/























































































































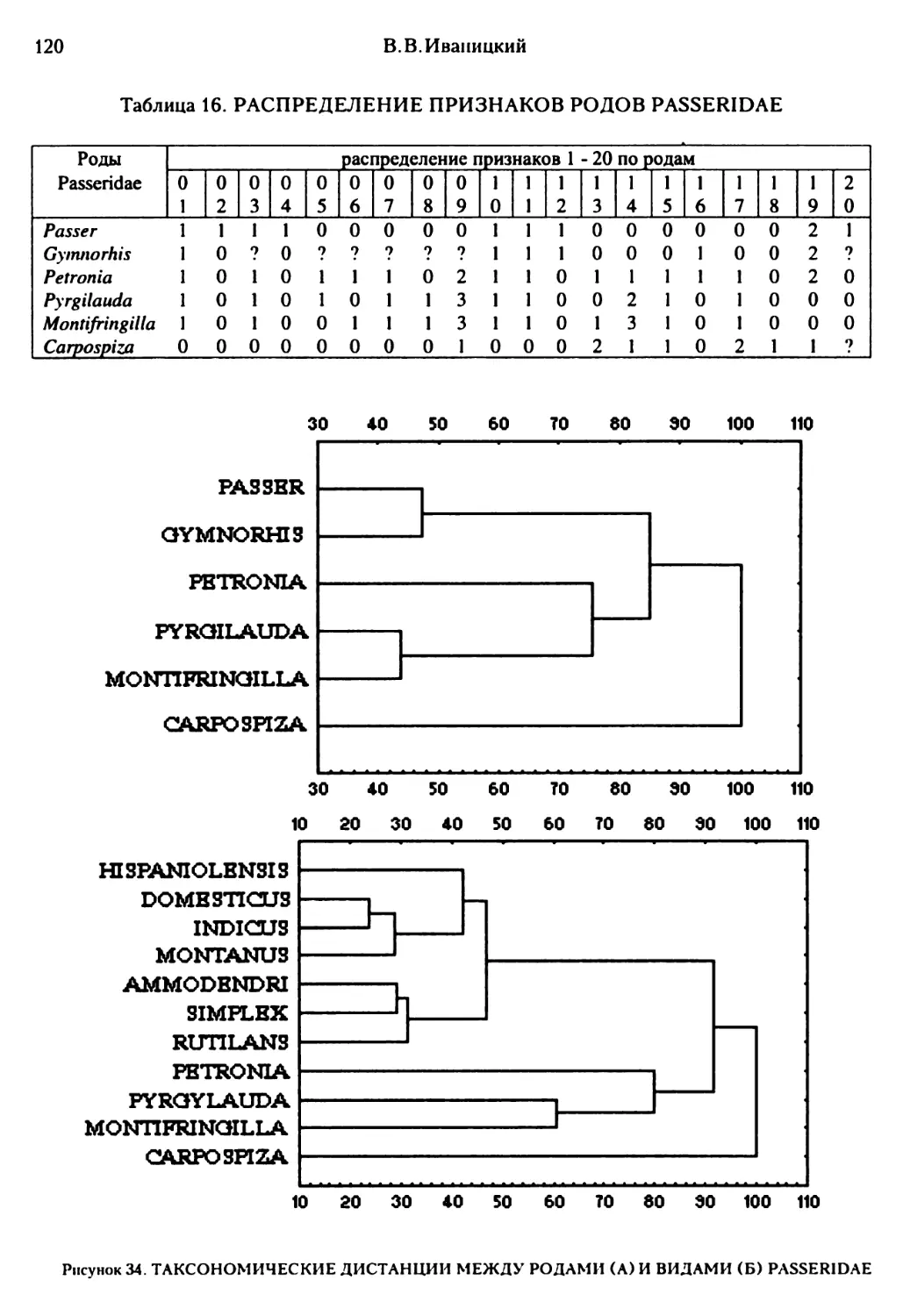
































Текст
В.В. ИВАНИЦКИЙ
ВОРОБЬИ
И РОДСТВЕННЫЕ ИМ ГРУППЫ
ЗЕРНОЯДНЫХ ПТИЦ:
ПОВЕДЕНИЕ, ЭКОЛОГИЯ, ЭВОЛЮЦИЯ
КМК SCIENTIFIC PRESS
МОСКВА 1997
В.В. ИВАНИЦКИЙ
ВОРОБЬИ
И РОДСТВЕННЫЕ ИМ ГРУППЫ
ЗЕРНОЯДНЫХ ПТИЦ:
ПОВЕДЕНИЕ, ЭКОЛОГИЯ, ЭВОЛЮЦИЯ
МОСКВА
1997
В.В.Иваницкий. ’’ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ
ПТИЦ: ПОВЕДЕНИЕ, ЭКОЛОГИЯ, ЭВОЛЮЦИЯ” Москва: КМ К Scientific Press, 1997.
АННОТАЦИЯ
Обобщены результаты многолетних исследований социальной организации, динамики
популяций, репродуктивного поведения, механизмов коммуникации и других аспектов обра-
за жизни 11 видов и 5 родов воробьев фауны России и сопредельных государств. В срав-
нительном аспекте в объеме мировой фауны рассмотрены литературные данные но поведе-
нию и экологии воробьев и родственных им групп зерноядных птиц. Подробно охарактери-
зованы область распространения, среда обитания и основные адаптивные тины воробьев,
пространственно-этологическая структура их поселений, гнездостроепие, взаимоотношения
иолов, способы заботы о потомстве, демонстративное поведение и акустическая сигнализа-
ция. Проведена таксономическая ревизия семейства и его положения в системе подотряда
певчих птиц. Предложен сценарии, трактующий основные этаны эволюции и адаптивной
радиации воробьев в природных и антропогенных экосистемах. Рассматриваются некото-
рые общие вопросы эволюции социального и репродуктивного поведения воробьиных птиц
на относительно высоких уровнях дивергенции.
Для биологов, этологов, зоологов, орнитологов, преподавателей вузов. 148 с.
Издано при финансовой поддержке
Российского фонда фундаментальных исследований
по проекту № 96-04-62148.
ISBN 5-87317—037-1
©—Иваницкий В.В., 1997.
©—КМ К Scientific Press,
оригинал-макет, 1997.
3
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ.................................*............................................5
ГЛАВА 1. СРЕДА ОБИТАНИЯ И АДАПТИВНЫЕ ТИПЫ................................................8
1.1. Область распространения.............................................................8
1.2. Места обитания и положение в ориктоценозах..........................................9
1.3. Адаптивные типы ................................................................... 12
1.4. Главные направления экогенеза...................................................... 14
ГЛАВА 2. ПРОСТРАНСТВЕННО-ЭТОЛОГИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИЙ.............................. 17
2.1. Общие принципы организации пространственных отношений............................. 17
2.2. Продвинутые формы колониальности ................................................ 17
2.2.1. Размеры, структура и динамика стай.............................................. 17
2.2.2. Социальная организация стай..................................................... 20
2.2.3. Пространственно-хронологическая структура и численность колоний................. 21
2.2.4. Поведение при формировании колоний.............................................. 22
2.2.5. Территориальное поведение в колониях ............................................23
2.3. Квазиколоннальность............................................................... 24
2.4. Изолированное гнездование......................................................... 25
2.5. Эволюция пространственной организации в популяциях воробьев и родственных нм птиц. 28
ГЛАВА 3. УСТРОЙСТВО И РАСПОЛОЖЕНИЕ ГНЕЗД................................................36
3.1. Расположение гнезд................................................................ 36
3.2. Воробьи в межвидовой конкуренции за гнездовые убежища............................. 40
3.3. Технология гнсздостросния..........................................................42
3.4. Гнезда воробьев в ряду гнездовых построек Ploccimorpha.............................46
ГЛАВА 4. РЕПРОДУКТИВНОЕ ПОВЕДЕНИЕ...................................................... 51
4.1. Образование пар................................................................... 51
4.2. Система спаривания и половое поведение............................................ 55
4.3. Внебрачные половые отношения.......................................................59
4.4. Половое поведение................................................................. 61
4.5. Гнсздостроенис.....................................................................62
4.5.1. Место гнездостроитслъного поведения в годовом и репродуктивном цикле.............62
4.5.2. Участие полов в постройке гнезд..................................................64
4.6. Насиживание ...................................................................... 70
4.7. Развитие птенцов, родительское поведение, полицикличность......................... 73
4.8. Основные направления эволюции репродуктивного поведения в разных группах зерноядных птиц. 76
ГЛАВА 5. СТРУКТУРА И ЭВОЛЮЦИЯ МОТОРНЫХ РЕПЕРТУАРОВ..................................... 83
5.1. Полет и передвижение по твердым субстратам.........................................83
5.2. Элементарные двигательные акты и их сочетания (позы) ..............................88
5.3. Структурно-функциональная дифференциация и эволюция визуальной сигнализации........96
ГЛАВА 6. СТРУКТУРА И ЭВОЛЮЦИЯ АКУСТИЧЕСКИХ РЕПЕРТУАРОВ .................................100
6.1. Общие принципы организации репертуаров............................................100
6.2. Пустынный и саксаульный воробьи ..................................................101
6.3. Рыжий воробей.....................................................................104
6.4. Полевой, домовый, индийский, испанский воробьи....................................105
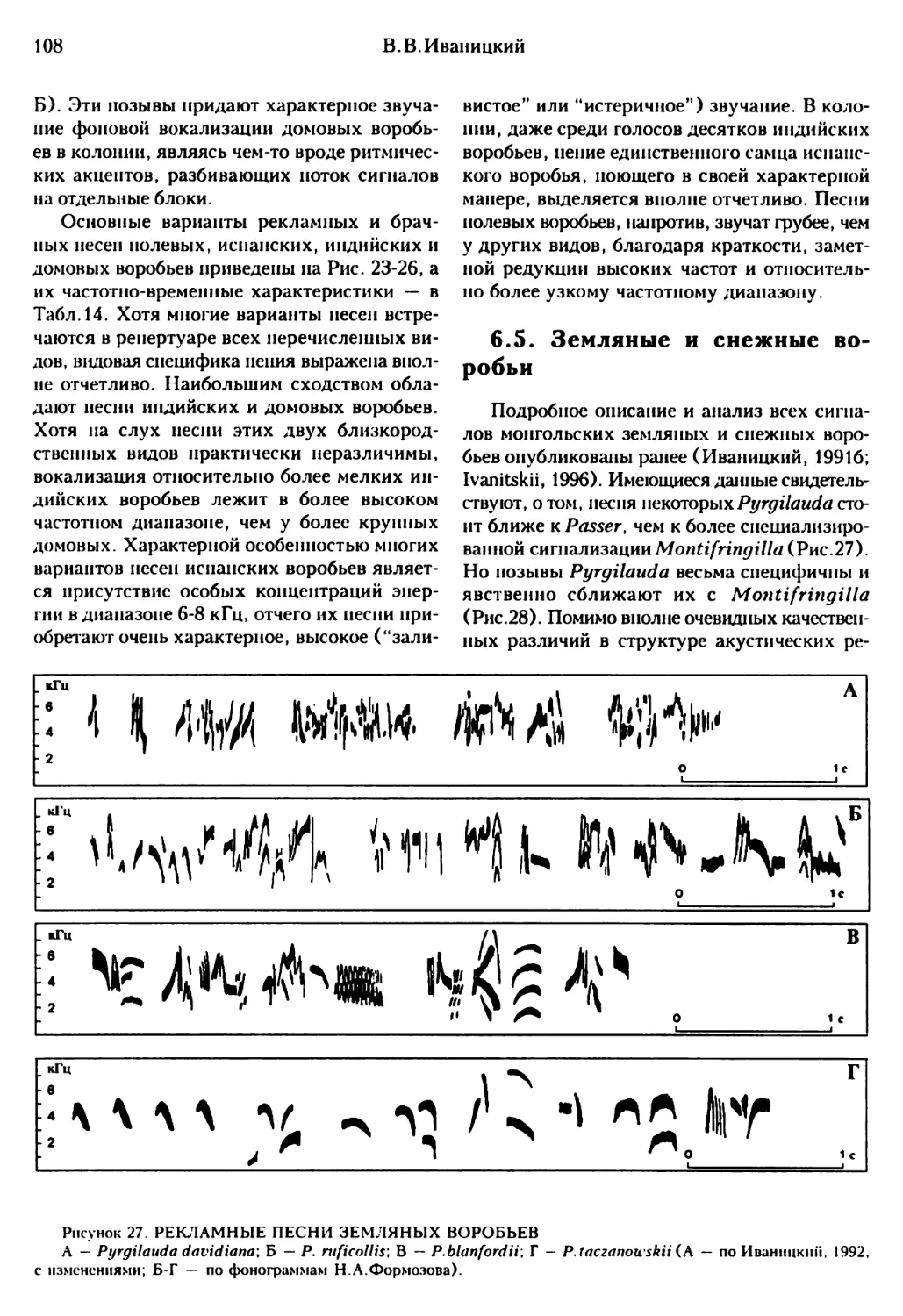
6.5. Земляные и снежные воробьи .......................................................108
6.6. Каменный воробей..................................................................109
6.7. Короткопалый воробей..............................................................111
6.8. Структурно-функциональная дифференциация и эволюция акустических репертуаров......112
ГЛАВА 7. КЛАССИФИКАЦИЯ И ЭВОЛЮЦИЯ......................................................117
7.1. Положение воробьев в системе отряда Passeriformes.................................117
7.2. Таксономическая структура семейства Passeridac....................................118
7.3. Гипотетический сценарий филогенеза Passeridac.....................................128
ЗАКЛЮЧЕНИЕ.............................................................................135
ЛИТЕРАТУРА .............................................................................137
SUMMARY................................................................................145
4
CONTENTS
PREFACE..............................................................................................5
Chapter 1. ENVIRONMENT AND ADAPTIVE TYPES.....................................................8
1.1. Geographic distribution.........................................................................8
1.2. Habitats and position within avian communities..................................................9
1.3. Adaptive types................................................................................. 12
1.4. Main directions of ccogencsis.................................................................. 14
Chapter 2. SPATIAL STRUCTURE........................................................................ 17
2.1. General patterns of spatial relations within the populations.................................. 17
2.2. Advanced formes of coloniality ............................................................... 17
2.2.1. Flocks size, structure, and dynamics........................................................ 17
2.2.2. Flocks social organization ................................................................. 20
2.2.3. Spatial structure and size of the colonics ................................................. 21
2.2.4. Behaviour during the colonics formation..................................................... 22
2.2.5. Territorial behaviour in colonics........................................................... 23
2.3. Quasicoloniality.............................................................................. 24
2.4. Solitary breeding............................................................................. 25
2.5. Evolution of the spatial structure............................................................ 28
Chapter 3. NEST DESIGH AND POSITIONS .............................................................. 36
3.1. Nest position................................................................................. 36
3.2. The sparrows in competition for nest shelters................................................. 40
3.3. Nest-building behaviour....................................................................... 42
3.4. Nest dcsighs in Passcridac and other Ploccimorpha..............................................46
Chapter 4. REPRODUCTIVE BEHAVIOUR.................................................................. 51
4.1. Pair formation................................................................................ 51
4.2. Mating systems................................................................................ 55
4.3. Extra-pair sexual relations................................................................... 59
4.4. Mating behaviour.............................................................................. 61
4.5. Nest-building behaviour....................................................................... 62
4.5.1. Nest-building witnin the annual and reproductive cycles......................................62
4.5.2. Male and female involmcnt in nest-building.................................................. 64
4.6. Incubation.................................................................................... 70
4.7. Nestlings development, parental behaviour, multi pic breeding................................. 73
4.8. Evolution of reproductive behaviour in different groups of granivorous birds.................. 76
Chapter 5. STRUCTURE AND EVULUTION OF MOVEMENTS AND POSTURES....................................... 83
5.1. Flighs diversity and hard substrata locomotion................................................ 83
5.2. Elementary movements and their compositions (postures)........................................ 88
5.3. Evolution of movements systems and locomotion................................................. 96
Chapter 6. STRUCTURE AND EVOLUTION OF ACOUSTIC REPERTOIRES.........................................100
6.1. General patterns of repertoire organization...................................................100
6.2. Desert and saxaul sparrows....................................................................101
6.3. Cinnamon sparrows.............................................................................104
6.4. Field, domestic, Indian, and willow sparrows..................................................105
6.5. Snow finches..................................................................................108
6.6. Rock sparrow..................................................................................109
6.7. Pale rock sparrow.............................................................................111
6.8. Evolution of acoustic repertoires.............................................................112
Chapter 7. CLASSIFICATION AND EVOLUTION............................................................117
7.1. Position of sparrows within the order Passeriformes...........................................117
7,2. Taxonomic structure of family Passcridac......................................................118
7.3. Supposed scenario of Passeridae phylogenesis..................................................128
CONCLUSION.........................................................................................135
LITERATURE CITED........................................A.........................................137
SUMMARY............................................................................................145
ПРЕДИСЛОВИЕ
Воробьи принадлежат к тем немногим
видам пернатых, которые стали нашими по-
стоянными соседями па городских и сельс-
ких улицах. Все события персональной и об-
щественной жизни воробьев день за днем
разворачиваются прямо па наших глазах, по-
рождая естественное желание узнать поболь-
ше о том, как живут эти птицы, откуда ведут
они свой род, как сложилась их история и
что в конце-копцов привело их к людям?
Ответы на эти вопросы мы попытались отыс-
кать на путях сравнительного анализа соци-
альной организации, репродуктивного пове-
дения, механизмов коммуникации и других
сторон образа жизни разных видов воробьев,
а также некоторых генеалогически и эколо-
гически близких к ним групп мелких зерно-
ядных птиц.
Вклад этологии в систематику птиц фун-
даментален и общепризнан. Вопросы клас-
сификации воробьев занимали в нашей ра-
боте важное, но не основное место и скорее
составляли ее прикладной аспект. Главную
задачу мы видели в том, чтобы па примере
воробьев и родственных им групп прояснить
некоторые общие закономерности эволюции
репродуктивного поведения певчих птиц, как
целостной, самоорганизующейся системы. Мы
попытались вскрыть те противоречия и кон-
фликты между отдельными структурными
блоками репродуктивного поведения, которые,
наряду с изменениями среды обитания, со-
ставляют важнейший внутренний источник
эволюционного развития. В соответствии с
традициями филогенетического направления
в сравнительной этологии программа наших
исследований была нацелена на конкретный
таксон и строилась так, чтобы возможно пол-
нее охватить все разнообразие составляющих
его видов и падвидовых группировок. Мы
исходили из того, что поведение теснейшим
образом связано с другими компонентами
образа жизни птиц и полагали полезным
рассмотреть синтетическую картину, включая
в анализ сведения но морфологии, экологии
и зоогеографии.
Для этолога воробьи интересны прежде
всего благодаря удивительному разнообразию
их поведения. У воробьев известны все типы
популяционных структур — от жесткой тер-
риториальности и дисперсного гнездования
до гипертрофированной колопиалыюсти. Учи-
тывая высокое разнообразие ландшафтных
связей воробьев, эту группу заманчиво исполь-
зовать для проверки гипотез, трактующих о
приспособительном значении разных типов
пространственных структур у птиц. Наряду с
этим, этологические исследования воробьев
способны пролить свет на закономерности
организации и функционирования систем
коммуникации в разных местообитаниях и в
разной социальной среде. Демонстративное
поведение и вокализация воробьев отличают-
ся значительным разнообразием, что позволя-
ет использовать коммуникативное поведение
как источник таксономических признаков.
Наконец, особенности таксономической струк-
туры семейства Passeridae, его место в системе
рядом с другими семействами мелких зерно-
ядных птиц, хорошо изученными в этологи-
ческом отношении, открывают возможность ис-
следования относительно высоких уровней
эволюции поведения.
По оценкам разных авторов мировая фа-
уна включает до 33 видов и до 6 родов воро-
бьев (Summers-Smith, 1988; Sibley, Monroe,
1992; Clement, Harris, Davis, 1993). Прежде
полагали, что воробьи близки к ткачихам и
придавали им ранг подсемейства в семействе
Ploceidae. В последнее время воробьев чаще
выделяют в самостоятельное семейство
Passeridae (Bock, Morony, 1978; Summers-Smith,
1988; Степанян, 1990; Ivanitskii, 1995,1996),
оставляя в Ploceidae настоящих ткачиков
Ploceinae, воробьиных ткачиков Plocepasse-
rinae, воловьих птиц Bubalornithinae и вдо-
вушек Viduinae. К более отдаленным род-
ственникам воробьев и ткачиков относят вос-
коклювых (вьюрковых) ткачиков Estrildidae.
Для краткости всех Passeridae, Ploceidae и
Estrildidae мы назовем ткачиконодобпыми
птицами Ploceimorpha.
По основным своим морфо-биологическим
свойствам воробьи — это типичные предста-
вители подотряда певчих птиц Oscines. Стро-
ение клюва и тип питания воробьев указы-
вает на их принадлежность к зерноядным
птицам, способным существовать на диете из
6
В.В.Иваницкий
созревших семян растений. Потребности в
белковой нище возрастают лишь в период
размножения, что побуждает птиц включать
в рацион насекомых, семена на стадии мо-
лочно-восковой спелости, генеративные орга-
ны растений (пыльники, завязи и т.н.). По-
мимо воробьев и большинства других
Ploceimorpha к зерноядным птицам в Старом
Свете относят жаворонков (Alaudidae), вьюр-
ковых (Fringillidae) и овсянок (Emberizidae).
Своеобразие воробьев во многом опреде-
ляется масштабами их адаптивной радиации,
почти не имеющими аналогов среди прочих
групп певчих птиц, сопоставимых с воробья-
ми по числу видов. Одни воробьи ведут пре-
имущественно древесный образ жизни и
строят массивные шаровидные гнезда среди
ветвей, другие обитают в абсолютно безлес-
ной местности — в горах или на равнинах,
где убежищами для гнезд служат щели в ска-
лах или норы мелких млекопитающих. Мно-
гие воробьи обитают в природных зонах с
крайне неблагоприятным климатом: в жар-
ких и сухих пустынях — Сахаре, Каракумах,
Гоби; на холодных и столь же засушливых
нагорьях Центральной Азии — Памире и Ти-
бете. В этих суровых местах птиц очень мало,
поэтому воробьи сплошь и рядом оказыва-
ются здесь очень характерными участника-
ми местных орнитокомплексов и вносят су-
щественный вклад в их совокупную числен-
ность и видовое разнообразие.
Глобальное антропогенное преобразование
природы глубоко повлияло на образ жизни
н эволюционную судьбу воробьев. На приме-
ре многих видов воробьев в разных регионах
Старого Света можно проследить все стадии
процессов синантропизации, начиная от фа-
культативных связей с разрозненными ант-
ропогенными элементами на стадиях первич-
ного изменения ландшафтов и кончая спе-
циализированным комменсализмом в искус-
ственных ландшафтах, отмеченных предель-
ными формами урбанизации. Это сообщает
семейству воробьев свойства упикалыю^эко-
логической и эволюционной модели. В ант-
ропогенных ландшафтах воробьи также вхо-
дят в состав крайне обедненных ориктоцено-
зов, но зато сами достигают колоссальной
численности и во многих регионах приоб-
ретают важное экономическое значение, при-
чиняя серьезный ущерб плодовым садам и по-
севам зерновых культур или же способствуя
распространению эктопаразитов и гельминтов
(Штегман, 1956а,б; Гаврилов, 1962; Ильенко,
1976; Костюнин, 1977; Кашкаров, Зинченко,
1986). Но практическая потребность изучения
воробьев диктуется не только важнейшим хо-
зяйственным значением массовых видов-ком-
менсалов, но и неблагополучным положением
редких видов. Многие воробьи имеют релик-
товые ареалы и крайне малочисленны. Пус-
тыш1ый, рыжий и монгольский воробьи вклю-
чены в Красные Книги СССР, России и со-
предельных государств. Мы считаем, что в обо-
их случаях незаменимы исследования поведе-
ния и экологии воробьев, ориентировашше
прежде всего на выявление механизмов орга-
низации их популяционных систем, способов
использования пространства, механизмов ком-
муникации и закономерностей динамики по-
пуляций, выполненные на основе тех теорети-
ческих принципов, приемов и методов, кото-
рые составляют суть популяционного подхо-
да к контролю численности животных (Ши-
лова, 1993).
Полные таксономические ревизии Passeridae
нам не известны, но вопросы классификации
и номенклатуры отдельных родов в литерату-
ре освещались неоднократно (Бианки, 1907;
Мауг, 1927; Stegman, 1932; Козлова, 1955; Abs,
1966; Нейфельдт, 1986; Summers-Smith, 1988).
Рассматривались также таксономические и эво-
люционные взаимоотношения воробьев с сест-
ринскими группами (Sushkin, 1927; Bock,
Могопу, 1978; Булатова, 1981; Sibley, Monroe,
1992). Мы полагаем, что усовершенствование
классификации воробьев и попытки реконст-
рукции их истории, наряду со всесторонним
учетом морфологических и биохимических
данных, должны базироваться и на комплекс-
ных исследованиях поведения и экологии пред-
ставителей возможно большего числа родов и
видов. В России, Казахстане, Средней Азии и
Закавказье встречаются 12 и гнездятся И ви-
дов воробьев, представляющие 5 родов: домо-
вый воробей Passer doniesticus (Linn., 1758);
индийский воробей Passer indicus(Jard. et Selby,
1831); испанский воробей Passer hispaniolensis
(Temm., 1820); саксаульный воробей Passer
aminodendri (Gould, 1872); рыжий воробей
Passer rutilans (Temm., 1820); пустынный во-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
7
робей Passer simplex (Licht., 1823); каменный
воробей Petronia petronia (Linn., 1766); мон-
гольский земляной воробей Pyrgilauda
davidiana (Verr., 1871); афганский земляной
воробей Pyrgilauda theresae (Mein., 1937)
(встречается только на зимовках); снежный
воробейMontifringilia nivalis (Linn., 1766); ко-
роткопалый воробей Carpospiza brachydactyla
(Bonap., 1850). Это треть мирового списка ви-
дов и почти все роды — за исключением кус-
тарниковых воробьев p.Gymnorhis, часто вмес-
те с P.petronia объединяемых в p.Petronia s.
lato. Мы полагали, что доступное нам разнооб-
разие воробьев вкупе с литературными дан-
ными может служить достаточным основани-
ем для суждений об эволюции, филогении и
классификации избранной группы.
Наши нолевые исследования охватили все
указанные виды воробьев (за исключением
афганского) и проходили в 1980-1995 гг. на
Украине, в Закавказье, Туркмении, Узбекис-
тане, Таджикистане, Казахстане, юго-восточ-
ном Алтае, юго-занадной Туве, в южных рай-
онах Сахалина, на Сейшельских островах и
на островах восточного шельфа Вьетнама.
Многолетние исследования динамики попу-
ляций домовых и полевых воробьев с при-
менением массового кольцевания и индиви-
дуального мечения проводили в Москве и
Подмосковье. По приблизительным подсчетам
мы провели в ноле около 20 месяцев и на-
блюдали за птицами в течение 2015 часов.
Численность и распределение воробьев, а так-
же сопутствующих им видов певчих птиц,
изучали по стандартной методике маршрут-
ных учетов с полосой регистрации от 50 до
100 м. Всего с учетами пройдено 390 км. Ал-
люмипиевыми и пластмассовыми кольцами
помечено 2780 воробья 8 видов. Из них по-
вторно встречено 719 особей. Голоса воробьев
записывали на магнитофоны “ Sanyo-М 101 А”
и “Веспа-207”. Общая нродолжителыюстышо-
лучениых фонограмм составила 860 мин. Сиг-
налы обрабатывали на соиографе Kay Electric
Со с применением узких и широких фильт-
ров. Всего получено 860 сонограмм с изобра-
жением 2034 песен и позывов.
Мы признательны руководству и коллек-
тиву кафедры зоологии позвоночных живот-
ных и общей экологии Биологического фа-
культета МГУ за те возможности, которые
были предоставлены для осуществления па-
шей программы, постоянную атмосферу доб-
рожелательной заинтересованности в ее ре-
зультатах. Многолетние исследования популя-
ционной биологии домовых и полевых воро-
бьев 6wj#i бы невозможны без активного, твор-
ческого участия А.В.Матюхина. С.А.Букреев
передал нам тотальный препарат короткопа-
лого воробья, а Л.П.Корзун по нашей просьбе
выполнил препаровку подъязычного аппара-
та этой птицы. Е.Н.Панов и Н.А.Формозов
любезно предоставили нам фонограммы не-
которых видов воробьев. При содействии
Н. А.Формозова ценные материалы получены
из фонотеки Британского музея естественной
истории. Работа на соиографе была продук-
тивной и приятной благодаря помощи
Н.К.Лысиковой и А.Б.Васильевой. И.М.Ма-
рова и Е.В.Шевченко делили с нами труд-
ности экспедиционной жизни, внесли свою
ленту в сбор материалов и помогли переве-
сти ряд статей с немецкого. Р.Л.Беме, В.М.О-
стапепко, Е.Н.Панов, А.С.Северцов, Г.Н.Сим-
кин, Л.С.Степанян и В.Е.Флинт взяли па
себя труд прочесть рукопись и высказали
критические замечания. Среди зарубежных
коллег, присылавших нам литературу и свои
комментарии, хотелось бы отметить доктора
D.Summers-Smith из Великобритании, про-
фессора университета в Йоханнесбурге
(ЮАР) D.Craig, профессора университета
Нью-Джерси (США) N.Collias, руководите-
ля Международной группы по зерноядным
птицам, профессора Варшавского универси-
тета J.Pinowski (Польша), профессора Ту-
ринского университета (Италия) D.Mala-
came. Всем коллегам мы выражаем свою глу-
бокую и сердечную благодарность. Автор
также бесконечно признателен Е.Г. Иваниц-
кой за неизменное понимание, терпение и
жизненную стойкость, позволившие ему со-
средоточиться па полевых исследованиях и
работе над рукописью.
ГЛАВА 1. СРЕДА ОБИТАНИЯ И АДАПТИВНЫЕ ТИПЫ
1.1. Область распространения
Благодаря широкой интродукции, домовых
воробьев ныне можно встретить на всех кон-
тинентах, кроме Антарктиды (см.Судиловс-
кая, 1952; Summers-Smith, 1988). Исконный же
ареал сем.Passeridae принадлежит исключи-
тельно Старому Свету. Но и здесь, лишь аре-
алы синантропных домовых и нолевых во-
робьев простираются выше 50 градуса с.ш.,
все прочие виды живут южнее. Максималь-
ного разнообразия воробьи достигают в Азии,
где представлены все роды: Pyrgilauda и
Carpospiza эндемичны, Petronia и Monti-
fringilla проникают в Европу, a Petronia — в
северную Африку. Passer и Gymnorhis рас-
пространены и в Азии, и в Африке, а общее
число всех видов воробьев на этих континен-
тах примерно одинаково (Табл.1). Однако,
среди африканских эндемиков из р.Passer
немало явно очень близких алло- или нара-
натрнчпых форм со спорным таксономичес-
ким статусом (комплекс “сероголовых”:
P.diffusus — P.griseus — et al. и “рыжес-
пинных”: P.motitensis — P.iagoensis — etal.
воробьев). Аргументы Summers-Smith (1988)
в пользу видовой самостоятельности боль-
шинства форм из указанных комплексов, по
природе своей более всего тяготеющих к ка-
тегории надвида, не всегда убедительны (Ива-
ницкий, 1989). Кроме названных надвидовых
групп в Эфиопской области обитают всего 4
бесспорных вида {luteus, eminibey, melanurus
и castanopterus). В противоположность это-
му, все азиатские эндемики Passer представ-
лены хорошо дифференцированными вида-
ми {aminodendri, flaveolus, montanus, pyrrho-
notus, rutilans, moabiticus). Есть основания
считать самостоятельными вид<&1и формы
indicus (Степанян, 1990) и italiae (Burhard,
1989; Massa, 1989). Ареалы еще двух пред-
ставителей рода {domesticus и hispaniolensis)
почти целиком лежат в Евразии, лишь незна-
чительно распространяясь в северные райо-
ны Африки.
В сущности, из 6 родов воробьев лишь
Gymnorhis бесспорно богаче представлен в
Африке, где обитают 3 нарапатричных вида
{superciliaris, dentatau pyrgita), чем в Азии,
где гнездится только xanthocollis. Основная
зона эндемизма и видового разнообразия во-
робьев приурочена к Центральной Азии. Здесь
много симпатричных и частично симбиото-
ничпых видов: так, в окрестностях оз.Куку-
Нор в юго-восточном Тибете живут бок о бок
4 вида Pyrgilauda и 2 Мontifringilla (Gebauer,
Kaiser, 1994). На окраине ареала семейства в
Юго-Восточной Азии живут 5 видов Passer, 3
из которых эндемичны {rutilans, pyrrhonotus,
flaveolus), тогда как на другой окраине в
южной Африке обитают 3 вида и только один
эндемик {melanurus). Таким образом, вопре-
ки версии об африканском происхождении
воробьев (Summers-Smith, 1988), зоогеографи-
ческие данные свидетельствуют в пользу древ-
них исторических связей воробьев с Азиатс-
ким континентом, где в настоящее время со-
средоточена самая богатая фауна этих птиц,
отмеченная .высоким уровнем эндемизма и, как
мы покажем далее, представленная здесь боль-
шим разнообразием адаптивных типов (жиз-
ненных форм), заселивших гораздо более
широкий спектр местообитаний, чем в Афри-
ке (см.Иваницкий, 1989).
Сравнивая распространение воробьев и
родственных им групп, отметим, что вопреки
ограниченному объему сем.Passeridae, его
ареал сопоставим но площади с ареалом весь-
Таблица 1. РАСПРОСТРАНЕНИЕ ВОРОБЬЕВ В АФРИКЕ И ЕВРАЗИИ
регион всего видов эндемичных видов всего родов эндемичных родов адаптивных типов
Евразия 19 15 6 3 8
Африка 16 12 3 - 7
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
9
ма многочисленного семейства Estrildidae
(107-115 видов) и значительно превосходит
область распространения другого, столь же
многочисленного семейства Ploceidae (97-103
вида). При этом видовые ареалы настоящих
и воскоклювых ткачиков за очень немноги-
ми исключениями не выходят за пределы
тропического пояса, тогда как воробьи, буду-
чи широко представленными в тропических
регионах, наряду с этим полностью освоили
субтропики и зону умеренного климата. По-
давляющее большинство Ploceidae приуроче-
ны к Эфиопской области, где они почти по-
всеместно принадлежат к числу ландшафт-
ных птиц. Лишь 5 представителей наиболее
многочисленного p.Ploceus (в Африке обита-
ет не менее 40 его представителей) гнездятся
в Индии и Юго-Восточной Азии, где во мно-
гих местах также достигают очень высокой
численности. Вероятно, прежде ткачики были
распространены значительно шире, чем теперь.
Что же касается Passeridae и Estrildidae, то
оба семейства имеют значительно более об-
ширные, сплошные ареалы в Африке, Южной
и Юго-Восточной Азии. Лишь воскоклювые
ткачики заселили Новую Гвинею, в Австра-
лию и Океанию, и только воробьи проник-
ли в северную Палеарктику. Отметим, что в
обширном ареала Estrildidae, в отличие от
других сестринских групп, имеется целых три
удаленных друг от друга очага эндемизма ро-
дового уровня: тропическая Африка, Юго-Во-
сточная Азия и Австралия (Goodwin, 1982),
что, как нам кажется, позволяет с биогеогра-
фической точки зрения рассматривать вос-
коклювых ткачиков в качестве наиболее древ-
него ствола ткачикоподобных птиц.
1.2. Места обитания и положение
в ориктоценозах
В своем распространении воробьи яв-
ственно тяготеют к аридным ландшафтам.
Судя но распространению биоклиматических
зон Африки и Евразии (Бабаев и др,, 1986),
они населяют экстраарндные районы (Саха-
ра, пустыни Аравии, Средней и Центральной
Азии, Тибетское нагорье), широко представ-
лены в аридных, семиаридных и субаридных
зонах, но лишь немногие виды проникают в
районы с более высокой влажностью. Тем са-
мым, воробьи должны быть причислены к
характерным обитателям пустынь, полупус-
тынь и саванн Старого Света, непрерывной
полосой протянувшихся от Калахари до Гоби.
Большинство воробьев живут там, где годо-
вые нормы осадков не превышают 200 мл и
преобладают весьма суровые, часто экстре-
мальные условия существования. В местах
обитания воробьев зарегистрирован абсолют-
ный минимум годовых осадков (10-15 мл),
абсолютный максимум температуры воздуха
(+59°С) и нагревание поверхности почвы до
80°С. Относительная влажность воздуха ле-
том опускается до 3%, характерны резкие су-
точные и сезонные колебания температуры
воздуха, которая зимой в материковых пус-
тынях надает иногда до -40°С.
Воробьи по преимуществу оседлы и в
полной мере испытывают на себе все преврат-
ности резко континентального климата. Так,
саксаульные и пустынные воробьи в Кара-
кумах размножаются в мае-июле, когда днев-
ные температуры редко опускаются ниже
+40°С. В конце сентября в Судане наблюда-
ли, как пустынные воробьи кормили птен-
цов при дневных температурах до +50°С, ког-
да отмечали массовую гибель мелких воро-
бьиных птиц — палеарктических мигрантов
(найдено 19 видов) (Ash, Nikolaus, 1991). В
противоположность этому, земляные, снежные
и каменные воробьи на холодных нагорьях
Центральной Азии даже летом подвержены
очень низким температурам, эпизодическим
снегопадам, непрерывным сильным и холод-
ным ветрам (Сушкин, 1938; Иваницкий, Шев-
ченко, 1991). Зимой же здесь, особенно в меж-
горных котловинах, стоит чрезвычайно холод-
ная погода; например, среднеянварские тем-
пературы в Чуйской степи па Алтае состав-
ляют ~32°С при абсолютном минимуме -62°С
(Ревякин, 1987).
Вполне естественно, что среда обитания
многих воробьев отличается не только клима-
тической агрессивностью, но и биоценотичес-
кой бедностью. Как показывают результаты
проведенных нами маршрутных учетов, мест-
ные сообщества певчих птиц имеют низкое
видовое разнообразие и численность (Табл.2).
Примечательно, что и в антропогенных ланд-
шафтах (урбоценозах, агроцеиозах и пр.) био-
тическая среда воробьев также крайне бедна
10
В.В.Иваницкий
Таблица 2. УЧАСТИЕ ВОРОБЬЕВ В ФАУНЕ И НАСЕЛЕНИИ ПЕВЧИХ ПТИЦ
Регион Ландшафт Гнездится видов певчих птиц Числен- ность пев- чих птиц экз/100 га Гнез- дится видов воробьев Численность воробьев экз/ 100 га Доля участия воробьев в%%
Южная Украина Овраги в меловых отложениях 7 162,5 1 67,6 41,6
Нахичевань Каменистые ушелья в предгорьях 15 97,2 2 15,0 15,4
Бадхыз Каменистые ушелья в предгорьях 16 137,0 3 55,5 40,5
Репетек Незакреп- ленные пес- ки 6 40,3 2 14,4 36,6
Репстек Черносакса- ульники 9 91,3 1 15,3 16,8
Ю.-В. Ал- тай Долины гор- ных рек 13 150,9 3 64,4 42,7
Ю.-З. Тува Остепненные плато 4 49,6 1 16,8 33,9
Ю.-З. Тува Пойменный леса 13 110,7 1 9,3 8,4
Южный Сахалин Пойменные леса 17 129,1 1 22,0 17,0
В среднем для всех местообитаний П,1 107,6 1,6 31,1 28,9
и часто вообще не включает других мелких
певчих птиц. Таким образом, воробьи повсе-
местно ведут себя, как типичные “цепофобы”,
избегающие условий, чреватых возникновени-
ем острой межвидовой конкуренции.
Воробьи тесно связаны с интразональпы-
ми местообитаниями, в то время как зональ-
ные ландшафты заселены этими птицами в
гораздо меньшей степени. Имеются региональ-
ные отличия. В тропической Африке преоб-
ладающим местом гнездования всех воробь-
ев (группаPasser motitensis-P.iagoensis, груп-
па P.griseus-P.diffusus, Р. eminibey, предста-
вители p.Gymnorhis) являются разнообраз-
ные тины зональных саванн на обширных
плоских водоразделах, причем воробьи пред-
почитают наиболее ксерофитпые варианты
пизкотравных саванн с очень разреженной и
бедной древесно-кустарниковой растительно-
стью (акациевые саванны) (Bannerman, 1948;
McLachlam, Liversidge, 1957; Archer, Goodman,
1961; Betts, 1966, Summers-Smith, 1988 ). 3a
пределами Африки, зональные ландшафты за-
селяются воробьями в песчаных пустынях
Средней и Центральной Азии: узко специа-
лизированный пустынный воробей гнездится
в незакрепленных, круппобугристых песках,
в то время как саксаульный воробей распро-
странен значительно шире и, будучи дуплог-
нездником, связан в основном с насаждения-
ми круппостволыюго белого саксаула и ту-
рапги на закрепленных песках. Наконец, пос-
ледний тин заселяемых воробьями ландшаф-
тов зонального типа представлен высокогор-
ными степями и полупустынями Централь-
ной Азии, где обитают глубоко специализиро-
ванные к жизни в открытых, безлесных био-
топах земляные воробьи рода Pyrgilauda.
Прочие виды воробьев гнездятся в интра-
зональных ландшафтах, среди которых вы-
деляются следующие основные тины:
1) Пойменные ландшафты с древесно-ку-
старниковой растительностью тугайного обли-
ка (галерейные леса). Эти ландшафты в той
или иной степени используют для гнездова-
ния 8 из 9 евроазиатских представителей
р.Passer. Единственное исключение — пус-
тынный воробей, однако африканская форма
вида (P.s.simplex) гнездится в характерном
для Сахары долинном ландшафте сухих ру-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
И
сел (вади), где растительность, питаемая под-
земными стоками, развивается лучше, чем па
безжизненных плакорах (Balzac de, 1929).
Только в тугайных лесах гнездятся Passer
moabiticus и P.pyrrhonotus, это преобладаю-
щее место обитания рыжих воробьев и очень
характерный гнездовой биотоп полевых, ин-
дийских и испанских воробьев, при том, что
в ряде мест его охотно используют домовые
и саксаульные воробьи, а также обитающий
в Юго-Восточной Азии P.flaveolus (Summers-
Smith, 1988).
2) Ландшафты с хорошо выраженными
элементами вертикального расчленения, сло-
жившиеся в результате разрушения интру-
зивных или осадочных толщ, при наличии
обрывистых обнажений и нагромождений
обломочного материала. К этим ландшафтам
приурочены прежде всего те виды воробьев,
которые нуждаются в закрытых убежищах
для гнездования. Это характерные гнездовые
стации трех видов p.Passer (indicus, mohtanus,
iagoensis), хотя здесь могут гнездится и дру-
гие виды, например domesticus и hispanio-
lensis. Наконец, это типичные гнездовые ста-
ции камешкых и снежных воробьев.
3) Антропогенные ландшафты, заселяемые
воробьями, отличаются огромным разнообра-
зием и образуют переход от первичных ста-
дий антропогенной трансформации естествен-
ных ландшафтов — через агроцеиозы — к
предельно урбанизированным территориям и
мегаполисам, где во множестве гнездятся до-
мовые и полевые воробьи. Среди И видов
воробьев Северной Палеарктики лишь пус-
тынный и короткопалый воробьи не связа-
ны с антропогенными ландшафтами, прочие
виды па определенных стадиях годового цикла
эпизодически или регулярно используют этот
тин ландшафтов (Иваницкий, 19846). По
Summers-Smith (1988), среди 20 рецептных
представителей р.Passer по крайней мере И
видов регулярно контактируют с антропоген-
ными ландшафтами.
Следует особо отметить явное тяготение
воробьев к речным долинам, где часто пред-
ставлены все три перечисленные типа интра-
зональных ландшафтов. Например, в Средней
Азии индийские, испанские и нолевые воро-
бьи особенно многочисленны в районах оазис-
ного земледелия в долинах Аму-Дарьи, Сыр-
Дарьи, Теджена, Мургаба и Атрека, с их не-
когда богатыми тугайными лесами и много-
числшшыми лессовыми обрывами. Индийс-
кие и испанские воробьи связаны прежде
всего с агроценозами, полевые воробьи гнез-
дятся в населенных пунктах. Рост численнос-
ти и широкая экспансия этих видов в после-
дние десятилетия обусловлены повсеместным
насаждением лесополос и садов, сменивших
сведенные тугаи, развитием зернового хозяй-
ства и ирригационных систем, что в общей
сложности создает идеальную среду обитания
для воробьев.
В Центральной Азии широкие, остепнеп-
пые долины ручьев в субальпийском поясе
населяют снежные, каменные и монгольские
земляные воробьи, а на расположенных здесь
кошарах и пастушьих домиках нередко гнез-
дятся домовые и полевые воробьи. Здесь легко
наблюдать и первые этапы синантронизации
у вышеупомянутых каменных, снежных и
монгольских земляных воробьев. Каменные
и снежные воробьи время от времени сами
гнездятся в щелях этих построек, сложенных
обычно из лиственничных бревен. Пастушьи
стоянки привлекают птиц обилием дефицит-
ных материалов для постройки гнезд, остат-
ками просыпанного овса и комбикормов, на-
личием минеральной подкормки. Особенно
важно присутствие корма у стоянок в зимнее
время, когда все горные воробьи, а с ними и
некоторые другие зерноядные птицы, кормят-
ся преимущественно или даже исключитель-
но возле стационарных пастушьих стоянок
или же в небольших поселках (Иваницкий,
19846; Зонов, Попов, 1987; Иваницкий, Шев-
ченко, 1992). Особенно подвержены сипапт-
ропизации снежные воробьи: они во множе-
стве гнездятся под крышами домов в посел-
ках, расположенных в соответствующих лан-
дшафтах, и в сущности их образ жизни здесь
ничем не отличается от образа жизни таких
классических синурбапистов, как домовые и
полевые воробьи (Тарасов, 1960; Степанян,
1974; наши наблюдения).
Помимо явного тяготения воробьев к до-
линным ландшафтам, которое, возможно, по-
служило одной из предпосылок их синаптро-
пизации, столь же характерно для воробьев по-
чти полное отсутствие лесных видов. Это тем
более удивительно, что многие представители
12
В. В. Иваницкий
рр.Passer и Gymnorhis в экологическом пла-
йе тесно связаны с древесно-кустарниковой
растительностью и в сущности являются пре-
имущественно древесными птицами.
Среди изученных нами видов в наиболь-
шей степени древесный образ жизни харак-
терен для рыжих воробьев, населяющих иво-
во-ольховые леса в поймах рек южной части
Сахалина и на Курильских островах. Мы
обнаружили, что рыжие воробьи ведут себя
как почти исключительно древесные птицы,
все формы жизнедеятельности которых со-
средоточены в кронах деревьев, причем зача-
стую па довольно большой высоте. На земле
этих воробьев нам почти вовсе видеть не
приходилось, хотя по данным других авто-
ров они кормятся на земле регулярно (Не-
чаев, 1973; Summers-Smith, 1988). Весь корм
птицы добывали в кронах, с большой ловко-
стью передвигаясь по самым топким ветвям,
легко подвешиваясь к ним в самых разнооб-
разных позах (вниз спиной, вниз головой и
т.д.). Тем не менее, хотя рыжие воробьи пре-
красно приспособлены к древесному образу
жизни, их нельзя причислить к лесным пти-
цам. Гнездовые участки пар и холостых сам-
цов тяготели к опушкам — не далее 50 м от
примыкающих нолей в тех случаях, когда
воробьи жили в высокоствольном лесу с сом-
кнутым пологом. По больше части рыжие во-
робьи гнездились вдоль самой окраины пой-
менного леса, в насаждениях паркового тина
и па отдельно стоящих деревьях.
Это довольно типичная картина для мно-
гих представителей p.Posser, которые, будучи
в трофическом отношении тесно связанными
с кронами деревьев и кустарников, тем не
менее отчетливо предпочитают разреженные,
светлые насаждения паркового или саванно-
вого тина или же самые окраины более гус-
тых насаждений, приближающихся по своему
облику к настоящему лесу.
1.3. Адаптивные типы
Мы предложили различать адаптивные
типы воробьев в зависимости от способа
распределения наиболее важных проявле-
ний их жизнедеятельности по основным
структурным элементам заселяемой ими ме-
стности (Иваницкий, 1991а; Ivanitskii, 1996).
Спектр адаптивных типов воробьев изоб-
ражен па Рис.1. Он построен с учетом 5 ос-
новных проявлений жизнедеятельности во-
робьев и 7 главных субстратов и убежищ
и включает, таким образом, 35 возможных
позиций, из которых каждый вид в зависи-
мости от широты своей экологической спе-
циализации занимает от 5 до 11. На одном
полюсе спектра сосредоточены преимуще-
ственно древесные формы, большая часть
активности которых протекает в кронах
деревьев или в их дуплах, на противопо-
ложном полюсе располагаются формы вов-
се не связанные с древесной растительнос-
тью и хорошо приспособленные к обитанию
в совершенно безлесных ландшафтах. Все-
го мы выделяем 8 адаптивных типов воро-
бьев, каждый из которых представляет со-
бой вполне универсальную модель исполь-
зования среды обитания.
При небольшом объеме семейства, разно-
образие адаптивных типов воробьев очень
велико. Среди них есть типичные дендрофи-
лы (адаптивные типы 1-4), дендро-нетрофи-
лы (5-6), типичные петрофилы (7) и геофи-
лы-норпики (8). Табл.З показывает распре-
деление адаптивных типов но родам воробь-
ев. Дендрофилы представлены лишь у Passer
и Gymnorhis, где к соответствующим адап-
тивным типам относится большинство видов.
Лишь некоторые представители Passer про-
являют определенную степень петрофильпо-
сти, которая наиболее ярко выражена у поле-
вых, домовых, индийских и, вероятно, близких
к последним двум — P.iagoensis, населяющих
аридные, скалистые побережья островов Зе-
леного Мыса (Summers-Smith, 1988). Опти-
мальные условия для гнездования все пере-
численные виды находят в тех местах, где
хорошо развитая древесно-кустарниковая ра-
стительность соседствует с элементами вер-
тикального расчленения и нагромождения-
ми обломочного материала, хотя в некоторых
случаях способны обходиться и без деревьев,
образуя как бы экологический переход к спе-
циализированным петрофилам из родов
Petronia и Montifring ilia, которые в процес-
се дальнейшей эволюции могли стать осно-
вой для формирования еще более продвину-
того адаптивного типа геофилов-норпиков из
р. Pyrgilauda.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
13
Рисунок 1. СПЕКТР АДАПТИВНЫХ ТИПОВ ВОРОБЬЕВ
1-8 — адаптивные типы; А — кроны деревьев; Б — скалы; В — воздух; Г — дупла; Д — поверхность почвы;
Е — полости в скалах; Ж — норы (по Ivanitskii, 1996 с изменениями).
Сравним направления адаптивной радиа-
ции воробьев и родственных им групп. Мно-
гочисленные Ploceidae и Estrildidae освоили
широкий набор местообитаний, по все они в
обязательном порядке связаны с древесно-ку-
старниковой растительностью и по нашей
классификации относятся к адаптивным ти-
пам 1 и 2. Лишь немногие амадины способны
гнездиться в закрытых убежищах (тип 3), а
один монотипичпый род африканских аст-
рильдов (Ortygospiza) приближается к адап-
тивному тину геофилов (тин 8). Судя но все-
му, адаптивная радиация настоящих и вос-
коклювых ткачиков протекала в основном
путем накопления относительно мелких меж-
видовых отличий в технике гнездостроения
и фуражировки на разнообразных раститель-
ных субстратах одновременно с прогресси-
рующим уменьшением размеров тела и все
более глубоким приспособлением к зериояд-
пости.
Многие ткачики целиком перешли к жиз-
ни среди травянистой растительности, пред-
почитая заросли относительно высокорослых,
жесткостебельных злаков или околоводных
осок, рогозов, тростников, нередко стоящих
прямо в воде. При этом сложился своеобраз-
ный адаптивный тин, ярко воплощенный в
представителях рода огненных ткачей Еир-
lectes, которые но свидетельству Алфреда
Брэма (цит. по Морозов, Остапенко, 1988) по
своим повадкам более всего напоминают ка-
мышевок p.Acrocephalus. Строго говоря, этот
адаптивный тип не имеет аналогов среди во-
робьев. Хотя испанские воробьи нередко гнез-
дятся в тростниковых зарослях, за кормом они
14
В.В.Иваницкий
Таблица 3. РАСПРЕДЕЛЕНИЕ АДАПТИВНЫХ ТИПОВ ПО РОДАМ ВОРОБЬЕВ
(по Ivanitskii, 1996)
Число видов рода, соответствующих адаптивным типам 1-8
Роды 1 2 3 4 5 6 7 8 Объём рода (видов)
Passer 2 5 1 4 8 - - - 20
Gymnorhis - - - 4 - - - - 4
Petronia - - - - - 1 - - 1
Pyrgilauda - - - - - - - 5 5
Montifringilla - - - - - - 2 - 2
Carpospiza - 1 - - - - - - 1
Всего 2 6 1 8 8 1 2 5 33
всегда улетают в другие места, нередко по-
крывая при этом большие расстояния, а гнез-
да укрепляют преимущественно на кустах
тамариска, тростниковых заломах или в стен-
ках гнезд цапель, бакланов и других круп-
ных птиц (Казаков, Ломадзе, 1984; Головано-
ва, 1985).
Другая важная черта экогенеза настоящих
и воскоклювых ткачиков состоит в том, что
они сумели утвердиться в лесных экосисте-
мах тропической биоты. Многих представи-
телей сем.Ploceidae и Estrildidae можно на-
звать настоящими лесными птицами, а тка-
чики p.M a limbus (10 видов) известны, как
специализированные обитатели верхнего яру-
са дождевых тропических лесов западной
Африки (Brosset, 1978).
1.4. Главные направления эко-
генеза
Таким образом, на всем протяжении эво-
люции ткачиков главной ареной их экогенеза
оставались ландшафты с хорошо развитой
древесно-кустарниковой растительностью или
же богатым высокотравьем. Возможность по-
стоянного использования объемных и плот-
ных растительных субстратов — древесных
крон, зарослей кустарников или высокотра-
вья, составляет одно из главных требований,
предъявляемых к среде обитания практичес-
ки всеми представителями Ploceidae и
Estrildidae. Что же касается экогенеза воробь-
ев, то он, как мы полагаем, проходил в целом
под знаком редукции их экологических и по-
веденческих связей с растительными субстра-
тами за счет приспособления ко все более пол-
ному использованию иных компонентов сре-
ды обитания и прежде всего разнообразных
скальных субстратов и поверхности почвы,
что открывало перед воробьями возможнос-
ти колонизации открытых, безлесных ланд-
шафтов. Это повлекло за собой глубокие пре-
образования внешнего облика, образа жизни
и поведения воробьев и в конечном итоге
привело к формированию широкого спектра
адаптивных типов, где легко отыскать приме-
ры конвергенции с представителями тех групп
зернвядных певчих птиц, которые по общему
мнению генеалогически удалены от воробьев
значительно дальше ткачиков.
Среди рецептных Passeridae мопотннич-
пый род Petronia благодаря своей удивитель-
ной экологической пластичности может слу-
жить наиболее полной иллюстрацией экоге-
неза всего семейства. На большей части сво-
его ареала каменные воробьи — настоящие
горные птицы. Будучи типичными нетрофн-
лами, они гнездится в глубоких ущельях с
обывистыми стенами и осыпями, не избе-
гая при этом и участков, покрытых доволь-
но густой древесной растительностью, кото-
рую охотно используют для отдыха и токо-
вания, а иногда и гнездятся в подходящих
дуплах и изредка даже открытым способом,
располагая гнезда среди густого переплете-
ния ветвей (Шульпин, 1956; Бельская, 1973).
Подобная обстановка составляет резкий кон-
траст с условиями обитания каменных во-
робьев в степях Северного Прикаспия, где
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
15
эти птицы живут среди совершенно плос-
кого, равнинного ландшафта, используя для
гнездования небольшие промоины и глав-
ным образом — разнообразные постройки
из саманного кирпича (Дубровский, 1961;
Гаврилов и др., 1968). В аналогичных усло-
виях нередко гнездятся каменные воробьи в
пустыне Гоби (Михайлов, 1992). Таким об-
разом, в некоторых частях своего ареала ка-
менные воробьи могут быть названы настоя-
щими степными и даже пустынными пти-
цами, распространение которых в открытых
ландшафтах, однако, жестко ограничивается
наличием подходящих гнездовых субстратов.
Более глубокое проникновение воробьев,
гнездящихся только в закрытых убежищах, в
стенные ландшафты стала возможным лишь
благодаря появлению совершешю нового адап-
тивного типа пориика, получившего широкое
распространение в р.Pyrgilauda. Осваивая рав-
нинные, степные ландшафты Азиатского кон-
тинента, воробьи приобрели отчетливые чер-
ты конвергентного сходства с жаворонками
(сем. Alaudidae). Отдельные этаны этого про-
цесса хорошо иллюстрируют представители
рода земляных воробьев {Pyrgilauda), принад-
лежащие к адаптивному тину 8. По способам
наземной локомоции, высокому разнообразию
токовых полетов, манере нения земляные во-
робьи также больше напоминают жаворонков,
хорошо отличаясь в этом отношении от боль-
шинства своих сородичей. В то же время, при-
спосабливаясь к жизни в степных ландшаф-
тах, земляные воробьи сохранили связи с
закрытыми убежищами для гнездования, что
обусловило их жесткую зависимость от рас-
пространения мелких млеконитающих-норни-
ков. Этим воробьи отличаются от жаворонков,
сооружающих открытые гнезда прямо па зем-
ле под кочкой, в куртине травы, в ямках, не-
редко выкапываемых специально для этой
цели самими птицами (Корелов, 1970). Зем-
ляные воробьи, также как и снежные, полнос-
тью расстались с растительными субстратами
и даже при наличии возможности никогда
не присаживаются на кусты или деревья, на-
поминая в этом отношении часто соседствую-
щих с ними рогатых жаворонков.
Снежные воробьи p.Montifringilla ио
внешнему облику и повседневному поведе-
нию, включая способы локомоции, местооби-
тания и характер их использования, очень
похожи на горных вьюрков p.Leucosticte из
ceM.Fringillidae и пупочек p.Plectrophenax из
ceM.Emberizidae. Все трое благодаря длинным
и острым крыльям в сочетании с длинным
хвостом обладают очень сильным, почти пря-
молинейным полетом и легко перемещаются
на большие расстояния как в горизонталь-
ной, так и в вертикальной плоскостях в усло-
виях беспрестанного сильного ветра. Все три
рода принадлежат к адаптивному тину 7 и
населяют сходные ландшафты альпийского
(аркто-алышйского) облика, где выходы скал,
обрывы и осыпи чередуются с выровненны-
ми пространствами на относительно поло-
гих склонах с пятнами альпийских лугов,
стенными и тундровыми растительными
ассоциациями. По окраске оперения в гнездо-
вом и нослеп[ездовом нарядах снежные во-
робьи и пуночки на удивление похожи друг
на друга, причем среди всех овсянковых, ио-
видимому, только у пупочек — как и у воро-
бьев родов Passer, Pyrgilauda и Montifringilia
— в брачный период клюв самцов чернеет
(Р.Л.Беме, личное сообщение). Не менее уди-
вительно, что пуночка является единственным
представителем овсянок с четко намеченной
тенденцией к синантропизации: если “эко-
логическую нишу” домового воробья — в от-
сутствие законного хозяина — в горах зани-
мают снежные воробьи, то в тундре ею владе-
ют пуночки (Портенко, 1973).
Особое направление экогенеза Passeridae
иллюстрирует монотнпичный род Carpospiza.
В облике и манерах короткопалых воробьев
угадываются черты представителей самых
разных семейств певчих птиц. Короткопалые
воробьи собирают корм на земле и при этом
еще более, чем земляные воробьи, напомина-
ют жаворонков или даже коньков но харак-
теру локомоции, нозам, постоянному склевы-
ванию насекомых с травинок, невзрачной се-
рой окраске. А во время нения самцы харак-
терной посадкой, манерой распушать опере-
ние па груди и широко раскрывать клюв, по
меткому наблюдению Е.Н.Панова (личное
сообщение), очень напоминают овсянок. В
матрице адаптивных типов короткопалый
воробей больше всего соответствует типу 2,
однако этот вид связан с крайне своеобраз-
ной растительностью — разбросанными но
16 В.В.Иваницкий
ксерофитным склонам колючими и нодуш-
ковидными кустарничками, высота которых
зачастую не превышает 20 см. Эти кустар-
нички воробьи используют как песенные
посты, места отдыха и — насколько это воз-
можно — прячут в них свои гнезда.
Судя по всему, главное направление экоге-
неза воробьев состояло, в постепенном перено-
се основных проявлений их жизнедеятельнос-
ти с растительных субстратов на иные компо-
ненты среды обитания, среди которых наиболь-
шее значение принадлежало поверхности по-
чвы и вертикальным элементам рельефа. По
мере все более полного освоения этих субстра-
тов воробьи постепенно утрачивали харак-
терные черты древесных птиц и приобрета-
ли черты адаптивных типов, свойствешшх пти-
цам открытых ландшафтов. По своему мор-
фо-биологическому облику воробьи все бо-
лее уклонялись от эволюционно родственных
групп ткачиков и эволюционировали по пути
конвергенции с представителями значитель-
но более удаленных в генеалогическом отно-
шении вьюрковых, жаворонков, и овсянок.
ГЛАВА 2. ПРОСТРАНСТВЕННО-ЭТОЛОГИЧЕСКАЯ СТРУКТУРА
ПОПУЛЯЦИЙ
2.1. Общие принципы организа-
ции пространственных отношений
Пространственная структура популяций
воробьев подвержена значительной видовой,
популяционной и сезонной изменчивости.
Наиболее существенные черты пространствен-
ной структуры определяются характером вза-
имного расположения гнезд и ночевок, нали-
чием или отсутствием стайности на данном
отрезке годового цикла. В свою очередь но
каждому из этих признаков можно выделить
несколько градаций (Табл.4).
2.2. Продвинутые формы колони-
альное™
Наиболее продвинутые формы колониаль-
ного гнездования в ceM.Passeridae свойствен-
ны индийским и испанским воробьям. В Сред-
ней Азии оба вида являются типичными миг-
рантами с относительно коротким репродук-
тивным периодом, па протяжение которого
отдельные пары все же успевают выкормить
но два выводка (Гаврилов, 1962; Семашко, 1962;
Кекилова, 1973). Большую часть годового цик-
ла занимают кочевки и миграции, когда пти-
цы держатся крупными стаями и много пе-
ремещаются в пространстве. Наряду с этим,
отчетливое тяготение к колониальному гнез-
дованию, хотя и не в столь яркой форме, свой-
ственно домовым и нолевым воробьям — пол-
ностью или преимущественно оседлым видам
с более продолжительным репродуктивным
сезоном и нолицикличпым размножением
(Благосклонов, 1950; Ильенко, 1976; Доржиев,
Доржиева, 1985; Hegner, Wingfield, 1986).
2.2.1. Размеры, структура и ди-
намика стай
В начале прилета к местам гнездования
индийские и испанские воробьи держатся
небольшими группами по 2-10 птиц, одиноч-
ные особи встречаются крайне редко. Со вре-
менем размер стай возрастает прямо пропор-
ционально общей численности птиц, мигри-
рующих через данный район. Иногда вдоль
дорог и лесополос почти непрерывно летят
сотенные и тысячные стаи, огромные скопле-
ния птиц кружатся над полями с поспеваю-
щими зерновыми культурами. Есть много
свидетельств очень высокой концентрации
воробьев в период их весеннпих миграций,
равно как и огромного ущерба, причиняемо-
го ими посевам озимых колосовых, достига-
ющих к этому времени молочно-восковой
спелости (Штегмап, 1956а,б; Гаврилов, 1962;
Голованова, 1965; Кашкаров, Зинченко, 1986).
Сплоченность стай, степень взаимного тя-
готения воробьев друг к другу зависят от сте-
пени обособленности от других скоплений и
в конечном итоге во многом определяются
общей численностью птиц в данном районе. В
период миграций воробьи неохотно покида-
ют стаю, если нет возможности тут же присо-
единиться к другой группе, по с готовностью
перемещаются из одной стаи в другую. Там
где воробьев немного, они держатся компакт-
ными группами. В местах массовой концент-
рации птиц, прежде всего на посевах и близ-
лежащих лесополосах, стаи ностояшю пере-
мешиваются, то распадаются, то формируются
вновь. Птицы то и дело перемещаются из од-
ного скопления в другое и непрерывно нахо-
дятся в состоянии повышенного возбуждения.
Чем Kpyrfliee стая, тем сильнее опа притягива-
ет воробьев. Особенно привлекательны летя-
щие стаи. С увеличением численности особей
в группировках растет площадь их рассосре-
доточепия и уменьшается синхронность их по-
ведения. Многотысячное скопление воробьев
па поле не реагирует как целое даже на выст-
рел. Ранее мы опубликовали подробный ана-
лиз стайного поведения мигрирующих воро-
бьев (Иваницкий, 1984а).
Размеры, структура и динамика стай осед-
лых домовых и нолевых воробьев подверже-
ны существенным изменениям на протяже-
ние годового цикла. Анализ стай этих видов
удобно начать с того момента, когда больший-
18
В.В.Иваницкий
Таблица 4.
ОСНОВНЫЕ ОСОБЕННОСТИ ПРОСТРАНСТВЕННОЙ СТРУКТУРЫ
В ПОПУЛЯЦИЯХ ВОРОБЬЕВ
Тип гнездования Тип ночевки Тип стайности
Изолированное гнездование. Гнездящиеся пары большую часть сезона размножения придерживаются своих обширных и обособленных участков, где сосредоточены все функциональные центры и зоны активности и которые используются относительно равномерно. Контакты соседей приурочены к нейтральным зонам или к стыкам охраняемых территорий. Индивидуальные ночевки. В одном убежище ночует только одна птица (две птицы противоположного пола) при полной акустической и визуальной изоляции от других особей Стаи как компактные группировки неродственных особей отсутствуют
Квазиколониальность. Расстояние между ближайшими гнездами варьирует от нескольких метров до нескольких десятков метров. Соседние пары на своих гнездовых участках постоянно видят и слышат друг друга, причем общее число таких пар обычно не превышает 10-ти. Квазикоммунальные ночевки Разлету на ночевку предшествует выраженный пик социальной активности в стаях, после чего птицы рассредоточиваются по индивидуальным убежищам обычно неподалеку друг от друга. Стаи наблюдаются преимущественно вне сезона размножения, а с его началом сохраняются лишь у негнездящихся особей.
Нормальная колониальность. Ближайшие гнезда расположены в нескольких метрах друг от друга . Поселение имеет компактный характер и достигает численности в несколько десятков пар. Коммунальные ночевки. В одном убежище (кроне дерева, пещере, здании)собирается на ночь множество птиц при возможности неограниченных визуальных и акустических контактов друг с другом. Участие самцов в коммунальных ночевках сохраняется и на протяжение репродуктивного цикла. Стайность сохраняется на всем протяжении годового цикла. Колония представляет собой осевшую на гнездование стаю. Молодые сбиваются в стаи сразу же по вылету из гнезд.
Суперколониальность. Расстояния между гнездами варьируют от нескольких метров до нуля (гнезда сливаются в конгломераты). Население колонии исчисляется сотнями и тысячами пар.
ство членов популяции занято размножени-
ем. В разгар первого репродуктивного цикла
— особенно с началом массового выкармли-
вания птенцов, стайность выражена в мини-
мальной степени. Этот единственный в годо-
вом цикле популяции и довольно непродол-
жительный период завершается с вылетом из
гнезд первых молодых, которые сразу но до-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
19
стижению относительной самостоятельности
сбиваются в многочисленные и сплоченные
стаи. С течением времени сюда вливаются
новые генерации молодых птиц и закончив-
шие размножение взрослые птицы. Стаи кон-
центрируются в местах с обильным кормом
и обязательным наличием деревьев или кус-
тарников, предоставляющих воробьям место
для дневного отдыха и ночевки.
По наблюдениям Pinowski (1971), в сель-
ской местности близ Варшавы стаи молодых
полевых воробьев насчитывают до 1000 осо-
бей и в течение всей второй половины лета
держатся довольно обособленно друг от дру-
га на относительно постоянных участках в ра-
диусе 1-2 км от колонии, откуда происходят
большинство (75%) членов стаи. В Подмос-
ковье наиболее постоянными центрами ак-
тивности таких стай являются места комму-
нальных ночевок в кронах, где часто объеди-
няются и полевые, и домовые воробьи. При
этом воробьи никогда не почуют в тех мес-
тах, где в основном кормятся и отдыхают днем,
по к вечеру всегда возвращаются в одно и то
же место па окраине поселка.
Ярко выраженное стремление скапливать-
ся на ночевку в постоянном месте вопреки
его подчас значительной удаленности от зон
фуражировки особенно характерно для до-
мовых воробьев. Количество одновременно
ночующих птиц к концу лета нередко исчис-
ляется тысячами особей, а их суточные пере-
леты на кормежку и обратно пролегают но
строго определенным маршрутам на расстоя-
ние до 6-8 км (Schmidt, 1954; Манучярян, 1963;
North, 1973 ). В Одессе в августе мы наблюда-
ли массовые суточные перелеты домовых
воробьев вдоль линии пляжей, составляющих
излюбленное место кормежки этих птиц. В
разгар вечернего перелета воробьи летят ста-
ями до 30 особей, причем численность стай
заметно возрастает вблизи места главной
ночевки, кда собиралось не менее 2000 осо-
бей. Общая картина при этом чрезвычайно
напоминала, на наш взгляд, сезонные мигра-
ции перелетных воробьев, описанные выше.
Как пишет North (1973), дальний разлет на
дневную кормежку характерен главным об-
разом для молодых воробьев, старые птицы
остаются днем на своих гнездовых участках
или ограничиваются незначительными ло-
кальными перемещениями: их доля в насе-
лении воробьев в центре города составила в
среднем 48%, на окраине — 28,3%, а на приле-
гающих сельскохозяйственных полях с посе-
вами зерновых — всего 7,5%.
К началу осени у домовых и нолевых
воробьев подходит к концу линька и активи-
зируется развитие гонад, что знаменует нача-
ло новой фазы годового цикла и влечет за
собой важные изменения пространственной
структуры популяций. Вначале взрослые, за-
тем молодые птицы (по порядку генераций)
все в большей степени концентрируют свою
активность вокруг гнездовых убежищ. Ста-
рые птицы возвращаются к своим гнездам,
молодые разыскивают убежища заново и
активно расселяются. Резко обостряется кон-
куренция за убежища, возрастает уровень со-
циальной активности и интенсивность вока-
лизации. Распадаются крупные коллективные
ночевки в кронах, и еще через некоторое
время большинство воробьев переходит к
индивидуальным ночевкам в закрытых убе-
жищах. Ранее опубликован детальный анализ
событий в популяциях нолевых воробьев в
период осеннего токования и расселения (так
называемого, абортивного цикла) (Pielowski,
Pinowski, 1962; Pinowski, 1971). В сущности
поведение полевых и домовых воробьев в это
время выглядит как формирование гнездо-
вых поселений (в том числе и колоний) и
будет подробно рассмотрено ниже.
Наступление холодов приостанавливает
репродуктивную активность, и домовые и по-
левые воробьи переходят в фазу зимовки,
хотя при некоторых специфических обстоя-
тельствах, например, при обитании домовых
воробьев в крупных животноводческих сель-
скохозяйственных фермах, развитие гонад и
сопутствующие ему формы поведения про-
грессируют, отчего первые нормальные клад-
ки, из которых вноследствие вылупляются
птенцы, гибнущие через несколько дней от
бескормицы, могут появиться уже в начале
января (Матюхин, Иваницкий, 1984). В тро-
пиках размножение домовых (индийских) и
нолевых воробьев в той или иной мере про-
исходит круглогодично, хотя большинство
репродуктивных циклов приурочено все же
к определенным сезонам (Ward, Poh, 1968;
Naik, Mistry, 1970).
20
В.В.Иваницкий
К зиме завершается расселение молодых
птиц, и они вместе со взрослыми особями
оседают до весны на относительно постоян-
ных участках, распределяя свою суточную ак-
тивность между местами ночевок (совпада-
ющими часто с местами гнездования и пе-
риодического зимнего токования).
Зимой свободное время полевые и домо-
вые воробьи проводят обычно в клубах —
местах коллективного отдыха и неспецифи-
ческого общения. По мерю увеличения про-
должительности светового для численность
и продолжительность прюбывания воробьев
в клубах возрастают, достигая наибольших
значений к началу марта, когда под воздей-
ствием активизации роста гонад (Ильенко,
1976) заметно возрастает социальная актив-
ность воробьев: они начинают петь в клубах,
устраивая характерные коллективные “кон-
церты” (Фетисов, 1981), и уже с середины
февраля при потеплениях самцы активно
ухаживают за самками.
2.2.2. Социальная организация стай
В зимних стаях у оседлых ворюбьев по
нашим наблюдениям преобладает индиффе-
рентное отношение друг к другу. Персонали-
зированные социальные взаимодействия слу-
чаются довольно редко, будучи связанными,
главным образом, с охраной индивидуальной
дистанции при кормежке и с эпизодически-
ми всплесками социо-сексуалыюй активно-
сти самцов.
Персональный состав зимних стай домо-
вых воробьев в местах кормежки па корот-
ких отрезках времени отличается непостоян-
ством. Птицы посещают зоны фуражировки
и перемещаются между ними поодиночке и
независимо друг от друга. Каждый воробей в
значительной мере следует собственной ин-
дивидуальной манере поисков корма в пре-
делах хорошо знакомого участка местности
и расценивает стаи других воробьев лишь
как одно из указаний на возможность при-
сутствия корма. В то же время, за счет того,
что контингент особей на данном участке ме-
стности почти всю зиму сохраняется относи-
тельно постоянным, в стаях в конечном итоге
встречаются одни и те же птицы (Barnard,
1980 а,b,с),
Данные о существовании внутривидовых
иерархических отношений в стаях домовых
ворюбьев противоречивы. В естественных ус-
ловиях обитания вида в Северной Америке
иерархии не обнаружено, в чем воробьи рез-
ко отличались от соседствующих с ними мек-
сиканских чечевичников Carpodacus mexi-
canus, взаимоотношения которых строились
на принципах четкой линейной иерархии
(Kalinoski, 1975). В неволе но наблюдениям
Уотсон (Watson, 1970) североамериканские
домовые воробьи также не поддерживают
четкой субординации: взаимоотношения осо-
бей отличались высокой изменчивостью и не-
стабильностью, а исход конфликтов между
•ними зависел от положения его участников
в пространстве вольеры.
В противоположность этому, Hegner и
Wingfield (1987b) обнаружили, что при со-
держании домовых воробьев группами по 6
птиц между ними примерно за 2 недели скла-
дывается довольно стабильная иерархия. В
62 % всех групп иерархия была линейной и
транзитивной, то есть реверсий доминирова-
ния не было. В других группах субодинация
была менее определенной, то есть взаимоот-
ношения некоторых особей не вписывались в
рамки простой линейной схемы. По данным
этих авторов, пол не оказывал существенного
влияния на социальные ранги особей. Если
в вольер помещали гнездовой ящик, первой
его всегда занимала одна из самок, но пару с
ней мог образовать самец любого ранга, и его
положение в группе при этом не изменялось.
Moller (1987b) экспериментально изучал,
как влияет величина черного пятна (маниш-
ки) на горле самцов домовых воробьев на их
социальный ранг в стаях (у самок пятна нет).
Обнаружена прямая зависимость между ве-
личиной манишки и частотой участия сам-
цов в агрессивных взаимодействиях как в
своей групне, так и после перемещения в дру-
гую группу. Искусственное увеличение маниш-
ки с помощью окрашивания также повысило
частоту участия самцов в стычках, однако пе
привело к повышению социальных рангов
обладателей искусственно увеличенных ма-
нишек. По мнению автора, иерархия у домо-
вых воробьев пе бывает жесткой и абсолют-
ной: относительно низкоранговые особи до-
вольно часто нападают на доминантов, несу-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
21
щих внешние признаки высокого ранга, вы-
ну ждая их постоянно подтверждать свое вы-
сокое иерархическое положение и осуществ-
ляя, тем самым, эффективный контроль за
реальными возможностями членов группы.
В целом, имеющиеся данные характеризу-
ют стаи воробьев как преимущественно ано-
нимные сообщества открытого тина с мини-
мальной степенью персонализации взаимоот-
ношений и аморфной социальной структурой.
Относительное постоянство персонального
состава оседлых стай обеспечивается главным
образом общностью жизненного пространства
сосуществующих бок о бок особей, а вовсе не
их индивидуальной привязанностью друг к
другу. В этом отношении стан воробьев со-
ставляют резкий контраст со стаями воробь-
иных и воскоклювых ткачиков, которые пред-
ставляют собой относительно замкнутые со-
общества с довольно постоянным составом и
чрезвычайно сложной социальной организа-
цией, целиком построенной па сугубо инди-
видуальных привязанностях и антипатиях,
имеющих долгосрочный характер (многолет-
нее постоянство брачных нар, четкая соци-
альная стратификация, гетеро- и гомосексу-
альные альянсы в борьбе за высокие соци-
альные ранги и т.н.) (Sparks, 1964; Collias,
Collias, 1978а, 1980а; Lewis, 1982).
2.2.3. Пространственно-хроноло-
гическая структура и численность
колоний
По данным Гаврилова (1962), колонии ис-
панских воробьев в юго-восточиом Казахста-
не насчитывают обычно но несколько десят-
ков тысяч гнезд, реже до 100000 и даже до
800000 гнезд. Колонии меньше 100 гнезд здесь
встречаются редко, и лишь как исключение
находили группы из 10-15 гнезд и совсем
редко — одиночно гнездящиеся пары. Коли-
чество гнезд на гектаре лесонасажден и й коле-
балось в крупных колониях от 5500 до 36000,
составляя в среднем около 13000 гнезд па 1 га.
Разумеется, однако, что совокупная плотность
популяций испанских воробьев даже в райо-
нах с их чрезвычайно высокой численностью
намного ниже. Например, здесь же в юго-вос-
точном Казахстане на территории одного из
совхозом общей площадью 10000 га распола-
гались 5 крупных колоний с общей численно-
стью около 75000 гнезд (Штегман, 1956а). Вы-
сокая степень колониальности присуща испан-
ским воробьям на всем протяжении их ареала
(Summers-Smith, 1988), хотя этот вид сохра-
нил все же способность к изолированному
гнездованию (Baumgart, 1984; наши наблюде-
ния в Бадхызе).
Индийские воробьи, которые в Средней
Азин являются ближайшими соседями испан-
ских воробьев и часто живут бок о бок с ними
в смешанных колониях, также способны фор-
мировать чрезвычайно многочисленные гнез-
довые поселения. Так, в южном Таджикистане
в обрывах вдоль дороги на протяжение 150 м
Голованова (1965) насчитала 10000 жилых нор
этих воробьев. Тем не менее, в отличие от пре-
дыдущего вида, индийские воробьи охотнее
гнездятся небольшими группами от 2 до 15
нар, не представляет редкости (а в ряде мест
являегся очень характерным) и изолирован-
ное гнездование отдельных пар.
Плотность гнезд в колониях испанских и
индийских воробьев очень высока, хотя и под-
вержена значительным вариациям даже в пре-
делах одной колонии. Иногда гнезда довольно
равномерно рассредоточены на расстоянии 1
— 5м друг от друга, другие гнезда сливаются
в массивные конгломераты, так что бывает
невозможно их отделить друг от друга (до 8
гнезд в одном конгломерате). Па одном дереве
бывает до 130 гнезд испанских воробьев (Гав-
рилов, 1962) и до 45 гнезд индийских воробь-
ев (Кекилова, 1973). Максимальная плотность
гнездования индийских воробьев в обрывах
отмечена нами в одной нз крупных колоний в
берегах канала близ Самарканда: на участке
ф<оло 2 кв. м гнездились 16 пар.
Численность и плотность колоний осед-
лых домовых и полевых воробьев в среднем
значительно уступает аналогичным показате-
лям для гнездовых поселений выше рассмот-
ренных мигрирующих видов, однако в ряде
мест бывают очень высокими.
Для домовых воробьев па большей части
их ареала характерны относительно немно-
гочисленные колонии (Summers-Smith, 1988),
однако там, где домовые воробьи по каким-
либо причинам переходят к открытому гнез-
дованию на деревьях, численность и плотность
их колоний приближается к аналогичным но-
22
В.В.Иваницкий
казателям в описанных выше колониях ин-
дийских воробьев. Так, в Ленкорани в неко-
торых колониях Р. domesticus hyrcanus на
пирамидальных тополях и акациях насчиты-
вали до 300 гнезд (Мустафаев, 1969). В Ка-
наде описана многочисленная колония инт-
родуцированных P.d.domesticus в кронах
елей: вдоль аллеи длиной 90 м найдено 146
гнезд — до 14 гнезд на одном дереве, отмече-
ны конгломераты гнезд (McGillvray, 1980а,b).
Во многих популяциях нолевых воробьев
явно преобладает изолированное гнездование.
В то же время, в подмосковных поселках, поле-
вые воробьи, часто соседствующие здесь с до-
мовыми воробьями, обнаруживают некоторую
тенденцию к квази колой иалыюсти. В Забайка-
лье на сельскохозяйственных животноводчес-
ких фермах нолевые воробьи гнездятся пре-
имущественно колониями но 4-5 пар, однако
некоторые колонии насчитывали до 25 нар
при средних расстояниях между ближайши-
ми гнездами 1,5-3,0 м (Доржнев, Доржиева,
1985). На одной из ферм в Пермской области в
козырьке вентиляционной трубы найдено 8
гнезд полевых воробьев (Болотников и др.,
1985). Однако наиболее многочисленные и
плотные колонии этот вид образует при гнез-
довании в обрывистых стейках оврагов в Вол-
гоградской области. Поданным Благосклонова
(1950,1991), плотность этих поселений дости-
гает 80 гнезд на 100 м обрыва (до 200 гнезд на
1 км), плотность доходит до 4 гнезд/м2, а об-
щая чмслеппость совместно гнездящихся нар
достигает нескольких сотен. Помимо оврагов, но
свндельству того же автора, нолевой воробей в
этом районе в массе гнездится в лесополосах,
где занимает практически все синичпнки, ‘‘не-
зависимо от плотности их установки”.
2.2.4. Поведение при формирова-
нии колоний
Поведение оседлых домовых и нолевых
воробьев, а также мигрирующих индийских
и испанских воробьев при формировании
гнездовых колоний выглядит очень сходно,
хотя, разумеется, у первых оно занимает зна-
чительно больше времени и по существу на-
чинается уже с осени. Тем не менее, в некото-
рых популяциях индийских воробьев но на-
шим данным период формирования поселе-
ний, исчисляемый от момента начала прилета
до начала массовой кладки, также может быть
очень растянутым — особенно при неблагоп-
риятных погодных условиях, достигая в от-
дельных случаях 3-4 недель. Колонии испан-
ских воробьев формируются значительно
быстрее и с поразительной синхронностью
(Гаврилов, 1962). В этом разделе мы рассмот-
рим главным образом поведение мигрирую-
щих воробьев, у которых все основные фор-
мы поведения особей, связанные с формиро-
ванием колоний, проявляются значительно
более рельефно, чем у оседлых видов.
Поведение воробьев на самых первых эта-
пах заселения колоний характеризуется зна-
чительной преемственностью но отношению к
предыдущему периоду зимовки или мигра-
ций с преобладанием стайного образа жизни.
Это проявляется в сохранении высокой сте-
пени коллективных действий, синхронизации
разных форм поведения, пониженном на пер-
вых норах уровне агрессивности. Только что
прилетевшие воробьи перемещаются по ко-
лонии группами по 5-10 особей, и если име-
ются прошлогодние гнезда (в кроне) или
поры (на обрыве), то активность таких групп
концентрируется возле них. Воробьи стара-
ются держаться поближе к гнезду, то и дело
залезают в пего, ведут себя очень возбужден-
но. Иногда можно наблюдать мимолетные
мимолетные агрессивные стычки, что не на-
рушает, однако, сплоченности таких микро-
групп. Птицы держатся у одного гнезда до 40
мин, после чего могут переместиться к друго-
му гнезду и далее.
В последующий дни, когда численность
птиц в формирующейся колонии возрастает,
коллективные инспекции гнезд происходят
уже в нескольких местах одновременно, а их
участники то и дело перемещаются из одной
группы в другую. В то же время, постепенно
усугубляется стремление самцов отделяться
от групп и оседать па постоянных участках,
где их агрессивность по отношению к другим
особям быстро увеличивается. В то же время
птицы еще долгое время сохраняют повышен-
ный интерес к инспектирующим стаям или
даже отдельным особям, демонстрирующим
характерный поведенческий стереотип осмот-
ра убежищ, которое в целом ряде случаев ско-
рее представляет собой чисто ритуальное но-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
23
ведение, нежели действительный поиск места
для гнезда. Важное свойство этого поведения
состоит в том, что оно оказывает мощное сти-
мулирующее воздействие на других особей, в
том числе и на тех самцов, которые уже име-
ют постоянные центры активности (убежи-
ща, гнезда, территории) в колонии. Мы много
раз наблюдали, как самцы в течение доволь-
но короткого времени их непрерывного про-
слеживания (30-40 мин) неоднократно (но 2-
5 раз) переходили от характерного режима
охраны и рекламирования определенного
центра активности к совершенно иному ре-
жиму коллективной инспекции убежищ. Воз-
можно, что устойчивое стремление к участию
в инспектирующих группах оказывает опре-
деленное влияние на формирование простран-
ственной структуры колонии, способствуя воз-
никновению характерных кластеров гнезд
(субколоний) за счет оседания членов таких
групп но-соседству друг с другом (см. выше).
Матюхин и др. (1990) детально изучили
формирования и структуры колоний индийс-
ких воробьев с использованием многолетнего
индивидуального мечения. Выявлена высокая
степень подвижности птиц не только в преде-
лах колонии, но и в пределах локального по-
селения, состоящего из нескольких близко рас-
положенных колоний. Многие воробьи без
видимых причин покидают обжитые убежи-
ща, других вытесняют более удачливые кон-
куренты. Наряду с этим, и у самцов, и у са-
мок этого вида хорошо выражена привязан-
ность к прошлогодним убежищам, которая ха-
рактерна и для близкородственных домовых
воробьев (Summers-Smith, 1988). По наблюде-
ниям Матюхина с соавторами, прошлогодние
владельцы убежищ, почему-либо запоздавшие
с прибытием в колонию, сразу же изгоняют со
своих территорий вновь обосновавшихся здесь
воробьев того же пола, что также служит од-
ной из причин обусловливающей постоянные
переселения особей с одного места на другое.
2.2.5. Территориальное поведение
в колониях
Всем колониальным воробьям свойствен-
но четко выраженное территориальное пове-
дение, которое проявляется в защите от дру-
гих особей определенного фрагмента кроны,
обрыва и пр. вокруг убежища, выбранного
владельцем территории для гнездования. В
смешанных колониях территория защищает-
ся от всех воробьев независимо от их видо-
вой принадлежности (Иваницкий, 1985в).
Территориальное поведение зависит от плот-
ности населения (Зыкова, 1980) и в ряде
случаев обладает хорошо выраженной видо-
вой спецификой.
У индийских воробьев при гнездовании
па обрывах территория представляет собой
фрагмент вертикальной плоскости с одной
или несколькими норами и присадами (не-
ровности грунта, торчащие корни и и т.д.),
которые служат в качестве наблюдательных
постов и мест нения. У изолированно живу-
щих самцов постоянно используемые нриса-
ды, где он проводит большую часть времени,
могут размещаться на расстояние до 8-10 м
от центрального убежища. В противополож-
ность этому, при максимальной плотности
населения в колониях территории и нриса-
ды но существу отсутствуют. Птицы все вре-
мя вынуждены сидеть у входа в свои норы,
которые иной раз расположены настолько
близко, что занимающие их воробьи способ-
ны дотянуться до соседа клювом, не покидая
своего убежища. Такую картину часто можно
видеть и в колониях домовых н полевых
воробьев под шиферными крышами домов,
когда из-под каждого изгиба шифера всего в
15-20 см друг от друга виднеются головы
оживленно чирикающих птиц.
В колониях индийских воробьев па дере-
вьях территория самца располагается вокруг
одной или нескольких мутовок, пригодных
для сооружения гнезд. Самец тщательно об-
следует территорию и много перемещается по
пей, то и дело вступая в пограничные конф-
ликты с соседями. Объем защищаемой терри-
тории при низкой плотности составляет око-
ло 50 м3, однако при возрастании плотности
резко уменьшается вплоть до того, что неко-
торые гнезда впоследствие входят в сопри-
косновение друг с другом.
В противоположность этому, территориаль-
ное поведение испанских воробьев выглядит
значительно проще. Пограничные конфликты
в том ваде, в котором они свойственны индий-
ским воробьям, в репертуаре этого вида отсут-
ствуют. Уже с самого начала освоения террито-
24
В.В.Иваницкий
рии внимание самца испанского воробья по-
чти целиком сосредоточивается на единствен-
ной мутовке, где он сразу же начинает строить
гнездо. Когда поблизости появляется другой
самец хозяин мутовки его атакует, однако дис-
танция проявления агрессивных реакций силь-
но меняется со временем даже у одного самца,
а конфигурация территорий отличается неопре-
деленностью. Даже при относительно разрежен-
ном гнездовании самцам испанского врробья
не свойственно стремление к расширению тер-
риторий. Также как и в колониях, одиночные
самцы поглощены главным образом строитель-
ством гнезда, а объем охраняемого простран-
ства вокруг него обычно не превышает 10 м3.
2.3. Квазиколониальность
Квазиколониальные поселения в сочета-
нии с довольно большим числом нар, гнездя-
щихся совершенно изолированно, характер-
ны для рыжих воробьев. Пространственные
отношения особей у этого вида устанавлива-
ются в процессе противоречивого взаимодей-
ствия двух побуждений: с одной стороны сам-
цы стремятся но возможности занять терри-
тории рядом с уже поселившимися нарами
или интенсивно поющими холостыми сам-
цами, но, с другой стороны, новые вселенцы
избегают и слишком близкого контакта, чему
препятствует и хорошо выраженное терри-
ториальное поведение резидентов, которое но
своим основным особенностям напоминает
территориальное поведение саксаульных во-
робьев (см. выше). Поданным Нечаева (1969),
на Курильских островах рыжие воробьи гнез-
дятся скоплениями по 7-8 пар в 30-50 м друг
от друга. Близкие по численности и плот-
ности поселения обнаружены нами в долине
р.Сусуя близ Ново-Николаевки па Южном
Сахалине (Табл.5), в этом же районе в долине
р.Шебупипки отмечено 12 пар воробьев на
15 км маршрута (Нечаев, 1973). Типичные ква-
зиколопиальпые поселения образуют рыжие
воробьи в Гималаях (Summers-Smith, 1988).
Стайность рыжих воробьев исчезает уже
па самых ранних стадиях формирования по-
пуляции. На Сахалине мы не встречали стай
или групп рыжих воробьев. Птицы вели ис-
ключительно древесный образ жизни и круг-
лосуточно оставались на своих участках, но-
чуя здесь же поодиночке в дуплах. Известно,
однако, что во время миграций и зимовок ры-
жие воробьи держатся довольно многочис-
ленными и сплоченными стаями (Нечаев,
1973; Summers-Smith, 1988).
Квазиколониальность очень характерна
для каменных воробьев. Пространственная
структура популяций этого вида отличается
особенно высокой изменчивостью и была под-
робно исследована нами в разных точках его
обширного ареала вида (P.p.exigua — Закав-
казье, P.p. in termedia — Туркмения нР.р.Ьге-
virostris — Алтай и Тува) (Иваницкий, 19856,
1986а). Изменчивость порождается отсутствию
жестких ограничений па взаимное размеще-
ние птиц в пространстве и их способностью
активно регулировать частоту и продолжи-
тельность социальных контактов. Если па
Алтае воробьи регулярно гнездятся в плот-
но заселенных колониях (нормальная коло-
пиалыюсть) и практикуют квазикоммупаль-
ные ночевки на традиционных местах, то в
Бадхызе отчетливо преобладают значитель-
но более аморфные гнездовые поселения (ква-
зиколониальность) и преимущественно ин-
дивидуальные ночевки, хотя в обоих этих
популяциях имеет место значительная инди-
видуальная изменчивость размещения гнез-
довых территорий и ночевок.
Квазиколониальные поселения при зна-
чительном числе одиночно гнездящихся нар
встречаются и у снежных воробьев. Взаим-
ное размещение гнездящихся пар и их плот-
ность па локальных участках сильно варьи-
рует. Так, Потапов (1966) сообщает об исклю-
чительно изолированном гнездовании вида па
Памире, однако в этом же районе Абдусаля-
мов (1977) нашел 5 гнезд на скале площадью
всего 10 кв. м. На юго-востоке Алтая Лоскот
(1986) па осыпи площадью 10 га насчитал 12
гнезд, наши данные но распределению гнез-
довых пар снежных воробьев в этом же рай-
оне приведены в Табл.5. Территориальность
снежных воробьев крайне аморфна. Актив-
ное территориальное поведение самцы про-
являют только в период образования пар, гнез-
достроепня и спаривания. В результате чис-
то агонистические реакции, непосредственно
направленные на защиту территорий, быва-
ют тесно переплетены с саморекламировани-
ем и брачным поведением (Иваницкий, 19916).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
25
2.4. Изолированное гнездование
Среди всех изученных нами видов мини-
мальной социальностью обладают пустынные
воробьи. И в Туркестане (Каракумы и Кы-
зылкумы), и в Северной Африке (Сахара)
отмечались преимущественно обособленные
пары с далеко отстоящими друг от друга гнез-
дами при общей крайне низкой плотности
популяций (Balzac de, 1929; Шестоперов, 1926;
Рустамов, 1958; Соныев, 1965, Пономарева,
1983; Третьяков, 1990). Тем не менее, имеется
свидетельство о коллективном гнездовании
пустынных воробьев: в Марокко на единствен-
ной пальме среди барханов примерно в мет-
ре друг от друга располагались жилые гнез-
да трех пар и держались три холостых сам-
ца (Densley, 1990а).
Мы подробно изучили структуру попу-
ляций пустынных воробьев в юго-восточных
Каракумах, где этот вид населяет крупнобуг-
ристые, подвижные, барханные пески с да-
леко расбросапными друг от друга деревья-
ми пустынной акации, кустами кандыма и
белого саксаула. Массивы незакрепленных
песков имеют форму довольно узких, вытя-
нутых гряд, простирающихся почти парал-
лельно друг другу и разделенных широки-
ми плоскими участками закрепленных и
иолузакренленных песков, покрытых значи-
тельно более богатой древесно-кустарнико-
вой и травянистой растительностью, где пу-
стынные воробьи не живут (Соныев, 1965).
В Табл.5 приведены данные о структуре
поселений пустынных и саксаульных воро-
бьев в Репетекском заповеднике. Судя о на-
шим наблюдениям в середине октября и в
конце февраля, пустынные воробьи круглый
год держатся парами или семьями па по-
стоянных и довольно обособленных друг от
друга участках, явно избегая контактов с со-
седями. Брачные партнеры очень привязаны
друг к другу. По данным мечения, пары со-
храняются круглогодично, причем не будучи
заняты насиживанием или выкармливанием
птенцов, самец и самка почти все время дер-
жатся вместе, перемещаясь в поисках корма
по своим обширным участкам. Гнездо в тече-
ние всего года служит главным функциональ-
ным центром участка обитания нары — оба
партнера неизменно почуют в своем гнезде и
периодически посещают его в дневное время.
В течение трех сезонов наблюдений (1988-
-1990 гг.) численность гнездящихся нар пус-
тынных воробьев и их распределение в про-
странстве оставались примерно одинаковы-
ми. Таким образом, паши наблюдения под-
твердили данные Сопыева (1965), о высоком
многолетнем постоянстве участков этого вида
в Репетеке. Значительную консервативность
проявляют пустынные воробьи и в отноше-
нии мест устройства своих гнезд: новые гнезда
птицы строят как правило неподалеку от ста-
рых, так что возле каждого жилого гнезда
почти всегда можно найти до 4-5 старых
гнезд. Гнезда одной нары могут располагать-
ся на одном дереве, сливаясь в довольно проч-
ный конгломерат.
В негнездовой период пустынные воробьи
большую часть дня проводят вдали от гнезд,
улетая в поисках корма на несколько сот мет-
ров, а иногда и намного дальше. Ранней вес-
Таблица 5. РАССТОЯНИЕ МЕЖДУ ГНЕЗДАМИ В НЕКОТОРЫХ ПОПУЛЯЦИЯХ
ВОРОБЬЕВ
расстояние между гнездами (м) плотность гнездования (гнезд/км^)
виды воробьев мини- мальное макси- мальное среднее число измерений
пустынный 200 1350 510,8±2,4 27 0,9
саксаульный 170 1100 419,4±33,6 42 1,2
рыжий 10 200 112,7±8,7 22 20,6
каменный 2 35 28,8±4,4 30 82,6
монгольский 50 350 216,0±15,1 *28 12,4
снежный 15 150 59,0 ± 6,2 34 37,8
26
В.В.Иваницкий
ной основные зоны фуражировки нар при-
урочены к участкам с наиболее густыми кус-
тами кандымов, где воробьи кормятся сами, а
впоследствие — собирают корм для птенцов.
Неоднократно мы наблюдали, как в течение
нескоьких дней пара неизменно кормилась на
одном и том же относительно небольшом уча-
стке, где ее можно было встретить регулярно.
Затем пара перемещается на другой участок
— иногда до 600 м от прежнего, а на освобо-
дившемся участке появляется соседняя пара.
В целом участки обитания соседних пар
широко перекрываются, однако, как уже ука-
зывалось, соседи явно избегают встречаться
друг с другом, предпочитая использовать при-
влекательные кормовые угодья поочередно. За
три сезона наблюдений мы зарегистрирова-
ли всего 8 встреч между соседними нарами
пустынных воробьев: в 6 случаях птицы про-
сто пе обратили внимания друг на друга и
только при 2 встречах зафиксировано пред-
намеренное сближение соседей и взаимные
демонстрации. Таким образом, пространствен-
ная структура популяции пустынных воро-
бьев базируется главным образом на их мно-
голетней привязанности к постоянным учас-
ткам обитания и избегании социальных кон-
тактов с соседями.
Иную картину мы видим у саксаульных
воробьев. В Каракумах оба вида часто сосед-
ствуют друг с другом, но саксаульный воро-
бей населяет более широкий спектр место-
обитаний в отличие от крайне специализиро-
ванного пустынного воробья. Будучи дуплог-
нездником, саксаульный воробей в гнездовой
период приурочен в основном к местам про-
израстания крупных кустов белого саксаула
вне зависимости от характера прочей расти-
тельности, рельефа и степени закрепленнос-
ти песков. Это типичный убиквист пустыни,
распространение которого определяется пре-
имущественно наличием пригодных для гнез-
дования убежищ. Другое важное отличие сак-
саульных воробьев состоит в том, что в их
годовом цикле отчетливо выражены перио-
ды миграций и кочевок (Потапов, 1959; Гав-
рилов, 1974).
По данным Соныева (1970) и нашим на-
блюдения, в Репетеке саксаульные воробьи,
гнездятся только отдельными парами, при-
чем соседние гнезда обычно располагаются
довольно далеко друг от друга (Табл.5). Толь-
ко изолированное гнездование отмечали для
этого вида в долине р.Или, где саксаульные
воробьи во множестве гнездятся в турапго-
вых рощах, но не ближе 100 м пара от пары
(Гаврилов, 1969). Тем больший интерес пред-
ставляют данные о возможности более плот-
ного гнездования саксаульных воробьев.
Шнитников (1949) наблюдал совместное гнез-
дование нескольких пар в парой, дуплистой
туранге в Прибалхашье. D.Summers-Smith
любезно сообщил нам о найденной им весной
1992 г смешанной колонии саксаульных (8
пар) и индийских (3 пары) воробьев, гнез-
дившихся в щелях казахского могильника в
пустыне Сары-Ишикограу (Прибалхашье).
Densley (1990) пишет о близком соседстве
двух пар, гнездящихся в щелях каменной за-
городки для скота в Монголии. Возможно,
коллективное гнездование саксаульных воро-
бьев не представляет редкости в пустыне Гобн,
где они гнездятся в безлесной местности в
обрывистых стенах характерных для этого
региона сухих русел — сайр (Курочкин, 1991).
В отличие от пустынных воробьев, струк-
тура популяций которых почти не меняется
на протяжение круглого года, у саксаульных
воробьев отчетливо выражен особый период
формирования гнездовой популяции сразу
после завершения весенней миграции, неиз-
менно сопровождающийся высокой социаль-
ной активностью птиц. Мы подробно изучи-
ли процесс формирования популяции сак-
саульных воробьев в Репетеке. Заметное уве-
личение численности птиц и рост их соци-
альной активности наблюдается в начале
марта. В это время самцы занимают и рекла-
мируют дупла, проявляя в то же время ак-
тивное территориальное поведение. С помо-
щью трансляции записанных на магнитофон
несен и визуальных наблюдений за естествен-
ными конфликтами установлено, что радиус
защищаемой территории может в отдельных
случаях достигать 40-50 м, хотя самое интен-
сивное территориальное поведение холостых
самцов наблюдается обычно в радиусе 15-20
м от дупла. Именно здесь в значительной мере
сосредоточена активность владельцев терри-
торий и располагаются несенные посты сам-
цов. Для периода формирования популяции
характерны многочисленные конфликты со-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
27
седиих территориальных самцов как друг с
другом (пограничные конфликты на стыках
территории), так и с многочисленными бро-
дячими особями или же оседло живущими
в этом районе птицами, не имеющими покуда
собственных регулярных центров активнос-
ти. Следует подчеркнуть, что акустическая
активность (пение) пришельцев служат важ-
нейшим стимулом, провоцирующим агрессив-
ность со стороны владельцев территорий, ко-
торые часто довольно миролюбиво относятся
даже к другим самцам, если те просто кор-
мятся и не поют.
На первых этапах формирования популя-
ции стремление к обособлению птиц па сво-
их территориях и активное рекламирование
последних наблюдается обычно в утренние
часы. Ближе к полудню у птиц все сильнее
проявляется тяготение к обществу себе по-
добных, и начинается формирование дневных
стай. Территориальные самцы все чаще при-
соединяются к другим воробьям, кормящим-
ся неподалеку в кронах саксаула, и проводят
в таких стайках все больше времени, возвра-
щаясь периодически па свои песенные по-
сты н возобновляя пение у дупел. Постепен-
но группы становятся все более многочислен-
ными и, наконец, стан воробьев покидают ок-
рестности гнездовых территорий в саксауль-
никах и перемещаются на более открытые
участки, где кормятся преимущественно на
земле среди редких кустов оставшуюся часть
дня вплоть до разлета на ночевку. Непосред-
ственно перед этим моментом все воробьи
собираются в плотную стаю па одном кусте и
в течение нескольких минут чистят оперение
или негромко ноют. Затем стайка взлетает и
устремляется в район гнездовий, причем сра-
зу же она веером распадается в воздухе, и
каждая птица летит ночевать к себе на учас-
ток. Коммунальных ночевок у саксаульных
воробьев мы не отмечали.
Таким образом, для саксаульных воробьев
в период формирования популяции харак-
терно закономерное чередование в их суточ-
ном цикле фазы индивидуальной обособлен-
ности, токования и активного террнториализ-
ма, фазы коллективной фуражировки в ста-
ях, куда собираются птицы, совсем недавно
оспаривавшие друг у друга убежища и тер-
ритории, и фазы индивидуальных ночевок в
постоянных местах или убежищах (в сфор-
мированных парах самец и самка почуют
вместе). Аналогичное чередование территори-
ализма и стайности у этого вида отмечал
Гаврилов (1969). По нашим наблюдениям,
саксаульные воробьи регулярно примыкают
к стаям вплоть до начала насиживания, после
чего приступившие к инкубации кладок пти-
цы придерживаются более компактных учас-
тков, ведут себя очень скрытно и явно избе-
гают встречаться друг с другом. На кормеж-
ку улетают от гнезд поодиночке или нарами
в густые саксауловые заросли, где в основ-
ном и собирают корм для птенцов. Каждая
пара имеет свои излюбленные места фура-
жировки и отдыха, которые, однако, могут на-
ходится в поочередном пользовании сосед-
них пар. На этой стадии размножения сакса-
ульные воробьи ведут себя точно также, как
и пустынные, явно избегая пользоваться теми
участками местности, где на данном отрезке
времени с высокой вероятностью можно встре-
тить других особей. Хотя зоны фуражиров-
ки соседних пар широко перекрываются, на-
блюдения за индивидуально помеченными
особями показывают, что наиболее привле-
кательные фрагменты таких зон всегда ис-
пользуются поочередно с продолжительнос-
тью периода монопольной утилизации от не-
скольких часов до нескольких дней. Уже сра-
зу после образования пар вокальная и тер-
риториальная активность самцов резко идет
па убыль и ко времени насиживания прак-
тически сходит на пет, что подтверждается, в
частности, прекращением реакции территори-
альных самцов на воспроизведение несен сво-
его вида.
Наиболее жесткие формы территориаль-
ности выявлены нами у монгольских земля-
ных воробьев. Хотя осенью и зимой они сби-
ваются на день в крупные стаи, па ночевку
отдельные особи разлетаются порознь и про-
водят ночи поодиночке в норах ннщух пли
в основании густых кустов караганы (Зоной,
1977; Зопов, Попов, 1987). В предгнездовой и
гнездовой периоды мы отмечали только ин-
дивидуальные ночевки этих воробьев в по-
рах пищух. По крайней мере с середины ап-
реля (т.е. примерно за месяц до начала клад-
ки) монгольские земляные воробьи уже дер-
жатся нарами на тщательно охраняемых тер-
28
В.В.Иваницкий
рнториях. Каждая пара перемещается ио тер-
ритории ио своим излюбленным маршрутам,
пролегающим обычно вдоль границ или по
самым крупным колониям пищух. Массивные
выбросы грунта из сурчиных нор служат сам-
цам в качестве постоянных песенных и на-
блюдательных постов. Лишь сильные снего-
пады и холода побуждают воробьев объеди-
няться в стайки и кочевать ио окрестностям
в поисках корма (Иваницкий, Шевченко, 1991).
На стыках соседних территорий во многих
местах отчетливо выражены постоянные зоны
пограничных конфликтов, протекающих до-
вольно интенсивно с пением, разнообразны-
ми демонстрациями и драками. Вне сферы
общения брачных партнеров пограничные
конфликты — это единственный регулярный
тип социальных контактов взрослых особей.
На горных плато Тувы и Алтая для монголь-
ских земляных воробьев типичны поселения
с довольно низко!! плотностью и относитель-
но равномерным распределением гнездящихся
нар в пределах пригодных битопов (Табл.5).
Самцы монгольских земляных воробьев
охраняют и рекламируют свои обширные
территории на протяжение почти всего реп-
родуктивного цикла. Заметная активизация
территориальной и токовой активности сам-
цов происходит с началом массового наси-
живания первой кладки, когда самцы, остав-
шись в одиночестве, много времени уделяют
пению, токовым полетам и патрулированию
границ своих территорий. С началом выкар-
мливания птенцов территориальная актив-
ность самцов падает, но вновь возрастает в
преддверии второго репродуктивного цикла.
Даже в период вождения выводков самцы
перемещаются главным образом в предела-
ми собственных территорий и крайне нега-
тивно реагируют друг на друга при эпизоди-
ческих встречах на границах. В целом же тер-
риториальное поведение и высокая агрессив-
ность монгольских земляных воробьев в со-
вокупности с хорошей обзорностью в зани-
маемых ими стациях очень существенно ог-
раничивают пространственные перемещения
особей и, возможно, существенно лимитируют
плотность локальных поселений этого вида
(Иваницкий, Шевченко, 1991).
Наши наблюдения за короткопалыми во-
робьями в Закавказье подтвердили данные
о его преимущественно изолированном гнез-
довании (Зарудный, 1896; Адамян, 1965). В
Закавказье, как и в других частях ареала,
короткопалые воробьи населяют довольно
резко пересеченный рельеф и нередко гнез-
дятся на довольно крутых склонах, покры-
тых разнообразными осыпями и поросших
редким кустарником. Структура поселений ба-
зируется па жесткой территориальности, хотя
границы территорий специально не патрули-
руются и выражены нечетко. Самцы занима-
ют территории сразу же по прилету и ин-
тенсивно рекламируют их с помощью нения
и мало ритуализированных трепещущих по-
летов. По нашим наблюдениям, самцы корот-
копалых воробьев на своих территориях от-
личаются высокой агрессивностью, часто на-
падая даже на своих самок и других мелких
птиц. Зарудный (1896) отмечает, однако, что
на водопое и во время кормежки эти воро-
бьи часто держатся стаями до 50 особей и
при этом ведут себя но отношению друг к дру-
гу вполне миролюбиво. Тем не менее, па но-
чевку устраиваются поодиночке в кронах ку-
старников на своих территориях.
2.5. Эволюция пространственной
организации в популяциях воробь-
ев и родственных им птиц
Пространственная структура популяций
воробьев очень пластична. Размах индивиду-
альной и популяционной изменчивости в
известной мере перекрывает даже видовые
различия. Это маскирует видовую специфику
популяционных структур и существенно зат-
рудияег эволюционный анализ. Например, го-
воря о суперколониалыюсти испанских н
индийских воробьев нельзя забывать о том,
что часть особен этого вида гнездиться все
же поодаль от сородичей. Наоборот, редкие
эпизоды колониального гнездования извест-
ны даже у сугубо одиночных саксаульных и
пустынных воробьев (Иваницкий, 1985а).
Градации “социальности” воробьев —
изолированное гнездование, квазиколониаль-
ность, нормальная колопиальность и сунер-
колониалыюсть — являются характеристи-
ками отдельных особей или поселений, по,
взятые поодиночке, они в большинстве слу-
чаев не охватывают всего спектра социо-нро-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
29
странственных стратегий того или иного вида,
включающего иногда все 4 градации (индий-
ский воробей). Поразительным разнообрази-
ем отличаются нространственые отношения
у каменных воробьев.
Кроме того, на базе изоморфных но своей
пространственной структуре популяционных
систем могут устанавливаться существенно
различные этологические отношения. Сравним
пустынных, саксаульных и монгольских зем-
ляных воробьев. У всех трех преобладает изо-
лированное гнездование, но видовые особен-
ности обозначены четко: у пустынных воро-
бьев социальные контакты вне семейной сфе-
ры сведены к минимуму, у саксаульных воро-
бьев они составляют характерную черту жиз-
ни социума на первых стадиях размножения,
у монгольских земляных воробьев — благо-
даря высокой социальной активности самцов
— поддерживаются па всем протяжении реп-
родуктивного цикла. Другой яркий пример это-
логических различий при очевидном сходстве
(изоморфизме) пространственных структур
дают испанские и индийские воробьи, тяготе-
ющие к противоположному краю спектра со-
цио-пространственных стратегий. Обоим видам
присущи продвинутые формы стайности и
колоииалыюсти, но испанские воробьи в сущ-
ности чрезвычайно индифферентны в соци-
альном плане и явно избегают вступать в пер-
сонализированные контакты с соседями но
стае или колонии, в то время как такие кон-
такты составляют существо социальной жиз-
ни индийских воробьев. И хотя но формаль-
ным признакам, обычно употребляемым при
оценках степени социальности видов (степе-
ни пространственной консолидации и обще-
му числу совместно гнездящихся пар), испан-
ский воробей превосходит индийского, все-таки
“более социальным” в этой паре следует, па
наш взгляд, считать индийского воробья.
Большинство видов воробьев тяготеют к
общественному образу жизни, причем стай-
ность распространена шире колониальное™:
многие воробьи, практикующие изолирован-
ное гнездование, сбиваются в стаи после окон-
чания размножения. Среди 20 видов Passer
исключительно или преимущественно изоли-
рованное гнездование известно лишь для
motitensis, diffusus, ammodendri и simplex,
хотя по крайней мере 2 последних вида эпи-
зодически размножаются в небольших коло-
ниях. Еще несколько видов сочетают преиму-
щественно изолированное гнездование с эпи-
зодической или регулярной квазиколоииаль-
ностью (gongoensis, rutilans, pyrrhonotus). Все
остальные представители рода во всяком слу-
чае достигают уровня нормальной колониаль-
ности (flaveolus, moabiticus, griseus, suachelicus,
castanopterus, eminibey, iagoensis) и по край-
ней мере 6 видов — уровня сунерколониаль-
ности (montanus, domesticus, indicus, hispani-
olensis, melanurus, luteus). Колониалыюсть весь-
ма характерна для каменных воробьев (моно-
типичный р.Petronia), а квазиколониалыюсть
не представляет редкости у 2 видов снежных
воробьев (pMontifringilla). В то же время, для
всех 4 видов Gymnorhis, 5 видов Pyrgilauda и
у монотипичиого Carpospiza достоверно изве-
стно лишь изолированное гнездование, однако
после окончания размножения эти воробьи
чаще всего встречаются стаями.
В родственных воробьям группах разнооб-
разие пространственных структур повторяет-
ся не только в целом, но и в мельчайших дета-
лях. Вот что мы видим у Ploceidae. Предста-
вители обширного рода Ploceus образуют пол-
ный ряд от изолированно гнездящихся, пре-
имущественно моногамных видов к высоко
колониальным (в том числе суперколониаль-
пым) преимущественно полигамным формам
(Crook, 1963а,с, 1964; Collias, Collias, 1969,1970).
Среди полигамных Euplectes, но-видимому, нет
видов, гнездящихся лишь дисперсным спосо-
бом, зато представлены все остальные града-
ции пространственных структур вплоть до су-
перколониалыюсти (Craig, 1980). Все Quelea
моногамны и суперколопиальпы (Ward, 1963;
Morel. More), 1978). Среди представителей
родов Foudia и Malimbus известны лишь изо-
лированно гнездящиеся моногамные формы
(Moreau, 1960b; Crook, 1961; Brosset, 1978).
Интересно, что у воробьев и ткачиков по-
вторяются не только варианты макрострук-
туры гнездовых сообществ, определяемые вза-
имным размещением и локальной плотнос-
тью гнезд, но и основные варианты микро-
структуры поселений. Например, у Euplectes
hordacea гнездовые территории варьируют
по площади от 400 до 1000 м2 и почти но
всему периметру очерчены довольно четки-
ми границами, которые усиленно натрулиру-
30
В.В.Иваницкий
ются самцами, в то время как у самцов
E.nigroventris, обитающих в тех же биотопах
бок о бок с предыдущим видом, территории
не имеют четких границ, а их площадь но мере
увеличения плотности может сокращаться
всего до 6 м2 (Moreau, 1938). В многочислен-
ных и плотных колониях Ploceus cucullatus,
где живут иногда сотни птиц, каждый самец,
тем не менее, защищает вокруг своих гнезд
строго определенную территорию, то и дело
вступая с соседями в пограничные конфлик-
ты, которые сопровождаются весьма специ-
фическим демонстративным поведением. В
противоположность этому, территории
P.nigerrimus, гнездование которых также про-
исходит в колониях, не имеют патрулируе-
мых границ и четкой конфигурации, а в ре-
пертуаре самцов отсутствуют и соответству-
ющие демонстративные позы (Crook, 1963с).
Среди сравнительно немногочисленных
представителей н/сем. Plocepasserinae так-
же отмечается значительное разнообразие
форм популяционною устройства. Например,
Plocepasser mahali живут нарами или груп-
пами до 9 особей на обособленных террито-
риях с четко обозначенными границами пло-
щадью до 12 га, но в ряде мест гнездятся со
значительно более высокой плотностью при
расстояниях между гнездами соседних групп
32-85 м, когда территориальное поведение
становится главным фактором, ограничива-
ющим дальнейший рост числа репродуктив-
ных группировок. В состав групп этого вида,
помимо пары размножающихся особей, час-
то входят так называемые помощники — до-
стигшие репродуктивного возраста и непо-
ловозрелые потомки данной пары, участву-
ющие в постройке гнезд, выкармливании
птенцов своих родителей и охране террито-
рии группы (Collias, Collias, 1978а,b; Levis,
1982; Wingfield et al., 1991).
Большей общественностью отличаются
Pseudonigritta arnaudii, образующие своеоб-
разные колонии. На одном дереве уживаются
до 5 групп, каждая из которых, подобно пре-
дыдущему виду, включают пару размножаю-
щихся особей и до 5 помощников. Члены ко-
лонии свободно используют все пространство
кроны. Впутриколоииальная (групповая) тер-
риториальность отсутствует, по в сообществе
четко выражена социальная иерархия, охва-
тывающая не только членов одной колонии,
по и жителей соседних колоний, расположен-
ных па расстоянии до 300 м друг от друга
(Collias, Collias, 1980а).
В колониях Philetairus socius насчитыва-
ются десятки (иногда более сотни) гнезд,
вплотную примыкающих друг к другу и при-
крытых сверху толстой и плотной массой гнез-
дового материала (общей крышей) (см.ниже).
Обращенные вертикально вниз входы в со-
седние гнезда могут располагаться всего в 10
см друг от друга, но гнездовые камеры всегда
полностью изолированы. В гпездостроепии и
выкармливании птенцов у этого вида также
участвуют помощники, однако их родственные
и социальные связи пока не изучены (MacLe-
an, 1973; Collias, Collias, 1978b, 1980b).
У Estrildidae изменчивость пространствен-
ных структур более ограничена. Отчетливо
преобладает изолированное гнездование, от-
дельные формы амадин (pp.Lonchura и
Taeniopygia) проявляют склонность к квази-
колониалыюсти и иногда селятся небольши-
ми колониями, а у амадипы Гульда {Chloebia
gouldiae) — во многих отношениях представ-
ляющую собой высоко специализированную,
уклоняющуюся форму, известно коллектив-
ное гнездование нескольких самок в одном
убежище. Вне сезона размножения все вос-
коклювые ткачики держаться сплоченными
стаями, социальная организация которых мо-
жет быть чрезвычайно сложной (Guttinger,
1970; Goodwin, 1982; Лукина, 1986).
Таким образом, во всех филогенетических
ветвях Ploceimorpha и на всех таксономичес-
ких уровнях от вида до семейства основные
варианты пространственных структур повто-
ряются (Иваницкий, 1985а). Эти наралеллиз-
мы свидетельствуют о монофилин Plocei-
morpha, ибо, как известно, сходные структуры
чаще возникают в близкородственных таксо-
нах. Если сравнить Ploceimorpha с другими,
генеалогически более отдаленными группа-
ми зерноядных воробьиных — жаворонко-
выми, вьюрковыми и овсянковыми, то мож-
но легко убедиться в том, что спектр изенчи-
вости пространственных структур последних
выглядит совершенно иначе. Он более узок
и в нем безусловно доминирует изолирован-
ное гнездование — лишь отдельные виды
вьюрков и овсянок достигают уровня квази-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
31
колониальное™. Отметим, что продвинутые
формы колониальное™ среди воробьиных
птиц в целом встречаются крайне редко, в то
время как правилом является рассосредото-
чешюе гнездование (см.Иваницкий, 19956).
В дополнение к высокой изменчивости
пространственных структур, разные виды во-
робьев резко отличаются но своей подвижно-
сти и общей стратегии использования про-
странства. Консервативная стратегия сопря-
жена с ярко выраженной, долгосрочной при-
вязанностью индивидов к постоянным участ-
кам обитания в форме оседлости (домовый
и пустынный воробьи) или гнездового кон-
серватизма (индийский воробей). Лабильная
стратегия, напротив, характеризуется перио-
дической смеиой участков обитания и непос-
тоянством мест гнездования (испанский и сак-
саульный воробьи, некоторые популяции по-
левого воробья). Конкретные данные о видо-
вых и популяционных особенностях динами-
ки персонального состава населения испанс-
ких, индийских, домовых, полевых, саксауль-
ных и пустынных воробьев приведены в
Табл.6 (см. также Иваницкий, Матюхин,
1990а,6, 1995). Оказалось, что приверженность
разных видов той или иной стратегии ис-
пользования пространства не зависит пи от
типа пространственной структуры, ни от сте-
пени оседлости. Весьма неоднозначна зави-
симость постоянства персонального состава
поселений и годовой динамики их числен-
ности. Примечательно, что резкие различия в
постоянстве населения обнаружены у видов
с очень сходной экологией, например у ин-
дийских и испанских (колониальные виды)
или пустынных и саксаульных (дисперсно
гнездящиеся виды) воробьев. Хотя различия
в образе жизни этих видов несомненно су-
ществуют, на наш взгляд, их трудно однознач-
но соотнести с глубокой дивергенцией их про-
странственных стратегий. Исчерпывающее
обсуждение проблемы дивергенции страте-
гий использования пространства птицами
содержится в монографиях Соколова (1991)
и Рябицева (1993).
Вопросу о происхождении колониально-
го гнездования у птиц посвящено колоссаль-
Таблица 6. ПОСТОЯНСТВО НАСЕЛЕНИЯ РАЗНЫХ ВИДОВ ВОРОБЬЕВ
(по Иваницкий, Матюхин, 1995)
Виды место контроля число мече- ных время наблюде- ний (лет) встречено повторно спустя год (в %%) макс ми ним. в сред. источник данных
Пустынный воробей у гнезд 15 3 55,5 33,3 44.4 наши данные
Саксаульный у гнезд 25 3 - - - наши
воробей на водопое 76 3 26,7 17,4 22,0 данные
Полевой у гнезд 207 3 6,4 5,0 5,8 Иваницкий, Матюхин, 1990
воробей на ночевке 196 3 6,1 4,9 5,2
Домовый воробей на ночевке 457 6 18,9 26,1 20,6 Иваницкий, Матюхин, 1990
Индийский у гнезд 198 3 20,6 28,2 24.4 Матюхин, неопубл.;
воробей у гнезд 9703 4 1,6 6,1 3,5 Остапенко и др., 1986
Испанский у гнезд 255 3 0 1,5 0,8 Гаврилов, 1962;
воробей у гнезд 3484 4 0 0,9 0,3 Остапенко и др., 1986
32
В. В. Иваницкий
ное число публикаций, которые были проана-
лизированы в ряде обзорных работ (Панов,
1983а,б; Burger, 1985; Wittenberger, Hunt,
1985). В фокусе обсуждения уже более полу-
века остаются три-четыре главные идеи, трак-
тующие колониалыюсть или как эпифено-
мен дефицита оптимальных для гнездования
стаций, или же как результат направленной
трансформации изолированного гнездования
в колонию под селектирующим воздействием
факторов внешней среды, в числе которых не-
изменно фигурируют хищники и пищевые
ресурсы. Утверждается, что птицы активно
стремиться устраивать свои гнезда рядом друг
с другом потому, что такой способ гнездова-
ния дает им определенные преимущества, но
сравиешно с особями, избравшими изолирован-
ное гнездование, которое в данной экологичес-
кой обстановке признается менее выгодным.
Современная среда обитания воробьев, на
наш взгляд, не содержит прямых указаний на
возможные пути становления колониальнос-
ти в это группе. Во-нервых, воробьи не спо-
собны ни к индивидуальной, ни к коллек-
тивной защите гнезд и колоний, представля-
ющих собой идеальное поле деятельности
хищников (рептилий, птиц, млекопитающих),
причиняющих тяжелый ущерб (Федянина,
1977; Голованова, 1985; Попов, 1962; паши
наблюдения). Во-вторых, в окрестностях круп-
ных колоний быстро истощаются пищевые
ресурсы, что вынуждает птиц летать за кор-
мом все дальше и ведет к возрастанию энер-
гетических затрат. Между тем, расчеты пока-
зывают, что в период выкармливания птен-
цов дистанция в 250 м является для индийс-
ких воробьев критической дальностью регу-
лярных полетов за кормом (Зинченко, 1991).
В-третьих, колония представляет собой бла-
гоприятную среду для массового распростра-
нения эктопаразитов (блох и клещей), бакте-
риальных и вирусных инфекций (Семашко,
1959, 1961 а,б). В-четвертых, перенаселение в
колониях воробьев ведет к многочисленным
эффектам дезорганизации и социо-ренродук-
тивным патологиям типа разрушения гнезд
при воровстве гнездового материала, повреж-
дения кладок при посещениях гнезд носто-
ропними особями, гнездового паразитизма, ин-
фантицида, клентопаразитизма слетков у гнезд
с маленькими птенцами, актов сексуальной
агрессии и общих высоких затрат на регу-
ляцию взаимоотношений в сообществе (Schi-
fferli, 1980: Иваницкий, 1984а, Ivanitskii, 1985,
1996; Cordero, Sanchez-Aquado, 1988; Kendra,
Roth, Tallamy, 1988; Yom-Tov, 1980; Veiga, 1990
a,b, 1992; Матюхин и др., 1990).
Вопрос о влиянии плотности гнездования
и тина социального окружения на успешность
размножения воробьев изучен пока недоста-
точно. Отрицательное влияние переуплотне-
ния на размножение индийских воробьев в
Казахстане было подтверждено Матюхиным
с соавторами (1992). Неоднозначные резуль-
таты получены Sasvari и Hegui (1993) при
изучении полицикличной популяции ноле-
вых воробьев в Венгрии. Первое гнездование
протекало более успешно в колониях, второе
— у одиночно гнездящихся нар, результаты
третьего цикла были одинаковыми. Авторы
считают, что успешность размножения вида
сильно зависит от погоды: при высокой влаж-
ности увеличивается вероятность заражения
кладок и птенцов патогенными микроорга-
низмами, распространение которых быстрее
идет в колониях. Эти данные, на наш взгляд,
представляют особый интерес потому, что, во-
нервых, убедительно подтверждают понижен-
ную устойчивость переуплотненных поселе-
ний птиц к патогенным инфекционным воз-
действиям, и, во-вторых, дают некоторую по-
чву для гипотез о природе изменчивости
пространственных отношений воробьев, ког-
да в одной популяции можно встретить и
многочисленные колонии, и одиночно гнез-
дящиеся пары.
В Табл.7 представлены наши данные ио
выживаемости гнезд индийских воробьев в
четырех популяциях. Гнезда располагались
в закрытых убежищах, н контролировать их
содержимое было невозможно. Поэтому, об
успешности размножения мы судили ио чис-
лу гнезд, выживших от начала инкубации
до вылупления птенцов. Для певчих птиц это
очень ответственный период, потому что реп-
родуктивные потери за время насиживания,
как правило, превышают потери в период
выкармливания птенцов (Паевский, 1985). Во
всех популяциях выживаемость одиночных
гнезд (> 50 м от других гнезд) была не-
сколько выше, чем у гнезд в колониях. Тем
не менее, эти различия недостоверны. Раз-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
33
личия по сумме наблюдений также не дос-
тигают порога достоверности, когда критерий
X2 применяется с поправкой па непрерыв-
ность, что в данном случае оправдано из-за
разных объемов выборок. Без поправки х2
достигает первого порога достоверности (Р
< 0,05). Таким образом, имеющиеся данные
не подтверждают более высокой успешнос-
ти размножения воробьев в колониях но
сравнению с одиночными парами. Скорее
можно говорить о противоположной тенден-
ции. И тем не менее, воробьи чаще выбира-
ют колонию. Так, в Бадхызе, где индийские
воробьи живут в естественном ландшафте
горной саванны, весной 1986 г. на площади
около 2,5 кв км 218 нар жили в одной круп-
ной колонии, 69 — в 7-и небольших колони-
ях разной величины и плотности и лишь
25 пар (8%) гнездились обособленно (не
ближе 50 м от других воробьев).
Пестрота биотопического спектра воробь-
ев, их оппортунистичность по отношению к
пищевым ресурсам, индивидуальная пластич-
ность при выборе социального окружения
оставляют мало шансов найти четкие корре-
ляции между экологическими особенностя-
ми разных видов и свойствами их популяци-
онной организации. В современной среде
обитания воробьев мы не видим явных ука-
заний на какие-либо особые преимущества
колониальное*™ по сравнению с дисперсным
гнездованием, которые можно было бы трак-
товать, как ее несомненное приспособитель-
ное значение. Парадокс состоит в том, что
очевидны как раз издержки колониальное™,
тогда как ее преимущества сугубо гипотетич-
ны и являются предметом беспрестанных
споров и дискуссий.
Иначе обстоит дело с дисперсным гнездо-
ванием, составляющим альтернативу колони-
альное™. Уже судя по абсолютному преобла-
данию дисперсного гнездования и продвину-
тых форм территориальности у рецентных
воробьиных птиц, можно предполагать, что
этот тип пространственной структуры обла-
дает широкими адаптивными возможностя-
ми и является чем-то вроде “оптимального
выбора для всех ситуаций”. Эти возможности
вполне убедительно подтверждаются и ре-
зультатами экспериментов. Твердо установ-
лена, например, роль территориальности в
регуляции плотности певчих птиц - по край-
ней мере на уровне локальных поселений
(Панов, 1983). Доказана высокая эффектив-
ность территориального поведения в борьбе
за пищевые ресурсы (см. обзоры на эту тему
Иваницкий, 19816, Ильина, 1996)). Наконец,
едва ли можно сомневаться в том, что рассос-
редоточененное гнездование затрудняет по-
иски гнезд для хищников, препятствует рас-
пространению инфекций и в известной сте-
пени гарантирует невмешательство чужих
особей в дела хозяев участка (Hinde, 1956;
Панов, 1983). Тем самым, дисперсное гнездо-
вание выглядит как типичная адаптация
широкого значения, отказ от которой в про-
цессе перехода к колониальное™ требует
очень веских оснований.
Таблица 7. ВЫЖИВАЕМОСТЬ ГНЕЗД ИНДИЙСКОГО ВОРОБЬЯ В КОЛОНИЯХ
И ПРИ ОДИНОЧНОМ ГНЕЗДОВАНИИ
Район одиночные гнезда гнезда в колонии достоверность
наблюдений наблю- дали гнезд вылу- пились птенцы наблю- дали гнезд вылу- пились птенцы различий
Бадхыз 18 15 62 47 Х2=0,12; Р=0,72
Репетек 8 6 34 26 Х2=0,14; Р=0,71
Самарканд 5 5 115 93 Х2=0,24; Р=0,62
Ашхабад 15 12 42 30 Х2=0.09; Р=0,76
Всего (в %%) 46 38 (82,6%) 253 196 (77,4%) Х2=0,34; Р=0.56
34
В.В.Иваницкий
В русле обсуждаемой проблемы лежит и
вопрос о соотношении колониальное™ и
стайности. Этологическая преемственность
между стаей и колонией несомненна, и для
большого числа видов птиц, приверженных
колониальному гнездованию, вполне спра-
ведливо замечание Симкина (1988) о том, что
колония в сущности представляет собой
“осевшую на гнездование стаю"’. Немногим
ранее аналогичную мысль высказал Литвин
(1985), отметивший, что: “сохранение груп-
повых [имеется е виду стайных - В.И.] свя-
зей, с относительно небольшими гнездовыми
территориями, и есть тот тип гнездования,
который обычно называют колониальным”.
Позиции упомянутых авторов обнаруживают
сходство и в том отношении, что оба рас-
сматривают взаимное стремление индивидов
к обществу себе подобных как имманентное
свойство всех птиц, составляющее первоос-
нову их стайного образа жизни и колониаль-
ного гнездования. Такая позиция подкреп-
ляется чрезвычайно широким распростране-
нием стайности в классе Aves, причем среди
зерноядных птиц этот тип организации соци-
ума в негнездовой период является практи-
чески универсальным. Заслуживает внима-
ния, на наш взгляд, и то обстоятельство, что
у многих птиц, в частности у всех Plocei-
morpha, стаи формируются непосредственно
в результате слияния выводков сразу после
вылета птенцов из гнезд, и, следовательно,
весь свой жизненный путь особи проходят в
условиях тесного социального контакта с
себе подобными.
Широкое распространение стайности и ее
раннее появление в онтогенезе у птиц позво-
ляет предполагать ее значительное превос-
ходство над одиночным существованием для
широкого диапазона экологических условий.
Это превосходство отчасти подтверждается
в цикле экспериментальных исследований
адаптивного значения стайности, выполнен-
ных в русле доминирующей ныне методоло-
гии оценки “преимуществ” и “недостатков”
разных поведенческих стратегий. Итоги этих
исследований, где в качестве одного из ос-
новных объектов использовали и домовых
воробьев, подведены недавно в обзоре Иль-
иной (1996). Как следует из приведенных
здесь источников, в стаях особи, полагаясь
друг на друга, меньше времени тратят на ска-
нирование окружающего пространства
(реже оглядываются) и в большей мере могут
сосредоточиться на кормежке. Однако, как
установлено в целом ряде исследований, это
преимущество полностью реализуется в
очень небольших стаях (менее 4-5 особей), и
дальнейший рост стаи не влияет на частоту и
продолжительность сканирования, зато вле-
чет за собой возрастание затрат времени и
энергии на агрессивное поведение и локомо-
торную активность, потребность в которой
возрастает под влиянием быстрого локально-
го исчерпания пиши. От себя добавим, что
важнейшей статьей расхода энергии и столь
же важным центром мобилизации внимания
особей в крупных стаях становятся социально
обусловленные всплески локомоции (коллек-
тивные взлеты, кружения и прочие воздуш-
ные маневры, описанные выше).
Экологический смысл преобразования
стаи в колонию мог бы состоять в том, что-
бы преимущества жизни в стае распростра-
нились бы и на репродуктивный период. Тем
не менее, это происходит крайне редко. У
подавляющего большинства птиц, живущих
в межсезонье крупными и сплоченными ста-
ями, с началом гнездового периода после-
дние быстро распадаются.
При обсуждении вопроса о происхожде-
ния колониального гнездования у
Ploceimorpha важно учитывать главные осо-
бенности их репродуктивного поведения.
Этому материалу посвящена отдельная глава,
и здесь мы отметим лишь наиболее суще-
ственные моменты, а именно то, что во всех
основных группах ткачикоподобных птиц
доминирующие позиции в гнездостроен и и
занимают самцы, что гнезда выполняют фун-
кции своеобразных брачных посредников, и
что само гнездостроительное поведение тесно
переплетается с токованием. В связи с упомя-
нутыми особенностями особого внимания
заслуживают, на наш взгляд, данные о том,
что интенсивность гнездостроительного по-
ведения самцов испанских воробьев и дере-
венских ткачей, а также успешность привле-
чения ими самок находится в прямой зависи-
мости от величины колоний (Collias, Collias,
1969; Collias et al., 1971; Metzmacher, 1989).
Еще ранее Crook (1960) обратил внимание на
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
35
то, что в колониях ткачиков Ploceus и Quelea
брачные демонстрации самцов часто вызыва-
ют аналогичные демонстрации у соседей (эф-
фект “социального облегчения”). Эти наблю-
дения создают определенную почву для пред-
положения о том, что преимущества коопе-
рации в деле привлечения самок (повышен-
ная заметность группировок токующих сам-
цов, взаимная стимуляция токового и гнездо-
строительного поведения) вполне могли стать
непосредственным селективным фактором,
действующим в направлении уменьшения
дистанций между центрами активности сам-
цов и сооружаемых ими гнезд.
Этот фактор, лежащий в сфере внутрипо-
пуляционных взаимоотношений, вполне мог
способствовать зарождению колониальное -
ти, однако дальнейшая ее эволюция неиз-
бежно должна оказаться под жестким кон-
тролем среды обитания. Продвинутые фор-
мы колониальности могли быть достигнуты
лишь в тех группах птиц, где еще ранее или
уже по ходу дела были “найдены” эффек-
тивные способы нейтрализации или компен-
сации отрицательных последствий переуп-
лотнения.
К числу таких “находок” можно отнести,
на наш взгляд, шаровидную форму гнезд
Ploceimorpha. Гнезда-камеры лучше защища-
ют кладку и птенцов от случайных поврежде-
ний, подбрасывания яиц и преднамеренных
насильственных действий со стороны соседей
по колонии, чем открытые сверху чашевид-
ные постройки. Кроме того, только шаровид-
ные гнезда допускают вентральное крепление
и могут подвешиваться к самым эфемерным
растительным субстратам, создающим серь-
езные проблемы для передвижения хищни-
ков (гнезда можно повесить над водой или
вокруг гнезд жалящих перепончатокрылых).
Колония обеспечивает своевременное опове-
щение о приближении хищников и тем са-
мым компенсирует один из главных недо-
статков шаровидных гнезд — невозможность
контроля за окружающим пространством, что
превращает гнездо в ловушку для наседки.
Ранее мы высказали предположение, что
высокий уровень смертности кладок и птен-
цов в колониях от хищников и социальных
помех может быть компенсирован увеличе-
нием выживаемости молодых и взрослых
птиц в межгнездовой период в благоприят-
ных условиях полной обеспеченности пищей
(Иваницкий, 19816). Примечательно в этой
связи, что уровня суперколониальности до-
стигают как раз те воробьи, которые теснее
всего связаны с агроценозами и неисчерпае-
мыми пищевыми ресурсами в виде посевов
культурных злаков (Passer hispaniolensis.
P.indicus, P.montanus. P.luteus, P.melanurus).
Среди Ploceidae наиболее многочисленные
колонии известны у деревенских (Ploceus
cucullatus) и красноклювых (Quelea quelea)
ткачиков, обладающих столь же тесными
связями с традиционными агроценозами тро-
пической Африки. Что же касается других
воробьев, обитающих зачастую в чрезвычай-
но суровых условиях, то на их примере, как
мы полагаем, скорее прослеживаются различ-
ные пути постепенной трансформации коло-
ниальности в разные варианты изолирован-
ного гнездования, составляющего основу
популяционного устройства наиболее про-
двинутых форм Passeridae и представляющего
собой господствующий тип пространствен-
ных структур во всем отряде Passeriformes.
ГЛАВА 3. УСТРОЙСТВО И РАСПОЛОЖЕНИЕ ГНЕЗД
3.1. Расположение гнезд
Воробьи, как и все другие Ploceimorpha
(короткопалый воробей составляет единствен-
ное исключение), являются типичными кла-
устрофилами, т.е. склонны гнездиться в отно-
сительно небольшом, замкнутом со всех сто-
рон пространстве, представляющем собой по-
лость естественного происхождения или же
специально построенное убежище (Иваниц-
кий, 19956). При клаустрофилии интерьер
гнезд, имеющих относительно узкий вход, ви-
зуально почти полностью изолирован от внеш-
него мира. У воробьев представлены все фор-
мы клаустрофилии: пассивные клаустрофи-
лы занимают уже готовые убежища, изготов-
ленные самой природой или же птицами
других видов, тогда как активные клаустро-
филы изготавливают убежища самостоятель-
но. В свою очередь виды, практикующие ак-
тивную клаустрофилию, подразделяются на
тех, кто выкапывает (или выщипывает) по-
лости в толще субстрата (поры в грунте или
дупла в древесине) (минирующий тин) и тех.
кто строит замкнутые гнезда-камеры из рас-
тительных материалов или грунта (конструк-
ционный тип). Таким образом, но тину распо-
ложения гнезда воробьев делятся на две ос-
новные группы. Одни гнезда располагаются
открыто в развилках ветвей древесных крон,
другие — в закрытых убежищах (дупла, тре-
щины в скалах, пустоты между камнями, норы
собственного изготовления и норы мелких
млекопитающих, укрытия антропогенного
происхождения, подходящие но конструкции
гнезда других птиц). Закрытое гнездование
распространено шире (Табл.8). Среди 20 ви-
дов Passer 4 вида практикуют только откры-
тое гнездование, 4 вида — только закрытое и
для 12 видов известны оба способа (Summers-
Smith, 1988).
Есть и переходные варианты. Среди них
особо упомянем расположение гнезд в кучах
плотно сложенных веток. Этот субстрат при-
влекает многие виды воробьев независимо от
того, который из двух типов гнездования яв-
ляется у них преобладающим. Иногда подоб-
ный субстрат имеет антропогенное происхож-
дение. Так, Голованова (1985) пишет, что ко-
лонии полевых воробьев в объемистых ку-
чах хвороста, оставшихся после лесотехничес-
ких работ, насчитывают десятки нар. Отме-
тим, что столь высокая степень колониалыю-
сти у этого вида встречается редко, что сви-
детельствует, видимо, о чрезвычайно высокой
привлекательности упомянутого субстрата. В
природе воробьи находят нагромождения
веток в виде гнезд хищных, голенастых и
вороновых птиц. Повсеместно это излюблен-
ные места гнездования домовых, индийских,
испанских, нолевых, саксаульных и пустын-
ных воробьев, поселяющихся здесь многочис-
ленными, часто смешанными колониями
(Ротшильд, 1959; Соныев, 1965; Фетисов и
др., 1981; Пономарева, 1983; Казаков, Ломад-
зе, 1984). Видимо, воробьев привлекает имен-
Таблица 8. РАСПРОСТРАНЕНИЕ ОТКРЫТОГО И ЗАКРЫТОГО ГНЕЗДОВАНИЯ
В РАЗНЫХ РОДАХ ВОРОБЬЕВ
Роды Закрытое гнездование Открытое гнездование Оба типа Объем рода (видов)
Passer 4 4 12 20
Gymnorhis 4 - - 4
Petronia 1 - - 1
Pyrgilauda 5 - - 5
Montifringilla 2 - - 2
Carpospiza - 1 - 1
Всего 16 5 12 33
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
37
но сам но себе субстрат, а не его изготовители,
потому что воробьи равно заселяют и жи-
лые, и пустые гнезда хищных птиц. Даже при
гнездовании па обрывах и очевидном избыт-
ке закрытых убежищ воробьи явно предпо-
читают расположенные здесь гнезда хищни-
ков, где формируются наиболее плотные суб-
колопии и долго не стихает ожесточенное
соперничество за “престижную жилплощадь”.
К числу видов с высокой пластичностью
устройства гнезд, принадлежит домовый во-
робей. На большей части своего громадного
ареала этот облигатный комменсал человека
гнездится в разнообразных закрытых убежи-
щах антропогенного происхождения (Ильен-
ко, 1976; Summers-Smith, 1988; Indykiewicz,
1991). Пока ничего не известно относительно
того, что побуждает этих птиц время от вре-
мени устраивать гнезда па деревьях в тех
популяциях, где безусловно преобладает зак-
рытое гнездование. Есть указания, что коло-
нии на деревьях возникают преимуществен-
но в местах высокой численности воробьев,
и что размножение здесь начинается значи-
тельно позже, чем у пар, гнездящихся в зак-
рытых убежищах (Ильенко, 1976; Храбрый,
1979; McGillivray, 1980а,b; Воробьев, 1991).
Понов (1962) и Воробьев (1991) обращают
внимание на многолетнюю приуроченность
эпизодов открытого гнездования домовых
воробьев к определенным локальным посе-
лениям.
На юге домовые воробьи гнездятся на де-
ревьях чаще, чем на севере (Келейников, 1953;
Мустафаев, 1969), но еще более широко от-
крытое гнездование распространено среди их
ближайших родственников — индийских во-
робьев. В Средней Азии крупные колонии
последних приурочены либо к древесным
насаждениям (тугаи, лесополосы, сады), либо
к глинистым, лессовым или скальным обры-
вам. Соотношение разных способов гнездова-
ния варьирует в разных популяциях (Шес-
топеров, 1926; Кашкаров, 1927; Ахмедов, 1957;
Понов, 1959,1962), но есть указания, что ин-
дийские воробьи охотнее гнездятся на обры-
вах (Семашко, 1962; Голованова, 1965, паши
данные). Крупные колонии индийских воро-
бьев на деревьях и на обрывах могут распо-
лагаться иногда всего в 2-3 км друг от друга,
причем возле колоний на обрывах нередко
отдельные пары воробьев гнездятся откры-
тым способом на ближайших деревьях. Воп-
рос о том, могут ли отдельные особи варьи-
ровать способ расположения гнезд, переходя
с обрывов на деревья и наоборот, нуждается
в специальном изучении. Мы располагаем на
этот счет лишь личным сообщением Т.Н.Зин-
ченко о том, что самка индийского воробья,
окольцованная птенцом в гнезде па обрыве,
через два года гнездилась в лесополосе в 2,5
км от места рождения. Учитывая высокий уро-
вень гнездового консерватизма этого вида и
важную роль запечатления в выборе убежищ
для гнездования у птиц (Вилкс, 1965;
Schaden,1992), в том числе и домовых воро-
бьев (Cink, 1976), можно ожидать известной
обособленности населения индийских воро-
бьев в колониях на разных субстратах, хотя, с
другой стороны, как будет показано ниже,
высокая социальная привлекательность коло-
ний может существенно модифицировать ин-
дивидуальные склонности.
В разных частях ареала индийских воро-
бьев хорошо выраженные отличия в манере
размещения гнезд. Например, в Гиссарской
долине (Таджикистан) нам не приходилось
наблюдать гнездование индийских воробьев
под крышами домов, где жили исключитель-
но нолевые воробьи, однако в Туркмении в
небольших аулах под крышами и в щелях
саманных построек бок о бок в большом чис-
ле селятся оба вида. Буквально на наших гла-
зах изменился стереотип гнездования индий-
ских воробьев в фисташниках горного Бад-
хыза. В 1976-1978 гг., мы не отмечали откры-
то расположенных гнезд этого вида — все
воробьи гнездились в скалах или в дуплах
фисташек. С начала 1980-х гг. в фисташковой
роще стали в массе гнездится испанские во-
робьи, которых прежде здесь не было. В 1986
г. мы наблюдали единичные случаи гнездо-
вания индийских воробьев в прошлогодних
открыто расположенных гнездах испанских
воробьев. В 1989 г. Л. В. Сим аки и (личное
сообщение) отмечал небольшие колонии ин-
дийских воробьев в кронах фисташек. В 1993
г. мы отметили несколько случаев гнездова-
ния индийских воробьев уже в самостоятель-
но построенных в развилках ветвей гнездах.
Скорее всего, открытое гнездование индийс-
ких воробьев было индуцировано именно по-
38
В.В.Иваницкий
явлением испанских воробьев, гнезда которых
во многих районах (см.ниже) охотно зани-
мают индийские воробьи. Интересен случай
нсевдосоциалыюй индукции открытого гнез-
дования домовых воробьев на высоком кусте,
в кроне которого с проехавшей вплотную под-
воды застряли клочки сена, отдаленно напо-
минавшие воробьиные гнезда (С.Винтер, лич-
ное сообщение). Что же касается настоящих,
открыто расположенных воробьиных гнезд
хорошей сохранности, то они могут привле-
кать даже специализированных дуплогнезд-
ников. Так, в Ренетеке мы отметили два слу-
чая гнездования саксаульных воробьев, отно-
сящихся именно к этой категории, в открыто
расположенных гнездах пустынных воробь-
ев. Несколько примеров подобного рода при-
ведены в нашем обзоре (Иваницкий, 19956).
Обратное направление социальной индук-
ции аберрантного способа гнездования ил-
люстрируют испанские воробьи. Oiui гнездят-
ся преимущественно открыто. Лишь отдель-
ные пары гнездятся в закрытых убежищах,
причем во всех известных случаях это были
пары, присоединявшиеся к колониям индийс-
ких, домовых или нолевых воробьев (Голо-
ванова, Попов, 1962; Кекилова, 1973; Konig-
stedt, Robel, 1978). По наблюдениям С.А.Бук-
реева (личное сообщение), в Западном Копет-
даге испанские воробьи изредка гнездятся в
скалах или на лессовых обрывах вдоль
р.Сумбар вместе с индийскими воробьями,
причем, в отличие от последних, гнезда испан-
ских воробьев всегда расположены более от-
крыто и хорошо видны снаружи. Таким об-
разом, воробьи способны быстро менять сте-
реотип гнездования, благодаря подражанию
близкородственным видам, использованию их
гнезд, ярко выраженному стремлению к гнез-
дованию в обществе себе подобных, включая
н особей близкородственных видов. Смена
стереотипа облегчается строением воробьиных
гнезд, представляющих собой закрытые со
всех сторон камеры, имитирующие естествен-
ного убежища.
Все 5 видов земляных воробьев
p.Pyrilauda принадлежат к крайне своеобраз-
ному адаптивному типу геофилов-норииков,
который редко встречается среди воробьи-
ных птиц. В ряду биологических особеннос-
тей этих птиц, обеспечивающих их существо-
вание в суровых климатических условиях
аридных и холодных нагорий Центральной
Азии, центральное место занимают их тесные
и разнообразные связи с норами мелких мле-
копитающих. Воробьи гнездятся и на протя-
жение всего года ночуют в норах. На Алтае
мы раскопали 9 пор пищух (Ochotona) с
гнездами воробьев. Длина галереи от входа
в нору до входа в гнездо варьировала от 120
до 306 см (в среднем 188,4±52,9 см), а глуби-
на гнезда — от 30 до 58 см (в среднем 46,2±8,9
см). Все найденные нами гнезда были устро-
ены в специальных камерах, сооружаемых пи-
щухами для хранения запасенного сена (Ива-
ницкий, Шевченко, 1992).
Хотя среди пернатых монгольский зем-
ляной воробей является одним из самых спе-
циализированных порников, нам ни разу не
удалось наблюдать самостоятельного рытья
нор. В то же время, рытье очень характерно
для индийских воробьев, гнездящихся в лес-
совых обрывах. Лесс легко поддается рытью
и обеспечивает многолетнюю сохранность нор.
Воробьи копают исключительно клювами,
откусывая небольшие кусочки грунта (до 25-
30 мм) и в клюве же вынося их наружу. По-
лость, вполне пригодную для гнездования, во-
робей выкапывает за 2 дня. Рытье — глубоко
укоренившаяся повадка индийских воробьев,
которая проявляется и у особей, занимаю-
щих готовые поры. Иногда это диктуется не-
обходимостью ремонта норы после частичных
обвалов, однако, по крайней мере в некото-
рых случаях рытье, также как и нсевдочист-
ка гнезд, явно представляет собой следствие
чисто внутреннего побуждения. Норы служат
ио много лет и их длина, увеличиваясь год от
года, может достигать 80 см (Иваницкий, 1996).
Предполагают, что основные черты воро-
бьиных гнезд, представляющих собой замк-
нутые со всех сторон шаровидные или ре-
тортовидпые камеры с узким входом сбоку
или сверху, сложились в условиях открыто-
го гнездования. После того, как воробьи вто-
рично освоили гнездование в закрытых убе-
жищах, надобность в дополнительной защите
исчезла, однако, в силу филогенетической
инерции характерный облик гнездовых по-
строек сохраняются и поныне (Collias, Collias,
1964). Представители рр.Passer и Pyrgilauda
даже в замкнутых, надежно изолированных
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
39
Рисунок 2. ПРЕДПОЛАГАЕМАЯ ЭВОЛЮЦИЯ ТИПОВ РАЗМЕЩЕНИЯ ГНЕЗД ВОРОБЬЕВ
А — типичное положение гнезд Passer hispaniolensis; Б — P.indicus; В — гнездо в дупле: Г — типичное
положение гнезда синицы Pants bokharensis в норе между корней саксаула (Рспстек) - возможный путь перехода
дуплогнездника к гнездованию в норах; Д — гнездо Pyrgilauda dauidiana в норе пищухи: Е — типичное располо-
жение гнезд Passer indicus, P.montanus, Petronia и Montifringilla на скальных стенках; Ж - один из вариантов
расположение гнезд Petronia и Montifringilla; 3 - гнездо Capospira brachydactila.
от внешнего мира естественных убежищах
сооружают типичные шаровидные гнезда
(Ivanitskii, 1996).
Видовые отличия в расположении гнезд
есть и у нетрофильных, и у дендрофильных
воробьев. Например, индийские воробьи от-
четливо тяготеют к высоким обрывистым кли-
фам (Рис.2, Е) и никогда не гнездятся под
лежащими на земле камнями. В противопо-
ложность этому, гнезда каменных или снеж-
ных воробьев можно обнаружить и на самой
вершине высокого утеса в десятках метров от
днища долины, и у подножия того же утеса
среди осыпей (Рис.2, Ж), и даже под неболь-
шим камнем, отдельно лежащим на ровной
почве.
При гнездовании в кронах у воробьев
прежде всего варьирует степень укрытости
гнезд. Виды, явно тяготеющие к убежищам
закрытого тина (домовый, полевой, каменный
воробьи), при эпизодическом гнездовании в
кронах всегда выбирают самые укрытые уча-
стки среди плотного переплетения ветвей и
густой листвы (Шульнин, 1956; Фетисов и
др., 1981; McGillvray, 1980а). Что же касается
видов, гнездящихся исключительно или пре-
имущественно открытым способом, то неко-
торые из них не только не стремятся спря-
тать свои гнезда в густой листве, по, даже при
наличии таковой возможности, выбирают под
постройку сухие деревья, засохшие и наиме-
нее облиственные ветви, благодаря чему мас-
сивные воробьиные гнезда, устроенные в столь
демаскирующей манере, бросаются в глаза с
далекого расстояния (Yom-Tov, 1980; Sum-
mers-Smith, 1988). В смешанных колониях ис-
панских и индийских воробьев на деревьях с
объемной кроной (белая акация, карагач) пер-
40
В. В. Иваницкий
вые, как правило, строят свои гнезда на пери-
ферии кроны (Рис.2, А), часто в конечных
мутовках ветвей, в то время как индийские
воробьи предпочитают центральные участки
кроны, а их гнезда отчетливо тяготеют к тол-
стым скелетным ветвям и часто располага-
ются у центрального ствола (Рис.2, Б) (Па-
нов, Раджабли, 1972; Иваницкий, 1984а).
Исходя из предположения о первичности
открытого гнездования, мы составили гипоте-
тическую схему эволюции способов располо-
жения гнезд у воробьев (Рис.2). Переход от
открытого гнездования в кронах к размноже-
нию в закрытых убежищах стал одним из клю-
чевых событий в эволюционной истории во-
робьев. После того, как воробьи преодолели
порог, отделяющий открыто гнездящихся птиц
от дуплогнездников, перед ними открылись
возможности быстрого освоения новых гнез-
довых субстратов и убежищ, а также колони-
зации новых ландшафтов. С высокой долей
вероятности можно утверждать, что предки
воробьев и других Ploceimorpha были дре-
весными птицами с открытым типом гнездо-
вания. По ряду причин, которые будут рассмот-
рены ниже, воробьи не смогли выдвинуться
на доминирующие позиции в лесных экосис-
темах. Адаптивная радиация воробьев прохо-
дила главным образом в интразональных мес-
тообитаниях и экотонах, где дуплогнездность,
несмотря на ряд связанных с нею ограничений,
с эволюционной точки зрения могла стать весь-
ма перспективным приобретением, благодаря
высокому разнообразию соседствующих друг
с другом гнездовых субстратов, когда, напри-
мер, недостаток дупел мог быть легко компен-
сирован обилием убежищ в скалах, обрывах jt
т.н. Все сказанное, с нашей точки зрения, дает
основания рассматривать переход к закрыто-
му гнездованию, как ведущую адаптацию воро-
бьев, хорошо отличающую их от всех Plocei-
morpha, равно как и от более удаленных групп
мелких зерноядных птиц (жаворонковых,
вьюрковых, овсянковых).
3.2. Воробьи в межвидовой кон-
куренции за гнездовые убежища
Перейдя к закрытому гнездованию в дуп-
лах, воробьи попали в жесткую зависимость
от наличия укрытий и вступили в конкурен-
цию за них. Имеется немало свидетельств того,
что воробьи вынуждены уступать обжитые
убежища более сильным конкурентам —
скворцам Stumus vulgaris, стрижам Apusapus
и др. (Ильенко, 1965; Lohrl, 1978; Фетисов и
др., 1981). Нов целом воробьи хорошо при-
способлены к борьбе за убежища, а их конку-
рентоспособность в этой сфере значительно
выше, чем у многих других птиц. Воробьи
чрезвычайно агрессивны, настойчивы и склон-
ны к насильственному захвату жилых гнезд
многих видов. Так, в Каракумах саксаульные
воробьи вытесняют из дупел бухарских си-
ниц (Соныев, 1967,1970; наши наблюдения),
па Сахалине рыжие воробьи изгоняют ост-
рокрылых дятлов (Нечаев, 1973), в Узбекис-
тане индийские воробьи захватывают норы
щурок Мег ops apiaster и М. superciliosus, ока-
зывая на этих птиц серьезное негативное воз-
действие (Косенко, 1992), множество приме-
ров захвата жилых дупел других певчих птиц
нолевыми воробьями приводят Елисеева
(1960) Фетисов и др. (1981). Гнезда ласточек
(Delichon urbica, Tachycineta bicolor) повсе-
местно служат излюбленным местом гнездо-
вания домовых воробьев, которые не только
изгоняют хозяев из принадлежащих им пост-
роек, выбрасывая их яйца и птенцов, но при
определенных обстоятельствах могут убить
и взрослых ласточек (Winkler, 1992). По на-
блюдениям Гаврилова (1962), в Казахстане
испанские воробьи, пользуясь своей многочис-
ленностью, воробьи вытесняют даже таких
крупных и агрессивных птиц, как черноло-
бые сорокопуты Lanius minor, что подтверди-
ли и наши наблюдения за взаимоотношени-
ями этих видов в Бадхызе.
Воробьи подолгу осаждают избранное
убежище. Один или оба члена воробьиной
пары почти непрерывно сидят у входа, вни-
мательно наблюдая за хозяевами и поджи-
дая момент для проникновения в убежище.
Такое поведение пе только в отношение дру-
гих видов, но присутствует и во взаимоот-
ношениях конспецифичных особей. Например,
у индийских воробьев в период формирова-
ния колоний тактику выжидания при захвате
чужих участков и убежищ применяют многие
самцы и самки (Матюхин и др., 1990).
Эффективность действий воробьев облег-
чается их численным превосходством, особен-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
41
но вначале формирования колоний, когда пре-
обладает коллективная инспекция убежищ, и
воробьи держатся преимущественно неболь-
шими группами. Пока хозяин гнезда гоняет-
ся за одним воробьем, в его гнездо проникают
другие претенденты. Мы не раз наблюдали,
как в период массового прилета индийских
воробьев каменные воробьи, скалистые попол-
зни, скворцы и щурки вынуждены были ос-
тавлять свои гнезда.
В Таджикистане мы наблюдали массовое
нашествие индийских воробьев в колонию
испанских воробьев накануне вылета птен-
цов последних. Многие гнезда оказались в
двойном владении — нары испанских воро-
бьев и одиночного самца или пары индийс-
ких воробьев. Интересно, что и хозяева и
пришельцы вели себя так, точно гнездо при-
надлежало только им. Испанские воробьи
почти не обращали внимания на конкурен-
тов, проявляя агрессию лишь в тех случаях,
когда последние пытались проникнуть в гнез-
до непосредственно в их присутствии. Индий-
ские воробьи защищали “свои” гнезда от кон-
специфичных особей, активно их рекламиро-
вали и даже образовывали нары. В отсут-
ствие законных хозяев индийские воробьи
свободно залезали в гнезда, но, судя но всему,
не причиняли заметного ущерба подросшим
птенцам испанских воробьев, которые к это-
му времени уже не уступали в размерам
индийским и с присущей птенцам этого вида
бурной экспрессией выпрашивали у них корм.
Всего в этой колонии гнездились 36 пар ис-
панских воробьев, и после вылета их птен-
цов 17 освободившихся, наконец, гнезд заня-
ли индийские воробьи.
К числу биологических черт воробьев,
увеличивающих их конкурентоспособность в
борьбе за убежища, принадлежат и некото-
рые особенности их гнездового поведения. Во-
первых, воробьи гнездятся в дуплах и дру-
гих убежищах вне зависимости от степени
их заполненности гнездовым материалом, в
чем состоит важное отличие воробьев от мно-
гих других дуплогнездников (синиц, мухоло-
вок, горихвосток), избегающих занимать убе-
жища с большим количеством прошлогодне-
го материала, из-за чего, как это хорошо изве-
стно в практике привлечения насекомоядных
птиц, существует необходимость ежегодной
очистки искусственных гнездовий от остат-
ков гнездового материала.
Вторая особенность гнездового поведения
воробьев состоит в том, что на первых эта-
пах освоения любого закрытого убежища и
у самцов, и у самок ярко выражено стрем-
ление к выносу из него разнообразных пред-
метов. По нашим наблюдениям, эта реакция
свойственная всем видам воробьев, практи-
кующим закрытое гнездование. Подробный
этологический анализ этого поведения мы
проведем в следующей главе, а здесь обсу-
дим лишь некоторые его экологические и
эволюционные аспекты.
Можно предполагать, что в нору ста-
новления этой реакции ее ведущая функ-
ция сводилась к очистке гнездовой полос-
ти от старого гнездового материала с мно-
гочисленными эктопаразитами, хотя сле-
дует заметить, что воробьи почти никог-
да не доводят очистку гнездовой полости
до конца и, как отмечено выше, проявляют
крайнюю терпимость к наличию старого
гнездового материала в любом количестве.
Вполне возможно, также, что в реакция
“выноса из гнезда” стала тем этологичес-
ким субстратом, на основе которого в про-
цессе последующей эволюции сформиро-
вались некоторые особые формы подго-
товки или даже специального сооруже-
ния гнездовых убежищ. Например, сакса-
ульные и нолевые воробьи расширяют
дупла, отщипывая кусочки гнилой древе-
сины и вынося их наружу (Гаврилов,
1962; Фетисов и др., 1981).
Захватывая убежища других видов, воро-
бьи выбрасывают яйца и птенцов хозяев —
реакция, которая также могла сложиться на
основе поведения “выноса из гнезда” (Ели-
сеева, 1960; Благосклонов, 1991; Winkler, 1992).
В Бадхызе мы дважды наблюдали успеш-
ные нападения индийских воробьев на лен-
ные гнезда большого скалистого поползня
Sitta tephronota. В первом случае воробьи
расклевали и выбросили яйца, во втором слу-
чае после длительной осады гнезда воробья-
ми мы нашли под ним четырех мертвых птен-
цов поползня, причем многочисленные гема-
томы в области головы указывали на то, что
птенцы вначале были убиты. В Репетеке под
одним из дупел, где активно строила гнездо
42
В. В. Иваницкий
пара саксаульных воробьев, мы нашли двух
сильно расклеванных птенцов бухарской си-
ницы. Интересно, что в Ренетекской популя-
ции бухарских синиц сложился весьма ха-
рактерный стереотип тревожного поведения
при виде воробьев (Соныев, 1967; 1970; наши
наблюдения).
Пиратские наклонности воробьев способ-
ствуют их успеху в межвидовой конкурен-
ции за убежища, но при определенных об-
стоятельствах распространяются на гнезда
своего вида и приводят к инфантициду. Так,
у домовых воробьев от инфантицида стра-
дают до 9-12% всех гнезд (Veiga, 1990
а,Ь, 1992). По наблюдениям Матюхина с со-
авторами (1990), инфантицид снижает ус-
пешность размножения индийских воробьев
в наиболее .плотных участках колонии. В
литературе появляется все больше сведений
о широком распространении инфантицида
среди воробьиных птиц с разным типом гнез-
дования, и в настоящее время некоторые
авторы рассматривают это явление как важ-
ный фактор внутривидовой и межвидовой
конкуренции (Belles-Isles, Pieman, 1986а,b;
Check, Robertson, 1991; Pieman, 1992).
Способности воробьев к захвату чужих
гнезд благоприятствуют популяционной или
даже видовой специализации к гнездовому
паразитизму. По свидетельству Благоеклопо-
ва (1991), в местах с высокой численностью
городских ласточек домовые воробьи прак-
тически полностью переходят к гнездованию
в их постройках. Глубокая специализация к
гнездовому паразитизму характерна для аф-
риканских Passer eminibey, использующих для
размножения исключительно только что по-
строенные, жилые гнезда сероголовых ткачи-
ков Pseudonigritta amaudii — примечатель-
но, что ареалы этих форм полностью совпада-
ют. Интересно, что ткачики заметно крупнее
и агрессивнее воробьев, и лишь беспрецеден-
тное упорство последних вынуждает закон-
ных хозяев уступать гнезда захватчикам
(Payne, 1969). Обобщив всю литературу,
Summers-Smith (1988) заключает все же, что
Р. eminibey, вопреки широкому распростра-
нению гнездового паразитизма, не утратил спо-
собности к самостоятельной постройке гнезд
типичной для открыто гнездящихся воробьев
шаровидной формы с боковым входом.
3.3. Технология гнездостроения
Будучи весьма разнообразными но раз-
мерам и употребляемым для их сооружения
материалам, все открыто расположенные гнез-
да воробьев характеризуются четко обозна-
ченным единством конструкции. Это всегда
закрытое со всех сторон, довольно массивное
сооружение, представляющее собой изолиро-
ванную от внешнего мира камеру шаровид-
ной, овальной или ретортовидной формы со
входом сбоку или сверху. От этого стандар-
та резко уклоняется лишь гнездо короткопа-
лых воробьев (Рис.2). Оно отличается значи-
тельно меньшими размерами и имеет типич-
ную для подавляющего большинства певчих
воробьиных форму открытой сверху полу-
сферы (“чашечки”), что уже сто лет служит
одной из главных причин сомнений в при-
надлежности этих птиц к I'pyiuie воробьев
(Зарудный, 1896; Адамян, 1966).
Размеры гнезд варьируют в широких пре-
делах. Самый мелкий представитель рода
(длина крыла 60 мм, вес 17,5 г), Passer
moabiticus, строит самые объемные и массив-
ные гнезда диаметром до 40 см и весом до 1
кг (Meinertzhagen, 1959). До 40 см в диамет-
ре и 470 г по весу достигают гнезда P.luteus
(длина крыла 63,5 мм, вес 14,5 г) (Morel,
Morel, 1973). Крупные и массивные гнезда с
толстыми стенками строят относительно мел-
кие пустынные воробьи Р.simplex (длина
крыла 76,5 мм, вес 19,5 г): длина гнезд дос-
тигает 250 мм, высота 180 мм (Соныев, 1965).
В то же время, гнезда испанского воробья —
одного из самых крупных видов рода (кры-
ло 80 мм, вес 28,1 г), не превышают в диамет-
ре 150-200 мм и весят от 70 до 295 г (в сред-
нем 146 г), причем весенние гнезда (163 г) тя-
желее летних (104 г) (Гаврилов, 1962). От-
крыто расположенные гнезда индийских во-
робьев(крыло 65-84 мм, вес 25 г), несколько
уступающих но размерам тела испанским во-
робьям, судя ио данным Зарудного (1896),
отличаются более крупными размерами (ди-
аметр большинства измеренных гнезд коле-
бался в интервале 210-270 мм). Открыто рас-
положенные гнезда близких по размеру до-
мовых воробьев бывают очень велики: масса
8 гнезд варьировала от 200 до 900 г, состав-
ляя в среднем 550 г (Воробьев, 1991). Самые
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
43
маленькие гнезда вьют, но-видимому, Р.руг-
rhonotus (крыло 67,4 мм) — диаметр пост-
роек не превышает 180 мм, а иногда составля-
ет всего 90 см (Summers-Smith, 1988).
Этот краткий обзор свидетельствует о
высокой изменчивости относительных разме-
ров воробьиных гнезд, размах которой в ли-
нейных измерениях достигает 4-кратной ве-
личины, в то время как размах изменчивости
размеров самих воробьев значительно мень-
ше — примерно в 2 раза но массе тела и
всего в 1,3 раза но длине крыла. Особой мас-
сивностью отличаются гнезда наиболее мел-
ких представителей Passer, населяющих са-
мые жаркие и сухие области Старого Света
— Сахару и Каракумы (Р.simplex), зону Са-
хеля и Аравию (P.luteus), котловину Мерт-
вого моря и прилегающие к ней районы
Ближнего Востока (Р. moabiticus). Самые
маленькие гнезда свойственны воробьям, на-
селяющим несколько более увлажненные лан-
дшафты (P.hispaniolensis, P.pyrrhonotus).
Технология постройки гнезд зависит от
свойств употребляемых материалов и распо-
ложения опорных элементов. При устройстве
первичного каркаса гнезда воробьи исполь-
зуют два основных приема — свивание мяг-
кого и гибкого материала (сухая или све-
жая трава, листья деревьев, гибкие прутики)
или же складывание в определенном поряд-
ке более жестких, не поддающихся сгибанию
ветвей или прутиков. Этот материал включа-
ется в конструкцию гнезда без изменения пер-
воначальной формы.
Так, основным материалом для гнезд
P.luteus (Рис.З, Л) служат толстые и длин-
ные ветки акаций с острыми шинами. На-
блюдения в природе (Morel, Morel, 1973) и в
неволе (Kunkel, 1959) показали, что манипу-
ляции со столь массивным гнездовым мате-
риалом явно затруднительны для птиц, а ост-
рые шины заставляют их действовать с осто-
рожностью. Гнездо складывается путем тща-
тельной и кропотливой пригонке веток друг
к другу и дополнительного сцепления колю-
чек. Вначале выкладывается основание, затем
стены, крыша и вестибюль (входная трубка).
Только через 10 дней самец завершает проч-
ный шаровидный каркас из ветвей, и затем
внутри самка выстилает лоток из мягких ма-
териалов.
По наблюдениям Гаврилова (1962), испан-
ские воробьи употребляют оба упомянутых
приема. Если гнездо закладывается в мутов-
ке с большим числом боковых огростков, да-
ющих опору для укрепления относительно
толстых прутиков и палочек, то строитель-
ство начинается с накопления и размещения
именно этих элементов. Если нет возможнос-
ти соорудить жесткий каркас, то употребля-
ются мягкие гибкие материалы. Тогда внача-
ле строится платформа, затем стенки, крыша
и в конце ко входу пристраивается входная
трубка-вестибюль.
Наши наблюдения за постройкой гнезд у
испанских, индийских и пустынных воробь-
ев позволяют детализировать эту схему. На
стадии первичного накопления материала он
укрепляется в мутовке в случайном порядке
и постепенно концентрируется вокруг того
места, где обычно сидит птица. Часто матери-
ал надает вниз, то и дело в отсутствие хозяи-
на его похищают соседи. Тем не менее, вскоре
устраивается базовая платформа — масса
материала, свободно лежащая среди ветвей в
мутовке. Всю последующую работу птица вы-
полняет, сидя па платформе. Важно, что, наря-
ду с типичными для всех воробьиных птиц
движениями опущенного вниз клюва, при
строительных манипуляциях у воробьев вы-
ражено инстинктивное стремление укреплять
материал не только "у себя под ногами”, по
и па боковых ветвях мутовки, дотягиваясь до
них с платформы. Воробей словно стремится
окружить себя со всех сторон гнездовым ма-
териалом. Одновременно птица наращивает
платформу, погружаясь в толщу материала
все глубже. Таким образом, сама птица пред-
ставляет собой центральное ядро, вокруг ко-
торого возводится оболочка.
Край гнезда является самым важным сти-
мулом, направляющим строительство. Клювом
птица то и дело подтягивает края па себя,
одновременно передвигая захваченный мате-
риал влево-вправо но краю гнезда и активно
формируя лоток корпусом и слегка отведен-
ными вбок крыльями. Тем самым стенка гнез-
да растет вверх, гнездовой материал равно-
мерно распределяется по краю гнезда, уплот-
няется и перемешивается за счет того, что
часть одной порции гнездового материала, зак-
репленной в каком либо месте, переносится в
44
В. В. Иваницкий
другое место и здесь также закрепляется в
общей массе материала.
Крыша гнезда образуется лишь после на-
копления относительно большой массы ма-
териала за счет того, что сидящая в гнезде
птица время от времени переносит свобод-
ные концы материала через спину, проделы-
вая характерные движения головой, и зак-
репляет их уже с другой стороны гнезда (см.
Рис. 19). Еще раз подчеркнем, что на всех
стадиях постройки птица ведет строитель-
ство “вокруг себя”, гнездовой материал все-
гда доставляется внутрь гнезда, и, таким об-
разом, стенки последнего всегда утолщают-
ся изнутри. При этом птица не только ут-
рамбовывает принесенный материал корпу-
сом и крыльями, но и клювом просовывает
его в стенки, что, по-видимому, значительно
повышает прочность всего сооружения. На-
пример, толщина стенок массивных гнезд
пустынных воробьев достигает, по нашим
данным, 4-5 см, и такое гнездо почти не де-
формируется под сидящим человеком.
На стадии формовки основного каркаса
гнезда вход, как таковой, отсутствует, и птица
иной раз в буквальном смысле слова выхо-
дит сквозь стену или крышу, придерживаясь,
однако, постоянного места, где но мере упро-
чения стенок и образуется входное отверстие,
и теперь уже его край становится важней-
шим источником стимуляции, контролирую-
щим дальнейшее поведение птицы. Входное
отверстие создает тот же самый набор сти-
мулов, что и край гнезда на предшествующей
стадии постройки. Теперь движения птицы
направлены преимущественно на край вход-
ного отверстия, чему в значительно степени
способствует и то обстоятельство, что к это-
му моменту гнездовая камера обычно уже
занята насиживающим и весьма агрессивно
настроенным партнером. Мы неоднократно
были свидетелями того, как самки испанс-
ких и индийских воробьев буквально вытал-
кивали из гнезд собственных самцов, принес-
ших очередную порцию материала (см.ниже).
В результате самец, часто желающий продол-
жать строительство вплоть до вылупления
птенцов, вынужден держаться как можно да-
лее от наседки, поэтому его деятельность ес-
тественным образом концентрируется у вхо-
да в гнездо. Здесь зачастую и скапливается
основная масса материала, которая по своему
объему и весу существенно превосходит мас-
су материала, употребленного для соружения
гнездовой камеры.
В литературе неоднократно отмечалась
чрезвычайно высокая внутривидовая измен-
чивость размеров и формы открыто распо-
ложенных воробьиных гнезд (Зарудный, 1896;
Гаврилов, 1962; Summers-Smith, 1988). Как
показали наши исследования, причины этой
изменчивости коренятся прежде всего в пер-
манентном характере гнездостроительной де-
ятельности воробьев и ее особой чувствитель-
ности к разнообразным социальным воздей-
ствиям, в ряду которых на первое место дол-
жны быть поставлены весьма сложные и про-
тиворечивые отношения брачных партнеров.
В результате и форма, и размеры воробьи-
ных гнезд испытывают непрерывные измене-
ния практически на всем протяжении реп-
родуктивного цикла, причем нередко кладка
начинается еще на стадии довольно эфемер-
ной базовой платформы, а все остальные час-
ти гнезда возводятся параллельно с насижи-
ванием (см.ниже). На Рис.З изображены не-
которые варианты формы гнезд, основные
отличия которых сводятся почти исключи-
тельно к тому, каким образом размещался
гнездовой материал после постройки гнездо-
вых камер.
В закрытых убежищах размеры и форма
гнезд во многом зависят от объема и конфи-
гурации выбранной для постройки полости.
У представителей р. Passer полнее всего спе-
цифические свойства гнезд-камер сохраняют-
ся при использовании относительно простор-
ных дупел или искусственных гнездовий.
Судя но нашим наблюдениям, последователь-
ность строительства здесь ничем не отличает-
ся от действий птиц при строительстве от-
крыто расположенных гнезд. Вначале на дне
убежища закладывается платформа, затем
возводятся стенки и крыша. Вход обычно
обращен ко входу в убежище — чаще всего
вверх и немного вбок.
Полости, занимаемые воробьями, сильно
варьируют по своему объему. В литературе
неоднократно отмечалось, что воробьи все-
гда стремятся заполнить убежище материа-
лом в максимальной степени (Ильенко, 1976;
Морозов, Остапенко, 1988; Indykiewicz, 1991).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
45
Во многих убежища материал накапливается
в течение многих лет почти круглогодично,
так что, несмотря на периодические чистки
убежищ, многие воробьиные гнезда представ-
ляют собой очень массивное и плотное скоп-
ление материала, состоящее из многих слоев,
причудливо перемешанных друг с другом,
внутри которого находится гнездовая каме-
ра, расположенная достаточно произвольно но
отношению к центру массы материала. Тем
интереснее, что наряду с чрезвычайно мас-
сивными гнездами, в некоторых убежищах,
занимаемых размножающимися воробьями,
гнездового материала может быть необычай-
но мало. Одно такое гнездо найдено нами у
саксаульных воробьев в очень маленьком ес-
тественном дупле черного саксаула в Репете-
ке, другое — на Сахалш1е, где пара рыжих
воробьев гнездилась в очень маленьком гнез-
де гаички Parus palustris, три других гнезда,
почти лишенные подстилки, принадлежали
каменным воробьям в Бадхызе и в Запад-
ном Коиетдаге. О гнездах с крайне ограни-
ченным количеством гнездового материала
известны у домовых воробьев (Indykievicz,
1991). Случаи откладки яиц и даже начала
насиживания в совершенно недостроенных, а
иногда и только что начатых гнездах, как по-
казывают наши наблюдения, отнюдь не пред-
ставляют редкости у всех изученных нами
представителей р.Passer, ио все же в боль-
шинстве случаев уже через несколько дней
гнездо приобретает законченную форму.
Наиболее ярким примером филогенетичес-
кой инерции базовых черт гнездовых конст-
рукций воробьев вопреки радикальным из-
менениям условий обитания могут служить
постройки земляных воробьев p.Pyrgilauda,
Судя но имеющимся данным, все 5 предста-
вителей рода гнездятся в норах мелких мле-
копитающих, где вьют типичные шаровидные
гнезда с боковым входом (Schaffer, 1938; Коз-
лова, 1950; Neithammer, 1967; Иваницкий,
Шевченко, 1991).
Возвращаясь к гнездам настоящих воробь-
ев (p.Awser), следует отметить, что, хотя под-
час их постройки в дуплах ничем не отлича-
ются от открыто расположенных гнезд, высо-
кая степень изоляции от внешнего мира внут-
ри замкнутых со всех сторон естественных
укрытий создает, по-видимому, некоторые пред-
посылки к редукции первоначальной формы
гнезд и ее постепенному приближению к ха-
рактерной для большинства певчих птиц ча-
шевидной форме. Среди всех изученных нами
представителей р.Passer, использующих для
гнездования закрытые убежища, глубже всего
первоначальная форма гнезда оказалось ре-
дуцированной у рыжих воробьев. Среди 7
гнезд, осмотренных нами в дуплах различных
деревьев на Сахалине, ни одно не имело кры-
ши и отчетливо выраженной задней стенки.
Во всех гнездах, кроме одного (см.выше), мы
нашли толстый слой подстилки из сухих стеб-
лей трав и листьев, не имевшей определенной
структуры, в верхней части которой был рас-
положен совершенно открытый сверху гнез-
довой лоток, свитый из травы и устланный
перьями и шестью. В сущности эти гнезда
мало чем отличались от гнезд прочих дуплог-
нездников — синиц, мухоловок и пр. Отсут-
ствие характерной крыши в гнездах рыжих
воробьев подчеркивают и другие авторы (Не-
чаев, 1973; Зембнцкая,1979).
Каменные воробьи строят гнезда преиму-
щественно в закрытых убежищах. Сообщение
Шулышпа (1956) остается пока единственным
указанием на возможность открытого гнездо-
вания этого вида. Найденные им гнезда были
расположены в густых кронах древовидной
арчи в Таласском Алатау и имели типичную
шаровидную форму, отличаясь от гнезд испанс-
ких и индийских воробьев несколько меньши-
ми размерами. Что же касается типичных для
каменных воробьев гнезд в закрытых убежи-
щах, то они ио своему устройству повторяют
некоторые варианты гнезд Passer, характери-
зующиеся отсутствием крыши. По данным Со-
колова (1986) в Забайкалье найденные им гнез-
да каменных воробьев заполняли обычно весь
объем полости, а если последняя была очень
велика, то имели вид полусферы. Осмотрев 12
гнезд каменных воробьев в Туве и на АЛтае,
мы также не обнаружили у них крыши. В про-
сторных убежищах, где птицы могли размещать
материал, не будучи стесненными ограничен-
ностью пространства, все гнезда были устрое-
ны довольно однотипно: в центре объемистой
платформе из сухих стеблей и травы диамет-
ром до 25 см и толщиной 3-4 см располагался
довольно небрежно свитый лоток диаметром SO-
75 мм, устланный шерстью и пухом.
46
В. В. Иваницкий
Гнезда снежных воробьев также далеко
удалились от исходной сферической формы.
Все известные гнезда этого вида с Памира
(Потапов, 1966) и Алтая (Сушкин, 1938;
Ирисова, 1990; наши данные) по своему уст-
ройству ничем не отличались от только что
описанных гнезд каменных воробьев, то есть
па платформе из относительно грубого рас-
тительного материала снежные воробьи со-
оружают лоточек из более топких травинок с
большим количеством шерсти и пуха, состав-
ляющих, но наблюдениям Ирисовой (1990),
до 63% всей массы гнезда.
3.4. Гнезда воробьев в ряду гнез-
довых построек Pfoceimorpha
По гнездам ткачиков и их гнездострои-
тельному поведению накоплен большой объем
данных (Crook, 1960а, 1963b; Collias, Collias,
1964,1969,1977,1978,1984; Collias, Victoria,
1978). В дополнение к литературным источ-
никам мы подробно ознакомились с гнезда-
ми мадагаскарского фу ди Foudia mada-
gascarensis — типичного представителя на-
стоящих ткачиков, интродуцированного на
Сейшельские и Амирантские острова с Ма-
дагаскара. Во время наших наблюдений тка-
чихи не гнездились, ио прекрасная сохран-
ность их многочисленных гнезд — как пол-
ностью завершенных, так и брошенных на
разных стадиях постройки, позволила не толь-
ко изучить конструкцию гнезд, но отчасти вос-
становить последовательность их сооружения.
Всего мы осмотрели 8 готовых и множество
недостроенных гнезд.
Мадагаскарский фуди принадлежит к
хорошо обособленному роду ceM.Ploceidae,
распространенному на тропических остро-
вах западной части Индийского океана.
Хотя все фуди обладают ярким половым
диморфизмом, это типично моногамные пти-
цы с равным участием полов во всех сфе-
рах гнездовой жизни, включая и постройку
гнезд (Moreau, 1960 a,b; Crook, 1961). Пред-
полагают, что технология гнездостроения
фуди отличается наибольшей простотой
среди всех родов настоящих ткачиков, хотя
но своим главным особенностям опа вполне
типичны для п/ceM.Ploceinae (Collias,
Collias, 1964).
Все найденные нами гнезда фуди при-
креплялись к впутренной (нижней) стороны
крупных перистых листьев кокосовых пальм.
Приступая к стройке, птицы сперва готовят
субстрат для крепления гнезда. Одни особи
расщепляют края листовых пластинок вдоль,
так что полоски шириной 2-3 мм и длиной
до 18 см образуют свисающий вниз пучок, к
которому и привязывается гнездо. Другие
птицы выгрызают вдоль края листа продол-
говатые щели, идущие параллельно друг дру-
гу, и уже к ним прикрепляют гнездо (Рис.З).
Интересно, что нам не удалось обнаружить
таких вариантов предварительной обработки
пальмовых листьев, которые можно было бы
рассматривать в качестве промежуточных,
хотя оба основных способа употребляются
птицами, живущими бок о бок друг с другом
па одном острове.
Хотя гнезда фуди внешне напоминают
воробьиные, это сходство поверхностно и не
распространяется далее того факта, что в обо-
их случаях постройка представляет замкну-
тую со всех сторон округлую камеру с боко-
вым входом. Наблюдателю, знакомому с мас-
сивными гнездами воробьев, прежде всего
бросается в глаза миниатюрность и внешняя
эфемерность гнезд ткачиков. Размеры гнезд
фуди варьировали от 100 до 125 мм, состав-
ляя в среднем 112,4 мм, при том, что по вели-
чине тела фуди лишь немного уступают во-
робьям. Гнезда относительно тонкостенны
(толщина стен около 3-5 мм), легки и нео-
бычайно прочны. Технология их изготовле-
ния и прикрепления базируется на уникаль-
ных для Ploceinae приемах сплетания и свя-
зывания длинных и узких полос гнездового
материала, которые детально описаны в пе-
речисленных выше источниках. Фуди соору-
жают свои гнезда исключительно из узких
полосок, отщепленных от пластинок пальмо-
вых листьев.
Касаясь отличий гнезд воробьев и ткачи-
ков, Collias и Collias (1964), отмечают, что для
сооружения похожих по форме конструкций
используются принципиально различные тех-
нологии. Выше мы подробно описали техно-
логию изготовления гнезд воробьями, в основе
которой лежат вполне ушшерсальные и отно-
сительно простые приемы манипуляций с ма-
териалом. Что же касается ткачиков, то они
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
Рисунок 3. НЕКОТОРЫЕ ТИПЫ ГНЕЗДОВЫХ ПОСТРОЕК PLOCEIMORPHA
А, Б — листья кокосовой пальмы, расклеванные ткачиками Foudia для изготовления отверстий или отщепле-
ния волокон, используемых для прикрепления гнезда; В — варианты расположения гнезд ткачиками Ploceus в
траве; Г-К — основные типы гнездовых камер: Л - гнездо Passer luteus; М — гнездо Bubalornis albirostris; Н —
коллективное гнездо Philetairus socius: 1 — крыша, 2 — гнездовая камера, заполненная материалом, 3 - жилая
гнездовая камера (А, Б — наши данные; В-М — по Crook, 1958,1963 b; Mac Lean, 1973; Morel, Morel, 1973 с
изменениями).
48
В. В. Иваницкий
пользуются чрезвычайно сложной и специа-
лизированной технологией строительства, не
имеющей аналогов среди прочих птиц. Это
отличие, будучи чрезвычайно важным, все же
не исчерпывает всего комплекса различий меж-
ду гнездами воробьев и ткачиков, в числе
которых укажем следующие:
1) Массивные гнезда воробьев всегда по-
коятся на субстрате, тогда как миниатюрные
и легкие гнезда ткачиков никогда не имеют
опоры снизу, имея лишь вентральные или
латеральные точки крепления к субстрату;
2) У воробьев полностью отсутствуют при-
емы подготовки места для гнезда или обра-
ботки гнездового материала, широко и раз-
нообразно представленные в гнездостроитель-
ном поведении ткачиков;
3) Воробьи на всех стадиях постройки
гнезда ведут строительство изнутри, тогда как
ткачики строят гнезда только снаружи, при-
ценившись к рабочему краю в том или ином
положении (часто вниз спиной). У полигам-
ных ткачиков самцы не участвуют в заботе о
потомстве и, судя но некоторым наблюдени-
ям, вообще никогда не залезают внутрь гнезд;
4) В разгар строительства воробьи но-
сят материал большими порциями, тогда как
ткачики всегда приносят по одной полоске
нли травинке, которую подолгу вплетают на
место;
5) Воробьиные гнезда плохо прикреплены
к субстрату, нз-за чего при сильном ветре мо-
гут упасть (Штегман, 19566; наши данные), в
то время как гнезда ткачиков “привязаны” к
субстрату необычайно прочно.
Таким образом, хотя гнезда воробьев и
настоящих ткачиков в целом ряде случаев
выглядят очень похоже, в сфере гнездострое-
ния эти группы характеризуются резкими,
системными отличиями.
Значительно больше общего в гнездостро-
ении воробьев и прочих групп Ploceimorpha.
Ближе всего к воробьям в этом отношении
стоят воскоклювые ткачики (Estrildidae),
которые, судя но литературным данным, строят,
в относительном измерении, еще более мас-
сивные гнезда с дорзальной точкой опоры и
пользуются традиционными приемами ук-
ладки материала. Спектры форм гнездовых
конструкций воробьев и астрильдовых в сущ-
ности одинаковы (Goodwin, 1982).
Гнезда буйволовых птиц (Bubalornithidae)
весьма своеобразны (Рис.З, М). По свидетель-
ству Crook (1958), вначале эти птицы, подоб-
но желтым воробьям, сооружают прочную
оболочку (’'скорлупу”) из довольно толстых,
длинных (15-75 см) и колючих веток. Обо-
лочка имеет примерно сферическую форму, и
в ее верней части время от времени возоб-
новляется накопление соответствующего ма-
териала, благодаря чему толщина оболочки
здесь может достигать 30 см. Внутрь оболоч-
ки птицы доставляют мягкий гнездовой ма-
териал — главным образом сухую траву —
для устройства гнездовых камер, число кото-
рых в одной оболочке может достигать шес-
ти, так как буйволовы птицы — типичные но-
лигамы, и каждое гнездо принадлежит семье
из одного самца и нескольких самок. Гнезда
служат в течение многих лет, непрерывно
достраиваются и перестраиваются всеми чле-
нами семьи, и в копце-концов внутри обо-
лочки образуется плотная, бесструктурная
масса сухой травы, где птицы но мере надоб-
ности сооружают камеры для гнездования
или ночевки. Таким образом, для буйволовых
птиц, также как и для некоторых воробьев,
характерно использование жестких и мягких
материалов для сооружения разных частей
своих построек.
Строительная деятельность воробьиных
ткачиков (n/ceM.Plocepasserinae) хорошо
изучена (Collias, Collias, 1964,1977,1978а, 1980;
MacLean, 1973). Судя но всему, технология
гнездостроения у Plocepasser и Pseudonigri-
tta похожа на воробьиную. Гнезда строятся
исключительно из сухой травы в мутовках
ветвей. Вход в гнезда воробьиных ткачиков
всегда снизу, а не сбоку или сверху, как у
воробьев. Кроме того, ткачики всегда вначале
строят гнездо с двумя входами, отчего пост-
ройка имеет характерный облик верхней по-
лусферы с двумя обращенными вниз отвер-
стиями. Такие гнезда круглый год употребля-
ются для ночевки, но перед началом кладки
одно из отверстий заделывается, а после вы-
лета птенцов — снова открывается.
Коллективные гнезда общественных тка-
чиков Philetairus socius, составляющих мо-
потипичпый род в n/ceM.Plocepasserinae,
принадлежат к числу самых удивительных
сооружений пернатых. Вся конструкция
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
49
представляет собой укрепленную па толстых
ветвях конусообразную спрессованную мас-
су сухой травы, прикрытую сверху довольно
толстым слоем плотно уложенных колючих
веток и грубых растительных стеблей, кото-
рый представляет собой своеобразную кры-
шу. Гнездовые камеры находятся в нижнем
слое мягкого материала, а их входы всегда
направлены вниз, так что при взгляде снизу
все сооружение напоминает пчелиные соты
(Рис.З). Толщина постройки достигает мет-
ра, длина по окружности — нескольких мет-
ров, общая численность населения доходит
до 500 птиц.
Доставкой и укладкой гнездового матери-
ала общественные ткачики занимаются по-
чти круглогодично. В негнездовой период они
предпочитают грубые материала, а основным
местом их накопления является крыша. По
мере активизации роста гонад птицы пере-
мещаются па нижнюю поверхность, где при-
ступают к постройке новых гнездовых камер,
поскольку старые помещения никогда пе ис-
пользуются повторно и быстро заполняются
сухой травой. Выбрав место, птица принима-
ется клювом аккуратно и методично засовы-
вать в толщу материала свисающие вниз кон-
цы травинок, отчего на нижней поверхности
мягкого слоя образуется ямка, которая рас-
ширяется и углубляется до тех нор, пока не
достигнет размеров гнездовой камеры. Покон-
чив с этим, птица начинает приносить новые
травинки и, действуя аналогичным образом,
вставляет их вокруг ямки, наращивая, тем са-
мым, слой мягкого материала, благодаря чему
гнездовая камера все глубже погружается в
его толщу. Таким образом, в пегнездовой пе-
риод коллективная постройка общественных
ткачиков растет вверх, а с началом размно-
жения — вниз. Что же касается самой первой
стадии “онтогенеза” коллективного гнезда, то,
как показали наблюдения в вольерах (Collias,
Collias, 1978b), она представляет собой накоп-
ление бесструктурной массы сухой травы до
тех пор, пока она пе станет достаточно боль-
шой для сооружения стандартным способом
первой гнездовой камеры.
Таким образом, гнезда Ploceimorpha край-
не разнообразны но форме, размерам, техно-
логии сооружения, подбору материалов, спо-
собам его транспортировки и укладки. Но
любое гнездо представляет собой замкнутую
со всех сторон камеру, сообщающуюся с вне-
шним миром через узкое отверстие. Многие
воробьи и некоторые амадины употребляют
для гнездования полости естественного про-
исхождения, однако большинство ткачикопо-
добных птиц сооружают гнездовые камеры
самостоятельно, употребляя для этого самые
разнообразные приемы и материалы. Напри-
мер, испанские воробьи строят шаровидные
гнезда исключительно из зеленой, только что
сорванной травы или листьев, пустынные
воробьи употребляют для постройки длин-
ные, сухие листья пустынного злака — сели-
па, желтые воробьи складывают гнезда из не-
гнущихся, толстых, колючих веток, а монголь-
ские земляные воробьи вьют гнезда из чис-
той шерсти и пуха.
Вопрос о происхождении шаровидных
гнезд у воробьиных птиц мы рассмотрели
ранее, в контексте филогенетического анали-
за основных типов гнездовых конструкций и
способов их расположения, взяв за основу
некоторые формы этолого-эволюционной
преемственности между разными типами
гнезд, социально-коммуникативные функции
гнездостроения, сложившиеся представления
о классификации и филогении Passeriformes
(по Карташев, 1974).
Большинство систематиков делит воробь-
инообразных на 4 подотряда : наиболее при-
митивные формы сводят в Eurilaimi, Tiranni,
Menurae (Sub-Oscines), наиболее совершен-
ные — в Oscines. Проведенный нами анализ
встречаемости и совстречаемости разных ти-
пов гнездования в подотрядах и семействах
Passeriformes показал, что в наиболее про-
грессивных группах певчих птиц безусловно
господствуют открыто расположенные чаше-
видные гнезда (Иваницкий, 1994, 1995 а,б).
Шаровидные гнезда у Oscines встречаются
намного реже и свойственны в основном
группам, известным своей трофической
(Hirundinidae, Sittidae, Cinclidae), социальной
(колониальное гнездования у Hirundinidae,
Stumidae, Icteridae) и репродуктивной (повы-
шенная плодовитость у Paridae и Aegithalidae)
специализацией. В противоположность это-
му, среди архаичных семейств Sub-Oscines,
включенных в подотряды рогоклювов, при-
митивных воробьиных и тиранновых, господ-
50
В.В.Иваницкнй
ствуют именно шаровидные гнезда и другие
формы клаустрофилии.
Клаустрофилия связана с рядом отрица-
тельных моментов. Во-первых, единственный
выход из гнезда в критический момент мо-
жет быть блокирован. Например, известны
случаи гибели больших синиц Parus major при
захвате их дупел вертишейками Jynx torquilla
(Левин, 1986), домовые воробьи, отбирая
гнезда у ласточек, нередко убивают застиг-
нутых дома хозяев (Winkler, 1992). Во-вторых,
гнезда-камеры, как правило, отличаются
чрезвычайной массивностью по сравнению
с размерами самих птиц, что повышает их
заметность для хищников и требует значи-
тельных затрат времени и энергии на стро-
ительство. Наконец, замкнутое пространство
затрудняет газообмен, причем кислородный
дефицит растет прямо пропорционально
температуре воздуха, что может замедлять
развитие птенцов (Erheldirung-Denk, Trill-
mich, 1990). Таким образом, по целому ряду
позиций массивные, трудоемкие, хорошо
заметные и плохо вентилируемые гнезда-
камеры безусловно уступают просто устро-
енным, маленьким чашевидным гнездам.
Шаровидная форма гнезд у воробьев, ви-
димо, также свидетельствует об их спениали-
зированности и относительной древности.
Мы относим воробьев (как и всех Ploce-
imorpha) к группе первичных клаустрофилов.
Эта группа включает в себя целый ряд се-
мейств воробьиных птиц (Hinindinidae, Pari-
dae и близкие к ним Sittidae, Troglodytidae и
близкие к ним Cinclidae и некоторые другие),
характеризующихся консервативной привер-
женностью к облигатной клаустрофилии при
сравнительной легкости переходов между ее
разными формами (прежде всего, между
шаровидными гнездами и естественными
убежищами — дуплами, полостями в скалах,
норами).
Деятельное, часто преобладающее участие
самцов в сооружении шаровидных гнезд дает
основание предполагать, что происхождение
этого типа построек, представляющих собой
имитацию естественных убежищ, было в зна-
чительной мере приурочено к сфере полового
отбора, где и само гнездо, и связанное с его
постройкой поведение выполняли функцию
привлечения самок. В пользу возможности
такого сценария говорят непропорционально
большие размеры многих типов шаровидных
гнезд, демаскирующая манера их расположе-
ния, использование гнезд в роли брчных
посредников или даже как места для спари-
вания (Hinindinidae, Estrildidae, Fumariidae),
серийное строительство множества гнезд или
смежных гнездовых камер, часто (но вовсе не
всегда) ведущее к полигинии (Remit,
Troglodytes, Ploceus и др.) (см.Иваницкий,
1995а).
ГЛАВА 4. РЕПРОДУКТИВНОЕ ПОВЕДЕНИЕ
4.1. Образование пар
У воробьев преобладает моногамия и рав-
ное участие полов в заботе о потомстве, хотя
конкретное распределение обязанностей ва-
рьирует у разных видов. Вне сезона размно-
жения воробьи как правило держатся стая-
ми, где в той или иной пропорции участвуют
и самцы, и самки. С приближением времени
размножения активизируется процесс форми-
рования гнездовых популяций, включающий
в себя постепенное распадение стай, распре-
деление и монополизацию центров репродук-
тивной активности, образование пар или по-
лигамных семейных групп.
У Ploceimorpha известны два основных
способа образования нар. У Estrildidae парт-
неры объединяются в стае, затем сообща по-
дыскивают участок для гнездования. Подоб-
ный тин отношений базируется па прочных,
часто пожизненных, глубоко персонализиро-
ванных связях брачных партнеров друг с
другом, поддерживаемых не только в период
размножения, но и в течение круглого года в
стаях. Известно, что у многих амадин подоб-
ные связи характерны не только для брач-
ных партнеров, но охватывают и многих дру-
гих особей из довольно постоянных по свое-
му составу стай (Sparks, 1962; Goodwin, 1982;
Лукина, 1986). Второй способ образования пар
вовсе не подразумевает необходимости ин-
дивидуального опознавания партнерами друг
друга и базируется главным образом на объе-
динении самца и самки возле общего цент-
ра активности. Образования нары здесь идет
не напрямую — путем непосредственного об-
ращения самца к самке, а через посредника,
т.е. через определенные элементы внешней
среды. Образование пар у воробьев целиком
базируется па системе посредников, построе-
ние которой отличается у разных видов. По-
средник первого уровня (гнездовая террито-
рия) имеется у всех воробьев, но ограничива-
ются им лишь короткопалые воробьи. У про-
чих видов самцы не только занимают терри-
торию, но и подыскивают место для гнезда
— развилку в кроне или убежище закрыто-
го типа (посредник второго уровня), которое
демонстрируют самкам в момент образования
пары. Наконец, некоторые воробьи и боль-
шинство представителей настоящих ткачиков
(Ploceinae) пользуются трехуровневой систе-
мой посредников, т.е. самцы занимают терри-
торию, находят мето для гнезда и начинают
его постройку еще до образования пары.
Временные рамки процесса образования
пар у воробьев часто могут быть очерчены
лишь с предельной условностью. В особенно-
сти это касается оседлых видов, особи кото-
рых на протяжение круглого года сохраняют
связь с постоянной системой функциональ-
ных центров и зон, некоторые из которых
одновременно в качестве составляющих час-
тей входят в присущую данному виду систе-
му брачных посредников.
Известно, что у домовых и нолевых воро-
бьев новый цикл роста гонад начинается сра-
зу же после завершения линьки в начале
осени и с малой скоростью протекает в тече-
ние всей зимы, испытывая, однако, мощное
ингибирующее воздействие низких темпера-
тур и короткого фотопериода (Ильенко, 1976;
Hegner, Wingfield, 1986b). Соответственно, ав-
торы, изучавшие поведение этих воробьев,
указывают на то, что пары у них могут фор-
мироваться вновь или возобновляться в пре-
жнем составе уже в период осеннего токова-
ния, когда и самцы, и самки попарно приоб-
ретают устойчивую привязанность к опреде-
ленному убежищу, которое зимой в период
спада социо-сексуальной активности может
использоваться для ночевки (Pielowski,
Pinowski, 1962; Pinowski, 1966; Summers-Smith,
1988).
Поведение воробьев вблизи посредников
в период латентного развития гонад в зимнее
время зависит главным образом от погоды, а
степень активизации социо-сексуальной ак-
тивности в благоприятные дни заметно на-
растает уже в конце января — начале февра-
ля. В это время начинают петь домовые и
полевые воробьи в Москве и Подмосковье.
В Великобритании домовые воробьи посе-
щают потенциальные места гнездования в
52
В.В.Иваницкий
течение всей зимы, а в конце января некото-
рые самцы начинают интенсивно рекламиро-
вать выбранные убежища (Summers-Smith,
1956). В Ленинградской области регулярное
пение нолевых воробьев возобновляется в
конце февраля (Фетисов, 1985). В 20-х чис-
лах февраля мы наблюдали пение возле убе-
жищ у каменных воробьев в Юго-Западном
Конет-Даге, саксаульных, нолевых и пустын-
ных воробьев в Реиетеке. Самки, хотя и в
значигелыюй меньшей степени, чем самцы, так-
же начинают посещать убежища в те же сро-
ки, что создает определенные предпосылки к
формированию нар уже на этой ранней ста-
дии за счет одновременного привыкания обо-
их партнеров к одному и тому же убежищу
(посреднику).
Активизация социо-сексуальной активно-
сти воробьев, включая пение и другие фор-
мы вокализации, пребывание у посредников,
их рекламирование, демонстративное поведе-
ние, образование нар и другие проявления,
происходит обыкновенно задолго до начала
кладки. Общая продолжительность этого пе-
реходного периода у оседлых видов состав-
ляет около 2-2,5 месяцев. В то же время, у
некоторых мигрирующих видов — в особен-
ности у испанских воробьев, переходный пе-
риод от прилета стаи ко времени начала
массовой кладки у сотен и тысяч самок иног-
да занимает всего несколько дней (Гаврилов,
1962). Генеалогически близкие к домовым во-
робьям, но относящиеся к мигрирующим пти-
цам индийские воробьи занимают ио продол-
жительности переходного периода промежу-
точное положение. На юго-востоке Туркмении
в Бадхызе по нашим наблюдениям они по-
являются у колоний в первых числах апреля
и при благоприятной, теплой погоде могут
отложить первые яйца уже через 8-10 дней
(Зыкова, 1980), но в холодную весну кладка
начинается лишь в начале мая.
У домовых и нолевых воробьев первыми
приступают к рекламированию посредников
самцы, уже имеющие опыт размножения, ко-
торые часто используют свои прошлогодние
убежища (Summers-Smith, 1988; Фетисов,
1985). В конце зимы и начале весны охрана
территорий даже у интенсивно поющих сам-
цов почти не выражена: при появлении по-
близости других воробьев самец замолкает
и внимательно следит за пришельцами, одна-
ко не предпринимает активных действий даже
в том случае, если они выказывают явный
интерес к имеющимся на его территории ук-
рытиям. Самки могут подолгу задерживать-
ся па таких территориях и обследовать убе-
жища, не обращая внимание на безучастно
сидящего поодаль самца. Кроме того, самки
домового воробья могут самостоятельно за-
нимать и удерживать за собой постоянные
убежища. Складывается впечатление, что на
этой стадии формирования популяции, когда
социальные процессы носят пока довольно
вяло текущий характер, образование нар мо-
жет происходить благодаря относительно не-
зависимому и случайному объединению осо-
бей разного пола, привлеченных в одно и то
же место специальными качествами после-
днего (в данном случае — пригодностью для
устройства гнезда).
Интересно, что аналогичный механизм от-
носительно случайного разбиения па пары
действует и в противоположных условиях, то
есть при относительно коротком и бурном
периоде формирования популяций у мигри-
рующих воробьев. У индийских воробьев это
происходит одновременно с формированием
гнездовой колонии и монополизацией убе-
жищ, приобрести которые стремятся и сам-
цы, и самки (Иваницкий, 1984а).
По-видимому, многие нары воробьев на
первых этапах освоения колонии формиру-
ются исподволь, в обход видоснецифйческих
ритуалов или взаимного антагонизма. Это
преимущественно стохастический процесс, в
основе которого лежит конгломерат противо-
речивых мотивов, руководящих поведением
птиц в этот переходный период. Временные
рамки этого процесса существенно отлича-
ются у оседлых и перелетных видов, однако,
и в том, и в другом случае выбранное убе-
жище с самого начала занимается и осваива-
ется (персонализируется) обоими партнера-
ми, каждый из которых расценивает другого,
как привычную деталь обстановки, не вызы-
вающую бурных эмоций с сопутствующими
им яркими проявлениями демонстративно-
го поведения.
Иначе протекает образование пар в тех
случаях, когда партнеры занимают убежище
не одновременно, а с определенным иптерва-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
53
лом, на протяжение которого убежище нахо-
дится в исключительной собственности од-
ного из них — как правило, самца. Такой сце-
нарий развития отношений может быть на-
зван типичным в том смысле, что поведение в
ходе подобных взаимодействий нередко от-
личается высокой экспрессивностью, а его
главные видоснецифические особенности
проявляются наиболее рельефно. Однако, при
любом сценарии образование пары у воро-
бьев, за исключением особых случаев (см.ни-
же), представляет собой довольно длительный,
растянутый во времени процесс, распадаю-
щийся на отдельные эпизоды взаимодействий
будущих партнеров у посредников. Частота
и продолжительность этих эпизодов, равно
как и разделяющих их периодов, сильно ва-
рьируют. Как показывают паши наблюдения
за меченными каменными и саксаульными
воробьями, в период образования пар самцы
возле своих посредников вступают во взаи-
модействия с разными самками, которые в
свою очередь посещают разных самцов. Мно-
жество изменений в составе нар в период
формирования колоний индийских воробьев
отмечено Матюхиным с соавторами (1990),
причем причины, побуждающие самцов или
самок перемещаться к другому убежищу и
менять партнеров, по большей части остают-
ся неизвестными.
Поведение холостых самцов при рекла-
мировании посредников имеет довольно чет-
кую ритмическую организацию. Интенсивное
пение на песенных постах, сосредоточенных
возле убежищ (рр. Passer и Petronia) или
разбросанных ио территории (p.Carpospiza),
а также в воздухе при токовых полетах
{рр.Pyrgilauda и Montifringilla), сменяются
осмотром и чисткой убежищ (у некоторых
Passer — доставкой и укладкой гнездового
материала), после чего самец может возобно-
вить пение и токование или же, наоборот,
перейти в состояние пониженной активнос-
ти и покинуть территорию, присоединившись
к стайке кормящихся воробьев. Если же он
остается в пределах территории постоянно,
то в эти периоды социальной инактивности
он ведет себя очень незаметно и в основном
кормится. Саморекламирование и инспекция
убежищ, помимо токовых полетов, часто со-
провождается и другими формами безадрес-
ного демонстративного поведения (см.ниже).
При появлении самки самец стремится
приблизиться к ней, чтобы напасть на нее,
предпринять попытку спаривания или без
определенной цели, но в то же время испыты-
вает и сильное побуждение приблизиться к
убежищу, а затем и забраться в него в соот-
ветствии с установившимся ранее стереоти-
пом. Как показали наши наблюдения, взаим-
ная агрессивность у воробьев является столь
же неотъемлемой частью взаимодействия при
образовании нар, что и показ убежищ. Суть
же всего взаимодействия состоит в том, что
самка пытается проникнуть в убежище воп-
реки активному или пассивному сопротив-
лению самца. Поэтому исход данного эпизода
определяется в конечном итоге степенью аг-
рессивности самца, настойчивостью и изоб-
ретательностью самки.
В начальный период формирования попу-
ляции самки часто ведут себя нерешительно,
быстро отступают даже при малейшей агрес-
сивности самцов и подолгу не залезают в
убежище даже при лояльном отношении со
стороны последних, хотя ведут себя очень
возбужденно, выказывая явный интерес к убе-
жищу, особенно когда туда залезает самец.
Иначе ведут себя самки на завершающих
стадиях формирования популяции. К этому
времени многие из них достигают состояния
экстренной репродуктивной готовности и
срочно нуждаются в убежище для откладки
созревающих яиц, развитие которых у воро-
бьев активизируется еще во время миграции,
отчего часть яиц откладывается в случайных
местах (главным образом на ночевках), а у
значительной части поздно Мигрирующих
самок даже развиваются наседные пятна (Гав-
рилов, 1962; Ковшарь, 1966; Брохович и др.,
1986). По-видимому, именно такие самки про-
являют чрезвычайную настойчивость, преодо-
левая противодействие самцов и часто всту-
пая с ними в ожесточенные схватки за пра-
во доступа в убежища, когда будущие партне-
ры, сцепившись когтями, надают на землю и с
остервенением колотят друг друга клювами.
Мы наблюдали три такие взаимодействия у
каменных воробьев и два — у саксаульных
воробьев: во всех случаях самки приступи-
ли к доставке строительного материала в день
образования нары, еще подвергаясь нападе-
54
В.В.Иваницкий
ниям со стороны самцов, и уже на следую-
щее утро отложили первые яйца на тонкую
подстилку и приступили к насиживанию. В
дальнейшем во взаимоотношениях партнеров
в этих парах и их гнездовом поведении мы
не заметили каких-либо отклонений от нор-
мы. Таким образом, даже самая жестокая и
бескомпромиссная агрессия при первой встре-
че пе мешает образованию пары.
Агрессия в этой ситуации может, по-види-
мому, утвердится и как неотъемлемая состав-
ляющая часть видовой нормы реакции. На-
пример, немедленная и жесткая агрессия от-
мечена как преобладающая и в сущности
единственная реакция холостых самцов ко-
роткопалых воробьев в ответ па появление
на их территории самок в тех двух эпизодах
образования пар, которые нам удалось наблю-
дать. Строго охраняемая территория служит
единственным брачным посредником у этого
вида, и, благодаря ее большим размерам, сам-
ка всегда может отыскать па ней укрытие от
постоянно преследующего ее самца. Брачные
отношения короткопалых воробьев выглядят
довольно своеобразно: в сущности самец и
самка всегда держатся порознь па общей тер-
ритории, поддерживая друг с другом главным
образом акустическую связь по типу дуэтной
переклички (см.выше). При этом самец, сидя
па верхушке куста, непрерывно следит за сам-
кой и время от времени бросается в погоню
за ней. Погоня может закончится дракой, спа-
риванием, по чаще всего самка попросту пря-
чется или улетает далеко за пределы терри-
тории. Судя по нашим наблюдениям, эти по-
гони исчерпывают весь спектр взаимоотно-
шений брачных партнеров у короткопалых
воробьев.
Поведение партнеров в ходе образования
пары зависит от социальной и предметной
обстановки на месте, где протекает взаимо-
действие, что можно иллюстрировать на при-
мере индийских воробьев, гнездящихся на
деревьях и па обрывах.
В момент появление самки действия сам-
ца, занявшего территорию в колонии в кроне
дерева, отличаются большим разнообразием,
однако с некоторой долей условности можно
наметить два основных варианта, соответству-
ющих преобладанию указанных выше тенден-
ций. Если поведение самца адресуется самке,
то он обычно бывает очень подвижен, стре-
мится приблизиться к самке и после корот-
ких демонстраций часто нападает па нее, пре-
следуя далеко за пределами территории. Дей-
ствия самца выглядят точно также, как и при
конфликтах с другими самцами, используются
те же демонстративные позы. Самка же пы-
тается проникнуть в центр территории, что
нередко приводит к стычкам и дракам. В дело
почти беспрестанно вмешиваются многочис-
ленные соседи, что постояшю отвлекает самца
и способствует успеху самки. Со временем
агрессивность самца уменьшается, и он позво-
ляет самке задерживаться па своей террито-
рии, которую она начинает тут же тщательно
обследовать, в свою очередь поминутно всту-
пая в конфликты с соседними самками.
Иногда же самец ведет себя совершенно
иначе. Прежде всего обращает на себя внима-
ние его ограниченная подвижность. Он впа-
дает в состояние своеобразного транса — все
движения его замедленны, он подолгу задер-
живается на одном месте, но тем пе менее не-
прерывно демонстрирует (позы Д,Е, Рис. 11)
и ноет. При этом внимание самца направле-
но в основном не па вновь прибывшую самку,
а на мутовки на своей территории, куда он то
и дело залезает, словно приглашая самку
проделать то же самое. Если самец избирает
подобную тактику, то пара часто образуется
без серьезных коллизий. Перемещаясь в ко-
лонии, любая холостая самка испытывает по-
стоянные нападения территориальных самок
и сексуальные домогательства самцов, поэто-
му, когда она попадает, наконец, на террито-
рию холостого самца, то, по-видимому, уже
одного лишь отсутствия беспокойства с его
стороны может быть достаточно для того, что-
бы она задержалась в данном месте, присту-
пила к его осмотру и к выяснению окружаю-
щей обстановки.
Особенно бескомпромиссной выглядит
половая конкуренция за убежища в плотных
колониях па обрывах, где многие самцы вы-
нуждены все время сидеть на входе в свою
нору, полностью закрывая его своим телом и
обратив клюв наружу. В этой позиции самец
неуязвим, и чтобы проникнуть в пору самке
приходится приложить немало усилий. Для
этого ей необходимо выманить самца из норы,
провоцируя его па проявление агрессивное-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
55
ти или сексуальности. Поведение самки в
этот момент поразительно напоминает пове-
дение собаки, которая пытается побудить к
игре партнера, к этому вовсе не расположен-
ного. Стоит самцу на мгновение оставить свой
пост, как самка стремительным обходным
маневром кидается ко входу и залезает
внутрь. Тут же за ней устремляется самец и
после короткой схватки, о перипетиях кото-
рой можно судить лишь но вылетающим на-
ружу перьям и крику, вытаскивает самку из
поры за хвост н восстанавливает прежнее
положение. Иногда, однако, н самка, проник-
нув в нору, успевает развернуться и занять
устойчивую позицию клювом наружу, что в
течение некоторого времени позволяет ей
успешно противостоять попыткам самца выг-
нать ее вон. Взаимная неприязнь может длит-
ся долго и проявляться в самых жестких фор-
мах. Образование нары можно считать состо-
явшимся, когда самка получает право относи-
тельно свободного доступа в убежище, хотя не-
редко ей (как, впрочем, и самцу) приходится
буквально протискиваться внутрь между стен-
кой норы и сидящим у входа партнером. За-
бегая вперед, отметим, что впоследствии —
особенно с началом насиживания, роль соци-
ального доминанта переходит к самкам.
Своеобразны проявления антагонизма при
образовании пар у испанских воробьев. При
виде самки самец сейчас же залезает в му-
товку, где обыкновенно уже лежит гнездовая
платформа, и принимается интенсивно демон-
стрировать (позы Д-М, Рис. 10). Самка под-
летает и тут же выказывает намерение заб-
раться в ту же самую мутовку. Вначале она
пытается вытеснить самца всеми доступны-
ми способами, включая сперва легкие, а затем
и полновесные удары клюва, что, однако, лишь
усиливает демонстрации самца, впадающего
в настоящий экстаз. Если самке никак не
удается вытеснить самца, она хватает его за
оперение и выбрасывает из мутовки, тут же
занимая его место. Самец мгновенно возвра-
щается и возобновляет интенсивные демон-
страции возле сидящей в мутовке самки, од-
нако более решительных действий не пред-
принимает. Всего мы отметили 12 случаев,
когда самки выбрасывали самцов из муто-
вок, в 7 из 10 наблюдавшихся нами эпизодов
образования нары, причем в 4 случаях, самки
удерживали самцов на вёсу несколько секунд
(до 12 с). Таким образом, у испанских воро-
бьев, в отличие от близкородственных индий-
ских воробьев, самки утверждают себя в роли
социальных доминантов с первых же часов
семейной жизни.
4.2. Система спаривания и поло-
вое поведение
В последние годы в литературе появляется
все больше данных, свидетельствующих о том,
что классическая моногамия — как наиболее
распространенная у птиц форма социальной
кооперации на почве выращивания потомства,
не всегда базируется на абсолютной супружес-
кой верности. У многих видов широко распро-
странены внебрачные копуляции, которые иног-
да фиксируются во время непосредственных
наблюдений, но чаще выявляются специаль-
ными методами анализа родственных отно-
шений (Birkhead et al., 1987).
Воробьи в этом смысле не представляют
исключения. Доминирующим типом взаимо-
отношений иолов у них является социальная
моногамия. Социальная полигиния, когда са-
мец состоит в относительно регулярных со-
цио-сексуальных отношениях с несколькими
(чаще — двумя) самками, обитающими на его
территории, встречается довольно редко (Ива-
ницкий, 19856,19866). По крайней мере у до-
мовых воробьев, судя но некоторым данным,
широко распространены внебрачные копуля-
ции (Wetton, Parkin, 1991).
В ходе многолетних наблюдений за десят-
ками нар полевых воробьев в Германии
Deckert (1973) наблюдала всего несколько
случаев полигинии. Все они касались овдо-
вевших самок, которые после исчезновения
своих партнеров остались в прежних гнездах
и вступили в брачный союз с одним из со-
седних самцов. Преобладала бигамия, лишь
один самец поддерживал регулярные отно-
шения с тремя самками, две из которых утра-
тили партнеров уже в ходе гнездования. Этот
самец насиживал кладки и выкармливал
птенцов во всех трех гнездах. В то же время,
Фетисов (1981) отмечает, что самки полево-
го воробья проявляют склонность к внебрач-
ным копуляциям даже при наличии у них
постоянных партнеров.
56
В.В.Иваницкий
Лишь три случая полигинии — все с уча-
стием овдовевших самок — зафиксированы
в детально исследованной в течение 5 лет
популяции домовых воробьев в Гуизборо (Ве-
ликобритания), где эти птицы гнездились в
своих обычных местах с относительно неболь-
шой численностью и низкой плотностью
(Summers-Smith, 1958).
По данным Veiga (1992) в Испании, где
домовые воробьи размножались в искусст-
венных гнездовьях при значительно более
высокой плотности, полигиния, воз1шкшая ес-
тественным путем, составляла регулярное яв-
ление и отмечалась в среднем у 9,6% самцов
(18 из них имели по 2 самки, у одного самца
было 3 самки). В то же время, склонность сам-
цов к нплигамии распространена широко .
Дополнительные брачные альянсы с сосед-
ними самками, оставшимися в одиночестве
после экспериментального устранения парт-
неров, заключают от 36,7 до 73,3% самцов.
Многие самцы захватывают и охраняют 2
или 3 близко примыкающих друг к другу
убежища, причем в большей степени это свой-
ственно самцам, имеющим более ярко выра-
женные вторично-половые признаки (вели-
чина черной манишки на груди). Тем не ме-
нее, вероятность приобретения второй самки
для самца не зависело от числа удерживае-
мых им гнездовий, будучи обусловленной
целым рядом случайных обстоятельств, спо-
собствующих утрате самками их партнеров
(Veiga, in press). По мнению автора, основным
фактором, противодействующим полигинии
у домовых ворюбьев, является агрессивность
самок в борьбе за поддержание моногамии, а
также широкое распространение инфантици-
да, хотя разрушение самцами содержимого
плохо охраняемых соседних гнезд при опре-
деленных обстоятельствах, наоборот, может
способствовать их спариванию с соседними
самками, возобновляющими размножение в
том же убежище (Veiga, 1990 а,Ь).
Как показали наши наблюдения, опреде-
ленные предпосылки к полигинии склады-
ваются и в плотных колониях индийских
воробьев. Как и у домовых ворюбьев, самцы
этого вида в период формирования колоний
часто занимают одновременно несколько нор
(иногда 5-6), расположенных обычно не да-
лее метра друг от друга в пределах единой
территории. Со своей стороны, самки способ-
ны самостоятельно занимать и долго удер-
живать за собой убежища. Такие одиночные
самки всегда привлекают к себе внимание
соседних самцов, уже имеющих собственных
самок, которые всеми способами пытаются
обозначить свое присутствие возле убежища,
занимаемого холостой самкой. Тем не менее,
нам не пришлось наблюдать перерастания
подобных временных альянсов в устойчивые
социо-сексуальные отношения. Главная при-
чина состоит в том, что в условиях высокой
плотности и постоянного пресса со стороны
претендентов на убежища, самцу трудно дол-
го сохранять контроль за многими убежища-
ми па большом расстоянии друг от друга.
Если же эти убежища расположены непода-
леку, то серьезным препятствием для полиги-
нии становится агрессивность самок, которые
проявляют крайнюю степень нетерпимости
друг к другу. Все это приводит к тому, что
полигиния у индийских воробьев отмечается
лишь как эпизодическое явление (Матюхин
и др., 1990; Иваницкий, 1996).
Постоянное стремление самцов к социо-
сексуальным взаимодействиям с соседками
но колонии составляет одну из наиболее ха-
рактерных черт социальной жизни испанс-
ких воробьев. Пик этих взаимодействий при-
ходится обычно на разгар спаривания в ка-
нун начала кладки, когда самки ведут мало-
подвижный образ жизни и подолгу проси-
живают возле своих гнезд, в то время как сам-
цы продолжают строить гнездо, то и дело
оставляя своих самок в одиночестве. Такие
самки очень привлекают чужих самцов. Мы
отметили 23 эпизода ухаживаний самцов за
чужими самками и все они протекали очень
интенсивно.
Как только самец приближается к сидя-
щей у гнезда в одиночестве самке, он сразу
начинает интенсивно демонстрировать в раз-
нообразных позах (Рис. 10, позы И,К), осо-
бенно интенсивность его демонстраций воз-
растает после того, как у самки включается
характерная для таких эпизодов агрессивная
реакция. Чем агрессивнее ведет себя самка,
чем с большим ожесточением клюет она сам-
ца и щиплет его за оперение, тем ярче его
демонстрации. Мы неоднократно наблюдали,
как самец продолжал демонстрировать даже
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
57
повиснув в клюве разъяренной самки, ухва-
тившей его за оперение и стащившей с ветви.
17 из 23 таких эпизодов были прерваны по-
явлением законного партнера вовлеченной во
взаимодействие самки, который немедленно
изгонял пришельца. В остальных случаях
последний сам покинул чужую территорию.
Во всех подобных эпизодов самка, демонст-
рируя незаурядные бойцовские качества, ни
разу не покинула своей территории, стойко
отражая домогательства пришельца (а иног-
да и двух-трех одновременно).
Интересно, что в 16 из 23 упомянутых эпи-
зодов в роли ухажеров фигурировали со-
седние самцы, нередко из ближайшей же пары,
имеющие и собственных самок. Часто мы
наблюдали такую картину: с одного из двух
соседних гнезд улетает самец, оставляя сам-
ку одну. Тут же сосед, сидевший прежде ря-
дом со своей самкой у своего гнезда, оставля-
ет ее и, переместившись на несколько десят-
ков сантиметров, начинает интенсивно демон-
стрировать перед соседкой. Взаимодействие
протекает но описанной выше схеме, причем
на протяжение всего эпизода самка демонст-
рирующего самца с безучастным видом си-
дит в прежней позиции. Возвращение улетев-
шего самца кладет конец ухаживанию, и де-
монстрировавший самец перемещается к сво-
ему гнезду. Однако, стоит улететь ему, как те-
перь уже его сосед может начать демонстра-
ции перед покинутой на время самкой.
Хотя мы не проводили индивидуального
мечения испанских воробьев, при наблюде-
ниях за их брачными отношениями у нас
сложилось твердое убеждение в том, что ко
времени начала спаривания самки персональ-
но опознают своих самцов. Самка замечает и
опознает самца еще на подлете в воздухе и
сразу же начинает трепетать крыльями в
предсовокупителыюй нозе (Рис.10,Н). Ее
поведение меняется мгновенно: то она с ос-
тервенением гоняет но территории ухажива-
ющего за пей соседа, а то вдруг замирает на
месте, приседает и сильно трепещет крылья-
ми, после чего сразу же па территорию опус-
кается ее партнер, и без всяких предваритель-
ных демонстраций может сразу же произой-
ти спаривание. Тем пе менее, вполне возмож-
но, что при гибели самца оставшаяся на тер-
ритории одинокая самка может в конце-кон-
цов проявить благосклонность к настойчиво-
му соседу, подобно тому, как это имеет место
у домовых или полевых воробьев.
Таким образом, полигиния у представите-
лей р.Passer — особенно в плотных колони-
ях — всегда возникает в результате развития
соседских отношений между моногамными
самцами и живущими неподалеку одиноч-
ными самками, утратившими своих партне-
ров, но сохранивших верность гнездовой тер-
ритории или убежищу. Вероятность такого
сценария зависит от многих случайных об-
стоятельств. К тому же, широкому распрост-
ранению полигинии у настоящих воробьев
противодействуют, как мы полагаем, вполне
определенные факторы: 1) затруднения с ох-
раной удаленных друг от друга убежищ при
высокой плотности населения в колониях; 2)
высокая агрессивность самок при охране тер-
ритории; 3) активное участие самцов в гпез-
достроепии и насиживании; 4) высокий уро-
вень социальных помех в колониях в форме
инспекций убежищ посторонними особями, во-
ровства гнездового материала и разрушения
гнезд, гнездового паразитизма и инфантици-
да, что обусловливает высокие требования к
охране гнезда и требует участия в этом деле
обоих партнеров.
Полигиния распространена у каменных
воробьев па юге Туркмении, однако взаимо-
отношения особей и их поведение выглядят
иначе (Иваницкий, 19856, 1986а). Как отмече-
но выше, многие самцы каменных воробьев
после успешного образования пары в первич-
ном центре активности часто занимают и
рекламируют вторичный центр активности,
одновременно поддерживая постоянные кон-
такты со своей первой самкой.
Динамика поведения в процессе образова-
ния полигамных ячеек отличается высокой
изменчивостью, но в общих чертах опа может
быть описана следующим образом. Когда пер-
вая самка (А) приступает к строительству
гнезда, самец, который сам гнезда пе строит,
всегда сопровождает ее при полетах за гнез-
довым материалом. Как только самка с мате-
риалом в клюве направляется к гнезду, самец
обгоняет ее и с интенсивными демонстраци-
ями усаживает у входа. Все время, пока сам-
ка укладывает материал внутри, самец не пре-
кращает пение и демонстрации, а затем вновь
58
В.В.Иваницкий
летит за самкой, направляющейся за новой
порцией материала. Возбуждение самца, его
вокальная активность и экспрессивность по-
ведения остаются на высоком уровне, поэто-
му сопровождая самку А, самец постоянно
сохраняет готовность вступить в контакт с
любой другой самкой, как в пределах своего
первичного центра активности, так и в лю-
бом другом месте.
Эти взаимодействия протекают в соответ-
ствии с типичной схемой “образования нары”:
завидев невдалеке самку, чье поведение свиде-
тельствует об отсутствии у нее постоянного
партнера и центра активности (см.выше), са-
мец тут же оставляет свою партнершу и на-
правляется к ближайшей скале, россыпи кам-
ней или отдельному камню, где принимается
громко иеть, демонстрировать характерные
позы и залезать в случайные отверстия. Заин-
тересовавшись действиями самца, новая сам-
ка нередко подлетает ближе и осматривает
предлагаемый ей участок и убежище. Хотя
большинство подобных взаимодействий пред-
ставляют собой лишь мимолетные эпизоды
(внешне они, однако, не отличимы от истинно-
го образования пары), ио крайней мере в двух
случаях нам установить, что именно эти места
случайных встреч с бродячими самками впос-
ледствие стали использоваться самцами в
качестве вторичных центров активности.
Всего нам удалось зарегистрировать 2 по-
лигамных трио каменных воробьев в юго-за-
падном Коиетдаге в 1981 г., 3 — в Бадхызе в
1982 г., 5 — в Бадхызе в 1983 г. В 1986 г. в
Бадхызе, несмотря на интенсивные и продол-
жительные наблюдения, полигинии не отме-
чено. Также не найдено полигинии у камен-
ных воробьев в горном Алтае в 1988 г. Таким
образом, полигиния у этого вида представля-
ет собой сугубо факультативное явление, как
впрочем и у подавляющего большинства ви-
дов мелких воробьиных птиц.
В Бадхызе нам удалось обнаружить два
достоверных случая нолитерриториалыюй по-
лигинии: расстояние между гнездами самки
А и второй самки (Б) составило 23 и 165 м. В
остальных случаях обе самки жили па общей
территории. Среднее расстояние между гнез-
дами самок одного самца составляет 26,5±4,1
м, максимальное — 40 м, минимальное — 8 м.
Расстояние между первичным и вторичным
центрами активности самца, ио-видимому, не
оказывает заметного влияния на вероятность
нолигинни, потому, что самка А обычно не про-
являет агрессии к самке Б, если последняя не
приближается вплотную к ее гнезду. Это об-
легчает вселение самки Б и заметно отличает
каменных воробьев от многих других птиц, у
которых формирование полигамных ячеек со-
провождается жестокими драками между
самками (см. Панов, 1983).
В 1983 г. в Бадхызе полигиния у камен-
ных воробьев была обычной, но в 1986 г. не
отмечено ни одного случая. В Табл.9 сумми-
ТаблицаЭ. ПАРАМЕТРЫ РАЗМНОЖЕНИЯ, СТРУКТУРЫ ПОПУЛЯЦИИ
И СРЕДЫ ОБИТАНИЯ КАМЕННЫХ ВОРОБЬЕВ В БАДХЫЗЕ
Параметры 1983 год 1986 год
Погода во 2-й половине марта сухая и теплая холодная и сырая
Начало массового насиживания конец марта- начало апреля середина апреля
Всего отмечено полигамных самцов 5 0
Полигамных самцов на площадке 3 0
Моногамных самцов на площадке 18 17
Холостых самцов на площадке 3 5
Число агрессивных взаимодействий 61 30
Число демонстраций самцов перед самками на своих территориях 22 12
Число демонстраций самцов перед самками вне территорий 18 12
Всего наблюдений (ч) 88 96
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
59
ровапы некоторые различия среды обитания
воробьев, фенологии их размножения, струк-
туры популяции и поведения в эти два се-
зона. Весна 1983 г. была сухой, ранней и теп-
лой, и каменные воробьи приступили к на-
сиживанию уже в конце марта-iгачале апреля.
В 1986 г. всю вторую половину марта стояла
исключительно сырая и холодная погода с
туманами, дождями и ночными снегопада-
ми, отчего насиживание у каменных воробь-
ев началось лишь в середине апреля, то есть
почти па две педели позже, чем обычно (см.
Бельская, 1973). Весной 1986 г. самцы вели
себя намного менее активно, чем в 1983 г. Хотя
их общая численность осталась па прежнем
уровне, не было полигинии, и больше стало
холостых самцов.
Общее число социальных взаимодействий в
популяции резко снизилось. По данным мече-
ния, большая часть этих взаимодействий про-
воцируется ХОЛОСТЫМИ особями, которые по-
стоянно держатся в районе наблюдений, посе-
щая время от времени территории разных сам-
цов. Эти особи в массе присутствовали в изу-
ченной популяции в 1983 г., внося постоянные
возмущения в сложившийся порядок жизни в
локальных поселениях воробьев. В 1986 г. час-
тота всех типов взаимодействий с участием этих
особей снизилась почти вдвое, а общий уровень
социальной активности самцов упал.
Половой дихроматизм у каменных воро-
бьев пе выражен, однако, скорее всего, боль-
шинство из этих особей, пе имеющих посто-
янных центров активности, представлено сам-
ками, потому что холостые самцы обычно
быстро оседают на постоянных территориях
и приступают к их рекламированию. В 1983
г. мы наблюдали за одной такой самкой, ко-
торая была впервые поймана 18 марта в цен-
тре активности одного из самцов и присту-
пила к постройке гнезда только 3 апреля на
другой территории в том же ущелье. В про-
межутке мы встречали ее 5 раз — трижды
она участвовала в эпизодах “образования
пары" с двумя другими самцами и дважды
ее видели во время кормежки прилегающей
к ущелью широкой долине, где воробьи не
гнездятся. В 1986 г. удалось проследить гнез-
достроепие еще у трех самок, которые были
помечены в центрах активности самцов за 8,
9 и 15 дней до окончательной стабилизации
собственных центров активности, па протяже-
ние которых они также неоднократно уча-
ствовали в брачных взаимодействиях с раз-
ными самцами.
Наши данные о годовых различиях соци-
альной организации и поведения каменных
воробьев в связи с климатическими услови-
ями, хорошо соответствуют выдвинутой
Wingfield (1984) гипотезе “социального вы-
зова", согласно которой интенсивность секре-
ции тестостерона, контролирующего брачное
и территориальное поведение птиц, у неко-
торых видов зависит от частоты и напряжен-
ности социальных взаимодействий в попу-
ляции, а не только от эндогенных факторов,
как полагали ранее. Эта гипотеза получила
подтверждение в исследованиях Silverin
(1990), которому удалось показать, что токо-
вание и захваты новых дупел (сииичников)
у нолигинпо-политерриториальных самцов
мухоловки-пеструшки Ficedula hypoleuca
стимулируются прежде всего холостыми сам-
ками, подыскивающими места гнездования.
4.3. Внебрачные половые отношения
Коллективные демонстрации самцов во-
робьев р. Passer перед самками составляют
одну из наиболее характерных особенностей
социо-сексуалыюго поведения этих птиц и
пе раз описаны в литературе (Summers-Smith,
1954; Blackbill, 1969; Moller, 1987а).
Вкратце, взаимодействие протекает так.
Начало ему может быть положено как на
гнездовой территории, так и в любом дру-
гом месте. Самец, испытывающий сексуаль-
ные побуждения, все ближе и ближе при-
ближается к своей или чужой самке, прыгая
вокруг нее и адресуя ей разнообразные де-
монстративные позы и интенсивную брач-
ную песню (см. ниже). Самка неизменно реа-
гирует на приближение самца (особенно чу-
жого) крайне агрессивно, пытается ударить
его клювом и даже гоняется за ним для этой
цели. Взаимодействие развивается под влия-
нием взаимной стимуляции: чем нетерпимее
реакция самки, чем в более жестких формах
проявляется ее агрессия, тем сильнее демон-
стрирует и громче поет самец и тем настой-
чивее сближается он с самкой, хотя, по-воз-
можности, и старается избегать ударов. Ког-
60
В.В.Иваницким
да самка пытается улететь, самец устремля-
ется в погоню, следуя за самкой и повторяя
все изломы ее траектории, и немедленно во-
зобновляет еще более интенсивные демонст-
рации при посадке.
Такое поведение сильно привлекает дру-
гих самцов, которые могут примкнуть к паре
на любой стадии взаимодействия, однако са-
мое сильным стимулом оказывается погоня,
после которой все примкнувшие самцы сооб-
ща принимаются демонстрировать перед сев-
шей куда-либо самкой. Каждый самец в неве-
роятной толчее пытается совершить копуля-
цию, приближаясь к самке со спины и пыта-
ясь забраться к пей на спину, что неизменно
встречает с ее стороны самый резкий отпор.
Одновременно самцы клюют друг к друга. В
копце-коицов взаимодействие перерастает во
всеобщую свалку и утрачивает какое-либо
подобие организации. Улучив момент, самка
улетает, и самцы тут же прекращают драку.
В коллективных демонстрациях у домо-
вых воробьев могут участвовать до 11 сам-
цов (Summers-Smith, 1954), в крупных коло-
ниях индийских воробьев, но нашим данным,
до 12, у нолевых воробьев — не более 5 сам-
цов. У гнездящихся нреимуществешю обособ-
ленными парами саксаульных, рыжих, камен-
ных, земляных и снежных воробьев коллек-
тивные демонстрации отмечаются редко и в
них принимают участие обычно не более 3-4
самцов.
Особенно характерны коллективные де-
монстрации для индийских воробьев, гнездя-
щихся на обрывах, где высокая численность
птиц сочетается с хорошим обзором, и пого-
ня тут же привлекает внимание всех самцов,
сидящих на обрыве у своих пор, и многие из
них включаются во взаимодействие. В круп-
ных колониях коллективные демонстрации
в разных частях поселения и с участием раз-
ных самцов происходят едва ли не поминут-
но и составляют фоновый вид социальной
активности. Взлетевшая со своей территории
самка часто стимулирует погоню соседних
самцов. Нередко коллективные погони сам-
цов за самками и коллективные демонстра-
ции совмещаются с коллективными спонтан-
ными взлетами всех воробьев (см.выше).
В колониях на деревьях коллективные
погони и демонстрации,также как и коллек-
тивные взлеты, происходят намного реже чем
на обрывах, вероятно потому, что в густой
кроне возможности для массовой мобилиза-
ции птиц для участия в каких-то коллектив-
ных действиях значительна затруднена огра-
ниченным обзором. Кроме того, в кроне сам-
кам значительно легче укрываться от пресле-
дующих их самцов. Для испанских воробьев,
гнездящихся огромными колониями в кро-
нах, коллективные демонстрации также не
характерны.
Есть все основания предполагать, что кол-
лективные демонстрации, будучи фоновым
элементом социо-сексуальной жизни воробьев,
требуют значительного расхода энергии, со-
пряжены с риском получить травму и стать
жертвой хищника, поскольку в разгар подоб-
ного рода взаимодействий, в чем мы неоднок-
ратно имели возможность убедиться, воробьи
в значительной степени теряют контроль за
внешней ситуацией. В этой связи возникает
вопрос о функциях н происхождении кол-
лективных демонстраций.
Поскольку подавляющее большинство сам-
цов в ходе коллективных взаимодействий,
демонстрируют перед чужими самками, мож-
но предполагать, что самцы стремятся к ко-
пуляции для увеличения своей совокупной
приспособленности (то есть собственного
вклада в генофонд следующих поколений).
Как показали наши наблюдения за испанс-
кими и индийскими воробьями, в колониях
привлекательность самок для соседних сам-
цов заметно возрастает по мере приближе-
ния времени кладки и увеличения вероят-
ности оплодотворения. Moller (1987,1990)
провоцировал коллективные погони и демон-
страции у домовых воробьев, выпуская са-
мок рядом со стайками сидящих на кустах
самцов ti обнаружил, что последние распоз-
нают рецептивных самок даже в полете.
На первый взгляд, коллективные демонст-
рации создают идеальную социальную среду
для внебрачных копуляций и промискуитета.
Об этом говорят данные, полученные для од-
ной из британских популяций домовых воро-
бьев с применением ДНК-дактилоскопии. В
разные годы от 10 до 19% птенцов не являют-
ся потомками воспитывающих их самцов. 28%
кладок содержали яйца, оплодотворенные при
внебрачных копуляциях, и 41,7% самцов вы-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
61
кармливали по крайней мере одного нерод-
ного птенца (Wettonetai., 1987, 1991).
Противоречие заключается в том, что ши-
рокое распространение внебрачных копуля-
ций у воробьев, на что недвусмысленно ука-
зывают молекулярно-генетические методы,
плохо подтверждается прямыми наблюдени-
ями. Мы подробно проследили 64 коллектив-
ные демонстрации в колониях индийских
воробьев, но добиться спаривания самцам не
удалось ни разу. Безрезультатность коллек-
тивных демонстраций у этого вида отмечает
и Зыкова (1980). Summers-Smith (1954) на-
блюдал более 200 коллективных демонстра-
ции у домовых воробьев, ио спаривания не
отметил. По другим данным, в 69 эпизодах у
этого вида было 3 внебрачные копуляции
(Moller, 1987, 1990). Поскольку в каждом из
них в среднем принимало участие по 3 самца,
то вероятность добиться успеха для каждого
нз них, с нашей точки зрения, выглядит исче-
зающе маленькой. Тем не менее, автор пола-
гает, что именно она побуждает самцов к уча-
стию в коллективных демонстрациях.
Иное объяснение предложили Wetton и
Parkin (1991). Указав на крайне низкую вероят-
ность оплодотворения при коллективных демон-
страциях, О1Ш предположили, что относительно
большая пропорция неродных птенцов объяс-
няется тем, что самки активно стремятся к тай-
ным (и потому трудно выявляемым при визу-
альных наблюдениях) спариваниям с чужими
самцами, чьи внешние признаки или поведе-
ние свидетельствуют об их более высоком гене-
тическом качестве, и тем, что копуляции с эти-
ми самцами приводят к более эффективному
оплодотворению, чем копуляции с собственны-
ми самцами, которые интересуют' самок, глав-
ным образом, не как производители генов, а как
воспитатели их потомства.
4.4. Половое поведение
Низкую вероятность оплодотворения при
коллективных демонстрациях воробьев сле-
дует ожидать еще и потому, что поведение
этих птиц при нормальном спаривании выг-
лядит в высшей степени своеобразно. Как
правило за один эпизод спаривания самец со-
вершает подряд несколько садок па спину
самке — как холостых (бесконтактных), так
и результативных, когда самец, удерживаясь
на спине партнерши, балансируя крыльями
и характерным образом подгибая хвост, со-
единяет свою клоаку с клоакой самки, благо-
даря чему становится возможным эффектив-
ная эйякуляция и оплодотворение. Спарива-
ние, протекающее в нормальных условиях,
представляет собой длительное и сложное
взаимодействие, требующее известной коор-
динации в действиях партнеров.
Спаривание у Passer длится иной раз до 2
мин. Интересно, что в те моменты, когда во
время спаривания самец сидит на спине сам-
ки, он часто бывает к ней крайне агрессивен
и с ожесточением клюет ее в спину, затылок
и темя. Чаще самец проявляет агрессивность
при бесконтактных садках. Иногда весь эпи-
зод включает только холостые садки, и тогда
самец клюет партнершу особенно сильно. Еще
раз подчеркнем, что эта агрессивность самцов,
как и вообще агрессивное поведение у воро-
бьев, носит весьма жесткие формы, далекие от
какой-бы то ни было ритуализации, хотя, ра-
зумеется, в ряду эпизодов можно проследить
полный переход от полновесных ударов до
легких прикосновений. Интересно, что во всех
прочих ситуациях (например, при залезании
в гнездо) ко времени начала спаривания во
взаимоотношениях партнеров устанавливает-
ся безоговорочное доминирование самок.
Помимо видов, указанных в Табл. 10, мно-
жественное спаривание достоверно известно у
Passer luteus (Morel, Morel, 1973). В ряду изу-
ченных нами видов существует прямая кор-
реляция между средней частотой спаривания
и количеством садок за одно взаимодействие
(иепараметрическнй коэффициент корреляции
Спирмена г = 0,735; Т = 3,25; Р < 0,01). При
этом намечается частота эпизодов спаривания
и числа садок в одном эпизоде уменьшается
при переходе от высоко колониальных к оди-
ночно гнездящимся видам воробьев. У Passer
спаривание представляет собой один из самых
распространенных способов взаимодействия
брачных партнеров, а его частота может быть
довольно высокой. Что же касается Petronia,
Pyrgilauda и Montifringilla, то у них множе-
ственное спаривание выражено вполне отчет-
ливо, хотя и частота копуляций и число садок
за один эпизод у них заметно меньше, чем у
Passer. Лишь у Carpospiza, по нашим иаблю-
62
В. В. Иваницкий
Таблица 10. ПОЛОВОЕ ПОВЕДЕНИЕ ВОРОБЬЕВ
(по Ivanitskii, 1996 с изменениями)
Виды Время наблю- дений в фазе спари-вания (час) Отмечено копуляций Средняя частота копуляций в час Максималь- ное число садок Среднее число садок
Испанский 39 86 2,23 ±0.15 17 6,5 ±0,3
Домовый 20 38 1,94 ±0,13 12 6,8 ± 0.6
Индийский 74 124 1,69 ±0,08 19 8,3 ±0,3
Полевой 56 45 0,81 ±0.06 16 7,9 ±0,5
Пустынный 29 32 1,15 ±0,04 9 5,8 ±0.4
Саксаульный 89 37 0,43 ±0,04 8 5,3 ±0,4
Рыжий 24 11 0,52 ±0,07 7 5,1 ±0,8
Каменный НО 28 0,29 ±0,05 3 1,8 ±0,1
Монгольский 29 7 0,21 ±0,08 5 3,1 ±0,7
Снежный 21 5 0,26 ±0,09 5 2.8 ±0.9
Короткопалый 10 5 0,50 ±0,02 1 1,0 ±0,0
дениям и данным Адамяна (1965), каждое спа-
ривание — как и у подавляющего большин-
ства прочих воробьиных птиц — включает
всего одну садку.
Таким образом, множественное спарива-
ние — характерный этологический признак
воробьев, который, как можно думать, имеет
весьма древнее происхождение. В литерату-
ре по поведению ткачикоподобиых птиц нам
пе встречались упоминания о множествен-
ном спаривании в других группах. Тем пе
менее, наблюдения за общественными тка-
чихами Philetairus socius дают основу для
гипотезы о происхождении множественно-
го спаривания (Collias, Collias, 1980b). Эти
ткачики живут многочисленными колония-
ми с элементами коммунального гнездост-
роепия и выкармливания птенцов. Гомо- и
гетеросексуальные садки и копуляции пред-
ставляют собой элемент повседневного об-
щения, не будучи жестко привязанными к
сезону размножения. Как полагают авторы,
спаривание у этого вида утвердилось в ка-
честве одной из форм проявления социаль-
ного доминирования: при спаривании до-
минирующий самец делает садку па подчи-
ненную особь, причем некоторые подобные
эпизоды завершаются типичным коитусом, ос-
новные признаки которого приведены в на-
чале этого раздела. Известно, что спарива-
ние (или садки), как проявление доминиро-
вания во взаимоотношениях самцов, встре-
чается у некоторых млекопитающих. Что же
касается птиц, то, по нашим наблюдениям,
типичное половое поведение (вплоть до ко-
итуса) составляет характерную деталь пове-
дения самцов горлиц {Streptopellia turtur,
S.senegalensis) при пограничных конфлик-
тах. Вполне возможно, что именно специфи-
ческая социальная среда коммунальных
групп была источником формирования мно-
жественного спаривания.
4.5. Гнездостроение
4.5.1. Место гнездостроительно-
го поведения в годовом и репродук-
тивном цикле
Гнездостроение у воробьев носит крайне
растянутый (пролонгированный) характер и
пе вписывается в четкие временные рамки,
приуроченные к определенным стадиям реп-
родуктивного цикла. Весной воробьи начи-
нают строить гнезда задолго до начала клад-
ки и продолжают приносить гнездовой ма-
териал вплоть до вылупления птенцов, а иног-
да и после этого перемежают доставку кор-
ма с отдельными приносами небольших пор-
ций гнездового материала (Благосклопов,
1950; Умрихина, 1955; Гаврилов, 1962; Поно-
марева, 1983).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
63
У оседлых домовых и полевых воробьев
гиездостроение носит почти перманентный ха-
рактер и может наблюдаться круглогодично
за исключением времени линьки (август) и
зимних месяцев с самым коротким фотопе-
риодом и ограниченным доступом к гнездо-
вому материалу (декабрь-январь). Начало
гпездостроения осенью в преддверии абортив-
ного цикла и в конце зимы — накануне на-
чала репродуктивного периода — всегда со-
впадает с началом обследования убежищ и
саморекламирования и является одним из
самых ранних свидетельств активизации
физиологической подготовки к размножению.
Осенью гиездостроение достигает макси-
мальной интенсивности в октябре, причем в
этот период воробьи могут доставлять гнез-
довой материал в разные убежища. В москов-
ском нарке Кусково в разгар абортивного
цикла наброски гнездового материала ноле-
вых воробьев, но нашим данным, присутству-
ют почти в 85% синичников, тогда как на но-
чевках птицы занимают не более 30% синич-
ников (Иваницкий, Матюхин, 19906). По на-
блюдениям Фетисова с соавторами (1981), в
Ленинградской области нолевые воробьи и
весной часто сооружают наброски во времен-
ных убежищах: брошенные постройки состав-
ляют 19% всех начатых гнезд, причем в 16%
случаев воробьи оставили гнезда после со-
оружения лотка, а в 9% - полностью закон-
ченные гнезда с крышами. Аналогичным об-
разом ведут себя, но нашим наблюдениям, сак-
саульные воробьи — к началу массовой клад-
ки в Ренетеке, которая в разные годы проис-
ходит в 20-35% развешенных синичников,
более или менее объемистые наброски гнез-
дового материала содержались уже в 60-75%
синичников.
Воробьи, обитающие зимой в животновод-
ческих помещениях, строят гнезда в течение
всей зимы, иногда уже в конце декабря здесь
появляются первые кладки (Матюхин, Ива-
ницкий, 1984). Домовые, нолевые и капские
воробьи Passer melanurus, обитающие в теп-
лом климате, заняты строительством гнезд
почти круглый год, используя их и для раз-
множения, и для ночевки (Siegfried, 1973;
Summers-Smith, 1988). Поданным Kendeigh
(1961), домовые воробьи, ночующие в закры-
том убежище при температуре воздуха — 30°
С, экономят за ночь около 13% энергии, рас-
ходуемой на поддержание постоянной темпе-
ратуры тела. Скорее всего, наличие в убежи-
ще плотной массы растительной ветоши, внут-
ри которой ночуют птицы, еще больше пони-
жает расход энергии, однако, с пашей точки
зрения, нет никаких оснований полагать, что
осеннее гиездостроение воробьев сформиро-
валось в процессе эволюции, именно как спе-
цифическая адаптация “направленная на эко-
номизацию энергетических ресурсов” (Наход-
кин, 1986). Пролонгированное гиездостроение
составляет базовую черту репродуктивного
поведения воробьев вне зависимости от кли-
матических условий их обитания. Более того,
оно свойственно всем прочим группам
Ploceimorpha, проявляется у них в разных
формах, и является, по-видимому, весьма древ-
ним признаком, становление которого проис-
ходило отнюдь не в ходе приспособления
нескольких видов воробьев к ночевкам в хо-
лодном климате, а шло в контексте фунда-
ментальных эволюционных перестроек реп-
родуктивного поведения воробьиных нтиц
(Иваницкий, 19956).
Весной оседлые воробьи иногда приступа-
ют к постройке гнезд почти за два месяца до
начала кладки, тогда как месячный разрыв
между этими событиями является вполне
типичным и при неблагоприятных погодных
условиях, отодвигающих начало кладки, иног-
да имеет место даже у мигрирующих индийс-
ких воробьев. При этом важно отметить, что
при определенных условиях гнездо может
быть построено всего за несколько дней. Как
пишет Гаврилов (1974), постройка гнезд у
мш-рирующих испанских воробьев занимает
от 2 до 7 дней, причем птицы, прибывающие в
колонию позднее, строят гнездо быстрее. Фети-
сов с соавторами (1981) также отмечают, что
ускоренное гиездостроение характерно для тех
пар нолевых воробьев, которые приступают к
стройке позже других. Таким образом, гиездо-
строение у многих пар растягивается на мно-
гие недели вопреки тому, что при концентра-
ции усилий вполне нормальное гнездо может
быть построено всего за 2-3 дня.
Особенно важен вопрос о согласованнос-
ти начала кладки с процессом постройки
гнезда. Его актуальность применительно к
воробьям обусловлена своеобразием протека-
64
В.В.Иваницкий
иия созревания гонад у самок, их зависимос-
тью в ряде случаев от самцов в деле построй-
ки гнезд или же, наоборот, возможностью от-
кладки яиц в закрытые убежища с минималь-
ным количеством гнездового материала или
даже вовсе без него. В некоторых случаях
самки вынуждены откладывать яйца в недо-
строенное гнездо (Голованова, 1985; С.А.Бук-
реев, личное сообщение; наши данные). Выше
мы уже упоминали о так называемых экст-
ренных кладках у воробьев. Многие самки,
особенно на поздних этапах формирования
популяции, появляются у мест гнездования
накануне откладки яиц, развитие которых у
воробьев — вплоть до овуляции и кладки
— контролируется социальной стимуляцией,
что ведет, в частности, к массовому отходу
яиц при их откладке в случайных местах
(Формозов, 1944; Гаврилов, 1962; Ковшарь,
1966; Брохович и др., 1986). В состоянии
экстренной репродуктивной готовности сам-
ки проявляют максимум настойчивости и
оперативности в розыске убежищ, образова-
нии пар и гнездостроепии (см.выше).
4.5.2. Участие полов в постройке
гнезд
Выделяются три главных способа разде-
ления обязанностей самцов и самок воробь-
ев в сфере гнездостроения:
1) Андролатералыюе гнездостроение. Ос-
новной вклад в строительство вносят самцы:
они начинают строить, они же выполняют и
основную работу, тогда как на долю самок при-
ходится в основном участие в выстилке гнез-
дового л очка.
2) Бинаренталыюе гнездостроение. Оба
партнера как правило вносят существенный
вклад в строительство, хотя конкретное рас-
пределение обязанностей сильно варьирует не
только у разных видов, но и в разных конс-
пецифичных парах.
3) Гннелатеральпое гнездостроение. Гнез-
до строит исключительно или преимуществен-
но самки. У самцов гнездостроительиые ре-
акции либо отсутствуют полностью, либо про-
являются в качестве атавистической (ритуа-
лизованной) формы поведения.
У всех Passer самцы обыкновенно участву-
ют в постройке гнезд и среди представите-
лей этого рода известны примеры как апдро-
латералыюго, так и бипаренталыюго гнездо-
строення (Summers-Smith, 1988; наши дан-
ные). Что же касается всех прочих родов
Passeridae {Petronia, Gymnorhis, Pyrgilauda,
Montifringilia и Carpospiza), то у них имеет
место строго гипелатералыюе гнездостроение
(Табл.И). Самцы каменных, снежных и ко-
роткопалых воробьев гнезд не строят, хотя
первые чистят гнездовые убежища и всегда
сопровождают самок при полетах за матери-
алов. Лишь у самцов монгольского земляно-
го воробья, сохранилось гнездостроительное
поведение, которое мы отмечали либо у хо-
лостых самцов, либо у самцов, рекламирую-
щих новые поры во время насиживания сам-
кой первой кладки (см.ниже).
Среди изученных нами видов андролате-
ральиое гнездостроение характерно для ис-
панских воробьев. Наши наблюдения в Тад-
жикистане, Узбекистане и Туркмении, а так-
же данные Гаврилова (1962) но Казахстану
свидетельствуют, что гнезда у испанских во-
робьев строят преимущественно самцы, хотя
Умрихина (1955) отмечает, что в Киргизии
самцы и самки вносят примерно равный
вклад в строительство.
Еще до образования нары самец испанско-
го воробья строит гнездовую платформу плос-
кой или чашевидной формы, перемежая дос-
тавку и укладку гнездового материала с пе-
нием и привлекающими демонстрациями.
Брачное и гнездостроителыюе поведение у
этого вида сливаются в единый конгломерат
(см. ниже). При этом холостому самцу то и
дело приходится ремонтировать платформу
или строить ее заново из-за расхищения ма-
териалов соседями (Иваницкий, 1985в). Но
даже при благоприятной обстановке самцы
приостанавливают строительство на стадии
платформы или чашечки, отдавая все время
саморекламированию и бесцельным манипу-
ляциям с накопленным гнездовым материалом.
Лишь после образования нары самец возоб-
новляет строительство и быстро доводит его
до конца. Интересно, что точно такая же двух-
фазная организация строительной деятельно-
сти самцов и стимулирующая роль вселения
самки типичны также и для других видов
певчих птиц с андролатеральпым гиездострое-
пием (настоящиеткачики, крапивники, ремезы).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
65
Таблица 11. УЧАСТИЕ ПОЛОВ В ДОСТАВКЕ ГНЕЗДОВОГО МАТЕРИАЛА
(ио Ivanitskii, 1996 с изменениями)
Виды Время на- блюдений в фазе гнездо- строения (ч) Число пар (самцов) под наблюде- нием Число приносов в час (самцы) Число приносов в час (самки) Число самцов, доставляющих материал (%%)
Испанский 98 18 8,4 ± 1,5 2,7 ± 0,8 18(100)
Домовый 27 6 6,3 ± 1,3 3,8 ± 1,0 4 (66,6)
Индийский 170 29 6,8 ± 1,1 5,1 ± 1,4 19(65,5)
Полевой 64 14 5,3 ± 1,1 6,2 ± 1,7 8(57,1)
Пустынный 50 7 6,7 ± 1,2 5,4 ± 1,8 3 (42,8)
Саксаульный 133 21 3,2 ±0,8 7,1 ± 1,2 7 (33,3)
Рыжий 44 8 2,9 ± 1,2 7,9 ± 2,2 3 (37,5)
Каменный 189 36 - 8,5 ± 1,4 -
Монгольский 44 16 0,3 ±0,1 6,9 ± 1,7 2(12,5)
Снежный 37 8 - 8,9 ± 1,0 -
Короткопалый 28 4 - 11.0 ±2,4 -
Сразу после образования нары самец ис-
панского воробья испытывает сильное побуж-
дение к строительству, однако на первых но-
рах получает доступ к гнездовой мутовке
лишь в моменты отсутствия самки, которая,
оккупируя мутовку, не позволяет самцу при-
ближаться слишком близко. Самец пе только
не сопровождает самку, покидающую колонию,
но в сущности ловит моменты ее отсутствия
для того, чтобы без помех запяться строитель-
ством. Возвращаясь па территорию, самка тут
же прогоняет самца и усаживается на плат-
форму.
Вскоре, однако, самка становится более тер-
пимой к партнеру, и в семье устанавливается
четкий ритм дежурства у гнезда. Завидев под-
летающего супруга, самка спешит покинуть
территорию, давая ему возможность занять-
ся строительством. В свою очередь, самец при
возвращении партнерши обычно сразу же
улетает за очередной порцией гнездового ма-
териала. В результате гнездо находится под
постоянной охраной, что резко снижает ско-
рость расхищения гнездового материала.
Среди других воробьев типичное андро-
латералыюе гнездостроение известно у Passer
luteus и P.moabiticus. Последовательность
работы такая же, как и у испанских воробь-
ев. Вначале самец строит в развилке платфор-
му и принимается ее рекламировать, затра-
чивая много усилий на ремонт и восстанов-
ление сооружения из-за набегов соседей. После
образования пары самец самостоятельно за
несколько дней складывает из колючих ве-
ток очень прочную, шаровидную оболочку с
боковым входом, внутри которой самка из
сухой травы и перьев выстилает лоток в
форме чашечки (Morel, Morel, 1973). В нево-
ле самка построила лоток гнезда вне оболоч-
ки (Kunkel, 1959).
Индийский воробей практикует типично
бипарепталыюе гнездостроение. Вначале мы
рассмотрим поведении этих воробьев при
гнездовании в готовых убежищах. При на-
блюдениях в Бадхызе (Туркмения) и на
Мальгузарском хребте (Узбекистан) ни нам,
ни Зыковой (1980) пе приходилось наблю-
дать доставку гнездового материала холос-
тыми самцами индийского воробья, за исклю-
чением тех самцов, которые долго не могли
образовать пару (Иваницкий, 1985в). Одна-
ко, Семашко (1962) и Матюхин с соавтора-
ми (1990) указывают па склонность некото-
рых холостых самцов индийских воробьев к
п 1ездострое1 шю.
Динамика гпездостроителыюго поведения
во всех обследованных нами парах индийс-
ких воробьев в общих чертах подчинялась
следующей схеме. Холостой самец, занимаю-
щий убежище, чередует саморекламирование
66
В.В.Иваницкий
(пенне, демонстрации) с удалением мусора из
полости. Следует, однако, отметить, что очист-
ка гнезда производится самцами в форме
одиночных выносов мусора с периодичнос-
тью раз в 15-20 мин и совпадает но времени
с окончанием очередного раунда саморекла-
мирования и часто предшествует кратковре-
менному вылету из колонии. На этой стадии
очистка полости никогда не выглядит как со-
средоточенная, целенаправленная деятель-
ность, ио следует повторить, что это поведе-
ние является очень характерным и в той или
иной степени его (в отличие от истинного
гиездостроения) можно наблюдать у холос-
тых самцов всех изученных нами воробьев.
Первое время после образования пары
внимание самца целиком обращено на самку,
и он перестает чистить убежище. Возобнов-
ление гиездостроения связано с началом рас-
чистки гнезда самкой, к чему она приступает
спустя 2-5 дней после образования нары. По-
ведение самок резко отличается от аналогич-
ного поведения самцов своей интенсивностью
(до 28 выносов в час). Самцы никогда не по-
могают интенсивно работающим самкам, од-
нако при длительном отсутствии партнер-
ши самец может возобновить расчистку гнез-
да, действуя при этом в своей обычной, нето-
ропливой манере. Общая продолжительность
очистки нор самками варьирует от 1-2 ча-
сов до 3-4 дней, однако, почти у всех самок
наиболее интенсивная деятельность бывает
приурочена к самому началу очистки полос-
ти и наблюдается обычно в течение всего
нескольких часов утром и в первой полови-
не дня.
Затем самки приступают к доставке гнез-
дового материала в убежище, тогда как сам-
цы вначале в этом не участвуют, но часто со-
провождают самок при полетах за гнездо-
вым материалом и через некоторое время в
свою очередь включаются в его доставку. Бли-
же к началу кладки самки больше времени
остаются у гнезд, их общая подвижность, а
также частота нриносов материала заметно на-
дают. Напротив, самец наращивает усилия и
строит очень интенсивно. Самка же в основ-
ном сидит у гнезда, неизменно встречая при-
летающего с материалом самца интенсивны-
ми предкопуляционными демонстрациями,
что часто стимулирует спаривание. Самец
продолжает строительство и во время отклад-
ки яиц, и во время инкубации, в которой он
также (как и у всех Passer) принимает оп-
ределенное участие. Что же касается самки,
то она тоже продолжает время от времени
доставлять гнездовой материал, однако мень-
шими порциями и лишь “попутно”, то есть
каждый раз возвращаясь к гнезду с кормеж-
ки с периодичностью 20-30 мин (Иваницкий,
1985 в).
Перейдем к более сложной ситуации, ког-
да прежде чем приступить к доставке мате-
риала, необходимо выкопать нору, как это
свойственно индийским воробьям. Самцы и
самки этого вида равно способны ко всем
работам но устройству гнезд, по конкретная
динамика их участия подвержена высокой из-
менчивости и имеет довольно сложный ха-
рактер. Инициатива постройки новой норы
принадлежит обыкновенно самцу. Занятый
раскопками воробей неизменно понадает в
фокус внимания многочисленных соседей и
расходует много сил и времени на то, чтобы
прогонять любопытных со своей территории.
Нетерпимость к присутствию посторонних в
непосредственной близости от сооружаемой
норы в полной мере распространяется и на
холостых самок, выказывающих повышенный
интерес к строительству и явно пытающих-
ся обосноваться в новостройке. Многие из
таких самок, будучи готовы к образованию
нары, буквально осаждают своих избранни-
ков, поджидая момент, когда улетевший на
кормежку самец освободит на время строи-
тельную площадку, и самка, получив доступ к
норе, сразу же со рвением берется за раскоп-
ку и копает до тех нор, пока ее не прогонит
самец и ей вновь приходится ждать нового
шанса для того, чтобы включиться в работу.
Спустя некоторое время самец привыкает
к присутствию самки, и она получает относи-
тельно свободный доступ к поре, после чего
довольно быстро происходит реверсия поло-
вых ролей. Самка включается в раскопку и
быстро наращивает интенсивность работы,
тогда как участие самца столь же быстро
уменьшается. Одновременно, к самке перехо-
дит и роль доминанта в сфере агрессивного
поведения. Она становится хозяйкой положе-
ния и теперь уже в свою очередь не допуска-
ет к норе самца в тех случаях, когда у пего
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
67
некоторое время сохраняется сильная моти-
вация к раскопкам, которая находит выход
лишь в периоды временного отсутствия в норе
самки.
Вскоре самцы утрачивают интерес к рас-
копкам и, предоставив их продолжение сам-
кам, берутся за доставку гнездового материа-
ла. С этого момента открывается особый этан
во взаимоотношениях партнеров, который ха-
рактеризуется относительной стабильностью и
значительной продолжительностью (до 3 не-
дель). Очень редко можно увидеть, как брач-
ные партнеры одновременно и с высокой ин-
тенсивностью копают нору или носят в нее
гнездовой материал (совместная раскопка норы
отмечена в 8, совместная доставка материала
— в 3 из 76 пятиминутных наблюдений). В
противоположность этому, высокий уровень
одновременной активности партнеров у норы
часто наблюдается в тех случаях, когда они
заняты разным делом, то есть самка копает нору,
а самец носит в нее гнездовой материал. В од-
ном случае самец с высокой частотой выносил
грунт, а самка была занята интенсивной лик-
видацией старого гнездового материала — за
все время наблюдений в этой популяции это
был единственный эпизод ярко выраженной
чистки гнездовой полости.
Выявлены заметные половые отличия в ча-
стоте эпизодов совмещения реакций выноса
грунта и доставки материала. У самцов такое
совмещение мы наблюдали всего в 2 из 37 на-
блюдений (5,4%), у самок — втрое чаше (16,2%).
Часто можно видеть, как самка, улетая за оче-
редной порцией гнездового материала, всякий
раз выносит в клюве из норы кусочек грунта,
чего почти никогда не делают самцы.
Частотный и корреляционный анализ выя-
вили обратную зависимость между интенсив-
ностью одноименных форм деятельности у
самцов и самок в парах индийских воробьев на
интервалах времени в 5 мин и 1 час., но не
обнаружили достоверной зависимости между
частотами разноименных форм активности,
выполняемых одновременно (Иваницкий,
1995а, 1996). Таким образом, брачные партне-
ры избегают вместе делать одно и то же, то есть
влияние социальных факторов на гнездостро-
ение в семье проявляется как взаимное тормо-
жение одноименных видов деятельности, что
соответствует гипотезе Гольцмана (1984).
Тем не менее, взаимные влияния партне-
ров в паре носят сложный и противоречи-
вый характер и отнюдь не исчерпываются
процессами торможения. Само ио себе при-
сутствие партнера оказывает сильно стиму-
лирующее воздействие, что можно показать
на примере наших наблюдений за полигам-
ным трио индийских воробьев. Полигамия у
этого вида встречается очень редко (Матю-
хин и др., 1990; Matuhin, 1994), поэтому каж-
дый такой случай представляет собой боль-
шой интерес, как своеобразный эксперимент,
иллюстрирующий поведение птиц в необыч-
ной социальной среде.
В момент начала наших наблюдений обе
самки почти непрерывно сидели у своих нор
всего в 15 см друг от друга. Большая часть
активности самца была сосредоточена у од-
ной норы (А), тогда как другую нору (Б) он
посещал лишь изредка. Самка А по отноше-
нию к самцу выступала в роли социального
доминанта, самка Б — явно занимала подчи-
ненное положение. Самец с высокой частотой
иосил гнездовой материал в нору А, самка А
столь же интенсивно занималась раскопкой
норы, тогда как самка Б неподвижно сидела
у своего входа. Картина резко менялась, как
только самец переключал свое внимание на
другую нору. Он переставал летать за гнездо-
вым материалом и принимался за раскопку
норы Б. Подвижность самки Б тут же резко
возрастала, и она также всеми способами
пыталась включиться в раскопку, наталкива-
ясь, однако на жесткую агрессивность самца.
Что же касается покинутой на время самки
А, то она немедленно останавливала раскоп-
ки, усаживалась у норы и погружалась в
полную неподвижность до тех нор, пока спус-
тя 5-10 мин сюда не возвращался самец, пос-
ле чего самка А сразу же возобновляла вы-
нос грунта. Такая ситуация сохранялась в
течение 2 дней, ио прошествии которых са-
мец переключил главное внимание на нору
Б, приступив к интенсивной доставке гнездо-
вого материала. Одновременно самка Б стала
раскапывать нору, а самка А приступила к
доставке гнездового материала в свою нору.
На основании этих наблюдений можно
сделать два вывода. Прежде всего удалось ус-
тановить, что в поведении самца одновремен-
но существуют две программы: доставки ма-
68
В.В.Иваницкий
Рисунок 4. ДИНАМИКА УЧАСТИЯ В ГНЕЗДОСТРОЕНИИ САМЦОВ И САМОК
I — испанский воробей; II — индийский воробей при гнездовании в скалах: III — саксаульный воробей; IV
- каменный воробей: V — индийский воробей при гнездовании в норах (доставка гнездового материала); VI -
индийский воробей при гнездовании в норах (рытье норы). По горизонтальной осп - стадии репродуктивного
цикла: А — холостой самец; Б, В, Г — первая, вторая и последняя трети отрезка времени от момента образования
пары до начала насиживания; Д — насиживание. По вертикальной оси — средняя частота приносов материала
или выноса грунта в час.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
69
тернала и раскопки норы. В зависимости от
обстоятельств он способен быстро переклю-
чаться па альтернативный вид деятельности,
хотя их совмещение в нормальных условиях
встречается очень редко (см.выше). Второй
вывод состоит в том, что у самок поведение,
связанное с раскопкой нор, явно стимулиру-
ется присутствием самца, однако, в дальней-
шем его одноименная деятельность либо тор-
мозит поведение самки, либо вызывает ее аг-
рессию.
Билатеральное гнездостроение свойствен-
но большинству представителей р.Passer. По
нашим наблюдениям, многие (хотя и не все)
холостые самцы саксаульных, полевых и до-
мовых воробьев регулярно приносят в рек-
ламируемые убежища гнездовой материал,
хотя частота доставки и величина порций
материала подвержены очень сильным и не-
предсказуемым изменениям даже у одного и
того же самца. Также, как и у испанских во-
робьев, холостые самцы упомянутых видов
строят в убежище плоскую или чашевидную
гнездовую платформу, после чего частота дос-
тавки материалов снижается, и самцы при-
нимаются интенсивно петь, то есть характер-
ная двухфазная организация гпездостроитель-
иой деятельности самцов Passer сохраняется
и у видов, гнездящихся в закрытых убежи-
щах. У рыжих воробьев, по нашим наблюде-
ниям, холостые самцы гнезд не строят. Пер-
вой носить материал в дупло начинает самка,
а к ней спустя некоторое время может присо-
единиться самец, хотя для некоторых самцов
нам не удалось зарегистрировать участия в
доставке материала (Табл. 11).
Хотя и самцы, и самки воробьев р.Passer
способны выполнять все работы но устрой-
ству гнезд, у всех видов прослеживается чет-
кая тенденция к разделению одноименной
деятельности партнеров во времени или же
к дивергенции видов деятельности. На Рис.4
суммированы наши данные об изменениях
частоты доставки гнездового материала у
самцов и самок трех видов воробьев в зави-
симости от стадий репродуктивного цикла. У
саксаульного воробья самцы наиболее склон-
ны строить гнездо до образования пары и в
начальный период насиживания, в промежу-
точный период строят в основном самки. Хо-
лостые самцы индийского воробья, распола-
гающие готовым убежищем, занимаются дос-
тавкой материала лишь в очень ограничен-
ной степени и часто вообще этим пренебре-
гают. По-настоящему строить гнезда начина-
ют самки, вскоре к ним присоединяются сам-
цы, которые быстро наращивают свое учас-
тие, тогда как активность самок постепенно
надает. Когда появляется возможность аль-
тернативного вида деятельности (рытье
норы), динамика участия партнеров услож-
няется, однако главная тенденция сохраняет-
ся. Если в строительстве активно участвуют
оба партнера, то один из них почти всегда
трудится намного интенсивнее другого. Если
же они работают с одинаково высокой ин-
тенсивностью, то, как правило, каждый из них
занимается своим делом вплоть до того, что
один может носить в убежище гнездовой ма-
териал, а другой в это же время выносит его
обратно, занимаясь “расчисткой” гнездовой
полости. Указанные закономерности харак-
терны для гнездостроительного поведения
всех изученных нами видов с билатеральным
п 1ездострое1 тем.
Совместная деятельность животных пред-
ставляет собой противоречивый процесс, вклю-
чающий скрытые и явные формы коопера-
ции и соперничества, взаимной стимуляции
(возбуждения) и подавления (торможения)
и построенный на обоюдных поведенческих
адаптациях и постоянных наблюдениях за
действиями партнера (или партнеров) (Голь-
цман, 1976). Примером может служить иссле-
дование билатерального гнездостроения у
корольков {Regulus sp.) (Thaler, 1976). Как
и у воробьев, постройка гнезда у этих моно-
гамных нтиц продолжается довольно долго
— до 18 дней. Гнездостроение тесно увязано
с динамикой социальных взаимоотношений
брачных партнеров, которые конкурируют
друг с другом за участие в строительстве. У
желтоголовых корольков R.regulus вначале
доминирует самец, он же начинает строить
гнездо. Некоторые самцы бывают крайне аг-
рессивностью н вообще не позволяют самкам
приближаться к своим гнездам, хотя без уча-
стия последних самцы могут построить лишь
очень массивные, но бесформенные сооруже-
ния. В большинстве нар, однако, примерно к
середине указанного срока роль социального
доминанта переходит к самке, которая актив-
70
В.В.Иваницкий
по вытесняет партнера со строительной пло-
щадки, крайне агрессивно реагируя па его
попытки внести свою ленту в постройку и
вынуждая его заниматься гнездом только в
свое отсутствие. У красноголовых корольков
R.ignicapillus на всех стадиях гнездострое-
ния доминируют самки, и самцы почти пол-
ностью исключаются из этой сферы. Но ин-
стинкт гиездостроения присущ самцамв пол-
ной мере, что легко выявляется в тех случаях,
когда самцы краспоголовых корольков обра-
зуют нары с более боязливыми на первых
норах самками желтоголового королька.
Таким образом, участие в постройке обще-
го гнезда способно порождать острую конку-
ренцию между партнерами даже в моногам-
ных нарах. У корольков она носит относи-
тельно простой характер и базируется пре-
имущественно на прямых агрессивных взаи-
модействиях между партнерами. Вследствие
конкуренции на каждом данном отрезке вре-
мени в строительстве преимущественно уча-
ствует либо самка, либо самец. У воробьев тот
же результат достигается, но-видимому, на
основе более тонких, опосредованных меха-
низмов психологической природы. Можно
предположить, что при дальнейших исследо-
ваниях совместного гиездостроения у птиц
особенно перспективной может оказаться мо-
дель социальной коммуникации, включающая
не только возможность взаимной стимуляции
но принципу “социального облегчения” (ими-
тации, подражания), но и возможность вза-
имного подавления (торможения, угнетения)
одноименных форм поведения (Гольцман,
1976, 1983). Помимо всего прочего, такая мо-
дель предсказывает широкую изменчивость
всех параметров гиездостроения и прежде
всего — относительного участия в нем сам-
цов и самок, что на примере воробьев хоро-
шо подтверждается нашими наблюдениями
и литературными данными.
4.6. Насиживание
Многое из того, что было сказано о гнез-
достроении воробьев характерно и для ин-
кубационного поведения этих птиц. Вне зави-
симости от участия полов в насиживании, оно
начинается сразу же после откладки перво-
го яйца и в соответствующее время плавно
переходит в необычайно длительное и плот-
ное обогревание птенцов.
Поданным круглосуточных наблюдений за
гнездами нолевых воробьев Фетисов и др.
(1981) обнаружили, что уже в день снесения
первого яйца партнеры находились в гнезде в
общей сложности 4 ч 40 мин, и плотность на-
сиживания составила 19,4%. Воробьи посети-
ли гнездо поочередно 22 раза, переворачивая
яйцо в среднем 5,1 раза в час. Максимальная
плотность насиживания у полевых воробьев
устанавливается на уровне 80% ко дню завер-
шения кладки, однако во вторых кладках мак-
симальная плотность насиживания достигает-
ся уже в день снесения 2 или 3-го яйца. Плот-
ность насиживания у домовых воробьев зави-
сит от температуры воздуха и варьирует от
58,7 до 87,0% (Summers-Smith, 1988).
Специальные исследования инкубационно-
го поведения Passer moabiticus, гнездящихся
в котловине Мертвого моря при экстремаль-
но высоких температурах, показали, что сам-
ки способны влиять па температуру кладки
пока температура воздуха не превышает 39°С,
после чего резко возрастает вероятность ги-
бели эмбрионов и птенцов (Yom-Tov,
Mendelssohn, 1978; Yom-Tov, Аг, 1980). Как
отмечают Соныев (1965) и Пономарева
(1983), изучавшие инкубацию пустынных во-
робьев в Каракумах, температура воздуха в
местах обитания этих птиц, часто бывает на-
столько высокой, что утрачивается необходи-
мость обогревания кладки. Как показали
наши наблюдения в Ренетеке, при прибли-
жении температуры воздуха к отметке 40°С
у пустынных, саксаульных и индийских во-
робьев наступает полная дезорганизация ин-
кубационного поведения: устанавливается
произвольный ритм посещения гнезда, про-
должительность пребывания в нем сильно ва-
рьирует, часто птица просто сидит на входе в
гнездо, не имея контакта с кладкой.
Данные но насиживанию у воробьев сумми-
рованы в Табл. 12. Как и в случае гнездострое-
ния, четко выделяются два поведенческих тина
инкубации: билатеральная и типе латеральная.
Первый тин характерен для большинства пред-
ставителей Passer. Исключение составляют
P.moabiticus и P.eminibey — самцы у них не
насиживают (Yom-Tov, Mendelssohn, 1978;
Summers-Smith, 1988). Во всех прочих родах
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
71
известна только пшелатеральиая инкубация.
Переходя к поведенческой характеристике
билатерального насиживания у воробьев, ука-
жем, что оно представляет собой совместную
деятельность двух индивидов, направленную на
общий объект интересов, что не только откры-
вает возможности для кооперации, по и со-
здает почву для конкуренции и взаимной
неопределенности. Наличие побуждения к на-
сиживанию у обоих партнеров легко приво-
дит к конфликту. У кольчатых горлиц
Streptopellia risoria ритм насиживания и рас-
писание участия в нем парнеров сильно отли-
чаются у разных пар — доля участи самцов
варьирует от 23 до 76%. Расписание насижива-
ния у каждой пары сохраняется в течешь ряда
репродуктивных циклов, однако смена парт-
нера влечет за собой его радикальную пере-
стройку, причем как самцы, так и самки стре-
мятся увеличивать свою долю в насиживании,
если новый партнер ведет себя менее иници-
ативно, чем прежний (Wallman et al., 1979).
Острая конкуренция за участие в насижива-
нии при однозначном доминировании более
крупных и агрессивных самок обнаружена у
хищных птиц — кондоров Gymnogyps
califoniicus (Harvey, Hart, Leet, 1994), сапсанов
Falco peregrinus и канюков Buteo buteo
(Hubert, Carlier, 1992). Авторы этих работ не-
зависимо друг от друга приходят к выводу о
том, что специфическое для каждой пары рас-
писание насиживания складывается в процес-
се взаимной адаптации партнеров. Каждый из
них стремится внести максимальный вклад в
общее дело, и это приводит к конфликту ин-
тересов, поскольку в каждый данный момент
насиживать может только одна птица.
Самцы Passer насиживают только днем,
ночью в гнездах сидят самки. У полевых во-
робьев доля самцов в насиживании состави-
ла около 30% дневного времени (Summers-
Smith, 1988), испанского и домового воробь-
ев — 40% (Гаврилов, 1962; Hegner, Wingfield,
1986а), у капских воробьев Passer melanurus
самки и самки днем насиживают поровну
(Immelmann, 1970), у пустынных воробьев
относительно участие иолов в насиживании
подвержено чрезвычайно высокой изменчи-
вости вплоть до того, что в одних нарах на-
сиживают преимущественно самцы, тогда как
в других — главным образом самки (Сопы-
ев, 1965; Пономарева, 1983). Высокую видо-
вую изменчивость поведения насиживания
подтверждают и наши наблюдения.
Как показали наблюдения Deckert (1968,
1973) за полевыми воробьями, а также наши
исследования индийских и испанских воро-
бьев, к моменту начала насиживания роль со-
Таблица 12. УЧАСТИЕ ПОЛОВ В НАСИЖИВАНИИ КЛАДКИ
(но Ivanitskii, 1996 с изменениями)
Виды Время наблюдений в фазе на- сиживания Число пар под наблю- дением Насиживают самцы (%% времени) Насиживают самки (%% времени) Не наси- живают (Я % вре- мени) Число на- сиживаю- щих сам- цов (%%)
Испанский 9 9 40.0 60.0 ? 9
Домовый 9 9 23.1 45.6 31.3 9
Индийский 65 12 15.2 50.8 34.0 ю (83.3)
Полевой 32 5 34.7 58.0 17.3 5(100,0)
Пустынный 22 4 34.8 40.1 25.1 4(100,0)
Саксаульный 40 7 12.3 48.9 38.8 4 (57.1)
Рыжий 21 5 15.7 51.3 33,0 3 (60.0)
Каменный 35 8 - 79.8 20,2 0
Монгольский 14 6 - 81.0 19,0 0
Снежный 8 5 - 75,4 24,6 0
Короткопалый 9 9 - 9 9 0
Источники данных: 1 — Гаврилов, 1962; 2 — Summers-Smith, 1988; 3-10 — наши наблюдения; 11 — Адамян,
1966.
72
В.В. Иваницкий
циалыюго доминанта в семье всегда принад-
лежит самке, которая во многом контролирует
ритм участия в насиживании партнера. Обыч-
но в начале насиживания самка очень агрес-
сивно относится к попыткам самца забраться
внутрь гнезда в ее присутствии, отчего, приле-
тев к гнезду самец часто вынужден дожидать-
ся, когда самка улетит, и лишь после этого про-
никает в него. Что же касается самки, то она
после кормежки сразу же устремляется в гнез-
до, откуда тотчас же вылетает самец. В то же
время, если самец не прилетает долго, самка в
свою очередь задерживается в гнезде, словно
“дожидаясь” смены. Напротив, самец никогда
не ждет самку и часто покидает гнездо до ее
появления, а эпизоды его присутствия на гнез-
де короче, чем у самки.
Экспериментальное устранение самцов до-
мовых воробьев в начале инкубации заменю
уменьшало успешность вылунле1шя птенцов
(Schifferli, 1978). Оценивая возможное термо-
регуляторное значение билатеральной инку-
бации у воробьев, приходится учитывать, что
самцы Passer — в отличие от самок — не име-
ют паседных пятен, поэтому их вклад в наси-
живание может состоять лишь в уменьшении
скорости остывания кладки при отсутствии
самки. Эта биологическая задача, па наш взгляд,
едва ли важна для воробьев, размножающих-
ся в гнездах с хорошими (а иногда — исклю-
чительно высокими) термостатическими свой-
ствами в условиях относительного теплого
или даже жаркого климата. Интересно, что ги-
нелатеральная инкубация утвердилась как раз
у тех воробьев, которые обитают в ландшаф-
тах с более холодным климатом (Petronia,
Pyrgilauda, Мontifringilia), в то время как у
живущих в жарком климате Passer кладку
всегда опекают оба партнера. Более вероятным
преимуществом билатеральной инкубации
представляется увеличение влажности в гнез-
довой камере из-за постоянного присутствия в
ней взрослых птиц, однако экспериментальные
данные на этот счет противоречивы (Кегп,
Knapic, 1991).
Возможно, поведение самцов воробьев
иногда представляет собой нсевдоинкубацню,
то есть пребывание в гнезде без контакта с
кладкой, что очень характерно, например, для
самцов городских ласточек Delichon urbica,
которые, помимо этого, часто сидят в гнезде
вместе со своими насиживающими самками
(Колоярцев, 1989).
Несомненно то, что, находясь в гнезде са-
мец вне зависимости от характера контакта с
кладкой охраняет его содержимое. Эта задача
особенно актуальна для воробьев, обитающих
в плотных и многочисленных колониях, в ус-
ловиях жесткого пресса социальной интерфе-
ренции в форме гнездового паразитизма, ин-
фантицида, постоянных инспекции убежищ
чужими особями, озабоченными поисками
гнезд (см. выше). У Passer melanurus ритм сме-
ны партнеров наиболее четко выражен в плот-
ных колониях, тогда как у одиночно гнездя-
щихся нар он носит значительно более сво-
бодный характер, и гнездо значительно чаще
остается без присмотра (Immelman, 1980). Ана-
логичная зависимость плотности насиживания
от социального окружения была выявлена и
у городских ласточек, которым также свой-
ственна билатеральная инкубация (Lind, 1960).
К числу очевидных недостатков билате-
рального насиживания следует отнести, на наш
взгляд, возрастание демаскирующего эффек-
та из-за присутствия у гнезда второй птицы, а
также повышение вдвое вероятности зараже-
ния кладки болезнетворными бактериями и
вирусами. Как показали исследования боль-
шого коллектива авторов в Польше, бактери-
альные и вирусные инфекции, поступающие
в гнезда с оперения взрослых птиц, составля-
ют одну из главных причин эмбриональной
смертности домовых п нолевых воробьев
(Pinowski, Kavanagh, Gorski, 1991).
У ближайших родствешшков воробьев пре-
обладает билатеральная инкубация. Это, но-
виднмому, единственный тин инкубации у всех
Estrildidae. У Ploceinae гинелатеральпая ин-
кубация известна лишь у полигамных пред-
ставителей p.Ploceus и всех Euplectes, в то
время как у многих моногамных Ploceus (см.
нанрймер Skead, 1953) и во всех прочих ро-
дах (Quelea, Foudia, Malimbus) в насижива-
нии участвуют и самцы, и самки (Crook, 1964,
Brosset, 1978). Эпизодическое участие самцов
в насиживании отмечено даже для некоторых
полигамных Ploceus (Ambedkar, 1972). Среди
Plocepasserinae у Plocepasser насиживают лишь
самки, по у Pseudonigritta и Philetairus —
оба пола. Таким образом, есть основания рас-
сматривать билатеральную инкубацию, как ар-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
73
хаичный признак для всех Ploceimorpha, кото-
рый, вероятно, был унаследован ими от еще
более далеких предковых форм. Более того, из-
вестно, что активное участие самцов в насижи-
вании кладки характерно для подавляющего
большинства представителей примитивных
воробьиных (Non-Oscines) а также отрядов, в
генеалогическом отношении наиболее близких
к Passeriformes. По мнению многих исследова-
телей, с эволюционной точки зрения, участие
самцов в насиживании в целом для птиц сле-
дует считать архаичным, примитивным при-
знаком (Kendeigh, 1952; Scutch, 1977; Аг, Yom-
Tov, 1978).
4.7. Развитие птенцов, родитель-
ское поведение, полицикличностъ
Одной из наиболее характерных особенно-
стей репродуктивной биологии воробьев яв-
ляется ярко выраженная гетерохронность эм-
брионального и постнаталыюго развития. Глав-
ная причина гетерохронности коренится, ви-
димо, в том, что инкубация, как сказано выше,
начинается сразу же после откладки первого
яйца и, благодаря участию обоих партнеров, яв-
ляется практически непрерывной. Оба эти
обстоятельства, как хорошо известно, резко ог-
раничивают возможности синхронизации раз-
вития потомства птиц на всех стадиях (Болот-
ников и др., 1985; Shmier, Evans, 1992).
Вылупление птенцов у испанских, палес-
тинских и нолевых воробьев растягивается на
1,5-2,0 суток (Гаврилов, 1962; Ильенко, 1976;
Yom-Tov, Аг, 1980; Фетисов, 1981). Вторые клад-
ки нолевых воробьев инкубируются в период
их формирования более плотно, поэтому пе-
риод вылупления птенцов иногда растягивает
доЗ суток (Доржиев, Доржиев, 1985). У пус-
тынных воробьев последовательность вылуп-
ления в ряде случаев практически повторяет
последовательность кладки яиц и занимает
до 4-5 суток (Соныев, 1965). У домовых воро-
бьев время вылупления птенцов в разных гнез-
дах подвержено сильной изменчивости и ва-
рьирует в пределах от 0 до 53,5 (в среднем
17,6± 10,9) часов, и находится в прямой зави-
симости от размера кладки и температуры
воздуха (Veiga, Vinuela, 1993).
В результате растянутого вылупления
птенцы в выводках воробьев всегда очень
резко отличаются друг от друга но размерам,
весу и и уровню общего развития. У ноле-
вых воробьев разница в размерах птенцов
одного выводка становится особенно заметна
па 4-5-й день жизни (Фетисов, 1981), причем
самые младшие птенцы бывают почти в 5
раз легче самых старших (Формозов, 1944).
Это оказывает самое неблагоприятное воздей-
ствие па судьбу младших птенцов. Прежде
всего, старшие птенцы способны нанести
физические повреждения младшим. По дан-
ным Фетисова (1977а,6), “затаптывание” слу-
жит причиной гибели 13% всех птенцов но-
левых воробьев и выступает в роли одного
нз главных факторов смертности. Соныев
(1965) пишет, что “затаптывание” ведет к
гибели 7% потомства у пустынных воробьев.
Гибель младших птенцов от “затаптывания”
— не редкость у домовых воробьев (Ильен-
ко, 1976; Морозов, Остапенко, 1988).
Помимо подверженности физическим по-
вреждениям, младшие птенцы проигрывают
в конкуренции за приносимый родителями
корм. Как полагают Елисеева (1961) и Фе-
тисов (1977), смертность птенцов нолевых
воробьев зависит от условий выкармливания
выводка, то есть от обилия и доступности
пищевых ресурсов, однако, 1-2 самых млад-
ших птенца обречены на гибель в любом слу-
чае. При этом, у домовых и полевых воробь-
ев при дефиците корма первыми гибнут са-
мые младшие птенцы, тогда как вероятность
гибели всех прочих птенцов от возраста не
зависит (Veiga, 1990),
Таким образом, воробьи несут ощутимые
репродуктивные потери в результате гетерох-
ронности развития птенцов, обусловленной
ранним началом насиживания. Судя но име-
ющимся данным, эти потери носят но суще-
ству облигатный характер и обусловлива-
ются главным образом физической несовмес-
тимостью разновозрастных птенцов. Анализи-
руя природу этих потерь, Veiga (1992) ука-
зывает на слабую зависимость смертности
младших птенцов домового воробья от уров-
ня обеспеченности пищей, что противоречит
известной гипотезе “редукции выводка”
(Lack, 1968) и полагает, что эта смертность
вызывается ранним началом инкубации, ко-
торое, как показали наблюдения этого авто-
ра, уменьшает эмбриональную смертность в
74
В. В.Иваницкий
период откладки яиц у домовых воробьев и
заметно увеличивает выживаемость птенцов
из первых яиц в ущерб самым младшим птен-
цам (см. также Veiga, Vinuela, 1993).
По литературным данным и нашим на-
блюдениям в выкармливании птенцов у всех
воробьев участвуют и самцы, и самки. Вмес-
те с тем, в этой сфере имеет место довольно
своеобразная динамика относительного учас-
тия партнеров в доставке корма, что обуслов-
ливает сложную временную организацию это-
го вида совместной деятельности и доволь-
но четкое разделение половых ролей на раз-
ных стадиях выкармливания птенцов.
Типичная картина изображена па Рис.5, где
суммированы наши наблюдения за поведени-
ем одной нары монгольских земляных воро-
бьев на Алтае. Самцы этого вида не участвуют
в насиживании, и их социальная активность
— как н многих других птиц (см.Панов, 1978),
резко активизируется после того, как самки
приступают к насиживанию. Возрастает общая
подвижность самцов, учащаются токовые по-
леты, пение, пограничные конфликты и пат-
рулирование границ. Многие самцы уже к се-
редине насиживания подыскивают нору под
вторую кладку и принимаются ее интенсивно
рекламировать с помощью пения и токовых
полетов, а иногда и приносить сюда гнездовой
материал. Обычно самец находит новую нору
неподалеку от прежней (в 10-15 м), то есть в
пределах видимости, и почти всегда сопро-
вождает самку во время ее периодических
вылетов из гнезда для кормежки. Характерно,
что самка всегда стремится улететь далеко, от-
правляясь на самую периферию территории,
нередко за 300-400 м от гнезда.
Самцы включаются в доставку корма сра-
зу же после вылупления птенцов, но в пер-
вые дни носят корм небольшими порциями
и довольно редко. Постепенно объемы при-
носимого корма и частота доставки увели-
чиваются. Самки в первые 5-6 дней подолгу
остаются в гнезде, обогревая птенцов, и в об-
щей сложности доставляют корм значитель-
но реже самцов. Примерно к 9-10 дню частота
приносов корма у обоих партнеров выравни-
вается, и почти сразу же вслед за этим учас-
тие самца начинает падать параллельно с
постепенной активизацией его демонстратив-
ного поведения, пения и токования. С 10-11
дня собственно родительское поведение сам-
ца сочетается с брачным поведением, интен-
сивным рекламированием второй норы и
ухаживанием за самкой.
Птенцы начинают вылезать из норы в воз-
расте 12-14 дней, но еще 2-3 дня избегают от-
даляться от входа и пользоваться соседни-
ми норами. Примерно в это время самки
приступает к постройке гнезд, спариванию и
кладке в указанных самцами новых норах.
Иногда самки начинают следующий репро-
дуктивный цикл еще до вылета первой гене-
рации птенцов. Мы наблюдали 3 нары, в ко-
торых самки уже насиживали, при том, что
самцы кормили гнездовых птенцов в возрас-
те 14-16 дней. Еще у двух пар, вторые гнезда
которых были раскопаны, кладка началась в
то время, когда птенцам первой генерации
исполнилось не более 17-18 дней.
Таким образом, выкармливание птенцов у
земляных воробьев состоит из нескольких ста-
дий, характеризующихся особым разделени-
ем ролей партнеров:
1) Возраст птенцов 1-10 дней. Самка подо-
лгу обогревает птенцов и на первых норах ее
поведение в сущности ничем не отличается от
инкубации. Доставка корма птенцам возложе-
на на самца, самка вначале приносит корм по-
путно, возвращаясь в гнездо с кормежки. Тем
не менее, к концу этого периода частота дос-
тавки корма у партнеров выравнивается и
достигает максимального уровня;
2) Возраст птенцов 10-15 дней. К концу
этого времени они начинают выходить из
норы Самец возобновляет интенсивное пение
и токование, резко снижая участие в достав-
ке корма, которая на этой стадии в основном
ложится на самку, у которой одновременно
возобновляется развитие гонад;
3) Возраст птенцов 15-20 дней. Птенцы
покидают гнездовую нору. Самка приступает
к откладке яиц и насиживанию, эпизодичес-
ки подкармливая слетков, основную заботу о
которых вновь берет па себя самец.
Эти стадии универсальны для всех изу-
ченных нами воробьев. Известно, что у домо-
вых воробьев самцы также резко ограничи-
вают участие в выкармливании птенцов при-
мерно к десятидневному возрасту, отдавая все
больше времени нению и токованию или воз-
ле ближайшего пригодного убежища (но,
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
75
Рисунок 5. НЕКОТОРЫЕ ОСОБЕННОСТИ ПОВЕДЕНИЯ МОНГОЛЬСКИХ ЗЕМЛЯНЫХ ВОРОБЬЕВ
ПРИ ВЫКАРМЛИВАНИИ ПТЕНЦОВ
1 — число приносов корма самкой в час; 2 — число приносов корма самцом в час; 3 — интенсивность
обогревания птенцов и насиживания кладки (%% от времени наблюдении); 4 — число токовых полетов в час; 5
- дни с момента вылупления птенцов первой генерации; 6 — начало насиживания второй кладки. По оси орди-
нат: левая шкала - число прилетов с кормом (в час) и интенсивность насиживания (®о°о); правая шкала — число
токовых полетов (в час)
обычно, не далее метра от гнезда), либо не-
посредственно рядом со своим гнездом, куда
самка постоянно носит корм (Ильенко, 1976;
Summers-Smith, 1956; Hegner, Wingfield, 1986).
Полицнкличность очень характерна для
размножения воробьев. Даже в условиях ко-
роткого и холодного горного лета Централь-
ной Азии монгольские земляные и снежные
воробьи успевают выкормить но два вывод-
ка (Нсйфельдт, 1986; Ирисова, 1990; Иваниц-
кий, Шевченко, 1991). С другой стороны, сак-
саульные и пустынные воробьи, гнездящиеся
в очень жарком климате, также выкармлива-
ют за сезон но два выводка (Соныев,
1965,1970; Пономарева, 1983). У рыжих во-
робьев даже на северном пределе распрост-
ранения этого вида на юге Сахалина и высо-
ко в горах (Гималаи) бывает но 2 кладки
(Нечаев, 1973; Summers-Smith, 1988), хотя
Зембицкая (1979) считает этот вид моноцнк-
личным.
На Кольском полуострове па усадьбе Лап-
ландского заповедника домовые воробьи ус-
певают выкормить до два, а в г.Мончегорске
76
В.В.Иваницкий
лапдского заповедника домовые воробьи ус-
певают выкормить до два, а в г.Мончегорске
— по 3-4 выводка за лето (Гилязов, Семепов-
Тяи-Шанский, 1990). В Болгарии, судя по не-
которым данным, у домовых воробьев также
предполагается до 4 выводков за сезон (Ива-
нов, 1987). Хотя эти данные, с нашей точки
зрения, нуждаются в подтверждении с приме-
нением методов индивидуального мечения, они
все же являются весьма впечатляющей иллю-
страцией способности воробьев к оперативно-
му, нолицикличному размножению, в особен-
ности, если учесть крайнюю тенлолюбивость
и в целом повсеместно довольно поздние сро-
ки начала гнездования этих птиц. Во всяком
случае в шт.Нью-Йорк (США), где наблюда-
ли за меченными птицами, многие пары до-
мовых воробьев в самом деле успевали вы-
кормить за сезон по 3 выводка, причем отдель-
ные самки откладывали за это время до 5
кладок (Hegner, Wingfield, 1986 а,Ь). Интерес-
но, что на той же широте в Испании, где прово-
дились столь же подробные наблюдения, у
домовых воробьев было только но 2 выводка
(Veiga, 1990, 1992). Обобщение большого ма-
териала по фенологии размножения нолевых
воробьев в разных точках обширного ареала
вида показало, что от Великобритании до Син-
гапура — за исключением самых северных
пределов распространения — этот вид спосо-
бен вырастить до 3 выводков за сезон (Гагин-
ская и др., 1981). Однако, н в Заполярье па
северном Урале нолевые воробьи выращива-
ют за лето по 2 выводка (Рябицев, 1987).
При нормальном размножении средняя
продолжительность интервала между выле-
том птенцов и началом следующей кладки у
домовых воробьев в Болгарии в разные годы
варьирует от 5,3 от 6,3 суток (Иванов, 1987).
В США этот интервал был больше при гнез-
довании в открыто расположенных гнездах,
чем при гнездовании в дуплянках, и досто-
верно возрастал в ходе сезона от 7,2 до 11,0
суток (McGillvray, 1983). Специальные экс-
перименты но индукции следующего цикла
путем удаления содержимого гнезд показали,
что домовые воробьи могут отложить первое
яйцо через 3-12 (в среднем 7,0) дней, а ноле-
вые воробьи — через 2-13 (в среднем 6,8)
дней, причем 65,5% из 29 повторных кладок
домовых и 66,7% повторных кладок поле-
вых воробьев были начаты в интервале 6-8
дней после начала эксперимента. Зависимость
скорости возобновления кладки от времени
была у этих видов совершенно разной: у до-
мовых воробьев она увеличивалась по мере
течения цикла, у нолевых — сокращалась, хотя
обе тенденции не обнаружили статистичес-
кой достоверности (Anderson, 1979).
Известно, что для формирования кладки
у домовых воробьев требуется примерно 8
дней (Krementz, Ankney, 1986). Таким обра-
зом, повторная активизация роста гонад у
многих самок воробьев начинается но суще-
ству в разгар выкармливания птенцов и со-
впадает по времени с резким ослаблением
родительского поведения самцов и нараста-
нием их социо-сексуальной активности, что,
как можно думать, и стимулирует переход
самок в состояние репродуктивной готовнос-
ти (см.ниже). В результате пауза между реп-
родуктивными циклами практически исче-
зает, и птицы находятся в состоянии “пер-
манентного” размножения.
Примечательно, что домовые воробьи —
подобно монгольским земляным воробьям —
способны отложить следующую кладку, не до-
жидаясь вылета птенцов предыдущей генера-
ции. При этом яйца могут быть отложены как
в новое гнездо, так и в прежнее убежище с
птенцами. Lowther (1979) проанализировал 8
случаев совмещения кладок и птенцов в од-
ном убежище. Самки приступали к следую-
щим кладкам при возрасте птенцов от 10 до
14 дней. Хотя многие эмбрионы в таких клад-
ках гибнут от повреждений, наносимых птен-
цами, некоторые все же развиваются нормаль-
но. Отмечается индивидуальная склонность
некоторых самок к перманентному размноже-
нию в постоянных убежищах и приуроченность
таких случаев к определенным колониям.
4.8. Основные направления эво-
люции репродуктивного поведения
в разных группах зерноядных птиц
Поведение, связанное с постройкой гнезд,
является одним из ключевых элементов реп-
родуктивного цикла всех Ploceimorpha. В од-
ном из разделов предыдущей главы мы про-
анализировали основные отличия в конструк-
циях и манере расположения гнезд у пред-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
77
ставителей разных групп ткачикоподобпых
птиц. Хотя эти различия, пе выходя все же за
рамки облигатной клаустрофилии, выраже-
ны очень рельефно, следует теперь обратить
внимание и на очень существенное сходство
но ряду базовых признаков, характеризую-
щих само ио себе гпездостроителыюе пове-
дение этих птиц:
1) За исключением вдовушек (Viduinae),
утративших навыки постройки гнезд в про-
цессе перехода к облигатному гнездовому
паразитизму, во всех остальных группах
Ploceimorpha существенную, а часто и доми-
нирующую роль в гнездостроении играют
самцы. Многим Ploceidae н Passeridae свой-
ственно андролатералыюе гиездостроение,
однако более широким распространением
пользуется бипарентальное гиездостроение,
которое характерно для всех групп (Passeridae,
Ploceinae, Plocepasserinae, Bubalornithinae,
Estrildidae), тогда как гнпелатералыюе гиез-
достроепие известно лишь у нескольких ро-
дов воробьев.
2) Во всех упомянутых группах гнездост-
роение не ограничено жесткими временны-
ми рамками, носит пролонгированный харак-
тер и в некоторых случаях представляет со-
бой в сущности перманентную форму актив-
ности, составляющую неотъемлемую часть ру-
тины повседневного существования в ходе
репродуктивного цикла или даже в течение
круглого года. Склонность ткачикоподобпых
птиц к пролонгированному, перманентному
гнездостроению является, на наш взгляд, од-
ной из главных причин широкого распрост-
ранения множественного гиездостроения, ког-
да птицы вскоре после завершения одного
гнезда приступают к постройке второго гнез-
да и так далее. Множественное гиездострое-
пие возникает параллельно во всех филоге-
нетических стволах Ploceimorpha, будучи
обусловлено перманентным характером соот-
ветствующего вида деятельности, но при этом
только у Ploceinae перманентное, множествен-
ное гиездостроение функционально связано
с полигинией.
3) Гиездостроение у Ploceimorpha сильней-
шим образом зависит от разнообразных соци-
альных регуляторов и в целом ряде случаев
сплетается в единый моторный конгломерат с
брачным поведением. При этом зависимость
гпездостроителыюго поведения от социальной
стимуляции проявляется на двух основных
уровнях. Во-первых, это уровень неснецифи-
ческого стимулирующего воздействия всего
социума на поведение отдельных индивидов
(эффект Дарлинга). Так, у деревенских тка-
чиков Ploceus с и си Hat us и испанских воробь-
ев интенсивность гиездостроения самцов пря-
мо пропорциональна величине колонии
(Collias, Brodman et al., 1971; Collias, Victoria
et al., 1971; Metzmacher, 1989b). Во-вторых, это
уровень репродуктивных ячеек — моногам-
ных нар, гаремов, коммунальных групп, гдегпез-
д встроите л ыюе поведение выполняет важные
коммуникативные функции и представляет
собой неотъемлемую составную часть происхо-
дящих здесь социальных процессов.
Мы полагаем, что все перечисленные при-
знаки в известной степени взаимообуслов-
лены. И с этологической, и с эволюционной
точек зрения они тесно связаны с андролате-
ральиым гнездостроен нем, которому, видимо,
с самого начала были присущи коммуника-
тивные и социальные функции (Иваницкий,
19956). Пролонгированное, андролатералыюе
гиездостроение можно считать наиболее древ-
ним, первичным типом, давшим начало всем
прочим типам гнездостроительного поведения,
обладающих отчетливой спецификой в раз-
ных стволах Ploceimorpha.
Так, у всех воскоклювых ткачиков Estri-
ldidae утвердилась строгая моногамия с
бипарентальиым гнездостроен нем, хотя конк-
ретное распределение обязанностей в сфере
гиездостроения у разных видов выглядит но-
разному (варьирует в основном доля учас-
тия самок, у некоторых видов, видимо, строят
только самцы). Постройка гнезда всегда на-
чинается после образования пары. В насижи-
вании кладок и выкармливании птенцов все-
гда участвуют оба партнера. Одно из основ-
ных направлений специализации гнездостро-
ителыюго поведения в этой группе, судя по
всему, состояло в своеобразном разделении
половых ролей: чаще всего самцы только
подносят гнездовой материал, показывают его
и передают самке в особой видоспецифич-
ной манере в сопровождении своеобразного
демонстративного поведения, тогда как основ-
ную работу по свиванию гнезда выполняет
самка. Однако, эта дифференциация имеет пре-
78
В.В.Иваницкий
имущественно социально-обусловленный ха-
рактер, потому что после того, как самка на-
чинает насиживание, самец, параллельно с
участием в этом процессе, уже целиком само-
стоятельно строит рядом еще одно гнездо или
же пристраивает дополнительную камеру
(иногда несколько камер) к действующему
гнезду (Goodwin, 1982; Лукина, 1986; Моро-
зов, Остапенко, 1988).
Бинаренталыюе гнездостроение и насижи-
вание при строгой моногамии свойственно
многим Ploceinae (все Amblyospiza, Foudia,
Quelea, многиеPloceus). У близких к Ploceus
моногамных лесных ткачей Malimbus состав
группировок па стадии постройки гнезд, от-
носительное участие самцов и самок в гнез-
достроепии и насиживании сильно варьи-
рует у разных видов (Brosset, 1978). По мне-
нию Crook (1963d, 1964), равное участие по-
лов во всех формах репродуктивного поведе-
ния является архаичным признаком. В ходе
эволюции оно сменялось специализацией
половых ролей. Ведущим обстоятельством,
повлиявшим на трансформацию репродуктив-
ного поведения настоящих ткачиков, Крук
считает эволюцию полигинии в наиболее мно-
гочисленных и широко распространенных рр.
Ploceus и Euplectes. У наиболее специализи-
рованных в этом направлении Ploceus сам-
цы практически непрерывно строят и рек-
ламируют гнезда, не участвуя ни в насижи-
вании кладок, ни в выкармливании птенцов.
Так, самцы P.cucullatus строят за сезон до 8
гнезд, причем интенсивность стройки прямо
пропорциональна величине колоний (Collias,
Collias, 1969,1970; Collias, Victoria, 1978). Что
же касается Euplectes, то у них полигиния
связана в основном с дифференцированной
способностью самцов удерживать и реклами-
ровать гнездовые территории в привлекатель-
ных для самок местообитаниях, тогда как роль
гнезд в общей системе брачных посредников,
равно как и относительно участие полов в
их сооружении отмечено значительной ви-
довой, популяционной и индивидуальной из-
менчивостью (Craig, 1980).
Вдовушки Viduinae являются облигатны-
ми гнездовыми паразитами. Мы полагаем, что
основные черты их поведения выводятся из
репродуктивного поведения огненных ткачей
р.Euplectes при допущении возможности
того, что на определенном этане эволюции у
предков вдовушек произошла полная эман-
сипации самцов из сферы гиездостроения с
параллельным усилением их территориаль-
ного и рекламного поведения. Эта тенденция
иллюстрируется рядом: Ploceus-Euplectes-
Vidua, где именно вдовушки отличаются са-
мыми продвинутыми формами территориа-
лизма, брачных демонстраций и полового ди-
морфизма, затронувшего не только размеры
тела и пигментацию оперения (как у
Euplectes), по и пластические признаки пос-
леднего. Понятно, что эмансипация самцов
могла осуществляться лишь в том случае, если
самки включались в гнездостроение сами
или же приучались откладывать яйца в гнез-
да других видов птиц (Fridmann, 1960). По
первому из этих путей, судя по всему, пошли
огненные ткачики, па примере которых мож-
но иллюстрировать все стадии эмансипации
самцов, по второму — вдовушки. Примеча-
тельно, что хозяевами вдовушек стали их
близкие родичи — воскоклювые ткачики
сем.Estrildidae, причем разные виды вдову-
шек паразитируют на строго определенных
видах астрильдов, копируя при этом их аку-
стические сигналы и специфические небные
структуры птенцов, выполняющие сигнальные
функции при выпрашивании корма у роди-
телей (Payne et al., 1992).
У воробьиных ткачиков Plocepasserinae
вопреки строгой моногамии имеет место мно-
жественное, перманентное, кооперативное гнез-
достроение. Хотя в группе размножается толь-
ко одна пара птиц в постройке многочислен-
ных гнезд на общей территории (или одного
большого общего гнезда) участвуют все чле-
ны группы, связанные друг с другом весьма
сложными кооперативно-конкурентными
отношениями. Хотя главным побудительным
мотивом, формирующим социальную страте-
гию индивидов, является постоянное соперни-
чество за удержание или приобретение реп-
родуктивного статуса, т.е. возможности непос-
редственно участвовать в размножении, вза-
имная стимуляция, кооперация и конкурен-
ция ярче всего проявляются именно па по-
прище ^прекращающегося строительства и
демонтажа гнезд, среди которых лишь отно-
сительно немногие используются для размно-
жения. При этом наиболее активно строят
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
79
гнезда особи, занимающие самые высокие со-
циальные ранги (Collias, Collias, 1978
a,b,1980а; Lewis, 1982а). Складывается впе-
чатление, что гнездостроение у воробьиных
ткачиков pp.Plocepasser, Pseudonigritta и
Philetairus представляет собой не только со-
оружение убежищ для выведения потомства
или ночевки, но и своеобразную форму со-
циального соперничества, настоящий краеу-
гольный камень социальных процессов, про-
текающих в замкнутых и относительно по-
стоянных по своему составу группировках.
У воробьев Passcridae моногамия утверди-
лась в качестве преобладающей системы спа-
ривания у всех видов. В процессе эволюции
этой группы произошли существенные изме-
нения в сфере разделения функций брачных
партнеров в сторону эмансипации самцов,
утративших навыки гнездостроепия и наси-
живания, но сохранивших за собой функции
фуражиров в наиболее ответственный пери-
од выкармливания потомства. Симметричная,
бинарентальная (бипареитальная) форма
организации репродуктивного поведения ус-
тупила место асимметричной, гипелатераль-
иой. Лабильная система регуляции совмест-
ной деятельности партнеров, построенная на
их взаимных адаптациях, стимуляции и кон-
куренции, сменилась внешне более простой и,
как можно думать, более эффективной систе-
мой жесткого распределения обязанностей в
моногамных парах. Отдельные стадии этого
процесса иллюстрируют представители
р.Passer если у пустынных воробьев отно-
сительное участие полов во всех сферах со-
вместной деятельности отличается предель-
ной оннортунистичностыо, то у рыжих воро-
бьев участие самцов в гнездостроении и на-
сиживании сведено к минимуму, тогда как у
остальных видов можно найти широкий
спектр переходных вариантов организации
репродуктивного поведения.
Оценивая основные тенденции в эволю-
ции репродуктивного поведения воробьев, мы
хотели бы обратить внимание па то, что наи-
более продвинутые его варианты, существен-
но отличаясь от свойственной в той или иной
мере всем ткачикоподобным птицам симмет-
ричной, бипаренталыюй системы, по многим
своим признакам приближаются к гинелате-
ральным системам репродуктивного поведе-
ния, присущим таким преуспевающим ныне
группам мелких зерноядных птиц, как вьюр-
ковые (Fringillidae), овсянковые (Emberizidae)
и жаворонковые (Alaudidae). Каждая из них
намного превосходит воробьев числом видов
и родов, значением в биоценозах и величи-
ной совокупного ареала, то есть но всем тем
признакам, которые характеризуют группы,
идущие но пути биологического прогресса
(Северное, 1939; Гентнер, 1965).
Остановимся вкратце па главных особен-
ностях репродуктивного поведения вьюрко-
вых, овсянок и жаворонков. Прежде всего,
обращает на себя внимание относительная
универсальность, “одинаковость” этих особен-
ностей во всех этих группах, в противовес
огромному разнообразию репродуктивного
поведения ткачикоподобных птиц. Имеются
и другие отличия.
Во-первых, пространственная структура
вьюрковых, овсянок и жаворонков базирует-
ся преимущественно на дисперсном гнездо-
вании и более или менее жесткой террито-
риальности. Аморфные агрегации гнезд не
выше уровня квазиколониалыюсти известны
лишь у немногих видов и возникают, как
преимущественно факультативное явление.
Во-вторых, у всех перечисленных групп
гнезда всегда имеют чашевидную форму и
невелики по размерам. Для них в любом ме-
стообитании легко найти места с благопри-
ятным микроклиматом и укрытые от глаз
хищников. Крышей служит сама наседка, ко-
торая имеет хороший обзор и способна ак-
тивно реагировать на происходящие вокруг
события. Подбор специальных материалов
(например, конского волоса, паутины и др.)
обеспечивает гнездам необходимую прочность
для того, чтобы служить надежным вмести-
лищем пе только для кладки, ио и для 5-6
подросших птенцов (Благосклонов, 1991).
В-1ретьих, репродуктивное поведение всех
этих птиц характеризуется практически пол-
ным устранением самцов из сферы гнездост-
роепия и насиживания. Участие самцов в этих
видах деятельности встречается у относитель-
но немногих видов, оно подвержено значитель-
ной индивидуальной изменчивости и, по-ви-
димому, носит сугубо факультативный харак-
тер (Долгушин и др., 1968; Доржиев,
Юмов, 1992).
80
В. В. Иваницкий
Интересное исключение составляют дар-
виновы вьюрки (ii/ceM.Geospizinae) с их
гинелатеральной инкубацией, но типичным ан-
дролатеральным, пролонгированным и мно-
жественным гнездостроением. Их гнезда от-
личаются массивностью, имеют шаровидную
форму с боковым входом и, судя но описа-
нию Лэка (1949), мало чем отличаются от во-
робьиных. По мнению Grant (1986), дарвино-
вы вьюрки представляют собой один из наи-
более древних стволов вьюрковых птиц, со-
хранившийся благодаря длительной изоляции
на Галапагосском архипелаге. Ближайшими
родственниками дарвиновых вьюрков этот
автор считает р.Гшгй, широко распростра-
ненный на островах Вест-Индии и в Цент-
ральной Америке, а также узкоареалыюго
вьюрка Melanospiza richardsonii с о.Санта-
Лючия, которые также строят массивные ша-
ровидные гнезда с боковым входом при том,
что самцы, выполняя основную работу по гнез-
достроению, активно участвуют и в насижи-
вании.
Обратимся теперь к роли гнездостроения
и социо-сексуальных факторов в регуляции
физиологических процессов, происходящих в
организме самок в начальные периоды реп-
родуктивного цикла. Судя по некоторым экс-
периментальным исследованиям и натурали-
стическим наблюдениям, в этой сфере также
намечаются существенные различия между
воробьями и вьюрковыми.
Так, Поликарпова (1940) провела экспе-
рименты но изучению влияния внешних фак-
торов па созревание фолликулов и кладку у
домовых воробьев. Установлено, что весенняя
активизация роста гонад у самок стимулиру-
ется фотопериодом, однако, в дальнейшем рост
продолжается только при социо-сексуальной
стимуляции со стороны токующих самцов.
Для полного развития гонад и овуляции
необходимо наличие гнезд, причем вполне до-
статочно того, чтобы самки имели возмож-
ность их видеть. Судя но этим данным, у во-
робьев визуальная и акустическая стимуля-
ция, исходящая со стороны токующих сам-
цов и построенных ими гнезд, является не-
обходимым и достаточным условием для ову-
ляции. Это подтверждается и данными по-
левых наблюдений, свидетельствующих о воз-
можности откладки яиц и развитии насед-
ных пятен у мигрирующих самок индийских
и испанских воробьев (Ковшарь, 1966; Бро-
хович и др., 1986). Многие самки появляют-
ся в гнездовых колониях с уже сформиро-
ванными яйцами и часто бывают вынужде-
ны откладывать их в прошлогодние, недостро-
енные, и подчас едва начатые самцами гнез-
да, что также ведет к отходу части яиц, ос-
тавшихся без присмотра, поврежденных при
гнездостроении или погребенных под слоем
гнездового материала (Гаврилов, 1962; Голо-
ванова, 1985; С.А. Букреев, личноесообщение,
наши наблюдения).
Таблица 13. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ПЛОЦЕИДНОГО
И ФРИНГИЛЛИДНОГО ТИПОВ РЕПРОДУКТИВНОГО ПОВЕДЕНИЯ
Этолого-физиологические признаки Тип социо-репродуктивного поведения
Плоцеидный Фрингиллидный
Пространственная структура колониальное гнездование дисперсное гнездование
Форма гнезд шаровидная чашевидная
Сроки гнездостроения растянутые, не приуроченные к началу кладки сжатые, приуроченные к началу кладки
Серийное (множественное) гнездостроение характерно отсутствует
Участие самцов в гнездостроении активное не участвуют
Участие самцов в насиживании активное не участвуют
Участие тактильных стимулов от гнезда в контроле овуляции факультативное (Passer domesticus - по Поликарпова, 1941) облигатное (Serinus canaria - по Hinde, 1966)
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
81
Регуляция процесса репродуктивного со-
зревания у самок канареек Serinus canaria
var. domestica — типичного представителя
вьюрковых, была детально изучена в экспе-
риментах Hinde (1958, 1967). Также, как и у
воробьев, начало роста гонад у самок канаре-
ек стимулируется изменениями фотоперио-
да, а затем поддерживается социо-сексуаль-
ной стимуляцией со стороны токующих сам-
цов. Тем не менее, система интегрирующих ме-
ханизмов у канареек выглядит более слож-
ной за счет того, что в нее встроена дополни-
тельная обратная связь, которая, по-видимо-
му, недостаточно развита у воробьев. Суть этой
обратной связи состоит в зависимости ско-
рости формирования яиц и овуляции от так-
тильных стимулов, исходящих от гнезда, а
также самой по себе гнездостроительной дея-
тельности самки. Если самкам не давали
возможность закончить постройку гнезд, на-
чало кладки сильно затягивалось. Известно,
что в природе при благоприятной погоде
постройка гнезд у вьюрковых, овсянок и
жаворонков продолжается всего несколько
дней и обрывается, как правило, за день-два
до откладки первого яйца. Наличие такой сво-
еобразной паузы (“просушка гнезд”) отмеча-
ли многие наблюдатели (Птушенко, Инозем-
цев, 1968; Благосклонов, 1991).
В Табл. 13 подытожены основные черты
двух различных типов репродуктивного по-
ведения у мелких зерноядных воробьиных.
Плоцеидный тип поведения пользуется ши-
роким распространением во всех основных
группах ткачикоподобпых птиц, где он, как
мы полагаем, является базовым типом для
эволюции более специализированных вари-
антов. Фрипгиллидный тип в наиболее пол-
ном варианте свойственней вьюрковым, овсян-
кам и жаворонкам, а также всем воробьям, за
исключением наиболее архаичного, как мы
полагаем (см. ниже), р.Passer, где сохранился
исходный, плоцеидный тип.
Мы считаем, что фрингиллидный тип со-
цио-репродуктивного поведения, свойствен-
ный в той или иной степени подавляющему
большинству рецентных воробьиных, по це-
лому ряду причин должен рассматриваться,
как более рациональный, надежный и эконо-
мичный способ организации взаимоотноше-
ний брачных парнеров в рамках моногамии.
Суть плоцеидного типа репродуктивного
поведения во многом состоит в том, что гнез-
до функционирует не только как вместили-
ще для кладки, но и как объект социальных
манипуляций. Примером последних могут
служить сложные взаимосвязи гнездострои-
тельного поведения и социального домини-
рования в репродуктивных группировках
некоторых Plocepasserinae (Collias, Collias,
1976, 1978). Использование гнезда в роли
брачного посредника также включает его в
сферу социальных взаимоотношений. Отсюда
преобладающая или даже исключительная
роль самцов в постройке гнезд и, соответ-
ственно, известная зависимость самок от
гнездостроительной деятельности партнеров,
что наряду с высокой восприимчивостью
самок к социальной стимуляции может выз-
вать преждевременное развитие гонад, насед-
ных пятен и отход яиц (Поликарпова, 1940;
Гаврилов. 1962; Ковшарь, 1966; Брохович и
др., 1986). При переходе к фрингиллидному
типу эта зависимость пропадает, и самки по-
лучают возможность точнее координировать
постройку гнезда и созревание гонад благо-
даря эффективной системе обратных связей
через наседное пятно (Hinde, 1965). Гнездо
теряет социально-коммуникативные функ-
ции, но включается в систему регуляции реп-
родуктивного созревания самок. Дополни-
тельные расходы энергии самок на самосто-
ятельное сооружение гнезд компенсируются
уменьшением размеров последних, а измене-
ние формы гнезд с шаровидной на чашевид-
ную повышает безопасность наседки и по-
зволяет ей активно реагировать на происхо-
дящие вокруг события. Возможно, что учас-
тие в насиживании только одной птицы
уменьшает риск привлечения к гнезду внима-
ния хищников и вероятность заноса инфек-
ции. Кроме того, уже сама возможность од-
новременного участия двух индивидов в од-
ноименной деятельности легко порождает
состояние неопределенности, физического и
психологического соперничества, чревата
утерей согласованности в действиях партне-
ров, стремящихся к наиболее полной реали-
зации собственных поведенческих программ
(Thaler, 1976; Иваницкий, 1996).Так, у реме-
зов Remit pendulinus* около 30% кладок гиб-
нет из-за несогласованности в действиях
82
В.В.Иваницкий
партнеров в момент начала инкубации. Хотя
после завершения кладки всю дальнейшую
работу может выполнять любой из партнеров
— как в одиночку, так и сообща, сильное по-
буждение самцов к строительству гнезд (пер-
манентное серийное гнездостроение) и сво-
еобразие контроля продукции яиц у самок
(серийные кладки) часто не позволяют им
сосредоточиться на насиживании (Franz,
Theid, 1982; Persson, Ohrstrom,1985).
Тем не менее, и плоцеидный тип социо-
репродуктивного поведения имеет свои пре-
имущества. Не будучи связанными необхо-
димостью самостоятельно строить гнездо,
самки имеют возможность переадресовать
соответствующие ресурсы на продукцию
кладки. Экономия энергии самками состав-
ляет не менее 50% даже у видов с минималь-
ными затратами на постройку гнезд (Доль-
ник, 1995, стр.297). Ключевым стимулом,
обеспечивающим формирование яиц вплоть
до момента овуляции, служит токование
самцов, что — в отсутствие блокировки га-
метогенеза со стороны тактильных стимулов
в процессе собственного участия в построй-
ке гнезда, может сокращать паузу между
кладками до минимума. При отсутствии
лимитирования со стороны пищевых ресур-
сов такая система этолого-физиологического
контроля репродуктивного состояния самок
значительно расширяет возможности для
полициклического размножения. Выше мы
привели целый ряд примеров, иллюстриру-
ющих насколько полно эти возможности
используют воробьи. Совершенно иная кар-
тина у зерноядных птиц с фрингиллидным
типом репродуктивного поведения. Так, в
зоне умеренного климата, где воробьи выра-
щивают за сезон по 2-3 выводка, большин-
ство вьюрковых и овсянок ограничиваются
одним репродуктивным циклом, и лишь
небольшое число пар имеют по 2 выводка
(Птушенко, Иноземцев, 1968).
Таким образом, плоцеидный тип в силу
внутренних конфликтов и противоречий
может привести к некоторым репродуктив-
ным потерям, зато в благоприятных условиях
он, видимо, обеспечивает быстрое наращива-
ние совокупного репродуктивного потенци-
ала. Фрингиллидный тип выглядит более
надежным, экономичным и пригодным для
значительно более широкого диапазона усло-
вий, но обеспечивает относительно умерен-
ные темпы воспроизводства.
ГЛАВА 5. СТРУКТУРА И ЭВОЛЮЦИЯ МОТОРНЫХ РЕПЕРТУАРОВ
5.1. Полет и передвижение по
твердым субстратам
У воробьев есть следующие типы полетов,
отличающиеся по режиму движений крыль-
ев, траекториям и целям:
1) Машущий режим с умеренно глубоки-
ми взмахами крыльев, частота которых от-
носительно невелика. У большинства мелких
воробьиных весом около 30 г она составляет
10-11 взмахов в секунду (Norberg, 1991);
2) Трепещущий режим с увеличенной, но
сравнению с предыдущим, частотой и умень-
шенной амплитудой взмахов;
3) Порхающий режим отличается от двух
предыдущих заметно меньшей частотой и
большей амплитудой (глубиной) взмахов;
4) Режим инерционного скольжения с не-
подвижными крыльями — сложенными или
приподнятыми над спиной в виде буквы V.
Таким образом, полет воробьев включает
три активных режима, когда птица может на-
бирать скорость и высоту, и один пассивный
режим, когда птица, двигаясь в воздухе но инер-
ции, теряет высоту и снижается. Повседнев-
ный транспортный полет у всех воробьев про-
текает но характерной дугообразной траекто-
рии: в активной фазе птица пользуется машу-
щим режимом, а затем скользит со сложенны-
ми или полусложенпыми крыльями.
Дугообразность транспортного полета осо-
бенно бросается в глаза у Passer, Petronia и
Pyrgilauda. У Montifringilla с их острыми
и длинными крыльями полет выглядит зна-
чительно более быстрым, сильным и прямо-
линейным. По-видимому, это достигается за
счет того, что снежные воробьи сильнее раз-
гоняются в активной и меньше снижаются в
инерционной фазе полета. Складывается впе-
чатление, что снежные воробьи больше дру-
гих видов приспособлены к полету на боль-
шие расстояния в горном ландшафте при
практически постоянном, сильном ветре.
На Рис.6 охарактеризовано присутствие
разных режимов полета в воздушной локо-
моции воробьев. У Passer полет исиользует-
название режима нормальный машущий трепещущий порхающий скольжение
крылья подняты крили прижаты
ПОЛОЖЕНИЕ КРЫЛЬЕВ Ж О
РОДЫ ПРИСУТСТВИЕ В ЛОКОМОЦИИ
Passer fa " "
Petronia ^ ^ ^
Pyrgilauda ^ ^ fa
Montifringilla ^ fa
Carpospiza ^ " a™ MM
Рисунок 6. ПРИСУТСТВИЕ РАЗНЫХ РЕЖИМОВ ПОЛЕТА В ВОЗДУШНОЙ ЛОКОМОЦИИ ВОРО-
БЬЕВ: 1 - да; 2 - нет.
84
В.В.Иваницкий
Рисунок 7. ТРАЕКТОРИИ НЕКОТОРЫХ ТИПОВ ПОЛЕТОВ ВОРОБЬЕВ
А — нсрптуализнрованный ориентировочный полет каменного воробья; Б - токовой, рекламирующий полет
монгольского земляного воробья; В — транспортный полет монгольского земляного воробья; Г — транспортный
полет снежного воробья; Д — токовой рекламирующий полет снежного воробья; Е — трепещущий полет монголь-
ского земляного и каменного воробьев, t — машущий режим; 2 — скольжение на поднятых крыльях; 3 —
скольжение со сложенными крыльями; 4 — трепещущий режим; 5 — порхающий режим.
ся только для транспортных целей и не не-
сет сигнальных функций. Некоторое возрас-
тание частоты взмахов и приближение к тре-
пещущему режиму все же удается наблюдать
в некоторых напряженных социальных кон-
текстах, например, при образовании пары или
копуляции (полевой и саксаульный воробьи).
У прочих родов воробьев режимы движений
крыльев отличаются большим разнообрази-
ем и входят в качестве составляющих частей
в различные типы воздушных эволюций, вы-
полняющих чисто коммуникативные функции.
Трепещущий режим активно используют
каменные и земляные воробьи в виде особо-
го демонстративного полета на расстояние до
20-30 м на высоте 0,5-1,5 м в социально зна-
чимых ситуациях (образование пары, рекла-
мирование территории, пограничные конфлик-
ты). Аналогичный, но значительно менее вы-
разительный полет, есть и у короткопалых
воробьев. Кроме того, трепещущий режим
употребляется в рекламирующих (токовых
полетах) земляных воробьев, а также в пред-
конуляционных воздушных демонстрациях
каменных и земляных воробьев (Рис.7,8).
Токовой полет монгольского земляного
воробья включает три режима. После старта
самец с песней поднимается на 10-20 м и пе-
ресекает территорию по прямой или летает
над ней кругами диаметром до 50 м в тече-
ние 1-2 мин, поразительно напоминая при
этом жаворонка и составляя резкий контраст
с привычными представлениями о воробьях.
Кроме того, почти во всех социальных кон-
текстах для земляных воробьев очень харак-
терны демонстративные взлеты свечой до 1-5
м с употреблением всех трех режимов. По-
видимому, это не что иное, как редуцирован-
ные формы полного токового полета при ус-
ловии, что исполнитель предпочитает не те-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
85
рять тесного контакта с социальным партне-
ром или оппонентом. Очень своеобразен по-
вседневный транспортный полет земляных
воробьев из-за необычайно крутых и ломан-
ных траекторий взлета и посадки. Эти воро-
бьи приземляются не по отлогой предсказуе-
мой территории, подобно другим птицам, а со-
скальзывают почти отвесно вниз с элемента-
ми скольжения на поднятых крыльях, благо-
даря чему положение птицы в воздухе меня-
ется самым неожиданным образом и ее в этот
момент легко потерять из виду.
Токовой полет снежных воробьев менее
выразителен, а его типичные формы контину-
ально связаны с повседневной локомоцией.
Главный демонстративный элемент полета —
порхающий режим. Глубокие, плавные взма-
хи крыльев чередуются со скольжением на
фиксированных и V-образно приподнятых
крыльях. После старта самец на машущем или
порхающем режиме, слегка фиксируя глубо-
кие взмахи, круто взлетает на 5-6 м и сразу же
соскальзывает полого вниз на поднятых на спи-
ной крыльях. Продолжительно полетов не пре-
вышает 20 с. Демонстративный эффект токо-
вого полета снежного воробья усиливается
яркими белыми пятнами на длинных кры-
льях и черно-белой окраской рулевых.
Судя но описанию Schaffer (1938), токо-
вые полеты тибетских M.adamsii выглядят
похоже, однако из-за более ярко выраженной
социальности этого вида, эти полеты часто
носят коллективный характер. Кроме того,
порхающий режим у М.adamsii выражен в
более яркой форме: взмахи крыльев столь
глубоки, что в нижней точке они с характер-
ным звуком сталкиваются друг с другом.
Предкопуляциоппый полет каменных во-
робьев представляет собой сложную мотор-
ную координацию (Рис.8,Б). В полном виде
поведение при спаривании выглядит так:
самец подлетает к самке замедленным, трепе-
щущим полетом с развернутым хвостом и
зависает у нее над спиной в порхающем по-
лете длительностью до 30 с. При этом самец
как бы комбинирует два режима движений
крыльев: замедленные глубокие взмахи, ко-
роткие трепещущие движения и мгновенные,
хотя визуально воспринимаемые вполне от-
четливо, фиксации высоко поднятых крыль-
ев. Тело самца в момент зависания ориенти-
ровано почти вертикально, благодаря чему
хорошо видны полностью развернутые руле-
вые с характерным рисунком (см. Рис.35), и
немного покачивается из стороны в сторону
за счет соскальзывания в воздухе в те момен-
ты, когда он фиксирует поднятые крылья и
как-бы слегка проваливается вниз. Затем са-
мец садится на спину самке, следует коитус,
после чего он взлетает и перелетает трепе-
щущим полетом на 2-4 м, садится в демонст-
ративной позе, продолжая интенсивно трепе-
тать крыльями, после чего тем же порядком
совершает новую садку.
У монгольских земляных и снежных во-
робьев предкопуляционные воздушные демон-
страции более динамичны (Рис.8,А). У обо-
их видов их схема примерно одинакова. Са-
мец, набрав скорость, стремительно подлетает
к самке и описывает в воздухе над ней до 5-
7 кругов диаметром до 3 м на постоянной
высоте или же, наоборот, резко меняя высоту
полета. При движении но кругу преоблада-
ет скольжение на поднятых крыльях, сопро-
вождаемое характерным шумом рассекаемо-
го воздуха. Для дополнительного разгона ис-
пользуется машущий или трепещущий (этот
последний — только у земляных воробьев)
режимы. В промежутках между садками сам-
цы земляных воробьев интенсивно демонст-
рируют (трепещущие полеты чередуются с
переползанием в позах И,К, Рис. 16). У снеж-
ных воробьев в промежутках между садка-
ми отмечаются лишь повседневные реакции
повышенной интенсивности.
Воробьи р.Passer передвигаются по твер-
дым субстратам преимущественно прыжка-
ми па обеих лапках, то есть отталкиваясь и
приземляясь сразу на обе конечности. Все
представители рода свободно чувствуют себя
на деревьях, прекрасно перемещаются в кро-
нах, включая периферические отделы с са-
мыми тонкими и гибкими ветвями. Часто
при кормежке воробьи демонстрируют не-
заурядные акробатические способности, лазая
но самым тонким ветвям и подвешиваясь в
самых разнообразных позах вниз головой и
спиной, как это делают синицы или некото-
рые мелкие вьюрковые,
Но далеко не у всех депдрофильных во-
робьев педальная локомоция осуществляет-
ся с помощью прыжков. Среди р.Passer из-
86
В. В. Иваницкий
Рисунок 8. ПРЕДКОПУЛЯЦИОННЫЕ ДЕМОНСТРАЦИИ ВОРОБЬЕВ
А — монгольский земляной воробей; Б — каменный воробей. 1 — трепещущий режим; 2 — машущий полет;
3 — скольжение на поднятых крыльях (по Ivanitskii, 1996 с изменениями).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
87
вестны виды, которые па твердом субстрате
передвигаются преимущественно шагами, то
есть ходят или бегают, переставляя лапки
поочередно. В той или иной степени это ха-
рактерно для многих представителей комп-
лекса близкородственных африканских “се-
роголовых” воробьев, причем наиболее ярко
эта повадка выражена у P.diffusus (Summers-
Smith, 1988). С другой стороны, чередование
прыжков и шага при наземной локомоции
свойственно P.luteus, которые генеалогичес-
ки, по-видимому, весьма удалены от “серого-
ловых” воробьев (Kunkel, 1959). Кустарни-
ковые воробьи p.Gymnorhis по данным це-
лого ряда наблюдателей также весьма при-
вержены к древесной растительности. Кро-
ны деревьев и кустарников используются
всеми видами, как основное место сбора кор-
ма, хотя иногда они кормятся и на земле,
где но крайней мере G.superciliaris передви-
гается шагом и может довольно быстро бе-
гать (McLacham, Liversidge, 1957).
Будучи связанными в трофическом от-
ношении преимущественно с древесными
субстратами, кустарниковые воробьи но это-
му признаку резко отличаются от каменных
воробьев (р.Petronia), которые кормятся ис-
ключительно на поверхности земли, употреб-
ляя древесно-кустарниковую растительность
исключительно для отдыха. Тем не менее, в
повседневной жизни и в кронах, и но земле
каменные воробьи перемещаются только
прыжками. Лишь при ухаживании за самка-
ми самцы способны перемещаться на рассто-
яние до 1,0-1,5 м с помощью своеобразных
мелких шажков. Самцы как-бы семенят по
субстрату в сопровождении характерных для
этой ситуации демонстраций и акустических
сигналов (см.ниже).
У земляных воробьев p.Pyrgilauda, глу-
боко приспособленных к обитанию в безлес-
ных, стенных ландшафтах и ведущих исклю-
чительно наземный образ жизни, на смену
прыжкам пришли иные аллюры, хотя и здесь
картина достаточно сложна. По нашим наблю-
дениям, P.davidiana никогда не прыгают и
способны к быстрому бегу и длительной
ходьбе на многие сотни метров. В ветрепные,
холодные дни, которые не представляют ред-
кости в суровых местах обитания этих птиц,
воробьи избегают пользоваться полетом, пред-
почитая подолгу ходить пешком от одной пи-
щучьей колонии до другой. В такие дни пти-
цы подолгу лежат у входов в поры, спасаясь
от пронизывающего ветра. В социальных кон-
текстах (образование нар, пограничные кон-
фликты) монгольские воробьи быстро бега-
ют в разнообразных позах, ползают и затаи-
ваются на земле (Рис. 16, позы Е,Ж,И,К).
Пробежки и переползания чередуются с тре-
пещущими полетами и образуют единую пос-
ледовательность, сопровождаясь почти непре-
рывным трепетанием крыльев или же серия-
ми резких отбрасываний их копчиков в сто-
роны. У тибетских представителей Pyrgilauda
Н.А.Формозов (личное сообщение) отметил
значительное разнообразие аллюров: Р.Ыап-
fordii — ходят и бегают, P.ruficollis — чере-
дуют бег и ходьбу с прыжками, a P.tacza-
nowskii, обладающие наиболее сильными зад-
ними конечностями среди всех воробьев, тем
не менее, преимущественно прыгают.
Снежные воробьи р. Montifringilla также
способны подолгу ходить в поисках корма, но,
в отличие от земляных воробьев, охотно пере-
двигаются прыжками и никогда не бегают. В
социальных контекстах педальная локомоция
сведена к минимуму. С ростом возбуждения
увеличивается частота перемещений птиц но
воздуху с использованием скольжешт вплоть
до типичных токовых полетов, растут частота
и амплитуда отбрасываний крыльев иногда с
короткой фиксацией их в приподнятом поло-
жении (Рис. 17,В).
Что же касается короткопалых воробьев
p.Carpospiza, то они собирают корм только
на земле и при этом преимущественно хо-
дят шагом или бегают, причем с такой легко-
стью, что чрезвычайно напоминают при этом
жаворонков или даже коньков (Рис. 18,Б).
Таким образом, эволюция воробьев сопро-
вождалась глубокой дивергенцией систем воз-
душной и педальной локомоции в процессе
приспособления этих птиц к чрезвычайно раз-
нообразным условиям существования парал-
лельно с формированием различных адаптив-
ных типов (жизненных форм). Физические
особенности среды обитания и прежде всего
свойства субстратов играли важную роль в
процессе этой дивергенции, однако, соответ-
ствие между локомоцией и субстратами но-
сит довольно сложный характер. Тенденция
88
В.В.Иваницкий
к смене прыжков на шаг или бег обозначи-
лась уже у дендрофильных форм (рр. Passer
и Gymnorhis). В то же время, прыжки сохра-
нились в качестве доминирующего типа пе-
дальной локомоции у такой чисто наземной
формы, как камешше воробьи, однако, в сфе-
ре социального поведения у них утвердился
альтернативный тин педальной локомоции
(шаг). В противоположность этому, коротко-
палые воробьи, часто живущие бок о бок с
каменными воробьями, используют передви-
жение шагом и быстрый бег в качестве до-
минирующего способа наземной локомоции.
Все это наводит на мысли о том, что эво-
люция педальной локомоции у воробьев про-
текала но сложному сценарию. В этой связи
следует иметь в виду возможность того, что
альтернативные способы локомоции могут
формироваться поначалу в сфере социаль-
ного поведения и выполнять на этом этапе
эволюции преимущественно коммуникатив-
ные функции, как это было показано выше па
примере каменных воробьев. Роль социальных
контекстов, как главной арены дивергенции
способов локомоции, особешю рельефно про-
является на примере полета воробьев. Имен-
но социальная среда послужила, как можно
предполагать, главным источником глубоких
преобразований воздушной локомоции, где со-
средоточено наибольшее разнообразие воз-
душных эволюций, функционирующих исклю-
чительно в коммуникативной сфере.
5.2. Элементарные двигательные
акты и их сочетания (позы)
Рассмотрим наиболее важные элементар-
ные двигательные акты (ЭДА) воробьев, обус-
ловливающие изменения формы тела, а так-
же наиболее характерные и устойчивые со-
четания этих движений, которые сохраняют-
ся в относительно неизменном состоянии в
течение определенного времени, достаточно-
го для того, чтобы опытный наблюдатель имел
возможность воспринимать такие сочетания,
как фиксированные позы, характерные для
дашюго вида. Описание поз неизбежно содер-
жит известную долю субъективизма, обуслов-
ленного трудностями вычленения фиксиро-
ванных моторных координаций из сплошно-
го потока поведения. Предложенный Пановым
(1978) и используемой нами комбинаторный
метод описания позволяет свести этот субъек-
тивизм к минимуму.
На Рис.9 изображены основные ЭДА во-
робьев. В соответствии с принятым нами
методом описания, ЭДА представляют собой
“нулевой” уровень интеграции — самые про-
стые движения, из которых строятся более
сложные координации — позы и их после-
довательности. Каждый ЭДА соответствует
изменению положения определенной части
тела или оперения и обозначается заглавной
буквой, а характер изменений — цифровым
индексом при букве, причем за нулевое зна-
чение индекса мы принимали то положение
органа, которое характерно для спокойного,
повседневного состояния птицы, тогда как
величина индекса (хотя и не во всех случа-
ях) прямо пропорциональна степени откло-
нения от этого состояния.
Всего в поведении воробьев мы выделяем
11 ЭДА: А — положение корпуса (от почти
вертикального до горизонтального); Б —
положение контурного оперения на груди и
брюшке (распушено или прижато); В — то
же на спине и пояснице; Г — то же на голо-
ве; Д — положение клюва (горизонтально,
опущен, приподнят); Е — положение хвоста
относительно оси тела ( параллельно, под
углом вверх или вниз); И — состояние ру-
левых перьев (собраны или развернуты); К
— положение крыльев (лежат в обычном
положении на спине, отведены в стороны,
приподняты вверх); Л — положение шеи
(вытянута или втянута); М — движение кры-
льев (неподвижность или вибрация); Н —
координированные движения корпуса и го-
ловы (поклоны, маятпикообразные движения
влево-вправо, повороты на месте и т.д.).
Остановимся теперь подробнее на основ-
ных межвидовых отличиях в ЭДА:
1) Вертикальная ориентация корпуса (А4)
присуща лишь монгольским земляным воро-
бьям, у которых это ЭДА составляет харак-
терную особенность ориентировочной позы
“столбик” (Рис. 16,В);
2) Комплекс разнообразных скоординиро-
ванных движений головы, шеи и корпуса
(Н1-3), употребляемых преимущественно при
брачных демонстрациях, очень характерен для
Passer и Petronia. У Pyrgilauda и Montifrin-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
89
gilla сохранились лишь отдельные следы этих
движений, причем у первых в несколько боль-
шем объеме;
3) Большой изменчивостью отличаются
движения крыльев (отведение в стороны, виб-
рация, фиксация в разных положениях). У
большинства Passer во время брачных демон-
страций крылья отводятся немного в сторо-
ны (в том числе и кистевые сгибы) и удер-
живаются в этом положении (К1) неподвиж-
но или же трепещут с разной интенсивнос-
тыо(М1).
Среди изученных нами видов Passer даль-
ше всего отводят крылья и сильнее всего тре-
пещут ими испанские воробьи, которые ха-
рактеризуюся очень интенсивными брачны-
ми демонстрациями (Рис.10,Е,Ж,К,Л). Тре-
пещущие крылья у этого вида не только ото-
двигаются от корпуса, но часто заметно при-
поднимаются вверх, что характерно также для
брачных демонстраций Р.eminibey, P.luteus
и P.moabiticusiKunkel, 1959; Harrison, 1965;
Payne, 1969) (Рис. 20, А). Помимо Passer, виб-
рация крыльев очень характерна для Petronia
и Pyrgilauda, но сведена к минимуму у
Montifringilia, у которых преобладают отбра-
сывания крыльев с отчетливо выраженной
склонностью к их фиксации в полностью под-
нятом V-образном положении (Рис. 17,В).
Вместе с тем, высокое поднимание крыльев с
их отчетливой фиксацией в этом положении
(как с вибрацией, так и без нее) составляет
очень характерную особенность брачных де-
монстраций Petronia, причем крылья могут
приподниматься как синхронно оба, так и но
одному (Рис.15,Д,Е; Рис.20,Б) Отличия в
употреблении этой реакции между снежны-
ми и каменными воробьями состоит в том,
что у последних она относится к числу высо-
конороговых и маркирует лишь состояния
90
В.В.Иваницкий
крайнего возбуждения самцов в напряжен-
ных социальных эпизодах (образование
нары), тогда как у первых (также как и у
земляных воробьев) высокое поднимание
крыльев отличается низкими порогами и со-
провождает малейшие всплески возбуждения
как у самцов, так и у самок;
4) Вибрация контурного оперения прида-
ет большое своебразие брачным демонстра-
циям испанских и каменных воробьев. Судя
по тому, что оперение вибрирует лишь одно-
временно с вибрацией крыльев, эти реакции
взаимосвязаны и обусловлены тем, что сам-
цы этих видов сильно распушают при демон-
страциях контурное оперение (БЗ), отчего кон-
чики перьев приподнимаются, и птица ста-
новиться взъерошенной.
5) Подгибание хвоста (ЕЗ) представляет
обычную реакцию самцов в момент копуля-
ции у всех видов. Вне этого контекста эта
реакция отчетливо выражена и составляет ха-
рактерную особенность демонстративного по-
ведения лишь у каменных воробьев, а также
у Passer eminibey (Payne, 1969).
Переходя к анализу следующего уровня
интеграции моторных актов — ноз, представ-
ляющих собой относительно устойчивые со-
четания ЭДА, следует сразу же отметить, что
здесь мы имеем дело уже со значительно бо-
лее глубокой дивергенцией вплоть до полной
утраты внешнего подобия отдельных поз
(Рис. 10-20).
На Рис. 19 изображена более детально одна
из наиболее распространенных демонстратив-
ных поз, разные варианты которой имеются
у многих Passer и у Petronia, Эта сложная
моторная координация включает следующие
ЭДА: 1) корпус расположен горизонтально
или слегка наклонен вперед (А2-3); 2) кры-
лья отведены в сторону и вибрируют (К1,
Ml); 3) хвост слегка приподнят, и рулевые
развернуты (Е2, И2); 4) ритмичные движе-
ния головы вверх-вниз и в стороны одновре-
менно с поворотами корпуса на месте (НЗ).
Контурное оперение при этом может быть
или сильно распушенным, или прижатым,
однако, наиболее характерным элементом этой
позы является распушение оперения на спи-
не и пояснице (В1-2).
Все перечисленные ЭДА могут использо-
ваться в комплексе с другими ЭДА и вхо-
дить в состав других ноз. Характерно, что
многие из них составляют неотъемлемую часть
гнездостроителыюго поведения: например, НЗ
употребляются при манипуляциях с гнездо-
вым материалом, в особенности при построй-
ке стенок и крыши гнезд, когда птица то и
дело крутит головой, перемещая клювом кон-
цы материала с одной стенки на другую. Столь
же характерны повороты корпуса и раздви-
гание крыльев, которые можно наблюдать в
процессе формовки лотка (см.выше). При-
мечательно, что эти движения птица, сидящая
в гнезде или в мутовке, иногда выполняет как-
бы “вхолостую”, то есть без захвата гнездово-
го материала и, таким образом, весь комплекс
перечисленных движений континуально свя-
зан с гнездостроительпым поведением как ио
своим структурным особенностям, так и по
некоторым функциональным (ситуационным)
характеристикам потому что самцы многих
Passer часто токуют и строят гнездо одно-
временно. В то же время, хотя самцы Petronia
гнезд не строят, в их демонстративном пове-
дении есть много сходства с Passer (особен-
но с испанскими воробьями).
Демонстративное поведение кустарнико-
вых воробьев (p.Gymnorhis) отчасти напо-
минает аналогичное поведение Passer. В мо-
мент брачных демонстраций G.dentata при-
нимает горизонтальную нозу (АЗ), плотно
прижимает все контурное оперение (Бо, Во,
Г1), отодвигает в стороны крылья и трепе-
щет ими (KI, Ml), импульсивно вздергивает
хвост (Е2) и приподнимает вверх клюв (ДЗ).
Демонстрации G.superciliaris отличаются от
только что описанной координации тем, что
производятся в почти вертикальной позе (А1)
и сопровождаются хорошо выраженным рас-
пушением оперения на груди и на брюшке
(Б2) (Abs,1966).
Демонстративное поведение земляных во-
робьев сильно изменилось под влиянием при-
способления к специфическим условиям оби-
тания, глубоких преобразований систем воз-
душной и наземной локомоции, изменений
репродуктивного поведения. У самцов полно-
стью исчезли ЭДА, имеющие связи с гнездос-
троепием (прежде всего Н2-3), что привело к
структурному упрощению демонстраций при
показе самцами убежищ в момент образова-
ния нар, где употребляются преимущественно
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
91
Рисунок 10. ОСНОВНЫЕ ПОЗЫ ИСПАНСКИХ ВОРОБЬЕВ (по Иваницкий, 1985в)
Рисунок 11. ОСНОВНЫЕ ПОЗЫ ИНДИЙСКИХ ВОРОБЬЕВ (по Иваницкий, 1985 в)
92
В. В. Иваницкий
Рисунок 13. ОСНОВНЫЕ ПОЗЫ САКСАУЛЬНЫХ ВОРОБЬЕВ
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
93
Рисунок 14. ОСНОВНЫЕ ПОЗЫ РЫЖИХ ВОРОБЬЕВ
Рисунок 15. ОСНОВНЫЕ ПОЗЫ КАМЕННЫХ ВОРОБЬЕВ (по Иваницкий, 1986а)
94
В.В.Иваницкий
Рисунок 16. ОСНОВНЫЕ ПОЗЫ МОНГОЛЬСКИХ ЗЕМЛЯНЫХ ВОРОБЬЕВ (по Иваницкий. 19916)
Рисунок 17. ОСНОВНЫЕ ПОЗЫ СНЕЖНЫХ ВОРОБЬЕВ (по Иваницкий. 19916)
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
95
Рисунок 19. ПОЗЫ ВОРОБЬЕВ ПРИ ГНЕЗДОСТРОЕНИИ (Л - В) И БРАЧНЫХ ДЕМОНСТРАЦИЯХ (Г Е)
96
В. В. Иваницкий
позы Г и Ж (Рис. 16). Некоторые позы мон-
гольских земляных воробьев сохранили по-
добие с позами Passer, однако они встречают-
ся крайне редко, и, как можно думать, их роль
в процессе коммуникации ограничена (позы
3,М, Рис. 16). Вместо этого у земляных воро-
бьев значительное распространение получили
позы, связанные с пробежками, переползанием,
затаиванием, характерным трепещущим поле-
том параллельно поверхности субстрата на
малой высоте (позы Д,Е,Ж,И,К, Рис. 16).
У снежных воробьев демонстративное по-
ведение в целом подверглось основательной
редукции (Рис. 17). Так же как и у земляных
воробьев, у них отсутствуют гнездостроитель-
ные ЭДА, по в отличие от последних, у них
нет и специфических ноз, связанных с назем-
ной локомоцией, потому что в социальных кон-
текстах снежные воробьи пользуются почти
исключительно полетом. Важную роль в сис-
теме визуальной коммуникации у снежных
воробьев играет приподнимание длшшых, ярко
окрашенных крыльев над спиной (К2). Эта низ-
копороговая реакция проявляется при малей-
шем возбуждении и неизменно сопровожда-
ет все типы социальных взаимодействий. Хотя
этот ЭДА пе меньше характерно и для земля-
ных воробьев, у снежных воробьев благодаря
контрастной окраске крыльев, оно имеет более
ярко выраженный сигнальный эффекта
Репертуар демонстративного поведения ко-
роткопалых воробьев в целом беден и почти
не содержит специализированных поз. Струк-
тура агонистических и социо-сексуальных вза-
имодействий у этого вида выглядит крайне
примитивной (см.выше). Нам всего дважды
удалось зарегистрировать достаточно сложные,
фиксированные моторные координации
(Рис. 18,Г), причем в обоих случаях короткие
демонстрации поз были отмечены у самцов
сразу же после копуляции. Больше всего, на наш
взгляд, эти позы напоминают демонстрации не-
которых Gymnorhis (Abs, 1966).
Изображенная на Рис. 19 поза Д и все со-
ставляющие ее ЭДА присутствуют в репертуа-
ре многих представителей настоящих ткачиков,
в особешюсти у p.Ploceus. Так, у деревенского
ткачика P.cucullatus похожая поза употребля-
ется самцами при пограничных конфликтах
на стыках их гнездовых территорий в колони-
ях (Рис.20,Е). Что же касается брачных демон-
страций самцов Ploceus при рекламировании
гнезд, то они очень похожи па аналогичные
демонстрации некоторых Passer, в ряду кото-
рых прежде всего должны быть перечислены
P.hispaniolensis P.emimibey и P.luteus). Раз-
личия касаются в основном положения токую-
щих самцов: воробьи демонстрируют, сидя на
ветке или на гнезде (платформе), тогда как
ткачики в момент интенсивных демонстраций
подвешиваются ко входу в гнездо или к его
рабочему краю вниз спиной (Рис.20,Г) и в та-
ком положении трепещут или размахивают
крыльями, причем частота и амплитуда этих
движений является во многих случаях видос-
пецифическим признаком (Crook, 1963с, 1964;
Collias, Collias, 1970). Красноклювые ткачики
Q.quelea (Рис.20, Д) так же как и воробьи, де-
монстрируют в обычном положении, причем в
момент демонстраций самцы приподнимают
слегка вибрирующие крылья настолько высо-
ко, что в верхнем положении они соприкаса-
ются своими наружными сторонами и удер-
живаются так несколько секунд (Crook, 1960с).
В целом, демонстративное поведение мно-
гих представителей Passer во многим напо-
минает поведение настоящих ткачиков
pp.Ploceus, Quelea (Harrison, 1965) и в не-
сколько меньшей степени — Foudia (Crook,
1961). Демонстрации огненных ткачиков
p.Euplectes отличается значительно большим
своеобразием (Craig, 1980).
Демонстративное поведение воскоклювых
ткачиков резко отличается от поведения воро-
бьев и настоящих ткачей потому, что при об-
разовании нар амадипы н астрильды не
пользуются посредниками, а гиездостроение у
этих птиц не связано с образованием нар. Роль
всевозможных моторных актов, направленных
на гнездо, уменьшилась, зато возросла доля ноз
и отдельных движений, адресованных непос-
редственнобрачному партнеру (Goodwin, 1982).
5.3. Структурно-функциональная
дифференциация и эволюция визу-
альной сигнализации
У многих видов воробьев патрулирование
границ территорий осуществляется целенап-
равленно, представляет собой одну из наибо-
лее обычных реакций и часто порождает фик-
сированные во времени и пространстве соци-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
97
Рисунок 20. НЕКОТОРЫЕ ДЕМОНСТРАТИВНЫЕ ПОЗЫ ВОРОБЬЕВ И ТКАЧИКОВ
А - Passer moabiticus; Б — Petronia petronia', В - Passer hispaniolensis, Г, Е — Ploceus cucullatus, Д —
Quelea quelea (Б, В - наши данные; А, Г, Е, Д — по Summers-Smith, 1988; Crook, 1960с; Collias, Collias, 1970; Payne,
1969 c изменениями).
альпых взаимодействий, главной особенностью
которых является отчетливое противостояние
социальных оппонентов (как правило — сам-
цов) друг другу. У других видов патрулиро-
вание периферии территории, как специфи-
ческая, особая форма активности, полностью
отсутствует, стычки за территории мимолет-
ны не связаны с ситуацией противостояния
соперников, которые можно было бы называть
пограничными конфликтами. Среди изучен-
ных нами видов воробьев патрулирование и
конфликты свойственны домовым, индийским,
полевым, саксаульным, рыжим и монгольским
земляным воробьям, тогда как у пустынных,
испанских, каменных, снежных и короткопа-
лых воробьев упомянутые формы поведения
отсутствуют. Межвидовые различия но этому
признаку универсальны, ибо их легко обнару-
жить у близкородственных видов в разных
группах птиц и млекопитающих (см.Иваниц-
кий, 1981; Гольцман, 1984).
Сравним поведение ненатрулирующих ис-
панских и патрулирующих индийских воро-
бьев. Испанские воробьи отличаются чрезвы-
чайно экспрессивными демонстрациями при
рекламировании мутовок и при образовании
пар (позы Д-М, Рис. 10) вплоть до того, что
самцы иногда теряют равновесие и вывалива-
ются из мутовок. У индийский воробьев брач-
ные демонстрации (нозы Д-Ж, Рис. 11) не
столь интенсивны. При стычках за террито-
рии самцы испанского воробья просто гоня-
ются друг за другом, присаживаясь время от
времени (при длительных конфликтах) в по-
вседневных или мало интенсивных позах А-Г
и 3 (Рис. 10). Алрессивные взаимодействия сам-
цов и самок индийских воробьев па стыках
территорий выглядят, как фиксированные вза-
имодействия, где соперники, сближаясь вплот-
ную, адресуют друг другу характерные покло-
ны (нозы К,Л, Рис. 11), отсутствующие в ре-
пертуаре испанских воробьев. Примечательно,
98
В.В.Иваницкий
что эти поклоны составляют заметный эле-
мент в демонстративном поведении всех пат-
рулирующих видов, по их пет у непатрулиру-
ющих. Позы, сопровождающие поклоны при
пограничных конфликтах, варьируют у раз-
ных видов: индийские воробьи кланяются в
сущности в обычных позах, рыжие — в позах
Д и 3 (Рис. 14), саксаульные — Г-Ж (Рис. 13),
полевые — П и Р (Рис. 12).
Проведенные нами исследования и име-
ющиеся литературные данные позволяют на-
метить несколько основных типов структур-
но-функциональной организации репертуаров
сигнальной моторики и высказать некоторые
суждения относительно их возможной дивер-
генции и эволюции.
Мы полагаем, что тесная структурная и
функциональная связь демонстративного и
гнездостроителыюго поведения является весь-
ма архаичным признаком, обусловленным до-
минирующей ролью самцов в сооружении
гнезд, утвердившейся еще до начала диверген-
ции главных стволов ткачикоподобных птиц.
Различные версии такой связи мы встречаем
практически во всех рецептных группах
Ploceimorpha. В пользу этого предположения
свидетельствует глубокое сходство основных
особенностей репродуктивного поведения и
брачных демонстраций воробьев р.Passer и
ткачиков р. Ploceus, которые по многим биоло-
гическим признакам представляются наиболее
древними ветвями семейств Passeridae и
Ploceidae. У многих представителей упомяну-
тых родов демонстративное и гнездостроитель-
ное поведение строятся из одинаковых дви-
жений и часто неотделимы друг от друга в
ситуационном отношении, поскольку и демон-
страции, и гнездостроение сплошь и рядом
выполняются самцами одновременно.
По-видимому, именно этот тин моторного
репертуара следует считать исходным в эво-
люции воробьев. Он характеризуется преж-
де всего высокой интенсивностью и разнооб-
разием демонстраций возле гнезда и пони-
женной склонностью к участию в персонали-
зированных социальных взаимодействиях.
Гнездо является главным фокусом внимания
и деятельности самца. При случайных или
навязанных агонистических взаимодействи-
ях употребляются те же демонстрации, что и
при рекламировании гнезд или убежищ. Этот
тип сигнальной моторики характерен для
ряда представителей р.Passer (hispaniolensis,
luteus, eminibey, nioabiticus). К этому же типу
тяготеет и поведение Petronia.
На примере остальных представителей
р.Passer можно проследить постепенную ре-
дукцию разнообразия и интенсивности де-
монстраций у гнезд и гнездовых убежищ. Так,
самцы индийских и домовых воробьев в мо-
мент образования нары обычно интенсивно
демонстрируют возле своих убежищ, у ры-
жих воробьев эти демонстрации относитель-
но менее интенсивны, тогда как у нолевых и,
в особенности, у саксаульных воробьев сколь-
ко-нибудь яркие демонстрации в момент об-
разования пары практически отсутствуют (по
крайней мере вне убежищ): самцы просто
показывают самкам дупла, то залетая в них,
то вылетая обратно и сопровождая свои дей-
ствия громким, непрерывным пением. Инте-
ресно, что в том же самом ряду видов парал-
лельно с редукцией экспрессивности брач-
ных демонстраций можно проследить посте-
пенное нарастание выразительности, интен-
сивности и специфичности демонстраций,
употребляемых в ситуациях, связанных с за-
щитой территории. Возможно, что становле-
ние этого тина сигнальной моторики проис-
ходило в результате постепенного перехода
воробьев от открытого гнездования в мутов-
ках к гнездованию в закрытых убежищах, а
также в результате постепенного усиления
территориальности и увеличения площади
охраняемых территорий.
Дальнейшее развитие упомянутых тенден-
ций привело к возникновению нового тина ре-
пертуара, который характеризуется прежде
всего почти полной утратой гнездостроитель-
ных движений и широким вовлечением в сиг-
нализацию разнообразных форм наземной и
воздушной локомоции, появлением всевозмож-
ных демонстративных и токовых полетов, ко-
торые используются не только как средство
широкого вещания при рекламировании об-
ширных территорий, но и как сугубо персона-
лизированные сигналы, употребляемые в са-
мых интимных сферах (нредкопуляциониые
демонстрации). Вместе с тем, связь демонстра-
ций самцов с заранее выбранными гнездовы-
ми убежищами сохранилась в полной мере,
однако при показе убежищ используются до-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
99
вольво простые по конструкции позы (мон-
гольский земляной и снежный воробьи).
Что же касается конечной стадии редук-
ции исходного сложного репертуара, то ее
иллюстрацией может, но-видимому, служить
крайне простой но своей структуре моторный
репертуар короткопалых воробьев. В нема-
лой степени эта простота обусловлена, как
можно думать, тем, что самцы этого вида не
выбирают, не рекламируют и не показывают
самкам конкретные убежища для гнезд, то есть
при образовании пар короткопалые воро-
бьи пользуются лишь посредниками перво-
го уровня (гнездовой территорией). Кроме
того, самцы короткопалого воробья, в отличие
от большинства других видов, никогда не со-
провождают своих самок при выборе места
для гнезд и их постройки.
Таким образом, эволюция демонстративно-
го поведения воробьев протекала, но-всей ви-
димости, в общем контексте исторических
преобразований репродуктивного поведения
и других аспектов образа жизни этих птиц.
Эти преобразования затронули ландшафт-
ную приуроченность воробьев, места располо-
жения гнезд и относительное участие полов
в их постройке, доминирующие типы локомо-
ции, пространственно-этологическую структу-
ру популяций. Будучи неотъемлемой частью
единого моторного репертуара видов, демон-
стративное поведение не только испытывало
на себя трансформирующее влияние всех этих
преобразований, но и служило, как можно
думать, своеобразной “экспериментальной
лабораторией”, где могли формироваться спе-
цифические моторные координации, которые
на первых порах выполняли преимуществен-
ной коммуникативные функции, однако, внос-
ледствие, вполне могли найти употребление
и в иных функциональных сферах.
ГЛАВА 6. СТРУКТУРА И ЭВОЛЮЦИЯ АКУСТИЧЕСКИХ
РЕПЕРТУАРОВ
6.1. Общие принципы организа-
ции репертуаров
Вокализация воробьев строится из отдель-
ных звуковых импульсов (йот). Они излуча-
ются поодиночке или группируются в серии
(секвенции), образуя более сложные акусти-
ческие конструкции — песни и позывы.
Остановимся вначале па характеристике наи-
более часто употребляемых нот и приемах
их группирования. Воробьи широко исполь-
зуют узкополосные поты с характерной /\-
образпой модуляцией и резкой сменой знака
изменения частоты. Они присутствуют в ре-
пертуаре всех изученных нами видов, а так-
же у воробьев р. Gymnorhis (Abs, 1966). Кон-
кретные характеристики пот — продолжи-
тельность, частотный диапазон, амплитудная
модуляция, относительное значение в репер-
туаре варьируют у разных видов.
Другой тип узкополосных сигналов отли-
чается большей длительностью и более уз-
ким частотным диапазоном. Что же касается
модуляции основной частоты, то опа отлича-
ется разнообразием и в разных сигналах ме-
няется от монотонного нарастания (или
уменьшения) до чрезвычайно сложной, пило-
образной формы. Обычно этот тип сигнала
характеризуется относительно более широкой
полосой заполнения, однако этот признак в
целом отличается высокой изменчивостью.
Узкополосные ноты с параболической мо-
дуляцией и плавной сменой знака изменения
частоты составляют характерный элемент
репертуара воробьев, но встречаются пе у всех
видов. Заметно варьирует характер энерге-
тического заполнения такого рода сигналов.
Здесь имеется множество переходных вари-
антов, так что даже в пределах одной фоног-
раммы в ряду сигналов можно видеть посте-
пенный переход шумовых сигналов в гармо-
нические или в узкополосные.
У воробьев выделяются два основных ре-
жима излучения нот. В одних случаях ноты
следуют друг за другом поодиночке с относи-
тельно постоянными интервалами, продолжи-
тельность которых значительно превышает
длительность самих нот. Этот режим харак-
терен для воспроизводства самых простых
(односложных) вариантов рекламных песен,
низкопороговых позывов и т.д. В других слу-
чаях поты отчетливо группируются в много-
сложные конструкции — фразы, длитель-
ность которых бывает обычно сопоставима
или же намного уступает продолжительнос-
ти разделяющих их пауз. Это наиболее ха-
рактерный тип организации акустических
рядов воробьев, который широко употребля-
ется и в песнях, и в позывах. Большинство
сигналов, воспринимаемых человеком, как от-
дельная песня или отдельный позыв, пред-
ставляют собой именно фразы, которые в свою
очередь образуют гомотипические или гете-
ротипические секвенции более высокого уров-
ня. Обращает на себя внимание ярко выра-
женная склонность многих видов воробьев к
попарному группированию нот в песнях и в
позывах.
Ключевую роль в коммуникативной систе-
ме всех воробьев, также как и у большинства
других певчих птиц, играет рекламная песня,
исполняемая самцами и направленная на
привлечение самок. Это типичный широкове-
щательный сигнал, исполняемый с большой
громкостью и имеющий преимущественно
эндогенную природу. Рекламные песни обыч-
но состоят из четко обозначенных фраз, раз-
деленных регулярными интервалами, причем
частота следования фраз маркирует уровень
возбуждения исполнителя. Количество пот во
фразах при рекламном пении заметно варь-
ирует и на примере нескольких видов здесь
легко построить ряды параллельной измен-
чивости от односложных нот до многочлен-
ных конструкций. Виесте с тем, этот признак
обладает и вполне отчетливой видовой спе-
цификой: у пустынных воробьев преоблада-
ют односложные песни, для саксаульных ти-
пичны двусложные конструкции или упот-
ребление сложной М-образной модуляции, пе-
сенные фразы рыжих воробьев включают до
8 пот и также, как и у саксаульных воробьев,
отдельные ноты могут иногда сливаться друг
с другом, образуя единый импульс с 3-4 ника-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
101
Рисунок 21. БРАЧНЫЕ ПЕСНИ ВОРОБЬЕВ
А - Passer ammodcndri; Б — P.montanus; В — P.indicus; Д — P.hispaniolensis; Е - P.rutilans.
ми. Максимальное число пот (до 20) харак-
терно для рекламных несен монгольских зем-
ляных воробьев.
Рекламная песня воробьев — их харак-
терное чириканье, представляет собой длин-
ную серию фраз, следующих друг за другом с
регулярными интервалами. Но воробьи спо-
собны петь и в иной манере, которая харак-
теризуется сокращением пауз между фраза-
ми и нотами и утратой их регулярности,
отчего песня приобретает своеобразное тороп-
ливое, слитное и как-бы сбивчивое звучание,
лишаясь четкой ритмической организации.
Такая свободная манера пения употребляет-
ся при нении вне посредников и, в особенно-
сти, при коллективном нении в клубах. Во-
вторых, слитная, непрерывная манера излуче-
ния нот и фраз употребляется в своеобраз-
ных брачных песнях самцов, озвучивающих
церемонии ухаживания, коллективных демон-
страций, и представляющих собой, в отличие
от двух предыдущих типов пения, строго ад-
ресованный акустический сигнал (Рис.21).
6.2. Пустынный и саксаульный
воробьи
Репертуары этих видов, живущих иногда
по-соседству друг с другом, крайне просты. У
пустынных воробьев вокализация в сущнос-
ти строится всего из двух базовых элемен-
тов. Преобладают узкополосные импульсы
длительностью 10-20 мс в диапазоне 2-7 кГц.
В зависимости от частоты следования они
звучат как отдельные щелчки, серии разре-
женных щелчков (потрескивание) (Рис.22,Б),
а в случае более плотных серий — как не-
громкое трещание или довольно мелодичная
трель, которая часто издается летящими во-
робьями (Рис.22,В). Другим характерным
элементом вокализации служат сигналы с
более широкой полосой заполнения, менее
глубокой, зато множественной (пилообразной)
модуляцией, воспринимаемые как мягкие и
тихие свисты (Рис.22,Г). Эти сигналы име-
ют длительность до 100 мс и частотный диа-
пазон от 4 до 5 кГц. В общем — крайне не-
102
В. В. Иваницкий
Рисунок 22. АКУСТИЧЕСКИЙ РЕПЕРТУАР ПУСТЫННЫХ (А-Г) И САКСАУЛЬНЫХ (Д-И) ВОРО-
БЬЕВ
А. Д — рекламные песни; Б, Е — отдельные щелчки и их серии (трещание); В — полетная трель; Г, И —
мягкие свисты; Ж — основной видовой позыв; 3 — храпение.
Таблица 14. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ РЕКЛАМНЫХ ПЕСЕН
Виды Воробьёв частотный диапазон (Гц) Длительность фраз (мс) Число песен
Верхняя граница Нижняя граница
Испанский 6,22 ±0,08 2,11 ±0,04 290.4 ± 18,1 35
Домовый 4,61 ±0,07 2,64 ± 0,05 129,7 ±9,2 33
Индийский 5,14 ± 0,11 2,75 ±0,04 161,7 ±20,1 23
Полевой 5.17 ±0,09 2,51 ±0,05 91,3 ±7,7 40
Пустынный 6,02 ±0,07 2,72 ± 0,09 57,8 ± 2,5 30
Саксаульный 5,35 ±0,06 2,69 ± 0,08 110,7 ±6,8 38
Рыжий 6,50 ± 0,09 2,64 ± 0,06 136,6 ± 14,9 27
Каменный 5,67 ± 0,06 2,73 ± 0,07 203,1 ±11,0 32
Монгольский 6,41 ±0,11 2,12 ±0,09 956,9 ±49,6 21
Снежный 5,80 ±0,08 2,70 ±0,07 178,0 ±31,4 52
Короткопалый 7,13 ±0,09 4,90 ± 0,05 650,3 ± 29,7 6
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
103
большом — объеме акустической продукции
пустынных воробьев свисты занимают зна-
чительно более скромное место, чем глубоко
модулированные, короткие, узкополосные
поты, комбинируясь с ними в гетеротипичес-
кие секвенции тревожной вокализации ро-
дителей у гнезд с большими птенцами. По-
мимо этого, свисты употребляются и как са-
мостоятельный сигнал при спаривании. Пес-
ни пустынных воробьев представляют собой
относительные простые конструкции из уз-
кополосных импульсов с длительностью фраз
40-70 мс в диапазоне 2,5-7,5 кГц (Рис.22,А,
Табл. 14).
Репертуар саксаульного воробья может
рассматриваться как более сложная версия
предыдущего репертуара. В отличие от пре-
имущественно одиночных, узкополосных им-
пульсов с относительно равномерным распре-
делением энергии по контуру сигнала, харак-
терных для пустынных воробьев, вокализа-
ция саксаульных воробьев строится из более
сложных нот длительностью 20-30 мс в диа-
пазоне 1-8 кГц с гармониками и отчетливы-
ми зонами концентрации энергии в диапа-
зоне 3,0-3,5 кГц (Рис.22,Е). Обычно хорошо
выражена 1-я гармоника, но основная энер-
гия сигнала сосредоточена во 2-й гармонике с
типичной ХЬбразной модуляцией, а также во
фронте 4-й гармоники. Эти сигналы образу-
ют гомотипические секвенции с интервала-
ми 50-100 мс, звучащие как трещание разной
интонации (звонкое или глухое, интонация
постоянна или плавно меняется). Саксауль-
ные воробьи явно предпочитают плотную
упаковку этих сигналов, в чем состоит замет-
ное отличие от более свободного режима их
использования пустынными воробьями. Тре-
щание саксаульных воробьев представляет
собой более высоконороговую реакцию и упот-
ребляется преимущественно в ситуациях, свя-
занных с нарушениями социальных дистан-
ций между особями или при тревоге у гнез-
да с птенцами. У пустынных воробьев щелч-
ки и трещание маркируют малейшее возра-
стание возбуждения птиц, а в полете, судя по
всему, излучаются спонтанно.
Сигнал саксаульных воробьев, который мы
условимся называть их основным видовым
позывом, имеет длительность 150-200 мс и
лежит в диапазоне 2,5-5,0 кГц. К концу сиг-
нала частота обычно плавно повышается па
0,5-1,0 кГц, хотя модуляция может быть и
более сложной (Рис.22,Ж). Сигнал звучит как
громкий и резкий свист (“иирри” или “при-
ррн”). Имеются дополнительные частотные
составляющие, идущие параллельно основной
частоте, которые видны лишь на сонофаммах,
выполненных с применением узких фильтров.
При использовании широких фильтров сиг-
нал выглядит широкополосным (полоса запол-
нения около 1 кГц) с крутым фронтом. Этот
позыв относится к числу высоконороговых и
издается лишь в ситуациях с сильной внеш-
ней стимуляцией: при образовании нары, в
начале гнездостроения, при сильном, внезап-
ном испуге. В социальных контекстах чаще
употребляется самцами: например, на первых
стадиях совместной постройки гнезда самцы
при эпизодических сближениях с самками у
гнезд издают именно этот крик, тогда как сам-
ки неизменно отвечают трещанием.
Особый позыв издают самцы при погра-
ничных конфликтах. Это очень короткие (око-
ло 50 мс) сигналы в диапазоне 2,0-4,0 кГц,
следующие с интервалами 500-800 мс
(Рис.22,3). Звучат как отрывистое, короткое
храпение, напоминающее известное “хорканье”
вальдшнепов. Этот позыв, судя по всему, не
имеет гомологов в репертуаре других видов.
Песни саксаульных воробьев хорошо от-
личаются от несен пустынного воробья боль-
шей длительностью фраз и более низкими
частотами (Табл. 14). Чаще всего фраза пес-
ни саксаульного воробья включает две /\ -
образные ноты, занимающие различные, по
обычно перекрывающиеся частотные диапа-
зоны (Рис.22,Д).
Особого упоминания заслуживает своеоб-
разный класс несен, лежащих в более высо-
ких отделах частотного диапазона и хорошо
отличающихся на слух от обычных песен. Вы-
сокочастотные песни лежат в дипазоие 3,5-
7,5 кГц, а их продолжительность достигает
300 мс. Примечательно, что такие песни, хотя
и отличающиеся но тонкой структуре состав-
ляющих импульсов, присутствуют в реперту-
аре практически всех изученных нами пред-
ставителей р.Passer (за исключением пустын-
ных воробьев) и являются проявлением па-
раллельной изменчивости песенных рядов
воробьев.
104
В.В.Иваницкий
Важно, что у саксаульных воробьев, как и
у других, отдельные варианты высокочастот-
ных рекламных песен по своей структуре
идентичны некоторым вариантам форсирован-
ной ювенильной вокализации, которую изда-
ют взятые в руки 10-12-дневные птенцы при
физическом воздействии (например, при по-
тряхивании) или при активном выпрашива-
нии корма. Ниже мы приведем еще несколь-
ко примеров того, как в “криках бедствия”
птенцов проявляются в готовом виде многие
сигналы, свойственные дефинитивным репер-
туарам (см.Рис.33). Интересно, что высоко-
частотные песни саксаульных воробьев но
физической структуре чрезвычайно напоми-
нают рекламные песни рыжих воробьев, хотя
обычные песни этих видов вполне опреде-
ленно отличаются (см.ниже).
6.3. Рыжий воробей
У рыжих воробьев, также как и у сакса-
ульных, имеется особый сигнал, который удоб-
но называть основным видовым позывом. Од-
нако у рыжих воробьев этот позыв характе-
ризуется очень низкими порогами и отра-
жает малейшие всплески возбуждения осо-
би. В период формирования гнездовой попу-
ляции позыв излучается практически при
любой стимуляции, а также, насколько можно
судить, и в режиме спонтанного воспроизве-
дения длинными сериями с интервалами не
менее 300 мс. Это фоновая вокализация сам-
цов, отчасти дублирующая функции песни.
Сигнал звучит как короткий, громкий и чис-
тый свист, напоминающий ниньканье зябли-
ков, но, в отличие от последнего, имеет слож-
ную структуру (Рпс.23,Б). Общая длитель-
ность позыва — 150-300 мс, диапазон — 2,5-
6,0 кГц. Позыв включает до 5 узкополосных
импульсов длительностью 20-100 мс и рез-
ким падением частоты но всему диапазону.
Глухие щелчки и трещание рыжие воро-
бьи употребляют редко. Ритмическая орга-
низация таких секвенций крайне вариабиль-
на. Это очень короткие (20-30 мс) н доволь-
Рисунок 23. АКУСТИЧЕСКИЙ РЕПЕРТУАР РЫЖИХ (А-Г) И ПОЛЕВЫХ (Д-З) ВОРОБЬЕВ
А. Д — трещание; Б - основной видовой позыв; Г, 3 — рекламные песни; В - мягкие свисты; Е - призыв-
ные и полетные позывы; Ж — позыв беспокойства.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
105
по тихие сигналы, основная энергия которых
сосредоточена в диапазоне от 3,0 до 8,0 кГц.
Тем не менее, некоторые сигналы включают
до 6 частотных составляющих, то есть имеют
довольно сложную энергетическую структуру
(Рис.23,Л).
Фразы в рекламных песнях рыжих воро-
бьев включают до 8 пот (Табл. 14, Рис.23,Г).
Составляют фразу узкополосные ноты в ди-
апазоне 2,0-7,0 кГц и длительностью 20-40
мс. Высокочастотные песни лежат в диапазо-
не 5,0-7,0 кГц. Иногда они выглядят на со-
нограммах как транспонированные вариан-
ты обычных песен, хотя могут обладать и
довольно своеобразной структурой. Брачные
песни отличаются крайне изменчивой ритми-
ческой организацией, частым употреблением
относительно высокочастотных нот, богат-
ством импровизаций, многие из которых име-
ют трелевую структуру (Рис.21). Для брач-
ных несен характерны короткие сильно мо-
дулированные поты длительностью от 20 до
50 мс в диапазоне 2,0-7,0 кГц.
6.4. Полевой, домовый, индийс-
кий, испанский воробьи
Акустические репертуары более соци-
альных видов устроены сложнее, чем у трех
предыдущих, гнездящихся преимущественно
одиночными нарами. Объем акустической
продукции в колониях социальных видов
очень высок и в этом отношении они замет-
но отличаются от более молчаливых сакса-
ульных, рыжих и в особенности пустынных
воробьев. В колониях существуют постоян-
ный акустический фон, который обладает хо-
рошо выраженной видовой спецификой. Из-
менчивость всех типов сигналов чрезвычай-
но велика, а границы между отдельными
структурными классами выражены нечетко,
благодаря обилию переходных вариантов. В
кратком конспекте вокализации испанских,
индийских и домовых воробьев Панов (1989)
рассмотрел некоторые, наиболее часто встре-
чающиеся акустические сигналы этих видов.
Рисунок 24. АКУСТИЧЕСКИЙ РЕПЕРТУАР ДОМОВОГО ВОРОБЬЯ
А - тревожный крик; Б - отрывистое трещание; В — позыв беспокойства; Г, Е - призывные крики; Ж -
трещание; И — рекламные песни.
106
В.В.Иваницкий
Рисунок 25. АКУСТИЧЕСКИЙ РЕПЕРТУАР ИНДИЙСКОГО ВОРОБЬЯ
А — тревожный крик; Б — трель при залезании в гнездо; В — позыв беспокойства; Г — призывный крик;
Д - крики у гнезда; Е — трещание; Ж — рекламные песни.
Рисунок 26. АКУСТИЧЕСКИЙ РЕПЕРТУАР ИСПАНСКОГО ВОРОБЬЯ
А — тревожный крик; Б — трель при залезании в гнездо; В — позыв беспокойства; Г — трещание; Д -
рекламные песни.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
107
Наши данные позволяют нарисовать более
полную и детализированную картину.
В отличие от предыдущих видов, испанс-
кие, индийские, домовые и полевые воробьи
широко используют ноты с относительно
плавной модуляцией и гармониками длитель-
ностью 40-150 мс. Энергетическое заполнение
этих сигналов подвержено высокой видовой
и ситуационной изменчивости. С известной
долей условности здесь можно, на наш взгляд,
выделить два основных класса в зависимос-
ти от характера заполнения центральных
частей сигналов и его периферии: у “кон-
турных” сигналов вся энергия сосредоточена
вдоль огибащей, у “шумовых” — равномерно
заполнен весь контур, у “гармонических” —
распределена но многим частотным состав-
ляющим (гармоникам).
“Контурные” сигналы у всех видов сви-
детельствуют о нарастании социальной деп-
ривации особи и часто издаются спонтанно.
Это громкие и звонкие крики, звучащие как
“цви” или “цви-ви” (Рис.24, Е; Рис.25, Г). В
противоположность этому, шумовые и гар-
монические сигналы с параболической оги-
бающей всегда свидетельствуют о появлении
внешнего источника тревоги вблизи особи:
при приближении хищника, вторжении кон-
курента на гнездовую территорию или нару-
шении индивидуальной дистанции на ночевке,
и звучат намного тише и глуше: “джи” или
“джи-жи” (Рис.23, Ж; Рис.24,25,26, В).
От испанских, индийских и домовых во-
робьев часто можно услышать довольно плот-
ные секвенции отдельных нот или фраз, сле-
дующих друг за другом с интервалами 50-
100 мс (Рис.25, 26, Б). На слух они восприни-
мается, как звонкое и громкое “ти-ти-ти-ти....”
продолжительностью до 2-2,5 с. Это один из
фоновых позывов испанских воробьев, свя-
занный с гнездостроительным поведением
самцов, которые неизменно издают его при
подлете к гнезду с гнездовым материалом вне
зависимости от прочих обстоятельств (вели-
чины колонии, присутствия самки, стадии
постройки и пр.). Это один из немногих по-
зывов воробьев, ситуационная специфика ко-
торого обозначена очень четко. Гомологичные
позывы индийских воробьев лежат в более
узком частотном диапазоне и хорошо отли-
чаются но структуре от сигналов испанских
воробьев, хотя на человеческий слух звучат
очень похоже. Индийские воробьи издают
трели примерно в той же ситуации, т.е. не-
посредственно перед проникновением в гнез-
до, однако у этого вида позыв пе имеет столь
четкой связи с приносом гнездового матери-
ала и излучается значительно реже, также, как
и у домовых воробьев.
Еще один специфический тин позывов
присущ репертуарам испанских, индийских и
домовых воробьев — это очень своеобразные,
трудно передаваемые словами звонкие, про-
тяжные посвисты с чуть хрипловатой инто-
нацией (“цииии”). Это типичные трели, и у
каждого из трех видов они заметно отлича-
ются (Рис.24-26, А). У индийских воробьев
эти сигналы в той или иной степени связа-
ны с беспокойством — внезапным появлени-
ем хищной птицы, приближением человека, со-
циальными конфликтами. Во всех этих ситу-
ациях свисты употребляются одновременно с
шумовыми и гармоническими сигналами, а
также с трещанием. У испанских и домовых
воробьев протяжные посвисты представляют
собой значительно более высоко!юроговые по-
зывы, но в целом приурочены к тем же ситуа-
циям и по своим функциям также должны
быть отнесены к сигналам тревоги.
Репертуар позывов домовых воробьев
наиболее сложен среди всех изученных нами
представителей рода. Два класса позывов до-
мовых воробьев не имеют очевидных гомо-
логов в репертуаре наиболее близких к ним
индийских воробьев. Во-первых, это громкий,
мелодичный свист, и но звучанию, и но струк-
туре напоминающий основной видовой позыв
рыжих воробьев. Этот позыв домовых воро-
бьев состоит из 2-3 импульсов длительностью
от 20 до 100 мс в диапазоне от 2 до 5 кГц,
имеет общую длительность 200-300 мс и зву-
чит как звонкое, громкое “цитью” или
“уитью” (Рис.24, Е). По характеру употреб-
ления эти сигналы не отличаются от выше-
описанных сигналов с периферическим запол-
нением и, вероятно, дублируют их функции,
выступая в роли широковещательного, при-
зывного сигнала. Другой своеобразный позыв
домовых воробьев слышится как короткий,
отрывистый треск и представляет собой сиг-
нал шумовой природы продолжительностью
50-130 мс в диапазоне от 2 до 5 кГц (Рис.24,
108
В.В.Иваницкий
Б). Эти позывы придают характерное звуча-
ние фоновой вокализации домовых воробь-
ев в колонии, являясь чем-то вроде ритмичес-
ких акцентов, разбивающих ноток сигналов
на отдельные блоки.
Основные варианты рекламных и брач-
ных песен полевых, испанских, индийских и
домовых воробьев приведены на Рис. 23-26, а
их частотно-временные характеристики — в
Табл. 14. Хотя многие варианты несен встре-
чаются в репертуаре всех перечисленных ви-
дов, видовая специфика нения выражена впол-
не отчетливо. Наибольшим сходством обла-
дают песни индийских и домовых воробьев.
Хотя па слух песни этих двух близкород-
ственных видов практически неразличимы,
вокализация относительно более мелких ин-
дийских воробьев лежит в более высоком
частотном диапазоне, чем у более крупных
домовых. Характерной особенностью многих
вариантов несен испанских воробьев являет-
ся присутствие особых концентраций энер-
гии в диапазоне 6-8 кГц, отчего их песни при-
обретают очень характерное, высокое (“зали-
вистое” или “истеричное”) звучание. В коло-
нии, даже среди голосов десятков индийских
воробьев, пение единственного самца испанс-
кого воробья, поющего в своей характерной
манере, выделяется вполне отчетливо. Песни
нолевых воробьев, напротив, звучат грубее, чем
у других видов, благодаря краткости, замет-
ной редукции высоких частот и относитель-
но более узкому частотному диапазону.
6.5. Земляные и снежные во-
робьи
Подробное описание и анализ всех сигна-
лов монгольских земляных и снежных воро-
бьев опубликованы ранее (Иваницкий, 19916;
Ivanitskii, 1996). Имеющиеся данные свидетель-
ствуют, о том, песня некоторыхPyrgilauda сто-
ит ближе к Passer, чем к более специализиро-
ванной сигнализации Monti fringilla (Рис.27).
Но позывы Pyrgilauda весьма специфичны и
явственно сближают их с Montifringilia
(Рис.28). Помимо вполне очевидных качествен-
ных различий в структуре акустических ре-
Рнсунок 27. РЕКЛАМНЫЕ ПЕСНИ ЗЕМЛЯНЫХ ВОРОБЬЕВ
А - Pyrgilauda davidiana; Б — Р. ruficollis', В — P.blanfordii\ Г - P.taczanouskii (А — по Иваницкий. 1992,
с изменениями; Б-Г — по фонограммам Н.А.Формозова).
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
109
Рисунок 28. ПОЗЫВЫ ЗЕМЛЯНЫХ И СНЕЖНЫХ ВОРОБЬЕВ
А-Г — позывы P.davidiana', Д.Е.З — позывы M.adamsiv, Ж — позывы P.blanfordi; И-Н — позывыМ.nivalis
("А-Г и И-Н — по Иваницкий. 1992. с изменениями; Д-З — по фонограммам Н.А.Формозова).
перту аров Pyrgilauda и Montifringilla, нам
представляется важным отметить два общих
отличия, имеющих свои проявления па уров-
не целого репертуара. Первое отличие состоит
в том, что репертуар Pyrgilauda складывает-
ся из нескольких вполне четко отграниченных
друг от друга классов позывов, которые по
своей структуре хорошо отличаются от песни,
тогда как репертуар Montifringilla построен
преимущественно по континуальному типу,
отдельные классы вычленимы с трудом, буду-
чи связанными переходными вариантами не
только друг с другом, но и с некоторыми не-
сенными строфами. Второе отличие состоит в
характере модулирования частот гомологич-
ных сигналов. У Pyrgilauda отчетливо преоб-
ладают резкие формы модуляции частоты,
нередко повторяющиеся на протяжение одно-
го сигнала, тогда как Montifringilla пользуют-
ся гораздо более плавной (медленно нараста-
ющей, убывающей, параболической) модуляци-
ей, которая лишь в некоторых несенных стро-
фах приобретает пилообразную форму.
6.6. Каменный воробей
На доступной нам территории известно гнез-
дование четырех подвидов каменного воробья:
Petronia petronia kirhizica Sushkin, 1925 (севе-
ро-западный Казахстан), P.p.exigua Hellmayr,
1902 (Кавказ и Закавказье), P.p.intermedia
Hartert, 1901 (Средняя Азия) YiP.p.brevirostris
Tachanowski, 1874 (Алтай, Тува, Забайкалье)
(Степанян, 1978). Нами установлено, что и по-
зывы, и песни центральноазиатских (алтайс-
ких) воробьев заметно отличается от вокализа-
ции кавказских, среднеазиатских и европейс-
ких птиц, причем эти различия хорошо выра-
жены и в позывах, и в песнях (Рис.29) (см.
также Bergmann, Helb, 1982).
Как и у других видов, одним из наиболее
распространенных и характерных позывов
в репертуаре каменных воробьев оказалось
трещание, порождаемое сильной экзогенной
стимуляцией в разнообразных конфликтных
ситуациях: тревоги, беспокойства, социальных
конфликтов (Рис.29,А,Е). Это плотная, гомо-
типическая секвенция сигналов, продолжи-
тельность которых варьирует в пределах 20-
40 мс, а продолжительность разделяющих
интервалов — в пределах 40-60 мс. Ритм сле-
дования нот отличается постоянством, однако
общая продолжительность трещания варьи-
рует в широких пределах, и при высоком
уровне возбуждения достигает почти 1,5 с.
Каждая нота представляет собой узкополос-
ный сигнал в диапазоне 2,5-4,2 кГц. Интона-
ция трещания сильно варьирует — оно мо-
жет быть глухим и звонким, грубым и мело-
дичным.
110
В. В. Иваницкий
Рисунок 29. АКУСТИЧЕСКИЙ РЕПЕРТУАР КАМЕННЫХ ВОРОБЬЕВ
А-Д — позывы P.p.exigua\ М — песни P.p.exigua\ Е-Л — позывы P.p.brevirostris\ Н — песни P.p.breuirostris
(А-Д и М — по фонограммам Е.Н.Панова, Е-Л и Н — наши данные).
Другой класс позывов также представлен
узкополосными сигналами продолжительно-
стью 22-70 мс в интервале 2-6 кГц (Рис.29,К).
По характеру модуляции позывы этого клас-
са отчетливо тяготеют к/\-образным сигна-
лам, однако, в отличие от предыдущего клас-
са, т.е. трещания, в зоне перегибы имеет место
резкий спад амплитуды, и основная энергия
сигнала здесь сосредоточена не на подъеме, а
на спаде, тогда как восходящая часть сигнала
может вообще отсутствовать. Хотя имеются
отдельные переходные варианты, весь класс
отчетливо распадается на два подкласса сиг-
налов, хорошо отличающихся и но структу-
ре, и по функциям. Позывы с восходящей
ветвью имеют несколько большую длитель-
ность и воспринимаются как громкое, мягкое
и несколько протяжное “пени” или “нси-пси”,
тогда как позывы без восходящей ветви обыч-
но короче и звучат значительно более резко
— “цит” или “ци-цит”, так как сигналы из-
лучаются либо поодиночке, либо попарно с
интервалом 20-30 мс.
Позывы “нсии” употребляются обыкновен-
но как широковещательные сигналы преиму-
щественно эндогенной природы, отражающие
нарастание социальной депривации. Часто их
можно слышать, когда воробьи после днев-
ной кормежки собираются к вечеру в коло-
нию или на ночевку, илн когда самец возвра-
щается на гнездовую территорию, где в это
время насиживает самка, которая неизменно
тут же отвечает на “пени” самца своим звон-
ким трещанием (см.выше). В противополож-
ность этому, позывы “цит” используются в
более напряженных ситуациях с сильной
внешней стимуляцией, где их можно часто
слышать вместе с трещанием. При сборе на
квазикоммунальные ночевки можно наблю-
дать очень характерную смену акустического
фона. Вначале, когда число птиц вблизи но-
чевки невелико и еще достаточно светло до-
минируют позывы “пси”, посредством кото-
рых воробьи, прилетевшие первыми, вступа-
ют в характерную для этой ситуации пере-
кличку. По мере возвращения воробьев рас-
тет частота стычек за убежища и других со-
циальных взаимодействий, сгущаются сумер-
ки, и теперь доминирующими сигналами
становятся позывы “цит” и трещание.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
111
У каменных воробьев имеется обширный
класс позывов, связанных с полетом (Рис.29,
Ж,3). В структурном отношении к полетным
позывам примыкают столь же изменчивые и
многочисленные позывы, излучаемые воробь-
ями в колониях н тоже, как показывают наши
наблюдения, связанные в основном с ситуаци-
ями прилета птиц — соседей или брачных
партнеров — в колонию (Рис.29, Б,Г,Л). Для
всех этих сигналов, излучаемых как поодиночке,
так и относительно разреженными гомоили
гетеротипическими секвенциями, можно вы-
делить пожалуй лишь две общие характерные
черты — относительно узкий частотный диа-
пазон в пределах 1,5-5,0 кГц при длительно-
сти 90-120 мс и отчетливо выраженную тен-
денцию к усложненной М-образной частотной
модуляции узкополосных импульсов, которая
при резких падениях амплитуды сигнала в
промежутках между пиками приводит к воз-
никновению двусложных сигналов.
По-видимому, именно присущая каменным
воробьям склонность к усложнению частот-
ной модуляции привела в конечном итоге к
формированию довольно специфических по-
зывов, главной отличительной особенностью
которых является наличие двух частотных со-
ставляющих с очень глубокой и множествен-
ной (пилообразной) частотной модуляцией
(Рис.29,И). Эти позывы имеют общую дли-
тельность 250-300 мс и располагаются в диа-
пазоне 1,2-6,5 кГц, т.е. по крайней мере часть
энергии этих сигналов располагается значи-
тельно выше, чем у сигналов предыдущей
группы. Нижняя частотная составляющая
выражена лучше верхней и может включать
в себя до 12 частотных пиков в области 3,0-
4,0 кГц и с промежутками между пиками в
30-40 мс. Позыв излучается поодиночке или
попарно как правило в одном ряду с М-об-
разными сигналами (см.выше) и звучит как
грубое “чррры” или “чррры-рры”. Позыв
отмечен только у P.p.brevirostris и только
при встречах брачных партнеров, принесших
корм к гнезду с уже подросшими птенцами
накануне их вылета. Акустические секвенции
при таких встречах отличаются высокой
сложностью, нередко включают в себя до де-
сяти сигналов, которые сами по себе имеют
сложную структуру, и могут служить одним
из самых впечатляющих примеров способно-
стей птиц к стереотипному воспроизведению
весьма сложных акустических конструкций.
Характерным элементом репертуаров закав-
казского Р. р. exigua, сред! (еазиатского Р. р. in-
termedia и европейского каменных воробьев
является позыв, звучащий как отрывистое
“жав” (“жав-жа” в спаренном варианте) и
употребляемый в самых различных ситуаци-
ях. Резкие отличия в характере модуляции со-
ставляющих указывают па то, что в формиро-
вании сигнала участвуют два независимых ис-
точника звука (Рис.29,В). У алтайских
P.p.brevirostris этот позыв не обнаружен, а в
целом репертуар этой формы заметно отли-
чается от репертуаров западных подвидов.
Песни каменных воробьев своеобразны по
звучанию и структуре (Рис.29,М,Н). Харак-
терно наличие нескольких частотных состав-
ляющих, которые тем или иным способом
располагаются вокруг типичных узкополос-
ных пот с очень крутой частотной модуля-
ций. Обычно песни лежат в диапазоне от 2,5
до 7,0 кГц, а продолжительность фраз варьи-
рует от 100 до 250 мс, хотя некоторые фразы
достигают почти 700 мс. Звучание песен за-
висит от ритма следования фраз и их струк-
туры: сигналы, где основная энергия сосре-
доточена преимущественно в центральном
импульсе воспринимаются как “чри” и весь-
ма напоминают чириканье домовых или по-
левых воробьев; сигналы с четко выражен-
ными гармониками звучат как очень гнуса-
вое “сини”, когда же в сигналах с тем же кон-
туром энергия раснределется более равномер-
но и гармоническая структура маскируется
шумовыми компонентами, песня каменного
воробья превращается в ритмизированное ши-
пение, имеющее на непосвященный слух мало
общего с птичьим пением. Примечательно, что
также, как и у некоторых представителей
р.Passer, специфические черты несен камен-
ных воробьев проявляются уже в ювениль-
ной вокализации и в совершенно очевидной
форме выражены уже в 10-12-дневном воз-
расте (Рис.33).
6.7. Короткопалый воробей
Акустический репертуар этого вида свое-
образен (Рис.30). Ведущую роль в коммуни-
кации короткопалых воробьев играет реклам-
112
В.В.Иваницкий
Рисунок 30. АКУСТИЧЕСКИЕ СИГНАЛЫ КОРОТКОПАЛЫХ ВОРОБЬЕВ И НЕКОТОРЫХ ТКАЧИКОВ
А - песня и Б — позывы короткопалого воробья (по фонограммам Е.Н.Панова); В - песня каменного
воробья при уменьшенной вдвое скорости вопроизведенпя; Г-Д - песни Ploccus cucullatus (по Collias, 1963); Е-
Л — фрагменты вокализации Philetairus socius (по Channing, Mac Lean, 1976).
пая песня самцов и очень звонкие и громкие
позывы самок, издаваемые ими в спонтанном
режиме чаще всего в тот момент, когда самка
летит к гнезду с очередной порцией материа-
ла, или же в ответ на песню самца. По Адамя-
ну (1965) и нашим наблюдениям, переклич-
ки брачных партнеров составляют одну из
наиболее характерных особенностей поведе-
ния вида. Обычно самка отвечает сразу же
после окончания песни самца, так что оба
сигнала на слух почти сливаются, т.е. имеет
место типичная дуэтная вокализация. Парт-
неры то и дело перекликаются, причем самец
избегает приближаться к самке и сопровож-
дать ее, предпочитая наблюдать за нею изда-
ли со своих несенных постов. Нам не уда-
лось записать голоса самок в природе, однако
на Рис.30 представлены сонограммы позы-
вов, записанных от особи неизвестного пола
в неволе, которые по нашему мнению очень
близки к вокализации самок.
Необычное звучание песни короткопалых
воробьев, напоминающее монотонное жужжа-
ние цикады, отмечалось многими наблюдате-
лями. На сонограмме песня выглядит как ши-
рокополосный сигнал с параметрами 800-850
мс и 5,0-7,5 кГц с тремя-четырьмя парал-
лельными частотными составляющими — от
начала к концу сигнала частоты монотонно
возрастают в среднем на 0,5 кГц при нали-
чии слабых дополнительных модуляций. В
другом варианте песня состоит из двух час-
тей, резко отличающихся по структуре и зву-
чанию: первая половина или треть песни
представляет собой плотную гомотопическую
секвенцию из 8-10 узкополосных импульсов
с глубокой модуляцией продолжительностью
в 20-30 мс в том же частотном диапазоне, пос-
ле чего почти без паузы следует только что
описанное жужжание.
6.8. Структурно-функциональная
дифференциация и эволюция акус-
тических репертуаров
Видовые акустические репертуары воро-
бьев резко отличаются по объему и разнооб-
разию составляющих их сигналов. Эти раз-
личия по большей части очевидны, хотя кон-
тинуальная природа сигналов сильно затруд-
няет количественную оценку репертуаров.
Последнее обстоятельство породило немало
острых дискуссий о соотношении дискрет-
ности и континуальности в акустической
коммуникации.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
113
Рисунок 31. СВЯЗЬ АКУСТИЧЕСКИХ РЕПЕРТУАРОВ И СОЦИАЛЬНОЙ ОРГАНИЗАЦИИ ВОРОБЬЕВ
1 — пустынный воробей; 2 — короткопалый воробей; 3 — рыжий воробей; 4 — саксаульный воробей; 5 -
монгольский земляной воробей; 6 — снежный воробей; 7 — полевой воробей; 8 — индийский воробей; 9 —
испанский воробей; 10 — домовый воробей; 11 — каменный воробей. По оси абсцисс — уровень социальности
(баллы), по осн ординат — число позывов в репертуаре.
Скорее всего, дискретность и континуаль-
ность не являются альтернативами. Они при-
сутствуют в репертуарах разных видов, но
проявляются неодинаково. Относительно бед-
ные репертуары саксаульных и рыжих во-
робьев распадаются на четко обозначенные
классы позывов. Промежуточных вариантов
между ними почти нет, но внутри классов
существует чрезвычайно высокая изменчи-
вость сугубо континуальной природы, так что
точное число позывов, составляющих репер-
туар вида, сосчитать невозможно. В противо-
положность этому, границы между разными
классами позывов, равно как между позыва-
ми и песнями, в богатых репертуарах испан-
ских, индийских и домовых воробьев выра-
жены значительно хуже, что неизбежно уве-
личивает долю субъективизма при оценке
числа таких классов. Аналогичные различия
существуют между вокализацией монгольс-
ких земляных и снежных воробьев.
Отдавая отчет в неизбежной субъективно-
сти разграничения акустических сигналов на
отдельные классы, мы все же попытались дать
количественную оценку объема репертуаров
изученных нами видов воробьев с использо-
ванием балльных оценок степени их “соци-
альности” в зависимости от тина ночевок, стай
и гнездовых поселений (см.Табл.4). Мини-
мальная социальность но этой шкале припи-
сывается пустынным воробьям (нет коллек-
тивных ночевок — 1 балл, нет стайности — 1
балл, нет колониального гнездования — 1 балл,
всего — 3 балла), максимальная — испанс-
ким, индийским, домовым и нолевым воробь-
ям (коллективные ночевки — 3 балла, круг-
логодичная стайность — 3 балла, сунерколо-
пиалыюсть — 4 балла, всего — 10 баллов).
Как видно из Рис.31, степень “социальности”
и богатство репертуара связаны сильной и
высоко достоверной корреляцией, что подтвер-
ждает гипотезу о том, что “высоко соци-
альные” виды птиц имеют в среднем более
обширные вокальные репертуары по сравне-
нию с “менее социальными” (Заблоцкая, 1982).
Касаясь функциональной организации
акустических репертуаров воробьев и ком-
муникативного значения составляющих его
элементов, мы полагаем, что наиболее важны
сигналы, регулирующие пространственные
114
В.В.Иваницкий
взаимоотношения особей. К их числу отно-
сится песня, обеспечивающая дистантную ком-
муникацию, и трещание, употребляемое для
регуляции межипдивидуальпых дистанций
при непосредственных социальных взаимо-
действиях. Трещание есть у всех видов, хотя
у каждого оно несет особые черты, благодаря
отличиям в тонкой структуре импульсов и
ритмической организации серий. Столь же
консервативны негромкие, мягкие свисты, ко-
торые воробьи издают во время спаривания
— они также есть у всех видов за исключе-
нием короткопалых воробьев.
Во всех прочих ситуациях, вокализация
обнаруживает резкие видовые отличия. Так,
при стычках за территорию рыжие и камен-
ные воробьи молчат, саксаульный воробьи
употребляют специализированный и стерео-
типный сигнал, который нигде помимо этой
ситуации не встречается, снежные воробьи
— наоборот, излучают при конфликтах пре-
дельно неспециализированные и крайне из-
менчивые позывы, у индийских воробьев в
этой ситуации в разных сочетаниях можно
слышать три разных позыва, излучаемых и
в иных случаях, тогда как земляные и корот-
копалые воробьи при пограничных конфлик-
тах вообще не употребляют позывов, зато
интенсивно поют. При появлении опаснос-
ти у гнезда с подросшими птенцами сакса-
ульные воробьи просто трещат — как и во
всех прочих случаях, пустынные воробьи к
трещанию прибавляют короткие посвисты,
близкие к тем, что можно слышать при спа-
ривании, домовые, индийские и нолевые и
испанские воробьи озвучивают ситуацию 2-3
типами неспециализированных позывов, на-
конец монгольский земляной и снежный во-
робьи в секвенциях тревожной вокализации
используют все имеющиеся в их репертуаре
позывы, причем у обоих видов имеется по два
типа специализированных позывов, которые
приурочены исключительно к этой ситуации.
Таким образом, даже у близких видов во-
кализация в однотипных ситуациях может
резко отличаться — одни виды ситуацию
вообще не озвучивают, другие употребляют
специализированные, третьи — неспециали-
зированные сигналы и т.д. Столь же высо-
кую видовую изменчивость мы наблюдаем и
в отношение ситуационной приуроченности
специализированных сигналов, которые мо-
гут озвучивать исключительно пограничные
конфликты (саксаульный воробей), встречи
брачных партнеров у гнезда с птенцами (ка-
менный воробей), спаривание (почти все
виды), окрикивание хищника у гнезда (зем-
ляные и снежные воробьи). Это наводит на
мысль о том, что в эволюции акустических
репертуаров существенную роль играли слу-
чайные факторы, отчего приуроченность сиг-
налов к определенным ситуациям была под-
вержена относительно быстрым и мало пред-
сказуемым изменениям.
В литературе, посвященной эволюции аку-
стических репертуаров птиц, простота репер-
туара часто рассматривается как доказатель-
ство его древности и примитивности. Тогда за
стартовый вариант в эволюции вокализации
рода Passer следовало бы принять репертуар
пустынных воробьев, а наиболее продвинутым
считать репертуары домовых воробьев. Тем не
менее, с позиций филогенетического подхода
такой ход событий выглядит маловероятным.
Во-первых, бедными репертуарами обладают
виды, которые по совокупности всех прочих
признаков (см. ниже) наиболее специализиро-
ваны. Кроме того, богатство репертуаров воро-
бьев во многом зависит от присутствия позы-
вов с параболической огибающей и многими
частотными составляющими. Эти сигналы ха-
рактерны для испанских, домовых, индийских
и полевых воробьев, по отсутствуют у пус-
тынных, саксаульных и рыжих. Куда направ-
лен в данном случае вектор эволюции? Для
ответа на этот вопрос необходимо учесть, что
сигналы с обертонами встречаются у всех
родов воробьев, кроме ^ Gymnorhis (Abs, 1966),
и широко распространены во всех прочих
rpymiax Ploceimorpha, где они не только ис-
пользуются в качестве позывов, по у многих
видов составляют основу их песенных конст-
рукций (Collias, 1963; Craig, 1976) (Рис.30).
Вероятно, обертоны свойственны как раз наи-
более древним акустическим репертуарам
Ploceimorpha, а их отсутствие есть результат
редукции.
На Рис.32 изображены гипотетические
этаны эволюции акустических сигналов во-
робьев. За исходный тип приняты плотные
пачки коротких узкополосных импульсов с
глубокой модуляцией. Подобная структура
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
115
Рисунок 32. ГИПОТЕТИЧЕСКИЕ ЭТАПЫ ЭВОЛЮЦИИ АКУСТИЧЕСКИХ СИГНАЛОВ ВОРОБЬЕВ
1 — низкоинтснснвный крик выпрашивания корма и маяковый сигнал Passer; 2 — форсированные крики
гнездовых птенцов Passer; 3 — крики интенсивного выпрашивания корма у птенцов Pyrgilauda; 4 — крики
бедствия птенцов Petronia; 5 — песня Passer indicus; 6 — песня Р. davidiana; 7,8 — песни Р. ruficollis; 9 — песня
P.taczanoivskii; 10 — песня P.blanfordii; 11,12 — песни Petronia; 13 — песня Carpospiza; 14 — основной видовой
позыв Р. rutilans; 15 — призывный крик Р.domesticus; 16 — тревожные крики (звонкие свисты) Р. davidiana; 17
— крики Carpospiza; 18 — трещание Р.davidiana; 19 — трещание Montifringilla nivalis; 20 - трещание
P.ammodendri.
видна уже в форсированных криках гнездо-
вых птенцов Passer, излучаемых, например,
при физическом раздражении или при ак-
тивном выпрашивании корма. У некоторых
представителей рода эти крики мало чем от-
личаются от базовых вариантов песни
(Рис.33). Песни Pyrgilauda davidiana еще
сохраняют много общего с исходным типом,
отличаясь, главным образом, большим числом
импульсов во фразах. У других земляных
воробьев заметно уменьшаются плотность
упаковки импульсов и глубина их частот-
ной модуляции. Эти тенденции достигают
максимального развития в песнях taczanow-
116
В.В.Иваницкий
Рисунок 33. ЭЛЕМЕНТЫ ЮВЕНИЛЬНОЙ ВОКАЛИЗАЦИИ ВОРОБЬЕВ
А — крики бедствия птенцов Petronia-, Б - маяковый сигнал птенцов Petronia, В - крики бедствия птенцов
Passer ammodendri; Г — крики выпрашивания птенцов P.montanus-, Д — тревожные крики слетков Pyrgilauda
davidiana.
skii и blanfordi, но если песня первого до-
вольно проста, то песня второго отличается
удивительной сложностью (см.Рис.27). Струк-
тура песни Petronia, вероятно, также’ берет
начало от плотно упакованных пачек глубо-
ко модулированных узкополосных импуль-
сов. Более специализированными варианта-
ми песни каменных воробьев, как и в па-
раллельной ветви, мы считаем фразы с наи-
более разреженными и наименее модулиро-
ванными импульсами. Специфическая тенден-
ция в эволюции песни Petronia состояла в
усилении дополнительных частотных состав-
ляющих, присутствующих и в ювенильной во-
кализации этого вида. В своей совокупности
все три тенденции вполне могли привести
к песне Carpospiza, которая представляет со-
бой относительно слабо модулированный по
частоте, длительный сигнал со многими час-
тотными составляющими.
ГЛАВА 7. КЛАССИФИКАЦИЯ И ЭВОЛЮЦИЯ
7.1. Положение воробьев в сис-
теме отряда Passeriformes
В конце прошлого и начале нынешнего
веков всех мелких зерноядных птиц Старого
Света, за исключением жаворонков, чаще все-
го объединяли в одно семейство Fringillidae,
куда, наряду с воробьями, включали ткачи-
ков, овсянок и собственно вьюрковых (см.Би-
анки, 1907). Эту традицию нарушил Chapin
(1917), который установил родство всех тка-
чикоподобпых птиц и выделил их в отдель-
ное семейство Ploceidae. Отметив определен-
ное сходство признаков скелета ткачиков и
воробьев, он все же оставил последних в сем.
Fringillidae.
Следующим шагом в развитии классифи-
кации мелких зерноядных стали исследова-
ния Sushkin (1927). Учитывая строение кост-
ного и рогового неба, особенности линьки и
конструкцию гнезд, Сушкин выделил воробь-
ев из Fringillidae и переместил их в Ploceidae
в ранге подсемейства Passerinae в составе рр.
Passer, Petronia и Montifringilia. Сушкин по-
лагал, что воробьи представляют собой наи-
более продвинутую ветвь семейства и проис-
ходят непосредственно от Plocepasserinae.
Точка зрения Сушкина господствовала,
пока Воск и Могопу (1978) не опубликова-
ли результаты своих исследований подъязыч-
ного аппарата воробьев и других мелких зер-
ноядных птиц. Изучив представителей
Passer, Petronia и Montifringilia, эти ав-
торы обнаружили, что подъязычный аппарат
воробьев состоит из двух окостенений:
preglossale и paraglossale, соединенных друг
с другом m.hypoglossus anterior. Наличие пос-
леднего признается этими авторами в каче-
стве необходимой предпосылки возникнове-
ния preglossale из-за тесной функциональной
связи между этими структурами. Между тем,
названная мышца отсутствует у всех ткачи-
ков, вьюрков и овсянок, у которых в языке
имеется только одно окостенение. Бок и Мо-
рони придают большое значение обнаружен-
ным отличиям и полагают невозможным
объединение воробьев с любой из названных
групп, выделяя их в особое семейство
Passeridae, не имеющее, по их мнению, близ-
ких родственников среди зерноядных птиц
Старого Света. Повышение ранга воробьев до
семейства было поддержано Summers-Smith
(1988) и Степаняном (1990).
Вместе с тем, Sibley, Ahlquist (1985) выс-
казали сомнения в том, что наличие preglossale
в самом деле кладет непреодолимый рубеж
между воробьями и прочими Ploceimorpha,
ссылаясь на возможность утраты этого при-
знака у Ploceidae и Estrildidae после отделе-
ния Passeridae. Опираясь па данные по гиб-
ридизации ДНК, эти авторы пришли к вы-
воду о моиофнлитичности всех ткачикопо-
добных птиц, которых они трактуют как еди-
ное семейство Ploceidae, где вместе с тради-
ционными членами — Ploceinae, Passerinae
и Estrildinae, неожиданно оказались трясо-
гузки Motacillinae и завирушки Prunellinae.
Эта же классификация приведена и в недав-
но опубликованном капитальном труде Sibley
и Monroe (1992) с той разницей, что но но-
менклатурным соображениям название семей-
ства изменено на Passeridae.
Наши данные подтверждают высокое био-
логическое своеобразие воробьев. Вероятно,
еще на самых ранних стадиях эволюции этой
группы ее представители освоили закрытое
гнездование, что в значительной мере предоп-
ределило эволюционную историю и характер
адаптивной радиации воробьев, которые сло-
жились существенно иначе, чем у других тка-
чикоподобпых птиц, сохранивших верность от-
крытому гнездованию и тесные экологичес-
кие связи с древесной растительностью. С
морфологической точки зрения воробьи ха-
рактеризуется уникальным строением подъя-
зычного аппарата — и этот признак сохра-
нялся в неизменном виде на всем протяже-
нии истории группы от самых архаичных
до наиболее специализированных форм. Что
же касается поведения, то некоторые этоло-
гические признаки также можно рассматри-
вать в качестве диагностических для всех во-
118
В. В. Иваницкий
робьев. Наиболее характерно множественное
спаривание, найденное у всех изученных ви-
дов за исключением короткопалых воробьев.
Последнее, впрочем, можно трактовать, как ко-
нечный результат отчетливо выраженной тен-
денции к редукции этого признака в эволю-
ции Passeridae. Множественное спаривание
— уникальный признак воробьев, резко отде-
ляющий их не только от других Ploceimorpha,
ио и от большинства других певчих птиц.
Другая своеобразная особенность всех воро-
бьев состоит в ярко выраженной склонности
к “купанию” в пыли, чего никогда не делают
ткачики (Moreau, 1967). Акустической “ви-
зитной карточкой” воробьев является их ха-
рактерное трещание.
Будучи действительно своеобразной груп-
пой, воробьи, тем не менее, обладают, как мы
полагаем, явственным поведенческим сход-
ством с другими ткачиконодобными птица-
ми. Все Ploceimorpha, включая воробьев, —
типичные клаустрофилы (т.е. гнездятся в
замкнутых камерах естественного происхож-
дения или собственного изготовления) с пло-
цеидным типом репродуктивного поведения
— сочетание, присущие наименее изменен-
ным ветвям Passeridae (Passer), Ploceinae
(Foudia, Quelea, Ploceus), а также всем
Bubalornithinae, Plocepasserinae и Estrildidae.
Что же касается наиболее уклонившихся
форм, то они подчас отличаются друг от
друга очень сильно. Трудно назвать общие
этологические признаки, сравнивая, например,
Ploceus и Montifringilla, однако, их связы-
вает полный ряд промежуточных форм (по-
лигамные Ploceus — моногамные Ploceus —
Passer hispaniolensis — Р. domesticus —
P.montanus — Pyrgilauda — Monti fringilla).
Интересно, что но данным электрофореза
яичных белков Montifringilla оказался бли-
же не к Passer, а к Ploceus (Sibley, 1970).
Все сказанное, с нашей точки зрения, дает
основания говорить об общности эволюци-
онной истории и монофилитичности
Ploceimorpha. Вместе с тем, главные ветви тка-
чнкоподобиых птиц к настоящему времени
претерпели достаточно глубокую диверген-
цию, что делает вполне приемлемым присвое-
ние каждой ветви одинакового таксономичес-
кого ранга (Воск, Могопу, 1978). Таким обра-
зом, группу Ploceimorpha составляют семей-
ства Ploceidae, Passeridae и Estrildidae. Ука-
зание Sibley па их близкое родство с зави-
рушками и трясогузками нуждается, на наш
взгляд, в дополнительном исследовании.
7.2. Таксономическая структура
семейства Passeridae
Разные авторы по-разному оценивают объем
и структуру семейства Passeridae. В современ-
ной литературе в качестве основных его под-
разделений чаще всего фигурируют следующие
роды: 1) настоящие воробьи Passer (Brisson,
1760), 2) снежные воробьи Montifringilla
(С.L.Brehm, 1828), 3) каменные воробьи
Petronia (Каир, 1829), 4) кустарниковые воро-
бьи Gymnorhis (Hodson, 1844), 5) короткопа-
лые воробьи Carpospiza (Muller, 1854), 6) зем-
ляные воробьи Pyrgilauda (Verreaux, 1871).
Более дробная классификация на этом уровне
не практикуется, наоборот, преобладает тенден-
ция к объединению родов: Gymnorhis и Carpo-
spiza сводят в синонимы Petronia, a Pyrgilauda
— в сшюнимы Montifringilla (Vaurie, 1956;
Гладков и др., 1970; Воск, Могопу, 1978;
Summers-Smith, 1988; Sibley, Monroe, 1992;
Clement, Harris, Davis, 1993).
Мы попытались представить взаимоотно-
шения шести перечисленных родов воробьев
с помощью кластерного анализа 20 морфоло-
гических и этологических признаков. Хотя
мы старались включать в анализ лишь те
признаки, наличие или отсутствие которых
было точно известно по нашим наблюдениям
или литературным данным, состояние неко-
торых из признаков у представителей наиме-
нее изученного р.Gymnorhis осталось невы-
ясненным. Описание признаков и их распре-
деление но родам приведены в Табл. 15 и 16.
Приняты Эвклидово расстояние и объедине-
ние но невзвешенпому парпо-грунновому
среднему. Как видно из Рис.34 наиболее близ-
ки Passer и Gymnorhis, образующие особый
кластер, противостоящий всем другим родам,
среди которых ближе всего стоят друг к дру-
гу Montifringilla и Pyrgilauda, к ним доволь-
но отчетливо примыкает Petronia, тогда как
Carpospiza занимает более обособленное по-
ложение.
С учетом результатов кластерного анали-
за и всей совокупности сведений мы нолага-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
119
Таблица 15. ПРИЗНАКИ ДЛЯ ГРУППИРОВКИ РОДОВ PASSERIDAE
NN Характеристика признаков Градации признаков
1 Выбор места для гнезда 0 - самка; 1 - самец
2 Самцы строят гнезда 0 - нет; 1 - да
3 Самцы сопровождают самку при сборе материала 0 - нет; 1 - да
4 Самцы насиживают кладку 0 - нет; 1 - да
5 Трепещущий режим в полете 0 - нет; 1 - есть
6 Порхающий режим в полете 0 - нет; 1 - есть
7 Скольжение на поднятых крыльях в полете 0 - нет; 1 - есть
8 Территориальный рекламирующий полет 0 - нет; 1 - есть
9 Предкопуляционное поведение 0 - неспецифическое; 1 - погоня за самкой; 2 - зависание над самкой; 3 - кружение над самкой
10 Число садок за одну копуляцию 0- 1; 1 - >1
И Гнездо в виде замкнутой полости 0 - нет; 1 - да
12 Кормятся в кронах деревьев 0 - нет; 1 - да
13 Структура песни 0 - только короткие узкополосные ноты; 1 - сильно модулированные ноты с гармониками; 2 - слабо модулированные ноты с обертонами
14 Окраска рулевых перьев 0 - однотонная (бурая или рыжеватая); 1 - темная с белыми пятнами на кончиках; 2 - темная со светлыми поперечными полосами; 3 - белая с черными кончиками
15 Форма обреза хвоста 0 - прямой или слегка вырезан; 1 - закруглен
16 Желтое пятно на груди 0 - нет; 1 - есть
17 Опушение новорожденных птенцов 0 - нет; 1 - пепельно-серое; 2 - желтое
18 Боковые движения головы у птенцов при выпрашивании корма (по Адамян, 1966) 0 - нет; 1 - есть
19 Окраска яичной скорлупы 0 - чисто белая; 1 - белая с маленькими четкими темными пятнами; 2 - охристо-бурая с размытыми пятнами
20 Хромосомные инверсии (по Булатова, 1981) 0 - нет; 1 - есть
ем, что сем.Passeridae распадается на три вет-
ви: одна включает Passer и Gymnorhis, дру-
гая — Petronia, Pyrgilauda и Montifringilla,
третья — лишь Carpospiza. Поскольку
Carpospiza биологически сильно отличается
от прочих воробьев, его целесообразно выде-
лить в монотипичпое подсемейство Сагро-
spizinae, а все другие роды объединить в под-
семейство Passerinae, которое в свою очередь
можно разбить на две трибы — Passerini (рр.
Passer и, предположительно, Gymnorhis) и
Montifringillini (рр. Petronia, Pyrgilauda и
120
В. В. Иваницкий
Таблица 16. РАСПРЕДЕЛЕНИЕ ПРИЗНАКОВ РОДОВ PASSERIDAE
Роды Passeridae распределение признаков 1 - 20 по родам
0 1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9 2 0
Passer 1 1110000011100000021 Gymnorhis 1 0 ? 0 ? ? ? ? ? 1 1 1 0 0 0 1 002? Petronia 1010111021101111 1020 Pyrgilauda 1010101 131100210 1000 Montifringilla 1010011 131101310 1000 Carpospiza 000000001000211021 1?
HI3PANIOLENSIS
DOME3TICLJ3
INDICUS
MONTANU3
AMMODENDRI
SIMPLEX
RUTILAN3
PETRONIA
PYRGYLAUDA
MONTIFRINGILLA
CARFOSFIZA
10 20 30 40 50 60 70 80 30 100 110
Рисунок 34. ТАКСОНОМИЧЕСКИЕ ДИСТАНЦИИ МЕЖДУ РОДАМИ (А) И ВИДАМИ (Б) PASSERIDAE
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
121
Montifringilla). Предлагаемая классификация
учитывает не раз звучавшие в литературе
высказывания о морфологических, биохими-
ческих и кариологических отличиях Passer
от всех прочих воробьев (Нейфельдт, 1970;
Sibley, 1970; Булатова, 1981). Тем не менее, но
этологическим признакам Passer обладает яв-
ственным сходством с другими родами
(Ivanitskii, 1996).
Перейдем теперь к отдельным родам, начав
с центрального рода Passer. Имеющиеся в ли-
тературе суждения о числе видов в нем расхо-
дятся. Summers-Smith (1988) приводит 20 ви-
дов Passer, Sibley и Morony (1990) — 23 вида,
Clements с соавторами (1993) — 21 вид.
Расмотрим классификацию Summers-Smith
— наиболее авторитетного специалиста по
роду Passer. Следуя давней традиции, он раз-
деляет его на 2 подрода. К Auripasser он от-
носит африканских luteus и eminibey, сход-
ных по своей склонности к номадизму, высо-
кой степени колониальное™, отсутствию по-
лового дихроматизма и характерного черно-
го пятна на горле, высокому подниманию кры-
льев при брачных демонстрациях. Прочие
формы объединены в номинальный подрод.
В их числе группа сероголовых воробьев, на-
селяющих всю тропическую Африку и также
лишенных полового дихроматизма, трактует-
ся как комплекс близких, нарапатрнчпых ви-
дов (надвил) в составе griseus, swainsonii,
gongoensis, suachelicus и diffusus. В группе
черпогорлых, рыжеспинных воробьев, также
широко распространенных в тропической Аф-
рике, по представленной относительно узко-
ареальными, аллопатричными формами, вы-
деляется моиотиничпый вид iagoensis (ост-
рова Зеленого Мыса) н политииичный вид
motitensis, куда в ранге полувидов отнесены
все материковые (motitensis, subsolanus,
cordofanicus, shelleyi,rufocinctus) и одна ос-
тровная (insularis) (о.Сокотра) формы ры-
жеспинных воробьев. Кроме того, в Африке
обитают хорошо дифференцированные виды
melanurus (Капская провинция) и casta-
nopterus (район Африканского Рога). Три
вида связывают северную Африку с Еврази-
ей — simplex (полувиды simplex в Африке и
zarudyni в Азии) domesticus (полувиды
domesticus и indicus) и hispaniolensis (по-
лувиды hispaniolensis и italiae). Все азиатс-
кие воробьи представляют собой хорошо диф-
ференцированные виды: moabiticus (Ближ-
ний Восток), pyrrhonotus, flaveolus, rutilans
(Восточная и Юго-Восточная Азия), ammo-
dendri (Средняя и Центральная Азия), monta-
nus (почти вся Евразия).
В монографии много внимания уделено
родственным связям разных видов. К сожа-
лению, рассуждения автора не подкреплены
анализом конкретных признаков, поэтому
мотивы многих решений неизвестны. Напри-
мер, форма italiae широко гибридизирует и
с domesticus, и с hispaniolensis, поэтому ее
объединение с любым из этих видов требует
дополнительной аргументации, тем более, что
высказывалось мнение о видовой самостоя-
тельности italiae (Stephan, 1986; Massa, 1989).
Очень иоверхпосто аргументировано выска-
зывание о близком родстве domesticus и
castanopterus, которое противоречит распро-
страненной точке зрения о том, что casta-
nopterus более близок к rutilans. Учитывая
большое морфологическое сходство этих ал-
лонатричиых форм их объединяют в надвид
(Hall, Moreau, 1970; Sibley, Monroe, 1990). Явно
ошибочно, утверждение о близком родстве
simplex и domesticus (см.ниже), а правомоч-
ность i 1адвида domesticus, включающего ши-
роко симнатричпые и, как мы полагаем, весь-
ма далекие друг от друга формы (domesticus,
hispaniolensis, simplex, castanopterus), вызы-
вает серьезные сомнения.
Переходя к анализу собственных данных,
отметим, что они касаются только 6 из 20 вы-
шеназванных видов Passer, но в соответствии
с традициями отечественной орнитологии
(Гаврилов, Корелов, 1968; Степанян, 1990;
Букреев и др., 1991), мы рассматриваем
domesticus и indicus как отдельные виды (в
зарубежной литературе они фигурируют как
подвиды пли полувиды). Краткая характе-
ристика признаков и их распределение по
видам представлены в Табл. 17 и 18, резуль-
таты кластерного анализа — на Рис.34. Бли-
же всего стоят друг к другу indicus и
domesticus, к которым отчетливо примыкает
montanus. Другой кластер образуют rutilans
и ammodendri, тогда как simplex и hispa-
niolensis занимают обособленное положение.
Таким образом, в выбранном пространстве
признаков hispaniolensis н domesticus разде-
122
В.В.Иваницкий
Таблица 17. ПРИЗНАКИ ДЛЯ ГРУППИРОВКИ ВИДОВ
NN Характеристика признаков Градации признаков
1 Выбор места для гнезда 0 - самка; 1 - самец
2 Самцы строят гнезда 0 - нет; 1 - да
3 Самцы сопровождают самку при сборе материала 0 - нет; 1 - да
4 Самцы насиживают кладку 0 - нет; 1 -да
5 Трепещущий режим в полете 0 - нет; 1 - есть
6 Порхающий режим в полете 0 - нет; 1 - есть
7 Скольжение на поднятых крыльях в полете 0 - нет; 1 - есть
8 Территориальный рекламирующий полет 0 - нет; 1 - есть
0 - неспецифическое;
9 Предкопуляционное поведение 1 - погоня за самкой; 2 - зависание над самкой; 3 - кружение над самкой
10 Число садок за одну копуляцию 0-7; 1 - >1
11 Гнездо в виде замкнутой полости 0 - нет; 1 - да
12 Кормятся в кронах деревьев 0 - нет; 1 - да
13 Склевывают с травинок, бегая по земле 0 - нет; 1 - да
14 Патрулирование границ и конфликты вдоль пограничной линии 0 - нет; 1 - есть
15 Поклоны при демонстрациях 0 - нет; 1 - есть
16 Синхронные движения головы и корпуса при демонстрациях 0 - нет; 1 - есть
17 Трепетание крыльев при демонстрациях 0 - нет; 1 - есть
18 Поднимание крыльев при демонстрациях 0 - нет; 1 - есть
19 Опускание рулевых при демонстрациях 0 - нет; 1 - есть
20 Вибрация оперения при демонстрациях 0 - нет; 1 - есть
21 Затаивание и переползание при демонстрациях 0 - нет; 1 - есть
22 Приподнимание оперения на пояснице при демонстрациях ("сгорбленная поза") 0 - нет; 1 - есть
0 - только короткие узкополосные ноты;
23 Структура песни 1 - сильно модулированные ноты с гармониками; 2 - слабо модулированные
ноты с обертонами
24 Трещание 0 - нет; 1 - естьс
25 Позывы с гармониками 0 - нет; 1 - есть
26 Полетная трель 0 - нет; 1 - есть
27 Трель при залезании в гнездо 0 - нет; 1 - есть
28 Тревожные крики 0 - нет; 1 - есть
29 Короткие свисты при тревоге у гнезда 0 - нет; 1 - есть
30 Призывные крики 0 - нет; 1 - есть
31 Основной видовой крик 0 - нет; 1 - есть
32 Короткое храпение 0 - нет; 1 - есть
33 Трели при тревоге у гнезда 0 - нет; 1 - есть
34 Сигналы с пилообразной модуляцией 0 - нет; 1 - есть
35 Одиночные шумовые импульсы (потрескивание) 0 - нет; 1 - есть
36 Шипение 0 - нет; 1 - есть
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
123
Таблица 17. ПРИЗНАКИ ДЛЯ ГРУППИРОВКИ ВИДОВ (Продолжение)
NN Характеристика признаков Градации признаков
37 Двусложные сигналы при тревоге у гнезда 0 - нет; 1 - есть
38 Мягкие свисты при спаривании 0 - нет; 1 - есть
39 Отрывистое циканье 0 - нет; 1 - есть
40 Дуэтная вокализация самцов и самок 0 - нет; 1 - есть 0 - однотонная (бурая или рыжеватая); 1 - темная с
41 Окраска рулевых перьев белыми пятнами на кончиках; 2 - темная со светлыми
поперечными полосами; 3 - белая с черными кончиками
42 Форма обреза хвоста 0 - прямой или слегка вырезан; 1 - закруглен
43 Желтое пятно на груди 0 - нет; 1 - есть
44 Опушение новорожденных птенцов 0 - нет; 1 - пепельно-серое; 2 - желтое
45 Боковые движения головы у птенцов при выпрашивании корма (по Адамян, 1966) 0 - нет; 1 - есть
0 - чисто белая; 1 - белая с
46 Окраска яичной скорлупы маленькими четкими темными пятнами; 2 - охристо-бурая с размытыми пятнами
47 При пении широко открывают клюв и распушают оперение на груди 0 - нет; 1 - да
48 Тип наземной локомоции 0 - прыгают; 1 - бегают
лены значительным расстоянием. Это хоро-
шо соответствует глубоким, системным отли-
чиям в их репродуктивном поведении, но явно
противоречит распространенной точке зрения
о том, что именно эти виды в пределах
р.Passer являются ближайшими родственни-
ками. Эта точка зрения нашла дальнейшее раз-
витие в монографии Summers-Smith, который
в дополнение к традиционному упоминанию
о гибридизации испанских и домовых воро-
бьев на юге Европы и на севере Африки —
как главном свидетельстве их генеалогичес-
кой близости — ссылается и на неопублико-
ванные данные D.Parkin по гибридизации
ДНК, из которых следует, в частности, что ис-
панский и домовый воробьи отделились друг
от друга всего лишь около 100000 лет тому
назад. С подобной оценкой нам согласиться
трудно. Домовый и испанский воробьи близ-
ки по экологическим требованиям, но резко
различаются по размерам, окраске, поведению
11 кариотипам (см.Панов, Раджабли, 1972).
Поэтому в вопросе о взаимоотношениях
domesticus и hispaniolensis более взвешенной
представляется нам позиция Stephan (1986),
который в результате исследований эволю-
ции окраски воробьев и критического пере-
осмысливания данных по гибридизации всех
представителей р.Passer пришел к выводу о
том, что домовый и испанский воробьи при-
надлежат к разным филетическим линиям
рода, и Козловой (1975), еще ранее не без
оснований полагавшей, что ближайшими ро-
дичами домового воробья следует считать
южноазиатского pyrrhonotus и африканско-
го iagoensis.
Отсутствие собственных наблюдений и
недостаточная изученность кустарниковых
воробьев рода Gymnorhis вынуждает нас ог-
раничиться самыми общими комментариями
по вопросу их классификации. Род включает
4 почти полностью аллонатричных вида:
pyrgita, dentata, superciliaris живут в тропи-
ческой Африке и xanthocollis — в южной
124
В.В.Иваницкий
Таблица 18. РАСПРЕДЕЛЕНИЕ ПРИЗНАКОВ ВИДОВ
изученные виды воробьев распределение признаков 1 - 48 по видам
01 1 02 | 03 1 04 | 05 1 06 | 07 | 08 | 09 | 10 111 1 12 1 13 1 I4 1 15 I 16
испанский 1 1 1 1 0 0 0 0 0 1 1 1 0 0 0 1
домовый 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 1
индийский 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 1
полевой 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 1
саксаульный 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 1
пустынный 1 1 1 1 0 0 0 0 0 1 1 1 0 0 1 1
рыжий 1 1 1 1 0 0 0 0 0 1 1 1 0 1 1 1
каменный 1 0 1 0 1 1 1 0 2 1 1 0 0 0 0 1
монгольский 1 0 1 0 1 0 1 1 3 1 1 0 0 1 1 0
снежный 1 0 1 0 0 1 1 1 3 1 1 0 0 0 0 0
короткопалый 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0
17 1 1 19 1 20 I 21 | 22 | 23 | 24 | 25 | 26 1 27 1 28 | 29 1 30 1 31 | 32
испанский 1 1 1 1 0 1 0 1 1 0 1 1 0 1 0 0
домовый 1 0 0 0 0 1 0 1 1 0 1 1 0 1 1 0
индийский 1 0 0 0 0 1 0 1 1 0 1 1 0 1 0 0
полевой 1 0 0 0 0 1 0 1 1 0 0 0 0 1 0 0
саксаульный 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1
пустынный 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0
рыжий 0 0 0 0 0 1 0 1 0 0 0 0 0 0 1 0
каменный 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0
монгольский 1 0 0 0 1 1 0 1 1 0 0 0 1 0 0 0
снежный 0 1 0 0 1 0 1 1 1 0 0 0 1 0 0 0
короткопалый 0 0 0 0 0 0 2 0 1 0 0 0 0 0 0 0
1 33 1 34 | 35 1 36 1 37 | 38 | 39 | 40 | 41 | 42 1 43 1 44 1 45 | 46 1 47 | 48
испанский 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
домовый 0 0 1 0 0 1 0 0 0 0 0 0 0 2 0 0
индийский 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
полевой 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
саксаульный 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
пустынный 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
рыжий 0 0 0 0 0 1 0 0 0 0 0 0 0 2 0 0
каменный 0 1 0 0 0 1 1 0 1 1 1 1 0 2 0 0
монгольский 0 0 0 0 1 1 0 0 2 1 0 1 0 0 0 1
снежный 1 0 0 1 0 1 0 0 3 1 0 1 0 0 0 0
короткопалый 0 0 0 0 0 0 0 1 1 1 0 2 1 1 1 1
Азии (Abs, 1966). Судя по опубликованному,
все эти виды очень близки экологически. Это
обитатели саванн на равнинах и в предгорь-
ях. Гнездятся в дуплах деревьев или трещи-
нах скал, хотя изредка могут, по-видимому, стро-
ить и открыто расположенные шаровидные
гнезда (Summers-Smith, 1988). Почти все на-
блюдатели отмечали тесные трофические свя-
зи всех видов с кронами деревьев и кустар-
ников, хотя эти птицы могут кормиться и па
земле (Shelly, 1902; Baker, 1926; McLacham,
Liverside, 1957; Archer, Godman; 1961, Chose,
1973).
В современной литературе преобладает
мнение о конгенеричности Gymnorhis и
Petronia. Между тем, Bannerman (1948) и
Козлова (1975) вполне убедительно, на наш
взгляд, показали искусственность такого объе-
динения. Оно основано на единственном об-
щем признаке — желтом пятне на груди, иг-
рающем, видимо, определенную роль во время
брачных демонстраций. Но даже но этому
признаку сходство не полное, потому что у
Petronia основания перьев, имеющих желтую
кайму, черные, а у Gymnorhis — белые (Коз-
лова, 1975). Внешне Petronia и Gymnorhis
сильно различаются: у Gymnorhis нет пест-
рин на контурных перьях мантии, рулевые
имеют однотонную бурую окраску, хвост об-
резан прямо или слегка выемчатый, птенцы
практически лишены эмбрионального опуше-
ния, тогда как у Petronia мантия покрыта от-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
125
четливыми пестринами, хвост закруглен, на
кончиках рулевых расположены белые пят-
на, а новорожденные птенцы покрыты доволь-
но густым пухом. Отметим, что Petronia кор-
мятся исключительно на земле, тогда как все
Gymnorhis — преимущественно в кронах.
Козлова (1975) предполагала близкое род-
ство Gymnorhis и Passer. Это предположе-
ние кажется нам не лишенным оснований. Со
своей стороны отметим, что по окраске опере-
ния лишенные полового дихроматизма, туск-
ло окрашенные кустарниковые воробьи не-
сколько напоминают тех представителей
Passer, которые в этом отношении, как мы по-
лагаем, наиболее специализированы (группа
форм griseus — diffusus). С экологической
точки зрения Passer и Gymnorhis очень по-
хожи. К сожалению данные о поведении
Gymnorhis неполны и фрагментарны, что зат-
рудняет проведение более содержательного
обсуждения. Судя по наблюдениям в неволе
(Abs, 1966), но своему демонстративному по-
ведению м вокализации Gymnorhis занимает
промежуточное положение между Passer и
Petronia. Вслед за Козловой мы считаем це-
лесообразным до проведения специальных
исследований рассматривать кустарниковых
воробьев как особый род Gymnoris, который,
по-видимому, лишь относительно недавно от-
делился от Passer.
Из 5 видов Pyrgilauda и 2 (возможно 3)
видов Montifringilla лишь ареал М.nivalis
простирается за пределы Центральной Азии,
с которой неразрывно связана история этих
своеобразных воробьев, составляющих ядро
авифауны Тибета и прилежащих к нему рай-
онов Монголии и Китая (Козлова, 1950).
В зарубежной литературе Pyrgilauda упо-
минается лишь как синоним Montifringilla
(Мауг, 1927; Vourie, 1956, 1972; Bock, Могопу,
1978; Summers-Smith, 1988) и только в при-
мечании к последнему нз опубликованных
списков птиц мира предполагается возмож-
ность существования Pyrgilauda в качестве
самостоятельного рода (Sibley, Monroe, 1992).
В противоположность этому, в отечественной
литературе принята более дробная класси-
фикация. Так, Бианки (1907) в первой реви-
зии снежных и земляных воробьев выде-
лил три рода в следующем составе: Monti-
fringilla (nivalis, adamsii, henrici), Pyrgilauda
(davidiana, ruficollis, blanfordi) и Onycho-
spiza (taczanowskii). Эта классификация с
незначительными изменениями повторялась
и в последующих ревизиях группы (Steg-
mann, 1932, Козлова 1950) и принимается в
сводках по фауне СССР (Гладков, 1970; Сте-
панян, 1990 и др.).
Как показали наши исследования, Monti-
fringilla и Pyrgilauda стоят ближе друг к
другу, нежели ко всем прочим родам. К чис-
лу наиболее характерных признаков, сближа-
ющих именно эти две группы, по дифферен-
цирующих их от всех прочих, мы относим
отсутствие пигментации на скорлупе яиц и
широкое использование специфических форм
воздушной локомоции в демонстративном
поведении (рекламирующий территориаль-
ный полет, предкопуляционный полет по ха-
рактерной траектории) и устройство их аку-
стических репертуаров (звонкие свисты в сек-
венциях тревожной вокализации, основные
видовые позывки). Вместе с тем, между снеж-
ными и земляными воробьями имеются не-
сомненные отличия во внешнем облике, пове-
дении и экологической специализации (Коз-
лова, 1950).
Schaffer (1938), изучавший образ жизни
представителей этих родов в Тибете, отмечал
большое сходство поведения и экологии
М. nivalis и М. adamsii по целому ряду при-
знаков (петрофилия, лоткообразпые гнезда в
трещинах скал, квазиколопиалыюе гнездова-
ние, тенденция к образованию стай при кор-
межке в гнездовой период, токовой полет со
скольжением и глубокими взмахами при от-
сутствии трепещущего режима, односложная
позывка). Косвенным подтверждением близ-
кого родства этих форм может служить, на
наш взгляд, их паранатрия с узкой зоной сим-
патрии в восточном Тибете, где они, по на-
блюдениям Шеффера, населяют разные мес-
тообитания: М,adamsii живет вдоль скалис-
тых бортов речных долин на высотах до 4000
м п.ур.м., а М.nivalis — преимущественно по
скальным гребням и осыпям в верхних час-
тях склонов и на плакорах в среднем на 500-
800 м выше. Интересно, что в Туве и на Ал-
тае, где отсутствует М.adamsii, М.nivalis, по
нашим наблюдениям, занимает оба этих типа
стаций, правда, значительно ниже (до 3000 м
н.ур.м).
126
В.В.Иваницкий
Что же касается трех других Тибетских
эндемиков, то, судя но описаниям Шеффера, а
позднее Gebauer и Kaiser (1994), они опре-
делению примыкают к монгольским земляным
воробьям: blanfordi, ruficollis и taczanowskii
тяготеют к мягкому субстрату и относитель-
но сглаженным формам рельефа, вьют шаро-
видные гнезда в норах нищух, гнездятся пре-
имущественно обособленными нарами и не
образуют стай в гнездовой период. В токовых
полетах отчетливо выражены скользящий и
трепещущий режимы, при наземных демон-
страциях воробьи, пригнувшись, бегают друг
возле друга с вытянутыми шеями, характер-
ны двусложные позывки и слитные песни.
Отметим, что все три вида широко симпат-
ричны почти но всему Тибету (cM.Vaurie,
1972), а их местообитания, по данным Шеф-
фера и Н.А.Формозова (личное сообщение),
здесь широко перекрываются. Данные по аф-
ганскому земляному воробью (Niethammer,
1967), будучи несколько более ограниченны-
ми, тем не менее со всей определенностью ука-
зывают на близость этой формы к другим зем-
ляным воробьям.
Как утверждает Майр (1971), присвоение
вновь отделившемуся таксону категории бо-
лее высокого ранга оправдано во всех случа-
ях, когда этот таксон осваивает новую адап-
тивную зону. У Montifringilla и Pyrgilauda
ситуация именно такова. Эти таксоны при-
урочены к разным адаптивным зонам, что, на-
ряду с четким хиатусом но морфологичес-
ким и этологическим признакам, дает весо-
мые аргументы в пользу присвоения им ста-
туса отдельных родов (Иваницкий, 19916;
Gebauer, Kaiser, 1994). Что же касается фор-
мы taczanowskii, то ее отличия от прочих
Pyrgilauda (иной тип песни, отсутствие пиг-
ментации клюва у самцов в брачную нору,
своеобразная окраска хвоста) кажутся нам
недостаточными для выделения ее в отдель-
ный род. Образ жизни, демонстративное по-
ведение и позывки taczanowskii близки к
другим Pyrgilauda (Gebauer, Kaiser, 1994).
Оптимальным решением, на наш взгляд, мо-
жет быть Pyrgilauda sensu lato в составе но-
минального подрода с тремя близкими алло-
патричными видами {davidiana, blanfordi,
theresae) и одним более удаленным видом
{ruficollis) и монотиничного Onychospiza,
единственный представитель которого принад-
лежит к числу наиболее специализированных
форм Passeridae.
Монотипичный род короткопалых воро-
бьев Carpospiza составляет одну из трудных
проблем в систематике мелких зерноядных
птиц Старого Света. Еще в 1896 г. Зарудный,
впервые изучивший образ жизни этих птиц
в природе, писал об их глубоких отличиях от
других воробьев и ткачиков, отмечая черты
сходства с некоторыми представителями
вьюрковых (напомним, что в то время воро-
бьев, ткачиков и вьюрковых еще не подраз-
деляли на отдельные семейства).
Однако, до недавнего времени Carpospiza
фигурировал в литературе именно как воро-
бей, причем его и поныне вместе с Gymnorhis
сводят в Petronia sensu lato (Clement et al.,
1993). Искусственность такого объединения
убедительно доказала Козлова (1975). Ука-
зав на ряд признаков, сближающих Carpospiza
с вьюрковыми (редукция наружного перво-
степенного махового, неоперенные ноздри,
конструкция гнезда), Козлова не высказалась
с определенностью относительно таксономи-
ческого положения короткопалых воробьев,
но, судя но всему, опа вовсе не видела необхо-
димости в перемещении их в сем.Fringillidae.
Впервые в новейшей литературе это предло-
жили Воск и Могопу (1978). Они исключи-
ли Carpospiza из Passeridae и поместили его
во Fringillidae с сохранением статуса моноти-
ничного рода. Позднее это решение поддер-
жал Summers-Smith (1988), однако Sibley и
Monroe (1990) в обзоре мировой авифауны
и Степанян (1990) в конспекте орнитологи-
ческой фауны СССР предпочли оставить
Carpospiza в Passeridae.
Поведение Carpospiza не дает твердых
оснований для сближения этого рода с ка-
кой-либо одной из известных групп мелких
зерноядных птиц. Трудно отрицать значитель-
ное сходство этих птиц с вьюрковыми (осо-
бенно в конструкции гнезд). Но хотелось бы
обратить внимание и на ряд моментов, про-
тиворечащих позиции Бока и Морони.
Прежде всего ни у кого из Палеарктичес-
ких Fringillidae или Emberizidae мы не встре-
чает хвостов с круглым обрезом — и у вьюр-
ковых, и у овсянок хвосты имеют вырезку,
т.е. центральные рулевые короче крайних.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
127
Рисунок 35. ОКРАСКА РУЛЕВЫХ ПЕРЬЕВ ВОРОБЬЕВ
А — Passer; Б — Petronia; В — Carpospiza; Г — Pyrgilauda dai idiana; Д — P.taczanoivsku; Е — Montifringilla
nivalis.
Между тем, у воробьев (за исключением
Passer и Gymnorhis) обрез хвоста круглый
(Бианки, 1907). Затем, у подавляющего боль-
шинства вьюрковых и овсянок рулевые не
имеют рисунка (исключением являются, на-
сколько нам известно лишь, два близких вида
щеглов из p.Carduelis). Что же касается во-
робьев, то в этой группе все роды за исключе-
нием Passer и Gymnorhis таким рисунком
обладают, причем у Carpospiza и Petronia
эти рисунки довольно сложны и практичес-
ки идентичны (Рис.35). Демонстративные
нозы короткопалых воробьев вполне типич-
ны для этой группы и заметно отличаются
от демонстраций вьюрковых (Hinde, 1956) и
овсянок (Andrew, 1957, 1961; Панов, 1989).
Песня короткопалых воробьев действитель-
но своеобразна, однако, на наш взгляд, опа
128
В.В.Иваницкий
вполне выводима из некоторых песен Petronia,
также включающих в себя несколько частот-
ных составляющих. Песни, звучащие как мо-
нотонное жужжание и напоминающие голос
сверчков или цикад, очень характерны для
настоящих ткачиков из pp.Ploceus и Euplectes
(Collias, 1963), хотя близкие по звучанию и
физической структуре песни изредка встре-
чаются у некоторых представителей Fringi-
llidae {Bucanethes githagineus — Панов, 1989)
и Emberizidae (Ammodramus savannarum —
Chandler et al., 1966). Отметим, что даже су-
губо наземные выорки и овсянки передви-
гаются исключительно прыжками, в то время
как Carpospiza в основном бегают — как и
многие другие воробьи.
Но самым серьезным свидетельством род-
ственных связей короткопалых воробьев вы-
ступают морфологические признаки. Как по-
казали результаты препаровки, проведенной
по нашей просьбе Л.П.Корзуном, подъязыч-
ный аппарат и язык Carpospiza весьма под-
вижны и по своему строению типичны для
Passeridae (см. Воск, Могопу, 1978), по отли-
чаются от прочих зерноядных птиц. Масш-
табные изменения привычной классификации
требуют серьезных оснований. В данном слу-
чае это кажется нам преждевременным. До-
полнительным аргументом, проясняющим си-
туацию, могли бы стать данные о биохими-
ческих или генотипических взаимоотношени-
ях Carpospiza с воробьями, вьюрковыми и
овсянками. Пока же, мы предпочитаем оставить
Carpospiza в Passeridae, но выделить его в
особое подсемейство Carpospizinae.
7.3. Гипотетический сценарий фи-
логенеза Passeridae
В отсутствие надежных сведений об иско-
паемых представителях таксона суждения о
его истории всегда ограничены рамками ги-
потез. Этот неоспоримый факт часто побуж-
дает к высказываниям о несостоятельности
и бесполезности исторических реконструк-
ций, построенных па неоптологическом мате-
риале. С подобной точкой зрения, па наш
взгляд, все же трудно согласиться целиком.
Эволюционный сценарий, трактующий клю-
чевые моменты истории группы, возникает как
закономерная стадия в ее исследовании и
может быть полезен, если учитывает все раз-
нообразие известных признаков, отвечает
принципу экономии, свободен от внутренних
противоречий и соответствует представлени-
ям об эволюции ландшафтов и материков,
принятых в исторической геологии и палео-
географии.
Рассмотрим с этих позиций версию об аф-
риканском происхождении рода Passer, пред-
ложенную Summers-Smith (1988). Он исходит
из того, что преимущественно монохромная
окраска оперения, отсутствие рисунка па го-
лове и полового дихроматизма характеризу-
ет аицестральиый тин окраски, носителем ко-
торого являются исключительно африканс-
кие виды. Именно таковы, но предположению
Summers-Smith, были предки современных
Passer, населявшие аридные тропические са-
ванны Африки. Позже здесь возникли виды
с более яркой окраской и сложным рисун-
ком оперения, наиболее характерной особен-
ностью которого стало черное пятно на горле.
Эти черпогорлые воробьи но долинам Аф-
риканского рифта и р.Нил вышли к Среди-
земному морю и начали колонизацию в за-
падном и восточном направлениях. По пред-
положению автора, это произошло в начале
плейстоцена и, следовательно, история воро-
бьев в Палеарктике насчитывает всего лишь
один миллион лет. Тем самым, основные эта-
ны формообразования палеарктических Pas-
ser приурочиваются к ледниковому периоду,
когда в рефугиумах южной, центральной и
юго-восточной Азии (прежде всего в доли-
нах крупнейших рек) обособились популя-
ции, давшие начало девяти современным ази-
атским видам.
На наш взгляд, изложенный сценарий со-
держит целый ряд уязвимых звеньев. Весьма
произвольно выбран временной рубеж для
проникновения воробьев в Евразию. Извест-
но, что фаунистические обмены между Аф-
рикой и Евразией, начиная по крайней мере
с миоцена, шли настолько интенсивно, что
временами состав фаун на обоих материках
сближался в высшей степени. Примером мо-
жет служить “фауна гиппариона”, ареал ко-
торой удивительно точно совпадает с облас-
тью максимального видового разнообразия
рецептных Passeridae (см.Пузанов, 1938,
стр. 137). Столь оживленные фаунистические
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
129
обмены были возможны благодаря преобла-
данию растительности саваннового типа на
большей части территории современной Са-
хары. Еще на рубеже третичного и четвер-
тичного периодов здесь текли многочислен-
ные реки, оставившие после себя “огромный
высохший скелет” сухих русел — вади (Ба-
баев и др. 1986). Скорее всего, ни миллион
лет тому назад, ни много ранее у воробьев не
было необходимости ограничивать пути сво-
его расселения на север долиной Нила. Судя
но всему, выбор автором момента проникно-
вения воробьев в Палеарктику мотивируется
главным образом его приверженностью к “лед-
никовой биогеографии” и подспудным же-
ланием приурочить происхождение азиатских
видов Passer к плейстоценовым рефугиумам.
Еще одно возражение касается темпов эво-
люции. При всей дискуссионности этой темы
и обескураживающей скудности паший зна-
ний ио этому предмету, особенно в его приме-
нении к певчим птицам, все же нелегко со-
гласиться с тем, что всего за один миллион
лет могли сформироваться 9 видов азиатс-
ких Passer, буквально во всех отношениях
весьма отличные друг от друга. К тому же,
если допустить, что черногорлые воробьи
попали в Азию всего один миллион лет тому
назад, то проблема нросхождепия трех энде-
мичных азиатских родов и, в частности рр.
Pyrgilauda и Montifringilla, где также пре-
обладают черногорлые виды, становится весь-
ма трудноразрешимой. В сущности единствен-
ная возможность примирить их существова-
ние с обсуждаемой гипотезой состоит в при-
знании африканского происхождения земля-
ных и снежных воробьев, их независимого
вселения в Евразию и последующего выми-
рания в Африке. С учетом данные налеогег-
рафии и экологичесой специфики Pyrgi-
lauda и Montifringilla, такая версия выгля-
дит неправдоподобной.
Сценарий Summers-Smith вызывает сомне-
ния в контексте истории всего семейства. Это
прежде всего касается тезиса о первичности
тусклой однотонной пигментации контурно-
го оперения и отсутствия полового дихрома-
тизма. Сочетание этих признаков характери-
зует как раз наиболее продвинутые формы
Passeridae, причем редукция пигментации, судя
но всему, шла в разных стволах семейства
параллельно, в соответствии с правилом Гло-
гера, т.е. в процессе приспособления воробьев
к прогрессирующей аридизации той геогра-
фической области, где протекала их эволю-
ция. Примером наиболее далеко зашедшей ре-
дукции окраски могут служить представи-
тели комплекса Passer griseus — Passer
diffusus, Pyrgilauda tazchanowskii, Monti-
fringilla adamsii, все Gymnorhis, монотипич-
ные Petronia и Carpospiza. Как полагал Вол-
чанецкий (1972), редукция пигментации и ут-
рата на этой почве полового дихроматизма у
птиц обычно сопровождает общее ухудшение
условий существования видов в результате
неблагоприятных изменений в их исконной
области обитания или быстрого расселения
в новые районы. Что же касается тезиса о
вторичпости “черногорлости” у воробьев, то
в его справедливости легко усомниться еще
и потому, что именно эта деталь окраски яв-
ляется одной из наиболее характерных прак-
тически для всех Ploceimorpha.
Зоогеографические основы наших пред-
ставлений об эволюции воробьев подробно
изложены выше. Мы полагаем, что данные о
современном распространении воробьев сви-
детельствуют в пользу древних связей воро-
бьев с территорией Азиатского материка. Ве-
роятно и первые этапы становления семей-
ства, и наиболее существенные стадии его
адаптивной радиации происходили именно
в Азин, тогда как Африка стала вторичным
очагом формообразования. К сожалению, све-
дения об ископаемых воробьях, способные
пролить свет на эволюционную историю этой
группы, отсутствуют. Опыты но гибридизации
ДНК позволяют предполагать, что Passeridae
и Ploceidae отделились друг от друга около
35 миллионов лет тому назад, что, вероятно,
можно в первом приближении рассматривать
как геологический возраст воробьев (Sibley,
Ahlquist, 1985). Судя потому, что точку рас-
хождения Passer и Petronia эти авторы да-
тируют более чем 10 миллионами лет, впол-
не можно допустить, что уже в миоценовых
саваннах Африки и Азии обитали птицы,
но многим признакам напоминавшие наибо-
лее архаичные формы современных воробьев.
В основу нашего сценария эволюции
сем.Passeridae положено предположение о том,
что анцестральные черты поведения полнее
130
В.В.Иваницкий
всего сохранились у представителей рода
Passer. Именно здесь мы находим максималь-
ное число экологических и этологических
признаков, сближающих воробьев с другими
филогенетическими стволами ткачиконодоб-
ных птиц. Это тесные связи с древесной рас-
тительностью, конструкции гнезд, активное
участие самцов в гнездостроении и насижи-
вании, специфические черты демонстративного
поведения, укказывающие на его тесные струк-
турные и функциональные связи с гнездос-
троительным поведением. Еще одно указание
па относительную древ!юсть Passer мы видим
в архаичности присущего им типа онтогене-
за песни, когда все основные физические чер-
ты дефинитивной вокализации отчетливо
проявляются уже в сигналах гнездовых птен-
цов. О древности Passer в какой-то мере го-
ворит обширность его ареала, сложность, “мно-
гослойпость” таксономической структуры,
обилие хорошо дифференцированных, укло-
няющихся н реликтовых форм.
Уже у Passer обозначились тенденции к
постепенной редукции упомянутых архаич-
ных, нлоцеидных черт поведения в процессе
перехода от колониалыюсти ко все более дис-
персным формам внутринонуляционной орга-
низации. Утрата нлоцеидных черт сопровож-
далась прогрессирующей перестройкой всей
сферы репродуктивного поведения в сторо-
ну конвергенции с Fringillidae. Этот процесс
удобно называть фрингиллизацией. После
того, как воробьи освоили закрытое гнездо-
вание, оставили древесные субстраты и рас-
селились в открытые ландшафты, процессы
фрингиллизации распространились и на вне-
шний облик продвинутых Passeridae, в ряду
которых можно видеть яркие примеры кон-
вергенции с вьюрками, овсянками (Monti-
fringilla, Carpospiza) и жаворонками (Pyrgi-
lauda).
Рассмотрим филетические отношения ви-
дов Passer. Здесь обращает на себя внимание
определенное сходство африканских luteus и
eminibey, образующих подрод Auripasser, с
ближневосточным moabiticus. По внешности
последний заметно отличается от лишенных
полового дихроматизма африканских воро-
бьев, однако, у всех трех есть, на наш взгляд,
немало общего: 1) это самые мелкие предста-
вители рода; 2) все гнездятся только откры-
тым способом; 3) гнезда luteus и moabiticus
очень массивны, а при их постройке широко
используются жесткие элементы; 4) при брач-
ных демонстрациях самцы всех трех видов
высоко поднимают крылья, что не характер-
но для прочих Passer, но типично для Petronia
и некоторых Ploceidae. Все три формы ал-
лоиатричны, имеют ограниченные ареалы, при-
чем moabiticus явно представляет собой ре-
ликтовую форму. У luteus и moabiticus стро-
ят гнезда почти исключительно самцы, у
eminibey гнездостроение, возможно, полностью
редуцировано в процессе перехода к парази-
тированию на гнездах ткачиков. Самцы moa-
biticus и eminibey в отличие от всех прочих
Passer не участвуют в насиживании. Вполне
возможно, что именно luteus, eminibey и
moabiticus представляют собой остатки пер-
вой волны дивергенции воробьев, уцелевшие
в узких экологических нишах на ограничен-
ной площади.
Ближе всего к рассмотренной группе сто-
ит, па наш взгляд, hispaniolensis с его супер-
колониалыюстыо, номадностью, преимуще-
ственно открытым гнездованием, апдролате-
ральным гнездостроением, отсутствим специ-
фических демонстраций при стычках за тер-
ритории, невыраженность границ последних,
тенденцией к приподниманию крыльев над
спиной при брачных демонстрациях. Эколо-
гически hispanioilensis близок к moabiticus
— исходно оба вида были связаны, как мож-
но думать, с зарослями тростников и тама-
рисков по берегам соленых водоемов — ре-
ликтов усыхающего Тетиса.
Мы видим в hispaniolensis звено, соединя-
ющее древний комплекс форм luteus-emini-
bey-moabiticus с более продвинутыми пред-
ставителями рода, среди которых наиболь-
шим сходством с hispaniolensis обладают ев-
разийские domesticus-indicus и, возможно, аф-
риканские iagoensis-motitensis. Как уже от-
мечалось, близкое родство iagoensis и domes-
ticus предполагала Козлова (1975). В подроб-
ном описании местообитаний, демонстратив-
ного поведения, социальной организации и
вокализации iagoensis (Summers-Smith, 1984
a,b ) мы нашли черты несомненного сход-
ства с domesticus, хотя по мнению самого ав-
тора, не нодтвердеиному, к сожалению, указа-
нием конкретных признаков, ближайшим род-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
131
ственником последнего в Африке является
castanopterus (Summers-Smith, 1988).
Этологически hispaniolensis все же суще-
ственно отличается от doinesticus-indicus, ко-
торые со своей стороны обнаруживают чер-
ты несомненного сходства с montanus. Это
сходство вызвано переходом к закрытому
гнездованию, увеличением роли самок в гнез-
достроепии, прогрессирующей редукцией ко-
лониальное™, становлением территориально-
сти и соответствующими изменениями аку-
стических репертуаров и демонстративного
поведения. Названные тенденции получили
развитие в наиболее продвинутой группе пред-
ставителей р. Passer, куда мы относим
ammodendri и rutilans. Это специализирован-
ные дуплогнездники с преобладанием дис-
персного гнездования и выраженной терри-
ториальностью. Экспрессивность брачных
демонстраций у них заметно меньше, чем у
описанных ранее видов, однако, четко выра-
жены специализированные демонстрации, а у
ammodendri — и акустические сигналы, об-
служивающие территориальное поведение.
Оба вида характеризуются еще большим со-
кращением участия самцов в гпездостроепии.
По данным фенетического анализа simplex
также примыкает к этой группе видов. Тем не
менее, эта форма обладает рядом специфичес-
ких особенностей. В настоящее время пустын-
ный воробей имеет явно реликтовый ареал и
крайне специализирован в отношении зани-
маемых стаций. Преобладает открытое гнез-
дование, однако в Сахаре нередко гнездится
и в щелях крутых каменистых бортов древ-
них речных русел (вади). Самцы активно
участвуют в постройке гнезда и нередко вы-
полняют основную работу но насиживанию
кладок. В целом, этот вид характеризуется
чертами глубокой специализации, но несет и
целый ряд архаичных черт.
Еще более продвинутые стадии эволюции
воробьев иллюстрируют роды Petronia и
Pyrgilauda. Оба они несомненно и примерно
в одинаковой степени родственны Passer, хотя
сходство сохранилось в разных сферах. У
Petronia и Passer оно затрагивает местооби-
тания, пигментацию яичной скорлупы, соци-
альную организацию (колопиалыюсть) и де-
монстративные позы, тогда как у Passer и
Pyrgilauda — внешний облик (окраску), струк-
туру песни, конструкцию гнезда, участие сам-
цов в гпездостроепии (правда, крайне ограни-
ченное у последних). Вместе с тем, \\Petronia,
и Pyrgilauda уже резко отличаются от Passer
отсутствием облигатных связей с древесной ра-
стительностью, сложными воздушными демон-
страциями, сокращением числа садок при спа-
ривании, редукцией гпездостроительного по-
ведения самцов, типично гинелатералыюй ин-
кубацией, хорошо развитым эмбриональным
пухом у птенцов, особешюстями кариотипов.
По всем этим признакам Petronia и Pyrgilauda
эволюционировали в сторону конвергенции
с Fringillidae и все больше уклонялись от ис-
ходного, плоцеидного тина.
Скорее всего Petronia и Pyrgilauda отде-
лились от предкового таксона (предположи-
тельно от Passer) в разное время и в даль-
нейшем эволюционировали параллельно и
независимо друг от друга, о чем свидетель-
ствуют прежде всего резкие отличия их аку-
стических репертуаров и предкопуляционных
воздушных демонстраций. Данные гибриди-
зации ДНК указывают на то, что Petronia
отделились от Passer около 10 миллионов лет
тому назад (Sibley, Aulquist, 1985), вероятно,
примерно тогда же отделились и Pyrgilauda.
Экологических обстоятельств этого отде-
ления в обоих случаях были скорее всего оди-
наковы. Становление Petronia и первые эта-
ны эволюции Pyrgilauda происходили, ви-
димо, в условиях нагорных саванн, подобных
современным фисташникам Бадхыза, где скла-
дывались благоприятные условия для пере-
хода от гнездования в дуплах к гнездованию
сначала в щелях скал, затем среди нагромож-
дений обломочных материалов и под отдель-
но лежащими камнями, откуда легко было
перейти к гнездованию в норах. По данным
палеогеографии подобные ландшафты в ми-
оцене были широко распространены в Цент-
ральной Азии, где простирались древние, силь-
но разрушенные нагорья времен герцинской
и каледонской складчатостей (Гвоздецкий,
Голубчиков, 1987).
Процессы формирования принципиально
нового биологического типа “воробья-жаво-
ронка” протекали в ходе эволюции Pyrgi-
lauda. По мнению Козловой (1955), становле-
ние этого рода могло происходить еще в ми-
оцепе в эпоху широкого распространения
132
В.В.Иваницкий
стенных ландшафтов в Центральной Азии.
Показательна геологическая история пищух
(Ochotonidae), норы которых представляют
собой основное убежище для гнезд и ноче-
вок всех земляных воробьев. Как пишет Ер-
баева (1988), миоцен был периодом расцвета
пищух. Второй аналогичный период пришел-
ся па средний и поздний плиоцен. Единствен-
ный рецептный род пищух (Ochotona) изве-
стен с позднего миоцена, тогда как современ-
ные виды — с конца плиоцена. Таким обра-
зом, версия о миоценовом возрасте Pyrgilauda
выглядит вполне правдоподобной. Позднее, с
началом нозднетретичпого орогенеза, прогрес-
сирующего похолодания и аридизации Цен-
тральной Азии Pyrgilauda все глубже при-
спосабливались к обитанию па густо заселен-
ных пищухами высокогорных плато с их эк-
стремальным климатом и крайне бедными
биоценозами.
Что же касается Montifringilla, то мы счи-
таем, что они отделились непосредственно от
Pyrgilauda, сохранив с предковой группой
немало общего в демонстративном поведении
и вокализации. Дивергенция Montifringilla
протекала в процессе освоения воробьями
совершенно новой экологической ниши, воз-
никновение которой оказалось возможным
лишь па продвинутых стадиях альпийского
горообразования. В настоящее время Monti-
fringilla, гнездящиеся в трещинах скал и сре-
ди осыпей, в трофическом отношении связа-
ны преимущественно с мозаично распределен-
ными и, как правило, небольшими по площа-
ди фрагментами альпийских лугов вокруг
водотоков и по периферии всевозможных
гляциалыю-нивальных образований, обеспе-
чивающих в течение лета постоянное увлаж-
нение соседних участков. Здесь развивается
богатая растительность, где кормятся многие
птицы. Таким образом, но сравнению с пред-
ковой формой Montifringilla осваивали в
ходе эволюции относительно более мозаич-
ную, гумидную и непредсказуемую среду.
Поистине впечатляющий успех синантроп-
ных видов воробьев, расселившихся вслед за
человеком но всему свету и достигших ко-
лоссальной численности, стал главной причи-
ной широкого распространения в литературе
представлений о воробьях, как о процветаю-
щей группе птиц (Судиловская, 1952; Келей-
ников, 1953; Ильенко, 1976; Summers-Smith,
1988). Результаты проведенного нами комп-
лексного этолого-экологического изучения
сем.Passeridae оказались в неожиданном кон-
трасте с этой точкой зрения. Многие экологи-
ческие и этологические характеристики во-
робьев указывают па их древность и архаич-
ность. Проигрывая в конкуренции с предста-
вителями более прогрессивных групп мел-
ких зерноядных птиц, воробьи вступили на
путь специализации к жизни в маргиналь-
ным местообитаниям с достаточно специфич-
ными и часто весьма суровыми условиями.
По всем показателям биологического про-
гресса, к числу которых относят количество
видов и родов в группе, роль ее представите-
лей в биоценозах, величину ее ареала (Север-
ное, 1939; Гентнер, 1965), воробьи сильно ус-
тупают всем прочим группам зерноядных
птиц Старого Света: вьюркам (Fringillidae),
овсянкам (Emberizidae), жаворонкам (Alau-
didae). Во всех трех семействах отчетливо
преобладает фрипгиллндпый тип репродук-
тивного поведения. Пространственная струк-
тура популяций базируется на дисперсном
гнездовании в сочетании с более или менее
жесткой территориальностью, гнезда с неболь-
шими вариациями повторяют форму откры-
той сверху полусферы, гиездостроение и на-
сиживание носит гипелатеральный характер,
роль социальных регуляторов в репродуктив-
ной сфере сведена к минимуму.
Вероятно, фрипгиллидпый тин поведения
оказался более надежным и эффективным
нежели плоцеидный тип, базирующийся на
сложной и противоречивой системе соци-
альных регуляторов, взаимозависимостей и
взаимовлияний социальных партнеров. Подоб-
ная система легко порождает состояние нео-
пределенности, физического и психологичес-
кого соперничества, чревата утерей согласо-
ванности в действиях парнеров, стремящих-
ся к наиболее полной реализации собствен-
ных поведенческих программ. Колониальное
гнездование добавляет свои издержки от
разрушения гнезд, гнездового паразитизма,
инфантицида, высоких затрат времени и
энергии на поддержание минимального про-
странственного суверенитета в условиях пе-
реуплотнения. Не случайно в эволюции
Passeridae мы видим неоднократные попыт-
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
133
ки перехода от плоцеидпого типа поведения,
наиболее полно представленного у некоторых
Passer, к фрипгиллидпому типу, к которому
ближе всего подошли Pyrgilauda, Montifri-
ngilla и Carpospiza. Интересно, что конвер-
генция воробьев с вьюрками, овсянками и
жаворонками затронула не только репродук-
тивное поведение, по и пропорции тела, ок-
раску оперения, что не раз вводило в заблуж-
дение систематиков (см. об этом Бианки, 1907;
Нейфельдт, 19866).
Мы упоминали о том, что отдельные чер-
ты плоцеидпого типа имеются у некоторых
архаичных групп Fringillidae и Emberizidae.
Это типично апдролатералыюе, множествен-
ное гнездостроение и шаровидные гнезда у
дарвиповых вьюрков (Geospizinae) и, вероят-
но, родственных им форм (рр. Turns и Mela-
nospiza) (Grant, 1986). Это ограниченное уча-
стие самцов в гнездостроении у некоторых
Carduelinae (pp.Serinus и Mycerobas) (Ков-
шарь, 1979; Elzen, Nemeschkal, 1991) или в
насиживании у некоторых, как полагают —
наиболее архаичных видов овсянок из
p.Emberiza s. lato (Шестакова, 1937; Юмов,
1985; Доржиев, Юмов, 1991). Вероятнее всего,
перечисленные особенности у вьюрков и
овсянок являются рудиментами предкового,
плоцеидпого типа репродуктивного поведения,
причем участие самцов в насиживании, судя
по всему, сохраняется в процессе эволюции
дольше, чем их участие в гнездостроении. Фун-
кциональным эквивалентом собственного уча-
стия в инкубации кладки является для сам-
цов доставка ими корма насиживающим сам-
кам, что свойственно многим Carduelinae.
По данным гибридизации ДНК, фрипгнл-
лидпая и плоцеидиая ветви зерноядных
птиц разделились около 40 миллионов лет
тому назад и развивались с тех пор парал-
лельно (Sibley, Aulquist, 1985). Наша гипоте-
за состоит в том, что обе ветви унаследова-
ли от общего предка плоцеидпый тип реп-
родуктивного поведения п затем параллель-
но, по с разной скоростью эволюционирова-
ли по пути фрипгиллизации. В стволах, ве-
дущих к современным вьюркам и овсянкам,
относительно быстро накапливались поведен-
ческие адаптации широкого значения, что и
составляет суть процесса фрипгиллизации.
Это непрерывно расширяло приспособитель-
ные возможности соответствующих групп и
привело в копце-копцов к появлению прак-
тически универсального по своих базовым
этолого-физиологических свойствам типа
зерноядной птицы, который с минимальны-
ми вариациями стал тиражироваться в сот-
нях видов, освоивших почти все местооби-
тания, достигших высокой численности и
расселившихся по всему Свету.
Что же касается плоцеидпой ветви, то про-
цессы фрипгиллизации здесь шли, вероятно,
более медленными темпами. Основным на-
правлением эволюции Ploceidae стала специ-
ализация, затронувшая технологию гпездост-
роепия, манеру расположения гнезд, соци-
альную организацию. Все эти адаптации по-
зволили ткачикам прочно утвердиться в со-
ставе тропической биоты Старого Света, где
они также достигают значительного видово-
го разнообразия и высокой численности.
В противоположность этому, воробьи и в
тропиках, и в умеренном поясе населяют наи-
менее благоприятные для жизни птиц арид-
ные и экстрааридные ландшафты с экстремаль-
но жарким или, наоборот, чрезвычайно холод-
ным климатом, по зато с ограниченным населе-
нием птиц и, как можно думать, низким уров-
нем межвидовой конкуренции. Другим убежи-
щем для воробьев послужили иптразональные
ландшафты и экотоны, существование в кото-
рых, по мнению ряда исследователей, связано с
необходимостью приспосабливаться к сильно-
му прессу хищников и гнездовых паразитов
(Gates, 1978; Brittingham, 1983; Moller, 1989;
Robinson, Suarez, Congdon, 1994).
Вероятно, уже к началу голоцена воробьи,
история которых к тому времени могла на-
считывать около 30-35 миллионов лет, пред-
ставляли собой глубоко специализированную
в экологическом отношении, маргинальную
группу, многие представители которой насе-
ляли реликтовые ареалы и были близки к
вымиранию. Ситуация резко изменилась все-
го за несколько тысячелетий — с точки зре-
ния биологической эволюции едва-ли не
мгновенно. Прогрессирующая антропогенная
трансформация коренным образом изменила
ландшафтный облик Старого Свела. Сложился
принципиально новый тип антропогенных
ландшафтов с чрезвычайно бедными (нена-
сыщенными) орнитоценозами, богатыми пи-
134
В. В. Иваницкий
щевыми ресурсами и пониженным прессом
хищников.
Отчего именно воробьи попали в число
пионеров освоения антропогенных ландшаф-
тов? Думается, здесь сыграли роль опреде-
ленные биологические черты этих птиц, став-
шие своего рода преадаптациями:
1) исконное тяготение воробьев к речным
долинам и другим интразональпым ланд-
шафтам предопределило быстрое расшире-
ние их первичных контактов с поселениями
человека;
2) способность гнездиться и ночевать в
закрытых убежищах открыла возможности
использования людских построек в качестве
экологического аналога привычных скальных
субстратов;
3) высокая степень социальности в тече-
ние всего года способствовала быстрому зак-
реплению и распространению этологических
адаптаций путем подражания и обучения
(см.Lefebvre, 1986; Monkkonen, 1993);
4) широкие трофические возможности
позволили воробьям использовать в пищу
семена культурных злаков и всевозможные
остатки со стола человека.
Вопреки распространенным представле-
ниям о деспециализации организмов, как не-
обходимой предпосылки их дальнейшей эво-
люции, Северцов (1984) обосновал мысль о
том, что новые адаптивные зоны могут ус-
пешно осваивать высокоспециализировапные
формы, ранее адаптированные к весьма спе-
цифическим условиям, которые выглядят
аберрантными по сравнению с экосистема-
ми, господствующими па данной территории.
Скорее всего, именно но такому сценарию
протекали завершающие этаны историчес-
кого развития сем.Passeridae. Как мы пола-
гаем, воробьи приступили к колонизации
антропогенных ландшафтов, будучи уже
явно маргинальной группой, вставшей на путь
специализации и оттесненной в аберрант-
ные местообитания биологически более про-
грессивными группами мелких зерноядных
птиц.
Ныне воробьи далеко продвинулись по
пути сипаптронизации. Разные представители
Passeridae независимо друг от друга в Африке,
Европе и Азии активно осваивают преобразо-
вашше человеком ландшафты, начиная от при-
митивных агроценозов и поселков аборигенов
и кончая предельно урбанизированными тер-
риториями современных мегаполисов (домо-
вый и полевой воробьи). При этом домовой
воробьей повсеместно но существу стал обли-
гатным комменсалом, нолевой воробей являет-
ся таковым на значительной части ареала, еще
несколько видов приближаются к этому {Passer
flaveolus в Малайзии, Р.castanopterus в Сома-
ли, P.iagoensis на островах Зеленого мыса,
P.melanurus в ЮАР). Среди И воробьев се-
верной Палеарктики лишь пустынный и ко-
роткопалый воробьи не связаны в той или иной
форме с антропогешшми ландшафтами — все
прочие виды более или менее регулярно ис-
пользуют их для гнездования, фуражировки,
ночевок и водопоя (Иваницкий, 19846). Рас-
ширение сельскохозяйственного использования
аридных зон, искусственное обводнение огром-
ных территорий, насаждение лесополос и са-
дов, посевы зерновых способствуют резкому
подъему численности и расселению многих ви-
дов (Summers-Smith, 1988).
ЗАКЛЮЧЕНИЕ
В настоящее время филогенетическое на-
правление в сравнительной этологии пере-
живает кризис и в значительной мере утра-
тило былую популярность. Приоритетное раз-
витие получили исследования этологических
изолирующих механизмов, рассматривающие
главным образом начальные стадии дивер-
генции поведения, или же исследования срав-
нительных аспектов поведенческих стратегий,
целиком сосредоточенные вокруг немногих
модельных видов. Что же касается филогене-
тического направления с его широким таксо-
номическим охватом, целостным взглядом на
поведение, плодотворными связями с систе-
матикой, зоогеографией и палеонтологией, то
оно оказалось едва ли не полностью забы-
тым (см. Панов, 1986; Rhijn, 1990, Panow, 1996).
Проблемы параллелизмов и конвергенции, го-
мологической изменчивости и нормы реак-
ции, целостности и мозаичности поведенчес-
кого облика видов редко обсуждаются в это-
логической литературе. Очень мало работ, рас-
сматривающих эволюцию поведения па отно-
сительно высоких уровнях дивергенции. Нам
видится в этом серьезная опасность элимина-
ции собственного предмета исследования
сравнительной этологии и ее превращения в
чисто прикладную дисциплину, предназначе-
ние которой сведется лишь к поиску новых
таксономических признаков.
Отсутствие "‘палеонтологического контро-
ля” в этологии определенным образом по-
влияло па структуру исследовательских про-
грамм, занимающих ныне доминирующие по-
зиции. Среди этологов распространилось
убеждение в том, что единственным надеж-
ным указанием на происхождение тех или
иных этологических признаков являются их
соответствие современным условиям обита-
ния видов. Что же касается вопроса о направ-
ленности вектора эволюционных изменений,
то его решению сплошь и рядом базируются
не на взвешенном анализе архаичности и
продвинутост и признаков, а на довольно про-
извольных, часто интуитивных и антропомор-
фных представлениях о “простоте” и “слож-
ности” изучаемых структур. Ограниченность
подобных представлений была убедительно
показана Пановым (1983), но и за минувшие
годы существенных сдвигов не произошло.
Представления об исторической преемствен-
ности структур, канализированности фило-
генеза, биологическом прогрессе, эволюции
функций, по-прежнему не находят употреб-
ления среди этологов, занятых скрупулезны-
ми подсчетами “достоинств” и “недостатков”
стратегий поведения.
Недавно Prum (1994) рассмотрел ситуа-
цию в сравнительной социоэтологии и при-
шел к выводу о том, что “большинство моде-
лей эволюции социального поведения, особен-
но позвоночных, совершенно антиисторичны”.
Это меткое замечание, на наш взгляд, не толь-
ко констатирует главную особенность совре-
менного состояния сравнительной этологии, по
н с определенностью указывает наиболее ве-
роятные пути дальнейшего прогресса этой
области. Возвращение к принципам историз-
ма представляется в этом смысле одним из
непременных условий.
При анализе главных тенденций в эволю-
ции воробьев мы стремились учесть все об-
стоятельства, способные пролить свет на ис-
торию группы, придавая особенное значение
тем из них, которые лежат за пределами соб-
ственно этологического материала (базовые
эколого-географические особенности, морфо-
логия, данные но экологически и таксономи-
чески близким группам и нр.). Вне конкрет-
но-исторического подхода и без учета всего
комплекса обстоятельств предложенные нами
трактовки могли бы выглядеть парадоксаль-
но, потому что практически но всем призна-
кам, в соответствии с нашими взглядами, век-
тор эволюции направлен от структур, интуи-
тивно воспринимаемых как “сложные”, к
структурам, воспринимаемым как “простые”.
Так, па смену колоннальности у воробьев
пришло дисперсное гнездование, бипаренталь-
ная система репродуктивного поведения сме-
нилась гинелатеральной, шаровидные гнезда
уступили место естественным убежищам и
чашевидным гнездам (короткопалый воробей),
у некоторых видов произошла глубокая ре-
136
В.В.Иваницкий
дукция акустических репертуаров, и даже в
эволюции окраски контурного оперения в
разных родах есть основания предполагать
параллельный переход от богатой пигмента-
ции и сложного рисунка к монохромной тус-
клой окраске.
Переходя от преимущественно древесно-
го образа жизни в саваннах к использованию
иных субстратов, местообитаний и ландшаф-
тов, воробьи приобрели сходство с предста-
вителями неродственных групп. Одним из его
источников могла стать конвергенция, обус-
ловленная приспособлением к одинаковым
экологических условиях и приведшая к по-
явлению ° гибридных" адаптивных типов “во-
робья-вьюрка” (^.Montifringilla) и “воробья-
жаворонка" (р. Pyrgilauda). Но конвергенция,
как всегда, затронула лишь “периферические"
признаки, которые прежде других реагируют
на изменения в условиях обитания. Это про-
порции тела, окраска, повседневное поведе-
ние, система локомоции и дистантной ком-
муникации. Между тем, сходство между про-
двинутыми формами воробьев и представи-
телями фрингиллидпой ветви зерноядных
птиц распространилось и на более интимные
сферы социо-репродуктивного поведения. Мы
предполагаем, что это сходство могло сложить-
ся в результате параллельной в обоих ветвях
трансформации исходного нлоцеидного типа
поведения во фрингиллидный тип. Главной
движущей силой этой перестройки стали, на
наш взгляд, уже не внешние условия, а внут-
ренние противоречия и конфликты, прису-
щие плоцеидной системе репродуктивного по-
ведения, которая была унаследована воробья-
ми от предковой формы.
Все же, вопреки конвергенции и парал-
лелизмам, воробьи в процессе эволюции в
полной мере сохранили свое своеобразие.
Наиболее устойчивыми оказались их склон-
ность к гнездованию в закрытых убежищах,
некоторые особенности полового и демон-
стративного поведения, акустической сигна-
лизации. Наблюдая за стайками снежных во-
робьев на кормежке или в полете, легко по-
нять, почему этих птиц называют снежны-
ми вьюрками. Но, как только птицы прини-
маются петь и токовать, их сходство с воро-
бьями и отличие от вьюрковых становятся
очевидными. Относительно полного “этоло-
гического разрыва" с предками достигли
лишь короткопалые воробьи. Об их принад-
лежности к группе Ploceimorpha и семей-
ству Passeridae уже невозможно судить толь-
ко по их поведению, и последнее слово здесь
остается за морфологическими и цитохими-
ческими методами.
ЛИТЕРАТУРА
Абдусалямов И.А. Фауна Таджикской ССР. T.XIX. Ч.З
Птицы. 1977. Душанбе: Изд-во Доншн. 273 с.
Адамян М.С. Об экологин короткопалого воробья
{Carpospiza brachydactyla) в Армянской ССР//
Зоол. жури. 1965. Т.44, вып. 4. С. 569-576.
Бабаев А.Г., Зонн И.С., Дроздов Н.Н., Фрейкин З.Г.
Пустыни. М.: Мысль, 1986. 316 с.
Бельская Г.С. Материалы к экологии туркестанского ка-
менного воробья в Туркмении// Фауна и экология
птиц Туркмении. 1973. Ашхабад: Ылым. С.34-54.
Бианки В.Л. Формы родов Montifringilla Brehm,
Pyrgilauda Verr, и Onychospiza Przev., ceM.Fringil-
lidac// Ежегодник Зоол. музея Академии Наук.
1907. Т. 12. С.555-597.
Благосклонов К.Н. Биология и сельскохозяйственное
значение палевого воробья в полезащитных насаж-
дениях юго-востока/ / Зоол. журн. 1950. Т.29. вып.З.
С.244-254.
Благосклонов К.Н. Гнездование и привлечение птиц в
сады п парки. М.: МГУ, 1991. 250 с.
Болотников А.М., Дьяконов Ю.В., Литвинов Н.А., Скры-
лсва Л.Ф. Происхождение и эволюция колониаль-
ного гнездования птиц / / Тсор. аспекты колониаль-
ное™ у птиц. М.: Наука, 1985. С. 11-13.
Болотников А.М., Печерский А.С. Насиживание, инкуба-
ция,// Полевой воробей. Л.: ЛГУ, 1981. С. 157-161.
Болотников А.М., Шураков А.И., Каменский Ю.И., Доб-
ринский А.Н. Экология раннего онтогенеза пищ.
Свердловск: УНЦ АН СССР, 1985. 226 с.
Брохович С.А., Гаврилов Э.И., Голощапов А.Б. О фор-
мировании нассдных пятен у самок черногрудого
и индийского воробьев на весеннем пролете в Ка-
захстане/ / Изучение птиц СССР, их охрана и ра-
циональное использование. 4.1. Л.: ЗИН АН СССР,
1986. С. 101-102.
Букреев С.А., Роздина О.Л., Леонович В.В. Взаимоот-
ношения домового и индийского воробьев в усло-
виях симбнотопии (юго-западный Копетдаг, Кара-
Кала) и отличительные черты их экологии / / Мат-
лы 10 Весе, орнптол. конф., Витебск, 17-20 сентября
1991 г. 4.2. кн.1. Минск, 1991. С.85-86.
Булатова Н.Ш. Положение вида в системе/ / Полевой
воробей. Л.: ЛГУ, 1981. С.6-8.
Вилке Е.К. Наши результаты изучения сложных форм
поведения птиц в естественных условиях / / Слож-
ные формы поведения. М., Л. 1965. С.130-133
Воробьев Г.П. Открытое гнездование домового воробья
{Passer domesticus) в городском ландшафте/ / Ор-
нитология. М.: МГУ, 1991. С. 182-183.
Волчанецкип И.Б. К изученino географической измен-
чивости рисунка и окраски птиц//Проблемы эво-
люции. Т.2. Новосибирск: Наука. 1972. С. 198-211.
Гаврилов Э.И. Биология испанского воробья {Passer
hispaniolensis Temm) и меры борьбы с ним в Ка-
захстане . Тр. НИИ зашиты растений Каз.ССР,
1962. вып.7. С.459-528.
Гаврилов Э.И. К экологии саксаульного воробья,// Ор-
нитология в СССР. Кн.2. Ашхабад. 1969. С. 155-158.
Гаврилов Э.И. СемействоТкачиковыс — Ploceidae/'/ Пти-
цы Казахстана. Т.5. Алма-Ата: Наука, 1974. С.363-406.
Гаврилов Э.И., Корелов М.Н. О видовой самостоятель-
ности индийского воробья//' Бюлл. МОИП. Отд.
биол. 1968. Т.73, вып.4. С. 115-122.
Таганская А.Р., Фетисов С.А., Шураков А.М., Болотни-
ков А.М., Нанкинов Д.Н. Сроки репродуктивного
цикла. Число циклов размножения / / Полевой во-
робей. Л.: ЛГУ. 1981. с. 133-138
Гвоздецкий Н.В., Голубчиков Ю.Н. Горы. М.: Мысль,
1987.399с.
Гептнер В.Г. Структура систематических групп и био-
логический прогресс/ / Зоол. журн. 1965. Т.44. Вып.9.
С.1291-1308.
Гилязов А.С., Семенов-Тян-Шанский О.И. Синантроп-
ные виды птиц Лапландского заповедника и его ок-
рестностей// Наземные позвоночные животные в
заповедниках севера европ. части РСФСР. М.:
ЦНИЛ Главохоты РСФСР, 1990. С.28-41.
Гладков Н.А., Дементьев Г.П., Птушенко Е.С., Судилов-
ская А.М. Определитель птиц СССР. М.: Высшая
школа, 1970.536 с.
Голованова Э.Н. Места гнездования и очаги вред-
ной деятельности перелетных воробьев в Тад-
жикистане// Орнитология. М.: МГУ, 1965. Вып.7.
С.464-466.
Голованова Э.Н. Мир птиц. Л.: Гидрометеоиздат, 1985.
176с.
Гаюванова Э.Н., Попов А.В. Наблюдения по естественной
гибридизации испанского и домового воробья/./ Мат-
лы по фауне наземных позвоночных животных Тад-
жикистана. Тр. Ин-та зоол. и паразитол. АН Тадж.ССР.
1962. Т.21. С.39-40
Гольцман М.Е. Социальное доминирование и социальная
стимуляция: частные проблемы и общие принци-
пы//' Системные принципы и этологические под-
ходы в изучении популяций. Пущино: НЦБИ АН
СССР. 1984. С. 108-134.
Датгупшн И.А., Гаврилов Э.И., Родионов Э.Ф. О гнездо-
вой биологии арчового дубоноса в Казахстане z / Тр.
Ин-та зоол. АН Каз.ССР. 1968. Т.29. С. 19-31.
Дольник В.Р. Ресурсы энергии и времени у птиц в
природе. Санкт-Петербург, Наука. 1995.359 с.
Доржиев Ц.З., Доржисва В.Д. Особенности гнездова-
ния полевого воробья в населенных пунктах Забай-
калья, / Фауна и экология позвоночных живот-
ных на территориях с разной степенью антропоген-
ного воздействия. М.. 1985. С. 21-34
Доржиев Ц.З., Юмов Б.О. Экология овсянковых птиц.
Улан-Удэ: Бурятское книжное издательство, 1991.
172 с.
Дубровский Ю.А. Заметки о распространении некото-
рых птиц в Актюбинских степях / / Тр. Ин-та зоол.
АН Каз. ССР. 1961. Т. 15. С. 192-197.
Елисеева В.И. Взаимоотношения между полевым воро-
бьем и мелкими дуплогнездниками при заселении
искусственных гнездовий/; Тр. Центрально-Чер-
ноземного зап-ка. 1960. Вып. 6. С. 321-331
138
В. В. Иваницкий
Елисеева В.И. О размножении полевого воробья в ис-
кусствсннных гнездовьях/ / Зоол. журн. 1961. Т.40.
№4. С. 583-591.
Ербасва М.А. Пищухи Кайнозоя. М.: Наука. 1988. 224с.
Заблоцкая М.М. Акустическая сигнализация коноплян-
ки Acanthis cannabina (L.). Препринт Ин-та бпол.
физики. Пущино: 1982. 41с.
Зарудный Н.А. Орнитологическая фауна Закаспийско-
го края. Мат-лы к познанию фауны и флоры Росс,
имп. Отдел зоол. 1896. Т.11. 555с.
Зембнцкая Л.В. Сравнительная характеристика биоло-
гии гнездования полевого Passer montan us и рыже-
го Passer rutilans воробьев на юге Сахалина// Био-
логия птиц юга Дальнего Востока СССР. Владивос-
ток. 1979. С. 87-89.
Зинченко Т.Н. Бюджет времени и энергии индийских
воробьев в репродуктивный период// Позвоноч-
ные животные Узбекистана. Ташкент: АН Узб. ССР,
1991. С. 103-111.
Зонов Г. Б. Методы изучения зимнего приспособитель-
ного поведения птиц/ / Методики исслсд. продук-
тивности и структуры видов птиц в пределах их
ареалов. Вильнюс: Мокслас, 1977. С. 49-55
Зонов Г.Б., Попов В.В. Зимующие птицы юго-западной
части Тувинской АССР/, Бюлл. МОИП. Отд. биат.
1987. Т.’ 92. №5. С. 55-63.
Зыкова Л.Ю. Поведение индийских воробьев {Passer
domes tic us indicus) в начальный период репродук-
тивного цикла/ / Зоат. журн. 1980. Т. 49. Вып. 10. С.
1500-1509.
Иваницкий В.В. О некоторых аспектах организации
внутрнпопуляционных отношений у воробьиных
птиц (Passeriformes)/ / Журн. Общ. Биат. 1981. Т.43.
№ 5. С. 708-720.
Иваницкий В.В. Поведение испанских {Passer
hispaniolensis) и индийских {Р. indicus) воробьев
при формировании колоний// Зоол. журн. 1984а.
Т. 55. Вып. 9. С. 1374-1387.
Иваницкий В.В. Филогенетические аспекты синантро-
пиэации и урбанизации воробьев (Passerinae) z /
Птицы и урбанизированный ландшафт (Сборник
кратких сообщений). Каунас: Ин-т зоол. и парази-
те л. АН Лит. ССР. 19846. С. 58-59.
Иваницкий В.В. О путях развития колониального гнез-
дования у ткачиков//' Тсор. аспекты колониально-
сти у птиц. Мат-лы совсрщ. по тсор. аспектам коло-
ииальности у птиц. М.: Наука, 1985а. С. 51-54.
Иваницкий В. В. Пространственная структура популя-
ции, территориальное поведение и полигиния у ка-
менных воробьев {Petronia petronia)// Биол. на-
уки. 19856. N 9. С. 50-56.
Иваницкий В.В. Социальная организация и демонстра-
тивное поведение испанских {Passer hispaniolensis)
н индийских {Р.indicus) воробьев/ / Зоол. журн.
1985в. Т. 64. Вып. 8. С. 1213-1223.
Иваницкий В.В. Сравнительный анализ поведения близ-
ких видов животных / / Методы исследований в эко-
логии и этологии. Пущино, 1986а. С. 141-156.
Иваницкий В.В. Структурно-функциональные и фило-
генетические аспекты репродуктивного поведения ка-
менных воробьев {Petroniapetronia)/ / Зоол. журн.
19866. Т. 65. Вып. 3. С. 387-397.
Иваницкий В.В. Рецензия на книгу D.J.Summers-Smith
“The sparrows. A study of the genus Passer”. 1988.
Calton: T and AD Poyscr// Новые книги за рубе-
жом. 1989. № 11. С. 32-38.
Иваницкий В.В. К изучению жизненных форм воробь-
ев// Мат-лы 10-й Всссоюзн. орнитол. конф. 4.1.
Минск: Навука i тэхжка, 1991а. С. 75-76.
Иваницкий В.В. Социальное поведение монгольских зем-
ляных Montifringilla [Pyrgilauda] и снежных
М. nivalis воробьев / / Зоат. журн. 19916. Т. 70. Вып.
9. С. 104-117.
Иваницкий В.В. О некоторых принципах координации
репродуктивной активности брачных партнеров у
моногамных птиц // Доклады РАН. 1995а. Т.343.
N? 1.С. 136-138.
Иваницкий В.В. Происхождение и распространение кла-
устрофилии воробьиных птиц: к вопросу об эво-
люции гнездостроения/ / Успехи современной био-
логии. 19956. T.115.NN». С.438-444.
Иваницкий В.В. Гнсздостроитслыюе поведение самцов
и самок индийского воробья {Passer indicus)/ /
Зоол. журн. 1996. Т.75. Вып. 2. С.249-255.
Иваницкий В.В., Матюхин А.В. О роли ночевок и свя-
занного с ними поведения в организации внутри-
популяцпонных отношений у разных видов воробь-
ев / / Тсор. аспекты колониальное™ у птиц. Мат-
лы совет, по тсор. аспектам колониальное™ v птиц.
М.: Наука. 1985. С. 54-59.
Иваницкий В.В., Матюхин А.В. К изучению коллектив-
ных ночевок и структуры популяций домовых во-
робьев {Passer domesticus) в сельской местности /
/ Биол. науки. 1990а. № 12. С. 50-56.
Иваницкий В.В., Матюхин А.В. К популяционной био-
логии полевых воробьев {Passer montanus) в го-
родских лесопарках / / Зоол. журн. 19906. Т. 69. Вып.
4. С. 78-89.
Иваницкий В.В., Матюхин А.В. Сравнительный анализ
постоянства персонального состава популяций раз-
ных видов воробьев (Passcridac) '/ Экология по-
пуляций: структура и динамика. М.: Типография
Росссльхозакадсмпи. 1995. С.546-560.
Иванинк1П1 В.В., Шевченко Е.В. К биологии монгольс-
кого земляного воробья в Туве и на Алтае/ Со-
временная орнитология. Т.2. М.: Наука, 1991. С. 30-47
Иванов Б.Е. Фенология на размножавансго на домаш-
него врабче {Passer domesticus (L.) в животновъд-
ни ферми край София/ / Екология (София). 1987.
Т. 20. С. 35-46.
Ильенко А.И. Конкуренция из-за мест гнездования в
популяции домовых воробьев/ / Зоол. журн. 1965.
Т. 44. Вып. 12. С. 1874-1878.
Ильенко А.И. Экология домовых воробьев и их экто-
паразитов. М.: Наука, 1976. 190 с.
Ильина Т.А. Роль энергетики в социальных взаимоот-
ношения птиц // Орнитология. М., МГУ, 1996.
Вып.27. С.98-126.
Ирисова Н.Л. Воробьиные птицы высокогорий Алтая
на пределе вертикального распространения. Авторсф.
дисс. насоиск. уч. степени канд. биол. наук. Новоси-
бирск: СО АН, 1990.26 с.
Ирисова Н.Л., Ирисов Э.А. Синантропизация птиц в вы-
сокогорье Алтая/' / Фауна, таксономия, экология мле-
копитающих и птиц. Новосибирск. 1987. С. 209-218
Казаков Б.А., Ломадзе Н.Х. О черногрудом воробьев
{Passer hispaniolensis Temm.) на Северном Кавка-
зе/ /' Орнитология. 1984. Т. 19. М.: МГУ. С. 179-180.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
139
Касссльтон П.Дж. Гнездящиеся птицы / / Сахара. М.:
Прогресс, 1990. С.266-280.
Кашкаров Д.Ю., Зинченко Т.Н. Проблема зерноядных
птиц в Средней Азми и некоторые пути ее реше-
ния// Изучение птиц СССР, их охрана н рацио-
нальное использование. Ч. 1. Л.: ЗИН АН СССР, 1986.
С.290-291.
Кскилова А.Ф. Материалы по экологии и сельскохо-
зяйственному значению некоторых видов воробьев
предгорий Копет-Дага// Фауна и экология птиц
Туркмении. Ашхабад: Ылым, 1973. С. 55-96.
Келейников А.А. Экология домового и патового воро-
бьев, как массовых вредителей зерновых культур в
южных районах СССР. Автореф. днсс. на соиск. учен,
степени канд. биол. наук. Москва: МГУ, 1953. 24 с.
Ковшарь А.Ф. Птицы Таласского Алатау. Алма-Ата:
Кайнар, 1966.433 с.
Ковшарь А.Ф. Певчие птицы в субвысокогорьс Тянь-
Шаня. Алма-Ата: Наука, 1979. 310 с.
Козлова Е.В. Филогенетические связи земляных и гор-
ных вьюрков родаMontifringilla / / Сборник, посвя-
щенный памяти П.П.Сушгата. М.-Л, 1950. С. 209-214.
Козлова Е.В. Авифауна Тибетского нагорья, ее родствен-
ные связи и история / / Тр. ЗИН АН СССР. 1952.
Т.9. Выл. 4. С.964-1028.
Козлова Е.В. Птицы зональных степей и пустынь Цент-
ральной Азии/ / Тр. ЗИН АН СССР. 1975. Т. 49.250 с.
Колоярцсв М.В. Ласточки. Л.: ЛГУ, 1989. 245 с.
Косенко С.М. Сравнительное изучение экологии золо-
тистой и зеленой щурок. Диес, на соиск. уч. степ. канд.
биол. наук. М., 1992. 188 с.
Костюнин В.М. О роли полевого воробья в поддержа-
нии природных очагов некоторых гсльминтозов в
окультуренных ландшафтах дельты р. Волги /VII
Всссоюз. орнитол. конф., Тезисы докладов. 4.2. Киев:
Наукова думка, 1977. С.77-78.
Курочкин Е.Н. К изучению птиц пустыни Гоби // Со-
временная орнитология. Т.2. М.: Наука, 1991. С. 25-29.
Левин А.С. Структура популяций большой синицы пой-
менных лесов долины реки Урал/ / Изучение птиц
СССР, их охрана и рациональное испапьэованпе. 4.2.
Л.: ЗИН АН СССР, 1986. С. 17-18.
Литвин К.Е. Взаимосвязь стайного поведения и колони-
альное™ у птиц / /Теоретические аспекты колони-
альности у птиц. М., Изд-во МОИП, 1985. С.83-86.
Лоскот В.М. Материалы по птицам окрестностей Та-
шанты// Распространение и биология птиц Алтая
и Дальнего Востока. Тр. ЗИН АН СССР, 1986. Т.
150. С. 44-56
Лукина Е.В. Экзопгческие птицы в нашем доме. Л.: ЛГУ,
1986.296 с.
Лэк Д. Дарвпновы вьюрки. М., Изд-во иностранной ли-
тературы, 1949. 200 с.
МайрЭ. Принципы зоологической систематики. М.: Мир,
1971.454 с.
Манучарян Г. Г. Биология кавказского домового воро-
бья // Изв. АН Армян. ССР. Биат. науки, 1963. Т. 16.
вып. 3. С. 77-84.
Матюхин А.В., Иваницкий В.В. О зимнем размноже-
нии домовых воробьев в Московской области//
Птицы и урбанизированный ландшафт (Сборник
кратких сообщений). Каунас, 1984. С. 95-96.
Матюхин А.В., Матюхина Т.Г., Любущенко С.Ю. К био-
логии гнездования индийского воробья в юго-вос-
точном Казахстане: социальная организация и ус-
пех размножения/, ' Современ. проблемы изучения
колониальное™ у птиц. 1990.
Михайлов К.Е. Некоторые случаи нетипичного гнездо-
вания птиц в Гоби / / Современная орнитология
1991. М.: Наука, 1992. С.125-144.
Морозов В.И., Остапенко В.А. Ткачихи. М.: Лесная про-
мышленность, 1988. 143 с.
Мустафаев Г.Т. К экологии трканского домового во-
робья в Талышс// Мат-лы 5-й Всссоюзн. орнитол.
конф. 1969. Вып.2. С.433-435.
Нейфельдт И.А. Пуховые птенцы некоторых азиатских
птиц/ / Орнитологический сборник к 100-летию со
дня рождения П.П.Сушкина. Тр. ЗИН АН СССР.
1970. Т.47. С.111-181.
Нейфельдт И. А. Из результатов орнитологической экс-
педиции на юго-восточный Алтай// Распростра-
нение и биология птиц Алтая и Дальнего Востока.
Тр. ЗИН АН СССР. 1986. Т.150. С.7-43.
Нейфельдт И.А. О неправомерности переименования
рода Pyrgilauda Verreaux, 1871 (Ploccidae, Aves)/
/ Там же. С. 171-173.
Нечаев В.А. Птицы южных Курильских островов. Л.:
Наука, 1969. 246 с.
Нечаев В. А. Рыжий воробей (Passer rut Hans) на Саха-
лине// Зоол. журн. 1973. Т.52. Вып. 7. С. 1033-1038.
Паевский В.А. Демография птиц. Л.: Наука, 1985. 285 с.
Панов Е.Н. Механизмы коммуникации у птиц. М.: На-
ука. 1978.303с.
Панов Е.Н. Поведение животных и этологическая струк-
тура популяций. М.: Наука, 1983. 423с.
Панов Е.Н. Гибридизация и этологическая изоляция у
ггпщ. М.: Наука, 1989. 510с.
Панов Е.Н., Раджабли С.И. Взаимоотношения домово-
го воробья (Passer domesticus) и черногрудого во-
робья (P.hispaniolensis) в Таджикистане и возмож-
ные механизмы изоляции// Проблемы эволюции.
1972. Т.2. Новосибирск: Наука. С.263-276.
Поликарпова Е.Ф. Влияние внешних факторов на раз-
витие половой железы воробья / / ДАН СССР. 1940.
T.26..N? 1. С.91-95.
Пономарева Т.С. Материалы по гнездовой биологии пу-
стынного воробья / / Орнитология. 1983. Вып. 18. М.:
МГУ. С.57-63.
Портенко Л.А. Птицы 4укотского полуострова и остро-
ва Врангеля. Л.,: Наука, 1973. Т.2. 323с.
Попов А. В. Воробьи — вредители зернового хозяйства
в Таджикистане и их истребление// Мат-лы по
фауне и экатогии позвоночных животных Таджи-
кистана. Тр. Ин-та зоол. и паразита!. Таджикистана.
1962. Т.22. С.26-38.
Потапов Р.Л. Очерк летней орнитофауны заповедника
“Тигровая балка” // Заповедник “Тигровая балка”.
Тр. Ин-та эоат и паразита!. Таджикистана. 1959. Т.15.
С.179-201.
Потапов Р.Л. Птицы Памира/ / Биология птиц. Тр. ЗИН
АН СССР. 1966. Т.31. Л. С. 1-119.
Птушенко Е.С., Иноземцев А.А. Биология и хозяйствен-
ное значение птиц Московской области и сопре-
дельных территорий. М.. Изд-во МГУ, 1968. 461 с.
Пузанов И.И. Зоогеография. М. Изд-во Наркомпроса
РСФСР. 1938.358 с.
Ревякин В.С. (ред.) Алтайский край. М.: Профнздат,
1987.259 с.
140
В. В. Иваницкий
Ротишльд Е.В. Совместное гнездование воробьев и хищ-
ных птиц в пустыне / / Зоол. журн. 1959. Т.38. Вып.
6. С.935-938.
Рустамов А.К. Птицы Туркменистана. Т.2. 1958. Ашха-
бад. 252 с.
Рябицсв В.К. Повторные кладки и бициклия у птиц
Ямала/ / Экология. 1987. №2. С.63-68.
Рябицев В.К. Территориальные отношения и динамика
сообществ птиц в Субарктике. Екатеринбург: Наука,
1993. 296 с.
Северное А.Н. Морфологические закономерности эво-
люции. М.: Изд-во АН СССР. 1939.
Северное А.С. Специализация как основа происхожде-
ния таксонов надвндового ранга// Журн. Общей
Биолопш. 1984. Т.45. № 5. С.586-595.
Семашко Л.Л. Полевой и домовый воробьи как носите-
ли клещей в Ашхабаде. Сообщ.1 / / Зоол. журн. 1959.
Т.38. Вып. 9. С. 985-991.
Семашко Л.Л. Полевой и домовый воробьи как носите-
ли клещей в Туркмении. Сообщ.2// Зоол. журн.
1961. Т.40. Вып. 10. С.1121-1129.
Семашко Л.Л. К изучению экологии домового воробья
в Южной Туркмении// Тр. Ашхабадского НИИ
эпидемиологии п гигиены. 1962. Т.5. С.225-233.
Сидоров Б.И. Материалы по размножению полевого во-
робья в Центральной Якутии / / Экология назем-
ных позвоночных таежной Якутии. Изд-во Якутско-
го ун-та. Якутск, 1984. С. 17-29.
Симкин Г.Н. О происхождении и эволюции колони-
альное™ у птиц / / Орнитология. 1988. Вып.23. Изд-
во МГУ. С.36-51.
Соколов Л.П. Филопатрия и дисперсия птиц. 1991. Л.,
Тр. ЗИН АН СССР. Т.230. 225 с.
Сопысв О. Пустынный воробей в Каракумах / / Орни-
тология. Вып.7. 1965. Изд-во МГУ. С. 134-140.
Сопысв О. О размножении некоторых птиц Караку-
мов? Орнитология. Вып.8. Изд-во МГУ, 1967.
С.221-235.
Сопысв О. О размножении саксаульного воробья в Во-
сточных Каракумах / / Животный мир Туркмении.
Ашхабад: Ылым, 1970. С.74-83.
Степанян Л.С. О зимней орнитофауне Восточного Па-
мира// Орнитология. Вып.11. 1974. Изд-во МГУ.
С.417.
Степанян Л.С. Конспект орнитологической фауны СССР.
М : Наука, 1990.726 с.
Судиловская А.М. О расселении домашнего воробья ?
/ Бюлл. МОИП. Отд. биол. 1952. Т.57. № 5. С.33-37
Сушкин П.П. Птицы Советского Алтая и прилежащих
частей северо-западной Монголии. М.-Л. 1938.
Тарасов М.П. Материалы по фауне птиц юго-восточной
части Монгольского Алтая и прилежащей Гоби//
Биологический сборник. Иркутск, 1960. С. 159-178.
Третьяков Г.П. К экологии пустынного воробья /,' Ред-
кие и малоизученные птицы Средней Азии. Мат-лы
3-й Рсспубл. орнитол. конф. Бухара, 1990. С.79-80.
Умрихина Г. Материалы о воробьях Чуйской долины / /
Уч. зап. Бполого-почвен. ф-та Киргиз, гос. ун-та. 1955.
Вып.5. С. 119-124.
Федянина Т.Ф. О взаимоотношениях обыкновенной гал-
ки и черногрудого воробья в летний период в Чуй-
ской долине/ / Тезисы докл. 7-й Всссоюзн. орнитол.
конф. 4.1. 1977. С.331-332.
Фетисов С.А. О степени постоянства брачных пар у
полевого воробья / / X Прибалт, орнитол. конф. Те-
зисы докл. Т. 2. 1981. С.192-194.
Фетисов С.А. Фенология размножения полевого воро-
бья в Ленинградской области// Экология птиц в
репродуктивный период. Л., 1985. С.39-51.
Фетисов С.А., Гагинская А.Р. и др. Гнсздостроснис ' '
Полевой воробей. Л.: ЛГУ, 1981. С.139-143.
Фетисов С.А., Носков Г.А. и др. Жилищные связи//
Полевой воробей. Л.: ЛГУ, 1981. С.246-251.
Фроиде М. Животные строят. М.: Мир, 1986. 210 с.
Формозов А.Н. Заметки об экологии и сельскохозяй-
ственном значении воробьев (Passer domesticus
bactrianus Zar. et Kudach, Passer montanus pallid us
Zar.) в Южной Туркмении/ / Зоол. журн. 1944. Т.23.
Вып. 6. С.342-350.
Храбрый В.М. О гнездовании домового воробья на де-
ревьях в Ленинграде/ / Экология гнездования птиц
и методы ее изучения. Самарканд, 1979. С.228-229.
Шестакова Г.С. К систематике и генетическим взаимо-
отношениям овсянок// Сборник памяти академи-
ка М.А.Мензбира. М.-Л.: Изд-во АН СССР, 1937.
С.597-627.
Шестоперов Е.Л. Воробьи Туркменистана и их эконо-
мическое значение / / Отчет о деятельности отдела
защиты растений за 1924/25 и 1925/26 гг. 1926. Аш-
хабад.
Шилова С.А. Популяционная экология как основа кон-
троля численности мелких млекопитающих. М.: На-
ука. 1993. 201 с.
Шилова С.А., Седых Э.Л. и др. Особенности биоцено-
зов горной щебнистой степи Тувы в связи с про-
блемой природопользования/ / Экология. 1977. №
3. С.55-62.
Шн1ггнпков В.Н. Птицы Семиречья. М.- Л.: АН СССР,
1949.665 с.
Штегман Б.К. Воробьи в Казахстане и изыскание мер
борьбы с ними z / Зоол. журн. 1956а. Т.35. Вып. 8.
С.1203-1213.
Штегман Б.К. Воробьи в Казахстане, их вредная дея-
тельность и меры борьбы с ними/ / Тр. рсспубл.
станнин запцггы раст. Казахст. фил. ВАСХНИЛ.
19566. Т.З. С.110-146.
Шульппн Л.М. Материалы по фауне птиц зхаповеднп-
ка Аксу-Джабаглы/ / Тр. Ин-та зоологии АН Каз.
ССР. 1956. Т.6.
Юмов Б.О. Сравнительная экология фоновых видов
овсянок западного Забайкалья в начальный период
розмножения / Экология и население птиц. 1985.
Иркутск. С. 142-162.
Abs М. Contribution to systematic problems and geograph-
ic variation within the genus Petronia (Arcs, Plo-
ccidac) / / Ostrich. 1966. Suppl.6. p.41-49.
Alvarez R.S., Gonzales P.J.A. Variacion intcranual de la
nidificacion de aves en cajas anidadcras en un cncinar
adehesado v en bosques de rebel lo ? / Ecologia. 1990.
N4. C.213-221.
Ambcdcar V.C. On the breeding biology of the black-throat-
ed (Ploceus be ng a lens is) and the streaked (Ploceiis
manyar) weaver birds in the Kumaon Tcrai//Journ.
Bombay Nat. Hist. Soc. 1972. Vol.69. P.268-282.
Anderson W. Experimental synchronization of sparrow repro-
duction / Wilson Bull. 1979. Vol.91. N 2. P.317-319.
Andrew R.J.The aggressive and courtship behaviour of cer-
tain Embcrizincs ' / Behaviour. 1957. Vol. 10. P.255-308.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
141
Andrew R.J. The displays given by Passerines in courths-
hip and reproductive fighting: a review. Part \//
Ibis. 1961. Vol. 103 (a). N3. P.315-348.
Ar A., Yom-Tov Y. The evolution of parental care in birds/
/ Evolution. 1978. Vol. 32. P.655-669.
Archer G., Godman E.M. The birds of British Somaliland
and the Gulf of Aden. Edinburg. London. 1961. Vol.4.
P. 1042-1570.
Ash J.S., Nikolaus G. Desert sparrows Passer simplex in
the Sudan/ / Bull. Brit. Ornithol. Club. 1991. Vol.111.
N 4. P.237-239.
Baker S.E.C. The fauna of British India. London. 1926.
Vol.3. Birds. 489 p.
Balzac H. de Remarques rur 1’cthologic de Passer sim-
plex// Alauda. 1929. Vol.1. P.68-77.
Bannerman D.A. The birds of tropical west Africa. London.
1948. Vol.6. 364 p.
Baumgardt W. Zur Charaktcrisicrung von Haus- und
Wcidcnspcrling, Passer domesticus und Passer hispan-
iolensis, als “zcitdiffcrcntc arten”// Bcitr. zur
Vogclkundc. 1984. Bd. 30. N 4/5. S.217-242.
Barnard C.J. Equilibrium flock size and factors affecting
arrival and departure in feeding house sparrows/ /
Anim. Bchav. 1980a. Vol. 28. N 2. P.503-511.
Barnard C.J. Factors affecting flock size mean and variance
in winter population of house sparrows (Passer domesti-
cus)// Behaviour. 1980b. Vol. 74. N 1-2. P.114-127.
Barnard C.J. Flock feeding and time budgets in the house
sparrow (Passer domesticus)/ / Anim. Bchav. 1980c.
Vol.28. P.295-309.
Bcllcs-Islcs J.C., Pieman J. House wren nest-destroying
behaviour/ / Condor. 1986a. Vol.88. P. 190-193.
Bcllcs-Islcs J.C., Pieman J. Destruction of hctcrospccific
eggs by the gray catbirds/ / Wilson Bull. 1986b. Vol.98.
P.603-605.
Bergmann H.H., Hclb H.W. Stimmcn dcr Vogel Euro-
pas// 1982. BLV Vcrlagcscllschaft Munchen, Wien,
Zurich. 415 Bd.
Betts F.N. Notes on some resident breeding birds of south
west Kenya// Ibis. 1966. Vol. 108. N 4. P.513-530.
Bock W.J.,Morony J. J. Relationships of the passerine finch-
es (Passeriformes: Passcridac)// Bonn. Zool. Bcitr.
1978. Bd.29. N 1-3. S.122-147.
Brittingham M.C., Temple S. A. Have cowbirds caused forest
songbirds decline?// Bioscicncc. 1983. Vol.33. P.31-35.
Brosset A. Social organization and nest building in the
forest weaver birds of the genus Malimbus (Plocein-
ae)// Ibis. 1978. Vol.120. N 1. P.27-37.
Bulatova N.S. A comparative karyological study of passe-
rine birds// Acta Sei. Nat. Brno. 1981. Vol. 15. N 3.
P.1-44.
Busse P., Gotzman J. Nesting competition and mixed clutch-
es among some birds inhabiting the nest-holes/ / Acta
ornithol. 1962. Vol.7. P.1-32.
Cink C.L. The influence of early learning on nest site se-
lection in the house sparrow (Passer domesticus)/ /
Condor. 1976. Vol.78. N 1. P.103-104.
Channing A., Mac Lean G.L. Analysis of vocalizations of
the sociable weaver/ / Ostrich. 1976. Vol. 47. N4. P.223-
225.
Chapin J.P. The classification of the weaver-birds/ / Bull.
Amer. Mus. Nat. Hist. 1917. Vol. 37. P.243-280 (цит. no
Bock ct Morony, 1978).
Chose R.K. Behaviour of the ycllowthroatcd sparrow Pet-
ronia xanthocollis/ / Journ. Zool. Soc. India. 1973.
Vol.25. N 1-2. P.141.
Clement R., Harris A., Davis J. Finches and sparrows. An
identification guide. A-C Black — London. 1993. 500 p.
Clutton-Brock Т.Н. The evolution of parental care. Princ-
eton Univ.Prcss, Princeton, New Jersey. 1991. 352p.
Collias E.C., Collias N.E. The development of nest build-
ing behaviour in a weaverbird// Auk. 1964. Vol.81.
N 1. P.42-52.
Collias N.E. A spectrographic analysis of the vocal reper-
toire of the african village weaverbird// Condor. 1963.
Vol.65. N6. P.517-527.
Collias N.E. Engineering aspects of nest building by
birds// Euro-Article, Endevour, New Series. 1986.
Vol. 10. N 1. P.9-16.
Collias N.E., Brodman M., Victoria J.K., Kiff L.F., Rischer
C.E. Social facilitation in weaverbirds: effect of vary-
ing sex ratio/ / Ecology. 1971. Vol. 52. N 5. P.829-836.
Collias N.E., Collias E.C. Evolution of nest-building in
the weaverbirds (Ploceidae)/ / Univ. Californ. Publ.
Zool. 1964. N 73. 199 p.
Collias N.E., Collias E.C. Size of breeding colony related
to attraction of mates in tropical passerine birds/ /
Ecology. 1969. Vol.50. P.481-488.
Collias N.E., Collias E.C. Behaviour of the west african vil-
lage weaver bird ' / Ibis. 1970. Vol. 112. N 4. P.457-480.
Collias N.E., Collias E.C. Weaverbird nest aggregation
and evolution of the compound nest//7 Auk. 1977.
Vol.94.N1. P.50-64.
Collias N.E., Collias E.C. Cooperative breeding behaviour
in the white-browed sparrow weaver// Auk. 1978 a.
Vol. 95. P.472-484.
Collias N.E., Collias E.C. Nest building and nesting be-
haviour of the sociable weaver Philetairus socius//
Ibis. 1978b b. Vol. 120. N 1. P.1-15.
Collias N.E., Collias E.C. Behaviour of the gray-capped
social weaver (Pseudonigritta amaudi) in Kenya//
Auk. 1980 a. Vol. 97. N 2. P.213-226.
Collias N.E., Collias E.C. Individual and sex differences
in behaviour of the sociable weaver Philetairus so-
cius/ / Proceed. 4-th Pan-African Ornithol. Congr.,
Seychelles 1976. 1980 b. P.243-251.
Collias N.E., Collias E.C. Nest building and bird behav-
iour. Princcnton, Princeton Univ. Press. 1984. 356 p.
Collias N.E., Victoria J.K. Nest and mate selection in the
village weaverbird (Plioccus cucullatus) z / Anim. Bc-
hav. 1978. Vol. 26. N 2. P.470-479.
Collias N.E., Victoria J.K., Schallcnberg R.J. Social facil-
itation in weaverbirds: importance of colony size//
Ecology’. 1971. Vol.52. N5. P.823-828.
Cordero P.J., Sanchez-Aquado F.J. Jovcncs de gorrion mo-
linero (Passer montanus) parasitando nidos des su pro-
piacspecic// Misccl. Zool. 1988. Vol. 12. P.385-386.
Franz D., Theib N. Brutbiologic und bestandsent wick lung
ci net farberingten population dcrbcutelmcise Remiz
pendulinus / / Verh. omitol. Ges. Bayern. 1982-1983.
Bd. 23. N 5-6. S.393-442.
Friedmann H. The parasitic weaverbirds/ / Bull. U.S. Nat.
Mus. 1960. Vol.223. p.1-196.
Craig A.J.F. Vocalization of male Euplectes in captivity/
/ Z. Ticrpsychol. 1976. Vol. 41. P.191-201.
Craig A.J.F. Behaviour and evolution in the genus
Euplectes / Journ. Ornithol. 1980. Vol. 121. N 2.
P.144-161.
142
В.В.Иваницкий
Crook J.H. Etudes sur lc comportment social de Bubalor-
nis a. albirostris (Vicillot)// Alauda. 1958. Vol. 26.
N 3. P.161-195.
Crook J.H. Nest form and construction in west african
weaver birds// Ibis. 1960 a. Vol. 102. N 1. P.1-25.
Crook J.H. Studies on the reproductive behaviour of the
baya weaver (Ploceus phillipinus)/ / Journ. Bombay
Nat. Hist. Soc. 1960 b. Vol. 57. N 1. P.1-44.
Crook J.H. Studies on the social behaviour of Quelea q.
quelea in French West Africa// Behaviour. 1960 c.
Vol. 16. P.1-55.
Crook J.H. The foudics (Ploccinac) of the Seychelles Is-
lands // Ibis. 1961. Vol. 103 a. N4. P.517-548.
Crook J.H. The Asian weaver birds: problems of coexist-
ence and evolution with particular reference to be-
haviour// Journ. Bombav Nat. Hist. Soc. 1963 a. Vol.
60.N1.P. 1-48.
Crook J.H. A comparative analysis of nest structure in the
weaver birds (Ploccinac)/ / Ibis. 1963 b. Vol. 105. N
2. P.238-262.
Crook J.H. Comparative studies on reproductive behav-
iour of two closely related weaver bird species (Plo-
ceus cucullatus and Ploceus nigerrimus) and their
races/ / Behaviour. 1963 c. Vol. 21. N 3-4, P. 177-232.
Crook J.H. Monogamy, polygamy, and food supply//
Discovery. 1963 d. Vol. 24. N 1. P.35-41.
Crook J.H. The evolution of social organization and visu-
al communication in the wcaverbirds Ploccidac// Be-
haviour. 1964. Suppl.10. 178 p.
Deckcrt G. Zu г Ethologic des feldsperlings (Passer monta-
nus) / /Journ. Omithol. 1962. Bd. 103. N4. S.428-486.
Deckcrt G. Dcr Fcldspcrling/ / DicNcue Brehm-Buchcrci 398.
1973. A. Ziemsen Vcrlag. Wittenberg Lutheistadt. 90 S.
Densley M. Desert sparrow in Marocco// Brit. Birds.
1990 a. Vol. 83. N 5. P.195-201.
Denslcv M. Saxaul sparrow in Mongolia// Dutch Bird-
ing. 1990 b. N 12. P.5-9.
Elzen R., Ncmesochkal H.L. Radiation in African canaries
(Carduelidac): a comparizon of different classifica-
tion approaches / / Acta 20 Congr. Internet. Omithol.,
Chrischurch, 2-9 Dee., 1990. Vol.l. Wellington. 1991.
P.459-467.
Erhcldirung-Denk, Trillmich, 1990 Das Mikroklima in Nist-
kasten und seine Auswirkungen auf die Ncstlingc heim
Star (Stumus vulgaris')// J. Omithol. 1990. Bd. 131.
S. 73-84.
Immclmann K. Be it rage zu cincr vcrglcichcndcn Biologic
australischcr Prachtfinkcn (Spermestidae)// Zool.
Jahrb. Abt. 3. 1962. Vol. 90. N 1. 196 S.
Immclmann K. Brutbiologischc Bcobachtungcn am Kapsper-
ling (Passer melanurus) im sudlichen Afrika/ / Bc-
itr. Vogclk. 1970. Bd.16. S. 195-203.
Indykicwicz P. Nest and nest-sites of the house sparrow
Passer domesticus (Linneus, 1758) in urban, suburban
an rural environments/ / Acta Zool. Cracov. 1991. Vol.
34. N 7. P.475-495.
Ivanitskii V.V. Social ethology of sparrows (Passeridae)
/ / Phvsiol. and General Biol. Rev. 1996. Harwood
Acad. Publ. P.1-65.
Ivanitskii V.V. Evolution of sociability, formation, and
structure of monospecific and mixed colonies of spar-
rows from the subfamily Passcrinae// Acta 18 Con-
gr. Intemat. Omithol., Moscow, August, 16-24,1982.1985.
Moscow, Nauka. P.900-904.
Ivanitskii V.V. Classification and phylogeny of the spar-
rows (Passeridae)/ / The Omithol. Notebook of the
21 Intcrnat. Omithol. Congr., Vienna, August, 20-25,
1994. 1994. P.580.
Ivanitskii V.V. Social ethology of sparrows (Passeridae)/
/ Gates J.E., Gyscl L.W. Avian nest dispersion and
fledging success in field-forest ecotones// Ecologv.
1978. Vol. 59. P.871-883.
Goodwin D. Estrildid finches of the world. Oxford Univ.
Press. 1982. 328 p.
Grant P.R. Ecology and evolution of Darwin’s finches.
Princcnton Univ. Press, 1986. New Jersey. 458 p.
Guttingcr H.R. Zur Evolution von Vcrhaltensweiscn und
Lotaubcruungen bci Prcchfinkcn (Estrildidae)/ / Z.
Ticrpsychol. 1970. Bd.27. N9. S.1011-1075.
Hall B.P., Moreau R.E. An atlas of speciation in African
passerine birds. London. 1970. Brit. Mus. Nat. Hist.
Harrison C.J.O. The nest advertisement display as a Pass-
er /Ploccidac link/ / Bull. Brith. Om. Club. 1965. Vol.
85. P.26-30.
Harvey N.C., Hart E.W., Lcctc A.J. Changes in incubation
sharing in one pair of captive califirnia condor (Gym-
nog у ps califomionus)/ / Zoo Biol. 1994. Vol. 13. N 2.
P. 157-165.
Hegner R.E., Wingfield J.C. Behavioural and endocrine
correlations of multiple breeding in the semicolonial
house sparrow Passer domesticus. Males / , z Hormones
and bchav. 1986 a. Vol. 20. N 3. P.294-312.
Hegner R.E., Wingfield J.C. Gonadal development dur-
ing autumn and water in house sparrows// Condor.
1986 b. Vol. 88. N 3. P.269-278.
Hegner R.E., Wingfield J.C. Effects of experimental ma-
nipulation on testosterone levels on parental inbvest-
ment and breeding success in male house sparrows /
Auk. 1987 a. Vol. 104. N 3. P.462-469.
Hegner R.E., Wingfield J.C. Social status and circulating
levels of hormones in flocks of house sparrows. Passer
domesticus // Ethology. 1987 b. Vol. 76. P.1-14.
Hinde R.A. The nest-building behaviour of domesti-
cated canaries/ / Proc. Zool. Soc. London. 1958.
Vol. 131. P.1-48.
Hinde R.A. Interaction of internal and external factors in
integration of canary reproduction// Sex and be-
haviour. Wiley, New York. 1965.
Hubert C., Carlier P. Etude comparative des relations
malc-fcmcllc chcz la buse variable Buteo buteo ct
chcz la faucon pelcrin Falco peregrinus au mo-
ment des eclosions/ / Cahiers d’Etol. Vol. 12. N
4. P.491-496.
Johnson R.F. Aggressive foraging behaviour in house spar-
rows / Auk. 1969. Vol. 86. P. 558-559.
Jones P.J., Ward P. A physiological basis for colony de-
sertion by red-billed quclcas (Quelea quelea) / ,
Journ. Zool.(London). 1979. Vol. 189. P.1-19.
Kalinoski R. Intra- and interspecific aggression in house
finches and house sparrows / / Condor. 1975. Vol. 77.
N 4. P.375-384.
Kcndcigh S.C. Parental care and its evolution in birds / /
Illinois Biol. Monogr. 1952. N 22. 356 p.
Kcndcigh S.C. Energy of birds conserved bv roosting in
cavities / / Wilson Bull. 1961. Vol. 73. P. 140-147.
Kendra P.E.R., Roth R., Tallamy D.W. Conspecific brood
parasitism in the house sparrow/ / Wilson Bull. 1988.
Vol. 100. P.80-90.
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
143
Kcm М., Knapic М. Short-term variations in water-vapor
pressure in nest of common canaries/ / Condor. 1991.
Vol. 93. N 3. P.768-773.
Krementz D.G., Ankney C.D. Bioenergetics of eggs pro-
duction bv female house sparrows / / Auk. 1986. Vol.
103. P.299-305.
Kunkel P. Allgemeeincs und soziales Vcrhaltcn des Braun-
rucken-Goldspcrling {Passer (Auri passer) luteus)/
Z. Ticrpsychol. 1959/61. Bd. 18. S.471-489.
Lefebre L. Cultural transmission of a novel food-finding
behaviour in urban pigeon: an experimental field test/
/ Ethology. 1986. Vol. 71. P.295-304.
Lewis D.M. Cooperative breeding in a population of white-
browed sparrow-weaver Plocepasser mahali/ / Ibis.
1982. Vol. 124. P.511-522.
Lohrl H. Hohlecnkonkurrenz und Hcrbst-Ncstbau bcim
Fcldspcrling (Passer montanus)// Vogelwclt. 1978.
Bd.99. S.121-131.
Lowther P.E. Overlap of the house sparrow’ broods in the same
nest / / Bird-Banding. 1979. Vol. 50. N 2. P.160-162.
Mac Lean G.L. The sociable w’cavcr/ / Ostrich. 1973. Vol.
44. N 3-4. P.176-261.
Massa R. Comments on Passer italiae (Viellot, 1817)//
Bull. Brith. Ornithol. Club. 1989. Vol. 109. P.196-198.
Мауг E. Die schnccfinkcn (Gattungen Montifringilla und
Leucosticte)/ / J.Ornithol. 1927. Bd. 75. S. 596-619.
McLacham and Liversidge. Roberts birds of South Africa.
London. 1957. 495 p.
Me Kitrick M.C. Phvlogcnetic analysis of avian parental
care // Auk. 1992. Vol. 109. N 4. P.828-846.
Me Gillvray B.W. Communal nesting in the house spar-
row’// Journ. Field Omitol. 1980 a. Vol. 51. N 4. P.371-
372.
Me Gillvray B.W. Nest grouping and productivity in the
house sparrow’/' / Auk. 1980 b. Vol. 97. N 4, P.396-399.
Me Gillvray B.W. Intraseasona) reproductive costs for the
house sparrow (Passer domesticus) / / Auk. 1983. Vol.
100.N1.P. 25-32.
Meinertzhagen C.R. Birds of Arabia. London. 1954. 624 p.
Mctzmachcr M. La densite des nichcurs influcncc-t-clle
Г activity des moincaux cspagnolc (Passer hispanio-
lensis) / / Aves. 1989. Vol. 26. N3-4. P.159-168.
Moller A.P. House sparrow, Passer domesticus, communal
displays z / Anim. Bchav. 1987 a. Vol. 35. P. 203-210.
Moller A.P. Social control of deception amon status sig-
nalling house sparrows Passer domesticus// Bchav.
Ecol. Sociobiol. 1987 b. Vol. 20. P.307-314.
Moller A.P. Nest site selection across ficld-w'oodland eco-
tones: the effect of nest predation / / Oikos. 1989. Vol.
56. P.240-246.
Moller A.P. Sexual behaviour is related to badge size in
the house sparrow Passer domesticus/ / Bchav. Ecol.
Sociobiol. 1990. Vol. 27. N 1. P.23-29.
Moller A.P. Clutch size, nest predation, and distribution
of avian unequal competitors in a patchv environ-
ment/ / Ecology. 1991. Vol. 72. N 4. P. 1336-1349.
Monkkoncn M., Koivula K. Ncophobia and social learning
of foraging skills in willow tits Parus montanus/ /
Ardca. 1993. Vol 81. N 1. P.43-45.
Moreau R.E. The comparative breeding ecology of two
species of Euplectes (bishop birds) in Usambaraz /
Journ. Anim. Ecol. 1938. Vol. 7. N 2. P.314-327
Moreau R.E. The ploccinc weavers of the Indian Ocean
islands // Journ. Omith. 1960 c. Bd. 101. S.29-49
Morel M.Y., Morel G. Premieres observations sur la reproduc-
tion du maineau dore Passer luteus cn zone semi-aride de
I’qucst Afrique// Oiscau. 1973. Vol. 43. P.96-118.
Naik R.M., Mistry L. Breeding season in tropical
population of the house sparrow’ // Journ. Bombay
Nat. Hist. Soc. 1970 (Suppl), p 1118-1142.
Niethammer G. On the breeding of Montifringilla there-
sae//Ibis. 1963. Vol. 109. N 1. P.117-118.
Norberg A.R. Mira fra rufocinnamomea doubles its
wingbeat rate to 24 Hz in wing-clap display flight: a
sexually selected feat / /J. Exper. Biol. 1991. Vol.
159. P.515-523.
North C.A. Movement patterns of the house sparrow’ in
Oklahoma // Omith. Monogr. 1973. N 14. P.79-91.
Payne R.B. Nest parasitism and display of chestnut spar-
rows in a colony of grey-capped social weaver/'/
Ibis. 1969. Vol. 111. P.300-307.
Payne R.B., Payne L.L., Nhlame M.E.D., Hustler K. Species
stutus and distribution of the parasitic indigo-birds
Vidua in East and Southern Africa/ / Proc. VIII Pan-
Afric. Ornith. Congr. 1992. P.40-52.
Pieman J. Egg destruction by eastern mcdow’larks/ / Wil-
son Bull. 1992. Vol. 104. N3. P.520-525.
Piclowski Z., Pinowski J. Autumnal sexual behaviour
of the tree sparrow/ / Bird Study. 1962. Vol. 9.
P.116-122.
Pinow’ski J. Dcr Jahrcszyklus dor Brutkolonic bcim Fcld-
sperling (Passer m.montanus)// Ekol. Pol. 1966. Ser.
A. Vol.14. P.145-174.
Pinowski J. Dispersal, habitat preferences and the regula-
tion of population numbers in tree sparrow s, Passer m.
montanus/ / Intern. Studies Sparrow’s. 1971. Vol. 5. N
1. P.21-39.
Rhijn J.G. Unidirectionality in the phylogeny of social
organization with special reference to birds/ / Be-
haviour. 1990. Vol. 115. N3-4. P.153-174.
Robinson S.K., Suarez A., Congdon K. Effect of edge types
on indigo buntings// The IllinoisNatur. Hist. Survey
Rep. 1994. Vol. 329. N8e. P.1-8.
Sandstrom U. Enhanced predation rates on cavity bird
nests at dccidious forest edges — an experimental
study/ / Ornis fenn. 1991. Vol.68. N 3. P.93-98.
Scutch J.L. The incubation patterns of birds// Parental
behaviour of birds. 1977 (repr. from 1957). P.176-200.
Shelley G.E. The birds of Africa, comprising all the species
which occur in the Ethiopian region. Vol. 3. London.
1902. 274 p.
Schaffer F. Ornithologischc Ergebnissc zweier Forschung-
sreisen nach Tibet// Journ. Ornithol. 1938. Bd. 88.
Sonderheft. 350 s.
Schiffcrli L. Juvcnil house sparrow’ parasiting nestling house
sparrows/, Brith. Birds. 1980. Vol. 73. P. 189-190.
Schiffcrli L. Die Rolle des Mannehens wahrend dcr Bc-
brutung dcr Eicr bcim Housspcrling Passer domesti-
cus/ / Ornith. Bcob. Vol. 75. S.44-47 (цнт. no Sum-
mers-Smith, 1988).
Schmidt G.A.J. Zur Sperlingsbekampfung// Omith. Mitt.
1954. Bd. 6. S. 147-153.
Sibley Ch.G. A comparative study of the egg-white pro-
teins of passerine birds// Bull. Pcabodv Mus. Nat.
Hist. Yale Univ. Vol. 32. P. 1-131.
Sibley Ch.G., Ahlquist J.E. The phylogeny and classifica-
tion of the passerine birds based on comparisons of
genetic material DNA// Acta XVIII Congr. Intern.
144
В.В.Иваницкий
Omithol., Moscow, August, 16-24, 1982. Vol.l. Moscow.
Nauka. 1985. P.83-121.
Sibley Ch.G., Monroe B.L. Distribution and taxonomy of
birds of the world. New Haven. 1990. London: Yale
Univ. Press. 1111 p.
Skead C.J. A study of the spectacled weaver (Ploceus ocu-
laris Smith/ / Ostrich. 1953. Vol. 24. N 2. P. 103-110.
Stegmann B. Untcrsuchungcniiber palacarktischc “Schncef-
inken/ / Joum. Omithol. 1932. Bd. 80. N 2. S. 99-114.
Stephan B. Die Evolutionthcoric und der Taxonomische
Status des italicn Sperlings/ / Mitt. Zool. Mus. Ber-
lin. 1986. 62, Supplement. S.25-68.
Summers-Smith D. The communal display of the house
sparrow Passer domesticus/ / Ibis. 1954 b. Vol. 96. N
1. P.116-128.
Summers-Smith D. Nest-site selection, pair formation and
territory in the house sparrow Passer domesticus/ /
Ibis. 1958. Vol. 100. N2. P.190-203.
Summers-Smith D. The rufous sparrows of the Cape Verde
Islands/ / Bull. Brith. Omith. Club. 1984 a. Vol. 104.
N4. P.138-142
Summers-Smith D. The sparrows of the Cape Verde Is-
lands// Ostrich. 1984 b. Vol. 55. P. 141-146.
Summers-Smith D. The sparrows. A study of the genus
Passer. T. AD Poyser. 1988. Calton. 342 p.
Sushkin P.P. On the anatomy and classification of weav-
er-birds/ / Bull. Am. Mus. Nat. Hist. 1927. Vol.57. art.1.
P.1-32.
Thaler E. Nest und Ncstbau von Winter- und Sommer-
goldhahnchen (Regulus regulus und R. ignicapillus) /
/ Joum. Omith. 1976. Vol. 117. (пит. по Фройдс, 1976).
Vaurie Ch. Systematic notes on Palaarctic birds. N 24.
Ploceidae: the genera Passer, Petronia, and Montifrin-
gilla/ / Amer. Mus. Novit. 1956. N 1814. 27 p.
Vaurie Ch. Tibet and its birds. H.F. and G.Wither Ltd.,
London.1972. 344 p.
Veiga J.P. Infanticide bv male and female house sparrows/
/ Anim. Bchav. 1990 a. Vol. 39. P.496-502.
Veiga J.P. Sexual conflict in the house sparrow: interfer-
ence between polygynously mated females versus asym-
metric male investment// Bchav. Ecol. Sociobiol. 1990
b. Vol. 27. P.345-350.
Volga J.P. Hatching asynchrony in the house sparrow: a
test of the eggs variabilitv hypothesis// Amer. Nat.
1992 a. Vol. 139. N 3. P.669-675.
Veiga J.P. Why are house sparrows predominantly monog-
amous? A test of hypothesis/ / Anim. Bchav. 1992 b.
Vol. 43. N3. P.361-370.
Veiga J.P., Vinucla J. Hatching asynchrony and hatching
success in the house sparrow: evidence for the egg via-
bility hvpothesis/ ,/ Ornis Scand. 1993. Vol. 24. N 3.
P.237-242.
Wallman J., Grabon M., Silver R. What determines the
patterns of sharing of incubation and brooding in ring
doves?// Journ. Comp, and Physiol. Psvchol. 1979.
Vol. 93. N3. P.481-492
Ward P. The breeding biology of the black-faced dioch Que-
leaquelea in Nigeria/ / Ibis. 1965. Vol. 107. P.326-349.
Ward P., Poh G.E. Seasonal breeding in an equatorial
population of the tree sparrow Passer montanus//
Ibis. 1968. Vol. 110. N 2. p.359-363.
Watson J.R. Dominance-subordination in caged groups of
house sparrows// Wilson Bull. 1970. Vol. 83. N 3.
P.268-278.
Wechsler B. The spread of food producing techniques in a
captive flock of jackdaw’s// Behaviour. 1989. Vol. 107.
P.267-277.
Wetton J.H., Carter R.E., Parkin D.T., Walters D. Demo-
graphic study of a wild house sparrow* population by
DNA fingerprinting/ / Nature. 1987. N327, P.147-149.
Wetton J.H., Parkin D.T. Sperm competition and fertility in
the house sparrow// Acta 20 Congr. Intern. Omithol.,
Vol. 4, 2-9 Dee., 1990, Wellington. 1991. P.2435-2441.
Wingfield J.C. Androgens and mating systems: testoster-
one-induced polvgyny in normalIv momogamous birs/
/ Auk. 1984. Vol. 101. P.665-67L
Winkler D.W. Causes and consequences of variation in
parental defence behaviour bv tree swallows/ / Con-
dor. 1992. Vol. 94. N 2. P.502-510.
Wittenberger J.F., Hunt G.L. The adaptive significance of
coloniality in birds// Avian Biol. Vol.8. New York
Acad. Press. 1985. P.1-79.
Yom-Tov Y. Intraspecific nest parasitism in the Dead Sea
sparrow*, Passer moabiticus/ • Ibis. 1980. Vol. 122.
P.234-237.
Yom-Tov Y., Ar A. On the breeding ecology of the Dead
Sea sparrow* Passer moabiticus/ / Israel Journ. Zool.
1980. Vol. 29. N 4. P.171-187.
Yom-Tov Y., Mcndclsson H. Incubation behaviour of the Dead
Sea sparrow* / / Condor. 1978. Vol 80. N 2. P.340-343.
THE SPARROWS AND RELATED GROUPS OF GRANIVOROUS
BIRDS: BEHAVIOUR, ECOLOGY AND EVOLUTION
Vladimir V. Ivanitskii
Department of Vertebrate Zoology and General Ecology
Biological Faculty of Moscow State University
The book represents main results of compara-
tive studies of social organization, reproductive
behaviour, displays, vocalization, population
ecology and general biology of house sparrows
(Passer domesticus), Indian sparrows (P. indicus),
Spanish sparrows (P.hispaniolensis), saxaul spar-
rows (P.ammodendri), desert sparrows (P.simp-
lex), cinnamon sparrows (P.rutilans), rock spar-
rows (Petronia petronia), snow finches (Monti-
fringilla nivalis), Pere David’s snow finches (Pyr-
gilauda davidiana), and pale rock sparrows (Car-
pospiza brachydactyla) m Russia and surrounding
countries. Taxonomic validity of the Passeridae
family is confirmed strongly. Montifringilla and
Pyrgilauda must be treated as separate genera as
well as Petronia and Gymnorhis. Petronia, Mon-
tifringilla and Pyrgilauda share many important
ethological characters to be well separated from
Passer and Gymnorhis which appear to be inter-
related closely. Carpospiza position is not clear
from behavioural data but their tongue sceleton
analisys lead to conclusion that it may be the
member of Passeridae.
We believe that ancestors of sparrows were
predominantly arboreal birds with their activities
proceeding mostly in crowns of bushes, trees and
large-stem grass weeds. Upon evolution, in ad-
dition to preserving mainly arboreal forms rep-
resented most completely in Passer genus, the
sparrows’ activity shifted gradually to soil surface
(foraging), tree holes (nesting and roosting), and
later to bedrock exposures, cliffs and small mam-
mals burrows. In the long run this permitted
sparrows to colonize successfully quite woodless
areas. Thus, the Pyrgilauda species have far more
in common with the larks rather than with other
sparrow species, distinguished from the latter long
and strong legs, ability to run quickly, and elabo-
rated display flights. On the other hand, upon
adaptation to conditions of highlands, Montifrin-
gilla species acquired a clear similarity with
mountain finches (Leucosticte). Switching from
open nesting in forks of tree and bush crowns to
hole nesting was one of the most important
events in the evolution of sparrows. Hole nesting
is common for the five sparrow genera, for 4
genera it being the only type known (all five Pyr-
gilauda species are known to be highly specialised
rodent burrow occupants).
Various types of spatial allocation of breeding
pairs can be detected in sparrows: (1) solitary
breeding pairs with tenacity to vast and isolated
breeding sites and without permanent social con-
tacts with other conspecifics; (2) pairs may be
virtually in constant acoustic and visual contact
with each other when the nests are dispersed at
a distance of tens of meters; (3) loose colonies
counting dozens of pairs when nests are scattered
at a distance of several meters; (4) supercolonies
counting more than a hundred of pairs with very
dense breeding and a plausible physical contact
of the neighbouring nests. Both nomadic and
site-tenacious strategies of using the space are
well represented within sparrows. The degree of
site tenacity does not correlate with characteris-
tics of the spatial structure (colonial vs solitary)
and seasonal movements (sedentary vs migratory)
of the species.
Monogamy was found to be predominant
type of mating system of sparrows. Both males
and females take part in nest building and incu-
bation in all Passer species studied but only fe-
males build and incubate in another genera be-
ing assusted by their males only under the time
of nestlings feeding.
Our scenario of the Passeridae evolution is
based on the assumption that ancestral forms of
behaviour have been sustained best of all in Pas-
sergenus, which is confirmed by its wide range,
complex taxonomic structure and existence of
numerous relict forms. We suppose that as far
back as in the Myocene, African and Asian sa-
146
В.В.Иваницкий
vannas were widely populated by avian species in
many characters resembling recent Passer. Most
likely, the Petronia species developed within eco-
logical boundaries of savannas with a broken low-
altitude relief which was very typical of the
Myocene in the Central Asia characterized by
predominating vast highlands of Hercynian and
Caledonian foldings. This lent a possibility to
colonize not only woodless but also purely steppe
landscapes lacking bed-rock outcrops. It is re-
markable that recent rock sparrows with an un-
usual broad ecological divergence can demon-
strate the whole range of ecological situations that
should occur in the sparrow evolution prior to
enhanced mountain formation in the Pliocene.
Although over a greater part of their spacious
breeding range, rock sparrows behave as typical
petrophilous species and prove to be dependent
on a broken relief, the P.p.kirgizika subspecies
breeding in the northwestern region near the
Caspian Sea can be classified as an entirely
steppe bird.
We assume that Pygrilauda were formed in
the Myocene when steppe became the prevailing
landscape in Central Asia. In the course of late
tertiary orogenesis and growing cooling, this spe-
cies became more adapted to high-altitude pla-
teau landscapes with marginal climatic conditions
and extremely poor biocenosis environment. It is
suggested that such conditions provided grounds
for reduction of all forms of coloniality and for-
mation of a quite rigorous system of territorial
interactions. This resulted in the maximal effi-
ciency in the utilization of rather scarce food
resources evenly ditributed over the area based on
a systematic foot inspection of spacious territo-
ries with flights reduced to the minimum. The
new life form became one of the most spread
adaptations and evoked corresponding transfor-
mations in communication behaviour.
Montifringilla could be suposed to originate in
the process of setting up in a new ecological
niche which had appeared upon Alpine orogen-
esis. At present these birds, utterly warranting
their name, are in most part dependent on rich
but small and widely dispersed beds of alpine and
subalpine vegetation near diverse glacial and
snow formations high in the mountains. As com-
pared to closely related Pere David’s snow
finches, in the process of evolution snow sparrows
inhabited a significantly more diverse, heteroge-
neous, humid and unpredicted environment.
Long-distance movements for picking food re-
quired, first of all, improvement of the flying
apparatus, caused diminishing of the territority,
renewal of quasicoloniality, prolongation of flock
behaviour in the breeding period. Parallel
changes occurred in the communication
behaviour as well.
The current concept on sparrows as a flour-
ishing avian group has appeared due to a virtu-
ally impressive breeding success of synantropic
sparrow species which have scattered over the
whole Earth following humans. Contrary to this
widespread view we suggest that most likely spar-
rows represent a distinctly marginal taxon whose
representatives are encumbered by numerous
biological characters which could hardly underlie
further development.
First, a typical sparrow domed nest is a rather
cumbersome bulky construction whose building
requires time and energy expenditures which can
be expedient only in rather specific conditions of
extremely hot or cold climatic conditions. Grow-
ing weight and size of the nest improve its ther-
mostatic properties, however enhancing predation
risk. Thus, many nests of sparrows are well seen
from a distance of hundred metres.
. Second, great costs are evoked by colonial
nesting which also increases predator vulnerabil-
ity, causes enhanced expenditures of food re-
sources near breeding sites and different forms of
social interference such as nest parasitism,
kleptoparasitism, infanticide, nest destruction,
and great time and energy expenditures due to
regulation of interaction between neighbouring
birds. So coloniality was gradually sustituted by
solitary breeding within the most advanced taxa
of sparrows. Colonial breeding have persisted and
even undergone the futher development in the
species occuring under the most favorable trophic
conditions in a human alterated environment.
Third, regulation and coordination of the
main phases in the reproductive cycle and the
corresponding forms of sparrow behaviour appear
to be rather imperfect. Mutual engagement of
males and females in nearly all the forms of re-
productive behaviour seems to be primitive. The
main phases of breeding cannot be distinctly
separated in time. Nest building starts long be-
fore breeding, in some cases prior to the final
choice of a shelter, and continues up to chicks
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ
147
hatch. Brooding patches may appear in females
and their first egg deposition may occur even
during migration, i.e. prior to the final choice of
the nesting site. As a result, having arrived to the
destination, females often have to lay eggs in
unaccomplished nests. They start brooding after
the first egg has been laid, therefore the nestlings
differ dramatically in size and in some cases the
youngest and smallest ones perish of hunger and
physical pressure of the elder.
The relative imperfection of nests, social
structure and breeding behaviour in sparrows can
be shown best of all by comparing them with such
currently prosperous birds as Fringillidae finches,
Emberizidae buntings and Alaudidae larks. Each of
them greatly exceeds sparrows in the number of
species and genera, the biocenosis value and the
dimensions of the total ranges. All these birds have
a typical simple cup-shaped nest, clear division of
sexual roles of male and female with a leading role
of the latter in nest building and incubation, clear
and regular switches between the main stages of
the nesting cycle, and mostly solitary breeding
based on more or less strong territoriality. On the
whole, the breeding behaviour of such a type
seems to be more simple, reliable and efficient
than that in sparrows.
Studies of habitat relations in recent sparrows
show that they apparently avoid rich communities
highly saturated with avian species and individu-
als. Sparrows have no forester species, however
many species breed in extreme landscapes, in the
first place mention should be done of arid and
extra-arid territories with extremely hot (the
Sahara and Kara-Kum deserts) or cold (the Tibet
and Pamirs mountain deserts) climatic condi-
tions. Adaptation to such severe conditions
evoked high specialization in shelter choice, nest-
building technology and reproductive behaviour,
which inevitably reduced the total evolutionary
perspectives of the taxon and its competitive
ability. Therefore it is not surprising that appar-
ently relict species inhabiting very small ranges
are abundant among sparrows. At present zonal
landscapes of Palearctics are occupied in most
part by finches, buntings and larks. The tropical
biota in Africa is occupied mainly by weavers,
waxbills and larks. Sparrows have been forced to
the driest acacia savannas and deserts, outskirts
of riparian and gallery forests at the boundaries
of open habitats, cold high-altitude steppes and
deserts.
It is most likely that at the onset of the
Holocoen sparrows were on the verge of their
extinction having bred for no less than between
30 or 35 million years. However, only for several
thousand years, i.e. almost instantaneously from
the point of view of evolution, the landscapes of
the Old World were dramatically changed due to
the developing human alterations in. natural
communities. A principally novel landscape ap-
peared virtually lacking avian species, and spar-
rows immediately took advantage of this. Their
synantropic development was likely to proceed
very rapidly, in various forms and with indepen-
dent involvement of various species in Africa, in
the region of the Mediterranean Sea, Asia Minor
and Southeastern Asia. Of 10 sparrow species
breeding in North Palearctics apparently only the
desert sparrow is not in any way dependent on
the man-altered landscape, while all the other
species use it for nesting, foraging or watering
places at different stages of their annual cycle.
Why have namely sparrows appeared to be best
adapted to breeding in man-altered landscapes?
It is assumed that in this case of vital importance
proved to be certain biological peculiarities of
these birds which can be regarded as specific
preadaptation. First, this is the ability to nest and
roost in hole shelters which allowed the sparrows
to breed directly in man-erected constructions.
Second, great trophic possibilities and relatively
large body sizes underlay efficient utilization of
remnants of human dieting and rather big grains
of principal cereals. Third, the high level of so-
ciality the year round lends support to rapid fixa-
tion and spread of novel types of ethological
adaptation by mimicking and learning.
148
В.В.ИВАНИЦКИЙ
ВОРОБЬИ И РОДСТВЕННЫЕ ИМ ГРУППЫ ЗЕРНОЯДНЫХ ПТИЦ: ПОВЕДЕ-
НИЕ, ЭКОЛОГИЯ, ЭВОЛЮЦИЯ
ISBN 5—87317—4)37—1
<^р»И
Сдано в набор 15.05.97. Подписано в печать 28.07.97.
Формат 70x100/8. Объём 18,5 уч. изд. листов.
Печать офсетная. Бумага офсетная №1. Гарнитура Кудряшёвская.
Тираж 300 экземпляров. Отпечатано фирмой "Подтеке".
"Товарищество научных изданий КМК”
111531. Москва. Шоссе Энтузиастов-100-5-56
ЯР № 070831 от 25.01.93