. 31-35. - Feldmann J. Une Nemalionale a carpotetraspores: Helmin-
thocladia hudsoni (C. Ag.) J. Ag. - Bull. Soc. hist, natur. Afr. Nord, 1939, t. 30, N 3,
p. 87-97. - Feldmann J. Les cycles de reproduction des Algues et leurs rapports avec
la phylogenie. - Rev. cytol. et biol. veg., 7952,1.13, fasc. 1 -2, p. 1 -49. - Friedmann J.
Structure, life history and sex determination of Praiola stipitata Suhr. - Ann. Bor., 7959,
vol. 25, N 92, p. 571 -594. - Fritsch F. E. The structure and reproduction of the algae.
Cambridge, 1935, vol. 1. 791 p. - Fritsch F. E. The structure and reproduction of the
algae. Cambridge, 1945, vol. 2. 936 p. - Gallagher J. C. Cell enlargement in Skeletonema
costatum (Bacillariophyceae). - J. Phycol., 1983, vol. 19, N 4,p. 539-542. - GayralP.,
Fresnel J. Description, sexualite et cycle de developpement d*une nouvelle Coccolitho-
phoracee (Prymnesiophyceae): Pleurochrysis pseudoroscoffensis sp.nov.— Protistologica,
1983, t. 19, fasc. 2, p. 245-261. - Gayral P., Haas C., Lepailleur H. Alternance morphol-
logique de generations et alternance de phases chez les Chrysophycees. - Mem. Soc. bot.
France, 1972, p. 215-230. - Goebel K. Morphologische und biologische Bemerkungen.
II. Ueber die Homologie in der Ent wicklung mannlicher und weiblicher Sexualorgane. -
Flora, 1902, Bd 90, S. 279-305. - Goebel K. Die Deutung der Characeen Antheridien:
Ein Versuch. - Flora, 1930, Bd 124, S. 491-498. - Goetz G. Ueber die Entwickelung
der Eiknospe bei den Characeen. - Bot. Ztg, 1899, Bd57,Nl,S.l-13.- Graham L. E.,
Wilcox L. W. The occurence and phylogenetic significance of patative placental transfer
cells in the green alga Coleochaete. - Amer. J. Bot., 1983, vol. 70, N 1, p. 113-120. -
Green B. R. Abnormal cells resulting from asexual reproduction in Acetabularia (Chloro-
phyceae, Siphonales). - Phycologia, 1976, vol. 15, N 2, p. 161-164. - Guiry M. D.,
West J. A. Life history and hybridization studies of Gigartina stellata and Petrocelis
cruenta (Rhodophyta) in the North Atlantic. - J. Phycol., 1983, vol. 19, N 4, p. 474-
494. - Guttenberg H. von. Lehrbuch der allgemeinen Botanik. Berlin. 1955. 708 S. -
Hafez A.-R. M„ Magne F. Existence d*une nouveau type de cycle de developpement chez
les Rhodophycees. - C. R. Acad. Sci., 1983, t. 296, N 14, p. 641 -644. - Hansgirg F. E.
Ueber der Polymorphysmus der Algen. - Bot. Centralbl., 1885, Bd 22, S. 246-251,
277-285, 343-352, 373-383, 385 -406. - Harada A., Yamagishi T. Meiosis in Spiro-
gyra (Chlorophyta). - Jap. J. Phycol., 1984, vol. 32, N 1, p. 10-18. - Henry E. C.
Syringodermatales ord. nov. and Syringoderma floridana sp. nov. (Phaeophyta). - Phy-
cologia, 1984, vol. 23, N 4, p. 419-426. - Herth W., Kuppel A., Franke W. W. Cellulose
in Acetabularia cyst walls. - J. Ultrastruct. Res., 1975, vol. 50, N 3, p. 289-292. -
Hofmeister W. Vergleichende Untersuchungen der Keimung, Entfaltung und Fruchtbil-
dung hoherer Kryptogamen und der Coniferen. Leipzig, 1851. 179 S. - Hopkins A. W.,
McBride G. E. The life history of Coleochaete scutata (Chlorophyceae) studied by
a feulgen microspectrophotometric analysis of the DNA cycle. - J. Phycol., 1976, vol. 12,
N 1, p. 29-35. - Hoxmark R. C. Experimental analysis of the life cycle of Ulva muta-
bilis. - Bot. Mar., 1975, vol. 18, fasc. 2,p. 123-129. - Hurdelbrink L., Schwantes H. O.
Sur le cycle de developpement de Batrachospermum. - Mem. Soc. bot. France, 1972,
p. 269-274. - Jensen J. B. Morphological studies in Cystoseiraceae and Sargassaceae
(Phaeophyceae) with special reference to apical organization. - Univ. Calif. Pubis Bot.,
1974, vol. 68, p. 3-61. - Jonsson S. Nouveau type de parth£nogen£se haploide chez
les algues: la parthenogenfese plasmogamique. - C. R. Acad. Sci., 1964, t. 258, N 7,
p. 2145-2148. - Jonsson S. Influence de 1’origine g&agraphique sur les cycles evolutifs
dans les chlorophycees marines du genre Acrosiphonia. - C. R. Acad. Sci., 1965, t. 261,
N 2, p. 499-502. — Jonsson S. Le cycle e'volutif de 1’Acrosiphonia sonderi (Kutz.) Kornm.
56
d’Islande et 1’origine de ses races asexue'es. - Rev. gen. bot., 1969, vol. 76, N 900—901,
p. 267-286. - Koeman R. P. T., Cortel-Breeman A. M. Observations on the life history
of Elachista fucicola (Veil.) Aresch. (Phaeophyceae) in culture. - Phycologia, 1976,
vol. 15, N 2, p. 107-117. - Koop HrU. Uber den Ort der Meiose bei Acetabularia medi-
terranea. - Protoplasma, 1975, vol. 85, N 1, p. 109-114. - Koop H. U. Observations
on the morphogenetic capacity of Residual nuclei” in Acetabularia. — Protoplasma, 1979,
vol. 101, N 4, p. 363-371. - Kornmann P. Die Entwicklung von Porphyra leucosticta im
Kulturversuch. - Helgoland. Wiss. Meeresuntersuch., 1961, Bd 8, N 1, S. 167-175. —
Kornmann P. Die Ulothrix-Arten von Helgoland. 1. - Helgoland. Wiss.Meeresuntersuch.,
1964, Bd 11, N 1, S. 27-38. - Kornmann P. Eine erbliche Variante von Derbesia ma-
rina. - Naturwissenschaften, 1966, Jg53,H. 6,S. 161,- Kornmann P. Eine Mutation
bei der siphonalen Griinalge Derbesia marina. - Helgoland. Wiss. Meeresuntersuch.,
1970, Bd 21, N 1, S. 1 -8. - Kornmann P. Ein Beitrag zur Taxonomie der Gattung Chae-
tomorpha (Cladophorales, Chlorophyta). - Helgoland. Wiss. Meeresuntersuch., 1972,
Bd 23, N 1, S. 1-31. - Kylin H. Ueber die Entwicklungsgeschichte der Florideen. -
Lunds Unuv. Arssk. N. F„ II, 1930, Bd 26, N 6,104 S. -Kylin H. Uber die Entwicfciung-
sgeschichte der Phaophyceen. - Lunds Univ. Arssk. Avd. 2,1933, Bd 29, N 7, 102 S. -
Kylin H. Zur Entwicklungsgeschichte der Ordnung Fucales. - Sven. bot. tidskr., 1940,
Bd 34, H. 3, S. 301 -314. - Lee R. E. Phycology. Cambridge, 1980. 478 p. - Lewin R. A.
The zygote of Chlamydomonas moewusii. - Can. J. Bot., 1957, vol. 35, N 6, p.795-804. -
Littler M. M„ Kanker B. J. Heterotrichy and survival strategies in the red alga Corallina
officinalis L. - Bot. Mar., 1984, vol. 27, fasc. 1, p. 37-44. - Littler M. M., Littler D. S.
Heteromorphic lifeJristory strategies in the brown alga Scytosiphon lomentaria (Lyngb.)
Link. - J. Phycol., 1983, vol. 19, N 5, p. 425—431. - Loiseaux S. Recherches sur les
cycles de developpement des Myrionematac^es (Pheophycdes). I-II. Hecatonematees
et Myrionematees. - Rev. gen. bot., 1967, t. 74, p. 529-576. - Lokhorst G. M.,
Trask B. J. Taxanomic studies on Urospora (Aerosiphonales, Chlorophyceae) in Western
Europe. - Acta bot. neer., 1981, vol. 30, N 5/6, p. 353-431. - Lokhorst G. M., Vro-
man M. Taxonomic study on three freshwater Ulothrix species. - Acta bot. neer., 1972,
vol. 21, N 5, p. 449-480. - Lombard E. H., Capon B. Observations on the tide pool
behavior of Peridinium gregarium. - J. Phycol., 1971, vol. 7, N 2, p. 188—194. -
Lubchenco J., Cubit J. Heteromorphic life histories of certain marine algae as adaptation
to variations in herbivory. - Ecology, 1980, vol. 61, N 3, p. 676-687. -Magne F. Sur
1’existence d*une reproduction sexuee chez le Rhodochaete parvula Thuret. - C. R. Acad.
Sci., 1960, t. 251, N 15, p. 1554-1555. - McCandless L., Vollmer С. M. The nemathe-
cium of Gymnogongrus chiton (Rhodophyceae, Gigartinales): immunochemical evidence
of meiosis. - Phycologia, 1984, vol. 23, N 2, p. 119-123. - Moe R. L., Henry E. C.
Reproduction and early development of Ascoseira mirabilis Skottsberg (Phaeophyta),
with notes on Ascoseirales Petrov. - Phycologia, 1982, vol. 21, N 1, p. 55-66. —
Mukai L. S., Craigie J. Brown R. G. Chemical composition and structure of the cell
walls of the Conchocelis and thallus phases of Porphyra tenera (Rhodophyceae). -
J. Phycol., 1981, vol. 17, N 2, p. 192-198. - Muller D. G. Generationswechsel, Kern-
phasenwechsel und Sexualitat der Braunalge Ectocarpus siliculosus im Kulturversuch. -
Planta, 1967, Bd 75, H. 1, S. 39-54. - Muller D. G. Life cycle of the brown alga Ecto-
carpus fasciculatus var. refractus (Kutz.) Ardis. (Phaeophyceae, Ectocarpales) in culture. -
Phycologia, 1972, vol. 11, N 1, p. 11-13. -Murray S. N., Scott J. L., Dixon P. S. The
life history of Porphyropsis coccinea var. dawsonii in culture. - Brit. Phycol. J., 1972,
vol. 7, p. 323 -333. - Nakahara H., Nakamura Y. Parthenogenesis,apogamy and apospory
in Alaria crassifolia (Laminariales). - Mar. Biol., 1973, vol. 18, p. 327-332. - Neuman K.
Beitrag zur Cytologie und Entwicklung der siphonalen Griinalge Derbesia marina. - Helgo-
land. Wiss. Meeresuntersuch., 1969, Bd 19, N 3-4, S. 355-375. -NewrothP.R. Studies
on life histories in the Phyllophoraceae. 1. Phyllophora truncata (Rhodophyceae, Gigarti-
nales). - Phycologia, 1971, vol. 10, N 4, p. 345-354. - Newroth P. R. Studies on life
histories in the Phyllophoraceae. II. Phyllophora pseudoceranoides and notes on P. crispa
and P. heredia (Rhodophyta, Gigartinales). - Phycologia, 1972, vol. 11, N 1, p. 99-107. -
Notoya M. The life history of Gloeophycus koreanum I. K. Lee et Yoo (Rhodophyta,
Gloiosiphoniaceae) in culture. - In: Abstr. Xlth Intern, seaweed symp. 19-25 june 1983.
Qingdao, China, 1983, p. 189. - Notoya M., Yabu H. Platythamnion yezoense Inagaki
(Rhodophyta, Ceramiales) in culture. - Jap. J. Phycol., 1980, vol. 29, N 1, p. 39—46. -
57
Pal В. P., Kundu В. C., Sundaralingam V. S., Venkataraman G. S. Charophyta. New
Delhi, 1962. 130 p. - Pedersen P. M. Culture studies on complanate and cylindrical
Scytosiphon (Fucophyceae, Scytosiphonales) from Greenland. - Brit. Phycol J., 1980,
vol. 15, p. 391-398. - Pedersen P. M. Phaeophyta: life histories. - In: The biology of
seaweeds. Oxford etc., 1981, p. 194-217 (Bot. monogr.; Vol. 17). - Perrot Y. Sur le
cycle de reproduction d’une cladophoracee marine de la r6gion de Roscoff: Lola implexa
(Harv.). - C. R. Acad. Sci., 1965, t. 261, N 2, p. 503-506. - Perrot Y. Sur le cycle de
deux formes d’Ulothrix flacca (Dillw.) Thuret de la region de Roscoff. - C. R. Acad.
Sci., 1968, t. 266, N 19, p. 1953-1955. - Pfiester L. A. Sexual reproduction of Peridi-
nium willei (Dinophyceae). - J. Phycol., 1976, vol. 12, N 2, p. 234-238. -Pfiester L. A..
Lynch A. R. Amoeboid stages and sexual reproduction of Cystodinium bataviense and its
similarity to Dinococcus (Dinophyceae). - Phycologia, 1980, vol. 19,N 3,p. 178-183. -
Popovsky J., Pfiester L. A. The life-histories of Stylodinium sphaera Pascher and Cystodi-
nedria inermis (Geitler) Pascher (Dinophyceae): Two freschwater facultative predator-
autotrophs. - Arch. Protistenk., 1982, Bd 125, H. 1-4, S. 115-127. - Richardson N.,
Dixon P. Life history of Bangia fuscopurpurea (Dillw.) Lyngb. in culture. - Nature,
1968, vol. 218, N 5140, p. 496-497. - Rietema H. A. A new type of life history in Bryo-
psis (Chlorophyceae, Caulerpales). - Acta bot. neer., 1969, vol. 18, N 5, p. 615-619. -
Rietema H. Life-history studies in the genus Bryopsis (Chlorophyceae). IV. Life-hystories
in Bryopsis hypnoides Lamx. from different points along the european coasts. - Acta
bot. neer., 1971, vol. 20, N 3, p. 291 -298. - Rieth A. Beitrage zur Kenntnis der Vau-
cheriaceae. XVI. Vaucheria hercyniana nov. sp. und ihre Entwicklung. - Arch. Protistenk.,
1974, Bd 116, H. 1/2, S. 201-209. - Rosowski J. R., Willey R. L. Colacium libellae sp.
nov. (Euglenophyceae) a photosynthetic inhabitant of the larval damselfly rectum. - J.
Phycol., 1975, vol. 11, N 3, p. 310-315. - Sandgren C. D. Characteristics of sexual and
asexual resting cyst (statospore) formation in Dinobryon cylindricum Imhof (Chryso-
phyta). - J. Phycol., 1981, vol. 17, N 2, p. 199-210. - Scherffel A. Kleiner Beitrag zur
Phylogenie einiger Gruppen niederer Organismen. - Bot. Ztg, 1901, Bd 59, Abt. 1,
S. 143—158. — Schussnig B. Handbuch der Protophy tenkunde. Jena,7960, Bd2. 1114S.-
Sears J. R., Wilce R. T. Reproduction and systematics of the marine alga Derbesia (Chlo-
rophyceae) in New England. - J. Phycol., 1970, vol.6,N 2,p. 381-392. - Skottsberg C.
Zur Kenntnis der subantarktischen und antarktischen Meeresalgen. I. Phaeophyceen. —
Wiss. Ergebn. Schwed. Siidpolar-Exped. 1901 -1903, Stockholm, 1907, Bd 4, S. 1 -172. -
South G. R., Hooper R. Stictyosiphon soriferus (Phaeophyta, Dictyosiphonales) from
Eastern North America. - J. Phycol., 1976, vol. 12, N 1, p. 24-29. - Spero H. J., Mo-
ree M. D. Phagotrophic feeding and its importance to the life cycle of the holozoic dino-
flagellate, Gymnodinium fungiforme. - J. Phycol., 1981, vol. 17, N 1, p. 43-51. -
Stosch H. A. von. Entwicklungsgeschichtliche Untersuchungen an zentrischen Diatomeen.
I. Die Auxosporenbildung von Melosira varians. - Arch. Microbiol., 1951, Bd 16, H. 2,
S. 101-135. - Stosch H. A. von. Die Oogamie von Biddulphia mobiliensis und die
bisher bekannten Auxosporenbildungen bei den Centrales. - In: 8 Congr. Intern. Bot.,
Paris, 1954, sect. 17, p. 58-68. - Stosch H. A. von. Entwicklungsgeschichtliche Unter-
suchungen an zentrischen Diatomeen. II. Geschlechtszellenreifung, Befruchtung und
Auxosporenbildung einiger grundbewohnender Biddulphiaceen der Nordsee. - Arch.
Microbiol., 1956, Bd 23, H. 4, S. 327-365. - Stosch H. A. von. Entwicklungsgeschicht-
liche Untersuchungen an zentrischen Diatomeen. III. Die Spermatogenese von Melosira
moniliformis Agardh. — Arch. Microbiol., 1958a, Bd 31, H. 1, S. 274-282. - Stosch H.A.
von. Kann die oogame Araphidee Rhabdonema adriaticum als Binderglied zwischen den
beiden grossen Diatomeengruppen angesehen werden? - Ber. Dtsch. bot. Ges., 1958b,
Bd 71, H. 4, S. 241 —249. — Stosch H. A. von. Manipulierung der Zellgrbsse von Diato-
meen im Experiment. - Phycologia, 1965, vol. 5, N 1, p. 21-44. - Stosch H. A. von.
La* signification cytologique de la „cyclose nucleate” dans le cycle de vie des Dinoflagel-
les. - Mem. Soc. bot. France, 1972, p. 201-212. - Stosch H. A. von. Observations on
vegetative reproduction and sexual life cycles of two freschwater dinoflagellates, Gymno-
dinium pseudopalustre Schiller and Woloszynskia apiculata sp.nov. - Brit. Phycol. J.,
1973, vol. 8, p. 105-134. - Stosch H. A. von, Theil G. A new mode of life history in
the freshwater red algal genus Batrachospermum. - Amer. J. Bot., 1979, vol. 66, N 1,
p. 105-107. - Stosch H. A. von, Theil G., Kowallik K. Entwicklungsgeschichtliche
Untersuchungen an zentrischen Diatomeen. V. Bau und Lebenszyklus von Chaetoceros
58
didymum, mit Beobachtungen uber einige andere Arten der Gattung. - Helgoland. Wiss.
Meeresuntersuch., 1973, Bd 25, N 2-3, S. 384-445. - Strasburger E. Zur Frage eines
Generationswechsels bei Phaeophyceen. - Bot. Ztg, 1906, Bd64,N 1,S. 1-7. -Tang Ti,
Loo Shih-wei. Culture of Acetabularia calyculus from the South China Sea. - In: Abstr.
Xlth Intern, seaweed symp. 19-25 june 1983, Qingdao, China, 1983, p. 231. - Tan-
ner С. E. Chlorophyta: life histories. - In: The biology of seaweeds. Oxford etc., 1981,
p. 218-247 (Bot. Monogr.; Vol. 17). - Tatewaki M. Life history and systematics in Mo-
nostroms. - In: Contributions to the systematics of benthic marine algae of the North
Pacific. Kobe, Japan, 1972, p. 1-15. - Taylor W. R. Marine algae of the eastern tropical
and subtropical coasts of the Americas. - Univ. Mich. Studies. Sci. sei., 1960, vol. 21,
870 p. - Thompson R. H. Schizochlamys gelatinosa and Placosphaera opaca. - Amer.
J. Bot., 1956, vol. 43, N 9, p. 665-672. - Tseng С. K., Chang T. J. Studies on the life
history of Porphyra tenera Kjellm. - Sci. Sinica, 1955, vol. 4, N 3, p. 375-398. -
Tuttle R. C. Crypthecodinium: sexual reproduction and mutagenesis. - In: Handb. phy-
cological methods: Developmental and cytological methods. Cambridge etc., 1980,
p. 143-151. - Umezaki I. The life histories of some Nemaliales whose tetrasporophytes
were unknown. - In: Contributions to the systematics of benthic marine algae of the
North Pacific. Kobe, Japan, 1972, p. 231-242. - Valet G. Contribution a 1’etude des
Dasycladales. 2. Cytologie et reproduction. - Nova Hedwigia, 1969, Bd 17, H. 1-4,
S. 551-573. - Van den Hoek C., Cortel-Breeman A. M„ Rietema H., Wanders J. B. W.
L Interpretation des donnees obtenues par des cultures unialgales, sur les cycles evolutifs
des algues. Quelques examples tirds des recherches conduites au laboratoire de Gronin-
gue. - Mem. Soc. bot. France, 1972, p. 45 -65. - Van der Meer J. P. Genetics of Graci-
laria sp. (Rhodophyceae, Gigartinales). IV. Mitotic recombination and its relationship to
mixed phases in the life history. - Can. J. Bot., 1977, vol. 55, N 22, p. 2810-2817. -
Van der Meer J. P., Todd E. R. The life history of Palmaria palmata in culture: A new
type for the Rhodophyta. - Can. J. Bot., 1980, vol. 58, N 11, p. 1250-1256. -
West J. A., HommersandM. H. Rhodophyta: life histories. - In: The biology of seaweeds.
Oxford etc., 1981, p. 133-193 (Bot. Monogr.; Vol. 17). - Wynne M. J., Loiseaux S.
Recent advances in life history studies of the Phaeophyta. - Phycologia, 1976, vol. 15,
N 3/4, p. 435-452. - Yabu H., Yasui H. The male gametophyte of Japanese Palmaria
palmata (Rhodophyta). - Jap. J. Phycol., 1984, vol. 32, N 3, p. 279-282. - Yotsui T.
The life 'cycle of Tinocladia crassa (Suringar) Kylin (Phaeophyta, Chordariales) without
a haploid gametophyte from Kuchinotsu, Kyushu, Japan. - Jap. J. Phycol., 1982, vol. 30,
N 2, p. 113-118. - Zakrys B. Aberrative divisions of Euglena and their phylogenetic
implications. - Nova Hedwigia, 1983, Bd 38, N 1-4, S. 471—476. - Zingmark R. G.
Sexual reproduction in the dinoflagellate Noctiluca miliaris Suriray. - J. Phycol., 1970,
vol.6,Nl,p.122-126.
59
ОГЛАВЛЕНИЕ
Введение .......................................................... 3
Циклы развития агамных водорослей ....................... 5
Циклы развития и смена форм развития эугамных
водорослей ...................................................... 8
Динофитовые водоросли (Dinophyta)............................... 8
Золотистые водоросли (Chrysophyta) .............................11
Цримнезиевые водоросли (Prymnesiophyta) ........................12
Диатомовые водоросли (Bacillariophyta) .........................13
Желтозеленые водоросли (Xanthophyta)............................16
Зеленые водоросли (Chlorophyta).................................17
Порядок Volvocales...........................................17
Порядок Ulotrichales.........................................21
Порядок Ulvales..............................................22
Порядок Schizogoniales ......................................23
Порядок Cladophorales........................................24
Порядок Oedogoniales.........................................25
Порядок Coleochaetales.......................................27
Порядок Dasycladales ........................................28
Порядок Siphonales...........................................31
Харовые водоросли ( Charophyta) ................................32
Красные водоросли (Rhodophyta) .................................34
Бурые водоросли (Phaeophyta)....................................42
Эволюция циклов развития и смены форм развития
у водорослей ....................................................50
Литература.........................................................55
Юрий Евгеньевич Петров
ЭВОЛЮЦИЯ ЦИКЛОВ РАЗВИТИЯ У ВОДОРОСЛЕЙ
Комаровские чтения. XXXV.
Утверждено к печати
Ботаническим институтом им. В. Л. Комарова
Академии наук СССР
Редактор издательства Е. А. Чекулаева
Технический редактор Н. А. Мяготина
Корректоры О. М. Бобылева и Э. Г. Рабинович
ИБ № 21356
Набор выполнен в издательстве на наборно-печатающем автомате. Подписано
к печати 14.02.86. М-18537. Формат 60 X 90 1/16 . Бумага офсетная № 1. Гарнитура
Пресс Роман. Усл. печ. л. 3.75. Усл. кр.-отт. 4.5. Уч.-изд. л. 4.52. Тираж 1000.
Тип. зак. № 786. Цена 70 к.
Ордена Трудового Красного Знамени
издательство „Наука”. Ленинградское отделение.
199164, Ленинград, В-164, Менделеевская лин., 1.
Ордена Трудового Красного Знамени
Первая типография издательства „Наука”.
199034, Ленинград, В-34, 9 линия, 12.