/
















































































































































































































































































































































































































































































































































































































Текст
Р. Саут, АУиттик
Основы
АЛЬГОЛОГИИ
Издательство «Мир»
Introduction to Phycology
G.Robin South
BSc PhD
Director
Huntsman Marine Science Centre
St Andrews
New Brunswick
Canada
and Professor of Biology
Memorial University of Newfoundland
St John’s, Newfoundland
Canada
Alan Whittick
BSc MSc PhD
Associate Professor of Biology
Memorial University of Newfoundland
St John’s, Newfoundland
Canada
Blackwell Scientific Publications
Oxford London Edinburgh
Boston Palo Alto Melbourne
Р. Саут, А. Уиттик
ОСНОВЫ АЛЬГОЛОГИИ
Перевод с английского
кавд. биол. наук К.Л. Тарасова
Москва "Мир" jagg
БЕК 28.591
C2I
УЖ 582.232/275
Саут Р.» Уиттик А.
C2I Основы альгологии: Пер. с англ. - М.: Мир, 1990. - 597 с.,
ил.
ISBN 5-03-00I522-I
Настоящее руководство по биологии водорослей, написан-
ное канадскими специалистами, освещает все аспекты этой дис-
циплины (систематику, морфологию, физиологию, биохимию,
экологию, эволюцию, филогению и хозяйственное использование
водорослей). По тематике книга не имеет аналогов в отечест-
венной литературе.
Для студентов и преподавателей биологических факульте-
тов, а также специалистов разного профиля, связанных с ку-
льтивированием и использованием водорослей.
Q 1906000000 — 248 Д09 — 90
056(01) - 90
ББК 28.591
Редакция литературы по биологии
ISBN
ISBN
5-03-00I522-I (русск.),
0-632-01726-0 (англ.)
(jo) 1987 by Blaokwell
Scientific Publica-
tions
© перевод на русский
язык, Тарасов К.Л.,
1990
От переводчика
Предлагаемая вниманию читателя книга Р. Саута и А.Уиттика
представляет собой развернутый обзор наиболее существенных совре-
менных работ (большинство - 1980-х гг.) в области альгологии, по-
нимаемой авторами достаточно широко: как наука не только о морфо-
логии, жизненных циклах и систематике водорослей, но и об основах
физиологии, биохимии, экологии и эволюции этих организмов. Подоб-
ной книги на русском языке нет. Наиболее существенным руководст-
вом по альгологии, изданным у нас, можно считать 3-й т. "Жизни
растений" под редакцией профессора М.М. Голлербаха (М., "Просве-
щение", 1977), но там подробно рассматриваются в основном строе-
ние и жизненные циклы различных видов водорослей. Поскольку в кни-
ге Р.Саута и А.УИттиха разбираются более общие вопросы, она и
этот том "Жизни растений" не "конкуренты”, а труды, хорошо допол-
няющие друг друга. Кроме того, в отечественных пособиях и учебни-
ках нет ссылок на соответствующую литературу, что не дает возмож-
ности более углубленно ознакомиться с отдельными деталями, а
здесь она насчитывает около 1500 названий! Таким образом, сведе-
ния, приводимые в книге, можно легко уточнить и изучить более
глубоко, обратившись к соответствующим первоисточникам.
Важно отметить, что именно в настоящее время водоросли во
воем мире привлекают все большее внимание исследователей самого
разного профиля, в частности, в связи с проблемой рационального
использования морских макрофитов (в нашей стране она чрезвычайно
актуальна из-за огромной протяженности береговой линии и недоста-
точно продуманного подхода к добыче этих ценных ресурсов: в од-
них местах они вообще не используются, а в других хищнически ис-
требляются, особенно агароносы) и возделывания их в марикультуре.
Водоросли вое шире используются как тест-объедты в физиоло-
гии, биохимии, генетике, биофизике и многих других областях фун-
даментальных биологических наук. Работающие в них ученые должны
иметь возможность на хорошем "своем" уровне ознакомиться о объек-
тами своих исследований.
Переведенная книга с этой задачей вполне справляется, при-
чем ее несомненное достоинство - обилие иллюстраций, в том чиоле
6
наглядных схем, которых зачастую явно не хватает нашим учебникам.
Некоторым недостатком книги можно, пожалуй, считать излиш-
нюю краткость при изложении систематики водорослей. Очень часто
по ходу рассмотрения особенностей размножения, экологии, биохимии
и т.д. авторы оперируют категорией "порядок", а в обзоре системы
дают характеристики только отделов и классов, просто перечисляют
порядки. Поэтому не всегда ясно, на основании каких критериев вы-
деляется та или иная крупная группа водорослей и какие семейства
и роды должны к ней относиться. Краткие характеристики пордцксв,
конечно, увеличили бы объем книги, но она от этого могла бы силь-
но выиграть.
Книга, несомненно, пригодится студентам при изучении ботани-
ки в целом и альгологии как спецкурса. Она хорошо дополнит не
только уже упоминавшуюся "Жизнь растений", но и основной совре-
менный учебник по этой теме - "Куро низших растений" (М., "Выс-
шая школа", 1981), послужив полезным справочником для преподава-
телей различного профиля при чтении лекций и проведении лабора-
торных и практических занятий, а также для специалистов в обла-
сти салих разнообразных фундаментальных и прикладных дисциплин.
К.Л.Тараоов
Посвящается нашим семьям: Барбаре,
Карен и Адриану, Джуди, Рейчел и
Кристсферу.
ПРЕДИСЛОВИЕ
На современной стадии развития науки вое труднее представить
обзор какого-либо ее раздела в рамках одного учебника для студен-
тов. Изучение водорослей, естественно, не является исключением,
и даже беглый просмотр биологической литературы за последнее де-
сятилетие показывает значительный рост наших знаний об этой груп-
пе растений^. Их эволюционное разнообразие, их важность как про-
дуцентов органики, их пригодность для многих физиологических и
экологических экспериментов, их ценность как индикаторов качест-
ва окружающей среды и их непосредственное экономическое значение
привлекли интерес ученых и инженеров, не считающих себя алъголв-
гами в классическом смысле. Продолжают появляться научные труды,
затрагивающие все более и более узкие разделы этого предмета, так
что с определенней точки зрения альгология как раздел биологии
уже прошла длительный путь развития. Однако мы все же вынуждены
представить предмет в целом в расчете на определенный срок его
преподавания.
"Введение в альгологию" - это наша попытка дать обобщенный
обзор альгологии для студентов. Здесь мы отошли от филетичеекого
подхода при изучении предмета, и в этом отличие предлагаемой кни-
ги от большинства современных учебников, в которых различные от-
делы водорослей обоуздаются индивидуально. Мы надеемся, что та-
ким образом дадим целостную картину мира водорослей и в результа-
те студенты воспримут предмет о точки зрения современных проблем
биологии, не занимаясь лишь таксономией и жизненными циклами.
"Введение в альгологию" мы начинаем с обзора классификации
водорослей, чтобы дать концептуальную основу для глав, которые
последуют дальше. Затем рассматриваются уровни организации с уль-
траструктурной, клеточной и морфологической точек зрения. После
этого идут обзоры жизненных циклов, особенностей размножения, фи-
зиологии, биохимии, экологии, филогении, эволюции и, наконец, ро-
ли водорослей в жизни человека.
Ту-------
ZB некоторых современных работах водоросли отделяют от растений,
включая в царство протиетов. - Прим, ред.
8
В эту книгу мы включили группу Cyanophycota , поскольку
считаем эти организмы шесте с Prochlorophycota особыми прока-
риотными водорослями, т.е. правомерным объектом альгологии, не
оспаривая, впрочем, их родства с бактериями. Каждый сравнительны!
анализ водорослей неизбежно должен обращаться к этим отделам,
иначе подробное рассмотрение структуры, физиологии, экологии и
эволюции водорослей будет неполным.
Мы полагаем, что студенты должны быть ознакомлены с основно!
литературой по предмету, поэтому даем многочисленные ссылки на
ключевые работы по соответствующим разделам. Цитируемая литератур
ра отражает наши собственные склонности, и ее не следует считать
исчерпывающей. В областях, где сейчас ведутся интенсивные иссле-
дования (таких, как классификация зеленых водорослей в широком
смысле), мы придерживались консервативного подхода, полностью от
давая себе отчет в том, что будущее неминуемо изменит картину,
которую мы попытались представить. (За короткое время, прошедшее
с момента окончания нашей рукописи, такие события уже произошли,)
Мы сознаем, что эта работа не лишена недостатков, в частно-
сти пропусков каких-либо интересных моментов, однако надеемся,
что достоинства все же перевесят. Беря на себя полную ответствен-
ность за книгу, мы сердечно благодарим многих друзей и коллег,
помогавших нам при ее создании. В частности, мы глубоко призна-
тельны тем, кто полностью или частично читал текст по ходу его
подготовки, - Эрику Хенри и Роберту Хуперу. Рой Фиккен предоста-
вил нам прекрасные фотографии, а Сью Мидс сделала ряд риоунков
к тексту. Марк Фавро и Ральф Кюленкамп помогли нам при цитирова-
нии французских и немецких источников, а специалисты Мемориально-
го университета при Ньюфаундлендской библиотеке королевы Елизаве-
ты К - при подборе ссылок. Мы особенно благодарны многим нашим
коллегам, охотно предоставившим нам копии оригинальных фотогра-
фий или разрешившим использовать их иллюстрации. Редакторский
коллектив издательства Blackwell Scientific Publications Ltd,
оказал нам неоценимую поддержку, помогая в работе, которая вре-
менами казалась невыполнимой. Наконец (но не в последнюю оче-
редь’), постоянным источником идей и энергии были для нас наши
прежние и нынешние студенты и аспиранты.
Г.Робин Саут
Элан УНттик
Глава I
ВВЕДЕНИЕ
Альгология и водоросли
Альгология (от латинского alga= водоросль) - наука о водо-
рослях^. Последние относятся к растительному царству и представ-
ляют собой очень разнообразную группу организмов, для которой с
древнейших времен не имелось четкого определения. Здесь мы прини-
маем их деление на шесть отделов (или типов), хотя в некоторых
работах признается 12 или 13 отделов; число таксонов более низко-
го ранга еще менее определенно. В книге рассмотрены как прокари-
отные, так и эукариотные формы. Ископаемые находки показывают,
что прокариотные водоросли были первыми фотосинтезирующими расте-
ниями; сейчас уже общепризнано, что именно от водорослей произош-
ли все прочие группы споровых растений и в конечном счете - цвет-
ковые.
Водоросли имеют простое строение: здесь можно встретить од-
ноклеточные формы, колонии клеток, нити и паренхиматозные талло-
мы. Многие одноклеточные формы подвижны, поэтому нельзя провести
четкой границы между ними и простейшими животными. Даже тело наи-
более сложных многоклеточных водорослей слабо дифференцировано
по сравнению о другими группами растений; только самые высокоор-
ганизованные формы обладают элементарными проводящими тканями.
Однако морфология крайне разнообразна, причем ряд бурых водорос-
лей (Phaeophycota ) может достигать размера небольшого дерева^.
Впрочем, относительная простота водорослей обманчива, так как на
клеточном уровне даже мельчайшие из них могут быть весьма сложно
устроены.
^В зарубежной литературе эта наука нооит название "фикология"
(по-гречеоки phykos = водоросль). Именно это слово (" phyco-
logy ") фигурирует в английском заглавии настоящей книги. В
отечественной литературе укоренился, однако, менее грамотный
(из-за смешения латинского и греческого корней) термин "альго-
логия", которым мы и вынуждены пользоваться, уступая традиции.
- Прим, перев.
^Некоторые бурые водоросли достигают длины 50 м и более. - Прим,
перев.
10
По типам воспроизведения водоросли столь же разнообразны,
как и по морфологии. У них известны вегетативный, бесполый и по-
ловой процессы, и у многих форм наблвдается чередование поколений.
Большинство образует подвижные зооспоры или гаметы. Половое раз-
множение эволюционировало независимо в разных отделах, его эволю-
ция включала развитие оогамии. Однако половой процесс отсутству-
ет у прокариот ( Cyanophyoota, Proohlorophycota ) И у
ряда эукариотных отделов (например, у Euglenophycota ). От
других споровых.растений водоросли отличаются отсутствием много-
клеточной стенки вокруг спорангиев и гаметангиев (исключение -
антеридии у Charophyoeae ). Половое воспроизведение у Rhodophy-
сеае , возможно, наиболее сложное в растительном царстве.
Биохимически и физиологически водоросли во многих аспектах
сходны с другими растениями. У них те же основные пути метаболиз-
ма; у всех есть хлорофилл а., а компоненты углеводов и белков срав-
нимы с присутствующими у высших растений. Многие водоросли - иде-
альные экспериментальные организмы, поскольку малый размер и про-
стота работы с ними в жидких средах позволяют изучать их в конт-
ролируемых лабораторных условиях.
Водоросли встречаются повсеместно, практически во всех при-
годных для жизни местообитаниях на суше - от жарких и холодных
пустынь до вечного льда и снежников - и в водных биотопах любого
типа. Они основные продуценты органических соединений. В водных
экосистемах им принадлежит важнейшая роль первого звена пищевой
цепи. Они могут непосредственно служить питанием для человека, а
экстракты из них используются при приготовлении пищевых и многих
других продуктов. Иногда водоросли засоряют системы водоснабже-
ния и водоеш в целом по мере их эвтрофизации. Некоторые вызываю-
щие "красные приливы" Dinophyoeae - причина отравлений. Одна-
ко лишь изредка водоросли бывают причиной серьезных заболеваний
человека. Вероятно, в дальнейшем эти растения будут все шире при-
меняться для получения энергии и пищи. Вероятно также, что они
будут одними из первых организмов, используемых человеком в кос-
мическом пространстве.
Развитие альгологии
Развитие альгологии связано с развитием западной культуры
в целом, особенно европейской (ом. Taylor , 1969), и может быть
разделено на четыре фазы. Первая началась о трудов древних гре-
ков (Теофраст, Диоскорид) и закончилась в конце ХУТИ в.; вторая
охватывает период с 1800 до примерно 1880 г.; третья - до начала
II
I950-x IT. (см. Prescott , 1951). Затем следует четвертая, т.е.
современная фаза.
Долгая ранняя история альгологии переплетается со становле-
нием ботаники в целом. Хотя первые письменные сведения о водо-
рослях обнаружены в древнекитайской классике (Porterfield ,
1922), именно греки заложили основы этой науки. Латинское слово
fucus , происходящее от греческого phykos , долго сохранялось
как название для недостаточно определенной группы примитивных ра-
стений , пока Линней ( Linnaeus , 1753) не описал род Fucus ,
положив тем самым начало формальной номенклатуре водороолей (гл.
2). Однако вплоть до конца ХУ1И в. они классифицировались грубо и
часто объединялись с грибами и лишайниками. Фон Цалузиан (Von Za-
lusian, 1592) включал их под названием "мхи" вместе с гриба-
ми, лишайниками и морскими травами в растения группы " Ruda et
Confusa " D, тогда как Боэн ( Bauhin , 1620) к водорослям при-
числял формы Muscus , Fucus , Conferva И Enuisetuta (= Chara).
Жгутиковые формы часто выделялись в так называемые "зоофиты".
Диллениус ( DiXXenius , 1741) писал о водорослях, но, как отмече-
но Преокоттом ( Prescott , 1951), трудно понять, какие растения
он имел в виду. К концу этого периода большинство макроскопиче-
ских водорослей было распределено между родами Fucus , Conferva.
Шуи. и Corallina , причем многие в современном понимании
разнообразные и неродственные виды были произвольно объединены
между собой.
Незрелая таксономия зтой ранней фазы в сочетании с непонима-
нием половых процессов, отсутствием подходящих микроскопов и гос-
подствовавшими неверными трактовками основных биологических прин-
ципов препятствовала возникновению альгологии как особой науки.
Например, среди ботаников тогда бытовало представление, что у во-
дорослей отсутствует половое размножение, хотя Реомюр возбудил
оживленные дебаты по этому поводу после своей публикации ( De
Reaumur , I7II) о половых органах Fucus . Много позже Гмелин
(Gmelin , 1768) заявлял, что водоросли развиваются партеногене-
тически.
По-видимому, своим превращением в настоящую науку альголо-
гия больше всего обязана Дж. Стэкхаусу, с которого начинается
вторая фаза ее развития. Возможно, он был первым, кто изучил про-
растание зиготы у Fucus И описал ( Stackhouse , 1801) процесс
«В переводе с латинского - "грубые и неясные". - Прим, ред.
12
оплодотворения; в результате были признаны несостоятельность оп-
ределения рода Fucus , данного Линнеем, и необходимость пере-
группировки многих включенных в него видов..
Совершенствование микроскопии и стимул, данный открытием
Стэкхауса, обеспечили бурное развитие таксономии. Революционные
преобразования систематики споровых растений длились с начала
XIX в. до 1880-х гг., - именно тогда были описаны многие извест-
ные сейчас роды водорослей. Среди первых здесь работали Досон Тер-
нер ( Dawson Turner » 1802), Воше ( Vaucher, 1803), Ламуру ( ia-
mouroux » 1813), Лингби ( l^ngbye г 1819), Гравий ( Greville»
1830) и УИльям Генри Харви (Harvey , I846-I85I, 1852-1858). Наи-
больший вклад внесли, однако, шведский альголог К.А. Агард (С.А.
Agardh ) и его сын Й.Г. Агард (j.G.Agardh ), обширные публика-
ции которых, появлявшиеся в течение трех четвертей столетия, до
сих пор остаются краеугольным камнем таксономии и номенклатуры
водорослей, а также немецкий альголог Ф.Т. Кютцинг (Kiitaing ,
1843, 1848). Для более полного ознакомления с этим важным перио-
дом, когда в альгологии были заложены основы современной таксоно-
мии и систематики, следует обратиться к обзорам Прескотта (Pres-
cott , 1951) И Тейлора ( Taylor,1969).
Когда большинство водорослей было описано и стало возможным
развитие их современной систематики, наступила третья фаза разви-
тия альгологии. В начале этого периода Де Тони (De Toni , 1889)
своей работой " Sylioge Algarum " ("Сводка водорослей"), завер-
шенной только через 35 лет (в 1924 г.), начал титанический труд
по обобщению таксономии и номенклатуры. Критические исследования
Шмица ( Sohmita > 1883, 1889) по красным водорослям, Куккука
(Kuckuck , 1912) и Чельмана ( Kjellman » 189?) по бурым водорос-
лям и Вилле ( wine > I897-I9II) по зеленым водорослям включали
уже новые таксономические обработки. Параллельно важные пионер-
ные исследования (например, whitяшн , 1897, 1898; Sauvageau
1899, 1915, Yamanouchi > 1906; Svedelius > 1908; Kylin • 1914)
способствовали пониманию жизненных циклов водорослей. Это привело
к совершенствованию их классификации и возникновению новых гипо-
тез о происхождении отдельных групп и родственных связях мевду
ними.
На протяжении третьей фазы было опубликовано много флор, мо-
нографий и общих таксономических обработок по микроскопическим и
макроскопическим, пресноводным и морским водорослям. Основными
вехами в становлении альгологии были классические сводки Ольтман-
13
оа (Oltmanns » 1904, 1922) и Фрича ( Fritsch » 2935, 1945); бу-
дучи не единственными обобщающими работами в этот период, они
тем не менее полнее всего представили самые различные аспекты
предмета. Для мощного взлета альгологии со времен Фрича характер-
но, что кому-то одному собрать все наши знания о водорослях в
двух томах теперь уже просто невозможно.
Четвертая, современная фаза, вероятно, началась публикацией
"Курса альгологии" Смита ( Smith. 1951, Manual of Phycology )•
Еде не включая электронномикроскопических данных, он тем не менее
знаменует собой конец "доэлектронной" эры развития предмета и,
многопланово освещая его постоянно растущую сложность, намечает
перспективы для более современных исследований в этой области.
Сейчас в альгологии происходят революционные изменения. Они
связаны с широким применением сканирующего и просвечивающего эле-
ктронных микроскопов, развитием тонких методов иоследований в
экологии, физиологии, биохимии, молекулярной биологии, а также
со значительно большей доступностью самих водорослей после изо-
бретения акваланга. Открывшиеся перспективы и не снились таким
классикам, как Ф.Э. Фрич. Может быть, будущие историки придут к
выводу, что происходящие сейчас изменения в альгологии глубже,
чем, например, вызванные в свое время изобретением светового ми-
кроскопа.
Современная техника значительно расширила наши представле-
ния о структуре и функциях водорослевых клеток и их органелл,
включая их микроморфологию, цитологию, физиологию, молекулярную
организацию и генетику. Альгологией сейчас занято множество ис-
следователей; читатели, интересующиеся конкретными вопросами,
могут найти ссылки на последние обзоры в списке литературы.
Изучение многих водорослей с помощью электронного микроско-
па и гораздо лучшее понимание их биохимии дали альгологам много-
численные дополнительные критерии, важные для таксономии. С их
учетом было предложено много новых систем классификации. Единст-
ва мнений по этощу вопросу не достигнуто, и разобраться в разных
таксономических построениях сложно даже для специалиста, не го-
воря уже о студенте. Особенно интенсивно обсуждайтся родственные
связи между разными группами зеленых водорослей и между этими
водорослями и другими зелеными растениями. По мере расширения
круга таксонов, изученных с помощью современных методов, многие
из современных разногласий, по-ввдимому, исчезнут.
Ряда альгологов росли параллельно развитию самого предмета,
и в начале "современной" фазы были основаны альгологические об-
14
щества США, Великобритании, Франции, Японии, а также Международ-
ное альгологическое общество. Взросление альгологии было отмечено
появлением журналов, посвященных исключительно водорослям: " Phy-
cologia ", " Journal of Thycology ", "British Thycological Bul-
letin " (ныне " British Phycological Journal "), " Phykos",
" Japanese Journal of Ihycology " И " Bulletin de la Sooi^te
Phycologique de France ", - а в последнее время и отдельным
группам водорослей (например, " Bacillaria" - о Bacillariophy-
cota ) и обзорам современного состояния знаний в этой области в
целом (например, " Advances in Thycology”), Ежегодно публикует-
ся несколько тысяч статей по альгологии, так что она вполне может
считаться сложившейся наукой.
ГОРИЗОНТЫ СОВРЕМЕННОЙ АЛЬГОЛОГИИ
Горизонты современной альгологии, как явствует из заголовков
сообщений наальгологических симпозиумах, поистине необъятны. Мож-
но назвать цитологию (включая электронную микроскопию), физиоло-
гию, биохимию, генетику, молекулярную биологию, экологию, таксо-
номию, систематику, прикладную биологию (например, аквакультуру,
сельское хозяйство, промышленное экстрагирование веществ и исполь-
зование фикоколлоидов), фармакологию, медицину, космическую биоло-
гию, планетологию и др. Если даже специалисты во многих из этих
областей не считают себя альгологами, они испольщуют водоросли в
своей работе; важна также;роль последних как крайне удобных эк-
спериментальных организмов.
Интересуетесь ли вы чисто, лабораторными исследованиями, по-
левой работой, или как-то их совмещаете - изучения вдет бесконеч-
ное множество проблем. Среди существенных достижений последних
10-15 лет - открытие двух различных путей цитокинеза (с фрагмо-
пластом или фикопластом), анализ и картирование пластвдных гено-
мов, осознание важности микротрабекулярных решеток как основы
ультраструктуры, цитоплазмы, обширные исследования глазков, кле-
точных мембран, жгутиков и цианобактериальных хлоропластен. Вы-
делять какую-либо область как наиболее актуальную было бы непра-
вомерным, хотя вряд ли многие станут отрицать выдающиеся успехи
высокоразрешающей электронной микроскопии или огромное поле дея-
тельности, ожидающее молекулярных биологов. Однако и классические
направления не теряют значимости, ибо таксономист по-прежнему ос-
тается в центре современных исследований: как молекулярный био-
лог опубликует свою работу, если он не знает названия организма,
с которым работает? Многие виды еще остаются неописанными, осо-
15
бенно среди мелких жгутиковых, а в случае относительно хорошо
известных групп применение новых методов приводит к пересмотру
прежних представлений.
Таким образом, альгология способна удовлетворить любые вку-
сы и интересы. Дополнительный плюс в том, что ее объекты - одни
из красивейших на планете организмов, если смотреть на них в ми-
кроскоп. Именно изучение водорослей помогло разрешить многие фун-
даментальные проблемы физиологии, и именно оно, возможно, даст
ответ на загадки происхождения жизни, эукариотной клетки (и, сле-
довательно, нас самих) и наземных растений.
Мы предполагаем в этой книге рассмотреть основы альгологии,
вполне сознавая, что это лишь беглый и поверхностный взгляд на
такой обширный предмет. Надеемся, многих наша книга побудит к
дальнейшим поискам: возможности у приобщившихся к альгологии не-
исчерпаемы.
Глава 2
КЛАССИФИКАЦИЯ ЮДОРОСЛЕЙ
Классификация растений и животных занимала лвдей со времен
Аристотеля. Шведский ботаник Линней ( Linnaeus , 1754) был пер-
вым, кто применил название Algae к одной из групп растений; од-
нако эта группа представляла собой смесь водорослей и печеночни-
ков. Трудность определения водорослей сохраняется и поныне; но
этому поводу Луин и Гиббс ( Lewin, Gibbs , 1982) отмечают: ”...
организмы, в настоящее время рассматриваемые как водоросли, в
сущности не имеют общих признаков, которые свидетельствовали бы
об их филогенетической однородности".
Большинство современных альгологов приняло бы в принципе,
если и не буквально, определение Фрича (Fritsch , 1935): ’’Пока
существуют чисто искусственные границы, понятие "водоросль" долж-
но включать все голофитные организмы (как и их многочисленные
бесцветные производные), которые не достигли уровня дифференци-
ровки архегониальных растений". Водоросли, таким образом, пред-
ставляют собой неоднородную группу продуцирующих кислород фото-
синтезирующих бессосудистых организмов с лишенными специальных
покровов репродуктивными структурами. Под определение подходят
как прокариотные, так и эукариотные формы. За исключением Cha-
rophyoeae (отдел Chlorophycota), водоросли отличаются от мохо-
образных отсутствием многоклеточных половых органов, имеющих по-
кровы из стерильных клеток, и тем, что опорофит не прикрепляется
внутри женского полового органа ( Silva , 1982).
Классификация водорослей, которой мы здесь пользуемся, взя-
та с некоторыми изменениями из Паркера ( Parker, 1982) (табл.
2.1). Они у него относятся к царству Protoctista ( Whittaker ,
1959, 1969; Whittaker, Margulis , 1978; Margulis , Schwartz ,
1982). Схема Паркера ( Parker. 1982) обобщает взгляды ряда веду-
щих альгологов. Признается разделение между прокариотными и эука-
риотными формами. У первых отсутствуют окруженные мембранами ор-
ганеллы. К ним ОТНОСЯТ бактерии, Cyanophycota (ИЛИ Cyanobac-
teria, "синезеленые водоросли"), и недавно описанные Prochloro-
phycota ( Lewin, 1976, 1977). Эукариотные формы включают водорос-
ли и все другие растения.
17
Подразделение водорослей долгой время было предметом споров.
Харви (Harvey , 1836) впервые выделил четыре основных отдела, ис-
ходя прежде всего из окраски (бурые, красные, зеленые и диатомо-
вые водоросли). Его схема служит основой и для наиболее современ-
ного подразделения водорослей на отделы. Хотя их ныне признается
больше четырех, основным фотосинтетическим пигментам и сопутствую-
щим биохимическим и структурным особенностям придается при клас-.
сифлкации водорослей огромное значение. Несомненно, остаются су-
щественные различия во взглядах альгологов на ее детали. Постоян-
ное поступление новой информации о тонкой структуре и биохимии
водорослей стимулирует новые подхода к их классификации, и каж-
дая схема, предлагаемая в настоящее время, неизбежно остается
приблизительной. К факторам, которыми обусловлены сложности клас-
сификации, относятся явно полифилетичеокое происхождение водорос-
лей, бедность (по-видимому) ископаемых остатков для большинства
групп и. тенденция делать далеко идущие выводы на основании изу-
чения относительно малого числа вадов.
Силва (Silva ,- 1982) выделяет 16 основных филетичееких ли-
ний, которым придается таксономический ранг класса. Классы разли-
чаются в основном пигментацией, запасными продуктами, особенно-
стями клеточной стенки и тонкой структурой таких органелл, как
жцутики, ядро, хлоропласты, пиреноиды и глазки. Эти различия сум-
мированы в табл. 2.2. Группировка классов в отделы в современной
альгологической литературе трактуется по-разному, и расхождения
являются в значительной степени результатом различного веса, при-
даваемого авторами тем или иным признакам. В этой книге принима-
ется шесть отделов: два из них прокариотные (Cyanophycota и
Irochlorophycota ) и четыре эукариотные ( Rhodophycota, Chromo-
phycota, Buglenophycota И Chlorophycota )• Встественно, мож-
но встретить и отклонения от иопользуемой здесь схема (см. Chap-
man , Chapman , 1973; Bold, Wynne , 1978, 1985; Trainor ,
1978; Round , 1973; Hoek , Jahns , 1978; lee , 1980; Christen-
sen , 1980; Soegel at al.,1982).
Классификация ниже уровня класса (или в отдельных случаях
подкласса, например, у Rhodophyceae ) производится последовате-
льно на порядки, семейства, роды и виды. Сходство меаду таксона-
ми возрастает по мере перехода на более низкие таксономические
уровни. Окончания названий таксонов определенного ранга постоян-
ны и соответствуют правилам Международного кодекса ботанической
номенклатуры ( Voss » 1983). Так, для бурой водоросли Fucua
vgalsalasua подучается следующая "леоенка":
18
Таблица 2.1. Общая классификация водорослей (по Parker , 1982)
Царство Отдел Plantae Класс Подкласс
Prokaryota Cyanophycota Prochlorophycota Eukaryota Rhodophycota Rhodophyceae Bangiophycidae Florideophyci- dae
Chromophyta Chrysophyceae Chrysophycidae Dictyoohophyci- dae Pr-ymnesiophyceae Xanthophyceae Eustigmatophyceae Bacillariophyceae Dinophyceae Dinophyoldae Ebriophycidae Ellobiophyoidae Syndiniophyci- dae Hiaeophyceae Raphidophyceae Cryptophyceae
Euglenophycota Chlorophycota Euglenophyceae Chlorophyceae Charophyoeae Prasinophyceae
Таблица 2.2. Обзор основных характеристик классов водорослей
Отдел/кла сс Основные пигменты Запасные вещества Особенности хлоропласта Клеточная стенка Жгутики
CZAJTOPHYCOTA Cyanophyceae PROCHLOROPHYCOTA RHODOPHYCOTA Hhodophyceae CHROMOPHYCOTA Chrysophyc eae Хлорофилл a, ft-Каротин с-Фикоэрит- рин Аллофикоциа- нин с-Фикоцианин Хлорофиллы £.>к Ь-Каротин Зеаксантин Криптоксан- тин Хлорофиллы 3.(1) г -фикоциа- нин Аялофикоциа- нин с-Фикоэрит- рин А- И £- Каротины Хлорофиллы а * сj, Со Фукоксантин Фикоцианино- вые гранулы (аргинин и аспарагино- вая кисло- та) Багрянковый крахмал Хризолами- нарин (J3- -I-3-глюко- пиранозид) Хлоропластов нет. Тила- коиды расположены сво- бодно в цитоплазме, не образу иг стопок Фикобилисомы Хлоропластов нет. Тила- коиды расположены сво- бодно в цитоплазме, не- которые стопками по 2 и более Оболочка хлоропласта 2-слойная, хлоропласт- ная эндоплазматическая сеть (ЭС) отсутствует. Тилакоиды не образуют стопок Две дополнительные мем- браны хлоропластной ЭС, внешняя из которых пере- ходит во внешнюю мем- брану ядерной оболочки Тилакоида в стопках по три 4-слойная, ос- нов ней компо- нент - пепти- догликан (му- реин) Как у Cyanophy- cota Целлюлозные (ксилановые у некоторых Bangiophycidae) полисахариды; некоторые ин- крустированы CaCOj Голые; у не- которых чешуй- ки или пан- цирь Отсутству- ют _и_ В норме 2; неравные: гладкий задний ко- роче; пери- стый перед- м ний длиннее «5
Продолжение табл. 2.2
Отд ел/к ла со Основные пигменты Запасные вещества Особенности хлоропласта Клеточная стенка Жгутики
Prynmesiophyceae (Haptophyoeae) Хлорофиллы a, S.J. а2 -Каротин ХризолаМи- нарин Две дополнительные мембра- ны хлоропластной ЭС, внеш- няя из которых переходит во внешнюю мембрану ядер- ной оболочки Тилакоады в стопках по три Клеточные покровы из органиче- ских чешуек, расположен- ных в один или несколь- ко слоев Обычно 2, неравные, гладкие Гаптонема
Xanthophyc еае Хлорофилл а (хлоро- филл £.У YauatieriLa) ь-каротин даатоксан- тин Диадино- ксантин Гетерок сан- гин Вошериакоан- тиновый эфир £-1,3-глю- кан Жиры и мас- ла Целлюлоза, глюкоза и уроновые кислоты У многих стенки из двух частей Обычно 2, неравные: передний длиннее, перистый; задний короче, гладкий
BustIgmatophycеае Хлорофиллы а, с 1-Каротин виолаксантин Вошериаксан- гин Масла Две дополнительные мем- браны хлоропластной ЭС, ни одна из которых не переходит в ядерную обо- лочку Тилакоиды в стопках по три Голые Единствен- ный перед- ний пеои- стнй жгу- тик и вто- рое базаль- ное тело
Продолжение тайл. 2.2
Отдёл/класс
Основные пиг- Запасные
мента вещества
Особенности хлоропласта
Клеточная Жгутики
стенка
Bacillariophyoeae
Хлорофиллы
Ja.c
Т>-Каротм
^ксксантин
иатсв сантин
иадиноксан-
тин
Хриэоламина- Как у Chrysophyceae
Липида
Dinophyceae
Хлорофиллы Крахмал
а,й2
Первдинин
Неоперидинин
J)-Каротин
Две дополнительные мем-
браны хлоропластной ЭС,
не связанные с яд ер но И
оболочкой. Тилакоиды
в стопках по три
Кремнезем- ный пан- Мужские гаметы е
цирь единст- венным перистым жгутиком без цент- ральных микротру- бочек
Целлюлоз- ная тека Чещуйки на клет- ках (ред- ко) Обычно 2, попереч- ный и про- дольный , гетеро- контные, резко раз- личающие- ся: попе- речный длиннее, спирально извитой, располо- жен в ка- навке, или пояс- ке; про- дольный короче, направлен назад
Отдел/класс Основные пиг- Запасные мента вещества
Phaeophyceae Хлорофиллы Ламинарии а, 2.2 Маннитол ^-Каротин Фукоксантин
Raphidophyceae Хлорофиллы ^иадиноксан- тин Динок сантин Гетероксан- тин ? Фукоксан- тин
Cryptophyceae Хлорофиллы Типа крахма- 8 t рг> ЛЭ оС-Каротин Диаток сантин Фикоэритрин Фикоцианин
Продолжение табл. 2.2
Особенности хлоропласта Клеточная стенка Жгутики
Две дополнительные мембра- ны хлоропластной ЭС, внеш- няя из которых переходит во внешнюю мембрану ядер- ной оболочки Тилакоиды в стопках по 2-6 Двуслойная, внутренний слой из целлюлозы, а внешний - из альги- новой кис- лоты и фукоидана Обычно 2, неравные: передний перистый длиннее; задний гладкий короче
Две дополнительные мембра- ны хлоропластной ЭС Тилакоиды в стопках по три Голые Обычно 2, неравные: передний перистый длиннее, задний гладкий короче
Две дополнительные мембра- Перипласт
ны хлоропластной ЭС
Тилакоиды парные
Два апи-
кальных
или боко-
вых, рав-
ных, пери-
стых
Отдел/класс Основные пиг- менты Запасные вещеотва
EUGLENOPHYCOTA Euglenophyceae Хлорофиллы “Ь^Каротии Астаксантин Антераксантин Диаданоксан- тин Неок сантин Парамилон (J5 -1,3- гликан)
СНЮВОРНУСОТА Chlorophyceae Хлорофиллы а> J и jp-Каротины Лютеин Сифоноксантин Сифоне ин Крахмал (амилоза, амилопек- тин)
Charophyoeae Хлорофиллы Крахмал а,к (оС-1,4-
Х-, р- и глюкан) Т-Каротины
Различные ксан-
тофиллы
Продолжение табл. 2.2
Особенности хлоропласта Клеточная стенка Жгутики
Одна дополнительная мем- Белковая брана хлоропластной ЭС, пелликула не связанная с ядерной мембраной Тилакоиды в стопках по три, иногда больше Обычно 2, неравные, перистые, один ча- сто не выступа- ет над поверхно- стью кле- тки
Хлоропластной ЭС нет Голые (не- Тилакоиды в стопках по которые) 2-6 или более Целлюлоза, Бывают граны гидрокои- пролин, гликозиды, к силаны, маннаны. У некоторых обызвест- вление В норме 2 (4), рав- ные, глад- кие (ино- гда о войлочным налетом), располо- жены на апикаль- ном конце
Хлоропластной ЭС нет ' Целлюлоза. Тилакоида в отопках по у многих 2-6 или более обызвест- Имеются граны влена Только у мужских гамет; 2 нерав- ных, суб- терминаль- ных; с чешуйка- ми
ОтделДласс Основные пиг- Запасные менты вещества
Prasinophycеае Хлорофилл Крахмал Л, Маннитол jS-Каротин Сифонеин Сифоноксантин
Продолжение табл. 2.2
Особенности хлоропласта Клеточная Жгутики стенка
Хлоропластной ЭС нет Тилакоиды в стопках по 2-6 Голые, I или 2 обычно о че- (нерав- щуйками вне) - или 4, более
ми менее
равные;
с чешуй-
ками
25
Отдел Chromophycota (-phycota)
Класс Phaeophyceae (-phyceae)
Порядок Fucales (-ales)
Семейство Fuoaceae (-aoeae)
РОД Fucus
ВИД vesioulosus
Принято также указывать фамилию автора, который описал род
или вад. Название Fucus vesioulosus должно писаться, следователь-
но , Fucus vesioulosus Linnaeus (или сокращенно Fuous vesioulo-
sus L.). Обиходные, или народные, названия (например, "лапуга"
для некоторых морских бурых водорослей) не имеют научного значе-
ния; преимущество формальной систе&ы в том, что она международна,
т.е. понимается учеными всего мира.
Названия рода и вида вместе представляют собой биномены и
уникальны для каадого вида. Биномиальная система номенклатуры
для большинства водорослей берет начало ст " Species Plantarurft
("Виды растений") Линнея (1-е издание - 1753 г.). Могут списыва-
ться и таксоны рангом ниже вида. В нисходящем порядке это подви-
ды, разновидности и формы.
Для каадого списанного вида водорослей должен быть типовой
образец, который неразрывно связан с присвоенным биноменом. Он
может храниться в гербарии, в форме микроскопического препарата
или во влажном виде. Для некоторых водорослей, особенно'для та-
ких микроскопических форм, как десмидиевые ( Chlorophyceae ), ти-
пом иногда служит рисунок (иконотип). Оригинальные описания каждо-
го ввда должны быть опубликованы, причем также и на латинском
языке (диагноз). Интересующимся вопросами номенклатуры можно по-
советовать обратиться к Международному кодексу ботанической но-
менклатуры ( Voss, 1983). Силва ( Silva, 1980) опубликовал хоро-
ший каталог названий классов и семейств ныне живущих водорослей,
а Фарр и др. ( Farr et al ., 1979) составили указатель родовых
названий растений, включающий детали публикации и типификации ро-
дов водорослей.
Ниже мы вкратце приводим признаки отделов и классов водоро-
слей. Для каждого класса дан список входящих в него порядков. В
последующих главах рассмотрены вопросы клеточной организации,
морфодогии, роста, физиологии и экологии.
26
Основные характеристики водорослей
рйокануота (прокариоты)
К прокариотам относятся Schizomycophyta (бактерии), Cyano-
phycota и Archaebacteria . У них ядерный материал не заключен в
особую оболочку, отсутствуют ядрышко и организованные хромосомы.
Нет также пластид, митохондрий, аппарата Гольджи и других окружен-
ных мембраной органелл. Осуществляющие аэробный фотосинтез прока-
риоты с хлорофиллом а относятся к Cyanophycota , а с хлорофиллами
а И b - к Prochlorophycota.
CYANOPHYCOTA
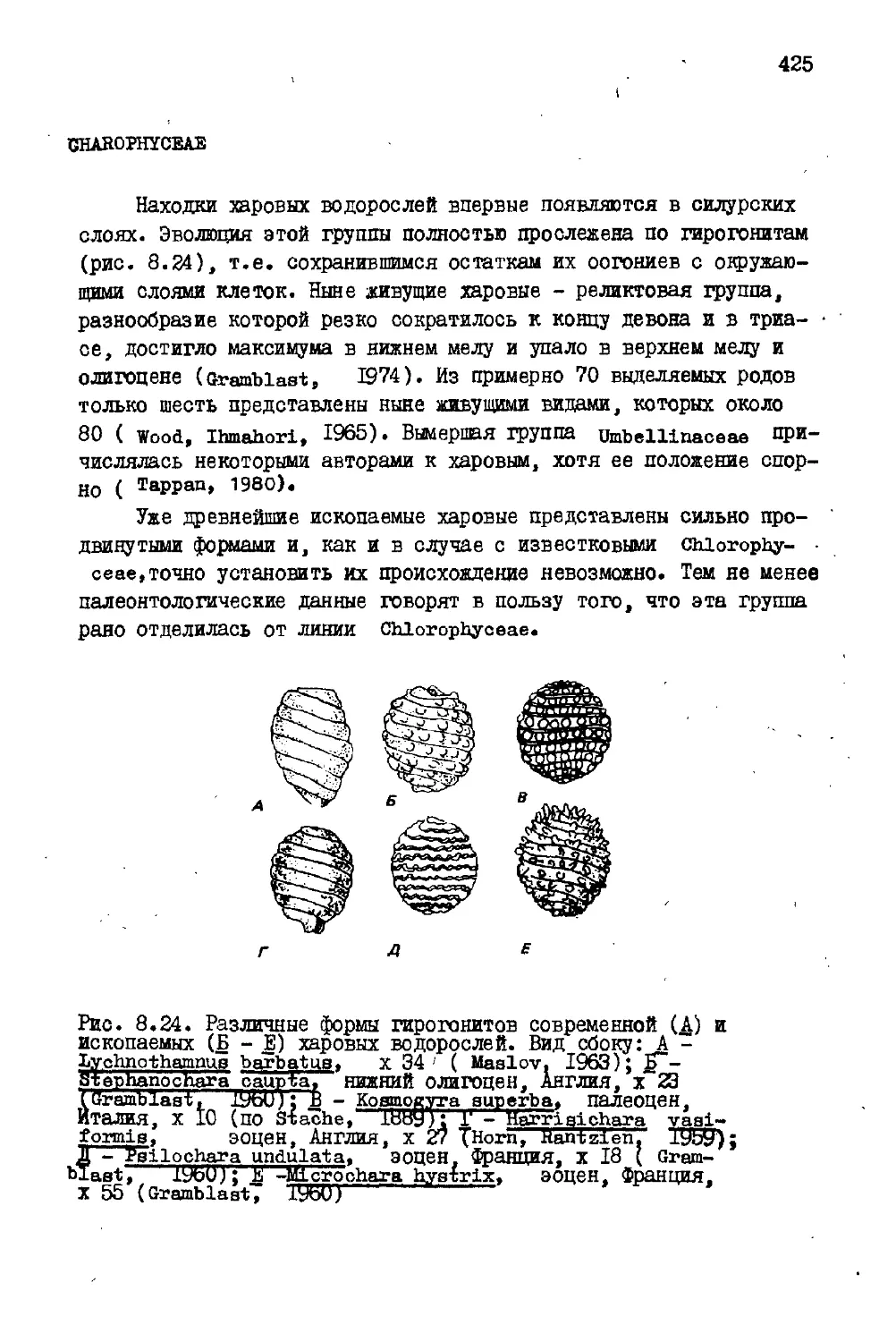
Cyanophyceae : синезеленые водоросли (рис. 2.1, 2.2)
Рис. 2.1. Chrooooccua ар.(Cyano-
phyceae, Ohroococcalee).
Планктонный пресноводный вид с
клетками, объединенными попар-
но и окруженными тонким бес-
цветным общим влагалищем. Мас-
штаб 10 мкм
)?ис. 2.2. Merismopedla ар.(Суа-
nophyceae,Chroococcales).
Многочисленные мелкие клетки
расположены в виде несколько-
искривленного слоя, который
увеличивается за счет их деле-
ния в двух направлениях, мас-
штаб 10 мкм
Имея в целом водорослевую морфологию, Cyanophycota обладают
и бактериальными чертами ( Friedmann , 1982). Название Cyanophy-
cota (=CyanochlorontajBold , 1973) сейчас, как правило, вытео-
27
няется названием Cyanobacteria (Stanier et al., 1978; ом.
-tead Walsby , 1982), но для однородности мы вслед за Паркером
( Parker , 1982) сохранили термин Cyanophyoeae sensu Sachs (1874).
у Cyanophyoota отсутствуют ядерная оболочка и митохондрии.
Их фотосинтетические ламеллы (тилакоиды) не собраны в отопки и
располагаются по периферии цитоплазмы, а не в окруженном мембра-
ной хлоропласте. Они содержат хлорофилл а и фикобилипротеины (фи-
коцианин, аллофикоцианин и фикоэритрин), а также полиглкжановые
гранулы (углеводы) и цианофицин (связанный азот) в качестве запас-
ных продуктов. Клеточная стенка четырехслойная, с основным струк-
турным слоем из пептидогликана (как у бактерий). Морфологическая
организация варьирует от одноклеточной до нитчатой; многие пред-
ставители окружены мощными слизистыми чехлами. К особым признакам
относятся способность фиксировать атмосферный азот в аэробных ус-
ловиях (это происходит в специализированных клетках - гетероцис-
тах) , наличие акинет и трихомов, т.е. особых нитчатых форм, у ко-
торых весь ряд клеток одет общим внешним слоем стенки.
Преобладает бесполое воспроизведение за счет деления клеток
пополам, множественного деления о образованием эндоспор (беоци-
тов) и о помощью гормогониев (коротких скользящих трихомов). Ис-
тинное половое размножение отсутствует, хотя имеются сведения об
изредка набладаемой генетической рекомбинации у некоторых видов
Б результате трансформации (Devilly, Houghton , 1977; Stevens,
Porter , 1980). Жгутики отсутствуют, однако многие виды способ-
ны к характерному скольжению.
Синезеленые водоросли, несомненно, одни из наиболее прими-
тивных автотрофов; их ископаемые остатки известны с докембрия
(3 млрд, лет назад) (Sohopf , 1976). Ныне живущие виды встречают-
ся повсеместно и могут обитать в таких экстремальных биотопах,
как горячие источники и каменистые пустыни. Весьма разнообразны
планктонные, бентосные и свободноживущие формы в морях, пресных
водоемах и почве; многие вступают в симбиотические отношения. Не-
которые виды могут вызвать токсичное "цветение” в эвтрофных место-
обитаниях, представляющее опасность для человека и домашнего ско-
та. Азотфиксирующие формы важны во воех биотопах, где они встре-
чаются, особенно в почвах тропиков, например на рисовых полях.
Таксономия и номенклатура этого отдела слабо разработаны, и
в настоящее время ореди специалистов здесь нет единогласия. Еде
Не решено, прибегать ли в данном случае к принципам, принятым
Для бактерий ( stanier et al. , 1978), или по традиции пользова-
28
тьоя правилами, применимыми к прочим водорослям (Drouet , 1968,
1973; Drouet , Daily , 1956). Концепция вида остается в крайне за-
путанном состоянии (lang , Walsby , 1982; Friedmann . 1982).
Порядки: Chroococcales, Pleurocapsales, Stigonematales,
Nostocales
PROCHLOROPHYCOTA
Prоchlorophyоeae
Этот отдел был недавно описан Луином (Lewin, 1976, 1977); он
включает один-единственный род Prochloron (lewin , 1981, 1984;
Chapman, Trench , 1982). Прокариотные, одноклеточные водоросли,
облигатные симбионты морских -асцвдий. К настоящему времени с до-
стоверностью описан только один вид; предполагается, что он обита-
ет совместно с различными видами асцвдий (Thinh , 1979; см. также
Stackebrandt et al. , 1982).
Prochlorophycota привлекли значительное внимание эволюциони-
стов. Будучи прокариотными по своей природе, клетки этих водорос-
лей содержат хлорофиллы а и (характерные для Chlorophyoota )•
ji-каротин, зеаксантин, криптоксантин, но не имеют фикобилипротеи-
новых пигментов Cyanophycota • В отличие от последних тилакоида
здесь собраны в стопки по два или более. Архитектура тилакоида
сходна с наблюдаемой у зеленых водорослей и высших растений (aid-
dings et al. , 1980; Сох , Dwarte, 1981), а структура стенки - С
наблюдаемой у синезеленых водорослей, причем подтверждено наличие
в ней мурамовсй кислоты ( Moriarty, 1979). Размножение посредст-
вом деления клеток пополам.
Луин ( Lewin, 1976, 1977) считает эти примитивные водоросли
связующим звеном между прокариотами и зелеными водорослями ( Ohlo-
rophycota ). Шадфо ( Chadefaud , 1978) не согласен с этим, счи-
тая, что хлорофиллы а и ъ_ могли возникать в ходе эволюции неодно-
кратно. Гиббс ( Gibbs, 1981) полагает, что напоминающие Prochio-
гоп эвдосимбиотические организмы дали начало хлоропластам Ohio- >
rophyceae , тогда как хлоропласта красных водорослей могли про-
изойти от коккоидной цианобактерии (синезеленой водоросли). Ряд
авторов (Antia > 1977; Stanier , Cohen-Baaire > 1977; Ragan,
Chapman, 1978; Trench , 1982) выступили против выделения само-
стоятельного отдела Prochlorophycota . Эволюционное значение
29
этой интереснейшей группы обсуждается более детально в гл. 8.
Достигнутый в настоящее время успех в лабораторном культиви-
ровании Trochloron (Patterson, Withers , 1982) и открытие овобод-
ноживущего прохлорофита (. Burger-Wiersma et al. , 1986), несом-
ненно, дают новый импульс исследованиям, в ходе которых будут вы-
яснены биохимия и физиология этих водорослей.
eukahiota (эукариоты)
Остальные водоросли наряду со всеми прочими организма™ от-
носятся к эукариотам. Их клетки имеют ядерную оболочку, хромосомы
и окруженные мембранами органеллы, в частности митохондрии, пла-
стиды и аппарат Гольджи.
вноооиясота : красные водоросли, или багрянки
(рис. 2.3 - 2.18)
RHODOPHYCEAE
Rhodophyceae • возможно, наиболее близкие к Cyanophycota
эукариотные водоросли, поскольку обладают простыми пластинами о
несобранными в стопки тилакоидами и фикобилипротвинами при отсут-
ствии хлоропластной эндоплазматической сети и жгутиков (Dixon
1982; Tforrls , Kurgens , 1982). Имеются также сведения о нали-
чии у них, как и у Cyanophycota , биосинтеза ненасыщенных жирных
кислот, отличного от наблюдаемого у зеленых водорослей ( Erwin,
Bloch , 1963).
У багрянок есть хлорофилл .а.'(наличие хлорофилла cl указыва-
лось только один раз; O'hEocha , 1971) наряду о фикобилипротеи-
нами (аллофикоцианин, фикоцианин, фикоэритрин) и рядом каротинои-
дов. Цвет клеток часто красный, но может быть фиолетовым, бурым,
черным или голубым. Запасной продукт - так называемый багрянко-
вый крахмал, который располагается свободно в цитоплазме,- клеточ-
ная стенка состоит из целлюлозы, а у некоторых видов - из ксила-
нов и галактанов, в частности из имеющих большое промышленное
значение агара и каррагинана. Слизь может составлять до 70$ сухо-
го веса клеточной стенки. Обызвествление последней встречается у
30
многих Cryptonemiales , некото-
рых Hemaliales , а также Б роде
Titanophora ИЗ Gigartinales
(см.Kraft > 1981). Специфиче-
ская особенность - поры (или
"поровые пробки", Puesohel .
1980), соединяющие клетки, у
всех Rhodophycota • за исклю-
чением нескольких порядков вап-
giophycidae (Brawley, Wether- ,
bee, , 1981). Относительно этой
уникальной структуры красных
водорослей остается еще много
неясного, но уже сейчас с нею
связывают межклеточный тран-
спорт веществ ( Wetherbee.
1979), упрочение структуры нит-
чатых и псевдопаренхиматозных
талломов ( Pueschel . 1980) и
взаимодействие паразит - хозяин
(Goff , 1979).
Морфологическое разнообра-
зие включает одноклеточные (ред-
ко), нитчатые и псевдопаренхи-
матозные формы; истинно парен-
химатозного развития нет, и у
большинства видов (за исключе-
нием некоторых Bangiophycidae)
наблюдается строго верху шечшй
(апикальный) рост нитей. У не-
которых представителей более
высокоорганизованных порядков
отмечен их интеркалярный рост
(например, среди Cryptonemia -
О
о
о
I
ёч
Рис. 2.3. Bangia atrqpurpurea (Rho-
dophyceae, Bangiales) . участ-
ки неразветвленного паренхиматозно-
го таллома. Растения образуют пло-
тные заросли на верхней литорали
в областях с прохладным климатом. .
Масштаб 50 мкм
les И Ceramiales )• Подавля-
ющее большинство из 4100 видов,
относящихся к 675 родам ( Kraft,
1981) багрянок, - обитатели морей, и лишь около 200 видов из 42
родов ( ott, Sommerfeld , 1982) известны из пресноводных место-
обитаний.
Половое воспроизведение оогамное и обладает рядом специфиче-
ских особенностей; обычно после оплодотворения происходит разви-
31
РИС. 2.4. Poruhyra umbilicales
(Rhodophyceae, JBangiales) Z
Таллом паренхиматозный, пласти-
нчатый, из двух слоев клеток.
Масштаб 2,0 см
Рис. 2.5. Audouinella infestans
(Rhodophyceae, Nernaliales).
Водоросль образует ползучие
Нити под поверхностью гидрои-
дов. Короткие вертикальные
ветви несут латеральные моно-
опоры. Масштаб 20 мкм (woel-
kerling, 1980)
Рис. 2.6. Erythro tri chia folii-
formis (Rhodophyceae, Bangiales).
Мелкие листовидные особи обра-
зуют дерновинки на бурой фуку-
совой водоросли Marginariella
boryana . Показана группа про-
стых Паренхиматозных пластин
с расположенными на них пятна-
ми моноспорангиальных сорусов
(М). Масштаб 500 мкм (South,
Adams, 1976)
Рис. 2.‘7. Brvthrocladia sr>. (Rhodophyceae, Bangiales).
Взрослый талломЖЖИЙТдйсковвдныи. с моностроматическим
краем и полистроматическим центром ( Nichols, Idssant,
1967)
Рис. 2.8. Beckerella scalaramosa (Rhodophyceae, Nemaliales).
[елое растение с двурядным двавды-перистым ветвлением,
’асштао 10 см ( Kraft • 1976)
33
тие in situ > приводящее к об-
разованию уникального диплоид-
ного поколения - карпоспорофи-
та прикрепленного к женскому
гаметофиту. Организмы в норме
раздельнополые, и в жизненный
цикл обычно входит последова-
тельное чередование одного
гаплоидного поколения (гаме-
тофита) и двух диплоидных -
карпоспорофита и тетраспоро-
фита. Гаметофит и тетраспоро-
фит обычно изоморфны, но из-
вестно и много примеров гете-
роморфной смены док еле ний.
Подразделение класса rho-
dcphyceae на два под-
класса вызывает постоянные
споры, особенно с тех пор,
как выяснилось, что многие
признаки, служившие для их
различения, для этого не го-
дятся. Таким образом, сущест-
вование приводимых здесь под-
классов в будущем может быть
окончательно опровергнуто
и багрянки станут считаться
единым классом.
Порядки: Bangiophycidae s
Porphyridiales, Bangiales,
Gompscpogomales, Rhodochaetales. Florideophycidae: Nemaliales,
Cryptonemiales, Gigartinales, Rhodymeniales, Palmariales, Cera-
aiales.
во. 2.9. Iiiagora tanakai (Rho-
dophyceae, Nemaliales ~).
Многоосевои таллом с собранны-
ми в центре осевыми нитями;
короткие боковые ветви ограни-
ченного роста образуют его
внешний слой. Без масштаба
(Abbott, 1967)
CHROMOPHYCOTA
Отдел Chromophycota включает все водоросли с хлорофиллами
2 и с, но без хлорофилла Ь. . Вспомогательные пигменты, преимущеот-
венно Jb-каротин и разнообразные ксантофиллы (последние специфич-
34
Рис. 2.10. Grateloupla stipitata (Rhodophyceae, Cryptonemiales).
Суживающиеся в черешковидные части листовидные
пластины полые внутри и очень слизистые; они отходят от бо-
лее жесткой и узкой многолетней стеблевидной оси. Морфоло-
гия сильно варьирует в зависимости от местообитания. Мас-
штаб 5 см (фотография: R.C.Ficken)
ны для каждого класса), придают хлоропластам буроватую или желтова-
тую окраску. Запасные продукты - углеводы (глюканы, крахмал) или
ЛИПИДЫ. У многих (за исключением Bacillarlophyceae ) встречаются
подвижные клетки, в норме - с двумя жгутиками, обычно неравными
по длине (за исключением Prymnesiophyceae , где они во многих
случаях равные) и гетеродинамичными: один чаще всего перистый (с
двумя рядами волосков^), другой - гладкий. Во всех клетках вокруг
В отечественной литературе волоски жгутиков чаще называют масти-
гонемами. - Прим, перев.
35
Рис. 2^11. Dumontia incrassata
(Rhodophyceae,Crypt onemialea ) . ’
Полые слизистые вет-
ки округлые в сечении, несколь-
ко уплощенные или вздутые. Мас-
штаб 2 см (фотография: R.C.Mc-
ken)
Рис. 2.12. btychodea marginife-
ra(Rhodophvceae. dlgartina-
Tea). Листовидный таллом
с лопатчатыми -выростами, от-
ходящими от края лопастей. Ма-
сштаб 5 см ( Kraft , 1978)
Рис. 2.13. Eucheuma gelatinae (Rhodophyceae, Gigartinales).
Целое растение (Tseng, 1981)
36
Рис. 2.14. Gymnogongrua sp.(Rhodophyceae, Gigartinales).
Русто расположенные, более или менее округлые в
сечении оси с дихотомическим ветвлением. Масштаб 5 см
(фотография: R.c.Ficken)
Риб. 2.15. Palmaria palmate
(Rhodophycеае, Palmarialea).
Целое растение с ли-
стовидной структурой и ложно-
дихотомическим ветвлением.
Масштаб 5 см (фотоградйя:
R.C.Ficken)
37
Рис2.16. Rhodomela о011-
fervoldes (Khodophyoeae,
Ceramiales).
Целое растение с поли-
сифоналнной структурой
и осями, покрытыми мощ-
ной корой. Масштаб 5 см
(фотография: R. C.Ficken)
Рир. 2.17. Cladhymenia
U^°^hyCeae ’
Уплощенные оси с много-
численными пролифериру-
ющими ветвями, несущими
Репродуктивные структу-
ры. Масштаб 5 см {Фото-
графия: R.C.Ficken)
38
Рис. 2..ТЯ~Д1усо1гу8 rubens (Rhodophyceae, Ceramiales).
Одноосевой таллом с листовидной структурой и выраженным
перистым жилкованием, напоминающий лист высшего растения.
Масштаб 2 см (фотография: R.c.Ficken)
хлоропласта находится цистерна (хлоропластная эндоплазматическая
сеть), а сами хлоропласты построены достаточно однообразно; ти-
лакоида в них собраны в стопки по три. Состав-клеточной стенки
различен в разных классах; в него могут входить кремнезем, пек-
тин или целлюлоза. Морфология крайне разнообразна: от одноклеточ-
ных жгутиковых до очень крупных паренхиматозных талломов, напри-
мер, у представителей порядка leminariales (Phaeophyceae) . Chro-
mophycota широко распространены как в морских, так и в пресно-
водных местообитаниях.
В настоящей книге отдел Chromophyccta включает девять клас-
сов и охватывает крайне неоднородную группу организмов. В литера-
туре некоторые из выделяемых здесь классов группируются в особые
отделы или даже сами считаются отделами (например, Xanthophyceae,
Bacillariophyceae, Dinophyceae И Phaeophyceae) . Так, В ОДНОМ
из учебников ( Bold, Wynne , 1978) рассматриваются четыре отде-
ла: Chrysophycophyta, Pyrrhophycophyta, Phaeophycophyta И '!
Cryptophycophyta , а Б другом ( Доек, Jahns , 1978) отдел Hete-
rocontophyta включает классы Chrysophyceae, Xianthophyceae, j
Bacillariophyceae, Phaeophyceae И Chloromonadophyceae , тогда
как классы Prymnesiophyceae (как Haptophyta), Bustigmatophyce-
ae, Cryptophyceae и Dinophyceaeсами получают статус отделов. ?
В принимаемой нами системе (согласно Kristiansen , 1982) опреде-
ляющим признаком отдела считается пигментация, тогда как в дру-
39
тих случаях ( Ноек, Jahns, 1978) за основу взята структура
ягутика. Учитывая неоднородность Chromophycota , вполне возможно,
что и в дальнейшем сохранится расхождение во взглядах на способы
группировки рассматриваемых ниже классов. Мы приводим основные
признаки каждого из них.
Chrysophyceae . золотистые водоросли
(рис. 2.19 - 2.24)
Детальное определение Chrysophyceae дает Хибберд ( Hibberd,
1976). Характерная группа здесь - подкласс Chrysophycidae •
подкласс Dictyochophyoidae объединен с ним только на основании
сходства пигментации и запасных продуктов ( Kristiansen, 1982).
Таксономия класса на порядковом, родовом И видовом уровнях ( Kris-
tiansen , Takahashi, 1982) сложна. Клетки голые, иногда покрыты
особой оболочкой или панцирем; в норме несут два неравных жгути-
ка, но гладкий жгутик иногда редуцирован или сохраняется внутри
клетки только как базальное тельце. Перистый жгутик покрыт трех-
частными волосками. Обычно имеется ассоциированный с хлоропластом
глазок. Клетки одноядерные и содержат от одного до нескольких
хлоропластов. Преобладающий вспомогательный пигмент - фукоксан-
тин, придающий клеткам золотисто-бурую окраску.Запасной продукт
- хризоламинарин. Воспроизведение преимущественно бесполое, хотя
для вое большего числа видов становятся известны половые стадии
( Kristiansen, Takahashi , 1982). Морфологическое разнообразие
включает одноклеточные жгутиковые (которые преобладают), коккоид-
ные, ризоподиальные, нитчатые и паренхиматозные (редко) формы.
Большинство Chrysophyceae- представители пресноводного планкто-
на, особенно в прохладных местообитаниях.
Порядки: Chrysophycidae: Ochromonadales, Chrysosphaerales,
Phaeothamniales, Sarcinochrysidales, Chrysamoebidales, Chryso-
oapsales. Dyotyochophycidae: Dictyocharles»
Prymnesiophyceae (Haptophyceae) : примнезиофиты
(рис. 2.25 - 2.29)
Prymmesiophyceae Hibberd (1976, 1980a = Haptophyceae ;
Christensen ’ представляют собой естественную группу
40
Рио. 2.19. Stiohogloea ар.(Chrysophyceae) . В жгутиковой
клетке (А, В) находится один хлоропласт со стигмой; в
клетке со стенкой (Б) заметны периферические вакуоли,
каждая из которых содержит градулы ( Norris, 1977)
Рис. 2.20. Ochromonas aphaerocvstia
< Chryaophyceaej . Удлиненная клетка
с гетеродинамичными жгутиками, хлоро-
пластом, ядром и периферическими дис-
коболоцистами. Масштаб 10 ом ( Ander-
sen, 1982)
41
Рио. 2.21. Chrysamoeba sp.
(Chrysophyceae). ме-
тка характерной формы с
четковидными ризоподиями
и сократительными ваку-
олями. Масштаб 10 мкм
( Bourelly , 1981)
Рио. 2.22. Sphaerldiothrix
Sompressa (Chrysophycеае ).
ветвистый,
сетчатый таллом (из
культуры), клетки кото-
рого содержат постенный
хлоропласт и несколько
капель запасного продук-
та, но лишены пиреноидов
Ui~J900) ( Andrews ,
<0)
42
Рис. 2.23. Phaeothaanion
ар, (Chrysophyceae).
Ветви с утолщенными попе-
речными перегородками. Мас-
штаб 10 мкм (из Geltier,
Schiman-Czeika , 1970,
с изменениями)
Рис. 2.24. - Chrvsococcus fPr-
catua (Chrysophycea^.Bere татив-
йая клетка с шипами, хлоропла-
стом (X), жгутиком (Ж), ядром
(Я) и капельками запасного
ламинариноподобного вещества
(Mchola , 1981)
(Boney,Green • 1982). Это преимущественно одноклеточные организмы
с двумя равными гладкими жгутиками (за исключением порядка Рат-
lovalea > где жгутики неравные, причем один несет чешуйки - см.
Green , I960). У многих видов имеется еще гаптонема - третий,
напоминающий жгутик придаток. Клетки одноядерные, с одним или
двумя хлоропластами, каждый из которых с одним-единственным пи-
реноидом; пигменты и запасные продукты те же, что и у Chrysophy-
ceae . Глазки имеются у Pavlovales - внутри оболочки хлоропла-
ста. Клетки обычно в один - несколько слоев покрыты органически-
ми (целлюлозными) чешуйками, образующимися внутри цистерн аппара-
та Голвджи, с которым может быть связана сократительная вакуоль.
Встречается половое размножение, а жизненный цикл обычно включа-
ет чередующиеся подвижную и неподвижвдю фазы. УCoccosphaerales
гаплоидная бентосная нитчатая фаза может чередоваться с диплоид-
ной одноклеточной подвижной.
Представители этого класса - в основном небольших размеров;
ныне живущие формы представлены примерно 75 родами с 300 вадами
43
Рис. 2.25. Chrvaochromulina
bireeri (Prynmeaiophyceae).
ФазевсГ-контрастная микрофото-
графия клетки, фиксированной в
растворе Люголя ( X 1000) (На1-
Ifors, Thomsen, 1979)
Рис. 2.26. Michaelaarsia elegans
(Prymnesionhyceae) . Све-
товая микрофотография клетки.
Обратите внимание на мутовку ди-
стальных лучей. Масштаб 10 мкм
(Heimdal, Gaarder, 1981)
Рис. 2.28. Неидентифицирован-
ный представитель кокколито-
Форид ( Prymnesiophyceae).
Микрофотография в сканирующем
электронном микроскопе целой
коккосферы. Масштаб I мкм
(Heimdal, Gaarder, 1981)
рИС. 2.27. Emiliania huxleyj
AHrymnesionhyceae) . Микро-
фотография в сканирующем элек-
тронном микроскопе целой кок-
косферы, показывающая детали
кокколитов. Масштаб I мкм
(Heimdal, Gaarder, 1981)
44
Norris, 1982), большинство
из которых - морские нанопланк-
тонные.. Установлено, что прим-
незиофиты могут составлять
около 45% общего фитопланктона
в средних широтах южной части
Атлантического океана. Их обыз-
вествленные чешуйки (кокколи-
ты) легко переходят в ископа-
емое состояние и могут быть
обильно представлены в захоро-
ненных илах. Очевидно, примне-
зиофиты ранее были многочислен-
нее, чем ныне, особенно в мезо-
зое (юрский и меловой периоды,
205 - 125 млн. лет.назад).
Порядки: Igochrysidales,
Cocoosphaerales, Prymnesiales,
Pavlovales.
Xanthophyceaes желтозеленые
водоросли
(рис. 2.30 - 2.32)
Рис. 2.29.Corysr>haera ягасШв
( Prymne s i ophyceae).
Микрофотография в сканирующем
электронном микроскопе целой
коккосферы. Масштаб I мкм
(Heimdal, Gaarder, 1980)
Эти организмы отличаются от других "зеленых" водорослей от-
сутствием хлорофилла ь , а от других Chromophycota-отсутствием
фукоксантина и преобладанием в качестве основного каротиноида ди-
атоксантина (Silva, 19795 Hibberd, 1980а, 1981, 1982as Ott 1982).
Подвижные клетки голые и несут два неравных жгутика, более круп-
ный из которых, перистый (о трубчатыми волосками), направлен впе-
ред по ходу движения, а более короткий, гладкий, - назад. Сложная
зооспора в роде Vaucheria покрыта многочисленными парами лишь
слегка неравных жгутиков, у каждого из которых волоски отсутству-
ют. Клеточные стенки коккоидных форм тонко скульптурированы; у
нитчатых и более крупных коккоидных видов отенка двустворчатая,
причем одна створка частично охватывает другую. Основное вещество
клеточной стенки у сифональных видов (например, у Vaucheria) -
целлюлоза; прочие компоненты - глюкоза и уроновые кислоты. Хлоро-
пласты обычно дисковидные, окружены двойной мембраной хлоропласт-
ной эндоплазматической сети и содержат один пиреноид. Глазок, ес-
45
Pur - RO - Vaucheria sp. (Xantho-
phyceae) “Плотные скопле-
ния на илиотой литорали. Нити
этой водоросли часто закрепля-
ют песок и ил б защищенных ча-
стях эстуариев. Примерным мас-
штаб 5 см
Рис. 2.RT.Vaucheria aHela (Xanthophyceae) . Таллом цено-
Цитный; к репродуктивным структурам относятся терминальные
антеридии на мужских ветвях, или андрофорах, и оогонии со
Зрелыми ооопорами (Ott, Honmersand., 1974)
46
ли имеется, расположен на
брюшной стороне внутри хлоро-
пласта; ему соответствуют уг-
лубление в клетке и базальное
вздутие короткого заднего жгу-
тика. Запасные продукты - глю-
кан С J3 - 1,3-связями (ср.
парамил® Euglenophycota),
жиры и маола; крахмал никогда
не встречается.
Бесполое размножение - за
счет деления клетки, фрагмен-
тации (у нитчатых форм), обра-
зования подвижных зооспор или
неподвижных апланоспор, акинет
и цист (статоцист - cp.Chryso-
phyceae). Половое размно-
жение известно оогамное в по-
рядке Vaucheriales(Botrvdium
И Vaucheria ) И изогамное у
Tribonematales (Tribonema).
Подразделение на порядки
основано на морфологии, причем
наблюдается эволюционно-морфо-
логическая последовательность,
сходная о представленной у зе-
леных водорослей. Организация
усложняется от одноклеточной
жгутиковой до коккоидной (боль-
шинство) , нитчатой (примерно
20%) и сифональной (примерно 10%,
Рис. 2.32. Onhiocvti-cun arbus-
culalXanthophvceae). Таллом
состоит из материнской" клет-
ки с дополнительными клетками,
отходящими от ее апекса. Клет-
ки многоядерные, содержат не-
сколько постенных хлоропластов.
Стрелкой показана апикальная
крышечка "материнской" клетки,
высвободившей споры. Масштаб
10 МКМ (Bourrelly, 1981),
преимущественно виды Vauche -
ria). Описано приблизительно 600 видов ( Hibberd , 1982 ),
большинство из которых пресноводные. Только 20 вадов - обитатели
морей и солоноватых вод (все - из рода Vauoheria ).
Порядки: Chloramoebales, Rhizoohloridales, Heterogloeales,
Mischococcaless Tribonematales, Vaucheriales.
Bustigmatоphyceae
Eustigmatophyceae - небольшой Класс, представленный толь-
47
л о 12 вадами, которые отличаются от желтозеленых водорослей уни-
кальной структурой подвижных клеток и преобладанием виолаксантина,
а не диатоксантина (Hibberd , 1982Ь , с). Зооспоры не имеют ана-
логов в растительном царстве: они несут единственный направленный .
вперед перистый жгутик; второй представлен лишь базальным телом
(однако у Ellinsoidion acuminatum И Eseudocharacionaia texensis
задний гладкий жтутик выступает наружу; см. Lee , Bold , 1973).
Крупный оранжево-красный глазок находится спереди, полностью не-
зависим от хлоропласта и связан с Т-образным вздутием у основания
перистого жгутика, которое тесно прижато к плазмалемме в области
глазка. Бесполое размножение с образованием автоспор или (редко)
зооспор; половое неизвестно. Все представители - голые одноклето-
чные формы, иногда прикрепленные; встречаются только в пресной
воде или в почве.
Baoillariophyceae : диатомовые водоросли, диатомеи
(рис. 2.33 - 2.36)
Эти микроскопические водоросли одноклеточные, иногда колони-
альные или образующие ложные нити ( Werner , 1977; Roas, 1982;
Amspoker, Czarnecki , 1982). Основные вспомогательные пигменты -
фукоксантин, Jd-каротин, диадиноксантин и диатоксантин, а основ-
ные запасные вещества - липиды и хризоламинарин; в этом отношении
диатомеи ближе всего к золотистым водорослям. Клетки одноядерные,
содержат один или несколько хлоропластов. Кремнеземный внешний
скелет (панцирь) состоит из двух половинок, каждая из которых в
свою очередь включает более или менее уплощенную пластинку (створ-
ку) и соединительный ободок, прикрепляющийся к ее краю. Два обод-
ка, накладываясь друг на друга, образуют так называемый поясок.
У некоторых пеннатных форм по створке проходит шов в ваде трещи-
ны; им обусловлено уникальное скользящее движение клеток. Беспо-
лое размножение, которое здесь преобладает, обычно проиохсдит за
счет деления клетки в плоскости, параллельной створке, о достраи-
ванием недостающей половинки панциря внутри доставшейся от роди-
тельской клетки; этот процесс в ряду поколений приводит ко все
большему измельчанию некоторого процента особей в популяции. По- >
левое размножение изо- или анизогамное посредством лишенных жгу-
тиков гамет или же оогамное (преимущественно у Centrales ) о од-
ножгутиковой мужской гаметой, уникальной в том плане, что у ее
48
Рио. 2.34. Cocconels pseudomar-
ginata (Bacillariophyceae).
Фотография в сканирующем эле-
ктронном микроскопе, показы-
вающая внутреннюю сторону
створки с. бахромчатым загибом
И швом. Масштаб I МКМ ( Holmes
et al., 1982)
Рис. 2.33. Thalassioaira oest-
Л1 (Bacillariophyceae).
_ рупная клетка co створкой,
на которой видны эксцентриче-
ские ареолы и два небольших
отверстия вблизи центра (х
1920). Фотография в скани-
рующем электронном микроскопе
поясковой зоны со зрелой эпи-
текой (внизу), состоящей из
створки, орнаментированного
загиба, вставочного ободка с
крупным кольцевым рядом пор
и трех соединительных обод-
ков. У незрелой гипотеки
(вверху) можно различить
створку, загиб и вставочный
ободок (х 2330) ( Fryxell,
Haalet 1980)
Рис. 2.35. Thalassiosira confu-
sa (Bacillariophyceae).
Фотография в сканирующем элек-
тронном микроскопе, показываю-
щая внешний вид теки. Видны
сквозные поры ( foramina ).
Масштаб 5 мкм ( Fryxell, Has-
le, 1979)
49
Рио. 2-3fi.Melqeira nummuloides (Baoillariophyceae).
Живая нить, образующая ауксоспоры (х 432)(Crawford, 1975)
жгутика нет центральных микротрубочек. После полового размножения
зигота (так называемая ауксоспора) дорастает до максимального для
данного вида размера клетки. У большинства бентосных и неритиче-
ских форм для переживания неблагоприятных условий формируются по-
коящиеся споры с сильно окремнелыми отенками, отличающиеся по мор-
фологии от обычных вегетативных клеток.
Диатомеи обильны в морских и пресноводных местообитаниях, в
бентосе или как эпифиты на более крупных водорослях или высших
растениях. Они легко сохраняются в ископаемом состоянии, и мощные
отложения их остатков (так называемый диатомит) известны с мелово-
го периода. Диатомит находит различное хозяйственное применение;
кроме того, его стратиграфический анализ помогает реконструиро-
вать прошлое водоемов (Round , 1957).
Два порядка диатомей выделяются по форме панциря: yBtddulphi-
alea его симметрия радиальная, а у Baoillar iales- билатераль-
ная. Электронная микроскопия позволяет существенно уточнить си-
стематику этого класса.
Порядки: Biddulphialea (aCentrales), Bacillariales
(=Pennales).
Dinophyceas : динофлагеллаты, динофиты
(РИС. 2.37 - 2.40)
В этом классе наиболее крупный по числу видов подкласс,
Dinophycidae , объединяет в основном планктонных фотосинтезирую-
щих одноклеточных весьма своеобразного строения (Dodge , 1979;
Podge, Steidinger , 7982; Loeblioh , 1982). Их подвижные клет-
ки несут на брюшной стороне два жгутика. Один располагается попе-
речно, обычно в бороздке, опоясывая клетку; он отличается наличи-
ем внешней аксонемы и внутреннего витого тяжа. Другой жгутик,
Рис. 2.37. Ceratium яр. (Dino-
phiceae) . Обычный предста-
витель морского фитопланктона.
Мощно вооруженная тека несет
три "рога , в свою очередь
орнаментированные шипами. Хо-
рошо заметен поясок. Масштаб
□О мкм
Рис. 2..38. Stylodinium Ij-ttora-
le (Ипорйусеае).При-
крепленная неподвижная клетка
( Horiguchi, Ch.ih.ara, 1983).
Рис. 2.39. Peridinium охпсШа
(Dinophyceae). фотография
в сканирующем электронном ми-
кроскопе вегетативной клетки
(Pfiester, Skvarla, 1980)
51
Рис 2.40. Pvrodiniuni bahamense (Dlnophyceae). фотогра-
фия'в сканирующем электронном микроскопе целой клетки со
стороны эпитеки. Видны пластинки и апикальная пора. Мас-
штаб 10 МКМ ( Dodge, Hermes, 1981).
гладкий, направлен назад. Клетки могут быть автотрофными или бес-
цветными и гетеротрофными; последние питаются сапро- или фаго-
трофно или ведут паразитический образ жизни. У фотосинтезирующих
видов хлоропласты окружены одной мембраной эндоплазматической се-
ти, которая не переходит во внешнюю мембрану ядерной оболочки.
Основные каротиноиды - перидинин и неоперидинин. Запасные продук-
ты - масло и крахмал, сходный с имеющимся у высших растений.
Клеточный покров - многослойный панцирь (амфиесма), состоящий из
верхней (эпикон) и нижней (гипокон) частей, каждая-из которых в
свою очередь разделена на ряд пластинок; перпендикулярно опоясы-
вающей бороздке располагается продольная. Клетки могут содержать
пиреноиды и глазки (менее чем у 5% видов). У ряда видов имеются
отстреливаемые образования - трихоцисты и книдоцисты, вероятно
имеющие защитную функцию. Ядро уникально по своей природе и назы-
вается динокарионом, или мезокарионом. Хромосомы в нем постоянно
конденсированы и прикреплены к мембране, ядерная оболочка сохра-
няется во время митоза. Бесполое размножение - посредством деле-
ния; вегетативные клетки обычно гаплоидные, единственная диплоид-
ная стадия - зигота. Часто встречаются покоящиеся споры, цисты
или гипноцисты; в ископаемом состоянии они известны как гистри-
хосфериды и относятся наряду с другими цистоподобными ископаемы-
ми к группе акритарх. Динофлагеллаты, веротно, доминировали ореди
52
продуцентов в палеозое ( Tasch, 1973), а древнейшие их остатки
известны из силура (435 - 460 млн. лет назад). Токсичные цветения
воды, вызываемые динофлагеллатами, - широко известное и присталь-
но изучаемое явление (Taylor, Seliger, 1979; Yentsch, Tay-
lor, 1982).
Из четырех подклассов наиболее крупный - Dinophycidae ; под-
класс Ebriophycidae представлен только двумя ныне живущими и
многочисленными’ ископаемыми видами. Ellobiophycidae - нефотосин-
тезирующие паразиты членистоногих (мизид) и полихет, a Syndinio-
phycidae - паразиты различных морских животных ( Loeblich,
1982).
Порядки: Dinophycidae : Blastodiniales, Brachydiniales,
Chytriodiniales, Desmocapsales, Dinemoebales, Dinocloniales,
Dinophysiales, Gloeodiniales, GymnodinialeS, Uoctiluoales,
Peridiniales, Phytodiniales, Prorocentrales, Pyrocystales, Zoo-
xanthellales. Ebriophyoidae: Ebriales. Ellobiophycidaei Thalas-
somycetales. Syndiniophycidae: Syndiniales. ,s
Phaeophyceae : бурые водоросли
(рис. 2.41 - 2.61) ’
Этот класс, насчитывающий около 265 родов и более 1500 ви-
дов, представлен почти исключительно бентосными морскими формам^;
( Wynne, 1981, 1982а,Ь ). Основной каротиноид - фукоксантин; в ;j
сочетании с различными таннинами (запасаемыми в физодах, или Й
фукозановых пузырьках) он придает этим растениям характерную бу-А
рую окраску. Обычно имеются две мембраны хлоропластной эндоплаз-*’
матической сети, из которых внешняя может переходить в наружную
мембрану ядерной оболочки. Клетки обычно одноядерные и содержат
от одного до многих хлоропластов; запасные продукты - ламинарии
и маннитол. Клеточная стенка двуслойная; внутренний слой из цел-
люлозы, а внешний - из альгиновой кислоты и фукоидина. Альгино-
вую кислоту, имеющую большое практическое значение, в изобилии
содержат представители порядков laminariales и Durvillaeales.
Карбонат кальция откладывается в стенках видов Padina (Dictyo-
tales ) б форме кристаллов арагонита. Одноклеточные виды неиз-
53
вестны. Зооспоры и гаметы в
норме несут два неравных жгу-
тика, прикрепляющихся сбоку
клетки; из них более длинный
- периотый (покрытый трехчаст-
ными волосками), а более ко-
роткий задний - гладкий (спе-
рмин Dictyotales , leminari-
ales, Desmarestiales И
некоторых представителей Fuca-
les отклоняются от этого ти-
па). Обычно имеются глазки,
сходные по расположению и
структуре с глазками золоти-
стых и желтозеленых водорослей.
Морфологическое разнооб-
разие охватывает от микроско-
пических разветвленных нитча-
тых форм до макроскопических
паренхиматозных, достигающих
наивысшего уровня морфологиче-
ской дифференцировки среди во-
дорослей (например, гигантские
морские представители порядка
laminariales ) • Бесполое раз-
множение - в основном посред-
ством подвижных зооспор, хотя
у некоторых видов в нем участ-
вуют и вегетативные структуры.
Половое размножение - от изо-
гамного до анизогамного и оога-
Рис. 2.41. Ectocarnus sp. (Pha-
eophyceae .Ectocarpales).А. Часть
таллома из одного ряда клеток
с многогнездными спорангиями;
масштаб 100 мкм. Б. внешний вид
растения с одногнездными спо-
рангиями; масштаб 1000 мкм (по
Cardinal ’ 1964)
много . Мейоз происходит в одногнездных (одноклеточных) споран-
гиях, где образуется от четырех до многих спор, обычно подвиж-
ных. Многоклеточные (многогнездные) структуры посредством мито-
тического давления дают начало бесполым спорам или гаметам. Ти-
пичный жизненный цикл включает чередование гаплоидной и диплоид-
ной фаз, которые могут быть изоморфными или гетероморфными. Осо-
бенности жизненного цикла учитываются при классификации бурых
Водорослей ( Wynne, Loiseaux, 1976; Pedersen, 1981). Для
многих видов он остается неизвестным; у некоторых представителей
54
Рис. 2.42. Elachista fuclcola (Phaeophyceae, Chordariales).
А. Разрез части взрослого растения; видны асси-
миляторы Та)> одногнездные спорангии (ос), парафизы (п),
вздутые клетки сердцевины (с) и ткань форофита Fucus
(х). Б. Вид расплющенного таллома, выращенного в культу-
ре; базальная зона с короткими ветвями, хорошо заметными
одногнездными спорангиями и ассимиляторами. Масштаб 100
МКМ ( Koeman, Cortel-Breeman, 1976).
имеет место его "прямой" тип без мейоза или сингамии, но иногда
с чередованием морфологических фаз.
Классификация бурых водорослей постоянно пересматривается;
на уровне порядков принимается во.внимание комбинация особенно-
стей жизненного цикла, морфологии и организации. Некоторые поряд-
ки четко разделить не удается, в частности Ectocarpales, Dictyo-
siphonales > Chordariales И Scytosiphonales (см. обзор Wynne,
1981). Педероен ( Pedersen, 1984) предлагает объединить поряд-
ки Dictyosiphonales и Tilopteri&les широком смысле), считая,
что двухфазный гетероморфный жизненный цикл, учитываемый при вы-
делении первого из них, не имеет диагностического значения. Хенри
( Henry, 1984) описал новый, состоящий всего из одного рода поря-
док Syringodermatales с гетероморфным жизненным циклом, включаю-
55
Рис. 2.43. Scytosinhon lomen-»
taria (Phaeophyceae, Soytosi-
рКопа1£з).Трубчатые неветвящие-
ся талломы псевдопаренхима-
тозного строения. Полый центр
местами сужается. Масштаб
5 см (фотография: R.С.Ficken)
Рис. 2.44. Sporochnus sp.(Pha-
eophyceae, SporoohnaJ.es).
Талломы с отчетливой осью
сильно ветвятся. Репродук-
тивные структуры располага-
ются пучками на верхушках
ветвей. Масштаб 5 см (фото-
графия: н.С.Ficken)
56
щим крайне редуцированную дву-
клеточную стадию гаметофита,
и вегетативной структурой,
отличной от так обой у Dictyota-
les И Sphacelariales.
Порядки: Ectocarpales,
Chordariales, Sporochnales,
Desmarestiales, Dictyosipho-
nales, Scytosiphonales, Tilo-
pteridales, Cutleriales,
Sphacelariales, Dictyotales,
Syringodermatales, la-mi nari -
ales, Fucales, Durvillaeales,
Asooseirales.
Raphidophyceae : рафидофиты
Это очень небольшой класс пре-
сноводных жгутиковых, иногда
включаемый в желтозеленые во-
доросли, с которыми здесь сход-
ны пигменты и жгутикование.
"Рис. 2.45, Punctaria plantagi-
nea (Phaeophyceae, Scytosipiho-
nales). Паренхиматозные
талломы листовидные, двуслой-
ные. Масштаб 2 см (фотография:
см (фотография:
Используемое иногда название
"хлоромонады"
R.C.Ficken)
Рис. 2.46. Dictyota sp.
(Phaeophyceae, Dictyota-
les) • Талломы парен-
химатозные с апикальным
ростом. Ветви уплощен-
ные, дихотомически вет-
вящиеся, оканчивающиеся
одиночными апикальными
клетками. Масштаб 5 см
(фотография: R.C.Ficken)
57
Рис. 2.47. Spatoglossum sp«( Phaeophyceae, Diotyotales).
Талломы лиотбЬаднае, йаренхиматозные с апикальным ростом.
Масштаб 5 см (фотография: R.C.Picken)
(или Chioromonadophyceae ) неприемлемо, так как основано на ро-
де Chioromonas , который не относится к рафидофитам. Имеются
значительные противоречия относительно того, какие водоросли от-
носятся к этоцу классу ( Heywood , 1982), хотя большинство ис-
следователей включает в него роды Chattonella. Gqnyostomum. Ме-
frotricha И Vacuolaria . Анализ пигментов (Fiksdahl et al.,
1984) показал, что у Gonyostomum semen И Vacuolaria virescens
их состав сходен с присутствующим у желтозеленых водорослей. Ос-
новные каротиноиды здесь диадиноксантин (50 - 60% всех каротинои-
дов) , диноксантин (8 - 17%), jb-каротин (7%) и гетероксантин
58
Рис. 2.48. Tilopteris mertensii
(Phaeоphyceae, Tilopteridales?.
Часть растения с глав-
ной осью и супротивным трихо-
таллическим двурядным ветвле-
нием. Масштаб 500 мкм ( South.
1975)
Рис. 2.49. Syringoderma phinneyi
(Phaeоphyсеае, Syringodermatales).
Целое растением
вееровидный таллом с каймой
апикальных клеток ( Henry, Muller,
1983)
Рис. 2.50. Halopteris sp.(Pha-
eophyceae, Sphacelariales).
Целое растение. Рост апикаль-
ный, основная ось покрыта
мощной корой. Масштаб 2 см
(фотография: R.C.Picken)
59
Рис. 2.51. Desmarestia firms.
(Phaeophyceae, Desmarestiales).
Внешний вид растения с боковы-
ми ответвлениями, на которые
приходится основной рост после
повреждения главной оси. Рост
трихоталлический, листовые оси
и ветви со средней жидкой. Мао-
штаб '5 см (фотография:
R. C.Ficken)
РйС. 2.,52. Chorda tomentosa
(Hiaeophyceae, leminariales).
Простой неветвящийся таллом
паренхиматозного строения по-
крыт волосками-ассимиляторами
и прикрепляется к субстрату
небольшой дисковидной ризои-
дальной структурой. Масштаб
5 см (фотография:
R. С. Sicken)
60
Рис. 2.53. laminaria oonroleuca (.Phaeophyceae, Laminariales).
Таллом состоит из основания с прикрепительной структурой, округ-
лой в сечении ножки, оканчивающейся меристемой (или переходной
зоной), и рассеченной на лопасти пластины
Рис. 2.54. Poatelaia palmaeformis (Phaeophyceae, laminariales).
Короткая твердая ножка несет многочисленные терминальные пласти-
ны со средней жилкой и зубцами по кромке. Растения встречаются
только в сильно открытых прибою местообитаниях на тихоокеанском
побережье Северной Америки
61
Рис. 2.55. Ecklonia sp.(Pha-
eophyceae, taminarlales).
Таллом уплощен в зоне средней
жилки и несет многочисленные
пролиферирующие боковые ответ-
вления
Рис. 2.56. Durvillaea.ant arc tic a (Phaeophyceae, Durvil-
laeales)J3 отличие от внешне сходных представителей лами-
нариевых Durviiiaea имеет пластины с апикальным ростом
Они могут быть глубоко рассечены на "языки"; их ткани за-
полнены воздухом, что придает растению плавучесть
Рис. 2.57.Aacoseira mirabIl-
lis (Phaeophyceae, Asoosei-
rales). Ратения внешне напо-
минают Durvillaea, но харак-
тер роста и размножения иной.
Масштаб 5 см (фотография:
Е. С, И.скеи)
Рис. 2.58 Fucus serratus
(Phaeophyceae, Fucalesj. Тал-
лом многолетний, паренхиматоз-
ный, дихотомически ветвящий-
ся. Оси со средней жилкой и
боковыми уплощенными "крыль-
ями" с пильчатым краем
(7%). В отличие от этого Chattonella .japonlca И Flbrocapsa ja-
ponica в качестве основных каротиноидов содержат фукоксантин
(40 - 68% общего состава), фукоксантиниол (30 - 42%), ^-каротин
(15 - 27%) и виолаксантин (I - 2%), что сближает их с золотистыми
водорослями. Фиксдаль и др.(И.кв1аЫ et al., 1984) делают на ос-
новании пигментного состава вывод, что рафидофиты родственны жел-
тозеленым и, возможно, золотистым водорослям. Более длинный, на-
правленный вперед жгутик - с микротубулярными волосками, а более
короткий, направленный назад, - гладкий. У рафидофитов отсутству-
ет клеточная стенка, а в качестве основного запасного продукта
указывается масло. Глазка также нет. К специализированным чертам
относится наличие слизистых телец (мукоцист), сходных с таковыми
у примнезиофитов и золотистых водорослей;у представителей рода
Gonyostomum имеются трихоцисты, гомологичные обнаруженным у
динофитов (Mignot, 1967). Половое размножение неизвестно.
Cryptophyceaes криптомонады, гриптофиты
Подавляющее большинство видов этого класса - типичные жгути- |
ковые морских и пресноводных местообитаний (Oakley, Santore, <
1982). У фотосинтезирующих форм основные каротиноиды - pi-каро- |
63
Рис. 2.59.HtoanthaJLift
elongata (Phaeophyceae,
Fucales) . Вегетативная
часть растения представ-
ляет собой мелкую "пуго-
вицу"» прикрепленную к
субстрату короткой нож-
кой. Видны языковидные
репродуктивные рецепта-
кулы, отходящие от "пу-
говицы"
Рис. 2.60.Cysi;ophora torulosa
(Phaeophyceae, Fucales;. Си-
льно ветвящийся таллом с воз-
душными пузырями, способствую-
щими плавучести (фотография:
R. С. Ficken.)
Рис. 2.61. Hormosira banks!!
(Ehaеophyсеае, Fucales) .
Этот южнополушарный вид зани-
мает местообитания, сходные с
заселяемыми видами Fucus в
Северном полушарии, чередова-
ние вздутых и суженных участ-
ков придает таллому четковид-
ную форму
64
тин и диатоксантин. Присутствуют также фикобилипротеины (фикоциа-
нин и фикоэритрин), которые отличаются ст имеющихся у синезеленых
и красных водорослей ( Stainer , 1974); они локализованы во внут-
ритилакоидных пространствах, а не в фикобилисомах, как в двух
этих отделах ( Gantt et al., 1971). Нефотосинтезирующие, бесцвет-
ные представители содержат лейкопласты, возможно являющиеся видо-
измененными фотосинтезирующими структурами предковых форм. Запас-
ной продукт крахмалоподобный; он находится между оболочкой хлоро-
пласта и хлоропластной эндоплазматической сетью. Может встречать-
ся глазок.
Клетки криптомонад овальной формы, дорсовентрально уплощены.
Предполагалось, что их сложная ультраструктура близка к предковой
для Chromophycota . Уникальным образованием является нуклеомор-
фа, находящаяся в перилластидном пространстве и окруженная двой-
ной мембраной; есть сведения, что она содержит дополнительный ге-
ном ( Giliot , Gibbs , 1980). Два расположенных апикально или
латерально несколько неравных жгутиков в норме отходят от дна уг-
лубления, или глотки. Они сужаются к вершине и несут микротубу-
лярные волоски. Снаружи клетка покрыта перипластом со спирально
расположенными белковыми пластинками (Hibberd et al. , 1971).
Имеются эжектосомы (трихоцисты) иной, чем у динофитов, структуры
и, возможно, наиболее близкие к R-тельцам каппа-частиц у инфу-
зорий ( Hovasse et al., 1967). Размножение - деление пополам; по-
ловое размножение неизвестно.
Порядок: Cryptomonadales.
EUGLENOPHYCOTA
Euglenophyceae : эвгленовые
(рис. 2.62 и 2.63)
К этому классу относится около 1000 видов из 40 - 50 родов
И 6 порядков ( Leedale , 1982а; Leedale et al. , 1982), но наи-
большее количество информации о нем получено при изучении предста-
вителей рода Euglena > особенно - в.gracilis (Buetow , 1968,
1982). Для эвгленовых характерно, как и для зеленых водорослей,
наличие хлорофиллов а. ик • Хлоропласты содержат также J5-каро-
тин, астаксантин, антераксантин, диадиноксантин и неоксантин,
65
Рис. 2.-62. Euglena gracilis (Eugleno-,
phyceae) . Вид живой клетки сбоку.
Гонкая испорченность пелликулы не по-
казана. Без масштаба (noieedale,
1967)
18 мкм
Рис. 2.63. Бесцветный двужгутиковый
представитель звгленовых из криофиль-
ного местообитания. Видны глотка,
гетеродинамичные
продукты ( Hoham
жгутики и запасные
, Blixm , 1979)
или пластин, окружены двойной мембраной и, кро-
g имеют вид дисков
ЦН' ме того, мембраной хлоропластной эндоплазматической сети, которая
не переходит в вдерную мембрану. Тилакоиды в стопках по три, две
тилакоидных ленты пересекают строму пиреноида. Запасной продукт -
парамилон ( J5-I,3-глюкан), образующий как бы экран вокруг пире-
ноида, но вне хлоропласта; он не окрашивается иодом. Может также
запасаться хризоламинарин. Все исследованные к настоящему време-
ни зеленые эвгленовые оказались фотоауксотрофными; они способны
расти фототрофно на свету и гетеротрофно в темноте. Известны так-
же гетеротрофы и паразиты. Все клетки имеют по два базальных те-
льца с отходящими от них одним или двумя жгутиками. Последние
сходны с таковыми у динофитов; по всей их длине проходит тяж, а
66
фибриллярные волоски расположены* вдоль жгутика в один спиральный
ряд. Жгутики выходят из передней камеры (резервуара). Обычно име-
ется глазок, независимый от хлоропласта, но связанный с околожгу-
тиковым вздутием. Клетка покрыта белковой пелликулой, состоящей
из нескольких спирально расположенных соединенных между собой по-
лосок. Пелликула может быть гибкой, что позволяет клетке совер-
шать характерное эвгленоидное движение. Ядро мезокариотное, о по-
стоянно конденсированными хромосомами; митоз протекает уникальным
способом. Размножение - только за счет деления клеток; половое
воспроизведение неизвестно.
Порядки: Eutreptiales, Euglenales, Rhabdomonadales, Spheno-
monadales, Heteronematales, Euglenamorphales.
CHLOROPHYCOTA : зеленые водоросли
Этот отдел включает все водоросли с хлорофиллами а и Ь , кро-
ме эвгленовых (Silva , 1982). Его характерные черты обобщены б
ряде работ ( Hoek , Jahns , 1978; Moestrup , 1978; Ноек > 1981;
Melkonian , Ichimura , 1982). Хлоропласта окружены только двой-
ной мембраной без дополнительной мембраны эндоплазматической се-
ти; тилакоиды в стопках по 2 - 6 и более. Пиреноиды (если имеют-
ся) находятся внутри хлоропласта; в них часто проникают тилакои-
ды, а вокруг них откладывается крахмал, являющийся основным запас-
ным продуктом. Глазок (если имеется) обычно расположен внутри
хлоропласта. Жгутики равные или неравные по длине, обычно апикаль-
ные; чаще всего их два. Волосков они не несут, но могут быть по-
крыты нежным "войлоком" или чешуйками. В зоне их выхода иэ клет-
ки находится девятиконечное звездчатое тело ( Moestrup , 1982).
Клеточные стенки состоят из целлюлозы, за исключением сифональ-
ных видов, у которых ее роль играют маннан или ксилан, и класса
Prasinophyoeae > гДе она содержит галактан и остатки уроновой
кислоты. Покровы многих зеленых водорослей обызвествлены, осо-
бенно у тропических морских сифональных форм и харовых.
Бесполое размножение - посредством фрагментации, образования
зооспор, апланоспор или автоспор. Половое размножение может быть
изогамным, анизогамным или оогамным. В целом особенности воспро-
изведения и жизненных циклов весьма разнообразны. За исключением
ризоподиальных одноклеточных и крупных многоклеточных талломов
67
сложного строения, здесь известны все морфологические типы, встре-
чающиеся у водорослей.
Современные исследования митоза, цитокинеза и структуры под-
вижных клеток вызвали предложения пересмотреть систему зеленых
водорослей (ср. Pickett-Heaps , 1975; Stewart, Mattox , 1975),
выделив несколько основных филетических линий. Одна из них ("хло-
рофитная", или "хламидомонадная") отличается наличием при цитоки-
незе фикопласта; для другой ("бриофитной", или "харофитной") ха-
рактерен фрагмопласт. Вторая линия считается предковой для высших
растений ( Steward, Mattox , 1975). Если такое деление обычно
и признается при построении современных систем, имеется еще много
расхождений в деталях классификации, и ее "тонкая настройка" про-
должается по мере публикации новых открытий (cM.Round , 1971,
1981; Bold , Wynne , 1978; Lee , 1980; Christensen , 1980;
Deason , 1984; Ettl , 1981; Ettl , Komarek , 1982; Hoek , 1981;
Moestrup , 1982; Melkonian , 1982; Melkonian , Ichimura , 1982;
Irvine , John , 1984)•
Нами приводятся здесь три класса (табл. 2.1). Prasinophyceae
иногда объединяются с Chlorophyceae (Hoek , 1981) ИЛИ получа-
ют статус отдела ( Round , 1981), однако чаще всего они рассмат-
риваются как самостоятельный класс. Раунд ( Round , 1981) выделя-
ет их в отдел в связи с очень характерной чешуйчатой орнаментаци-
ей клеточной стенки. Хук ( Hoek, 1981) использует эту же особен-
ность, чтобы показать, как "празинофитный" тип мог служить связу-
ющим звеном между покрытым чешуйками жгутиковым предком зеленых
водорослей и современными представителями этого отдела с целлю-
лозными стенками; при этом он включает Prasinophyceae в порядок
Volvocales • Харофиш занимают изолированное положение мезду зе-
леными водорослями, с одной стороны, и мхами и печеночниками (бри-
офитами) - с другой, являясь, возможно, тупиковой ветвью эволю-
ции. Обычно их выделяют в самостоятельный отдел, признавая его
раннее отделение от основных эволюционных линий зеленых водорос-
лей.
Большинство Chlorophycota - пресноводные формы (90%), ос-
тальные - морские. Морская альгофлора их весьма однообразна в
тропиках и становится богаче в более прохладных регионах. Многие
пресноводные виды - космополиты. Ископаемые зеленые водоросли из-
вестны еще из палеозойских отложений: харовые - из силура (435 -
460 млн. лет назад), а морские известковые роды - из ордовика
(500 - 530 млн. лет назад). Приводим ниже краткий обзор трех клас-
сов этого отдела.
68
Chlorophyceae
(риб. 2.64 - 2.75)
Этот класс в широком смысле включает по одним источникам
около 560 родов и 860 видов (Silva, 1982), по другим - свыше
20 000 видов ( Melkomian, Ishimura, 1982). Наряду С хлорофилла-
вегетативной клетки. Ж - жгути-
ки, Г - глазок, X - хлоропласт,
П - пиреноид, СВ - сократитель-
ная вакуоль.Без масштаба
ГИС. 2.65. Pandorlna unlcocca
(Chlorophyceae, Volvocales),
32-клеточная колония. Замет-
но отсутствие дифференциров-
ки клеток ( Nozaki, 1981)
Рис. 2.66. Pvrobotrye (Chloro-
phyceae). Структура
отдельной колонии, показываю-
щая дифференцировку (на самом
верху - передние клетки мень-
шего размера); у клеток по
два гомодинамичных жгутика
(Sarma , Shyam, 1974)
69
Рис. 2.fi7. Ulothrix apeciosa (Chlorophyceae, Ulotrichales).
Схема апекса (А). базальной части с ризоидами (Б) и зрелого
участка нити (В). Каждая клетка содержит постенный хлоропласт с
одним или двумя пиреноидами ( Berger-Perrot, 1981)
ми _а и ъ_ хлоропласты содержат различные ксантофиллы, в том чис-
ле виолаксантин, зеаксантин, антераксантин и неоксантин; сифонок-
сантин и сифонеин характерны для порядка Bryopsidales.Qco6eHHoo-
ти хлоропластов, пиреноидов и запасания крахмала - как описано
выше для отдела в целом. Подвижные клетки обычно несут два или
четыре равных гладких жгутика, расположенных апикально и.не выхо-
дящих из какого-либо углубления. Чешуйки и волоски на жгутиках
редки. Клеточная стенка, если имеется, состоит в основном из цел-
люлозы. Она может обызвествляться, а также содержать в больших
количествах гликозиды гидроксипролина, ксилан и маннан.
У Chlorophyceae имеется полный набор морфологических ти-
пов, за исключением ризоподиального или крупных, сложно устроен-
ных многоклеточных форм. Силва ( siiva,1982) на основе современ-
ных данных по ультраструктуре описывает три эволюционные линии
зеленых водорослей. Согласно этому автору, к Chlorophyceae
относятся представители отдела, "характеризующиеся разрушением
70
Рис. 2.68. Hosenvingiella •ро-
lyrhiza (Chlorophyceae, Pra-
siolales). Вегетативная нить с
однорядными и многорядными
участками. Масштаб 50 мкм ( Ho-
oper, South 1977)
Рис. 2.69. ^raparanaldia sp.
(Chlorophyceae, Chaetophora-
ceae). Часть нити; видны клет-
ки главной оси с поясковидными
хлоропластами и боковые ветви
ограниченного роста, оканчиваю-
щиеся щетинками. Масштаб 50 мкм
Рис. 2.70. Xanthidium sp. (Chlo-
rophyceae, Conjugales).
Клетка этой десмидиевой водо-
росли представляет собой две
равные половины, соединенные
узким перешейком, в котором на-
ходится единственное ядро.
Утолщенные стенки орнаментиро-
ваны шипами. Масштаб 25 мкм
71
Рис. 2.7I.Buastrum insi-
gne (Chlorophyceae, Con-
Jugales). Стенки этой
десмидиевой водоросли, хо-
тя и не несут шипов, ха-
рактерно вздутые и разрос-
шиеся у полюсов. Масштаб
25 мкм
Рис. 2.72. Caulerna brownlKChlo-
rophyoeae, Caulerpales).
Ценоцитныи таллом состоит из сте-
лющегося "корневища" (здесь уда-
лено), которое дает начало иду-
щим вверх выростам и прикрепи-
тельным ризоидам снизу. Масштаб
5 см (фотография:Н.С.Р1скеп)
Рис. 2.73. Penis,Шив sapitatus
(Chlorophyoeae. Cai<erpales).
Обызвествленный таллом из ножки
с ветвящимися терминальными си-
фональными выростами закрепляет-
ся в мягком субстрате, предпочи-
таемом этим тропическим видом,
плотной массой погруженных ризо-
идов (Meinesz, 1980)
72
Рис. 2.74. Udotea petiolata (Chlo-
rophyceae , Caulerpalea).
Обызвествленный сифональный тал-
лом из вееровидной верхней час-
ти на ножке, отходящей от ризо-
идального основания, погруженно-
го в мягкий субстрат (ifeinesz,
1980)
Рио. 2.75.Cladonhora
rupestris (Chlorophy-
ceae, Cladophorales).
Целое растение с обиль-
ным ветвлением. Струк-
тура семиценоцитная;
клетки содержат много-
численные одра. Мас-
штаб 2 см (фотография:
R.С.Ficken)
митотического веретена в телофазе, наличием фикопласта во время
цитокинеза и подвижными клетками, у которых два или четыре апи-
кальных жгутика имеют базальные тельца, связанные о четырьмя или
более относительно узкими пучками микротрубочек". Такому опреде-
лению соответствуют традиционно признаваемые вольвоксовая, тетра-
споровая и хлорококковая линии эволюции (Bold, Wynne, 1978),
а также порядки Chlorosarcinales, Ittcrosporales, Oedogoniales
и Chaetophorales в узком смысле ( Silva, 1982). Вторая филетиче-
ская линия включает Ulvophyceae (“Ulvaphyceae по Sluiman et al.
73
1980; см, O’Kelly, Floyd, .198$ Floyd, O’Kelly, 1984). Силва
( Silva, 1982) не придает формального ранга этой группе, у пред-
ставителей которой отсутствует фикопласт. Локхорст и Стар (Lok- ,
horst, Star, 1983) также ставят под сомнение независимый статус
этого класса. Другие возможные диагностические признаки uivophy-
сеае включают: центральную симметрию жгутикового аппарата, абсо-
лютную ориентацию его компонентов по часовой стрелке, если смот-
реть сверху, и терминальные двулопастные колпачки, прикрывающие
проксимальные концы базальных телец ( O'Kelly, Floyd , 1983). По-
нимаемые так Ulvophyceae должны включать порядки Ulotriohales,
Ctenooladales, Ulvales, Acrosiphoniales, Cladophorales, Siphono-
oladales, Bryopsldales и Sphaeropleales ( Silva, 1982).
Третью линию составляет класс Charophyceae в понимании некоторых
авторов ( Stewart, Mattox, 1975), т.е. включающий наряду с ха-
рами порядки Chlorokybales, Klebsormidiales, Zygnematales, Tren-
tepohliales и Coleochaetales.Cифoнaльныe зеленые водоросли
могут считаться четвертой линией, рано отделившейся от линии Ш-
vophyoeae. Здесь мы придерживаемся консервативного подхода Сил-
вы ( Silva, 1982), признавая, что для упорядочения накапливаемой
информации по ультраструктуре рано или поздно потребуется сущест-
венный пересмотр высших подразделений этого отдела.
Порядки: Volvocales, Tetrasporales, Chlorococcales, Chloro-
saroinales, Microsporales, Oedogoniales, Chaetophorales, Ulotri-
chales, Ctenooladales, Ulvales, Prasiolales, Sphaeropleales,
Acrosiphonales, Cladophorales, Siphonocladales, Bryopsldales,
Dasyoladales, Chlorokybales, Klebsormidiales, Zygnematales, Tren-
tepohliales, Coleoohaetales.
Charophyoeae : харовые, или лучицы
(рис. 2.76)
В настоящее время этот класс представлен единственным поряд-
ком с семью родами ( Daily, 1982; Grant, Sawa, 1982), Клетки од-
ноядерные и обычно содержат многочисленные эллипсоидные хлороплас-
ты, расположенные окрученными продольными параллельными друг дру-
гу рядами. Пигменты и запасные продукты - как описано выше для
отдела в целом. Клеточные стенки состоят из целлюлозы и часто
сильно обызвествлены; это приводит к отложению мергеля (СаСОд +
MgCOj ) в пресноводных местообитаниях, где в изобилии встречают-
74
Рис. 2.76. Chara vulgaris
(Charophyoeae) А.Целое рас-
тение с главной осью и мутов-
ками простых веточек в узлах,
разделенных междоузлиями.
Б. Оогоний, или "глобула",
женский репродуктивный орган.
В Участок покрытой корой оси
и зона ^?л|. Без масштаба (по
ся харовые. Обызвествленные
остатки легко переходят в иско-
паемое состояние, особенно в
случае женских половых органов,
которые при этом образуют так
называемые гирогониты. Клеточ-
ное деление включает образова-
ние фрагмопласта. Из особеннос-
тей этой группы, сближающих ее
с бриофитами, можно отметить
репродуктивные структуры, за-
ключенные в стерильный покров
(уникальный среди водорослей
случай), и спермин с двумя не-
сколько неравными жгутиками,
прикрепленными субапикально и
покрытыми чешуйками. Имеется
только один жгутиковый корень,
обычно связанный с многослой-
ной структурой (МСС), сходной
с обнаруженной в спермиях архе-
гениальных растений (Moestrup9
Ichimura, 1982; Moestrup,
1982). Тело спермин удлиненное
и состоит из трех частей, чего
никогда не наблюдается у Chlo-
rophyceae. Бесполые зооспо-
ры у харовых не образуются, а
вегетативное размножение осуще-
ствляется посредством заполнен-
ных крахмалом структур в форме
звездочек, луковичек на ризои-
дах и выростов в виде прото-
нем, отходящих от узлов.
Рост апикальный, начинаю-
щийся со стадии протонемы (ср.
с бриофитами) и приводящий к
структуре с чередующимися узлами и междоузлиями. В узлах формиру-
ются мутовки ветвей, а междоузлия состоят из одной крупной клет-
ки, которая иногда покрыта нитями коры (например, в роде Chara).
75
Харовые - главным образом пресноводные организмы, хотя не-
сколько видов может встречаться и в солоноватой воде. То, что эта
линия очень рано дивергировала от Chlorophyceae, подтвержда-
ется ископаемыми находками: харовые известны уже из силурских от-
ложений {435 - 460 млн. лет назад).
Порядок:Charales (с одним семейством, обычно подразделяе-
мым на две трибы:Nitelleae И Chareае)*.
Prasinophyceae : празинофиты
К этому классу относятся коккоидные или жгутиковые однокле-
точные; у видов со жгутиками последние, как и тело клетки, покры-
ты одним или несколькими слоями чешуек нескольких типов. Клетки
одноядерные, с крупным лопастным пристенным хлоропластом. Среди
пигментов может встречаться сифоноксантин (кроме этого известный
только у Chlorophyceae). Как правило, присутствует пиреноид, в
который у этих водорослей проникают тилакоиды, а у некоторых ви-
дов - и лопасти цитоплазмы (Norris, 1982а, ъ). Основные запас-
ные продукты - крахмал и маннитол. Обычно в клетках имеется гла-
зок, а в некоторых - трихоцисты (Pearson, Norris, 19755 Norris,
1980, 1982Ъ), напоминающие эжектосомы криптофитов. Жгутиков -
один, два, четыре, шеоть или восемь. У одно- и двужгутиковых ви-
дов они прикрепляются сбоку, тогда как у четырехжгутиковых они
примерно одинаковой длины и выходят из углубления в апикальной
части сердцевидной клетки. Некоторые виды (например, Halosphae-
ra viridis.gM.Parke. Adams, 1961) имеют чередующиеся подвижную и
неподвижную стадии. Клетка может быть покрыта панцирем (например,
У родов Platymonas и Prasinccladus). Бесполое размножение про-
исходит с образованием зооспор или путем простого деления; поло-
вое размножение неизвестно.
Большинство Prasinophyceae - свободноживущие обитатели мо-
рей, солоноватых и пресных вод. Некоторые - симбионты, например
Р°Д ^etraselmis, вступающий в хорошо известный симбиоз с морским
плоским червем Convoluta.
Относительно тонкой структуры представителей класса еще мно-
гое неясно; в настоящее время его таксономия приблизительна, так
же как И сам статус Prasinophyceae (Parke, Green, В76). Мёст-
РУП (Moestrup, 1982) выделйет poUHMcnomastix, Pedinomonas и
Soourfjpid-iH (и Micromonas) в класс Loxophyceae, Не признавае-
Мый Боррисом ( Norris, 1980, 1982b, с).
76
Порядки: Pedinomonadales, Monomastigales, Pyramimonadales,
Pterospormatales, Chlorodendrales.
"Glaucophyta" , ИЛИ "Glaucophyoeae"
Паркер ( Parker, 1982) не включает этот отдел или класс в
свою систему водорослей, хотя последние работы показывают, что
признание его может оказаться необходимым. В отдел Glauoophyta
выделяли ряд жгутиковых форм, содержащих эндосимбиотические сине-
зеленые водоросли ( Skuja, 1954). Впоследствии из него были ис-
ключены динофлагелляты с эндосимбиотическими синезелеными "фео-
сомами" (Taylor, 1973). Эти комплексные организмы (известные
под названием синцианозы), представителей которых ныне сохрани-
лось немного, возможно, иллюстрируют важные этапы эволюции содер-
жащих хлоропласты эукариотных клеток. Среди дошедших до нас ви-
дов есть формы с голыми цианидами (клетками хозяина) и их произ-
водными, окруженными клеточными стенками ( Geitler, 1959). Сине-
зеленый компонент (цианелла), как и Cyanophycota, содержит хло-
рофилл а и фикобилипротеины.
Статус этой группы остается под вопросом, хотя Кис ( Kies,
1980) полагает, что она представляет собой естественную совокуп-'
ность форм, несколько родственных зеленым водорослям. Этот вывод
подкрепляется данными по структуре жгутиков ( Moestrup, 1982).
Glaucophyoeae (в понимании Moestrup, 1982) можно сблизить с
эвгленовыми и зелеными водорослями. Более раннее отнесение Glau-
cocystis К багрянкам (Robinson, Preston, 1971) сейчас опро-
вергнуто ( Kremer et al., 1979).
Глава 3
СТРУКТУРА КЛЕТКИ
Общие особенности
Клетки водорослей, как и любых других организмов, состоят
из протопласта, который либо голый, либо окружен клеточной стен-
кой или иным покровом. Внешняя клеточная мембрана называется
плазмалеммой. У прокариотных водорослей ( Cyanophycota И Proch-
lorophyoota, рис. 3.1) внутри протопласта отсутствуют окружен-
Рис. 3.1. Prochloron sp.(Prochlorophycota) . Полученная
методом замораживания - скалывания реплика клетки, показы-
вающая ее общую организацию. Видны тилакоиды, просвет кото-
рых (и) можно отличить от цитоплазмы (Ц) по более гладкой
текстуре (х 11000) ( Giddings et al., 1980)
ные мембранами органеллы, однако сами клетки разделены на зоны
таким образом, что ядерный материал сосредоточен в центре (цент-
роплазма), а фотосинтетические ламеллы - по периферии (хромоплаз-
ма). У эукариотных форм имеется ряд внутриклеточных органелл,
окруженных мембранами: ядро, хлоропласты, митохондрии и тельца
Гольджи. Эндоплазматическая сеть (ЭС) представляет собой особую
систему внутриклеточных мембран или цистерн. К клеточным включе-
78
ниям относятся отложения углеводов (часто связанные с пиреноида-
ми) и различные кристаллические, липидные и другие образования.
У многих водорослей подвижность обусловлена наличием жгутиков-би-
чевидных придатков, окруженных мембраной, составляющей единое це-
лое с плазмалеммой. Внутри клеток здесь встречаются сложные ко-
решковые структуры, а у многих подвижных форм присутствует глазок
(стигма), часто связанный с базальной системой жгутиков и хлоро-
пластом.
Широко распространена точка зрения (эндосимбиотическая тео-
рия), что пластиды и митохондрии эукариот возникли в ходе эндосим-
биоза из прокариотных фотосинтезирующих форм (например, цианелл)
и бактерий (см. гл. 8, а также Margulis, 1981; Giddins et al.,
1980; Brawley, Wetherbee, 1981). Эта теория выстраивает непре-
рывные эволюционные ряды от прокариот к эукариотам и между отдель-
ными группами эукариот. Эндосимбиотическая теория признается не
всеми (см. обсуждение в гл. 3 и Dillon, 1981); ее слабостью
является доступность для доказательств только ныне живущих видов.
Возможно, дальнейшее изучение очень древних ископаемых эукариот
со временем прольет некоторый свет на этот интересный вопрос (по-
пулярное обсуждение см. Vidal, 1984). Хотя клеточное и субклеточ-
ное строение эукариотных водорослей и других эукариот во многом
сходно, здесь наблюдается также много специфических и уникальных
черт. Именно последние и будут рассмотрены ниже.
Клеточные особенности классов водорослей суммированы в гл. 2.
Предполагается, что у большинства классов в основе организации
клетки лежит строение подвижного одноклеточного "предка" (за ис-
ключением прокариот и красных водорослей - см. гл. 4 и 8). Хотя
структура и развитие клеток сильно варьируют, дифференцировка
тканей у водорослей развита незначительно. В ходе роста, созрева-
ния и размножения могут происходить значительные изменения кле-
точной и субклеточной организации ( Pickett-Heaps,I975), динамич-
ность которой всегда следует иметь в виду.
Клеточные покровы: клеточная стенка
Хотя многие жгутиковые формы, а также споры и гаметы водорос-
лей "голые", большинство водорослевых клеток покрыто одним или не-
сколькими слоями относительно инертного материала (в основном уг-
леводной природы), который иногда пропитан или прослоен неоргани-
79.
ческими веществами» например карбонатом кальцин или кремнеземом. У
многих видов стенки по строению и составу близки к имеющимся у
высших растений, однако в некоторых группах встречаются специфи-
ческие покровы (в частности, пелликула и панцирь). О формирова-
нии химическом составе и структуре стенки многих водорослей сей-
час имеется детальная информация. Как ни удивительно, хотя эти
особенности близки в пределах ряда хорошо очерченных групп, у дру-
гих групп в структуре и химии стенки на разных стадиях роста или
одного жизненного цикла наблюдаются сильные различия.
Додж (Dodge, 1973) предложил классификацию, которая до сих
пор полезна при обсуждении покровов водорослевых клеток. В
табл. 3.1 дан обзор основных типов этих покровов по разным клас-
сам водорослей.
Голые клетки бывают основным вегетативным состоянием, напри-
мер, у некоторых Bangiophycidae, Chrysophyceae (рис. 3.2) И
Chlorophyceae, а также у всех Raphidophyceae. Они мотут пред-
ставлять собой переходную фазу в форме бесполых зооспор ( Eustig-
matophyceae), гамет (особенно Мужских, например у Bacillariophy-
сеае) пли как зооспор, так и гамет (Chlorophyceae, Xanthophyceae
и Phaeophyceae, рис. 3.3). Голые клетки окружены только плазма-
тической мембраной (плазмалеммой). У многих из них внешний покров
гибкий, что позволяет им совершать амебоидное, или ризоподиаль- _
ное, движение. Поверх плазмалеммы иногда откладывается белковый
материал, а у Porphyridium (Bangiophycidae) она может быть
окружена полисахаридной капсулой ( Нашив, 1972). У подвижных од-
ноклеточных представителей золотистых водорослей и рафидофитов
имеются различные "отстреливаемые" органеллы1. У первых они двух
типов: слизистые тельца и дискоболоцисты;первые сходны со слизис-
тыми тельцами, встречающимися у примнезиофитов, рафидофитов и ди-
нофлагеллат (Lee, 1980). И те и другие возникают внутри аппара-
та Гольджи. Слизистые тельца окружены одной мембраной и при выбро-
се формируют вне клетки волокнистую сеть. Дискоболоцисты детально
описаны у Oohromcnas tuberoulatua (Hibberd , 1970). Они располо-
жены в наружном слое цитоплазмы и представляют собой окруженные
одной мембраной пузырьки с полым диском на стороне, обращенной к
~~ "Г~Такие органеллы одноклеточных (трихоцисты, эжектосомы,
и^‘стн> токсицисты, рабдоцисты, дискоболоцисты, нематоцисты
“Др.) объединяют под названием экструсомы. - Прим, ред.
80
Таблица 3.1. Обзор типов клеточных покровов у водорослей (по
Dodge » 1973, с изменениями)
Голые Модифике ЦИИ ПОД плазма- леммой - чешуйки снаружи плазма- леммы А а _ В клеточ- ная стенка
промежуточные i сталии нормальное состояние Перипласт Пелликула Тека Органические Известковые Кремнеземные 1 Кремнеземный панг неполная (домик) Полная
Cyanophyoeae +
Prcchlorophyceae +
Rhodophyceae + +
Chrysophyceae + + +
Prynmesiophyceae + +
Xanthophyceae + +
BustIgmatophyceae + +
Bacillariophyceae + + j
Dinophyceae + +
Phaeophyceae + +
Raphidophyceae +
Cryptophyceae *
Buglenophyc eae + +
Chlorophyceae + + +
Charophyceae + +
Prasinophyceae
81
Рис. 3.2. Chryaamoeba radicans (Chrysophyceae). Схемати-
ческий разрез голой амебоидной стадии. В - волосок жгути-
ка; Г - тельце Гольджи; Жт - главный жгутик; Ж?— второй
жгутик; Л - пузырьки лейкозина; М - митохондрий; МГ - мас-
ляные глобулы; Ох - оболочка хлоропласта; П - пиреноид;
ПГ - плотные глобулы; ПК - пиреноидные каналы; ПС.- пери-
пластидная сеть; Р - ризоподии; СВ - сократительная ваку-
оль; X - хлоропласт; ХЭС - хлоропластная эндоплазматичес-
кая сеть; Я - ядро (Hibberd, 1971)
внешней части клетки. При их взрывообразном отстреливании высво-
бовдается тонкая нить с диском на конце. Слизистые тельца обиль-
ны в наружной цитоплазме рафидофита Vacuolaria; трихоцисты
GonyoBtomum из того же класса сходны с наблюдаемыми у динофи-
т°в (Mignot, 1967).
В случае переходных фаз голое состояние обычно быстро утра-
чивается, как только зооспоры или гаметы прекращают плавать и
прикрепляются к субстрату; за этим быстро следует образование
стенки.
82
Рис. 3.3. Обобщенная схема зооспоры ламинариевой водорос-
ли ( Phaeophyceae, leminariales); прикрепление жгутиков,
цитоскелетные микротрубочки и основные органеллы. ЗЖ -
задний жгутик; М - митохондрия; Ма - волоски (мастигоне®-
мы); П - пузырек типа "В"; Ш - передний жгутик; X - хло-
ропласт; Я - ядро (Henry, Cole, 1982)
Покровами клетки, имеющими различные модификации и обычно
находящимися под плазмалеммой, являются перипласт, пелликула и
панцирь (тека). Они характерны для криптофитов, эвгленовых и ди-
нофитов соответственно. В некоторых работах (см., например, Ro-
und, 1973; Bold, Wynne, 1978) водоросли с клеточными покрова-
ми этих типов называются "голыми".
Перипласт криптофитов состоит из внешнего волокнистого слоя
с белковыми пластинками под плазмалеммой. Они располагаются по-
разному в зависимости от видовой принадлежности водоросли (San-
tore, 1982 а, Ъ; рис. 3.4), могут быть шестиугольными ( Crvutomo-
nas) или прямоугольными ( Chroomonas). Трихоцисты ассоциированы
с пластинками.
Пелликула эвгленовых (рис. 3.5) ( Leedale, 1967, 1982b )
сходна с перипластом белковой природой материала, расположенного
под плазмалеммой (80% белка, остальное - липиды и углеводы) (Wal-
пе, 1980). Предполагается ( West et al., 1980), что эволю-
ция пелликулы шла в двух разных направлениях: I) по пути усложне-
ния; 2) по пути упрощения и редукции. У Egglena поверхность клет-
ки на поперечном срезе имеет волнистый контур из-за наличия бел-
83
Рис. 3.4. Электронная микрофотография поперечного среза
Hemiselmis brunnescens (Cryptophyceae) У переднего
конца ядра, видны оелковые пластинки перипласта и общие
детали клетки. Ли - лизосомы; Нм - нуклеоморфа; Хп - хло-
ропласт; Я -ядро (х 178000) ( Santore, 1982b)
ковых пелликулярных полосок, расположенных спирально или продоль-
но и смыкающихся или соединяющихся между собой. У Trachelomonas
такая волнистость отсутствует и клетка одета минерализованным
панцирем, состоящим из слизистых тяжей (рис. 3.6; West, Walne
1980), содержащих большое количество (20-60% массы) марганца
( West et al., 198ОЪ).
У многих эвгленовых пелликула гибкая и способна сокращаться
и растягиваться (метаболийное движение), когда организм не плава-
ет. Под белковым слоем находятся сеть микротрубочек и часто при-
мыкающие к углублениям пелликулы слизистые тельца, которые сооб-
щаются через узкие поры с внешней средой.
Тека динофитов (рис. 3.7) ( Dodge, Crawfold, 1970), или ам-
фиеома ( boeblich, 1970), состоит из нескольких слоев. Единства
мнений относительно того, находится ли плазматическая мембрана
под ней или снаружи, не достигнуто ( Bold, Wynne, 1978), хотя,
поскольку самая внешняя мембрана переходит в мембрану жгутиков,
возможно, она и является плазмалеммой. Динофлагеллат подразделя-
ли на "голые" и "вооруженные", причем последние формы характери-
зовались наличием микрофибриллярных пластинок теки, которые от-
кладывались вместе со слоем уплощенных пузырьков под внешней мем-
браной. Такие пузырьки присутствуют в теках обоих типов, и Лёб-
ли* ( Loeblich, 1970) полагает, что структура их в основных чер-
тах одинакова; тонкие пластинки могут встречаться и у некоторых
84
Рис. 3.5. Электронная микрофо-
тография продольного среза Еи-
alena granulata (Euglenophy-
оеае). йидны передний резерву-
ар, ядро с хромосомами, хлоро-
пласты и белковая пелликула с
многочисленными поверхностны-
ми складками (х 5400) (walne,
Arnott, 1967)
Рис. 3.6. Пустая клеточная
обмотка TrachelomonaB lefev-
rei (Euglenophyceae),состо-
ящая из анастомозирующих мине-
рализованных слизистых тяжей.
Видно окруженное валиком от-
верстие для выхода жгутиков
(х 2800) (фотография любезно
предоставлена P.L.Walne)
видов, считающихся "голыми".
Число и расположение пласти-
нок имеют большое таксономиче-
ское значение, и таксономисты
разработали системы однознач-
ной их нумерации для каждого
вида (рис. 3.8 и 3.9). Форма
и морфология пластинок также
важны для таксономии. Они мо-
гут быть орнаментированы греб-
нями, нести сетчатый рисунок,
шипики или какие-либо другие
выросты, иногда окремнелые.
У многих видов пластинки перфорированы, что обеспечивает выброс
трихоцист.
Клетки представителей золотистых водорослей (рис. 3.10),
примнезиофитов, празинофитов и динофитов (изредка) бывают покры-
85
ж
Рис. 3.7. Вертикальные срезы динофлагеллатных тек различ-
~ QKmfhte». Ё-" -М11Щда> В. - некоторые
рЯДЫ Gynmodinium. Г - Katodinium.Д - Woloszamakla.
Ofitarocspsa» • IL - , некото-
рое виды Peridinlum.У - Prorocentrum (Dodge, Crawford,
1970) ------- ~ ----------------
86
Рис. 3-8.Cerati.uia brachyceros
(Dinophyceae).
Фотография клетки в сканирую-
щем электронном микроскопе,
показывающая общее расположе-
ние и орнаментацию пластинок
теки и поясковую бороздку „
(х 1040) ( Bourrelly, Coute,
1976)
Рис. 3.9. Примеры расположения и нумерации пластинок теки
у динофлагеллят. A. Ceratium brachyceros; схематический
вид клетки с вентральной стороны, показывающий последова-
тельность нумерации пластинок теки: I' - апикальный ряд;
I' - перицингулярный ряд; Г" - постцингулярный ряд;!'*' -
антапикальный ряд. С?, Cg, CL - цингулярные пластинки
( Bourrelly. Coute, 1976). Peridinium greeatum С вен-
тральной (Б) и дорсальной (В) сторон (обозначения см.
рис. 3.9,А). ПБ - продольная борозда ( Lombard, Capon,
87
Рис. 3.10. Syncrypta glomifera (Chrysophyceae).
Схематический разрез клетки с покровом из чешуек. Г - гла-
зок; ДЖ - длинный жгутик; ЖУ - жгутиковое углубление; КЖ -
короткий жгутик; М - митохондрия; П - пиреноид; ПГ - пиг-
ментные гранулы; ПМ - пузырек мастигонем (жгутиковых во-
лосков); ТГ - тельце Гольджи; X - хлоропласт; ЦКУ - цистер-
на жгутикового углубления; Ч - чешуйки; ЭПМ - злектроно-
плотный материал; Я - ядро ( Clarke, Pennick, 1975)
ты чешуйками, иногда нескольких типов,расположенными на поверхно-
сти плазмалеммы. Их образование - интересный процесс, тесно свя-
занный с пузырьками Гольджи (впервые зто показали Manton, Parke,
88
I960). У некоторых водорослей (например, у празинофита Pyramimo-
nas) описан специальный "резервуар чешуек" (рис. 3.II). Морфоло-
гия чешуек, как показывают тонкие исследования с помощью просве-
чивающего и сканирующего электронного микроскопа, сильно варьиру-
ет. В основном признается, что если у водорослей они покрывают и
тело, и жгутики (например, у празинофитов), то относятся к раз-
ным типам. Однако они формируются из одинаковых пузырьков Голь-
джи, и остается загадкой, каким образом один тип откладывается
на плазмалемме жгутиков, а другой - на теле. У Pyramimonas orien-
talis и Р,parkeae , например, чешуйки шести различных
типов проходят через резервуар чешуек, а затем каждый тип зани-
мает специфическое положение на жтутиках или на теле (Moestrup,
1982). Выход чешуек на поверхность у других водорослей также
обычно наблюдается у основания жгутиков. У динофита Heterocansa
наблюдали (Morrill, Loeblich, 1983), как мембраны особых пузырь-
ков Гольджи выполняют роль "литейной формы" для развивающихся че-
шуек; предполагается, что у этого рода их образование идет непре-
рывно. Причины столь крупных различий чешуек по химизму и морфо-
логии неясны, хотя в числе их предполагаемых функций можно на-
звать защиту клетки, увеличение ее размеров и плавучести.
На основании химического состава описано три основных типа
чешуек: органические, известковые и кремнеземные (табл. 3.1). Ни-
же мы рассмотрим их по очереди. Другие образуемые аппаратом Голь-
джи структуры жгутиков или стенки включают так называемые "шиш-
ковидные" чешуйки в порядке Pavlovales (примнезиофиты; Green,
1980), представляющие собой электроноплотные тельца,и цилиндри-
ческие чешуйки у Chrysochromullna (примиезиофиты; beadbeater,
1972; Manton et al., 1981).
Органические чешуйки, обычно разных типов, встречаются на
теле и на жгутиках празинофитов ( Moestrup, 1982), а также сос-
тавляют нижний слой чешуек у образующих кокколиты примнезиофи-
тов. У празинофитов (рис. 3.12) на клетке может присутствовать
до пяти разных типов чешуек по морфологии; Норрис ( Norris, 1980)
предполагает, что эволюционная сложность клетки возрастает по
мере увеличения числа слоев чешуек. У высокоорганизованной водо-
росли Tetraselmis (рис. 3.13) эти слои объединены между собой
в жесткую оболочку (Norris, 1982). У органических чешуек фибрил-
лярная структура, но их химический состав известен недостаточно.
89
Рис. З.П. Схематический продольный разрез Pyramimonas lu-
nata (Prasinophyceae), показывающий особенности ультра-
структуры (кроме чешуек на жгутиках). Г - глазок; К - жгу-
тики; м - микротрубочковые корни жгутиков: КП - слои
крахмала на пиреноиде; ЛК - латеральное фиброзное кольцо;
М - митохондрии; МТ - микротельце; МТУ - микротрубочки
углубления; П - пиреноид; ПЧК - промежуточные чешуйки
клетки; Р - ризопласты; РЧ - резервуар чешуек; С - сини-
стосома; СВЧК - самые внешние чешуйки клетки; СНЧК - самый
нижний слой чешуек клетки; X - хлоропласт; ЦП - цилиндри-
ческие пузырьки; Я - ядро. Без масштаба (Inouye et al.,
1983)
В них обнаружены пектиноподобные вещества; в органических чешуй-
ках нижнего слоя у Apistonema (примнезиофит) отмечен целлюлоз-
ный гликопротеин.
Известковые чешуйки, или кокколиты, характерны для кокколи-
тофорид (порядок Coccoaphaerales) из примнезиофитов (Norris,
90
Рис. 3.12. Чешуйки на клетке Pyramimonas gelidicola
( Prasinophyceae). Показаны коробчатые чешуйки ( box
scales), чешуйки в виде отпечатка ноги ( footprint sca-
les) между ними и корончатые чешуйки ( crown scales)
(стрелки), сцепляющие углы коробчатых чешуек. Масштаб
0,2 МКМ ( McFadden et al., 1982)
1982). Эти образования были первоначально описаны из меловых от-
ложений, причем их водорослевая природа тогда не была понята.
Они состоят из органической чешуйки с карбонатом кальция (реже
арагонитом), откладывающимся на одной из поверхностей характер- '
ним .для каждого вида образом. Морфология чешуек очень сложна и
необязательно одинакова для каждой стадии одного и того же жиз-
ненного цикла; часто имеются шипы. Число чешуек изменяется с
возрастом клетки. Известковые чешуйки, как и органические, фор-
мируются в аппарате Гольджи.
На теле золотистых водорослей кремнеземные чешуйки могут
откладываться в тонкой пектиновой оболочке снаружи плазмалеммы.
Они бывают очень сложно устроены (например, у Mallomonas
рис. 3.14) и иногда относятся к нескольким разным типам на одной
и той же клетке. Такие чешуйки отличаются от органических и из-
вестковых чешуек тем, что обычно образуются не в аппарате Голь-
джи, а в пузырьках, связанных с эндоплазматической сетью, на по-
верхности хлоропласта. Однако Мёструп ( Moestrup, 1982) описы-
вает второй тип чешуек золотистых водорослей (например. Snhale-
91
Рис. 3.14. Чешуйки тела и аии-
кальные чешуйки со щетинками
Mallomonas harrisea (Chryso-
phyceae) (* 5500) (Takahashi,
1975)
Рис. 3.13. Tetraselmis asti-
ematica (Prasinophyceae).
Показаны твердая оболочка (или
тека, Т) и общие особенности на
срезе через середину клетки. БТ-
базальное тельце; В - вакуоль;
Г - аппарат Гольджи; М - мито-
хондрия; П - матрикс пиреноида;
X - хлоропласт; ЭС - эндоплазма-
тическая сеть; Я - ядро ( Hori
et al.,1982)
rnrmntiM- НахОДЯЩИХСЯ На П0-
верхности как жгутиков, так и
самой клетки и возникающих в
аппарате Гольджи.
Клеточная стенка, или пан-
цирь, диатомей состоит из крем-
незема с органическим покровом.
Эта структура интенсивно изуча-
лась с помощью световой и элек-
тронной микроскопии (особенно
с использованием угольных реп-
лик и СЭМ). Структура и орна-
ментация панциря диатомей ле-
жит в основе их классификации
и лучше всего наблюдается при удалении протопласта; таким обра-
зом , зти водоросли обычно идентифицируют в неживом состоянии.
Панцирь состоит из двух почти равных по размеру половинок,
смыкающихся между собой, как в чашке Петри (рис. 3.15«А); внеш-
Рис. 3.15. А. Обобщенный вид панциря диатомеи со створки (вверху) и с пояска (внизу). Г - гипотека; ПУ-
полярный узелок; X - хлоропласт; ЦУ - центральный узелок; Ш - шов; Э - эпитека. Б. Детали структуры
панциря в районе сочленения створки и пояскового кольца у диатомеи Tricerattum favus Д - дендритная
структура; КВ - краевой вырост; КК краевой канал, ПОт о - поясковые ооодки; шК - шестиугольная каме-
ра ( Miller, Collier, 1978) L"°
3
v
о
93
ние, более или менее уплощенные их поверхности, противоположные
друг другу, называются створками,более крупная - эпивальва, бо-
лее мелкая - гиповальва. Между собой их соединяет поясок
(рис. 3.15,Б), состоящий из двух перекрывающих частей, верхнего
пояскового кольца ( epicingulum) и нижнего пояскового кольца
(hypooingulum). Эпивальва и верхнее кольцо составляют зпитеку,
гиповальва и нижнее кольцо - гипотеку. У более сложно устроенных
диатомей поясок может быть расширен за счет дополнительных, ин-
теркалярных (вставочных), поясковых колец. Клетки, если смотреть
со створки, обычно имеют радиальную (центрические диатомеи, Bid-
dulphiales) или билатеральную (пеннатные, или перистые, диато-
меи, Bacillariales) симметрию. Хенди ( Hendey, 1964) выделяет
два дополнительных типа (рис. 3.16): треллисоидный, когда струк-
туры створки располагаются по радиусам и дугам концентрических
окружностей, центр которых расположен вне клетки (например, у
Eunotia) , и гоноидный, т.е. с угловатой створкой (например, у
Triceratium). Многие перистые диатомеи имеют шов, т.е. централь-
ную s-образную щель, проходящую вдоль одной или обеих створок,
разделенную центральным узелком и оканчивающуюся полярными узел-
ками. Формы со швом способны к скользящему движению за счет выде-
ления слизистого материала, возникающего из кристаллоидных телец
(Dirum, Hopkins, 1966). У бесшовных перистых диатомей на
створке имеется центральная неорнаментированная область, называе-
мая ложным швом.
Кроме шва панцирь несет разнообразную и сложную орнамента-
цию и перфорации, придающие клеткам диатомовых свойственную им
красоту и индивидуальность. Перфорации могут быть простыми пора-
ми или же сложными камерами, часто расположенными в виде линий
или штрихов. Поры бывают усилены окремнелыми утолщениями, называ-
емыми ребрышками. Камеры (рис. 3.17 и 3.18) представляют собой
полости, заполняющие стенку наподобие сот. С одной их стороны
располагается перфорированный слой (velum), с другой - отверстие
( foramen). Поверхности створок часто покрыты выростами, а их
края - лучами или щетинками. Большой интерес представляют двугу-
бые выросты, встречающиеся у центрических и перистых форм. Хасл
(Hasie, 1973) предполагает, что они могут быть предшественника-
ми шва, поскольку отсутствуют у перистых видов с хорошо развитым
швом. Различные выросты значительно увеличивают отношение поверх-
94
Рис. 3.16. Типы клеток диатомей (Hendey, 1964). Объясне-
ния см. в тексте
ности клетки к ее объему и могут способствовать как плавучести,
так и сцеплению клеток у колониальных форм.
Степень окремнения панциря различна (до 95,6%); только в не-
которых редких случаях кремнезем отсутствует (например, у ihaeo-
dactvlum tricornuturn: Lewin et al., 1958). С химической
точки зрения кремнезем - зто кварцит (зю2 • пн^о), но здесь он
включает также следовые элементы, среди которых - алюминий, маг-
ний, железо и титан (Mehta et al., 1961, Levin, 1962). "Кожа",
покрывающая панцирь, состоит из аминокислот и сахаров ( Coombs,
Volcani, 1968, Heokey et al., 1973); описан также ряд вне-
клеточных органических выделений ( Werner, 1977). Кремнистый ма-
териал откладывается упорядоченным образом; пузырьки, отделяющие-
95
А Б
Рис. 3.17. Типы локул в стенках панциря диатомей. А. Отвер-
стие, или пора ( Chaetoeегоз dldvmus var.aaglica) Б. Не-
локула (Sviiedra tq.-bulata). В. локула, откры-
пая мембрана; ГС - гуочатая поровая структура; Ж - жилка;
ЛИ - латеральная, или проходная, пора; 0 - отверстие; ОП -
опорная пластинка; ПМ - покровная мембрана; Ш - покровная
пора; СМ - ситовидная мембрана; СП - ситовидная пора ( Неп-
<*еу, 1971)
96
Рис. 3.18. Вид центральных
ВЫРОСТОВ у ДИаТОМеИ ThelaSBi-
ов1та incerta. ,
Масштаб! МКМ (Hasle, 1978)
ся от аппарата Гольджи, собираются под плазмалеммой и сливаются
в так называемые силикалеммы ( Reinmann et al., 1966), по од-
ной для каждой половинки панциря. Следует быстрое отложение крем-
незема, и материал приобретает морфологию, характерную для данно-
го вида. У Hantzschia amphloxvs микротрубочковый центр (МЦ) игра-
ет главную организующую роль в развитии новых половинок панциря
после митоза ( Piokett-Heaps, Kowalski, 1981). Удивительно, что
новая створка возникает здесь в силикалемме вблизи центра клетки
и лишь позже перемещается к месту своего окончательного положения
в ходе существенной внутриклеточной перестройки под контролем МЦ.
После ее образования формируются новые пузырьки Гольджи, которые
сливаются с образованием пояскового кольца ( Dawson, 1973). Во
время отложения кремнезема кремний поглоп&ется клеткой в виде
кремниевой кислоты посредством активного транспорта (lewin,Chen,
1968). Этот процесс ингибируется в присутствии двуокиси германия
(lewin, 1966).
Клеточная стенка водорослей всегда формируется вне плазма-
леммы (как у высших растений). Ее структура весьма разнообразна,
но в целом можно выделить два типа: неполный и полный. Первый ха-
рактерен для ряда жгутиковых, а второй - для всех прокариотных
водорослей и большинства представителей Chlorophyceae, Xanthophy-
ceae, Phaeophyceae И Rhodophyceae (табл. 3.1).
Неполную клеточную стенку обычно называют панцирем или доми-
ком. Это бутылевидная или чашевидная структура, внутри которой
находится клетка со жгутиками. Примеры можно найти среди prasi-
nophyceae, Chrysophyceae (рис. 3.19) И Chlorophyceae.
97
у празинофита Platvmonaa панцирь (иногда неправильно называемый
такой) состоит из полисахаридов; он формируется из звездчатых
частиц, продуцируемых пузырьками Гольджи. Микрофибриллярные цел-
люлозно-пектиновые панцири обычны у. золотистых водорослей; их об-
разование, возможно, происходит ±n situ, вне клетки (Dodge,
1978). Иногда наблюдается их импрегнация кремнеземом, а у неко-
торых родов - обызвествление. Панцири золотистых водорослей край-
не разнообразны по форме, что используется в таксономии. Среди
Chlorophyceae панцирь характерен для видов семейства Phacota-
ceae (Volvocales).y ИХ клеток со структурой, типичной для других
представителей порядка, он импрегнирован соединениями железа. У
Dysmorphococcus globosus незрелый панцирь включает фибрилляр-
ные (внешние) и гранулярные '(внутренние) компоненты. На началь-
ном этапе развития происходит биоминерализация железом и форми-
руются поры. В конечном счете через поры выделяется слизистый
материал, и они становятся незаметными (Porcella, Walne, 1980).
Клеточные стенки Cyanophyoota и Prochlorophycota обладают
прокариотными чертами. У первых стенка, или оболочка, очень сход-
на с таковой у грамотрицательных бактерий (рис. 3.20; Allen,
1968). Она располагается между плазмалеммой и образованиями, на-
зываемыми слизистым влагалищем, капсулой или чехлом. Ранее обо-
лочка описывалась как четырехслойная, а слои обозначались И, ЫТ,
Ш1И L1V (от центра к периферии): Ы и Ш1- электронопро-
зрачные, а их и liv - электроноплотные. Электронопрозрачные
слои ныне считаются пространствами между Ы1 и ЫУ; приписывав-
шаяся им структура сейчас признается артефактом фиксации ( Drews,
Weckesser, 1982). Самый внутренний слой (Ы1) содержит основную
часть пептидогликанов (до 50% сухой массы), включая муреин, гли-
копептид и мукоцептид. Он отделен от плазмалеммы электронопро-
зрачным пространством ы . Внешняя клеточная мембрана (ЫУ)
выглядит на ультратонких срезах под электронным микроскопом как
двухрядная структура ( Butler, Allsopp, 1972), отделенная от
пептидогликанового слоя электронопрозрачным пространством ЫН.
У нитчатых форм через поперечные стенки могут проходить тон-
кие плазмодесмы. Влагалище предохраняет клетки от высыхания и у
многих представителей важно при скольжении. Единства мнений отно-
сительно его химической природы нет, но возможно, что оно состоит
из пектиновых кислот и мукополисахаридов. Предполагаются ( Martin,
98
-Рис,. 3.19. Схематический разрез панцирной золотистой водо-
росли Chrvsococcus rufescens. В - ветвистый цитоплазмати-
ческий вырост; Г - тельце Гольджи; ПГ - пузырьки Гольджи;
ДЕ - длинный жгутик; КЖ - короткий жгутик; Ии - клеточная
мембрана; М - митохондрия; МГ - масляная глобула; П - пан-
цирь; ПК - пигментные камеры; ПЛ - пузырьки лейкозина;
ПЦ - перипластидные цистерны; ПЭС - перипластидная эндо-
плазматическая сеть; Р - рибосомы; СВ - сократительная ва-
куоль; X - хлоропласт; У1Г -уплощенные пузырьки; ФП - фиб-
риллярные пучки; Я - ядро; ЯО - ядерная оболочка ( Belcher,
99
Рис. 3.20. Организация клеточных покровов у синезеленых во-
дорослей по данным электронной микроскопии. В - влагалище;
КС - клеточная стенка; Мр - муреин; ПЛ - плазмалемма; ПС -
пластичные слои; т,т _ ыу - слои клеточной стенки (на ос-
нове Carr, Whitton, no Jost)
Wyatt, 1974) три типа структур: влагалище, видимое без окраски,
слизистый чехол с неясными границами и сходный чехол с хорошо
очерченным контуром. Влагалище часто пигментировано. Толщина его
варьирует в зависимости от внешней среды и роста.
Клеточная стенка Prochiorophycota напоминает стенку Суа-
ncphycota (Whatley, 1977); в ней обнаружена мурамовая кисло-
та (Moriarty, 1979).
Истинные клеточные стенки, во многих отношениях сходные с
таковыми у высших растений, встречаются у Rhodophyceae, Phaeophy-
ceae, Xanthophyceae, Chlorophyceae И Charophyceae;
кроме того, истинная стенка окружает цисты некоторых Dinophyceae
и вегетативные клетки представителей Eustigmatophyceae . Как
правило, эти стенки построены из микрофибриллярного каркаса
(рис. 3.21), погруженного в аморфный слизистый материал. Моцут
также присутствовать инкрустирующие вещества, например кремнезем,
карбонат кальция или спорополленин (см. обзоры B.Siegel, S.Sie-
8el 1973; Mackie, Preston, 1974). За исключением отдель-
ных Rhodophyceae И Phaeophyceae , особенно тех, ИЗ стенок кото-
рых получают экономически важные вещества, и ряда Chlorophyceae
(RickettaHeaps, 1975), У подавляющего большинства водорослей
клеточные стенки подробно не изучались. В целом, однако, считает-
100
Рис. 3.21. Стенка Valonia
(Chlorophyceae). Видны два
последовательных слоя фиб-
рилл, ориентированных друг
относительно_друга под уг-
лом около 90 и (х 14 000)
(Dodge, 1973)
тропиках. Хорошо сохраняющиеся
ся, что, как и у высших расте-
ний, в образовании их микро-
фибриллярного материала участ-
вуют тельца Гольджи. Этот ма-
териал собирается в пузырьках
Гольджи под плазмалеммой и за-
тем выходит наружу, где из на-
го синтезируются микрофибрил-
лярные компоненты. Электронная
микроскопия (особенно исполь-
зование методов замораживания-
скалывания) внесла огромный
вклад в понимание микроархи-
тектуры стенки, однако точный
механизм, определяющий ориен-
тацию микрофибрилл, до сих
пор выяснен слабо. Накопление
и синтез аморфного материала
стенки, возможно, также связа-
ны с тельцами Гольджи, посколь-
ку его формированию часто со-
ответствует повышенная актив-
ность аппарата Гольджи. Пред-
полагается однако, что место
синтеза спорополленина - едер-
ная оболочка (Atkinson et al.,
1972). Активно изучалось накоп-
ление инкрустирующих веществ,
в частности процесс обызвест-
вления у Rhodophyceae и
Chlorophyceae (см. гл. 6),
крайне важный и с экологичес-
кий точки зрения, особенно в
ископаемые остатки известковых и
окремнелых форм широко используются в стратиграфических исследо-
ваниях.
Ниже мы рассмотрим разнообразие истинных клеточных стенок
по основным классам водорослей.
101
Bhodophyceae
У Bangiophycidae стенка макроскопической фазы (например, у
Кяпс1а. Porphyra) содержит микрофибриллярный каркас из ксилана
с J5-1,3-связями; недавно показано (Gratz et al., 1980;
Iftikai et al., 1981), что на стадии Conchooells этих водорос-
лей в стенке присутствует целлюлоза. У некоторых бангиевых также
отмечалась внешняя кутикула из маннана с J3-1,4-связями
(Frei, Preston, 1964) или белка (Hanic,Craigie,I969)• У Flori-
deophycidae каркас стенки из целлюлозы, причем его фибриллы рас-
положены беспорядочно. Аморфные слизи, составляющие остальную
часть стенки (дс 70%, Dixon, 1973), обычно образованы водораст-
воримыми сульфатированными галактанами ( Percival, McDowell,1967).
Среди слизей имеются экономически важные агары и каррагинаны, ко-
торые при определенных условиях образуют гели. У всех представи-
телей Corallinaceae (Cryptonemiales ) и некоторых Nemaliales
(например, Падога. Захаига ) откладывается карбонат кальция
(кальцит у Corallinaceae и арагонит у Nemaliales ) и обычно на-
ряду с ним, но в меньших количествах, карбонаты магния и строн-
ция. Процесс обызвествления связывали с фотосинтезом (Pearse,
1972), хотя он может происходить и в темноте ( Okazaki, 1970).
Имеется несколько теорий относительно его биохимического механиз-
ма (см.Digby, 1977а,Ъ; Lee, 1980);
Поровая пробка, или поровое соединение (рис. 3.22, Puesohel,
1980), - структура стенки Florideophycidae и фазы Conohocelis
представителей родов Bangia и Porphyra (Bangiophycidae). Она
отмечена и у одной морской зеленой водоросли ( Brawley, Sears,
1980). У багрянок первичные поровые пробки формируются между дву-
мя клетками во время деления, а вторичные развиваются между клет-
ками, примыкающими друг к другу латерально. Эти пробки обычно
Двояковыпуклые, со срединной перетяжкой ( Pueschel, 1977). Обра-
зование их происходит путем неполного цитокинеза, с накоплением
эндоплазматической сети в отверстии между дочерними клетками и
последующим сосредоточением здесь материала пробки ( Norris, Kug-
rens, 1982). Ее матрикс содержит белок ( Rames, 1971, Pues-
chel, 1980), а также, возможно,липиды и полисахариды. Не исклю-
чено, что поровые пробки обеспечивают структурную прочность тела
водоросли ( Kugrens, West, 1973; Pueschel, 1980), межклеточный ,
транспорт(wetherbee, 1979) или связи паразит-хозяин (Golf, 1979).
102
Рис. 3.22. Поровая пробка у багрянок (A) Galaxaura sp. (х 30 400)
и (Б) Pseuaogloiophloea sp. (х 64 ООП1 < электронные микрофото-
графии любезно предоставлены д-ром С.М. Pueschel)
При развитии после оплодотворения гонимобласта межклеточный тран-
спорт метаболитов облегчается за счет удаления пробок с образова-
нием клеток слияния ( Turner, Evans, 1978).
Phaeophyceae
Клеточные стенки этих водорослей по крайней мере двуслойные,
с внутренним целлюлозным микрофибриллярным окелетом. Микрофибрил-
лы обычно расположены беспорядочно, хотя иногда ориентированы в
продольном или горизонтальном направлениях (Dodge, 1973). Аморф-
ные компоненты - в основном альгиновая кислота и фукоидан (суль-
103
фатированные полисахариды; Mackie, Preston, 1974). Слизь и кути-
кула состоят главным образом из альгиновой кислоты, полимера
пятиуглеродных кислот( D-маннуроновой и L—гулуроновой кислоты),
относительная доля которых варьирует (Hang et al., 1969). На
альгиновую кислоту иногда приходится белее 30% сухой массы во-
доросли. Альгинаты (соли этой кислоты), обладающие ценными эмуль-
гирующими и стабилизирующими свойствами, широко используются в
промышленности (гл. 9). (0 биосинтезе альгиновых кислот см.
barsen, 1981; Percival, 1979; McCandless, 1981.)
У паренхиматозных бурых водорослей между большинством клеток
находятся плазмодесмы или поры (рис. 3.23), а уLaminariales,
Fucales и некоторых Dictyotales поры группируются в первичные
поровые поля. Ситовидные элементы (рис. 3.24), сходные с имеющи-
мися у высших растений, встречаются у laminariales , обеспечи-
вая быстрое перемещение продуктов фотосинтеза (Schmitz, 1981).
Xanthophyceae
Стенка отдельных желтозеленых водорослей (Tribonema aequale;
Clear, Percival, 1972; Vaucheria spp.; Parker et al.,1963)
состоит из беспорядочно расположенных целлюлозных микрофибрилл
(до 90% у Vaucheria ). У нитчатых форм она образована двумя час-
тично перекрывающими одна другую половинами. Эта особенность не-
заметна под световым микроскопом, пока клетки не обработаны соот-
ветствующим реактивом, например концентрированной гидроокисью ка-
лия. У Ophiccytium majus (Hibberd. Leedale, 1971) половинки
представлены колпачком постоянного размера, посаженным на труб-
чатую базальную часть,которая удлиняется во время роста. У нит-
чатой водоросли Tribonema стенки состоят из Н-образных частей
более или менее равного размера, поочередно перекрывающих друг
друга.
104
Рис. 3.23. Обобщенная схема ультраструктурних особенностей клет-
ки Scytosiphon (Phaeophyceae). В - вакуоль; Г - аппарат Голь-
джи ; КП - Колпачок пиреноида; М - митохондрия; ОХ - оболочка хло-
ропласта; П - плазмодесмальное поровое поле; Пи - пиреноид; и -
стенка; Ф - физода; X - хлоропласт; ХЭС - хлоропластная эндоплаз-
матическая сеть; Ц - центриоль; 30 - шероховатая эндоплазматичес-
кая сеть; Я - ядро; ад - ядрышко ( Clayton, Beakes, 1983)
105
Рис. 3.24. Продольный разрез Alarla marginala (Phaeo-
phyceae). Видны коровые клетки (К) и внутренняя сердце-
вина из ситовидных (С) и гифальных (Г) клеток (х 170)
( Schmitz, Srlvastava, 1975)
Chlorophyceae
Хотя клеточная стенка подробно изучена у относительно не-
многих представителей этого класса, в его пределах обнаружено
большое разнообразие ее структуры и состава. У многих видов она
двухслойная, с более компактным внутренним слоем и капсулой или
слизистым слоем снаружи. Додж ( Dodge, 1973) выделяет три основ-
ные категории стенок: I) состоящие преимущественно из целлюлоз-
ных микрофибрилл (некоторые Cladophcrales, Slphcnocladales);
2) с менее четкой организацией целлюлозных микрофибрилл и гораз-
до большей долей аморфного материала (некоторые uivales; Oedc-
goniales; многие десмидиевые и коккоидные виды); 3) не содержа-
щие целлюлозного микрофибриллярного каркаса (многие сифональные
виды и некоторые Volvccalea ). У сифональных форм стенки состо-
ят из микрофибрилл полимеров ксилозы (например, у Caulerpa; Ma-
ckie, Percival, 1959) или маннозы ( Acetabularia. Codium: Frei,
Freston, 1961), погруженных в матрикс, по-видимсиу состоящий
106
в основном из гемицеллюлоз. У многих сифональных морских видов
стенки пропитаны соединениями кальция; процесс обызвествления и
отложения карбоната кальция у ризофитовых водорослей, в частно-
сти у няНшоНя , играет важную роль в формировании структур рых-
лого дна мелководных тропических местообитаний. Хитин обнаружен
в стенках Fifho-phora oedogoniana (Cladophorales) (Pearlmutter,
Lembi, 1978). Стенки представителей Oedogonialee уникальны в
том плане, что разрываются в верхней части при клеточном делении
с образованием рубцов, или "колец" (апикальных колпачков) (Hill,
Machlis, 1968; Pickett-Heaps, 1975). Этот процесс схематично
показан на рис. 3.25.
Иногда Число слоев стенки достигает шести или семи (напри-
мер, у Chlamydomonae reinhardtil; Robert et al., 1972). У
Valonia она заметно упрочняется за счет чередования слоев, мик-
рофибриллы в которых располагаются перпендикулярно микрофибрил-
лам соседнего слоя (рис. 3.21; ср. Roelofsen,1966).
Интересно, что у некоторых Chlorophyceae по составу кле-
точной стенки происходит биохимическая смена поколений (ср.щс_
Candless, 1981). Наиболее известный пример - Bryopsis , У гаме-
тофитов которого она состоит из ксилана и глюкана, а у спорофи-
тов - ИЗ маннанов ( Huizing, Rietema, 1975).
Charcphyceae
Стенки харовых состоят из целлюлозных микрофибрилл и в
большинстве случаев сильно обызвествлены. В ризоидах Kttella
( Chen,1980) эти микрофибриллы ориентированы преимущественно
параллельно оси клетки, а в клетках междоузлий - перпендикулярно
ей. Для внутренних продольных стенок Chara описаны сложные мем-
бранные структуры, харасомы; их функция неясна, но, по-видимому,
они не связаны с транспортом бикарбоната (НСО~)( Franceschi,
Lucas, 1980).
4
107
?ис. 3.25- Схема клеточного деления Oedogonium (Chlorophyceae),
показывающая разрыв стенки и образование колпачков. А. Телофаза
митоза. Б. Цитокинез. В, Г, Д. Разрыв стенки и образование кол-
пачка пс“мере удлинения протопласта (по Pickett-Heaps, 1975)
Жгутики
Жгутики встречаются в большинстве отделов водорослей. Эти
локомоторные структуры чрезвычайно важны для выяснения филогении;
электронная микроскопия произвела настоящую революцию в нашем по-
нимании их структуры и функции (исчерпывающий обзор см. Moestrup,
1982).
Обладая в значительной степени однообразней внутренней мик-
ротубулярной структурой, жгутики сильно различаются по внешней
морфологии, способу прикрепления и корневым системам.
Относительно происхождения жгутиков в ходе эволюции до сих
пор имеются только предположения (см. Margulis, /981). Так, Ка-
валье-Смит ( Cavalier-Smith, 1978) утверждает, что микротрубочки
жгутиков или их предшественники возникли вместе с ядром, тогда
как Стьюарт и Маттокс ( stewart, Mattcx, 1980) полагают, что
жгутики возникли первжи.а митотическое веретено эукариот эволю-
ционировало из жгутикового аппарата или его предшественника. Как
бы то ни было, жгутики должны рассматриваться как составные части
клетки, поскольку их мембрана составляет единое целое с плазмалем-
мой, а их внешние структуры являются клеточными производными. Их
роль в обеспечении подвижности клетки несомненна, однако только
сейчас цитологи берутся за решение сложных вопросов, касающихся
связи между морфологией и функцией жгутиков.
108
Число, характер прикрепления и организация жгутиков иногда
довольно постоянны для отдела; схематично основные формы жгутико-
вания отражены на рис. 3.26. Подвижные клетки водорослей чаще
всего двужгутиковые, хотя у зеленых водорослей нередок и четырех-
жгутиковый тип; обычно считается, что последний возник из перво-
го (ср. Melkonian, 1982а). Одножгутиковые формы, возможно, про-
изошли от двужгутиковых, тогда как уникальны? трехжгутиковые
зооспоры.зеленой водоросли Entocladia wittrockii.возможно. име-
ли четырехжгутикового предка. У многожгутиковых (стефаноконтных)
типов известны различные сложные модификации базальной системы
жгутиков или их внутренней структуры (обсуждение см. Moestrup,
1982).
Формы с одинаковыми по длине и облику жгутиками называют
изоконтными; у гетероконтных форм жгутики различны (например,
гладкий и перистый), равной или неравной длины ( Moestrup, 1982).
Формы со жгутиками одного типа, но разной длины называют анизо-
контными. Егутики на клетке могут двигаться координированно (гомо-
динамично) или независимо друг от друга (гетеродинамично). Абер-
рантные, или необычные, жгутики встречаются у диатомей, криптофи-
тов, у некоторых бурых водорослей, динофитов, эвгленовых и прим-
незиофитов ( Pavlovales).
Истинные жгутики состоят из наружной аксонемы, расположенной
под плазмалеммой и включающей девять дублетов микротрубочек, окру-
жающих две центральные микротрубочки (9+2)-схема (рис. 3.27). Мо-
дификации микротубулярной (9+2)-архитектуры редки, но известны
для спермиев некоторых диатомовых (9+0) и зеленой водоросли
Golenkinia minutisaima (9+1; Moestrup, 19?2); сюда же можно от-
нести гаптонему примнезиофитов (рис. 3.28) (семь микротрубочек,
окруженных тремя концентрическими мембранами; Manton, 1964а) и
"псевдоцилии" тетраспоровых Chlorophyceae (9 одиночных + 0),
например у Tetraspcra (Lembi, Walne, 1971).
Морфология кончика жгутика вызывает определенный филогене-
тический интерес. Установлено (Melkonian, 1982а), что у многих
изученных форм периферические дублеты исчезают ниже кончика, а
центральные микротрубочки, заходят дальше их, хотя эта картина,
по-видимому, меняется во время движения за счет скольжения микро-
трубочек относительно друг друга (рис.’ 3.29). У Friedmannlя
israe.1.6?3,13 описано (Melkonian, 1982 ъ ) следующее изменение ор-
109
Рис. 3.26. Некоторые типы жгутиков и их расположения у во-
дорослей. А. Изоконтные, гомодинамичные, гладкие, прикреп-
ленные апикально (например, у Chlamydomonas ). Б. Гетеро-
контные, неравные, прикрепленные апикально ;например, у
желтозеленой водоросли Tribonema). В. Изоконтные, гомоди-
намичные, прикрепленные апикально, с центральной гаптонемой
(например, у примнезиофита Chrysochromullna )• Г. Изоконт-
ные, гомодинамичные, прикрепленйые аййкально. Д.“Изоконтные,
выходящие из жгутикового углубления (например, у празинофи-
та Pyramimonas ). Е. Многожгутиковый, или стефаноконтныи,
тип (жгутики расположены субапикальным венцом) (например, у
Oedogonium из Chlorophyceae ). Ж. Гетероконтные, гетероди-
намичные, с боковым прикреплением (например, у бурой водо-
росли Ectocarpus). 3. Поперечный и продольный гетероконт-
ные., прикрепленные базально (например, у динофита GymnodiM-
“ч>. И,Одиночный апикальный, двусторонне-перистый. К. Сда-
ночный“апикальнцй однорядно-перистый. Л. Одиночный субапи-
кальный
ганизации микротрубочек от основания к кончику жгутика: 9+2>4+2>
2+2>0+2. Суживающаяся вершина (акрснема) содержит продолжение
центральной пары микротрубочек (ср. Jonsson, Chesnoy, 1974) или
одну-единственную микротрубочку.
no
Рис. 3.2?. Схематическая реконструкция жгутиковой аксоне-
мы, переходной зоны, базального тела, элементов корневых
соединительных волокон и т.Д. у Chlamydomonaa reinhardtii
(Chlorophyceae) (Cavalier^-Smith ?., 1974)
Ill
Рис. 3.28. Pavlova oaloeolata
(Phynmesiophyoeae). Полусхе-
матическое изображение co жгу-
тиковыми придатками, показанны-
ми только там, где они не закры-
вают собой других клеточных
структур. Вентральные лопасти
хлоропласта удалены, чтобы пока-
зать внутри него ламеллы. В -
вакуоль, содержащая полифосфаты
(ПФ) и парамилон (П); Г - аппа-
рат Гольджи; ГЛ - глазок; ГН -
гаптонема; М - митохондрия; ПЦ -
поверхностные цистерны; У - уг-
лубление; X - хлоропласт; Я -
ядро. Масштаб I мкм ( van der
Vee*. 1976)
Апикальный
колпачок
Центральная
трубочка
Дистальные
нити
Пробка
Компонент А
Компонент В
Рис. 3.29. Реконструкция конца жгутика на основе не-
гативного окрашивания И ультратонких срезов жгутиков Chlamy-
domonaa u Tetrahymena ( no Dentler;oM.Oillon, 1981)
112
Акронемы’(рис. 3.30) обычны у Prymnesiophyceae, Chlorophy-
ceae И части Prasinophyceae (Loxophyceae aensu Moestrup,
1982). Тупые вершины с (9+2)-организацией в норме встречаются у
криптофитов, данофитов, эвгленовых, празинофитов (за исключением
Loxophyoeae sensu Moestrup, 1982) и харовых. Перистый жгутик
всегда тупой у гетероконтных форм, за исключением бурых водорос-
лей. У последних он часто несет крайне тонкую акронему, тогда как
у гладкого жгутика она двутяжевая. Предполагалось, что у мужской
гаметы (рис. 3.31) она служит для спаривания( Muller, Falk,
1973), а у зооспор, где акронема может первой вступать в контакт
с субстратом, она клейкая (ср.Chorda tomentosa; Toth, 1976).
Наружная аксонема растет за счет удлинения; этот процесс
описан у Chlamydomonas spp. и Ochromonas danica (Rosenbaum,
Child, 1967;Rosenbaum et al., 1969). По-видимому, новый мате-
риал добавляется дистально к девяти периферическим парам микро-
трубочек, тогда как центральные, прикрепленные к терминальной мем-
бране жгутика, увеличиваются в ходе проксимального добавления ком-
понентов.
О формировании и функционировании жгутиковой мембраны извест-
но мало. У многих водорослей она гладкая, хотя встречаются различ-
ные типы орнаментации, включая волоски и чешуйки. Эта орнамента-
ция по структуре и образованию бывает сходна с орнаментацией те-
ла клетки или отличается от нее. Она увеличивает диаметр жгутика,
но ее функция с точки зрения динамики жгутика точно не выяснена.
Жгутиковые чешуйки встречаются только у водорослей и некото-
рых, возможно родственных им, нефотосинтезирующих организмов
(Moestrup, 1982). Они отмечены у празинофитов, харовых и изредка
у, криптофитов и динофитов. Образование чешуек обычно связано с
цистернами и пузырьками Гольджи (рис. 3.32), хотя у Rickertia
sagittifera (Moestrup, 1982) они формируются в пузырьках на по-
верхности митохондрий. Жгутиковые чешуйки празинофитов откладыва-
ются из особого резервуара. На одном и том же жгутике иногда
встречается несколько типов чешуек, которые могут располагаться
слоями (рис. 3.33, А,Б).
Относительно жгутиковых волосков существует значительная пу-
таница, и от терминологии, сложившейся до широкого использования
электронного микроскопа, следует отказаться ( Manton, 1965). Они
обнаружены во всех классах chromophyoota и зеленых водорослей,
хотя у большинства Prymnesiophyceae и Chlorophyceae ЖГУ тики, как
из
Рис. 3.30. Фотография с отте-
ненного электронномикроскопи-
ческого препарата спермин
Golenkinia minutissima;
видно два жгутика с акронема-
ми (х 5250) ( Moestrup, 1982)
Рис. 3.31. Схематическая обобщенная реконструкция спермия лами-
нариевой водоросли (Phaeophyceae ), показывающая расположение
основных органелл. ЗЖ - задний жгутик; М - митохондрия; Ма - во-
лоски (мастигонемы); П - пузырек типа "В"; Ш - передний жгу-
тик; х - хлоропласт; Я - ядро ( Henry, Cole, 1982Т
8—121
114
Рис. 3.32. Электронная микрофотография примиезиофита
Chrysochromuilna.показывающая расположенное над ядром
крупное тельцегольджи, продуцирующее чешуйки (х 37100)
(любезно предоставлено д-ром J.D.Dodge)
правило, голые. Функция жгутиковых волосков выяснена недостаточ-
но, хотя предполагается, что,увеличивая диаметр жтутина, они уси-
ливают его удар ( Moestrup, 1982). Недавние исследования позво-
лили предположить, что эти волоски могут формироваться двумя пу-
тями: в аппарате Гольджи (например, у празинофитов; Моеstrap,
Thomsen, 1974; Melkonian, 1982с) И эндоплазматической сети
(включая околоядерное пространство) ( Moestrup, 1982). Выделяют
( Moestrup, 1982) два типа волосков: I) трубчатые и 2) простые
(нетрубчатые). Первые подразделяются на криптофитовые, трехчаст-
ные и празинофитовые.
Простые волоски тонкие и очень нежные; возможно, они состо-
ят из одного ряда субъединиц. Трубчатые волоски включают две или
более областей, из которых проксимальная обычно толстая и трубча-
тая, а дистальные напоминают простые волоски.
Криптофитовые трубчатые волоски (рис. 3.34) расположены в
115
A
Б
'4 ^"здал.'Л-ай*- л л *> и » ' w -
Рис. 3.33. А. Чешуйки празинофита Mamiella(Mephroaelmia)
gilva, расположенные восемью рядами в один слой, покры-
ВЙЙ1ЦЙЙ жгутик (х 75 000). Б. Чешуйки Nephroeelmia. oliva-
сеа, расположенные в три слоя; здесь же видны толстые
вЗлоски (х 60 000). В. Егутиковые чешуйки празинофита
Dolichomaatrix nummuTifera (х 40 000) (А - Moeatrup,
ТШЗ; Б - Moeatrup, Ettl? 1979; £- Manton, 1977)
два супротивных ряда на более длинном жгутике и в один ряд на бо-
лее коротком. В отличие от трехчастных у них отсутствует четкое
подразделение на основание и ствол. Только у празинофитов с хлоро-
филлом Ь. я некоторых харовых трубчатые волоски присутствуют бо-
лее чем на одном, жгутике. Трехчастные (гетероконтяые) волоски об-
наружены у рафидофитов, золотистых, диатомовых, желтозеленых, бу-
рых (рис. 3.35) водорослей И Euatigmatophyoeae, а также у неко-
торых низших грибов ( Oomycetea и Hypliochytridiomycetes; Moest*-
rup 1982). Они прикрепляются к жтутику двумя супротивными ряда-
ми, возможно в плоскости центральных трубочек. Выделены ( Bouck,
1969) три зоны таких волосков: базальная часть, полый отвод и ди-
116
Рис. 3.34. Трубчатые жгутиковые волоски крип-
тофита (врезка: х 750), расположенные в два
супротивных ряда на более длинном жгутике и
в один ряд на более коротком (х 14 625)
(Moestrup, 1982)
стальные части, очень нежные и часто утрачиваемые при изготовле-
нии препаратов для электронной микроскопии. Морфология празинофи-
товых волосков, по-видимому, разнообразна, и на одном и том же
жгутике иногда встречается несколько их типов (например, у
Mantoniella; Manton, Parke,I960). У харовых жгутиковые волоски
обнаружены на зооспорах Chaetоsphaeridimn (Moestrup, 1974), а
также на зооспорах и спермиях Coleochaete scutata (Marchant in
Pickett-Heaps, 1975; Marchant in Moestrup, 1978; Graham, Mc-
Bride,1979). У них есть родственные черты с волооками празинофитов
и других зеленых водорослей.
Проотые, или нетрубчатые, волоски имеются у представителей
разных групп. У эвгленовых и динофитов они уникальны, хотя еще
мало изучены. В отличие от трубчатых простые волоски не дифферен-
цированы на области. У эвглеяовых они расположены в один ряд пуч-
ками (ср.воцсь et al., 1978), у Eutreptiella gymnastics - ПО
четыре (Throndsen in Moestrup, 1982),a у Euglena gracilis ~ ОТ
трех до семи в пучке (Moestrup, В82). Эти волоски у Eugle-
na, проходя через мембрану, прикрепляются к параксиальному
117
Рис. 3.35. Детали трехчастных волосков на
переднем КГУТИКе.СПерМИЯ Laminaria ^igita-
ta (Phaeophyceae;. QB - основание вбЗоока;
СВ - ствол волоока; И - терминальная нить
(х 47 600) ( Maier, Muller, 1982)
тяжу (см. ниже) и к одному из периферических дублетов аксонемы.
На жгутиках некоторых эвгленовых присутствует, кроме того, плот-
ный войлок из более коротких волосков, расположенных продольны-
ми, поперечными или спиральными рядами. Детали строения этого
войлока крайне сложны (Bouck et al., 1978) (рис. 3.36).
Оба жгутика динофитов мохут нести волоски (рис. 3.37), но
разных типов. У некоторых видов на поперечном жгутике волоски на-
ходятся с одной стороны и, видимо, сгруппированы в пучки (clarke,
Pennick, 1972; Leadbeater, Dodge, 1967), в которых ОНИ
имеют разные размеры. Сведения о волосках продольного жгутика ме-
нее определенны; у некоторых видов они,- возможно, отсутствуют.
Простые волоски, описанные у других групп, не собраны пучка-
ми. ОНИ известны у Pedinomonas (Loxophyceae, выделяемые из Рга-
sinophyceae; Manton, 1965), некоторых видов Chlamydomonaa
(Chlorophyceae; Ringo, 1967;McLean et al., 1974), Pteromonaa
(Chlorophyceae; Belcher, Swale,1967), Purcilia (Chlorophyceae;
Belcher, 1968), Qloeochaete(Kiea. 1976) И Cyanophora
118
Рис. 3.36. Реконструкция сложного войлока из
коротких волосков на жгутике Ецк1епа кгасШа
( Euglenophyceae) (из Moestrup, 1У82, 6 Измене-
ниями; ПО Bouck et al., 1978)
( Glaucophyta*, Thompson, 1973). Очень тонкие волоски образуют
плотный покров на переднем жгутике Pavlovales (примнезиофиты) и
у некоторых золотистых водорослей (см. Moestrup, 1982).
Отличный от чешуек или волосков покров, описываемый как
"войлок", обнаружен у ряда групп в результате электронномикро-
скопических исследований. У некоторых chlorophyceae и Prymne-
siophyceae его имеют жгутики, ранее считавшиеся гладкими. У жаро-
вых он находится под внешним слоем чешуек. Среди функций этого
покрова, состоящего, вероятно, из гликопротеинов, предполагают
слипание при спаривании ( Cavalier - Smith, 1975), защиту от фаго-
цитоза и прикрепление (Moestrup, 1982).
Дополнительные детали внешнего жгутика - параксиальные вклю-
119
Рис. 3.37. Фотография в просвечивающем
электронном микроскопе поперечного жгутика
динофита Prorocentgum rainimiua » Видны ак—
сонема (широкий тяжТ7"11опёречно полосатый
тяж (парафлагеллярное тело) и жгутиковые
волоски (х 5600) ( Dodge, 1983)
чения. Эти структуры находятся под его мембраной и могут быть
подразделены на жгутиковые вздутия, параксиальные тяжи и жгути-
ковые ШИПИКИ ( Moestrup, 1982).
Большинство вздутий встречается у основания жгутика и свя-
зано с глазками (см. ниже). Их ламеллярная или кристаллическая
структура наводит на мысль, что они функционируют как часть фо-
торецепторного аппарата (см. Robenek, Melkonian, 1983). У гете-
роконтных групп кроме Rustigmatophyoeae.T.e. у золотистых, жел-
тозеленых и бурых водорослей, вздутие находится у основания
120
гладкого жгутика, а у Euetigmatophyoeae (уникальная черта клас-
са) - у основания перистого. Жгутиковое вздутие присутствует и у
одного не реагирующего на свет примнезиофита Diacronema (Green,
Hibberd, 1977). У эвгленовых оно (парафлагеллярное тельце,
PFB Т; ср. Robenek, Melkonian, не такое, как у гетероконт-
ных водорослей, и в большинстве случаев овязано с более длинным
жгутиком, находясь на некотором расстоянии от его основания меж-
ду переходной зоной и каналом резервуара. Очень необычные жгути-
ковые вздутия имеются у Urospora (Chlorophyceae), нескольких
золотистых водорослей и представителей криптофитов. У Uroapora
(рис. 3.38) проксимальные части каждого из четырех жгутиков зоо-
споры несут вздутия, или "крылья", которые прикрепляются изнутри
параллельными плоскими пластинками к периферическим дублетам
( Kristiansen, 19745 Sluiman et al., 1981).
Параксиальные тяжи, по-видимому, механически укрепляют жгу-
тики динофитов (рис. 3.37), эвгленовых и некоторых золотистых
водорослей ( Bedinellales sensu Moestrup, 1982).
Они обнаружены также в жгутиках бодоноидов и трипанооом
(простейшие). Обычно такие тяжи проходят вдоль всей или большей
части длины внешнего жгутика и ммут быть поперечно исчерченными
(например, у поперечного жгутика динофитов) или полыми. Наличие
АТРазы вдоль параксиального тяжа у Euglena (Piccini et al.,
1975) говорит о том, что он как-то связан с потреблением энергии.
Жгутиковые шипики встречаются нечасто и известны только у
мужских гамет оогамных бурых водорослей. У Biotyota имеется
ряд из 12 коротких шпиков на перистом жгутике; они прочно при-
крепляются к одному из периферических дублетов аксонемы (Manton,
1959). У некоторых PUcales (Pucu.st H-imnnthaH а; Xiphophora и
Hormoairatcp. Moestrup, 1982) присутствует один-единственный
птипик длиной до 1,0 мкм (например, у mrnanthalia; рис. 3.39),
хорошо видимый уже в световом микроскопе.
Входя в клетку, аксонема оканчивается базальным телом. Дета-
ли строения переходной зоны (рис. 3.27, 3.40 и 3.41) между этими
структурами очень сложны и являются одним из наиболее широко ис-
пользуемых показателей филогенетических отношений ( KLbberd,I972;
Moestrup, 1982). Мёструп ( Moestrup, 1982) различает шесть типов
От английского paraflagellar body _ Прим. ред.
121
Рис. 3.38. Схематическая модель жгутикового аппарата
зооспоры grospora penicilliformis (Chlcrophyceaej.
ВВ - восходяще'ё,"5олдйИйд" ЬВ’ - бййовое волокно; КП -
колпачковая пластинка; К - корни с полосатыми волок-
нами I (Р1) и II (Р2)
Показана также внутренняя структура крылатой части
жгутиковой аксонемы (Sluiman et al.f 1982)
переходной зоны: I) криптофитовнй; 2) динофитовый; 3) гетероконт-
ный (или тип Oohromonas ); 4) гаптофитовый (или тип Ргуипеsl-
um) ; 5) эвгленоидный (или тип Entosiphon) и 6) тип зеленых водо-
рослей (см. рис. 3.41). К существенным различительным признакам
относятся: один или более дисков на конце центральных трубочек,
переходная улитка (TH) , обнаруженная у некоторых гетероконтных
классов (Chryscphyceae, Xanthcphyceae И Eustigmatophyceae ),
но отсутствуювдя у бурых водорослей, рафидофитов и диатомей,
звездчатые структуры (примнезиофиты, за исключением Pavlovales,
и все водоросли о хлорофиллом^. ). На продольном срезе звездча-
тая зона выглядит Н-образной, а на поперечном видно, что ее лучи
соединяются с А.-трубочками периферических дублетов аксонемы. Род
Pvraminionaa (празинофиты) уникален в том отношении, что у него
имеется и "спиральное волокно", или переходная улитка (напоминаю-
щая признак гетероконтных форм), и звездчатая структура (харак- .
От английского transitional helix. - Прим, ред.
122
Рис. 3.39. Спермий Hlman-
thalia (Phaeophyceae);на жгу-
WKe"xopomo заметен шипик,
длина которого может превы-
шать 1,0 мкм (х 5400)
(Moestrup, 1982)
Рис. 3.40. Внутренняя структура базального тела и жгути-
ковой переходной зоны празинофита Kephroselmis clivacea.
СВ - соединительное волокно; ТК - ’’тележное колесо'П '
К - микротрубочковые жгутиковые корни; 3 - звездчатая
структура (х 48000) ( Moestrup, Bttl, 1979)
123
Рис. 3.41. Схематическая реконструкция жгутиковой переход-
ной зоны у Cryptophyceae (А - Cryptomonaa cryophila:
£ - Amphidinium вр>). Chryacphyceae □ - ПгояГёпа).
EuglenophyceaeQ? - Entcaiphcn sulcatum). ЗВ’ушйёаГо-
phyceae (£ - Prymneaium parvum). Ргав1ПопЬусеяа (E -
flephrcaelmis clivaoea), Chlorophyceae (Ж - Chlmnydgnnnna
reinhardtii),а также Monomaatrix minima (Tcxcphyceae
eengu Moeatrup, 1982). Масштаб 0,2 мкм ( Moeatrup, 1982:
£ -ПО Taylcr, Lee; Б - Dodge; В - Hibberd; Г - Mig-
nct> S - Manton; IS - Moestrup, Ettl- Cavalier-Smith)
терная для водорослей с хлорофиллом ъ ;
У эвгленовых, за исключением Entcaiphcn,
м°Му, пустая.
Moeatrup, Thomeen,1974).
переходная зона, по-види-
124
Недавние исследования (Floyd et al., 1980; Melkonian,
I981a,b ; Melkcnian, Berns, 1983; O’Kelly, FlcydJ983; Flcyd,
O’Kelly, 1984) показали, что понимание абсолютной ориентации
жгутикового аппарата (рис. 3.42 и 3.43) имеет фундаментальное зна-
чение для разделения ТИПОВ Uivophyceae И chlamydomonasсреди
зеленых водорослей. Ориентация основания жгутика у бурых водорос-
лей ( O’Kelly, Floyd, 1984а) отличает их в этом плане от других
классов, содержащих хлорофилл с. Можно ожидать, что дальнейшие
исследования выяснят взаимоотношения между этими группами.
Базальное тело служит завершением каждого жгутика внутри
клетки. В дистальной части его оканчиваются периферические микро-
трубочки. Основные структуры базального тела мало варьируют в за-
висимости от группы водорослей. Ниже переходной зоны к перифери-
ческим дублетам добавляется третья С-трубочка.Эти периферические
триплеты располагаются под углом примерно 130° к радиусам. В ди-
Рис. 3.42. Схема типов абсолютной ориентации жгутиковых
структур с центральной симметрией. А. Строго крестовид-
ное расположение базальных тел и микротрубочковых кореш-
ков. Б. Вращение против часовой стрелки, как у Entocla-
dia viridls и Uivophyceae sensu 0’Ке11у,Р1оусЦВ. ВрАщёШё
ПО часовой стрелке, типичное Для Chaetophoraceae sensu Н
stricto Chlorophyceae (О’Kelly,Floyd, 1983) -----------
IZ5
Рис. 3.43. Обобщенная схема жгутикового аппарата четырех-
жгутиковой зооспоры представителя Ulotrichales или U1-
vales(Chlcrcphyceae),BHH сверху. К9 q 4 - микротрубочко-
вые корешки (с числом микротрубочек>в’корешке ) ;ПН - по-
лосатая лента, связывающая корешки с нижней парой базаль-
ных тел; ПВ - полосатая лента, связывающая корешки с
верхней парой базальных тел. Базальные тела пронумерова-
ны 1-4 (KLcyd, O’Kelly, 1984)
стальной части они обычно прикреплены тонкими нитями к одной цен-
тральной нити - такую конфигурацию называют "тележным колесом"
(рис. 3.27). В проксимальной части просвет базального тела может
содержать аморфный материал или несколько обособленных электроно-
плотных телец, бывает пуст (у большинства эвгленовых) или запол-
нен рибосомами ( Moestrup, 1982). Соединительные волокна могут
связывать между собой базальные тела, но, по-видимому, отсутству-
ют у базальных тел, которые не соответствуют функциональным жгу-
тикам.
Все жгутики водорослей, вероятно, имеют жгутиковые корни,
прикрепленные к базальным телам и распространяющиеся пел поверх-
ностью клетки или вглубь цитоплазмы. Современные электронномик-
Роскопичеокие исследования показали, что эти системы жгутиковых
126
корней крайне сложны. Внутренние корни могут, контактировать с яд-
ром, митохондриями, аппаратом Гольджи или хлоропластом. Однако
овязь между жгутиками и ядром, обычно утверждавшаяся в более ран-
них работах, часто не подтверждается. Корни представляют собой
микротубулярные или фибриллярные, часто поперечнополосатые струк-
туры (рис. 3.44). В последнем случае их нередко называют ризопла-
стами, хотя Мёструп (Moestrup, 1982) предостерегает против упо-
требления этого термина. Етутиковым корням приписывали разнообраз-
ные функции, включая заякоривание, сенсорное преобразование, пере-
распределение нагрузок, а также скелетную или организующую роль.
Сейчас о них известно достаточно для того, чтобы делать обобщения
филогенетического плана.
Микротубулярные корни обнаружены у всех изученных к настоя-
щему времени водорослей. Они могут быть необычными, как у диато-
мей, или однотипными, как у бурых водорослей, примнезиофитов,
"глаукофитов" (sensu Moestrup. 1982), харовых, мохообразных и
сосудистых растений. Возможно, наиболее сложны они у динофитов,
где связаны с рядом вспомогательных структур ( Dodge, Crauford,
1971), и у рафидофитов (Mignot, 1967, 1976), где иногда сочета-
ются с электроноплотной пластинкой и многослойной структурой. Осо-
бым расположением микротубулярных корней у спермиев Fucaceae (бу-
рые водоросли) поддерживается хоботок - уникальный вырост в перед-
ней части клетки. РЛикрофибриллярная крестовидная система жгутико-
вых корней известна у большинства зеленых водорослей (за исключе-
нием Oedcgoniales; см. Moestrup, 1978; Melkonian, 1980), неко-
торых локсофитов (в смысле Moestrup, 1982), празинофитов и "глау-
кофитов". Она состоит из четырех корней, расходящихся от базаль-
ного тела; расположенные друг против друга корни обычно имеют
одинаковое число микротрубочек. Крестовидные корни относятся к
наиболее широко изучавшимся. Они оказались полезны для разграни-
чения классов "хлорофитов". В отличие от других водорослей с хло-
рофиллом Ь эвгленовые, по-видимому, обладают одним микротубуляр-
ным корнем против каждого базального тела и одним корнем между ни-
ми ( Moestrup, 1982).
Поперечнополосатые волокнистые корни обнаружены у Cryptcphy-
oeae, Dinophyceae,RaphXdophyceae, Chrysophyceae, Xanthophyceae,
Eustigmatcphyceae, Euglenophyceae, "Loxophyceae" (sensu Moes*
trap, 1982), Prasinophyceae (рис. 3.44) И Chlorophyceae
( Moestrup, 1982). Они различаются по размеру и положению и в не-
127
Рис. 3.44. Упрощенная схема комплекса базального тела у
празинофита Tetraselmis.Б - базальное тело; $ - жгутик;
ТК - "тележное колесо"; МТ - микротрубочки, связанные с
половинными десмосомами; ПД - половинная десмосома; Р -
ризопласт; С - синистосома; ОФ ~ связующие фибриллы;
Я - ядро (Norris et al., 1980)
котором отношении (по виду и, возможно, по функции) напоминают
мышечные волокна. У золотистых водорослей они связаны с ядром,
аппаратом Гольджи или хлоропластом; эти типы корней описывались
как.ризостиль. Волокнистые корни без поперечной полосатости ука-
зывались у примнезиофитов. Поперечнополосатые корни обычно встре-
чаются в сочетании с микротубулярными; у желтозеленых водорослей
расположение корней уникально в том смысле, что представляет со-
бой комбинацию нисходящего, поперечнополосатого волокнистого и
нескольких микротубулярных корней ( Hibberd, Leedale, 1971; Hib-
berd, 1980а).
128
„ I
Хлоропласта
У фотосинтезирующих водорослей воспринимающие свет пигменты
локализованы внутри или на поверхности мембран, образующих упло-
щенные пузырьки, называемые тилакоидами. У прокариот (Cyanophy-
cota и Prcchlorophycota) последние свободно расположены в ци-
топлазме, а у эукариот они окружены мембранами, составляя вместе
с ними хлоропласт. Матрикс хлоропласта называют стромой. Хлоро-
пласты содержат хлоропластную ДНК и хлоропластные рибосомы, одна-
ко неполностью автономны от ядерного генома (обзор синтеза белка
и нуклеиновых кислот в хлоропластах см. Ellis, 1976). Развитие
и деление хлоропласта и клетки мотут происходить синхронно (осо-
бенно у клеток, содержащих только один хлоропласт) или независи-
мо друг от друга (особенно у клеток со многими хлоропластами)
(обзор сложных вопросов наследования и возобновления хлороплас-
те в см. Whatley, 1982, а также гл. 5).
Имеются общие обзоры, посвященные хлоропластам водорослей
(например, Dodge, 1973; Coombs, Greenwood, 1976). Любое знаком-
ство с хлоропластами обязательно включает изучение пигментов и
запасных продуктов (часто связанных с пиреноидами), а также глаз-
ков. Последние мы рассмотрим отдельно. Следует заметить, что, ес-
ли у сосудистых растений хлоропласты крайне однообразны по своей
структуре, у водорослей их особенности имеют диагностическое зна-
чение на уровне класса. Общие их признаки приведены в табл. 3.2
и 3.3.
Прокариоты
У прокариот тилакоиды расположены по периферии клетки и
окружены цитоплазматическим матриксом (рис. 3.1). После фиксации
глутаральдегидом для электронной микроскопии фикобилиновые пиг-
менты выглядят собранными в фикобилисомы, напоминающие одноимен-
ные структуры в тилакоидах красных водорослей, но отличающиеся
от тех, что присутствуют у содержащих фикобилины криптофитов. В
гетероцистах азотфиксирующих синезеленых водорослей тилакоиды ре-
I В отечественной литературе в качестве синонима часто
встречается постепенно выходящий из употребления термин "хро-
матофоры". - Прим, перев.
129
Таблица 3.2. Некоторые особенности хлоропластов эукариотных
водорослей
Число тилакои- дов на ламеллу Имеет- ся опо- ясываю- щая ла- мелла Форми- руются граны Число мембран вокруг хлоро- пласта крах- мал в хло- ропла- сте
Rhodophyceae I 2 —
Chryeophyoeae 3 — — 4 —
Prymnesiophyceae 3 — 4 —
Xanthophyceae 3 + — 4 —
Eustigmat ophyc eae 3 — — 4 —
Bacillariophyceae 3 + (?) — 4 —
Dincphyoeae (I) - 3 — — 3 —
Phaeophyceae 3 + — 4 —
Raphidophyceae 3 — 4 —
Cryptophyceae (I) - 2 — — 4 —
Euglenophyceae 3 — — • 3 —
Chlorophyceae 2 - мно- го — + 2 +
Charophyceae 2 - мно- го — + 2 +
Prasinophyceae 2-4 + (?) 2 +
130
Таблица 3.3. Общая классификация хлоропластов, фотосинтетических пигментов и запасных продуктов водо-
рослей (по Coombs, Greenwood, 1976, с изменениями)
Прокариоты Эукариоты
Локализация Тилакоиды в цитоплазме Тилакоиды в хлоропластах
Мембраны Отсутствуют Хлоропласты в цитоплазме Оболочка из 2 мембран Хлоропласты в особой части цитоплазмы Оболочка + цистерны ЭС = 4 мембраны (у некоторых 3 - см.*)
Вторичные Фикобилисомы на мембране пигменты Хлорофилль или с и каротиноиды в мембране
Группировка Стопок нет. тилакоида фор- тилакоидов мируют простые ламеллы Отопки в ци- топлазме. Сложные ла- меллы и граны Сложные ламеллы или ленты из 2-3 тила- коидов. Часто очень правильные 3-тила- коидные ленты. 4 1 = опоясывающие ла- меллы L J
Классификация chlorophycota chromophycota ( I I a) PROCHLORO- jChlorophyceae ных пигментов ( 1 МУТОТА ’Prasinophyceae ! [ jCharophyceae q (а) ФИК06ИЛИНЫ । CYAKOPHYCOTA RHODOPHYCOTA 1 1 EpDinophyceae* (б) Хлорофилл Ь 1 1 j ' 1 1 (в) Хлорофиллу j ' . ^Cyanophyceae^ ^Rhodophyceae [_ _Cryptqphyceae|_ и euglenophycota) 1 Euglenophyceae ♦ (б) 1 Prymnesiophyceае Eustigmatophyceae Raphydophyceae Л Chrysophyceae, PhaeophyJ ceae, Baoillariophyceae! _<XaHtkophyceay
Типичные запас- "Гликоген" ные углеводы Амилопектин Амилопектин и амилозы Ламинарии, парамилон и т.д.
вС-(1,4)-Глюканы "Крахмалы" >Д-(1,3)-Глюканы Крахмала нет
131
организованы в извилистую сетчатую систему, фикобилисомы утраче-
ны но хлорофилл а_ сохраняется. "Упаковка" тилакоидов у синезе-
леных водорослей отсутствует, но у содержащих хлорофилл ь_ тила-
коидов Proohlorophycota уже намечается, напоминая имеющуюся у
всех' других водорослей, кроме багрянок.
Rhodophycota
Структура и организация тилакоидов красных и синезеленых во-
дорослей (рис. 3.45) в некоторой степени гомологичны, хотя имеют-
ся существенные различия в функции вследствие отграничения их у
багрянок от цитоплазматического матрикса особой оболочкой хлоро-
пласта, представленной двумя параллельно идущими мембранами. Ла-
меллы не контактируют друг с другом (не собраны в стопки) и о
окружающей хлоропласт мембраной. Фикобилисомы находятся на обра-
щенных к строме поверхностях ламелл. У более примитивных багря-
нок ( Bangiophycidae) часто имеется пиреноид (например, у г0Г-
phyridium; Coombs, Greenwood, 1976), внутри которого ламеллы
образуют сеть. Пиреноиды встречаются только у самых примитивных
представителей KLorideophycidae, причем ламеллы здесь проходят
параллельно друг другу от одного края пиреноида к другому ( Gibbs,
1962а,ь ); крахмал (если он запасается) содержится в цитоплазме
вне хлоропласта. У этих более высокоорганизованных красных водо-
рослей под мембранами хлоропласта может лежать периферическая ла-
мелла или тилакоид, однако гомологизировать эту "опоясывающую ла-
меллу", как ее иногда называют, с одноименной структурой у Chro-
mophyoota следует с осторожностью ( Coombs, Greenwood, 1976).
Chromophyoota
У Chromophyoota структура хлоропласта весьма разнообразна
(см. табл. 3.2 и 3.3). Хлоропласты представителей этого отдела
отграничены от цитоплазмы клетки двумя дополнительными мембрана-
ми (у динофитов - одной, как и у эвгленовых). Внешняя мембрана
иногда бывает продолжением ядерной. Околохлороплаотное простран-
ство имеется во всех классах, кроме криптофитов и пинпфитоп
(Coombs,Greenwood, 1976). У динофитов связь хлоропласта с ядром
Не обнаружена. Внутренняя структура хлоропласта в этом отделе
Уникальна среди эукариот. Тилакоиды сложно и неодинаково сгруппи-
132
Рис. 3.45. Электронная микрофотография среза красной кок-
коидной водоросли Rhodella reticulata, показывающая
расположение органелл, в том числе хлоропласта и пиренои-
да. В - вакуоль; Г - аппарат Гольджи; К - зерна крахмала;
2 - радиально расходящиеся пластидные лопасти; Ог - ос-
миофильные глобулы; П - пиреноид; Я - ядра. Масштаб
2,0 мкм ( Deason et al., 1983)
рованы у разных классов; описано несколько типов их "упаковки',
но наиболее обычны ленты из двух (рио. 3.46) или трех тилакоидов.
Опоясывающие ламеллы отмечены у пяти классов, указанных в
табл. 3.3. Хотя возможно, что в этом отделе ламеллы внутри хлоро-
пласта образуют трехмерную сеть, как у высших растений, их мем-
браны не организованы в виде гран ( Coombs, Greenwood, 1976).
Криптофиты стоят особняком по своей пигментации, а также
структуре и организации тилакоидов (рис. 3.46). Их фикобилиновые
пигменты, отличающиеся химически от таких же пигментов у Суапо-
phyoota, Proohlorophyoota и Rhodophyoota, не образуют фикобили-
сом, а просто находятся на внутренней стороне тилакоидов, которые
собраны в "стопки" попарно. В классе динофитов упаковка тилакои-
133
Рис. 3.46. Хлоропласт криптофита Chroomonas meaoatigma-
tlca. Видны парные тилакоида. ИкНВдбЬё® МбЯНШги за-
полнен фикобилиновым питментом (х 81 000) ( Dodge, 1969b)
дов разнообразнее, чем у всех прочихгводорослей. Ламеллы часто
встречаются парами или даже поодиночке.
В большинстве классов Chromophycota отмечены внутриплас-
тидные пиреноида, часто на своего рода стебельке (общий признак
бурых водорослей; см. рис. 3.23 и Dcdge, 1973). Ламеллы тила-
коидов могут пересекать матрикс пиреноида у некоторых примнезио-
фитов, криптофитов и динофитов (рис. 3.47). У последних организа-
ция пиреноидов самая разнообразная в отделе (см. обзорDcdge,
1973). Крахмал у Chromophyccta образуется нев хлоропласте,
а в цитоплазме или, у некоторых представителей, между мембраной
хлоропласта и его эндоплазматической сетью.
Chlorophyccta
Хлоропласты зеленых водорослей не связаны с мембранами эндо-
плазматической сети или ядра; эта особенность напоминает строение
клетки багрянок. В отличие от Chromophycota крахмал запасается
в строме хлоропласта, часто в виде слоя вокруг пиреноида. Тила-
134
Рис. 3.47. Пиреноид на
ножке у динофита Amphi-
dinium carterae
(х 22 ЬОО) (любезно пре-
доставлено J.D.Dodge)
коиды зеленых водорослей собраны стопками по 2 - 6 или более; их
многослойное расположение может выглядеть как граны у высших рас-
тений, связанные ламеллярными перемычками. В этом смысле особен-
но много общего с высшими растениями у харовых. Пиреноиды отмече-
ны у большинства зеленых водорослей, обычно внутри хлоропласта.
На их поверхности образуются крупные зерна крахмала. В пиреноиды,
которые никогда не имеют стебелька, как у некоторых Chromophy-
cota, может проникать от нескольких до многих тилакоидов.
Глазки (стигмы)
Глазок (стигма) встречается у многих подвижных водорослей
или у гамет и зооспор неподвижных форм. Он состоит в основном из
осмиофильных каротиноидных липидных глобул (пластоглобул), часто
связанных со светочувствительной палочковидной или кристалличес-
_кой зоной вблизи аксонемы жгутика. Как фоторецептор глазок обус-
ловливает фототаксис.
135
У подвижных клеток рафидофитов и цримнезиофитов глазки обыч-
но отсутствуют; относительно редко они встречаются у динофитов
(особые типы) и криптофитов, однако известны у большинства жгути-
ковых клеток Chlorophyceae, Prasincphyoeae, Euglenophyceae,
Xanthophyceae, Chrysophyceae И Phaeophyceae (за исключением
зооспор большинства Laminariales и спермиев многих оогамных ви-
дов; Henry, Cole, 1982а, Ъ).
Додж (Dodge, 1969а, 1973) различает три основных типа глаз-
ков и особо выделяет несколько их вариантов, встречающихся у дино-
фитов.
I. Chlorophyceae, Prasinophyceae И некоторые Chrysophy-
ceae : глазок является частью хлоропласта, но не показывает явной
связи со жгутиками.
2. Cryptophyceae (рис. 3.48), Xanthophyceae, мужские гаме-
ты и зооопоры большинства Phaeophyceae (за исключением зооспор
большинства I<amlnariales и спермиев многих оогамных видов;
Henry, Cole,1982b): глазок является частью хлоропласта и явно
связан со жгутиками.
3. Euglenophyceae, некоторые Xanthophyceae И Eustigma-
tophyceae: глазок независим от хлоропласта, но связан со жгутика-
ми.
У динофитов глазки разнообразные: от скопления лишенных мем-,
бран липидных глобул, содержащих каротиноиды, до более сложных
комплексов аналогичных глобул, окруженных мембранными оболочками
(cp.Bold, Wynne 1978). У Glenodinium foliaceum глазок связан
с органеллой, представляющей собой стопку уплощенных пузырьков,
число которых доходит до 50 ( Dodge, Crawford, 1969). Додж
( Dodge, 1984) описывает три типа глазка динофлагеллат: независи-
мый и не окруженный мембранами; независимый и окруженный тремя
мембранами; расположенный на периферии хлоропласта. Все они нахо-
дятся позади продольной борозды, и между глазком и текой лежит пу-
чок микротрубочек; предполагается, что он участвует в передаче ди-
рекциональных стимулов от глазка к жгутику. Додж (Dodge, 1984)
считает, что по структуре глазка динофлагеллат можно сблизить с
эвгленовыми и золотистыми водорослями. Наиболее сложная светочув-
ствительная органелла водорослей - глазок некоторых морских дино-
фитов (например.Wamowia ). Он представляет собой камеру, обра-
зованную впячиванием плазмалеммы, закрытую светопреломляющей лин-
зой из плотноупакованных вздутых пузырьков. Экспериментально по-
136
Рис. 3.48. Глазок криптофита Chrcomonas mesostigmatica.
состоящий из единственного слоя пластоглобул, расположен-
ных на конце короткого выроста хлоропласта, отходящего за
пиреноидом (справа) (х 44 100) ( Dodge, 1973)
казано (Francis, 1967), что падающий на клетку свет фокусирует-
ся на выстилающем камеру слое, названном ретиноид. Такой глазок
наиболее близок к светочувствительным органам животных.
Большое внимание уделялось вопросу о связи между глазками,
хлоропластом, жгутиками и фототаксисом. Особенно это касается эв-
гленовых, относительно которых преобладают два взгляда. Одни счи-
тают, что глазок и является собственно фоторецептором, другие -
что свет воспринимается в жгутиковом вздутии, или околожгутиковом
теле(РГВ). Современные данные позволяют связать фоторецепцию с
флавйноподобным пигментом, локализованным bpfb (Colombetti et al.,
1982). Предполагается, что в систему фотосенсорного преобразова-
ния входят две самостоятельные фоторецепторные системы (реагирую-
щие на повышение и понижение освещенности), сопряженные с натрие-
во-калиевой транспортной системой в мембране грв (Robenek, Melko-
nian, 1983).
Для Chlorophyceae показано (Hartshorne, 1953), что не име-
ющие глазка мутанты Chlamydomonas. хотя и способны к положитель-
ному фототаксису, реагируют значительно медленнее, чем клетки ди-
кого типа с нормальным глазком. Изучая находящийся внутри единст-
венного хлоропласта глазок спермия Pucus , Баук (Bouck, 1970) вы-
сказал предположение, что парабазальное тело и глазок передают
137
световое воздействие на фоторецептор, связанный с плазмалеммой.
у золотистой водоросли Dlnobryon возможный механизм фоторецеп-
ции И реакции на свет описан более детально ( Kristiansen, Walne,
1976, 1977). Два базальных тела соединены здесь мостиком, причем
то из них, которое относится к гладкому жгутику, связано с зоной
глазка в хлоропласте некой структурой, отходящей от двух трипле-
тов. Такая система, возможно, служит для передачи импульсов от
глазка к одному или обоим жгутикам.
Строение глазка - одна из характернейших особенностей Eus-
tigmatophyoeae. У зооспор здесь он находится вне хлоропласта
(как у эвгленовых)и представляет собой группу глобул у переднего
конца клетки, примыкающих к жгутиковому основанию и не окружен-
ных мембраной (см.Lee, Bold, 1973).
Ядро, деление ядра и цитокинез
Интерфазное ядро
Ядро (рис. 3.49) - основное хранилище генетической информа-
ции (генома) в клетке и центр, контролирующий ее селективную экс-
прессию (как упоминалось ранее, генетический материал содер-
жится й в хлоропласте, а кроме того, в митохондриях). Исключение
составляют криптофиты, обладающие нуклеоморфой. Предполагалось,
что эта структура содержит дополнительный геном, и недавние ис-
следования ( Gillott, Gibbs, 1980), похоже, подтверждают это.
Как у всех про- и эукариот генетическая информация водорослей
"записана" последовательностями триплетов нуклеотидных оснований
в цепочках молекул дезоксирибонуклеиновой кислоты (ДНК). Репли-
кация ДНК и процессы транскрипции и трансляции в ходе экспрессии
этой информации у водорослей аналогичны таковым у других про- и
эукариот и детально здесь не рассматриваются.
У прокариотных водорослей ДНК кольцевая; она располагается
в центре клетки и не окружена системой мембран. Репликация здесь
происходит во время деления клетки, как у эукариот, а процессы
транскрипции и трансляции тесно сопряжены. У эукариотных водорос-
лей ДНК находится в хромосомах, которые кроме ДНК содержат ядер-
белки и небольшие количества РНК в комплексе, называемом хро-
иатином. Хромосомы располагаются внутри ядра, отграниченного от
Цитоплазмы двойной мембраной (ядерной оболочкой), составляющей
единое целое с эндоплазматической сетью клетки. Транскрипция про-
138
исходит внутри, ядра,, а трансля-
ция - вне его.
Интерфазное ядро водорос-
лей в целом относится к обычно-
Рис. 3.49. Интерфазная апикаль-
ная клетка Sphacelaria tribu-
loides (Phaeopnyсеae)(на
врезке клетка целиком). В -
вакуоли: Ф - физоды; X - хлоро-
пласт; Яд - ядрышко: Я - ядро
(х 2125: врезка х 2о5) (Katsa-
ros et al., 1983)
му для эукариот типу, встречаю-
щемуся у большинства животных и
растений (общий обзор см.Dodge,
1973; обсуждение некоторых спе-
цифических черт см., например,
Dillon, 1981). Исключение сос-
тавляют динофиты, эвгленовые и
криптофиты, у которых наблюдают-
ся уникальные особенности строе-
ния ядра, его деления и цитоки-
неза (см. обзоры beedale, 1967,
1982; Dodge, 1973). Эти типы
ядер названы мезокариотными
(рис. 3.50), поскольку наряду с
другими особенностями они отли-
чаются хромосомами, остающимися
конденсированными на протяжении
всего клеточного цикла. К дру-
гим характерным признакам отно-
сятся сохранение ядерной мембра-
ны (у динофитов и эвгленовых) и
ядрышка (или эндосомы) при деле-
нии (у эвгленовых).
Мембраны ядерной оболочки
имеют толщину 7 - 8 нм и разде-
лены перинуклеарной полостью,
которая в некоторых случаях,
возможно, выполняет специфичес-
кие функции, например участвует
в образовании жгутиковых волос-
ков. Оболочка пронизана много-
численными порами ( Dillon,
1981), иногда располагающимися
в определенном порядке ( Dodge,
1973). Кроме хромосом ядро содержит основное вещество, или нуклео-
плазму, и одно или более ядрышек, которые представляют собой скоп-
139
ления базофильного материала, богатого рибонуклеопротеином. Они
тесно связаны со специфическим участком хромосомной ДНК, кодирую-
щим рибосомную РНК. Транскрипция рибосомной РНК обеспечивается
особой РНК-полимеразой, обнаруженной только в ядрышке (cp.Hojh-
1978). Другие возможные функции этих структур до сих пор
полностью не выяснены. Размер и число ядрышек важны при оценке
различий между ядрами водорослей (Dodge, 1973).
Большинство клеток эукариотных водорослей одноядерно. Одна-
ко многоядерность типична для вегетативных клеток большинства
^lorideophycidae (багрянки) и является нормой для сифональных
Xanthophyceae(Vaucheria) и Chlorophyceae. За Некоторыми ис-
ключениями (например, Динофиты, эвгленовые) ядра и хромосомы во-
дорослей мелкие, вследствие чего их трудно изучать в световом
микроскопе (обзор более ранней информации о хромосомах водорослей
См. Godward, 1966).
Деление ядра
Важнейшее свойство клеток - их способность к самовоспроизве-
дению. Во время вегетативного роста эукариотных водорослей с этим
140
связаны два процесса: ядерное деление (митоз) и цитокинез (деле-
ние цитоплазмы). Хотя этапы митоза у водорослей широко изучались
с помощью световой микроскопии, о нем накоплено сравнительно ма-
ло ультраструктурных данных. Вместе с тем значительное внимание
было уделено особенностям ядерного деления yDindphyceae, Eu-
glenophyceae, Cryptophyceae И Chlorophyceae. В случае по-
следней группы эти исследования дали много нового для понимания
родственных связей между водорослями и наземными растениями
(Pickett-Heaps, Marchant, 1972; Pickett-Heaps, 1975, 1976,19795
Stewart, Mattox, 1975, 1980).
За исключением мезокариотных форм, ядерное деление у водо-
рослей во многом сходно с митозом у животных, хотя кинетохор,
универсальный признак хроматид животных, и центриоли встречаются
не всегда.
Сейчас уже достаточно известны (см., например, обзоры
Dodge, 19735 Brawley, Wetherbee,1981) особенности ядерного деле-
ния, отличающие разные классы водорослей друг от друга и от дру-
гих эукариот (табл. 3.4). Сюда относятся поведение при митозе
ядерной оболочки (остается ли она интактной, возникают ли в ней
разрывы и отверстия или она полностью разрушается), полярная
структура и ее связь с организацией мшдэотрубочек, а также приро-
да веретена. К этому можно добавить наличие или отсутствие кине-
тохоров, центриолей и специализированных образований.
Информация о тонких структурных перестройках при митозе у
красных водорослей получена в основном при исследованиях предста-
вителей Ceramiales (McDonald, 1972; Scott et al., 1981s Phil-
lips, Scott, 1981). Ядра и хромосомы очень мелкие; известны хро-
мосомные числа от п = 2 до п = 90. Наиболее характерная особен-
ность - наличие полярных колец,»которые возникают в ранней профа-
зе и сохраняются на протяжении всего митоза; предположительно они
функционируют как организующие микротрубочки центры, хотя не мо-
гут считаться истинными центриолями, поскольку никаких следов
жгутиков у багрянок нет.
, У золотистых водорослей организующую роль при митозе играет
ризопласт ( Slankis, Gibbs, 1972). Во время препрофазы базальные
тела двух жгутиков и аппарат Гольджи реплицируются. С каждым по-
люсом связывается ризопласт, и микротрубочки веретена, по-видимо-
му, прикрепляются к ризопластам. Ядерная оболочка иногда разруша-
ется В профазе ( Ochromonas danicas Slankis, Gibbs, 1972), HO
может оставаться и относительно интактной ( Hydrurus foetidus:
Таблица 3.4. Отличительные особенности митоза и цитокинеза у эукариотных водорослей
Хрома- сомы Кинето- Яцрыш- Тип веретена Полярная Цитокинез хор ко2 та-додаа.-. структура 5 СП ф CD g g К &Я f>> g its & g sa £
Rhodophyceae - + — + Закрытый адер-- Полярные коль- ца; настоящих Кольцевидная
ный и цитоплаз- стенка и по-
матический центриолей нет ровая пробка
Chrysophyceae - — - + Ядерный и цито- плазматический Ризопласт Расщепление
Prymnesiophyceae - - + п Центриолей нет _ И _
Xanthophyceae — *• . — + Обширный внут- риядерный Центриоли _ м
Eustigmatophyceae - - + Ядерный и цито- плазматический Основания жгу- тиков действуют _ м _
как центриоли
Bacillariophyceae - - — + Цитоплазматиче- ский ? "Предшествен- ник" веретена Борозда де- ления
Dinophyceae + - — + Цитоплазматиче- Расщепление
прикреп- ленные к ядер- ной обо- лочке ские туннели
Phaeophyceae — + - (+) + Внутриядерный Внеядерные центриоли Клеточная пластинка
Хромо- Кинето- Яцрыш- Яцерная
сомы! хор ко2 оболочка
Тип веретена Полярная
структура
Цитокинез
Raphidophyceae Cryptophyceae + + Внутриядерный + Ядерный и цито- Центриоли Основания хгу- Расщепление Цитокинети-
и нук- леомор-. фа + (прос- той) круп- ное плазматический тиков; истинных центриолей нет ческое коль- цо
Euglenophyceae (эндо- сома) + Внутриядерный и подаеретена ? Расщепление
Chlorophyceae — - или + (+) + (+) Ядерный и цито- плазматический или внутриядер- ный Центриоли Фикопласт и фрагмопласт
Charophyceae - и - + Ядерный и цито- плазматический Фрагмопласт
Prasinophyceae — — t — ° — Фрагмопласт или фрагмо- пласт и бо- розда деле- ния
+: постоянно сконденсированы; сконденсированы при митозе
%) +: сохраняется при митозе; не сохраняется при митозе.
143
yee»t et al», 1984)»
Среди желтозеЛеных водорослей Vaucheria во время митоза име-
ет очень длинное интерзональное веретено, тогда как ядерная мем-
брана остается интактной ( ott, Brown, 1972); особенности ядерно-
го деления здесь, возможно, более типичны для:различных фикомице-
тов, чем для других водорослей ( Dodge, 1973).
Своеобразно происходит формирование веретена диатомей; Ман-
тон и др. ( Manton et al., 1969) описали у Lithodeamium (Biddul-
phialea) его предшественник, представляющий собой прямоугольное
тельце, образованное рядами параллельных пластинок и играющее
определенную роль в организации веретена до разрушения ядерной
оболочки. Структура, сходная с центросомой, описана у Surirella
( Drum, Pankratz, 1963)? хотя ее роль в организации веретена не-
ясна.
У динофитов ядрышко и ядерная мембрана во время митоза сохра-
няются. Хромосомы, которые, по-видимому, прикреплены к ядерной
мембране, лишены кинетохора. Цитоплазматические микротрубочки,
проникающие в ядро, создают видимость веретена, но не участвуют
в разделении хромосом и как будто не связаны с какой-либо органи-
зующей структурой. В конце митоза ядерная оболочка перешнуровыва-
ется вслед за разделением хроматид.
Веретено у бурых водорослей, по имеющимся сведениям, внутри-
ядерное, с внеядерными центриолями. Ядерная оболочка во время
митоза сохраняется интактной, если не считать полярных разрывов,
через которые проникают микротрубочки веретена ( Katsaros et al.,
1983).
Митоз изучен только у двух криптофитов (см. Oakley, Santore,
1982). Центриоли здесь, по-видимому, отсутствуют, а микротрубочки
веретена возникают до профазы у оснований жгутиков, от которых
они отделяются при разрушении ядерной оболочки. Кинетохор - прос-
того типа, и в анафазе трудно различить в массе хроматина индиви-
дуальные хромосомы. Образование трубочек веретена до профазы и
разрушение ядерной оболочки - особенности, которые роднят эти во-
доросли С примнезиофитами ( Manton, 1964Ь).
Митоз эвгленовых имеет ряд необычных черт (Leedale, 1982).
Ядерная мембрана остается интактной, а эндосома (ядрышко) делит-
ся надвое.Затем хромосомы располагаются вдоль оси деления эндосо-
мы. Разделение на хроматиды может происходить до, во время или
после этого процесса. Расхождение хроматид в метафазе несинхрон-
но> и некоторые из них уже могут достигать полюсов, тогда как
144
другие остаются у экватора. Нуклеоплазматические микротрубочки
формируются во время профазы и ввязаны с хромосомами; они оканчи-
ваются у полярных областей под ядерной оболочкой, цитоплазматиче-
ские микротрубочки в процессе не участвуют. Более ранние сообще-
ния об отсутствии кинетохора (Leedale, 1968s Dodge, 1973)
не подтвердились. Он отмечен в митотических хромосомах Euglena
gracilis (Pickett Heaps, Weike, 1977) , причем Лидейл ( Leedale,
1982), заново проводя свои исследования, также его обнаружил.
Примечательно, что каждая хроматида, по-видимому, имеет свое соб-
ственное миниатюрное "субверетено", включающее четыре центромер-
ные и 8 - 12 более крупных микротрубочек.
Обширная информация о митозе у зеленых водорослей (особенно
Chlorophyceae) собрана в ряде обзоров ( Pic^ett-Heaps, 1975,
1979s Brawley, Wetherbee, 1981). В этом отделе ядерная оболочка
может при делении сохраняться интактной (рис. 3.51 и 3.52), обра-
Рис. 3.51. Митоз у Cladophora (Chlorophyceae)s ВИДНО
полностью внутриядерное веретено в средней анафазе
(х 15 720) ( Scott, Bullock, 1976)
145
Рис. 3.52. Митоз у Cladophora (Chlorophyceae); поздняя
телофаза (х 9000). В - вакуоли; 0В - остатки веретена
(Scott, Bullock, 1976)
зовывать полярные отверстия или разрушаться в профазе (например,
у десмидиевых). Веретено - как ядерное, так и цитоплазматическое,
хотя у Oedogonlum оно полностью внутриядерное. Этот род отлича-
ется также наличием сложных кинетохоров.
Поведение веретена зеленых водорослей после ядерного деле-
ния представляет огромный филогенетический интерес. Оно обсужда-
ется ниже.
Цитокинез
Следуя за ядерным делением, цитокинез обычно идет одним из
Двух путей: за счет роста новой стенки или перешнуровки клетки.
Микротрубочки могут играть определенную роль в управлении этим
процессом, а в формировании клеточной пластинки (структуры, кото-
рая типична также для клеток высших растений) участвует аппарат
Гольджи.
Ю-121
Цитокинез, или деление надвое, прокариот (например, синезе-
леннх водорослей) - относительно простой процесс, обычно являю-
щийся следствием роста клетки. В этой отделе описано три типа ци-
токинеза (Drews, Weckesser, 1982)s с перешнуровкой; с формирова-
нием септы; почкование. В первом случае происходит одновременное
впячивание стенки по окружности (Fogg et al., 1973). .Септа форми-
руется за счет впячивания цитоплазматической мембраны и пептидо-
гликанового слоя до начала впячивания внешней мембраны. Тилакои-
ды выталкиваются к центру клетки и как бы разрезаются поперек
при завершении септирования. Особенность цитокинеза синезеленых
водорослей в том, что образование септы является непрерывным про-
цессом и последовательно могут закладываться три или более септ.
Их образование происходит, таким образом, независимо от цикла
репликации в противоположность типичному цитокинезу эукариот, ко-
торый всегда происходит в конце митотического цикла (Dillon,
1981). Почкование показано v Chamaesiphon (Rippka et al., 1979).
Наиболее примитивная форма цитокинеза эукариотных водорос-
лей - продольное расщепление - известна у жгутиковых; оно начина-
ется от переднего (у большинства) или заднего (например, у неко-
торых динофитов) конца клетки. Интересная модификация этого про-
цесса наблюдается у Chlamydononas, где расщепление происходит
только после того, как весь протопласт повернется на четверть
оборота внутри клеточной стенки; это рассматривается как шаг на
пути к поперечному делению, типичному для нитчатых зеленых водо-
рослей (Dillon, 1981).
Поведение митотического веретена в ходе цитокинеза широко
изучалось у зеленых водорослей (Pickett-Heaps, 1975, 1979). Если
оно исчезает, ядра остаются вблизи друг друга и новый набор мик-
ротрубочек, перпендикулярный прежнему веретену, действует как ор-
ганизатор стенки между дочерними клетками; эти микротрубочки на-
зываются фикопластом, а сам тип цитокинеза - фикопластным. Если
же веретено сохраняется, дочерние ядра расходятся на большое рас-
стояние и новая стенка формируется фрагмопластаи (слияние пузырь-
ков диктиосом, содержащих компоненты стенки, между микротрубочка-
ми веретена) или путем комбинации фрапиопласта и впячивания плаз-
малеммы клетки. Фрагмопластный тип цитокинеза характерен для выс-
ших растений, а его наличие у зеленых водорослей (некоторых Chlo-
rophyceae, Prasinophyceae и всех Charophyceae) легло В
основу новых подходов к их классификации (stewart, Mattox, 1975s
Pickett-Heaps, 1975).
147
Рис. 3.53. Апикальная клетка
gflhacelaria tribuloiaes (Jha-
eophyceae) на ранней стадии
Цитокинеза; видны развивающая-
ся борозда (стрелки) и дочернее
ЯДРО (Я) (х 900) ( Katsaros
et al.,- 1983)
У багрянок на ранних ста-
диях цитокинеза происходит не-
полное разделение дочерних кле-
ток, так что их цитоплазма ос-
тается единым целым. После это-
го образуется поровая пробка
(см. выше).
У бурых водорослей (рис.
3.53) клеточная пластинка раз-
вивается от центра к периферии
и утолщается за счет фибрилляр-
ного материала, поставляемого
пузырьками Гольджи; этот про-
цесс очень напоминает образова~
ние клеточной пластинки у выс-
ших растений ( Markey, Wilee,
1975).
Глава 4
УРОВНИ ОРГАНИЗАЦИИ
Разнообразие морфологии
Понимание организации крайне важно при рассмотрении эволюции
и классификации водорослей. Современная микроскопия значительно
расширила возможности их описательно-морфологического изучения, а
дополнительные физиологические и ультраструктурные детали значи-
тельно уточнили таксономию.
Многие водоросли отличаются высокой морфогенетической плас-
тичностью, проявляющейся в ответ на факторы внешней среды (гл. 6).
У многих морфология совершенно различна на разных стадиях жизнен-
ного цикла (гл. 5). Однако уровень организации водорослей остает-
ся сравнительно цростш, и, за несколькими исключениями (среди
крупных бурых водорослей), сложной тканевой дифференцировки у них
нет.
Морфологическая эволюция имела тенденцию идти в разных отде-
лах водорослей параллельно, что подробно обсуждалось еще Фричем
( Fritsch, 1935). Предполагается, что от одноклеточных предков
сходные ряды этапов могли привести к колониальным, нитчатым парен-
химатозным и в некоторых случаях сифональным типам в каждой из фи-
логенетических линий, которые сейчас в общих чертах представлены
разными отделами. В одних отделах это развитие далеко не зашло
(или более "продвинутые" формы утрачены), в других весь морфологи-
ческий спектр представлен ныне живущими видами (например, среди
зеленых водорослей), а в некоторых (например, у бурых водорослей)
наиболее примитивные формы уже не существуют или рассматриваются
современными альгологами как представители других отделов. Если
между видами в отделе существует значительное сходство, предпо-
лагается единая линия его эволюционного развития (монофилия); в
других случаях ясно, что таких линий несколько (полифилия, напри-
мер, у зеленых водорослей). Эволюционные аспекты еще будут обсуж-
даться в гл. 8, однако они служат полезной основой для последую-
щего рассмотрения морфологического разнообразия.
Водоросли бывают одноклеточными, колониальными, нитчатыми,
псевдопаренхиматозными, паренхиматозными, сифонокладальными (полу-
ценоцитными) и сифональными (ценоцитными) (табл. 4.1).
Одноклеточные формы могут быть неподвижными, или коккоидными
(протококкоидными по Round, 1973), амебоидными, или ризоподиаль-
149
Таблица 4.1. Обзор уровней организации у водорослей
Однокле- точные с Колони- альные Нитчатые | Псевдопаренхима- тозные
§
1 ь « S i §
m isi о s®
НЫЙ нал вый So . « « й о я „ « СЦ *3 о ь «3 8 § i | §
S *=[ о « Pt Ен ф о S и М
я ом ФО я оно оом о се
9 о « Жсб о Е-* g О< оря я л
« Ofc S со £>» и?1 2 О Рч Ф о а § о Я § >§<
о я Ь Мн Рч N ЛФ ф Ден ст Л © & ИМ Рч 5 S S 3 3
Прокариоты
Cyanophyceae + + + + +
»ig^ycots +
Rhodophyceae + + + + + +
Chrysophyceae + + + + j + + 4*
Prymnesiophyceae + + !
Xanthophyceae + + + + ! + + +
Eustigmatophyceae + i
Baoillariophyceae +
Dinophyceae + + + +
Phaeophyceae i I + + + + +
Raphidophyc eae + +
Cryptohyceae + + +
Euglenophyo eae + +
Ch lorophyceae + + + + + + + + + + + + +
Charophyceae 4-
Prasinophyc eae + + +
150
ними, и подвижными, или жгутиковыми. Колониальные формы варьиру-
ют от пальмеллоидной (часто переходная стадия в результате потери
подвижности) до тетраспоровой, более устойчивого состояния, когда
продукты клеточного деления остаются агрегированными в слизистой
массе. У ценобиальных видов число клеток неподвижных или подвиж-
ных колоний после их образования не изменяется; клетки здесь рас-
полагаются определенным образом.
Простое нитчатое строение становится возможным, когда клетки
после деления остаются соединенными между собой. Дополнительные
плоскости деления позволяют развиваться ветвящимся нитям, а когда
деления пропорционально более часты в нижних частях растения, мо-
гут формироваться отличающиеся друг от друга вертикальная и про-
стертая части водоросли, давая начало гетеротрихальным типам.
Псевдопаренхиматозные талломы образуются за счет ветвления одной
(одноосевой тип) или нескольких (многоосевой тип) центральных ни-
тей, находящихся в общем матриксе. Деление нити в двух или более
плоскостях приводит к истинно паренхиматозному росту. В последнем
случае клеточные деления могут происходить в разных местах талло-
ма или локализоваться в апикальной или интеркалярной меристемах.
Ценоцитные талломы возникают при неоднократных ядерннх делениях с
относительно малым количеством возникающих клеточных стенок (сифо-
нокладальный тип; Hoek , 1981) или вообще без них (сифональный
тип). Как и у нитчатых форм, здесь могут развиваться сложные цено-
цитные талломы за счет соединения нескольких сифонов в многоосе-
вое образование.
Одноклеточная организация - ризоподиальные формы
(рис. 4.2)
Одноклеточные (или неклеточные) виды известны во всех клас-
сах, за исключением бурых и харовых водорослей. Независимо от то-
го, самые это примитивные формы или нет, все они обладают харак-
терными цитологическими и физиологическими особенностями соответ-
ствующей группы.
Одноклеточные, у которых отсутствуют (или временно исчезают)
жгутики, а голый протопласт движется амебоидно, можно считать ри-
зоподиальными. Они образуют цитоплазматические выросты, известные
как псевдоподии (иногда способствующие захвату пищевых частиц) и
ризоподии. Такие виды наиболее многочисленны среди золотистых и
желтозеленых водорослей, но встречаются также среди динофитов и
в некоторых других классах (табл. 4.1). К этому же типу можно от-
151
нести и представителей Chrysophyceae и Xanthophyceae с голыми
клетками, находящимися внутри теки, из которой выходят цитоплаз-
матические нити.
Золотистые водоросли с доминирующей амебоидной фазой отно-
сятся к порядку Chrysamoebidales . Семейство Chrysamoebidaoeae
включает род Chrvsamoeba с жгутиковой стадией, у которой вто-
ричный жгутик редуцирован до базального тела. У других амебоид-
ных форм из этого порядка жцутиковая стадия отсутствует (семей-
ство Khizoohrysidaoeae по Bourrelly , 1981)а Сетевидное обра-
зование из клеток, соединенных между собой ризспсдиями, образует-
ся у Chrysostephenosphaera . Виды с панцирем встречаются в семей-
стве stylococoaceae 5 их репродуктивные стадии преимущественно
амебоидные, а не жгутиковые. Род Myxochrysis в этом порядке
примечателен тем, что в вегетативном состоянии он представляет
собой многоядерный плазмодий.. Его жизненный цикл, включающий ста-
дию подвижных зооспор, очень сходен с таковым у слизевиков (ify-
xomyoota).Высказывалось предположение, что начало последней груп-
пе дали именно такие типы золотистых водорослей ( Hollande ,
Enguinet, 1955).
Спектр форм ризоподиальных желтозеленых водорослей такой же,
что и у Chrysophyceae, т.е. ст свсбодноживущих амебоидных типов
с псевдоподиями или филоподиями (например, Heterochlcris) до пан-
цирных (например, Rhizoohloris) и плазмодиальных представителей
(например,Myxochloris).
У динофитов в качестве примера ризоподиальнсго вида можно
привести Dinamoeba varians (порядок Dinamoebales ). Преоблада-
ет амебоидная стадия, а гимнодинисидная подвижная фаза кратковре-
менна.
Одноклеточная организация - коккоидные формы
х (рис. 4.1 и 4.2)
Под ксккоидным типом понимается строение одноклеточных водо-
рослей, у которых преобладает неподвижная стадия, а подвижность
либо полностью отсутствует (например, азооспоровые зеленые водо-
росли, в частности Chlorella), либо ограничена репродуктивными
Фазами (Chloroooooum из Chlorophyceae). Коккоидные формы име-
ется в большинстве классов. В некоторых на их долю приходится
основное число видов (70% представителей желтозеленых водорослей;
152
Рис. 4.1. Уровни организации синезеленых водорослей: А -
Glceccapsa ralfsiana - КОККОИДНЫЙ; Б - Pleurocaosa
ccmuana - пальмеллоидный с тенденцией к соЬазованйб'
простых нйтей; В - Stigonema mamillosum И Г - Pisohe-
rella major - нитчатый с истийныМ ветвлением; Д - Soy-
tonema mllei - нитчатый с ложным ветвлением; Jg - Os-
olllatoria margaritifera - простая неветвящаяся нить;
А - Ncstcc piaoinale - колониальный с трихомами и мно-
го численными гетероцистами, погруженными в слизистую мас-
су; 3 - Anabaena flos-aquae - планктонные трихомы с
крупными интеркалярными гетероцистами; И - Calothrix
crslniana - колониальный; трихомы с оазальяымй гетеро-
цййтами и хорошо различимым влагалищем (по Bourrelly.
1970)
Ott, 1982), иногда - все (Eustigmatophyoeae; Hibberd, 1982с)«,
У Prochiorophycota единственный род Prochloron - коккоид-
ный. Кскксидными являются многие синезеленые водоросли (напри-
мер, Aphanocapsa. Synechcooccus). особенно В порядке Chro-
153
Рис. 4.2. Коккоидная (А - Ж), ризоподиальная (3) и,паль-
меллоидная (И - Л) организация: А - Miorasterias dentiou -
latus (Chlorophyceae'); Б - Staurbdesmus validus(Chlo-
rophyceae)} В - Euastrum didelta (Chlorophyceae); Г - Sai-
rotaenia ooncTensaia (Chlorophyceae) j Д - Pinnularia
sp. (Bacillariophyceae); E - Staarastrum anatinum (Ohio -
rophyoeae) (вид сбоку); Г- S.&natinum (ВИД с полюса):
3 - Bhygoohrysia sp. (Chrysophyceae)» Z ~ Tetraspora sp.
(Chlorophyceae ) ; К - Gloeooystis naegeliana (Cnloro-
Phyoeae)j ~
JJ - Sphaerooystis sohroeteri (Chloj-ophyoeae); M - Dicty-
osphaerium pulohellum (ChloTOPhvceae) . Без масштаба (по
West, Fritsch, Doflein, Reinke, Artari, Chodat, G.M. Smith)
ocoooales (Chroococoaoeaej Bold, Wynne,1978)7 многих его пред-
ставителей (например. Chroooccous Gloeooapsa) имеется тенденция
к частичному соединению между собой дочерних клеток после деле-
ния или к расположению их в общем слизистом матриксе. Число ви-
154
дов коккоидных синезеленых водорослей крайне неопределенно. Ис-
ходя в основном из морфологических признаков, число таких родов
сокращали до шести ( Grcuet, Daily, 1956) , а на основании био-
химических особенностей выделяли несколько различных и далеко
отстоящих друг от. Друга групп ( Stanley et al., 1971; Rippka
et al., 1979).
Среди багрянок коккоидный облик редок и ограничен примерно
восемью родами порядка Pcrphyridiales (Bangiophycidae sensu Bold,
Wynne, 1978) .Наиболее известный пример - Pcrhvridium (Gantt,
Conti, 1965; Gantt et al., 1968),сферические особи которого су-
ществуют поодиночке или образуют скопления в слизи. Амебоидное
скольжение описано у Petrovanella (Pringshelm, 1968) и Pcrphy-
ridium (Pin et al., 1975; Sommerfeld, Nichols, 1970). Как родо-
вой признак коккоидных красных водорослей используется форма хлс-
ропластов (одиночные и звездчатые у Porphyridium ; многочислен-
ные дисковидные у Rhodcspora; чашевидные с 3 - 7 лопастями у
Rhodosorus). У других багрянок коккоидный облик если и описывал-
ся, то как артефакт, связанный с условиями культивирования; так
у морской нитчатой водоросли Asterccytis таллом может распа-
даться в культуре при низкой солености на отдельные клетки ( Le-
win, Robertson, 1971) , напоминающие особей Chrootheoe (Porphy-
ridiales).
Среди золотистых водорослей коккоидные формы встречаются у
некоторых Chrysccapsales и большинства Chrysosphaera (Kri-
stiansen, 1982 ). У первых клетки (например, Chrysocaosat
Phaesaster ) погружены в слизь, и тогда как некоторые из них кск-
коидные, наблюдается более сильная тенденция к пальмеллоидному
состоянию; подвижные зооспоры напоминают собой chromulina (Och-
romcnadai.es). У Chrysosphaerales спектр коккоидных форм сходен
С отмечаемым В порядке Chloroccccales (Chlorophyceae)(Fritsch,
1935). Клетки Chryscsnhaera сферические, имеют отчетливую стен-
ку и два постепенных хлоропласта. После деления дочерние клетки
могут оставаться сгруппированными по четыре. Имеются сднсжгути-
ковые зооспоры (Starmach, 1972). Epichrysis , эпифит нитчатых
пресноводных водорослей, образует плотные гроздья из отдельных
клеток.
Большинство коккоидных видов желтозеленых водорослей поме-
щают в порядок Mlschocccoales (Hibberd, 1982а). Вегетативные
клетки содержат от одного (например, у Characlcpsis) до многих
155
(например, у Onhiocytlum) ядер, лишены сократительных вакуолей
и глазков. У Heterogloeales клетки, одиночные или чаще в паль-
меллоидных слизистых колрниях, сохраняют сократительные вакуоли
и глазки (Hibberd, 1982). Клеточная стенка может быть цельной
(jBotrvdiopsis) или состоять из двух частей (например, Onhiooytl-
ща), а поверхность бывает гладкой или скульптурированнсй. Фср-
ма"клетки варьирует от сферической до эллипсоидной. .Подвижные
зооспоры, если образуются, несут два гетерсконтных, гетеродина-
мичных жгутика.
Вегетативная стадия всех Eustigmatopbyoeae представлена
кокксидными одноклеточными организмами (Hibberd, 1982). Форш
их варьирует от сферической до многогранной, звездчатой или яй-
цевидной, а размер - от 2 до 18 мкм (Hibberd, 1982b). Присутст-
вует един глубокслопастнсй хлоропласт, придающий клеткам типич-
ную для них желто-зеленую окраску. Зооспоры, если формируются,
имеют крайне характерную для класса структуру (гл. 2).
Ксккоидные формы динофитов могут быть либо свободноживущими'
( Phytodiniales, 13 родов), либо симбионтами (Zooxanthellales,
три морских рода) ( Loeblioh, 1982). Другие одноклеточные непо-
движные представители класса крайне специализированы, и многие
из них паразиты. У Phytodiniales вегетативные клетки одноядер-
ны и содержат многочисленные мелкие хлоропласты. Это - эпифиты с
разными типами прикрепления или свободноживущие виды ст сферичес-
кой до серповидной формы. Вегетативные клетки не мотут делиться
и воспроизводятся неподвижными автоспорами, гимнодиниоидными или
панцирными зооспорами (например. stvlodi. Роды и виды раз-
личаются между собой преимущественно морфологией и способом раз-
множения. Представители порядка Zooxanthellales - внутриклеточ-
ные симбионты некоторых морских беспозвоночных; у большинства из
них свободноживущие формы отсутствуют. Клетки - от сферических
до удлиненных; вегетативное размножение - путем деления надвое.
Зооспоры, если формируются, - двужгутиковые и гимнодинисидные
(Ideblich, 1982).
Строго коккоидные формы у рафидофитов отсутствуют., хотя мо-
жет встречаться пальмеллоидное состояние ( Heywood, 1982).Боль-
шинство криптофитов - жгутиковые формы, но некоторые могут быть
кокксидными или пальмеллоидными; форма клетки варьирует ст (обыч-
н°) яйцевидной дорсовентральней до сферической в неподвижном со-
стоянии (Santore, 1977). Эвгленовые - исходно жгутиковые, но мо-
156
гут инцистироваться или становиться пальмеллоидными (Bold, Wyn-
ne, 1978) 5 коккоидные формы здесь не встречаются.
Очень разнообразны коккоидные виды среди Chlorophyceae
(в основном порядки Chlorocoooales и Chloroaarcinales , боль-
шинство представителей которых обитает в почве). Коккоидные
Chlorocoooales и Chlorosarcinalea - зооспоровые (способные об-
разовывать подвижные зооспоры) или азооспоровые (неспособные к
этому) организмы. Некоторые авторы (например, Bold, Wynne, 1978)
помещают азооспоровые типы в порядок Chlorellalea. Chlorocoooa-
les и Chlorosarcinalea отличаются друг ст друга способом раз-
множения. У первых родительские клетки не образуют потомства не-
посредственно путем деления, тогда как именно это происходит у
Chlorosarcinalea.У последних тип цитокинеза может, по термино-
логии Фрича (Fritsch, 1935), рассматриваться как вегетативное
деление клетки; впоследствии этот процесс был назван десмосхизис
(Groover, Bold, 1969)u Основная его особенность в том, что до-
черние клетки частично получают родительскую клеточную стенку.
Как показано Фричем ( Fritsch, 1935), такой способ деления - от-
правной пункт эволюции истинно нитчатого строения; он характерен
для всех высокоорганизованных водорослей и высших растений. При
альтернативном типе размножения, как показано у Chlorococcales,
потомство либо голое, либо строит целиком новую стенку. Этот про-
цесс был назван (Groover, Bold, 1969) элевтеросхизисом.
Chlcrococoum (около 38 видов; Archibald, Bold, 1970 ) И
Trabmivi а - хорошо известные зооспоровые коккоидные представите-
ли порядка Chlorocoooales . У Chloroooocum клетки сферические
или эллипсоидные, одноядерные, с одним постенным, иногда чашевид-
ным хлоропластом с одним или несколькими пиреноидами. Клетки не
поляризованы и лишены глазков и сократительных вакуолей, за ис-
ключением случаев, когда формируются подвижные стадии. Виды это-
го рода можно различать на основе биохимических особенностей
(Archibald, Bold, 1970). Trebouxia - фикобионт (водорослевый
симбионт) лишайников (Ahmajian, яЯ1Д>1973). Клетки содержат
крупный хлоропласт и один пиреноид. Повсеместно обитающая сыо-
rella _ наиболее известная азооспоровая водоросль этого поряд-
ка. Ее мелкие клетки (2-12 мкм) имеют сферическую или эллипсо-
идную форму и отличаются от особей Chlorooocoum своей азооспо-
ровой природой и наличием в стенке спорополленина (Atkinson et
al., 1972). Таксономия родаChlorella сложная, и число его ви-
157
дов спорно. Для разделения их использовались (с разными результа-
ми) физиологические, биохимические (Shlhira, Krauss, 1964; VI-
nayakumar, Kessler, 1985) и морфологические (Fott, Nbvakova,
1969) признаки. Бесцветный родственник Chlcrella, Prototheca,
вызывает болезнь прототекоз у животных и человека (ст. Klin-
tworth et al.,1968 и гл. 9).
Высокоспециализированные кокксидные одноклеточные организмы
встречаются среди десмидиевых (Chlorophyceae, zygnematales ) И
диатомовых (рис. 4.2). Группа десмидиевых объединяет многочислен-
ные пресноводные виды, обычно относимые к двум семействам (поряд-
кам богласно Mix, 1975). В одном из них (Mesotaeniaoeae), харак-
теризующемся так называемым саккодермным типом строения, одиноч-
ные клетки палочковидные или удлиненные, без перетяжки посереди-
не. Единственное ядро расположено в центре. Могут быть два спира-
левидных (например, у Spirotaenia ), два звездчатых хлоропласта
(например, у Cylindrccystis ) или хлоропласт в виде пластинки
(например, у Mesct^enium ). Стенка цельная, гладкая, без пор. У
представителей другого семейства ( Desmidiaoeae), отличающегося
так называемым плаксдермным типом, клетки чаще всего уплощенные
в поперечном сечении и имеют три плоскости симметрии, расположен-
ные под прямыми углами друг к другу. Стенка состоит из двух поло-
винок, тесно примыкающих друг к другу (иногда различить их, ис-
пользуя световой микроскоп, крайне трудно). Перетяжка (синус) у
клетки может отсутствовать (например, у Closterium ) или (у по-
давляющего большинства плаксдермных видов) быть хорошо выражен-
ной между двумя ее половинами (пслуклетками), соединенными пере-
шейком (например, у Cosmarium, Euastrum, Micrasterias). Единст-
венное ядро обычно располагается в зоне перешейка. Стенки орна-
ментированы складками, шипиками, бородавками и выростами, имеют
многочисленные поры. Осевые хлсрспласты, обычно по одному на каж-
дую полукДетку, часто причудливо лопастные или рассеченные и со-
держат пиреноиды. Жгутиковых стадий у десмидиевых нет, а воспро-
изведение посредством вегетативного клеточного деления усложняет-
ся двучастной природой клетки; оно детально описано Пиккетт-Хип-
сом ( Pickett-Heaps, 1975). Вслед за митозом и цитокинезом две
полуклетки разделяются, и каждая отпочковывает от себя недостаю-
щую полуклетку, которая дорастает до размера взрослой как зер-
кальное отражение родительской полуклетки.
Кремнеземные стенки диатомовых также состоят из двух полови-
аок, их структура и морфология описаны в гл. 3. Как примеры цен-
158
трических (радиально симметричных) диатомей (Biddulphiales )
можно привести Biddulphia. Ogclotella и Melosira , а билате-
рально симметричных (Baoillariales) - Miischia, Navicula и
Pinnularia.
Основной способ размножения у диатомовых - деление клетки,
которое, как у десмидиевых, приводит к сохранению половины роди-
тельской стенки у каждого потомка. Вследствие этого у большинст-
ва диатомей в отличие от десмидиевых постепенно размер клетки
уменьшается (гл. 5).
Как десмидиевые, так и диатомовые способны к скользящим дви-
жениям; первые - выделяя обильную слизь из пор, а вторые - за
счет системы шва у перистых видов.
Одноклеточная организация - жгутиковые
(рис. 4.3)
Вегетативная фаза у многих водорослей - подвижные, снабжен-
ные жгутиками одиночные клетки. Такие одноклеточные организмы
можно назвать фитофлагеллатами, хотя отмечалась сложность исполь-
зования этого термина с альгологической точки зрения ( Dodge,
1979). Фитофлагеллаты встречаются во всех классах эукариотных во-
дорослей, кроме Bhodophyoeae, Bacillariophyceae, Eustigmatcphy-
сеае и Charophyoeae (в последних трех, однако, жгутиковыми
являются гаметы и/или зооспоры). Жгутиковые одиночные клетки -
основная вегетативная фаза динофитов, рафидофитов, криптофитов,
эвгленовых и празинофитов. Признаки таких клеток (как вегетатив-
ных, так и генеративных) довольно однообразны внутри многих клас-
сов; общие особенности их обобщены в гл. 2, а цитологические де-
тали обсуждаются в гл. 3.
Подвижные одноклеточные формы обычно сферические, удлинен-
ные или яйцевидные, округлые в поперечном сечении. Иногда они
уплощены, что приводит к дорсовентральному строению. Содержимое
клетки обычно пслфизсвано: ее передний конец, как правило, слу-
жит местом прикрепления жгутиков, а также локализации световос-
принимающих органелл и сократительных вакуолей. У большинства
одно ядро и ст одного до нескольких хлоропластов, располагающих-
ся по бекам или в задней части клетки. Встречаются различные за-
пасные продукты. Жгутиковые одноклеточные могут быть голыми,
иметь различные покровы под плазмалеммой или настоящую стенку
(гл. 3). Впрочем, часты и отклонения от этой картины. Жгутиковые
159
Рис. 4.3. Одноклеточная жгутиковая организация: А - Phacua pie-
uroneotes (Euglenophyceae )s Б - Ceratium hirundi “
nella(~Dinophyceae ) ; В - Euglena polymorpha (Bug -
lenophyceae )s T - Chlamydomonasspl (Chlorophyceae );
Л _ Phacotua sp. (Chlorophyceae); E - Dinodinium oonradi
( 1 Dtnophyceae ); Ж - Chloromeson agile (Xanthophy-
ceae 3 - Uroglena volvox (OhnBophyceae )t И - Heterochloris
mutabilis (Chrysophyceae )5 К - Pyramimonaa
delicatua (Prasinophyceae); 5 - Pyramimonaa tetrarhynchus
(Prasinophyceae). Без масштаоа (ПО Keen, Schilling,
Dangeard, West, Pascher, Peterson, Griffiths, Dill)
одноклеточные обладают уникальными комбинациями морфологических
и биохимических особенностей, характерными для соответствующего
класса.
Ряд одноклеточных типов встречается среди золотистых водо-
160
рослей ( Bourrelly, 1981j Keistiansen, 1982). Здесь известны
виды с двумя гетероконтными гетеродинамичными жгутиками (напри-
мер, Ochromonas). единственным жгутиком, выходящим наружу, и
вторым, редуцированным до короткого выроста (например, Chroma-
11да ), единственным наружным жгутиком, имеющим укрепляющую лен-
ту и волнообразную конфигурацию (типа поперечного жгутика дино-
фитов) ( Ре dine Hales), ИЛИ с ОДНИМ наружным жгутиком в сочета-
нии с цитоплазматическим окремнелым скелетом ( Diotyoohales =
силикофлагеллаты). Если жгутика два, то один из них перистый, а
второй гладкий, причем перистый обычно несет трехчастные волоски
(с. П5); если из двух жгутиков один редуцирован, то сохраняется
перистый. Клетки могут быть голыми (например, у Oohromonas ),
покрытыми окремнелыми чешуйками (например, у Mallomonas). орга-
ническими чешуйками (например, у Apedinella). панцирем (напри-
мер, у Chrysocooous) или плотно прилегающей стенкой. К золотис-
тым водорослям относили своеобразных, обычно бесцветных хоанофда™
геллат, напоминающих хоаноциты губок ( Bourrelly,1981), однако
большинство авторов считает их все же животными (Leadbeater,
1982;Leadbeater, Manton, 1974» Kristiansen,1982; Kristiansen,
Takahashi, 1982). Большинство золотистых водорослей (не из поряд-
ка Pedinellales) образует цисты (статоцисты или стоматоцисты),
состоящие из двух неравных частей: бутылевидной, более гфупной,
и подогнанной к ней более мелкой "пробки". Стенка их всегда окрем-
нелая независимо от природы плавающей стадии ( Bourrelly, 1957).
Признаки одноклеточных жгутиковых примнезиофитов (составляю-
щих подавляющее большинство представителей этой группы) типичны
для класса в целом (с. 39). Виды с редуцированной гаптонемой
или вовсе без нее помещаются в порядок Isoohrysidales (Green,
Pienaar, 1977), У представителей которого равные или почти рав-
ные жгутики и клеточный покров из органических чешуек. У некото-
рых видев этого порядка (семейство Prinsiaceae) чешуйки играют
роль "матриц" для обызвествления и образования кокколитов (напри-
мер, у Gephryocansa). Однако большинство образующих кокколиты
видов (группа коккслитофорид) относится к порядку Coooosphaerales
( Norris, 1982а ). Описано более 150 родов кокколитофорид
(Loeblioh, Tappan, 1966; Tappan, 1980), многие из которых - ис-
копаемые формы. Известно два основных типа кокколитов: голококко-
литы, когда поверх органических матриц откладываются ромбоэдриче-
ские кристаллы кальцита, и гетерококколиты, когда кристаллы
161
(обычно кальцита, но иногда и арагонита) морфологически различ-
ны. Кокколиты могут плотно прилегать друг к другу, образуя кок-
косферу (например, у Calcidiscus). Предложена их сложная клас-
сификация ПО форме (Braarud et al., 1955? Tappan, 1980), хотя
искусственность такого подхода очевидна из результатов все воз-
растающего числа работ, показывающих наличие стадий с кокколита-
ми и без них в одном и том же жизненном цикле (например, Parke,
Adams, I960, von Stosch, 1967? Leadbeater, 1970; Klaveness ,
1973). Очень мало известно о живых клетках большинства кокколи-
тофорид, но имеется обширная литература,детально рассматривающая
тонкую структуру их чешуек (см. Boney, Creen, 1982).
Гаптонема (с. 108) хорошо развита в семействе Prymnesia-
oeae (Prynmesiales); иногда в ответ на механическое воздействие
она сворачивается. Клетки здесь покрыты одним или несколькими
слоями неминерализованных чешуек (например, у Prvmnesium). Поря-
док Pavlovales, характеризующийся наличием двух неравных жгути-
ков, один из которых покрыт чешуйками (с. 108), выделяется среди
примнезиофитов присутствием стигмы. Большое внимание уделялось
их тонкой структуре (Green, 1980.). К дополнительным особенно-
стям этой естественной группы относятся шишковидные чешуйки на
более длинном жгутике и канал, входящий в клетку рядом с местом
прикрепления жгутиков (Boney, Green, 1982).
Представители Желтозеленых водорослей, существующие в веге-
тативном состоянии в виде жгутиковых клеток, помещаются в поря-
док Chloramoebidales (Ettl, 1978, Hibberd, 1982а). Не исключе-
но, что на самом деле они относятся к другим классам, например
золотистым водорослям, примнезиофитам и празинофитам (Hibberd,
1982а).
Большинство динофитов - двужгутиковые одноклеточные. Они от-
личаются структурой жгутиков (с. 117, 49 ), текой (с. 51 ), ме-
зокариотным ядром (о. 138), наличием трихоцист, пузул и специфи-
ческого глазка. Трихоцисты состоят из шейки, прикрепленной к те-
ке, и тела, содержащего паракристаллический белковый материал
(Dodge, 1979). Они отличаются от эжектосом криптофитов тем, что
выбрасывают длинную поперечнополосатую нить квадратного или ром-
бического сечения (Bouck, Sweeney, 1966, Dodge, 1973).yNemato-
Ainium и Polykrikoa обнаружены нематоцисты, напоминающие ана-
логичные структуры у кишечнополостных ( Mornin, Francis, 1967;
®reuet, 1972). Глазок (с. 134) отмечен у некоторых крупных пред-
162
ставителей класса, в частности у Wamowia (Grenet, 1977). Пузу-
ла - постоянная осморегуляторная органелла; у динофитов описано
несколько ее типов ( Dodge, 1972). Она представляет собой впячи-
вание плазмалеммы у основания жгутикового канала, открытое нару-
жу и имеющее толстую стенку из двух мембран.
В то время как для большинства динофлагеллат характерна
структура теки с вентрально прикрепленными жгутиками (например,
Gymnodinium, Peridinium). У некоторых форм (например, Ртого-
centrum) жгутики отходят спереди, а тека разделена на равные
правую и левую половины. Среди первых некоторые имеют асимметрич-
ную теку с поперечной бороздой ( cingulum), сдвинутой вперед (на-
пример, Amphidinlum pacificum). или о относительно мелким гипо-
коном (например, Katodinium го tundatum).
Для панцирных динофлагеллат характерны пластинки теки (до
100 штук). Их число и расположение использовались как таксономи-
ческие признаки для описания родов и видов. Пластинки зпитеки и
гипотеки (соответствующих зцикону и гипоксну беспанцирных форм
по некоторым классификациям) обозначаются определенным образом,
так что таксон можно охарактеризовать однозначной формой панциря
(рис. 3.9). Используя такую систему, можно выделить три порядка:
Peridinialea (различное число пластинок; cM.ioeblioh, 1970),
Dinophysialea( 18 пластинок, гипотека крупнее эцитеки;коГо1й,
Skogsberg, 1928; Dodge, 1979) и Prorocentralea (две латераль-
ные пластинки, прикрепленные спереди жгутики; loeblich, 1982).
Большинство плавающих клеток динофлагеллат гаплоидные. Обыч-
ный для них способ вегетативного размножения - деление надвое,
которое после цитокинеза может происходить в поперечной, продоль-
ной ИЛИ КОСОЙ ПЛОСКОСТИ (Bold, Wynne, 1978). У голых форм при
этом образуется перетяжка; иногда новая амфиесма формируется во-
,круг каждой половинки, иногда старая сбрасывается (экдизис) еще
до деления. У большинства панцирных форм две половины родитель-
ской теки вслед за цитокинезом разделяются и каждая из дочерних
клеток образует недостающую половину.
У динофлагеллат обычны цисты, устойчивые к неблагоприятным
факторам внешней среды. Их два типа: напоминающие вегетативные
клетки (например, уPeridinium ) или сильно отличающиеся от них,
обычно сферические и покрытые полыми выростами (например, у Go-
nyaulax). Цисты вскрываются с образованием отверстия (археопи-
ле), особенности которого учитываются в таксономии. Многие из-
163
вестны только в ископаемом состоянии (так, называемые гистрихоспо-
ры, Sarjeant, 1974, Tappan, 1980); они будут обсуждаться ниже
(с. 420).
Некоторые рафидофиты (или Chloromonadophyceae, см. Bold,
Wynne, 1978; Dodge, 1979 - жгутиковые одноклеточные очень свое-
образной структуры (с. 029). Клетки Gonyoatornum крупные, покры-
тые тонким перипластом, дорсовентэально уплощенные, с двумя гете-
родинамичныминеравными жгутиками. Они содержат многочисленные
дисковидные хлоропласты, много трихоцист, сократительные вакуоли
( Heywood, 1982). Воспроизведение - путем продольного деления еще
движущейся клетки.
Большинство криптофитов - жгутиковые с уникальной ультра-
структурой хлоропластов, ждутиков и эжектосом ( Dode, 1979). К
хорошо известным родам относятся Crytomonaa. Chilomonaa и
Hgmiaeimia.Клетки несут два передних жгутика, которые выходят из
глотки, разделенной на вестибулярный отдел и выстланный эжектосо-
мами канал, задним концом впадающий в клетку (Oakley, Santore,
1982). Жгутики - более или менее одинаковой длины; у Cryotomonas
на одном из них - два ряда трубчатых волосков и вздутие у основа-
ния, а на другом - один ряд .волосков (Hibberd et al., 1971).
Таксоны традиционно разделялись по числу рядов эжектосом в глотке
и пигментации, хотя оба этих признака ненадежны (см. Bold, Wynne,
1978; Oakley, Santore, 1982). Клетки покрыты тонким перипластом;
пластинчатые структуры под плазмалеммой в разных родах имеют раз-
ную форму (например, шестиугольные у Cryptomonaa; Hibberd et
al,, 1971; прямоугольные у Chroomonaa; Grant, 1971). Особенно-
стям пластинок может соответствовать специфическое расположение
эжектосом.
Эжектосомы (трихоцисты, или тениоболоцисты, у некоторых ав-
торов; Dodge, 1979) встречаются у всех криптофитов, кроме Hil-
las. Они аналогичны таковым у динофитов и инфузорий, но отлича-
ются по структуре. В отстрелянном виде это утончающиеся к концу
нити, а внутри клетки - смотанный в "рулетку" мембранный матери-
ал.
Криптофиты обычно размножаются продольным делением, во вре-
мя которого клетки иногда продолжают двигаться. В пальмеллоидной
фазе жгутики сохраняются, плотно закручиваясь вокруг находящихся
в слизистых колониях клеток.
Большинство эвгленовых - жгутиковые одноклеточные, объединя-
емые в хорошо очерченный класс на основании ряда уникальных цито-
164
логических и биохимических особенностей (с. 64). Все они имеют
два базальных тела и один (например, Euglena ) или два (напри-
мер, Eutreptta ) выходящих наружу жгутика со специфической струк-
турой ( beedale, 1967). Встречаются бесцветные (например. Гегапе-
та) и пигментированные виды; Лидейл ( LeedaLe, 1982Ъ) считает,
что последние скорее всего произошли от первых за счет эндосимби-
оза с хлоропластами зеленых водорослей (последовательность, обрат-
ная предполагавшейся во многих более ранних источниках). Некото-
рые бесцветные виды (например, Paranema triohophorum) фагоцити-
руют другие клетки и детрит. Их "ротовое отверстие", или цито-
стом, снабжено параллельными крючковидными палочками, прикреплен-
ными к его жесткому краю. Лучше всего изучен род Euglena (leeda-
le, 1967; Buetow, 19б8, 1982), разделяемый на виды по размеру и
форме клеток, длине и характеру движения жгутиков, числу и разме-
ру хлоропластов и пиреноидов, жесткости пелликулы, слизистым
тельцам и форме парамилоновых гранул ( Leedale, 1982).
Выделяются ( Boveey 1982) четыре типа движения эвгленовых:
плавание, сокращение, ползание и скольжение. С тремя последними
связаны разные формы метаболии.
Мукоцисты, или слизистые тела, располагаются рядами под пел-
ликулой и выделяют растворимый в воде полисахарид. Клетки посто-
янно покрыты тонким слоем слизи ( Rosowskl, 1977). Она может об-
разовывать ножки (например, у Colacium). сложные покровы клеток
(например, у Trachelomonas) и оболочку цист. Последние у Eugle-
na и Dtatigma покрыты утолщенным слизистым слоем, хотя и у не-
которых других родов особый слизистый чехол служит временной за-
щитой от высыхания. Под таким слоем может происходить деление
(например, у Eutreptia) с образованием объемистых пальмеллоид-
ных колоний ( Leedale, 1967). У Euglena клетки иногда стано-
вятся неподвижными, значительно увеличиваются с возрастом в раз-
мере и накапливают оранжевые и черные пигментные частицы.
Размножение у эвгленовых только бесполое, путем деления.
После потери жгутиков происходят митоз и репликация базальных
тел; формируются два дочерних канала, и передний конец клетки
между ними разделяется. Ленты пелликулы удваиваются, и расщепле-
ние клетки идет спереди назад, следуя их спиральному расположе-
нию. У форм с упругой пелликулой во время деления наблюдается ин-
тенсивная метаболия.
Одноклеточные жгутиковые Chlorophyceae и их колониальные
производные относят к порядку вольвоксовых (Volrocales). У одно-
165
клеточных видов по два, четыре или восемь изоконтных гладких го-
модинамичных жгутиков. Клетки голые (семейство Polyblepharida_
ееае, например, Punaliella) или в большинстве случаев окруже-
ны стенкой, иногда сложной структуры (например, уChlamydomonas )•
Кгутики обычно отходят спереди через каналы в клеточной стенке, у
двужгутиковых видов - под углом 180° друг к другу по обе стороны
"носика". Клетки бывают заключены в панцирь, состоящий у гнасо-
tus из двух частей, которые при делении расходятся. У Haemato-
сосctis стенка сильно ослизнена и пронизана радиально расходящи-
мися тяжами цитоплазмы; у Brachlomonas на ней находятся высту-
пающие базальные лопасти. Представители своеобразного рода Medu-
sochloria CPascher, 1917) напоминают медузу; их клетка пред-
ставляет собой выпуклую полусферу с четырьмя выростами, каждый
из которых несет у своего нижнего края жгутик.
Клетки вольвоксовых обычно одноядерны и лишены крупной цент-
ральной вакуоли; вместо этого у них одна или несколько осморегу-
ляторных сократительных вакуолей (отсутствующих у морских видов),
расположенных непосредственно под носиком (например, у Chiamy-
domonas ), у оснований жгутиков или рассеянных по всей клетке
(например, у Chiorogonlum). Хорошо заметный хлоропласт бывает
чашевидным, постенным звездчатым, сетчатым, пластиновидным,
Н-образным и т.д. Обычно внутри его находятся глазок и один или
более пиреноидов. Не все вольвоксовые пигментированы; имеются
как двужгутиковые (Polytoma). так и четырехжгутиковые (Poiyto-
mella) бесцветные виды.
Бесполое воспроизведение является результатом митоза и деле-
ния клетки. При этом у голых видов (например, Punaliella) клетка
может сохранять подвижность, но большинство форм со стенкой (на-
пример, Chlamydomonas) на время митоза и цитокенеза становятся
неподвижными. При делении происходит удвоение хлоропластов и пи-
реноидов, хотя некоторые органеллы (например, сократительные ва-
куоли, глазки), по-видимому, возникают у дочерних клеток de novo
(Bold, Wynne, 1978). Деления часто происходят в темноте и могут
Достаточно быстро следовать друг за другом, так что внутри роди-
тельской стенки возникают две, четыре, восемь или более дочерних
клеток. Обычно они меньше размером, чем взрослая особь, но после
выхода наружу постепенно дорастают до величины, типичной для со-
ответствующего вида, прежде чем переходят к вегетативному деле-
нию.
Жгутиковые одноклеточные празинофиты (большинство предста-
166
вителей этого класса) принципиально отличаются от Chlorophyceae
чешуйчатым покровом жгутиков и клеток. У видов с одним (напри-
мер, redinomonas) и двумя жгутиками последние прикреплены в пе-
реднебоковой части клетки с углублением, или канавкой. У четы-
рехжгутиковых (или восьмижтутиковых) видов (например. Platymo _
пая) жгутики выходят спереди из углубления в виде поры. По пиг-
ментации и внутренней структуре празинофиты сходны с Chlawydo-
дюп-аа (Chlorophyceae);здесь имеются чашевидный хлоропласт, гла-
зок и крупный пиреноид. Уникальная особенность некоторых пред-
ставителей класса (например, Byramimonas. Haloaphaera. Mono-
mastix) - наличие трихоцист ( Norris, 1982), очень сходных с та-
ковыми у криптофитов ( Dodge, 1979).
Воспроизводятся празинофиты исключительно путем вегетатив-
ного деления клетки. Согласно Норрису ( Norris, 1982b), наибо-
лее примитивные виды сохраняют во время деления подвижность,
тогда как более высокоорганизованные теряют жгутики. В начале ми-
тоза происходит последовательное (у более примитивных) или син-
хронизованное (у более продвинутых) удвоение органелл ( Norris,
1982 b). У некоторых видов (например, Pterosperma. Dachvsr>haera)
может встречаться устойчивая к неблагоприятным воздействиям, со-
держащая спорополленин стадия фикомы, важная для изучения отно-
симых к празинофитам ископаемых находок ( Norris, 1980, 1982b).
Колониальная организация - пальмеллоидный, тетраспоровый
и дендроидный типы
(рис. 4.1, 4.2 и 4.4)
Большинство обычных жгутиковых или коккоидных одноклеточных
водорослей может переходить (часто временно) в стадию пальмеллы -
состояние, когда жтутики теряются (если в норме имелись) и клет-
ки, погруженные в общий слизистый матрикс, последовательно веге-
тативно делятся. Этот термин, образованный от названия рода
Palmella (вольвоксовые), можно в узком смысле применять к водо-
рослям, клетки которых способны легко восстанавливать подвиж-
ность ( Bold, Wynne, 1978), или распространять и на все формы
с относительно устойчивым пальмеллоидным обликом (Fritsch ,
1935). Однако некоторые авторы (например, Round , 1973) называют
последние виды тетраспоровыми (по роду Tetrasnora из Chloro-
phyceae J . При таком типе организации подвижные стадии (если они
167
•ceae );
Рис. 4.4. sartularl.a
простые нити: Д
toon ар. 1^Уа??^севК - ТЙ nnthrixрвдае«> - с
4^wTChior°pMyeaeJi .- Xemmermaim, Meitler,
да^е). Без ®сш*дба p&acHerJ
ter, Cienkowski, «еаь,
168
вообще имеются) ограничены репродуктивными клетками. Пальмелло-
идные и тетраспоровые формы характеризуются отсутствием опреде-
ленного облика, дифференцировки и специфического предельного
размера колонии.
К тетраспоровому типу относят и некоторые более правильные
колонии синезеленых водорослей (например, представляющие собой
плоские пластинки из клеток у Meriamopedia и Kalopedia. или
кубические-скопления у Eucapaia). У одних видов этого отдела ко-
лонии состоят всего из нескольких клеток в общем матриксе (на-
пример, yghropcoccuB.Gloeooapaa ) и мало отличаются от коккоид-
ных форм; в других случаях они выглядят как крупные уплощенные
слизистые массы.
У некоторых специализированных водорослей слизь выделяется
только основанием клетки, так что образуются древовидные (дендро-
идные) колонии, напоминающие обычные нити (например, у Prasino-
cladua из празинофитов, Dinobryon из золотистых водорослей,
Cdlaclwn из эвгленовых); клетки здесь легко переходят в подвиж-
ное состояние.
Кроме синезеленых водорослей тетраспоровые формы (по Round,
1973) встречаются у золотистых водорослей (например, sticho -
gloea ). примнезиофитов (например, Phaeocyatis. Saroinochryaia).
желтозеленых водорослей (например, Qioeochloria). динофитов
(например, Gloeodinum montanum) и в порядке Tetrasporalea
из Chlorophyceae (нацример, Gloeocyatia. Palmodlctyon. Tetra-
spora). Особенность Tetraapora - наличие слизистых псевдоци-
лий.
Крупные колонии встречаются у диатомей; они мотут быть по
природе капсулярными (например, некоторые виды Mavicuin ) или
иметь форму слизистой трубки (например, у Amnhipieura). Индиви-
дуальные клетки продолжают двигаться и делиться внутри слизи, а
трубка, как правило, прикрепляется к твердому субстрату; она мо-
жет достигать нескольких сантиметров в длину. В этом классе (осо-
бенно в порядке Biddulphialea) известен ряд специфических коло-
ний, образующихся за счет соединения клеток слизью после их де-
ления. Они бывают нитчатыми (например, у Meloaira. Pragilaria.
EunotialnpH нежестком соединении панцири могут скользить друг
относительно друга (например, у Baciilaria) или образовывать
зигзагообразную нить (например, у Grammatophora. Tabeliaria).
169
Для некоторых видов Tabellaria и Diatoms характерны звездча-
тые колонии. У ряда эпифитных диатомей (например, Licmoohora)
клетки могут соединяться между собой и прикрепляться к субстрату
слизистыми ножками, формируя ветвящиеся древовидные (дендроид-
ные) колонии.
Многие эукариотные тетраспоровые виды иногда размножаются
путем простой фрагментации колонии, но при определенных условиях
(например, резком изменении температуры) образуют подвижные зоо-
споры.
Колониальная организация - ценобии
(рис. 4.5)
Колониальные формы с упорядоченной организацией, или цено-
бии, состоят из жгутиковых или неподвижных клеток. Первый случай
встречается в нескольких эукариотных классах (табл. 4.1), тогда
как последний - только среди Ohiorcphyceae Подвижные ценобии •
отличаются от пальмеллоидных, тетраспоровых или дендроидных форм
в основном тем, что число особей в них довольно постоянно для
данного вида, колонии имеет определенную форму, а между особями
часто наблюдаются координация деятельности и некоторая степень
дифференцировки.
Наиболее полный спектр подвижных колониальных форм встреча-
ется в порядке вольвоксовых (Chlorophyceae). Ценобии здесь сос-
тоят из клеток, число которых кратно двум; каждая клетка напоми-
нает особь Chlamydomonaa j все они находятся в,общей слизистой
оболочке. Количество клеток в колонии колеблется от относительно
небольшого (например, у Gonium 4 - 32) до довольно существенно-
го (например, у Eudor-j^n 16 - 128) и достигает кульминации у
Volvox (500 - 40 000). Внутри колонии клетки особым образом ори-
ентированы друг относительно друга; форма колоний плоская ( Go-
aium; Blatvdorina). шаровидная (Bandorina) или сферическая, с
однослойным расположением клеток ( Eadorina; VolvoxIIIo мере уве-
личения размера колонии возрастает тенденция к развитию ее поляр-
ности. Появляются ориентация клеток относительно направления дви-
жения и переднезадний градиент размеров их самих и глазков. По-
лярность проявляется также в появлении специализированных клеток,
осуществляющих половое и бесполое размножение. Подвижные цено-
170
Рис. 4.5. Колониальная организация - ценобии: & - gonium
pectorals (Chlorophyceae); Б - Chrygoaphaerella longj^"
spina (flErysophyceae): В - "Scenedesmus quadricauda
( Chlorophyceae)$ J - Synuraadanaonii (Chrysophyceae)
£ - Chlorodesmus hiapidus (Chlorophyceae): E - Pediaat-
rwn duplex ( Chlorophyceae). Без масштаба (no Hart-
mann, Lauterborn, G.M, Smith, Conrad, Philipps)
бии воспроизводятся бесполым путем, формируя дочерние колонии,
число клеток в которых такое же, как у исходной. Размер родитель-
ского ценобия достигается в ходе последующего роста клеток. У бо-
лее примитивных видов (например. Pandorina) формировать дочер-
ние колонии способны все клетки, а у более высокоорганизованных
(например, Voivox) - только специализированные (гонидии), кото-
рые лишены жгутиков и выделяются крупными размерами. Образование
дочерних колоний у Voivox схематически представлено на рис. 5.3.
171
Неподвижные ценобии у зеленых водорослей встречаются в се-
мействах Hydrodictyaceae (например, Pediastrum), Serastrum.
Hvdrodictyon J И Scenede emaceae (например, Scenedesmu,6.
Ccelastrum. Dictyosr'h^giMiiinl порядка Chlorococcales. У Pedlast—
rum и Borastrum они представляют собой пластинки из одного
слоя расположенных концентрическими окружностями одноядерных
клеток, крайние из которых (а иногда и внутренние) снабжены ро-
говидными выростами. Такие ценобии размножаются бесполо, двужгу-
тиковыми зооспорами, формирующимися в каждой из клеток в резуль-
тате синхронных митозов. Родительская клеточная стенка затем раз-
рывается, высвобождая активные зооспоры, остающиеся внутри обще-
го пузыря. После короткого периода подвижности они сбрасывают
жгутики и слипаются вместе в колонию характерного для данного ви-
да облика. Затем клетки образуют стенки, а пузырь разрушается.
У Hydrodlctyon ценобий представляет собой цилиндрическую сете-
видную структуру до I м длины. Он состоит из ценоцитных трубко-
видных клеток с крупной центральной вакуолью и множеством мелких
ядер. Клетки смыкаются концами, образуя многоугольные (обычно
шестиугольные) ячеи сети. Ценобий размножается бесполо многочис-
ленными одноядерными двужгутиковыми зооспорами, которые образу-
ются в каждой его клетке. Зооспоры собираются тонким слоем между
вакуолью и клеточной стенкой. После периода активной подвижности
они сбрасывают жгутики и устойчиво соединяются в характерную для.
данного вида структуру. Новые ценобии высвобождаются при разруше-
нии родительской клеточной стенки ( Pickett-Heaps, 1975).
Ценобии Scenedesmus состоят из цилиндрических клеток, сое-
диненных боковыми сторонами в группы по 4, 8 или 16. Внешние (а
иногда и внутренние) клетки несут один или более шипиков. Вегета-
тивное размножение представляет собой модифицированную форму об-
разования автоспор (Pickett-Heaps, Staehelln , 19?5). Изредка
встречаются подвижные зооспоры, не связанные с образованием до-
черних колоний. Ценобии Со elastrum- полые сферы из 4 - 128 кле-
ток ( Bold, Wynne , 1978). У Dictrosphaerium сферические сли-
зистые ценобии организованы совершенно иначе. Здесь клетки, напо-
минающие особи Chlorella. сгруппированы по четыре на ножках,
которые могут формировать обширную дендроидную систему в преде-
лах сферической колонии.
172
Нитчатая организация - простые нити
(рис. 4.1 и 4.4)
Простая нить состоит из одного неветвящегося ряда плотно
скрепленных друг с другом клеток. Она представляет собой исход-
ную форму многоклеточного водорослевого таллома ( Fritsch, 1938).
Простые нити встречаются только у яескольких классов (табл. 4.1);
их можно вывести из подвижных или неподвижных одноклеточных форм,
у которых (среди эукариот) модифицировались ориентация веретена,
цитокенез (например, возник десмосхизис) и образование стенки.
Обычно одноядерные виды (например .Ulothrix из Chlorophyceae)
формируют нити длиной от всего нескольких (например,Stichocoo -
cus из Chlorophyceae ) ДО многих клеток (например, Osoillato -
ria из синезеленых водорослей). Встречаются как свободноживущие
(например, uiothrix, Osoillatoria ), так и прикрепленные формы.
У синезеленых водорослей простая нить называется трихомом.
Отдельные клетки могут быть крайне мелкими (меньше I мкм в диа-
метре у lamgbya). Ширина их равная по всей длине нити (например,
У Osoillatoria) с только слабым изменением у конечной клетки
или заметно уменьшается от основания к вершине трихома, так что
его терминальная часть волосовидная (например, у Soytoneaa. Ri-
vularia). а базальная клетка является гетероцистой. Сужающиеся
трихомы могут группироваться, создавая подобие ветвления (напри-
мер, у Rivularia). расходиться из общего центра (например, у
Sloeсtrichina) или образовывать пленчатую массу. Колонии корот-
ких трихомов в общем слизистом матрикое характерны для Arhani-
zomenon и Nostoo.y некоторых более мелких планктонных видов (на-
пример, Spiralina) трихом спирально скручен. Большинство прос-
тых нитей синезеленых водорослей способны к скользящему движению,
которое может быть плавным, скачкообразным или колебательным
( Fogg et al., 1973) и связано с порами и выделением слизи.
Трихомы синезеленых водорослей легко распадаются на гормого-
нии либо за счет естественных разрывов, либо по специальным двоя-
ковогнутым клеткам, называемым разделительными дисками, или яе-
кридиями. Гормогонии обычно высвобождаются из родительского вла-
галища, но могут и оставаться внутри его (например, у Microco-
leus).
Tribonema - наиболее известный нитчатый представитель жел-
тозеленых водорослей. Ее неприкрепленные нити состоят из обычно
одноядерных одинаковых цилиндрических или бочковидных клеток,
173
стенки которых образованы двумя равными и слегка перекрывающими
друг друга половинками. В результате на концах нити остаются по-
ловинки стенок, и вся она может распадаться на Н-образные фраг-
менты. Бесполое воспроизведение включает образование одной-двух
гетероконтных зооспор в неспециализированных клетках, апланоспор
или толстостенных гипноспор.
Большое разнообразие простых нитчатых форм встречается сре-
ди Chlorophyceae в порядках Micrcspcrales(например, Microspo-
ra), Oedogoniales (например, Oedogoniun). Ulotrichales (напри-
мер ,Ulothrix), Sphaeropleales (например, Sphaeroolea). Klebsc-
rmidiales (например, Klebsormidium)И Zygnematales (например,
Spirogyra. Zygnema. Desmidium).
Нити могут быть свободноплавающими (например, у Micrisnora.
Sphaeroolea. Klebsormidium. большинства Zygnematales) или при-
крепленными. Прикрепление обычно осуществляется за счет простой
модификации базальной клетки, стенка которой разрастается в "при-
соску" на конце (например, у Oedogonium). У одноядерных клеток
стенки сплошные (за исключением Microsnora. где они состоят из
Н-образных частей, и Oedogoniales, где стенки во время деления
клетки разрываются с образованием апикальных колпачков; с. 106).
Часто имеется слизистый чехол (особенно у Zygnematales). Если
не считать прикрепительной базальной клетки, дифференцировка ни-
тей в целом незначительна. У Desmidium они спирально закручены.
Идентификация многих простых нитчатых Chlorophyceae часто
трудна (например, в роде Oedogonium выделяют несколько сотен
видов, у Spirogyra - свыше 275 видов; Bold, Wynne, 1978). Одна-
ко различение на уровне рода упрощается благодаря особенностям
хлоропласта и пиреноидов.
У большинства простых нитей в этом классе рост диффузный,
т.е. клеточные деления происходят в любом месте. Фрагментация
или распад нити - обычнейший способ вегетативного размножения; у
некоторых форм (например, Klebsormidium) разделение может дохо-
дить до отдельных клеток. Широко распространено образование по-
движных зооспор (за исключением Zygnematales), возможное в лю-
бой клетке, кроме базальной, служащей для прикрепления. Также ча-
сто образуются апланоспоры или толстостенные гипноспоры, послед-
Дие как средство выживания при неблагоприятных условиях.
174
Нитчатая организация - ветвящиеся нити
(рис. 4.1, 4.4 и 4.6 - 4.13)
Имеются три принципиальные модификации ветвящихся нитчатых
талломов. В первом случае прямостоячий таллом прикрепляется к
субстрату простым диском, образованным базальной клеткой; во вто-
ром (так называемый гетеротрихальный тип) - таллом состоит из
двух частей - стелющегося основания и прямостоячей ветвящейся
системы, в третьем - одна (одноосевой тип) или несколько (много-
осевой тип) ветвящихся нитей образуют поевдопаренхиматозный тал-
лом. Наряду с этими модификациями наблюдается усиливающаяся тен-
Рис. 4.6. Нитчатые (А), гетеротрихальные (Б) и сифонокла-
дальные (В, Г)Chlorophyceae. A.Draparnaldia ivenaa-
rii, часть таллома с крупными клбттаяи ТОШВИОЙ ЙИТЙ и
расположенными в мотовках боковыми ветвями со щетинками.
Б.Pritschiella tuberosa. гетеротрихальная форма С парен-
химатозной оазалйГОй"Системой, образующей ризоиды, и нит-
чатыми вертикальными выростами. В. Cladophora ар.,
ветвящаяся сифонокладальная форма. Г. Chaetomcrpha dar-
те1п11гневетвяшаяся сифонокладальная форма. Молодой экзем-
пляр (слева) с базальной прикрепительной клеткой и нити
(справа) с клетками, вытягивающимися после деления.
А, Б, В без масштаба (по Tiwari, Pandey и Iyengar,
ffoek, Sigula)
175
Рис. 4.7. Псевдопаренхиматозные (гаплостихальные) и парен-
химатозные (полистихальные) бурые водоросли. A.Wrionema
strangulans: видны двуслойная базальная система, гетеро-
трихальные вертикальные выросты с беспорядочными попереч-
ными перегородками и волоски. Б.СЬаоозвога, obtusifolia:
апикальная зона паренхиматозного таллома с аликеЬкьйой ме-
ристемой из мелких клеток. В. Вид с поверхности на апекс
дорсовентрально уплощенното“таллома Dictyola dichotoma:
заметны единственная апикальная клетка и развитие псевдо-
паренхимы. Г. Срез через апекс Carpomitra costata: вид-
ны меристематические волоски, формирующие многоосевой
псевдопаренхиматозный таллом. £. Продольный срез через
апекс Dioty.osiphon. foeniculaceus: видны единственная апи-
кальная клетка и ее производные, из которых развивается
паренхиматозный таллом. Е. Вершина Sphacelaria pltumda
с апикальными клетками основной и боковых осей; слева вни-
зу - поперечные срезы паренхиматозных осей. Ж. Продольный
срез через середину апикальной зоны паренхиматозного про-
ростка споры Fucus sp. .• видна единственная апикальная
клетка (затушевана) в апикальной выемке. 3. Продольный
срез через середину более взрослого проростка Fucus sp.
И» Апикальная зона Acrothrix gracilis: видно развитие
гаплостихального таллома. R. Вершина таллома Desmarestia
aculeata; гаплостихальный таллом развивается в результате
трихоталлического роста и имеет единственную осевую нить.
Боковые ветви и последующие покровные веточки дают либо
билатеральную псевдопаренхиматозную структуру, как показа-
но для Desmarestia ligulata (Л), либо, как у Arthrocla-
dia villosa (М, радиальную структуру с осевой нитью,
остающейся хорошо различимой на поперечном срезе. Н. Про-
дольный срез апр.кс.а MvriQKloia sciurus: видна гаплости-
хальная структура, развившаяся из многоосевой трихоталли-
ческой меристемы. Без масштаба (по Sauvageau, Kuckuck,
Cohn, Murbeck, Fritsch, Reinke, Jonsson)
176
Рис. 4.8. Одноосевые нитчатые багрянки. A. Antithmimion
defectums видна основная ось неограничённоТб' ДООТА И рас-
поДокёИйые в два супротивных ряда ветви ограниченного рос-
та с молодыми тетраспорангиями. Б. Апикальная зона
ria elegana с осевыми клетками (затушеваны) и ветвяМЙ
бГр'аИИчейИОТо роста. В. Delesseria' deoipiena. фотография
верхушки с единственной апикальной клеткой. Осевые нити
маскируются коровыми клетками, образующими утолщенную сред-
нюю жилку (см. j). Г. Общий вид Bonnemaiaonia asparagoi-
des, показывающий характер ветвления, и деталь W ашГ-
"йайьной зоны. Е. Общий вид листовидного таллома РеХевве-
ria вапяи1пеа со средней и боковыми жилками. Ж. Вид о по-
верхности апекса Phycodrys rubens с главнойГосевой нитью
и отходящими от нее боковыми ветвями ограниченного роста
(затушеваны). Без масштаба (по Kylin, Newton)
денция к локализации клеточного деления, специализации клеток
(т.е. дифференцировка) и переходу репродуктивных функций к спе-
циальным структурам (например, спорангиям и гаметангиям).
Ветвящиеся нитчатые системы встречаются в восьми из рассмат-
риваемых в этой книге классов (табл. 4.1). У синезеленых водорос-
177
лей известны два типа ветвления. Ложное, более примитивное, про-
исходит за счет разрывов нити (например, по гетероцисте или раз-
делительному диску). Одна (например, у Toiypothrix) или обе (на-
пример, у Scytonema) части разделившегося трихома выталкивают-
ся из влагалища как ветви. Новой плоскости деления здесь не воз-
никает. При более продвинутом истинном ветвлении ветви отходят в
результате деления клеток во второй плоскости (например, у нара-
losiphon.).
Ветвящиеся нити, редки у динофитов (например, Dinothrix).
желтозеленых (например, Heterodendron) и золотистых (например,
Рис. 4.9. Одноосевые багрянки. A. Audow-nella seoundata:
видны волоски, простые боковые ветви и моноспоры. Б, В. Ча-
сти таллома Sirodotia gueoioa , состоящего из крупных
осевых клеток и нитеи ограниченного роста, дающих либо
свободные ветви, либо кору. Г.Griffithsia corallina :
апикальная зона и (£) крупные бочонковидные клетки, несу-
Щие редуцированные или более длинные ветви ограниченного
роста. Е. Dumontia incrassata , продольный срез, показы-
вающий осевую нить с сердцевинным и коровым слоями, обра-
зованными ветвями ограниченного роста. Ж. Поперечный срез
gloeosinhonia capillaris с единственным осевым рядом и
четырьмя радиально расположенными пучками ограниченного
Роста, придающими таллому округлую в сечении форму. 3. По-
перечный срез Sphaerocoocus coronopifolia с одноосевым
строением (осевая клетка затушевана) и уплощенным талломом.
*ез масштаба (по Kylin, Bornet и Thuret)
I?8
Рис. 4.10. Одноосевые багрянки. А. Апекс Plocamium carti-
.,2agl^gum„_,; показан характер ветвления (1 - 4); каждый по-
следова те явный участок таллома включает два ветвящихся бо-
ковых выроста и один неветвящяйся. Каждая веточка оканчи-
вается единственной апикальной клеткой. Б. Фотография це-
лого растения P.cartilagineum (по Haegeli)
Рис. 4. II. Многоосевые багрянки. A. Chylocladia эр., фо-
тография целого растения с уплощенным полям талломом, вну-
тренняя часть которого полностью или неполностью разделена
диафрагмами. Б. Продольный срез через середину апикальной
зоны 0Ку1ос1ёй1а reniformla , показывающий апикальные
клетки (затушеваны) и диафрагму, формируемую поперечно
растущими боковыми выростами (Б - по Kylin)
179
Рис. 4.12. Полисифональные и коровые одноосевые багрянки.
А. Апикальная зона Polysiphonia nigresoens: показаны
апикальные клетки и развитие полос '‘сифонов” из каждой
осевой клетки. Б, В, Г. Поперечные срезыPolysiphonia sp.,
показывающие последовательность развития перицентральных
клеток ("сифонов”) (I - 7). £. Вершина С eramium deslong-
champii с апикальными клетками и псевдодихотомическим
ветвлением. Е. Ceramium sp., участок продольного среза
ветви ограниченного роста: показано образование частичной
коры в виде полос у мест сочленения осевых клеток (вокруг
средней части осевых клеток коры нет). Без масштаба. (По
Kylin, W, Н. Taylor, Cramer)
Phaeothamnion) водорослей. В каждом из этих классов встречаются
и гетеротрихальные представители (например, Dinocladium. Hetero-
coccus , •phaeodermatium соответственно).
Гетеротрихальные нити - наиболее простая форма таллома у бу-
рых водорослей. Тогда как у более примитивных форм (например. Во-
tooarpus ) рост диффузный, с повышением уровня организации уси-
ливается тенденция к интеркалярному делению клеток (например, у
pylaiella) или трихоталлическому росту (когда деления сконцент-
рированы в субтерминальной части апикальных волосков, например,
У Tilopteridales). У многих мелких представителей класса преобла-
дает стелющаяся система (например, у Streblonema). Гетеротрихаль-
180
Рис. 4.13. Схематическое изо-
бражение апекса (А) членистой
кораллиновой водоросли. Необыз-
вествленные членики (Ч) состо-
ят из необызвествленных осевых
нитей, а чередующиеся с ними
обызвествленные участки - из
сердцевины (С) и коры (К) во-
круг этих нитей
ные виды и их псевдопаренхиматозные производные обычно называют
гаплостиховыми бурыми водорослями. Среди них имеются одноосевые
представители типа Acrothrix , где рост начинается одиночной
трихоталлической нитью и переходит в апикальный, или Desmares-
tia. у которой из первичной трихоталлической оси развиваются
как цилиндрические, так и листовидные талломы (chapman, 1972а,&).
Многоосевые типы бурых водорослей встречаются чаще (напри-
мер, представители порядков Chordariales, Cutleriales и Sporo-
chnales). Они могут бить слизистыми и аморфными (например, Leat-
hesia). ветвящимися ( Eudesme). корковыми (например, Haif a la
или листовидными (например, Cutieria). Значительная дифференци-
ровка клеток может приводить к выделению тканей (например, коры,
сердцевины, ассимиляционного слоя), имеющих в любом случае стро-
го нитчатое происхождение. Ветвление многоосевых типов подразуме-
вает, что группа нитей и связанных с ними ветвей отходит от ос-
новной оси. Ветвящиеся нитчатые стадии могут являться частью жиз-
ненного цикла бурых водорослей, включающего также паренхиматоз-
ную фазу (см. ниже). Так устроены, например, микроскопические
гаметофиты представителей laminariales.
Ветвящееся нитчатое строение характерно для большинства баг-
рянок; только среди Bangiophjroidae имеются истинно паренхиматоз-
ные представители. Конструкция таллома уPlorideophycidae основа-
на полностью на нитчатой организации ( Dixon, 1973), выводимой
181
из первично гетеротрихального состояния (Fritsch, 1945). Как и
у бурых водорослей, встречаются одно- и многоосевые типы роста,
дающие весьма разнообразную внешнюю морфологию. Однако внутрен-
няя дифференцировка ограничена и развита значительно менее, чем
у бурых водорослей. Нити багрянок растут строго верхушечно, т.е.
развиваются из одной (одноосевые типы) или нескольких (многоосе-
вые типы) апикальных клеток. Эти клетки обычно куполообразные;
плоскость деления проходит параллельно основанию, что дает нача-
ло осевым клеткам. Интеркалярное деление у Florideophycidae
встречается редко - только у некоторых Cryptonemiales и Ceramia-
les. Осевые клетки растут, удлиняясь и расширяясь; отделяют две
или более перицентральных клеток, которые (за исключением Rhodo-
melaceae из ПатяиНяЛоя) выступают в роли потенциальных апикаль-
ных клеток боковых ветвей. Такая же картина может наблюдаться и
у боковых ветвей второго, третьего и т.д. порядков. В семействе
Hhodomelaceae (например, у Folyaiphonia ) перицентральные клет-
ки (называемые сифоны) образуются последовательно и удлиняются
вместе с осевой клеткой, от которой они произошли, окружая ее.
Образование обширной коры из сифонов встречается у многих Rhodo-
melaceae (например.Rhodomela. laurencia). причем возникают
как листовидные (например, у Symphocladia). так и в основном про-
стертые (например, у Placo.nhora) талломы.
Нити обычно обильно моноподиально ветвятся; истинно дихото-
мическое ветвление у одноосевых форм не встречается, поскольку
апикальные клетки не делятся вертикально (ср. с верхушечным рос-
том перенхиматозных бурых водорослей). Ложная дихотомия имеет
место, когда боковые ветви у верхушки нити и основная ось разви-
ваются одинаково. Истинная дихотомия может встречаться у много-
осевых форм с равным числом осевых нитей, расходящихся при вет-
влении. Однако в целом считают, что у главной оси рост неограни-
ченный (недетерминированный), а у боковых он в разной степени
ограничен (детерминирован); эти термины указывают на наличие апи-
кального доминирования (см. гл. 7). Окончательная морфология тал-
лома определяется относительным развитием первичной и боковых ни-
тей. Осевые клетки и их производные образуются в строгой последо-
вательности, которая у некоторых багрянок детально проанализиро-
вана (ихоп, 1973). Наличие первичных и вторичных поровых пробоя
облегчает такой анализ, поскольку позволяет проследить за отдель-
ными линиями клеток, начиная от апикальной зоны.
182
Среди наиболее простых гетеротрихальныхFlcrideophycidae
типичен род Audoulnella (Woelherling , 1983). Базальная прикре-
пительная структура варьирует здесь от единственной клетки-ножки
до ползучей или пластинчатой стелющейся системы. Ba-fcrachoaper-
mum - пример одноосевого типа строения таллома, осевые клетки
которого образуют мутовки ветвей ограниченного роста. Кроме то-
го, перицентральные клетки могут давать нисходящие ризоидальные
нити, формирующие покров из коровых клеток, в свою очередь способ-
ных образовать вторичные мутовки ветвей (Fritsch, 1945). Уgloio-
aiahonia одноосевой таллом значительно компактнее. Боковые вет-
ви образуют вторичные ветви, дающие начало все меньшим по размеру
клеткам, которые в конечном счете остаются соединенными вместе
в сплошной поверхностный слой. Внутренние клетки дифференцируют-
ся в кору. УDvuncntia они преобразуются в гифы - узкие удлинен-
ные клетки, проходящие по таллому вертикально и формирующие вто-
ричные поровые соединения с другими клетками. Невероятное разно-
образие листовидных талломов одноосевого типа встречается в се-
мействе Delesseriaceae (Ceramiales)$ здесь даже наблюдается
поверхностное жилкование, напоминающее таковое у листьев высших
растений (например, уDelesseria. Mebrancptera. Phvcodrys.
Bolyneura). У большинства видов этого семейства апикальные
клетки заметны на протяжении всего периода роста (например, у
Delesseria). но у некоторых (например, Hitophyllum) они у более
старых экземпляров неразличимы. Образуются четыре перицентральные.
клетки, боковая пара которых продолжает делиться, что приводит к
листовидному разрастанию таллома. Он обычно моностроматический,
хотя у некоторых видов включает до пяти слоев клеток (например, у
Hemineurai Kylfn, 1956). Более подавленный рост дорсальной и
вентральной перицентральных клеток приводит к образованию "жилок".
В отличие от большинства Florideophycidae у ряда представителей
Delesseriacea происходят многочисленные интер калярные деления.
Подавление прямостоячей системы привело к широкому набору
корковых видов, включай такие, у которых редуцированные простые
и ветвящиеся вертикальные нити расположены в слизистом матриксе
(например. Betrocelis). плотно агрегированы (например, Hidden-
brandia) или сильно обызвествлены (например. Clathromorphwn).
В последнем случае талломы бывают мелкими, с правильными очерта-
ниями (например, Fosliella) или широко расходящимися, с неопре-
деленным контуром (например. Ltthothamnion); поверхность их ли-
бо довольно гладкая (например, Ehytomalithon). либо несет корот-
183
кие ветвящиеся или неветвящиеся вертикальные выросты (например,
Tjithothamnion glaciale). Корки прикрепляются к субстрату корот-
кими ризоидами.
Множество одноосевых видов встречается в семействеCerami-
aceae (Ceramiales)j все ОНИ обильно ветвятся и варьируют от
форм совершенно без коры (например, Antlthamnion) до тех, у ко-
торых осевые нити из крупных клеток частично или целиком одеты
полосами коровых нитей с ограниченным ростом (например, Cerami-
urn), а также видов, полностью покрытых мощной корой (например,
Ptilota).
Типичная простая многоосевая организация наблюдается у ае-
mallon с мягкими дихотомически ветвящимися студенистыми таллома-
ми. Центральные бесцветные осевые нити легко разделяются при сла-
бом нажиме; многочисленные ветви ограниченного роста одевают тал-
лом слоем мелких фотосинтезирующих клеток, большинство из кото-
рых оканчивается гиалиновым волоском. Сходная структура наблюда-
ется у Galaxaura. только здесь апикальные клетки осевых нитей
сгруппированы в верхушечном понижении. Таллом сильно обызвеств-
ленный и жесткий.
Более выражена многоосевая специализация у сильно пропитан-
ных известью членистых кораллиновых водорослей (Cryptonemiales),
в частности, в родах Corallina. Amphipoa. Jania и Calliar- .
thron. Ветви ограниченного роста развиваются здесь вдоль таллома
прерывистыми рядами, между которыми слабообызвествленные осевые
нити образуют "суставные" промежутки.
К значительно более сложным многоосевым типам относятся ци-
линдрические, мясистые или листовидные плотные талломы; многоосе-
вая природа и наличие апикальных клеток здесь легкоразличимы
только на ранних стадиях развития. Из. цилиндрических форм можно
назвать Ahnfeltia. Gymnogongrus и Agardhiella. а из крупных
листовидных - Gigartina (ДО I м в длину), Iridaea и Rhodo -
glossum.y некоторых Rhodymeniales отмечен специфический тип
роста, при котором полые трубчатые талломы разделяются на камеры
поперечными сердцевинными гифами (диафрагмами) (например, Gas-
trooIonium, bomentaria. Champia).
Дифференцировка у даже наиболее высокоорганизованных и.о-
rideophycidae не приводит к появлению сложных тканей. Цилиндри-
ческие и листовидные талломы (как слизистые, так и обызвествлен-
ные) обычно дифференцированы на поверхностные, коровые j>l сердце-
винные клетки; внутрь таллома к сердцевине или между коровыми
184
клетками часто развиваются гифы к специализированным структу-
рам относятся волоски, усики (например, у Нурпеа) и железистые
клетки (например, у Antithantnion. Tumerella).
Ветвящаяся нитчатая структура широко распространена у Chia-
rophyceae и типична для некоторых Oedagoniales, всех Chaeto-
phorales И Ctenocladialee. Гетеротрихальность - отличительная
черта двух последних порядков, объединяющих весьма разнообразные
формы.
Талломы Bulbochaete (Oedogoniales) обильно ветвятся, При-
чем ветви возникают последовательно с одной стороны. Большинство
клеток несет бесцветную щетинку с луковицеобразно расширенным ос-
нованием. Клеточное деление базальное и интеркалярное. Среди ге-
терртрихальных Chaetophorales и CtenocladialeB имеются виды
с развитыми как стелющейся, так и вертикальной системами (напри-
мер, Stigeoclonlum, Trentepohlia). со слаборазвитой стелющей-
ся и доминирующей относительно сложной вертикальной системой (на-
пример, Chaetophora, Draparnaldia). ® доминирующей стелющейся
системой дисковидной структуры (например, Chaetopeltla. Prlng-
sheimiella. Baeudulvella. Ochlochaete) ИЛИ в виде ползучей ни-
ти (например, Aphanochaete. Acrochaete. Bulbocoleonk у стелю-
щихся видов вертикальная часть может быть либо редуцирована до
одной-двух клеток (например, Aorochaete). либо ограничена во-
лосками или щетинками (например, Bulbocoleon. Coleoehaete).
У харовых (например, Chara, Mtella) строго организованные
преимущественно вертикальные талломы заканчиваются куполообраз-
ной апикальной клеткой неограниченного роста. От нее поочередно
отделяются клетки узлов и междоузлий. Первые дают начало 6-20
перицентральным клеткам, образующим мутовки ветвей ограниченного
роста, тогда как вторые больше не делятся, но сильно удлиняются
(до нескольких сантиметров у некоторых видов), становясь много-
ядерными в результате амитотического деления ( Shen, 1967). У не-
которых видов от базальных клеток некоторых ветвей ограниченного
роста развиваются нити, покрывающие междоузлия наподобие коры.
Талломы прикрепляются к субстрату многоклеточными ризоидами. Раз-
витие новых растений начинается с протонемы - стелющейся нитча-
той фазы, сходной с одноименной структурой у мохообразных. Харо-
вые легко воспроизводятся бесполым путем - образуя либо верти-
кальные отростки от протонемальной базальной системы, либо спе-
циальные структуры типа выводковых почек на базальной или верти-
кальной системе. Бесполые зооспоры не формируются.
185
Паренхиматозная организация
(рис. 4.14 и 4.15)
О паренхиматозной организации говорят тогда, когда клетки
первичной нити делятся во всех направлениях, в результате чего
нитчатая организация рано утрачивается. Такое строение характер-
но для многих бурых водорослей, хотя некоторые более примитив-
ные случаи его известны среди багрянок ( Bangiophycidae, напри-
мер, Porphyra. Smithora). золотистых водорослей (нацример,
Phaeosacclon) И Chlorophyceae (например, Ulva. Enteromorpha).
В большинстве случаев клетки паренхиматозных талломов одноядерны;
у эволюционно продвинутых видов бурых водорослей может наблюдаться
высокий уровень тканевой дифференцировки (например, у представи-
телей Fucales, Xaminariales и Durvillaeales). Рост более при-
митивных представителей диффузный (например, у Deamotrichum из
бурых водорослей, Porphyra, Ulva). иногда приводящий к образова-
нию листовидных талломов из одного слоя клеток (моностроматичес-
КИХ, например У Desmotrichum. Monoairoma ИЗ Chlorophyceae, неко-
торых видов Porphyra ) или двух их слоев (дистроматических, на-
цример уUlva. бурой водоросли Punctaria . некоторых видов рог _
phyra). В других простых случаях встречаются трубчатые (например,
у Enteromorpha) или исходно мешковидные (например, Omphalophy-
llum из бурых водорослей, Monoatroma из Chlorophyceae)тал-
ломы. У таких листовидных или трубчатых водорослей слабая клеточ-
ная дифференцировка ограничивается образованием в нижней части '
ризоидов, формирующих "присоску".,Однако клетки бывают организо-
ваны в правильные группы (например, у Prasiola из Chlorophy-
ceae) или ряды (например, у некоторых видов Blidingia и Ente-
romornha из Chlorophyceae).У многих более примитивных паренхи-
матозных форм отсутствует дифференцировка репродуктивных струк-
тур; почти любая клетка может образовывать бесполые споры, хотя
У листовидных видов спорогенез иногда ограничен краями таллома
(например, у Щуа).
Верхушечный рост от единственных апикальных клеток, находя-
щихся на концах ветвей, дает разнообразные цилиндрические или
листовидные типы строения бурых водорослей, в том числе дихотоми-
чески ветвящиеся талломы у Dictyotales (например, Dictvota)
м Scales (например, Eucus).B последнем порядке апикальная
клетка (или клетки, см. McCully, 1966), расположенная в верху-
®ечной бороздке или ямке, в сечении треугольная или четырехуголь-
Рис. 4.14. Паренхиматозная и псевдопаренхиматозная организация. A. Wereocystis luekeana (Phaeo-
phyceae). Б. Punctaria plantaginea (Phaeophyceae ) - целое растение И срез (Jjj. i'. Palmaria
palmata (Rhodophyceae), HeЛое растение и срез (Е), показывающий крупные сердцевинные ТЫё^кй'и
тштоотую из мелких клеток кору с тетраспорангиями. В отличие от представителей Bangiophycidae
виды Plorideophycidae, кажущиеся паренхиматозными, развиваются из нитей, и их, таким образом,
правильнее считать псевдопаренхиматозными. Е. Багрянка Chondrus crispus ; целое растение и срез (Ж),
показывающий толстостенные гифальные нити сердцевины и пЖШоуПЭТЙ5ЭнПВе мелкие клетки коры. 3. Це-
лый экземпляр Viva lactuca (Chlorophyceae ), вид сверху участка его таллома (И) и поперечный срез,
показывающий двуслойное строение (К). Без масштаба (по Postels и Ruprecht, Rosenvinge, W.R.Taylor)
187
Рис. 4.15. Паренхиматоз-
ное строение. Талломы /
видов Fucus (вверху 4
F.yesiculosus , внизу -
F,diatichus ssd. 1
edentatus). демонстри-
рующие один из высших
уровней дифференцировки4
и-организации у водорос-
лей
ная и имеет три (у более примитивных форм; Jensen, 1974) или че-
тыре боковые плоскости деления и одну базальную. Примечательно,
что апикальная клетка у Fucus в эмбриогенезе закладывается у
основания трихоталлически растущего волоока (см. Fritsch, 1945).
Медианные, перпендикулярные деления апикальной клетки приводят к
истинно дихотомическому ветвлению, типичному для многих родов это-
го порядка. Талломы билатерально или радиально ветвятся и могут
быть уплощенными или цилиндрическими.
Представители порядка Durvillaeales (например, Durvil-
laea) имеют крупные талломы, напоминающие талломы laminaria и
Формируемые краевой полосой апикальных клеток. У большинства вы-
сокоорганизованных паренхиматозных бурых водорослей развитие пла-
стинки, ножки и ризоидов или "присосок" обусловлено интеркалярны-
’«и меристемами из активно делящейся меристематической ткани (на-
пример, у laminaria, Alaria). Эта меристема (или переходная
зона) может иметь сезонную активность у многолетних видов типа
188
laminaria hyperborea. Классификация laminariales на уровне се-
мейств основана на природе переходной зоны (например, расщеплен-
ной у Lessanlaceae, нерасщепленной у taminarlaceae И А1а-
riaoeae). Поверхностные слои клеток у этих эволюционно продви-
нутых видов продолжают делиться на протяжении всего роста талло-
ма и называются меристодерма. У высокодифференцированных lamina-
riales и Pucales развиты коровые и сердцевинные ткани. У пер-
вых (например, Macrocystis длиной до 100 м) транспорт метабо-
литов осуществляется через специализированные (трубчатые) гифы,
которые имеют ситовидные поля и аналогичны флоэме высших расте-
ний ( Schmitz, 1981).
Сифонокладальная организация
(рис. 4.6 и 4.16)
Сифонокладальная организация ( Hoek, 1981) встречается у
представителей класса Chlorophyceae с неветвящимися (например,
Urospora из Acrosiphonales, Chaetomorpha из Cladophorales)
или ветвящимися (например, AcrosiphonlaH3 Acrosiphonales, Cla-
donhora из Cladophorales) нитями, состоящими из многоядерных
(семиценоцитных) клеток, а также в порядке stphonocladales (иног-
да объединяемом с Cladophorales; см. Hoek,1981), например у
Volonia. Slphonocladus. Anadyomene и Mlcrodlotyon, Некоторые
Stphonocladalesотличаются также наличием сегрегационного деле-
ния клеток, при котором протопласт распадается на одетые обо-
лочкой округлые части, дающие начало новым сегментам таллома
(например, Dictyosphaeria, Slphonocladus),
Valonla существует в виде одиночного сферического пузыря
диаметром до 10 см, описывавшегося как самая крупная раститель-
ная клетка, хотя это не совсем так, поскольку по его периферии
сегрегационное деление дает много мелких клеток ( Egerod, 1952);
те из них, что образуются в нижней части таллома, выдаются над
поверхностью в виде прикрепительных ризоидов.
У более простых неветвящихся сифонокладальных видов (напри-
мер, Urospora) дифференцировка нити незначительна, хотя базаль-
ные клетки образуют ризоиды, растущие вниз и объединяющиеся в
"присоску". Деления имеют тенденцию происходить чаще в базальной
зоне, так что клетки дистальной части нити крупнее. В любых клет-
ках, за исключением находящихся у самого основания, может идти
Рис. 4.16. бифокальная организация У Chlorophyceae И Xanthophy-
ceae: А - Protosiphon botryoldes (Chlorophyceae): — ~ Botry-
granulatum (Xanthophyceae): 2 ~ Rhizocephalua phoenix
п,251°^°рКуоёаё)” Hallmeda incrasaata (Chlorophyceae).
звестковый вид: Д - Bryopais corymbose,» (Cfilorophyceae):
s - вертикальный срез 'гтвий извнсткивой водоросли леотеГ1В annu-
iSia.(Chlorophyceae)5 £ ~ Valonia utrlcularis (Chlorophy-
“ Acetabularia mediterranea (Chlorophyceae): разрез
шляпки" фертйЛШУГО'ТЦСташЯ' V” 'герминальными волоскам и ради-
(съ?° Расположенными гаметангиями; И - Caulerpa racemosa
Беч oroPhyceae)t Д “ Dlchotomoaiphon tuberoaua (Xanthophyceae )•
яс масштаба (ПО Kleba. Roatafinski Я Woronin. Gepp, Boerge-
se»» Cramer, Schmitz, Erast)
190
зооспорогенез. Рост у ветвящейся Cladophora бывает апикальным,
интеркалярным или обоих этих типов ( Hoek, 1982), а организация
таллома акропетальная, неправильная или изредка дорсовентральная.
УAcroalphonia (Spongomorpha) канатовидное сплетение нижних
осей образовано растущими вниз ризоидами, отходящими от клеток
вблизи основания, или особыми изогнутыми боковыми ветвями.
Удивительно красивые листовидные талломы Anadyomene иМ1с-
rodictyon построены из ветвящихся многоядерных семиценоцитных
нитей, анастомозирующих с образованием моностроматической струк-
туры. Под микроскопом клетки Anadyomene выглядят тесно прижаты-
ми друг к другу, причем нижние из них значительно крупнее крае-
вых, тогда как уMjcrodictyon все они примерно одинакового раз-
мера и цилиндрические. 1 "
Сифональная организация
(рис. 4.16)
У ряда морских Chlorophyceae (порядки Siphonales, Вгуор-
sidales, Dasycladales) И некоторых желтозеленых водорослей (на-
пример, Botrydium. Vaucheria) увеличение и усложнение таллома
происходит в отсутствие септ. Ядерные деления (так называемые
свободные) не сопровождаются цитокинезом, и в результате возни-
кают ценоцитный многоядерный таллом и сифональная организация.
Последняя, однако, не ограничила возможности морфогенеза, по-
скольку известны самые разнообразные ценоцитные формы. Палеонто-
логические данные говорят о том, что ныне живущие представители
некоторых порядков (например, Dasycladales) - только остатки
некогда гораздо более многочисленных групп.
Сифональная организация включает мешковидные (например. Bot-
rydium). одноосевые (например. Vaucheria. Bryopsis ) и мно- '
гоосевые (например, Codium) формы. Сложные радиально ветвящиеся
талломы отмечены среди представителейDasycladales (например,
Heomeris). а листовидные встречаются у некоторых Caulerpales.
У многих видов жесткость несептированным талломам придают отложе-
ния извести или трабекулы, т.е. перегородки, которые (например,
У Caulerpa) представляют собой обширные внутренние разраста- '
яия клеточной стенки. За исключением некоторых одноосевых видов
и представителей желтозеленых водорослей, большинство сифональ-
ных форм не образует зооспор. У Chlorophyceae сифональные виды
191
отличаются специфическими ксантофилловыми пигментами. В этом
классе такая организация, вполне возможно, произошла от форм ти-
па мешковидного Protosiphon (порядок Chlorococcales), у кото-
рого за счет свободного ядерного деления также возникает ценоцит-
ное состояние.
Сифональное строение среди желтозеленых водорослей ограниче-
но мешковидным Botrydium И нитевидной Vaucheria. обитающими
на почве и иле. У Vaucheria септы образуются только для отделе-
ния гаметангиев или зооспорангиев от основного таллома. Непра-
вильно ветвящиеся нити удлиняются на концах, где заметна зональ-
ность цитоплазмы ( Ott, Brown, 1974).
У Bryopsis нежные одноосевые талломы несут акропетально
развивающиеся свободные ветви, перисто и радиально расположенные
на основной оси. Вертикальные части отходят от стелющегося корне-
вищеподобного основания, прикрепленного к субстрату ризоидами.
Обнаружена интересная реакция на повреждение: белковые тельца в
цитоплазме мигрируют к его месту и образуют пробку, которая пред-
отвращает дальнейшую потерю цитоплазмы (Burr, West, 1971). У
Derbesia одноосевые талломы ветвятся более беспорядочно; здесь
сифональная спорофитная фаза чередуется с мешковидной ( Halicys-
tis) половой’*’. Более примитивные виды широко распространенного
тропического рода Caulerpa по основной структуре напоминают
Bryopsis (например, C.fastlgiata. С, verticillata). хотя у
большинства таллом гораздо сильнее дифференцирован. Базальная
система в виде столонов дает начало вертикальным выростам, по
форме сходным с побегами высших растений ( Fritsch, 1935). Они
могут быть мутовчатыми (С.verticillata). листовидными (например,
C.prolifera). нести ветви, расположенные в два ряда (например,
с.taxifolia) или радиально (например, C.racemosa). Хотя
стенки у видов Caulerpa не обызвествлены, значительную проч-
ность несептированным талломам придают хорошо развитые трабекулы.
У Dasycladales пропитанные известью талломы радиально сим-
- метричные, одноосевые, с расположенными в мутовках ветвями. К
Другим отличительным особенностям относятся воспроизведение по-
средством оперкулятных (т.е. открывающихся крышечкой) цист и на-
личие средних частей таллома (диафизов), рост которых происходит
° Разрывом стенки и образованием рубцов ( Bold, Wynne, 1978).
I
и у некоторых видов Derbesia половая фаза по строению на-
сминает описанный выше рол Brvonsis. - Прим, перев.
192
Виды Dasycladales по существу одноклеточны, поскольку остаются
одноядерными вплоть до момента размножения. Единственное, так на-
зываемое "гигантское", или первичное, ядро расположено в базаль-
ном ризоиде, когда растение находится в вегетативном состоянии.
Мутовки ветвей могут располагаться свободно (например, у Неоте-
ris. Batophora, Cymopolia) или расширяясь, плотно прижиматься
друг к другу, создавая компактный поверхностный слой. У Aoetabu-
laria мутовки ветвей взрослого растения находятся только на
верхнем конце относительно длинной ножки, которая на остальном
своем протяжении ничем не покрыта. Первичное ядро остается в ба-
зальной зоне ниже уровня субстрата. Acetabularia широко исполь-
зовалась в морфогенетических исследованиях с применением экспери-
ментальной меротомии; при этом ядро удалялось и энуклеированное
растение изучалось в контролируемых условиях. Эти работы способ-
ствовали выяснению роли ядра в морфогенезе (см. гл. 7).
Многоосевые сифональные формы встречаются в семействах Со-
diaceae (например, Сodium) и Udoteaoeae (например, Avrain-
villea. Udotea. Beaicillus. Rhipocephalus. Halimeda). Губча-
тые, не пропитанные известью талломы Codium состоят из сердце-
вины, образованной бесцветными дихотомически ветвящимися сифона-
ми, и коры из вздутых, булавовидных пузырей (утрикул) с хлоро-
пластами. Утрикулами оканчиваются разветвления апикально расту-
щих сердцевинных нитей. К роду Codium относятся виды с корко-
выми, .сферическими, ветвящимися прямостоячими или пластинчатыми
талломами, сднакс их важнейший диагностический признак - форма
утрикул ( Silva, 1951). Кольцевидные утолщения сифонов, иногда
почти замыкающие их просвет придают таллому'определенную проч-
ность. Репродуктивные структуры располагаются у Codiaeeae по
бокам утрикул.
Представители семейства Udoteaoeae отличаются от видов
Codiaeeae двумя особенностями: специализированными гаметангия-
ми и наличием как хлоропластов, так и амилопластов (гетероплас-
тидностью). У Avrainvillea и Udotea обызвествленные многоосе-
вые талломы представляют собой веерообразную пластинку на прос-
той или ветвящейся ножке, прикрепляющейся к субстрату ризоидаль-
ной массой или несущим столоны основанием. Внутри талломы диффе-
ренцированы на сердцевину из переплетенных сифонов со слегка
вздутыми перпендикулярно отходящими боковыми ответвлениями, которые
тесно прижаты друг к другу, образуя кору. У своеобразного рода
193
рыnocenhalus неветвящаяся ножка несет напоминающую щетку мас-
су многочисленных скученных веерообразных структур, тогда как у
-РятНсШиз сходная щетковидная верхушка образована свободными
ветвящимися пропитанными известью нитями. Предствители рода на-
Ijmeda состоят из ветвящихся рядов обычно уплощенных, сильно
обызвествленных сегментов, разделенных не пропитанными известью
перетяжками.
Глава 5
РАЗМНОЖЕНИЕ И ЦИКЛЫ РАЗВИТИЯ
Введение
У водорослей с воспроизведением помимо увеличения числа осо-
бей связан еще ряд функций. К ним относятся механизмы, способст-
вующие расселению, выживанию при неблагоприятных условиях, а так-
же повышению приспособленности видов за счет генетической реком-
бинации. Многие жизненные циклы обязательно включают как беспо-
лое, так и половое размножение.
У многих одноклеточных видов деление клетки надвое - единст-
венный механизм бесполого воспроизведения. Такой его тип может
распространяться и на многоклеточные формы, размножающиеся либо
путем фрагментации таллома, либо за счет образования специализи-
рованных многоклеточных структур, называемых геммами или пропагу-
лами. Наиболее сложный характер имеет этот процесс у пенобиаль-
ных и колониальных форм, воспроизводящих бесполым путем с форми-
рованием автоколоний. Часто это называют вегетативным размножени-
ем, ограничивая понятие "бесполое размножение" механизмами,
включающими образование специализированных одноклеточных спор^.
Споры бесполого размножения функционально отличаются от га-
мет тем, что развиваются без сингамии, т.е. не сливаются с обра-
зованием зиготы. Подвижные жгутиковые споры называют зооспорами;
это отражает традиционное представление о том, что подвижность -
отличительный признак животных. Неподвижными гомологами зооспор
являются апланоспоры, по существу отличающиеся от них только тем,
что "пропускают" подвижный период, и автоспоры, онтогенетически
безжгутиковые и обычно представляющие собой уменьшенную копию
клетки, из которой произошли. Некоторые авторы не проводят тако-
го различия и называют все неподвижные споры апланоспорами. Экзо-
споры и эндоспоры встречаются у синезеленых водорослей; они фор-
мируются соответственно за счет поверхностного отделения части
клеточного содержимого или по нескольку внутри родительской клет-
ки. Сходным образом все содержимое клетки некоторых эукариотных
^"именно так трактуется бесполое размножение в отечественной
литературе. - Прим, перев.
1Й5
водорослей может высвобождаться из оболочки как одна неподвижйая
моноспора. Известны многие другие типы спрр, связанные с половым
процессом, например тетраспоры у бурых и красных водорослей или
ауксоспоры у диатомей. Неслившиеся гаметы могут развиваться пар-
теногенетически и,'таким образом, функционировать как споры.
Споры или специализированные вегетативные клетки ряда видов
обеспечивают юл выживание при условиях, неблагоприятных для веге-
тативного роста. Они имеют утолщенные или как-то иначе модифици-
рованные клеточные стенки и пониженную метаболическую активность.
Сюда относятся акинеты,образующиеся из вегетативных клеток, а
также покоящиеся споры, цисты, статоспоры и гипноспоры, которые
онтогенетически являются устойчивыми к внешним воздействиям cnof
рами. Зиготы в некоторых группах водорослей могут также оставать-
ся покоящимися и называются тогда гипнозиготами или зигоспорами^
Многие жгутиковые (табл. 4.1) способны образовывать временные не\-
подвижные ("пальмеллоидные") стадии, представляющие собой скопле-
ния клеток, погруженных в слизистый матрикс. Это может быть реак-
цией на неблагоприятные условия, но, поскольку в таких пальмел-
лах иногда происходит деление клеток и образование спор, их нель-
зя считать покоящимися стадиями.
Половое воспроизведение включает два функционально противо-
положных процесса:
I) сингамию, т.е. двустадийный процесс, включающий цитоплаз-
матическое слияние гамет (плазмогамию), за которым следует вос-
становление двойного набора хромосом (кариогамия);
2) мейоз, т.е. обмен генетической информацией (в синапсисе)
и независимое расхождение хромосом (по одной из пары) в дочерние
клетки.
Сингамию называют изогамной (изогамией), когда гаметы морфо-
логически идентичны, и анизогамной, если одна из них (мужская)
меньше по размеру и более подвижна, чем другая (женская). При
изогамии гаметы разного пола чаете отличаются друг от друга осо-
бенностями поведения или физиологии, как, например у Eotocarpus ,
Где женская гамета первой оседает на дно и выделяет феромон, при-
вклекающий мужские гаметы (Muller , 1981). В случае безжгутико-
вых гамет, например у zygnematales, одна из них может амебообраз-
но передвигаться, тогда как другая остается внутри клетки, где
она сформировалась. Между морфологическими изогаметами бывают
196
также ультраструктурные различия, например у Chlamvdomonas rein-
h«rd~fcii (Triemer, Brown * 1975) • Под оогамией понимают ели—
яние мужской гаметы (спермия) с лишенной жгутиков женской гаметой
(яйцеклеткой). Оогамны размножающиеся половым путем красные водо-
росли; их безжгутиковые мужокие гаметы называют спермациями. Чет-
вертый тип сингамии, который не так легко наблюдать, - автогамия,
т.е. слияние двух гаметичерких ядер внутри одной особи.
Один из наиболее важных аспектов полового воспроизведения -
место мейоза в жизненном цикле по отношению к сингамии. У водо-
рослей он может происходить:
I) при прорастании зиготы, являющейся единственной диплоид-
ной фазой (зиготический мейоз);
2) при образовании спор, прорастающих затем в гаплоидное
растение, которое в свою очередь образует гаметы путем .митоза
(спорический мейоз);
3) при образовании гамет, которые являются единственной гап-
лоидной фазой (гаметический мейоз);
4) при вегетативно^ делении клеток, за которым не следует
немедленного образования спор или гамет. В результате одна часть
таллома является гаплоидной, а другая - диплоидной (соматический
мейоз).
Цитологическую и морфологическую последовательность событий,
происходящих при половом воспроизведении, обычно называют жизнен-
ным циклом. (Иногда этим термином обозначают более широкое поня-
тие, которое может быть определено как сумма адаптаций организма
к выживанию и размножению. Сюда относятся физиологические и эко-
логические моменты, например распределение ресурсов между ростом
и размножением.) Полные жизненные циклы многих водорослей воспро-
изведены в культуре, но только для некоторых из них получены ци-
тологические данные, включая уровень плоидности и место мейоза.
Хромосомы водорослей обычно мельче, чем у цветковых растений, и
точный подсчет их сложен. Прямые количественные определения ядер-
ной ДНК с помощью микроспектрофотометрии и особенно использова-
ние ДНК-специфичных флуорохромов - весьма перспективные методы,
уже использовавшиеся для регистрации изменений в уровне плоидно-
сти ( Golf, Coleman, 1984).
Жизненные циклы водорослей классифицируются в соответствии
с местом мейоза, т.е. подразделяются на зиготический, споричес-
кий, гаметический и соматический (рис. 5.1). Представленность их
197
Рис. 5.1. Жизненные циклы водорослей: I - зиготический:
И - спорический; БГ - гаметический; 1У - соматический. До-
минирующая фаза в случаях I и Ш бывает многоклеточной, а
если она одноклеточная, то наиболее долговечная и способ-
ная к митотическому воспроизведению
Диплоидная фаза
в разных классах и типы сингамии отражены в табл. 5.1. Как и в
отношении морфологии (см. табл. 4.1), Chlorophyceae наиболее
разнообразны с точки зрения репродуктивных механизмов.
Зиготические жизненные циклы обычно считают примитивными;
чаще всего они встречаются среди одноклеточных таксонов и их ко-
лониальных или пальмеллоидных сородичей.
Спорический жизненный цикл включает обычно диплоидную спо-
рообразующую фазу, спорофит, и, как правило, гаплоидную гамето-
образующую, гаметофит. Периодическое их повторение .друг за дру-
гом называют чередованием поколений. Фазы изоморфны, если иден-
тичны по вегетативной морфологии, и гетероморфны, если внешне
198
Таблица 5.1. Половое размножение и жизненные циклы у водорослей
£орма сингамж
Место мейоза
Cyanophyceae
Prochlorophycota
Rhodophyceae
Chrys о phyо eae
Prymnesicphyoeae
Xanthophyceae
Eus tugmat ophyc e ae
Bacillar!ophyc eae
Dinophyceae
Phaeophyceae
Raphidophyc eae
Cryptcphyceae
Euglenophyceae
Chlorophyceae
Charophyce ae
Prasinophyceae
Включая поведенческую или биохимическую анизогамию,
о
Показан в отдельных случаях, например у Hymenomonaa carterae.
но в других случаях может быть плеиоморфизм оез чередования пло-
идности.
3 Могут образовываться планозиготы, а мейоз часто постзиготичес-
кий, т.е. происходит после прорастания цисты.
Указан для Jfoctiluca millarls (Zingmark, 1970).
5 Встречается только yiiicales и родственных порядков, однако
некоторые исследователи рассматривают оогоний как редуцированный
гаметофит и, таким образом, считают мейоз спорическим, а не гаме-
тическим.
® Микроденсиометрическое измерение ДНК показывает одинаковые ее
уровни в спермиях и соматических клетках, так что мейоз не может
быть гаметическим.
7 Обнаружены (Huber, lewin, 1986) электрофоретические фенотипы
Tetraselmis. характерные для гетерозигот. Предполагается, что
они диплоидны, а это может свидетельствовать о наличии полового
размножения.
199
различны. Известны и водоросли с несходными морфологическими фа-
зами, имеющими одинаковый набор хромосом (т.е. между ними нет
сингамии илимейоза); это явление называют плейоморфизмом. Эволю-
цию и успех таких гетероморфных/плейоморфных видов связывают с
использованием различных физиологических возможностей разных форм
(см. гл. 7), в частности фотосинтетических и ростовых (так называ-
емая "гипотеза функциональной формы"; м, Littler, D.Littler,
1980). Кроме того, морфологически редуцированные фазы менее под-
вержены выеданию растительноядными животными, чем вертикальные
мясистые талломы ( Lubchenco, Cubit, 1980).
Возможно, лучше избегать термина "чередование поколений"
при обсуждении жизненных циклов водорослей, поскольку строгое че-/
редование происходит далеко не всегда. Многие виды способны раз-;
множиться в одной или обеих фазах бесполыми спорами, у других на-
блюдаются плейоморфные смены. Также неоправданно использование
этого термина в отношении тех багрянок, у которых в обычную гап-
ловдно-диплоидную последовательность вклинивается вторичная ди-,
плоидная фаза карпоспорофита.
Гаметические жизненные циклы, которые присущи также живот-
ным и оомицетам, встречаются у водорослей нечасто и, по-видимому,
возникли в разных их группах независимо (табл. 5.1). Соматичес-
кий мейоз еще более редок и достоверно известен только у Prasi-
olales (Chlorophyceae) и некоторых пресноводных представителей
Batrachospermales (Rhodophyceae).
Вегетативное размножение
Размножение путем простого митотического деления отмечено
во всех отделах водорослей, где есть одноклеточные представите-
ли. Однако примитивные одноклеточные Chlorophyceae размножают-
ся бесполым путем исключительно с образованием спор. Такой про-
цесс, называемый элевтеросхизисом (Groover, Bold, 1969), отли-
чается от истинного клеточного деления (десмосхизиса) тем, что
клеточные стенки формируются de novo и не включают в себя части
родительской стенки.
В некоторых группах водорослей возникли специфические формы
цитокинеза, позволяющие потомству сохранять характерную для так-
сона морфологию.
У диатомей две половинки панциря при цитокинезе расходятся,
и в борозде деления формируются новые половинки. Как эпитека,
200
так и гипотека родительской клетки становятся у потомства эпите-
ками, поэтому в активно увеличивающихся популяциях размеры клеток
постепенно уменьшаются (рис. 5.2). Обычно максимальный размер
восстанавливается путем полового размножения и образования ауксо-
спор, хотя отмечались отдельные случаи выхода цитоплазмы из пан-
циря с регенерацией более крупных его половинок (Stosch, 1965а).
У панцирных динофитов отмечено несколько ферм цитокинеза
(Walker, 1984). В некоторых родах (например.Peridinium и
GlanodinJiim ) тека накануне деления может сбрасываться, что тре-
бует ее полного восстановления у потомства. У других видов, в
частности Ceratium spp. И Pyrodinium bahamense. части роди-
тельской теки распределяются между дочерними клетками и заново
формируются только недостающие пластинки.
Плакодермные, или истинные, десмидиевые состоят из полукле-
ток, и цитокинез здесь происходит в зоне перешейка, после чего
каждая полуклетка отпочковывает недостающую.
Колонии вольвоксовых размножаются с образованием автоколо-
ний путем деления воех или только некоторых (у более высокоорга-
низованных родов) клеток. Дочерняя колония отличается от роди-
тельской размером клеток, но не их числом; после ее высвобожде-
ния в окружающую среду они дорастают до величины родительских
(рис. 5.3). У Voivox (Starr, 1969; Kam et al., 1974) делиться
способны лишь некоторые клетки, образующие внутри родительской
колонии полые сферы, клетки которых с кавдым митозом становятся
мельче. Затем новая колония выворачивается так, что концы кле-
ток, прежде смотревшие внутрь, оказываются снаружи, на них форми-
руются жгутики, а сама колония высвобождается в среду за счет
разрыва родительской сферы. Образование автоколоний встречается
также у колониальных Tetrasporales, например Hydrodictyon и
Hediastrum 1 ( Marchant, 1974). Протопласт некоторых или всех
клеток колонии делится с образованием двужгутиковых'зооспор. Они
не высвобождаются, а агрегируют между собой в новую колонию под
стенкой родительской клетки (рис. 5.4).
Вегетативное размножение фрагментацией таллома хорошо из-
вестно во всех отделах как пресноводных, так и морских водорос-
лей и особенно распространено у нитчатых видов. По-видимому, оно
важно для сохранения популяций в таких местообитаниях, как эстуа-
отечественной литературе роды Hydrodictyon и Pediastrwn
относят к nop.Chlorococcaleg . - Прим, перев. ’
201
Родительская Первое поколение
клетка (а, в) (г, 6}
(а, б)
Второе поколение
(а, д) (е, в) (г, з) (ж, б)
Число и относитель-
ные размеры створок
во втором
поколении
Рис. 5.2. Схема постепенного уменьшения размеров диатомей
(вид с пояска) при клеточных делениях (по Hendey, 1964)
рии ( Nierihuis, 1974) или соленые марши ( Norton, Mathieson,
1983), а также на границах ареалов ( Asoaragopsis; Dixon, 1965;
Callithamnion8 Whittick, 1978; Codium: Fralick, Mathieson,
1972). Одна из планктонных популяций Pyiaiella littoralis под-
держивается фрагментацией, которой способствует снижение прочно-
сти нитей из-за заражения хитридиомицетом Eurychasma dicksonii
(Wilce et al., 1972).
Некоторые водоросли образуют специфические вегетативные
пропагулы. Гормогонии (рис. 5.5,В), представляющие собой корот-
кие нити из недифференцированных клеток, возникают путем распа-
да трихома некоторых синезеленых водорослей (Nichols, Adams,
1982). Многоклеточные бесполые структуры, геммы, описаны у прес-
новодной багрянки Hildenhrandia rivularis (Nichols, 1965), а
пропагулы, напоминающие сперматангиальные веточки, встречаются
на мужских, женских и тетраспорангиальных экземплярах Polysipho-
nia ferulaoea (Kapraun, 1977). Среди бурых водорослей неко-
торые представители порядка Sphacelariales формируют булавовид-
ные, дву- или трехлучевые пропагулы из модифицированных ветвей
(рис. 5.6).Fucus может размножаться, образуя придаточные заро-
дыши на ризовдах ( Mclachlan, Chen, 1972). У харовых на ризои-
дах встречаются клубеньки, способные перезимовывать и прорас-
тать в протонему, из которой развивается прямостоячая фаза
( Grant, Sawa, 1982).
202
Рис. 5.3. Жизненный цикл Voivox . А. Бесполое размножение
путем образования дочерних колоний? Б Половое размножение
(ПО G.M.Smith, 1955)
Рис. 5.4. Формирование автоколоний у Pediastrum.
А. Взрослая вегетативная оообь, образующая зооспоры.
П. Подвижные зооспоры. В, Г. Зооспоры прекращают движе-
ние и формируют новую колонию (по G.M.Smith, 1955)
203
Рис. 5.5. Репродуктивные структуры синезеленых водорослей.
A. Gloeotrichia: акинета развивается рядом с базальной
Тет'ёроЦйстой.'"Б. Wollea.B. Calothrix: образование гормо-
гониев. Г. Chamaesiphon;’* образование экзоспор почковани-
ем от свободных концов нитей. Д. Деипосаграэндоспоры
внутри родительской клетки (по Pogg et al., 1973)
Рис. 5.6. Вегетативные пропагулы
Sphacelaria den Ноек»
Flinterman,
204
Бесполое размножение спорами
Экзоспоры и эндоспоры
Экзоспоры и эндоспоры (рис. 5.5.Е) характерны для порядков
Chamaesiphonales И Pleurocapsales из синезеленых водорослей
( Nichols, Adams, 1982). Первые развиваются экзогенно, отделяясь
от концов вегетативных клеток, тогда как вторые образуются при
дроблении вегетативных йлеток увеличенного размера.
Моноспоры, биспоры и параспоры
Моноспоры (рис. 5.7) - наиболее обычные беополые споры багря-
нок. Они отличаются от спермациев обычно большим размером и нали-
чием пластвд ( Dixon, 1973). Особенно ваашы они yBangiales, где
являются единственными известными репродуктивными клетками более
примитивных нитчатых видов ( Dixon, 1973),. У Bangia и Porphyra
моноспоры встречаются как на стадии Conohocelis , так и в макро-
скопической фазе; в последнем случае они в сущности представляют
собой отшелушивающиеся вегетативные клетки. Сходние клетки отме-
чались также уUlva из Chlorophyceae (Bonneau, 1978).
Среди Nemaliales (багрянки) некоторые виды Audouinella раз-
множаются исключительно моноспорами, тогда как другие - половым
путем; например, у Pseudogloiouhloea confusa могут самовозоб-
новляться как гаметофит, так и спорофит (Ramus, 1969). Моноспо-
ры описаны в культурах ряда морских высокоорганизованных багря-
нок ( Dixon, 1973), но их роль в природе неясна. Сейчас известно,
что моноспоры, указывавшиеся у Tilopteridales (бурые водоросли),
являются на самом деле партеногенетически развивающимися яйце-
клетками или четырехъядерными апомейотическими спорангиями (Kuh-
lenkamp, Muller, 1985). Моноопоры могут также формироваться
на диплоидных частях таллилов Prasiola spp. (Chlorophyceae)в
возобновляя эту фазу ( Friedmann, 1959).
У багрянок на тетраспорофитах часто отмечаются биспорангии
(рис. 5.7,g), которые в некоторых случаях могут быть стадиями
развития тетраспорангиев ( Dixon, 1973). Они известны у корковых
представителей Corallinales, а также у Lj-tophyllum litorale и
L.corailinae. Установлено ( Sunescn, 1950), что такие двуядер-
ные спорангии являются апомейотическими.Считалось, что у
205
Рис. 5.7. А. Выход моноспор у багрянки Goniotrichum (по Abbott,
Hollenberg^ 1976). £. БиспоранЩ Gallithamnion.
В. Пара спорангии Callithamnion (по Rosenvinge; 1У2'4Т.1
£. Полиспорангии ptilothamnion (по Wollaston, 1984)
206
Corallinales жизненный цикл такой же, как v Polysiphonia (West,
Hommersand, 1981), поэтому биспорангии, возможно, связаны с бес-
полым воспроизведением тетраспорофитной фазы.
Параспоры (рис. 5.7,3), обнаруженные у некоторых багрянок,
можно рассматривать как неупорядоченные массы моноспор. Лучше
всего они известны у Plumaria elegans (Diew, WSiWhittick^g??)
И Ceramium strieturn (Rueness,1973), *№ отмечены на бесполых
триплоидных поколениях в северной части ареала видов.
Зооспоры, апланоспоры и автоспоры
Зооспоры и апланоспоры возникают путем деления протопласта
родительской клетки и известны у Chlorophyceae, Prasinophyceae,
Xanthophyseae, Phaeophyceae, Eustigmatophyceae, Chrysophyceae,
Dinophyceae, Prymnesiophyceae.
Среди Chlorophyceae У Chlamydomonas образуются два, че-
тыре или восемь дочерних протопластов, которые формируют свои
клеточные стенки внутри родительской (рис. 5.8,4) и высвобожда-
ются за счет разрыва или лизиса последней ( Schlosser, 1976).
Представители порядков Chloroccccales и Chlorosarcinales, ли-
шенные в вегетативном состоянии жгутиков, способны размножаться
двужгутиковыми или четырехжгутиковыми зооспорами, напоминающими
особи Chlamydomonas. В отличие от этого также безжгутиковые
вида Chlorellales не образуют зооспор и размножаются бесполым
путем автоспорами (рис. 5.8,£).
Описан ( Belcher, 1966) коккоидный празинофит Prasinochlo-
rie, формирующий четырехжгутиковые покрытые чешуйками подвиж-
ные клетки, напоминающие особи Pyramimonas . Свободноживущие
стадии цистообразующих родов здесь можно рассматривать как зоо-
споры ( Norris, 1980).
У одноклеточных желтозеленых водорослей пальмеллоидные фор-
мы могут давать зооспоры и апланоспоры, но многие коккоидные ви-
да образуют автоспоры ( Hibberd, 1980а). Сходные варианты спор
известны и у Eustigmatophyceae (Hibberd, 1980). Ценоцитная
желтозеленая водоросль Vaucheria образует спорангии, в которых
развивается одна многоядерная зооспора с многочисленными парами
жгутиков (рис. 5.8, В, Г) ( ott, Brown, 1974в). После ее оседа-
ния на субстрат жгутики отбрасываются, формируется клеточная
стенка и спора развивается непосредственно в ценоцитную нить.
207
Рис. 5.8. А. Образование зооспор у Chlamydomonas eugametos
Б. Вегетативная клетка и образование автоспор у Ohio-
rella (по Bold, 1973). В, Г. Бесполое размножение Vau-
bheria • В. Развитие зооспорангия и выход зооспоры.
Г. Зооспора (по G.M.Smith, 1955)
Многоклеточные представители золотистых водорослей, дино-
фитов и примнезиофитов образуют зооспоры, сходные со свебодножи-
вущими одноклеточными формами соответствующих классов. У phaeo-
208
ЬИяттН on (золотистые ВОДОРОСЛИ) ИЗ ОТДвЛЬНЫХ КЛвТОК ВОЗНИКНЕТ
4-8 зооспор, напоминающих свободноживущие особи Ochromonas.
Нитчатый представитель динофлагеллат Dinothrtx высвобождает
зооспоры, напоминающие Gymnodinium, но при последующих делени-
ях эти клетки не расходятся. Ложнонитчатая стадия "Apisthonema"
примнезиофита Hymenomonas carterae образует зооспоры без кок-
колитов (рис. 5.9) (Stosch, 1967).
Зооспоры часто встречаются у бурых водорослей и многоклеточ-
ных Chlorophyceae как часть полового жизненного цикла. Обычно
только у более примитивных таксонов этих классов зооспоры могут
возникать вне связи с половым размножением. У бурых водорослей
они образуются в многогнездных или одногнездных спорангиях
(рис. 5.10). В первом случае клетка увеличивается в размере и
делится митотически на несколько более мелких клеток (гнезд),
каждая из которых дает одну зооспору. В одногнездном спорангии
первое деление может быть либо мейотическим, либо митотическим;
последовательные деления не связаны с образованием стенки, и зоо-
споры созревают внутри одной клетки (гнезда). У многих предста-
вителей Ectocarpales зооспоры из многогнездных спорангиев
воспроизводят спорофитное поколение (например, у pyialella;
Knight, 1923; Ectocarpus; Muller, 1967; laminariocolax
tcmentosoides; Russel, 1964; Giffordia mitchelliae; Edwards
1969). Много раз указывалось, особенно в отношении представите-
лей Ectocarpales, Chordariales и Scytosiphonales, на отсутст-
вие в одногнездном спорангии мейоза и следующее за спорогенезом
повторение родительской фазы (Wynne, Loiseaux, 1976; Pedersen
1981).
Размножение бесполыми зооспорами - обычное явление для при-
митивных нитчатых Chlorophyceae. Во многих родах Ulotriohales
они дву- или четырех»гутиковые. Как у пресноводных, так и у мор-
ских видов uiothrix четырехмгутиковые зооспоры развиваются не-
посредственно в новые нити, идентичные родительским ( Lokhorst,
1978). Сходные процессы наблюдаются среди Chaetophorales у
Stigeoclonium, Chaetophora И Coleochaete ( Bold, Wynne
1985). Двужгутиковые зооспоры, воспроизводящие родительскую фа-
зу, указывались среди Cladophorales у Cladophora и Rhizo-
clonium; у последнего рода это, по-видамаиу, обычный метод
размножения спорофитной фазы в эстуариях (Nienhttis, 1974). Сход-
ное бесполое размножение с помощью дву- или четырехжгутиковых
209
Рис. 5.9. Жизненный ЦИКЛ Hymenomonas carterae (ПО Lee,
1980; из von Stosoh, 1967; Leabeater, 1У70)
зооспор отмечено у нитчатого гаметофита Urospora в частности,
при низких температурах ( Kornmann , 1961). Некоторые ввды Еп-
teromorpha (Innes, Yarish, 1984 И Blidingia (Tatewaki;
lima, 1984) размножаются зооспорами, но у Ulva размножение
спорофита четырехжгутиковыми зооспорами неизвестно (Tanner
1981). Такие диплоидные зооспоры (рис. 5.II) отмечались у uiva
matabilis (Bjeld, Lovlie, 1976), но они развиваются в диплоид-
ные гаметофиты.
Воспроизводящие родительское поколение (рис. .’5.12) стефано-
Контные зооспоры С кольцом из жгутиков образуются у Oedogonium
(Hoffman, Manton, 1962) И других представителей Oedogoniales
(Boid, Wynne, 1985). Кроме них, среди зеленых водорослей такие
210
Рис. 5.ТО.Ectocarpus.
Многогнездные (А) и одногнезд-
ные (Б) репродуктивные струк-
туры ?по Miiller, 1972)
зооспоры дают только Derbesia
и близкие рода. У многих из
них зооспорогенез сопровождает-
ся мейозом и является только
частью полового жизненного цик-
ла ( Tanner, 1981), однако из-
вестен бесполый изолят Derbe-
sia marina, воспроизводящийся
непосредственно апомейотически-
ми стефаноконтными зооспорами
(рис. 5.13,Г) ( Kornmann
1970а).
Покоящиеся стадии
Специализированные покоя-
щиеся стадии обнаружены во мно-
гих группах водорослей ( Егу-
xell, 1983). Они особенно час-
ты в пресных водах, где могут
выдерживать полное высыхание
или гораздо более сильные коле-
бания температуры и концентра-
ции питательных веществ, чем'
их морские аналоги. Наиболее
примечательны среди морских
форм гипнозиготы динофлагеллат
(Dale, 1983), встречающиеся в прибрежном бентосе и иногда выно-
симые к поверхности апвеллингом (см. гл. 9). У многих морских
центрических диатомей в жизненном цикле также присутствуют поко-
ящиеся споры (рис. 5.14) (Hargraves, Trench, 1983).
У морских макрофитов споры обычно прорастают сразу же, хо-
тя у багрянки Uemalion helminthoides указывались толстостенные
зимующие карпоспоры (Martin, 1969). Acetabularia (Chlorophy-
ceae) также образует толстостенные цисты (Tanner, 1981).
Акинеты (рис. 5.5,A, J>) - это одиночные клетки, предназна-
ченные для переживания неблагоприятных условий, у представителей
нескольких порядков синезеленых водорослей, включая некоторые од-
ноклеточные формы (Nichols, Adams, 1982). Они обычно’крупнее,
211
Рис. 5.II. Жизненный цикл Ulva . Во внутреннем прямоуголь-
нике можно видеть основной1 тип чередования изоморфных по-
колений: диплоидного спорофита и гаплоидного гаметофита.
Вокруг показаны альтернативные репродуктивные варианты.
Гаметофиты могут развиваться партеногенетически в гамето-
фиты или спорофиты. Партеноспорофиты гаплоидны, диплоид-
ны или частично гаплоидны и частично диплоидны. Процент-
ные соотношения указаны для Ulva mutabills. М! - мейоз
(по Tanner, отчасти из Fjeld, lovlie, 1975)
чем вегетативные клетки, имеют утолщенные стенки и содержат обиль-
ные запасы азота в цианофициновых тельцах. У некоторых родов (на-
пример, Cylindroapermum: Walk, 1982) соседство гетероцист,
по-видимсму, стимулирует образование акинет, но это не общее яв-
ление.
Факторы внешней среды, стимулирующие образование акинет,
недостаточно ясны, но иногда оно связано с недостатком питатель-
ных веществ. В периодических культурах акинеты могут появляться
212
Рис. 5.12. Жизненный никл Oedogonium А. Прорастание зиго-
ты (мейотическое) и выход дтефанокойтных зооспор. Б. Раз-
витие нити. В. Бесполое размножение зооопорами. £. Разви-
тие оогония.~Д. Сперматогенез у макрандрового вида.
Е. Нанандровый вид: андроспоры оседают на оогоний и раз-
виваются в карликовые мужские особи. Спермий. 3. Яйце-
клетка и спермий проникающий через стенку оогония?
И. .Развитие и высвобождение зиготы (по Esser, 1982)
в конце экспоненциальной фазы роста (Sutherland et al.., 1979),
а прорастание их удается индуцировать обновлением культуральной
среды (Miller, leng, 1968). В природе играет роль также увели-
чение освещенности и долготы дня ( Wildman et al., 1975). Миллер
и Ланг ( Miller, lang, 1968) дали детальный обзор прорастания
акинет Cyiindrospeiraun. показав, что клетка способна делиться
еще до выхода через пору в клеточной стенке.
213
Рис. 5.13. Жизненный цикл комплекса Derbesia-Halicystis -
Bryopsis (Chlorophyceae) . А. Чередование гаметофи-
Фа HeXi'cystis и спорофита Derbesia . Б. Чередование
Bryppsia со спорофитом типаЬегЪёаТа .~В. Чередование
гамйТофйФа Bryopsis hypnoIdes с редуцированной споро-
фитной стадией протонемы. Г. Бесполое размножение Derbe-
sia стесбаноконтными зооспорами. М! - мейоз (по Tanner,
1581) ч
При неблагоприятных условиях, особенно при высыхании, многие
Chlorophyceae образуют толстостенные покоящиеся клетки
(рис. 5.14,В) (Coleman, 1983). Акинеты представляют собой моди-
фицированные вегетативные клетки с утолщенной стенкой. Гипноспо-
Ры и гипнозиготы также имеют утолщенные стенки, но последние об-
разуются de novo протопластом, который предварительно отделяет-
ся от стенок родительской клетки. В лаборатории такие клетки мо-
гут сохранять жизнеспособность в течение белее 20 лет ( Coleman,
1983). Многие из одноклеточных вольвоксовых образуют толстостен-
214
Эпициста
Гипоциста
Циста
Недоразвитая
поперечная
бороздка
Недоразвитая
продольная
бороздка
Выросты
Апекс
Пожвижная стадия
Эпитека
Поперечная
бороздка
Г ипотека
- Жгути к
Б
А
Рио» 5.14. А. Сравнение морфологии покоящейся цисты и по-
движной клетки панцирного динофита (по Dale 1983).
Б. Покоящиеся споры и вегетативные клетки в нитевидной
колонии диатомеи Chaetoceroa (по Hendey, 1964).
В. Гипноспоры Chlorococcum hypnoaporum (Chlorophyceae)
fno Coleman, 1983) --------------
215
ные покоящиеся клетки (Lembi, 1980). Среди пресноводных нитча-
тых зеленых водорослей акинеты встречаются у stigeoclgnium
(Chaetophorales Сох, Bold, 1966) И Zygnema (Zygnematales ),
но, по-видимому, отсутствуют у Spjrogyra которая переносит не-
благоприятные условия исключительно в виде покоящихся зигот
( Bold, Wynne, 1985). Апланоспоры пресноводной водоросли Ulo-
-t-h-rix fimbriata могут развиваться в гипноспоры. Бесполые цисты
пальмеллоидных стадий, возможно, встречаются у большинства прес-
новодных и некоторых морских празинофитов (Norris, 1980).
Одна из характерных особенностей золотистых водорослей - их
способность образовывать окремнелые статоспоры, или покоящиеся
цисты ( Pienaar 1980; Sandgren, 1983). В процессе их формиро-
вания цитоплазма родительской клетки мигрирует в эту цисту, кото-
рая затем отделяется пробкой (рис. 5.15). При прорастании пробка
растворяется, и наружу выходят одна или несколько подвижных кле-
ток. У Dinobryon cylindricum ядерное деление происходит до об-
разования стенки, и в возникающей двуядерной клетке, вероятно,
имеет место автогамная рекомбинация (Sandgren, 1980, 1981). Вис-
ты примнезиофитов изучены недостаточно и, возможно, не образуют-
ся у видов, в жизненном цикле которых присутствует бентосная фа-
за (Hibberd, 1980с). У Prymnesium это клетки овальной формы С.
широкой порой, которая, по-видимому, не закрыта пробкой. Стато-
споры образуются также у некоторых подвижных желтозеленых водо-
рослей и имеют стенки из двух налегающих друг на друга частей
(Hibberd, 1980а).
У диатомей известны покоящиеся толстостенные споры и покоя-
щиеся клетки (рис. 5.14,Б), которые отличаются от вегетативных
прежде всего овоей более темной пигментацией. У Stephanopyxis
palaeriana образование покоящихся спор вызывается недостатком
фосфатов (Droebes, 1966). Такие споры могут длительное время
выживать в темноте, у Melosira italics subsp. antarctica - до
трех лет ( Lund, 1954).
Многие свободноживущие динофиты инциотируются в ответ на
неблагоприятные условия (Dale, 1983), причем у одних видов эти
стадии напоминают родительские клетки, а у других отличны от них
(Рис. 5.14,А). Ныне известно (Evitt, 1963), что к последнему ти-
РУ относятся гистрихосферы, которые в изобилии известны в иско-
паемом состоянии. Покоящиеся цисты известны также у паразитичес-
к°Го рода Djasodinium (Drebes, 1981).
216
Рис. 5.15. Половое размножение Dinobryon cvlindricum.
А. Контакт гамет. J>. Плазмогамид. В. Двувдерная зигота.
Т. Начало образования цисты; жгутики исчезли; Д. Выделе-
ние инцистирующего дузырька (ИП) и выход из панциря (П).
Е. Незрелая статоспора перед отложением кремнезема.
Ж. Зрелая гипнозигота с окремнелой стенкой цисты (СЦ) и
порой, запечатанно!? пробкой (ПП). ВЗХ - вакуоль, где за-
пасается хризоламинарин; Пл - пластида (по Sandren,
1981)
Половое размножение
Половое размножение отсутствует или не установлено досто-
верно у Prochlorophycota, Eustigmatophyceae, Raphidophyceae,
Cryptophyceae И Euglenophyceae(табл. 5.1). Оно не встречается
также у Cyanophyceae, хотя здесь доказан перенос генов между
особями. В частности, установлено ( Bazin, 1968), что штаммы
21?
cnvstis НйчЛяпа. устойчивые либо к полимиксину, либо к стреп-
^щину, давали при совместней выращивании шталм, устойчивый к
обоим антибиотикам. Это связано с генетической трансформацией,
при которой выделяемая в среду ДНК клетки-донора захватывается
оеткой-ренипиентом. Последующие эксперименты по инкубации сине-
зеленых водорослей вместе с экстрагированной ДНК ( Devilly, Hough-
toiij 1977» Stevens, Porter, 1980) подтвердили передачу
функциональных генов. Хотя хорошо известны цианофаги ( Safferman
1973). трансдукция в этсм отделе окончательно не установлена,
как неизвестна и конъюгация, хотя образование гетероклонов за
счет анастомозов нитей указывалось у Noatoc muacorum (Delaney
etal., 1976).
Зиготические жизненные циклы
Среди золотистых водорослей половое размножение лучше всего
известно у панцирных родов благодаря хорошо наблюдаемому образо-
ванию зигот (Pienaar, 1980). Клетки здесь прикрепляются друг к
другу зоной отверстия в панцире, и их протопласты сливаются, да-
вая зиготу (рис. 5.15). У Dinobryon cylindricum сингамия анизо-
гамная, хотя известна и автогамия в двуядерных покоящихся цистах
(Sandgren, 1980, 1981).
Половое размножение желтозеленых водорослей отмечалось ред-
ко и, по-видимому, ограничено в основном филогенетически продви-
нутыми сифональными формами, нацример Vaucheria (Hibberd, 1987).
Многоядерный антеридий образует большое число бесцветных сперми-
ев ( Moestrup, 1970), проникающих в одноядерный оогоний через
пору в его стенке (рис. 5.16). Формируется толстостенная покоя-
щаяся зигота, при прорастании которой происходит мейоз с образо-
ванием сифональной нити.
Уокер (Walker, 1984) перечисляет 22 вида динофитов, имею-
щих половой цикл; у всех, за исключением Noctiluca miliaris
он,по-видимому, зиготический. Большинство видов изогамно; гаме-
ты мельче вегетативных клеток, а у панцирных видов могут быть
голыми (рис. 5.17). В результате сингамии возникает подвижная
планозигота, которая иногда делится мейотически, но в большинст-
во случаев развивается в толстостенную покоящуюся гипнозиготу.
Период покоя последней длится от нескольких часов у Peridinium
218
Рис. 5.16. Антеридий и оогоний Vaucheria (по G.M.Smith,
1955) ----------
Рис. 5.17. Жизненный никл свободноживущей динофлагеллаты
(по Walker, 1984)
gatunenae (Pfiester, 1977) до нескольких месяцев у Protogonya-
ulax tamarenaia (Anderson > 1980). Мейоз обнаруживается при на-
блюдении циклоза, когда ядро, по-видимему, вращается. Это явле-
ние, связываемое со спариванием хромосом ( stosch, 1973), мо-
жет отмечаться внутри зиготы или при выходе из цисты. Обычно
219
образуются две клетки, Но у Gymnodinium pseudonalustre высво-
бождается только одна двужгутиковая, которая впоследствии делит-
ся мейотически, давая четыре дочерние клетки ( stosch, 1973).
Ни в какой группе водорослей половое размножение не иссле-
довано так подробно, как в роде Chlaaydomonas и у родственных
ему одноклеточныхаяусиковых Chlorophyceae (рис. 5.18). Большин-
ство видов здесь изогамно, хотя известны также анизогамные и
оогамные ( Tschermak-Woess, 1954, 1962). Несмотря на морфологи-
ческое сходство гамет, изогамвые вида биохимически анизогамны и
сливаются только комплементарные типы спаривания. В зависимости
от вида клональные популяции, образуемые одной особью, могут
быть гомоталличными, т.е. дающими совместимые гаметы, или гете-
роталличными, нуждающимися для спаривания в присутствии другого
клона. Половое размножение у Chlaaydomonas reirihardtii удает-
ся индуцировать недостатком в среде азота; два митотических де-
ления приводят к образованию четырех гамет на клетку. Соедине-
ние и последующее слияние гамет обеспечиваются высокомолекуляр-
ными сульфатированными гликопротеинами, гамонами, на концах жгу-
тиков ( Wiese, 1974). После агглютинации этих концов происходит
лизис стенки гамет ( Goodenough, Weiss,1975), и их протопласты
сливаются. В течение 24 ч зигота увеличивается в размере и фор-
мирует утолщенную клеточную стенку, превращаясь в зигоспору
( Cavalier-Smith, 1976), которая после периода покоя прорастает
с мейотическим делением, давая 4-8 гаплоидных подвижных вегета-
тивных клеток ( Buffaloe, 1958). Хорошо изучена генетика Chla-
туНотппяя , особенно в отношении питания, фотосинтеза и по-
движности ( Lewin, 1976а). Исследование гибридизации между С.
moewusii и c.eugametos показало наличие рекомбинации в хлоро-
пластной ДНК и жесткого отбора, благоприятствующего хлороплас-
там, наиболее совместимым о гибридным ядром ( Lemieux et al.,
1981). Это может говорить о тем, что ядерно-пластвдная несовме-
стимость является важным источником гибели потомства от таких
скрещиваний.
Колман ( Coleman, 1979) и Старр ( Starr, 1980) опублико-
вали обзоры половых жизненных циклов у колониальных вольвоксо-
вых. Gonium И Pandorina ИЗОГЭМНЫ, Eudorina (Goldstein,1964)
анизогамна, a Voivox оогамен (рис. 5.3) (Starr, i960). Таким
220
Рис. 5.18. Зиготический жизненный цикл обитающей в снегу
зеленой водоросли Chloromonas brevlaoina_ .А. Вегетатив-
ная клетка. Б - Г. ОТраыиШНИа 900бЦЩГ.~^. Зооспора.
Е. Сингамия.”! Разные стадии развития покоящейся
зигоспоры. Эти стадии часто определялись как разные виды.
0. Мейоз при прорастании зигоспоры (по Hoham et al.,
1979)
образом, в этой группе эволюция вегетативного строения шла па-
раллельно эволюции процессов размножения. У Volvox известны
как гомоталличяые штаммы, образующие спермин и яйцеклетки в пре-
делах одной колонии, так и гетероталличные виды, у которых муж-
ские и женские гаметы формируются разными колониями. Мелкие бес-
цветные спермин возникают в материнских клетках пакетами по 16-
512 штук. У гетероталличных видов эти пакеты, высвобождаясь,
плывут к женской колонии; после растворения отверстия в матрик-
се спермин разделяются, поодиночке оплодотворяя яйцеклетки. У
221
гомоталличных видов пакеты распадаются и зиготы формируют утол-
щенные клеточные стенки внутри родительской колонии, которая со
временем разрушается, высвобождая их. Цитологически и генетичес-
ки показано (Starr, 1975), что мейоз у Voivox происходит при
прорастании зиготы. Образующийся двужгутиковый протопласт перво-
начально окружен ее стенкой и образует новую колонию путем деле-
ния клеток и последующего их роста. Зиготические жизненные цик-
лы также являются правилом у Tetrasporales, Chlorococcales,
Chlorosarcinalea И Chlorellales (Bold, Wynne,1985) . У ценобиаль-
ных форм, например Pediastrum , половое размножение изогамное, с
немощью двужгутиковых гамет. При прорастании образуется зооспора,
щротопласт которой делится внутри стенки зигоспоры, давая новую
колонию ( Davis, 1967).
Зиготические жизненные циклы предполагаются для многих нит-
чатых Ulotriohales И Chaetophorales, особенно пресноводных, хо-
тя цитологические подтверждения этого обычно отсутствуют. У рода
Coleochaete« который ТО помещают В порядок Chaetophorales (Bold,
Wynne, 1985), то Сближают с харовыми (stewart, Mattox,
1975), половое размножение оогамное, а яйцеклетки напоминают ве-
гетативные клетки или имеют небольшую воспринимающую трихогину.
Двужгутиковые спермин формируются внутри антеридиев. Зигота оста-
ется внутри родительской особи и обрастает вегетативными клетка-
ми ( Graham, 1985) •
В двух других группах зеленых водорослей, Oedogoniales и
Zygnematales, зиготический жизненный цикл более специализиро-
ванный. У Oedogonium (рис. 5.12) половое размножение оогамное,
причем каждый оогоний образует одну клетку. Антеридии мельче,
каждый из них дает 2-4 многожгутиковых стефаноконтных спермин.
Некоторые виды двуполые, с антеридиями и оогониями на одном и
том же талломе, тогда как другие (называемые макрандровыми) име-
ют вегетативно изоморфные разнополые нити. У нанандровых видов
наблюдается резкий половой диморфизм: карликовая мужская нить
Растет эпифитно на женской. Такие мужские экземпляры возникают
из андроепбр, которые по размеру занимают промежуточное положе-
ние между спермиями и нормальными зооспорами. Они образуются в
андроспорангиях, находящихся у гинандроспоровых видов на той же
нити,что и оогонии,а у идиоандроспоровых - на отдельных нитях. В
последнем случае андроспора химически привлекается к женской нита
222
(Raurtscher-Kunkel, Machlis, 1962). Высвобождающиеся спермин
макрандроных видов плывут к оогониям за счет хемотакоиса ( Mach-
lis et al., 1974), тогда как у нанандровых форм развитие карли-
кового мужского экземпляра стимулирует образование в непосредст-
венной близости от него оогония со слизистым чехлом, обволакиваю-
щим вершину мужской нити. Спермий проникает в оогоний через спе-
циальный сосочек, а стенка зиготы утолщается, и возникает покоя-
щаяся зигоспора, при прорастании которой происходит мейоз
(Hoffman, 1965).
Подвижные клетки в порядке Zygnematales отсутствуют. У
Sptrogyra выросты двух соседних нитей соприкасаются, и стенки
их в местах контакта растворяются с образованием конъюгационного
канала (рис. 5.19). Одно ядро мигрирует и сливается с другим, не-
подвижным ядром, а возникающая зигота окружается утолщенной стен-
кой. У S.crassa мейоз происходит до прорастания, и из образую-
щихся ядер три дегенерируют ( Godward, 1961). Механизм размноже-
ния у Mougeotia сходный, за исключением того, что имеет место
изогамия и зигота возникает в конъюгационном канале; у sirogoni,
um наблюдается прямое слияние нитий и конъюгационных каналов
не образуется ( Hoshaw, 1965).
Описаны ( Starr, 1955) методы изоляции среди десмидиевых по-
тенциально половых штаммов. Они часто встречаются в мочажинах на
торфяниках, но половое размножение у них наблюдалось редко. Пред-
полагается ( Coesel, Texeira, 1974), что это не следствие от-
рицательного воздействия факторов среды, а наследственная особен-
ность многих штаммов. Нитчатые представители саккодермных десми-
диевых, в частности Desmidium И Hyalotheca. образуют конъюга-
Изогамия
Анизогамия
'Изогамия Анизогамия
Боковая
Лестничная
Рис. 5.19. Конъюгация у Zygnemataceae (по Esser,
223
ционные каналы, и в зависимости от вада зигота формируется внут-
ри их или в одной из родительских клеток. Такие же каналы извест-
ны и у некоторых плакодермннх родов. У Micrasteriaa papillifera
зигота формируется внутри конъюгационного канала (Coessel, Те-
xeira,1974)Пары особей совместимых типов спаривания у Cloate-
rium образуют слизистый матрикс, внутри которого оболочки обеих
клеток расходятся у перешейка, высвобождая протопласты, сливаю-
щиеся с образованием зигоспоры (рис. 5.20). При ее прорастании
мейотическое деление дает две клетки противоположных типов спари-
вания. Половое размножение у саккодермных десмвдиевых протекает
сходным образом, с развитием широких конъюгационных каналов. У
Ifeaotaenlum (Starr, Rayburn, 1964) И Cylindrooyatia (Biebel,
1973) прорастание зиготы также сопровождается мейозом, но в отли-
чие от плакодермных форм при этом могут возникать четыре клетки.
Кариологические наблюдения и количественные исследования
ДНК ( Sawa , 1965; Shen , 1967) свидетельствуют о наличии зиготи-
ческого жизненного цикла у харовых. Однако в морфологическом пла-
не половое размножение представителей этого класса, возможно,
наиболее эволюционно продвинутое среди всех водорослей. Как муж-
ские, так и женские структуры окружены здесь специализированны-
ми вегетативными клетками, что сближает их с репродуктивными ор-
ганами архегениальных растений (Graham, 1985). У однодомных или
двудомных (Proctor, 1971) видов Chara антеридии и оогонии
развиваются в несущих ветви узлах (рис. 5.21,А). Антеридии сидят
на колонкообразной клетке-ножке и окружены клетками-щитками
(рис. 5.21,Б). На апексе клетки-ножки находятся восемь так назы-
ваемых клеток-головок, образующих вторичные и третичные головки,
от которых отходят сперматогенные нити, т.е. ряды тонкостенных
клеток, дающих каждая один спермий (рис. 5.21,В). При формирова-
нии оогония от исходной клетки узла развивается ряд клеток, из
которых дистальная дает единственную яйцеклетку (оосферу) и не-
сколько стерильных клеток (Sawa, Frame, 1974). Из базальной
клетки развивается прикрепительная ножка с клеткой, в конечном
счете дающей начало пяти спиральным покровным клеткам, окружаю-
щим развивающуюся яйцеклетку (рис. 5.21.Г). Каждая покровная
Клетка отделяет мелкую апикальную клетку, в результате чего
224
Рис. 5.20. Конъюгация у Closterium roatratum . А. По-
парное сближение клеток. Выдвижение сосочков (U) по
мере расхождения полуклеток по средней линии (СЛ).
В. Контакт сосочков с вакуолизированными кончиками (В).
J. Слияние протопластов в конъюгационном канале.
, Е. Развитие зигоспоры (3). (По Brook, 1981)
формируется коронка оогония. При созревании под коронкой между по
кровными клетками образуется щель, через которую проникает спер-
мий. После оплодотворения стенки покровных клеток и зиготы утол-
щаются и образуется темная ооспора (рис. 5.21, £, Е). При ее про-
растании развивается нитевидная протонема, которая дифференциру-
ется, давая у каждого конца клетки узлов; у базального полюса они
формируют ризоиды, а у противоположного закладывается апикальная
клетка. Не считая некоторых различий в расположении клеток репро-
в
Е
Рис. 5.21. Развитие репродуктивных структур у Chara • А. Узел (У), несущий оогоний й антеридий.
Б. Зрелый антеридий в разрезе: Н - ножка: Эп - ёгВДермальная клетка; ОН - сперматогенная нить.
В. Конец сперматогенной нити и спермий (С). Г. Оогоний в разрезе: К - коронка; Я - яйцеклетка;
ПК - покровная клетка. Д. Разрез оогония с зиготой (3) и ее утолщающейся стенкой. Е. Зрелая зй-
гота с утолщенной стенкой (по G.M. Smith, 1955) “
226
дуктивных органов, размножение и жизненные циклы у представите-
лей этого класса по сути своей идентичны.
Спорические жизненные циклы
Почти у всех размножающихся половым путем красных водорос-
лей отмечены спорические жизненные циклы и оогамия (West, Hommer-
sand, 1981). У большинства Blorideophycidae цикл трехфаз-
ный, типа известного для Polysinhonia (рис. 5.22). Двудомные
гаметофиты и тетраспорофиты чаще всего изоморфны, тогда как кар-
поспорофит редуцирован и остается прикрепленным к женокому гаме-
тофиту. Этот цикл встречается во всех порядках Plorideophyoidae,
признаваемых Кюлином (Kylin, 1956), но отсутствует у Palmaria-
les (van der Meer, Todd, 1980) и Bonnemaisoniales (Chihara
Yoshizaki, 1972). Мужские гаметы без жгутиков и клеточной стен-
ки, называемые спермациями, образуются по одной в сперматангиях,
хотя после их высвобождения сперматангии могут регенерировать
(Dixon, 1973). Апекс оогония (карпогона) несет волосовидный вы-
рост, трихогину, воспринимающий спермаций при оплодотворении.
Расположение карпогона и характер развития карпоспорофита
после оплодотворения положены в основу выделения порядков бч0-
rideophycidae (Kylin, 1956). У Hemaliales карпоспорофит обра-
зуется непосредственно из оплодотворенного капогона и формирует
массу карпоспор. В других порядках зиготическое ядро переходит в
так называемую ауксиллярную клетду (рис. 5.23); иногда это сопро-
вождается развитием длинной связующей нити, или ообласта. От
ауксиллярной клетки могут отходить вторичные нити ообласта, сли-
вающиеся с другими ауксиллярными клетками, те в свою очередь
способны сливаться с другими вегетативными клетками, образуя в
конечном счете нити гонимобласта, которые уже и дают карпоспоры.
Ауксилдарная и производные от нее клетки составляют карпоспоро-
фит. Часто как синоним карпоспорофита используется термин цисто-
карпий, но в это понятие иногда включают сопутствующую ткань
женского гаметофита, в частности бутылевидные перикарпы, окружаю-
щие карпоспорофиты у некоторых представителей Bonnemaisoniales
И Ceramiales.
227
Рис. 5.22. Жизненный цикл Polyslphonia. Обратите внимание
на тетраэдральное деление tfe^padnoparitiiH (по West, Hommer-
sand, 1981)
Высвободившиеся карпоспоры прорастают в диплоидную тетраспо-
Рофитную фазу, которая, созрев, Дает в ходе мейотического деле-
ния внутри тетраспорангия тетраспоры. При этом с помощью элек-
тронного микроскопа установлено наличие синаптонемальных комплек-
са, рассматриваемых как доказательство мейоза (Kugrens, West,
1972). Встречаются три основных типа деления тетраспорангия: тет-
раэдрический (рис. 5.22), крестообразный и зональный
(Рис. 5.24.А.Б). Обычно каждому виду свойственна только одна
228
50 мкм
Рио. 5.23. Развитие карпоспорсфитного поколения у Fimbrio-
nhvllum. А. Карпогониальная ветвь: КВ - клетка карпого-
нйальнои ветви, К - карпогон о удлиненной трихогиной.
Б. Развитие ауксиллярной (Аукс) клетки и переход диплоид-
ного ядра из оплодотворенного карпогона. В. Ауксиллярная
клетка, образующая нити гонимобласта. £. Развивающийся
карпоспорофит с цепочками карпоспорангиев (КВ) (по Hansen.
1980) ’
форма деления ( Guiry, 1977). Тетрадный анализ ряда видов показы-
вает генотипическое определение пола при мейозе; это было подтвер-
ждено на Gracilaria в ходе изучения пола и пигментный мутантов.
Есть данные, что пол у багрянок определяет пара аллелей ,с непол-
229
I____________।
50 мкм
A
I______________I
50 мкм
Б
Рис. 5.24. А, Зональные теграопораргии Haematocelis.
2. Крестовидные тетраспорангии Petrocelia (no Abbott.
Hollenberg, 1976)
ним доминированием; диплоидные клетки тетраспорофитов должны
быть, таким образом, гетерозиготны и нейтральны в смысле пола
(van der Meer, Todd , 1977). Может происходить митотическая ре-
комбинация с образованием парных островков гомозиготной ткани,
которая, будучи диплоидной, является мужской и женской; этим мо- ,
жет отчасти объясняться наблюдаемое иногда появление у тетраспо-
Рафитов гаметофитных репродуктивных органов. Griffithaia pacifi-
£а имеет крупные многоядерные клетки, у которых можно индуциро-
вать соматическое слияние. Когда таким образом сливаются клетки
Мужских и женских особей, возникающий экземпляр несет структуры
типа тетраспорангиев, хотя слияние ядер и не доказано. Таким об-
230
разом, уровень плоидности багрянок - не главный фактор, определя-
ющий, является растение гаметофитом или спорофитом. То же самое
можно сказать и о бурых водорослях (Nakahara, Nakamura, 1973;
Henry, Muller, 1983) И Chlorophyceae (Field, Lovlie, 1976).
Кроме жизненного цикла типа известного для Polysiphonia у
красных водорослей обнаружен ряд гетероморфных циклов. В них
обычно чередуются крупный прямостоячий гаметофит и редуцирован-
ный нитчатый или корковый тетраспорофит. Часто эти фазы ранее
рассматривались как разные виды.
Для Audouinella pectinatum характерна гетероморфная смена
тетраспорофитов высотой 1-2 см и карликовых нитчатых двудомных .
гаметофитов высотой менее 0,5 мм (West , 1968). У других видов
порядка Nemaliales, Nemalion multifidium (Fries, 1969), Id. agora
farinosa'. (Stosch, 1965), Pseudogloiophloea confusa (Ramus,
1969) гаметофиты более крупные, а тетраспорофиты типа отмечаемо-
го у Audouinella. У представителей Bonnemaisoniaceae (Chiha-
ra, Yoshizaki, 1972) также Гетероморфная смена поколений с нит-
чатым тетраспорофитом, редуцированным либо до сильно ветвящейся
стелющейся стадии (Hymenoclonium serpens), либо до менее про-
стертой ползучей фазы Trailliella intricate ИЛИ Falkenbergia.
К Cryptonemiales и Gigartinales относится ряд родов, у ко-
торых тетраспорофит обычно редуцирован до корковой фазы; сюда от-
носятся ВИДЫ Farlowia (DeCow, West, 1982а), Pikea (Scott, Dixon,
1971)и Gloiosiphonia(Edelstein, McLachlan, 1971).Карпоспоры Acro-
symphyton purpuriferum развиваются в фазу HymenocloniuBua у Halyme-
nia floresia—в фазу типа Audouinella(Cortel-Breeman,Hoeh, 1970).
У других представителей Gigartinales отмечены разные гетеро-
морфные жизненные циклы, которые лучше всего проиллюстрировать
на примере видов Mastocarpus. ранее выделявшихся в секцию рода
Gigartina (Guiry et al., 1984), тогда как у видов Gigartina В
узком смысле жизненные циклы в целом типа цикла Polysiphonia .
Тетраспоры корковой формы Petrocelis развиваются в Gigartina
papillata и G, jardinii (West. 1972); сходная взаимосвязь пока-
зана между Mastocarpus stellata И Petrocelis cruenta (Chen
et al., 1974). Жизненный цикл w»gtocarpus усложнен в некоторых
популяциях таи, что карпоспоры могут развиваться непосредствен-
но в женские гаметофиты, несущие карпоспорофиты (Polanshek,
231
West, 1977; Rueness, 1978).
У представителей Rhyllophoraoeae жизненные циклы весьма
разнообразны. Корковые фазы отмечены в цикле Gymnogrongus lep-
tophyllus (PeCew, West 1981). У Phyllophora pseudoceranoides
( Newroth, 1972) цикл типа Roly sinh onia. a у P.truncata он
тетраспоробластический ( Newroth , 1971). У последнего вида и у
Gymnogrongus griffithsiae (Cordeiro-Marino, Poza, 1981) карпо-
спорангии, образующиеся внутри женского гаметофита, дают начало
нитям, которые вынооят тетраспорангии на поверхность таллома. У
родственного вида Gvmnogrongus chiton иммунологически (на осно-
ве распределения типов каррагинанов) показано, что мейоз происхо-
дит при тетраспорогенезе (McCandless, Vollmer, 1984). Предпо-
лагается ( Guiry, in McCandless, Vollmer, 1984), что эти реду-
цированные фазы следует считать тетраспорофитами, а не карпотет-
распорофитами, обнаруженными у некоторых видов Ыаяога и Не1-
mlnthocladla. Карпотетраспорофиты, например у Ijiagora tetra-
sporifera. напоминают карпоспорофиты образующей карпоспоры фа-
зы 1. farlnosa (Stoschf, 1965), но дают мейотические тетраспо-
рангии на вершинах нитей гонимобласта.
У Palmarlales (рис. 5.25) женский гаметофит редуцирован,
а карпоспорофит отсутствует. Тетраспорофиты и мужские особи у
Palmaria palmata отмечались, а женские растения были неизвест-
ны. Цветовые мутанты, используемые для различения гаметофитной
и спорофитной тканей, показали, что тетрады спор из тетраспоро-
фитов прорастали в отношении 2:2 в крупнолопастные мужские гаме-
тофиты, изоморфные с тетраспорофитами, и карликовые дисковидные
женские особи диаметром в зрелом состоянии 0,1 мм (van der Meer,
Todd, 1980). На женских гаметофитах образуются многочислен-
ные карпогоны с удлиненными трихогинами, которые после оплодо-
творения развиваются непосредственно в лопастной тетраспорофит.
В жизненном цикле коркового вида Rhodophysema elegans
'наблюдается дальнейшая редукция: здесь отсутствует тетраспорофит-
ное поколение. Корковый таллом представляет собой однодомный га-
метофит, а оплодотворенные карпогоны развиваются непосредствен-
но в мейотические тетраспорангии, споры из которых воспроизво-
дят корковую фазу (DeCew, West, 1982).
232
Рис. 5.25. Жизненный цикл Falmaria palmate (
Todd, 1980)
van der Meer,
У представителей порядков Rhodymeniales И Ceramiales в ос-
нове жизненного цикла лежит тип Foivsiphonia. но имеется много
незначительных отклонений от него, обычно в результате отсутствия
мейоза в те траспорангии. Как примеры можно привести виды Lomen-
taria orcadensia (Svedelius, 1937) И Antlthamnlon boreale
( Sundene, 1962), у которых происходит непосредственное возобнов-
ление тетраспорофитного поколения. В этой группе на одном и том
же талломе часто отмечались тетраспорангии и гаметангии ( К1п-
aggs, 1969), хотя мало что известно о лежащих в основе этого
цитологических механизмах. Несколько родов, например Пеопоаро-
rum и Tiffanleiia. образуют полиспорангии (рис. 5.7,Г), гомоло-
гичные тетраопорангиям тем, что в них протекает мейоз, а возника- х
ющие споры делятся внутри спорангия митотически ( Drew, 1937). )
233
• У представителей Bangiaies репродуктивный цикл может вклю-
чать сверлящую раковины моллюсков водоросль Conchocelis rosea
(рис. 5.26). Это было впервые показано для Pornhyra umbilicalis
( Drew,, 1949) и впоследствии также для Erythrotrichia (Нееге-
bout, 1968), Bangla (Richardaon, Dixon, 1968), Smithora. (Riohar-
dson, Dixon, 1969) и Pornhyronsis(Murray et al., 1972). Ны-
не общепризнано, что жизненный цикл Porphyra и Bangia по суще-
ству половой, причем фаза Conchocelia предположительно соответ-
ствует здесь диплоидному спорофитному поколению ( Cole, Conway,
1980). Однако у многих видов указаны идентичные хромосомные чис-
ла листовидной стадии и фазы Conchocelia , причем оплодотворе-
ние и мейоз не наблюдаются. Фаза Conchocelia образует в конхо-
спорангии цепочки конхоспор, поочередно высвобождающихся на его
вершине и прорастающих в макроталломную фазу. В зависимости от
вида в клетках на поверхности таллома формируется от 16 до 256
опермациев, называемых также -спорами. Высвободившись, они
сливаются с небольшими выростами на талломе, прототрихогинами.
Клетка, несущая их, считается карпогоном. Она делится с образо-
ванием 4-16 карпоспор, называемых dt-споры; последние прораста-
ют в фазу Conchocelis. Изучение хромосом позволяет предпола-
гать, что мейоз происходит при развитии конхоспор. Изящное уль-
траструктурное исследование (Hawkes, 1978) однозначно продемон-
стрировало сингамию. Следовало бы, однако, подчеркнуть, что по-
ловое размножение показано лишь у немногих видов этого порядка.
Жизненные циклы примнезиофитов плохо понятны. У большинст-
ва, если не всех, кокколитофорид в жизненном цикле больше одно-
го типа клеток (Hibberd, 1980с), и поэтому можно считать его
спорическим, но место мейоза, как правило, четко не установлено
и возможен плейоморфизм. Отмечено чередование диплоидной свобод-
ноживущей кокколитофорида (рис. 5.9) Hymenomonas carterae И
гаплоидной бентосной ложнонитчатой стадии Anisthcnema (Stosoh,
1967; Leadbeater, 1970). Hymenomonas воспроизводится клеточ-
ным делением, но каждая клетка может также образовать четыре
меньшие по размеру зооспоры без кокколитов, которые вырастают в
бентосную фазу. Последняя способна воспроизводиться зооспорами
или формировать более мелкие гаметы, сливающиеся с образованием
зиготы, восстанавливающей фазу с кокколитами. У Ochrosphaera -
234
Рис. 5.26. Жизненный цикл Pornhvra (по West, Hommer-
aand, 1981)
рода, представленного неподвижными формами с кокколитами, - жиз-
ненный цикл такой »е ( Gayral, Presnel-Morange, 1971) и предпо-
лагает плейоморфизм. Описано ( Parke, Adams, I960) чередование
ДВУХ ОВОбОДНОЖИВУШИХ КОККОЛИТОфОрИД - НеПОДВИЖНОГО Coccolithum
pelaglcus И подвижного Crystallollthus hvallnus ; мейоз И
сингамия не наблюдались, и неясно, является ли этот цикл поло-
вым. У Bnliiania huTievi доминирующая фаза неподвижна, хотя
235
может давать подвижные, чешуйчатые, но лишенные кокколитов клет-
ки, в которых, по-видимому, меньше ДНК, чем в фазе с кокколитами
( Paasche, Klaveness, 1970).
Ьпорический жизненный цикл с чередованием свободноживущих
гаплоидного гаметофитного поколения и морфологически сходного
или несходного диплоидного спорофита характерен доя всех воспро-
изводящихся половым путем бурых водорослей, за исключением Fuca-
1еа и родственных порядков. Это было установлено свыше .100 лет
назад среди Cutleriales у Zanardinia(Reinke. 1878) с изоморф-
ной сменой и у Cutleria (Falkenberg,1879) С гетероморфным цик-
лом, включающая стелющееся спорофитное поколение, ранее тракто-
вавшееся как самостоятельный род Aglaozonia.
Мейоз происходит в одногнездных спорангиях на спорофитах,
причем синапсис установлен с помощью электронного микроскопа у
Pylaiella и Chorda (Forth, Markey, 1973). В большинстве поряд-
ков образующиеся споры многочисленны и подвижны. При прорастании
они дают изогамные, анизогамные или оогамные гаметофиты. Выражен-
ные анизогамия или оогамия среди бурых водорослей характерны для
порядков Cutleriales, Sphacelariales, Dictyotales, Desmaresti-
ales и Laminariales, тогда как виды Ectocarpales, Dictyosi-
phonales и Chordariales преимущественно изогамны. В поряд-
ках Ectocarpales, Tilopteridales, Dictyotales, Sphacelariales
и Cutleriales (частично) наблюдается в основном изоморфная
смена поколений, тогда как в других порядках отмечено чередование
гетероморфных фаз. У изогамных и анизогамных видов гаметангии
многогнездные; их гнезда возникают в результате последовательных
делений и содержат в зрелом виде по одной гамете каждое. Сходные
многогнездные спорангии могут встречаться на спорофитах некото-
рых видов, давая споры, а не гаметы. У оогамных водорослей в каж-
дом оогонии образуется одна яйцеклетка, а мужские гаметы форми-
руются либо в большом количестве многогнездными антеридияии, ли-
бо поодиночке одногнездными, доторые можно рассматривать как ре-
дуцированные многогнездные структуры ( Henry, Cole, 1982b).
Отклонения от характерного чередования спорофита и гамето-
фита встречаются во многих порядках ( Wynne, Doiseaux, 1976) И
чаще всего связаны с возобновлением гаплоидной или диплоидной
фазы с помощью нейтральных в половом отношении подвижных клеток
(зооидов), возникающих в многогнездных спорангиях; одна из фаз
может также подавляться в результате апомейоза или партеногене-
за. Имеется также ряд сообщений (см. обзор Carат, 1972) о слия-
236
нии продуктов одногнездных спорангиев, т.е. зооспор, функциони-
рующих как гаметы, что дает гаметический жизненный цикл. Это бы-
ло впервые описано у Ectocarpus silioulosus (Knight, 1929), Но
затем для данного вида опровергнуто (Mailer, 1975); по-видимо-
му, убедительных цитологических данных й пользу такого цикла у
бурых водорослей нет.
Показано (Knight, 1929) изоморфное чередование поколений
у Ectocarpus ailiculosua. Многогнездные структуры встречаются
здесь как в гаплоидной, так и в диплоидной фазе, но одногнездные
спорангии присущи только диплоидным спорофитам. Впоследствии
(Muller, 1967, 1972) эти данные были подтверждены, причем ока-
залось, что гаметофиты могут быть гапловдными или диплоидными, а
спорофиты - гаплоидными, диплоидными или тетраплоидными (рис.
5.27). Гаплоидные спорофиты развиваются при партеногенетическом
прорастании неслившихся мужоких или женских гамет и образуют апс-
мейотические одногнездные спорангии. Тетраплоидные спорофиты
иногда развиваются спонтанно из зооидов, высвобождаемых много-
гнездными спорангиями этих гаплоидных спорофитов. Зооиды из однс-
гнездных спорангиев тетраплоидов дают как диплоидные опорофиты,
так и диплоидные гаметофиты. Спорофиты и гаметофиты обычно раз-
личимы морфологически; при внешней изогамии наблюдается выражен-
ная физиологическая анизогамия: женская гамета оседает на суб-
страт и выделяет феромон, привлекающий мужскую гамету (см. гл.6).
У многих других представителей этого порядка, особенно у корко-
вых форм, половой процесс, по-видимому, отсутствует (Wynne,
loiseaux, 1976).
Среди представителей Dictyotales у Pictyota dichotoma
И Dilophus ligulatua (Gaillard, 1972) отмечен характерный
изоморфный цикл: антеридии и оогонии развиваются из коровых кле-
ток мужских и женских гаметофитов, образуя сорусы. Антеридии
многогнездны (рис. 5.28,А); каждое из тысяч гнезд дает единст-
венный одножгутиковый спермий, тогда как в каждом оогонии раз-
вивается одна яйцеклетка (рис. 5.28,Б). Спорофиты неоут мейоти-
ческие тетраспорангии (рис. 5.28,В), хотя апомейоз отмечался у
Padina pavonica и Dilophua faaicola. Изоморфный жизненный цикл
характерен также ДЛЯ Sphacelariales, хотя у Sphacelaria rigidu-
la (Hoek, Flinterman, 1968, как S, furcigera) он слабо гете-
роморфный, причем спорофиты здесь мощнее двудомных гаметофитов. \
Женские гаметы могут развиваться партеногенетически в женские га-
метофиты или гаплоидные спорофиты.
237
Рис. 5.27. Схема жизненного
никла Ectocarpus siliculo-
sus с гаплоидной, дйпло-
идной и тетраплоидной фаза-
ми. О - одногнездные струк-
туры; М - многогнездные
структуры; М! - мейоз (по Wy-
nne, Lois ваше, 1976,
ИЗ ffiiller, 1967)
В истинно гетероморфных циклах два поколения заметно разли-
чаются по форме. Это характерно для Порядка Laminariales, ЧТО
впервые продемонстрировал Соважо (Sauvageau, 1915) ДЛЯ Saccorhj__
ga polvschides (рис. 5.29). Мейотические одногнездные споран-
гии в сорусах располагаются либо на вегетативной пластине, либо
на специализированных спорофиллах макроскопического спорофита.
Зооспоры каждого спорангия дают начало равному числу микроскопи-
ческих мужских и женских гаметофитов (Schreiber, 1930). Показа-
но (Evans, 1965), что пол у этого вида определяется генотипиче-
ски наличием крупной Х-хромоссмы у женских особей и мелкой
Y-хромосомы - у мужских. Наблюдалось спаривание этих хромосом
при мейозе. При прорастании протоплазма обычно выходит из споры
в проростковую трубку; образующаяся стенка обособляет первую
клетку гаметофита. У мужского гаметофита клетки мельче, и ветвит-
238
Антеридии
I_________1
50 мкм
Рис. 5.28. Репродуктивные структуры Dictyota: А - анте-
ридии; Б - оогонии; J. - тетраспорангий; Un - спорангий,
б - спора (по Scagel et al., 1982)
ся он сильнее, нем*женский; на нем располагаются антеридии, в
каждом из которых образуется по одному спермию. У женских особей
каждый оогоний высвобождает единственную яйцеклетку, и после
оплодотворения зигота развивается в спорофит, который быстро
увеличивается в размерах и дифференцируется. Яйцеклетки
могут развиваться партеногенетически в гаплоидные спорофиты,
морфологически неотличимые от диплоидных ( Nakahara, Nakamura,
1973). Представители одинаковых морфологических секцией рода
Tam-inerta ПО-ВИДИЛОМУ, ЛЭ ГКО ДЭЮТ ГИбрИДЫ ( liming et al. ,
1978); указывались также межродовые гибриды между Macrocystis
и Pelagonhycus (Sanbonsuga, Neushul, 1978). Процессы развития,
связанные с жизненными циклами представителей laminariales J
ХОРОШО прослежены у laminaria (Kain, 1979).
239
Рис. 5.29. Еизненный цикл laminaria . А. Макроскопичес-
кий спорофит. Б. Срез пласТМы 'бйброфита с одногнездными
спорангиями (Ос), местом мейоза. В. Гаплоидные зооспоры.
J. Микроскопический мунской гаметофит с антервдиями, об-
разующими спермин, g. Микроскопический женский гаметофит
с оогониями, образующими яйцеклетки. Е. Зигота. 2. Моло-
дой, спорофит (по Bold, 1973) ~ ~
Сходные жизненные циклы известны у Sporocnnalesи Desma-
rest iaies.y Sporochnus pedunculatus крупный спорофит с мейотичес-
" кими одногнездными спорангиями чередуется с микроскопическим га-
t метофитом ( Caram, 1965). Чередование поколений по типу 1япн_
L narla происходит И у Desmarestiales: У Desmarestia viridis
гаметофиты однодомные ( Kornmann, 1962), а у D, aculeata - дву-
И домные ( Chapman, Burrows, 1971). Однодомные гаметофиты встреча-
11 ются также у Arthrocladia ; оплодотворение не наблюдалось, а
ж сходные хромосомные числа отмечены у обеих фаз, что предполагает
наличие как партеногенеза, так и апомейоза (Ifiiller, Мее1,1982).
240
Выраженное чередование морфологических фаз встречается в не-
давно описанном порядке Syringodermatales (рис. 5.30) (Нему,
1984). У Svringoderma nhinnevi (Henry, Mdller, 1983) мейоспоры
из одногнездных спорангиев дают начало микроскопическим двудом-
ным изогамным стелющимся гаметофитам; из зиготы восстанавливает-
ся опорофитное поколение. У S. floridana (Henry, 1984) зооспо-
ры из одногнездных спорангиев слабоподвижны, а гаметофитное поко-
ление, развивающееся in aitu , редуцировано до двух клеток, каж-
дая из которых дифференцируется в изогамету. Считается, что у
s, abvasicola имеются похожие четырехклеточные гаметофиты
( Walker, Henry, 1978); морфологически это наиболее редуцирован-
ные свободноживущие гаметофиты бурых водорослей.
Рис. 5.30. Схемы жизненных
циклов четырех видов Syrtn-
Roderma-.ro - гаметофит; Г -
гамета; С - спора; 3 - зиго-
та (по Henry, 1984)
S. phinneyi S. australa
241
ПРЯМОСТОЯЧАЯ ФАЗА РАЛЬФСИОИДНАЯ ФАЗА
О’ Гаметофит (п) -->М—► о" Гаметы -----------------------
И!__________________________
(Партеногенез)
I—Зооиды*— О *— Корки/дернины (п)--------------
Ml v
---Зооиды-*— О Корки/дернины *— Зигота -4-Слияние
(2п)
г—Зооиды*— О *— Корки/дернины ----------------
(л)
I (Партеногенез)
Геметофит (п) —М—► (р Гаметы —-----------------------
Рис. 5.31. Схема жизненного цикла Scytosiphon lomentaria:
О - одногнездные структуры: М - многогнездные структуры, М' -
мейоз (по Wynne, Loiseaux,1976> из Nakamura, Tatewaki, 1975)
Жизненные циклы Chordariales, Diсtyosiphonales 0 Scyto-
siphonales относятся к числу наиболее- запутанных в этом отделе;
в основном они гетероморфны, но во многих случаях встречается
плейоморфизм без указаний на смену плоидности. Редуцированные фа-
зы могут быть представлены нитчатыми "плетизмоталломами" или кор-
ками с особенностями морфологии и размножения, сближающими зги
порядки с Ectocarpales в широком смысле. Жизненные циклы еще
более усложнены у некоторых видов тем, что, по-видимому, меняют-
ся в зависимости от географического положения. Истинная смена по-
колений известна У Panenfussiella calUtri.cha (Peters, 1984),
Sphaeorotrichia divaricata (Ajisaki, Umezaki, 1978), Hecato-
nema streblonematoides (Loiseaux, 1970a), Soranthera ulvoidea
(Wynne,1969) И Stictyosiphon adriaticus (Caram, 1965).
У Scytosiphon lomentaria описано (Tatewaki, 1966) чере-
дование прямостоячего гаметофита и коркового спорофита типа
Ralfsia, образующего одногнездные спорангии (рис. 5.31). Это че-
редование морфологических фаз подтверждено в других местообитани-
ях вида и указывалось также для родственного таксона Petalonia
fascia (Wynne, 1969); Hsiao, 1969s Edelstein et al.,1970). Сооб-
щалось также о наличии в этом комплексе жизненных циклов фаз
Microspongium (Lund, 1966), Streblonema и Compsonema
(Loiseaux, 1970b) . В отличие от Татеваки (Tatewaki, 1966)
последние авторы не получили данных в пользу полового
процесса, хотя позже он был вновь подтвер-
16— 1 91
242
жден ДЛЯ японских экземпляров ( Nakamura, Tatewaki, 1975) и про-
демонстрирован ДЛЯ Colnonema bullosa. Petalonia fascia и Endo-
rachne binghamiae из этого же порядка. Показано ( Clayton,
1981), что половая активность у Scytosinhon зависит как от окру-
жающей среды, так и от возраста особей и сильнее всего выражена
у более старых водорослей в зимние месяцы. Гетероморфные чередо-
вания без явных изменений плоидности указывались также в порядке
Chordariales у Haplogloia andersonii (Winne, 1969) И Splach-
nidium rugosum (Prioe, Duoher, 1966), а кроме того, среди
Dictyosiphonales Г Hiaeostrophion irregulars (Mathieson,I967),
Coliodesme californica (Wynne, 1972) И Striaria attenuate
(Nygren, 1975) из Швеции. Последние данные представляют особый
интерес, поскольку этот жизненный цикл в Средиземноморье описы-
вался как половой (Caram, 1965).
Чередование особей, выглядящих как гаметофиты и спорофиты,
но без доказательств мейоза или сингамии, встречается не только
у гетероморфных видов. С морфологической точки зрения Tilopteris
mertensii И Haplospora globosa (Tilopteridales) размножа-
ются половым путал, поскольку у них образуются яйцеклетки и спер-
мин, однако последние нефункциональны и мейоза в спорангиях не
происходит (Kuhlenkamp, Miller, 1985). У Haplospora (рис.
5.32) неоплодотворенные яйцеклетки однодомного гаметофита разви-
ваются в изоморфные спорообразующие спорофиты. Ситуация еще про-
ще у Tiiopteris. поскольку здесь неоплодотворенные яйцеклетки
дают непосредственно гаметофит, а спорофитное поколение, видимо,
отсутствует. Представляет особый интерес то, что, хотя сингамии
и не происходит, яйцеклетки все-таки образуют феромон, привлека-
ющий спермин. Это подразумевает позднейшую утрату полового про-
цесса и возможность обнаружения популяций, воспроизводящихся по-
ловым путем.
Другое, до сих пор плохо понятное явление у бурых водорос-
лей - гетеробластия, при которой зооиды независимо от внешних
условий и пола прорастают двумя разными*путями. Исходно яйцевид-
ные проростки становятся нитчатыми,а амебоидные - обычно звезд-
чатыми и позже развиваются в дисковидные образования. Гетеробла-
стия особенно распространена у Myrionemataceae (Chordariales)
(Pedersen, 1981).
Сейчас в целом признается, что жизненные циклы нельзя ис-
пользовать как основной критерий при классификаций^ Chlorophy-
ceae на уровне порядков, поскольку каждый их тип.упо-видимсму,
243
Рис. 5.32. Жизненный цикл Tilonteris mertensiit
О - оогоний; А - антеридий; ГГ - гипертрофированное гнез-
до; Я - яйцеклетка; С - спермий; Ср - спора из гипертро-
фированного гнезда; П - проросток; Г - гаметофит; Р - ри-
зоид; Н - нить; ОФ - спорофит; Си - спорангий; 0/ - четы-
рехъядерная спора; Ст - одноядерная спора (по KuKLenkamp,
Mailer, 1985)
возникал независимо в разных группах и неоднократно (Hoek, 1981).
Тем не менее изоморфные спорические циклы свойственны Cladopho-
rales и Ulvales, а гетероморфные - Acrosiphonales и некоторым
представителям Caulerpales.
У морских Cladophorales чередуются спорофитное поколение,
дающее четырехжгутиковые зооспоры, и однодомное гаметофитное, об-
244
разующее двужгутиковые гаметы. Это подтверждено как исследования-
ми в культуре, так и цитологическими данными .для Chaetomornha
(Patel, 1971, 1972; Kornmann, 1972), Cladonhora (Wik-Sjostedt.
1970) и Rhizoclonium (Bidding 1957). Однако имеются сведения,
что у пресноводного вида Cladonhora crispata жизненный цикл
зиготический ( Siddeque, Paridi, 1977), а у С. glomerata- гаме-
тический (list, 1930). Гаметы Cladonhora могут развиваться
партеногенетически и спонтанно диплоидизироваться; встречается
также полиплоидия (Wik-Sjostedt, 1970).
Среди Ulvales у родов Percursaria. Enteromomha. Ulva,
Ulvaria и Blidingia изоморфное чередование поколений, сходное с
указывавшимся для Cladophorales (Kornmann, I956;Foyn, 1934;
Sliding, 1963; Dube, 1967; Tatewaki, lima, 1984). Наиболее
детально исследован вид Ulva mutabilis (рис. 5.II) (Field, Lov-
lie, 1976; Lovlie, Bryhni 1978), у которого 12 морфологичес-
ки идентичных форм различаются особенностями цитологии и размно-
жения. Типы спаривания определяются генотипически одной парой ал-
лелей, и спорофиты при мейозе дают равное число зооспор каждого
типа. Развивающиеся из них гаплоидные гаметофиты образуют гаме-
ты, которые сливаются, если принадлежат к комплементарная типам
спаривания, в результате чего восстанавливается диплоидный спо-
рофит. Встречается партеногенетическое развитие гамет, когда в
1-2% случаях возобновляется родительский гаметофит, а в осталь-
ных возникают гаплоидные спорофиты. Может происходить диплоиди-
зация гаплоидных спорофитов ( Hoxmark, Nordby, 1974), иногда не-
полная, так что известны гаплоидные, диплоидные и гаплодиплоид-
ные партеноспорофиты. Вдобавок два спаривающихся штамма различа-
ются по своей способности к диплоидизации. Как диплоидные, так и
гаплоидные партеноспорофиты дают путем мейоза зооспоры; в послед-
нем случае случайное расхождение хромосом приводит к образованию
спор с очень низкой жизнеспособностью. Зооспоры таких партеноспо-
рофитов всегда относятся к одному типу спаривания, что еще раз
демонстрирует аллельную природу определения пола. Партеногенети-
ческое развитие гамет наблюдалось и у других видов Ulva (Sliding,
1968; Tanner, 1981), а также в родах Enteromorpha(Kapraun,I97Q)
11 Percursaria (Kornmann, 1956), причем у некоторых штаммов,
по-видимому, отсутствует нормальное половое воспроизведение.
Половое размножение продемонстрировано^ двух родах порядка
Chaetophorales. У Entocladia (O’Kelly, Yariph, 1981) изоморф-
ная смена поколений; она отмечена и у Eugomontia (Kornmann ,
245
I960), хотя для этого рода указывалась также стадия типа
Codiolum (Wilkinson, Burrows, 1970).
Гетероморфное чередование макроскопического гаметофита и
редуцированного спорофита типа Codiolum или Chlorochytrium
указывалось для целого ряда зеленых водорослей, например
Ulothrix (Bokhorst, 1978), Monostroma (Kornmann, Sahling 1962),
Urospora (РИС. 5.33,A) (Kornmann, 1961) И Spongomorpha
(рис. 5.33,£>, В, Г)(= Acrosiphonia) (Kornmann, 1964), Эти ро-
ды имеют неясное таксономическое положение на уровне порядка, и
их относят то к Ulotrichales, то кAcrosiphonales, го (в слу-
чае талломов сложного строения) К семейству Monostromataceae
из Ulvales (Hoek, 1981; Tanner, 1981). Для них был также пред-
ложен специальный класс Codiolophyceae (Kornmann, 1973). Толсто-
стенные одноклеточные фазы типа Codiolum можно рассматривать
как гипнозиготы на ножках о прикрепительными дисками ( Bold,
Wynne, 1985). Мейоз происходит при зооспорогенезе, так что жиз-
ненный цикл здесь как бы зиготический. С другой стороны, эти фа-
зы можно считать редуцированным одноклеточным спорофитом (Tanner ,
1981), тогда цикл представляется спсрическим. Полагают, что раз-
личия в морфологии между прикрепленной фазой ' Codiolumи коккоид-
ной фазой Chlorochytrium контролируются условиями внешней сре-
ды (Kornmann, 1973).
На примере Aorosjphonia прослеживается постепенный переход
от гетероморфного спорического цикла к зиготическому(Когптапп>
1970, 1973). A.spinescens - типично гетероморфный вид (рис.
5.33,5). У A. grandis отсутствует зооспорогенез (рис. 5.33,В),
однако ядро фазы Codiolum перед развитием гаметофита делится не-
сколько раз; у a.arcta с Гельголанда (рис. 5.33,В) в зиготе до
прорастания в гаметофит происходят два ядерных деления (возмож-
но, мейотические).
Для представителей порядка Caulerpales в целом характерен
гаметический жизненный цикл, однако у видов комплекса Halycystis-
Derbesia—Bryopsis и родственных родов отмечено гетероморфное че-
редование поколений; впрочем, разные варианты встречаются даже в
пределах одного вида (Rietema 1970; Tanner, 1981). Нитчатая фа-
за Derbesia считается спорофитом: мейоз происходит при образова-
нии стефаноконтных зооспор, которые в зависимости от вида прорас-
тают либо в многоядерную мешковидную форму Halicystis (рис.
5.13,А), либо в ветвящиеся ценоцитные нити Bryopsis. (рис.5.13,Б).
246
Рис. 5.33. Жизненные циклы Uroapora (А) и Acrosiphonia (£ - Г), показывающие последователь-
ную редукцию спорофита типа Cpdiolum . М! - мейоз < по Tanner, 1981)
247
Гаметофиты анизогамны; гаметы у Bryopsis образуются в вегета-
тивных ветвях, превращающихся в гаметангии ( Burr, West, 1970);
У Ня1icvatis в гаметы преобразуется часть вегетативной прото-
плазмы ( Hollenberg, 1935).
У некоторых изоляторов Bryopsis полная стация Derbesia
(рис. 5.13,В) отсутствует и гаметы развиваются в одноядерную
протонему, которая образует споры или развивается непосредствен-
но в гаметофит ( Rietema, 1971; Bartlett, South, 1973). Неодно-
кратно сообщалось также ( Tanner, 1981) об апомейотических штам-
мах и различных добавочных бесполых процессах размножения. Убеди-
тельные цитологические данные об уровнях плоидности и наличии
мейоза для этой группы почти полностью отсутствуют.
Паметические жизненные циклы
Noctiluca miliaris (рис. 5.34) - динофлагеллата с гамети-
ческим жизненным циклом ( Zingmark,I970). Этот вид выделяется
среди своих сородичей внешне эукариотным вегетативным ядром, хо-
тя у его гамет оно типично динофитное. При гаметогенезе ядро де-
лится мейотически, а затем митотически, с образованием свыше
2000 одножгутиковых изогамет. Пары гамет сливаются, и зигота
после периода покоя развивается непосредственно в вегетативную
клетку.
У диатомей половое размножение гаметическое. У большинства
видов, кроме генетической рекомбинации, при половом воспроизве-
дении происходит восстановление максимального размера клетки
(Drebes, 1977). Перистые диатомеи изогамны (рис. 5.35), центри-
ческие - оогамны (рис. 5.36). У последних мужские клетки в ре-
зультате серии митозов могут формировать сперматогонии, остаю-
щиеся внутри родительских панцирей, а затаи делящиеся мейотичес-
ки с образованием одножгутиковых спермиев. Детали сперматогене-
за различаются в зависимости от родовой принадлежности; так, у
Cyciotella как сперматогонии, по-видимому, функционируют веге-
тативные клетки (Schultz, Trainor, 1968); в других таксонах,
например Stephanophvxis (Drebes, 1966), сперматогонии накануне
мейоза окружаются тонкими кремнезаиными оболочками. Оогонии у
Диатомей развиваются непосредственно из вегетативных клеток,
причем цитокинез при оогенезе отсутствует. После каждого мейоти-
248
Рис. 5.34. Жизненный цикл цинофлагеллаты Noctiluca mi-
liaria (по Lee , 1980; Из Zingmark, 19/0}
ческого целения сохраняется только одно функциональное ядро. Не-
которые ВИЦЫ, В частности Lithodesmwn undulatum и Biddulphia
mobiliensis , дают две функциональные яйцеклетки (Stosch, 1954).
После оплодотворения зигота увеличивается в размере, формируя
ауксоспору, под оболочкой которой развивается вегетативная клет-
ка исходного типа. Образованию каждой створки предшествует аци-
токинетический митоз с возникновением дополнительных ядер. Обо-
лочка ауксоспоры в конечном счете разрывается, высвобождая новую
укрупненную клетку ( Drebes, 1977).
У центрических диатомей отмечалась и автогамия. У Cyclotel-
la menegh-i.niana (Schultz, Trainor, 1968) спермин известны, НО
оплодотворение не наблюдалось. Вместо этого после мейоза в мате-
ринской клетке ауксоспоры сливаются два ядра. )
249
Юмкм
Рис. 5.35. Половое размножение перистой диатомеи Rhoieo-
s-phenia curvata. A, Б. Ламетангиальные клетки знуТрГТли-
з истой оболочки (CU); более активный партнер располагает-
ся выше. В. Четыре гаметы непосредственно перед слиянием.
£. Две зиготы между пустыми панцирями гаметангиев. Д. Рас-
хождение зигот. Е. Ранние стадии образования аукоспор.
Д. Достигшие максимального размера ауксоспоры с пластида-
ми, приуроченными к центральной части. 3. Ауксоспоры пос-
ле отложения створок; пластиды увеличились и заншают поч-
ти всю дайну клетки. Левая клетка частично вышла из свое-
го перизония (по Mann, 1982)
В отличие от представителей Biddulphlales перистые диато-
меи размножаются половым путем с образованием безжгутиковых физи-
ологически анизогамных изогамет. В геологической истории центри-
ческие диатомеи появляются раньше перистых (Drebes, 1977) и счи-
таются более примитивными, т.е. у предположительно производных
от них форм наблюдается эволюционно более древняя изогамия. Одна-
ко изогаметы здесь лишены жгутиков, что указывает на их удаление
от исходного типа. У большинства перистых диатомей изоморфные
250
Рис. 5.36. Жизненный цикл центрической диатомеи Chaetoce-
ros didymum; I - 4 - деление вегетативных клетой; К - W
- ПОЛТОой Цикл; 5 - II - образование спермиев; 20 - оого-
ний; 21 - 27 - последовательные этапы сингамии; 28 - 30 -
развитие ауксоспор; 31 - 37 - цикл бесполых покоящихся
СПОр(по Stosch et al., 1973)
251
стимулированные к половому размножению клетки сближаются попарно
и выделяют популяционную слизь. Первое деление мейоза нормальное,
но1 при втором цитокинеза не происходит, так что образуются две
гаметы, каждая с одним гаплоидным и одним дегенерирующим допол-
нительным ядром. Известна изогамия, но более распространена фи-
зиологическая анизогамия, при которой каждая материнская клетка
дает одну активную и одну пассивную гамету. Слияние их дает две
зиготы, развивающиеся в ауксоспоры. Последние растут, и их внеш-
няя мембрана заменяется окремнелым перизонием; как и у центриче-
ских форм, образование створок сопровождается митозом и ядерной
дегенерацией. У таких перистых диатомей, как Eunotia и Соссо-
neia материнская клетка образует только одну гамету; в этой
группе также встречается автогамия (Drebea, 1977).
Гаметические жизненные циклы предпрлагаются и yFucalea
из бурых водорослей (рис. 5.37), где диплоидны только свободно-
живущие экземпляры, а мейоз тесно связан с гаметогенезом. Размно-
жение в этом порядке лучше всего изучено у рода Pucua. к которо-
му относятся однодомные (например, spiralis) и двудомные (на-
пример, F, serratua) виды. При достижении половой зрелости вер-
шины вегетативных ветвей превращаются в репептакулы с погружен-
ными в них концептакулами^, где формируются антеридии и оогонии
шесте со стерильными парафизами. Не отделяющиеся стенками
( Berkaloff, Rouaaeau, 1979) антеридии располагаются на ветвя-
щихся парафизах и образуют многочисленные гаплоидные спермин.
Оогонии сидят на клетке-ножке; в них происходит мейоз, за кото-
рым следует митоз, так что образуется восемь яйцеклеток. При со-
зревании внешняя стенка оогония разрушается и яйцеклетки высво-
бождаются все имеете, пакетом. После выхода из концептакула оста-
вавшаяся вокруг яйцеклеток внутренняя стенка оогония также раз-
рушается, высвобождая яйцеклетки, хотя спермин способны прони-
кать в оогониальный мешок До этого. Прорастание зигот и физиоло-
гия раннего эмбрионального развития интенсивно изучались ( Evans
et al., 1982). Роды фукусовых различаются по числу яйцеклеток
на оогоний: у Aacophyllum их четыре, у Helvetia две, а у Суа-
toaeira и Sargasaum одна. Во всех случаях сначала образуется во-
семь ядер, но лишние удаляются или дегенерируют. У Xiphophora
В отечественной литературе концептакулы называют также
скафидиями. - Прим, перев.
252
Рио. 5.37. Жизненный цикл однодомного вида Fucus . А,. Зре-
лый спорофит. Б. У концов ветвей образуются рецептакулы.
В. Концептакулы внутри рецептакул содержат оогонии, анте-
ридии и стерильные многоклеточные волоски (парафизы).
£. Оогоний. Ц. Мейотическое, затем митотическое деления
внутри оогония с образованием яйцеклеток. Ё. Выход яйце-
клеток. Ж. Развитие антеридиальной ветви и антеридиев;
мейоз с последующим митозом внутри антеридия приводят к
образованию спермиев. £. Сингамия. Д. Зигота. К. Развитие
зародыша спорофита (по Scagel et al., 1982)
(Naylor, 1954) и Bifurcarionais (Jensen,1974) в развивающемся
оогонии предположительно на восьмиядерной стадии формируются
внутренние стенки, изолируя конечные продукты деления в четырех
двуядерных камерах, где к зрелости сохраняется только одно ядро.
Молодой оогоний может, таким образом, рассматриваться как одно-
гнездный спорангий, мейотически дающий четыре споры, каждая из
которых делится зндоспорически с образованием гаметофитов. Эти
наблюдения подтвердили теорию, впервые развитую Страсбургером
( Strasburger, 1906), который предположил, что у Fucales проис-
ходит смена поколений с крайне редуцированным гаметофитом.
Эта теория использовалась дая объяснения жизненного цикла
в родственном порядке Ascoseirales (Мое,Henry,1982). Свободно-
253
живущий таллом. Aacoaeira рассматривается как спорофит, на кото-
ром в концептакулах развиваются цепочки одногнездных спорангиев.
Каждая спора делится с образованием двуклеточного гаметофита;
одна его клетка дегенерирует, а другая дает единственную подвиж-
ную гамету. Сингамии не наблюдалось, и предполагается, что гаме-
ты развиваются партеногенетически; мейоза не отмечено.
Представители Basycladales - единственные Chlorophyceae,
для которых гаметический жизненный цикл продемонстрирован цито-
логически и генетически (рис. 5.38) ( Коор, 1979}. В вегетатив-
ном состоянии еифональный таллом Acetabularia содержит одно
крупное ядро, расположенное в одном из базальных ризоидов.
Рис. 5.38. Жизненный цикл Acеtabularia mediterranea:
I - прорастающая зигота; 2 - 5 - развитие спорофита; 6 -
мейотическое деление первичного ядра и миграция вторичных
ядер в шляпку; 7 - развитие цист в шляпке; 8 - выход га-
мет из цист; у - изогаметы; 10 - сингамия. М! - мейоз
(по Tanner, 1981; из Bonotto, 1975)
254
Гаметогенез начинается с мейотического деления этого ядра с после-
дующими митозами, что приводит х большому числу гаплоидных дочер-
них ядер, многие из которых мигрируют в "шляпку" и включаются
поодиночке в гаметангиальные цисты. Покоящиеся цисты высвобожда-
ются из шляпки, и митотическое Деление внутри них приводит к вы-
ходу в среду двужгутиковых изогамет. Зигота, образующаяся при
слиянии гамет, развивается непосредственно в новый таллом Aceta-
bularia (Bonotto, 1975). Ядра Цист содержат то же количество
ДНК, что и гаметы, но вдвое меньше, чем зигота, т.е. мейоз про-
исходит до образования гамет (Коор, 1979).
Считается, что, за исключением комплекса Derbesia-Bryopsis
у представителей Caulerpales гаметической жизненный цикл
(рис. 5.39), но цитологические доказательства этого, как прави-
ло, отсутствуют. У Codium дифференцированные гаметангии отделяют-
ся ст сифонального таллома септой ( Borden, Stein, 1969), а
У Udotea (Meinesz, I960) И Caulerpa (Goldstein, Morrall, 1970)
такой дифференцировки не происходит и гаметы формируются путем
деления соматической протоплазмы. Большинство видов дает анизо-
гаметы, хотя известны и изсгамные формы. У Caulerpa (Price,
1972) зигота развивается непосредственно в новый взрослый тал-
лом, но у Udoteaceae (например, Udotea и Halimeda) при прора-
стании возникает сферическая одноядерная структура, протосфера,
которая в свою очередь дает нитчатую стадию, а уже она - взрос-
лый таллом (Meinesz, 1980). Протосфера и нитчатая стадия могут
быть аналогичны одноядерным нитчатым стадиям в жизненных циклах
некоторых видов Bryopsis. способным формировать зооспоры или не-
посредственно взрослые талломы (Hoek, 1981). Такая интерпрета-
ция предполагает наличие спорического жизненного цикла с крайне
редуцированным спорофитом и соматическим мейозом. Этого не наблю-
далось у Udoteaceae (Meinesz, 1980'. Однако интересна гипотеза
о том (Rietema, 1971, 1972), что в северных широтах спорофит
Bryopsis утрачивает способность давать споры и происходит сома-
тический мейоз. В подтверждение этого Сирс и Уиле (Sears, Wilce,
1970) показали, что в спорангиях спорофитнсй Derbesia отсут-
ствует цитокинез и они развиваются непосредственно в гаметофит
Bryopsis.
255
5 мм
Рис. 5.39. Хизненный цикл Udotea petiolata А. Фертиль-
ный таллом. Б: I - мужская fAMASAt 2'жбйОКая гамета;
3 - сингамия; 4 - зигота через 10 дней. £. Протосфера.
£: I - апикальный рост протосферы; 2 - развитие ризовдов.
, Ветвящаяся стадия с ризоидами и вертикальными нитями,
которые агрегируют в зрелый сифональный таллом (помез-
nesz, 1980)
256
Соматические жизненные циклы
Жизненные циклы с мейозом, происходящим в вегетативной
клетке и непосредственно не ведущим к образованию спор или га-
мет, по-видимому, встречаются в нескольких группах водорослей,
но достоверно продемонстрированы только у Praslolales из СШо-
rophyceae И у некоторых багрянок.
У Prasiola meridionalis и Р.stipitata (рис. 5.40) клетки
верхней части пластины могут делиться мейотически, а затем ми-
тотически, давая мозаичные участки парных мужских и женских гаме-
тангиальных зон. Это считалось свидетельством генотипического
определения пола при мейозе (Friedmann, 1959; Cole, Akintobe,
1963). Однако у P.meridionalia показано ( Bravo, 1965), что лю-
бая клетка, возникающая при мейозе, способна дать как мужские,
так и женские гаметангии. Высвобождение гамет облегчается разру-
шением гаметангиальных стенок. Сингамия оогамная: двужгутиковый
спермий оплодотворяет яйцеклетку. Один жгутик утрачивается, а
другой сохраняется и перемещает зиготу перед ее оседанием и про-
растанием в новый таллом. К другим жизненным циклам, указывавшим-
ся для этого рода, относятся зиготический у пресноводной р.japo-
nica (Fujiyama, 1955)и гетероморфный спорический у морской Р.саг-
lophvlla (Kornmann, Sahling, 1974).
Рис. 5.40. Жизненный цикл Prasiola с соматическим мейо-
зом. Вегетативные зоны таллома моностроматические, споро-
генные - дистроматические, а гаметогенные - полистромати-
ческие. М! - мейоз (по Tanner, 1981; из Friedmann,1959)
257
У пресноводных красных водорослей жизненные циклы обычно ге-
тероморфные (Sheath , 1984). У Batrachospermales они включают
макроскопический гаметофит и редуцированную нитчатую диплоидную
фазу, называемую "шантранзия". Единственные образующиеся споры -
карпоспоры, даваемые карпоспорофитом в результате оплодотворения
карпогона спермацием. Тетраспоры неизвестны, и гаметофитное поко-
ление возникает непосредственно из шантранзии. У Lemanea mamil-
losa (РИС. 5.41) (Magne , 1967) и Batrachoapermwn mahabaleahwa-
renaia (Balakrlshnan, Chagule, 1980) мейоз происходит в апикаль-
ной клетке шантранзии, после чего лишние ядра выталкиваются в бо-
ковые выросты, в дальнейшем не развивающиеся. У некоторых видов,
10 мкм
Б
---------------1
Ю мкм
В
Рис. 5.4Т. Lemanea mamilloaa . А. Жизненный цикл: К - кар-
поспор офит; С - диплоидная "спорофитная часть таллома":
Г - гаплоидная гаметофитная часть таллома; М! - мейоз (по
Dixon, 1973). £. В результате мейотического деления воз-
никают гаплоидная апикальная клетка и латеральные остаточ-
ные ядра. В. Развитие гаплоидного таллома; стрелкой пока-
заны остаточные ядра (по Magne, 1967)
258
например Batrachospermum ар.(Stosch, Tiel, 1979) и Lemanea
fluviatilis (Huth, 1981), мейоз, по-видимому, задерживается и
идет не в шантранзии, а в апикальной клетке того, что морфологи-
чески является гаметофитной фазой.
•У других красных водорослей с редуцированными тетраспорофи-
тами тетраспорогенез, вероятно, может подавляться, что ведет к
непосредственному развитию гаметофита за счет соматического мей-
оза. Некоторые такие случаи рассмотрены Хоксом (Hawkea, 1983),
который описывает возможное-наличие этого феномена уHummbrella
hydra. представителя Gigartinales из Новой Зеландии. Одна-
ко окончательное цитологическое подтверждение жизненного цикла
типа lemanea отсутствует у всех таксонов, кроме Batrachosper-
males.
Глава 6
ФИЗИОЛОГИЯ И БИОХИМИЯ
Введение
Хотя многие физиологические и биохимические процессы, харак-
терные для высших растений, очень сходны с происходящими у водо-
рослей, между ними имеется и ряд заметных отличий. Группы водо-
рослей далеко отстоят друг ст друга в эволюционном плане, обычно
проще, чем высшие растения, устроены анатомически и в .большинст-
ве являются водными организмами. Эволюционное разнообразие отра-
жается на составе их фотосинтетических пигментов и запасных пита-
тельных веществ, тогда как структурные и экологические различия
можно видеть в механизмах поглощения и транспорта питательных ве-
ществ. Эта глава посвящена вопросам физиологии, которые либо уни-
кальны для тех или иных групп водорослей, либо особо важны для
их существования.
Свет и его измерение
Фотосинтез у водорослей протекает преимущественно в видимой
области спектра (400 - 700 нм), а фотсморфогенетические реакции
иногда расширяют область физиологического действия света в даль-
нюю красную область (от 700 до 760 нм). Спектральная чувствитель-
ность этих реакций сильно отличается от свойственной человеческо-
му глазу или фотографическим эмульсиям, поэтому единицы измере-
ния, принятые для регистрации освещенности- люксы или фут-свечи, -
неудобны для большинства биологических измерений. Альгологов в
основном интересуют плотность потока фотонов (1ШФ) и число кван-
тов (фотонов в световой области электромагнитного спектра). Эти
единицы приемлемы для характеристики большинства фотобиологичес-
ких явлений, которые зависят от числа поглощаемых квантов, а не
от их энергии. В системе СИ единица такого квантового излучения -
микромоль на квадратный метр в секунду (мкмоль/м2. с) ( Incoll
et al., 1977); часто также используется идентичная, но не входя-
щая в систему СИ единица микроэйнштейн на квадратный метр в се-
кунду (мкЭ/м .с). Излучение может также измеряться и выражаться
энергетически в ваттах на квадратный метр (Вт/м2). Для монохрома-
тического света излучение и плотность потока фотонов связаны
260
соотношением
I мкмоль/м2 . с = (119,7/Д )• Вт/м2,
где Я - длина волны света в нанометрах.
Для света с более широким спектральным диапазоном переход
от излучения к ППФ требует интегрирования значений ППФ, получен-
ных по всей интересующей области спектра. Это легко сделать, ис-
пользуя спектрорадиометр для измерения энергии узких спектраль-
ных полос с последующим переводом ее в ППФ, построением кривой
зависимости ППФ от длины волны и определением площади под кривой.
Хотя в целом невозможно непосредственно соотнести освещен-
ность в люксах с энергией света или плотностью потока фотонов,
уравнение
I Вт/м2 = 5 мкмоль/м2 . с = 250 лк
является хорошим приближением для большинства источников белого
света. Известны также переводные коэффициенты для большинства
обычных источников света (Luning, 1981); они применимы там,
где не требуется абсолютная точность.
Фотосинтетические пихменты и поглощение света
Фотосинтез у водорослей, как и у всех фотоавтотрофных орга-
низмов, зависит от поглощения световой энергии и преращения ее в
химическую путем образования АТР и восстановительного агента
nadp*h. Первая стадия этого процесса - взаимодействие света с
фотосинтетическими пипаентами. Распределение различных хлорофил-
лов, билипротеинов и каротиноидов по определенным таксономичес-
ким группам водорослей представляет интерес с эволюционной точ-
ки зрения и обсуждается в гл. 2 и 8.
Молекула хлорофилла (рис. 6.1,А) состоит из четырех связан-
ных между собой пиррольных колец с хелатированным атомом магния
в ее центре; in vivo пихмент связан с белками ( Thornber, Bar-
ber, 1979). Хлорофиллы а и Ъ. имеют также длинную фитольную
цепь, отсутствующую у хлорофилла с. Хлорофилл а есть у всех эука
риотных фотосинтезирующих водорослей и у прокариотных Cyanophy-
cota и Prochlorophycota. В спектре поглощения содержащих его
ацетоновых экстрактов два максимума примерно одинаковой интеноив
ности: полоса с°ре в синей области (430 нм) и другой пик - в
красной (660 нм).
261
(I) Ri = CH3
(II) R, = CH3
(I) Хлорофил a
R2 = CH3
R2 = CHO
(II) Хлорофилл b
(I) R = CH2CH3
(II) R = CH=CH2
(I) Хлорофилл c, (||) Хлорофилл c2
0-Каротин
Фикоцианобилин
Фи коэритробилин
Рис. 6.1. Химическая структура трех основных групп фото-
синтетических пигментов (по Ragan, 1981)
262
Зеленые водоросли содержат хлорофилл ъ_ , отличающийся от
хлорофилла а тем, что- метильная группа молекулы замещена альде-
гидной. В тех же условиях экстракции он показывает максимум по-
глощения в полосе Соре (435 нм), примерно втрое более интенсив-
ный, чем в красной области при 643 нм ( Prezlin, 1981). У Chromo-
phycota хлорофиллъ. отсутствует, но есть хлорофилл с, для кото-
рого известны две основные формы: хлорофилл Cj и хлорофилл £2;
как и хлорофилл ъ., они интенсивно поглощают синий свет.
Билипротеины (фикоэритрины и фикоцианины) содержат билино-
вые пигменты фикоэритробилин и фикоцианобилин (рис. 6.1,В), яв-
ляющиеся не замкнутыми, а линейными тетрапирролами и в отличие
от хлорофиллов прочно связанные ковалентной связью с белком.
Красные фикоэритрины и сине-зеленые фикоцианины существуют в раз-
ных формах, известных у синезеленых, красных водорослей и крипто-
фитов. Эти формы, различающиеся организацией полипептидных компо-
нентов, неодинаковы и по своим спектральным характеристикам
( Prezlin, 1981).
Каротиноиды (рис. 6.1,Б) - это изопреноидные полиеновые пиг-
менты, присутствующие у многих организмов, но образуемые de novo
только растениями и фотосинтезирующими бактериями. К ним относят-
ся каротины и их гидроксилиройанные производные ксантофиллы обыч-
но красного, бурого или желтого цвета, поглощающие свет в сине-
зеленой области спектра (обзор сведений о каротиноидах различных
групп водорослей см. Ragan, 1981). Роль многих встречающихся
в небольших количествах каротиноидов не светоулавливающая, а све-
тозащитная, поскольку они эффективно поглощают потенциально опас-
ное синее и ближнее ультрафиолетовое излучение ( Prezlin, 1981).
Многие зеленые водоросли по составу каротиноидов напоминают
высшие растения; здесь отмечаются J3-каротин и ксантофиллы люте-
ин, виолаксантин, антераксантин, зеаксантин и неоксантин. Боль-
шинство сифональных представителей этого отдела содержит dC-каро-
тин,сифоноксантин и его жирнокиолотный эфир сифонеин. В линиях
Chromophycota преобладают ксантофиллы; у бурых, диатомовых и зо-
лотистых водорослей среди них наиболее обилен фукоксантин, а дино-
флагелляты вместо него содержат перидинин. Эти два ксантофилла
сходны по структуре и отличаются от большинства других относи-
тельно высоким содержанием кислорода. Зеаксантин и лютеин - са-
мые обычные ксантофиллы багрянок, тогда как у синезеленых водо-
рослей присутствуют миксоксантин (эхиненон) и максоксантофилл.
263
Поглощение света
Фотосинтетическое превращение световой энергии в химическую
описывается так называемой Z-схемой ( Hill, Bendall, I960)
(рис. 6.2). Существуют две фотосистемы - ФС1 и ФСП - каждая со
своей энергетической ловушкой P?qq и PgQ8 соответственно. Эти
фотосистемы различаются набором пишентов, не всегда встречаются
в соотношении 1:1 и могут быть физически разделены путем обработ-
ки мягкими детергентами. Донор электронов для ФСП - вода, а ко-
нечный их акцептор в ФС1 - окисленный никотинамидадениндинуклео-
тидфосфат ( nadp).
Многие виды синезеленых водорослей способны к анокоигенному
фотосинтезу с использованием в качестве донора электронов сульфи-
да ( Padan, Cohen, 1982). Этот факультативный путь указывает на
их связи как со строго анаэробными фототрофными бактериями, так
и с облигатно аэробными эукариотными водорослями.
Комплекс Pr,QQ выделен у нескольких классов водорослей
(Phaeophyceae; Anderson, Barrett, 1979; Rhodophyceae; Gantt,1981;
Chlorophyceae; Nakamura et al., 1976; Dinophyoeae; Prezlin,
Alberts, 1978). Он включает 40 - 50 молекул светособираю-
щих единиц“хлорофилла - белок"на каждую молекулу реакционного
центра Р700’ К0Т°Рая сама является особой формой хлорофилла а.
Природа ловушки P68q в ФСП менее ясна; по-видимому, сюда входят
формы хлорофилла а, поглощающие свет меньшей, чем в случае P?qq,
длины волны, а также хлорофилл Ь и вспомогательные пигменты.
Процесс улавливания света описан по аналогии с системой
антенна - приемник - преобразователь (Ramus, 1981): сигналы
(фотоны) различного качества (длины волны) улавливаются антенной
приемника (пигментным комплексом) и направляются в ловушку, где
преобразуются (превращаются в химическую энергию). Когда пихмент-
ная молекула антенны поглощает фотон, ее электрон переходит на
более высокий энергетический уровень и молекула становится менее
стабильной. Без сопряжения с другой системой такой возбужденный
электрон вернулся бы в исходное состояние с высвобождением энер-
гии в виде флуоресценции. Поскольку энергия теряется на повыше-
ние энтропии системы, излучался бы свет большей, чем у поглощен-
ного, длины волны. Пигментные молекулы антенны расположены таким
образом, что энергия электрона, возвращающегося в исходное состо-
яние, не тратится на флуоресценцию, а передается другой пигмент-
ной молекуле и в конечном счете - реакционным центрам фотосисте-
264
Фотосистема 11
Рис. 6.2. Z-схема Хилла - Бендолла потока электронов в
световых реакциях фотосинтеза. Направленные вверх стрелки
означают фотохимические, или световые, реакции. Показано
только нециклическое фотофосфорилирование. Q и х -
первичные акцепторы электронов ФС1Г и ФС1, соответственно;
Цит (цитохром), ПХ (пластохинон), ПЦ (пластоцианин),
ФД ферредоксин) - переносчики электронов (по Ramus,
мы (рис. 6.3). Чем короче длина волны поглощаемого света, тем
больше участвует в этом процессе вспомогательных питментов. Так
у красных водорослей коротковолновый диапазон поглощается фико-
эритрином, а затем энергия передается фикоцианину, аллофикоциа-
нину и хлорофиллу а ( Ramus, 1981); такая цепочка соответствует
пространственному расположению пигментов внутри фикобилисомы
(рис. 6.4) ( Gantt, 1981).
265
Рис. 6.3. Гипотетическое расположение пихментов в свето-
улавливающих антеннах ФС1Г и ФИ (по Govindjee, Braun,
Спектры действия
Роль конкретных пипаентов в фотосинтезе продемонстрирована
с помощью спектров действия, когда регистрируется фотосинтетиче-
ская реакция на постоянную ППФ при различных длинах волн
(рис. 6.5). У Chlorophycota и Chromophycota спектры действия
и поглощения in vivo хорошо коррелируют между собой, что го-
ворит о важности как хлорой^ллов, так и вспомогательных пигмен-
тов для улавливания света. Однако у красных водорослей картина
существенно отличается: спектр поглощения здесь шире спектра
действия. Это предполагает, что основная роль в улавливании
света принадлежит билипротеинам, а хлорофиллы участвуют в этом
процессе в меньшей степени.
266
Рис. 6.4. Структура фикобилисом. А. Полудисковидный тип,
обнаруженный у синезеленых водорослей Rhodella и Pseu-
danabaena. £• Полусферический тип у оагряйййвогрЬугхД
сНшп черное пятно - белок, предположительно прикрепляю-
ЦЙификобилисому к тилакоиду (по Gannt, 1981)
Рис. 6.5. Спектры действия фотосинтеза (•) и спектры
поглощения (—). А. Хлоррфит tilva_ lactuca . Б. Хромофит
laminaria aaccharina \ бурая водоросль). В. Багрянка
Beieaaeria sanguinea (по Dring, 1982: неопубликован-
таа 'ДАЙЙые xidning)
267
Хроматическая адаптация
Широко известно, что в клетках водорослей, выращенных при
низкой освещенности, концентрации пигментов выше, чем в тех, ко-
торые развивались при ярком свете (Ramus, 1981; Yen-tach,
1980), т.е. поглощение доступного света в первом случае идет эф-
фективнее, хотя энергетический выход на квант остается без изме-
нений. У диатомей это наблюдается при различной освещенности в
синем конце спектра (Vesk, Jeffrey, 1977); соотношение различ-
ных пигментов в клетке остается постоянным, т.е. комплементарной
спектральной адаптации не происходит. Последняя обнаружена у си-
незеленых водорослей, способных к синтезу с-фикоэритрина, причем
отношение фикоцианин/фикоэритрин наибольшее при красном свете и
наименьшее при зеленом (Tandeau de Marsac, 1977). Аналогич-
ной пигментной адаптации у багрянок неизвестно ( Waaland et al.,
1974; Ramus et al. ,1976), но, возможно, она встречается у крип-
тофитов (Vesk, Jeffrey, 1977).
Комплементарная хроматическая адаптация и вертикальное
распределение водорослей
Зигельман (Engelmann, 1693) выяснил общий спектр действия
фотосинтеза у ряда водорослей, используя бактериальный аэротак-
сис как биотест на выделение кислорода. Он отметил важность зеле-
ного света для багрянок и предположил, что состав пигментов в
основных труппах морских водорослей является их адаптацией к
длинам волн света, преобладающего на тех глубинам, где они оби-
тают в океане. Поэтому Chlorophyceae наиболее обычны на мелко-
водье, а багрянки - на существенных сублиторальных глубинах. Эта
теория комплементарной хроматической адаптации используется в ли-
тературе для демонстрации приспособления растений к условиям
обитания, но сейчас создается впечатление, что она не подтверж-
дается сведениями по физиологии и распространению водорослей
( Ramus, 1981; Dring,I98I). На континентальном шельфе шт. Север-
ная и Южная Каролина ( Schneider, 1976) 25% зеленых водорос-
лей обитает на глубине более 50 м; на Гавайях ( Doty et al.,1974)
их видовое богатство также увеличивается с глубиной: на 90 м их
обилие выше, чем у багрянок, а зеленая водоросль Johnson-sea-
linkia profunda на ^7 м ( et ai,> 1985) ставит ре-
268
корд глубины для форм с мясистым талломом. Некоторые красные и
зеленые водоросли встречаются и глубже, но это уже корковые или
иные редуцированные формы, и их выживание обусловлено способно-
стью не только расти при низкой освещенности, но', по-видимому,
также противостоять выеданию. Опыты по пересадке на глубину и
затенению мелководий показывают, что любая известная фенотипиче-
ская адаптация обусловлена скорее общим снижением освещенности,
чем изменением спектрального состава света ( Ramus et al., 1976,
1977).
Фотосинтез и освещенность
Взаимосвязь между интенсивностью фотосинтеза (Р) и осве-
щенностью (I) описывается кривыми зависимости между этими вели-
чинами. При низкой освещенности интенсивность фотосинтеза лими-
тирована световыми реакциями и линейно растет с увеличением
освещенности (не завися от температуры), пока не достигнет пла-
то ( Ртах) при так называемом уровне светового насыщения. Даль-
нейшее небольшое увеличение освещенности не влияет на фотосин-
тез, хотя очень сильный свет действует на него подавляюще
(рис. 6.6). Максимальная интенсивность фотосинтеза при уровне
светового насыщения определяется скоростью фиксации углерода,
которая часто может быть увеличена путем добавления в среду пи-
тательных веществ или повышения температуры.
Изменение содержания пиплентов в ответ на рост освещеннос-
ти отражается на фотосинтетической активности. Возрастание их
концентрации в типичном случае приводит к увеличению начально-
го наклона кривой зависимости Р от I и к уменьшению ftnax-
Точка световой компенсации, когда фотосинтез и катаболические
процессы уравновешивают друг друга, также снижается.
•
Долгота дня и фотопериодические реакции
Фотопериодизм определяется как "контроль некоторых аспек-
тов жизненного цикла соотношением по времени света и темноты"
(Dring, 1984). Важно различать реакции на количество света и
на длительность его действия (рис. 6.7). Фотопериодические реак-
ции, по-видимому, особенно распространены у представителей
Chlorophyceae, Phaeophyceae и Rhodophyceae С гетероморфными
фазами в жизненном цикле (bring, 1984). Истинная фотопериоди-
269
Рис. 6.6. Кривая светового насыщения фотосинтеза: Р -фото-
синтез; Ршлт - максимальный фотосинтез;Pg - валовой фо-
тосинтез; Рп - чистый фотосинтез; R - дыхание; I - плот-
ность падающего потока фотонов; К - насыщающая плот-
ность потока фотонов (по Ramus, ±981)
Рис. 6.7. Развитие при разной долготе дня у двух бурых
водорослей при постоянных освещенности и температуре:
О - процент особей 3c:rt о siphon loment aria образующих
вертикальные^талломы; • - процент особей Sphacelaria
rigidula . образующих гаметангии (поDring—198Г)—
270
ческая реакция должна протекать по закону "всё или ничего", со-
гласно которому ночь, прерываемая вспышкой света, равнозначна
по действию долгому дню (Vince-Prue, 1975). Такие реакции из-
вестны для многих водорослей-макрофитов (Dring, 1984); напри-
мер, они участвуют в образовании конхоспор у багрянок Porphyra
(Dring, 1967; Rentschler, 1967) и Bangia (Richardson, 1970).
Известны также случаи влияния долготы дня, не удовлетворяющие
принципу "всё или ничего" или критерию прерывания темноты. Тет-
распорогенез у багрянки Acrosymphyton purpuriferum (Cortel-
Breeman, ten Hoopen, 1978), требующий малой долготы дня, не
подавляется прерыванием темноты. У багрянки Halymenia latifolia
наблюдается частичная фотопериодическая реакция: прерывание тем-
ноты на вид действует, но спорогенез при большой долготе дня
все-таки происходит, хотя и с пониженной интенсивностью ( Maggs,
Guiry, 1982).
Среди бурых водорослей фотопериодические реакции наиболее
детально изучены у Sc.vtosiphon lomentaria. При большой долготе
дня зооспоры этого вида образуют корковые талломы, но последую-
щее помещение их в условия короткого дня индуцирует развитие
вертикальных структур ( Dring, Luning, 1975). Прерывание темно-
ты меньше чем на одну минуту подавляет эту реакцию, причем эф-
фективно действует здесь только синий свет.
В отличие от высших растений физиологическим основам фото-
периодических реакций у водорослей уделялось мало внимания. По-
лагают, что все такие реакции у высших растений связаны с фито-
хромом. Красный свет (660 нм) наиболее эффективно стимулирует
опосредованный фитохромом ответ, который затем может быть инги-
бирован дальним красным светом (730 нм). Известно, что фитохром
содержат все основные группы зеленых растений ( Dring, 1984).
Связанные с ним движения хлоропластов подробно описаны у зеле-
ных водорослей Mougeotia и Mesotaenium ( Haupt,1983). Среди
багрянок типичные для фитохрома реакции на свет показывает
?orph.vra(Dring. 1967; Rentschler, 1967), а сходный с ним рецеп-
торный пигмент обнаружен в экстрактах из Acrochaetium daviesii
(van der Velde, Hemrika-Wagner, 1978). Синий свет вызывает
аналогичные реакции у некоторых красных и бурых водорослей, и
Дринг ( Dring, 1984) предполагает, что в этих случаях ответ на
долготу дня может обеспечиваться альтернативным пигментом -
"криптохромом".
271
Показана ( Abdel-Rahman, 1982) связь между общей продолжи-
тельностью цикла свет-темнота и временем темнового периода при
тетраспорогенезе у Acrcchaetium asparagopsis (рис. 6.8). В нор-
ме для этого требуется короткий день, но только при длине цикла
в целом 24 или 48 ч. Эти наблюдения свидетельствуют о связи цир-
кадианных ритмов с фотопериодизмом.
Свет другого спектрального состава может вызывать фотомор-
фогенетические реакции, не относящиеся к фотопериодизму. Рост
зародышей Fucus резко подавляется длинами волн 575 - 625 нм,
причем это влияние может быть аннулировано добавочным более длин-
новолновые или более коротковолновым освещением (McLachlan, Bid-
well, 1983). Реакции на синий свет у бурых водорослей вклю-
чают образование волосков и двумерных корковых талломов у 5су-
tosiphon lomentaria (Dring, Liining, 1975), а также индукцию фор-
мирования яйцеклеток у гаметофитов ламинариевых ( Liming, Dring,
1975).
Рис. 6.8. Влияние цикла свет-темнота на образование тет-
распорангиев у Acrochaetium asparagopsis» За фотопери-
одами в 6 ч (свёЖё" иЛй 8 ч (Затушеванные круж-
ки) следовали темновые периоды различной длительности.
Обратите внимание на пики фертильности при 24- или 44-
чаоовом циклах и почти полное ее подавление в остальных
случаях (по Adel-Rahman, 1982).
272
Метаболизм углерода
Фиксация углерода
Пути метаболизма, связанные с фотосинтетической фиксацией
углерода, широко изучались у одноклеточных зеленых водорослей.
Кратковременное предоставление chlorella и Scenedesmus мечен-
ной 14С двуокиси углерода показало, что фиксация идет путем кар-
боксилирования рибулозо-I,5-бисфосфата (РуБФ) с участием фермен-
та рибулозобисфосфаткарбоксилазы (РуБФК), а первый стабильный
продукт этих реакций - фосфоглицериновая кислота (ФГК). Много-
численные эксперименты подтвердили универсальность И важность
этого фотосинтетического, механизма карбоксилирования. Он отме-
чен у таких неродственных групп, как синезеленые (Pelroy, Bass-
ham, 1972), красные (Bean, Hassid, 1955) и бурые водоросли
(Bidwell et al.,. 1958).
Показано ( Hatch, Slack, 1966), что у сахарного тростника
имеется альтернативный путь фотосинтетического усвоения углеро-
да, включающий карбоксилирование фосфоенолпирувата (ФЕЛ) с обра-
зованием оксалоацетата. В этом потенциально участвуют два фермен-
та - фосфоенолпируваткарбоксилаза (ФЕПК) и фосфоенолпируваткар-
боксикиназа (ФЕПКК); первый использует бикарбонат-ион, а послед-
ний - С02- Ныне известно, что такой механизм широко распростра-
нен среди высших наземных растений. Однако он является вспомога-
тельным, облегчающим восстановительную ассимиляцию углерода, и
все растения,у которых он обнаружен, полностью зависят от РуБФК-
механизма. У водорослей показано наличие ряда -кислот, которые
быстро включают радиоактивный углерод в опытах по поглощению
^COg, что предполагает важную роль у них -метаболического пу-
ти (Joshi et al., 1974). Однако активность ФЕПК у chlorophy-
ceae, Phaeophyceae, Hhodophyceae (Kremer, Kuppera, 1977) и
Baoillariophyceae (Kremer, Berks, 1978) крайне низка. И напро-
тив, активность ФЕПКК, почти нулевая у Chlorophyceae, соответ-
ствует у багрянок 10%, а у некоторых бурых водорослей - 70% ак-
тивности РуБФК ( Kremer, Kuppers, 1977). Ее высокая активность
указывалась также для диатомей ( Kremer, Berks, 1978). Однако у
этих групп первый продукт кратковременной фотосинтетической фик-
сации. углерода - фосфоглицеральдегид (ФГАЛ), который, очевидно,
используется для образования ФЕП для дальнейшего кар боксилирова-
ния ( Kremer, 1981а,Ь ). Сведений о фотосинтетическом С^-пути,
273
сходном с известным у высших растений, нет. Активность ФЕПКК у
Laminarialee выше всего в меристематических зонах и, по-видимо-
му, важна для сохранения углерода, обеспечивая повторную.фикса-
цию С02, возникающей при метаболизме маннитола ( Weidner, Kup-
pers, 1982). Такое темновое усвоение С02 с участием ФЕПКК мо-
жет иметь большое экологическое значение.
Создается впечатление, что, хотя водоросли.биохимически
являются Cg-растениями, они физиологически напоминают С^-виды
низкими значениями компенсационных точек С02, отмеченными для
некоторых видов ( Birmingham, Colman, 1979; Coughlan, Tatters-
field, 1977). Эти наблюдения могут быть объяснены неким механиз-
мом концентрации С02 У водорослей (Raven, Beardall, 1981). Точ-
ное происхождение углерода, используемого при водорослевом фото-
синтезе, определить трудно, поскольку в отличие от наземных рас-
тений, которым доступна атмосферная С02, водные организмы имеют
в распоряжении также угольную кислоту (Н2С0д) и ее ионы - бикар-
бонат (НСОд“) и карбонат (СОд^-). Общие количества и соотношения
этих соединений в воде зависят от pH, сЛтености, парциального
давления С02 в атмосфере и температуры. В кислых водах доминиру-
ет свободная двуокись углерода, но ее содержание резко падает
при нейтральном значении pH, при pH 7 - 9 доминирует НСОд-, а
при более щелочной реакции становится ощутимым присутствие ионов
СОд^” (рис. 6.9). По-видимому, все водоросли способны поглощать
свободную С02 (Raven, 1974), используемую исключительно как
субстрат для РуБФК. Для рядайодорослей показана также способ-
ность усваивать НСОд" ( Beardall et al., 1976; Jolliffe, Tre-
gunna, 1970), возможно, в результате образования С02 за счет
дегидратационной активности карбоангидразы (Reed Graham,
1981). Бидуэлл и Мак-Лохлан (Bidwell, McLachlan, 1985) изучали
фотосинтез ряда морских водорослей, находящихся во влажном воз-
духе, а не в водной культуральной среде, и пришли к заключению,
что они обычно поглощают из морской воды бикарбонат, а не С02.
Фотосинтетическое поглощение углерода - преобладающий меха-
низм углеродного питания, однако известна и гетеротрофная фикса-
ция С02,и некоторые водоросли способны использовать органический
углерод путем зависящего от фотосинтеза его усвоения или гетеро-
трофно (Droop, 1974а). Виды, лишенные фотосинтетических пигмен-
тов, известны в большинстве классов водорослей; они по необходи-
мости должны быть гетеротрофами; другие виды, сохраняя фотосин-
тетический аппарат, могут утилизировать простые органические суб-
274
Рис. 6.9. Связь между pH и долями различных форм неоргани-
ческого углерода в пресной воде (по Goiterman et al., 1978)
страты (например, ацетат) с участием АТР и восстановителя, полу-
чаемых при фотосинтезе. Экологические аспекты гетеротрофности
водорослей обсуждаются в гл. 7.
Темновое дыхание
Основные особенности темнового дыхания водорослей в целом
идентичны таковым у других аэробных растений ( Lloyd, 1974;
Raven, Beardall, 1981). Гликолиз, или путь ЭМП (Эмдена - Мейер-
гофа - Парнаса), осуществляется параллельно гексозомонофосфатно-
му шунту (1МФ), а цикл трикарбоновых кислот (ЦТК), или цикл
Кребса, следует за этими процессами. Эта система дает клетке ,
АТР, но еще большие его количества синтезируются в аэробных ус-
ловиях в ходе связанного с мембранами окислительного фосфорили-
рования при использовании восстановителя, получаемого путем воз-
действия дегидрогеназ на органические субстраты. Восстановитель,
преимущественно NADP-н, образуется также в 1МФ.
Локализуются эти процессы в клетке в основном так же, как
у высших растений. Ферменты гликолиза и 1МФ находятся в цитозо-
ле, но встречаются и в хлоропластах, содержащих крахмал (Raven,
275
Beardall, 1981). Ферменты ЦТК локализованы в митохондриальном
матриксе, тогда как процессы окислительного фосфорилирования свя-
заны с внутренней митохондриальной мембраной. Отличия от этой об-
щей схемы обнаружены у синезеленых водорослей, где ферменты гли-
колиза, ГМФ и ЦТК, находятся в цитозоле, а окислительное фосфори-
лирование связано с мембранами.
Значительная часть АТР и nadp»h, продуцируемых при фотосин-
тезе, идет на восстановление С02 до углевода, но эти соединения
могут использоваться и в других клеточных процессах. Определение
роли темнового дыхания в энергетике фотосинтезирующей клетки -
задача нелегкая. Для сравнительных целей дыхательная способность
водорослевой клетки может быть выражена в виде отношения макси-
мальной продукции двуокиси углерода в темноте (ur тах) к макси-
мальной скорости роста ( ug). При фототрофном росте это отноше-
ние доходит до 0,3 - 0,5, тогда как при гетеротрофном оно состав-
ляет 0,5 - 1,2. Это значит, что дыхательная способность при данной
максимальной скорости роста у фототрофов ниже, чем у гетеротро-
фов, т.е. существенная доля АТР и восстановителя образуется у
них непосредственно при световых реакциях фотосинтеза. Как пока-
зывают данные по Anaoystis nidulans (Doolittle, Singer, 1974),
это отношение у синезеленых водорослей может быть меньше 0,5,
что позволяет предполагать почти полную зависимость от фотосин-
тетически продуцируемых АТР и восстановителя. И напротив, дино-
флагеллаты, по-видимому, единственная группа водорослей с интен-
сивным темновым дыханием ( ц„ = 1,1; Prezlin, Sweeney,
1978). ,
Фотодыхание, гликолатный метаболизм и
потеря органического
углерода
Выполняя функцию карбоксилирования, РуБФК является также ок-
сидазой, катализирующей окисление РуБФ до фосфоглицерата и фос-
фогликолата ( Raven, Beardall, 1981). У сосудистых растений,
бриофитов и харовых гликолатоксидаза катализирует дальнейшее
окисление гликолата в глиоксилат с участием кислорода. У большин-
ства водорослей это окисление обеспечивается другим ферментом -
гликолатдегидрогеназой ( Raven, Beardall, 1981). Две молекулы
глиоксилата могут в конечном счете преврапдться в одну молекулу
глицерата с потерей двуо иси углерода. Это светозависимое погло-
276
щение кислорода с образованием COg У высших растений называют фо-
тодыханием; на него иногда расходуется свыше 50% фиксированной
при фотосинтезе двуокиси углерода без чистого прироста АТР (Bid-
well, 1983). Хотя четко установлено, что у водорослей есть гли-
колатный путь, относительно его роли в углеродном обмене их попу-
ляций в природе остаются значительные противоречия. Цри изучении
широкого круга водорослей (Lloyd et al., 1977) установить фосфо-
респираторный газообмен не удалось. Однако гликолат является вне-
клеточным продуктом многих водорослей ( Raven, Beardall, 1981;
Bidwell, 1983), причем его выделяемые количества варьируют от
крайне малых до соответствующих 70% фиксированного углерода. В
лаборатории максимальная экскреция гликолата наблюдается, если
выращивать водоросли при высоких концентрациях углекислоты (бо-
лее 0,2%), а затем переносить их в условия низкого ее содержания
(Ingle, Colman, 1976). Предполагается, что относительно большие
количества РуБФ, образующиеся при высоком содержании COg, впо-
следствии окисляются до гликолата. При более близких к естествен-
ным условиях выделение гликолата иногда стимулируется ограничен-
ной доступностью питательных веществ, и не исключено (Harris,
1980), что посредством этого механизма водорослевая клетка избав-
ляется от избытка углерода, когда скорость его поглощения времен-
но превышает скорость, с которой'он может включаться в обмен ве-
ществ или запасаться.
Водоросли способны выделять и ряд других органических ве-
ществ, в том числе углеводы, аминокислоты, липиды, фенолы и фер-
менты (Hellebust, 1974; Kremer, 1981а). Известно, что более
крупные представители бурых водорослей экскретируют большие коли-
чества фенолов (Kremer, 1981а), которыми, как считается, обус-
ловлено наличие желтых гумусовых веществ в прибрежных морских во-
дах. Выделение гликолата наблюдается у некоторых морских водорос-
лей-макрофитов, но оно незначительно по сравнению с происходящим
у фитопланктона. Растворенные углеводы экскретируются многими
морскими водорослями, но данные о деталях этого процесса и выде-
ляемых количествах противоречивы (Kremer, 1981). У Aacophyllum
nodosum такое выделение очень незначительно ( Moebus, et al.,
1974), тогда как у laminaria в виде углеводов и фенолов теряет-
ся 30 - 35% фиксированного углерода ( Johnston et al., 1977;
Hatcher et al., 1977).
277
Накопление и запасание продуктов фотосинтеза
Идентичный в целом механизм фиксации углерода во всех груп-
пах водорослей не отражается на составе их запасных веществ. За-
пасаемые продукты фотосинтеза у эукариотных водорослей - углево-
ды (Craigie, 1974; Kremer, 1981а); хотя более ранняя литерату-
ра содержит указания на запасание жиров и масел, основная роль
водорослевых липидов - вхождение в состав мембран (Wood, 1974).
Кроме запасных полисахаридов водоросли образуют различные саха-
ра и сахароспирты ( Craigie, 1974; Kremer, 1981а). Один из по-
следних, многоатомный спирт маннитол, может составлять до 20 -
30% сухой массы некоторых бурых водорослей. Эти низкомолекуляр-
ные соединения играют осморегуляторцую роль ( Kauss, 1978; Reed
et а2«, 1985) и используются непосредственно в обмене веществ.
Истинные запасные углеводы ( Craigie, 1974; McCandles, 1981) -
более крупные, относительно нерастворимые молекулы, являющиеся
преимущественно полимерами глюкозы. Chlorophyceae образуют ти-
пичные для высших растений крахмалы, состоящие из двух полиглю-
канов: линейных цепочек амилозы с <Z-1,4-связями и разветвлен-
ных молекул амилопектина с оС-1,4- и аС-1,6-связями; они да-
ют характерное сине-черное окрашивание при реакции с иодом.
Крахмалы багрянок и криптофитов примерно такие же, но содержат
больше амилопектина; с иодом они дают красное окрашивание. Крах-
мал синезеленых водорослей по размеру молекул и продуктам гидро-
лиза напоминает скорее гликоген, чем крахмал высших растений.
Ламинарии бурых водорослей - линейный полиглюкан из 16 - 32 мо-
номеров с Ji-1,3-связями, который может иметь на концах остат-
ки маннитола и несколько 1,6-ответвлений. Хризоламинарин золоти-
стых водорослей сходен с ним по структуре, но не содержит манни-
тола, а у парамилона эвгленовых, указываемого также у примнези-
офитов ( Kreger, van der Veen, 1971), более крупная молекула.
У многих водорослей, особенно у одноклеточных, перенос про-
дуктов фотосинтеза по телу не составляет проблемы, поскольку
большинство их используется in situ. Вместе с тем этот перенос
иногда осуществляется между симбиотическими партнерами - водо-
рослями и гетеротрофами (гл. 7). Внутри водорослевого таллома
он изучался главным образом у Phaeophyceae, в частности у
Scales и Laminariales (рис. 6.10) ( Buggeln, 1983). Тран-
спорт ассимилятов практически у всех представителей Laminari-
ales идет по ситовидным трубкам, причем у Macrocystis послед-
278
Высвобождение НСО3
д МОРСКАЯ ВОДА
----------------f МЕРИСТОДЕРМА
I
Полифенолы « I - Фотоассимипяты Запас (ламинарии)
(физоды) (остаточный пул)
__________► Альгинат
___________► Белок, липид и т. д.
Перенос ____________
---------------------------Г КОРАи СЕРДЦЕВИНА
Выпадение на дно
Остаточный пул
---► Альгинат, каллоза
Ламинарии
’«--► Белок, липид и т. д.
Рис. 6.10. Предполагаемые пути преобразования продуктов
фотосинтеза в тканях пластины ламинариевой водоросли (по
Buggeln, 1983)
ние сходны с известными для высших растений. У laminaria они бо-
лее заполнены цитоплазматическими органеллами, а поры в ситовид-
ных полях мельче и многочисленнее. У Saocorhiza ситовидные труб-
ки отсутствуют, а их транспортная функция выполняется соленоци-
тами (рис. 6.II) ( Emerson et al., 1982). Последние представ-
ляют собой удлиненные клетки, находящиеся в сердцевине и соеди-
ненные поперечными перемычками с другими клетками, аллелоцитами.
Результаты наблюдений за передвижением фотоассимилятов у Lamina-
riales совпадают с гипотезой массового потока ( Munch),хотя
подтверждается это только косвенно ( Buggeln, 1983). Показано
( Buggeln et al., 1985), что скорость движения продуктов фото-
синтеза неодинакова, массовому потоку предшествует и на него
накладывается пульсация (рис. 6.12). Известно, что сходные явле-
ния встречаются у высших растений и, возможно, указывают на ме-
ханизм транспорта с затратой энергии.
279
Минуты
6. II. Импульсы меченного 11С продукта фотосинтеза
Рис
проходящего детекторы, расположенные последовательно с
интервалом в I см на пластине Macrocystls pyrifera.
Предположительно связанные между собой пики радиоактив-
ности показаны пронумерованными стрелками. Радиоактивная
метка удалена в Т = 0 (по Buggeln et al., 1985)
Неорганические питательные вещества (биогены)
К Всем водорослям требуются углерод, водород, кислород, фос-
» фор, азот,магний, железо, медь, маганец, цинк, молибден, сера,
калий и кальций, а некоторым также натрий, иод, бром, бор, вана-
дий, хлор и кремний (табл. 6.1) (О1 Kelly, 1974). Неорганические
биогены типа азота и фосфора, которых нужно сравнительно много,
называются макроэлементами. Другие, например медь и молибден,
требуются обычно в относительно малых количествах и называются .
микроэлементами, они
мах или используются
играют роль кофакторов в ферментных систе-
в электронотранспортных цепях.
Азот
После углерода
кислорода и водорода азот - наиболее обиль-
. ный элемент у водорослей без минерализованных стенок. Если его
поступление неограниченно, он может составлять 10%их сухой мас-
сы ( Syrett, 1981). Азот доступен водорослям в трех основных
формах: как газообразный Ng или в виде неорганических и органи-
ческих соединений.
280
Фиксация азота
Рис. 6.12. Радиоавтография
среза через сердцевину плас-
тины Saccorhiza dermatodea,
в которую введена метка ^С-
бикарбоната, сконцентрировав-
шаяся в соленоцитах; вероятно,
их функция заключается в пере-
носе веществ на большие рас-
стояния (любезно предоставле-
но Carolyn Emerson)
кие билипротеины (Wolk, 1982).
Способность восстанавли-
вать, или фиксировать, газооб-
разный азот обнаружена только у
прокариот, а среди водорослей -
исключительно у Cyanophyceae.
Для этого требуется нитрогеназа -
чувствительный к кислороду фер-
ментный комплекс, содержащий же-
лезо, серу и молибден (Bothe,
1982), который способен восста-
навливать другие субстраты с
тройными ковалентными связями
(окись азота, цианиды, изоциани-
ды, циклопропен и ацетилен).
Восстановление ацетилена до эти-
лена и простое разделение суб-
страта и продукта с помощью га-
зовой хроматографии применяются
как стандартная проба на фикса-
цию азота.
Большинство азотфиксирую-
щих синезеленых водорослей пред-
ставлено нитями, в состав кото-
рых входят крупные бледные тол-
стостенные клетки, называемые
гетероцистами (рис. 6.13). Они
образуются в отсутствие доступ-
ного связанного азота, лишены
продуцирующей кислород фотосис-
темы К, РуБФК, не содержат во-
все или содержат лишь в неболь-
ших количествах фотосинтетичес-
В стенках гетероцист находятся
связывающие кислород гликолипиды ( lambein, Wolk, 1973), которые
Q Q м г< М
наряду с респираторным потреблением кислорода поддерживают ана-
эробные условия, необходимые для фиксации азота ( Bothe, 1982).
281
ГЕТЕРОЦИСТА
ВЕГЕТАТИВНАЯ КЛЕТКА
Рис. 6.13. Схема потока углерода и азота между гетероцис-
тами и вегетативными клетками азотфиксирующих синезеленых
водорослей: ГС - глутаминсинтетаза; РОГАТ - глутаминоксо-
глутаратаминотрансфераза; Глу - глутаминовая кислота;
Глн - глутамин. Слоистая стенка гетероцисты непроницаема
для азота, COg и кислорода (по Haselkorn, 1978)
Для нитрогеназной активности необходимы АТР и восстановлен-
ный ферредоксин (Bothe, 1982). АТР и часть восстановленного фер-
редоксина образуются в гетероцистах путем циклического фотофос-
форилирования фотосистемы I. Однако большая часть ферредоксина,
по-видимому, восстанавливается с участием NADP«H, получен-
ного в основном за счет гексозомонофосфатногр шунта с использо-
ванием углеводов, поступающих из соседних клеток. Азот восста-
навливается до кн4+ , который перед переносом из гетероцист в
другие клетки нити превращается в глутамин.
Фиксация азота отмечена и у не имеющих гетероцист планктон-
ных колониальных видов Osoillatoria (Trichodesmium)«OHa проте-
кает здесь в клетках, расположенных в центре колонии, которые
не образуют фотосинтетически 0g и лишены в отличие от гетеро-
цист специализированных утолщенных клеточных стенок ( Carpenter,
Price, 1976). Азотфиксация морской планктонной водоросли Qsci-
llatoria erythraea связана со способностью нитей группи-
роваться в пучки ( Bryoeson, Рау, 1981).
282
Таблица 6.I-. Роль неорганических биогенов в метаболизме водо-
рослей(по Deboer, 1981)
Элемент Возможные функции Примеры соединений
Азот В соединениях имеет основ- ное значение для метабо- лизма Аминокислоты, пурины, пи- римидины, порфирины, ами- носахара, амины
Фосфор Структурная функция, пере- нос энергии АТР, GTP, нуклеиновые кислоты, фосфолипиды, ко- ферменты, кофермент А, фо сфо енолпиру ват
Калий Осмотическая регуляция, контроль pH, конформация и стабильность белков Возможно, встречается пре- имущественно в ионной форме
Кальций Структурная функция, ак- тивация ферментов, ионный транспорт Пектат кальция, карбонат кальция
Магний Фотосинтетические пигмен- ты, активация ферментов, ионный транспорт, стабиль- ность рибосом Хлорофилл
Сера Активные группы в фермен- тах и коферментах, струк- турная функция Метионин, цистин, цисте- ин, глутатион, агар, кар- рагинан, сульфолипиды, кофермент А
Железо Активные группы порфири- новых молекул и (ферментов Ферредоксин, цитохромы, нитратредуктаза, нитрит- редуктаза, ферритин, ка- талаза
Марганец Электронный транспорт в фо- тосистеме Н, поддержание структуры мембраны хлоро- пласта Манганин
Медь Электронный транспорт (фо- тосинтез), ферменты Пластоцианин, аминокси- даза
Цинк Ферменты, метаболизм аукси- нов (?), структура рибо- сом (?) Карбоангидраза
Молибден Восстановление нитратов, абсорбция ионов Нитратредуктаза
Натрий Активация ферментов, вод- ный баланс, ферменты Нитратредуктаза
Хлор Фотосистема Н Терпены
Бор Регуляция потребления угле- рода (?), метаболизм РНК(?)
Кобальт Компонент витаминов Вто, С4> путь фотосинтеза Витамин Bj2
283
Продолжение табл. 6.1.
Эдемент Возможные функции Примеры соединений
Бром^ Иод1^ ? Широкий набор галогени- рованных соединений у ? красных водорослей
‘''Возможно, незаменимый элемент для некоторых морских водо-
рослей-макрофитов .
284
Неорганический связанный азот
Почти все содержащие хлорофилл водоросли мохут расти на
ионах нитрата ( NOj), нитрита ( ^°2 ) или аммония ( NH4+), причем
в большинстве случаев их развитие на этих источниках азота не
сильно различается. У клеток, не испытывающих недостатка азота,
при добавлении в среду №4* поглощение uoj прекращается, хо-
тя при сильном азотном дефиците происходит одновременное поглоще-
ние этих ионов ( Syrett, 1981).
Каков бы ни был источник неорганического азота, в целом при-
знано, для включения в клеточные органические соединения требует-
ся его превращение в NH4+ (Syrett, 1981). Оно представляет со-
бой двустадийный процесс:
- Нитратредуктаза Нитритредуктаза
Н°3 ------2?=--------* К02 ------6Г-------~№4 *
У эукариотных водорослей нитратредуктазный комплекс сходен
с таковым у высших растений и состоит из молибденового, флавино-
вого и цитохромного компонентов; восстановителем служит NAD»H
(иногда HADP«H). Прокариотная нитратредуктаза проще; она также
содержит молибден, но использует как источник электронов восста-
новленный ферредоксин. Второе восстановление до аммония катализи-
руется феррелоксиннитритредуктазой; ни NAD.H , ни МАЗР.Н как
доноры электронов в процессе не участвуют ( Syrett, 1981).
Наиболее распространенный путь включения иона аммония в кле-
точные органические соединения (Syrett, 1981; De Boer, 1981)
предполагает катализируемую глутаминсинтетазой реакцию NH*
с глутаматом и образование глутамина. Одна аминогруппа затем
переносится на оксиглутарат, и возникают две молекулы глутамата.
Одна из них опять включается в цикл, а другая может использовать-
ся для образования аминокислот за счет реакций трансаминирова-
ния (рис. 6.14 и 6.15).
В целом процесс поглощения и превращения нитрата в амино-
кислоты требует 10 электронов на молекулу, а восстановление COg
до углевода - только четырех. Поэтому неудивительны многократные
сообщения о том, что ассимиляция и кнд+ , и но^~ у нормально
растущих водорослей зависит от фотосинтеза и не происходит или
идет очень слабо в темноте ( Syrett, 1981). Однако это не обяза-
285
COOH CONH2
сн2 сн2
сн2 СН2 .
I < I
CHNHj со
I I
соон соон
Глутаминовая / -оксо глутаровая
кислота кислота
Рис. 6.14. Пути включения аммония в органические соедине-
ния с использованием РОГАТ (глутаминоксоглутаратамино-;
трансферазы) (по Syrett, 1981)
Рис, 6.15. Основные особенности ассимиляции и распределе-,
ния азота в водорослевой клетке (по Syrett, 1981)
286
тельно, и при дефиците азота клетки Chlamydomonas с запасом уг-
леводов ассимилируют какмо~, так и 1Ш^+ даже в темноте,
хотя на свету делают это с еще большей скоростью ( Thacker,
Syrett, 1972).
Органические источники азота
Многие водоросли способны использовать органические соеди-
нения (особенно аминокислоты, мочевину и пурины) как единствен-
ный источник азота ( Neilson, Larsson, 1980). Из 26 исследован-
ных морских планктонных видов ( Antia et al., 1975) 23 могли ус-
ваивать мочевину, а 17 - пурин гипоксантин. Другие водоросли ис-
пользуют аминокислоты, в частности глицин, серин, аланин, глута-
миновую и аспарагиновую кислоты ( Wheeler et al., 1974).
Запасание азота
Ассимилированный азот может сохраняться в виде ионов нитра-
та и аммония или низкомолекулярных органических соединений, а
затем использоваться для будущего роста. Ряд морских водорослей,
В том числе Ulva lactuca (Chlorophyceae), багрянка Gracilaria
folicera (Rosenberg, Ramus,1982) И бурые ВОЛОрОСЛИ Alaria escu-
lent a (Buggeln, 1978) И Laminaria longicruris (Chapman, Crai-
gie, 1977), запасают азот, что позволяет им продолжать рост
при ограниченных концентрациях этого биогена в океане (гл. 7).
Синезеленые водоросли, по-видимому, уникальны в своей способнос-
ти накапливать азот в виде структурированных гранул цианофицина
( Lang et al., 1972) - полипептида из аргинина и аспарагиновой
кислоты. Синтезируется он необычно и, по-видимому, не на рибосо-
мах (Simon, 1973).
Фосфор
Фосфор нужен клеткам для образования нуклеиновых кислот,
фосфолипидов и различных сложных эфиров фосфорной кислоты, напри-
мер фосфорилированных сахаров, АТР и NADP.
Ортофосфаты - единственный существенный источник неорганиче-
ского фосфора для водорослей, хотя, по-видимому, у них широко
287
распространена способность усваивать полифосфаты и различные ор-
ганические фосфаты (Kuenzler, 1965). Последние гидролизуются
фосфатазами, локализованными на клеточной мембране. Щелочная фос-
фатаза индуцируется дефицитом фосфора (Healey, 1973) и может
высвобождаться из клеток в окружающую среду (Reichardt et al.,
1967; berry, 1972). Способность использовать органические фос-
фаты выгодна в водных местообитаниях, где такие сложные соедине-
ния присутствуют в более высоких концентрациях, чем свободные
фосфаты.
Большинство водорослей запасает избыток фосфата в виде по-
лифосфатных гранул диаметром 30 - 500 нм (Harold, 1966), хотя
некоторые виды моцут накапливать фосфат-ион в вакуолях (Healey,
1973). Поглощение и запасание*питательного вещества сверх непо-
средственных метаболических нужд, возможно, обеспечивают водо-
росли биогеном на тот период, когда его внешняя концентрация
становится недостаточной. Предполагается, однако, что запасная
функция полифосфатов только побочная, а основная их роль заклю-
чается в регуляции уровня свободных фосфат-ионов в клетке
(Kulaev, 1975).
Отношения азот:фосфор
У фитопланктонных клеток, растущих в условиях неограничен-
ной доступности питательных веществ, атомное отношение С:К:Р
обычно равно 106:16:1; оно называется отношением Редфилда
(Redfield et al.,1963). Анализ, при котором клетки отделялись от
сопутствующего детритного материала, показал, что отношение N=P
для "нормального фитопланктона" находится в пределах от 5:1 до
15:1 ( Ryther, Dunstan, 1971). Однако изучение Scenedesmus
в хемостате (Rhee, 1978) позволяет предполагать, что клетки ис-
пытывают недостаток азота вплоть до отношения 1Т:Р около 30 и
недостаток фосфора, если этот уровень выше (рис. 6.16). Бентос-
ные морские водоросли больше, чем фитопланктон, ограничены по
фосфору и меньше по азоту относительно углерода; у них соотноше-
ние С:И:Р равно в среднем 550:30:1 (Atkinson, Smith, 1983).
Значительные отклонения от этих цифр указывают на дефицит пита-
тельных веществ, причем общей реакцией является снижение доли
ЛИИ1 тирующего биогена в клетке. Влияние недостатка фосфора и
азота подробно изучалось у морской .диатомеи Thalaasiosira
288
Рис. 6.16. Реакция Scenedesmug ня
отношение N :Р в хемостате. Кон-
центрация фосфора поддержива-
лась постоянной, а концентоация
азота менялась. А. Кривая рос-
та. Б. Концентрации азота
(светлые кружки) и фосфора (за-
тушеванные кружки) в клетке
(по Rhee, 1978)
•paeudonana (Eppley, Render,
1974; Perry, 1976; McCarthy,
Goldman, "1979). Дефицит азо-
та вызывает снижение скорости
роста (выражаемой временем ге-
нерации) и интенсивности фото-
синтеза. Содержание азота,
фосфора и хлорофилла а в клет-
ке снижается быстрее, чем со-
держание углерода, что повыша-
ет его долю относительно дру-
гих клеточных компонентов. Не-
достаток фосфора также увели-
чивает время генерации, но
при падении доли фосфора азот
остается на прежнем уровне, а
доля углерода в клетке возрас-
тает. ;• •
Кальций
Обызвествление у водорос-
лей предполагает отложение
СаСО3 вокруг или внутри кле-
ток. Оно встречается у Суапо-
phyceae, Rhodophyceae, Dino-
phyceae, Prymnesiophyceae,
Phaeophyceae, Chlorophyceae
И Charophyceae (Borowitzka,
1982). Фотосинтетическое no-
личению pH и содержания ионов C0|
Са^+, образуя осаждающийся СаСО,. У
требление СО 2 ПРИВ°ДИТ к Уве-
личению pH и содержания ионов COj ; последние соединяются с
Са^+, образуя осаждающийся СаСОд. У водорослей встречаются две
кристаллические формы СаСОд - кальцит и арагонит, хотя каждое
семейство известковых водорослей содержит только одну из них.
Арагонит чаще всего обнаруживается у морских форм, а кальцит - у
пресноводных видов. Исключение составляют морские кораллиновые
багрянки, образующие кальцит.
Процесс обызвествления лучше всего прослежен у тропической
зеленой водоросли Halimeda, откладывающей игловидные кристаллы
289
арагонита вне клеточных стенок в межклеточном пространстве, отде-
ленном от внешней морской воды слоем плотно спрессованных утри-
кул (рис. 6.17). Такая система позволяет быстро использовать С©2
и HcoJ , высвобождать ОН- и увеличивать pH, что приводит к от-
ложению СаСОд. Эту модель можно применить и к другим откладываю-
щим арагонит морским водорослям, у которых плотно переплетенные
нити создают длинный диффузионный путь от внешней среды до места
осаждения карбоната (Borowitzka, 1982).
Представители Corallinaceae (багрянки) откладывают каль-
цит между фибриллами клеточной стенки ( Oiraud, Cabioch, 1979).
И здесь этот процесс связан с фотосинтезом ( Borowitzka, 1982).
Механизмы отложения выяснены недостаточно хорошо, а предложенная
электрохимическая модель, не основанная на использовании COg
(Digby, 1979), оспаривается ( Borowitzka, 1982
Рис. 6.17. Схема ионных потоков углерода, ведущих к отло-
жению карбоната кальция в стенках известковой зеленой во-
доросли Halimeda (по Borowitzka, 1982)
I9 121
290
Представители примнезиофитов из группы кокколитофорид -
единственные водоросли с внутриклеточным обызвествлением
(рис. 6.18). У большинства видов отложение кальцита протекает на
органической основе внутри цистерн аппарата Гольда (Klaveness,
Paasche, 1979) с формированием тонко скульптурированных извест-
ковых пластин, или кокколитов; Последние содержат полисахариды и
белковый материал, способный связывать ионы Са^+ и предположи-
тельно действующий как центр осаждения (Borowitzka, 1982). Про-
цесс идет на свету, и у Emiliania huxleyj со2 используется
при фотосинтезе, тогда как НС03“ дает углерод для обызвествления
(Sikes et al., 1980). В условиях дефицита азота этот вид откла-
дывает арагонит, а не кальцит (Wibur, Watanabe, 1963); это дает
основание полагать, что именно белковый компонент определяет от-
кладываемую кристаллическую форму карбоната.
Важно понять, почему одни водоросли остаются необызвествлен-
ными в тех условиях, когда другие обызвествляются. Согласно од-
ной из теорий, они образуют на поверхности клеток определенные
Рис. 6.18. Схема ионных потоков углерода, ведущих к обыз-
вествлению и образованию кокколитов у Emiliania huxleyi.
Карбонат кальция откладывается внутри клетки"' 'в цибт'ёр-
нах аппарата Гольда (по Borowitzka, 1982)
291
вещества, например полифенолы, которые подавляют образование кри-
сталлов СаС03 ( Reynolds, 1978),
•
Кремний
Диатомеи со своими кремнеземными панцирями нуждаются в крем-
нии в виде ортокремниевой кислоты. Этот элемент обнаружен также
у бурых водорослей и Chlorophyceae (Parker, 1969), золотистых
водорослей, образующих окремнелые чешуйки или цисты ( Paasche,
1980), и празинофита Plat.ymonas (Fuhrman et al., 1978). Крем-
ний транспортируется в клетку с участием фермента-переносчика,
локализованного на клеточной мембране ( Sullivan, 1976, 1977).
Его поглощение требует потребления энергии, которая по-видимому,
обеспечивается процессом дыхания, поскольку скорость поглощения
на свету заметно не увеличивается (Nelson et al., 1976).
Опыты с синхронизированными культурами диатомей показали,
что поглощение кремниевой кислоты происходит только во время ци-
токинеза И образования новой стенки ( Darley et al., 1976; Chi-
sholm et al., 1978). Некоторые из крупных морских планктон- ,
ных диатомей могут, вероятно, непрерывно поглощать и запасать
кремний в своих вакуолях ( Chisholm et al.,1978). Длительный
дефицит кремния приводит к накоплению дочерних клеток, которые
остаются соединенными, поскольку не мотут образовывать стенки
(Coombs et al., 1978).
Диатомеи часто бывают нежелательными примесями в культурах
водорослей-макрофитов, и их абсолютную потребность в кремнии мож-
но использовать в борьбе с ними путем добавки к среде германия,
конкурентного ингибитора его поглощения ( Lewin, 1966). Двуокись
германия задерживает рост некоторых бурых водорослей ( Parker,
1969; Markham, Hagmeir, 1982), что указывает на их возможную по-
требность в кремнии. Однако избыток силикатов в среде не устра-
няет токсичности германия для Fucus (McLachlan et al.,1971),
позволяя полагать, что ее природа не овязана с конкурентным ин-
гибированием поглощения кремния.
Калий и сера
Калий и сера - биогены, необходимые в количествах, превышаю-
щих следовые. Концентрация калия у многих водорослей выше, чем в
окружающей среде. К его функциям относятся осмотическая регуля-
292
ция ( Kauss, 1978) и поддержание электрохимических свойств кле-
ток. Сера входит в состав аминокислот метионина и цистеина, а
также ряда других соединений; некоторые из них, по-видимому,
встречаются только у водорослей. Сульфатированные полисахариды -
важные компоненты стенок у морских водорослей-макрофитов (McCand-
less, 1981, см. гл. 9); серосодержащие соединения используются
некоторыми из них для осморегуляции (Reed, 1983).
Микроэлементы
Потребность во многих микроэлементах достоверно показана у
относительно немногих водорослей. Опыты по обогащению среды пока-
зали, что продуктивность возрастает при добавке следовых коли-
честв ряда металлов (Axler et al., 1980), которые включаются в
большинство искусственных культуральных сред (Provasoli et al.,
1957).
В количественном'отношении железо - наиболее важный из та-
ких элементов для фитопланктона ( Huntsman, Sunda, 1980). Оно
необходимо для разнообразных окислительно-восстановительных ре-
акций и синтеза хлорофилла (O’Kelly, 1974). Многие водорооли
накапливают цинк в более высоких, чем в окружающей среде, концен-
трациях (гл. 9); значительный интерес альгологов к этому элемен-
ту был связан с механизмом его поглощения (O’Kelly, 1974). Недо-
статок цинка коррелирует с исчезновением цитоплазматических рибо-
сом у Chlorella (Prask, Plocke, 1971) и со снижением интенсив-
ности синтеза хлорофилла и белка у Porphyra tenera (Noda, Hori-
guchi, 1971). Молибден требуется для метаболизма азота и свя-
зан с восстановлением нитрата. У синезеленых водорослей он наря-
ду с железом входит в нитрогеназный ферментный комплекс (Bothe,
1982). Медь незаменима как составная часть пластоцианина (белка,
участвующего в фотосинтетическом транспорте электронов; Katoh,
I960) и как кофактор для нескольких ферментов. Многие водоросли
крайне чувствительны к избытку меди (гл. 9), и это свойство на-
шло применение в разработке составов альгицидов и стойких к об-
растанию красок. Кобальт, входящий в молекулу витамина В^,
также является незаменимым микроэлементом (Bunt, 1970). Для фо-
тосинтетических реакций фотосистемы I необходим марганец (O’Kel-
ly, 1974). Предполагается, что доступность марганца и кобальта -
один из факторов, контролирующих рост Macrocystis pyrifera в
южной Калифорнии ( Kiwabara, 1982). Потребность в боре продемон-
293
стрирована у зародышей Fucus (McLachlan, 1977) и у ряда водорос-
лей-микрофитов (Lewin, 1976). Установлена ( stosch, 1964) по-
требность в мышьяке у багрянки Asparagopsis armata.
Многие морские водоросли - активные накопители галогенов.
По-видимому, хлорида необходимы для реакций фотофосфорилирования
(O’Kelly, 1974), а багрянка Polysiphonia urceolata ( Fries,
1975) и бурая водоросль Fucus edentatus (Pedersen. 1969) нужда-
ются в броме. Иод требуется Polysiphonia urceolata ( Rries,
1966) и, вероятно, необходим для роста и морфогенеза Petalonia
fascia из бурых водорослей (Hsiao, 1969). Галогенированные ор-
ганические соединения присутствуют у многих морских макрофитов
(гл. 9) и, возможно, эффективно отпугивают растительноядных жи-
вотных.
Витамины и ростовые факторы
Помимо неорганических биогенов многим водорослям нужны по-
ступающие извне витамины: цианокобаламин (В^), тиамин и биотин
(Swift, I960). Из исследованных 388 видов водорослей (Provaso-
li, Carlucci?I974) 203 были гетеротрофны по этим витаминам; 85%
нуждались в Bjg,
40% - в тиамине и 7% - в биотине. За некоторы-
ми исключениями, в биотине не нуждаются Xanthophyceae, Phaeophy-
ceae, Cyanophyceae и Chlorophyceae,а большинство Cyanophy-
сеае и Rhodophyceae аэтотрофны по тиамину. У золотистых во-
дорослей, примнезиофитов, эвгленовых и динофитов тенденция к
ауксотрофности по витаминам значительно сильнее. Многие виды, не
нуждающиеся в поступлении витаминов извне, могут синтезировать и
высвобождать их в количествах, достаточных для поддержания роста
тех водорослей, которым витамины требуются (Carlucci , Bowes,
1970). Однако некоторые виды образуют внеклеточные продукты, ко-
торые хелатируют свободный В^, делая его недоступным для других
водорослей ( Pintner, Altmyer, 1979).
Другие органические соединения также могут быть необходимы
для роста водорослей или действовать на него ингибирующе. Много-
клеточные виды часто не развивают нормальных морфологических
структур при выращивании в аксеничных условиях ( Provasoli, Car-
lucci, 1974; provasoli et al., 1977), но восстанавливают нор-
мальный рост при добавке в среду живых или автоклавированных мор-
ских бактерий. Известны многочисленные аллелопатические взаимо-
294
отношения фитопланктонных водорослей ( Maestrini, Bonin, 1981а).
Рост корковой багрянки Rhodonhysema elegang подавляется вблизи
корковой бурой водоросли Ralfsia apongiocarpa (Fletcher,1975),
т.е. диффундирующие в воду соединения, образуемые бентосными фор-
мами, по-видимому, также могут служить экологически важными ре-
гуляторами роста.
Поглощение биогенов
Кислород и двуокись углерода обычно поглощаются путем пас-
сивной диффузии, хотя при высоких значениях pH для поглощения би-
карбоната со скоростью, достаточной для поддержания чистого фото-
синтеза, необходим активный транспорт (Raven, 1970). Другие не-
органические биогены, как правило, поглощаются в ионной форме за
счет активного транспорта, связанного с фотосинтезом, который
обеспечивает необходимую для этого процесса энергию.
Механизмы и кинетика активного поглощения биогенов у водо-
рослей интенсивно исследовались на протяжении двух последних де-
сятилетий (De Boer, 1981; McCarthy, 1981). Основной стимул та-
ким исследованиям дали две статри ( Dugdale, 1981; Eppley, Coat-
worth, 1968), в которых оценивалась роль этой кинетики в рос-
те фитопланктона. Было показано, что скорость поглощения биогена
связана с его внеклеточной концентрацией по формуле (рис. 6.19)
где v - скорость поглощения, vmav - максимальная скорость по-
глощения, S - концентрация биогена,Kg - константа полунасыще-
ния. т.е. концентрация субстрата, при которой v = v /2.
Это означает, что при низких концентрациях скорость погло-
щения возрастает с увеличением внешней концентрации биогена бы-
стро, а при более высоких концентрациях рост скорости замедляет-
ся, пока не достигается ее максимальное значение (v ). уже не
зависящее от дальнейшего повышения концентрации. Сходство между
зависимостью .для поглощения биогенов и уравнением Михаэлиса -
Ментен для ферментативного катализа позволяет предполагать нали-
чие сходных механизмов, т.е. специфических энергоемких фермент-
ных систем поглощения, связанных с мембранами, с биогеном в ка-
честве субстрата.
295
Применение этого уравнения
Концентрация биогена {S)
Рис. 6.19. Связь между концен-
трацией биогена в субстрате
и скоростью его поглощения.
Утят - максимальная скорость
поглощения. Кд - концентрация
в субстрате, при которой ско-
рость поглощения равна полови-
не максимальной (константа по-
лунасыщения) (по Parley, 1982)
подразумевает присутствие только
одного биогена или постоянные
концентрации других биогенов,
измерение для каждой концентра-
ции только исходных скоростей
поглощения и достаточную переме-
шанность среды, чтобы предотвра-
тить ее истощение по данному био-
гену у поверхности водорослевых
клеток.
Скорость роста водоросли
будет снижаться, если внешняя
концентрация биогенов упадет ни-
же необходимой для поддержания
интенсивности поглощения, доста-
точной для обеспечения прежних
темпов роста. При таких условиях
окорость роста и концентрация
связаны соотношением, аналогич-
ным уравнению Михаэлиоа - Ментен
для поглощения биогенов. Это - уравнение Моно ( Monod,I942), пер-
воначально примененное для описания роста бактерий в хемостате.
но описывающее также рост водорослей с постоянной скоростью при
лимитированном питании (Droop, 1974-а):
( S \
4- S J ’
где JA - удельная скорость роста, J4 ^ах - максимальная удель-
ная скорость роста, з - концентрация субстрата и кд - конотанта
полунасыщения для роста.
Другие исследования (Droop, 1968) показали, что скорость ро-
ста связана также с внутриклеточной концентрацией лимитирующего
биогена (рис. 6.20). Это отражает уравнение Друпа (Droop, 1968):
/ It Q А
= (Д- Q
где q - количество питательного вещества в водорослевых клет-
ках, q0 - самый низкий уровень q, при котором водоросль способна
расти, и р'тад “ скорость роста при бесконечно большом значе-
нии Q . J
При стабильных условиях для описания роста при ограниченно-
сти биогенов применимы уравнения как Моно, так и Друпа. Использо-
вать последнее из них часто более удобно, поскольку более высокие
296
Скорость роста (/с )
Рис. 6.20. Связь между ско-
ростью роста (JU ) и концентра-
цией биогена внутри клетки (Q):
Q - минимальное содержание;
Лп>ах “ максимальная скорость
роста (поRhee, 1980)
уровни биогена в клетке изме-
рить легче, чем его очень низ-
кие внешние концентрации. Урав-
нение Друпа - теоретическое
приближение, поскольку <5 не
может быть бесконечным, а ис-
тинная максимальная скорость
jUmaxnpz меньше, чем
ее теоретический максимум; та-
ким образом, и , HjMmax
должны определяться эксперимен-
тально (Rhee, 1980).
Принцип поглощения пита-
тельных веществ макроскопичес-
кими водорослями в целом тот
же, что и у микроскопических
( DeBoer, 1981). Однако здесь
его механизм усложнен в связи
с размером таллома и увеличе-
нием свободного апопластного
пространства; это следует учи-
тывать во всех опытах по погло-
щению при измерении доли со-
держания в клетке, соответствую-
щих веществ.
Рост
и размножение
феромоны
Половые аттрактанты известны у многих животных, у оомицетов
и некоторых пресноводных зеленых водорослей, в том числе Chla-
mydomonasCTsubo. 1961) и Oedogonium. (van den Ende, 1976). Гаме-
тогенез у ряда видов Voivox также контролируется феромонами
( Starr -et al., 1980); у двудомных форм развитие половых сферо-
идов индуцируется гликопротеином, а у однодомных критические
уровни L-глутаминовой кислоты, концентрация которой возрастает
при увеличении плотности популяции, стимулируют переход к полово-
му размножению.
Среди водорослей строение и действие таких феромонов лучше
297
Таблица 6.2. Химическая структура и распространение феромонов
бурых водорослей (Maier, Mailer;1986)
Структура
Тривиальное название Таксоны
Десмарестен
Диктиоптерен
Ламоксирен
Ectocarpus siliculosus
Е. fasciculatus
Sphaceiaria rigidula
Adenocystis utricularis
Desmarestia aculeata
D. viridis
Dictyota dichotoma
Мупьтифидем
Lammariaceae
Alariaceae
Lessoniaceae
(29 видов)
Cutleria multifida
Chorda tomentose
Виридиен
Фукосерратеи
Syringoderma phinneyi
Fucus serratus
F. vesiculosus
F. spiralis
Финаваррен
AscophyHum nodosum
Dictyosiphon foenicula -
ceus
Цистофорен
Cystophora sUiquosa
Гормозирвн
Hormosira banksii
Xiphophora chondrophylla
X. gladiate
Durvilaea potatorum
D. antarctica
D. willana
Scytosiphon lomentaria
Colpomenia peregrins
298
всего известны для Phaeophyceae) (табл. 6.2). Они активны уже
при концентрациях I0”? моль/л и на расстоянии ло 0,5 - I мм
( Maier, Muller, 1986).
Ectocarpug ailiculosus - номинально изогамный вид, но его
гаметы, которые можно считать женскими, отличаются более корот-
ким периодом подвижности, оседают на субстрат и отбрасывают жгу-
тики. Вскоре такая гамета начинает привлекать мужские гаметы, ре-
агирующие на образуемое ею крайне летучее соединение. Запах его,
отдаленно напоминающий апельсиновый, обнаружен в клональных куль-
турах женских гаметофитов. Это вещество было очищено, идентифици-
ровано как циклической Сц-олефин, названный эктокарпе ном, и син-
тезировано. Другие низкомолекулярные олефиновые феромоны сходной
структуры известны у Cutleriales,Syringodermatales, Chordariales,
Dictyotales, Larainarialea, Desmarestiales, Sporochnales, Sphace-
lariales И Tilopteridales (Muller, 1981; Maier, Muller,
1986). Показано, что одинаковые феромоны вызывают высвобождение
спермиев у Laminariales, Desmarestiales и Sporachnalea(Maier,
1982; Maier, Muller 1986). Представители Fucales и Durvilla-
ealesобразуют линейные или содержащие циклопропановое кольцо оле-
финовые феромоны. У iVcus обнаружено Cg-соеданение фукосерратен,
а у других - Cjj-соединения. С j-[-феромон Ascophyllum nodosum
финаваррен отмечен также у Dictyosiphonales, тогда как Cjj-гормо-
сирен, типичный для нескольких австрало-азиатских видов, встреча-
ется у Scytosiphonales (Maier, Miller, 1986).
Оплодотворение и прорастание
Хотя имеются многочисленные данные об анатомических и морфо-
логических процессах, характеризующих прорастание спор и зигот
водорослей, почти все сведения о его физиологии получены на яйце-
клетках Fucus. Две особенности делают этот объект привлекатель-
ным для физиологов, занимающихся развитием, - крупные размеры
(70-80 мкм в диаметре) и отсутствие клеточной стенки в момент
высвобождения, хотя выделение альгиновой кислоты для ее образо-
вания начинается непосредственно после оплодотворения. На нем
изучались механизмы распознавания клетки клеткой, половые аттрак-
танты и полярность ( Evans et al., 1982); система яйцеклетка/зи-
гота Fucus, несомненно, является важной моделью, способной дать
ответ на фундаментальные вопросы теории развития в целом.
299
У Fucus слияние гамет в отличие от действия феромонов про-
исходит высоковидоспецифично ( Bolwell et al., 1977). С яйце-
клеткой сливаются лишь несколько спермиев (Evans, et al., 1982),
возиозно,в связи с изменениями в ее мембране, индуцируемыми опло-
дотворением, или же вследствие малого числа воспринимающих участ-
ков. Специфические белки, лектины, участвуют в этом распознава-
нии, связывая, как показано, остатки фукозы, маннозы, галактозы
и глюкозы (Bolwell et al., 1979). Лектины подавляют оплодотво-
рение предположительно путем блокировки воспринимающих спермин
участков на поверхности яйцеклетки Fucus. Белки, виде специфично
подавляющие оплодотворение при добавлении извне, выделены из мем-
• браны яйцеклетки, а связывающие углеводы - из спермиев Fucus
( Bolwell et al., 1980).
Полярность
Полярность - это наличие .любого асимметричного состояния
внутри живей системы. Она обычно выражается в различиях между
двумя концами клетки или органа. Только что высвободившиеся яйце-
клетки Fucus неполярны, и первые внешние признаки поляризации
появляются через несколько часов после оплодотворения. При этом
развивается вырост, который удлиняется, формируя прикрепительный
ризоид. На этой стадии зародыш становится резко полярным, с не-
равным распределением органелл и отчетливыми химическими градиен-
тами. Сильное одностороннее освещение индуцирует формирование ри-
зоидального выроста с затененной стороны, а более слабое застав-
ляет его отходить под углом 90 - 135° к источнику света ( Jaffe,
1958).
Интенсивность дыхания после оплодотворения возрастает, что,
возможно, коррелирует с уменьшением запаса питательных веществ
(Quatrano, Stevens, 1976). Наиболее заметные из изменений в
этот период связаны с образованием клеточной стенки. В течение
нескольких минут после оплодотворения происходит секреция альги-'
новой кислоты, а через 60 мин клеточные стенки содержат равные
количества альгинатов и целлюлозы (Quatrano, Stevens, 1976).
За этот период еще до ядерного деления и цитокинеза внутренние
органеллы распределяются асимметрично fcrawley et al., 1977).
Взрослый таллом всех многоклеточных водорослей имеет своего
Рода апикально-базальную полярность, предположительно возникаю-
щую уже при первом делении споры или зиготы. Она может быть за—
300
заметной в морфологии или анатомии или проявляться более тонко в
физиологических градиентах таких параметров, как содержание хло-
рофилла, фотосинтетическая активность, интенсивность дыхания и
темновая фиксация СО2 вдоль, например, пластины Laminaria sac-
charins (Johnston et al., 1977). Наличие такой полярности в от-
дельных клетках ясно продемонстрировано при регенерации багрян-
ки Griffiths la (Duffield et al.,1972), когда базальные полюса изо-
лированных осевых клеток образуют ризоиды, а апикальные - новые
осевые клетки (рис. 6.21).
Полярность отдельных клеток, возможно, является одним из
проявлений тотипотентности, при которой клетка сохраняет способ-
ность делиться, дифференцироваться и в конечном итоге развивать-
ся в целый организм. Тотипотентны изолированные протопласты це-
НОЦИТНОЙ водоросли Bryopeie plumosa из Chlorophyceae (Tatewaki,
Hagata, 1970), ризоидальные клетки бурой водоросли Pelvetia
wrightll (Зада et al., 1978) и сердцевинные клетки багрянки
Chondrus crlspus (Chen, Taylor, 1978). Возможно, наиболее ярко
тотипотентность продемонстрирована у Laminaria angustatat здесь
отдельные клетки формируют каллусовую ткань, которая при добав-
лении бактеризованного экстракта родительской особи развивается
в спорофиты (Saga et al., 1978).
Имеется много других данных по апикально-базальным градиен-
там, проявляющимся в многоклеточных фрагментах макроскопических
водорослей и указывающим на физиологические различия между верх-
ней и нижней частями исходного таллома. У Enteromorpha (Chloro-
phyceae) больше ризоидов образуется от базальных сегментов, чем
от апикальных (Eaton et al., 1966), а у Caulerpa prolifera
из того же класса отрезанные горизонтальные части ("корневища")
с большей частотой регенерируют ризоида у апикального конца
( Jacobs, 1964).
Апикальное доминирование
Явления апикального доминирования указывались у ряда водо- -
рослей, особенно у имеющих хорошо развитые апикальные клетки
или меристематические зоны. Этот эффект предполагает прежде все-
го подавление роста боковых ветвей. У Fucus встречается доба-
вочная эмбриония. На поврежденном талломе возникают молодые осо-
би, число и ориентация которых коррелируют с их положением отно-
сительно апикально-базального градиента родительской водоросли
301
Рис. 6.21. Экспериментальная система восстановления
повреждений с использованием Griffithsia spp. Д. Нить
Griffithsla разрезают на базальный (ЕС) и апикальный
(АС) сегменты и помещают в цилиндр, представляющий со-
бой свободную от цитоплазмы клеточную стенку Hitella
(10 (см. Б). Регенерация ризоида (Р) на апикальйоМ cet-
менте индуцирует образование восстановительной клетки
(ВК) на базальном сегменте; происходит их слияние (В).
Когда апикальный и базальный сегменты принадлежат раз-
ным видам (Г), на последнем чаще регенерирует апикаль-
ная (АК), а~не восстановительная клетка и слияния не
происходит (по Buggeln, 1981, из Waaland, 1975)
(Moss, 1964, 1966). Сходные проявления полярности можно наблю-
дать у багрянки Schottera nicaeensls (рис. 6.22) ( Perrone,Fe-
licini, 1972). Удаление апикальных клеток способствует росту
боковых ветвей у красных водорослей Apoglossum ruscifolium
( Abelard, L’Hardy-Halos, 1975) и Antithemnion plumula (L’Hardy-
Halos, 1971a), а также бурой водоросли Sphacelaria cirrhosa
( Ducreux, 1977). Среди Fucales удаление верхушки приводит к
развитию примордиев боковых ветвей у Ascophyllum nodosum (Moss,
1970) и вторичных боковых ответвлений у Sargassum muticum
( Chamberlain et al., 1979); их наличие в свою очередь контро-
лирует развитие третичных ответвлений.
302
Рис. 6.22. Способы регенерации после экспериментального
повреждения или удаления части пластины красной водо-
росли Schottera nicae&nse. в каждом варианте слева по-
казано исходное состояние; линии - надоезы, штриховка -
удаленные части. Справа показана добавочная эмбоиония,
наблюдавшаяся через 30 - 90 дней (по Perrone, Fellaini’,
1972)
Взаимодействия между клетками таллома
Морфогенетические взаимодействия между клетками интактных
водорослей отмечались, в частности, у багрянок, апикальный рост
и волокнистое псевдопаренхиматозное строение которых делают их
идеальными системами .для таких исследований. Клетки простых нит-
чатых красных водорослей могут по мере развития значительно уве-
личиваться в размерах (в 40 000 раз у An-tithamnion plumula;
Dixon, 1973. Для сравнения отметим, что среднее увеличение кле-
точного объема у цветковых растений - в 100 - 200 раз). Такие
клетки легко окрашиваются прижизненными флуоресцентными красите-
лями для определения скорости роста; наблюдения могут сочетаться
с простой визуальной оценкой скорости деления клеток ( Waaland,
Waaland, 1975). Эти два компонента - скорости деления и увеличе-
ния объема - нетрудно соотнести с возрастом клетки и ее положе-
нием в талломе. У ряда видов Fiorideophyceae окончательный раз-
зоз
Апекс 20 40 60 80 1 00 120 1 40Основание
Число сегментов
Рис. 6.23. Гистограмма длин клеток вдоль главной оси
Ceramium rubrum. Стрелки указывают места основных ди-
хотомических разветвлений (по Dixon, 1979)
мер клеток, по-видимому, зависит не только от их возраста и
расстояния до апикального конца водоросли, но и от расположения
по отношению к боковым ветвям ( Dixon, 1971). Осевые клетки у
оснований последних часто значительно мельче, чем расположенные
ближе к вершине особи.
Гормоны и развитие водорослей
У сосудистых растений хорошо известны пять групп гормонов:
ауксины, гиббереллины, цитокинины, абсцизовая кислота и этилен,
однако судить об их наличии и действии у водорослей приходится
в основном по косвенным данным ( Buggeln, 1981).
Ауксин (индолил-3-уксусная кислота, ИУК) - эндогенный регу-
лятор увеличения клеток у высших растений. По-видимому, в кле-
точной мембране Valonia (Chlorophyceae) он регулирует тран-
спорт К+ ( Zimmermann et al., 1976), не влияя на эластичность
304
стенки ( Zimmermann, 1977). Он обнаружен в полевых сборах бурой
водоросли Undaria pinnatlfida (Abe et al., 1972), но вполне
возможно, что образован здесь эпифитными микроорганизмами. У
Alaria esculents ростовые реакции на ауксин и его неактивные
аналоги различить не удалось ( Buggeln, 1976). Рост пластин по-
давлялся, что коррелировало с ингибированием фотосинтеза и струк-
турными изменениями хлоропластов (Buggeln, Bal, 1977). Имеются
сведения, что апикальное доминирование у декапитированных водо-
рослей восстанавливается при добавлении ауксина извне ( Mobs,
1965; Chamberlain et al., 1979). Росту бактеризованных культур
Codium fragile (Chlorophyceae) способствовало добавление аукси-
на, но не его синтетических аналогов ( Hanisak, 1979). ИУК обна-
ружена также у Caulerpa (Chlorophyceae) с помощью высокочувстви-
тельной жидкостной хроматографии и масс-спектроскопии; ее концен-
трация, составляющая примерно I млн-'2’ овежего материала, сходна
с приводимой для высших растений ( Jocobs et al., 1985).
Гиббереллиноподобная активность показана в экстрактах из
морских водорослей ( Augier, 1976) и морской воды (Fries, 1973).
Подавление бокового роста у бурой водоросли Sargassum muticum
частично снималось добавлением экзогенной гиббереллиновой кисло-
ты ( Gorham, 1977), но у других видов этого эффекта не отмечено
(Buggeln, 1981). Обработка крупной ценоцитной зеленой водоросли
Caulerpa prolifera гормональными дозами этой кислоты стимулиро-
вала рост "корневища" и более чаотое заложение ризоидов, но ника-
кого влияния на пластины не наблюдалось (Jacobs, Davis, 1983).
Цитокининоподобная активность показана в экстрактах из мор-
ских водорослей (Pedersen, 1973), а экзогенный кинетин требует-
ся для формирования вертикальных нитей в аксеничных культурах бу-
рой водоросли Ectocarpus faBoiculatus (Pedersen, 1968). Сообща-
лось, что’ этилен образуется у обработанных ауксином багрянки
Porphyra tenera, Codium latum ИЗ Chlorophyceae И бурой водорос-
ЛИ Padina arborescens (Watanabe, Kondo, 1976), хотя вполне
возможно, что его продуцируют эпифитные микроорганизмы. Он не
влияет на удлинение пластин Alaria esculenta (Buggeln, 1981).
Наилучшее свидетельство образования специфичного для водо-
рослей эндогенного регулятора роста - существование диффундирую-
щего гликопротеина "родоморфина" (Watson, Waaland, 1983). Он
возникает в ризоидах багрянки Griffithsia spp. (Waaland, 1975)
и индуцирует формирование на апикальном конце поврежденной клет-
305
ки выроста, который растет в сторону ризоидальной клетки и в ко-
нечном счете сливается с ней.
Тропизмы
Тропизмы - это движение в определенную сторону в зависимос-
ти от того, откуда воздействует тот или иной внешний фактор. У
высших растений известно много типов подобных реакций, а у водо-
рослей лучше всего изучены фототропизм и геотропизм. Фототропизм
указывался для нитчатых представителей Xanthophyceae, Chloro-
phyceae, Phaeophyceae И Rhodophyceae (Buggeln, 1981). Синий
свет вызывает при этом максимальную реакцию, которая обычно поло-
жительна у вертикальных частей таллома и отрицательна у ризоидов
и "присосок".
Показано (Kataoka, 1975а, Ъ), что положительный фототро-
пизм апикальных частей таллома ценоцитной желтозеленой водоросли
Vaucheria geminata - многостадийный процесс, подавляемый силь-
ным освещением. Он связан с ростом, точка которого перемещается
на светлую сторону. Ветвление также можно индуцировать односто-
ронним освещением таллома. Катаока (Kataoka, 1977) делает вывод,
что с процессом искривления связан циклический АМР, а ауксин в
нем не участвует. Разделение восприятия стимула и реакции на не-
го наблюдается в ризоидах Boergesenia forbesii ИЗ Chlorophyceae
( Ishizawa, Wada, 1979a, b) при I5°C: роста не происходит и соот-
ветственно нет реакции на одностороннее освещение. Когда стимули-
рованные растения переносятся в темноту при 25°С, рост возобнов-
ляется и наблюдается изгиб ризоидов в сторону, противоположную
исходному направлению на стимул. Более крупные многоклеточные
структуры, например "присоски" бурой водоросли Alaria esculenta.
также показывают отрицательный .Тототропизм (Buggeln, 1974). Все
такие реакции максимально стимулируются синим светом (430 -
450 нм), а свет с длиной волны более 500 нм в этом смысле влияет
слабо. Предполагается, что рецепторный пигмент может быть кароти-
ноидом или рибофлавинам.
Положительный геотропизм у водорослей достоверно продемон-
стрирован на ризоидах Chara , в которых обнаружены статолиты
( Sievers et al., 1979). Опыты с использованием двустороннего
освещения наводят на мысль, что земное притяжение может влиять
На ориентацию ризоидов Caulerpa racemosa (Chlorophyceae), при-
чем. его восприятие связано с накоплением содержащих крахмал ами-
306
лопластов у нижних поверхностей ризоидов ( MatПеку, Jacobs,
1983). Предполагается также, что в реакциях геотропизма участ-
вует эндогенный гиббереллин ( Jacobs, Davis, 1983).
Контактные явления наблюдались у талломов красных водорос-
лей (L-*Hardy-Halos, 1971 bj; образование ими прикрепительных ри-
зоидов может говорить о тигмотропизме.
Эндогенные ритмы
Эволюция водорослей проходила в среде с суточной периодич-
ностью поступления энергии, и регулярное чередование света и
темноты требует координации процессов синтеза и воспроизведения
их клеток ( Chisholm, 1981). Циркадианные ритмы с периодом при-
мерно 24 ч отсутствуют у синезеленых, но имеются у всех эукари-
отных водорослей (рис. 6.24) (Sweeney, 1983). Точная длитель-
ность таких циклов зависит от эндогенных часов, которые в приро-
де подстраиваются под 24-часовой режим смены света и темноты.
Когда водоросль помещается в усложя постоянного света, ритм со-
храняется, т.е. циркадианные часы не зависят от внешнего стиму-
ла. Исчезновение ритма в популяции при постоянных условиях обус-
ловлено потерей синхронности в функционировании особей, а не са-
мих циркадианных часов ( Chisholm, 1981).
Циркадианным ритмам подчинены многие физиологические и эко-
логические явления. Два наиболее изученных в этом плане вида -
Euglena gracilis И Gonyaulax polyedra (Chisholm, 1981). Цир-
кадианные ритмы у многоклеточных водорослей проявляются в кле-
точном делении у багрянки Griffithsia Pacifica (Waaland, Cle-
land, 1972), фотосинтетической и ферментативной активности у
бурой водоросли Spatoglossum pacificum (Yamada et al., 1979),
размножении представителей Chlorophyceae Oedogonium cardiacum
(Huddat, 1961), Pseudobryonsis so. (Okuda, Tatewaki, 1982) И
багрянки Acrochaetium asparagopsis (Abdel-Rahman , 1982).
У Gonyaulax все ритмические процессы, по-видимому, контро-
лируются единым часовым механизмом ( Sweeney, 1983). Периодич-
ность люминесценции связана с ритмом концентрации белка люцифди-
зы ( Johnson et al., 1984), возможно, со скоростью обновления
его клеточного пула.
Полевые наблюдения за литоральными морскими водорослями
продемонстрировали у некоторых из них периодичность размножения,
которая совпадает со специфическими периодами лунного приливно-
307
Рис. 6.24. Циркадианный ритм биолюминесценции Qonyau-
lax polyedra при постоянном освещении. Первый 24-часо-
вой цикл' из 12 ч света и 12 ч темноты является настро-
ечным (по Sweeney, 1979)
Лунный месячный цикл
Рис. 6.25. Образование яйцеклеток в лабораторных культу-
рах Dictyota dichotoma. выращиваемых в режиме 14-часо-
вогосветового дня. Темновой период заменялся тусклым
освещением через каждые 28 дней (стрелки) чтобы имити-
ровать условия полнолуния (по Muller, 1962)
308
го цикла (Tanner, 1981). Эндогенный лунный ритм показан у бу-
рой водоросли Dictyota dichotoma (Mfiller, 1962), высвобождаю-
щей в природе гаметы с примерно 14-дневными интервалами в соот-
ветствии с сизигийными приливами (рис. 6.25). В культуре этот
ритм может быть задан помещением особей в условия слабой осве-
щенности на то время, когда в норме должен быть темновой период.
Высвобождение гамет происходит после этого примерно с двухне-
дельными интервалами, подразумевая, что в природе настройка
цикла определяется светом полной луны.
Глава 7
ЭКОЛОГИЯ
Введение
Водоросли освоили практически все местообитания, где фото-
синтез может идти со скоростью, достаточной для образования чис-
той продукции, т.е. роста, несмотря на потери на метаболизм и вы-
едание. Они встречаются от полярных районов до жарких пустынь,
от вершин гор до границ проникновения света в океанах и озерах,
обитают почти на всех неорганических субстратах, вступают в раз-
личные симбиотические ассоциации с други® организмами, свободно
плавают в планктоне всех водоемов, за исключением тех, которые
образуются на очень короткое ьремя.
Раунд (Round, 1981) был первым альгологом, попытавшимся
дать обобщающий обзор экологии водорослей в одной книге; такую
работу можно сравнить разве что с проведенной на 50 лет раньше
Фричем (Fritsch, 1935, 1945), собравшим сведения о структуре и
размножении этих организмов. Здесь мы применяем функциональный
подход и описываем некоторые процессы, связанные с экологически-
ми взаимодействия!®. В результате выделяются три основные эколо-
гические группы водорослей: I) планктонные, 2) бентосные, в част-
ности эпилитные морские и приуроченные к относительно рыхлым
осадкам и почвам, 3) симбиотические. Вдобавок к этому будут упо-
мянуты водоросли, обитающие в экстремальных температурных усло-
виях: в горячих источниках, снежных и ледяных биотопах. Многие
альгоэкологи составили детальные списки видов, обнаруженных в
самых разнообразных местообитаниях и районах планеты. Объем на-
шей книги не позволяет включить в нее такие данные, но это не
значит, что мы против подобного метода; наоборот, как отмечено
Раундом ( Round, 1981), игнорировать списки водорослей - значит
отрицать сложность их сообществ. Для понимания экологии водорос-
лей необходимы как экспериментальный, так и описательный подхо-
ды, а сочетание их должно особенно приветствоваться.
Общий и чрезвычайно важный признак подавляющего большинства
водорослей - их роль продуцентов; путем фотосинтеза они превра-
щают физическую энергию солнечного излучения в химическую. Как
продуценты, они составляют основу пищевой цепи в водных место-
обитаниях, потребляясь либо непосредственно при выедании, либо
310
через детритные системы. Важность этой продукции легко оценить,
вспомнив, что свыше 70% поверхности нашей планеты покрыто водой.
Многие водорослевые сообщества чрезвычайно продуктивны и соперни-
чают в этом смысле со многими наземными экосистемами (рис. 7.1).
Фитопланктон
Фитопланктон - наиболее распространенная и хорошо изученная
из всех экологических трупп водорослей. Он встречается практичес-
ки во всех водоемах, где плавает свободно и в значительной степе-
ни пассивно. Большинство планктонных видов не связано в своем су-
ществовании с берегом и дном, однако жизненный цикл некоторых
мелководных форм включает бентосные покоящиеся стадии (Fryxell,
1983). Во всех классах, за исключением багрянок, харовых и бу-
ры^ водорослей, есть планктонные представители, а в прибрежной
зоне хотя бы временно пополнять планктон могут зооспоры бентос-
ных бурых водорослей и Chlorophyceae. В последнее время появил-
ся ряд прекрасных книг и обзоров, посвященных экологии фитопланк-
тона (например, Sournia, 1978; Morris, 1980; Platt, 1981; Round,
1981; Reynolds, 1984).
Фитопланктонные водоросли преимущественно одноклеточные, хо-
тя среди них много колониальных и нитчатых форм, особенно в прес-
ных водах. Размеры составляют от менее 5 мкм у мелких жгутиковых
и коккоидных видов до 500 мкм у колониальных, напримерVoivox<.
Выделяются две группы: сетной планктон, улавливаемый сетями о
размером ячеи примерно 64 мкм, и более мелкий, или нанопланктон.
Согласия относительно того, какой размер считать переходным от
одной группы к другой, пока не достигнуто. Одни ( Zeitzschei,
1978) относят к нанопланктону формы диаметром менее 20 мкм, дру-
гие (Manny, 1972) - менее 10 мкм. Размер клетки и отношение пло-
щади ее поверхности к объему очень важны с точки зрения плавуче-
сти, поглощения питательных веществ и устойчивости к выеданию.
Крупные свободноплавающие водоросли-макрофиты, например Sargas-
sura в Саргассовом море, по определению тоже являются планктон-
ными, но обычно не рассматриваются как таковые из-за своей вели-
чины и родства с бентосными видами.
В океанах сетной фитопланктон представлен прежде всего диа-
томеями и динофлагеллятами, а нанопланктон - синезелеными водоро-
слями и мелкими жгутиковыми, в основном примнезиофитами, многие
из которых относятся к группе кокколитофорид. Диатомеи и дино-
зп
МОРСКИЕ ПРОДУЦЕНТЫ
Кораллы
Донные макрофиты
Травы соленых маршей
Морские травы
Мангры
Бентосные микроскопические
водоросли
Прибрежный фитопланктон
Фитопланктон открытого моря
ПРЕСНОВОДНЫЕ ПРОДУЦЕНТЫ
Макрофиты
Фитопланктон (богатая биогенами
среда)
Фитопланктон (бедная биогенами
среда)
НАЗЕМНЫЕ ПРОДУЦЕНТЫ
Дождевые леса
Леса умеренного климата
Тревяные формации
Прерии, стели
Пустыни, тундра
Рис. 7.1. Сравнение ежегодной чистой первичной продукции
в морских, пресноводных и наземных местообитаниях (noVa-
liela, 1984)
флагеллаты обильны также в пресных водах, но там мотут домини-
ровать и представители других классов, в частности Cyanophyceae,'
Chlorophyceae и Chrysophyceae. В олиготрофных (бедных бИОГена-
ми) условиях преобладают золотистые и десмидиевые водоросли,
тогда как в эвтрофных (богатых биогенами) обычны оинезеленые и
диатомовые; эвгленовые бывают особенно обильны в мелких богатых
питательными веществами лужах ( Round, 1981).
Методологая
Изучение фитопланктона идет в основном по двум направлени-
ям: в первом случае исследуют все сообщество, во втором - от-
312
дельные виды, которые определяют и рассматривают порознь при
дальнейшем анализе. Первое направление особенно полезно при
определении наличных запасов планктона, которые выражаются в био-
массе, общем углероде, органическом азоте, хлорофилле, АТР и мно-
жестве других показателей. Продукцию всего фитопланктонного сооб-
щества можно оценить путем измерений вышеуказанных параметров
или методами, основанными на определении интенсивности фотосин-
теза с использованием 14 С (parsons et al., 1984).
Когда требуется детальная информация о роли отдельных видов,
следует применять методы микроскопического анализа и подсчета
(sournia, 1978). Радиоавтографические методы позволяют оцени-
вать продуктивность индивидуальных клеток и таким образом опре-
делять вклад различных видов в продукцию фитопланктонных сооб-
ществ ( Knoechel, Kalff, 1976).
Использование широкого набора методов значительно способст-
вовало пониманию экологии фитопланктона, но привело и к опреде-
ленным трудностям: отсутствие стандартизации часто осложняет
сравнение результатов разных исследований или делает его вообще
невозможным.
Плавучесть
Чтобы выжить, фитопланктонная популяция должна удерживаться
в той части водной толщи, где освещенность обеспечивает уровень
фотосинтеза, достаточный для роста и размножения. Однако не сле-
дует однозначно рассматривать тенденцию к опусканию ниже как от-
рицательную, поскольку вертикальное перемещение способствует по-
глощению питательных веществ и может быть также механизмом избе-
гания консументов и сильного, потенциально вредного солнечного
излучения у поверхности воды.
Плотность всех водорослей, за исключением синезеленых, об-
разующих газовые вакуоли, больше, чем у воды, т.е. они неизбежно
должны погружаться в глубину. Для подвижных клеток это не пробле-
ма, поскольку они могут активно плыть на свет, но многие другие
организмы, например диатомовые или коккоидные формы из разных
отделов, к этому не способны. Плотность большинства компонентов
цитоплазмы выше, чем у воды (1,0008 г/мл у пресной и 1,027 г/мл
у морской). Кремнеземные стенки диатомей, одной из наиболее про-
цветающих фитопланктонных групп, имеют плотность 2,6 г/см3. Толь-
ко у липидов плотность меньше, чем у воды (0,86 г/см3), что сни-
313
жает общую плотность клетки, но не может обеспечить нейтральной
плавучести. Подсчитано (Smayda, 1970), что увеличение содержа-
ния липидов у морской диатомеи с 9 до 40% сократило бы общую
плотность клетки на 3,5%, т.е. до 1,15 г/см3, а значит, она все
равно осталась бы значительно выше плотности морской воды, хотя
это и привело бы к снижению на 25% скорости погружения. Обычно
липиды составляют 2-20% сухой массы водорослей, причем накапли-
ваются в сходных концентрациях и у бентосных видов, следователь-
но, если они и'регулируют распределение клеток в толще воды, то
лишь в исключительных случаях (Reynolds, 1984). Ионная регуля-
ция, при которой более тяжелые ионы замещаются более легкими,
возможно, как-то влияет на плотность у морских видов. Показано
(Anderson, Sweeney, 1978), что плотность вакуолярного сока диа-
томеи Ditylum brightwelli снижается за счет селективного накоп-
ления ионов к+ и Иа+, замещающих более тяжелые двухвалентные
ионы. Динофлагеллата Pyrocystts noctiluca также способна сни-
жать концентрацию тяжелых ионов so2- Са2* и Mg2+ до уровня,
обеспечивающего всплывание (каьп, Swift, 1978).
Планктонные синезеленые водоросли обладают специальными
плавательными приспособлениями, так называемыми газовыми вакуо-
лями (Walsby, 1978), которые состоят из нескольких тесно упако-
ванных цилиндров, или пузырьков, длиной до I мкм при диаметре
70 нм, окруженных белковой мембраной. При достаточном внешнем
давлении, в природе вызываемом клеточным тургором, пузырьки раз-
рушаются и клетка теряет плавучесть.
КлеткиAnabaena fios-aquae при низкой освещенности обра-
зуют газовые вакуоли и становятся плавучими, но на ярком свету
их вакуолизация снижается и плавучесть теряется. В ходе фотосин-
теза клеточное тургорное давление может увеличиваться до несколь-
ких атмосфер вследствие накопления фотоассимилятов и поглощения
Ионов K+(Dinsdale, Walsby, 1972; Allison, Walsby, 1981).
Таким способом планктонные синезеленые водоросли, по-видимому,
Регулируют свое положение в столбе воды, удерживаясь на глуби-
нах, наиболее благоприятных для роста ( Reynolds, Walsby, 1975).
Это дает им значительное преимущество перед другими планктонны-
ми видами и возможность вызывать "цветение" в стабильном водном
слое. Виды с газовыми вакуолями нечасты в море, где планктон
обычно адаптирован к турбулентной среде ( Margalef, 1978) и эф-
фективность рассмотренного выше плавательного приспособления по-
нижена. Заметное исключение - колониальная нитчатая водоросль
314
Trichodesmium, вызывающая цветение в тропических морях (Fogg.
1982). Давление, необходимое для разрушения ее газовых пузырь-
ков, порядка 37 атм, т.е. в 8 - 10 раз выше требуемого для Апа-
baena и намного больше обеспечиваемого любым изменением клеточ- ,
ного тургора. Таким образом, Trichodesmium сохраняет положитель-
ную плавучесть, но не способен ее регулировать. Он пассивно под-
нимается в столбе воды и нуждается в ее турбулентном перемешива-
нии для погружения в более богатые питательными веществами слои,
где его прочные газовые пузырьки могут противостоять внешнему
гидростатическому давлению. Эта стратегия по сути дела противо-
положна стратегии неплавучего фитопланктона, использующего тур-
булентное перемешивание, чтобы удерживаться в верхней части вод-
ного столба.
Таким образом, большинство фитопланктонных клеток не погру-
жается вниз благодаря течениям. Конвекционные вихри, т.е. вызы-
ваемое ветром вращательное движение воды у поверхности (рис. 7.2)
(Langmuir, 1938), возможно, играют при этом наиболее важную
роль, создавая даунвеллинг (движение воды вниз) в местах схожде-
ния (конвергенции) потоков и апвеллинг (движение воды вверх) в
местах их дивергенции (расхождения).
Скорость погружения фитопланктонной клетки непосредственно
зависит от ее размера; при прочих равных условиях крупная-клетка
будет погружаться быстрее мелкой (рис. 7.3). Микроскопические
размеры в сочетании с механизмами уменьшения плотности - важней-
шая адаптация, уменьшающая скорость опускания. При постоянных
объеме и массе клетки любые морфологические изменения, увеличи-
вающие площадь ее поверхности, повысят трение и возможность увле-
чения турбулентным водным потоком. Это продемонстрировано экспе-
риментально (Walsby, Xypolyta, 1977): после ферментативного
удаления хитиновых волокон с диатомеи Thalasaioaira fluviatilis
скорость ее погружения удваивалась. Физиологическое состояние
диатомей также может влиять на скорость опускания: мертвые или
даже стареющие клетки движутся вниз примерно вдвое быстрее, чем
живые. Не исключено, что это - приспособление, которое позволяет
голодающим клеткам войти в более богатые глубинные вода, но ме-
ханизм его не выяснен (Reynolds, 1984). Оно не зависит от обра-
ботки клеток до гибели, а доказательств образования прижизненных
приспособлений к плавучести типа газовых вакуолей нет.
315
Подповерхностный поток
Рис. 7.2. Схема вызываемого ветром поверхностного потока,
приводящего К циркуляции Ленгмюра (по Reynolds, 1984)
Рис. 7.3. Взаимоотношение между скоростью погружения и
диаметром клетки у морских диатомей, формирующих цепочки
клеток (по Reynolds, 1984)
Свет
Многие фитопланктонные виды, возможно, способны изменять
свою фотосинтетическую реакцию на свет. Так, клетка, находящая-
ся в нижней части освещенной (фотической) зоны, увеличивает кон-
центрацию фотосинтетических пигментов и снижает уровень светово-
го насыщения фотосинтеза. Такое увеличение концентрации хлоро-
филла связано с увеличением числа тилакоидов на клетку; с умень-
316
пением освещенности возрастает размер фотосинтетической единицы
(Perry et al., 1981). Хотя не исключено, что теневыносливые
водоросли эффективнее поглощают свет, они не могут повысить эф-
фективность его использования и выход фотосинтеза на фотон погло-
щенного излучения всегда одинаков. У поверхностного или светолю-
бивого планктона содержание хлорофилла ниже, а ртах (см.
рис. 6.6) выше (об этом говорит возрастание уровней карбоксили-
рующих ферментов). Изменения в концентрации хлорофилла, возможно,
вызываются увеличением доли синего света с глубиной, а не просто
уменьшением общей освещенности (Vesk, Jeffrey, 1977). За исключе-
нием ряда синезеленых водорослей и, возможно, некоторых крипто-
фитов, никакой комплементарной хроматической адаптации не наблю-
далось (гл. 6). Многие синезеленые водоросли вызывают мощное по-
верхностное цветение, и у Microcystis увеличивается концентра-
ция каротиноидов, что способствует улавливанию света и отфильтро-
выванию потенциально опасных уровней ультрафиолета ( Paerl et
al., 1983).
Один из интересных вопросов экологии.фитопланктона: встре-
чаются ли постоянно затененные формы в более глубоких слоях оке-
анов? Сообщалось, что многие виды обитают предпочтительно или
исключительно на глубинах 100 м или более. Составленный их спи-
сок ( Sournia, 1982) включает порядка 20 видов из 12 родов и пя-
ти классов и охватывает по сути дела весь спектр размеров и форм
фитопланктона. Хотя, по-видимому, в этих условиях слабой освещен-
ности существует сложившееся водорослевое сообщество, общих мор-
фологических адаптаций не обнаружено. Все еще остается открытым
вопрос, действительно ли водоросли растут или только выживают
на этих глубинах. Показана выживаемость микроскопических водо-
рослей без света в лаборатории ( Antia, 1976); наблюдается также
физиологическая адаптация к слабому свету ( Falkowski, 1981;
Geider et al., 1986). Phaeodactylum tricomutum. Диатомея НО ИЗ
числа теневыносливых, способна существовать, сохраняя основной
метаболизм при освещенности ниже I мкмоль-см^.с--1-, и быстро уве-
личивает интенсивность фотосинтеза при повышении доступности све-
та.
Кроме фотосинтетически активной области спектра, солнечный
свет содержит инфракрасные лучи с длиной волны около 700 нм,
которые поглощаются водой и нагревают ее. Интенсивность нагрева
зависит от объема воды и окружающей температуры. Продолжительное
нагревание поверхности воды снижает ее плотность и приводит к
317
появлению двух (или более) слоев: теплого сверху, более прохлад-
ного под ним (рис. 7.4). Зона между двумя этими слоями называет-
ся термоклином; в ней температура резко падает, и обусловленные
этим различия в плотности препятствуют обмену воды между глубин-
ным (гиполимнион) и поверхностным (эпилимнион) слоями, хотя цир-
куляция в пределах их происходит (рис. 7.5). В прибрежных зонах
расслоение по плотности может также вызываться снижением солено-
сти на поверхности из-за притока пресной воды с суши или таяния
морского льда. Границы между такими обусловленными плотностью
или соленостью слоями воды называются соответственно пикноклином
или галоклином. Расслоение (стратификация) имеет важные экологи-
ческие последствия, поскольку поверхностные воды мотут оказаться
обедненными питательными веществами, а более глубокие будут бога-
ты биогенами, недостижимыми для водорослей, находящихся выше тер-
моклина. Термоклины на мелководье развиваются только в спокойных
условиях, когда не происходит турбулентного перемешивания ветром.
Если же они оказываются на глубине, недоступной для влияния вет-
ра, то могут сохраняться до тех пор, пока поверхностные воды не
охладятся достаточно дая того, чтобы опуститься на глубину и пе-
ремешаться. В умеренных регионах они в значительной степени се-
зонны, т.е. встречаются летом. В более высоких широтах, по край-
ней мере в океанах, поверхностные воды бывают холодными на протя-
жении всего года, термоклин развивается здесь очень слабо. И на-
оборот, в тропиках расслоение воды существует круглогодично.
Биогенные элементы
Фитопланктон нуждается в ряде как неорганических, так и ор-
ганических биогенов, представленных разбавленными растворами,
концентрации которых варьируют в пространстве и времени. Их до-
ступность имеет огромное экологическое значение, поскольку все
водоросли нуждаются в одних и тех же макроэлементах (исключение
составляет потребность диатомовых и некоторых жгутиковых в сили-
катах) .
Фосфаты, неорганические соединения азота и силикаты (для
Диатомей)требуются в наибольшем количестве (гл. б), и их влияние
Да фитопланктонные популяции изучалось наиболее интенсивно. В
Морской среде азот, по-видимому, является макроэлементом, чаще
всего лимитирующим рост ( Ryther, Dunstan, 1971), тогда как опы-
318
Глубина, м
Рис. 7.4. Схема положения температурных слоев в зависимо-
сти от глубины и времени в озере.умеренной зоны, показы-
вающая влияние летнего термоклина (по Jenkin, 1942)
Рис. 7.5. Схема вертикальной
циркуляции выше и ниже термо-
клина. Т - изменение темпера-
туры в завис илости от глубины
(по Reynolds, 1984)
319
ты по удобрению озера позволяют приписать такую же роль фосфору
В пресных водах (рис. 7.6) ( Schindler, 1974, 1977; см. также
гл. 9).
Обмен энергией и биогенами происходит через поверхность фи-
топланктонной клетки, а для клетки любой формы отношение площади
к объему (S/V) падает с увеличением последнего. Следовало бы,
таким образом, ожидать, что скорость роста фитопланктона и отно-
шение s/v положительно коррелируют; это было показано как в
природе, так и на лабораторных культурах ( Sournia, 1982). Пар-
сонс и Такахаси ( Parsons, Takahashi, 1973) сравнили влияние раз-
мера клетки на скорости роста диатомеи Ditylum brightwelli
и нанопланктонной кокколитофориды Emiliania huxleyi. Скорость
роста крупных клеток была выше, чем у мелких, только при сильном
освещении, высоких концентрациях биогенов и нестабильности вод-
ного столба. Это, возможно, объясняет, почему наноцланктои коли-
чественно превосходит сетной планктон в большинстве морских эко-
систем. Показано, что отношение s/v возрастает в средах с дефи-
цитом биогенов (Harrison et al., 1977). Это дает основание по-
лагать, что мелкие водоросли с высоким отношением s/v получат
преимущество в олиготрофных водах. Для пресных вод подтверждено
( Watson, Kalff, 1981) увеличение доли нанопланктона вдоль сни-
жающегося градиента концентрации биогенов. Однако крупные водо-
росли способны запасать большие их количества. Планктонные формы
типа динофлагеллят, мигрирующие ночью в богатые биогенами воды
гиполимниона и поднимающиеся к освещенной поверхности днем, име-
ют явные экологические преимущества ( Dugdale, Goering, 1967).
Добавление питательных веществ может повышать скорость рос-
та и конечную биомассу фитопланктонной популяции, однако некото-
рые данные говорят о том, что биогены не всегда лимитируют ее
рост, даже когда их концентрация и биомасса водорослей малы. При-
родные фитопланктонные популяции не показывают резких отклонений
от отношения Редфилда с:Н:Р , типичного для сильного ограниче-
ния питательных веществ (гл. 6), и рост их, по-видимому, близок
к потенциально максимальному ( Goldman et al., 1979). Выращенный
при дефиците азота фитопланктон быстро поглощал ионы бикарбона-'
та в темноте, когда получал ионы аммония ( Yentsch et al., 1977).
Эта методика использована для проверки азотного голода у популя-
ций прибрежного фитопланктона в заливе Мэн. Наблюдались три цве-
тения: Chaetoceros в апреле, Skeletonema в июле и Gonyanlax
в сентябре; недостаток азота обнаружен только в периоды цвете-
320
Рис. 7.6. Влияние экспериментального внесения в озеро удоб-
рений на биомассу фитопланктона (количество хлорофиллах, В
1269 г. удобрение не проводилось; в 197£ г. вносили NH,,
ррХ- и сахарозу, а в 1973 г. - NH. и сахарозу.
Пик 1971 г. показывает, что из трех вносимых веществ лими-
тирующим является фосфат (по Schindler, 1974)
ния; это дает основание полагать, что размер популяций между ни-
ми уравновешен круговоротом азота, а цветения связаны с апвеллин-
гом, приносящим дополнительный азот в поверхностные воды.
Таким образом, наблюдается парадокс: популяция в целом, су-
дя по конечной биомассе, лимитирована питанием, а индивидуальные
клетки, судя по концентрациям внутри их биогенов, - нет. Полага-
ют ( Maestrini, Bonm, 1981 b ), что концепция нелимитированно-
го питанием роста должна ограничиваться видами, способными к бы-
строй ассимиляции и получающими преимущество в краткие периоды
поступления биогенов. Такие богатые питательными веществами мик-
ронити, вероятно, связаны с экскретами животных или реминерализа-
цией мертвых организмов ( McCarthy, Goldman, 1979).
Многие представители фитопланктона способны поглощать орга-
нические соединения в темноте ( Droop, 1974). Сообщалось, что во-
доросли с нижней поверхности морского льда в Антарктике ассимили-
руют аминокислоты при их внешних концентрациях, но это дает ме-
нее 1% углерода, получаемого за счет фотосинтетической фиксации
( Palmisano et al., 1985). Однако наиболее убедительно экологиче-
ское значение поглощения органических веществ показано .для фото-
321
гетеротрофности, а не для темнового поглощения. Свет стимулиро-
вал поглощение ацетата фитопланктоном в нижних слоях фотической
зоны озера Тахо (граница Калифорнии и Невада), но не влиял на та-
кое поглощение в более высоких частях водного столба. В этом фо-
тогетеротрофном сообществе отношение потребления углерода из аце-
тата и неорганических источников составляет от 0,25 до 2,5 ( Vin-
cent, Goldman, 1980). Сообщалось также о поглощении органичес-
ких частиц потенциально фотосинтезирующими водорослями, а неко-
торые пресноводные золотистые водоросли из родов Dinobryon и
Uroglena способны поедать бактерии со скоростями, близкими к
указывавшимся для нефотосинтезирующих жгутиковых (Bird, Kaiff,
1986). В более глубоких водах Dinobryon получает по крайней ме-
ре 50% углерода за счет бактерий и извлекает их из вода больше,
чем ракообразные, коловратки и инфузории, вместе взятые.
v
Выедание животными
Потери в фитопланктонных популяциях количественно оценить
еще труднее, чем их прирост. Клетки мотут непрерывно опускаться
в глубину, страдать от паразитизма или нападения микробов ( Shi-
1о, 1971; Youngman et al., 1976), но, по-видимому, общепризнано,
что потери от выедания (рис. 7.7) наиболее существенны ( Frost,
1980; Crumpton, Wetzel, 1982). Ведущую роль в этом плане играет
зоопланктон. В океанах наиболее широко изучалось выедание водо-
рослей веслоногими ракообразными. Другие потребители юрского
фитопланктона относятся к простейшим,преимущественно инфузориям
( Penchel, 1980) и тинтиннидам ( Capriulo, Carpenter, 1980), а
также аппендикуляриям (Alldreage, 1981), сальпам и крылоногим
моллюскам (Silver, Bruiand, 1981); в пресных водах это преимуще-
ственно ракообразные и коловратки, хотя простейшие (Canter, 1973)
и хитридиомицеты (canter, Jaworski, 1978) могут существенно
влиять на популяции диатомей.
Любые обобщения относительно влияния размера водорослевой
клетки на возможность ее поедания представляются рискованными.
Полагают (Kaiff, Knoechel, 1978), что в олиготрофных пресных
водах меньше всего страдают крупные планктонные формы типа дино-
флагеллат и колониальных видов. Часто указывается, что более мел-
кие водоросли чувствительнее к выеданию. Портер (Porter, 1973)
показал, что зоопланктон снижает долю мелких водорослевых клеток
и позволяет развиваться более крупным, таким, как десмидиевые,
322
г Юг-
Май Июнь Июль Авг Сен Окт
Рис. 7.7. А. Популяцион-
ные кривые для Cyclotel-
la michiganiana (...).
Cyclotella comehsist---)
и Sphaerocystie sohroe-
terl (----b. про-
зрачность (измеренная по
диску 1!екиь В. Скорости
выедания. Первый из каж-
дой пары вертикальных
столбиков - для .дневного
времени, второй - для
ночного. Г. Биомасса
зоопланктона в дневное
(...) и ночное (----)
время. Видна корреляция
в середине августа между
уменьшением биомассы во-
дорослей и увеличением
прозрачности, скорости
фильтрации и биомассы
зоопланктона (по Crump-
ton, Wetzel, 1982)
323
диатомеи и динофлагелляты. Подчеркивалось также ( Prost, 1980;
Sournia, 1981), что водоросли любого размера не застрахованы от
выедания и значительная часть биомассы любого фитопланктона до-
ступна растительноядным животным, хотя отдельные синезеленые ви-
ды могут быть менее съедобны, чем представители других трупп
( Arnold, 1971; Porter, 1973). Давно известны отрицательная кор-
реляция между плотностью популяций фитопланктона и зоопланктона
и неравномерное распределение планктонных видов; растительнояд-
ные животные - один из основных факторов, обусловливающих эту
пятнистость. Наблюдение за популяцией веслоногого рачка Calanus
finmarchicus в течение трех месяцев ( Cushing, Vucetik, 1963)
показало, что численность фитопланктона сокращалась, в то время
как неорганические биогены оставались в изобилии.
Не следует полагать, что фитофаги усваивают весь съеденный
органический материал. У веслоногих-каланид примерно 20-30% орга-
нического углерода фитопланктона выделяется с фекалиями ( Eppley,
Peterson, 1979). Некоторые тропические пресноводные рыбы, питаю-
щиеся водорослями, усваивают в среднем только 43% поглощенного
углерода ( D.Moriarty, С,Moriarty, 1973). Кроме того, выеда-
ние животными позволяет органическому материалу перейти на более
высокие трофические уровни, являясь первой ступенью в реминерали-
зации неорганических биогенов, позволяющей расти другим водорос-
лям. Выедание может быть благоприятным для некоторых видов. Так,
колонии пресноводной зеленой водоросли Sphaerocystis schroeteri
только частично разрушаются, проходя через кишечник Daphnia тая-
ла, а основная часть выделяется наружу неповрежденной и в жизне-
способном состоянии. При этом они усваивают из пищеварительного
тракта рачка фосфаты, стимулирующие фотосинтез и деление дефеци-
рованных клеток.Это улучшение роста может компенсировать незначи-
тельные потери популяции вследствие выедания (Porter, 1976).
Динамика популяций
Фитопланктон редко распределен равномерно по водоему; зна-
чительная горизонтальная пятнистость - скорее правило, чем исклю-
чение, причем выражена она в разном масштабе; пятна достигают раз-
меров от менее миллиметра до десятков или даже сотен километров
( Platt, Denham, 1980). Наблюдаются также различия в вертикаль-
ном распределении, причем оно,как и горизонтальное, может менять-
ся со временем. Изменения происходят с разным временным масшта-
324
бом: они бывают суточными, сезонными или долговременными, зани-
мающими много лет. Сообщество фитопланктона должно, таким обра-
зом, рассматриваться как трехмерное пятно, границы и видовой сос-
тав которого меняются со временем.
Хорошо известны суточные флуктуации в биологии водорослевых
клеток (Chisholm, 1981; Sweeney, 1983); многие из них обуслов-
лены сменой дня и ночи и являются эндогенными (гл. 6). Такой цик-
личности подчинены клеточное деление, вертикальная миграция, фо-
тосинтетическая активность, биолюминесценция и поглощение биоге-
нов. Эти циркадианные ритмы гарантируют оптимальное физиологиче-
ское состояние клетки в условиях среды, изменяющейся с периодом
24 ч. Поскольку не у всех видов ритмы одинаковы, это может быть
механизмом, снижающим интенсивность межвидовой конкуренции.
Исследованы ( Happey-Wood, 1976) вертикальные миграции не-
скольких фитофлагеллат (включая ОоЪгошопав, Chroomonas, Mallomo-
nas. Cryptomonas)за 24 ч в пресноводном пруду (рис. 7.8). Виды
перемещались вниз в темноте и возвращались вверх в течение дня.
При ясном небе они скапливались немного ниже поверхности, а в
пасмурную погоду - практически на ней самой. Неподвижная зеленая
водоросль Oocystis служила контролем, показывая, что эти пере-
мещения были активными, не являясь только следствием циркуляции
воды.
Сезонные изменения, особенно в озерах умеренных широт, от-
носятся к числу наиболее изученных аспектов экологии фитопланкто-
на. Как температура, так и освещенность зимой низкие, и хотя био-
гены могут встречаться в изобилии, биомасса фитопланктона и его
продуктивность понижаются. Возрастание освещенности весной стиму-
лирует рост и в зависимости от стабильности водного столба может
приводить к цветению. В поверхностных слоях этому способствует
также развитие термоклина. Цветение продолжается до тех пор, по-
ка выедание водорослей и истощение ими запаса биогенов не приво-
дят к летнему спаду обилия. Видовой состав в любой момент време-
ни зависит от многих факторов и, возможно, более всего от богат-
ства воды основными биогенами. В целом протекает сукцессия от
мелких видов к крупным, которую можно предсказать исходя из тео-
ретических предпосылок ( Margalef, 1978). В весенних цветениях
доминируют диатомовые, они выедаются фитофагами и сменяются ле-
том более крупными динофлагеллатами и зелеными жгутиковыми. В
эвтрофных водах в более теплые периода позднего лета может проис-
ходить цветение синезелеными водорослями. Осенью, когда поверх-
325
Время, ч
Рис. 7.8. Вертикальное распределение фитопланктона в пруду
за 24 ч. Сверху показана облачность несоответствующее время
(ПО Happey-Wood, 1976)
костные воды охлаждаются и исчезновение термоклина делает возмож-
ным их смешение с более глубокими, богатыми биогенами слоями воды,
возможно второе диатомовое цветение (рис. 7.9).
Высокоширотные озера отличаются от озер умеренной зоны бо-
лее длительным сохранением ледяного покрова. В полярных регионах
известны постоянно замерзшие озера с альгофлорой (Baker, 1967).
Однако в большинстве из них максимальный рост водорослей наблюда-
ется непосредственно после таяния льда, а видовой состав таких
озер крайне изменчив (Round, 1981). Сукцессия видов, возможно,
коррелирует с длительностью свободного от льда периода, и иногда
бывает только одно летнее цветение (рис. 7.10).
326
Рис. 7.9. Сезонные изменения биомассы водорослей (концент-
рации хлорофилла) в озере умеренных широт. Тонкая линия -
1961 г., жирная линия - 1963 г. Заметны весенний и осенний
пики (по Заоиерапвку, 1966)
Рис. 7.10. Сезонные изменения биомассы в высокоширотном
озере, которое в течение пяти месяцев в году покрыто
льдом (по Ilmavirta, Kotimaa, 1974)
327
И наоборот, в тропических озерах сезонная изменчивость
меньше, чем в озерах умеренных широт. Многие из них эвтрофны
вследствие быстрой реминерализации при более высоких температу-
рах воды. Одна из интересных особенностей планктона таких озер -
наличие в нем перистых диатомей, например Nitaschia. которые яв-
ляются преимущественно бентосными в озерах умеренных широт
( Richardson, 1968). В тропических озерах биомасса водорослей,
по-видимому, в целом больше, а видовое разнообразие меньше, чем
в умеренной зоне. Луис (Lewis, 1978) отмечает незначительные
сезонные изменения и доминирование представителей Chlorophyceae
в озерах Филиппин. В озере Джордж в Уганде - одном из наиболее
интенсивно изучавшихся тропических водоемов - преобладают сине-
зеленые водоросли, составляющие до 70 - 80% общей биомассы. Хо-
рошо представлены здесь и зеленые водоросли, а диатомеи относи-
тельно редки. Колебания биомассы в течение года незначительны:
минимум от максимума отличается всего вдвое. Это было определе-
но по уровням хлорофилла со слабыми пиками, приходящимися на пе-
риоды максимальных осадков (рис. 7.II) (Ganf, 1974-)»
Если взять обзоры распространения фитопланктона и сукцес-
сии видов в морских местообитаниях (Smayda, 1980), то окажется,
что, как и в случае пресных вод, больше всего исследований про-
ведено в зоне умеренного климата, особенно у побережья. В целом
диатомеи доминируют в более холодных, богатых биогенами водах,
кокколитофориды - в более теплых олиготрофных, а динофлагеллаты,
по-видимому, предпочитают промежуточные условия. Основное эколо-
гическое различие обычно наблюдается между фитопланктонными фло-
рами прибрежных (неритических) и океанических вод, хотя экологи-
ческие факторы, которыми обусловлены их различия, остаются неяс-
ными. Прибрежные воды, несомненно, испытывают влияние стока с
суши, возможно более богатого, чем море, питательными вещества-
ми. Вдобавок к этому у прибрежного планктона вероятнее, чем у
обитающего в открытом море, присутствие бентосных стадий в жиз-
ненном цикле.
В крупных водоемах важно различать истинную сукцессию, пред-
ставляющую собой изменение видового состава в пределах данной
водной массы, и смены, вызванные замещением одного объема воды
с характерной для него альгофлорой другим, с иным набором видов.
Хороший пример истинной сукцессии приводился для прибрежных вод
Норвегии ( Braarud et al., 1958), где весеннее диатомовое цве-
тение сменялось меньшей по видовому составу и обилию флорой эв-
328
Месяц
Рис. 7.II. Сезонные изменения биомассы водорослей (кон-
центрации хлорофилла) в тропическом озере Джордж. Каждая
линия - данные за один год между 1967 и 1971 гг.; заме-
тим, что сезонные изменения биомассы незначительны. Для
контраста приведены максимальная и минимальная концент-
рации хлорофилла за тот же период для озера умеренных
широт (Лох-Ливен) (по Ganf, 1974)
гленовых, а та в свою очередь сменялась летним максимумом дино-
флагеллат и кокколитофорид (рис. 7.12). Однако у побережий измен-
чивость видового состава фитопланктона очень велика. В проли-
ве Лонг-Айленд и заливе Наррагансетт у восточных берегов США из-
вестны сезонные сукцессии, отличающиеся от описанных для Норве-
гии и даже для соседнего залива Мэн ( Smayda, 1981). Популяции
фитопланктона здесь крупнее и "цветут" чаще, так что четкие пики
весеннего и летне-осеннего цветений смазываются (рис. 7.13).
Здесь меньше особенно обильных видов, а диатомеи, в частности
Skeletonema coetatum . доминируют постоянно, тогда как динофла-
геллаты повышают численность летом. Однако в отличие от прибреж-
ных вод Норвегии динофлагеллаты, в частности Ceratium. и кокко-
литофориды почти полностью отсутствуют в летней альгофлоре. Сук-
цессия, таким образом, сведена прежде всего к серии сезонных ко-
лебаний обилия наиболее обычных видов.
В открытых океанах значительны широтные различия видового
состава альгофлоры и ее сезонные изменения. При низких темпе-
329
Рис. 7.12. Сезонные изменения основных компонентов фито-
планктона на двух станциях (Скрова и Эггум) в прибрежных
водах Норвегии. В каждом случае максимальная численность
принимается за 100% (по Smayda, 1980; данные из Braarud
et al., 1958)
Месяц
Рис. 7.13. Сезонная встречаемость и частота важнейших фи-
топланктонных видов в проливе Лонг-Айленд (по Smayda,
1980; данные из Riley, 1967)
ратурах моря Лабрадор (летний максимум ниже 10°С) в годичном
цикле доминируют диатомеи, причем Fragilaria папа- дважды, в
июне и в сентябре-октябре ( Holmes, 1956). За весенним цветени-
ем не следовало преобладания динофлагеллат, хотя Exnviaella
baltica была относительно обильна с июня по конец августа. В бо-
лее умеренных водах ближе к Исландии (летний максимум примерно
15°С) за мартовским кокколитофоридным цветением Emiliania hux-
leyi в мае следовал пик крупных диатомей с доминированием
Rhizosoienia врр. К июлю он сменялся максимумом динофлагеллат,
представленных несколькими видами Ceratium и Peridinium .которым
в свою очередь в сентябре приходили на смену виды Exuviaella
(динофлагеллаты) и Thalassioeira (диатомеи).
331
В тропиках у берегов часто находятся зоны апвеллинга глубин-
ных вод, которые прохладнее и богаче биогенами, чем поверхност-
ные. Здесь значительны различия между прибрежными и более морис-
тыми популяциями фитопланктона, а поскольку апвеллинг может
иметь сезонный характер, встречаются хорошо выраженные годичные
никлы планктона. У берегов Западной Африки холсдноводная альго-
флора с доминированием диатомей, в основном Chaetoceros, Rhizo-
aolenia, Coscinodiscus, развивается между июлем и октябрем. В
остальное время года альгофлора более тепловодная, в ней преоб-
ладают динофлагеллаты ( Reyssac, Roux, 1972).
Тропические океанические воды бедны биогенами, и эта их де-
фицитность подтверждена опытами по обогащению среды ( Giooschen-
ko, Curl, 1971). Численность фитопланктонных клеток обычно неве-
лика, диатсмовые и кокколитофориды являются наиболее обильными
группами ( Round, 1981). В некоторых тропических морях довольно
обычна и может давать обширные цветения синезеленая водоросль
Trichodesmium thiebaudii (Fogg, 1982). Сезонная сукцессия отме-
чена (Hulbert et al., I960) в субтропических водах Саргассова
моря (рис. 7.14). Кокколитофориды встречаются на протяжении все-
го года, причем доминирует единственный их вид Emiliania hux-
leyi. Летом, с мая по октябрь, альгофлора изреживалась и домини-
рование переходило от кокколитофсрид к динофлагеллатам. В ноябре
исчезал летний термоклин и доминантой становилась Emiliania
huxleyi. В апреле кратковременно доминировала диатомея Rhizoso-
lenia, что совпадало с началом температурного расслоения
воды.
Парадокс планктона
Хатчинсон (Hutchinson, 1961) описал феномен, который он
назвал "парадоксом планктона", заключающийся в том, что 10 - 50
видов водорослей, по-вицимсму, сосуществуют в как будто однород-
ных по условиям водоемах. Это на первый взгляд противоречит прин-
ципу конкурентного исключения, утверждающему, что два или более
вида не могут занимать одну и ту же экологическую нишу (Hardin,
I960). Хатчинсон и другие авторы ( Richerson et al.,1970; Piatt,
Denham, 1980) полагают, что взаимоподавление отсутствует здесь
из-за неоднородности местообитания вследствие временных и про-
зтранственных флуктуаций условий среды.
. Альтернативное объяснение этсму явлению - возможность сосу-
332
Месяц
Рио. 7.14. Сезонные изменения основных фитопланктонных
групп в поверхностных водах Саргассова моря. Концентра-
ции показаны в процентах от максимальных, наблюдавшихся
для каждой группы. Жирная линия для кокколитофорид озна-
чает процент, представленный Emiitania huxleyi (по
Smayda, 1980; данные из Hilbert et al., 19ЬО)
шествования нескольких видов в равновесии в стабильных условиях
среды, если каждый из них лимитирован специфическим ресурсом.
Это было проверено (Tilman, 1977) при выращивании в полунепре-
рывной культуре двух пресноводных диатомей со сходными максималь-
ными скоростями роста - Aaterionella Тоипоаа И Cyclotella mene-
ghiniana - при разных соотношениях концентраций силиката и
фосфата (рис. 7.15). Aaterionella способна поглощать фосфор эф-
фективнее, чем Cyclotella, при низких его концентрациях; для
силикатов ситуация обратная. Таким образом, при высоких отноше-
ниях Si:Р конкурентное преимущество получает Aaterionella,
а при низких в более благоприятных условиях оказывается Cyclo-
tella, При промежуточных отношениях виды будут стабильно сосу-
ществовать, поскольку лимитированы разными биогенами. Это объяс-
нение распространено на многовидовые системы ( Sommer, 1983).
Однако показано также ( Sommer, 1984), что колебания уровня
333
Si:P Si:P
Рис. 7.15. Конкуренция между Asterionella formoaa и
?yElo.te-Lla иепея;Мп1апа вдоль градиента концентраций
31:р. а. Связанная о источником питания кинетика роста
каждого вида в чистой культуре, g. Результаты опытов
по изучению конкуренции в зависимости от скорости роста
и отношения si :Р.светлые кружки - доминирует Aeterloneiia;
затушеванные кружки - доминирует c^cloteiia; светлые тре-
угольники - оба вида сосуществуют. В. Относительное оби-
лие Cyclotell а как процент общего числа клеток обоих ви-
дов в пробах из озера Мичиган и отношение si.-p в ?дх же
Пробах (по Parley, 1982; данные из Tilman, 1977)
единственного биогена повышают видовое разнообразие, позволяя
сосуществовать конкурирующим за это вещество видам.
Эпилитные водоросли-макрофиты
Эпилитные водоросли встречаются на твердых субстратах в
биотопах, где движение воды препятствует накоплению осадков;
к таким местообитаниям относятся ложа рек и прибрежные зоны озер
334
t
и океанов. Морские эпилитные макрофиты привлекли наибольшее вни-
мание экологов. Сравнительная структурная простота сообществ с их
доминированием в сочетании с быстрым ростом видов делает такие
экосистемы идеальным полигоном для экспериментов с перемещением
видов, в которых проверяются экологические гипотезы. За послед-
ние два десятилетия такие исследования значительно повлияли на
общеэкологические теории, особенно на связанные с конкуренцией
и выеданием.
Жизненные формы и адаптивная морфология
Морские макрофиты относятся к трем основным таксономическим
группам: Chlorophyceae, Phaeophyceae И Rhodophyceae(Lobbап,
Wynne, 1981), хотя местами в изобилии встречаются и важны в эко-
логическом плане колониальные или нитчатые формы синезеленых,
желтозеленых, золотистых и диатомовых водорослей. Морские макро-
фиты - самые крупные и наиболее структурно сложные из водорос-
лей, причем в трех первых классах наблюдается примечательный па-
раллелизм морфологии (гл. 4), хотя с эволюционной точки зрения
они представляют далеко отстоящие друг от друга линии (гл. 8).
По всей видимости, этот параллелизм имеет адаптивную природу и
обусловлен приспособленностью филетически неродственных организ-
мов к общему местообитанию. Это предположение подтверждается
тем, что на общую морфологию макрофитов условия среды могут вли-
ять сходным образом независимо от систематического положения
( Morton et al., 1981).
Признание важности морфологии привело к попыткам экологиче-
ских классификаций макрофитов на основе их жизненных форм, как
это было сделано Раункиером ( Raunkiaer, 1934) для наземной рас-
тительности. При этом стремятся объединить под одним названием
морфологию таллома, длительность жизни и жизненный цикл; такой
подход наиболее пригоден для описания стратегий переживания в
биогеографическом контексте ( Garbary, 1976). Имеются и другие
классификации жизненных форм (Feldmann, 1951; V.Chapman, D.Chap-
man, 1976; Ruaaell,I977). Многие исследователи пользуются та-
кими терминами, как "однолетний", "многолетний" и "ложномноголет-
ний" (например, Seara, Wilce, 1975), которые имеют некоторые
аналоги в схемах жизненных форм.
Предложена ( «.Littler, D.Littler, 1980) гипотеза функ-
циональной формы, согласно которой экологическая классификация
335
водорослей учитывает адаптивные особенности структуры и функции
в виде баланса издеркек/преимуществ (табл. 7.1 и 7.2). Проводит-
ся различие между оппортунистическими формами с быстрым ростом,
малой биомассой и высоким репродуктивным потенциалом (прошедшими
г-отбор) и позднесукцессионными видами с белее медленным ростом,
более высокой конечной биомассой и меньшим репродуктивным потен-
циалом (прошедшими К-отбор). Литтлер и Арнолд ( littier, Arnold,
1982) анализируют фотосинтетическую активность шести таких функ-
циональных форм; она сильнее всего у тонкотрубковидных и пластин-
чатых вадов и слабее всего у корковых. Экологически это отража-
лось в том, что быстрорастущие г-вида характеризовали ранние
стации сукцессии в колеблющихся или нестабильных условиях, тогда
как К-виды встречались в более устойчивых местообитаниях. Следует
ожидать, что К-виды устойчивее к выеданию. Все говорит за то,
что развитие медленно растущих корковых талломов будет подавлять-
ся эпифитами, если последние не уничтожаются растительноядными
животными или какими-либо физическими факторами (Jackson, 1977;
Wanders, 1977). Предположено, что гетероморфные жизненные циклы
водорослей, в которые входит корковый таллом, могли развиться в
ответ на выедание ( Lubchenco, Cubit, 1980; Slocum, 1980).
Сукцессия в понимании экологов наземных растений подразуме-
вает изменение среды каждой из последовательных стадий таким об-
разом, что облегчается развитие серии до формирования климаксово-
го сообщества. Многие внешне сукцессионные закономерности наблю-
дались и в морском бентосе. Лодж (Lodge, 1948) указывает четыре
сукцессионные стадии на расчищенной полосе литорали острова Мэн
(Великобритания): I) диатомовая слизь; 2) нитчатые эфемеры, та-
кие, как Ulothrixs 3) Плотный ковер Enteromomha-.4) климакоо-
вне заросли Pucus. Другие исследования показали, что сукцессион-
ные стадии бывают менее преданазуемиии, причем к климаксу может
вести несколько путей ( Poster, 1975; Sutherland, Karlson, 1977).
Мало подтверждается теория, согласно которой ранние стадии гото-
вят условия для последующих ( Connell, Slayter, 1977); наблюдае-
мая сукцеооия, возможно, просто обусловлена быстрым ростом и ре-
яродуктивнши способностями эфемерных пионерных водорослей.
Диогены
В отличие от ситуации с фитопланктоном нам мало известно о
Роли биогенов в контроле роста и структуры сообществ морских мак-
336
Таблица 7,1. Гипотетические (a priori) стратегии выживания оп-
портунистических макроскопических водорослей (представителей
нарушенных!'сообществ) и макроскопических водорослей ненарушен-
ных* сообществ (по Littler, Littler, 1980)
Оппортунистические формы Позднесукцессионные формы
I. Быстро заселяют свободную
поверхность
2. Эфемеры, однолетники или
многолетники с недолгой веге-
тативной фазой в жизненном
цикле
3. Форма таллома относительно
простая (недифференцированная);
мелкий размер; биомасса неболь-
шая; высокое отношение площади
поверхности таллома к его объе-
му
4. Быстрый рост и высокая чис-
тая первичная продуктивность
целого таллома; практически
все ткани фотосинтезирующие
I. Не заселяют быстро субстрата
(присутствуют преимущественно
на поздних стадиях сукцессии);
внедряются в пионерные сообщест-
ва в определенный сезон
2. Более сложные и длительные
жизненные циклы; размножение в
оптимальный сезон
3. Таллом структурно и функцио-
нально дифференцирован (круп-
ные талломы с большой биомас-
сой) ; низкое отношение площади
поверхности таллома к его объе-
му
4. Медленный рост и низкая чис-
тая продуктивность палого тал-
лома вследствие дыхания нефото-
синтезирующей ткани и меньшего
количества протоплазмы на еди-
ницу тела водоросли
5. Высокий общий репродуктив-
ный потенциал, почти все клет-
ки в принципе способны к вос-
произведению; много диаспор,
но энергия, вкладываемая в об-
разование каждой из них, низка;
диаспоры высвобождаются круг-
лый год
6. Калорийность высока и рав-
номерно распределена по тал-
лому
5. Низкий общий репродуктивный
потенциал; имеются специализи-
рованные репродуктивные ткани;
на отдельную диаспору приходит-
ся относительно много энергии
7. Разные звенья жизненного
цикла имеют сходную оппортуни-
стическую жизненную стратегию;
изоморфная смена поколений; мо-
лодые талломы представляют со-
бой лишь уменьшенные варианты
более старых
6. Калорийность низка в некото-
рых структурных компонентах и
неоднородно распределена по
частям таллома; могут запасать-
ся высококалорийные соединения
для переживания вполне опреде-
ленных неблагоприятных сезонов
7. Разные звенья жизненного
цикла могут проявлять разные
жизненные стратегии; гетероморф-
ная смена поколений; стратегии
молодых талломов бывают близки
к известным для оппортунистиче-
ских форм
337
Продолжение табл. 7.1
Оппортунистические формы Позднесукцессионные формы
8. Избегают выедания за счет несовпадения с фитофагами по встречаемости во времени или пространстве или же благода- ря быстрому росту (фитофаги не успевают их уничтожить) 8. Избегают выедания фитофага- ми благодаря комплексу структур- ных и химических механизмов за- щиты
Молодые или флуктуирующие во времени.
2 Зрелые, устойчивые во времени.
338
Таблица 7.2. Гипотетические издержи и выгоды стратегий выжива-
ния, рассмотренных в табл. 7.1 для оппортунистических и поздне-
сукцессионных водорослей-макрофитов (по Littler, Littler, 1980)
Оппортунистические формы
Позднесукцессионные формы
Издержки
I. Высокая смертность диаспор
2. Мелкие и просто устроенные
талломы легко вытесняются при
затенении их более крупными
формами
3. Нежные талломы легко сми-
наются и повреждаются более
грубыми формами
I. Медленный рост; низкая чис-
тая продуктивность целого тал-
лома приводит к длительному на-
чальному периоду развития
2. Образование диаспор редкое
и в небольших количествах
4. Таллом относительно легко
доступен для фитофагов и чув-
ствителен к выеданию ими
5. Нежные талломы легко выры-
ваются прибоем и повреждаются
твердыми частицами осадков
6. Высокое отношение площади
поверхности к объему приво-
дит к большему высыханию на
воздухе
7. Меньший выбор экологичес-
ких условий из-за сходства
фаз жизненного цикла
3. Низкое отношение площади по-
верхности к объему делает отно-
сительно неэффективным поглоще-
ние биогенов, присутствующих в
низких концентрациях
4. Случаи массовой гибели бо-
лее опасны из-за медленного
возобновления и общей меньшей
плотности популяций
5. Требуется относительно мно-
го энергии и материалов для за-
щиты долгоживущих структур
(эта энергия становится недо-
ступной для роста и размноже-
ния)
6. Физиологическая специализа-
ция чревата стенотопностыо
7. Большие издержки на дыхание
из-за необходимости обеспечи-
вать существование структурных
тканей (особенно при неблаго-
приятных для роста условиях)
Выгоды
I. Высокая продуктивность и
быстрый рост позволяют быстро
осваивать незанятый субстрат
I. Большая жизнеспособность
диаспор (больше энергии на диа-
спору) снижает их смертность
339
Продолжение табл. 7.2
Оппортунистические формы
Позднесукцессионные формы
2. Образование диаспор в изо-
билии и постоянно
3. Высокое отношение площади
поверхности к объему благо-
приятствует быстрому поглоще-
нию биогенов
4. Быстрое восстановление тка-
ней может снижать эффект выеда-
ния и компенсировать смертность
5. Могут избегать выедания
из-за несовпадения с фитофа-
гами во времени или простран-
стве
6. Физиологически неспециа-
лизированы и потому более эв-
ритопны
2. Дифференцировка (например,
длинная ножка) и больший раз-
мер повышают конкурентоспособ-
ность в борьбе за свет
3. Структурная специализация
повышает прочность и конкурен-
тоспособность в борьбе за про-
странство
4. Фотосинтезирующие и репро-
дуктивные структуры относи-
тельно недоступны и устойчивы
к выеданию эпилитными фитофа-
гами
5. Устойчивы к физическим
стрессам типа вырывания волна-
ми и механического повреждения
частицами осадка
6. Низкое отношение площади по-
верхности к объему снижает по-
терю воды на воздухе
7. Больший выбор экологических
ниш из-за неодинаковой страте-
гии разных фаз жизненных цик-
лов
8. Механизмы запасания пита-
тельных веществ, сбрасывание
"дорогостоящих" частей тела и
изменчивая физиология позволя-
ют выживать в неблагоприятные,
но наступающие в строго опре-
деленное время сезоны
340
рофитов. Наиболее детальная информация накоплена об азотном бюд-
жете представителей Laminarialea. Чапман и Крейги (Chapman, Сга-
gie. 1977) связывали рост Laminaria, longicrur1зВ бухте
Сент-Маргарет (Новая Шотландия, Канада) как с окружающим уров-
нем; так и с внутренними запасами азота (рис. 7.16). Быстрый
рост начинается в декабре, что коррелирует с увеличением концен-
трации нитрата в морской воде. Скорость роста продолжает возрас-
тать, достигая пика в начале мая - июне, примерно через два ме-
сяца после резкого снижения в море концентрации нитрата. Этот
рост поддерживается нитратом, запасенным в ткани Laminaria. Пик
фотосинтеза наблюдается в шале, когда рост медленнее всего; это
приводит к накоплению углеводов (лаиинарина), вероятно исполь-
зуемых для роста в начале зимы, когда освещенность падает. Таким
образом, Laminaria longicruria благодаря своей способности за-
пасать как нитрат, так и углеводы, поглощает азот в условиях эго
максимального обилия осенью и зимой и использует его для роста
весной, когда освещение наиболее благоприятно для фотосинтеза.
В другой месте у берегов Новой Шотландии, где апвеллинг поддер-
живал относительно высокие уровни азота в воде круглый год, этой
стратегии роста не наблюдалось; запасы как углеводов, так и не-
органического азота были незначительны, и рост коррелировал с
сезонный характером освещенности ( Gagne et al., 1982). У дру-
гих родов лимитирующая роль азота не столь выражена, на скорость
летнего роста Alaria (Buggeln, 1978), Laminaria hyperborea и
L.aaccharina (Liining, 1979) внесение в среду дополнительного
нитрата не влияло.
Сходные примеры с углеродом и азотом продемонстрированы
для макрофитов меньшего размера, в частности багрянки Gracila-
ria и Ulva из Chlorophyceae (Rosenberg,Ramus, 1982). Пики тка-
невого содержания азота следуют за пиками его концентрации в
окружающей морской воде; обе водоросли накапливают этот элемент
зимой и расходуют весной пропорционально скорости роста. Ни од-
на из них не зимует со значительными запасами углеводов, и нача-
ло весеннего роста не зависит от запасов крахмала, созданных в
предыдущий вегетационный период.
Гетеротрофность у морских макрофитов не продемонстрирована
так же четко, как у фитопланктона. Имеются некоторые данные ла-
бораторных исследований, согласно которым может происходить по-
глощение ацетата, сахаров и аминокислот (гл. 6), но его значе-
ние в природе еще не установлено. Уиле ( wilce, 1967) предпола-
341
Время года
Рис. 7.16. Сезонное взаимоотношение между скоростью роста
пластины, внутренними запасами углерода и азота и концен-
трацией нитрата в морской воде ДЛЯ Laminaria longicruris
(по Parley, 1982; данные из Chapman, Craigie,1977J
Ганг, что гетеротрофность встречается у некоторых арктических
макрофитов, обнаруженных на большой глубине, где освещенность
зимой для роста недостаточна. Однако Чапман и Линдли ( chapman,
Bindley, 1980) показали, что окружающего овета хватает арктиче-
ской водоросли Laminaria solidungula. Иопользующей для зимне-
го роста накопленные запасы углеводов.
342
Конкуренция
В морском бентосе конкуренция за пространство происходит не
только между видами водорослей, но и между ними и сидячими бес-
позвоночными. Площадь субстрата для прикрепления во многих слу-
чаях, по-видимому, является основным лимитирующим ресурсом, а
скученность особей лишь вторично ведет к затенению и конкуренции
за биогены. Результат борьбы за пространство зависит также от
наличия или отсутствия фитофагов и хищников, выедающих сидячих
животных. Обильное оседание на субстрат водорослевых опор может
приводить к конкуренции между сородичами, причем наблюдается об-
ратная зависимость между густотой зарослей и индивидуальным разме-
ром (рис. 7.17) (Cousens, Hutchings, 1983). Иногда затрагивает-
ся и размножение: Fucus vesiculosus в условиях скученности рас-
ходует на него ресурсов меньше, а на вегетативный рост - больше
( Russell, 1976).
Межвидовая конкуренция морских водорослей-макрофитов изуча-
лась в культуре (Russell, Fielding, 1974). Три вида - зеленая
ВОДОРОСЛЬ Ulothrix flacca, бурая водоросль Ectocarpus siliculo-
sus и багрянка Erythrotrichia,carnea - выращивались порознь и
попарно при разных температурах, соленостях и режимах освещения.
Все вида показали больший диапазон толерантности в монокультуре,
чем в двухвидовых сообществах; встречались условия, в которых
вид при парнем культивировании погибал, но выживал в изоляции.
Большинство опытов по конкуренции проведено в полевых усло-
виях и включало изменение плотностей популяций предполагаемых
конкурентов. Бурая водоросль Egregia laevigata на юге Калифор-
нии вытесняется произрастающими в нижнем ярусе дерновинными баг-
рянками, прежде всего Gigartiпа, laurencia и Gastroclonium.
Последние, размножаясь вегетативно, захватывают доступное про-
странство, а песок, скапливающийся между их талломами, препятст-
вует развитию спор Egregia (Sousa et al., 1981). Возобновление
этого вида возможно только при освобождении участка дна под дей-
ствием каких-либо внешних нарушений. Сходные конкурентные отно-
шения наблюдались между бурой водорослью Postelsia palmaeformis
и мидией Mytilus califomianus на незащищенных участках тихо-
океанского побережья США (Dayton, 1973).
В серии опытов о удалением бурой водоросли Hedophylium
sessile (Dayton, 1975) показана роль крупного доминирующего
макрофита в контролировании сопутствующей флоры посредством ме-
343
Десятичный логарифм средней массы пластины (г)
Saccorhiza polyschides
Chordaria flagelliformis
Ecklonia radiata
Sargassum sinclairii
Fucus vesiculosus
Ascophyllum nodosum (средняя арифме-'
. . , тическая масса)
A. nodosum (средняя геометрическая
масса)
Рис. 7.17. Зависимость между средней массой пластин и
густотой их размещения для природных одновидовых за-
рослей некоторых морских водорослей-макрофитов (по сои-
sens, Hutchings)
344
ханизмов, отличных от конкурентного исключения. Удаление приво-
дило к выпадению обычных видов нижнего яруса и способствовало
росту эфемерных водорослей. Сходные изменения отмечали Хокинс и
Харкин (Hawkins, Harkin, 1985), удалявшие виды верхнего яруса
на сублиторали острова Мэн.
Неумышленная интродукция новых видов также дает возможность
изучить конкурентное исключение. Конкуренция между представите-
лем ламинариевых Macrocystis pyrifera и занесенным из Японии
видом Sargassum muticum из Fucales отмечалась у острова Санта-
Каталина (США, Калифорния) ( Ambrose, Nelson, 1982). Sargassum
появился здесь в 1971 г. и после отмирания м.pyrifera в 1976 г.
захватил часть освободившегося пространства, препятствуя возоб-
новлению ламинариевого макрофита. Таким образом, распространение
здесь Sargassum, по-видимому, сдерживается конкуренцией с
м.pyrifera, но, поселившись, первый вид способен вытеснять по-
следний.
Выедание и хищничество
Любченко и Гейнс (Lubchenco, Gaines, 1981) составили вели-
колепный обзор данных о взаимодействиях между морскими водоросля-
ми-макрофитами и фитофагами, ясно показывающий, что питание водо-
роолями и хищничество - мощные биотические факторы, контролирую-
щие видовой состав и биомассу водорослевых сообществ. Основные
группы животных, потребляющих водоросли, - моллюски (особенно
брюхоногие и панцирные), ракообразные (в частности, бокоплавы),
морские ежи и рыбы. Растительноядные рыбы редки в умеренных во-
дах, но в тропиках 15 - 25% рыб питаются водорослями, и значи-
тельная доля здесь токсичных видов последних (гл. 9), возможно,
следствие адаптации к интенсивному выеданию.
Для многих фитофагов характерна высокая избирательнбсть в
отношении поедаемых видов, которые можно связать с морфологией
водорослей. Описано (steneck, Watling, 1982) семь функциональ-
ных групп последних пс мере увеличения трудности их выедания: от
микрофитов через нитчатые формы и покрытые корой макрофиты до
хрящеватых и, наконец, корковых кораллиновых водороолей
(рис. 7.18). Размеры или стадии развития могут также иметь важ-
ное значение: улитка Littorina ест проростки спор Chondrus
crispus, а взрослые особи этого вида почти не потребляет (.Che-
ney, 1982). Питательная ценность водорослей влияет на общую при-
345
функциональная
группа
Представители
Микроскопимес
кие водоросли
Диатомеи
Синезеленые
водоросли
Нитчатые
водоросли
Пластинчатые
водоросли
Claduphora
Ecrocarpns
Acrochaehum
U/va
Porphyra
Коровые
макрофиты
Bryothamnium
Chondna
Acanthophora
Кожистые
макрофиты
Laminaria
Fucus
Неизвесткован
корка
Членистые извест
ковые водоросли
Coralhna
Корковые корал -
линовые водоросли
Корковидные
корал линовые
Мо|Х|)рлогия
Анатомия на
поперечном срезе
Т РУДНОСТЬ
выедания
Рио. 7.18. функциональные группы бентосных морских водо-
рослей. Затрудненность для выедания связана оо структур-
ной жесткостью талломов (ПО Steneck, Watling, 1982)
способленность фитофагов ( Vadas, 1977). Съеденное количество
определенного вида не обязательно отражает важность его для жи-
вотного. Littorina littoreа предпочитает Enteromorpha. но лучше
всего существует на смешанном рационе из Chondrus и уцсиз (Che-
ney, 1982).
Любченко ( lubohenco, 1978) изменял плотность популяции
bittorina littorea в литоральных лужах на скалах Новой Англии
(рис. 7,19). Все улитки удалялись из луж с доминированием Chon-
dru3 и переносились туда, где доминировала Enteromorpha. Вслед-
ствие усиленного выедания обилие последней водоросли снижалось,
а другие эфемерные виды, приходившие ей на смену, в конце концов
тоже потреблялись. В лужах с доминированием Chondrus уменьше-
ние выедания позволяло развиваться Enteromorpha. которая быст-
ро вытесняла другие вида. В контрольных лужах со средний уровнем
выедания состав альгофлоры оставался без изменений. Таким обра-
346
Процент покрытия
А- < Эфемеры
в-----в Enteromorpha
•.....• Chondrus
Рис. 7.19. Влияние Littorina littorea на видовой сос-
тав водорослей в литоральных лужах Новой Англии: А -
контроль; Б - внесение littorina в удаление Jattori-
На • Представлена средняя численность Littorina в лу-
жах (по Lubchenoo, 1978)
зом, при интенсивной выедании все виды, кроме самых устойчивых,
исчезают, тогда как при слабом воздействии фитофагов конкурент-
ный доминант, который в норме был бы съеден, захватывает все до-
ступное пространство. В результате видовое разнообразие водорос-
лей достигает максимума при средних уровнях выедания.
Сходный эффект отмечен в зарослях бурых водорослей, где
347
наиболее активные фитофаги - морские ежи ( Lawrence, 1975). При
удалении Strongylocentrotus franciscanus (Paine, Vadas, 1969) в
районе Фрайди-Харбор обнаружено увеличение видового разнообразия
и доминирование Laminaria, тогда как на контрольных площадках
эстречались только известковые кораллины и несколько эфемерных
водорослей. Удаление Echinus esculentus в Великобритании ( Jones,
Kain, 1967) И Strongylocentrotus droebachiensis в ВОСТОЧ-
НОЙ Канаде ( Breen, Mann, 1976) привело к сходному увеличению
бисмассы и разнообразия водорослей.
Животные, питающиеся водорослями и конкурирующие с ними за
пространство, в свою очередь взаимодействуют с хищниками, что
существенно влияет на соответствующие водорослевые сообщества.
Мидия Mytilus califomianus местами конкурентно доминирует на
литорали тихоокеанского побережья Северной Америки. Основной хищ-
ник, питающийся этим моллюском, - морская звезда Pisaster ochra-
сеив» Удаление ее вызывает изменение богатого видами литораль-
ного сообщества в сторону доминирования мидий ( Paine, 1974).
Пейн ( Paine, 1969) называет таких хищников, истребляющих конку-
рентных доминантов, "краеугольными видами".
Отмечалась ( Lubchenco, Menge, 1978) конкуренция между мор-
ским желудем Balanus balanoides, мидией Mytilus edulie и баг-
рянкой Chondrus на литорали Новой Англии, сдерживаемая хищ-
ным брюхоногим моллюском Thais и морскими звездами. Опыты по уда-
лению хищников показали, что беспозвоночные были более конкурен-
тоспособны, но хищничество приводило к возникновению свободного
пространства, которое могло быть занято водорослью Chondrus .
На открытых учаотках, где хищники уничтожались прибоем, домини-
ровала мидия.
Калан Enhydra lutris ранее занимал широкий ареал вдоль Ти-
хоокеанского побережья от севера Японии до Нижней Калифорнии
(Мексика). Он был почти полностью уничтожен в результате охоты
и сохранился в изобилии только местами на Алеутском архипелаге.
Этот хищник питается морскими беспозвоночными, в частности мор-
скими ежами, и увеличение численности последних наряду с сопут-
ствующим уменьшением биомассы бурых водорослей можно отчасти
приписать уничтожению калана в том или ином регионе. У острова
Амчитка, где каланы встречаются, в сублиторальной зоне имеется
обильный покров водорослей, а у острова Шемья в 400 км к западу
°ба этих компонента экосистемы отсутствуют, зато морские ежи
крупнее и Обильнее (рис. 7.20) ( Ester, Palmisano, 1974). В рай-
346
д Плотность морских ежей
(число особей на 1Мм21
20 40 60 80 100
Покрытие растительноти, "о
Диаметр особей, мм
Рис. 7.20. Каланы, морские ежи и бентосная водорослевая
растительность. А. Растительный покров и популяционная
плотность морских ежей в зависимости от глубины; график
для растительного покрова острова Шемья совпадает с осью
ординат. £. Остров Шемья (каланы отсутствуют). В. Остров
Амчитка (каланы имеются). Классы размера и биомасса мор-
ских ежей. Пунктирная линия на рис. £ приведена для срав-
нения с рис. В и показывает наибольший размерный класс
морских ежей на Амчиткв (ПО Estes, Palmisano, 1974)
онах Аляски, куда каланы реинродуцированы, снова восстанавливают-
ся заросли бурых водорослей ( Duggins, 1980). Сходная роль крае-
угольного камня предполагается для омаров в Новой Шотландии
( Breen, Mann, 1976); здесь очмечена корреляция между снижением
обилия омаров и увеличением численности морских ежей с одновре-
менным разрушением водорослевого покрова. Эту гипотезу еще сле-
дует доказать, поскольку другие беспозвоночные и рыбы могут так-
349
we активно поедать морских врой ( Miller, 1985).
Демографические и фенологические исследования
В отличие от ситуации с бентосными животными имеется относи-
тельно мало демографических данных о морских водорослях; еще мень-
ше известно об их продолжительности жизни. Наиболее детальное ис-
следование было проведено на многолетнем представителе Pucales
Pelvetia fastigiate из Калифорнии (Gunnill, 1980); его макси-
мальная продолжительность жизни была 2 5 года, причем 80% экзем-
пляров погибало в первый год, а еще 10 - 15% - во второй
(рис. 7.21). Описаны ( Rice, Chapman, 1982) изменения биомассы
и выживаемости за вегетационный период двух когорт однолетней
бурой водоросли Chordaria flagelliformis; эти данные были исполь-
зованы для подсчета первичной продукции. Вид появлялся в июне-
июле и исчезал в середине сентября; на протяжении большей части
сезона наблюдалось снижение численности популяции, коррелирующее
с увеличением биомассы отдельных экземпляров.
Большинство морских водорослей-макрофитов показывает сезон-
ные различия в росте, являющиеся ответом на изменения факторов
окружающей среда - освещенности,фотопериода, температуры, доступ-
ности биогенов, интенсивности выедания. Эти фенологические раз-
личия иногда выражаются в изменениях морфологии или репродуктив-
ного статуса, а у гетероморфных или имеющих криптические фазы
форм - в кажущемся исчезновении на часть года. Хупер с соавт.
(Hooper et al., 1980) детально изучили альгофлору фиорда Бонн-
Бей в западном Ньюфаундленде, проанализировав кластерный методом
сезонную встречаемость и репродуктивный статус видов. Данные по
вегетативному состоянию показывают, что сезонные эфемеры можно
подразделить на зимне-весенние и летне-осенние (рис. 7.22), а
Данные по размножению демонстрируют существование трех основных
групп - с зимней, весенней и летне-осенней фертильностью
(Рис. 7.23). Такие исследования говорят о необходимости провер-
ки в культуре гипотез относительно сезонных факторов, способных
контролировать рост и размножение видов.
350
Рис. 7.21. Кривые выживаемости для пополнения популяции
Pelvetia faatlsiata в южной Калифорнии. Затушеванные
кружки -" прбТСенТ"Выживаемости 314 особей; светлые круж-
ки - десятичный логарифм выживаемости 314 особей, отне-
сенный к популяции из 1000 особей (по Gunnill, 1980)
Monostroma undulatum
Chorda tomentosa
Giffordia granulosa
Eudesme virescens
Delamarea attenuate
Myriodadia lovemi
Monostroma grevillei
Urospore wormskiofdii
Dumontia mcrassata
Audouinella alariae
Petalonia fascia
Bangia atropurpurea
Porphyra hneans
Cladophora sericea
Acrothnx novae-anghae
Chorda filum
Asperococcus fistulosus
Capsosiphon fulvescens
Sphaerotnchta divaricate
Spongomorpha aeruginosa
Stictyosiphon soriferus
Derbesia manna
Calhthammon corymbosum
Dictyosiphon foemculaceus
Chordana flagelliformis
Ectocarpus sihculosus
Saccorhiza dermatodea
Petalonia zosterifoha
Porphyra miniata
40 60 80 100
Процент сходства
Рис. 7.22. Кластерный анализ, показывающий наличие ви-
дов с выраженной вегетативной сезонностью в заливе
Бонн-Бей, Ньюфаундленд. Затушеванные кружки - присутст-
вие в определенный месяц (по Hooper et al., 1980)
351
Delamerea attenuate
Eudesme virescens
Myriocladia lovenii
Chorda tomentosa
Giffordta granulosa
Urospora wormsktoldii
Monostroma grevillei
Bangia atropurpurea
Fucus vesioulosus
Ascophyltum nodosum
Monostroma undulatum
Fucus edentatus
Callophyllis cnstata
Agarum cribrosum
Phycodrys rubens
Dictyos/phon foeniculaceus
Saccorhiza dermatodea
Chorda filum
Spongomorpha aeruginosa
Chordaria flagelhformts
Phymatolithon rugulosum
Ectocarpus siliculosus
Stictyosiphon soriferus
Plumaria elegans
Derbesia marina
Asperococcus fistulosus
Acrothrix novae-angliae
Sphaerotrichia divaricate
Callithammon corymbosum
Phymatolithon laevigatum
Capsosiphon fulvescens
Cladophora sericea
Phymatolithon lenormandii
Petalonia zosterifolia
Cystoclonium purpureum
Polysiphonia nigrescens
Porphyra miniata
Antithamnion floccosum
Clathromorphum compactum
Petalonia fascia
Audouinella purpurea
Alaria esculenta
Antithamnion pylaisaei
Halosaccion ramentaceum
Leptophytum laeve
Uthothamnium ierroineae
Porphyra linearis
Odonthaha dentate
Membranoptera alata
Rhodophysema elegans
Sphacelaria plumose
Ahnfeltia plicate
Laminaria solidungula
Neodifsea integra
Pantoneura baerii
Turnerelfa penny!
•---1__, _1_____________I__I
40 60 80 100
Процент сходства
mbS’ 7.23. Кластерный анализ, показывающий фенологию раз-
ния отделкных бентосных водорослей из залива Бонн-
na?J Ньюфаундленд. Затушеванные кружки - наличие функцио-
нальных репродуктивных структур (по Hooper et al., 1900)
352
Вертикальное распределение макрофитов
Большинство морских водорослей-макрофитов ограничено в рас-
пространении прибрежными участками океана, где субстрат достаточ-
но стабилен, чтобы обеспечить их прикрепление. Они занимают уз-
кую полосу от верхних границ заплеска волн через литораль и суб-
литораль к нижним пределам фотической зоны. Рекордная к настоя-
щему времени глубина для прикрепленных водорослей указана для
неописанной кораллиновой багрянки у Багамских островов - 268 м
( Littler et al., 1985). Высыхание у верхних границ и низкая осве-
щенность у нижних, по-видимому, основные абиотические факторы,
лимитирующие вертикальное распространение, но их влияние заметно
модифицируется конкуренцией и выеданием. Водоросли распределены
вдоль этого вертикального градиента не беспорядочно; многие за-
нимают определенные зоны с четкими верхними и нижними границами,
которые особенно хороню различимы на литорали.
При наиболее простом подходе прибрежную полосу морей делят
на ряд горизонтальных зон в соответствии с' границами встречае-
мости диагностических видов. Одна из наиболее широко принятых
схем ( Т.Stephenson, A.Stephenson, 1949; Lewis, 1961; T.Stephen-
son, A.Stephenson, 1972) признает три основные зоны
(рис. 7.24). На умеренно защищенных побережьях эулиторальная зо-
на составляет большую часть литорали, от верхней границы сизигий-
ных приливов до нижней границы сизигийных отливов. Выше находит-
ся литоральная кайма"’’, а ниже - сублиторальная зона. В северной
умеренной зоне переход между сублиторалью и эулиторалью характе-
ризует верхняя граница ламинариевых, а между эулиторалью и лито-
ральной каймой - верхняя граница морских желудей. Чем незащищен-
нее побережье, тем эти зоны поднимаются выше и становятся шире
(рис. 7.25) ( Burrows et al., 1954; Lewis, 1961; Seapy,Littler,
1979). Хотя диагностические виды неодинаковы в разных регионах,
эта "универсальная система зональности" широко принята и приме-
няется для береговой линии всего мирового океана (Г.Stephenson,А.
Stephenson, 1972). В пределах каждой из этих широких зон водо-
росли могут также занимать определенные пояса. На многих северо-
европейских побережьях присутствует верхняя полоса эфемерных зе-
леных и синезеленых водорослей, часто с доминированием Entero-
morpha ;ниже ее доминируют фукусовые, обычно в последовательно-
I
Эту зону иногда называют супралиторалью. - Прим, ред.
353
Супралитораль
Верхняя граница
Litton па
Верхняя граница
(мпример) ламинариевых
ЭСО
Верхняя граница морских
Ж ВЛ У Д8Й
Инфралитораль
сти Pelvetia canaliculata - Fu-
cus spiralis - F.vesiculosus -
F, serratus; еще ниже на-
ходится узкая полоса из lamina-
ria digitata, за которой сле-
дует сублиторальная зона с 1.
hyperborea. Описания бере-
говых линий в других частях
света показывают существование
сходных горизонтальных полос
(TStephenson,A.Stephenson, 1972;
Round, 1981).
Наиболее очевидный из фак-
торов, способных влиять на зо-
нальность, - амплитуда и перио-
дичность приливов. Колман
(Colman, 1933) предположил,
что приливы и отливы в Плимуте
(Великобритания) заливают и об-
нажают литораль с неодинаковой
скоростью, причем наиболее рез-
кие ее изменения происходят в
трех точках цикла, что коррели-
рует с границами между поясами
литоральной растительности.
Эта теория критических прилив-
Рис. 7.24. Общая схема зональ-
ности каменистой литорали. ЭСП -
экстремальный сизигийный при-
лив; ЭСО - экстремальный сизи-
гийный отлив (ПО Stephenson,
Stephenson, 1949)
ных уровней' была развита Доти
(Doty, 1946), который в более
сложных приливных циклах тихо-
океанского северо-запада США
(два прилива неравной высоты в
день) выделил семь критических
Уровней. Подразумевается, что водоросли в зонах между критически-
ми приливными уровнями точно физиологически адаптированы к соот-
ветствующим периодам осушения и погружения под воду и не могут
выживать в соседних зонах. Существование критических приливных
УРовней вообще и их влияние на зональность литорали многими оспа-
риваются. Точные измерения времени пребывания субстрата под во-
Дой там же, где работал Колман ( Underwood, 1978), показали плав-
НСе непрерывное изменение. И напротив, измерения по трем трансек-
354
Рис. 7.25. Влияние прибоя на расширение и поднятие лито-
ральных зон. ЭСП - экстремальный сизигийный прилив; ЭСО -
экстремальный сизигийныи отлив (по Lewis, 1Э64)
там на литорали Британской Колумбии (Druehl, Green, 1982) про-
демонстрировали неравные периоды затопления и выхода из-под во-
да. Эти измерения, учитывавшие как действие прибоя, так и при-
ливно-отливный цикл, использовались для построения кривых погру-
женного - осушенного состояния. Волны вызывают значительные из-
менения в цикле обнажения из-под воды - погружения под воду по
сравнению с тем, что можно было бы предсказывать, исходя только
из приливно-отливного цикла. Распределение водорослей по тран-
сектам показывало некоторые достоверные корреляции с этими резки-
ми сменами режима выхода из-под воды - погружения.
Чапман ( Chapman, 1973) придерживается другой крайней точки
зрения, согласно которой существование резких границ между лито-
ральными зонами на фоне непрерывного средового градиента может
целиком объясняться межвидовой конкуренцией без всякого привлече-
ния приливно-отливного цикла.
За последнее десятилетие проведен ряд опытов с целью опре-
деления факторов, влияющих на вертикальное распределение водорос-
лей. Шонбек и Нортон ( Schonbeck, Norton, 1978) показали, что
отливы в сочетании с иссушающими условиями вызывали наибольшее
повреждение литоральных фукусовых, причем фукусовые нижней лито-
рали (Fucus vesiculosus и ?« eexvatue) менее устойчивы к выси-
355
Ханию, чем F,gpilalis и Helvetia canaliculata. приуроченные
К верхней литорали. Дринг и Браун (Dring, Brown, 1982) также
приводят данные в пользу физиологической основы зональности. Их
исследования показали, что потеря воды зависит от отношения по-
верхности к объему, а не от положения на литорали. Восстановле-
ние после сильного высыхания занимало около двух часов у каждого
вида, но масштабы восстановления от данного уровня высыхания
больше у видов верхней литорали. У Pelvetia и. г, spiralis оно бы-
ло полным после потери 80 - 90% воды, у F.veaiculoaug - при-
мерно 70%, а у. serratus - 60%. Таким образом, способность к
восстановлению, измеренному по фотосинтезу, наиболее четко кор-
релирует с характером зональности.
На нижней литорали Калифорнии показана (Hodgson, 1980) важ-
ность для поддержания зональности межвидовой конкуренции. Крас-
ная водоросль Gastroclonium coulter! занимает полосу шириной
примерно 0,3 м над средней линией отлива. Выше находится зона
других багрянок, в основном Gigartina papillata и Rhodoglossum
affine. а ниже - морского покрытосеменного растения Phyllos-
padix. После удаления Gastroclonium Gigartina распространились
ниже, захватив свободное пространство; когда же удаляли ату водо-
росль, соответствующего расширения зоны Gastroclonium вверх не
было. Сходным образом при удалении Phyllospadix зона Gastroclo-
nium расширялась вниз, но обратный вариант не приводил к распро-
странению вверх Phyllospadix. Установлено, что перемещению
вверх йо литорали препятствует высыхание, тогда как нижний пре-
дел распространения определяется конкуренцией. Сходные опыты с
верхнелиторальными фукусовыми Великобритании свидетельствуют,
что в отсутствие конкурентов зона их произрастания может расши-
ряться вниз ( Schnonbeck, Norton, 1980; Hawkins, Hartnell, 1985).
Любченко ( Lubchenco, 1980) также показывает, что конкуренция а
выедание - лимитирующее факторы, определяющие нижние границы
встречаемости видов водорослей на литорали Новой Англии.
Таким образом, стало общепризнанным, что верхние пределы
Распространения водорослей-макрофитов определяются физислогичес-
кими реакциями на высыхание, а нижние - биологическими взаимодей-
ствиями. Правда, Андервуд и Денли ( Underwood, Denley, 1984)
Упорно считают, что доказательств для такого обобщения недоста-
т°чно, а имеющиеся данные неубедительны. Кьюбит ( Cubit, ©84)
отмечает, что на самых верхних уровнях скалистого побережья, где
обильны фитофаги, состав водорослей показывает четкую сезонность.
356
Быстрорастущие их виды обильны в холодные зимние месяцы, но ис-
чезают теплым летом, что согласуется с гипотезой высыхания. Уда-
ление моллюсков блюдечек позволяет этим водорослям сохраняться
в летний сезон и увеличивать свое обилие и разнообразие зимой.
Этот автор предполагает, что приживание здесь многолетних видов
лимитируется интенсивностью выедания летом и конкуренцией с ко-
роткоживущими водорослями зимой.
Большинство экологических экспериментов на- сублиторали было
связано с установлением влияния света,конкуренции и выедания на
структуру водорослевых сообществ, а не- с выяснением, какой из
этих факторов определяет зональность и влияют ли они на нее вооб-
ще. Исключение составляет проведенная недавно серия опытов по
удалению водорослей на нижней литорали ( Fucus serratus) и субли-
торали ( Laminaria digitata И L.hyperborea) острова МэН в Вели-
кобритании ( Hawkins, Harkin, 1985). Сделан вывод, что зональ-
ность в значительной степени определяется конкуренцией за про-
странство, причем взаимодействия между макрофитами на границах
зон важнее, чем выедание. Уиттик ( whittick, 1983) исследовал
распространение четырех доминирующих эпифитов на "черешках" Lami-
naria hyperborea. показав наличие четких зон и конкуренции за
пространство между видами.
Биогеография морских макрофитов
Некоторые таксономические группы водорослей особенно хороно
представлена в определенных географических областях, например
Laminariales в Северной Пацифике фгиеЫ, 1981), a Fucales -
в Австралазии Qlizamuddin, 1962). Dictyotales и сифональные
зеленые водоросли особенно обычны в тропических водах, где в це-
лом отсутствуют виды с крупными талломами, зато много известко-
вых форм. Любченко и Гейнс (Lubchenco, Gaines, 1981) считают,
что в тропиках такая альгофлора обусловлена выеданием и что изме-
нения в растительноядной фауне с увеличением широты, в частности
исчезновение раб-фитофагов, объясняют смену господствующих жиз-
ненных форм водорослей. Северная Атлантика, по-видимому, обедне-
на крупными бурыми водорослями: например, вид Macrocystis pyri-
fera здесь отсутствует, но встречается во многих других холод-
ных водах и в принципе способен расти в этом регионе ( Boalch,
1981). Laminariales не встречаются в Антарктике, и их ниша там
занята представителями Desmarestiales (Мое, Silva, 1977).
357
Общепризнано также, что относительное обилие бурых водорослей по
сравнению с красными возрастает в Северном полушарии с увеличе-
нием широты. Этого не наблюдается в Южном полушарии, и багрянки,
по-видимому, особенно хорошо представлены б Субантарктике.
В целом признается, что наличие большого числа видов опреде-
ленной таксономической группы, включая эндемики, свойственно для
центра ее происхождения, но без ясного представления о предковых
формах или наличия палеонтологических находок с равным успехом
можно считать, что наблюдаемая картина отражает быстрое видообра-
зование после расселения группы в новую для нее область. Оконча-
тельные выводы требуют дальнейших палеонтологических исследований,
особенно чтобы увязать современноераспространение, морской кли-
мат в прошлом и геологические события.
Фитогеографический анализ опирается на сведения о локальных
флорах. В этом отношении флоры Средней и Северной Атлантики из-
вестны особенно хорошо, и их нумерический анализ проведен соот-
ветственно Ван ден Хутом (van den Hoek, 1975) и Лосоном ( Lawson,
1978) (рис. 7.26). Ван ден Хук разграничивает пять фитогеографи-
ческих областей с характерными температурами воды у поверхности.
В 1915 г. Сетчелл разделил океаны на 9 биогеографических зон в
соответствии с распространением водорослей. Внутри их изменчи-
вость температуры у поверхности воды составляла 5°С, за исключе-
нием полярных регионов, где она была Ю°С. Хатчинс (Hutchins,
1947) предполагает наличие четырех критических температур, влияю-
щих на распространение морских водорослей-макрофитов: I) мини-
мальная для выживания, 2) минимальная для размножения, 3) макси-
мальная для размножения, 4) максимальная для выживания. Ван ден
Хук ( van den Hoek, 1982 b ) добавляет еще две - верхнюю и ниж-
нюю предельные для роста - и рассматривает распространение бен-
тосных морских водорослей в связи с регуляцией их жизненных цик-
лов температурой и долготой дня ( van den Hoek, 1982b ,c). Такой
анализ требует детальной получаемой в культуре информации о фак-
торах, ответственных за рост и размножение (см., например, Yarish
et al., 1984), но зато дает возможность объяснить потенциаль-
ные пределы географического распространения. Например, гаметофит-
иая фаза гетероморфной водоросли Bonnemaisonia hamlfera отсутст-
вует у Ньюфаундленда, тогда как ее вегетативно размножающаяся фа-
За £railiella там обычна. Тетраспорогенез у этого вида требует
с°нетания высоких температур воды и короткого дня ( Luning, 1980),
а это условие у Ньюфаундленда не соблюдается, хотя местные изо-
358
Рис. 7.26. Распределение флор макрофитов в Северной
(А - Н), а также Центральной и Южной (1-7) Атлантике
(ПО Druehl, 1981; данные из van den Hoek, 1975 И Law-
son, 1978)
ляты генетически способны к тетраспорогенезу в культуре. Сходным
образом багрянки Ceramium фертильны на юге Великобритании и
стерильны в Шотландии (Edwards, 1973).
У некоторых водорослей известны физиологические расы. Уэст
(West, Т972) показал существование генотипически различных штам-
мов красной водоросли Rhodochorton purpureum в отношении спору-
ляции, а Люнинг ( Liining, 1980) продемонстрировал широтные раз-
личия в требованиях к долготе дня для образования вертикальной
359
фазы У бурой водоросли Scytosiphon lomentaria (рис. 7.27).
Болтон (Bolton, 1983) установил оптимальные температуры роста и
максимальные температуры длительного выживания нескольких изоля-
тов Ectocarpus ailiculoaua, собранных из разных местообитаний
от Арктики до теплой умеренной зоны. Они поддерживались в куль-
туре до 20 лет, и для многих была показана способность к интер-
бридингу. Между изолятами наблюдались генотипические и фенотипи-
ческие различия в реакции на температуру, но четких экотипов не
установлено.
Таким образом, температура и долгота дня, по-видимому, яв-
Рйс. 7.27. Влияние долготы дня на образование вертикально-
£0 таллома различными географическими линиями Scytosiphon
-^entaria при 10°С (светлые кружки) и 15°С (затушеванные
кРУжки). Каждое значение основано на просмотре 250 экзем-
пляров (noLiining, 1980)
360
ляются основными абиотическими факторами, определяющими распро-
странение морских водорослей-макрофитов по планете. Соленость
имеет некоторое региональное значение. Установлено (Druehl,
1978), что устойчивость вида Macrocystie integrifolia к низкой
солености при высоких температурах понижена, и это обусловило
его отсутствие в ряде мест Британской Колумбии. Влияние биологи-
ческих взаимодействий, столь важное в локальном экологическом
масштабе, почти неизвестно на биогеографическом уровне. Примеча-
тельное исключение составляют две работы (Pielou, 1977, 1978),
в которых сравнивается распространение видов одного рода по ши-
роте вдоль восточного и западного побережий Северной Америки.
Ареалы их в значительной мере перекрываются, причем у близкород-
ственных видов - больше, чем у более далеко отстоящих друг от
друга в эволюционном плане. Отсюда делается вывод, что на геогра-
фические ареалы морских водорослей-макрофитов межвидовая конку-
ренция не влияет.
Водоросли рыхлых осадков
Эпипелитные водоросли
Эпипелитные водоросли развиваются как в морских, так и в
пресных водах на поверхности отложений, варьирующих от тонких
илов до грубых песков. В этой альгофлоре доминируют перистые диа-
томеи, синезеленые водоросли и подвижные формы из других групп;
в пресных водах бывают обильными также десмидиевые. Хорошо про-
слежена вертикальная миграция в нескольких верхних миллиметрах
литоральных отложений; сборы здесь можно сделать, положив на по-
верхность осадков покровное стекло микроскопа.
В целом клетки движутся к поверхности на свету и возвраща-
ются вглубь осадка ночью, хотя некоторые могут уходить туда, что-
бы избежать интенсивного облучения в середине дня. У водорослей
литоральных и эстуарных морских отложений известны ритмы, согла-
сованные с приливно-отливными циклами. Предполагается (Palmer,
Round, 1965), что такой ритм у диатомей и Euglena в эстуарии
Эйвона выработался из циркадианного в связи с периодическим за-
темнением осадков очень мутной водой. В некоторых более прозрач-
ных эстуарных водах водоросли остаются на поверхности на свету
при всех приливах, кроме самых высоких ( Perkins, I960). Эта
361
вертикальная миграция подчинена эндогенному ритму, сохраняющему-
ся в лаборатории в течение нескольких суток при постоянном осве-
щении. Euglena сохраняла циркадианный ритм, основанный на цик-
личной смене света и темноты в день сбора, тогда как движения
диатомеи Hantzehia virgata оставались скоррелированными с при-
ливно-отливным режимом в природе (рис. 7.28). Эта корреляция про-
является в суточном сдвиге миграции на 50 - 60 мин до того, как
отлив придется.на вечер; затем цикл меняет фазу, чтобы захватить
отлив утром (Palmer, Round, 1967). У речных диатомей миграция
вверх начинается перед рассветом; это дает основание полагать,
что если цикл свет - темнота и важен для синхронизации ритма с
природными процессами, свет сам по себе не является пусковым фак-
тором миграции.
Экологическое значение этих вертикальных перемещений выясне-
но недостаточно хорошо; возможно, это механизм, позволяющий избе-
гать выедания или расширять доступность заключенных в осадке био-
генов. Опыты по обогащению среды показали, что рост эпипелитных
водорослей соленых маршей может быть лимитирован доступностью
азота (Barley et al., 1981).
Эписаммйтные водоросли
♦
В отличие от эпипелитных водорослей, эписаммитные прикреп-
лены к частицам осадков. Как в морских, так и в пресных водах
эта альгофлора состоит отчасти из мелких коккоидных зеленых и
синезеленых водорослей, занимающих углубления в частицах. Однако
наиболее заметные ее элементы - диатомеи, которые либо прочно
прилипают к частицам, либо прикрепляются к ним короткими слизис-
тыми ножками; в отличие от эпипелитной флоры здесь преобладают
неподвижные формы (Round, 1981). Мосс (Моээ, 1977) отмечает,
что эпиоаммитные виды устойчивее эпипелитных к длительным перио-
дам темноты и анаэробным условиям. В текучих водах они могут от-
рываться от субстрата и пребывать во взвешенном состоянии, осо-
бенно около полудня. Предполагают (Itieller-Haekel, 1973),
что это следствие периодического клеточного деления, при котором
одна дочерняя клетка остается на субстрате, а другая уносится
токами воды, чтобы прикрепиться в другом месте.
362
Время суток
Рис. 7.28. Сохранение вертикаль-
ной миграции (сплошная линия)
при постоянном освещении в лабо-
ратории у диатомеи Hantzechia
virgata. Верхняя схема показы-
вает чередование приливов и отли-
вов в день полевых сборов, а схе-
мы I - 5 - в следующие дни; точ-
ки означают темновой период. За-
метен сдвиг фазы вертикальной
миграции между I-м и 3-м днями
(ПО Palmer, Round, 1967)
Макрофиты и рыхлые осадки
Представители харовых -
наиболее обильные ризобентос-
ные водоросли пресных вод,
иногда образующие обширные
сообщества в щелочной среде
на мелководьях озер и медлен-
но текущих рек. Их прикрепи-
тельные ризоиды связывают
частицы и повышают стабиль-
ность осадка (Round> 1981).
В морской среде к ризо-
бентосным видам относятся
преимущественно тропические
сифональные зеленые водорос-
ли ИЗ родов Cauls д-р a, Halims-
da и Udotea,которые обра-
зуют обширные скрепляющие
осадки ризоидальные системы,
дающие им возможность проти-
востоять умеренному движению
воды (рис. 7.29).
Другие нитчатые морские
водоросли, такие, как виды
Audouinella, также могут иг-
рать важную экологическую
роль в связывании минераль-
ных отложений и песка. Это
несколько напоминает образо-
вание синезелеными водоросля-
ми в тропиках строматолитов
(см. рис. 8.4) - структур ти-
па плотных горных пород.
Классическое место современ-
ного накопления строматоли-
тов - залив Марк в Западной
Австралии, где они формируют-
ся в защищенных от волнения
гиперсалинных водах (Logan,
1961).
363
Рис. 7.29. Лленкообразующие нитчатые водоросли, связываю-
щие частицы песка (по Scoffin, 1970)
Неприкрепленные или слабо прикрепленные морские макрофиты
также встречаются в защищенных местообитаниях; 87 их видов отме-
чено в заливе Порт-Ирин на острове Мэн ( Burrows, 1958), около
60 - в небольших бухтах Флориды ( Нити, Нитт, 1976). Другие ви-
ды, по-видимому, способны сохраняться в более открытых песчаных
биотопах, где они могут свободно плавать в толще воды, как,напри-
мер, бурая водоросль Pylaiella(Wilce et al., 1982) и багрянка
Anoenothamnicon planctonicum (Womersley, Norris, 1959).
Соленые марши, на которых доминируют в основном сосудистые
Растения, - еще одно местообитание, где можно встретить свобод-
но лежащие на субстрате или неприкрепленные водоросли (Chapman,
1974; Ranwell, 1972). Некоторые нитчатые формы, например Entero-
BPfrpha prolifera и Rhizoclonium sp., растут, оплетаясь вокруг
основания стеблей высших растений. Другие (например, Catenella
£ggens. Bostrychia scorpoides) являютоя эпифитами, а формы типа
Xspcheria из желтозеленых водорослей прикрепляются к отложениям.
Синезеленые водоросли ( Byngbya, Oscillatoria, Phormidium)оообен-
364
но обычны на осадках соленых маршей, где они вместе с бактерия-
ми фиксируют значительные количества азота ( Patriquin, McLung,
1978). Сходные ассоциации водорослей с участием тропических ро-
дов, например Caulerpa, встречаются также в мангровых сообщест-
вах, замещающих соленые марши в безморозных областях.
Важным компонентом флоры соленых маршей северной умеренной
зоны являются фукусовые. Некоторые из них растут неприкрепленны-
ми, другие могут непрочно удерживаться на меоте, как бы "укоре-
няясь" в осадке. Те же самые виды встречаются как эпилиты на ска-
листых побережьях, но здесь они показывают значительные модифи-
кации в морфологии и размножении ( Norton, Mathieson, 1983) И ча-
сто считаются особыми разновидностями или, точнее, экадами. Обыч-
но у них карликовые, скрученные талломы без особых прикрепитель-
ных приспособлений, а в крайнем варианте талломы преобразуются в
сферические структуры, наиболее известные у Ascophyllum nodosum
ecad makaii (South, Hill,1970). Такие фукусовые редко становятся
фертильными, рецептакулы у них редки, а если и формируются, то
образуют мало жизнеспособных гамет или таковых не возникает во-
все; размножение, таким образом, только вегетативное. Это наличие
исключительно вегетативного размножения - характерная черта не-
прикрепленных или слабо прикрепленных водорослей, встречающаяся
независимо от того, являются ли их репродуктивные структуры слож-
ными (типа рецептакул фукусовых) или маломодифицированными веге-
тативными клетками, как у зеленых водорослей вроде видов Cladopho-
га.
Почвенные водоросли
Водоросли широко встречаются в почвах всех континентов, от
полярных областей до жарких пустынь (Starks et al., 1981). Поч-
венная альгофлора состоит в основном из представителей Chlorophy-
ceae, Baoillariophyceae И Xanthophyceae И достаточно специфич-
на для того, чтобы не считать ее просто частью водной биоты, жи-
вущей в неблагоприятных условиях ( Round, 1981).
Таксономически эта группа трудна, и определение видов требу-
ет их выделения в культуре. Такие методы,несомненно, должны быть
высокоселективными, и чтобы свести к минимуму ошибки, необходимо
использовать широкий набор сред я условий культивирования. Этот
подход был разработан в основном Харолдом Бодцом ( Harold Bold)
и его учениками (обзоры, включающие систематику и экологию поч-
365
веяных водорослей, CM. netting, 1981; Starks elal., 1981).
Еще сложнее получить количественные данные. Шараби и Прей-
мер ( Sharabi, Pramer, 1973) сравнили методы, основанные на пря-
мом наблюдении, культуре и уровнях хлорофилла, и сдежли вывод,
что максимальные оценки обилия водорослей дает последний из них.
Ясно, что для обоснованного сопоставления данных по почвенной
альгофлоре необходима стандартизация методов сбора и культивиро-
вания этих водорослей.
Определение 685 таксонов почвенных водорослей, приводивших-
ся в литературе ( Feher, 1948), позволило говорить о невозможности
установления каких-либо закономерностей в их географическом рас-
пространении или какой-либо корреляции с типами почв. Однако
создается впечатление, что щелочные почвы благоприятны для сине-
зеленых водорослей; последние не обнаружены в почвах с pH менее
5 (Brock, 1973; Starks et al., 1981). И напротив, представите-
ли Chlorophyceae часто встречаются на кислых почвах. Отмечено
( Schields, Durrel, 1964), что многие аридные местообитания ха-
рактеризуются щелочной реакцией, а влажные - кислой, т.е. при
анализе распределения почвенной альгофлоры можно принимать во
внимание и влажность почвы. Нитчатые синезеленые водоросли часто
указываются как стабилизаторы обнаженных и эродированных почв,
тогда как зеленые водоросли мотут также образовывать слизи, спо-
собствующие гумификации. Имеются данные в пользу положительной
корреляции разнообразия и обилия водорослей о возрастом некото-
рых датированных вулканических почв и содержанием в них органи-
ческого вещества и биогенов (Carson, Brown, 1978). Флористиче-
ское разнообразие водорослей также наибольшее в ненарушенных
почвах ( King, Ward, 1977). Почвенные водоросли растут лучше в
частично сухих почвах ( Hunt et al., 1979), а поскольку мно-
гие их виды образуют толстостенные акинеты или гипнозиготы, они
таким образом хорошо приспособлены к перенесению засушливых усло-
вий. В сухом состоянии некоторые виды способны выдерживать темпе-
ратуру 140°с в течение I ч и 40°С до трех месяцев ( Trainor,
1983).
Многие представители альгофлоры встречаются на поверхности
яочвы, другие обитают на лишенной света глубине, предположитель-
Ro попав туда с просочившейся водой. Такие водоросли бывают пред-
ставлены покоящимися стадиями, но некоторые из них, вероятно,
способны и к гетеротрофному росту (Droop, 1974а). Показано
Sheath, Hellebust, 19?4), что Bracteococcus minor (Chlorophy-
366
сеае) может расти гетеротрофно на глюкозе; предполагается, что
такой механизм поглощения углеводов у почвенной водоросли не
возник бы, если бы не способствовал выживанию. Возникает мысль,
что такие виды могут по крайней мере поддерживать основной об-
мен, вымываясь в почве за пределы фотической зоны.
Симбиотические ассоциации
Водоросли часто растут в тесной связи с другими организма-
ми, включая водоросли, грибы, сосудистые растения и беспозвоноч-
ных. Симбиозом сейчас называют "ассоциацию в течение значитель-
ной части жизненных циклов особей, являющихся представителями
разных видов" (Margulis, 1981), включая сюда все степени пара-
зитизма, а также мутуалистические и комменсалистские отношения
(Goff, 1982а).
Ассоциации водорослей с беспозвоночными
Эти ассоциации привлекли внимание как зоологов, так и аль-
гологов, и им посвящено много обборов (см., например, Glider,
Pardy, 1982; Goff, 1983; Taylor, 1983; Trench, 1979).
Тренч (Trench, 1975, 1982) рассматривает ассоциации с уча-
стием хлоропластов и цианелл. Среди наиболее известных из них -
те, что встречаются у мешкоязычных моллюсков, питающихся путем
высасывания цитоплазмы таких сифональных зеленых водорослей, как
Codium и Caulerpa. Хлоропласта накапливаются в клетках гепатопан-
креаса, где сначала остаются неповрежденными (Trench, 1980;
Hinde, 1983) и продолжают фотосинтезировать, но неспособны к
делению и синтезу хлорофилла или рибулозобисфосфаткарбоксилазы,
т.е. к действиям, отчасти контролирующимся ядерным геномом. Плас-
тиды Codium fragile обычно теряют in vitro 15 _ 20% только ЧТО
фиксированного углерода преимущественно в виде гликолата. Эта
цифра может быть увеличена до 74% обработкой гомогенатами гепа-
топанкреаса (Gallop, 1974). Высвобожденные продукты фотосинте-
за используются в обмене веществ хозяина ( Trench et al., 1973),
и моллюск Elysia в отсутствие света и пищи теряет в веое вдвое
быстрее, чем при лишении только пищи ( Hinde, Smith, 1974). в ко-
нечном счете хлоропласты перевариваются, и при хорошем питании
у Elysia viridis еженедельно сменяется от трети до половины их
количества ( Gallop et al., 1980).
367
Мутуалистический характер этой ассоциации с достоверностью
не установлен. Моллюск питается водорослью, и, возможно, единст-
венная выгода для нее в том, что благодаря "подкормке" за счет
фотосинтеза внутри животного последнее снижает интенсивность вые-
дания ( Hinde, 1983).
Представители Cyanophyceae, Prochlorophyceae, Chlorophy-
ceae, Era sinophyceae, Bacillariophyceae, Prymnesiophyceae,
Dinophyceae и Hhodophyceae указывались как эндосимбионты ряда
пресноводных и морских хозяев. Наиболее изучены три типа ассоциа-
ций-
I. ПразИНОфИТ Tetraselmis (= Platymonas) В морском ПЛОС-
КОМ черве Convoluta.
2. Преимущественно пресноводные ассоциации с участием зеле-
ных водорослей, по ультраструктуре напоминающих chlorella и из-
вестных, особенно в зоологической литературе, как зоохлореллы.
3. Морские симбиозы динофитов, называемых зооксантеллами,
наиболее распространенный представитель которых - Symbiodinium
m-in-rowd-r-intimim показывающий значительную морфологическую из-
менчивость. В животной ткани он представлен коккоидной формой, а
в свободноживущем состоянии может быть подвижен и по признакам
приближается к Gymnodinium (Loebiich, Sherley, 1974). Предполо-
жительно ( Taylor, 1974) он обитает по меньшей мере в 80 видах
морских беспозвоночных из трех типов (простейшие, кишечнополост-
ные, моллюски). Особенности биохимии и заселения хозяина позво-
ляют предполагать наличие нескольких генетически различных форм
водоросли ( Schoenberg, Trench, 1980а, Ъ,с).
Ассоциации водорослей с беспозвоночными можно рассматривать
как стабильные сложные организмы ( Glider, Pardy, 1982) с при-
знаками, обеспечивающими им занятие ниш, недоступных для каждого
из компонентов по отдельности. К этим особенностям относятся ме-
ханизмы сохранения симбиоза при размножении и переноса веществ
между симбионтами.
Водоросли передаются потомству хозяина различным путем; у
гидры они внедряются в оплодотворенную яйцеклетку ( Muscatine,
McAuley, 1982) или приобретаются de novo личинками из окружаю-
щей среды. Свободноплавающие особи Platymonas convolutae хемо-
тактически привлекаются к защитным оболочкам яйца Convoluta
( Holiigan, Gooday, 1975). Некоторые голожаберные моллюски при-
обретают своих симбионтов-динофлагеллат, поедая кишечнополостных
с зооксантеллами и впоследствии включая поглощенные клетки в
свои ткани.
368
От водорослей-симбионтов хозяин получает значительную часть
питательных веществ. На свету апосимбиотические актинии исполь-
зуют запасы липидов, а симбиотические особи - углевода, предпо-
ложительно водорослевого происхождения ( Pitt, Pardy, 1981).
Взрослые экземпляры Convoluta полностью зависят от своих симби-
онтов и голодают в темноте. В то же время Hydra viridis меньше
зависит от водоросли chlorella и, хотя и не встречается в при-
роде в апосимбиотическом состоянии, гложет быть получена и под-
держана в таком виде в лаборатории ( Pardy, 1981). У рифообра-
зующих кораллов обызвествление и рост коррелируют с обилием сим-
бионтов; свыше 85% органического углерода они получают от водо-
рослей (Muscatine, Porter, 1977). Концентрация фотосинтетичес-
ких пигментов у этих зооксантелл возрастает с глубиной (Dunstan,
1979), что, возможно, компенсирует снижение освещенности, как у
растений, обитающих в тени. Обмен метаболитами между водоросля-
ми и хозяином продемонстрирован во многих ассоциациях, причем
большинство исследований было посвящено переходу углерода к жи-
вотному. В кораллах эта величина может достигать 50% фиксирован-
ного углерода, который в животной клетке встречается преимущест-
венно в виде белка или липида, хотя перелается в основном в ви-
де глицерина (Muscatine, 1973). В случае симбиозов с участием
Chlorella переносимый метаболит - мальтоза ( Ziesenisz et al.,
1981), тогда как Piatymonas convolutae продуцирует аминокисло-
ты (рис. 7.30) (Boyle, Smith, 1975).
Внутренний цикл углерода показан у коралла Porillopora
capitata, где зооксантеллы фотохимически перерабатывают ацетат,
получаемый от хозяина, образуя жирные кислоты. Вдобавок к этому
кораллы с симбионтами активно поглощают неорганические азот и
фосфор, в то время как апосимбиотические особи теряют их в окру-
жающую среду. Различие обусловлено переходом биогенов к зооксан-
теллам от хозяина, т.е. симбиотической адаптацией к запасанию
питательных веществ в бедных ими тропических водах (Muscatine,
1980). Piatymonas получает свой азот oTConvoluta в виде моче-
вой кислоты, и последняя накапливается в хозяине при подавлении
фотосинтеза.
С эволюционной точки зрения одной из наиболее интересных
ассоциаций между водорослью и беспозвоночными является симбиоз
прокариоты Prochloron и тропических асцидий (Dewin, 1984).
Водоросль обитает в тунике этих беспозвоночных, не проникая ни
в клетки, ни в органы. Prochloron способен к фотосинтезу внут-
369
Рис. 7.30. Предполагаемый обмен биогенами между плоским
червем Convoluta и его симбиотической водорослью (по
Boyle, Smith, 1975)
ри хозяина и in vitro (Fisher, 1980), но ауксотрофен по трипто-
фану ( Patterson, Withers, 1982). Изучению биологии Prochloron
мешает невозможность выращивания этой водоросли длительное время
вне животного-хозяина. Открытие свободноживущего протохлорофита
из одной эвтрофной лагуны в Нидерландах имеет, таким образом,
огромное значение ( Burger-Wiersma et al., 1986).
Взаимодействия водорослей с простейшими
Взаимоотношения фотосинтезирующих телец с простейшими, явля-
ющимися во всем, кроме наличия эндосимбионтов, гетеротрофами, ши-
роко изучались с эволюционной точки зрения. Некоторые из этих те-
лец явно родственны свободноживущим водорослям, например эндосим-
бионты Peridinium balticum (Thomas, Сох, 1973), другие, В част-
ности цианеллы, в криптомонадоподобном организме Cyanophora име-
Ют геном пласти,иного типа ( Herdman, Stanier, 1977), причем не-
которые аспекты их жизнедеятельности, например синтез фотосинте-
Т0ческих пигментов, находятся под частичным контролем ядерного
^ННома Хозяина ( Trench, Siebens, 1978).
370
Современные фораминиферы являются хозяевами представителей
Chlorophyceae, Dinophyceae, Bacillariophyceae и Rhodophyceae
ф<ее, McEnergy, 1983). Для ряда их видов указывался обычный ди-
нофлагеллатный симбионт кораллов Symbiodinium microadriaticum.
Эти простейшие особенно обильны в мелких тропических и субтропи-
ческих морях. Пески с фораминиферами могут быть в 50 раз продук-
тивнее планктона в том же местообитании ( Sournia, 1976) и обес-
печивают интенсивный круговорот азота и фосфора между водорослью
и ее хозяином (bee, 1980). Для роста некоторых видов необходим
свет. Рост Amphistegina lessonnii идет со скоростью, пропорци-
ональной освещенности, и прекращается в темноте даже при нали-
чии пищи ( Rottger et al., 1980); при ее отсутствии Globigeri-
noideB sacculifer медленно растет и иногда неспособен к размно-
жению ( в® et al., 1981). Водоросли играют определенную роль в
обызвествлении; около 20% фотосинтетически фиксированного угле-
рода впоследствии появляется в раковине фораминифер ( Smith,
Wiebe, 1977). Механизм получения животным соединений углерода
от водоросли неясен. Кример и др. ( Kremer et al., 1980) полага-
ют, что фотоассимилят высвобождается ее живыми клетками, а не в
ходе переваривания водорослевого симбионта, но некоторые авторы
( Lee, McEnergy, 1983) относятся к этому утверждению скептически
и, основываясь на ультраструктурных данных, утверждают, что уг-
лерод, фиксированный водорослью, поступает к хозяину после гибе-
ли последней.
Кроме симбиоза с животными водоросли образуют тесные и час-
то облигатные ассоциации с другими водорослями, высшими расте-
ниями и грибами.
Ассоциации водорослей с грибами
Среди всех симбиозов с участием водорослей их ассоциации с
грибами в виде лишайников уникальны тем, что имеют морфологичес-
кие, физиологические и экологические особенности, отсутствующие
и у грибного, и у водорослевого компонентов в отдельности
( Ahmadjian, 1981). Все лишайники, кроме самых примитивных, от-
личаются четкой внутренней структурой, причем водоросли распола-
гаются в подповерхностной зоне слоевища (рис. 7.31, А, Б). Со-
временные описания их биологии можно найти в монографиях Лори
( Lawrey, 1984) и Кершо ( Kershaw, 1985).
371
Объемные доли таллома
Гриб 43.3% ± 0.9
Водоросль 6.7% ± 0.3
Внеклеточный матрикс 34.1% ± 1.4
Воздушные пространства 18.2% ± 1.1
Процент стенки водоросли в контакте:
со стенкой гриба 21.3 ± 2.5
с неклеточным веществом 35.1 ± 3.6
Рис. 7.31. Морфология и анатомия лишайника Xanthoria
Eartetina: А - внешний вид; Б - поперечный ргАйрей талло-
водоросли располагаются непосредственно под верхней
"°Рой; В - грибные гифы, тесно контактирующие с поверх-
ностью водорослевых клеток (по Ahmajian, 1981)
Как фикобионты, так и микобионты способны к независимому
Росту Б аксеничной культуре, т.е. их взаимосвязь не облигатна
Abaadjian, 1981). Лишайники часто приводят как пример мутуализ-
372
ма, но, если они занимают ниши, недоступные для фикобионта в
свободноживущем состоянии, трудно говорить о каком-либо преиму-
ществе от этого для индивидуальных водорослевых клеток.
Б большинстве лишайников микобионт составляет более 90%
биомассы. Симбиоз образует почти 30% всех видов грибов. Преиму-
щественно это аскомицеты, иногда - базидиомицеты ( Smith, 1978),
Фикобионт представлен зелеными или синезелеными водорослями при-
близительно из 30 родов, причем в более 50% случаев это зеленые
водоросли Trebouxia И Paeudotrebouxia (Ahmadjian, 1981).
Известны химероидные лишайники, например Peltigera, с зеле-
ными водорослями внутри слоевища и синезелеными - в цефалодиях,
выростах на его поверхности.
Все имеющиеся данные свидетельствуют об одностороннем пото-
ке питательных веществ от водоросли к грибу - углеводов в глав-
ном талломе и аммония в цефалодиях. До 90% всего фотосинтетичес-
ки фиксированного углерода передается микобионту. Зеленые водо-
росли выделяют при этом многоатомные спирты эритритол, рибитол
или сорбитол ( Farrar, 1978), а синезеленые - глюкозу ( Hill,
1976); эти водорослевые продукты превращаются в грибном талломе
в маннит (рис. 7.32).Выделение фикобионтом углеводов прекращает-
ся почти сразу же после его перевода в культуру ( Staith, 1980).
Поступление этих веществ во много раз больше, чем требуется гри-
бу для роста. Избыток их, по-видимому, используется при таких
связанных со стрессами процессах, как усиленное дыхание во вре-
мя регидратации сухого лишайникового слоевища и регуляция осмо-
са при высыхании (Farrar, Smith, 1976). Хотя симбионты могут
быть тесно связаны грибными гаусториями или специализированными
контактами клеточных стенок в местах примыкания аппрессориев
(рис. 7.31,В), доказательств передачи через такие "мостики" уг-
лерода нет.
Способность лишайников фиксировать атмосферный азот зави-
сит от наличия в них синезеленых водорослей в качестве главного
фикобионта или компонента цефалодиев ( Ahmadjian, 1981). Азот-
фиксация происходит в гетероцистах, которые у фикобионтов встре-
чаются чаще (21% объема, 4% клеток), чем у тех же видов в сво-
бодноживущем состоянии. Фотосинтез в цефалодиях идет, по-видимо-
му, относительно слабо, и предполагается ( Hitch, Millbank,
1975), что зеленые водоросли главного слоевища поставляют угле-
воды синезеленым. Активность глутаматсинтетазы, с участием кото-
рой аммоний превращается в глутамат, в этих сияезеленых водорос-
373
Рис. 7.32. Поток углерода в лишайнике (по Parley, 1982)
лях подавлена, и нн^+ передается микобионту. Интересно, что
у водоросли могут наблюдаться симптомы азотного голодания fete-
wart, Rowell, 1977). По-видимому, активность глутаминсинтетазы
регулируется микобионтом, поскольку у выделенного из лишайника
фикобионта восстанавливается нормальная активность этого фермен-
та ( Sampaio et al., 1979).
Многие виды лишайников размножаются вегетативным путем с
образованием структур, обеспечивающих совместное распростране-
ние микобионта и фикобионта. Простейшие из этих вегетативных
Диаспор, соредии, представляют собой шары .диаметром 25 - 100 мкм
из нескольких клеток водоросли, окруженных грибными гифами. Ког-
да микобионт размножается половим путем, для образования лишай-
ника необходимо, чтобы его споры оказались возле водорослевого
партнера. Некоторые обычные фикобионты, особенно из синезеленых
водорослей, часто встречаются рядом с лишайниками, но самые ти-
пичные из них, например виды Trebouxia, в свободноживущем сос-
тоянии крайне редки ( Tschermak-Moess, 1978). Хотя Trebouxia
размножается в составе лишайника, как правило, апланоспорами,
она способна также образовывать и высвобождать зооспоры ( Slocura
et al., 1980). Вероятно, последние дают новый лишайник, всту-
пая в контакт с прорастающими грибными спорами.
374
В аксеничной культуре недавно удалось синтезировать лишай-
ник Cladonia cristatella (Ahmadjina, Jacobs, 1981). Методика
включала смешивание комков водорослей и гиф микобионта на толь-
ко что разделенных пластинках слюды, смоченных культуральной сре-
дой для водорослей и инкубированных при относительной влажности
примерно 95%. При этом показано, что отношения гриба к водоросли
носят агрессивный характер; лишь некоторые виды становятся фико-
бионтами лишайников, на остальных гриб паразитировал и убивал их.
Существуют симбиотические ассоциации морских водорослей и
морских грибов, когда грибной компонент растет в межклеточном
пространстве водоросли, но не изменяет ее морфологию и не приво-
дит к образованию специфических для этого симбиоза химических
соединений. Такие симбиозы, хотя они, по-видимому, и являются об-
лигатными, нельзя рассматривать как истинные лишайники; они наз-
ваны микофикобиозами ( J.Kohlmeyer, Е.Kohlmeyer, 1972). Наибо-
лее известно присутствие гриба Mycosphaerella ascophylli в бУ“
рых водорослях Ascophyllum nodosum И Pelvetia canaliculata.^н
растет между клетками водоросли, не проникая в них, и образует
в рецептакулах хозяина аскокарпы и спермогонии, которые могут вы-
глядеть на поверхности таллома черными точками. Показано, что в
аксеничной культуре этот гриб использует маннитол и ламинарии;
оба этих вещества вырабатываются хозяином ( Fries, 1979).
Ассоциации с бриофитами и сосудистыми растениями
Гетероцистные представители семейства Nostocaceae встреча-
ются в ассоциации с некоторыми бриофитами, например антоцеросом
и печеночником Blasia, водным разноспоровым папоротником Azolla;
несколькими саговниковыми и покрытосеменным растением Gannera
( Peters, Calvert, 1983). В гаметофитных талломах антоцероса и
Biasia находятся содержащие Nostoc полости; частота гетеро-
цист у водоросли может достигать 60% вегетативных клеток. Азот
выделяется в виде аммония и поглощается бриофитом-хозяином, кото-
рый, возможно, снабжает водоросль фиксированным углеродом (Ste-
wart, Rodgers, 1977). Как и в цефалодиях лишайников, активность
глутаминсинтетазы здесь подавлена, но восстанавливается при изо-
ляции- водор'осли.
В полостях верхних лопастей плавающих листьев Azolla нахо-
дится Anabaena azollae (рис. 7.33). Эта водоросль отличается
от других синезеленнх симбионтов сосудистых растений сложностью
375
Рис. 7.33. Продольный разрез через ось папоротника Azolla,
показывающий различные стадии формирования полостей на
листьях и захвата симбиотической синезеленой водоросли
Anabaena azollae. А.К. - аминокислоты (по Peters, 1978)
выделения, причем большинство ее клеток, вероятно, неспособно к
свободному существованию ( Peters, Calvert, 1983). Было изолиро-
вано несколько штаммов с гетеротрофным ростом в темноте и пони-
женной интенсивностью азотфиксации по сравнению с симбиотичес-
кими формами ( Newton, Herman, 1979).
В развитии этой ассоциации наблюдается выраженная синхрон-
ность. Нити Anabaena вступают в контакт с листовыми примордия-
ми апикальной меристемы папоротника. На нижней стороне листа
сначала образуется углубление, которое затем замыкается, захва-
тывая водоросль; деление водоросли и формирование гетероцист
происходят уже внутри полости. В зрелом растении частота гетеро-
цист варьирует от 20 до 60%; на водорослевый симбионт приходит-
ся 15% биомассы ассоциации; он способен обеспечить всю потреб-
Ность Azolla в азоте (рис. 7.34) (Peters, Calvert, 1983).
Ассоциации синезеленых водорослей с саговниковыми формируют-
ся внутри специализированных коралловидных корневых клубеньков,
а У Ounnera - в основаниях листьев (whitton, 1973). Симбиоз
S°stoc о Gunnera - исключение среди фотосинтезирующих рас-
тений в том плане, что нити водоросли мохут проникать в межкле-
точное пространство покрытосеменного вида (Schaeda, 1951).
376
Листья
Рис. 7.34. Относительная нитрогеназная активность, процент
сухого вещества, отношение углерода к азоту и процентное
отношение азота к сухой массе в разных участках главных
стеблей Azolla, несущих последовательные группы листьев
(по Calvert, 1983)
Паразитические водоросли
В семействе Chroolepidaceae (Chlorophyceae) и среди багря-
нок известны водоросли, ставшие паразитами и в разной степени за-
висящие в своем питании от растения-хозяина.
Лучше всего известные паразиты среди Chroolepidaceae - это
виды ceohaieuros. распространенные в тропиках и субтропиках.
Водоросли растут под кутикулой хозяина на стеблях, листьях, цвет-
ках и плодах. Иногда они образуют в своих выходящих на поверх-
ность репродуктивных структурах оранжево-красные гематохромные
пигменты, придающие поражению облик, напоминающий грибную ржавчи-
ну. Заражаться может широкий круг сосудистых растений, в том чис-
ле чайный куст, кофейное дерево, цитрусовые и многие другие эко-
номически важные виды. Только для одной Луизианы указывается 116
видов, восприимчивых к c.virescens (Holcomb. 1975). Хотя за-
377
ражаются многие виды, наблюдаются значительные различия в устой-
чивости к инфекции даже в пределах одного вида.
На специальных фертильных ветвях, поднимающихся над поверхно-
стью листа, развиваются зооспорангии. Они толстостенные, но на
ранней стадии в них формируется выводная пора, закрытая пробкой
из белкового материала. Зрелые зооспорангии отпадают и разносят-
ся ветром, водой, насекомыми и паукообразными. При наличии доста-
точного количества воды пробка растворяется и высвобождаются зоо-
споры. Чтобы выжить, они должны быстро проникнуть через кутикулу
растения, и в этом им помогают животные-переносчики, которые, пи-
таясь, повреждают поверхность листьев.
Эксперименты по культивированию показывают, что виды Cepha-
leuroe .потенциально способны получить питание от хозяина, по-
скольку добавление глюкозы или дрожжевого экстракта к среде
вызывало заметную стимуляцию их роста (Joubert et al., 1975).
Наблюдался также гетеротрофный рост в полной темноте на ряде гек-
соз, хотя восцроизведение отсутствовало, если не добавлялось низ-
ких концентраций ауксинов (JOse, Chowdray, 1978). Это дает осно-
вание полагать, что in vivo размножение водоросли зависит от
гормонов, продуцируемых хозяином.
Паразиты-багрянки - таксономически неоднородная группа,
включающая более 100 видов из 50 ролов. Они распадаются на две
четкие категории: большинство - адельфопаразиты, родственные сво-
им хозяевам, например относящиеся к тому же семейству или той же
трибе; остальные - аллопаразиты, неродственные своим хозяевам.
Паразитические водоросли в основном мелкие (мельче 0,5 см), с
ослабленной или отсутствующей пигментацией; морфологически они
устроены просто: состоят из ветвящихся нитей,’ распространяющихся
между клетками хозяина. Репродуктивные структуры закладываются в
массе ткани, выступающей над поверхностью хозяина ( Evane et al.,
Goff, 1982b).
Багрянки паразитируют только на других красных водорослях
по-видимому, высокоспецифичны в отношении хозяев. Janczew-
skia mortmotoi в природе, вероятно, встречается только на
kaurencia nipponica. хотя есть данные о ее способности заражать
Другие вида Laurencia, но не представителей каких-либо других
РОДОВ Rhodomelaceae (Nonomura, West, J98I). Тесты ПО пере-
крестной инокуляции водорослью Asterocolax Rardneri, паразити—
РУЮщей на хозяевах из семейства Deleaseiiaceae (Wienburgia anders-
goniar^. PhycodrVB setohellii и folyneura latissima).
378
показывают, что, хотя речь как будто идет об одном и том же так-
соне, паразит, похоже, способен заражать только тот вид, из кото-
рого выделен (Goff, 1982b).
Характер питания изучен лишь у немногих паразитических ба-
грянок-, но для большинства, особенно с ослабленной или отсутству-
ющей пигментацией, предполагается, по крайней мере частичная, за-
висимость ОТ хозяев. Hclmsella pachydexma. паразит Gracilaria
verrucosa, передает хозяину флорэдозид, тот превращает его в
маннитол, а затем в крахмал, что создает градиент концентрации,
обеспечивающий движение фотоассимилятов посредством диффузии
(Evans et al., 1973). Harveyella airabilia получает углерод
в виде дигенеазида и, возможно, нескольких кислот (рис. 7.35)
(Kremer, 1983). Более крупные, сильнее пигментированные парази-
ты способны фотосинтезировать сахара и аминокислоты, и свиде-
тельств их передачи от хозяина к паразиту нет (court 1980). Не-
которые из таких фотосинтезирующих паразитов, например Erythro-
cygtig aaccata, нормально развиваются только в присутствии хозяи-
на, что предполагает получение от него регуляторов роста ( Goff,
1982b).
Вторичные поровые соединения, по-видимому, обязательный при-
знак любых ассоциаций хозяина с паразитом среди красных водорос-
лей. Ядра и другие цитоплазматические компоненты Choreocolax
polygiphoniae переходят через пору В клетку Polysiphonia con—
fusa. Ядра остаются при этом интактными и стимулируют синтез
ящерной ДНК хозяина. Происходят также пролиферация клеточных ор-
ганелл последнего и накопление крахмала. По-видимому, генетичес-
кая информация паразита изменяет в его пользу физиологию хозяи-
на ( Goff, Colleman, 1985).
Эпифиты
Эпифиты, встречающиеся на других водорослях или сосудистых
растениях, - наименее специализированные из симбиотических водо-
рослей. Многие такие виды с равным успехом могут быть найдены на
неорганических субстратах и не показывают какой-либо специфично-
сти в выборе форофита. Токида ( Tokida, I960) перечисляет почти
300 видов водорослей-макрофитов, встречающихся эпифитно на пред-
ставителях Laminariales. Харлин ( Harlin, 1980) приводит СПИСОК
свыше 400 макроскопических и более 150 микроскопических эпифит-
ных водорослей (преимущественно диатомей), обнаруженных на мор-
379
Rhodo meta
Рис. 7.35. Ежедневный баланс углерода в тропических отно-
шениях между Rhodomela confervoides И паразитом Harvey-
ella mirabilis^ иырй.я масса хозяина принята за“2ии мг,
паразита - КТО ,05 мг. РОВ - растворенное органическое ве-
щество (ПО Kremer, 1983)
ских растениях. По-видимому, эпифитизм у большинства видов -
следствие конкуренции за субстрат, свет и биогены в цусто засе-
ленной среде.
Известны, по крайней мере в природе, и облигатные эпифиты,
произрастающие на очень небольшом числе видов-форофитов. В от-
личие от паразитизма здесь нет явно перемещения веа'еств от "хо-
зяина". Наиболее хороню изучена такая, видимо, облигатная ассо-
циация между красной водорослью Polysiphonia lanosa и фукусовым
Ascophyllum nodosum (Turner, Evans, 1977). Эпифит образует
ризоиды, проникающие в ткань форофита, вероятно, за счет фермен-
тативного переваривания (Rawlence, 1972), однако переноса мета-
болитов не наблюдается (Barlin, Craigie, 1975; Turner, Evens,
1978). В пресных водах не обнаружено ( Cattaneo, Kalff, 1979)
Различий в биомассе и продукции видов, зпифитно произрастающих
Яа рдесте и выращенных на пластиковых пластинах. Однако в первом
случае у них была понижена активность кислотной фосфатазы, т.е.
380
не исключено, что органические соединения фосфора они получают
от сосудистого растения (рис. 7.36).
Отрицательное влияние эпифитов на форофиты - следствие за-
тенения и механической нагрузки. Сообщалось (D’Antonio, 1985) о
подавлении роста и увеличении травматизма таллома у сильно оброс-
ших эпифитами экземпляров Rhodomela larix у побережья Орегона
(рис. 7.37).
Некоторые растения развили механизмы, которые могут ограни-
чивать развитие на них эпифитов. Известно, что у ряда бурых водо-
рослей, например Halidrys вШдиова (Moes, 1982) И Hlmanthalia
(Russell, Veltkamp, 1982), слущивается поверхность эпидер-
мы; рост эпифитов может подавляться полифенолами, выделяемыми
морскими растениями ( McLachlan, Craigie, 1964).
Водоросли при экстремальных температурах
Снежные водоросли
Такие водоросли встречаются повсюду, где имеется постоян-
ный или почти постоянный снеговой погфов. Чаще всего это хламидо-
монады, но отмечались также эвгленовые, золотистые, синезеленые,
желтозеленые и диатомовые водоросли, динофлагелляты и криптофиты
( НоЬаш, 1980). Их таксономия разработана плохо; многие могут
быть стадиями в жизненном цикле других видов, а большое число
форм, считавшихся хлорококковыми, оказалось зиготами видов Chla-
mydomonas (рис. 5.18). Полагают ( Hoham, 1975а,ъ ), что
истинно снежные водоросли оптимально растут при температурах ни-
же Ю°С. Chloromonas pichinchae в более теплых условиях не рас-
тет и существует в пальмеллоидном состоянии; подвижная фаза на-
блюдается только между I и 5°с.
Хламидомопадные цветения начинаются тогда, когда температу-
ра воздуха остается достаточной для таяния несколько дней подряд
и в массе снега появляется вода. В течение этого периода свет,
биогены и растворенные газы становятся доступными для водорослей.
Покоящиеся зиготы могут прорастать в снегу или под ним у поверх-
ности почвы, подвижные клетки за счет фототаксиса плывут к по-
верхности и размножаются зооспорами, chloromonas pichinchae
перемещается вверх на рассвете и в сумерках, но, вероятно, избе-
гает сильного света в полдень, заглубляясь на 10 - 15 см в снег.
381
Рис. 7.36. Активность вислой фосйатазы слабо прикреплен-
ных эпифитных сообществ, развивающихся на высшем водном
растении Potamogeton и на пластиковых лентах; для срав-
нения взят обитающий вокруг фитопланктон(по Cattaneo,
Kalff, 1979)
Рис. 7.37. Выживание индивидуально меченных осей Rhodo-
pela larix С эпифитами и без НИХ (по d’Antonio, 1985)
382
Через несколько дней образуются гаметы, а возникающие при их
слиянии зиготы засыпаются снегом или при его таянии попадают в
почву. В любом случае вегетативная фаза снежных водорослей длит-
ся примерно неделю в год (Hoham, 1980).
Эти виды иногда образуют вспомогательные пигменты, которые
в условиях цветения придают снегу красный или желтый цвет. Крас-
ный пигмент Chlatnydomonas nivalis - ксантофилл астаксантин;
его синтез, возможно, следствие недостатка азота (Czygan, 1970).
Другие вторичные каротиноиды - желтые, розовые и оранжевые - мо-
гут встречаться в зиготах. Распространение снежных водорослей
связано с экспозицией, затенением, высотой над уровнем моря и
доступностью биогенов. На юго-западе США зеленые водоросли мож-
но обнаружить в нескольких сантиметрах под поверхностью снега в
лесах; в оранжевый цвет бывают окрашены его верхние слои близ
деревьев; красный цвет снега характерен для открытых пространств
в горах выше лесного пояса (Hohan, Biinn, 1979).
Водоросли морского льда
Такие водоросли встречаются в морском паковом льду полярных
областей; они растут в канальцах с соленой водой между крис-
таллами льда и в его рыхлом слое на нижней стороне льдин (Hor-
ner, Alexander, 1972). Наиболее обычны здесь диатомеи, но
указывались также динофлагеллаты, зеленые жгутиковые и криптофи-
ты. Большинство из них - непланктонные формы, и их группировки
отличаются от планктонных (Bunt, Wood, 1963). Оценки биомассы
для сообществ донного льда в Антарктике достигают 300 мг хлоро-
филла на I ( РаЛлДвапо, Sullivan, 1983).
Эти водоросли адаптированы к росту при низких освещенности
и температуре; сообщалось, что ffragilaria sublinearis способ-
на развиваться ниже 0°С при 50 - ЮО лк (Bunt et al., 1966).
Предполагалось, что в зимние месяцы здесь возможна факультатив-
ная гетеротрофность (Allen, 1971), хотя Хорнер и Александер
( Homer, Alexander, 1972) сделали вывод о пренебрежимо малой
интенсивности гетеротрофного метаболизма глицина, глюкозы и аце-
тата у микроскопических водорослей морского льда в Арктике. За-
то темновая гетеротрофность обнаружена у трех видов диатомей ан-
тарктического морского льда ( Palmisano, Sullivan,1983), причем
у таких водорослей отмечено ( Palmisano et al., 1985) поглоще-
ние серина при концентрациях субстрата, близких присутствующим
383
в окружающей среде, хотя это дает менее 0,3% углерода, фиксиро-
ванного фотосинтетическим путем.
Водоросли горячих источников
На другом конце биотического температурного спектра находят-
ся водоросли горячих источников, обычные в районах активной вул-
канической деятельности. Кроме повышенной температуры, такие во-
ды отличаются в 10 - 15 раз более высоким содержанием растворен-
ных солей, чем в обычных водоемах; они особенно богаты натрием,
калием, карбонатом, силикатом, хлоридом и сульфидами. У некото-
рых значение pH низкое; они могут иметь высокие концентрации тя-
желых металлов (гл. 9). Эти экстремальные условия постоянны в
течение года, а водоемы обычно мелкие и прозрачные. Обитающим
здесь водорослям нет необходимости адаптироваться к сезонным
изменениям среды, кроме связанных с освещенностью. Обзор основ-
ной информации об этих термофильных организмах дает Брок (Brock,
1978).
По-видимому, существует обратная взаимосвязь между сложно-
стью клеточной организации и способностью выживать при повышен-
ных температурах. Некоторые гетеротрофные бактерии устойчивы к
кипящей воде; фотосинтезирующие синезеленые водоросли, например
Synechococcua lividus (Brock, 1967), могут встречаться при
+?0°С, а вид неясного систематического положения Cyanidium cal-
darium растет при температуре до 57°С ( Doemel,Brock, 1970).
Между 45 и 40°С отмечаются уже эукариоты - .диатомеи ( Achnanthes,
Bavicula ) и зеленые водоросли (например, Ulothrix ) ( Kullberg,
1971). Вид Synechococcus lividus наблюдался в интервале темпера-
тур от 53 до 7Э°С ( Fair child, Sheridan, 1971); изолировано его
четыре генетически различных штамма, каждый - со своим темпера-
турным оптимумом. Установлено ( Sheridan, 1979) существование
генетически различных световых и теневых Форм Plectonema nota-
tum , встр ечающихся на разной глубине внутри водорослевой пленки.
Многие водоросли горячих источников (например, Cyanidium
caldarum, MaatiRocladus врр.) имеют широкое географическое рас-
пространение, что трудно объяснить с помощью любой из гипотез
Распространения на большие расстояния. Брок ( Brock, 1969) пред-
полагает, что такие термофильные виды представлены реликтовыми
популяциями, сохранившимися с раннего периода истории Земли, ког-
384
да вулканические условия встречались чаще, чем сейчас. Интерес-
но, что на Исландии, сравнительно молодом вулканическом острове,
найдено только 6-8 видов синезеленых водорослей горячих источ-
ников (Castenholz, 1969), т.е. в несколько раз меньше, чем мож-
но ожидать в таких биотопах на континентах.
Глава 8
ЭВОЛЮЦИЯ И ФИЛОГЕНИЯ
Основные этапы эволюции водорослей
Разгадка происхождения водорослей - центральная тема изуче-
ния их эволюции. Процесс развития этой группы охватывает большую
часть известного периода существования жизни на Земле - с появ-
ления пержх прокариот до нашего времени. Он связан с рядом глав-
ных событий ботанической эволюции: появлением фотоавтотрофных
прокариот, эукариот, митоза, оингамии и мейоза, чередования поко-
лений, а также с адаптацией к наземной жизни.
Возраст прокариот приближается к возрасту древнейших извест-
ных нам горных пород, т.е. примерно к 3,8 млрд, лет (рис. 8.1).
Первые автотрофные организмы, синезеленые водоросли, появились
примерно в середине архея (3,8 - 2,6 млрд, лет назад), т.е. в
раннем докембрии (Schopf, Walter, 1982а). В позднем докембрии,
Рис. 8.1.Примерная последовательность событий в эволюции
биосферы и гипотетические уровни кислорода (в процентах
к современному). Масштабы логарифмические, а кривая край-
не обобщенная; возможно, происходили отклонения от сред-
них цифр (по Cloud, 1976, с изменениями)
386
протерозое (2,6 - 0,58 млрд, лет назад), они были распростране-
ны повсеместно (рис. 8.2), причем в ископаемых остатках этого
периода, часто называемого веком цианобактерий, известны предста-
вители большинства их современных порядков (Schopf, Walter,
1982а).
Клетки с оформленным ядром (эукариоты), возможно, появились
около 1,4 млрд, лет назад (schopf, 1978 ). мейоз и эукариотное
разделение полов - по крайней мере 800 млн. лет назад (Нойпап,
Atiken, 1979), но не исключено, что и на 530 млн. лет раньше
(waiter et al., 1976). Последующее формирование жизненных цик-
лов с чередованием диплоидного и гаплоидного поколений происхо-
дило по четырем основный направлениям (гл. 5); сведений о точнсм
времени зарождения каждого из них нет, хотя последовательность,
в которой они развивались, может быть выведена на основе жизнен-
ных циклов современных растений (Scagel et al., 1982). Чередо-
вание поколений оказалось одним из приспособлений, существенных
при жизни на суше и в конечном итоге сыгравшем важную роль в эво-
люции наземных растений (эмбриофитов). Широко распространено мне-
ГЕОЛОГИЧЕСКОЕ РАСПРЕ
ДЕЛЕНИЕ ИСКОПАЕМЫХ
СИНЕЗЕЛЕНЫХ
ВОДОРОСЛЕЙ
"Коккоидные "
Chroococcales
Chroococcaceae
Entophysalidaceae
Pleurocapsales
Hyellaceae
"Нитчатые"
Nostocales
Oscillatoriaceae
Nostocaceae
Scytonemataceae
Rivulariareae
Stigonematales
Stigonemataceae
"Строматолиты"
• — Достоверные находки © Спорные данные
Рис. 8.2. Гистограмма появления семейств синезеленых во-
дорослей, построенная по ископаемым находкам (по schopf,
1970, с изменениями)
387
ние, что последним дали начало предки зеленых водорослей, возмоя-
но, где-то между 700 и 400 млн. лет назад ( Cloud, 1976).
Происхождение прокариот и фотоавтотрофов
Основные события ранней эволюции Земли и жизненных форм
обобщены на рис. 8.3. Предполагается, что возраст нашей планеты -
4-6 млрд, лет; на протяжении примерно половины ее существова-
ния в атмосфере отсутствовал молекулярный кислород ( Cloud, 1976).
Прокариоты развивались в течение биогенного периода 4,5 -
3,5 млрд, лет назад, когда атмосфера содержала н2, со2, л2,
SO2,HC1 и была практически лишена свободного02 (Walter et.
al., 1982). Первые прокариоты являлись предположительно водными
анаэробами. Их ископаемые остатки встречаются в различных слан-
цах и стрсматолитах. Строматолиты - это плоские, выпуклые, колон-
чатые или куполообразные осадочные структуры с внутренней слоис-
тостью, возникшие в результате послойного захватывания, связыва-
ния и/или осаждения материала микробными сообществами (в основ-
ном синезеленых водорослей), называемыми водорослевыми пленками.
Сходные сообщества формируют строматолиты или подобные им образо-
вания и в настоящее время (рис. 8.4), но с конца докембрия их
деятельность значительно сократила свои масштабы. Строматолиты
были наиболее широко распространены и обильны в позднем архее и
в протерозое, когда на протяжении примерно 2,5 млрд, лет прока-
риоты были доминирующей формой жизни на Земле (век прокариот).
Неясно, когда появились первые прокариоты и в какой после-
довательности; как бактерии, так и синезеленые водоросли встре-
чаются в наиболее ранних строматолитах. Предполагают, что сине-
зеленые водоросли возникли от эубактерий, одной из трех древних
эволюционных линий прокариот (архебактерии, уркариоты и эубакте-
рии), отделинпихся в свою очередь от единого предка ( Woese,
1981; рис. 8.5). Даже лучше всего сохранившиеся микроскопичес-
кие ископаемые из архейских отложений дают о морфологии клетки
информацию, недостаточную для различения примитивных сине зеленых
водорослей и других прокариот (например, Beggiatoaceae из отде-
лаSchizomycophycota). На основании имеющихся к настоящему вре-
мени данных невозможно также определить, были или нет первые си-
чезеленые водоросли автотрофами, а если были, то использовали ли
обе фотосистемы - I и П - т.е, высвобождали ли как побочный про-
дукт фотосинтеза кислород (Schopf, Walter, 1982а). Пока этот
388
ОСНОВНЫЕ
ОБСУЖДАЕМЫЕ
МИКРОБИОТЫ
СОБЫТИЯ И ТЕНДЕНЦИИ
ЛИТОЛОГО-
ИСТОРИЧЕСКИЕ
ПОДРАЗДЕЛЕНИЯ
Саблино
(пирофиты)
Хектор (эндоспоры)
Биттер-Спрингс
(мейоз?)
Бек-Спрингс (митоз)
Парадайз-Крик
Ганфлит (гетероцисты)
Покегама (синезеленые
водоросли)
Булавайские стромато-
литы (синезеленые
водоросли?)
Строматолиты
Понголы (синезе-
леные водоросли?)
Примерный
возраст, млрд, лет
0.7
МНОГОКЛЕ-
ТОЧНЫЕ
Оледенение
Обычны
отло-
жении
CaSO4
___t___
_ Обычны
Увеличение содержания О2 СаМд (ср («„гаи
Снижение содержания СО2 СаСО3
ЭУКАРИОТЫ
02 в атмосфере * , г
Оледенение Главный этап ।
ипж ।
2,5 ТОЛЬКО ПРОКАРИОТЫ
Биогенный О2 в гидросфере
в равновесии с Fe2*
4Fe2+ + О2 + ВОН 2Fe2O3 + 4Н2О
6Fe2O3 + С 4Fe3O4 + СО2
3,4--------Древнейшие хорошо
сохранившиеся осадки
и океанические лавы
2 g____Сильно метаморфизованные
' гнейсы и метаосадки
Неок пелен ные
эпикратонные
осадки, насто-
ящие граниты
(протерофит/
Окисленные
эпикратонные
осадки, много
клеточные
(фанерозой/
Окисленные
эпикратонные
осадки, много-
клеточных нет
(палеофит/
। (Зулусский '
ЭЕ I клин)
ё
Зеленокаменные
породы, граувакки,
< натриевые II
“граниты”,
серые гнейсы
(архей/
Первые автотрофы?
Химическая эволюция биогенез?
Главная дегазация (свободного О_ нет)
Происхождение концентрической структуры
Земли
Гравитационное нагревание
4,65 Возраст метеоритного РЬ , происхождение
Земли
Достоверных
наземных
находок нет
(хэд)
Рис. 8.3. Основные этапы (I - V) истории Земли с соответ-
ствующими аспектами биологической и геологической эволю-
ции (по Cloud, 1976, с изменениями). 01Ш - образование
полосчатых железистых пород
вопрос не решен, невозможно в деталях проследить их происхожде-
ние от предположительно фотосинтезировавших, но не высвобождав-
ших кислород предков. Фокс и др. ( Рох et al., 1980), деталь-
но анализируя последовательность 16S-рибосомной РНК бактерий,
-показали, что бактериальные фотосинтезирующие фенотипы крайне
389
Рис. 8.4. Синезеленая водоросль Aphanothece sp. -
живая "мозговидная" пленка с морской литорали в заливе
Акаба. Такие современные формы, вполне возможно, напо-
минают те, что дали начало ископаемым строматолитам.
Масштаб 10,0 CM (Potts, 1980)
древние и многие нефотосинтезируюшие группы, по-видимому, воз-
никли от фотосинтезирующих предков; это ставит под сомнение дол-
го бытовавшее мнение об анаэробной гетеротрофности первых бакте-
рий. Предками синезеленых водорослей могли быть анаэробные фото-
синтезирующие бактерии, представленные тремя основными ныне жи-
вущими группами: зелеными и пурпурными серными и пурпурными не-
серными (Margulis, Schwartz, 1982). Эти прокариоты не исполь-
зуют воду в качестве донора водорода при фотосинтезе и высвобож-
дают не кислород, а ряд других продуктов, в частности серу.
Однако по косвенным признакам можно предположить, когда в
архее начался кислородный фотосинтез. Вероятно, кислород прежде
всего высвобождался в гидросферу, где процессы окисления могли
затрагивать донные осадки. В период 3,5 - 2,0 млрд, лет назад
Происходило широкомасштабное отложение полосчатых железистых
формаций. Они состоят из мощных последовательностей сланцев,
390
Рис. 8.5. Гипотетическое филетическое дерево, построен-
ное исходя из гипотезы трех первичных царств и отделе-
ния архебактерий от всех остальных бактерий. Предполага-
ется, что архебактерии, эубактерии и уркариоты происхо-
дят от обще го предка (прогеноты), который был значитель-
но примитивнее самых просто устроенных современных про-
кариот. Эукариоты, происшедшие от уркариоты, стали "хо-
зяевами" бактериальных эндосимбионтов, развившихся в хо-
де эукариогенеза в митохондрии и хлоропласты (по woese,
1981, с изменениям)
слои которых поочередно то богаты, то бедны железом, причем в
первом случае оно находится в окисленном, трехвалентном состоя-
391
нии. В окислении и осаждении двухвалентного железа, растворенно-
го в морской воде, должен был участвовать высвобожденный при фо-
тосинтезе кислород. Следовательно, происхождение кислородного
фотосинтеза или предшествовало, или совпадало по времени (свыше
3,5 млрд, лет назад) с появлением первых полосчатых железистых
формаций. Отложение красноцветных пластов сменило накопление
этих осадков около 2,0 млрд, лет назад. Следовательно, к тому
времени кислород уже накопился в атмосфере в достаточных количе-
ствах для окисления обнажающихся пород в ходе их выветривания.
Когда его содержание в атмосфере достигло примерно 1% от совре-
менного уровня, у верхних ее границ сформировался слой, богатый
озоном. Это событие предопределило появление эукариот. Предпо-
лагается ( Schopf, Walter, 1982а), что кислород накапливался
в атмосфере относительно быстро, если исходить из геологических
масштабов времени.
Таким образом, косвенные свидетельства позволяют предполо-
жить, что синезеленые водоросли возникли примерно 2,5 млрд, лет
назад. Однако интерпретация имеющихся палеонтологических нахо-
док чревата сложностями. В древнейших микроскопических остатках
(3,5 млрд, лет) синезеленые водоросли не определены (Schopf,
Walter, 1982а). Строматолиты Инсузи из Южной Африки (Мавоп,
Van Brunn, 1977) возрастом 3,1 млрд, лет чрезвычайно сходны с
образующимися ныне в Хот-Спрингс в Йеллоустонском национальном
парке (США) и, возможно, откладывались фототаксическими нитча-
тыми фотоавтотрофами, однако микроорганизмов в них не найдено.
Первые достаточно достоверно определенные синезеленые водоросли
обнаруживаются только в позднеархейских породах возрастом при-
мерно 2,8 - 2,5 млрд, лет (группа Фортескью в Западной Австра-
лии; Schopf, Walter, 1982b). По морфологии они напоминают
Oscillatoria И Lyngbya.
"Век цианобактерий" (синезеленых водорослей)
Синезаленые водоросли были преобладающими на Земле орга-
низмами в период примерно 3,0 - 1,0 млрд, лет назад (Schopf,
1974а,Ь ). В это время они существенно влияли на эволюцию атмо-
392
сферы, которая постепенно изменялась от бескислородной к кисло-
родной в результате фотосинтеза; в ответ на это среди синезеле-
ных водорослей появились фотоавтотрофные формы. Вдобавок нитча-
тые виды развили гетероцисты, которые "защищают" их чувстви-
тельный к кислороду нитрогеназный ферментный комплекс.
Протерозойские синезеленые водоросли встречались повсемест-
но, обычно образуя пленчатые сообщества, дававшие начало строма-
толитам, хотя занимали такие любое доступное местообитание, ку-
да попадал свет, особенно в более поздний, кислородный период.
Появление эукариотных известковых и необызвествленных водорос-
лей, а также сверлящих И грызущих Metazoa (Schopf.Walter, 1982)
в конце докембрия положило конец веку синезеленых водорослей.
Однако эволюционный консерватизм и развитая способность к адап-
тации способствовали сохранению этой группы до нынешнего време-
ни.
Происхождение эукариот
Наибольший эволюционный скачок среди живых организмов -
это переход от прокариот, у которых отсутствуют окруженные мем-
бранами органеллы, нет жгутиков, организованных по схеме "9+2",
и ядерной оболочки, к эукариотам, обладающий, всеми этими при-
знаками (кроме багрянок, лишенных "9+2"-структур). Однако
вплоть до 1960-х гг. фундаментальные различия между про- и эука-
риотами четко не осознавались (Stanier, 1961). Скачок оказался
не только морфологическим, но и биохимическим ( Ragan,Chapman,
1978), и кандидатов на роль промежуточных форм нет, кроме, воз-
можно, Prochlorophycota (Lewin, 1976, 1977). Однако отмечено
( Chapman, Trench, 1982), что, если не считать такого существен-
ного отличия, как присутствие хлорофилла ъ, Prochloron - это
"хорошая" цианобактерия.
В последние 20 лет достигнут большой прогресс в поисках от-
вета на вопрос, когда и как появились эукариоты. Предполагают
(Cloud, 1976), что это произошло после примерно 2,0 млрд, лет
назад, т.е. когда земная атмосфера содержала достаточно свобод-
393
него кислорода. По-видимому, они уже были достаточно распростра-
нены к 1,3 млрд, лет назад, так как описаны ( Walter et al.,
1976) макроскопические, возможно эукариотные, остатки водорослей
этого возраста из глинистых сланцев Грейстон супергруппы Белт в
[Лонтане (например, Proterotainia, Lanceoforma) Этапы их эволю-
ции связаны с развитием окруженных мембранами органелл, вклкяая
ядро, хлоропласты, митохондрии и "9+2"-жгутики (за исключением
багрянск), а также линейных хромосом и веретена в сочетании с
митозом.
Имеются дне научные школы, резко различающиеся по взглядам
на детали-этого процесса. Одна предполагает экзогенное происхож-
дение окруженных мембранами органелл через серию эндосимбиозов
( Mereschkowsky, 1905; Margulis, 1970, 1981), например, мито-
хондрий - от бактерий, хлоропластов - от синезеленых водорослей,
жгутиков - от спирохет (рис. 8.6). Эта теория очень привлекатель-
на тем, что хорошо известны симбиозы одних ныне живущих водорос-
лей с другими (нагример, Glaucophyta, см. с. 76) и водорослей
с животными. Однако она требует признания до пяти отдельных эн-
досимбиотических событий и 27 независимых этапов перехода от ми-
тотического клеточного деления к мейотическому ( Margulis, 1970;
Cloud, 1976). Еще один недостаток в том. что для успеха процес-
са требуется серия совпадений. Постулировано (McQuade, 1983),
что митоз, фаготрофность, митохондрия, жгутик, половое размно-
жение и хлоропласт - слишком сложные явления и структуры, и
трудно предположить их неоднократное появление de novo. Экзо-
генная теория еще должна удовлетворительно объяснить происхожде-
ние ядра, а предложенный ею эндосимбиотический путь приобрете-
ния "9+2"-жгутиков не получил общего признания.
Другая теория предполагает постепенную эволюцию окруженных
мембранами органелл от неограниченных мембранами систем прокари-
отного предка в ходе аутогенного процесса ( Aiisopp, 1969; Cava-
lier-Smith, 1975). Кавалье-Смит ( Cavalier-Smith, 1975, 1978)
считает, что это наиболее правдоподобная альтернатива эндосим-
биозу. Важнейшим этапом здесь была эволюция эндоцитоза (фагоци-
тоза и пиноцитоза, рис. 8.7), обеспечившего разделение клетки
внутренними мембранами. Возможно, эндосимбиоз был бы одним из
394
Базидии
Аски
Соредии
ГРИБЫ
♦
Таннин Кость
Лигнин Раковина
Целлюлоза Эмбрионы Мышца
РАСТЕНИЯ ЖИВОТНЫЕ
\ Бластулы । Дикариоз i
' Сложные соединения ’ Спорогенез путем !
\ клеток зиготического I
I Гаметический мейоз 1 мейоза /
\ \ /
Г кетогенез
протоктисты
(водоросли, оомицеты,
слизевики, простейшие)
t
л
Конъюгация
Митоз
Мейоз
Центриоли
Ундулилодии
Зооспоры
Фагоцитоз \
Внутриклеточная '
подвижность
♦
Эндомембраны
Споры
\ Многоклеточность
Фотосинтез
Дыхание
Брожение \
1\
Коккоидные синезе-
леиые водоросли
Хлорокси бактерии
Паракокки
Виды
Bdellovibrio
Устойчивость к Подвижность
высоким темпера- Спирохеты
турам и кислотам Спироллазмы
Термоплазмы
Рис. 8.6. Модель экзогенного Формирования эукариотной
клетки посредством симбиоза. Прослежено происхождение
растений, животных и грибов. Предполагаемое здесь появ-
ление жгутиков из спирохет не нашло широкого признания
(по Margulis, 1981)
неминуемых следствий фагоцитоза, но не причиной эукариотного
состояния. Способность к фагоцитозу дала предку эукариотных во-
дорослей селективное преимущество перед другими синезелеными
формами (от которых он возник), позволив фотосинтезировать на
свету и фагоцитировать в темноте. Аутогенная теория предлагает
механизмы эволюции ядра, митоза, мейоза, веретена, митохондрий
и микротрубочек без привлечения эндосимбиоза. Кавалье-Смит
(Cavalier-Smith, 1975) допускает, однако, что симбиотическая
природа митохондрий и хлоропластов совместима с аутогенным про-
395
Рис. 8,7. Предполагаемая эволюция эукариотной клетки в хо-
де аутогенного процесса, включающего две стадии зндоцито-
за или фагоцитоз и пиноцитоз), давшие начало хлоропласту,
митохондриям, ядру и эндоплазматической сети. Модель не
показывает происхождения жгутиков. А. Первичные впячива-
ния. Б. Вторичные впячивания. X - хлоропласт, ДНК - де-
зоксирибонуклеиновая кислота, цЩЖ - пластидная ДНК;
мДНК - митохондриальная ДНК; ЭС - эндоплазматическая
сеть; Я - ядро; М - митохондрия; Т - тилакоиды ( chade-
faud, 1974).
похождением ядра и митотического аппарата, предполагая ( Cavali-
lier-smith, 1983а), что митохондрии и хлоропласты появились
значительно позже (около 700 млн. лет назад), чем сами эукарио-
ты (около 1400 млн. лет назад). Он включает эту концепцию коэво-
люции в предложенную им схему унифицированной филогении (Cavali-
396
er-Smith, 1983b; рис. 8.8), однако твердо уверен, что экзоген-
ная теория не дает удовлетворительного объяснения происхождению
ядра, митотического аппарата и жгутиков. Он полагает ( Cavalier-
Smi'th 1978), что последние возникли в ходе сложного ступенча-
того процесса, начинающегося образованием филоподиальных вырос-
тов с микротрубочками у простых планктонных прокариот. Однако,
как отмечалось (Taylor, 1974;Ragan, Chapman, Г978), свои до-
стоинства имеют обе школы. В любом случае вряд ли мы когда-ни-
будь точно узнаем, как возникли эукариоты.
Рис. 8.8. Возможные филогенетические взаимоотношения
между 14 подцарствами, признаваемыми Кавалье-Смитом. Ди-
вергенции эукариот предшествует приобретение хлороплас-
тов И митохондрий ( Calaviei-Smith, 1983)
397
Эволюция митоза была сопряжена с развитием ядра, хотя име-
ются фундаментальные различия между экзогенной и аутогенной схе-
мами, объясняющими эти процессы. Первая ( Margulia, 1981) пред-
полагает, что митоз предшествовал мейозу и формирование ядра,
no-видимому, зависело от симбиотического события, приведшего к
эволюции митохондрий. Хит (Heath, 1980) предлагает II этапов
эволюции митоза. Однако Кавалье-Смит ( Cavalier-Smith, 1975,
1978) придерживается мнения о параллельном развитии митоза, кле-
точного деления и мейоза (т.е. микротрубочек веретена, ядерной
оболочки и борозды деления), считая, что половой процесс появил-
ся в эволюции эукариотных клеток очень рано, еще до завершения
развития ядра.
Ранние эукариоты были, судя по всему, амебоидными или
планктонными, способными к фагоцитозу автотрофами, не имевшими
клеточной стенки. Независимо от того, возникло эукариотное сос-
тояние экзогенным, аутогенным или смешанным путем, полагают,
что произошло это в масштабах всей эволюции относительно быстро,
вероятно, не белее чем за 100 млн. лет.
Эволюция мейоза, сингамии и чередования поколений
Развивалась ли раздельнополость (мейоз и сингамия) посте-
пенно или более или менее одновременно с эукариотным состоянием
( Cavalier-Smith, 1975), ее появление было основным этапом на
цути к сосудистым растениям. Эволюция различных эукариот стала
возможной за счет процессов расхождения хром осой, рекомбинации
и естественного отбора. Неизвестно, когда возникла раздельнопо-
лость, но биогеологические данные свидетельствуют, что она су-
ществовала уже 700 млн. лет назад, когда в ископаемых остатках
появляются первые Metazoa (Cloud, 1976). Обсуждение конкрет-
ного возраста этого явления возвращает нас к дебатам относитель-
но экзогенной и аутогенной теорий эволюции эукариот. Учитывая
палеонтологические данные и сведения о цитологии, молекулярной
биологии и физиологии ныне живущих организмов, Клауд ( cloud,
1976) представляет аргументы в поддержку взглядов Кавалье-Сми-
та ( Cavalier-Smith, 1975, 1978) о более или менее одновремен-
398
нем возникновении митоза и раздельнополости. Такое заключение со-
гласуется с биогеологическими данными о том, что митоз и мейоз
существовали уже 1,3 млрд, лет назад (см. Walter et al., 1976),
а также с выводом Шопфа ( Schopf, 1974 ) о возможном появлении
раздельнополости ранее 1000 млн. лет назад.
Эукариотная раздельнополость обеспечила прогрессивное разви-
тие от клеток к тканям и органам (Cloud, 1976). Хотя нет аде-
кватных палеонтологических находок для большинства современных
классов водорослей, возможно, они обособились в ходе эволюции
эукариотного состояния; однако разделились ли основные группы до
или после появления раздельнополости, неясно. Например, эвглено-
вые, у которых полностью отсутствует половое размножение, могли
либо возникнуть до его появления, либо утратить его впоследствии.
Половое размножение, несомненно, привело к увеличению разнообра-
зия организмов и в целом ускорило процесс эволюции.
Среди древнейших многоклеточных остатков водорослей, у кото-
рых не исключена раздельнополость, - Papillomembrana. напоминаю-
щая современных Dasycladales (Spjeldnaes, 1963), предположи-
тельно бурые водоросли Vendotaenia и Tryrasotaenia (Гниловская,
1971) и отпечатки макрофитов из Австралии возрастом около
760 млн. лет ( Milton, 1966).
Следствием развития раздельнополости было сохранение у ран-
них водорослей как диплоидной, так и гаплоидной фаз; возможно,
первоначально обе они были одноклеточными ( Scagel et al., 1982).
Последующая эволюция в направлении усовершенствования обеих фаз
или только одной из них привела к крайне разнообразным жизненным
циклам и у наиболее продвинутых форм к чередованию поколений:
гаплоидных гаметофитов, дающих гаметы, и диплоидного спорофита,
образующего споры (гл. 5). Четких ископаемых свидетельств того,
как и когда происходило это развитие, нет, хотя анализ ныне жи-
вущих форм показывает четыре основных пути эволюции в зависимо-
сти от момента мейоза в жизненном цикле (гл. 5).
Предложена схема этапов эволюции водорослей (Scagel et al.,
1982; рис. 8.9); все эти стадии в разной степени сохраняются у
современных видов. В первую очередь, возможно, возник зиготичес-
кий мейоз, давший начало двум другим его типам: гаметическому и
399
ПРОКАРИОТЫ
ПРОКАРИОТА
ЭУКАРИОТА
Эволюция мейозе и
сиигемии
п + 2п, оба одноклеточные
I
Эволюция способов
чередования поколений
Зиготический мейоз
(преобладает гаплоидная фаза)
ДИКАРИОН
Чередование
моиокериотного
и дикариотного
ми цели ее
(некоторые грибы)
Л- * •
СПОРИЧЕСКИЙ МЕЙОЗ ГАМЕТИЧЕСКИЙ МЕЙОЗ
(п + 2п, многоклеточные) (гаплоидная фазе
I преобладает)
| (несколько групп
водорослей и грибов)
ИЗОМОРФНОЕ ЧЕРЕДОВАНИЕ ГЕТЕРОМОРФНОЕ ЧЕРЕДОВАНИЕ
ПОКОЛЕНИЙ ПОКОЛЕНИЙ
(некоторые водоросли,
изредка у грибов)
Некоторые водоросли
Наземные растения
Преобладает Преобладет
гемеофит спорофит
Преобладает Преобладает
гаметофит спорофит
Рис. 8.9. Этапы эволюции основных типов чередования поко-
лений (ПО Scagel et al., 1982)
спорическому. Гаметический путь привел к диплоидным организмам,
а спорический - к чередованию поколений. Дальнейшая эволюция че-
редования поколений дала его изоморфный и гетероморфный типы. В
последнем случае была возможность усовершенствования либо гаме-
тофитного, либо спорофитного поколения. Оба этих пути привели к
появлению наземных растений, но именно водоросли (и животные) с
Доминированием диплоидной фазы приобрели относительно сложные
ткани, органы и физиологические процессы, необходимые для адап-
тации к суронм экологическим требованиям наземного существова-
ния.
400
Четвертый вариант смены ядерных фаз, дикарион, не встреча-
ется у водорослей и известен только у некоторых грибов. Он пред-
ставляет собой задержку кариогамии после плазмогамии или сомато-
гамии (слияния талломов), в результате чего совместимые ядра по-
парно располагаются в цитоплазме , не сливаясь до образования
спор. Происхождение этого механизма неясно.
Эволюция наземных растений
Эволюция наземных растений (эмбриофитов) затрагивается на-
ми, поскольку поиск эволюционных связей между ними и водоросля-
ми - одна из основных проблем современной эволюционной биологии.
Хотя точное их происхождение неизвестно, в целом существует
единство мнений относительно того, что наземные растения - по-
томки фрагмопластных зеленых водорослей ( Scagel et al.,1982).
Возможных предков можно искать среди нескольких группу но скорее
всего - среди Ulotrichales. Этапы, необходимые для перехода к
неблагоприятным наземным условиям, обобщены Джеффри (Jeffrey,.
1962). Как упоминалось выше, основное расхождение, происшедшее
на этол пути, - между формами с наиболее развитой гиплоидной га-
метофитной фазой и с доминированием диплоидного спорофита. Пер-
вые дали начало бриофитам, ставшим в целом тупиковой ветвью эво-
люции и ограниченным в своем распространении сырыми местообита-
ниями, поскольку для оплодотворения им нужна вода. От вторых
произошли все остальные высшие растения, успешно приспособившие-
ся к наземному существованию.
Гипотетические предковые водоросли скорее всего были нит-
чатыми и гетеротрихальными, с чередованием двух многоклеточных
поколений и, возможно, оогамным половым размножением, причем
оплодотворение происходило в оогонии. Одной из самых ранних
адаптаций могло быть то, что зигота оставалась на гаплоидном
растении (как у Coleochaete; Graham, 1985) под защитой стериль-
-----j—
И могут многократно строго синхронно митотически делить-
ся. - Прим, перев.
401
нах клеток. Бриофитная линия взяла начало от форм с диплоидным
спорофитным поколением, развивающимся в прикрепленном к гамето-
фиту состоянии, а эмбриофитная - от тех, у которых зигота в ко-
нечной счете отделялась от гаметофита и продолжала развитие как
независимое растение, дающее споры. В ходе приспособления к на-
земному существованию у спорофита произошла кутинизапия надзем-
ных частей, появились водопоглощающие структуры, проводящие тка-
ни, специализированные фотосинтезирующие органы (листья) и усть-
ица; гаметофитное поколение постепенно редуцировалось.
Четкой границы между зелеными водорослями, бриофитами и эм-
бриофитами нет. Наземные отделы просто объединяют наиболее высо-
коорганизованные из зеленых растений и во многих схемах (обзор
см. Edwards, 1976) включаются вместе со своими водными пред-
ками в единое царство.
Филогения водорослей
Выяснение эволюционной истории водорослей требует изучения
как ныне живущих, так и вымерших их представителей. Ход эволю-
ции может быть представлен в виде филогенетического дерева, пер-
вый вариант которого опубликовал Геккель ( Haeckel, 1896). Ес-
тественную систему ныне живущих видов (гл. 2) можно рассматри-
вать как совокупность конечных ответвлений такого дерева, при-
чем группировка таксонов проводится на основе признаков общей
предковой формы. Такая классификация основана на филогенетичес-
кой систематике, или кладистике. Однако идентичная или очень
сходная схема может быть получена путем группировки современных
видов по уровню их общего сходства. Этот подход называется фоне-
тическим. Классификация водорослей, принятая в этой книге и мно-
гих других современных работах, является результатом комбинации
обоих подходов. Идеальной классификации не существует, и здесь
остается широкое поле для умозрительных построений из-за отсут-
ствия соответствующих палеонтологических данных для большинства
групп. В целом признано, что некоторые из выделяемых отделов яв-
ляются полифилетичными, т.е. имеют несколько предковых групп,
тогда как другие монофилетичны (например, эвгленовые).
402
Предложено множество филогенетических схем для водорослей
(наиболее примечательные см. Copeland, 1938; Cronquist, I960;
Christensen, 1964; Sagan, 1967; Whittaker, 1969; Leedale,
1974; Cavalier-Smith, 1975; Scagel et al., 1982). Практически
все они учитывают биохимические признаки (например, пигментацию)
в сочетании с цитологическими и морфологическими особенностями,
хотя некоторые (например, Dodge, 1974) опираются лишь на морфо-
логические и ультраструктурные характеристики или (например,
Ragan, Chapman, 1978) только на биохимические данные. Клейн и
Кронквист ( Klein,Cronquist, 1967) дали исчерпывающий сравни-
тельный обзор биохимических, микроморфологических и физиологиче-
ских аспектов филогении водорослей, построив полный набор филе-
тических схем, основанных на индивидуальных признаках. Филоге-
нию можно также выводить, исходя только из одного признака (на-
пример, митоза: Heath, 1980; жгутикового annaparazMoestrup,
1982; Melkonian, 1982а; фоточувствительного аппарата: Kivic,
Walne, 1983). Построения могут быть ограничены объемом отдель-
ной группы (например, эвгленовыми: Leedale, 1978).
Хотя многие схемы различаются в деталях, в целом сложилось
единство мнений относительно наиболее существенных аспектов эво-
люции водорослей; так, признаются отдаленность багрянок от
остальных эукариот, изолированность эвгленовых, динофитов и
криптофитов, четкая граница между хрсмофитами и хлорофитами и
происхождение наземных растений от древних зеленых водорослей.
Данные биохимии и молекулярной биологии в значительной степени
подтверждают выводы, основанные преимущественно на результатах
исследований морфологии, цитологии и ультраструктуры. Новая ин-
формация по мере поступления постепенно включается в филогене-
тические схемы.
Общие взаимосвязи между классами водорослей показаны на
рис. 8.10. Они основаны прежде всего на сходстве в пигментном
составе, типе жгутикового аппарата, клеточных покровах и основ-
ных запасных продуктах. Все классы, за исключением золотистых
водорослей, по-видимому, представляют собой четкие эволюционные
линии, хотя следует помнить, что построения эти в значительной
степени фонетические. Из сказанного выше (гл. 2, 3 и 6) очевид-
но, что кроме учтенных различий, основанных на пигментации, име-
403
ХРОМОФИТЫ
Phaeophyceae /
Chlorophyceae
Bacillariophyceae f
Л
• Зеленая
Мрокари-
I ova
'с '
с \
\
ЭВГЛЕНОВЫЕ
•t
. Raphidophyceae
Xanthophyceae х
Charophyceae
Prasinophyceae
Oinophyceae
Eustlgmatophyceae
Ч
\ Prymnesiophyceae
С
Chrysophyceae у
Cryptophyceae
I
/C\
БАГРЯНКИ
| Поли мастигмиы
Трипаносомы*-^ \ ।
ПРЕДКОВЫЙ
ЖГУТИКОНОСЕЦ
СИНЕЗЕЛЕНЫЕ
ВОДОРОСЛИ
♦
ПРЕДКОВЫЙ
ЖГУТИКОНОСЕЦ
---ПРЕДКОВАЯ —
ПРОКАРИОТА
Рис. 8.10. Обобщенное филетическое дерево, показывающее
гипотетические родственные связи между классами водорос-
лей и учитывающее 5 симбиотических комбинаций (С). Пред-
полагается две исходные линии жгутиковых; одна из них да-
ла начало хлорофитному ряду, развитие которого сопровож-
далось эндосимбиозом с зеленой прокариотой (возможно, на-
поминающей Pro chi огор). а другая - хромофитному ряду,
за счет двуЯ"ЭВДСХ!ИМбйозов с участием предшественников
синезеленых и красных водорослей. Происхождение динофи-
тов проблематично (по Dodge, 1979)
404
имеется много других морфологических, цитологических и физиоло-
гических расхождений между классами, которые позволяют с боль-
шим основанием полагать, что построенная схема действительно от-
ражает важнейшие точки дивергенции в эволюции водорослей. Одна-
ко интерпретировать такую в основном фонетическую схему следует
о осторожностью, поскольку она не может служить доказательством
того, что эволюционный процесс происходил во времени именно
так, а не иначе. Тем не менее общие направления, по которым шла
эволюция водорослей, определены ею довольно четко.
Сведения об известных ископаемых остатках водорослей обсуж-
даются ниже (с. 0253). Для большинства классов количество их
очень мало, и хорошо представлены только известковые (например,
Dasycladales из Chlorophyceae ) Или окремнелые (например, диа-
томеи) формы, а остальные группы известны главным образом по
фрагментам очень немногих вадов. Поэтому построение филогенети-
ческого дерева водорослей, правильно отражающего их эволюцион-
ное прошлое, крайне затруднено. Тем не менее полезно рассмот-
реть состояние наших знаний о филогении основных классов этих
организмов.
Прокариоты: Cyanophyceae И Prochlorophyceae
Широко распространена гипотеза, согласно которой примитив-
ные коккоидные синезеленне водоросли возникли параллельно с "со-
временными" бактериями от архаичных бактериальных предков
( Klein, Cronquist, 1967). Действительно, у этих водорослей
много общих черт с бактериями. Родство между ними было впервые
предложено Коном (Cohn, 1853) и обсуждалось в деталях многи-
ми современными исследователями (например, Echlin, Morris,
1965; Hall 1971; Fogg et al., 1973; Ragan Chapman, 1978;
Fox et al., 1980; Starr et al., 1981; Woese, 1981; Schopf ,
Walter, 1982a). Значительный интерес представляет вывод (Fox
et al., 1980; woese, 1981) о наличии трех основных эволюцион-
ных линий живых организмов, выводимых от общего предка, или
прогеноты ( Woese, 1981): архебактерии, настоящие бактерии и
эукариоты. Синезеленые водоросли, согласно такой схеме, произо-
405
шли от настоящих бактерий (эубактерий), однако обладают некото-
рыми фундаментальными отличиями, что позволяет предполагать
очень раннее расховдение двух этих эволюционных линий. Структур-
но они разнообразнее бактерий (встречаются, например, многоклет
точные талломы, ветвящиеся нити, колонии), имеют цитологические
особенности (например, нет бактериальных эндоспор, но есть гете-
роцисты) и никогда не образуют жгутиков бактериального типа. На-
блюдаются также существеннейшие различия в фотохимическом процес-
се и составе жирных кислот ( Fogg et al., 1973).
Эволюцию синезеленых водорослей можно проследить с опреде-
ленной точностью по ископаемым находкам (см. рис. 8.2). Самыми
ранними были коккоидные одноклеточные формы, которые достигли
некоторого разнообразия примерно 1,7 млрд, лет назад, когда на-
метился ряд современных семейств. Нитчатые виды, возможно, воз-
никли от коккоидных; представители нескольких современных се-
мейств имеют возраст более 2,0 млрд, лет (Schopf, 1974 ). Опи-
сано (Hoffmann, 1976) 18 родов и 24 вида коккоидных и нитчатых
синезеленых водорослей из докембрийской (1,9 млрд, лет назад)
микрофлоры островов Белчер (Канада). Фрич (Fritsch, 1945) пола-
гал, что разнообразие нитчатых форм говорит о наличии нескольких
независимых эволюционных линий, которые, возможно, рано разо-
шлись от общего нитчатого предка. Время и последовательность
эволюционных событий, приведших к дивергенции синезеленых водо-
рослей, точно установить трудно, хотя вполне вероятно, что sti-
gonemataies - самый молодой порядок среди нитчатых форм
(Schopf, Walter, 1982а), поскольку он единственный не пред-
ставлен в ископаемых находках докембрийского возраста.
Прохлорофиты, по своим особенностям занимающие промежуточ-
ное положение между синезелеными и зелеными водорослями ( Lewin,
1981а, ; рис. 8.II), послужили основой для ряда интересных фи-
логенетических гипотез (Lewin, 1983а). Поскольку маловероятно,
что предковые формы этой группы могут быть обнаружены в ископае-
мых находках, их происхождение неизбежно придется обосновывать,
исходя из фонетических данных. Если признать экзогенное проис-
хождение эукариот, прохлорофиты, по-видимому, будут филогенети-
чески близки к предшественникам хлоропластов, а с аутогенной
точки зрения допустимо мнение ( Lewin, 1983а), что они родствен-
406
ны предковым эукариотным хлорофитам. Луин ( Lewin, 1983а) обоб-
щает эти взгляды в виде нескольких простых филогенетических де-
ревьев (рис. 8.12). Он излагает, что происхождение прохлорофи-
тов от синезеленых водорослей потребовало бы утраты фикоцианина
и приобретения способности к синтезу хлорофила ъ.
Зеленые
водоросли
Синезеленые
водоросли Ргоспюгоп
Э у кариогенез
Фикобилины
Хлорофилл b
Хлорофилл а
Рис. 8.II. Микрофотография
тонкого среза Prochioron
(Prochlorophycota). централь-
ная зона, по-видим ему, огра-
ничена мембранами, а тилакои-
ды и электроноплотная цито-
плазма сосредоточены в пери-
ферической зоне (х 7500)
(Giddings et al., 1980)
Рис. 8.12. Схемы филогенетиче-
ских деревьев, показывающих
возможное происхождение Proch-
loron, и хлорофитов от прока-
риотных водорослей. И хлорофи-
ты, и Prochioron синтезируют
хлорофиллы а и ь и не содер-
жат фикобилинов!" Возможно,
Prochiогоп возник от синезе-
леных водорослей путем утраты
фикобилинов и приобретения спо-
собности к синтезу хлорофил-
ла к (Б - й). но он может
быть филогенетически родствен
и предковым хл^орофитам(Д)
407
Rhodophyceae
Багрянки, несомненно, представляют собой древнюю естествен-
ную группу водорослей, достоверные ископаемые остатки которых из-
вестны еще из кембрийских отложений (Schopf, 1970). Исходя из
особенностей биохимии, физиологии и строения, можно с большим ос-
нованием предположить, что они произошли от синезеленых водорос-
лей (Klein, Crounquist, 1967). Водоросль типа Cyanidium счита-
ют при этом промежуточной формой. Однако нет единства мнений от-
носительно того, является ли Cyanidium caldarium организмом,
содержащим цианеллы, гомологичным, например, Cyanophora (Kremer.
1982), или это истинная багрянка (Bisalputra, 1974; Ford, 1984-
Сложность в вопросе происхождения красных водорослей от синезеле-
ных связана с проблемой эволюции эукариот: уникальные особенно-
сти багрянок, несомненно, говорят об очень ранней независимой ди-
вергенции их от остальных водорослей, что нашло отражение в раз-
личных филетических схемах.
Возможно, первые красные водоросли были одноклеточными, т.е.
от них уже на ранней стадии эволюции возникли простые нитчатые
формы. При отсутствии надежных палеонтологических данных любые
построения, представляющие собой попытку проследить происхожде-
ние отдельных эволюционных линий, остаются в значительной степе-
ни спекулятивными. Нет единства мнений относительно того, пред-
ставляют ли собой Два подкласса, Bangiophycidae И Florideophy—
cldae, естественные группы, соответствующие двум разным направ-
лениям эволюции (как предполагает р,ее, 1980). Современные от-
крытия показали значительно меньшее несходство между ними, чем
предполагалось ранее (см., например, Fritsch, 1945). Очень ин-
тересно предположение (Scagel et al., 1982) о том, что стадия
Conchocelis видов Porphyra и Bang!a (Bangiophycidae) прототип
ЭВОЛЮЦИОННОЙ ЛИНИИ багрянок, ОТ которой ВОЗНИКЛИ Florideophy-
cidae , a Bangiophycidae остались тупиковой ветвью.
Ряд исследователей (в том числе Klein, Cronquist, 1967;
Cavalier-Smith, I978;Kohlmeyer, Kohlmeyer, 1979) полагает,
что красные водоросли, потеряв пластиды, дали начало высшим гри-
бам. Отсутствие подвижных клеток и сходство процесса оплодотво-
рения и последующего развития часто считают основанием для сбли-
40В
жвния двух этих групп ( Scagel et al.J982). Согласно другой точ-
ке зрения, грибы возникли от примитивных Protozoa, а высшие
грибы - от зоомицетов, и любое их сходство с багрянками - резуль-
тат параллельной эволюции. Эти предположения вынужденно остаются
в значительной мере спекулятивными, поскольку о происхождении
каждой из групп известно очень мало.
Хромофитная и хлорофитная линии
Как было показано выше, классификационные (и филвтические)
схемы для водорослей зависят от значения, придаваемого различиям
в пигментации или особенностям жгутиков. Схема, принятая нами,
предполагает раннюю дивергенцию между формами, синтезировавшими
хлорофилл £ (хром офитами), с одной стороны, и хлорофилл Ъ (хло-
рофитами) - с другой. Однако это расхождение должно было произой-
ти после появления жгутиков - ключевого события в ранней эволю-
ции эукариот. Как указывает Кавалье-Смит (Cavalier-Smith , 1978),
основная адаптивная радиация, связанная с появлением главных
групп эукариот (растений и животных), произошла на стадии одно-
клеточных жгутиковых. Егутик - достаточно сложная и при этом
стабильная структура (особенно в строении базального тельца и
9+2-аксонемы), чтобы служить надежным признаком родства ( Manton,
1965; Taylor, 1976; Moestrup, 1982). Другие характеристики жгу-
тиков (например, корни, внешние особенности, расположение и чис-
ло) часто объединяют между собой несколько крупных групп ( Moes-
trup, 1982) и указывают на их связь при наличии общих модифика-
ций. <
И напротив, Кавалье-Смит (Cevalier-Smith, 1978) полага-
ет, что признаки, относящиеся к питанию (например, хлоропласта
и связанные с ними цитологические изменения), часто более полез-
ны для выяснения различий между группами. Как пигменты, так и
особенности жгутикового аппарата, по-видимому, мало изменились
при переходе от одноклеточных к многоклеточным растениям, и мож-
но с большими основаниями считать, что многоклеточные формы воз-
никли в ходе эволюции неоднократно (Cavalier-Smith, 1978). Та-
ким образом, при рассмотрении филогении водорослей (кроме багря-
нок) особенности жгутиков и пигментов (с сопутствующЕМи им цито-
409
логическими и биохимическими признаками) чрезвычайно важна при
установлении связей и различий на уровне классов и в конечном
счете при построении филогенетических деревьев.
| Додя (Dodge, 1979; см. рис. 8.10), исходя из эндосимбиоти-
| ческой гипотезы, предлагает три или четыре отдельные линии эука-
риот, идущие от предковой прокариоты. Хромофитную и хлорофитную
5? линию он сохраняет: симбиоз красных водорослей и криптофитов дал
в хромофитную линию, а прохлорофитоподобный предок, став эндосим-
Й бионтомпредкового жгутиконосца, - хлорофитную. Только динофиты
остаются в стороне, и для их образования требуется "поздняя" эн-
I' досимбиотическая интеграция примнезиофита, золотистой водоросли
I' и предкового жгутикового. Реальной хронологической последователь-
?? ности событий не предложено, и в этой схеме столько же неяснос-
тей, сколько и в строго аутогенной; однако она демонстрирует,
насколько различны могут быть взгляды на филогению водорослей в
зависимости от исходных предпосылок.
Родственные связи хрсмофитной и хлорофитной линий реконстру-
; ируются главным образом на фонетической основе (хромофиты; Н1Ъ-
( herd, 1979; хлорофиты: Moeetrup, Ettl, 1979; Melkonian, 1982а).
' В этих схемах не удается найти бесспорного места для динофитов и
эвгленовых, эндосимбиотическое происхождение хлоропластов кото-
рых вызывает серьезные споры ( Gibbs, 1978; Leedale, 1978; What-
ley , 1981). Динофиты не связаны с другими хрсмофитными группами,
если исходить из особенностей жгутикования ( Moestrup, 1982) или
некоторых биохимических признаков ( Ragan, Chapman, 1978). Вызыва-
ет разногласия также место криптофитов, хотя Додж (Dodge, 1979)
и находит возможным включить их в хромофитную линию (см. также
Gillot, Gibbs, 1980); они стоят особняком по ряду фундамен-
тальных черт, включая уникальное расположение жгутиковых волос-
ков ( Moestrup, 1982).
Если оставить в стороне эвгленовые, в целом общепризнано,
что хлорофитная линия монофилетична. Когда разошлись Chlorophy-
ceae, Prasinophyceae И Charophyceae неясно; Мёструп (Moesta '
nip, 1982) предполагает, что "Loxophyceae" (не рассматриваемые
здесь) представляют собой примитивный элемент хлорофитной линии,
больше всего родственный празинофитам, но обладающий рядом проме-
жуточных празинофитно-хлорофитных признаков. Однако места для
Loxophyceae в предложенной им филетической схеме этот автор не
। нашел.
Все в большем числе работ при выяснении эволюционного родст-
410
ва между хлорофитными группами (Melkonian, 1982Ь) или внутри
их (например, O’Kelly, Floyd, 1982а, Ь) придается основное зна-
чение консервативным (например, 9+2-структуре аксонемы), а не ва-
риабельным (например, переходной зоне) особенностям жгутиков.
О’Келли и Флойд (O'Kelly, Floyd, 1984) подчеркивают важность
абсолютной ориентации жгутикового аппарата для филогении зеленых
водорослей; по их схеме ориентация против часовой стрелки (прими-
тивная) встречается во всех классах, кроме Chlorophyceae, У ко-
торых жгутики ориентированы по часовой стрелке (более продвину-
тая). В предложенной ими филетической схеме три эволюционные ли-
нии (Chlorophyceae, Ulvophyceae И Pleuraatrma, Pleuraatrophy-
сеае) исходят из общего предка, напсминающего современный прази-
нсфит Pyramimonas; ответвление к Charophyceae предполагается
еще более ранним. При этом клетка зеленого жгутикового предка об-
ладала такими примитивными признаками, как ориентация жгутиково-
го аппарата против часовой стрелки и ромбовидными чешуйками на
теле. Мелкониан (Melkonian, 1982Ъ ) предлагает детальную гипо-
тетическую филетическую схему (рис. 8.13), которая интерпретиру-
ет накопленную к настоящему времени информацию об особенностях
жгутиковых корней. Предполагаются четыре эндосимбиоза: внедре-
ние в гетеротрофного жгутиконосца эукариотной зеленой водоросли
(приведшее к эвгленовым), зеленой прокариоты типа Prochioron
(приведшее к Chlorophyceae), синезеленой прокариоты (приведшее
К Glaucophyceae^ И красной ВОДОРОСЛИ (приведшее К Cryptophy-
ceae). Эта схема, как и все прочие, ставит много вопросов, исхо-
дит из ряда недоказанных предположений и лишена какой-либо вре-
менной шкалы. Она не отрицает обычно признаваемых родственных
связей между современными группами, хотя отличается в деталях
их трактовки.
Филетическая изоляция эвгленовых обсуждалась многими авто-
рами. Лидейл ( Leedale, 1978) приводит длинный список отличи-
тельных признаков этой группы, крайне затрудняющих определение
ее места в рамках любой схемы. Давно ведутся споры, зеленую или
бесцветную линию эвгленовых считать исходной для всех современ-
ных родов. Дидейл (Leedale, 1978) И Додж ( Dodge, 1979) В сво-
их филетических схемах отдают предпочтение бесцветным формам.
Их гипотеза, согласно которой эвгленовые только недавно приобре-
ли хлоропласты за счет эндосимбиоза, по-видимому, заслуживает
внимания, если учесть возможную эволюционную изоляцию группы.
Положение "Giaucophyceae" (с. 76) подробно обсуждалось
Мёструпом (jfoeetx’up, 1982). Филогенетические взаимоотношения
411
Chroolepidaceae
Chlorophyceae
Trebouxia
Глазок
Система II
Pleurastrum
Chlamydomonas
Система 11
Friedmannia
-Чешуйки
MCC
Zygnematales
Утрата двух
жгутиков
Утрата двух
жгутиков
Подвижные
клетки
Чешуйки
Hafniomonas
Развитие MMC
MCC редуцируется
. Х-2-Х-2
Дубли-
рование
жгутиков
Чешуики
Co/eochaete
Characeae
Chlorokybus
Односторонний
орень
Г лазок
редковый зеленый
жгутиконосец -
ЭС (багрямкаЬ- Cryptophyceae
Апикальное
углубление
Система II
Bryopsldales
Глазок
Microthamnion
Преобразование МСС
Клеточная стенка
Чешуйки
Фиколласт
Клеточная
стенка у
подвижной стадии
Ulvaohvceae I Клеточная
Дублирование
системы II
Наземные
растения
.стенка МСС
Неполосатые
соединительные
волокна
Перекрывание
базального тела
К/ей$огтй/итчешуйкиК5-енОк?ая
Englenophyceae
Mesostigma Система к
ЭС(аеленая I
прокариота)
ЭС (хлоропласты) xi
Чешуйки МСС Система ||
Tetraselmis
Тека
Pyramimonas
МСС
Апикальное углубление
МСС редуцируется
Nephroselmis
Чешуйки Глазок
Предковая зоофпагеллата
G laucophyceae
ЭС (синезеленая
' водоросль)
Рис. 8.13. Предполагаемая эволюция хлорофитсв и наземных
растений. Показаны четыре эндосимбиотических события
(изогнутые стрелки ЭС): в эволюции эвгленовых это включе-
ние хлороплаотов из эукариотных зеленых водорослей, у зе-
леных водорослей - зеленой прокариоты типа Prochlorcn,
у криптофитов - красной водоросли, у "Glaucophyceae"-
синезеленой прокариоты. Стрелки, указывающие на линии
или от линий, означают' соответственно приобретение или
утрату каких-либо структур или функций: МСС - многослой-
ная структура; система Ц - волокнистый корень: Х-2-Х-2 -
крестовидная система с двумя типами микротубулярных жгутико-
вых корней ( Melkonian, 1982а)
412
этих водорослей, содержащих цианеллы, неясны, хотя некоторые от-
носят их к отдельной монофилетичной линии эукариот, содержащих
хлорофилл ъ_ • Ли Сьее 1980), напротив, полагает, что глауко-
фйты - живые примеры предковых жгутиковых, доказывающие эндосим-
биотическое происхождение хлоропластов.
Ископаемые находки
Возраст известных ископаемых находок представителей клас-
сов хромофитной и хлорофитной линий показан на рис. 8.14. Инфор-
мация имеется преимущественно о водорослях со стенками или дру-
гими скелетными структурами, легко переходящими в окаменелое
состояние; вследствие этого четкая филогенетическая картина мо-
жет быть прослежена только дая нескольких групп.
RHODOPHYCEAE
Построение фонетических рядов дая Elorideophyoidae край-
не затруднительно, поскольку палеонтологические данные неполны
и взаимосвязи порядков, особенно Nemaliales, неопределенны
( Scagel et al., -1982). Однако имеются отличные окаменелые ос-
татки известковых кораллиновых водорослей, которым посвящена об-
ширная литература, к сожалению плохо известная альгологаы
Johansen, 1981). К настоящему времени большинство признает, что
ископаемые представители относятся к четырем семействам извест-
ковых багрянок ( догау, 1977): Solenoporaoeae, Gymnooodiaoeae,
Squamariaceae, Coralliiraoeae, которых первые два полностью
вымерли. Solenoporaoeae впервые появились в кембрии и были
обильны В ордовике (например, Solenopora, Parachaetetes).
Две отчетливые линии стали заметны к среднему силуру; одна из
них, по-видимому, привела к современным Corallinaceae, имею-
щим клетки меньшего размера ( Wray 1977; Johansen, 1981). К
концу мезозойской эры-, начиная с мела, вполне сформировались
многие современные роды (например, lithothamnion. Idthophyllum).
Выдвигались разные предположения относительно того, как возник-
ли членистые и нечленистые формы. Согласно современным морфоло-
413
Рис. 8.14. Представленность в различные геологические
эпохи бактерий и основных отделов водорослей (включая
акритарх). Прерывистые линии означают сомнительные или
непроверенные остатки (по Scagel et al., 1982, с из-
менениями)
гическим и культуральным исследованиям (Scagel et al., 1982),
деление современных кораллин на членистые и нечленистые искус-
ственно, поскольку фонетические данные показывают, что членис-
тые роды могли возникнуть от двух разных групп нечленистых пред-
ков, а не от одной линии, как часто предполагалось ( Johansen,
414
1981).
Нечленистые кораллиновые водоросли в виде свободноживущих
узловатых структур известны в ископаемом и современном состоя-
нии как родолиты (рис. 8.15 - шары Lithothamnion, или родолит,
но не мергель; см. Adey, MacIntyre, 1973; Johanaen, 1981). Они
широко распространены на океаническом дне в интервале глубин
50-200 м, растут очень медленно и за ВОО и более лет могут до-
стигать диаметра 30 см ( Adey, MacIntyre, 1973). Возраст родо-
литов с банки Агульяс в районе Южной Африки, определенный радио-
углеродным методом, составляет до 13 760 лет ( Sleager, 1972).
Их ископаемые остатки указывались из эоценовых, олигоценовых
и миоценовых отложений.
Мягкоталломные представители багрянок в окаменелостях пред-
ставлены плохо, хотя известна ( Campbell, 1980) интересная на-
ходка Palaeooonchocelia etarmachii из верхнего силура Польши
(425 млн. лет назад). Водоросль отлично сохранилась благодаря
эндолитнсму образу жизни (рис. 8.16). Ископаемые нити морфологи-
чески идентичны фазе Conchocelia современных- Bankiaи Porphy-
ra. которую описывают как "живое ископаемое" (Campbell, 1980).
Предпринята попытка ( Tappan, 1976) отнести к бангиевым гораз-
до более ранние протерозойские остатки (возрастом порядка
1,9 млрд, лет), датируя таким образом происхождение этой группы
примерно тем же временем, Зто и возникновение эукариотной клет-
ки.
PHAEOPHYCEAE
Хотя предполагалась возможность докембрийского происхожде-
ния бурых водорослей (более 570 млн. лет назад), ранние ископае-
мые находки здесь немногочисленны, поскольку, за исключением со-
временных видов Padina. представители этого класса неизвестко-
вые. По имеющимся сведениям, некоторые окаменелости силурского
и девонского возраста сходны с современными представителями
Pucaleg, Dictyotalea, Chordarialea И Sphacelariales (Scagel
et al»,. 1982); сообщалось о находке некоторых других бурых во-
дорослей в триасовых отложениях (возраст около 250 млн. лет).
415
Рио. 8.15. Родолит, живая ко-
раллиноюя водоросль ( Litho-
thamnion glaciale, Rhodo-
phycota, Corallinaceae),
дающая роцолитные отложения '
на океаническом мелководье.
Масштаб 2,0 см
Рис. 8.16. Palaeoconchocelia starmachii (Rhodonhycota.
Bangiophycidae). ЭТб РнДЬЛИФйоё йскбйаемое из верхнего
силура (около 425 млн. лет назад) очень напоминает фазу
Conchocelia совремeHHHXBangiqphycidae. А. Петрогра-
фйчёбкЬй тонкий срез in situ. Б. Конхоспорангиальная
ветвь, извлеченная из породы. В. Стадия Conohooelis
современной водоросли Porphyra nereocyatis. flit — пИрит
внутри клетки; к - конЯЬйПОра (СашрЪедТ;—1980)
416
Лучше всего сохранились остатки бурых водорослей из миоцена
( Parker, Dawson, 1965), включающие роды, отнесенные к Lamina-
rialea (Julescraneia) И Fucales (Palaeohalidryg. Cystoseirites.
Palaeocystophora; рис. 8.17). Некоторые совсем недавние
(примерно 6450 лет назад) неизвестковые ископаемые бурые водо-
росли с севера Канады сохранились исключительно хорошо ( Ulman
et al., 1972; рис. 8.18).
Немночисленные остатки бурых водорослей дают мало пищи для
филогенетических построений. Одноклеточные представители в насто-
ящее время неизвестны. По цитологии и биохимии бурые водоросли
ближе всего к золотистым, и предполагают (Christenaen, 1980),
что их можно считать четко дифференцированными многоклеточными
представителями большой группы, охватывающей оба этих класса. То
же самое следует из биохимии (Ragan, Chapman, 1978); вполне воз-
можно, что бурые водоросли - наиболее продвинутая группа в линии
водорослей с хлорофиллом с, или хромофитов. Педерсен ( Pederaen,
1984) склоняется в пользу прямой связи между золотистыми и буры-
ми водорослями (= Pucophyceae), полагая, что предковая форма по-
следних произошла от золотистой водоросли с прямостоячим парен-
химатозным талломом.
BACILLARIOPHYCEAE
Кремнеземные стенки диатомей хорошо сохраняются в виде ис-
копаемых остатков, самые ранние из которых относятся к нижнему
мелу ( Tappan, 1980). Причисление более ранних находок (напри-
мер, из докембрия) к диатомеям было признано необоснованным, но
в обширном обзоре о возможном происхождении этого класса Раунд
и Крофорд ( Round, Crawford, 1981) считают, что эти данные сле-
дует еще раз проверить. Древнейшие формы были центрическими (на-
пример, Stephanophyxis). Изучение обширного ископаемого матери-
ала (см. Tappan, 1980) в сочетании с известными физиологически-
ми и морфологическими особенностями современных диатомей позво-
ляет полагать ( Round, Crawford, 1981), что предок этих центри-
ческих форм должен был иметь эукариотные органеллы и кремнезем-
ные чешуйки, из которых развился зачаточный панцирь; построен
гипотетический эволюционный ряд от клеток с чешуйками к настоя-
щей диатомее со створками и пояском. Считают (Tappan, 1974),
что предковый организм был жхутиковнм и ( Round Crawford, 1981),
вполне вероятно, размножался половым путем. От центрического
417
Рис. 8.17. Palaeocystophora
acuminata (Phaeophyciae,
Fucales?. Плиоценовая бу-
рая водоросль, найденная в
мягком белом диатомите в Ка-
лифорнии. Тринадцать ископа-
емых видов семейства Cysto-
seiraoeae, к которому при-
надлежит вид, обнаружено в
богатых калифорнийских отло-
жениях, т.е. это семейство -
древняя и долго дивергиро-
вавшая группа (х 1,16) (Par-
ker, Dawson, 1965)
предка легко можно проследить последующую эволюцию диатомей по
,трем классическим линиям (центрические, бесшовные и шовные),
представленным современными вилами.
Все имеющиеся данные говорят за то, что диатомеи возникли
в мелководных бентосных местообитаниях, но произошло это в соле-
ной воде или нет - неясно (Round, Crawford, 1981). Центричес-
кие виды преобладают в морских биотопах, а перистые - в пресно-
водных; обширные отложения ископаемых диатомей (диатомит) сфор-
мировались как в морских, так и в пресноводных средах. Все 60
родов, известных из верхнего мела, существуют и сейчас, не изме-
нив существенно отличительные родовые признаки ( Johnson, 1951).
Положение диатомей в пределах хромофитной линии вряд ли
удастся определить, пока среди ископаемых находок не будут рас-
познаны их предшественники. Однако возможно, что они возникли
как четкая группа от эукариотного жгутикового предка с кремне-
418
Рис. 8.18. Sphacelaria plumo-
sa (Phaeophyceae, Sphacelaria-
les). Послеледниковая (около
8000 лет назад) ископаемая на-
ходка ныне живущего вида из от-
ложений восточной части Канад-
ской Арктики, показывающая за-
мечательную сохранность. Терми-
нальная ветвь с супротивно-пе-
ристым ветвлением. Масштаб
0,5 мм (Ulman et al., 1972)
земными чешуйками на очень раннем этапе эволюции хромофитов.
CHRYSOPHYCEAE
Вегетативные стадии золотистых водорослей редко попадаются
в ископаемом состоянии; исключение составляют силикофшагеллаты,
обычные в морских отложениях. Хорошо сохранились также окремне-
лые цисты морских и пресноводных форм. Морские цисты относят к
ископаемому семейству Archaeomonadaceae, известному от верхнего
мела до голоцена ( Tappan, 1980). В верхнем эоцене Австрии они
были описаны как почти чистый археомонадит (Tynan, 1971). Остат-
ки силикофпагеллат встречаются в силикатных породах (особенно
диатомитах), где они мотут составлять до 40-60% по массе (силико-
флагеллит; см.Таррап, 1980). Впервые они появляются в среднем
мелу (в юре согласно рис. I у Taylor, 1978), и с тех пор их раз-
нообразие несколько раз колебалось, дав пик в миоцене (Tappan,
loeblich, 1972). Попытки проследить эволюцию силикофлагеллат по
данным палеонтологии приводят к признанию двух возможных тенден-
ций - к упрощению скелета и к его усложнению (Tappan, 1980).
419
Такие тенденции наблюдаются и в современных популяциях, т.е. мо-
гут быть выведены фенетически. Следовательно, ископаемые находки
проливают мало света на происхождение и эволюцию золотистых во-
дорослей.
PRYMNESIOPHYCEAE
Цримнезиофиты хорошо известны в ископаемом состоянии как
кокколитофориды (с. 44; рис. 8.19). Имеется обзор очень обшир-
ной литературы по этой группе Tappan, 1980). Известковые чешуй-
ки (кокколиты) прекрасно сохраняются, и для описания их морфоло-
гии разработана сложная и детальная терминология. Ископаемые кок-
колиты известны из мезозоя и были обильны на протяжении большей
части юры. Меловые отложения мелового периода в значительной
степени состоят из кокколитов и их фрагментов, а максимального
разнообразия группа достигла в позднем мелу, причем многие виды
существовали 5-10 млн. лет ( Tappan, 1980). Затем последовало
резкое вымирание, а в третичном периоде возникли новые формы,
большинство из которых сохранилось до настоящего времени.
Удовлетворительной схемы филогении примнезиофитов пока нет,
поскольку палеонтологические данные не дают достаточной информа-
ции для построения филетических рядов. До сих пор плохо понятны
в этом плане ныне живущие виды, что затрудняет интерпретацию ис-
копаемых находок. Следует упомянуть недавно открытую группу
холодноводных примнезиофитов ( Manton et al., 1977), роль которой
Рис. 8.19. Вид в сканирующем
электронном микроскопе ископае-
мого представителя кокколитофо-
рид (примнезиофиты) Bidigcus
xgnotus. Целая коккосфера,
мел, Испания (х 7000) Csrun,
Allemann, 1975)
420
в установлении связей между ископаемыми- формами еще требует вы-
яснения. Вдобавок современные исследования показали (см. Ме1-
konianj;982b ), что кокколиты в отложениях мотут изменяться под
воздействием внешней среды; последствия этого для таксономии мо-
гут быть очень существенны. Примнезиофиты, будучи внешне сходны-
ми с золотистыми водорослями, отличаются от других хромофитов
по большинству основных признаков ( Hibberd, 1976), хотя мохут
быть и общим предком обоих классов. Их положение в любой филоге-
нетической схеме в настоящее время проблематично.
DINOPHYCEAE
Несмотря на свое как будто древнее происхождение, динофиты
(рис. 8.20) недостаточно хорошо известны в ископаемом состоянии
вплоть до мезозоя и кайнозоя. Полагают ( Tappan, 1980), что их
могли не распознать в более ранних отложениях, ошибочно приняв
за предотавителей Arcitaroha (Evitt, 1963) - неоднородной
труппы микроскопических ископаемых. С другой стороны, не исклю-
чена их длительная эволюция до появления прочных, хорошо сохра-
няющихся структур. Допускается (Loeblich, 1974) возможность
возникновения динофитов от предковых хромофитов в позднем докем-
брии. Широко обсуждался вопрос о том, рассматривать Prorocen-
ralea как примитивную или как продвинутую труппу (см. Dodge,
1983) и не относить ли ее к акритархам, которые доминировали
в раннепалеозойской микрофлоре (Loeblich, 1976). Додж (Dodge,
1983) делает вывод, что это продвинутая группа с относительно
недавней историей; противоположную точку зрения высказывают
Лёблих (Loeblich, 1976) и Тейлор (Taylor, 1980).
Силурский род Arpylorue,возможно, самый ранний ископаемый
представитель динофлагеллат; развитию этой цисты с прочной стен-
кой могла предшествовать более ранняя дивергенция ( Tappan,
1980; Dodge, 1983), хотя фенетические данные и не говорят о
бесспорности такой гипотезы. Динофлагеллаты - прекрасные руко-
водящие ископаемые, и их находки от перми до настоящего време-
ни хорошо документированы (Tappan, Loeblich, 1973). Их разно-
образие быстро возрастало на протяжении юры и мела, после чего
началось продолжающееся сокращение числа цистообразующих видов.
Таксономия почти полностью ископаемых Ebryophycidae, или
эбридий ( Tappan, 1980), вызывает споры, хотя мы относим их к
динофлагеллятам. Живые клетки, лишенные теки и несущие апикаль-
421
Рис. 8.20. Ископаемые динофлагеллаты. A. Phthanoperidini-
um amoenum; олигоцен, Миссисипи. Вид с дорсальной сто-
роны (X 1280) ( Drugg, Loeblich, 196?). Б. Palaeoperi-
dinium pyrophorym; палеоцен, Алабама.“Целая клетка
Гх Ь40) ( Drugg, 1969)
но прикрепленные жгутики, внешне напоминают виды Deemocapsalea
и Proroceritraies. Эбридии сохраняются в ископаемом состоянии в
виде внутреннего окременелого скелета, тек или цист, неизвестны
до кайнозоя, но в палеоцене представлены уже восемью родами и
становятся еще разнообразнее в эоцене и миоцене ( Tappan, Loeb-
Hchi972). Их родство с динофлагеллятами ныне широко признает-
ся, хотя классификация этой группы запутана из-за отнесения ее
то к растениям, то к животным. Филогения эбридий неясна, но в
будущем может иметь решающее значение для понимания эволюции ди-
нофлагеллат.
Хлорофитная линия
Геологические данные по хлорофитам - самые древние (от до-
кембрия до наших дней) и полные по сравнению с имеющимися по лю-
бым другим группам водорослей. Однако эвгленовые представлены в
остатках слабо, и многие более ранние сообщения об их находках,
как оказалось, относятся к цистам золотистых водорослей. Хорошо
сохранившийся TrachelomonaB найден в плейстоценовых битуминоз-
ных сланцах Мадагаскара (Deflandre, Lenoble, 1948); имеются
сведения о находках форм, напоминающих Phacus. в эоценовых от-
422
ложениях ( Bradley, 1929). Однако сохранившиеся в породах остат-
ки мало говорят об эволюции эвгленовых. С другой стороны, прази-
нофиты известны по крайней мере с ордовика (около 500 млн. лет
назад); они сохранились в виде цистоподобных фиком (рис. 8.21),
толстостенных тасманитов и тонкостенных лейосфер ( Tappan, 1980).
Род Tasmanites, широко распространенный в палеозое, напоминает
современный род Pterospeima (Scagel et al., 1982), а некоторые
из докембрийских и палеозойских лейосфер (например, beiosphaeri-
dia ) сходны С представителями современных Halosphaeraceae
( Wall, 1962). Подчеркивался эволюционный консерватизм фазы фи-
комы в жизненном цикле празинофитов ( Colbath, 1983). Ископаемых
представителей этого класса можно, по-видимому, считать "видами
катастроф”, пережившими периоды массовых вымираний, уничтожившие
большинство акритарх в среднем палеозое (Tappan, 1980). Они ста-
ли менее обильными, когда возросло разнообразие динофлагеллат и
других групп водорослей. Пермские отложения тасманитов в Тасма-
нии, образующие протяженные слои, свидетельствуют о многочислен-
ности празинофитов в прошлом.
Рис. 8.21. Ископаемый празинофит Cymatosphaera reticulosa;
силур. А. Вид в сканирующем электронном микроскопе слабо
орнаментированной фикомы (цисты) с местом разрыва (стрел-
ка). Б. Вид в сканирующем электронном микроскопе фикомы
с умеренно развитой орнаментацией (х 2500) ( Colbath,
1983)
423
CHLOROPHYCEAE
Немногочисленные ископаемые находки большинства Chlorophy-
ceae компенсируются остаткам# множества сигнальных видов, воз-
раст которых достигает докембрия ( Р1а, 1920). Эти формы, откла-
дывающие арагонит, особенно важны как породообразователи. Пред-
полагают, что они, как и их современные сородичи (например, на-
limeda, Udotea, Penicillus). предпочитали теплые мелководья,
где насыщенность СаСО3 благоприятствовала отложению извести. В
развитии и распространении ископаемых зеленых водорослей наблю-
даются значительные флуктуации: максимального разнообразия они
достигли в среднем мелу, и с тех пор оно более или менее посто-
янно сокращалось.
Современные Dasycladaceae (Dasycladales)представлены только
небольшим числом довольно сложно устроенных остатков некогда
значительно более обширной группы (рис. 8.22 и 8.23). Это поис-
тине реликтовое семейство, в котором примерно 95% описанных ро-
дов вымерло (Tappan., 1980). Начиная с докембрия (1200-1400 млн.
лет назад; например.Timanella, Templuma) и на протяжении палео-
зоя дазикладовые все более усложнялись по строению, а основного
разнообразия достигли в мезозое. Пиа ( Р1а, 1920 и более поздние
работы; см. Tappan, 1980) разбирал ископаемые находки предста-
вителей этой группы в своих классических монографиях, а Вале
( Valet, 1969) провел их более современный филогенетический ана-
лиз, включающий ныне живущие и вымершие формы. Сведения об этой
группе достаточно полные, и для нее можно проследить эволюцию
структуры таллома, однако происхождение давикладовых на основа-
нии палеонтологических находок установить невозможно.
Ископаемые остатки Chlorophyceae отнюдь не исчерпываются
известковыми видами. Коккоидные и нитчатые формы, которые можно
отнести К Chlorococcales и Ulotrichalea,встречаются В не менее
древних отложениях (Tappan, 1980; Scagel et al.,1982). Остатки
представителей большинства других порядков впервые появляются в
породах от девона до нашего времени. Ископаемые сифональные фор-
мы древнее нитчатых примерно на 230 млн. лет, и неясно, какой
тип строения возник раньше.
424
Рис. 8.22. Palaeodasycladus
mediterraneus (.Chlorophyceae.
basycladales);нижняя юра, Ита-
лия. А. Реконструкция продоль-
ного среза обызвествленного
(слева) и лишенного извести
(справа; некоторые ветви уда-
лены) таллома. Б, В. Попереч-
ные срезы частично_обызвест-
вленного и частично лишенного
извести таллома. В апикальной
(Б) части таллома много вет-
вей, в нижней (В) - неветвя-
щих боковых выростов (х 18)
( Pia, 1920)
Рис. 8.23. Cymonolia paktia
(ehlorophyceae, Dasycladales);
эоцен. Афганистан. Продольный
срез (X 30) (Kaever, 1969) '
425
OHAROPHYCEAE
Находки харовых водорослей впервые появляются в силурских
слоях. Эволюция этой группы полностью прослежена по гирогонитам
(рис. 8.24), т.е. сохранившимся остаткам их оогониев с окружаю-
щими слоями клеток. Ныне живущие харовые - реликтовая группа,
разнообразие которой резко сократилось к концу девона и в триа-
се, достигло максимума в нижнем мелу и упало в верхнем мелу и
олигоцене (Gramblast, 1974). Из примерно 70 выделяемых родов
только шесть представлены ныне живущими видами, которых около
80 ( Wood, Ihmahori, 1965). Вымершая труппа Umbellinaceae при-
числялась некоторыми авторами к даровым, хотя ее положение спор-
но ( Tappan, 1980).
Уже древнейшие ископаемые харовые представлены сильно про-
двинутыми формами и, как и в случае с известковыми Chlorophy- •
сеае,точно установить их происхождение невозможно. Тем не менее
палеонтологические данные говорят в пользу того, что эта группа
рано отделилась от линии Chlorophyceae.
Рис. 8.24. Различные формы гирогонитов современной (А) и
ископаемых (Б - Е) харовых водорослей. Вид сбоку: А -
lychnothamnus barbatив. х 34 ' ( Maslov. 1963); Б~-
stephanochara саирта, нижний олигоцен, Англия, х~2Э
7 Gramblast, I960): В - Kosmogyra superba, палеоцен,
Италия, X 10 (по St ache, 1В8У): I1 - Harris! chara vasi-
formis, эоцен, Англия, X 27 ТН°ГГЧ Hantzien, 1УЬУ);
Д - Psilochara undulata, эоцен, Франция, X 18 ( Gram-
blast j 1960): hi -biter о chara hystrix. эоцен, Франция,
X 55 (Gramblast? I960)
Глава 9
ВОДОРОСЛИ, ЧЕЛОВЕК И ОКРУЖАЮЩАЯ СРЕДА
Введение '
Водоросли играют очень незначительную роль как возбудители
болезней, но могут создавать ряд серьезных проблем, связанных с
отравлением водоемов и нарушением работы гидротехнических соору-
жений. Они, несомненно, являются важнейшими продуцентами (гл. 7)
и на протяжении геологической истории участвовали в образовании
известковых и силикатных пород, а также углеводородных отложе-
ний. Водоросли мотут служить источниками пищи и полезных химиче-
ских соединений; развитие современных лабораторных и культиваци-
онных методик дает широкие возможности их использования сейчас и
в будущем (Lewin, 1983 Ъ).
Водоросли и болезни человека .
Роль водорослей и их метаболитов как возбудителей заболева-
ний человека рассмотрена Стейном и Борденом ( stein, Borden,
1984). Поскольку автотрофы вообще плохо приспособлены к существо-
ванию в качестве патогенов, имеются сведения только о заражении
человека и животных лишенной хлорофилла, напоминающей представи-
телей рода Chlorella водорослью Prototheca (Chlorophyceae;
Chlorococcales)(Kessler, 1977, 1982). Виды Prototheca встреча-
ются в различных как морских, так и пресноводных местообитаниях,
и установить связь между дикими типами и болезнетворными форма-
ми сложно. Они могут быть выделены в культуру и из больных, и из
здоровых людей. Клетки размножаются бесполым путем, образуя до
20 эндоспор ( Cooke, 1968 _а,Ъ).
После подтверждения инфекционной природы водоросли Proto-
theca (Davies et al., 1964) (болезнь обычно называют прототеко-
зом) сообщалось о заражении ею человека. Болезнь обычно проявля-
ется в поражениях кожи, подкожных инфильтратах, иногда затраги-
ваются суставы ( Ahbel et al*, 1980).Prototheca редко вызыва-
ет системное заражение, но встречаются и рассеянные инфекции
( Сох et al., 1974). Из животных прототекозы указывались у со-
бак; у крупного рогатого скота эта водоросль приводит к маститу
(Frank et al., 1969). Есть мнение (Munday, Peel, 1983), что
427
Prototheca возбуждает язвенный дерматит у утконоса (ornitho -
rhynchus anatinua), т.е. такие инфекции, по-видимому, могут
бкть у широкого круга животных. Борьба с прототекозом сложна, по-
скольку Prototheca устойчива ко многим антибиотикам, но неко-
торые из них все же эффективны, например амфотерицин В или его
смесь с тетрациклином ( Stein, Borden, 1984).
В Швеции отмечен силикоз из-за вдыхания диатомовой земли
( Beskow, 1978). В судебной медицине диатомеи использовались при
диагностике утопления ( Peabody, 1980), но здесь в выводах нуж-
на осторожность, поскольку эти водоросли встречаются повсеместно
и часто обнаруживаются в нормальных человеческих тканях.
Токсичные водоросли
Токсины, выделяемые многими как пресноводными, так и морски-
ми водорослями, попадая в организм человека и животных, мохут на-
носить вред здоровью или даже вызывать смерть. Наиболее известны
в этом плане некоторые синезеленые водоросли, один вид примнезио-
фитов, различные динофлагелляты и несколько морских макрофитов.
Токсичные синезеленые водоросли
Цветение воды, вызываемое синезелеными водорослями, давно
связывали с отравлениями животных (Francis, 1878). Для видов
Microcystis aeruginosa, Anabaana flos-aquae И Aphanizomenon
flos-aquae подтверждено образование токсинов. Цветения о учас-
тием Anabaena дают наиболее тяжелые последствия, Microcystis
наиболее распространен, aAphanizomenon указывается реже всех
( Carmichael, 1981).
Токсины Anabaena, так называемые анатоксины ( Carmichael,
Gorham, 1978), по-видимому, бывают трех типов: алкалоиды, пеп-
тиды и птеридины. Алкалоиды дают наиболее острые отравления с
неврологическими симптомами через несколько минут после попада-
ния в организм, тогда как дейстше пептидов требует от одного до
нескольких часов; они вызывают некроз печени. Токсины Microcys-
tis - преимущественно пептиды (Carmichael, Gorman, 1980), а у
Aphanizomenon они близки к сакситоксинам динофлагеллат (Alam
et al., 1978). Токсины образуются не при всех цветениях, и да-
же в рамках одного цветения существует мозаика токсичности
(Carmichael, Gorham, 1981). С определенной степенью достовер-
428
ности можно предположить, что для образования токсинов Microcys-
tis требуется наличие в клетках плазмид ( Hauman, 1981).
Острых отравлений людей обычно не возникает, поскольку явно за-
грязненная вода для питья не используется. Однако наличие токси-
нов синезеленых водорослей в питьевой воде может приводить к
гастроэнтериту ( Keleti et al., 1979).
Токсины морских видов Lyngbya . возможно, вызывают контакт-
ный дерматит у пловцов на Гавайях ( Grauer, Arnold, 1961) и Оки-
наве ( Hashimoto st al., 1976). Активное вещество здесь - поли-
фенол дебромоаплизиатоксин ( Mynderse st al., 1977).
Токсичность Prymnesium parvum
Prynmesium parvum _ широкораспространный в морских и соло-
новатых водах вид, токсины которого избирательно поражают жабро-
дышащие организмы. Он способен вызывать массовую гибель рыбы,
нанося серьезный ущёрб аквакультуре. Токсичные вещества пред-
ставляют собой смесь протеофосфолипидов ( Shilo,. 1981). Они тре-
буют активации катионными кофакторами (ионами натрия, магния или
кальция) и изменяют проницаемость жаберных мембран. Природа ток-
сичности у Prynmesium сложная, и корреляция между обилием кле-
ток водоросли и смертностью рыб незначительна.
Красные приливы и токсичные динофлагеллаты
"Красные приливы" на самом деле не всегда красные и не всег-
да приливы; они обычно представляют собой цветение с участием
одного из видов динофлагеллат, образующего токсины, которые вызы-
вают паралитическое или нейротоксическое отравление моллюсками у
питяющится ими людей, рыб и морских птиц. Другие нетоксичные
морские организмы, изменяющие окраску воды, могут приводить к
гибели морских животных от асфиксии, заооряя поверхность их
Жабер ( Steidinger, Haddad, 1981).
Динофдагеллатные красные приливы наблюдаются в прибрежной
зоне и эстуариях морей как тропических, так и умеренных широт.
Из более 1000 видов динофлагеллат приблизительно для 20 показано
образование паралитических токсинов ( steidinger, 1983), потен-
циально смертельных для человека. Хотя сведения о таких отравле-
ниях имеются с ХИГ в. ( Steidinger, Haddad, 1981), их связь с ди-
нофлагеллатами установлена только в 1937 г. ( Sommer et al.,
1937). Экстракты из Protogonyaulax (dGonyaulax) catenella
429
вызывали у мышей те же симптомы, что и экстракты из токсичных
моллюсков; нетоксичные моллюски становились токсичными, если их
кормили этими динофлагеллятами.
Protogonyaulax tamarensis (включая Р. excavata ) описан
как возбудитель паралитических токсикозов в Северной Атлантике
(Needier, 1949), а р,catenella - в Британской Колумбии (Pra-
kash, Taylor, 1966). Предполагали (Schmidt, Loeblich, 1979),
что подобные отравления связаны только с труппой видов "catenel-
la", но установлена причастность к ним и других родов динофитов
(Steidinger, 1983). Токсины Protogonyaulax, концентрируясь в
фильтрующем зоопланктоне, мотут вызывать гибель рыб (White,
1981).
Паралитические отравления моллюсками вызываются двенадцатью
близкими водорастворимыми токсинами (steidinger, 1983), из кото-
рых лучше всего известен сакситоксин. Все они - сильные нейроток-
сины, подавляющие передачу нервного импульса путем блокировки
натриевых каналов в аксоне (Schantz, 1981).
Цветения Protogonyaulax связаны с половым циклом. У P.ta-
marenais бентосные покоящиеся цисты накапливаются в удаленных
от берега осадках ( Lewis et al., 1979). Апвеллинг переносит их
в прибрежные воды, где они прорастают (Hartnell, 1975). Начало
прорастания, возможно, обусловлено повышением температуры (Ander-
son, 1980); затем происходит быстрое увеличение бесполым путем
численности планктона ( Anderson et al., 1983), которая зависит
как от температуры, так и от солености (watrass et al., 1982).
Наиболее впечатляющие красные приливы встречаются у запад-
ных берегов Флориды; они вызываются в основном Ptychodiacus
( =Gymnodinium) brevis возбудителем нейротоксического моллюс-
кового отравления. Эти приливы могут приводит к массовой гибели
рыб. Нирорастворимый нейротоксин отличается по структуре и дей-
ствию от рассмотренных выше паралитических токсинов. У людей,
которые съели накопивших это вещество моллюсков, может наблюдать-
ся оцепенение наряду с.симптомами пищевого отравления, но парали-
ча и смертельных случаев не зарегистрировано ( steidinger, 1983).
Полагают, что цветение р,brevis начинается в особой зоне
( steidinger, 1975 а, ъ),удаленной на 18 - 74 км от берега, при-
чем его развитие зависит от насыщения поверхностных вод донными
цистами. Прибрежные красные приливы, по-видимому, коррелируют с
притоком вод открытого моря, приносящих динофлагеллат в область
Шельфа ( Haddad, Carter, 1979).
430
Механизмы развития красных приливов в эстуариях лучше всего
Известны ДЛЯ вида Prorocentrum minimum fcExuviaella mariae-lebo-
uriae), приводящего к отравлениям двустворчатыми в Японии
( Nakajima, 1965). Ежегодные цветения Р.minimum наблюдаются в
отдельных частях Чесапикского залива (США); они начинаются в зо-
не фронтальной конвергенции и перемещаются в залив под пикнокли-
ном подповерхностным течением, вызываемым стоком полых вод вес-
ной. Фототаксис и граница солености концентрируют водоросли во
фронтальной области. Летом эти границы исчезают, и Р.minimum
проникает в поверхностные воды внутренней части залива ( Tyler,
Seliger, 1978, 1981).
Бентосный представитель динофлагеллат Gambierdiscus toxi-
cus образует токсин сицуатеры ( Bagnia et al., 1979), отравле-
ния, вызываемого поеданием внутренних органов некоторых рыб ко-
ралловых рифов. Его симптомы - тошнота, мышечная слабость и судо-
роги, но смертность невелика. Другие бентосные динофлагелляты, в
том числе космополитный вид Prorocentrum lima , также могут
быть причиной сигуатеры ( Yasumoto et al., 1980).
Токсичные макрофиты
Хорошо известны ядовитые высшие растения; представители мно-
гих их семейств образуют вторичные метаболиты, которые служат
для защиты от фитофагов и патогенных микроорганизмов. Такие сое-
динения могут быть токсичными и для человека; многие из них ис-
пользуются в медицине.
Сходные вещества встречаются и у морских водорослей-макро-
фитов (обзоры CM.Penical, 1980; Norris, Fenical 1982), веро-
ятно предохраняя их от поедания беспозвоночными и рыбами. Они об-
наружены у Chlorophyceae, Phaeophyceae и Rhodophyceae и пред-
ставляют собой ацетиленовые производные, терпеноиды, галогенотер-
пеноиды, галогенометаны, полифенолы и алкалоиды. Водоросли, об-
разующие эти вещеотва, распространены почти исключительно в тро-
пических водах; не исключено, что синтез подобных соединений -
реакция на интенсивное выедание (особенно рыбами) в таких место-
обитаниях ( Hiat, Strasburger, i960).
В нескольких семействах тропических зеленых водорослей име-
ются виды, не поедаемые обычными растительноядными животными.
Среди них Caulerpa образует производное индола каулерпин ( Hai-
ti et al.,1978) и токсичные для рыб терпеноиды (Sun, Penical,
1979).
431
Общая особенность бурых водорослей - синтез фенольных сое-
динений (Glombitza, 1977), обычно продуктов полимеризации флоро-
глуцинола (1,3,4-тригидроксибензола), обладающих антибактериаль-
ным действием ( Conover, Siebur'th, 1964) И способных также отпу-
гивать беспозвоночных. Ряд тропических видов из семейств Dictyo-
taceae и Cystoseiraceae образует терпеноиды, которые у неко-
торых видов DictyotaMorvT составлять дс 5% сухой массы.
У багрянок известно больше всего токсичных вторичных метабо-
литов. Ядовитые виды обитают прежде всего в тропиках, хотя пред-
ставители Bonnemaisoniaceae из умеренной зоны также синтезиру-
ют галогенированные токсины. Asparagopsis taxiformis, считаю-
щийся лакомством у местных жителей Гавайских островов (Abbott,
Williamson, 1974), образует канцерогенный бромоформ, галогени-
рованные ацетаты, акрилаты и кетоны, а Liagora farinosa (Nema-
liales, Helminthocladiaceae) - ацетиленовый липид, токсичный
ДЛЯ рыб. Laurenica obtusa (Ceramiales, Rhodomelaceae) - ВОДО-
росль, избегаемая большинством растительноядных животных, в том
числе морским ежом Diadema (Ogden, 1976)-синтезирует бронирован-
ные соединения, исключительно токсичные для оплодотворенных яйце-
клеток морских ежей.
Однако токсичные морские водоросли-макрофиты поедаются неко-
торыми фитофагами. Некоторые мешксязычные заднежаберные моллюски
выборочно потребляют Caulerpa; они концентрируют водорослевые
токсины ( Doty, Aguilar-Santos, 1970 ) и выделяют их в ядовитой
для рыб слизи ( Lewin, 1970). Морские зайцы ( Ар1уа1а~) питаются
токсичной водорослью Laurencia, также концентрируя ее токсины
как средство защиты от хищников ( Stallard, Faulkner, 1974).
Большинство морских макрофитов, используемых в пищу челове-
ком, не образует токсичных соединений или содержит их в относи-
тельно малых, легко обезвреживаемых количествах ( Fenical, 1980).
Однако некоторые таксоны, например бурая водоросль Hizikia
(Pucales), потенциально токсичны, поскольку накапливают мышьяк
(Watanabe et al., 1980).
Загрязнение и водоросли
Загрязнение возникает в результате человеческой деятельнос-
ти, когда либо повышаются концентрации природных веществ, либо
не встречающиеся в природе синтетические соединения (ксенобиоти-
ки) попадают в окружающую среду. Они могут как подавлять, так и
432
стимулировать рост водорослей, причем в последнем случае сами
водоросли становятся источником беспокойства.
Органическое загрязнение
Этот вид загрязнения дают органические вещества, попадающие
в окружающую среду с бытовыми, сельскохозяйственными и промышлен-
ными отходами. Основную опасность для окружающей среды представ-
ляют канализационные стоки, углеводороды и галогенированные угле-
водороды.
Канализационные стоки создают значительный недостаток кисло-
рода, а многие водоросли чувствительны к его содержанию в воде.
Описаны (Palmer, 1977; Round, 1981) водорослевые сообщества,
обычно встречающиеся в загрязненных пресных водах- В наиболее за-
грязненных местах водоросли могут отсутствовать или быть пред-
ставленными только Cyanophyceae и несколькими жгутиковыми, осо-
бенно эвгленовыми. По мере уменьшения загрязненности сначала по-
являются диатомеи и нитчатые зеленые водоросли, а затем развива-
ется обильная альгофлора, типичная для эвтрофных условий. Значи-
тельная часть неочищенных стоков сбрасывается в море, где они
обычно быстро разводятся водой до низкой концентрации. На юге
Калифорнии такие стоки привели к гибели зарослей Macrocyatla из
ламинариевых (North et al., 1964, 1971). Реколонизация подав-
лялась накоплением осадков и увеличением численности растительно-
ядных иглокожих. Однако стоки могут и способствовать росту бен-
тосных морских водорослей (Kindig, Littler, 1980).
Углеводороды, в том числе из природных выходов нефти (Straug-
han, 1982), попадают в водную среду и без участия человека. Мор-
ской фитопланктон образует углеводороды в количествах, которые
мотут достигать 5 т/км^-год ( Smith, 1954). Из них ароматические
более токсичны, чем алифатические, а более растворимые низкомо-
лекулярные фракции - самые вредные, в значительной степени из-за
того, что присутствуют в наибольших концентрациях. При одинако-
вых концентрациях менее растворимые соединения о более высоким
коэффициентом распределения в системе липид/вода обладают наибо-
лее острым токсичным действием на водоросли (Hutchinson et al.,
1978}, так, нафталин в концентрации 3 млн токсичнее более раст-
воримого бензола при такой же его концентрации.
В лаборатории общая реакция водорослевых культур на углево-
дороды заключается в стимуляции роста при низких концентрациях и
433
его подавлении при высоких (Davenport, 1982). Показано (Gordon,
Prouse, 1973), что уровни углеводородов ниже 3-5 млн"1 стиму-
лируют фотосинтез, но при дальнейшем повышении их наблюдается
его постепенное подавление. Нафталин в концентрации I млн~" сни-
жал скорость (фотосинтеза у Fragilaria (Bacillariophyceae), Mono-
chrysie lutheri(Chrysophyceae) и Dunaliella ep. (Chlorophyceae)
на 40%, а в концентрации 20 млн"! - на 80% ( Vandermuelen, Ahern,
1976). Это вещество не влияло на содержание у водорослей хлоро-
филла, но снижало количество АТР в клетке. Нафталин подавляет
также фотосинтез И ПОДВИЖНОСТЬ у Chlamydomonas angulosa (Chloro-
phyceae; Kauss et al., 1973). В свежих культурах Chlorella .его
добавление сокращало длительность лаг-фазы, но снижало и конеч-
ную концентрацию клеток. Низкие концентрации углеводородов усили-
вают деление клеток у ряда одноклеточных зеленых водорослей, при-
чем уровень стимуляции коррелирует с канцерогенностью этих ве-
ществ. Сходная стимуляция наблюдалась у проростков Antithamnion
plumula и представителей близких родов бентосных морских багря-
нок, обработанных бензантраценом; реакция зависела от дозы, а
рост ускорялся за счет усиленного деления клеток, а не увеличе-
ния их размеров (Boney, Corner, 1962; Boney, 1974).
Полихлорированные бифенилы способствуют фотосинтезу и кле-
точному целению при низких концентрациях, а при белее высоких
оказывают обратное действие ( Gorham, Rhee, 1982). Накопление
их у водорослей представляет собой двустадийный процесс: за ад-
сорбцией на поверхности следует поглощение клеткой за счет диф-
фузии, а не активного транспорта. Десорбция происходит медленнее
абсорбции, т.е. восстановление нормального состояния идет дольше,
чем загрязнение ( Lederman, Rhee, 1981). Скорость накопления и
влияние в разведенных водорослевых суспензиях больше, чем в гус-
тых (cole, Plapp, 1974). Многие полициклические углеводороды
разрушаются солнечным светом, и тлеются сведения, что это проис-
ходит активнее, когда вещества связаны с поверхностью водоросле-
вых клеток ( Zeppa, Schlotzhauer, 1993).
Эксперименты в условиях мезскссма дали информацию о влиянии
углеводородов на смесь видов фитопланктона в присутствии расти-
тельного зоопланктона. В Канаде в изолированный участок залива
Саанич объемом 60 000 л спустили богатую нафталином сырую нефть
в концентрации 40 млн"! (ьее, Takahashi, 1977). Продукция фито-
планктона увеличивалась, причем наблюдалось снижение обилия .диа-
томеи Ceraulina hergonii и массовое развитие мелкого жгутиксво-
434
ГО Chrysochromulina kappa из золотистых водорослей, а ЯВНОГО вли-
яния на зоопланктон не отмечалось. В огороженные 300 000 л залива
Лох-Ю (Шотландия) закачали сырую нефть из Северного моря до кон-
центрации 100 млн~1 (Davies et al., 1980). Через пять недель
концентрация нефти упала в четыре раза; половина потерянных 75%
выпала в осадок, другая половина приходится на микробный обмен
веществ и испарение. Никаких изменений биомасоы планктона или
структуры сообщества не отмечено. В 13 000-литровых объемах, за-
грязненных нефтяным топливом, отмечен тот же видовой состав фи-
топланктона, что и в исходной воде, и увеличение его обилия, свя-
зываемое с меньшим выеданием ( Vargo et al., 1982).
Разливы нефти в природе, по-видимому, мало влияют на фито-
планктонные популяции. После крушения танкера "Торри-Каньон" от-
мечено нормальное обилие динофлагеллат и диатомовых, хотя и на-
блюдалось большое число мертвых цист празинофитов (Smith, 1968).
Нефть из танкера "Цесис" в Балтийском море ( Johansson et al.,
1980) вызвала кратковременное увеличение на прилегающей аквато-
рии биомассы фитопланктона, что можно объяснить снижением обилия
зоопланктона; однако обычная численность восстановилась в тече-
ние пяти дней.
Широко изучалось влияние нефти в природе на бентосные мор-
ские водоросли, особенно литоральные. Разнообразие литоральной
альгофлоры в Кол-Пойнте (Калифорния), где имеются природные выхо-
ды нефти, меньше по сравнению со сходными местообитаниями, где
нефти нет ( Nicholson, Cimberg, 1971). Посадка на мель судна
"Тампико-Мару" у берегов Нижней Калифорнии привела к попаданию
около 9500 м3 дизельного топлива в небольшую бухточку за период
8 месяцев (North, 1967). Непосредственным результатом этого бы-
ли исчезновение бентосных фитофагов и последующее увеличение чис-
ленности альгофлоры. Оценка влияния нефти с "Терри-Каньон" была
осложнена использованием токсичных детергентов при очистке моря.
Ряд литоральных видов был поражен непосредственно ( O'Brien,Di-
xon, 1976), а на залитых нефтью участках до обработки поврежде-
ние биоты часто было меньше, чем после применения эмульгаторов.
В местах, где испарение удаляло более легкие растворимые фракции
нефти до выброса ее на берег, основной эффект был механический:
литоральные фукусовые страдали от удушья и ударов "утяжеленных"
волн ( Nelson-Smith, 1973). Важнейшим результатом воздействия
-нефти и детергентов в случае "Торри-Каньон" явилось уничтожение
поедающих водоросли животных ( Bellamy et al., 1967; Smith,
435
1968), что наряду с освобождением пространства из-за гибели бо-
лее крупных водорослей позволяло оппортунистическим видам, напри-
мер Enteromorpha (Chlorophyceae),захватывать новые места и доми-
нировать там. При крушении танкера "Амоко-Кадис" у побережья
Франции детергенты широко не использовались и отмирание водорос-
лей-макрофитов коррелировало с длительностью воздействия на них
нефти. Больше всего пострадали фукусовые верхней литорали fucus
spiralis И Pelvetia canaliculate (Floc’h Diouris, 1981). Спус-
тя четырнадцать месяцев в них все еще можно было обнаружить остат-
ки нефти, нс на скорость роста это не влияло ( Topinka, Tucker,
1931).
Культурная эвтрофизация
Под культурной эвтрофизацией (Hutchinson, 1969) понимается
обогащение воды питательными веществами за счет деятельности че-
ловека; увеличение содержания доступных азота, фосфора и углеро-
да способствует росту продуктивности водорослей и изменению их
видового состава. Озера, по-видимому, не подвержены постепенной
эвтрофизации в ходе своего естественного развития (Beeton, Edmon-
ston, 1972; Round, 1981). Уайтсайд (whiteside, 1983) счита-
ет, что эвтрофизация - неподходящий термин для описания процес-
сов обогащения воды вследствие старения озер и что биотические
взаимодействия и/или морфометрия озера могли бы равным образом
объяснить палеолимнологические изменения. Полностью проблему оце-
нить трудно, но установлено, что в США 50% поверхностных вод в
той или иной мере эвтрофизировано ( Jewell, McCarthy,1968). Ос-
новные трудности создает массовый рост водорослей, физически пре-
пятствующих водному транспорту и усиливающих недостаток кислоро-
да при гниении. Сельскохозяйственные стоки - основной источник
азота; в Англии вода, выносимая с пахотных земель, содержит в
среднем 10 мг/л нитратов (Cooke, Williams, 1970), а иногда и на
порядок выше. Канализационные стоки - основной источник фосфатов
и органического углерода, но в отличие от азота они обычно попа-
дают в воду локально.
На примере озера Вашингтон, возможно, наиболее детально изу-
чены ( Edmonston, 1970, 1972, 1979; Edmonston, Lehman,1981), вли-
яние стоков и последующее восстановление водоема после соответст-
вующей обработки. Отвод стоков всзвраирл уровень фосфатов к быв-
шему до эвтрофизации, чему сопутствовало уменьшение биомассы во-
дорослей. Однако такое ее уменьшение не сопровождало снижения
436
уровня нитратов или углерода; это позволяет предполагать ведущую
роль фосфора в процессе эвтрофизации. Годфри (Godfrey, 1982) по-
казал, что за последние 60 лет биомасса водорослей в озере Каюга
(США) удвоилась в летние месяцы и выросла в 30 раз весной. Эти
изменения коррелировали с использованием детергентов на основе
фосфатов.
Хотя фосфор явно причастен к процессу эвтрофизации, относи-
тельная роль в нем различных бисгенов долго дебатировалась. Ис-
следования в небольших бутылях последствий искусственного обога-
щения озерной воды (Schindler, 1971) позволяли полагать, что
большое значение имеет при этом также.углерод. Он редко бывает
лимитирующим фактором в природе, и в озере, экспериментально обо-
гащенном фосфором и азотом ( Schindler et al., 1973), его содер-
жание увеличивалось вследствие роста фитопланктона, хотя естест-
венная концентрация углерода в воде была низкой. Флетт с сотр.
( Flett et al., 1980) показали, что азотфиксирующие синезеленые
водоросли мотут повышать содержание азота в озерной воде, если
другие биогены поступают в избытке. Это еще раз подтверждает ве-
дущую роль фосфора в эвтрофизации, заставляя полагать, что конт-
роль за поступлением в-экосистему азота и углерода не имеет смыс-
ла без одновременного контроля фосфора.
Эвтрофизация влияет также на видовой состав альгофлоры и ее
сезонные изменения. В необогащенных водах умеренных широт у попу-
ляций водорослей пики численности обычно приходятся на весну и
осень. В сильно эвтрофных водах они становятся менее заметными
из-за дополнительных цветений, происходящих круглый год ( Davie,
1964). В Великих озерах Северной Америки умеренное увеличение
содержания фосфатов вызывало быстрое истощение запасов силикатов
и обусловило переход доминирования от диатомей к зеленым и сине-
зеленым водорослям (Makarewicz, Baybutt, 1981). Еще большее по-
ступление фосфатов вело к уменьшению содержания доступного азота
и способствовало развитию азотфиксирующих синезеленых водорослей
( Bierman et al., 1980).
Наиболее бросающееся в глаза следствие культурной эвтрофи-
зации - усиление роста нитчатых видов. Лучше всего изучена в
этом плане Cladophora glomerata (Chlorophyceae), но усиленно
развиваться и причинять беспокойство могут и другие роды, в част-
ности Spirogyra, Rhizoclonium, Enteromorpha, (Chlorophyceae),
Compsopogon и Thorea (Rhodophyceae) (Palmer, 1977; Whitton,
1971). Засорение рек длинными тяжами Cladophora может быть
437
крупной проблемой в умеренных регионах ( Lund, 1972), и драмати-
ческие последствия этого наблюдаются в наиболее эвтрофных из Ве-
ликих озер.
Cladophora glomerata была впервые обнаружена в озере Эри
в 1847 г. и разрасталась там все сильнее по мере эвтрсфизации
(Miller, Sweeney, 1982). Цветения происходят весной и осенью, и
биомасса превышает 400 г/м^ сухого вещества на глубинах 0,5 - 3 м.
Водоросль отрывается от места прикрепления и может попадать на бе-
рег, засоряя пляжи (Niell, Jackson, 1982). Показано (Graham et.
al., 1982), что оптимальный рост С. glomerat а происходит между
13 и Г7°С; когда температура воды превышает 20°С, он прекращает-
ся. Реактивный фосфор в скоплениях Cladophora при этом ниже опре-
деляемого уровня, и его добавление не стимулирует роста водорос-
ли, пока температура не падает ниже 20°С.
Использован ряд методов уничтожения С.glomerata, включая
обработку сульфатом меди, арсенитом натрия и несколькими продаж-
ными гербицидами. Однако успешное решение проблемы заключается в
борьбе с эвтрофизацией, а не с водорослью, т.е. в устранении при-
чины, а не следствия.
Загрязнение металлами и кислотами
В последнее время появились обзоры работ о влиянии на водо-
росли тяжелых металлов (например, Rai et al., 1981;stokes, 1983; .
Davies, 1983). Эти металлы можно использовать как альгициды, на-
пример добавляя в водоемы растворы соединений меди, хотя некото-
рые виды весьма устойчивы к такого рода загрязнению. Данные мно-
гих исследований противоречивы, и до сих пор нельзя сделать обще-
го вывода относительно влияния тяжелых металлов на отдельные во-
доросли или их сообщества.
Изучение альгофлор в местах влияния рудничных вод показыва-
ет, что металлы снижают как разнообразие, так и продуктивность
сообществ, причем синезеленые и диатомовые водоросли в целом ме-
нее устойчивы, чем представителиChlorophyceae. Фостер (Foster,
1982а) сравнивает участки рек, загрязненные медью и свинцом, и
приходит к заключению, что видовой состав определяется уровнем
загрязнения, а не конкретным металлом, причем uiothrix и Micro-
зрога (Chlorophyceae) особенно толерантны, а представители zyg-
nematales (Chlorophyceae), как правило, встречаются в водах с
низкой концентрацией металлов. Гейл с соавт. (lale et al., 1973),
438
Напротив, указывают, ЧТО Mougeotia, Zygnema HSpirogyra (Zygnema-
tales) обильны в загрязненных металлами водотоках шт. Миссури
(США). Сей и Уиттон (Say, Whitton, 1978) относят виды Natrium,
Fleetonema, Nitzschia, Hormidium, Ulothrix и Mougeotiaic ТИПИЧ-
НЫМ для альгофлоры водотоков, загрязненных цинком. Ряд данных
свидетельствует о котолерантности к металлам. Фостер ( Foster,
1982Ъ) показывает, что отбор водорослей на устойчивость к меди
повышает их толерантность и к никелю. И напротив, при отборе
штаммов Anacyatis midulans( Cyanophyсеае)на устойчивость К ко-
бальту, меди, никелю и кадмию обнаружены признаки понижения толе-
рантности ко второму металлу (Whitton, Shehata, 1992).
Информация о механизмах толерантности, включающих непроница-
емость мембран, внеклеточное хелатирование или внутреннюю деток-
сикацию, скудна и часто противоречива (stokes, 1983). При сход-
ных внешних уровнях меди толерантные клетки Chlorella sp-содер-
жат ее в тех же концентрациях, что и нетолерантные, что предпола-
гает отсутствие механизма исключения. По мнению Батлера и др.
(Butler et al., 1980), исключение отчасти обусловлено образова-
нием комплексов тяжелых металлов с внеклеточными водорослевыми
продуктами, причем возникающий комплекс более стабилен у толе-
рантных штаммов. Однако у Scenede3iaUB (chlorophyceae) толерант-
ность' к меди, по-видимому, связана не с внеклеточным хелатирова-
нием ( Mierle, Stokes, 1976), а со снижением проницаемости мем-
браны. Эксперименты в течение более длительного времени показали,
что поглощение не является первичным механизмом толерантности
( Stokes, 1981). Исследования с помощью электронного микроскопа
позволили выявить медные комплексы в ядрах толерантных водорос-
лей ( Silverherg et al., 1976). Нетолерантные формы накапливают
медь сходным образом, но это сопровождается обширным повреждени-
ем мембраны. У чувствительных к металлам штаммов .диатомеи Di ato-
ms tenue свинец и медь включались в полифосфатные тельца, но
также происходило повреждение мембран митохондрий и вакуолей
( Sicko-Goad, Stoermer, 1979; Sicko-Goad, 1992).
В местах многих горнорудных разработок под действием окисля-
ющих серу бактерий возникают сильнокислые воды. Таким образом, с
загрязнением тяжелыми металлами бывает связана кислотность и вли-
яние этих двух факторов часто трудно разграничить. При исследова-
нии в Великобритании биотопов со значением pH ниже 3,0 и анализе
литературы по кислым ведам в Северной Америке ( Hargreaves
et al.1975) выявлено 40 видов водорослей, обитающих в таких уело-
439 '
ВИЯХ. Показано, BTgChlamydomonaB acidophila может расти при pH
2,0 (Cassin, 1974), a Chlamydomonas applanataH Euglena mutabi-
lis - при pH 1,3 (Hargreaves, Whitton, 1976). При низких значе-
ниях pH большинство тяжелых металлов находится в доступной для
водорослей форме, поэтому они обычно токсичнее в кислой среде
( Rai et al., 1981). Однако показано (Hargreaves, 1976), что ток-
сичность меди и цинка для синезеленой водоросли Hormidium rivu-
lare при pH 3,5 меньше, чем при pH 6,0.
Имеется много современных обзоров о кислотных дождях или
кислотных осадках в целом (Haines, 1981). Кроме снижения pH в
озерах, они могут повышать концентрации алюминия и тяжелых метал-
лов, в частности марганца, цинка и железа, которые вымываются из
отложений озер и зоны их питания. Сообщалось также об увеличении
концентрации свинца, никеля, меди и кобальта, но это объяснялось
осаждением твердых частиц, а не локальным вымыванием. В озерах с
pH 4,1-4,3, загрязненных медью и никелем, состав фитопланктона
не такой, как в расположенных рядом незагрязненных озерах, и схо-
ден с отмечаемым для кислых озер на расстоянии 80 км, в которых
отсутствуют тяжелые металлы ( Yan, 1979).
Подкисление озер вызывает снижение числа и разнообразия ви-
дов ( Schindler, 1980; Hendrey,1982; Eriksson et al.,1983), ХОТЯ
биомасса и первичная продукция могут остаться прежними. В трех
озерах гор Адирондак в США ( Hendrey, 1982) начинали доминировать
жгутиковые меныпего размера, что отчасти объясняется меньшей до-
ступностью фосфата. Изменения в видовом составе отмечались также
для подкисленных озер Швеции ( Eriksson et al., 1983), причем
основная часть биомассы фитопланктона приходилась здесь на кисло-
тостойкие и олиготрофные динофлагеллаты Gymnodinium uberrima и
Feridinium inconspicuum. Известкование озер повышало pH, но ха-
рактер фитопланктона в первый год оставался ацидофильным вслед-
ствие малого содержания биогенов. В последующие годы его видовой
состав начинал напоминать известный для соседних олиготрофных
озер с pH 6,0, где доминировали диатомеи и золотистые водоросли.
Увеличение биомассы бентооных и эпифитных водорослей в соче-
тании с уменьшением видового разнообразия наблюдалось при подкис-
лении многих озер ( Lazarek, 1982); это результат снижения интен-
сивности микробного разложения и выедания беспозвоночными.
Если кислотность не создает проблем в морской среде, в при-
брежных и эстуарных водах может происходить локальное повышение
концентрации тяжелых металлов. В ряде широкомасштабных экспери-
440
ментов в мезокосмах исследовано влияние этих металлов на морской
фитопланктон. Добавление меди в дозах 10 мкг/л и 50 мкг/л ( Tho-
mae et al., 1977а) через три дня снижало скорость фотосинтеза
до 12 и 2% от контроля соответственно, но наблюдался постепенный
возврат к исходному состоянию вследствие развития толерантных к
меди микроскопических жгутиковых. Сходные результаты получены с
ртутью в1 концентрациях I мкг/л и 5 мкг/л: продуктивность падала,
после чего восстанавливалась в ходе изменения видового состава
( Thomas et al., 1977b). Томас С соавт. ( Thomas et al. 1980) ис-
следовали влияние смесей металлов при относительных концентраци-
ях, сходных с отмеченными в умеренно-загрязненных эстуариях. Су-
щественных изменений не наблюдалось, но,когда общая концентрация
повышалась, снижалась скорость фотосинтеза. Удаление отдельных
металлов из смеси показало, что ингибирующее действие обусловле-
но медью и ртутью.
Морские бентосные водоросли также способны поглощать тяже-
лые металлы. Цинк абсорбируется Fucus vesiculosus и'может концен-
трироваться более чем в 60 000 раз, причем более высокое его со-
держание наблюдается в более старых тканях. Он прочно связан и
не может легко вымываться из водорослей (Bryan, 1969; Bryan,
Hummerstone, 1973). Концентрации меди, мышьяка, свинца, цинка и
серебра (но не других тяжелых металлов) в тканях р.vesiculosus
( Luoma et al., 1982) коррелируют с их концентрациями в донных
отложениях, а не в морской воде. Металлы могут проходить через
пищевую цепь; так, свинец из Egre^is 1аеу1жаЬаобнаружея в расти-
тельноядном моллюске морское ушко (Stewart, Schitz-Baldes, 1976).
Много экспериментов по выяснению устойчивости бентосных водо-
рослей к тяжелым металлам проведено на обрастающих суда и толе-
рантных К меди штаммах Ectocarpus siliculosusf Phaeophyceae)
(Russell, Morris 1970). Толерантность, по-видимому, обусловле-
на исключением ( Hall et al., 1979; Hall, 1981), а не детоксика-
цией, поскольку чувствительные штаммы аккумулируют в несколько
(до семи) раз больше меди, чем толерантные. Внеклеточные хелаты
не образуются, и медь не связывается с клеточной стенкой, т.е.
толерантность является следствием снижения мембранной проницаемо-
сти. И напротив, ряд устойчивых видов бентосных диатомей (Daniel ,
Chamberlain, 1981) поглощает и детоксицирует медь, локализуя ее
в клетке в виде нерастворимых медно-полифосфатных телец.
441
Тепловое загрязнение
Загрязнение нагретыми стоками встречается в реках, озерах,
эстуариях и в меньшей, но все возрастающей мере - в морских при-
брежных водах. Хотя многие предприятия возвращают в окружающую
среду воду с повышенной темперачурой, основной источник ее - теп-
лоэлектростанции, использующие воду для охлаждения конденсаторов
и обычно сбрасывающие ее в водоемы при температуре на 10 - 15°0
выше окружающей.
Разнообразие фитопланктона, как правило, уменьшается в во-
дах, нагретых выше 30°С, причем там, где температура длительное
время превышает это значение, доминируют синезеленые водоросли.
В диапазоне от 20 до 30°0 могут увеличиваться как разнообразие,
так и первичная продуктивность. В морских прибрежных водах какое-
либо долговременное воздействие теплых стоков на фитопланктон-
ные сообщества практически не прослеживалось ( Langford, 1983).
Механическое повреждение фитопланктона при проходе через си-
стемы охлаждения ( Kreh, Dewort, 1976), по-видимому, минимальное;
большинство исследований этих вод показывает незначительные изме-
нения плотности клеток и концентрации пигментов ( Langford, 1983).
Однако окружающая температура иногда отрицательно влияет на фото-
синтез засасываемого в трубы планктона ( Seilner et al., 1984).
Весной его интенсивность при выходе из системы была выше, чем
при входе в нее, а летом наблюдалась обратная картина. Охлаждаю-
щие воды часто хлорируют для предотвращения обрастания труб бес-
позвоночными и образования бактериальной слизи; это приводит к
снижению фотосинтетической активности засасываемого фитопланкто-
на ( Davis, Coughlan, 1978). Она сокращается на 30% при концентра-
ции остаточного хлора 0,4 мг/л и полностью подавляется при
0,8 мг/л. Между температурой и хлором наблюдается синергизм
(Eppley et al., 1976).
Пресноводные бентосные и эпифитные водоросли по своей реак-
ции близки к фитопланктону. В целом умеренное увеличение темпера-
туры приводит к росту продуктивности и биомассы, хотя видовое
разнообразие может снижаться ( langford, 198З). В одном из озер
в Канаде нагретые стоки обеспечивали существование свободной ото
льда акватории, позволяя диатомеям, эпифитно растущим на Scirpus
validus. развиваться на протяжении всей зимы и весны; летом они
замещались нитчатыми зелеными водорослями. Эти стоки были впос-
ледствии отведены от озера, и за год в прежней зоне их влияния
442
водорослевое сообщество вернулось к исходному состоянию ( Hick-
man, 1982): возвратились ранее вытесненные виды диатомей, а
обилие нитчатых зеленых водорослей Oedogonium, Spirogyra и
Cladophora уменьшилось»
Крайний пример теплового загрязнения ( Tison et al., 1981) -
неравномерный сброс воды температурой до 45°С.При этом развива-
лась водорослевая пленка, состоящая прежде всего из синезеленых
водорослей из родов Fiacherella И PhormidiumH сохранявшаяся,
когда температура выравнивалась с окружающей. Оптимальная для
роста температура оставалась у водорослей близкой к верхней гра-
нице температур, воздействию которых они подвергались, - реакция,
сходная с указывавшейся для альгофлоры горячих источников ( Воу-
len, Brock, 1973).
Морские водоросли-макрофиты также местами испытывают влия-
ние теплых стоков. В Терки-Пойнт (Флорида) отмечено ( Thorlaug
et al., 1973) Совращение численности Halimeda И Panicillua
(chlorophyceae) там, где температура зимой на 3-5°С превышала
окружающую. Освободившийся субстрат впоследствии-заселялся сине-
зелеными водорослями. У подверженного воздействию теплых стоков
побережья шт. Мэн (США) обнаружены незначительные общие измене-
ния сообществ литоральных водорослей-макролитов ( Vadas et al.,
1976). Сначала рост фукусовых продолжался в некоторых умеренно
подогретых местах, но в некоторых других пунктах апикальный рост
Agcophyilum nodosum не восстановился через год после начала
прогревания. Базальные части таллома выживали, но прикрепление
их ослаблялось. В еще более холодном регионе, у Ньюфаундленда
(Канада), стоки пиковой электростанции, функционирующей лишь в
зимние месяцы, влияли на альгофлору только непосредственно в зо-
не сброса воды ( whittick, Hooper, 1979). Они способствовали
росту фукусовых и багрянки chondrug, а типичная зимняя флора с
Monogtroma, Uroapora И Bangia отсутствовала.
Механические помехи
Чрезмерный рост пресноводных водорослей иногда ограничивает
движение воды в реках и искусственных каналах. Морские водоросли
могут препятствовать прохождению воды через фильтры и обрастать
корпуса судов.
443
Помехи движению воды
Забивающие фильтры водоросли затрудняют работу систем водо-
снабжения ( Palmer, 1977). Вода обычно проходит через песчаные
фильтры для задержки твердых и коллоидных примесей. Во многих
местах она предварительно обрабатывается коагуляторами и из нее
до фильтрования удаляются осаждаемые вещества, однако фильтры
все равно засоряются и должны периодически промываться. Забиваю-
щие их водоросли обычно имеют жесткие стенки, нитчатую или коло-
ниальную организацию и могут образовывать слизь. Наибольшие про-
блемы создают диатомеи, прежде всего ВИДЫ Asterionella, Tabel-
laria И Synedra.
Морские макрофиты иногда мешают работе систем охлаждения
электростанций. На атомной электростанции Уилфа (остров Англси,
Великобритания) талломы бурой водоросли laminaria , оторванные
от субстрата во время осенних штормов, могут серьезно блокиро-
вать забор воды в такую систему (langford, 1983). Японская бу-
рая водоросль ^argae^s^^U^H^i^lJTl еабивает мелкие гавани и водные
пути на тихоокеанском побережье Северной Америки, куда она была
занесена с японскими устрицами (Soagel, 1956). Сейчас этот вид
уже встречается в проливе Па-де-Кале, где возникла угроза конку-
рентного вытеснения им местных макрофитов (parnhaffl et al. 1973).
Codium fragile (Chlorophyceae) появился на атлантическом побе-
режье Северной Америки в 1950-х гг. Предполагают, что он занесен
на раковинах устриц из Европы (Churchill, Moeller, 1972). Ныне
вид распространился от Мэна до Нью-,Вжерси (Malinowski, Ramus,
1973), нанося ущерб устричным банкам к юту от полуострова Кейп-
Код; он предпочтительно поселяется на раковинах моллюсков и при-
водит к их отрыванию от субстрата.
Оба последних вида - примеры интродукции в районы, где, по-
видимому, было слишком низким давление конкуренции и выедания.
Они служат предостережением о последствиях непродуманного заво-
за в новые биотопы "экзотических" форм; следует, например,
очень осторожно отнестись к предложению об интродукции гигант-
ской ламинариевой водоросли масгосузИа pyrifera в европей-
ские ВОДЫ (Boalch, 1981).
Обрастание судов
На современных судах водоросли - основная групп» организ-
444
мов-обрастателей ( Evans, 1981). Увеличение трения о воду, кото-
рое снижает скорость супертанкера всего на доли узла, может повы-
сить ежегодные эксплуатационные расходы на сумму свыше 100 000
долл. США (Evans, 1981). Основные растительные обрастатали -
диатомеи, нитчатые зеленые и бурые водоросли, чаще всего виды
Enteromorpha (Chlorophyceae) и Entocarpus (Phaeophyceae), способ-
ные развиваться в широком диапазоне солености и температуры. Они
также обладают эффективными механизмами прикрепления спор и спо-
собностью к регенерации таллома из базальной части после неполно-
го удаления.
Стандартные предохраняющие от обрастания краски содержат
купорос или другие соединения меди и обычно составляются таким
образом, чтобы ежедневно из них выделялось 10 мкг ионов меди на
I см^ поверхности. Толерантность к меди у водорослей-обрастателей
разная (Evans, 1981). Устойчивые к меди виды Enteromorpha ста-
ли основной цроблемой при эксплуатации крупных танкеров в 1950-х
гг., но добавление в краски оловоорганических соединений позволи-
ло ее решить. Другие виды, например uiothrix и Ectocarpus,
не столь чувствительны к этим веществам ( Miller, Evans, 1980).
По-видимому, современные сополимерные краски с токсичными соеди-
нениями, включенными в водорастворимую метакрилатную основу, име-
ют большое будущее, поскольку кроме выделения токсичных ионов
происходит постоянная эрозия самой поверхности, т.е. покрытие эф-
фективно самополируется.
Неподвижные конструкции: нефтяные платформы и преобразователи
тепловой энергии в открытом море
Беспокойство может причинять и рост водорослей на нефтяных
платформах в открытом море (Moss et al., 1981), особенно в бо-
лее холодных водах с достаточным количеством питательных веществ
и слабой конкуренцией со стороны сидячих беспозвоночных. Водорос-
ли-макрофиты увеличивают гидродинамическую нагрузку на структур-
ные компоненты и метают осматривать критические зоны, в частнос-
ти сварные швы. Они способствуют коррозии стали, особенно там,
где их толстые слои создают бескислородные условия. И напротив,
тонкие слои фотосинтезирующих водорослей могут придавать некото-
рую устойчивость к коррозии ( Terry, Edyvean, 19В1).
Один из возможных методов получения электричества в тропи-
ках - преобразование тепловой энергии в открытом море благодаря
445
различию температур между поверхностным и расположенными ниже
Золее холодными слоями воды. Основная проблема при эксплуатации
таких систем - обрастание поверхностей теплообменников водорос-
ЛЯЫИ ( Thorhaug, Marcus, ^981).
Использование водорослей
Питание человека
Морские водоросли - постоянный элемент пищевого рациона на
побережье Китая с 850 г. до н.э. ( Waaland, 1981); сейчас они
используются в пищу преимущественно на Востоке и островах Тихого
океана. Известно (Chapman, chapman, 1980) 160 видов съедобных
морских макрофитов (25 Chlorophyceae,54 Phaeophyceae и 81 Rhodo-
phyoeae). Наибольшее значение имеют представители родов:
Chlorophyceae: Monostroma, Caulerpa, Enteromorpha, Ulva;
Paeophyoeae: Laminaria, Undaria, Alaria, Eisenia, Ecklonia;
Rhodophyceae: Porphyra, Palmaria, Gracilaria, Gelidium.Eucheuma.
Доти ( Doty 1979) оценивает ежегодную стоимость пищевых во-
дорослей более чем в I млрд. долл. США; преимущественно это виды
трех родов: Porphyra (500 млн. Долл.), Laminaria И Undaria.
В Северной Атлантике основными потребляемыми водорослями помимо
багрянок Gig art ina и Chondrus, местами используемых для приго-
товления желе, являются Palmaria на востоке Канады и в Ирландии
и Porphyra на юге Уэльса. Обе последние водоросли считаются ла- .
комством, а не одним из главных пищевых продуктов.
Основанием для использования в пищу могут служить питатель-
ность, вкус, цвет и консистенция. Структурные углеводы морских
макрофитов в основном не усваиваются, но некоторые растворимые
углеводы включаются в обмен веществ. Содержание белка во многих
съедобных водорослях составляет 20 - 25% сухой массы. Морские
макрофиты - прекрасный источник витаминов (Kanazawa, 1963), в
том числе витамина С на уровне, эквивалентном его содержанию в
плодах цитрусовых, витаминов A, D , Bj, В-^, К, рибофлавина, ниа-
цина, пантотеновой и фолиевой кислот. В них содержатся также все
необходимые для человека микроэлементы ( Yamamoto et al., 1979).
Приятный вкус Porphyra связывают с наличием изофлоридиозидов и
свободных аминокислот (McLachlan et al., 1972).
В пищу человеком используются и водоросли-микрофиты. Qhio-
yeiia - единственная микроскопическая зеленая водоросль, регу-
446
лярно включаемая в рацион ( Boeder, 1980), но потребляется и не-
сколько видов синезеленых водорослей. Наземные виды Uostoc едят
в Китае и Юкной Америке (Johnston, 1970). Предполагается (i'arrer
1966), что основную пищу ацтеков в доколумбовские времена сос-
тавляли лепешки из высушенной синезеленой водоросли Snirulina.
Ее собирают и едят также в районе озера Чад в Северной Африке
(Leonhard, Compere, 1967); содержание белка в ней более 60%,
сейчас она в широких масштабах разводится.
Корма для животных
Отмечалось, что дикие животные (лисы, олени, кролики и мед-
веди) едят морские водоросли; в Европе и Северной Америке круп-
ный рогатый скот, овцы и лошади часто пасутся на литорали (chap-
man, chapman, 1980). Морские макрофиты можно непосредственно
давать скоту как корм ( Hailsson, 1964), но при промышленном под-
ходе их собирают, высушивают и измельчают в муку, которую приме-
няют в виде добавки к кормам. В основном используются бурые водо-
росли laminaria, Ascophyllum и Fgcug ; ежегодно их стоимость
оценивается в 10 млн. долл. США (Jensen, 1979 )ч. Питательность
морских водорослей для животных в целом такая же, как и для чело-
века. Особенно важны микроэлементы, в частности для скота, пасу-
щегося на бедных лугах. При кормлении домашней птицы водорослями
их каротиноиды придают более темную окраску яичному желтку.
Фикоколлоиды
Издавна морские макрофиты использовались для получения со-
ды и иода, но впоследствии им на смену пришли минеральные ресурсы
и химические процессы. В настоящее время основными химическими
соединениями, выделяемыми из водорослей в промышленном масштабе,
являются фикоколлоиды.
Фикоколлоиды широко используются в пищевой, косметической и
фармацевтической отраслях как эмульгаторы и желирующие агенты.
Как и их аналоги, экстрагируемые из высших растений, пектины, они
представляют собой компоненты клеточной стенки, растворимые в го-
рячей воде и относительно нерастворимые в холодной. В последнее
время появилось несколько обзоров о структуре, биосинтезе и функ-
ции фикоколлоидов ( McCandless, Craigie, 1979; Percival, 1979;
Larsen, 1981; McCandless, 1981). Промышленным путем производят-
447
ся три основных типа фикоколлоидов: альгинаты, каррагинаны и ага-
ры. Общая стоимость их ежегодно превышает 200 млн. долл. США.
Альгиновую кислоту и ее соли (альгинаты) получают из бурых
водорослей, в основном относящихся к родам MacrocyatiB. Lamina-
ria и Ascophyllum. Важными потенциальными источниками их являют-
ся также представители родов Ecklonia и Durvillaea. Общее миро-
вое производство составляет 15-17 тыс. т при стоимости свыше
100 млн. долл. США ( Jensen, 1979).
Альгиновая кислота - сополимер маннуроновой и гулуроновой
кислот. Показано (Haug et al., 1966, 1967), что встречаются три
типа сочетания этих мономеров: блоки полиманнуроновой кислоты,
блоки политулуроновой кислоты и блоки из чередующихся маннуроно-
вой и гулуроновой кислот. Эти полимеры имеют различные структур-
ные свойства (McCandless, 1981) и встречаются в разных частях
растения. Концептакулы и растущие верхушки Agcophyllum богаты
полиманнуроновыми альгинатами, а в более зрелых тканях преоблада-
ют полигулуроновые блоки ( Haug et al., 1974). Общее содержание
альгинатов варьирует в зависимости от части растения и сезона. В
восточной Канаде показано (Cardinal, Breton-Provencher, 1977),
что максимум его у Laminaria longicruris .приходится на сентябрь.
Для L.longicruris установлено, что генетическая составляющая
дисперсии в изменчивости альгинатов незначительна ( chapman, Doy-
le, 1978).
Laminaria и Ascophyllum собирают вручную с берега или с
лодок. Гораздо более крупный Macrocystis с плавающими пластина-
ми убирается механически с помощью вращающихся или движущихся
взад-вперед ножей, установленных на баржах.
Основные этапы промышленного производства альгинатов ( Chap-
man, chapman, 1980) - вымывание кислотой, разрушение щелочью,
промывание, отбеливание, сушка и измельчение для получения неочи-
щенного экстракта альгината кальция, который в свою очередь пре-
вращают в свободную кислоту или другие соли. Соли щелочных метал-
лов растворимы в воде, а .двух- и трехвалентных металлов - относи-
тельно нерастворимы. Так, альгинаты натрия и калия дают растворы
с высокой вязкостью, а соли поливалентных ионов имеют тенденцию
к образованию гелей. Растворимые соли используются в текстильной
и бумажной промышленности для придания глянца и проклеивания; в
производстве красок, пищи, косметики и лекарств - как обеспечи-
вающие вязкость агенты для суспендирования несмешивающихся с во-
дой веществ. Приблизительно половина их в США применяется для
448
стабилизации молочных продуктов. Желирующие альгинаты, в частно-
сти альгинат кальция, используются в производстве гелей, целлофа-
ноподобных пленок и резины, в зубоврачебной практике - для приго-
товления форм, а в мелицине - как безжировая смазка. Из высушен-
ных альгинатов можно изготовить пряжу, а из нее выткать специаль-
ные ткачи. Они растворимы в щелочах, а волокна альгината натрия -
в воле, однако такие материалы устойчивы к обычному микробному
разрушению и достаточно огнеупорны.
Каррагинаны - фикоколлоиды, получаемые из представителей се-
мейств красных водорослей Hypneaceae, Phyllophoraceae, Solieria-
сеае и Gigartinaoeae. В промышленном масштабе используют два
основных источника: chondrue crispua в Северной Атлантике (в основ-
ном шт. Мэн, США,и приморские провинции Канады) и различные виды
Eucheuma на Филиппинах. Меньшее значение имеют виды Gigartina,
Iridaea и Hypnea, а Purcellaria и Phyllophora дают структур-
но и функционально сходные соединения. Ежегодное производство
каррагинанов составляет 10 000 т (Моза, 1978).
Каррагинаны - полимеры сульфатированной или пируватирован-
ной галактозы и 3,6-ангидрогалактозы (McCandieaa, 1981). Описа-
но два основных их вида; растворы ЗЕ-каррагинана образуют гель в
присутствии калийных солей, а растворы ^-каррагинана - нет. Ос-
новное химическое различие между ними в том, что желирующие типы
несут сульфатные группы в положении С-4 галактозных остатков,
тогда как нежелирующие их не имеют (McCandieaa, Craigie, 1979).
Показано (McCandieaa et al., 1973; Chen et al., 1973), ЧТО 3E-
каррагинан образуют в основном гаметофиты, а 'Л-каррагинан наи-
более обилен у спорофитов. Такое чередование наблюдается у пред-
ставителей Gigartinaoeae (McCandieaa, 1978), но, видимо, отсут-
ствует у Eucheuma (Doty, Santoa, 1978) И Hypnea (McCandieaa,
1978).
Сырье получают, собирая выброшенные на берег талломы, сры-
вая их вручную на литорали или с лодок на сублиторали. Eucheuma
на Филиппинах в основном культивируется, а не собирается в при-
родных местообитаниях.
Каррагинан используется в пищевой промышленности как эмуль-
гатор, особенно молочных продуктов, как клеящее вещество в тек-
стильном и кожевенном производствах и как эмульгатор в фармацев-
тике.
Агар - малайское слово, означающее желирующее вещество, эк-
страгируемое из Eucheuma. которое по иронии судьбы оказалось
449
каррагинаном. Агар в современном понимании получают из багрянок
Gelidium и в меньшей мере из Gracilaria, Pterocladia, Acantho-
peltie И Ahnfeltia (Chapman, Chapman, 1980). Ежегодное цроИЗВОД-
ство - 10 000 т стоимостью примерно 50 млн. долл. США (Моав,
1978).
Подобно каррагинану, агар в основном состоит из сульфатиро-
ванной и пируватированной галактозы. Выделены (Araki, 1966) два
его компонента: нейтральная желирующая фракция, агароза,и анион-
ная фракция, содержащая сульфатную и пируватную группы, агаропек-
тин. По-видимому, речь идет о сложном спектре соединений от не-
сульфатированных до сильно сульфатированных и пируватированных
полигалактанов (McCandless, 1981).
Водные растворы 1-2% лучших по качеству агаров образуют
цри комнатной температуре плотные гели, которые разжижаются цри
нагревании. В основном они используются в бактериологии и гисто-
логии для приготовления культуральных сред. Чистая агароза в на-
стоящее время применяется как гель для электрофореза и хромато-
графии (Pluzek, 1981).
Водоросли как удобрения
Использованию водорослей в качестве удобрений посвящен це-
лый ряд обзоров (см., например, Chapman, Chapman, 198O). Их тал-
ломы, выброшенные прибоем, применялись для этой цели во всех рай-
онах с прибрежными сельскохозяйственными угодьями (Booth, 1965).
Морские макрофиты содержат достаточные количества калия и азота,
но бедны фосфатами. Для сравнения можно указать, что в навозе
втрое меньше калия, примерно столько же азота и втрое больше фос-
фатов. Морские макрофиты богаты также микроэлементами ( Stephen-
son, 1974) и мотут содержать гормоны и регуляторы роста (Blun-
den, 1977; wildgoose et al., 1978). Удобрения из водорослей име-
ют то преимущество, что в нем нет диаспор сорняков и патогенных
грибов. В Европе и Северной Америке отложения известковых водорос-
лей (в основном из родов Lithothamnion И Phymatolithon) применя-
ются для снижения кислотности почвы ( Biundon et al., 1975). Азот-
фиксирующие синезеленые водоросли широко используются на рисовых
полях в тропиках вместо азотных удобрений ( Watanabe, 1975).
Выпускается ряд продажных жидких удобрений, полностью или
частично производимых из водорослей и предназначенных для исполь-
зования в интенсивном садоводстве и тепличном хозяйстве (Stephen-
450
sen, 1981; Povolny, 1981), где они, как сообщалось, повы-
шают всхожесть семян, урожайность, выносливость, устойчивость к
болезням, а также срок хранения продукции.
Водоросли в медицине
Водоросли широко использовались в народной медицине примор-
ских стран как глистогонные и анестезирующие средства, .для изго-
товления мазей, а также лечения кашля, ран, подагры, зоба, гипер-
тонии, венерических заболеваний, рака и ряда других болезней
(Норре 1979; Chapman, Chapman, 1980; Stein, Borden, 1984).
Как характерно вообще для народных средств, некоторые из них бес-
полезны, но основу других действительно составляют биологически
активные соединения. К последнему случаю относятся применение
багрянки Digenia simplex , содержащей сильнодействующее глисто-
гонное вещество, каининовую кислоту, а также йода из видов Lami-
naria для предотвращения зоба. Витамины и минеральные вещества
морских, водорослей потенциально важны и в профилактике других бо-
лезней, связанных с неполноценным рационом.
Неощищенные экстракты из водорослей многих видов содержат
вещества с антибиотическими свойствами по отношению к грибам,
бактериям и вирусам (Glombitza., 1979). К противовирусным соеди-
нениям, обнаруженным у багрянок, относятся полисахариды, содержа-
щие В-гликозильные группы, которые, по-видимому, эффективны в
борьбе с вирусами герпеса ( Ehresmann et al., 1979). Они действу-
ют путем блокировки точек прикрепления вируса к клеточной мембра-
не. Многие морские макрофиты содержат стеролы и родственные им
соединения, являющиеся антагонистами холестерола в организме мле-
копитающих; они могут снижать повышенное кровяное давление,
связанное с атеросклерозом (chapman, Chapman, 1980). Сделан вы-
вод ( Weinheimer, Karns, 1974), что многие водорослевые экст-
ракты "удивительно неэффективны" в опытах по выяснению противора-
ковой активности,, однако Ямамото и др. (Yamamoto et al., 1982)
показали, что экстракты из Sargassum и Ъат1паг1аподавляют рост
саркомы и лейкемических клеток у мышей.
Ряд специфических соединений водорослевого происхождения
был очищен, охарактеризован и экспериментально используется в ме-
дицине. Сакситоксины, образуемые динофлагеллятами, вызывающими
паралитическое отравление моллюсками, используются в нейробиоло-
гических исследованиях (Schantz, 1981). Каининовая кислота также
451
нейротоксична и вызывает разрушение дендритов; она полезна в свя-
зи со своей способностью имитировать симптомы хореи Хантингтона
(Coyle et al., 1979) и используется также при изучении эпилеп-
сии ( Pisa et al., 1980). Морские водоросли образуют широкий на-
бор гемагглютининов ( Rogers et al., I960); один из них, получен-
ный из Ptilota piumosa, специфичен по отношению к группе В кро-
ви человека ( Rogers, Blundon, 1980), следовательно применим в
диагностике.
Каррагинан можно использовать, чтобы вызывать язву желудка
и кишечника у животных для последующего испытания противовоспали-
тельных соединений (Di Rosa, 1972). Сообщалось, что он не дейст-
вует таким образом на человека ( Grasso, Sharrat, 1973) и исполь- .
зовался при лечении пептических язв ( MacPherson, Pfeiffer, 1976).
Способность каррагинанов и альгинатов образовывать соли с метал-
лами предполагает возможность их применения как нетоксичных хела-
тирующих агентов при лечении отравлений тяжелыми металлами и ра-
дионуклидами ( Tanaka, Stara, 1979).
Высушенные "стебли" Laminaria (называемые в литературе ла-
минариевыми тампонами) используются как расширители шейки матки
в акушерстве ( Feochari 1979; stein, Borden, 1984). Введенные в
шейку матки, они медленно набухают, впитывая окружающую жидкость
и давая требуемое расширение.
Возможности использования водорослей в медицине еще требуют
своего изучения. Широкий набор соединений, обнаруженных у тропи-
ческих морских макрофитов (Morris, Penical, 1982), открывает в
этом плане особенно широкие перспективы.
Культивирование водорослей
Крупномасштабное промышленное культивирование водорослей для
употребления в пищу особенно развито на Востоке, в частности в
Японии и Китае (современные обзоры см. Mathieson, 1982; Tseng,
1981). Разводятся зеленые водоросли Enteromorpha, Monostroma
И Ulva, бурые водоросли Laminaria, Undaria, Nemacystis, Clado-
siphon и Endarachne (Saito, 1982) и многочисленные багрянки, из
которых экономически наиболее важны виды Porphyra (Tseng, 1981;
Kite, 1982).
Имеется хороший обзор основной части работ по массовому куль-
тивированию ОДНОКЛеТОЧНЫХ Водорослей ( Shelef, Boeder, 1980). К
микроскопическим видам, выращиваемым для потребления человеком,
452
относятся Chlorella (Chlorophyceae) и Spirulina (Cyanophyceae ),
a Scenedeamua используетоя на корм домашним животным. Аквакуль-
тура моллюсков и рыб также зависит от постоянного широкомасштаб-
ного снабжения водорослями-микрофитами; чаще всего здесь исполь-
зуются роды Chlorella. а также Monochrysis и Isochrysis ИЗ
Chrysophyceae, Chaetoceros И Skeletonema Из Bacillariophyceae .
Культивирование водорослей-макролитов
Несколько видов Porphyra выращивается на продажу в Шонии
и Китае; из них наиболее важен P.yezoensis, но P.tenera име-
ет более высокое качество, хотя и меньшие размеры. Разведение на-
чалось в ХУЛ в., когда пучки талломов размещали на литорали, да-
вая возможность водорослям прикрепляться естественным путем. В
начале нашего столетия стали применять плавающие бамбуковые .маты
и сети. Когда было впервые показано ( Drew, 1949), что сверлящая
раковины нитчатая водоросль Conohocells - фаза жизненного цикла
Porphyra, стало возможным развитие коммерческого культивирова-
ния с использованием искусственно засеянных сетей. Производство
Conchocelis ведется в емкостях с раковинами устриц; затем засе-
янные раковины прикрепляются к сетям. В настоящее время наблюда-
ется тенденция к посеву на сети непосредственно конхоспор, обра-
зуемых свободноживущими особями Conchocelis (Ktto, 1982). Сети
затем помещают в океан. При литоральном разведении они прикреп-
ляются к шестам либо на постоянней высоте, либо так, что могут
подниматься и опускаться в соответствии о уровнем воды. Это пред-
отвращает высыхание при отливе, но позволяет сетям оставаться на
поверхности воды во время прилива. Шире всего применяется метод
культивирования на сетях, прикрепленных к плотам, которые мотут
периодически подниматься над поверхностью воды (талломы растут
быстрее без периодического всплытия, но тогда они восприимчивее
к болезням и имеют более низкое качество). Когда сети покрывают-
ся молодыми пластинами Porphyra, их можно извлечь из моря, час-
тично высушить и хранить при температуре -20°С; оттаяв, водоросль
возобновляет рост. Хранение в замороженном состоянии позволяет
получать несколько урожаев за оезон и подстраховывает от неурожая.
Зрелые водоросли механически удаляются и отмываются от сетей, а
затем измельчаются, прессуются в пластины и высушиваются.
Другие водоросли-макрофиты, выращиваемые в широких масштабах
для непосредственного потребления человеком, - Laminaria и Unda-
453
ria(Tseng, 1981). Хотя Laminaria .japonica используется в
Китае в пищу свыше 100 лет, она не является там местным видом
и импортировалась из Японии и Кореи. Случайно с морокими суда-
ми она была занесена в северную часть Желтого моря в 1927 г. и
после этого стала сознательно вводиться в соотав морской альго-
флоры Китая. В настоящее время плоты, на которых выращивается
L. japonic а. покрывают общую площадь более 18 000 га
и дают ежегодно свыше миллиона тонн сырой продукции.
В Китае гаметофиты и молодые споровые проростки ламинарий
получают в питомниках с искусственным охлаждением воды до 8-
Ю°С. Зооспоры, образуемые родительскими пластинами, оседают на
шкурах, натянутых на рамах, в июле и остаются в питомнике до ок-
тября, когда температура воды в море падает ниже Ю°С. Рамы с
молодыми спорофитами высотой 1-2 см продаются фермерам для вы-
садки в море. Через 1-2 месяца, когда водоросли достигают высо-
ты 10-15 см, их снимают с рамок и прикрепляют к тросам, спускае-
мым с плотов. Талломы T«minaria собирают через 6-7 меся-
цев при длине 3-4 м. Для роста водоросль нуждается в удобрени-
ях. В период быстрого ее развития вокруг плотов разбрызгивают
5-10%-ный раствор сульфата аммония. За сезон вносится пример-
но 2 т удобрений на гектар; это позволяет культивировать Lami-
naria .japonica на ранее неплодородных акватори-
ях.
Undaria pinnatifida - более теплолюбивый ВИД,
выращиваемый примерно так же, как L. japonica. причем
часто в смешанной культуре. У Undaria вегетационный
период короче, и ее собирают в марте, до созревания
L. .japonica.
Тропическая багрянка Eucheuma широко используется
в пищу, а сейчас является и важным источником каррагинана. Ее
активно разводят на Филиппинах (Eucheuma spinosum и
E.cottonii; Doty, 1977; Tseng, 1981); свыше
Й00 ферм производят более 10 000 т сухой массы ежегодно. Водо-
росль размножается вегетативно и растет прикрепленной к шнурам
или сетям.
Способы культивирования водорослей в Северной Америке
(см. обзор Mathieson, 1932) сильно отличаются от при-
меняемых на Востоке. Здесь это производство находится в основ-
ном на стадии "опытных предприятий"; будучи технологически
454
более прогрессивным и менее трудоемким, оно еще не освоено в
промышленном масштабе. Хорошие перспективы имеет разведение
продуцента каррагинана iridaea (Jtaford., 1979).
В североамериканской аквакультуре морских макрофитов
главней упор делается на красные водоросли, дающие фикоколлои-
ды; ЭТО В ОСНОВНОМ Chondrus crispus И ВИДЫ Eucheuma.
Gracilaria. Hypnea. Gigartina. Gelidium и Pterocladia ,
Основной объем работ в Канаде и Новой Англии связан с Chondrus
crispus (Harvey,McLachlan, 1973). Вид успешно выращивает-
ся в сосудах в закрытых помещениях (Neish et al., 1977),
хотя этот способ в промышленном отношении считается менее пер-
спективным, чем разведение в приморских лагунах под открытым
небом ( Mathieeon 1982). Основная проблема при-культиви-
ровании водорослей - развитие сорной водорослевой растительно-
сти; новый подход к ее решению в аквакультуре Chondrus
контролируемое использование фитофагов ( Stacklock, Croft,
1981), избирательно выедающих нежелательные' виды. Химический
состав Chondrus зависит от доступности биогенов.
При больших дозах азота раст^ниярастут быстро и содержат мало
каррагинана, а при переносе их в необогащенную морскую воду
снижают концентрацию азота внутри таллома, но увеличивают про-
цент каррагинана. Это может иметь значение для обработки воды
после сбора урожая и дальнейшего культивирования в лагу-
нах.
Известны попытки генетического изменения водорослей
(van der Meer, 1983), но в основном улучшение культур
основано на селекции более перспективных клонов из диких попу-
ляций. Т4-И30ЛЯТ Chondrus (Neish, Fox, 1971) показал
шестикратное увеличение ежемесячного прироста, но для поддер-
жания максимальной скорости развития нуждался в удобрениях.
Получены (van dor Meer, 1983) многочисленные му-
танты продуцента агара Gracilaria tikvahlae , в том
числе с ускоренным ростом и лучшим качеством агара по сравне-
нию С ДИКИМИ типами ( Patwary, van der Meer, 1983).
кие генетические исследования весьма перспективны с точки зре-
ния выведения промышленных штаммов морских макрофитов.
455
Культивирование одноклеточных водорослей
Важность водорослей как корма в аквакультуре беспозвоноч-
ных и рыб часто недооценивается из-за уделения основного вни-
мания разводимым в этом случае животным (Shaw, 1982). Одшко
часто производство водорослей - наиболее дорогостоящая и трудо-
емкая часть таких систем. Наиболее широко применяются водоросли
в моллюсковых садках для кормления личинок. Существуют два ос-
новных метода: с использованием одновидовых культур, выращивае-
мых при определенных условиях, и с использованием морской воды,
которая фильтруется для удаления более крупных частиц и зоо-
планктона. Спонтанно развивающиеся водорослевые цветения обес-
печивают моллюсков кормом. Первый метод обеспечивает более
контролируемые условия, но второй значительно дешевле. Вариант
его использовали при разведении двустворчатых моллюсков на Вир-
гинских островах, где богатая питательными веществами глубин-
ная морская вода закачивалась насосами в мелкие бассейны и за-
севалась фитопланктоном, который массово размножался и скармли-
вался моллюскам»
Сходные культуры в бассейнах с удобрением или без него
используются и в других тропических областях. На Гавайях так
получают фитопланктон для выращивания устриц. Аналогичные сис-
темы дают корм при разведении креветок и других ракообразных.
Водоросли важны и в рыбоводстве. В этих случаях они используют-
ся как корм для зоопланктона, которым в свою очередь питаются
мальки.
Одноклеточные ВОДОРОСЛИ, например Chlorella И Spirulina
в широких масштабах культивируются в Мексике и Юго-Восточной
Азии для внутреннего потребления и экспорта, особенно в Япо-
нию. В Мексике Spirulina разводится в открытых пру-
дах общей площадью 900 та в Караколе близ озера Тескоко
(Durand-Chastel, 1980). Эту водоросль и Chlorella
выращивают в промышленных масштабах также на Тайване. В 1977 г.
заводы по производству Chlorella давали свыше 1000 т в
год продукции, преимущественно для диетических целей ( Soong,
1980).
456
Будущее разведения водорослей
В идеальной системе разведения водорослей в качестве удобре-
ния используются органические отходы и образуются различные ком-
мерческие продукты без (или почти без) возвращения в среду побоч-
ных соединений. Водоросли уже играют важную роль в системах очист-
ки сточных вод (Palmer, 1977). Экспериментальные поликультуры,
основанные на сбросах установок по вторичной обработке бытовых
стоков, созданы в Вудс-Холе (США.) (Ryther et al., 1979). Эти
сбросы смешиваются с морской водой и используются для выращива-
ния одноклеточных водорослей в прудах; развивавшиеся в результа-
те цветения Skeletonema costatum зимой И Phaeodact lum tricor-
nutum летом непрерывно обеспечивали кормом моллюсков. Затем
сточные воды поступают в контейнеры с образующими фикоколлоида
макрофитами Gracilaria tikvahiae и Agardhiella tenera , которые
извлекают оставшиеся питательные вещества. Проблемы возникают
из-за колебаний температуры, необходимости коррелировать рост
моллюсков с поступлением биогенов и загрязнения.моллюсков нежела-
тельными веществами, поступающими из канализации. Такие многови-
довые системы лучше приспособлены к климату с меньшими перепада-
ми температуры по сезонам. В настоящее время организовано эконо-
мически рентабельное производство ракообразных, рыб и Gracilaria
в изолированных от моря емкостях на Тайване (Chiang, 1981).
Исследуются также возможности культивирования водорослей
как энергетического сырья с целью получения метана путем фермен-
тации талломов ламинариевых (Flowers, Bryce, 1977). Для этого
предложено использовать крупные плоты, засеваемые Macrocystis
В небольшом масштабе ряд водорослей может выращиваться для полу-
чения специфических веществ, например глицерина и J5-каротина
(из Punaliella) (Ben-Amotz, Avron, 1980).
ЛИТЕРАТУРА
Гниловская М.Б. (1971). Ископаемые водоросли венда с Русской плат-
формы. Палеонтол. ж., 3, ТОТ-107.
Маслов В.П. (1963). Введение в изучение ископаемых харовых водо-
рослей. Труды Геологического института АН СССР, вып. 82, М.,
изд-во АН СССР, 104.
Abbott I.A.(1967). Liagora tanakai, a new species from southern
Japan.Bull. Jap. Soc.Phycol., 15. 32-37.
Abbott I.A., Hollenberg G.J.(1976). Marine Algae of California
Stanford University Press, Stanford.
Abbott I.A., Williamson E. (1974). Lima: An Ethnobotanical Study
of Some Edible Hawaiian Seaweeds. Pacific Tropical Botanical
Garden, Lawai, Kauai, Hawaii.
Abdel-Rahman M.H. (1982). The involvement of an endogenous cir-
cadian rhythm in photoperiodic timing in Acrochaetium aspara-
gopsis (Rhodophyta, Acrochaetiales). Br. Phycol. J., 17,
389-392.
Abe H., Uchiyama M., Sato R.(1972). Isolation and identification
of native auxins in marine algae. Agr.Biol.Chem., 36, 2259-
2260.
Abelard C., L Hardy-Halos MjT. (1975). Correlationsmorphogenes
entre les differents elements d’un meme cladome chez I1Apog-
lossum ruscifolium (Turner), J.Ag. (Delesseriaceae, Cerami-
ales). Bull.Soc. Phycol.Fr., 22, 111-119.
Adey W.H., Macintyer I.G. (1973)- Crustose coraline algae: a re-
evaluation in the geological sciences. Geol.Soc.Am.Bull., 84
883-904.
Ahbel D.E., Alexander A.H., Klein M.L. (1980). Protothecal ole-
cranon bursitis - a case and review of the literature. J. Bone
Jt.Surg. Am., 62. 835-836.
Ahmadjian V. (1981). Algal/fungal symbioses. Prog. Phycol,Res.,
1, 179-233.
458
Ahmadjian V., Hale M.E. (1973)- The lichens. Academic Press*
New York.
Ahmadjian V., Jacobs J.B. (1981). Relationship between fungus and
alga in the lichen Cladonia criststella Tuck. Nature, 289,
162-172.
Ajisaki T., Umezaki I. (1978). The life history of Sphaerotrichia
divaricata (Ag.) Kylin (Phaeophyta, Chordariales) in culture.
Jap. J.Phycol., 26, 53-59.
Alam M., Shimizu M. , Ikawa M. i Sasner J.J. (1978). Re-investiga-
tion of the toxins from the blue green alga, Aphanizomenon
flos-aquae, by a high performance chromatographic method. J.
Environ.Sci. Health, A13 (7), 493-499.
Alldredge A.L.(1981). The impact of appendicularian grazing on
natural food concentration in situ. Limnol. Oceanogr., 26,
247-257-
Allen M.B. (1971). High latitude phytoplankton. Ann.Rev. Ecol.
Systematics, 2., 261-276.
Allen M.M. (1968). Ultrastructure of the cell wall and cell di-
vision of unicellular blue-green algae. J.Bact., 96, 842-852.
Allison E.M., Walsby A.E. (1981). Potassium in the control of
turgor pressure in a gas-vacuolate blue-green alga. J.Exp.
Bot., 32, 241-249.
Allsopp A. (1969). Phylogenetic relationships of the Prokaryota
and the origin of the eukaryotic cell. New Phytol., 68, 591-
612.
Ambrose R.P., Nelson B.V. (1982). Inhibition of giant kelp rec-
ruitment by an introduced brown alga. Bot.Mar., 25, 265-267.
Amspoker M.G., Czarnecki D.B. (1982). Baoillariophyceae: intro-
duction and bibliography. In: Selected Papers in Phycology II
(eds. Rosowski J.R., Parker B.C.), pp. 712-718. Phycological
Society of America, Allen Press, Lawrence, Kansas.
Andersen R.A. (1982). A light and electron microscopical inves-
tigation of Ochromonaa sphaerocastis Matvienko (Chrysophy-
ceae): the statopsore, vegetative cell and its peripheral
vesicles. Phycologia, 21, 390-398.
Anderson D.M. (1980). Effects of temperature conditioning on de-
velopment and germination of Gonyaulax tamerensis (Dinophyce-
ae) hypnozygotes. J. Phycol., 16, 166-172.
Anderson D.M., Chisholm S.W., Watrass C.J. (1983). Importance of
life cycle events in the population dynamics of Gonyaulax
tamerensis. Mar. Biol., 76,179—202.
459
Anderson J.M., Barrett J. (1979). Chlorophyll-protein complexes ’
of brown algae: P700 reaction centre and light harvesting
complexes. CIBA Foundation Symposium, 61, 81-96.
Anderson L.W.J., Sweeney B.M. (1978). Role of inorganic ions in
controllung sedimentation rate of a marine centric diatom,
Ditylum brightwelli. J. Phycol., 14, 204-214.
Andrews H.T. (1970). Morphology and taxonomy of the euryhaline
chrysophyte Sphaeridiothrix compressa. J. Phycol., j5, 133-
136.
Antia N.J. (1976). Effects of temperature on the darkness survival
of marine microplanktonic algae. Microb. Ecol., 3, 41-54.
Antia N.J. (1977). A critical appraisal of Lewin’s prochlorophy-
ta. Brit. Phycol. J., 12, 271-276.
Antia N.J., Berland B.R., Bonin D. J., Maestrini S. Y. (1975). Com-
parative evolution of certain organic and inorganic sources
of nitrogen for phototrophic growth of marine microalgae. J.
Mar. Biol.Ass. U.K., 55, 519-539.
Araki 0. (1966). Some recent studies on the polysaccharides of
agarophytes. Proc. Tnt. Seaweed Symp., 5, 3-7.
Archibald Р.Л., Bold H. C. (1970). Phycological Studies XI. The
genus Chlorococcum Meneghini. Univ. Texas Pub., 70 15, 1-115»
Austin.
Arnold D.E. (1971). Ingestion, assimilation, survival and repro-
duction by Daphnia pulex fed seven species of blue-green al-
gae. Limnol. Oceanogr., 16, 906-920.
Atkinson A.W.Jr., Gunning B.E.S. , John P.C.C.(1972). Sporopolle-
nin in the cell wall Chlorella and other algae: ultrastruc-
ture, chemistry and incorporation of1^C acetate, studied in
synchronous cultures. Planta, 107, 1-32.
Atkinson M.J., Smith S.V. (1983). C:N:P rations of benthic marine
plants. Limnol. Oceanogr., 28, 568-574»
Augier H. (1976). Les hormones des algues. Etat actuel des connais-
sances. II. Recherche et tentatives d'identification des gib-
berellines, des cytokinines et de diverses autres substances
de nature hormonale. Bot. Mar., 19, 245-254.
Axler R.P., Gersberg R.M., Goldman C.R. (1980). Stimulation of
nitrate uptake and photosynthesis by molybdenum in Castle Lake,
California. Can.J.Fish.Aquat.Sci., 37, 707-712.
Bagnis R., Hurtel J;M., Chanteau S., Chungyem E., Inoue A.,
Yasumoto T. (1979). Le dinoflagelle Gambierdiscus toxicus
Adachi et Fukuyo: agent causal probable de la ciguatera. C.R.
Acad.Sci. Paris-D, 289. 671-674.
460
Baker A.N. (1967)» Algae from lake Miers, а .solar heated Antarctic
lake. N.Z.J.Bot., _5, 453-468.
Balakrishnan M.S., Chagule B.B. (1980). Cytology and life history
of Batrachospermum mahabaleehwarenais Balakrishnan et Chagule.
• Crypogam.Algol., .1., 83-97.
Bartlett R.B., South G.R. (1973). Observations of the life history
of Bryopsis hypnoides Lamour from Newfoundland. Acte Bot.
Neer., 22, 1-15.
Bauhin C. (1620). Prodromus theatri botanici. Basileae. (not seen
by the authors; quoted from Prescott, 1951).
Bazin M.J. (1968). Sexuality in a blue green alga: genetic recom-
bination in Anacystis nidulans. Nature, 218,282-283.
Be A., Caron D.A., Anderson O.R. (1981). Effects of feeding fre-
quency on life processes of the planktonic fonaminifer Globi-
gerinoides sacculefer in laboratory culture. J. Mar. Biol.
Asb. U.K., 61, 257-277.
Bean R.C., Hassid W.Z. (1955). Assimilation of 1^COg by a photosy-
nthesizing red alga, Iridophycus flaccidum. J. Biol.Chem.,
212. 411-425.
Beardall J.D., Mukerji D., Glover H.E. , Morris I. (1976). The
path of carbon in photosynthesis by marine phytoplankton. J.
Phycol., 17, 134-141.
Beeton A.M., Edmonston W.T. (1972). The eutrophication problem.
J.Fish.Res.Bd.Can., 29, 673-682.
Belchei' J.H. (1966). Prasinochloris веввШа gen. et вр. nov.,
a coccoid member of the Prasinophyceae, with some remarks of
cyst formation In Pvramiтопая. Br. Phycol.Bull., 3, 43-51.
Belcher J.H. (1968). The fine structure of Purcilia stigamastopho-
ra (Skuja) Korehikov. Arch. Mikrobiol., 60, 84-94.
Belcher J.H. (1969). A morphological study of the phytoflagellate
ChryBococcus rufescenc Klebs in culture. Brit. Phycol.J., 4,
105-117.
Belcher J.H., Swale E.M.P. (1967). Observations on Pteromonas
tenuis sp. nov. and P.angusta (Carter) Lemmermann (Chloro-
phyceae, Volvocales) by light and electron microscopy. Nova
Hedwiga, 1 3, 353-359»
Bellamy D.J., Clarke P.H., John D.M., Jones D., Whittick A., Dar-
ke T. (1967). Effects of pollution from the Torrey Canyon on
littoral and sublittoral ecosystems. Nature, 216. 1170-1173»
Ben-Amotz A., Avron M. (1980). Glycerol, JJ-carotene and dry meal
production by commercial cultivation of Dunaliella. In: Alga
461
Biomass: Production and Use (edg. Shelef G. , Boeder C.J.),
pp. 603-610. Elsevier/North-Holland, Amsterdam.
Berger-Perrot Y. (1981). Mise au point ear le problems concer-
nant I’Ulothrix epecioga (Carm, ex Harvey) Kutzing et 1'Upos-
-pora kornmannii Berger-Perrot. Etude comparee de la reproduc-
tion et du cycle de developpement des deux especes sur les
cotes de Bretange. Phycologia, 20, 147-164.
Berkaloff C., Rousseau B. (1979)» Ultrastructure of male gameto-
genesis in Fucus serratus (Phaeophyceae). J.Phycol., 15, 163-
173.
Beskow R. (1978). Silicosis in diatomaceous earth factory workers
in Sweden.Scand. J.Resp.Dis., .59, 216-221.
Bidwell R.G.S. (1983). Carbon nutrition of plants: photosynthe-
sis and respiration. In: Plant Physiology: A treatise, Vol.
VII; Energy and Carbon Metabolism (ed. Stewerd F.C.), pp.287-
457. Academic Press, Hew York.
Bidwell R.G.S., McLachlan J. (1985). Carbon nutrition of seaweeds:
photosynthesis, photorespiration and respiration. J. Exp.Mar.
Biol.Ecol., 86. 15-46.
Bidwell R.G.S., Craigie J.S., Krotkov G. (1958). Photosynthesis
and metabolism in marine algae. III. Distribution of photosyn-
thesis carbon from ^со in Fucus vesiculosus. Can.J.Bot.,
36, 581-590.
Biebel P. (1973). Morphology and life cycles of saccoderm desmids
in culture. Nova Hedwiga, 42, 39-47»
Bierman V.J.Jr. , Dolan D.M. , Stoermer E.F. , Gannon J.E., Smith
V.E. (1980). The development and calibration of a spatially
simplified multi-class phytoplankton model for Saginav Bay,
Lake Huron.Great Lakes Environ. Planning Study Contribution No.
33., Great Lakes Basin Commission.
Bird D.F., Kalff J. (1986). Bacterial grazing by planktonic lake
algae. Science, 231, 493-495.
Birmingham B.C., Colman B. (1979). Measurement of carbon dioxide
compensation points of freshwater algae. Plant. Physiol., 64,
892-895.
Bisalputra T. (1974). Plastids. Tn: Algal Physiology and Bioche-
mistry (ed. Stewart W.D.P.), pp. 124-160. Blackwell Scientific
Publications, Oxford.
Bliding C. (1957), Studies in Rhizoclonium. I. Life history of two
species. Bot. Not., IIP. 271-275.
Sliding C. (1963). A critical survey of European taxa in Ulvales.
462
Part I. Capsosiphon, Percursaria, Blidingia, Enteromorpha.
Opera Bot., 8, 1-60.
Sliding C. (1968). A critical survey of European taxa in Ulvales.
Part II. Ulva, Ulvarja, Monostroma, Kornmannia. Bot. Not., 121,
535-629.
Blunden G. (1977). Cytokinin activity of seaweed extracts. In:
Marine Natural Products Chemistry (ede. Faulkner D.J., Fenical
W.H.), pp. 337-343. Plenum Press, New York.
Blunden G. , Binns W.W., Perks F. (1975). Commercial collection
and utilization of maerl. Econ. Bot., 29, 140-145*
Boalch G.T. (1981). Do we really need to grow Macrocystis in
Europe? Proc. Int^eaweed . Symp., 10, 657-667.
Bold H.C. (1973). Morphology of Plante, 3rd Edition. Harper and
Bow, New York.
Bold H.C., Wynne M.J. (1978). Introduction to the Algae. Struc-
ture and Reproduction. Prentice-Hall, Englewood Cliffs, New
Jersey.
Bold H.C. , Wynne M.J. (1985). Introduction to the Algae: Struc-
ture and Reproduction, 2nd Edition. Prentice-Hall, Englewood
Cliffs, New Jersey.
Bolton J.J. (1983). Ecoclinal variation in Ectocarpus siliculosus
(Phaeophyceae) with respect to temperature growth optima and
survival limits. Mar. Biol., 73, 131-138.
Bolwell G.P., Callow J.A., Callow M.E. (1977). Cross-fertiliza-
tion in fucoid seaweeds. Nature, 268, 626-627.
Bolwell G.P., Callow J.A., Callow M.E., Evans L.V. (1979). Fer-
tilization in brown algae. II. Evidence for lectin-sensitive
complimentary receptors involved in gamete recognition in
Fucus serratus. J.Cell Sci., 36 , 19-30.
Bolwell G.P., Callow J.A., Evans L.V. (1980). Fertilization in
brown algae. III. Preliminary characterization of putative
gamete receptors from eggs and sperm of Fucus serratus. J.
Cell Sci., 43, 209-224.
Boney A.D. (1974). Aromatic hydrocarbons and the growth of mari-
ne algae. Mar. Poll.Bull., _5, 579-585.
Boney A.D., Corner E.D.S. (1962). On the effects of some carci-
nogenic hydrocarbons on the growth of sporelings of marine
red algae. J.Mar.Biol.Ass. U.K., 42, 579-585.
Boney A.D., Green J.C. (1982). Prymnesiophyceae (Haptophyceae):
introduction and bibliography. In: Selected Papers in Phyco-
463
logy II (ede. Rosowski J.R., Parker В.C.), pp. 705-711» Phyco-
logical Society of America, Allen Press, Lawrence, Kansas.
Bonneau E.R. (1978). Asexual reproduction capabilities in Ulva
lactuca L. (Chlorophyceae). Bot. Mar., 21, 117-121.
Bonotto S. (1975). Morphogenesis in normal branched and irradiated
Acetabularia mediterrania. Publ. Staz.Zool. Napoli, 39, 96-107.
Booth E. (1965). The manurial value of seaweed. Bot. Mar., _8, 138-
U3.
Borden C.A., Stein J.R. (1969). Reproduction and early develop-
ment in Podium fragile (Suringar) Hariot: Chlorophyceae. Phy-
cologia, 8, 91-99.
Borowitzka M.J. (1982). Mechanisms in algal calcification. Prog.
Phycol.Res. , 1, 137-178.
Bothe H. (1982). Nitrogen fixation. In: The Biology of Cyanobac-
teria (ede. Carr. N. G. , Whitton B.A.), pp. 87-104. Blackwell
Scientific Publications, Oxford.
Bouck G.B. (1969). Extracellular microtubules. The origin, struc-
ture, and attachment of flagellar hairs in Fucus and Ascophy-
llum antherozoids. J.Cell Biol., 40. 446-46O.
Bouck G.B. (1970). The development and post-fertilization fate
of the eyespot and the apparent photoreceptor in Fucus sperm.
Ann. N.Y. Acad.Sci., 175, 673-685.
Bouck G.B., Sweeney B.M. (1966). The fine structure and ontogeny
of marine diniflagellates. Protoplasma, 61. 205-223.
Bouok G.B., Rogalski A., Valaitis A. (1978). Surface organization
and compisition of Euglena. II. Flagellar mastigonemee. J.
Cell Biol., 77, 805-826.
Bourrelly P. (1957). Recherches sur les Chrysophycееs. Morpholo-
gie, phylogenie, systematique. Rev.Algol. Mem. Hors Ser., 1,
412 pp.
Bourrelly P. (1970). Les Algues d’Eau Douce. Initiation & la
Systematique. III. Les Algues Bleues et Rouges, les Eugleni-
ens, Peridiniens et Cryptomonadines. N. Boubee et Cie, Paris.
Bourrelly P. (1981). Les Algues d'Eau Douce. Algues Jaunes et
Brunes. N.Boubee et Cie, Paris.
Bourrelly P., Coute A. (1976). Observations en microscopie elec-
tronique a balayage des Ceratium d?eau douce (Dinophycees).
Phycologia, 15, 329-338.
Bovee E.0. (1982). Movement and locomotion of Euglena. In: The
Biology of Euglena,. Vol. Ill, Physiology (ed. Buetow D.E.)
464
pp. 143-168. Academic Press, New York.
Boyle J.E., Smith D.C. (1975). Biochemical interactions between
the symbionts of Convoluta roscoffensis. Proc. Roy.Soc. Bond.
B., 189, 121-135-
Boylen C.W., Brock T.D. (1973). Effects of thermal additions from
the Yellowstone Geyser Basins on the algae of the Firehole
river. Ecology, 54, 1283-1291. ,
Braarud T., Deflandre G., Halldal P., Kamptner E.K. (1955). Ter-
minology, nomenclature and systematics of the Coccolithopho-
ridae. Micropaleontology, 1., 157-159-
Braarud T., Gaarder K.R. , Nordii 0. (195^, Seasonal changes in
the phytoplankton at various points off the Norwegian west
coast (observations at the permanent oceanographic stations,
1954-1946). Rep. Norweg. Fish. Mar. Invest., 12 (3), 1-77.
Bradley W.H. (1929). Freshwater algae from the Green River For-
mation of Colorado. Bull. Torrey Bot. Club, 56, 421-428.
Bravo L. (1965). Studies on the life history of Prasiola meridi-
onalis. Phycologia, 4_, 177-194.
Brawley S.H. , Sears J.R. (1980). Septal plugs in a marine green
alga. J.Cell Biol., 8£, 62a.
Brawley S., Wetherbee R. (1981). Cytology and ultrastructure. In:
The Biology of Seaweeds (eds. Lobban 0.S., Wynne M.J.), pp.
248-299. Blackwell Scientific Publications, Oxford.
Brawley S., Quatrano R.S., Wetherbee R. (197?). Pine structural
studies of the gametes and embryos of Fucus vesiculosus L.
(Phaeophyta). J.Cell Sci., 87, 275-294-
Breen P.A., Mann K.H.(1976). Destructive grazing of kelp by sea
urchins in Eastern Canada. J.Pish Res.Bd.Can., 33, 1278-1283»
Brock T.D. (1967). Life at high temperatures. Science, 158, 1012-
1019.
Brock T.D. (1969). Microbial growth under extreme environments.
Symp.Soc.Gen. Microbiol., 19, 15-41.
Brock T.D. (1973). Lower pH limit for the existence of blue green
algae: Evolution and ecological implications. Science, 179,
480-483.
Brock T.D. (1978). Thermophilic Micro-organisms and Life at High
Temperatures. Springer-Verlag, New York-
Brook A.J. (1981). The Biology of Desmids. Blackwell Scientific
Publications, Oxford.
Brown R.M., Johnson C., Bold H.C. (1968). Electron and phase-
contrast microscopy of sexual reproduction in Chlamydomonas
465
moewusii. J. Phycol., _4, 100-120.
Bryan G.W. (1969). The absorption of zinc and. other metals by
brown seaweed Laminaria digitata. J. Mar.Biol.Ass.U.K., 49, '
225-245.
Bryan G.W., Hummerstone L.G. (1973). Brown seaweed as an indicator
of heavy metals in estuaries in south-west England. J. Mar.
Biol. Ass. U.K., 52, 705-720.
Bryceson I., Fay P. (1981). Nitrogen fixation in Osoillatoria
(Trichodesmium) erythraea in relation to bundle formation
and differentiation. Idar. Biol., 61, 159-166.
Buetow D.E. (ed.) (1968). The Biology of Euglena, Vols I, II.
Academic Press, New York.
Buetow D.E. (ed.) (1982). The Biology of Euglena, Vols III, IV.
Academic Press, New York.
Buffaloe N. (1958). A comparative cytological study of four spe-
cies of Chlamydomonas. Bull.Torrey Bot.Club., 85. 157-178.
Buggeln R. G. (1974). Negative phototropism of the haptera of
Alaria esculenta (Laminariales). J.Phycol., .10, 80-82.
Buggeln R. G. (1976). Auxin, an endogenous regulator of growth in
algae? J. Phycol., 12., 355-358.
Buggeln R.G. (1978). Physiological investigations on Alaria es-
culenta (Laminariales, Phaeophyceae). IV. Inorganic and
organic nitrogen in the blade. J.Phycol., 14., 156-160.
Buggeln R.G. (1981). Morphogenesis and growth regulators. In:
The Biology of Seaweeds (eds. Lobban C.S., Wynne M.J.), pp.
627-660. Blackwell Scientific Publications, Oxford.
Buggeln R.G. (1983). Photоassimilate translocation in brown al-
gae. Prog. Phycol.Res., 2, 283-332.
Buggeln R.G., Bal A.K. (1977). Effects of auxins and chemically
related non-auxins on photosynthesis and chloroplast ultras-
tructure in Alaria esculenta (Laminariales). Can.J. Bot.,
55, 2098-2105.
Buggeln R.G., Fensom D.S., Itaerson C.J. (1985). Translocation of
1C—photoassimilate in the blade of Macrocystis pyrifera
(Phaeophyciae). J. Phycol», 21, 35-40.
Bunt J.S. (1970). Uptake of cobalt and vitamin B^ by tropical
marine macroalgae. J. Phycol., _6, 339-343.
Bunt J.S., Wood E.J.C. (1963). Microbiology and antarctic sea
ice. Nature, 199. 1254-1255.
Bunt J.S., Owens 0., Van H., Hoch G. (1966). Expolatory studies
on the physiology and ecology of a psychrophilic marine diatom.
466
J. Phycol., 2, 96-100.
Burger-Wiersma T., Veenhuis M., Korthals H.J., Van der Wiel C.C.
M., Mur L.R. (1966). A new prokaryote containing chlorophylls
a and b. Nature, 320, 262-264-
Burr F.A., West J.A. (1970). Light and electron microscope obser-
vations on the vegetative and reproductive structures of
Bryopsis hypnoides. Phycologia, 9, 17-37.
Burr F.A., West J.A. (1971). Protein bodies in Bryopsis hypnoides:
their relationship to wound-healing and branch septum develop-
ment. J.Ultrastr. Res., 35, 476-498.
Burrows E.M. (1958). Sublittoral algal population in Port Erin Bay,
Isle of Man. J.Mar.Biol.Asse. U.K., 37, 687-703»
Burrows E.M., Conway E., Lodge S.M., Powell H.T. (1954). The rai-
sing of intertidal algal zones in Fair Isle. J.Ecol., 42, 283-
288.
Butler M., Haskew A.E.J., Young Ш.М. (1980). Copper tolerance in
the green alga, Chlorella vulgaris. Plant Cell Environ., 3,
119-126.
Butler R.D., Allsopp A. (1972). Ultrastructural investigations in
the Stigonemataceae. Arch. Mikrobiol., 82. 283-299.
Campbell S.E. (1980). Palaeoconchocelis starmachii, a carbonate
boring microfossil from the Upper Silurian of Polant (425
million years old): implications for the evolutions for the
evolution of the Bangiaceae (Rhodophyta). Phycologia, 19,
25-36.
Canter H.M. (1973). A new primitive protozoan devouring centric
diatoms in the plankton. J.Linn.Soc.Lond. Zool., 52, 63-83.
Canter H.M., Jaworski G.H.M. (1978). The isolation maintenance and
host range studies of a chytrid Rhizophidium planktonicum
Canter emend., parasitic on Asterionella formosa Hassall. Ann.
Bot., 4_2, 967-979.
Capriulo G.M., Carpenter E.J. (1980). Grazing by 35 to 202 mm
microzooplankton in Long Island Sound. Mar. Biol., 56, 319-
326.
Caram B. (1965). Rechercher sur la reproduction et le cycle sexu-
el de quelques Pheophycees. Vie et Milieu, 16, 21-226.
Caram B. (1972). Le cycle de reproduction des Pheophycees-Pheos-
porees et ses modifications. Soc. Bot.Fr.Memoires, 1972, 151-
160.
Cardinal A. (1964). Etude sur les Ectocarpacees de la Manche.
Beih. Nova Hedwiga, IS, 1-86.
МП
Cardinal A., Breton-Provencher M. (1977)» Variations de la teneur
en acide alginique des Laminariales de I'eetuaire du Saint-
Laurent (Quebec). Bot. Mar., 20, 243-252.
Carlucci A.F., Bowes P.M. (1970). Vitamin production and utili-
zation by phytoplankton in mixed culture. J. Phycol., 6, 393-
400.
Carmichael W.W. (ed.) (1981). Water Environment. Algal Toxins and
Health. Environmental Science Research, Vol. 20: International
Conference on Toxic Algae. Plenum Press, New York.
Carmichael W.W., Gorham P.R. (1978). Anatoxins from clones of
Anabaena flos-aquae isolated from lakes of western Canada.
Mitt. Int. Ver.Limnol. , 21_, 285-295.
Carmichael W.W., Gorham P.R. (1980). Freshwater cyanophyte toxins:
types and their effects on the use of micro algae biomass. In:
Algal Biomass: Production and Use (eds. Shelef G., Boeder C.J.),
pp. 437-448. Elsevier/North-Holland, Amsterdam,,
Carmichael W.W., Gorham P.R. (1981). The moxaic nature of toxic
blooms of Cyanobacteria. In: Water Environment. Algal Toxins
and Health, (ed. Carmichael W.W.). Environmental Science Re-
search, Vol. 20, pp.161-172. International Conference on Toxic
algae. Plenum Press, New York.
Carpenter E.J., Price С.C. (1976). Marine Oscillatoria (Tricho-
desmium): explantation for aerobic nitrogen fixation without
heterocysts. Science, 191. 1278-1280.
Carson J.I., Brown R.M. (1978). Studies of Hawaiian freshwater
and soil algae. II. Algal colonization and succession on a
dated volcanic substrate J. Phycol., 24, 171-178.
Cassin P.E. (1974). Isolation growth and phyciology of acidophi-
lic Chlamydomonads. J.Phycol., 10, 439-447.
Castenholz R.W. (1969). Thermophilic cyanophytes of Iceland and
the upper temperature limit. J.Phycol., 5., 360-368.
Cattaneo A., Kalff J. (1979). Primary production of algae growing
on natural and artificial aquatic plants: a study of interac-
tions between epiphytes and their substrate. Limnol. Oceanogr.,
24, 1031-1037.
Cavalier-Smith T. (1974). Basal body and flagellar development
during the vegetative cell cycle and the sexual cycle of
Chlamydomonas reinhardii. J. Cell Sci., 16, 529-556.
Cavalier-Smith T. (1975). Electron and light microscopy of gameto-
genesis and gamete fusion in Chlamydomonas reinhardii. Proto-
plasma, 86, 1-18.
468
Cavalier-Smith T. (1976). Electron microscopy of zygospore forma-
tion in Chlamydomonas reinhardii. Protoplasma, 87, 1-18.
Cavalier-Smith T. (1978). The evolutionary origin in phylogeny of
microtubules, mitotic spindles and eucaryote flagella. Biosys-
tems, 10, 93-114.
Cavalier-Smith T. (1983a). Endosymbiotic origin of the mitochon-
drial envelope. Endocytology, 2, 265-279.
Cavalier-Smith T. (1983b). A six-kingdom classification and a
unified phylogeny. Endocytology, 2, 1027-1034.
Chadefaud M. (1974). Possibilityd’une origine non symbiotique de
la cellule des Eucaryotes. C.R. Acad. Sci.Paris D, 278. 3079-3081.
Chadefaud M. (1978). Sur la notation de prochiorophytes. Rev.
Algol. N.S. , 12, 203-206.
Chamberlain A.H.L., Gorham J., Kane D.F. , Lewey S.A. (1979). la-
boratory growth studies on Sargassum muticum (Yendo) Pensholt.
Bot. Mar.,, 22, 11-19.
Chapman A.R.0. (1972a). Morphological variation and its taxonomic
implications in the legulate members of the genus Desmarestia
occurring on the west coast of North America. Syesis, 1-20.
Chapman A.R.O. (1972b). Species delimination in the filiform,
oppositely branched members of the genus Desmarestia Lamour.
(Phaeophyceae, Desmarestiales) in the northern hemisphere.
Phycologia, 11, 225-231•
Chapman A.R.O. (1973). A critique of prevailing attitudes towards
the control of seaweed zonation on the sea shore. Bot. Mar.,
16, 80-82.
Chapman A.R. 0., Burrows E.M. (1971). Field and culture studies
of Desmarestia aculeata. Phycologia, 10, 63-76.
Chapman A.R.O., Craigie J.S. (1977). Seasonal growth in Laminaria
longicrusis: relation with dissolved inorganic nutrients and
internal reserves of nitrogen. Mar.Biol., 40, 197-205.
Chapman A.R.O., Doyle R.W. (1978). Genetic analysis of alginate
content in Laminaria longicruris (Phaeophyceae). Proc. Int.
Seaweed Symp., 9_, 125-132.
Chapman A.R.O., Lindley J.E. (1980). Seasonal growth of Lamina-
ria solidungula in the Canadian High Arctic in relation to
irradiance and dissolved nutrient concentrations. Mar. Biol.,
57, 1-5.
Chapman D. J. , Trench R.K. (1982). Prochlorophyceae: introduction
and bibliography. In: Selected papers in Phycology II (eds.
Rosowski J.H., Parker B. 0.), pp. 656-658. Phycological So-
469
ciety of America, Allen Press. Lawrence, Kansas.
Chapman V.J. (1974). Salt Marshes and Salt Deserts of the World,
2nd Edition. Cramer, Lehre.
Chapman V.J., Chapman D.J. (1973). The Algae, 2nd Edition. Mac-
Millan, London.
Chapman V.J., Chapman D.J. (1976). Life forms in the algae. Bot.
Mar., 19, 65-74.
Chapman V.J., Chapman D.J. (1980). Seaweeds and their Uses. Chap-
man and Hall, London and New York.
Chen J.C.W. (1980). Longitudinal microfibrillar alignment in the
wall of cylindrical Nitella rhizoidal cells - observation
under polarizing and interference microscopes. Am.J.Bot.,
67, 859-865.
Chen L.C.-M., Taylor A.R.A. (1978). Medullary tissue culture of
the red alga, Chondrus crispus. Can J.Bot., 56, 883-886.
Chen L. C.M., McLachlan J., Neish A.C., Shacklock P. P, (1973). The
ratio of kappa-to lambda-carrageenan in nuclear phases of the
rhodophycean algae Chondrus crispus and Gigartina stellata.
J.Mar. Biol.Ass. U.K., 53, 11-16.
Chen L.C.-M., Edelstein T., McLachlan J. (1974). The life history
of Gigartina stellata in culture. Phycologia, 13, 287-294.
Cheney D.P. (1982). The determining effects of snail herbivore
density of intertidal algal recruitment and composition.
Abstr. 1st. Int. Phycol.Congr., p. 48. St. Johns, Nev.'f oundland
Chiang Y.-M. (1981). Cultivation of Gracilaria (Rhodophycophyta,
Gigartinales) in Taiwan. Proc.Int.Seaweed Symp., 10, 657-667.
Chihara M., Yoshizaki M. (1972). Bonnemaisoniaceae: their goni-
moblast development, life history and systematics. In: Contri-
butions to the Systematics of Benthic Marine Algae of the
North Pacific (eds. Abbott I., Kurogi M.). Japanese Society
of Phycology, Kobe, Japan.
Chisholm S.W. (1981). Temporal patterns of cell division in
unicellular algae. Can. Bull.Pish. Aquat.Sci., 210, 150-181.
Chisholm S.W., Azam P., Eppley R.W. (1978). Silicic acid incorpo-
ration in marine diatoms on light-cycles: use as an assay for
phased cell division. Limnol. Oceanogr., 23. 518-529.
Christensen T. (1962). Alger. In: Botanik, II, Nr. 2; Systematisk
Botanik (eds. Bocher T.W., Lange G., Srensen T.). Munksgaard,
Copenhagen.
Christensen T. (1964). The gross classification of the algae.
In: Algae and Man (ed. Jackson D.p.), pp. 59-64. Plenum Press,
New York.
470
Christensen T. (1980). Algae, A Taxonomic Survey. Раве. I. AiO
Tryk, Odense.
Churchill A.C. , Moeller H.W. (1972). Seasonal patterns of repro-
duction in hew York populations of Codium fragile (Sur.)
Hariot ssp. tomentosoides (van Goor) Silva. J. Phycol.,
147-152.
Clarke K.J,, Pennick IJ.C. (1972). Flagellar scales in Oxyrrhis
marina Dujardin. Brit. Phycol. J., J7, 357-360.
Clarke K.J., Pennick IJ.C. (1975). Syncrypta glomerifera sp. nov.,
marine member of the Chrysophyceae bearing a new form of scale.
Br. Phycol. J., 10, 363-370.
Clayton M.N. (1981). Correlated studies on seasonal changes in
the sexuality, growth rate longevity of complanate Scytosiphon
(Scytosiphonaceae: Phaephyta) from southern Australia, growing
in situ. J.Exp. Mar.Biol. Ecol., 51, 87-96.
Clayton M.N., Beakes G.Y'. (1983). Effects cf fixatives on the ul-
trastructure of physodes in vegetative cells of Scytosiphon
lomentaria (Scytosiphonales, Phaeophyta). J.Phycol., 19,
4-16.
Cleare M., Percival E. (1972). Carbohydrates of the freshwater
alga Tribonema aequale I. Low molecular weight polysaccharides.
Brit. Phycol. J., 7, 185-193.
Clocchiatti M. (1971). Sur I’existence de coccospheres portant
des coccolithes de Gephyrocapsa oceanica et de Emiliana hux-
leyi (Coccolithophorides). C.R. Acad.Sci. Paris D, 273,
318-321.
Cloud P. (1976). Beginnings of biospheric evolution and their
biogeochemical consequences. Paleobiology, _2_, 351-387.
Coesel P.F.M., Texeira H.M.V. (1974). Notes on sexual reproduc-
tion in desmids. I. Zygospore formation in nature (with spe-
cial reference to some unusual records of zygotes). Acta Bot.
Neer., 23, 361-368.
Cohn F. (1853). Untersuchungen uber die Entwicklungsgeschichte
mikroskopischer Algen und Pilze. Nov. Act. Acad.Leop. Carol.,
24, 103-256.
Colbath G.K. (1983). Fossil prasinophycean phycomata (Chlorophyta
from the Silurian Bainbridge Formation, Missouri, U.S.A.
Phycologia, 22, 249-265.
Sole D.K., Plapp F.W. (1974). Inhibition of growth and photosyn-
thesis in Chlorella pyrenoidosa by a chlorinated biphenyl and
several incecticides. Environ. Entomol., 3, 217-220.
lole K., Akintobe S. (1963). The life cycle of Prasiola meridi-
471
onalis Setchel and Gardner.Can.J.Bot., 41. 661-668.
Cole K.> Conway E. (1980). Studies in the Bangiaceae: reproduc-
tive modes. Bot.Mar. , 23, 545-553.
Coleman A.W. (1979). Sexuality in the colonial flagellates. In:
Physiology and Biochemistry of the Protozoa, Vol. 1 (eds.
Hunter S.H. , Lewsndowsky M. ), pp.307-340. Academic Press,
Nev/ York.
Coleman A.W. (1983). The role of resting spores and akinetes in
chiorophyte survival. In: Survival strategies of the Algae
(ed. Eryxell G.A.), pp. 1-21. Cambridge University Press,
Cambridge.
Colman J.S. (1933). The nature of the interidal zonation of
planta and animals. J.Mar.Biol.Ass. U.K., 18, 435-476.
Colombetti G., Lend F., Diehn B. (1982). Responses to photic,
chemical and mechanical stimuli. In: The Biology of Euglena.
Vol. Ill, Physiology (ed. Buetow D.E.), pp.169-195. Academic
Press, Nev, York.
Conell J.H., Slayter R.O. (1977). Mechanisms of succession in
natural communities and their rde in community stability
and organization. Am.Nat., Ill, 1119-1144.
Conover J.T., Sieburth J. McN. (1964). Effects of Sargassum
distribution on its epibiota and antibacterial activity.
Bor. Mar., 6, 147-157.
Cooke G.W. , Williams R.J.B. (1970). Losses of nitrogen and phos-
phorus from agricultural land. Water Treat.Exam., 19, 253-276.
Cooke W.B. (1968a). Studies in the genus Prototheca. I.Literature
review. J.Elisha Mitchell Sci.Soc., 84, 213-216.
Cooke W.B. (1968b). Studies in the genus Prototheca. II. Taxono-
my. J.Elisha Mitchell Sci.Soc., 84, 217-220.
Coombs J., Greenwood A.D. (1976). Compartmentation in the photo-
synthetic apparatus. In: The Intact Chloroplast. Topics in
Photosynthesis Vol. I (ed. Barber J. ), pp. 1-51. Elsevier/
Horth-Holland, Amsterdam.
Coombs J., Volcani B.E. (1968). Studies on the biochemistry and
fine structure of silica shell formation in diatoms. Chemi-
cal changes in the wall of Navicula pelliculosa during its
formation. Planta, 82, ,280-292.
Coombs J., Halicki P.J., Holm-Hansen 0., Volcani B.E. (1967).
Studies on the fine structure and silca shell formation in
diatoms. Exp.Cell Res., 47, 315-328.
Copeland H.P. (1938). The kingdoms of organisms. Quart.Rev.
472
Biol., 12» 383-420.
Cordeiro-Marino M. , Poza A. (1981). Life history of Gymnogrongus
griffitheiac (C.Ag.) J.Ag. (Phyllophoraceae, Gigartinales).
Proc. Int. Seaweed Symp., 10, 155-161.
Cortel-Breeman A.M., Ten Hoopen A. (1978). The short day response
in Acrosymphyton purpuriferum (J.Ag.) Sjost. (Rhodophyceae:
Cryptonemiales). Phycologia, 17. 125-132.
Cortel-Breeman A.M., Hoek C. van den (1970). Life history studies
on Rhodophyceae. I. Acrosymphyton purpuriferum (J.Ag.) Kyi.
Acta Bot. Neer., 19, 265-284.
Goughian S., Tattersfield D. (1977). Photorespiration in larger
littoral algae. Bot.Mar. , 20, 265-266.
Court G.J. (1980). Photosynthesis and translocation studies of
Laurencia spectabilis (Rhodophyceae). J. Phycol., 16. 270-279.
Cousens R., Hutchings M.J. (1983). The relationship between densi-
ty and mean frond weight in monospecific seaweed stands. Natu-
re, 301. 240-241.
Cox E.R., Bold H.C. (1966). Taxonomic investigations of Stigeo-
clonium. Univ. Tex.Publ., 6612, 167 pp.
Cox G., Dwarte D.M. (1981). Freeze-etch ultrastructure of a
Prochloron species - the symbiont of Didemnum mo lie. New
Phytol., 88, 427-438.
Cox G.E., Wilson J.D., Brown P. (1974). Protothecosis: a case of
disseminated algal infection. Lancet, ii, 379-382.
Coyle J.T., London E.D. , Biziere K., Zaczek R. (1979). Kainic
acid neurotoxicity: insights into the pathophysiology of
Huntington's disease. Adv. Neurol., 23» 593-608.
Craigie J.S. (1974). Storage products. In: Algal Physiology and
Biochemistry (ed. Stewart W.D.T.), pp. 206-235. Blackwell
Scientifics Publications, Oxford.
Crawford R.M. (1975). The taxonomy and classification of the
diatom genus Melosira C. Ag. The type species M. nummuloides
C.Ag. Br. Phycol. J., 10, 323-338.
Cronquist A. (I960). The divisions and classes of plants. Bot.
Rev. (London), 26, 425-482.
Crumpton W.G., Wetzel R.G. (1982). Effects of differential growth
and mortality in the seasonal succession of phytpplankton
populations. Ecology, 63, 729-739.
Cubit J.D. (1984). Herbivory and the seasonal abundance of algae
on a high intertidal rocky shore. Ecology, 65, 1904-1917.
Cushing D.H., Vucetic T. (1963). Studies on a Calanus patch. Щ,
473
The quantity of food eaten by Calanus finmarchicus.J. Mar.
Biol. Ass. U.K., ££, 349-371.
Czygan FrC. (1970). Blutregen und Blutschnee: Stickstoffmangel-
zellen von Haematococcus pluviales und Chlamydomonas nivalis.
Arch. Microbiol., 24» 69-76.
Daily F.K. (1932). Charophyceae. In: Synopsis and Classification
of Living Organisms, Vol. I (ed. Parker S.P.), pp.161-162.
McGraw-Hill, New York.
Dale B. (1983).'Dinoflagellate resting cysts: "benthic plankton",
In: Survival Strategies of the Algae (ed. Fryxell G.A.), pp.
69-136. Cambridge University Press, Cambridge.
Daniel G.F., Chamberlain A.H.L. (1981). Copper immobilization
in fouling diatoms. Bot.Mar., 24. 229-243.
D’Antonio C. (1985). Epiphytes on the rocky intertidal red alga
Rhodomella larix (Turner) C.Ag.: negative effects on the
host and food for herbivores. J.Exp.Mar.Biol.Eool., 86, 197-
218.
Darley W.M. (1982). Algal Biology: A Physiological Approach.
Blackwell Scientific Publications, Oxford.
Darley W.M., Sullivan C.W., Volcani В.E. (1976). Studies on the
biochemistry and fine structure of silica shell formation in
diatoms. Division cycle and chemical composition of Navicula
pelliculosa during light dark synchronized growth. Planta,
130. 159-167.
Darley VI.M., Montague C.L., Plumley F.G. , Sage W.W., Psalidas
A.T. (1981). Factors limiting edaphic algal biomass and
productivity in a Georgia salt-marsh. J.Phycol., 17, 122-128.
Davenport J. (1982). Oil and planktonic ecosystems. Trans. Roy.
Soc.Lond. B, 297. 369-381.
Davies A.G. (1983). The effects of heavy metals upon natural
marine phytoplankton populations. Prog. Phycol.Res., 2, ИЗ-
145.
Davies J.M., Baird I.E., Massie L.C., Hay S.J., Ward A.P. (1980).
Some effects of oil-derived hydrocarbons on pelagic food webs
from observations in an enriched ecosystem and considerations
of their implications for monitoring. Rapp. P-v. Reun.Cons.
Int. Explor. Mer., 179, 201-211.
Davies R.R., Spencer H., Wakelin P.O. (1964). A case of human
protothecosis. Trans. Roy.Soc.Trop.Med.Hyg., 58, 448.
Davis C.C. (1964). Evidence for the eutrophication of Lake Erie
474
from phytoplankton records. Limnol. Oceanogr., 275-283.
Davie J.S. (1967). The life cycle of Pediastrum simplex. J. Phy-
col., 2» 95-103.
Davis M.H., Goughian J. (1978). Response of entrained phytoplank-
ton to low level chlorination at a coastal power station. Tn:
Water Chlorination: Environmental Impacts and Health Effects,
Vol. 2. (ed. Jolley R.L.), pp. 369-376. Ann Arbor Science,
Michigan.
Dawson P.A. (1973). Observations on the structure of some forms
of Gomphonema parvulum Kutz. ITT. Frustule foimation. J. Phy-
col., 9, 353-364.
Dayton P.K. (1973). Dispersion, dispersal and persistence of the
annual intertidal alga Postelsia palmaeformis Ruprecht. Eco-
logy» 54 . 433-438.
Dayton Г.К. (1975). Experimental evaluation of ecological domi-
nance in a rocky intertidal algal community. Ecol. Monogr.,
_45, 137-159.
Deason T.R. (1984). A discussion ofthe classes Chlaraydophyciae
and Chlorophyceae and their subordinate taxa. Pl. Syet. Evol.,
146. 75-86.
Deason T.R., Butler G.L., Rhyne C. (1983). Rhodella reticulata
sp. nov., a new coccoid rhodophytan alga (Porphyridiales).
J. Phycol., 19, 104-111.
DeBoer J.A. (1981). Nutrients. In: The Biology of Seaweeds (eds.
Lobban C.S., Wynne M.J.), pp. 356-392. Blackwell Scientific.
Publications, Oxford.
DeCew T.C., West J.A. (1981). Life histories in the Phyllophora-
ceae (Rhodophyta: Gigartinales) from the Pacific coast of
North American. I. Gymnogrongus linearis and G. leptophyllus.
J. Phycol., IJ, 240-250.
DeCew T.C., West J.A. (1982a). Investigations on the life his-
tories of three Farlowia spp. (Rhodophyta, Cryponemiales,
Dumontiaceae) from Pacific North America. Phycologia, 20,
342-351.
DeCew T.C., West J.A. (1982b). A sexual life history in Rhodo-
physema (Rhodophyceae); a re-interpretation. Phycologia, 21,
67-74.
Deflandre G., Lenoble A. (1948). Sur la presence d'eugleniens
fossiles du genre Trachelomonas Ehr. dans un schiste pliocene
de^ Madagascar. C.R. Acad.Sci. Paris D, 226, 509-511.
Delaney S.F., Herdman M., Carr N.G. (1976). Genetics of blue-green
475
algae. Tn: The Genetics of Algae (ed. Lewin R.A.), pp. 7-28.
Blackwell Scientific Publications, Oxford.
De Reaumur R.A. (7711). Description des fleurs et des graines des
divers Fucus, et quelques autres observations physiques sur
ces mfeies plantes. Mem.Acad.Sci. Paris, 1711, 383.
De Toni G.B. (1889). Sylloge algarum omnium hucusque cognitarum.
1 (Sylloge Chlorophycearum). 1-12, I-CXXXIX, 1-1315. Patavii.
De Toni G.B. (1924). Sylloge algarum omnium hucusque cognitrum.
6 (Sylloge Ploridearum 5, Additamenta). I-XI, 1-767. Patavii.
Devilly C.I., Houghton J.A. (1977). A study of genetic transfor-
mation in Gleocapsa alpicola. J. Gen. Microbiol., 98, 277-280.
Digby P.S.B. (1979). Reducing activity and the formation of base
in thecoralline algae: an electrochemical model. J. Mar. Biol.
Ass. U.K., 59, 455-477.
Dillenius J.J. (1741). Histori Muscorum in qua circiter sexcen-
tae veteres et novae ad sua genera Telatae describuntus et
inconibur genuimis......Oxonii (not seen by the authors; quo-
ted from Prescott, 1951).
Dillon L.S.(1981). Ultrastructure, Macromolecules and Evolution.
Plenum Press, Hew York and London.
Dinsdale M.T., Walsby A.E. (1972). The interrelations of cell
turgor pressure, gas vacuolations, and buoyancy in a blue
green alga. J.Exp.Bot., 23, 561-570.
Di Rosa M (1972). Biological properties of carrageenan. J. Pharm.
Pharmacol., 24, 89-102.
Dixon P.S. (1965). Perennation, vegetative propagation and algal
life histories, with special reference to Asparagopsis and
other Rhodophyta. Bot.Gothoburg., 67-74.
Dixon P.S. (1971). Cell enlargement in relation to the develop-
ment of thallus form in Plorideophyceae. Brit. Phycol, J.,
6, 195-205.
Dixon P.S. (1973). The Biology of the Rhodophyta. Oliver and
Boyd, Edinburgh.
Dixon P.S. (1982). Rhodophycota. In: Synopsis and Classifica-
tion of Living Organisms, Vol. I (ed. Parker S.P.), pp.61-79.
McGraw-Hill, Her: York.
Dodge J.D. (1969a). A review of the fine structure of algal
eyespots. Brit. Phycol. J., _£, 199-210.
Dodge J.D. (1969b). The ultrastructure of Chroomopas mesostigma-
tica Butcher (Cryptophyceae). Arch. Mikrobiol., 69, 266-280.
476
Dodge J.D. (1971)» A dinoflagellate with both a mesocaryotic and
a eucaryotic nucleus. I. Fine structure of the nuclei.
Protoplasma, 73. 145-157.
Dodge J.D. (1972). The ultrastructure of the dinoflagellate pu-
sule : a unique osmo-regulatory organelle. Protoplasma, 75.
285-302.
Dodge J.D. (1973). The Fine Structure of algal cells. Academic
Press, New York.
Dodge J.D. (1974)» Fine structure and phylogeny in the algae.
Sci. Prog. (Oxford), 61., 257-274.
Dodge J.D. (1979). The phytoflagellates: fine structure and phy-
logeny. In: Biochemistry and Physiology of the Protozoa (eds.
Hunter S.H., Levandarsky M. ), pp. 7-57. Academic Press, New
York.
Dodge J.D. (1983). Dinoflagellates: investigation and phylogene-
tic speculation. Brit. Phycol. J., 18, 335-356.
Dodge J.D. (1984). The functional and phylogenetic significance
of dinoflagellate eyespots. Biosystems, 16, 259-267.
Dodge J.D., Crawford R.M. (1969). Observations on the fine
structure of the eyespot and associated organelles in the
dinoflagellate Glenodinium foliaceum. J.Cell Sci., _5, 479-
493.
Dodge J.D., Crawford R.M. (1970). A survey of thecal fine struc-
ture in the Dinophyceae. Bot. J.Linn.Soc., 63. 53-67.
Dodge J.D. , Crawford R.M. (1971). Fine structure of the dinofla-
gellate Oxyrrhis marina. II. The flagellar system. Protisto?.
logica, 7, 399-409.
Dodge J.D., Hermes H.B. (1981). A scanning electron microscopial
study of the apical pores of marine dinoflagellates (Dinophy-
ceae). Phycologia, 20, 424-430.
Dodge J.D., Steidinger K.A. (1982). Dinophyceae: introduction
and bibliography. In: Selected Papers in Phycology II (eds.
Rosowski J.R., Parker B.C.), pp. 691-697. Phycological Socie-
ty of America, Allen Press, Lawrence, Kansas.
Doemel W.N., Brock T.D.(1970). The upper temperature limit of
Cyanidium caldareuin. Arch. Mikrobiol., 72, 326-332.
Doolittle W.F., Singer R.A. (1974). Mutation analysis of dark
endogenous metabolism of the blue green bacterium Anaoystis
nidulans. J. Bacteriol., 119, 677-683.
Dop A.J. (1980). The genera Phaeothamnion Lagerheim. Tetrachry-
sis, gen. nov. and Sphaeridiothrix Pascher et Vlk (Chrysophy-
4Т1
ceae). Acta Bot. Neerl., 29, 65-86.
Doty M.S. (1946). Critical tide factors that are correlated with
the vertical distribution of marine algae and other organisms
along the Pacific coast. Ecology, 27, 315-328.
Doty M.S. (1977). Eucheuma current marine agronomy. In: The Ma-
rine Plant Biomaes of the Pacific North-West Coast. A Potenti-
al Economic Resource (ed. Kraus R.W.), pp.203-214. Oregon Sta-
te University Press, Corvallis.
Doty M.C. (1979). Status of marine agronomy, with special referen-
ce to the tropics. Proc. Int. Seaweed Symp., 9_> 35-58.
Doty M.S., Aguilar-Santos G. (1970). Transfer of toxic algal sub-
stances in marine food chains. Pacific Science, 24, 351-355.
Doty M.S., Santos G. (1978). Carrageenans from tetrasporic and
oystocarpio Eucheuma species. Aquatic Botany, 4 143-149.
Doty M.S., Gilbert W.J.» Abbott I.A. (1974). Hawaiian narine al-
gae from seaward of the algal ridge. Phycologia, 12, 345-357.
Drebes G. (1966). Of the life history of the marine plankton dia-
tom Stephanopyxis palmeriana (Grev.) Grunow. Helgol. Wise.
Meeresunters., 13 101-114.
Drebes G. (1977). Sexuality. In: The Biology of Diatoms (ed. Wer-
ner D.). University of California Press, Berkeley.
Drebes G. (1981). Possible resting spores of Dissodinium pseudo-
lunula Dinophyta and their relation to other taxa. Brit. Phy-
col. J., 16, 207-215.
Drew K.M. (1937). Spermothamnicn snyderae Parlow, a floridean
alga bearing polysporangia. Ann. Bot. N.S. , 2, 463-476.
Drew K.M. (1939). An investigation of Plumaria elegans (Bonnem.)
Schmitz with special reference to triploid plants bearing
parasporangia. Ann. Bot. N.S., 2, 347-367.
Drew K.M. (1949). Conchocellis phase in the life history of Por-
phyra umbilical!s (L.) Kutz. Nature, 164, 748-749.
Drew G«, Weckesser J. (1982). Eunction, structure and composition
of cells walls and extremal layers. In: The Biology of Cyano-
bacteria (eds. Carr N.G., Whitton B.A.), pp. 333-357. Black-
well Scientific Publications, Oxford.
Dring M.J, (1967). Effects of daylength on growth and reproduc-
tion of the Conchocelis-phase of Porphyra tenera. J. Mar.
Biol. Ass. U.K., Д7, 501-510.
Dring M.J. (1981). Chromatic adaptation of photosynthesis in
benthic marine algae: an examination if its ecological signi-
478
ficance using a theoretical model. Limnol. Oceanogr., 26,
271-284.
Dring M.J. (1982). The Biology of Marine Plante. Edward Arnold,
London.
Dring M.J. (1984). Photoperiodism and phycology. Prog. Phycol.
Res., 3, 159-192.
Dring M.J., Brown P.A. (1982). Photosynthesis of interidal brown
algae during and after periods of emersion: a renewed search
for physiological causes of zonation. Mar. Ecol. Prog.Ser.,
8, 301-308.
Dring M.J., Liining K. (1975). A photoperiodic effect mediated by
blue light in the brown alga Scytosiphon lomentaria. Planta,
125. 25-32.
Droop M.R. (1968). Vitamin B12 and marine ecology. IV. The kine-
tics of uptake, growth and inhibition in Monochrysis lutheri.
J. Mar. Biol.Assoc. U.K., 48, 689-733.
Droop M.R. (1974a). Heterotrophy of carbon. In: Algal Physiolo-
gy and Biochemistry (ed. Stewart W.D.P.), pp.530-559. Black-
well Scientific Publications, Oxford.
Droop M.R. (1974b). The nutrient status of algal cells in conti-
nuous culture. J. Mar. Biol.Ass. U.K., 54. 825-855.
Drouet F. (1968). Revision of the classification of Oscillato-
riaceae. Monogr. Acad.Kat.Sci. Philad., 12» 370 p.
Drouet (1973). Revision of the Nostocaoeae with cylindrical
trichomes. Hafner Press, New York.
Drouet P., Daily W. (1956). Revision of the ooccoid Myxophyceae.
Butler Univ. Bot.Stud., 12, 1-218.
Druehl L.D. (1978). The distribution of Macrocystis integrlfo-
lia in British Columbia as related to environmental parame-
ters. Can.J. Bot., 56, 69-79.
Druehl L.D. (1981). Geographical distribution. In: The Biology
of Seaweeds (eds. Lobban C.S., Wynne M.J.), pp. 306-325.
Blackwell Scientific Publications, Oxford.
Druehl L.D., Green J.M. (1982). Vertical distribution of inter-
tidal seaweeds as related to patterns of submersion and
emersion. Mar. Ecol. Prog.Ser., 9, 163-170.
DruggW.S. (1969). Some new genera, species and combinations of
phytoplankton from the Lower Tertiary of the Gulf Coast,
U.S.A, in ultra mioroplankton. Proc. K.Am.Paleontol. Conven-
tion, Chicago, pp. 809-843» Allen Press, Lawrence, Kansas.
479
Drujn R.W., Hopkins J.T. (1966). Diatom locomotion: an explanation.
Protoplasma, 62, 1-33.
Drum R.W., Pankratz H.S. (1963). Fine structure of a diatom centro-
some. Science (N.Y.), 142,61-63.
Dube M.A. (1967). On the life history of Monostroma fuscum (Pos-
tels et Rup?.). Mttrock. J. Phycol,, 2» 64-73.
Ducreux P.(1977). Etude experimentale des correlation et des pos-
sibilites de regeneration au niveau de l'apex de Sphacelaria
cirrhosa Ag. Ann.Sci. Kat.Bot., 18, 163-184.
Duffield E. C. S., Waaland S.D., Cleland R. (1972). Morphogenesis
in the red alga Griffithsia pacifica: regeneration from single
cells. Planta, 105, 185-195.
Dugdale R.C. (1967). Nutrient limitation in the sea: dynamics,
identification, and significance. Limnol. Oceanogr., 12.
685-695.
Dugdale R.C., Goering J.J. (1967). Uptake of new and regenerated
forms nitrogen in primary productivity. Limnol. Oceanogr., 12.
685-695.
Duggins D. 0. (1980). Kelp beds and sea otters: an experimental
approach. Ecology, 61, 447-453.
Dunstan P. (1979). Distribution of zooxanthellae and photosynthe-
tic chloroplast pigments of the reef building coral Motastrea
annularis Ellis and Solander in relation to depth on a West
Indian coral reef. Bull. Mar. Sci., 29. 79-95.
Dura dChastel H. (1980). Production and use of Spirulina in Me-
xico. In: Algal Biomass: Production and Use (eds. Shelef G. ,
Boeder C.J.), pp. 51-64. Elsevier/ North Holland, Amsterdam.
Eaton J.W., Brown J.G., Bound F.E. (1966). Some observations on
polarity and regeneration in Enteromorpha. Brit, Phycol. J.,
3, 53-62.
Echlin P., Morris I. (1965). The relationship between blue-green
algae and bacteria. Biol.Rev., 40, 143-187.
Edelstein T., McLachlan J. (1971). Further observations on Gloio-
siphonia capillaris (Hudson) Carmichael in culture. Phycolo-
gia, 10, 215-219.
Edelstein T. , Chen L., McLachlan J. (1970). The life cycle of
Ralfsia clavata and R.bometii. Can. J.Bot., 48, 527-531.
Edmonston W.T. (1970). Phosphorus, nitrogen and algae in Lake
Washington after diversion of sewage. Science, 169, 690-691.
Edmonston W.T. (1972). Nutrients and phytoplankton in Lake Washin-
gton. Limnol. Oceanogr., Special Syrnp., I., 172-193.
480
Edmonston W.T. (1979)* Lake Washington and the predictability
of limnological events. Arch. Hydrobiol., 13. 234-241.
Edmonston W. T., Lehman J.T. (1981). The effect of changes in
the nutrient income on the condition of Lake Washington. Lim-
nol. Oceanogr., 26. 1-29.
Edwards P. (1969). Field and cultural studies on the season pe-
riodicity of growth and reproduction of selected Texas benthic
marine algae. Contrib. Mar. Sci., 14. 59-114.
Edwards P. (1973). Life history studies of selected British Ge-
rami um species J. Phycol., _9, 181-184.
Edwards P. (1976). A classification of plants into higher taxa
based on cytological and biochemical criteria. Taxon, 25.
529-542.
Egerod L.E. (1952). An analysis of the siphonous Chlorophycophyta.
Univ.Calif. Publ. Bot., 2£, 325-454.
Ehresmann D.W., Dieg E.F., Hatch M.T. (1979). Antiviral properties
of algal polysaccharides and related compounds. In: Marine Al-
gae in Pharmaceutical Science (eds. Hoppe H.A., Levringe T.,
Tanaka Y. ), pp. 294-302. Walter de Gruyter, Berlin.
Ellis R.J. (1976). Protein and nucleic acid synthesis by chloro-
plasts. In: The Intact Chloroplast (ed. Barber J.), pp. 335-
364. Topics in Photosynthesis, Vol. I. Elsevier/North-Holland,
Amsterdam.
Emerson C.J., Buggeln R. G., Bal A. K. (1982). Translocation in
Saccorhiza dermatodea (Laminariales, Phaeophyceae): anatomy
and physiology. Can.J.Bot., 60, 2164-2184.
Engelman T.W. (1893). Farbe und Assimilation. Botanische Zeitung,
Ц, 1-29.
Eppley R.W., Coatsworth J.L. (1968). Nitrate and nitrite uptake
by Bitylum brightwellii. Kinetics and mechanisms. J. Phycol.,
£, 151-156.
Eppley R.W., Peterson B.J. (1979). Particulate organic matter
flux and planktonic new production in the deep ocean. Nature,
282 , 677-680.
Eppl®r R.W., Renger E.H. (1974). Nitrogen assimilation of an
oceanic diatom in nitrogen-limited continuous culture. J.Phy-
col. , 10, 15-23.
Eppley R.W., Renger E.H., Williams P.M. (1976). Chlorine reactions
with seawater constituents and the inhibition of photosyntheis
of natural marine phytoplankton, Est.Coast. Mar.Sci., £, 147-
161.
481
Eriksson F., Homstrom E., Mossberg P., Nyberg P. (1983). Ecolo-
gical treatment of acidified lakes and rivers in Sweden. Hyd-
robiologia, 101, 145-164.
Erwin J., Bloch K. (1963). Polyunsaturated fatty acids in some
photosynthetic organisms. Biochem.Z., 388, 496-511.
Esser K. (1982). Cryptogams: Cyanobacteria, Algae, Fungi, Lichens.
Cambridge University Press, Cambridge.
Estes J.A., Palmisano J.F. (1974). Sea otters: their role in
structuring nearshore communities. Science, 185. 1058-1060.
Ettl H. (1978). Xanthophyceae. In: SiJsswasserflora von Mitteleuro-
pa, Bd. 3, 1 Teilf (eds. Ettl H., Gerloff J., Heynig H.).
Gustav Fischer, Stuttgart.
Ettl H. (1981). Die neue Klaese Chlamydophyceae, eine naturliche
Gruppe der Grunalgen (Chlorophyta). Pl.Syst. Evol., 137, 107-
126.
Ettl H., Komarek J. (1982). Was verstehl man unter dem Begriff
"coccale Grunalgen"? Arch.Hydrobiol.(Suppl.), 60, 345-374.
Evens L.V. (1965). Cytological studies in the Laminariales.
Ann.Bot.N.S. , 22, 541-562.
Evans L.V. (1981). Marine algar and fouling: a review with par-
ticular reference te ship fouling.Bot.Mar. , 24. 167-171.
Evans L.V., Callow J.A., Callow M.E.(1973). Structural and phasio-
logical studies of the parasitic red alga Holmsella. New Phy-
tol., 22, 393-402.
Evans L.V., Callow J.A., Callow M.E. (1978). Parasitic red algae:
an appraisal. In: Modern approaches to the Taxonomy of Red and
Brown algae (eds. Irvine D.E.G., Price J.H. ), pp.87-140. Aca-
demic Press, New York.
Evans L.V., Callow J.A., Callow M.E. (1982). The biology and bio-
chemistry of early development in Fucus, Prog. Phyool.Res.,
1, 67-110.
Evitt W.R. (1963). A discussion and proposals concerning fossil
dinoflagellates, hystrichospheres and acritarchs. Proc. Natl.
Acad. Sci. U.S.A., 49, 158-164.
Fairchild E., Sheridan R.P. (1971). A physiological investigation
of the hot spring diatom Achnanthes exigua Grun.J. Phycol.,
10, 1-4.
Palkenberg P. (1879). Die Befruchtung und der Generationswechsel
▼on Cutleria. Mit. Zcol.Stat.Neapel., 1, 420-447.
Falkowski P.J. (1981). Light-shade adaptation and assimilation
482
numbers. J. Plankton Res., £, 203-216.
Farnham W.F., Fletcher R.L., Irvine L.M. (1973). Attached Sarga-
SBum found in Britan. Nature, 243. 231-232.
Farr E.R., Leussink J.A., Stafleu F.A. (1979). (eds.). Index Nomi-
num.Genericorum (Plantarum). Bohn, Scheltema and Holkema, Utre-
cht.
Farrar J.F. (1978). Symbiosis between fungi and algae. In: CRC
Handbook Series in Nutrition and Food (ed.Reichigl M.), pp.
121-139. CRC Press, Cleveland;
Farrar J.F., Smith D.C. (1976). Ecological physiology of the li-
chen Hypogymnia physodes. III. The importance of the rewetting
phase. New Phytol., 72. 115-125.
Farrer W.V. (1966). Tecuitlatl, a glimpse of Aztec food technolo-
gy. Nature, 211, 341-342.
Feher D. (1948). Researches on the geographical distribution of
soil microflora. Part II. The geographical distribution of so-
il algae. Communications of the Botanical Institute of the
Hungarian University of Technical and Economic Sciences, Sop-
ron (Hungary), 21. 1-37.
Feldmann J. (1951). Ecology of marine algae. In: Manual of Phy-
oology - an Introduction to the Algae and their Biology (ed.
Smith G.M.), pp.313-334. Waltham, Massachusetts.
Fenchel T. (1980). Suspension feeding in ciliated protozoa: feed-
ing rates and ecological significance. Microb. Eool., 6, 13-25»
Fenical W. (1980). Distributional and taxonomic features of toxin
producing macroalgae. In: Pacific Seaweed Aquaculture (eds.
Abbott I.A., Foster S. , Eklund L.F. ), pp. 144-151. Institute
of Marine Resources, University of California, La Jolla.
Feochari C. (1979). Use of Laminaria tent in obstetrical practi-
ce In: Marine Algae in Pharmaceutical Scicence (eds. Hoppe H.A.,
Levring T., Tanaka Y.), pp. 663-673. Walter de Gruyter, Berlin.
Fjeld A., Lovlie A. (1976). Genetics of multicellular algae. In:
Genetics of Algae (ed. Lewin R.A.), pp. 219-235. University
of California Press, Berkeley.
Fiksdahl A., Withers N., Guillard R.R.L., Liaaen-Jensen S. (1984).
Garotenoids of the Raphidophyceae - a chemosynthetio contri-
bution. Comp.Biochem.Physiol., 78, B(1), 265-271.
Fisher C.R., Trench R.K.* (1980). In vitro carbon fixation by
Prochloron sp. isolated from Diplosoma virens. Biol.Bull.,
159, 636-648.
Fitt W.K., Pardy R.L. (1981). Effects of starvation, and light
483
and dark on the energy metabolism of symbiotic and aposymbio-
tic sea anemones, Anthopleura elegantissima.Mar. Biol., 61,
199-205.
Fletcher R.L. (1975). Heteroantagonism observed in mixed algal
cultures. Nature, 253. 534-535.
Flett R.J., Schindler D.W., Hamilton R.D., Campbell N.E.P. (1980).
Nitrogen fixation in Canadian pre-cambrian Shield lakes. Can.
J. Fish Aquat.Sci., 21» 494-505-
Floc'h J.Y., Diouris M. (1981). Impact du petrole de I'Amoco Cadiz
sur les algues de Portsail: suivi ecologique dans une anse
tree polluee. Tn: Amoco Cadiz: Consequences d'une pollution
aooidentelle par les hydrooarbures, pp.381-391. Actes du Col-
loque International Publ. Centre National pour 1’Exploration
des Oceans, Paris.
Flowers A., Bryce A.J. (1977). Energy from marine biomass. Sea
Technol., Oct., 1977, 18-21.
Floyd G.L., O'Kelly C.J. (1984). Motile cell ultrastructure and
the circumscription of the orders Ulotriohales and Ulvales
(Ulvophyceae, Chlorophyta). Am.J.Bot., 71» 111-120.
Floyd G.L., Hoops H.J., Swanson J.A. (1980). Fine structure of
the zoospore of Ulothrix belkae with emphasis on the flagel-
lar apparatus. Protoplasms, 104. 17-31.
Fogg G.E. (1967). Observation on the snow algae of the South Or-
kney Islands. Phil.Trans.Roy.Soc.Bond. B., 252. 279-287.
Fogg G.E. (1982). Marine plankton. In: The Biology of the Cyano-
bacteria (eds. Whitton B.A., Carr N.G.), pp. 491-513. Blackwell
Scientific Publications, Oxford.
Fogg G.E., Stewsrt W.D.P., Fay P., Welsby A.E. (1973). The Blue-
Green Algae, Academic Press, New York.
Ford T.W. (1984). A comparative ultrastructural study 6f Cyani-
dium caldarium and the unicellular red alga Rhodosorus mari-
nus. Ann. Bot., 22. 285-294.
Foster M.S. (1975). Algal succession in a Macrocystis pyrifera
forest. Mar.Biol., 22, 313-329.
Foster P.L. (1982a). Species associations and metal contents of
algae from rivers polluted by heavy metals. Freshwater Biol.,
22, 17-39.
Foster P.L. (1982b). Metal resistances of Chlorophyta from rivers
polluted by heavy metals. Freshwater Biol., 12, 41-46,
Fott B., Novakova M. (1969). A monograph of the genus Chlorella.
The fresh water species. In: Studies in Phyoology (ed. Fott B.),
484
pp.10-70. Academia, Prague.
Fox G.E., Stackebrandt E., HespellR.B., Gibeon J., Maniloff J.,
Dyer T.A., Wolfe R.S., Balch W.E., Tanner R.S., Magrum L.J.,
Zablen L.B., Blakemore R., Gupta R., Bohen L., Lewis B.J.,
Stahl D.A., Luenrsen K.R., Chen K.N., Woese C.R. (1980). The
phylogeny of prokaryotes. Science, 209, 457-463.
Foyn B. (1934). Lebenszyklus und Sexualitjlt der Chlorophyceae
Ulva lactuca L. Arch. Protistenk., 83, 154-177.
Pralick R.A., Mathieson A.C. (1972). Winter fragmentation of
Codium fragile (Suringer) Hariot spp. tomentosoidea (van Goor)
Silva (Chlorophyceae, Siphonales) in New England. Phycologia,
11, 67-70.
Pranceschi V.R., Lucas W.J. (1980). Structure and possible fun-
ctions) of charasomes complex plasmalemma - cell wall ela-
borations present in some Characean species. Protoplasma,
104. 253-272.
Francis D. (1967). On the eyespot of the dinoflagellate Nemato-
todinum. J.Exp.Biol., 47, 495-502.
Francis G. (187B). Poisonous Australian lake. Nature, IB, 11-12.
Frank N., Fergusson L.C., Cross R.F., Redman D.R. (1969). Proto-
theca, a cause of bovine mastitis. Am.J.Vet.Res., 22» 1785-
1794.
Frei E., Preston R.D. (1961). Variants in the structural poly-
saccharides of algal oell walls. Nature, 192, 939-943»
Frei E., Preston R.D. (1964). Non-cellulosis structural polysac-
charides in algal cell walls. II. Association of xylan mannan
in Porphyra umbilicalis. Proc. Roy.Soc.Lond. B., 160, 314-327.
Friedmann I. (1959). Structure, life history and sex determina-
tion of Prasiola stipitata Suhr.Ann.Bot. N.S., 23, 571-594.
Friedmann E.I. (1982). Cyanophycota. In: Synopsis and Classifi-
cation of Living Organisms, Vol. 1 (ed. Parker S.P.), pp.
45-52. McGraw-Hill, New York.
Fries L. (1966). Influence of iodine and bromine on growth of
some red algae in axenic culture. Physiol. Plant., 19, 800-
B08.
Fries L. (1969). The sporophyte of Nemalion multifidum (Weber
and Mohr) J.Ag. found on the Swedish West Coast. Svensk Bot.
Tidskr. , 61., 457-462.
Fries L. (1973). Requirements for organic substances in seaweeds.
Bot.Mar. , 1.6, 19-31.
485
Fries L. (1975). Requirement of bromine in a red alga. Z. Pflan-
zenphysiol., 76, 366-368.
Fries N. (1979). Physiological characteristic of Mycosphaerella
ascophylli, a fungal endophyte of the marine brown alga As-
cophyllum nodosum. Physiol. Plant., 45., 117-121.
Fritsch F.E. (1935). Structure and Reproduction of the Algae,
Vol. I. Cambridge University Press, Cambridge.
Fritsch F.E. (1945). Structure and Reproduction of the Algae,
Vol. II. Cambridge University Press, Cambridge.
Frost B.W. (19B0). Crazing. In: The Physiological Ecology of
Phytoplankton (ed. Morris I. ), pp. 465-491. University "'f
California Press, Berkeley.
Fryxell G.A. (ed.) (1983). Survival Strategies of the Algae.
Cambridge University Press, Cambridge.
Fryxell G.A., Hasle G.R. (1979). The genus Thalassiosira: spe-
cies with internal extensions of the strutted processes.
Phycologia, 18, 378-393.
Fryxell G.A., Hasle G.R. (1980). The marine diatom Thalassiosira
oestrupii; structure, taxonomy and distribution. Am.J.Bot.,
67, 804-814.
Fuhrman J.A., Chisholm S.W., Guillard R.R.L. (1978). Marine al-
gae Platymonas sp. accumalates silicon without apparent
; requirement. Nature, 272, 244-246.
' Fujiyama T. (1955). On the life history of Prasiola japonica
' Yatabe. J.Fac.Fish.Hiroshima Univ., 1_, 15-45.
Gagne J.A., Mann K.H., Chapman A.R.O. (1982). Seasonal patterns
of growth and storage in Laminaria longicruris in relation
j!.*,- to patterns of availability of nitrogen in the water. Mar.
Biol., 69, 91-101.
A* Gaillard J. (1972). Quelques remarques sur le cycle reproducteur
des Dictyotales et sur see variations. Soc. Bot.Fr, Mein.,
1972, 145-150.
Gale N.I. , Hardie M.G., Jennet J.C*, Aleti A. (1973). Transport
of trace pollutants in lead mining waters. In: Trace Sub-
stances and Environmental Health, Vol. VI (ed. Hemphell D.D.),
pp.95-106. University of Missouri, Columbia, Missouri.
Gallop A. (1974). Evidence for the presence of a "factor" in
Elysia viridis which stimulates photosynthate release from
its symbiotic chloroplasts. New Phytol., 73, 1111-1117.
Gallop A., Bartrop J., Smith D.C. (1980). The biology of chloro-
plast acquisition by Elysia viridis. Proc. Roy.Soc.bond. B.,
486
207, 335-349.
Ganf G. G. (1974). Phytoplankton biomass and distribution in a
shallow eutrophic lake (Lake George, Uganda). Oecologia, 16,
9-29.
Gantt E. (1971). Mlcromorphology of the periplast of Chroomonas
sp. (Cyanophyceae). J. Phycol., 7, 177-184.
Gantt E. (1981). Phycobilisomes. Ann.Rev. Plant Physiol., 32,
327-347.
Gantt E., Conti S.P. (1965). The ultrastructure of Porphyridium
cruentum. J.Cell Biol., 26, 365-381.
Gantt E., Edwards M.R., Conti S.P. (1968). Ultrastructure of
Porphyridium aeruginosum, a blue-green colored Rhodophytan.
J. Phycol., 4, 65-71.
Gantt E., Edwards M.R., Provaeoli L. (1971). Chloroplast struc-
ture of the Cryptophyceae. Evidence for phycobilisomes within
intrathylakoidal space. J. Cell.Biol. , 4jB, 280-290.
Garbary D.J. (1976). Life forms of algae and their distribution.
Bot. Mar., 12, 97-106.
Gayral P., Presnel-Morange J. (1971). Resultats preliminaires
sur la structure et la biologie de la coccolithacee Ochro-
sphaera neopolitana Schussnig C.R. Acad.Sci. Paris D., 273.
1683-1686.
Geider R.J., Osborne B.A., Raven J.A. (1986). Growth photosyn-
thesis and maintenance metabolio cost in the diatom Phaeodac-
tylum tricornutum at very low light levels. J. Phycol., 22,
39-48.
Geitler L. (1959). Syncyanosen. In: Handbuch der Pflansenphysio-
logie II, pp.530-536. Springer-Verlag, Berlin.
Geitler L., Schiman-Cseika H. (1970). Ueber das sogennante Pal-
mellastadium von Phaeothamnion confervicola. Oesterr. Bot. Z.,
118, 393-296.
Gibbs S.P. (1962a). The ultrastructure of the pyrenoids of algae,
exclusive of green algae. J.Ultrastr. Res., 7, 247-261.
Gibbs S.P. (1962b). The ultrastructura of the chloroplasts of
algae. J.Ultrastr.Res., 7, 41B-435.
Gibbs S.P. (1978). The chloroplasts of Euglena may have evolved
from symbiotic green algae. Can.J.Bot., 56, 28B3-28B9.
Gibbs S.P. (1981). The chloroplasts of some algal groups may
have evolved from endosymbiotic eukaryotic algae.Ann.N.Y.
Acad.Sci., 361, 193-207.
487
Giddings T.R.Jr., Withers N.W., Staehelin L.A. (1980). Supramo-
lecular structure of stacked and unstacked regions of the
photosynthetic membranes of Prochloron sp., a prokaryote.
; Proc. Natl Acad.Sci. U.S.A., 77, 352-356.
.Gillott M.A., Gibbs S.P. (1980). The cryptomonad nucleomorPh:
its ultrastructure and evolutionary significance. J.Phycol.,
1_6, 558-568.
Giraud G., Gabioch J. (1979). Ultrastructure and elaboration of
calcified cell walls in the coralline algae (Rhodophyta, Cryp-
tonemailes). Biol.Cell., 36, 81-86.
Glider W.V., Pardy R.L.(1982). Algal endozoic symbioses: an
introduction and bibliography. In: Selected Papers in Phyco-
logy II (eds. Rosowski J.R., Parker B.C.), pp.761-772.
Phycological Society of America, Lawrence, Kansas.
Glombitza KrW. (1977). Highly hydroxylated phenols of the Phae-
ophyceae. In: Marine Natural Products Chemistry (eds. Faulk-
ner D.J., Fenical W.H.). pp. 191-204. Plenum Press, New York.
Glombitza KrW. (1979). Antibiotics from algae. In: Marine Algae
and Pharmaceutical Science (eds. Hoppe H.A., Levring T.,
Tanaka Y.), pp. 303- 342. Walter de Gruyter, Berlin.
Glooschenko W.A., Curl H.Jr. (1971). Influence of nutrient en-
richment on photosynthesis and assimilation rations in na-
tural North Pacific phytoplankton communities. J.Pish.Res.Bd.
Can., 28, 790-793.
Gmelin S.G. (176B). Historia Fucorum. Petropol.
Godward M.B.E. (1961). Meiosis in Spirogyra crassa. Heredity,
16, 53-62.
Godward M.B.E. (1966). The Chromosomes of the Algae. Edward
Arnold, London.
Godfrey P.J. (1982). The eutrophycation of Cayuga Lake: a his-
torical analysis of the phytoplankton’s response to phospha-
te detergents. Freshwater Biol., 12, 149-166.
Goff L.J. (1979). The biology of Harveyella mirabilis (Crypto-
nemiales, Rhodophyceae). VI. Translocation of photoassimila-
te UC. J. Phycol., 1_5, 82-87.
Goff L.J. (1982a). Symbiosis and parasitism: another viewpoint.
BioScience., 32, 255-256.
Goff L.J. (1982b). The biology of parasitic red algae. Prog.
PhycoIl Res., 1, 239-369.
Goff L.J. (ed.) (1983). Algal Symbiosis: A Continuum of Interac-
tion Strategies. Cambridge University Press, Cambridge.
Goff L.J., Coleman A.W. (1984). Elucidation of fertilization
488
and. development in a red alga by quantitative DNA microspec-
trophotometry.Dev.Biol., 102, 173-194»
Goff L.J., Coleman A.W. (1985)» The role of secondary pit connec-
tions in red algal parasitism. J.Phycol. , 21, 483-508.
Goldman J.C., McCarthy J. J., Peavey D.G. (1979). Growth rate
influence on the themical composition of phytoplankton in
oceanic waters. Nature, 279, 210-215.
Goldstein M. (1964). Speciation and mating behaviour in Eudorina
J.Protozool., 11, 317-344.
Goldstein M., Marrall S. (1970). Gametogenesis and fertilization
in Caulerpa. Ann.N.Y. Acad.Sci., 175, 660-672.
Goiterman H.L., Clymo R.S., Ohmstad M.A.M. (eds.) (1978). Methods
for Physical and Chemical Analysis of Freshwaters, 2nd Edition
IBP Handbook No. 8. Blackwell Scientific Publications, Oxford.
Goodenough U.W., Weiss R.L. (1975). Gametic differentiation in
Chlamydomonas reinhardtii, III. Cell wall lysis and micro-
filament=associated mating structure activation in wild-type
mutant strains.J.Cell Biol., 67. 623-637.
Gordon D.C.Jr. , Prouse N.J. (1973). The effect of three oils on
marine phytoplankton photosynthesis. Mar.Biol., 22, 329-333»
Gorham J. (1977). Plant growth regulators in Sargassum muticum.
J. Phycol., 13 (suppl.), 25.
Gotham I.J., Rhee GrY. (1982). Effects of hexachlorobiphenyl
and pentachlorophenol on growth and photosynthesis of phyto-
plankton. J.Great Lakes Res., 8, 328-335»
Govindjee, Braun B.Z. (1974)» Light absorption, emission and
photosynthesis. In:Algal Physiology and Biochemistry (ed.
Stewart W.D. F.), pp. 346-390. Blackwell Scientific Publica-
tions, Oxford.
Graham J»M., Auer W.R., Canale R.P., Hoffman J.P. (1982).
Ecological studies and mathematical modelling of Cladophora
in lake Huron.4. Photosynthesis and respiration as functions
of light and temperature. J.Great Lakes Res., 8, 100-111.
Graham L.E. (1985). The origin of the life cycle of land plants.
Am.Sci., 7J, 178-186.
Graham L.E., McBride G.E.(1979)» The occurrence and phylogenetic
significance of a multilayered structure in Coleochaete врез>-
matozoids. Am.J.Bot., 66, 8B7-924.
Gramblast L.J. (1960). Extension Chronologique des generes chez
les Charoideae.Soc.Edit.Technip., Paris.
489
Gramblast L.J. (1974)» Phylogeny of the Charophyta.Taxon, 23.
463-481.
Grant M.C., Sawa T. (19B2). Charophyceae: introduction and biblio-
graphy. In: Selected Papers in Phycology II (eds. Rosowski J.R.,
Parker B.C. ), pp.754-759. Phycological Society of America,
Lawrence,Kansas.
Grasso P., Sharrat M. (1973). Studies on carrageenan and large
bowel ulceration in mammals. Pood Cosmet.Toxicol., 11, 555-564.
Grauer F.H., Arnold H.L.Jr., (1961). Seaweed dermatitis: first
report of a dermatitis^producing marine alga. Arch.Dermatol.,
84, 720-732.
Green J.C. (1980). The fine structure of Pavlova pinguis Green
and a preliminary survey of the order Pavlovales (Prymnesio-
phyceae). Brit. PhycolJ., 15. 151-191.
Green J.C., Hibberd D.J. (1977). The ultrastructure and taxonomy
of Diacronema vlkianum (Prymnesiophyceae) with special refe-
rence to the haptonema and flagellar apparatus. J.Mar.Biol.
Ass.U.K., 5X, 1125-1136.
Green J.C., Pienaar R.N. (1977). The taxonomy of the order Isochry-
sidales (Prymnesiophyceae) with special reference to the ge-
nera Isochrysis Parke, Dicrateria Parke, and Imantonia
Reynolds. J.Mar.Biol.Ass. U.K., 57, 7-17.
Greta M.R. , Aronson J.M., Sommerfeld M.R. (1980). Cellulose in
the cell walls of the Bangiophyceae (Rhodophyta). Science,
207. 779-780.
Greuet C. (1972). La nature trichocystaire du cnidoplaste dans le
complexe cnidoplaste-nematocyste de Polykrikos schwartai But-
schli.C.R.Acad.Sci. Paris D., 275, 1239-1242.
Greuet C. (1977). Evolution structurale et ultrastruoturale de
I'ooellofde d'Erythropsidinium pavillardi Kofoid et Swezy
(Peridinien Warnowiidae Lindemann) au Cours des divisions bi-
naire et palintomiques. Protistologica, 1 3. 127-143.
Greville R.K. (1830). Algae Britannicae.Edinburgh.
Groover R.D., Bold H.C. (1969). Phycological studies. VIII. The
taxonomy and comparative physiology of the Chlorosaroinales
and certain other edaphio algae. Univ.Texas.Publ., 6907.
1-165.
Grim W., Allemann P. (1975). The lower cretaceous of Caravaca
(Spain): Berriasian calcareius nannoplankton of the Mlrauetes
section (Subbetic Zone, Prov. of Murcia). Ecologae Geol.Helv.,
490
68, 147-211.
Guiry M.D. (1977). The importance of sporangia in the classifi-
cation of the Florideophyceae. In: Modern Approaches to the
Taxonomy of Red and Brown Algae (edB. Irvine D.E.G., Price
J.H.), pp. 111-144. Academic Press, London.
Guiry M.D., West J. A., Kim D.H., Masuda M. (1984). Re-instate-
ment of the genus Mastocarpus Kiitzing (Rhodophyta). Taxon,
33, 53-63.
Gunnill P.O. (1980). Demography of the intertidal brown alga
Pelvetia fastigiata in southern California, U.S.A. Mar. Biol.,
59, 169-179.
Haddad K., Carter K. (1979). Oceanic intrusion: one posible ini-
tiation mechanism of red tide blooms on the west coast of
Florida. In: Toxic Dinoflagellate Blooms (eds. Taylor D.L.,
Seliger H.H.), pp. 269-274. Developments in Marine Biology 1.
Elsevier/North-Holland, New York and Amsterdam.
Haeckel E. (1B96). Systematische Phylogenie, Entwurf eines na-
tiirlichen Systems der Organismen auf Grund ihrer Stammenges-
ohiohte. Zweiter Theil. Systematische Phylogenie der Wirbello-
sen Thiere (Invertebrata). Verlag V. Georg Reiser, Berlin,
8 vols.
Haines T. (1981). Acid precipitation and its consequences for
aquatic ecosystems: a review. Trans.Am.Fish,Soc.,110,669-707.
Hall A. (1981). Copper accumulation in copper tolerant and non-
tolerant species populations of the marine fouling alga,
Ectocarpus siliculosus (Dillw.) Lyngbye. Bot. Mar., 24,
223-228.
Hall A., Fielding A.H., Butler A.H. (1979). Mechanisms of cop-
per tolerance in the marine fouling alga Ectocarpus silicu-
losus - evidence for an exclusion mechanism Mar. Biol., 54,
195-199.
Hall J.B. (1971). Evolution of the Prokaryotes. J. Theor. Biol.,
30, 429-454.
Hallfors G., Thomsen H.A. (1979). Further observations in Chry-
sochromulina birgeri (Prymnesiophyceae) from the Tvarminne
archipelago, SW coast of Finland, Acta Bot. Fennica, 110,
41-46.
Hallsson S.V. (1964). The uses of seaweed in Iceland. Proc. Int.
Seaweed Symp., 4_, 398-405.
Hamm D., Humm H.J. (1976). Benthic algae of the Anclote Estuary.
II. Bottom-dwelling species. Florida Sci., 39, 209-229.
491
Hanic L.A., Craigie J.S. (1969)» Studies on the algal cuticle.
J.Phycol., 89-102.
Hanisak M.D. (1979). Effect of indole-3-acetic acid on growth of
Codium fragile subsp. tomentosoides (Chlorophyceae) in cul-
ture. J.Phycol., 15. 124-127.
Haneen G.I. (1980). A morphological study of Fibriofolium, a new
genus in the Cystocloniaceae (Gigartinales, Rhodophyta), J.
Phycol., 16, 207-217.
Happey-Wood C.M. (1976). Vertical migration patterns in phyto-
plankton of mixed species composition. Brit. Phycol. J., 11,
355-369.
Hardin G. (I960). The competitive exclusion principle. Science,
131. 1292-1298.
Hargraves P.E., French F.W. (1983). Diatom resting spores: signi-
ficance and strategies. In: Survival strategies of the Algae
(ed. Fryxell G.A.), pp.49-68. Cambridge University Press,
Cambridge.
Hargreaves J.W. (1976). Effects of pH on tolerance of Hormidium
rivulare to zinc and copper. Oecologia, 26, 235-243.
Hargreaves J.W. , Lloyd E.J.H., Whitton B.A. (1975). Chemistry
and vegetation of highly acidic streams. Freshwater Biol.,
5, 563-576.
Hargreaves J.W. , Whitton B.A. (1976). Effect of pH on growth
of acid stream algae. Brit. Phycol. J., 11, 215-223.
Harlin M.M. (1980). Seagrass epiphytes. In: Handbook of Seagrass
Biology. An Ecosystem Perspective (ed. McRoy C.P.), pp. 117-
152. Garland STPM Press, New York.
Harlin M.M., Craigie J.S. (1975). The distribution of photosyn-
thate in Ascophyllum nodosum as it relates to epiphytic
Polysiphonia lanosa. J.Phycol., 11, 109-113.
Harold F.M. (1966). Inorganic polyposphates in biology: structure
metabolism and function. Bacteriol.Rev., ДО» 772-794.
Harris G.P. (1980). The measurement of photosynthesis in natural
populations of phytoplankton. In: The Phycological Ecology
of Phytoplankton (ed. Morris I.), pp.129-187. Blackwell
Scientific Publications, Oxford.
Harrison P.J., Conway H.L., Holmes R.W., Davis C.O. (1977). Ma-
rine diatoms growth in chemostat under silicate of ammonium
limitation. III. Cellular chemical composition and morphology
of Chaetoceros debilis, Skeletonema costatum. and Thalassio-
sira gravida. Mar. Biol., 43, 19-31.
492
Hartnell A.D. (1975). Hydrographic factors affecting the distri-
bution and movement of toxic dinoflagellates in the western
Gulf of Marine. Ins Proceedings of the First Conferenoe on
Toxic Dinoflagellate Blooms, (ed. LoCicero V.R.), pp.47-68.
Massachusetts Scientific and Technical Foundation, Wakefield,
Mass.
Hartshorne J.N. (1953). The function of the eyespot in Chla-
mydomonas. New Phytol., 52, 292-297.
Harwey M.J., McLachla J. (eds.) (1973). Chondrus crispus. Nova
Scotia Institute of Science, Halifax, Nova Scotia.
Harvey W.H. (1836). Algae. InzFlora Hfbernica. Mackay J.T., Dub-
lin.
Harvey W.H. (1846-51). Phycologia Britannica, 4 Vols. London.
Harvey W.H. (1852-58). Nereis Boreali-Americana. Pt. I. Melano-
spermae. Smithson. Contrib. Knowl., 3 (Art.4), 1-150, 12 pls.
Pt. II. Rhodospermae. ibid., 4, (Art. 5), 1-258, 24 pls. Pt.
III. Chiorospermae.ibid., 10 (Art.2), 1-140, 14 pls.
Hashimoto Y., Kamiya H., Yamazato K., Nozawa K. (1975). Occurren-
ce of a toxic blue green alga inducing skin dermatitis in
Okinawa. Taxicon, 1_2,, 95—96.
Hasle G.R. (1973). The "mucilage pore" of pennate diatoms. Nova
Hedwiga, 45. 167-194.
Hasle G.R. (1978). Some freshwater and brackish water species
thediatom genus Thalassiosira Cleve. Phycologia, 17, 263-
292.
Haselkorn R. (1978). Heterocysts. Ann.Rev.Plant Physiol., 29,
319-344.
Hatch M.D., Slack C.R. (1966). Photosynthesis by sugar cane
leaves. Biochem., J., 101, 103—111.
Hatcher B. G., Chapman A.R.O., Mann K.H, (1977). An annual carbon
budget for the kelp Laminaria longicruris. Mar. Biol., 44.
85-96.
Haug A., Larsen B., Smidsrod 0. (1966). A study of the consti-
tution of alginic acids by partial acid hydrolysis. Acta Chem.
Scand., 20, 183-190.
Haug A., Larsen B., Smidsrod 0. (1967). Studies on the sequence
of uronic acid residues in alginic acid. Acta Chem.Scand., 21,
691-704.
Haug A., Larsen B., Baardseth E. (1969). Comparison of the consti-
tution of alginates from different sources. Proc. Int. Sea-
weed Symp., 6, 443-451.
493
Haug A., Larsen B., Smidsrod 0. (1974). Uronic acid sequence in
alginate from different sources. Carbohyd.Res., 32, 217-225.
Hauman J.H. (1981). Is a plasmid(s) involved in the toxicity of
Microcystis aeruginosa? In: Algal Toxins and Health. Environ-
mental 'Science Research. Vol. 20 (ed. Carmichael W.W.), pp.97-
102. Plenum Press, New York.
Haupt W. (1983). Movement of chloroplasts under the control of
light. Prog. Phycol.Res., 2, 227-282.
Hawkes M.W. (1978). Sexual reproduction in Porphyra gardneri
Smi th et Hollenberg) Hawkes (Bangiales, Rhodophyta). Phycolo-
gia, 17, 326-350.
Hawkes M.W. (1983). Hummbrella hydra Earle (Rhodophyta, Gigarti-
nales) 4 seasonality, distribution and developnent in labora-
tory culture. Phycologia, 22, 403-413.
Hawkins S.J., Harkin E. (1985). Preliminary canopy removal experi-
ments in algal dominated communities low on the shore and in
the shallow subtidal on the Isle of Man. Bot.Mar., 28., 223-230.
Hawkins S.J., Hartholl R.G. (1985). Factors determining the upper
limits of intertidal canopy forming algae. Mar.Ecol. Prog.Ser.,
20, 265-271.
Healey F.P, (1973). Inorganic nutrient uptake and deficiency in
algae. Crit.Rev. Microbiol., 3, 69-113»
Heath I.B. (1980). Variant mitoses in lower eukaryotes: indicators
of the evolution of mitosis. Int.Rev.Cytol., 64, 1-80.
Hecky R.E., Kilham P. (1974). Environmental control of phytoplank-
ton cell size. Limnol. Oceanogr., 19. 361-366.
Heckey R. F,, Mopper K., Kilham P., Degens E. T. (1973). The amino
acid and sugar composition of diatom cell walls. Mar.Biol., 19.
323-331.
Heerebout G.R. (1968). Studies on the Erythropeltidaceae (Rhodo-
phyceae, Bangiophycidae). Blumea, 16, 139-157.
Heimdal B.R., Gaarder K.R. (1980). Coccolithophorids from the nor-
thern part of the eastern central Atlantic I. Holococcolithopho-
rids. "Meteor" Forsch. - Ergebnisse D, 32, 1-14.
Heimdal B.R., Gaarder K.R. (1981). Coccolithophorids from the
northern part of the eastern central Atlantic II. Heteroroccoli-
thophorids. "Meteor" Forsch, - Ergebnisse D, 33. 37-69,
Hellebust J.A. (1974). Extracellular products. In: Algal Physio-
logy and Biochemistry (ed. Stewart W.D. P.), pp. 838-8 63.
Blackwell Scientific Publications, Oxford.
^endey N.I. (1964). An introductory account of the smaller algae
of British waters. Part V. Bacillariophyceae (Diatoms). Ministry
494
of Agriculture, Fisheries and Food Fisheries Investigation
Series IV, pp. xii + 317. HMSO, London.
Hendey N.I. (1971). Electronmicroscope studies and the classifi-
cation of diatoms. In: The Micropaleontology of Oceans (eds.
Funnell B.M,, Riede R. W. ), pp. 625-631. Cambridge University
Press, Cambridge.
Hendrey G.R. (1982). Acid precipitation effects on algal produc-
tivity and biomass in Adirondak lakes. NTIS publ. PB83-173203.
Henry E.C. (1984). Syringodermatales ord. nov. and Syringoderma
floridana sp. nov. (Phaeophyceae).Phycologia, 23, 419-426.
Henry E.C., Cole K.M. (1982a). Ultrastructure of swarmers in
the Laminariales (Phaeophyceae). I. Zoospores. J.Phycol., 18,
550-569.
Henry E.C., Cole K.M. (1982b) Ultrastructure of swarmers in the
Laminariales (Phaeophyceae). II. Sperm.J.Phyco 1., 18, 570-579.
Henry E.C., Muller D. G. (1983). Studies on the life history of
Syringoderma phinneyi sp. nov. (Phaeophyceae). Phycologia, 22,
387-393.
Herdman M., Stanier R.V. (1977). The cyanelle: chloroplast of
endosymbiotic prokaryote. FEMS Letters 1, 7-12»
Heywood P. (1982). Raphidophyaeae (Chloromonadophyceae)• In:
Selected Papers in Phycology II (eds. Rosowski J.R., Parker
В.C.), pp.719-722. Phycological Society of America, Allen
Press, Lawrence, Kansas.
Hiat R.W., Strasburg D.W. (I960). Ecological relationships of
the fish fauna of the Marshall Islands, Ecol. Monogr., 30,
67-127.
Hibberd D.J, (1970). Observations on the cytology and ultrastruc-
ture of Ochromonas tuberculatus sp. nov. (Chrysophyceae),
with special reference to the discobolocysts. Brit. Phycol.
J., 5, 119-143.
Hibberd D.J. (1971). Observations on the cytology and ultrastruc-
ture of Chrysamoeba radians Klebs (Chrysophyceae). Br. Phycol.
J., 6, 207-223.
Hibberd D.J. (1976). The ultrastructure and taxonomy of the
Chrysophyceae and Prymnesiophyceae (Haptophyceae), a survey
with some new observations on the ultrastructure of the
Chrysophyceae. Bot. J. Linn.Soc., 7.2, 55-80.
Hibberd D.J. (1979). The structure and phylogenetic significance
of the flagellar transition region in the chlorophyll c_-con-
taining algae. Biosystems, 11 , 243-261.
495
Hibberd D.J. (1980a). Xanthophytes. In: Phytoflagellates (ed.
Cox E.R. ), pp.243-272. Elsevier/ North-Holland, New York.
Hibberd D.J. (1980 b). Eumastigiophytes. In: Phytoflagellates
(ed, Cox E.R.), pp.319-334. Elsevier/North-Holland, New York.
Hibberd D.J. (1980p). Prymnesiophytes (=Haptophytes). In: Phyto-
flagellates (ed. Cox E.R.), pp.273-318. Elsevier/North-Holland,
New York.
Hibberd D.J. (1981). Notes on the taxonomy and nomenclature of
the algal class Eustigmatophyceae and Tribophyceae (synonym
Xanthophyceae). Bot. J.Linn.Soc., 82, 93-119.
Hibberd D.J. (1982a). Xanthophyceae. In: Synopsis and Classifi-
cation of Living Orgnaisms, Vol. 1 (ed. Parker S.P.), pp.
91-94- McGraw-Hill, New York.
Hibberd D.J. (1982b). Eustigmatophyceae: introduction and bib-
liography. In: Selected Papers in Phycology II (eds. Rosowski
J.R., Parker B.C.), pp.728-730. Phycological Society of Ame-
rica. Allen Press, Lawrence, Kansas.
Hibberd D.J. (1982c) Eustigmatophytes. In: Synopsis and Classi-
fication of Living Organisms, Vol. 1 (ed. Parker S.P.),
p. 95. McGraw-Hill, New York.
Hibberd D.J., Leedale G.E, (1971). Cytology and ultrastructure
of the Xanthophyceae. II. The zoospore and vegetative cells
of coccoid forms, withspecial reference to Ophiocytium majus
Naegeli. Brit. Phycol. J. , 6, 1-23.
Hibberd D.J., Greenwood A.D., Griffiths H.B.(1971). Observations
on the ultrastructureof the flagella and periplast in Crypto-
phyceae. Brit. Phycol. J. , _6, 61-72.
Hickman M. (1982). The removal of a healted discharge from a
lake and the effect upon an epiphytic algal community. Hydro-
biologia, 82, 21-32.
Hill D.J. (1976). The physiology of lichen symbiosis. In: Liche-
nology (Progress and Problems) (eds. Brown D.H., Hawksworth
D.L,, Bailey R.H.), pp. 457-496. Academic Press, New York.
Hill G.J.C., Machlis L. (1968). An ultrastructural study of
vegetative cell division in Oedogonium bonsianum.J. Phycol.,
4, 261-271.
Hill R., Bendall F. (i960). Function of two cytochrome compo-
nents: a working hypothesis. Nature, 186, 136-137.
496
Hinde R. (1983). Retention of algal chloroplasts by molluscs.
In: Algal Symbiosis: A Continuum of Interaction Strategies
(ed. Goff L. J. ), pp.97-107. Cambridge University Press,
Cambridge
Hinde R,, Smith D.C. (1974). Chloroplast "symbiosis" and the
extent to which it occurs in Sacoglossa (Gastropoda, Mollus-
cs). Biol. J.Linn.Soc., 6, 349-356.
Hitch C.J.B., Millbank J.W. (1975). Nitrogen metabolism in li-
chens. VII. Nitrogenase activity and heterocyst frequency
in lichens with blue-green phycobionts. New Phytol., 75. 239-
244.
Hodgson L.M. (1980). Control of the intertidal distribution of
Gastroclonium coulter! (Harvey) Kylin, in Monterey Bay, Cali-
fornia. Mar. Biol,, 57. 121-126.
Hoek C. van den (1975). Phytogeographic provinces along the coast
of the northern North Atlantic Ocean. Phycologia, 14. 317-330.
Hoek C, van den (1981). Chlorophyta: morphology and classifica-
tion. In: The Biology of Seaweeds (eds. Lobban C.S. , Wynne
M.J,), pp.86-132. Blackwell Scientific Publications, Oxford.
Hoek C. van den (1982a). A Taxonomic Revision of the American
Species of Cladophora (Chlorophyceae) in the North Atlantic
Ocean and their Geographic Distribution. North-Holland, Am-
sterdam.
Hoek C, van den (1982b). Phytogeographic distribution of benthic
marine algae in the North Atlantic Ocean. A review of experi-
mental evidence from life history studies. Helgol. Will. Meere-
sunters., 25» 153-214.
Hoek C. van den (1982c). The distribution of benthic marine algae
in relation to the temperature regulation of their life his-
tories. Biol. J.Linn.Soc., 18. 81-144.
Hoek C. van den, Flinterman A. (1968), The life history of Spha-
celaria furcigera Kutz (Phaeophyceae). Blumea, 16. 193-242.
Hoek C. van den, Jahns H.M. (1978). Einfuhrung in die Phykologie.
Georg Thieme Verlag, Stuttgart.
Hoek C. van den, Colijn F. , Cortel-Breeman A.M., Wanders J.B.W.
(1972). Algal vegetation-type along the shores of inner bays
and lagoons of Curacao and the lagoon (Bonaire) Netherlands
Antilles. Ver.K.Ned.Akad.Wet. , 61. 1-72.
Hoffman L.R. (1965). Cytological studies of Oedogonium I. Oospo-
re germination in 0,foveolatum. Am.J.Bot., 52. 173-181,
Hoffman L.R,, Manton I. (1962). Observations on the fine struc-
497
ture of the zoospore of Oedogonium cardiacum with special
reference to the flagellar apparatus. J.Exp.Bot., 13. 443-
449.
Hoffmann H.J. (1976). Precambrian microflora, Belcher Islands,
Canada: significance and systematics. J.Paleontol., 52, 1040-
1073.
Hoffmann H.J., Aitken J.P. (1979). Precambrian biota from the
Little Gal Group, McKenzie Mountains, northwestern Canada.
Can.J, Earth Sci., 16. 150-166.
Hoham R.W. (1975a). Optimum temperatures and temperature ranges
for growth of snow algae.Arct.Alp.Res., 2, 13-24.
Hoham R.W. (1975b). The life history and ecology of the snow
alga Chloromonas pichinchae (Chlorophyta, Volvocales). Phy-
cologia, 16, 53-68.
Hoham R.W. (1980), Unicellular chiorophytes - snow algae. In:
Thytoflagellates (ed. Cox E.R.), pp. 61-84. Elsevier/North-
Holland, New York.
Hoham R.W,, Blinn D.W. (1979). The distribution of algae in an
arid region. Phycologia, 18. 133-145.
Hoham R.W., Roemer S.C., Mullet J.E. (1979). The life history
and ecology of the snow alga Chloromonas breviaspina comb,
nov. (Chlorophyta, Volvocales). Phycologia, 18. 55-70.
Holcomb G.E. (1975). Hosts of the alga Cephaleuros virescens
in Louisiana. Proc.Am.Phytopath.Soc,, 2, 134.
Hollande A., Enguinet M. (1955). Sur I'evolution et la syste-
matique des Labyrinthulidae; Etude de Labyrinthula algenensis
nov. sp. Ann.Soi.Nat.Zool. II ser., 17. 357-368.
Hollenberg G.J. (1935). A study of Halicystis ovalis. I. Morpho-
logy and reproduction. Am,J.Bot,, 22. 782-812.
Holligan P.M. , Gooday G.W. (1975). Symbiosin in Convoluta ros-
cbffensis. Symp.Soc.Exp.Bio1., 29. 205-227.
Holmes R.W. (1956). The annual phytoplankton in the Labrador
Sea, 1950-1951. Bull.Bingham Oceanogr.Col., 16. 1-74.
Holmes R.W., Crawford R.M., Round P.E. (1982). Variability in
the structure of the genus Cocconeis Ehr. (Bacillariophyta)
with special reference to the cingulum. Phycologia, 21. 370-
381.
Hooper R., South G.R. (1977). Additions to the benthic algal
. flora of Newfoundland III, with observations on species
new to eastern Canada and North America. Naturaliste Can.
(Qu6.), 104 383-394.
498
Hooper R.G., South G.R.,Whittick A. (1980). Ecological and phe-
nological aspects of the marine phytobenthos of the Island
of Newfoundland. In: The Shore environment. Vol. 2; Ecosys-
tfems (eds. Price J.H., Irvine D.E.G,, Farnham W. F.) ,PP«395-423.
Systematics Association Special Volume 17b Academic Press,
London and New York.
Hopkins G.R, (1978). Structure and Function of Cells. W.B. Saun-
ders , London.
Hoppe H.A. (1979). Marine algae and their products and constitu-
ents in pharmacy. In: Marine algae in Pharmaceutical Scienoe
(eds. Hoppe H.A., Levring T. , Tanaka Y.), pp.25-119. Walter
de Gruyter, Berlin.
Hori T,, Norris R.E,, Chihara M. (1982). Studies on the ultras-
tructure and taxonomy of the genus Tetraselmis (Frasinophy-
ceae). I. Subgenus Tetraselmis. Bot. Mar.Tokyo, 22» 49-61.
Horiguchi T., Chihara M. (19B3). Stylodinium littorale. a new
marine dinococcalean alga (Pyrrhophyta), Phycologia, 22. 23-
28.
Horn af Rantzien H. (1959). Morphological types and organ-genera
of Tertiary charophyte fructifications. Stockh. Contr. Geol.,
4, 45-197.
Horner R., Alexander V. (1972). Algal populations in arctic sea
ice. An investigation of heterotrophy. Limnol. Oceanogr., 17.
454-458.
Hoshaw R.W. (1965). A cultural study of sexuality in Sirogonium
melanosporum J. Phycol., 2» 134-138.
Hovasse R., Mignot J.P,, Lyon L. (1967). Nouvelles observations
sur les trichocystes des Crypotomonadines et les "R boches"
des particules Kappa de Paramecium aurelia Killer. Protisto-
logica, 2» 241-255.
Hoxmark R.C., Nordby 0. (1974). Haploid meiosis as a regular
phenomenon in the life cycle of Ulva mutabilis. Hereditas,
26, 239-250.
Hsiao S. (1969). Life history and iodine nutrition of the mari-
ne brown alga Petalonia fascia (O.F.Miill.) 0. Kuntze.Can.
J.Bot., 1611-1616.
Huber M.E., Lewin R.A. (1986), An electrophoretic survey of
the genus Tetraselmis (Chlorophyta, Prasinophyceae). Phyco-
logia, 22, 205-209.
Huizing H.J., Rietema H. (1975). Xylan and mannan as cell wall
constituents of different stages in the life histories of
499
some siphoneous green algae.Brit.Phycol. J., 10, 13-16.
HUlbert E.M. , Ryther J.H. , Guillard R.R.L. (I960). Hie phyto-
plankton of the Sargasso Sea of Bermuda. J,Cons, Int, Ex-
plor. Mer., Hi, 115-118.
Hunt M.E. , Floyd G.C., Stout В,B. (1979). Soil algae in field
and forest environments. Ecology, 60, 362-375.
Huntsman S.A.,, Sunda W.G. (1980). The role of trace metals in
regulating phytoplankton growth. In: The Physiological Eco-
logy of Phytoplankton (ed. Morris I.), pp.285-328. Blackwell
Scientific Publications, Oxford.
Hutchins L.W, (1974). The basis for temperature zonation in
geographical distribution. Ecol.Mongr., 17. 81-148.
Hutchinson G.E. (1961). The paradox of the plankton. Am.Nat,,
21, 137-145.
Hutchinson G.E. (1969), Eutrophication past and present. In: Eu-
trophication: Causes, Consequences, Correctives, pp.17-26.
Academy of Science, Washington D.C,
Hutchinson T.C., Hellebust J.A., Tam D., Mackay D., Mascarenhas
R.S., Shiu W.Y. (1978). The correlation of the toxicity of
algae of hydrocarbons with their physical-chemical properties.
In: Hydrocarbons and Halogenated Hydrocarbons in the Aquatic
Environment (eds.Afghan B.K., Mackay D. ). pp.577-586, Envi-
ronmental Science Research, Vol. 16. Plenum Press, New York,
Huth K. (1981). Der Generationswechsel von lemanea fluviatilis
C.Ag. in Kultur. Nova Hedwiga., 34, 177-189.
Ulman W.I., McLachlan J., Edelstein T. (1972). Two assemblages
of marine algae from post-glacial deposits in the eastern
Canadian arctic.Can, J.Earth Sci., 2., 109-115.
Ilmavirta K., KotimahA.L. (1974). Spatial and seasonal varia-
tions in phytoplanktonic primary production and biomass in
the oligotrorhic lake Paajarvi, southern Finland. Ann.Bot.
Fennici, 11, 112-120.
Incoll L.D., Long S.P., Ashmore M.R. (1977). SI units in pub-
lications in plant science,Curr.Adv.Plant Sci., 28, 331-343.
Ingle R.K., Colman B. (1976). The relationship between carbonic
anhydrase activity and glycolate excretion in Coccochloris
peniocystis. Planta, 128, 217-223.
limes D.J., Yarish C. (1984). Genetic evidence for the occur-
rence of asexual reproduction in populations of Enteromorpha
linza (L.) J.Ag. (Chorophyta, Ulvales). Phycologia, 23, 311-
320.
500
Inouye I., Hori T,, Chihara M. (1983). Ultrastructure and taxo-
nomy of Pyramimonaa Lunata. a new marine species of the class
Prasinophyceae. Jap,J.Phycol., 31. 238-249.
Irvine D.E.G., John D.M. (eds.) (1984). Systematics of Green
Algae. Academic Press, London.
Ishizawa K., Wada S. (1979 a). Growth and phototropic bending
in Boergesenia rhizoids. Plant Cell Physiol., 20, 973-982.
Ishizawa K., Wada S. (1979b). Action spectrum of negative photo-
tropism in Boergesenia forbesii. Plant Cell Physiol,, 20.
983-987.
Jackson J. В. C. (1977). Competition on marine hard substrata:
the adaptive significance of solitary and colonial structu-
res. Am.Nat., Ill, 743-767.
Jacobs W.P. (1964). Rhizoid phoduction and regeneration of
Caulerpa prolifera. Publ.Staz.Zool. Napoli, 34. 185—196.
Jacobs W.P., Davis W. (1983). Effects of gibberellic acid on
the rhizome and rhizoids of the algal coenocyte Caulerpa
prolifera in culture. Ann.Bot., 52, 39-41.
Jacobs W.P,, Falkenstein K., Hamilton R.H. (1985). Nature and
amount of auxin in algae. Plant Physiol., 78. 844—848.
Jaffe L.P. (1958). Tropistic responses of zygotes of the Fuca-
ceae to polarized light.Exp.Cell Res., 1 5. 282-299.
Jeffrey C. (1962). The origin and differentiation of the arohe-
goniate landplants. Bot. Not., 115. 446—454.
Jenkin P.M, (1942). Seasonal changes in the temperature of lake
. Windermere (English Lake District). J.Animal Ecol., 11, 248-
269.
Jensen A. (1979). Industrial utilization of seaweeds in the
past, present and future. Proc.Int.Seaweed Symp., 2» 17-34.
Jensen J.B. (1974). Morphological studies in Cystoseiraceae
and Sargassaceae (Phaeophyceae) with special reference to
apical organization. Univ.Calif. Publ.Bot., 68, 1-61.
Jewell W.J., McCarty D.L. (1968). Aerobic decomposition of
algae and nutrient regeneration. Tech.Rep.Fed. Water Pollut.
Control Admin. USA., No. 91.
Johansen H.W, (1981). Coralline Algae, First Synthesis. CRC
Press, Boca Raton, Florida.
Johansson S., Larsson U,, Boehm P, (1980). The Tsesis oil
spill Impact on the pelagic ecosystem. Ma.j> Poll,Bull., 11.
284-293.
501
Johnson C.H., Roeber J.F., Hastings J.W. (1984). Circadian chan-
ges in enzyme concentration account for rhythm of enzyme
activity in Gonyaulax. Science, 223» 1428-1430.
Johnson J.H. (1951). Fossil Algae. In: Manual of Phycology. An
Introduction to the Algae and their Biology, (ed. Smith G.M.),
pp. 193-202. Ronald Press Co., New York.
Johnson C.S., Jones R.G., Hunt R.T. (1977). A seasonal carbon
budget for a laminarian population in a Scottish sea loch.
Helgol.Wiss.Meeresunters., 30, 527-545.
Johnston H.W. (1970). The biological and economic importance of
algae. 3. Edible algae of fresh and brackish waters. Tuatara,
18, 19-35.
Jolliffe E.A., Tregunna E.B. (1970). Studies of HCO^ ion uptake
during photosynthesis in benthic marine algae. Phycologia, 9,
293-303.
Jones N.S., Kain J.M. (1967). Subtidal algal colonization follow-
ing the removal of Echinus.Helg.№i-ss.Iteeresimt., £0, 611-621.
Jonsson S., Chesnoy L. (1974). Etude ultrastructurale de I’inbor-
poration des axonemes flagellaires dans les zygotes in Mono-
stroma grevillei (Thuret) Wittr. Chlorophycee marine. C.R.
Acad.Sci. Paris D. 278. 1557-1560.
Jose G., Chowdray Y.B.K. (1978). Effect of some growth substan-
ces on Trentepohlia efuusa (Kremp) Harlot. Indian J. Plant.
Physiol., 21, 6-33.
Joshi G.V., Karekar M. D., Gowda C.A., Bhosale L. (1974). Photo-
synthetic carbon metabolism and carboxylating enzymes in al-
gae and mangrove under saline conditions. Photosynthetica, 8,
51-52.
Joubert J.J., Rijkenberg F.H.J., Steyn P.I>. (1975). Studies on
the physiology of a parasitic green alga Cephaleuros sp.
Phytopath. Z., 84, 147-152.
Kaever M. (1969). Neue Dasycladaoeen - Afghanopolia fragills
Nogen., Nosp. aus dem Mittel - Eozan von Ost-Afghanistan.
Arguments Paleobotanies, 2» 15-42.
Kahn N., Swift E. (1978). Positive buyoyancy through ionic con-
trol in the non-motile marine dinoflagellate Pyrocystis noti-
luca Murray ex Schuett. Limnol. Oceanogr., 23. 649-658.
Kain J.M. (1979). A view of the genus Laminaria. Oceanogr. Mar.
Biol. Ann.Rev., Г£, 101-161.
Kalff J., Knoechel R. (1978). Phytoplankton and their dynamics
502
in oligotrophic and trophic lakes. Ann.Rev.Ecol.System,
9, 475-495.
Kanazawa A. (1963). Vitamins in algae. Bull.Jap.Soc.Sci. Fish.,
29, 713-731.
. Ketpraun D.F. (1970). Field and cultural studies of Ulva and
Enteromorpha in the vicinity of Port Aransas, Texas. Contrib.
Mar.Sci., 1£, 205-283.
Kapraun D.F. (1977). The genus Polysiphonia in North Carolina,
U.S.A. Bot.Mar., 20, 313-331.
Karn R.C., Starr R.C., Hudock G.A. (1974). Sexual and asexual
differentiation in Volvox obversus (Shaw) Printz, Strains
WD3 and TO7. Arch. Protistenk, 116, 142-148.
Kataoka H. (1975a). Phototropism in Vaucheria geminata. I. The
action spectrum. Plant Cell Physiol., 16, 427-437.
Kataoka H. (1975b). Phototropism in Vaucheria geminata. II. The
mechanism of bending and branching. Plant Cell Physiol.,
16, 439-448.
Kataoka H. (1977). Phototropic sensitivity in Vaucheria gemina-
ta regulated by 3Z , 5Z cyclic AMP. Plant Cell Physiol., 18,
431-440.
Katoh S. (i960). A new copper protein from Chlorella ellipsoideac
Nature, 186, 533-534.
Katsaros C., Galatis B., Mitrakos K. (1983). Fine structural
studies on the interphase and dividing apical cells of
Sphacelaria tribulpides (Phaeophyta). J.Phycol., 19. 16-30.
Kauss H. (1978). Osmotic regulation in algae. Prog. Phytochem.,
5, 1-28.
Kauss P., Hutchinson C.S., Hellebust J., Griffiths M. (1973).
The toxicity of crude oil and its components to fresh water
algae. In: Proceedings of the 1973 Conference on Prevention
and Control of Oil Spills, pp. 703-714. American Petroleum
Institute, Washington D.C.
Keleti G., Sykora J.L., Lippy E.C., Shapiro M.A. (1979). Com-
position and biological propertiosof lipopolysaccharides
isolated from Schizothrix calciola (Ag.) Gomont (Cyanobac-
teria). Appl.Environ. Microbiol., 38, 471-477.
Kershaw K.A. (1985). Physiological Ecology of Lichens (Cambridge
Studies in Ecology). Cambridge University Press, Cambridge.
Kessler E. (1977). Physiological and biochemical contributions
to the taxonomy of the genus Prototheca. I. Hydrogenase,
acid tolerance thermophily, and liquefaction of gelatin.
503
Arch. Mikrobiol., 113. 139-144.
Kessler E. (1962). Physiological and biochemical contributions
to the taxonomy of the genus Prototheca III. Utilization of
organic carbon and nitrogen compounds. Arch. Mikrobiol., 132,
103-106.
Kies L. (1976). Untersuchungen zur Feinstruktur und taxonomischen
Einordung von Gloeoohaete wittrockiana. einer apoplastidialen
capsalen Alge mit blaugriinen Endosymbioten (Cyanellen). Pro-
toplasma, 87. 419-446.
.Kies L. (1980). Morphology and systematic position of some endo-
cyanomes. In: Endocytology. Endosymbiosis and Cell Biology,
Vol. 1 (eds. Schwemmler W., Schenk H.E.A.), pp. 7-10. Walter
de Gruyter, Co., Berlin, New York.
i Kinding A.C., Littler M.M. (1980). Growth and primary productivi-
ty of marine macrophytes exposed to domestic sewage effluents.
Mat. Environ. Res., 2» 81-100.
King J.M., Ward C.H. (1977). Distribution of edaphic algae as
related to land . usage. Phycologia, 16. 23-30.
Kito H. (1982). Recent problems of nori (Porphyra) culture in
Japan. In: Proceedings of the 6th U.S. - Japan Meeting of
Aquaculture (ed. Sinderman C.J.), pp. 7-12. NOAA Technical
Report NMFC Circ. 442.
Kivic P.A., Walne P.L. (1983). Algal photosensory apparatus pro-
1 bably represents multiple parallel evolutions, Biosystems, 1_6,
\ 31-38.
Kjellman F.R. (1897). Phaeophyceae (Fucoideae). In: Die natur-
lichen Pflanzenfamilien, Vol. 1, Ab. 2 (eds. Engler A., Prantl
K.A.E.), 121 pp., 62 figs. Leipzig.
Klaveness D. (1973). The microanatorcy of Calyptrosphaera sphaeroi-
dea, with some supplementary observations on the mfttile stage
of Coccoluthus pelagica. Norw. J.Bot., 20, 151-162.
Klaveness D., Paasche E. (1979). Physiology of coccolithophorids.
In: Biochemistry and Physiology of Protozoa, Vol. 1, 2nd
f Edition (eds. Lewandowsky M., Hunter S.H.), pp.191-215.
Academic Press, New York.
:Klein R.M., Gronquist A. (1967). A consideration of the evolutio-
nary and taxonomic significance of some biochemical, micromor-
phological, and physiological characters in the thallophytes.
Quart.Rev. Biol., 42, 105-296.
KLintworth G.K., Fetter B.F., Nielsen H.S. (1968). Protothecosis,
an algal infection: report of a case in man. J.Med. Microbiol.,
504
1, 211-216.
Knaggs F. (1969). A review of Florideophycidean life histories
and the culture techniques employed in their investigation.
Nova Hedwiga, 18, 293-330.
Knight M. (1923). Studies in the Ectooarpaceae. I. The life
history and cytology of Pylaiella littoralis (L.) Kjellm.
Trans.Roy.Soc. Edinb., 53» 343-361.
Knight M, (1929). Studies in the Ectooarpaceae. II. The life
history and cytology of Ectocarpus siliculosus Dillw. Trans.
Roy.Soc. Edinb., 56. 307-332.
Knoechel R., Kalff J. (1976). Track autoradiography, a metod
for the determination of phytoplankton species productivity.
Limnol. Oceanogr. , 21, 59<^-596.
Koeman R.P.T., Cortel-Breeman A.M. (1976). Observations on the
life history of Elachista fucicola (Veil.)Aresch. (Phaeophy-
ceae) in culture, Phycologia, 15. 107-117.
Kofoid C.A., Skogsberg T. (1928). The Dinoflagellate: the Di-
nophysoidae. Met.Mils. Comp. Zool. Harv., 51. 1-766.
Kohlmeyer J., Kohlmeyer E. (1972). Is AscophyHum lichenized?
Bot. Mar., 1£, 109-112.
Kohlmeyer J., Kohlmeyer E. (1979). Marine Ideology. The Higher
Fungi.Academic Press, New York.
Koop Hs-U. (1979). The life cycle of Acetabularia (Dasyoladales,
Chlorophyceae): a compilation of evidence for meiosis in
the primary nucleus. Protoplasma, 100 , 353-366.
Kornmann P. (1956). Zur Morphologie und Entwicklung von Per-
cursaria percursa. Helgol. Wiss. Meeresunters., £, 259-272.
Kornmann P. (I960). Die heterogene Gattung Gomontia II. Der
fadige Anteil, Eugomonita sacculata nov. gen et. nov.spec.
Helgol. Wiss. Meeresunters., 7, 59-71.
Kornmann P. (1961). Uber Codiolum und Urospora. Helgol. Wiss.
Meeresunters., 8, 42-57.
Kornmann P. (1962). Der Lebenszyklus von Desmarestia viridis.
Helgol. Wiss Meeresunters., 8, 287-292.
Kornmann P. (1964). Der Lebenszyklus von Acrosiphonia arcta.
Helgol. Wiss. Meeresunters., 11. 110-117.
Kornmann P. (1970a). Mutation in the siphonaceous green alga
Derbesia marina. Helgol.Wiss. Meeresunters., 21, 1-8.
Kornmann P« (1970b). Phylogenetische Beziehungen in der Grunal-
gengattung Acrosiphonia. Helgol. Wiss. Meeresunters., 21,
292-304.
505
Kornmann P. (1972). Ein Beitrag zur Taxonomie der Gattung
Chaetomorpha (Cladophorales, Chlorophyta). Helgol. Wise.
Meeresunters., 23. 1-31.
Kornmann P. (1973). Codiolophyceae, a new class of Chlorophyta.
Helgol. Wise. Meeresunters., 25, 1-13.
Kornmann P., Sahling PrH. (1962). Zur Taxonomie und Entwicklung
der Monostroma-Arten von Helgoland. Helgol.Wiss. Meeresunters
8, 302-320.
Kornmann P., Sahling P.H. (1974). Prasiolales (Chlorophyta) von
Helgoland. Helgol.Wise.Meeresunters., 8, 320.
Kraft G.T. (1976). The morphology of Beckerella scalaramosa.
a new specie of Gelidiales (Rhodophyta) from the Philippines.
Phycologia, 15, 85-91.
Kraft G.T.(1978). Studies on marine algae in the lesser-known
families of the Gigartinales (Rhodophyta). III. The Mychodea-
ceae and Itychodeophyllaceae. Aust. J. Bot., 26, 515-610.
Kraft G.T. (1981). Rhodophyta: morphology and classification.
In: The Biology of Seaweeds (eds. Lobban C.S., Wynne M.J.),
pp. 6-51. Blackwell Scientific Publications, Oxford.
Kreger D.R., van der Veer J. (1971). Paramylon in a Chrysophyte.
Acta.Bot. Neer., 19, 401-402.
Kreh T.V., Dewort J.E, (1976). Effects of entrainment through
Oconee Nuclear Station on carbon-14 assimilation rates
of phytoplankton. In: Thermal Ecology II (eds. Esch G.W.,
McFarlane R.W.), pp. 331-335. ERDA. Symposium Series (Conf.
750425), Springfield, Mass., U.S.A.
Kremer B.P. (1981a) Carbon metabolism. In: The Biology of Sea-
weeds (eds. Lobban C.S., Wynne M.J.), pp.493-533, Blackwell
Scientific Publications, Oxford.
Kremer B.P, (1981b). Dark reactions of photosynthesis. Can.Bulli
Fish. Aquat.Sci., 210, 55-82.
Kremer B.P. (1982).'Cyanidium oaldarium: a discussion of bio-
chemical features and taxonomic problems. Brit.Phycol. J.,
11, 51-61.
Kremer B.P^ (1983). Carbon economy and nutrition of the allo-
parasitic red alga Harveyella mirabilis. Mar. Biol., 76,
231-239.
Kremer B.P., Berks R. (1978). Photosynthesis and carbon meta-
bolism in marine and freshwater diatoms. Z.Pflanzenphysiol.,
j. §1. 149-167-
e^er B.P., Kuppers U. (1977). Carboxylating enzymes and path-
506
way of photosynthetic carbon assimilation in different marine
algae - evidence for the pathway? Planta, 1^3» 191-196.
Kremer B.P., Kies L., Rostami-Rabet A. (1979). Photosynthetic
performance of cyanellea in theendogamones Cyanophora,
Glaucosphaera. Gloeochaete. and Glaucocystis. Z. Pflanzenphy-
siol., 22, 303-317.
KremPr B.P., Schmaljohann R., Rottger R. (1980). Features and
nutritional significance of photosynthetates produced by
unicellular algae symbiotic with larger foraminifera. Mar.
Eool. Prog.Ser., 2, 225-228.
Kristiansen J. (1974). The fine structure of the zoospore of
Urospora penicilliformis, with special reference to the
flagellar apparatus. Brit.Phycol. J., 2, 201-213.
Kristiansen J. (1982). Chromophyoota and Chrysophyceae. In: Sy-
nopsis and Classification of Living Organisms, Vol. 1 (ed.
Parker S.P.), pp. 81-86. McGraw-Hill. New York.
Kristiansen J., Takahashi E. (1982). Chrysophyceae: introduction
and bibliography. In: Selected Papers in Phycology II (eds.
Rosowski J.R., Parker В. C.), pp. 698-704.. Phycological Socie-
ty of America, Allen Press, Lawrence, Kansas.
Kristiansen J., Walne P.L.(1976). Structural connections
between flagellar base and stigma in Dinobryon. Protoplasms,
99, 371-374.
Kristiansen J., Walne P.L. (1977). Fine structure of photokine-
tic systems in Dinobryon cylindricum var. alpinum (Chryso-
phyceae). Brit. Phycol. J., 12. 329-341.
Kuckuck P. (1912). Die Fortpflanzung der Phaeosporeen. Wise.
Meeresunters. N.P. (Abt. Helgoland), 2, 153-186.
Kuenzler E.J. (1965). Glucose-6-phosphate utilization by marine
algae. J.Phycol., 1, 156-164.
Kugrens P., West J.A. (1972). Ultrastructure of tetrasporogenesis
in the parasitic red alga Levringiella gardneri (Setchell)
Kylin. J.Phycol., 8, 370-383.
Kugrens P., West J.A* (1973). The ultrastructure of an allopa-
rasitic red alga Choreocolax polysiphoniae. Phycologia, 12.
175-186.
Kuhlenkamp R., Muller D.G. (1985). Culture studies of the life
history of Haplospora globosa and Tilopoteris mertensii
(Tilopteridales, Phaeophyceae). Brit. Phycol. J., 20. 301-312.
Kulaev I.S. (1975). Biochemistry of inorganic polyphosphates.Rev.
Physiol. Biochem.Pharmacol., 22» 133-157.
507
gullberg R.G. (1971). Algal distribution in six thermal stream
effluents. Trans. Am.Microscop.Soc., £0, 412-434.
Kutaing F.T. (1843). Phycologia generalis. Leipzig.
Kutzing F.T. (1849). Species algarum.Leipzig.
Kuwabara J.S. (1982). Micronutrients and kelp cultures: evidence
for cobalt deficiency insouthern California depp seawater.
Science, 216, 1219-1221.
Kylin H. (1914). Studien uber die Entwicklungsgeschichte von
Rhodomela virgata Kjellm.Svensk. Bot.Tidsskr., 8, 33-70.
Kylin H. (1956). Die Gattungen der Rhodophyceen. C.W.K. Gleerup,
Lund.
Lambein F., Wolk C.P. (1973). Structural studies on the glycoli-
pids from the envelope of the heterocyst of Anabaena cylindri-
ca. Biochemistry, 12, 791-798.
Lamouroux J.V.F. (1813). Essai sur les genres de la famille des
thalassiophytes non articulees. Mem.Hus.Nat.Hist. Paris, 20.
21-47, 115-139, 267-293.
Lang N.J., Walsby A.E. (1982). Cyanophyceae: introduction and
bibliography. In: Selected Papers in Phycology II (eds.
Rosowski J.R., Parker B.C.), pp. 647-655. Phyoological Socie-
ty of America, Allen Press, Lawrence,Kansas.
Lang N.J., Simon R.D., Wolk C.P. (1972). Correspondence of cya-
nophyoean granules with structured granules in Anabaena cy-
lindrica. Arch. Mikrobiol., 83. 313-320.
Langmuir I. (1938). Surface motion of water induced by wind.
Science, 87, 119-123.
Langford T.E. (1983). Electricity Generation and the Ecology
of Natural Waters. Liverpool University Press, Liverpool.
Larsen B. (1981). Biosynthesis of alginate. Proc.Int.Seaweed
Symp., 10, 7-34.
Lawrence J.M. (1975). On the relationship between marine plants
and sea-urchins. Oceanogr. Mar.Biol. Ann.Rev., 13. 213-286.
Lawrey J.D. (1984). Biology of Lichenized Fungi. Praeger, New
York.
bawson G.W. (1978). The distribution of seaweeds in the tropical
and subtropical Atlantic Ocean: a quantitative approach. Bot.
J. Linn.Soc., 26, 177-193.
lazarek S. (1982). Structure and productivity of epiphytic algal
communities on Lobelia dortmanna in acidified and limed lakes.
Water Air Soil Pollut,, 18, 333-342.
beadbeater B.S.C. (1970). Preliminary observations on differen-
508
ces of scale morphology at various stages in the life cycle of
"Apistonema-Syracosphaera" sensu von Stosch.Brit.Phycol. J.,
2, 57-69.
Leadbeater B.S.C. (1972). Fine structural observations on six
new species of Chryeochromulina (Haptophyceae) from Norway,
with preliminary observations on scale production in C.micro-
cylindrica sp. nov.Sarsia, 49. 65-80.
Leadbeater B.S.C., Dodge J.D. (1967). An electron microscope stu-
dy of dinoflagellate flagella. J.Gen.Microbiol., 46. 305-314.
Leadbeater B.S.C., Manton I. (1974). Preliminary observations on
the chemistry and biology of the lorica in a collared flagel-
late (Stenhanoeca diplocostata). J.Mar. Biol. Ass.U.K., 54.
269-279.
Lideiraan T.C., Rhee GrY. (1981). Bioconcentration of hexaohoro-
biphenyl in Great Lakes planktonic algae. Can.J.Fish.Aquat.
Sci., 39, 380-387.
Lee J.J. (1980). Nutrition and physiology of the foraminifera.
In: Biochemistry and Physiology of the Protozoa, Vol. 3 (eds.
M. Levandowsky and S.H. Hunter), pp. 43-66, Academic Press,
New York.
Lee J.J., McEnery M.E. (1983). Symbiosis in foraminifera. In:
Algal symbiosis: A Continuum of Interaction Strategies
(ed.L.J.Goff), pp. 37-68. Cambridge University Press, Cant-
bridge.
Lee K.W., Bold H.C. (1973). Pseudocharaciopsis texensis gen et
sp. nov., a new member of the Eustigmatophyceae. Brit. Phycol.
J., 8, 31-37.
Lee R.E. (1980). Phycology. Cambridge University Press, Cambridge.
Lee R.F., Takahashi M. (1977). The fate and effect of petroleum
in controlled ecosystem enclosures. Rapp. P.v. Reun. Cons.
Int. Explor. Met., 171, 150-156.
Leedale G.F. (1967). Euglenoid Flagellates. Prentice-Hall Inc.,
Englewood Cliffs, New Jersey.
Leedale G.F. (1968). The nucleus in Euglena. In: The Biology
of Euglena. Vol. 1 (ed. Buetow D.E.), pp. 185-242. Academic
Press, New York.
Leedale G.F. (1974). How many are the kingdoms of organisms?
Taxon, 2£, 261-270.
Leedale G.F. (1978). Phylogenetic criteria in euglenoid flagel-
lates. Biosystems, 10, 183-187.
Leedale G.F. (1982a). Euglenophycota. In: Synopsis and Classifi-
509
| cation of Living Organisms, Vol. 1 (ed. Parker S.P.), pp.129-
£ 131. McGraw-Hill, New York.
feedale G.P. (1982b). Ultrastructure. Tn: The Biology of Euglena
Vol. ITT, Physiology (ed. Buetow D.E.), pp.1-27, Academic
Press, New York.
Leedale
G.P. ,
Schiff J.A.,
Buetow D.E. (1982). Euglenophyceae:
introduction and bibliography, In: Selected Papers in Phycolo-
gy TI (eds. Rosowski J.R. and Parker В. C.), pp. 687-690).
Phycological Society of America, Allen Press, Lawrence, Kansas.
Lembi C.A.
(1980). Unicellular chiorophytes. In: Phytoflagella-
tes (ed. Cox E.R,), pp. 5-60. Elsevier/North-Holland, New
York.
Lembi C.A., Walne P.L. (1971). Ultrastructure of pseudocilia in
Tetraspora lubrica (Roth). Ag.J.Cell Sci., 569-579.
Tunnel M., Lee R.W. (1981). Physical evidence for
Lemieux
C.,
recombination of chloroplast DNA in hybrid progeny of Chlamy-
domonas eugametos and C. moewusii. Current Genetics, 2» 97-
ЮЗ.
Leonhard J., Compere P. (1967). Spirulina platensis (Gom.) Geitl.,
algue bleue de grande valeur alimentaire par sa richesse en
proteines. Bull.Jard.Bot. Bruxelles, 37. Suppl. 1.
Lewin J.C. (1962). Silicification. Tn: Physiology and Biochemis-
try of the Algae (ed. Lewin R.A.), pp. 445-455. Academic Press,
New York.
Lewin J.C. (1966). Silicon metabolism in diatoms. V. Germanium
dioxide a specific inhibitor of diatom growth. Phycologia, 6,
1-12.
Lewin J.C. (1976). Effects of boron deficiency on chemical compo-
sition of a mirine diatom. J.Expl. Bot, 27. 916-921.
Lewin J.C., Chen CrH. (1968). Silicon metabolism in diatoms.
VI. Silicic acid uptake by a colourless marine diatom, Nitz-
schia alba Lewin and Lewin.J.Phycol., £, 161-168.
Lewin J.C., Lewin R.A., Philpott D.E. (1958). Observations on
Phaeodactylum tricornutum. J.Gen.Microbiol., 18. 418-426.
Lewin R.A. (1970). Toxin secretion and tail autonomy by irrita-
ted Oxynoe panamensis. Pacific Sci, 24. 356-359.
Lewin R.A. (1976a). The Genetics of Algae. Blackwell Scientific
Publications, Oxford.
Lewin R.A. (1976b). Prochlorophyta as a proposed new division
of algae. Nature, 261, 697-698.
Lewin R.A. (1977). Prochloron-type genus of the Prochlorophyta.
Phycologia, 16. 217.
510
Lewin R.A. (1981a). Prochloron and the theory of symbiosis.
Ann. N.Y. Acad.Sci.. 361, 325-329.
Lewin R.A. (1981b). The prochlorphytes. In: The Prokaryotes.
A Handbook on Habitat, Isolation, and Identification of Bac-
teria (eds. Starr M.P. , Stolp H., Truper H.G., Balows A. and
Schlegel H.G. ), pp. 257-266, Springer-Verlag, Berlin.
Lewin R.A. (1983a). The problems of Prochloron. Ann. Microbiol.
(Inst. Pasteur), 134 B, 37-41.
Lewin R.A. (1983b). Phycotechnology - how microbial geneticists
might help. BioScience, 22» 177-179.
Lewin R.A. (1984). Prochloron - a status report. Phycologia, 23,
203-208.
Lewin R.A., Gibbs S.P. (1982). Algae of uncertain position:
introduction and bibliography. In: Selected Papers in Phyco-
logy II (eds. Rosowski J.R. and Parker B.C.), pp. 659-662.
Phycological Society of America. Allen Press, Lawrence, Kan-
sas.
Lewin R.A., Robertson J.A. (1971). Influence of salinity on
the form of Asterocytis in pure culture. J. Phycol., 7., 236-
238.
Lewis C.M., Yentsch C.M., Dale B. (1979). Distribution of Gony-
aulax excavata resting cysts in the sediments of the Gulf of
Maine. In: Toxic Dinoflagellate Blooms. Proceedings of the
2nd International Conference on Toxic Dinoflagellate Blooms
(eds. Taylor D.L. and Seliger H.H,), pp.235-238. Developments
in Marine Biology 1. Elsevier/North-Holland, Amsterdam.
Lewis J.R. (1961). The littoral zone on rocky shores - a biolo-
gical or physical entity? Oikos, 12, 280-281.
Lewis J.R. (1964). The Ecology of Rocky Shores. English Univer-
sities Press, London.
Lewis W.M. (1978). A compositional phytogeographical and elemen-
tary structural analysis of the phytoplankton in a tropical
lake, Lake Lanao, Philippines. J.Ecol., 66, 213-226.
L’hardy-Halos MrTh. (1971a). Manifestation d'une dominance
apicale chez les algues a structure cladomienne du genre
Antithamnion (Rhodophycees, Ceramiales). C.R.Acad.Sci. Paris
D 272, 2301-2304.
L'hardy-Halos M.Th. (1971b). Recherches sur les Ceramiacees
(Rhodophycees-Ceramiales) et leur morphogenese. III. Observa-
tions et recherches experimentales sur la polarite cellu-
laire et sur la hierarchisation des Elements de la fronde.
511
Rev.Gen.Bot., 78. 407-491.
Lin H., Sommerfeld M.R., Swafford J.R. (1975). Light and elect-
ron microscope observations on motile cells of Porphyridium
purpurearn (Rhodophyta). J. Phycol., 11, 452-457»
Linnaeus C. (1753). Species plantarum, Ed. 1. Holtniae .
Linnaeus C. (1754). Genera plantarum., Ed. 5. Holtniae.
List H. (1930). Die Entwicklungsgeschichte von Cladophora glo-
merate Kutz.Arch. Protistenk., 72, 453-481.
Littler Ы.М., Arnold K.E. (1982). Primary productivity of mari-
ne macroalgal functional form groups from south-western
North America. J. Phycol. , 18, 307-311.
Littler M.M,, Littler D.S. (1980). The evolution of thallus form
and survival strategies in benthic marine macroalgae: field
and laboratory tests of a functional form model.Am.Nat., 116,
25-42.
Littler M.M., Littler D.S., Blair S.M., Norris J.N. (1985). Dee-
pest known plant life discovered on an uncharted seamount.
Science, 227. 57-59.
Lloyd D. (1974). Dark respiration. In: Algal Physiology and Bio-
chemistry (ed. Stewart W.D.P.), pp. 505-529. Blackwell Scienti-
fic Publications, Oxford.
Lloyd N.D.H, , Canvin D.T. , Culver D.A. (1977). Photosynthesis
and photorespiration in algae. Plant Physiol., 59. 936-940.
Lobban C.S., Wynne M.J. (eds.) (1981). The Biology of Seaweeds.
Blackwell Scientific Publications, Oxford.
Lodge S.M. (1948). Algal growth in the absence of Patella on an
experimental strip of foreshore, Port St. Mary, Isle of man.
Proc. Trans.Liverpool Biol.Soc. , 56, 78-83.
Loeblich A.R. Jr (1974). Protistan phylogeny as indicated by
the fossil record. Taxon, 23. 277-290.
Loeblich A.R. Ill (1970). The amphiesma or dinoflagellate cell
covering. North American Paleontological Convention, Chicago,
1969. Proc. G, 867-929.
Loeblich A.R. III. (1976). Dinoflagellate evolution: speculation
and evidence. J. Protozool., 23. 13-28.
Loeblich A.R. III (1982). Dinophyceae. In: Synopsis and Clasifi-
cation of Living Organisms, Vol. 1 (ed. Parker S.P.), pp. 104-
115. McGraw-Hill, Nev: York.
Loeblich A.R. III, Loeblich L.A. (1978). Division Eustigmatophy-
ta. In: CRC Handbook of Microbiology, 2nd Edition. Vol. II,
Pungi, Algae, Protozoa and Viruses (eds. Laskin A.I. and
512
and Lechevalier H.A.), pp. 484-487. CRC Press, West Palm
Beach.
Loeblich A.R. III., Sherley J.L. (1979). Observations on the
theca of the motile phase of freeliving and symbiotic isola-
tes of Zooxanthella microadriatica (Freudenthal) comb. nov.
J. Mar.Biol., Ass. U.K., £9, 195-205.
Loeblich A.R., Jr., Tappan H. (1966). Annotated index and bibli-
ography of the calcareous nanoplankton. Phycologia, 2» 81-216.
Logan B.W. (1961). Cryptozoon and associated stromatolities
from the Recent, Shark Bay, Western Australia. J. Geol., 69.
517-533.
Loiseaux 3. (1970a). Rotes on several Hyrionemataceae from
California using culture studies. J. Phycol., 6, 248-260.
Loiseaux S. (1970b). Streblonema anomalum S. et G. and Compso-
nema sporangiiferum S. et G. stages in the life history of
a minute Scytosiphon. Phyoologia, 2» 185-191.
Lokhorst G.M. (1978). Taxonomic studies on tie marine and brack-
ish-water species of Ulothrix (Ulotrichales, Chlorophyceae)
in western Europe. Blumea, 24. 191-299.
Lokhorst G.M., Star W. (1983). Fine structure of mitosis and
cytokinesis in Urospora (Acrosiphoniales, Chlorophyta).
Protoplasma, 117» 142-153.
Lombard E.H., Capon B. (1971). Peridinium gregarium, a new spe-
cies of dinoflagellate. J. Phycol., 7, 184-187.
Lovlie A., Bryhni E. (1978). On the relation between sexual and
parthenogenic reproduction in haplodiplontic algae. Bot.Mar.,
21., 155-163.
Lubchenco J. (1978). Plant species diversity in amarine interi-
dal community: importance of herbivore food preference and
algal competitive ability. Am.Nat., 112'» 23-39.
Lubchenco J. (1980). Algal zonation in the New England rocky
intertidal community: an experimental analysis. Ecology, 61.
333-344.
Lubchenco J,, Cubit J. (1980). Heteromorphic life histories of
certain marine algae as adaptations to variations in herbivory
Ecology, 61. 676-687.
Lubchenco J., Gaines S.D. (1981). A unified approach to marine
plant-herbivore interactions. I. Populations and communities.
Ann.Rev.Ecol., Syst., 12, 405-437.
Lubchenco J., Menge B.A. (1978). Community development and per-
513
sigtence in a low rocky interitdal zone. Ecol.Monogr., 48.
67-94.
bund J.W. G. (1954). The seasonal cycle of the planktonic diatom
Melosira italics subsp. subarctica Mull. J.Ecol., 42, 151-179.
bund J.W.G. (1972). Eutrophication. Proc.Roy.Soc.Lond. B, 180,
371-382.
bund S. (1966). On a sporangia-bearing microthallue of Scytosiph-
on lomentaria from nature. Phycologia, 6, 67-78.
buning K. (1979). Growth strategiesof three Laminaria species
(Phaeophyceae) inhabiting different depth zones in the sub-
littoral region of Helgoland (North Sea). Mar.Ecol. Prog.
Ser., 1, 195-207.
buning K. (1980). Control of life histories by daylength and
temperature. In: The Shore Environment . Vol. 2: Ecosystems
(eds. Price J.H., Irvine D.E.G. and Farnham W.F.), pp.915-
945, Systematics Association Special Volume 17b. Academic
Press, London and New York.
Luning K. (1981). Light. In: Biology of Seaweeds (eds. Lobban
C.S. and Wynne M.J.), pp. 326-355. Blackwell Scientific Pub-
lications, Oxford.
Luning K., Dring M.J. (1975). Reproduction, growth and photosyn-
thesis of gametophytes of Laminaria saccharins grown in blue
and rod light. Mar.Biol., 29, 195-200.
Luning K., Chapman A.R. 0., Mann K.H. (1978). Crossing experiments
in the non-digitate complex of Laminaria from both sides of
the Atlantic. Phycologia, 17., 293-298,
Luoma S.N., Bryan G.W. , Langston W.J. (1982). Scavenging of heavy
metals from particulates by brown seaweed.. Mar.Poll.Bull.,
12, 394-396.
Lyngbye H.C. (1819). Tentamen Hydrophytologiae Danicae. Copen-
hagen.
McCandless E.L. (1978). The importance of cell wall constituents
in algal taxonomy. In: Modern Approaches to the Taxonomy
of Red and Brown Algae (eds. Irvine D.E.G. and Price J.H.)»
pp.63-85. Academic Press, London and New York.
McCandless E.L. (1981). Polysaccharides of seaweeds. In: The Bio-
logy of Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp.559-
588. Blackwell Scientific Publications, Oxford.
McCandless E.L., Craigie J.S. (1979). Sulphated polysaccharides
in red and brown algae. Ann.Rev. Plant Physiol., 20, 41-53.
McCandless E.L., Craigie J.S., Walter J.A. (1973). Carrageenans
33-121
514
in the gametophytic and sporophytic stages of Chondrus cris-
pus« Can. J.Bot. , 55 2053-2064.
McCandless E.L., Vollmer C.M. (1984). The nemathecium of Gymno-
grongus chiton (Rhodophyceae, Gigartinalea): immunochemical
evidence of meiosis. Phycologia, 23, 119-123.
McCarthy J.J. (1981). The kinetics of nutrient utilization.Can.
Bull.Fish Aquat.Sci., 210, 211-233.
McCarthy J.J., Goldman J.C. (1979). Nitrogenous nutrition of
marine phytoplankton in nutrient depleted waters.Science,
203, 670-672.
McCully M.E. (1966). Histological studies on the genus Fucus.
I. light microscopy of the mature vegetative plant. Proto-
plasma, 62, 287-305.
McDonald K. (1972). The ultrastructure of mitosis in the marine
red alga Membranoptera platyphylla. J. Phycol., 8, 156-166.
McFadden G.I., Moestrup 0., Wetherbee R. (1982). Pyramimonaa
gelidicola sp.nov. (Prasinophyceae), a new species isolated
from Antarctic sea ice. Phycologia, 21, 103-111.
Machlis 1., Hill G.G.C., Steinbeck K.E., Reed W. (1974). Some
characteristics of the speztn attractant in Oedogonium cardia-
cum.J. Phycol., 10, 199-204.
Mackie I.M., Percival E. (1959). The constitution of xylan from
the green seaweed Caulerpa filiformis. J.Chem.Soc., 30, 10-15.
Mackie W., Preston R.D. (1974). Cell wall and intercellular
region polysaccharides. In: Algal Physiology and Biochemistry
(ed. Stewart W.D.P.), pp. 40-85. Blackwell Scientific Publi-
cations, Oxford.
McLachlan J. (1977). Effects of nutrients on growth and develop-
ment of embryos of Fucus adentatus Pyl. (Phaecophyceae;
Fucales). Phycologia, 16, 329-338.
McLachlan J., Bidwell R.G.S. (1983). Effects of colored light
on the growth and metabolism of Fucus embryos and apices
in culture. Can.J.Bot., 61, 1993-2003.
McLachlan J., Chen LiC. (1972). Formation of adventive emtryos
from rhizoidal filaments in sporelings of four species of
Fucus. Can.J.Bot., 22» 1841-1844.
McLachlan J., Chen L.CrM., Edilstein T. (1971). The culture of
four species of Fucus under laboratory conditions.Can.J.
Bot., 1463-1469.
McLachlan J., Craigie J.S. (1964). Algal inhibition by yellow
ultraviolet absorbing substances from Fucus vesioulosus.
515
Can.J.Bot., £2, 287-292.
McLachlan J., Craigie J.3., Chen L.CrM., Ogetze E. (1972). Por-
phyra linearis Grev.: an adible species of nori from Nova
Scotia. Proc. Int. Seaweed Symp., 7, 473-476.
McLean R.J., Laurendi C.J., Brown R.M., Jr. (1974). The relation-
ship of gamone to the mating reaction in Chlamydomonas moe-
wusii. Proc. Natl.Acad.Sci. , 71. 2610-2613.
MacPherson B., Pfeiffer C.J. (1976). Experimental colitis. Diges-
tion, 1J., 424-452.
McQuade A.B. (1933). Origins of the nucleate organisms. II.
Biosystems, 16, 39-55.
Maestrini S.Y., Bonn D.J. (1981a). Allelopathic relationships
between phytoplankton species.Can.Bull.Fish Aquat.Sci., 210,
323-346.
Maestrini S.Y., Bonn D.J. (1981b). Competition among phytoplank-
ton based on inorganic macronutrients.Can.Bull. Fish Aquat.
Sci., 210, 264-278.
Maiti В.C., Thomson R.H., Mahendran M. (1978). The structure of
caulerpin , a pigment from Caulerpa algae.J.Chem.Res., £,
126-127.
Magg C.A., Guiry M.D. (1982). Morphology, phenology and photo-
periodism in Halymenia latifolia Kutz (Rhodophyta) from
Ireland.Bot. Mar., 25, 589-599.
Magne F. (1967). Sur le deroulement et le lieu de meiose chez
Lemaneacees (Rhodophycees, Nemalionales). C.R. Acad. Sci. Paris
D, 265, 670-673.
Maier I. (1982). New aspects of pheromone-triggered spezmatozoid
release in Laminaria digitata (Phaeophyta). Protoplasms,
113. 137-143.
\ Maier I., Iffiller D.G. (1982). Antheridium fine structure and
spermatozoid release in Laminaria digitata (Phaeophyceae).
Phycologia, 21, 1-8.
Maier I., Muller D.G. (1986). Sexual pheromones in algae. Biol.
Bull., 170, 145-175.
Makarewicz J.C., Baybutt R.I. (1981). Longterm (1927-1978)
changes in the phytoplankton community of Lake Michigan at
Chicago, Bull. Torrey Bot.Club., 108, 240-254.
Malinowski K.C., Ramus J. (1973). Growth of the green alga
Coldium fragile, in a Connecticut estuary. J. Phycol., 2.,
102-110.
Mann D.G. (1982). Structure, life history and systematics of
516
Rhoicosphenia (Bacillariophyta). II. Auxospore formation
and perizonium structure of Rh.gurvata. J. Phycol., 18.» 264-
274.
Manny B.A. (1972). Seasonal changes in organic nitrogen content
of net and nanoplankton in two hardwater lakes. Arch.Hydro-
biol.f Zl, 103-123.
Mantofil. (1959). Observations on the internal structure of the
sperraatozoid of Dictyota J.Exp. Bot., 10, 448-461.
Mantoal. (1964a) Further observations on the fine structure of
the haptonema in Prymnesium parvum. Arch Mikrobiol., 49» 315-
330.
Manton I. (1964b). Observations with the electron microscope on
the division cycle in the flagellate Prymnesium parvum Car-
ter. J. Roy.Microsc.Soc.Ser. 3, 83» 317-325.
Manton I. (1965). Some phyletic implications of flagellar struc-
ture in plants. Ins. Advances in Botanical Research, Vol. 2
(ed. Preston R.D.), pp. 1-34. Academic Press, New York.
Manton I. (1977). Dolichomastix (Prasinophyceae) from arctic
Canada, Alaska, and South Africa: a new genus of flagellates
with scaly flagella. Phycologia, 16, 427-438.
Manton I., Parke M. (I960). Further observation on small green
flagellates with special reference to possible relatives of
Chromulina pusilia Butcher. J. Mar.Biol.Ass. U.K., 39,
275-298.
Manton I., Kowallik K., Stosch H.A., von (1969). Observations
on the fine structure and development of the spindle at mi-
tosis and meiosis in a marine centric diatom (Lithodesmium
undulatum). I. Preliminary survey of mitosis in spennatogonia
J. Microscop., §2, 295-320.
Manton I., Sutherland J., Oates K. (1977). Arctic coccolithopho-
rids: Wigwamma arctica gen.et sp. nov. from Greenland and
Arctic Canada, W«annulifera sp. nov. from South Africa and
S.Alaska and Calciarus alaskensis gen.et sp. nov. from S.
Alaska, Proc.Roy. Soc.Lond. В 197, 145-168,
Manton I., Oates K., Sutherland J. (1981). Cylinder-scales in
marine flagellates from the genus Chrysochromulina (Hapto-
phyceae-Prymnesiophyceae): further observations from C« micro-
cylindrica Leadbeater and C«cyathophora Thomsen. J.Mar.Biol.
Ass.U.K., 61, 27-33.
Marchant H.J. (1974). Mitosis, cytokinesis and colony formation
in Pediastrum borvanum.Ann. Bot., 38, 883-888.
517
Margalef R. (1978). Life forms of phytoplankton as survival alter-
natives in an unstable environment. Oceanologica Acta, 1,
493-509.
Margulis L. (1970). Origin of Eukaryotic Celle. Yale University
Press, New Haven.
Margulis L. (1981). Symbiosis in Cell Evolution. Life and its
Environment on the Early Earth. W.H. Freeman and Co., San
Francisco.
Margulis L. (1982). Phyla of the Fine Kingdoms. An Illustrated
guide to the Kinds of Life on Earth. W.H. Freeman and Co.,
San Francisco.
Margulis L., Schwartz K.V. (1982). Five Kingdoms. An Illustrated
Guide to the Phyla of Life on Earth. W.H. Freeman and Co.,
San Francisco.
Markey D.R., Wilce R.T. (1975). The ultrastructure of reproduc-
tion in the brown alga Pylaiella littoralis. I. Mitosis and
cytokinesis in the plurilocular gametangia. Protoplasma, 85.
219-241.
Markham J. W., Hagmeier E. (1982). Observations on the effects of
germanium dioxide 'On the growth of macroalgae and diatoms.
Phycologia, 21, 125-130.
Martin M.T. (19^9). A rewiew of life histories in the Nemalio-
nales and some allied genera. Brit. Phycol. J., 4, 145-158.
Martin T.C., Wyatt J.T. (1974). Extracellular investments in
blue-green algae with particular emphasis on the genus Nostoc.
J. Phycol., 10, 204-210.
Mason T.R., Van Brunn V. (1977). 3-Gyr-old stomatolites from
South Africa. Nature, 266. 47-49.
Mathieson A.C. (1967). Morphology and life history of Phaestro-
phion irregulare S. et G. Nova Hedwiga. 13, 293-318.
Mathieson A.C. (1982). Seaweed cultivation: a review. In: Pro-
ceedings of the 6th U.S. - Japan Meeting on Aquaculture
(ed. Sindeiraan C.J.), pp.25-66, NOAA Technical Report NMFS
Circ. 442.
Matilsky M.B., Jacobs W.P. (1983). Accumulation of amyloplasts
on the bottom of normal and inverted rhizome tips of
Caulerpa prolifera .(Forsskal) Lamouroux. Planta, 159. 189-
192.
Mehta S.C., Venkataraman G.S., Das S.C. (1961). The fine struc-
ture and the cell wall nature of Diatoma hiemale var.
mesodon (Ehr.) Grun.Rev.Algol. U.S., 6, 49-52.
518
Meinesz A. (1980). Connaiseances actuelles et contribution a 1
etude de la reproduction et du cycle des Udoteac^es (Cauler-
pales, Chiorophytes). Phycologia, 19., 110-138.
Melkonian M. (1980). Ultrastructural aspects of basal body asso-
ciated fibrous structures in green algae: a critical review.
Biosystems, 12, 85-104.
Melkonian M. (1981a) Structure and significance of cruciate fla-
gellar root systems in green algae: female gametes of Bryop-
sis lyngbyei (Bryopsidales). Helgol. Wiss. Meeresunters,
2£, 355-369.
Melkonian И, (1981b). The flagellar apparatus of the scaly green
flagellate Pyramimonas obovata: absolute configuration. Proto-
plasma, 108, 341-355.
Melkonian M. (1982a). Structural and evolutionary aspects of
the flagellar apparatus in green algae and land plants. Taxon.,
21, 255-365.
Melkonian M. (1982a). The functional analysis of the flagellar
apparatus in green algae. In: Prokaryotic and Eukaryotic Fla-
gella (eds. Amos W.B. and Duckett J.G.). Cambridge University
Press, Cambridge.
Melkonian M. (1982c) Effect of divalent cations on flagellar
scalles in the green flagellate Tetraselmis cordifonais.
Protoplasma, 111, 221-233»
Melkonian M. (1982d). Systematics and evolution in the algae.
Prog. Bot., 44, 315-344.
Melkonian M., Berns B. (1983). Zoospore ultrastructure in the
green alga Friedmannia israelensis: an absolute configuration
analysis. Protoplasma, 114, 67-84.
Melkonian M., Ichimura T. (1982). Chlorophyceae: introduction
and bibliography. In: Selected Papers in Phycology II (eds.
Rosowski J.R. and Parker В.C.), pp. 747-753. Phycological
Society of America, Allen Press, Lawrence, Kansas.
Mereschowsky C. (1905). Uber Natur und Ursprung der Chromato-
phoren im Pflanzenreiche. Biol.Centr., 25, 593-604.
Wetting B. (1981). The systematics and ecology of soil algae.
Bot. Rev., Ц, 195-312.
Mierle G.M., Stokes P.M. (1976). Heavy metal tolerance and metal
accumulation by planktonic algae. In: Trace substances and
Environmental Health Vol. XI (ed. Hemphill D.D.), pp. ИЗ-
122. University of Missouri, Columbia, Missouri.
Mignot JtP. (1967). Structure et ultrastructure de quelques
519
Chloromonadines. Protistologica, 2» 5-23.
Mignot JrP. (1976). Complements a I'etude dee Chloromonadines.
Ultrastructure ds Chattonella subsalsa Biecheler. Flagelle
d’eaux saumatres. Protistologica, 12, 279-293»
Miller M.M., Lang N.J. (1968). The fine structure of akinete
formations and germination in Cylindrospermum. Arch. Mikro-
biol., 60, 303-313.
Miller R.J. (1985). Seaweeds, sea-urchins and lobsters: a re-
appraisal. Can. J.Fish. Aquat.Sci. , 42, 2061-2072.
Miller W.I., III, Collier A. (1978). Ultrastructure of the frus-
tule of Triceratium favus (Bacillariophyceae). J. Phycol.,
14, 56-62.
Millner G. C., Sweeney R.A. (1982). Lake Erie Cladophora in per-
spective, J.Great Lakes Res., 8, 27-29»
Millner P.A., Evans L.V. (1980). The effects of triphenyltin
chloride on respiration and photosynthesis of the green algae
Enteromorpha intestinalis and Ulothrix flacca. Plant Cell
Environ., 2» 339-348.
Milton D.J. (1966). Drifting organisms in the Precambrian sea.
Science, 153. 293-294»
Mix M. (1975). Die Feinstruktur der Zellwande der Conjugaten
und ihre systematische Bedeutung. Nova Hedwiga, 42, 179-194»
Moe R.L.,Henry E.C. (1982). Reproduction and early development
of Ascoseira mirabilis Skottsberg (Phaeophyta), with notes
on Ascoseirales Petrov. Phycologia, 21, 55-66.
Moe R.L., Silva P.C. (1977). Antarctic marine flora: uniquely
devoid of kelps. Science, 196, 1206-1208.
Moebus K., Johnson K.M., Sieburth J. McN (1974). Re-hydration of
desiccated intertidal brown algae: release of dissolved or-
ganic carbon and water uptake. Mar.Biol., 26, 127-134»
Moestrup 0 (1970). On the fine structure of the speiraatozoids
of Vaucheria sescuplicaria and on the later stages in sper-
matogenesis. J. Mar.Biol.Ass. U.K., 50, 513-523.
Moestrup 0. (1972). Observations on the fine structure of sperma-
tozoids and vegetative cells of the green alga Golenkinia.
Brit. Phycol. J., 7, 169-183.
Moestrup 0 (1974). Ultrastructure of the scale-covered zoos-
pores of the green alga Chaetosphaeridium. a possible ances-
tor of the higher plants or bryophytes. Biol. J.Linn.Soc.,
6, 111-125.
Moestrup 0 (1978). On the phylogenetic validity of the fla-
520
gellar apparatus in green algae and other chlorophyll a and
b containing plants. Biceystems, 10, 117-144.
Moestrup 0. (19S2). Flagellar structure in algae: a review, with
new observations particularly on the Chrysophyceae, Phaeophy-
ceae (Fucophyceae). Euglenophyceae, and Reckertia. Phycologia,
21_, 427-528.
Moestrup #., Ettl H. (1979). A light and electron microscopical
study of the flagellate Nephroselmis olivacea (Prasinophy-
ceae). Opera Bot., 49, 1-39.
Moestrup J0., Thomsen H.A, (1974). An ultrastructural study of
the flagellate Pyramimonas orientalis with particular empha-
sis on Golgi apparatus activity and the flagellar apparatus.
Protoplasma, 81, 247-269.
Monod J. (1942). Recherches sur la Croissance des Cultures Bac-
teriennes, Hermann and Cie, Paris.
Moriarty D.J.W. (1979). Muramic acid in the cell walls of Prochlo-
ron. Arch. Mikrobiol., 210, 191-194.
Moriarty D.J.W., Moriarty C.M. (1973). The assimilaiion of carbon
froml phytoplankton by two herbivorous fishes: Tilapia niloti-
ca and Haplochromis nigripinnis. J.Zool., 171, 41-55.
Mornin L., Francis D« (1967). The fine structure of Nematodinium
armatum.a naked dinoflagellate, J. Microscop., 6, 759-772.
Morril L.C., Loeblich A.R. Ill (1983). Formation and release of
body scales in the dinoflagellate genus Heterocapsa. J.Mar.
Biol. Ass. U.K., 62, 905-913.
Morris I. (ed.). (1980). The Physiological Ecology of Phytoplank-
ton. Blackwell Scientific Publications, Oxford.
Moss B. (1977). Adaptations of epipelic and episammic freshwa-
ter algae. Oecologia, 28, 103-108.
Moss B.L. (1964). Growth and regeneration of Fucus vesiculosus
in culture. Brit. Phycol.Bull., 2, 377-380.
Moss B.L. (1965). Apical dominance in Fucus vesiculosus.New
Phytol., 6£, 387-392.
Moss B.L. (1966). Polarity and apical dominance in Fucus
vesiculosus.Brit. Phycol.Bull., 2» 209-212.
Moss B.L. (1970). Meristems and growth control in Ascophyllum
nodosum (L) Le Jolis New Phytol., 69. 253-260.
Moss B.L. (1982). The control of epiphytes on Halidrys siliquo-
sa (L.) Lyngb. (Phaecophyta, Cystoseiraceae). Phycologia,
21_, 185-191.
521
Moss B.L., Tovey D., Court P. (1981). Kelps as fouling organisms
on North Sea oil platfonns. Bot.Mar., 24. 207.
Moss J.R. (1978). Essential considerations for establishing sea-
weed extraction factories. In: The Marine Plant Biomass of
the Pacific North-west Coast (ed. Kraus R.W.), pp. 301-314.
5 Oregon state University Press, Corvallis.
Mueller-Haekel A. (1973). Experiments zum Bewegungsverhalten von
einzelligen Eliesswasseraigen, Hydrobiology, 41, 221-246.
Mukai L.S., Craigie J.S., Brown R.0. (19S1). Chemical composition
j.j«f and structure of the cell walls of the Conchocelis and thallus
phases of Porphyra tenera (Rhodophyceae). J. Phycol., 17,
Й 192-198.
Miller D.G. (1962). Uber jahres-und lunarperiodische Erscheinun-
gen bei einigen Braunalgen. Bot.Mar., £, 140-155.
Muller D.G. (1967). Generationswechsel, Kernphasenwechsel und
Sexualitat der Braunalge Ectocarpus im Kulturversuch. Planta,
Z2, 39-54.
Muller D.G. (1972). Life cycle of the brown alga Ectocarpus fasci-
culatus var. refractus (Kutz)Ardis. (Phaeophyceae, Ectocarpa-
les) in culture. Phycologia, 11, 11-13.
Muller D.G. (1975). Experimental evidence aginst sexual fusion
of spores from unilocular sporangia of Ectocarpus siliculosus
(Phaeophyta). Brit. Phycol. J., 10, 315-320.
Muller D.G. (1981). Sexuality and sex attraction. In: The Biolo-
gy of Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp.661-674.
Blackwell Scientific Publications, Oxford.
Muller D.G., Falk H. (1973). Flagellar structure of the gametes
of Ectocarpus siliculosus (Phaeophyta) as revealed by nega-
tive staining.Arch. Mikrobiol., 91. 313-322.
Muller D.G., Meel H. (1982). Culture studies on the life history
of Arthrocladia villosa (Desmarestiales, Phaeophyceae). Brit.
Phycol. J., £, 419-425.
Mumford T.F. (1979). Pield and laboratory experiments with Iri-
daea cordata (Florideophyceae) grown on nylon netting. Proc.
Int. Seaweed Symp., g_» 515-523.
Munday B.L., Peel B.F. (1983). Severe ulcerative dermatitis in
Platypus (Ornithorphynchus anatinus). J. Wildlife Diseases,
19, 363-365.
Murray S., Dixon P.S., Scott J.L. (1972). The life history of
Porphyropsis coccininea var. dawsonii in culture. Brit.
Phycol. J., 7, 323-333.
522
Muscatine L. (1973)* Nutrition of corals. In: Biology and Geo-
logy of Coral Reefs.Vol. II, Biology I (eds.Jones O.A. and
Endean R. ), pp. 77-115. Academic Press, New York.
Muscatine L. (1930). Productivity of zooxanthellae. In: Primary
Productivity in the Sea (ed. Falkowski P.G.), pp.381-402,
Plenum Press, New York.
Muscatine L., McAuley P.J. (1982). Transmission of symbiotic
algae to eggs of green hydra. Cytobios, 22» 111-123.
Muscatine L., Porter J.W. (1977). Reef corals} mutualistic sym-
bioses adapted to nutrient-poor environments. Bioscience
2£, 454-460.
itynderse J.S., Moore R.E., Kashiwagi M., Norton T.R. (1977).
Antileukemia activity in the osoillatoriaceae, isolation of
debromoaplysiatoxin from Lyngbya. Science, 196, 538-540.
Nakahara H., Nakamura Y. (1973). Parthenogenesis, apogamy and
apospory in Alaria crassifolia (Laminariales). Mar. Biol.,
18, 327-332.
Nakajima M. (1965). Studies on the source of shellfish poison
in Lake Hamana. III. Poisonous effects of shellfishes feeding
on Prorocentrum sp. Bull.Jap.Soc.Sci.Fish,, 31. 282-285.
Nakamura K., Ogawa T., Shibata K. (1976). Chlorophyll and pepti-
de composition in the two photosystems of marine green algae.
Biochim. Biophys.Acta, 423, 227-236.
Nakamura Y., Tatewaki M. (1975). The life history of some spe-
cies of Scytosiphonales. Sci.Pap. Inst. Algol.Res. Hokkaido
Univ., 6, 57-93.
Naylor M. (1954). A note on Xiphophora chondrophylla var. maxi-
ma J. Ag. HewPhytol., 53. 155-159.
Needier A.B. (1949). Paralytic shell fish poisoning and Gonyaulax
tamarensis. J. Fish.Res.Bd Can., X» 490-504.
Neilson A.H., Larsson T. (1980). The utilization of organic
nitrogen for growth of algae: physiological aspects. Physiol.
Plant, 48, 542-553.
Neish A.C., Fox C.H.(1971). Greenhouse experiments on the vege-
tative propagation of Chondrus crispus Irish Moss. Natl Res.
Coun.Can. Atl.Reg.Lab.Tech. Rep., 14. 25p.
Neish A, C., Shacklock P.F., Fox C.H., Simpson F.J. (1977). The
cultivation of Chondrus crispus. Factors affecting growth
under greenhouse conditions. Can.J.Bot., 55. 2263-2271.
Nelson D.M., Goering J.J., Kilham S.S., Guillard R.R.L. (1976).
Kinetics of silicic acid uptake and rates of silica disso-
523
lution in the marine diatom Thalassioaira paeudonana. J. Phy-
col., 12, 246-252.
jjeig°n-Smith A. (1973). Oil Pollution and Marine Ecology. Plenum
Press, New York.
jjewroth P. (197D. Studiea on life hiatoriea in the Phyllophora-
ceae. I. Phyllophora truncata. (Rhodophyceaea, Gigartinales).
Phycologia, 10, 345-354.
Bewroth P. (1972). Studiea on life histories in the Phyllophora-
oeae II. Phyllophora paeudoceranoides and notes on P.criapa
and P, heridia (Rhodophyta, Gigatinalea). Phycologia, 11.
99-Ю7.
Newton J.W., Herman A. I. (1979). Isolation of cyanobacteria from
the aquatic fern Agolla. Arch. Mikrobiol., 120 161-165.
Nichole K.H. (1981). Chryaococcus furcatus (Dolg.) comb, nov.:
a new for Chryaaatrella furcata (Dolg.) Defl. baaed on the
discovery of the vegetative stage. Phycologia, 20, 16-21.
Nichols H.W. (1965). Culture and development Hildenbrandia rivu-
laria from Denmark and North America. Am.J.Bot., 51. 180-188.
Nichols H.W., Liasant E.K. (1967). Developmental atudiea of
Erythrocladia Rosenvinge in culture. J. Phycol., 2» 6-18.
Nichole J.M., Adame D.G. (1982). Akinetea. In: The Biology of
Cyanobacteria (eds. Whitton B.A. and Carr N.G.), pp.389-412.
Blackwell Scientific Publications, Oxford.
Nicholson N.L., Cimberg R.L. (1971). The Santa Barbara oil spilla
of 1969» a post spill survey of the rocky interidal. In: Bio-
logical and Oceanografic Survey of the Santa Barbara Channel
Oil Spills of 1969-1970. Vol. 1, pp.325-400. University of
Southern California.
Niell J.H., Jackson M.B. (1982). Monitoring Cladophora growth
conditions and the effect of phosphorus additions at a shore-
line site in North-eastern Lake Erie. J.Great Lakes Res., 8,
30-34.
Nienhuis P.H. (1974). Variability in the life cycle of RhizoclonL
urn ripariurn (Roth) Harv. (Chlorophyceae: Cladophoralea) under
Diitch estuarine conditions. Hydrobiol. Bull., 8, 172-178.
Niaamuddin M. (1962). Classification and distribution ofthe
Pucales. Bot.Mar., £, 191-203.
Noda H., Horiguchi Y. (1971). The significance of zinc as a nut-
rient for the red alga Porphyta tenera. Proc. Int. Seaweed
Symp., 7 , 368- 372.
Nonomgra A.M., West J.A. (1981). Host speoifity of Janczewakia
524
(Ceramiales, Rhodophyta). Phycologia, 20, 251-258.
Norris J.N., Fenical W. (1982). Chemical defense in tropical
marine algae. In: The Atlantic Barrier Reef Ecosystem at
Carrie Bow Cay.Belize.I. Structure and Communities (eds.
Rutzler K. and Macintyre I.S.), pp.417-432. Smithsonian
Institute Press, Washington.
Norris J.N., Kugrens P. (1982). Marine Rhodophyceae: introduc-
tion and biblography. In: Selected Paper in Phycology II (eds.
Rosowski J.R. and Parker B.C.), pp. 663-670. Phycological
Society of America, Allen Press, Lawrence, Kansas.
Norris R.E. (1977). Flagellate cells in the life-history of Sti-
chogloea (Chrysophyceae). Phycologia, 16, 75-78.
Norris R.E. (1980). Prasinophytes. In: Phytoflagellates (ed. Cox
E.R.), pp.85-146. Elsevier/North-Holland, New York.
Norris R.E. (1982a). Prymnesiophyceae. In: Synopsis and Classi-
fication of Living Organismsyol. 1 (ed. Parker S.P.), pp.
86-91. McGraw-Hill, New York.
Norris R.E. (1982b). Prasinophyceae. In: Synopsis and Classifica-
tion of Living Organisms, Vol. 1 (ed. Parker S.P.), pp. 163—
164. McGraw-Hill, New York.
Norris R.E. (1982c). Prasinophyceae: introduction and bibliogra-
phy. In: Selected Papers in Phycology II (eds. Rosowski J.R.
and Parker В.C.), pp. 740-746. Phycological Society of Ame-
rica, Allen Press, Lawrence, Kansas.
Norris R.E., Hori T., Chihara M. (1980). Revision of the genus
Tetraselmis (Class Prasinophyceae). Bot.Mag.Tokyo, 93» 317-
339.
North W.J. (1964). Ecology of the rocky near shore environment
in southern California and possible influence of discharged
wastes. Adv.Water Poll.Res., 2» 247-274.
North W.J. (1967). Tampico: a study of destruction and restora-
tion.Sea Frontiers, 13. 212-217.
North W.J. (ed.) (1971). The biology of giant kelp beds in
California. Nova Hedwiga, 32, 1-600.
Norton T.A., Mathieson A.C. (1983). The biology of unattached
seaweeds. Prog. Phycol.Res., 2, 333-386.
Norton T.A., Mathieson A.C., Neushul M. (1981). Morphology and
environment. In: The Biology of Seaweeds (eds. Lobban C.S.
and Wynne M.J.), pp.421-451. Blackwell Scientific Publica-
tions, Oxford.
Nozaki H. (1981). The life history of Japonese Pandorina uni-
525
росса (Chlorophyta, Volvocales). J.Jap.Bot., 56, 65-72.
jlygren S. (1975). Life history of some Phaeophyceae from Sweden.
Bot. Mar., 18, 131-141.
Oakley B.R., Santore U.J. (1982). Cryptophyceae: introduction and
bibliography. In: Selected Papers in Phycology II (eds. Ro-
aowski J.R. and Parker B.C.), pp. 682-686. Phycological So-
ciety of America, Allen Press, Lawrence, Kansas.
O'Brien P.Y., Dixon P.S. (1976). The effects of oils and oil
components on algae: a review. Brit. Phycol. J., 11, 115-142.
Ogden J.C,(1976). Some aspects of herbivore - plant relationships
on Caribbean reefs and seagrass beds. Aquatic Botany, 2, ЮЗ-
106.
O'hEocha C. (1971). Pigments of the red algae. Oceanogr. Mar,
Biol.Ann.Rev., 9, 61-82.
Okazaki M.Ikawa T., Ригцуа K., Nisizawa K., Miwa T. (1970).
Studies on calcium carbonate deposition of a calcareous red
alga Serraticardia maxina. Bot. Mag. Tokyo, 83, 193-201.
O'Kelly C.J., Floyd G.L. (1983). The flagellar apparatus of
Entocladia viridis motile cells, and the taxonomic position
of the resurrected family Ulvellaceae (Ulvales, Chlorophyta).
J.Phycol. , 12., 153-164.
O'Kelly C.J., Floyd G.L. (1984a). The absolute configuration
of the flagellar apparatus in zoospores from two species
of Laminariales (Phaseophyceae). Protoplasma, 123, 18-25.
O'Kelly C.J., Floyd G.L. (1984b). Flagellar apparatus absolute
orientations and the phylogeny of the green algae. Biosys-
tems, 16, 227-251.
O'Kelly C.J., Yarish C. (1981). On the circumscription of the
genus Entocladia Reinke, Phycologia, 20, 32-45»
O'Kelly J.C. (1974). Inorganic nutrients, In:Algal Physiology
and Biochemistry (ed. Stewart W.D.P.), pp.610-634. Blackwell
Scientific Publications, Oxford.
Okuda K., Tatewaki M. (1982). A circadian rhythm of gametangium
formation in Pseudobryopsis sp. (Chlorophyta, Codiales). Jap.
Soc.Phycol ДО, 171-180.
Oltmanns F. (1904). Morphologie and Biologie der Algen, Volg I
and II. Jena.
Oltmanns I. (1922). Morphologie und Biologie der Algen, Vols I
and II, 2nd Edition, Jena.
Ott D.W. (1982). Tribophyceae (Xanthophyceae): introduction and
bibliography. In: Selected Papers in Phycology II (eds.
526
Rogowski J.R. and Parker B.C.), pp. 723-727. Phycological
Society of America, Allen Press, Lawrence, Kansas.
Ott D.W., Brown R.M. , Jr. (1972). Light and electon microscopic
cal observations on mitosis in Vaucheria litirea Hofman ex
C.Agardh. Brit. Phycol. J., 7, 361-374.
Ott D.W., Brown R.M., Jr. (1974a). Developmental cytology of
the genus Vaucheria. II. Sporogensis in V.fontinalis (L.)
Christensen. Brit. Phycol. J., 2» 333-351.
Ott D.W., Brown R.M. (1974b). Developmental cytology of the
genus Vaucheria. I. Organization of the vegetative filament.
Brit. Phycol. J., 2» 111-126.
Ott D.W., Honimersand M.H. (1974). Vaucheriae of North Carolina.
I. Marine and brackish water species. J. Phycol*., 10, 373-
385.
Ott P.D., Sommerfeld M.R. (1982). Freshwater Rhodophyceae :in-
troduction and bibliography. In: Selected . Papers in Phyco-
logy II (eds. Rosowski J.R. and Parker B.C.), pp. 671-681.
Phycological Society of America, Allen Press, Lawrence,
Kansas.
Paasche E. (1980). Silicon. In: The Physiological Ecology of
Phytoplankton (ed. Morris I.), pp. 259-284. Blackwell Scien-
tific Publications, Oxford.
Paasche E., Klaveness D. (1970). A physiological comparison
of coccolith forming and naked cells of Coccolithus huxleyi.
Arch. Mikrobiol., 73. 143-152.
Padan E., Cohen Y. (1982). Anoxogenic photosynthesis. In: Bio-
logy of Cyanobacteria (eds. Carr N. G. and Whitton B.A.),
p. 215-236. Blackwell Scientific Publications, Oxford.
Paerl H.W., Tucker J., Bland P.T. (1983). Carotenoid enhancement
and its role in maintaining blue green algal (Microcystis
aeruginosa)surface blooms. Limnol. Oceanogr., 28, 847-857.
Paine R.T. (1969). A noteon trophic complexity and community
stability.Am.Nat., 103. 91-93.
Paine R.T. (1974). Intertidal community structure: experimental
strudies on the relationship between a dominant competitor
and its principal predator. Oecologia, 15, 93-120.
Paine R.T. , Vadae R.L. (1969). The effects of grazing by sea
urchins Strongylocentrotus spp. on benthic algal populations»
Tdmnol. Oceanogr., 16., 86-98.
Palmer C.M. (1977). Algae and Water Pollution: An Illustrated
Manual on the Identification, Significance and Control of
527
Algae in Water Supplies and Polluted Water. ЕРА-6ОО/9-77-ОЗ6.
US Environmental Protection Agency, Cincinnati, U.S.A.
palmer J.D., Round R.E. (1965). Persistent, vertical migration
rhythms in benthic microflora. I. The effect of light and
temperature on rhythmic behaviour of Euglena obtusa.J. Mar.
Biol. Ass.U.K., 45, 567-532.
Palmer J.D., Round R.E. (1967). Persistent, vertical migration
rhythms in benthic microflora. VI. The tidal and diurnal
nature of the rhythm in the diatom Hantzschia virgata.Biol.
Bull*, 132 , 44-55.
Palmisano A.C., Kottmeier S.T., Moe R.L., Sullivan C.W. (1985).
Sea ice microbial communities. IV. The effect of light pertur-
bation on microalgae at the ice seawater interface in McMurdo
Sound, Antarctica. Mar.Ecol. Prog.Ser. 21, 37-45.
Palmisano A.C., Sullivan C.W. (1933). Sea ice microbial communi-
ties (SIMCOs). I. Distribution, abundance and primary produc-
tion of ice microalgae in McMurdo Sound in 1980. Polar Biol.,
2, 171-177.
Pardy R.L. (1981). Cell size distribution of green symbionts
from Hydra viridis. Cytobios, 32, 71-77.
Pardy R.L. , Lewin R.A., Lee K. (1983). The Prochloron symbiosis.
Ins Algal Symbiosis: A Continuum of Interaction Strategies
(ed. L.J. Goff), pp. 91-96. Cambridge University Press, Cam-
bridge.
Parke M., Adams I. (I960). The motile (Crystallolithus hyalinus
Gaarder and Markali) and non motile phases in the life histo-
ry of Coccolithus pelagicus (Wallich) Schiller. J. Mar.Biol.
Ass. U.K., 3g, 263-274.
Parke M., Adams I. (1961). The Pyramimonas-Iike motile stage of
Haloshaera viridis Schmitz. Bull.Res.Coun.Israel, 10D, 94-100
Parke M., Green J.C. (1976). Chlorpphyta, Prasinophyceae. Ins
Parke M. and Dixon P.S. Check-list of British marine algae -
third revision, 564-566. J.'Mar.Biol.Ass. U.K., 56, 527-594.
Parker B.C. (1969). The occurrence of silica in brown and green
algae. Can. J.Bot. , 47, 537-540.
Parker B.C., Dawson E.Y. (1965). Non-calcareous marine algae
from California Mioccene deposits. Nova Hedwiga, 10, 273-295.
Parker B.C., Preston R.D., Fogg G.E. (1963). Studies on the
: structure and chemical composition of the cell walls of
f' Vaucheriaceae and Saprolegniaceae. Proc.Roy.Soc.Lond. B, 158,
Й' 435-445.
528
Parker S.P. (ed.). (1982). Synopsis and Classification of living
Organisms, Vols 1 and 2. McGraw-Hill, New York.
Parsons T.R., Takahashi M. (1973). Environmental control of
phytoplankton cell size. Limnol. Oceanogr., 18.,, 511-515.
Parsons T.R., Malta Y., Lalli C.M, (1984). A Manual of Chemical
and Biological Methods for Seawater Analysis. Pergamon Press,
Oxford.
Pascher A. (1917). Von der merkwurdigen Bewegungsweise einiger
Flagellaten. Biol. Centralbl., 37, 421-429.
Patel R.J. (1971). Cytotaxonomical studies of British species
of Chaetomorpha I. Chaetomorpha linum Kutz, and Chaetomorpha
a ere a Kutz. Phykos, 10, 127-136.
Patel R.J. (1972). Cytotaxonomical studies of British species of
Chaetomorpha II. Chaetomorpha melagonium Kutz. Phykos, 11,
17-22.
Patriquin D.G. , McClung C.R. (1978). Nitrogen accretion, and
the nature and possible significance of Ng fixation (acetyle-
ne reduction) in a Nova Scotian Spartina alterniflora stand.
Mar. Biol., 16, 49-58.
Patterson G.M.L. , Withers N.W. (1982). Laboratory cultivation
of Prochloron, a trypophan auxotroph. Science, 217, 1934-1935.
Patwary M.U., Van der Meer J.P. (1983). Genetics of Gracilaria
tikvahiae (Rhodophyceae). IX. Some properties of agar extrac-
ted from morphological mutants. Bot.Mar., 26, 295-299.
Peabody A.J. (1980). Diatoms and drowning-a review. Med.Sci.
Law, 20, 254-264.
Pearlmutter N.L. , Lembi C.A. (1978). Localization of chitin in
algal and fungal cell walls by light and electron microscopy.
J. Histochem.Cytochem., 26, 782-791.
Pearse V.B, (1972). Radioisotope study of calcification in the
articulated coralline alga Bossiella orbigniana.J. Phycol.,
8, 88-97.
Pearson B.R., Norris R.E. (1975). Fine structure of cell divi-
sion in Pyramimonas parkeae Norris and Pearson (Chlorophyta,
Prasinophyceae). J. Phycol., 11, 113-124.
Pedersen M. (1968). Ectocarpus fasciculatus: marine brown alga
requiring kinetin. Nature, 218, 776.
Pedersen M. (1969). The demand for iodine and bromine of three
marine brown algae grown in bacteria-free cultures. Physiol.
Plant., 22, 680-685.
Pedersen M. (1973). Identification of a cytokinin, 6-(3-methyl-
529
2-butenylamino) purine, in seawater and the effects of cytoki-
nins on brown algae, physiol. Plant., 2B, 101-105.
Pedersen P.M. (1981). Phaeophyta: Life histories. In: The Bio-
logy of Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp.
194-217. Blackwell Scientific Publications, Oxford.
Pedersen P.M. (1984). Studies on primitive brown algae (Eucophy-
ceae). Opera Bot., 74. 1-76.
Pelroy R.A., Bassham J.A. (1972). Photosynthetic and dark car-
bon metabolism in unicellular blue-green algae. Arch. Mikro-
biol., 86, 25-38.
Percival E. (1979). The polysaccharides of green, red, and brown
seaweeds: their basic structure, biosynthesis and function.
Brit. Phycol. J., 1£, 103-117.
Percival E., McDowell R.H. (1967). Chemistry and Enzymology
of Marine Algal Polysaccharides. Academic Press, New York.
Perkins E.J. (I960). The diurnal rhythm of the littoral diatoms
of the River Eden estuary. J.Ecol., 48, 103-108.
Perrone G., Feiicini G.P. (1972). Sur les bourgeons adventifs
de Petroglossum nicaeese (Duby)Schotter (Rhodophycees,
Gigartinales) en culture. Phycologia, 11. 87-95.
Perry M.J. (1972). Alkaline phosphatase activity in subtropical
central North Pacific waters using a sensitive fluorometric
method. Mar.Biol., 15. 113-119.
Perry M.J. (1976). Phosphate utilization by an oceanic diatom
in phosphorus-limited chemostat culture and in oligotrophic
waters of the central North Pacific. Limnol. Oceanogr., 21,
88-107.
Perry M.J., Tolbert M.C., Alberts R.S. (1981). Photoadaptation
in marine phytoplankton: response of the photosynthetic unit.
Mat. Biol., 62, 91-101.
Peters A.F. (1984). Observation on the life history of Papenfus-
siella calliricha (Phaeophyceae, Chordariales) in culture.
J. Phycol., 20,. 409-414.
Peters G.A. (1978). Blue-green algae and algal associations.
Biosciehce, 28, 580-585.
Peters G.A., Calvert H.E. (1983). The Azolla- Anabaena azollae
symbiosis. In Algal symbiosis: A Continuum of Interaction
Strategies (ed.Goff L.J.), pp.109-146. Cambridge University
Press, Cambridge.
Pfiester L.A.(1977). Sexual reproduction on Peridinium gatunense
(Dinophyceae). J.Phycol., 13. 92-95.
530
Pfiester L.A., Skvarla J.J. (1980). Comparative ultrastructure
of vegetative and sexual thecae of Peridinium limbatum and
Peridinium cinctum (Dinophyceae). Am.J.Bot., 67, 955-958.
Phillips D., Scott J. (1981). Ultrastructure of cell division
and reproductive differentiation of male plants in the Flori-
deophycidae (Rhodophyta). Mitosis in Dasya baillouviana.
Protoplasms, 106, 329-341.
Pia J. (1920) . Die Siphoneae verticillatae vom Karbon bis zur
Kreide. Abh.Zool.Bot.Ges.Wien, 11(2), 1-263.
Piccinni E., Albergoni V., Coppellotti 0. (1975). ATP-ase activi-
ty in flagella from Euglena gracilis.Localization of the enzy-
me and effects of detergents. J. Protozool., 22, 331-335.
Pickett-Heaps J.D. (1975). Green Algae. Sinauer Association,
Sunderland, Mass.
Pickett-Heaps J.D. (1976). Cell division in eukaryotic algae.
Bioscience, 26, 445-450.
Pickett-Heaps J.D. (1979). Electron microscopy and the phylogeny
of green algae and land plants. Am. Zool., 19, 545-554.
Pickett-Heaps J., Kowalski S.E. (1981). Valve morphogenesis
and the microtubule center of the diatom Hantzschia amphioxys.
Eur. J. Cell Biol., 25, 150-170.
Pickett-Heaps J., Marchant H.J. (1972). The phylogeny of the
green algae: a new proposal. Cytobios, .6, 255-264.
Pickett-Heaps J.D,, Staehelin L.A. (1975). The ultrastructure
of Scenedesmus (Chlorophyceae). II.Cell division and colony
formation. J.Phycol., 11, 186-202.
Pickett-Heaps J.D., Weik K. (1977). Cell division in Euglena
and Phacus. I. Mitosis. In: Mechanisms of Control of Cell
Division (eds. Post L. and Gifford E.M.), pp. 308-336. Dow-
den, Hutchinson and Ross, Stroudsburg, Pennsylvania.
Pielou E.C. (1977). The latitudinal spans of seaweed species
and their patterns of overlap. J.Biogeogr., 4, 299-311.
Pielou E.C. (1978). Latitudinal overlap of seaweed species:
evidence of quasisympatric speciation. J.Biogeogr., 5, 227-
238.
Pienaar Н.П. (1980). Chrysophytes. In: Phytoflagellates (ed.
Cox E.R.), pp.213-242. Elsevier/North Holland, New York.
Pintner I.J., Altmyer V.L. (1979). Vitamin binder and
other algal inhibitors. J. Phycol., 15. 391-398.
Pisa M., SanbergP.R., Corcoran M.E., Pibiger H.C. (1980). Spon-
taneously recurrent seizures after intracerebral injections
531
of kainic acid in rat: a possible model of human, temporal
lobe epilepsy Brain Res., 200, 481-487.
, 1 Platt T. (ed.) (1981). Physiological bases of phytoplankton
ecology. Can.Bull.Pish.Aquat.Sci., 210, 1-346.
Platt T., Denham K. (1980). Patchiness in phytoplankton distribu-
tion. In: The Physiological Ecology of Phytoplankton (ed.
Morrisl.), pp. 413-431. Blackwell Scientific Publications,
Oxford.
J
?Pluzeck K. (1981). Applications of agarose in biomedical tech-
niques in relation to chemical and physical properties. Proc.
Int.Seaweed Symp., 10, 711-718.
Polanshek A.R., West J.A.(1977). Culture and hybridization stu-
dies on Gigartina papillata. J. Phycol., 13. 141-149.
Porcella R.A., WalneP.L. (1980). Microarchitecture and envelope
, . in Dysmorphocoocus globobue (Phacotaceae, Chlorophyceae).
J. Phycol., 16, 280-290.
Porter K.G. (1973). Selective grazing and differential digestion
of algae by zooplankton. Nature, 244, 179-180.
Porter K.G. (1976). Enhancement of algal growth and productivity
by grazing zooplankton.Science, 192, 1332-1334.
Porterfield W.M. (1922). References to the algae in the Chinese
classics. Bull. Torrey Bot.Club., 49, 297-300.
Potts M. (1980). Blue-green algae (Cyanophyta) in marine coastal
environments of the Sinai Peninsula; distribution, zonation,
stratification and taxonomic diversity. Phycologia, 19, 60-73.
Povolny M. (1981). The effect of steeping of peat-cellulose
flower pots (Jiffypots) in extracts of seaweeds on the quali-
ty of tomato seedlings. Proc.Int.Seaweed Symp., 8, 730-733.
Prakash A., Taylor P.J.R. (1966). A red water bloom of Gonyaulax
aoatenella in the strait of Georgia and its relation to
paralytic shell fish toxicity. J.Pish.Res. Bd.Can., 23,
1256-1270.
Prask J.A., Plocke D.J. (1971). A role for zinc in the structural
integrity of the cytoplasmic ribosomes of Euglena gracilis.
Plant. Physiol. Lancaster, 37, 428-433.
Prescott G.W. (1951). History of phychlogy. In: Mannual of Phy-
cology (ed. Smith G.M.), pp. 1-11. The Ronald Ргевв Company,
New York.
Prezlin B.B. (1981). Light reactions in photosynthesis. Can.
Bull.Pish.Aquat.Sci. , 210, 1-43»
Prezlin B.B., Alberte R.S. (1978). Photosynthetic characteristics
532
and organization of chlorophyll in marine dinoflagellates.
Proc. Natl.Acad.Sci., 75, 1801-1804.
Prezlin B.B., Sweeney B.M. (1978). Photoadaptation of photosyn-
thesis in Gonyaulax polyhedra. Mar. Biol., 48. 27-35.
Price I.A. (1972). Zygote development in Caulerpa (Chlorophyta,
Caulerpales). Phycologia, 11, 217-218.
Price I.R., Ducker S.C. (1966). The life history of the brown
alga Splachnidium rigosum. Phycologia, 261-273.
Pringsheim E. G. (1968). Klein Mitteilungen uber Flagellaten
und Algen. XV. Zur Kenntnis der Gattung Pqphyridium.- Arch.
Mikrobiol., 61, 169-180.
Proctor V.W. (1971). Taxonomic significance of monoecism and
dioecism in the genus Chara.Phycologia. 10, 299-307.
Provasoli L., Carlucci A.P. (1974). Vitamins and growth regu-
lators. Ins Algal Physiology and Biochemistry (ed. Stewart
W.D.P.), pp.741-787. Blackwell Scientific Publications, Ox-
ford.
Provasoli L., McLaughlin J.J.A., Droop M.R. (1957). The develop-
ment of artifical media for marine algae. Arch. Mikrobiol.,
2£, 392-428.
Provasoli L., Pintner I.J., Sampathkumar S. (1977). Morphogene-
tic substances for Monostroma oxyspenaum from marine bacteria.
J. Phycol., 13 (suppl.), 56.
Pueechel C.M. (1977). A freeze-etch study of the ultrastructure
of red algal pit plugs. Protoplasma, 91. 15-30.
Pueechel C.M. (1980). A re-appraisal of the cytochemical pro-
perties of rhodophycean pit plugs. Phycologia, 19. 210-217.
Quatrano R.3., Stevens P.T. (1976). Cell wall assembly in Fucus
zygotes. Plant Physiol., 58, 224-231.
Ragan M.A., (1981). Chemical constituents of seaweeds. In: The
Biology of Seaweeds (eds. Lobban C.S. and Wynne M.J. ), pp.
589-626. Blackwell Scientific Publications, Oxford.
Ragan M.A., Chapman D.J. (1978). A Biochemical Phylogeny of the
Protistв.Academic Press, New York.
Rai L.C., Gaur J.P., Kumar H.D. (1981). Phycology and heavy
metal pollution, Biol.Rev., 56, 99-151.
Ramus J. (1969). The developmental sequence of the marine red
alga Pseudogloiphloea in culture. Univ.Calif. Publ. Bot., 52.
1-42.
Rsmus J. (1971). Properties of septal plugs from the red alga
GriffithBia pacifica. Phycologia, 10, 99-103.
Ramus J. (1972). The production of extracellular polysaccharide
533
by the unicellular red alga Porphyridium aerugineum J. Phy-
col. , 8, 97-111.
Ramus J. (1981). The capture and transduction of light energy.
In: The Biology of Seaweeds (eds. Lobban C.S. and Wynne M.J.),
pp. 458-492. Blackwell Scientific Publications, Oxford.
Ramus J., Beale S.X., Mauzerall D., Howard K.L. (1976). Changes
in photosynthetic pigment concentration in seaweeds as a
function of water depth. Mar.Biol., 37. 223-229»
Ramus J., Lemons F., Zimmerman C. (1977). Adaptation of light-
harvesting pigments to downwelling light and consequent
photosynthetic performance of the eulittoral rockweeds Asco-
phyllum nodosum and Fucus vesiculosus. Mar.Biol., 42. 293-303»
Ranwell D.S. (1972). Ecology of Salt Marshes and Sand Dunes.
Chapman and Hall, London.
Raunkiaer C. (1934). The Life Forms of Plants and Statistical
Plant Geography. Clarendon Press, Oxford.
Raven J.A.(1970). Exogenous inorganic carbon sources in plant
photosynthesis. Biol.Rev., 45. 167-221.
Raven J.A. (1974). Carbon dioxide fixation. In:Algal Physiology
and Biochemistry (ed. Stewart W.D.P. ), pp. 434-455. Black-
well Scientific Publications, Oxford.
Raven J.A., Beardall J. (1981). Respiration and photorespiration.
Can.Bull.Fieh.Aquat.Sci., 210, 55-82.
Rawitscher-Kunkel E., Machlis L. (1962). The hormonal integra-
tion of sexual reproduction • in Oedogonium. Am.J.Bot., 49.
177-183.
Rawlence D.J. (1972). An ultrastructural study of the relation-
ship between the rhizoides of Polysiphonia lanosa (L). Tandy
(Rhodophyceae) and the tissue of Ascophyllum nodosum (L.)
Le Jolis (Phaeophyceae). Phycologia, 11, 279-290.
Redfield A.C., Ketchum B.H., Richards F.A. (1963). The influence
of organisms on the composition of seawater. In: The Sea
(ed. Hill M.N.), pp.26-77. Wiley-Interscience, New York.
Reed M.L., Graham D. (1981). Carbonic anhydrase in plants:
distribution, properties and possible physiological function.
Prog. Phytochem., 7, 47-96.
Reed R.H. (1983). Measurement and osmotic significance of J5~
dimenthylsulphonioproponate in marine macroalgae. Mar.Biol.
Lett., £, '173-181.
Reed R.H., Davision I.R., Chudek J.A., Foster R. (1985). The
osmotic role of mannitol in the Paeophyta: an appraisal.
534
Phycologia, 24, 35-47»
Richardt W., Overheck J., Stenting L. (1967)» Free dissolved
ansymes in lake waters. Nature, 216, 1345-1347.
Reimann B.E.F., Lewin J.C., Volcani B.E. (1966)» Studies on the
biochemistry and fine structure of silica shell formation in
diatoms II. The structure of the cell wall of Naviculla pelli-
culosa (Breb.)Hilse. J.Phycol., 2, 74-84»
Reinke J. (1878). Entwicklungsgeschichtliche Untersuchungen
uber die Cutleriaceen des Golfs von Neapel. Nova Acta K»Leopj-
Carol. Deutsch. Akad. Naturforsch., 40. 59-96.
Rentschler HrG. (1967). Photoperiodische Induktion der Monospo-
renbild.ing bei Porphyra tenera Kjellm. (Phodophyta-Bangiophy-
ceae). Planta, 76, 65-74»
Reynolds C.S. (1984)» The Ecology of Freshwater Phytoplankton.
Cambridge University Press, Cambridge.
Reynolds C. S. , Walsby A.E. (1975). Water Uooms.Biol.Rev. , 50,
437-481.
Reynolds R.C., Jr. (1978). Polyphenol inhibition Of caloite pre-
cipitation in lake Powel, Idmnol. Oceanogr., 23.589-997.
Reyssac J., Roux M. (1972). Communautes phytoplanctoniques dans
les eaux de la Cote d'Ivoire. Groupes d'especes associees.
Mar.Biol., 1J, 14-33.
Rhee GrY. (1980). Continuous culture in phytoplankton ecology.
Advances Aquatio Microbiol., 2, 151-203.
Rice E.L., Chapman A.R.O. (1982). Net productivity of two cohorts
of Chordaria flagelliformis (Phaeophyta) in Nova Scotia,
Canada. Mar. Biol., 71, 107-111.
Richardson J.L. (1968). Diatoms and lake typology in East and
Central Africa. Int.Rev.Ges.Hydrobiol.Hydrogr., 53, 299-338.
Richardson N. (1970). Studies on the photobiology of Bang!a fus-
copurpurea. J. Phycol., 6, 215-219.
Richardson N., Dixon P.S.(1968). Life history of Bangia fuscopur-
purrea (Dillw.) Lynb. in culture. Nature, 218, 496-497.
Richardson N., Dixon P. (1969). The Conchocelis phase of Smithora
naidum (Anders.) Hollenb. Brit. Phycol. J., 7, 49-51.
Richerson P., Armstrong R., Goldman C.R. (1970). Contempoeraneous
disequilibrium, a new hypothesis to explain the"paradox of
the plankton", Proc.Natl.Acad.Sci., 67» 1710-1714.
Rietema H. (1970). Life histories of Bryopsis plumosa. Csulerpa-
les, Chlorophyceae from European coasts, Acta Bot.Neer., 19,
859-866.
535
Bietema H. (1971). Ute history studies in the genus Bryopsis.
Chlorophyceae. Part 4, life histories in Bryopsis hypnoides,
from different parts of the European coasts. Acta Bot. Neer.,
20, 291-298.
Hietema H. (1972). A morphological, developmental and caryolo-
gical study oh the life history of Bryopsis halymeniae (Chlo-
rophyceae). Neth.J.Sea Res., 2» 445-457.
Riley G.A. (1967). The plankton of estuaries. In: Estuaries
(ed. lauff G.H.), pp. 316-326. A.A.A.S. Publ. 83, Washington,
D.C.
Ringo D.l. (1967). Flagellar motion and fine structure of the
flagellar apparatus in Chiamydomonas.J.Cell Biol», 33. 543-
571.
Rippka R., Deruelles J., Waterbury J.B., Herdman М», Stanier
R.Y. (1979). Generic assignments, strain histories and pro-
perties of pure cultures of Cyanobacteria. J.Gen. Microbiol.,
111, 1-61.
Robenek H., Melkonian M. (1983). Structural specialization of
the paraflagellar body membranes of Euglena.Protoplasma.
117.154-157.
Roberts K., Gurney-Smith M., Hills G.J. (1972). Structure,
composition and morphogenesis of the cell wall of Chiamydo-
monas reinhardii I. Ultrastructure and preliminary chemical
analyses. J.Ultrastr. Res., 40» 599-613.
Robinson D.G., Preston R.D. (1971). Studies on the fine struc-
ture of Glaucocystis nostochinearum Itzigs. II. Membrane
morphology and taxonomy. Brit. Phycol. J., 6, 113-128.
Roelofsen P.A. (1965). Ultrastructure of the wall in growing
cells and its relation to the direction of growth.Adv.Bot.
Res., 2, 69-149.
Rogers D.J., Blunden G. (1980). Structural properties of the anti-
B lectin from the red alga Ptilota plumosa (Huds.) G.Ag.
Bot.Mat. 23, 459-462.
Rogers D.J., Blunden G., Topliss J.A., Guiry M.D. (1980). A
survey of some marine organisms for haemagglutinins. Bot.
Mar*» 22» 569-577.
RoseribaumJ.Ii., Child F.M. (1967). Flagellar regeneration in
protozoan flagellates. J.Cell Biol., 34» 345-364.
Rosenbaum J.L., Moulder J.E., Ringo D.L. (1969). Flagellar,
elongation and shortening in Chlamyd omonas. The use of
cycloheximide and colchicine to study the synthesis and
536
assembly of flagellar proteins. J.Cell Biol., 41» 600-619.
Bosenberg G., Ramus J. (1982). Ecological growth strategies in
the seaweeds Gracilaria foliifera (Rhodophyceae) and Ulva
sp. (Chlorophyceae): soluble nitrogen and reserve carbohyd-
rates. Mar.Biol., 66. 251-259.
Rosenvinge 1.K. (1924). The marine algae of Denmark, contribu-
tions to their natural history. Pt. Ill. Rhodophyceae
(Ceramiales). Det. Kgl. Vidensk, Selsk. Skrift., 7(3),
287-486.
Rosowski J.R. (1977). Development of mucilaginous surfaces in
euglencids. II. Flagellated, creeping and palmelloid cells
of Euglena. J. Phycol., 13. 323-328.
Ross R. (1982). Bacillariophyceae. In: Synopsis and Classifica-
tion of Living 0т.еЯ'п1ятя, Vol. 1 (ed. Parker S.P.), pp.95-
101. McGraw-Hill, New York.
Rcttger R., Irwan A., Schmaljohann R., Pranziket L. (1980).
Growth of the endosymbiont-bearing forminifer Amphistegina
lessonii d’Orbigny and Heterosteglna depressa d’Orbigny
(Protozoa). In: Endocytobiology, Vol. 1 (eds. Schwemmler
W. and Schenk H.E.A.), pp.155-162. Walter de Gruyter, Berlin.
Round P.E.(1957). The late-glacial and post-glacial diatom su-
oession in the Kentmere valley deposit.I. Introduction,
methods and flora. New Phytol., 58. 98-126.
Round P.E. (1971). The taxonomy of the Chlorophyta. II. Brit.
Phycol. J., 6, 235-264.
Round P.E. (1973). , The Biology of the Algae, 2nd EditioruSt.
Martin’s Press, New York.
Round P.E. (1981). The Ecology of the Algae. Cambridge Univer-
sity Press, Cambridge.
Round P.E., Crawford R.M. (1981). The lines of evolution of
the Bacillariophyceae. I. Origin. Proc.Rpy.Soc.Lond. В 211.
237-260.
Ruddat M* (1961). Versuche Zur Beeinflussung und Auslosung
der Endogen tagesrhythmik bei Oedogonium cardiaceum Wittr.
Z. Bot., 42, 23-46.
Rueness J. (1973).Culture and field observations on growth
and reproduction of Ceramium atrictum. Harv. from the Oslof"
jord, Norway. Norw. J.Bot., 20. 61-65.
Rueness J. (1978). A note on development and reproduction in
Giagartina stellata (Rhodophyceae, Gigartinales from Norway»
Brit. Phycol. J., 22» 87-90.
537
Bussell G. (1964). Laminariocolax tomentosoides on the Isle
of Man.J.Mar. Biol. Ass. U.K., ££, 601-612.
Bussell G. (1977). Vegetation on rocky shores at some north
Irish sea sites. J.Ecol., 65. 485-495»
Bussell G. (1979). Heavy receptacles in estuarine Fucus vesica-
loans L. Est.Cstl Mar.Sci., 2.» 659-661.
Bussell G., Frieding A.H. (1974). The competitive properties
of marine algae in culture. J.Ecol., 62. 689-698.
Russell G., Morris O.P. (1970). Copper tolerance in the marine
foulding alga Ectocarpus siliculosua» Nature, 228, 288-289.
Ruasell G., Veltkamp C.J. (1982). Epiphytes and anti-fouling
characteristics of Himanthalia (brown algae). Br. Phycol.J.,
17, 239.
Ryther J.H., Dunstan W.M. (1971). Nitrogen, phosphorus and eutro-
phication in the coastal marine environment. Science, 171,
1008-1013*
Ryther J.H., Deboer J.A., Lapointe B«E.(1979)« Cultivation of
seaweeds for hydrocolloids, waste treatment and energy con-
version. Proc. Int.Seaweed Symp., £, 1-16.
Sachs J. (1874). Lehrbuch der Botanik, 4th Edition. Leipzig.
Safferman R.S. (1973). Phycoviruses. Ins The Biology of the Blue
Green Algae (eds. Carr N.G. and Whitton B.A.), pp.214-237.
Blackwell Scientific Publications, Oxford.
Saga N», Uchida T., Sakai Y. (1978). Clone Laminaria from sinop-
le isolated cell. Jap.Soc.Sci. Fish., 44. 87.
Sagan L. (Margulis L.) (1967). On the origin of mitosing cells.
J. Theor. Biol., 14, 225-275.
Saito Y. (1982). Information on the culture of phytoplankton for
aquacultural needs in Japan. Ins Proceedings of the 6th
U.S.-Japan Meeting on Aquaculture (ed. Sinderman C.J.),
pp. 1-6, NOAA Technical Report NMP3 Circ, 442.
Sampaio M.J.A.M., Rai A.N., Rowell P., Stewart W. D. Г. (1979).
Occurrence, synthesis and activity of glutamine synthetase
in Nj fixing lichens. FEMS Microbiol. Lett., 6, 107-110.
Sanbonsuga Y., Neushul M. (1978). Hybridization of Macrocystis
(Phaeophyta) with other float bearing kelps.J. Phycol., 14.
214-224.
Sandgren C.D. (1980). An Ultrastructural investigation of rest-
ing cyst formation in Dinobryon cylindricum Imhoff (Chry-
sophyceae, Chrysophycota). Protistologica, 16. 259-276.
Sandgren C.D. (1981). Characteristics of sexual and asexual
resting cyst (statospore) formation in Dinobryon cylindri-
538
cum. J. Phycol., 17. 199-210.
Sandgren C.D. (1983). Survival strategies of chrysophycean fla-
gellates: reproduction and the formation of resistant cysts.
In: Survival Strategies of the Algae (ed. Fryxell G.A. ),
pp. 23-49. Cambridge University Press, Cambridge.
Santore U.J. (1977). Scanning electron microscopy and comparative
micromorphology of the periplast of Hemiselmis rufescens.
Chroomonas sp. and members of the genus Cryptomonas (Crypto-
phyceae). Brit.Phycol. J., 12, 255-270.
Santore U.J. (1982a). Comparative ultrastructure of two members
of the Cryptophyceae assigned to the genus Chroomonas - with
comments on their taxonomy.Arch, Protistenk., 125, 5-29.
Santore U.J. (1982b). The ultrastructure of Jemiselmia brunnes-
cens and Hemiselmis virescens with additional observations
on Hemiselmis rufescens and comments about the Hemiselmidaceae
as a natural group of the Cryptophyceae. Brit. Phycol. J.,
17, 81-99.
Sarjeant N.A.S. (1974K Fossil and Living Dinoflagellates. Aca-
demic Press, New York.
Sarma Y. S.R.K., Shyam R. (1974). A new member of colonial Vol-
vocales (Chlolrophyceae), Pyrobotrys acuminata sp. nov. from
India. Phycologia, 13, 121-124.
Sauvageau C. (1899). Les Culteriacees et leur alternance de ge-
nerations. Ann.Sci. Nat.Bot.Ser. 8, 10. 265-362.
Sauvageau C. (1915). Sur la sexualite heterogamique d'une Lami-
naire (Saocorhiza bulbosa). C.R. Acad.Sci. Paris D, 161. 769-
799.
Sawa T. (1965). Cytotaxonomy of the Charaoeae: karyotype analysis
of Njtella opaca and Nitella flexilis. Am.J.Bot., 52. 962-
970.
Sawa T., Frame P. W. (1974). Comparative anatomy of Charophyta. I.
Oogonia and oospores of Tolypella with special reference
to the sterile oogonial cell.Bull.Torrey Bot.Club, 101, 136-
144-
Say P.J., Whitton B.A. (1978 ). Chemistry and benthic algae of
a zinc polluted stream in the northern Pennines. Brit. Phycol.
J., 1£, 206.
Scagel R.P. (1956). Introduction of a Japanese alga. Sargassum
muticum, into the North west Pacific. Fish. Res.Pap. Wash.
Dept. Fish, 1, 49-58.
Scagel R. F. , Bandoni R. J., Maze J.R., Rouse G.E., Schofield W.G. ,
539
: Stein J.R. (1982). Konvaacular Plants.An Evolutionary Survey,
!’ Wadsworth, Belmont.
Schaede R. (1951). Uber die Blaualgensymbiose von Gunneria.Planta.
39, 154-170.
Schantz E.J. (1981). Poisons produced by dinoflagellates - a re-
view. In: The Water Environment: Algal Toxins and Health (ed.
Carmichael W.W.), pp.25-36, Plenum Press, Mew York.
Schindler D.W. (1971). Carbon, nitrogen and phosphorus and the
eutrophication of freshwater lakes. J.Phycol., 7, 321-329.
Schindler D.W.(1974).Eutrophication and recovery in experimental
lakes:implications for lake management, Science.184. 897-898.
Schindler D.W.(1977). Evolution of phosphorus limitation in lakes
Science, 195, 260-267.
Schindler D.W. (1980). Experimental acidification of a whole lake,
> . a test of the oligotrophioation hypothesis. In: Ecological Im-
;; pact of Acid Precipitation (eds. Drablos B. and Tollan A.), pp.
'370-374, SNFS, Oslo.
Schindler D.W. , Kling H., Schmidt R.V., Prokopowich J., Frost V.E. ,
Reid R.L., Capel M. (1973). Eutrophication of Lake 227 by addi-
tion of phosphate and nitrate: the second, third and forth years
of enrichment 1970,1971,1972, J.Fish Res.Bd Can..30.1415-1446.
Schlosser U.(1976). Enzymatisch gesteuerte Freisetzung von Zoospo-
ren bei Chlamydomonas reinhardtii Dangeard in Sybchronkultur
Arch.Mikrobiol., 54.129-154.
Schmidt R.J., Loeblich A.R.III (1979) Distribution of paralytic
shell fish poison among Pyrrophyta.J.Mar.Biol.Ass.U.K..59.479-487
Schmitz Fr.(1883). Untersuchungen uber die Befruchtung der Flori-
deen, Sitzbr.Akad.Wiss.Berlin, 883. 215-258.
Schmitz Fr. (1889). Systematische Ubersicht der bisher bekannten
Gattungen der Florideen.Flora (Jena), 72. 435-456.
Schmitz K. (1981). Translocation. In: The Biology of Seaweeds(eds.
Lobban C.S. and Wynne M.J.), pp.534-558. Blackwell Scientific
Publications, Oxford.
Schmitz K., Strivastava L.M. (1975). On the fine structure of sea-
ve tubes and the physiology of assimilate transport in Alaria
marginata. Can. J. Bot., 53, 861-876.
Schneider C.W.(1976). Spatial and temporal distributions of ben-
thic marine algae on the continental shelf of the Carolinas.
Bull.Mar.Sci., 26, 133-151.
Schoenberg D.A., Trenoh R.K. (1980a). Genetic variation in
Symbiodinium (=Gymnodinium) microadriaticum Freudenthal,
540
and specificity in its symbiosis with marine invertebrates. I.
Isoenzyme and soluble protein patterns of axenic cultures
of Symbiodinium microadriaticum. Proc. Roy.Soc.Lond. B, 207,
405-427.
Schoenberg D.A., Trench R.K. (1980b). Genetic variation in Sym-
biodinium (»Gymnodinium) microadriaticum Freudenthal, and
specificity in the symbiosis with marine invertebrates, II.
Morphological variation in Symbiodinium microadriaticum.Proc.
Roy.Soc.Lond. B, 207, 429-444.
Schoenberg D.A., Trench R.K. (1980c). Genetic variation in Sym-
biodinium (=Gymnodinium) microadriaticum Freudenthal, and
specificity in its symbiosis with marine invertebrates. III.
Specificity and infectivity of Symbiodinium microadriaticum.
Proc.Roy.Soc. Lond. B, 207, 445-460.
Schonbeck M., Norton T.A. (1978). Factors controlling the upper
limit of fucoid algae on the shore. J.Exp. Mar. Biol.Ecol.,
21» 303-330.
Schonbeck M., Norton T.A. (1980). Factors controlling the lower
limits of fucoid algae on the shore. J.Exp. Mar.Biol.Ecol., 43.
131-150.
Schopf J.W. (1970). Precambrian micro-organisms and evolutionary
events prior to the origin of vascular plants. Biol.Rev., 45.
319-352.
Schopf J.W. (1974a). Paleobiology of the Precambrian: the age
of blue-green algae.Ins Evolutionary Biology(eds, Dobzhansky
T., Hecht M.K. and Steere W.C,), pp.1-43. Plenum Press, New
York.
Schopf J.W. (1974b). The development and diversification of
Precambrian life. Origins of Life, 2» 119-135.
Schopf J.W. (1976). Are the oldest "fossils" fossils? Origins
of Life, 2» 19-36.
Schopf J.W. (1978). The evolution of the earliest cells.Sci.
Am., 239. 110-134.
Schopf J.W., Walter M.R. (1982a). Origin and early evolution of
Cyanobacteria: the geological evidence. In: The Biology of
Cyanobacteria (eds. Carr N.G. and Whitton B.A.), pp.543-564.
Blackwell Scientific Publications, Oxford.
Schopf J.W,, Walter M.R, (1982b). Archean microfossils: new evi-
dence of amcient microbes. In: Origin and Evolution of Earth's
Earliest Biosphere: An Interdisciplinary Study (ed. Schopf
J.W.). Princeton University Press, Princeton, New Jersey.
Schreiber E. (1930). Untersuchungen fiber Parthenogenesis Ge -
541
gchlechtsbestimmung und Bastardierungs vermogen bei leminari-
en Planta, 12, 331-351.
Schultz M.E. , Trainor E.R. (1968). Production of male gametes
and auxospores in the centric diatoms Cyclotella meneghiniana
and C. crypta.J. Phycol., Д, 85-88»
Scoffin T.P. (1970). The trapping and binding of aubtidal carbo-
nate sediments by marine vegetation in Bimini lagoon, Bahama,
J. Sedim.Petrol., ДО, 249-273.
Scott J., Bullock K.W. (1976). Ultrastructure of cell division
in Cladophora. Pregametangial cell division inthe haploid
generation of Cladophora flexuosa. Can. J.Bot., 54. 1546-1560.
Scott J.L., Dixon P.3.(1971). The life history of Pikea califor-
nica Harv. J. Hiycol., 2, 295-300.
Scott J.L., Bosco C. , Schornstein K., Thomas J. (1980). Ultra -
structure of cell division and reproductive differentiation of
male plants in the Plorideophyceae (Rhodophyta): cell divi-
sion in Polysiphonia. J. Phycol., 16. 507-524.
Seapy R.R., Idttler M.M. (1978). The distribution, abundance,
community structure and primary productivity of macro orgas.
nisms from two central California rocky intertidal habitats.
Pacific Sci., 22, 293-314.
Sears J.R., Wilce R.T. (1970). Reproduction and systematics
of the marine alga Derbesia (Chlorophyceae) in New England.
J. Phycol., 6, 381-392.
3ears J.R., Wilce R.T. (1975). Sublittoral, benthic algae of
southern Cape Cod and adjacent islands: seasonal periodicity,
associations, diversity and floristic composition. Ecol.
Monogr., 45. 337-365.
Sellner K.G.,Kachur M.E., Lyons L. (1984). Alterations in car-
bon fixation during power plant entrainment of estuarine
phytoplankton. Water Air Soil Poll., 21. 359-374.
Setchell W.A. (1915). The law of temperature connected with
the distribution of the marine algae.Aim, Mo.Bot.Gard,, 2»
287-305.
Shacklock P.P., Croft G.B. (1981). Effect of grazers on Chon-
drus crispus in culture. Aquaculture, 22. 331-342.
Sharabi N.E., Pramer D. (1973). A speotrophotofluorometric
method for studying algae in the soil.Bull.Ecol.Res.Comm.
(Stockholm), JJX, 77-84.
Shaw W.N. (1982). The use of phytoplankton for aquaculture
needs - a status report. In: Proceedings of the 6th U.S. -
Japan Meeting on Aquaculture (ed. Sinderman C.J,), pp.
542
19-24» NOAA Technical Report NMFS Clrc., 442.
Sheath R.G. (1984). The biology of freshwater red algae. Prog.
Phycol.Res., 2» 89-157.
Sheath R.G., Hellebust J.A. (1974). Glucose transport systems and
growth characteristics of Bracteococcus minor.J. Phycol.,
12, 43-41.
Shelef G., Soeder C.J. (eds.) (1980). Algal Biomass: Production
and Use Elsevier/North Holland, Amsterdam.
Shen E.Y.F, (1967). Amitosis in Chara. Cytologia, 32. 481-488.
Shen E.Y.F. (1967). Microspeotrophotometric analysis of nuclear
DNA in Chara zeylanica.J.Cell Biol., 22» 377-384.
Sheridan R.P. (1979). Seasonal variation in sun-shade ecotypes
of Plectonema notatum (Cyanophyta). J. Phycol., 22» 223-226.
Shields L.M., Burrell L.W. (1964). Algae in relation to soil fer-
tility. Bot. Rev. , 22» 92-128.
Shihara I», Krauss R.W, (1964). Chlorella: Physiology and Taxo-
nomy of Forty-one Isolates. University of Maryland, College
Park.
Shilo M. (1971). Biological agents which cause lysis of blue-
green algae. Mitt. Int.Verein.Limnol., 19. 206-213-
Shilo M. (1981). The toxic priciples of Prymnesium parvum.In:
Water Environment. Algal Toxins and Health. Environment Scien-
ce Research Vcl. 20 (ed, Carmichael W.W.), pp.34-47. Plenum
Press, New York.
Sicko-Goad L. (1982). A morphometric analysis of algal response
to low dose, short term heavy metal exposure, ProtOpiaeaa
110. 75-86.
Sicko-Goad L., Stoermer E.F. (1979). A morphometric study of
j lead and ocpper effects on Diatoma tenue var, elongatum
(Bacillariophyta). J. Phycol., 15. 316-321.
Siddeque M., Faridi M.A.F. (1977). The life history of Cladopho-
ra crispata (Roth). Ag. Pak.J.Bot., 2» 159-162.
Siegel B.T., Siegel S.M. (1973). The chemical composition of
algal cell walls.Crit.Rev.Microbiol., 2» 1-26,
Siesser W.G.(1972). Relief algal nodules (rhodolites) from the
South African continental shelf, J.Geol., 80, 611.
Sievers A., Heinemann B., Rodriguez- Garcia M.I, (1979). Nachwei
des subapikalen differentiellen Flankenwaohstums in Chara-
Rhizoid wahrend der Graviresponse, Z.Pflanzenphysiol., 91.
435-442.
Sikes C.S., Roer R.D., Wilbur K.M. (1980). Hiotosynthesis and
coccolith formation: inorganic carbon sources and net inor-
543
ganio reaction of deposition. limnol. Oceanogr., 25. 248-261.
Silva P.C. (1951). The genus Codium in California with observa-
tions cn the structure of the walls of the utricles. Univ.
Calif.Pub. Bot., 22, 79-114.
(1979). Review of the taxonomic history and nomen-
of the yellow-green algae.Arch.Protistenkd., 121.
(1980). Names and classes of living algae. Regnum
Silva P.O.
clature
20-63.
Silva P.O.
Vegetabile, 103. 1-156.
Silva P.O. (1982). Thallobionta. In: Synopsis and Classification
of living Organisms, Vol. 1 (ed. Parker S.P.), pp.59-60.
McGraw-Hill, New York.
Silver M.W,, Bruland K.W. (1981). Differential feeding and fe-
cal pellet composition of salps and pteropods, and the pos-
sible origin of the deep water flora and olive green cells.
Mar.Biol., 62, 263-273.
^Silverberg B.A., Stokes P.M. ,Ferstenberg L.B. (1976). Intranu-
clear complexes in a copper-tolerant green alga. J.Cell Biol.
62, 210-214.
imon R.D. (1973). The effect of chloramphenicol on the produc-
tion of cyanophycin granule polypeptide in the blue green
alga Anabaena cylindrica. Mikrobiol., 92. 115-122.
Skuja H. (1954). Glaucophyta. In:Engler A., Syllabus der Pflan-
zenfamilien, 12, I (eds. Melohoir H. and Nerdermann E.),
pp. 56-57. Bomtraeger,Berlin.
Slankis T,, Gibbs S.P. (1972). The fine struotureof mitosis and
cell division in the Chrysophycean alga Ochromonas danica.
J. Phycol., 8, 243-256.
Slocum C.J. (1980). Differential susceptibility .to grazers in
two phases of an intertidal alga: advantagesof heteromorphic
generations. J.Exp.Mar.Biol.Ecol., 46, 99-110.
Slocum R.D., Ahmadjian V., Hildreth K.C. (19S0). Zoosporogenesis
in Trebouxia gelatinosa: ultrastructure, potential for zoos-
pore release and implications for thelichen association.
Idchenologist, 12, 173-187.
Sluiman H.J., Roberts K.R., Stewart K.D., Mattox K.R. (1980).
Comparative cytology and taxonomy of the Ulvaphyoeae. I.
The zoospore of Ulotrhix zonata (Chlorophyta). J. Phycol.,
16, 537-545.
Sluiman H.J., Roberts K.R., Stewart K.D,, Mattox K.R., Lokhorst
G.M. (1981). A reinvestigation of theflaggellar apparatus
544
of the zoospore of Urospora (Chlorophyta). Brit. Phycol. J.,
16, 140.
Sluiman H.J., Roberts K.R., Stewart K.D., Mattox K.R. (1982).
The flagellar apparatus of the zoospore of Urospora penicilli-
formisCChlorophyta). J. Phycol., 18, 1-12.
Smajfda T.J. (1970). The suspension and sinking of phytoplankton
in the sea. Ann.Rev.Oceanogr. Mar.Biol., 8, 353-414.
Smayda T.J. (I960). Phytoplanktqn species succession. In: The
Physiological Ecology of Phytoplankton (ed. Morris I.), pp.
493-570. Blackwell Scientific Publications, Oxford.
Smith D.C. (1978). What can lichens tell us about real fungi?
Mycologia, 70. 915-934.
Smith D.C. (1980). Mechanisms of nutrient movements between li-
chen symbionts. In: Cellular Interactions in Symbiosis and
Parasitism (eds. Cook C.B., Pappas P.W. and Rudolph E.D.),
pp.197-227. Ohio State University Press, Columbus.
Smith D., Wiebe W. (1977). Rates of carbon fixation, organic
carbon release and translocation in a reef building foramini-
fer Maringopora vertebralis. Aust. J. Mar. Freshwater Res.,28.
311-319.
Smith G.M. (ed.) (1951). Manual of Phycology. An Introduction
to the Algae and their Biology. The Ronald Press Company,
New York.
Smith G.M. (1955). Cryptogamic Botany. Vol. I, Algae and Fungi,
2nd Edition McGraw-Hill, New York.
Smith J.E. (ed.) (1968). "Torrey Canyon" Pollution and Marine
life. Cambridge University Press, Cambridge.
Smith P.V. (1954). Studies on the origin of petroleum; occurence
of hydrocarbons in recent sediments. Bull.Am.Ass. Petrol.
Geol., 3.8, 377-404.
Soeder C.J. (1980). The scope of microalgae for food and feed.
In: Algal Biomass: Production and use (eds. Shelef G. and
Soeder C. J.), pp. 9-20. Elsevier/North-Holland, Amsterdam.
Sommer H., Whedon W.F., Kofoid C.A., Stohler R. (1937). Relation
of paralytic shellfish poison to certain plankton organisms
of the genus Gonyaulax.A.M.A.Arch. Pathol., 24. 537-546.
Sommer U. (1983). Nutrient competition between phytoplankton
species in multispecies chemostat experiments.Arch.Hydrobiol.,
26 , 399-416.
Sommer U. (1984). The paradox of the plankton: fluctuations of
phosphorusavailability maintain diversity of phytoplankton
545
( in flow-through cultures. Limnol.Oceanogr. , 29. 633-636»
। Sommerfeld M.R., Nichols H.W. (1970). Comparative studies in
! the genus Porphyridium Naeg. J.Phycol., 6, 67-78.
Joong P. (1980). Production and development of Chlorella and Spi-
rulina in Taiwan.In: Algal Biomass: Production and Use (eds.
I Shelef G. and Soeder C.J.), pp.97-114. Elsevier/North-Holland,
Amsterdam.
Soumia A. (1976). Primary production of sands in the lagoon of
| an atoll and the role of foraminiferan symbionts. Mar.Biol.,
27, 29-32.
Soumia A. (ed.) (1978). Phytoplankton Manual. UNESCO, Paris.
I Soumia A. (1981). Morphological bases of competition and succes-
sion. Can. Bull.Fish.Aquatic Sci., 210, 339-346.
Soumia A. (1982). Is there a shade flora in the marine plankton?
J. Plankton Res., 4, 391-399.
Sousa W.P., Schroeter S.C., Gaines S.D. (1981). Latitudinal vari-
ation intertidal algal community structure: the influence of
grazing and vegetative propagation. Oecologia, 48, 297-307.
.South G.H. (1975). Contribution to the flora of marine algae of
eastern Canada. III. Tilopteridales. Naturalists Can. (Que),
102, 693-702.
South G.R •» Adams M.M. (1976). Erythrotrichia foliifonnis sp.
nov. (Rhodophyta, Erythropeltidaceae) from New Zealand, J.
I Roy.Soc. N.Z., 6, 399-405.
; South G.R., Hill R.D. (1970). Studies on marine algae of Newfound-
land. 1. Occurence and distribution of free-living Ascophyllum
nodosum in Newfoundland. Can.J.Bot., 48. 1697-1701.
Spjeldnaes N. (1963). A new fossil (Papillomembrana sp.) from the
upper pre-Cambrian of Norway. Nature, 200. 63-65.
Stackebrandt E., Seewaldt E., Fowler V.J., Schleifer KrH. (1982).
The relatedness of Prochloron sp. isolated from different
didemnid ascidian hosts. Arch.Mikrobiol., 132. 216-217.
Stackhouse J. (1801). Nereis Britannica, Bath.
Stache K.H.H.G. (1889). Die Libumische Stufe und deren Grenz-Ho-
rizonte. Eine Studie liber die Schichtenfolgen der Cretacisch-
Eucanen Oder Protocanen Landbildung speriode im Bereiche
der Kustenlander von Oesterreich-Ungarn, Wien,Geol.Abh., 22
(1), 1-170.
; Stallard M.O., Faulkner D.J. (1974). Chemical constituents of
К the digestive gland of Aplysia californica. Importance of diet
к Comp.Biochem.Physiol., 49B, 25-36.
546
Stanley R.Y. (1961). La place dee bacteries dans le monde vivant.
Annl.Inst. Pasteur Paris, 101. 297-312.
Stanier R.Y. (1974). The origins o-f photosynthesis in eukaryotes.
Symp.Soc.Gen. Microbiol., 24, 219-240.
Stanier R.Y., Cohen-Bazire G. (1977). Phototrophic prokaryotes:
cyanobacteria. Ann.Rev.Microbicl., 31. 225-270.
Stanier R.Y., Kunisawa R., Mandel M., Cohen-Bazire G. (1971).
Purification and properties of unicellular blue-green algae
(order Chroococcales). Bacteriol.Rev., 35. 171-205.
Stanier R.Y., Sistrom W.R., Hansen T.A., Whitton B.A., Casten-
holz R.W., Pfennig B.A., Gorlenko V.N., Kondratieva E.N.,
Eibihjellen K.E., Whittenbury R., Gherna R.L., Truper H.G. (1978).
Proposal to place the nomenclature of the cyanobacteria (blue-
green algae) under the rules of the international code of
nomenclature of bacteria. Int.J.Syst. Bacteriol., 28, 335-336.
Starks T.L., Shubert L.E., Trainor F.R. (1981). Ecology of soil
algae: a review. Phycologia, 20. 65-80.
Starmach K. (1972). Chrysosphaera stigmatica n. sp. (Chrysophy-
ceae). Bull.Acad. Polon.Sci.Ser.Biol., 20, 577-579.
Starr R.C.(1955). Isolation of sexual strains of placodexra des-
mids. Bull.Torrey Bot.Club, 82, 261-265.
Starr R.C. (1969). Structure reproduction and differentiation
in Voivox carter! f. nagarensis. Iyengar, strains HK9 and 10.
Arch. Protistenk., Ill, 204-222.
Starr R. C. (1975). Meiosis in Voivox carter! f. nagariensis.
Arch. Protistenk., 117, 187-191. '
Starr R.C. (1980). Colonial chiorophytes. Ins Phytoflagellates
(ed. CoxE.R»), pp.147-164. Elsevier/North-Holland, New York.
Starr R.C., Rayburn W.R. (1964). Sexual reproduction in Mesotae-
neum kramstai. Phycologia, 4, 23-46.
Starr R.C., O'Neil R.M., Miller III C.E. (1980). L-glutamic acid
as a mediator of morphogenesis in Voivox capensis. Proc. Natl.
Acad. Sci., 77, 1025-1028.
Starr M.P., Stolp H., Truper H.G,, Balows. A., Schlegel H. G. (eds.)
(1981). The Prokaryotes, Vols 1 and 2. Springer-Verlag, Berlin.
Steidinger K.A. (1975a). Implications of dinoflagellate life cyc-
les on initiation of Gymnodinium breve red tides. Environ.
Lett., £, 129-139.
Steidinger K.A. (1975b). Basic factors influencing red tides.
In: Proceedings of the 1st International Conference cn Toxic
Dinoflagellate Blooms (ed. LoCicero V.R.), pp.153-162. Massa-
547
chusetts Science and Technical Foundation, Wakefield, Maas.
Steidinger K.A. (1983). A re-evaluation of toxic dinoflagellate
biology and ecology.Prog.Phycol.Res., 2, 147-188.
Steidinger K.A., Haddad K. (1981). Biologic and hydrographic as-
pects of red tides. BioScience, 31, 814-819.
Stein J., Borden C.A. (1984). Causative and beneficial algae in
human disease conditions: a review. Phycologia, 23, 485-501.
Steneck R.S., Watling 1. (1982). Feeding capabilities and limi-
tation of herbivorous molluscs: a functional approach. Mar.
Biol., 68, 299-312.
Stephensen J.W. (1981). The effects of a seaweed extract on the
yield of a field and glasshouse crops.Proc. Int.Seaweed Symp.,
8, 740-744.
Stephenson T.A., Stephenson A. (1949). The universal features of
zonation between tide marks on rocky coasts. J.Ecol., 37, 289-
305.
Stephenson T.A., Stephenson A. (1972). Life Between Tide Marks
on Rocky Shores. W.H.Freeman, San Francisco.
Stephenson W,G.(1974). Seaweed In Agriculture and Horticulture,
3rd Edition. Rateaver Publ.Valley, California.
Stevens S.E., Porter R.D. (1980). Transformation in Agmenellum
quadruplicatum. Proc. Natl.Acad.Sci. U.S.A., 77, 6052-6056.
Stewart K.D., Mattox K.R. (1975). Comparative cytology, evolu-
tion and classification of the green algae with some consi-
deration of the origin of other organisms with chlorophylls
a and b. Bot.Rev., 41. 104-135. ,
Stewart K.D., Mattox K.R. (1980). Phylogeny of phytoflagellates.
In: Phytoflagellates (ed. CoxE.R.), pp.433-462. Elsevier/
North-Holland, New York.
Steward J.G., Schultz-Baldes M. (1976). Long term lead accumula-
tion in the abalone (Haliotus sp.) fed on lead treated brown
algae (Egregia laevigata) Mar.Biol., 36. 19-24.
Stewart W.D.P., Rodgers G.A. (1977). The cyanophyte hepatic sym-
biosis. II. Nitrogen fixation and the interchange of nitrogen
and carbon. New Phytol., 78, 459-471.
Stewart W.D.P., Rowell P. (1977). Modifications of nitrogen fix-
ing algae in lichen symbioses. Nature, 265. 371-372.
Stokes P.M. (1981). Multiple metal tolerance in copper tolerant
green algae. J.Plant Nutrit., 2» 667-678.
Stokes P.M. (1983). Responses of freshwater algae to metals. Prog
Phycol.Res., 2, 87-112.
548
Stosch H.A., von (1954)» Die Oogamie von Bidduiphia mobiliensis
und die bisher bekannten Auxosporenbildungen bei den Centra-
les. Int. Bot.Congr. 8, Rap. Coram.Sec. , 17 . 58-68.
Stosch H.A. von (1964). Wirkungen von Jod und Arsenit auf Meer-
salgaen in Kultur'. Proc. Int.Seaweed Sytnp., 4, 151-156.
Stosch H.A. von (1965a). Manipulierung der Zellgrosse von Diato-
meen im Experiment. Phycologia, 2» 21-44.
Stosch H.A. von (1965b). The sporophyte of Liagora farjnosa La-
mour. Brit. Phycol.Bull., 2, 486-496»
Stosch H.A. von (196?). Haptophyceae. ins Vegetative Fortplan-
zung, Parthenogenese und Apogamie bei Algen. (ed. Rohland W. ),
pp. 646-656. Encyclopedia of Plant Physiology, Vol. 18.
Stosch H.A. von (1973). Observations on vegetative reproduction
and sexual life cycles of two freshwater dinoflagellates, Gym-
nodinium pseudopalustre Schiller and Woloszynskia apiculata
sp. nov.Brit. Phycol. J., 8, 105-134.
Stosch H.A. von, Tiel G. (1979). A new mode of life history in
the freshwater red algal genus Batrachospenaum. Am.J.Bot.,
66, 105-107.
Stosch H.A., von Theil G., Kowallik K.V. (1973). Entwicklungsge-
schichtliche Untersuchungen an zentrisohen Diatomeen. V. Bau
und Lebenszyklus von Chaetoceros didymum mit Beobachtungen
fiber einige andere Arten der Gattung. Helgol.Wise Meeresunters
22, 384-445.
Strasburger E. (1906). Zur Frage eines Generationswechsels bei
Phaeophyceen, Bot.Zeit., 64, 1-7.
Straughan D. (1982). Observations on the effects of oil seeps
in the Coal Oil Point area. Phil.Trans.Roy.Soc.Lond. B, 297.
269-281.
Sullivan C.W. (1976). Diatom mineralization of silicic acid. I.
Si(OH)4 transport characteristics in Navicula pelliculosa. J.
Phycol., 12, 390-396.
Sullivan C.W. (1977). Diatom mineralization of silicic acid. II.
Regulation of Si(OH)^ transport rates during the cell cycle
of Havicula pelliculosa. J. Phycol., 13. 86-91.
Sun H., Fenioal W. (1979). Rhipocephenal and rhipocephalin, no-
vel linear sesquiterpenoids from the tropical green alga
Rhipocephallus phoenix. Tetrahed. Lett., 1979 (8), 84—88.
Sundene 0. (1962). Reproduction and morphology in strains of
Antithamnion boreale originating from Spitzbergen and Scandi-
navia. Nor.Vidensk.Akad.Oslo. I.Marrnat.KT. N.S., 2» 1-19.
549
Suneson S. (1950)» The cytology of bispore fonnation in two spe-
cies of Lithophyllum and the significance of the bispores
in the Corallinaceae. Bot.Not., 1950, 429-450.
Sutherland J.M., Herdman M., Stewart W.D.P. (1979). Akinetes of
the Cyanobacterium Nostoc PCC 7524: macromolecular composition,
structure and control of differentiation. J. Gen.Microbiol.,
115,( 273-287.
Sutherland J.P., Karlson R.H. (1977). Development and stability
of the fouling community at Beaufort, North Carolina. Ecol.
Mongr. , 47, 425-466.
Svedelius N. (1908). Uber den ваи und die Entwicklung der Blori-
deengattung Martensia. Vet.Akad.handl.Bd., 43, Stockholm.
Svedelius N. (1937). The apomeiotic tetrad division in Lomentaria
rosea in comparison with the normal development in Lomentaria
clavellosa. Symb. Bot.Uppsala, 2, 1-53.
Sweeney B.M. (1979). The bioluminiscence of dinoflagellates. In:
Biochemistry and Physiology of Protozoa, 2nd Edition, Vol. I.
(eds. Lewandrowsky M. and Hunter G.H.), pp.287-306. Academic
Press, London.
Sweeney B.M. (1983). Circadian time keeping in eukaryotic cells,
models and hypotheses. Prog. Phycol.Res., 2, 189-225.
Swift D.G. (198(J|, Vitamins and phytoplankton growth. Ins The
Physiological Ecology of Phytoplankton (ed. Morris I»), pp.
329-368. Blackwell Scientific Publications, Oxford.
Syrett P.J. (1981). Nitrogen metabolism of microalgae.Can.Bull.
Fish.Aquat.Sci., 210, 182-210.
Szczepanski A. (1966). Bermerkungen iiber die Primarproduction
des Pelagials. Verb.Int.Ver.Limnol., 16, 364-371.
Takahashi E. (1975). Studies on genera Mallomonas and Synura, and
other plankton in the fresh water with the electron microscope
IX. Mallomonas harrisae sp. nov. (Chrysophyceae). Phycologia,
Ц, 41-44.
TanakaY., Stara J.P. (1979). Algal polysaccharides, their poten-
tial use to prevent chronic metal poisoning. In: Marine Algae
in Pharmaceutical Science (eds. Hoppe H.A., Levring T. and
Tanaka Y.), pp. 525-543. Walter de Gruyter, Berlin.
Tandeau de Marsac N.T. (1977). Occurrence and nature of chroma-
tic adaptation in cyanobacteria. J.Bact., 130, 82-91.
Tanner C.E. (1981). Chlorophyta life histories. In: The Biology
of Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp.218-247.
Blackwell Scientific Publications, Oxford.
550
Tgppan H. (1974)» Protistan phylogeny: multiple working hypothe-
ses. Taxon, 32, 271-276.
Tappan H. (1976). Possible eukaryotic algae (Bangiophycidae) among
early Proterozoic miorofossils. Geol.Soc.Am.Bull., 87, 633-
639.
Tappan H. (1980). The Paleobiology of Plant Protists. W.H. Free-
man and Co., San Francisco.
Tappan H., Leoblich A.R., Jr. (1972). Fluctuating rates of pro-
tistan evolution, diversification and extinction. XXVI Int.
Geol.Congr., Sec. 7 Paleont, 205-213.
Tappan H., Loeblich A.R., Jr. (1973). Evolution of the oceanic
plankton. Earth-Sci. Rev., 9.» 207-240.
Tatewaki M. (1966). Formation of a crustose sporophyte with uni-
locular sporangia in Scytosiphon lomentaria. Phycologia, 6,
62-66.
Tatewaki M., lima M. (1984). Life histories of Blidingia minima
(Chlorophyceae) especially sexual reproduction. J. Phycol.,
20, 368-376.
Tatewaki M., Nagata N. (1970). Surviving protoplasts in vitro
and their development in Bryopsis. J. Phycol., 6, 401-403.
Tasch P. (1973). Paleobiology of the Invertebrates. John Wiley
and Sons, New York.
Taylor D.L. (1973). The cellular interaction of algal-inverte-
brate symbiosis.Adv.Mat.Biol., 11, 1-56.
Taylor D.L. (1974). Symbiotic marine algae: taxonomy and biolo-
gical fitness.
In: Symbiosis in the Sea (ed.Verriberg W.B.), pp.245-262. Univer-
sity of South Caroline Press, Columbia.
Taylor D.L., Seliger H.H. (eds.) (1979). Toxic Dinoflagellate
Blooms. Elsevier/North-Holland, New York.
Taylor F.J.R. (1974). Implications and extensions of the serial
endosymbiosis theory of the origin eukariotes. Taxon, 23,
229-258.
Taylor F.J.R. (1976). Autogenesis theories for the origin of
eukaryotes. Taxon, 25. 377-390.
Taylor F.J.R. (1978). Problems in the development of an explicit
hypothetical phylogeny of the lower eukaryotes. Biosystems,
10, 67-89.
Taylor F.J.R. (1980). On dinoflagellate evolution. Biosystems,
1_3, 65-108.
Taylor W.R. (1969). Phycology. In: A Short History of Botany
551
in the United States (ed. Ewan J.), pp.74-81. Hafner Publi-
shing Co., New York and London.
Terry L.A., Edyvean R.G.T. (1981). Microalgae and corrosion.Bot.
Mar., 24, 177-183.
Thacker A., Syrett P.J. (1972). The assimilation of nitrate and
ammonium by Chlamydomonas reinhardii. New Phytol., 71. 423-
433.
Thinh L.V. (1979). Prochloron (prochlorophyta) associated with
the ascidian Tridemnum cyclops. Phycologia, 18, 77-82.
Thomas R.N. , Cox E.R. (1973). Observation on the symbiosis of
Peridinium halticum and its intracellular alga. I. Ultra-
structure. J. Phycol., 2» 304-323.
Thomas W.H., Holm-Hansen 0., Seibert B.L.R., Azam P., Hodson R.,
Takahashi M. (1977a). Effects of copper on the dominance and
the diversity of algae: controlled ecosystem pollution expe-
riment. Bull. Mar.Sci., 27, 19-33.
Thomas W.H., Seibert D.L.R., Takahashi M. (1977b). Controlled
ecosystem pollution experiment: effect of mercury of enclosed
water columns III. Phytoplankton dynamics and production. Mar.
Sci, Commun., 2» 331-354.
Thomas W.H. , Hollibaugh J.T. , Seibert D.L.R. , Wallace G.T., Jr.
(1980). Toxicity of mixture of ten metals to- phytoplankton.
Mar. Ecol.Prog.Ser., 2, 213-220.
Thompson A. (1973). The flagella of Cyanophora paradoxa Korsch.
J. South Afr.Bot., 29, 35-39.
Thorhaug A., Marcus J.H. (1981). The effects of temperature
and light on attached forms of tropical and semi-tropical
macroalgae potentially associated with OTEC (ocean thermal
energy conversion) machine operation. Bot.Mar., 24, 393-398.
Thorhaug A., Segar D., Roessler M.A. (1973). Impact of a power
plant on a subtropical estuarine environment. Mar. Poll.Bull.
4, 166-169.
Thornber J.P., Barber J. (1979). Photosynthetic pigments and
models for their organization in vivo. In: Photosynthesis in
Relation to Model Systems (ed. Barber J.), pp.27-70. Elsevier,
New York.
Tilman D. (1977). Resource competition between planktonic algae:
an experimental and theoretical approach. Ecology, 58, 338-348
Tison D.L., Wilde E.W., Pope D.H., Poiermans C.B. (1981). Produc-
tivity and species composition of algal matcommunities expo-
sed to a fluctuating thermal regime. Microbiol.Ecol., 7,
552
151-165.
Tokida J. (I960). Marine algae epiphytic on Laminariales plants.
Bull.Fac.Fish.Hokkaido. Univ., 11, 73-105.
Topinka J.A., Tucker L.R. (1981). Long-term oil contamination of
fucoid macroalgae following the Amoco Cadiz oil spill. Ins
Amoco Cadiz: Consequences d'une Pollution Accidentelle par
les Hydrocarbures, pp.381-391. Publ.Centre national pour
lexploration des oceans, Faris.
Toth R. (1976). The release, settlement and germination of zoospo-
res in Chorda tomentosa (Phaeophyceae, Laminariales). J. Phy-
col., 12, 222-233.
Toth R., Markey D.R. (1973). Synaptonemal complexes in brown
algae. Nature, 243, 236-237.
Trainor F.R. (1978). Introductory Phycology, Wiley, New York.
Trainor F.R. (1983). Survival of algae in soil after high tem-
perature treatment. Phycologia, 22, 201-202.
Trench R.K. (1975). Of "leaves that crawl": functional chloro-
plasts in animal cells.Symp.Soc.Exp.Biol., 29. 229-265.
Trench R.K. (1979). The cell biology of plant - animal symbiosis.
Ann.Rev.Plant Physiol., 30. 485-531.
Trench R.K. (1980). Uptake, retention and function of chloro-
plasts in animal cells. In: Endocytobiology: Endosymbiosis
and cell Biology, a Synthesis of Recent Research (eds.
Schwemmler W. and Schenk H.E.A. ), pp. 703-727. Walter de
Gruyter, Berlin.
Trench R.K. (1982). Physiology, ultrastructure and biochemistry
of cyanellae. Prog. Phycol.Res., 1, 257-287.
Trench R.K. .Siebens H.B. (1978). Aspects of the relation between
Cyanonhora paradoxa (Korschikoff) and its endosymbiotic
cyanelles Cyanocyta korschikoffiana (Hall and Claus). IV.
The effects of rifampicin, chloramphenicol and cycloheximide
on the synthesis of ribosomal ribonucleic acids abd chloro-
phyll. Proc. Roy.Soc. Lond. B, 202, 473-483.
Trench R.K., Boyle J.E., Smith D.C. (1973). The association
between chloroplasts of Codium fragile and the mollusc
Elysia viridis. II. Chloroplast ultrastructure and photosyn-
thetic carbon fixation in E.viridis. Proc.Roy.Soc.Lond. B,
184, 63-81.
Triemer R.E., Brown R.M., Jr. (1975). Ultrastructure of ferti-
lization in Chlamydomonas reinhardtii. Brit. Phycol. J.,
12, 23-44.
553
Tschermak-Woess E. (1959). Extreme Anisogamie und ein bemerkene-
werter Pall der Geschlechtsbestimmung bei einer neuen
Chlamydomonas-Art. Planta, 52» 606-622.
Tschermak-Woess E. (1962). Zur Kenntnis von Chlamydomonas suboo-
gama. Planta, 59. 68-76.
Tschermak-Woess E. (1978). Myrmecia reticulata as a phycobiont
and free living Trebouxia - the problem of Stenocybe gepatata.
Lichenologigt, 10, 69-79.
Tseng C.K. (1981). Commercial cultivation. In: The Biology of
the Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp. 680-741.
Blackwell Scientific Publications, Oxford.
Tsubo Y. (1961). Chemotaxis and sexual behaviour in Chlamydomonas.
J. Protozool., 8, 114-121.
Turner C.H., Evans L.V. (1977). Physiological studies on the re-
lationships between Asoophyllum nodosum and Polysiphonia
lanosa. New Phytol., 79, 363-371.
Turner C.H., Evans L.V. (1978). Translocation of photoassimilated
1^C in the red alga Polysiphonia lanosa.Brit. Phycol. J., 13.
51-55.
Turner D. (1802). A Synopsis of the British Fuci.London.
Tyller M.A., Seliger H.H. (1978). Annual subsurface transport of
a red tide dinoflagellate in its bloom area: water circula-
tion patterns and organism distribution in the Chesapeake
Bay.Limnol. Oceanogr., 23, 227-246.
Tyler M.A., Seliger H.H. (1981). Selection for a red tide orga-
. nism: physiological responses to the physical environment.
1 Limnol. Oceanogr., 26, 310-324.
, Tynan E.J. (1971). Geologic occurrence of the'archaeomonads.
In: Proceedings II. Planktonic Conference, Roma, 1970, Vol. 2
(ed. Parinacci A.) pp.1225-1230. Edizioni Tecnoscienza, Rome.
Underwood A.J. (1978). A refutation of critical tide leves as
determinants of the structure of intertidal communities on
British Shores. J.Exp.Mar.Biol.Ecol., 22» 261-276. z
Underwood A.J., Denley E.J. (1984). Paradigms, explanation, and
generalizations in models for the structure of intertidal
communities on rocky shores. In: Ecological Communities:
Conceptual Issues and the Evidence (eds. D.R^Jr., Simberloff
D., Abele L. G. and Thistle A.B.). pp. 151-180. Princeton
University Press, Princeton.
: Vadas R.L. (1977). Preferential feedings: an optimization strate-
gy in sea urchins. Ecol. Monogr., 47, 337-371.
554
Vadas R.L., Keser M., Rusanowski P.O. (1976). Influence of ther-
mal loading on the ecology of intertidal algae. In: Thermal
Ecology II. (eds. Esch G.W. and McFarlane R. W.), pp. 331-335*
E.R.D.A. Symposium Series, Conf. 750425 Springfield, Mass,
U.S.A.
Valet G. (1969). Contribution a l'etude des Dasycladales, 2.
Cytologie et reproduction. 3. Revision systematique. Nova
Hedwiga, 1/, 551-644, pls 133-157.
Valiela I. (1984). Marine Ecologioal Proceases. Springer-Verlag,
New York.
Van den Ende H. (1976). Sexual Interactions in Plants. Academic
Press, London.
Van der Meer J.P. (1983). 'The domestication of seaweeds. Bio-
Science, 22» 172-176.
Van der Meer J.P., Todd E.R. (1977). Genetics of Gracilaria sp.
(Rhodophyceae, Gigartinales). IV. Mitotic recombination and
its relationship to mixed phases in the life history. Can.
J.Bot., 22» 2810-2817.
Van der Meer J.P., Todd E.R. (1980). The life history of Pal-
maria palmata in culture. A new type for the Rhodophyceae.
Can.J.Bot., 58, 1250-1256.
Vandermeulen J.H. , Ahern T.P. (1976). Effect of petroleum hydro-
carbons on algal physiology: review and progress report. In:
Effects of Pollutants on Aquatic Organisms (ed. Lockwood A.P.
M. )| pp. 107-126. Cambridge University Press, Cambridge.
Van der Veer J. (1976). Pavlova calceolata (Haptophyceae), a
new species from the Tamar Estuary, Cornwall, England, J.
Mar.Biol. Ass. U.K., 56., 21-30.
Van der Velde H.H., Hemrika-Wagner A.M. (1978). The detection
of phytochrome in thered alga Achrocheaetium daviesii. Plant.
Sci. Lett., 11, 145-149.
Vargo S.A., Hutchins M., Almquist G« (1982). The effect of low,
chronic levels of No.2 fuel oil on natural phytoplankton as-
semblages in microcosms: 1. Species composition and seasonal
succession. Mar. Environ.Res., 6, 245-246.
Vaucher J.P. (1803). Historic des Conferves d'eau douce. Geneve.
Vesk M., Jeffrey S.W. (1977). Effect of blue-green light on
photosynthetic pigments and chloroplast structure in unicel-
lular marine algae in six classes. J. Phycol., 13. 280-288.
Vidal G. (1984). The oldest eukaryotic cells.Sci.Am., 250. 48-57»
Vinayakumar M., Kessler E. (1975). Physiological and Mnr.Wn»!
555
contributions to the taxonomy of the genus Chlorella. Arch.
Mikrobiol., 103. 13-19.
Vince-Prue D. (1975). Photomorphogenesis and flowering. In:
Encyclopedia of Plant Physiology.Vol. 16B, Photomorphogenesis
(eds. Shropshire W. and Mohr H. ), pp.457-490. Springer, Heidel
berg.
Vincent W.P., Goldman C.R. (1980). Evidence for algal heteroptro-
phy in Lake Tahoe, California - Nevada. Limnol. Oceanogr,, 25,
89-99.
Von Zaluzian Z. (1592). Methodi herbariae libri tres. Prague
(not seen by the authors) quoted from Prescott, 1951).
Voss V.G. (1983). International Code of Botanical Nomenclature.
Bohn.Scheltema and Holtema, Utrecht and Antwerpen.W Junk,
The Hague and Boston.
Waaland J.R. (1981). Commercial cultivation. In: The Biology of
the Seaweeds (eds. Lobban C.S. and Wynne M.J.), pp. 726-741.
Blackwell Scientific Publications, Oxford.
Waaland J.R., Waaland S.D., Bates G. (1974). Chloroplast struc-
ture and pigment composition in the red alga Griffithsia
pacifica: regulation by light intensity. J. Phycol., 10, 193-
198.
Waaland S.D. (1975). Evidence for a species-specific cell fusion
hormone in red algae. Protoplasma, 86 253-263»
Waaland S.D., Cleland R. (1972). Development in the red alga
Griffithsia pacifica: control by internal and external fac-
tors. Planta, 105, 196-204.
Waaland S.D., Waaland J.R. (1975). Analysis of cell elongation
in red algae by fluorescent labelling. Planta, 126. 127-138.
Walker D.C., Henry E.C. (1978). Unusual reproductive structures
in Syringoderma abyssicola (S. and G.) Levring. New Phytol.,
80, 193-197.
Walker J.C.G., Klein C., Schidlowski M., Schopf J.W., Stevenson
D.J., Walter M. R. (1982). Environmental evolution of the
Archean - early Proterozoic earth. In: Origin and Evolution
of Earth’s Earliest Biosphere: An Interdisciplinary Study
(ed.Schopf J.W.). Princeton University Press, Princeton,
New Jersey.
Walker L.M.( 1984). Life histories, dispersal and survival in
marine, planktonic dinoflagellates. In: Marine Plankton Life
Cycle Strategies (eds. Steidinger K.A. and Walker L.M.), pp.
19-34.
556
CHC press, Воса Raton,
Wall D. (1962). Evidence from recent plankton regarding the bio-
logical affinities of Tasmanites Newton 1875 and beiosphaeri-
dta Eisenack 1958. Geol.Mag., 22.» 353-362.
Walne R.L. (1980). Euglenoid flagellates. In: Phytoflagellates
(ed. Cox E.R.), pp.165-212. Elsevier/North-Holland, New York.
Waine P.L., Arnott H.J. (1967). The comparative ultrastructure
and possible function of eyespots: Euglena granulata and
Chlamydomonas eugametos.Planta. 77. 325-353.
Walsby A.E. (1978). The gas vesicles of aquatic prokaryotes. In:
Relations Between Structure and Function in the Prokaryotic
Cell (eds.Stanier R.Y. et al.), pp.327-357. Cambridge Univer-
sity Press, Cambridge.
Walsby A.E., Xypolyta A. (1977). The form resistance of chitin
fibres attached to the celles of Thalassiosira fluviatilis
Hustedt. Brit.Phycol. J., 12., 215-223.
Walter M.R., Oehler' J.H., Oehler D.Z. (1976). Megascopic algae
1300 million years old from the Belt Supergroup. Montana:
a reinterpretation of Walcott's helminthoidichnites. J. Pale-
ontol., 50, 872-881.
Wanders J.B.W. (1977). The role of benthic algae in the shallow
reef of Curacao (Netherlands Antilles). III. The significance
of grazing. Aquatic Bot., 357-390.
Watanabe A. (1975)» Practical significance of algae in Japan.
Nitrogen fixation by algae. In: Advance of Phycology in Japan
(eds. Tokida J. and Hirose H.W.), pp.255-272. Junk, The Hague.
Watanabe T., Kondo N. (1976). Ethylene evolution in marine algae
and a proteinaceous inhibitor of ethylene biosynthesis from a
red alga. Plant Cell Physiol., 1J. 1159-1166.
Watanabe T., Hirayama T., Takahasi T., Kokubu T., Ikedam M. (1980)
Toxicological evaluation of arsenic in edible seaweed Hizikia
species. Toxicology, 14. 1-22.
Watrass C.J., Chisholm S.W., Anderson D.M. (1982). Regulation
of growth in an estuarine clone of Gonyaulax tamarensis
Lebour: salinity dependent temperature responses. J.Exp.Mar.
Biol.Ecol., 62, 25-37.
Watson B.A., Waaland S.D.(1983). Partial purification of a gly-
copritein cell fusion hormone from Griffithsia pacifica,
a red alga. Plant Physiol., 71. 327-332.
Watson S., Kalff J. (1981). Relationships between nannoplankton
and lake trophic status. Can. J. Fish. Aquat.Sci. , 38 , 960-967.
Weidner M., Kuppers U. (1902). Metabolic conversion ofUC=aspar-
557
tate, ^C-malate and ^C-mannitol by tissue disks of Laminaria
hyperborea: role of phosphoenolpyruvate carboxykinase. Z.
Pflanzenphysiol., 108, 353-364»
Weinheimer A.J., Karns T.K.B. (1974). A search for anti-cancer
and cardiovascular agents in marine organisms. In: Food-Drugs
fromthe Sea (eds. Webber H.H. and Ruggieri R. ), pp.491-496.
Marine Science Center, Puerto Rico.
Werner D. (ed.) (1977). The Biology of Diatoms, Blackwell Scien-
tific Publications, Oxford.
West J.A. (1968). Morphology and reproduction of the red alga
Acrochaetium pectination.J. Phycol., 4, 89-99.
West J.A. (1972). The life history of Petrocells franciscana.
Brit. Phycol. J., 7, 299-308.
WestJ.A. (1972). Environmental regulation of reproduction in
Rhodochorton purpureum. In: Contributions to the Systematics
of the Benthic Marine Algae of the North Pacific (eds. Abbott
I.A. and Kurogi M.), pp.213-230. Japanese Society of Phycolo-
gy, Kobe, Japan.
West J.A., Hommersand M.H.(1981). Rhodophyta: life hosieries. In:
The Biology of Seaweeds (eds. Lobban C.S. and Wynne M.J.),
pp. 133-193. Blackwell Scientific Publications, Oxford.
West L.K., Walne P.L. (1980). Trachelomonas hispida var. coronata
(Euglenophyceae): II. Envelope substructure. J.Phycol., 16.
498-506.
West L.K., Walne P.L. , Rosowski R.J. (1980a). Trachelomonas hispi-
da var. coronata (Euglenophyceae): I. Ultrastructure of cytos-
keletal and flagellar systems. J. Phycol., 1_6, 489-497.
West L.K., Walne P.L., Bentley J. (1980b). Trachelomonas hispida
var. coronata (Euglenophyceae): III. Envelope elemental compo-
sition and mineralization. J.'Phycol., 16, 582-591.
Wetherbee R. (1979). "Transfer connection": specialized pathways
for nutrient translocation in a red alga? Science, 204. 858-
859.
Whatley J.M. (1977). The fine structure of Prochloron. New Phy-
tol., 79, 369-373.
Whatley J.M. (1982). Ultrastructure of plastid inheritance:
green algae to andiosperms. Biol.Rev., 57. 527-569.
Whatley J.M., Whatley P.R. (1981). Chloroplast evolution. New
Phytol., §2, 233-247.
Wheeler P.A., North B.B., Stephens C.G. (1974). Aminoacid uptake
by marine phytcplanktere. Lomnol.Oceanogr., 19, 249-295.
558
White A.W. (1981). Marine zooplankton can accumulate and retain
dinoflagellate toxins and cause fish kills. Limnol.Oceanogr.,
26, 103-107.
Whiteside M.C. (1983). The mythical concept of eutophication.
Hydrobiologia, 103, 107-111.
Whittaker R.H. (1959). On the broad classification of organisms.
Quart. Rev. Biol., 34, 210-226.
Whittaker R.H. (1969). New concepts of kingdoms of organisms.
Science, 163. 150-160.
Whittaker R.H., Margulis L. (1978). Protist classification and
the kingdoms of organisms. Biosystems, 10, 3-18.
Whittick A. (1977). The reproductive ecology of Plumaria elegans
(Bonnem) Schmitz (Ceramiaceae, Rhodophyta) at its northern
limits in the western Atlantic J. Exp.Mar.Biol.Ecol., 29.
223-230.
Whittick A. (1978). The life history and phenology of Callitham-
nion corymb os um (Rhodophyta: Ceramiaceae) in Newfoundland.
Can.J. Bot., 56, 2497-2499.
Whittick A. (1983). Spatial and temporal distributions of domi-
nant epiphytes on the stipes of Laminaria hyperborea,(Gunn.)
Fosl. (Phaeophyta: Laminariales) in S.E,Scotland. J.Exp.
Mar.Biol. Ecol., 7J3, 1-10.
Whittick A., Hooper R.G. (1979). Seasonal changes in the phyto-
benthos of Conception Bay, Newfoundland, in the vicinity of
a thermal outfall. Brit. Phycol. J., 14, 128-129.
Whitton B.A. (1971). Filamentous algae as weeds Ln: 3rd Euro-
pean Weed Res.Council Symposium, pp. 249-263. Oxford.
Whitton B.A. (1973). Interactions with other organisms. In: The
Biology of the Blue Green Algae (eds.Carr N.D. and Whitton
B.A.), pp. 415-433» University of California Press, Berkeley
and Los Angeles.
Whitton B.A., Shehata P.H.A. (1982). Influence of cobalt, nicket,
copper and cadmium on the bluegreen alga Anacytis nidulans.
Environ. Pollute (Ser. A), 27, 275-281.
Wiese L. (1974). Nature of sex specific glycoprotein agglutinins
in Chlamydomonas. Ann. N.Y. Acad.Sci., 234 , 383-395.
Wik-Sjostedt A. (1970). Cytogenetic investigations in Cladophora.
Hereditas, 66, 233-262.
Wilbur K.M., Watabe N. (1963). Experimental studies of calcifi-
cation in molluscs and the alga Coccolithus huxleyi. Ann.
N.Y.Acad. Sci., 109, 82-112.
559
Wilce R.T. (1967)» Heterotrophy in Arctic sublittoral seaweeds:
an hypothesis. Bot.Mar., 10, 185-197.
Wilce R.T., Schneider C.W.Quinlan A.V., van den Bosch K. (1982).
The life history and morphology of a free living Pilayella lit-
toral! s (1.) Kjellrn. (Ectooarpaceae, Ectocarpales) in Nahant
Bay, Massachusetts, Phycologia, 21. 336-354»
, Wildgoose P.B., Blunden G., Jewers K. (1978). Seasonal variations
in gibberellin activity of some species of Fucacaeae and
Laminariaceae. Bot. Mar., 21, 63-65.
Wildman R.B., Loescher J.H., Winger C.L. (1975). Development and
germination of akinetes of Aphanizomenon flos-aquae. J. Phy-
col., 11, 96-104.
Wilkinson M., Burrows E.M. (1970). Eugomontia sacculata Kornm.
in Britain and North America.Brit.Phycol., J., 5_, 235-238.
Wille J.N.F. (1897<-1911). Conjugatae. Chlorophyceae. Characeae.
In: Die naturalichen Pflanzenfamilien, Teil 1, Abt. 2 (1897)
(eds. Engler A. and Prantl K.A.E. ), 175 pp; 128 figs; ibid.
(1909), 134 pp., 70 figs; ibid. (1911), 136 pp., 70 figs,
Leipzig.
Williams J.L. (1897). The antherozoids of Dictyota and Taonia.
Ann.Bot., 11, 545-553.
Williams J.L. (1898). Reproduction in Dictyota dichotoma. Ann.
Bot., 12, 559-560.
Woelkerling W.J. (1983). The Audouinella (Acroohaetium - Rhodo-
chorton) complex (Rhodophyta): present perspectives. Phycolo-
gia, 22, 59-92.
Woese G.R. (1981). Archaebacteria.Sci.Am., 299. 98-122.
Wolk C.P. (1982). Heterocysts. In: The Biology of Cyanobacteria
(eds. CarftN.G. and Whitton B.A.), pp. 359-386. Blackwell
Scientific Publications, Oxford.
Wollaston E. (1984). Species of Ceramiaceae (Rhodophyta) recor-
ded from the International Indian Ocean Expedition, 1962.
Phycologia, 23, 281-299*
Womersley H.B.S., Norris R.E. (1959). A free floating marine
red alga. Nature, 811, 828.
Wood B.J.B. (1974). Fatty acids and saponifiable lipids. In:
Algal Physiology and Biochemistry (ed. Stewart W.D.P.), pp.
236-265. Blackwell Scientific Publications, Oxford.
Wood R.D. (1967). Charophytes of North America, Stella’s Prin -
ting, West Kingston, Rhode Island.
Wood R.D., Imahori K. (1965). A Revision of the Characeae.Vol.
560
1, Monograph of the Characeae»Cramer, Weinheim.
Wray J.L. (1977). Late Paleozoic calcareous red algae. In: Fossil
Algae,Recent Results and Development (ed.Flugel E.), Springer-
Verlag, Berlin.
Wynne M.J. (1969). Life history and systematic studies of some
Pacific North American Phaeophyceae (brown algae). Univ. Calif.
Publ.Bot., 50, 1-88.
Wynne M.J. (1972). Culture Studies of Pacific coast Phaeophyceae.
Soo.Bot.Fr.Memoires, 1972. 129-144.
Wynne M.J. (1981). Phaeophyta: morphology and classification.
In: The Biology of Seaweeds (eds. Lobban C.S. and Wynne M.J.),
pp. 52-85» Blackwell Scientific Publications, Oxford.
Wynne M.J. (1982a). Phaeophyceae. In: Synopsis and Classification
of Living Organisms, Vol. 1 (ed. Parker S.P.), pp.115-125.
McGraw-Hill, New York.
Wynne M.J. (1982b). Phaeophyceae: introduction and bibliography.
In: Selected Papers in Phycology II (eds. Rosowski J.R. and
Parker B.C.), pp.731-739. Phycological Society of America,
Allen Press, Lawrence, Kansas.
Wynne M.J., Loiseaux S. (1976). Recent advances in life history
studies of the Phaeophyta, Phycologia, 15. 435-453.
Yamada T., Ikawa T., Nisizawa K. (1979). Carcadian rhythm of the
enzymes participating in the COg photoassimilation of the
brown alga Spatoglossum pacificum.Bot. Mar., 22. 203-209.
Yamamoto I., Takahashi M., Tamura E., Maruyama H. (1982). Anti-
tumor activity of crude extracts from edible marine algae
against L1210 leukemia. Bot.Mar., 25. 455-457.
Yamamoto T., Yamaoka T., Tuno S., Tokura R., Nishimura T., Hi-
rose H. (1979). Microconstituents in seaweeds. Proc. Int.
Seaweed Symp., 2» 445-450.
Yamanouchi S. (1906). The life history of Polysiphonia violacea.
Bot.Gaz., 42. 401-449.
Yan N.D. (1979). Phytoplankton community of an acidified heavy
metal-contaminated lake near Sudbury, Ontario: 1973-1977.
Water Soil Air Pollut., 11, 43-55.
Yarish C., Breeman A.M., Hoek C. van den (1984). Temperature,
light and photoperiod responses of some Northeast American abd
West European endemic rhodophytes in relation to their geo-
graphic distribution, Helgol.Wiss.Merresunters., 38, 273-304.
Yasumoto T., Inoue A., Ochi T., Fujimoto K., Oshima Y., Fukuyo
Y., Adachi R., Bagnis R. (1980). Environmental studies on a
561
toxic dinoflagellate responsible for ciguatera*Bull»Jap.
Soc.Sci. Pish., 4j5, 14O5-U11.
Yentsch C.S. (1980). Light attenuation and phytoplankton phos-
tosynthesis. In: The Physiological Ecology of Phytoplankton
(ed. Morris I.), pp.95-128. University of California Press,
Berkeley and Los Angeles.
Yentsch C.M., Yentsch C.S., Strube L.R. (1977). Variations
in ammonium enhancement, an indication of nitrogen deficien-
cy in New England coastal phytoplankton populations. J.Mar.
Res., 22» 539-555.
Yentsch C.M., Taylor D.L. (1982). Red Tides.Harvard University
Press, Boston.
Youngman R.E., Johnson D., Parley M.R. (1976). Factors influen-
cing phytoplankton growth and succession in Farmoor Reservoir
Fresh water Biol., 6, 253-263.
Zeitzschel B. (1978). Why study phytoplankton? In: Phytoplank-
ton Manual (ed. Sournia A.), pp. 1-6. UNESCO, Paris.
Zeppa R.G., Schlotzhauer P.F. (1983). Influence of algae on
photolysis rates of chemicals in water.Environ.Sci.Technol.,
17, 426-468.
Ziesenisz E., Reisser W., Wiessner W. (1981). Evidence of de
novo synthesis of maltose excreted by the endosymbiotie
Chlorella from Paramecium bursaria. Planta, 153, 481-485.
Zimmermann U. (1977). Cell turgor pressure regulation and
turgor pressure mediated transport processes. In: Integra-
tion of Activity in the Higher Plant (ed. Jennings D.H.),
pp. 117-154. Cambridge University Press, Cambridge.
Zimmermann U., Steudle E., Lelkes P.I. (1976). Turgor pressure
regulation in Valonia utricularis, effect of oell wall elas-
ticity and auxin. Plant Physiol., £8, 608-613.
Zingmark R.G. (1970). Sexual reproduction in the dinoflagellate
Noctiluca miliaris Suriray. J.Phycol., 6, 122-126.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Автогамия 248
Автоколонии, образование 200, 202
Автоспоры 206
Агар, свойства и использование 448-449
Аденозинтрифосфат, образование 275
Азот, фиксация 280-281
— у лишайников 372-373
- запасание 286-287
- метаболизм 272-278
- неорганический связанный 284
- органические источники 286
- соотношение с фосфором 287-288
Акинеты 203, 2I0-2II
Акронема, у жгутиков 109, 112, ИЗ
Аксонема, у жгутиков 108, 112, 121
Альгинаты 103
Альгиновая кислота, промышленное получение'447
— соли - см. Альгинаты
Альгология, задачи 14-15
- история и развитие 9-14
Аммоний, включение 286
Андроопоры 221
Анизогамия 251
Апикальное доминирование 300
Апланоспоры 194 , 206
Архабактерии 387, 390
Атлантический океан, распространение водорослей 356, 357
Ауксины 303-304
Багрянки - см. Красные водоросли
Базальное тело жгутиков 124, 127
Бактерии, архе- 387, 390
- циано— см. Синезеленые водоросли
Бентосные водоросли
— влияние загрязнения 434, 440, 442
— функциональные группы 345
563
Бесполое/вегетативное размножение 194, 199-215
Биогены, значение для фитопланктона 317-321
- поглощение 294-296
- роль в сообществах мороких макрофитов 335-341
Биогеография морских водорослей 356-360
Биохимия 10, 256-296
- различия между классами 26-76
Биспорангии 204
Биспоры 205
Болезни человека, вызываемые водорослями 426-427
Бриофиты, ассоциации водорослей с ними 374
Бурые водоросли, гетеробластия 242
— зооспоры 208
— клеточная стенка 102
— митоз 143
— нитчатые, ветвящиеся 179-180
— общая характеристика 22, 52-56
— паренхиматозные 175
— псевдопаренхиматозные 175
— токсичные 431
— фотопериодизм 268-270
— цитокинез 147
— эволюция и филогения 414-416
Вегетативное/басполое размножение 194, 199-201
Ветвящиеся нити 174-184
Виды 25
- краеугольные 347, 348
- состав, сукцессия 335
---- влияние подкисления 439
---- влияние эвтрофизации 436
---- сезонные явления 324-331
- сосуществование 331-333
Витамины, потребность в них 293-294
Влагалище, слизистое 97
Вода, влияние потери у водороолей 354
- помехи ее движению из-за водороолей 442-443
564
"Войлок" (покров жгутика) 118
Волны, влияние подверженности им 354
Волоски, у жту тиков II2-II9
Выедание, морских водорослей-макрофитов 344-349, 355-356
- фитопланктона 321-323
Выживаемости кривые - см. Кривые выживаемости
Выживания стратегии у морских водорослей-макрофитов - см.
Стратегии выживания у морских водороолей-макрофитов
Высыхание, влияние на водоросли 355
Галогены, потребности в них 293
Гаметические жизненные циклы 196, 247 - 254
Гаметофита - спорофита чередование - см. Чередование поколений
Гаметические жизненные циклы 196, 247-254
Гаметы, мужские, акронема жцутика у них 109, 112, 113
- образование, лунный ритм 306, 308
Гексозомонофосфатный щунт 274
Генетическая манипуляция у морских водорослей-макрофитов 454
Гены, перенос их у синезеленых водорослей 216-217
Географическое распространение водорослей 356-360
Геотропизм 305-306
Гетеробластия 242
Гетеротрихальные формы 179-180
Гетеротрофность у морских водорослей-макрофитов 340-341
Гиббереллины 303, 304
Гирогониты, зеленых водорослей 425
"Глаукофиты", общая характеристика 76
- эволюция и филогения 410-412
Глазки (стигмы) 134-137
- классификация 135
Гликолат, метаболизм 275-276
Гликолиз (путь ЭМП) 274-275
Глутаминоксиглутаратаминотрансфераза (ГОГАТ) 281
Голые клетки 79-81
— классы, в которых они встречаются 80
Гольджи тельца, участие в синтезе клеточной стенки 100
Гормоны и развитие 303-305
565
Горячие источники, водоросли в них 383-384
Грибы, ассоциации с ними водорослей 370-374
- эволюция 407-408
Дезоксирибонуклеиновая кислота (ДНК), общие свойства 137-138
— хлоропластов, рекомбинация и отбор 219
Деление цитоплазмы (=цитокинез) 14I, 145-147
- ядра 134-139
Демографические исследования морских водорослей 349
Дендроидные формы - см. Древовидные формы
Десмидиевые, коккоидные одноклеточные 157
- половое размножение 222-223
Диатомеи (см. тж. Фитопланктон), жизненный цикл 247-251
- коккоидные одноклеточные 157-158 -
- колониальные 168-169
- метаболизм кремния 291
- митоз 141, 143
- обпря характеристика 21, 47-49
- периотые 247-251
- размножение вегетативное 199-200, 201
- - половое 247-251
- споры, покоящиеся 215
- структура панциря 91-96
- типы клеток 94
- центрические 247-250
- эволюция и филогения 416-418
Динофлагелляты (см. тж. Фитопланктон), глазки 135-136
- жгутиковые I6I-I63
- жизненный цикл 247, 248
- коккоидные 155
- митоз 141, 143
- нитчатые, ветвящиеся 177
- общая характеристика 21, 24-25
- особенности клеточной стенки 83-84, 85, 86, I6I-I62
- покоящиеся цисты 214, 215
- размножение вегетативное 200
— половое 217-219
566
- структура хлоропласта 1Э0, 131
- токсичные 428-430
- эволюция и филогения 420-421
Динофиты - см. Динофлагеллаты
ДНК - см. Дезоксирибонуклеиновая кислота
Долгота дня, взаимосвязь с распространением морских водорос
лей-макрофитов 357, 358, 359-360
- и фотопериодические реакции 268-271
Древовидные формы 168
Друпа уравнение 295-296
Дыхание темновое 274 -275
- фото- 275-276
Жгутики, акронема 109, 112, 113
- аксонема 108, 112, 121
- анизоконтные 108
- аппарат, абсолютная ориентация 124
- базальное тело 124-125
- вздутия H9-I20
- волоски П2-П9
— простые/не трубчатые П4, 116
— типы 114
— трехчастные II5-II6
— трубчатые 114, 116
- гетероконтные 108
- движение 108, 114
- изоконтные 108
- кончик 108-I12
- корни 125-127
- мастигонемы - см. волоски
- мембрана 107, 112
- параксиальные тяжи 120
- переходная зона НО, 120-123
- перистые 112
- различия в зависимости от отдела/клаооа 19-24
- стефаноконтные 108
- схема структуры НО
567
- типы и расположение 109
- чешуйки 112, 114, 115
- шипики 120, 122
- эволюция 408-413
Жгутиковые 158-166
Железо, метаболизм 282, 292
Желтозеленые водоросли, классификация 44-46
— клеточная стенка 103
— коккоидные 154-155
--- митоз 143
— нитчатые 172-173
--- общая характеристика 20, 44-46
— половое размножение 217, 218
— сифональные 189, 190
— споры, бесполые 206, 207
Жизненные циклы 194, 196, 197, 217-258
Журналы, поовященные водорослям 14
Загрязнение 431-442
Запасные продукты - см. Виогены
Зеленые водоросли, глазок 135, 136
— жизненный цикл гаметический 253-255
------- соматический 256
— — спорический 242-247
--- классификация 18, 67, 72-73, 75
— клеточная стенка 105-106
--- колониальные I69-I7I
--- митоз 142, 144-145
— нитчатые ветвящиеся 174, 184
----- простые 172-173
— общая характеристика 23-24, 66-76
— одноклеточные жгутиковые 164-166
----- коккоидные 153, 156-157
— покоящиеся цисты 215
— сифональные 189, I9O-I93
— сифонокладальные 188-190
--- споры бесполые 206, 208-210
568
--- покоящиеся 213-215
— сигма - см. глазок
— структура хлоропласта 129, 133-134
— ценобии I69-I7I
— цитокинез 146
— эволюция и филогения 408-412, 421-425
Зиготические жизненные циклы 217-226
Золотистые водоросли амебоидные 151
— коккоидные 154
--- половое размножение 217
— митоз 140
— общая характеристика 19, 39
— эволюция и филогения 418-419
Зональность, на литорали Э52-356
- факторы, влияющие на нее 354-356
Зоопланктон, внедание им водорослей 321-323
Зооспоры 194, 206-210
- жгутиковый аппарат 121
- образование 206-208
- стефаноконтные 209-210
- структура 82
Изогамия 247, 248
Индолил-З-уксусная кислота (ИУК) 303-304
Ископаемые находки 386, 405, 407, 412-425
Использование водорослей 445-451
История альгологии 10-14
ИУК - см. Индолил-З-уксусная киолота
Калан, влияние хищничества 347-348
Калий, метаболизм 282, 291-292
Камеры - см. Локулы
Кальций, отложенные (обызвествление) 288-291
Капсула, слизистая 97
Каротиноиды 262
Карповой, оплодотворение 226, 227, 218
569
Карпоспорофит, развитие после оплодотворения 226, 228
Каррагинаны, использование и свойства 448
Кислотное загрязнение 438-439
Кислород, эволюция водорослей в зависимости от его содержа-
ния 385, 391
Кладистика 401
Классификация/таксономия и систематика 9, 16-76
Классы 16-17, 18
- жизненные циклы, различия 198
- общие характеристики 16-24
- родственные связи 401-404
Клетка, покровы 78-106
— типы 80
- стенка - см. Клеточная стенка
- структура и организация 77-147
Клеточная стенка неполная 96-97
— особенности 98-107
— полная (истинная) 97-100
Кокколиты, виды, у которых они встречаются - см. Кокколито-
фориды
- типы I60-I6I
Кокколитофориды 160
- ископаемые 419
- морфология 43, 44
- обызвествление у них 290
Колониальные формы 166-171
Конкуренция межвидовая 342, 355
- у морских макрофитов 342-344, 347
- у фитопланктона, межвидовая 332, 333
Кораллины, клеточная стенка 101
- обызвествление 289
- эволюция 412-414
Корковая фаза, имеющие ее виды 182-183, 230-231
Красные водоросли
— жизненный цикл соматический 257-258
------- спорический 226-233
— классификация 18
--клеточная стенка I0I-I02
570
— коккоидные 154
— кора, имеющие ее формы 179
— митоз 140, 141
- - нитчатые многоосевые 178
-----одноосевые 178, 180-182
— общая характеристика 19, 29-33
— паразитические 377-378
— пол, определение 228-230
— полисифональные 179
- - споры бесполые 204-206
----- половые 226-233
— токсичные 431 ,
— хлоропласты 131
— цитокинез 141, 147
Красные приливы 428-430
Кремний, метаболизм 291
Кривые выживаемости 350
Криптомонады (криптофиты) жгутиковые 163
- жгутиковые волоски II4-II5, 116
- клеточная стенка 82, 83
- митоз 143
- / общая характеристика 22, 62-64
- хлоропласт, структура 132-133
Культивирование водорослей 451-456
— дальнейшие перспективы 456
Лекарства из водорослей 450-451
Ленгмюра циркуляция 314, 315
Лишайники 370-374
Локулы 93, 95
Лунные ритмы 306-308
Макроскопические водоросли, культивирование 451-456
----- токсичные 430-431
— экология 333-360, 362-364
Макрофиты и рыхлые осадки 362-364
Марши, водоросли в них 363-364
571
Мель, загрязнение ей и устойчивость к нему 438, 440
Медицина, применение в ней водорослей 450-451
Мейоз 195,<196
- место в жизненном цикле 196
----- классификация по нему жизненных циклов 196
----- связь с половым размножением 196
Металлы, загрязнение 437-440
Миграция, вертикальная у динофлагеллат 324
— у эпипелитных водорослей 360-361
Микофикобиозы 374
Микроскоп, электронный, эра его 13
Микроэлементы, метаболизм 292-293
Митоз, различия между классами 140-145
- эволюция 397
Митохондрии, эволюция 393, 395
Многоосевые нитчатые формы 178, 180, 183
Моллюски, ассоциации с ними водорослей 366-368
Моно уравнение 295
Моноспора 204
Море, распространение в нем фитопланктона 323-331
Морские водоросли-макрофиты -см. Макроскопические водоросли
Морской лед, обитание в нем водорослей 382-383
Морфология 9
- адаптивная у морских макрофитов 334-335
- разнообразие 148-150
- особенности в разных отделах и классах 26-76
- эволюция ее 148
Моря и океаны, распространение фитопланктона 323-331
Мохообразные - см. Бриофиты
Мукоцисты 164
Наземные растения, эволюция 140-401
Нанопланктон 3I0-3II
Нефть, загрязнение ей 432-435
Нефтяные платформы, обрастание водорослями 444
Нитчатая организация 152, 167, 172-184
— образование пленок из нитчатых форм 363
572
Одноклеточные водоросли 150-166
Одноосевые нитчатые формы 174-184
Озера, динамика популяций фитопланктона 324-327
- подкисление 439
- эвтрофизация 435-437
Океаны, Атлантический, распространение водорослей 356, 357
Окружающая среда 426-445
Оогоний, развитие 223-226
Оплодотворение - см. Сингамия - . .
Организация клеточная/ультраструктурная 77-147
- уровни 148-193
Органическое загрязнение 266-267
Особенности, общие, классификация в связи с ними 26-76
Отделы 17, 18
Пальмеллоидные формы 153, 166-168
Панцирь 96-97
Паразитические водоросли 376-378
Параспоры 205, 206
Параксиальные тяжи 120
Паренхиматозные (полистиховые) формы 175, 185-188
Пелликула, эвгленовых 82-83, 164
Переживание неблагоприятных условий I3I-I33
Песок, живущие на нем водоросли 360-364
Пигменты фотосинтетические 260-262
- - особенности в разных отделах/классах 19-24, 25-76
Пиреноиды 131, 133
Питательные вещества - см. Биогены
Пища - см. тж. Биогены
- животных 446
- человека 445-446
Плавучесть фитопланктона 312-314
Плаэмодесмы - см. Поры
Планктон - см. Зоопланктон; Фитопланктон
Плотность, у фитопланктона 312-314
Побережье, зональная классификация 352-353
Поверхность, отношение ее к объему, взаимосвязь с биогена-
ми 319
573
Погружение, скорости 312-314, 315
Подклассы 17
Поколения, чередование - см. Чередование поколений
Покоящиеся стадии 210-215
Пол, определение 237-238
— аллели, ответственные за 228-229
Полифосфаты, функция 287
Половое размножение 10,. 216-217
— жизненные циклы, его включающие 217-258
— характеристики в зависимости от класса 9-74, 197
--- эволюция 397-399, 400-401
Половые аттрактанты 296-298
Популяции, динамика 323-331
Поровая пробка (поровое соединение) 101-102
Поры между клетками (= плазмодесмы) 97, 103
Порядки зеленых водорослей 73, 75, 76
- красных водорослей 33
- синезеленых водорослей 28
- хромофитов 39, 44, 46, 49, 52, 56
- эвгленовых 66
Почвенные водоросли 364-366
Преобразователи тепловой энергии в открытом море, обрастание
водорослями 444-445
Приливы, величина и периодичность 353-354
- красные 428-430
Примнеэиофиты жтутиковые I60-I6I
- жизненные циклы 233-235
- общая характеристика 20, 39-44
- эволюция и филогения 419-420
Происхождение прокариотных водорослей 78, 385-386, 387-391
- эукариотных водорослей 386, 392-397
Прокариоты, классификация 16, 18, 19, 26-29
- клеточная организация 77-78
- происхождение и эволюция 385, 387-392, 404-406
Прорастание 298
Прохлорофиты коккоидные 152
- общая организация 28-29
- общая характеристика 19, 28-29
- эволюция 28, 405-406
574
pH (см. тж. Кислотное загрязнение), доступность неорганическо-
го углерода в связи с ним 273 , 274
Развитие, роль гормонов 303-305
Размер, взаимосвязь с густотой зарослей 342, 343
Размножение 10, 194-217
- бесполое 194, 199-215
- половое - см. Половое размножение
- рост и 296-308
- у лишайников 373
Распределение водорослей, вертикальное 352-856
— факторы, влияющие на него 354-356
Рафидофиты жгутиковые 163
- общая характеристика 22, 56-62
Рдест, эпифиты на нем 379-380, 381
Регенерация/восстановление, системы 300, 301, 302
Ризоподиальные формы I50-I5I, 153
Ритмы, эндогенные 306-308
Родолиты 414
Рубцы при образовании клеточной стенки 106
Саргассово море, сезонные изменения фитопланктона 331, 332
Свет, измерение 259-260
- улавливание/поглощение 263-264, 265, 266
- фитопланктон и 315-317, 318
Сезонная динамика популяций 323-331
Сера, метаболизм 291-292
Симбиоз, участие в нем водорослей 366-380
- эволюция эукариотных форм посредством него 393, 403, 409
Сингамия (оплодотворение) 195-196, 296-299
- карпогона 226, 229
- изогамная 195-196
- оогамная 196
- формы 195-196, 197
- эволюция 397-400
Синезеленые водоросли, бесполые споры 204
- - "век" их 386, 391-392
575
— классификация 28
— клеточная стенка 97-99
— колониальные 167, 168
- - нитчатые 172
— общая характеристика 19, 26-28
— перенесение неблагоприятных условий 210-212
--- перенос генов 216-217
— термофильные 383-384
--- токсичные 427-428
— уровни организации 152
— фиксация азота 280-281, 286, 372-373
--- эволюция 385-392, 404-406
Систематика - см. Классификация
Сифональные формы 189, 190-193
Сифонокладальные формы 174, 188-190
Снежные водоросли 380-382
Соленость, обусловленная ей стратификация 317
Соматические жизненные циклы 196, 256-258
Спектры действия фотосинтеза 265, 266
Спермий, акронема жгутика 112, ИЗ
Спорические жизненные циклы 196, 226-247
Спорофит-гаметофит, чередование - см. Чередование поколений
Споры бесполые 194, 204-215
- покоящиеся, связь' с выживанием 210-215
- половые 226-247
Ститмы - см. Глазки
Стратегии выживания у морских водорослей-макрофитов 335
Строматолиты 387, 389
Сукцессии видов - см. Виды
Таксономия - см. Классификация
Талломы ветвящиеся нитчатые 174-184
- клетки, взаимодействия между ними 302-303
- образование, влияние долготы дня 358-359
- у сифональных форм 190-193
Тека, у динофлагеллат 83-84, 85, 86, 162
Текальные пластинки 84, 86, 162
576
Температура воды, биогеографические зоны, определяемые ей 356
358
-- стратификация 317, 318
- как фактор загрязнения 441-442
- экстремальная, водоросли при ней 380-384
Тепловое загрязнение 441-442
Термоклин 317, 318
Термофильные водоросли 383-384
Тетраспоровые формы 166-168
Тетраспорангий, деление, типы 227, 229
Тетраспорогенеэ, необходимые для него условия 357-358
Тетраспорофитное поколение 230-233
Тилакоиды, расположение 77, I28-I3I, 133
Типовой образец 25
Токсины, водорослевые 427-431
Трансаминирования реакции 284, 285, 286
Трикарбоновых кислот цикл 274 .
Трихоцисты 16I
Тропиэмы 305-306
Тяжелые металлы, загрязнение ими 437-440
Углеводородное загрязнение 432-434
Углеводы, запасание 277-278
- поступление у лишайников 372, 373
Углерод, метаболизм 272-278
- органический, потери 276
- поток в лишайниках 372, 373
- фиксация 272-274
--гетеротрофная 273-274
-- фотосинтетическая 272-273
Удобрения, влияние внесения их в озера на фитопланктон 3J7-
319, 320
- использование водорослей как 449-450
Уркариоты 387, 390
Фенетика 401
Фенологическое изучение морских водорослей-макрофитов 349
: 577
Феромоны 296-298
Фибриллы, клеточной стенки 99-100
Физиология 259-308
- особенности в зависимости от класса/отдела 26-76
Фикобилисомы, структура 264, 266
Фикоколлоиды, промышленное использование 446-449,451
Фикология - см. Альгология
Филетические деревья 387, 390
Филогенетические связи, строение переходной зоны жцутика как
из показатель 120-123
Филогения водорослей 401-425
Фильтры, забивание их водорослями 443
Фитогеографические области, Атлантический океан 356-358
Фитопланктон, парадокс его 331-333
- разнообразие, влияние теплового загрязнения 441-442
- экология 310-333
Фораминиферы, ассоциации с ними водорослей 370
Фосфоенолпируват, метаболизм 272
Фосфор 282 , 286-288
- метаболизм 286-287
- роль в эвтрофизации 436
- соотношение с азотом 287-288
Фотоавтотрофная эволюция 387—391
Фотодыхание 275-276
Фотопериодизм, долгота дня 268-271
Фотосинтез, кривая светового насыщения 269
- освещенность и 268
- продукты, накопление и запасание 277-278
- спектры действия 265 , 266
- у фитопланктона 315-317
- фиксация углерода фотосинтетическая 272-274
- эволюция 389-391
Фотосинтетические пигменты - см. Пигменты фотосинтетические
Фотосистемы I/II 263, 264
Фототропизм 305
Фрагментация, вегетативное размножение посредством нее 200-201
Функциональные группы бентосных морских водорослей 345
578
Характеристики классов водорослей 26-76
Харовые,ветвящиеся нитчатые 184
- клеточная стенка 106
- половое размножение 223-226
- общая характеристика 23, 73-75
- эволюция и филогения 425
Хилла - Бендолла 2-схема 263-264
Хищничество 347-349
Хламидомонады (см. тж.Chlanydomonaa ) обитающие на снегу 380
Хлоропласты, различия в зависимости от отдела/класса 19-24,
130, 131-134
- структура/особенности 128-134
- эволюция 392-396
Хлорофилл 260-262
Хлорофитная линия эволюции 408-412, 421-422
Холод, живущие при экстремальных условиях водоросли 380-383
Хризамёбы, одноклеточные 151
Хроматическая адаптация 267
-- комплементарная 267-268
Хроматофоры - см. Хлоропласты
Хромофиты - см. Chromophycota
Цветения синезелеными водорослями 427-428
- фитопланктонные 3244325, 427, 428
Ценобии 169-171
Цианобактерии - см. Синезеленые водоросли
Цинк, загрязнение им 440
Циркадианные ритмы 306, 307
Цисты динофлагеллат 162-163
- золотистых водорослей 160, 215
- муко- 164
- покоящиеся 215
Цитокинез, различия в зависимости от класса I4I-I42, 145-147
Цитокинины 304-305
Цитоплазма при делении (цитокинезе), различия в зависимости
от класса I4I-I42, 145-147
579
Человек и водоросли 426-456
Чередование поколений 226-247
— изоморфное 236
— характерные особенности 226-235
— эволюция 385-387, 398
Чешуйки на жгутиках 112, 114, П5
- на клетке 84-91
Чехол, слизистый 97
Шипик, на жгутике 120, 122
Шов 93
Эбридии - см. Ebriophycidae
Эвгленовые, глазки 135
- жгутиковые 163-164
- митоз 143-144
- общая характеристика 23, 64-66
- эволюция и филогения 410-412
Эволюция 3854387
- морфологическая 148
- основные этапы 385-387
- особенности в зависимости от класса 26-76
- от прокариот к эукариотам 78
- симбиоз и 368-369
- филогения и 396, 401-425
Эвтрофизация культурная 435-437
Эжектосомы 163
Экзоспоры 203, 204
Экология 10, 309-384, 426-445
- особенности в зависимости от класса/отдела 26-76
Электронный микроскоп, эра его 13
Эндогенные ритмы 306-308
Эндосимбиоэа теория 78
Эндоспоры 203, 204
Эпилитные водоросли - см. Макроскопические водоросли
Эпипелитные водоросли 360-361
-- влияние загрязнения 442
580
Эписаммитные водоросли 361
Эпифитные водоросли 378-380
— влияние загрязнения 439, 441-442
ЭухЗактерии 387, 390
Эукариоты, классификация 16, 18, 19-24, 29-76
- клеточная организация 77-78
- происхождение и эволюция 78, 386-387, 392-397
Ядро 137-139
- деление 139-145
- интерфазное 137-139
- мезокариотическое 138
- число на клетку 139
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Acetabularia. жизненные циклы 253-254
- морфогенетические исследования 192
Aoroohaetum aaparagopsis. фотопериодизм 271
Aoroaiphonia. жизненные циклы 245, 246
Alaria marginata. особенности клеточной стенки 105
Amphidinium carterae. пиреноиды 134
Anabaena. водорослевые ассоциации с 374-375
- плавучесть 313
Aphanothece ар.. образование пленки 389
Asccseira mirabilia. морфология 62
Ascoseirales, жизненный цикл 252-253
Aaterionella formoaa. конкурентные отношения 332-333
Audouinella infeatans. морфология 31
Agolla. водорослевые ассоциации 374-375
Bangia atropurpurea. морфология 30
Benglales, жизненный цикл 233
Bangiophycidae, клеточная стенка 101
- эволюция 407
Beokerella sealaramosa. морфология 32
Bidlacus ignotua. ископаемое 419
Bryopaia ар., клеточная стенка 106
- комплекс Derbesia - Halocystia. жизненные циклы 213, 245-247
Caulerpa brownii. морфология 71
Caulerpales, жизненные циклы гаметичеокие 254, 255
-----споричвские 245-247
Cephaleuros. паразитизм 376-377 '
Ceremiaceae, одноосевые нитчатые 183
Ceremiales, жизненные циклы 232
Ceramium rub ram. взаимодействия между клетками 303
Ceratlum ар., морфология 50
- brachyceros. особенности клетки 86
Chaetocercs didymum. жизненный цикл 250
Chaetophorales, половое размножение
Chara, репродуктивные структуры, развитие 223 - 226
“ vulgaris. морфология 74
Charophyceae - см. Харовые
582
Chiamvdomonas (см. тж. Хламидомонады), морфология 68
- половое размножение 219
- hreviB-pina. зиготический жизненный цикл 220
- reinhardtii. аксонема жгутика ИО
Chlorococcales, одноклеточные коккоидные 156-157
Chlorophyceae - см. Зеленые водоросли
Chlorophycota - см. Зеленые водоросли
Chondrus. выедание 344-346
- конкурентные взаимоотношения 347
~ культивирование 454
Chorda tomentosa. морфология 59
Chordariales, жизненные циклы 241-242
Choreooolax polysIphonlae. паразитизм 378
Chromophycota, классификация 18, 19-22, 33-39, 44, 46, 49, 52,56
- общая характеристика 19-22, 33-56
- структура хлоропласта I3I-I33
- эволюция 408—110
Chroococcus sp.. морфология 26 ’
Chroolepidaceae, паразитические 376-377
Chroomonas mesostigmatica. глазок 136
— хлоропласт 133
Chrysamoeba sp., клеточная организация 81
- морфология 41
Chrysamoebidales, одноклеточные 151
Chrysoohromullna. образование чешуек на жгутиках 114
“ blrgeri, морфология 43
Chrysoeoccus furoatus. морфология 42
- rufescens, клеточная организация 98
Chrysophyceae - см. Золотистые водоросли
Chylocladia sp., ветвящиеся нитчатые формы 178
Cladhymenia oblongifolia. морфология 37
Cladophora, митоз 144, 145
- glomeratа. влияние эвтрофизации 436-437
- rupestris. морфология 72
Cocconeis pseudomarginata. морфология 48
Codiaeeae, сифональные 192-193
Conchocelis. культивирование 452
Convoluta. взаимоотношения с водорослями 367-368
Corallinaceae - см. Кораллины
Corisphaera gracilis, морфология 44
583
Cryptophyceae - см. Криптофиты
Cryptonemialea, спорические жизненные циклы 230
Cyanobacteria - см. Синезеленые водоросли
Cyanopliyceae - см. Сине зеленые водоросли
Cyanophycota - см. Синезеленые водорооли
Cyclotella , выедание 322
- конкурентные взаимоотношения 332-333
Cymatiosphaera reticulosa. ископаемое 422 •
Cymopolia paktia. ископаемое 424
Cyatophora toruloaa. морфология 63
Daaycladalea, сифональные I9I-I92
- эволюция и филогения 423
Derbeaia, зооспоры 209-210
- комплекс Halicystis - Bryopsla. жизненные циклы 213,245-270
Desmarestia firma. морфология 59
Deamarestiales, жизненные циклы 239
Deamidialea - см. Десмидиевые
Dlotyotales, изоморфные жизненные циклы 236, 238
Dictyota sp,. лунные ритмы при образовании яйцеклеток 307, 308
-морфология 56
Dinobryon cylindricum. половое размножение 216, 217
Dinophyceae - см. Динофлагеллаты
Dolichomastix numniulifera. чешуйки на жгутиках 115
Draparnaldia. в.р., морфология 70 ,
Dumontia incrasaata. морфология 35
Durvillaea antarctica. морфология 61
Dysmorphococcua globoaus. развитие панциря 97
Ebriophycidae, эволюция и филогения 420-421
Ecklonia ар., морфология 61
Ectocarpua ар., морфология 53
- репродуктивные структуры 210
- ailiculosua
-- жизненный цикл 236, 237
—обраотание судов 440, 444
— устойчивость к меди 440
Elachifata fucicola. морфология 54
Emiliania huxleyi. морфология 43
— сезонные изменения популяций 331, 332
•НпЪуНгя int-r-ig - см. Калан
584
Enteromorpha. выедание ее 345-346
Erythrooladia ар,. морфология 32
Erythrotriohia folilformis. морфология 31
Euaatrum insigne, морфология 71
Eucheuma, культивирование 453
- gelatinae. морфология 35
Euglena gracilis, морфология 65
— чешуйки на жгутиках 117, 118
- granulata. организация клетки 84
Euglenophyceae - см. Эвгленовые
Euglenophycota - ом. Эвгленовые
Eukaryote - см. Эукариоты
Eustigmatophyceae, глазок 137
- коккоидные 155
-общая характеристика 20 , 46-47
Eimbriophyllum, развитие карпоспорофита 228
Florideophycidae, клеточная стенка 10I-102
-нитчатые, ветвящиеся 175-184
- спорические жизненные циклы 226-232
-эволюция 407, 412-414
Foraminifera - см. Фораминиферы
Fucales, жизненные циклы 251-252
Fucus, жизненный цикл 251, 252
-модельная система яйцеклетка/зигота, изучение оплодотворения
и прорастания 298-299
. - паренхиматозные формы 185-187
distichus edentatus. морфология 187
serratua. морфология 62
- vesioulosus, классификация 17-25
---морфология 187
I.
Galexaura sp. поровая пробка 102
Glaucophyceae - см. Глаукофиты
Glaucophyta - см. Глаукофиты
Golenklnia mlnutissima, акронемы жгутиков 113
Gonyaulax . цветения 428-429
-циркадианные ритмы 306, 307
Grateloupia stipitata. морфология 34
Griffithsia регенерация 300, 301
- pacifica. спорический жизненный цикл 229
585
Gymnodinium. цветения 429
Gymnogongrus ар.. морфология 36
Halicystis - Derbesia - Bryopaia комплекс, низменные одклы 213,
245-247
Halimeda. обызвествление 288-289
Halopteris ар. морфология 58
Hantzschia amphioxys. отложение кремнезема 96
- virgata, вертикальная миграция 361, 362
Haplospora globosa. жизненный цикл 242
Haptophyceae - ом. Примнезиофиты
Harveyella mirabilia, паразитизм 378
Hemiselmis brunnescens. организация клетки 83
Heterocapsa. ядро, мезокариотное 139
Himanthalia, шипик на жгутике 120, 122
- elongata. морфология 63
Hormosira banks!!, морфология 63
Hydrpdiotjpa. ценобии 171
Hymenomonas carterae, жизненный цикл 208, 209
Johnson-sea-linkia profunda, обитание на большой глубине 267 •
laminaria, жизненный цикл 237-238, 239
- japonicа. культивирование 453
- longicruris. питание 340
- ochroleuca. морфология 60
laminariales, гетероморфные жизненные циклы 237-238
- паренхиматозные 186, 187-188
Lemanea mamillosa. жизненный цикл 257-258
Id-agora tanakai. морфология 33
Idthothamnion. ЭВОлиция 414 '
- filaciale, морфология 415
Id.ttorina, выедание водорослей ею 344-346
Maprocystis pyrlfgrs, транслокация продуктов фотосинтеза 279
Mallomonas harrisea. чешуйки 91
Mamiella - см, Nephroselmis
Melosira nununuloides. морфология 49
Merismopedia ар., морфология 26
Michaelsarsia elegans. морфология 43 '
586
Mychodea marginifera. морфология 35
Mytilus califomianus. конкуренция 347
Nemaliales, клеточная стенка 101
Nephroselmis (Mamiella) gilva. чешуйки на жгутике 115
-olivacea. жгутики 115, 122
Noctiluca miliaria. жизненный цикл 247, 248
Ochromonas aphaerooyatia. морфология 40
Oedogoniales, половое размножение 212, 221-222
Oedogonium sp., жизненный цикл 212
- клеточная стенка при делении клетки 106, 107
- размножение 212, 221-222
Ophiocytium. морфология 46
Oacillatoria. фиксация азота 281
Palaeoconchocelis starmaohii. ископаемое 414, 415
Palaeodasycladus mediterraneus, ископаемое 424
Palaeoperidinium pyrophorum. ископаемое 421
x'alaeocystophora acuminata, ископаемое 417
Palmarialea, спорический жизненный цикл 231
Palmaria palmata, жизненный цикл 232
— морфология 36
Pandorina unicocoa, морфология 68
Pavlova calceolata. жгутики III
Pelvetia faatigiata. выживание 349, 350
Penicillus capitatus. морфология 71
Peridinium cinctum. морфология 50
Phaeophyceae - см. Бурые водоросли
Phaeothamnion borzianum. морфология 42
Phthanoperidinium amoenum. ископаемое 421
Phycodrys rubens. морфология 38
Phyllophoraceae, спорические жизненные циклы 231
Pisaster. эффект выедания 347
Piatymonas - см. Tetraaelmis
Plocarnium cartilagineum. характер ветвления 178
Polysiphonia. жизненный цикл 226-227
Porphyra. жизненный цикл 234
- культивирование 452
- umbilicalia. морфология 31
587
Poatelaia palmaeformis. морфология 60
Potamogeton, - см. Рдест
Praeinophyceae - см. Празинофиты
Prasiolalea, жизненные циклы 256
Prasiola sp.. жизненные циклы 256
Prochloron ар,, ассоциации с беспозвоночными 368-369
- общая характеристика 28
- организация клетки 77
-----филогения и 405-406
Prochlorophyceae - см, Прохлорофиты
Prochlorophycota - СМ. Прохлорофиты
Prorocentrum minimum, жгутики 119
Protogonyaulax - см. Gonyaulax
Prototheca. связанные С ней болезни 426-427
Prymnesiophyceae (Haptophyceae) - см. ПримнезиофйШ
Prymnesium parvum. токсичность 428
Pseudogloiophlcea sp,. поровая пробка 102
Punctaria plantaginea , морфология 56
Pyramimonaa gelidicola. чешуйки на клетке 90
- lunata, особенности клетки 89
Pyrobotrya, морфология 68
Pyrodinium bahamense. морфология 51
Raphidophyceae - см. РафИДофИТЫ
Rhodella reticulata, структура хлоропласта/пиреноида 132
Rhodomela confervoidea. морфология 37
- larix. заселение эпифитами 380, 381
Rhodomelaceae, нитчатые ветвящиеся 18I
Rhodophyceae - см. Красные водоросли
Rhodophycota - см. Красные водоросли
Rhodymeniales, жизненные циклы 232
Rhoicosphenia curvata. половое размножение 249
Roaenvingiella polyrhiza. морфология 70
Saocorhiza dermatodea. транслокация продуктов фотосинтеза 280
- polyschides, определение пола 237-238
Schottera nicaeensia. регенерация 302
Soenedeamua, реакция на отношение Ь1:Р 287 , 288
Scytosiphcnalea, жизненные циклы 241-242
Scytoaipho.n sp., ультраструктура клетки 104
- J,omentaria. жизненный цикл 241-242
588
-- морфология 55
- - образование таллома, влияние долготы дня 358-359
Spatogloesum а-р., морфология 57
Sphaoelaria, вегетативные пропахали 203 -
- plumoaa, ископаемое 418
- tribuloidea
: : «SW6™1 138
Sphaeridiothrix comoreasa. морфология 41
Sphaerocyatis sohroeteri. выедание 322, 323
Spiroдуга, зиготический жизненней цикл 222
Sporoehnales, жизненные циклы 239
ЗрогооЬлца ар.. морфология 55
Stichogloea ар., Морфология 40
Stylodinium littorale. морфология 50
Synorypta elomifera. особенности клетки 87
Syringoderma. жизненные циклы 240
- phiiuieyi. морфология 58
Syringodermatales, жизненные циклы 240
Tetraselmia ( я Piatymonas). ассоциации с беспозвоночными 367,368
- жгутиковые корни 127
Thalaasjoaira confuaa. морфология 48
- incerta. центральные выросты 96
- oeatrupii. морфология 48
Tilooteria mertenaii. жизненный цикл 242
— морфология 58
Trachelomonaa lefevrei. клеточный покров 84
Triceratium favua. структура панциря 92
Udoteaceae, сифональные 192-193
Udotea petiolata. жизненный цикл 254
-- морфология 72
Ulothrix apecloaa. морфология 69
Viva, жизненный цикл 211
Ulvales, жизненные циклы 244
Uivophyceae (Ulvaphyoeae) 72-73
Urospora penicilliformis, жгутики зооспор 120, 121
yalpnia. клеточная стенка 100
- сифонокладальное развитие 188, 189
Vauoheria ар., морфология 45
- половое размножение 217
589
Volvocales, жгутиковые колониальные 169-170
— одноклеточные 164 ,
- размножение, половое 200, 219-221
Volvox, размножение бесполое 200, 202
— половое 202, 219-221
Xanthidium sp., морфология 70
Xanthophyceae - см. Желтозеленые водоросли
Xanthorla partetins, морфология и анатомия 371
Zygnematales, стратегии конъюгации 222
ОГЛАВЛЕНИЕ
От переводчика .............................................. 5
Предисловие ................................................... 7
Глава I. Введение ............................................. 9
Альгология и водоросли ................................... 9
Развитие альгологии ..................................... 10
Горизонты современной альгологии ........................ 14
Глава 2. Классификация водорослей ............................ 16
Основные характеристики водорослей ...................... 26
Prokaryota (прокариоты) ........................... 26
Cyanophycota....................................... 26
Cyanophyceae: синезеленые водоросли ............. 26
Prochlorophycota.................................... 28
Prochlorophyceae............................... 28
Eukaryota (эукариоты) ............................. 29
Rhodophycota: красные водоросли, или багрянки 29
Rhodophyceae ..................... 29
Chromophycota ..................................... 33
Chrysophyceae: ЗОЛОТИСТые ВОДОРОСЛИ ... 39
Prymnesiophyceae (Haptophyceae): примне-
зиофиты.................................. 39
Xanthophyceae: желтозеленые водоросли ... 44
Eustigmatophyceae ........................... 46
Bacillariophyceae: ДИЭТОМОВЫе ВОДОРОСЛИ,
диатомеи .................................... 47
Dinophyceae; динофлагеллаты, динофиты .... 49
Phaeophyceae: бурые водоросли ........... 52
Raphidophyceae: рафИДОфИТЫ ................ 56
Cryptophyceae: криптомонады, криптофиты 62
Euglenophycota .................................... 64
Euglenophyceae: эвгленовые ........................ 64
Chlorophyccta: зеленые водоросли •••••......... 66
Chlorophyceae ............................ 68
Charophyceae : харовые, ИЛИ лучицы......... 73
Prasinophyceae: празинофиты ....... 75
"Glaucophyta", или "Glaucophyoeae1’ .......
591
Глава 3. Структура клетки ..............
Общие особенности .................
Клеточные покровы: клеточная стенка
Rhodophyceae ......................
Phaeophyceae .................
Xanthophyceae ................
Chlorophyceae ................
Charophyceae .................
Жгутики .......... ................
Хлоропласты .......................
Прокариоты ...................
Rhodophycota .................
Chromophycota ................
Chlorophycota ................
Глазки (стигмы) ...................
Ядро, деление ядра и цитокинез •••
Интерфазное ядро ..................
Деление ядра .................
Цитокенез ....................
77
77
78
101
102
ЮЗ
105
106
107
128
128
ХЗЕ
I3X
133
134
137 '
137
139
145
Глава 4. Уровни организации ................................ 148
Разнообразие морфологии .............................. 148
Одноклеточная организация - ризоподиальные формы ...... 150
Одноклеточная организация - коккоидные формы ......... 151
Одноклеточная организация - жтутиковые ............... 158
Колониальная организация - пальмеллоидный, тетраспоро-
вый и дендроидный типы ................................ 166
Колониальная организация - ценобии ................... 169
Нитчатая организация - простые нити .................. 172
Нитчатая организация - ветвящиеся нити ............... 174
Паренхиматозная организация .......................... 185
Сифонокладальная организация ......................... 188
Сифональная организация .............................. 190
Глава 5. Размножение и циклы развития...................... 194
Введение ............................................. 194
Вегетативное размножение ............................. 199
Бесполое размножение спорами ......................... 204
Экзоспоры и эндоспоры ........................... 204
592
Моноспоры, биспоры и параспоры ..................
Зооспоры, апланоспоры и автоспоры ............... 206
Покоящиеся стадии .................................... 210
Половое размножение .................................. 216
Зиготические жизненные циклы ....................
Спорические жизненные циклы ..................... 226
Гаметические жизненные циклы ............ 247
Соматические жизненные циклы .................... 256
Глава 6. Физиология и биохимия ............................ 259
Введение ............................................. 259
Свет и его измерение ................................. 259
Фотосинтетические пигменты и поглощение света ........ 260
Поглощение света ................................. 263
Спектры действия................................. 265
Хроматическая адаптация.......................... 267
Комплементарная хроматическая адаптация и вертикаль-
ное распределение водорослей.........;............ 267
Фотосинтез и освещенность ....................... 268
Долгота дня и фотопериодические реакции .............. 268
Метаболизм углерода ................................ 272
Фиксация углерода ............................... 272
Темновое дыхание ................................ 274
Фотодыхание, гликолатный метаболизм и потеря ор-
ганического углерода ............................. 275
Накопление и запасание продуктов фотосинтеза .... 277
Неорганические питательные вещества (биогены) ........ 279
Азот ............................................ 279
Фиксация азота ............................. 280
Неорганический связанный азот .............. 284
Органические источники азота ............... 286
Запасание азота........... ................. 286
Фосфор .......................................... 286
Отношение азот:фосфор ........................... 287
Кальций ......................................... 288
Кремний ......................................... 291
Калий и сера..................................... 291
Микроэлементы ................................... 292
Витамины и ростовые факторы .......................... 293
Поглощение биогенов ..................................
593
Рост и размножение ................................... 296
Феромоны ........................................ 296
Оплодотворение и прорастание .................... 298
Полярность ..................................... 299
Апикальное доминирование ........................ 300
Взаимодействия между клетками таллома ........... 302
Гормоны и развитие водорослей ................... 303
Тропизмы ........................................ 305
Эндогенные ритмы ............................... 306
Глава 7. Экология.......................................... 309
Введение ............................................. 309
Фитопланктон ......................................... 310
Методология...................................... 311
Плавучесть ...................................... 312
Свет ............................................ 315
Биогенные элементы ............................ 317
Выедание животными .............................. 321
Динамика популяций .............................. 323
Парадокс планктона ............................... 331
Эпилитные водоросли-макрофиты ........................ 333
Жизненные формы и адаптивная морфология ....... 334
Биогены.......................................... 335
Конкуренция...................................... 342
Выедание и хищничество .......................... 344
Демографические и фенологические исследования... 349
Вертикальное распределение макрофитов ........... 352
Биогеография морских макрофитов ................. 356
Водоросли рыхлых осадков ............................. 360
Эпипелитные водоросли.............'.............. 360
Эписаммитные водоросли .......................... 361
Макрофиты и рыхлые осадки ....................... 362
Почвенные водоросли .................................. 364
Симбиотические ассоциации ............................ 366
Ассоциации водорослей с беспозвоночными ......... 366
Взаимодействия водорослей с простейшими ......... 369
Ассоциации водорослей с грибами ................. 370
Ассоциации с бриофитами и сосудистыми растениями 374
Паразитические водоросли ........................ 376
Эпифиты ......................................... 378
Водоросли при экстремальных температурах ............. 380
Снежные водоросли ............................... 380
594
Водоросли морского льда ........................ 382
Водоросли горячих источников .................... 383
Глава 8. Эволюция и филогения............................... 385
Основные этапы эволюции водорослей .................... 388
Происхождение прокариот и фотоавтотрофов .............. 387
"Век цианобактерий" (синезеленых водорослей) .......... 3®^
Происхождение эукариот ................................ 392
Эволюция мейоза, сингамии и чередования поколений ... 397
Эволюция наземных растений ........................... 400
Филогения водорослей .................................. 401
Прокариоты: Cyanophyceae и Prochlorophyceae .., 404
Hhodophyceae ..................................... 407
Хромофитная и хлорофитная линии .................. 408
Ископаемые находки .................................... 412
Rhodophyceae..................................... 412
Phaeophyceae..................................... 414
Bacillariophyceae................................ 416
Chrysophyceae*.................................. 418
Prymnesiophyceae.................................. 419
Dinophyceae..................................... 420
Хлорофитная линия ................................ 421
Chlorophyceae..................................... 423
Charophyceae...................................... 425
Глава 9. Водоросли, человек и окружающая среда ............. 426
Введение ............................................. 426
Водоросли и болезни человека .......................... 426
Токсичные водоросли ................................... 427
Токсичные синезеленые водоросли................... 427
Токсичность Prymnesium parvum.................... 428
Красные приливы и токсичные динофлагеллаты ....... 428
Токсичные макрофиты............................... 430
Загрязнение и водоросли ............................... 431
Органическое загрязнение ......................... 432
Культурная эвтрофизация .......................... 435
Загрязнение металлами и кислотами ................ 437
Тепловое загрязнение ............................. 441
Механические помехи ................................... 442
Помехи движению воды .............................. 44 я
595
Обрастание судов ................................. 443
Неподвижные конструкции: нефтяные платформы и пре-
образователи тепловой энергии в открытом море... 444
Использование водорослей .............................. 445
Питание человека.................................. 445
Керма дая животных ............................... 446
Фикоколлоида ..................................... 446
Водоросли как удобрения .......................... 449
Водоросли в медицине ............................. 450
Культивирование водорослей ............................ 451
Культивирование водорослей-макрофитов ............ 452
Культивирование одноклеточных водорослей ......... 455
Будущее разведения водорослей ................. 456
Литература ........................................... 457
Предметный указатель .................................. 562
Указатель латинских названий ...........................
Уважаемый читатель!
Ваши замечания о содержании книги,
ее оформлении, качестве перевода и
другие просим прислать по адресу:
129820, Москва,
I-й Рижский пер., д. 2,
издательство "Мир"
Научное издание
Р. Саут, А. Уиттик
ОСНОВЫ АЛЬГОЛОГИИ
Заведующий редакцией чл.-корр. АН СССР Т.М. Турлаев
Зам. зав. редакцией М.Д. Гроздова
Научн. редактор Ю.Л. Амченков
Мл. редактор М.С. Карнюшина
Художник А.В. Евстратов
Художественные редакторы А.Я. Мусин, Л.М. Аленичева
Технический редактор Г.А. Мирошина
Корректор В.И. Киселева
ИВ * 7245
Сдано в набор 18.08.89 г. Подписано к печати 5.02.90 г.
Формат 60x90 I/I6. Бумага офсетная М 2.
Печать офсетная. Объем 18,75 бум.л. Усл.печ.л. 37,50.
Усл.кр.-от. 37,50. Уч. изд. л. 36,90.
Изд. Л 4/6773. Тираж 2950 экз. Зак. 121. Цена 5р.60к.
Издательство "Мир"
В/0 "Совэкспорткнига" Государственного комитета СССР
по печати
129820, ГСП, Москва, I-й Рижский пер., 2
Тульская типография Государственной) комитета СССР
по печати. 300600, ЮТ, г.Тула, пр. Ленина, 109.