/







































































































































































































































































































































Текст
Állattan
Szerkesztette Bakonyi Gábor
allattan
Szerkesztette
dr. Bakonyi Gábor
Második, átdolgozott, bővített kiadás
MEZŐ
GAZDA
Az ábrákat készítette
Bagosi Zoltán
dr. Bakonyi Gábor
Hencz Helga
dr. Kiss István
Soproni Emese
Szabóky Csaba
Szunyoghy András
Varga Sándor
Lektorálta
dr. Altbacker Vilmos (21. fejezet)
dr. Kordos László (22. fejezet)
dr. Majer József (15. és 17. fejezet)
dr. Matskási István (6-14. és 16. fekezet)
dr. Nősek János (19. és 20. fejezet)
dr. Réz Gábor (1-3. fejezet)
dr. Széky Pál (18. fejezet)
dr. Vásárhelyi Tamás (5. fejezet)
dr. Zboray Géza (4. fejezet)
© dr. Bakonyi Gábor, dr. Juhász Lajos, dr. Kiss István, dr. Palotás Gábor, 2003
Minden jog fenntartva. A könyv egészének vagy bármely részletének, ábráinak elektronikus vagy bármi más
módon való tárolása, sokszorosítása, felhasználása középiskolai, főiskolai, egyetemi jegyzetek, tankönyvek
készítése során vagy bármi más célból, kizárólag a szerkesztő és az érintett fejezetek szerzőinek előzetes írás-
beli engedélyével és a munkába való bevonásával lehetséges.
cit.r Bakonyi G., Juhász L., Kiss I. és Palotás G. (szerk. Bakonyi G. (1995): Állattan.
Mezőgazda Kiadó, pp. 720
ISBN 963 286 044 6
Mezőgazda Kiadó - az 1795-ben alapított
Magyar Könyvkiadók és Könyvterjesztők
Egyesülésének a tagja -
1165 Budapest, Koronafürt u. 44.
Felelős kiadó: dr. Lelkes Lajos
Felelős szerkesztő: Wenszky Ágnes
Műszaki vezető: Gerlóci Judit
Műszaki szerkesztő: Berkes Tamás
A borító Kiss István sorozatterve alapján készült
Boritófotó: Szelényi Gábor/Farkasalmalepke (Zerinthia polyxena)
Megjelent 64,3 (A/5) ív terjedelemben, 457 ábrával
DTP: TiMac Bt.
MGK 712 044/2003
TARTALOM
1. AZ ÁLLATI SEJTEK FELÉPÍTÉSE ÉS MŰKÖDÉSE
(dr. Kiss István)....................................................... 19
1.1. Sejtbiológia....................................................... 19
1.2. A sejt membránrendszerei........................................... 20
1.2.1. A plazmamembrán.................................................. 20
1.2.2. A sejt belső membránrendszerei és membránnal határolt sejtszervecskéi... 27
1.3. A sejt alapállománya és vázrendszere............................... 30
1.4. A sejtközpont és a sejtek mozgási organellumai..................... 33
1.5. A sejtek mozgása................................................... 33
1.6. A sejtek osztódása................................................. 38
1.6.1. Kromoszómaszám-tartó osztódás.................................... 38
1.6.2. Kromoszómaszám-felező osztódás................................... 41
1.7. Rákos sejtek....................................................... 43
1.8. A sejtek szerepe a szervezet immunfolyamataiban.................... 43
2. AZ ÁLLATOK EGYEDFEJLŐDÉSE (dr. Kiss István)......................... 46
2.1. Az ivarsejtek felépítése és kialakulása............................ 46
2.1.1. A petesejt kialakulása és felépítése............................. 47
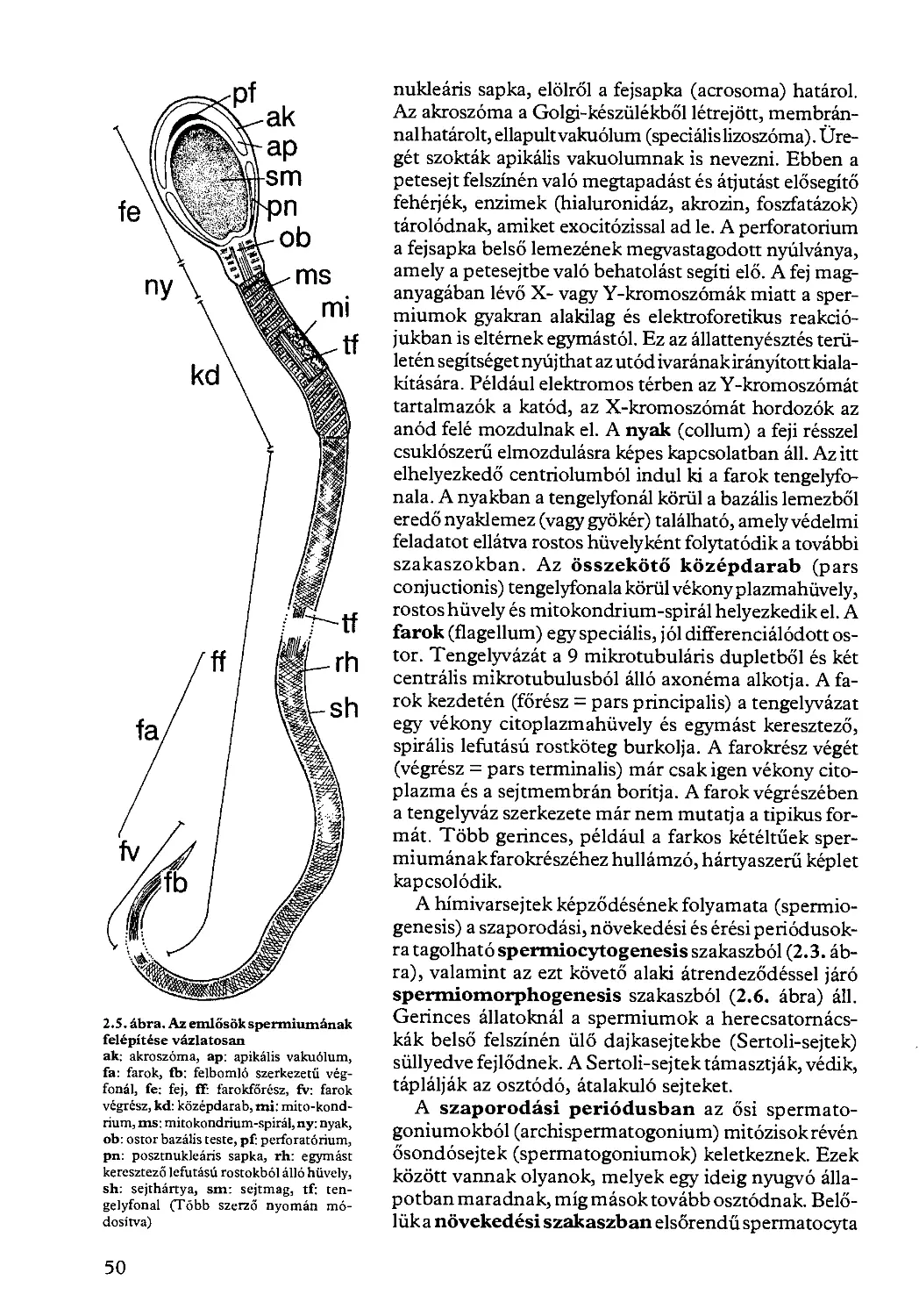
2.1.2. A hímivarsejt kialakulása és felépítése.......................... 49
2.2. Megtermékenyítés................................................... 51
2.3. Az embrionális fejlődés citotipikus szakasza....................... 54
2.3.1. Barázdálódás..................................................... 54
2.3.2. Szedercsíra állapot.............................................. 58
2.3.3. Hólyagcsíra állapot.............................................. 58
2.4. Az embrionális fejlődés organotipikus szakasza..................... 60
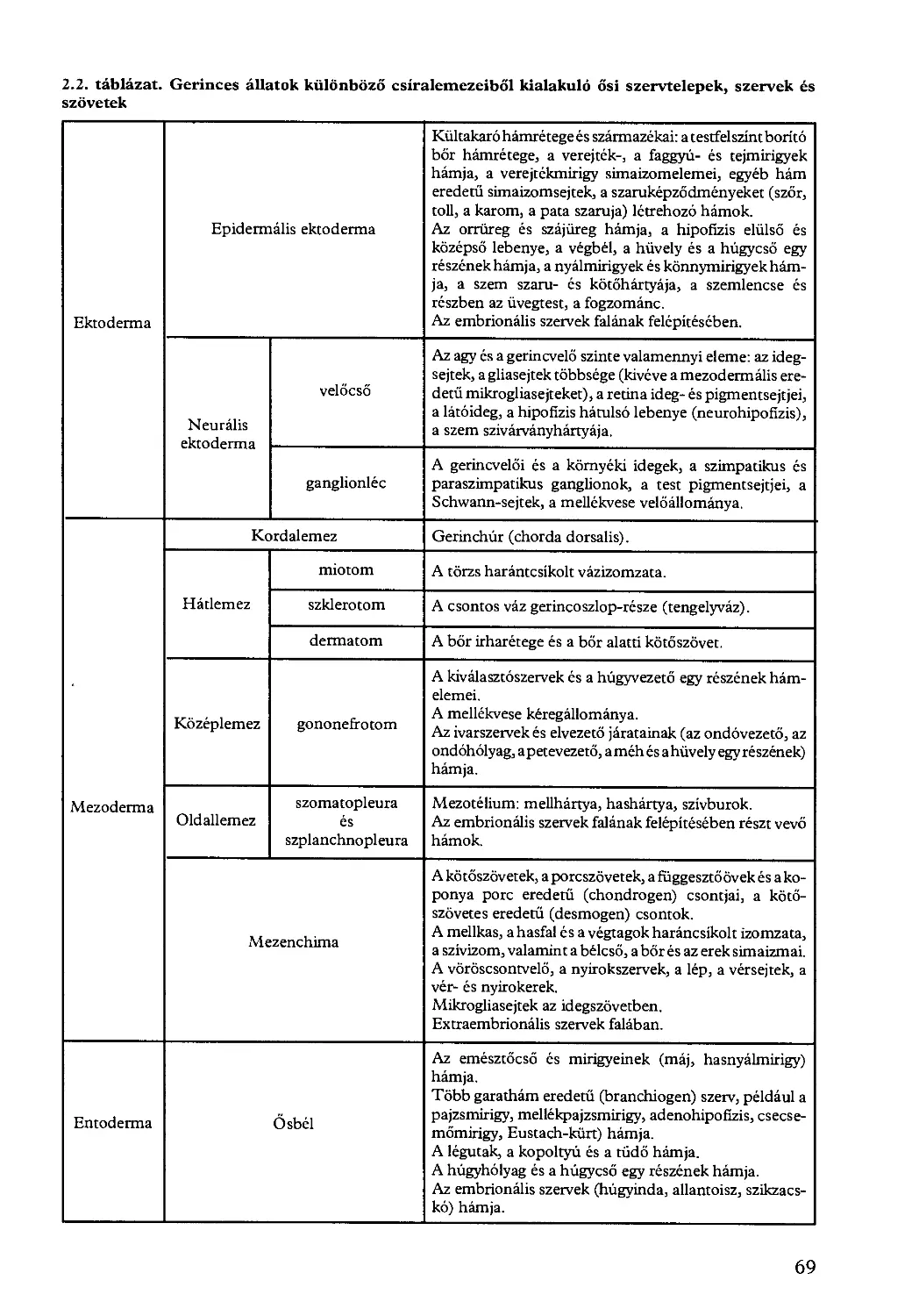
2.4.1. Csíralemezek kialakulása......................................... 60
2.4.2. Szervfejlődés és szöveti differenciálódás........................ 67
2.5. Embrionális szervek................................................ 70
2.5.1. Szikzacskó....................................................... 70
2.5.2. Magzatburkok..................................................... 71
2.6. Az egyedfejlődés szabályozása ..................................... 75
2.7. Posztembrionális fejlődés.......................................... 78
2.8. Regeneráció ....................................................... 79
2.9. Az állati szervezet öregedése, a halál............................. 80
3. AZ ÁLLATI TESTET FELÉPÍTŐ SZÖVETEK (dr. Kiss István) ............... 82
3.1. Hámszövetek........................................................ 82
3.1.1. A hámok csoportosítása morfológiai jellemzők alapján............. 83
3.1.2. A hámok csoportosítása funkciók alapján.......................... 85
5
3.2. Kötőszövetek és támasztószövetek................................... 88
3.2.1. Kötőszövetek...................................................... 89
3.2.2. Vér és egyéb testfolyadékok....................................... 92
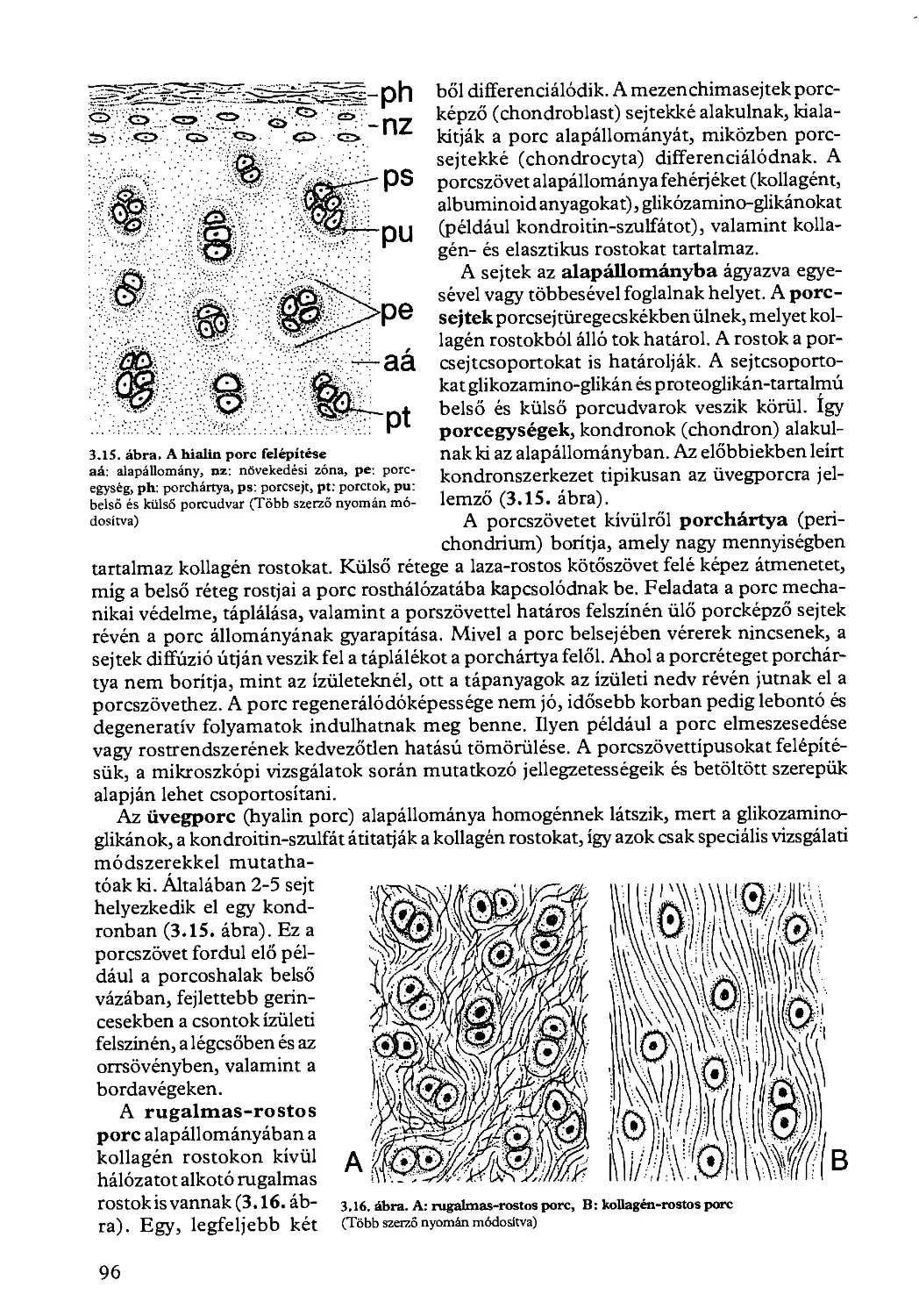
3.2.3. Támasztó szövetek................................................. 95
3.3. Izomszövetek...................................................... 101
3.3.1. Simaizomszövet................................................... 101
3.3.2. Harántcsíkolt izomszövetek....................................... 101
3.4. Idegszövet........................................................ 103
4. AZ ÁLLATOK SZERVRENDSZEREINEK FŐ TÍPUSAI
(dr. Bakonyi Gábor) ................................................. 107
4.1. Kültakaró és mozgásszervek........................................ 108
4.1.1. Az egységes kültakaró és mozgásrendszer.......................... 109
4.1.2. Az önálló kültakaró.............................................. 110
4.1.3. Az önálló mozgásrendszer......................................... 112
4.1.4. A kültakaró funkciói ............................................ 114
4.2. Táplálkozás és emésztés........................................... 116
4.2.1. Kétszakaszos bélcsatoma ......................................... 116
4.2.2. Háromszakaszos bélcsatorna....................................... 117
4.2.3. Intracelluláris és extracelluláris emésztés...................... 119
4.3. Anyagszállítás.................................................... 120
4.3.1. Az anyagszállítás szervrendszerei ............................... 120
4.3.2. Nyílt keringési rendszer......................................... 120
4.3.3. Zárt keringési rendszer.......................................... 121
4.3.4. A gázok szállítása............................................... 122
4.3.5. Egyéb anyagok szállítása ........................................ 122
4.4. Légzés............................................................ 123
4.4.1. A légzőszervek típusai........................................... 124
4.4.2. A külső gázcsere mechanizmusa.................................... 128
4.5. Kiválasztás és ozmoreguláció...................................... 128
4.5.1. A kiválasztás szervrendszerei ................................... 129
4.5.2. Kiválasztás egyéb módon.......................................... 133
4.5.3. A kiválasztott anyagok raktározása............................... 134
4.5.4. Ozmoreguláció.................................................... 134
4.6. Szaporodás........................................................ 134
4.6.1. Ivartalan szaporodás ............................................ 135
4.6.2. Szűznemzés....................................................... 135
4.6.3. Ivaros szaporodás ............................................... 136
4.6.4. Nemzedékváltakozás............................................... 138
4.7. Endokrin szervek rendszere........................................ 138
4.7.1. Hatásmechanizmusok a célsejteknél................................ 140
4.7.2. Endokrin szervek és hatásmechanizmusok........................... 141
4.7.3. Feromonok ....................................................... 145
4.8. Idegrendszer...................................................... 146
4.8.1. Az idegrendszer típusai ......................................... 146
4.8.2. A reflexek....................................................... 150
4.8.3. Evolúciós trendek................................................ 150
4.9. Érzékszervek...................................................... 151
4.9.1. Mechanikai érzékszervek.......................................... 152
6
4.9.2. Látószervek................................................... 156
4.9.3. Kémiai érzékszervek........................................... 158
4.9.4. Egyéb érzékszervek............................................ 159
5. RENDSZERTANI ALAPISMERETEK (dr. Bakonyi Gábor) ................... 161
5.1. Klasszifikáció ................................................. 161
5.2. Az élővilág fő csoportjai....................................... 165
5.3. A heterotróf protiszta élőlények rendszere...................... 167
5.4. Az állatvilág áttekintő rendszertana............................ 167
6. ÁLLÁBAS-OSTOROS EGYSEJTŰEK (SARCOMASTIGOPHORA)
(dr. Kiss István).................................................... 177
6.1. Állati ostorosok (Zoomastigophorea)............................. 178
6.1.1. Parabazális-testes ostorok (Kinetoplastida) .................. 178
6.1.2. Tükrözött-testű ostorosok (Diplomonadida)..................... 179
6.1.3. Tengelytálcás ostorosok (Trichomonadida) ..................... 179
6.2. Gyökérlábúak (Rhizopoda) ....................................... 180
6.2.1. Csupaszamőbák (Amoebida)...................................... 180
6.2.2. Házasamőbák (Arcellinida)..................................... 180
6.3. Tengelyeslábúak (Actinopoda).................................... 181
6.3.1. Napállatocskák (Heliozoea).................................... 181
7. APLIKÁLIS-SZERVŰ EGYSEJTŰEK (APICOMPLEXA)
(dr. Kiss István).................................................... 182
7.1. Sejtélősködő spórások (Eimeriina)............................... 184
7.2. Vérélősködő spórások (Haemosporina)............................. 185
7.3. Vérélősködő spórátlanok (Piroplasmida).......................... 186
8. APRÓSPÓRÁSOK (MICROSPORA) (dr. Kiss István) ...................... 187
8.1. Apróspórás alakúak (Microsporida)............................... 187
9. CSILLÓSOK (CILIOPHORA) (dr. Kiss István)......................... 188
9.1. Egyforma csillójúak (Kinetofragminophorea)...................... 192
9.1.1. Csupasz-szájúak (Gymnostomatia)............................... 192
9.1.2. Szájtölcséresek (Vestibulifera) .............................. 192
9.1.3. Hasi szájmezősök (Hypostomatia)............................... 193
9.1.4. Szívókások (Suctoria) ........................................ 193
9.2. Kevés-csillószervesek (Olygohymenophorea) ...................... 193
9.2.1. Hasi szájgödrösök (Hymenostomatia)............................ 193
9.2.2. Csillókoszorúsok (Peritrichia)................................ 194
9.3. Sok-csillószervesek (Polyhymenophorea).......................... 194
9.3.1. Örvényszervesek (Spirotrichia)................................ 195
7
10. LAPOSFÉRGEK (PLATYHELMINTHES) (dr. Kiss István).................. 196
10.1. Mételyek (Trematoda).......................................... 198
10.1.1. Villásfarkú-cerkáriás (Strigeatida).......................... 202
10.1.2. Májmétely-alakúak (Echinostomatida).......................... 203
10.1.3. Szájszuronyos-cerkáriájú mételyek (Plagiorchiida)............ 204
10.2. Csáklyásférgek (Monogenea).................................... 205
10.2.1. Egyszerű-tapadószervű (Monopisthocotylea).................... 206
10.2.2. Összetett-tapadószervűek (Polyopisthocotylea)................ 207
10.3. Galandférgek (Cestoda)........................................ 207
10.3.1. Tagolatlan galandférgek (Caryophyllidea)..................... 209
10.3.2. Szívógödrös galandférgek (Pseudophyllidea)................... 209
10.3.3. Szívókás galandférgek (Cyclophyllidea)....................... 211
11. FONÁLFÉRGEK (NEMATODA) (dr. Kiss István)......................... 215
11.1. Pórusszerű-oldalszervűek (Secementia) ........................ 223
11.1.1. Szabadonélő fonálférgek (Rhabditida) ........................ 223
11.1.2. Burzás fonálférgek (Strongylida)............................. 223
11.1.3. Kevésajkú fonálférgek (Ascaridida)........................... 224
11.1.4. Szájszuronyos fonálférgek (Tylenchida) ...................... 226
11.2. Betüremkedett oldalszervűek (Penetrantia)..................... 228
11.2.1. Hordószájú fonálférgek (Mononchida) ......................... 228
11.2.2. Tüskefogú fonálférgek (Dorylaimida) ......................... 228
11.2.3. Páratlan ivarszervű fonálférgek (Trichocephalida) ........... 228
11.2.4. Raktárbelű fonálférgek (Mermithida).......................... 229
12. KEREKESFÉRGEK (ROTATORIA) (dr. Kiss István)...................... 231
12.1. Araszoló kerekesférgek (Bdelloidea)........................... 236
12.2. Egypetefészkűek (Monogononta) ................................ 236
13. BUZOGÁNYFEJŰEK (ACANTHOCEPHALA) (dr. Kiss István)................ 238
13.1. Spiráltüskés buzogányfejűek (Archiacanthocephala)............. 239
13.2. Sortüskés buzogányfejűek (Palaecanthocephala) ................ 240
14. PUHATESTŰEK (MOLLUSCA) (dr. Kiss István)......................... 241
14.1. Csigák (Gastropoda)........................................... 243
14.1.1. Pajzskopoltyúsok (Aspidobranchiata).......................... 249
14.1.2. Fésűkopoltyúsok (Pectinibranchia)............................ 249
14.1.3. Ülőszemű tüdőscsigák (Basommatophora)........................ 250
14.1.4. Nyelesszemű tüdőscsigák (Stylommatophora).................... 251
14.2. Kagylók (Bivalvia) ........................................... 253
14.2.1. Lemezkopoltyúsok (Eulamellibranchiata) ...................... 258
15. GYŰRŰSFÉRGEK (ANNELIDA) (dr. Kiss István)........................ 259
15.1. Kevéssertéjűek (Oligochaeta) ................................. 260
15.1.1. Egypár-heréjű kevésszertéjűek (Tubificida)................... 267
8
15.1.2. Lumbriculida .................................................... 268
15.1.3. Kétpár-heréjű giliszták (Diplotesticulata)....................... 268
15.2. Piócák (Hirudinoidea) ............................................ 269
15.2.1. Ormányos nadályok (Rhynchobdellidea)............................. 273
15.2.2. Állkapcsos nadályok (Gnathobdellidea) ........................... 273
15.2.3. Garatos nadályok (Pharyngobdellidea)............................. 274
16. ÍZELTLÁBÚAK (ARTHROPODA) (dr. Bakonyi Gábor)........................ 275
16.1. Rákok (Crustacea)................................................. 278
16.1.1. Csupasz levéllábúrákok (Anostraca)............................... 282
16.1.2. Pajzsos levéllábúrákok (Notostraca) ............................. 283
16.1.3. Ágascsápúrákok (Cladocera)....................................... 283
16.1.4. Haltetvek (Branchiura) .......................................... 284
16.1.5. Féregatka-alakúak (Pentastomida)................................. 284
16.1.6. Kagylósrákok (Ostracoda)......................................... 285
16.1.7. Evezőlábú rákok (Copepoda)....................................... 285
16.1.8. Tízlábú rákok (Decapoda) ........................................ 286
16.1.9. Hasadtlábú rákok (Mysidacea)..................................... 288
16.1.10. Ászkarákok (Isopoda)............................................ 288
16.1.11. Felemáslábú rákok (Amphipoda)................................... 289
16.2. Százlábúak (Chilopoda)............................................ 289
16.3. Ikerszelvényesek (Diplopoda)...................................... 290
16.4. Hatlábúak (Hexapoda) ............................................. 291
16.4.1. Előrovarok (Protura)............................................. 310
16.4.2. Lábaspotrohúak (Diplura)......................................... 310
16.4.3. Ugróvillások (Collembola)........................................ 310
16.4.4. Ugró ősrovarok (Archeognatha) ................................... 311
16.4.5. Pikkelykék (Zygentoma)........................................... 311
16.4.6. Kérészek (Ephemeroptera)......................................... 312
16.4.7. Szitakötők (Odonata)............................................. 313
16.4.8. Álkérészek (Plecoptera).......................................... 314
16.4.9. Fülbemászók (Dermaptera)......................................... 315
16.4.10. Fogólábúak (Mantodea)........................................... 315
16.4.11. Csótányok (Blattodea)........................................... 316
16.4.12. Sáskák (Caelifera).............................................. 317
16.4.13. Tojócsöves egyenesszárnyúak (Ensifera) ......................... 319
16.4.14. Fatetvek (Psocoptera)........................................... 320
16.4.15. Tetvek (Phthiraptera) .......................................... 320
16.4.16. Tripszek (Thysanoptera)......................................... 321
16.4.17. Poloskák (Heteroptera).......................................... 322
16.4.18. Színkabócák (Auchenorrhyncha) .................................. 328
16.4.19. Növénytetvek (Stemorrchyncha)................................... 330
16.4.20. Vízifátyolkák (Megaloptera) .................................... 334
16.4.21. Tevenyakú fátyolkák (Raphidiop tér a) .......................... 335
16.4.22. Recésfátyolkák (Planipennia).................................... 335
16.4.23. Bogarak (Coleoptera)............................................ 335
16.4.24. Legyezőszámyúak (Strepsiptera).................................. 345
16.4.25. Hártyásszámyúak (Hymenoptera)................................... 346
16.4.26. Tegzesek (Trichoptera).......................................... 351
9
16.4.27. Lepkék (Lepidoptera)............................................ 352
16.4.28. Csőrös rovarok (Mecoptera)...................................... 362
16.4.29. Kétszárnyúak (Diptera).......................................... 363
16.4.30. Bolhák (Siphonaptera)............................................ 372
16.5. Pókszabásúak (Arachnida) .......................................... 372
16.5.1. Álskorpiók (Pseudoscorpiones)..................................... 374
16.5.2. Szálfarkúak (Palpigradi).......................................... 375
16.5.3. Kaszáspókok (Opiliones)........................................... 375
16.5.4. Valódi pókok (Araneae) ........................................... 376
16.5.5. Atkák (Acari)..................................................... 379
17. GERINCESEK (VERTEBRATA).............................................. 385
17.1. Ingolák (Cephalaspidomorphi) (dr. Kiss István)..................... 386
17.1.1. Ingolaalakúak (Petromyzontiformes) ............................... 386
17.2. Sugarasúszójú halak (Actinopterygii) (dr. Kiss István)............. 387
17.2.1. Tokalakúak (Acipenseriformes)..................................... 401
17.2.2. Angolnaalakúak (Anguilliformes)................................... 401
17.2.3. Heringalakúak (Clupeiformes) ..................................... 402
17.2.4. Pontyalakúak (Cypriniformes)...................................... 402
17.2.5. Harcsaalakúak (Siluriformes) ..................................... 405
17.2.6. Lazacalakúak (Salmoniformes)...................................... 405
17.2.7. Tőkehal-alakúak (Gadiformes)...................................... 406
17.2.8. Fogasponty-alakúak (Cyprinodontiformes)........................... 407
17.2.9. Pikóhal-alakúak (Gasterosteiformes)............................... 407
17.2.10. Sárkányfejűhal-alakúak (Scorpaeniformes)........................ 407
17.2.11. Sügéralakúak (Perciformes)...................................... 408
17.3. Kétéltűek (Amphibia) (dr. Kiss István)............................. 409
17.3.1. Farkos kétéltűek (Caudata)........................................ 410
17.3.2. Békák (Anura)..................................................... 412
17.4. Halántékablak-nélküliek (Anapsida) (dr. Kiss István)............. 424
17.4.1. Teknősök (Testudines)............................................. 424
17.5. Kettős-halántékablakúak (Diapsida)................................. 426
17.5.1. Pikkelyes hüllők )Squamata) ...................................... 427
17.5.2. Lábatlangyík-alakúak (Anguimorpha)................................ 432
17.5.3. Vakondgyík-alakúak (Scincomorpha)................................. 433
17.5.4. Kígyók (Serpentes)................................................ 434
17.6. Madarak (Avés) (dr. Juhász Lajos) ................................. 437
17.6.1. Struccalakúak (Struthioniformes).................................. 459
17.6.2. Tyúkalakúak (Galliformes)......................................... 460
17.6.3. Lúdalakúak (Anseriformes)......................................... 461
17.6.4. Harkályalakúak (Piciformes) ...................................... 463
17.6.5. Bankaalakúak (Upupiformes)........................................ 464
17.6.6. Szalakótaalakúak (Coraciiformes).................................. 464
17.6.7. Kakukkalakúak (Cuculiformes)...................................... 465
17.6.8. Sarlósfecske-alakúak (Apodiformes) ............................... 465
17.6.9. Bagolyalakúak (Strigiformes)...................................... 466
17.6.10. Galambalakúak (Columbiformes).................................... 467
17.6.11. Darualakúak (Gruiformes)......................................... 468
17.6.12. Lilealakúak (Charadriiformes).................................... 469
10
17.6.13. Sólyomalakúak (Falconiformes).................................. 473
17.6.14. Gólyaalakúak (Ciconiiformes)................................... 475
17.6.15. Enekesmadár-alakúak (Passeriformes) ........................... 477
17.7. Emlősök (Mammalia) (dr. Palotás Gábor)........................... 490
17.7.1. Rovarevők (Insectivora)......................................... 519
17.7.2. Denevérek (Chiroptera).......................................... 521
17.7.3. Nyúlalakúk (Lagomorpha)......................................... 523
17.7.4. Rágcsálók (Rodentia)............................................ 524
17.7.5. Ragadozók (Camivora)............................................ 528
17.7.6. Páratlanujjú patások (Perissodactyla)........................... 531
17.7.7. Párosujjú patások (Artiodactyla)................................ 532
18. SZÜNBIOLÓGIA (dr. Bakonyi Gábor)................................... 536
18.1. A szupraindividuális szerveződés................................. 537
18.2. Populációk....................................................... 539
18.2.1. A populációk térbeli eloszlása.................................. 540
18.2.2. Koreloszlás..................................................... 542
18.2.3. A populációk struktúrájának egyéb jellemzői..................... 544
18.2.4. Populációk létszámváltozásai.................................... 545
18.2.5. Populációk kölcsönhatásai....................................... 548
18.2.6. Élettörténetek.................................................. 560
18.2.7. Koevolúció...................................................... 562
18.2.8. A populációk létszámának szabályozása .......................... 563
18.3. Életközösségek................................................... 566
18.3.1. Az életközösségek szerkezete ................................... 567
18.3.2. Energiaáramlás.................................................. 574
18.3.3. Biogeokémiai ciklusok........................................... 579
18.3.4. Szukcesszió .................................................... 581
18.4. A bioszféra...................................................... 583
19. TERMÉSZETVÉDELEM (dr. Juhász Lajos)................................ 585
20. VISELKEDÉSTAN (dr. Bakonyi Gábor).................................. 598
20.1. Egyszerű viselkedésformák........................................ 600
20.2. A tanulás........................................................ 603
20.3. A viselkedés idegi és hormonális irányítása...................... 606
20.4. A viselkedés irányításának hierarchiája.......................... 609
20.5. A viselkedés genetikai alapjai................................... 610
20.6. A viselkedés ontogenezise és filogenezise........................ 613
20.7. A domesztikáció hatása a viselkedésre............................ 616
20.8. Szociobiológia................................................... 617
20.9. Az állatok viselkedése csoportokban.............................. 619
20.10. Kommunikáció az állatok között.................................. 624
20.11. A viselkedés optimalizálása..................................... 626
11
21. AZ EVOLÚCIÓELMÉLET ALAPJAI ÉS AZ ÁLLATOK
FILOGENEZISE (dr. Kiss István) ...................................... 630
21.1. Korai evolúciós gondolatok.................................... 631
21.2. Darwin evolúcióelmélete....................................... 631
21.2.1. Az elmélet alaptézisei....................................... 631
21.2.2. Az evolúció bizonyítékai Darwin szerint...................... 632
21.3. Szintetikus evolúcióelmélet................................... 634
21.4. Megközelítési szintek és vizsgálati lehetőségek............... 634
21.5. Mikroevolúció................................................. 635
21.5.1. Az evolúció genetikai alapjai................................ 636
21.5.2. Adaptáció.................................................... 639
21.5.3. Természetes szelekció........................................ 640
21.5.4. Evolúciós ökológia........................................... 643
21.6. A fajok evolúciója............................................ 644
21.6.1. A faj fogalmának értelmezése................................. 644
21.6.2. Izolációs mechanizmusok...................................... 645
21.6.3. Fajon belüli változatok kialakulása földrajzi okok miatt..... 646
21.6.4. A fajképződés típusai........................................ 647
21.6.5. Az evolúciós változások sebessége............................ 650
21.6.6. A fajok kihalása............................................. 651
21.7. Makroevolúció................................................. 651
21.7.1. Makroevolúciós események..................................... 652
21.7.2. A makroevolúció vizsgálatának lehetőségei.................... 655
21.7.3. Globális változások a fauna összetételében................... 658
21.8. Az ember szerepe az evolúció folyamatában..................... 668
21.8.1. A modem emberi populációk evolúciója......................... 668
21.8.2. Az ember hatása más élőlényekre.............................. 669
21.8.3. Evolúció a fogságban tartott állatoknál...................... 670
23. VÁLOGATOTT IRODALOM.............................................. 675
24. TÁRGYMUTATÓ...................................................... 683
12
Előszó
Az állattan (zoológia) fontos alapozó tantárgy az agrár-felsőoktatásban. Az állattani
ismeretek jelentősége napjainkban a környezet állapotának romlása, a biodiverzitás
csökkenése, az új mezőgazdasági technológiák kialakulása miatt egyre nő. A tárgy ok-
tatásához jelenleg új szemléletű, magyar nyelvű tankönyv nem áll rendelkezésre, mert a
Fábián Gyula szerkesztette „Állattan, mezőgazdasági mérnökök részére” című tankönyv
utolsó kiadása 1977-ben jelent meg és régen elfogyott. Jelen könyv az említett munka
szellemi hagyományait követve kívánja összefoglalni az agráregyetemi hallgatóknak a
zoológia tantárgy elsajátításához szükséges korszerű ismereteket. A tankönyv tematiká-
jának kidolgozásában dr. Bakonyi Gábor (GATE), dr. Havasi András (GATE), dr.
Juhász Lajos (DATE), dr. Kiss István (GATE), dr. Palotás Gábor (DATE) és dr.
Sey Ottó (PATE) vett részt. A tankönyv általános alapelveket tárgyal. Részletekben el
nem merülve, áttekintést ad nagy, ma már önálló tudományterületekről (sejttan, szö-
vettan, szervezettan, rendszertan, ökológia, etológia, evolúció). A képzés céljának meg-
felelően szintézisre, az általánosan jellemző jelenségek, elvek bemutatására törekszik.
Napjainkban kiemelt fontosságúnak tűnik az állatvilág és az állatokkal kapcsolatos
jelenségek sokféleségének bemutatása. A mezőgazdasági termelésben az elmúlt években
világszerte elterjedt szabványosított, a helyi sajátosságokat figyelmen kívül hagyó tech-
nológiák katasztrofális következményekhez, a termőterületek leromlásához, elszennye-
ződéséhez vezettek. Világosan látható, hogy a jövőben világszerte a komplex biotóphá-
lózatokon, fajgazdag életközösségeken alapuló termelés az egyetlen lehetséges alterna-
tíva. Új megközelítési elvet jelent e könyvben az, hogy a mezőgazdasági jelentőség mellett
az ökológiai jelentőségre is hangsúlyt helyezünk. Ezért a hazánkban általánosan elterj edt,
gyakori fajokkal, a ritka, védelemre szoruló fajokkal is foglalkozunk. Figyelmet fordítunk
arra, hogy a mezőgazdasági termelésben is speciális, új igények jelentkeznek, amelyek a
hagyományostól eltérő zoológiái ismereteket igényelnek. A biotechnikai módszerek
elterjedése a mezőgazdaságban (ivarsejt és embriómanipulációk), csigák, rákok, békák,
orvosi pióca tenyésztése, szerves hulladék komposztálása, biológiai növényvédelem, nö-
vények tápanyagellátásának segítése a talajfauna kezelésével mind példa arra, hol lehet
az alapozó állattani ismereteket a gyakorlatban hasznosítani. Ezért is szükséges, hogy a
jelenleg közvetlen mezőgazdasági jelentőséggel bíró fajok, csoportok ismertetése mellett
szélesebb kitekintést adjunk a többi csoportokra is.
A tankönyv anyagának összeállítása során figyelembe vettük a középiskolai biológiai
törzsanyagot, elsősorban a gimnáziumokban tanított ismereteket. Amennyiben az le-
hetséges, használjuk az ott bevett magyar szakkifejezéseket, terminológiát és az anyagot
továbbfejlesztve építünk rá. Reményeink szerint azonban a könyv önmagában is érthető
a némi biológiai előképzettséggel rendelkező olvasó számára.
1995. július 16.
A szerkesztő
Előszó a második kiadáshoz
Tankönyvünk második, javított kiadása kerül most az olvasó kezébe. Az első kiadáshoz
képest a rendszertani fejezet változott meg jelentősen. Ebben a kiadásban a Papp L.
(szerk.) (1996): Zootaxonómia jegyzetben található rendszerezést követjük, mint jelen-
leg a legtöbb egyetemen és főiskolán alkalmazott beosztást. Lényegesen átdolgoztuk és
korszerűsítettük a természetvédelmi fejezetet. Az elmúlt évek pedagógiai tapasztalatait
figyelembe véve egy tanulási útmutatóval egészítettük ki a könyvet. Szintén didaktikai
célok miatt illesztettünk minden jelentősebb fejezet elé egy rövid összefoglalót és ezét
tettünk fel a tanulást segítő kérdéseket is a fejezetek végén. Javítottuk az apróbb,
értelemzavaró, vagy technikai hibákat. Ezekre számos kollégánk és hallgatónk hívta fel
a figyelmet. Tanácsaikat itt is köszönöm.
2003. április 2. A szerkesztő
Bevezetés
Az agráregyetemi állattan oktatása keretében régebben csupán azokkal az állatfajokkal
vagy egyéb állatcsoportokkal foglalkoztak, amelyek hasznuk vagy kártételük miatt me-
zőgazdasági szempontból fontosaknak tekinthetők. Az ilyen kategorizálás azonban: ez
hasznos faj, amaz viszont káros, már elavult szemléletmódot tükröz. Napjainkban tel-
jesen egyértelművé vált, hogy a biocönózisokat egységes egésznek kell tekinteni, olyan
rendszernek, ahol az egyes tagok egyformán fontosak a teljes rendszer működése szem-
pontjából, egyik sem hasznos vagy káros, hanem egyaránt szükséges. Világosan látható,
hogy a hagyományosan kártevőnek vagy haszonállatnak számító fajok mellett további
fajok ezrei élnek egy mezőgazdaságilag művelt területen is. Ezen állatok szerepe ma még
sokszor kevéssé ismert. Ugyanakkor számos példát lehet látni arra, hogy a korábban
jelentéktelennek tartott állatok (ragadozó rovarlárvák, trágyagiliszta, szabadon élő
fonálférgek stb.) fontos gyakorlati szerepét felismerik, tevékenységüket tudatosan kezdik
hasznosítani. További szempontot jelent az is, hogy sok agrármérnök fog dolgozni más,
esetleg távoli földrészeken található országokban. Helyes, ha legalább bepillantást kap-
hatnak abba a változatos állatvilágba, amit meg kell majd ismerniük. Mindezek indokol-
ják, hogy ha csupán egy rövid áttekintés erejéig is, de foglalkozzunk olyan törzsekkel,
amelyeknek képviselői nem vagy csupán kis számban élnek hazánkban. Emiatt igen
rövid, átfogó képet adunk a ma ismert állattörzsekről. így egyértelművé válik, hogy a
mezőgazdasági szempontból kiemelkedő jelentőségű állattörzsek csupán egy kisebb
részét képezik a Földön élő összes állatcsoportnak. A könyv súlyponti részét természe-
tesen azok a csoportok jelentik, melyek mezőgazdasági jelentőséggel bírnak.
A könyv írásakor különös nehézséget jelentett, hogy Magyarországon jelenleg nincsen
általánosan elfogadott állatrendszertani beosztás. A Dudich E.-Loksa L: Állatrendszer-
tan című könyvében található tagolás ma már túlhaladott. Ezért arra törekedtük, hogy
a legáltalánosabban elfogadott rendszerekből állítsunk össze egy modem, koherens, nem
túl bonyolult, áttekinthető, az agráregyetemi képzési célnak megfelelő beosztást. A kér-
désről konzultáltunk az egyes csoportok specialistáival, valamint tekintetbe vettük a más
hazai szaktanszékeken oktatott rendszertani beosztásokat. A szakirodalomban természe-
tesen számos más rendszertan is létezik. Hangsúlyozni kell, hogy a csoportok jellemzé-
sénél található fajszámok csak nagyságrendet jelentenek. Azt teszik lehetővé, hogy az
egyes csoportok relatív nagyságát meg lehessen állapítani. A nevek helyesírása és a
magyar nevek használata során Gozmány L.: A magyar állatnevek helyesírási szabályai
című dolgozata és Gozmány L. (1979): Európa állatvilága + hétnyelvű szótár című műve
volt általában irányadó.
A rendszertani kérdések tárgyalása során a csoportjellemzésekre helyeztük a hang-
súlyt. Úgy gondoljuk, hogy ma már számos könyv hozzáférhető, amely a mi rendelke-
zésünkre álló kereteket jóval meghaladóan, bővebb szöveggel, részletesebb rendszertani
beosztást ismertetve, színes ábrákkal kigészítve a fajismeret elsajátítására kiválóan
felhasználható. Ennek következtében a tisztán taxonómiai ismeretek rovására bővítettük
a többi anyagrészt. így a könyvben bemutatásra kerülő fajszám csekély és óhatatlanul
szubjektív. Az előbbi gondolatmenet fokozottan vonatkozik a fajok habitusképeire. A
15
mezőgazdasági gyakorlatban közönséges fajokkal csak érintőlegesen foglalkozunk, mivel
ezekről a hallgatók más tárgyak keretében (állategészségtan, növényvédelemtan, vad-
biológia stb.) részletesen hallhatnak.
Néhány technikai megjegyzés a könyv használatához
Az érthetőség kedvéért az idegen szavakat és kifejezéseket igyekeztünk a lehetőségekhez
mérten csökkenteni. A szövegben a szakkifejezéseket, fajneveket magyarul vagy magyaros
átírásban használjuk, ahol csak lehet. A latin szavak átírásához Brencsán J. (1993):
Orvosi szótár című könyvét használtuk elsősorban. Az első említés alkalmával a magyar
név mellett a latin neveket zárójelbe raktuk és eredeti írásmóddal írtuk. Ezután viszont
általában a magyar vagy magyaros kifejezéseket használjuk. A magyar és latin kifejezé-
seket számban egyeztettük, tehát ahol a magyar kifejezés egyes, illetve többes számban
áll, ott a zárójeles latin kifejezés is annak megfelelő.
Mivel a legtöbb hallgató az egyetemi tanulmányok megkezdése előtt nem tanul
latinul, ezért a következőkben ismertetünk néhány egyszerű szabályt, melyek segítségé-
vel a legtöbb latin kifejezés helyes kiejtése megtanulható. Betűk kiejtése:
ae = é, eh = k, oe = ő, qu = kv, gu = gv, ph = f, s = sz, c = c (e, i, ae, oe és y előtt),
egyébként k, i = j (ha utána magánhangzó következik, egyébként i), ti = ci, ha utána
magánhangzó következik és nem áll előtte s, t, vagy x (egyébként ti-nek kell ejteni) y = i
(görög eredetű nevekben ü).
Felhívjuk a figyelmet arra, hogy a tartalomjegyzék decimális beosztása nem fedi az
egyes csoportok rendszertani besorolását. A rendszertani áttekintés a megfelelő fejeze-
tekben bekeretezve található.
Köszönetnyilvánítások
Külön köszönettel tartozunk lektorainknak gondos munkájukért. Tanácsaik, megjegy-
zéseik nagyban segítették a kézirat színvonalának emelését, a hibák kijavítását.
A tankönyv témaköreit egyeztettük azokkal az agráregyetemi tárgyfelelős kollégákkal,
akiknek a tananyaga szorosan kapcsolódik a könyvben foglalt fejezetekhez, és figyelembe
vettük az általuk javasolt és igényelt módosításokat. A Gödöllői Agrártudományi
Egyetemen a következő kollégákkal konzultáltunk: dr. Bárdos László (sejttan, egyed-
fejlődéstan, szövettan, szervezettan), dr. Heszky László (evolúció), dr. Keszthelyi
Tibor (etológia), dr. Kovács Margit (szünbiológia), dr. Szalai-Marzsó László
(ízeltlábúak, szünbiológia), dr. Péczeli Péter (sejttan, egyedfejlődéstan, szövettan).
Egyes fejezeteket a témában szakértő kollégáink is átolvastak. Fáradságukért, hasznos
tanácsaikért ezúton is köszönetét mondunk a következőknek: dr. Bárdos László (sejttan,
egyedfejlődéstan, szövettan, szervezettan), dr. Bozsik András (Neuropteroidea), dr.
Csányi Sándor (szünbiológia), Csorba Gábor (Mammalia), Csutomé dr. Bereczky
Magdolna (Sarcomastigophora, Apicomplexa, Microspora, Myxozoa, Ciliophora), dr.
Csúzdi Csaba (Annelida), Fábián Miklós (szünbiológia, etológia), dr. Forró László
(Crustacea), dr. Gulyás Pál (Rotatoria), dr. Heszky László (evolúció genetikai
vonatkozásai), dr. Hollósi Gábor (Mammalia), dr. Horváth László (Cyclostomata,
Osteichthyes), dr. Korsós Zoltán (Diplopoda, Chilopoda, Amphibia, Reptilia), dr.
Kovács Gábor (Aves), dr. Kovács Margit (szünbiológia), Kovácsné dr. Murai Éva
(Platyhelminthes, Acanthocephala), László Gyula (Lepidoptera), dr. Mahunka Sán-
dor (Acari), Nagy Péter (Nematoda), dr. Péczeli Péter (sejttan, egyedfejlődéstan,
szövettan), dr. Pintér László (Mollusca), dr. Rózsa Lajos (evolúció), Sinkovitsné dr.
Hlubik Ilona (sejttan, egyedfejlődéstan, szövettan), dr. Sréter Tamás (Sarcomasti-
gophora, Apicomplexa, Microspora, Myxozoa, Ciliophora), Szendrei László (Aves), dr.
Szovátay György (Aves, Mammalia), dr. Turcsányi Gábor (sejttan), Sárospataki
Miklós (szünbiológia), dr. Vásárhelyi Tamás (Heteroptera).
16
Több kolléga segített a rendszertannal kapcsolatos problémáink megoldásában. A
konzultációkért köszönetünket fejezzük ki a következőknek: dr. Kalotás Zsolt (Aves),
dr. Mahunka Sándor (Arachnoidea), dr. Merkl Ottó (Coleoptera), dr. Mészáros
Ferenc (Platyhelminthes, Nematoda), dr. Papp Jenő (Hymenoptera), dr. Papp
László (Diptera), dr. Ronkay László (Lepidoptera), dr. Szalay-Marzsó László
(Sternorrchyncha), Vida Antal (Cyclostomata, Osteichthyes).
A könyv kéziratának technikai előkészítésében munkatársaink Dányi Sándomé, dr.
Menyhért Zoltánná, Tóth Béláné és Fábián Miklós voltak. Segítségüket ezúton is
köszönjük.
Köszönetünket fejezzük ki a Mezőgazda Kiadó és TiMac Bt. munkatársainak
segítőkész munkájukért.
Családtagjaink semmivel nem pótolható segítséget nyújtottak azzal, hogy nyugodt
feltételeket teremtettek munkánkhoz. Türelmük és áldozatos segítségvállalásuk nélkül
ez a könyv nem készült volna el.
1995. június 16.
A szerzők
Hogyan tanuljunk a könyvből?
A tanulás módszertanával sokan foglakoztak már, tanárok, pszihológusok, oktatáskuta-
tók egyaránt. Számos eljárást dolgoztak ki annak érdekében, hogy a befogadók, a
hallgatók ki tudják szűrni a leglényegesebb információkat az olvasottak közül, és azokra
jobban emlékezzenek, mint a módszer alkalmazása nélkül. A következőkben röviden
összefoglalom egy olyan módszer lényegét, amelyet egyetemi tankönyvek anyagának
elsajátítására, tehát viszonylag nagy mennyiségű ismeret befogadására nagyon hatékony-
nak találtak. Mielőtt nekifognak a könyv anyagának tanulásához, érdemes alaposan
áttanulmányozni, majd alkalmazni a következőket!
A tankönyvet 21 fő fejezetre osztottuk. Minden fő fejezetet tekintsen egy oktatási
egységnek. Minden egységet a következő elvek szerint, négy egymást követő lépésben
dolgozzon fel és az itt megadott sorrendben próbáljon elsajátítani. A lépések és azok
sorrendje a következő:
1. Áttekintés; 2. Olvasás; 3. Felmondás; 4. Ellenőrzés.
1. Áttekintés
Futólag nézze át az egész fejezetet. Olvassa el az alfejezetek címeit, próbálja meg
megérteni a fejezet felépítését, nézze meg az ábrákat és táblázatokat, de ne mélyüljön el
a szöveg tanulásában. Ezután a szöveg elején található összefoglalást nagyon alaposan
olvassa el, tanulmányozza át. Ne feledje: itt sűrítve találhatók a fejezet legfontosabb
információi és minden szónak fontos szerepe van. Itt nem fejtjük ki a kérdéseket, csak
felhívjuk a figyelmet a súlyponti, legfontosabb tananyag részekre, ráirányítjuk a figyel-
mét, hogy mit tanuljon meg különösen alaposan a fejezet anyagából. A bevezetés
megértése, átgondolása és alapos elsajátítása nagyon fontos. Gondolkozzon el azon,
hogy miért pont az ott bemutatott ismereteket emeltük ki a többiek közül. Ha valamit
nem ért a fejezetben leírtak közül, azt jelölje meg és a későbbi tananyagrészekben a
vonatkozó ismereteket a többinél alaposabban tanulja meg.
17
2. Olvasás
Olvassa el a fejezetet az alfejezetek sorrendjében. Elsősorban a vastag betűvel kiemelt
szavak, és kifejezések jelentésének megtanulására koncentráljon. Ha szükségesnek látja,
jelöljön ki további szövegrészeket, hogy ismétlésnél már csak ezekre figyeljen. Ne jelöljön
ki többet, mint legfeljebb a szöveg 10-15%-át. Több jelöléssel ugyanis ismétléskor már
nem találja meg a lényeget. Csak azután kezdje el a kijelölést, miután egy alfejezet teljes
szövegét gondosan elolvasta!
3. Felmondás
Miután elolvasott egy alfejezetet, próbáljon meg emlékezni a legfontosabb információk-
ra. Lehetőleg hangosan (ha erre nincs mód, akkor magában) mondja el, hogy mire
emlékezik a fejezetből, majd ellenőrizze a szövegben, hogy valóban jól emlékezik-e? Az
önálló felmondás rendkívül fontos, semmiképpen se hagyja ki ezt a lépést! Elemzések
szerint jó, ha a tanulásra szánt idő 80%-át erre a fázisra, az aktív ismétlésre szánja. A - le-
hetőleg- hangos felmondás tehát a tanulás egyik legfontosabb fázisa. Nem véletlen, hogy
sok előadó, színész, az otthonában, tükör előtt hangosan gyakorolja a szövegét.
4. Ellenőrzés
Amikor egy fő fejezet feldolgozásával és megtanulásával végzett, újra át kell tekinteni az
egész (^pyagot. Olvassa el megint a fő fejezet összefoglalóját és bővítse ki saját maga
számára azokkal az információkkal, amiket a fejezetből megtanult. Nézze át megint a
jegyzeteit, és a fejezetben található, vastag betűkkel kiemelt anyagrészeket. A fejezet
végén található kérdésekre adjon válaszokat. Válaszai legyenek minden részletre kiterje-
dők.
Ha mindezekkel végzett, kezdje el a következő fő fejezet tanulását a bemutatott
sorrend szerint. Ha olyan anyagrészekre, információkra bukkan, amelyeket a tankönyv-
ben már máshol is olvasott, akkor az azt jelenti, hogy a bemutatott információt,
jelenséget, ismeretet különösen fontosnak tartjuk, tehát feltétlenül tanulja meg. Figyel-
jen a határidőkre. Jusson ideje mindegyik fejezet tanulására. A félévközi beszámolók és
zárthelyi dolgozatok segítik abban, hogy helyes ütemben sajátítsa el a tananyagot.
Sok sikert a módszer használatához!
18
1. AZ ÁLLATI SEJTEK FELÉPÍTÉSE
ÉS MŰKÖDÉSE
Az állati szervezet sejtekből áll. A sejtes testfelépítés az élővilág egységének egyik bizonyítéka. Sejtek
építik fel a szöveteket, amelyek a szerveket, a szervezetet alakítják ki. A szöveti kötelékben élő sejtek
saját anyagcseréjükön túl általában speciális, a szövet, a szerv vagy a szervezet számára nélkülöz-
hetetlenfeladatot is végeznek. Alakjuk, nagyságuk, felépítésük ennek megfelelően módosul.
Az állati sejt felszínét, a sejtorganellumok egy részét membrán borija vagy építi fel. A memb-
ránok kettős lipidrétegből épülnek fel. Ebbe a rétegbe különböző arányban eltérő összetételű
fehérjék merülnek, illetve különböző szénhidrátok kapcsolódnak hozzá. A folyékony mozaik-
membránnnak nevezett sejthártya fontos szerepet tölt be a sejt életében. Elhatárol és összeköttetést
biztosit a környezettel, a szomszédos sejtekkel. Különböző membránstruktúrák létesítenek kap-
csolatot a szöveti kötelékben élő sejtek között. A sejthártyán keresztül zajló aktív vagy passzív
transzportfolyamatok biztosítják a sejt anyagcseréjét. A felszínén lévő jelrendszer alapján a sejtek
azonosítani tudják egymást. Az információk felvételében és továbbításában a sejtmembránnak
kiemelt fontossága van. Igen kifinomult rendszer alakult ki az idegsejtekben. A sejt alkotói az
alapállománybanülnek,asejtvázrendszerétfilamentumhálózatokképezik. Az endoplazmatikus
retikulum és a Golgi-készülék membránrendszerekből épül fel. Az endoplazma tikus retikulumnak
a zsírok és fehérjék különböző formáinak szintézisében, a Golgi-készüléknek pedig főleg a fehérjék
tárolásában, átalakításában van szerepe. A lizoszómák és peroxiszómák membránnal határolt
testek, amelyek a sejt működéséhez szükséges enzimeket tartalmaznak. A sejtmag és a mitokond-
riumok membránnal határolt sejtszervek. A mitokondriumok a sejt energiatermelő, raktározó
telepei, a sejtmag pedig a legfőbb koordináló egység, amely a sejt DNS állományát tartalmazza.
A sejtközpontot nem határolja membrán. Szerepe a sejten belüli mozgások és a mozgási sejtor-
ganellumok működésének irányításában van. A sejtek mozgásának különböző formái vannak,
legdifferenciáltabb az izomsejtek működése. A sejtek osztódása a szervezetre jellemző sejtszám
elérése és fenntartása, illetve az ivaros szaporodáshoz szükséges sejtek létrehozása érdekében
zajlik. A testi sejtekre a kromoszómaszám-tartó, az ivarsejtek képződésére a kromoszómaszám-
felező osztódás jellemző.
1.1. Sejtbiológia
A sejtbiológia már régen kinőtt akár a botanikai, akár a zoológiái tudományok közül,
és ma mint önálló tudományág működik. Még így sem nevezhetjük egységesnek, mert
például a sejt biokémiai folyamatainak vizsgálata és a sejtgenetikai kutatások újabb
irányokat jelöltek ki. Éppen ezért jelen állattani könyv keretein belül kizárólag az állati
sejt felépítését mutatjuk be olyan szinten, amely véleményünk szerint a könyv további
fejezeteiben leírtak megértéséhez szükséges. A biokémiai, genetikai, speciális sejt-élet-
tani folyamatok bemutatása más szakkönyvek és tantárgyak keretein belül történik.
Az eukarióta sejtek számos vonásban alapvetően különböznek a prokariótáktól.
Ezek közül a membránpárral határolt, elkülönült sejtmag, a mitokondriumok, valamint
más citoplazmatikus membránból felépülő organellumok megléte a legismertebb. Az
19
eukarióta sejtek nagyobbak (átmérőjük átlagosan 10-100 pm) a prokariótáknál és velük
ellentétben képesek mitózissal vagy meiózissal osztódni. Az állatok, valamint aheterotróf
táplálkozáséi egysejtűek az eukarióták közé tartoznak. A továbbiakban elsősorban a sok-
sejtű állatok sejtjeinek felépítéséről lesz szó.
Az eukarióta sejtek kialakulására, evolúciójára vonatkozóan több elképzelés született,
amelyek közül ma leginkább az endoszimbionta teória elfogadott. Eszerint a fejlett
eukarióta sejt egy ősi eukarióta és prokarióták szimbiózisából alakult ki. Ezt a feltételezést
állati sejtek esetében leginkább a mitokondrium felépítése és működése, saját DNS-
tartalma támasztja alá. A mitokondrium valaha prokarióta sejt lehetett, amely endo-
szimbiózisba került egy másik sejttel. A csiliók és az ostorok is származhattak szimbionta
prokariótáktól. Egy másik elképzelés szerint az eukarióta sejtek az ősi állapotból fo-
lyamatos fejlődés, közvetlen leszármazás útján jöttek létre.
1.2. A sejt membránrendszerei
Valamennyi biológiai membrán lipidekből és fehérjékből áll, de azok aránya igen változó,
így a sejtmembrán és a különböző sejtőrganellumok membránjainak felépítése eltérése-
ket mutat. A lipidek legnagyobb részét különböző foszfolipidek alkotják, bár a glikolipidek,
és a koleszterol előfordulása is jelentős lehet. A szénhidrátokglikoproteidekés glikolipidek
kialakításában vesznek részt. Elsősorban a plazmamembrán külső rétegében fordulnak
elő, míg hiányoznak például a mitokondrium belső membránjából. A membrán lipid-
szerkezete a biológiai membránokban egységes. A kettős lipidrétegben az egyes
lipidmolekuláknak van egy hidrofil része, amely a membrán valamelyik felszíne felé néz,
és van egy hidrofób része. Ez utóbbiak zsírsavak vagy alkoholok szénhidrogénláncai,
amelyek a két réteg egymás felé eső oldalán helyezkednek el és egymással szembe fordul-
va összeköttetést létesítenek.
1.2.1. A plazmamembrán
A felületet borító plazmamembrán elhatárolja a sejteket a külvilágtól, de összeköttetést
is létesít vele. Szabályozza a sejt és a környezete közötti anyagkicserélődést. A plazma-
membrán vastagsága általában 6-12 nm között változik. Felépítésére vonatkozóan szá-
mos elképzelés született, amelyek közül a folyékony mozaikmembrán elmélet a leg-
korszerűbb (1.1. ábra).
A kettős lipidrétegben a lipidmolekulák oldalirányú, laterális diffúziója, azaz egy-
más közötti helycseréje igen gyors folyamat. A nagy mozgásszabadság azonban csak a
saját rétegében igaz, mert a membrán külső és belső lipidrétege közötti helycsere csak
nagyon ritkán megy végbe. Egyes lipidmolekulák nagyfokú hajlékonyságot is mutatnak.
A kettős lipidréteg folyékonysága (fluiditása) függ például a membránlipidek zsírsav-
összetételétől és a plazmamembrán felépítésében részt vevő koleszterolmolekuláktól.
A magasabb telítetlenzsírsav-szint a membrán fluiditását növeli, míg a koleszterol-
molekulák csökkentik. A plazmamembrán külső és belső lipidrétegének összetételében
különbség van. A glikolipidek például csak a külső rétegben fordulnak elő, szénhidrát-
láncaik a sejten kívüli tér felé néznek. A glikolipidek hozzájárulnak a plazmamembrán
külső felszíne negatív töltésének kialakításához.
A membránt felépítő glikoproteideknek két formája figyelhető meg. Vannak globuláris
és a-helix szerkezetűek. A globuláris fehérjék vagy a külső, vagy a belső membránré-
tegbe ágyazottan helyezkedhetnek el, de vannak olyanok is, amelyek az egész memb-
ránt átérik. A membránt átérő fehérjemolekuláknak a különböző anyagok transzport-
folyamataiban van katalizátor szerepük. A lipidrétegbe ágyazott fehérjemolekulák több-
20
1.1. ábra. A sejthártya felépítése, a sejtek kapcsolódási formái és a sejt belső vázrendszerének alkotói
af: aktín filamentumok, ah: alfa helix szerkezetű fehérje, al: alaphártya, fd: féldezmoszóma, fi: foszfolipid molekulák,
gf: globuláris fehérje, gl: glíkolípid, if: intermedier filamentum, ko: koleszterol, mb: mikrobolyhok, mt: míkrotubulus,
ol: oligoszacharid oldallánc, öd: övszerű dezmoszóma, pb: plazmamembrán-betüremkedés, pd: pontszerű dezmoszóma,
pm: plazmamembrán, rk: réskapcsolat, sb: sejtburok, sv: sejtváz, th: terminális hálózat, tr: transzmembrán fehérje, sz: szo-
ros kapcsolódás (Damell-Lodish-Baltimor és más szerzők nyomán módosítva)
21
sége korlátozottan képes oldalirányú elmozdulásra. A plazmamembrán külső felszíni
fehérjéihez szénhidrátláncok kapcsolódnak. A glikoproteidek közül számos receptor
funkciót tölt be, azaz megköti és érzékeli a speciális kémiai hírvivő anyagokat, például
a hormonokat.
A membránfehérjékhez és lipidekhez kapcsolódó oligoszacharid oldalláncok a sejt
felszínén kialakítják a sejtburkot (glycocalix) (1.1. ábra). A glikokalix speciális össze-
tétele révén az adott sejttípusra jellemző azonosító jelet (marker) képez a külvilág, a
környező sejtek számára. A markerek egy másik sejt receptorai számára jelentenek infor-
mációt. A markerek egy része faj-, más része egyedspecifikus. Ezeken túl vannak olyan
markerek, melyek az egyedfejlődés során töltenek be fontos szerepet. A sejt-sejt kapcso-
latot közvetítő markerek az úgynevezett celluláris adhéziós molekulák. A vörösvértestek
felszínén lévő glikokalix fehérjéi hozzák létre például a vércsoportok különbségét. Ugyan-
csak ilyen fehérjék biztosítják a szervezet immunrendszerének működési alapját, amikor
a védekező sejtek felismerik a testidegen sejtet, illetve anyagot. A specifitáson túl a
szénhidrát-összetevők meghatározzák a sejt felületi töltésviszonyait is. A glikokalix sze-
repet játszik a különböző anyagok felvételében, valamint mechanikai védelmet is nyújt
a sejt számára.
A plazmamembránból a sejt citoplazmája felé benyúló fehérjék kapcsolódnak a sejt
belső vázrendszeréhez. A plazmamembrán alatti filamentumok sejtkérget kialakítva
erősítik a sejthártyát.
A szöveti kötelékben élő sejtek egymáshoz való kapcsolódását különböző membrán-
struktúrák teszik lehetővé (1.1. ábra). Alakja szerint ez lehet sejten körbefutó „övecske”
(zonula), illetve kis területre kiterjedő, szabályos alakú, „szeplő” (macula) típusú kap-
csolódás. A sejtkapcsoló struktúrák hámsejtek esetében a legdifferenciáltabbak.
A hámsejtek szabad felszínéhez közel, övszerűen körbefutó szoros kapcsolódás
(zonula occludens) figyelhető meg. Itt az érintkező sejtek szorosan összeérnek. A sejt-
membrán fehérj éit C a2+- és Mg2+-ionok kapcsolj ák össze. Ez a kapcsolódási forma megaka-
dályozza a szabad diffúziót a két sejt közötti résben. A szoros kapcsolódás a hámmal
határolt terek között teremti meg a sejten keresztül végbemenő (transzcelluláris) transz-
portot, míg a szomszédos sejtek közötti (intercelluláris) diffúziót nem teszi lehetővé. Ez
valamennyi hámban fontos, így például a bélbolyhok felszínét borító tápanyagfelszívó
sejtekben és az emésztőenzimeket termelő mirigyekben.
A különböző típusú dezmoszomális kapcsolódásokban a sejtek membránjai között kis
tér van.
Az övszerű dezmoszóma (zonula adherens) területén a sejtek között 15-20 nm-es
távolság van, amit amorf anyag tölt ki. A két sejt találkozási felülete közötti összeköttetés
kevéssé ismert. A szomszédos sejtek citoplazmatikus felszínéhez aktintartalmú fila-
mentumok csatlakoznak. Az aktin filamentumok ebben a szintben más sejtvázfehérjék-
kel kapcsolódva az úgynevezett terminális hálózatot alakítják ki.
A pontszerű dezmoszóma (macula adherens) típusnál a két sejt azonos pontjain,
a membránhoz illeszkedő, ellipszoid alakú korongokból kiinduló, a membránon is átnyúló
glikoproteid szálak kapcsolják össze a sejteket. A két sejt között mintegy 30 nm a távolság.
A sejten belüli, korongszerű plazmatömörülési helyekről speciális fehérjeszálak, tonofila-
mentumok indulnak ki, amelyek a sejt belső vázrendszeréhez kapcsolódnak. Ilyen sejt-
kapcsolódás a hámsejtek között tipikus.
A féldezmoszóma vagy hemidezmoszóma az előző típushoz hasonló felépítésű, de
nem a sejtek közötti kapcsolatot, hanem a sejtek alaphártyához való rögzülését teszi
lehetővé. Ez fordul elő például az egy rétegben álló hámsejtekben, illetve a többrétegű
hámok legalsó sorát alkotó sejtjeiben.
A réskapcsolat (macula communicans) a sejtek közötti információcserét teszi lehe-
tővé. Az információ átadás a fehérjemolekulák által kialakított csatornákon keresztül
22
megy végbe a két sejt citoplazmája között. A csatornák felépítésében hat fehérjemolekula
egység vesz részt. Kis molekulájú anyagok, ionok, aminosavak, valamint az ATP juthat-
nak át egyik sejtből a másikba. Fontos szerepe van a Ca2+-ioneloszlás kialakításában.
Lehetővé teszi az elektromos impulzusok gyors továbbítását. Ilyen kapcsolat jellemző az
elektromos szinapszisban, illetve például a hámsejtek, a szívizomsejtek, valamint a csont-
sejtek között.
Számos, a szervezet belső üregrendszerét határoló sejt felszínén speciális nyúlványok,
mikrobolyhok (microvilli) találhatóak. Mikrobolyhok elsősorban a felszívóhámok (pél-
dául bélhám, vesetubulushám) sejtjein figyelhetőek meg. A bélhámsejtek felszínét nagy
számbanborító mikrobolyhok átmérője 100 nm körüli. Óriási felületet hoznak létre, ami
a tápanyagok felszívásában jelentős tényező. Membránjuk számos transzportfehérjét
tartalmaz, melyek a tápanyag-molekulák sejtbe való bejutását tesziklehetővé. A bélhám-
sejt plazmamembránja enzimeket (peptidázok és a glikozidázok) is tartalmaz, melyek a
bélcsőben történő enzimatikus bontást befejezik. A mikrobolyhok állandó átmérőjét,
alakját a főleg aktin filamentumokból felépülő tengelyvázuk adja meg, melyeka sejt belső
vázrendszerével vannak kapcsolatban. A mikrobolyhokban található aktinszálakkal több
fehérjemolekula (például viliin, fimbrin) is kapcsolatot létesít. Az aktin filamentumköteg
a mikrobolyhok membránjához és az alapjuknál lévő aktinmiozin filamentumokból álló
terminális hálózathoz kapcsolódik (1.1. ábra és 4.5. ábra). Ez a hálózat kapcsolatban
van az övszerű dezmoszómákkal is. A mikrobolyhok a bennük lévő filamentumok révén
képesek megrövidülni, illetve megnyúlni.
A sejtek alapján felületnövelő képződményeket, membránbetüremkedéseket lehet
találni (1.1. ábra). Ezek egyrészt a rögzítés lehetőségét növelik, másrészt az anyagfelvé-
telt segítik elő. Ilyen képződmények vannakpéldául a mirigyhámsejteken és a vesecsatomák
hámsejtjein.
Transzportfolyamatok a sejthártyán keresztül
A plazmamembrán egyes anyagok bejutását vagy távozását megkönnyíti, míg másokét
megnehezíti vagy teljes mértékben megakadályozza.
A passzív transzport során a sejt nem fektet be energiát a plazmamembránon ke-
resztül végbemenő anyagforgalomba. A membrán - kettős lipidrétegének szerkezete
miatt - a vízoldékony anyagok számára átjárhatatlan (impermeábilis). Egyes apoláris és
töltés nélküli kicsi molekulák (például az oxigén, a szén-dioxid, a víz) azonban szabad
diffúzióval jutnak át rajta. A gyorsított passzív transzporthoz sem kell külön ener-
giabefektetés a sejt részéről. Ilyenkor az anyagok (például a glükóz) bejutását hordozó
fehérjemolekulák segítik elő. Ezek a fehérjék megkötik a transzportálandó anyagot, majd
konformációváltozásuk révén csatornát képezve bejuttatják azokat a sejtbe.
Az aktív transzport energiát igénylő folyamat, melynek során a magasabb koncentráció
irányába mozdulnak el az anyagok. A hordozó fehérjemolekulák általában igen specifi-
kusak, csak meghatározott anyagokat, szubsztrátumot szállítanak. Energiát igénylő fo-
lyamatok révén valósul meg a makromolekulák felvétele is. A Na+- és K+-pumpa a
legáltalánosabban előforduló aktív transzportfolyamat, amely bizonyos sejtek energia-
termelésének 10-40%-át is felemészti. A legtöbb állati sejt magas sejten belüli K+-kon-
centrációt igényel. Ez a riboszómákon végbemenő fehérjeszintézishez és bizonyos enzi-
matikus funkciókhoz nélkülözhetetlen. A Na+-ionok sejten belüli koncentrációja viszont
csak mintegy tizede a sejten kívülinek. Ez a nagy különbség csak aktív transzport révén
tartható fenn. Az aktív transzportot mindkét ion számára ugyanaz a transzport-fehérje-
komplex is lehetővé teheti, mint például a Na+-K+-csere esetében. Különösen intenzíven
megy végbe ez a folyamat például az idegsejtek esetében, ahol szerepe van a membrán
elektromos potenciáljának, polarizáltságának fenntartásában, illetve megváltoztatásá-
ban (1.2. ábra).
23
1,2. ábra. Elektromos potenciálváltozások az idegsejt plazmamembránján, valamint az ingerület terjedésének és
átadásának folyamata
am: axon membránja, ap: akciós potenciál, ax: axon, de: dendrit, EL: elektromos szinapszis, hp: hiperpolarizáció,
iá: ingerületátvivő anyagok, IC: ingerületvezetés a csupasz axonon, if: intermedier filamentum, ih: ingerület haladásának
iránya, IS: idegsejt, IV: ingerületvezetés a velőhüvelyes axonon, K: K*-ionok kiáramlása, KE: kémiai szinapszis, mt: mik-
rotubulus, Na: Na*-ionok beáramlása, ny: nyugalmi potenciál, pm: posztszinaptikus membrán, pr: preszinaptikus membrán,
Ra: Ranvier-féle befuződés, re: repolarizáció, ré: szinaptikus rés, rk: réskapcsolat, sm: sejtmag, vf: végfácska, vh: velőshüvely
(Damell-Lodish-Baltimor, Dórit és más szerzők nyomán módosítva)
24
A passzív és az aktív transzport során vízmolekulák is áramlanak. A víz az ozmotikus
viszonyoknak megfelelően áramolhat és helyezkedik el a membrán által határolt két
térben.
A transzportfolyamatok lehetnek egy-, illetve kétirányúak. Előfordul, hogy az egyik
molekula bejutása egyben egy másikét is elősegíti, amit kotranszportnak nevezünk.
Az aktív transzportfolyamatok speciális esete az, amikor a sejt képes szilárd vagy
folyékony anyagok felvételére. Ennek során azokat plazmamembránba csomagolja, így
kerülnek a felhasználási helyükre. A folyamat összefoglaló neve az endocitózis. Az
endocitózis lehet fagocitózis, pinocitózis és receptorközvetítésű endocitózis. A fagocitózis
adszorpcióval kezdődik, majd az így plazmamembránhoz kötött konzisztens szemcsét a
sejt betűri magába. Az így képződött hólyag leválik a sejthártyáról és bekerül a citoplaz-
mába (1.3. ábra). Itt az enzimeket tartalmazó lizoszómák segítségével végbemegy az
emésztés. Nagyméretű táplálékrészecskék esetén a membrán nyúlványokat növeszt, ezek-
kel körülveszi azokat, míg a kis szemcsék észlelésekor csak betüremkedés történik. A pi-
nocitózis a fagocitózishoz hasonlóan megy végbe azzal a különbséggel, hogy ebben az
esetben folyadékot vesz fel a sejt a besüllyedő és hólyaggá záródó plazmamembránrész
segítségével. A folyadék felvétele egyben az abban oldott anyagok bejutását is jelenti.
A receptor közvetítette endocitózis során a sejt membránjának egyes receptorfehérjéi
az általuk megkötött anyagokkal (szabályozó vagy tápanyag-molekulákkal) együtt kerül-
nek endocitotikus felvételre, majd feldolgozásra. Az exocitózis az endocitózissal ellen-
tétes irányú folyamat, amikor a sejt kilöki magából a káros anyagcsere-végtermékeket
(excretio) vagy éppen a szervezet számára fontos anyagokat, például hormonokat és
végső metabolitokat (secretio) (1.3. ábra).
A plazmamembrán szerepe az információcserében
Minden sejt ad jelet a környezetének és minden sejt egyben célsejtje is általában több
jelnek. A többsejtű állati szervezetekben a sejtek egy része közvetlen, sejt-sejt közötti
j elcserét folytat. A már említett réskapcsolat révén lehetőség van a koordináló vegyületek
egy részének átadására is. Az egymástól távolabb lévő sejtek között ilyen formában
viszont már nem valósítható meg az információátadás, ezért speciális jeladó sejtek
jöttek létre, amelyek például hormont termelnek. A jelek felfogása, feldolgozása és a
válaszreakció a legtöbb sejtben alapvetően hasonló módon megy végbe. A folyamatban
részt vevő enzimrendszerek, közvetítő anyagok jórészt azonosak vagy nagyon hasonlóak.
Az extracelluláris jelek felvételére a fogadósejtek membránjába épült, kizárólag az adott
vegyület megkötésére alkalmas speciális receptorfehérjék képesek. Az információt hor-
dozó molekulák hidrofób és hidrofil tulajdonságaik alapján csoportosíthatóak. A hidro-
fóbok, mint például a szteroid hormonok, a vérplazma fehérjéihez kötődve jutnak el a
sejtekhez. A hordozó fehérjékről leválva átjutnak a membránon és az intracelluláris
receptorfehérjékhez kapcsolódnak. Ez utóbbiak tulajdonságai megváltoznak, képesek a
DNS-hez kötődni, így annak működését befolyásolni. A polipeptid hormonok, amino-
sav-származékok és biogén aminok hidrofil jelhordozók, amelyek a sejtmembrán felszí-
nére kinyúló speciális receptorfehérjékhez kötődnek, a membránon nem tudnak
átjutni (lásd még 4.7.1. fejezet).
A sejt felszínét ért elektromos vagy kémiai inger újabb, sejten belüli hírvivő rend-
szer működését indítja be. Ilyen inger például a kalciumion-kon centráció gyors meg-
változása, vagy a ciklikus adenozin-monofoszfát hatása. A sejt kalciumkoncentráció-
ját elsősorban a sejthártyába épült fehérjék (kalciumcsatomák) szabályozzák. Ezek a
fehérjék a membrán egyik oldalán lévő kalciumot megkötik és a másik oldalra viszik át.
A sejtekbelső terükben alacsony kalciumszintet tartanakfenn, amelynek mértéke mintegy
tízezred része a sejten kívüli térben mérhető szintnek. A kalciumszint kismértékű válto-
zása igen sok enzim működését szabályozza. A kalcium rövid idő alatt és nagyobb mennyi-
25
ségben az akciós potenciál kialakulása során megnyíló kalcium csatornákon keresztül jut
be. A kalciumionok sejten belüli aktivitását speciális fehérjék szabályozzák. Ezek meg-
kötik, így inaktiválják, tárolják a kalciumot, illetve inger hatására felszabadítják azt. Az
aktívvá vált kalcium ilyenkor képes információ továbbításra. A sejten belüli kalciumkötő
fehérjék kötést létesítve vagy közvetlenül fejtik ki hatásukat, vagy más enzimeket hoznak
működésbe. A sejten belüli térben a szabad kalciumszint fő szabályozói az endoplazmatikus
retikulum és a mitokondriumok membránrendszeréhez kapcsolódnak, illetve a citoplaz-
ma alapállományában vannak. Az intracelluláris kalciumkötő fehérjék közé tartozik pél-
dául a troponin-C, amely az izom-összehúzódás kiváltásában játszik szerepet. A sejten
belüli információ továbbításában leggyakrabban a kalmodulin nevű kalciumkötő fehér-
je kap szerepet. Számos enzim működését szabályozza, így például az izomműködéshez
szükséges energia felszabadítását végzőkét is. Az endoplazmatikus retikulum a kalcium
sejten belüli elosztásának, koncentrációjának gyors szabályozását biztosítja. Ez különö-
sen jól megfigyelhető például az izomrostok szarkoplazmatikus hálózata által alkotott
tubulusrendszer működésében.
Az idegsejtek membránja speciális feladat ellátását végzi, részt vesz az intercelluláris
kommunikációban. A sejtek (neuron) felépítése ennek megfelelően erősen módosult
(1.2. ábra). A fajra jellemző idegsejtszám elérésekor elvesztik mozgási és osztódási ké-
pességüket, megtelepedve nyúlványokat növesztenek. A sejttestben és a nyúlványokban
található fehérjeszálak, neurofilamentumok és csövek, mikrotubulusok elősegítik a mak-
romolekulák, fehérjék, hólyagocskák (vesiculae), mitokondriumok mozgatását nemcsak
a sejttest területén, hanem az axon teljes hosszában is egészen a végfácskáig (telodendron).
Ez az úgynevezett axoplazmatikus transzport mindkét irányban folyik, a végfácska
működéséhez feltétlenül fontos.
A különböző állatfajok idegsejtjeiben az idegi impulzus létrejötte és terjedése alap-
vetően egyforma. Az idegsejtek - más sejtekhez hasonlóan - aktív, energiát igénylő transz-
portfolyamat eredményeként a sejten belüli és sejten kívüli tér között átlagosan mintegy -
70 mV értékű feszültségkülönbséget tartanak fenn. Ez a sejt nyugalmi potenciál ér-
téke. A sejten belül mintegy 30-40-szer több K+-ion van, mint kívül, illetve a sejten kívüli
térben 10-szer nagyobb a Na+-ionok mennyisége, mint belül. Intracellulárisan jelentős
mennyiségű és kulcsszerepet játszó negatív töltésű fehérjemolekula, valamint különböző
anionok találhatók. A membránt ért helyi inger hatására megváltozik a membrán áteresz-
tőképessége. Nátriumionok hatolnak be, csökkentve a sejt külső térhez viszonyítva negatív
töltését. Egy küszöbértéket (ingerküszöb) elérve a nátriumcsatomák megnyílnak, rövid idő
(a millisecundum egyötöd része) alatt igen nagy mennyiségű Na+-ion hatol be a sejtbe. Az
ingerlés helyén bekövetkezik az előzőleg polarizált membrán depolarizációja, létrejön az
akciós potenciál. Ez azt jelenti, hogy a sejt belső tere a külső térhez viszonyítva átmene-
tileg pozitív töltésűvé válik. Értéke mintegy +30, +40 m V csúcspotenciált is elér. A küszöb-
érték alatti erősségű inger nem vált ki akciós potenciált, míg felette, az inger erősségétől
függetlenül kialakul a csúcspotenciál. Ez a minden vagy semmi elvének érvényesülése.
Az akciós potenciál a mérés pontján nem tart tovább mint fél milliszekundum, majd a sejt
az energiaigényes K+-Na+-pumpa segítségével elkezdi visszaállítani az eredeti, polarizált
membránállapotot. A nátriumcsatomák becsukódnak, de a káliumcsatomák kezdetben
még nyitottak. így káliumionok jutnak ki a sejtből, ami a sejten kívüli tér pozitív töltését
emeli. A membrán- eredeti polarizáltsága visszanyerésének (repolarizáció) ideje alatt- az
adott területen nem képes ingerek felvételére, úgynevezett refrakter állapotba kerül. Az
akciós potenciált követően a membrán rövid időre hiperpolarizálódik (értéke a nyugalmi
potenciál alá csökken), mert a sejtet igen nagy mennyiségű kálium hagyta el. A sejt nát-
rium-kálium-pumpája végül is visszaállítja az eredeti nyugalmi potenciált. A membrán egy
pontján létrejött akciós potenciál a sejt szomszédos területei számára jelent elektromos
ingert, így az ingerület a membrán felületén végigfut. A csupasz axon membránján az
26
ingerület pontról pontra terjed. Ebben az esetben a vezetés gyorsasága elsősorban az axon
vastagságától függ. Ennek oka az, hogy a beáramló és a membrán belső felszínén előre
áramló Na+-ionok megnövelik a depolarizáció sebességét. Minél nagyobb az axon átmé-
rője, annál több Na+ áramlik be azonos időegység alatt, és így az előre terjedő depolarizáció
sebessége felgyorsul. Ilyen óriásrostok elsősorban a gerinctelenekben fordulnak elő (gyűrűs-
férgek, rákok, tintahal), de megfigyelhetők egyes halak és kétéltűek idegrendszerében is.
Az ingerületvezetés gyorsításának másik lehetősége az axon körüli, elektromosan szigetelő
védőburkok kialakulása (3.4. fejezet). A speciális gliasejtek többszörösen feltekeredett
plazmamembránjából kialakult mielinhüvely nem folytonos, a Ranvier-féle befűződések-
nél az axon membránja kis területen csupasz. Az itt keletkezett helyi áramkör a mielin-
hüvelynek az axonmembránnál nagyobb elektromos ellenállása miatt nem tud a szomszé-
dos membránrészletre átterjedni, hanem a következő befűződéshez átugorva hozza inge-
rületbe a membránt. így a membrán csak kis területen depolarizálódik, a Na+-K+-pumpa
működéséhez kevesebb energia szükséges, és az ingerületvezetés is gyorsabb, mint a csu-
pasz axon esetében.
Az ingerület egyik idegsejtről a másikra vagy a célsejtre történő átadásának helye a
szinapszis. Két formája ismert: az elektromos és a kémiai. Az elektromos szinapszis
nem más, mint a már említett réskapcsolat, ahol a két sejt között az ionáram szabadon
áthalad. Akémiai szinapszisban az ingerületet ingerületátvivő anyagok közvetítik a két
sejt között. Az ingerületátadó, preszinaptikus neuron által hozott impulzus a végfácska
végződésében kiváltja a szinaptikus hólyagokban tárolt ingerületátvivő anyag (például
acetilkolin) exocitotikus leadását. Az ingerületátvivő anyag bejut a 20-30 nm-es szinaptikus
résbe, majd kapcsolódik a fogadó, posztszinaptikus membrán receptor molekuláihoz.
Ha elegendő receptorra kapcsolódik acetilkolin, akkor az ingerületet vált ki, amely a
membránon továbbhalad. Az acetilkolin az acetilkolin-észteráz révén hamar lebontódik
és ezt követően visszajut a preszinaptikus idegsejtbe. Ha az enzim munkája gátolt (pl.
szervesfoszfát-tartalmú inszekticidek, egyes harci gázok által), akkor az acetilkolin folya-
matosan fejti ki stimuláló hatását, ami igen veszélyes a szervezet számára. Az acetil-
kolinon kívül a noradrenalin az előbb említett serkentő, míg a gamma-aminovajsav a
gátló szinapszisokban fordul elő. Ez utóbbi hiperpolarizációt vált ki, ami meggátolja
a membránt az újabb ingerek felvételében. Az egyes idegsejtek membránján serkentő és
gátló szinapszisok egyaránt előfordulnak. Ezek együttes hatása dönti el az idegsejt reak-
cióját. A serkentő szinapszisokfőleg az axon-dendrit kapcsolatokra jellemzőek. Az ismert
ingerületátvivő (neurotranszmitter) anyagok száma már az ötvenet is meghaladja. Az
eddig felsoroltakon túl ismertebb még a szerotonin, a dopaminok, az enkefalinok és az
endorfinok. Az intemeuronális szinapszisokon túl vannak ideg-izom ingerületátadási
helyek, azaz neuromuszkuláris szinapszisok is.
1.2.2. A sejt belső membránrendszerei és membránnal határolt
sejtszervecskéi
A sejten belüli térben található membránok burkolják a belső szervecskéket és ezen túl
óriási felül etet hoznak létre, amely az enzimatikus folyamatok színtere. Becslések szerint,
egy gramm élő szövetben lévő valamennyi membránfelületet egymás mellé helyezve,
azok összfelülete elérné a harminc négyzetmétert is.
Az endoplazmatikus retikulum bonyolult, lemezekből, hólyagocskákból, csövek-
ből álló membránrendszert alakít ki a sejten belül (1.3. ábra). Felületi struktúrája alapján
két típusa van. Főleg a sima felszínű endoplazmatikus retikulumon szintetizálódnak azok
a zsírsavak, a foszfolipidek és a koleszterol, melyek a membrán felépítéséhez szükségesek.
A sima felszínű endoplazmatikus retikulumnak szerepe van a detoxikálásban is,
amely különösen a májsejtekben kifejezett. A szemcsés felszínű endoplazmatikus
27
1.3. ábra. Az állati sejtet felépítő néhány sejtorganellum
cs: csöves szerkezetű mitokondrium, di: diktioszóma, en: endocitózis, el: enzimtároló lizoszóma, ex: exocitózis, hm:
harántlemezes mitokondrium, ho: hosszanti lemezes mitokondrium, lm: lebontandó mitokondrium, mb: membrán-
betüremkedés, mh: maghártya, ml: táplálékszemcsét bontó másodlagos lizoszóma, mt: maradványtest, öe: önemésztő
másodlagos lizoszóma, se: sima endoplazmatikus retikulum, sh: sejthártya, sk: sejtközpont, sm: sejtmag, sv: sejtmagvacska,
sz: szemcsés endoplazmatikus retikulum (Több szerző nyomán módosítva)
28
retikulumon riboszómák ülnek. A szemcsés endoplazmatikus retikulum elsősorban
azoknak a fehérjéknek a szintézisében vesz részt, melyek a sejtből leadásra kerülnek,
illetve amelyek a membránok felépítéséhez szükségesek. Az üregrendszerébe bekerült
polipeptidekre oligoszacharid oldalláncok épülnek rá. A plazmában szabadon lévő
riboszómák pedig főleg a sejt saját felhasználására kerülő fehérjéit termelik. Az
endoplazmatikus retikulumon szintetizálódott fehérjékhólyagocskákba, vezikulákba ke-
rülnek, amelyek kapcsolatba lépnek a Golgi-készülék membránrendszerével (1.3. ábra).
A riboszómák a fehérjeszintézis helyei. Itt kapcsolódnak össze a szállító RNS-ek által
hozott aminosavak a hírvivő RNS-ek által meghatározott sorrendben. Egy részük az
endoplazmatikus retikulum membránrendszeréhez kapcsolódik, míg mások a citoplaz-
mában szabadon vannak.
Szinte valamennyi állati sejtben a Golgi-apparátus több Golgi-halmaz (dictyosoma)
összekapcsolódásából, hálózatából áll (1.3. ábra). Az egyes diktioszómák sima felszínű,
középen összesimuló, a végükön kiszélesedő zsákocskákból (cistema) állnak. A ciszter-
nák átmérője egy mikrométer körüli. A fehérjék tárolásában, átalakításában, csomago-
lásában és osztályozásában játszanak szerepet. Működésük főleg a szekréciós sejtekben
kifejezett. Az endoplazmatikus retikulum felől vezikulákba zárva érkező fehérjék a Golgi-
készülékben feldolgozódnak és újabb szénhidrátokkal egészülnek ki. Ezzel befejeződik
a glikoproteid-szintézis. A Golgi-készülék szerkezetük alapján válogatja a fehérjemole-
kulákat, például a szekréciós fehérjéket elkülöníti a lizoszomális fehérjéktől. Az elkészült
termék egy része szekrétumként (szekréciós vakuólákban) a sejtből leadásra kerülhet,
más részük a sejtben marad és elsődleges lizoszóma formájában ismert.
A lizoszómák (lysosoma) membránnal határolt, emésztőenzimeket (hidrolázokat)
tartalmazó testek. Az újonnan képződött elsődleges lizoszómák a Golgi-készülékről
fűződnek le, általában apró vezikulák formájában. Az enzimek potenciális veszélyforrást
jelentenének a sejt számára. Egészséges sejtekben azonban ezek az enzimek a lizoszómák
membránjával körülvéve, biztonságosan tárolódnak. Működésükhöz savas közeg (pH 3-
5) szükséges, ami a lizoszómán belül rendelkezésre áll. Ezt a körülményt tf lizoszómák
folyamatosan működő protonpumpája teszi lehetővé, amely a belső térben a H+-ionok
felhalmozását eredményezi.
A fagoszómák plazmamembránnal körülvett vakuólák vagy vezikulák, amelyek az
endocitózissal felvett anyagokat tartalmazzák. Az autofág vakuólák a sejt saját, le-
bontandó alkotórészeit tartalmazzák. Ezek membránj át feltételezhetően a Golgi-készülék
vagy az endoplazmatikus retikulum képezi. Az enzimhordozó primér lizoszómákösszeol-
vadnak a fentiekben említett emésztendő anyagot tartalmazó testekkel, és megindul a
bennük foglalt anyag lebontása. Az így kialakuló hólyagok a másodlagos lizoszómák.
Az emészthetetlen anyagok az úgynevezett harmadlagos lizoszómákban halmozód-
nak fel. Az itt lévő anyagok exocitózissal leadásra kerülhetnek, vagy a sejten belül ma-
radványtestekként (reziduális testekként) tartósan megmaradhatnak (1.3. ábra).
A peroxiszómák membránnal határolt vezikulák, amelyek nagyszámú és sejttípu-
son-ként különféle enzimet tartalmazhatnak. Enzimfehérjéik a szabad riboszómákon
szintetizálódnak, és azokat utólagos importtal veszik fel a peroxiszómák. Oxidázaik sok-
féle szubsztrátumot oxidálnak, többnyire dehidrogénezéssel, hidrogén-peroxidok kép-
ződése közben. Ehhez légzési oxigént használnak fel. Peroxidáz enzimük a hidrogén-
peroxidot vízzé redukálja, így a sejt számára nem jelent veszélyt. A peroxiszómák emlős-
állatoknál különösen a máj és vese sejtjeiben gyakoriak, ahol méregtelenítő (detoxiká-
ciós) szerepük a legkifejezettebb. Részt vesznek a hőtermelésben, mert ellentétben a
mitokondriumok hasonló működésével, az oxidációs folyamataik kémiai energiája nem
épül be ATP-be, hanem teljes egészében hővé alakul.
A mitokondriumok méretben, alakban és számukat tekintve igen változatosak
lehetnek (1.3. ábra). Vannak pálcikaszerűek, de gömbölydedek is. A sejtben ott
29
helyezkednek el, ahol nagy a metabolikus aktivitás. A mitokondrium kettős falú: a külső
membrán sima, míg a belső a felületet jelentősen megnövelő lemezeket, vagy ujjszerű
nyúlványokat képez. E képletek alapján beszélhetünk például hosszanti és harántlemezes,
valamint csöves szerkezetű mitokondriumokról. A belső membránban található az
energiafelszabadító, aerob metabolitikus folyamatokhoz szükséges enzimkészlet. Itt kép-
ződik és tárolódik az adenozin-trifoszfát (ATP), ezért a mitokondriumot a sejt „energia-
termelő telepének” szokták nevezni. A mitokondrium részleges önreprodukcióra képes,
mivel van saját DNS-e. Ez a prokariótákra jellemző gyűrű alakú DNS megkettőződik
minden mitokondrium-osztódás előtt. Az egyed szaporodásakor is átadásra kerül a
mitokondriális DNS. Csak a petesejtben lévő jut tovább a következő generációba, míg a
hímivarsejté nem, mivel az a megtermékenyítéskor nem jut be a petesejtbe. A mitokond-
rium fehérjéinek szintézisét részben a saját, részben a sejtmagban lévő DNS kódolja.
A sejtmag (nucleus) két membránnal határolt, általában gömb vagy tojásdad alakú
szervecske (1.3. ábra). A sejtmaghártyája pórusokkal átjárt, amelyen keresztül anyagok
átjutása könnyen lehetséges. A póruskomplexeket globuláris és fibrilláris fehérjék alakít-
ják ki, és irányítják az anyagok átjutását. A sejtmag tartalmazza a kromatinállományt, ami
a genetikai információt magában hordó DNS-ből, valamint a bázikus hiszton és savanyú
nonhiszton fehérjék komplexéből áll. A hisztonok feladata a DNS-molekula védelme,
struktúrájának kialakítása, az RNS-átírás megakadályozása. A nonhiszton fehérjék kö-
zött vannak regulátorfehérj ék és enzimek, amelyek a génállomány működésének szabá-
lyozását végzik. A DNS aktív szakaszán fordulnak elő, felszabadítják őket a hisztonok
gátló hatása alól. A nonhiszton fehérjék a plazmában betöltött vázalkotó szerepük mellett
a kromatinállomány kondenzálásában is részt vesznek. A kromoszómák a kromatin-
állomány transzportformái, a kromatint szállítható formába szervezve, erősen konden-
zált formában tartalmazzák. A sejtmagban található az egy vagy több sejtmagvacska
(nucleolus), amely egyes kromoszómák specializált része. A riboszomális RNS szintézi-
séhez szükséges gének másolatait foglalja magába. Az itt átíródó (szintetizálódó) ribo-
szomális RNS-fehérjékkel kapcsolódva riboszóma-alegységeket alkot, amelyek a sejt-
mag pórusain át kijutnak a citoplazmába és ott funkcióképessé válnak.
1.3. A sejt alapállománya és vázrendszere
A membránorganellumok a citoplazma alapállományába (cytoplasma mátrix) van-
nak beágyazva. Ez főleg fehérj ék által alkotott, gél állapotú állomány, de benne találhatjuk
a riboszómákat és a különböző RNS-molekulákat is. Az alapállomány része a sejtnedv
is, amelynek egy részét a víz és az abban oldott kis molekulájú anyagok képezik. A víznek
oldó, hidratáló, transzport-elősegítő képessége révén fontos szerepe van a sejt anyag- és
eneriaforgalmának szinte minden területén. Az alapállományban található az anyagcsere
köztes és végtermékeinek egy része is. Jelentős a fehérjetartalma, amely elérheti a sejt
összes fehérjetartalmának 20-30%-átis. Az itt lévő fehérjék legnagyobb részét a reverzibilis
szerkezetváltozásra képes, úgynevezett strukturfehérjék képezik. Kialakítják a sejt belső
vázrendszerét, részt vesznek az összehúzódásra képes filamentumkomplexek és a sejtosz-
tódás járulékos képleteinek (1.6. fejezet) kialakításában.
Az eukarióta sejtek jellegzetessége a filamentumokból és tubulusokból álló belső
sejtváz (cytosceleton) (1.1. ábra). Szerepe van a sejt alakjának létrehozásában, a sejt-
alkotók sejten belüli elmozdulásában, a sejt aljzathoz való rögzítésében, illetve önálló
mozgásra képes sejtekben a helyváltoztatás kivitelezésében. Afilamentumokés tubulusok
fehérjemonomerekből jönnek létre polimerizációval.
A mikrofilamentumok közé tartoznak az aktin- és a miozinfilamentumok.
A spirális alakú (hélix) aktmfilamentumot globuláris monomerek építik fel. Ezek-
30
nek mindegyike az adott szálon azonos polaritást mutat, ami meghatározza az egész
filamentum polaritását. Az aktinfilamentum felépítése az átlagos testi sejtekben és az
izomsejtekben alig különbözik egymástól, ennek ellenére a madarak és az emlősök sejt-
jeiben legalább hatféle típusa különíthető el, melyek az adott előfordulási helyre jellem-
zőek. így eltér egymástól a vázizomban, a szívizomban, a bélcső és a véredények sima-
izomsejtjeiben található aktinfilamentum. Az egyes aktintípusok között - a több száz
közül - mindössze néhány aminosav tekintetében van eltérés. Ez azt bizonyítja, hogy az
eukarióta sejtekben az aktin felépítését meghatározó géneknek közös eredete volt. Az
aktinfilamentumok a hozzá kapcsolódó fehérjemolekulákkal (tropomiozin, troponin
komplex egységei) együtt a 7-8 nm átmérőjű vékony filamentumot képezik.
A miozinmolekulának van egy hosszú, fibrilláris típusú farok- vagy nyélrésze és egy
feji vége, amely fibrilláris és globuláris szerkezetet egyaránt mutat. Számos, a hosszú
farokrészekkel összekapcsolódó miozinmonomer alkotja a körülbelül 12 nm átmérőjű
vastag filamentumot. A kiálló feji részek képesek kapcsolódni a vékony filamentumhoz,
illetve elmozdulhatnak. A vékony filamentumok a vastag körül helyezkednek el az izom-
sejt típusának megfelelő számban és rendezettségben. Aktinfilamentum jelenlétében a
miozinmolekuláknak jelentős mértékű ATP-hidrolizáló képessége van, képesek a vé-
kony filamentumhoz kötődni.
A mikrofilamentumok nemcsak az izomsejtekben képeznek hálózatot. Kapcsolódnak
a sejtmembránt átérő fehérjékhez, így tudják a sejt alakját változtatni. A filamentumoknak
szerepük van a nagyobb fehérjemolekulák, a sejtorganellumok mozgatásában, a citokinézis
során a sejtek elkülönítésében, valamint a plazmaáramlás és az amőboid mozgás kivite-
lezésében.
Az átmeneti vagy köztes (intermedier) filamentumok (általában 10 nm-es átmérő-
jűek) a vékony filamentumoknál vastagabbak, de a vastag filamentumoknál vékonyabb
struktúrák. A sejt alapplazmájában és a sejtmagban is hálózatot képeznek, a sejt vázát
alkotják. Az intermedier filamentumok egyik fajtája az idegsejtekben található neuro-
filamentum. A neurofilamentumok a gerincesek központi és környéki idegrendszerében
egyaránt előfordulnak a neuronok axonjaiban. A hámsejtekben az intermedier filamen-
tumok másik típusa, a keratinfilamentumok fordulnak elő. Keratinfilamentumok alkot-
ják például a tonofilamentumokat, amelyek a dezmoszómákat kötik a sejtvázhoz. A ke-
ratinfilamentumok igen nagy mennyiségben vannak jelen az elszarusodó többrétegű
laphámban és módosulatainak (karom, köröm, tülök, szőrszál) szaruképleteiben. A bőr
hámrétegében a felszín felé haladva a sejtekben egyre nagyobb mennyiségben halmozó-
dik fel a keratin, a felszíni rétegben lévő elhalt sejtek szinte csak ezt tartalmazzák. A de-
zminrost elsősorban az izomrostok Z-vonalában (lásd 1.5. fejezet) fordul elő, rögzíti a
filamentumokat a plazmamembránhoz és más sejtszervecskékhez, valamint a rostok
hossztengelyére merőleges irányban álló hálózatot képez. A vimentin tartalmú filamen-
tumok a sejtorganellumok rögzítésében játszanak szerepet.
A mikrotubulusok a filamentumoknál vastagabbak (22-24 nm átmérőjűek), és spi-
rálisan feltekeredett a- és 0-tubulin nevű globuláris fehérjékből épülnek fel. Keresztmet-
szetben a 12-15 alegységből álló mikrotubulus közepén üreg látszik. A mikrotubulusok
általában kötegekbe rendeződnek, ilyenkor kereszthidak kötik össze őket. Fontos szere-
pük van a sejt belső szerkezetének kialakításában, egyes szervecskék, például a sejtosz-
tódás során a kromoszómák sejten belüli mozgatásában. A sejtosztódás során a mitotikus
orsó a mikrotubulus-rendszerből alakul ki. Megfigyelhetők az idegsejtek axonjában is,
ahol az intermedier filamentumok közbeiktatásával egészen a végfácskáig húzódnak és
részt vesznek a transzportfolyamatokban. Az ingerületátvivő anyagokat tartalmazó
vezikulákat a sejttestből a végkészülékbe szállítják. A halak, kétéltűek, hüllők gyors szín-
változtatásában is szerepet kapnak, mivel a pigmentsejtekben lévő pigmentszemcsék
sejten belüli mozgatását végzik.
31
1.4. ábra. A: a csilló és az ostor felépítésének és működésének vázlata. B: a spermium és C: az ostoros egysejtű
ostorának csapásai, D: egy csilló működése. A D ábrarészen a számok a csilló csapássorozatának egymás utáni fázisait
jelzik (1-3: erőteljes csapás, 4-6: a csilló helyzetének visszaállítása)
at: alapi test, bl: bazális lemez, ct: centrális mikrotubuluspár, di: külső és belső dinéin karok, fe: csilló, illetve ostor felépítése
és elhajlása, hi: sejt haladásának iránya, km: külső mikrotubuluspár, kü: küllő, me: mikrotubulus-duplet tagjainak elcsúszása,
mh: az ostor mozgási hullámainak iránya, pm: plazmamembrán, pr: protofilamentumok keresztmetszete (Damell-Lodish-
Baltimor, Flock és Duvall, Ruthmann és Hauser nyomán módosítva)
32
1.4. A sejtközpont és a sejtek mozgási organellumai
A sejtközpont (cytocentrum) két henger formájú központocskából (centriolum) áll. A
két központocska hossztengelye egymással 90 fokos szöget zár be. Utóbbiak közelében
erednek a sugárirányban kiinduló mikrotubulusok. Ezt a centriólumok körüli, sejt-
organellumokat alig tartalmazó területet centroszférának (centrosphaera) nevezik.
A centriolum membránnal nem burkolt organellum, amely csövecskékből áll (1.3. ábra).
A henger alakú képlet palástját mikrotubulustripletek alkotják.
Az alapi vagy bazális test (kinetoszóma) a centriólumból alakul ki, a felépítésük főbb
vonásaiban megegyező. A légutak csillés hámsejtjeinek kialakulása időszakában például
a centriólumok a sejt csúcsi, apikális membránjához húzódnak és számos centriólumszerű
képletet hoznak létre. Ezek alapi testekké válnak, melyek kialakítják a csillókat. Csillés
egysejtűek osztódását követően a már meglévő bazális testek hozzák létre a még hiány-
zókat, amelyekből azután a csillék kialakulnak.
A hámsejtek apikális felszínét, és számos egysejtű sejtfelszínét csillék boríthatják, míg
egyes egysejtűek és a hímivarsejtek többsége esetében ostor fordul elő. Az egysejtűek és
a kevés sejtből álló többsejtűek egész testének mozgatását ezek az organellumok végzik.
A többsejtűek egyes hámsejtjeinek (például a légutakban) csiliói tisztító szerepet játsza-
nak. A petevezető belső felszínét borító csillós sejtek a petesejt mozgatását végzik.
A csillék általában 5-10 pm hosszúak, míg az ostor elérheti a 150 pm-t is. A csillék
a sejt felszínével, míg az ostorok a sejtre merőlegesen, saját tengelyükkel párhuzamos
irányban mozgatják a környezetükben lévő folyadékot. Az ostor és a csilló is az alapi testből
indul ki, amiben a részt vevő mikrotubuláris tripletek a centriólumhoz hasonló felépítést
mutatnak (1.4. ábra). Az alapi test a sejt belső vázrendszeréhez kötött. A csilló és az ostor
palástját 9 duplet alkotja, középen pedig mikrotubuluspár van. Együttesen úgynevezett
„axonémát” képeznek. A duplet az „A” és „B” tubuláris egység közös képlete. A centrális
mikrotubuluspárt magába foglaló hüvelyhez az „A” tubuláris egységből küllők nyúlnak be,
valamint ugyanezekből külső és belső karok indulnak a szomszédos „B” tubuláris egység-
hez. A mikrotubulusok tubulin, az összekötő karok dinéin fehérjéből állnak. A szomszédos
„A” és „B” tubuláris egységeket rugalmas, nexinfehérje-tartalmú szálak is összekapcsol-
ják. Az ostor és a csilló felszínét sejtmembrán határolja. A dupletek egymáshoz képest
történő elcsúszása a csilló vagy ostor elhajlásához vezet, mivel tubuluspárok alapjukon
rögzítve vannak. A dinéin hidak működése az izomsejtekben lévő vékony és vastag
filamentumokéhoz hasonló. Számos csillós egysejtű képes arra, hogy a kívánt mozgási
iránynak megfelelően megváltoztassa a csillók csapásának irányát.
Az ostor felépítése és működése a csillóéhoz igen hasonló. Ostorral mozoghatnak az
egysejtűek és az állatok többségénél a hímivarsejtek. Utóbbiak felépítését a hímivarsejt
jellemzésénél (2.1.2. fejezet) mutatjuk be.
1.5. A sejtek mozgása
A soksejtűek több sejttípusa, például az embrionális sejtek, a gerinctelen állatok test-
folyadékának számos sejtje, a fibroblaszt sejtek, a fehérvérsejtek néhány típusa, valamint
sok egysejtű lény (amőbák) állábak (pseudopodiumok) segítségével amőboid mozgás-
ra képes. A képződő állábak egyaránt szerepet kapnak a mozgásban, az aljzathoz való
rögzülésben és a táplálékfelvételben. A sejt meghatározott pontjain, a tapadási helyeken
rögzül az aljzathoz. Alláb a sejt bármely pontján képződhet. Kialakulásának helyén a szol
állapotú endoplazma előrepréselődik, a képződés irányában a gél állapotú ektoplazma
elvékonyodik. Ennek révén az endoplazma a membránt maga előtt tolva kitüremkedést
33
1.5. ábra. A vázizomzatot alkotó harántcsíkolt izomrost felépítése és a miofibrillumok működésének (1-4)
alapelve (a folyamat magyarázata a szövegben található)
A: anizotrop csík, ak: aktinfilamentum, am: aktinmonomer, Ca: kalcium ion, fi: miofibrillum, H: H-sáv, I: izotróp esik, ih:
izomrost hártyája, ik: ideg-izom kapcsolat, lt: longitudinális tubulus, M: M-vonal, mf: miozinfej, mi: miozinfilamentum, mo:
motoros ideg velőshüvelyu axonja, sm: sejtmag, sz: szarkoméra felépítése nyugalmi és összehúzódott állapotban, te:
terminális ciszterna, tm: tropomiozin, tr: troponin komplex, tt: transzverzális tubulus, Z: Z-vonal (Több szerző nyomán
módosítva)
34
hoz létre. Aszol állapotú endoplazmában az aktin diffúz, nem polimerizálódott globuláris
állapotban van, míg az álláb kialakulási helyén polimerizálódik és más fehérjemolekulák-
kal összekapcsolódik. A sejt belső szervecskéi részben az aktinmiozin rendszer működése
révén a haladás irányában a nyúlványba folynak. A haladási iránnyal ellentétes végen a
sejt nyúlványai fokozatosan elvékonyodnak, belső anyaguk a sejttestbe áramlik. A sejt
membránjának tapadóképessége a képződő állábakon a legerősebb, míg a régebbieken,
illetve a mozgás irányával ellentétes állásúakon fokozatosan megszűnik. Ez lehetővé
teszi, hogy mindig a haladási iránynak megfelelően rögzüljenek a legerősebben.
Egyes egysejtűek (például Vorticella spp.) sejtizmocska segítségével változtatják
helyzetüket. A sejt alapi részéről kiinduló összhúzékony fehérjefílamentumokat tartal-
mazó rost biztosítja a sejt rögzítését, valamint elmozdulását.
A soksejtű állatok izomsejtjei a többi sejthez viszonyítva sokkal nagyobb mennyiség-
ben tartalmaznak összehúzódásra képes aktinmiozin komplexet. Ahhoz, hogy a kontrak-
ció végbemehessen, az aktinfilamentum egyik végének rögzített állapotban kell lennie.
Az izomsejtek sejthártyáját szarkolemmának, míg plazmáját szarkoplazmának nevezik.
A harántcsíkolt izmok miofibrillumaiban a kettős fénytörés mértéke szakaszonként
váltakozva eltérést mutat. A kevésbé kettősen fénytörő csíkot nevezték izotróp (I), míg
az erősebben fénytörőt anizotrop (A) csíknak. A vékony és vastag fílamentumok
(miofilamentumok) összekapcsolódva a rost egész hosszában futó miofibrillumokat al-
kotnak. A miofibrillumok működési egysége a nyugvó sejtekben a szarkoméra
(sarcoméra), amely egy összehúzódásra képes aktinmiozin komplexet jelent. A szarkoméra
határait a „Z” vonalak (Krause-féle lemezek) jelentik (1.5. és 1.6. ábrák). A „Z” vonalak
az „I” csíkok közepén húzódnak, azokat kétfelé tagolják. A vékony fílamentumok egyik
végükkel a „Z” lemezhez kapcsolódnak, míg a másik végük a miozinfílamentumok közé
nyúlik be, a szarkoméra közepe felé. Aharántcsíkolatot az „I” és a „A” csíkok váltakozó,
a szomszédos szarkomérákban megismétlődő elhelyezkedése alakítja ki. Az „A” csík
közepén világos „H” sáv különíthető el, amelynek közepén a sötét „M” vonal látszik. Az
„M” vonal a miozinmolekulákat összekötő fehérjemolekulákból álló középlemez. Az
izomrost összehúzódásakor a szarkoméra határait képező „Z” vonalak közelítenek egy-
máshoz, az „I” csík elvékonyodik, a „H” sáv pedig szinte eltűnik.
A harántcsíkolt izomzatnak két típusa van. Az egyik a test mozgatását végző vázizom-
szövetet, a másik a szívizomszövetet építi fel.
A vázizomzatot sok sejtmagvú óriás rostok (fiber) alkotják (1.5. ábra). Ezek az
embrionális élet során a mezodermális eredetű embrionális izomsejtek (myoblastok)
összeolvadásából jöttek létre. A sejtmagvak száma egy rostban akár több ezer is lehet. A
rostok hosszúsága néhány millimétertől több tíz centiméterig terjedhet. A sejtmagok
közvetlenül az izomrost plazmamembránja (szarkolemma) alatt, tehát a rost perifériáján
helyezkednek el. A harántcsíkolt izomrostok fő tömegét a miofibrillumok teszik ki, a
szarkoplazma aránya általában jóval kisebb.
A szívizomsejtek egy sejtmagvúak. Az ovális sejtmag a sejt közepén helyezkedik el.
A szívizomsejtek hosszúsága 50-150 pm közötti, és a vázizomrostokhoz viszonyítva vé-
konyabbak. A sejtek közötti réskapcsolat lehetővé teszi az ingerület gyors terjedését. A
szívizomrost össze nem olvadt sejtekből áll, ezért a rost el is ágazhat. Az egymás után
következő sejtek membránjainak találkozási helyein az ujjszerű betüremkedések révén
jellegzetes, fénymikroszkóppal is látható, úgynevezett Eberth-féle vonalak (intercalaris
korongok) jönnek létre (1.6. ábra). Az interkaláris korongban az aktinfílamentumok
rögzítését többek között az aktinin- és vinkulin-fehérjék végzik. A szívizomsejtekben a
harántcsíkolat a vázizomhoz képest fénymikroszkóppal vizsgálva gyengébben figyelhető
meg, ugyanis kevesebb miofíbrillumot tartalmaz.
Az izom-összehúzódás során a miofibrillumokat alkotó aktin- és miozin-fila-
mentumok elcsúsznak egymás mellett. A folyamatot a vázizomzat ábrájához (1.5. ábra)
35
mi Zv Mv rf
1.6. ábra. A: a szívizomsejt felépítésének részlete,
valamint B: a két sejt találkozásának térbeli képe az
Eberth-féle vonalban
ak: aktinfilamentum, Ev: Eberth-féle vonal, mi: miozinfila-
mentum, mk: mitokondrium, Mv: M-vonal, ny: membrán-
nyúlványok, rf: rögzítő fehérjeszálak, sh: sejthártya, Zv: Z-
vonal (Geier és Lindner nyomán módosítva)
kapcsoltan mutatjuk be, de az alapvetően megegyezik a szívizom működési elvével is. A
harántcsíkolt izom rostjaiban a vékony filamentum aktinból és a troponin-tropomiozin
alegységekből polimerizálódott komplex. A troponin egyik alegysége kalcium megköté-
sére képes. A kalcium az ingerület hatására az izomrost sima endoplazmatikus reti-
kulumából (szarkoplazmatikus retikulum) szabadul fel. A kalcium megkötése megvál-
toztatja a troponinkomplex, rajta keresztül az egész vékony filamentum szerkezetét (kon-
formációját), így szabaddá válnak az eddig fedett kötési helyek. A vastag filamentum
miozinfejeiben ekkor végbemegy az ATP hidrolízise, és a fejek kötődnek a vékony fila-
mentumhoz. Az ATP-molekulákbontásából származó energia a miozinfejek elfordulá-
sát („bólintását”) váltja ki. Az ATP hidrolíziséből keletkezett kémiai energia ekkor alakul
át mechanikai energiává. A miozinfejek az aktinszálon „lépegetnek”, így a két filamentum
egymás mellett elcsúszik. A vékony filamentumok az „M” vonal felé közelítenek. Mindad-
dig, amíg újabb ATP nem jut a miozin feji részéhez, a kötés fennáll. Ha ATP kapcsolódik
a miozin feji részéhez, akkor a két filamentum közötti kapcsolat oldódik, visszaáll a nyu-
galmi állapotban jellemző helyzet, azaz a miozinfejek lazán vagy egyáltalán nem kötődnek
a vékony filamentumokhoz. Addig, amig az izom ingerületet kap és elegendő ATP áll
rendelkezésre, ez a működési ciklus ismétlődik, másodpercenként akár 50-100-szor is.
Amikor az izomrost szarkolemmájához kapcsolódó idegvégződésben (1.5. ábra) az inge-
rület hatására felszabadul az átvivő anyag (általában acetilkolin), az megváltoztatja a memb-
rán permeábilitását. A gyorsan beáramló Na+-ionok kiváltják az akciós potenciált, ami vé-
gigfut a rost felszínén és a rost belsejébe mélyen behatoló, keresztirányú transzverzális
36
B
1.7. ábra. A: helikális izomsejt, B: egy kiemelt részlet
térbeli felépítése
ak: aktinfilamentum, fi: fonálféreg izomsejtjének metszete,
mi: miozin, sh: sejthártya, sm: sejtmag, sz: szarkoplazma-
tikus retikulum tubulusa, Zp: Z-pont (Rosenbluth, Heumann
és Zebe nyomán módosítva)
tubulusokon. Ez utóbbiak a szarkolemma
betűrődései, általában a Z- vonal szintjé-
ben futnak. Az impulzust a transzverzális
tubulusok vezetik be a rost belsejébe, ahol
az áttevődik a szarkoplazmatikus retiku-
lumra. A szarkoplazmatikus retikulumból
kialakult terminális ciszternákban és a
hosszanti, longitudinális tubulusaiban
tárolt Ca2+ felszabadul. Mivel a szarko-
plazmatikus retikulumnak szarkomérákat
borító részei is vannak, ezért annak teljes
hosszában biztosítja a Ca2+ hozzáférhető-
ségét (1.5. ábra). Tekintve, hogy az in-
gerületi folyamat továbbterjedése millisze-
kundumokon belül megy végbe, az összes
szarkoméra gyakorlatilag egy időben húzó-
dik össze. Ha az összehúzódás megtörtént,
akkor a kalciumionok újból a ciszternákba
kerülnek, az acetilkolin enzimatikusan le-
bomlik. A Ca2+-pumpa működéséhez és az
izom relaxációjához ATP szükséges. Ha az
izomrost újabb ingerületet nem kap, a kal-
ciumszint olyan mértékben lecsökken, hogy
a rost összehúzódása leáll. A szívizomsejtek
plazmájában lévő szarkoplazmatikus reti-
kulum tubulusrendszere a vázizomhoz ké-
pest kevésbé fejlett, különösen a longitudi-
nális tubulusok fejletlenek.
Az izomsejteknek természetesen nyugalmi állapotban is folyamatosan szükségük van
ATP-re, amely lehetővé teszi az aktív iontranszportot, a membránpotenciál és a sejt bel-
ső környezetének állandóságát. Az izom-összehúzódás alkalmával az ATP-felhasználás
a többszörösére nő.
A klasszikus értelemben vett simaizomsejtek alakja általában hosszúkás, megnyúlt,
a két végén kihegyesedő, ritkábban elágazó formájuk is előfordul. Hosszúságuk 15 pm és
1 mm között változik. Sejtmagjuk pálcika alakú, a sejt középvonalában helyezkedik el. Az
összehúzódott sejtekben a sej tmagok hullámos vagy spirálisan feltekert képet mutathatnak.
Az összehúzódást ezekben a sejtekben is az aktinmiozin komplexek hozzák létre, de a
vékony és vastag fílamentumok nem alkotnak szarkomérákat, illetve miofibrillumokat. Az
aktinfilamentumok nem rendeződnek el szabályos alakzatban a vastag fílamentumok kö-
rül, mint a harántcsíkolt izomrostokban. A miozin fílamentumok körül nagyobb számban
fordulnak elő aktinfilamentumok, mint a harántcsíkolt izomban. Az aktin fílamentumok
egy része az intermedier filamentumokkal közösen a plazmamembránhoz is kapcsolódik
(sötét foltok). A többletben lévő aktinfilamentumok között is kialakulnak kötési helyek
(sötéttestek). A simaizmokvékonyfilamentumai tartalmaznaktropomiozint, de a troponin-
komplex hiányzikbelőlük. A simaizomsejtek összehúzódó képessége, erőkifejtése a haránt-
csíkolt izomrostokhoz képest kisebb mértékű, de tartós működésre képesek.
A helikális izom sejtjeiben az egymással párhuzamosan álló szomszédos mio-
fibrillumok azonos felépítésű szakaszai egymáshoz képest elcsúszva, a térben spirális
(helikális) felcsavarodást mutatnak (1.7. ábra). Metszeti képükön ebből eredően ferde
harántcsíkolat figyelhető meg. A sejtek összehúzódó képessége jó. Ilyen izomtípus fordul
elő például a fonálférgek és a gyűrűsférgek bőrizomtömlőjében.
37
A paramiozin típusú izom sejtjeiben a miofibrillumokhosszanti lefutásúak, a miozin-
hoz paramiozin is kapcsolódik. Az ilyen izom igen kis energiafelhasználással hosszú ideig
tartó kontrakciót tesz lehetővé. Ez az izomféleség található a kagylók záróizmában.
1.6. A sejtek osztódása
A sejtciklus az osztódóképes sejt egyik osztódásának kezdetétől a következőig zajló
életciklusa. A szöveti kötelékben élő sejtek egy része és az egysejtűek a fajra jellemző
méretet elérve, megfelelő körülmények között osztódnak. A sejt osztódása két fő részre
tagolható: először végbemegy a magosztódás (karyokinesis), amit a sejt osztódása követ
(cytokinesis).
Két osztódás között rövidebb-hosszabb ideig tartó interfázis különíthető el (1.8.
ábra). A sejt ciklusában ez a leghosszabb szakasz. Ekkor a sejt növekszik és létrejönnek
a későbbi osztódása során nélkülözhetetlen fehérjék, ribonukleinsavak. Az S-fázisban
kettőződik meg a DNS-állomány. Ezt megelőzi a Gj-, illetve követi a G2-fázis. A Gt-
fázisban fontos előkészületi folyamatok zajlanak, például szállító RNS, hírvivő RNS,
riboszómák és enzimek szintézise. A G2-fázisban főként a mitotikus apparátus fehérjéi
képződnek. Az interfázist alkotó szakaszok időtartamának aránya a különböző funkciójú
sejtekben eltérő. A gyorsan növekvő, osztódó sejtekben a G^fázis igen rövid, míg a lassan
növekvő sejtekben ugyanez a szakasz igen hosszú lehet. Az osztódóképességüket elvesz-
tett sejtek, például az idegsejtek a ciklusból kilépve (szokták G0-fázisnak nevezni) diffe-
renciált sejtekké válnak, a ciklusba visszalépni nem képesek.
Az osztódás során a sejtek kromatinállományában és bizonyos sejtszervecskékben je-
lentős változások zajlanak le. A magosztódások egyik típusa a mitózis, amelynek vég-
eredményeként az utódsejtek kromoszómaszáma az anyasejtével azonos lesz, míg a
meiózisban feleződik.
A sejtciklus szabályozottságára vonatkozóan számos elképzelés született, de a folya-
mat minden részletét ma sem ismerjük. Megállapítást nyert, hogy valamennyi eukarióta
sejtben a ciklus szabályozását fehérjemolekulák aktivitása határozza meg.
A sejtciklus menetét nagymértékben serkentik a sejt membránjához kapcsolódó nö-
vekedést szabályozó anyagok. Ezek kötődnek a receptoraikhoz. Hatásuk befolyásolja a
sejten belüli enzimatikus láncreakciókat, így a sejtosztódás menetét és gyakoriságát is.
1.6.1. Kromoszómaszám-tartó osztódás
A kromoszómaszám-tartó osztódás (mitózis) az egysejtűekben és a soksejtű szervezetek
testi (szomatikus) sejtjeiben fordul elő. Az aszexuálisan szaporodó egysejtűeknél ez az
osztódás teszi lehetővé, hogy a létrejövő utódok azonos genetikai állományúak legyenek.
A soksejtű szervezetek testi sejtjeinek mindegyike rendelkezik a szervezet létrehozásáért
felelős teljes genetikai állománnyal. A differenciálódás során azonban ez a képességük
beszűkül, és általában csak a speciális feladatuknak megfelelő funkciót képesek ellátni.
A mitózis biztosítja a DNS-állomány folytonosságát a sejtgenerációk során.
Egy bizonyos faj testi sejtjeiben jellemző számú kromoszóma található, melyek együtt
a kromoszómagarnitúrát alkotják. A kromoszómaszám és az adott faj törzsfejlődésben
elfoglalt helye között nincs összefüggés, mint ahogy a szám nem mutatja egyértelműen
a rokoni kapcsolatokat sem. A kétszeres (diploid) kromoszómaszámú sejtekben a kro-
moszómák egyik fele anyai, míg a másik apai eredetű. A két különböző eredetű sorozat
egymásnak megfelelő tagjai a homológ kromoszómák. A homológ kromoszómák mellett
még előforduló ivari kromoszómák azonos vagy különböző alakúak és eltérő méretűek
lehetnek. Általában az őspetesejtek azonos tulajdonságú (XX), míg az ős-hímivarsejtek
38
eltérő (XY) ivari kromoszómákat tartalmaznak. Egyes rovarokban (lepkék), halakban,
kétéltűekben, hüllőkben és madarakban a helyzet fordított, tehát ezeknél a hím-ivarsej-
tek ivari kromoszómái hordoznak azonos tulajdonságot meghatározó géneket. Több
rovarfajban (például egyes szöcskékben) a hímivarsejtekfelében nincs ivari kromoszóma,
míg a többiben egy női jellegű X kromoszóma van.
1.8. ábra. Az állati sejt mitózisának folyamata
AN: anafázis, cc: citocentrum, cm: centroméra, ek: egy kromatidás, kevéssé kondenzálódott kromoszóma, et: egy kro-
matidás, pólus felé húzódó kromoszóma, fs: felbomló sejtmaghártya, Gj-S-Gji az interfázis alfázisai, hf: húzófonalak, IF:
interfázis, ki: kinetokor, kk: két kromatidás, kevéssé kondenzálódott kromoszóma, kr: kromatida, kt: két kromatidás
kromoszóma transzportformája, ME: metafázis, MI: mitózis, mt: osztódási orsó mikrotubulusai, oc: osztódási csillag, öm:
összehúzódó filamentumgyurú, PR: profazis, SO: sejtosztódás, TC: telofazis (TE) és a citokinézis (Cl) kezdete, tk: tömörülő kro-
matinállomány, ús: újraképzödö sejtmaghártya, ut: utódsejtek (Miller és Harley, valamint más szerzők nyomán módosítva)
39
Abban az esetben tehát, ha az X kromoszómájú hímivarsejt termékenyíti meg az X
kromoszómát tartalmazó petesejtet, akkor mindig nőivarú utódok fejlődnek. Ezzel szem-
ben az ivari kromoszómát nem tartalmazó hímivarsejttel egyesülő petesejtből hímek
(XO) alakulnak ki.
A szervezetet felépítő sejtek élettartama jelentősen eltérhet egymástól. A bélbolyhok
felszínét borító sejtek és egyes fehérvérsejttípusok például csak néhány napig élnek. Az
egészséges máj sejtjei ritkán pusztulnak el, és az idegsejtek jelentős része is igen hosszú
életű. A szervezetet alkotó legtöbb sejt pusztulása és szaporodása folyamatosan megy
végbe, míg például az idegsejtek egy idő után elveszítik osztódóképességüket, számuk
csak csökkenhet. Egyes sejtek a posztembrionális élet kezdetekor vagy később veszítik el
osztódóképességüket. Más sejtek viszont az egyed egész élete során rendelkeznek ezzel
a képességgel. Az osztódások a sejtek funkciójától függően végbemehetnek óránkénti,
naponkénti, hónaponkénti vagy akár éves ritmusban is.
A sejtállandóságot nem mutató soksejtű szervezetekben egyidőben általában a sejtek-
nek csak mintegy öt százaléka osztódik, a többi interfázisban van (1.8. ábra).
A profázis elej én befej eződik a centriolumok megkettőződése, máj d a két citocentrum
egymástól elválva a sejtmag ellentétes oldalára húzódik (1.8. ábra). Körülöttük osztódási
csillag, míg közöttük magorsó alakul ki. A magorsót alkotó rostok tubulinfehérjetartalmú
mikrotubulusok. Egyrészt a két citocentrum között (pólusrostok), másrészt a sejtköz-
pontok és a kromoszómák között (kromoszóma- vagy húzórostok) létesítenek kapcsola-
tot. Mindezek együttesen a mitotikus apparátust alkotják. A kromatinállomány fokoza-
tos tömörülése, kondenzálódása révén kezdetben fonál alakú, két kromatidából álló
kromoszómák jönnek létre. A két kromatida a DNS-replikációban keletkezett 1-1 új
DNS-molekulát tartalmazza. A sejtmaghártya eltűnik, a kromoszómafonalak kezdetben
gomolyformát mutatnak. Ez az egygomolyállapot (monospirema). A profázis végén a
kromoszómák fokozatosan elnyerik transzportformájukat.
A metafázisban a kromoszómák a sejt központi síkjába rendeződnek (1.8. ábra).
A két kromatida meghatározott szakaszon (centromera) kapcsolatban áll egymással.
A centrom érákhoz fehérjéből álló korongok (kinethochor) tartoznak, melyekhez a húzó-
rostok kapcsolódnak. Egy kinetokorhoz rendszerint 15-30 mikrotubulus is kapcsolódik,
melyek másik vége a sejtközponthoz tartozó centroszférához kötődik. A kromoszómák
karjaikkal a központtól kifelé néznek, így egycsillag alakzatot (monaster) alkotnak.
A kromoszómák ebben a fázisban mutatják a tipikus transzportformát. A metafázis végén
a testvérkromatidák elválnak egymástól.
Az anafázisban a kromatidák a saját kinetokorjaikhoz kapcsolódó húzófonalak révén
a sejt két pólusára húzódnak. így a sejten belül kétcsillag alakzat (diaster) jön létre (1.8.
ábra).
A telofázisban a kromoszómák dekondenzálódnak, újra fonal alakúvá válnak (1.8.
ábra). A mitotikus apparátust alkotó mikrotubulusok szétesnek, belőlük alakul ki az
utódsejtek belső váza. A sejt két pólusán a fonál alakú kromoszómák kétgomoly alakot
(dispirema) hoznak létre. Kialakul a két leendő sejtmag körül a maghártya. A sejt-
magvacska igen hamar megjelenik, megindul a riboszómaképzés. Ezután általában végbe-
megy a citoplazma kettéválása (cytokinesis) is (1.8. ábra). A szétváló sejtek határán
a sejtmembrán befűződik, és az itt lévő aktinmiozin komplexek összehúzódása segítség-
ével a két sejtfél közötti plazmahíd fokozatosan szűkül. A membránok összeérnek, majd
összeolvadnak és a határfelületen elválnak egymástól, így a két sejt elkülönül.
Ha a mitózist nem követi a citokinézis, akkor soksejtmagvú sejtek (plasmodium)
jönnek létre. Sokmagvú sejtek keletkezhetnek akkor is, ha a sejtek összeolvadnak
(syncytium). Ez utóbbi történik a harántcsíkolt izomrostok esetében, amikor a még
embrionális izomsejtek olvadnak össze.
40
1.6.2. Kromoszómaszám-felező osztódás
A mitózissal ellentétben a kromoszómaszám-felező osztódás (meiózis) során az anya-
sejtből haploid, azaz fele kromoszómaszámú utódsejtek keletkeznek. A meiózisnak két
típusát különíthetjük el, a zigogén és a gametogén meiózist. A zigogén meiózis az egy-
sejtűek közül egyes ostorosokra és a spórásokra jellemző, amikor az ivaros alakok összeol-
vadását követően az első osztódás meiózissal történik. Ezt követően mitózisok sorozata
mehet végbe, így a következő ivaros szaporodás bekövetkeztéig valamennyi sejt haploid.
A napállatocskák, csillósok és egyes ostorosok ivaros alakjai gametogén meiózis
eredményeként jönnek létre. Gametogén meiózis játszódikle az állatokivarsejtjeinek kiala-
kulásakor is. Ennek során két egymást követő osztódás (a meiózis két főszakasza) zajlik le,
amelyek közül csak az első előtt megy végbe a DNS-állomány megkettőződése. Az első
főszakaszban a kromoszómaszám megfeleződik, de mivel a kromatidák nem váltak szét
egymástól, ezért minden DNS-molekulából két (az átkereszteződés miatt nem mindig
azonos) kópia kerül az utódsejtbe. A DNS-állomány az első főszakasz végén keletkezett
sejtekben ugyanannyi, mint a DNS-replikáció előtti anyasejtben volt. A kromoszómaszám
a második osztódásban már nem változik, de a DNS-állomány a felére csökken, mert a
kromatidák ekkor elválnak egymástól és a haploid utódsejtekbe kerülnek (1.9. ábra).
Az első meiotikus osztódás előtti interfázisban végbemegy a DNS-állomány dup-
likációja. A profázis több alfázisra osztható. A leptotén szakaszban a még vékony kro-
moszómák szalag alakúak, a sejtmaghártyához csokorszerűen kapcsolódva rendeződnek
el. A zigotén szakaszban a homológ kromoszómák hosszukban összekapcsolódnak.
A pachitén szakaszban a kromoszómák megrövidülnek, megvastagodnak, és láthatóvá
válik, hogy mindkét homológ kromoszóma két-két kromatidából áll. A két összekapcso-
lódott homológ kromoszóma ily módon úgynevezett tetrádot alkot. A diplotén szakasz-
ban a homológ kromoszómák között a kromatidák átkereszteződései révén génkicseré-
lődés megy végbe. Az új génkombinációkat tartalmazó rekombináns kromoszómák kiala-
kulása az élőlények genetikai változékonyságát növeli meg. A diplotén szakaszban megin-
dul, majd a diakinezisben folytatódik az összetapadt homológ kromoszómák szétválása.
A sejtmaghártya ciszternákra tagolódik, a kromoszómák pedig a sejt egyenlítői síkja felé
mozdulnak el.
Az első meiotikus osztódás metafázisában kialakul az osztódási orsó és a kromoszó-
mák a sejt egyenlítői síkjába rendeződnek (1.9. ábra).
Az anafázisban a homológ kromoszómapárok szétvált tagjai, tehát kétkromatidás
kromoszómák (diádok) húzódnak véletlenszerűen a pólusok felé. Ez újból növeli az
utódsejtek genetikai változékonyságát. így haploid, fele kromoszómaszámú, de kromo-
szómánként még kétszeres DNS-állományú sejtek jönnek létre.
A telofázisban a sejtmaghártya és a sejtmagvacska újból kialakul, és az osztódási orsó
eltűnik. Végbemegy a citokinézis, a sejtek elválnak egymástól. Bizonyos esetekben a
telofázis kromoszómákat érintő változásai nem mennek végbe, de a citokinézis lezajlik.
Az első főszakasz eddig ismertetett fázisai után általában nem következik interfázis, és
a második főszakasz profázisa is elmaradhat. A második meiotikus osztódás felé való
átmenetet interkinézisnek nevezzük. Ennek során a DNS-állomány nem kettőződik
meg. Az első osztódással létrejött két sejt vagy közvetlenül a második főszakasz meta-
fázisába lép be vagy egy profázis is beiktatódik, amikor az osztódási orsó újból kialakul.
A második meiotikus osztódás metafázisában a citocentrum elkülönült centriólumai
a sejt két pólusára húzódnak. A kromoszómák a sejt egyenlítői síkjába állnak be (1.9.
ábra).
Az anafázisban a húzófonalak a sejt két pólusa felé húzzák el az egymástól elvált
testvérkromatidákat. így az utódsejtekben a DNS-állomány az előző állapothoz képest
a felére csökken.
41
A telofázisban a kromoszómák elveszítik transzportformájukat, dekondenzálódnak.
Kialakul a sejtmaghártya, majd bekövetkezik a citokinézis, azaz a sejtek citoplazmája is
kettéválik (1.9. ábra).
A két egymást követő osztódás eredményeként négy haploid (ivar)sejt alakul ki. A
testi sejtekre jellemző diploid kromoszóma szám a megtermékenyítés alkalmával áll vissza.
1.9. ábra. Az állati sejt meiózisának folyamata
AN-1: anafázis-1., AN-2: anafázis-2., cc: citocentrum, ce: centriolum, ek: egy kromatidás, kevéssé kondenzálódott kromoszó-
ma, et: egy kromatidás, pólus felé húzódó kromoszóma, IF: interfázis, kk: két kromatidás, kevéssé kondenzálódott kromoszóma,
kt: két kromatidás kromoszóma transzportformája, ME-1: metafázis-1., ME-2: metafázis-2., PR-1: profázis-1., PR-2:
profázis-2., TE-1: telofázis-1., TE-2: telofázis-2., us-1: utódsejtek a meiózis első főszakaszának végén, us-2: utódsejtek a
meiózis második főszakaszának végén (Miller és Harley, valamint más szerzők nyomán módosítva)
42
1.7. Rákos sejtek
A sejtek osztódása szabályozott folyamat. Általában annyi sejt keletkezik, amennyire a
szervezetnek az adott helyen szüksége van, ami pótolja az elpusztult sejteket. Az osztódást
szabályozó sejtspecifikus faktoranyagoknak egy része már ismert, de többsége valószínű-
leg még ismeretlen. Ezek a faktoranyagok jeleket adnak az egyes sejtek számára, hogy
osztódjanak-e vagy maradjanak nyugalmi állapotban. Ha ez a szabályozási folyamat
zavart szenved, akkor a sejtek növekszenek és osztódnak anélkül, hogy a szervezetnek ez
hasznos lenne. Ilyenkor a létrejövő sejttömeg tumort alkot. A daganatos elfajulás
növekedési és differenciálódási zavart jelent. A daganatsejtek differenciálatlanok marad-
nak. A rákos daganatok képződését a sejtek genetikai állományában bekövetkezett mu-
táció okozza, amit a rákkeltő, karcinogén hatások, például vírusok, kemikáliák, káros
sugárzások hoznak létre. A sejtek információs jelpályáinak működéséhez szükséges fe-
hérjék szintézisét gének irányítják. Azokat a géneket, amelyek hibás működésük révén
nemkívánatos sejtnövekedést és osztódást válthatnak ki, onkogéneknek nevezzük. A
létrejövő daganatok egy része „jóindulatú”, lokalizált, a szervezet számára általában
kisebb veszéllyel jár. Az ilyen kötőszövetes tokba zárt sejttömeg akkor okoz gondot, ha
például a szervezet működését befolyásoló hormonhatású vegyületeket juttat a vérpályá-
ba vagy létfontosságú eret, ideget, agyrészt nyom el.
Ha a sejt az említett mutációk következtében olyan változáson megy át, hogy invazíwá
válik, azaz a szövetben helyét változtatja és eközben korlátlanul osztódó, az eredetinél
differenciálatlanabb sejttömeget hoz létre, akkor „rosszindulatúan átalakult” sejtről be-
szélünk. A rosszindulatú sejt kötőszövetes tokkal nem határolt úgynevezett primer da-
ganatot képez. Ebből további változások után olyan sejtek is kialakulhatnak, melyek
képesek hámokon és érfalakon is áthatolni. Ezek a vérárammal eljutnak más szövetekbe,
szervekbe, ahol osztódásuk, növekedésük tovább folyhat, és ezáltal új tumort hoznak
létre. Ezt a folyamatot nevezik áttételnek (metasztázis). A ráksejtekkromoszómaállománya
abnormális, nem stabil. Gyorsan növekszenek és gyakran osztódnak. Eredeti speciális
funkciójukat elvesztik. Az ektodermális és entodermális eredetű sejtekből származó rossz-
indulatú daganatokat karcinómának nevezik, míg a mezodermális eredetűeket szarkó-
mának. Ez utóbbi speciális esete a fehérvérűség tünetével járó leukémia, amikor a rákos
sejtek nem képeznek a szó eredeti értelmében vett tumort.
1.8. A sejtek szerepe a szervezet immunfolyamataiban
A szervezetet felépítő sejteknek fel kell ismerniük egymást, illetve meg kell tudni
különböztetniük a saját anyagokat az idegentől.
Védekező immunrendszer a gerinctelen és a gerinces állatokban egyaránt előfordul,
de annak felépítése, működése nagymértékben eltér egymástól. A szivacsokban és
csalánozókban amőboid sejtek látják el ezt a feladatot. A puhatestűekben, gyűrűsférgekben
és az ízeltlábúakban a cölomaciták és a hemociták védekeznek. E sejtek közös jellemzője,
hogy fagocitózisra képesek. A limfoid sejtek, illetve az általuk termelt anyagok révén
megvalósuló immunrendszer a gerincesek szervezetében jelenik meg. Az állkapocs-
nélküliekben, a porcos és a csontos halakban nyiroksejtek és nyirokcsomók még nem
találhatók. Az immunrendszer elemeiként a vörös csontvelő és a nyirokcsomók egyes
kétéltűektől kezdődően fejtik ki hatásukat. Madarakban és emlősökben további speciális
nyirokszervek is vannak.
A védekezés feladatainak egy részét a retikuloendotéliális rendszer (RÉS) sejtjei látják
el. A májban (Kupffer-sejtek), a lépben, más nyirokszervekben (retikulumsejtek), a
43
kötőszövetekben (hisztiociták) és a vérben fagocitózisra képes sejtek vannak. A vérben
főleg a neutrofil granulociták (mikrofágok), míg a környező szövetekben az oda kilépő
monocitákból átalakuló szöveti makrofágok szerepe jelentős. A gerinces állatok közül az
emlősök immunfolyamatai a legismertebbek.
A természetes immunitás működésének alapelve egy sejtszintű tanulási folyamat.
Ennek során a szervezetbe került idegen anyagok vagy kóros folyamat miatt idegenné vált
anyagok indukálják a védekezősejtek felismerő képességét. Az immunrendszer működé-
sének alapvető kritériuma az, hogy a védekező sejtek fel tudják ismerni a testidegen anya-
gokat. Ezt követően az idegen jelrendszerű fehérjemolekulát, vírust, baktériumot, gombát
vagy a fajazonos, de másik egyed bármely sejtjét elpusztítják vagy a szervezetből kilökik.
Antigénnek minősül minden olyan, elsősorban fehérjetermészetű anyag, amellyel az
immunsejtek az embrionális fejlődés időszakában nem kerültek kapcsolatba és ezért a
későbbiekben védekezési reakciót váltanak ki. A posztembrionális fejlődés kezdeti idő-
szakában különösen nagy intenzitással folyik az immunsejtek antigénfelismerést szolgáló
tanulási folyamata.
A védekezés két módon mehet végbe. Ez történhet a testfolyadékban szabadon lévő
immunfehérjék, immunglobulinok (humorális vagy antitestes immunitás) vagy az immun-
sejtek közvetlen beavatkozása (sejtes immunitás) révén. A testidegen anyagokat mindkét
esetben immunfehérjék kötik meg.
Az immunválasz kialakításában a limfociták két típusa, a B- és a T-sejtek játszanak
döntő szerepet. Ezek a sejtek alakilag nem különböznek egymástól, de eltérnek a memb-
ránjukba beépült fehérjéik összetételében és funkciójukban.
Az immunfolyamatokban és a vér egyéb alakos elemeinek képzésében részt vevő sejtek
valamennyien egy közös leszármazási vonalba tartoznak. A pluripotens haemopoietikus
őssejtekből keletkező limfoid őssejtek az elsődleges vérképző- és nyirokszervekben, így
az emlősökben a vörös csontvelőben és a tímuszban, madarakbanpedig a bursa Fabriciiban
termelődnek és fejlődnek tovább. Az őssejtek osztódások sorozatán keresztül olyan utód-
sejteket hoznak létre, amelyeknek a legkülönbözőbb antitesteik vannak. Differenciáló-
dásuk kezdetben tehát még antigéntől függetlenül megy végbe. Antigéntől függő diffe-
renciálódásuk a másodlagos nyirokszervekben zajlik le. Az immunológiai szempontból
érett B- és T-limfociták a vérben és a nyirokban keringenek, illetve a perifériális, másod-
lagos nyirokszervekben (lép, nyirokcsomók, mandulák, bélcsatoma nyiroksejtes képle-
tei, féregnyúlvány) tárolódnak. A csontvelő és a bursa Fabricii állandó limfocita-utánpótlást
nyújt a szervezet számára.
Ellenőrző kérdések az 1. fejezet témaköreiből
1. Mit jelent a biológiai membránok egységes felépítése?
2. Mi jellemző a plazmamembrán kettős lipidrétegére?
3. Mi a feladata a sejtburoknak?
4. Milyen membrán kapcsolódási formákat ismer?
5. Mi a feladata a mikrobolyhoknak?
6. Milyen típusú transzportfolyamatok mennek végbe a membránon keresztül?
7. Sorolja fel a sejt membránnal határolt sejtszervecskéit!
8. Mi a feladata az endoplazmatikus retíkulumnak?
9. Mi a feladata a Golgi-apparátusnak?
10. Mi a feladata a lizoszómáknak és a peroxiszómáknak?
11. Milyen speciális tulajdonságai vannak a mitokondriumoknak?
12. Mi a feladata a sejtmagnak?
13. Mi alkotja a sejt alapállományát?
44
14. Miből épül fel a sejtváz?
15. Milyen felépítésű és működésű a sejtközpont?
16. Jellemezze a sejtek mozgásformáit!
17. Mi a különbség a mitózis és a meiózis között?
18. Ismertesse a mitózis során lezajló eseményeket!
19. Ismertesse a meiózis során lezajló eseményeket!
20. Milyen szerepet töltenek be egyes sejtek a szervezet immunfolyamataiban?
45
2. AZ ÁLLATOK EGYEDFEJLODESE
Az állatok egyedfejlődése (ontogenesis) a megtermékenyítéskor veszi kezdetét, és mindaddig tart,
amíg az életfolyamatok meg nem szűnnek. Az embrionális fejlődés a megtermékenyítéstől a
peteburok elhagyásáig, illetve a megszületésig tart, mig a posztembrionális szakasz ezt követően
a szervezet kifejlődésén keresztül a halálig terjed.
Az ivarsejtek kialakulásának folyamata három fő szakaszra osztható. Először az ősi ivarsej-
tek kromoszómaszám-tartó osztódással szaporodnak, majd az így keletkezett sejtek a növekedési
periódusban tartaléktápanyagokat halmoznak fel. Az érési periódusban történik a kromoszóma-
szám felezödés, haploid sejtek alakulnak ki a meiózis eredményeként. A petesejt és a himivarsejt
kialakulásának folyamata azonos alapelven zajlik, de a kimenetele eltéréseket mutat. A hímivar-
sejt speciális formáját az osztódásokat követő alaki átrendeződéssel éri el. A petesejt az állati
szervezet egyik legértékesebb sejtje, amit védőburkok vesznek körül. Ezek képződhetnek az ivar-
szervben vagy az elvezető ivari járatokban. A petesejt egyik, a testi sejtektől eltérő tulajdonsága,
hogy plazmájában, az utód táplálása céljából szikanyagot tartalmaz. A hímivarsejtek legálta-
lánosabb formája ostoros. Végső alakjának elnyerésekor csupán a mozgáshoz, a genetikai állo-
mány védelméhez, a petesejtbe való behatoláshoz legszükségesebb sejtorganellumok maradnak a
sejtben, a többitől megszabadul.
A megtermékenyítés lehet külső vagy belső. Az állat élőhelyétől, élet- és szaporodásmódjától,
utódgondozási formájától függően egyaránt sikeresek lehetnek. A megtermékenyülés folyamata
bonyolult kölcsönhatások eredménye. A megtermékenyült petesejt osztódásnak indul, amit baráz-
dálódásnak nevezünk. A petesejtben lévő szikanyag mennyisége és sejten belüli eloszlása megha-
tározza a barázdálódás, ezen keresztül pedig a csíralemezek kialakulásának és további differen-
ciálódásának menetét. A fejlődő utód védelmét és táplálását embrionális szervek (magzatburkok,
szikzacsó) látják el, amelyek különösen jól differenciáltak a gerincesek körében.
A posztembrionális fejlődés során számos állatcsoportnál lárvaformák jelennek meg, így fejlő-
désük közvetett. A kifejlettekrejellemző testformát és belső felépítést jellegzetes átalakulás során érik
el. A fejlődő, növekvő egyed élete során zajló élettani változások alapján életszakaszokat lehet
elkülöníteni.
Ebben a fejezetben az ivarosán szaporodó állatok egyedfejlődésének általános és összehasonlító
vonásait tárgyaljuk, kiegészítve az ontogenezist megelőzőfolyamatok (proontogenesisj, vagyis az
ivarsejtek kialakulásának bemutatásával.
2.1. Az ivarsejtek kialakulása és felépítése
A legtöbb állatcsoportban a petesejt (oviim) és a hímivarsejt (spermium) kialakulása
(gametogenesis) háromfő periódusra osztható. A szaporodási szakaszban az ősivarsejtek
mitózis révén nagy számban hoznak létre utódsejteket. Ezt követi a növekedési szakasz,
amelyben méretnövekedés következik be. Az érési szakaszban meiózissal osztódnak,
ekkor alakul ki a haploid kromoszómagarnitúra.
46
2.1.1. A petesejt kialakulása és felépítése
A különböző állatfajok petesejtjei általában gömb vagy gömbölyded formájúak, míg egyes
rovarokban hosszúkás, megnyúlt alakúak. A szivacsok és a csalánozók petesejtjei változó
alakúak, amőboid mozgásra képesek. Méretük általában az átlagos testi sejtekének sokszo-
rosa, átmérőjük akár több centiméteres is lehet, mint például a madarak esetében. Méretét
elsősorban az szabja meg, hogy mennyi szikanyagot tartalmaz a fejlődő utód táplálására.
Egyes állatcsoportokban (például igen sok laposféregben) a kevés szikanyagot tartalma-
zó petesejtet kívülről veszi körül szikanyag, ilyenkor ektolecitális (ectolecithalis) pete-
sejtről beszélünk, megkülönböztetve a következőkben felsoroltaktól, ahol a szik-anyag a
petesejten belül (endolecithalis) található (2.1. ábra).
Azendolecitális petesejteknél a felhalmozódott szikanyagmennyisége alapján meg-
különböztetünk szik nélküli (alecithalis), kevés szikű (oligolecithalis), közepes szikű
(mesolecithalis) és sok szikanyagot tartalmazó (polylecithalis) petéket. A sziktartalom
eloszlása alapján beszélhetünk egyenletes (isolecithalis) és egyenlőtlen (anisolecithalis)
szikeloszlásról. Ha a szikanyag a petesejt egyik pólusán tömörül, akkor véghelyzetű
(telolecithalis), míg ha középen helyezkedik el, akkor középszikű (centrolecithalis) típus-
ról beszélünk (2.1. táblázat). Az egyenlőtlen szikeloszlású petesejtekben elkülöníthető
egy szikmentes vagy kevés szikanyagot tartalmazó animális és egy több szikanyagot
magába foglaló vegetatív pólus. A sejtmag az animális póluson helyezkedik el.
A szivacsok és néhány csalánozó kivételével a petesejteket védőburkok veszik körül,
amelyek elsődleges szerepe a mecha-
nikai, hő- és vegyi hatások kivédése.
Szerepük lehet még a petesejt táplálá-
sában, helyzetének rögzítésében, illet-
ve a vízbe lerakottaknál a lebegtetés
kialakításában.
Az elsődleges burkot a petefészek-
ben való fejlődés során a petesejt hozza
létre maga körül. Elsődleges buroknak
tekinthető mindenekelőtt a sejthártya
(ovolemma). A petesejt felszíne mikro-
bolyhokkal borított, amelyek között a
teret a sejt által leadott anyagok töltik ki.
Fénymikroszkópos vizsgálatok ezt a ré-
teget sugarasan csikóitnak mutatj ák, így
kapta a sugaras zóna (zóna radiata)
elnevezést. Megtalálható a gyűrűsférgek,
a puhatestűek, egyes tüskésbőrűek, a
halak, a kétéltűek és a hüllők körében,
de megfigyelhető a madarak között és
az emlősök egy részénél is.
A másodlagos burkokat a petesej-
2.1. ábra. A petesejtek típusai a szikanyaggal való ellátottság
és a szik elhelyezkedésétől függően. (A pontozás sűrűsége a
szikanyag mennyiségével arányos)
ektolecitális (A), endolecitális (B), ml: mezolecitális és telolecitális,
ol: oligolecitális és izolecitális, pb: peteburok, pc: polilecitális és
centrolecitális, pe: petesejt, pt: polilecitális és telolecitális, sz:
sziksejtek (Több szerző nyomán módosítva)
tét körülvevő tüszőhámsejtek képezik, vagy e sejtek átalakulásából jönnek létre. Másodlagos
burkok jönnek létre a puhatestűek, egyes tüskésbőrűek körében, valamint a legtöbb gerin-
cesben. Emlősök esetében a tüszőhámsejtek által létrehozott másodlagos burkot fénylő
hártyának (zóna pellucida) nevezik (2.2. ábra). A rovarok, fejlábúak, tüskésbőrűek és a
legtöbb csontoshal petesejtjének másodlagos burkán a spermium behatolását lehetővé tevő
nyílás (mikropyle) van. Amikor a halikra vízbe kerül, ez a burok megduzzad, megerősödik.
Madarakban a zóna radiata és a tüszőhámsejtek által létrehozott másodlagos burok (membrana
perivitellina), valamint a petevezető tölcsérében e fölé felrakódó extravitellinális membrán
47
2.2. ábra. Az emlősállatok petesejtjének felépítése
cr: corona radiata, dl: diktioszóma, mi: mitokondrium, ny:
tüszőhámsej-tek nyúlványa, sh: sejthártya, sm: sejtmag, sv:
sejtmagvacska, th: tüszőhámsejtek, zp: zóna pellucida (TörŐ-
Csaba, Kovács-Fehér nyomán módosítva)
együttesen kapta a szikhártya (membrana
vitellina) elnevezést. Az extravitellináris memb-
rán a hímivarsejtek behatolásával közel egy
időben jön létre. Madarak esetében a tüsző-
hámsejtek csak a petefészek tüszőiben borít-
ják apetesej tét, az ovulációt követően már nem.
A méhlepényes emlősök petesejtjének s ejthár-
tyáját kívülről borító zóna pellucida rétegen a
tüszőhámsejteknyúlványai áthatolnak, és táp-
lálják, védik, illetve támasztják a petesejtet. A
tüszőhámsejtek másodlagos burkot (corona
radiata) alkotnak, amely még az ovulációt
követően is megfigyelhető a petesejt körül (2.2.
ábra).
Az elsődleges és másodlagos burkok az
utód keléséig megmaradnak a halak és a két-
éltűek esetében.
A megtermékenyítési vagy fertilizációs
membrán a megtermékenyítést követően jön
létre általában a már meglévő védőburkokból.
Kialakulása a kortikális reakció révén követ-
kezik be. A halak és a kétéltűek körében ez a folyamat gyorsan megy végbe, a megterméke-
nyítési membrán itt a zóna radiata rétegből jön létre. Ezt követően a felette lévő másodlagos
burok is fokozatosan megduzzad, megkeményedik. Lassan alakul ki a fertilizációs membrán
a hüllők és a madarak esetében,
ahol a szikhártya alakul át. Vala-
mivel gyorsabban megy végbe a
méhlepényes emlősök fertilizá-
ciós membránjának kialakulása,
ahol a zóna pellucida válik el a
petesejt membránjától. Fertili-
zációs membrán a gerinctelen
állatok körében is létrejön.
A harnaadlagos burkok
azután képződnek, miután a
petesejt kikerül a petefészek-
ből és végighalad a kivezető
járatrendszeren vagy már el-
hagyja a szervezetet. A harmad-
lagos burkok létrehozásában
tehát részt vehet a petevezető,
a méh, a héjmirigyek (például
laposférgek esetében) vagy a
bőr váladéka (például gyűrűs-
férgek körében). Ez a burok ro-
varoknál kitintartalmú, a bé-
2.3. ábra. A spermio- és ovogenezis folyamatának vázlata
A: spermíogenezis, B: ovogenezis, eo: elsőrendű oocita, es: elsőrendű sperma-
tocita, ér: érési szakasz, mo: másodrendű oocita, ms: másodrendű spermatoc-
ita, nö: növekedési szakasz, oo: oogoniumok, ov: ovum, po: polocita, sd:
spermida, sg: spermatogoniumok, sm: spermiomorfogenezis, sp: spermium,
sz: szaporodási szakasz (Több szerző nyomán módosítva)
kák petesejtjén többrétegű, víz-
ben megduzzadó. A hüllők és
a madarak petesejtjét a pete-
vezető öbölben képződő fe-
hérjeburkok veszik körül. Ezt
48
a hüllőknél elsősorban pergamen- vagy bőrszerű (ritkábban meszes) héj, míg madaraknál
a petevezető-szorosban képződő kettős héjhártya és az „uterus” nevű petevezetői sza-
kaszban termelődő meszes héj is határolja. A hüllők tojásában egyes csoportok (például
teknősök) kivételével nem jön létre számottevő vastagságú fehérjeburok, nem különül el
jégzsinór, valamint nem alakul ki légkamra. A méhlepényes emlősök petesejtje körül nem
alakulnak ki harmadlagos burkok.
A kokon egy vagy több petesejt és az ezeket körülvevő sziksejtek közös burokban való
elhelyezkedését jelenti. Előfordul a laposférgek, a gyűrűsférgek, a csigák körében és egyes
rovarok (például a csótányok) között is.
Néhány állatcsoport kivételével (szivacsok, egyes csalánozók) a petesejtek a petefészekben
(ovarium) jönnek létre. A petesejt kialakulásának folyamata (ovogenesis) három fő sza-
kaszra osztható.
A szaporodási periódusban az embrionális fejlődés korai szakaszában elkülönült ős-
csírasejtekből differenciálódó ősi oogoniumokból (archioogoniumok), több egymást követő
mitózis révén őspetesejtek (oogoniumok) keletkeznek (2.3. ábra). Anövekedési periódus-
ban az oogonium mérete nagyobbodik, jelentős mennyiségű szikanyagot halmoz fel, első-
rendű oocyta képződik belőle. Az érési periódusban az első érési osztódás, vagyis a meiózis
első főszakasza végén egy elsőrendű oocitából egy másodrendű oocyta és egy sarki sejt
(polocyta) alakul ki. A második érési osztódás (meiózis második főszakasza) alkalmával
mindkét sejt tovább osztódik, így a másodrendű oocitából keletkezik egy érett petesejt (ovum)
és egy sarki sejt, illetve az előzőekben kialakult sarki sejt tovább osztódva két sarki sejtet hoz
létre. A petesejt kialakulása során tehát egy elsőrendű oocitából egy érett, megtermékenyít-
hető petesejt és három, jóval kisebb méretű, megtermékenyülésre nem alkalmas sarki sejt
keletkezik. A sarki sejtek általában elpusztulnak. Az érési periódus a tüskésbőrűeknél a pe-
tefészekben megy végbe, az emlősöknél a petevezetőben fejeződik be, míg például a fonál-
férgeknél teljes mértékben az ivarszerven kívül zajlik.
A petesejt petefészekből való kiszabadulásának folyamata az ovuláció. A tüsző felrepe-
désének folyamata, a petesejt kilökődése neurohormonális szabályozás alatt áll. Egyes
állatoknál (görények, macskák, nyulak) az ovulá-
ciót közvetlenül a párzás váltja ki.
Ha a megtermékenyítés elmarad, akkor a pete-
sejt élettartama a legtöbb állatcsoportnál néhány
óra vagy nap.
2.1.2. A hímivarsejt kialakulása
és felépítése
A spermiumok lehetnek amőboid típusúak, mint
például a fonálférgek és egyes rákfajok esetében.
Számos tízlábú rák explóziós típusú spermiu-
mán rugószerű képlet van (2.4. ábra). Az állábai-
val a petesejten megtapadt spermium kitines tok-
ját ez a rugó- mechanizmus lelöki, miközben az
ellenerő a petesejtbe löveli a spermium maganya-
gát.
A legáltalánosabban elterjedt spermiumforma
az ostoros (flagellospermium). A gerinces állatok
flagellospermiuma jól differenciált, közülük is legin-
kább az emlősöké (2.5. ábra). Ennek feji részében
(caput) van a DNS-állományt magába foglaló sejt-
mag, melyet hátulról a citoplazma eredetű poszt-
2.4. ábra. Különböző spermiumformák
a: amőboid típus, b: explóziós típus, c-g: ostoros
típus, c: rovar, d: pisztráng, e: gőte, f: gyík, g:
madár, h: egér (Nelsen és más szerzők nyomán
kissé módosítva)
49
2.5. ábra. Az emlősök spermiumának
felépítése vázlatosan
ak: akroszóma, ap: apikális vakuólum,
fa: farok, fb: felbomló szerkezetű vég-
fonál, fe: fej, ff: farokfőrész, fv: farok
végrész, kd: középdarab, mi: mito-kond-
rium, ms: mitokondrium-spirál, ny: nyak,
ob: ostor bazális teste, pf: perforatórium,
pn: posztnukleáris sapka, rh: egymást
keresztező lefutású rostokból álló hüvely,
sh: sejthártya, sm: sejtmag, tf: ten-
gelyfonal (Több szerző nyomán mó-
dosítva)
nukleáris sapka, elölről a fejsapka (acrosoma) határol.
Az akroszóma a Golgi-készülékből létrejött, membrán-
nal határolt, ellapultvakuólum (speciálislizoszóma). Üre-
gét szokták apikális vakuolumnak is nevezni. Ebben a
petesejt felszínén való megtapadást és átjutást elősegítő
fehérjék, enzimek (hialuronidáz, akrozin, foszfatázok)
tárolódnak, amiket exocitózissal ad le. A perforatórium
a fejsapka belső lemezének megvastagodott nyúlványa,
amely a petesejtbe való behatolást segíti elő. A fej mag-
anyagában lévő X- vagy Y-kromoszómák miatt a sper-
miumok gyakran alakilag és elektroforetikus reakció-
jukban is eltérnek egymástól. Ez az állattenyésztés terü-
letén segítséget nyújthat az utód ivarénak irányított kiala-
kítására. Például elektromos térben az Y-kromoszómát
tartalmazók a katód, az X-kromoszómát hordozók az
anód felé mozdulnak el. A nyak (collum) a feji résszel
csuklószerű elmozdulásra képes kapcsolatban áll. Az itt
elhelyezkedő centriolumból indul ki a farok tengelyfo-
nala. A nyakban a tengelyfonál körül a bazális lemezből
eredő nyaklemez (vagy gyökér) található, amely védelmi
feladatot ellátva rostos hüvelyként folytatódik a további
szakaszokban. Az összekötő középdarab (pars
conjuctionis) tengelyfonala körül vékony plazmahüvely,
rostos hüvely és mitokondrium-spirál helyezkedik el. A
farok (flagellum) egy speciális, jól differenciálódott os-
tor. Tengelyvázát a 9 mikrotubuláris dupletből és két
centrális mikrotubulusból álló axonéma alkotja. A fa-
rok kezdetén (főrész = pars principális) a tengelyvázat
egy vékony citoplazmahüvely és egymást keresztező,
spirális lefutású rostköteg burkolja. A farokrész végét
(végrész = pars terminális) már csak igen vékony cito-
plazma és a sejtmembrán borítja. A farok végrészében
a tengelyváz szerkezete már nem mutatja a tipikus for-
mát. Több gerinces, például a farkos kétéltűek sper-
miumának farokrészéhez hullámzó, hártyaszerű képlet
kapcsolódik.
A hímivarsejtek képződésének folyamata (spermio-
genesis) a szaporodási, növekedési és érési periódusok-
ra tagolható spermiocytogenesis szakaszból (2.3. áb-
ra), valamint az ezt követő alaki átrendeződéssel járó
spermiomorphogenesis szakaszból (2.6. ábra) áll.
Gerinces állatoknál a spermiumok a herecsatomács-
kák belső felszínén ülő dajkasejtekbe (Sertoli-sejtek)
süllyedve fejlődnek. A Sertoli-sejtek támasztják, védik,
táplálják az osztódó, átalakuló sejteket.
A szaporodási periódusban az ősi spermato-
goniumokból (archispermatogonium) mitózisokrévén
ősondósejtek (spermatogoniumok) keletkeznek. Ezek
között vannak olyanok, melyek egy ideig nyugvó álla-
potban maradnak, míg mások tovább osztódnak. Belő-
lük a növekedési szakaszban elsőrendű spermatocyta
50
alakok jönnek létre. Az érési periódus-
ban az első érési osztódás (a meiózis első
főszakasza) során másodrendű sperma-
tocyták, míg a második érési osztódás (a
meiózis második főszakasza) végére a
spermidák jönnek létre. A petesejt kiala-
kulása során megismert méret és funkció-
beli különbségekkel bíró utódsejtekkel
szemben a spermium esetében egy első-
rendű spermatocitából négy egyenrangú
spermida keletkezik. A spermidák az os-
toros spermiumok esetén még egy alaki,
felépítésbeni átalakuláson mennek keresz-
tül. Ez a spermiomorfogenezis, amely so-
rán a citoplazma döntő hányada lefűző-
dik, a sejt belső szervecskéi átrendeződ-
nek, kialakul a hosszú ostor (2.6. ábra).
Az átalakulás kezdetén a Golgi-készülékről
leváló, megnövekedett méretű vakuólum
alakítja ki a fejsapkát. A Golgi-készülék
többi része részben a nyak úgynevezett
bazális lemezét képezi, részben a későb-
biekben leváló citoplazmacseppel együtt
távozik. Az akroszómában felhalmozód-
nak a petesejt védőburkát feloldó enzimek.
A sejtmag kromatinállománya kondenzá-
lódik. A centriólum bazális testté alakul,
belőle indul ki a mikrotubulusokból álló
2.6. ábra. A spermiomorfogenezis folyamatának váz-
lata
ak: akroszóma, ce: centriólum, cp: citoplazma, di: diktioszó-
ma, Ip: lefüződő citoplazmarész, mi: mitokondrium, sm:
sejtmag, sd: spermida, sp: majdnem érett spermium (Több
szerző nyomán módosítva)
tengelyfonal. A mitokondriumok a tengelyfonál köré rendeződnek. Az így létrejött hím-
ivarsejtek a herecsatomák üregébe kerülnek, bejutnak a mellékherébe, ahol érésük be-
fejeződik, a citoplazma maradványa csepp formájában lefűződik róluk. Membránjuk
felszíni elektromos töltése negatív. A csatornarendszerben képződő burkuk révén nyu-
galmi állapotba kerülnek, életműködéseik a minimumra csökkennek, így hosszabb ideig
megtartják életképességüket.
A spermiumok általában a járulékos nemi mirigyek váladékával együtt ürülnek ki
ondó formájában. Az ondó plazmája biztosítja a megfelelő pH-jú közeget, a szállítást és
a táplálást. Számos állatfaj kocsonyás tokban (spermatophora) rakja le spermiumait,
melyek onnan később szabadulnak ki. Ez jellemző például a laposférgekre, a piócákra,
egyes rákokra, rovarokra, pókokra, csigákra és a farkos kétéltűekre.
A spermiumok alakjának, mozgásképességének vizsgálata igen fontos tényező az ondó
minősítésekor, ha mesterséges termékenyítést alkalmaznak.
2.2. Megtermékenyítés
A megtermékenyítés (fecundatio vagy fertilisatio) során a női és a hímivarsejtek plaz-
ma- és maganyagai összeolvadnak, amely visszaállítja asejtekrejellemző kétszeres, diploid
(2n) kromoszómaszámot, és lehetővé válik az embrionális fejlődés megindulása. A meg-
termékenyítés két részmozzanata a hímivarsejt behatolása a petesejtbe (impregnatio),
valamint maganyagaik összeolvadása (conjugatio). Maga a folyamat igen bonyolult és
számos eltérés mutatkozik az egyes állatcsoportok között.
51
Külső megtermékenyítés esetén az ivarsejtek találkozása az állatot körülvevő kö-
zegben, leggyakrabban vízben történik meg. Mivel ennek biztonsága, eredményessége,
valamint a közegben többnyire védelem nélkül fejlődő utódok életbenmaradása gyakran
igen kis mértékű, ezért az így szaporodó állatok általában nagyon sok ivarsejtet adnak le.
Ez figyelhető meg például a halak, kétéltűek többségénél.
Belső megtermékenyítés esetén a peték az anya testén belül találkoznak a hímivarsej-
tekkel. A valódi értelemben vett belső megtermékenyítésnél a hímek párzószervének
segítségével jutnak be a spermiumok a női ivari járatokba. Ez a tipikus belső megtermé-
kenyítés előfordul például egyes laposférgekben és fonálférgekben, valamint a kerekes-
férgek, a szárazföldi csigák, a rovarok, a hüllők, a madarak és az emlősök körében. Ezzel
szemben a farkos kétéltűek hímjei által lerakott spermatofórát a nőstények kloákájukkal
veszik fel. A kagylók esetében a víz szállítja a spermiumokat a nőstény köpenyüregében
lévő petesejtekhez, míg a pókszabásúak hímjei tapogatólábaik segítségével juttatják be
ivarsejtcsomóikat a nőstény ivarnyílásába.
A belső megtermékenyí-
tésű állatokban a női ivari jára-
tokba bekerült hímivarsejtek
petesejthez való eljutását po-
zitív rheotaxisuk (a folyadék
áramlásával szemben történő
mozgás), valamint a méh és a
petevezető összehúzódásai se-
gítik. A női ivarjáratokba ke-
rült spermiumok termékenyí-
tőképessége fajonként változó,
lehet néhány óráigvagy akár he-
tekig tartó is. Azt az elképze-
lést, miszerint a spermiumok
kemotaxis révén jutnak el cél-
jukhoz a petesejt által kibocsá-
tott kémiai anyagok hatására,
ez ideig csak néhány faj (egyes
csalánozók, tüskésbőrűek, ha-
lak) esetében sikerült bizonyí-
tani. A két ivarsejt találkozása,
közvetlen érintkezése bonyolult
membrán- és sejtszerkezeti vál-
tozásokat vált ki mindkét ivar-
sejtben.
A megtermékenyítés bekö-
vetkezhet akkor, amikor a pe-
tesejt még nem ment keresztül
az érési osztódáson, de ebben
az esetben a spermium beha-
tolt feji és nyaki része mindad-
dig várakozik, amíg a petesejt
éretté nem válik. Ilyen például
a fonálférgek között fordul elő.
A spermium behatolásakor a
petesejt az első érési osztódás
közben van például a rovarok
2.7. ábra. Az akroszóma reakció lezajlása. A: külső és B: belső meg-
termékenyítésű állatoknál
af: aktinfilamentumok, av: apikális vakuólum, en: enzim, ami a védőburkot
feloldja, kg: kortikális granulum, kh: kötőhelyek, kt: kortikális granulum
kiömlő tartalma, pc: petesejt citoplazmája, sf: spermium farki részének kezdeti
szakasza, sh: petesejt sejthártyája, sm: spermium sejtmagja, vb: védőburok a
petesejt körül (Jungermann, Möhler és Darnell nyomán módosítva)
52
és a kagylók körében. A kétéltűek, a madarak és az emlősök petesejtje az első érési
osztódáson már keresztüljutott, amikor a hímivarsejt behatol. A tüskésbőrűeknél a sper-
mium a már érett petesejtbe jut be.
A nőstény emlősállatok ivari útjaiba került spermiumoknak bizonyos időt kell ott
eltölteniük, amíg elnyerik termékenyítőképességüket. Ez a kapacitáció (capacitatio)
folyamata, amely alatt megváltozik a hímivarsejtek membránjában lévő fehérjék össze-
tétele és elhelyezkedése, ezzel együtt a felületi töltésviszonya is. A felszíni negatív töltés
mértéke csökken. A kapacitáció alatt a spermiumról fokozatosan leválik az a védőburok,
amely még a hím ivari utakban borította. A respiráció foka emelkedik, jelentősen megnő
a Ca2+-felvétel. Ez utóbbi az akroszómareakció végbemeneteléhez szükséges kalciumfel-
halmozást eredményezi. A kapacitáció végén a spermium membránja destabilizálódik.
Mindezeket a változásokat a női ivari utak ionösszetétele, pH-ja és hormonális hatások
váltják ki.
Az akroszómareakció során a petesejt közelébe került spermium csúcsi plazma-
membránja összeolvad az akroszomális membránnal. így az akroszóma felnyílik, tartal-
ma a közegbe kerül (speciális exocitózis). A kiszabaduló enzimek, melyek közül a hi-
aluronidáz és az akrozin a legjelentősebb, a petét körülvevő burkok feloldását végzik.
A folyamat másik jelentősége abban van, hogy szabaddá válnak az eddig külvilágtól
elzárt, fajspecifikus kötőhelyek, és így a spermium számára lehetővé válik a petesejthez
való kötődés. Az akroszómareakció a külső és a belső megtermékenyítéssel szaporodó
állatok körében eltérő módon következik be (2.7. ábra). A külső megtermékenyítésűek
(például a tengeri sün és a halak közül a tokfélék) esetében a spermium akroszomális
membránjának és a sejt membránjának fúziója mindig az akroszómasapka alapi részén
kezdődik el. Az akroszóma alatti területről, a vakuólum alapi membránjával határolva
aktin-filamentumköteg indul ki. Ez az akroszomális nyúlvány a petesejthez való kapcso-
lódásban és a behatolásban játszik szerepet. A belső megtermékenyítéssel szaporodó
állatok esetében az akroszóma membránja több ponton fuzionál a plazmamembránnal,
és nem képződik aktinszálakból álló nyúlvány. Az akroszomális reakciót a petesejt bur-
kának glikoproteid részei, illetve más hormonhatású, fajspecifikus induktoranyagok vált-
ják ki. Ennek hatására megváltozik a spermium membránjának kationtranszportja, fo-
kozott mértékben jut be kalcium a sejtbe, ami a membránfúziók közvetlen kiváltó oka.
A spermium petesejtbe való behatolásának folyamata az impregnáció. Az ivarsejtek
membránjainak felszínén lévő specifikus azonosító glikoproteid molekulák - kötőhelyek
- általában biztosítják, hogy csak az azonos fajba tartozók ivarsejtjei egyesülhetnek. Sper-
miumokon a kötőhelyek a volt akroszóma megnyílt területein vannak, míg petesejteknél
ez változó lehet. A membránok fúzióját követően a spermium sejtmagja és a centriólum
a petesejt citoplazmájába kerül. A petesejt egész felületén vannak kötőhelyek például a
tengeri sünnél, így a spermium behatolása bárhol végbemehet. A kétéltűekpetesejtjében
ez például csak az animális póluson történhet meg. A petesejtbe általában csak a sper-
miumfeji és nyaki része (a centriólummal) hatol be, a többi leválva kívül marad. Amennyi-
ben a petesejtet keményebb védőburok veszi körül, akkor azon kis nyílás (micropyle)
található, ahol a spermium be tud hatolni (például a halaknál).
Az impregnáció a petesejt membránján kortikális reakciót vált ki. Ezt a spermium-
ban lévő nagyobb mennyiségű kalciumnak a petesejtbe való bejutása okozza. Kezdetben,
néhány másodpercen keresztül megváltozik a petesejt membránpotenciálja, akciós po-
tenciál alakul ki. A felszíni töltésviszonyok megváltozása megakadályozza, hogy további
spermiumok a membránhoz kötődjenek. Ezzel egy időben a kortikális granulumokmemb-
ránja összeolvad a sejtmembránnal és tartalmuk a felszínre ömlik (2.7. ábra). A felsza-
baduló proteolitikus enzimek a petesejt felszínén a kötőhelyek fehérjemolekuláit részle-
gesen bontják, ami a kötőképesség elvesztéséhez vezet. A spermium behatolási helyétől
kiindulva ez a folyamat a petesejt teljes felszínére rövid idő alatt kiterjed. A petesejt
53
membránszerkezetének megváltozása csak egy hímivarsejt behatolását (monospermia)
teszi lehetővé. A kortikális granulumokból peroxidázenzim is felszabadul, ami a szik-
hártya megduzzadásához, szilárdságának növekedéséhez, az úgynevezett megterméke-
nyítési burok kialakításához vezet. A főleg glikozamino-glikánokból álló szikhártya vízzel
érintkezve abból jelentős mennyiséget vesz fel, megduzzad. A megtermékenyített pete-
sejt kissé összezsugorodik, így a megtermékenyítési membrán alatt perivitellináris tér
(spatium perivitellinum) jön létre. Ezek a változások együttesen megakadályozzák to-
vábbi spermiumok behatolását, mert azok enzimjei már nem képesek a megtermékenyí-
tési membránt feloldani.
A polyspermia, tehát több hímivarsejt petesejtbe való behatolása nem rendellenesség.
Több állatcsoportnál, például egyes rovaroknál, a porcoshalaknál, egyes kétéltűeknél, a
hüllőknél és a madaraknál is előfordul. Ilyenkor a szikhártyán számos spermium hatol
be, azonban ebben az esetben is csak a leendő csírakorong, illetve az animális pólus
területén behatoló termékenyíti meg a petesejtet. Ha nemcsak egy spermium kromoszó-
maállománya vesz részt a további folyamatokban, akkor rendellenes kromoszóma-kom-
binációk jöhetnek létre, ami abnormális fejlődést okozhat.
A spermium petesejtbe való behatolását követően az emlősök esetében a sejtmagokból
létrejönnek a hím és a női előmagvak (pronucleusok). A két pronukleusz kromatin-
állományából kialakulnak a kromoszómák transzportformái. A maghártyák ciszternákra
tagolódnak, kialakul a mitotikus orsó, a két centriolum az orsó két végére kerül és
megindul a mitózis.
Az állattenyésztési kutatásokban és a gyakorlatban széles körben használják ki azt a
lehetőséget, hogy az ondó (sperma) mélyhűtve nagyon hosszú ideig eltartható, és benne
a spermiumok megtartják életképességüket. A spermahűtés lehetősége számos állat-
fajnál mindennapi gyakorlattá tette a művi ondóbevitelt (inszemináció - mesterséges
termékenyítés).
2.3. Az embrionális fejlődés citotipikus szakasza
2.3.1. Barázdálódás
A barázdálódás (segmentatio) a zigóta osztódási folyamata, amelynek során számos
utódsejt (blastomera) jön létre. Az újonnan képződött sejtek kezdetben általában még
igen hasonló tulajdonságokkal és nagyfokú önállósággal bírnak.
A szűznemzés (parthenogenesis) során a petesejt barázdálódása megtermékenyítés
nélkül is végbemegy. Ez előfordul például egyes kerekesféigek, rákok, rovarok, halak, két-
éltűekés gyíkok körében (4.6.2. fejezet). A meg nem termékenyített petékből az adott állatfaj
ivari kromoszómáitól függően rendszerint nőstények, ritkábban hímek (például a méhek
esetében), vagy akár mindkét ivar egyedei (például egyes rák- és levéltetű-fajok esetében) is
kifejlődhetnek. Természetes parthenogenezis esetén az utódok diploidok lesznek abban az
esetben, ha a petesejt barázdálódása az érési osztódás (meiózis) előtt végbemegy. Ha a
meiózis már lezajlott, akkor az utódoknak haploid kromoszómagamitúrája lesz.
Ginogenezis (gynogenesis) alkalmával a petesejtbe behatoló inaktív vagy nem faj-
azonos spermium csak elindítja a barázdálódást, de genetikai állománya a további folya-
matokban nem vesz részt. Előfordul például számos fonálféreg és az ezüstkárász esetében.
Mesterségesen mind a szűznemzés, mind a ginogenezis kiváltható bizonyos vegyi
anyagokkal, extrém hő- és fényhatással, mechanikai beavatkozással, sugárzó anyagok
segítségével.
A barázdálódás során a sejtosztódások típusa mitózis, a blasztomérák száma többnyire
mértani haladvány szerint nő (2,4,8,16, 32 ... sejt), de ettől eltérő módon is alakulhat.
54
A képződő sejtek osztódásai között növekedési periódus nincs, egyre kisebb sejtek jönnek
létre, melyek össztérfogata a petesejt eredeti térfogatával egyezik meg. A blasztomérák
a tartalék tápanyagaik felhasználásával igen intenzív DNS-szintézist folytatnak.
Az utódsejtek létrejötte történhet egyszerre (szinkron) vagy különböző (aszinkron)
időritmusban.
A barázdálódás végbemenetele nagymértékben függ a petesejt szikanyagtartalmától
és eloszlásától (2.1. táblázat, 2.9. ábra). A nagyobb mennyiségű szik lassítja vagy gátolja
a blasztomérák osztódását. Ahol a zigótán belül a szikanyag koncentrálódik, ott nagyobb
méretű sejtek (macromera) jönnek létre, míg a szikben szegényebb területen kismére-
tűek (micromera) keletkeznek. így elkülönül a szikdús sejteket tartalmazó vegetatív
pólus és a szikben szegény animális pólus.
Ha az osztódás a teljes térfogatra kiterjed, akkor a barázdálódás teljes (segmentatio
totális). Ellenkező esetben a barázdálódás részleges (segmentatio partialis). A teljes
barázdálódást mutató (holoblasticus) zigóta utódsejtjeinek nagyságbeli eltérése alapján
a szegmentáció lehet aequalis, amikor a blasztomérák egyformák, adaequalis, amikor
2.1. táblázat. Összefüggés a petesejt sziktartalma és eloszlása, valamint a barázdálódás módja
között
Sziktartalom Szikeloszlás Barázdálódás típusa Állatcsoport
ENDOLECITÁLIS PETÉK:
alecitális- oligolecitális izolecitális totális ekvális adekválís radiális csalánozók
bilaterális zsákállatok
aszimmetrikus emlősök*
mezolecitális anizolecitális inekvális radiális tüskésbőrűek
spirális puhatestűek gyűrűsférgek
diszimmetrikus bordásmedűzák
bilaterális tüdőshalak kétéltűek
polilecitális anizolecitális telolecitális parciális inekvális diszkoidális skorpiók fejlábúak porcos halak csontos halak hüllők madarak
centrolecitális szuperficiális ízeltlábúak (kivétel: skorpiók)
EKTOLECITÁLIS PETÉK:
totális ekvális anarchikus laposférgek
‘Megjegyzés: Az emlősök teljes barázdálódása másodlagosan alakult ki, amit az is jelez, hogy a gasztrulációs
folyamatok a sok szikanyagot tartalmazó petesejtü hüllőkéhez és madarakéhoz igen hasonlóan mennek végbe.
55
közel egyformák és inaequalis, amikor kü-
lönbözőek.
A teljes barázdálódású zigóta animális és
vegetatív pólusát képzeletben összekötő ten-
gelyhez viszonyítva a képződő sejtek helyze-
te lehet radiális, spirális, bilaterális, diszim-
metrikus, aszimmetrikus és anarchikus. A
radiális típusú barázdálódás esetében a
blasztomérák a tengely körül sugárszimmet-
rikusan, egymás feletti sorokban, rétegek-
ben helyezkednek el (2.8. ábra). Ennek so-
rán az első osztódási sík meridionális, vagyis
az animális és vegetatív póluson megy át. A
második osztódási sík is meridionális, de az
2.8. ábra. A teljes barázdálódás típusai a képződő
blasztomérák helyzete és a szimmetriaviszonyokszerint
bi: bilaterális, dí: diszimmetrikus, ra: radiális, sp: spirális
(Siewing, Conglin, Stewing és Speknyomán kissé módosítva)
előzőre merőleges helyzetű. A harmadik sík
egyenlítői helyzetű. Ha a petesejt eredeti szik-
anyagtartalma egyenletes eloszlású volt, az
animális és a vegetatív fél egyforma, míg
egyenlőtlen szikeloszlás esetén az első egyen-
lítői barázda az animális pólushoz esik közelebb. A további osztódások során függőleges,
vízszintes és a belső sejtrétegben a felszínnel párhuzamos barázdák alakulnak ki. Az
osztódások kezdetben szinkron módon, később ettől eltérően alakulhatnak. Ez a baráz-
dálódási mód tipikus formában jellemző a szivacsokra, csalánozókra és a tüskésbőrűekre.
A kétéltűek és az emlősök zigótájának barázdálódása csak a kezdeti szakaszban mutat
radiális típust, a későbbiekben már nem. A spirális barázdálódáskor az egyenlőtlen
osztódással létrejövő sejtek nem egymás felett szabályosan helyezkednek el, hanem egy-
máshoz képest 45 fokkal elcsúszva (2.8. ábra). Egy felső mikroméra így két alatta lévő
makromérához kapcsolódik. A blasztomérák a képzeletbeli tengelyhez képest eltolód-
hatnak egy azonos vagy váltakozó irányban, így az animális pólus felől nézve a barázdá-
lódás spirálisnak látszik. Ez a soksertéjű gyűrűsférgekben és a puhatestűekben fordul elő.
Bilaterális barázdálódás esetében már néhány sejtes állapotban szimmetriasík jelenik
meg, az egyik oldalon lévő sejteknek a másik oldalon megtalálhatók a szimmetrikusan
elhelyezkedő megfelelőik (2.8. ábra). Ilyen fordul elő például a fonálférgek, kerekes-
férgek, fejlábúak, zsákállatok körében. A kétéltűek kezdeti radiális barázdálódása később
bilaterális vonásokat mutat. Diszimmetrikus barázdálódáskor a négysejtes állapot-
tól kezdődően két szimmetriasík alakul ki (2.8. ábra). Ez jellemző egyes csalánozókra és
a bordásmedúzákra. Anarchikus barázdálódás esetében a blasztomérák látszólag tel-
jesen rendezetlen állapotban helyezkednek el, mint például egyes csalánozók hidrome-
dúza-alakjaiban, egyes laposférgekben.
A teljes barázdálódást mutató petesejttel rendelkező fajok körében a leggyakoribb két
barázdálódási típus a radiális és a spirális. Ez jól jellemzi a bilaterális, többsejtű álla-
tok között meglévő két ágat, az ősszájúakat (Protostomia) és az újszájúakat (Deuterosto-
mia). A radiális barázdálódás általában az újszájúakra, a spirális pedig az ősszájúakra
jellemző.
A részleges barázdálódású (meroblasticus) zigóta fejlődése során csak a szikmentes
vagy szikben szegény területen megy végbe a szegmentáció (2.9. ábra). Ez megvalósulhat
az egyik póluson, csírakorongot létrehozva (segmentatio partialis discoidalis) vagy
a felszíni rétegben (segmentatio partialis superficialis). Diszkoidális barázdálódás
jellemző például a skorpiókra, a fejlábúakra, a csontos halakra, a hüllőkre, a madarakra
és a kloákás emlősökre. Szuperficiális barázdálódás fordul elő az ízeltlábúak körében (a
skorpiók kivételével). Ennél a barázdálódási típusnál a sejtmagok osztódnak, a felszíni
56
2.9. ábra. A barázdálódás típusai
1.: első osztódások, 2.: morula állapot, 3.: blasztula metszetben, A: segmentatio totális adaequalis, B: segmentatio totális
(emlősök esetében), C: segmentatio totális inaequalis, D: segmentatio partialis discoidalis, E: segmentatio partialis
superficialis (Több szerző nyomán módosítva)
57
rétegbe húzódnak, majd ezt követően jönnek létre a sejthatárok a plazma feldarabolódása
során.
A korai blasztomérák képessége alapján meg lehet különböztetni regulációs és mozaik
típusú barázdálódást, illetve embriókat.
A regulációs képességű embriók blasztomérái képesek korrigálni a bekövetkezett
sérüléseket, sejtkieséseket. A blasztomérákat egymástól elválasztva, azok normális egye-
dekké fejlődnek. Ez azt jelenti, hogy ezek a sejtek totipotenciájukat bizonyos számú
osztódásig nem vesztik el. Ilyen képességgel rendelkeznek az elsősorban radiális baráz-
dálódást mutató újszájú állatok, így a tüskésbőrűek, valamint a gerincesek embriói a
korai fejlődés időszakában. Az emlősállatok in vitro történt megtermékenyítése esetén
(az embernél is) például a nyolcsejtes állapotban lehetőség van egy blasztoméra eltávo-
lítására az esetleges genetikai eredetű betegségek tesztelése céljából. A megmaradt sejtek
normális utóddá fejlődnek.
A mozaik típusú embriók blasztomérái eleve determináltakká válnak, azaz beszű-
kül a fejlődési potenciájuk. Ha az ilyen sejteket szétválasztjuk, akkor azokból nem fejlőd-
nek normális egyedek, mindegyik blasztoméra azt a fejlődési irányt folytatja, amelyiken
már elindult. A zigótán már előre meg lehet határozni azokat a területeket, amelyekből
az egyes csíralemezek, ősi szervtelepek ki fognak alakulni, így már az első osztódás során
létrejövő sejtek meghatározott fejlődési irányúak. Igen gyakori, hogy már a petesejt is
determinált régiókra oszlik, azaz elkülöníthetőek a leendő egyed elülső és hátulsó végé-
nek, illetve a háti és hasi oldalának megfelelő területek. Ez a determinisztikus fejlődési
típus az ősszájú, főleg spirális barázdálódást mutató állatokra, például az örvényférgekre,
a fonálférgekre, a kerekesférgekre, a gyűrűsférgekre, a legtöbb puhatestűre és számos
rovarfajra jellemző.
2.3.2. Szedercsíra állapot
A barázdálódás eredményeként egyre több blasztoméra keletkezik. A tizedik osztódás
után általában már több mint 1000 sejt jön létre, kialakul a szedercsíra (morula) ál-
lapot. Az egyre csökkenő méretű blasztomérák elhelyezkedése a petesejt szikanyag-
tartalmától és annak eloszlásától függ (2.9. ábra). A barázdálódás végére például az
ízeltlábúakban mintegy 6000, a lándzsahalban 9000, a békákban közel 700 000 blaszto-
méra alakul ki.
Az állattenyésztési kutatásokban és gyakorlatban felhasználják az emlősmorulát alko-
tó blasztomérák azon képességét, hogy néhány sejtes állapotban még totipotensek. A
morulát alkotó sejteket szétválasztva és továbbnevelve (klónozás) elérhető, hogy gene-
tikailag teljesen azonos egyedeket hozzanak létre. Kiméra (chymera) előállítása esetén
két különböző morula blasztoméráit helyezik egy közös peteburokba. A kiváló képességű
anyaállatokból (donor) a morula állapotban lévő utódokat ki lehet mosni, és azok más,
kevésbé értékes nőstények (fogadó-recipiens) méhébe helyezhetők be. A morula mély-
hűthető, és ilyen formában a felhasználási helyre szállítható.
2.3.3. Hólyagcsíra állapot
A fejlődő csíra belsejében, a felszíni rétegbe rendeződő blasztomérák alatt általában
barázdálódási üreg (blastocoel) jön létre. Ezt az állapotot hólyagcsírának (blastula)
nevezik. A barázdálódás típusától függően a barázdálódási üreg alakja, mérete, elhelyez-
kedése különbözőképpen alakul (2.9. ábra).
Ha a barázdálódási üreg igen erősen redukálódott vagy teljesen hiányzik, akkor
tömött blasztula (sterroblastula) jön létre (például egyes csalánozók és soksertéjű
gyűrűsférgek).
58
Teljes barázdálódás esetén a blasztocöl nagy kiterjedésű üreg (coeloblastula) lehet.
Szikmentes vagy kevés szikű peték teljes barázdálódása terjedelmes, általában központi
elhelyezkedésű blasztocölt hoz létre. Közepes szikanyagtartalmú petéknél (például a
kétéltűeknél) a vegetatív póluson lévő, nagyobb méretű makromérák miatt a blasztocöl
excentrikus elhelyezkedésű (amphiblastula), vagyis az animális pólushoz közelebb toló-
dott és alakja nem szabályos gömb.
Részleges barázdálódásnál, ha az diszkoidális módon ment végbe, a korong alak-
ban rendeződött blasztomérák (madaraknál megközelítőleg 60 ezer sejt) blasztodermát
alakítanak ki. A blasztoderma és az alatta lévő nagy mennyiségű szik között keskeny üreg
(subgerminalis üreg) jön létre. Halaknál a blasztoderma alatt, a szikanyag felszínén
szincíciális periblast sejtréteg alakul ki. A leendő embrió farki, kaudális végén a
blasztodermasejtek felszaporodása megvastagodáshoz vezet. Az itt lévő hypoblast sejtek
az entodermát fogják kialakítani (2.13. ábra). Hüllőknél ősmadaraknál a csírapajzs széli
része a szikanyaggal létesít kapcsolatot (area opaca), míg a centrális helyzetűek elvesz-
tették a közvetlen érintkezést (area pellucida) a szikanyaggal, alattuk jön létre a
szubgerminális üreg. A sok sejtrétegű blasztoderma szikanyag felé néző alsó sejtjei elvál-
nak a felette lévőktől, kialakítanak egy alsó sejtréteget (hypoblast) (2.14. ábra). A felszíni
réteg (epiblast) és a hipoblaszt között a blasztocöl jön létre (discoblastula).
A centrolecitális peték szuperficiális barázdálódása során a felszíni vékony periplazma
rétegbe húzódott, kevés plazmával határolt sejtmagok kezdetben sejthatár nélküli
szincíciumot alkotnak. Később sejtekre tagolódnak, blasztodermát alkotnak. Előfor-
dul, hogy a blasztoderma kezdetben csak bizonyos területen alakul ki, majd később
szétterjedve borítják be a teljes felszínt (2.16. ábra). Barázdálódási üreg nem képződik,
periblastula alakul ki.
Annak ellenére, hogy az emlősállatok zigótája másodlagosan oligolecitális és izolecitális
típusú, mégis teljes barázdálódást mutat. A kialakuló blasztula (blastocysta) felépítése,
további differenciálódása viszont a diszkoidális barázdálódásúakra jellemző, valamint más
speciális vonásokat is mutat. A
külső elhelyeződésű sejtek (tro-
phoblast) rétegéből a blaszto-
cölbe türemkedik az embrió-
csomó (embryoblast). Később
az embriócsomó alsó sejtjei el-
különülve az elsődleges ento-
dermával határolt szikzacskót
alakítják ki (2.15. ábra). A felső
helyzetű sejtekből az amnion jön
létre. Az embriócsomó többi sej t-
jei az ektodermából, mezo-
dermából és az entodermából ál-
ló, lapos, korong formájú csíra-
pajzsot alkotják (2.15. ábra). A
blasztociszta a trofoblaszt sejtek
által termelt enzimek aktivitása
révén a méh nyálkahártyájába süllyed, beágyazódik (implantatio). A trofoblaszt sejtek egy
része az anyai placentával való szoros kapcsolatot alakítja ki (syncytiotrophoblast sejtek),
míg a többiek kívülről határolják a fejlődő embriót (cytotrophoblast sejtek).
Hólyagcsíra állapotban a blasztodermán speciális módszerekkel meg lehet különböztet-
ni azokat a területeket (praesumptiv areák), amelyekből a leendő csíralemezek, ősi szerv-
telepek, illetve szervek majd kialakulnak (2.10. ábra), így megrajzolható a fejlődő embrió
sejtjeinek „sorstérképe”.
B
2.10. ábra. A preszumptiv areák
Iáján. A: oldalnézetben, B: az őss
ee: epídermális ektoderma, en: entc
mezoderma, ne: neurális ektoderma, <
módosítva)
elhelyezkedése a békák blasztu-
táj felől nézve
derma, ko: korda mezoderma, me:
>s: összájnyílás (Torrey nyomán kissé
59
2.4. Az embrionális fejlődés organotipikus szakasza
C2
D2
Dl
Dl
A
B
C
2.4.1. Csíralemezek kialakulása
A gasztruláció (gastrulatio) eredeti értelmezésben az ősbéllel bíró csíra, azaz a gasztrula
(gastrula) képződését jelenti. Mivel ennek során a csíralemezek is kialakulnak, valamint
a cöloma- (coeloma) formálódás is lezajlik, ezért a gasztruláció fogalmának tágabb ér-
telmezését használjuk.
Az embrionális fejlődés során a blasztomérák származásuknál és helyzetüknél fogva
meghatározott kiterjedésű területeket,
sejtrétegeket, azaz csíralemezeket
alkotnak. A folyamat során a sejtek
száma tovább nő, a morfogenetikus
mozgások, a sejtrendeződéssel járó el-
mozdulások létrehozzák a két vagy há-
rom csíralemezt. A sejtképződés, dif-
ferenciálódás során a fejlődő csíra
morfológiai, méretbeni változásával
párhuzamosan egyes helyeken a sej-
tek elhalhatnak. A csíralemezeket al-
kotó sejtek fejlődési potenciája megha-
tározott, belőlük alakulnak ki a szer-
vezetet felépítő szövetek, szervek. A
gasztruláció során kialakul a szerve-
zet felépítésének alapja.
A gasztruláció alkalmával megva-
lósuló sejtmozgások, átrendeződések
különböző típusait lehet elkülöníte-
ni. Ezek gyakran egymással párhuza-
mosan, egyidőben zajlanak, különbö-
ző mértékben módosulhatnak, illetve
egy állatcsoporton belül több típus is
jellemző lehet.
A külső csíralemezt (ektoderma)
alkotó sejtek a felszíni rétegben nagy
kiterjedésben helyezkednek el.
A belső csíralemez (entoderma)
kialakulásának különböző típusai van-
nak. A betűrődés (invaginatio) so-
rán a hólyagcsíra egy területén a külső
sejtekrétegebehajlik, benyomul a bel-
ső üregbe és mozgását tovább folytat-
va a külső sejtréteg alatt kialakít egy
belső réteget (2.11. ábra). A betűrő-
désben részt nem vevő sejtek hozzák
létre az ektodermát, míg a többi az
entodermát. Ez tipikusan a kevés és
egyenletes eloszlású szik-anyagot tar-
talmazó petesejtekből fejlődő állatok-
ban (szivacsok, tengeri sün, lándzsa-
D
2.11. ábra. Az entoderma kialakulásának gyakoribb típusai
A: betürödés (invaginatio), B: körülnövés (epibolia), C: bevándor-
lás (immigratio), C2: poláris immigratio, C2: multipolaris immigra-
tio, D: leválás (delaminatio), az osztódással létrejött belső
sejtréteg leválása, D2: a belső sejttömeg leválása (Plfugfelder és
Siewing nyomán módosítva)
60
2.12. ábra. A kétéltűek gasztrulációjának fontosabb lépései (At-A2: agasztruláció folyamata, B: az embrió térbeni
metszete a neuruláció kezdetén, C: az embrió testének metszete a neurulációt követően, a csíralemezek további
differenciálódásának kezdetén)
bl: blasztocöl, ee: epidermális ektoderma, ek: ektoderma, en. entoderma, gh: gerinchúr, me: mezoderma, őb: ősbél ürege,
ős: ősszájnyílás, őv: ősbélvályú, ve: velőcső, vé: vérér, vl: velőlemez, vt: valódi testüreg (Torrey és Siewing nyomán kissé
módosítva)
hal) fordul elő, de megfigyelhető a gerincesekben (például a kétéltűekben) is (2.12.
ábra). A kétéltűekben a szikdús sejtekből álló entoderma az ősbélvályút alakitja ki. Ennek
szélei később felhajlanak, csővé zárulnak, létrehozzák az ősbelet. A betüremkedés révén
keletkező üreg az ősbélüreg (archenteron), nyílása az ősszáj (blastoporus). Az ektoderma
és entoderma közötti üreg (elsődleges testüreg) a blasztocöl maradványa.
A rátüremkedés (epibolia) során az ektoderma gyorsabban osztódó sejtjei beborít-
j ák, körülnövik a többi sej tét, így azok az ektoderma alá kerülnek (2.11. ábra). A folyamat
az embrió fej-farok irányú megnyúlásával és a másik két csíralemez sejtjeinek az ektoderma
alá való bekerülésével is együttjár. Ez fordul elő a szivacsokban, az örvényférgekben, a
kerekesférgekben, a gyűrűsférgekben, a csigákban, egyes ízeltlábúakban, a halakban és
a kétéltűekben is. Halakban a csírapajzs kraniális és oldalsó (laterális) szélén a sejtek igen
intenzíven osztódnak, gyorsan körülnövik a szikanyagot (2.13. ábra). A fejlődő embriót
beborító animális eredetű sejtek a csírapajzs kaudális végén, az embrió farki régiójánál
találkozva fejezik be mozgásukat. Ezt követően a csírapajzs leendő testvég felé néző,
kaudális szélénél lévő, a felszínnel nem érintkező hipoblaszt sejtek fokozott osztódás
révén a fej felé, kraniális irányban a külső sejtréteg (ilyenkor már epiblaszt a neve) alatt
betüremkednek és előrefelé húzódnak. E folyamatot a mezodermasejtek befordulása
(involutio) is követi (2.13. ábra). A sejtek befordulási helyén résszerű ősszáj alakul ki.
A bevándorlás (immigratio) során a felszíni rétegből egy (unipoláris) vagy több
pontból (multipoláris) kiindulva sejtek válnak le, és alatta önálló sejtréteget, entodermát
alkotnak (2.11. ábra). Ilyen fordul elő egyes csalánozókban és puhatestűekben.
A leválás (delaminatio) alkalmával legegyszerűbb esetben a hólyagcsíra falát alkotó
több sejtréteg belső sora leválik a külsőről (2.11. ábra). A blasztula külső sejtrétege
például osztódással válhat kétrétegűvé, majd a belső sor leválik. Szterroblasztula esetén
61
2.13. ábra. A valódi csontoshalak gasztrulációjának fontosabb lépései (A: a gasztruláció kezdete, B: embrió az
epibólia befejeződése előtt, C: az embrió testének keresztmetszete a csíralemezek kialakulásakor, D: az embrió
teste a testszelvények ki alakulásának időszakában, E: az embrió testének keresztmetszete a neurulációtkövetően,
a csíralemezek további differenciálódása kezdetén)
em: ento-mezoderma, en: entoderma, ep: epiblaszt, gh: gerinchúr, hi: hipoblaszt, me: mezoderma, mh: mezoderma
hátlemeze, mk: mezoderma középlemeze, mo: mezoderma oldallemeze, ne: neurális ektoderma, Őb: ősbél, pe: periblaszt,
sz: szik, ts: testszelvények, ve: velőcso, vt: valódi testüreg (Lagler, Portmann, Siewing és Wilson nyomán kissé módosítva)
a belső sejtcsomó elkülönül a külső rétegtől. Ezek az entoderma-kialakulási módok tipikus
formában jellemzőek például a csalánozókra. Halakban a befordult entodermális, kordális
és mezodermális sejtek kezdetben egymáshoz szorosan illeszkedő sejt-rétegeket alkotnak,
melyek később delaminációval különülnek el (2.13. ábra). A hüllők és a madarak blaszto-
diszkuszának szubgerminális üreggel érintkező sejtjei a felső rétegről leválva kialakítják a
hipoblaszt réteget, amely részt vesz az entoderma létrehozásában (2.14. ábra). Az entoderma
végleges formájának kialakításában azonban a primitív csík területén beforduló és a hipoblaszt
rétegbe beépülő sejtek is részt vesznek. Emlősökben a csírapajzsot (embriópajzsot) alkotó
sejtek igen korán elkülönülnek ekto- és entodermarétegekre, amelyek később, a mezo-
derma differenciálódásakor egymástól elválnak (2.15. ábra).
A rovarok szuperficiális barázdálódású zigótájából kialakuló periblasztuláján, a leen-
dő szervezet hasi oldalán embriócsík jön létre a blasztoderma sejtjeinek tömörülése
folytán (2.16. ábra). Az embriócsík felszínén maradó sejtek alkotják az ektodermát.
Ennek középvonalában különül el a neurális ektoderma. Az ősbél entodermarétege
részben a szikanyagba ágyazott, kezdetben nem összefüggő vitellofág sejtekből, illetve az
embriócsík leváló sejtjeiből alakul ki.
A középső csíralemez sejtjei a külső és a belső csíralemez között fejlődnek. Kiala-
kulásuk alapvetően két típus szerint mehet végbe, de ennek lehetnek módosulásai is
(2.17. ábra).
62
Az enterocöl (enterocoel) eredetűnél a mezodermasejtek az ősbélről kitüremkedő,
majd leváló és egymással összeolvadó zsákokat formálva jelennek meg. A mezoderma
által határolt üreg (coeloma) tehát valójában az ősbélüregről fűződik le. Ez a forma az
újszájú állatok közül a tüskésbőrűekre jellemző.
A másik típus a szkizocöl (schizocoel) eredetű, amelynél a leendő mezoderma sejtjei
az ősszájnyílás mellett betüremkedéssel (invaginatio), befordulással (involutio), illetve
bevándorlással (immigratio) indulnak fejlődésnek a külső és belső csíralemez között. A
formálódó gasztrulában a leendő mezoderma sejtek szétterjednek (divergencia) a végle-
ges helyükre. A leendő mezodermasejtek már a barázdálódás kezdeti szakaszában elkü-
lönülnek a többi sejttípustól. A fejlődő, kezdetben tömör, több sejtrétegből álló mezo-
dermában hasadással alakul ki a cöloma. Ez elsősorban az ősszájú állatok (például a
gyűrűsférgek, puhatestűek, ízeltlábúak) jellemzője, de a gerinceseknél is előfordul.
Csontoshalaknál az entodermasejteket követve a mezoderma is befordul. Az entoderma-
és mezodermasejtek kezdetben szorosan egymáson fekszenek, de később elválnak egy-
mástól (2.13. ábra). A kétéltűeknél a kordamezoderma lemezének kezdeménye az
entoderma invaginációját követve befordulással (involutio) alakul ki. Kezdetben az
ősbélvályú felett húzódik, majd laterális irányban is kiterjed (2.12. ábra). Középső részé-
ből alakul ki a gerinchúr, míg az oldalsó lemezek a mezodermát képezik. Hüllőknél,
madaraknál (2.14. ábra) és az emlősöknél (2.15. ábra) a primitív csík (közepén a baráz-
B
2.14. ábra. A madarak gasztrulációjának fontosabb lépései és a magzatburkok kialakulásának kezdete (A:
jellegzetes képletek a blasztodiszkusz területén felülnézetben, B: sejtmozgások a gasztruláció és a neuruláció
folyamán felülnézetben, C-D: blasztodiszkusz hosszmetszete a gasztruláció kezdetén, E: sejtmozgások a primitív
barázda területén, F: embrió testének keresztmetszete a neuruláció során, G: a magzatburkok kialakulásának
kezdeti szakasza)
ao: area opaca, ap: arca pelhicida, ar: amnionredő, bl: blasztocöl, ee; epidermális ektoderma, en: entoderma, ep: epiblaszt,
fe: fejnyúlvány, gh: gerinchúr, hi: hipoblaszt, me: mezoderma, ne: neurális ektoderma, őb: kialakuló ősbél, pc: primitívcsík,
pg: primitív gödör, sz: szik, ve: velőcső, vr: velőredő, vt: valódi testüreg (Geiler, Huettner, Pattén, Portmann, Starck és
Torrey nyomán)
63
da) és a Hensen-féle primitív csomó (közepén a gödör) területén lévő összetömörült
sejtek befordulnak és sejtvándorlással kialakítják a mezodermaréteget a külső és belső
csíralemez között. A cöloma a mezoderma sejtrétegei között létrejövő hasadás következ-
tében jön létre. A primitív gödör és a primitív csík funkcionálisan az ősszájnak felel meg,
ahol nyílás kialakulása nélkül az eredetileg külső elhelyezkedésű sejtek befordulnak a
csíra belsejébe. A primitív gödörnél befordulva a gerinchúr (chorda dorsalis) előtelepe
és a testszelvényekre, szomitákra tagolódó mezoderma fejnyúlvány formájában kraniális
irányban indul fejlődésnek. A primitív csíknál kraniálisan és laterálisán zajlik a mezo-
derma befordulása. A többirányú, fokozatos kiterjedés eredményeként kialakul az extra-
embrionális szervek mezodermális rétege. Emlősöknél az extraembrionális mezoderma
előbb alakul ki, mint az embrionális. Amikor az emlősembrió teste még csak két csíra-
lemezből áll, a citotrofoblasztból leváló sejtek létrehozzák a laza hálózatot alkotó extra-
embrionális mezodermát (2.15. ábra). Ez körülveszi a már meglévő képleteket, kibéleli
2.15. ábra. Az emlősök gasztrulációjának fontosabb lépései és a magzatburkok kialakulásának típusai (A-B: az
amnionüreg hasadás útján való kialakulása, C: embriópajzs metszete, D: az amnionüreg kialakulása az amnion-
redők felhajlása révén, E: ragadozó embriójának magzatburkai)
aa: amnioallantoisz, al: allantoisz ürege, aü: amnion ürege, ar: amnionredő, ct: citotrofoblaszt, ee: epidermális ektoderma,
ek: ektoderma, en: entoderma, ex: extraembrionális mezoderma, gh: gerinchúr, in: intraembrionális mezoderma, ka:
korioallantoisz, kb: korionbolyhok, kö: köldökzsinór, me: mezoderma, ne: neurális ektoderma, őb: kialakuló ősbél, pa:
embriópajzs, pl: korioallantoisz placenta, sp: sziktömlöplacenta, st: szincíciotrofoblaszt, sz: szikzacskó, vt: valódi testüreg
(Huttner, Portmann és Zietschmann nyomán kissé módosítva)
64
a blasztociszta üregét és kialakítja az extraembrionális cölomát. A fejlődő embriót (emb-
riópajzs, amnion, szikzacskó) a testnyél köti a trofoblaszt réteghez. Az extraembrionális
mezodermából kialakuló testnyél kötőszövete a később kialakuló köldökzsinór alapállo-
mányát képezi. Az embriópajzs területén zajló csíralemez-elkülönülések révén kialakul
az intraembrionális mezoderma is, amely összeköttetésbe lép az extraembrionális mezo-
dermával. A rovarok mezodermája általában az embriócsík középvonalában kialakuló
bemélyedés, betűrődés révén, speciális módon alakul ki (2.16. ábra). Létrejötte után
szelvényesen cöloma zsákok alakulnak ki benne. Az embriócsík szelvényekre tagolódása
után megjelennek a végtagok kezdeményei. Az embrió kifejlődése végbemehet a szik
felszínén vagy a szikanyagba besüllyedve, azzal teljesen körülvéve (16.4. fejezet).
A csíralemezek kialakulásával és kiterjedésükkel (extensio) a fejlődő embrió a bél-
csíra (gastrula) fejlődési állapotot éri el. Ekkor elkülöníthető rajta az ősszáj, amelyen
keresztül az ősbélüreg közlekedik a külvilággal. Az ősszájú állatoknál ez a nyílás megma-
rad szájnyílásnak, míg az újszájúaknái ebből a végbélnyílás jön létre (vagy elzáródik és új
végbélnyílás nyílik). Gerinces állatoknál a gerinchúr kezdeménye már elkülönül a mezo-
dermától. Az embrió fej-farok irányban megnyúlik. Az ősszáj dorzális ajkának bizo-
nyos sejtjei fontos szerepet töltenek be az embrió fejlődésének irányításában. Az embrió
a gasztruláció végén a szervképződés, a sejtdifferenciálódás szakaszába lép.
2.16. ábra. A rovarok gasztrulációjának fontosabb lépései és a testi szerveződés kialakulása (A: az embrió
hosszmetszete a gasztruláció megindulása előtt, B: a dorzális blasztodermasejtek kiterjedése az embrió kereszt*
metszetében, C: az amnion és a szeroza kialakulása - keresztmetszetben, D-E: a csíralemezek differenciálódása
- keresztmetszetben, F: az embrió hosszmetszete a testi szerveződés fázisában, G: az embrió képe a hasi oldal
felől, az embrionális fejlődés késői szakaszában)
ad: agydúc, am: amnion, bk: blasztoderma-kettőzet, cs: embriócsík, db: dorzális blasztodermasejtek, fe: fej, ek: ektoderma,
em: entomezoderma, en: leendő entodermasejtek, ev: entoderma-betüremkedés a leendő végbélnyílás területén, ko: korion,
Is: leendő szájnyílás, me: mezoderma, ne: neurális ektodermasejtek, po: potroh, se: szeroza, sm: sziksejt magja, sz: szikanyag,
to: tor (Johansen-Butt, Wehner-Gehring nyomán kissé módosítva)
65
ACOELOMATA
I
PSEUDOCOELOMATA
2.17. ábra. A három csíralemezből felépülő testű állatok testüregviszonyai és a valódi testüregesek mezoder-
májának kialakulási lehetőségei (A: schizocoel, B: enterocoel)
át: áltestüreg, bé: bélcsö, bl: blasztocöl, ek: ektoderma, en: entoderma, me: mezoderma, mi: mezodermális eredetű izomzat,
mp: mezodermális parenchima, mt: mezenterium, őb: ősbél, sz: mezodermális eredetű szerv, tb. testüreget bélelő
mezodermális hám, vt: valódi testüreg (Hickman és Dórit nyomán kissé módosítva)
A csíralemezek kialakulása, a mezoderma hiánya vagy megléte, illetve sejtjeinek elren-
deződése megszabja a kifejlett állat testüregviszonyait. így megkülönböztetünk testüreg
nélküli, áltestüreges és valódi testüreges állatokat (2.17. ábra).
A testüreg nélküli (acoelomata) többsejtű állatok egy részében (szivacsok, csalá-
nozók, bordásmedúzák) az ektoderma és entoderma között nincs üreg. Alaposférgekben
a két csíralemez közötti teret ugyan mezodermális eredetű sejtek töltik ki, de a teret
mezodermális hámfal nem béleli.
Az áltestüreges (pseudocoelomata) állatoknál a külső és a belső csíralemez közötti
blasztocöl eredetű testüreget csak részben határolhatják mezodermális eredetű sejtek.
Ilyen testüregűek például a kerekesférgek és a fonálférgek.
Valódi, másodlagos testüregű (eucoelomata) állatoknak azokat tekintjük, ame-
lyeknél az ektoderma és az entoderma között mezodermális eredetű hámmal bélelt üreg
(coeloma, deuterocoeloma) jönlétre. A valódi testüreg megjelenése evolúciós szempont-
ból fontos lépés volt, mert megnövelte a test mozgékonyságát, egyes esetekben hidroszta-
tikai vázat alkot, elősegíti a mozgást, az üreg- és járatásás képességét. A mezodermális
hámmal bélelt üreg a zsigeri szervek számára megfelelő belső környezetet biztosít, nagy
felületet képez a mezodermális sejtek számára, lehetővé teszi az anyagáramlás nagyobb
hatásfokát.
ízeltlábú állatokban a valódi testüreg a mezodermális szervek differenciálódása során ösz-
szeolvad az elsődleges testüreggel, így kevert, harmadlagos testüreg (mixocoel) alakul ki.
66
2.4.2. Szervfejlődés és szöveti differenciálódás
A gasztruláció során kialakult két vagy három csíralemez további fejlődését cső- vagy
zsákszerű képletekké alakulásuk jellemzi. A csíralemezekből szervtelepek, majd ezekből
szervek, szövetek alakulnak ki. A szervfejlődés (organogenesis) magába foglalja nem-
csak a maradandó, hanem az embrionális életben működő szervek kialakulását is. A
szöveti differenciálódás (histogenesis) során a sejtek fokozatosan unipotenssé válnak,
azaz speciális feladatokat ellátó sejtekké fejlődnek.
A gerinces állatok neurulációjának folyamán
(2.18. ábra) az ektoderma neurális területének, a
velőlemeznek (lamina neurális) szélei felgyűrődnek,
kialakulnak a velőredők (plica neurális), amelyek a
velőbarázdát (sulcus neurális) határolják. A velőre-
dők a barázda felett áthajolva összeérnek, csővé zá-
rulnak, amely az epidermisz alá lesüllyedve az elemi
velőcsövet (canalis neurális) hozzalétre. Ennekürege
az idegrendszer központi csatornájának (canalis
centrális) felel meg. Ez a fejlődési állapot az idegcsí-
ra (neurula). Az összehajlási széleken futó gang-
lionléc (velőléc) sejtjei mozgásra képesek, leválnak
és eljutnak a fejlődő szervezet különböző pontjaira,
ahol részben idegsejtekké, részben pigmentsejtekké
differenciálódnak.
A csíralemezek további differenciálódása a kü-
lönböző gerincesek esetében hasonlóságot mutat.
Ennek szemléltetésére a 2.19. ábra szolgál, amelyet
a 2.2. táblázatban foglaltakkal kell összevetni.
Amikor az állati testet felépítő szerveket eredetük
szerint csoportosítjuk, akkor elsődleges szempont-
ként hámszövetének kialakulását tekintjük, de tudni
kell, hogy felépítésében más csíralemezek is (illetve a
mezoderma szinte mindig) szerepetkapnak(2.2. táb-
lázat).
Az ősi ivarsejtek (primordiális ivarsejtek) a fejlődés korai szakaszában elkülönülnek
a többi sejttől. A csíralemezekkialakulása során kezdetben az entodermasejtekkel együtt
figyelhetők meg, majd bevándorolnak a mezoderma által létrehozott ivarszervkezdemé-
nyekbe.
A neurális ektodermából alakul ki az összes idegsejt és a gliasejtek döntő többsége.
Az epidermális ektodermából a kültakaró hámrétege és származékai jönnek létre.
Amezodermából a gerinces állatok esetében hamar elkülönül a kordalemez, amely-
ből a gerinchúr (chorda dorsalis) fejlődik ki. Ez az embrionális életben a szilárdító váz
szerepét tölti be, de kifejlett korban már csak maradványai figyelhetők meg a csigolyák
között. Amezodermatömlők az embrió testének két oldalán hosszanti irányban növek-
szenek, majd ős-szelvényekre (somita) tagolódnak. A gerinces állatok embrióiban a
szemiták száma több, mint a kifejlett állat csigolyáinak száma, mivel a test hátulsó végén
lévők az embrionális fejlődés végéig visszafejlődnek. A szemiták centrális része az embrió
hasoldala felé növekszik, majd a középvonalban össze is érnek. Elkülönül a hátlemez
(epimer), a középlemez (mesomer) és az oldallemez (hypomer). A hátlemez megtartja
szelvényezettségét, míg az oldallemez nem. A hátlemez velőcsőre és gerinchúrra fekvő
részéből (myotom) alakul ki a törzs szelvényezetten fejlődésnek induló harántcsíkolt
izomzata. A miotomsejtek mioblasztsejtekké alakulnak át, intenzíven osztódnak, összeol-
2.18. ábra. A neuruláció folyamatának ál-
talánosított vázlata (Az embrió hátoldalán
az ektodermának csak kis részlete van
feltüntetve.)
ee: epidermális ektoderma, ev: elemi velőcső
ürege, gl: ganglionléc, nb: migráló neuroblaszt
sejtek, vb: velőbarázda, vl: velőlemez, vr:
velőredő (Több szerző nyomán módosítva)
67
2.19. ábra. Gerinces állatok csíralemezeinek differenciálódása a
neurulációt követően. A: egy korai és B: egy későbbi fejlődési
állapotban (embrió keresztmetszetének általánosított, egyszerűsített
vázlata)
de: dermatom, ee: epidermális ektoderma, ev; elemi velőcső ürege, gh:
gerinchúr, gl: ganglionléc, hl: hátlemez, iv: ivarszerv kezdeménye az ősi
ivarsejtekkel, ki: kiválasztószerv járata, ki: középlemez, me: mezenchi-
masejtek, mi: miotom, ol: oldallemez, őb: ősbél, so: somatopleura, sp:
splanchnopleura, sz: szklerotom, vé: vérér, vt: valódi testüreg (Portmann
nyomán kissé módosítva)
vadva izomrostot képeznek, amely-
ben kialakulnak a miofibrillumok. A
hátlemez gerinchúrra fekvő részéből
(sclerotom) jön létre a gerincoszlop,
míg az epidermális ektodermára fek-
vő területéből (dermatom) a bőr ir-
harétege, a bőr alatti kötőszövet és a
simaizmok alakulnak ki. A középle-
mez gononephrotom része a húgy-
ivarszervek és elvezető járataik hám-
ját alakítja ki. Az oldallemezek
laterális (somatopleura) és mediális
(splanchnopleura) részekre különül-
nek el. A pleuralemezek a magzat-
burkos gerincesek embrióinak széli
részein az extraembrionális pleura-
lemezekben folytatódnak. Az oldal-
lemezek hám jellegű részeiből fejlő-
dik a szervezet savóshártya-rendsze-
re. A savóshártyák által körülvett üreg
a másodlagos vagy valódi testüreg. A
mezoderma egyes sejtjei amőboid
mozgással igen hamar elkülönülnek
a többiektől, és az embrionális kötő-
szövetet, a mezenchimát (mesen-
chyma) hozzák létre. Sokféle sejttí-
pus differenciálódhat belőlük. A me-
zenchima az üvegtest és a szemlen-
cse kivételével valamennyi szövet és
szerv felépítésében részt vesz. A fej-
lődő szervezet legkülönbözőbb he-
lyein jönnek létre belőlük más kö-
tőszövetféleségek, a porc és a csont-
szövet, a sima és harántcsíkolt izom-
szövetek (anem miotom eredetűek),
a vér- és nyirokerek, a vér- és nyirok-
sejtek, a szív és a vérképző szervek.
Az entodermából az ősbél (archenteron) formálódik, amelyből az emésztőcsatorna
hámja és származékai, a máj, a hasnyálmirigy és több belső elválasztású mirigy hámja,
a tüdő hámrétege jön létre.
Az organogenezis során a kialakult szervkezdemények növekszenek, differenciálód-
nak, a posztembrionális élet megkezdéséig elérik a fajra jellemző méretet.
A differenciálódás során számos sejt elveszti osztódóképességét, mint például az ideg-
sejtek, a vörösvértestek, az érett ivarsejtek. A vese csatornarendszerét bélelő hámsejtek
és a májsejtek bizonyos körülmények között még képesek osztódásra. Vannak olyan
sejtek, melyek állandóan osztódásra képesek, mint például a többrétegű elszarusodó lap-
hám stratum germinativum rétegét alkotók.
Néhány állatcsoportban az utódok testi sejtjeinek körében szinte nem is figyelhető
meg osztódás a posztembrionális élet megkezdését követően. Ilyen állandó sejtszám
fordul elő a kerekesférgekben és a fonálférgekben.
68
2.2. táblázat. Gerinces állatok különböző csíralemezeiből kialakuló ősi szervtelepek, szervek és
szövetek
Ektoderma Epidermális ektoderma Kültakaró hámrétege és származékai: a testfelszínt borító bőr hámrétege, a verejték-, a faggyú- és tejmirigyek hámja, a verejtékmirigy simaizomelemei, egyéb hám eredetű simaizomsejtek, a szaruképződményeket (szőr, toll, a karom, a pata szaruja) létrehozó hámok. Az orrüreg és szájüreg hámja, a hipofízis elülső és középső lebenye, a végbél, a hüvely és a húgycső egy részének hámja, a nyálmirigyek és könnymirigyek hám- ja, a szem szaru- és kötőhártyája, a szemlencse és részben az üvegtest, a fogzománc. Az embrionális szervek falának felépítésében.
Neurális ektoderma velőcső Az agy és a gerincvelő szinte valamennyi eleme: az ideg- sejtek, a gliasejtek többsége (kivéve a mezodermális ere- detű mikrogliasejteket), a retina ideg- és pigmentsejtjei, a látóideg, a hipofízis hátúlsó lebenye (neurohipofízis), a szem szivárványhártyája.
ganglionléc A gerincvelői és a környéki idegek, a szimpatikus és paraszimpatikus ganglionok, a test pigmentsejtjei, a Schwann-sejtek, a mellékvese velőállománya.
Mezoderma Kordalemez Gerinchúr (chorda dorsalis).
Hátlemez miotom A törzs harántcsíkolt vázizomzata.
szklerotom A csontos váz gerincoszlop-része (tengelyváz).
dermatom A bőr irharétege és a bőr alatti kötőszövet.
Középlemez gononefrotom A kiválasztószervek és a húgyvezető egy részének hám- elemei. A mellékvese kéregállománya. Az ivarszervek és elvezető járatainak (az ondóvezető, az ondóhólyag, apetevezető, a méh és a hüvely egy részének) hámja.
Oldallemez szomatopleura és szplanchnopleura Mezotélium: mellhártya, hashártya, szívburok. Az embrionális szervek falának felépítésében részt vevő hámok.
Mezenchima A kötőszövetek, a porcszövetek, a függesztőövek és a ko- ponya porc eredetű (chondrogen) csontjai, a kötő- szövetes eredetű (desmogen) csontok. A mellkas, a hasfal és a végtagok haráncsíkolt izomzata, a szívizom, valamint a bélcső, a bőr és az erek simaizmai. A vöröscsontvelő, a nyirokszervek, a lép, a vérsejtek, a vér- és nyirokerek. Mikrogliasejtek az idegszövetben. Extraembrionális szervek falában.
Entoderma Ősbél Az emésztőcső és mirigyeinek (máj, hasnyálmirigy) hámja. Több garathám eredetű (branchiogen) szerv, például a pajzsmirigy, mellékpajzsmirigy, adenohipofízis, csecse- mőmirigy, Eustach-kürt) hámja. A légutak, a kopoltyú és a tüdő hámja. A húgyhólyag és a húgycső egy részének hámja. Az embrionális szervek (húgyinda, allantoisz, szikzacs- kó) hámja.
69
2.5. Embrionális szervek
Az organogenezis folyamatában keletkeznek olyan szervek, amelyek csak az embrionális
fejlődés alatt funkcionálnak, tehát ideiglenes képződmények. A gerincesek körében két
fő formájuk különböztethető meg, az egyik a szikzacskó, a másik a magzatburok.
2.5.1. Szikzacskó
A szikzacskó vagy sziktömlő (saccus vitellinus) a polilecitális petesejtű gerinces állatok
(illetve az ilyen csoportból származtathatóak) embrióinál alakul ki. Extraembrionális
képződmény, amely a fejlődő embrió tartalék tápanyagait foglalja magába, és lehetővé
teszi annak folyamatos felhasználását. Tipikus formában előfordul a halakban, hüllők-
ben és madarakban. Emlősökben az embrionális fejlődés folyamán csökevényes, nem
funkcionáló szikzacskó formálódik. Ez is egy bizonyítéka annak, hogy az emlősök
polilecitális ősöktől (hüllők) származnak.
A halak, a hüllők és a madarak zigótája csak az animális póluson, egy viszonylag kis
kiterjedésű, szikben szegény területen barázdálódik. A hüllők és a madarak szikzacskója
falának felépítésében mindhárom csíralemez részt vesz, mert sejtjeik a csírakorong szé-
léről kiindulva osztódásuk révén körülnövik a szikanyagot (2.14. ábra).
A szikzacskó a szikvezeték (ductus vitellointestinalis) segítségével az embrió hasol-
dalán, a bélcsőhöz kapcsolódik a hüllőkben, a madarakban és az emlősökben. Halakban
a szikjárat hiányzik, mert a fejlődés korai szakaszában a szikzacskó elkülönül a bél csőtől
(2.13. ábra).
A szikzacskó falánakbelső rétegét alkotó entodermális sejtekkezdetbenfagocitózissal
veszik fel a szik alkotórészeit. A későbbiekben enzimatikus úton bontják, majd a táp-
anyagokat bejuttatják a szikvérkeringésbe. Madarakban a szikzacskó az embrióval ellen-
tétes oldalán nem záródik teljesen, egy kis nyíláson (sziktömlőköldök) át kapcsolatban
marad a fehérjezsákkal (2.21. ábra). A fejlődő embrió testébe a szikraktár tápanyagait a
szikvérkeringés szállítja. Az elsődleges szikerek a szikzacskó falának felépítésében részt
vevő középső, mezodermális sejtrétegből differenciálódnak. A fejlődés során az embrió
testéből is kiindulnak vérerek, amelyek a másodlagos szikvérkeringést alakítják ki. Ez
utóbbi érrendszer szerepe egyre nagyobb lesz, az elsődleges szikkeringés ereivel a kapil-
lárisok révén kapcsolatba is kerülnek. Az embrionális fejlődés végén megindul a szikerek
felszívódása, amely a tojásból való kikelést követő napokban fejeződik be. Madaraknál
a szikzacskó maradványa még a hasfal záródása előtt a hasüregbe kerül. A benne lévő
szikanyag ilyenkor a bélcsővel kapcsolatot létesítő csatornán keresztül közvetlenül beke-
rülhet az emésztőszervrendszerbe.
Emlősállatoknál is kialakul a szikzacskó vagy sziktömlő, de az embrió táplálásában
gyakorlatilag nem játszik szerepet. Az embriópajzs centrális oldalát alkotó entoderma
sejtek osztódása és a citotrofoblaszt-rétegből leváló, szplanchnopleurát kialakító sejtek
együtt alkotják a falát (2.15. ábra). Kialakul a szikzacskó falában a vérkeringés is, de
feladata az egyes csoportoknál eltérő lesz. A patások, ragadozók, rágcsálók és rovarevők
egy részének esetében a szikvérkeringésen kívül a sziktömlőplacenta (omphaloplacenta)
is kialakul. A többi emlősállatnál a sziktömlő-vérkeringés szerepe nem jelentős, igen
hamar elsorvad, a fejlődő embrió anyagcseréjét a placentán keresztül megvalósuló vér-
keringés látja el. A placentális cirkuláció beindulása után már magzatnak nevezzük a
fejlődő utódot. A szikzacskó a fejlődés során redukálódik, a létrejövő köldökzsinór bel-
sejébe húzódik be, majd teljesen elcsökevényesedik.
70
2.5.2. Magzatburkok
A magzatburkok (velamenta foeti) embrionális sejtekből képződő olyan testfüggelé-
kek, amelyeknek szerepe elsődlegesen a mechanikai és a kiszáradástól való védelem,
állandó környezet és a gázcsere biztosítása, de ezen kívül más, speciális funkciójuk is
lehet.
A gerinctelen állatoknál (például rovaroknál és skorpióknál) magzatburoknak neve-
zett, általában csak néhány sejtrétegből álló védőburkok nem homológok a gerinces
állatokmagzatburkaival. Rovarokesetében a hátoldal felől kiinduló blasztodermakettőzet,
az amnionredő a hasoldalon összeér. A belső sejtsor az amniont, míg a külsők a szerozát
alakítják ki (2.16. ábra).
A gerinces állatok közül a hüllőknek, a madaraknak és az emlősöknek van magzatbur-
ka. Ezeket az osztályokat együttesen magzatburkosoknak (Amniota) is nevezik, míg a
többi gerinces a magzatburok nélküliekhez (Anamnia) tartozik (17. fejezet).
A magzatburkos gerincesekben három magzatburok, a magzating (amnion), savós
burok (membrana serosa) és a húgytömlő (allantois) jön létre. Ezek a kezdeti, részben
hasonló kialakulást követően különböző mértékben fejlődtek ki, illetve módosultak a
speciális feladat ellátására (2.20. ábra).
2.20. ábra. A hüllők, a madarak és az emlősök magzatburkainak kialakulása vázlatosan, leegyszerűsítve. A; az
embrió testének keresztmetszete a három csíralemezes állapotban, B: az amnion és a szeroza ki alakulása az ekto-
és mezoderma felhajlásával, C: embrionális szervek helyzete és felépítésük
al: allantoisz, am: amnion, an: amnion ürege, ar: amnionredő, aü: allantoisz ürege, ek: ektoderma, em: embrió, en:
entoderma, ex: extraembrionáliscöloma,me: mezoderma, se: szeroza, za: szikzacskó (Több szerző nyomán kissé módosítva)
A hüllők és a madarak embriójának teste körül az ektoderma és a mezoderma fali le-
meze (somatopleura) a sejtek fokozottabb osztódás révén redőt képez. A felemelkedő
redő az embrió hátoldala fölött lassan összeér és egy varrat formájában látható. A redő
külső és belső rétege között a kapcsolat megszűnik, elválnak egymástól. A belső réteg az
71
2.21. ábra. A madárembrió magzatburkai kialakulásának egyszerűsített vázlata (Az ábrán nincs feltüntetve, hogy a
szeroza-allantoisz közös burok nagy területen összenő a héjhártyával.)
al: allantoisz, am: amnion, ek: ektoderma, em: embrió, en: entoderma, ex: extraembrionális cöloma, fe: fehérje, fz:
fehérjezsák széle, hh: kettős héjhártya, lu: légudvar, me: mezoderma, sa: szeroza-allantoisz összenövése, sh: szeroza-
allantoisz összenövése a héjhártyával, sk: sziktömlőköldök, sz: szikzacskó, th: tojáshéj (Goerttler, Schauinsland-Grosser és
Torrey nyomán módosítva)
amniont alkotja, míg a külső a szerozát. Az embrió így közös eredetű, kettős burokba
kerül (2.21. ábra).
Az amnion és a szeroza között lévő extraembrionális testüreget (exocoeloma) folyadék
tölti ki, amelybe az utóbélből kiindulva a húgytömlő türemkedik be.
Emlősállatoknál az amnion és a szeroza ettől eltérő módon is kialakulhat (2.15. ábra).
A magzatburkok a blasztociszta anyai méhfalba való beágyazódását (implantáció) követően
alakulnak ki. Az implantáció úgy jön létre, hogy a trofoblaszt sejtek által termelt enzimek
emésztik a méh nyálkahártyáját, így a blasztociszta lassan besüllyed, beágyazódik. Az
amnion ürege a ragadozók és a patások esetében a felhajló redő-amnion révén keletkezik.
Ez utóbbi folyamat a madarakéhoz hasonlóan megy végbe. A többi emlősnél a trofoblaszt-
sejtek és az embriókezdemény közötti rés szélesedik ki amnionüreggé. A külső magzat-
burok az emlősöknél irhahártyává (chorion) alakul. Formálódása a trofoblasztsejtekből
indul ki. Az embriócsomó feletti trofoblasztsejtek többsége a sejthártyák felszívódásával
szincíciumot (syncytiotrophoblast) alkot, amiben üregek jelennek meg. Ezek az üregek
később felnyílnak, belső faluk részt vesz a korionbolyhok felszíni rétegének kialakításá-
ban. A rágcsálókban nem jön létre összefüggő korion. A sejtes szerkezetüket megtartó
trofoblasztsejtek (cytotrophoblast) a somatopleura sejtjeivel együtt a szerozát hozzák
létre. Számos ragadozó, rágcsáló, patás és rovarevő méhlepényes emlős esetében a szik-
zacskó a szerozával kisebb-nagyobb felületen összenő (choriovitellin), sziktömiőplacen-
tát alakíthat ki. A szikplacenta fejletlen például a ló, közepesen fejlett a ragadozók,
erősebben fejlett a rágcsálók és a nyulak esetében. Egyeseknél ez a burok addig tartja fenn
a magzati placenta vérkeringését, amíg a kezdetben lassan fejlődő allantoisz érrendszere
ezt át nem veszi. Másoknál a sziktömlő az allantoisz mellett biztosítja a korion érrend-
szerét. A legtöbb méhlepényes emlősnél a szikzacskó visszafejlődik és csak az allantoisz
72
biztosítja a vérkeringést. A korion az allantoisszal összeolvadva (allantochorion) a mag-
zati placentát hozza létre. Az embriópajzs ektodermális sejtjeiről levált sejtek a
szomatopleurával együtt az amniont alakítják ki. Az amnion egyes emlősöknél (például
a főemlősöknél) jelentősen megnő, végül összeolvad a korionnal, így egy közös magzat-
burok (amnochorion) alakul ki. Növekedése közben körülveszi a szik-vezetéket és szik-
zacskót, valamint az allantoisztömlőt, így a folyamat végén kialakul a köldökzsinór. Az
extraembrionális cöloma ezzel párhuzamosan fokozatosan beszűkül.
Az amnion falát két csíralemez sejtjei alkotják, az ektoderma az embrió felé eső
oldalon, a mezoderma a külső felületén helyezkedik el. Az embriót kezdetben szorosan
veszi körül, később, a falát alkotó sejtek által termelt folyadék, a magzatvíz (liquor amnii)
felhalmozódásával ürege egyre tágul, fala az embriótól eltávolodik. Az amnion az emb-
riót (és az emlősöknél a köldökzsinórt) állandó nyomás alatt tartja, védi a mechanikai
behatásoktól, hozzájárul a vízszükséglet kielégítéséhez. A leadott tápanyagok és a leváló
hámsejtek révén kismértékben táplálékul is szolgál. Emlősöknél a szülés során szerepe
van a szülőút tágításában, majd az amnion felrepedését követően annak sikamlóssá
tételében is.
A szeroza falában az ektodermális sejtek a külső felszínen, míg a mezodermálisak a
belsőn találhatóak. A mechanikai védelem funkcióján túl madaraknál kezdetben önál-
lóan, később az allantoisszal összenőve részt vesz az embrió gázcseréjében.
Az embrionális bélcső kaudális szakaszának falából kitüremkedéssel keletkező
allantoisz az embrió hasi területéhez kapcsolódik. Falát entodermális sejtek és a mezo-
derma zsigeri lemezének (splanchnopleura) sejtjei alkotják. Az entodermális sejtek az
üreg belső, a mezodermális eredetűek a külső felszínén vannak. Az allantoisz ürege a
húgyinda (urachus) révén madaraknál a kloákához, emlősöknél a köldök közelébe kap-
csolódik. A húgytömlő feladata elsősorban a vesék által termelt vizelet összegyűjtése,
tárolása. A húgytömlő az amnion és a szeroza között növekedve terjedelmes üreggé
alakul, amelynek fala összeolvad a másik két magzatburokéval is. így kialakul az
amnioallantoisz és a szerozaallantoisz membrán. A húgyhólyag az allantoiszzsák marad-
ványából alakul ki.
Madarakban az amnioallantoisz folyadékfelvételre képes, melyet a magzati vérkeringés-
be juttat. A szerozaallantoisz, mivel a tojáshéj belső felszíne alatt fut, gazdag érhálózata
miatt fontos szerepet játszik az embrió gázcseréjének lebonyolitásában, valamint a kal-
cium- és a vízfelvételben. A húgytömlő allantoamnion-területének egy része a szikzacskó-
val is összeér, de azzal nem vagy legfeljebb csak a sziktömlőköldök szegélyénél nő össze.
Más területén a szerozaallantoisz membrán a tojásfehérjét is körülöleli, redője kialakítja a
szikzacskóval közvetlen kapcsolatban álló fehérjezsákot (saccus albuminis). A zsák falát
alkotó sejtek felveszik a fehérjéket, és a magzati vérkeringésbe juttatják. Mivel a fehérjezsák
a fejlődés későbbi szakaszában az amnionüreggel is összeköttetésbe kerül, a fehérje oda
közvetlenül bekerülhet, így az embrió a magzatvízzel együtt a fehérjéket is lenyelheti,
bélcsatomájában hasznosíthatja (2.21. ábra). Madaraknál az allantoiszfolyadék víztartal-
ma a kelést megelőzően felszívódik, a húgyanyagok szilárd formában maradnak vissza. Az
allantoisz membránrendszerei a kelés idejére fokozatosan elsorvadnak, elhalnak.
Emlősök esetében az allantoisz nem vesz részt sem a húgyanyagok tárolásában, sem
a légzésben. A legtöbb fajban körülnövi az embriót. Az emlősök többségében az allantoisz
fala összenő a külső savós burokkal, vérereivel behatol annak felszíni bolyhaiba is, együt-
tesen kialakítják az irhahártyát (chorion). A korion kezdetben többnyire az egész fel-
színén bolyhos, de azok a későbbiekben axonokra jellemzően csak meghatározott terü-
leteken fejlődnek tovább. Az amnionburokban fejlődő magzat a köldökzsinór (funiculus
umbilicalis) révén van kapcsolatban a méhlepénnyel (placenta). A köldökzsinór kocso-
nyás kötőszöveti alapállományába ágyazva két artéria, egy vagy két véna és a húgyinda
teszi lehetővé a megfelelő anyagcserét. A vérerek az allantoiszkeringés ereiből maradnak
73
meg. Amíg a sziktömlő funkcionál, a köldökzsinórban megfigyelhető annak egy szaka-
sza, a szikbéljárat is. A méhlepény rögzíti a fejlődő magzatot, biztosítja az anyagcseréjét
(szabad diffúzióval: oxigén, széndioxid, víz, ásványi anyagok nagy része, aktív transz-
porttal: glükóz, aminósavak, vitaminok, endocitózissal: egyes makromolekulák) és az
anyai ellenanyagokat átjuttatja a magzati vérkeringésbe. A méhlepény termeli a terhesség
fennmaradásáért felelős hormonokat.
A megerősödő, elágazó bolyhokból kialakuló magzati placenta (placenta foetalis) az
anyaméh falával (placenta materna) a keringési rendszerük révén különböző formá-
ban kerülhet kapcsolatba (2.22. ábra). Az epitheliochoriális placentában a kapcso-
lat a legkevésbé közvetlen, mert az anyai vérerek endotheliumsejtjeiből kilépő tápanya-
goknak át kell jutniuk a méhnyálkahártya kötőszöveti és hámrétegén, a korionboholy
hámrétegén és kötőszövetén, amíg eljut a magzati vérerek endotheliumsejtjeihez. Ez
a típus például a cetekre, a lovakra, a sertésekre, egyes félmajmokra jellemző. A szülés
alkalmával a két placentafél vérzés nélkül, könnyen elválik egymástól, de az anyai
placenta egy része (decidua) nem válik le. Az ilyen placentát félplacentának (se-
miplacenta), azokat az állatokat pedig, ahol ez előfordul, hullóhártyanélkülieknek (Ade-
2.22. ábra. Az emlősállatok placentatípusainak (A: epi-
theliochoríalis, B: syndesmochorialis, C: endotheliocho-
ríalis, D: haemochorialis) szöveti felépítése
ah: anyai placenta hámrétege, ak: anyai placenta kötőszöveti
állománya, kk: korionbolyhokkötőszöveti állománya, mh: mag-
zati placenta korionbolyhainakhámrétege, sn: sejthatárok nélküli
hámréteg, vé: vérér, vö: véröböl (Torrey és más szerzők nyomán
módosítva)
ciduata) nevezzük. A syndesmocho-
riális placentában a fenti kapcsolat-
rendszerből a méhnyálkahártya hámré-
tege hiányzik, vagyis a korionbolyhok
közvetlenebb kapcsolatba kerültek az
anyai placentával. Ilyen jellemző a laj-
hárra és egyes palásokra (szarvasmarha,
juh, kecske). Szüléskor az ilyen piacénták
kisebb vérzés kíséretében válnak el egy-
mástól. Az endotheliochoriális placen-
tában az anyai vérerek hámrétege köz-
vetlenül a korionbolyhok szincíciális
hámrétegéhez kapcsolódik, itt tehát a
méhnyálkahártyának sem a hám, sem a
kötőszövetes rétege nem képez akadályt
az anyagáramlás útjában. Előfordul pél-
dául egyes denevérfajok és a ragadozók
között is. A szülés alkalmával jelentősebb
vérzés alakul ki, mert a decidua is leválik.
A haemochoriális placentában az
anyai szövetekből származó határoló hár-
tya nincs, a korionbolyhok szincíciális
hámrétege az anyai placentában kiala-
kult véröbölben végződik. Ez jellemző a
rovarevőkre, a denevérekre, a rágcsálók-
ra, a nyúlalakúakra és a főemlősökre. Szü-
léskor a decidua itt is erősebb vérzés kí-
séretében válik le. Azokat az állatokat,
amelyeket az utóbbi placentatípusok jel-
lemeznek, hullóhártyás méhlepényesek-
nek (Deciduata) nevezzük.
A fent említett csoportosítás csak főbb
kategóriákat ad meg, de számos átmene-
ti forma is megfigyelhető. A főemlősök
kezdetben epithelochorialis placentája
74
például később haemochorialis for-
mát ölt.
Attól függően, hogy a korion fel-
színén hol találhatóak a bolyhok,
különböző placentatípusok is-
mertek (2.23. ábra). A szétszórt,
diffúz placentán (placenta dif-
fusa) a bolyhok egyenletes eloszlá-
súak (sertés, ló). A lebenyes pla-
centán (placenta cotyledonaria) a
bolyhok számos helyen, csoportok-
ba tömörülve találhatóak (juh, szar-
vasmarha). Az öves placentán
(placenta zonaria) a bolyhok övsze-
rűen helyezkednek el, mint például
a ragadozók esetében. A mosómed-
ve placentáján az öv nem teljes. A
bolyhok egy korongot képeznek
(placenta discoidea simplex) egyes
rágcsálók, a rovarevők, a denevé-
rek körében és a főemlősök egy ré-
szében, közöttük az ember eseté-
ben is. Kétkorongos placentája
(placenta discoidea duplex) van
számos főemlősnek.
2.23. ábra. Az emlősállatok placentatípusai a korionbolyhok
eloszlása szerint
A: szétszórt placenta, B: lebenyes placenta , C: öves placenta, D: nem
teljesen öves placenta, E: kétkorongos placenta, F: egykorongos placenta
(Több szerző nyomán kissé módosítva)
2.6. Az egyedfejlődés szabályozása
A soksejtű állatok egyedfejlődése során egyetlen sejtből, a zigótából jön létre az egész
szervezet, amelynek sejtjei ugyan összehangoltan működnek, de egyedileg igen eltérő
feladatokat látnak el. A zigóta totipotens, azaz képes az egész szervezet kialakítására,
ezzel szemben a kifejlett állat osztódó testi sejtjeinek döntő részéből csakis egy sejttipus
differenciálódhat, tehát unipotens. Az embrionális fejlődés során az eredetileg totipotens
sejt differenciálódóképességefokozatosan beszűkül. A csíralemezek sejtjei multipotensek,
belőlük már csak meghatározott szervtelepek, szervek alakulhatnak ki. Az igy létrejött
szervtelepek sejtjei még pluripotensek. Szinte valamennyi testi sejt (kivételt az immun-
rendszer sejtjei jelentenek) genetikai állománya rendelkezik a teljes szervezet kialakítá-
sához szükséges génekkel. Mivel azonban a gének egy része a differenciálódás során
irreverzibilisen inaktiválódik, ezért a lehetőség nem használható ki.
Egyes állatoknál a testi sejtek totipotensek maradnak. Ha egy édesvízi hidrát sejtekre
szedünk szét, akkor azok bármelyike az egész szervezetet létre tudja hozni (szomatikus
embriogenezis).
A determináció a differenciálódási potencia beszűkülésének folyamata, ami azért
jön létre, mert a differenciáció során egyes gének irreverzibilisen kikapcsolódnak. Az,
hogy egy adott sejt a benne rejlő differenciálódási potenciának mely részét valósítja meg,
attól függ, hogy génjei közül melyek vannak „bekapcsolt”, illetve „kikapcsolt” állapot-
ban. Valamennyi sejtre jellemző az aktív és inaktív gének mintázata, amely az egyedfej-
lődés során megváltozik. Az elnyomott differenciálódási potencia egy része bizonyos
sejtekben a regeneráció során a gátlás alól felszabadulhat. A zigóta totipotenciája azt
jelenti, hogy genetikai állománya nincs véglegesen blokkolt állapotban. A fejlődés során
75
azonban a sejtek génállományának egyre nagyobb része kerül véglegesen blokkolt álla-
potba, így a sejtek potenciája beszűkül. A teljesen differenciált sejtben a genetikai állo-
mány döntő hányada nem fejtheti ki hatását. Az embrionális fejlődés során az embrió
testének különböző helyein lévő sejtek génállománya egymástól eltérő működést mutat,
ami a sejtek és szövetek differenciálódásához vezet.
Az egyedfejlődés szabályozásának pontos feltárása mind ez ideig a biológia megoldat-
lan problémái közé tartozik. A jelenleg folyó molekuláris szintű kísérletek azonban egyre
több támpontot adnak a folyamat megértéséhez.
A klasszikus embriológiai vizsgálatok is rámutattak már arra, hogy a barázdálódás
menetének, az embrió kifejlődésének irányításában a sejtmag hatásán kívül döntő jelen-
tősége van a citoplazmának is.
A kétéltűek megtermékenyített petesejtjének membránhoz közeli plazmájában megfi-
gyelhető szürke félhold területe a fejlődés további irányításában meghatározó szerepű.
Determinálja a kifejlődő állat bilaterális szimmetriasíkját, és amennyiben az utódsejtekbe
nem kerül e területnek egy-egy részlete, akkor ott a fejlődés nem következik be. A szürke
félhold területén a citoplazmában olyan anyagok vannak, melyek a normális fejlődés fel-
tételei. A néhány blasztomérából álló embrióban a sejtmagok teljesen azonos fejlődési
potenciájú génállománnyal rendelkeznek, de a citoplazmájuk már nem egyforma. Az
ovogenezis során kialakuló petesejt citoplazmájában kémiailag egymástól eltérő területek
jönnek létre, amelyek az első néhány barázdálódás során egymástól is elkülönülhetnek. A
citoplazmában található anyagok („anyai hatás”) determinálják, hogy a génkészlet mely
részei juttathatják érvényre a hatásukat. Kísérletek rámutattak arra, hogy a sejtmagban
található gének a fejlődés kezdetén túlnyomórészt inaktívak, csak a gasztruláció kezdetén
vagy röviddel előtte válnak aktívvá. Ezt jelzi, hogy ilyenkor a mRNS mennyisége ugrássze-
rűen megemelkedik, új citoplazmatikus fehérjék szintézise indul be. Egy aktivált, de meg
nem termékenyített petesejtből a sejtmagot eltávolítva az többé-kevésbé normálisan kezd
osztódni. Ez azt jelenti, hogy a barázdálódó zigóta számára szükséges fehérjék, illetve
hírvivő RNS-molekulák már a petesejtben szintetizálódtak. Ez az anyai hatás magyarázata.
A további fejlődés azonban leáll a gasztruláció kezdetén, amikor az újabb genetikai infor-
mációkra már szükség van. Közvetlenül a megtermékenyítést követően, a sejtmagfúzió
előtt a női előmagot eltávolítva a zigóta ablastula állapotignormálisanfejlődik. Agasztruláció
végbemenetele azonban attól függ, hogy a spermium haploid génkészlete mennyire kompa-
tibilis a petesejt citoplazmájával. Ha a megtermékenyítés ugyanazon faj hímjének spermiu-
mával történt, akkor haploid, de normális utód alakul ki. Közel rokon faj spermiumával
történt megtermékenyítésnél a fejlődés a neurula stádiumig is eljut, de ott leáll. Távoli
rokon spermiumával megtermékenyítve a fejlődés már a blasztula állapotban vagy a
gasztruláció kezdetén leáll. Ezek a különbségek az immunológiai inkompatibilitás külön-
böző mértékével magyarázhatóak. Ez akkor jelentkezik, amikor a spermiumból származó
gének az embrió fehérjéit kezdik kódolni. Minél távolabbi a rokonság, az inkompatibilitás
annál nagyobb, a fejlődés annál korábbi állapotokban áll le.
Az anyai hatást a közönséges muslicával (Drosophila melanogaster) folytatott kísér-
letek is igazolták. Ha petéjének hátulsó végéből egy kevés citoplazmát eltávolítottak,
akkor a fejlődő embriónak nem alakultak ki a potrohszelvényei. Ha az eltávolított cito-
plazma helyére egy másik pete elülső csúcsából juttattak be plazmát, akkor az embrió
mindkét testvégén fej alakult ki. Ennek magyarázata a petesejt citoplazmájában lévő
hírvivő RNS-molekulák hatásában és elhelyezkedésében keresendő.
Az oogenezis során a petesejt DNS-állományának meghatározott szakaszain képző-
dött mRNS-ek a citoplazmában, specifikus helyeken a sejtvázhoz kapcsoltan kötődnek.
Ezért a barázdálódás során meghatározott sejtekbe kerülnek, ahol az általuk irányítottan
létrehozott fehérjék mint morfogének (morphogen) egyes géneket bekapcsolnak, míg
másokat működésükben gátolnak. A citoplazmában található anyagok (speciális anyai
76
hírvivő RNS-ek) tehát meghatározzák az embrió polaritását, az egyes sejtek differenciá-
lódási képességét.
Az egyedfejlődés korai szakaszában, a gasztrulációs folyamat szabályozását például a
kétéltűekben az ősszáj dorzális ajkának bizonyos sejtcsoportja, mint elsődleges orga-
nizátor centrum látja el. Kialakulását tekintve visszavezethető a megtermékenyített, de
még nem barázdálódott békapetén megfigyelhető szürke félhold területére. Ezeket a
sejteket átültetve más, a gasztruláció kezdetén álló embrió külső sejtrétegébe, azok ott
gasztrulációs folyamatot indítanak el, másodlagos embrió kialakulásához vezetnek. Ha
azonban a befogadó (recipiens) embrió már túljutott e fejlődési szakaszon, az átültetett
sejtek nem tudják kifejteni hatásukat. Nemcsak a kétéltűekben, hanem számos más ál-
lat-csoportban is kimutattak különböző organizációs központokat. A korongos barázdá-
lódást mutató állatok és az emlősök esetében az őscsomó és a fejnyúlvány, míg az ízelt-
lábúakban a mikropyle közelében lévő sejtek funkcionálnak elsődleges organizációs cent-
rumként. Az eddigi vizsgálatok alapján úgy tűnik, hogy az organizátor centrum sejtjei
kémiai anyagaik révén nem közvetlenül szabályozzák a differenciálódást, hanem speciá-
lis környezeti feltételeket teremtve a környező sejtekben lévő képességet engedik ér-
vényre jutni. Ezek a genetikai képességet felszabadító vegyületek vagy sejtről sejtre ter-
jedve, vagy a sejt közötti térben szétáramolva fejtik ki hatásukat. Általában egy már
differenciálódott szövet mint induktor hat a körülötte lévő még nem differenciálódott
sejtekre, szövetekre. így egyre szőkébb hatáskörű másodlagos, harmadlagos és további
centrumok jönnek létre. Az idő- és a helytényező nagyon fontos. Ez azt jelenti, hogy
bizonyos gének csak egyes szövetekben és a fejlődésnek csak meghatározott szakaszában
képesek érvényre juttatni képességeiket. Ha egy elsődleges induktor kivált egy differen-
ciálódási folyamatot egy szövetben, akkor azt számos másodlagos indukció követi. A fej-
lődés folyamatát azonban nemcsak az induktorok határozzák meg, hanem fontos szerepe
van a sejtmozgásoknak, a sejtek adhéziós tulajdonsága megváltozásának és a sejtek osz-
tódási képességének. A sejtek a sejthártya jelzőmolekulái révén képesek egymást felis-
merni, így ha az elrendeződésükben valamilyen oknál fogva zavar támadna, képesek
egymást megkeresve újra összekapcsolódni. A sejtek felismerik az embrió testén belüli
helyzetüket is. Egy adott testrészt vagy szervet alkotó, különböző sejtek által leadott
vegyületek, morfogének segítenek behatárolni egy adott sejttípus térbeli helyzetét. Ez a
helyzetfelismerés az egyedfejlődés során egyre pontosabb, kisebb területekre vonatkozik.
Kimutatták, hogy közvetlenül a megtermékenyítést követően a petesejt plazmájában
hirtelen megnő a fehérjeszintézis intenzitása, annak ellenére, hogy a mRNS-képzés csak
a diploid zigóta kialakulását követően, később kezdődik meg. A petesejt plazmájában
még a megtermékenyítést megelőzően felhalmozódnak bizonyos mRNS-molekulák, me-
lyek az állatcsoporttól függően az első néhány sejtes állapot kialakulása alatt irányító
szerepet játszanak. Eloszlásuk a zigóta citoplazmájában nem egyenletes, hatásuk befo-
lyásolja a létrejövő blasztomérák képességeit, megszabják az embrió polaritását.
Rovarok egyedfejlődésének vizsgálata során megfigyelték, hogy az alapvető polaritás
kialakulását követően a szegmentációért felelős gének kezdik kifejteni hatásukat. A két-
szárnyúak körében több mint húsz ilyen gént azonosítottak. Az egyes szelvények felépí-
tését, különbségeiket a homeotikus gének szabályozzák. A homeotikus gének további
végrehajó gének működését indítják be.
Homeotikus géneket a gyűrűférgektől a gerincesekig számos állatcsoportban sikerült
már kimutatni. Ezeknek a géneknek egyes szakaszai (úgynevezett homeobox) felépíté-
süket tekintve nagyfokú hasonlóságot mutatnak. A homeoboxok által irányított amino-
sav-sorrend az evolúció során meglepően jól megőrizte eredeti jellemzőit. Ahomeoboxokat
megtalálták nemcsak a zigóta első néhány osztódását szabályozó, még a petesejtben
működő, de a szegmentációért felelős génekben is. Úgy tűnik, hogy a homeoboxok az
általuk irányítottan előállított fehérjék révén az embrionális fejlődés kapcsolói, melyek
77
ellenőrzik a különböző génekről történő átírást. A regulációs proteinek lehetnek aktiváló
és gátló hatásúak, amelyek a DNS adott génjéhez kapcsolódhatnak. A homeobox gének
a gerinctelen és gerinces állatok kromoszómáiban egyaránt géncsoportokba tömörülnek.
A kromoszóma e szakaszán a homeoboxok szigorúan meghatározott sorrendben következ-
nek egymás után. A homeobox gének ugyanolyan sorrendben rendeződnek el a DNS-
szálon, mint ahogyan a kialakítandó szervezet fej-farok tengelye mentén hatásuk megjele-
nik. Aktiválódásuk időbeli sorrendje is megegyezik a DNS-szálon elfoglalt helyzetükkel.
Az egyedfejlődési események időzítésének szabályozása a különböző állatcsoportokban
eltéréseket mutat. Egyik végletként a sejtszám-konstanciával jellemezhető fonálférgek em-
lithetőek. Náluk a sejtek sorsa már a legelső osztódást követően szigorúan meghatározott,
állandó sejtvonalakon alapul. A másik véglet az, ha az egyedfejlődés kezdetén létrejövő
sejtek sorsát az embrió testében elfoglalt helyzetük szabja meg, fejlődési potenciájuk csak
jóval később szűkül be. A legtöbb állatfaj e két véglet között helyezkedik el. A közönséges
muslica esetében például a hólyagcsíra állapot kialakulásáig az egyes sejtek megváltoztat-
hatják helyzetüket, sejtmagjaik ekkor még igen nagyfokú fejlődési potenciával bírnak.
2.7. Posztembrionális fejlődés
A peteburok elhagyását vagy a megszületést követően a fiatal állat megkezdi posztemb-
ríonális életet. Fejlődése lehet közvetlen, amikor lárvaállapotok kialakulása nélkül fej-
lődik ki. Közvetett fejlődés során a kifejlett egyedtől alakilag és testfelépítésben is eltérő
egy vagy több fejlődési forma, lárvaalak iktatódik be.
A lárvaformák kialakulása főleg a vízben élő, rögzült életmódú vagy lassan mozgó
fajok számára jelentős tényező az adaptáció és az elterjedés szempontjából. Hasonló
fontossággal bír azonban a szárazföldi fajoknál is. Amozgásra képes és eltérő táplálkozást
folytató lárvák nem jelentenek konkurenciát a kifejlett egyedek számára. Új területeket
hódíthatnak meg és eltérő táplálékbázisokat használhatnak ki. Az evolúció során, a
természetes szelekció révén egyes fajok lárvái olyan mértékben adaptálódtak speciális
környezeti feltételekhez, hogy a kifejlett állatokra egyáltalán nem, vagy csak kis mérték-
ben hasonlítanak. Egy fejlődési alak sikeressége meghatározza a soron következőekét is.
Jellegzetes lárvaforma például a planula (csalánozók), a Müller-féle lárva (örvényfér-
gek), a trochophora (puhatestűek), valamint a nauplius és a zoea (rákok). A mezőgaz-
dasági szempontból is fontosabb állatcsoportoknál megfigyelhető, de itt fel nem sorolt
lárva-formákat, valamint az átalakulási folyamat jellegzetességeit az adott rendszertani
egység tárgyalásakor mutatjuk be.
A lárvaalakok közbeiktatásával fejlődő fajok életciklusa általában összetett. A fajra
jellemző tulajdonságok eléréséhez gyakran bonyolult morfológiai, fiziológiai és életmód-
beli átalakulásoknak kell végbemennie. Ha a lárva és a kifejlett állat között testfelépítés-
ben, táplálkozásmódban és az élőhely-adaptációban nagy eltérés mutatkozik, akkor ezt
a folyamatot metamorphosisnak nevezzük. Minél nagyobbak ezek a különbségek,
annál drámaibb a metamorfózis folyamata. Az átalakulás elsősorban a szervezet hormo-
nális szabályozása alatt áll.
Az esetlegesen lárvaformákat is magába foglaló kezdeti, fiatalkori fejlődési szakasz
után az egyed eléri ivarérettségét, majd ezt követően teljes kifejlettségét. Az örege-
dési periódusban bekövetkező változások végül az egyed halálával végződnek.
Az állatok élete során lehetnek olyan szakaszok, amikor anyagcseréje, így a fejlődése
és növekedése lelassul, esetleg szinte teljesen leáll. Az ilyen nyugalmi szakaszokat
(diapausa) elsődlegesen az időjárás évszakos változása, a rossz táplálkozási feltételek
hozzák létre. Az állat részéről fiziológiai adaptációt jelent, amelynek segítségével képes
átvészelni a kedvezőtlen körülményeket. Ilyen például egyes egysejtűek ciszta állapota,
78
számos édesvízi szervezet „téli petéje”. A rovarok diapauzája végbemehet akkor, ha a
körülmények kedvezőtlenné válnak, vagy ettől függetlenül is, ha egy adott fejlődési állapo-
tot elért az egyed. A diapauza bármely fejlődési alakjukat érintheti, legyen az pete, lárva,
báb vagy imágó. Rovaroknál a diapauza kiváltásában leginkább a fotoperiódúsnak van
szerepe. A fény közvetlen hatása az agyra befolyásolja a neuroszekréciós rendszert: leállít-
hatja például a vedlést kiváltó hormon termelését. A nyári álom olyan élőhelyen előfor-
duló fajoknál jellemző, ahol van egy igen száraz periódus, ami az állat kiszáradását okoz-
hatná. Tipikus formában jellemző például a tüdőshalakra, több kétéltűre, vagy a mérsékelt
övben a szárazföldi tüdőscsigák egy részére is. Ahibemáció előtt az állat jelentős energia-
tartalékokat halmoz fel szervezetében, majd anyagcseréjének intenzitását jelentősen csök-
kenti és álomba merülve vészeli át a kedvezőtlen hideg időszakot. A valódi hibemáló állatok
(például denevérek, rágcsálók és rovarevők) testhőmérséklete ilyenkor oly mértékben le-
csökken, hogy megközelítheti a kömyezetéét. Alapanyagcseréjük a normálisnak néhány
százalékára csökken le. Testük átfagyása ellen úgy védekeznek, hogy fagymentes helyre
húzódnak el, összegömbölyödnek vagy több állat összebújik, hogy a lehető legkisebb test-
felületet hagyják szabadon. A medve a rágcsálóktól és rovarevőktől eltérően végigalussza
téli álmát, közben nem táplálkozik. Ha a rágcsálók és a rovarevők testének hőmérséklete
a veszélyes szintet közelítené meg, illetve ha átmeneti enyhülés következik be, akkor álmuk-
ból felébrednek. Ilyenkor az elraktározott táplálékukból fogyasztanak, ürüléket, vizeletet
adnak le. Anyagcseréjüket, testhőmérsékletüket ilyenkor átmenetileg megemelik, majd
ismét álomba merülnek. A kétéltűek és hüllők ilyen szabályozásra képtelenek. Téli nyugal-
mi állapotukat, anyagcseréjüket a környezet hőmérséklete döntően meghatározza. Számos
fajuk képes akár a fagypont alatti hőmérsékletet is elviselni, mert szervezetében fagyásgátló
anyagok vannak. Ezek bizonyos határon belül megakadályozzák a szervezet víztartalmának
jégkristályokká alakulását, a sejtek szétroncsolását.
Az embrionális és posztembrionális életszakaszban az állatok fejlődési és növekedési
folyamatai egymástól el nem választhatóan, egymást feltételezve mennek végbe. A fej-
lődés elsősorban új tulajdonságok megjelenését, minőségi változásokat jelent. A növe-
kedés az élő szervezetek alapvető sajátossága, amelynek során a már meglévő sejttípu-
sok, szövetek és szervek méretükben, tömegükben gyarapodnak. Ez mindaddig tart,
amíg el nem érik a fajra jellemző mértéket. Egyes állatfajok egyedei egész életük folyamán
növekednek (például tízlábú rákok, hüllők).
A fejlődést és a növekedést elsősorban a genetikai faktorok és a szervezet fiziológiai
hatótényezői szabják meg. Érvényre jutásukat azonban a környezeti tényezők jelentősen
befolyásolhatják.
Az állat fejlődése és növekedése során lezajló változásokat legegyszerűbb módon a tömeg,
a méret- és a formaváltozások vizsgálatával lehet nyomon követni. A fajra jellemző
testméretek eléréséig az állati szervezet különböző organizációs szintjei meghatározott fizio-
lógiai változásokon mennek keresztül. Ezeket az adott szinten általában morfológiai és tö-
megváltozások kísérik, amelyek mérhetőek. Az így nyert adatokból az állattenyésztési kuta-
tások vagy akár a közvetlen gyakorlati élet számára is következtetések vonhatók le.
Egy állatfaj fiatal és kifejlett egyedeinek testformái legtöbbször nem egybevágóak
geometriailag. A gerincesek közül a halaknál viszonylag kisebb, míg a szárazföldieknél
jelentősebb forma- és arányváltozások mennek végbe.
2.8. Regeneráció
Az állati szervezetet felépítő sejtek élettartama eltérő. A szöveteket, szerveket alkotó
sejtek elpusztulását normális esetben kompenzálják az osztódó sejtek. Ez a fiziológiai
regeneráció, amely általában folyamatos utánpótlását jelenti az elpusztult sejteknek.
79
Az idő előtt elpusztult szövetek, szervek, testrészek pótlása a reparatív regeneráció.
Ez általában valamilyen külső behatásra bekövetkezett sérülés kijavítására szolgál. Ekkor
a sérülés közelében lévő sejtek időlegesen visszanyerik differenciálódási potenciájukat.
Az egyes állatcsoportokban a regenerációs képesség eltérő mértékű. Igen jó regenerációs
képességgel rendelkeznek például a csalánozók és az örvényférgek, de a gyűrűsférgek,
tüskésbőrűek is jól, míg a puhatestűek és az ízeltlábúak kevésbé jól pótolják elvesztett
testrészeiket. A halak sérült úszóikat tudják regenerálni, a békák ebihalai és a farkos
kétéltűek kifejlett alakjai pedig képesek kopoltyújuk, végtagjuk vagy farkuk pótlására. A
hüllők körében az öncsonkítás (autotomia) képességével találkozunk, ami azt jelenti,
hogy veszélyhelyzetben képesek egyes testrészeiket (elsősorban a farkukat) a testről
ledobni. Ez igen fontos védekezési reakció, segítségével a ragadozót átmenetileg meg-
tévesztheti, ami lehetőséget ad a menekülésre. A farok később regenerálódik.
2.9. Az állati szervezet öregedése, a halál
Valamennyi állatfajnak jellemző biológiai élettartama van. Ez általában szoros össze-
függésben van például a testtömeggel, hiszen a nagy testű állatok többnyire hosszabb
életűek. Az emlősök között például az indiai elefánt 70, az orrszarvú 50, a gorilla 25, a
kutya 15, a szürke mókus 10, a házi egér 2, a vízicickány 1 évig él. Hasonló vonás
figyelhető meg a gerincesek más osztályainál is. A hüllők közül például az óriás galapagosi
teknősök évszázadokat is megélhetnek.
A többsejtű állatok egyedi életének vége felé közeledve szembetűnővé válik az örege-
dés folyamata. E jelenség okainak feltárása régóta foglalkoztatja a kutatókat. Egyes
jelenlegi elképzelések szerint a folyamat végbemenetele ugyanúgy a genetikai programra
vezethető vissza, mint a fajra jellemző maximális élettartam. Az öregedés jelei megmu-
tatkoznak a makromolekulák, a sejtorganellumok, a sejtek, a szövetek, a szervek, illetve
az egész szervezet szintjén is. A sejtekben az öregedés során a DNS felépítésében egyre
több mutációs eredetű változás jöhet létre, annak ellenére, hogy egy folyamatos javítási
mechanizmus zajlik. Összefüggést találtak a javító mechanizmus működésének haté-
konysága és az adott faj biológiailag lehetséges élettartama között.
Az egyes egyedek halállal (exitus) végződő öregedési folyamata populációs szinten
megakadályozza a túlnépesedést. A természetes halálon kívül az egyed életének végét
okozhatják például a ragadozók, a betegségek, a táplálékhiány és a kedvezőtlen klimati-
kus hatások. Valójában az állatok általában nem érik el a lehetséges maximális, poten-
ciális élettartamukat, mert már az előtt elpusztulnak.
Ellenőrző kérdések a 2. fejezet témaköreiből
1. Miben tér el egymástól egy átlagos testi sejt és a petesejt felépítése?
2. Az endolecitális petesejtek típusai hogyan határozzák meg az embrionális folyamatok
menetét?
3. Mi a jelentősége a petesejtet körbevevő burkoknak?
4. Miben különbözik egymástól az ovogenezis és a spermiogenezis?
5. A hímivarsejt felépítése milyen speciális vonásokat mutat?
6. A himivarsejt kialakulása során miért van szükség a spermiomorfogenezisre?
7. Értékelje a külső és a belső megtermékenyítés jelentőségét, a közöttük lévő különbséget!
8. Mi határozza meg a barázdálódás menetét?
9. A barázdálódás milyen típusait ismeri?
10. Mi jellemző a szedercsíra állapotra?
80
11. Mi jellemző a hólyagcsira állapotra?
12. Az embrionális fejlődés során mikor lehet elkülöníteni egymástól a leendő csíralemezeket?
13. Milyen csíralemezek alakulnak ki az embrionális fejlődés organotipikus szakaszában?
14. Sorolja fel az ektodermából kialakuló ősi szervtelepeket, szerveket, szöveteket!
15. Sorolja fel a mezodermából kialakuló ősi szervtelepeket, szerveket, szöveteket!
16. Sorolja fel az entodermából kialakuló ősi szervtelepeket, szerveket, szöveteket!
17. Milyen embrionális szervek alakulnak ki az organogenezis során?
18. Milyen állatcsoportokra jellemző a szikzacskó kialakulása?
19. Milyen állatcsoportokra jellemző a magzatburkok kialakulása?
20. Mi a jelentősége az egyes magzatburkoknak?
21. Milyen kapcsolatot jelent a méhlepény az anya és utóda között?
22. Mi a különbség fiziológiai és reparativ regeneráció között?
23. Mutassa be az autotomia képességének jelentőségét!
24. Mit értünk az öregedés folyamata alatt?
81
3. AZ ALLATI TESTET FELEPITO
SZÖVETEK
Az embrionális fejlődés során kialakul a három csíralemez. A csíralemezek további differenciá-
lódása során szervtelepek, szervek jönnek létre. Az egyes szerveket túlnyomórészt többféle szövet
építi fel. A szövet (tela) olyan sejtek egységétjelenti, melyek hasonló alakúak és azonos feladatot
töltenek be, valamint az egyedfejlődés során általában azonos csíralemezből származnak. A
szövetfelépítésében a sejtes állományon kívül a sejt közötti állománynak (substantia intercellularis)
is fontos szerepe van, mert részt vesz a szövet tulajdonságainak kialakításában.
Tekintettel arra, hogy egy adott alapszövetféleség több csíralemezből is kialakulhat, ezért
csoportosításuk nem származásuk, hanem elsődlegesen morfológiai tulajdonságaik és funkció-
juk alapján történik. Ennek alapján megkülönböztetünk hámszövetet (tela epithelialis), kötő-
és támasztószövetet (tela connectiva), izomszövetet (tela muscularis) és idegszövetet (tela
nervosa).
Hámszövetek mind a három csíralemezből kialakulhatnak, a testfelszínt és a belső üregek
falát borítják. A felépítő sejtek szorosan illeszkednek egymáshoz, táplálásuk az alaphártya felől
megy végbe. A hám lehet egy- vagy többrétegű, elszarusodó vagy el nem szarusodó. A hámokat
funkciójuk szerint a fedő-, a felszívó-, a pigment-, az érzék- és a mirigyhámok közé sorolhatjuk.
A kötő- és támasztószövetek mezodermális eredetűek. Felépítő sejtjeik és a sejtközötti állomány-
ban lévő rostok (enyvadó-, rugalmas- és rácsrostok) megléte és arányaik meghatározzák a
tulajdonságaikat. Szerepük főleg a szövetek és szervek összekapcsolásában, támasztásában,
táplálásában, és a hőszigetelésben van. A kötőszövetek (embrionális-, laza-rostos-, retíkuláris-,
lemezes-, tömött-rostos kötőszövet, zsírszövet, vér és egyéb testfolyadékok) szinte valamennyi
szerv felépítésében résztvesznek. A támasztószövetek (kondroid-, korda-, porc- és csontszövet)
sejtközötti állománya rugalmas vagy igen szilárd lehet. A csontszövet az állati szervezet leg-
keményebb szövetfélesége. Az izomszövetek (sima- és harántcsíkolt izomszövetek) jórészt
mezodermális eredetűek. Felépítő sejtjeik Összehúzódásra képesek. Az idegszövet alkotásában
szerepet játszó idegsejtek más testi sejteknél sokkal jobban differenciálódtak ingerfelvételre,
ingerület továbbításra, feldolgozásra.
Ebben a fejezetben csak az általános szövettan bemutatására szorítkozunk, tehát az alap-
szövetek típusainak jellemzését adjuk meg. Néhány szövet esetében az azokat felépítő speciális
sejteket a sejttani fejezetben ismertettük. A szervszövettan az egyes szervek szöveti felépítésével
foglalkozik, és más tantárgyak anatómiai, élettani témaköreiben kerül kifejtésre.
3.1. Hámszövetek
Hámok mind a három csíralemezből kialakulhatnak. Az ektodermális eredetűek a test-
felszínt borítják, a mezodermából származók a testüregeket bélelik, a húgy-ivari szervek
felépítésében vesznek részt, míg az entodermálisak a bélcső és a légzőszerv hámját alkot-
ják (lásd még 2.2. táblázat).
A hámszövetet alkotó sejtek szorosan illeszkednek egymáshoz, a sejt közötti
(intercelluláris) állományuk igen kevés vagy hiányzik. Valamennyi hámszövetre jellem-
82
ző, hogy alatta poliszacharidokból, speciális kollagénrostokból és proteoglikánokból álló
alaphártya (membrana basalis) van. Az alaphártyával érintkező sejtek hemidez-
moszómákkal és adhéziós molekulákkal kapcsolódnak hozzá, ami jó rögzülést tesz lehe-
tővé számukra. Ez az alaphártya elválasztja a hámot az alatta lévő kötőszövettől, de rostjai
révén egyben össze is fűzi azzal. A hámsejtek tápanyagokkal való ellátása a kötőszövet
felől az alaphártyáig futó kapillárisok segítségével valósul meg. Innen a tápanyag diffú-
zióval jut tovább a távolabbi sejtekhez. A hámsejtek rétegében vérereket nem találunk.
A többrétegű hámokban a kötőszövetes alap kiemelkedései (papillák) növelik a hám és
a kötőszövet határfelületét. Ez szorosabb kapcsolódást, a nagyobb felület révén pedig
intenzívebb anyagáramlást és nagyobb számú osztódóképes sejt elhelyezkedését teszi
lehetővé. A hámrétegben érzőideg-végződések, valamint idegrostok fordulnak elő.
A hámszövet regenerálódóképessége a többi szövethez viszonyítva jó. A folyamatosan
elpusztuló sejtek pótlásán túl képes a kisebb-nagyobb sérülések kijavítására is.
A hámokat csoportosítani lehet morfológiai bélyegeik és funkciójuk szerint. Morfo-
lógiai jellemzőik közül a sejtek alakja és a sejtrétegek száma az elkülönítés alapja.
A hámsejteken megkülönböztetünk alapi (basalis) és csúcsi (apicalis) pólust. A kettő
közötti tengely a sejt magassági méretét adja meg, míg erre merőlegesen a szélességi és
hosszúsági méretek vehetők fel.
3.1.1. A hámok csoportosítása morfológiai jellemzők alapján
Az egyrétegű hám valamennyi sejtje érintkezik az alaphártyával, míg a többrétegűek
esetében csak az alsó sejtsor.
Az egyrétegű laphámot felépítő sejtek erősen lapítottak, sokszögletűek (polygonalisak)
és egymáshoz igen szorosan kapcsolódnak (3.1. ábra). Lapított vagy gömbölyded sejt-
magjuk van. Különösen vékony laphámsejtek közepén lévő kidomborodás a sejtmag
helyét jelzi. Tipikus formában fordul elő például az utóvesében (metanephros), ahol a
Bowman-tok fali (parietális) lemezét képezi. A laphám speciális formája az érfalhám
(endothelium) és a testüreghám (mesothelium).
3.1. ábra. Egyrétegű hámokat felépítő sejtek alakjai
A: laphámsejtek, B: köbhámsejtek, C: hengerhámsejtek (Több szerző nyomán)
Az egyrétegű köbhámsejtek szélességi, hosszúsági és magassági méretei közel azo-
nosak (3.1. ábra). A gömbölyű sejtmagok a sejt közepén helyezkednek el. Tipikus for-
mában fordulnak elő a vese kanyarulatos csatomácskáiban, mirigyvégkamrákban és
azok kivezetőcsatomáiban, valamint a szemlencse elülső felszínén. A vese elsőrendű
kanyarulatos csatornácskáinak kezdeti szakaszán a sejtek apikális felszínén lévő mikro-
bolyhok úgynevezett kefeszegélyt alkotnak, mely a felszívófelületet növeli meg.
83
3.2. ábra. Speciális egyrétegű (átmeneti) hám, az urothehum
A: üres húgyhólyag falában, B: telt húgyhólyag falában, ah: alaphártya, es: emyősejtek
(Több szerző nyomán)
Az egyrétegű hen-
gerhámsejtek ma-
gassági mérete felül-
múlja a szélességi és
hosszúsági méreteket
(3.1. ábra). Az általá-
ban ugyanezen irány-
ban megnyúlt sejtmag
a sejt alapi részéhez kö-
zel helyezkedik el. Ti-
pikus formában talál-
ható számos gerincte-
len állat kültakarójá-
ban és például az utó-
vese gyűjtőcsatornái-
ban, az epeutak falában. A bélcső belső felületét borító hámsejtek aplikális felszínén ún.
pálcikaszegélyt alkotnak a hosszú mikrobolyhok. A légutakban csillós (kinociliumos),
többsejtmagsoros hengerhám található. Ez utóbbinál valamennyi hámsejt kisebb-na-
gyobb felületen érintkezik az alaphártyával, de sejtmagj aik különböző szintekben helyez-
kednek el.
Az egyrétegű hámok speciális formája a gyakran átmeneti hámnakis nevezett urothelium.
Fénymikroszkópban vizsgálva úgy tűnik, mintha a többnyire szabálytalan alakú sejtek több
rétegben helyezkednének el (3.2. ábra). A látszólag felszíni réteget alkotó sejtek (emyősej-
tek) beborítják ugyan az alattuk lévőket, azonban egy-egy vékony nyúlványuk segítségével
kapcsolatot létesítenek az alaphártyával. A sejtek kapcsolódó membránjai erősen redő-
zöttek, ami lehetővé teszi, hogy megnyúláskor a sejtek egy-máshoz viszonyított helyzete
jelentősen megváltozhasson. Ez a hámféleség a vesemedence, a húgyhólyag és a húgyve-
zeték falában található. A húgyhólyag telített állapotában a hám erősen elvékonyodik, a
sejtek rugalmas alakváltoztatás révén követik a tágulást.
A többrétegű hámot mindig a legkülső
3.3. ábra. El nem szarusodó többrétegű laphám
réteg sejttípusa alapján nevezzük el.
A többrétegű laphámban a fel-
színi rétegekben lévő sejtek mutatnak
laphám formát, míg az alsó sorokban
g| ülők nem. Legalul osztódásra képes,
henger alakú, középen pedig átme-
neti köbhámformát mutató sejtek ta-
lálhatók. Attól függően, hogy a felső
réteg elszarusodik-e vagy nem, meg-
különböztetünk el nem szarusodó és
tS elszarusodó többrétegű laphámot. Az
el nem szarusodó többrétegű laphá-
mot legalul az osztódó sejtek rétege
(stratum germinativum), felette a tüs-
Qg kés réteg (str. spinosum), a felszínen
pedig az ellapult sejtek rétege (str. pla-
, nocellulare) építi fel (3.3. ábra). A
3 n tüskés sejtek nevüket a szövettani pre-
parálás során alkalmazott fixálási el-
ah: alaphártya (membrana basalis), el: ellapult sejtek rétege (str.
planocellulare), os: osztódó sejtek rétege (str. germinativum), ts:
tüskés sejtek rétege (str. spinosum) (Katznelson-Ríchter nyomán
kissé módosítva)
járás által kialakított műtermékről
kapták. Az ilyenkor zsugorodó sejtek
az erős dezmoszomális kapcsolat ré-
84
vén egymással helyenként érintkezésben marad-
nak, így az egymástól részlegesen elváló sejtek kö-
zött tüskeszerű nyúlványok mutatkoznak. A leg-
felső ellapult sejtek fokozatosan elhalnak, a sejt-
szervecskék lebomlanak. A halak kültakarójának
felszínét ez a hám alkotja. A fejlettebb gerincesek-
nél ilyen hám fordul elő a mindig nedves szájgarat
üregben, a nyelőcsőben, a hüvely egy részében, a
szem szaruhártyájának elülső felszínén.
A többrétegű elszarusodó laphám felszínén
elhalt sejtekből álló szaruréteg van (3.4. ábra).
A kétéltűek, hüllők és madarak kültakarójában ez
a hám három rétegből áll. Legalul van az alapi
réteg (str. germinativum), felette a közbülső (str.
intermedium) és a felszínen az elszarusodott ré-
teg (str. comeum). A kétéltűek hámja csak gyen-
gén elszarusodó. Az emlősök epidermiszében öt
réteget lehet elkülöníteni. A hám alapi részén he-
lyezkednek el az osztódó sejtek (str. germina-
tivum), majd ezt borítja a tüskés sejtek (str. spino-
sum) rétege. A tüskés réteg sejtjeiben jelentős
mennyiségű keratinfii amentum képződik. Felet-
tük van a szemcsés réteg (str. granulosum), ahol
a sejtekben szemcsék vannak. A fénylő rétegben
(str. lucidum) a szemcsék eltűnnek, megváltozik
a réteg törésmutatója, ezért látszik fénylőnek. A
sejtek ebben a rétegben már degenerálódnak, a
sejtorganellumok gyakran már nem ismerhetők
3.4. ábra. Többrétegű elszarusodó laphám
ah: alaphártya (membrana basalis), fr: fénylő réteg
(str. lucidum), os: osztódó sejtek rétege (str. germi-
nativum), se: szemcsés sejtek rétege (str.
granulosum), sz: szaruréteg (str. comeum), ts: tüskés
sejtek rétege (str. spinosum) (Katznelson-Richter
nyomán kissé módosítva)
fel, mert a lizoszómák lebontják azokat. A legfelső szaruréteg (str. corneum) elhalt
sejtjeiben nagy mennyiségben halmozódik fel keratin. A keratin vízben nem oldódó
fehérje, amely felhalmozódva a kültakaró felszíni rétegében védelmet nyújt a kiszáradás-
sal szemben, és gátat szab a káros anyagok és kórokozók bejutásának. Az elszarusodás
folyamata egy speciálisan szabályozott, programozott sejthalál. A felszíni réteg fokoza-
tosan kopik. Ez a hám alkotja az emlősök bőrének hámrétegét és hozza létre származékait
(szőr, karom, köröm stb.). A többrétegű hengerhám felső és alsó rétegében tipikus
hengerhámsejtek ülnek, míg közöttük több rétegben ettől kissé eltérő alakú sejtek van-
nak. Ez a hám nem gyakori. A nagyobb mirigyek kivezetőcsöveiben, a szem kötőhártya-
boltozatán, a kétéltűek nyelvén és garatján található.
3.1.2. A hámok csoportosítása funkciók alapján
A fedőhámok elsődleges feladata a védelem. Tágabb értelemben szinte valamennyi eddig
bemutatott hám besorolható lenne ebbe a csoportba, de tipikusan a testfelszínt borító
epidermisz, valamint az endothelium és a mesothelium tartozik ide. Az endothelium és
mesothelium a test belső üregrendszereit fedi. Az endothelium a vér- és nyirokerek, vala-
mint a szív belső felszínét borítja. Az endotélium sejtjeinek kapcsolata olyan is lehet, hogy a
közöttük képződő réseken át akár a vérsejtek is áthatolhatnak. Az endotélium kétirányú
anyagáramlást tesz lehetővé a vérplazma és a vérereket körülvevő szövetek között. Az
endotéliumsejtek átalakulhatnak kötőszöveti sejtekké. A mesothelium a savóshártyák
(mellhártya, szivburok, hashártya) felszínén található. A sejtek által termelt folyadék bevonj a
a felületét, lehetővé teszi, hogy a zsigeri szervek súrlódásmentesen elcsússzanak egymáson.
85
kh
3.5. ábra. Endoepiteliális mirigyek
A: váladékkal (vá) telt kehelysejt (kh) a
bélhámsejtek kozott, B: csoportot alkotó
mirigysejtek (Welsch-Storch nyomán mó-
dosítva)
A mezotélium felszínét mikrobolyhok növelik, így nagy
felülete révén jelentős felszívóképessége van. Különleges
fedőhám-módosulat a számos hüllő és madár pislogóhár-
tyájának belső felszínét borító tollepithelium. Ennél a
hámsejtek szabad felszínén tollszerűen elágazó nyúlvá-
nyok vannak, ami a szem kötőhártyájának tisztántartására
szolgál.
A felszívóhám a tápanyagokfelvételét és továbbadá-
sát végzi. Jellegzetes formája a vékonybél bélbolyhait
borító, az apikális felszínén mikrobolyhokat viselő hen-
gerhám. Természetesen bizonyos fokig minden hámsejt
képes anyagok felszívására, de ezek a bélhámsejtek pél-
dául felületi struktúrájuk révén specializálódtak erre a
feladatra.
A pigmenthám sejtjeiben nagy mennyiségben talál-
hatóak festékanyagok, amelyeknek szerepe a fény útjá-
nak elzárásában van. Tipikus formában fordul elő pél-
dául a gerincesek szemében a retina mögött.
Az érzékhám az érzékszervek felszínét alkotja. Az ér-
zéksejtek támasztósejtekkel együtt fordulnak elő benne.
A csillós hám speciális feladatokat láthat el. A lég-
utak hámját alkotva elősegíti, hogy a szennyeződések a
csiliók csapkodása révén a testből kikerüljenek. A pete-
vezetőben ezek a hámsejtek továbbítják a petesejtet.
A mirigyhám a mirigyek (glandulae) váladéktermelő részét alkotja. A belső elválasz-
tású (endocrín) mirigyeknek nincs kivezetőjáratuk, váladékukat a véráramba juttatják. A
külső elválasztású (exocrín) mirigyek vagy a test-
felszínre, vagy szervek üregeibe öntik váladé-
kukat, kivezetőjáratukon keresztül. Ha az exo-
krin mirigy a hámrétegbe ágyazott, akkor endo-
epithelialis mirigyről beszélünk. A nyálkaanya-
got, méreg- és illatanyagokat termelő egysejtű
exokrin mirigyek nagy számban fordulnak elő a
gerinctelenek (példáulférgek, puhatestűek) kül-
takarójában. A körszájúak bőrében a nyálkater-
melő mirígysejtek szorosan egymás mellett el-
helyezkedve, csoportokat alkotnak (3.5. ábra).
A gerinces állatokbélcsatomájánakbelső felszí-
nét borító hámrétegben az önemésztéstől védő,
mucinozus váladékot termelő kehelysejtek van-
nak (3.5. ábra). Exoepithelialis típusú a mirigy
akkor, ha a hám alatti kötőszövetbe süllyedve
fekszik (például az emlősök verejtékmirigyei).
A mirigyek lehetnek egy vagy több sejtből
állóak. Ez utóbbi esetben a mirígyhámsejtek
többnyire a vázalkotó kötőszöveti részekkel
(interstitium), vérerekkel és idegekkel együtt
alkotnak mirigyet. A végkamra a mirigyek vá-
ladéktermelő sejtekkel bélelt ürege, amely a
kivezetőj áraihoz csatlakozik.
A soksejtű exokrin mirigyeket a végkamra
3.6. ábra. Mirigyek csoportosítása felépítésük
alapján
A: egyszerű csöves, B: egyszerű bogyós, C: egyszerű
csöves-bogyós, D; elágazó csöves, E: elágazó bogyós, F:
elágazó csöves-bogyós, G: összetett csöves, H: összetett
bogyós, I: összetett csöves-bogyós (Több szerző nyomán)
86
alakja, száma és a kivezetőcsövek száma szerint
csoportosítják (3.6. ábra). A végkamra alakja sze-
rint csöves, bogyós és csöves-bogyós miri-
gyeket különböztetünk meg. Az egyszerű miri-
gyekben egy kivezetőcsőhöz egy végkamra tarto-
zik. Egyszerű csöves mirigy például a verejtékmi-
rigy és a bélcsőben lévő Lieberkuehn-mirigy. Egy-
szerű bogyós mirigyek vannak a békák bőrében.
Az elágazó mirigyekben több végkamrának van
egy közös kivezetőcsöve. A nagyobb faggyúmiri-
gyek elágazó bogyós felépítésűek. Az összetett
mirigyeknél a kivezetőcső több mellékágra osz-
lik és ezek végein vannak a végkamrák. Összetett
csöves-bogyós mirigy például a hasnyálmirigy, a
tejmirigy és a nyálmirigyek. Attól függően, hogy
a mirigyhámsejtek milyen módon adják le a vála-
dékot, megkülönböztetünk merokrin, apokrin és
holokrin működésűeket (3.7. ábra).
Merokrin működés esetén a mirigysejtből
exocitózissal csak a váladék távozik el. így műkö-
dik a mirigyek többsége, például a nyálmirigyek
is. A merokrin működés egy speciális esetének
tartják az ekkrin szekréciót. Ebben az esetben a
szekrétum (elektrolit, szteroid) a membránon ke-
resztül zajló transzporttal kerül leadásra. Például
a verejtékmirigyek működnek ilyen módon.
A
3.7. ábra. A mirigyhámsejtek váladék-
leadásának különböző típusai
A: merokrin, B: apokrin, C: holokrin, vá: váladék,
1-4: a holokrin működésű sejt átalakulásának folya-
mata (Kühn nyomán)
Apokrin módon működő sejteknél a váladékkal együtt a plazma egy része is távozik,
körülvéve a plazmamembrán egy részével. Ez jellemző a tejmirigy zsírszekréciójára és
egyes illatmirigyekre.
3.8. ábra. Különböző végkamratípusok egy exokrin mirigyben
er: endoplazmatikus retikulum, Ga: Golgi-apparátus, kj: kivezetőjárat, KV; kevert
végkamra, MU: mucinozus végkamra, SE; szerozus végkamra, sm. sejtmag, vá;
váladék (Welsch-Storch nyomán kissé módosítva)
Holokrin típusú sej-
teknél a váladéktermelés
során az egész sejt vála-
dékká alakul át, az egész
sejt kiürül. Az elpusztult
sejteket a végkamra alap-
hártyáján ülő osztódó
sejtekpótolják. Ilyen mű-
ködésforma jellemző a
faggyúmirigyekre.
Előfordul, hogy egy
mirigy két módon is ad-
hat le váladékot. A tej-
mirigyben apokrin mó-
don kerülnek leadásra a
lipidcseppek, merokrin
módon pedig a fehérjék
és a tejcukor.
A váladék összetétele,
termelésének és leadá-
sánakmódja szerint a me-
rokrin működésű nyál-
87
mirigyekben három végkamra típust lehet elkülöníteni (3.8. ábra). A savós (serosus) mi-
rigyvégkamra sejtjei fehérjeváladékot termelnek. A sejtek endoplazmatikus retikuluma
ezért különösen jól fejlett, a sejt alapi részén és a sejtmag körül helyezkedik el. Az apikális
póluson halmozódnak fel a váladékszemcsék, melyek exocitózissal ürülnek. A mirigyvég-
kamra ürege viszonylag szűk. Szerózus mirigy például a fültőmirigy és a könnymirigy. A
nyákos (mucinosus) mirigyvégkamra sejtjeinek endoplazmatikus retikuluma kevésbé
fejlett, mert csak kevés szekréciós fehérjét szintetizál. A Golgi-készülék viszont igen jól
fejlett, sok szénhidrátot szintetizál. A terjedelmes váladékgranulumok a szerózus váladék-
hoz képest nagyobb víztartalmúak, méretük is nagyobb. A váladék sűrű, viszkózus, a
termelt fehérjéhez sok szénhidrát kapcsolódik. A váladék leadása történhet egyszerű
exocitózissal is, de gyakran úgynevezett sorozatexocitózis figyelhető meg. Ebben az eset-
ben egy, az apikális plazmamembránnal összeolvadt granulum membránjába beleolvad a
következő, majd sorban még továbbiak is. Sokszor az egész granulumrészlet így ürül ki
egyszerre, ami a fénymikroszkópos vizsgálatokban azt a benyomást keltette, hogy a felgyü-
lemlett váladék a sejthártyát átszakítva ürül ki. Mucinózus típusúak például a nyelőcső és
a szájpadlás nyálka termelő mirigyei. A kevert (seromucinosus) mirigyvégkamrát mucinózus
sejtek borítják, míg a szerózus sejtek félholdszerűen veszik körül őket. Utóbbi sejtek vála-
déka a mucinózus sejtek közötti réseken át jut be a végkamra üregébe. Az állkapocs alatti
és a nyelv alatti nyálmirigyek végkamrái ilyen kevert típusúak.
3.2. Kötőszövetek és támasztószövetek
A kötő- és támasztószövetek mezodermális eredetűek. Elsődleges feladatuk a sejtek, szö-
vetek, szervek összekapcsolása, a hézagok kitöltése, a támasztás. A kötő- és támasztószö-
vetek megadják a szervek és a test külső formáját, biztosítják más szövetek táp-anyagellá-
tását, egyes sejtjeik regenerációs képessége révén képesek az elvesztett részek pótlására. A
szervezetben majdnem mindenütt előfordulnak, szinte valamennyi szerv felépítésében
szerepük van. A kötő- és támasztószövetekben - a porcszövet kivételével - mindig találha-
tóak vér- és nyirokerek, valamint idegek. Sejtes állományuk igen változatos (bővebben
lásd a kötőszövetek ismertetésénél), de általában a sejt közötti állomány dominál. Az
intercelluláris állomány rostokból és alapállományból áll, összetétele meghatározza a szö-
vet tulajdonságait. Az enyvadó (collagen) rostok szkleroproteidek, melyek nem nyújtha-
tóak, de szakítási szilárdságuk igen nagy. A rostok kollagénből épülnek fel. A kollagén
molekulák polaritása és a rost felépítőegységeinek szabályos ismétlődése miatt a kollagén
rostok elektronmikroszkóppal vizsgálva enyhe harántcsíkoltságot mutatnak. Avastag, hul-
lámos lefutású, elemi rostokból álló rostnyalábok gyakran elágaznak. Az elemi rostokat
glikozamino-glikán (régebbi nevén mukopoliszacharid) tartalmú anyag köti össze. A kol-
lagén rostok valamennyi többsejtű állatban megtalálhatóak. A rugalmas (elasticus) ros-
tok vékonyak, elasztin- és glikoproteid-tartalmúak, nem alkotnak kötegeket. A rostok jól
nyújthatóak, az erőhatás után pedig visszanyerik eredeti méretüket. Sok rugalmas rost van
az irhában, az elasztikus típusú artériák falában, a rugalmas-rostos porcban és a tüdőben.
A rácsrostok (reticularis rostok) igen vékonyak, elágazóak, egymással hálózatot képez-
nek. A rácsrost a kollagén egyik változatának tekinthető fehérjékből és glikozamino-glikánból
épül fel. Szinte valamennyi szervben megtalálhatóak, sűrű hálózatot alkotnak például a
hámok alatti alaphártyában és a nyirokszervekben. Körülveszik a zsírsejteket és a vérereket.
Az intercelluláris állomány kolloid jellegét a fehérje- és a glikozamino-glikán tartalom,
viszkozitását a hialuronsav (egy speciális, savanyú glikozamino-glikán) mennyisége szabja
meg. Az alapállománynak fontos szerepe van az anyagforgalomban és a vízháztartásban.
A kötőszövetek és támasztószövetek csoportosítása a celluláris és intercelluláris állo-
mány összetétele alapján történik:
88
Kötőszövetek: - embrionális kötőszövet: mezenchima
kocsonyás kötőszövet
- laza-rostos kötőszövet
- retikuláris kötőszövet
- lemezes kötőszövet
- tömött-rostos kötőszövet: rugalmas-rostos kötőszövet
kollagén-rostos kötőszövet (ín)
- zsírszövet: fehér zsírszövet
barna zsírszövet
- vér és egyéb testfolyadékok
Támasztószövetek: - kondroid- és kordaszövet
- porcszövet: üvegporc
rugalmas-rostos porc
kollagén-rostos porc
- csontszerű szövetek
- csontszövet
3.2.1. Kötőszövetek
A kötőszöveteket a sejtes és sejt közötti állományuk összetétele, arányai alapján lehet
csoportosítani. A mezodermából, illetve az arról korán elkülönülő mezenchimasejtekből
különböző sejtek differenciálódnak. A mezenchimasejtek egy része (megtartva nagyfokú
differenciálódóképességét) a kifejlett állatok laza-rostos kötőszövetében is megtalálható.
A kötőszövetekben vannak rögzített (fixált) és szabad vagy vándorsejtek.
A helytülő, rögzített sejtek azt követően, hogy egy adott helyen megtelepedtek,
esetleg nyúlványokat növesztettek, általában már nem mozognak. A kezdetben mozgás-
ra képes fibroblast sejtekből képződő fibrociták (fibrocyta) rostokat szintetizálnak, részt
vesznek az alapállomány glikozamino-glikán anyagainak képzésében. A kollagén mole-
kulák az endoplazmatikus retikulumban képződnek, ezeket a Golgi-apparátus csoma-
golja, majd merokrin szekrécióval leadásra kerülnek. A kollagén molekulák a sejten kívül
rendeződnek rosttá. A másik két kötőszöveti rosttípus is hasonló módon képződik a
fibrociták révén. A vér monocitáiból (monocyta) származó hisztiociták (hystiocyta) nyugvó
vándorsejtek, mert gyulladásos folyamatok esetén állábakat növesztenek és mozognak,
valamint fagocitózissal mikroorganizmusokat fogyasztanak. Ez utóbbi képességük miatt
nevezik őket makrofág sejteknek is. Az immunrendszerben szerepet játszó retikulum-
sejtek (reticulocyta) is a rögzített sejtekhez tartoznak, de szükség esetén szintén képesek
amőboid mozgásra ésfagocitózisra. Azínsejtekazínszövetbenfordulnakelő. Azsírsejtek
számos kötőszövetben megtalálhatóak.
Aszabad vagy vándorsejtek közé tartoznak a plazmasejtek, melyek aB-limfocitákból
alakulnak ki. Fő feladatuk a gamma-globulin előállítása, amely a szervezet védekező
folyamataiban fontos. A hízósejtek heparint, hisztamint, szerotonint szintetizálnak. A
heparin a véralvadást gátolja. A hisztamin tágítja a vérereket, fokozza azokpermeábilitását,
míg a szerotonin szűkíti a kapillárisokat. A pigmentsejtek (melanocyták) a ganglion-
lécből származnak. Kezdetben mozgásra képesek, a szervezet számos pontjára elvándo-
rolnak, majd megtelepednek. Nagyobb számban fordulnak elő a hám alatti kötőszövet-
ben és a pigmentes kötőszövetben.
A kötőszövetek egyes típusaiban ezeken kívül előfordulhatnak idevándorolt fehérvérsej-
tek, melyek a humorális (antitestes) és sejtes immunitás kialakításában vesznek részt.
A fentiekben felsorolt sejtes elemek és rostok elsődlegesen a gerinces állatokra jellem-
zőek. Gerinctelen állatoknál más, speciális feladatot ellátó sejtek is előfordulhatnak.
89
3.9. ábra. Embrionális kötőszövetek
A: mezenchima, B: kocsonyás kötőszövet (Geiler és Törő nyomán kissé módosítva)
Mezenchima
A mezenchima (mesenchyma) az embrionális élet során alakul ki a mezodermából.
Ebből differenciálódnak a többi kötő- és támasztószövetek. A nagy mennyiségű folyadé-
kot tartalmazó sejt közötti állomány nyomása feszességet ad e szövetféleségnek. A sejtek
nyúlványaik révén képeznek hálózatot. A sejt közötti állomány rostokat nem tartalmaz
(3.9. ábra). Kifejlett állatokban önálló szövetként már nem fordul elő, de a mezenchima-
sejtek a kötőszövetekben még megtalálhatóak.
Kocsonyás kötőszövet
Warthon-féle kocsonyának is nevezik ezt az embrionális élet során kialakuló kötőszöve-
tet. A nyúlványos sejtek között gél állapotú alapállomány van, amiben már kollagén
rostok találhatók (3.9. ábra). Emlősállatokban a fejlődő embrió testében és a köldök-
zsinór alapállományában figyelhető meg.
3.10. ábra. A: laza-rostos kötőszövet, B: retikuláris kötőszövet (Törő és Krölling-Grau nyomán kissé módosítva)
90
Laza-rostos kötőszövet
Az állati szervezet leggyakoribb kötőszövet-félesége, amely szinte valamennyi szerv fel-
építésében részt vesz (3.10. ábra). Összeköti a különböző szöveteket, kitölti a hézagokat,
vázat alkot és fontos szerepe van a környező szövetek anyagforgalmában. Szinte vala-
mennyi kötőszöveti sejttípus előfordul benne. Az alapállományban kollagén, elasztikus
és rácsrostok vannak. Attól függően, hogy a szövet hol helyezkedik el, a sejttípusok és a
sejtközötti állomány aránya eltérő lehet. A bőralatti kötőszövetben például legnagyobb
arányban a fibroblasztok, hisztiociták, hízósejtek és fehérvérsejtek fordulnak elő. A ros-
toklaza hálózatot alkotnak, közülük a kollagénrostok dominálnak. Vérerekkel és idegek-
kel gazdagon ellátott.
Retikuláris kötőszövet
Nyúlványos sejtjei (úgynevezett re tikul ocyták) hálózatot képeznek, amelyet a retikuláris
rostok is megerősítenek (3.10. ábra). A rostokat a sejtek nyúlványai körülölelik. A vér-
képző szervek (vöröscsontvelő, lép, nyirokszervek) alapvázát alkotják, ahol a kötőszövet-
ben lévő szabad terekben foglalnak helyet a fejlődő vérsejtek. Sejtjei képesek arra, hogy
legömbölyödve vérsejtekké és nyiroksejtekké, valamint zsírsejtekké alakuljanak át.
Lemezes kötőszövet
Felépítésében elsősorban kollagén, kisebb arányban elasztikus rostok és sejtes elemek
vesznek részt. Mechanikai erőknek igen ellenálló szövetféleség. Ilyen kötőszövet képezi
például az ízületi tok külső rétegét, de megtalálható a bőr alatti kötőszövetben is.
Tömött-rostos kötőszövet
A tömött-rostos kötőszövetben a rostok dominálnak, a sejtek és az alapállomány aránya
kisebb. Aránylag kevés vérér fordul elő benne. Két formáját különböztetik meg, a rugal-
mas-rostos és a kollagén-rostos kötőszövetet.
A rugalmas-rostos kötőszövetben az elasztikus rostok adjákmeg a szövet tulajdon-
ságait. Olyan helyeken fordul elő a szervezetben, ahol a gyakori nyújtó hatások után
izommunka nélkül áll vissza az eredeti állapot. Ilyen rugalmas-rostos kötőszövet van a
tarkószalagban, a csigolyákat összekötő szalagokban, az artériák falában, a mirigyvég-
kamrák körül.
A kollagén-rostos kötőszövet tipikus
megjelenési formája az ín (tendo). Az insej-
tek (tendocyták), amelyek valójában fibro-
citák, a kötegekbe rendeződött, szorosan ál-
ló rostok között helyezkednek el (3.11. áb-
ra) . Alakjuk megnyúlt, keresztmetszetben ál-
talában „Y” vagy „X” alakúak. Az ínben más
sejttípus csak elvétve fordul elő. A rostnya-
lábok között még vérereket tartalmazó laza-
rostos kötőszövet fordul elő. Az int kívülről
kötőszöveti lemez burkolja. A csonthártyá-
hoz kapcsolódó inak az izmok segítő szervei,
szilárdan rögzítik azokat a csontokhoz.
Zsírszövet
A zsírszövetben legnagyobb mennyiségben
zsírsejtek vannak. Ezek a sejtek vagy a me-
zenchimából vagy retikulumsejtekből, hisz-
ti oci iákból, esedegfibroblasztokból alakulhat-
3.11. ábra. ínszövet felépítése. A: keresztmetszet-
ben, B: hosszmetszetben
ír; ínrost, is; ínsejt (Törő, valamint Grau és Bucher
nyomán módosítva)
91
nak ki. Szerepük van többek között a táp-
anyag-raktározásban, a mechanikai vé-
delemben és a hőszabályozásban. A zsír-
sejtek zsírfelhalmozási típusa és funkció-
ja alapján fehér és barna zsírszövetet lehet
elkülöníteni.
A fehér zsírszövetet alkotó sejtek-
ben a fokozatosan felhalmozódó zsir-
cseppek végül összeolvadva szinte telje-
sen kitöltik a belső teret, kiszorítva a sejt-
magot és egy kevés plazmát a sejt egyik
szélére (3.12. ábra). A szövettani prepa-
rátumok készítésekor alkalmazott zsír-
elvonó szerek hatására a készítmények-
ben a zsírsejtekben nagy üres teret lehet
látni. Ha a sejtmag is a metszési síkban
van, akkor a sejtek pecsétgyűrű alakot
mutatnak. Minden egyes zsírsejtet reti-
kuláris rostok hálózata vesz körül. A zsír-
szövet vérerekben gazdag. Kötőszövetes sövények tagolják lebenyekre. A zsírsejtek egye-
sével vagy kisebb csoportokban, számos kötőszövet-féleséghez társultan is előfordulnak.
Nagyobb kiterjedésű zsírpárnák jönnek létre például a bőr alatti kötőszövetben, az ujj-
pámákban, a vese körül, a tejmirigyben. A fehér zsírszövet sejtjeiben lévő, elsősorban
neutrális trigliceridekből álló zsír állandó lebontás és felépülés alatt áll. A szervezet
számára energiában gazdag tápanyagot szolgáltat, oxidatív lebontása során víz szabadul
fel, így ez a szövetféleség éhezés és szomjazás alkalmával fontos szerephez jut. A zsírszö-
vetnek hézagkitöltő, támasztó, formát és rugalmasságot adó, mechanikai védelmet és
hőszigetelést nyújtó szerepe is van. Az úgynevezett struktúr-zsírszövetek (például az ujj-
pámákban) még tartós éhezés esetén sem bomlanak le.
A barna zsírszövet kisebb sejtekből áll, melyek sejtmagja nem szorul ki a széli részre,
mert jóval kevesebb zsircsepp raktározódik a sejtben. A plazmában sok mitokondrium
fordul elő. Jelentős mennyiségű glikogént is tárolhat. A RÉS sejtjeiből átalakult sejteket
retikuláris rostok, vérerek veszik körül, idegvégződések kapcsolódnak hozzájuk. A barna
zsírszövet gazdag vérellátású, és elsődleges feladata a hőtermelés. Különösen jól fejlett
a téli álmot alvó állatoknál, ahol a zsírpárna a tarkó, a nyak és a lapocka tájékán helyez-
kedik el. A barna zsírszövetben a hőtermeléssel járó lebontási folyamatok felmelegítik az
átáramló vért. Ez fontos szerepet játszik a téli álomból ébredő állatok testhőmérsékleté-
nek emelésében.
3.2.2. Vér és egyéb testfolyadékok
A vér (sanguis) a zárt keringési rendszerben áramló testfolyadék. Nem tipikus kötőszö-
vet. Sejtjeit mezodermális eredetű vérsejtképző őssejtek osztódással hozzák létre. A ge-
rinces állatok esetében a vöröscsontvelőben és az egyedfejlődés korai szakaszában a
nyirokszervekben képződnek.
Gerinctelen állatok közül zárt keringési rendszerük van például a gyűrűférgeknek.
Vérükben amöbociták (amoebocyta), sőt a soksertéjű gyűrűférgekében vörösvértestekis
vannak. A rákok és a puhatestűek testfolyadékában fehérvérsejtek találhatók. A rovarok
testfolyadékában hemociták (haemocyta) fordulnak elő.
A gerinces állatok vére komplex folyékony szövet, amely vérplazmából és alakos elemek-
ből áll. A plazma mintegy kilencven százalékban vizet tartalmaz, amelyben többek között
92
fehérjék (például albumin, globulinok, fibrinogén), glükóz, aminosavak, szervetlen ionok,
enzimek, hormonok, vitaminok, anyagcsere-bomlástermékek és még számos más szervet-
len és szerves anyag található. A plazmában oxigén, szén-dioxid és nitrogéngázok is oldód-
nak. A gerincesekben a vér alakos elemeit a vörösvérsejtek (erythrocyták), a fehérvér-
sejtek (leukocyták) és a vérlemezkék (thrombocyták) alkotják (3.13. ábra). A vérben
főleg a vörösvértesteknek és a vérlemezkéknek van fontos szerepük, mig a fehérvérsejtek
elsősorban a kapillárisokat elhagyva (extravasculárisan) működnek. A vér alapvető felada-
tai közé tartozik a gázok, tápanyagok és bomlástermékek szállítása, az immunfolyamatokban
való részvétel, a szervezet sav-bázis egyensúlyának fenntartásában és a hőszabályozásban
való közreműködés. A vér mennyisége és összetétele fajtól függően eltérő, de az egyedek
között is változhat. E tekintetben különbség lehet például a hímek és a nőstények között,
változhat az életkor, a testméret, az egészségi állapot és az évszakok szerint is.
A vörösvérsejtek az emlősök kivételével ovális alakúak, sejtmagot tartalmaznak. Az
emlősök eritrocitája korong alakú és sejtmagot nem tartalmaz, ezért vörösvértest a neve.
A tevefélék vörösvértestjei ovális alakúak. Az eritrociták membránja a felületén lévő
3.13. ábra. Vérsejtek kialakulása és típusai (egyszerűsített vázlat)
Bl: B-limfocita, eb: eritroblaszt, er: eritrocita, e^: sejtmagvas eritrocita, er2: sejtmag nélküli eritrocita, ív: fehérvérsejtek, gr:
granulocíta, grt: neutrofil granulocita, gr2: eozinofil granulocita, gr3: bazofil granulocita, hp: hemopoietikus őssejt, K: limfoid
őssejt, mb: mieloblaszt, mc: mielocita, me: megakarioblaszt, mi: mieloid őssejt, mk: megakariocita, mo: monocita, TI: T-
limfocíta, tr: trombocita (Reman, Storch, Welsch nyomán módosítva)
tr
ív
93
különféle glikoproteidek révén a vércsoport-tulajdonságokat alakítja ki. A vércsoport-
rendszerek közül legismertebb az emberi ABO vércsoport és az Rh-faktor előfordu-
lása. A vörösvérsejtek feladata az oxigén szállítása, amit hemoglobintartalmuk tesz lehe-
tővé. A vörösvérsejtek nagysága a gerinces állatok között jelentős eltéréseket mutathat.
A halak ellipszoid alakú vörösvérsejtjei például átlagosan 13x9 pm méretűek, az em-
lősök korong alakú vörösvértestei 6-7 pm átmérőjűek. Egy köbmilliméter vérben az
eritrociták száma halaknál 1-2 millió, az emlősöknél 4-8 millió között mozog. Az eritrociták
a vörös-csontvelőben keletkeznek az eritroblaszt (erythroblast) sejtekből. A sejtek soro-
zatosan osztódnak, közben megtörténik a hemoglobin szintézise. Az emlősállatokban a
vörösvérsejt fejlődése során kettéválik. A sejtmembránnal körülvett sejtmagot tartalma-
zó részt fagociták emésztik meg. A mag nélküli részből alakul ki a vörösvértest, amely
végül minden organellumát lebontja. A plazmamembránnal határolt teret speciális cito-
plazma-állomány (stroma) tölti ki.
A színtelen fehérvérsejtek a vörösvérsejtekhez viszonyítva jóval kisebb számban for-
dulnak elő. A legtöbb gerincesnél egy köbmilliméter vérben a számuk 10 000 alatti, ma-
daraknál közel 20 000. A fehérvérsejtek között megkülönböztetünk agranulocita típusú
limfocitákat (lymphocyta) és monocitákat (monocyta), valamint granulocitákat (granulocyta).
A limfocitáknak nagy, 5-7 pm átmérőjű, kerekded sejtmagjuk van. A nyirokszervekben
válnak éretté, innen kerülnek ki a véráramba, és a szervezet immunrendszerében töltenekbe
fontos szerepet. Alakilag megkülönböztethető kis és nagy limfocita. A nagy limfocita korai
fejlődési állapotú (sejtmagja körül vastagabb plazmaszegély figyelhető meg, mint a kis
limfocitánál), a kis limfocita érettebb. A differenciálódás során belőlük T és B sejtek alakul-
nak ki, amelyek szerepe sokrétű. A monociták nagyméretűek, emlősöknél elérhetik a 20 pm
átmérőt is. Sejtmagjuk bab vagy patkó alakú, a sejt egyik felére tolódva helyeződik el.
Fagocitózisra képesek, bekebelezik a kórokozó baktériumokat. A vérereket elhagyva makrofág
sejtekké alakulnak. Képződési helyük elsősorban a vöröscsontvelő és a nyirokszervek. Élet-
tartamuk az emlősöknél 4-5 nap. A vöröscsontvelőben képződő granulociták sejtmagja
karéjozott, lebenyezett, plazmájuk pedig szemcsézett. Átmérőjük 10-15 pm között változik.
A granulumok festődése alapján meg lehet különböztetni neutrofil, bazofil és acidofil
granulocitákat. A neutrofil granulociták sejtmagja több lebenyre tagolt, plazmájukban a
szemcsék kisméretűek. Élénk amőboid mozgásra és fagocitózisra képesek. Ez utóbbi képes-
ségük miatt mikrofágoknak is nevezik őket. Igen fontos szerepet töltenek be a szervezet
védekező folyamataiban. A bazofil granulociták plazmájában nagyméretű szemcsék vannak.
A sejtekben heparin és hisztamin található, amit a sejtek a sérült, vagy gyulladt helyeken
leadnak. A heparin részint gyulladásgátló, részint kemotaktikusan vonzza például a makro-
fágokat. Mivel a hisztamin növeli az erek permeábilitását, ezért a sérült szövetrészekbe több
neutrofil granulocita tud kijutni. A bazofil granulociták élettartama mindössze néhány nap.
Az acidofil granulociták plazmájában nagyméretű szemcsék vannak, a sejtmag általában
kétlebenyű. Amőboid mozgásra és fagocitózisra képesek.
A vérlemezkék az emlősökben nem tartalmaznak sejtmagot, míg a többi gerinces
állatban igen. Utóbbi esetben a sejtek ovális, orsó vagy gömbölyded alakúak. Emlősöknél
a vöröscsontvelő óriássejtjeiből (megakaryocyta) keletkeznek, azok citoplazmájának fel-
darabolódása során. Méretük emlősöknél 1-4 pm közötti. A trombocitáknak a véralva-
dás folyamatában van szerepük.
A vér alakos elemeinek képződése (haemopoiesis) elsősorban a vöröscsontvelőben
és a nyirokszervekben történik. A vöröscsontvelő a kétéltűektől kezdve tölt be kiemelke-
dő szerepet. Az említetteken kívül a máj, a lép és más szervek is részt vesznek a vérsejtek
előállításában. A vöröscsontvelőben a nagy differenciálódási képességgel rendelkező
pluripotens őssejtek differenciálódása két fő irányban indul meg (3.13. ábra). Elkülönül-
nek a mieloid sejtek és a limfoid őssejtek. A mieloid őssejtekből alakulnak ki a mieloblaszt
(myeloblast), az eritroblaszt (erythroblast) sejtek, valamint a megakariociták előfutárai
94
(megakaryoblast). A mieloblasztokból a mielociták, míg az eritroblasztokból az eritrociták
alakulnak ki. A mielociták a granulociták, illetve a monociták közös előfutárai. A limfoblaszt
sejtekből alakulnak ki a T- és B-limfociták különböző formái. A limfociták és a monociták
a nyirokszervekben differenciálódnak.
A nyirok (lympha) sejt közötti folyadék, szövetnedv, amely a nyirokkapillárisokba
szivárog. Sejtes állományát leukociták, túlnyomórészt limfociták alkotják. Plazmájában
fibrinogén található. A nyirokrendszert az ereken kívül még a nyirokszervek építik fel.
A nyirokfolyadéknak szerepe van az anyagszállításban, így az anyagcserében, valamint
a limfociták révén a kórokozók elleni védelemben (4.3.2. fejezet).
Az ízületi nedv, szinovia (synovia) az ízületi üregeket, ínhüvelyeket kitöltő folyadék,
amelyben fehérvérsejtek, levált kötőszöveti sejtek fordulnak elő.
Az agyi-gerincvelői folyadék (liquor cerebrospinalis) az agykamrát és a gerincvelő
központi csatornájának üregét tölti ki. Kevés fehérje, fehérvérsejt és levált ependimasejt
lehet benne.
A savós testüregek folyadéka kevés fehérjét, fehérvérsejtet és levált mezotélium-
sejtet tartalmaz.
3.2.3. Támasztószövetek
A támasztószövetek intercelluláris állománya a kötőszövetekhez képest sokkal szilár-
dabb, de gyakran igen rugalmas is.
Kondroidszövet
A kondroid- (chondroid) szövet egyes gyűrűsférgekben, rákokban és a csigákban fordul
elő. Celluláris állományát összetömörült hólyagos kötőszöveti sejtek alkotják, míg az inter-
celluláris térben kollagén rostok vannak. Nyomásszilárdsága a kordaszövetnél kisebb. A
kondroidszövet a csigák reszelőnyelvében rugal-
mas pamat kepez, illetve a testükben különböző
szöveteket választ el egymástól.
Kordaszövet
Előfordul az előgerinchúrosoknál, a fejgerinchú-
rosoknál, valamint a gerinces állatok egyedfejlő-
désének kezdeti szakaszában. Nyomással szem-
ben ellenálló szövetféleség. A kordaszövetből álló
gerinchúr (chorda dorsalis) felépítése a gerinchú-
rosoknál elég változatos képet mutat. Gerince-
sekben a gerinchúrt felépítő kordaszövetet kolla-
gén és elasztikus rostokból álló hüvely veszi körül.
A felszíni rétegében lévő kordoblaszt (chordoblast)
sejtek hozzák létre a kordasejteket, amelyekben
vakuolumok képződnek. Az egymással dezmo-
szómákkal kapcsolódó, egymásnak feszülő folya-
dékkal telt sejtek nyomása és a külső kordahüvely
alakítja ki a rugalmas gerinchúrt (3.14. ábra).
Porcszövet
A porcszövet (tela cartilaginea) támasztószere-
pét a sejtközötti állomány biztosítja, amelynek
minősége megszabja a porcszövet tulajdonságait.
A porcszövet a mezoderma mezenchima sejtjei-
3.14. ábra. A kordaszövet felépítése (részlet az
ingolák gerinchúrjának keresztmetszetéből)
fc: fíbrocita, kb: kordoblaszt sejt, ko: kollagén rostok,
ks: kordasejtek nagy sejten belüli üreggel (Welsch-
Storch nyomán kissé módosítva)
95
3.15. ábra. A hialin porc felépítése
aá: alapállomány, nz: növekedési zóna, pe: porc-
egység, ph: porchártya, ps: porcsejt, pt: porctok, pu:
belső és külső porcudvar (Több szerző nyomán mó-
dosítva)
bői differenciálódik. A mezenchimasejtek porc-
képző (chondroblast) sejtekké alakulnak, kiala-
kítják a porc alapállományát, miközben porc-
sejtekké (chondrocyta) differenciálódnak. A
porcszövet alapállománya fehérjéket (kollagént,
albuminoid anyagokat), glikózamino-glikánokat
(például kondroitin-szulfátot), valamint kolla-
gén- és elasztikus rostokat tartalmaz.
A sejtek az alapállományba ágyazva egye-
sével vagy többesével foglalnak helyet. A porc-
sejtek porcsejtüregecskékben ülnek, melyet kol-
lagén rostokból álló tok határol. A rostok a por-
csejtcsoportokat is határolják. A sejtcsoporto-
kat glikozamino-glikán és proteoglikán-tartalmú
belső és külső porcudvarok veszik körül. így
porcegységek, kondronok (chondron) alakul-
nak ki az alapállományban. Az előbbiekben leírt
kondronszerkezet tipikusan az üvegporcra jel-
lemző (3.15. ábra).
A porcszövetet kívülről porchártya (peri-
chondrium) borítja, amely nagy mennyiségben
tartalmaz kollagén rostokat. Külső rétege a laza-rostos kötőszövet felé képez átmenetet,
míg a belső réteg rostjai a porc rosthálózatába kapcsolódnak be. Feladata a porc mecha-
nikai védelme, táplálása, valamint a porszövettel határos felszínén ülő porcképző sejtek
révén a porc állományának gyarapítása. Mivel a porc belsejében vérerek nincsenek, a
sejtek diffúzió útján veszik fel a táplálékot a porchártya felől. Ahol a porcréteget porchár-
tya nem borítja, mint az ízületeknél, ott a tápanyagok az ízületi nedv révén jutnak el a
porcszövethez. A porc regenerálódóképessége nem jó, idősebb korban pedig lebontó és
degenerativ folyamatok indulhatnak meg benne. Ilyen például a porc elmeszesedése
vagy rostrendszerének kedveződen hatású tömörülése. A porcszövettipusokat felépíté-
sük, a mikroszkópi vizsgálatok során mutatkozó jellegzetességeik és betöltött szerepük
alapján lehet csoportosítani.
Az üvegporc (hyalin porc) alapállománya homogénnek látszik, mert a glikozamino-
glikánok, a kondroitin-szulfát átitatják a kollagén rostokat, így azok csak speciális vizsgálati
módszerekkel mutatha-
tóak ki. Általában 2-5 sejt
helyezkedik el egy kond-
ronban (3.15. ábra). Ez a
porcszövet fordul elő pél-
dául a porcoshalak belső
vázában, fejlettebb gerin-
cesekben a csontok ízületi
felszínén, a légcsőben és az
orrsövényben, valamint a
bordavégeken.
A rugalmas-rostos
porc alapállományában a
kollagén rostokon kívül
hálózatot alkotó rugalmas
rostok is vannak (3.16. áb-
ra). Egy, legfeljebb két
3.16. ábra. A: rugalmas-rostos porc, B:
(Több szerző nyomán módosítva)
kollagén-rostos porc
96
porcsejt található együtt. Ez a porcféleség rugalmas alakváltozást tesz lehetővé. Emlő-
sökben fordul elő például a fülkagylóban és a gége-fedőben.
A kollagén-rostos porcban a nagy mennyiségben előforduló és az erőhatásoknak
megfelelően rendeződött kollagén rostkötegek között a porcsejtek többnyire egyesével
helyezkednek el (3.16. ábra). Nagy mechanikai igénybe vételnek kitett helyeken előfor-
duló szövet, amely tipikus formában található meg például a csigolyák közötti porcko-
rongban és a pataporcban.
Csontszerű szövetek
A dentin felépítése nagyon hasonlit a csontéhoz, bár szerveden állománya jóval magasabb
(75%) és ezért sokkal szilárdabb. A dentin belső állománya nem tartalmaz sejteket vagy
nagyobb üregeket. A képzésben részt vevő odontoblaszt (odontoblast) sejtek a leendő
dentinállomány belső határán helyezkednek el és csak nyúlványaik hatolnak be az állo-
mányba. Az általuklétrehozottcsatomácskákban vérerek és idegekfutnak. Az odontoblaszt
sejtek kollagén rostokat képeznek az alapállomány felé, amely apatitkristályokkal való
impregnálás útján megkeményedik. A sejtek a felszínhez közel maradnak, egymással kis
csatomacskak ré-
vén nem létesíte-
nek kapcsolatot, de
nyúlványaik beha-
tolnak a dentin bel-
ső állományába
(3.17. ábra). A
dentin a fog zo-
máncrétege és a
pulpaüreg között
található. Az ele-
fántok agyarában
(„elefántcsont”)
nagy mennyiség-
ben található den-
tin. Megtalálható a
porcoshalakpikke-
lyében is, amelynek
felépítése a gerin-
cesek fogáéval ho-
A B
3.17. ábra. A: dentinállomány, B: csontszövet kialakulása
(1: első fázis, 2: második fázis) ck: csontszövet kiindulási állománya, cs: csontszövet alapállománya,
da: dentin alapállománya, kf: kiindulási felszín, ny: sejtnyúlvány, ob: oszteoblaszt sejt, od:
odontoblaszt sejt, pu: pulpa (Currey nyomán erősen módosítva)
mológ.
A gerincesek fogának cementállománya a csontszövethez nagyon hasonló felépíté-
sű, de a Havers-féle csatornák legfeljebb idős korban alakulnak ki benne.
A csontoshalak többségének pikkelyeit csontszerű szövet alkotja. Képzését csontépí-
tő oszteoblaszt (osteoblast) sejtek végzik. Ezek a pikkelytasak falában ülő sejtek változó
intenzitással termelik a pikkely kemény állományát, de abba nem épülnek be. A pikkely
csontszerű anyagának képzése elsődlegesen a széli részen megy végbe.
A halak egy részének (például tokfélék) a pikkelyeit részben ganoin alkotja. A ganoin
elmeszesedő kollagén rostnyalábokból áll.
Csontszövet
A csontszövet (tela ossea) a csontszerű szövetekhez hasonlóan igen kemény szövetféle-
ség. A csontok fő tömegét alkotja. A csontsejtek, oszteociták (osteocyták) üregecskék-
ben ülnek és csatomácskákban futó nyúlványaik révén kapcsolatot létesítenek egymással
(3.18. ábra). A csontsejtek hozzák létre az intercelluláris állományt és mobilizálj ák annak
97
3.18. ábra. A: csontszövet felépítése egy csövescsont (B) kompakt állományában
cs: csontsejt a csontsejtüregben, eh: csonthártya, He: Havers-féle csatorna, Hl: Havers-féle lemezrendszer, id: ideg, ká: külső
általános lemezrendszer, ki: különböző irányban futó rostok rétegei, ko: kollagén rost apatitkristályokkal körülvéve, öl-
összekötő lemezrendszer, sz: szivacsos állomány, tá; tömött kéregállomány, Ve: Volkmann-féle csatorna, vé: vérér (Becker-
Hoegen és Pfeffer, Miller, Törö és más szerzők nyomán összeállítva)
98
ásványi anyagait. A sejtközötti állomány adja a csont jellegzetes tulajdonságait, a szilárd-
ságot, keménységet, rugalmasságot. Szerves anyagai: a kollagén fehérjék, a glikoproteidek
és a citrátok a szárazanyag 30-35%-át alkotják. A 65-70%-ot kitevő szerveden anyagok
közül fontosabbak a kalcium- és magnézium-foszfát, a kalcium-karbonát, a magnézium és
a kálium sói, valamint a fluoridok. A főleg kalcium- és foszfáttartalmú hidroxiapatit, va-
lamint a fluort is tartalmazó fluoro-apatit kristályok a kollagén rostok mentén, felületén
helyezkednek el. Térrácsaikba számos kation és anion épül be.
A rostos vagy fonadékos csontszövet a csontoshalak és a kétéltűek csontos vázát építi
fel. A nyúlványos, orsó alakú csontsejtek körül a kollagén rostok párhuzamos vagy csavart
helyzetben állnak. A sejtek és a rostok nem képeznek lemezrendszereket. A hüllők, a
madarak és az emlősök fejlődésének embrionális szakaszában is megjelenik ez a típusú
csontszövet, azonban később átalakul lemezes szerkezetű csontszövetté.
A lemezes szerkezetű csont tömött kéregállományában (substantia compacta) a
szövet jellegzetes szerkezetet mutat, lemezrendszereket alakít ki (3.18. ábra). A csont
hossztengelyével párhuzamosan futó, keresztmetszetben kör alakú Havers-féle csatornák
körül a sejtek koncentrikus gyűrűkbe rendeződnek. Az egymást követő rétegekben a spi-
rális lefutású kollagén rostok mintegy 90 fokban keresztezik egymást. Az így kialakult,
jellegzetes formát csontegységnek (osteon) nevezik. Elsősorban a statikailag jobban igénybe
vett csontok mutatják az említett tipikus szerkezetet. A Havers-féle csatornákat haránt
és ferde irányban haladó Volkmann-féle csatornák kötik össze. Ezekben a csatornákban
futnak a csontsejtek táplálását szolgáló vérerek és az idegek, melyek a csonthártyából
ágaznak le. A Havers-féle csatornák körül kialakult speciális lemezrendszert (laminae
speciales) a köztes lemezrendszerek (laminae intercalares) kötik össze. Ez utóbbiakban
a sejtek nem mutatnak gyűrűszerű elrendeződést. A csont kompakt állományát a felszí-
nen és csöves csont esetén a velőüreg felé eső oldalán külső és belső alap lem ezek határolják.
Ezek szerkezete réteges. A csont szivacsos állományát (substantia spongiosa) csontge-
rendák és lemezek épitik fel, melyek köztes tereit a vöröscsontvelő tölti ki. A csont lemez-
rendszerei az egyedfejlődés során a megváltozott erőviszonyok hatására átépülnek. A cson-
tot kívülről csonthártya (periosteum) borítja. Legbelül a csontsejtképző és mezenchima-
sejtek ülnek (stratum osteoblasticum), középen kollagén és elasztikus rostokban, vérerek-
ben gazdag, kötőszövetes réteg (stratum fibroelasticum), míg kívül erősen rostos és vérerek-
ben igen gazdag réteg (stratum fibrovasculosum) található. Csöves csontoknál a velőüre-
get csontbelhártya (endosteum) határolja. A csontosodás (ossificatio) végbemehet nem
valódi és valódi módon.
A nem valódi (metaplasztikus) csontosodás folyamán a kötőszövetbe vagy porcszö-
vetbe szervetlen sók épülnek be, így az megkeményedik. így következik be például az inak
elcsontosodása egyes madarak lábában.
A valódi (neoplasztikus) csontosodás során a kiindulási szövet átalakulása révén
jön létre csontszövet. A valódi csontosodásnak két formája különböztethető meg. El-
sődleges (primer) csontosodás során a csontszövet a mezenchimából közvetlenül
alakul ki. Nem gyakori csontképződési forma. A koponyacsontok varratainál vagy az el-
tört csont érintkező felei közötti szűk térben figyelhető meg. Ezeken a helyeken a kötő-
szöveti rostrendszerek előzetes kialakulására, majd későbbi elcsontosodására nincs mód.
A mezenchimasejtek közvetlenül csontsejtekké alakulnak át és termelik a csont alapál-
lományát. A másodlagos (secunder) csontosodás során egy már meglévő szövetfé-
leség fokozatos átépülése zajlik le. Egyik típusa a dezmális (desmalis) csontosodás, ami-
kor a mezenchimasejtek a bőséges vérellátású, kollagén rostokban gazdag kötőszövetes
telepekben (például kötőszöveti hártyákban) csontosodási magokat alakítanak ki. Átala-
kulnak oszteoblaszt, azaz csontképző sejtekké, melyek létrehozzák a még el nem mesze-
sedéit csontszerű alapállományt (osteoid szövet). Az ásványi anyagok lerakódásával
párhuzamosan az osteoblasztsejtek csontsejtekké, oszteocitákká fejlődnek. A csontképző
99
A
B
oszteoblaszt és a csontfaló, oszteoklaszt (osteoclast)
sejtek együttes tevékenysége révén az erőhatásoknak
megfelelően kialakul a csont szerkezete. így jönnek
létre a bőrcsontok (például a hüllőknél). Ez a csonto-
sodási forma jellemző egyes koponyacsontok fejlődé-
sének kezdeti szakaszára és egyéb laposcsontok kiala-
kulására is. Akondrális csontosodásnál a csontszövet
másodlagosan, közvetve alakul ki a mezenchimából
létrejöttporctelep helyén (3.19. ábra). A csontoktöbb-
sége ilyen módon alakul ki. A csöves csontoknál a fo-
lyamat először a középső szakaszon (diaphysis), a porc
felszínén, perichondrálisan kezdődik, majd ezt
követően a porc belső állományában, enchondrálisan
is folytatódik. A porctelep felszínét boritó porchártya
csonthártyává alakul, a belső rétegében lévő oszteo-
blasztok fokozatosan vastagítják a felszíni réteget. így
a perikondrális csontosodás révén a diafízis területén
vastagabb, a csontvégek (epiphysis) felé közeledve fo-
kozatosan vékonyodó csontszövetréteg alakul ki. Ez a
réteg alkotja majd a csont külső általános lemezrend-
szerét. A folyamat a porcfelszínt borító első csontos
réteg kialakulását követően már nem tekinthető valódi
kondrális csontosodásnak. A perikondrális csontoso-
dás az embrionálisán már kialakult csontok vastago-
dásában játszik szerepet. A külső rétegben zajló folya-
mattal párhuzamosan a csonthártyából vérerek, velük
együtt kötőszöveti sejtek tömekbe az időközben elme-
szesedett alapállományú porc belső terébe. A bejutott
mezenchimasejtekporcfaló (chondroclast), csontkép-
ző, csontfaló és a későbbiekben kialakuló velőüregben
funkcionáló vérsejtképző sejtekké alakulnak. Enkond-
rálisan, a porc belsejében porcfaló sejtek az alapállo-
mányt és a sejteket enzimatikusan bontják, míg a meg-
maradt, elmeszesedettporcgerendákfelszínéna csont-
3.19. ábra. A kondrális csontosodás
folyamata (A: korai, B: későbbi állapot)
áp: átépülő porcállomány, eh: csonthártya,
cü: csontüreg, di: diafízis, en: enkondrális
csontszövetképződés, ef: epifízis, ep: epifízis
porc, ic; irányító csontgerendák, ip: irányító
porcgerendák, pk: perikondrális csontvasta-
godás, po: porckorong, vé: vérér (Wumbach
nyomán erősen módosítva)
építő sejtek az oszteoid szövetet, majd a csontszövetet
hozzák létre. A dialízisben a csontfaló sejtek a képző-
dött csontgerendákat is lebontják, kialakítják a velő-
üreget. Az epifízis csontosodása hasonló módon megy
végbe az irányító porcgerendák mentén, de az ízületi
felszínig nem terjed ki. Ott megmarad az üvegporc-
állomány. Az epifízis és a diafízis határán fajonként
eltérő életkorigporckorong marad fenn és intenzívporc-
képzés révén teszi lehetővé a csont hosszanti növekedését. A porcsejtoszlopok közötti
elmeszesedett gerendákra csontképző sejtek telepszenek. A porcsejteket a porcfaló sejtek
fokozatosan lebontják. A csontosodással párhuzamosan vagy azt követően zajlik a csont
szerkezetének kialakulása. Az embrionális életben először a filogenetikailag is ősibb rostos
vagy fonadékos csontszövet jön létre, amelynek elmeszesedett alapállományában kollagén
rostok hálózata és a csontsejtek találhatók. A fonadékos csontszövet a csontfaló és -képző
sejtek összehangolt munkájának eredményeként átalakul lemezes szerkezetűvé.
A csontképzés és a csont szerkezetének kialakulása, folyamatos átépülése a belső
elválasztású mirigyek szabályozása alatt áll.
100
3.3. Izomszövetek
Az izomszövet mezodermális eredetű, kivéve a gerincesek szemének belső izmait, amelyek
ektodermálisak. Az izomszövet az állati szervezetben a zsigeri szervek működésében, a test
helyzet- és helyváltoztatásában játszik fontos szerepet. Legjellemzőbb tulajdonsága az
összehúzódó képesség, de rugalmassága és ingerlékenysége is fontos a működésében. A
nagy mennyiségű izomzat elhelyezkedése meghatározza az állat alakját is. Egyes gerinctelen
állatcsoportokban előforduló speciális izmok jellemzése a sejttani fejezetben található.
3.3.1. Simaizomszövet
A simaizomsejtek egyesével, kisebb
csoportokban kötőszövetbe ágyazva
vagy szövetet alkotva fordulhatnak
elő. Szöveti kötelékben a sejtek szo-
rosan kapcsolódnak egymáshoz, ré-
tegeket, lemezeket, nyalábokat, kö-
tegeket alkotnak az adott szervben
betöltött funkciónak megfelelően
(3.20. ábra). A sejteket retikuláris ros-
tok hálózata borítja. A sejtek közötti
kötőszövetben kollagén és elasztikus
rostokvannak, valamint vérerek, nyi-
rokerek és idegek futnak. A sima-
izomzat vegetatív beidegzés alatt áll,
főleg akarattól függetlenül működik.
Bár beidegzést nem minden izom-
sejt kap, a közöttük lévő nyitott kap-
csolódások révén az ingerület gyor-
san terjed a sejtek között. A gerinces
állatok zsigeri szervein kívül a tipikus
simaizomszövet számos gerinctelen
állatban is megfigyelhető, mint pél-
dául a laposférgekben.
3.20. ábra. Simaizomszövet felépítése. A: hosszmetszetben, B:
keresztmetszetben
si: simaizomsejt, sm: sejtmag, sö: kötöszöveti sövény (Voss nyomán
módosítva)
3.3.2. Harántcsíkolt izomszövetek
A harántcsíkolt izomrostokban lévő miofibrillumok felépitésének és működési elvének
leírása az 1.6. fejezetben található meg. Harántcsíkolt izomszövet a vázizom és a szív-
izom.
V ázizoms zö vet
A vázizomszövet előfordul például a fonálférgek nyelőcsövében, az ízeltlábúak és a
gerincesek testében, ahol az utóbbiaknál a test mozgatásának szolgálatában áll. Ez az
izomszövetféleség tartós működésre nem, de nagy erőkifejtésre képes. Gerinces állatok-
nál sokmagvú rostokból épül fel (3.21. ábra). A sejtmagok az emlősökben az embrionális
kor után a szarkolemma alatt, a rost perifériáján helyezkednek el, míg a többi gerincesben
a rost belső területén. A rostok átmérője 15-150 pm között mozog. A halak harántcsíkolt
izomrostjai a legnagyobb átmérőjűek, ezt követik a kétéltűek, a hüllők és az emlősök
izomrostjai. A legkisebb átmérő a madarak harántcsíkolt izomrostjain mérhető. A rostok
101
3.21. ábra. Harántcsíkolt vázizomszövet felépítése. A: hosszmetszet-
ben, B: keresztmetszetben, C: egy rost keresztmetszete
em: endomízium, kő: kötőszövet, mf: miofibrillumok, sl: szarkolemma,
sm: sejtmag, sp: szarkoplazma, vé: vérér (Voss és Törő nyomán módosítva)
lizmusa révén állítják elő.
körüli kötőszöveti burokban, az
endomiziumban (endomysium)
erek és idegek futnak. Több ros-
tot együtt a perimizium (perimy-
sium) köti össze nyalábbá. A nya-
lábokat epimizium (epimysium)
kapcsolja össze izomköteggé. Ha
az izomrostban a szarkoplazma
aránya kisebb mint a miofibril-
lumoké, akkor fehér izomrostról,
hanagyobb, akkorvörösizomrost-
ról van szó. A fehér izomrostok
a gyors mozgások, míg a vörösek a
lassú mozgások kivitelezésében j al-
szanak szerepet. A két rosttípus a
mozgás energiaszükségletének biz-
tosításában is eltér egymástól. A
gyors összehúzódásra képes fehér
izomrostok az ATP-t glikolízisből
nyerik. A lassúbb összehúzódás-
ra képes vörös izomrostokban
nagyobb számban található mi-
tokondrium és oxigén megköté-
sére képes pigmentanyag, a mioglobin. Az ATP-t a glükóz és a zsírsavak aerob katabo-
Szívizomszövet
A szívizomszövet szintén ha-
rántcsíkolt, de felépítése több vo-
násban különbözik a vázizométól.
A vázizomtól eltérően sejtes szer-
kezetű (3.22. ábra). Az ovális sejt-
mag a sejt közepén helyezkedik el.
A sejtekből felépülő rostok elága-
zóak. A sejtek találkozási helyei-
ken Eberth-féle vonal (1.6. feje-
zet) látszik. Ezen a területen az
érintkező sejtek dezmoszómák és
réskapcsolódások segítségével lé-
tesítenek összeköttetést egymás-
sal. Az utóbbi kapcsolódási for-
ma az ingerület igen gyors terje-
dését teszi lehetővé a szívizomsej-
tek között.
A szívizom hosszú ideig képes
kitartóan működni. Az ingerület-
vezetést emlősökben speciális, mó-
dosult szívizomrostok (Purkinje-
féle rostok) végzik. A sejtek körüli
kötőszövetben gazdag kapilláris-
hálózat van.
3.22. ábra. Szívizomszövet, A: hosszmetszetben, B: keresztmet-
szetben
Ev: Eberth-féle vonal, kö: kötőszövet, sm: sejtmag (Voss nyomán
módosítva)
102
3.4. Idegszövet
Az állati szervezet valamennyi sejtje képes ingerek felvételére, azonban az idegszövetet
alkotó idegsejtek különösen alkalmasak ingerfelvételre, annak továbbítására és feldolgo-
zására. Az idegszövet speciális elemei az idegsejtek és a gliasejtek. Nem specifikus elemei
a kötőszövetbe ágyazott vérerek. Az ideg-
sejtek felépítését és működését a sejttani
fejezetben már jórészt bemutattuk, így itt
csak az eddig nem ismertetett jellemzők-
re térünk ki. Az idegsejtek speciális funk-
ciójuknak megfelelően eltérő alakúak le-
hetnek, nyúlványaik száma és elágazódá-
suk különbözhet egymástól (3.23. ábra).
Unipoláris, tehát mindössze egy axon-
nal rendelkező idegsejtek főleg a gerinc-
telenekben fordulnak elő, de ritkábban
megtalálhatóak a gerincesekben is, pél-
dául a szaglóhámban és a retinában. Az
érző agyidegek ganglionjaiban, csigolyák
közötti ganglionokban előforduló pszeu-
dounipoláris idegsejtek axonja az ere-
dés után rögtön elágazik. A bipoláris
idegsejteknek tipikus esetben két axonja
van. Többnyire a receptorsejtek és az el-
vezető ideg között létesítenek kapcsola-
tot. Megfigyelhető például a retinában,
ahol intermedier neuron szerepét töltik
be. A csap- és a pálcikasejtektől veszik át
az ingerületet és a központ felé továbbít-
ják azt. Amultípoláris idegsejteknek egy
axonja van és számos dendritje. Ilyenek
például a gerincvelőben lévő motorikus
idegsejtek vagy a kisagyban lévő Purkinj e-
sejtek. A Golgi-I. típusú multipoláris ideg-
sejteknek hosszú, kevéssé elágazó, míg a
Golgi-II. típusúaknak rövid, gazdagon
3.23. ábra. Gyakoribb idegsejttípusok (az ábrán a dendritek
folyamatos, az axonok pontozott vonallal vannak ábrázolva)
bi: bipoláris idegsejt, cs: csapsejt, G-I.: Golgi-I. típusú multi-
poláris idegsejt, G-II.: Golgi-II. típusú multípoláris idegsejt,
pá: pálcíkasejt, pi: piramíssejt, Pu: Purkínje-féle idegsejt, sz:
szemcsesejt (Törő és más szerzők nyomán módosítva)
elágazó axonjuk van. A központi idegrendszerben a legnagyobb mennyiségben a Golgi-
I. típusú idegsejtek fordulnak elő. A Golgi-II. típusú sejtek is a központi idegrendszer
felépítésében vesznek részt, de rövidebb nyúlványaik nem hagyják el azt. A piramissejtek
a nagyagykéreg jellemző sejtjei, axonjuk közvetlenül a végrehajtó izomhoz fut. A három-
szög alakú sejtek leghegyesebb csúcsa a kéreg felszíne felé néz. A kisagyban található
Purkinje-sejt multipoláris neuron, amelynek jellegzetesen elágazó dendritnyúlványa van,
míg az axon igen vékony. A gömb alakú szemcsesejtek is a kisagyban fordulnak elő. Dend-
ritnyúlványaik minden irányban kiindulnak, az axon a felszín felé halad, majd két hori-
zontálisan futó ágra válik szét.
A környéki, perifériás idegrendszerben a Schwann-sejtek (lemnocyta) a központi
idegrendszerben az oligodendroglia-sejtek velős (myelin) hüvelyt alakítanak ki (3.24.
ábra). Ilyenkor ezeknek a sejteknek a membránja sokszorosan, akár 50-100 rétegben is
feltekeredik az axon körül. A feltekeredő, egymáshoz tapadó sejtmembránrétegek alkot-
ják a velős hüvelyt. Az oligodendroglia-sejtek nyúlványai a Schwann-sejtektől eltérően
103
3.24. ábra. A neuritnyúlvány körül kialakuló velős hüvelyek felépítése. A: oligodendroglia-sejtek által kialakított
velőshüvely, B: Schwann-sejtek által kialakított velős hüvely, C: velőhüvely nélküli axonok
ax; axon, bl: bazális lamella, cs: csupasz axon, is: idegsejt, mi: mielinhüvely, ol: oligodendroglia-sejt, Rb: Ranvier-féle
befüzödés, Se: Schwann-sejt, sm: sejtmag, vf: axonvégfácska (Voss és más szerzők nyomán módosítva)
104
egyszerre több axon szigetelésében is részt vehetnek. A hüvely nem folytonos, a gliasejtek
találkozásánál az axon 1-2 milliméterenként kis távon csupasz (Ranvier-féle befűződés).
Itt mehetnek végbe az elektromos potenciálváltozások. Az elektromos impulzus gyor-
sabban, ugrálva terjed. Ha a Schwann-sejtek vagy a központi idegrendszerben az oli-
godendroglia-sejtek csak egy rétegben veszik körül az axont, akkor velőhüvely nélküli
axonról beszélünk. A velőhüvely nélküli szigetelésnek egy változata az, amikor több
axon ágyazódik be egy Schwann-sejt plazmájába. Az így kialakult szürke (Remák-féle)
rostok a szimpatikus idegrendszer idegdúcokat követő, posztganglionáris rostjait alkot-
ják. A hüvellyel borított axont idegrostnak nevezik. Az idegrostok kezdeti és végszakasza
hüvellyel nem borított, így ezek a szakaszok csupaszok.
Az idegrendszerben az idegsejtek egymással és más szövetekkel létesítenek kapcsola-
tot. Az intercalaris idegvégződések az idegsejtek között, szinapszisok révén jönnek
létre. Az idegsejtek kapcsolódhatnak egymáshoz divergens vagy konvergens módon.
Előbbi esetben egy idegsejt axonja több idegsejthez, utóbbinál számos idegsejt egy neu-
ronhoz kapcsolódik. Konvergens kapcsolódásnál egy mozgató idegsejten akár több ezer
szinapszis is lehet. A terminális idegvégződéseknél az idegsejtek nyúlványa más szö-
vethez kapcsolódik. Funkcióját tekintve lehet ingerleadó (effektorikus) vagy érző (re-
ceptorikus) idegvégződés. Az effektorikus idegvégződés lehet mozgató vagy szekretorikus
típusú. A mozgató idegvégződés az ideg és az izom kapcsolatát jelenti. A szekretorikus
idegek végfácskája a mirigyhámsejtek közé hatol be. A külvilágból érkező ingereket exte-
roreceptorok, a szervezet belsejében keletkezőket az interoreceptorok veszik fel. Kü-
lönleges receptorforma a proprioceptor, melynek ingerfelvevő és -leadó végkészüléke
ugyanabban a szervben helyezkedik el. Ez a típus jellemző az izomorsóra.
Felépítésük alapján a receptorok lehetnek szabad idegvégződések vagy idegvég-
testek. A szabad idegvégződésnél a végfácska csupasz, kötőszövetes tok nem borítja.
Ilyen például a fajdalomérzekelo idegvegzodes a borben. Az idegvegtestek közös jellem-
zője, hogy a végfácska körül kötőszövetes tok található. Fontosabb típusainak felépítését
a 4.9.1. fejezetben mutatjuk be.
A gliasejtek az idegsejtek működéséhez szükséges feladatokat látnak el. Támasztják,
más szövetektől izolálják azokat, és szállítják a tápanyagokat is. Kitöltik az idegsejtek
közötti hézagokat, és mivel képesek osztódásra, az elpusztult idegsejtek helyét is pótolják.
A gliasejtek a mikrogliasejtek kivételével
ektodermális eredetűek, a velőcsőból és
a ganglionlécből származnak. A mikro-
gliasejtek mezodermális eredetűek. A köz-
ponti idegrendszerben találhatók az oli-
godendroglia sejtek, a makrogliasejteknek
is nevezett asztrociták (astrocyta), a mik-
rogliasejtek és az ependima -(ependyma)
sejtek (3.25. ábra). Azoligodendroglia-
sejtek a központi idegrendszerben a ve-
lőshüvelyt alakítják ki az axon körül, így
főleg a fehér állományban találhatóak. A
térkitöltő, támasztó, tápláló, elektromo-
san izoláló szerepű asztrocitáknak két tí-
pusa van, aplazmás és a rostos asztrocita.
A plazmás asztrociták nyúlványai rö-
videbbek, vaskosabbak, főleg a központi
idegrendszer szürke állományára jellem-
zőek. Hálószerűén veszik körül az ideg-
sejteket, kitöltik a köztük lévő hézagokat,
3.25. ábra. Gyakoribb gliasejttípusok
ep: ependimasejt, mg: mikroglia, ol: oligodendroglia-sejt, pa:
plazmás asztrocita, ra: rostos asztrocita (Törő nyomán kissé
módosítva)
105
valamint az erek körül alakítanak ki hártyahüvelyt. A rostos asztrociták nyúlványai
vékonyabbak és hosszabbak, a plazmájukban rostok figyelhetőek meg. Főleg a központi
idegrendszer fehér állományában jellemzők. Itt rostjaik szeptumokat képezhetnek, míg
a koponya és a gerincagy felszínén hártyát alakítanak ki. A mikrogliasejtek amőboid
mozgásra és fagocitózisra képesek, az idegszövetben a RÉS tagjaiként funkcionálnak.
Kisméretű sejtek, kevés elágazó nyúlványuk van. Megtalálhatók az idegsejtek és a vérerek
körül. Az ependimasejtek az agykamrákat és a gerincvelő központi csatornáját (canalis
centrális) bélelik. Hám jellegű feladatokat látnak el, valamint részt vesznek az agyfolya-
dék anyagforgalmában. Felszínüket hosszúra nyúlt sztereociliumok növelik. A perifériás
idegrendszerben a Schwann-sejtek és a satellitasejtek fordulnak elő. A Schwann-sejtek
az idegrostok velőhüvelyes és velős hüvely nélküli szigetelését alakítják ki. A satellita
(amphicyta) sejtek az idegdúcok neuronjai körül helyezkednek el.
A központi idegrendszer területén található idegsejtek csoportosulását idegmagvak-
nak (nucleus), a környéki idegrendszerben idegdúcoknak (ganglion) nevezik. A perifé-
riás idegrendszer ganglionjait az idegsejteken és rostjaikon kívül gliasejtek, valamint laza-
rostos kötőszövetbe ágyazott vérerek alkotják. A dúcokat kívülről kötőszöveti tok hatá-
rolja.
Ellenőrző kérdések a 3. fejezet témaköreiből
1. Melyek az állati testet felépítő alapvető szövet-típusok?
2. Milyen speciális tulajdonságai vannak a hámszöveteknek?
3. Hogyan lehet csoportosítani a hámszöveteket morfológiai jellemzők és funkciók alapján?
4. Jellemezze az egyes hámszövet típusokat!
5. Sorolja fel a többrétegű elszarusodó laphám rétegeit!
6. Milyen szöveti felépítést mutatnak a mirigyek?
7. Milyen feladatokat látnak el a kötőszövetek?
8. Mi jellemző a kötőszövetek sejtes és sejtközötti állományára?
9. Sorolja fel a kötőszövetek típusait!
10. Melyik az állati szervezet leggyakoribb kötőszövete?
11. Milyen felépitésbeni és funkcionális eltérés van a fehér és barna zsírszövet között?
12. Milyen feladatokat látnak el a vér alakos elemei?
13. A porcszövetek milyen típusait ismeri?
14. Mutassa be részletesebben az üvegporc felépítését!
15. Milyen tulajdonságai alapján kapta az állati szervezet legkeményebb szövete elnevezést
a csontszövet?
16. Az izomszövetek milyen típusait ismeri?
17. Milyen speciális feladatokat látnak el az egyes izomszövet-típusok?
18. Milyen sejtekből épül fel az idegszövet?
106
4. AZ ÁLLATOK SZERVRENDSZEREINEK
FŐ TÍPUSAI
Ebben a fejezetben a legelterjedtebb állati szervek, szervrendszerek felépítésének és - bizonyos
mértékig-működésének alapvonásait ismertetjük. Az evolúció során ugyanazon, vagy hasonló
feladat elvégzésére különböző szervek, szervrendszerek jöttek létre. Csupán a fő típusok legfon-
tosabb jellemzőit mutatjuk be, hiszen az élővilágban rengeteg különböző szervforma fordul elő.
Mivel a specialitások megismertetésére a rendszertani fejezetekben, az állatcsoportok jellemzése
során rendszeresen visszatérünk, ezért ebben a fejezetben ritkán fejtünk ki részletesen egy-egy
kérdést. Ha nem értenek valamit, lapozzák fel a későbbi fejezeteket. A szervrendszerek struk-
túráinak bemutatása mellett röviden kitérünk a működés néhány fontosabb jellegzetességére is,
anélkül azonban, hogy a folyamatok mélyebb élettani magyarázatát adnánk. Ez más tantár-
gyak feladata. Figyeljen továbbá arra, hogy azonos elnevezés különböző embrionális szervből
eredő és egymástól eltérő felépítésű szervet jelenthet.
A különféle féregfajok bőrizomtömlőjében egy, két, vagy három izomréteg kapcsolódik egy
külső sejtréteghez, amiben gyakran mirigysejtek vannak, és kívülről kutikula is boríthatja. Az
ízeltlábúak kültakarójában az élő sejtréteg a kitines kutikula alatt, a test belseje felé helyezkedik
el. Ez a kültakaró kemény, kissé rugalmas, néha mészsók még keményebbé teszik. A gerincesek
epidermiszéből indulnak ki a tollak és a szőrök. Az irha táplálja az epidermiszt. A színek a
sejtekben lévő színanyagok miatt, vagy a fényvisszaverődés következtében alakulnak ki. A
kültakaró az a szerv, amely talán a legtöbb feladatot látja el a szervezetben. Védi a szervezetet
fizikai és kémiai módon is. Szoros kapcsolatban áll a mozgásszervekkel. Ezen keresztül jutnak
az anyagok a testbe, sokszor az anyagcsere végtermékek a külvilágba. Részt vehet a gázcsere
lebonyolításában. Része van a színek kialakításában. Számos érzékszerv helyezkedik el benne.
A test vázát jelentheti a testfolyadéknak a kültakaróra gyakorolt nyomása (hidrosztatikus
váz), vagy valamilyen vázrendszer. Az izeldábúaknak külső, a gerinceseknek belső vázrend-
szerük van. A vázhoz különféle szövettani felépítésű izmok kapcsolódnak.
A tápanyagokat változatos szájszervekkel és a körülötte lévő segédszervekkel szerzik meg az
állatok. Leggyakrabban az elő-, közép-, és utóbélből álló bélcsatomában termelődő enzimek
segítségével bontják le a tápanyagokat. A rákok és puhatestűek bélcsatomájához középbéli
mirigy csatlakozhat. Az emésztés történhet a testen kívül, vagy a testen belül. A sejten belüli
emésztés a fagocitózis. Ritkán fordul elő az állatok körében. A sejten kívüli emésztés a bélcsa-
toma üregében történik. A lebontott tápanyagokat a keringési rendszer szállítja a felhasználás
helyére, a sejtekhez, majd az elhasznált anyagokat a kiválasztás helyére. Vannak állatok,
amelyeknek nincsenek ereik, hanem a testüregben kering a testfolyadék. Máskor nyílt, vagy
zárt keringési rendszer, érrendszer figyelhető meg. Csak a gerinceseknek van nyirokrendszerük.
A légzés történhet a testfelületen. Ez kistestü, el nem szarusodott kültakarójú állatok esetében
megfelelő gázcsere-módszer. A legtöbb állat kopoltyúval, trachea rendszerrel, vagy tüdővel
lélegzik. A légzéshez szükséges gázok szállítását transzportfehérjék segíthetik. Az anyagcsere
folyamatok végtermékei a széndioxid, víz, ammónia, karbamid és húgysav. Ezek eltávolítása
a szervezetből a kültakarón, a légzőszerveken és a kiválasztó szerveken keresztül történik. A
fejezetben az elövesécske, vesécske, Malpighi-edény és vese típusú kiválasztó szervrendszereket
tárgyaljuk. Nitrogén tartalmú anyagcsere végtermékek kiválasztása történhet a kopoltyún
keresztül, kloridsejtekkel és más itt nem tárgyalt módokon is.
107
Az állatok szaporodhatnak ivarsejtek nélkül (hasadás, bimbózás), vagy ivarsejtekkel. Szűz-
nemzésről akkor beszélünk, ha megtermékenyülés nélkül indul fejlődésnek a petesejt. Ivaros
szaporodás esetén a petesejt és a hím ivarsejt egyesül. Az állatok lehetnek hímnősek, vagy vált-
ivarúak. Bár az ivarszervek egyes részeinek elnevezései egyformák nagyon különböző állatcso-
portokban (nőstény ivarkészülék: petefészek, petevezető, méh, hüvely, hím ivarkészülék: here,
kivezető csatorna, ondóvezetö, kilövellő járat), azok felépítése jelentősen eltérhet egymástól. A
heterogónia során a szűznemzéssel szaporodó és a hímeket és nőstényeket egyaránt tartalmazó
populációk követik egymást. A metagenezis során viszont az ivarosán és az ivartalanul sza-
porodó populációk váltakozásáról van szó.
A hormonok aminosav származékok, proteinek és proteidek, szteroidok, arachidonsav-
származékok és juvenilis hormonok. Termelődhetnek szöveti sejtekben, idegsejtekben, vagy
belső elválasztású mirigyekben. A közvetlen hatású hormonok átjutnak a sejthártyán, majd a
citoplazma receptorfehérjéihez kötődnek. A közvetett hatású hormonok a sejthártya receptor
fehérjéihez kötődve váltanak ki egy reakciósorozatot a sejten belül. Ha összehasonlítjuk a
gerinctelenek és gerincesek hormonrendszerének és idegrendszerének jelentőségét az életfolyama-
tok szabályozásában, akkor a hormonrendszer a gerinctelenek, az idegrendszer pedig a gerin-
cesek esetében valamivel fontosabb az életfolyamatok szabályozásában. Az ízeltlábúak köré-
ben a vedlés hormonális szabályozása különösen jelentős.
A fejezet címe azt kívánja hangsúlyozni, hogy nem különböző állatcsoportok szervrend-
szereinek összehasonlítása a cél, hanem az, hogy a legelterjedtebb szervek, szervrendszerek
felépítésének és - bizonyos mértékig - működésének alapjait ismertessük. A szervek,
szervrendszerek fő típusainak legfontosabb jellemzőit kívánjuk bemutatni, hiszen az
élővilágban rengeteg különböző forma fordul elő. Mivel a specialitások megismertetésére
a rendszertani fejezetekben, minden egyes állatcsoport jellemzése során, a szervek,
szervrendszerek leírásakor rendszeresen visszatérünk, ezért ebben a fejezetben csak a
legszükségesebb esetekben utalunk a részletesebb kifejtésre és az ott található példákra. A
heterotróf egysejtüekkel és a hazánkban nem élő csoportok testfelépítésével ebben a
fejezetben nem foglalkozunk. A szervrendszerek struktúráinak bemutatása mellett röviden
kitérünk a működés néhány fontosabb jellegzetességére is, anélkül azonban, hogy a
folyamatok mélyebb élettani magyarázatát megadnánk. Ez más tárgyak feladata. A
szervek, szervrendszerek embrionális fejlődésének alapvonásai a 2. fejezetben találhatók.
4.1. Kültakaró és mozgásszervek
Az állatok testét borító kültakaró a szervezet és a környezete közötti határfelület.
Ugyanúgy, ahogy a sejtek sejthártyája, az egyedek kültakarója is bonyolult felépítésű
szerv, amely határfelületet képez a szervezet és környezete között. Részben tehát el-
választja a szervezetet a külvilágtól, ugyanakkor azonban össze is köti, hiszen a kültaka-
rón keresztül anyagáramlás, energiaáramlás és információáramlás is történik. Nagyon
sok állatfaj ezen keresztül veszi fel a tápanyagokat és az oxigént, és itt adja le a felesleges
anyagokat, a szén-dioxidot. A kültakaró számos egyéb funkcióval is rendelkezik. Védi
a szervezetet a különböző fizikai, kémiai és biológiai hatások ellen, megakadályozza
a fertőző mikroorganizmusok bejutását a testbe, és hatásos lehet a ragadozók ellen is.
Itt találhatók az állat színezetét kialakító anyagok, struktúrák: ezek számos viselkedés-
formát befolyásolnak. Végül, nem kevésbé fontos, hogy a kültakaróban a külvilág in-
gereinek felfogására alkalmas érzékszervek sokasága található, ami miatt a kültakaró
maga is érzékszervnek tekinthető. Az érzékszervek különös jelentőségűek a környezethez
történő adaptáció során, mert működésük döntően befolyásolja az egyedek túlélési
esélyeit.
108
Az állatokra az aktív, helyváltoztató mozgás és az ehhez szükséges számos mozgás-
szerv különösen jellemző. Ez teszi lehetővé, hogy az egyedek gyorsan eljussanak egyik
helyről a másikra. Az aktív mozgás képessége valószínűleg nagy szerepet játszott az
állatok evolúciójában, mert segítette a gyors szétterjedést, új élőhelyek felfedezését,
benépesítését, a versengés csökkenését. A gyorsabb egyedek könnyebben menekültek
meg ragadozóik elől, mint a lassúak.
A mozgásszervek a vázrendszerből és az izomrendszerből állnak. A vázrendszer
meghatározza a test alakját, valamint többnyire ez az izmok eredési és tapadási helye is.
A testalak, a testforma összefüggésben áll az élőhellyel, ahol az adott populáció él, mint
ezt a halak példája mutatja. A gyors úszáshoz az áramvonalas testforma alapkövetel-
mény. A sebes folyású hegyi patakokban, folyókban hengeres testű fajok élnek, míg az
állóvizek lakóinak teste oldalról lapított. A vázrendszer fontos a testnagyság szempont-
jából is, hiszen a kis test fenntartható gyenge vázzal, viszont a nagy testnek erős tartó-
rendszerre van szüksége. A tengeri emlősök azért lehetnek nagyobbak, mint a szárazföl-
diek, mert a test tömegének csak egy részét viseli a vázrendszer. Őslénytani vizsgálatok
nyomán úgy látszik, hogy a 30 tonnánál nehezebb állatok nem tudtak a szárazföldön
megfelelően mozogni. A nagy testű hüllők például a vízi életmódra tértek át. A jelenleg
élő legnagyobb tömegű állatok, a cetek is vízben élnek. A váz lehet külső, ekkor az izmok
belülről tapadnak rá. Ha a vázrendszer a test belsejében található, akkor az izmok kívülről
veszik körül.
Az izmok és izomrendszerek sokféleképpen alakulhatnak. Molekuláris alapjaiban
azonban mindegyik rendszer rendkívülien hasonló, működésük lényegében az aktin- és
a miozin-fehérjék tulajdonságaira vezethető vissza (1.5. fejezet).
4.1.1. Az egységes kültakaró és mozgásrendszer
A kültakaró és a mozgásszervek integrációja sok esetben nagyon szoros, működésük
egymással összehangoltan, egységben történik. Ilyen egységes kültakaró és mozgásrend-
szer a bőrizomtömlő és az ízeltlábúak kitinváza az izmokkal együtt. A bőrizomtömlő
több rétegre tagolható. Kívülről egyrétegű hám fedi. A hengeresférgek hámjában (hypo-
dermis) a sejtek gyakran összeolvadnak és így sokmagvú plazma (syncytium) jön létre.
A hámrétegben lehetnek csillós sejtek (örvényférgek), előfordulhat számos mirigysejt
(pl. gyűrűsférgek, puhatestűek) és boríthatja vékonyabb (szívóférgek, galandférgek)
vagy vastagabb (fonálférgek, kerekesférgek) kutikula. Különösen az endoparazita fonál-
férgek kutikulája vastag, mert ez védi az állatot az emésztőnedvek hatásaitól. A kutikula
mukopoliszacharidokból, fehérjékből és kollagénből áll, így nagyfokú mechanikai védel-
met és szilárdságot nyújt. A hámréteget alapi hártya (membrana basalis) választja el az
alatta található kötőszöveti rétegtől. A kötőszövet lehet igen vékony (fonálférgek, ke-
véssertéjű gyűrűsférgek), de tekintélyes vastagságú is (nadályok). A kötőszövet alatt ta-
lálhatók az izomrétegek. A fonálférgek bőrizomtömlőjében csupán hosszanti lefutású,
a gyűrűsférgekében külső körkörös és belső hosszanti lefutású, a laposférgekében pedig
az említett két rétegen kívül ferde lefutásúak is találhatók. A rétegek mellett gyakran hát-
hasi irányban húzódó izomkötegek is felismerhetőek (laposférgek, nadályok). A puha-
testűek bőrizomtömlőjében az izomrostok a hám alatt található kötőszövetbe ágyazód-
nak. Izomréteget csak ritkán alkotnak.
Az izmok támasztórendszer nélkül nem képesek a testet megfelelő irányba mozgatni.
Szükséges valamilyen vázrendszer (sceleton), hogy az izmok megtapadhassanak és a
testet célra orientáltan mozgathassák. Ősi, különleges forma a hidrosztatikus váz, a
férgek jellemző vázrendszere. Ez annyiban váz, hogy a testüreget teljesen kitöltő testfo-
lyadék kifeszíti a bőrizomtömlőt, ezzel kialakítja a test alakját és feszesen tartja a
kültakarót. Mivel a folyadékok összenyomhatatlanok, és a test, illetve egy szelvény tér-
109
fogatának nagysága nem változik lényegesen, ha egy izomköteg megrövidül, a test
ellentétes oldalán lévő izmoknak meg kell nyúlniuk. A fonálférgek testürege egységes és
csak hosszanti izomkötegeik vannak. Hol az egyik, hol a másik oldali izomkötegek
húzódnak össze. A viszonylag kemény kutikula miatt a test nem tud összehúzódni és
megnyúlni. A fonálférgek ezért tekeregve, jobbra-balra kígyózva mozognak. Ezzel
szemben a gyűrüsférgek teste szelvényezett. A szelvényeket hártyák választják el egymás-
tól, amelyek gátolják a testfolyadék szabad áramlását. Továbbá körkörös és hosszanti
izomrétegeik is vannak. így minden szelvény külön-külön képes összehúzódásra és
megnyúlásra, de az előrehaladó mozgás során a szelvények mozgása meghatározott
rendben követi egymást.
Az ízeltlábúak kitinváza a kültakaró és a vázrendszer is egyidejűleg. Legnagyobb
tömegét a vastag kutikula teszi ki. Alatta egy rétegben epidermiszsejtek találhatók. A
kutikulát három réteg (endo-, exo- és epicuticula) alkotja. Ezekben sejtek nem találha-
tók. A rétegek elsősorban összetett fehérjékből, szénhidrátokból és szervetlen sókból
épülnek fel. Jellemző anyaguk a kitin, ami egy nitrogéntartalmú poliszacharid (acetil-
glükózamin) és a szklerotin, ami kínon- vagy acetildopamin-molekulák hidjaival erősí-
tett fehérje. A szervetlen anyagok közül jellemzőek a kálciumsók. Ezek gyakoriak a
magasabb rendű rákok (Malacostraca) és ikerszelvényesek (Diplopoda) kültakarójában.
A kitinváz fizikai és kémiai behatásokkal szemben igen ellenálló. Bár rugalmas, de
kevéssé nyújtható, ezért a növekedő állatoknak időről időre le kell vedleniük, különben
gátolná a test növekedését. A vedlés ideje alatt az állatok kültakarója puha, mozogni csak
korlátozott mértékben tudnak, ezért ez idő alatt könnyebben eshetnek áldozatul a
ragadozóknak.
A kitinváznak a testet fedő része a külső váz (exosceleton). Innen nyúlványok in-
dulnak a test belsejébe, amiket ugyan belső váznak (endosceleton) hívnak, de az el-
nevezés megtévesztő. A test belseje felé irányuló nyúlványok ugyanis nem homológok a
valódi belső vázzal (pl. a gerincesek csontvázával), mert ektodermális eredetűek, a
kültakaróból származnak és azzal mindig kapcsolatban maradnak. A két említett váz-
rendszer együttesen nagy tapadási felületet jelent az izmok számára: relatíve nagyobbat,
mint a gerincesek belső váza. Mindez sok izomrost megtapadását, számos önálló, finom
mozgást tesz lehetővé, ahogy ez például a szájszervek, tapogatók, ivarszervek működése
során látható.
4.1.2. Az önálló kültakaró
A gerincesek körébe tartozó állatfajok kültakarója (bőr) a mozgás szervrendszerétől
felépítés és működés szempontjából többé-kevésbé elkülönül. A kétéltűek kültakarója a
nyirokzacskók miatt könnyen elválik az izomzattól, a bőrük felemelhető. A többi gerinces
kültakarója viszont az izomrendszerrel erősebb-gyengébb kapcsolatban áll. A bőr (in-
tegumentum commune) három rétegből áll (4.1. ábra). Kívülről többé-kevésbé elsza-
rusodó többrétegű laphám (epidermis) borítja, amely egyedfejlődési szempontból nézve
ektodermális eredetű. Alatta a jóval vastagabb irharéteg (cutis, corium, dermis) találha-
tó. Ez mezodermális eredetű. A harmadik, szintén a mezodermából származó, legbelső
réteget a bőralja (subcutis) képezi.
Az epidermisz elszarusodása jelentős lépés a gerincesek evolúciója szempontjából.
A halak és a kétéltűek epidermisze csupán kismértékben szarusodott el, ezért életük a
vízhez kötött, hiszen a szárazföldön kiszáradnának. A halak pikkelye az irhát és az
epidermiszt maga előtt nyomva fejlődik. A pikkelyeket tehát irharéteg és epidermisz is
borítja. A hüllők, madarak és emlősök kültakarójában a test felszínét borító többrétegű
laphám szemcsés rétegének sejtjei (stratum granulosum) egy vízben oldhatatlan
struktúrfehérjét, keratint képeznek, majd elpusztulnak. így jön létre a szaruréteg (stra-
110
4.1. ábra. A: emlősbőr és -szőr felépítése, Bl: a szőrszál és a szőrtüsző hosszmetszete, B2: ránézeti képe, B3: a
szőrszál részei
af: apokrin faggyúmirigy, ba:bőralja, fin: faggyúmirigy, há: hám, ir: irha, iz: szőrmerevítő izom, ka: kapilláris ér, ké: a szőr
kérge, ku: kutikula, lk: laza-rostos kötőszövet, or: orsósejtek, pi: pigmentszemcsék, sg: stratum germinativum, tt: tapintótest
(nyomásérzékelő), tü: szőrtüsző, ve: a szőr velőállománya, vm: verejtékmirigy, zs: zsírszövet (Több szerző nyomán)
111
tűm comeum) (3.1.1. fejezet). Ebből azután a test felszínét teljesen befedő szilárd, de
rugalmas réteg keletkezik, ami megakadályozza a párolgást és amit rendszerint vetítések-
kel távolítanak el az állatok magukról.
Az önálló kültakaró epidermiszéből képződik a kültakaró függelékeinek (tollak,
szőrök, körmök, karmok, agancsok stb.) legnagyobb tömege. A madarak tollai a hüllők
pikkelyeiből alakultak ki az evolúció során, tehát a kültakaró kitüremkedésének tekint-
hetők. Az egyedfejlődés során először a tolikezdemény kiemelkedése (papilla) látható a
bőrön, majd ezután jön tétre az irhába besüllyedő toll. Az emlősök szőre törzsfejlődéstani
szempontból nem homológ a toliakkal. Úgy képződik, hogy az epidermisz az irhába
betüremkedve, rövid, vakon végződő csövet képez. A cső végébe az irhából erek nyo-
mulnak be. Itt kis tágulat jön tétre. így alakul ki a szőrhagyma (bulbus pili). A gazdag
tápanyagellátás miatt a hám e helyen fokozott mértékben képez szarut. A szaru fonal
alakban nő ki a betűrődési csatornából és közben átalakul szőrszállá, felveszi a szőrre
jellemző szerkezetet.
Az irharéteg alapja kötőszövet. A hám alatt közvetlenül általában laza-rostos kötő-
szövet, a test belseje felé pedig tömött-rostos kötőszövet található. Ezen kívül sok haj-
szálér, különféle mirigyek, idegvégződések és receptorok találhatók még ebben a
rétegben. A mirigyek szintén az epidermiszből képződnek, de mélyen behúzódnak az
irhába. A halak és a kétéltű lárvák bőrében csak egysejtű mirigyek találhatók. A kifejlett
kétéltűek bőrében azonban jól fejlett, soksejtű méregmirigyek (holokrin mirigy) és nyál-
kamirigyek (merokrin mirigy) alakulnak ki (17. 25. ábra). A hüllők és a madarak bőre
ezzel szemben mirigyekben igen szegény. Hüllőknél legfeljebb néhány feromontermelő
mirigy, madaraknál pedig a fartőmirigy (glandula uropygii) található a bőrben. Az em-
lősök bőre mirigyekben rendkívülien gazdag. Számos verejtékmirigyük és faggyúmiri-
gyük van. A faggyúmirigyek váladéka teszi a szőrt vízállóvá és ezek a mirigyek termelik
a feromonokat is. A szőrtüszőhöz kapcsolódó apokrin mirigyekből alakultak ki az evo-
lúció során a tejmirigyek. A bőralja laza-rostos kötőszövet, melyben zsírsejtek találha-
tók. Ez a zsírréteg gyakran vastag lehet, egyben mechanikai védelmül szolgál és tartalék
tápanyagot jelent.
4.1.3. Az önálló mozgásrendszer
A puhatestűek és a gerincesek vázrendszere és izomrendszere a kültakarótól többé-
kevésbé elkülönül. A kettő együtt önálló strukturális és funkcionális egységet képez. A
vázrendszert a mozgás passzív szervének, az izomrendszert pedig a mozgás aktív szer-
vének nevezik.
A legtöbb puhatestű köpenye (pallium) házat vagy héjat választ ki, amely az állat
számára menedéket ad, az izomrendszernek pedig eredési helyül szolgál, tehát ez a külső
váz. Kívülről a héjhártya (periostracum) borítja. Alatta vastag, kemény oszlopos,
kristályos réteg (ostracum) helyezkedik el. A kagylókhéján legbelül (eltérően a csigáktól)
még egy csillogó gyöngyházréteg (hypostracum) is van, amely szintén kristályos (14.10.
ábra). A puhatestűek izmai többnyire a váz belső felszínéről erednek és az egyes
szerveken tapadnak. A kagylók héjának erős záróizmai gyors összehúzódásra képesek,
ezzel szemben a láb a testet csak lassan tolja előre. A láb nyújtását a beléáramló test-
folyadék okozza. A csigák hasi oldalán izmos láb található. Az itt végigfutó izomhullámok
viszik lassan előre az állatot.
A csontos vázrendszer a gerincesekre jellemző képződmény, ami csontokból, por-
cokból, valamint a csontok összeköttetéseiből áll. A csontvázrendszert három fő részre,
a koponyára, a törzs vázára és a végtagok vázára lehet felosztani. A koponya (cranium)
a fej váza. Két nagy tájéka az agykoponya (neurocranium) és az arckoponya
(viscerocranium). A két nagy tájék viszonyában az agykoponya fokozatos növekedése, az
112
arckoponyához viszonyított relatív túlsúlya jelenti az evolúciós
trendet a halaktól az emlősök felé haladva. A törzs vázának
központja a gerincoszlop (columna vertebralis), mely csigo-
lyákból (vertebrae) áll. A csigolyák háti oldalán a csigolyaív
(arcus vertebrae) öleli körül a gerincvelőt. A hasi oldalon a
halaknál és a farkos kétéltűeknél vérív (haemapophysis) találha-
tó, ami a farokereket átfogó, keskeny csontív a farokcsigolyá-
kon. Aszerint, hogy a csigolya testén hol van homorú bemélye-
dés, ízületi vápa, a következő típusokat különböztetik meg: elöl-
hátul vájt (amphicoelicus), elöl vájt és hátul kidomborodó
(procoelicus), hátul vájt és elöl ki domborodó (opisthocoelicus),
nyereg formájú (heterocoelicus) vagy lapos felszínű (acoelicus)
csigolya. A csigolyák száma az egyes osztályokon belül is rend-
kívül variábilis. Egyedül az emlősöknél viszonylag állandó, ahol
hét nyakcsigolya, 12 hátcsigolya, 6-7 ágyékcsigolya, 2-5 ke-
resztcsigolya és 1-47 farokcsigolya a jellemző. A gerincoszlop-
hoz bordák (costae) csatlakoznak. A bordák egyik típusa nem
kapcsolódik a szegycsonthoz (lengőbordák: costae fluctuantes).
Más esetben több borda egy közös porccal (álbordák: costae
spuriae) vagy külön-külön (valódi bordák: costae verae) kap-
csolódik a szegycsonthoz. A halak, békák és kígyók kivételével
a gerincoszlop, a bordák és a szegycsont (sternum) együttese
zárt mellkast képez. A szárazföldi gerincesek ötsugaras
(pentadactyl) végtagja (4.2. ábra) a csontoshalak soksugarú,
páros úszóiból alakult ki. Függesztőövből és végtagcsontokból
áll. Az első végtag függesztőöve a vállöv (cingulum membri
thoracici) három csontból: lapocka (scapula), kulcscsont
(clavicula) és hollócsőrcsont (coracoideum) áll. Ugyanennyi
csont alkotja a hátsó végtag függesztőövét, a medenceövet
ev hv
4.2. ábra. Gerincesek ötsu-
garú végtagjának alapsémá-
ja
co: combcsont, ev: elülső vég-
tag, fk: felksrcsont, hv: hstulsó
végtag, kk: kézközépcsontok,
kt: kéztőcsontok, Ik: lábközép-
csontok, It: lábtőcsontok, or:
orsócsont, sí: singcsont, sí: síp-
csont, sz: szárkapocscsont, uj:
uj jak (Wehner és Gehring nyo-
mán)
(cingulum extremitatis pelvinae) is. Ezek: csípőcsont (ilium), ülőcsont (ischium) és
szeméremcsont (pubis). A mellső végtag a felkarcsonttal (humerus) kezdődik. Két
csont, a singcsont (ulna) és az orsócsont (radius) kapcsolódik hozzá (alkarcsontok).
Ezután a kéztőcsontok (carpalia), kézközépcsontok (metacarpalia) és az ujjak (digiti)
következnek. Az ujjak ujjpercekre (phalanges digitorum) tagolódnak. A hátulsó végtag
a combcsonttal (femur) kezdődik, a sípcsonttal (tibia) és szárkapocscsonttal (fibula)
folytatódik (ezeka lábszárcsontok), végül a lábtőcsontokkal (tarsalia), láb középcsontokkal
(metatarsalia) és szintén az ujjakkal ér véget. A végtagok csontjai az életmódnak
megfelelően rendkívül változatosan alakultak. így egyes csontok részben, vagy teljesen
visszafejlődhettek, vagy rudimentálódhattak (pl. emlősök hollócsőrcsontja, ami a lapoc-
ka hollócsőmyúlványát, a processus coracoides-t képezi), mások pedig hatalmas fejlett-
séget érhettek el (pl. sok madár szegycsontja).
A gerincesek izomrendszere (systema musculorum) a vázrendszer elemeihez
kívülről csatlakozik. Ez az elrendezés pontosan az ellentéte annak, ami az ízeltlábúak
testfelépítésében látható, ahol a külső vázhoz az izmok belülről tapadnak (4.3. ábra).
A gerincesek izomzata a test tömegének felét-harmadát teszi ki. Az izmok izomszövet-
ből épülnek fel (3.3. fejezet). A végtagok és a törzs izmai mozgatják a testet. A zsigeri
izmok a szervek falában találhatók és azok mozgatásáért (bélperisztaltika, húgyhólyag
ürítése stb.) felelősek. A halaknál jól láthatóan szelvényezett törzsizmok figyelhetőek
meg. A szelvények száma megegyezik a csigolyákéval. Az izomszelvényeket (myomer)
kötőszövetes hártyák választják el egymástól. A kétéltűek lárváin az izomrendszer
szelvényezettsége még jól látszik. Kifejlett korban és a további osztályokban a szelvé-
113
4.3. ábra. A: külső vázhoz belülről tapadó izmok, B: és belső vázhoz kívülről csatlakozó izmok
fe: feszítőizom, ha: hajlítóizom, hi: hosszanti izmok (Több szerző nyomán)
nyezett izmok mennyisége csökken. Ezért tekintik ősi bélyegnek például az emlősök
esetében a gerincoszlop izmainak szelvényezett voltát. A röpképes madaraknak - ért-
hetően - a mellizmai a legtömegesebbek. A nagy mellizmok a szárnyat lefelé, a kis
mellizmok felfelé mozgatják. Az emlősök jellemző, speciális izma a rekeszizom (dia-
phragma), ami a mellüreget választja el a hasüregtől. Ez az egyik legfontosabb légző-
izom.
4.1.4. A kültakaró funkciói
A következőkben két, nagymértékben a kültakaróhoz köthető feladatot, a színezet
kialakulását és a hőszabályozást tárgyaljuk.
Az állatok színezete
Az állatok jó része képes arra, hogy színezetével, színváltoztatással az élőhelyébe be-
olvadjon, az ott jellemző színekhez, mintákhoz idomuljon (mimikri). Ez a képesség
mindazon fajok esetében előnyös tulajdonság, amelyeket ragadozói optikai úton találnak
meg. Azoknak az egyedeknek nagyobb a túlélési esélyük, amelyek mintázatukkal,
színeikkel jobban illeszkednek a környezetükbe, mint társaik. Hasonlóan fontos szerepet
játszhat a színezet a párválasztás során. Azon túl, hogy a fajra jellemző színezet és min-
tázat fontos a nemek egymásra találása miatt, a feltűnőbb állat szaporodási esélye meg-
nőhet, mivel a másik nem egyedeinek figyelmét jobban felkelti.
A színek alapvetően kétféle módon alakulhatnak ki. Lehetnek kémiai színek és struk-
túrszínek. A kémiai színek a kültakaróban található színanyagok következtében jönnek
létre. A színanyagokat tartalmazó sejtek (pigmentsejtek, kromatofor sejtek) citoplazmá-
jában színes szemcsék, lapocskák halmozódnak fel. A következő sejttípusok különböz-
tethetőek meg ebből a szempontból:
a) barna és fekete pigmenteket, melanint tartalmazó (melanophor),
b) sárga, karotinoid pigmenteket tartalmazó (xantophor),
c) piros, pteridin pigmenteket tartalmazó (erythrophor) és
d) ezüstszínű, guanint tartalmazó (guanophor) sejtek.
A természetben igen elterjedt színek, mint a zöld és a kék - ritka kivételektől eltekintve
- nem a sejtekben lévő színanyagoktól származnak. Ezek struktúrszínek, vagyis a
kültakaró felépítésének következtében alakulnak ki. A testre vetődő fehér fény visszave-
rődik, a hosszabb hullámhosszúságú sugarai interferálnak, kioltják egymást vagy elnye-
114
lödnek, a rövidebb tartományba esők viszont nem, így különféle kék és zöld színek
alakulnak ki. Természetesen a kémiai és a struktúrszínek keveredhetnek egymással és így
rendkívülien változatos, árnyalt színpompa jön létre. Színek másféle módon is kialakul-
hatnak. Az erek falán átpirosló vér színesítheti a csupasz bőrfelületet. Ha a szőrök belső
üregét levegő tölti ki, akkor a szőr fehér színű (ősz) lesz.
A színváltozás lehet lassú. Ez annak a következménye, hogy egyes pigmentsejtek
elpusztulnak és mások jönnek létre, vagy az idő előrehaladtával megváltozik a kültakaró
struktúrája. A jelenséget morfológiai színváltozásnak nevezik. ízeltlábúak, madarak
és emlősök esetében gyakori (pl. a hermelin fehér téli és barna nyári bundája). Számos
gerinctelen állat, hal, kétéltű és hüllő kültakarójában a fiziológiai színváltoztatás jel-
lemző. Ez azt jelenti, hogy ha a pigmentszemcsék a nyúlványos kromatofór sejtek
citoplazmájában szétterjednek, akkor sötétebb lesz az állat, ha pedig a sejtmag közelébe
húzódnak össze, akkor világosabb. A pigmentszemcsék mozgatását a sejten belül a
citoszkeleton végzi. A színváltoztatást a hormonrendszer (rákok, kétéltűek, sok hüllőfaj)
vagy az idegrendszer és a hormonrendszer együttesen (halak és hüllők) szabályozza.
Hőszabályozás
A hőfelvétel és hőleadás, a szervezet hőegyensúlyának fenntartása komplex folyamat.
Vannak olyan állatok, amelyeknek testhőmérséklete a környezet hőmérséklet-ingadozá-
sait követve változik (gerinctelenek, halak, kétéltűek, hüllők). Ezeket nevezik változó
testhőmérsékletű (hidegvérű, poikilotherm, ektotherm) állatoknak. Mások (madarak,
emlősök) többé-kevésbé függetleníteni tudják testhőmérsékletüket a környezetétől,
tehát viszonylag állandó testhőmérsékletnek (melegvérű, homoiotherm, endotherm
fajok). A változó testhőmérsékletű állatok anyagcsere-intenzitása (az életet megengedő
hőmérsékleti határok között) tíz fok hőmérséklet-növekedés esetén közel megduplázó-
dik (van’t Hoff-törvény). Az állandó testhőmérsékletűeké viszont növekvő hőmérséklet
mellett fokozatosan csökken, egészen egy kritikus pontig, hiszen ebben a hőmérsékleti
tartományban egyre kevesebb energiára van szükségük a testhőmérséklet fenntartására.
Ezután viszont újból emelkedik az anyagcsere intenzitása, mert a nagy meleg miatt
fokozott hőleadásra (verejtékezés, lihegés) van szükség.
Az állandó testhőmérséklet fenntartása sok energiát igényel. Az oxidatív anyagcseré-
nek mintegy 80%-át kell ezeknek az állatoknak erre a folyamatra fordítaniuk. A test-
nagyság csökkenésével az energiafelhasználás fokozódik, mivel a testfelület/testtömeg
aránynő. Némelyik változó testhőmérsékletű állatfaj az izommozgás révén annyihőttud
termelni, hogy testhőmérsékletét tartósan és jóval a környezet hőmérséklete felett képes
tartani. Számos rovar (méhek, dongók, lepkék) is képes arra, hogy intenzív izommozgás
segítségével a testhőmérsékletét szabályozza. Az ilyen állatokat vegyes testhőmérsék-
letűeknek (heterotherm, fakultatív homoiotherm) nevezik.
A külső hőmérséklet változásaihoz az állatok különféle fiziológiai mechanizmusok
segítségével is alkalmazkodnak. Rovarok és pókok testfolyadékában a hőmérséklet csök-
kenésével párhuzamosan fagyásgátló anyagok (glicerin, szorbit, trehalóz) jelennek meg,
amelyek a környezeti hőmérséklet jelentős csökkenésének átvészelését lehetővé teszik.
Vándormadarak, emlősök körében közismert a tollváltás, szőrváltás, valamint az, hogy téli
időben a bőr alatt zsír halmozódik fel. Mindez a jobb hőszigetelést szolgálja. A téli álmot
alvó fajok szívműködése, anyagcsere-intenzitása és testhőmérséklete csökken. Egyes kis
testű madár- (pl. kolibrik) és emlősfajok (pl. cickányok, hörcsög), ha éheznek, éjszakára
csökkentik a testhőmérsékletüket, így gazdálkodnak az energiával. Ennek a jelenségnek
torpor a neve. Nagy testű fajok esetében ez nem lehetséges, mert a testhőmérséklet
felemelése napkeltekor több energiát emésztene fel, mint amennyit éjjel megtakarítot-
tak.
115
4.2. Táplálkozás és emésztés
Az állatok heterotóf módon táplálkoznak. Vizet, ásványi sókat és különböző szerves
anyagokat kell felvenniük ahhoz, hogy testüket felépítsék, anyagcseréjüket fenntartsák és
szaporodni tudjanak. A legtöbb állatnak van a táplálék felvételére, lebontására és fel-
szívására szolgáló elkülönült emésztőkészüléke. A bélcsatomában a táplálék fizikailag
aprózódik és az emésztőenzimek a nagymolekulájú anyagokat lebontják. Olyan kis-
molekulájú anyagok (monoszacharidok, aminosavak, glicerin stb.) jönneklétre, amelyek
a bél falán keresztül fel tudnak szívódni, majd a testfolyadék áramlása vagy a keringési
rendszer segítségével a felhasználás helyére és a tápanyagraktárakba (máj, izmok) ke-
rülnek. Az emésztés során le nem bontott, a szervezet által nem asszimilált anyagok az
ürülékbe kerülnek. Az itt található anyagok tehát (legalábbis a döntő többségük) -
ellentétben a vizeletben található anyagokkal - nem kerültek felhasználásra, nem vettek
részt a szervezet anyagcsere-folyamataiban. Ha a bélcsatoma hiányzik (galandférgek),
akkor a tápanyagok a test falán keresztül jutnak a testfolyadékba és onnan a felhasználás
helyére.
A táplálékkal különféle anyagok és energia jut az állat szervezetébe. Az anyagok a test
felépítéséhez, a bioszintézis folyamataihoz, a normális anyagcseréhez szükségesek, a
felvett kémiai energia pedig a szervezet működése, a biokémiai reakciók során keletkező,
majd hő formájában eltávozó energiát pótolja.
Az állatok minden lehetséges táplálékforrást kihasználnak. A növényevők (herbivora)
elfogyaszthatják az egész növényt vagy annak csupán egy részét. Leharaphatnak darabo-
kat (pl. csipkézőbogarak) vagy kiszívhatják a növények nedveit (pl. levéltetvek), ehetik
a terméseket (pl. magevő madarak). A növényevők között gyakoriak a táplálékspecia-
lista fajok. Állati táplálékon élnek a ragadozó (camivora), a parazita (pl. az állatokban
élősködő laposférgek és fonálférgek) és a parazitoid életmódú fajok (pl. számos hár-
tyásszárnyú rovar). A ragadozók kevésbé válogatnak a táplálékban, mint a növényevők.
Rendszerint sokféle állatot fogyasztanak. A paraziták gyakran csupán meghatározott
gazdában vagy gazdán élhetnek. A vegyes táplálkozású (omnivora) állatok növényi és
állati eredetű táplálékot egyaránt fogyasztanak. A vörös róka (Vulpes vulpes) rovarok
és kisemlősök mellett, különösen az őszi időszakban sok növényi táplálékot, gyömölcsöt
is fogyaszt. Sok állatfaj elhalt szerves anyagokon él (detritivora). Számukra nem csupán
az elpusztult szervezetek, hanem az azokat bontó mikroorganizmusok is táplálékforrásul
szolgálnak. (A táplálkozási kapcsolatokról a 18.3.1. fejezetben lesz részletesebben szó.)
Nincs minden állatnak elkülönült emésztőszerve. Agalandférgekbélcsatomájahiány-
zik, ezért a táplálék diffúzió útján kerül a szervezetükbe. A bélcsatoma hiánya másodla-
gos jelenség: az endoparazita életmód következtében rudimentálódott a bélrendszer.
Mivel a galandférgek környezetében, a gazda bélcsatomájában bőven található lebontott
tápanyag, ez átdiffundálhat a kültakarón, vagy a hámsejtek veszik fel és a lapos test
bármely sejtjéhez - rövid úton - eljuthat.
4.2.1. Kétszakaszos bélcsatorna
A laposférgek egyes csoportjaiban (örvényférgek, szívóférgek) kétszakaszos bélcsatoma
található. Ez az emésztőszervrendszer szájnyílással kezdődik, melyet a szívóféregfajok
esetében szívókorong vesz körül. Az örvényférgeknél a garat a szájnyíláson át kiölthető,
a zsákmány testét ezen keresztül szívják ki az állatok. A szájnyílás után az izmos garat
következik. Ezután a bélrendszer elágazik. Néha csupán kétágú csatorna látható, máskor
az elágazás a fa gyökérzetéhez hasonlóan dús. Minél nagyobb az állat, annál gazdagabban
ágazik el a bélcsatomája. A bélcsövek vakon végződnek. Ebben a szervrendszerben tehát
116
nem csupán emésztés történik, de a részben vagy teljesen lebontott anyagok szállítása is,
ezért szokták gasztrovaszkuláris rendszernek nevezni. Ez lehet az egyik oka annak is, amiért
a laposférgeknek nincs fejlett keringési rendszerük. Mivel a csőrendszer falában mindenütt
vannak emésztőenzimeket termelő sejtek, a lebontás és a felszívás bármely bélszakaszon
megtörténhet. A bél falában kétféle sejttípus található. A bélüreg felé néző felszínükön
mindkettő csillókkal borított. A nagyobb sejtek fagocitózisra képesek, a kisebbek pedig
minden valószínűség szerint az emészőenzimeket állítják elő és szekretálják. A táplálék
maradványai a szájnyíláson át távoznak a testből, mivel a végbélnyílás hiányzik.
A kétszakaszos bélcsatomában a friss és az emésztett táplálék keveredik egymással, a
lebontás, az energia kihasználása ezért nem hatékony. Az egész rendszer kevéssé spe-
cializált. Ezzel szemben a háromszakaszos bélcsatorna lehetővé teszi, hogy a táplálék egy
irányba mozogjon. Meghatározott feladatot ellátó bélszakaszok alakulhatnak ki, és így
az energiakihasználás fokozódik.
4.2.2. Háromszakaszos bélcsatorna
A bélcsatorna egyes részeit több állatcsoportban is hasonlóan nevezik. (Nem szabad
azonban elfelejteni, hogy a közös név ellenére az egyes szakaszok eredete, felépítése és
működése eltérő lehet). A legtöbb állat bélcsatornája három szakaszra tagolható. E
szakaszokat előbélnek, középbélnek és utóbélnek nevezik. Az előbél a szájnyílással,
más nevén a szájréssel (rima oris) kezdődik, amit a szájüreg (cavum oris) követ. A
szájnyílás és környéke igen változatosan alakulhat és elsősorban a minél hatékonyabb
táplálékszerzést segíti (pl. fonálférgek szájszuronya, ízeltlábúak szájszervei, madarak
csőre). A szájüregben a táplálék megragadására és/vagy megőrlésére szolgáló fogak
(dentes) helyezkedhetnek el és igen gyakran nyálmirigyek nyílnak ide. Néhol a nyálmi-
rigyek részben méregmirigyekké alakultak át és a csáprágókhoz (csáprágós ízeltlábúak)
vagy a méregfogakhoz (kígyók) csatlakoznak. A szájban gyakori a nyelv (lingua) is. A
szájüregben a táplálék fizikai aprózása folyik és megkezdődik a kémiai bontás is.
A garat (pharynx) a szájüreget követő tágulat a bélcsatornán. Gyakran igen izmos
szerv. Számos állat ennek segítségével szívja fel szájüregébe a táplálékát (fonálférgek,
rovarok, pókszabásúak). A nyelőcső (oesophagus) rövidebb-hosszabb, vékonyfalú cső.
Alsó része kitágulhat és ekkor begyet (ingluvies) alkot. A begy a táplálék raktározására
szolgál (gyűrűsférgek, rákok, rovarok, madarak), de néha emésztő és erjedési folyamatok
is történhetnek benne (pl. méhek, madarak). Speciális szakasz a bélcsatomán egyes fajok
előgyomra (proventriculus). Egészen más eredetű és felépítésű bélszakaszokat nevez-
nek előgyomomak különböző állatcsoportokban. A közös bennük az, hogy mindegyik
közvetlenül a gyomor előtt helyezkedik el. Előgyomor fordul elő a gyűrűsférgek és a ro-
varok bélcsatomáján. A kérődző emlősök bendő-recés-százrétű gyomoregyüttesét szin-
tén előgyomomak nevezik.
Az előbél utolsó szakasza a gyomor (ventriculus). Állhat egy üregből (gyűrűsférgek,
kétéltűek, hüllők, számos emlős) vagy több üregből (madarak, egyes emlősök). A kérőd-
zők többüregű gyomra előgyomorra és valódi gyomorra tagolható. Ha a gyomor nyálka-
hártyái a mindenütt egyforma szöveti felépítésű, akkor egyszerű gyomorról van szó, de ha
többféle szövet alkotja, akkor összetett gyomornak nevezik. A gyomorban lebontó folya-
matok játszódnak le. Itt elsősorban a fehérje emésztése zajlik. A gyomorhoz felületna-
gyobbító kitüremkedések, függelékek (pylorusfüggelék) kapcsolódhat (pl. ragadozó
halak). A kérődzők nagy, többüregű, összetett gyomra valószínűleg azért alakulhatott ki
az evolúció során, mert jelentős mennyiségű (de alacsony tápértékű) táplálék befogadá-
sát tette lehetővé, és azt, hogy a hosszú ideig tartó emésztés már ragadozóktól védett
helyen, vagy szintén védelmet jelentő csoportokban történjen.
A középbél (intestinum) hosszú, gyakran kanyargós bélszakasz. Az emésztés kémiai
117
4.4. ábra. Gerincesek középbelének felépítése. A: a bél keresztmetszete, B: bélbolyhok, C: egy bélboholy, D: a
felszívóhám egy sejtje és E: a mikrovillik
al: alaphártya, ak: aktinszálak, ar: artéria, bü: bél ürege, de: dezmoszóma, gl: glikokalix, há: egyrétegű kutikuláris hengerhám,
ho: hosszanti simaizmok rétege, id: ideg, kö: körkörös simaizmokrétege, kt: kötőszövet, mirmikrovilli, ny: nyirokér, sr: sejt
közötti rés, vé: véna (Wehner és Gehring nyomán)
folyamatainak döntő többsége itt történik. Ebben a szakaszban szívódik fel a tápanyagok
zöme. Gerinctelen állatoknál általában kevéssé differenciálódott, morfológiai és szövettani
szempontból egyaránt viszonylag egységes rész. Puhatestűekben, rákokban középbéli
mirigy (hepatopancreas) kapcsolódhat ide. A középbéli mirigy feladata az emésztőenzi-
mek termelése, a felszívott tápanyagok raktározása, esetenként pedig méregtelenítés is.
Gerinces állatoknál a hasnyálmirigy és a máj is ebbe a bélszakaszba juttatja váladékát. A
halak, a kétéltűek és a hüllők középbele kevéssé tagolt. A madarak és az emlősök középbele
további három szakaszra osztható: patkóbél (duodenum), éhbél (jejunum) és csípőbél
(ileum). E csoportokban a középbél falát hosszanti és körkörös elhelyezkedésű simaizom-
réteg, ezen belül kötőszövet, majd a felszívást végző egyrétegű hengerhám borítja (4.4.
ábra). A felületet nem csupán a bél falának betüremkedései, a bélbolyhok, hanem a
hámsejtek felszínét borító citoplazmanyúlványok (microvilli) is növelik.
A középbélhez számos gerinctelen csoportban (piócák, haltetvek, rovarok) több nagy,
zsákszerű kitüremkedés csatlakozik. Ezek a táplálék raktározására szolgálnak, de gyakran
emésztés is folyik bennük. A középbél és utóbél határán található a vakbél (caecum) a
madarakra és emlősökre jellemző szerv. Lehet egyetlen zsákszerű képződmény (emlő-
sök), de páros is (madarak). A vakbél a növényevő madarakban és emlősökben jól fejlett.
Ezen a helyen szimbionta baktériumok és egysejtűek segítségével cellulózbontás törté-
118
nik. A ragadozók vakbele fejletlen. A vakbél végéhez egy nyiroktüszőket tartalmazó,
zsákszerű szerv, a féregnyúlvány (appendix vermiformis) csatlakozhat.
Az utóbél a bélcsatoma végső szakasza. Általában rövid cső. Ha hosszabb, akkor
rendszerint két szakasz, a remese (colon) és a végbél (rectum) különíthető el. Az utó-
bélben formálódik az ürülék, a bélsár. A rovaroknál igen fontos kiválasztási funkciót is
ellát (16.4. fejezet). A háromszakaszos emésztőszervrendszer utolsó pontja a végbélnyí-
lás (anus). Ha az utóbélnek közös kivezetőnyílása van az urogenitális rendszerrel, akkor
a nyílást kloákának nevezik (halak, kétéltűek, hüllők, madarak).
4.2.3. Intracelluláris és extracelluláris emésztés
Az emésztés folyamatában a nagy molekulájú tápanyagok enzimatikus hidrolízis során
kis molekulákra (aminosavak, monoszacharidok, zsírsavak, glicerin stb.) bomlanak.
Eközben kevés energia szabadul fel, ellentétben a sejtekben lezajló bontó folyamatokkal,
ahol az eredetileg a táplálékban rejlő energia nagy része ATP-kötésekbe megy át.
Külső emésztésről van szó, ha a szájnyíláson keresztül az állat emésztőenzimeket
bocsájt a táplálékára és ezért az emésztés már a testen kívül megkezdődik (egyes rovarok,
pókszabásúak). A legtöbb esetben azonban az emésztés az állat testén belül, a bélcsator-
nában történik. Ezt a folyamatot nevezik belső emésztésnek.
Lehet az emésztés sejten belüli (intracelluláris). Ez azt jelenti, hogy endocitózissal
kerül a táplálékdarab az emésztést végző sejt belsejébe, ahol emésztő vakuóla jön létre
körülötte. A lebontás ebben történik. Hasonló emésztési mód figyelhető meg a szivacsok
galléros ostoros sejtjeiben (choanocyta), valamint a csalánozók és örvényférgek bélfa-
lának sejtjeiben is.
A háromszakaszos bélcsatomával rendelkező állatok csaknem teljes mértékben sejten
kívüli (extracelluláris) emésztést folytatnak, vagyis az emésztőenzimeket a bélcsator-
na üregébe bocsájtják. Itt történik a lebontás, majd a kis molekulájú vegyületek fel-
szívása.
A gerinctelenek bélcsatomájában az emésztőenzimek szekréciója elsősorban a közép-
bélben (rovarok) vagy a középbéli mirigyben (puhatestűek, rákok) történik. A bélcsator-
na egyes szakaszainak a pH-ja nem tér el olyan nagymértékben, mint a gerincesek
esetében. Ezzel szemben a gerincesek bélcsatomájának szakaszai az emésztés és a kém-
hatás szempontjából határozottan elkülönülnek. A gyomor kémhatása erősen savas, a
pH 1,5-2,5 között van. A többi szakasz lúgos, esetleg semleges kémhatású. A kémhatás
mellett a detergensek jelenléte is igen fontos. Ilyen szerepet játszik a gerincesek epéje.
Egyes rovarok középbelében a redoxpotenciál erősen negatív. Ennek következtében a
fehérjék diszulfidhídjai fellazulnak és a szokásos endopeptidázok számára bonthatóvá
válnak. így képesek a ruhamoly (Tineola biselliella) lárvái, továbbá a csótányok a
selymet, a kollagént és a keratint is megemészteni.
Az endoszimbionta mikroorganizmusok egészen speciális és igen fontos szerepet
játszanak az emésztésben. Az orvosi pióca (Hirudo medicinalis) belében egy baktériumfaj
(Pseudomonas hirudinis) szinte minden emésztési funkciót betölt. Rendszerint azonban az
endoszimbiontáknak speciális feladatuk van. Mivel cellulózbontó enzimeket az állatok
általában nem képesek termelni (a kivételek közé tartoznak például a Lepisma spp. rovarok),
ezért a cellulóznak, ennek az igen fontos és a szervezetbe nagymennyiségben bejutó anyagnak
a bontását baktériumok és egysejtűek (legtöbbször ostoros és csillés egysejtűek) végzik. A
kérődzők bendőjében élő csillós egysejtűek elsősorban a cellulóz bontásában vesznek részt,
míg a bendő baktériumai a nyálba kiválasztott karbamidot, a vérből a bendőbe átszűrődő
karbamidot, a fehérjék hidrolíziséből származó nitrogént és a takarmánnyal felvett nem
fehérje nitrogént alakítják saját testanyagaikká. A mikrobiális emésztés jelentőségét mutatja,
hogy a kérődzők ürülékének esetleg a felét is az elhalt baktériumok tehetik ki.
119
4.3. Anyagszállítás
A szervezetben szükség van az anyagok térbeli mozgatására. A tápanyagok, salak-
anyagok, hormonok, légzési gázok stb. szállítása nélkül az anyagcsere nem valósulhatna
meg. A keringési rendszer segítségével jut az oxigén a légzőszervtől a szövetekig és a
széndioxid a szövetektől visszafelé, a légzőszervhez. E szervrendszer útján kerülnek a
tápanyagok az emésztőszervrendszertől a szövetekig, a hormonok a belső elválasztású
mirigyektől a hatás helyére, és az anyagcsere-végtermékek a kiválasztószervekhez. Már
e néhány, felsorolt feladat is mutatja a keringési rendszer jelentőségét a szervezetben.
Legegyszerűbben diffúzióval történik anyagszállítás. A légzési gázok felvétele és le-
adása például a többsejtűek légzőrendszerében ilyen módon megy végbe. Gyakran
azonban valamilyen szállítóanyag végzi ezt a feladatot. Ez az anyag lehet testfolyadék,
vér, illetve nyirok. A testfolyadék (haemolympha) a testüregben vagy nyílt csőrendszer-
ben kering. Sejtjei az oxigén szállítására alkalmas transzportfehérjéket nem tartalmaz-
nak. A vér (sanguis) jellemzője, hogy zárt csőrendszerben keringés oxigénszállítást vég-
ző transzportfehérjéket tartalmaz. A nyirokban (lympha) oxigénszállításhoz szükséges
transzportfehérje nincs. A szövetektől a szív felé haladó csőrendszerben áramlik. Az
említett folyadékok másodlagos feladatként számos egyéb funkciót is elláthatnak: fenn-
tarthatják a test alakját, részt vehetnek a testhőmérséklet szabályozásában, illetve a
szervezet immunrendszerének egy részét alkothatják.
4.3.1. Az anyagszállítás szervrendszerei
Számos állatcsoportnak nincs elkülönült keringési szervrendszere. Szivacsokban és csa-
lánozókban olyan sejtek (vándorsejtek) találhatók, melyek - több más feladat mellett
- anyagszállítási funkciót is ellátnak. Számos féregfajból szintén hiányoznak a keringés
szervei. A testüregben, a szervek között levő folyadék azonban viszonylag szabályos utat
tesz meg és ezért szerepe van az anyagszállításban. A testfolyadék áramlása ebben az
esetben az állatok mozgásának következménye.
Törzsfejlődéstani tanulmányok alapján úgy látszik, hogy kezdetben zárt keringési
rendszer alakult ki az állatvilágban. Ebből az ősi típusból fejlődhetett ki a jelenleg ismert
nyílt és zárt keringési rendszerek sokasága. Mindkét rendszernek vannak előnyei és
hátrányai, tehát nem mondható az, hogy a zárt keringési rendszer specializáltabb, mint a
nyílt. A zárt rendszerben a vér gyorsabban kering, az anyagszállítás sebessége nagyobb, ami
előnyös a gyorsabb mozgás szempontjából. A puhatestűeken belül a zárt keringési
rendszerű lábasfejűek például sokkal gyorsabb mozgásra képesek, mint a nyílt keringésű
csigák vagy kagylók. A zárt keringési rendszerrel sokkal pontosabban lehet irányítani az
anyagok áramlását és a szöveteket sokkal pontosabban és célzottabban lehet ellátni a
szükséges mennyiségű tápanyaggal vagy oxigénnel. Kisebb mennyiségű anyagot kell egy
időben mozgatni, ami energiatakarékos eljárás. Ezzel szemben a nyílt keringési rend-
szerrel bíró állatok szervezetében a tápanyagok eloszlása sokkal egyenletesebb lehet. A
testüregben keringő folyadék részt vehet a test alakjának fenntartásában (hidrosztatikus
váz), vedlés után a test alakjának kialakításában, a szárnyak kifeszítésében (rovarok), sőt
az ugrásban is, azáltal, hogy az állat a testfolyadékát hirtelen a lábakba préseli (ugró pókok).
4.3.2. Nyílt keringési rendszer
A puhatestűek és az ízeltlábúak körében elterjedt rendszer. Szívből (cor), bevezető
(afferens) és kivezető (efferens) erekből áll. Abevezető ereket vénáknak, a kivezető ereket
artériáknak nevezik. Az erek nem alkotnak zárt csőhálózatot, így a testfolyadék a
120
keringési rendszerből a testüregbe jut, majd innen vissza az erekbe. A testfolyadék aránya
az említett gerinctelenek esetében a testtömeg 50%-nál is több lehet.
A rákok és a csigák légzőszervében a testfolyadék oxigénben dúsul és úgy kerül tovább
a szívbe, majd innen ereken keresztül a szervek irányába. A rovarok esetében a légző- és
a keringési rendszer alapjában véve elkülönül. A testfolyadék csupán kis mértékben vesz
részt a szén-dioxid szállításában. A szívből kiinduló aorta a testfolyadékot a fej felé
továbbítja. A pókokra jellemző általános vonás, hogy a keringési rendszer annál fejlet-
tebb, minél fejletlenebb a légzőrendszer. A nyílt keringési rendszerrel bíró állatok szí-
vében aránylag magas nyomás uralkodik (az éti csigánál például 1,6 kPa), viszont az
aortából kiáramló testfolyadék nyomása gyorsan csökken és a testüregben a nyomás már
igen alacsony. Ennek következtében itt az áramlás is lassú. Gyors izomösszehúzódások
(kontrakció) segítségével az áramlás gyorsítható (rovarok).
A nyílt keringési rendszer speciális változata a nyirokrendszer, amely csak a ge-
rincesek szervezetében fordul elő. A gerinceseknél ugyanis a szövetnedveket, az erek
falain átpréselődő nedveket egy sajátos csőrendszer, a nyírokérrendszer gyűjti össze. Ez
a szövetektől indul és egyre nagyobb átmérőjű csöveket képezve végül a fővénákba nyílik.
A nyirokfolyadék nyomása a nyírokerekben sokkal kisebb, mint a vérnyomás. Halak,
kétéltűek és hüllők testében különleges, pulzáló nyirokszivek találhatók a nyirokerek
mentén.
A gerincesek nyirokerei mellett nyíroktüszők (lymphonoduli) helyezkednek el. Ilyen
nyiroktüszők például a mandulák. Ha a nyíroktüszők egy csoportját kötőszöveti tok veszi
körül, akkor nyirokcsomóról (lymphonodus) van szó. Nyirokcsomók az emlősökben
fordulnak elő. A nyiroktüszők és nyirokcsomók a többi nyírokszerwel együtt (lép,
csecsemőmirigy, a csontvelő öblei, a madarak kloákájának falában lévő bursa fabricii) a
szervezet immunrendszerének tagjai. A nyírokszervek alapját retikuláris kötőszövet
(3.2.1. fejezet) alkotja és nagy mennyiségű nyiroksejtet is tartalmaznak. A nyírok, össze-
tételét tekintve, hasonlít a vérhez, de eritrociták, trombociták és nagy molekulájú
fehérjék nem találhatók benne. Jellemző sejtjei a limfociták.
A nyirokrendszemek amellett, hogy a szövetnedveket szállítja a keringési rendszerbe,
egyéb funkciói is ismertek. Fenntartja a sejtekközötti folyadék ionegyensúlyát, részt vesz
a bélcső falán át felszívódó zsírok továbbításában és a sejtekben keletkező fehérjéket a
vérbe szállítja. Elsősorban ebben a rendszerben érnek meg a limfociták, valamint a
leukociták lebontása is itt történik. A nyírokrendszer különlegesen fontos szerepet tölt
be a szervezetben azzal, hogy a testbe bejutó baktériumokat, testidegen fehérjéket vagy
egyéb szerves anyagokat különböző immunfolyamatok révén kiszűri.
4.33. Zárt keringési rendszer
Több állatcsoportban (gyűrűsférgek, gerincesek) a vér zárt csőrendszerben, az érrend-
szerben áramlik. Ennek központja a szív (cor) vagy az érrendszer egyes izmos falú
szakaszai. E szakaszok feladata a vér mozgatása. Ha kitágulnak (diastole), viszonylag
nagy mennyiségű vért fogadnak magukba, majd összehúzódva (systole) az artériákba
préselik azt. A gerincesek szíve egy kötőszövetes burokban (pericardium) foglal helyet.
Két (halak), három (kétéltűek) vagy négy (hüllők, madarak, emlősök) üregből áll. Az
üregeket pitvarnak (átrium) és kamrának (ventriculus) nevezik. A vér a testből vagy a
légzőszervből először a pitvarba érkezik, majd innen jut a kamrába. A szívben zseb alakú
és vitorla alakú billentyűk (valva) találhatók. Ezek irányítják a vér áramlását. A szív
intenzív működését segíti az, hogy saját érrendszere (koszorús erek) látja el tápanyaggal
és igy a nagy oxigénigény könnyebben kielégíthető. A saját ingerképző központok (szi-
nuszcsomó, pitvar-kamrai csomó) biztosítják, hogy a szív az idegrendszertől viszonylag
függetlenül működjék.
121
Az artériák fala izmos és egyben rugalmas. Ez teszi lehetővé, hogy a szívből hirtelen
kiáramló vért folyamatosan, de nagy erővel juttassák a kapillárisokba. Az artériák között
nevezetes ér a főütőér vagy aorta, a kopoltyú artéria, vagy a test felé futó erek első
szakasza. Lehet egy (madarak, emlősök) vagy több (kétéltűek, hüllők) aorta is. Az aorta
gyakran izmos falú kiöblösödéssel, az aortahagymával (bulbus aortae) kezdődik. Az
artériákból szétágazó kapillárisokban a vér nyomása lecsökken, mivel az egy időben
nyitva levő kapillárisok keresztmetszete nagyobb az artériákénál. Ebből következően a
vér áramlási sebessége kisebb lesz és így megfelelő idő áll rendelkezésre az anyagcseréhez,
a gázcseréhez a sejtek és a vér között. A vért ezután a vénák gyűjtik össze. Falukban
kevesebb izomrost van, mint az artériákéban, viszont üregükben a vér visszaáramlását
megakadályozó billentyűk találhatók. A halak esetében a testből érkező erek több gyűj-
tőéren (venae cardinales) keresztül vezetik a vért a vénás öbölbe (sinus venosus). A többi
gerincesben a vénák az üres vénákba (venae cavae) egyesülnek és így jut vissza a vér a
vénás öbölbe vagy a jobb pitvarba. A két vénás rendszer különböző eredetű.
A gerincesek keringési rendszere egy vagy két vérkörből állhat. A halaknak egy
vérkörük alakult ki, a kétéltűek, hüllők, madarak és emlősök körében viszont a kis vérkör
(a szív és a tüdő közötti véráramlás) és a nagy vérkör (a szív és a test egyéb részei közötti
véráramlás) együttesen alkotja a keringési rendszert. Kétéltűek és a legtöbb hüllő szívé-
ben az artériás és a vénás vér még többé-kevésbé keveredik egymással. Lényeges lépés
a kis vérkör és a nagy vérkör teljes elkülönülése egymástól az evolúció során. Ez növeli
az oxigénszállítás hatékonyságát és lehetővé teszi a madarak és az emlősök intenzív
anyagcseréjét.
4.3.4. A gázok szállítása
A gázokat speciális fehérjék (transzportfehérjék) szállítják. Ezek közül a legelterjedtebb
a hemoglobin (gyűrűsférgek, gerincesek vérében található). A hemoglobin négy alegység-
ből álló összetett fehérje. Részben polipeptid (globin) alkotja. Mindegyik polipeptidhez
egy-egy hem-gyűrűrendszer (központjában egy Fe2+-ion) kapcsolódik. Igen alkalmas ez a
felépítés arra, hogy az emlős tüdő léghólyagocskáiban (alveolus), ahol az oxigén parciális
nyomása 13 kPa-nál nagyobb, a hemoglobin oxigént kössön meg, majd a szövetek
közelében, ahol az oxigén parciális nyomása 5 kPa-nál kisebb, azt leadja. Ha a vér pH-
értéke csökken (ez a helyzet a szövetek közötti folyadékban, amikor növekszik a CO2-
koncentráció), a hemoglobin-molekuláról könnyebben válik le az oxigénmolekula.
Gerinctelen állatok körében (különösen puhatestűek, rákok, pókok testfolyadékában)
elterjedt, oxigént szállító vegyület a hemocianin. Felépítése hasonló a hemoglobinéhoz,
de a vasion helyett itt rézion található. Ennek következtében a testfolyadék színe is kékes.
Hasonló feladatú, de kevésbé elterjedt transzportfehérje az egyes gyűrűsférgek vérében
előforduló hemeritrin és klorokruorin is.
Az oxigénnel ellentétben a szén-dioxid szállításában a hemoglobin csak kismérték-
ben vesz részt. A szén-dioxid elsősorban hidrogén-karbonát formában kerül a szövetek-
től a tüdőbe. A hidrogén-karbonát kialakulását a karboanhidratáz enzim gyorsítja. Kis-
emlősökben különösen nagy ennek az enzimnek az aktivitása, összhangban az intenzív
anyagcserével.
4.3.5. Egyéb anyagok szállítása
A keringési rendszer a gázok mellett számos egyéb anyagot (tápanyagokat, egyszerű
vegyületeket, ionokat, hormonokat, fehérjéket stb.) is szállít. Ezek az anyagok a zárt
keringési rendszerben a kapillárisok vékony hámján, valamint keskeny, sejt közötti
réseken, pórusokon keresztül kerülnek felhasználási helyükre, a sejtekhez. A vér alakos
122
elemeinek és a nagy molekulájú fehérjéknek a kivételével az összes többi anyag ezen a
módon jut ki az erekből. Különösen gazdag például a vese ereinek pórusrendszere a
vesetestecskék érgomolyagaiban. Az erekből kilépett anyagok útja ezután rövid: maxi-
mum két-három sejtnyi távolságot kell diffúzió vagy aktív transzport segítségével
megtenniük a felhasználás helyéig, a célsejtig.
Az anyagáramlás iránya a kapilláris erek falán keresztül a nyomásviszonyoktól függ.
Az emlősök esetében például a kapillárisokban a hidrosztatikus nyomás az artériás
szakasztól a vénás felé csökken (4,5 kPa-ról 2,0 kPa-ra) a kolloidozmotikus nyomás
viszont állandó marad (3,3 kPa). Ezért az első (artériás) szakaszon a folyadékok
kiáramlása, a hátsó (vénás) szakaszon pedig a beáramlásuk figyelhető meg.
4.4. Légzés
Az anyagcsere oxidatív folyamatai miatt a sejtek állandó gázcserét bonyolítanak le a
környezetükkel. Oxigént vesznek fel és szén-dioxidot adnak le. A felvett oxigén segítsé-
gével égetik el a szőlőcukrot szén-dioxiddá és vízzé a glikolízis, a citrátciklus és az oxidatív
4.5. ábra. A külső gázcsere főbb módozatai. A: testfelszínen keresztül vízi állatok esetében, B: testfelszí-
nen keresztül szárazföldi állatok esetében, C: tracheakopoltyúval, D: trachearendszerrel, E: kopoltyúval, F:
tüdővel
ka: kapilláris ér, ko: kopoltyú, kü: kültakaró, lé: léghólyagocska, ta: tracheola, te: testi sejtek, tr: tracheacső (Calow nyo-
mán)
123
foszforilálás során. Majdnem mindegyik állatfaj - beleértve az endoparazitákat is -
oxigént vesz fel a környezetéből, és így nyeri az életfenntartásához, a szaporodáshoz
szükséges energiát (aerob anyagcsere). Csupán a fonálférgek egy kicsi (tengerek
mélyén élő) csoportja az, amelyik állandóan oxigénmentes környezetben él és ezért
erjedési folyamatok során jut energiához (anaerob anyagcsere).
A levegőben 21%, a vízben 8% (0 °C-on) az oxigén aránya. Ezekből a közegekből
veszik fel az állatok az oxigént különböző módon, különböző szervekkel, de lényegében
mindig a diffúziós törvények alapján. A gázcsere mértékét a légzési hányados (respi-
rációs kvóciens, rövidítve: RQ) adja meg, ami a leadott szén-dioxid és felvett oxigén
mennyiségének a hányadosa. A gázcsere első lépése a külső légzés, ami a közeg és a
légzőszerv közötti gázcserét jelenti. Ez történhet a kültakarón keresztül, kitüremkedő
kopoltyúval, betüremkedett tüdővel, trachearendszerrel, vagy az említett lehetőségek
kombinációjával (4.5. ábra). A külső légzés után a gázok szállítása következik a fel-
használás helyére, a szövetekhez, ezeken belül a sejtekhez és a sejtorganellumokhoz.
Legegyszerűbben diffúzió útján valósul meg a szállítás, de történhet a keringési szerv-
rendszer segítségével a testfolyadék, illetve a vér által is. Rovaroknál direkt gázszállításról
beszélnek, mert itt a trachearendszeren keresztül közvetlenül juthatnak a gázok a
sejtekig. A belső légzés (más néven sejtlégzés) az oxigén felvételét jelenti a sejt
citoplazmájába, illetve a keletkező szén-dioxid és egyéb gázok leadását a sejt környeze-
tébe.
A következőkben a külső légzés elterjedtebb szervtípusait, a külső gázcsere jellegzetes
módjait ismertetjük (4.5. ábra). Meg kell j egyezni, hogy számos esetben egyidejűleg több
szerv is funkcionálhat (pl. halaknál a kopoltyú, a kültakaró és az úszóhólyag) légzőszerv-
ként.
4.4.1. A légzőszervek típusai
A kültakaró mint légzőszerv
Számos élőlény testfelület/testtömeg aránya olyan nagy lehet, hogy a gázcsere egyszerű
diffúzió útján is megvalósulhat. A Paramecium spp. (csillós egysejtű) fajoknál ez az érték
1000 cm2/g körüli, míg az embernél átlagosan 0,5 cm2/g. Mindez világosan mutatja, hogy
ahol a testfelület/testtömeg arány nagy, nincs feltétlenül szükség elkülönült légzőszervre
a külső légzés lebonyolításához. A laposférgeknek, fonálférgeknek, kerekesférgeknek,
buzogányfejűeknek, számos gyűrűsféregnek többek között ezért nincs légzőszerve. Más
esetekben a bőrlégzés igen jelentős annak ellenére, hogy az állatnak egyéb légzőszerve is
van. A gőtefajok (Triturus spp.) gázcseréjük 70%-át, az angolna (Anguilla anguilla)
légzésének 60%-át képes a kültakarón keresztül lebonyolítani.
Kopoltyúlégzés
A vízi állatok igen elterjedt légzőszerve a kopoltyú. A kopoltyúk lehetnek lemezes,
fonalas, lebenyes, nagy felületű szervek, de felépítésük és működésük meglehetősen
egységes. Két vékony laphámréteg (légzőhám) között találhatók a légzési gázokat
szállító kis átmérőjű, vékony falú erek, bennük a gázcserében részt vevő folyadék. Ha
tracheacsövek hálózzák be a kopoltyút, akkor tracheakopoltyúról, ha vérerek, akkor
vérkopoltyúról van szó. A kopoltyúk kiemelkedhetnek a test felszínéről (rovarlárvák,
ebihal) vagy belső üregekben lehetnek elrejtve (puhatestűek, rákok, halak). A gázcsere
az ellenáram elve alapján történik. A szén-dioxidban dús vér vagy testfolyadék ta-
lálkozik először a friss, oxigénnel telített vízzel, így igen nagyfokú, hatékony gázcsere
lehetséges. Jellemző, hogy a csontos halak a vízben található oxigén 80-90%-át képesek
felvenni, míg az emlősök a tüdőbe került levegő oxigénjének körülbelül 25%-át ve-
szik fel.
124
4.6. ábra. Rovarok trachearendszere. A: a csótány főbb tracheacsövei, B: légzőnyílás felépítése, C: tracheavég-
sejt, D: a tracheolák végződése az izomrostban
en: endokutikula, ft: folyadékot tartalmazó tracheola, ho: hosszanti tracheatörzs, ke: keresztirányú tracheatörzs, ki: kitinléc,
lé: légzőnyílás, It: levegőt tartalmazó tracheola, ma: izomrost magja, mi: mitokondrium, mo: miofibrillum, sl: szarkolemma,
ta: tracheola, tr: tracheacső, vs: tracheavégsejt (Több szerző nyomán)
125
Trachearendszer
A szárazföldi ízeltlábúak, elsősorban a rovarok tipikus légzőszervrendszere. A keringési
rendszertől teljesen elkülönül, így a két rendszer funkciói különválnak. A kültakaró
betüremkedése során keletkezett, amit bizonyít, hogy a vedlés során az ízeltlábúak a
trachearendszer egy részét is levetik.
A trachearendszer elágazó csövek szövevénye (4.6. ábra). A csövek falát kívülről, a
testüreg felől egyrétegű hám borítja, belülről pedig spirális lefutású kitinlécek (taenidia)
merevítik és egyben nagyfokú rugalmasságot is adnak. A trachearendszer a test felületén
található számos nyílással, légzőnyílással (stigma) kezdődik. Ezek a tor és a potroh
szelvényeinek két oldalán találhatók, tehát szelvényenként egy pár figyelhető meg.
Gyakran bonyolult, kitinszőrökből, lemezekből álló záróberendezés található itt, melyet
izmok mozgatnak. A légzőnyílásokból induló csövek a fejlettebb ízeltlábúak testében a
hosszanti lefutású fő tracheatörzsekbe jutnak. A tracheatörzseket harántágak kötik össze,
valamint kiöblösödések, léghólyagok is megjelennek, különösen a jól repülő rovaroknál.
A tracheacsövek azután egyre kisebb átmérőjű csövecskékre oszlanak és gazdagon
behálózzák az egész testet. A csövecskék végén egy sajátos tracheavégsejt ül. Ezen át
futnak a legfinomabb, vékony hajszálcsövek (tracheolae), melyek más sejtek közé vagy
a sejtekbe nyúlnak. Átmérőjük mindössze 0,1-0,5 pm. A bél és a nyálmirigy szöveteiben
a traheolák nem hatolnak be a sejtekbe, hanem a sejt közötti térben végződnek. A zsír-
sejtekbe és a végbél papilláinak sejtjeibe egy-egy csövecske hatol. Kimutatták, hogy az
izomrostokban elágazó rendszerük látszik és a mitokondriumok közelében végződnek a
tracheolák. Ezzel intenzívebb ATP-szintézis válik lehetővé, egységnyi idő alatt több ATP
keletkezik. Különleges a méhek és egyes lepkefajok azon teljesítménye, hogy a nyugalmi
anyagcseréjüket 150-szeresére képesek növelni anélkül, hogy szervezetben oxigénhiány
lépne fel.
A légcserét az izmok összehúzódása, a potroh mozgása idézi elő. A trachearendszert,
a csöveket hosszukban összenyomva a teljes belső térfogat csökken, a gázok egy része a
külvilágba áramlik. Ha visszaáll az eredeti helyzet, megnövekszik a teljes rendszer tér-
fogata, nyomáscsökkenés lép fel benne és a friss levegő beáramlik. Mivel a térfogatvál-
tozás nem nagy, egyszerre kevés levegő cserélődik ki, a gázcsere csekély. (Meg kell je-
gyezni, hogy a légzőmozgások pontos mechanizmusát ma még kevéssé ismerjük.) Ez a
módszer nem teszi lehetővé, hogy a tracheákon keresztül a sejtekhez gyorsan eljusson az
oxigén. A szén-dioxid eltávolítása csak részben történik a trachearendszeren keresztül.
Jelentős szerepet játszik ebben a folyamatban a kültakaró is. Különösen sok szén-dioxid
távozik a testből a szelvényeket összekötő vékony hártyákon (interszegmentális hártyák)
keresztül. Valószínűleg a lassú gázcsere az egyik oka annak, hogy a rovarok nem lehetnek
túlságosan nagy testűek, mivel ekkor a test belsejében levő szervek nem kapnának
megfelelő oxigénellátást.
Tüdőlégzés
A gerincesek tüdeje az előbél kitüremkedése során jött létre, entodermális eredetű,
ellentétben a trachearendszerrel vagy a kopoltyúval, amelyek az ektodermából képződ-
nek. A szárazföldi gerincesek légzőkészüléke a felső légutakból (ormyílás, orr-üreg, száj-
üreg, garat) az alsó légutakból (gége, légcső) és a tüdőből áll. Jellegzetes a tüdő felü-
letének változása az evolúció folyamán. A kétéltűek tüdeje vékony falú, kevéssé tagolt,
nem nagy felületű szerv. A szervezet számára szükséges légcsere lebonyolítására az ilyen
felépítésű tüdő önmagában nem elegendő, ezért annyira fontos a bőrlégzés és a száj
nyálkahártyáján keresztül történő gázcsere szerepe. A hüllők, de kiváltképpen a madarak
és az emlősök tüdejének belső felszíne egyre kiterjedtebbé és differenciálódottabbá válik.
Felülete megnő, mivel kiemelkedések, lécek, csövek jönnek létre benne.
A levegő a felső légutakon és a gégén keresztül jut a légcsőbe. A változatos hosszúságú
126
légcső (trachea) először kettéágazik, majd fagyökérhez hasonlóan szétágazik egyre ki-
sebb átmérőjű csövekre, a hörgőkre (bronchi). Végül a kétéltűek, a hüllők és az emlősök
esetében a légutak a tüdőhólyagocskákban (alveoli) végződnek (4.7. ábra). így a
tüdőbe áramló friss és a tüdőből kiáramló elhasznált levegő bizonyos mértékig keveredik
egymással és a tüdőhólyagocskákban is marad valamennyi elhasznált levegő. Mindez
4.7. ábra. Emlősök tüdejének felépítése. A: a tüdő hosszmetszete, B: a léghólyagocskák, C: a léghólyagocskák
metszete, D: a léghólyagocska falának finomszerkezete
ar: artéria, en: endotelsejt, ep: epitelsejtek, hö: hörgő, ka: kapilláris, kő: kötőszövet, lh: egy léghólyagocska, lé: légcső, Is:
léghólyagocska falának sejtjei, si: simaizom, vé: véna, w: vörösvértest (Több szerző nyomán)
127
4.8. ábra. A madárlégzőszerv vázlata és a légcsere főbb mozzanatai.
A: belégzés, B: kilégzés
el: elülső légzsákok (elhasznált levegőt tartalmaz), fo. főhörgő, ha: tüdőből
származó, használt levegő, há: hátsó légzsákok (friss levegőt tartalmaz),
ki: külső levegő, lé; légcső, tü; tüdő (Kühn és Starck nyomán)
rontja a gázcsere hatásfokát. A
gázcsere hatékonysága ezzel
szemben a madarak tüdejében
jóval nagyobb, mint a többi ge-
rinces tüdejében. Itt a tüdő állo-
mányát nagyrészt a para-
bronchiális egységek alkotják
(4.8. ábra). Ezek rövid csövek,
melyek falán kiöblösödések (at-
ria) találhatók. A kiöblösödé-
sekből indulnak ki a légka-
pillárisok (canaliculi aeripheri),
melyek mindkét végükön nyi-
tott csövek, a gázcsere színhe-
lyei. Mivel a légkapillárisok a
harmadlagos és a másodlagos
bronchusok útján a légzsákokba
nyílnak, egy speciális áramlási
rendszer segítségével belégzés-
kor és kilégzéskor is friss levegő
áramlik rajtuk keresztül (kettős
légzés). A gázcserét a mellkas, a mellkasi izmok, a hasizmok és - az emlősök esetében-
a rekeszizom mozgása teszi lehetővé. Ahol a mellkas hiányzik (pl. békák), az állatok
lenyelik a levegőt.
4.4.2. A külső gázcsere mechanizmusa
A légzéshez nagy felületű légzőhám szükséges. Ez azért van így, mert a gázcsere passzív
módon bonyolódik, a diffúzió törvényeit követi. A gázcsere gyorsaságát a légzőfelület
nagysága, a gázok parciális nyomáskülönbségei, a vérkeringés intenzitása és a légzőhám
vastagsága szabja meg. A légzőfelület/testtömeg arány növelése tette lehetővé, hogy a
madarak és az emlősök anyagcseréje intenzívebb lehet, mint a többi állatfajé. A nyo-
máskülönbség a légzőfelület két oldalán azáltal marad fenn, hogy a légzőhám külső
felszínén a levegő vagy a víz, a belső felszínén pedig a testfolyadék vagy a vér ellentétes
irányban áramlik (ellenáram elve). Tehát a kicserélt gázok eltávoznak és az eredeti
nyomáskülönbségek folyamatosan helyreállnak. A halak kopoltyúinak légzőhámja nagy-
jából 20 pm vastag. Az emlősök léghólyagjainak falában található légzőhám ennél vas-
tagabb. A madaraké viszont csak 0,1 pm, ami segíti, hogy ezek az állatok alacsony
parciális oxigénnyomású viszonyok között is (például nagy magasságokban repülve) ki
tudják elégíteni a szervezet oxigénszükségletét. Egy veréb nagyságú madár 6000 méter
magasan, ahol a levegő oxigénjének parciális nyomása a tengerszinthez viszonyítva
mindössze 50%, még mindig jól repül. Ilyen körülmények között egy hasonló tömegű
emlősállat már nem tudna mozogni.
4.5. Kiválasztás és ozmoreguláció
Az állati szervezetben lezajló anyagcsere-folyamatok végtermékei a szén-dioxid, a
víz és különböző nitrogéntartalmú anyagok. A legfontosabb nitrogéntartalmú anyagok
az ammónia (NH3), a karbamid (NH2)2CO és a húgysav (2,6,8-trihidroxi-purin). A vízi
állatok nitrogéntartalmú anyagcsere-végterméke elsősorban az ammónia, míg a száraz-
128
földieké a karbamid és a húgysav. Kivételek az ászkarákok, mivel elsősorban ammóniát
választanak ki, ezzel is bizonyítva szoros kapcsolatukat vízi rokonaikkal. A kétéltűek és
az emlősök nitrogéntartalmú anyagcsere-végterméke főleg a karbamid, míg a csigáké, a
rovaroké, a hüllőké, a madaraké a húgysav.
A szén-dioxidot és a nitrogéntartalmú anyagokat el kell távolítani a szervezetből,
mert mérgezőek. A szén-dioxid a légzőközpontokat bénítja meg. Eltávolítása a légzés
során könnyen megvalósul. Az ammónia mérgező anyag, különösen a gerincesekre
veszélyes. A gerinctelenek sokkal nagyobb koncentrációban képesek elviselni, mint a
gerincesek. A vízben könnyen oldódik, így eltávolítása a szervezetből egyszerű. A kar-
bamid még az ammóniánál is jobban oldódik a vízben, de nehezen diffundál. Kevésbé
mérgező, mint az ammónia. A húgysav nehezen oldódik vízben, de kevéssé mérgező
anyag. Ezért a szervezetben raktározódhat, nem szükséges rövid időn belül a szervezetből
eltávolítani. Húgysav raktározódik például a rovarok és a százlábúak zsírtestjeiben.
A víz kiválasztását úgy kell szabályozni, hogy a szervezet víz-ion egyensúlya, a
sejtekben és a sejt közötti terekben található ozmotikus nyomás a szükséges határértékek
között maradjon. Az ellentmondás, különösen a szárazföldi állatok esetében az, hogy az
anyagcsere-végtermékeket rendszerint vízben oldott formájukban lehet csak kiválaszta-
ni. Ekkor viszont a mérgező anyagok eltávolítása során vízveszteség következik be, amely
az ozmózisos viszonyokat befolyásolja. Túl sok víz nem adható le, mert kiszárad a szer-
vezet. A vázolt probléma különféle kiválasztószervekkel oldható meg. A legtöbb állatfaj-
ban jellegzetes felépítésű szervek alakultak ki a fölösleges vagy káros anyagok kiválasz-
tására.
A kiválasztási mechanizmus alapjai néhány fő lépésben foglalhatók össze. A
testfolyadékból vagy a vérből szűrés útján (filtráció), vagy aktív transzport, esetleg
szekréció segítségével különböző anyagok kerülnek a kiválasztó szervrendszerbe. Ha
nyomáskülönbség van a kiválasztó szerv valamely belső ürege és a vér, illetve testfolyadék
között, akkor szűrés történik, egyébként (mint például a rovaroknál) aktív transzportfo-
lyamatokra van szükség. Ezután a kiválasztó szerv csatornáiból szelektíven szívódnak
vissza az anyagok. A visszaszívásra hosszú csőrendszer és megfelelően hosszú idő áll
rendelkezésre. Attól függően, hogy a szervezetnek az adott pillanatban mire van szük-
sége, igen finom differenciálásra van lehetőség.
A kiválasztó szervek csatornáinak falában sokszor speciális felépítésű sejtek találhatók.
Rajtuk keresztül jutnak a kiválasztandó anyagok a kiválasztó szervekbe, illetve a vissza-
szívás is segítségükkel történik (transzportfolyamatok). Előfordulnak a Malpighi-edények
falában (rovarok) csakúgy, mint az utóvesében (hüllők, madarak, emlősök) lévő kanya-
rulatos csatornák falában (4.11. ábra). Ezek egymáshoz szorosan illeszkedő hámsejtek.
Apikális végüket citoplazma-kitüremkedések (mikrovilli) borítják, amik igen nagy
mértékben megnövelik a felületet. A sejtek bazális részén sejthártyakettőzetek nyomul-
nak a citoplazma belsejébe (szintén jelentősen megnövelik a felületet). Itt nagyszámú
mitokondrium található. A sejt nagy felülete sok ion és molekula számára teszi lehetővé
egy időben a diffúziót, a mitokondriumok pedig az energiaigényes aktív transzport-
folyamatokhoz szükséges energiát adják.
4.5.1. Akiválasztás szervrendszerei
Elővesécske (protonephridium)
A gerinctelenek körében igen elterjedt kiválasztó szerv. Megtalálható a laposférgek, a
kerekesférgek mindegyik fajában, a fonálférgek és a gyűrűsférgek némelyik csoportjában,
valamint a gyűrűsférgek és a puhatestűek lárváiban. Gazdagon elágazó csőrendszerről
van szó (4.9. ábra). Minden cső végén egy végsejt (cyrtocyta) található. Ez zárja le a
csövecskét. A cső üregébe egy vagy több ostor nyúlik. Ezek mozgásának látványa
129
4.9. ábra. A: elovesécske, B: végsejtje
cs: csilló, gs: gyűjtőcsatorna falának sejtje, gy: gyűjtőcsatorna, ré: sejtek
közötti rés, sm: sejtmag, vs: végsejt (Wehner és Gehring nyomán)
mikroszkópban megfigyelve
emlékeztethet a lángokéra, in-
nen ered a lángsejt elnevezés.
Az ostorok mozgása miatt a
csövecske belsejében található
folyadék kifelé áramlik. Ennek
következtében viszont nyomás-
csökkenés lép fel a csőben, így
a végsejten keresztül, a test-
üregben áramló testfolyadék-
ból a felesleges anyagok (min-
denekelőtt a víz) kiszűrődnek.
A szűrést segítik a sejten vagy a
végsejt és a szomszédos sejt
nyúlványos kapcsolódásánál le-
vő, a két sejthártya között talál-
ható rések. A réseket sejthártya
fedi. Ez biztosítja az ultraszű-
rést. A lángsejtek üregéből az
elsődleges vizelet gyűjtőcsövek-
ben halad tovább. Eközben
visszaszívás és további kiválasz-
tás is történik. A vizelet a kivá-
lasztó pórusokon keresztül jut
a külvilágba. Míg az állatok szer-
vezetéből a víz elsősorban az elovesécske segítségével távozik el, addig a nitrogéntartalmú
bomlástermékek a test felszínén keresztül, diffúzióval távoznak a testből.
Vesécske (nephridium, metanephridium)
Elsősorban a gyűrűsférgek és a puhatestűek jellemző
kiválasztó szerve. Típusos formájában (gyűrűsférgek)
egy csillós tölcsérrel (nephrostoma) kezdődik, majd
hosszú, több szakaszból álló, kanyargós, nem elágazó
csőrendszer következik (4.10. ábra). Ezen több he-
lyen, a végén rendszerint, kiöblösödés, hólyag találha-
tó, majd a kiválasztó póruson keresztül jutnak a külvi-
lágra a fölösleges anyagok. A tölcsér csilléi hajtják a
test üregfolyadékot a cső belsejébe. A hosszú áramlás
során a cső falát alkotó sejtek aktív transzport segítsé-
gével kivonják a szervezet számára még szükséges
anyagokat és visszajuttatják azokat a csövet körülvevő
erekbe vagy a testfolyadékba. Ugyanakkor visszaszí-
vódik a szervezet ozmotikus nyomásának fenntartását
lehetővé tevő vízmennyiség is.
A vesécskének több, erősen módosult változata is-
mert. Ezek felépítése jelentősen eltérő lehet, de emb-
rionális fejlődésük analógiája folytán mégis közös cso-
portba sorolhatók. A puhatestűek kiválasztó szervé-
ben a csillós tölcsér a szívet körülvevő üregbe (peri-
cardium) nyílik és innen folytatódik kanyargó csövek-
ben, kiöblösödésekben. Az ízeltlábúak esetében a mó-
4.10. ábra. Vesécske felépítése
am: amőboid sejteket tartalmazó hólyag,
cs: csillós tölcsér, hh: húgyhólyag, kö: közepes
vastagságú elvezető csatorna, Ös: összhú-
zékony végszakasz, va: vastag elvezető csa-
torna, vé: vékony elvezető csatorna (Geiler
nyomán)
130
dosult vesécskék aszerint kapták a nevüket, hogy a test melyik tájékán helyezkednek el.
így a rákoknál csápmirigyekről és állkapcsi mirigyekről, a pókoknál csípőmirigyekről
beszélnek. Kérdéses, hogy ezek a kiválasztó szervek valóban módosult vesécskék vagy
másféle kiválasztó szervek, mert nem csillós tölcsérrel, hanem üregekkel gazdagon
behálózott, zárt zsákkal (sacculus) kezdődnek. A zsák falát kiválasztásra specializálódott
sejtek, podociták alkotják.
Malpighi-edények
A legtöbb szárazföldi ízeltlábú állat kiválasztószervei a Malpighi-edények. Felépítésük-
ben és működésükben jelentősen eltérnek az összes többi kiválasztó szervtől. Gyakran
a vesécskékkel együtt fordulnak elő (pókok). Vékony, hosszú csövekről van szó, amelyek
a közép- és utóbél határától indulnak ki és vakon végződnek a testüregben. Számuk változó,
a fajokra jellemző, néha több száz is lehet. A csövek falát a béltől indulóan egyre laposodó
hámsejtek bélelik. Eltérően az eddig tárgyalt kiválasztószerv-típusoktól, a Malpighi-
edények falán keresztül csak kiválasztás (excretio) történik, visszaszívás nem. Az
anyagok nem szűréssel (filtratio) kerülnek a csövekbe, hanem diffúzió, aktív transzport és
szekréció segítségével. Ez azért van így, mert a hemolimfa hidrosztatikai nyomása alacsony,
és a Malpighi-edények csöveiben nincsenek csillék vagy ostorok, lassú a folyadékáramlás,
tehát nyomáscsökkentés a csövek üregein belül nem lehetséges. Ennek következtében
viszont a testfolyadék (haemolympha) és a csövekben található folyadék között nincs
olyan nagy nyomáskülönbség, amekkora a szűréshez szükséges lenne.
Különleges szerepük van a káliumionoknak, mivel ezek szekrécióval kerülnek a
Malpighi-edények üregébe, a legvégső, leglaposabb sejtek régiójában. Következéskép-
pen ezen a területen igen magas ozmotikus nyomás alakul ki. Ezért áramlanak ide a víz-
molekulák, immáron passzív módon a testüregfolyadékból. Az alsóbb szakaszokon aktív
vagy passzív módon választódnak ki az ionok, a cukrok, a karbamid és a legfontosabb,
legjellegzetesebb bomlástermék, a húgysav. A kiválasztott húgysav teljes mennyisége az
ürülékbe kerül.
Vese (ren, nephros)
A gerinces állatok kiválasztó szerve a vese. A vese szelvényezett vagy szelvényezetlen
voltát és az elsődleges szűrletet előállító működési egység (nephron) felépítését figyelem-
be véve elővesét (pronephros), ősvesét (mesonephros) és utóvesét (metanephros) lehet
megkülönböztetni. Az elnevezések onnan származnak, hogy az elővese az embrionális
fejlődés során a test elülső részén található szelvényekből, az ősvese a középsőkből, míg
az utóvese a hátulsókból fejlődik ki. Az elővese szelvényezett. A nefron itt egy, az
aortából kiágazó érgomolyból (glomerulus) és a közeli, ezzel szemben nyíló csillós töl-
csérből áll. A tölcsérek vesecsatomákban folytatódnak. Ez utóbbiak az elővese-vezeték-
ben (Wolff-féle vezeték) egyesülnek, ami végül a kloákába torkollik. Az ősvese az
embrionális fejlődés kezdetén szintén szelvényezett, de a szelvényezettség később eltű-
nik. A viszonylag kevés csillós tölcsér mellett számos vesetestecske (Malpighi-testecske)
is megjelenik. Egy vesetestecske egy érgomolyagból és az azt körülvevő, kettős falú tokból
(Bowman-féle tok) áll. Ebből ered a vesecsatomácska. Az utóvese szelvényezetlen
szerv. Benne csillós tölcsérek nincsenek. A nefron itt is a Malpighi-testecske (4.11. ábra).
A Malpighi-testecskék felépítése hasonló, mint az ősvesében, de számuk (relatíve)
lényegesen nagyobb. Ebből következően megnövekszik a szűrőfelület is. így az utóvese
nagyobb mennyiségű vér szűrésére és hatékonyabb kiválasztásra képes, mint az ősvese.
(Hangsúlyozni kell, hogy az előbbi jellemzés csupán néhány általános vonást emel ki,
mert mindhárom vesetípusnak számos változata ismert.)
Elővese található a csontos halak és a kétéltűek embrióiban, valamint a körszájúak
ivarérett egyedeiben is. Kifejlett korban a csontos halak és a kétéltűek veséjének fel-
131
4.11. ábra. Utóvese felépítése. A: sertés veséjének hosszmetszete, B: a metszet egy része nagyítva, C: az elsődleges
kanyarulatos csatorna falának egy sejtje, D: a Malpighi-testecske, E: a Malpighi-testecskében levő ér egy kis
részlete
al: alaphártya, ar: artéria, bo: Bowman-féle tok, ea: elvezető arteriola, el: elsődleges kanyarulatos csatorna, en: a kapilláris
endothel hámja, go: érgomoly, gy: gyűjtőcső, ha: határállomány, ht: hajtűkanyar, hú: húgyvezető, ke: kefeszegély, ké:
kéregállomány, kő: kötőszövetes tok, mi: mitokondrium, oa: odavezető arteriola, po: podocita, ré: a sejtbe húzódó rések, ve:
velöállomány, vé: véna, vm: vesemedence (Több szerző nyomán)
132
építése az ősvesének felel meg. Mivel azonban kialakulásában nem csupán az ősvese,
hanem az utóvese létrejöttéért felelős embrionális szelvények is részt vesznek, azért
speciális ősvesének (opistonephros) tekinthető. A hüllők, madarak és emlősök embrio-
nális korában az elővese és az ősvese is megjelenik, de kifejlett egyedeinek jellemző
vesetípusa az utóvese.
A gerincesek veséje páros szerv, a gerincoszlop két oldalán található. Néha igen
szoros összeköttetésben áll a szaporodás szervrendszerével, azzal anatómiai és működési
egységet képez. Ekkor húgy-ivar készülékről (urogenitális rendszer) van szó (4.12. és
4.13. ábra). A kétéltűek heréje például közvetlenül a vese háti oldalához simul. Az
ondóvezető csatornák a vese fej felé eső területein keresztülhatolnak és ezután csatlakoz-
nak a húgy-ivar vezetékhez (Wolff-cső).
A csontos halak, a kétéltűek és a hüllők veséjében kéregállomány még nem látható, ez
a madaraknál és az emlősöknél jelenik meg. Az emlősök utóveséje kéregállományból
(cortex), határállományból és velőállományból (medulla) áll (4.11. ábra). A Malpighi-
testek a kéregállományban találhatók, míg az ebből kiinduló csatornák mindhárom
rétegen áthaladnak. A velőállomány kicsúcsosodó részeit nevezik vesepiramisoknak. A
csatornák itt nyílnak a vesemedencébe. A vesemedencéből a vizelet a húgyvezetőbe
(urether), majd a húgyhólyagba (vesica urinaria) kerül. Végül a húgycsövön keresztül jut
a vizelet a külvilágba.
Az utóvesében a kiválasztás az érgomolyoknál kezdődik (4.11. ábra). A kapillárisok-
ban nagy nyomás uralkodik, többek között azért, mert az érgomolyból kilépő (efferens)
ér keresztmetszete kisebb, mint az oda belépő (afferens) éré. A szűrés a pórusokkal áttört
endotél sejteken, a rákövetkező alaphártyán és a kapillárisokat borító nyúlványos sejtek
(podociták) közötti réseken keresztül történik. A vérplazma fehérjementes szűrlete
(ultrafiltrátum) kerül a csillós tölcsérbe vagy a Bowman-féle tokba. Ebben az elsődleges
szűrletben az alakos elemeket és a vérben lévő fehérjéket kivéve minden egyéb véralkotó
megtalálható.
Az elsődleges szűrlet egy hosszú, kanyargós csőrendszerben (elsődleges kanyarulatos
csatorna) folyik tovább, amit gazdag érhálózat ölel körül. Már a csőrendszer kezdeti
szakaszán visszaszívódik az erekbe a kiválasztott glükóz és az aminosavak majdnem teljes
mennyisége, a kálium- és nátriumionok, valamint a vízmolekulák nagyobbik része.
Ugyanitt különféle anyagok, például gyógyszerek kerülhetnek az erekből aktív transzport
útján a vese csatornáiba. A vese velőállománya felé futó egyenes csatornaszakaszon víz-
és nátriumionok lépnek ki. A hajtűkanyar után a kéregállomány felé visszaforduló
szakasz fala azonban a víz számára nem átjárható (impermeábilis). Ezért itt az ionok aktív
transzport segítségével jutnak vissza a véráramba. Végül, mivel a vesepiramisok csúcsán,
a gyűjtőcsatornákat körülvevő szövetekben alacsony ozmotikus nyomás uralkodik,
újabb vízvisszaszívás lehetséges és a vérnél sűrűbb (hiperozmotikus) vizelet keletkezhet
(madarak, emlősök). A halak, a kétéltűek és a hüllők veséjében ez a vizeletkoncentráció
nem lehetséges.
4.5.2. Kiválasztás egyéb módon
Az említett kiválasztó szervek mellett más szervekkel és más módokon is lehetséges a
káros vagy fölösleges anyagok eltávolítása. Példa volt már erre a rovarok utóbele, amely
a Malpighi-edényekkel közösen végzi a kiválasztás feladatát. A rákok és a csontoshalak
az ammóniát a kopoltyún keresztül adják le, ahol egyszerű diffúzió útján kerül az
ammónia a vízbe. Vízi rovarok testfelszínén és tengeri halak kopoltyúján fordulnak elő
a kloridsejtek. Mitokondriumokban gazdag sejtekről van szó, amelyek aktív transzport
útján képesek a kloridionokot az ozmotikus nyomással ellentétesen mozgatni, a szerve-
zetből kiválasztani.
133
4.5.3. A kiválasztott anyagok raktározása
Az állatok az anyagcsere végtermékeit sok esetben időszakosan tárolni képesek a test
különböző helyein. Ez a növények világában megszokott jelenség, az állatoknál azonban
már egyáltalán nem olyan általános. A rovarok és a százlábúak zsírtestjeiben húgysav és
a húgysav különböző sói (urátok) halmozódhatnak fel. Ezek egy része később még
visszaszivódhat és az aminosavak felépítésében vehet részt. Viszonylag gyakori, hogy
ezüstszínű anyagcsere-végtermék, guanin halmozódik fel a bél falában (pókok) vagy a
bőrben (halak). Ekkor az állat színének kialakításában, a környezethez való alkalmazko-
dásban fontos szerepet játszik.
4.5.4. Ozmoreguláció
A tengeri állatok testfolyadékának ionösszetétele, sőt az ionok koncentrációja is
feltűnő módon megegyezik a tengervízével. Ezzel szemben a szárazföldi állatok szerve-
zetében az ionkoncentráció némileg megváltozott, de ennek ellenére a mai napig em-
lékeztet arra, hogy őseik a tengerekből származnak. Világosan kiderül ez abból, hogy a
vér és a sejtek közötti (intercelluláris) térben levő folyadék ionösszetétele messzemenően
hasonlít a tenger vízéhez: magas a nátrium- és a kloridionok koncentrációja, viszont ala-
csony a káliumionoké. A sejt citoplazmájában ezzel szemben pontosan fordított a hely-
zet. Annak ellenére azonban, hogy az ionösszetétel különböző a sejthártya két oldalán,
az ozmotikus nyomás rendszerint nem tér el lényegesen egymástól. Ha mégis, akkor
ionregulációs mechanizmusok (ioncsatomák, ionpumpa) működnek és fenntartják a
megfelelő nyomásviszonyokat.
Ha az állatok testfolyadékának és a környezetnek az ozmotikus nyomása közel egyenlő,
izozmotikus fajokról van szó. Ezek tengeri állatok. Az édesvizeket és szárazföldet
azonban csupán úgy tudták meghódítani az állatok, hogy testfolyadékuk ozmotikus nyo-
mását fenntartják (ozmoreguláció). Bizonyos fajok (tengeri rákok és kagylók) a környezet
ozmotikus nyomásváltozásait követik (poikilozmotikus fajok), mások függetlenül a
környezettől állandó nyomást tartanak fenn a szervezetükben (homoiozmotikus fajok).
Ez utóbbi mechanizmus függetleníti legjobban az állatokat a környezet ozmotikus nyomás-
viszonyaitól és segíti elterjedésüket különféle biotópokban. Az édesvízi állatok szempont-
jából a legnagyobb probléma az, hogy a víz ozmotikus nyomása lényegesen alacsonyabb,
mint a testfolyadéké, így folyamatos vízdiffúzió történik a test belseje felé. Ezért a kiválasztó
szerveknek állandóan nagy mennyiségű vizet kell a szervezetből eltávolítaniuk. Előnyös, ha
a szervezet sókoncentrációja olyan alacsony szintű, amennyire csak lehet, mert így
kevesebb energiára van szükség a sóvíz háztartás fenntartásához. A szárazföldi állatok
esetében a legnagyobb veszély a túlzott vízvesztés. Számos stratégia figyelhető meg
ennek megakadályozására. A légzéssel történő vízvesztést csökkenti a belső légzőszerv, a
nedves környezet vagy az éjszakai aktivitás. A kültakaró elszarusodása is hasonlóan
előnyös. A vízben oldhatatlan húgysawal, tömény vizelettel, száraz ürülékkel alig távozik
víz a szervezetből. A sejtlégzés során, számos folyamat végeredményeképpen víz szabadul
fel. Ez a metabolitikus víz is felhasználható. Mindezek a mechanizmusok hozzájárulnak
ahhoz, hogy a fajok elterjedjenek a Földön.
4.6. Szaporodás
A szaporodás során az élőlények önmagukhoz hasonló utódokat hoznak létre. Ez a
folyamat ivarsejtek, ivarszervek nélkül (ivartalan szaporodás) és azok segítségével (ivaros
szaporodás) is megvalósulhat. Az ivartalan szaporodás során létrejövő utódok a szülőhöz
134
genetikai szempontból nagymértékben hasonlítanak. Ezzel szemben az ivaros szaporodás
folyamán a két szülő egyed genetikai állománya keveredik, ezért az utódok genetikai
szempontból változatosak lesznek. Amíg a növényekre az ivartalan, addig az állatokra az
ivaros szaporodásmód túlsúlya jellemző. Az ivaros és az ivartalan szaporodási módok
váltakozhatnak egymással.
4.6.1. Ivartalan szaporodás
Ivarsejtek nélkül történő szaporodás (monogonia) során egyes testi sejtek osztódni
kezdenek, speciális egyedfejlődés kezdődik, végül létrejön az utód. Genotípusukat
tekintve az utódok szüleikhez nagymértékben vagy teljesen hasonlítanak, hiszen az új
sejtek mitózissal keletkeznek. Az egysejtűek körében különösen gyakori az ivartalan
szaporodás, de előfordul az állatoknál is. Az ivartalan szaporodás legfontosabb típusai az
állatvilágban a hasadás és a bimbózás. A hasadás során az állat harántirányban vagy
hosszirányban befíiződik, kettéválik és két, közel egyforma utód jön létre. A harántirányban
történő hasadás a csalánozók, laposférgek és gyűrűsférgek körében is előfordul. A
bimbózás a csalánozókra és egyes gyűrűsférgekre jellemző. Lényege, hogy a test fel-
színének egyes pontjain gyors sejtosztódásokindulnakmeg és egy még kevéssé szervezett
sejttömeg keletkezik. Ez azután vagy leválik az anyaegyedről és úgy fejlődik önálló
egyeddé, vagy együttmarad az anyával és ilyenkor telepek (kolóniák) jönnek létre.
Bimbózás figyelhető meg a galandférgek végleges lárvaformáinál is, amikor hólyagszerű
képződmények belső felszínén jön létre egy vagy több szkolex. Ivartalan szaporodás csak
ott fordul elő, ahol egyes szövetek sejtjei megőrizték totipotenciájukat. Az ivartalan
szaporodás jelenségét használják fel a biotechnológiában akkor, amikor egy vagy néhány
sejtből mesterséges körülmények között genetikailag egyforma, identikus egyedeket
állítanak elő, vagyis klónoznak.
Az élőlények a biológiai evolúció első, leghosszabb szakaszában minden bizonnyal
ivartalan úton szaporodtak. Ez előnyös volt abból a szempontból, hogy gyorsan nagy-
számú utód jöhetett létre egyetlen egyedből is. Ugyanakkor a hatékony szelekcióhoz
szükséges genetikai változatosságot csak a mutációk jelentették. Ezért az evolúció lassú
lehetett, hiszen a mutációk nagy része közömbös, vagy igen gyakran halálos. A jelen
korban azokban a biotópokban lehet előnyös az ivartalan szaporodásmód, amelyekben
a környezeti tényezők értékei kevéssé változnak. Nem meglepő, hogy a viszonylag
állandó környezetet jelentő tengerekben fordulnak elő olyan nagy számban azok az
állatfajok, amelyek életciklusaiban az ivartalan szaporodási mód a döntő.
4.6.2. Szűznemzés
Azt a jelenséget, amikor az ivarsejtek megtermékenyítés nélkül indulnak fejlődésnek,
szűznemzésnek (parthenogenesis) nevezik. Előfordulhat, hogy hirtelen megváltozik a
petesejt környezetének hőmérséklete, pH-ja, sókoncentrációja vagy a petét mechanikus-
inger éri. Ilyen esetekben elkezdődhet a barázdálódás anélkül, hogy megtermékenyítés
történt volna. A partenogenetikus úton létrejövő egyedek rendszerint azért diploidok,
mivel diploid petéből fejlődnek ki, vagy pedig azért, mert az első barázdálódási osztódás
után létrejövő blasztomérák összeolvadnak. Más esetekben (ágascsápú rákok, levéltet-
vek) ahaploidnőstényekhaploidutódokathoznaklétre. Ezek egy részéből (szintén egyes
környezeti tényezők változásainak hatására) hímek alakulnak ki. így a megtermékenyí-
tett pete már diploid lesz.
Sok állatfaj (például kétéltűek, sőt nyulak) petesejtjeit stimulálni lehet, hogy osztódni
kezdjenek. Az így létrejövő aduit egyedek azonban rendszerint nem csupán kisebbek,
gyengébbek, mint az ivarosán keletkező társaik, hanem terméketlenek is.
135
Némely fajnál aszerintlesznekhímekvagynőstényekazutódok,hogymegtermékenyített,
vagy megtermékenyitetlen petéből fejlődnek-e ki. A mézelő méh királynője rajzás után
számos hímmel párzik és 5-6 millió spermiumot raktároz el az ivarszervében. Ha pete-
rakáskor nem termékenyülnek meg a peték, akkor haploid hímek (herék) bújnak ki a
petéből, ellenkező esetben viszont diploid nőstények (királynők, dolgozók).
4.6.3. Ivaros szaporodás
Az ivaros szaporodás (amphigonia) lényege nem a közvetlen egyedszám-növekedés,
hanem az, hogy az utódok genetikai állománya a szülőkétől bizonyosan eltérő lesz. A két,
ellentétes nemi jelleget hordozó ivarsejt lehet egyforma (isogameta) vagy különböző
felépítésű (anisogameta). Ha az ivarsejtek nagysága, felépítése eltérő, akkor a kicsi, a
hímnemre jellemző tulajdonságokat hordozó sejt neve: mikrogaméta, a nagyobb, a női
nemre jellemző tulajdonságokat hordozó sejt neve pedig: makrogaméta.
Az állatok ivarszerveiben alakulnak ki az ivarsejtek. A hímnős (hermafrodita) állatok
testében mindkét ivarkészülék megtalálható, míg a váltivarú állatoknál hím és női
egyedek is léteznek. A hím és a női egyedek közötti külső morfológiai különbségek néha
jelentéktelenek (számos ízeltlábú, kisemlősök), máskor viszont nagyon feltűnők, test-
nagyságban, színezetben is megnyilvánulnak (madarak). Ismert az a jelenség is, amikor
több, ivarhoz kötött forma fordul elő egy fajon belül (pl. méhek).
Az ivarszervek rendkívül változatos morfológiai képet mutatnak. Elhelyezkedhetnek
ízenként ismétlődve (galandférgek), alakjuk hasonlíthat hosszú, fokozatosan elkeske-
nyedő fonalhoz (fonálférgek), lehet páros (ízeltlábúak, gerincesek) vagy páratlan (női-
varú madarak). E nagy formagazdagság ellenére, funkcionális szempontból az ivarszer-
vek egységes szakaszokra tagolhatok és ezeknek a szakaszoknak hasonló a nevük a
legkülönbözőbb állatcsoportokban.
A női ivarszerv a petefészekkel (ovarium) kezdődik (4.12. ábra). Ez az oogenezis
helye. Lehet egységes szerv, de tagolódhat csövecskékre is (rovarok). Az oogenezis
4.12. ábra. Női urogenitális rendszerek. A: kétéltű, B: hüllő, C: emlős
em: embrió, ev: elővese, hú: húgyhólyag, hu: hüvely, iv:ivamyílás, ki: kloáka, mé: méh, mü: Müller-féle vezeték, őv: ősvese,
pf: petefészek, pv: petevezető, to: tojás,ub: utóbél, uv: utóvese, vb: végbél, wo: Wolf-féle vezeték (Gerber nyomán)
136
szaporodási, növekedési és - legalább részben - az érési szakasza itt megy végbe (2.1.1.
fejezet). A legtöbb gerinctelen esetében a petevezető (oviductus) a petefészekhez köz-
vetlenül csatlakozik. A gerincesek körében (a halakat kivéve) az jellemző, hogy a
petefészekhez a petevezető funkcionálisan kapcsolódik a petetranszportot végző mecha-
nizmusok által, de nincsen összenőve azzal. A petevezető rendszerint hosszú, izmos falú
cső. Belsejében a pete passzívan mozog: csiliók és a petevezető falának összehúzódásai
hajtják a méh felé. A méh (uterus) gyakran igen terjedelmes (laposférgek) és a meg-
termékenyített petéket raktározza. Máskor az embrionális fejlődés színhelye (emlősök).
A méhhez párzótáska (bursa copulatrix) csatlakozhat (laposférgek, rovarok). Ide jut a
sperma a párzás során, majd itt raktározódnak a spermiumok. A női ivarszerv utolsó
szakasza a hüvely (vagina), a kloákába, illetve közvetlenül a külvilágba nyílik. Az
ivarnyílás környékén tojócső lehetséges (rovarok, halak). Ennek segítségével az állatok
petéiket rejtett helyekre, kövek alá, résekbe helyezhetik, így a petéből kikelő fiatal állatok
túlélési esélyei jelentősen megnőhetnek.
A hímivarszerv a herével (testis) kezdődik (4.13. ábra). Ez a spermiogenezis helye
(2.1.2. fejezet). Előfordulhat, hogy egy állatnak több száz heréje van (laposférgek), de
lehet viszonylag egyszerű, páros szerv is (gerincesek). A herékből kivezető csatorna
(ductus efferens) az ondóvezetőben folytatódik (ductus deferens), ami az ondóhólyagba
(vesicula seminalis) torkollik. Az ondóhólyag hiányozhat is (fonálférgek, puhatestűek,
rákok). Innen az ondókilövellő járat (ductus ejaculatorius) vezeti tovább a spermát
rendszerint a párzókészüléken (penis) keresztül a külvilágba. Azoknak az állatoknak
nincs párzókészülékük, amelyeknél külső megtermékenyítés figyelhető meg (gyűrűsférgek,
halak, kétéltűek). A rovaroknál igen komplikált felépítésű a párzószerv. A rákok és a
pókszabásúak lábai úgy módosultak, hogy a spermát rejtő tokokat a nőstények ivar-
készülékébe tudják juttatni. Számos állat ivarkészülékéhez különböző járulékos szervek
és mirigyek kapcsolódhatnak. Az emlősök hímivarszerveiben ezek a következők: a
mellékhere (epididymis), a már említett ondóhólyag (vesicula seminalis), a dülmirigy
(prostata), a Cowper-féle mirigy (glandula bulbourethalis) és az ampulláris mirigyek. A
mirigyek váladéka táplálja, aktiválja és szállítja a spermiumokat.
4.13. ábra. Hím urogenitális rendszerek. A: kétéltű, B: hüllő, C: emlős
ev: elővese, he: here, hű: húgyhólyag, ki; kivezető csatorna, ki: kloáka, me: mellékhere, mü: Müller-féle vezeték, oh: ondó-
hólyag, ov: ondóvezető, őv:ősvese, pr: prosztata, uv: utóvese, vb: végbél, wo: Wolf-féle vezeték (Gerber nyomán)
137
Az ivaros szaporodásmód bonyolult és energiaigényes folyamat. Nagyszámú
ivarsejtet kell létrehozni ahhoz, hogy a viszonylag kevés utód létrejöhessen. Rengeteg
pete, spermium és fiatal állat pusztul el, amíg az új, szaporodásra képes egyedek
kialakulnak. Egy ponty nőstény több millió petét termel egy szaporodási ciklusban, de
ezekből - természetes viszonyok között - legfeljebb néhány száz utód lehet. A peték
sziktartalma nagy, tehát sok anyag és energia szükséges a képzésükhöz. Az emlősök
ugyan kevesebb utódot szülnek, viszont a tejtermelés, a fiatalok védelme ugyancsak sok
energiát kíván, elsősorban az anyától.
Mint látható, az ivaros szaporodás energiaigénye igen magas, mégis fennmaradt az
evolúció során, tehát számottevő előnyének is kell lenni, ami ellensúlyozza a ráfordítá-
sokat. Ilyen előny az utódpopuláció fokozott genetikai variabilitása. Változó környe-
zetben a változatos génállomány a populáció (faj) fennmaradásának biztosítéka. Bizo-
nyítékok vannak arra vonatkozóan, hogy a meiozis során megsérülő DNS eredeti
bázisszekvenciájának helyreállítása a zigótában megtörténhet. Ez fontos előnye ennek a
szaporodási módnak.
Az ivaros szaporodásmód különösen előnyös lehetett a biológiai evolúció azon
szakaszaiban, amikor a környezeti változások gyorsak voltak, nem kis mértékben azért,
mert a kialakuló, egyre változatosabb életmódú élőlények is alakították saját élőhelyüket,
a bioszférát. Egyes kutatók azt állítják, hogy az ivaros szaporodásmód már régen kialakult
és rögzült úgy, hogy ma már ez stabilan létező szaporodási forma. Mások szerint az ivaros
szaporodás által létrejövő genetikai változatosság annyira fontos napjainkban is, hogy a
ráfordítások (energiafelhasználás, nagyobb kockázata annak, hogy zsákmányul esik az
állat stb.) még így is kisebbek, mint a várható haszon, és ezért maradt fenn ez a
szaporodásmód. A kérdés továbbra is nyitott.
4.6.4. Nemzedékváltakozás
Számos gerinctelen állatcsoportban megfigyelhető, hogy egyes fajok egyedei ivaros és
ivartalan módon egyaránt szaporodhatnak. Ez a képességük igen előnyös a következők
miatt: állandó környezetben ivartalan módon, gyorsan, nagyszámú, de genetikailag
hasonló utódot képesek létrehozni. Ugyanakkor, ha a környezeti tényezők számukra
kedvezőtlenre fordulnak, ivaros szaporodás történik. Ekkor az utódok genetikai szem-
pontból változatosak lesznek, tehát nagyobb a valószínűsége annak, hogy a populáció
számára kedvezőtlen körülményeket egyes egyedek átvészeljék.
A nemzedékváltakozásnak két alapvető formáját szokták megkülönböztetni, a
heterogoniát és a metagenezist. A heterogonia azt jelenti, hogy az egyféle ivarból álló
és a kétféle ivarból álló generációk (nemzedékek) váltják egymást. A szűznemzéssel
szaporodó, csak nőstényekből álló generációkat ivarosán szaporodó, mindkét nemű
állatokból álló generáció váltja fel. Ez a jelenség gyakori a kerekesférgek (Rotatoria), az
ágascsápú rákok (Cladocera) és a növénytetvek (Stemorrchyncha) körében. A metage-
nezis során az ivartalanul és az ivarosán szaporodó generációk váltják egymást. Ez akkor
lehetséges, ha a faj valamelyik formája ivartalan szaporodásra (hasadás, bimbózás)
képes. Ilyen rendszer figyelhető meg a csalánozók (Cnidaria), némely galandféreg
(Cestodes) és soksertéjű gyűrűsféreg (Polychaeta) esetében.
4.7. Endokrin szervek rendszere
A szervezetben a sejtek kémiai kommunikációja, a szervek, szervrendszerek vegyi
szabályozása a hormonok segítségével valósul meg. A hormonok egyes sejtek, sejtcso-
portok vagy a belső elválasztású mirigyek olyan szekrétumai, amelyek speciális anyag-
138
csere-folyamatok eredményeképpen keletkeznek, a termelő sejt vagy más sejtek specifi-
kus működését váltják ki. Leggyakrabban a keringési rendszer segítségével jutnak el a cél-
sejtekhez. A célsejt felszínén vagy a sejt citoplazmájában csak az adott hormont fogadó
kötőhely, receptor található. A hormonok kémiai szerkezete igen változatos, és moleku-
latömegük lényegesen eltér egymástól. Egy adott hormon szerkezetében történő csekély
változás következtében hatása lényegesen megváltozik. Már igen kis koncentrációban
(10 6-10~12 mol/liter) hatásosak. Kémiai szempontból a következő fő hormoncsoportok
különböztethetők meg.
Lehetnek:
a) aminosav-származékok (pl. tiroxin, adrenalin, noradrenalin, melatonin),
b) különböző proteinek és proteidek (pl. a hipofízis, mellékpajzsmirigy és hasnyál-
mirigy hormonjai, a szöveti hormonok),
c) szteroidok (pl. a mellékvese kéregállományának, az ivarszerveknek a hormonjai),
d) arachidonsav-származékok (pl. a minden szövetben termelődő prosztaglan-
dinok), valamint
e) juvenilis hormonok.
Megjegyzendő, hogy kellően általános és pontos hormondefiníciót nehéz megfogal-
mazni. A noradrenalint például szinaptikus ingerületátvivő anyagnak kell tekinteni, ha
az axonban termelődik, de a hormonok közé sorolandó, ha a mellékvesevelőből szabadul
fel.
A hormonok az érzékszervekkel és az idegrendszerrel együtt a szervezet működését
koordinálják. Az információ átvitel hormonális módja lényegesen eltér az idegrendszer
működésétől. Mivel a hormonok a keringési rendszer útján jutnak el egyik sejttől vagy
szervtől a másikig, hatásuk viszonylag lassú, és az idegrendszeri hatásokhoz képest tartós.
Szállítás közben egyes hormonok (szteroidok, pajzsmirigy hormonjai) speciális transz-
portfehérjékhez kötődnek. Ezek megakadályozzák a hormonok elbomlását és hatásukat
is fokozhatják. Ugyancsak a terjedés, szállítás módjából következik, hogy a hormonok
kémiai struktúrájának, természetének igen specifikusnak kell lenni, mert a testfolyadék-
ban található számos vegyület közül a sejtek specifikus receptorai csak ebben az esetben
ismerhetik fel a hormonokat. Ugyanez az idegrendszer esetében nem ennyire fontos,
hiszen ott az idegsejtek felépítése és működése, az ingerületvezetés sajátos rendszere (és
nem az átvivő anyagok kémiai típusa) teszi lehetővé, hogy az információk kellően
változatosak legyenek.
A sejtek közötti kémiai kommunikációnak többféle módja ismert aszerint, hogy hol
termelődik a hormon és milyen úton kerül a célsejtekhez.
1. A helyi hatású hormonok az egymás mellett lévő sejtek fiziológiai állapotát
változtatják meg, de nem kerülnek a véráramba. így működnek a zsírsavakból
keletkező prosztaglandinok. Ezeknek a vegyületeknek számos hatásuk van. Befo-
lyásolhatják a simaizmok összehúzódását és elernyedését. Részt vesznek az ivari
ciklusok kialakításában, a táplálékok okozta allergiás reakciók megjelenésében. A
lázas állapot kialakításában is szerepük van. (Az Aspyrin úgy csökkenti a lázat, hogy
a prosztaglandin-szintézist gátolja.)
2. A neurohormonok speciális idegsejtekben (a neuroszekréciós sejtekben) terme-
lődnek. Innen a testfolyadékba vagy a véráramba kerülve jutnak el a célsejtekig. A
hormont termelő idegsejtek nyúlványai végződhetnek a keringési szervek falában
vagy egy erekkel dúsan átszőtt szervben, a neurohemális szervben is. Ekkor a
hormonok itt kerülnek a véráramba vagy a testfolyadékba. Neuroszekréciós sejtek
termelik például a rovarokban az előtöri, protorakális mirigy működését szabályozó
hormont (prothoracotroph hormon), a vedlési mozgások stimulálását végző
hormont (eclosion), a vedlés után a kültakaró keményedését gyorsító hormont
(bursicon), a gerincesek méhizmainak összehúzódását és a tejleadást serkentő
139
hormont (oxitocin), valamint a vizelet koncentrációját szabályozó hormont
(antidiuretikus hormon).
3. Egyes hormonok elkülönült szervekben, a belső elválasztású mirigyekben
képződnek és a keringési rendszer segítségével jutnak el a célsejtekhez. A gerinctelenek
körében aránylag kevés csoport rendelkezik elkülönült belső elválasztású miriggyel,
legtöbbször kisebb-nagyobb, más szövetek közé beágyazódott sejtcsoportok látják
el ezt a feladatot. A gerincesekben viszont 7-8 jól felismerhető szerv, belső
elválasztású mirigy figyelhető meg.
4.7.1. Hatásmechanizmusok a célsejteknél
A hormonok hatásukat csak a célsejteknél váltják ki. Célsejteknek nevezik azokat a
sejteket, amelyek az adott hormon felismerésére, megkötésére alkalmas helyekkel
4.14. ábra. Közvetlen hatású hormonok hatásmechanizmusa
F: szállító fehérje, H; szteroid hormon, K: kromoszóma, M: messenger-RNS,
R: receptor, ak: aktivált hormon-receptor komplex, sh: sejthártya, sm:
sejtmag
4.15. ábra. Közvetett hatású hormonok hatásmechanizmusa
A: adenil-cikláz, H: N-tartalmú hormon, R: receptor
rendelkeznek a sejthártyán vagy
a citoplazmában, és amelyek-
ben a hormon meghatározott
biokémiai reakciókat indít el.
A hormonok alapjában véve
kétféle módon, közvetlenül
vagy közvetve irányítják a sej-
tek egyes biokémiai reakcióit.
A közvetlen hatású hormonok
a citoplazmába, majd a sejt-
magba jutva az mRNS-szin-
tézist befolyásolják (szteroi-
dok). A közvetett hatású hor-
monok a sejthártya speciális
receptoraihoz kötődnek (de
nem jutnak a sejt belsejébe) és
ezzel egy folyamatot indukál-
nak. E folyamat végeredmé-
nyére a hormonok közvetlenül
már nem hatnak (proteinek és
proteidek, aminosav-szárma-
zékok).
A közvetlen hatású, zsír-
ban oldódó hormonok (ép-
pen e tulajdonságuknál fogva)
átjutnak a sejthártyán és a ci-
toplazma speciális receptorfe-
hérjéihez kötődnek (4.14. áb-
ra) . Ez a komplex azután meg-
határozott DNS-részlethez kö-
tődik és ott mRNS-képződést
vált ki. Következménye a spe-
ciális fehérjék képzése. Először
árvaszúnyog- (Chironomus
spp.) lárvák kromoszómáin
mutatták ki, hogy az ekdizon
hormon hatására egyes génhe-
lyek aktiválódnak. A közve-
140
tett hatású hormonok a sejthártya felszínén található receptorfehérjékhez kötődnek
(4.15. ábra). Ennek következményeképpen a receptorfehérjék aktiválnak valamilyen
közvetítő molekulát (gyakran a GTP-áz fehérjét). A közvetítő molekula aktiválja azután
a sejthártya belső felszínén található adenilát-cikláz enzimet, aminek hatására az ATP-
ből ciklikus AMP keletkezik. (Ismert, hogy a sejthártya közvetítő molekulái az adenilát-
ciklázon kívül hathatnak más enzimekre is és így egyéb sejten belüli folyamatokat
indukálhatnak.) A ciklikus AMP hatására a citoplazma enzimjei (leggyakrabban pro-
teinkináz) aktiválódnak és ezért megváltoznak az anyagcserében résztvevő biokémiai
reakciók.
4.7.2. Endokrin szervek és hatásmechanizmusok
Az ősibb gerinctelenek (laposférgek, fonálférgek, gyűrűsférgek, kerekesférgek, puha-
testűek) hormonjait legtöbbször az idegrendszerben, elsősorban a különböző idegdú-
cokban található speciális idegsejtcsoportok termelik (neuroszekréció). Egy meztelen
csiga faj (Aplysia sp.) hasi idegdúcaiban az idegsejtek olyan hormonokat termelnek,
amelyek részben a környező idegsejtek működését befolyásolják (hatás az idegrend-
szerre), részben a petevezető izmainak összehúzódását serkentik. A két hatáscsoport
együttesen váltja ki a peterakást. Mesterségesen, e sejtek kivonatának befecskendezésé-
vel elérhető, hogy az állatok lerakják a petéjüket még akkor is, ha sem párzás, sem
megtermékenyülés nem történt.
Az ízeltlábúak körében gyakori, elkülönült neuroendokrin szervek a következők:
az X-szerv-szinuszmirigy komplexum és az Y-szerv a rákokban, az előtöri mirigy, a
corpus cardiacum és a corpus allatum a rovarokban.
Az X-szerv-szinuszmirigy komplexum számos magasabb rendű rák (Mala-
costraca) szemnyelében fordul elő. Az X-szerv neuroszekréciós sejtek csoportja. Ezek
a sejtek váladékukat a szinuszmirigybe juttatják, ahol az raktározódik. A komplexum
hormonjai elsősorban a vedlés szabályozásában játszanak fontos szerepet, de részt
vesznek a nemi érés, a szénhidrát-anyagcsere, az ozmoreguláció és a színváltoztatás
szabályozásában is.
Az Y-szerv a rágok töve mögött található (16.3. ábra). Nem idegi eredetű szerv,
hanem a kültakaróból származik. A rákok vedlési hormonját (crustecdyson) termeli.
Ennek hatására megindul a vedlés. Az Y-szervnek része van az ivari működések szabá-
lyozásában is.
A rovarok előtöri, protorakális mirigye lapos, néhány sejtsorból álló, lebenyes fel-
építésű szerv. Hormonja a vedlési hormon vagy más néven ekdizon. Az előtöri mirigy
csak a lárvákban létezik, a báb állapot alatt szövete szétesik, a szerv eltűnik a szervezetből.
A corpus cardiacum páros, neurohemális szerv. Az agy mögött, a nyelőcső háti
oldalán helyezkedik el. Ide futnak az előagy ún. pars intercerebralis területén található
idegsejtek nyúlványai, melyeken keresztül az előtöri mirigy működését serkentő hormon
(prothoracotroph hormon) jut a corpus cardiacumba. Itt raktározódik, illetve innen jut
a testfolyadékba. A corpus cardiacum találhatók még a vízürítést növelő, az anyagcserére
ható, a szív összehúzódását és a bélperisztaltikát fokozó hormonok is.
A corpus cardiacum mögött helyezkedik el a corpus allatum. A két mirigy egymással
idegi összeköttetésben áll. Önálló hormontermelése is van, nevezetesen ebben a szerv-
ben képződik a juvenilis hormon, ami a normális lárvakori fejlődéshez szükséges. Ivari
működést serkentő hormonokat is mutattak ki ebből a szervből.
Az ízeltlábúak körében a neurohormonális szabályozás bonyolult rendszerei isme-
retesek. Gyakorlati jelentősége miatt a vedlés szabályozásának mechanizmusát különös
alaposságai tanulmányozták. Kiderült, hogy ugyanazon jelenség, a vedlés, egészen más
hormonális szabályozás alatt áll a rákok és a rovarok szervezetében. A legfontosabb
141
4.16. ábra. Rákok ved-
lését és ivarérését irá-
nyító belső elválasztásig
mirigyek és hatásaik
(Több szerző nyomán)
mechanizmusok a következők. A tízlábú rákok vedlése (4.16.
ábra) külső és belső tényezők együttes hatására indul meg. AzX-
szerv neuroszekréciós sejtjeinek aktivitása csökken, ezért a szi-
nuszmirigyben egyre kevesebb hormon lesz, egyre kevesebb
juthat a testfolyadékba. A vedlés tehát akkor kezdődik, amikor
az X-szerv nem termel hormont, tehát nem blokkolja az Y-szerv
működését. Ha a szemnyeleket eltávolítják, az állatok folyama-
tos vedlésekbe kezdenek. A rovarokban ezzel szemben akkor
következik be a vedlés, amikor a testfolyadékban a vedlés sza-
bályozásában részt vevő hormonok koncentrációja és az egy-
máshoz viszonyított arányuk megváltozik (4.17. ábra). A me-
chanizmus vázlata a következő: különböző ingerek (pl. a nappa-
lok és éjszakák hosszának változása miatt, táplálékfelvétel követ-
keztében feszülő potroh miatt) a pars intercerebralisban levő
sejtek intenzív hormontermelésbe kezdenek. A protorakotróf
hormon a corpus cardiacumon át a testfolyadékba kerül. Eljut az
előtorba és serkenti az előtöri mirigy ekdizontermelését. Az
ekdizon hatására indul a vedlés. Ellentétes hatást vált ki a corpus
allatum hormonja (juvenilis hormon), mivel gátolja a vedlést,
viszont serkenti a lárvakori növekedést. A juvenilis hormon a
fiatal lárvák testfolyadékában egy lipoproteinhez kötve találha-
tó, ezért nem bomlik le. Idős lárvákban, majd a bábban a corpus
allatum működése, a juvenilis hormon termelése fokozatosan
csökken, majd az imágóban teljesen megszűnik.
A rovarok vedlésének kezdetekor a kutikula elválik az epidermiszsejtektől és a sejtek
inaktív emésztőenzimeket választanak ki a kutikula felőli, apikális végükön (4.18. ábra).
A sejtek, valamint a kutikula közötti térben vedlési
folyadék halmozódik fel. Alatta kialakul az új kutikula
első rétege (kutikulinlemez). Ez aktiválja a régi kutiku-
lát bontó enzimeket és megakadályozza az epidermisz-
sejtek önemésztését. Ezzel párhuzamosan kicsi lapocs-
kák (membránplakkok) alakulnak ki a sejtek felszí-
nén, amelyek a keletkező kutikula fehérjehálózatai-
nak kialakításáért felelősek. A kutikulin-lemeztől ki-
felé a lebontófolyamatok, a test felőli térben pedig az
építő folyamatok mennek végbe. A régi endokutikula
anyagának nagy része visszaszívódik és részt vesz az új
kutikula felépítésében. A jelenség példa arra, hogy
egy gazdaságos, anyagtakarékos fiziológiai folyamat
elősegítheti egy állatcsoport, jelen esetben az ízeltlá-
búak sikerességét az evolúció során. Az epikutikula és
az exokutikula a fajokra jellemző helyeken felhasad.
Ezeken a nyílásokon keresztül, jellegzetes mozgások
sorozata segítségével bújik ki az állat. A kibújáshoz
szükséges mozgások az eklozion hormon hatására
kezdődnek meg. Ez a hormon az agyban termelődik,
de a corpus cardiacumban raktározódik. Az új kutikula
megkeményedéséért a burzikon nevű hormon felelős.
4.17. ábra. Rovarok vedlését és ivar-
érését irányító belső elválasztású miri-
gyek és hatásaik (Több szerző nyomán)
A gerincesek belső elválasztású (endokrin) miri-
gyei az agyalapi mirigy (hypophysis), a pajzsmirigy
(glandula thyroidea), a mellékpajzsmirigy (glandula
142
4.18. ábra. Rovarok vedlésének mechanizmusa
ak: aktív kutikulabontó enzimek, ek: az új kutikula endokutikulája, en: endokutikula, er: endoplazmatikus retikulum, ep: új
epikutikula, in: inaktív kutikulabontó enzimek, ku: kutikulinlemez, li: lizoszóma, Ik: lebomló kutikula, me:membránplakk,
pr: prokutikula, rk: a régi kutikula visszaszívott anyagait tartalmazó vakuólum, sm: sejtmag (Darvas nyomán)
parathyroidea), az ultimobranchiális szerv, a hasnyálmirigy Langerhans-féle sejtcso-
portjai, a mellékvese (glandula suprarenalis), a csecsemőmirigy (thymus) és a tobozmi-
rigy (corpus pineale). A következőkben ezeket tárgyaljuk.
A hormonrendszer és idegrendszer kapcsolatának legismertebb példája a gerincesek
hipotalamusz-hipofízis rendszere. Az agyalapi mirigy vagy hipofízis egy rövid
nyélen keresztül összeköttetésben áll a hipotalamusszal, azzal egységes hipotalamusz-
hipofízis rendszert alkot. Ez szövettani és működési kapcsolatot is jelent. A hipotalamusz
egyes idegsejtjeinek nyúlványai (az ún. nagysejtes magok sejtjeinek axonjai) ugyanis az
agyalapi mirigy hátul só lebenyében végződnek és hormonjaikat neuritjaikon át közvetlenül
ide juttatják. A hipofízis más idegsejtjeinek (ún. kissejtes magok) váladéka egy speciális
érhálózaton, a hipofízis portális érrendszerén keresztül jut az elülső és középső lebenyek-
be, ahol az ott termelődő hormonok szintézisét fokozza vagy gátolja. A hipofízis elülső
és középső lebenye az egyedfejlődés során az embrionális garatüreg kitüremkedéséből
jön létre, a hátulsó lebeny viszont a köztiagy (diencephalon) nyúlványából alakul ki.
Az elülső lebeny (adenohypophysis) hatféle hormont termel. Ezek a következők:
növekedést serkentő, szomatotrop hormon (STH), a mellékvesekéreg működését ser-
kentő, adrenokortikotrop hormon (ACTH), a pajzsmirigy működését serkentő tireotrop-
hormon (TSH), a tüszőérést stimuláló hormon (FSH), az ovulációt, illetve a tesztoszteron-
szintézist serkentő luteinizáló hormon (LH), valamint a tej elválasztást serkentő prolaktin
(LTH). Az elülső lebeny hormonjai hathatnak közvetlenül az anyagcserére, vagy közvetet-
ten más szervek, belső elválasztású mirigyek működését szabályozva. Közvetlen hatású a
növekedést serkentő hormon, a többi viszont közvetetten hat más belső elválasztású
mirigyekre, az ivarszervekre, a pajzsmirigyre és a mellékvese kéregállományára. A középső
lebeny (pars intermedia) hormonja, a melanotropin (MSH, más néven: intermedin) a sötét
színanyagok termelését fokozza. Hatására terjednek szét a pigmentszemcsék a melanofor
sejtek citoplazmájában. A madarak és az emlősök agyalapi mirigyéből ez a rész hiányzik.
A hátulsó lebenyben (neurohypophysis) a hipotalamusz neuroszekréciós sejtjei által
termelt két hormon raktározódik. Az antidiuretikus hormon (ADH) a vizelet koncentrá-
cióját határozza meg, az oxitocin pedig a méh simaizmainak összehúzódását stimulálja.
A pajzsmirigy tiroxint és trijód-tironint termel. A legtöbb sejt alapanyagcseréjét
fokozza, de különösen az izmok, a szív és a máj működését befolyásolja. Hat a szervezet
növekedésére és fejlődésére is. A kétéltűek metamorfózisát gyorsítja.
143
A mellékpajzsmirigyben jön létre aparathormon. A szervezet kalciumanyagcseréjét
szabályozza. A vér kalciumszintjét növeli, mivel fokozza a kalcium felszívódását a bél
falán keresztül, mobilizálását a csontokból, visszaszívódását a szűrletből. Mellék-
pajzsmirigy halakban nincsen.
Az ultimobranchiális szerv működés szempontjából a mellékpajzsmirigy antago-
nistája. Az itt termelődő kalcitonin hatására a vér kalciumszintje csökken, mert fokozó-
dik a kiválasztás és a csontokba történő beépülés. Emlősökben ez a szerv nem található
meg. Itt a pajzsmirigy parafollikuláris sejtjei felelnek meg a többi gerinces ultimobranchiális
szervének. Ezek a sejtek tireokalcitonint termelnek.
A hasnyálmirigyben elszórtan kishormontermelő sejtcsoportok, azún. Langerhans-
féle szigetek találhatók. A sejtek többfélék. Az alfasejtek termelik a glukagonhormont,
amely a máj működésére hat. Serkenti a glikogén lebontását, a glükóz felszabadulását a
májból. A bétasejtek az inzulint termelik. Az inzulin minden szövetféleségben hathat. A
glukagonnal ellentétes hatású. Serkenti a glikogén felépítését glükózból, az izomsejtek
glükózfelvételét fokozza, de befolyásolja a fehérjék és a zsírok szintézisét is.
A mellékvese a halakban még nem egységes szerv. Itt a vesében található kétféle
sejtcsoportból, az adrenális és az interrenális sejtek csoportjából áll. A mellékvese elkü-
lönült szervként a farkos kétéltűeknél jelenik meg, de az adrenális és interrenális sejtcso-
portok még keveredve helyezkednek el. Kéreg- és velőállomány pedig csak az emlősöknél
figyelhető meg. Az interrenális sejtek a kéregállományban, az adrenális sejtek a velőál-
lományban csoportosulnak. Az interrenális sejtek glükokortikoidokat (pl. cortison, cortisol,
corticosteron) termelnek. Ezek a hormonok a szénhidrát-anyagcserében vesznek részt. A
legtöbb szövetben hatásosak. Segítik a szénhidrátok előállítását fehérjékből és zsírokból.
Csökkentik a szervezetben a gyulladásos reakciókat. Ugyancsak az interrenális sejtek
termékei a mineralkortikoidok (pl. aldosteron). Avese működésére hatnak. Azt szabályoz-
zák, hogy a vese kanyarulatos csatornáiból mennyi nátriumion, és ezáltal mennyi víz
szívódjon vissza a szervezetbe. Az adrenális sejtek képezik az adrenalint és a noradrenalint.
Részt vesznek a vércukorszint regulációjában. Fokozzák a májban a glükogén bontását és
így növelik a vér cukortartalmát. Az adrenalin fokozza a szív működését, a vérkeringést és
a hipofízis ACTH-termelését. A noradrenalin hatására a környéki erek összehúzódnak, de
a szív erei tágulnak.
A tobozmirigy a melatonin nevű hormont termeli. Ez hat a pigmentsejtekre, mert a
pigmentszemcsék koncentrációját idézi elő. Ellentétes hatású az agyalapi mirigy középső
lebenyének melanotropin hormonjával. így termelődésének következtében világosabb lesz
a halak, kétéltűek és hüllők testszíne. Szabályozza ezenkívül a napi ritmust is, valamint a
FSH, LH szekrécióját a hipotalamuszon keresztül.
A csecsemőmirigy nem tipikus belső elválasztású mirigy, hanem elsősorban a
szervezet sejtes immunitásának kialakításáért felelős szerv. Emellett azonban hormont is
termel. Hormonja - fiatal korban - az ugyanezen szervben képződő T-limfociták ter-
melődését fokozza. Az állatok élete során a csecsemőmirigy lassan elsorvad és elzsírosodik.
Az elkülönült belső elválasztású mirigyek mellett számos egyéb szervben is jöhetnek
létre hormonok. A herében termelődnek az androgének, a petefészekben pedig az
ösztrogének. Az ivarszervekben termelődő hormonok az ivarszervek fejlődését és normá-
lis működését szabályozzák, valamint a másodlagos nemi jellegek kialakulásáért felelősek.
Kiemelendők a prosztaglandinok is, amelyek sokféle szövetben keletkezhetnek. Különö-
sen sok termelődik a hímek dülmirigyében és az ondóhólyagban. Hatásukat rendszerint
lokálisan fejtik ki. Nőstényekben fokozzák a simaizmok összehúzódását és szabályozzák a
női nemi ciklust.
Számos emlős bélcsatomájának falában, veséjében és méhlepényében is termelődnek
hormonok. A gyomorkapu tájékának egyes speciális sejtjei állítják elő a gasztrint. Ennek
a hormonnak a hatására a gyomor falának sejtjei pepszint és sósavat termelnek. A
144
szekretin nevű hormon a patkóbél falában lévő sejtekben jön létre. Fokozza a hasnyálmi-
rigy szekrécióját. Gátolja a sósav termelődését a gyomorban. A vesében termelődő
eritrogenin hatására képződik a vérplazmában az eritrociták fejlődését segítő eritropoietin.
A méhlepényben termelődő koriongonadotropin hatása megegyezik a luteinizáló hor-
monéval.
A következő példa, a kétéltűek lárvakori metamorfózisának hormonális sza-
bályozása. A legfontosabb folyamatok leírásával mutatjuk be a hormonok szabályozó
hatásait az ebihalak átalakulására. A hipofízisben termelődik egy pajzsmirigyserkentő
hormon (tirotropin, TSH), amely glikoprotein. ATSH-szekréciót a hipotalamusz serkentő
hormonja (TRH, tirotropin-releasing-hormon) fokozza. A kétéltűeknél TSH-termelést
gátló hormon nem ismert. A TSH hatására a pajzsmirigy fokozza a trijód-tironin és tiroxin
kibocsájtását. Ha megnő e két hormon mennyisége a vérben, akkor az egyrészt gátló hatású
a TSH- és valószínűleg a TRH-képzésre is (negatív visszacsatolás). Másfelől fokozódik a
sejtek anyagcseréje és ezzel párhuzamosan az egyedfejlődés sebessége is megnő. Az
ebihalak átalakulásának idejében a vérben 3-4-szer több pajzsmirigyhormon található,
mint más időszakokban, és ezért a szervezetben a lebontó és felépítő folyamatok gyorsasága
jelentősen fokozódik. Normális esetben gyorsan kialakulnak az elülső végtagok, a farok
visszafejlődik (lehetővé válik a szárazföldi mozgás), megváltozik a hemoglobin minősége
(mivel légköri oxigént kell megkötni), létrejönnek a karbamid szintéziséért felelős enzimek
(a húgysav mellett ez a szárazföldi állatok nitrogéntartalmú anyagcsere-végterméke, 4.5.
fejezet.) stb. Ha a pajzsmirigy nem működik megfelelően, akkor az ebihalak nem tudnak
átalakulni, vagy a kifejlett állatok csököttek maradnak, mivel a pajzsmirigy hormonjainak
hiányában számos szervük nem tud kifejlődni. (Megjegyzendő, hogy a pajzsmirigy csök-
kent működése minden gerinces állat fejlődését gátolja.) Az átalakulás után a kétéltűek
növekedését elsősorban a növekedési hormon szabályozza.
Általában úgy vélik, hogy törzsfejlődéstanilag a hormonrendszer ősibb szabályozója a
szervezeteknek, mint az idegrendszer. Elképzelhető, hogy a jelenleg ismert hormonok
olyan molekulákból alakultak ki az evolúció során, amelyek lassan jutottak el a termelődés
helyétől a célsejtekhez, de ott hatásukat hosszú időn át fejtették ki. Azok a molekulák pedig,
amelyek a testfolyadékban vagy a szövetek közötti folyadékban gyorsabb mozgásra voltak
képesek, de rövid ideig hatottak a célsejtekben, az idegsejtek átvivő anyagaivá
(neurotransmitter) váltak.
4.7.3. Feromonok
A feromonok olyan vegyül etek, amelyek egy faj egyedei közötti kommunikációt szolgálják.
A legtöbb állatcsoportban megtalálhatók, de a madarakból hiányoznak. Különösen
ismertek a rovarok és az emlősök feromonjai. Rendszerint fajspecifikusak, ritkábban több
rokon faj számára is jelenthetnek információt. Többek között lehetnek ivari viselkedést
befolyásoló anyagok, kijelölhetik a territóriumok határait, jelezhetik a veszélyt vagy a
fajtársak nyomait is (20.10. fejezet). A feromonok exokrin mirigyekben termelődnek, a
külvilágba kerülnek, majd egy másik egyed érzékszerveire jutnak és ezután speciális
élettani, viselkedési hatást váltanak ki. Igen kis mennyiségben hatásosak. Kémiai termé-
szetüket tekintve a szárazföldi állatok feromonjai általában hosszú láncú alkoholok, ezek
aldehidjei és észterei, valamint szteroidok és terpenoidok. A vízi állatok feromonjai
elsősorban szteroidok és különböző proteinek. A rovarok szexferomonjai között gyakoriak
a 10-20 szénatomot tartalmazó szénhidrátok. Ilyen feromonok termelődnek a méhkirály-
nő rágómirigyében. Nászrepülés alkalmával az egyik feromon (transz-9-keto-2-decensav)
vonzó hatást gyakorol a hímekre. A kaptárban egy vegyületkeverék gátolja a többi nőstény
ivari érését (20.9. fejezet). A feromonok rendszerint többféle vegyületből állnak. Hatásukat
csak akkor fejtik ki, ha a komponens vegyületek relatív koncentrációja a fajra jellemző
145
értéket mutatja. A hormonokhoz hasonlóan a molekulastruktúra csekély változása komoly
hatásváltozást okoz. Példa erre egyes kevéssertéjű gyűrűsférgek (Platynereis sppj
szexferomonja (5-metil-3-heptanon). A vegyület R-izomerje a nőstényeket, az S-izomerje
ezzel szemben a hímeket vonzza.
4.8. Idegrendszer
A hormonrendszer mellett az idegrendszer vesz részt a szervezet működésének szabályo-
zásában, a folyamatok összehangolásában. Hatása sokkal gyorsabb, mint a hormonrend-
szeré. Az állatok életében ez a szervrendszer igen fontos szerepet játszik, mert általánosan
lehetővé teszi az aktív helyváltoztató mozgást és azt, hogy az egyedek gyors választ ad-
hassanak a környezet ingereire. Ezek a tulajdonságok nagyban segítették az állatok el-
terjedését a Földön, és azt is, hogy szoros szociális szerveződés jöhessen létre. Az ideg-
rendszernek kiemelkedő szerepe volt a fajok evolúciója során, mert a környezet ingereire
való egyre gyorsabb és pontosabb reagálást tette lehetővé.
Az idegrendszer olyan bonyolult felépítésű és működésű lehet, mint egyetlen más szerv
sem. Mindez azon múlik, hogy az idegsejtek nyúlványaik segítségével, rendkívül komplex
struktúrát hozhatnak létre. Az idegrendszer kiemelkedő szerepe élettani paraméterek
alapján is látható. Például az emberi agy a szervezet tömegének mindössze 2%-át teszi ki.
Ugyanakkor a teljes energiafelhasználás 20%-a szükséges az agy működéséhez.
Az egyes állatcsoportok idegrendszerének felépítésében és a rendszer működésében
hatalmas különbségek vannak, olyan nagyok, mint talán egyetlen más szerv esetében sem.
A szivacsoktól és néhány erősen specializált parazitától eltekintve, minden állatnak van
ilyen szervrendszere. A csalánozók diffúz idegrendszerétől az emlősök agyáig azonban több
fejlődési fokozat, számos átmenet és igen nagy változatosság figyelhető meg.
4.8.1. Az idegrendszer típusai
Egyszerű idegi szerveződések
A szivacsok testében elszórtan felismerhetők egymással és más sejtekkel összeköttetésben
álló nyúlványos sejtek. Kimutattak olyan anyagokat is a szivacsokból, amelyek más
állatokban, mint neurotranszmitterek ismertek (pl. 5-hidroxi-triptamin). A szivacsok idegi
szerveződéséről azonban egyenlőre kevés biztos információ áll rendelkezésre.
A csalánozóknak diffúz idegrendszerük van. A test külső és belső rétegében is
megfigyelhető az egymással kapcsolódó idegsejtek hálózata. A háló nem egyenletesen sűrű:
a szájnyílás környékén lényegesen több idegsejt található, mint máshol. Idegdúcok nem
fordulnak elő, ezért központi irányítás sincs. A szinapszisok is átjárhatók mind a két
irányban (Hydra sp.), tehát dendritek és axonok (legalábbis funkcionális szempontból)
nem különíthetők el. A felépítés következménye, hogy a csalánozók válaszai az ingerekre
kevéssé differenciáltak. Ha inger éri az állatot, az ingerület minden irányban tovaterjed és
az állat az egész test mozgásával reagál.
Gerinctelenek dúcidegrendszerei
Az idegsejtek (1.2. ábra) a legtöbb állat testében jellegzetes módon rendeződnek.
Tömörülésükből a test meghatározott helyein idegdúc (ganglion) jöhet létre. Idegdúcok
először a laposférgekben jelennek meg. A dúcok később egyre komplikáltabb felépítésűek
és működésűek lesznek. Ennek a tömörülésnek az egyik előnye, hogy az információ gyor-
san átjuthat egyik idegsejtből a másikba. A másik, még fontosabb előny, hogy igen sok
kapcsolat alakulhat ki az idegsejtek között, így bonyolult struktúrák jöhetnek létre. Ezek
azután komplex idegi működéseket tesznek lehetővé. A gerinctelen állatok idegdúcaiban
146
az idegsejtek sajátosan helyezkednek el. A sejtek teste (perikarion) a dúcok felszínén
található, a nyúlványaik pedig a dúc belsejében vannak. Az idegsejtek nyúlványai a dúcban
szigetszerűen tömörülhetnek, kis területen egymással igen sok szinaptikus kapcsolatot
létesíthetnek és így speciális feladatok irányítására alkalmas központokat (neuropilema)
hozhatnak létre.
A dúcok mellett az idegek és az idegkötegek kialakulása ugyanilyen fontos. Az idegeket
az idegrostok kisebb-nagyobb nyalábjai alkotják. Idegek az idegrendszeri központokon
kívül találhatók. Az idegkötegek komplikáltabb felépítésűek, nem csupán idegrostokból,
hanem idegsejtekből is állnak. Az idegek és az idegkötegek kis átmérőjű, hosszú képletek.
Rajtuk keresztül nagy mennyiségű információ áramlik, viszonylag kis helyen, nagy
távolságokra a testen belül. Az idegkötegek további előnye az is, hogy az ingerületet gyorsan
vezetik. Könnyen belátható, hogy evolúciós szempontból előnyt élveznek azok az egyedek,
amelyek nagyobb sebességű ingerülettovábbításra, ezért gyorsabb reagálásra képesek.
A dúcok kialakulása az állatok feji részén kezdődött. Érthetően, hiszen a legtöbb inger
itt éri a testet. Megfordítva is igaz: ott alakulhatott ki a fej, ahová a legtöbb inger érkezett.
A feji dúc a test szimmetriaviszonyainak megfelelően szerveződik. Általában ez a leg-
nagyobb dúc a szervezetben, ezért - még a gerinctelen csoportokban is - gyakran agy-
dúcnak nevezik. Egyszerűbb esetben (laposférgek) a fejben található dúcokból hoszszanti
idegtörzsek indulnak ki. Ezek a test hosszában futnak, szétágaznak, összeköttetésbe lépnek
egymással, behálózzák az egész testet, beidegzik az érzékszerveket. A fonálférgek feji dúca
nem egységes tömb, hanem gyűrűre emlékeztető szerv, amely a garatot veszi körül
(comissura cephalica). A fő idegkötegek már - harántágak révén - egymással szorosabb
összeköttetésben állnak. A találkozási pontokon sokszor kisebb dúcok láthatók. A puha-
testűeknek a legtöbb esetben meghatározott számú (5 pár) idegdúcuk van, amelyek a
fontosabb testtájakon helyezkednek el. Az idegrendszer felépítése ebben a törzsben igen
változatos.
A szelvényezett testfelépítés következtében a gyűrűsférgek és az ízeltlábúak idegrend-
szere igen jellegzetes felépítésű, hasdúclánc idegrendszer. Nevét központi részének
kötélhágcsóhoz hasonlítható felépítéséről kapta. A hasdúclánc szelvényenként elhelyezke-
dő idegdúcpárokból áll, melyek a test hosszában futó és a dúcpárok tagjai között lévő
idegkötegekkel állnak kapcsolatban egymással. Egy szelvényen belül a dúcok igen közel
kerülhetnek egymáshoz. A test első szelvényének dúcpárja vagy néhány első szelvény dúc-
párjai összeolvadhatnak egymással, és egységes agydúcot alkothatnak. így jön létre a garat
feletti dúc (ganglion supraoesophageale). A többi idegdúcpár a test hasi oldalán, a bélcsa-
torna alatt található. A garat feletti dúcot a nyelőcső két oldalát körülölelő idegköteg (con-
nectivum) köti össze a következő idegdúcokkal. A garat alatti dúc (ganglion inphrao-
esophageale) gyakran szintén több dúcpár összeolvadásával alakul ki. A vázlatosan be-
mutatott központi részekből számos ideg fut az izmokhoz, a mozgásszervekhez, zsigeri
szervekhez, érzéksejtekhez, illetve vissza a központi területekhez. Igen jellegzetes tenden-
cia, különösen az ízeltlábúak körében, hogy a hasi idegdúcpárok összeolvadásából egységes
dúctömeg jön létre. A dúcok összeolvadása jellemző a törzsfejlődés szerint fiatalabb
rovarrendekre (kétszárnyúak, hártyásszámyúak) és különösen kifejezett a pókok körében,
ahol fejtorba tömörült hatalmas, egységes idegdúc látható.
Csőidegrendszer
E helyen nem feladatunk a különböző gerinces állatfajok rendkívül változatos ideg-rendsze-
rének részletes ismertetése. Csupán annyira érintjük ezt a kérdéskört, amennyire a gerincesek
- csontos halak, kétéltűek, hüllők, madarak és emlősök- idegrendszerének összehasonlítása
során felismerhető főbb evolúciós tendenciák bemutatása ezt szükségessé teszi.
A gerincesek idegrendszerét csőidegrendszemek nevezik. Felépítés szempontjából
központi és környéki idegrendszer különböztethető meg. A központi idegrendszert az
147
agyvelő (encephalon) és a gerincvelő (medulla spinalis) alkotja, a környéki idegrendszer
pedig az agyidegekből, gerincvelői idegekből és idegdúcokból áll. A csőidegrendszer
működés szerint az akarattól függő működést végző, szomatikus és akarattól független
működésű, vegetatív részre osztható. A vegetatív idegrendszer a szervezet belső egyensú-
lyának fenntartásáért felelős. Működését a szomatikus idegrendszer befolyásolja.
A gerincesek központi idegrendszerében az agyvelő a következő öt részre tagolható:
előagy, köztiagy, középagy, nyúltagy és kisagy. Az előagy (telencephalon) igen változatos
nagyságú a többi agyrészlethez viszonyítva (4.19. ábra). A szaglószerva páros szaglóhagymán
(bulbus olfactorius) és - gyakran - az ebből kilépő axonkötegen (tractus olfactorius)
keresztül kapcsolódik az előagyhoz. A csontos halak előagya relatíve igen kicsi. Legfőbb
feladata a szaglás szabályozása. Később, a többi gerinces osztályban az előagyhoz egyéb,
fontos fünkciók kapcsolódnak. A szaglás központja a legősibb kérgi részben, a paleo-
cortexben lokalizálódik. Ennek területe - különösen a gyenge szaglású csoportok, pl. a
madarak, egyes emlősök esetében- csekély lesz. A hüllőknél megjelenik a neocortex, mely
nagy jelentőségre tesz szert az emlősök körében. A legfejlettebb emlősök és az emberi
agykéreg felületének legnagyobb részét ez borítja. Itt találhatók a legfelsőbb érző- és
mozgatóközpontok, ez a legbonyolultabb idegi szabályozás helye. A hüllők és különösen
a madarak legmagasabb rendű idegtevékenységei ezzel szemben az előagy legnagyobb
tömegét kitevő corpus striatumhoz kötődnek. A legfontosabb idegi irányítási fünkciók
is ehhez a területhez kapcsolódnak. Evolúciós trend, hogy az előagy relatíve, a többi
agyrészlethez viszonyítva egyre nagyobb tömegűvé válik. Az emlősök előagya a legnagyobb
a többi agyrészlethez viszonyítva.
A köztiagy (diencephalon) két legfontosabb része a talamusz és a hipotalamusz (4.19.
ábra). A terjedelmesebb terület a talamusz. A hipotalamusz a talamusz alatt helyezkedik
el. A hipofízis kap csolódik hozzá. A hipotalamusz a vegetatív működések legfelső központ-
ja. Szerepe van a testhőmérséklet szabályozásában, a vegetatív idegrendszer szimpatikus-
paraszimpatikus működésében, a szervezet homeosztázisának fenntartásában. A köztiagy
relatív nagysága, differenciáltsága többé-kevésbé az előagyéval párhuzamosan alakul a
különböző gerinces osztályokban.
A középagy (mesencephalon) a legtöbb gerinces agyában jól fejlett terület. Fontos
érző- és mozgatóközpont. Felső lemeze a tektum, alatta található a tegmentum. A közép-
agy a csontoshalak és kétéltűek látásában különösen fontos szerepet játszik, mivel a
tektumban találhatók a legfelső látó központok (4.19. ábra). Jelentős a középagy részvétele
a mozgás (szemmozgás, helyváltoztató mozgás koordinációja) szabályozásában is. A
tegmentum irányítása alatt áll a csontos halak és kétéltűek legtöbb helyváltoztató mozgása,
különösen az összetett viselkedési mozgásformák. A hüllőkben itt jelenik meg az a fontos
idegsejtcsoport (nucleus ruber), amelyik a madarakban és emlősökben a mozgatórendszer
(ezen belül az extrapiramidális rendszer) igen fontos része lesz.
A nyúltagy (medulla oblongata) létfontosságú vegetatív központok helye. Az agyvelő-
nek a gerincvelővel kapcsolódó része. A halak és a kétéltűek V-X. agyidege, a többi gerinces
V-XII.-ig számozott agyidege ered belőle. A nyúltagy ezeken keresztül számos fontos
működést (keringés, légzés stb.) befolyásol. Ugyancsak lényeges a mozgást, testtartást és
az izomtónust szabályozó szerepe.
A kisagy (cerebellum) az izomrendszer legfinomabb összerendezettséget kívánó mű-
ködésének szabályozója, koordinálója (4.19. ábra). A kisagy kérge rendszerint három
rétegből (stratum moleculare, stratum gangliosum, stratum granulosum) áll. A normális
testtartásért és mozgásformákért, az ehhez szükséges izommozgások összehangolásáért a
kisagy a felelős. E fünkció következtében különösen azoknál az állatoknál fejlett, amelyek
gyors, bonyolult mozgásformákra képesek (halak, madarak, emlősök).
A gerincvelő (medulla spinalis) hosszú, hengeres szerv. Belső részét a szürkeállomány
(substancia grisea) alkotja. Főleg idegsejttestek, asszociációs neuronok találhatók itt.
148
Közepén húzódik végig a központi csatorna (canalis centrális). A külső, fehérállomány
(substancia alba) elsősorban az idegsejtek nyúlványait, a velős hüvellyel körülvett axonokat
tartalmazza. A gerincvelő felépítésére és működésére a szelvényezettség jellemző. Ennek
következtében egy szelvény önállóan is szabályozhat egy egyszerű idegi működést (pl.
gerincvelői reflex). Az egymás mellett található szelvények működése egymással összehan-
golt lehet. Ilyen szabályozású a vakaródzási reflex. Általános tendencia, hogy a gerincvelő
tömege az agy tömegéhez képest a gerincesek evolúciója során csökkent. A gerincvelő
kapcsolata az előaggyal egyre fontosabb lesz a halaktól az emlősökig tartó fejlődés során.
4.19. ábra. Gerincesek agyának hosszmetszete. A: csontos hal, B: béka, C: gyík, D: madár, E: emlős
am: agyalapi mirigy, ea: előagy, hm: szaglóhagyma, ha: harmadik agykamra, hí: híd, kai kisagy, ké: agykéreg, ki: közti-
agy, kp: középagy, It: látótető (tectum opticum), ne: negyedik agykamra, ny: nyúltagy, pa: parietális szem (Wurmbach
nyomán)
149
Ezzel párhuzamosan a gerincvelő önálló (autonóm) működése csökken, tevékenységét
egyre jobban befolyásolja az agyvelő. Ezt mutatja, hogy amíg a halak, kétéltűek vagy hüllők
agyának eltávolítása a gerincvelőtől (dekapitálás) a mozgásokat nem vagy csupán kismér-
tékben befolyásolja, addig az emlősöknél mozgás azután már nem lehetséges.
A környéki idegrendszert alkotó idegek a központi idegrendszerből indulnak ki és az
egész szervezetre kiterjednek. Az agyidegek és a gerincvelői idegek is párosak. Az agy-
idegek az agyvelő alapjáról erednek. Számuk a halak és a kétéltűek osztályában 10, a
többiben 12. A gerincvelői idegek szelvényenként lépnek ki a gerincvelőből. A háti
oldalhoz futnak az érzőidegek, a hasi oldalról indulnak a mozgatóidegek. A környéki
idegrendszerhez tartozó idegdúcok lehetnek a gerincoszlop mellett, egyes zsigeri szervek
mellett vagy azok falában is. A perifériás dúcokat és a zsigeri szerveket is összekötik idegek.
Funkcionális szempontból érző- (afferens) idegek azok, amelyek az érzékszervekből a köz-
ponti idegrendszerbe vezetik az ingerületet és mozgató- (efferens) idegek azok, melyek a
központi idegrendszerből elsősorban az izmokhoz, de az érzékszervekhez is futnak és oda
továbbítják az ingerületet. A zsigereket a vegetatív idegrendszer szimpatikus és paraszim-
patikus idegei együttesen idegzik be. Kölcsönös hatásuk szabja meg, hogy a szerv mű-
ködése fokozódik, vagy csökken.
4.8.2. A reflexek
Az idegsejtek működéséről, az elemi idegi jelenségekről már volt szó az 1. fejezetben. Most
az idegrendszer működésével kapcsolatban csupán a reflexekről beszélünk, mert ez
szükséges a viselkedéstannal foglalkozó 20. fejezet jobb megértéséhez.
Az idegrendszer működésének legegyszerűbb megnyilvánulási formái a reflexek. Már a
kezdetleges testfelépítésű állatok, mint a laposférgek, számos reflexmozgást végeznek. A
gerinces állatok mozgásainak jelentős része is reflexmozgás. Ha inger éri az állatot, pédául egy
bögöly megszúr egy legelő szarvasmarhát, akkor a bőrben levő fájdalomérzékelő receptorok
ingerületbe jönnek. Az ingerület az érző idegpályákon keresztül a gerincvelőbe jut. Ott
átkapcsolódik, vagyis az asszociációs neuronok segítségével azokra a mozgató pályákra
tevődik át, amelyek az adekvát válaszhoz szükséges izmokat idegzik be. Bizonyos izmok
összehúzódásának és elernyedésének következménye, hogy a szarvasmarha a farkával a
bögölyhöz csap. Fontos megjegyezni, hogy a receptorok ingerküszöbét az idegrendszeri
központok növelni vagy csökkenteni képesek, ami az idegrendszer működésének optimali-
zációját segíti elő. Az izmok tónusa a központi idegrendszer állandó ellenőrzése alatt áll. A
reflex tehát gyorsan és a központi idegrendszer pillanatnyi befolyása nélkül, de mégis azzal
összhangban működik. Megkülönböztetnek feltétlen és feltételes reflexeket. A feltétlen
reflexek veleszületettek, öröklődnek, megfelelő inger hatására, egyéb feltétel nélkül is
kiváltódnak (szopás, nyál elválasztás, bélnedvszekréció stb.). A reflexet kiváltó inger hatására
változatlan formájú válasz következik. Ha a borjú szájnyílása körüli mechanoreceptorokat
ingerük, a szopómozgások elkezdődnek. A feltételes reflexek az állat élete során alakulnak
ki. Ügy jönnek létre, hogy egy feltétlen reflexet kiváltó ingerrel egy időben egy másik, semleges
inger is éri az állatot. Amennyiben ez az ingertársítás rövid időn belül többször ismétlődik,
a kezdetben semleges inger is kiváltja azt a választ, amit kezdetben csak a feltétlen inger váltott
ki. A feltételes reflexek nem öröklődnek, és ha tartósan nem történik megerősítés, akkor meg
is szűnnek. Igen fontos szerepet játszanak a tanulási folyamatokban (20. fejezet).
4.8.3. Evolúciós trendek
Egyetlen más szervrendszer felépítése sem mutat olyan világos, egy irányba (a komplexitás
irányába) mutató változást, mint az idegrendszer. Megfigyelhető, hogy minél bonyolultabb
felépítésű egy állat szervezete, annál összetettebb, komplikáltabb felépítésű és működésű
150
az idegrendszere. Az idegsejtek egyre szorosabb kapcsolatba kerülnek egymással.
Létrejönnek azok a speciális helyek, a szinapszisok, ahol az ingerület gyorsan átjut az
egyik sejtről a másikra. Ezek a legősibb, idegrendszerrel bíró állatcsoportokban mindkét
irányban átjárhatók, később azonban csak egy irányú ingerületvezetést tesznek lehetővé.
Az ősi idegsejtek nem egyformák. Különböznek nyúlványaikban., az ingerületvezetés
gyorsaságában és nem teljesen egyenletesen oszlanak el a testben. így a test azon részei,
ahol az idegsejtek idegdúcokba tömörülnek, gyorsabb és adekvátabb válaszreakcióra
képesek. Ez a jelenség különösen a táplálékszerzés és a fajtársak felismerése, a párválasz-
tás szempontjából fontos. A feji dúc arányának növekedése (kefalizáció) szintén igen
feltűnő jelenség. A dúcokban rövid, soknyúlványú (multipoláris) idegsejtek alakulnak ki.
Ezek száma egyre nő, így a kapcsolódási rendszer is egyre bonyolultabb lehet, ami
komplikáltabb működésű szervrendszer irányítását, folyamatainak összehangolását teszi
lehetővé. Ugyanakkor a bonyolultabb viselkedésformák nagyobb adaptív értéke az
összetettebb, nagyobb teljesítményű idegrendszerrel bíró egyedeknek kedvez. Ezek
ugyanis hatékonyabban tudják kihasználni a környezetben található forrásokat, minek
következtében nő az evolúciós értelemben vett rátermettségük (21.8. fejezet). így génjeik
jobban elterjedhetnek a populációban, mint azoké az egyedeké, amelyek erre - éppen a
kisebb teljesítő képességű idegrendszerük miatt - nem képesek.
4.9. Érzékszervek
Az állatok környezetükkel állandó, szoros információcserében állnak. Az információkat
az érzékszervek veszikfel és továbbítják az ingerületek feldolgozásának színhelyére, rend-
szerint az idegrendszer valamelyik központjába. Az érzékszervek tehát döntő szerepet
játszanak abban, hogy egy egyed különbséget tudjon tenni a rázúduló ingerek között,
adekvát (az ingernek megfelelő) választ adhasson és környezetében élni tudjon.
Egy adott élőhelyen található állat számára a környéknek csupán meghatározott ingerei
lényegesek. Ezért az érzékszervek az evolúció során erőteljesen specializálódtak. Kifino-
multak azok, amelyek az egyedek számára fontos ingerek felfogását végzik és csöke-
vényesedtek azok, amelyek a kevésbé létfontosságú ingerek érzékelésére alkalmasak. A
kígyók különleges hőérzékelő szerve például lehetővé teszi, hogy teljes sötétségben is ész-
revegyék táplálékukat, a meleg vérű emlősállatokat. Nem csupán a jelentős (releváns)
ingerek köre, hanem a hatás spektruma is fontos. Az emlősök és a rovarok életében egyaránt
fontos tényező a fény. Sok rovar az ultraibolya fényt is látja. Ezzel lehetővé válik számukra,
hogy olyan táplálékforrást is észrevegyenek, amelyet más rovarok nem.
Mielőtt egy inger az érzékelő idegsejtet (receptor sejt) elérné, egy speciális ingervezető
rendszeren halad át. Ez a rendszer érzékszervenként igen különböző lehet. Az emlősök
hallásakor például a dobhártya rezgéseit a hallócsontocskák veszik át, majd továbbítják a
belső fülhöz. A belső fülben levő folyadék mozgása ingerli a receptorsejteket. A szaglás során
is először a szaglóhámot borító nyálkahártyába jutnak be a vegyületek és csupán ezután
ingerük az idegsejteket. Amíg az inger az említett rendszeren keresztüljut, jelentősen
módosulhat. Az emberi fül ovális ablakának (fenestra ovális) egységnyi területére a kengyel
mintegy 20-szor akkora energiát fejt ki, mint amekkorát a kalapács a dobhártyáról átvett.
Az idegsejtek inger hatására ingerületbe jönnek. Az ingert elektromos ingerületté
alakítják át (transzdukció) azáltal, hogy a membrán áteresztőképességét, így az ion-
egyensúlyt megváltoztatják. Létrejön a receptorpotenciál. A receptorpotenciál - a sejt
evolúciója során kialakult határok között - az inger erősségével arányos. Az arány nem
lineáris. Az érzéksejtek rendelkeznek azzal a képességgel, hogy felerősíthetik az ingereket.
Az emlősök szemében található egyes érzéksejtek (pálcikák) már egyetlen foton hatására
ingerületbe jöhetnek, és közben az ingert körülbelül egymilliószorosára erősítik.
151
Az érzékszervek érzékenysége különböző lehet, az inger szélesebb vagy szűkebb
spektrumát foghatja át. Ezen a spektrumon belül az érzékenység csúcspontja változhat, az
érzékszerv alkalmazkodni képes. Alkalmazkodás az is, ha az ingerküszöb változik meg. Ha
csökken, akkor a szerv érzékenyebb lesz, ha növekszik, akkor pedig érzéketlenebb.
Különbséget kell tenni egyetlen érzéksejt és az érzékszerv érzékenysége között. Előfordul,
hogy egy vagy néhány sejt ingerületbe kerül, de az egész szerv nem. A selyemlepke (Bom-
byx móri) hímjének egy szaglósejtje reagál a nőstény egyetlen szexferomon-molekulájára,
ingerületbe jön attól. Az állat azonban csupán akkor indul a nőstény keresésére, ha legalább
200 szaglósejtben keletkezik egyszerre ingerület.
A külvilágból származó ingerek felfogását az exteroreceptorok, a test belsejéből
származó ingerek érzékelését pedig az interoreceptorok végzik. Ugyanaz az érzék-
szervtípus (pl. Vater-Paccini-féle test) lehet exteroreceptor és interoreceptor is, attól füg-
gően, hogy az inger a külvilágból vagy a test belsejéből éri.
Az érzékszerveket aszerint szokták csoportosítani, hogy milyen típusú ingerek felfogá-
sára alkalmasak. így mechanikai, fényérzékelő, vegyi és egyéb érzékszervekről lehet
beszélni.
4.9.1. Mechanikai érzékszervek
Ezekben a szervekben mechanikai hatásokra (nyomás, áramlás, rezgés, hullámzás stb.) jön
létre az ingerület. Nyomást, érintést már a legősibb állatok is érzékelnek. A lágy köz-
takarójú gerinctelen állatok tapintószervei egyszerű felépítésűek. Itt szabad idegvégződé-
sek, egyszerű érzéksejtek, érzékbimbók szolgálnak ingerfelvételre. Az egyszerűbb vagy
komplikáltabb felépítésű érzékelőszőrök (sensilla trichodea) az állatvilágban sok cso-
portban elterjedtek (4.20. ábra). Ezek egy, a kültakaróból kiemelkedő szőrszálból és a
hozzá kapcsolódó érzéksejtből állnak. A szőr mozgatható, rövidebb, hosszabb, néha
elágazó nyúlvány. Már igen csekély erő hatására elmozdul és ingerületet kelt a csatlakozó
4.20. ábra. A: selyemlepke csápjának részlete, B: egy érzékszőr felépítése
ep: epidermiszsejtek, ér: érzékszőr, és: érzéksejt, há: kötőszövetes hártya, ír: idegrost, js: járulékos sejt, ku: kutikula, ta: tra-
cheola, ti: trichogén sejt (a szőrt hozza létre), to: tormogén sejt, (a szőrt körülölelő üreg falát hozza létre), tr: trachea (Több
szerző nyomán)
152
érzéksejtben, tehát igen érzékeny szerv. A káposzta bagolylepke (Mamestra brassicae)
hernyójának testén számos ilyen receptor található. Az érzékelőszőrök félfokos kilengése
(150-1000 Hz között) már ingerületet kelt. Elmozdulás következik be a légmozgás
hatására, például akkor is, ha egy másik rovar repülve közeledik. Ilyen szőrökkel rendelkeznek
a laposférgek, fonálférgek, gyűrűsférgek, kerekesférgek,
de ezek az érzékszervek az ízeltlábúakra különösen
jellemzőek.
Az érzékelőkupola (sensilla campaniformia) első-
sorban a rovarok szárnyán fordul elő és a légnyomás-
változás, a szélerősség, tehát a repülés gyorsaságának
mérésére szolgál (4.21. ábra). Működési elve, hogy az
érzéktest kitinvázba illeszkedő kupolája a légnyomás
hatására benyomódik és ingerli az alatta elhelyezkedő
érzéksejtet. Ilyen érzékszerv található a kétszárnyúak
(Diptera) billérein. A kitinképződményekkel kapcsola-
tos érzékszervek közé tartoznak a hasadékérzékszer-
vek is. Ahogy az elnevezés jelzi, a kitinvázat finom
hasadék járja át, amelyben érzéksejtek nyúlványai vég-
ződnek a réseket elzáró vékony kitinlemezeken. Ez az
4.21. ábra. Érzékkupola vázlata
ep: epidermiszsejtek, és: érzéksejt, kh: kitin-
hártya, ku: kutikula, ti: trichogén sejt, to: tor-
mogén sejt (Davies nyomán)
érzékszerv a méhek testén sokfelé megtalálható. A
pókok kültakaróján szintén számos ilyen szerv van. Gyakran egymás mellett helyezkednek
el. A rések sorozatának alakjáról kapta a lantképű (lyriform) szerv elnevezést.
A szkolopidiális szervek az ízeltlábúak különleges mechanoreceptorai. Alapegységei
4.22. ábra. Szkolopidium váz-
lata
ep: epidermiszsejtek, hu: hüvely-
sejt, id: idegsejt, ku: kutikula, ny:
idegsejt nyúlványa, sk: szkolopidi-
ális sejt, ss: süvegsejt, sü: süveg
a szkolopídiumok (4.22. ábra). Minden szkolopídium három-
féle egységből, egy bipoláris érzékelősejtből, a süvegsejtből és a
járulékos sejtekből áll. A két utóbbi csoportba tartozó sejttípus
az érzékelősejtet köpenyszerűen burkolja. A járulékos sejtek
(hüvelysejt és süvegsejt) rögzítik a szervet a kitinvázhoz. A
legegyszerűbb ilyen szerv a csótányok, sáskák, tücskök térd
alatti (subgenualis) szerve. Ezzel képesek a földfelszín legcse-
kélyebb rezgéseit is érzékelni a 30-5000 Hz tartományban. Igen
egyszerű felépítésű az ún. kordotonális szerv is. Szinte vala-
mennyi rovarrendnél előfordul. Rendszerint két, egymással
mozgathatóan összekötött kitinlemez között feszül ki (erre utal
a szerv nevében szereplő „tónus” kifejezés) és akkor jön ingerü-
letbe, ha a két lemez egymáshoz viszonyított helyzete, tehát a
szerv feszessége, tónusa megváltozik. Helyzetétől függően al-
kalmas a testrészek elmozdulásának érzékelésére vagy a közeg-
ben terjedő rezgések felvételére. Mindenütt előfordulhat a
testben. Egyes speciális szkolopidiális szerveket (tümpanális
szerv) később tárgyalunk (16.4. fejezet).
A gerincesek körében számos tapintószerv ismeretes (4.23.
ábra). Gyakoriak az elágazó vagy el nem ágazó, Schwann-
sejtekkel körülvett idegvégződések. Ezek körül másféle sejtek
nem találhatók. Az irhában, a szőrök gyökerénél vagy az
epidermiszsejtek között végződnek. Az éles fájdalom érzékelé-
sére a csupasz idegvégződések alkalmasak. A bőrben, a
szőrökön és aspeciális érzékszőrökben (pl. emlősökbajuszsző-
rei) fordulnak elő a Merkel-féle tapintótestek. Az érzékelő
idegkötegek nyúlványai kiszélesednek és kapcsolatba léphet-
nek más érzékelősejtekkel. A Meissner-féle testek az irhá-
153
4.23. ábra. Különböző érzékideg-végkészülékek. A: szabad idegvégződés, B: Vater-Paccini-féle végtest,
C: Merkel-féle tapintótest, D: Meissner-féle tapintótest, E: izomorsó
ep: epidermisz, id: idegrost, le: lemezes kötőszövet, mi: miofibrillum, sg: stratum germinativum, sp: spirális idegvégződés,
ve: idegrost lemezszerűén kiszélesedő vége, vé: vérér (Több szerző nyomán)
bán találhatók, közvetlenül az epidermisz alatt. Az érzékelő idegrost végződése
felcsavarodik, körülfogja a Schwann-sejtek kis csoportját. Innen és az egész testet
körülvevő burok sejtjeiből kollagén rostokfutnak az epidermiszsejtek tonofibrillumaihoz.
A Vater-Paccini-féle testek a bőr alatti kötőszövetben, az erek közelében, az iz-
mokban és a csonthártyában találhatók nagyobb számban, ennek következtében a
belső ingerek felvételére is alkalmas tapintókészülékek. A harántcsíkolt izmok mozgá-
sának szabályozásában játszik fontos szerepet az izomorsó. Az emlősök izomorsója
néhány izomrostból, az ide csatlakozó idegvégződésekből és az ezeket körülvevő
kötőszövetes tokból áll. Egyes idegvégződések spirálisan rácsavarodnak az izomorsón
belül levő izomrostokra. Ezek az annulospirális végződések. Más idegvégződések
szétágaznak az izomorsóban. Ezeket szekunder végződésekneknevezik. Az annulospirális
végződéseken akkor jön létre ingerület, amikor az izomrostok megnyúlnak, a szekun-
der végződéseken pedig akkor, amikor a rostok megfeszülnek. Az izomorsó receptoraiból
fontos gerincvelői reflexek indulnak ki. Elsősorban a normális testtartás fennmaradását
szolgálják.
A csontos halak és egyes farkos kétéltűek oldalvonalszerve szintén mechanoreceptor.
Ennek felépítése és működése az említett csoportok jellemzésénél található (17.2. és
17.3. fejezetek).
A gravitáció érzékelése létfontosságú az állatok számára, hiszen csak ennek segítségével
képesek a helyes testtartásra és a térbeli orientációra. A nehézségi erő érzékelésére
154
szolgáló, egyensúly-érzékelő szervek (statocysta) alap-
jában véve azonos elven épülnek fel és működnek. Ez az
elv a következő: az egyensúly-érzékelő szerv üregét fo-
lyadék tölti ki. Falában érzéksejtek találhatók. Az üreg-
ben egy vagy több speciális kristály (statolith) van (4.24.
ábra). Az állat testének elmozdulása következtében, a
tehetetlenség miatt, a kristály is elmozdul a helyéről és
ingerli az érzéksejteket. Az állat ennek alapján módosít-
hatja a test helyzetét. Már a csalánozókban, a különböző
féregtörzsekben is találhatók ilyen szervek. Különösen
fontos szerepet látnak el a puhatestűek és a magasabb
rendű rákok életében. A gerincesek egyensúlyérzékelő
szerve (4.25. ábra) a három félkörös ívjárat (ductus
semicircularis), melyek végén kiöblösödés (ampulla)
található, valamint a csatlakozó két, kisebb üreg, a
zsákocska (sacculus) és a tömlőcske (utriculus). A fél-
körös ívjáratok a fej gyorsuló vagy lassuló mozgását, míg
4.24. ábra. Kagyló helyzetérzékelő
szerve
ér: érzéksejt, id: idegköteg, kr: kris-
tályszemcse (statolith), tá: támasztósejt
(Wurmbach nyomán)
a zsákocska és a tömlőcske a fej térbeli helyzetét, tehát a statikus állapotát érzékeli.
Evolúcióját tekintve a belső fül a halak oldalvonalából származtatható. Ellátja a moz-
gásérzékelés (félkörös ívjáratok), a helyzetérzékelés (zsákocska) és a hanghullámok
érzékelésének funkcióját (csiga) egyaránt.
A gerinctelenek körében a hallószervek a rovarokra jellemzőek. A hallószervként is
funkcionáló szkolopidiális szervek közül a Johnston-féle szerv (16.40. ábra) minden
kifejlett rovar második csápízében (pedicellus) megtalálható. A tümpanális szerv
(16.39. ábra) elhelyezkedhet a torban (egyes lepkék), a potrohban (kabócák, poloskák,
egyenesszárnyúak) vagy a lábban (szöcskék, tücskök). (E szervek részletes jellemzése a
16.4. fejezetben található.)
A gerincesek hallószervének felépítését tekintve, csupán a belső fül az, amely
mindegyik osztályában megtalálható. A hallósejtek a csontoshalak és a kétéltűek esetében
csupán mint speciális sejtcsoportok fordulnak elő a tömlőcske, illetve egy másik kitürem-
kedés (papilla amphibiorum) falában. A madarak zsákocskája megnyúlik, hosszabb-
rövidebb tömlőt képez, míg az emlősök esetében még hosszabbra nyúlik és felcsavarodva
a csigát (cochlea) alkotja. A madaraknak még nincsen, de az emlősöknek már van Corti-
4.25. ábra. Gerincesek helyzetérzékelő szervei. A: csontos hal, B: madár, C: emlős
ca: csiga, cs: csatorna, melyen keresztül agyvelőfolyadék (endolympha) jut a szervbe, fé: félkörös ívjárat, tá: tágulat (ampulla)
a csavaró mozgás érzékelésének helye, tö: tömlőcske, zs: zsákocska (Gerber nyomán)
155
féle szervük. A halaknak nincsen külső és középső füle. A víz rezgéseit vagy az úszóhólyag
egymaga vagy az úszóhólyag és az ehhez kapcsolódó Weber-féle csontok továbbítják a
belső fülhöz. A kétéltűektől kezdve megjelenik a dobhártya és a középfül. Ebben először
egyetlen hallócsont, az oszlop (columella) található (kétéltűek, hüllők, madarak), majd az
emlősöknél három ilyen csont, a kalapács (malleus), az üllő (incus) és a kengyel (stapes)
található. Annak ellenére, hogy bőrredők, illetve tolikoszorúval kombinált bőrredők már
a hüllők és a madarak esetében is fokozzák a hang erősségét és segítik a hangforrás helyének
meghatározását, valódi külső fülről csak az emlősök esetében beszélnek.
4.9.2. Látószervek
Az összes érzékszerv közül a fejlett látószervek közvetítik a legámyaltabb, legrészletesebb
információt a környezetből az állat számára. Ahhoz, hogy érzékelhető kép alakuljon ki,
fényérzékeny receptorra van szükség. Ezek a sejtek az állatvilágban alapvetően két
csoportra, a csöves sejtek és a csillós sejtek típusaira oszthatók. A csöves látósejtekre
az jellemző, hogy a fényelnyelő fehérje (rhodopsin) a sejtekből kitüremkedő nagyszámú
nyúlványban (microviílus) található. Ilyen sejtek vannak a kerekesférgek, laposférgek,
fonálférgek, gyűrűsférgek és ízeltlábúak szemében. A csillós sejtekben ezzel szemben a
szabályos felépítésű (9+2 tubulusrendszer) csiliók tubulusaiban vagy az azokat körülve-
vő lemezrendszerekben található ez a fehérje. A puhatestűek és a gerincesek szemére
jellemző ez a lemezes látósejttípus.
A receptorsejtek működése nagymértékben hasonlít egymáshoz. A fény fotonjait
elnyeli egy speciális anyagból (retinái) és fehérjéből (opsin) összetett membránfehérje
(rhodopsin). A fehérje struktúrájától függ az, hogy melyik hullámhosszúságú fénysuga-
rat nyeli el legnagyobb mértékben. A rodopszin hatására az érzéksejtek hártyáján
található ion csatornák vagy kinyílnak (gerinctelenek) vagy bezáródnak (gerincesek). A
fényérzékenység mellett a képfelbontás mértéke jellemzi a látószervet. A képfelbontás
attól függ, hogy az ideghártya (retina) egységnyi felületén hány érzéksejt található.
Az egyszerű felépítésű látószervek a fény erősségét és irányát érzékelik. Ez a
fényérzékelés legegyszerűbb módja. Erre már a látószervvel nem rendelkező egysejtűek
is képesek. Számos csoportban (csigák, rákok, egyes gerincesek) a valódi szemek mellett
még a bőrben is találhatók fényérzékelő sejtek. Ezek működéséről még nagyon kevés
információ áll rendelkezésre. Amennyiben a látósejteket pigmentszemcsék vagy pig-
menttartalmú sejtek veszik körül és árnyékolják azokat, akkor csupán a meghatározott
irányból érkező fénysugarakat képes érzékelni a sejt. Ezzel lehetővé válik az iránylátás.
Legegyszerűbb formájában egy síkban, a test felszínén elhelyezkedő sejtcsoportról van
szó (halpiócák). Ha a sejtcsoport félgömb formában besüllyed a testbe, létrejön a
kehelyszem (férgek, puhatestűek) (4.26. ábra). A kehelyszem igen nagy nyílású, tehát
fény szegény környezetben alkalmas érzékszerv, mivel sok fényt képes befogni. Érzékeli
a fény irányát is abból, hogy a kehelyszerű szerv melyik falán levő sejtek kerülnek
ingerületbe. A kehelyszem lehetővé teszi az iránylátást, hiszen a kehely falában lévő sejtek
és a hozzájuk kapcsolódó idegek, központok, a test tengelyéhez viszonyított, adott irányú
fény felfogására specializálódhatnak. Ez azonban az iránylátásnak nagyon durva módja,
hiszen a fénynyaláb széle nem éles. A problémát a lencsével ellátott szemekkel lehet
megoldani. A lencse ugyanis a fényt a retina meghatározott helyére koncentrálja.
A kehely belső sejtjei a test mélyebb területeibe süllyedhetnek, ezért a szem testfelü-
leten lévő nyílása csökken és létrejön a hólyagszem (4.26. ábra). A hólyagszem
egymástól függetlenül, párhuzamosan alakult ki az evolúció során a lábasfejűek
(Cephalopoda) és a gerincesek (Vertebrata) csoportjaiban. Ebben a szem típusban a beeső
fény mennyiségét a szivárványhártya szűkülésével vagy tágulásával a pupillarés szabá-
lyozza, míg a felbontóképességet a nagyon pontosan irányított és koncentrált fénysuga-
156
4.26, ábra. A: puhatestű kehely szeme, B: gyík hólyagszeme
ak: akkomodációs izmok, cp: conus papillaris, cs: szemcsont, ep: epidermisz, ér: érhártya, id: idegsejt, ih: ideghártya,
ik: idegköteg, ín: ínhártya, sá; sárga folt, su: sugártest, üv: üvegtest, vá: váladék (Melhom és Stark nyomán)
rak fokozzák. Javítja a fényérzékenységet és a felbontóképességet az is, hogy az ideghár-
tyában (4.27. ábra) található egyes érzéksejtek között feladatmegosztás jött létre. A
pálcikák elsősorban a fény mennyiségére, a csapok a fény színére váltak érzékennyé. Éjjeli
állatok, mint a denevérek, sünök, egerek szemében szinte csak pálcikák találhatók, míg
a nappal aktív állatok (gyíkok, madarak, mókus)
fordulnak elő a csapok. A hólyagszemet belülről
viszkózus folyadék tölti ki. Ha a hólyagszem nyílása
elég kicsi, akkor - lencse nélkül is - kép jön létre a
hólyag hátsó falán. Egyes puhatestűek szeme mu-
tatja ezt a fejlődési fokot. Hátránya ennek a megol-
dásnak, hogy a szembe lépő fény mennyisége erő-
sen csökken, mire az ideghártyához ér. A hólyag-
szemek legfejlettebb típusában lencse is található,
ami a képélességet fokozza.
A rákok és a rovarok összetett szeme másféle
módon, de hasonlóan jó hatásfokkal gyűjti a fényt
és bontja fel a képet, mint az említett hólyagszem.
Itt a receptorok nem süllyednek a testbe, ellenke-
zőleg: kiemelkednek abból. így nagyobb mennyi-
ségű fény juthat a szembe. A képfelbontás javítását
pedig az alapegységek számának növelése teszi
lehetővé. Minél több egyszerű szem (ommatidium)
alkotja az összetett szemet, annál finomabb lehet a
felbontás.
A fénymennyiség szabályozása és a képéles-
ség beállítása a gerincesek hólyagszemében a
következő. A szivárványhártya (iris) közepén talál-
ható nyílás, a pupilla szabályozza a beeső fény
mennyiségét, egy optikai rendszer (comea, csar-
nokvíz, lencse) pedig a képfelbontás, képélesség
mértékét. A halak és a farkos kétéltűek a levegőnél
retinájában nagy mennyiségben
ÜV
4.27. ábra. Hólyagszem ideghártyája (afény
az üvegtest felől érkezik)
am: amakrin sejt, bp: bipoláris sejtek, cs: csap
ga: ganglionsejt, ho: horizontális asszociációs
sejtek, li: látóideg, pá: pálcika, pi: pigmentsejt,
üv: üvegtest (Wehner és Gehring nyomán)
157
sűrűbb közegben, vízben élnek, ezért közeire látnak jól. A szemlencséjük erősen dom-
ború, gyakran gömb alakú. A lencse alakját nem tudják változtatni, de a lencse távolságát
a retinától képesek növelni vagy csökkenteni egy izom (musculus retractor lentis)
segítségével. A szárazföldi állatok messzire látnak. A szembe belépő fény legjobban a sza-
ruhártyán (cornea) törik meg. Ezután a lencse gyűjti össze és vetíti az ideghártya meg-
felelő pontjaira. A lencse domborúságát, tehát a képtávolságot a kétéltűek, a hüllők, a
madarak és az emlősök is képesek változtatni. Elsősorban a madarak (kisebb mértékben
a kétéltűek és a hüllők is) távolítani, illetve közelíteni tudják a szemlencsét az ideghár-
tyához. Ez a madarak kiváló látásának egyik oka.
A legtöbb látószerv a látható fény tartományában érzékeny. A halak, a madarak és az
ízeltlábúak azonban kiválóan érzékelik az UV-sugarakat és a polarizált fényt is. (A méhek
UV-receptorai a 350 nm hullámhosszra a legérzékenyebbek.) Mindkét ingerforrás segíti
a térbeli orientációt és a táplálékválasztást. A látható fény hullámspektrumának másik
végén található infravörös sugarak érzékelésére is képesek egyes állatok.
4.9.3. Kémiai érzékszervek
A kémiai érzékszervek képviselik a legelterjedtebb receptortípust az állatvilágban, hiszen
ez az érzékelésmód igen ősi, jellemző már a legegyszerűbb élőlényekre is. Ezért a
következőkben csupán két olyan receptorcsoportról
lesz szó, amelyek a külvilágból származó vegyületek
felfogására specializálódtak. Ezek a következők: a
szaglószervek és az ízérzékelő szervek.
A szaglószervek távoli ingerek felfogására alkal-
mas, igen érzékeny készülékek. A szél vagy a víz
segítségével érkező vegyületeket már igen kis kon-
centrációban képesek felfogni. A rovarok szagló-
szervei elsősorban a csápon (antenna), kisebb rész-
ben a szájszervi tapogatón (palpus) találhatóak. Két
legfontosabb típusuk a testből kiemelkedő szag-
lószőrök (sensilla trichodea) (4.28. ábra) és a kuti-
kulába besüllyedő szagtól emezek (sensilla placodea).
Mindkét szaglószerv esetében a levegőből kis póru-
sokon keresztül jut a vegyület a receptorsejtet körül-
vevő üregben található testfolyadékba, majd onnan
az idegsejt membránjára. Az ingerület a csápidegen
keresztül fut a garat feletti dúc területén (a deuteroce-
rebrumban) található központba. A szaglás egyik
legfontosabb funkciója a rovarokban, hogy segítségé-
vel találják meg a hímek a nőstényeket. Ennek meg-
felelően a hímeken lényegesen több szaglószerv
található, mint a nőstényeken. A háziméh (Apis
mellifera) hímjei hatszor annyi (18000) kemoreceptort
viselnek a csápjaikon, mint a nőstények.
A gerincesek szaglószervei a fejen találhatók.
A halaknak páros szaglógödrük van, a többi gerinces
szaglószerve az arckoponyába mélyebben süllyedt
és alul nyitott, egy csövön keresztül a szájüreggel
közlekedik. A kiváló szaglással rendelkező halak a
vízben található molekulákat érzékelik, a szárazföldi
4.28. ábra. Rovarok szaglószőre
de: dendritnyúlvány, id: idegsejt, fo: folyadék,
ku: kutikula, ny: nyílás, ti: trichogén sejt, to:
tormogén sejt
158
állatok ezzel szemben a belélegzett levegőben található molekulákat érzékelik. A szaglóhám-
ban az érzéksejtek között támasztósejtek vannak. A hám felületét nyálka borítja, amit
nagyrészt a hám alatti nyálkasejtek, kisebb részben a támasztósejtek termelnek. A
szaganyagok először ebben a nyálkarétegben kötődnek meg (adsorptio), majd az
érzéksejtek csillóihoz jutnak, ahol cAMP képződését váltják ki. A gerincesek szagló
receptorsejtjei kevéssé képesek a szaganyagokat megkülönböztetni egymástól. Az elkü-
lönítés feladata az agyi központokra hárul.
Különböző állatfajok szaglása jelentősen eltérő egymástól. A selyemlepke hímje
például a nőstény feromonjára (bombykol), az angolna a béta-feniletanolra, a kutya a
vajsavra igen érzékeny.
A rovarok ízlelőszervei főleg a szájszerveken, néha a lábfejen (legyek) találhatók.
Felépítésük nagymértékben hasonlít a szaglószervéhez. A gerincesek ízlelősejtjei kis
csoportokban állnak (a sejtek száma 80-ig térj edhet). Az apikális végükön levő mikrovillik
egy póruson keresztül közlekednek a külvilággal. A receptorsejtek csoportját hámsejtek
veszik körül és támogatják azokat. Az ízlelősejtek a halak testén elszórva, mindenütt
megtalálhatóak, néha igen nagy számban. Egyes gerinceseknek nincs ízlelőszervük
(kígyók), míg másoknál igen fejletlen (legtöbb madár). Az emlősök ízérzékelése fejlett.
A különböző állatcsoportok az eltérő ízeket különböző mértékben érzékelik. A ro-
varok táplálkozásuk során rendszerint az egyszerű cukrok, aminosavak minősége és
mennyisége alapján válogatják ki a számukra megfelelő táplálékot. A kinin erősen keserű
az emlősök számára, de a halak és a kétéltűek kísérletekben nem reagálnak rá. A békák,
macskák, kutyák esetében a víz is ízérzetet vált ki. Az ízlelőszervek lényegesen kevésbé
érzékenyek, mint a szaglószervek. Nagy általánosságban az mondható, hogy mintegy
kétszer nagyobb koncentráció szükséges az ízérzékelő szervek ingerléséhez, mint a
szaglószervekéhez.
4.9.4. Egyéb érzékszervek
A Föld mágneses hatása is befolyásolja az állatok életét. Bár olyan receptorokat eddig még
nem mutattak ki, amelyekkel az állatok a mágnesességet érzékelnék, számos kísérlet
eredménye szerint az ízeltlábúak és a madarak (különösen a nagy távolságokra vándorló
fajok) érzékelik a földmágnesesség hatását. A kismértékű hőmérsékletváltozás érzé-
kelésére képes termoreceptorokkal rendelkeznek különösen az ízeltlábúak és az emlősök.
Parazita rovarok hőoptimuma a gazda testhőmérsékletén van. Ha magasabb hőmérsékletű
(lázas) lesz a gazdaállat vagy kihűl (elpusztul), akkor a paraziták elhagyják. A termorecep-
torokfeladata a hőmérséklet-szabályozás és a viselkedés befolyásolása. Hőmérsékletváltozás
következménye lehet a vándorlás vagy a téli és/vagy nyári álom (diapausa).
Ellenőrző kérdések a 4. fejezet témaköreiből
1. Keressen példákat arra vonatkozóan, amikor két szerv neve azonos, felépítése, működése
és egyedfejlődési eredete viszont eltérő!
2. Hasonlítsa össze a gerincesek kültakaróit abból a szempontból, hogy milyen módon nyúj-
tanak védelmet a külvilág mechanikai ingerei ellen!
3. Mutassa be, hogy a kültakaró funkciói miképpen valósultak meg a rovaroknál!
4. Magyarázza el, hogy a hidrosztatikus váz miért jelent támaszt az izmok számára!
Milyen típusú mozgásformákat tesz lehetővé egy ilyen vázrendszer?
5. Milyen előnyei vannak a három szakaszos bélcsatomának a két szakaszos bélcsatomával
szemben?
6. Gyűjtsön példákat a sejten belüli és a testen kívüli emésztésre!
159
7. Hasonlítsa össze a nyílt és a zárt keringési rendszer előnyeit és hátrányait!
8. Milyen légzésformához, milyen keringési rendszer társul a laposférgek, ízeltlábúak és a
gerincesek esetében? Miért nem alakulhatott ki a madaraknak nyílt keringési rendsze-
rük?
9. Hasonlítsa össze a madarak és az emlősök tüdejét! Miért hatékonyabb a gázcsere a
madarak tüdejében?
10. A kerekesférgek általában nyáron szűznemzéssel szaporodnak, ősszel viszont ivarosán.
Mik ennek a szaporodásmódnak az előnyei?
11. Hasonlítsa össze az emlősök endokrin rendszerét és az idegrendszerét felépítés és működés
szempontjából!
12. Milyen módon lehetne felhasználni a rovarok hormonrendszeréről tanultakat növényvé-
delmi célokra, a kártevő rovarok elleni védekezésre?
13. Sorolja fel a gerincesek hormonjait!
14. Milyen hasonlóságokat és különbségeket ismer a hormonok és a feromonok között?
15. Milyen evolúciós trendek ismerhetők fel a gerinces agyfelépítésében a halaktól az emlősökig
terjedő osztályokban?
16. Vázlatosan rajzoljon le egy szkolopidiális szervet!
17. Mi történik akkor, ha egy tízlábú rák egyensúly érzékelő szervébe vasport juttatunk és
mágnest tartunk fölé? Miért?
18. Hasonlítsa össze az állatok szemtípusait abból a szempontból, hogy milyen mértékben
alkalmas a fény, irány és színlátásra?
160
5. RENDSZERTANI ALAPISMERETEK
Az állatvilág fajszámát senki sem ismeri pontosan, csupán durva becslések vannak arra
vonatkozóan, hogy hány faj él a földön. A már leirt fajok száma több mint 1,5 millió. A
fajoknak kettős nevük van. Az első név jelzi a nemet, a második a faj saját neve. A fajta kifejezés
nem rendszertani kategória, hanem tenyésztett állatok esetében használt kifejezés. A rendszertani
beosztásokat a fajok hasonlósága alapján alkotják. Régebben elsősorban morfológiai és anatómiai
bélyegek alapján csoportosították a fajokat. A klasszikus taxonómia, numerikus taxonómia és
a kladisztikus módszer alkalmazása terjedt el legjobban. A molekuláris biológiai eljárások
térhódításával egyre több adatunk van a fajok genetikai (DNS, RNS) hasonlóságáról, tehát
a tényleges rokonságról. A legújabb rendszerek már így készülnek. Egyelőre csupán néhány
taxonra van ilyen adatsor. Ebben a fejezetben az élővilágot öt ország szerint csoportosítva
mutatjuk be (természetesen más csoportosítás is létezik).
5.1. Klasszifikáció
Ahogyan az emberek a tárgyaikat, fogalmaikat megnevezik, ugyanúgy nevet adnak az őket
körülvevő természetben élő lényeknek is. Ez az elnevezés nyelvtől, népcsoporttól, földrajzi
helytől függő. Hermán Ottó például a század elején 21 különböző magyar nevet gyűjtött
össze a búbos banka (Upupa epops) megnevezésére. Ez a változatosság a nyelvi gazdagság
szempontjából igen fontos. Ugyanakkor rendkívül zavaró a szaknyelvi, tudományos
kommunikáció szempontjából, mivel félreértésekhez vezethet. Előfordulhat a másik véglet
is. Vannak olyan fajok, melyeknek nincs magyar nevük. Ekkor a név hiánya okoz gondot.
Olyan egyértelmű neveket kell tehát adni az élőlényeknek, amelyek mindenkinek ugyanazt
jelentik.
Az elnevezés során abból indulnak ki, hogy léteznek és felismerhetők a természetben
olyan csoportok, melyek egyedei genetikai, morfológiai, anatómiai, élettani, ökológiai,
viselkedésbeli stb. szempontok szerint jobban hasonlítanak egymáshoz, mint más csopor-
tok egyedeihez. Ez annak a következménye, hogy a csoportok tagjai egymással képesek
ivarosán szaporodni és termékeny utódokat létrehozni, más csoportok tagj aival pedig nem.
Ezeket az elkülöníthető csoportokat nevezik fajoknak (species). A faj a legfontosabb
rendszertani kategória (taxon). A faj definíciója (mint minden más definíció) megálla-
podás kérdése. Az említett, sokak által elfogadott meghatározás alkalmazása azonban
például a mikroorganizmusok körében különösen sok nehézséggel jár.
A különböző fajok egyedei néha igen hasonlóak egymáshoz, mégsem párosodnak
egymással. Egy vízparti nádasban több, szemre szinte egyforma nádiposzáta faj (Acro-
cephalus spp.) élhet egymás mellett anélkül, hogy a különböző fajok egyedei párzanának,
szaporodnának egymással. Ugyanakkor léteznek olyan fajok is, amelyek rendkívül nagy
területeken terjedtek el a Földön és igen változatossá váltak, a távoli területekről származó
egyedek mégis képesek termékeny utódokat létrehozni. A mezei nyúl (Lepus europaeus)
Eszak-Európát kivéve az egész kontinensen előfordul. A különböző populációk egyedeinek
testtömege, testmérete, viselkedése stb. eltérhet egymástól. Ennek ellenére a Nagy-
161
5.1. táblázat. Akülönböző állattörzsekbe tartozó máig leírt fajokhozzávetőleges
száma (Melhorn nyomán, kiegészítve)
Sejthalmazosok (Mesozoa) 50
Lapszerűek (Placozoa) 1
Szivacsok (Porifera) 10 000
Csalánozók (Cnidaria) 10 000
Bordás medúzák (Ctenophora) 80
Laposférgek (Platyhelminthes) 18 000
Állkapcsos férgecskék (Gnathostomulida) 80
Zsinórférgek (Nemertini) 900
Fonálférgek (Nematoda) 30 000
Húrférgek (Nematomorpha) 230
Buzogányfejűek (Acanthocephala) 1 200
Kerekesférgek (Rotatoria) 1 800
Csillóshasúak (Gastrotricha) 400
Öves férgecskék (Kinorhyncha) 100
Nyelesférgek (Kamptozoa) 300
Vértesek (Loricifera) 30
Farkosférgek (Priapulida) 16
Fecskendőférgek (Sipunculida) 300
Ormányosférgek (Echiurida) 150
Gyűrűsférgek (Annelida) 12 000
Puhatestűek (Mollusca) 130 300
Karmosak (Onychophora) 100
Medveállatkák (Tardigrada) 600
Féregatkák (Pentastomida) 100
ízeltlábúak (Arthropoda) több mint 1 000 000
Tapogatókoszorúsok (Tentaculata) 4 300
Serteállkapcsúak (Chaetognata) 80
Tapogató szakállasok (Pogonophora) 120
Tüskésbőrűek (Echinodermata) 6 000
Félgerinchúrosok (Hemichordata, Branchiotremata) 80
Előgerinchúrosok (Prochordata, Tunicata) 2 100
Fejgerinchúrosok (Cephalochordata, Acrania) 30
Gerincesek (Vertebrata) 53 000
Britanniából származó példányok minden további nélkül termékeny utódokat hozhatnak
létre tadzsikisztáni fajtársaikkal. Egy faj domesztikált formáinak körében (szarvasmarha,
házityúk, kutya stb.) még kifejezettebb példák láthatók. Kutyafajták körében gyakori
például, hogy a nagy testméret-különbség ellenére termékeny utódok jönnek létre.
Napjainkra már igen sok fajt írtak le. Az ismert fajok száma 1,3-1,8 millió között lehet
(5.1. táblázat), de a pontos értéket senki sem tudja, mivel központi világkatalógus, ahol a
neveket összegyűjtenék, nem létezik. Egy-egy rendszertani csoport specialistái a saját
területükhöz tartozó fajok számáról aránylag jó becsléseket tudnak adni, de teljes, minden
fajra kiterjedő összesítést még nem készítettek. Érthető, hogyha a leírt fajok száma sem
ismert pontosan, még nehezebb becsülni a Földön található összes faj számát. Még a
nagyságrendet sem tudjuk, mivel van, aki 3 millió, más pedig 30 millió vagy még ennél is
több faj létét feltételezi. Legtöbben mégis 3-5 millió közötti értéket tartanak valószínűnek.
A fajszám nem állandó, hiszen a fajok keletkezése lassú, de folyamatos. Figyelembe kell
venni azt is, hogy az ember természetpusztító hatására (erdőirtás, levegő-, víz- és talaj-
szennyezés stb.) gyors ütemben pusztulnak a fajok anélkül, hogy létüket tudomásul
lehetett volna venni (19. fejezet). Az ember kömyezetpusztító hatásának következtében
kihalt fajok számát sem lehet megmondani, hiszen a tudomány számára ismeretlen lények
pusztultak ki. Csupán az ismert fajok eltűnésének arányából lehet következtemi a kihalás
mértékére. További problémát jelent, hogy különösen a mikroorganizmusok körében (de
162
más csoportoknál is) nagyon nehéz meghatározni az egy fajhoz tartozó egyedek körét. A
morfológiai, élettani, ökológiai szempontok szerint egy fajnak tartott Legionellapneumophilia
baktérium különböző vonalainak nukleotid sorrendjét elemezve megállapították, hogy
közöttük a hasonlóság 50%-nál kisebb. Ez akkora genetikai különbséget jelent, amekkora
a halak és az emlősök között van. Várható tehát, hogy a mikroorganizmusok fajszáma a
molekuláris biológiai módszerek fejlődésével párhuzamosan nőni fog.
A fajokat tudományosan kettős latin névvel illetik (binominális nómenklatúra). Az
első név a nem (genus) neve, amelybe az adott faj tartozik. A második név a faj (species,
rövidítése: sp., több faj: spp.) saját megkülönböztető neve, hiszen egy nembe több faj is tar-
tozhat. Tudományos közleményekben a faj nevét kiegészítik még egy névvel és évszámmal
is. A harcsa neve például Sílurus glanis L. 1758. A két latin szó után tett „L. ” betű Linné
(Carolus Linnaeus 1707-1778) nevének rövidítése. Az igen ismert szerzők esetében csak
a kezdőbetűt vagy a név rövidítését írják ki. Kevésbé ismert szerzők teljes nevét ki szokták
írni. Az évszám a fajleírás megjelenésének évét jelöli. Fajokon belül különböztetik meg az
alfajt (subspecies, rövidítése: ssp.) és a változatot (varietas, rövidítése: var.).
A háziállatok latin nevét jelenleg különféle szabályok szerint írják. Elterjedt rendszer
az, amikor a faj latin neveként a „domesticus” vagy „domestica” szó szerepel (Gallus
domesticus, Felis domesticus stb.J. Ez az elnevezés azonban helytelen, mivel a háziállatok
esetében nem az ősi fajtól elkülönült új fajról van szó, hanem egy vadon élő (esetleg már
kihalt) ős háziasított formájáról. A vadon élő egyedek termékenyen szaporodhatnak a
háziasított formákkal. Ezért a helyes írásmód szerint a faj kettős latin neve után következik
a „forma domestica” (f. domestica) megjelölés. Abban az esetben, amikor a fajnév már
erősen rögzült a köztudatban, használható az is a forma neveként a „domesticus” szó
helyett (21.1. táblázat).
A mezőgazdasági szakemberek alfaj vagy változat helyett a fajta fogalmát használják. Ez
nem rendszertani kategória, hanem gyakorlati megnevezés az egy fajhoz tartozó egyedek
kisebb tenyésztett csoportjaira. A fajtákat mesterséges szelekció során hozták létre, vala-
melyik tulajdonságuk (rezisztencia, terméshozam, hústermelő képesség, tojástermelő
képesség, forma, szín stb.) erőteljes fejlesztésével. Szarvasmarhafajta például a magyar
szürke, a magyar tarka, az osztrák tarka, a fekete-tarka lapálymarha, a hereford stb.
A központi rendszertani kategória (taxon; többes száma: taxa) a faj (species). A fajnál
magasabb, főbb rendszertani kategóriák a következők: nem (genus), család (família), rend
(ordo), osztály (classis), törzs (phylum). Mindegyik fő kategória több alkategóriára bont-
ható, vagy nagyobb csoportba vonható össze. (Ezeket az áttekinthetőség kedvéért nem
tárgyaljuk, és ahol lehet, a későbbiekben is kerüljük.) A harcsa (Sílurus glanis) például a
Sílurus nembe, a harcsafélék (Siluridae) családjába, a harcsaalakúak (Süuriformes) rend-
jébe, a sugarasúszójú halak (Actinoptergyii) osztályába és a gerincesek (Venebrata)
törzsébe tartozik.
A fajok feletti rendszertani kategóriákban igen nagy különbségek lehetnek a fajszámot
tekintve az egyes csoportok között. Van olyan törzs, ahova a jelenlegi ismeretek szerint
mindössze egyetlen faj tartozik (Placozoa: Trichoplax adhaerens), míg az ízeltlábúak
(Arthropoda) törzsébe sorolják a ma élő fajok mintegy háromnegyed részét. Hasonló helyzet
látható a például madarak rendszerezésében is, ahol a fajok mintegy fele tartozik az
énekesmadár-alakúak (Passeriformes) rendjébe, a másik fele pedig a többi rendbe. Elvben
az egymástól távolra helyezett rendszertani egységekbe tartozó fajok származás szempontjá-
ból távol esnek egymástól, a közös csoportba soroltak pedig közelebbi rokonok. A legtöbb
esetben a fajok tényleges rokonsági kapcsolatait nem ismerik. A hagyományos rendszerek
általában a fajok morfológiai leírásán, azok összehasonlításán alapultak. Az utóbbi években
sokasodó molekuláris genetikai kutatások lehetővé tették a genetikai alapokon nyugvó
rendszertanok megjelenését. Ezek számos faj rendszertani helyét, egyes taxonok rokonsá-
gát a hagyományos elképzelésekhez viszonyítva jelentősen megváltoztatják.
163
Rendszertani kategóriákat úgy hoznak létre, hogy egyedek, illetve csoportok tulajdon-
ságait összehasonlítják. Rokonságot a homológ tulajdonságok alapján állapítanak meg. A
homológra azt jelenti, hogy két vagy több hasonló szerv, tulajdonság (aminosav-sorrend
a DNS-ben, végtag csontos váza, viselkedéssor stb.) egy közös, ősibb formából alakult ki.
Az emlősök végtagjai például homológ szervek, függetlenül attól, hogy a denevérek szár-
nya, a lajhárok kapaszkodó lába vagy a kenguruk kicsi mellső lába kevéssé hasonlít egy-
máshoz. Az említett változás a szétválás (divergencia) folyamatának következménye. Az
ősi forma a környezet hatására, a szelekció következtében, ritkábban mutációk miatt
megváltozik és a létrejövő új formák nemzedékről nemzedékre egyre jobban különböznek
egymástól. Ezért a közös őstől származó csoportok rokonsági vizsgálatát a homológ
bélyegek vizsgálata alapján el lehet végezni. Ehhez azonban különbséget kell tenni törzs-
fejlődési értelemben fiatalabb, leszármaztatott (apomorf) és törzsfejlődési értelemben
ősibb (pleziomorf) bélyeg között. Egy apomorf bélyeg megléte két csoportban közeli
rokonságot jelez, és arra utal, hogy a két csoport közös őstől származik (monofiletikus
csoport). A pleziomorf bélyegek a rokonsági kör kijelölésében nem használhatók. Elő-
fordulhat ugyanis, hogy ősi bélyegek egyes fajokon visszafejlődnek, eltűnnek, ezért úgy
látszódik, mintha a rokonság hiányozna. A pleziomorf bélyegek ősiségük folytán a fajok
túlságosan nagy körében láthatók. Rendszertani értékük ezért is kicsi. Az említett eljárás
természetesen körültekintéssel kezelendő, hiszen a tulajdonságok evolúciója történhet
párhuzamosan és az apomorf bélyegek visszafejlődése, eltűnése is lehetséges.
Előfordulhat az, hogy hasonló fünkció ellátására egészen különböző eredetű szervek
egyforma vagy nagyon hasonló felépítésűvé válnak az evolúció során. Ezt a folyamatot
konvergenciának nevezik. Konvergens fejlődés egymástól nagyon távol eső fajok eseté-
ben is megfigyelhető. A fejlődés során létrejövő különböző eredetű, de hasonló felépítésű
szerveket, tulajdonságokat analóg szerveknek, analóg tulajdonságoknak nevezik. Analó-
gia figyelhető meg a vakondok és a lótücsök ásólába között. Ez a jelenség a rokonság
megállapítására természetesen nem alkalmas.
Amikor a taxonokat valamely rendszer szerint csoportosítják, klasszifikációt végez-
nek. Ennek az eljárásnak többféle módja lehet, melyek közül az alábbiakban ismertetjük
a leggyakrabban alkalmazottakat.
1. Klasszikus taxonómia: a fajok szemügyre vétele után különböző, elsősorban morfo-
lógiai tulajdonságok alapján
történik a csoportok, rend-
szertani kategóriák kialakí-
tása. A leírások elsősorban a
fajokról készült rajzok, fény-
képek, méretfelvételek alap-
ján történnek. Nagy szere-
pet játszik a vizsgáló személy
tapasztalata, szintetizálóké-
pessége, szubjektuma.
2. Numerikus taxonómia:
ebben az esetben igyekez-
nekminél több adatot össze-
gyűjteni a besorolni kívánt
taxonokról. Ezek az adatok
lehetnek genetikai, morfo-
lógiai, élettani, viselkedési
stb. bélyegek. Ezután, anél-
kül, hogy az egyes tulajdon-
ságokat súlyoznák, a közös
5.1. ábra. Négy taxon (A, B, C, D) hipotetikus kladogramja. Legkö-
zelebbi rokonság a C és D taxon között áll fenn. A Ti, T2 és T3 jelek
a szétválás idejére utalnak (Mayr nyomán)
164
tulajdonságok megléte, illetve hiánya alapján vonnak le következtetéseket a rokon-
ságról. Lehetőség nyílik arra, hogy számszerűen kifejezhető adatok alapján a több
közös tulajdonsággal rendelkező fajt közelebbi rokonságba, a kevesebb közös
jellemzővel rendelkezőeket pedig távolabbi rokonságba állítsák.
3. Kladisztikus módszer: itt a bélyegeket apomorf és pleziomorf kategóriákba
sorolják. Ezután az apomorf bélyegek összehasonlítása segítségével keresik meg
a közel rokon csoportokat. Nehézséget jelent, hogy az apomorf bélyegektől meg
kell különböztetni a konvergens evolúció során létrejött tulajdonságokat is. A ro-
konsági kapcsolatokat ábrán mutatják be. Ezt az ábrát kladogramnak nevezik (S.l.
ábra).
Világosan kell látni azt a tényt, hogy az élővilág egyféle módon alakult ki, evolválódott
a törzsfejlődés során az elmúlt mintegy 3,5-4 milliárd év alatt. Egymással közelebbi és
távolabbi rokonságban lévő fajok jöttek létre és haltak ki. Egy természetes, a valóságban
lezajlott történéseket tükröző rendszertannak ezt a leszármazási és rokonsági kapcsolathá-
lózatot kellene bemutatnia. Mivel azonban az evolúció lefolyását, az élőlények tényleges
rokonsági viszonyait senki sem ismeri, számos rendszert alkottak a kutatók különböző
elképzelések szerint. Egységes, mindenki által elfogadott rendszertan sem hazánkban, sem
a világon másutt nincs. Ezért ebben a könyvben egy olyan, több forrásból öszszeállított
rendszert használunk, amely alkalmas az élővilág diverzitásának bemutatására, figyelembe
veszi a rendszertani kutatások újabb, már széleskörűen elfogadott eredményeit, ugyanak-
kor konzervatív abban az értelemben, hogy a Magyarországon ismert és elterjedt nevek,
kategóriák megőrzésére törekszik.
5.2. Az élővilág fő csoportjai
Az élőlényeket kezdetben két nagy csoportba, a növények országába (Regnum Plantarum)
és az állatok országába (Regnum Animalium) sorolták. E. Haeckel (1834-1919) a múlt
században a mikroorganizmusok, növények és állatok csoportjait tekintette egy-egy
országnak. Később, századunk harmincas, negyvenes éveiben négy országot különböztet-
tek meg: a monérákat (Monera) (baktériumok és kékeszöld algák), a protoktisztákat
(Protoctista) (egysejtűek, vörös- és bamamoszatok, gombák), a növényeket és az állatokat.
Az utóbbi évtizedekben azonban annyi új ismeret halmozódott fel, különösen a moleku-
láris biológia és a mikrobiológia területén, hogy ez a felosztás elavult. Az újabb ismeretek
alapján 1969-ben Whittaker megalkotott egy olyan csoportosítást, melyet ma már
általánosan használnak. Ennek alapján az élőlények következő öt fő országát különböztetik
meg: monérák (Monera), protiszták (Protista), gombák (Fungi), növények (Plantae),
állatok (Animalia). Ezt a beosztást követjük ebben a könyvben.
Megjegyzendő, hogy a rendszertani felosztások számos egyéb lehetősége ismert még. A
hagyományos, elsősorban a fajok morfológiájára, anatómiájára alapozott rendszerek
mellett a molekuláris szintű struktúrák is lehetőséget és új szempontokat adnak a ka-
tegorizáláshoz. Lehetőség nyílt arra, hogy ezeknek az információknak az alapján alakítsa-
nak ki csoportokat. A riboszomális RNS 500. és 545. pozíciója között található nuk-
leotidok száma alapján például a fajokat 12 országba sorolták. A legnagyobb különbséget
ennek a bélyegnek az alapján a baktériumok és az összes többi élőlény között találták.
Látható tehát, hogy a fajok csoportosításának, rendszerbe sorolásának, vagyis a klasszi-
fikációnak számos lehetősége és módja van.
A következőkben röviden összefoglaljuk a Whittaker által kidolgozott rendszer szerint
az országok legfontosabb jellemzőit. (Megjegyezzük, hogy a vírusokat most nem tárgyal-
juk, mivel önálló életre nem képesek, a gazdasejtjük nélkül élettelen, anabiotikus állapotot
vesznek fel és ezért nem tekinthetjük élőlényeknek.)
165
A legegyszerűbb élőlényeket soroljuk a monérák (Monera = Procariota) közé. Ebbe a
csoportba tartoznak többek között a cianobaktériumok (Cyanobacteria), nyálkabakté-
riumok (Myxobacteriae), sugárgombák (Actinomycota), spirochéták (Spirochaetae) és
valódi baktériumok (Eubacteriae). Sejtmagjuk (cárion) nincs, azaz prokarioták. Ez a
tulajdonság különbözteti meg egyértelműen az ide tartozó élőlényeket a következő
csoportok tagjaitól. DNS-ükzárt, körkörös. A citoplazma aránylag homogén felépítésű,
membránnal körülvett sejtszervecskék (pl. kloroplasztisz vagy mitokondrium) nem talál-
hatók benne. Kemoszintézissel vagy heterotróf módon táplálkoznak. Az osztódás kromo-
szómák kialakulása nélkül történik. Lehetnek egy sejtből álló vagy kolóniákban élő lények.
A prokarioták rendszerezése és jellemzése nem tartozik a könyv tárgykörébe.
A következő négy csoport tagjainak mindegyike eukarióta, mert elkülönült sejtmaggal
rendelkezik. A sejtmagot sejtmaghártya veszi körül és benne található a DNS. Ezért a DNS-
replikáció, az RNS-képzés és a fehérjeszintézis folyamata térben elkülönül egymástól.
A protiszta (Protista) ország, egysejtű, sejtkolóniákból álló és igen kezdetleges test-
felépítésű soksejtű élőlényeket foglal magába. A sejtekben számos, különböző típusú
sejtszervecske található. Gyakran láthatók ostorok és csiliók is. Jellemző a nemzedékvál-
takozással történő szaporodás.
Ide tartoznak többek között az ostoros moszatok (Euglenophyta), sárgás moszatok
(Heterokontophyta), nyálkagombák (Myxomycota) egyes csoportjai és az alább felsorolt,
heterotróf táplálkozásmódú hat törzs:
1. Sarcomastigophora, 2. Labyrinthomorpha, 3. Apicomplexa, 4. Microspora,
5. Acetospora, 6. Gsillósok (Ciliophora).
Mint látható, a jelenleg alkalmazott rendszer szerint a protiszták nem állatok. Is-
mertetésük tehát - szigorúan véve - nem tartozik a zoológia tárgykörébe. Néhány, ebbe az
országba sorolt egysejtű (Protozoa) törzset azonban a legújabb zoológiái szakkönyvek is
tárgyalnak. Tekintettel erre a hagyományra az említett hét törzs ismertetését felvettük e
könyv anyagába és a későbbiekben tárgyaljuk.
A növények (Plantae) olyan soksejtű élőlények, amelyek sejtjeit sejtfal veszi körül.
Autotróf módon táplálkoznak, fotoszintézissel állítják elő a számukra szükséges szerves
anyagokat. Helyváltoztató mozgásra általában nem képesek. Ismert csoportjaik a vörös-
moszatok (Rhodophyta), a barnamoszatok (Phaeophyceae), a zöldmoszatok
(Chlorophyceae) és a többi, klasszikusan ismert soksejtű növénytörzs.
Gombák (Fungi) közé tartoznak azok a soksejtű, nem állati élőlények, melyek hete-
rotróf módon táplálkoznak. A fotoszintézishez szükséges színanyagok a sejtjeikből hiá-
nyoznak. Az elhalt szerves anyagokat sejten kívül emésztik. Egy sejtben gyakran több
sejtmag is található (syncycium). A szöveti differenciáció hiányzik vagy gyengén fejlett. A
növényekhez hasonlítanak abban, hogy gyakran sejtfallal rendelkeznek, de annak legna-
gyobb tömegét nem cellulóz, hanem kitin alkotja. Helyváltoztató mozgásra csak egyes
csoportjaik képesek. Szaporodásuk során rendszerint nemzedékváltakozás figyelhető meg.
A kezdetlegesebb fajok micéliumai haploidok, a fejlettebbeké diploidok. Ebbe a csoportba
tartoznaktöbbekközöttanyálkagombák (Myxophyta), a tömlősgombák (Ascomycotina),
a bazídiumos gombák (Basidiomycotina), a konídiumos gombák (Deuteromycetes) és
a valódi gombák (Eumycota).
Az állatok (Animalia) soksejtű élőlények. Sejtfaluk nincs. Sejtjeiket sejtmembrán veszi
körül. Heterotróf módon táplálkoznak. Rendszerint aktív mozgással könnyen változtatják
a helyüket. Jellemző az idegi szabályozás megjelenése. Az ivaros szaporodás gyakoribb, az
ivartalan kevésbé jellemző. Könyvünkben a következő állattörzsekkel foglalkozunk:
1. Sejthalmazosok (Mesozoa), 2. Lapszerűek (Hacozoa), 3. Szivacsok (Porifera),
4. Csalánozók (Cnidaria), 5. Bordás medúzák (Ctenophora), 6. Laposférgek (Plathy-
helminthes), 7. Zsinórférgek (Nemertini), 8. Fonálférgek (Nematoda), 9. Húrférgek
(Nematomorpha), 10. Buzogányfejűek (Acanthocephala), 11. Kerekesférgek (Rotatoria),
166
12. Csillóshasúak (Gastrotricha), 13. Nyelesférgek (Kamptozoa), 14. Öves férgecskék
(Kinorhyncha), 15. Vértesek (Loricifera), 16. Farkosférgek (Priapulida), 17. Csillagfér-
gek (Echiurida), 18. Fecskendőférgek (Sipunculida), 19. Gyűrűsférgek (Annelida), 20.
Puhatestűek (Mollusca), 21. Karmos féreglábúak (Onychophora), 22. Medveállatkák
(Tardigrada), 23. ízeltlábúak (Arthropoda), 24. Tapogatókoszorúsok (Tentaculata), 25.
Serteállkapcsúak (Chaetognatha), 26. Tapogatószakállasok (Pogonophora), 27. Tüs-
késbőrűek (Echinodermata), 28. Félgerinchúrosok (Hemichordata), 29. Előgerinchú-
rosok (Prochordata), 30. Fejgerinchúrosok (Cephalochordata), 31. Gerincesek (Vertebrata).
Meg kell jegyezni a következőt. Az ismertetett rendszer szerint a Mesozoa, Placozoa és
Porifera törzsek az állatok országába sorolandók. Egy szigorúbb filogenetikai megfontolás
alapján azonban csupán azok az élőlények tekinthetők állatoknak, amelyek egyedfejlődése
során a bélcsíra (gastrula) állapot megfigyelhető. Mivel az említett három törzsbe tartozó
fajok egyedfejlődése során ez a fejlődési stádium nem jelenik meg, ez arra utal, hogy
törzsfejlődésük különálló, a többi állattörzsétől eltérő, sajátos úton valósult meg.
5.3. A heterotróf pro tiszta élőlények rendszere
A következőkben azt a két heterotróf protiszta törzset jellemezzük, amelyeknek nincs
mezőgazdasági jelentősége. A többi törzs részletes bemutatása a következő fejezetben
található.
Labyrinthomorpha
Tengeri egysejtűek. Rendszerint csoportokban, kolóniákban élnek. Spórákat képeznek. Algák
parazitái lehetnek.
Acetospora
Bizonytalan rendszertani helyzetű csoport. Fajai egy vagy több sejtből állnak. Spórákat képeznek, de
ezen belül sem azApicomplexa törzs fajaira jellemző kisebb tok, sem a Micro spóra törzs fajaira jellemző
hosszú fonal nem figyelhető meg. Különböző állatfajok (pl. halak) parazitái.
5.4. Az állatvilág áttekintő rendszertana
Az állatvilág törzseit, különböző szempontok szerint, nagyobb csoportokba szokták so-
rolni. Anélkül, hogy e csoportosítás szempontjainakrészletes tárgyalásába bocsátkoznánk,
a következőkben ismertetjük a legfontosabb, széleskörűen használt fogalmakat. Kézenfek-
vő, ezért elterjedt a sejtszám figyelembevétele. Az egysejtűek (Protozoa) testalakulása a
soksejtűektől (Metazoa) sok egyéb jellemző mellett a sejtek számában is eltér egymástól.
Az egysejtű élőlények mindazokat az életjelenségeket (szaporodás, fejlődés, növekedés,
mozgás stb.) mutatják, mint a soksejtűek. A soksejtűek között régebben elkülönítették a
valódi szövetekkel nem rendelkező, álszövetes állatokat (Parazoa) a többi állattörzset
összefogó szövetesektől (Eumetazoa). Az álszövetesek közé a szivacsokat sorolták. Ma
már úgy vélik hogy a szivacsok testében is található szövet. A szövetes állatokon belül
további csoportosítás lehetséges a csíralemezek száma és a közöttük kialakuló üregek
figyelembevétele alapján. így megkülönböztetnek a) űrbelű, b) testüreg nélküli és c)
testüreges állatokat. Az űrbelű állatok (Coelenterata) teste az ősbél üregét körülvevő külső
és belső csíralemezből áll. A két csíralemez szorosan összesimul, közötte üreg nincsen.
Ebbe a csoportba sorolják a csalánozókat és a bordásmedúzákat. Fejlődéstörténetileg
fontos bélyeg az, hogy az entoderma és ektoderma közötti üreget egyes állatok testében
parenchimaszövet tölti ki, mint a testüreg nélkülieknél (Acoelomata), vagy már megje-
167
lenik a másodlagos testüreg, a mezodermával határolt tér is, amit a testüregeseknél
(Coelomata) lehet megfigyelni. A testüreges állatok egyedfejlődésének jellegzetességei
szerint különbség tehető az ősszájúak (Protostomia) és az újszájúak (Deuterostomia)
között is. Az ősszáj a bélcsíra nyílása. Ez képezi az emésztőcsatorna kezdeti szakaszát a
kifejlett állaton. Az ősszájúak közé a laposférgektől (Platyhdminthes) az ízeltlábúakig
(Arthropoda) terjedő törzseket sorolják. Az újszájúak esetében az ősszájból a bélcsatoma
végső szakasza lesz, az újszáj pedig a bélcsíra más területéből alakul ki. Az újszájúak a
tapogatókoszorúsoktól (Tentaculata) a gerincesekig (Venebrata) található törzsek az
általunk használt rendszertanban. További különbségek tehetők a szövetes állatok között
a szimmetriaviszonyok alapján. Léteznek sugaras, több síkra nézve szimmetrikus
testfelépítésű csoportok (Radiata), a legtöbb állat azonban kétoldalian részarányos, a
test függőleges középsíkjára nézve szimmetrikus (Bilateria).
A következőkben röviden ismertetjük azokat a törzseket, amelyeknek nincs hazánkban
mezőgazdasági jelentőségük. Nem törekszünk teljességre, nem vesszük rendszeresen sorra
a törzseket jellemző összes tulajdonságot. A leírások során csupán azokra a morfológiai,
fiziológiai, ökológiai bélyegekre mutatunk rá, melyek az adott csoportot legjobban
megkülönböztetik a többitől. Igen rövid leírást adunk azokról a törzsekről, melyek tagjai
hazánkban egyáltalán nem fordulnak elő és valamivel hosszabbat azokról, melyekben
mezőgazdasági jelentőség nélküli hazai fajok is találhatók.
Sejthalmazosok (Mesozoa)
Bizonytalan rendszertani helyzetű fajok tartoznak ebbe a törzsbe. Származásukról ellentétes nézetek
léteznek. Egyesek szerint az egysejtűek és a soksejtűek közötti átmeneti formákról, tehát egyszerűbb-
től a bonyolultabb felé történő változást tükröző szervezetekről van szó. Erre mutat laposféregszerű,
kevés sejtből álló testük. Kívül egy gyakran csillós sejtréteg (somatoderma) található. Alatta
láthatók a hossztengely mellett elhelyezkedő axiális sejtek. Mások szerint ezek az állatok leegysze-
rűsödött testfelépítésű paraziták. Erre utal komplikált szaporodási ciklusuk, a férgekéhez hasonló
lárvatípusaik és az a tény, hogy különböző férgek, kagylók, lábasfejűek, tengeri csillagok testében
élősködnek.
Lapszerűek (Placozoa)
Csupán egyetlen faj (Trichoplax adhaerens, Schulze, 1883J tartozik ebbe a törzsbe. Testfelépitésének
sajátosságai azonban annyira különlegesek, hogy ilyen magas szinten kellett elkülöníteni a többi
fajtól. E sajátos testszerveződésű élőlény léte valószínűsíti a következő evolúciós elméletet: a soksejtű
állatok létrejöhettek olyan ostoros egysejtű kolóniákból, melyek kezdetben egy sejtréteget alkottak,
majd a sejtek osztódása után létrejövő leánysejtek nem váltak le az anyasejtekről, hanem egy újabb,
hasi réteget képeztek. Ezt a primitív, két sejtrétegből álló lényt nevezik piacúidnak (vö. a törzs
nevével). Később a két réteg eltávolodott egymástól. így kialakulhatott egy ősi testüreg, majd egy
újabb, közbenső sejtréteg.
Ez a lapított testű állat változtatni tudja az alakját. Teste három sejtrétegből épül fel. A hasi oldalon
hengeres, a háti oldalon pedig lapított sejtek szorosan egymás mellé simuló rétege található. Minden
testfedő sejtből egy ostor indul ki. A hasi oldal sejtjei közé mirigysejtek is ékelődnek. Az említett két
réteg között található rostos sejtek hálózata a mezenchimaszövethez hasonlít. E sejtek citoszkeletonja
jól fejlett, segíti a viszonylag gyors és erőteljes alakváltoztatást. A soksejtű állatok között eleddig
ezekben a sejtekben találták a legalacsonyabb DNS-tartalmat. Meleg tengerek parti zónáiban élnek.
Algákkal táplálkoznak.
Szivacsok (Porifera)
A szivacsok valódi soksejtű, egyszerű szövettel (hámszövet, epithelium), sőt primitív szervvel,
ostoroskamrákkal rendelkező állatok. Testüket kívülről és belülről egy igen lapos sejtekből álló
hámszövet borítja. A két réteg között alapállomány található, amely kollagénből, egy jódtartalmú
168
fehérjéből (spongin), mész- és kovatűkből, valamint számos, különböző
felépítésű és működésű sejtből áll. Az amőbákhoz hasonló sejt (archeocyta)
totipotens, vagyis bármely másik sejttípussá átalakulhat. A spongint a
spongioblasztok hozzák létre, a tűket a szkleroblasztok (kísérősejtek) képezik,
a kollagénfonalak nyalábjait a lofociták termelik.
A szivacsegyedek testén számos apró nyílás fedezhető fel. Ezeken keresztül
áramlik a víz az állat testében található üregrendszerbe, az űrbél (coelenteron)
járataiba, majd innen egy nagyobb nyíláson át (osculum) a külvilágba. Az ősi
szivacsfajok esetében az űrbélüreg falát galléros-ostoros sejtek (choanocytae)
rétege alkotja (aszkonoid típusú szivacs). Ezek a megnyúlt, egymáshoz lazán
kapcsolódó sejtek, egy hosszú ostorral rendelkeznek. Az ostort rövid cito-
plazmakitüremkedések veszik körül egy sorban, mint egy gallér. Ez a felépítés
igen alkalmas arra, hogy a vízáramból kiszűrje a táplálékszemcséket. Egy vi-
szonylag kis csoportnál (szüconoid típusú szivacsok) a test falában csőszerű nyílásokat találunk, ahol
a galléros-ostoros sejtek ezek falát bélelik. Végül a harmadik nagy csoportnál (leukonoid típusú
szivacs) igen nagy mennyiségű az alapállomány. Az ősbélüreg elágazik, az ostoroskamrák tömege
szabálytalanul helyezkedik el.
A szivacsok álló vagy lassan folyó vizek aljzatán élnek. Aktívhelyváltoztató mozgásra nem képesek.
Táplálékukat, a baktériumokat, elhalt növénydarabkákat, kisebb lebegő élőlényeket a vízből szűrik
ki. Ha tömegesen fordulnak elő, ezzel a tevékenységükkel jelentősen elősegíthetik a vizek öntisztu-
lását. A szivacsok hímnősek. Ivartalanul, bimbózással szaporodnak. Ivaros szaporodásuk során az
archeocitákból ivarsejtek, majd a zigótából csillós lárvák jönnek létre. Kedvezőtlen környezetben
gyöngysarjakat (gemmulae) hoznak létre. Ez lényegében archeociták tömege, amelyet kettős burok
vesz körül. A gemmulának mind a szétterjedésben, mind a kedvezőtlen időszakok átvészelésében
fontos szerepe van.
Szivacsok tengerekben és édesvizekben is élnek. A Balatonban gyakori a balatoni szivacs
(Spongilla carteri). Fürdéshez ősidők óta használják a fürdőszivacsot (Euspongia officinalis). Egyes
szivacsfajok antibiotikumokat termelnek. Vannak ehető fajok (Chondrosia spp.J is.
Csalánozók (Cnidaria)
Testük két hámréteg közé zárt középrétegből (mesogloea) áll, amely körülveszi az űrbélüreget.
Ellentétben a szivacsokkal, a csalánozók űrbélürege a bélcsíra ősbélüregének (coeloma) felel meg.
Ennél a csoportnál már megfigyelhető, hogy a külső hámréteg (epidermis) az ektodermából a belső
hámréteg (gastrodermis) pedig az entodermából fejlődik ki. A középrétegben, a két csíralamez
közötti térben sejtek ritkán találhatók. Ennek a területnek az állományát fehérjék, összetett
szénhidrátok és gyakran igen nagy mennyiségű víz (medúzák) alkotja.
A csalánozók jellegzetes képződménye a csalánsejt (nematocyta, cnidocyta). A test felületén
mindenütt előfordulnak csalán-sejtek, de elsősorban a karokon
gyakoriak. A sejtek külső felszínén egy merev serte található.
Ha ezt megérintik, a sejt belsejéből egy fonal csapódik ki. Attól
függően, hogy milyen típusú sejtről van szó, a fonal lehet
ragadós, injekciós fecskendő módjára juttathat mérgező anyagot
az áldozatba vagy egyszerűen összeszoríthatja azt. A kisülés
után a csalánsejt elpusztul, majd a helyén újabb képződik.
A táplálékot részben az ősbélüregben emésztik (extracel-
luláris emésztés), részben ún. abszorpciós sejtek veszik fel és
fejezik be a lebontást (intracelluláris emésztés). A csalánozók
vegyestáplálkozásúak. Nem válogatósak, mindent megesznek,
amit meg tudnak fogni és ami a testüregnyílásukon befér.
Aránylag fejlett, diffúz idegrendszerrel rendelkeznek. Idegsejt-
jeik hálózatot alkotnak, központosulás alig figyelhető meg. Az
169
ingerekre ezért kevéssé differenciált válaszokat adnak. Külön sejtcsoportokkal érzékelik a fényt, a
helyzetüket, a mechanikai és kémiai ingereket. A legtöbb faj esetében generációváltás figyelhető meg.
Az aljzaton ülő, kevéssé mozgékony hidraalak ivartalanul, bimbózással vagy egyszerű lehasadással
hozza létre a vízben szabadon lebegő medúzaformát. A medúzák termelik azután az ivarsejteket.
Megtermékenyülésután azigótából csillós lárva (planula) fejlődik ki, majd megtelepszik az aljzaton,
köveken, növényeken, más állatokon (csigák, kagylók, rákok stb.) és újra hidra formát vesz fel
(metagenesis).
A csalánozók elsősorban tengeri állatok, bár a közönséges hidra (Hydra vulgáris) hazai
vizeinkben is gyakori. A tengerekben néha hatalmas telepeket alkotnak. Ide tartoznak a koraitok,
melyek testében felhalmozódhat a mész. Hatalmas tömegükből sok helyen szigetek jönnek létre. A
koraitok a legproduktívabb állatok a Földön, vagyis egységnyi területen és idő alatt ezek képezik a
legtöbb szerves anyagot. Egyes fajok prosztaglandin hormonokat és a szívműködésre, valamint az
idegrendszerre ható összetett fehérjéket állítanak elő.
Bordás medúzák (Ctenophora)
E gömbszerű vagy vastag szalaghoz hasonló állatok
testének középrétege a legtömegesebb. Igen sok vizet
tartalmazó kocsonyás kötőszövetből áll, melyben
már simaizomsejtek is megjelennek. A testfelszínt
borító hám helyenként megvastagodik és nyolc, rend-
szerint a test hosszában futó, kiemelkedő bordát hoz
létre. A bordákon csillókból összenőtt finom lemezek
ülnek és segítik az állatok helyváltoztató mozgását. A
test belsejében egy speciális csőrendszerben folyik az
emésztés. Ragadozó fajok tartoznak ebbe a törzsbe.
Posztembrionálisan, kifejtéssel fejlődnek. Jellegzetes
azonban, hogy kezdetben minden faj utódja hasonló
(cydippe-stádium), még akkor is, ha kifejlett korban egészen eltérő testalakot vesznek fel.
Tengerekben sokfelé előfordulnak, néha nagyobb tömegekben is.
Zsinórférgek (Nemertini)
Testük hengeres vagy szalagszerű, rendszerint néhány centiméter hosszú. Az óriás zsinórféreg
(Lineus longissimus) testhossza azonban eléri a 30 métert. Ez a tengeri faj a leghosszabb gerinctelen
állat a Földön. A zsinórférgek testét bőrizomtömlő fedi. Legkívül csillós hámréteg található, melybe
számos mirigy sejt ékelődik. A hám alatt elhelyezkedő izomkötegek lefutása jellegzetes, felhasználha-
tó a fajok rendszerezésében. Kültakarójuk gyakran színes. Tipikus
testrészük az igen hosszúra nyúlt ormány (proboscis). Az ektodermális
eredetű ormány rendszerint nem áll összeköttetésben a bélcsatomával.
Nyugalmi állapotában a testbe visszagyűrve található. Ha az állat
zsákmányt fog, az ormányával körülöleli, összeszorítja és gyakran mér-
get is fecskendez bele szúrósertéje (stylet) segítségével. Bélcsatomájuk
háromszakaszos, a középbélhez számos kitüremkedés csatlakozik. Álta-
lában ragadozó fajok tartoznak ebbe a törzsbe. Gyűrűsférgeket, rákokat
esznek. Páros elővesécskék szolgálják a kiválasztást. Összeköttetésben
állnak az igen fejlett, zárt keringési rendszerrel azáltal, hogy lángsejtjeik
a hosszanti véredények falába ágyazódva helyezkednek el. A testfolya-
dék színtelen, kivételesen azonban hemoglobin is előfordulhat benne.
Elkülönült légzőszervrendszerük nincs, diffúz úton lélegeznek. Ideg-
rendszerük a laposférgekéhez hasonlóan négy feji dúcból, garatideg-
gyűrűből és 2-4 hosszanti idegtörzsből áll. Látó- és helyzetérzékelő
szerveik vannak. Váltivarúak, ivarszerveik felépítése igen egyszerű.
170
Külső megtermékenyítéssel szaporodnak. A posztembrionális fejlődés rendszerint közvetlen, egyes
csoportoknál azonban csillós (pilidium) lárvák figyelhetők meg.
Legtöbb fajuk tengerekben él, de ritkán szárazföldön is előfordulnak. Egyes fajaik parazita
életmódra tértek át. Ezek rákok kopoltyúüregében vagy kagylók köpenyüregében élnek. A szabadon
élő formák gyakran találhatók kövek alatt vagy talajba ásott járataikban. A gráci zsinórféreg
(Prostoma graecense) a Balatonban és a Tiszában is megtalálható.
Húrférgek (Nematomorpha)
Vékony, hosszú, több tíz centiméteres állatok, testük átmérője mindössze 1-2 milliméter. A vi-
szonylag kemény kutikula alatt egyrétegű hám (hypodermis) és a hozzá kapcsolódó hosszanti izmok
helyezkednek el. Bélcsatomájuk redukálódott, tápanyagfelvételre és emésztésre már nem alkalmas.
A fiatal állatok parazita életmódúak, táplálékukat a test kültakaróján keresztül veszik fel. A kifejlett
állatok nem táplálkoznak. Kiválasztó-, légző- és keringési szervrendszerük hiányzik. Idegrendszerük
központja a test első felében lévő idegdúc, melyből a hasi oldalon végig húzódó idegköteg indul ki.
Csak a mechanikai érzékszerveik fejlettek. Váltivarú állatok. Mindkét nem páros ivarszervekkel
rendelkezik. A vízbe lerakott petékből kikelő első stádiumú lárvák erőteljes, kiölthető szájszervvel
bírnak. Ennek segítségével hatolnak a köztigazdába, valamilyen ízeltlábú állatba.
Leggyakrabban rákok és vízi rovarok endoparazitái. A táplálékláncokon keresztül azután más
rovarokba is átkerülhetnek. Az utolsó stádiumú lárvák kijutnak a gazdából, vizes környezetben
fejeződik be a fejlődésük. A kifejlett állatok édesvizekben vagy tengerekben élnek. A közönséges
húrféreg (Gordius aquaticus) forrásokban, hegyvidéki patakokban, kisebb folyóvizekben hazánkban
is sokfelé előfordul.
Csillóshasúak (Gastrotricha)
Rendszerint egy milliméternél kisebb testnagyságú fajok tartoznak ebbe a tözsbe. Fejükön rövidebb-
hosszabb csili ók ülnek, amelyek a táplálékot sodorják a szájnyíláshoz. Csili ók találhatók a hasoldalon
is, de ezek a mozgást szolgálják. A kültakarót kívülről egy kutikularéteg borítja. Ebből jellegzetes,
lapos pikkelyek nyúlnak ki. Felépítésük, formájuk fajonként eltérő, így a határozásnál fontos bélyeg.
A test két oldalán csövek találhatók, melyek segítségével az állatok időlegesen rögzíteni tudják
magukat az aljzaton. Bélcsatomájuk hosszú, egyenes, kevéssé tagolt. Elsősorban szerves törmelék-
kel, baktériumokkal, egysejtűekkel táplálkoznak, a ragadozó fajok ritkák. Egy pár elővesécskéjük
végzi a kiválasztást. Idegrendszerük a garat környékén elhelyezkedő páros idegdúcból és az innen
induló hosszanti idegkötegekből áll. Rendszerint hímnősek, ritkán váltivarúak. Az édesvízi fajok
kétféle petét termelnek: az egyik vastag burokkal körülvett, a kedvezőtlen környezeti tényezőknek
ellenálló, tartós pete, a másik burka vékony. A petéből néhány nap alatt kikelnek a fiatal állatok.
Posztembrionális korban kifejtéssel fejlődnek. Rövid ideig, mindössze néhány napig élnek. A
nőstények ez alatt az idő alatt 4-5 petét képeznek. Tengerekben, illetve édesvizekben előforduló fajaik
külön rendszertani csoportba tartoznak. Az iszapot, növényi törmeléket népesítik be, vagy növények,
állatok testfelszínén élnek.
Ritkábbak a talajban, avarban, mohában élő fajok. Magyarországon gyakori a Chaetonotus
maximus.
Nyelesférgek (Kamptozoa)
E néhány milliméter hosszú állatkák testalakja hosszú nyelű kehelyhez hasonlít. A kehelyben (calyx)
találhatók a belső szervek, míg a nyélben erős izomzat van. A testüreget a mezenchimához hasonló
kötőszövet tölti ki, melyben fixált és szabadon mozgó sejtek egyaránt találhatók. A nyél végén
elhelyezkedő ragasztómirigy váladéka a megtapadást segíti. A kehely szélén 8-40, csillókkal belepett
tapogató ül. Ezekkel segítik a táplálékot (baktériumokat, algákat, egysejtűeket) az előtérbe (átrium),
ami nem más, mint a karok tövénél található bemélyedt üreg. Itt található a szájnyílás, és nem messze
a hajlott bélcsatoma végbélnyílása is. A gyomor nagy kiterjedésű üreg, a bélrendszer legterjedelme-
sebb része. Kiválasztórendszerük páros elővesécske. A bélcsatoma belső íve mentén található az
171
idegrendszer központja, egyetlen nagy dúc formájában. Innen indulnak az idegek a kémiai és
mechanikai érzékszervekhez, valamint a tapogatókhoz. Ivartalanul, bimbózással szaporodnak. Az
együtt maradó egyedek gyakran képeznek nagyobb telepeket. Ivarosán is szaporodnak. Lehetnek
hímnősek és váltivarúak is. A peték már a nőstény petefészkében megtermékenyülnek és egy, az
ivarnyílás közelében elhelyezkedő költőüregben fejlődnek tovább. Lárváik koszorúslárvák
(trochophora típus), hasonlóak a gyűrűsférgekéhez.
Szinte kizárólag tengerek parti régióiban élnek, gyakran kövekhez, csigák, kagylók héjához, rákok
páncéljához tapadva. Szivacsokkal, gyűrűsférgekkel, virágállatokkal élhetnek együtt. Egyetlen
édesvízi nemük egyik faja, az édesvízi nyelecske (Umatella gracüis) hazánkban is előfordul. Még
csapvízből is kimutatták.
Öves férgecskék (Kinorhyncha)
Egy milliméternél kisebb állatok. Ellentétben az előző két törzs tagjaival, külta-
karójukon csillók nincsenek. Igen jellegzetes sajátosságuk, hogy testüket 13
gyűrű (zonit) építi fel.
Kutikulájuk vastag, helyenként lemezeket képez. Háromszakaszos bélcsator-
nájuk egyenes lefutású, izmos garattal ellátott. Főleg kovamoszatokat és elhalt
növénydarabkákat esznek. Idegrendszerük garatideggyűrűből és a hasi oldalon
futó idegtözsből áll. Törzsfejlődéstani szempontból tekintve fejlett bélyeg, hogy
a 3-13. gyűrű kültakarójában idegdúc is található. Az idegdúcokat idegsejtek
hálózata köti össze. Váltivarú állatok tartoznak ebbe a csoportba. Posztemb-
rionális fejlődésük kifejlés. A fiatal lárvák hat vedlés után érik el az ivarérett
állapotot. Kizárólag tengeri fajok, melyek a partközeli területek iszapjában
mászkálnak.
Vértesek (Loricifera)
A legutoljára, 1983-ban leírt állattörzs. Fajai milliméternél kisebbek.
Testük legnagyobb táját az altest (abdomen) képezi. Ezt négy oldalról, a
tulipán szirmaihoz hasonló lemezek veszik körül. Ezekről a lemezekről
kapták a nevüket is (lorica = bőrpáncél, mellvért).
Kétoldalian szimmetrikus állatok. A fej és a tor a lemezek közé vissza-
húzható. A szájnyílás körül nyolc szúrótövis található. Bélcsatomájuk
terjedelmes. Agyuk jól fejlett, idegrendszerük a hasdúclánchoz hasonló.
Váltivarú állatok. Lárváik a kifejlett egyedekhez hasonlóak. Biológiájuk
még kevéssé ismert. Tengerfenék iszapjában élnek. 8000 m mélységben
is előfordulnak.
Farkosférgek (Priapulida)
Néhány centiméter hosszúságú tengeri állatok. Testük 25 hosszanti bordá val díszített
előtörzsre és az ehhez csatlakozó törzsre tagolódik. Ez utóbbi lebenyes szerkezetű
farokfüggelékben végződik. Szelvényezettséget nem mutatnak.
Nincs véredényrendszerük, de testfolyadékukban gáz szállítására alkalmas szína-
nyag (hemerythrin) előfordulhat. Posztembrionális fejlődésük kifejlés. A lárvák
sokszor évek alatt, számos vedlés után érik el a kifejlett, ivarérett álla potot. Tengerek
parti zónájában élnek. Igen érzékenyek a viz oxigéntartal mának változására, ezért
ebből a szempontból indikátorszervezeteknek tekinthetők.
Csillagférgek (Echiurida)
Hengeres testű állatok, hosszú megnyúlt, ámbár a testnél rendszerint rövidebb ormányt (proboscis)
viselnek. Kivételes esetben azonban testarányaik jelentősen eltérőek lehetnek, a 15 centiméteres
testhez még 2 m hosszúságú ormány is csatlakozhat (Boniella spp.J. Igen hosszú bélcsatornájukból
172
a középbélnél egy oldalág indul ki. Ormányukat a hasi oldalon felcsavarva tartják. Az így kialakuló
cső belső felszínén található csiliók vizáramot létesítenek és az algákból és elhalt növényi részekből
álló táplálékot a szájnyíláshoz hajtják. Zsákszerű kiválasztószerveik (metanephridium) a test külön-
böző részein találhatóak. Zárt keringési rendszerük hasonlít a gyűrűsférgekéhez, de testfolyadékuk-
ban színanyag nem található. Váltivarú állatok, gyakran nagyon kifejezett ivari különbségekkel
(sexualis dimorphismus). Néha a törpe himek a nőstények testében élnek. Csillókoszorús lárváik
vannak. Tengeri állatok.
Fecskendőférgek (Sipunculida)
Hengeres testük kevéssé tagolt. Mindössze a kiölthető és visszahúzható, tapogatókkal r jP
ellátott előtörzs különül el a test többi részétől. Kutikulával ellá tott bőrizomtömlőjük Cj
igen gazdag mirigyekben. Bélcsatomájuk lefutása különleges: csavarodottan fut a
szájnyílástól a test végéig, ott visszakanyarodik és az előző szakasszal összesimulva kettős / \
spirált képez. A végbélnyílás az előtörzs végén, az állatok hátoldalán található. A /
testfolyadékban az előző törzshöz hasonlóan, itt is színanyag található. Lárváik a
gyűrűsférgek csillókoszorús lárváihoz (trochophor lárva) hasonlítanak. Tengeri, az \
aljzaton, homokban, iszapban élő állatok. \ /
Karmos féreglábúak (Onychophora)
Megnyúlt, gyűrűs testű állatok, melyeknek fején egy pár, nem ízelt tapogató látható. Jellemzőjük,
hogy a száj felett egy szemölcs (papilla orális) található, melynek ragadós váladéka segíti a zsák-
mányolást.
Idegrendszerük nem hasdúclánc típusú, hanem egymástól távol futó idegtörzsekből áll, amiket
gyűrűnként 9-10 harántidegköteg köt össze. Igen érdekes csoport, ahol az említett sajátos bélyegek
mellett a gyűrűsférgekre és az ízeltlábúakra jellemző tulajdonságok egyaránt felismerhetők. Az
egyforma testszelvények, a bőrizomtömlő és a szelvényenként előforduló, páros metanefridiumok a
gyűrűsférgekhez teszik hasonlóvá a testet. Ezzel szemben az ízeltlábúakhoz hasonló tulajdonságuk,
hogy kitines kutikulájuk, valamint nyílt keringési rendszerük van, vedlenek, és az embrionális
fejlődésük is az utóbbi csoportéhoz hasonlít. Általában trópusi erdők avarjában élnek.
Medveállatkák (Tardigrada)
Másodlagosan leegyszerűsödött testfelépítésű állatok, vagyis őseik szervezete bonyolultabb felépíté-
sű volt, mint a ma élő fajoké. Tömzsi testük fejből és négy testszelvényből áll. Ritkán hosszabbak egy
milliméternél.
Kültakarójuk kitines kutikula, amit gyakran szép mintázat díszít. A kutikula néhol lemezeket
alkot. Kültakarójukat rendszeresen vedlik. Egy állat élete során rendszerint 12 vedlés következik be.
Négy pár lábuk végén karmok vagy tapadókorongok figyelhetők meg. Szájukban mészből lévő
szúróserte (stylet) található. Segítségével szúrjákmeg
növényi (algák, mohalevelek), vagy állati (fonálférgek,
kerekesférgek, más medveállatkák) táplálékukat, majd
izmos garatjuk pumpáló mozgásával szívják ki a folyé-
konynedveket. A bélcsatoma legterjedelmesebb része
a középbél, ahol a táplálék lebontása és felszívása
történik. Kiválasztószervük a rovarokéhoz hasonló Mal-
pighi-edény. Elkülönült keringési és légzőszervrend-
szerük nincs. Hasdúclánc idegrendszerrel rendelkez-
nek. Néha primitív, egyetlen pár, egy-egy sejtből álló
szem is megjelenik. A legtöbb faj váltivarú.
Egyes fajok hímjei még nem ismertek. Posztembrionális fejlődésük epimorfózis. Nagyon gyorsan
fejlődnek, egy-két hét alatt elérik a kifejlett állapotot. Gyakori a szűznemzés (parthenogenesis) is.
Tengeri, édesvízi, többségükben pedig szárazföldi fajok. A szárazföldiek igen nedves környezetben,
173
a talajokban sokfelé élnek. Ha élőhelyük kiszárad, testük víztartalma lecsökken, összetöppednek,
életfolyamataik rendkívül alacsony szintre csökkennek és így vészelik át a száraz időszakot. Ezt
nevezik anabiotikus vagy kriptobiotikus állapotnak. Ilyen formában egyes fajok elviselik az
abszolút nulla fokhoz közelítő hőmérsékletet, vagy azt is, ha tiszta szeszbe helyezik őket. Ezért extrém,
rendkívüli élőhelyek lakói lehetnek. Amikor a kedvező életfeltételek újra helyreállnak, néhány óra
alatt visszanyerik eredeti, aktív állapotukat. Életmódjuk rendkívül specializált. Sok faj a mohák és
páfrányok felszínét bevonó vízhártyában él. A lajhárállatka (Arctíscon tardigradum) hazánkban
mindenfelé gyakori.
Tapogatókoszorúsok (Tentaculata)
Viziállatok, rendszerint az aljzathoz tapadva élnek, gyakran nagy telepeket is alkotnak. Testük alig
tagolt, fejük redukálódott. Kitintartalmú kutikulájukba néha mészsók is rakódnak. A kültakaróhoz
kapcsolódó izomrétegek hiányoznak. A szilárd külső test-
fal és a bélcsatoma között nagyméretű testüreg található.
Szájnyílásukat változó számban csillós tapogatók veszik
körül (lophophor). Ezek körben vagy patkó alakban he-
lyezkedhetnek el. Segítségükkel szűrikki a vízből az állatok
a táplálékukat. Bélcsatomájuk visszahajlik, a végbélnyílás
a szájnyílás közelében, a test hátoldalán található. A plank-
tont alkotó növényeket és állatokat fogyasztják. Keringési,
légzési és kiválasztó szervrendszerük hiányzik. Idegrend-
szerüket a garat környékén lévő idegdúcok, a tapogatókba
futó ideg-kötegek és a testet behálózó idegek alkotják.
Specializált érzékszervekkel nem rendelkeznek. Átalaku-
lással fejlődnek, lárváik a gyűrűsférgek csillókoszorús lár-
váihoz hasonlítanak.
Igen heterogén csoport, amelyet ezért egyes rendszere-
zők több törzsre bontanak. Édesvízi fajai közül hazánkban
sokfelé megtalálható az ágbogas mohaállat (Fredericella
sultana).
Serteállkapcsúak (Chaetognata)
Nagyon gyakori tengeri állatok. Gyors mozgásuk és átlátszó, hengeres testük miatt
nyílférgeknek is nevezik az idetartozó fajokat. Ragadozó életmódot folytatnak, zsákmányu-
kat a fejen lévő hegyes, iveit tövisekkel ragadják meg. Központi idegrendszerüket garat-
ideggyűrű és egy hatalmas hasi idegdúc képezi. Himnős állatok. A petesejtek érése csupán
a megtermékenyülés után következik be. Lárváik nincsenek.
Tapogatószakállasok (Pogonophora)
A tengerek mélyebb rétegeiben (gyakran több
ezer méter mélységben) találhatók. A testük
átmérője ahosszukhoz képest igen csekély. Rend-
szerint kitines csövekben élnek.
A test elülső végéről néha több száz tapogató
indul ki. Bélcsatomájuk hiányzik, tehát a táp-
anyagokat valószínűleg diffúzió útján veszik fel.
Váltivarúak, de ivari dimorfizmus nem figyelhe-
tő meg náluk. Regenerációs képességük kifeje-
zetten nagy. Rendszertani helyzetük bizonytalan. Ezt a
törzset egyesek az összájúakhoz, mások az újszájúakhoz
sorolják.
174
Tüskésbőrűek (Echinodermata)
Eredetileg kétoldalian szimmetrikus állatok, melyek
testfelépítése másodlagosan (a törzsfejlődés során) vált
álötsugaras szimmetriájúvá. Testük szilárdságát a hám
alatt elhelyezkedő mészváz adja. Innen némely cso-
portnál mozgatható mésztüskék indulnak ki. Speciális
szervük a csövekből és hólyagokból álló vízedény-
rendszer (hydrocoel), melyből a hasoldalon található
zsákszerű nyúlványokba (ambulakrális lábak) folya-
dékot tudnak préselni. így, magukat lassan tologatva
képesek helyváltoztatásra. Központi idegrendszerük és
érzékszerveik gyengén fejlettek. Váltivarúak, átalaku-
lással fejlődnek. Lárváik kétoldalúan szimmetrikusak.
Egyes fajok mérget termelnek.
Testük magas mész- és nitrogéntartalma miatt az állatok testéből készített őrleményt némely
vidékeken trágyaként is használják.
Félgerinchúrosok (Hemichordata)
Magányosan vagy csoportokban élő tengeri állatok. Bélcsatomá-
juk kezdeti szakaszából a test eleje felé kraniálisan rövid nyúlvány
irányul, amelyet a gerinchúrral homológnak tartottak (illetve
tartanak). Az előbélből kitüremkedések indulnak ki, melyek kes-
keny nyílásokon keresztül kapcsolatban állnak a külvilággal. A
szájon keresztül vízáram jut beléjük és ekkor történik a gázcsere
(kopoltyúbél). Nemcsak a hasoldalukon, de a hátoldalukon is
húzódik egy idegtörzs. Váltivarúak. Lárváik hasonlóak némely
tüskésbőrű lárváihoz (tomarialárva).
Számos faj egyedei a tengerek fenekén csöveket vájnak maguk-
nak és ezekben élnek.
Előgerinchúrosok (Prochordata)
Kizárólag tengeri állatok. Lárvakorban farokkal rendelkeznek, s ebben megjelenik a gerinchúr
(chorda dorsalis). Kifejlett korban számos faj helybenülő életmódra tér át, ami testük módosult
felépítésén is tükröződik.
Egyrétegű hámból álló kültakarójuk egy cellulózhoz hasonló anyagot, tunicint választ el. Ez
alkotja rugalmas, és szilárd külső vázukat. Kopoltyúbéllel rendelkeznek, ami nem csupán a légzés,
de a kiválasztás funkcióját is ellátja. Keringési rendszerük nyílt. A kifejlett állatok idegrendszerének
központját egyetlen idegdúc alkotja. Hímnős állatok, amelyeknél az ivaros szaporodásmód mellett
bimbózás is megfigyelhető. Egyes fajok világítanak, mert fénykibocsájtó baktériumokkal élnek
szimbiózisban.
Fejgerinchúrosok (Cephalochordata)
Halformájú tengeri állatok. Belső szerveik legtöbbje szelvényezett. Testük hátoldalán a velőcső alatt
húzódik végig a gerinchúr. Vékony kültakarójuk alatt az izomzat jellegzetes „V” formájú kötegeket
alkot. Kopoltyúbelük egy üregben található. Keringési rendszerük zárt. Valódi agyuk nincs, de a feji
részen nagyobb idegdúc található. Ebből nyúlik hátrafelé a gerincvelő. Váltivarúak. A kopoltyúbél
üregében mintegy 25 pár ivarszerv található. Legismertebb képviselőjük a lándzsahal (Branchiostoma
lanceolatum).
175
Ellenőrző kérdések a 5. fejezet témaköreiből
1. Mit jelent a binominális nómenklatúra kifejezés? Mondjon példákat (legalább hármat) az
alkalmazására.
2. Sorolja be a házi verebet a fő rendszertani kategóriákba!
3. Hasonlítsa össze a klasszikus taxonómia, numerikus taxonómia és a kladisztikus módszer
alkalmazásának előnyeit és hátrányait!
4. Táblázatosán foglalja össze, hogy miben hasonlítanak, és miben különböznek az élővilág
öt országának képviselői!
5. Milyen bélyegek alapján lehet az állatfajokat az országnál kisebb, de a törzseknél nagyobb
kategóriákba sorolni?
6. Jellemezze a szivacsokat, a csalánozókat, a húrférgeket és a medveállatkákat!
7. Jellemezze a szivacsokat!
8. Jellemezze a csalánozókat!
9. Jellemezze a húrférgeket!
10. Jellemezze a medveállatkákat!
176
6. ALLABAS-OSTOROS EGYSEJTŰEK
(SARCOMASTIGOPHORA)
A törzsbe tartozó fajok ostorral vagy állábakkal, esetleg mindkettővel rendelkező protiszták.
Egy vagy több sejtmagjuk van. A több sejtmagvú fajoknál magdimorfizmus nincs, ezért egy-
félemagvúáknak is hívják őket. Az előbbi megállapítás alól a likacsoshéjúak kivételt képeznek,
mert sejtmagjaik eltérőek, tehát ezek az egysejtűek heterokarióták. A protiszták között ez a törzs
a legnagyobb fajszámú csoport. A következőkben részletesebben bemutatásra kerülő három
altörzsbe tartozó ismert, ma is élő fajok száma közel húszezer.
A törzsbe tartozó fajok közös tulajdonsága, hogy ivartalanul osztódással szaporodnak, míg
ivaros szaporodásuk az ivari jelleggel bíró egyedek összeolvadása (syngamia). Az ivaros sza-
porodás során kialakuló formák alakilag eltérhetnek az ivartalanul osztódóaktól. A szabadon
mozgó alakokat ilyenkor trofozoitáknak (trophozoita) nevezik. Heterotróf táplálkozásúak, a
fajok jelentős része parazita, így állat- és emberegészségügyi szempontbólfontosak. Táplálékukat
a sejthártyán keresztül vagy sejtszájon át veszik fel. Egyes fajok védőburkot képeznek maguk
körül. A szabadon, vízben élőknek szerepük van a talaj és a víz szerves anyag forgalmában.
Törzs: Állábas-ostoros egysejtűek (Sarcomastigophora)
Altörzs: Ostorosok (Mastígophora)
Osztály: Állati ostorosok (Zoomastigophorea) 6.1. fejezet
Rend: Parabazális-testes ostorosok (Kinetoplastida) 6.1.1. fejezet
Rend: Tükrözött-testű ostorosok (Diplomonadida) 6.1.2. fejezet
Rend: Tengelypálcás ostorosok (Trichomonadida) 6.1.3. fejezet
Altörzs: Gyöngyállatkák (Opalinata)
Altörzs: Állábasok (Sarcodina)
Főosztály: Gyökérlábúak (Rhizopoda) 6.2. fejezet
Osztály: Lebenyes-lábúak (Lobosea)
Rend: Csupaszamőbák (Amoebida) 6.2.1. fejezet
Rend: Házasamőbák (Arcellinida) 6.2.2. fejezet
Osztály: Hálózatos-lábúak (Granuloreticulosea)
Rend: Likacsoshéjúak (Foraminiferida)
Főosztály: Tengelyeslábúak (Actinopoda) 6.3. fejezet
Osztály: Napállatocskák (Heliozoea) 6.3.1. fejezet
Megjegyzés: A rendszertani felosztást Levin és munkatársai (1980) munkája alapján állítottuk össze.
Az ostorosok altörzsbe tartozó fajok trofozoita alakjainak mozgási sejtorganelluma általában az
ostor (flagellum). Mintegy 7000 fajuk ismert. Két osztályuk közül az egyik az autotróf táplálkozást!
növényi ostorosokat (Phytomastigophorea) foglalja magába.
A gyöngy állatkák altörzsbe tartozó fajok kettő vagy több sejtmagja egyféle típusú. Sejtszájuk és
lüktetőűröcskéjük nincs. Valamennyi faj belső élősködő, főleg kétéltűekben, hüllőkben és halakban
fordulnak elő. Ide tartozik például az Opalina ranarum, amely a kétéltűek kloákájában él.
177
Az állábasok altörzsébe tartozó fajok kifejlett alakjai állábakkal (pseudopodium) vagy citoplaz-
ma-áramlással mozognak. Néhány faj korai fejlődési szakaszában ostor fordulhat elő. Vannak
közöttük vízi vagy nedves körülmények között szabadon élők és paraziták is. Az ide tartozó taxonok
egy része ősi, eukarióta fejlettségi állapotból vezethető le, míg más részük az ostorosokkal mutat
rokonságot. A körülbelül 41 600 leírt fajból közel 30 000 fosszilis, a ma élők közül pedig mintegy 250
parazita. A Lobosea osztályba tartozó fajok állábai lebenyszerűek, karéjosak vagy többé-kevésbé vé-
konyak, ujjszerűek. A Granuloreticulosea osztály képviselői finoman szemcsézett vagy áttetsző állábaik
nyúlványai (reticulopodium) hálózatot képeznek. Ebbe az osztályba tartoznak a likacsoshéjúak
melyek döntő többsége tengerben él. Főleg kalcium-karbonát tartalmú, kamrákra tagolt vázat hoz-
nak létre. Közel 30 000 fosszilis, illetve 4000 ma is élő fajuk van. Ismertebbek a Nummulites nembe
tartozó fajok (21.8.3. fejezet).
6.1. Állati ostorosok (Zoomastigophorea)
Az osztályba tartozó fajok minden bizonnyal polifiletikus eredetűek. Valószínűleg a
növényi ostorosok különböző csoportjaitól származtak, melynek során elvesztették
színtesteiket, áttértek a heterotróf táplálkozásmódra. A közel 4300 leírt, ma élő fajnak
mintegy a fele parazita.
Testük alakját a rugalmas alakváltozást lehetővé tevő bőrke (pellicula) szabja meg.
Igen sok protiszta kültakarója a pellikula, amely a sejtmembránból és az alatta megha-
tározott elrendeződésű vezikulumok, mikrotubulusok és belső membránok rendszeré-
ből áll. Mozgásszervük lehet egy vagy több ostor, de néhány csoportnál az ostoros
mellett előfordulhat állábas mozgás is. Az ostor mozoghat két dimenzióban vagy spirális
pályán. Az utóbbi a sűrűbb közegen való áthaladást is lehetővé teszi. A fajok egy részénél
a sejt teste és az ostor között a pellikulából kialakult hullámzó hártya (membrana
undulans) van, amely a mozgásukat segíti. Az ostor alapján a bazális test (kinetosoma)
található, amely még egy nagyobb méretű és DNS-tartalmú parabazális testtel (kinetoplast)
is kapcsolatban állhat (Kinetoplastida rend).
Az állati ostorosok, a többi heterotróf protisztához hasonlóan táplálékukat a pelli-
kulán keresztül aktív transzport révén vagy endocitózissal vehetik fel. A protiszták egy
része, így az állati ostorosok is rendelkezhetnek sejtszájjal (cytostoma). Ez a sejtga-
ratban (cytopharynx) folytatódik, amelynek végéről a táplálékszemcsék a vakuólákba
zártan fűződnek le. Az emészthetetlen részecskéket vagy exocitózissal, vagy a sejt egy
meghatározott helyén kialakult sejtalrésen (cytopyge) keresztül adják le a külvilágra.
A lebontó folyamatok során keletkezett nitrogéntartalmú bomlástermékeket elsősor-
ban ammónia formájában távolítják el a sejthártyán keresztül.
Az ostorosok általában ivartalanul, hosszanti kettéosztódással szaporodnak,
amelynek során két egyforma utódsejt jön létre. Az ostor bazális teste és az egyéb
szervek megkettőződése általában már az osztódás előtt végbemegy. Ivaros szaporodá-
suk során a két egyed összeolvad (syngamia). Ezt a fajok egy részénél a gametikus
meiózis előzi meg, míg a többieknél ezt követően zigotikus meiózis zajlik le. A gametikus
meiózis során ivari pronukleuszok jönnek létre. A szingámia után zajlik le a két egyed
pronukleuszainak összeolvadása.
6.1.1. Parabazális-testes ostorosok (Kinetoplastida)
Egy vagy két ostorral rendelkeznek. A fajok többsége parazita, a szabadon élők száma
kevés.
A Cryptobia fajoknak két ostora van (6.1. ábra). A 10-30 pm testhosszúságú Cryptobia cyprini a
halak álomkórjának okozója. Halpiócák terjesztik, amelyeknek testében az ivaros szaporodás során
178
a mozgó alakoktól morfológiailag eltérő fejlődési fonnák
jönnek létre. A halpióca a vérszivások alkalmával fertőzi
meg a halakat. Az ostoros alakok a vérben élnek, a vér-
plazma összetevőit fogyasztják és avérsejteket károsítják.
Az álomkórt okozó egyostoros Trypanosoma fajok szá-
mos állatfaj, így a háziállatok, valamint az ember para-
zitái lehetnek, elsősorban a trópusi területeken. Testhosz-
szúságuk általában 10-30 |um közötti. A gerincesek véré-
ben, egyes szerveikben élnek. A sejtekben előforduló
fejlődési alakjuk nem rendelkezik ostorral, de az extra-
celluláris forma igen. Egy ostoruk a megnyúlt test teljes
hosszában végighúzódik és hullámzó hártyát alkot (6.1.
ábra).
A Trypanosoma equiperdum a lovakban a tenyészbéna-
ság nevű fertőző betegséget okozza. Terjedése a párzás
útján történik. Kezdetben a nemi szervek hámrétegét
károsítja, később a vérárammal eljut egyes idegekhez,
valamint a bőrbe, ahol gyulladásos folyamatokat vált ki.
Hazánkban az utóbbi évtizedekben nem fordult elő ez a
parazita.
Emberegészségügyi szempontból is jelentős kórokozó a Trypanosoma brucei, amelynek három
alfaját különítik el. A T. b. brucei az afrikai emlősállatok parazitája, míg a T. b. gambiense és a T.
b. rhodesiense az ember álomkór betegségét okozza. Mindhárom alfaj terjesztői a cecelegyek
(Glossinapalpalis), amelyek a vérszívás során juttatják újabb gazdába a parazitát. Az ember vérébe
bekerült Typanosoma alakok többszörös ivartalan osztódással szaporodnak, majd elárasztják az
egész szervezetet. Előfordulnak a vérben és a nyirokban, a lépben, a központi idegrendszerben és
az agy-gerincvelői folyadékban. A cecelégyben a parazita ivartalanul és ivarosán is szaporodik, és
a vérszíváskor a nyállal jut be a gerinces gazdaállatokba és az emberbe is.
6.1. ábra. Állati ostorosok (Zoomastigophorea):
A; Trypanosoma sp., B: Cryptobia sp., C:
Giardia sp., D: Trichomonas sp. (Mehlhom és
más szerzők nyomán)
6.1.2. Tükrözött-testű ostorosok (Diplomonadida)
Testük kétoldali részarányosságot mutat. Egy-négy pár ostorral rendelkezhetnek. Több-
ségük parazita életmódú. Cisztaképzés megfigyelhető.
A Giardia fajok az egész Földön elterjedtek. A gerincesek bélcsatomájában élnek (6.1. ábra).
Ide tartozik a sejtszájnyílás nélküli parazita Giardia intestinalis, mely 10-20 |jm hosszú. Az ember,
elsősorban a gyerekek vékonybelében fordul elő és hasmenéses megbetegedést okoz. A fertőződés
a bélsárral szervezetből kikerülő cisztákkal történik.
6.1.3. Tengelypálcás ostorosok (Trichomonadida)
Általában 4-6 ostoruk van, amelyek közül az egyik hullámzó hártyával van kapcsolatban.
Valamennyi ide tartozó faj parazita.
A Trichomonas fajoknál három ostor előre néz, míg a negyedik a test oldalához kapcsolódik hul-
lámzóhártya segítségével (6.1. ábra). Ahullámzó hártya alapjánál asejttest szegélyén egy kiemelkedő
megvastagodás (costa) húzódik. A sejtet a tengelypálca (axostyl) támasztja. Az emlősállatok, az
ember bélcsatomájában és az ivari kivezetőjáratokban élnek. A Trichomonas foetus 10-18 |Jm hosszú,
a szarvasmarha nemi szerveinek nyálkahártyáján élősködik. A paraziták a fedeztetés során terjednek.
A fertőződés a teheneknél vetélést, időszakos meddőséget, ivarzási zavarokat okoz. A Trichomonas
gallinae különböző madárfajokban él. Az erős fertőzöttség akár elhullással is járhat. Főleg a szájüreg,
a garat, a nyelőcső és a begy nyálkahártyáját támadja meg, de más belső szervekben is előfordulhat.
A Trichomonas vaginalis az ember parazitája, a nemi szervek nyálkahártyáján élősködik.
179
6.2. Gyökérlábúak (Rhizopoda)
Mozgásuk elsősorban állábakkal történik. Az állábak különböző alakúak lehetnek, így
elkülöníthető a lobopodium, filopodium, reticulopodium és axopodium típus. Alobo-
podiumok a mozgást és a táplálékrészecskék bekebelezését szolgáló lebenyszerű nyúl-
ványok. Bennük az ekto- és endoplazma egyaránt megtalálható. A filopodiumok csak
ektoplazmát magukba foglaló vékony nyúlványok. A retikulopodiumok az előző típus-
hoz hasonlóak, de a nyúlványok elágazásai hálószerűén összekapcsolódnak. Az axopo-
diumok igen vékony fílamentumok, amelyeket mikrotubulusokból álló tengely merevít.
Testük többnyire csupasz, de külső vázuk is lehet.
Sejtszájuk nincs, a táplálékrészecskéket fagocitózissal veszik fel. Azoknál a fajoknál,
ahol az állábak igen vékonyak, a bekebelezés a sejt álláb nélküli részén történik.
Édesvízben élő fajaik az ozmoregulációt biztosító lüktető vakuólával rendelkeznek,
amelynek azonban nincs állandó helye a sejtben. Ott jön létre, ahol a felgyülemlett víz
és a benne oldott ionok eltávolítására szükség van. A vakuólák a sejtmembránnal fu-
zionálva időnként kilökik a bomlástermékeket.
Ivartalanul kettéosztódással szaporodnak. A házzal rendelkező fajoknál ilyenkor a ház
is kétfelé válik, vagy a létrejött leánysejt újat épít magának. Parazita fajoknál a cisztán
belül többszörös osztódás megy végbe. Ivarosán szingámiával szaporodnak.
6.2.1. Csupaszamőbák (Amoebida)
Az amőbafélék legismertebb képviselői az édesvízi, tavakban, lassú folyású patakokban gyakori
fajok, mint például az óriás amőba (Amoeba proteus). Az áttetsző sejt mérete körülbelül 250-600
pm között változik (6.2. ábra). Táplálékát szerves törmelék és más protiszták képezik.
Avérhasamőba (Entamoeba hystolytica) az ember dizentériás megbetegedését okozó, 10-40 pm
nagyságú patogén amőba (6.2. ábra). A trofozoita alakok a bélcső üregében szabadon mozognak,
táplálkoznak. A bél szöveteit proteolitikus enzimjeikkel oldják, fekélyeket hoznak létre. A véráram-
mal különböző szervekbe (például a májba) is eljutnak, ahol tályogok képződnek. A környezeti
hatásoknak ellenálló ciszták a gazdaállat bélsarával kerülnek ki a külvilágra. Az ember a cisztát
tartalmazó táplálékkal vagy vízzel fertőződik, elsősorban a trópusi, szubtrópusi területeken.
Az Entamoeba coli az ember bélcsatomá-
6.2. ábra. Gyökérlábúak (Rhizopoda). A: óriás amőba
(Amoeba proteus), B: vérhasamőba (Entamoeba hysto-
litica) mozgásra képes (trophozoita) és kerekded ciszta-
alakja, C: bárkaállatka (Arcella spj, D: zománcállatka
(Difflugia sp.)t E: sugaras napállatocska (Actinosphaeri-
um eichhomi) (Deflandre, Stemm, Smidt-Roberts nyomán)
jában él, de nem parazita, hanem baktériu-
mokkal, béltartalommal táplálkozik.
Számos más, nem patogén amőbafaj is él
a gyomor- és béltraktusban.
6.2.2. Házasamőbák
(Arcellinida)
Az ide tartozó amőbák maguk körül
védőburkot, héjat, házat hoznak létre.
A héj anyaga lehet kalcium-karbonát,
szilicium-dioxid, kitinszerű, vagy állhat
fehérjéből. A ház megerősítéséhez né-
hány faj homokszemcséket vagy egyéb
törmeléket használ fel, amit az általa
elválasztott alapanyagba ágyaz be.
Édesvizekben közönségesek a bárkaál-
latkák (Arcella spp.), melyeknek sárga szí-
180
nű háza az egyik felén lapított, a másikon domború (6.2. ábra). Állábaik a lapított oldalon lévő
nyíláson át nyúlnak ki. Testük mérete 50-150 pm közötti.
A zománcállatkák (Difflugia spp.J szintén gyakoriak, testük 180-500 pm nagyságú. Házuk
váza alakú, létrehozásához ásványi részecskéket használnak fel (6.2. ábra).
6.3. Tengely eslábúak (Actinopoda)
Gyakran gömbölyded alakúak, az állábaik tűszerű nyúlványok (axopodium), amelyeket
mikrotubuláris struktúrák támasztanak.
Ide tartoznak a régebben összefoglaló néven sugár-állatkáknak (Radiolaria) nevezett, ma több
osztályba sorolt tengeri, planktonikus életmódú, finoman mintázott házú egysejtűek is.
6.3.1. Napállatocskák (Heliozoea)
A napállatocskák vékony, tűszerű állábai, az axopodiumok sugárirányban állnak. Ha van
vázképződményük, akkor az szilíciumtartalmú, vagy idegen testek felhasználásával
készül. Ivaros szaporodásuk során gametikus meiózis zajlik le. Legtöbb fajuk édesvízben
él, de a tengerekben is előfordulnak. A napállatocskák főleg planktonikusak, de vannak
rögzült fajok is. Az axopodiumokon megtapadó táplálék a membrán továbbító tevékeny-
sége folytán jut el a sejttesthez, ahol azt fagocitózissal veszi fel.
Ide tartozik például az édesvízi sugaras napállatocska (Actinosphaerium eichomi), amelynek
átmérője elérheti az egy millimétert (6.2. ábra).
Ellenőrző kérdések a 6. fejezet témaköreiből
1. Melyek a legfontosabb közös bélyegek, amelyek az állábas-ostoros egysejtűek (Sarcomasti-
gophora) törzsbe tartozó fajokra jellemzők?
2. Foglalja össze az állati ostorosok fontosabb jellemzőit!
3. Jellemezze a parabazális-testes ostorosok (Kinetoplastida) rendbe tartozó fajok biológiáját
és mezőgazdasági szerepét!
4. Jellemezze tükrözött-testü ostorosok (Diplomonadida) rendbe tartozó fajok biológiáját és
mezőgazdasági szerepét!
5. Foglalja össze az állábasok (Sarcodina) fontosabb jellemzőit!
6. Jellemezze a gyökérlábúak (Rhizopoda) főosztályába tartozó fajok biológiáját és mezőgaz-
dasági szerepét!
181
7. APIKALIS-SZERVU EGYSEJTŰEK
(APICOMPLEXA)
Nevüket arról kapták, hogy bizonyos fejlődési állapotukban a test elülső (apikális) végén a gaz-
daállat sejtjeibe való behatolást segítő speciális sejtorganellumok, képletek találhatók. Emésztő- és
lüktetővakuólumaik nincsenek, a tápanyagokat a sejthártyán keresztül veszik fel. Egy vagy
több sejtmaggal rendelkeznek, magdimorfizmusuk nincs. Mozgási sejtorganellumaik kevéssé
differenciáltak, nem jellemzők. Kedveződen körülmények között védőburkot hoznak létre maguk
körül, cisztát képeznek. Nemzedékváltakozást mutatnak: ivaros szaporodásuk a gamogónia,
amit többnyire védőburkon belül zajló mitotíkus osztódások (sporogónia) követnek. Az így ke-
letkezett alakok fertőzik mega gazdaállat sejtjeit, azokban többszörösen osztódnak (merogónia).
Az Apicomplexa törzsbe számos olyan faj tartozik, amelyik a mezőgazdasági szempontból
káros gerinctelen állatokban, férgekben, rovarokban élősködik. A jelenleg folyó kutatások
várhatóan lehetővé teszik több faj felhasználását a biológiai védekezésben. Valamennyi, közel
4600 ez ideig leirt, ma élő faj parazita. Állat- és emberegészségügyi szempontból igen veszélyesek
lehetnek. A még le nem irt fajok számát az ismertek tízszeresére becsülik. A jelenleg ismert fajokat
több mint 60 családba sorolják, a törzsön belüli rendszertani felosztásra számos elképzelés van.
7.1. ábra. Az Apicomplexatörzsbe tartozó
fajok infekciós alakja
di: diktioszoma, ep: elülső poláris gyűrű, er:
endoplazmatikus retikulum, hp: hátulsó poláris
gyűrű, ko: konoid szerv, mi: mitokondrium,
mn: mikronéma, mp: mikropórus, rh:
rhoptria, sm: sejtmag, sp: szubpellikulárismik-
rotubulus (Stamm nyomán kissé módosítva)
A törzsbe tartozó fajok néhány fejlődési állapotában,
például a merozoitákban és a sporozoitákban, a test
elülső végén gyűrűszerű, csöves, fonalas szerkezetű
apikális komplex fordul elő (7.1. ábra). Az apikális
komplex magába foglalja a poláris gyűrűt, amely nem-
csak a sejt elülső, hanem a hátulsó végén is előfordul,
mint a pellikula belső membránjának megvastagodá-
sa. A sejt apikális végén számos, spirálisan csavarodott
mikrotubulusból felépülő gyűrű (conoid) a gazdaállat
sejtjeibe való behatolást segíti elő mechanikus úton.
Ennek működését támogatják a valószínűleg enzim-
tartalmú, hosszú nyakú zsákok (rhoptria), valamint
körülöttük a plazmában lévő, rövid pálcika alakú kép-
letek (micronema). Az elülső poláris gyűrűből a sejt
hátulsó vége felé szubpellikuláris mikrotubulusok fut-
nak, melyek szerepe a mozgásban és a támasztásban
van. Mindezekhez a pellikulán egy vagy néhány mik-
ropórus társul, amelyek feltehetően a táplálékfelvétel-
ben játszanak szerepet. Emésztő- és lüktetővakuólu-
maik nincsenek, a tápanyagokat a sejthártyán keresz-
tül veszik fel.
Egy vagy több sejtmaggal rendelkeznek, magdi-
morfizmusuk nincs. A törzsbe tartozó fajok mozgási
sejtorganellumai kevéssé differenciáltak, nem jellem-
zők. A csillék teljesen hiányoznak, míg egyes csopor-
182
toknál ostoros mikrogamétákat lehet megfigyelni. A gazdaállat sejtjein belül fejlődő ala-
koknál állábak lehetnek. A folyékony közegben történő mozgásukat a plazmában, a pel-
likula alatt lévő összehúzódásra képes fíbrillumok biztosítják. Testalakjukat elsősorban a
pellikula szabja meg. Kedvezőtlen körülmények között védőburkot hoznak létre maguk
körül, cisztát képeznek.
7.2. ábra. Az Eitneria fajok életciklusának vázlata
A: merogonia (schizogonia), gamogonia, C: sporogonia, ma: makrogametocita, me: merozoita, mg: mikrogaméta,
mi: mikrogametocita, mk: makrogaméta, oo: oociszta, se: sporociszta, si: sizonta (meronta), sm: gazdasejt sejtmagja,
sz: sporozoita, zi: zigóta (Noble nyomán kissé módosítva)
Életciklusuk alapvető jellegzetessége, hogy az ivaros és ivartalan szakaszok váltakozva
követik egymást. Ennek a nemzedékváltakozásnak azonban több csoportnál speciális
esetei lehetnek, egyes fázisok akár hiányozhatnak is. Bizonyos általánosításra mégis van
lehetőség (7.2. ábra). Életciklusuk három fő szakaszra osztható. Ezek a gamogonia, a spo-
rogonia, és a merogonia. Az ivaros szaporodási fázis (gamogonia) kezdetén a merozoi-
tákból gamonták jönnek létre. A mikrogamontákból (mikrogametocitákból) kettéosztó-
dással vagy többszörös osztódás útján kialakulnak az önálló mozgásra képes mikrogamé-
ták. A makrogamontákból (makrogametocitákból) általában közvetlenül makrogaméták
jönnek létre, de többszörös osztódással is keletkezhetnek. A mikro- és makrogaméták
kialakulhatnak az előbb említett úton egymástól függetlenül, vagy egyes csoportoknál a
fejlődés során összetapadhatnak (syzygia), illetve közös burokban helyezkednek el. A
mikro- és makrogaméták összeolvadásából (syngamia) zigóta keletkezik. Egyes fajoknál a
zigóta kezdetben mozgékony (ookineta), másoknál a kialakulását követően tok veszi körül
183
(oocysta). A zigóta meiózissal osztódik (zigotikus meiózis), majd az így létrejövő sejtek
ismételt mitózisokkal tovább osztódnak. Ez a folyamat a sporogonia, amelynek során
számos pálcika alakú sporozoita alakul ki. Az oocisztán belül lévő sporozoiták egyesével,
kettesével vagy többesével külön burokban lehetnek (sporocysta vagy „spóra”). A fajok
egy részénél a sporogonia végén az oocisztában vagy a sporocisztában tartalék tápanyag-
ból álló maradéktest keletkezik. Egyes fajoknál a fertőzőképes sporozoiták a gazdaállat
sejtjeibe bekerülve előbb lekerekednek, trofozoitákká válnak, majd megindul a sejtmag
osztódása. Ekkor már meronta (schizonta) a nevük. Többszörös osztódással (merogonia,
schizogonia) merozoitákat hoznak létre. Fajra jellemző módon a merontáknak és a
merozoitáknak egy vagy több nemzedéke fejlődik ki. Több generáció esetén a merozoiták
új sejtekbe hatolnak be, és ott merontákat képeznek, amelyekből újból merozoiták lesz-
nek. Végül néhány merozoita makro- és mikrogamontává alakul, és kezdetét veszi az
ivaros szaporodási fázis. A törzsbe tartozó fajok egyedei a zigóta állapot kivételével egész
életükön át haploidok.
Törzs: Apikális-szervű egysejtűek (Apicomplexa)
Osztály: Spórások (Sporozoea)
Alosztály: Üregélősködők (Gregarinia)
Alosztály: Sejtélősködők (Coccidia)
Rend: Valódi sejtélősködők (Eucoccidida)
Alrend: Sejtélősködő spórások (Eimeriina) 7.1. fejezet
Alrend: Vérélősködő spórások (Haemosporina) 7.2. fejezet
Osztály: Spóra nélküliek (Piroplasmia)
Rend: Vérélősködő spórátlanok (Piroplasmida) 7.3. fejezet
Megjegyzés: Jelen könyvben Levin és munkatársai (1980) felosztását követjük, kizárólag a
jelentősebb, hazai előfordulású taxonokat feltüntetve.
A spórások osztályába tartozó fajok az ivaros szaporodás végén mozgékony ookinétát vagy
burokkal körülvett oocisztát, benne sporocisztákat (spórákat) hoznak létre. A sporocisztában a spo-
rogónia eredményeként fertőzőképes sporozoiták jönnek létre. Az állábak többnyire hiányoznak, de
ha mégis lennének, akkor azoknak nem a mozgásban, hanem a táplálkozásban van szerepük. A
kifejlett alakok mozgása a test hajlítgatásával, csúsztatásával, hosszanti redők hullámoztatásával
történik. Ostor csak egyes fajok mikrogamétáinál fordul elő. Az üregélősködők alosztályába tartozó
egyes fajok életciklusából a merogonia hiányozhat, míg a gamogonia és a sporogonia általánosan
megfigyelhető. A fajok egy részénél az anizogaméták bimbózással jönnek létre. Hazánkban a gyű-
rűsférgek és rovarok testüregeiben élősködő Monocystis és a Gregarina fajokat találták meg. A sejt-
élősködők alosztálya képviselőinek életciklusában a merogonia, a gamogonia, és a sporogonia is
végbemegy.
A spóra nélküliek osztályának képviselői nem képeznek oocisztát, illetve sporocisztát. Sejt-
élősködők, főleg a vörösvérsejtekben fordulnak elő.
7.1. Sejtélősködő spórások (Eimeriina)
A mikro- és makrogamonták egymástól függetlenül fejlődnek, a sizigia nem fordul elő.
A mikrogamonták általában igen sok, aktívan mozgó mikrogamétát hoznak létre. A
zigóta mozgásra nem képes. A ciszta átmérője általában 10-40 pm közötti.
Az Eimeria fajok egy oocisztájában négy sporociszta van, mindegyikében két sporozoitával
184
(7.3. ábra). A merogonia és a gamogonia a gazdaál-
lat sejtjeiben, míg a sporogonia a sejteken kívül zajlik
(7.2. ábra). Főleg gerincesekben, azok bélcsator-
nájának nyálkahártyájában fordulnak elő. Az álta-
luk előidézett kórképet kokcidiózisnak nevezzük,
amely a gazdaállatban általában különféle súlyossá-
gú hasmenés formájában jelentkezik. Fejlődésük
során köztigazdára nincs szükségük. Az állatok
fertőződése a bélsárral kiürülő sporulált oocisztákkal
történik. Szinte valamennyi vadon élő és háziasított
gerinces állatnak van többgazdás vagy gazdaspecifi-
kus Eimeria élősködője. A háziállatok fertőződését a
zsúfolt tartás, a nem kielégítő takarmányozási és
higiéniai viszonyok segítik elő. Súlyos fertőződés
esetén elhullás is bekövetkezhet.
A házityúk kokcidiózisának előidézésében pél-
dául kilenc Eimeria faj játszhat szerepet. Az egyes
fajoka bélcsatoma különböző szakaszaiban élnek. A
legfontosabb kórokozó faj az Eimeria tenella, amely
7.3. ábra. Eimeria fajok oocisztájának felépítése
bu: kettős burok, ku: kupak, ma: maradéktest, se: spo-
rociszta, sm: sejtmag, sz: sporozoita (Több szerző
nyomán kissé módosítva)
a vakbélben él. A merogonia, majd ezt követően a gamogonia a vakbél hámrétegében zajlik. Súlyos
fertőzés esetén vérzéses bélgyulladás alakulhat ki, amelynek következtében a gazdaállat elpusztul-
hat. Az oociszták a bélsárral kikerülnek a külvilágra, ahol végbemegy a sporogonia, létrejönnek a
sporozoiták. A fertőződés akkor következik be, amikor a gazdaállat a táplálékkal együtt felveszi a
sporulált oocisztát, amelyből az emésztőenzimek hatására kiszabadulnak a sporozoiták. Igen ve-
szélyes lehet még például az Eimeria necatrix és az E. maxima, amelyek a vékonybélben károsítanak.
A számos faj közül gyakoribb előfordulású például az Eimeria bovis és az E. zümii, amelyek a
szarvasmarhák, az Eimeria magna és az E. irresidua, amelyek a nyulak, valamint az Eimeria carpelli,
amely a ponty bélkokcidiózisát okozza.
A Sarcocystis fajok fejlődése során az ivartalan szaporodás a köztigazda izomzatában zajlik, az
utolsó meronta generáció tömlőszerű egyedei (sarcocysta) kerülnek be a végleges gazda (ragado-
zók, ember) szervezetébe, a bélcsatornába, ahol a gamogonia megy végbe. Az oociszta a végleges
gazda sejtjeiben sporulálódik, majd a bélsárral ürül ki. Gyakoribb előfordulású a Sarcocystis
suicanis, amelynek végleges gazdái a kutyafélék, köztigazdái a disznófélék.
A Toxoplasma fajok életciklusa során az ivartalan szaporodással létrejött fejlődési alakok egyik
köztigazda fajból átkerülhetnek egy másikba is, mielőtt a végleges gazdába jutnának. A toxo-
plazmózis az emlősökben, így az emberben is előfordulhat. A Toxoplasma gondii kozmopolita faj,
ivaros szaporodása a macskafélékben zajlik, míg köztigazdája közel 200 emlős- és madárfaj lehet.
A 11-14 pm átmérőjű oociszták a végleges gazda bélsarával kerülnek a külvilágba. Az emberbe
leggyakrabban cisztaállapotban, vagy a táplálékul szolgáló köztigazda húsának nyersen történő
fogyasztásával juthatnak be a fejlődési alakok. Terhes nők fertőződése esetén vetélést, halvaszü-
letést, súlyos magzatkárosodást (például vízfejűséget) okozhatnak.
7.2. Vérélősködő spórások (Haemosporina)
A makro- és mikrogaméták egymástól függetlenül alakulnak ki, sizigia nem történik. A
mikrogamonták ostoros mikrogamétát hoznak létre. A zigóta mozgásra képes (ookineta).
Fejlődésük köztigazda segítségével megy végbe. A merogonia és a gamogonia első fele
a köztigazda gerincesek bizonyos sejtjeiben, főleg a vér vörösvérsejtjeiben zajlik. A ga-
mogonia további része, valamint a sporogonia a végleges gazda, és egyben terjesztő vérszívó
rovarokban játszódik le. Az ookineta átlagosan 75 pm, a sporozoita 10-15 pm átmérőjű.
185
Az ember maláriás megbetegedését négy Plasmodium faj okozhatja (P. vivax, P. falciparum, P.
malariae, P. ovale). A merogonia kezdetben az ember májsejtjeiben, később a vörösvértestekben
megy végbe. A kiszabaduló merozoiták újabb vörösvértestekbe hatolnak, ahol először trofozoitákká
alakulnak, majd meronták, ezekből merozoiták képződnek. A betegség jellegzetes, fajra jellemző,
időszakonként fellépő lázas tüneteit a merozoiták kiszabadulása, illetve a hemoglobin emésztésé-
ből származó bomlástermékek vérplazmába kerülése váltja ki. A gamogonia is az eritrocitákban
indul meg. A gametociták vérszíváskor a maláriaszúnyog (Anopheles sp.) bélcsatornájába kerülnek,
ahol gamétákká alakulnak, majd összeolvadva létrehozzák a zigótát. Ez a kezdetben mozgásra
képes alak a szúnyog bélhámsejtjeibe hatol, ahol oocisztává alakul. A sporogonia eredményeként
kialakuló haploid sporozoiták a szúnyog nyálmirigyeibe vándorolnak, majd vérszíváskor a nyállal
kerülnek az emberbe. Hazánkban régebben számos helyi megbetegedést észleltek, de ma már csak
az ide látogató, a trópusi területeken fertőződött emberek gyógykezelése jelent feladatot.
Más Plasmodium fajok hüllőket, madarakat és emlősöket támadnak meg. A merogonia és a
gamonták kialakulása a köztigazda gerincesek bizonyos sejtjeiben, főleg az eritrocitákban zajlik. A
madarak között a fertőzést főleg a vérszívó Cidex szúnyogok terjesztik.
7.3. Vérélősködő spórátlanok (Piroplasmida)
Az ide tartozó fajok változatos alakúak lehetnek. Nem képeznek oocisztát, spórát. Az
ivaros alakok axopodiumszerű nyúlvánnyal mozoghatnak. A kifejlett alakok a test haj-
lítgatásával, csúsztatásával mozognak. Fejlődésük során a merogonia a gerinces állatok
vérsejtjeiben, míg a gamogonia és a sporogonia főleg a fertőzést terjesztő és egyben
végleges gazda kullancsokban játszódik le.
ABabesia fajok általában kerek vagy ovális alakúak, mindössze néhány mikrométer átmérőjűek.
Apikális komplexük erősen redukálódott, a konoid szerv minden fajnál hiányzik. A Babesia fajok
végleges gazdái és terjesztői a kullancsok. A köztigazda hüllők, madarak és emlősök testében az
egyes fejlődési alakjaik főleg a vérsejtekben, leginkább az eritrocitákban találhatók. Leggyakoribb
ivartalan szaporodási formájuk a kettéosztódás. A kullancs vérszívása során jut be a sporozoita a
köztigazda emlősök vérébe, ahol kettéválással többször egymás után osztódik. A köztigazda vérével
a kullancs szervezetébe kerülnek az így létrejött alakok, ahol a bél falában végbemegy a gamogonia.
Az itt kialakult formák behatolnak a petesejtekbe, majd a fejlődő kullancsokban lezajlik a spo-
rogonia. Köztigazdában történő károsításuk a vérfestékvizelés nevű megbetegedést okozza, amely
súlyos esetben elhullással is járhat. A kullancs vérszívása révén az ember is fertőződhet.
A Babesia divergens, és a B. bigemina az őzek, szarvasok, szarvasmarhák megbetegedését okozza.
A Babesia canis a kutyákat, míg a B. caballi a lovakat károsítja.
Ellenőrző kérdések a 7. fejezet témaköreiből
1. Melyek azok a tulajdonságok, amelyek az apikális-szervü egysejtűek (Apicomplexa) törzsbe
tartozó fajok közös jellemzői?
2. Milyen részekből áll az apikális komplex?
3. Ismertesse a spórások életciklusát!
4. Jellemezze a spórások osztályába tartozó fajok biológiáját és mezőgazdasági jelentőségét!
5. Jellemezze a vérélősködő spórádanok (Piroplasmia) osztályába tartozó fajok biológiáját és
mezőgazdasági jelentőségét!
186
8. APROSPORASOK (MICROSPORA)
Számos állatcsoportban, elsősorban az ízeltlábúakban fordulnak elő. Oocisztájuk egy sejtből
jön létre. Egy vagy több sejtmaggal rendelkeznek, de magdimorfizmus nincs. Nincs apikális
komplexük. A gazdaállatban fonálszerü képlettel rögzülnek. A kifordult fonal üregén keresztül
jut ki az amőboid mozgású sporoplazma (sporozoita) a tokból. Körülbelül 550 leírt fajuk van,
melyek sejtélősködö paraziták. Ivaros szaporodásuk a szingámia, amelynek során két egyed
olvad össze. Fejlődésük köztigazda nélkül megy végbe.
Törzs: Apróspórások (Microspora)
Osztály: Apróspóraképzők (Microsporea)
Rend: Apróspórás alakúak (Microsporida) 8.1. fejezet
Megjegyzés: A Microsporea osztályba tartozó fajok oocisztájában csak egy sarki tok és spirális
rögzítő fonal található (8.1. ábra). Az oociszta fala három rétegből áll. A legkülső fehérjéből, a
középső kitinből épül fel, míg a belső egy vékony membrán.
8.1. Apróspórás alakúak
(Microsporida)
A rendbe tartozik a Nosema apis, amely a méhek
gyomorvészét, hasmenését okozza. A fertőző, átla-
gosan 5 pm hosszú és 3 pm széles oociszták a bél-
sárral ürülnek. Életképességük 1-2 évig is megma-
rad. A szaporodás során kialakult alakok felélik a
gazdasejtet, a sporogonia végén új fertőző oociszták
kerülnek ki a bél üregébe.
A Nosema bombycis a selyemlepke parazitája. A
hernyók a táplálékkal veszik fel az oocisztákat. Ha a
fertőzés nem súlyos, a hernyó átalakulása végbe-
megy, de az imágó testében, a képződő petesejtekbe
is behatolnak a Nosema fejlődési alakjai. így a peték-
ből kikelő hernyók már fertőzöttek.
8.1. ábra. Microspora fajok oocisztájának felépítése
és a sporozoita kijutásának módja
bu: kettős burok, sf: spirális filamentum, sm: sejtmag, sp:
sporozoita, st: sarki tok (Kudo és Daniels, Dórit nyomán
kissé módosítva)
Ellenőrző kérdések a 8. fejezet témaköreiből
1. Melyek azok a tulajdonságok, amelyek az apróspórások (Microspora) törzsbe tartozó fajok
közös jellemzői?
2. Jellemezze az apróspórás alakúak (Microsporea) osztályába tartozó fajok biológiáját és
mezőgazdasági jelentőségét!
187
9. CSILLOSOK (CILIOPHORA)
Életüknek legalább egy szakaszában csülókat mseinek. Kétféle magvűak, tehát a sejtmagjaik
alakilag és funkciójukat tekintve is különböznek. Valamennyi fajnak van egy vagy több
nagyméretű poliploid sejtmagja (macronucleus), amely DNS-t és RNS-t egyaránt tartalmaz.
A makronukleuszok mérete és alakja igen változatos lehet a különböző fajoknál. A makro-
nukleuszok irányítják a sejt alapvető anyagcsere-folyamatait, a differenciálódást és a regene-
rációt. Egy vagy több kisméretű sejtmagjuk (micronucleus) is van. A mikronukleuszok száma
általában jóval meghaladja a makronukleuszokét. Egyes fajokban előfordulhat akár több mint
80 mikronukleusz is. A mikronukleuszok az ivaros szaporodáshoz szükségesek. A legtöbb
protisztával ellentétben a csillósok diploidok, és a meiózis megelőzi az ivaros szaporodást. A sejt
felszínén lévő csillák száma redukálódhat, eloszlásuk lehet egyenlőtlen, speciális képződménye-
ket (csillótüske, hullámzó- és sodróhártyák) alakíthatnak ki. Sejtorganellumaik, a többi euka-
rióta egysejtűhöz képest jól differenciáltak. Egyes fajok a körülmények kedvezőtlenné válásakor
cisztát képeznek.
A körülbelül 7500 leírt faj egyaránt előfordul tengerekben, édesvizekben és a nedves talajban.
Legtöbbjük mikroszkopikus nagyságú, méretük 10 mikrométertől 3 milliméterig terjed.
A fajok mintegy kétharmada szabadon élő, de vannak más állatok szervezetében elő-forduló
mutualista, kommenzalista fajok is, ülewe körülbelül 2200 faj parazita. Magas fokon
specializált szimbionta fajok élnek a kérődző állatok előgyomraiban. Lehetnekplanktonikusak,
míg egyesek aljzathoz rögzülnek. Többnyire egyesével élnek, de vannak telepet alkotók is. A
környezeti tényezők kedvezőtlen változásaira érzékenyen reagálnak, így indikátorszervezetek-
ként is fel lehet őket használni.
A csillósok testét borító bőrke (pellicula) különböző erősségű lehet. A bőrkét a plaz-
malemma alatt lévő megnyúlt, lapított hólyagok (alveolus), és egy belső, az endo-
plazmatikus retikulumból kialakult további membránréteg erősíti (9.1. ábra).
A sejt felszínén látható csillók felépítését az 1.4. fejezetben már bemutattuk. A csillók
koordináltan működnek. A sorokba rendeződött csillók egymáshoz képest kissé eltérő
ütemben zajló működése a csillósoron hullámok végigfutását eredményezi. Számos
csilló hirtelen eltérő csapásiránya irányváltoztatást eredményez. Egy csilló elhajlása az
extracelluláris mechanikai inger hatására kiváltja a szomszédos csilló megmozdulását. A
csillók működésének koordinálását valószínűleg a membránon végigfutó depolarizációs
hullámok végzik. A szomszédos csillók tövében lévő alapi (bazális) testek (kinetosoma)
hosszanti sorokba rendeződve, finom rostok révén összekapcsolódva biztosítják a kap-
csolatot. Biztos rögzítést adnak a csili óknak, és a pellikula merevítésével hozzájárulnak
a sejt alakjának kialakításához. Az evolúció során az eredetileg egyforma csillók egyes
fajoknál specializálódtak, mind alakilag, mind működésben. Egymással szoros kapcsolat-
ban állva, nagyobb méretű csillónyalábot, csillótüskét (cirrhus) képezhetnek. Ezek egyes
fajoknál „járólábakként” is működhetnek. A csillósok hullámzó hártyáját (membrana
undulans) lemezszerűén összetapadt csillók alkotják. A sodróhártyák (membranella)
kialakításában több sorban álló, viszonylag kevés csilló vesz részt, három- vagy négy-
188
szögletü, evezőszerűen csapko-
dó lemezkéket létrehozva. A kü-
lönböző csillómódosulások a
mozgás jobb kivitelezésében, a
táplálék sejtszáj felé terelésében
játszanak szerepet. Egyes fajok
esetében a csillók a sej t bizonyos
területeiről hiányozhatnak. Né-
hány faj elkeskenyedő testvé-
gével rögzül az aljzathoz. Apelli-
kula összehúzódásra képes fib-
rillumokat tartalmaz, amelyek a
test mozgatását, helyzetének vál-
toztatását teszik lehetővé. Kü-
lönösen jól fejlett a Stentor, még
inkább a Vorticella fajok rögzítő-
nyúlványa, amiben az összehú-
zódásra képes spasmonema ta-
lálható (9.8. és 9.9. ábrák).
A plazma kéregrészében szá-
mos (körülbelül tízféle) zsák-
szerű sejtalkotó elem található,
amelyek gyűjtőneve extruso-
ma. Legrégebben ismert a pa-
pucsállatkák orsó-trichocystája
(9.1. ábra). Sok, igen vékony,
tűszerű képletet tartalmaznak,
9.1. ábra. Ciliophora fajok bőrkéjének (pellicula) felépítése
al: alveoláris lemez, at: alapi test, ci: citoplazma, cs: csilló, ki: kinetodezmá-
lis rost, mi: longitudinális és transzverzális mikrotubulusok, pa: paraszomá-
lis zsák, pe: pelükula, re: redő, sh: sejthártya, tr: trihociszta (Wehner-
Gehring nyomán)
amelyeket ki tudnak lökni magukból. Szerepük még nem teljesen tisztázott. Valószínűleg
a támadók elleni védekezésben, a zsákmány elbódításában, elmenekülésének megaka-
dályozásában, illetve egyes elképzelések szerint az aljzathoz való rögzülésben is funkció-
juk van. Ez utóbbi esetben a tűk a kilökődés után ragadós fonál segítségével kapcsolatban
maradnak a pellikulával.
A csillések többsége jól fejlett sejtszájjal, sejtgarattal rendelkezik. Táplálékukat a
szabadonélőknél főleg baktériumok, algák és kisebb testű csillósok képezik. Csili óik mé-
rete és szájuk tágulási képessége szabja meg, hogy milyen méretű táplálékot képesek
megfogni. A kisméretű csillókkal rendelkezők kicsi táplálékrészecskéket vesznek fel. A
membranellák elősegítik a táplálék továbbítását a sejtgarat felé. A hosszabb csillék lehe-
tővé teszik, hogy a vízáramból kiszűrjék a táplálékot. Egyes ragadozó fajok speciális, a
zsákmány megölésére, elfogyasztására alkalmas sejtorganellummal rendelkezhetnek. Az
emészthetetlen részecskék az alrésen (cytopyge) keresztül kilökődnek.
Mikroszkopikus méretük miatt nincs speciális sejtorganellumuk a nitrogéntartalmú
bomlástermékek eltávolítására, sem pedig a gázcseréhez. A test víztartalma szabályo-
zásának, az ozmoregulációnak elkülönült szerve a lüktetőűröcske. Ez a csillósoknál
meghatározott alakú és helyzetű. Számos fajnál a központi gyűjtőhólyaghoz sugárirányú
csatornák kapcsolódnak, amelyek felveszik, majd a központi hólyagba ürítik a vizet.
Innen a pellikula nyílásán át távozik el az összegyűjtött folyadék.
A csillósok ivartalan szaporodása általában harántosztódással, a Peritrichia
alosztályba tartozó fajoknál hosszirányban, vagy ritkán, mint például a Suctoria fajoknál
bimbózással történik. Ez utóbbi esetben kisméretű csillós utódsejtek jönnek létre, ame-
lyek a megfelelő szubsztrátumhoz tapadva kifejlődnek. A harántosztódás a környezeti
feltételektől függően 3-20 óránként következhet be. A makronukleusz és a mikronukleuszok
189
kettéosztódása egyaránt végbemegy, majd az osztódást követően a többi sejtorganellum is
kiegészül. A makronukleuszok az egyszerű befúződéssel való kettéválást megelőzően ösz-
szeolvadhatnak. A mikronukleuszok mitózissal osztódnak. Az ivartalan szaporodás során
létrejövő egyedek azonos genetikai állományúak az őket létrehozó egyedekkel, kiónt al-
kotnak. Amennyiben hosszú időn keresztül csak ivartalanul szaporodnak, az a populációt
alkotó egyedek életképessége szempontjából kedvezőtlen.
Az ivaros szaporodásuk lényege az, hogy a gametogén meiózissal létrejött ivari
pronukleuszok összeolvadnak (conjugatio). Ez a forma valamennyi más protisztától
eltérően tehát nem egész sejtek összeolvadását (syngamia), hanem csak a nukleuszok
egyesülését jelenti. A csillósok ivaros szaporodása leggyakrabban időleges egyesülés révén
történik (9.2. ábra). A két egyed ilyenkor a fajokra jellemző helyzetben egymáshoz tapad
a pellikula által kibocsátott ragadós váladék segítségével. A plazmamembránok a tapadási
helyen összeolvadnak. A poliploid makronukleusz nem vesz részt a genetikai anyag
kicserélődésében, hanem felszívódik, és újraképződése a későbbiekben a mikronukleuszokból
történik. Azoknál a fajoknál, ahol csak egy diploid mikronukleusz van, a gametikus
meiózist követően négy haploid pronukleusz alakul ki, amelyek közül három degenerá-
lódik, felszívódik. Amennyiben több mikronukleuszból indult ki a meiózis, akkor is csak
egy utódmag marad meg. A haploid pronukleusz mitózissal osztódik, és az így létrejöttek
közül egy-egy a két egyed között kicserélődik. A két különböző egyedből származó
pronukleuszok összeolvadnak, diploid zigotikus nukleuszt (synkaryon) létrehozva. Az
ideiglenesen összekapcsolódott egyedek ezek után elválnak egymástól. A kialakult
diploid synkaryon mitózissal történő osztódások sorozatán megy végig, amíg el nem éri
J K
9.2. ábra. A közönséges papucsállatka (Pavamecium caudatum) ivaros szaporodása
A: két, különböző ivari típusba tartozó egyed, B: az összekapcsolódott egyedek mikronukleuszai meiózissal osztódnak, C: a mak-
ronukleuszok felszívódnak, a mikronukleuszok meiózis révén pronukleuszokat hoznak létre, D: három pronukleusz felszívódik,
a megmaradt mitózissal kettéosztódik, E: egy-egy pronukleusz a két egyed között kicserélődik, F: pronukleuszok összeolvadása,
G: az egyedek elkülönülnek, synkarionjuk mitózissal mikronukleuszokat hoz létre, H: 4 mikro- és 4 makronukleusz kialakulása,
I: a mikronukleuszok egy része felszívódik, a megmaradt mitózissal osztódik, J: a sejtmagok osztódását a sejtek elkülönülése
követi, K: újabb sejtosztódások zajlanak, ma: makronukleusz, mi: mikronukleusz (Pechenik nyomán kissé módosítva)
190
a fajra jellemző mikronukleusz-számot, il-
letve ki nem alakítja a makronukleuszokat.
A Paramecium caudatum esetében a mitózis
eredményeként 8 mikronukleusz jön létre,
amelyből 4 a makronukleuszokat hozza
létre. A többi négy közül 3 felszívódik.
Ezután a Paramecium kétszer egymás után
kettéosztódik, így négy utódsejt jön létre.
Az ivartalan szaporodás tovább folyik, az
utódsejtekben egy makro- és egy mikro-
nukleusz marad meg. A csillósok között
számos faj esetén, például bizonyos pa-
pucsállatkánál is, a populációt alkotó egye-
dek genetikailag eltérőek a szaporodást
illetően, így csak ezek nukleuszai képesek
egymással konjugálni. A konjugáció a po-
pulációt alkotó egyedek életképességének
fennmaradását biztosítja.
Egyes fajoknál időnként az egy egyed-
ben lévő mikronukleuszok meiózissal osz-
tódnak (pronukleoszok képződnek), ezt
követően pedig ezek a haploid magok
összeolvadhatnak (autogamia). A kez-
detben szétesett makronukleuszok újra-
képződnek. Eredményét tekintve ez a fo-
lyamat hasonló az egyedek közötti génki-
cserélődéshez (9.3. ábra).
9.3. ábra. Az autogámia folyamata egy papucsállat-
kánál (Paramecium aurelia)
A: makronukleusz felszívódása, B és C: mikronukleuszok
meiózissal osztódnak, D:apronukleuszoktöbbsége felszívódik,
E: a megmaradt két pronukleusz összeolvad, F és G; a
synkarion mitózissal osztódik, H és I: kialakulnak a
makronukleuszok és a mikronukleuszok,}: osztódás révén két
egyed keletkezik (Raikov nyomán)
Igen sokfaj képes maga körül cisztát létrehozni (betokozódás, encisztálódás), amikor
a környezeti tényezők kedvezőtlenné válnak. Ilyenkor maguk körül védőburkot válasz-
tanak el, elvesztik víztartalmuk egy részét, életműködésük erősen lecsökken. Ennek a
képességnek nagy jelentősége van a fajok fennmaradásában és elterjedésében. Egyes
fajok ilyen állapotban akár évekig is életképesek maradhatnak. A papucsállatka nem
képes ciszta képzésére.
Törzs: Csillósok (Ciliophora)
Osztály: Egyforma csillójúak (Kinetofragminophorea) 9.1. fejezet
Alosztály: Csupasz-szájúak (Gymnostomatia) 9.1.1. fejezet
Alosztály: Szájtölcséresek (Vestibulifera) 9.1.2. fejezet
Alosztály: Hasi szájmezősök (Hypostomatia) 9.1.3. fejezet
Alosztály: Szívókások (Suctoria) 9.1.4. fejezet
Osztály: Kevés-csillószervesek (Olygohymenophorea) 9.2. fejezet
Alosztály: Hasi szájgödrösök (Hymenostomatia) 9.2.1. fejezet
Alosztály: Csillókoszorúsok (Peritrichia) 9.2.2. fejezet
Osztály: Sok-csillószervesek (Polyhymenophorea) 9.3. fejezet
Alosztály: Orvényszervesek (Spirotrichia) 9.3.1. fejezet
Megjegyzés: A csillés egysejtűek rendszertani csoportosítása ez ideig már igen sok elven
valósult meg. Jelen könyvben Levin és munkatársai (1980) alapján adjuk meg a fontosabb hazai
fajok besorolását.
191
9.1. Egyforma csillójúak (Kinetofragminophorea)
Az osztályba tartozó fajok sejtszája közelében lévő csillók alig különböznek a test más
területein lévőktől. A csillókból kialakult képletek általában hiányoznak. A sejtszáj
többnyire apikális helyzetű, vagy a hasoldal közepén, illetve a sejt felszínén, vagy egy
kisebb mélyedésben (átrium, vestibulum) található.
9.1.1. Csupasz-szájúak
(Gymnostomatia)
Csilióik általában rövidek és közel egyfor-
ma hosszúak. Eloszlásuk lehet egyenletes,
de egyes fajoknál csak bizonyos területekre
korlátozódnak. A sejtszáj területe a pelli-
kulával egy szintben van, felszíni elhelyez-
kedésű. A sejtszáj-mezőben (peristoma) a
csillózat legfeljebb annyiban módosul, hogy
a bazális testek (kinetoszómák) egymáshoz
közelebb állnak.
A pellikulához toxiciszták kapcsolód-
nak, amelyek kilökhető, alapjukon széle-
sebb, tüskeszerű képletek.
Az ormányos csillós (Didinium nasutum)
a nála nagyobb méretű papucsállatkát támadja
meg, és annak citoplazmáját szívja ki ormány-
szerű nyúlványával (9.4. ábra). Mérete 80-150
pm közötti.
A lúdnyakú csillós (Dileptus anser) 250-
600 pm hosszú teste elöl ormányszerűen meg-
nyúlt (9.4. ábra).
9.4. ábra. Gymnostomatia és Vestíbuliferia fajok. or-
mányos csillós (Didinium nasutum), A2: ormányos csilló-
sok papucsállatkát támadnak meg, B: lúdnyakú csillós
(Dileptus anser), C: Colpoda sp. (Hell, Mackinnon és
Hawes, Mást nyomán)
9.1.2. Szájtölcséresek (Vestíbulifera)
9.5. ábra. Bendőben élő csillósok. A: Entodi-
niumcaudatum, B: Ophryoscolexpurkinjei
(Bütschli nyomán)
A sejtszáj nyílása általában apikális vagy ehhez közeli
helyzetű, és a csillósorokkal, esetleg ostorszerű cirru-
szokkal ellátott bemélyedő szájtölcsérből (vestibu-
lum) nyílik. Vannak közöttük szabadon élők, szim-
bionták és paraziták. Ide tartoznak például a talajban
gyakori szabadon élő Colpoda fajok (9.4. ábra).
Az emlősök bélcsatomájában, gyomrában számos
olyan „bendőprotozoa” faj él, amely a növényi táplálék
rostos, cellulóztartalmú, nehezen emészthető részeinek
feldolgozását végzi. A lebontás általában tejsavas, ecetsa-
vas, vajsavas erjedés révén valósul meg. Különösen nagy
számban fordulnak elő a kérődzők előgyomraiban. Több-
ségük testét a pellikulában található glikogéntartalmú
lemezkék szilárdítják. Páncélzatuk jellegzetes nyúlványo-
kat is képezhet. A testüket borító csillók csökevényesek,
vagy hiányozhatnak is.
192
A kérődzők előgyomraiban gyakoriak az Isotricha, Diplodinium, Entodinium, Ophryoscolex fajok
(9.5. ábra). A táplálkozásuk során a bontásra felhasznált szubsztrátum anyaga különböző lehet. Az
egyes fajok testmérete igen változatos, de a legtöbb faj hosszúsága 100-200 pm közé esik.
NBalantidium coli az emlősállatok és az ember vastagbelében élő kozmopolita faj, amely általában
nem okoz problémát, de néha enzimjei a bélhámsejteket károsítják. A fertőzőképes ciszták a bélsárral
ürülnek.
9.1.3. Hasi szájmezősök (Hypostomatia)
Az ide tartozó fajok sejtszája a hasoldalon nyílik. Testük orsó
alakú, vagy hát-hasi irányban lapított. A test csillózata gyak-
ran redukálódott. A bemélyedő szájmező csillókkal borított.
Sok faj szabadon élő, de vannak köztük parazita fajok is.
Ide tartoznak például az édesvizekben, elsősorban az állóvizek-
ben gyakran előforduló varsaállatkák (Nasstda spp.J, melyek teste
többnyire henger alakú, csilléik nem redukálódtak..
Aparazita ChilodoneUa cyprini szájnyílása a hasoldalán van. Test-
hosszúsága 35 pm körüli. Cisztát is tud képezni. Csilléi a test két
szélén és a hasoldalon találhatók. Főleg a pontyfélék kopoltyúján és
bőrén okoz hámkárosodást. Károsítása a zsúfoltan tartott, legyen-
gült ivadékállományban és a telelő halak között lép fel.
9.1.4. Szivókások (Suctoria)
A kifejlett alakok testét csiliók nem borítják. Táplálékukat,
például más csillósok plazmáját a. számos karszerű szívócső
(suctellum) segítségével veszik fel (9.6. ábra).
Bimbózással is szaporodhatnak. Édesvizekben és tengerekben
mindenütt gyakoriak. Hazai vizeinkben is él az 50 pm nagyságú
csupasz szívókás (Podophrya fixa).
9.2. Kevés-csillószervesek
(Olygohymenophorea)
Az osztály képviselőinek sejtszája közelében kis számban,
összetett csillószervek (például membranellák) találhatók.
A szájgödör alján nyíló sejtszáj általában ventrális helyzetű,
vagy a sejt apikális vége felé elhelyezkedő.
9.2.1. Hasi szájgödrösök
(Hymenostomatía)
A test nagyszámú, közel egyforma csillókkal borított. A száj-
gödör ventrális helyzetű. Elsősorban édesvizekben fordul-
nak elő.
A Tetrahymena pyriformis szájnyílása körül hullámzó membrán
és membranellák egyaránt előfordulnak. Gyakori, 50pmtesthosz-
szúságot is elérő faj, amelyet laboratóriumi vizsgálatokban is
kiterjedten használnak (9.7. ábra).
9.6. ábra. A szivókások (Sucto-
ria) egyik képviselője a Po-
dophrya fixa (Móczár nyomán)
9.7. ábra. Hymenostomatia fa-
jok. A: Tetrahymena pyriformis,
B: Ichthyophthirius multifiliis
(Mackinnon és Hawes nyomán)
193
A veseállatkák (Colpidium sppj poshadó vizekben gyakoriak. Nevüket az emlősvesére
emlékeztető alakjukról kapták.
Az Ichthyophthirius multifiliis a halak darakórját okozza (9.7. ábra). A hal hámsejtjei közé
befurakodott rajzó alakok növekszenek, gömb alakúakká válnak, károsítják a sejteket. A hámsejtek
burjánzása fehéres göb kialakulását eredményezi. A megfelelő nagyságot (0,5-1,5 mm) elérve
elhagyják a hámfelületet, és vízinövényekre, szilárd aljzatra tapadva nyálkás tokot hoznak létre
maguk körül. A cisztán belül osztódnak, nagy számban hoznak létre rajzó, nauplius alakokat. Ezek
felkeresik a halat, megkapaszkodnak rajta, és befúrják magukat a hámsejtek közé.
Aközönségespapucsállatka (Paramecium caudatum) orálisredőjét (szájmező) borító csillók,
a módosult hullámzó membrán és a memb-
B
9.8. ábra. Csillókoszorúsok (Perítrichia):
A: harangállatka (Vorticella sp.), B: Trí-
chodinaspp. testfelépítésének vázlata (a
részben megnyitott sejtben látszik a központi
horogkoszorú és a körülötte lévő módosult
csillók) (Sleigh, Hausmann nyomán)
ranellák sodorják a táplálékszemcséket a sejt-
szájhoz. Álló- és folyóvizekben gyakori faj, mely-
nek hosszúsága 120-330 pm közötti.
9.2.2. Csillókoszorúsok
(Peritrichia)
Az orális helyzetű csillók jól fejlettek, a
sejtszáj körüli korong peremén több sort
alkothatnak. Ezzel szemben a testet borító
csillók redukálódhatnak. A legtöbb faj rög-
zülést segítő nyéllel rendelkezik, amely-
ben úgynevezett spasmonema, összehú-
zódásra képes miofibrillumokból álló szál
van. A fajok többsége szabadon élő, de
vannak közöttük paraziták is.
Ide tartoznak az összehúzódásra képes rögzí-
tőnyelű harangállatkák (Vorticella spp.) (9.8.
ábra) és az összehúzódásra nem képes nyéllel
rendelkező, telepeket alkotó Epistylis fajok, ame-
lyek édesvizeinkben gyakoriak. A Vorticella fa-
jok nyél nélküli teste 40-80 pm közötti.
A Trichodina fajok teste sapkaszerű, 70-90
pm nagyságú. A hasi oldalon lévő horogkoszorú
(corona) biztosítja a gazdaállaton való megtele-
pedést (9.8. ábra). A tapadókorong széléről indul ki az erősebb, helyváltoztatást szolgáló csillósor,
valamint a befelé álló, finom csülökből összetapadt hullámzó hártya. A test többi részén további
csillósorok lehetnek. A halivadék kopoltyúján megtelepedve komolyabb károsodást is okozhatnak a
légzőfelületet alkotó hámsejtek roncsolásával.
9.3. Sok-csillószervesek (Polyhymenophorea)
Az osztályba tartozó fajok sejtszája közelében nagy számban találhatók feltűnő, összetett
csillószervek. A testet borító csillók vagy megtalálhatók, vagy redukálódtak, esetleg
cirruszok formájában vannak jelen. A sejtszáj a szájgödör alján nyilik. Egyesek kemé-
nyebb páncéllal rendelkeznek.
194
9.3.1. Örvényszervesek (Spirotrichia)
A közönséges kürtállatka
(Stentor polymorphus) vi-
zeinkben gyakori, tipikusan
szerves törmeléket szűrő
szervezet (9.9. ábra). A száj-
nyílás körül számos memb-
ranellahelyezkedik el. Nagy-
sága 0,5-1,0 mm közötti.
A kagylóállatka (Stylo-
nichia mytilus) erősen lapí-
tott testű, a hátoldalán lévő
sörték a víz áramlását érzé-
kelik, míg a hasoldali, pálca
alakú csillómódosulások
(cirrhus) a mozgás szolgála-
tában állnak (9.9. ábra). Fel-
színivizekben gyakori, rend-
kívül nagy alkalmazkodóké-
pességű, 100-400 pm nagy-
ságú faj.
Ellenőrző kérdések a 9. fejezet témaköreiből
1. Melyek azok a tulajdonságok, amelyek a csillósok (Ciliophora) törzsbe tartozó fajok közös
jellemzői?
2. Mit jelent a csillósok törzsében a magdimorfizmus?
3. Jellemezze az egyforma csillójúak (Kinetofragminophorea) osztályába tartozó fajok bioló-
giáját és mezőgazdasági jelentőségét!
4. Jellemezze a kevés-csillószervesek (Olygohymenophorea) osztályába tartozó fajok bioló-
giáját és mezőgazdasági jelentőségét!
195
10. LAPOSFÉRGEK (PLATYHELMINTHES)
A laposférgek testét a három csíralemezből létrejött szövetek és szervek alkotják. Az ektoderma
és entoderma közötti teret azonban a mezodermából származó parenchyma és az izomsejtek
teljesen kitöltik. Testüregük nincs. Testük kétoldalion részarányos. Az aktív úszás, a mászás
az érzékszervek, fejlettebb idegrendszer, koordináció létrejöttét feltételezi. Ezek a szervek az
elkülönült feji végen találhatók, hiszen ez a testrész találkozik legelőször a környezet ingereivel.
A feji vég, az agy dúc kialakulása nagy evolúciós lépést jelent.
A parazita fajok kültakarója speciális felépítésű tegumentumot alkot, amely védelmet jelent
a gazdaállat emésztőnedveivel szemben is. Bélcsatomájuk kétszakaszos, elágazó, de egyes
parazitáknál hiányozhat. Nincs elkülönült légzési és keringési szervrendszerük. Kiválasztó-
szervük elovesécske típusú. A fajok többsége hímnős, az ivarszervek igen jól differenciáltak. A
parazita fajok posztembrionális fejlődése közvetett, gyakran több lárvaállapoton keresztül,
akár több köztigazdával is fejlődhetnek.
Közel 18 000 fajuk ismert. Vannak közöttük szabadon élők és paraziták is. A parazita
fajok növényi vagy állati kártevők lehetnek. Az emberben is számos laposféreg élhet.
A laposférgeknek a csalánozókkal való közeli rokonságát testfelépítésük több bélyege is
tükrözi. Egyes vonásaik, így például a mezodermális parenchima megléte viszont
sokkal fejlettebb tulajdonság. Néhány laposféregnél azonban - jól felismerhetően - a
csalánozóknál jellemző izomsejtstruktúra, a hámizomsejt-típus fordul elő. Ennek a
sejtnek egyik része a plazma nagy részét és a sejtmagot tartalmazza, míg a másik, meg-
nyúlt része az összehúzódásra képes fibrillumokat foglalja magába.
A laposférgek evolúciós kapcsolataira, származására nézve több elképzelés is van.
Sok csiliót viselő sejtjeik, a petesejtjük spirális barázdálódása azt jelzi, hogy a kétoldalian
részarányos testfelépítésű állatok fejlődési szintjének nem a legalsó lépcsőfokán állnak.
Egyes elképzelések szerint a laposférgek ősei testüregnélküliek voltak, és bélcsatomájuk
fejletlen volt, tehát ezek a tulajdonságaik elsődleges bélyegek. Mivel van egy olyan
gyűrűsféreg csoport, amelynek testfelépítése számos vonásban a laposférgekére emlé-
keztet, elképzelhető az is, hogy a laposférgek a testüreggel rendelkező állatokból ve-
zethetők le, amennyiben a testüregnélküliségük csak másodlagos, leegyszerűsödési fo-
lyamat eredménye. így a laposférgek egy olyan oldalágnak tekinthetők, melyek speciális
életmódjuk kialakulása során elvesztették testüregüket.
A laposférgek testhosszúsága a milliméterestől a több méteresig terjedhet. Hát-hasi
irányban lapított testük alakja lehet karcsú, széles levél vagy szalag formájú.
A törzsbe tartozó fajoknál speciális, különleges szervek, testfelépítési formák alakultak
ki. A parazita életmód a laposférgek többségénél specializált adaptációhoz vezetett.
Testükön szívókák, tapadókorongok és rögzítőhorgok alakultak ki. Ezek szolgálnak a
megkap aszkodásr a.
A fajok egy része nem rendelkezik bélcsatomával. Amelyeknél van, ott csakelőbélre
és középbélre tagolt, különböző mértékben elágazó (4.2.1. fejezet). A bélcső falát
egyrétegű hengerhám alkotja.
196
Elkülönült légzőszervük nincs, így a test belső elhelyezkedésű sejtjeinek ellátása
kritikus tényező. Ennek megfelelően a legtöbb laposféreg kis testű, vagy ha nagyobb is,
a test erősen lapított.
Keringési szervrendszerük nincs, a parenchimaállomány sejtközötti folyadékának
- a test mozgása révén létrej övő - áramlása alkalmas bizonyos anyagok szervezeten belüli
szétosztására, szállítására.
Valamennyi faj kiválasztószervei az elővesécskék (protonephridium), amelyek elve-
zető csőrendszerük felépítésében, a kivezető pórus helyzetében mutatnak eltérést az
egyes fajoknál (4.5.1. fejezet). Ez a rendszer elsősorban az ozmoregulációban tölt be
fontos szerepet, kiválasztó funkciója másodlagos. Az anyagcsere végtermékeit, például
az ammóniát főleg a testfalon át, diffúzióval távoh'tják el.
Idegrendszerük központja a feji végben lévő idegsejt-tömörülés, illetve a fejlettebbek-
nél általában kétlebenyű agydúc. Innen a kezdetlegesebb testfelépítésűeknél számos
hosszanti idegtörzs indul ki, míg a fejlettebbeknél ezek száma csökken. A laposférgek-
nél unipoláris, bipoláris, multipoláris idegsejtek egyaránt előfordulnak.
Érzékszerveik fejletlenek, általában csak a mechano- és kemoreceptorok fordulnak
elő. A szabadon élő fajoknak és egyes ektoparazitáknak látószervei lehetnek. Az örvény-
férgek és egyes ektoparazita fajok látószervei pigmentkehely típusúak. Ez a szem
elsődlegesen a fény erősségét és irányát képes érzékelni. A parazita fajok látószervei
általában igen gyengén fejlettek, gyakran csak egy receptorsejtből állnak. A kemo-
receptoroknak fontos szerepük van a gazdaállat, illetve a megfelelő táplálék azonosítá-
sában. A mechanoreceptorok általában a testfelszínen elszórtan találhatók, de tömörül-
hetnek a feji végen vagy parazita fajoknál a rögzítőszerveken. A receptorsejtekhez
többnyire érzékelőserték kapcsolódnak, melyek a testfelszínről nyúlnak ki. A galandfér-
geknek általában nincsenek specializált érzéksejtjeik. Az ingereket szabad idegvégződé-
sek veszik fel.
A legtöbb faj hímnős. Az ivarszerveik jól fejlettek, sokszor igen bonyolult felépítésűek
(10.3., 10.8., 10.14. ábrák). Ivarszerveik általában hasonlók, de egyes csoportokban,
fajokban a petefészkek és herék száma, az egyes szakaszok alakja változhat, vagy a
járulékos részek hiányozhatnak. A hím ivarszervrendszer lebenyezett, páratlan vagy
páros heréből (testis), a kivezető csövekből (ductus efferens), a gyűjtő szerepű ondóve-
zetőből (ductus deferens) áll. A közös ondóvezetőn páratlan ondóhólyag (vesicula
seminalis) is lehet. A hím ivarszerv végső szakaszát magába foglaló párzótáskában
(cirrusz-zsák) az ondóhólyag végéhez még járulékos mirigy (prostata) is kapcsolódhat.
A cirrusz-zsák vége az ivamyíláson át kitolható párzószerv (cirrus) szerepét is betölti. Az
ondókilövelő járat (ductus ejaculatorius) a párzószervben húzódik. Utóbbi gyakran a női
ivarszerv kivezető járatával együtt a közös ivarpitvaron (átrium genitale), ivamyíláson át
érintkezik a külvilággal.
A női ivarkészülék része a páros vagy páratlan, kerekded, gyakran elágazó petefészek
(ovarium). A petevezetőhöz (oviductus), vagy utolsó tágultabb üregéhez (ootype)
csatlakoznak a szikmirigyek (vitellinea). Ezek a petesejteket kívülről látják el szikdús
sejtekkel. A termelt szikanyag átmeneti tárolására szikraktár is elkülönülhet. Az ootype
ürege körül található a Mehlis-féle mirigy, amelynek váladéka az ektolecitális petesejt
védőburok anyagának összetapasztásában játszik szerepet. A petevezető felső szakaszá-
ból vagy az ootype üregéből nyílhat a párzótárs ondójának tárolására szolgáló ondótar-
tály (receptaculum seminis). A hím ivarsejtek felhatolnak az ootype üregéig, itt megy
végbe a megtermékenyítés. Az ootype üregéből a sziksejtekkel és védőburokkal körülvett
peték az innen kiinduló, kanyargó lefútású, tágult üregű méhbe (uterus) kerülnek. Ez a
megtermékenyített peték tárolására szolgál. Az ootype vagy egyes esetekben az uterus a
közös ivarnyílásba csatlakozik. A fajok egy részénél a petevezető felső szakaszából indul
ki a Laurer- féle vezeték, amely a test hátoldalán a külvilágra nyílik. A hüvely szerepét
197
töltheti be a párzás során, illetve a felesleges mennyiségben lévő sziksejtek vagy sper-
miumok kiürítésére szolgál.
A legtöbb laposféreg hímivarsejtje vékony, fonalszerű és mozgékony farokrésze lehet.
Számos örvényféregnek és más laposféregnek két ostorral rendelkező spermiuma van,
amelynek finom szerkezete is eltér az átlagostól, ugyanis 9+1 mikrotubulus építi fel a 9+2
helyett. A laposférgek többségénél, szemben az állatvilágban általánosan jellemző
endolecitális típussal, a petesejt ektolecitális, ami azt jelenti, hogy a szikanyag a petesejt
körül helyezkedik el. A megtermékenyített, a sziksejtekkel körülvett egy vagy néhány
petét a laposférgek petetokba (kokon) zárva vagy kocsonyás burokban rakják le. Ezt a
petetokot a gyakorlatban egyszerűen petének nevezik. A laposférgek megtermékenyített
petesejtje általában teljes, spirális barázdálódást mutat, amelynek során mikromérák és
makromérák jönnek létre. A gasztruláció epiboliával történik. A parazita fajok ektolecitális
petesejtjének fejlődése igen erősen módosult formában megy végbe.
A szabadon élő fajok posztembrionális fejlődése általában közvetlen, míg a
parazitáké közvetett. Ez utóbbiak életciklusa gyakran bonyolult, több köztigazdában és
számos lárvaállapot közbeiktatásával zajlik. Köztigazdának azt az állatot nevezzük,
amelyikben a féreg lárvaformái fordulnak elő, míg a végleges gazda az, ahol az ivaros
szaporodás történik.
Törzs: Laposférgek (Platyhelminthes)
Osztály: Örvényférgek (Turbellaria)
Osztály: Mételyek (Trematoda) 10.1. fejezet
Rend: Villásfarkú-cerkáriás mételyek (Strigeatida) 10.1.1. fejezet
Rend: Májmétely-alakúak (Echinostomatida) 10.1.2. fejezet
Rend: Szájszuronyos-cerkáriájú mételyek (Plagiorchiida) 10.1.3. fejezet
Osztály: Csáklyásférgek (Monogenea) 10.2. fejezet
Rend: Egyszerű-tapadószervűek (Monopisthocotylea) 10.2.1. fejezet
Rend: Összetett-tapadószervűek (Polyopisthocotylea) 10.2.2. fejezet
Osztály: Gálándférgek (Cestoda) 10.3. fejezet
Rend: Tagolatlan galandférgek (Caryophyllidea) 10.3.1. fejezet
Rend: Szívógödrös galandférgek (Pseudophyllidea) 10.3.2. fejezet
Rend: Szívókás galandférgek (Cyclophyllidea) 10.3.3. fejezet
Megjegyzés: A parazita laposférgek rendszertani felosztása Mehlhom és Walldorf (1988) és
Papp szerk. (1996) szerint került összeállításra.
Az örvényférgeknek több mint 3000 faja ismert, és újabban több osztályukat különítik el.
Többségük tengerben él, de megtalálhatók az édesvizekben és a talajban is. Testméretük a néhány
millimétertől a több tíz centiméterig terjedhet. Főleg ragadozók. Regenerálódóképességük igen
nagy. Hazánkban a gyakoribb fajok közé tartozik az állóvizekben élő gyászplanária (Planaria
lugubris) és a tejfehér planária (Dendrocoelum lactewn), valamint gyors folyású vizekben a füles
planária (Euplanaria gonocephala).
10.1. Mételyek (Trematoda)
Valamennyi ide tartozó, közel 11 000 faj parazita, és kifejletten szinte valamennyien
gerinces állatok belső élősködői. A végleges gazdában elsősorban a bélcsatomában
élnek, ritkábban a vérerekben vagy más zsigeri szervekben is előfordulnak.
198
10.1. ábra. A: Trematoda, B: Cestoda fajok kültakarójának felépítése
ah: alaphártya, íz: különböző irányban futó izomrétegek, mb: mikroboholy, pi: pikkely vagy tüskeszeríi képlet, pk: perikarion-
sejtek, ps: parenchimasejtek, te: tegumentum (Storch, Threadgold, Welsch nyomán módosítva)
Színük általában áttetsző vagy fehéres. Lapított testük megnyúlt vagy kiszélesedett,
levél alakú. Testhosszúságuk általában 50 mm-ig terjed, de előfordulnak ennél jóval
nagyobbak is.
Kültakarójuk az örvényférgektől eltérően csillós sejteket nem tartalmaz, különleges
felépítésű (10.1. ábra). A felső réteg (tegumentum) a mélyebben ülő, testfelszínt képző
subtegumentális (perikarion) sejtek plazmanyúlványainak összeolvadásából kialakult
szinciciális védőburok. Felszíne mikrobolyhokkal borított, rajta a megkapaszkodást
segítő, tüske- és pikkelyszerű képletek lehetnek. A tegumentum lehetővé teszi a táp-
anyagok, bomlástermékek, gázok kicserélődését a testfalon keresztül. Mitokondriumok
is előfordulnak benne. Védi a parazitát a gazdaállat enzimjeitől, immunrendszerének
védekező sejtjeitől. A tegumentum alatt alaphártya van, ez alatt pedig az izom- és pa-
renchimasejtek, valamint a külső felszíni réteget létrehozó perikarionsejtek ülnek.
Testfelépítésük következőekben bemutatásra kerülő jellegzetességei elsősorban a
májmételyre vonatkoznak, a különböző fajoknál ettől kisebb eltérések lehetnek (10.2.
ábra).
Jellegzetességük a szájnyílás körüli feji szívóka, valamint egyes fajoknál a hasi
oldalon, a test középső szakaszán vagy a test végén kialakult tapadószerv, a hasi szívóka
(acetabulum).
Emésztőszervrendszerük kétszakaszos, elő- és kőzépbélre tagolt, és különböző
mértékben elágazó lehet. A legtöbb ide tartozó fajnak izmos garatja, rövid nyelőcsöve és
vakon végződő, kétágú vagy elágazó bélrendszere van. Táplálékukat elsősorban a
sejttörmelékek, a szövetnedv, a nyálkaanyagok és a vér képezik.
Kiválasztó szervrendszerüket (elovesécske) igen sok lángsejt, és a hozzájuk tartozó
elvezető csatornák alkotják, a közös kivezető pórus a test végén található. A lángsejtek
száma, az elvezető csatornák elrendeződése faji jellegzetesség.
199
10.2. ábra. A májmétely (Fasciola hepaticá) testfelépítése. A: a test belső szervei, B: a béledényrendszer, C: a
kiválasztó szervrendszer
ad: agydúc, be: béledényrendszer, bé: bélág, fe: feji szívóka, ga: garat, ha: hasi szívóka, he: here, ip: ivarpitvar, it: idegtörzs,
kp: kiválasztópórus, mé: méh, Mm: Mehlis-féle mirigy, ny: nyelőcső, ok: ondókivezetőcső, oo: ootyp, ov: ondóvezető, pe:
pete, pf: petefészek, pt: párzótáska, pv: petevezető, sz: szikmirigy (Sommer, Landois és más szerzők nyomán módosítva)
200
10.3. ábra. A Trematoda fajok ivarszervrendszere fel-
építésének általánosított vázlata
he: here; ki: közös ivarpitvar, La: Laurer-féle vezeték, mé:
méh, Mm: Mehlis-féle mirigy, oh: ondóhólyag, ok: kivezető-
cső, oo: ootyp, ot: ondótartály, ov: ondóvezető, pá: párzószerv,
pf: petefészek, pr: prosztata, pt: párzótáska, pv: petevezető, ra:
szikraktár, sz: szikmirigy (Edelényi nyomán)
A parazita szívóférgek idegrendsze-
re általában az agydúcokból kiinduló,
páros, dorzális, ventrális, laterális ideg-
törzsekből, az azokat összekötő haránt-
ágakból (comissurák) és a kilépő környé-
ki idegekből áll. Az érzékszervek gyengén
fejlettek.
A szívóférgek általában himnős álla-
tok. Szaporodásmódjuk, egyedfejlődésük
sokféle. A kifejlett állatok ivarszervei igen
jól fejlettek, a laposférgek általános jel-
lemzésénél leírtakkal megegyeznek (10.3.
ábra). A páros, olykor gazdagon elágazó
herék általában a test középső részén he-
lyezkednek el. A páratlan, sokszor elága-
zó szerkezetű petefészek változatos elhe-
lyezkedésű. A test két szélén igen sok, a
petesejteket sziksejtekkel ellátó szikmirigy
található. Az uterusz a fajok többségénél
tömve van a már barázdálódásnak indult
petékkel, melyek a közös ivamyíláson át
kerülnek leadásra. A közös ivarnyílás
általában a test elülső vége felé, a hasol-
dalon található. A peteképzés és -leadás
folyamatosan történik. A megterméke-
nyítés általában kölcsönös, de önmegter-
mékenyítés is előfordulhat.
Az ide tartozó fajok fejlődése bonyolult. Legalább egy köztigazdájuk van, amely egy
csigafaj, de egyes férgeknek kettő, ritkábban három köztigazdája is lehet. A közvetett
fejlődésű szívóférgek fejlődésének részleteiben számos eltérés van, mégis valamennyinek
alapvetően ugyanolyan lárvaformái jönnek létre (10.4. ábra). A pete általában a végleges
gazda bélsarával kerül a külvilágra, ahol a további fejlődéséhez többnyire nedves közegre
van szüksége. Kedvezőtlen körülmények között a pete igen hosszú ideig, a májmételynél
például akár évekig is életképes maradhat. A petetokon kupak (operculum) található,
amelynek kinyílása lehetővé teszi a csillós lárvakibújását. A petéből szabadon úszó csillés
lárva (miracidium) alakul ki, amely felkeresi a köztigazda csigát és behatol a testébe.
Előfordulhat, hogy a csiga a petét még a lárva kibújása előtt táplálkozása során felveszi. A
miracidiumnak4-5 sejtrétegből álló epidermisze van, amelynek felszíni sejtsora csillós. Feji
vége kihegyezett, van két szemfoltja, és két lángsejtje. Az izmokat és érzékszerveket a
kezdetleges agy idegzi be. A miracidium folyamatosan úszik, míg véletlenszerűen vagy
kemoreceptorai segítségével rá nem talál a köztigazdára. Mirigyei által termelt enzimek
segítségével behatol a köztigazda csiga testébe, ahol csilióit elvesztve átalakul csíratöm-
lővé (sporocysta). Ezzsákszerűen megnő, benne olyan embrionális sejtek vannak, amelyek
leánysporocisztává fejlődhetnek, vagy közvetlenül dajka- (redia) alakokat képezhetnek.
Fajoktól függően egy sporociszta tartalmazhat leánysporocisztákat, rediákat vagy már
cerkáriákat is. A rediáknak szájnyílása, rövid bélszakasza, a lángsejtekhez kapcsolódó
nefridiális csatornája és kezdetleges idegrendszere van. Testük laterális vagy ventrális
részén függelékeik lehetnek. Egyes fajoknál a redia újabb leányrediákat képezhet vagy
cerkáriákat hoz létre. A farkos lárva- (cercaria) alaknak kialakulnak a szívókéi, nefridiális
csatornái gyűjtőhólyagban egyesülnek, valamint mozgékony farokrésze van. A cerkária
elhagyja a csiga testét, és fajtól függően vagy a második köztigazda testébe kerül, vagy a
201
10.4. ábra. Trematoda fajok fejlődésének vázlata. A: pete a petetokban (op: operculum, pe: petesejt, sz: sziksejtek),
B: csillós lárva (miracidiuxn), amelynek valóságban a teljes testfelszínét csillók borítják, C: csíratömlő (sporo-
ciszta), D: dajka (redia), E: farkos lárva (cercaria), F: betokozódott cerkária (metacercaria), G: kifejlett féreg
(Mattes nyomán módosítva)
szabadban betokozódik (encisztálódik), és átalakul metacerkária-alakká. Ha ez a táp-
lálékfelvétel közben bekerül a végleges gazdába, akkor ott kifejlődik a szívóféreg és sza-
porodni kezd. Egy miracidiumból kiindulva fajoktól függően tehát akár több száz sporo-
ciszta, illetve azok mindegyikéből több száz redia, illetve cerkária fejlődhet. Ez egyes
fajoknál akár 200 000 cerkária létrejöttét is jelentheti. Nem valódi polyembriogonia zajlik
le (amikor egy petesejtből számos embrió fejlődik), hanem itt egy speciális esetről van szó.
Az első lárvaállapotú alak testében lévő embrionális sejtek differenciálódhatnak, de egyben
az új alakokban is lesznek e képességgel rendelkező utódsejtek.
10.1.1 . Villásfarkú-cerkáriás mételyek (Strigeatida)
A miracidiumoknak két pár elővesécskéjük van. A cerkáriák farka elágazó.
Az Alaria alata ragadozó állatok (kutya, macska, róka) vékonybelében élősködő,
gyakori faj (10.5. ábra). Testhosszúsága 3-6 mm közötti. Két részre tagolt testének elülső
része kétszer hosszabb, mint a hátulsó. A feji vég két oldalán fülszerű nyúlványok vannak.
Első köztigazdája vízicsiga, míg a második ebihal. A cerkáriaalakok a vivőgazda kétél-
tűek, hüllők izomzatában, bőr alatti kötőszövetében telepszenek meg és encisztálódnak.
202
A halak feketepettyességét okozza a Posto-
diplostamum cuticola metacerkária alakja. A kifej-
lett férgek a halevő vízimadarak bélcsatomájában
élnek. Az 1,2-2,0 mm hosszú testük két részre
tagolódik, amelyek közül az elülső erősen kiszéle-
sedő. A bélsárral vízbe kerülő petéből először a mi-
racidium alakul ki, majd ez bekerül a Planorbis,
Planorbarius csigafajok testébe, ahol a sporociszta,
redia, majd cerkária alakok jönnek létre. A cerká-
riák felkeresik a halivadékot, a fiatal halakat, és
azok bőrébe befúrják magukat. Körülöttük a ha-
lak kötőszöveti sejtjei pigmentanyagokban gaz-
dag, fekete színű tokokat alakítanak ki, amelyben
a metacerkáriák életben maradnak, amíg a halat
egy madár el nem fogyasztja.
A Diplostomum spathaceum kifejlett alakjai a
vízimadarakban, főleg a sirályok bélcsatomájá-
ban élnek (10.5. ábra). Testük 1,2-4,4 mm hosszú
és szintén két fő részre különül el. A vízbe került
petéből kibúvó miracidiumok a Lymnaea, Radix,
Stagnicola csigafajok testébe kerülnek, ahol to-
vábbi lárvaformák alakulnak ki. A cerkária felke-
10.5. ábra. Strigeatida fajok. A: Alaria alata, B:
Diplostomum spathaceum (Edelényi nyomán)
resi a pontyfélék közé tartozó halakat, befúrja magát a bőrbe, a vérereken át eljut a szemgolyóba,
ahol a szemlencsében telepszik meg. Itt a mozgásra képes metacerkária, az úgynevezett diplosto-
mulum betokozódás nélkül évekig életképes marad. A fertőzött hal látása, így táplálékkereső
képessége erősen legyengülhet, gyakran úszik a vízfelszín közelében, ahol könnyen a sirályok
zsákmányává válhat. Igen gyakori faj.
A Schistosoma fajok a trópusi területeken az ember legelterjedtebb és egyik legveszélyesebb
megbetegedését okozzák, a kifejlett paraziták a vérerekben telepszenek meg.
A Bilharziella polonica a víziszámyasok (például kacsafélék, hattyúk, gémek) véredényrendsze-
rében, főleg a vénáiban élősködik. Köztigazdái a Planorbis és Lymnaea csigafajok, amelyekben
cerkáriák alakulnak ki. A kifejlett, 2-4 mm-es alakok váltivarúak, de hasonló alakúak. A két fő
bélcsőágon további leágazások nincsenek. Az ágak a test hátulsó felében újra egyesülnek.
A ponty vérében élősködik az egy milliméternél nem nagyobb Sanguinicola inermis, amely az
úgynevezett vérmételykórt okozza. A vérerekben, szívben megtelepedő kifejlett férgek szívókák-
kal nem rendelkeznek, nyelőcsövük rövid, vakon végződő bélcsőágakra ágazik el. Ez a faj is
váltivarú. A lerakott peték a kapillárisokban a test különböző szerveibe sodródnak, és ha ott nagy
számban megakadnak, károsítják az adott szervet. A kopoltyúkapillárisokba jutott, petéből ki-
bújt miracidiumok a hámrétegen átjutva a vízbe kerülnek. További fejlődésükhöz köztigazdára,
a Lymnaea csigafajokra van szükségük. Ezekben jön létre a sporociszta, redia, majd cerkária
alak. Az utóbbi elhagyja a csiga testét. Felkeresi a halat, befúrja magát a bőrbe, így kerül a vér-
áramba.
10.1.2 . Májmétely-alakúak (Echinostomatida)
A miracidiumoknak csak egy pár protonefridiuma van. A cerkáriák farka egyszerű, elágazás
nélküli.
A leggyakoribb szívóférgek közé tartozik a májmétely (Fasciola hepatica), amely Földünkön
rendkívül elterjedt. A kifejlett féreg elsősorban a kérődzők, ezek közül is főleg a juhok és a szarvasmarhák
májában él, az epeutak falát károsítja. Testhossza elérheti, sőt meghaladhatja a három centimétert.
Élettartama több év lehet. A szájnyüás körüli szívókához közel helyezkedik el az acetábulum. Az
203
10.6. ábra. Echinostomatida fajok.
A: Echinostoma revolutum, B:
Calicophoron daubneyi (Dinnik és
Edelényi nyomán)
ivamyílás a két szivóka között van. Bélcsatomája gazdagon elága-
zó béledényrendszert alkot. Fejlődéséhez nedves, vizes terület
szükséges. Köztigazdája az elterjedési területétől függően más-
más csigafaj lehet. Európában a törpe iszapcsiga (Lymnaea
truncatida) a köztigazda. A csigába bekerült miracidiumból kiala-
kul a sporociszta, ebben a rediák. A rediákban létrejönnek a
cerkáriák. A csigát elhagyó, vízben mozgó farkos lárva az ivóvízzel
kerülhet be a végleges gazdába. Ha ez néhány órán belül nem
következik be, akkor a lárva a növényekre felkapaszkodva en-
cisztálódik. A gazdaállat ilyenkor a legelés során veszi fel a lárvákat.
A kifejlett féreg élete során akár félmillió petét is rakhat, egyetlen
petéből pedig közel 300 fárkos lárva alakulhat ki. Földünk egyes
területein, ahol a nedves területen termesztett növényeket élelem-
ként nyersen fogyasztják, ott az ember, valamint a sertés és a
rágcsálók is fertőződhetnek a féreg növényre telepedett, betokozó-
dott lárvájával.
Az Echinostoma revolutum főleg a víziszámyasok (hidak, ka-
csák) bélcsatomájában él, de előfordul más vadon élő és házia-
sított madarakban, valamint a rágcsáló, a mindenevő és a raga-
dozó emlősökben is (10.6. ábra). Kozmopolita faj, amelynek első
köztigazdája csiga, míg a második csiga, ebihal vagy esetleg rovar
lehet. Feji vége különleges felépitésű, mert gallérszerű peremén
tüskék sorakoznak. Az 1-2 cm testhosszúságot elérő féreg hasi
szívókája igen jól fejlett. Erős fertőződés esetén a gazdaállat le-
gyengül, véres hasmenés jelzi a paraziták kártételét.
A szarvasmarhák, juhok, szarvasfélék
bendőmételykórját okozó gyakoribb fajok
közé tartozik a Calicophoron daubneyi (10.6.
10.7. ábra.
Lándzsásmétely
(Dicrocoelium
dendriticum)
(Edelényi nyomán)
ábra) és a Paramphistomum cervi. Testhosszúságuk 6-12 mm közötti. Fejlett
acetábulumuk a test végén helyezkedik el. Fejlődésük a májmételyéhez hason-
lóan megy végbe. Köztigazdaként Lymnaea és Planorbis csigafajok szerepelnek.
A legelés során felvett metacerkáriák a bélcsatomába kerülnek. A fiatal mételyek
először a vékonybélben telepszenek meg, majd visszavándorolnak a bendőbe.
10.1.3. Száj szúr ony o s- cerkáriáj ú mételyek
(Plagiorchiida)
A cerkária alakoknak szájszuronya van és a farokrészben az elovesécske
gyakran hiányzik.
A megnyúlt, 6-10 mm hosszú testű lándzsásmétely (Dicrocoelium dend-
riticum) fejlődése száraz területeken megy végbe (10.7. ábra). A szabadba
került petéket felveszik például a kórócsigák (Helicella obvia) és a zebracsi-
gák (Zebrina detrita), amelyekben az egyes lárvaformák létrejönnek. A csigák
testéből nyálkás csomókba ágyazva kerülnek ki a farkos lárvák. A cerkáriákat
hangyák (Formica spp.) veszik fel táplálkozásuk során, bennük jön létre a
metacerkária. A garat alatti dúcban megtelepedő cerkáriák károsító hatására
megváltozik a hangyák viselkedése. A hűvösebb esti órákban felkapaszkodnak
a növényekre és nem mennek vissza a bolyba, mint a nem fertőzött társaik. A
végleges gazdák (főleg a juh, a kecske, a szarvasmarha és más vadon élő
kérődzők) a fűszálakra kapaszkodó hangyákkal együtt, a legelés során veszik fel
a parazitát.
204
10.2. Csáklyásférgek (Monogenea)
A valamivel több, mint 1000 ismert faj parazita.
Elsősorban az édesvízi és tengeri halak kopoltyú-
ján és testfelszínén élnek, de néhány faj a halak,
kétéltűek és a teknősök húgyhólyagjában élőskö-
dik. Megnyúlt, lapított, mozgékony testük igen
kicsi. Általában csak 1-2 mm hosszúak.
Mivel ektoparaziták, ahhoz, hogy jól meg
tudjanak kapaszkodni a gazdaállaton és ellenáll-
janak az erős vízáramnak is, testvégükön külön-
leges tapadószervük (opisthaptor) alakult ki
(10.8. ábra). Ez viselhet kisebb vagy nagyobb ki-
tines horgokat, kampókat, csappantyúkat, eset-
leg szívókákat. Az egyes fajok elkülönítését lehe-
tővé teszi a központi és szegélyhorgok számá-
nak, alakjának és nagyságának különbözősége.
Kültakarójuk felépítése a Trematoda osz-
tálynál leírtakkal megegyező.
Bélcsatornájuk a gyakran fajra jellemző,
hosszúkás alakú, izmos garattal kezdődik, majd a
nyelőcső kétfelé ágazik (10.8. ábra). A két bélcső-
ág nem ágazik tovább, a test vége felé vakon vég-
ződik, esetleg a két ág egyesülhet. Táplálékukat a
gazdaállat sejtjei, testfolyadéka képezi, amit az
izmos garat szívóhatása révén vesznek fel.
A kiválasztó szervrendszer két nefridiopó-
rusa a feji vég közelében nyílik a külvilágra.
Idegrendszerük a Trematoda taxonnál leír-
takhoz képest módosult. Nincsenek dorzális ideg-
törzsek és a laterálisak is hiányozhatnak, így álta-
lában csak a két ventrális ág található meg.
Érzékszerveik gyengén fejlettek, mechano-
és kemoreceptorokon kívül egyes fajoknál szem-
folt, pigmentkehelyszem fordul elő.
Valamennyi faj hímnős. A legtöbb fajnál pár-
zószerv (cirrus) is előfordul. A petefészkük min-
dig páratlan. A petevezetőn gyakran ondótartály
(receptaculum seminis) van. A fajok egy részénél
előforduló méh (uterus) rövid és kevés petét tar-
talmaz.
A csáklyásférgek életciklusa egyszerű, egy
gazdaállatuk van, fejlődésükhöz köztigazdára
nincs szükség. A megtermékenyítés általában
10.8. ábra. Monogenea fajok testfelépítésének
vázlata
ad: agydúc, bé: bélcsőág, fin: fejmirigyek, ga: garat,
he: here, ho: szegélyhorgok, ip: ívarpítvar, it: idegtörzs,
iz: izom, ka: kapaszkodószerv, kh: központi horgok,
La: Laurer-féle vezeték, If: látófoltok, Mm: Mehlis-
féle mirigy, oh: ondóhólyag, oo: ootyp petével, ov: on-
dóvezető, pá: párzószerv, pf: petefészek, pv: pete-
vezető, sz: szikmirigy (Kollmann nyomán módosítva)
kölcsönös. A petéből horgokat is viselő csillós lárva (oncomiracidium) bújik ki,
amelynek látószerve lehet, és testfelszínét haránt csillókötegek boríthatják. Felkeresi a
gazdaállatot, és a testvégen lévő horgainak segítségével megkapaszkodik azon. A
metamorfózis során elveszti csillóit, gyakran látószerveit is, illetve kialakul a fajra
jellemző felépítésű tapadószerve. A kopoltyúférgek gazdaspecifikusak, általában csak
egy fajt vagy annak közeli rokonait képesek megtámadni.
205
Bár széles elterjedésűek és gyakoriak, természetes viszonyok között általában nem
jelentenek komoly veszélyt a gazdaállat számára. Ezzel szemben zsúfolt tartás mellett, a
halgazdaságokban tömeges elszaporodásuk esetén komolyabb károkat okozhatnak. A
paraziták általában a kopoltyún telepszenek meg, akadályozzák a gázcserét, megkapasz-
kodásuk és táplálkozásuk révén roncsolják a hámsejteket. A megtámadott területen a
hámsejtek burjánzani kezdenek, a szervezet igyekszik a behatolót tokkal körülvéve el-
szigetelni. A sérült kopoltyúlemezek letöredezése a férgek egy részének eltávolítását
eredményezi. Ez a folyamat a hámréteg további károsodásával jár, csökken a hasznos
légzőfelület. A paraziták toxikus anyagokat is termelnek. Jelenlétük fokozza a hámréteg
nyálkaleadását, amely így, vastagabb rétegben, csökkenti a gázcsere hatékonyságát.
Kevés számú parazita megtelepedése is kaput nyithat kórokozó mikroorganizmusok
számára.
10.2.1. Egyszerű-tapadószervűek (Monopisthocotylea)
A gazdaállaton való megkapaszkodásra a tapadószerv horgai, horgocskái szolgálnak.
A pontyfélék kopoltyúférgességét elsősorban a Dactylogyrus fajok okozzák. A megnyúlt testű
férgek testhossza ritkán éri el az egy millimétert. Bélcsatornájuk a garatszakaszon kettéágazik, majd
a testvégén a két bélcsőág egyesül. Feji végükön négy pigmentes látószervük van. A hím ivarszerv
kivezető járatának végén párzószerv található. A test végén lévő tapadókészüléken egy pár központi
és 14 darab szegélyhorgot viselnek. A megtermékenyített petéket egyesével adják le a vízbe,
amelyek az aljzatra lesüllyedve indulnak fejlődésnek. Az onkomiracidiumok megtelepszenek a
halon, majd további fejlődésük alatt elvesztik csilióikat. Ha néhány óra alatt nem képesek a
gazdaállatra kapaszkodni, akkor elpusztulnak. Különbség figyelhető meg a fajok között abban,
hogy a kopoltyún hol telepszenek meg.
A Dactylogyrus vastator a ponty- és a kárászivadékot támadja meg (10.9. ábra). A kopoltyúlemezek
végein telepszik meg. A Dactylogyrus extensus szintén gyakori pontykárosító, ivadék és idősebb korban
10.9. ábra. Monopisthocotylea fajok. A: Dactylogyrus vastator,
YizAncylodiscoidesvistulensis, C: Gyrodactylushatharineri (Bihovszkij
és Gussev nyomán kissé módosítva)
egyaránt. Ez a faj a kopoltyúleme-
zek középső területein telepszik
meg. A különböző Dactylogyrus fa-
jok nemcsak az őshonos, hanem a
betelepített pontyféléken is előfor-
dulnak.
PizAncylodiscoides vistulensis 1-2
mm hosszúságú féreg, amely
morfológiailag és biológiája tekin-
tetében a Dactylogyrus fajokra ha-
sonlít (10.9. ábra). Kizárólag a
harcsa kopoltyúján élősködik, az
ivadékon jelentős hámkárosodást,
így gázcsere problémákat okoz. Ta-
padókorongján nem kettő, hanem
négy központi horog van.
A hámférgeknek nevezett Gy-
rodactylus fajok apró, általában egy
milliméternél kisebb testű álla-
tok, melyek a halak testfelületén,
az úszóikon, a kopoltyúikon élős-
ködnek. Feji végükön látószerv
nincs, viszont két mirigyet tartal-
mazó nyúlványt viselnek. Izmos
206
garatjuk kétfelé ágazik, a bélágak vakon végződnek. A farki végükön lévő tapadókorongban két
középhorog és 16 szegélyhorog található. Petéiket netn rakják le, a kifejlett állat testében több,
egymás testében kialakuló egyed fejlődik (polyetnbriogonia), majd a legfejlettebb elevenen jön a
világra.
A hazai halgazdas ágokban a Gyrodactylus katharineri
és a Gyrodactylus medius a fiatal pontyok gyakori kár-
tevője (10.9. ábra). Az előbbi faj főleg a kültakarón,
míg az utóbbi inkább a kopoltyún megtelepedő para-
zita.
10.2.2. Összetett-tapadószervűek
(Polyopisthocotylea)
Jellegzetességük az, hogy a tapadószervükön több,
bonyolult felépítésű, kitinvázas csappantyú van,
amelyek harapófogószerűen működnek.
Az ikerféreg (Diplozoon paradoxuni) nevét onnan
kapta, hogy a lárvaszakasz után, az ivarérés idején két
diporpa fejlődési alak a hasi szivókái és háti szemölcsei
révén összetapad, ivarszerveik egymással összekötte-
tésbe kerülnek. A pontyfélék kopoltyúján élnek és
károsítanak ezek az 5-6 milliméteres nagyságú, vérszí-
vó férgek (10.10. ábra). A kifejlett alakok évekig élet-
ben maradnak, ürítik a petéiket. A petéből kibújó on-
comiracidium megtelepszik a kopoltyún és átalakul
diporpa lárvává.
10.10. ábra. Ikerféreg (Diplozoon pavadoxum)
(Gussev nyomán)
10.3. Galandférgek (Cestoda)
Közel három és fél ezer, az élősködő életmódhoz magasfokúan specializálódott fé-
regfaj tartozik ebbe az osztályba. Testük általában fehér, sárgás vagy szürkés árnyalatú.
A galandférgek testfelépítése sok szempontból eltérhet az előzőekben tárgyalt két
osztályétól. A gazdaállat speciális, de viszonylag állandó belső környezetéhez adap-
tálódtak. A galandférgeknek több olyan szerve visszafejlődött, eltűnt, amely a feltéte-
lezett örvényféregszerű ősöknél még megvolt. Leegyszerűsödött testfelépítésük jó
példa arra, hogy az evolúció során nemcsak egyre bonyolultabb, hanem egyszerű szer-
vezetű fajok is kialakulnak. Testük általában három részre tagolható. Speciális testré-
szük a fej (scolex), amely a megkapaszkodást szolgálja. Rajta szívógödrök, szívókák,
horgok, tüskeszerű nyúlványok lehetnek. Tipikus esetben a scolex után van egy nyaki
(germinatív) zóna, amely az új ízeket (proglottis) hozza létre (10.14. ábra). Hosszú,
lapított testük kevés kivételtől eltekintve ízekre tagolt. Az egyes ízek találkozási helyét
egy felszíni barázda, és a vékonyabb izomzat jelzi. Az ízekre tagolt test, az ízek lán-
colatának neve a sztrobila. Minden új proglottisz megjelenésével párhuzamosan a
régebbiek hátrébb kerülnek, és a bennük található ivarszervek fokozatosan fejletté
válnak.
Testük védőburka (tegumentum) az előző taxonoknál megismert alapfelépítéshez
hasonló (10.1. ábra). A felszíni szinciciális plazmaréteg számos nyúlványt visel. Ennek
elsődleges szerepe feltehetően a felszívó felület növelése. A tegumentum megvédi a
férget a gazdaállat lúgos kémhatású emésztőnedveitől, de a vizet és a tápanyagokat át-
207
engedi. A galandférgek a pH szempontjából jó tűrőképességnek. Közvetlenül a tegu-
mentum alatt vékony, körkörös és egy vagy két hosszanti izomréteg van. A kültakaró
alatti parenchimát külső és belső rétegre tagolják az ott lévő erősebb hosszanti és kör-
körös izomkötegek. Néhány dorzoventrális izomköteg is megfigyelhető. A különböző
irányban futó izmoknak különösen fontos szerepük van a szkolexben, ahol a megkapasz-
kodást biztosító szívókák, horgok mozgatását végzik.
Bélcsatornájuk valamennyi fejlődési állapotukban hiányzik, a tápanyagokat teljes
mértékben a kültakarón át veszik fel. Lehetséges, hogy a gazdaállat bélcsövének belső
falába befuródó szkolex olyan anyagok felvételére is képes, melyre a proglottiszok nem.
A tápanyagok felvételét elősegíti, hogy testük belső ozmotikus koncentrációja alacso-
nyabb a gazdaállat bélcsatomájában lévő anyagoknál, testfolyadékainál. Testüknek ma-
gas a glikogéntartalma. Ez száraz tömegüknek akár 60%-át is kiteheti, míg zsírokból és
különösen fehérjékből igen keveset tartalmaznak. A zsírsavtartalom a glikogén anaerob
bontásából származik. A gazdaállat bélcsatomájában lévő szénhidrátok fokozzák a
növekedésüket és a petetermelést. A fehérjék esetleges hiányát jól elviselik.
Gázcseréjük a kültakarójukon keresztül megy végbe. Az oxigént felhasználják, ha
rendelkezésre áll, de az anaerob légzés dominál. A glikogén anaerob lebontásakor szén-
dioxid és különböző szerves savak keletkeznek.
A kiválasztó szervrendszer gyűjtőcsatornái a test két szélén, dorzo- és ventrolate-
rálisan futnak végig, hozzájuk csatlakoznak a lángsejtekben végződő csatomácskák. Min-
den ízben, annak poszterior végén harántágak kötik össze a ventrális helyzetű hosszanti
gyűjtőcsatornákat (10.14. ábra). Egyes fajoknál ez az alaptípus leegyszerűsödhet, míg
másoknál - például egyes szívókás galandférgeknél - a főcsatornákon kívül számos
további, de kevésbé fejlett csatornák is előfordulhatnak.
Idegrendszerük központja, a szkolexben lévő ideggyűrű- és idegsejtcsoport. Ebből
a testbe két hosszanti, jól fejlett laterális, és általában további két dorzális és két ventrális
helyzetű, vékonyabb mellékidegtörzs indul ki az egyes ízeken keresztül hátrafelé. Min-
den íz legalább egy harántirányú összekötő ideggyűrűt tartalmaz. A szkolexben a kétle-
benyű agydúc a transzverzális komisszúrák révén a legfejlettebb laterális törzsekhez és az
ott lévő ideggyűrűhöz csatlakozik. A rosztellumban szintén megfigyelhető egy kevésbé
fejlett ideggyűrű (10.14. ábra). Speciális érzékszerveik nincsenek, de a kültakaróban
módosult érzékelőcsillók ülnek.
Az érett ízek fejlett izomzatot, parenchimát és ivarszerveket tartalmaznak. Az utolsó,
legérettebb, esetleg már leváló ízekben viszont gyakorlatilag csak a pete tárolására szol-
gáló, megnövekedett uteruszt és a parenchimát lehet megtalálni.
Szinte valamennyi galandféreg hímnős. Minden egyes proglottiszban van ivarszerv
(10.14. ábra). A herék elszórtan helyezkednek el az ízen belül. A cirrusz alakja, tüské-
zettsége faji jellegzetesség. A petefészek többnyire páros, elágazó felépítésű. Az ootype
üregéből nyíló uterusz a legtöbb fajnál igen nagy, alakja fajra jellemző alakú, zsákszerű,
elágazó vagy hálózatos felépítésű. A belőle kiinduló vagina az ivarnyíláshoz vezet, rajta
a párzás során felvett spermiumok tárolására szolgáló ondótartály lehet. Egyes galand-
férgeknél (Bothriocephalus, Dipylidium, Moniezia) egy ízben két ivarszerv-garnitúra van,
így a proglottisz mindkét oldalán van ivarpitvar. A hím ivarkészülék általában előbb válik
éretté, így a petesejtek megtermékenyítése többnyire egy ízen belül nem megy végbe.
Általában két egyed termékenyíti meg egymást kölcsönösen, vagy ugyanazon egyed két
íze között történik megtermékenyítés. A megtermékenyített peték az ivamyíláson át
jutnak ki a külvilágra, vagy a testvégen lévő petékkel telt íz válik le a sztrobiláról. Azok
a fajok, melyek uteruszukból folyamatosan adják le a petesejteket, és így az ízek nem
válnak le rendszeresen, hosszúra megnőtt sztrobilával rendelkeznek. A peték általában
igen ellenállók a környezeti hatásokkal szemben, gyakran hónapokig fertőzőképesek
maradnak. Kevés kivételtől eltekintve valamennyi galandféreg fejlődéséhez legalább egy,
208
és általában gerinctelen köztigazdára van szükség. A kifejlett állatok a
gerincesek minden osztályának képviselőit károsíthatíják. Az emberek is
fertőződhetnek. Erre főleg a trópusi területeken, a paraziták fejlődésé-
hez kedvező táplálkozási szokások és a higiéniás körülmények miatt
kerülhet sor.
10.3.1. Tagolatlan galandférgek (Caryophyllidea)
Az ide tartozó fajok teste a többi galandféregtől eltérően nem ízekből áll,
hanem tagolatlan, szalag alakú. Fejükön valódi értelemben vett szívó-
gödröket, szívókákat nem viselnek. Az elülső testvég formája legyező-
szerűen kiszélesedő, gyakran csipkézett. Főleg édesvízi halak élőskö-
dői.
Hazánkban őshonos a szegfűféreg (Caryophyllaeus fimbriceps), míg a Khawia
sinensis (10.11. ábra) nevű fajt a Távol-Keletről betelepített növényevő halakkal
hurcoltákbe hazánkba. A két említett faj morfológiailag igen hasonló. AK. sinensis
kifejlett alakjainak mérete meghaladja a 80 mm-t, míg a C. fimbriceps csak 25 mm
hosszú. Kiszélesedő feji végük fodrozott, virágszirmokra emlékeztető nyúlványo-
kat visel. A jellegzetes, „H” alakú petefészek a test hátulsó vége felé található. A
szikmirigyek a test szélein, az uteruszban tárolt peték a középső régióban helyez-
kednek el. Köztigazdával fejlődnek. A férgek a halak bélcsatomájában élnek,
ahonnan a petéik a bélsárral jutnak ki a vízbe. A kialakuló lárva (coracidium) a
csővájó férgek, leggyakrabban a Tubifex tubifex testébe kerül, ahol belőle a
procercoid, majd a plerocercoid lárva jön létre. A hal testébe a fertőzött Tubifex
elfogyasztásával kerül be a plerocerkoid lárva. A féreg a hal bélcsatomájában éri
el az ivarérettségét. A bélben hámsérüléseket, bélgyulladást okoz, a halak leso-
ványodnak, súlyosabb esetben a fiatal halak elhullanak.
10.11. ábra. Kha-
wia sinensis (Bi-
hovszkij nyomán)
10.3.2. Szívógödrös galandférgek (Pseudophyllidea)
Az ide tartozó fajoknak a szkolexén általában kettő, ritkán több ventrális és dorzális hely-
zetű tapadóbarázda vagy szívógödör (bothrium) található. A szívógödör falában nincs
jól fejlett, erős, elkülönült izomzat, az csak a bőrizomtömlő kismértékben módosult
része. A sztrobilát felépítő ízek többsége azonos fejlettségi állapotban van. Egyes fajoknál
a test tagolatlan, az ízek határa elmosódott. Az uteruszuknak általában a ventrális oldal
középvonalában külön kivezető nyílása van.
Fejlődési ciklusuk során petéik a végleges gazda bélsarával kerülnek a vízbe. Itt kialakul
a koracidium lárva, amely csiliókkal rendelkezik, de hat kapaszkodóhorga is van (10.12.
ábra). Az első köztigazda, általában alacsonyabbrendű rák testüregében ebből procerkoid
lárva jön létre, míg a második köztigazdában, vagy ennek hiányában a végleges gazdában a
plerocerkoid fejlődik ki. A procerkoid lárva megnyúlt testű, farki részén kapaszkodó-
horgokat visel. A második köztigazdába vagy a végleges gazdába kerülve átfúrja magát a bél
falán. Ebben a folyamatban valószínűleg az igen fejlett, úgynevezett frontális mirigy váladéka
segíti. Megtelepszikazizomzatbanvagyatestüregben.Átalakulmegnyúlttestűplerocerkoiddá,
amelynek feji végén már kialakul a szkolex. Az ivarszervek kivételével valamennyi, a kifejlett
állatban is meglévő belső szerv kezdeménye megtalálható benne. Az ivarszervek csak a
végleges gazdába kerülve alakulnak ki. A végleges gazda az egyetlen vagy a második
köztigazda elfogyasztásával fertőződik, ahol a plerocerkoid kifejlett féreggé alakul.
A Bothriocephalus acheilognathi a Távol-Keletről behurcolt parazita (10.13. ábra). Valamennyi
hazai pontyfélében megtalálja életfeltételeit, de elsősorban a pontyot károsítja. Jellegzetes formájú
209
10.12. ábra. A szívógödrös galandférgek (Pseudophyllidea) fejlődési ciklusának egyszerűsített vázlata. A: fejlő-
désnek indult zigóta, B: koracidium, C: procerkoid, D: plerocerkoid, E: kifejlett féreg (Vogel, Rosen és más szerzők
nyomán módosítva)
szkolexén két sekély üregű, megnyúlt alakú szívógödör van, amelyek segítségével megtapad a hal
belében. A kifejlett alakok teste 15-20 cm hosszú lehet. Mechanikusan károsítják a bél nyálkahár-
tyáját, gyulladásos folyamatot okoznak. Jelentős mennyiségű táplálékot vonnak el, ezért a halak
10.13. ábra. Szívógödrös galandférgek. A:
szíjgalandféreg (Ligula intestinalis)^ B:
Bothriocephalus acheilognathi (Bihovszkij,
Edelényi és Molnár nyomán kissé módosítva)
lesoványodnak. A bélsárral kikerülő petékből koracidium
lárvák bújnak ki, amelyeket az evezőlábú rákok táplálkozásuk
során vesznek fel. Testüregükben a lárva továbbfejlődik
procerkoiddá. A halak a rákok elfogyasztásával fertőződnek,
bélcsatomájukban kialakul a plerocerkoid, majd ebből a
kifejlett féreg.
A szíjgalandféreg (Ligula intestinalis) kifejlett alakjai
(10.13. ábra) a vizimadarak bélcsatomájában élősködnek,
ahol a peterakást követően rövidesen elpusztulnak. Abélsárral
kiürült petékből kialakuló koracidiumot a köztigazda evezőlá-
bú rákok veszik fel, bennük kialakul a procerkoid. A rákokat
elfogyasztó pontyfélékben, azok hasüregében több mint egy
évig fejlődnek a plerocerkoid alakok. Testméretük több tíz
centiméteres is lehet. Szinte valamennyi belső szervre igen
káros hatással van a hatalmasra növő lárva, így akár már 1-2
példány megtelepedése is a hal elhullásához vezethet. A
fertőzés következtében legyengült, a vízfelszín közelében las-
san mozgó halakat elfogyasztó madarakban kialakul a kifejlett
féreg. Ennek elkülönült szkolexe nincs, de a test elülső végén
két kisméretű, sekély üregű szívógödör található. Testük
külsőleg tagolatlan, rajta harántirányú, felületi beíűződések
figyelhetők meg. A hosszanti sorba rendezett ivarszervek
kivezető nyílásai a test hasi oldalán egymás után következnek.
210
10.3.3. Szívókás galandférgek (Cyclophyllidea)
A galandférgek fajokban leggazdagabb rendje. Szkolexükön négy jól fejlett, erős,
elkülönült izomzattal bíró szívóka van, amelyek segítségével erősen meg tudnak tapadni
a gazdaállat bélcsatomájánakfalán (10.14. ábra). Egyes fajok szkolexének csúcsán lévő
10.14. ábra. A szívókás galandférgek (Taenia sp.) testfelépítésének vázlata. A: fej, B: idegrendszer a feji végben,
C: ízelt test, D: íz, E: érett íz, amelyben jól látszik a petékkel teli, elágazó méh,
ad: agydúc, ge: germinatív zóna, he: here, hi: haránt ideggyürü, ho: horog, hü: hüvely, id: ideggyűrű, ip: ivarpitvar, it:
idegtörzs, ki: kiválasztószerv csatornája, mé: méh petékkel, Mm: Mehlis-féle mirigy, oo: ootyp, ot: ondótartály, pf:
petefészek, pt: párzótáska, pv: petevezetö, ri: rosztellum ideggyűrűje, sí: szívóka, sz: szikmirigy (Meglitsh, Braun és más
szerzők nyomán módosítva)
211
nyúlványon, ormányon (rostellum) horogkoszorú található, amely még biztonságosabb
megkapaszkodást eredményez. A sztrobilát alkotó ízek különböző fejlettségi állapotban
vannak. A szikmirigy kompakt és az íz hátulsó felében helyezkedik el. Az ivamyílás
általában az íz laterális szélén van. Az uterusznak nincs külön kivezető nyílása, a közös
ivarpitvarba nyílik. Fejlődési ciklusuk a szívógödrös galandférgekhez képest egyszerűbb.
A pete a végleges gazda bélsarával kerül ki a külvilágra. A petét a köztigazda táplálkozása
közben veszi fel. Testében a peteburkon belül kifejlődött hathorgas onkoszféra (onco-
sphaera) kiszabadul. Kezdetben a gazdaállat szervezetében vándorol, majd egy bizonyos
szervben megtelepedve kialakul belőle a végleges lárvaforma (10.15. ábra). A fertőzőké-
pes, végleges lárvaformáknak több típusa van, amelyeket felépítésük alapján csopor-
tosítani lehet. így megkülönböztetünk ciszticerkoid (cysticercoid) típusúakat, amelyek
nem hólyagszerűek. A szkolexet ebben az esetben a test középső részéből kialakult
párénchimális védőtok veszi körül, a test végén csak a farokrész szabad. Igen változatos
alakúak lehetnek, számos formájuk ismert. Ilyen típusú lárva például a kriptocisztisz
(cryptocystis), amely a Dipylidium caninum fertőzőképes fejlődési alakja. Ciszticerkoid
típusú lárvája van a Moniezia fajoknak is. A másik fő csoportba a ciszticerkusz (cys-
ticercus) típusú lárvákat lehet sorolni, amelyeknél egy vagy több betüremkedett szkolex
van egy folyadékkal telt, a test többi részéből kialakult hólyagban. Ide tartozik a valódi
ciszticerkusz, népies nevén borsóka. Ebben a hólyagban egy betüremkedett szkolex van.
Ilyen végleges lárvája van például a Taenia saginata fajnak. A cönurusz (coenurus) lár-
vában egy hólyagban több betüremkedett szkolex fejlődik ki. Ez a lárvatípus a Taenia
multiceps fajnál fordul elő. A harmadik ciszticerkusz lárvatípus az echinokokkusz (echi-
nococcus), egy terjedelmes nagy hólyag (rívó ka tömlő), amelyben a hólyag belső, ger-
10.15. ábra. A szívókás galandférgek (Cyclophyllidea) lárvaformái. A: hathorgas onkoszféra, B-E: végleges
lárvaformák: B: kriptocisztisz, C: ciszticerkusz, D: cönurusz, E: echinokokkusz
ho: horog, kb: külső sejtmentes védőburok, kh: költőhólyag, se: betüremkedett scolex, te: tegumentum, amelynek belső
felszínén differenciálódásra képes sejtek ülnek (Belding és Mehlhom nyomán módosítva)
212
minatív rétegéről sarjadzással további utódciszták különülnek el, és
tartalmazzák a betüremkedett szkolexeket. Ilyen lárvája van az
Echinococcus granulosus fajnak.
Elsősorban a juhok galandférgességét okozza a Moniezia expansa, amely
Földünkön igen gyakori előfordulású. A bélcsatomában élő kifejlett férgek
testéről leváló, duzzadt rizsszemre hasonlító ízek a bélsárral kerülnek ki a
külvilágra. A petéket a talajban élő páncélosatkák (Oribatida) veszik fel.
Bennük alakul ki a hathorgas onkoszféra, majd a ciszticerkoid típusú lárva.
A végleges gazda legelés közben, a fűszálakra felkapaszkodott atkákkal veszi
fel a féreg lárváit. A kifejlett féreg 1-5 m hosszúságot érhet el.
A baromfiak és a vadon élő madarak gyakoribb galandférgei a Davainea
és Raillietina fajok. A bél nyálkahártyáján megkapaszkodó férgek helyi elha-
lásokat okoznak. A fiatal baromfi erős fertőződése elhulláshoz vezethet. A
Davainea proglottina kis méretű (1,5-4 mm), és teste kevés számú (4-9 db)
proglottiszból áll (10.16. ábra). A rostellumon és a szívókákon is apró horgok
vannak. A férgek köztigazdái csigák (Cepaea sp., Limax spj. Ezekben jönnek
létre a ciszticerkoid típusú lárvák. A Raillietina cesticillus gyakori parazita.
Testhossza a 13 centimétert is elérheti, a sztrobilát több száz íz alkotja. Köz-
tigazdaként számos bogárfaj (például az alombogár -Alphitobius diaperinus)
lehet, amelyekben a ciszticerkoid lárvák alakulnak ki. A baromfi az ilyen bo-
garak elfogyasztásával fertőződik.
Az uborkamagképű galandféreg (Dipylidium caninum) végleges gazdái
a kutyák, macskák és néha az ember (főleg a gyerekek) is. Köztigazdái a bol-
hák, amelyekben a hathorgas onkoszférából kialakul a ciszticerkoid típusú
kriptocisztiszlárva. A kutyák és macskák a bolhászkodás közben szétharapott
és lenyelt bolhákkal fertőződnek. Simogatás közben az ember kezére, innen
a szájába kerülhetnek a szőrzetre tapadt, a bolhászkodás során szabaddá vált
lárvák. A kifejlett féreg testhosszúsága 15-40 cm között lehet, a proglottiszok
száma nem több, mint 200 darab.
A simafejű galandféreg (Taenia saginata) kifejlett alakjai az ember
bélcsatomájában élnek, ahol testhosszúságuk eléri a három, néha több métert
is. A sztrobila akár 2000 proglottiszból is állhat, egy-egy íz pedig 80 000 petét
is tartalmazhat. A bél falához a szkolexen található négy szívókával tapadnak
hozzá. A bélsárral külvilágra kerülő levált ízekből kiszabadult petéket a szarvas-
marha legelés közben veheti fel. A hathorgas onkoszféra a köztigazda belében
kiszabadul, az izomzatba vándorol, ahol kialakul belőle a borsóka. Az ember
a kellően meg nem sütött vagy nyersen fogyasztott borsókás marhahús révén
fertőződik. Bélcsatornájában a lárva szabaddá válik, megkapaszkodik a bél
falában, sztrobilát fejleszt és szaporodóképessé válik.
A macskafélék leggyakoribb galandférge a 15-25 cm hosszú Taenia tae-
niaeformis. A kifejlett alak a fején horogkoszorút visel. Köztigazdái az egerek,
patkányok és pockok, amelyekben a ciszticerkusz lárvák találhatóak.
A Taenia pisiformis végleges gazdái a kutyák, vadon élő húsevők, míg a
köztigazdái a nyulak. A ciszticerkusz lárvák a nyulak hasűri szerveinek sa-
vóshártyáján telepszenek meg. A kifejlett alakok testhosszúsága elérheti az
egy métert is.
A kergeféreg (Taenia multiceps) végleges gazdája a kutya, amelynek bél-
sarával kikerülnek a peték a legelőre. A legelő juhok táplálkozás közben veszik
fel a petéket, majd a bélcsatornájukban kiszabadulnak a hathorgas onkoszférák.
Átfúrva a bél falát, a vérárammal eljutnak az agyvelőt védő hártyákhoz, ahol
megtelepszenek. Itt kialakul a cönurusz lárvaforma, amely károsítja az agyve-
10.16. ábra. Davai-
nea proglottina
(Kotlán nyomán)
10.17. ábra. Rívó**
kaféreg (Echinococ-
cus granulosus)
(Mehlhorn és más
szerzők nyomán kissé
módosítva)
213
löt, kiváltva a juhok jellegzetes körbe forgó mozgását. A kutyák fertőződése a kényszervágott juhok
agyának elfogyasztásakor következik be. A kifejlett férgek 0,4-1,0 méter hosszúságot érhetnek el.
A 3-6 mm testhosszúságú háromízü galandféreg vagy rivókaféreg (Echinococcus granulosus) a
kutyafélékben éri el kifejlettségét (10.17. ábra). Köztigazdája számos emlősfaj (főleg a patások)
lehet, többek között az emberis. A kutya bélsarával ürülő petéket felvehetik a legelő állatok, például
a sertés, a juh vagy simogató keze révén az ember is. A hathorgas onkoszféra a bélcsatomából a vér-
áramba kerül, és különböző szervekben, leggyakrabban a májban és a tüdőben telepszik meg. Ott
hosszú ideig fejlődik. Akár ököl nagyságú hólyaggá, echinokokkusz-tömlővé is válhat. A gazdaállat
vagy az ember a rivókaféreg fertőződése nyomán súlyosan megbetegszik.
Ellenőrző kérdések a 10. fejezet témaköreiből
1. Sorolja fel azokat a tulajdonságokat, amelyek a laposférgek parazita életmódját tükrözik!
2. Ismertesse a mételyek testfelépítését!
3. Jellemezze a mételyek fejlődésének vázlatát, az egyes lárvaformákat!
4. Jellemezze a mételyek osztályába tartozó fajok biológiáját és mezőgazdasági jelentőségét!
5. Ismertesse a csáklyásférgek testfelépítését!
6. Jellemezze a sáklyásférgek fejlődésének vázlatát, az egyes lárvaformákat!
7. Jellemezze a sáklyásférgek osztályába tartozó fajok biológiáját és mezőgazdasági jelentő-
ségét!
8. Ismertesse a galandférgek testfelépítését!
9. Jellemezze a galandférgek fejlődésének vázlatát, az egyes lárvaformákat!
10. Jellemezze a galandférgek osztályába tartozó fajok biológiáját és mezőgazdasági jelentő-
ségét!
214
11. FONÁLFÉRGEK (NEMATODA)
A törzsbe tartozó fajok száma pontosan nem ismert, mivel a közel 20 000 leirt fajon túl a még
ismeretlenekkel együtt számukat több százezerre becsülik. Egyedszámuk az ízeltlábúakhoz és
puhatestüekhez hasonlóan igen magas.
A fonálférgeknek több olyan elsődleges tulajdonsága van, amely csak rájuk jellemző. Ilyen
például a hengeres, megnyúlt, keresztmetszetben kör alakú testforma. Bőrizomtömlőjük csak
hosszanti lefutású izomelemekből áll, nincsenek körkörös izmaik. Kiválasztószervük különle-
ges felépítésű. A fajok egy része párzóhoroggal rendelkezik. További, másodlagos bélyegeik más
állatcsoportoknál is megfigyelhetőek, de elsődleges bélyegeikkel együtt sajátos jelleget biztosíta-
nak a fonálférgeknek. Testüket felépítő sejtek száma nagy állandóságot mutat. Ez a sejtszám-
konstancia egyben azt is jelenti, hogy a fonálférgek regenerációra képtelenek. Kutikulájukat
vedlés útján tudják eltávolítani. Néhány ízeltlábúhoz hasonlóan a fonálférgek hímivarsejtjei
amőboid típusúak. Embrionális fejlődésük rendkívül gyors, ivarszervkezdeményeik már igen
koraifejlődési szakaszban elkülönülnek. Egyes szerveik sugaras szimmetria szerint épülnek fel.
Képesek anabiotikus állapot kialakítására. Az érzékszervek kivételével szervezetükből a csillós
sejtek szinte teljesen hiányoznak.
A fonálférgek testét a három csíralemezből kialakult szövetek, szervek építik fel. Emésztő
szervrendszerük három szakaszos. Szájszerveik felépítése jól mutatja életmódjukat. Elkülönült
légzési és keringési szervük nincs. A legtöbb faj váltivarú, ivari dimorfizmus is megfigyelhető.
Több lárvaállapot közbeiktatásával érik el kifejlettségüket.
A fonálférgek óriási alkalmazkodási képességük révén meghódították a legkülönbözőbb
élőhelyeket, ott is képesek fennmaradni, ahol egyetlen más állatfaj sem. Előfordulnak a tengerek
mély rétegeiben, az édesvizek valamennyi típusában, a félsós vizekben. Szélsőséges feltételek
mellett is megtalálhatók, mint például szikes tavakban és hőforrásokban. A talaj többméteres
mélységeibe is lehatolnak, de élnek még a sivatagokban is.
Nagyrészük szabadon élő faj, és fontos szerepet tölt be a szervesanyag-lebontás folyamatá-
t bán. Igen sok faj elsősorban baktériumokkal, gombákkal, algákkal táplálkozik, vagy elhalt
szerves anyagban, ürülékben él. Vannak ragadozók is, amelyek főleg kerekesférgekkel,
medveállatkákkal, kis testű gyürűsférgekkel vagy más fonálférgekkel táplálkoznak. Számos faj
növényi nedvet szív. Szinte valamennyi élőlénynek van gazdaspecifikus fonálférge. A termesz-
tett növényeken a gyökér, a szár, a gumó és a levél károsítása révén jelentős veszteséget okoz-
hatnak. Vírus-, baktérium- és gombavektor szerepük sem elhanyagolható. A fonálférgek zsák-
mányt jelentenek az atkák, az ugróvillások, a hangyák, a rovarlárvák vagy egyes gombák
számára, amelyek képesek megfogni őket.
Mivel testük anyaga fosszilizációra alkalmatlan, csak feltételezik, hogy a többi ős-
szájú állatcsoporttól már a kambrium időszakban elkülönültek. Lenyomataik 50 millió
évesek, míg borostyánkőbe zártan 30 millió évvel ezelőtt ék fajok maradtak meg. A
néhány fellelhető fosszília azonban nem ad elegendő információt a csoport törzsfejlődé-
séről. Ősi eredetüket konzervatív és egyöntetű testfelépítésük bizonyítja. Nem
valamilyen magasabb fejlettségű csoport leegyszerűsödött formáját, hanem egy egyenes
215
irányban, progresszíven fejlett taxont képeznek. Fejlettségüket jelzi a magasfokú élet-
módbeli differenciáltság. Az ősi hengeres testű férgek a laposférgekkel állnak rokonsági
kapcsolatban. Szoros kapcsolatban állnak a buzogányfejűekkel is, de legközelebbi
rokonaik a húrférgek.
Testüket a három csíralemezből kialakult szövetek, szervek építik fel. Testhosszú-
ságuk igen változó lehet, a legkisebbek az egy millimétert sem érik el, míg a legnagyobbak
akár több méteresek is lehetnek. A talajban szabadon élők általában mikroszkopikus
méretűek, míg az állatokban élősködők ennél jóval nagyobbak. Az egy fajhoz tartozó
egyedek közül a nőstények mindig vastagabbak. Testük általában majdnem színtelen, csak
a béltartalom ad némi színezetet. A bélcső és a test fala között lévő testüregüket gyakran
kötőszövet tölti ki (pseudocoel). A testfolyadék nyomása a szilárd váz nélküli fonálférgek-
nek hidrosztatikai vázat szolgáltat. Testük tagolatlan, szelvényezetlen. A test egyes
szervei sugaras, míg mások bilaterális szimmetriát mutatnak. Testük végén ragadós
anyagot termelő mirigy van.
Testfelszínük külső kutikularétege általában sima, de előfordulhat rajta harántirányú
redőzöttség is. Ez egyes fajoknál olyan kifejezett, hogy szelvényezettségnek tűnik. A
kutikula kinövései tehetnek serték, hólyagszerű duzzanatok vagy szemölcsök, az élőskö-
dőknél kapaszkodóhorgok. A serték elsősorban a vízben élőkre jellemzők. A Torquentia és
Secementia osztályoknál a test két oldalán hosszanti kiemelkedések (oldalmezők vagy
oldallemezek) futnak végig. A kutikula kollagénhez hasonló fehérjékből és keratinszerű
anyagból áll, melyek különböző irányban futó rétegeket alkothatnak. Az Ascaris fajok
kutikulájában például kilenc réteget tehet elkülöníteni. A kutikula funkciója a belső
hidrosztatikai nyomás fenntartása, a mechanikai behatásokkal és endoparazitáknál a gaz-
daállat emésztőnedveivel szembeni védetem. Fejlődésük során négyszer vedlenek. Vedlés
11.1. ábra. Fonálféreg (Ascaris sp.) testének keresztmetszete
bf: bélcsö fala, hl: hipodermisz, hl: hipodermiszléc, ik: izomsejt karja, ír:
idegrostok, íz: izomsejt, ki: kiválasztócsatoma, ku: kutikula, mé: méh petékkel,
pf: petefészek, pv: petevezető (Alexander és számos más szerző nyomán
módosítva)
alkalmával a kutikula régi, fel-
ső rétegét eltávolítják az álla-
tok. A rákoktól és rovaroktól
eltérően a fonálférgek kutiku-
lája a vedlések közötti időszak-
ban is növekszik. A kifejtett
állatok nem vedlenek, de a ku-
tikula tovább nő.
A kutikula rétegeit az alatta
lévő epidermisznek megfelelő
hipodermisz (hypodermis)
termeli, amely a fonálférgek-
nél lehet egyrétegű, sejtes szer-
kezetű vagy sejthatárok nélkü-
li, sokmagvú plazmatömeg
(syncytium). A hipodermisz he-
lyenként a test hosszában meg-
vastagodhat, léceket képezhet
és az izomréteget hosszanti kö-
tegeltre tagolja. Általában dor-
zális, ventrális és laterális léce-
ket tehet elkülöníteni (11.1.
ábra). A lécekben futnak az
idegtörzsek, illetve a két oldal-
sóban a kiválasztószerv. A hi-
podermiszsejtek magjai is itt, a
lécekben találhatók.
216
A hipodermiszréteghez kapcsolódnak a
hosszanti lefutású izomrostok. Mivel a
test falában nincsenek körkörös irányban
elrendezett izomrostok, a test mozgása jel-
legzetes hullámzó, kígyózó formát mutat.
Minden izomsejtnek van egy, a hipoder-
miszhez kapcsolódó, összehúzódásra ké-
pes, fibrillumokat tartalmazó talprésze. A
fibrillumok aktin és miozin filamentu-
mokból állnak. Az izomsejt teste kontrak-
cióra nem képes, mert a miofibrillumok
nélküli szarkoplazmát és a sejtmagot foglal-
ja magába (11.2. ábra). A testüregbe nyúló
sejttest plazmája jelentős mennyiségű tar-
talék tápanyagot, glikogént tárol, és a sejt-
mag is itt van. Minden izomsejt rendelkezik
egy karral, amely a legközelebbi idegpályá-
hoz kapcsolódik. Ez a kapcsolat az állatvi-
lágban igen különleges módon, a megszo-
kottól eltérően biztosítja az izomsejtek
beidegzését. Nem az idegek viszik az im-
pulzusokat az izomsejtekhez, hanem fordít-
va, az izomsejtek karjai veszik át az idegros-
toktól.
11.2. ábra. Fonálféreg (Ascaris sp.) kültakarójánakés
izomzatának részlete
hl: hipodermisz, hl: hipodermiszléc, ik: izomsejt karja, ir:
idegrostok, it: izomsejt talpa, íz; izomsejt teste, ku: kutikula
(Rosenbluth nyomán kissé módosítva)
A folyadékkal telt testüreg (pseudocoel) az állat hidrosztatikai vázát adja, valamint
ebben helyezkednek el a belső szervek. A testüregben meghatározott helyeken és
számban rögzült sejtek (coelomacyta) lehetnek, melyeknek szerepe a testbe bekerült
idegen anyagok bekebelezése. A feji végben található cölomaciták szoros kapcsolatban
állhatnak a kiválasztást végző sejtekkel. Az Ascaris fajokban a feji végen találhatók a
fagocitózisra képes rögzült cölomaciták.
11.3. ábra. A fonálférgek gyakoribb szájszervtípusainak felépítése vázlatosan. A: mikroorganizmust fogyasztó
(Rhabditis típus), B: növényi nedvet szívó (Tylenchus típus), C: vegyes táplálkozást! (Dorylaimus típus), D:
ragadozó (Mononchus típus)
fin: fogmódosulásból alakult szurony, fo: fog, go: szájszuronygomb, sz: szájszurony, to: szájtok (Andrássy és más szerzők
nyomán kissé módosítva)
217
11.4. ábra. A fonálférgek testfelépítése vázlatosan. A: nőstény, B: hím
aj: ajak, as: ajakserte, fa: fazmidium, fk: farok, fin: farokmirigy, he: here, hü: hüvely, id: ideggyuru, kb: középbulbusz, kö:
középbél, kv: kiválasztószerv kivezetőnyílása, mé: méh, ni: női ivamyílás, ny: nyelőcső, oh: ondóhólyag, ok: ondókilövellő cső,
ot: ondótartály, ov: ondóvezető, pá: párzótáska, pf: petefészek, pt: párzótüske, pv: petevezető, sz: szájszurony, ub: utóbél, vb:
végbulbusz, vé: végbélnyílás, vi: végbél és ivamyílás, vp: vezetőpálca (Andrássy és más szerzők nyomán módosítva)
218
Háromszakaszos emésztő szervrendszerük a szájnyílásból, az izmos garatból, a
nyelőcsőből és a nem izmolt bélcsőszakaszokból áll. A szájnyílás körül a fajok egy
részénél, például az Ascaris fajoknál ajkak, míg másoknál tapintószemölcsök (papilla) és
sertékvannak. A bélcsatomájuk kezdeti szakaszánakfelépítésében meglévő különbségek
fontos támpontot adnak a rendszertani egységek és a fajok elkülönítéséhez (11.3. ábra).
A szájüreg kutikulával bélelt, ha ez erős, vastag, akkor szájtok a neve. A mikroorganiz-
musokat fogyasztó fonálférgek szájürege ilyen alakulású. A szájüregben a ragadozó fajok
többségénél fogszerű képletek lehetnek. A szájszurony (stylus) azoknál a fajoknál for-
dul elő, melyek áldozatukat vagy a növényi részeket megtámadva, annak védőrétegét
átszúrva a lágy részeket szívják ki. A szájszurony a legtöbb növényi nedvet szívó fajnál az
alapján lévő gombokhoz kapcsolódó izmokkal mozgatható, az állat a szájnyílásán át ki
is tudja tolni. A vegyes táplálkozáséak szájszuronya az előző típushoz képest tágasabb
üregű, és fogmódosulásnak tekinthető. A fonálférgek a táplálékot elsősorban folyékony
formában veszik fel. Ha szükséges, akkor a felszívás előtt enzimtartalmú váladékot bo-
csátanak a táplálékba. A megnyúlt nyelőcső lehet egyszerű, tagolatlan cső. Számos fajnál
egy vagy két izmos falú tágulat (bulbus) figyelhető meg rajta (11.4. ábra). Ezeknek
alakja, elhelyezkedése bizonyos mértékig a fajok elkülönítését is lehetővé teszi. A bul-
buszok zárókészüléke megakadályozza a táplálék visszaáramlását. A garatüreg és a
bulbuszok izmainak gyors összehúzódása tágítja az üreget, amely a táplálék felszívását
eredményezi. Az elemyedő izmok - a bulbusz üregét szűkítve - a bélcső felé préselik
tovább a táplá-lékot. Az emésztőnedvet termelő mirigyek váladékukat vagy a bulbuszokba,
vagy a nye-lőcső és a középbél határán a bélcsőbe öntik. A bélcső fala egyrétegű
hengerhámsejtekből áll, belső felszínét mikrobolyhok borítják. A kis testű fonálférgeknél
a bélcsövet viszonylag kevés sejt alkotja. így például a Rhabditis fajoknál 18-64, a
Tylenchidea fajoknál 16-24 darab, míg a nagy testűeknél több száz vagy ezer sejtből épül
fel a bélcső. A táplálékot a test mozgása és az újonnan felvett táplálék tolóhatása
továbbítja a bélcsőben. Az Ascaris fajok utóbele többé-kevésbé lapított. A hámréteg alatt
igen vékony izomréteg van. Az ürülék az izomgyűrűvel nyitható-zárható végbélnyíláson
keresztül távozik, leadását a testüregfolyadék nyomása is elősegíti. A végbélnyílás a farki
vég közelében van, a szaba-don élő fajoknál általában a tapadást elősegítő ragadós
váladékot termelő mirigy kísére-tében. Ide torkollik a hímek ivari kivezető járata is.
A fonálférgeknek nincs elkülönült légzőszervük, a gázcsere a kültakarón át megy
végbe. Az endoparazita fajok elsődlegesen anaerobok, de képesek hasznosítani az oxi-
gént, ha az rendelkezésükre áll. Számos kifejlett endoparazitának csak anaerob méta-
hólizmusa van, az aerob hiányzik. Az energiát glikolízisből és a ma még kevéssé ismert
elektrontranszport-folyamatokból nyerik. A szabadon élő fonálférgek és a paraziták sza-
badon élő lárvaformái obiigát aerob szervezetek. Ez a túlélési stratégiájuk szempont-
jából jelentős tényező. A legkülönbözőbb élőhelyeken fordulnak elő, alacsony és magas
oxigéntartalom mellett egyaránt. A legtöbb szabadon élő fonálféreg a speciális légző-
rendszer hiányában kisméretű. A nedves talajban, a vízfenékiszaprétegében, a gazdaállat
bélcsatornájában élő fajok általában speciális módon alkalmazkodtak az alacsony oxi-
génellátottsághoz. Egyes fajoknál hemoglobin található a testfolyadékban vagy a test
falában. Csekély mennyisége révén általában nincs jelentős szerepe az oxigén szállításá-
ban. Ha a környezetben igen alacsony az oxigéntartalom, akkor a hemoglobin által meg-
kötött oxigén rövid időre biztosíthatja a szervezet oxigénszükségletét. Azok a paraziták,
melyek a gazdaállat vérével kapcsolatba kerülhetnek, felszívják azt, és a bélcsatomájukon
keresztül hasznosítják a vérben lévő oxigént.
A biokémiai specializáció az anatómiainál is fontosabb. A citrátkör és a terminális
oxidáció egyes fajoknál (például a Trichinella esetében) megtalálható, míg másoknál,
például az Ascaris fajoknál hiányzik. Ez utóbbiakban helyette egy hasonló rendszer
működik. Anaerob szénhidrátbontás esetén számos végtermék keletkezhet. A környe-
219
11.5. ábra. Fonálférgek kiválasztószerveinek alaptípusai. A: renett-
sejtekkel (re), B: hipodermiszlécben futó csatornával (cs)
hl: hipodermiszléc, ki: kiválasztó csatorna ürege, kp: kiválasztószerv kivezető
pórusa, ku: kutikula (Maggenti, Schneiderés másszerzőknyomán módosítva)
zet oxigéntartalmának emelke-
désekor többnyire csökken a
leadott szerves savak mennyisé-
ge. Ascaris és Trichinella eseté-
ben ez a hatás csekély mértékű.
Ezek a fajok elsősorban a gli-
kolízisből nyerik a szükséges
energiát, így a táplálék szénhid-
ráttartalma döntő tényező szá-
mukra. Képesek nagy mennyi-
ségben tárolni a glikogént, hogy
a lebontó folyamatokhoz min-
dig rendelkezésre álljon a szük-
séges alapanyag. A fonálférgek
csak kis hányadát képesek hasz-
nosítani a táplálék energiatartal-
mának, ami megmagyarázza fo-
kozott táplálékfelvételi igényü-
ket.
A fonálférgeknek nincs elkü-
lönült keringési rendszerük. A testüregfolyadék igen sok szervetlen és szerves vegyületet
tartalmaz. A test mozgása révén, a folyadék áramlása eljuttatja azokat a test minden
részébe. A sejtek számára lehetővé válik a szükséges anyagok felvétele, vagy a feleslegesek
leadása. Az Ascaris fajok testüregfolyadéka anyagcseretermékeket, például valeriánsavat
tartalmaz.
A fonálférgek kiválasztó működése sokrétűen zajlik. Az elkülönült kiválasztószerv
más áltestüreges (pseudocoelomata) állatokhozhasonlóan elsődlegesen a szervezet vízház-
tartását és kisebb mértékben a bomlástermékek eltávolítását szolgálj a. A vízben élő fajoknál
a garattájék mögött, ventrálisan speciális mirigysejtek, úgynevezett renettsejtek vannak
(11.5. ábra). Ezek a sejtek a testüregfolyadékból felveszik a bomlástermékeket, majd a
hasoldali kivezető póruson át adják le a külvilágba. Parazita fajoknál e sejtek összeolvadása
révén egy vagy két hosszú csőrendszer alakult ki. Ha a kivá-lasztószerv páros, akkor
csatornái a test két oldalán lévő hipodermiszlécekben fúrnak, majd a hasoldali közös
kivezető járatban egyesülnek. A legtöbb fonálféreg nitrogéntartalmú bomlásterméke az
ammónia és a karbamid. Az Ascaris fajok például a teljes nitrogénmennyiség 52%-át
képesek karbamid formájában leadni akkor, ha kevés viz áll rendelkezésükre. Normális
körülmények között a karbamidürítés mindössze 7%-os mértéket ér el. A kiválasztás
módjának ilyen megváltoztatása nagyfokú alkalmazkodást tesz lehetővé, különösen olyan
környezetben, ahol a víztartalom szélsőséges határok között változhat, mint például
kiszáradó talajokban és elhalt szerves anyagban. A re-nettsejtek felépítése azt jelzi, hogy
kisebb mértékű szerepük lehet a nitrogéntartalmú bomlástermékek eltávolításában is. A
fonálférgek kutikulája mindkét irányban átjárható a víz és a benne oldott sók számára. Igen
sok fonálféreg jól alkalmazkodott a szél-sőséges sótartalomhoz is. Az ozmoreguláció
részben az epidermiszsejtek tevékenységének tudható be. A legtöbb viz a bélcsatomán
keresztül távozik, de a kiválasztó szerv-rendszer működése is felgyorsul, ha a környezet
sótartalmának megváltozása miatt nagy mennyiségű vizet kell a szervezetnek leadni.
Idegrendszerük kevés, meghatározott számú idegsejtből áll. A garat körüli ideggyű-
rűből a feji vég felé rövidebb idegek erednek, míg a test vége felé legalább két, jól fejlett,
hosszanti törzs indul ki, a háti és a hasi hipodermiszlécben húzódva. Hosszanti idegágak
előfordulhatnak az oldalsó lécekben és a hipodermisz más területein is. A hosszanti törzsek
között aszimmetrikusan elhelyezkedő harántágak (comissura) létesítenek kapcsolatot. A
220
növekedés, vedlés, új kutikulaképzés és a metamorfózis folyamataiban fontos szerepet
játszanak a garatideggyűrűben lévő neuroendokrin sejtek által termelt hormonok.
Az érzékelő papillák és serték legsűrűbben a feji végen fordulnak elő, de a papillák
száma a farki végen is számottevő. Szerepük elsősorban a mechanikai hatások érzékelése.
A fej két szélén bemélyedések, úgynevezett oldalszervek (amphidium) vannak, ame-
lyekben módosult csilló kkal bíró érzékelősejtek ülnek. Az amfidiumokkemoreceptorként
működnek, és a garatideggyűrűvel állnak kapcsolatban. Az üregben lévő folyadék
kicserélődését az ide kapcsolódó mirigy váladékának folyamatos termelődése biztosítja.
Az állatokban élő parazitáknál az oldalszervek általában redukálódtak. A Secementia
osztályba tartozó legtöbb parazita a farki végén hasonló struktúrájú és funkciójú faz-
midiumokkal (phasmidium) rendelkezik.
A legtöbb fonálféreg váltivarú. A női ivarkészülék a többnyire páros vagy ritkábban
(Trichinella, Capíllaria^ Trichuris) páratlan, igen vékony, feltekeredett petefészekkel kez-
dődik. Az ivarsejtek a petefészek középső, sejtmag nélküli plazmatengelyéhez (rachis)
kapcsolódva fejlődnek. A petevezető egy szakasza ondótartállyá módosulhat. A peteveze-
tők a tágultabb üregű méhben (uterus) folytatódnak, majd egy közös, erősen izmolt, rövid
hüvely (vagina) következik. Ez a hasoldal középvonalában lévő önálló ivamyíláson (vulva)
keresztül nyílik a külvilágra. Az uterus falát vékony izomréteggel határolt laphámsejtek
alkotj ák. A hímek a nőstényeknél kisebbek. A hím ivarszerv a páratlan vagy páros heréből
és folytatásaként az ondóvezetőből (ductus deferens) áll, amelyen ondóhólyag (vesicula
seminalis) lehet (11.4. ábra). Ezután még egy izmos szakasz, az ondókilövellő cső (ductus
ejaculatorius) található. Végső szakaszához járulékos mirigyek csadakozhatnak. A hímek
ivarvezetéke a végbéllel közös nyíláson át kapcsolódik a külvilágba. A hímek farki végén
gyakran párzótáska (bursa copulatrix) fordul elő, amely izomelemeket tartalmazó, suga-
rakkal merevíthető kutikularedőzet. Szerepe a párzótárshoz való kapcsolódásban van.
Ehhez a ragadós váladékot termelő mirigyek is hozzájárulnak. A hímek a farki végükön
általában páros, ritkábban páratian párzótüskét (spiculum) viselnek (11.4. ábra). A
hajlott, izmokkal mozgatható párzótüskék a párzás alkalmával elősegítik a férgek egymás-
hoz való kapcsolódását és a spermiumok bejuttatását a női ivamyílásba. A szpikulumot egy
vezetőpálca (gubemaculum) támaszthatja. Aburza, aszpikulumésagubemákulumalakja
fontos lehet a fajok elkülönítésében. A hímek farki vége az Ascaris fajoknál élesen felhajló.
A megtermékenyítés belső. A párzásra készülő egyedek egymásra találásában elsősorban
a kemoreceptoroknak van szerepük. A párzás általában rövid ideig tart, az amőboid
mozgásra képes spermiumok átpréselődnek a női ivarjáratba, ahol az ondótartályban
raktározódhatnak. A megtermékenyítést követően a petesejtek körül védőburok jön létre.
A védőburok felszíni rétegének képzésében az uterusz mirigysejtjei játszanak szerepet.
Egyes fajok peteburkán kis nyílás van, ahol a lárva ki tud bújni. A megtermékenyített pete-
sejtek a testüregfolyadék nyomásának eredményeként az ivamyíláshoz kerülnek. Sok
parazita és néhány szabadon élő fonálféreg a petét lerakáskor rögzíti a szilárd aljzathoz. A
nőstények által termelt peték száma a fajtól függően eltérő lehet. Egyes fajok csak néhány
száz petét raknak, míg mások naponta akár több százezer petét is termelhetnek. Lehetnek
úgynevezett peterakók (ovipar), amikor zigóta állapotban rakják le az utódokat, vagy
ovoviviparok, amikor a peteburkon belül a fejlődés már egy bizonyos állapotig eljutott.
Vannak viviparok is. E fajok nőstényei már mozgásra képes lárvákat raknak le a külvilágra.
A zigóta fejlődését elsősorban a környezet hőmérséklete és nedvességtartalma befolyásolja.
A fonálférgek megtermékenyített petesejtjei módosult spirális barázdálódást mutatnak.
Embrionális fejlődésük rövid ideig tart. A lárvák nagymértékben hasonlítanak a kifejlett
állatokra. Az ivarszervek kialakulása csak a negyedik, utolsó vedlés előtt kezdődik meg. A
lárva általában négy vedlésen megy keresztül kifejlődéséig, de előfordul, hogy az első és a
második vedlés még a peteburkon belül végbemegy. A harmadik stádiumú lárvák
kutikulája már ellenáll az igen szélsőséges körülményeknek is. Számos faj petéi és bizonyos
221
fejlettségi állapotú lárvái hosszú ideig, évekig életképesek, fertőzőképesek maradhatnak.
Egyes talajlakó hímnős fonálférgek önmegtermékenyítéssel vagy mások nőstényei
partenogenetikus úton szaporodhatnak, mivel hímjeik rendkívül ritkák vagy ismeretlenek.
A parazita fonálférgek az evolúció során számos tulajdonságban adaptálódtak speciális
életmódjukhoz. Ezek közé tartozik a magas szaporodási ráta, speciális életciklusuk, amely
lehetővé teszi, hogy fejlődésük során egyik gazdaszervezetből átkerüljenek a másikba.
Továbbá jellemző vonásuk a gazdaállat emésztőnedveinek is ellenálló kutikula, valamint
a környezeti feltételekkel szemben igen ellenálló pete, és a ciszta kialakítására képes lárva.
Életciklusaik nem olyan bonyolultak, mint a laposférgeké, mert általában csak egy, leg-
feljebb két gazdaállatban fordulnak elő. A szabadon élő fonálférgek életciklusa még
egyszerűbb, itt gazdaszervezetekre általában nincs szükség.
A fonálférgek egy része képes arra, hogy kedvezőtlen környezeti feltételek esetén -
elvesztve teste víztartalmának legnagyobb részét - összezsugorodjon, és életműködéseit
szüneteltetve hosszú évekig megtartsa életképességét. Ez az anabiózis jelensége, amely
kozmopolita elterjedésüknek is egyik magyarázata.
Törzs: Fonálférgek (Nematoda)
Osztály: Felcsavarodott oldalszervűek (Torquentia)
Osztály: Pórusszervű oldalszervűek (Secernentia) 11.1. fejezet
Rend: Szabadonélő fonálférgek (Rhabditida) 11.1.1. fejezet
Rend: Burzás fonálférgek (Strongylida) 11.1.2. fejezet
Család: Gyomor- és bél-fonálférgek (Trichostrongylidae)
Család: Nagy tüdőférgek (Dictyocaulidae)
Család: Strongylidák (Strongylidae)
Rend: Kevésajkú fonálférgek (Ascaridida) 11.1.3. fejezet
Család: Hegyesfarkú férgek (Oxyuridae)
Család: Orsóférgek (Ascarididae)
Család: Lebbencsajkú férgek (Toxocaridae)
Rend: Szájszuronyos fonálférgek (Tylenchida) 11.1.4. fejezet
Család: Levélfonálférgek (Aphelenchoididae)
Család: Szívogató fonálférgek (Anguinidae)
Család: Gyökérgubacsképző fonálférgek (Meloidogynidae)
Család: Cisztaképző fonálférgek (Heteroderidae)
Osztály: Betüremkedett oldalszerűvek (Penetrantia) 11.2. fejezet
Rend: Hordószájú fonálférgek (Mononchida) 11.2.1. fejezet
Rend: Tüskefogú fonálférgek (Dorylaimida) 11.2.2. fejezet
Rend: Páratlan ivarszervű fonálférgek (Trichocephalida) 11.2.3.fejezet
Család: Ostorférgek (Trichuridae)
Család: Hajszálférgek (Capillaridae)
Család: Trihinellák (Trichinellidae)
Rend: Raktárbelű fonálférgek (Mermithida) 11.2.4. fejezet
Megjegyzés: A fonálférgek rendszertana, részben az evolúciójuk ismeretének hiányossága
miatt, nem nevezhető lezártnak. Jelen könyvben egy viszonylag egyszerűsített felosztást követünk
Maggenti (1981) és Andrássy (1988) nyomán.
A felcsavarodott oldalszervűek (Torquentia) osztály a ma élő fonálférgek legősibb csoportja
oldalszervük felületi elhelyezkedésű, felcsavarodott képlet. Feji és testi sertéik egyaránt vannak.
Szájüregük változatos alakulásé, szájszuronyuk nincs. A nyelőcső általában bulbuszban végződik.
222
A kiválasztószerv egyetlen, ventrális helyzetű sejtből áll. Fazmidiumaiknincsenek. Farokmirigyeik
vannak. A hímeknek bursája nincs, a gubemákulum megtalálható. Kizárólag szabadon élők, főleg
a tengerekben, kisebbrészt az édesvizekben és a talajban fordulnak elő.
11.1. Pórusszerű-oldalszervűek (Secernentía)
A Secernentía osztály képviselői igen egységes testfelépítést mutatnak. A feji és testi
serték hiányoznak. Az amfidium általában kicsi, pórusszerű. A fazmidiumok általában
előfordulnak, a farokmirigyek viszont hiányoznak. A fajok egy részénél kiölthető száj-
szurony van. A nyelőcsövön egy vagy két bulbusz általában megtalálható. A kiválasztó-
szerv a páros laterális csatornákból és a közös kivezető járatból áll. A cölomaciták száma
kevés, általában csak 4-6 darab. A hímek heréi közül csak az anterior van meg, tehát
páratlan szerv, a gubernákulum megtalálható és gyakran burza is előfordul. Többségük
szárazföldön él, kevés fajuk fordul elő édesvízben. Vannak köztük szabadon élők, vala-
mint obiigát növényi nedvet szívó és állati paraziták is.
11.1.1. Szabadonélő fonálférgek (Rhabditida)
A szájüreg változatos felépítésű lehet. A nyelőcsövükön egy vagy két bulbusz található.
Az amfidiális szerv kicsi, zsebszerű. A szájszurony hiányzik. A fazmidiumok mindig
megtalálhatóak. A hímeknél burza előfordulhat. A fajok többsége szárazföldi, csak kevés
él édesvízben.
Ide tartoznak a Rhabditis fajok, melyek főleg ürülékben, elhalt szerves anyagban élnek. Terje-
désükben a rovarok játszhatnak szerepet.
Az ecetféreg (Turbatrix aceti) bomló, rothadó szerves anyagban, erjedő folyadékokban, állott
és nem tisztán kezelt ecetben él. Elsődlegesen baktériumokkal táplálkozik. Elevenszülő,
testhosszúsága 1,2-2,3 mm közötti.
A Panagrellus redivivus szintén elevenszülő. Erjedő anyagokban, például a földre hullott gyü-
mölcsben fordulhat elő nagyobb mennyiségben. Testének hossza egy milliméter körüli.
11.1.2. Burzás fonálférgek (Strongylida)
A szájnyílásuk körül ajkak nem találhatók, viszont szájtokvan. Nyelőcsövükön bulbuszok
nincsenek. A nőstényeknek általában a pete lerakását segítő nyúlványuk van az ivar-
nyílásnál. A hímeknek páros szpikuluma és általában burzája is van, utóbbit izmos
sugarak támasztják.
Gyomor- és bél-fonálférgek (Trichostrongylidae)
A Haemonchus contortus a kérődző állatok 20-30 mm hosszú, ovovivipar gyomorférge. Hazánkban
leggyakrabban a juhokban fordul elő, de más vadon élő és tenyésztett kérődzőben is megtalálható.
A himek burzája két jól fejlett oldalsó és egy apró háti lebenyből áll. A nőstények ivamyílását
vulvafedő takarja. A lárvák és a kifejlett állatok is vérszívók. Erős fertőzöttség esetén az állatok
lesoványodnak, jelentős lehet a vérveszteségük. A fertőződés a legelőn, a bélsárral kikerült és fűre
tapadt peték felvételével következik be.
Nagy tüdőférgek (Dictyocaulidae)
A Dictyocaulus fajok a gazdaállat légcsövében, tüdejében élősködnek. Erős fertőzés
esetén komoly légúti károsodást, légzési nehézséget és allergiás tüneteket okoznak,
amely akár az állat elhullásával is járhat.
223
11.6. ábra. Dictyocaulus filaria hímjének burzája. A: oldal-
nézetben, B: a hasoldal felől
bt: burzatámasztó pálca, bu: burza, pt: párzótüske (Gerichter
nyomán)
Az ovovivipar Dictyocaulus filaria a ju-
hók és kecskék légcső- és tüdőférgességét
okozza (11.6. ábra). T esthossza 30-100mm
közötti. A talajra kikerült 1. stádiumé lár-
vák kétszer vedlenek, így kialakul a 3. stá-
diumé lárva. Ez kettős védőburokban vé-
szeli át a kedvezőtlen körülményeket, majd
a legelő állat felveheti. A bélcsatomában e
fertőzöképes lárva elhagyja a burkot és a nyi-
rokerekbe behatolva eljut a nyirokcsomók-
ba. Itt vedlik, így kialakul a 4. stádiumé
lárva. Ez eljut a légutakba és ott kifejlődik.
A Dictyocaulus viviparus a páros-
ujjú patások (leggyakrabban a szar-
vasmarhák, szarvasok, őzek) tüdőfér-
gességét okozza. Testhosszúsága az
előző fajhoz hasonló, 35-80 mm közötti. Viviparfaj, amelynek fejlődése az előző fajéhoz
hasonlóan megy végbe.
Strongylidák (Strongylidae)
A Strongylus vulgáris a páratlanujjú patások vastagbélférgességét okozó ovovivi-
par faj. A nőstények 20-22, a hímek 14-15 mm hosszúak. Köztigazda nélkül
fejlődnek. A szabadba kijutott petékből kifejlődő, kétszeri vedlés után létrejövő
harmadik stádiumé lárvák a fertőzőképesek. A táplálékkal a bélcsatomába
bekerülő lárvák a vastagbélben újra vedlenek. A negyedik stádiumé lárvák a
bélfodor lemezei közé, majd a véráramba kerülnek. Visszajutnak a vastagbélbe,
itt újra vedlenek. Kifejlődésüket követően párosodnak, lerakják a petéiket. A
bélhámsejteken való megkapaszkodást a szájtokban lévő fogszeru képletek
biztosítják. Hámsejtekkel és vérrel táplálkoznak, ami komoly károsodást okoz
a gazda szervezetében, akár az állat elhullását is okozhatja.
A Syngatnus trachea a madarak légcsőférgességét okozza. A nőstények 15-
30, a hímek 2-6 mm hosszúak. A kifejlett hímek és nőstények állandó kopu-
lációban, a légcsőben megtelepedve élnek (11.7. ábra). A peték a felköhögött
váladékkal közvetlenül, vagy lenyelve, a bélsárral kerülnek ki a külvilágra. A
peteburkon belül létrejön a harmadik stádiumú lárva, amit vagy a végleges
gazda, vagy a vivőgazdák, például a földigiliszták, csigák vesznek fel. A ma-
darak ezek elfogyasztásával is fertőződhetnek. A lárvák a bélcsatomából a
vérárammal eljutnak a tüdőbe, a légcsőbe. Hámsejtekkel és vérrel táplálkoz-
nak, gyulladást, légzési elégtelenséget okoznak.
11.7. ábra. Synga-
tnus trachea (Kotlán
nyomán)
11.1.3. Kevésajkú fonálférgek (Ascaridida)
Szájnyílásukat általában három, ritkábban hat ajak veszi körül. A páros amfidium pó-
russzerű. Nyelőcsövükön bulbusz előfordulhat. A kiválasztószervnek vannak laterális
helyzetű csatornái és közös kivezető járata, ezért alakja „H” formájú. Általában nincs
bur-zájuk. A gerincesek parazitái.
Hegyesfarkú férgek (Oxyuridae)
Az ajkak erősen redukálódtak vagy hiányoznak. Nyelőcsövükön a hátulsó bulbusz van
meg, az elülső hiányzik. A hímeknek páros vagy páratlan szpikuluma van, de erősen
redukálódhat is. A burza előfordul. Általában kétéltűek, hüllők, emlősök parazitái.
224
A hegyesfarkú bélgiliszta vagy cérnagiliszta (Enterobius vermicularis') az ember egyik leggya-
koribb férge, elsősorban a gyermekkorban. A kifejlett alakok a vastagbélben élnek. A nőstények 8-
13, a hímek 3-5 mm hosszúak. A nőstények ovoviviparok, petéiket a végbélnyílás közelébe rakják.
A faj köztigazda nélkül fejlődik. Az ember önmagát fertőzi meg például akkor, amikor a viszkető
érzést kiváltó férgek tartózkodási helyéhez nyúlva kezére, majd innen a szájába kerülnek a peték
vagy lárvák. Elsősorban a mérsékelt és a hideg égövi területeken gyakori parazita.
Orsóférgek (Ascarididae)
Kutikulájuk erős, felszínén gyűrűzött. A szájnyílás körül három ajak található. A nős-
tények ivarszerve páros, a hímeké páratlan. Számos gerinces állatfaj orsóférgességét
okozzák. A kifejlett férgek a bélcsatoma hámját károsítják, toxikus anyagokat, valamint
a gazdaállat emésztőenzimjeinek hatását gátló vegyületeket termelnek. A lárvák vándor-
lásuk során szöveti gyulladásokat okozhatnak.
Az Ascaris és Parascaris fajok kifejlett alakjai a bélcsatornában élnek. Megtermékenyített petéik
a bélsárral kerülnek ki a külvilágra. Ott a peteburkokon belül kialakulnak a fertőzőképes lárvák.
Bekerülve ugyanazon vagy más gazdaállat szájüregébe, majd bélcsatomájába, annak érrendszerén
keresztül eljutnak a májba, innen a tüdőbe, a légcsőbe. A lárvák vándorlásuk során szöveti káro-
sodást, gyulladásos folyamatokat okoznak. A kifejlett férgek a bélcsatomában károsítanak és sza-
porodnak. Az Ascaris és Parascaris fajok nőstényei naponta 50-100 ezer petét is rakhatnak. A
szabadba került peték igen ellenállóak, életképességüket hosszú évekig megtarthatják.
A sertés-orsóféreg (Ascaris suum) a sertésfélék parazitája (11.8. ábra). A kifejlett nőstények
20-30 cm, a hímek 15-20 cm hosszúak. A szabadba került, lárvát tartalmazó petét a gazdaállat a
szennyezett táplálékkal veszi fel. A peték a gilisztákbélcsatomájába is bejuthatnak, és a kiszabaduló
lárvák a testben hosszú ideig (évekig is) életképesek maradhatnak. A giliszták így vivőgazdák le-
hetnek. Az A. suum petéi az ember bélcsatomájába is bejuthatnak. A lárvák vándorolnak a szer-
vezetben és gyulladásos folyamatokat idéznek elő.
Az ember-orsóféreg (Ascaris lumbricoides) az ember parazitája. Morfológiája megegyezik a
sertés orsóférgével. Fejlődése köztigazda nélkül történik. A gazda a petéket tartalmazó táplálékkal
vagy ivóvízzel fertőződik.
A ló-orsóféreg (Parascaris equorum) a lovak parazitája. A kifejlett nőstények elérhetik a 40 cm-
es, a hímek a 28 cm-es testhosszúságot.
Amadár-orsóféreg (Ascaridia gaUi) a madarak parazitája. A kifejlett nőstények 8 cm, ahímek
5-7 cm hosszúak. Ennek a fajnak is köztigazda nélkül történik a fejlődése, de az előző fajoktól
eltérően a lárvák nem vándorolnak a gazda szervezetében, csak a bélcsatomában élnek. A bélsárral
11.8. ábra. Ascaridida fajok: A: Ascaris suum, B: Toxocara canis feji vége
aj: ajak, fv: feji vég, hf: hím Ascaris farki vége, nő: nőstény Ascaris, sz: oldalszárnyak (Kotlán, Mehlhom és Storer nyomán
kissé módosítva)
225
kikerülő peték a szabadban fejlődésnek indulnak. A második stádiumú lárvák a peteburkon belül
maradnak, így veszik fel azokat táplálkozásuk során a madarak. A megtelepedett férgek roncsolják
a bél falát, gyulladásos folyamatokat okoznak.
Lebbencsajkú férgek (Toxocaridae)
Feji végükön nyílhegy formát kialakító kutikuláris oldalszárnyak vannak. A nyelőcső
vége duzzadt.
A Toxocara canis a kutyaféléket (kutya, farkas, róka) és macskaféléket károsító ovipar faj (11.8.
ábra). A hímek 11, a nőstények 18 centiméter hosszúak is lehetnek. Főleg a vékonybélben telep-
szenek meg. A fertőződés a bélsárral ürülő peték révén történik. A féreg vivőgazdái rágcsálók le-
hetnek. Ez a faj az ember számára is veszélyes, mert a bélcsatomába bekerült, peteburkukat elhagyó
lárvák a szervezetben vándorolnak (akár az agyba is eljuthatnak), és súlyos károsodást okozhatnak.
11.1.4. Szájszuronyos fonálférgek (Tylenchida)
A szájüregben szájszurony van, melynek végén általában három gomb található. Az
amfidium kicsi. A nyelőcsövön egy vagy két bulbusz van. Egyes fajoknál a fazmidiumok
hiányoznak. A kiválasztószerv csatornája a testnek csak az egyik oldalán van meg. A pe-
tefészkeknem tekerednekfel, legfeljebb visszahajlanak. A hímek burzája ritkán hiányzik.
Az ide tartozó fajok többsége élő növények, illetve gombák sejtjeinek nedveivel táplál-
kozik. Egyes fajok rovarokban élnek.
Levélfonálférgek (Aphelenchoididae)
Karcsú, egy milliméternél kisebb, mozgékony állatok. A szájszuronyuk vékony, gombjai
gyengén fejlettek vagy hiányoznak. Valamennyi nyelőcsői mirigy a középbulbuszba csat-
lakozik. Kutikulájuk finoman gyűrűzött. A nőstények páratlan petefészke hosszú, nem haj-
lik vissza. A hímek burzája fejletlen vagy hiányzik. Vannak köztük rovarokban élősködők
és növénykárosítók is. Az utóbbiakhoz tartozó fajok levélkárosítók. A lárvák kívülről beha-
tolnak a levél szöveteibe, ahol kifejlődnek, szaporodnak. Évente több nemzedékük lehet.
A megtámadott levelek ráncosodnak, összesodródnak, részben vagy teljesen elszáradnak.
A gyakoribb fajok közé tartozik a szamócafonálféreg (Aphelenchoides fragariae), a rizsfonál-
féreg (Aphelenchoides bessey) és a krizantémfonálféreg (Aphelenchoides ritzemabosi).
Szívogató fonálférgek (Anguinidae)
A szájszurony kicsi, az alapj án lévő gombokis fejletlenek. Kutikulájuk többnyire finoman
gyűrűzött. Közép és végbulbuszuk egyaránt megtalálható. A női ivarkészülék páratlan.
A hímeknek van burzája. Többségük növénykárosító életmódú.
A búzafonálféreg (Anguina tritici) kozmopolita faj, hazánkban is előfordul. A nőstény 3-5, a
hím 2,0-2,5 mm hosszú. A termesztett növények közül elsősorban a búzát, zabot, rozsot támadja
meg. Évente egy nemzedéke alakul ki. A nedves talajba kerülő magban mozgóképessé válnak a
lárvák, amelyek a növény gyökerén, szárán keresztül a kalászban fejlődő magokba hatolnak be. Itt
zajlik le a kifejlett egyedek párzása, majd a nőstények lerakják petéiket. A kikelő lárvák vedlenek,
és a második stádiumban lévők anabiotikus állapotba kerülnek a beszáradó gabonaszemben. Ilyen
állapotban igen hosszú ideig, akár évtizedekig is megtarthatják életképességüket. Az idő előtt
beszáradó magvak jellegzetes formája alapján a golyóüszög elnevezést kapta ez a kártétel.
A szárféreg (Ditylenchus dipsaci) számos tápnövényben (lucerna, hagymások, zab) megtalálja
életfeltételeit. A nőstény 1,0-2,2, a hím 1,0-1,9 mm hosszúságú. Évente több nemzedéke van. A
talajban vagy a növényi maradványokban telel át lárva alakban. Amikor a körülmények kedvezővé
válnak, akkor a lárvák megtámadják a növényeket, és a szárban vagy a levélben megtelepedve
torzulásokat okoznak. A nőstények petéiket a növényekbe rakják le.
Hasonlóan veszélyes kártevő a gumórontó fonálféreg (Ditylenchus destructor), amely főleg a
burgonya és a komló kártevője. Évente több nemzedéke is lehet.
226
Gyökérgubacsképző fonálférgek
(Meloidogynidae)
Az ide tartozó fonálférgek nőstényeinek teste
érett állapotban megduzzadva körte vagy gömb
alakú petezsákká válik (11.9. ábra). A hímek
megnyúlt testűek maradnak. Színük fehér, sár-
gás. Testük mindig a gyökér belsejében marad,
körülöttük a növény gubacsot képez. Kutikulá-
juk lágy marad, nem keményedik meg úgy, mint
a Heteroderidae család tagjainál. Petéiket kocso-
nyás anyagba ágyazva a talajba adják le. A ku-
tikula csak a hímeknél és a lárváknál gyűrűzött.
A lárvák és a hímek szájszuronya jól fejlett. A
hímeknek jól fejlett szpikulumai vannak, de
burzájuk nincs. A Meloidogyne fajok életciklusa
11.9. ábra. Meloidogyne sp. kifejlett nősténye
(Maggenti nyomán kissé módosítva)
a Heterodera fajokétól főleg abban tér el, hogy a nőstények a megtermékenyítést követően
nem pusztulnak el rövid időn belül, hanem folyamatosan képeznek petéket. Szaporodá-
suk hímek hiányában parthenogenetikus módon történhet. Testhosszúságuk általában
egy milliméter alatt marad.
Agyökérgubacsképzőkközül ismertebb a szabadföldi gubacsfonálféreg (Meloidogyne hapla),
melynek több száz tápnövénye ismert. Leggyakrabban a zöldségféléket és a szőlőt károsítja.
A kertészeti gubacsfonálféreg (Meloidogyne incognita) üvegházakban és szántófoldön egyaránt
előforduló polifág kártevő. Leggyakrabban a zöldség- és dísznövényféléken károsít.
Cisztaképző fonálférgek (Heteroderidae)
A Heterodera fajok lárvái kezdetben a gazdanövény
gyökérsejtjeit szívogatják, majd a hímek és a nőstények
további fejlődése, életmódja eltérő lesz. A nőstények
teste fokozatosan megrövidül, kezdetben tömlőszerű,
majd egyre gömbölydedebb lesz. Duzzadó testük fel-
repeszti a gyökér szöveteit, majd nagyobb része a nö-
vényen kívülre kerül, csak a feji végével kapaszkodik a
gyökérhez. A hímek az utolsó vedléskor nem hagyják
el a kutikulaburkot, hanem azon belül fejlődnek to-
vább, válnak ivaréretté. Ezt követően elhagyj ák a tokot,
és újból táplálkozni kezdenek. Felkeresik a nősténye-
ket, megtermékenyítik azokat, majd elpusztulnak. A
nőstények ezután jelentős átalakuláson mennek ke-
resztül. Néhány pete kezdeti lerakása után (11.10. áb-
ra) a belső szerveik fokozatosan felszívódnak, kutiku-
lájuk megkeményedik, erős védőtokot, fajra jellemző
alakú cisztát képeznek. Közben a megtermékenyített
petesejtek fejlődésnek indulnak, a lárvák a tokon belül
kikelnek. A cisztában hosszú ideig megtartják életké-
pességüket, majd kedvező feltételek esetén elhagyják
azt.
A répafonálféreg (Heterodera schachtii) egész Európá-
ban előfordul, így hazánkban is (11.10. ábra). Átlagos test-
hosszúsága egy milliméter alatt marad. A megtámadott nö-
vény a tápanyagfelvétel növelése érdekében járulékos gyöke-
reket fejleszt, a répagyökér „szakállassá” válik. Évente két
11.10. ábra. A répafonálféreg (He-
terodera schachti) kifejlett nősténye az
első néhány pete lerakását (A) követően
cisztává (B) alakul át (Thome nyomán
kissé módosítva)
227
nemzedékük lehet. A lárvák a talajban mélyre lehúzódva telelnek át, anabiotikus állapotban sokáig
fertőzőképesek maradnak. A lárvák károsítása miatt a cukorrépa gyengén fejlődik, jelentős lehet
a terméskiesés.
A közönséges burgonyafonálféreg (dobodéra rostochiensis) a burgonyafélék családjába
tartozó növények gyökerét szívogatja. Komoly károkat okoz a termesztett fajoknál. A faj elterjedt
egész Európában. Ahímek testhosszúsága meghaladja az egy millimétert, míg a nőstényeké ez alatt
marad.
11.2. Betüremkedett oldalszervűek (Penetrantia)
A Penetrantia osztályba tartozó fajok amfidiumai változatos felépítésűek, üregük a
testfelszín alá hatol. A szájüreg alakulása sokféle lehet. Egyes fajoknak kiölthető száj-
szuronyuk van. A nyelőcsövön bulbusz nincs. Fazmidiumuk nincs és a farokmirigyek is
hiányozhatnak. A kiválasztó csatornák csökevényesek vagy hiányoznak. Kutikulájuk fel-
színe általában sima. A hímeknek nincs burájuk. Heréjük általában páros, a guberná-
kulum többnyire hiányzik. Az ide tartozó fajok többsége szabadon élő, de növényi
táplálkozású és állatokban élősködő paraziták is vannak köztük.
11.2.1. Hordószájú fonálférgek (Mononchida)
Fejsertéik hiányoznak. Tágas szájüregük kutikulája vastag, rajta egy vagy több fog ta-
lálható. Valamennyi faj ragadozó. Kis testű, mezőgazdasági szempontból káros fonálfér-
geket is fogyasztanak, így szerepük lehet a biológiai növényvédelemben. Talajban és
édesvízben fordulnak elő, mozgékonyak. A hímeknek szpikuluma van, de burzája nincs.
A farokrészen ragadós váladékot termelő mirigy lehet. Ennek váladéka az állat rögzülését
segíti.
Gyakori fajok közé tartozik a ClarkuspapiUatus. Testhosszúsága 2,5 mm alatt marad. Talajban,
mohapámákban él.
11.2.2. Tüskefogú fonálférgek (Dorylaimida)
Fejsertéik hiányoznak. Kiölthető, tüskeszerű foguk van. Bélcsatomájukban nincsenek
izomsejtek, így bulbuszok sem figyelhetők meg. Nincsenek farokmirigyeik. A hímek
szpikuluma megtalálható, de a burza hiányzik.
A Dorylaimus fajok talajban, édesvizekben és félsós vízben fordulnak elő. A Dorylaimus
stagnalis a hazai édesvizekben gyakori.
Az óriási tűfonálféreg (Paralongidorus maximus) a talajban élő fonálférgek óriása, mert
testhossza eléri az 1,0-1,2 centimétert is. Külső élősködő. Főleg a fás szárú növények, de számos
haszonnövény gyökérzetét is károsítja.
A Longidorus tűfonálféregfajok kártétele elsősorban abban nyilvánul meg, hogy a növényeket
vírusokkal fertőzik meg. Gyakoribb a vírusoltó tűfonálféreg (Longidorus elongatus), az európai
bökőfonálféreg (Trichodorusprimitivus) és a szőlőszívó tűfonálféreg (Xiphinema index).
11.2.3. Páratlan ivarszervű fonálférgek (Trichocephalida)
Lárvaállapotban kiölthető szájszuronyuk van, ami a kifejletteknél hiányzik. A nőstények-
nek és a hímeknek egyaránt páratlan ivarszerve van, és különlegessége az, hogy a ger-
minális zóna az ivarjárat teljes hosszára kiterjed. A hímek szpikuluma páratlan, vagy
hiányzik.
228
Ostorférgek (Trichuridae)
A Trichuris fajok testének elülső kéthar-
mada hajszálvékony, míg a hátulsó har-
mad jóval vastagabb. Ovipar fajok, me-
lyek fejlődése közvetlen, köztigazda nél-
küli. A fertőződés a petéket tartalmazó
bélsárral szennyezett táplálékkal törté-
nik. Súlyos esetben gyulladásos folya-
matok jelentkeznek a bélben, amelyek
halálos kimenetelűek is lehetnek. Szá-
mos emlősfajnak és az embernek is van
specifikus ostorféreg-parazitája.
Az ostorféreg (Trichuris trichtura) 3-5
11.11. ábra. Az ostorféreg (Trichuris trichiura).
a: kifejlett nőstény, b: him (Jeffrey és Leach nyomán kissé módosítva)
cm hosszú, a sertés és az ember vakbelében és vastagbelében élősködik (11.11. ábra).
Hajszálférgek (Capillaridae)
A Capillaria fajok a gerinces állatok, főként madarak és emlősök élősködői. Leginkább a májban
és a bélcsatornában károsítanak, a hámfelületet teszik tönkre. A férgek hajszálvékonyak, néhány
centiméter hosszúak. Valamennyi faj ovipar. A szabadba kikerült peték embrionális fejlődése a
talajon zajlik. Az egyes fajok posztembrionális fejlődése eltérő módon megy végbe. A fertőződés
történhet az első stádiumú lárvák közvetlen felvételével vagy köztigazda (például földigiliszta)
közbeiktatásával.
Trichinellák (Trichinellidae)
A trichinella (Trichinella spirális) kifej-
lett alakjai 1,5-4,0 mm-es testhosszúsá-
gúak, főleg a rágcsálók, disznófélék és
ragadozók bélcsatomájában fordulnak elő
(béltrichinella). A nőstények viviparok. A
lárvák a bélfal ereibe fúrják be magukat, és
eljutnak az izomzatba. Itt megtelepszenek
(izomtrichinella), körülöttük meszes tok
jön létre, amelyben igen hosszú ideig, akár
több mint öt évig is életben maradnak
(11.12. ábra).
Az ember a fertőzött, kellően meg nem
főzött vagy sütött vaddisznó és házisertés
húsának elfogyasztásával válhat a féreg
11.12. ábra. Trichinella spirális betokozódott lárvái az izom-
zatban (Brown, Mehlhom nyomán kissé módosítva)
gazdájává. A gyomorsavak hatására a meszes tok feloldódik, a lárvák kiszabadulnak, és a bélben rövid
idő alatt szaporodóképessé válnak.
11.2.4. Raktárbelű fonálférgek (Mermithida)
Hosszú, igen karcsú állatok. A szájszurony csak a lárváknál van meg. A kifejlett alakok
bélcsatornája degenerálódik, a nyelőcső a bélből kialakult raktározószervbe nyílik. Az
ivarszervek általában mindkét ivarnál párosak.
A Mermis fajok lárvái szárazföldi vagy vízirovarokban és rákokban élősködnek, míg a kifejlettek
szabadon élnek és nem táplálkoznak. A Mermis nigrescens nősténye petéit a füfélék levelére rakja,
ahol azt a táplálkozó rovarok, főleg az egyenesszárnyúak veszik fel. Ennél a fajnál a kifejlett nős-
tény testhosszúsága elérheti a tíz centimétert is.
229
Ellenőrző kérdések all. fejezet témaköreiből
1. Melyek azok az elsődleges tulajdonságok, amelyek csak a fonálférgekre jellemzőek?
2. Sorolja fel a fonálférgek óriási alkalmazkodó képességének néhány példáját!
3. Miért különleges felépítésű a fonálférgek izomzata?
4. Jellemezze a fonálférgek testfelépítését!
5. Jellemezze a pórusszerű oldalszervűek (Secementia) osztályába tartozó fajok biológiáját és
mezőgazdasági jelentőségét!
6. Jellemezze a betüremkedett oldalszervűek (Penetrantia) osztályába tartozó fajok biológiá-
ját és mezőgazdasági jelentőségét!
230
12. KEREKESFERGEK (ROTATORIA)
A kerekesférgek nevüket a fejükön található kerékszervről (rota) kapták, amely a mozgás és a
táplálkozás szolgálatában áll. Földünkön körülbelül 1800, hazánkban mintegy 600fajuk ismert.
A kerekesférgek az egész Földön elterjedtek. Többségük az édesvízi életterekben él, de a
tengerekben is előfordulnak Megtalálhatók a nedves talajban, avarban, a mohapárnákban, a
fák odvának vizterében is. A fajok többsége szabadon élő, mások aljzathoz rögzültek, valódi
paraziták ritkán találhatók közöttük. Kedvező életfeltételek között gyakran igen nagy faj- és
egyedszámban elszaporodhatnak. Előfordulhatnak egész évben folyamatosan vagy csak bizonyos
évszakokban. Viszonylag könnyen terjednek, mert a beszáradt egyedeket, a védőburokkal kö-
rülvett petéket a szél és az állatok széthurcolhatják, és ha az új élettérbe^ a körülmények kedve-
zőek, ott gyorsan elszaporodnak. Változatos élőhelyeiken elhalt szerves anyaggal, algákkal,
gombákkal, baktériumokkal, kisebb egysejtűekkel táplálkoznak, ami nagymértékben hozzájá-
rulhat a vizek természetes tisztulásához. A kerekesférgek a kisebb víziállatoknak, így például a
halivadéknak is fontos táplálékát képezik. Az ivadéknevelő halgazdaságok a tavakat megfelelő
előkészítéssel, trágyázással mintegy „kerekesféreg-tenyészetté” változtatják a halivadék kihe-
lyezésének kezdeti időszakában. így biztosítják az ivadék számára a jó minőségű táplálékot.
A kerekesférgek a legkisebb méretű többsejtűek. A legtöbb faj testhossza 100 és 500 mikrométer
közé esik, de vannak ettől kisebbek (30-40 pm) és nagyobbak (2 mm-ig) is. A törzsbe tartozó
fajok közös jellemzője, hogy testük szelvényezetlen, nincs bőrizomtömlőjük és elkülönült légzőszer-
vük, valamint keringési rendszerük sem alakult ki. Jellegzetes tulajdonságuk az alakváltozás. A
tes-tüket felépítő sejtek száma állandó. Bélcsatomájuk háromszakaszos, de számos fajnál ettől
jelentős eltérések lehetnek. Kiválasztószervük elővesécske típusú. A kerekesférgeket ugyan
váltivarú állatoknak tartják, de a hímek csupán a fajok közel tíz százalékánál ismertek. Szinte
mindig csak a nőstényekkel találkozhatunk ott is, ahol a hímek előfordulnak, mert a hímek
többnyire csak a szaporodás rövid periódusában jelennek meg. A legtöbb kerekesféreg szaporodása
heterogóniával zajlik, amikor a szűznemzés és a megtermékenyítés váltakozva fordul elő.
Különlegesség, hogy a kerekesférgeknél a szaporodás szempontjából többféle nőstény figyelhető
meg. A hímek testfelépítése jelentősen eltér a nőstényekétől, így a következőkben sorra kerülő
alaktani és szervezettani jellemzés elsősorban a nőstényekre vontakozik.
A nagy fajszám és alakgazdagság azt jelentheti, hogy ez a csoport hosszú földtörténeti
múltra tekint vissza. Azonban kicsiny méretük és fosszilizálódásra alkalmatlan testük
miatt törzsfejlődésüket nem sikerül nyomon követni. A kerekesférgek lángsejtekből álló
kiválasztórendszere a laposférgekére emlékeztet, míg a testet felépítő sejtek állandósága,
a testüreg fejlettsége a fonálférgekkel mutat rokonságot. Néhány faj kifejlett egyedeinek
testalakja egyes puhatestűek és gyűrűsférgek trochophora lárvájára emlékeztet. Ez a
hasonlóság az esetleges rokonsági kapcsolatokat is tükrözheti, bár számos kutató inkább
a primitív örvényférgekkel hozza őket kapcsolatba.
Testükön általában elkülöníthető a háti és hasi oldal. Kétoldalian részarányos testük
alakja rendkívül változatos lehet. Találunk köztük megnyúlt orsó alakú, kerekded,
szögletes vagy zsákszerű formát is.
231
Kül takar ójuk (tegument) hámrétege igen vékony, sejtjei szinciciális réteget (hypo-
dermis) alkotnak. Ennek felszíni rétegébe ágyazódva átlátszó, keratinszerű réteg találha-
tó, amely egyes fajoknál jellegzetes páncélzatot (lorica), különböző nyúlványokat ké-
pez. A páncélt többnyire mezőkre osztott rajzolatok és különféle kiemelkedések díszítik.
A testükön esetleg megfigyelhető gyűrűszerű bemélyedések csupán a kültakaró befűző-
dései, a belső szervek nem mutatnak szelvényes elrendeződést. Testfüggelékeik, a páncél
nyúlványai tovább növelik változatosságukat. Jellegzetes tulajdonságuk az egymást
követő generációk során bekövetkező alakváltozás (cyclomorphosis) képessége, ame-
lyet elsősorban a víz fizikai-kémiai paramétereinek megváltozása idéz elő, ezért évszakos
alaki különbségek is megfigyelhetők. Bekövetkezhet ez a változás a kedvezőtlen táplál-
kozási viszonyok, sőt a ragadozó kerekesféreg fajok jelenlétében is. Ilyenkor a megnöve-
kedett hosszúságú testfüggelékek, a páncél nyúlványai többnyire megakadályozzák azt,
hogy a ragadozó elnyelje a kisebb testű zsákmányszervezetet. A páncél nélküli fajok
testfüggelékei elsősorban az úszás és lebegtetés szolgálatában állnak, mert a testhez
mozgékonyán kapcsolódó hosszú tüskék vagy lapátszerű nyúlványok hirtelen elmozdítva
az állat helyváltoztatását eredményezik.
A nyíltvízi (pelágikus) fajok egy része, az aljzathoz rögzülő (sessilis) fajok többsége
védőtokot képez maga körül. Ez lehet átlátszó kocsonyás burok vagy kitinszerű, ke-
ményebb váladéktok is. Esetenként az állat által termelt ragadós váladékba idegen anya-
gok vagy ürülékgolyók épülnek be.
Áltestüregüket (pseudocoeloma) színtelen folyadék tölti ki. Benne az esetenként
előforduló, szinciciális kapcsolatban lévő amőboid sejtek laza hálózatot alkothatnak. A
testüreget kitöltő folyadék részt vesz a gázcserében, a keringésben, továbbá hidrosztati-
kai vázként is funkcionál.
A fejen található a jellegzetes alakulású kerékszerv, ezenkívül a szájnyílás, az ér-
zékszervek egy része, mint pl. a látószervek és a tapogatók (12.1. ábra). Ez utóbbiak a test
más pontjain is előfordulnak.
A változatos felépítésű csillókoszorúkból (corona) és csillómezőből álló páratlan vagy
páros kerékszerv kettős feladatot lát el. Részben biztosítja az állat mozgását, részben
pedig elősegíti a táplálék szájnyílásba juttatását. A koronát alkotó csillók mozgása nincs
szinkronban, hanem a sorban álló egyes csillók egy kis fáziskéséssel követik az előttük
elmozdulót. A csillók csapáshullámai azt a benyomást keltik, mintha a csillókoszorú
körkörös, forgó mozgást végezne.
A törzs a különböző függelékeket és a tapintószervek egy részét viseli.
A fajok egy részénél előforduló láb általában gyűrűszerű ízekre tagolt, jól mozgatható.
Teleszkópszerűen visszahúzható a testbe, illetve a páncélzatba (12.1. ábra). Elsődleges
funkciója alapján megkülönböztetünk fajokra jellemző felépítésű araszoló-, úszó-, ugró-
és rögzítőlábat. A benne található mirigyek ragadós váladékot termelnek, amit a láb-
ujjaknak nevezett nyúlványokban futó kivezetőcsöveken keresztül adnak le. Ez a ragadós
anyag biztosítja az aljzathoz való tapadást. Egyes planktonikus fajok (például Asplanchna
spp., Keratella spp., Polyarthra spp.) lábai redukálódtak, vagy hiányoznak. A fajok egy
része úszik, mások a piócákra emlékeztető módon mozognak az aljzaton.
A testüket felépítő sejtek kicsik és számuk állandó. Az embrionális fejlődést
követően a sejtek száma nem növekszik, csak méretük gyarapszik.
A kerekesférgeknek nem bőrizomtömlőjük van, hanem testtájanként elkülönült
izomkötegeik vannak (12.1. ábra).
Mozgásuk sebessége a testméretüktől függ. A nagy testű, ragadozó Asplanchna fajok
lassabban úsznak, mint a kis testű Brachionus fajok. Megállapították, hogy egyes Bra-
chionus fajok teljes energiafelhasználásuk 62%-át fordítják a mozgásra.
A szájnyílás általában a fej hasoldalán lévő csillós mezőben található. Alegtöbb fajnál
a bélcsatoma belső felszínét teljes hosszában csillós hám borítja. A szájnyílás szájcsőben
232
12.1. ábra. Egy Monogononta kerekesféreg (Brachionus calyciflorus) testfelépítése
A: testtájanként elkülönült izomelemek, B: a nőstény szervezete, C: a hím csökevényes szervezete
aa: agydúc alatti mirigy, ad: agydúc, cs: kerékszerv csilléi, ev: elővesécske, gy: gyomor, he: here, hh: húgyhólyag, it: idegtörzs,
iz: izom, kb: középbél, ki: kloáka, Iá: láb, lm: lábmirigy, It: látószerv, lu: lábujjak, mi: gyomormirigy, pá: páncél, pe: pete,
pf: petefészek, pr: párzószerv, rá: rágógyomor, re: retrocerebrális szerv, rk: rágókészülék, sm: szikmirigy, sp: spermium, sz:
szájnyílás, ta: tapintószerv, tü: tüske, ub: utóbél (Beaucham, Donner, Voight nyomán módosítva)
233
folytatódik, amely a táplálékot a garatban elhelyezkedő speciális rágógyomorba (mas-
tax) juttatja (12.1. ábra). A masztaxban lévő fogak, lécek, pálcikák, lemezkék mérete és
alakja attól függ, hogy a faj milyen táplálkozásmódot folytat. A rágókészülék (trophi)
egyes elemeihez izmok kapcsolódnak, amelyek mozgatják azokat. Az algákat fogyasztó
fajoknál a zúzó, szívó, őrlő típusúak, míg ragadozó fajoknál a tépő vagy a szájnyíláson át
kitolható hegyes, harapófogóra emlékeztető fogó és csípő felépítésűek a jellemzőek. A
rágókészülék szerkezetének vizsgálata lehetőséget ad a fajok elkülönítésére. A rágógyo-
morban a táplálék apró részekre őrlődik, majd a gyomorba jut. A gyomorhoz emésztő-
nedvet termelő páros mirigy kapcsolódik. Több fajnál a bélcsatoma e szakasza egy sej-
tekre el nem különült citoplazmatömeg, amelynek közepén egy cső fut, míg más fajoknál
még ez a cső is hiányzik. Ezeknél a bekerülő táplálékrészecske lassan süllyed tovább, és
közben emésztődik. A bélcső a kloákában végződik, amely a testvégen, a hátoldal kö-
zépvonalában nyílik a külvilágra. Egyes fajok (pl. Asplanchna) gyomra vakon végződik,
ezért az emészthetetlen részek a szájnyíláson át távoznak.
Az úszó és araszoló kerekesférgek oxigénszükséglete nagy. Légzőszerv hiányában az
oxigén felvétele a kültakarón át történik. Előfordul, hogy igen kicsi oxigéntartalmú
vizekben is megtalálhatók. Ekkor a szükséges energiát anaerob lebontó folyamatok révén
nyerik. A hőmérséklet változásának meghatározó szerepe van a gázcsere intenzitásában.
A Brachionus fajoknál megfigyelték, hogy +20 és +28 °C között a metabolitikus folya-
matok rátája nem változik, míg ez alatt és felett jelentősen megemelkedik.
Kiválasztószervük a páros elővesécske (protonephridium) (12.1. ábra). A csatornák
közös kivezető járattal a húgyhólyagba, vagy annak hiányában közvetlenül a kloákába nyíl-
nak. Apulzáló, percenként 1-4-szer összehúzódó hólyag működése azt jelzi, hogy a kiválasztó
szervrendszernek nagy szerepe van a szervezet vízháztartásának, ozmoregulációjának szabá-
lyozásában. Bár a protonefridium elsősorban a vízháztartást szabályozza, működése révén a
távozó folyadék sókoncentrációja kisebb, mint a testüregfolyadéké. A kerekesférgek a nit-
rogéntartalmú anyagcsere-végtermékeiket a testfelületen át is képesek leadni.
A nőstények ivarszervei az araszoló kerekesférgeknél párosak, míg az egypetefész-
kűeknél páratlanok. A kloákába nyíló, vékony hártyával burkolt ivarszerv (germovitel-
larium) felépítése különleges, mert mind a petefészek (germarium), mind a szikmirigy
(vitellarium) egy szinciciális sejthalmaz (12.1. ábra). A germáriumból leváló petesejtet
a szikmirigy szikdús plazmával gazdagítja, majd a pete a kloákán át távozik.
A Monogononta csoport hímjeinek ivarszerve a csökevényes testfelépítés ellenére is
terjedelmes méretű, csaknem az egész testüreget kitölti. A herében kialakuló spermiu-
mokhoz a prosztatamirigyek öntik váladékukat, amely az ondóvezetőn és ennek végén
esetenként kialakult cirrusz-zsákon keresztül jut el az ivamyílásig. Számos faj hímjének
van párzószerve. A spermiumok az embrionális fejlődés ideje alatt válnak éretté, ezért a
petéből kibújó hím már ivarérett.
A legtöbb faj peterakó (ovipar), de néhányuk elevenszülő (vivipar), mint például az
araszoló kerekesférgek közé tartozó Rotaria rotatoria, vagy az egypetefészkű Asplanchna
fajok. Az ovipar fajok petéiket a vízbe juttatják, vagy az aljzathoz rögzítik, míg a pelágikus
fajok gyakran a fiatalok kikeléséig a testvégükön hordozzák azokat.
Egyes tengeri fajok kizárólag megtermékenyített petékkel, az araszoló kerekes-
férgek pedig (ahol a hímek ismeretlenek) mindig megtermékenyítés nélkül (parthe-
nogenetikus úton) szaporodnak. A legtöbb fajnál ez a két szaporodási típus a ciklusos
szaporodás során váltakozva fordul elő (heterogonia) (12.2. ábra).
Heterogonia esetében a parthenogenetikus szakaszban a diploid, úgynevezett amiktikus
nőstények megtermékenyítés nélkül rakjákle a „nyári” vagy amiktikus petéket. Ezek vékony
héjúak, nagy méretűek, és belőlük újabb amiktikus nőstények fejlődnek ki rövid idő alatt.
Mindaddig, amíg a víztér kedvező életfeltételeket nyújt, ilyen módon hoznak létre számos
generációt. A kedvezőtlen körülmények (pl. hőmérséklet-csökkenés, táplálékhiány) hatására
234
a petékből az amiktikus nőstényekkel általában azonos alakú (kivéve Asplanchna fajok)
diploid, miktikus nőstények fejlődnek ki. A kiváltó tényezők közül legfontosabb és legálta-
lánosabb a hőmérséklet csökkenése. Apopuláció nagy egyedsűrusége, a fellépő táplálékhiány
és egyes fajoknál még egyéb speciális környezeti tényezők bekövetkezése is erre az eredmény-
re vezet. A miktikus nőstények - a peték érése során lezajló meiózis miatt - haploid kromo-
szómaszámú petéket képeznek. Ezekből a petékből, amennyiben a miktikus nőstényben nem
termékenyülnek meg, hímek fejlődnek ki. A gyors mozgású hímek megtermékenyítik a hap-
loid petesejtet termelő, miktikus nőstényeket. Az amiktikus nőstényekkel a hímek nem pár-
zanak. Maga a párzás igen gyors lefolyású. Történhet a nőstény kloákanyílásán át, vagy a
páncél nélküli fajoknál a test más pontján (pl. nyaktájékon) is, mivel a hím párzószerve,
vagy az erre a célra szolgáló testvége átfúrja a kültakarót és így a hím ivarsejtek bejutnak
a test-üregbe. Ha a miktikus nőstény megtermékenyül, akkor úgynevezett tartós vagy
„téli” pete képződik. Ez már diploid, vastag védőburokkal ellátott pete, amelyik képes
átvészelni a kiszáradást, vagy a hideg időszakot. A kedvező körülmények beköszöntével
ezekből amiktikus nőstények fejlődnek ki, és újból elindul a parthenogenetikus szaporodási
szakasz (12.2. ábra).
A nyári petékből kibúvó fiatalok néhány óra vagy nap alatt befejezik növekedésüket.
A kerekesférgek életkora általában 10-20 nap.
Kétoldalian részarányos idegrendszerük egy központi kétlebenyű agyból és a belőle
kiinduló két ventrolaterális helyzetű hosszanti törzsből áll (12.1. ábra). Utóbbiak a test
különböző részein lévő dúcokkal állnak összeköttetésben. Ezek a dúcok látják el ide-
gekkel az egyes szerveket.
Érzékszerveik többsége a feji területen fordul elő. Ezek elsősorban mechanore-
ceptorok és kemoreceptorok, amelyek gyakran csillókkal állnak kapcsolatban. Tapoga-
tók találhatóak az állatok hátán és oldalán, a páncél nyílásainál és a lábon is. Látószervük
12.2. ábra. Monogononta kerekesférgek heterogénjával történő szaporodása
am: amiktikus nőstény, hí: hím, hp: hímpetés nőstény, kk: kedvezőtlen környezeti tényezők hatása, me: megtermékenyítés,
mi: miktikus nőstény, mm: miktikus, megtermékenyíthető nőstény, tn: tartós petés nőstény, tp: tartós pete (Több szerző
nyomán módosítva)
235
(szemfolt) egyszerű, de nem szokványos felépítésű, mert az egyes látósejteken belül meg-
található a rendszerint vörös pigmentszemcsék halmaza és a fénytörő, parányi, gömb
alakú lencse. Az 1 -5 darab szem elhelyezkedhet az agydúcon, a fej két oldalán és a fejtetőn
is (12.1. ábra).
Víztartalmukat fokozatosan elvesztve, anabiotikus állapotban képesek átvészelni a
kedvezőtlen időszakot.
Törzs: Kerekesférgek (Rotatoria)
Osztály: Araszoló kerekesférgek (Bdelloidea) 12.1. fejezet
Osztály: Egypetefészkűek (Monogononta) 12.2. fejezet
12.1. Araszoló kerekesférgek (Bdelloidea)
Az araszoló kerekesférgekre az összehúzódásra képes test, az úszás mellett az araszoló
mozgásmód, valamint a páros germovitellarium megléte jellemző. Általában két, jól
fejlett kerékszervük van. Lábukon gyakran kettőnél több ujj található, és a lábmirigyek
száma is több, mint kettő.
A hímek ennél a csoportnál ez ideig ismeretlenek, így feltételezhető, hogy szaporo-
dásuk mindig parthenogenetikus úton megy végbe.
Számos faj a szárazföldi nedves élőhelyeken, a talajban fordul elő. Legtöbbjük om-
nivora, így elsősorban szerves törmelékkel és mikroorganizmusokkal táplálkoznak. A
rágókészülékük főleg őrlő típusú. A vizekben és a talajban hozzájárulnak a szerves
anyagok lebontásához.
Különleges képességük, hogy testük víztartalmának jó részét fokozatosan elvesztve,
maguk körül idővel megkeményedő, kezdetben zselatinszerű tokot képeznek, miáltal
anabiotikus állapotba kerülnek. így igen hosszú ideig, hónapokig, sőt évekig is életben
maradhatnak.
Vizeinkben leggyakrabban előforduló két faj a 320-400 |tm testhosszúságú rózsaszínű fér-
gecske (Philodina roseola) (12.3. ábra) és a közönséges férgecske (Rotaria rotatoria).
12.2. Egypetefészkűek (Monogononta)
Az egypetefészkűek nőstényeinek germovitellariuma páratlan. Az alkalmanként meg-
jelenő, mindössze néhány napig élő hímek többé-kevésbé csökevényes testfelépítésűek.
Nem táplálkoznak, általában hiányzik a száj- és a végbélnyílásuk. Heréjük páratlan, nagy
tömegben termel spermiumokat. Igen gyorsan tudnak úszni, a petéből való kibújásukat
követően akár már egy órán belül képesek a miktikus nőstényekkel párzani.
Szaporodásuk tipikus heterogónia, tehát a parthenogenetikus szakaszt időnként fel-
váltja a megtermékenyítéssel járó folyamat. Fajoktól függően évente általában egy vagy
néhány miktikus generációjuk van. Egyes fajoknál a populációk igen kis részét (kevesebb,
mint 0,5%-át) amfoterikus nőstények alkotják, amelyek egyaránt képesek diploid és
haploid peték előállítására.
A fajok egy része aljzathoz rögzülve él. Ezek kerékszerve módosult felépítésű lehet,
esedeg gyengén fejlett vagy csillók nélküli. Egyes fajoknál a szájnyílás körül hosszú,
tűszerű szálak szolgálnak a táplálékszervezet csapdába ejtésére és a szájnyílásba való
terelésére. Több szesszilis faj védőtokot, csövet hoz létre maga körül. A zselatinszerű
alapanyag szilárdításához idegen részecskéket vagy saját ürüléküket használják fel.
236
A planktonikus, szabadon úszó fajok teste lehet egészen lágy, mérsékelten formatartó,
vagy merev páncél is boríthatja. Lábuk gyakran hiányzik.
A háromsertéjű férgecske (Filinia longiseta) planktonikus életmódú. Teste 70-140 pm hosz-
szú. Három hosszú, serteszerű nyúlványa van (12.3. ábra). A Collotheca omata helytülő faj, amely
kocsonyás, cső alakú házat képez (12.3. ábra). Feji vége erősen módosult, a rágókészülék tépő
típusú. Testhosszúsága 500-1500 pm közötti.
A továbbiakban bemutatásra kerülő fajok főleg szabadon úszók, amelyeknek a kerékszerve jól
fejlett. Testük alakja igen változatos (12.3. ábra).
12.3. ábra. Gyakoribb előfordulású kerekesférgek. A: rózsaszínű férgecske (Phylodina roseola), B: Collotheca
omata, C: háromsertéjű férgecske (Filinia longiseta), D: kardos férgecske (Polyarthra vulgáris), E: négytüskés
férgecske (Brachionus calyciflorus), F: kockaalakú férgecske (Keratella quadrata), G: zsákbelű férgecske
(Asplanchna priodonta) (Bancsi és más szerzők nyomán)
A gyakoribb fajok közé tartozik például a 250-300 pm-es négytüskés férgecske (Brachionus
calyciflorus). A zsákbelű férgecske (Asplanchnapriodonta) ragadozó, más kis testű kerekesférgekkel
is táplálkozik. Testhosszúsága 500-1500 pm lehet, ami jóval meghaladja az itt bemutatásra kerülő
fajok általában 100-500 pm közötti testméretét. A kardos férgecske (Polyarthra vulgáris) jellegzetes
evezőlapát-szerű testfüggelékei a lebegést, a mozgást segítik. A 150-230 pm-es kockaalakú fér-
gecske (Keratella quadrata) páncélja mezőkre osztott és finom mintázottsággal díszített.
Ellenőrző kérdések a 12. fejezet témaköreiből
1. Milyen képességei teszik lehetővé a kerekesférgeknek, hogy a legkülönbözőbb élőhelyeket
rövid idő alatt benépesítsék?
2. Jellemezze a kerekesférgek testfelépítését!
3. Ismertesse a kerekesférgek különleges szaporodásmódját!
4. Jellemezze a gyakoribb kerekesférgek biológiáját és mezőgazdasági jelentőségét!
237
13. BUZOGANYFEJUEK
(ACANTHOCEPHALA)
A törzsbe tartozó fajok jellegzetes tulajdonsága a feji végen lévő, visszahúzható ormány (probos-
cis), amelyen horgok találhatók. A visszahajló végű horgok segítségével kapaszkodnak meg a
végleges gazda bélcsatomájának belső felszínén. Jelentős károsodást idézhetnek elő a hámréteg-
ben, de a legnagyobb kárt akkor okozzák a gazdaállatnak, ha teljesen átfúrják a bélfalát. A
buzogányfejű férgeknek nincs elkülönült bélcsatomájuk, légző- és keringési rendszerük, kiválasz-
tószervük is hiányozhat. Váltivarú állatok.
Közel 1200fajuk ismert, amelyek Földünkön mindenütt előfordulnak. Evolúciójuk menete,
pontos rokonsági kapcsolataik nem ismertek.
Valamennyi ide tartozó faj endoparazita, kifejlett állapotban gerincesekben, így főleg halak-
ban, madarakban, emlősökben élnek. Lárváik a fajoktól függően rákokban, vagy rovarokban
élnek. Bár az embernek nincs specifikus buzogányfejü férge, fertőződése ritkán mégis előfordulhat.
Testméretük két millimétertől egy méterig terjedhet. Egy fajon belül a nőstények na-
gyobbak, mint a hímek.
Testük általában két oldalról kissé lapított, rajta esetleg számos haránt irányú redő
figyelhető meg. Színük általában fehéres, világos sárgásbarna.
Kültakarójukat a tegumentum alkotja, amelynek vastagságát a benne lévő csatoma-
és üregrendszer megnöveli. A test felszínén tüskék lehetnek. A testfal izomzatában a
külső körkörös és a belső hosszanti izomrétegek egyaránt megtalálhatók. Az izomréte-
13.1. ábra. Avchiacanthocephala fajok testfelépítésének vázlata. A: nőstény, B: hím
ad: agydúc, ev: elővesécske, he: here, hs: üregrendszer hasoldali zsákja, ht: üregrendszer hátoldali zsákja, hü: hüvely, iv: ivar-
nyílás, mé: méh, or: ormány, pá; párzószerv, pe: pete, pf: petefészek, rv: ragadós váladékot termelő mirigy, tb: tegumentum
betüremkedése (lemniszkusz), te: tegumentum és a testfal izomzata (Brumpt, Reman nyomán módosítva)
238
get is csatornák rendszere hálózza be, amely a tegument üregrendszerével áll kapcso-
latban. A benne áramló folyadék elősegíti a tápanyagok, bomlástermékek szállítását. A
glikogénben és lipidekben gazdag folyadékot a testfal izomzatának működése mozgatja.
A proboszcisz a fejben lévő ormányüregbe külön izmok segítségével visszahúzható. A
nyaki részen, az ormányhoz két megnyúlt lemniszkusz kapcsolódik, amelyek az üreg-
rendszerrel rendelkező tegumentumbetüremkedései (13.1. ábra). Aproboszcisz vissza-
húzásakor magukba fogadják az ormány merevítését szolgáló folyadékot.
A buzogányfejű férgeknek nincs bélcsatornájuk, a tápanyagokat a csatornákkal át-
járt kültakarón keresztül veszik fel. Egyes molekulák speciális membrántranszport-
folyamatok révén, mások pinocitózis segítségével jutnak át a kültakarón. A glükózt és a
glikogént anaerob úton bontják le.
Elkülönült légzőszervük nincs. Kiválasztószervük hiányozhat. Ha megtalálható,
akkor elővesécske típusú, két lángsejtből és a hozzájuk tartozó elvezető csőből áll.
Idegrendszerük központja a proboszcisz tövében lévő ganglion, amelyből az ormány
területére és a test vége felé indulnak ki idegek (13.1. ábra). Érzékelő idegvégződések van-
nak az ormányon és az ivamyílás környékén.
Váltivarú állatok, az ivari dimorfizmus a testméret különbségében kifejezett. A hí-
mek mindig rövidebb testűek. Ivarszervük a páros heréből áll (13.1. ábra). Az ivarsejtek
a párzótáskából kitüremkedő párzószerven át kerülnek be a nőstény ivarjárataiba. A
nőstény petefészkéből levált petesejtek a megtermékenyülést követően egy speciális üreg-
rendszerben fejlődnek, amelynek van egy háti és egy hasi zsákja. A hátoldali üregből a
még kevéssé fejlettek a hasoldali üregbe kerülve folytatják fejlődésüket (13.1. ábra). A
már fejlettebb, embriót tartalmazók a hátoldali zsákból a méhbe, majd a hüvelybe, innen
pedig a külvilágra jutnak (ovoviviparia).
A gazdaállat bélsarával kijutott petéket köztigazdaként szárazföldi rovarok vagy vízben
élő rákok vehetik fel. Ha a különböző lárvaalakokat hordozó köztigazdákat a halak, a
madarak és az emlősök elfogyasztják, akkor bél csatornájukban kifejlődik az ivarérett féreg.
Törzs: Buzogányfejűek (Acanthocephala)
Osztály: Spiráltüskés buzogányfejűek (Archiacanthocephala) 13.1. fejezet
Osztály: Sortüskés buzogányfejűek (Palaeacanthocephala) 13.2. fejezet
13.1. Spiráltüskés buzogányfejűek (Archiacanthocephala)
Az ide tartozó fajok fejlődése
szárazföldön zajlik. Végső gaz-
dáik az emlősök és a madarak,
míg köztigazdáik a rovarok.
A buzogányfejű féreg (Mac-
racanthorhynchus hirudinaceus) fő-
leg a legeltetett sertés és a vaddisz-
nó parazitája (13.2. ábra). Ahímek
testhossza elérheti a 10, mig a nős-
tényeké akár a 60 centimétert is.
Fejlődésnek indult petéi a sertés
bélsarával kerülnek ki a külvilágra,
ahol további átalakulásukhoz közti-
gazdára van szükségük. Leggyak-
13.2. ábra. A buzogányfejű féreg (Macracanthorchynchus hirudina-
ceus). A: acanthor lárva, B: kifejlett alak
(Meyer és más szerzők nyomán módosítva)
239
rabban a talajban élő bogárlárvák, például a cserebogarak pajorjai veszik fel őket, ahol a petéből
kikikelő acanthor nevű lárva (13.2. ábra) átfúrja magát a bélcső falán, és a hemocölben először
acanthella, majd fertőzőképes cystacanth lárvává fejlődik. A sertés turkálás közben fertőzött
rovarlárvákat fogyaszt, így fertőződik. Ha sok példány telepszik meg a bélcsatomájában, az igen
komoly károsodást okozhat. A kifejlett nőstény férgek testüregében akár tízmillió lárva is fejlődhet
egy időben.
13.2. Sortüskés buzogányfejűek (Palaeacanthocephala)
Fejlődésük általában vízben zajlik. Végleges gazdáik halak, vízimadarak és vízben élő
emlősök lehetnek, míg köztigazdáik főleg rákok.
Ahalak bélcsatomájában viszonylag gyakoriak az 1 -3 cm-es Echinorhynchus és Acanthocephalus
fajok. Köztigazdáik a Gammarus és Asellus rákok lehetnek.
Ellenőrző kérdések a 13. fejezet témaköreiből
1. Jellemezze a buzogányfejűek testfelépítését!
2. Ismertesse a buzogányfejűek szaporodásmódját, posztembrionális fejlődését!
3. Jellemezze a gyakoribb buzogányfejűek biológiáját és állategészségügyi jelentőségét!
240
14. PUHATESTŰEK (MOLLUSCA)
A puhatestűek az ízeltlábúak után az egyik fajokban leggazdagabb és legváltozatosabb állat-
csoport. Mintegy 130 000 ma élő és közel ugyanennyi kihalt fajt tartanak számon. Hazánkban
mintegy 240 puhatestű faj él.
A puhatestűek Földünkön szinte mindenütt megtalálhatók. így előfordulnak a sarkköri és trópusi
tengerekben, az édesvízi patakokban, folyókban, tavakban és mocsarakban egyaránt. Többségük
a tengerekben, főleg a parti sávban él. Fajszámuk elsősorban a meleg trópusi területeken magas.
Testük a legősibb csoportok kivételével szelvényezetlen. Lehetnek kétoldalion részarányosak, de
aszimmetriát is mutathatnak. Testfaluk a hasoldalon megvastagodott, és általában jól fejlett izmos
lábatalkot. A testhátoldalán létrejött redőzet, a köpeny (pallium) a héj anyagátválasztja el. A visz-
szahajló része hozza létre a köpenyüreget. Itt helyezkedik el a kopoltyú vagy a tüdő. A legtöbb faj
külső vázzal rendelkezik. A héj kezdeménye már az embrionális fejlődés végén, illetve a lárvastá-
diumban megjelenik, és növekedése legtöbbször az ivarérettség eléréséig tart. A héj előállításához
szükséges kalciumot a vízből vagy a talajból, illetve a táplálékból veszik fel. A héj egyes csoportoknál
csökevényes lehet, vagy szinte teljesen eltűnhet. Testtájaik a fej, alábésazsigerzacskó. A fejen található
a szájnyílás és a főbb érzékszervek. Dúcidegrendszerük van. Egyes csoportoknál a fej hiányozhat. A
belső szervek többségét a vékonyfalú zsigerzacskó zárja magába. Kiválasztószervük vesécske típusú,
keringési rendszerük nyitott. Kültakarójukfelszíne mindignyálkás, mert a hámrétegben, vagy az alatt
igen sok nyálkaanyagot termelő mirigy van. A puhatestűek váltivarúak vagy himnősek.
A puhatestűek jelentősége igen sokrétű. A geológiai, paleontológiái kutatásokban kitűnően
felhasználhatók, mint a földtörténeti korokjelzői. Számos fajuk emberi fogyasztásra alkalmas.
A külső váz sokrétű kézműipari feldolgozásra használható. Néhány faj igazgyöngy előállítá-
sára képes. Mezőgazdasági szempontból nem elhanyagolhatók, mint jó néhány féreg közti-
gazdái, illetve mint növényi kártevők.
Testméretük változó, az alig fél milliméterestől az óriási, több méteresig is találunk példákat.
Legtöbbjük, a fajok mintegy háromnegyed része azonban nem éri el az 5 centimétert.
Másodlagos testüregük csak a szívburok üregére, valamint az ivarszervek és a
kiválasztószervek üregeinek egy részére korlátozódik.
Kültakarójuk felszíne mindig nyálkás, mert a hámrétegben, vagy az alatt igen sok
nyálkaanyagot termelő mirigy van.
A puhatestűek váltivarúak vagy himnősek. Ivarszerveik párosak vagy páratlanok.
Parthenogenezis csak néhány fajnál ismert. A megtermékenyítés a hímnős fajoknál min-
dig kölcsönös. Megtermékenyített petéjük teljes, egyenlőtlen, spirális barázdálódást
mutat. Egyes fajok fejlődése során először egy úgynevezett trokofora (trochophora) lárva
jelenik meg. Ebből a lárvából vagy a fiatal állat jön létre metamorfózis révén, vagy a
trokofora után egy szabadon úszó vitorlás (veliger) lárva alakul ki. Ez a lárva csiliói se-
gítségével úszásra képes, testét vékony héj borítja. A legtöbb puhatestű trokofora lárvája
csak a peteburkon belül jön létre és a veliger lárva hagyja el a peteburkot. A fejlábúak,
néhány tengeri kagyló, az édesvízi kagylók jó része, valamint az édesvízi és szárazföldi
csigák átalakulás nélkül, közvetlenül fejlődnek.
241
A puhatestűek szelvényezett testű gyűrűsférgekkel való rokonságát a pete spirális
barázdálódása és a trokoforalárva megjelenése jelzi. A laposférgekkel való kapcsola-
tot a néhány puhatestűnél még megfigyelhető kötélhágcsószerű hasdúclánc-idegrend-
szer, valamint az ősi puhatestű osztályoknál meglévő mészpikkelyek mutatják. Valószí-
nű, hogy egy laposféregszerű ősből két irányban indult meg az ősszájúak fejlődése. Az
egyik ágon a szelvényezetien testű puhatestűek, míg a másik ágon a szelvényezett testű
gyűrűsférgek és ízeltlábúak felé.
Törzs: Puhatestűek (Mollusca)
Osztály: Csigák (Gastropoda) 14.1. fejezet
Alosztály: Elölkopoltyúsok (Prosobranchia)
Rend: Pajzskopoltyúsok (Aspidobranchiata) 14.1.1. fejezet
Család: Bödöncsigák (Neritidae)
Rend: Fésűskopoltyúsok (Pectinibranchia) 14.1.2. fejezet
Család: Fialócsigák (Viviparidae)
Család: Kerekszájú vízicsigák (Valvatidae)
Család: Változékony vízicsigák (Hydrobiidae)
Család: Felfújt vízicsigák (Bithyniidae)
Család: Tüskés vízicsigák (Melanopsidae)
Alosztály: Tüdőscsigák (Pulmonata)
Rend: Ülőszemű tüdőscsigák (Basommatophora) 14.1.3. fejezet
Család: Mocsári csigák (Lymnaeidae)
Család: Ellobiidae
Család: Balogcsigák (Physidae)
Család: Tányércsigák (Planorbidae)
Család: Sapkacsigák (Ancylidae)
Rend: Nyelesszemű tüdőscsigák (Stylommatophora) 14.1.4. fejezet
Család: Borostyánkőcsigák (Succineidae)
Család: Bábcsigák (Pupillidae)
Család: Tonnacsigák (Chondrinidae)
Család: Csavarcsigák (Enidae)
Család: Orsócsigák (Clausiliidae)
Család: Zonitidae
Család: Félmeztelen csigák (Daudebardia)
Család: Tarajos csupaszcsigák (Limacidae)
Család: Tarajoshátú csupaszcsigák (Milacidae)
Család: Simatestű csupaszcsigák (Arionidae)
Család: Eticsigafélék (Helicidae)
Osztály: Kagylók (Bivalvia) 14.2. fejezet
Rend: Lemezkopoltyúsok (Eulamellibranchiata) 14.2.1. fejezet
Család: Festőkagylók (Unionidae)
Család: Vándorkagylók (Dreissenidae)
Család: Gömbkagylók (Sphaeriidae)
Család: Borsókagylók (Pisidiidae)
Megjegyzés: A hazánkban élő puhatestűek rendszertani felosztása Kemey-Cameron (1979),
Meglitsch-Schram (1991), Richnovszky-Pintér (1979) munkái alapján került összeállításra.
Az elölkopoltyúsok a fajokban leggazdagabb alosztályt alkotják, mert körülbelül 60 000 ma
élő fajuk ismert. Főleg tengerben és édesvízben élnek, de van szárazföldi fajuk is. A torzió vala-
242
mennyi fajnál végbemegy, míg a detorzió egyiknél sem. Idegrendszerükben a zsiger- és köpeny-
dúcok közötti konnektívumok kereszteződtek egymással. Fésűskopoltyújuk a szív előtt helyezke-
dik el. Egy tapogatópárjuk van. Általában váltivarúak.
A körülbelül 50 000 ma élő fajt számláló tüdőscsigák alosztályának tagjai főleg szárazföldön
élnek. A detorzió jeleit elsősorban a szárazföldi és az édesvízi fajok mutatják. Légzőszervük a
köpenyüregből kialakult úgynevezett „tüdő”. A fésűskopoltyú tehát hiányzik. A köpenyüreg
nyílása kerek lyukká szűkült le. A végbélnyílás és a kiválasztószerv kivezető járata a köpenyüregbe
nyílik. Hímnősek, fejlődésük közvetlen. Házuk főként jobbra csavarodó.
14.1. Csigák (Gastropoda)
A puhatestűeken belül a csigák messze a legnagyobb faj- és egyedszámú, a legkülönbözőbb
élőhelyekhez sikeresen alkalmazkodó csoport. Fajszámuk a ma élőket figyelembe véve
körülbelül 110 000.
Megtalálhatók a tengerek parti (litorális) övében, a nagyobb mélységekben is, de
vannak a nyílt víztérben úszó (pelágikus) fajok is. Egyesek alkalmazkodtak a félsós
(brack) vizekhez, míg mások az édesvizekhez. A szárazföldön való elterjedésüket nagy-
mértékben meghatározza a talaj ásványianyag-tartalma, kémhatása, az előforduló szél-
sőséges hőmérsékleti értékek, a száraz periódusok. A szárazföldiek a talajban, az avar-
rétegben, a mohapámákban, a kövek alatt, a növényekre felmászva vagy más állatok
testére kapaszkodva fordulnak elő.
Az ősi csigák bilaterális szimmetriájúak voltak. Ama élők szimmetriaviszonyai azon-
ban jelentősen megváltoztak. Ezek a változások az embrionális fejlődés során, a veliger
állapotban mennek végbe. Testfelépítésük az evolúció során a csavarodás (torsio) és a
visszacsavarodás (detorsio), másrészt a ház feltekeredése folytán erősen módosult, így
változatos testfelépítést eredményezett (14.1. ábra). A torziót megelőzően az embrió
szájnyílása a fejlett feji végfelé irányult, míg a végbélnyílás és a köpenyüreg kivezető járata
az ellenkező testvégen nyílt. A torzió alkalmával a zsigerzacskó szervei és a köpeny 180
fokkal elfordultak, így a jobb és bal oldali szervek helyzete felcserélődött. Ennek során
a páros szervek egyik oldali része általában elcsökevényesedett, és kialakult a keresztezett
idegűség (streptoneuria) (14.1. ábra). A végbél és a köpenyüreg a fejen lévő szájnyílás
felett nyílt a külvilágra. Ilyen állapotot tükröz a ma élő ősibb csigák testfelépítése. A torzió
a ma élő valamennyi csigánál végbement. E folyamat evolúciós jelentőségére vonatko-
zóan több elképzelés is született, azonban kielégítő magyarázatot egyik sem tud adni.
Abban azért egyetértés van, hogy a köpenyüregben lévő érzékszervek, mint például az
oszfradium feji végfelé történt áthelyeződése az állat szempontjából előnyt jelent, mert
így a haladás irányában képes érzékelni környezete kémiai összetételének változását.
A köpenyüreg nyílásának a haladási irányban való elhelyezkedése biztosítja a légző-
szervek hatékonyabb működését, és a végbélnyíláson át a salakanyagok mielőbbi tá-
vozását. Azoknál a taxonoknál, ahol a detorzió is végbement, ott az egyes szervek visz-
szakerültek a torzió előtti helyükre, de az egyszer már elcsökevényesedett szervek nem
fejlődtek ki újra (14.1. ábra). Az idegrendszer felépítésében visszaállt egy másodlagos
egyenesidegűség (másodlagos orthoneuria). Ilyen folyamat ment végbe például a tü-
dőscsigáknál is.
A zsigerzacskó és a ház spirális feltekeredése nem tévesztendő össze a torzió fo-
lyamatával. A feltekeredés evolúciós szempontból a torziótól függetien folyamat. Előbb
következett be, mint a csavarodás. Valamennyi ma élő csiga ilyen feltekeredett zsiger-
zacskójú csoporttól származik, azonban a fejlődés során ezt a tulajdonságot egyes cso-
portok többé-kevésbé elvesztették. A ma élő fajoknál a torzió és a feltekeredés akár egy
időben is végbemehet, a lárvaállapot során lezajló változások alkalmával. A ház felte-
243
14.1. ábra. A csigák szimmetriaviszonyainak és testfelépítésének fontosabb változásai
A: ősi csigák feltételezett testfelépítése, B: torzió kezdete, C: primitív elölkopoltyús csiga, a torzió végbemenetele
után, D: fejlett elölkopoltyús csiga, a torziót követően, E: a tüdőscsigáknál detorzió révén másodlagos egyenesideguség
alakul ki
ad: agydúc, fd: fali dúc, kd: köpenydúc, ki: kiválasztószerv kivezető nyílása, ko: kopoltyú, kö: középbéli mirigy, li: lábidegfonat,
os: osphradium, sz: szájnyílás, si: szív, tü: tüdő, vé: végbélnyílás, zs: zsigerdúc (Stempell, Storer-Usinger-Hennignyomán kissé
módosítva)
keredésének evolúciós jelentősége a szilárd, ellenálló védőburokkal ellátott és kevéssé
terjedelmes, ugyanakkor hosszú test kialakításában volt.
A csigák teste fejre, lábra és zsigerzacskóra tagolódik. A fej viszonylag jól differenciált.
Rajta található a szájnyílás, a tapogatók és a látószervek. A láb általában mászásra, csú-
szásra, illetve úszásra szolgál. A zsigerzacskó a belső szerveket foglalja magába. A kö-
peny képezi a héjat, illetve a köpenyüreget hozza létre. A köpenyüreg nyílása közvedenül
a fej mögött, annak jobb oldalán található.
A kültakaró felszíni rétegét egyrétegű hengerhámsejtek alkotják. Az epidermisz alatt
elkülönült irharéteg nincs, mivel a kötőszöveti és izomelemek egymással keveredve
fordulnak elő. A mirigyek által termelt nyálka megakadályozza a testfelszín kiszáradását,
244
megkönnyíti a mászótalp mozgását, valamint mechanikai védelmet is biztosít az állatnak.
Különösen jól fejlett a lábmirigy, amely a talp felszínére önti váladékát.
A fajok többségének külső váza van, amely az ősi formáknál csak sapkaszerű, a fej-
lettebb alakoknál spirálisan feltekert. A csigák héja a test gyarapodásával párhuzamosan
növekszik. A csúcstól (apex), a legidősebb résztől kiindulva, a kanyarulatok egyre na-
gyobbak lesznek (14.2. ábra). A legelső kanyarulatok még a peteburkon belül kialakulnak,
ezért azokat embrionális héjnak nevezik. A kanyarulatok belső találkozási felszíne a köz-
ponti tengelyben, oszlopban (columella) forr össze, míg a külső felszínen is látható érint-
kezési vonaluk a varrat. Ha a kanyarulatok illeszkedése nem szoros, akkor a ventrális ol-
dalról kiindulva köldökrés (umbilicus) indul ki. A csavarodás irányától függően a ház lehet
jobbra vagy balra csavarodó. Leggyakoribb a jobbra csavarodás. A csavarodás iránya úgy
állapítható meg, hogy a házat a csúcsával felfelé és szájadékával felénk tartva megnézzük,
hogy a szájadék a képzeletbeli tengelytől balra vagy jobbra helyezkedik-e el. A jobbra csa-
varodóknál ez a jobb oldalon található. A héj felszíne fajra jellemzően lehet sima, hosszanti
vagy haránt irányban bordázott, tüskékkel vagy szőrszerű képződményekkel ellátott. A héj
színe, mintázottsága (sávok, foltok) a színanyagot termelő mirigyek jellegétől és elhelyezke-
désétől függ. Az utolsó kanyarulat nyitott vége a szájadék. Ennek széle lehet éles szegély vagy
megvastagodó ajakduzzanat, esetieg kihajló perem. A ház alakja, a kanyarulatok elhelyezke-
dése igen változatos lehet. Ha a kanyarulatok egy síkban kapcsolódnak egymáshoz, akkor a
ház lapított is lehet. A ház lehet gömbölyded, kúpos, tojásdad vagy éppen orsó alakú. A héj
növekedése nem egyenletes, így rajta növekedési vonalak figyelhetők meg, melyek jelzik pél-
dául a kedvezőtlen táplálkozási feltételeket, a nyári szárazság miatt bekövetkezett nyugalmi
periódust is. A héj felszínén található a pigmentált, föleg szaruszerű szerves anyagból, konhi-
olinból álló héjhártya (periostracum). Ez védi az alatta lévő réteget a kopástól, illetve más
állatok behatolásával szemben. A periosztrákum-réteget a köpeny szegélyrésze termeli. így
növekedése csak a héj szélén zajlik. A héj idősebb részein ez a réteg gyakran lekopik. A külső
réteg alatt fehérjékből álló alapanyagba ágyazott, szorosan egymás mellett álló aragonit- és
kaldtkristályok által létrehozott prizmás, oszlopos réteg (ostracum) van. A köpenyszegé-
lyen termelődő oszlopos réteg az állat növekedésével együtt vastagszik. A harmadik, legbelső
gyöngyházréteg (hypostracum) a csigák többségénél gyengén fejlett vagy hiányzik, míg
egyes tengeri fajoknál igen jól fejlett. Az édesvízi puhatestűek héját általában vastag peri-
osztrákum-réteg borítja, amely némi védelmet ad a bomló szerves anyagokból felszabaduló
savakkal szemben. A ház nyílását, a szájadékot a fajok egy része állandó képződménnyel
képes lezárni. Ilyen az elölkopoltyús vízicsigáknál lévő héjfedő (operculum). Az állat lá-
bának hátoldalán ülő operkulum gyakran szaruszerű, néha pedig konhiolinból és kalcium-
karbonátból áll. Az orsócsigák zárólemeze (clausilium) nem a szájadék pereménél,
hanem annál mélyebben helyezkedik el. A legtöbb mérsékelt övben élő szárazföldi csigánál
a nyugalmi időszakban ideiglenes, a nyálkaanyag megszilárdulásából kialakuló, néha mész-
tartalmú képződmény (epiphragma) figyelhető meg, amely a szájadékot bezárja, és meg-
védi az állatot a kiszáradástól. A ház ugyan védelmet jelent az állat számára, de megnöveli
a testtömegét, megnehezíti mozgását. Ezért például a szárazföldi csigák héja vékonyabb,
vagy teljesen eltűnik. A csupasz csigák héja oly mértékben csökevényes, hogy azt a köpeny
el is takarja.
A csigák többsége lassú, csúszó mozgással halad, mások képesek mászásra, úszásra,
ásásra is. Mozgásukat az igen erősen módosult bőrizomtömlő, a kötőszövetes állomány-
ban lévő izomelemek biztosítják. A mászótalp izomzatán az egyes fajoknál eltérő gya-
korisággal és terjedési iránnyal kontrakciós hullámok haladnak végig, amelyek a testet
igen kicsi távolsággal előre csúsztatják. A talp mozgásához feltétlenül szükség van a láb-
mirigyek által termelt nyálkára. A mozgás irányításáért elsődlegesen az idegrendszer
lábdúca (ganglion pedale) felelős, de az agydúcnak is van hatása, ami a mozgás gátlá-
sában nyilvánul meg.
245
Táplálkozásmódjuk változatos. A többségük növényevő, vannak detritusfogy asztok,
mindenevők, de előfordulnak ragadozók is. Az ajkak által határolt szájnyílás a terjedel-
mes szájüregbe nyílik. Itt helyezkedik el a reszelőnyelv (radula) és vele szemben a száj-
üreg boltozatán egy meszes-kitines állkapocslemez (14.2. ábra). A radula egy izmokkal
mozgatható és kordoid szövetből álló, támasztópálcával ellátott duzzanaton helyezkedik
el. Rajta, a fajra jellemző számban, sorokba rendeződve változatos alakú fogak ülnek. A
fogak száma néhány darabtól akár a több százezerig is terjedhet. A csiga a táplálékot a
radulával az állkapocslemezhez szorítja és így harap le apró darabokat. A radula az apró
táplálékrészecskéket a nyelőcső felé továbbítja. A garat egyes ragadozó fajoknál kiölthe-
tő, a többieknél rövid, izmos falú. A növényi táplálékot fogyasztó csigák nyálmirigyeinek
váladéka lúgos kémhatású, benne keményítőbontó amiláz van. A ragadozók nyála savas,
benne fehérjebontó enzimek, néha méreganyagok is lehetnek.
A nyelőcső a gyomorba juttatja a táplálékot. Ide csatlakozik a középbéli mirigy (he-
patopancreas) (14.2. ábra). A táplálék a gyomorban osztályozásra és részben emésztésre
kerül. Az osztályozást segíti elő egy kitüremkedés (vakzsák), ahol a táplálék összetorló-
pi vé ub kk vb lm id re ák
14.2. ábra. A házas csigák testfelépítésének vázlata. A: külső jellemzők, B: a tüdőlégzés módja, C: belső szervek
(az ivarszerveknek csak a kivezetőszakaszai, az idegrendszernek csak a feji dúcai vannak feltüntetve)
ar: artéria, ák: állkapocslemez, be: begy, cs: csúcs, gy: gyomor, hé: héj, ht: hosszabbik tapogató, id: idegdúc, iv: ivamyílás,
ka: kanyarulat, kb: középbél, kk: kiválasztószerv kivezető nyílása, km: középbéli mirigy, kn: köpenyüreg nyílása - légzőnyílás,
kö: köpenyüreg, kr: kamra, lm: lábmirigy, ná: nyálmirigy, ny: nyelőcső, pi: pitvar, re: reszelőnyelv, rt: rövidebbik tapogató,
tü: tüdő, ub: utóbél, va: varrat, vb: végbélnyílás, ve: vesezsák, vé: véna (Hereter, Runham, Stiasny, Storer-Usinger-Hennig
nyomán módosítva)
246
dik, majd izmok segítségével visszajut a gyomorba. Itt a redőzet és az izomkontrakciók
révén a pépes táplálék a középbéli mirigybe kerül, illetve részben visszajut a gyomorba.
Ez a folyamat többször ismétiődik, a szemcsés táplálékrészecskék végül a középbélbe
kerülnek. A középbéli mirigy egy terjedelmes csöves mirigy, amelynek feladata az emész-
tőnedvtermelés, a lebontott tápanyagok felszívása és részben a raktározás. A közép-
bélben az emésztési folyamat a fagocitózisra képes sejtekben, tehát intracellulárisan
zajlik. Növényevő csigákban a fehérjeemésztés szinte teljes mértékben a középbéli mi-
rigyben történik, míg ragadozóknál a gyomorban extracellulárisan is. A középbéli mi-
rigyben raktározódnak a zsírok, a glikogén és a kalciumsók. A középbél hosszú és
kanyargós. Belső felszínét csillós hám borítja, míg külső falában hosszanti és körkörös
izomrétegek fúrnak. Ezen a szakaszon szívódik vissza a víz, a szénhidrátok és a zsírok egy
része. Az utóbél izomrétege a középbélénél erősebb. A salakanyagok a végbélen át
kerülnek ki a külvilágra.
Légzőszervük változatos felépítést mutat. Az ősi formáknál két fésűkopoltyú (cte-
nidium) van a köpenyüregben, de a legtöbb faj esetében a test torziója miatt csak a bal
oldali maradt meg. A víz áramlását a köpenyüreg és a kopoltyúk felszínén lévő csillózat
biztosítja. A szárazföldi csigáknál a köpenyüreg nedves hámrétegén át történik a gáz-
csere. A levegő a köpenyüregbe a légzőnyíláson (pneumostoma) át jut be, cseréjét
részben a fenéklemez mozgása, részben az állat házba való be-, illetve kibújása eredmé-
nyezi (14.2. ábra). A vízben élő tüdőscsigák levegőért feljönnek a felszínre, vagy rö-
videbb-hosszabb légzőcső (sipho) segítségével töltik fel levegővel a tüdejüket. Egyes
fajok köpenyüregének hámja közvetlenül a vízből veszi fel az oxigént.
Keringési rendszerük nyílt típusú. Központja a szívburokban lévő szív. A csigák
többségénél a torzió miatt az eredetileg két kamrából csak egy maradt meg. A légzőszervben
felfrissült testfolyadék a szívpitvarba, majd a kamrába, innen pedig az elágazó artériák
révén a testhézagokba kerül (14.2. ábra). A szervek, sejtek innen veszik fel az oxigént és a
tápanyagokat, illetve ide adják le a szén-dioxidot. A hemolimfa a testüregekből a légző-
szervbe, majd a szívbe jut. A testfolyadékban oldott hemocianin segíti az oxigén szállítását.
Az amőboid sejtek a szervezet védekezési reakcióiban, a tápanyagok raktározásában, szál-
lításában vesznek részt. A testfolyadék szerepet játszik a láb mozgásában is.
Kiválasztószervük vesécske (metanephridium) típusú. Kezdeti szakasza a szívburok
üregével van kapcsolatban (14.2. ábra). Innen egy csillós hámmal bélelt csatorna vezet
a vesezsákba. Ennek fala gazdagon redőzött. A hámrétegben nefrociták ülnek. A ve-
sezsákból a húgyvezető indul ki és létesít kapcsolatot a köpenyüregen keresztül a
külvilággal. A húgyvezető szakaszán jelentős a só- és a vízvisszaszívás. Száraz időszakban
a víz szinte teljesen visszaszivódik. A vízben élő fajok jelentős mennyiségű ammóniát
távolítanak el a kiválasztószerven, a testfelszínen és a kopoltyúkon keresztül. A száraz-
földi fajok húgysavak formájában ürítik nitrogén tartalmú anyagcsere-végtermékeiket. A
csigák többsége képes túlélni szervezetének jelentős vízveszteségét is. Ennek során a
Helix fajok testsúlyuk 50%-át, a Limax fajok testsúlyuk 80%-át is képesek elveszíteni
anélkül, hogy elpusztulnának.
Az elölkopoltyús csigák (Prosobranchia) általában váltivarúak, míg például a tüdős-
csigák (Pulmonata) hímnősek. Ez utóbbiak legtöbbször kölcsönös megtermékenyítéssel
szaporodnak. Hímnős mirigyük (glandula hermaphroditica) páratlan szerv, amely
mindkét ivarsejtféleséget termeli (14.3. ábra). A hímnős vezeték (spermoviductus) belső
tere általában egy barázdával két félre osztott, majd kétfelé ágazik, egy ondóvezetőt
(ductus deferens) és egy petevezetéket (oviductus) képezve. Elágazásuknál található a
megtermékenyítési kamra, ahol a párzótárs felfelé hatoló spermiumai a lefelé haladó
petesejteket megtermékenyítik. Az ide kapcsolódó fehérjemirigy (glandula albuginea)
fehérjeburokkal veszi körül a megtermékenyített petesejteket. Az ondóvezetőhöz csatiako-
zik az ostor (flagellum), amelyben az ondótok (spermatophora) jön létre. Ez a párzószervbe
247
14.3. ábra. Az éticsiga ivarszervrendszerének felépítése
fin: fehérjemirigy, hn: himnős mirigy, hu: hüvely, iv: ivar-
nyilás, mk: megtermékenyítés! kamra, ny: nyílkamra, os:
ostor, ot: ondótarrály, ov: ondóvezető, pá: párzószerv, pv:
petevezető, uj: ujj alakú mirigy, ve: hímnős vezeték, vi:
visszahúzó izom (Baudelot nyomán kissé módosítva)
14.4. ábra. Az éticsiga erősen központosodott idegrend-
szerének felépítése
ad: agydúc, fz: fali- és zsigerdúc, kő: kőpenydúc, Iá: lábdúc,
po: pofadúc, sz: szemideg (BŐhmig nyomán kissé módosítva)
(penis) jut, és az erekció csökkenésével
kerül be a párzótárs ondótartálya (re-
ceptaculum seminis) felé vezető járatba. A
spermiumok a párzást követően ebből az
ondótartályból kerülnek a petevezetőbe. A
párzó társak egymáshoz tapadását segíti elő
az ivamyíláshoz kapcsolódó, ragadós vála-
dékot termelő, ujj alakú mirigy (glandula
mucosae). Számos szárazföldi csigánál a
párzási kedvet fokozzák a szerelemtáská-
ból vagy más néven nyílkamrából (bursa
teloris) kiszabaduló és egymás testébe lövelt
mésztűk. A kopuláció után a szárazföldi
fajok kisebb üregekbe rakják le petéiket,
ahol a fiatal állatok kifejlődnek. Vízben élő
fajok kocsonyás csomókban rögzítik petéi-
ket valamilyen aljzatra, például a vízinövé-
nyek leveleire. Néhány tengeri és édesvízi
faj ovovivipar. Ezek petéi a petevezetőben
indulnak fejlődésnek, és a fiatalok nem
sokkal a pete lerakása után hagyják el a
burkot. A szárazföldi csigák petéi nagyok,
szikanyagban gazdagok. A megterméke-
nyített peték teljes, egyenlődén és spirális
barázdálódást mutatnak. Számos tengeri
és valamennyi édesvízi, valamint szárazföl-
di fajnál a fejlődés közvetien, nincs lárvaál-
lapot. A peteburkot fiatal kis csigák hagy-
ják el.
A csigák erősen központosodott ideg-
rendszerének felépítése, az egyes dúcok
megléte, fejlettsége vagy hiánya az egyes
rendszertani egységeknél igen változatos
képet nyújt. Az elölkopoltyús csigák ke-
resztezett idegűek. A tüdőscsigák egyenes
idegűek, ami a detorzió révén alakult ki. A
páros agydúc (ganglion cerebrale) általá-
ban a legfejlettebb dúc, amely kisebb-
nagyobb mértékben irányítja a többi dú-
cot. Ezenkívül a látószervből, a tapogató-
szervből, a helyzetérző szervből jövő inge-
rületek feldolgozását is végzi (14.4. ábra).
Fejlett még a lábdúc (ganglion pedale),
amely a láb izmainak működését irányít-
ja. Az általában páratian zsigerdúc (gang-
lion viscerale) az ivarszerveket, a kiválasz-
tószervet, a szívet, és a bélcsatoma egy
részét idegzi be. A fali dúc (ganglion pa-
rietale) a testfalat és a köpeny egy részét
idegzi be. A köpenydúc (ganglion ple-
urale) beidegzi a köpenyt, szabályozza a
248
légzőmozgásokat. A pofadúc (ganglion buccale) a bélcsatoma kezdeti szakaszát idegzi be.
Ez a vegetatív idegrendszer központ]aként is működik. A testben lévő dúcokat idegek kötik
össze.
A hormonrendszer elsősorban a kiválasztószerv, az ivarszervek és az anyagcsere
szerveinek működésére hat. A neuroszekréciós sejtek az idegdúcokban ülnek. A látószerv
közelében lévő és azzal funkcionális kapcsolatban álló endokrin mirigy peteérést
szabályozó hormont termel.
Érzékszerveik közül a látószervek igen változatosak. A gödörszemek fénylátásra
szolgálnak. A hólyagszem alak- és színlátásra is képes. Hólyagszemük van például a
tüdőscsigáknak. A szaglószerv a tüdőscsigák hosszabbik tapogatóján található. A vízi-
csigák köpenyüregének anterior részén vagy a kopoltyúk alapjánál receptorsejtekkel bíró
bemélyedés, egy kemoreceptor (osphradium) van, amely a beáramló víz kémiai össze-
tételét érzékeli (14.1. ábra). Különösen jól fejlett a ragadozó fajok kemoreceptora. A
zsákmányállat felderítésében van jelentős szerepe. Mechanoreceptorok találhatók a ta-
pogatókon, a fejen, a talpon, a köpenyszegélyen. Az elölkopoltyús csigáknak (Pro-
sobranchia) egy pár, a tüdőscsigáknak (Pulmonata) egy, vagy két pár tapogatójuk van. A
lábdúc közelében lévő sztatociszták érzékelik a test helyzetét.
14.1.1. Pajzskopoltyúsok (Aspidobranchiata)
Házfedőjük vastag, belső oldalán fogszerű nyúlvány van. Túlnyomórészt tengeri fajok
tartoznak ide, de hazánkban is előfordulnak.
Bödöncsigák (Neritidae)
Félgömb vagy lapított házuk fala vastag, porcelánszerű. Lehetnek egyszínű feketék, lilák
vagy jellegzetesen zegzugos keresztrajzolatúak is. Növényevő csigák, melyek a vizek parti
sávjában a szilárd aljzatra, kövekre tapadva ülnek vagy másznak.
Gyakori, védett képviselőjük a rajzos csiga (Theodoxus danubialis), amely hazánkban elsősorban
a Duna vízrendszerében található. A szájadékára fektetett csigahossza 9-15 mm közötti (14.5. ábra).
14.1.2. Fésűkopoltyúsok (Pectinibranchia)
Fésűkopoltyújuk páratlan, a kö-
penyüreg falához teljesen hozzá-
nőtt. Szívük pitvara és vesécskéjük
is páratlan.
Fialócsigák (Viviparidae)
Váltivarú, elevenszülő, nagy ter-
metű vízicsigák, házkanyarulataik
domborúak. Az utódok fejlődésé-
nek kezdeti időszaka a petevezető
tágult, utolsó szakaszában megy
végbe.
Hazánkban a folyókban, holtágak-
ban, tavakban és mocsarakban egy-
aránt előfordulhatnak. Gyakori a he-
gyes fialócsiga (Viviparus acerosus)
(14.5. ábra). Házának magassága 30-
55 mm közötti. Elsősorban az iszap-
ban tartózkodik.
14.5. ábra. Pajzskopoltyús és fésűkopoltyús csigák. A: rajzos csiga
(Theodoxus danubialis), B: közönséges vízicsiga (Bithyna tentac-
ulata), C: pettyes csiga (Fagotía esperi), D: hegyes fialócsiga
(Viviparus acerosus)
(Krolopp és más szerzők nyomán kissé módosítva)
249
Kerekszájú vízicsigák (Valvatidae)
Házuk szélessége általában nagyobb, mint a magassága. A szájadék kerek, a házfedő
vékony, hajlékony, szaruszerű.
Hazánkban gyakori a 6,0-7,5 mm házmagasságú kerekszájú csiga (Valvata piscinalis) és az
1,2-3,0 mm-es lapos kerekszájú csiga (Valvata cristata). Mindkét faj előfordul álló és lassú
folyású vizekben, de az utóbbi forrásokban és mocsaras helyeken is megtalálható.
Változékony vízicsigák (Hydrobiidae)
Házuk kúp alakú, hengeres vagy gömbölyű, az utolsó kanyarulat kitáguló.
Hazánkban a folyók lassúbb folyású szakaszaiban, csatornákban, tavakban fordul elő a vastag
héjú gyöngycsiga (Lithoglyphus naticoides). Háza 7-13 mm magas, gömbölyded-kúpos.
Felfújt vízicsigák (Bithyniidae)
Házuk kanyarulatai meredeken emelkednek.
Hazánkban álló- és folyóvizekben általánosan elterjedt a közönséges vízicsiga (Bithynia ten-
taculata) (14.5. ábra). Háza vékony, áttetsző, 7,7-13,7 mm magas.
Tüskés vízicsigák (Melanopsidae)
Házuk magas, sok kanyarulatból áll, és a szájadék is általában megnyúlt.
A rozsdabarna foltokkal tarkított pettyes csiga (Fagotia esperi) hazánkban leggyakrabban
a Dunában fordul elő. Védett faj. Házának magassága 14,5-22,0 mm között változik (14.5.
ábra).
14.1.3. Ülőszemű tüdőscsigák (Basommatophora)
Szemük az egy pár, vissza nem húzható tapogató tövében ül. Házfedőjük nincs.
Mocsári csigák (Lymnaeidae)
14.6. ábra. Ülőszemű tüdőscsigák. A: nagy mocsárcsiga (Lymnaea
stagnalis), B: májmételyes csiga (Lymnaea truncatula)J C: pocso-
lyacsiga (Lymnaea peregra), D: nagy tányércsiga (Planorbarius
corneus), E: sapkacsiga (Ancylusfluviatilis) (Kemey, Krolopp,Needon
és Pintér nyomán kissé módosítva)
Házuk többnyire magas, kúpos.
A héj többnyire vékony, áttetsző,
törékeny. Házfedőjük nincs. A
legtöbb faj igen változékony.
Hazánkban álló- és folyóvizekben
egyaránt gyakori faj a nagy mocsári-
csiga (Lymnaea stagnalis) (14.6. áb-
ra) és a karcsú mocsáricsiga (Lym-
naea palustris). Előbbi fajnak 42-67
mm, utóbbinak 20-24 mm magas a
háza. Táplálékukat elsősorban élő és
korhadó növényi részek alkotják. Kü-
lönböző mételyeknek (Trematoda) le-
hetnek köztigazdái. A kisebb vizek-
ben, nedves helyeken előforduló máj-
mételyescsiga (Lymnaea truncatula)
a májmétely köztigazdája (14.6. áb-
ra). Házának magassága 7-12 mm
közötti. A pocsolyacsiga (Lymnaea
peregra) házának szájadéka igen nagy
(14.6. ábra). Háza 11-22 mm magas.
Ez a faj rendkivül változékony, szá-
mos változata ismert. A legtöbb víztí-
pusban igen közönséges.
250
Ellobiidae
Szárazföldön, a nyirkos talajban élő csigák. A hazai fajok igen apró testűek.
A kétéltű csiga (Carychium minimum) hazánkban gyakori, de apró, a 2 mm-t el nem érő
termete és rejtett életmódja miatt ritkán lehet észrevenni.
Balogcsigák (Physidae)
Házuk a legtöbb csigával ellentétben balra csavarodó. Vízinövényekkel jól benőtt álló-
és folyóvizekben egyaránt előfordulnak.
Hazánkban a hólyagcsiga (Physa fontinalis) gyakori. Háza általában 10-12 mm magas.
Tányércsigák (Planorbidae)
Házuk egy síkban balra csavarodott, korong alakú. Házukat nem a mászótalpra me-
rőlegesen állítva, hanem mindig balra billentve tartják. Házfedőjük nincs.
Gyakori, gyakran tömeges előfordulású a nagy tányércsiga (Planorbarius comeus) (14.6. ábra)
és az éles csiga (Planorbis planorbis). Mindkét faj elsősorban az állóvizek, a lassú folyású vizek
lakója. Előbbi faj házának szélessége 20-30 mm, utóbbinak 14-21 mm.
Sapkacsigák (Ancylidae)
Házuk kanyarulatok nélküli, egyszerű kúp alakú, általában vékony falú.
Ide tartozik a sapkacsiga (Ancylus fluviatilis), amely főleg hegyi patakokban, de folyókban és
tavakban is előfordul (14.6. ábra). Szájadékának hossza 4,5-9,0 mm közötti.
14.1.4. Nyelesszemű tüdőscsigák (Stylommatophora)
Két pár visszahúzható tapogatójuk közül a hátulsón ül a látószerv. Ez utóbbi tapogatót
ezért inkább szemtartó nyélnek (ommatophor) nevezik. Valamennyi faj szárazföldi, de
legtöbbje nedvességkedvelő. A fajok egy része a termesztett növények fogyasztásával,
megrágásával közvetlen kárt okoz. Mások
közvetve okozhatnak kárt, mert nyálkájuk,
ürülékük bevonja a növény levelét, továbbá
kórokozókat is terjeszthetnek.
Borostyánkőcsigák (Succineidae)
Kihegyesedő, tojásdad alakú házuk igen tö-
rékeny. Házfedőjük nincs. A szárazföldi csi-
gák közül a leginkább vízhez kötött fajok, így
többnyire vízparti vagy vízben gyökerező
növényeken élnek.
Általánosan elterjedt faj a kis borostyánkő-
csiga (Succinea oblonga) és a karcsú boros-
tyánkőcsiga (Oxyloma elegáns) (14.7. ábra). Az
előbbi faj háza 7,5 mm, míg az utóbbié 12-16 mm
magas.
Bábcsigák (Pupillidae)
Házuk általában kicsi, szájadékában legtöbb-
ször fogazat található. Sziklákon, kövek alatt,
avarban és növényekre kapaszkodva egyaránt
előfordulhatnak.
Gyakoribb a bábcsiga (Pupilla muscorum),
amelynek háza 2-3 mm magas lehet.
14.7. ábra. Nyelesszemű tüdőscsigák 1. A: karcsú
borostyánkőcsiga (Oxyloma elegáns), B: sokfogú csi-
ga (Gvanaria frumentum), C: zebracsiga (Zebrina
detrita), D: csavarcsiga (Ena obscura), E: redős-
csiga (Laciniaria plicata)
(Kemey, Krolopp és más szerzőknyomán kissé módosítva)
251
Tonnacsigák (Chondrinidae)
A sokfogú csiga (Granaria frumentum) háza 6-10 mm magas, hegyes, kúpos. Meleg, napos
helyeken közönséges faj (14.7. ábra).
Csavarcsigák (Enidae)
Ide tartozik a száraz, napsütötte, meszes talajú területeken előforduló zebracsiga (Zebrina
detrita), amely a lándzsásmétely köztigazdája (14.7. ábra). Erős héjú, 15-28 mm magas háza fehér,
vagy többnyire barnán sávozott. Bár elsősorban elhalt növényi részekkel táplálkozik, nagy tö-
megben elszaporodva a pillangós virágú növényeket károsíthatja.
A csavarcsiga (Ena obscura) főleg a domb és hegyvidéki erdőkben, az avarban, vagy sziklákra
kapaszkodva található (14.7. ábra). Házának magassága 11 mm alatti.
Orsócsigák (Clausiliidae)
Házuk általában igen erősen megnyúlt, orsó alakú. A ház nyílása többnyire zárólemezzel
(clausilium) ellátott. Főként domb- és hegyvidéki erdőkben az avarban, fatörzseken,
sziklákon fordul elő.
Gyakori a erdei orsócsiga (Balea biplicata), valamint a redőscsiga (Laciniariaplicata) (14.7.
ábra). Házuk magassága 12-22 mm közötti.
Zonitidae
A népes családba a növényevő fajokon kívül ragadozók is tartoznak.
Ilyen például az emberi települések közelében avarban, kövek alatt, pincében, barlangokban
előforduló gyors mozgású csillogó csiga (Oxychilus drapamaudi). Még a nagyobb testű csigákat
is megtámadja. A pincében tárolt zöldségfélék fogyasztásával kárt okozhat. Házának magassága
11-16 mm közötti.
Félmeztelen csigák (Daudebardia)
Házuk igen csökevényes, az állat méretéhez képest rendkívül kicsi.
Domb- és hegyvidéki erdőkben a nedves avarban, a talajban, kövek között, barlangokban él a
félmeztelen csiga (Daudebardia rufa) (14.8. ábra). Ragadozó faj, gilisztákat, rovarlárvákat,
ászkákat, más csigákat fogyaszt.
Tarajos csupaszcsigák (Limacidae)
Megnyúlt testük csupasz, és héjuk annyira visszafejlődött, hogy a pajzsnak nevezett
köpeny eltakarja. Testük hátoldalán vagy csak a testvégen taraj húzódik. Főleg növényi
táplálékot fogyasztanak, és mivel táplálékhasznosításuk igen rossz, ezért hatalmas meny-
nyiséget vesznek fel.
14.8. ábra. Nyelesszemű tüdőscsigák II. A: félmeztelen csiga
(Daudebardia rufa), B: pincelakó házatlancsiga (Limax fia-
vus) (Kemey és más szerzők nyomán kissé módosítva)
A termesztett növények fogyasztásával
nagy kárt okozhat például a 3,5-5,0 cm-es
recés meztelencsiga (Deroceras reticula-
tum) és a 10-20 cm-t is elérő nagy há-
zatlancsiga (Limax maximus).
A pincelakó házatlancsiga (Limax
flavus) vermekben, pincékben, üregekben
él, ahol a tárolt növényeket károsítja (14.8.
ábra). Testhossza a 7,5-10,0 cm is lehet.
Tarajoshátú csupaszcsigák
(Milacidae)
Hátoldalukon, a köpenytől a test végéig,
kissé kiemelkedő taraj húzódik.
252
A szemcsés meztelencsiga (Tandonia budapestensis) gyakori, mindenütt előforduló faj.
Testhossza elérheti az 5-6 cm-t. Termesztett növényeken kárt okozhat.
Simatestű csupaszcsigák (Arionidae)
A nagy testű 10-15 cm-es vörös csupaszcsiga (Árion rufus) és a 7-10 cm-es Árion lusitanicus fajok
az utóbbi években kerültek be hazánk területére. Ahol nagy tömegben fordulnak elő, ott a nö-
vényzetben jelentős pusztítást végezhetnek.
Éticsigafélék (Helicidae)
Igen sok faj tartozik ide. Gyakori faj a napsütötte, száraz helyeken élő kórócsiga (Helicella obvia).
amely a lándzsás métely köztigazdája (14.9. ábra). Házának magassága 7-10 mm. Kártételét több
pillangós virágú termesztett növényen
is megfigyelték.
Erdőszéleken, ligetekben, kertekben
fordul elő a gyakori pannoncsiga (Ce-
paeavindobonensis) (14.9. ábra), alige-
ti csiga (Cepaea nemoralis) és a kerti
csiga (Cepaea hortensis). Előbbi két faj
házának magassága 21-22 mm-t, míg
az utóbbié a 17 mm-t érhet el.
Az éticsiga (Helix pomatia) ha-
zánkban a legismertebb nagy testű faj,
amelynek háza 30-50 mm magas lehet
(14.9. ábra). Védettsége ellenére nagy
mennyiségben gyűjtik étkezési célok-
ra, ezért állománya egyre csökken.
A pettyes éticsigát (Helix aspersa)
tenyésztési célból is tartják (14.9. áb-
ra). Házának magassága 25-35 mm
közötti. Ahol természetes körülmények
között nagyobb létszámban fordul elő,
ott jelentős kárt okozhat.
14.9. ábra. Nyelesszemű tüdőscsigák III. A: kórócsiga (Helicella
obvia), B: pannoncsiga (Cepaea vindobonensis), C: éticsiga (Helix
pomatia), D: pettyes éticsiga (Helix aspersa) (Kemey és Krolopp
nyomán kissé módosítva)
14.2. Kagylók (Bivalvia)
A puhatestűek törzsén belül fajszámot tekintve a második legnagyobb osztályt alkotják.
Közel 20 000 faj tartozik ide. Hazánkban jelenleg 23 fajuk ismert. Tengerben és édes-
vízben egyaránt elterjedtek. A csigáktól eltérően azonban nem figyelhető meg nagymér-
vű adaptív radiációjuk. Többnyire a vízfenéken lassan mozgó, valamilyen aljzathoz
rögzülő, vagy maguknak üreget kialakító életmódú fajok jöttek létre. Az életmódban
megjelenő nagy hasonlóság a testfelépítésük igen egységes voltában is megnyilvánul.
Testük általában kétoldalian részarányos és két oldalról lapított. Testüket két
oldalról a nagy kiterjedésű köpeny és az általa termelt héj (valve) borítja. Testükön feji
részt elkülöníteni nem lehet. A kagylók hátulsó testvégén a köpeny módosulásából lét-
rejött egy vízkivezető és egy vízbevezető készülék (sipho). Testméretük lehet 1-2 mm-
es, de az egy métert is meghaladhatja.
Kültakarójuk hámrétegét csillós hengerhám alkotja. Az alatta lévő kötőszövetes
réteg simaizomelemeket és sok, nyálkaanyagot termelő mirigyet tartalmaz.
A külső köpenyhám termeli a héj anyagát (14.10. ábra). A héjat kívülről a fehérje-
tartalmú héjhártya (periostracum) borítja, amely konhiolinból áll. Alatta van a porce-
253
14.10. ábra. A kagylók köpenyszegélyének és
héjának metszete
bk: belső köpenyredő, gy: gyöngyházréteg,
hh: héjhártya, íz: izomnyaláb, kh: köpenyhám,
kk: külső köpenyredő, kö: középső köpenyredő,
ny: nyálkatermelő mirigy, or: oszlopos réteg
(Kennedy nyomán kissé módosítva)
lánszerű prizmás vagy oszlopos réteg (ostracum),
amely a konhinba ágyazott aragonit- és kalcitegy-
ségeket tartalmazza. A legbelső gyöngyházréteg
(hypostracum) igen vékony lemezekből álló szer-
kezetű, itt a konhinba aragonit rakódik be. Számos
fajnál ennek a rétegnek egy centiméter vastagságú
része akár 450-5000 finom, párhuzamosan álló
lemezből is állhat.
A kétoldali héjat fajra jellemző kialakítású fogak-
ból, lécekből álló, egymásba illeszkedő zárpár-
kányzat (14.11. ábra) kapcsolja össze, a felszínen
lévő sarokpánt (ligamentum) segítségével. Ezutób-
bi rugalmassága révén a héj izommunka nélküli
nyitását teszi lehetővé. A búb (umbo) a héj leg-
idősebb része. A növekedési gyűrűk e körül kép-
ződnek és koncentrikusan helyezkednek el. Az állat
növekedése során újabb és újabb rétegek képződ-
nek a héj széli részén, így a növekedési vonalak ál-
talában jól felismerhetők.
A kétoldali héj zárását egy vagy kettő igen erős
összehúzódásra képes záró-(abduktor) izom bizto-
sítja. Ezek és még más, speciális feladatot végző
izmok tapadási helyei jellegzetes izomlenyomato-
kat hoznak létre a héj belső felszínén. A sarokpánt
és az abduktorizmok ellentétesen működnek. Az
abduktorizmok kétféle működést ellátó fíbrillu-
mokból állnak. Egy részük a kétoldali héj igen gyors összezárását biztosítja, míg a másik
típusúak a lassú összehúzódást követően igen erősen zárva tartják a héj két felét. Igen
kevés energiát felhasználva, kifáradás nélkül hosszú időn keresztül képesek ezt a feladatot
ellátni. Ennek a képességnek rendkívül nagy jelentősége van a kagylók ragadozók elleni
védekezésében.
A gyöngyképződés valójában az állat védekezési reakciója a külső köpenyhám és a
gyöngyházréteg közé került testidegen anyaggal szemben. Ez lehet például egy homok-
szemcse, egy parazita valamely fejlődési alakja vagy bármi más idegen test. Mesterséges
körülmények között rendszerint kagylók héjdarabkáját helyezik a köpeny és a héj közé.
A köpenyhám a bekerült idegen test köré számos gyöngyházréteget választ el, így hozza
létre a gyöngyöt.
A kagylók a kétoldali héjból kinyúló, visszahúzható izmos lábukkal mozognak.
Miután kinyúlt a láb, a szöveteibe préselt testfolyadék révén megduzzad, megkapaszko-
dik az aljzaton. A longitudinális izmok összehúzódása megrövidíti a lábat, így a test a láb
után húzódik. A mozgás igen lassú, és jellegzetes barázdaszerű mászásnyomot hagy az
aljzaton. Számos tengeri faj és a hazánkban is előforduló vándorkagyló (Dreissenapoly-
morpha) más kagylókra tapadva viteti magát, vagy szilárd aljzathoz rögzül. Lábukban
fehérjefonalakat előállító mirigyek (byssus) találhatók. Ezekkel a vízben rugalmas szá-
lakká alakuló fonalakkal rögzítik magukat az aljzathoz.
Táplálkozásuk során kevés kivételtől eltekintve szerves törmeléket, baktériumokat,
egysejtű lényeket szűrnek ki a vízből. A kétoldali héj által határolt testbe, a köpenyüregbe
a víz a bevezető nyíláson keresztül jut be (14.11. ábra). A táplálékrészecskék vagy köz-
vetlenül jutnak el a szájvitorlákkal határolt szájnyíláshoz, vagy a kopoltyúlemezeken lévő
nyálkarétegben fennakadva, a csillós hám segítségével a táplálékvezető barázdákba so-
dorva kerülnek a megfelelő helyre. A kopoltyúlemezek csillós hámja főleg a kopoltyúk
254
tv xcs
14.11. ábra. A kagyló kopoltyúsának és köpenyüreg-rendszerének felépítése
be: bevezető szifó, bú: búb, cs: csillók, ei: előrehúzó izom, el: emelőizom, hé: héj, kf: a kopoltyúlemezek feletti vízvezető
csatorna, ki: kivezető szifó, ko: kopoltyúlemezek, kö; köpeny, ks: köpenyszegély, kz: közelítőizom, Iá: láb, Öi: összehúzó izom,
sv: páros szájvitorla, sz: szájnyílás, tv: táplálékvezető barázda, vb: vízbevezető nyílás, vé: vérér, ví: vízcsatorna, zp:
zárpárkányzat (Hickman, Russel-Hunter nyomán kissé módosítva)
ventrális vége felé továbbítja az apró táplálékrészecskéket, ahonnan a láb oldalán lévő
nyálkás barázdákban jutnak el a szájnyíláshoz. A nagyobb részecskék nem akadnak fenn
a kopoltyúk felszínét borító nyálkában, hanem a gravitáció révén arról leesnek, távoznak
a szervezetből. A bevezető szifó és a köpenyszegély hámsejtjei között receptorsejtek
ülnek, amelyek a táplálékfelvételhez szükséges vegyi és mechanikai érzékelést végzik. A
tápcsatorna bevezető nyílása után egy rövid nyelőcső, majd a gyomor következik (14.12.
ábra). Ide kapcsolódik a középbéli mirigy, amelynek számos funkciója van: emésztő-
enzimeket termel, sejtjei intracellulárisan emésztenek, illetve tápanyag felszívására és
raktározására is képesek. A gyomorhoz egy zsákszerű üreg kapcsolódik, amelyben egy
elsősorban emésztőnedveket tartalmazó kocsonyás állagú, úgynevezett kristálynyél
képződik, amely benyúlik a gyomorba. A zsákszerű üreg csillós hámja azt folyamatosan
forgatja, így a kristálynyél lassan leadja a magába foglalt anyagokat, a szénhidrát- és zsír-
bontó enzimeket. Forgása egyben a nyelőcső felől érkező, nyálkával bevont tápálék-
részecskéket is mozgatja a gyomor üregében. A gyomor üregében lévő terelőlebeny és
redőzet a táplálékszemcséket méretük alapján osztályozza. Az emésztés a gyomor-
ban indul meg, de jórészt a középbéli mirigyben fejeződik be. A középbéli mirigybe
kerülnek a legkisebb táplálékrészecskék. A nagyobb táplálékrészecskék a középbélbe
jutnak, ahol folytatódik az emésztés és a felszívás. Ezen a szakaszon a béltartalom a csillós
255
hám segítségével jut tovább. A bélcső belső felszínét hosszanti lefutású redő növelheti.
Az utóbél áthúzódik a szívkamrán. A végbélnyílás a kivezető szifó közelében nyílik
(14.12. ábra). Az emésztő szervrendszer nagy része mélyen benyúlik a lábba.
A kagylók életben maradásához a folyamatos vízáram létfontosságú. A kopoltyúk a
tágas köpenyüregben, a láb két oldalán húzódnak (14.11. ábra). A víz mozgatását a
kopoltyúk és a köpenyüreg hámjának csillós sejtjei végzik. A kagylók ősi formáinál fé-
sűkopoltyú fordul elő. A fonalas, majd lemezes kopoltyúk kialakulásával nőtt a légző-
felület. A szomszédos, érintkező filamentumok egymással kapcsolódva lemezes, lamel-
lás szerkezetet hoztak létre. A visszahajlott lemezek a lábhoz és a köpenyüreg hámjához
nőttek hozzá. A kopoltyúlemezek felszínét a másodlagos redők tovább növelhetik. Az
egymással kapcsolatban lévő fonalas egységek között nyílások (ostia) biztosítják a víz
áramlását a kopoltyúk belső üregrendszerébe. Az üregrendszer vízszállító csatornái a
kopoltyú dorzális részén lévő hosszanti csatornával (suprabranchialis csatornával) van-
nak kapcsolatban. Ez a csatorna összegyűjti és a kivezető szifóhoz vezeti a kopoltyúleme-
zek között már átáramlott vizet. A kopoltyúlemezek kapillárisokban gazdagok. A gázok
kicserélődése diffúzió útján valósul meg. A gázcsere nemcsak a kopoltyú, hanem a kö-
peny belső hámrétegén keresztül is végbemegy.
Egyes kagylóknál a kopoltyúlemezek a fejlődő petéket magukba foglaló költőüreggé
módosulhatnak a szaporodási időszakban. Az iszapban maguknak mélyebb csatornát,
üreget kialakító fajok szifói megnyúlt, gyakran hosszú csövet formálhatnak, amely a
szabad víztérbe nyúlva biztosítja a gázcserét. A kopoltyúlemezek ventrális és dorzális
hajlatában a vízzel besodort táplálékrészecskék elvezetésére és összegyűjtésére szolgáló,
nyálkával borított barázdák futnak.
Nyílt keringési rendszerük van, amelyben a hemolimfa részben az erekben, részben
a szövet közötti hézagokban áramlik. Központja a szívburokban lévő szív, amely két
14.12. ábra. A kagyló belső szervei
ao: sorra, be: bevezető szifó, bu: szívburok, ei: előrehúzó izom, el: emelőizom, gy: gyomor, hé: héj, id: idegdúc, iv: ivarszerv,
kb: középbél, ki: kivezető szifó, km: középbéli mirigy, ko: kopoltyúlemezek, kő: köpenyszegély, kv: kiválasztószerv, kz: kö-
zelítőizom, Iá: láb, ny: nyelőcső, öi: összehúzó izom, si: szív, sz: szájnyílás, ub: utóbél, vb: végbélnyílás, vi: páros szájvitorla,
vz: vakzsák (Hickman, Russel-Hunter nyomán kissé módosítva)
256
pitvarból és egy kamrából áll (14.12. ábra). A két pitvar a kopoltyúkból érkező, oxigén-
ben dúsult hemolimfát a kamrába löki, majd innen az aorták viszik a testbe. A szö-
vetközötti hézagokból a vénákon át jut a kopoltyú felé a testfolyadék. Szívük miogén
beidegzésű. Működését a zsigerdúc gátolja, és a víz hőmérséklete nagymértékben be-
folyásolja. A szív működése általában igen lassú, percenként körülbelül hatszor húzódik
össze. A köpenyhámban részben oxigénnel gazdagodott testfolyadék a pitvarokon ke-
resztül a kamrába jut. A testfolyadék másik része a test üregeiből a kiválasztószerv vé-
náiba jut, ahonnan a kopoltyúkba kerül. Itt oxigént vesz fel, majd a szívbe kerül. A test-
folyadékban fagocitózisra képes amőboid sejtek találhatóak.
Kiválasztószervük a szívburok üregével csillós tölcsér révén kapcsolatban álló, erő-
sen módosult vesécske (14.12. ábra). Az elsődleges szűrlet a szívburok üregében
keletkezik, majd a tágas vesezsákba kerülve lezajlik a visszaszívás, illetve az egyes anyagok
kiválasztása. A főként ammóniát és kevesebb húgysavat tartalmazó vizelet a köpenyüreg-
be kerül, majd a kivezető szifon át távozik.
A kagylók többsége váltivarú, azonban ivari kétalakúság nem figyelhető meg. Egy-
szerű felépítésű ivarszerveik a lábban helyezkednek el (14.12. ábra). A spermiumok
a szuprabranchiális csatornába kerülnek, ahonnan a vízárammal jutnak ki a külvilágra.
Számos tengeri faj és a hazánkban is előforduló vándorkagyló (Dreissena polymorpha)
esetében a megtermékenyítés külső, míg az édesvízieknél belső. Belső megterméke-
nyítéskor a hím ivarsejtek az áramló vízzel a nőstények köpenyüregébe, kopoltyúüre-
gébe kerülnek. A petesejtek itt termékenyülnek meg, és további fejlődésük is itt foly-
tatódik.
A kikelő kajmacsos lárvák (glochidium) egy ideig itt, a kopoltyúüregből alakult köl-
tőüregben maradnak, majd a vízárammal elhagyják a kagyló testét. Amelyeknek sikerül
a vízfenéken keresgélő halak bőrére, kopoltyújára tapadni, azok néhány hétig mint ekto-
paraziták fejlődnek tovább. A gazdaállaton való megkapaszkodásukat a kétoldali vékony
héj szélén lévő erős, hajlott fogak segítik elő (14.13. ábra). A gazdaállat testnedveivel
táplálkozó lárvákat körülvevő hámsejtek tokot képeznek. Bizonyos fejlettséget elérve a
vízfenéken folytatják átalakulásukat, növekedésüket. Kártételük az ivadéknevelő halgaz-
daságokban esetenként jelentős lehet. A halakon való megtelepedésük révén nagyobb
távolságra is eljuthatnak. Az Unió fajoknál a szaporodás és lárvafejlődés a nyár folyamán
megy végbe, míg azAnodonta fajok késő nyáron megtermékenyült petéiből fejlődő lárvák
csak a következő év tavaszán hagyják el a nőstény tes-
tét. A vándorkagyló megtermékenyített petéiből ki-
alakuló, szabadon úszó csillós lárvák a plankton tagjai-
ként fejlődnek tovább.
A kagylók idegrendszere viszonylag fejletlen dúc-
idegrendszer, mert csak három pár ganglionjuk van,
amelyeket idegkötegek kötnek össze. Az agydúcnak
(ganglion cerebrale) koordináló szerepe van. A lábdúc
(ganglion pedale) elsősorban a mozgást irányítja. A
zsigerdúc (ganglion viscerale) (14.12. ábra) a legfejlet-
tebb, mert ez látja el a legtöbb belső szerv irányítás át. Az
idegdúcokból lépnek ki a szerveket ellátó idegek.
Érzékszerveik a speciális életmód következtében
kevéssé differenciálódtak. Helyzetérzékelő szervük
(statocysta) a lábdúc közelében fekszik. A kemore-
ceptorként funkcionáló oszfradium a köpenyüregben,
a láb alapjánál ül. Kemo- és mechanoreceptor sejtjeik
a köpenyszegélyen, a szifók és a szájvitorlák hámjában
találhatók.
14.13. ábra. A kagylók kajmacsos lár-
vája
fo: fog, hé: héj, kö: közelítőizom, rö: rög-
zítőszál, sz: érzékelőszőrök (Pennák nyo-
mán kissé módosítva)
257
14.2.1. Lemezkopoltyúsok (Eulamellibranchiata)
Kopoltyújuk a szomszédos fílamentumok összenövése révén lemezes szerkezetű.
Festőkagylók (Unionidae)
Alakjuk hosszúkás, tojásdad. A héjon zárószerkezet van.
Hazánkban gyakori a festőkagyló (Unió pictorum), a tompa folyamikagyló (Unió crassus) és a
tavikagyló (Anodonta cygnea) (14.14. ábra). A festőkagyló és a tompa folyami kagyló átlagosan 5-6
cm, a tavi kagyló pedig 10-15 cm hosszú. Az utóbbi években egyre több helyen fordul elő a tekintélyes
hosszúságot (20 cm) elérő amuri kagyló (Anodonta woodiana). A kagylók a jelentős mennyiségű víz
átszurése révén fontos szerepet töltenek be a víztisztításban. Egyesek a vízminőség romlására
érzékenyen reagálnak, ezek felhasználhatók a biológiai vízminősítésben. Testüket számos állat fo-
gyasztja (halak, madarak, emlősök). Héjukat iparilag ma is feldolgozzák. Régebben gombgyártáshoz
alkalmazták, ma elsősorban a dísztárgyak és ékszerek készítéséhez használják.
14.14. ábra. Lemezkopoltyús kagylók. A: tavikagyló
(Anodonta cygnea), B: vándorkagyló (Dreissena
polymorpha), C: gömbkagyló (Sphaerium corne-
um) (Krolopp és más szerzők nyomán kissé módo-
sítva)
Vándorkagylók (Dreissenidae)
Héjuk szögletes, általában háromszög alakú,
egyik végén kihegyezett. Bisszuszmirigyeik ál-
tal termelt rögzítőfonalaikkal szilárd aljzatra
vagy más kagylókra kapaszkodnak.
Ide tartozik az átlagosan 35 mm hosszú vándor-
kagyló (Dreissena polymorpha) (14.14. ábra).
Gömbkagylók (Sphaeriidae)
A gömbkagyló (Sphaerium comeum) szerves anyag-
ban gazdag vizekben fordul elő (14.14. ábra). Vé-
kony falú héja majdnem gömb alakú, átlagosan 12 mm
hosszú.
Borsókagylók (Pisidiidae)
Hazánkban gyakori a nagy borsókagyló (Pisidium
amnicum), amely álló- és folyóvizekben egyaránt elő-
fordul. A borsókagylók közül a legnagyobb, hosszú-
sága 8-11 mm közötti.
Ellenőrző kérdések a 14. fejezet témaköreiből
1. Milyen változások következtek be a csigák testfelépítésében evolúciójuk során?
2. Jellemezze a csigák testfelépítését!
3. Hogyan szaporodnak a csigák?
4. Mutassa be az elölkopoltyúsok (Prosobranchia) és a tüdőscsigák (Pulmonata) fontosabb
családjait és a gyakoribb fajokat!
5. Jellemezze a kagylók testfelépítését!
6. Hogyan szaporodnak a kagylók?
7. Mutassa be a kagylók fontosabb családjait és a gyakoribb fajokat!
258
15. GYURUSFERGEK (ANNELIDA)
Földünkön mintegy 12 000gyürűsféregfaj él. Hazánkban kevesebb mint 100fajt mutattak ki.
Legjellegzetesebb vonásuk a test szelvényezettsége és a másodlagos testüreg megléte.
A valódi szelvényezettség (metameria) egyedfejlődéstani alapja az, hogy a mezodermában
a fej felöl kiindulva, egymás után következve párosával jönnek létre a testüregszakaszok
(coelomazsákok). Ez egyben az adott szakaszon kialakuló szervek szelvényes elhelyezkedését
is maga után vonja. Az egyedfejlődés során létrejövő szelvények közül tehát a legidősebbek a
feji végen, míg a fiatalabbak a farki végen lesznek. A legtöbb fajnál a szelvényezettség egynemű
(homonom metameria), vagyis a szelvények nagy része egyforma, vagy igen hasonló egymás-
hoz. Ritkán fordul elő, hogy a szelvények nagymértékben különböznek egymástól, és ezáltal
testtájak jönnek létre (heteronom metameria).
A kültakaró és függelékei külső, míg az egyes szervek és a testüreg belső szelvényezettséget
mutatnak. Bélcsatomájuk három szakaszosból tagolt. Keringési rendszerük a kevéssertéjűeknél
zárt, a piócáknál erősen módosult. Elkülönült légzőszervük nincs. Kiválasztószervük vesécske
típusú. Dúcidegrendszerük van, amelynek központja a feji végben lévő agydúc, a szelvényen-
ként! dúcok helyi reflexközpontokként működnek. Ivartalan és ivaros szaporodás egyaránt
előfordul. Himnős állatok, a megtermékenyítés kölcsönös.
A gyűrűsférgek közé tartozó kevéssertéjűek a vizek és a talaj szervesanyagforgalmában
fontos szerepet töltenek be. A giliszták élettevékenységeinek termőtalajt javító hatása régóta
ismert. A vérszivó piócák gyakran a vérélősködő paraziták terjesztői.
A testük mutathat elsődleges vagy alapgyűrűzöttséget (primer annulatio) amely a
szelvénybarázdáknak felel meg, valamint másodlagos gyűrűzöttséget (secunder an-
nulatio) is, amikor a szelvénybarázdák között további gyűrűk (annulum) figyelhetők
meg. Az elsődleges gyűrűzöttség a kevéssertéjű gyűrűsférgek többségére jellemző, míg
a másodlagos gyűrűzöttség néhány, a Tubificidae családba tartozó fajnál, valamint a
nadályoknál fordul elő.
A kifejlett állat testüregét borító hám a peritoneum, amelynek van egy testfalhoz
simuló fali (parietalis) és a bélfalhoz fekvő zsigeri (visceralis) lemeze. A jobb és bal oldali
cöloma fala a középvonalban a bélcső felett és alatt, mint dorsalis és ventrális bélfodor
(mesenterium) találkozhat, de üregeik gyakran egybeolvadnak. Az egymást követő cölo-
maüregeket a szeptális peritoneumból kialakuló haránt válaszfalak (septum) különítik
el egymástól. Egyes csoportoknál ez a válaszfal hiányozhat.
A lágy testű gyűrűsférgek hidrosztatikai vázát a folyadékkal teli testüreg képezi.
Mivel a cöloma üregei egymástól elkülönülnek, az egyes egységekben, illetve egyes
testtájakon a testfolyadék nyomása a többitől eltérő lehet. Ez a rendszer kifinomultabb
mozgásokat is lehetővé tesz.
A testüreg és a cöloma tagolódása biztosítja, hogy az egyes szelvények a többiektől
eltérő fázisban legyenek képesek összehúzódni (a szelvényt megrövidíteni), illetve
elernyedni (a szelvényt megnyújtani). A testen végigfutó izomkontrakciós hullám
- a váltakozva összehúzódó és elernyedő szelvények idegrendszer által összehangolt
259
munkája - erőteljes mozgást tesz lehetővé. A gyűrűsférgek többsége cső alakú járatokat
hoz létre táplálkozása során, később ez lesz a lakóhelye is.
A féregszerű állattörzsek képviselői közül a legsikeresebben alkalmazkodtak az édes-
vizekhez, a talajbeli élethez, de a legnagyobb adaptív radiációt és fajszámot a tengerekben
érték el.
Számos elképzelés van a gyűrűsférgek származására vonatkozóan. Legvalószínűbb,
hogy a gyűrűsférgek olyan ősi, lapított testű férgekre vezethetők vissza, amelyeknek egyes
szervei szelvényezett elhelyezkedést mutattak. A gyűrűsférgek testfelépítésének néhány vo-
nása az ízeltlábúakra emlékeztet, mint például a mezodermasejtek eredete, a kültakaró és az
idegrendszer felépítése tekintetében. Úgy tartják, hogy e két törzs szorosabb származástani
kapcsolatban van egymással. A gyűrűsférgek a puhatestűekre is hasonlítanak az embrionális
fejlődés több vonatkozásában, mint például a spirális barázdálódási típusban, a mezoderma
kialakulási formájában, valamint a tengeri soksertéjű gyűrűsférgeknél megjelenő koszorús
lárva (trochophora) révén. Ez a hasonlóság is a közös őstől való származást jelzi.
Törzs: Gyűrűsférgek (Annelida)
Osztály: Olajosgiliszták (Aelosomatida)
Osztály: Nyeregképzők (Clitellata)
Alosztály: Kevéssertéjűek (Oligochaeta) 15.1. fejezet
Rend: Egypár-heréjű kevéssertéjűek (Tubificida) 15.1.1. fejezet
Család: Csővájóférgek (Tubificidae)
Család: Televényférgek (Enchytraeidae)
Rend: Lumbriculida 15.1.2. fejezet
Család: Mocsári giliszták (Lumbriculidae)
Rend: Kétpár-heréjű giliszták (Opisthopora) 15.1.3. fejezet
Család: Földigiliszták (Lumbricidae)
Alosztály: Piócák (Hirudinea) 15.2. fejezet
Rend: Ormányos nadályok (Rhynchobdellida) 15.2.1. fejezet
Család: Lapospiócák (Glossiphoniidae)
Család: Halpiócák (Piscicolidae)
Rend: Allkapcsos nadályok (Gnathobdellida) 15.2.2. fejezet
Család: Valódi nadályok (Hirudinidae)
Rend: Garatos nadályok (Pharyngobdellida) 15.2.3. fejezet
Család: Erpobdellidae
Megjegyzés: A kevéssertéjűek osztályát Papp (1996) szerint rendszerezzük. A gyűrűsférgek
tengerben élő csoportját alkotja a számos tulajdonság tekintetében heterogén soksertéjűek (Poly-
chaeta) osztálya. Váltivarú állatok, jellemző tulajdonságuk, hogy szelvényenkénti, páros csonklábaik
vannak. Az olajosgiliszták néhány mm nagyságú, többnyire ivartalanul szaporodó, édesvízben élő
állatok. A nyeregképzőkhimnős, kifejlett példányain legalább a szaporodási időszakban megtalálható
a mirigyes megvastagodás, csonklábaik hiányoznak, sertéik száma redukálódott vagy hiányzik.
15.1. Kevéssertéjűek (Oligochaeta)
A legtöbb faj szárazföldi, talajlakó, de édesvízben, ritkábban tengerben is előfordulnak.
A szárazföldiek szinte mindenütt, de elsősorban a mérsékelt éghajlati övben, nyirkos,
szerves anyagban gazdag talajban élnek. A vízben élők főleg a vízfenéken élnek, a szá-
razföldieknél mozgékonyabbak, érzékszerveik jól fejlettek, sertéik hosszabbak. A további
jellemzés elsősorban a földigilisztákra vonatkozik.
260
Testük alakja általában hosszan megnyúlt, a hasoldalon egyes fajoknál kissé, má-
soknál erősebben lapított.
Testhosszúságuk igen változó. Előfordulnak alig 1 mm és több 10 cm nagyságú faj ok
is. A szárazföldi fajok a vízben élőknél nagyobbak, trópusi területeken előfordulnak több
méter hosszúak is.
Testük színét vörös, barna, bíbor és zöld pigmentek is adhatják, de vannak fajok, ahol
ez nem fordul elő. A végső szín a pigmentek kevert hatásából, illetve az interferencia jelen-
ségéből áll össze. Sok fajnál a vöröses szín a vér hemoglobintartalmából ered, vagy a táp-
lálék lebontása során keletkező termékek adhatnak jellegzetes színezetet. Hátoldaluk söté-
tebb árnyalatú, mint a hasoldaluk.
A kifejlett állatok teste általában 30-300 gyűrűszerű szelvényre osztott. Testükön az el-
sődleges és másodlagos gyűrűzöttség egyaránt előfordulhat. Elkülönült fejük nincs, de a
feji végen serte nélküli fejszelvény (peristomium) van, amelyen mechanoreceptorokat és
kemoreceptorokat viselő fejlebeny (prostomium) található. Ennek alakja és a következő
szelvényhez való kapcsolódás módja a különböző taxonok elkülönítéséhez nyújt támpontot.
A feji és a farki vég szelvényeinek kivételével testüket rendkívül változatos alakulásban
serték boríthatják. Rendeződhetnek háti és hasi sorokba, vagy gyűrűszerűén körbeve-
hetik a szelvényt. Számuk változó, lehet akár 50-100 serte is egy szelvényen, de általában
ennél kevesebb fordul elő, sőt egyes fajoknál hiányozhatnak is. A serték tokokban ülnek,
melyeknek saját izomzatúk van. A serték fontos szerepet töltenek be a helyváltoztatás-
ban. A serték alakja, elhelyezkedése és száma jellegzetes taxonómiai bélyeg.
A farokszelvény (pigidium) az utolsó, sertéket nem viselő szelvény, amelyen a vég-
bélnyílás található.
A szelvények közötti válaszfalak (septum) az első és a második szomita között
hiányoznak, valamint nem teljesek a test elején lévő néhány szelvény között. A dorzális
mezentérium csak a szeptumoknál ismerhető fel, míg a ventrális mezentérium nem éri el
a test falát, így a jobb és bal oldali testüregfél egymással kapcsolatban van. A szeptumok
rugalmas alakváltozása a moz-
gás során lehetővé teszi a test-
folyadék elmozdulását.
A fajok egy részénél a test-
üreg a háti póruson keresz-
tül kapcsolatban van a kül-
világgal. Ezen keresztül cölo-
mafolyadékot préselhet ki ma-
gából az állat. A kibocsátott
folyadék nedves en tartj a a test-
felszínt, dezinficiáló és riasztó
hatású. Valószínűleg szerepe
van a hibemáció alkalmával
maguk körül kialakított védő-
tök falának szilárdításában,
keményítősében is.
Bőrizomtömlőjük felszí-
ni rétegét az epidermisz alkot-
ja, amely egyrétegű henger-
hámsejtekből áll (15.1. ábra).
Felszínükön általában nagyon
vékony, rugalmas, albumoid
anyagú kutikularéteg van. A
hámsejtek között egysejtű mi-
15.1. ábra. A földigiliszta testének keresztmetszete
bh: bélcső hámrétege, bt: betűrődés a középbél hátoldalán, da: hasdúclánc alatti
ér, ha: hasi ér, há: hámréteg, hé: háti ér, hi: hosszanti izomréteg, hl: hasdúclánc,
ki: körkörös izomréteg, kk: kiválaszt ószerv kivezető nyilasa, ki: kloragogén sejtek,
ku: kutikularéteg, se: serte, tb: testüreget bélelő hám, vk: vesécske kanyargós
csatornája, vt: vesécske tölcsére (Pechenik és más szerzők nyomán módosítva)
261
rigyek termelik azt a nyálkaanyagot, amely nedvesen tartja a hámsejtek felszínét, lehetővé
téve a gázcserét és elősegítve a mozgást. A hámrétegben számos érzékelősejt van.
A testfal izomzata a külső, vékony, körkörös és a belső, vastag - a szárazföldi fajoknál
több egységre tagolt -, hosszanti lefutású izomsejtek rétegéből áll (15.1. ábra). A kör-
körös izomrétegben pigmentsejtek, kötőszöveti sejtek, kapillárisok fordulnak elő. Perio-
dikus és ellentétes fázisban zajló összehúzódásuk eredményeként testükön jellegzetes
perisztaltikus mozgás fut végig. Mozgáskor az állat a megnyúlt szakaszok szelvényein
lévő sertékkel az aljzaton megkapaszkodik, és a hátrább lévő szelvényeket megrövidítve
maga után húzza a test többi részét.
Bár a táplálkozásmódjuk változatos lehet, azonban jól tagolt emésztőkészülékük
felépítése a különböző csoportoknál nagy hasonlóságot mutat. Ennek részei a szájnyílás,
a szájüreg, a garat, a nyelőcső, a begy, a gyomor, a középbél, az utóbél és a végbélnyílás
(15.4. ábra). A garat erős izomzatú, esetleg kiölthető. A földigiliszták garatjának kitü-
remkedését a hozzá kapcsolódó, ferdén álló szeptumok összehúzódása váltja ki. A garat
fala mirigyekben igen gazdag. Az itt termelődött nyálka csúszóssá teszi a táplálékot, ami
a talajlakó, többnyire kevéssé nedves táplálékot fogyasztó giliszták számára rendkívül
fontos. A Lumbricus fajoknál proteáztermelést is kimutattak ezen a szakaszon. A nye-
lőcső kevésbé izmolt. A szárazföldi fajok egy részénél járulékos szervként ide kapcsolód-
nak a Morren-féle mészmirigyek (15.4. ábra). Számuk és elhelyezkedésük fontos
rendszertani bélyeg. A mirigyhámsejtek képesek felvenni a vérből a táplálékkal bejutott
kalcium-ionokat, és azokat kalcitkristályokká alakítva visszajuttatják a bélcsőbe. A kalcit-
kristályok a bélcsatomából már nem szívódnak vissza. A táplálékkal felvett szerves savak
közömbösítése is a kalciumionok segítségével valósul meg. így nem borul fel a szervezet
sav-bázis egyensúlya. Az ürülékkel távozó kalcium hozzájárul a föld alatti járatok falának
szilárdításához. A nyelőcső és a begy belső felszínét vékony kutikularéteg borítja. A begy-
nek raktározó szerepe van. Innen a táplálék a gyomorba kerül. A gyomor belsejét borító
vastagabb kutikularéteg és az erőteljes izommozgás segíti a táplálék szétdarabolását. A
középbél belső felszíne tipikus esetben egyrétegű, csillós hámmal borított. Alatta kapillá-
risokat tartalmazó laza rostos kötőszövet, majd körkörös és hosszanti izomrétegek és
legkívül a peritoneum viszcerális lemeze található. A bélcső hámrétegében egysejtű
enzim- és nyálkatermelő mirigyek vannak. Az emésztés jórészt extracellularis. Proteáz
(pepszin, tripszin), lipáz, amiláz, és még nem egyértelműen eldöntötten celluláz enzim-
termelést mutattak ki. A gyűrűsférgek jó része rendelkezik azzal a képességgel, hogy
kültakaróján keresztül képes aminosavakat, zsírsavakat és glükózt felvenni. A kültakaró
felszínére leadott proteáz enzim nemcsak a kórokozóktól védi meg, hanem a külső
emésztés révén a hámsejtek a tápanyagot is fel tudják venni. A középbél háti oldalán
hosszanti betűrődés (typhlosolis redő) futhat végig, amely a felszívó felületet növeli. A
bélcső külső felületén peritoneumsejtekből módosult, sárgás színű kloragogén
(chloragogen) sejtek ülnek (15.1. ábra). A glikogén és zsír szintézisére, raktározására,
szállítására, valamint a karbamid kiválasztására, aminosavak bontására és káros vagy fe-
lesleges anyagcsere-végtermékek tárolására is szolgálnak. Az utóbélszakaszon redő
nincs, kloragogén sejtek nem borítják. Az ürülék a végbélnyíláson át távozik. A vízi ke-
véssertéjűek alapvetően detritusz- és algaevők, míg a szárazföldiek elhalt növényi része-
ket fogyasztanak. A talajlakó giliszták egy része táplálkozás céljából általában éjszaka jön
fel a talaj felszínére. A növényi részekkel együtt felvett talajszemcsék, ásványi részek
segítik a táplálék őrlését, aprítását.
Elkülönült légzőszervük nincs. A testfelszínüket borító nyálkarétegből az oxigén
diffúzióval jut be a hámsejtekbe, majd innen a kapilláris erek vérplazmájába. Az oxigén
szállítását a vérplazmában oldott hemoglobin segíti elő. A keletkezett szén-dioxid dif-
fúzió útján távozik. A földigiliszták az alacsony oxigénellátottsághoz igazítják anyag-
csere-intenzitásukat, és meglepő módon a hemoglobin elsősorban magasabb oxigéntenzió
262
15.2. ábra. A földigiliszták keringési szervrendszere egy szelvény
keresztmetszetében
bé: bélcső érrendszere, ha: hasi ér, hé: háti ér, ho: hasdúclánc oldalsó
hosszanti ere, da: hasdúclánc alatti ér, ki: kiválasztószerv érrendszere,
la: laterális ér, tf: testfal érrendszere, ti: tiflozólisz redő hosszanti ere
(Meglitsh nyomán kissé módosítva)
esetén szolgál az oxigén szállítására. A földigiliszták érzékenyen reagálnak az oxigén-
tartalom hirtelen csökkenésére vagy hiányára, igyekeznek az adott helyről elmenekülni.
Ezért jönnek fel esőzés után a talajfelszínre is. Az Enchytraeidae család egyes képviselői-
nek nincsen az oxigén szállítására szolgáló légzési pigmentjük. Egyes vízi életmódú
fajoknál (pl. Tubifex) a kültakarón keresztül zajló gázcserén kívül fontos szerepe van a
végbéltájéki béllégzésnek is. A friss víz bejutását a végbél pumpáló mozgása biztosítja. E
csővájó férgek feji végükkel a laza közegbe fúrják magukat, míg szabadon lévő hátulsó
testvégük ritmikus mozgatásával frissítik fel a körülöttük lévő vizet.
Keringési rendszerük zárt típusú, az erekben vér kering. A folyékony plazmában
színtelen sejtek, amőbociták és a fajok többségénél oldott állapotban lévő hemoglobin
található. A kis kapillárisok falát csak a vékony endotélium alkotja, a nagyobb és nem
lüktető ereket még egy erősebb alaphártya is körülveszi, kevés izomelemmel kiegészítve.
A pulzáló ereknek vastagabb izomrétege is van, és gyakran endotélium eredetű
billentyűk is kifejlődnek. A bélcső
feletti és alatti hosszanti erek külö-
nösen jól fejlettek. A vér a háti érben
előre, a hasi érben hátrafelé áram-
lik. Ezt a két főeret szelvényenként
oldalsó erek kötik össze. Ezek közé
tartoznak a testfalhoz futó érpályák,
amelyeknek elsősorban a légzési gá-
zok, kisebb részben a tápanyagok és
anyagcsere-végtermékek szállításá-
ban van szerepük (15.2. ábra).
Ugyancsak az oldalsó erekhez tar-
tozik a kiválasztószervek érrend-
szere. A kis testű kevéssertéjű gyű-
rűsférgeknél hiányozhat az epider-
misz alatti kapillárishálózat. A háti
ér kapcsolatban van a hasdúclánc
alatt futó hosszanti érpályával is,
így az idegrendszer mindig friss,
oxigénben gazdag vért kap. Továb-
bi hosszanti erekfordulhatnak elő a
hasdúclánc két oldalán, illetve a
tiflozóliszredőben is. A vér mozga-
tásában szerepet játszhat a háti
hosszanti ér és a hozzá kapcsolódó haránterek is. A vér mozgatásában legfontosabb
szerepük a nyelőcső két oldalán lévő aortaíveknek („szívek”) van (15.4. ábra). Erő-
teljes pulzálásuk a vért a háti edényből a hasi edénybe löki. Számuk változó, a vízben
élőknél általában egy pár, a Lumbricus fajoknál öt pár. A lüktető érszakasz működését,
a falában lévő izomsejtek összehúzódását részben az erek falában lévő ganglionsejtek,
másrészt az erek tágulása válthatja ki. A hosszanti háti ér és a pulzáló haránterek
működése nem koordinált, és az egymást követő szelvényekben lévő szívek is eltérő
ritmusban húzódnak össze.
Kiválasztószervük vesécske (metanephridium) típusú. Ez az első és utolsó néhány
szelvény kivételével minden szelvényben párosán előforduló szerv. Csillós tölcsérrel
(nephrostoma) kezdődik az egyik szelvényben, ami egy igen rövid csőszakaszban folytató-
dik és átvezet a mögötte lévő szelvénybe. Itt hosszú, kanyargós, kapillárishálózattal kö-
rülvett csatornarendszert alkot, amelynek kezdeti szűkebb és tágasabb középszakaszát
csillós hám béleli. Ezt követően egy kiszélesedett ampulla, majd egy hosszabb tágult
263
csőszakasz, végül egy izmos rész (terminális vesiculum) következik. Ez utóbbi, húgyhó-
lyagként is szolgáló rész megléte, alakja vagy hiánya fontos rendszertani bélyeg. A ki-
vezető nyílások (nephridioporus) tehát a nephrostomát követő szelvényben nyílnak a
külvilágra. Fejlett keringési rendszerrel bíró fajoknál (pl. földigilisztáknál) a kanyargó
csatornát körülvevő kapillárishálózat teszi lehetővé, hogy a bomlástermékek ne csak a
cölomából, hanem a vérből is a kiválasztószervbe kerülhessenek. A gyűrűsférgek többsé-
ge, a vízben élők elsődlegesen ammóniát adnak le kiválasztó működésük során. A szá-
razföldi élettérhez való alkalmazkodás során ez többnyire megváltozott, megnőtt az urea
ürítésaránya. A gilisztáknál még így is jelentős az ammónialeadás.
Idegrendszerük központja a garat feletti dúcpár összeolvadásából létrejött agydúc,
amely a garatideggyűrű révén a garatalatti dúcpárral létesít kapcsolatot (15.3. ábra). Ez
utóbbi a hasdúcláncban folytatódik, amely a hasoldali izomzat felett, a testüregben
helyezkedik el. A gyűrűsférgek fejletlenebb formáinál az agydúc a prosztomiumban van,
míg a fejlettebbeknél fokozatosan egyre hátrébb került, így például a Lumbricus fajoknál az
agydúc és a garat alatti dúc a 3-4. szelvényben található. Valamennyi szelvényben dúc-
párok vannak, amelyek a Lumbricus fajoknál egymással összeolvadtak. A garat alatti dúc
általában fejlettebb a soron következőknél. Az egymást követő szelvényekben lévő dúcokat
connectivumok kötik össze. Perifériális idegrendszerüket a dúcokból mindkét oldalra
kiinduló 4 pár (vízi kevéssertéjűeknél), illetve 3 pár (szárazföldieknél) elágazó ideg alkotja
(15.3. ábra). Ezek látják el afferens és efferens idegágakkal a belső szerveket, a testfal
15.3. ábra. A: a földigiliszták idegrendszerének felépítése, B: egy összeolvadt dúcpár metszeti képe egy kilépő ideg
síkjában
ad: agydúc, da: hasdúclánc alatti ér, di: dúcból kílképő ideg, ga: garat alatti dúc, gi: garatideggyűrű, ha: hasdúclánc, id: ideg-
sejtek, ór: óriás rostok (Hess és Schneider nyomán)
izomzatát, az érzékszerveket. A szelvényenként így kialakult neuroszomita egységek helyi
reflexválasz adására képesek. Neuroszekréciós sejtek elsősorban az agydúcban fordulnak
elő. Szerepük van a szaporodás regulációjában és a regenerációban.
A szárazföldi fajok hasdúcláncában óriásrost-rendszer alakult ki (15.3. ábra). Az
impulzus szinaptikus átkapcsolások nélkül fut végig a test hosszában. A vastagabb axon
264
15.4. ábra. A földigiliszta ivarszervrendszerének és az elülső testvég más szerveinek elhelyezkedése, felépítése.
A: a giliszta testének elülső vége a hasoldalról, B: az ivarszervek elhelyezkedése a testüregben a hátoldalról nézve,
C: az ivarszervek helyzete oldalnézetben
ad: agydúc, ao: aortaív, be: begy, bt: bőrizomtömlő, ga: garat, gy: gyomor, há: hátoldali hosszanti ér, he: here, hi: hím
ivamyílás, ib: ivari barázda (spermaredő), is: ivari serték, ki: kiválasztószerv, Mm: Morren-féle mészmirigyek, ne: nyelőcső,
ni: női ivamyílás, ny: nyereg, oh: ondóhólyag, on: ondótartály nyílása, ot: ondótartály, ov: ondóvezető, pf: petefészek, pö:
petevezető tölcsére, pt: petetartó, pv: petevezető, sd: serdülési dudor, sv: szelvényválaszfal, tő: ondóvezető tölcsére (Bames,
Meglitsh és más szerzők nyomán kissé módosítva)
265
gyorsabb vezetést tesz lehetővé. Tipikus esetben a hasdúclánc hátoldalán három, a has-
oldalán két (a hátoldaliakhoz képest jóval kisebb) óriás rost fut. A középső háti ág az
agyból visz mozgató impulzusokat a végrehajtó izomzathoz. Ebből az ágból minden
szelvényhez kétoldalt leágazás lép ki. Az oldalsó háti ágak a szelvényekben lévő ér-
zékszervekből viszik az ingerületet a fej felé. Az említett két ág az egyes szelvényekben
egymással is kapcsolatot létesít. A Lumbricus középső dorzális óriás rostja 1600-szor
gyorsabban (45 m/s) vezeti az impulzusokat, mint a kis neuronok.
Érzékszerveik az epidermiszrétegbe ágyazottak, általában egyszerű felépítésűek,
mert csak néhány, támasztósejtekkel körülvett érzékelősejt alkotja őket. Sűrűbben
fordulnak elő a feji és farki végen, a szájüregben, az egyes szelvények legvastagabb részén.
Minden érzékelősejt apikális csúcsán szőrszerű nyúlvány van, amely a kutikula pórusán
át érintkezik a külvilággal. Szabad idegvégződések is találhatók az epidermiszsejtek
között. A fényérzékelő sejtek a test különböző pontjain elszórtan találhatók, de különösen
sok van a feji végen a prosztomiumban és az anális szelvényben. Ezek a test fényre
legérzékenyebb részei. A legtöbb kevéssertéjű gyűrűsféreg a gyenge fénnyel szemben
pozitív fototaxist, míg erős fénnyel szemben negatív fototaxist mutat. Számos, több inger
felvételére is alkalmas általános érzékelősejt is megfigyelhető a hámrétegben. A tapintó
idegvégződések a sertékhez vagy speciális érzékelő szőrökhöz kapcsolódnak. Különösen
a feji végen fordulnak elő nagy számban. A földigiliszták rendelkeznek ízérzékelő recep-
torokkal a szájnyílás környékén és a szájüregben. Jól meg tudják különböztetni a szá-
mukra kedvező táplálékot a többitől.
A kevéssertéjű gyűrűsférgek között előfordul az ivartalan és ivaros szaporodás is.
Ivartalan szaporodás, fragmentáció többek között előfordul az Enchytraeidae család
egyes képviselőinél is. Testük meghatározott szelvénye képes arra, hogy az előtte lévő feji
részt viselő tesztrészt a testvég szelvényeivel kiegészítse, illetve az utána következő farki
vég elé feji véget alakítson ki.
Az ivarszervek felépítése a kevéssertéjűek között nagy változatosságot mutat, így
csak a Lumbricus fajoké kerül bemutatásra (15.4. ábra). A giliszták hímnős állatok. A női
ivarszerv részei a páros petefészek (ovarium), a testüregbe leadott petesejtekbefogadására
szolgáló tölcsér és a petevezető (oviductus). A tölcsér közelében esetleg egy petetartó
tágulat (ostium) is kialakul. A hím ivarszerv része a két pár here (testis), amelyek az egy-
mással kapcsolatban lévő 3 pár ondóhólyag (vesicula seminalis) belső üregében találha-
tók. A heréből kikerülő még éretlen ivarsejtek itt nyerik el teljes fejlettségüket, és itt rak-
tározódnak a lerakásig. Innen indul csillós tölcsérrel a két pár kivezetőcső (ductus
efferens), amely közös ondóvezetővel (ductus deferens) nyílik a külvilágra. A női ivar-
készülékhez tartozik még a párzótárs ondójának befogadására szolgáló ondótartály (re-
ceptaculum seminis). Az ivarszervek valamennyi kivezető járata a hasoldalon nyílik a
külvilágra. A hím ivamyílások és az ondótartályok nyílásai körül a testet borító sertéktől
eltérő nagyságú és alakú, úgynevezett ivarserték lehetnek.
A legtöbb kevéssertéjű gyűrűsféreg ivaros szaporodása jellegzetes módon, kölcsönös
megtermékenyítéssel zajlik (15.5. ábra). Ilyenkor a párba álló állatok hasi oldalukkal
úgy tapadnak egymáshoz, hogy feji végeik ellentétes irányba néznek, így a saját ondót
kölcsönösen egymás ondótartályába tudják üríteni. Ha az ivamyílás és az ondótartály
nem illeszkedik pontosan egymással szemben, mint például a Lumbricus fajoknál, akkor
egy felszíni ivari barázda vezeti a spermiumokat a megfelelő helyre. A bőséges nyálkater-
melés elősegíti az állatok összetapadását a kopuláció ideje alatt, valamint a spermiumok
ivari barázdában való továbbítását. A peték lerakása nem következik be feltétlenül
azonnal a párzás után. A nyereg (clitellum) fontos szerepet tölt be a szaporodás során.
A has oldalán lévő mirigyes megvastagodások a serdülési dudorok. A peték lerakásakor
a nyereg mirigyhámsejtjei által termelt váladékgyűrűből az állat lassan hátrafelé kicsú-
szik, így az a női ivamyíláshoz érve először a petesejteket, majd az ondótartályhoz érve
266
15.5. ábra. A: a giliszták kölcsönös megtermékenyítése, B: a kokon képzésének folyamata
gy: nyálkagyűrű, hi: hím ivamyílás, ib: ivari barázda, ko: kokon, ni: női ivamyílás, ny: nyereg, ot: ondótartály (Grove és
Cowley, Tembe és Dubash nyomán kissé módosítva)
a párzótárs ondóját fogadja magába (15.5. ábra). Ebben a kocsonyás gyűrűben történik
meg a megtermékenyítés. Ezután a gyűrű a feji végről lecsúszva bezárul, felszíni rétege
megszilárdul és egy gubó (cocon) jön létre belőle. A szárazföldi fajok a talajba rakják le
a kokonokat. A fiatal giliszták közvetlen fejlődéssel alakulnak ki. Első táplálékukat a
körülöttük lévő fehérjedús váladék képezi. A Lumbricus terrestris kokonjában több
petesejt is van, de azok közül csak egy-kettő fejlődik ki. Az Eisenia fetida 2-10 kokont rak
3-5 napos intervallumokban. Egy egy kokonban 1-8 petesejt van, amelyekből átlagosan
csak 3-4 fejlődik ki.
A barázdálódás holoblasztikus, inekvális, spirális típusú. Az embrionális fejlődéshez
általában néhány hét szükséges, amely a faji különbségeken túl a környezeti feltételektől
is függ.
A kifejlett gyűrűsférgek képesek regenerálni a testvégeken leszakadt szelvényeket.
A feji végen az első hat-kilenc, míg a hátulsó testvégen ennél több szelvény elvesztését
is pótolni tudják. A nyári száraz és a téli fagyos időszak alatt a földigiliszták egy része
mélyebb talaj-rétegbe (1,5-2 méter) húzódik le, és ott vészeli át a kedvezőtlen körülmé-
nyeket. Mások nem húzódnak ilyen mélyre, hanem inaktív állapotba kerülnek. Ilyenkor
nyáron testük víztartalmának akár 70%-át is elveszíthetik.
15.1.1. Egypár-heréjű kevéssertéjűek (Tubificida)
A kevéssertéjűek közé tartozó Tubificata alosztály képviselőinek sertekötegeiben általá-
ban kettő vagy több, gyakran szőrszerű serte is van. Az egy pár herét a következő
szelvényben egy pár petefészek követi. A klitellum hámrétege egy sejtrétegű.
Csővájóférgek (Tubificidae)
Vízben élő, bentikus férgek. A víz kiszáradásakor mélyebbre húzódnak az iszapba, vagy
betokozódnak. A halak előszeretettel fogyasztják őket. A férgek a Khawia sinensis parazita
laposféreg köztigazdái is lehetnek. Oxigénben szegény és szennyezett vizekben is
előfordulnak.
267
15.6. ábra. Tubificata fajok. A: cső-
vájóféreg (Tubifex tubifex), B: kö-
zönséges televényféreg (Enchytrae-
us albidus) (Több szerző nyomán kissé
módosítva)
Legismertebb faj a csővájóféreg (Tubifex tubifex), amely a
szerves anyagban gazdag vizekben fordul elő nagyobb mennyi-
ségben (16.6. ábra). Akváriumi halak táplálására is használják.
Televényférgek (Enchytraeidae)
Kis testű, általában 5-15 mm hosszú állatok. Az egyes
szelvényeken lévő serték csomókban állnak, de hiányoz-
hatnak is. Vérük színtelen is lehet. Szaporodásuk főleg
ivarosán történik, de egyes fajoknál ivartalan mód is
előfordul. Fontos szerepük van az elhalt növényi anya-
gok feldolgozásában. Nyálmirigyeik erősen lúgos vála-
dékot termelnek, amely jól fellazítja a növényi sejtfala-
kat, és egyben a kisebb, káros növényi fonálférgeket is
elpusztítja. Ezért egyes televényféreg-fajok a biológiai
védekezésben is szerepet játszhatnak. Nedves talajban és
vízben egyaránt előfordulhatnak.
Ismertebb faj a közönséges televényféreg (Enchytraeus
albidus), amely tápanyagban gazdag talajban, trágyában, mo-
csaras területen él. Teste 2-4 cm hosszú, fehér színű (15.6. áb-
ra). Kertészetekben esetenként kárt is tehet. Akváriumi halak
tápláléka is lehet.
15.1.2. Lumbriculida
Mocsári giliszták (Lumbriculidae)
A mocsári gilisztáknak egy-két pár heréjük és 1-3 pár petefészkük van. Sertekötegenként 2-2
sértőjük van. Vízben, a bentoszban élnek, detritusszal táplálkoznak. Képesek harántosztódással
szaporodni.
Ismertebb faj a Lumbriculus variegatus, amely mocsarakban, tavakban, folyókban fordul elő.
15.1.3. Kétpár-heréjű giliszták (Opisthopora)
Az alosztályba tartozó fajok egy részénél ősi bélyegként két pár here és két pár petefészek
figyelhető meg négy egymást követő szelvényben. A törzsfejlődés során később kialakult
többi fajnál ennek az alaptípusnak módosulásai, redukciói jöttek létre.
Földigiliszták (Lumbricidae)
Szelvényenként négy pár serte található. Ivarszerveiket általában két pár here és egy pár
petefészek alkotja.
A legtöbb talajféleségben nagy egyedszámban fordulnak elő. A mezőgazdaságilag
művelt, valamint a gyorsan száradó, homokos talajokban kevesebb fordul elő, mint
például erdős területen. Szerepük igen sokrétű, mert táplálkozásuk és járatkészítő moz-
gásuk révén forgatják a talajrétegeket, javítják a talaj szeliőzöttségét, vízháztartását és
szerkezetét. Jelentős mennyiségű szerves növényi maradványt fogyasztanak, gyorsítják a
szerves anyagok lebontását. Termésfokozó hatásuk számszerűleg is kimutatható. Ürü-
lékük igen kedvező hatású a mikroorganizmusok elszaporodásához. Szárazság vagy fagy
közeledtével a mélyebb talajrétegekbe húzódnak. A fagyra különösen érzékenyek, így
mezőgazdasági területeken igen sok elpusztul a túl későn végrehajtott mélyszántások
következményeképpen. A giliszták köztigazdái lehetnek egyes laposférgeknek és fonál-
férgeknek, illetve vivőgazdái a Syngamus trachea nevű fonálféregnek.
Hazánkban jelenleg mintegy hatvan fajuk ismert.
268
Mocsaras helyeken gyakori a 2-5 cm hosszú simanyergű giliszta (Dendrabaena rubida) és a
2-5 cm hosszú kétéltű giliszta (Eiseniella tetraedra). Mocsaras területeken is előfordul, de inkább
szárazföldi faj a 4-16 cm hosszú tejfehér giliszta (Octolasium lacteum).
Legismertebb a közönséges földigiliszta (Lumbricus terrestris), amely főleg emberi települések
körül, parkokban fordul elő. Testhosszúsága 9-30 cm közötti. Táplálékát a talajfelszínről szerzi be.
Az erdei giliszta (Lumbricus polyphemus) a legnagyobb termetű hazai gilisztafaj, mert test-
hossza általában 15-35 cm közötti, de az ötven centimétert is elérheti. A 3-13 cm hosszú ró-
zsaszínű giliszta (Allobophora rosea) a talaj felső rétegében él. Az egyik leggyakoribb faj a
bűzgiliszta (Allobophora caliginosa). Majdnem minden biotópban előfordul, mérete 5-10 cm
között változik. A horgászok előszeretettel gyűjtik a nádikukacot (Allobophora dubiosa), amely
vízpartokon, nádasokban, vízzel időlegesen borított területeken él. Nagysága a 15-25 cm-t is
elérheti. Trágyában, komposztban fordul elő a 3-8 cm-es trágyagiliszta (Eisenia fetida). A vilá-
gító giliszta (Eisenia lucens) a Bükk és a Zemplén területén korhadó fában, fakéreg alatt él. Kézben
tartva, ingerlésre biolumineszkáló cölomafolyadékot bocsát ki magából, ami sötétben jól megfi-
gyelhető. Alaposfarkú giliszta (Fitzingeriaplatyura) erdővel borított területeken él. Testhosszú-
sága 7-17 cm közötti.
15.2. Piócák (Hirudinoidea)
Földünkön közel 500, hazánkban mintegy húsz faja ismert ennek a magas fokon
specializálódott csoportnak. Előfordulnak a legkülönbözőbb víztípusokban, elsősorban
édesvizekben, kisebb számban tengerekben, de megtalálhatók a szárazföldön is.
Vannak közöttük szabadon élő, elhalt növényi maradványokat, detrituszt evők vagy
ragadozók, amelyek rovarlárvákat, férgeket, puhatestűeket fogyasztanak. Egyesek vér-
szívók, amelyek puhatestűeket, rákokat vagy gerinceseket támadnak meg. Lehetnek
állandó vagy csak ideiglenes paraziták. Táplálkozásmódjuk a különböző fejlődési álla-
potokban eltérő lehet.
Testük alakja megnyúlt, általában hát-hasi irányban lapított, csak néha hengeres.
Testhosszúságuk változó, az alig egy centiméterestől akár több tíz centiméterig is ter-
jedhet. Általában mindkét, de legalább a hátulsó testvégen jól fejlett tapadókorongjuk van.
Színüket a hám alatti kötőszövetben elhelyezkedő pigmentsejtek barna, fekete, zöld
színanyagai okozzák.
Testfelszínükön a szelvénybarázdák között általában három másodlagos gyűrű
(secunder annulus) látható, de lehet ennél kevesebb, vagy további tagolódás révén jóval
több (15.8. ábra). A piócák (vagy nadályok) teste tipikus esetben 34 szelvényből áll. Az
orvosi pióca egy-egy szelvényén 5-5 másodlagos gyűrű van. Bár szelvényválaszfalaik
nincsenek, valódi szegmentumaik többsége tipikus szelvényszerveket tartalmaz.
Bőrizomtömlőjük külső rétegét a vékony kutikulával borított epidermisz alkotja
(15.8. ábra). Testük csupasz, sertéik nincsenek. A hámréteg alatti kötőszövet rostokban
és pigmentsejtekben gazdag. Itt helyezkednek el a fejlett nyálkatermelő mirigyek is. A
tapadókorongban lévő mirigyek ragadós váladékot termelnek, míg a klitellumban ülők
váladékából képződik a kokon. Izomrétegük jól fejlett, vastag, benne körkörös, hosszanti
és átlós lefutású rétegek húzódnak (15.8. ábra). A hát- és a hasoldalt dorzoventrális
izomnyalábok kapcsolják össze. Mozgásuk lehet a tapadókorongok segítségével araszoló,
vagy haladhatnak úszva - a test hullámzó mozgatásával. Ezeket a mozgásmódokat a jól
fejlett hoszszanti izmok biztosítják.
A testüreget mezodermális eredetű bothrioid sejtek töltik ki, így csak szűk cölo-
majáratok ismerhetők fel (15.8. ábra). A bothrioid sejtek hasonló feladatokat láthatnak
el, mint a kevéssertéjű gyűrűsférgek kloragogén sejtjei.
Emésztőszervrendszerük felépítése a különböző táplálkozásmódhoz igazodva
269
15.7. ábra. A: állkapcsos nadályok és B: ormányos nadályok feji
végének felépítése
ák: állkapocs, ga: garat, gi: garat izmai, hd: hasdúclánc, or: ormány, ov:
ormányvisszahúzó izom (Herter, Pfúrtscheller és Scribín nyomán)
differenciált felépítésű. A szájnyí-
lás az elülső szívókorongon talál-
ható. Szájüregükben kitines, fo-
gazott állkapcsok lehetnek, mint
pl. az állkapcsos nadályoknál
(Gnathobdellidea) (15.7. ábra).
Ezekkel ejtik a sebet a gazdaállat
kültakaróján, majd a vérét szív-
ják. A garat (pharynx) különö-
sen a garatos nadályoknál (Pha-
ryngobdellidea) igen erősen izmolt.
Ezek a fajok zsákmányukat általá-
ban egészben nyelik le. Az ormá-
nyos nadályok (Rhynchobdel-
lidea) lehetnek ragadozók, elhalt
szerves anyagot fogyasztók, illet-
ve vérszívók. Garatukban kiölt-
hető ormány (proboscis) helyez-
kedik el (15.7. ábra). A vérszívó
piócáknál a garat falában véralva-
dásgátló, hirudint termelő mirigyek vannak, amelyek váladékukat közvetlenül az
állkapcsok közelébe ürítik. Ugyanitt egysejtű nyáltermelő mirigyek is találhatók. Nyelő-
csövük rövid, és begyszakaszban folytatódik. A ragadozó fajoknál ezen vagy nincsenek,
vagy csak kis számban találhatók kétoldali kitüremkedések. A vérszívók többségénél a jól
fejlett, terjedelmes, hátrafelé haladva egyre nagyobb vakzsákok (diverticulumok) a
szívott vér akár hónapokig történő tárolására szolgálnak (15.8. ábra). Vérszíváskor
testtömegüknek négyszeresét, néha akár tízszeresét is képesek felvenni. A vakzsákokba
kerülő vérből a kiválasztószervek folyamatosan eltávolítják a vizet és a kloridokat, így a
szívott vér besűrűsödik. A vérsejtek hosszú hónapokig, néha több mint egy évig is vál-
tozatlan állapotban maradnak. A középbélben az emésztés igen lassan megy végbe. A
vérszívóknál a vér hemoglobinjának fehérjerészét elsősorban a Pseudomonas hirudinis
baktérium emészti. A ragadozó fajok csekély mértékben termelődő fehérjebontó enzi-
mekkel is rendelkeznek. A csillós hámmal bélelt rektális hólyag után következő végbél
rövid, kutikulával bélelt szakasz. A végbélnyílás a hátulsó tapadókorong előtt, a hátolda-
lon található.
A hazai fajoknak elkülönült légzőszervük nincs, a gázcsere a hámrétegen keresztül
történik. A gázok szállítását a hemolimfa végzi, amely légzőpigmentet is tartalmazhat. A
piócák testük ritmikus mozgatásával frissítik fel a körülöttük lévő vizet.
Keringési rendszerűk a kevéssertéjű gyűrűsférgekhez képest jelentősen módosult.
Ennek egyik oka az egységes testüreg. A cölomazsákok válaszfalai az egyedfejlődés során
redukálódnak. A testüreget a fejlett izomzat és a botrioid sejtek tömege erősen beszűkíti,
így a cöloma csak szűk járatok, öblök formájában maradt meg. Ezekben áramlik a test-
folyadék, esetleg nagyobb vérerek futnak benne. Egyes ormányos nadályoknál néhány
ér még felismerhető. Más nadályoknál az eredeti keringési rendszer telj esen eltűnt, helyét
és szerepét a cöloma üregrendszere vette át. Legfejlettebb üregek a háti és a hasi öböl,
melyekben a testfolyadék elölről hátrafelé áramlik, valamint az oldaledények, amelyek
pulzálásra képesek és a testfolyadékot hátulról előre mozgatják (15.8. ábra). Ahosszanti
járatokat haránt irányúak kötik össze, amelyek elágazva cölomakapillárisokban végződ-
nek. Az ormányos nadályoknak nincs légzőpigmentjük, míg az állkapcsos és garatos
nadályoknak van. Az orvosi pióca testfolyadéka vörös színű a benne oldott állapotban
lévő erithrokruorintól.
270
15.8. ábra. A nadályok testfelépítésének vázlata. A: a feltárt testüreg (az áttekinthetőség érdekében a kiválasztószervek
csak az ábra bal oldalán, az idegrendszer részei csak a test elején vannak feltüntetve), B: az orvosi pióca ivarszerve, C: a
test keresztmetszete (A római számok testszelvényeket jelölnek.)
bé: gyomorbél, bi: bőrizomtömlő, bo: botrioid szövet, dü: dülmirigy, et: elülső tapadókorong, ha: hasi öböl, benne a
hasdúclánc, há: hámréteg, hd: hasdúclánc, he: here, hh: hát-hasi izomköteg, ho: hosszanti lefutású izomréteg, hő: háti öböl,
ht: hátulsó tapadókorong, hü; hüvely, kb: középső bélág, ki: körkörös lefutású izomréteg, oe: oldalsó edény, ok: ondókilövellő
cső, on: ondóvezető, pá: párzószerv, pf: petefészek, pv: petevezető, re; rektális hólyag, se: szelvény, sm; szikmirigy, sz:
szájnyílás, va: vakzsákok, ve: vesécske (Kükenthal-Mathes, Leuckart és Brandes, Mann, Storer és Usinger nyomán
módosítva)
271
Kiválasztószervük általában erősen módosult metanefridium. Leginkább az ormá-
nyos nadályoké emlékeztet a kevéssertéjűekére. Előfordul, hogy nincs csillós tölcsére,
vagy ha az megtalálható, akkor nem a testüregbe, hanem a hasi öbölbe vagy az oldal-
edénybe nyúlik be. A kezdeti szakaszon lévő hólyag fagocitáló sejteket tartalmaz,
amelyek a botrioid sejtekkel és a cöloma amőboid sejtjeivel együtt részt vesznek az exkré-
tum kialakításában. A hólyag egy csatornában folytatódik, amelynek végén, a kiürítő
nyílás előtt, egy tágulat, húgyhólyag lehet. A nadályok nitrogén-anyagcseretermékeik
háromnegyed részét ammónia formájában adjákle, amelyben fontos szerepet kaphatnak
a szimbionta baktériumok is. Nem elhanyagolható azonban a botrioid sejtek kiválasztás-
ban betöltött szerepe sem, mert jelentős mennyiségű bomlástermék tárolására képesek.
Az édesvízi fajok hiperozmotikus vizeletet képeznek, jelezve, hogy a nefridiumoknak
elsődleges szerepe a víztartalom szabályozásában van.
Idegrendszerükben az agydúc kevéssé differenciálódott, kisebb méretű, a hasdúc-
lánccal garatideggyűrű köti össze (15.8. ábra). A tapadókorongokban a dúcpárok egy-
mással összeolvadtak. Tipikus esetben a garat alatti dúcot 6, míg a test végén kialakult
összetett dúcot 7 dúcpár alkotja. A szelvényes elhelyezkedésű dúcpárok között komisz-
szúrák nincsenek, egymással látszólag összeolvadtak. Az egyes szelvényeket a test fe-
lületén tovább tagoló gyűrűk közül az egyik az úgynevezett érzékelő annulus. Az itt el-
helyezkedő érzékelősejtek kapcsolatban állnak az alattuk lévő szubepidermális ideg-
gyűrűvel, amely az ingerületet a hosszanti elhelyezkedésű, asszociációs neuronoknak
adja át. Az egyes testszelvények mint neuroszomita egységek funkcionálnak. A szel-
vényenkénti dúcokból általában két vagy három pár ideg lép ki. A piócákban az em-
lősökéhez hasonló ingerületátvivő anyagok vannak. A táplálékkal teli test falának fe-
szülése a szerotonint termelő sejtek működését gátolja. A piócák táplálkozását, táplál-
kozási viselkedését csupán néhány sejt, és az általuk előállított szerotonin szabályozza.
A szerotoninról kiderült, hogy fontos szerepe van a piócák nyálmirigyei működésének
serkentésében, a testfal izmainak elernyesztése révén pedig lehetővé válik a nagyobb
mennyiségű táplálék felvétele. Fokozott mozgásaktivítást is kivált, ami a táplálékforrás
gyorsabb felkeresését eredményezi. Annak ellenére, hogy viselkedésformáikat kevés
számú idegsejt irányítja, sikerrel képesek megtámadni még az emlősöket is. A garat feletti
és alatti dúcokban neuroszekréciós sejtek vannak, amelyek elsősorban az ivari működésre
hatnak.
Érzékszerveik fejlettebbek, mint a kevéssertéjűeké, mivel sokkal aktívabb életet
élnek. A testfelületen elszórtan előforduló látószervek felépítése viszonylag egyszerű,
pigmentkehely típusú. A látóidegekhez kapcsolódó, a hámrétegben elhelyezkedő látósejtek
kis csoportját a pigmentsejtek kehelyszerűen veszik körül. A fejen lévő szemek több látó-
sejtet tartalmaznak. Kemoreceptoraikkal a zsákmány- vagy gazdaállat testnedveit érzé-
kelik messzebbről, vagy már a szájüregben. Az orvosi piócának jól fejlett hőérzékelése
van, amely segíti a melegvérű zsákmányállat testének felkeresését.
A nadályok hímnős állatok. Az orvosi pióca hím ivarszerveinek része a kilenc pár here
(testis), a rövid kivezetőcsövek (ductus efferens), a közös ondóvezetők (ductus defferens),
amelyek ondóhólyagban (vesicula seminalis) és az izmos falú ondókilövellő járatokban
(ductus ejaculatorius) folytatódnak (15.8. ábra). Ez az ivarpitvarba (átrium) nyílik,
ahová a dülmirigy (prostata) is önti a váladékát, illetve innen tolható ki a párzószerv
(penis). A női ivarszerv egy pár petefészekből (ovarium), a rövid petevezetőkből (ovi-
ductus) és a közös petevezetőből, illetve hüvelyből (vagina) áll. A petevezetőbe a szik-
vagy fehérjemirigy önti a váladékát. A hím ivamyílás a 10., a női all. szelvényben ta-
lálható.
Szaporodáskor a párba állt egyedek párzószervük segítségével kölcsönösen egymás
hüvelyébe juttatják hím ivarsejtjeiket. A spermiumok az állkapcsos nadályok kivételével
kocsonyás csomóban (spermatophora) kerülnek leadásra. A szaporodási időszakban a 9-
272
11. szelvények mirigyeinek fokozott működése eredményeként láthatóvá válik a nyereg
(clitellum). Az itt lévő mirigyek által termelt váladék a fej felé csúszva magába zárja a
kipréselt, megtermékenyített petéket, majd a testről lecsúszva összezárul, kokont képez.
Az ormányos nadályok közé tartozó Glossiphoniidae család fajai gondozzák utódaikat. A
kokont testük hasoldalára tapasztják, a kibúvó fiatalok a testvégi tapadókorongjukkal is
ide kapaszkodnak és itt folytatják fejlődésüket. A szülő összetekeredve védi őket, ha az
oxigénellátottság csökkenne, akkor teste ritmikus mozgatásával frissíti maga körül a
vizet. Petéik teljes, egyenlőtlen, spirális módon barázdálódnak. A kokonból a fiatal ál-
latok 4-6 hét múlva bújnak ki, teljes fejlettségüket pedig 3-4 éves korukban érik el. Növe-
kedésük, fejlődésük során kezdetben gerinctelen állatok, például rovarok vérét szívják,
míg később a gerinceseket is felkeresik.
A nadályok a kevéssertéjű gyűrűsférgekkel ellentétben alig képesek regenerációra.
15.2.1. Ormányos nadályok (Rhynchobdellidea)
Az ide tartozó fajok kiölthető ormánnyal (proboscis) rendelkeznek (15.7. ábra). A feji
tapadókorong a fajok egy részénél hiányzik. Vérük színtelen.
Lapospiócák (Glossiphoniidae)
Lapított testű édesvízi piócák, kevéssé fejlett elülső tapadó-
koronggal. A ragadozók aktív mozgással keresik fel a zsák-
mányul szolgáló férgeket, csigákat. Vannakhulladékevőkis.
A vérszívók csigákat, ízeltlábúakat, hidegvérű gerinceseket
támadnak meg. Ivadékgondozók.
Ismertebb faj az 5-12 mm hosszú íiahordó nadály (HelobdeUa
stagnalis). Nevét onnan kapta, hogy petéit és rövidebb ideig a
fiatalokat is a hasoldalán hordozza (15.9. ábra).
A csigapióca (Glossiphonia complanata) gyakori, 1-3 cm test-
hosszúságú faj. Elsősorban a vízicsigákat támadja meg, azok
testnedvét szívja.
Halpiócák (Piscicolidae)
Megnyúlt testük feji végén jól fejlett szívókorong van.
Rákok és halak ektoparazitái. A petetokokat egyesek vala-
milyen aljzathoz tapasztják, míg mások közvetlenül a gaz-
daállatra helyezik.
Legismertebb faj a 2-5 cm hosszú halpióca (Piscicolageometra),
amely időszakosan ahalak vérét szívja (15.9. ábra). Emellett mint
vérélősködő, a Cryptobia fajok vivőgazdájaként a halgazdaságok-
ban okoz kárt. Testhosszúsága körülbelül 3 cm. A kokonokat a
vízinövényekre, vagy egyéb tárgyakra rakja. A kikelő fiatal piócák
azonnal képesek vérszívásra. Ha sok pióca telepszik meg a halon,
jelentős mennyiségű vért szívhatnak, legyengíthetik a gazdaálla-
tot. A hal testéről levált pióca szívásnyoma sokáig nem gyógyul
be, a vér a sebből lassan tovább szivárog, mivel a véralvadást gátló
hirudin hatása még tart.
15.9. ábra. Gyakoribb nadályok.
A: orvosi pióca (Hirudo medicina-
lis), B: halpióca (Piscicolageomet-
ra), C: fiahordó nadály (Helabdel-
la stagnalis) (Több szerző nyomán
kissé módosítva)
15.2.2. Allkapcsos nadályok (Gnathobdellidea)
Van feji szívókorongjuk is. Fogazott, három pár kitines állkapcsukkal ejtenek sebet az
áldozaton (15.8. ábra). Vérük piros.
273
Valódi nadályok (Hirudinidae)
Legismertebb faj az orvosi pióca (Hirudo medicinalis), amelyet régebben, elsősorban a XVIII.
század végén és a XIX. század elején kiterjedten használtak gyógyászati célokra. Ma a modem
orvostudományban ismét szerepet kaptak egyes speciális sebészeti műtéteknél. Sikerült már
klónozni a hirudin előállításáért felelős gént. Az orvosi pióca a mérsékelt égövi és szubtrópusi
területek sekély vizeiben fordul elő (15.9. ábra). Színezete, a hátoldal mintázottsága változó lehet.
Testhossza akár a 20 cm-t is meghaladhatja.
Hazánkban előfordul még a ragadozó lópióca (Haemopis sanguisuga). Kis testű férgekkel,
csigákkal, rovarlárvákkal táplálkozik. Álló és folyóvizekpartján gyakori faj, melynek testhosszúsága
10 cm-nél is nagyobb lehet.
15.2.3. Garatos nadályok (Pharyngobdellidea)
Állkapcsuk, ormányuk nincs. Kiölthető garatjuk segítségével nyelik le kisebb testű
áldozataikat, főként a férgeket. Vérük piros. Nedves talajban és vízben fordulnak elő.
Erpobdellidae
Legismertebb fajuk a nyolcszemű nadály (Erpobdella octoculatd), amely a hazai vizekben igen
gyakori. Főleg rovarlárvákkal és kis testű férgekkel táplálkozik. Változó színű és rajzolatú, 2-6 cm
testhosszúságú faj.
Ellenőrző kérdések a 15. fejezet témaköreiből
1. Ismertesse a gyürüsférgek (Annelida) valódi szelvényezettségét és a másodlagos testüreg
kialakulását!
2. Hogyan mozognak a gyürüsférgek?
3. Ismertesse a kevéssertéjü gyürüsférgek (Oligochaeta) testfelépítését!
4. Hogyan táplálkoznak a földigiliszták (Lumbricidae) ?
5. Mutassa be a giliszták ivarszerveinek felépítését és szaporodásukat!
6. Ismertesse a kevéssertéjü gyürüsférgek (Oligochaeta) rendszertani felosztását és jellemezze
a gyakoribb hazai fajokat!
7. Mi a földigiliszták jelentősége a talaj szervesanyag-forgalmában?
8. Miben tér el a piócák testfelépítése a kevéssertéjü gyürüsférgekétől?
9. Ismertesse a piócák rendszertani felosztását és jellemezze a gyakoribb hazai fajokat!
274
16. ÍZELTLÁBÚAK (ARTHROPODA)
Ebben a fejezetben bemutatjuk, hogy melyek azok a legfontosabb tulajdonságok, amelyek az
ízeltlábúakat a Föld legsikeresebb, fajokban leggazdagabb csoportjává tették.
A rákok rendkívül sokfélék. A hazánkban élő rákok általában a vizekben lebegve, vagy azok
fenekénéinek, de vannak szárazföldiek (Isopoda) és paraziták (Branchiura, Pentastomida) is.
Tes felépítésükben a többi ízeltlábúhoz viszonyítva az a legfontosabb különbség, hogy két pár
csápjuk van. Ezek a csápok sokféle feladat elvégzésére módosulhattak. Lábaik hasadtlábak: egy
alapízből két, további ízekből álló ág, a belső ág (endopodit) és a külső ág (exopodit) indul ki.
A lábak szerepe rendkívül sokrétű (a táplálék megszerzése, mozgás, párzás, a párzótárs rögzítése,
a petéket védelme, légzőviz mozgatása). Testüket, vagy annak egy részét gyakran a ff hátulsó
részéből kiinduló kettősfalú héj vagy pajzs fedi. A két fal között testfolyadék található, ezért a héj
légzőszervként isfunkcionálhat. A kültakaróba mészsók épülhetnek be, keményítve azt. A legtöbb
rák váltivarú. Jellemző lárvájuk a ff lárva (nauplius) és a tövises lárva (zoea). Fontos
haltáplálék szervezetek is vannak közöttük.
A soklábúak teste megnyúlt, nagyjából hasonló szelvényekből áll. A szelvények egyes csopor-
tokban összeolvadhatnak, ekkor „ikerszelvények” jönnek létre, amelyeken két pár lábat látunk.
Szemeik egyszerű szemek laza halmaza. A ffen, a csápok mögötti területen jellemző alakú kémiai
érzékszerv (temporális szerv) található. A százlábúak ragadozók, a legtöbb ikerszelvényes elhalt
növényi anyagokkal táplálkozik. Szárazföldi állatok.
A rovarok teste kicsi, könnyű. Lábaik és gyakran szárnyaik is vannak, ezért passzív és aktív
módon is képesek gyors helyváltoztatásra. Ilyen módon sikeresen vándorolhatnak el, ha a kör-
nyezeti viszonyok kedvezőtlenre fordulnak. Kitines kültakarójuk szilárd, de a rugalmas interszer-
mentális hártyák mégis sokféle mozgásformát tesznek lehetővé. A belső vázhoz kapcsolódó
speciális harántcsíkolt izmok óriás mitokondriumokat tartalmaznak. Öt ízből álló lábaik
felépítése alkalmazkodott az élőhelyhez, illetve az életmódhoz. Az ősi rágó szájszerv számos
irányba módosult. Ezekkel az új típusú szájszervekkel egyre hatékonyabban lehet a táplálékot
megszerezni. A hosszanti tracheatörzseket harántágak kötik össze, ezzel is segítve a gyorsabb
gázcserét. Erre szükség is van, hiszen a gázáramlás a tracheában lassú. Lehet, hogy a rovarok
teste éppen ezért nem lehet egy bizonyos méretnél nagyobb. A testben keletkező széndioxid mintegy
negyede nem a trachearendszeren, hanem a kültakarón keresztül kerül a külvilágba. A rovarok
váltivarúak. A hímek külső, kitines ivarkészüléke legtöbbször bonyolult és jellegzetes felépítésű,
állandó struktúrájú képlet, ami éppen ezért a fajok rendszerezéséhez kiválóan felhasználható. A
posztembrionális fejlődés sokfélesége szintén nem véletlen: különböző élőhelyek benépesítése, az ott
található források hatékony kiaknázását teszi lehetővé. Posztembrionális fejlődésüket elsősorban
az előagy dúcok közötti területén található neuroszekréciós sftek, az előtöri mirigy és a corpus
allatum által termelt hormonok szabályozzák. A corpus cardiacum hormonokat raktároz. Az
előagy (a gombatestekkel), a középagy és az utóagy alkotja a garat feletti dúcot. Számos
érzékszervük, mint például az érzékelőszőrök, érzékelőkupola, hasadékszerv, tümpanális szerv,
Johnston-féle hallószerv, kordotonális szerv, szaglószerv, hőérzékelő receptorok, pontszem és -
appoziciós vagy szuperpoziciós egyszerű szemekből álló-összetett szem szolgálja tájékozódásukat
a környezetben.
ősi bélyegnek tekintik a rovarok körében a fonalas csápot, rágó szájszervet, járó lábat,
275
gazdagon erezett szárnyat, a kifejlést, mint posztembrionális fejlődési módot. A számyerezet
redukciója jellemző a fiatalabb rovarrendekre, mint például a kétszámyúakra, vagy a hártyás-
számyúakra. A rendekfajszáma nagymértékben különbözik egymástól. A legtöbb faj apoloskák,
bogarak, lepkék, kétszárnyúak és hártyásszámyúak rendjeibe tartozik.
A rovarok és az emberek között rengeteg kapcsolat van. A házi méhet és a selyemlepkét
tenyésztjük, mint akármelyik más házi állatot. Számos faj növényi kártevő, ellenük rendszeresen
védekeznek a mezőgazdák. Nem csupán a szántóföldeken, de üvegházakban, kertészetekben is
okozhatnak károkat. A háztartásokban élő fajok kéretlen, nemszeretem lakótársaink. Vannak
beporzók. Mások ragadozók, parazitoidok. Külső, belső élősködők is akadnak közöttük. Számos
faj az elhalt növényi maradványok lebontásában nélkülözhetetlen.
A pókszabásúak legtöbbje a szárazföldön él, bár vannak vízi fajok is. Jellemző testfüggelék a
tapogató. Ezen igen sok mechanikai és kémiai érzékszerv található, és párzószervvé is módosul-
hat. A csáprágó a táplálékszerzés fontos szerve. Méregmirigy kapcsolódik hozzá. Szájnyílásuk
szűk, ezen keresztül szívják fel a testen kívül részben megemésztett táplálékot. Tracheatüdővel és!
vagy trachea rendszerrel lélegeznek. Nincsenek összetett szemeik. A valódi pókokra igen jellemző
a szövőmirigyek jelenléte. A valódi pókok átalakulás nélkül, az atkák átalakulássalfejlődnek. A
valódi pókok hasznos állatok, mert pusztítják a mezőgazdaságban kártevő rovarokat. Sok
atkafaj szívogalja a növények leveleit, gyökereit, lehet raktári kártevő, ektoparazita is. A páncélos
atkák viszont nélkülözhetetlenek a normális szervesanyag-lebontás során.
Az ízeltlábúak származásával kapcsolatos nézetek egymásnak ellentmondóak. Egyesek
szerint az ízeltlábúakra jellemző bélyegek egymástól függetlenül fejlődtek ki a különböző
csoportokban. így jöhettek létre többek között a rákok, a pókszabásúak és az egy pár, el
nem ágazó csáppal rendelkező ízeltlábúak, az Uniramia csoport tagjai. Ez utóbbiak közé
tartoznak a rovarok és az ezerlábúak. Mások elfogadják azt a nézetet, mely szerint az
ízeltlábúak és a gyűrűsférgek közös őstől származnak, és az ízeltlábúak törzse is egységes,
vagyis monofiletikus törzsnek tekinthető. Egyes újabb molekuláris biológiai adatok sze-
rint viszont az ízeltlábúak polifiletikus származásúak. A kérdés, hogy az ízeltlábúak mo-
nofiletikus vagy polifiletikus eredetűek-e, tehát továbbra is nyitott.
Az ízeltlábúak az állatvilág fajokban leggazdagabb, legnépesebb csoportja. Az eddig
leírt fajok száma már jóval egymillió felett van. A törzsbe tartozó állatok testfelépítése,
élettani folyamatai, életmódjuk, viselkedésük, ökológiai jellemzőik igen változatosak.
Ennek ellenére néhány, nagyon jellegzetes tulajdonság hasonló az ízeltlábúak közé sorolt
fajoknál. Ezek a következők:
A. A zigóta részleges, felületi barázdálódása. Az ízeltlábúakpetéje sok szikanyagot
tartalmaz. A sziktömeg a petesejt közepén helyezkedik el. A barázdálódás, a blasz-
tomérák kialakulása a sziktömeg felszínén következik be.
B. Szelvényekből felépülő test. A gyűrűsférgek teste is szelvényezett, azonban a test
többé-kevésbé hasonló egységekből, gyűrűkből épül fel (homonóm szelvényezettség),
míg az ízeltlábúak esetében a szelvények felépítése különböző (heteronóm szel-
vényezettség) . Az ősibb ízeltlábúak szelvényszáma változó, míg az újabban kialakultaké
állandó. Ezeknél megfigyelhető, hogy állandó számú szelvényből egységes testrészek
(tagmata) alakulnak ki. A rovaroknál például a fej, a tor és a potroh ilyen testrész. To-
vábbi jellegzetesség, hogy a szelvények összeolvadhatnak (tipikusan például az iker-
szelvényeseknél), esetenként a kívülről látható szelvényezettség el is tűnik (például a
pókok; atkák testén).
C. ízelt láb. A névadó testrész, az ízelt láb, rövidebb-hosszabb csövekből áll. Ezeket
puha kitines hártyák kötik össze egymással. Belül komplikált izomrendszer található. Ősi
bélyeg, ha szelvényenként egy pár láb látható. A lábak az evolúció során átalakultak,
módosultak. A lábak száma redukálódott, bizonyos szelvényekről a lábak teljesen el-
276
tűntek. Elágazó, hasadt lábak figyelhetők meg egyes rákoknál. Ezeknél szájszervekké,
párzószerwé, úszószerwé is változtak a járólábak. A pókszabásúak jellegzetes csáprágói
vagy a rovarok szájszerveinek egyes részei szintén lábakból alakultak ki.
D. Kitines kutikula. A férgeknél megismert, puha, kollagén tartalmú kutikulával
borított bőrizomtömlő helyett az ízeltlábúaknál kemény, kitint tartalmazó kültakaró
jelent meg. A kitin az élővilágban rendkívül elterjedt anyag. Előfordul például a gombák
sejtfalában, a gyűrűsférgek és a puhatestűek testében is. Azonban sehol sem nyer annyira
fontos szerepet, mint az ízeltlábúak esetében, ahol a kültakaró egyben a külső váz is.
E. Trachearendszer. A légzőszerv nagyon sok fajnál a testfelszínéről induló és egyre
jobban elágazó csőrendszerből, üregekből, a trachearendszerből áll. A legvékonyabb
hajszálcsövek (tracheolae) tracheavégsejtekben végződnek. E sejtek nyúlványai a testi
sejtekbe vagy azok közé nyúlnak. A trachearendszer tipikus formája a rovaroknál és a
százlábúaknál figyelhető meg. Több kutató véleménye szerint ez a légzésmód az evolúció
során többször és több csoportnál, egymástól függetlenül jött létre.
F. Koncentrálódó hasdúclánc-idegrendszer. Az ízeltlábúak idegrendszere alap-
vonásait tekintve a gyűrűsférgek hasdúclánc-idegrendszeréhez hasonló. Az ősi fajoknál
a szelvényezettség még jól felismerhető, az idegdúcok (ganglionok) különállóak. Jelleg-
zetes változás a törzsfejlődés folyamán az, hogy az egymás mellett található dúcok ösz-
szeolvadnak egymással. A garat feletti dúcokból egységes agy jön létre, mely az ideg-
rendszer legfőbb irányító központjává válik. Morfológiai és működési szempontból is
szoros kapcsolatban áll a garat alatti dúcok egységével. A hasi dúcok összeolvadnak,
egységes idegtömeget képeznek, és a járólábak tövének környékére helyeződnek át.
G. Pontszernek és összetett szemek. Az ízeltlábúak látószerve különleges felépí-
tésű, kitinlencsével ellátott szem. Állhat önmagában, ekkor pontszemnek (ocellus)
nevezik. A pókszabásúaknak csak pontszemeik vannak. A legtöbb csoportban azonban
összetett szemek vannak, melyek egyszerű szemekből épülnek fel. Összetett szemei
vannak a rákoknak, a rovaroknak, az ikerszelvényeseknek és a százlábúaknak. Sok
csoport fajainak pontszemei és összetett szemei egyaránt vannak.
A felsorolt jellegzetességek nem külön-külön, hanem együttesen teszik az ízeltlábúa-
kat igen jellegzetes, másokkal össze nem téveszthető csoporttá.
Törzs: ízeltlábúak (Arthropoda)
Altörzs: Rákok (Crustacea) 16.1. fejezet
Altörzs: Soklábúak (Myriapoda)
Osztály: Százlábúak (Chilopoda) 16.2. fejezet
Osztály: Ikerszelvényesek (Diplopoda) 16.3. fejezet
Altörzs: Hatlábúak (Hexapoda) 16.4. fejezet
Altörzs: Csáprágósok (Chelicerata)
Osztály: Pókszabásúak (Arachnida) 16.5. fejezet
Megjegyzés: A soklábúak altörzsébe tartozó fajokat homonóm szelvényezettség, egy pár
tapogató a fejen, függelék nélküli mandibula, és érzékszervként szolgáló temporális szerv, a csáp
mögött jellemez. A csáprágósok szájnyílásának közelében lévő egyik lábpár csáprágóvá alakult át.
Ehhez méregmirigy is csatlakozik.
277
16.1. Rákok (Crustacea)
16.1. ábra. A folyami rák morfológiája
el: első csáp, ex: exopodit, hp: hátpajzs, jr: járólábak, má: második
csáp, ok olló, pl: párzóláb, po: potrohlábak, rá: rágó, te: telson, vb:
végbélnyílás (Hatsek és Cori nyomán)
A rákok teste jól láthatóan szelvénye-
zett. A szelvények száma változó, álta-
lában 10-20 szelvény figyelhető meg,
de lehet ennél több is. A test felépítése
változatosan alakult. A fej (cephalon),
a tor (thorax) és a potroh (abdomen,
pleon) különböző mértékben össze-
nőhet egymással. A testtájak (tagmata)
azonban a lábak elhelyezkedése és fel-
építése alapján mégis elhatárolhatók
egymástól (16.1. ábra). A rákok tora és
potroha nem homológ a rovarok ha-
sonló testtájaival.
A testfüggelékek (az első pár csáp ki-
vételével) a rákokra jellemző alapvető
lábtípusból, a hasadt lábból vezethe-
tők le. A hasadt láb alapízéhez (basi-
podit) egy - rendszerint rövidebb -
belső ág (endopodit) és egy külső ág
(exopodit) kapcsolódik. Gyakori, hogy
a láb alap-ízéhez egy vékony falú, elága-
zó, lapos kitüremkedés (epipodit) csat-
lakozik. Ez a légzőszerv a kopoltyú. A
hasadt láb két utolsó íze ollóvá (chela
vagy subchela) alakulhat.
A szájszervek is a hasadt lábból ve-
zethetők le (16.2. ábra). Némely eset-
ben a lábakról a külső ág hiányzik. Ek-
kor nem elágazó, pálcalábakról beszé-
lünk. Ha a láb lapos, széles lemezzé,
levéllábbá (phyllopodium) alakult, ak-
kor a légzés feladatát látja el, szűrő-
szervként működik és/vagy úszásra is
szolgál.
A fej hat szelvény összeolvadásából
jött létre, és öt pár függelék található
rajta. Ezen két pár csáp látható. A
csápok mozgásszervek vagy érzékszer-
vek (mechanikai és kémiai érzékelés). Az első csáp egy, a második rendszerint két ágból
áll. Ez utóbbi szervet a csáprágósok tapogatólábával tekintik rokon szervnek, mivel
mindkettőt az utóagy (tritocerebrum) idegzi be. A két pár csáp a rákok jellegzetessége,
más ízeltlábú csoportnál ez nem fordul elő. A szájszerv (gnathopodium) három
lábpárból alakult ki. Változatos felépítésű lehet, bár nem annyira sokféle, mint a rovarok
szájszervei. Egy pár rágó (mandibula) és két pár állkapocs (1. és 2. maxilla) alkotja ezt
a szervet. A torszelvények részben vagy egészben összeolvadhatnak a fejjel, így jön létre
a fejtor (cephalothorax). A tor lábai járásra szolgáló pálcalábak, vagy az úszás szol-
gálatában álló levéllábak és hasadt lábak. Az első tor-lábak gyakran részt vesznek a táp-
lálék vagy a nőstények megragadásában és fogva tartásában. Felépítésükben az állkap-
278
csokhoz hasonlítanak. A potroh jól elkülönülő ízekből áll. Utolsó
szelvényén (telson) gyakran egy pár ízelt függelék (furca) található.
A rákok testét sok esetben a kültakaró kettőzetéből kialakuló
burok fedi. Előfordul, hogy a fej hátsó szegélyéről indul ki, ráborul
a fejre és a torra is. Ekkor fejpajzsnak (carapax) nevezik (Decapoda).
Máskor kétoldalról veszi körül a testet. Ez esetben héj (testa) a neve
(Ostracoda, Cladocera). A kettős fal külső rétegébe sokszor mészsók
rakódnak le, így a fejpajzs vagy a héj kiváló védelmet nyújt az állatnak.
A belső réteg ezzel szemben vékony, hártyás és légzőszervként mű-
ködik.
A kültakaró külső rétege kitines kutikula. A kutikula alatt talál-
ható a prokutikula. A mészsók ebbe a rétegbe rakódnak be. A kül-
takaró legbelső rétegét a hipodermisz képezi. A kültakaróban pig-
16.2. ábra. Egy ágas-
csápú rák (Calanus
sp.) hasadt lábból ki-
alakult rágója
en: endopodit, ex: exo-
podit (Hennig nyomán)
mentsejtek is előfordulnak, a prokutikula külső felszínén.
A rákok bélcsatornája a száj szervekkel kezdődik (16.3. ábra). Ezeket két kitines
lemez határolja: felülről a felső ajak (labrum), alulról pedig az alsó ajak (labium). Az
előbél több esetben rágógyomorrá (mastrix) alakult át, melyben kitinfogak, kitinleme-
zek vagy mészszemcsék is találhatók. Az emésztés fő helye a változó nagyságú középbél.
Egy vagy több pár kitüremkedés van itt, melyek közül az első átalakult középbéli
miriggyé (hepatopancreas). A középbéli mirigy néha alig észrevehető, máskor terjedel-
mes. Igen összetett feladatot lát el: emésztőnedveket termel, felszívja a lebontott
16.3. ábra. Tízlábú rák (Decapoda) fontosabb belső szervei. A: a test hosszmetszete, B: a fej hosszmetszete
ag: agydúc, cs: csáp, er: erek, gy: gyomor, hd: hasdúclánc, he: here, ki: kiválasztószetv, km: középbéli mirigy, kü: kopoltyúüreg,
ny: nyelőcső, ov: ondóvezető, ra; rágó, ri: rágóizom, si: szív, sz: szem, yo: Y-szetv, vb: vékonybél, vn: végbélnyílás (Bames
és Burghause nyomán)
279
tápanyagokat, raktározza azokat, sőt még méregtelenítésre is alkalmas. A középbéli
mirigy glikogént, zsírt és kalciumot is raktároz. A tápanyagok felszívása a középbél falán
és a középbéli mirigyet átszövő csöveken keresztül történik. A végbél az utolsó potroh-
szelvényen nyílik. A rákok körében sokféle táplálkozási típus fordul elő. Néhány példát
sorolunk csak fel: a legelterjedtebbek azok a növényevő fajok, amelyek szűrik a vízben
lebegő növényeket (fitoplanktont). Az ágascsápúrákok (Cladocera) szűrőkészülékéta
3. és a 4. lábpár alkotja, az evezőlábú rákok (Copepoda) esetében pedig a szájszervek
szűrik a táplálékot. A szűrés az ágascsápúaknál inkább passzív folyamat. A lábakról
kiinduló kitinszőrök meghatározott lyukbőségű szitát alkotnak, amelyen az áramló
vízben úszó táplálékdarabok (algák, egysejtűek, elhalt növényi részek stb.) fennakadnak.
Az evezőlábúak elsősorban szájszerveik aktív mozgása segítségével veszik fel a tápláléku-
kat. Vannak vegyestáplálkozású rákok, melyek elhalt növényi és állati anyagokat is esznek
(Decapoda). Előfordulnak körükben ragadozók (egyes Copepoda fajok), de ektoparaziták
(Branchiura) is.
A planktonikus rákok elsősorban a kültakarón keresztül, külön légzőszerv nélkül
bonyolítják a gázcserét. A legtöbb faj azonban kopoltyúkkal rendelkezik. A kopoltyúk
vagy a torlábak függelékei, vagy a test oldallemezeiből emelkednek ki. A lábak mozgása
biztosítja, hogy a kopoltyúkhoz mindig friss, oxigénben dús víz áramoljék. A szárazföldi
rákok (egyes Isopoda fajok) a trachearendszerhez hasonló csőhálózattal rendelkeznek.
Ennek segítségével lélegeznek.
A rákok keringési rendszere igen különböző módokon alakult. Vannak zárt kerin-
gési rendszerű fajok, de olyanok is, amelyeknek testéből ez a szervrendszer teljesen
hiányzik. Rendszerint azonban nyílt keringési rendszerük van (16.3. ábra). Ennek köz-
pontja a hosszú, megnyúlt, zsákszerű szív, melyhez előre és hátra irányuló, elágazó erek
csatlakoznak. A szív oldalán nyílások (ostia) találhatók. Számuk 1-18 pár. A testfolya-
dékban színanyagok (hemocianin, néha hemoglobin) is előfordulhatnak, de meglehető-
sen ritkán. Alakos elemek a testfolyadékban az amőboid, fagocitáló sejtek. Sérüléskor a
testfolyadék alvadását segítik.
Kiválasztószerveik a vesécske (nephridium) típusba sorolhatók, bár erőteljesen
módosultak (16.3. ábra). Felépítésüket és eredetüket tekintve igen hasonlítanak a csáp-
rágósok csípőmirigyeihez. Nevüket elhelyezkedésükről kapták. A 2. csáp tövében lévő
vesécskét csápmirigynek, a 2. állkapocs tövében találhatót állkapcsi mirigynek ne-
vezik. A lárvákban mindkét típus megtalálható, a kifejlett állatokban azonban együttesen
ritkán fordulnak elő. Meg kell említeni, hogy a kopoltyúk (miként a halaknál is) fontos
szerepet játszanak az ammónia kiválasztásában. A rákok testfolyadékuk ionegyensúlyát
elsősorban a kopoltyúkon keresztül szabályozzák. A vesécskék jelentősége az ioncseré-
ben csekély, feladatuk elsősorban a szervezet vízháztartásának szabályozása. Eltérően a
rovaroktól, a rákoknál Malpighi-edények sohasem fordulnak elő. Viszont - hasonlóan a
többi ízeltl ábúhoz- az anyagcsere szilárd végtermékeit fagocitáló sejttípus (nephrocyta)
itt is megtalálható.
A rákok ivarszervei nagyon egyszerű felépítésűek (16.3. ábra). Mind a hímek, mind
a nőstényekivarszerve páros, hosszúkás, zsákszerű szerv. Az ivarszerveka test hátoldalán,
a bélcsatoma mellett helyezkednek el. A legtöbb rákfaj váltivarú, de az ivari dimorfizmus
nem feltűnő, a nemeket nehéz első látásra megkülönböztetni. A hímek párzószerwel
(penis) rendelkeznek, vagy valamelyik lábukkal (gonopodium) juttatják spermájukat a
nőstény ivarnyílásába. A spermiumoknak rendszerint nincs farkuk, mozgásra nem ké-
pesek. Zigótáik teljes vagy felületi barázdálódással fejlődnek tovább. Embrionális fejlő-
désük néha a gyűrűsférgekére emlékeztet. Posztembrionális fejlődésük lehet közvetlen
(epimorphosis) vagy közvetett (metamorphosis). A közvetlen fejlődés másodlagos, le-
származtatott jelleg. Tipikus a közvetett fejlődés, annak is az a módja, amikor a petéből
kikelő lárva szelvényszáma kevesebb, mint a kifejlett állaté, és a lárvák testfelépítése,
280
életmódja eltér a kifejlett állatokétól
(anamorphosis). A rákoknak számos,
rendkívül eltérő lárvatípusa van, ezért
a következőkben csupán a hazánk-
ban leggyakoribb formákról lesz szó
(16.4. ábra). A legősibb lárvatípus a
naupliusz. Tojás formájú kis állat,
melyen a két pár csáp, az egy pár rá-
gó és a naupliusz szem jól felismer-
hető. A naupliuszokból fejlődnek ki
fokozatos vedlések után a már több
szelvénnyel és lábbal rendelkező me-
tanaupÜuszok. A naupliusz lárvák
egy speciális, gyakori típusa a ciprisz
(cypris) lárva. A magasabb rendű
rákok jellemző lárvája a tövises lár-
va (zoea). Fejpajzsáról hosszú, erős tövis nyúlik ki. A torán 2-3 hasadt láb található. A
tövises lárvákhoz hasonlítanak a mizisz (mysis) lárvák. Ezeken azonban már az összes
torláb megtalálható. A rákok körében előfordul az ivadékgondozás is. A nőstények a
potrohlábaikhoz ragasztva, azok védelmében, vagy a héj alatti költőüregben hordják
petéiket egészen a kikelésig. Egyesekre jellemző a szűznemzés, valamint a nemzedékvál-
takozás is.
A rákok hasdúclánc idegrendszerrel rendelkeznek (16.3. ábra). Az ősi felépítés
azonban sok csoportban módosult, és összeolvadások is megfigyelhetők. Kivételesen az
összes idegdúc egyetlen hatalmas garat alatti dúcot alkot. Igen jellemzőek a központi
idegrendszerben a gyors ingerületvezetésre képes óriás idegsejtek. A rákok garat feletti
dúcában számos neuroszekréciós sejt található. Ezeken kívül belső elválasztású mirigyek
is találhatók a szervezetükben (16.3. ábra). Ezek a következők: szinuszmirigy, X-szerv
(neurohaemalis szervek), petefészek, androgén mirigy, Y-szerv. A rákok vedlésének
mechanizmusa alaposan kutatott. A szemnyélben található X-szerv a vedlést gátoló hor-
mont (MIH) termeli. Innen a hormon a szinuszmirigybe, majd a vedlés alatt a test-
folyadékba kerül. Ezzel megakadályozza, hogy az Y-szerv által termelt vedlési hormon
(crustecdyson) kifejtse hatását. A színváltozást két hormon szabályozza. Az egyik hor-
mon (PDH) hatására a pigmentsejtekben levő színanyagok a sejtben szétoszlanak, ha
pedig a másik hormon (ECH) kerül túlsúlyba, akkor a citoplazmában koncentrálódnak.
Érdekes, hogy a színváltozás gyakran napi ritmust követ, nappal sötétek (így védekeznek
az UV-sugárzás ellen), éjjel világosak az állatok. Ez a ritmus akkor is megmarad, ha
állandó sötétségben tartják őket. Az ivarszervekben termelődő hormonok hatására
fejlődnek ki a másodlagos nemi jellegek (vö. 4.7. fejezet).
A rákok érzékszervei jól fejlettek. A lárvakorban jellemző naupliusz szem páratlan,
egyszerű felépítésű, rendszerint a fej közepén található. Általában három, néha négy
egyszerű szem (ocellus) szorosabb-lazább kapcsolatából áll. Valószínűleg a fény irányá-
nak érzékelésére szolgál. A rákok fejének két oldalán legtöbbször jól fejlett összetett
szemek vannak, melyek számos esetben rövidebb, hosszabb nyélen (peduncle) ülnek.
A szemek képlátásra, sőt sokszor a színek megkülönböztetésére is alkalmasak. A me-
chanikai érzékszervek elsősorban a csápokra és a lábakra koncentrálódtak. Ugyanitt
találhatók a kémiai érzékszervek sorba rendezett érzékszőrei is. A magasabb rendű rákok
a csápok tövében, a potrohlábak eredésénél vagy az utolsó potrohszelvényben található
helyzetérzékelő szervvel (statocysta) rendelkeznek.
281
Osztály: Phyllopoda
Rend: Csupasz levéllábúrákok (Anostraca) 16.1.1. fejezet
Rend: Pajzsos levéllábúrákok (Notostraca) 16.1.2. fejezet
Rend: Ágascsápúrákok (Cladocera) 16.1.3. fejezet
Osztály: Maxillopoda
Rend: Haltetvek (Arguloida) 16.1.4. fejezet
Rend: Féregatka-alakúak (Pentastomida) 16.1.5. fejezet
Alosztály: Kagylósrákok (Ostracoda) 16.1.6. fejezet
Alosztály: Evezőlábú rákok (Copepoda) 16.1.7. fejezet
Osztály: Magasabbrendű rákok (Malacostraca)
Rend: Tízlábú rákok (Decapoda) 16.1.8. fejezet
Rend: Hasadtlábú rákok (Mysidacea) 16.1.9. fejezet
Rend: Edriophthalma
Alrend:Aszkarákok (Isopoda) 16.1.10. fejezet
Alrend:Felemáslábú rákok (Ámphipoda) 16.1.11. fejezet
Megjegyzés: A Phyllopoda osztály fajainak testszelvény száma változó, a testet boríthatja héj, a
toron legtöbbször levéllábak, az utolsó testszelvényen villás függelék található. A Maxillopoda
osztály jellemzője, hogy az első csáp nem ágazik el, hat torszelvényük van és az öt potrohszel vényen
nincsenek végtagok. A magasabbrendű rákok teste majdnem mindig 19 szelvényből áll (fej: 5,
tor: 8, potroh: 6 szelvény). Jellemző a hátpajzs. Lárvájuk rendszerint zoea típusú. Az Edriophthal-
ma rend fajainak nincs hátpajzsuk, a torlábak egy ágúak, az összetett szemnek pedig nincsen
nyele.
16.1.1. Csupasz levéllábúrákok (Anostraca)
Körülbelül kétszáz faj tartozik a csupasz
levéllábú rákok közé. Megnyúlt testű, 1-
10 cm hosszú állatok (16.5. ábra). Hát-
pajzsuk hiányzik. A hímek második csáp-
ja erős fogókészülékké alakult át, mellyel
szilárdan tartják a nőstényeket párzás
közben. A tor 11 (ritkán 17, vagy 19)
szelvényből áll. Mindegyik szelvényhez
egy pár levélláb csatlakozik. Ezek moz-
gásra, légzésre és a táplálék kiszűrésére
egyaránt alkalmas szervek. Hasdúclánc-
idegrendszerük ősi felépítésű, a dúcok
összeolvadása nem figyelhető meg. A hí-
mek gyakran csupán elenyésző számban
fordulnak elő. Kisebb édesvizű vagy
sósvizű tavak lakói, de a tengerekben
nem élnek.
A sóféreg (Artemia salina) különböző jel-
legű, elsősorban erősen sós vizekben él. A legnagyobb sótűrésű rákfaj. Elterjedt hazánkban a
közönséges tócsarák (Branchipus stagnalis). Teste nyúlánk. Összetett szemei nyélen ülnek. A
nőstény petéit költőtáskában hordja. Haltáplálék.
282
16.1.2. Pajzsos levéllábúrákok
(Notostraca)
A pajzsos levéllábúrákok mintegy húsz fajt
magába foglaló csoportot alkotnak. Az álla-
tok 3-6 cm hosszúak. Nevüket a hatalmas
fejpajzsról (carapax) és lapos, széles lá-
baikról kapták (16.6. ábra). A lábak száma
egészen 70 párig terjedhet. Jellemző, hogy a
potroh hátulsó szelvényein több pár láb is
lehet. A hatalmas, lapos fejpajzs szinte az
egész testet befedi. Az első pár torláb vége
hármasán elágazik, hosszú, ízelt nyúlványok-
ban végződik.
Az érzékelésben, a mechanikai ingerek
felfogásában játszik szerepet. A második pár
csáp redukálódott. Fejükön két összetett
szem és négy pontszem található. A potroh
végéről két igen hosszú, sok ízből álló nyúl-
vány (furca) indul ki. Erős rágó szájszer-
vükkel virágos növényeket rágnak meg.
Hazánkban a rizsföldeken mindenfelé gyakori
kártevő a fiatal rizsnövények gyökerét elrágó, a
palántákat a talajból kitúró, 2-4 centiméter hosszú
16.6. ábra. Nyári pajzsosrák (Triops cancriformis)
testfelépítésének vázlata (Kaestner nyomán)
nyári pajzsosrák (Triops cancriformis). Hasán
és hátára fordulva egyaránt jól úszik. Csápjai rövidek, a hátpajzs alól az elülső lábakról kiinduló
elágazó ostorok nyúlnak ki.
16.7. ábra. A: a kis vízibolha szervezetének
vázlata. B: efippiuma, C: ciklomorfózisa
(a: nyári forma, b: tavaszi és őszi forma).
bé: bélcsatoma, el: első csáp, hé: héj, iz: izmok,
kő: költőüreg, má: második csáp, pe: peték,
pv: potrohvég, si: szív, sz: szem (Sars nyomán)
B
16.1.3. Ágascsápúrákok
(Cladocera)
Több száz fajt sorolnak ebbe a csoportba. Testnagy-
ságuk néhány tizedmillimétertől egy centiméterig
terjed. Testüket jobbról és balról egy-egy héj veszi
körül, ezek rendszerint a teljes testet elfedik. A két
héjat erős izom tartja össze. A nőstények testének
hátoldala és a héj között található a költőüreg, amely-
ben a peték fejlődnek. A 2. pár csáp igen erősen
fejlett, szőrökkel gazdagon ellátott és elágazó szerv
(16.7. ábra). A lebegő állat lefelé csapva ezzel az
evezővel felemelkedik, majd a következő csapásig
lefelé süllyed, így ugráló mozgás jön létre (vízibol-
hák). A potroh vége jellegzetes alakulású, fontos
határozóbélyeg. Bélcsatomájuk kevéssé tagolt, eny-
hén kanyarulatos cső. Táplálékukat a vízből szűrik
ki. Mivel sok algát fogyasztanak, gátolják a vizek
eutrofizálódását. Szívük (ha van) egyszerű, rövid, a
hátoldalon található. A lábakon levő kopoltyúkkal és
a héj belső falán keresztül lélegeznek. Kiválasztó-
szervük az állkapcsi mirigy. Kedvező körülmények
között szűznemzéssel szaporodnak, ezért rövid idő
283
alatt nagy egyedsűrűséget érhetnek el. Posztembrionális fejlődésükkifejlés (epimorfózis).
Ha a környezeti tényezők kedvezőtlenné fordulnak, megjelennek a hímek, megterméke-
nyítik a nőstényeket. Ezek tartós petéket termelnek, amelyek egy jellegzetes alakú tokban
(ephippium) (16.7. ábra) találhatók. Az efippiumban levő peték (számuk általában 1-
2 darab) képesek átvészelni a vizek kiszáradását, a szárazságot, a hideget, sőt a halak és
a madarak bélcsatomáján is áthaladhatnak anélkül, hogy a védőburkot az emésztőenzi-
mek feloldanák. A kagylós ágascsápú rákok testalakja az egymás után következő gene-
rációkban lassan változhat (cyclomorphosis), és szabályos évszakos ciklust mutathat
(16.7. ábra). Hasonló változásokat a kerekesférgeknél és más állatoknál is megfigyeltek.
Bár többféle magyarázat van a jelenségre, pontos okát nem ismerik.
A legtöbb faj az ágascsápúrákok (Cladocera) alrendjébe tartozik. Igen elterjedt a tapadó
lencserák (Limnadia lenticularis) és számos vízibolhafaj (Daphnia spp.). Ez utóbbiak fontos
haltáplálékot jelentenek a halgazdaságokban és nélkülözhetetlenek az ivadéknevelésben. Nagy
mennyiségű algát fogyasztanak, ezáltal az eutrofizációt csökkentik.
16.1.4. Haltetvek (Branchiura)
Megközelítően nyolcvan faj tartozik a haltetvek közé. Ezeknek a lapított testű, ovális alakú
állatoknak 0,5-3 cm a testhosszúságuk (16.8. ábra). Testfelépítésük az ektoparazita élet-
módhoz alkalmazkodott. Fejpajzsuk széles, hosszú. Csápjaik rövidek, az 1. csápon erős
fog látható, amely a kapaszkodást segíti. Szúró-szívó szájszerveik a felső és alsó ajakból, va-
lamint a szúrósertévé alakult rágóból épülnek fel. Az első pár állkapocs alapízei tapadó-
koronggá módosultak. A tapadókorong segítségével szívja rá magát a haltetű a táplálékul
szolgáló hal kültakarójára. Páros összetett szemeik és két csoportban álló 3-3 naupliusz
szemük van. A torról négy pár elágazó láb indul ki. Ezekkel úszik az állat. Bélcsatomájuk
terjedelmes, üregét a középbélből kiinduló
zsákok növelik. Mivel a halakon csak a
vérszívás idejéig tartózkodnak, ezekben a
kitüremkedő üregekben raktározzák a táp-
lálékot. A szívből a fej felé hosszú aorta
indul ki. A véráramlást a potroh mozgása is
segíti. A nőstényeket a hímek rendszerint
vérszívás közben termékenyítik meg. A pe-
ték a vizek aljzatára hullanak. A naupliusz
lárvák már a peteburkon belül kialakulnak,
ezért a kikelő állatok a kifejlett alakokhoz
hasonlítanak. Elsősorban halakat károsíta-
nak, de a gőtéket is megtámadják.
Halastavainkban gyakori kártevő a ponty-
tetű (Argulusfoliaceus). Vérszívása során a ha-
lak kültakaróját felsérti, és utat nyit a másodla-
gos fertőzések (baktériumok, gombák) számá-
ra. További károkat okoz azzal, hogy a halak
16.8. ábra. Haltetű (Argulu, sp.) szervezetének vázlata vérében é10 vírusokat és baktériumokat is tér-
(Kaestner nyomán) jCSZti.
16.1.5. Féregatka-alakúak (Pentastomida)
Féregszerűen kinéző, de embrionális fejlődésük alapján az ízeltlábúakhoz közelebb álló
fajokat egybefogó csoport. Néhány centiméteres testnagyságú állatok tartoznak ebbe a
törzsbe. Kivétel nélkül paraziták, rendszerint gerincesekben, mindenekelőtt hüllőkben
284
élősködnek, de az emlősöket is gyakran fertőzik. Különösen sok faj él a kígyók légutaiban.
Kültakarójuk gyűrűzött, kitines kutikulával borított bőrizomtömlő. Szájnyílásuk kör-
nyékén két pár csökevényes, karmokkal ellátott lábpár található, mellyel a gazda légző-
szervében megkapaszkodnak, de szükség esetén lassan mászni is képesek. Bélcsatomá-
juk egyszerű felépítésű, kevéssé tagolt, egyenes cső. Élősködő életmódjuk következtében
számos szervük visszafejlődött (keringési szervrendszer, szemek). Kiválasztó szervrend-
szerüket speciális sejtek, kloridsejtek alkotják. Idegrendszerük erősen redukálódott has-
dúclánc idegrendszer, mindössze az előtestben található néhány idegdúcpár. Ezek ideg-
zik be az érzékszerveket. Az endoparazita életmódnak megfelelően fejlett mechanikai és
kémiai érzékszervekkel rendelkeznek. Váltivarúak, többféle lárvatípus figyelhető meg a
posztembrionális fejlődés során. Gyakori a köztigazdával (főleg halak, emlősök) történő
fejlődés is.
A kutyák orrférge (Linguatula serrata) a szakirodalomban sűrűn emlegetett, kozmopolita faj,
amelyet azonban hazánkban már évtizedek óta nem mutattak ki. A sirályok orrférge (Reighardia
stemae) egész Európában elterjedt.
16.1.6. Kagylósrákok (Ostracoda)
Mintegy tizenötezer kagylósrakfaj ismert napjainkban. Mivel aranylag jól fosszilizalód-
nak, a kihalt fajokról is sok információ áll rendelkezésre. Igen kicsi állatok, testnagyságuk
egy milliméter körüli (16.9. ábra). Testüket két oldalról a kagylók alakjához hasonló két
héj veszi körül. Ezeket záróizmok tartják össze.
Nyugalmi állapotban a héjak zárva vannak és az egész testet beborítják. Ilyen
állapotban a szárazságot is átvészelik. Törzsük (a tor és a potroh együtt) igen rövid, a tor
legfeljebb két pár lábat visel. Szervezetük erősen leegyszerűsödött. Jellemző a bőrlég-
zés, a csápmirigyek és az állkapcsi miri-
gyek megléte és az, hogy a hasdúclánc-
idegrendszer csökevényes. Lárváik cip-
risz lárvák. Rendszerint tengerek vagy
édesvizek fenekén élnek. Különféle táp-
lálékot fogyaszthatnak. Szűrő tevékeny-
ségükkel hozzájárulnak a vizek tisztítá-
sához. Különös jelentőségük, hogy se-
gítségükkel a földkéreg egyes rétegeinek
korát meg lehet állapítani. Ezért a kő-
olajkutatások során kagylósrákh átárazá-
sokat is végeznek.
A legnagyobb édesvízi faj a magyar
kagylósrák (Hungarocypris madará-
szi).
16.9. ábra. ábra. Kagylósrák (Cypris sp.) szervezetének
vázlata (Zenker és Gerstácker nyomán)
16.1.7. Evezőlábú rákok (Copepoda)
Fajgazdag, több ezer fajt magába foglaló, általában 1-2 mm testhosszúságú állatok
csoportja. Testszelvényeik jól láthatók, mert a fejpajzs vagy héj hiányzik (16.10. ábra).
A fejtor a fej szelvények és az első vagy első két torszelvény összeolvadásából jött létre.
Az első csáp rendkívül hosszú. Mint lebegtető szerv játszik szerepet. A második csáp
rövid. A toron öt pár rövid, elágazó úszóláb látható. A potroh végén egy pár jellemző
alakú függelék (furca) helyezkedik el, de a lábak hiányoznak (16.10. ábra). A szabadon
élő fajok általában algaevők: a vízből szűrikki táplálékukat. Számos ragadozó, endoparazita
és ektoparazita evezőlábú rákot is ismerünk. A paraziták szájszerve erősen módosult,
285
szúró-szívó típusú. Rendszerint a kültakarójukon ke-
resztül lélegeznek és a szívük hiányzik. Csak egyetlen
naupliusz szemük van, az összetett szemek hiányoz-
nak. Jelentős az ivari dimorfizmus. A hímek a nősté-
nyekhez viszonyítva sokszor igen kicsinyek. A megter-
mékenyített petéket a nőstények egy vagy két pete-
zsákban a potroh végéhez ragasztva hurcolják maguk-
kal. A posztembrionális fejlődés során számos naupli-
usz, metanaupliusz és copepodit stádiumon mennek
keresztül, mire az ivarérettséget elérik. Tengerekben
és édesvizekben egyaránt nagy tömegben fordulnak
elő, ezért fontos haltáplálékot jelentenek, különösen
az ivadékok számára. Kártételt jelent, hogy külön-
böző férgek (pl. Ligula intestinalis) köztigazdái lehet-
nek.
Egyes parazita fajok (Lemea cyprinacea) kifejlett egyedei
olyan nagymérvű morfológiai változásokon estek át az evo-
lúció során, hogy evezőlábú rák voltukat csupán poszt-
embrionális fejlődésük alatt megjelenő lárvaformáik igazol-
ják. Hazánkban az állóvizekben mindenütt gyakoriak a
Cyclops és a Diaptomus nemzetség fajai.
16.1.8. Tízlábú rákok (Decapoda)
16.10. ábra. A: evezőlábú rák (Diaptomus
sp.) testfelépítése,B.*egyCyclopsfajpot-
rohvége
el: első csáp, ev; evezőláb, iv: ivamyílás, má:
második csáp (Marshall és Orr nyomán)
A legismertebb rend, mintegy tízezer rákfaj tartozikide. A test hosszúsága 1 millimétertől
60 centiméterig terjed. Jellemző a fejből és a torszelvényekből kialakult, a fejtort beborító
egységes fejpajzs. A fejpajzs oldalsó lemezei befedik a kopoltyúüreget. A csápok igen
hosszúak, elsősorban tapintószervek. Az első csáp tövében található a páros helyzet-
érzékelő szerv (statocysta). A második pár csáp belső ága (endopodit) fejlett, sok ízből
áll. A fej toron levő lábak közül az első három pár az állkapcsot (maxilla) alkotja, a hátulsó
öt pár pedig járólábbá alakult. Innen ered a rend neve. A járólábak közül az első pár
gyakran hatalmas ollót visel, de a következő lábak is lehetnek ollósok (16.11. ábra). A
potroh szelvényei világosan láthatók. A potroh lehet hosszú, izmos, ekkor az utolsó
szelvényhez több kiszélesedő kitinlemez (uropodit) csatlakozik. Ha a potroh rövid, csö-
kevényes, akkor ezek a lemezek is hiányoznak. A nőstények potrohlábaik között
hordozzák és védik a petéket. A tízlábú rákok potrohlábak segítségével hajtják a kopoltyú
felé az oxigénben dús vizet. Kutikulájuk vastag, kemény a berakódott kalciumsóktól.
Bélcsatomájuk viszonylag tagolt. Jól fejlett rágógyomor és egy kitinszőrőkkel borított
szűrőgyomorszakasz is található rajta. Középbéli mirigyük hatalmas. Vegyes táplálékon
élnek, a dögöket is megeszik. Keringési rendszerük központja a rövid, izmos szív,
ahonnan a kopoltyúk irányába jól fejlett artériarendszer indul ki. Vérükben hemocianin
található. Kopoltyúkkal lélegeznek, de mivel a kopoltyúüreget borító kitinlemez belső
fala vékony, hányás, ezért ezen keresztül is lehetséges a gázcsere. Kiválasztószerveik a
csápmirigyek és az állkapcsi mirigyek (metanephridium). Érzékszerveik közül a már
említett tapintószervek mellett fejlettek az összetett szemek, melyek gyakran rövid nyélen
ülnek. Váltivarú állatok. A petéből vagy tövises lárva (zoea) bújik ki vagy a lárvakori
átalakulások már a peteburkon belül megtörténnek. Ekkor a kikelő fiatal rák a szülőkhöz
hasonlít. Gyakori az ivadékgondozás. Egyes tízlábú rákok hasznosak, mivel eltakarítják
az elpusztult növények és állatok maradékát. Húsuk emberi táplálkozásra alkalmas, ezért
az édesvízi fajok mesterséges tenyésztésével is próbálkoznak. A természetes vizekben a
betegségek és a vízszennyezés következtében populációik száma megcsappant.
286
16.11/ ábra. A folyami rák anatómiája
bé: bél, el: elülső csáp, gy: gyomor, iz: izom, km: középbéli mirigy, ko. kopoltyúlemezek, má: második csáp, sir szív, sz: szem,
vé: végbél (Kükenthal-Matthes nyomán)
Hazánkban elterjedt a folyami rák (Astacus astacus). Lassú folyású vizekben a part közelében él.
Elhalt növényi és állati részeket, puhatestűeket, rovarlárvákat egyaránt fogyaszt. A nőstények a
petéket a potroh hasoldalára ragasztva hordozzák, ahol azok védett helyen vannak és a lábak
mozgásnak következtében a friss oxigénellátás biztosított. A kecskerák (Astacus leptodactylus) a
síkvidéki vizekben, tavakban, holtágakban fordul elő elsősorban. Jellegzetessége a hoszszú, keskeny
olló. Újabban terjed a cifrarák (Orconectes limosus). Napjainkban komoly erőfeszítéseket tesznek
több (külföldről származó) tízlábú rákfaj nagyüzemi tenyésztésének kidolgozására.
287
16.12. ábra. Hasadtlábú rák testfelépítése
(Dudich és Loksa nyomán)
16.1.9. Hasadtlábú rákok
(Mysidacea)
Mintegy félezer, 0,5-3,5 cm hosszúságú faj
tartozik ebbe a csoportba. Hátpajzsuk rövid,
nem nőtt össze az utolsó négy torszelvénnyel.
Potrohúk vékony, hosszú és véglemezben
végződik (16.12. ábra). Az első csáp két hosz-
szú ágra válik szét. A torlábak meglehetősen
egyforma hasadtlábak. Szemük nagy, moz-
gatható nyélen ülő összetett szem. Lárváik
mizisz lárvák. Általában tengeri állatok.
A pontusi tanúrák (Litnnotnysis benedeni) a Dunában és a Balatonban is él. Jó haltáplálék.
16.1.10. Ászkarákok (Isopoda)
Számos, mintegy ötezer faj tartozik az ászkarákok rendjébe. A néhány milliméteres vagy
centiméteres állatok teste ovális, hát-hasi irányban kissé lapított. Hátpajzsuk nincs, a test
szelvényezettsége igen jól látszik (16.13. ábra) A szelvényekről oldalirányban gyakran
széles kitinlemezek indulnak ki. A torszelvények közül csak azelső (ritkán a másodikis) olvad
össze a fejjel fejtorrá. A csápok nem ágaznak el. Az első pár csáp rövidebb, mint a második
pár. Lábaik - az első torláb kivételével - nagyjából hasonló felépítésűek. Rágó szájszervük
felépítésében gyakran két állkapocs vesz részt. A tapogató (palpus) viszont hiányzik. A hat
potrohszelvény részben összeolvad, ilyenkor rövid. A potrohon öt pár hasadt láb található.
A láb (exopodit) külső ága légzőszerv, a belső pedig ezt védi. A külső ágon található a
tracheákkal átszőtt légzőszerv (fehér test). A szárazföldi fajokon a külső ágban bemélyedő,
elágazó, vékony falú csöveket találunk, melyek a légzés feladatát látják el. A kültakaró
bőrszerű. Rendszerint növényevők vagy avart fogyasztanak, de paraziták is lehetnek.
má
16.13. ábra.Ászkarák testfelépí-
tése hasi oldalról nézve
lé: légzőszerv, má: második csáp,
pe: peték (Eisenbeín és Wichard
nyomán)
Szimbionta baktériumaik segítségével bontják a cellulózt. Az
emésztés fő helye az utóbél. A szívük részben-egészben a pot-
rohban található. Összetett szemeik a fej tetején helyezkednek el.
Általában váltivarúak. Posztembrionális fejlődésük (a parazita
fajokat kivéve) epimorfózis. Az egyetlen rákrend, amelyben a
vízi fajok mellett vannak szárazföldiek is. Ezek a fajok rendsze-
rint éjszakai életmódot élnek, és nedves helyeken (avar között,
pincékben, kövek alatt) fordulnak elő. Ellentétben a rovarok-
kal és a pókokkal, az ászkarákoknak nincs viaszt tartalmazó
epikutikulájuk, ezért könnyen kiszáradnak.
Hazánkban minden víztípusban gyakori és helyenként tömeges a
közönséges víziászka (Asellus aquaticus). A növényi maradványo-
kat fogyasztja, így fontos a dekompozíciós folyamatokban. Az ász-
karákok fontos szerepet játszanak az elhalt növényi maradványok
lebontásában. Üvegházakban, kertészetekbenmegrághatjákanövé-
nyeket, tehát kártevők is lehetnek.
Az érdes pinceászka (Porcellio scaber) igen sokfelé, erdőkben,
kertészetekben is megtalálható. Elsősorban üvegházakban okoz káro-
kat. A rokon fajokkal együtt népiesen „pincebogámak” nevezik.
Tökéletesen össze tud gömbölyödni a közönséges gömbászka
(ArmadiUidium vulgare). Alföldi és dombvidék! területeken minde-
nütt előfordul. Kedveli a napos, száraz területeket, ahol kövek alatt
húzódik meg. Ez a faj helyenként a cukorrépa kártevője lehet.
288
16.1.11. Felemáslábú rákok
(Amphipoda)
Több ezer fajt foglal magába ez a rend. A
testnagyság változó, 0,2-14 centiméterig
terjed. Testük oldalról lapított (16.14.
ábra). Hiányzik a fejpajzs. A második
csáp sohasem ágazik el. Első pár torlábuk
állkapcsi lábbá alakult, a további négy pár
fogókészülékké. Összetett szemeik kiemel-
kednek a fejből. Szájszerveik rágó típu-
súak, elhalt növényi részeket esznek vagy
ragadozók. A torlábak csípőit széles kitin-
lemezek borítják. A hat pár potrohláb kö-
zül az első három pár hosszú, elágazó és
úszásra is alkalmas. A hátulsó három pár
pedig erős, rövid ugróláb. Szívük a pot-
rohban található. Kopoltyúik a torlábak
16.14. ábra. Felemáslábú rák testfelépítésének vázlata
el: első csáp, fo: fogóláb, já: járóláb, má: második csáp, rá: rágó,
sz: szem, ul: úszóláb (Meglitsch és Schram nyomán)
csípőízeiről kiinduló lemezek vagy hólyagok. Posztembrionális fejlődésük során lárvafor-
mák nem jelennek meg (epimorfózis). A felemáslábú rákok folyóvizekben és állóvizek-
ben is sokszor tömegesen fordulnak elő, ezért fontos, jó minőségű haltáplálékot jelen-
tenek.
Hazánk kisebb-nagyobb folyóvizeiben mindenfelé előfordul a tüskés bolharák (Gammarus
roeseli). Elhalt növényi szerves anyaggal (detritusz) táplálkozik. Kiváló haltáplálék. Föld alatti
vizekben, kutakban, barlangokban sokfelé előfordulnak a vakrákok (Niphargus spp.J. Testük
világos vajsárga vagy fehér, szemük hiányzik.
16.2. Százlábúak (Chilopoda)
Hosszú, lapított testű, rendszerint több centiméteres állatok (16.15. ábra). Mintegy há-
romezer fajuk ismert. Testükön az egyes szelvények jól elkülönülnek egymástól. Fejük
határozottan elválik a törzstől. Egy pár csápjuk és lábaik néha igen hosszúak. Fürge ál-
latok. Szájszervük egy pár rágóból (mandibula) és két pár állkapocsból (maxilla) áll. Az
első törzsszelvényen található lábpár hatalmas, sarlószerűen hajlott, hegyes fogóké-
szülékké (állkapcsi lábbá) alakult át, melynek tőíze méregmirigyet tartalmaz (16.15.
ábra). Ezzel ragadják meg és pusztítják el áldozataikat, majd rágóikkal aprítják fel és
CS
16.15. ábra. A: százlábú (Scolopendra) testfelépítésének, B: egy Scutigera sp. fejének vázlata
ál; állkapcsi láb, cs: csáp, eá: első állkapocs, má: második állkapocs, ml: második járóláb csípője (Meglitsch és Schram
nyomán)
289
továbbítják a szájnyílásba a táplálékot. Külső emésztés is előfordul, amikor a zsákmányra
emésztőnedveket bocsátanak, majd a folyós táplálékot felszívják. Egyenes, kevéssé tagolt
bélcsatornájukhoz egy pár igen hosszú Malpighi-edény csatlakozik. Tapintószerveik és
kémiai érzékszerveik jól fejlettek. Sok faj a ragadozók ellen védekezésül ártalmas anya-
gokat termel. Egyesek rendkívülien mérgező, a sejtlégzést gátló hidrogén-cianidot (HCN)
is képesek előállítani. A hímek spermájukat egy fonalból készített gubóba helyezik. A
nőstények ezt a gubót veszik fel az ivamyílásukba. Posztembrionális fejlődésük történhet
szelvényszámnövekedéssel (anamorphosis) vagy kifejtéssel (epimorphosis). Annak elle-
nére, hogy a rovarokhoz hasonlónak tűnnek, semmiképpen sem tekinthetők ezek őse-
inek. Az avarban, kövek alatt, talajban, komposztban élnek és vadásznak. Rendszerint
éjjel járnak zsákmány után. Mint ragadozók, fontos szerepet játszanak a mezőgazdasági
szempontból káros talajállatok létszámának szabályozásában, de a biocönózisok struk-
túrájának fenntartásában is.
Hazánkban gyakori százlábú a sárgás színű földirinya (Pachymerium ferrugineum). Karcsú,
hosszú testű, rövid lábú, ezért aránylag lassú mozgású állat. Avarban, talajban, kövek alatt fordul
elő. A barna százlábú (Lithobius forficatus) teste csillogó vörösbarna. Pontszemei csoportosan
állnak. Éjszakai állat, ragadozó. Talajlakó rovarokat, ászkarákokat, férgeket eszik. Mindenütt
gyakori.
A százlábúakat az ikerszelvényesekkel, valamint a könyvünkben nem tárgyalt villásé sápúakkal
(Pauropoda) és szövőcsévésekkel (Symphyla) együttsoroljákasoklábúak (Myriapoda) altörzsébe.
16.3. Ikerszelvényesek (Diplopoda)
Testük hosszúkás, rendszerint henger alakú, de lehet lapított is. A szelvényzettség igen
feltűnő (16.16. ábra). Némelyik faj össze tud gömbölyödni vagy tekeredni. Fejük jól
elkülönül a törzstől, rövid, 7-9 ízből álló csápot visel. Szájszervüket csupán egy pár rágó
(mandibula) és egy pár állkapocs (maxilla, gnathochilarium) alkotja. Az első négy törzs-
szelvényen 1-1 pár, az ez után következőkön 2-2 pár láb található. Ezekben a szelvények-
ben a belső szervek (szívkamrák, idegdúcok, tracheanyílások) is duplán fordulnak elő,
bizonyítva, hogy a törzsfejlődés során a szomszédos szelvények összeolvadtak. így alakult
ki az ikerszelvény (diplosomit).
Lábaik rövidek, lassú mozgásra alkalmasak. Kültakarójuk kemény, rendszerint
vm
17.16. ábra. A: ikerszelvényes testfelépítésének, B: fejénekvázlata
cs: csáp, ga: első szelvény (gallér), sz: pontszemhalmaz, vm: védekező
mirigy nyílása, (a rajzokon csupán a jobb oldali lábsor látható) (Meglitsch
és Schram nyomán)
mésztartalmú, ezért alkalmasak a
kövületképződésre, sokfosszíliájuk
maradt meg, elsősorban a karbon-
és a permkorból. Védekezőmiri-
gyeik vál adékát ragadozóikra fröcs-
kölik. Bélcsatomájuklegfejlettebb,
nagy üregű szakasza a középbél.
Két pár Malpighi-edényük van.
Idegrendszerük fejletlen. Vannak
vak fajok is. A hímek egyes lábai
párzószerwé alakultak át. Ezzel
juttatják spermájukat a nőstények
ivamyílásába. A petéből kikelő fia-
tal állatok hét szelvénnyel és há-
rom lábpárral rendelkeznek. Poszt-
embrionális fejlődésük anamor-
fózis.
290
A talajban, a talaj felszínén, a komposztban gyakoriak. Fontos szerepet játszanak az
erdei avar lebontásában, a humusz előállításában. Néha kertészeti kártevők lehetnek.
Ezerlábúaknak is nevezik ezeket az állatokat, ámbár ezer lábuk soha nincs. Van olyan faj,
melynek 20 lábpára van (pl. Polydesmus spp.), de olyan is, mely 340 lábpárral rendelkezik
(Siphonophorella progressor).
A világon élő majdnem tízezer fajuk közül hazánkban mindenfelé gyakori a karimás
ezerlábú (Polydesmus complanatus), melynek testszelvényei a hátoldalon lapos, számy-
szerűen kiszélesedő kutikulalemezeket viselnek. Erdeinkben a fák kérge alatt és az avar-
ban található. A nőstény petéi köré földkupacot hord, amin egy szellőzőnyílást készít. Az
egyik leggyakoribb ikerszelvényes faj a korhadó fákban gyakori, sárgásbarna, hengeres
testű gombaevő vaspondró (Cylindroiulus boleti). Az alföldi száraz réteken néha
tömegesen elszaporodik a vonalas vaspondró (Megaphyllum unilineatum). Fényes
fekete, hengeres testén világossárga vagy rozsdaszínű háti középvonal van. Helyenként
és időszakosan cukorrépa-kártevő is lehet.
16.4. Hatlábúak (Hexapoda)
A hatlábúak teste három nagy, rendszerint egymástól jól elkülöníthető tájékra (tagmata)
tagolható (16.17. ábra). Ezek: a fej (caput), a tor (thorax) és a potroh (abdomen). A fej
hat (egyes vélemények szerint öt) szelvény összeolvadásából keletkezett. A torszelvények
száma három. A potroh legfeljebb húsz szelvényből állhat, de a szelvények igen gyakran
összeolvadtak a törzsfejlődés
során, ezért ma ennél keve-
sebb szelvény látható. A szel-
vények eredetileg négy kitin-
lemezzel határoltak. Felülről
a hátlemez (tergit), alulról a
haslemez (stemit), két oldal-
ról pedig egy-egy oldallemez
(pleurit) veszi körül a belső
szerveket. A potrohszelvények
között vékony, puha hártyás
kitinlemez található, ezért a
szelvények könnyen mozgat-
hatók.
A fej a többi testtájékhoz
viszonyítva lehet kicsi (pl. po-
loskák, bogarak), lehet nagy
(pl. szitakötők), de hiányoz-
hat is, mint például számos
légyfaj lárvájánál. A fejet egy-
séges kitinváz borítja (fejtok),
melyről nyúlványok, lécek in-
dulnak a fej belső ürege felé
(16.18. ábra). Szelvényezett-
ség nem ismerhető fel a fejen,
de különböző árokszerű be-
mélyedések, varratok igen.
Ezek a varratok azonban nem
az egykori szelvények határai.
16.17. ábra. Egy hatlábú testfelépítésének vázlata
aa: alsó ajak, ál: állkapocs, cs: csáp, et: első torszelvény, fa: felső ajak,
fc: fsrcsuta, iv: ivarszerv, ny: légzőnyílás, ös: összetett szem, ps: pontszem,
rá: rágó (Több szerző nyomán módosítva)
291
ft
16.18. ábra. A fej vázlata. A: oldalnézetből, B: metszetben, C: hátulnézetből
aa:alsó ajak, ál: állkapocs, bv: a fej belső kitinváza, cs: csáp, fa: felső ajak, fp:
fejpajzs, ft: fejtető, fii: fej ürege, ga: garat, ho: homlok, íz: a garatot és a szájszerveket
mozgató izmok, nk: nyálmirigy kivezető csöve, ny: nyakszirt, ős: összetett szem,
po: pofa, ps: pontszem, rá: rágó (Davíes és Weber nyomán)
csápok a homlokról (frons) indulnak ki. Mindig párosán fordulnak elő, kivéve az
előrovarokat (Protura), amelyeknél a csápok hiányoznak.
A szájszervek is igen változatos
felépítésűek, attól függően, hogy mi-
vel táplálkozik, illetve hogyan szerzi
meg táplálékát az állat. Legősibb és
legelterjedtebb típusa a rágó szájszerv.
A ma élő hatlábúak szájszerve az evo-
lúció során három lábpárból és a fej-
tok néhány részéből jött létre. Végtag
eredetű a páros rágó (mandibula), az
állkapocs (maxilla) és az alsó ajak (la-
bium). A fejtokból jött létre a felső
ajak (labrum) és a hypopharynx, mely-
nek testét nyelvnek (lingua) nevezik.
A felső ajakon gyakran ízérzékelő re-
ceptorok helyezkednek el. A rágok
egységes, tömör szervek, fogas élű ol-
lóhoz hasonlóan működnek. A rágó
típusú szájszervben belső élük karé-
lyozott, az ehhez egyébként igen ha-
sonló ragadozó szájszervben pedig
kiemelkedő kitinfogakkal, szőrökkel
ellátott. A poloskák és a szúnyogok
rágója hosszú, erős szúrósertévé ala-
kult át. A legyek és lepkék rágói csö-
kevényesek, vagy teljesen el is tűntek.
A fejen két nyílás találha-
tó: a szájnyílás, melyet a
szájszerv részei vesznek kö-
rül, és a nyakszirti nyílás,
amely az előtorba nyílik. A
fej különböző irányokba
állhat. Leggyakrabban füg-
gőleges állású (hypog-
nath), ekkor a szájszerv le-
felé néz (pl. csótányok, egye-
nesszárnyúak, legyek),
vagy vízszintesen helyez-
kedik el (prognath), ek-
kor a száj szerv előre irá-
nyul (pl. bogarak).
A fej függelékei a csá-
pok (antenna) és a szájszer-
vek (16.19. ábra). Acsápok
mechanikai érzékszervek és
szaglószervek egyszerre.
Több-kevesebb ízből áll-
nak. Az ízek száma és alak-
ja, a csáp formája gyakran
fontos határozó bélyeg. A
16.19. ábra. A rágó szájszerv vázlatos felépítése
at: alsó ajki tapogató, áÖ: álltő, ár: állrész, át: állkspcsi tapogató, bk:
belső karéj, cs: csáp, fa: felső ajak, fh: fióknyelv, fp: fejpajzs, ft:
fejtető, hí: hipofarinx, ho: homlok, kk: külső karéj, nv: nyelv, ny:
nyél, ős: Összetett szem, po: pofa, ps: pontszem, rá: rágó, sa: sarokíz
(Több szerző nyomán módosítva)
292
16.20. ábra. A torszelvényt határoló lemezek és a láb vázlata
co: comb, cs: csípő, fo: forgató, hl: hátlemez, hs: haslemez, ka: karom, Iá: lábszár,
If: lábfej, It: lábtő, ny: légzőnyílás, ol: oldallemez, sz: szárny (Több szerző nyomán
módosítva)
Az állkapcsok a rágó szájszerven a táplálék megragadását, rögzítését segítik. A poloskák-
nál hosszú nyálvezető és táplálékot szívó csővé alakultak. A szúnyogoknál szúrósertévé
változtak, a méheknél a két karéj összenőtt, a lepkéknél pedig hosszú, feltekeredő csövet
(pödömyelv) képeznek. Az állkapcsokhoz sokszor tapogatók (palpi) kapcsolódnak. Az
alsó ajkak összenőttek. Terjedelmesek a csótányok, bogarak esetében, csővé alakultak a
poloskáknál, szúnyogoknál, méheknél. Az alsó ajak is visel tapogatót. A tapogatókon
igen sok mechanikai érzékszerv található.
A fejen ezen kívül szemek is láthatók. A szemek lehetnek összetett szemek és pont-
szernek. A fej két oldalán (a vak fajok kivételével) mindig található két összetett szem.
Ezek néha igen nagyok (pl. szitakötők), máskor viszont gyengén fejlettek, mindössze
néhány egyszerű szemből állók (pl. tetvek). Fejlettségük az életmóddal áll összefüggésben.
Pontszernek gyakran, de nem mindig láthatók. Hiányoznak például az egyenesszár-
nyúaknál, terveknél, bogaraknál. A pontszernek száma rendszerint három. Kettő a
homlok oldalán, egy a közepén helyezkedik el.
A tor három szelvényből, az előtorból (prothorax), a középtorból (mesothorax) és az
utókorból (metathorax) áll.
Mindegyik lemezt négy ki-
tinlap, a hátlemez (tergit),
a haslemez (sternit) és két
oldallemez (pleurit) hatá-
rol (16.20. ábra). A három
szelvény egymáshoz viszo-
nyított aránya különböző le-
het. A szárnyatlan csopor-
tokban a három torszelvény
nagyjából egyformán fejlett.
Több csoportban az első
szelvény igen fejlett, és a
hátlemez egy pajzsszerű
képletet alkot, amit elő-
hátnak neveznek (pl. csó-
tányok, egyenesszárnyúak,
poloskák, bogarak). Mások-
nál (pl. kétszárnyúak, hártyásszámyúak) az előtör mindössze egy keskeny gyűrű. Ahol
a két pár szárny egyformán fejlett, a közép- és utótor is hasonló nagyságú (pl. szitakötők).
A kétszámyúaknál pedig, ahol a második számypár hiányzik, a középtor igen megerő-
södött.
A tor viseli a mozgásszerveket, a lábakat és a szárnyakat. A legtöbb hatlábú mindegyik
torszelvényén egy pár láb található. Egyes lárvatípusok (pl. kukac, nyű, szúnyoglárva)
egyáltalán nem rendelkeznek lábbal. A láb öt ízből áll (16.20. ábra). Az utolsó ízhez a
lábfej (praetarsus) csatlakozik. Ezen lehet egy páratlan vagy páros karom, de kiegészülhet
további kitinlapokkal, nyúlványokkal, kapaszkodó- és tapadókészülékekkel is. A lábak az
életmód függvényében módosulhatnak kiszélesedő ásólábbá (pl. lótücsök), megnyúlt,
erős ragadozólábbá (pl. imádkozósáska), hosszú felületnövelő szőrökkel ellátott úszó-
lábbá (pl. vízipoloskák), izmos combú ugrólábbá (pl. sáskák, bolhák), hosszú futólábbá
(pl. futóbogarak), erős karmokkal, fejlett izomzattal rendelkező kapaszkodólábbá (pl.
tetvek), de előfordul az is, hogy egy fajon belül a hímek elülső lába erőteljes fogólábbá
alakul, amivel párzás közben szilárdan átfoghatják a nőstényt (pl. csíkbogarak).
A legtöbb hatlábúnak kifejlett állapotában két pár, jellemző alakú és erezettségű
szárnya van (16.21. ábra). Az erek száma és lefutása állandó, a fajokra jellemző bélyeg.
Arovarszerűek (Parainsecta) azonban elsőrendűen szárnyatlanok, vagyis őseiknek sem
293
16.21. ábra. Szárny általános vázlata. A fo erek elágaznak. Ezután az adott ér
kezdőbetűjével és sorszámmal jelölik azokat
an: anális erek, ho: hónaljtőér, ju: jugális erek, kk: a szárnyat a torral összekapcsoló
készülék, kö: középér, me: mellékér, mh: a középső eret és a hónaljtőeret összekötő
harántér, su: sugárér, sk: a sugáreret és a középső eret összekötő harántér, sz: szegélyér
(Meglitsch és Schram nyomán)
volt szárnyuk. Mások,
mint a tetvek, a bolhák,
a hangyák dolgozói stb.
másodlagosan szár-
nyatlanok, mert őseik
szárnyakkal rendelkez-
tek. A szárnyak kezde-
ményei ilyen esetben az
embrionális korban ma
is megfigyelhetők. A
szárnyak lehetnek kár-
tyásak és többé-kevésbé
egyformák (pl. egyes
szitakötők, álkérészek),
vagy különbözők (pl.
kérészek), lehet az első
pár kemény szárnyfedő
és a hátsó pár hártyás (pl. bogarak). Egy pár szárnyuk van a kétszámyúaknak, míg a
fülbemászók első pár szárnya kicsiny, pikkelyszerű, de a hátsó hatalmas, hártyás. A
ki
szárnyak repülés közben mozoghatnak külön-külön (pl.
szitakötők), de különféle kapcsolórendszerek, kitinhor-
gok, tüskék, lapok össze is kapcsolhatják azokat, ilyenkor
a két szárny egységes repülőfelületet képez (pl. hártyás-
számyúak, lepkék) (16.22. ábra).
A szárnyak valószínűleg úgy jöttek létre az evolúció
során, hogy a hátlemez oldalirányba kiterjedt, léceket,
lapokat alkotott. Ezek a nyúlványok előnyt jelenthettek
a párválasztásban, a testhőmérséklet szabályozásában és
mint siklófelület, a mozgásban is, ami erős szelekciós
16.22. ábra. A szárny kapcsolódásának
egyik módja
es: első szárny, hs: hátsó szárny, ki:
kitinléc, le: kitinlemez
nyomást jelenthetett a szárnyak kifejlődésének irányába. Van olyan elképzelés is, hogy
a szárnyak a tracheakopoltyúkból alakultak ki (Id. például a kérészek vagy álkérészek
lárváit), de ezt a teóriát kevés adat támasztja alá. A szárnyak kettős kitinlemezből és az
általuk közrefogott erekből állnak. A lárvabőrből vagy bábból való kibújás után rövid
ideig puhák, máj d az erekbe préselt testfolyadékhatásár a kifeszülnek és megszilárdulnak.
Az erekben tracheacsövek és idegek találhatók. Igen jellemző, állandó és gyakran faji
bélyeg, hogy milyen erek talál-
hatók a szárnyon. Ezért az erezet
alakulását felhasználják a hat-
lábúak rendszerezésében. Az ősi
csoportok szárnya gazdagon ere-
zett, míg az újabban kialakultak
szárnyain az erezet redukciója,
egyes erek eltűnése figyelhető
meg. A kevesebb, de erősebb ér
fokozza a szárny szilárdságát. A
szárnyak felszínén ritkább vagy
sűrűbb elhelyezkedésben kitin-
szőrök (pl. poloskák, kétszár-
nyúak, hártyásszámyúak), más-
korpikkelyek (pl. lepkék) figyel-
hetők meg.
16.23. ábra. A szárny mozgatásánakmódja. A: háthasi izmok össze-
húzódása esetén a szárny felemelkedik, B: elernyedésükkor a hosz-
szanti és a direkt szárnymozgató izmok húzódnak össze és a szárny
lesüllyed
di: direkt számymozgató izmok, ha: haslemez, hh: háthasi indirekt izmok,
hl: hátlemez, ho: hosszanti indirekt izmok, ol: oldallemez, sz: szárny
(Davies nyomán)
294
A szárnyak mozgatását a torlemezekhez és a szárnyhoz közvetlenül kapcsolódó
(direkt) és a csak torlemezekhez kapcsolódó (indirekt) izmok végzik (16.23. ábra). A
szárnyak emelkedése úgy jön létre, hogy a háti, valamint a hasi torlemezhez kapcsolódó
izmok összehúzódnak, ezáltal a hátlemez lenyomja a szárny test belsejében levő végét.
A lefelé történő mozgásban mind a di-rekt, mind az indirekt izmok részt vehetnek, attól
függően, hogy melyik rovarrendről van szó. Emellett a szárnyak csavarodó, elliptikus
vagy nyolcashoz hasonló pályát is leírnak. A számycsapás sebessége igen különböző:
egyes sáskák, lepkék mindössze 4-20 csapást tesznek másodpercenként, míg szúnyogok-
nál ez az érték 1000/sec is lehet.
A hatlábúak potrohún embrionális korban szelvényenként egy pár lábkezdemény
alakul ki, de ezek a lábak rendszerint visszafejlődnek, mire az állat kibújik a petéből.
Kivételek vannak, mivel az előrovarok és a lábaspotrohúak kifejlett korukban is viselnek
csökevényes potrohlábakat. Jellegzetes végtagcsökevények a páros farcsuták (styli) és
fartoldalékok (cerci). A faroktoldalékok a fejlettebb rovarcsoportokban rendszerint
hiányoznak. A potroh végén nem végtag eredetű végfonal (filum terminálé) is lehet.
16.24. ábra. A: nőivarú, B: hímivarú rovar potrohvége
fa: farcsuta, ft: fartoldalék, iv: ivamyílás, ny: légzőnyilás, pa: paramer, pá: párzószerv, ps: tizedik potrohszelvény, ti:
titillatorium, to: tojócső, us: utolsó potrohszelvény hátlemeze, va: virga, vb: végbélnyílás (Webet nyomán)
A potroh legjellegzetesebb képletei a külső ivarkészülékek (gonapophyses). Első-
sorban a hím, de gyakran a nőstény külső ivarkészüléke is komplikált felépítésű, számos
kitinlemezzel, nyúlvánnyal kigészült szerv. A hímek külső ivari készüléke a kilencedik
potrohszelvényen található (16.24. ábra). Legteljesebb formájában két kitinlemezből áll,
melyek a nőstény megragadását szolgálják. Ezek között rendszerint páratlan párzószerv
(aedeagus vagy penis) helyezkedik el. Páros párzószervük van a kérészeknek. A párzó-
szerv végén érzékszervek (virga és titillatorium) találhatók. Végül újabb kétoldali kitin-
lemezek (paramerek) csatlakoznak és teszik teljessé a külső ivarszervet. Mivel a felsorolt
struktúrák fajokra jellemzőek és felépítésük igen állandó, ezért a rovarok rendszerezésé-
nek talán legfontosabb bélyegei. A nőstények külső ivarkészüléke a tojócső (ovipositor)
és a kapcsolódó kitinlemezek (16.24. ábra). A tojócső a nyolcadik és a kilencedik szelvény
hasi oldaláról kiinduló, három pár, hosszú kitinlemezből áll. Ezek fogják körül azt az
üreget, melyen keresztül a peték a külvilágba jutnak. A méhek és darazsak tojócsöve
fullánkká alakult át. Számos rovarrendben kifejezett tojócső nem található, de a potroh
vége rendszerint ilyenkor is vékony, kitűrhető csőben végződik, amely funkcionálisan
hasonló szerepet játszik, mint a tojócső (pl. kérészek, álkérészek, lepkék, bogarak, két-
szárnyúak).
295
ön ms is to
16.25. ábra. A kültakaró felépítése
al: alaphártya, en: endokutikula, ep: epikutikula, ex: exokutikula, ha: hámréteg, is:
idegsejt, kn: kitinnyúlvány, mn: mirigysejtek kivezetőjáratánaknyilása, ms: mirigysejt
(trichogen sejt), ön: önocita, pi: kitinpikkely, sz: szőr, to: tormogén sejt (Davies
nyomán)
A hatlábúak kültaka-
ró ja bonyolult felépítésű
szerv(16.25. ábra). Az egy-
rétegű hámsejtek (epi-
dermis) rétegét kívülről
borító vastag, háromréte-
gű kutikulából és a csatla-
kozó speciális sejtek, ér-
zékszervek, felszíni struk-
túrák együtteséből áll. Ezt
a struktúrát nevezik külső
váznak (exosceleton). A
hámsejtek hozzák létre a
kutikulát a vedlések so-
rán. A kutikula külső réte-
ge az igen vékony epi-
kutikula. Elsősorban de-
naturált fehérjékből és
viaszból áll. Megakadá-
lyozza a szervezet vízvesztését a kutikulán keresztül, valamint a kémiai hatások ellen is
véd. A kutikula középső rétege, az exokutikula, sokkal vastagabb. Főként kitinből
(nitrogéntartalmú poliszacharidból) és fehérjéből képződik. A fehérjék között cserző
hatású anyagok (fenolok) hatására kötések keletkeznek, így térhálós szerkezetű, szilár-
dabb, kevésbé nyúlékony anyag (sklerotin) jön létre. Ahol hajlékony, puha kitinhártyák
láthatók, ott ez a réteg hiányzik a kültakaróból. A kutikula legbelső, egyben legvastagabb
rétege az endokutikula. Ez is elsősorban kitinből és fehérjékből épül fel, de a fehérjék
cserzése elmarad, ezért ez a réteg hajlékony. A kültakarónak 25-60%-a kitin.
A kültakaró felszínéből rövid szőrök, lapos pikkelyek, lemezek indulhatnak ki. Ezek
a képletek mozdulatlanul, szilárdan kapcsolódnak a felszínhez. Vannak hosszú, mozgé-
kony szőrök, amelyek belül üregesek. Ezeket hámsejtek (trichogen sejtek) hozzák létre.
Alapjuk rugalmas hártyával kapcsolódik a kültakaróhoz. Ez a hártya másik hámsejttípus
(tormogén sejtek) terméke. Ilyen szőrök borítják például a poszméhek testét. Ha
kiszélesednek, pikkelyeket alkotnak (ugróvillások, lepkék, bogarak, kétszárnyúak). Néha
mirigyek kivezetőcsövei nyílnak a szőrök végén, amint ez számos hernyón megfigyelhető
(pl. aranyfarú szövőlepke - Euproctis chrysorrhoea). Mechanikai ingerek felfogására is
alkalmas lehet a szőr, ha alapjához idegsejt csatlakozik.
A hámsejtek között különleges sejtek is találhatók. Egyik részük a viasz előállításában
vesz részt. Másik csoportjuk feromonokat termel.
A hatlábúak között számos gyönyörűen színezett faj fordul elő. A színek ugyanúgy
jönnek létre, mint a többi állatcsoportnál. A színek eredhetnek a kültakaróban előforduló
színanyagoktól, a kültakaró struktúrájából fakadó fényinterferenciából, illetve e két ok
kölcsönhatásából.
A kültakaró belső felszínéről kitinnyúlványok, lemezek, lécek, csövek indulnak a test
belseje felé. Ezt a rendszert belső váznak (endosceleton) nevezik. A belső váz biztosítja
az egyes testtájak szilárdságát (különösen a fej belső vázrendszere fejlett), valamint
biztos, szilárd alapot nyújt az izmok tapadásához.
A hatlábúak vázizmai és a zsigeri szervekben található izmok egyaránt speciális
harántcsíkolt izmok. Ezek felépítése eltér a gerincesek testében megfigyelhető harántcsí-
kolt izmoktól. A miofibrillumok száma izomrostonként kevés, de nagy miofibrillumok
láthatók. Óriásiak a mitokondriumok is, melyek a gyors és nagy mennyiségű ATP
előállítását biztosítják. A hatlábúak és a gerincesek harántcsíkolt izmainak működése
296
fiziológiai szempontból hasonló. A hatlábúaknál azonban szokatlanul gyors és nagymér-
tékű ATP-termelés figyelhető meg, ami a számos, nagy mitokondrium jelenlétének
köszönhető. A szárnyak mozgatásához kismértékű izomösszehúzódás szükséges, ezért
lehetséges, hogy egyes fajok má-sodpercenként száz vagy annál is több számycsapásra
képesek. A szükséges energia a rövid távon intenzíven repülő csoportok esetében (legyek,
méhek) gyorsan mobilizálható szén-hidrátokból, a kitartóan, hosszan repülő csoportoknál
(sáskák, vándorló lepkék) pedig elsősorban zsírból származik. A tápanyagok teljes lebon-
tásához (elégetéséhez) szükséges oxigént az izmok gazdag tracheahálózata biztosítja.
A hatlábúak bélcsatornája háromszakaszos (16.26. ábra). Az elő- és az utóbél az
ektodermából ered. Ennek megfelelően ezeken a szakaszokon a bél falának szövettani
felépítése hasonlít a kültakaróéhoz többek között abban, hogy belülről kutikula béleli. A
középbél viszont az entodermából alakul ki. A bélcsatoma a szájnyílást körülvevő
szájszervekkel kezdődik. Legősibb típusa a rágó szájszerv. Az életmódnak, táplálkozási
módnak megfelelően, számos módosulás során szúró-szívó, nyaló, nyaló-szívó típusok
jöttek létre a különféle táplálékforrások kiaknázására. A szájüregbe nyílik a három pár
nyálmirigy. A rágóhoz kapcsolódó nyálmirigyben a méheknél feromon is termelődik.
Az állkapcsi nyálmirigy a poloskáknál méregmiriggyé alakult. Az alsó ajakhoz kapcsoló-
dó nyálmirigy váladéka szénhidrátbontó enzimeket (amiláz) tartalmaz. A lepkék her-
nyóinál és egyes hártyásszámyúak álhernyóinál az alsó ajki nyálmirigyek választják el a
selymet, melyből a báb gubója készül. A nyálban lehet pektináz (a növényi sejtfalban
található nagy mennyiségű pektin bontására), továbbá mérgező anyagok a zsákmány
elpusztítására és véralvadást gátló anyagok is a vérszívó fajoknál. A szájüreg a garatban
(pharynx) folytatódik. A lepkék és a hártyásszámyúak garatja részt vesz a táplálék
felszívásában. A garat után vékony, hosszú nyelőcső (oesophagus) következik. Ennek
alsó vége kitágul és esetenként begyet (ingluvies) alkot (16.27. ábra). A nyelőcső után
némely csoportnál (csótányok, egyenesszárnyúak, bogarak) a rövid, de igen izmos falú
SZ
16.26. ábra. A belső szervek elhelyezkedése a testben
ag: garat feletti idegdúc (agydúc), an: alsó ajak nyálmirigye, ao: az aorta testüregbe nyíló vége, án: az állkapocs nyálmirigye,
be: begy, ga: garat, hd: hasdúclánc, iv: ivarszerv, kb: középbél, ma: Malpighi-edények, nő: nyelőcső, rn: a rágó nyálmirigye,
sí: szív, sn: szájnyílás, ti: a tor izomzata, sz: szárny, vb: végbél, vn: végbélnyílás (Kaestner nyomán)
297
előgyomor (proventriculus) található. Belsejébe
a táplálékot megőrlő kitinfogak, -lécek nyúlnak. A
folyékony táplálékon élő rovarok nyelőcsövének
végén előgyomor helyett begy található (méhek).
A lepkék és a dongólégyfélék nyelőcsövéhez vi-
szont oldalról csatlakozik egy terjedelmes kiöblö-
södés (diverticulum). Abélcsatománakezeka sza-
kaszai rendszerint mint raktárak játszanak szere-
pet, ahonnan kis csomagokban kerül tovább a táp-
lálék a középbélbe. Kisebb mérvű emésztés azon-
ban itt is előfordul. Az előbelet a középbéltől egy
izmos szelep választja el. A középbelet kívülről hosz-
szanti és körkörös izmok határolják. Alatta egyré-
tegű kutikuláris hengerhám található. Itt két alap-
vető sejttípus látható: egyrészt emésztőenzimeket
termelő mirigysejtek, másrészt a felszívást végző
sejtek. E sejtekfelszínén található nyúlványok (mic-
rovilli) a felület nagyobbítására szolgálnak. Ugyanez
a funkciójuka zsákszerű kitüremkedéseknek (coeca)
is, melyek a bélszakasz elején helyezkednek el.
Rendszerint - ahogy a szövettani felépítésből is
látható - a kitüremkedő bélszakaszok és a közép-
bél első szakasza a tápanyagok felszívásának leg-
fontosabb helye. A béltartalmat közvetlenül egy
vékony hártya (peritrophicus membrán) veszi
körül. Ez az emésztőenzimek és tápanyagok szá-
mára átjárható, viszont megóvja a bél falát a me-
16.27. ábra. A középbél és utóbél találkozása
(általános vázlat)
be: begy, bk: bélkitüremkedések, iz: hosszanti és
körkörös izomkötegek, kő: körkörös izmok, ki:
kitinlécek, ks: középbél sejtjei, pm: peritrofikus
hártya, rá: rágógyomor, se: a táplálékáthaladást
szabályozó izomcsoport, mely szelepként működik
(Több szerző nyomán módosítva)
chanikai sérülésektől. A részben emésztett anyagok - átlépve a peritrofikus membránon -
még felszívásuk előtt további bontáson mennek keresztül a membrán és a bél fala közötti
térben található enzimek segítségével. A középbél és (ha van) a begy az emésztés színtere.
A nyálmirigy és a középbél amiláz enzimjei a keményítőt bontják, míg a középbél glikozidáz
enzimjei a maltóz, a laktóz és a szukróz bontásáért felelősek. A nektárt fogyasztó fajok
(pillangók, molylepkék) bélcsatomájában az invertázon kívül, mely a nádcukrot bontja,
szinte nincs más emésztőenzim. Cellulózbontó enzimek ritkán fordulnak elő (pikkelykék,
cincérek lárvái), viszont endoszimbionta baktériumok (ganéjtúró bogarak lárvái) vagy
egysejtűek (csótányok, termeszek) segítségével a cellulóz emésztése is lehetséges. E
szimbionta lényeket a petéből frissen kikelő állatok gyakran a fajtársak végbélnyílásának
vagy ürülékének nyalogatásával veszik fel. A zsírok bontását lipázok végzik. A rovarok
zsíremésztése gyenge. A fehérjéket az endopeptidázok polipeptidekre, az exopeptidázok
pedig aminosavakra hasítják. Vérszívó szúnyogokból a szénhidrátbontó enzimek hiányoz-
nak és a fehérjéket hasítok dominálnak. A rovaroknak számos fontos aminosavat a táp-
lálékkal kell felvenniük. Ugyancsak életfontosságúak a szterolok és a B-vitaminok. Ezeket
több esetben különleges sejttípusban (mycetocyta) előforduló szimbionta baktériumok
termelik.
Az utóbelet két szakaszra, a vastagbélre (ileum) és az ennél is vastagabb végbélre (rec-
tum) lehet osztani. Sok rovar végbelében hat hosszanti kiemelkedés vagy sok betürem-
kedő szemölcs (papillae rectalis) látható. Ez a felületnövelő berendezés a végbél kivá-
lasztó funkciója miatt alakult ki, hiszen nem csupán a víz, hanem a sók (ionok) vissza-
szívása is itt történik.
A testüreg különböző helyein a kültakaróhoz vagy a bélcsatomához rögzítve speciális
sejtcsoportok, zsírtestek találhatók. A zsírtesteket alkotó sejtek, a trofociták központi
298
szerepet játszanak a szervezet anyagcseréjében. Ezek a tápanyagok raktározása szem-
pontjából hasonlóan működnek, mint a gerincesek májának sejtjei. Képesek glikogént,
fehérjét és zsírt szintetizálni és raktározni, valamint innen kerülnek az említett anyagok
a felhasználás helyére az egyes szövetek szükségleteinek megfelelően. Azok a fajok, ahol
az imágók nem táplálkoznak, a lárvakori fejlődés során a zsírtestekben felhalmozott
tápanyagokat használják fel. A zsírsejtek elraktározhatják a keletkező méreganyagokat,
és bennük szimbionta baktériumok is élhetnek.
A hatlábúak elsősorban trachearendszerrel (16.28. ábra), ritkábban kopoltyúval,
tracheakopoltyúval, vagy egyéb módon lélegeznek. A trachearendszer légzőnyílása
(stigma) a közép- és az utó tor, valamint a potrohszelvényekoldalán található. N éha állandó
nyílások (legyek), máskor viszont nyithatók és csukhatok. A nyílást fedheti két, mozgatható
kitinlapocska, amely a levegő kiáramlásakor passzívan nyílik ki, zárni viszont izmok se-
gítségével zárja az állat (sáskák). Más esetben a légzőnyílás mögötti tracheaszakasz fala
mozgatható. A szemben lévő két fal összeszorítása gátolja meg a levegő áramlását (her-
nyók). Vízirovarok testén a légzőnyílások teljesen hiányozhatnak. A primitív hatlábúak
körében minden egyes légzőnyílástól elágazó, önálló trachearendszer indul ki, közöttük
összeköttetés nincsen. Legtöbbször azonban nagy, hosszanti tracheatörzsek futnak végig
a testen és kapcsolatot létesítenek az egyes szelvények trachearendszerei között. A tra-
cheákból induló jelentős nagyságú kitüremkedések, a légzsákok kapcsolódhatnak a nagy
tracheatörzsekhez (kétszárnyúak, méhek) vagy a tracheákhoz (cserebogarak, sáskák). A
légnyomás kiegyenlítésében játszanak szerepet. Különösen repülés közben fontosak, mert
segítik, hogy a friss levegő megfelelő gyorsasággal és nyomással jusson az izmokhoz.
A kis testű vagy kevéssé aktív hatlábúak gázcseréje különösebb légzőmozgások nél-
kül, a szokásos izommozgások következtében megy végbe. Ha a tracheolákban csökken
az oxigén parciális nyomása, a külvilágból újabb molekulák áramlanak oda. A keletkező
szén-dioxid nagy része a trachearendszeren keresztül távozik, de egy jelentős mennyiség
(mintegy 25%) a kültakarón keresztül hagyja el a testet. Ennek az az oka, hogy a szén-
dioxid könnyebben nyelődik el a szövetnedvekben, mint az oxigén. A nagy testű vagy
intenzív mozgást végző hatlábúak gázcseréjük mértékét a légzőnyílások nyitásával és
zárásával, valamint a légzőmozgások gyakoriságának változtatásával tudj ák b efolyásolni.
A légzőmozgások a potrohon figyelhetők meg. Ha a potroh izmai összehúzódnak, a
trachearendszer térfogata csökken, ezáltal levegő préselődik ki. Ellentétes esetben, a ru-
galmas kültakaró visszaállítja az eredeti térfogatot, és a levegő beáramlik a tracheákba.
Repülés közben a szárny mozgatása hasonló következményekkel jár. így biztosított a re-
pülés fokozott oxigénszükséglete. A légzés idegi szabályozása két szinten történik: egy-
részt az adott szelvény hasi dúcaiban, másrészt pedig a tor (a légzőmozgásokintenzitásának
változtatásával) és az agy idegdúcaiban (a diffúzió mértékének befolyásolásával).
16.28. ábra. A rovarok trachearendszerének vázlata
ht: hosszanti tracheatörzs, Íz: légzsák, ny: légzőnyilás, tr: tracheacsövek (Jannone nyomán)
299
A vízirovarok lélegezhetnek tracheakopoltyúkkal (szitakötők, kérészek, álkérészek,
tegzesek lárvái) is. A kopoltyút ekkor tracheacsövek hálózzák be. A gázcsere ilyenkor a
víz és a tracheacsövekben levő gázkeverék között történik. Máskor a kopoltyúban test-
folyadék kering, melyben esetenként hemoglobin is található (árvaszúnyogok). A gáz-
csere tehát a víz és a testfolyadék között megy végbe. A nyitott trachearendszerrel bíró
vízirovarok (vízibogarak, vízipoloskák) testét kitinszőrzet fedi. A szárnyfedők alá és a
szőrök közé levegőbuborékot gyűjtenek az állatok, mely az egész testet befedi. A bu-
borékból az állat légzése során felvett oxigén helyére a vízből diffúndálnak újabb mole-
kulák. Egyes bogarak és légy ek lárvái a víz alá merült növények levegőt tartalmazó üregeit
szúrják meg, és innen biztosítják a szükséges oxigént. A bagócsok (Diptera) lárvái
emlősök bőrében élnek. Átfúrják az epidermiszt és a külvilágból jutnak friss levegőhöz.
A hatlábúak nyílt keringési rendszerrel bírnak (16.26. ábra). A keringési szerv-
rendszert-a központi szervek elhelyezkedése miatt-háti edényrendszemekhívják. Aháti
edényrendszer két részre, a szívre (cor) és a kapcsolódó erekre osztható. A szív a potrohban
helyezkedik el (16.29. ábra). Testszelvényenként egy-egy kamra alkotja, de ezek száma
általában kevesebb, mint a potrohszelvényeké. A kamrák két oldalán réshez hasonló,
szelepekkel ellátott nyílások (ostia) találhatók. A szelepek csak a testfolyadék beáramlását
teszik lehetővé, a visszaáramlást nem. Minden kamrához számyszerű izmok, a legyezőiz-
mok kapcsolódnak. Ha összehúzódnak, a kamrák térfogata megnő, és a szív körüli térből
(pericardium) a testfolyadék a szívbe áramlik. A kamrák falában található izmok összehú-
zódása csökkenti a kamra üregét.
Az egymást követő kamrák felvált-
va húzódnak össze. A testfolyadék
a szívből a fej irányába áramlik. A
szív pumpáló mozgása tartja egy
irányú mozgásban a testfolyadé-
kot. A szívet egy hártyás zsák, a
szívburok veszi körül. Az első szív-
kamra folytatása az aorta. Az agy
mögött a testüregbe nyitottan vég-
ződik. Ritkán (csótányok, méhek)
a csápokbanis vannakerek, és ezek-
ben a testfolyadékot a csáp tövénél
elhelyezkedő izmos szerv (járulé-
kos szív) tartja mozgásban. A test-
folyadék alapja a plazma, melyben
számos különböző sejt (haemocyta)
található. A sejtek száma 1-100
ezer/mm3. Négy fő típusukat kü-
lönböztetik meg: a) prohemociták:
ezekből alakulnak ki a többi sejtek, b) fagociták: ezek pusztítják el a kórokozókat, elhalt
szövetdarabkákat, c) önocitoidok: ezek funkciója nem ismert és d) cisztociták: a testfolya-
dék alvadásában játszanak szerepet.
A Malpighi-edények a hatlábúak legfontosabb kiválasztószervei (16.30. ábra).
Számuk jellemző a fajokra, ezért a rendszerezésben felhasználható jellemző. AMalpighi-
edények felépítését és működését tekintve feltűnő különbségek ismerhetők fel az egyes
csoportok között. Az egyenesszárnyúak, a recésszárnyúak és egyes bogarak Malpighi-
edényeiben csupán folyadék található. A húgysav a végbélben kristályosodik ki, ahol a
vízvisszaszívás is történik. A rablópoloskáknál a húgysav már a csöveknek a bélhez kö-
zelebb eső szakaszán kialakul. Bogarak és hernyók testében a Malpighi-edények disztális
vége a végbél fala közelében, vagy azzal kötőszövetek segítségével összekapcsolva látható
16.29. ábra. A rovarok szívének felépítése
ep: epidermisz, há; szívet felfüggesztő hártya, hd: háti szövetlemez
(diaphragma), ku: kutikulasejtek, le: legyezőizmok, ne: nefrociták, ny:
szív oldalán lévő nyílás, sf: szív fala (Weber nyomán, módosítva)
300
(kriptonefridiális rendszer). Ennek kö-
vetkeztében a Malpighi-csövek és a végbél
közösen még hatékonyabban szívják vissza
a vizet. Ellentétben a szárazföldi fajokkal,
ahol a test víztartalmának fenntartása lét-
fontosságú az állat számára, a vízrovarok
nem rendelkeznek olyan fiziológiai rend-
szerrel, mely a vízvisszaszívást lehetővé ten-
né. A kiválasztás végterméke sem húgysav,
hanem ammónia. A Malpighi-edényekmel-
lett kiválasztó funkcióval rendelkeznek a
zsírtestek, a nefrociták és az önociták is. A
zsírtestek az embrionális testüreget bélelő
mezodermából keletkeznek. Speciális sejt-
jeik, a trofocitákhúgysavat tartalmaznak. A
bábozódás során ezek a sejtek nem kerül-
nek a kifejlett állatba (imago). A nefrociták
a szív és a nyálmirigyek két oldalán helyez-
kednek el. Pinocitózissal képesek a felesle-
es
16.30. ábra. A: rovarok Malpighi-edényei, B: rova-
rok kriptonefridiális rendszere
es: a Malpighi-csövek első szakasza a lapos kiválasztó
sejtekkel, ms: a Malpighi-edények második szakasza a
magas kiválasztó sejtekkel, vb: végbél, vn: végbélnyílás
(Wall nyomán)
ges anyagokat felvenni. Az önociták a test
legnagyobb sejtjei közé tartoznak. A kültakaró alatt vagy a zsírsejtek közelében találha-
tók. A vedlés idején különösen aktívak, amikor a kutikula képzéséhez szükséges anyago-
kat állítják elő.
A hatlábúak váltivarú állatok. A nőstények páros petefészke rendszerint egyenként
4-8 petecsövecskéből (ovariolum) tevődik össze (16.31. ábra). Ezeket a végfonal (filum
terminálé) függeszti a potroh hátlemezéhez. A petecsövecske végén (germarium) talál-
hatók az őspetesejtek. Az oogenezis szaporodási, növekedési szakasza és az érési szakasz
kezdete a petecsövekben zajlik, ahogy a fejlődő pete a petevezető felé halad, de az érési
szakasz általában csaka peterakáskor fejeződikbe. Kezdetleges rendekben (pl. szi-takötők,
egyenesszárnyúak) a peték tápanyagaikat a petecsövet bélelő hámsejtektől kapják (panoisticus
ovariolum). A fejlettebbeknél (legtöbb rovar) minden érőfélben levő petesejthez tápláló-
sejtek csoportja csatlakozik (politroph ovariolum). Ezek a sejtek hozzák létre a petesejt
szikállományát. A poloskák, kabócák és bogarak ováriolumainak végén találhatók a
táplálósejtek. Innen hosszú csöveken keresztül jut a szikanyag a petékbe (acrotroph
ovariolum). A petecsövecskékből a pete a jobb és bal oldali petevezetőkbe, majd azok
egyesülése után a közös petevezetőbe kerül. Ez a cső az izmos falú hüvelybe torkollik.
Ugyancsak ide nyílik a spermatartó (spermatheca) kivezető csöve is. Párzás után a sper-
miumok a spermatartóba kerülnek és itt raktározódnak. A megtermékenyítés akkor tör-
ténik, amikor a pete a vaginába csúszik és elhalad a spermatartó kivezető csövének nyílása
előtt. A hüvelyhez járulékos mirigyek kapcsolódnak, melyek ragasztóanyagokat, vagy a
petéket burkoló, a kokont felépítő anyagokat termelnek. Alepkéknőstényein két ivamyílást
találunk. Itt a spermiumok a párzótáska (bursa copulatrix) nyílásán kerülnek a nőstény
testébe. Innen egy csövön keresztül jutnak a vaginába, ahol a megtermékenyítés történik.
A megtermékenyített petéket a hüvely nyílásán át rakja le az állat.
A páros herék finom csövecskékből állnak (16.31. ábra). A herecsöveket végfonal
függeszti fel a hátlemezhez. A herecsövekben történik a spermiogenezis. A herékhez páros
elvezető járat (vas deferens) kapcsolódik. Az elvezető járatok az ondóhólyagban (vesicula
seminalis) folytatódnak, majd a kilövellő járatban (ductus ejaculatorius) egyesülnek. Számos
járulékos mirigy kapcsolódhat a hím ivarszervhez. Ezek tokot képeznek (spermatophor) az
ejakulátum körül. A párzás legtöbbször úgy történik, hogy a hím bevezeti a párzószervét a
nőstény hüvelyébe vagy párzótáskájába. Ez a mechanikai inger váltja ki a pete lerakásához
301
16.31. ábra. Rovarok belső ivarszerveinek vázlata. A: nőstény rovar, B: hím rovar, Cl: panoisztikus petecső, C2:
politrof ováriolum, C3: akrotróf ováriolum
cs: szikanyagot vezető cső, ej: elvezető járat, és: érőfélben lévő petesejt, ge: germarium, he: here, hl: hátlemez, hü: hüvely,
jm: járulékos mirigy, kj: kilövellő járat, mi: mirigy, pa: paramer, pc: petecső, pf: petefészek, ps: párzószerv, pv: petevezető,
st: spermatartó, oh: ondóhólyag, vi: virga, ti: titillatorium, vf: végfonal, ts: táplálósejt (Davies nyomán)
302
szükséges izommozgásokat. A petéket a nőstények tehát akkor is lerakják, ha steril hímekkel
párzanak. A megtermékenyülés ekkor természetesen elmarad. Ezt a jelenséget felhasználják
a biológiai növényvédelemben. Az ugróvillások hímjei által lerakott spermacsomókat a
nőstények aktívan veszik fel. Az agyi poloskák (Cimex spp.) hímjei a nőstények szelvények
közötti (intersegmentalis) hártyáit átszúrva, a testüregbe juttatják spermiumaikat, melyek
innen jutnak a petefészekbe, ahol a megtermékenyítés történik.
A szűznemzés sok rendben gyakori. A hártyásszámyúak haploid, meg nem termé-
kenyített petéiből hímek kelnek ki, míg a megtermékenyített, teljes kromoszómaszámú
petékből nőstények. Levéltetű-populációk szaporodása során több generáció jön létre
igen gyorsan egymás utáni szűznemzéssel. Levéltetvek és legyek esetében élveszülés
(viviparia) is megfigyelhető. Ekkor az embrionális fejlődés és a posztembrionális fejlődés
egy része is a nőstény hüvelyében történik. Előfordul, hogy a megtermékenyített peték
a hüvely egy tágulatában maradnak mindaddig, amíg az embrionális fejlődés végbe nem
megy. A lerakott, érett embriókat tartalmazó petékből azonnal kikelnek a fiatal állatok.
Ez a folyamat az álelevenszülés (ovoviviparia). Egyes legyek és bogarak lárváiban
petesejtek jönnek létre, majd szűznemzéssel fejlődésnek indulnak. Újabb lárvák alakul-
nak ki, melyek felfalják a szülő lárvát. Ezt a jelenséget lárvánemzésnek (paedogenesis)
nevezik. Hártyásszámyúak és legyezőszámyúak körében fordul elő, hogy a zigótából
néha egy, néha több utód lesz (poliembrionia). Előfordult, hogy a parazita Litomastix
truncatella darázs egyetlen petét rakott a gammalepke (Autographagamma) hernyójába
és ebből 1000 utódja kelt ki.
A hatlábúak petéje változatos alakú, színű, felépítésű lehet (16.32. ábra). A pete típusát
tekintve általában sokszikű, és a zigóta barázdálódása a felületen indul meg. A zigóta
sejtmagja a szikanyag felületén többszörösen osztódik, így hozza létre a blasztomérákat. A
blasztomérák a has középvonalában gyűlnek össze és az embriócsíkot alkotják. Később
minden szerv ebből az embriócsíkból jön létre. Az embriócsík preszumptív areái egysé-
gesek az egész rovarvilágban, csupán a lábaspotrohúak és ugróvillások alkotnak külön
csoportot ebből a szempontból. A pete elülső részéből (aktivációs központ) vegyületek
áramlanak az embriócsík felé, és meghatározzák annak kialakulását. Ekkor a blasztomérák
még omnipotensek. Miután az említett vegyületek termelése megszűnik, egy másik, a
jövendő tor területére eső szabályozó központ, a differenciációs központ lép működésbe.
Kialakulnak az ősi szervtelepek, majd a szervek. Közben az embrió a szikanyagba süllyed
(immersio), vagy oda betüremkedik (invaginatio). Az embriót egy belső (amnion) és
egy külső (serosa) embrionális burok veszi körül. A szel-
vényezettség már az embrionális fejlődés elején, a hólyag-
csíra állapotban megjelenik.
A posztembrionális fejlődés a petéből való kibújás
után kezdődik. A hatlábúak sorozatos vedlések után érik
el a kifejlett (imágó) állapotot. Testhosszuk nem fokoza-
tosan, hanem minden vedlés után ugrásszerűen nő. A
hatlábúak posztembrionális fejlődése rendkívül változa-
tos. A főbb jelenségek jobb áttekintésére, a fejlődési típu-
sok bemutatására többféle csoportosítási lehetőség kínál-
kozik. A következőkben egy olyan rendszert ismertetünk,
amelyben a testben lezajló szöveti, szervi átalakulások,
tehát a fejlődési jelenségek mértéke szerint sorolják cso-
portokba a rovarokat.
Az előrovarok (Protura) posztembrionális fejlődése
során szelvényszámnövekedés történik (anamorphosis).
A petéből kikelő fiatal állatok potroha 8 szelvényből áll.
Három vedlés után, folyamatos növekedéssel jön létre a
17.32. ábra. Rovarok néhány pete-
formája
bő: böglyökhalomba tett petéi, fá: fátyol-
kák nyélen ülő petéje, le: lepkék fiizére-
sen elhelyezett petéi, Ik: lepke hagy-
maformájú petéje, me: méhek petéje, te:
tetvek serkéje hajszálhoz ragasztva (Több
szerző nyomán módosítva)
303
16.33. ábra. Hatlábúak gyakori lárvatípusai
A: fiatal (juvenilis) ugróvillás, B: fiatal és idősebb poloskalárva, C: szitakötőlárva, Dl-3: teljes átalakulással fejlődő
rovaroklárvái, D1: lemezescsápú rovar fiatal lárvája, D2: lemezescsápú rovar idős lárvája, D3: lemezescsápú rovar
belső szervei
kb: középbél, ma: Malpighi-edények, sz: szájnyílás, vb: végbél, vn: végbélnyílás (Több szerző nyomán módosítva)
16.34. ábra. Kevéslábú és lábatlan lárvatípusok
A: futóka (kampodeoid) lárva, B: pondró, C: drótféreg, D: áldrót-
féreg, E: pajor, F: futókaszerű (kampodeoidszerű) lárva, G: nyű, H:
árvaszúnyog lárvája, I: kukac (Több szerző nyomán módosítva)
kifejlett állatokra jellemző 11 potrohszelvény. A többi rovarfaj esetében a petéből kikelő
fiatal állat szelvényszáma megegyezik az imágóéval (holomorphosis) (16.33. ábra). A
holomorfózissal fejlődő rovarok csoportja további három, jelentősen különböző
posztembrionális fejlődésű alcsoportra bontható. A kifejtéssel (epimorphosis) fejlődő
rovarok lárváinak semmiféle lárvakori szerve nincs. Ennek következtében az utolsó
lárvastádium során sem történnek jelentős szöveti átalakulások, vagy szervek lebontása és
újabb szervek felépítése. Az állatok testnagysága megnő, a petéből való kibújás után a már
meglévő szárnyak, valamint az ivarszervek kifejlődnek (16.33. ábra). Az epimorfózis során
a következő fejlődési sor figyelhető meg: embrió, elsődleges (primer) lárva, imágó.
[Megjegyzendő, hogy az elsőd-
leges lárvát gyakran fiatal (ju-
venilis) állatnak nevezik.]
A következő nagy csoportba
tartozó fajok átváltozással (he-
mimetamorphosis) fejlődnek.
Ennek során az imágótól jelen-
tősen eltérő testfelépítésű lárvák
bújnak ki a petéből. Lárvakori
szervekkel rendelkeznek és élet-
módjuk is eltér a kifejlett állato-
kétól. Ebben különböznekaz elő-
ző csoport elsődleges lárváitól.
A lárvakori struktúrák jól lát-
hatóak (16.33. ábra). Legkésőbb
az utolsó vedlés során a lárvakori
szervek leépülnek, és kialakul-
nak az imágókra jellemző szer-
vek. Bábállapot azonban nem
jön tétre. A fejlődési sor ebben
az esetben: embrió, másodlagos
(secunder) lárva, imágó.
Ahatlábúakposztembrionális
fejlődésének harmadik típusa a
teljes átalakulás (holometa-
304
morphosis). A teljes átalaku-
lással fejlődő rovarok lárvái nagy-
mértékben különböznek a kifej-
lett állatoktól. Más a testfelépíté-
sük, és számos lárvakori szervvel
is rendelkeznek (16.33. ábra). A
lábak száma alapj án három nagy
csoportba sorolhatók, (16.34.
ábra) a kevéslábú, lábatlan és a
soklábú lárvák közé (16.35. áb-
ra) . A lárvakori fejlődés után egy
nyugvó stádium a báb- (pupa)
16.35. ábra. Soklábú lárvatípusok
A: hernyó, B: álhernyó, C: araszoló hernyó. A különbség az egyes
típusok között abban áll, hogy a potrohlábak száma pontosan 4 pár
(hernyó), annál több (álhernyó), vagy annál kevesebb (araszoló
hernyó) (Több szerző nyomán módosítva)
17.36. ábra. A báb felépítése és néhány jellegzetes
bábtípus
A: szabad báb oldalnézetből, B: szabad láb elöl-
nézetből, C: hangyaleső (szabad) bábja, D: lepke
(fedett) bábja, E: csípőszúnyog (fedett) bábja, F:
légy (tonna) bábja.
cs: csáp, eh: előhát, fe. fej, kb: kitinburok, Iá: láb, sz:
szárny (Több szerző nyomán módosítva)
állapot következik. A báb lehet szabad báb (pumpa libera), fedett báb (pupa obtecta) és
tonna báb (pupa coarctata) (16.36. ábra). A bábállapot alatt egy, az egész szervezetet érintő
nagy átalakulás következik be. Hangsúlyozni kell azonban, hogy ámbár a legtöbb lárvakori
szerv lebomlik és átalakul a kifejlett állatokra jellemző szervvé, ez az átalakulás a szívet, a
belső ivarszerveket és az idegrendszert nem érinti. A teljes átalakulással fejlődő fajok
végleges szervei az imaginális korongokból jönnek létre. Az imaginális korongok kis
sejtcsoportok, amelyek a lárvakor alatt a kültakaróhoz kapcsolódva találhatók meg. A sejtek
differenciálódási képessége nagymértékű. Miközben a lárvakori szervek leépülnek, ezekből
a sejtekből fejlődnek ki az imágó szervei. A teljes átalakulás során a következő stádiumok
figyelhetők meg: embrió, harmadlagos (tertier) lárva, báb, imágó. A teljes átalakulás stra-
tégiai előnyöket nyújt a faj számára. Különböző forrásokat használhat ki a lárva és az imágó,
a báb pedig kellő védelem alatt vészelheti át a kedvezőtlen környezeti körülményeket. A
posztembrionális fejlődés során nyugalmi szakaszok (lárvakori diapauza) is megfigyelhe-
tők. Számos rovarfaj esetében a nappalok rövidülése vagy a hőmérséklet csökkenése
vezet diapauzához. Ez igen fontos jelenség, mert segíti a lárvákat, hogy a kedvezőtlen
körülményeket táplálkozás, mozgás nélkül át-
vészeljék. A diapauza hozzájárulhat ahhoz is,
hogy a populáció egyedeinek aktivitási idejét
szinkronizálja, ami a források hatékonyabb
kihasználását segítheti.
A hatlábúak hormonrendszere fejlett. A
hormonok számos életműködés szabályozá-
sában vesznek részt, de a legfontosabb szere-
pet a vedlések során játszák. A hormonrend-
szer legfontosabb tagj ai az agy neuroszekréciós
sejtjei, a corpus cardiacum, a corpus allatum
és az előtöri vagy protorakális mirigy (16.37.
ábra). Az előagy dúcok közötti területe (pars
intercerebralis) neuroszekréciós sejtjeinek
egy csoportja torakotrop hormont termel
(16.38. ábra). Az agyban található másik sejt-
csoport képzi a burzikont, a kutikula kemény-
ségéért és színezetéért felelős hormont. A cor-
pus cardiacum egy neurohemális szerv. Ben-
ne raktározódik a neuroszekréciós sejtek által
termelt torakotrop hormon. Itt termelődnek a
testfolyadék összetételét szabályozó hormo-
nok is. Amennyiben a corpus cardiacum több
torakotrop hormont juttat a testfolyadékba,
305
16.37. ábra. Az idegrendszer felépítése egy hangya (Formica sp.) fejében.
(A rágó és az állkapocs az áttekinthetőség kedvéért hiányzik az ábráról)
ai: alsó ajkat beidegző ideg, ao: aorta, ái: állkapcsot beidegző ideg, ca: corpus
allatum, cc: corpus cardiacum, ci: csápokat beidegző idegkötegek, ea: előagy, ei:
előtöri idegdúc, fc: frontális idegdúc, fi: felső ajkat beidegző ideg, ga: garat alatti
dúc, gl: glossa, gt: gombatest, iz: izomkötegek, ka: középagy, ns: neuroszekréciós
sejtek, ny: nyelőcső, ös: összetett szem, pm: protorakális mirigy, ps: pontszem,
ri: rágót beidegző ideg, sn: szájnyílás, ua: utóagy (Goll nyomán).
akkor fokozódik az előtöri
mirigy hormontermelése.
Több vedlési hormon (ecdy-
son, illetve származékai) ter-
melődik, és megkezdődik a
vedlés. Ezzel a folyamattal el-
lentétes hatást váltanak ki a
corpus allatum hormonjai,
a juvenilis hormonok. Hatá-
sukra a normális lárvakori nö-
vekedés következik be. Ha az
előtöri mirigy hormonterme-
lése csökken, akkor óriási lár-
vák, ha a corpus allatum mű-
ködése csökken, akkor törpe
imágók jönnek létre. Az álla-
tok egyik esetben sem képe-
sek szaporodásra. Ezért nap-
jainkban rendkívül intenzíven
kutatják a kártevő rovarok
hormonrendszerének műkö-
dését, hogy természetbarát
inszekticideket állítsanak elő.
Az előtöri mirigy az imágók-
ban eltűnik, kivéve azokat a
csoportokat, amelyek egész
életük során vedlenek (szá-
mos elsődlegesen szárnyatlan
16.38. ábra. Légy előagyának néhány fontosabb részlete (a fej homlokrészéről a kültakarót eltávolították)
cd: csápok irányítását végző idegdúc, cs: csáp, fp: fejpajzs, gt: gombatest, kt: központi test, ns: neuroszekréciós sejtek,
ös: összetett szem, pi: pars intercerebralis (Heisenberg nyomán)
306
rovar [Apterygota], mint például az ugróvillások). A corpus allatum az imágókban is aktív
marad addig, amíg az ováriumok és az ivarszervek járulékos mirigyei ki nem fejlődnek.
Idegrendszerük hasdúclánc idegrendszer (16.26. ábra). Ez központi és környéki
részekből áll. A központi idegrendszert az agy és a hasdúclánc, a környéki idegrendszert
a kültakaró alatt futó idegek alkotják. Az agy (ganglion supraoesophageale) a garat felett
helyezkedik el. Az első három fejszelvényben található idegdúcpár összeolvadásából jött
létre az evolúció során. Az előagy (protocerebrum) a legterjedelmesebb része (16.38.
ábra). Ezidegzibe a szemeket. Fejlettsége egyenes arányban álla látószervek fejlettségével.
Benne találhatók a gombatestek, melyekben igen sok asszociációs neuron található. Ez
a képlet számos viselkedésformáért felelős. A középagy (deuterocerebrum) a csápokat
és a csápizmokat látja el idegekkel. Az utóagy (tritocerebrum) viszonylag gyengén fejlett,
mivel a rovarok második pár csápja (ellentétben a rákokkal) hiányzik. Az utóagy végzi a
felső ajak (labrum) és az előbél beidegzését. Az agyból a garat két oldalát körülölelő
idegköteg indul ki, és a garat alatti dúchoz (ganglion infraoesophageale) csatlakozik.
A garat alatti dúc idegzi be a rágókat, az állkapcsokat és az alsó ajkat. A tor dúcaiból
fúrnak az idegek a szárnyakba és a lábakba, illetve a végtagokat mozgató izmokhoz. A
potroh szelvényei nagyfokú autonómiával rendelkeznek, önállóan irányítják a szelvény
számos mozgását. Kísérleti körülmények között a torról levágott potroh szabályosan
lerakta a petéket az agy irányítása nélkül is. Az agy előtt található dúc a testfolyadék
ionkoncentrációjának szabályozását végzi. Az agy mögötti, hipocerebrális ganglionnal
együttműködve szabályozza az előbél és a szív mozgását is. A hasi dúcokból kiinduló
idegek szabályozzák a légzőnyílások zárórendszereinek mozgásait, míg az utolsó szel-
vényből kiinduló idegek az utóbél és az ivarszervek beidegzésében vesznek részt. A
környéki idegrendszer nem más, mint a kültakaró alatt futó idegsejtek hálózata. Ezek
kötik össze az érzékszerveket a központi idegrendszerrel.
Az érzékszervek igen változatos felépítésűek és működésűek a hatlábúak körében. A
legelterjedtebb mechanikai érzékszervek az érzékelőszőrök és a szkolopidiális szervek, de
sokfelé található érzékelőkupola és hasadékszerv is. A hallószervek közül a tümpanális
(tympanalis) szerv a legjellegzetesebb (16.39. ábra). Számos csoportban, így az egyenes-
szárnyúak, fogólábúak, poloskák, kabócák, recés fátyolkák, molylepkék és kétszárnyúak
rendjében fordul elő. Felépítése mindegyik csoportban messzemenően egységes. A
16.39. ábra. A: szöcske lába, a lábszáron lévő hallónyílásokkal, B: a lábszár keresztmetszete a hallószerv
magasságában, C: tümpanális hallószerv
do: dobhártya, ha: hallónyilás, hv: hallószerv, id: idegköteg, ku: kutikula, sz: szkolopidium, te: testüreg, tr: trachea
(Davies és Weber nyomán)
307
16.40. ábra. Szúnyog Johnston-féle hallószerve
es: első csápíz, id: idegköteg, ms: második csápíz, se: szkolopi-
díum, sm: idegsejt magja, ső: szőr, tr: trachea (Weber nyomán)
állkapcsi tapogatón és a lábfejen fordulnak
rendelkezésre.
trachearendszemek egy széles üregét a
testfelszín közelében feszes kitinhártya zárj a
le. A levegő hullámai ezt rezegtetik meg, és
az alatta elhelyezkedő idegsejtek ennek
megfelelően jönnek ingerületbe. Az egye-
nesszárnyúak az 5-20 ezer Hz frekvenciára
érzékenyek, míg a molylepkék hallószerve
a magasabb frekvenciatartományokban
(ultrahangok) is érzékeny. A Johnston-
féle hallószerv sok rovar csoporban megta-
lálható (16.40. ábra). A hím szúnyogok
második csápízében (pedicellus) helyez-
kedik el, és a csápostor (flagellum) mozgá-
sa ingerli. így érzékelik a szúnyogok a
nőstények számycsapásai által keltett faj-
specifikus hullámokat, de a saját repülé-
sük sebességét is. A térd alatti szerv az
egyenesszárnyúak, álkérészek, poloskák,
lepkék és hártyásszámyúak körében ter-
jedt el. A kordotonális szerv, amely halló-
szervként vagy a testrészek egymáshoz va-
ló helyzetének érzékelésére szolgáló szerv-
ként funkcionál, mindegyik rovarcsoport-
ban megtalálható. A jól fejlett szaglószer-
vek mellett, amelyekkel a feromonokat
érzékelik, a rovarok rendelkeznek hőér-
zékelő receptorokkal is. Ezek a csápon, az
elő. Működésükről kevés információ áll
A hatlábúak legfejlettebb érzékszerve a szem. Pontszemeket (ocelli) (16.41. ábra) és spe-
ciális egyszerű szemekből álló,
összetett szemeket (oculi com-
positi) különböztetnek meg.
Vannakfajok, amelyeken mind-
két szemtípus előfordul, máso-
kon csupán az egyik vagy másik
szemtípus található meg. A
pontszernek elhelyezkedhetnek
a fej tetején (lárvák és imágók)
vagy a fej oldalán. A fejtetőn
egy háromszög sarkainak meg-
felelően három pontszem ül-
het. A középső is páros volt a
törzsfejlődés során valamikor,
mivel két idegrost idegzi be. A
háti pontszem egy nagy szaru-
lencsével gyűjti a fényt. Alatta
átlátszó sejtek rétege (komagén
sejtek) található, mely befedi az
egymás mellett álló érzéksejtek
(retinula) felszínét. Kívülről és
16.41. ábra. Rovarok szemének felépítése. A*, oldalsó pontszem,
B: appozíciós egyszerű szem, C: szuperpozíciós egyszerű szem
ili: ideghánya, ip: az idegsejteket körülvevő pigmentsejtek, is: idegsejt,
kk: kristálykúp, ki: kitinlemez, kp: kristálykúpot körülvevő sejtek, ks: komagén
sejtek, sz: szarulencse (Davies nyomán)
308
alulról sötét pigmentsejtek veszik körül (árnyékolják) az említett rendszert. Az oldalsó
pontszernek struktúrája nem egységes. Ez a szem legfőképpen a közeli tárgyak mozgását
érzékeli, de a fény irányára és egynémely színekre is reagál. Az összetett szemek állhatnak
néhány tucat egyszerű szemből, ommatidiumból (hangyadolgozók), de több tízezerből
(szitakötők) is. Az átlátszó kutikulából alakul ki a lencse (kitinlencse), melyet a komagén
sejtek hoznak létre. Ezt a réteget az állat vedléskor leveti. A komagén sejtek alatt kristálykúp
található. Ezen keresztül vetül a fény az alatta található retinulára, amely hét sejt együtteséből
áll. Az említett egységet többé-kevésbé zártan pigmentsejtek veszik körül és szigetelik el
egymástól. Ha az egyes egyszerű szemeket a pigmentsejtek egymástól teljesen elszigetelik
(appozíciós szem), akkor erős fényben (nappal) éles képet lát az állat. Ha viszont a szigetelés
nem teljes, több lencse is vetítheti a fényt ugyanarra a receptorsejtre (szuperpozíciós szem)
(16.41. ábra). Ezzel a szemmel kevesebb fényben (éjjel) is jól lehet látni. Vannak esetek,
amikor a pigmentsejtekben a színanyag mennyisége változik, így lehetővé válik, hogy nappal
és éjjel is élesen lásson az állat.
Osztály: Rovarszerűek (Parainsecta)
Rend: Előrovarok (Protura) 16.4.1. fejezet
Rend; Lábaspotrohúak (Diplura) 16.4.2. fejezet
Rend: Ugróvillások (Collembola) 16.4.3. fejezet
Osztály: Rovarok (Insecta)
Rend: Ugró ősrovarok (Archeognatha) 16.4.4. fejezet
Rend: Pikkelykék (Zygentoma) 16.4.5. fejezet
Rend: Kérészek (Ephemeroptera) 16.4.6. fejezet
Rend: Szitakötők (Odonata) 16.4.7. fejezet
Rend: Álkérészek (Plecoptera) 16.4.8. fejezet
Rend: Fülbemászók (Dermaptera) 16.4.9. fejezet
Rend: Fogólábúak (Mantodea) 16.4.10. fejezet
Rend: Csótányok (Blattodea) 16.4.11. fejezet
Rend: Sáskák (Caelifera) 16.4.12. fejezet
Rend: Tojócsöves egyenesszárnyúak (Ensifera) 16.4.13. fejezet
Rend: Fatetvek (Psocoptera) 16.4.14. fejezet
Rend: Tetvek (Phthiraptera) 16.4.15. fejezet
Rend: Tripszek (Thysanoptera) 16.4.16. fejezet
Rend: Poloskák (Heteroptera) 16.4.17. fejezet
Rend: Színkabócák (Auchenorrhyncha) 16.4.18. fejezet
Rend: Növénytetvek (Sternorrhyncha) 16.4.19. fejezet
Rend: Vízifátyolkák (Megaloptera) 16.4.20. fejezet
Rend: Tevenyakú fatyolkák (Raphidioptera) 16.4.21. fejezet
Rend: Recésfátyolkák (Plannipennia) 16.4.22. fejezet
Rend: Bogarak (Coleoptera) 16.4.23. fejezet
Rend: Legyezőszárnyúak (Strepsiptera) 16.4.24. fejezet
Rend: Hártyásszámyúak (Hymenoptera) 16.4.25. fejezet
Rend: Tegzesek (Trichoptera) 16.4.26. fejezet
Rend: Lepkék (Lepidoptera) 16.4.27. fejezet
Rend: Csőrös rovarok (Mecoptera) 16.4.28. fejezet
Rend: Kétszárnyúak (Diptera) 16.4.29. fejezet
Rend: Bolhák (Siphonaptera) 16.4.30. fejezet
Megjegyzés: A rovarszerűek szájszerve a fejtokba, visszahúzva található. A tapogató, a szem
és a Malpighi edények gyengén fejlettek. Az ide tartozó rendeket másokkal együtt régebben az
alacsonyabbrendű rovarok (Apterygota) taxonba sorolták. A rovarok szájszerve kívülről is jól
látható. Csak a csápok alapízeiben vannak izmok.
309
16.4.1. Előrovarok (Protura)
Apró, fél-két milliméter hosszú állatok. Több száz fajukat ismerik. Tes-
tük viszonylag egyforma szelvényekből áll (homonom szelvénye-
zettség). Igen feltűnő különlegességük, hogy a csápjaik hiányoznak.
Ezek helyett a kissé megnyúlt első lábpár szolgál tapogatószervként
(16.42. ábra). Szúró-szívó szájszervük a fejbe visszahúzódva helyezkedik
el (entognath szájszerv). Elsődlegesen szárnyatlanok, tehát őseiknek sem
volt szárnyuk. Az első három potrohszelvényen egy-egy pár csökevényes
láb van. Külső ivarszerveik jól fejlettek. A petéből kikelő lárvák potroh-
szelvényeinek száma 8, a posztembrionális fejlődés során további három
alakul ki (anamorphosis).
A hajnalrovar (Eosentomon transitorium) a csoport jellemző képviselője. A
potrohún három pár, két ízből álló csökevényes lábat visel. Avarban, talajban
sokfelé előfordul. A többi előrovarhoz hasonlóan a gombák hifáit szivogatja.
16.42. ábra. Elő-
rovar testfelépí-
tésének vázlata
át: állkapcsi tapo-
gató, el: első láb,
pl: csökevényes
potrohlábak (We-
ber nyomán)
16.4.2. Lábaspotrohúak (Diplura)
Mintegy félezer faj tartozik ebbe az ősi rovarrendbe (16.43. ábra).
Testhosszuk néhány millimétertől több centiméterig terjed. Rágó száj-
szervvel rendelkeznek, amely a fejtokba süllyedt üregbe visszahúzható.
Ebben az állapotban a rágónak és az állkapocsnak csak a vége emelkedik
ki az üregből. Szárnyuk hiányzik, elsődlegesen szárnyatlanok. Potrohúk tizenegy szel-
vényből áll. Az 1 .(2.)-7. szelvényeken csökevényes lábak láthatók. Ez igen ősi bélyeg.
Vannak farcsutáik (styli), de a faroktoldalékok (cerci) jellemzőek igazán. Ezek lehetnek
16.43. ábra Lábaspotrohú rovar testfelépítésének vázlata
át: állkapcsi tapogató, cs: csáp, el: első láb, ft: faroktoldalék, pl:
csökevényes potrohlábak (Weber nyomán)
hosszúak, fonalasak (Campodeidae),
vagy rövidek, hajlottak, erősek (Japy-
gidae). Már a petéből kikelő lárvák
ugyanannyi szelvénnyel rendelkeznek,
mint az ivarérett állatok. Életük végéig
rendszeresen vedlenek. A lábaspotro-
húak fehér vagy sárga színűek. A talaj-
ban, a talaj felszínén élnek. Növény-
evők, vegyestáplálkozásúak és ritkán
ragadozók. Rendszerint meleg, nedves
helyeken fordulnak elő.
Sokfelé gyakori a fonálfarkú ősrovar
(Campodea staphylinus).
16.4.3. Ugróvillások (Collembola)
Néhány ezer fajuk ismert ezeknek a pár milliméteres állatoknak (16.44. ábra). Fejükön
rövid, négyízű csáp és a fejtokba visszahúzott rágó (ritkán szúró-szívó) szájszerv találha-
tó. Elsődlegesen szárnyatlanok. Az első torszelvény kisebb a többinél, gyakran a második
alá húzódik, így nem látható. Potrohúk mindössze hat szelvényből áll. Az első potrohszel-
vényen található a hasi tömlő (centrális tubulus), egy nagy, kitüremkedett hólyag, aminek
a funkciója még nem tisztázott. Valószínűleg a légzésben és a vízfelvétel-vízleadás bizto-
sításában van szerepe. A harmadik szelvényen van az akasztó (retinaculum), amibe a ne-
gyedik szelvényről előre hajló ugróvilla (furca) kapcsolható. Ilyen állapotban az ugróvilla
a has alá simul, és olyan állapotban van, mint egy felhúzott rugó. Ha az állatot megzavarják,
az akasztó elengedi az ugróvillát és a test előrepattan. Trachearendszerük hiányzik vagy
310
erősen redukálódott. Hiányoznak
a Malpighi-edények is. Helyettük
az alsó ajaki mirigy lát el kiválasztási
funkciót a hasi tömlővel és a zsír-
mirigyekkel együtt. Gyakran va-
kok, vagy legfeljebb 8 szem laza
halmaza alkotja a szemet a fej két
oldalán. A csáp mögötti szervük
kémiai érzékelésre szolgál. Az aduit
állatok is vedlenek. A talajban élő
fajok fehérek, míg a talaj felszínén
találhatók sárga, barna, fekete szí-
nűek. Vegyestáplálkozásúak, nö-
16.44. ábra. Ugróvillás testfelépítésének vázlata
ak: akasztó, cs: csáp, ht: hasi tömlő, sz: szem, uv: ugróvilla (Davies
nyomán)
vényeket, avart, algát, baktériumokat, de mindenekelőttgombahifákat esznek. Igen nagy
egyedszámban fordulnak elő (a mezőgazdasági területek talajában 2-3000 db/m2).
Általában hasznosak, mert segítik a szerves anyagok lebontását, a tápanyagok minerali-
zációját. Néhány fajuk (pl. Onychiurus fimatus) kártevő is lehet.
A Folsomia candida sokfelé előforduló faj, melyet gyakran használnak laboratóriumi kísérletek-
ben. Mindennapos a kertészetekben és az üvegházakban, valamint a lakások virágcserepeiben is.
Hazánkban a zárt erdők kivételével mindenütt megtalálható a zöld gömböcugróka (Sminthurus
viridis). Ellentétben az előbb említett nyúlánk, hengeres potrohú fajokkal, ennek a fajnak és
rokonainak is ráncos, gömb alakú a potroha. A lucerna és a lóhere kártevője lehet.
CS
át fc_
16.45. ábra. Osiszájszervu testfelépítésének vázlata
át: állkapcsi tapogató, cs: csáp, fc: farcsuta, ft: faroktoldalék,
ös: összetett szem, vf: végfonal (Weber nyomán)
ös
16.4.4. Ugró ősrovarok
(Archeognatha)
Pár száz fajt számláló csoport. A barnás
színű állatok testhossza fél-két és fél
centiméter között változik. Fejük kicsi,
toruk erőteljes
(16.45. ábra).
Fonalas csáp-
juk, melyet
nyugalomban
a testhez simítva viselnek, feltűnően hosszú. Rágó szájszervük
van. Az állkapcsi tapogató (palpus maxillaris) is hosszú. A tor
hátlemezei oldal irányban peremszerűen kiterjednek (para-
notum). Elsődlegesen szárnyatlanok. A potroh végén fartol-
dalékok és végfonal található. Életük végéig vedlenek. Extrém
élőhelyeken is elterjedtek (sziklák, gleccserek), mivel a hőmér-
séklet-változásra aránylag kevéssé érzékenyek. Ismert csoport-
juk a pattanók (Machilidea). Hazánkban sokfelé, különböző,
főleg száraz élőhelyeken megtalálható az ugró ősrovar
(Lepismachilis notata) nevű faj.
16.4.5. Pikkelykék (Zygentoma)
Mind fajszámukat, mind testnagyságukat, testalakulásukat
tekintve hasonlítanak a pattanókhoz. Testük lapított, a széles
előtör után fokozatosan elkeskenyedő (16.46. ábra), minde-
nütt ezüstösen csillogó pikkelyekkel borított. Csápjaik hosszúak,
CS
fc
ft
vf
16.46. ábra. Pikkelyke testfel-
építésének vázlata
cs: csáp, fc: farcsuta, ft: fartol-
dalék, ös: összetett szem,vf: vég-
fonal (Weber nyomán)
311
fonalasak. Összetett szemeik közepes méretűek. Szárnyaik nincsenek. Faroktoldalékok
és végfonal is látható a potroh végén. Az aduit állatok is vedlenek. Kimondottan
melegkedvelők, ezért a szárazságot is jól bírják.
Jól ismert csoportjuk a pikkelykék (Lepismatidea). Ezek közül a legismertebb faj a lakásokban,
raktárakban élő, centiméteres testhosszú ezüstös pikkelyke (Lepisma saccharina). Minden édes,
szénhidráttartalmú táplálékot megrág, de az enyvet, tapétaragasztót, könyveket sem kíméli.
Raktárakban a magvakat károsíthatja. Hasonlóan a többi pikkelykéhez, képes a cellulóz emészté-
sére is.
16.4.6. Kérészek (Ephemeroptera)
A kérészek ősi állatok. Ez a rend már a permkorban is népes csoport volt. Mintegy kétezer
faj tartozik ebbe a jellegzetes, törékeny testű rovarokat összefogó csoportba. Testnagy-
ságuk 0,3-5 cm között változik. Az imágók feje viszonylag kicsi, rövid csápokat visel
(16.47. ábra). Rágó szájszervvel rendelkeznek, de az imágókon ez csökevényes.
Lábaik vékonyak, gyengék, de kapaszkodásra alkalmasak. A hímek első pár lába
megnyúlt, párzáskor ez segíti az állatot a nőstény átkarolásában. Két pár gyenge, hártyás
szárnyuk közül az első pár lényegesen nagyobb, mint a második, amely teljesen re-
dukálódhat is. Igen feltűnő jellegzetességük, hogy szárnyaikat nem tudják a hátukra
fektetni, ezért azok nyugalmi állapotban függőleges irányban állnak. A legtöbb faj szár-
nya átlátszó, ritkábban szürkés foltokkal díszített, füstös vagy tejszerűen fehér. Hosszú
potrohúk végén általában egy pár faroktoldalék és egy végfonal található. Ennek a
bélyegnek a segítségével könnyen elkülöníthetők az álkérészektől lárva korban is, mert
ott végfonal nem található. Összetett szemeik és három pontszemük is jól fejlett. Az
imágók nem táplálkoznak, bélcsatomájuk vékony falú, fejletlen. Ismert jellegzetességük,
hogy a kifejlett állatok - az elevenszülők kivételével - csak rövid ideig élnek. A hímek
repülés közben termékenyítik meg a nőstényeket, majd elpusztulnak. A nőstények pe-
téiketfolyó- és állóvizekbe rakják,
és szintén elpusztulnak. A lárvák
több évig (1-3 év) fejlődnek az
iszapban. Rendszerint algával és
elpusztult növényi részekkel táp-
lálkoznak, de vannak közöttük ra-
gadozók is. Trachearendszerük
zárt, tracheakopoltyújuk van
(16.47. ábra), de a végbelükbe
szivattyúzott vízből is fel tudják
venni az oxigént. Többszöri ved-
lés után - a lárvafejlődés végén -
a lárvából egy szárnyas, de még
nem ivarérett szubimágó bújik ki.
Ez rövid időn belül újra vedlik,
így jelenik meg a szaporodásra
képes imágó.
A kérészek főleg a mérsékelt és a
hideg égövek lakói. Állóvizektől a se-
besen folyó patakokig mindenütt elő-
fordulnak. Testalakjuk is ennek meg-
felelően alakult. A lárvák halaink fon-
tos táplálékállatai. Lárváik érzéke-
nyek a vizek szennyezettségére, első-
cs: csáp, eh: első torszelvény hátlemeze, esi első szárny, ft: fartoldalék,
ms: második szárny, ös: összetett szem, pe: peték, ps: pontszem, sk:
számykezdemény, tk: tracheakopoltyú, vf: végfonal (Weber nyomán)
312
sorban az oxigéntartalom csökkenésére, ezért a biológiai vízminősítés során mint indikátorállatok
kerülnek szóba. Jellegzetes, nagy termetű faj a tiszavirág (Palingenia longicauda). Fartoldaléka
nagyon hosszú, végfonala viszont csökevényes. Sokfelé megtalálható, de a Tisza egyes szakaszain
néha megint tömeges.
16.4.7. Szitakötők (Odonata)
A szitakötők is igen ősi rovarok. Napjainkban mintegy ötezer faját ismerik ezeknek a
2-15 cm hosszú állatoknak. A lárvák és az imágók testfelépítése és életmódja eltér egy-
mástól. A lárvák teste lehet rövid, tömzsi, de megnyúlt is (16.48. ábra). Ragadozó száj-
szervük igen jellegzetes fogókészülékkel (álarc) egészült ki. Az alsó ajak egyes részei
(submentum, mentum) megnyúltak, végükön egymás felé mozgatható, fogazott leme-
zek találhatók. Az egész készülék hirtelen előre nyújtható, így igen alkalmas az áldozatok
megragadására. A légzés történhet tracheakopoltyúkkal (Zygoptera) (16.48. ábra) vagy
a végbélbe felszívott víz segítségével (Anisoptera). A lárvakor végén báb nem alakul ki,
de az utolsó vedlés előtt rövidebb-hosszabb nyugalmi periódus figyelhető meg. A
kifejlett állatok feje nagy, a csáp igen rövid (16.48. ábra). A könnyen mozgatható fej két
oldalán hatalmas összetett szemek ülnek, a fejtetőn pedig három pontszem található,
ezért a levegőben vadászva, gyors repülés közben is megtalálják zsákmányukat. Lábaik
a zsákmány vagy a pihenőként szolgáló nádszál átkarolására is alkalmasak. Szárnyaik
többé-kevésbé egyforma, gazdagon erezett hártyás szárnyak. Repülés közben az első és
a hátsó pár egymással ellentétesen mozog. Potrohúk hosszú, a végén jól fejlett külső
ivarszervekkel. Az imágók tipikus repülő állatok. A felemásszámyúakra (Anisoptera) kü-
lönösen jellemző, hogy a levegőben ejtik el áldozataikat, ott párzanak, repülésükkel jelzik
territóriumaik határait. A szitakötők viszonylag ritka számycsapásokkal (20/sec) előre és
hátrafelé is képesek repülni, vagy egy helyben lebegni a levegőben. Mindenütt előfordul-
nak, ahol a szaporodá-
sukhoz szükséges vize-
ket megtalálják. Kifej-
lett korban messze el-
távolodhatnak a vizek-
től. A mezőgazdaság
szempontjából szere-
pük kettős. A lárvák
halgazdasági kártevők.
Ivadéknevelő tavakban
sok kis halat elpusztít-
hatnak. A kifejlett álla-
tok viszont számos kár-
tevő rovart, vérszivó
böglyöket foghatnak
meg, amivel hasznot
hajtanak.
Vizek partján minden-
felé elterjedt, igen gya-
kori faj a sávos szitakö-
tő (Agroion splendens). Fé-
mes kék vagy zöld teste
karcsú, szárnyain kereszt-
ben széles sötét sáv hú-
zódik. Kedvelt élőhelyei
16.48. ábra. A: szitakötőlárva, B: imágó, C: Zygoptera lárva potrohvégén lévő
lemezes tracheakopoltyúk, D: szitakötőlárva-fej
aa: alsó ajak, cs: csáp, et: előtör, ft: faroktoldalék, kt: középtor, ős: összetett szem, rá:
rágó, ut: utótor (Több szerző nyomán módosítva)
313
a lassú folyású patakok. Főleg a síkvidéki és dombvidéki vizekben fordul elő. A hímek territóriumot
tartanak, ahonnan a többi hímet elűzik. Gyakori faj a közönséges szitakötő (Sympetrum
vulgatum) is.
16.4.8. Álkérészek (Plecoptera)
Bár a rend neve hasonlóságot sejtet, törzsfejlődésükben és testfelépítésükben sem
hasonlítanak a kérészekhez. Néhány ezer fajt magába foglaló rend. Az állatok testhossza
fél-négy centiméter. A lárvák teste hát-hasi irányban lapított (16.49. ábra). Színezetükkel,
mintázatukkal beolvadnak a vizek aljzatának színébe, így nehezen vehetők észre. Lábaik
erősek, gyors mozgásra alkalmasak. Rágó szájszerveikkel elhalt növényi részeket fogyasz-
tanak. A ragadozó fajok ritkák. Az oxigént legtöbbször a test teljes felületén, diffúzióval
veszik fel. A nagyobb testű fajok torán, néha a potrohán is tracheakopoltyúk találhatók.
A tiszta, oxigénben gazdag vizeket kedvelik, ezért leggyakrabban a sebes folyású patakok
kövei alatt bukkanunk rájuk. A vizek szennyeződésekor gyorsan elpusztulnak, ezért a
biológiai vízminősítés során figyelembe veszik az álkérészlárvák fajösszetételének válto-
zásait. Az imágók testformája hasonlít a lárvákéhoz, szintén kissé lapított (16.49. ábra).
Csápjaik hosszúak, vékonyak. Két pár hárty ás szárnyat viselnek. A második pár szárny
kissé nagyobb, nint az első. Nyugalomban szárnyaikat a test hátoldalára fektetve hordják.
A szárnyak gazdagon erezettek. Lábaik erősek. Három pontszemük és egy pár, igen jól
fejlett összetett szemük van. Rágó szájszervük nem, vagy gyengén működik, mivel az
imágók alig táplálkoznak. Rendszerint a vizek környékének növényzetén üldögélnek. A
lárvák és az imágók potrohának végén egy pár fartoldalék (cercus) található, de vég-
fonaluk nincs. A fartoldalék lehet csökevényes, de hiányozhat is. Az imágók rövid
életűek, néhány hétig élnek, a párzás után pár órával elpusztulnak. A legtöbb faj teljes
életciklusa egy év vagy annál rövidebb. A lárvák a halak táplálékaként fontosak lehetnek.
Hegyi patakokban különösen gyakran fordul elő az óriás álkérész (Perla maxima). A
közönséges álkérész (Nemoura cinerea) az egyik leggyakoribb faj.
16.49. ábra. A: álkérészlárva, B: álkérész-imágó
at: alsóajki tapogató, cs: csáp, ft: fartoldalék, ős: összetett szem, ps: pontszem, sk: számykezdemény (Steinmann nyomán)
314
16.50. ábra. Fülbemászó testfelépítésének vázlata
cs: csáp, es: elülső szárny, et: előtör hátlemeze, ft:
faroktoldalék, há: hátulsó szárny, hs: hátulsó szárny
kilógó vége, ös: összetett szem, su: sugárerek (Webet
nyomán)
16.4.9. Fülbemászók (Dermaptera)
Egységes testű, jellegzetes felépítésű, mintegy másfél ezer faj tartozik ide. Testük
nyúlánk, hát-hasi irányban lapított (16.50. ábra). Testhosszuk 0,5-5 cm közötti. A fej
előre áll (prognath), jól mozgatható, hosszú, fonalas csápokat, két nagy összetett szemet
és rágó szájszervet visel. A pontszernek hiányoznak. A csápokon található mechanikai
érzékszervek elsődlegesen fontosak az állatok életében, mivel rejtett életmódú fajokról
van szó. Az első pár szárny kicsi, pikkelyhez
hasonló, bőrszerű képződmény (tegmina).
Az erek szinte teljesen hiányoznak róla. A
hátsó szárnyak nagyok, hártyásak. Nyugalom-
ban többszörösen összehajtva az első szárnyak
alatt helyezkednek el. A lábak viszonylag rövid
járólábak. A potroh végén a fajokra jellemző
alakú, hatalmas, fogószerű fartoldalékok
(cerci) találhatók. Ezekkel bontják ki, illetve
hajtják össze hártyás szárnyaikat, de a fartolda-
lékok a táplálék megragadására is alkalmasak.
A hímek fartoldaléka jóval erősebb, mint a
nőstényeké. Petéiket a talajban levő gödörbe
rakják, ahol a nőstény őrzi a petéket. Meleget
és nedvességet kedvelő állatok. A talaj felszí-
nén, repedésekben, a fák kérge alatt találhatók.
Általában éjszaka aktívak. Elsősorban növény-
evők vagy vegyestáplálkozásúak, gyümölcsö-
ket, virágokat, növényi részeket, más rovarokat
fogyasztanak. Élő és elhalt szerves anyagokkal
egyaránt táplálkoznak. Kertészetekben, üveg-
házakban néha tömegesen lépnek fel, ekkor
kártevők lehetnek.
Mindenfelé elterjedt, ismert faj a közönséges
fülbemászó (Forficulaauricularia). Fák kérge alatt
gyakran fellelhető. Vegyestáplálkozású. Ritkán lép
fel akkora tömegben, hogy konyhakertekben kártevővé váljon. A törpe fülbemászó (Labia minőt)
hátsó szárnya fejlett, jó repülő. Nem csupán erdőkben, mezőkön, hanem komposztban és
trágyadombokon is megtalálható.
16.51. ábra. Fogólábú testfelépítésének vázlata
cs: csáp, es: elülső szárny, et: előtör, ft: fartoldalék, Ös: összetett szem
(Csiby nyomán)
16.4.10. Fogólábúak
(Mantodea)
Mintegy kétezer fajuk a meleg,
trópusi területeken mindenütt
elterjedt. Hosszú, megnyúlt tes-
tük 1-16 centiméteres. Fej ük ki-
csi, igen jól mozgatható (16.51.
ábra). Ez szükséges is, hiszen
nagy, összetett szemeikkel loka-
lizálják zsákmányukat. Szájszer-
veik erős, ragadozó száj szervek.
Az előtör (prothorax) hosszan
megnyúlt és két hatalmasan
315
megerősödött fogólábat visel. Számos erős kitintövis borítja a lábat. Hasonlóan pl. a
szitakötők lárváihoz, ezek is lesben állnak, majd hirtelen mozdulattal ragadják meg
áldozatukat. A 2. és 3. pár lábak gyenge járólábak. Első pár szárnyuk pergamenszerű, a
második hártyás. Rossz repülők. Az idegrendszer dúcainak nagyfokú az autonómiájuk,
ezért a párzás folytatódik akkor is, ha pl. a nőstény közben leharapta a hím fejét. Petéiket
jellegzetes tokba (ootheca) rakják és a növényekre ragasztják.
Hazánk füves területein, a dombvidékek rétjein fordul elő az egyetlen hazai faj, az imádkozó-
sáska (Mantis religiosa). A száraz, meleg, napos területeket részesíti előnyben. Rendszerint
mozdulatlanul ül és így várja zsákmányát, de ha megzavarják, gyorsan fut vagy nehézkesen, rövid
távra repül. Védett faj.
16.4.11. Csótányok (Blattodea)
Viszonylag sok, több ezer fajt magába foglaló csoport. Ovális, hát-hasi irányban erő-
teljesen lapított testük 0,2-10 cm hosszú. Testfelépítésük a rovarok ősi típusára
emlékeztet (16.52. ábra). Mozgatható fejük aránylag kicsi. Csápjaik igen hosszúak,
fonalasak, a rejtett, éjszakai életmódhoz szükséges tapogató funkciónak kiválóan meg-
felelnek. Az első torszelvény hátlemeze lapos, széles, jellegzetes formájú előháttá
(pronotum) alakult, ami a fejet többé-kevésbé befedi. Lábaik erős futólábak, az állatok
gyors mozgásúak. Két pár szárnyuk közül az első bőrszerű, a második hártyás, nyugalom-
ban egymás tetején, a potrohon fekszenek. A nőstények szárnya visszafejlődhet. Egyesek
tudnak repülni, ámde rosszul, inkább ugrásaikat hosszabbítják meg a szárnyak segítsé-
gével. A potroh végén rövid, ízelt fartoldalékok (cerci), a hímeken ezen túl még far-
csuták (styli) is láthatók. A csótányok rágó szájszervvel rendelkeznek. Vegyestáplálko-
16.52. ábra. A közönséges csótány testfelépítése. A: hím, B: nőstény, C: kitintok
cs: csáp, es: elülső szárny, et: előtör, fc: farcsuta, fe: fejlődő embriók, ft: fartoldalék, kt: kitintok, ös: összetett szem (Ziegler
nyomán)
316
zásúak, növényeket, állatokat, szerves maradványokat egyaránt elfogyasztanak. A táplá-
lékhasznosítás foka magas, képesek nehezen bontható anyagok (szaru, cellulóz, keratin
stb.) emésztésére is, elsősorban szimbionta baktériumok és egysejtűek segítségével. Kö-
rükben gyakori a kannibalizmus is. Petéiket jellegzetes alakú kitintokba (ootheca) he-
lyezik (16.52. ábra). Ezeket azután a nőstények a potroh végén, részben a testbe süllyeszt-
ve magukkal hordozzák. A csótányok erdőkben, mezőkön élnek, de néhány fajuk kivá-
lóan alkalmazkodott az emberi környezethez. Ezek lakásokban, élelmiszerboltokban,
középületekben, ahol viszonylag állandó, magas a hőmérséklet, elégséges búvóhely és
táplálék akad, hatalmas egyedsűrűséget érhetnek el. Kártételük, hogy megrágják, ma-
radványaikkal szennyezik az élelmiszereket, és kórokozókat terjeszthetnek. Számos em-
ber allergiás a csótányokra. Mezőgazdasági raktárakban vagy kertészetekben kártevők is
lehetnek. Rendszerint éjjel aktívak.
Az emberek közelében élő legismertebb hazai faj a német csótány (Blattela germanica) és a
közönséges csótány (Blatta orientalis). A német csótány karcsú, szalmasárga állat. Főleg
élelmiszereket rág meg. Lakótelepeken gyakori. A közönséges csótány széles testű, sötét barnás-
vörös színű rovar. Vegyestáplálkozású, az alacsony tápértékű anyagokat (szaru, kitin) is jól
hasznosítja. Kórokozó baktériumokat, férgek petéit terjesztheti, ezért közegészségügyi jelentőséggel
bír. Nem csupán régi bérházakban, de sütödékben, zöldségesboltokban, kórházakban is elterjedt.
A lapp erdeicsótány (Ectobius lapponicus) különböző erdőkben, napos helyeken, erdő szélén a
bokrokon tanyázik, repülni is tud.
Ősi testfelépítésű, fajgazdag (több mint tizenötezer faj) csoport az egyenesszár-
nyúaké (Orthoptera). Mozgékony fejükön nagy összetett szemek, erős rágó szájszervek
találhatók. Előtoruk nagy, oldalról és felülről nyeregszerűen alakult, egységes kitinlemez
veszi körbe. Az 1. és 2. pár láb egyszerű járóláb, a harmadik pár viszont többé-kevésbé
erős ugrólábbá módosult. Különösen a combok (femur) erősek, izmosak. Az első
szárnyak pergamenszerűek, a hátsók hártyásak, mindkettő gazdagon erezett. Általában
gyenge repülők (kivéve a vándorsáskákat), a szárnyak az ugrásokban segítenek, néha
redukálódhatnak is. A potroh végén egy pár fartoldalék (cercus) és a nőstényeken
rendszerint egy jellegzetes alakú tojócső található. A peték lerakása után a nőstények
rendszerint elpusztulnak. A lárvák kifejtéssel fejlődnek. Hangokat adnak (ciripelnek), és
hallószerveik is fejlettek. Elsősorban növényeket esznek, de vannak vegyestáplálkozású
fajok is. Nagy tömegeik komoly károkat okozhatnak a kertészetekben vagy a szántóföldi
növénykultúrákban. Újabban külön rendként tárgyalják a sáskákat (Caelifera), vala-
mint a tojócsöves egyenesszámyúakat (Ensifera). Ezért a következő jellemzés során
ezt a beosztást követjük.
16.4.12. Sáskák (Caelifera)
Testük kétoldalról kissé összenyomott. Csápjaik rövidek, nem haladják meg a test
hosszánakfelét, legfeljebb 30 ízből állnak (16.53. ábra). Rágó szájszerveik erősek. Fejlett
előhátukat nyereg formájú, jellegzetes alakú, egységes kitinlemez határolja felülről és
kétoldalról. A harmadik lábpár nagy, erős ugróláb. A hímek párzókészüléke komplikált
felépítésű. A nőstények tojókészüléke rövid, külső ivarszerveikkel lyukat fúrnak a talajba,
így rakják le petéiket. Számos faj képes a színezetét megváltoztatni, így alkalmazkodik a
háttér mintázatához. Hangot úgy adnak, hogy az ugróláb combjának belső felén talál-
ható kitincsapsort hozzádörzsölik a nyugalomban fekvő elülső szárny kiemelkedő eré-
hez. A hang frekvenciája, ritmusa, a ciripelés hosszának a szünetekhez viszonyított
aránya a fajra jellemző. A ciripelés a párválasztás fontos eszköze. A nőstények hangadó
szerve gyengébben fejlett, mint a hímeké. Megjegyzendő, hogy egyes sáskafajok kommu-
nikációjában fontos szerepet játszik az összedörzsölt rágok, a szárnyak mozgatása, vagy
317
a talaj ütögetése által keltett hang is. Számos faj szárnya redukálódott. Az utolsó vedlés
után még hosszabb időnek kell eltelni, amíg az állatok párzása elkezdődhet. Az ivari érést
feromonok gyorsíthatják. A tümpanális hallószerv az első potrohszelvény két oldalán
található.
A sáskákra jellemző, hogy ugyanazon faj magányosan élő alakjai, a szoliter egyedek
morfológiai, fiziológiai és ökológiai szempontok szerint is eltérnek a csoportosan élő
alakoktól, a gregariner egyedektől. Általában a nyílt biotópok, mezők, rétek, legelők,
tisztások lakói.
A sáskák jellegzetesen növényevő állatok. Néha tömegesen lépnek fel és szántóföldi növényeket
is károsíthatnak a sokféle növényt fogyasztó (polifág) rövidnyakú sáskák (Dociostaurus brevicollis).
Könnyen összetéveszthetők a marokkói
sáskával (Dociostaurus maroccanus). Mind-
két faj hajlamos a tömeges elszaporodásra
(gradáció). A két faj összetéveszthetősége
miatt a sáskajárásokra vonatkozó régebbi
adatok között nehéz meghatározni, hogy
mikor károsított a rövidnyakú és mikor a
marokkói sáska. A robusztus olasz sáska
(Calliptamus italicus) színe szürkésbamán
foltozott, a háttérbe illő. Hátulsó szárnya
részben rózsaszínű, ami repüléskor jól lát-
szik. Homokos vidékek kivételével minde-
nütt előfordul. Majdnem mindegyik ter-
mesztett növényünket károsíthatja. A Kár-
B
16.53. ábra. A: sáska testfelépítésének vázlata, B: hím ivarszerv, C: női ivarszerv
cs: csáp, ej: elvezető járat, et: előtör, he: here, hü: hüvely, jm: járulékos mirigy, kj: kilövellő járat, ős: összetett szem,
pc: petecsövecske, pv: petevezető, ts: tümpanális szerv, vf: végfonal (Weber és Jannone nyomán)
318
pát-medence jellegzetes sáskafaja a védett sisakos sáska (Acrida hungarica). Feje csúcsosan
megnyúlt, csápjai laposak, kard formájúak, harmadik pár lába gyenge. Száraz, meleg homokpusz-
tákon fordul elő.
16.4.13. Tojócsöves egyenesszárnyúak (Ensifera)
A tojócsöves egyenesszárnyúak két nagy csoport) át a szöcskék (Tettigoniidae) és a tücskök
(Gryllidae) alkotják. Mindkét csoport fajaira jellemző, hogy csápjuk igen hosszú, rend-
szerint jelentősen túllépi a potroh végét (16.54. ábra). Rágó szájszerveik - hasonlóan az
előző csoporthoz-jól fejlettek. Az első pár lábaikon levő kitintövisek a zsákmány rögzítését
segítik. Harmadik pár lábuk rendszerint erős ugróláb. A hímek párzó készüléke egyszerű,
a nőstények tojócsöve fejlett, hosszú, egyenes, vékony vagy rövid, tömzsi, erős (16.54.
ábra). A hímek hangadó szervei az első pár szárnyon találhatók. Ahogy a két szárny lapj ával
egymáson fekszik, az alul levő szárnyról kiemelkedő kitinlemezsort dörzsölik a felső szárny
egyik vastag eréhez és így keltenek hangot (16.54. ábra). A legtöbb esetben a nőstények
nem rendelkeznek hangadó szervvel. Tümpanális hallószervük az első lábpár lábszárá-
nak (tibia) két oldalán található. Általában vegyestáplálkozású állatok, ritkábban növény-
evők vagy ragadozók.
Kertészetekben gyakori kártevő alótücsök (Gryllotalpa gryllotalpa). Nem annyira rágásával
okoz kárt (mivel sok gerinctelen állatot is eszik), mint azzal, hogy járatokat ás a talajban és ezzel
a palántákat kitúrja. A barnás színű állat testét bársonyos szőrök borítják. Első lábai jellegzetesen
kiszélesedő ásólábak.
Füves területeken mindenfelé közönséges a vegyestáplálkozású mezei tücsök (Gryllus campestris).
Nagy testű, három-négy centiméteres faj a zöld lombszöcske (Tettigonia viridissima). A ma-
gasabb hegyvidékeket kivéve az egész országban elterjedt. Ragadozó állat, növényt ritkán fogyaszt.
A tömzsi testű, vegyestáplálkozású homokpusztai szöcske (Montana montana) az alföldi és
hegyvidéki sovány réteken gyakori.
felső felülete, D: jobb szárnytöve alsó
felülete
cé: cirpelő ér, cs: csáp, et: előtör, kh: ki-
tinhártya, ki: kitinlemezek sora, amit a cir-
pelö érrel penget az állat, ös: összetett
szem, ts: tümpanális szerv (Weber és
Knetsch nyomán)
319
16.4.14. Fatetvek (Psocoptera)
Ezeknek a néhány milliméteres állatoknak mintegy
kétezer fajuk ismert. Testük zömök, tömzsi, erősen
szőrözött, fejük jól mozgatható és hosszú csápokat
visel (16.55. ábra). Szájszervük érdekes felépítésű,
mert alapvetően rágó típusú, de az állkapocs belső
karélya (lacinia) árszerű szúrósertévé alakult. Ezzel
tudnak kisebb faforgácsokat kivésni a fából. Az alsó
aj ak (labium) mellett nyílnak a nyál- és szövőmirigyek
kivezető csövei a külvilágra. A fejlett összetett sze-
mek mellett 3 pontszemük is van. A szárnyak gyak-
ran hiányoznak, vagy csak a hímek viselnek szárnya-
kat. Ha rendelkeznek szárnyakkal, akkor is rosszul
repülnek. Lábaik erős járólábak. A hímek általában
kisebbek a nőstényeknél. Egyes fajok hímjei nem is
16.55. ábra. Fatetű testfelépítésének vázlata
cs: csáp, et: előtör, ös: összetett szem (Weber
nyomán)
ismertek. Szűznemzéssel és ivaros úton szaporodnak. A petéket a nőstények hálószövedék-
be helyezik. Általában 5-8 vedlés figyelhető meg, míg az állatok kifejlődnek. Jellegzetes-
ségük, hogy han^adásra is képesek. A potroh végét ütögetik az aljzathoz és ezzel kopogó
hangot adnak. Általában fakéreg alatt élnek, de madárfészkekben és méhkasokban is
megtalálhatók. Elsősorban gombákkal, algákkal, zuzmókkal, virágporral táplálkoznak.
Élelmiszer raktárakban az elpusztult fatetvek maradványai szennyezhetik az élelmet.
Lakásokban, könyvtárakban, raktárakban, méhkasokban gyakori, néha tömeges a kopogó por-
tetű (Trogium pulsatorium). Az ugró könyvtetű (Troctes divinatorius) egy milliméteres állat. Me-
16.56. ábra. A: egy vérszívó tetű (Anophtra) testfelépítése, B: a fej
hosszmetszete, C: serkéje, D: egy bunkóscsápú tetű (Amblycera)
testfelépítése
cs: csáp, et: előtör, ga: garat, ha: hajszál, íz: izom, ka: fejlett karom, nm:
nyálmirigy, ny: nyelőcső kivezetőcsöve, ös: összetett szem, ss: szúróserte,
sz: szájnyílás (Weber és Kaestner nyomán)
neküléskor gyorsan fut és nagyokat
ugrik. Könnyen összetéveszthető a ro-
kon fajokkal. Az egész világon előfor-
dul az ember környezetében, laká-
sokban, könyvtárakban, gyűjtemé-
nyekben, ahol kártevő is lehet.
16.4.15. Tetvek
(Phthiraptera)
Fajgazdag, több ezer fajt magába
foglaló, kizárólag ektoparazita ál-
latokat tartalmazó rend. Kicsik,
rendszerint egy milliméternél is
kisebbek vagy néhány milliméte-
resek. Az életmódjuknak megfe-
lelően testük hát-hasi irányban
erőtelj esen lapított, kerekded vagy
hosszúkás alakú (16.56. ábra).
Kutikulájuk erős, kemény, miri-
gyekben szegény képződmény.
Csápjaik rövidek, legfeljebb 5
ízből állnak. Száj szerveik rágó vagy
szúró-szívó típusúak. Szemeik
gyengén fejlettek, néha hiányoz-
nak. Lábaik igen erősek, izmo-
sak, számos kitintüskével és szé-
320
les, hajlott karommal ellátottak. Szárnyaik hiányoznak, de másodlagosan szárnyatlan
állatok. A tracheanyílások a test hátoldalán találhatók. Petéiket (serke) a gazdaállatok
tollára vagy szőrére ragasztják (16.56. ábra). A kikelő kis lárvák az imágókra hasonlíta-
nak, kifejtéssel fejlődnek. Három lárvastádium után érik el az ivarérett, aduit állapotot.
Az ivaros szaporodás mellett előfordulhat szűznemzés (parthenogenesis) is. Erőteljesen
kötődnek a gazdafajhoz. Egész életüket ugyanazon a gazdán töltik. Néha megfigyelhető,
hogy ezen belül meghatározott testtájakon élnek. Kizárólag madarakon vagy emlősökön
találhatók. Jelentős kártevők lehetnek, mivel a gazda kültakaróját mechanikusan sértik
és fertőzéseket is terjeszthetnek.
Három alrendjüket különböztetik meg.
A bunkóscsápú tetvek (Amblycera) alrendjébe sorolt fajok ötízű, bunkós csápot viselnek
(16.56. ábra). A csáp egy gödörbe részben vagy teljesen visszahúzható. Házimadarainkon a
közönséges tyúktetű (Menopon gallinae) gyakran megtalálható. A toll ágai között lapul meg, így
védekezik a tollászkodó madár ellen. A tolikezdemények rágásával okoz kárt.
A fonalascsápú tetvek (Ischnocera) alrendjébe tartozó fajok csápjai 3-5 ízűek, fonalasak,
szabadon helyezkednek el. Rágó szájszerveik fejlettek, erősek. Ide tartozó faj a madarakon gyakori
karcsú galambtetű (Columbicola columbae) és a szarvasmarhán élősködő Damalinia bovis.
A szívótetvek (Anoplura) alrendjének fajai a fejbe visszahúzható szúró-szívó szájszervvel
rendelkeznek (16.56. ábra). Fejük keskenyebb, mint a tor szélessége. Csápjaik rövidek, szemeik
hiányozhatnak. Kizárólag emlősökön élnek, a gazda szőrzetébe kapaszkodva. Karmaik felépítése a
szőrön történő sikeres megkapaszkodást segiti, a karmok nagyok, erősek, sarlószerűen hajlottak.
Vérszívók, de vírusokat és baktériumokat is terjesztenek. A sertéseken gyakori a disznótetű
(Haematopinus suis). Az emberen két faj, a ruhatetű (Pediculus humánus) és a lapostetű (Phthirus
pubis) élhet. Az előbbi faj egyedei a fejen vagy a ruházatban élnek, míg az utóbbié a szeméremszőr-
zetben.
16.4.16. Tripszek (Thysanoptera)
Egy-két milliméteres rovarok tartoznak ebbe a több ezer fajt összefoglaló csoportba. Kis
fejükről rövid csápok irányulnak előre (16.57. ábra). Összetett szemeik mellett rendsze-
rint három pontszem is található. Szúró-szívó szájszervük aszimmetrikus, három szú-
rósertét tartalmaz. A szúróserték az állkapocs belső karélyaiból és a bal oldali rágóból
alakultak ki. A jobb oldali rágó visszafejlődött. Lábaik járólábak, végükön két karommal.
A karmok küzül egy hólyag (arolium) türemkedhet ki, mely a tapadást segíti (16.57.
ábra). A lábszár végén található mirigy váladéka tovább fokozza a lábak tapadását a nö-
vények felületén. A két pár, egyforma szárny igen jellegzetes felépítésű. Karcsú, meg-
nyúlt képletek erősen redukálódott, de jellegzetes lefutású erezettel. A szárnyak széléről
hosszú kitinszőrök indulnak ki, ezek megnövelik a repülőfelületet (16.57. ábra). Jól
repülnek, néha nagy csapataik láthatók a levegőben. Petéiket a növények felületére, vagy
ha tojócsővel rendelkeznek, akkor a növény szöveteibe egyesével rakják le. A lárvák poszt-
embrionális fejlődése során először két lárvastádium, majd egy előnimfa-stádium,
végül egy vagy két nimfastádium figyelhető meg. A nimfastádiumú állatok nyugalomban
vannak, akár több hónapig is. Ez a fejlődési forma a holometamorfózis egy sajátos
változata. Körükben szűznemzés is előfordul. Termesztett növényeinken néha nagy
tömegben jelennek meg. Ekkor szívásukkal károsítják a növényeket. Egyes fajok sokféle
tápnövényt fogyasztanak, mások tápnövényköre viszont korlátozott. A ragadozó tripszek
elsősorban levéltetveket esznek, de a kannibalizmus sem ritka. Gyakran károsítanak
üvegházakban. Jellegzetes kártételük, hogy vírusokat és baktériumokat vihetnek át egyik
növény egy edről a másikra (vektorok).
A hazánkba behurcolt közönséges tripszet (Frankliniella intonsa) már százötven növény
virágjában találták meg. Pillangós növényeken rendszeresen nagy tömegekben léphet fel. Évente
321
több nemzedéke lehetséges. Hasonlóan gyakori, elterjedt, sok tápnövényű faj a dohánytripsz
(Thrips tabaci). Főleg a dohányt, hagymát, uborkát és néhány dísznövényt károsítja. Gyakori
kártevő a búzatripsz (Haplothrips tritici). A faj által megtámadott búzaszemek tömege csökken,
és gyengébb lesz a csírázóképességük is. A szántóföldi növények tarlójának talajába húzódva telel
át a lárva. A monokultúra különösen kedvező elszaporodásához.
16.4.17. Poloskák (Heteroptera)
Több tízezer fajt sorolnak ebbe a rendbe. Testnagyságuk néhány millimétertől tpbb,
mint^űxaiti^terig^^ei Többnyire ovális, felülről kissé lapított testű állatok (16.58.
ábrá). KicsvSnUszérinJ,háromszög alakú fejűit illeszkedik a tőrhöz. Csápjaik fonalas
csápok, általában 4 ízből állnak. Szájszeryűk,.^mit szipókának (rostrum) neveznek,
szurok-szívó típusú (16.58. ábfá).'?i szipóka nyugalmi állapotban sokszor a hasi oldalon
húzódó vályúba simul. Aszúróserték a rágókból, a nyálvezető és a táplálékot vezető" cső
az állkapocs bélső karelyából alakult ki. Mindezeket egy három-négy ízből álló, vastag
falú cső veszi körül, mély az alsó ajakból alakult ki. Az állkapcsokhoz és az alsó ajakhoz
kapcsolódó tapogatók hiányoznak. Nem túl nagy összetett szemek ülnek a fej két oldalán,
a fejtetőn pedig legtöbbször két pontszem található. Az első torszelvény hátlemeze, az
előhát (pronotum) erőteljesen fejlett. Mögötte, a hátoldalon rendszerint kisebb, néha
nagyobb háromszög alakú lemez, a pajzsocska (scutellum) található. Ez a középtorról
indul ki. A lábak járólábak. Az első pár szárny, az ún. félfedő (hemelytron), bőrszerű
fedőszámy, amely hártyában (membrán) végződik. A második pár hártyás szárny erezete
erősen redukálódott (16.58. ábra). Szárnyatlan csoportjaik is vannak. Egyes fajoknak
teljesszámyú (macropter) és rövidszárnyú (micropter, brachypter) alakjaik is vannak. A
322
16.58. ábra. A: poloskalárva, B: kifejlett poloska, C: elülső szárny, D: hátulsó szárny, E: szájszerve, F: szú-
rósertéje
aa: alsó ajak, an; anális ér, ál: állkapocs, cs: csáp, et: előtör, fa: felső ajak, há: hártya, hí: hipofarinx, ho: hónaljtőér, ös:
összetett szem, ny: nyálvezető cső, pa: pajzsocska, ra: rágó, sk; számykezdemény, su: sugárér, sz: szegélyér, tá: táplálékvezető
cső (Több szerző nyomán)
rövidszámyú formák nem tudnak
repülni. A szárnyas fajok jól re-
pülnek. Táplálékukat elsősorban
növényilíedvek'képezik. Vannak
véfsZívó fajaik is. Nyáluk, a han-
gyasav-tartalom miatt viszkető ér-
zést okoz. Gyakori a bűzmirigy
(16.59. ábra). A lárvák bűzmiri-
gye a 3.-5. potrohszelvény hátol-
dalán, az imágóké az utótor két
oldalán nyílik a külvilágra. Vála-
déka mérget és detergenst tartal-
maz. Az utótor felszínén finoman
eloszlatják a mirigyváladékot.
Vannak fajok, amelyek a váladé-
kot a támadóra lövellik. Bűzmi-
rigyeik segítségével riasztják el
vagy tartják távol az ellenséget.
Számos faj hangot is ad. Általá-
ban öt, igen ritkán csak négy lár-
vastádiumuk van.
17.59. ábra. A: poloska elülső testfele, B: bűzmirigye
bv: a bűzmirigy váladéka, es: elülső szárny kezdeti része, et: előtör, há:
hámsejtek rétege, iz: izom, ku: kutikula, mh: mirigyhám, ny: bűzmirigy
nyílása, sz: szájszerv, ut: utótor (Weber, Eisenbeis és Wichard nyomán)
323
Rend: Poloskák (Heteroptera)
Alrend: Vízipoloskák (Nepomorpha)
Család: Búvárpoloskák (Corixidae)
Család: Hanyattúszó-poloskák (Notonectidae)
Alrend: Kétéltűpoloskák (Gerromorpha)
Család: Molnárpoloskák (Gerridae)
Alrend: Címerespoloska-alkatúak (Pentatomomorpha)
Család: Földipoloskák (Cydnidae)
Család: Pajzsospoloskák (Scutelleridae)
Család: Cimerespoloskák (Pentatomidae)
Család: Karimáspoloskák (Coreidae)
Család: Verőköltő poloskák (Pyrrhocoridae)
Család: Bodobácsok (Lygeidae)
Család: Kéregpoloskák (Aradidae)
Alrend: Vérszívópoloska-alkatúak (Cimicomorpha)
Család: Csipkéspoloskák (Tingitidae)
Család: Vérszívópoloskák (Cimicidae)
Család: Rablópoloskák (Reduviidae)
Család: Mezeipoloskák (Miridae)
Megjegyzés: a poloskák áttekintő rendszere Kerzhner és Stys nyomán.
Búvárpoloskák (Corixidae)
Testnagyságuk néhány milliméter és másfél centiméter között mozog. Szárnyfedőiken
sötétebb és világosabb csíkok szép mintázata látható. Szájszervük a fejtokhoz széles
alapon ízesül, ezért kevéssé mozgatható. A hímek első lábainak lábfeje (tarsus) kiszéle-
sedik. Harmadik pár lábaik hosszú szőrökkel borított úszólábak. Jól repülnek, fénycsap-
dával tömegesen gyűjthetők. Vegyestáplálkozású állatok. Halak táplálékai lehetnek.
Egyes fajok jól hallható hangot adnak.
Hazánk mindenféle víztípusában, álló- és folyóvizekben egyaránt általánosan elterjedt a csíkos bú-
várpoloska (Sigarafalleni) (16.60 ábra). Különösen a szerves anyagokban dús, eutrófvizekben tömeges.
Hanyattúszó-poloskák (Notonectidae)
Másfél centiméteres testük zömök, erőteljes. Száj szervük erőteljes, szabadon mozgatható.
Szúrásuk fájdalmas, mivel nyálmirigyükben hangyasav található. A vízben hassal felfelé
tartózkodnak és így is úsznak. Harmadik pár lábuk erőteljes úszóláb. Ragadozó fajok,
amelyek más rovarokat, halivadékot zsákmányolnak és szájszervükkel kiszívják azokat.
Sokfelé, álló- és folyóvizekben egyaránt gyakori a közönséges hanyattúszó-poloska (Notonecta
glauca). Ha az ivadéknevelő halastavakban elszaporodik, akkor komoly károkat okozhat.
Molnárpoloskák (Gerridae)
A molnárpoloskák teste karcsú, megnyúlt, igen könnyű. Második és harmadik pár lábuk
hosszú. Lábfejeiken legyezőszerűen elhelyezkedő szőrök találhatók, ezért a víz felszínén
nem süllyednek el. A víz felületi feszültségének segítségével a víz felszínén futkároznak.
Ragadozók, a vízfelszínre hulló rovarokat első pár lábaikkal ragadják meg. Akár sebes
folyású patakok felszínén is képesek a vízáramlással szemben futva egyhelyben maradni.
Jellemző állat a mintegy centiméter hosszúságú rozsdáshátú molnárpoloska (Gerris thora-
cicus). Az álló- vagy lassan folyó vizeket kedveli (16.60. ábra).
324
16.60.ábra. Jellegzetes poloskafajok
habitusképei
A: csíkos búvárpoloska (Sigara falle-
ni) (Soós nyomán), B: rozsdáshátú mól**
nárpoloska (Gerris thoracicus) (Bene-
dek nyomán)
Földipoloskák (Cydnidae)
Kicsi, sötét színezetű, ovális testalakú állatok. Lábaik erőteljesek, a lábszárakon erős
tüskék találhatók. Ezekkel ássák magukat a talajba, mely szokásos élőhelyük. Rendsze-
rint a növények gyökereit szívogatják.
A foltos foldipoloska (Tritomegas bicolor) (16.61. ábra) nedves élőhelyeken sokfelé gyakori.
Pajzsospoloskák (Scutelleridae)
Viszonylag nagy, domború testű, nehézkes állatok. Pajzsuk igen nagy, szinte az egész
potrohot befedi, csupán a potroh szélén körbefutó, széles kitinlemezekből álló oldalsze-
gély (connexivum) marad szabadon.
A nagyüzemi gabonatermesztés elterjedésével párhuzamosan terjedtek el a gabonapoloskák
(Eurygaster spp.) és váltak rendkívül jelentős kártevőkké. Az osztrák poloska (Eurygaster aust-
riaca) pofái elől körülzárják a fejpajzsot. Főleg a búza, az árpa és a rozs kalászait károsítja. A lárvák
kifejlődéséhez fűfélék szükségesek. A nagyon hasonló mórpoloska (Eurygaster maura) egyedein
a fejpajzsot a pofák nem ölelik körül. Életmódja az előző fajéhoz hasonló, de gyakrabban található
az alföldi területeken.
Címerespoloskák (Pentatomidae)
Közepes testnagyságú, ovális, megnyúlt testű poloskák. Szemük gyakran kis nyélen ül.
Szájszervük részben vagy egészben a fej alján található vályúban fekszik. Pajzsocskájuk
elég nagy, a potroh felét is meghaladhatja. Lábaik nem túl erős futólábak. Gyakran szép,
élénk csillogó színekkel tarkítottak. Petéik hordó alakúak, rendszerint jellegzetes csíkos
mintázattal díszesek. Legtöbb fajuk növényevő, de néhány ragadozó faj is található
ebben a családban.
A közönséges szipolypoloska (Aelia acuminata) feje megnyúlt, hátsó lábainak belső oldalán
két fekete folt van. Mezőgazdasági területeken és természetes növényközösségekben egyaránt
gyakori. Tápnövényeit a gabona- és pázsitfűfélék képezik. A bogyómászó-poloska (Dolycoris
baccarum) az egyik leggyakoribb poloskafajunk. Pajzsocskájának vége világos krémszínű. Lárvái-
nak testét rövid, sűrű szőrök borítják. Sokféle növényen megél, elsősorban az éretlen magvakat és
terméseket fogyasztja. A bencepoloska (Rhaphigaster nebulosa) nagy testű, másfél centiméteres
325
állat. Igen elterjedt, közismert faj, ősszel a lakásokba is behúzódik. Ha megfogják, büzmirigyének
váladékát nehéz a kézről lemosni. Igen gyakori, feltűnő faj a csíkos pajzsospoloska (Graphosoma
lineatum). Elénkpiros alapszínű testét az előháton hat, a pajzson négy széles, fekete, hosszanti
szalag díszíti. Elsősorban az emyősvirágzatú növényeken (Umbelliferae) fordul elő. Gyakori
kártevő a káposztapoloska (Eurydema ventrale) és a paréjpoloska (Eurydema oleraceum).
Karimáspoloskák (Coreidae)
Változatos testnagyságú és alakú poloskák. Fejük négyszögletes, az előhátnál jóval
keskenyebb. Jellemzőjük, hogy a potroh szélső területei ellaposodnak, az oldalszegély
feltűnően széles, a szárnyak alól kiálló.
A közönséges karimáspoloska (Coreus marginatus) (16.61. ábra) elsősorban a nedvesebb
erdő széli terüle-teken, ni trotil növényekben gazdag élőhelyeken gyakori.
Verőköltő poloskák (Pyrrhocoridae)
Testük lapított, feji irányban megnyúlt, ovális. Pontszemeik nincsenek. Rendszerint
rövid számyúak. Növényevők, magvakat, terméseket szívogatnak.
Ide tartozik a legismertebb hazai poloskafaj, a fekete-piros mintázatú verőköltő bodobács
(Pyrrhocoris apterus). A meleget rendkívül kedveli, ezért napsütötte helyeken hatalmas tömegekben
található. Amagasabbhegyvidékeketkivéve mindenütt rendkívül közönséges. Elterjedt laboratóriumi
állat, elsősorban a rovarok hormonrendszerének jellegzetességeit tanulmányozzák rajta.
Bodobácsok (Lygeidae)
Kis és közepes testű, hosszúkás fajok tartoznak ebbe a családba. Általában élénk színűek,
tarkák. Szemük határozottan kidomborodik a fej oldalából. Pontszemeik is megtalálha-
tók. Növényevők, a rétek, legelők növényeit szívogatják.
Jellegzetes a fekete alapon piros mintás lovagbodobács (Lygeus equestris), amely faj félfedőjé-
nek hártyáját nagy, szabályos fehér folt díszíti. Dombvidéki réteken, bokros területeken min-
denütt gyakori.
16.61.ábra. Jellegzetes poloskafajok habitusképei
A: foltos földipoloska (Tritomegas bicolor) (Cs. Halászfy nyomán), B: közönséges karimáspoloska (Coreus
marginatus ) (Vásárhelyi nyomán), C: közönséges kéregpoloska (Aradus betulae) (Vásárhelyi nyomán)
326
írta
dr. Bakonyi Gábor
dr. Juhász Lajos
dr. Kiss István
dr. Palotás Gábor
©M
Oktatási
Minisztérium
A könyv az Oktatási Minisztérium támogatásával,
a Felsőoktatási Pályázatok Irodája által lebonyolított Felsőoktatási
Tankönyv- és Szakkönyvtámogatási Pályázat keretében jelent meg.
A Földművelésügyi és Vidékfejlesztési Minisztérium Intézményközi
Tankönyvkiadási Szakértő Bizottsága támogatásával készült.
Az egyetemek és a főiskolák agrártudományi karain javasolt tankönyv