/





























































































































































































































































































































































































































Автор: Шамшинов А.М. Волков В.В.
Теги: офтальмология глазные болезни и травмы медицина оптика
ISBN: 5-225-04516-2
Год: 1999




Текст
А.М.Шамшинова, В.В.Волков
функциональные
методы
исследования
в офтальмологии
УДК 617.7-072.7
ББК 56.7
Ш19
Рецензент
О. В. Груша, докт. мед. наук, профессор
Шамшинова А. М., Волков В. В.
Ш19 Функциональные методы исследования в офтальмологии: —
М.: Медицина, 1999. — 416 с.: ил. — ISBN 5-225-04516-2.
В монографии представлены современные функциональные методы иссле-
дования, используемые в офтальмологии. В 1-ю главу включены электрофизи-
ологические и психофизические методы исследования сенсорной зрительной
системы, во 2-ю — придаточного аппарата глаза, его гемо- и гидродинамики.
Изложены новые данные о механизмах нарушения зрительных функций, кото-
рые обусловливают выбор метода офтальмологического исследования, его
нейро-, биофизические и анатомофункциональные основы. Показана роль этих
методов в диагностике заболеваний зрительной системы.
Для офтальмологов, невропатологов, врачей кабинетов функциональной
диагностики, психиатров, эндокринологов, а также профпатологов.
ББК 56.7
ISBN 5-225-04516-2
© Издательство «Медицина»,
Москва, 1999
Все права авторов защищены. Ни одна часть этого издания не может
быть занесена в память компьютера либо воспроизведена любым спосо-
бом без предварительного письменного разрешения издателя.
ОГЛАВЛЕНИЕ
Предисловие.................................................. 8
Часть I
Глава 1. Анатомия и нейрофизиология сетчатки и зритель-
ных путей (А.М.Шамшинова) ...................... 11
1.1. Строение сетчатки...................................... 11
1.2. Нейрофизиология сетчатки .............................. 14
1.3. Нейромедиаторы....."................................... 21
1.4. Клетки Мюллера и их взаимоотношения с нейронами ....... 24
1.5. Анатомо-физиологические особенности зрительного пути *. 26
Глава 2. Острота зрения, контрастная чувствительность и
устойчивость к слепящим засветим (В. В. Волков) 32
2.1. Оптико-анатомо-физиологические основы функции.......... 32
2.1.1. Острота зрения .................................. 32
2.1.2. Контрастная чувствительность..................... 40
2.1.3. Устойчивость к слепящим засветим................. 43
2.2. Основные методы функциональных исследований ........... 45
2.2.1. Методы оценки остроты зрения..................... 45
2.2.2. Методы исследования контрастной чувствительности. 52
2.2.3. Методы оценки устойчивости к слепящим засветим... 57
2.3. Клинические проявления нарушений остроты зрения, контраст-
ной чувствительности и устойчивости к ослеплению с учетом
особенностей их этиологии и патогенеза ................... 61
2.3.1. Основные варианты и причины нарушений остроты зрения
в клинической практике ............................ 61
2.3.2. Варианты и возможные причины изменения пространствен-
ной контрастной чувствительности при заболеваниях
зрительной системы.................................... 62
2.3.3. Клинические примеры нарушений устойчивости зрения к
слепящим засветим..................................... 66
Глава З.Светоощущение и зрительная адаптация (В, В. Вол-
ков, А. М. Шамшинова)..................................... 67
3.1. Основные фотометрические понятия и единицы............. 67
3.2. О механизмах зрительного восприятия и физиологических воз-
можностях зрения ......................................... 69
3.2.1. Физиологические регуляторы светового потока, поступаю-
щего в глаз ...................................... 71
3
3.2.2. Морфологические основы и субъективные проявления фото-
и скотопического зрения ............................... 72
3.2.3. Пространственные аспекты световой чувствительности .... 73
3.2.4. Временные аспекты светочувствительности......... 77
3.3. Клинические проявления нарушений светоощущения и зритель-
ной адаптации ............................................ 82
3.4. Клинические методы тестирования светочувствительности. 86
3.4.1. Темновая адаптометрия........................... 86
3.4.2. Световая адаптометрия .......................... 88
3.4.3. Исследование критической частоты слияния мельканий ... 88
3.4.4. Периметрия...................................... 89
Глава 4.Цветоощущение (В.В.Волков, А.М.Шамъиинова) .. 106
4.1. Анатомо-физиологические основы цветового зрения...... 106
4.1.1. Трихроматичность цветового зрения ............. 106
4.1.2. Цветовой треугольник и координаты цветов ...... 107
4.1.3. Основные параметры цвета ...................... 109
4.1.4. Об одновременном и последовательном цветовом контрасте 111
4.2. Основные причины и признаки нарушений цветового зрения .... 113
4.3. Функциональные тесты на цветовое зрение.............. 115
4.4. О классификации нарушений цветового зрения .......... 125
Глава 5. Электроретинография (А.М. Шамшинова).............. 133
5.1. Общая электроретинограмма ........................... 133
5.1.1. Современные представления о происхождении ЭРГ... 133
5.1.2. Факторы, влияющие на ЭРГ....................... 137
5.1.3. Классификация ЭРГ.............................. 141
5.1.4. Метод регистрации ЭРГ (рекомендации ISCEV)..... 147
5.1.5. Роль ЭРГ в диагностике заболеваний сетчатки и зритель-
ного нерва ........................................... 154
5.2. Локальная ЭРГ ....................................... 167
5.2.1. Происхождение локальной ЭРГ.................... 167
5.2.2. Методы регистраций локальной (макулярной) ЭРГ . 167
5.2.3. Локальная ЭРГ в диагностике заболеваний макулярной
области сетчатки ..................................... 174
5.3. Паттерн-ЭРГ ......................................... 178
5.3.1. Происхождение паттерн-ЭРГ ..................... 178
5.3.2. Методологические основы регистрации паттерн-ЭРГ (ре-
комендации ISCEV) .................................... 181
5.3.3. Паттерн-ЭРГ в диагностике заболеваний сетчатки и зри-
тельного нерва .. .•.................................. 185
5.4. Ритмическая ЭРГ...................................... 187
5.4.1. Происхождение ритмической ЭРГ.................. 187
5.4.2. Методологические основы регистрации ритмической ЭРГ 189
5.4.3. Роль ритмической ЭРГ в диагностике заболеваний сетчатки 189
5.5. Электроретинография при катаракте.................... 192
Глава 6. Электроокулография (А.М,Шамшинова, В,Я,Эскин) 196
6.1. Постоянный потенциал глаза ......................... 1'96
6.2. Физиологические основы и происхождение ЭОГ............197
4
6.3. Методы регистрации ЭОГ ............................. 199
6.3.1. Стандартная методика регистрации ЭОГ ...........199
6.3.2. Импульсная электроокулография ................ 206
6.3.3. Другие методические подходы в электроокулографии .... 206
6.3.4. Методологические основы регистрации ЭОГ (рекомен-
дации ISCEV) ......................................... 208
6.4. Роль электроокулографии в диагностике заболеваний пигмент-
ного эпителия сетчатки и хороидеи ....................... 212
Глава 7. Зрительные вызванные корковые потенциалы
(А.М.Шамшинова) ......................................... 222
7.1. Происхождение зрительных вызванных корковых потенциалов . 222
7.2. Виды зрительных вызванных корковых потенциалов........228
7.3. Метод регистрации зрительных вызванных корковых потенциа-
лов .......................................................229
7.4. Картирование зрительных вызванных корковых потенциалов ... 236
7.5. Зрительные вызванные корковые потенциалы и острота зрения . 243
7.6. Роль зрительных вызванных корковых потенциалов в диагнос-
тике заболеваний зрительных путей ....................... 244
Глава 8. Энтоптические феномены в диагностике функцио-
нальных нарушений (В. В. Волков)......................... 254
8.1. Роль особенностей структуры слезной пленки, роговицы и хру-
сталика ................................................. 254
8.2. Роль особенностей структуры стекловидного тела ..... 256
8.3. Роль особенностей витреоретинальных отношений....... 257
8.4. Фактор наличия непрозрачных кровеносных сосудов в нормально
функционирующей прозрачной сетчатке ..................... 257
8.5. Фактор наличия дугообразности в нормальной топографии
нервных волокон сетчатки ................................ 258
8.6. Фактор наличия в норме строго определенной (относительно
центра макулы) ориентации молекул ксантофильного пигмента
в наружном плексиформном слое сетчатки................... 259
8.7. Роль функционального состояния фоторецепторов сетчатки в про-
исхождении эктопических явлений ......................... 260
Часть П
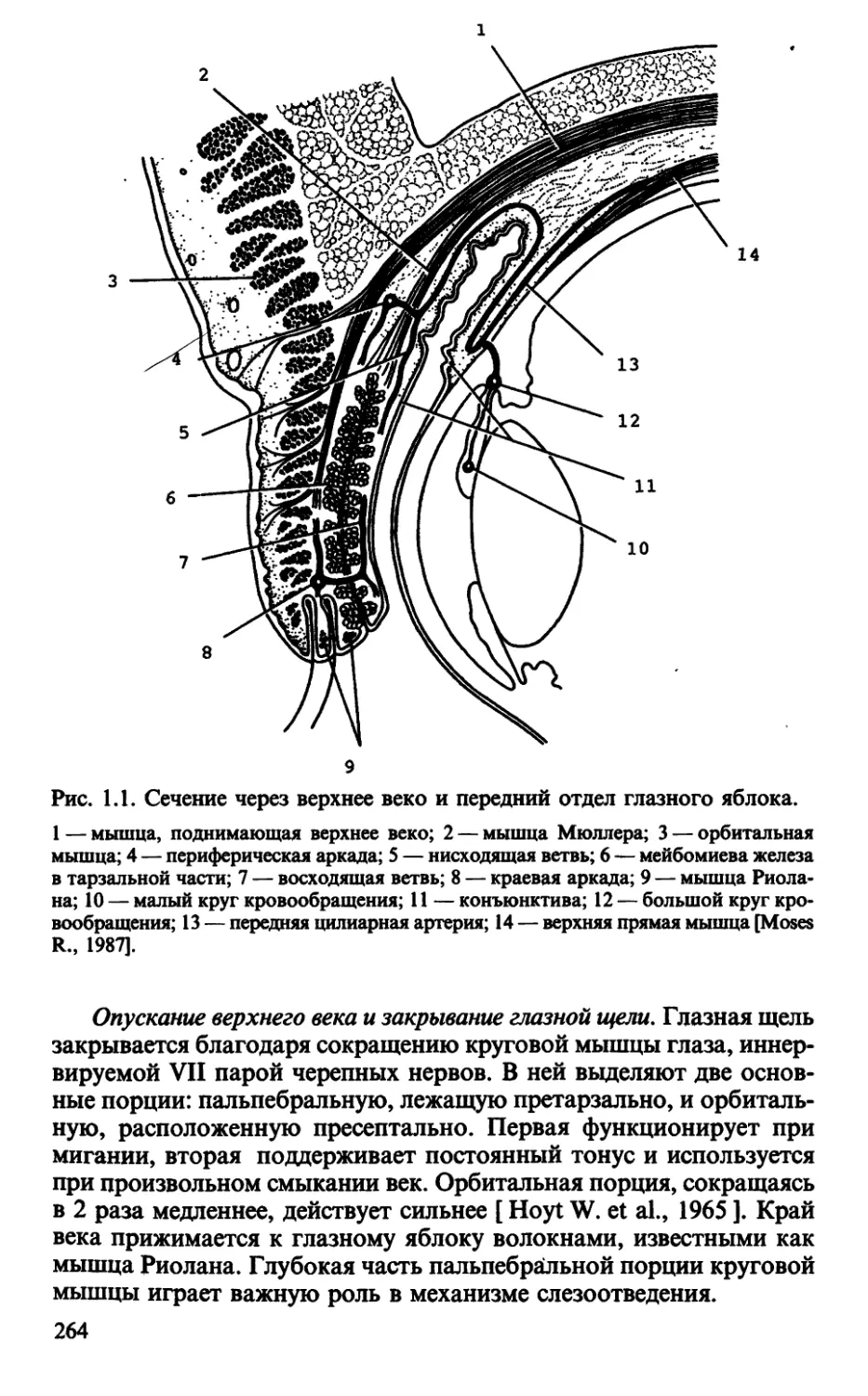
Глава 1. Веки как защитный аппарат глаза (В.В.Волков) ... 263
1.1. Строение и функция век в норме ..................... 263
1.2. Клинические варианты нарушения функции век ......... 266
1.3. Приемы функциональной диагностики и трактовка результатов . 267
1.3.1. Тестирование мигательного рефлекса ........... 267
1.3.2. Функциональная оценка края век................ 268
1.3.3. Тестирование функции открывания глазной щели.... 269
1.3.4. Тестирование функции закрывания глазйой щели ..272
1.3.5. Тестирование ассоциированных движений ........ 273
5
Глава 2. Слезопродуцирующий и слезоотводящий аппарат
(В.В.Волков) ............................................ 274
2.1. Строение и функции слезного аппарата в норме ........ 274
2.1.1. Слезная пленка ................................ 274
2.1.2. Слезопродукция ................................ 274
2.1.3. Слезоотведение................................. 276
2.2. Клинические варианты нарушений функции увлажнения глаза ... 277
2.3. Приемы функциональной диагностики и трактовка результатов . 278
2.3.1. ’’Сухой” глаз, тестирование слезопродукции и качества
слезной пленки ....................................... 278
2.3.2. ’’Переувлажненный” глаз, тестирование слезоотведения ... 281
Глава 3. Циркуляция крови и тканевых жидкостей в глазу
(В.В.Волков).............................................. 284
3.1. Прикладная анатомия сосудистой системы глаза......... 284
3.2. Структура и функции гематоофтальмических барьеров ... 286
3.3. Глазной кровоток в норме. Механизмы регулирования крово-
тока в глазу и клинические методы его исследования ...... 289
3.3.1. Артериальное давление и методы исследования активной
(моторной) части глазного кровотока ............. 290
3.3.2. Венозное давление и методы исследования ’’стопорной” ча-
сти глазного кровотока ............................... 293
3.3.3. Ауторегуляция кровотока в глазу и методы его исследо-
вания ................................................ 295
3.4. Этиология, патогенез и клинические варианты нарушения
глазной гемоциркуляции................................... 298
3.4.1. Нарушения артериального кровотока.............. 298
3.4.2. Нарушения венозного кровотока.................. 299
3.4.3. Нарушения капиллярного кровотока .............. 301
3.5. Тестирование нарушений внутриглазной гемодинамики в кли-
нической практике (по данным флюоресцентной ангиографии
и других методик)........................................ 301
3.5.1. Оценка кровотока при симптомах гипофлюоресценции ... 301
3.5.2. Оценка тематических барьеров при симптомах гиперфлюо-
ресценции ....................................... 303
Глава 4. Аппарат, регулирующий гидродинамику и опти-
ческую установку глаза (В.В.Волков) ...................... 311
4.1. Анатомо-физиологическая характеристика иридохрусталико-
вой диафрагмы, ресничного тела, камерного угла и решетчатой
мембраны диска зрительного нерва......................... 311
4.1.1. Продукция и отток камерной влаги, офтальмотонус. 311
4.1.2. Взаимоотношения офтальмотонуса с артериальным и вну-
тричерепным давлением, а также с тканевым давлением
в зрительном нерве.................................... 315
4.1.3. Зрачковые реакции ............................. 321
4.1.4. Структурно-функциональные элементы аккомодацион-
ного аппарата ........................................ 322
4.1.5. Аккомодация и оптическая установка глаза....... 324
6
4.1.6. 0 механизмах аккомодации .......................... 328
4.1.7. Попытки биомеханического моделирования одновремен-
ного действия внутренних мышц глаза на оптическую
установку и дренажную систему.......................... 330
4.2. Основные причины, клинические признаки и методики иссле-
дования нарушений глазной гидродинамики, функции зрачка,
аккомодации и оптической установки глаза ................. 332
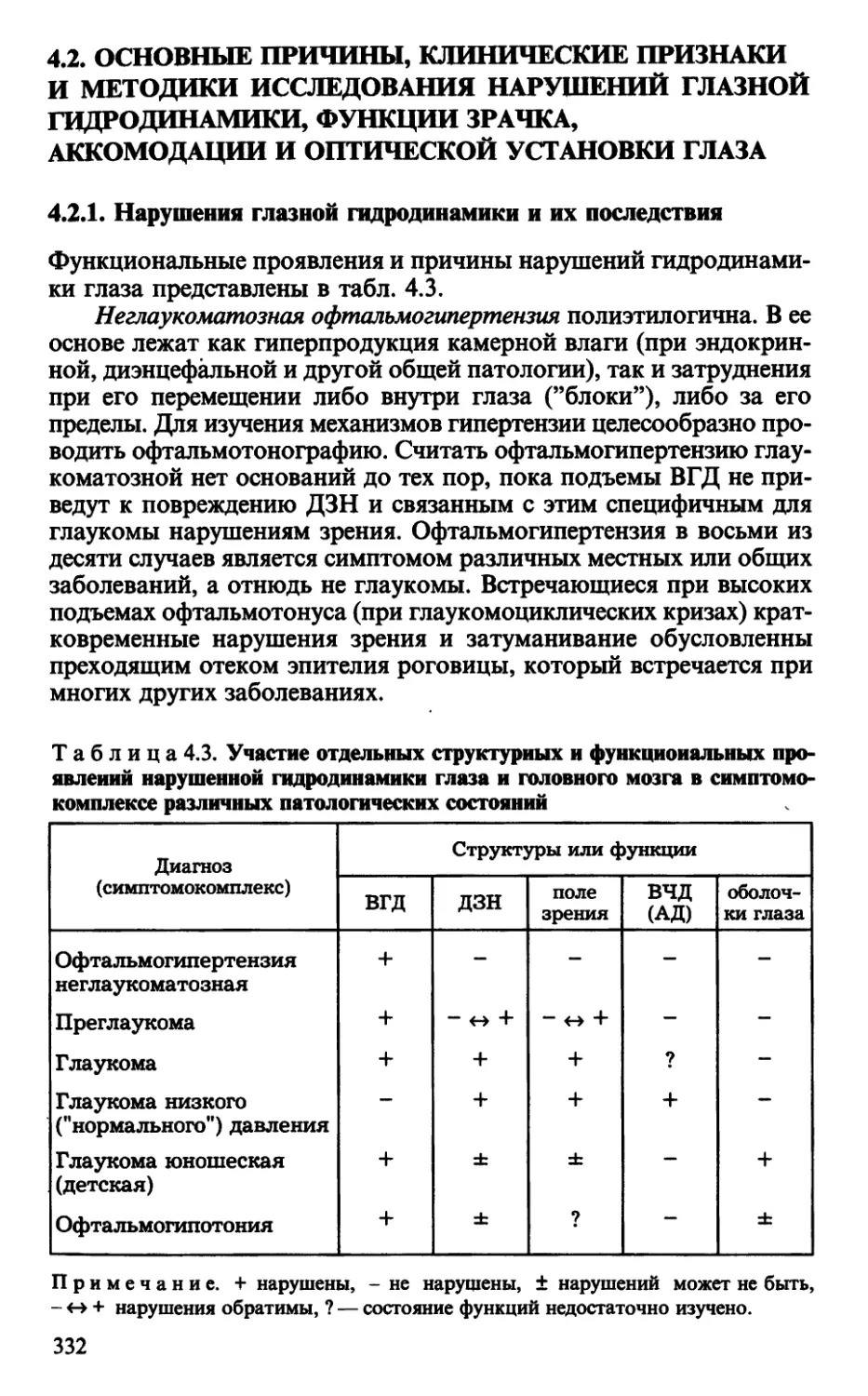
4.2.1. Нарушения глазной гидродинамики и их последствия .... 332
4.2.2. Методики и устройства для исследования офтальмотонуса
и компонентов глазной гидродинамики ................. 334
4.2.3. Нарушения зрачковых реакций, их диагностика и послед-
ствия ................................................. 338
4.2.4. Нарушения аккомодации, их связь с ’’неудачными” вари-
антами оптической установки глаза. Основные признаки
и причины нарушений ................................... 342
4.3. Методы и устройства для функциональных исследований акко-
модации и оптической установки глаза...................... 344
4.4. Сводный материал об ответной реакции механизмов регуляции
офтальмотонуса зрачка и ресничной мышцы на фармакологиче-
ские воздействия ......................................... 358
Глава 5. Глазодвигательный аппарат и бинокулярное
зрение (В.В.Волков)........................................ 361
5.1. Устройство и работа глазодвигательного аппарата в норме . 361
5.1.1. Глазницы и биомеханика глазодвигательных мышц... 361
5.1.2. Виды произвольных движений здоровых глаз .......... 363
5.1.3. Виды непроизвольных движений здоровых глаз ........ 365
5.2. Бинокулярное зрение ..................................... 367
5.2.1. Совместное одиночное видение ...................... 367
5.2.2. Совместное раздвоенное видение..................... 374
5.3. Клинические проявления нарушений функции глазодвигатель-
ного аппарата и бинокулярного зрения с учетом различий
этиологии и патогенеза ................................... 377
5.4. Методики функциональных исследований глазодвигательного
аппарата и бинокулярного зрения........................... 379
5.4.1. Измерение угла косоглазия.......................... 379
5.4.2. Измерение мышечного баланса, в том числе конвер-
гентно-аккомодационных отношений .................... 382
5.4.3. Определение слабофункционирующей (парализованной)
мышцы и оценка ее силы ................................ 384
5.4.4. Выявление сенсорной аномалии, в том числе аномальной
корреспонденции сетчаток .............................. 387
5.4.5. Оценка глубинного зрения и стереопсиса............. 389
5.5. О тенденциях в развитии представлений о механизмах бино-
кулярного зрения и выборе адекватныхметодов для его исследо-
вания .................................................... 391
Список основной литературы.................................... 395
ПРЕДИСЛОВИЕ
За последнее десятилетие функциональные исследования в офталь-
мологии поднялись на качественно новую ступень, в связи с чем не-
обходимы теоретическое осмысление и все более широкая клиниче-
ская апробация многочисленных методик.
Сложность структурной организации сетчатки глаза, проводящих
путей и обширных областей центральной нервной системы послужи-
ла основанием для создания разных научных направлений, призван-
ных решать общую задачу — поиск методов, позволяющих адекват- ч
но оценивать зрительное восприятие и качество переработки зритель-
ной информации.
Электрофизиологические и психофизические исследования, про-
веденные в последние десятилетия, подтвердили существование па-
раллельных подсистем, или каналов, в зрительной системе у прима-
тов. В экспериментах на клеточном уровне получены доказательства
того, что большинство зрительных нейронов избирательно активи-
руются специфическими стимулами различного цвета, контраста,
яркости, ориентации, движения и т.д. и относительно нечувствитель-
ны к диффузному освещению. Функциональная организация зритель-
ной системы и определяет уровень наиболее выраженного ответа ней-
ронов на тот или иной раздражитель.
Значительные успехи, достигнутые в изучении химических меди-
аторов (нейротрансмиттеров) зрительных сигналов, свидетельству-
ют о том, что именно нарушения синаптической передачи являются
одним из главных звеньев патогенеза при ряде заболеваний сетчат-
ки. Поскольку изменения межнейрональных контактов неизбежно
отражаются на суммарном биоэлектрическом ответе сетчатки, кли-
ническая электроретинография обладает большими потенциальны-
ми возможностями в изучении тонких патофизиологических меха-
низмов при различной офтальмопатологии органа зрения.
Вопросы клинической физиологии зрения относятся к “главным
проблемам офтальмологии”, о чем свидетельствуют монографии
Н.Н.Зислиной (1993), А.А.Шпак (1993), J.H.Jacobson (1961), G.EJayl
(1965), A.E.Krill (1970), J.Babel (1978), N.R.Galloway (1981), R.E.Carr
и I.M.Siegel (1990), Heckenlively (1991), A.M.Halliday (1993) и др., по-
священные электрофизиологическим методам исследования и их роли
в клинике глазных болезней, а также многочисленные работы
А. И. Богословского, О. И. Щербатовой и др.
Теоретические вопросы, связанные с психофизиологией зрения,
представлены в работах Н.Д.Нюберга (1928,1933,1971), С.В.Кравко-
8
ва (1945,1951), М.М.Бонгард (1955,1956,1958), В.И.Говардовского
(1975,1985), О.Ю.Орлова(1971,1984), В.В.Максимова (1984), В.В.Вол-
кова (1983,1988), Ю.Е.Шелепина (1985,1992) и др.
В то же время развитие электрофизиологических методов иссле-
дования, таких, как электроретинография, исследование вызванных
зрительных кортикальных потенциалов головного мозга, электро-
окулография, а также психофизических методов, включающих пери-
метрию, метод исследования контрастной и цветовой чувствитель-
ности глаза, позволило не только оценивать функцию зрительного
анализатора, определять локализацию процесса в различных слоях
сетчатки, зрительном нерве и в вышележащих отделах зрительной
системы, но также осуществлять раннюю диагностику заболеваний
и интоксикаций, выявлять побочные действия лекарственных препа-
ратов, прогнозировать восстановление зрительных функций под вли-
янием консервативного и хирургического лечения и в процессе реа-
билитации. Указанные методы исследования позволяют оценивать
яркостные, цветовые, контрастные каналы зрительной системы, ис-
точники нарушения восприятия, функциональное состояние колбоч-
ковой и палочковой систем, наружных и внутренних слоев сетчатки,
а также передачу информации в центральные отделы зрительного
анализатора, первичные и вторичные зрительные центры.
Во второй части монографии отражены традиционные современ-
ные представления о методах исследования век и защитного, слезо-
продуцирующего и слезоотводящего аппаратов глаза, гемодинами-
ки и гидродинамики глаза, а также глазодвигательного аппарата и
бинокулярного зрения. Эти вопросы освещены в работах ведущих
ученых нашей страны — А.Я.Бунина, А.П.Нестерова, Э.С. Аветисо-
ва, Т.П.Кащенко, Ю.З.Розенблюма и др. В каждом из разделов изло-
жены анатомические и физиологические основы рассматриваемого
метода, а также современные представления о механизмах наруше-
ния тех или иных функций зрительного аппарата, предопределяю-
щих выбор метода исследования. Так, в главе, посвященной цирку-
ляции крови и тканевых жидкостей в глазу, описываются не только
прикладная анатомия, структура и функция гематоофтальмических
барьеров, но также этиология, патогенез и клинические варианты
нарушений гемодинамики. В описание аппарата, регулирующего гид-
родинамику, включены вопросы, связанные с оптической установ-
кой глаза. Авторы попытались не только донести до читателя совре-
менные данные по рассматриваемым проблемам, но и передать свой
клинический опыт, накопленный в течение многих десятилетий. Объе-
динение в одном издании функциональных исследований различных
отделов зрительной системы определено необходимостью руковод-
ства по этой проблеме.
Бурный технический прогресс, триумфальное шествие компью-
терной техники, большие достижения в биологии, генетике, иммуно-
логии, биохимии обусловили необходимость на новом методическом
9
уровне пересмотреть патогенез некоторых, далеко еще не раскрытых
процессов в зрительной системе, представления о механизмах нару-
шения зрительных функций. Это позволит выработать более адек-
ватные, патогенетически обоснованные подходы к лечению, разра-
ботке методов реабилитации слепых и инвалидов от рождения и в
результате перенесенных заболеваний, травм, интоксикаций, различ-
ных экологических воздействий, приводящих к мутациям, дистрофи-
ям, дегенерациям, атрофиям и прочим патофизиологическим процес-
сам, ведущим к потере зрительных функций и снижению работо-
способности.
Мечтой офтальмологов является возрождение терапевтической
школы; основой этого может быть осмысление и понимание приро-
ды зрительных нарушений при заболеваниях сетчатки различного
происхождения, в чем должны помочь современные функциональ-
ные методы исследования. Описание методов функционального ис-
следования световой, цветовой и контрастной чувствительности, элек-
троретинографии, электроокулографии, зрительных вызванных кор-
ковых биопотенциалов включает представление о современных
нейрофизиологических механизмах, лежащих в основе этих методов.
В монографии описаны методические приемы, применяемые при ис-
следовании, и кратко изложена роль функциональных методов ис-
следования в оценке функции зрительной системы при некоторых
заболеваниях сетчатки ц зрительного нерва.
Авторы руководства — А.М.Шамшинова, доктор медицинских
наук, член Международного общества клинических электрофизио-
логов зрения (ISCEV), главный научный сотрудник Московского
научно-исследовательского института глазных болезней им.Гельм-
гольца, и В.В.Волков, академик Российской медико-технической и
Российской лазерной академий, профессор, доктор медицинских наук,
заслуженный деятель науки РФ, член Международного общества
исследователей глаза (ISER).
Настоящая книга предназначена для широкого круга читателей,
врачей — офтальмологов, невропатологов, нейрофизиологов и мо-
жет быть использована как методическое руководство в медицинс-
ких вузах и клиниках.
Авторы выражают благодарность за предоставленный материал
и помощь в создании и оформлении монографии В.Я.Эскину,
Е. Н. Юстовой, Ю.Е.Шелепину, М.В.Зуевой, а также, В.В.Кобилец-
кой, А.Ж.Смирновой, И.В.Зольниковой, С.А.Коскину, В.П.Никола-
енко, И.В.Рубцовой, И. Л.Симаковой, А.А.Кривошееву.
Авторы были бы очень признательны за отзывы о настоящей
монографии, а также за возможные пожелания для переработки и
добавления материала, необходимого для офтальмологической прак-
тики.
ЧАСТЬ I
ГЛАВА 1
АНАТОМИЯ И НЕЙРОФИЗИОЛОГИЯ СЕТЧАТКИ
И ЗРИТЕЛЬНЫХ ПУТЕЙ
1.1. СТРОЕНИЕ СЕТЧАТКИ
Сетчатка эмбриологически является частью мозга. Она состоит из 10
слоев: внутренней пограничной мембраны, слоя волокон зрительно-
го нерва, слоя ганглиозных клеток, внутреннего плексиформного
слоя, внутреннего нуклеарного слоя, наружного плексиформного
слоя, наружного нуклеарного слоя, наружной пограничной мембра-
ны, слоя палочек и колбочек и пигментного эпителия. Таким обра-
зом, в сетчатке различают три иерархически организованных клеточ-
ных слоя (рис. 1.1): наружный нуклеарный слой, представленный
ядрами фоторецепторов, внутренний слой, состоящий из биполяр-
ных клеток (нейроцитов), и слой ганглиозных нейроцитов. Из отрост-
ков (аксонов) ганглиозных нейроцитов формируется зрительный
нерв, который направляется в мозг. В структуре зрительного пути,
включающей фоторецепторы, биполярные и ганглиозные нейроци-
ты, имеются два типа интернейронов: горизонтальные клетки в на-
ружном плексиформном слое и амакриновые клетки во внутреннем
плексиформном слое. Горизонтальные клетки имеют синаптические
контакты друг с другом и биполярными нейроцитами, а также об-
ратную связь с фоторецепторами. Амакриновые клетки, богатые ней-
ромедиаторами, имеют синапсы с другими амакриновыми и гангли-
озными клетками, а по системе обратной связи — с биполярными
нейроцитами.
Внеклеточное пространство занимают большие глиальные клет-
ки Мюллера, проходящие радиально через все слои от фоторецепто-
ров до стекловидной мембраны сетчатки. Известно, что глиальные
клетки Мюллера, выполняя опорную, буферную и трофическую функ-
ции, играют важную роль в поддержании стабильности всех нейро-
нальных функций в центральной нервной системе. Метаболически
клетки Мюллера объединены с нейронами в единую функциональ-
но-биохимическую систему, что определяет их участие в модулиро-
вании специфической функциональной активности нейронов.
11
Рис. 1.1. Схема нейрональной организации периферической и макулярной
областей сетчатки и слои генерации светового подъема ЭОГ, общей ЭРГ,
ПЭРГиЗВКП.
1 — хороидея; 2 — пигментный эпителий; 3 — фоторецепторы (палочки и колбочки);
4 — биополярные клетки (нейроциты); 5 — ганглиозные нейроциты; 6 — зрительный
нерв; 7 — зрительная область коры.
Ганглиозные клетки сетчатки являются началом афферентных
зрительных путей; в них продуцируются спайки, потенциал действия.
По аксонам ганглиозных клеток — волокнам зрительного нерва —
импульсные сигналы передаются в мозг. Количество аксонов в зри-
тельном нерве от 564 776 до 1 200 000. В сетчатке имеются ганглиоз-
ные клетки разных типов: те, которые деполяризуются и генерируют
спайки в ответ на включение света (“on’’-центр клетки), и те, кото-
рые гиперполяризуются на свету и генерируют спайки в ответ на
его выключение (“off’-центр клетки).
Распределение и синаптическая организация клеточных элементов
сетчатки неодинаковы, так как плотность фоторецепторов меняется от
центра к периферии. При толщине сетчатки менее 0,4 мм она содержит
около ПО—125 млн палочек и 6,3—6,8 млн колбочек и только 1 млн
ганглиозных клеток. Наибольшая плотность колбочек 147 000,
178 000—238 000 на 1 мм2 в центральной зоне (фовеа) размером
50х 50 ммк (5°). Дальше от центра плотность колбочек уменьшается, в
12
парафовеа (8,6°) она составляет 9500 на 1 мм2, а в перифовеа (20°) —
10 000 на 1 мм2 [Osterberg G., 1935].
Различие физиологических свойств палочек и колбочек обуслов-
ливает двойственную природу зрения, скотопического (ночного) и
фотопического (дневного), которую описал еще в XIX столетии
M.Schultze. Палочки, содержащие родопсин (520 нм), определяют
ночное видение, функционируют при низком освещении, высокочув-
ствительны к слабому свету, имеют низкую пространственную и вре-
менную разрешающую способность, максимальную чувствительность
к сине-зеленой части спектра, слабую дирекциональную чувствитель-
ность и способность медленной адаптации к темноте. Колбочки, со-
держащие пигменты с максимумом чувствительности в красной, зе-
леной и синей частях спектра, определяют функцию цветового зре-
ния, контрастной чувствительности, пространственного и временного
разрешения, функционируют при дневном свете, обладают слабой чув-
ствительностью к свету, высокой пространственной и временной раз-
решающей способностью, максимальной чувствительностью к желто-
зеленой части спектра, высокой дирекциональной чувствительностью,
быстрой фазой адаптации в темноте.
Центральная зона диаметром 250—750 ммк, что соответствует
зрительному углу в 53 дуговых мин, свободна от палочек. Плотность
палочек максимальна в кольце вокруг фовеа (10—18° от центра) —
150 000—160 000 на 1 мм2, затем их количество уменьшается к край-
ней периферии, где имеется около 60 000 палочек на 1 мм2. Средняя
плотность палочек — 80 000—100 000 на 1 мм2.
Каждый из основных типов нейронов делится на множество под-
типов. В сетчатке приматов различают три класса цветочувствитель-
ных колбочек в зависимости от содержащихся в их наружных сег-
ментах зрительных пигментов с разной спектральной чувствитель-
ностью, у которых пик абсорбции проявляется в различных областях
спектра (560, 550 и 440 нм). Распределение различных типов колбо-
чек в гексагональной мозаике фовеа до конца не изучено. К настоя-
щему времени известно, что более чувствительные к длинноволновой
и средневолновой части спектра колбочки человека распределены
случайным образом и имеют одинаковое число, а не соотношение
1:2, как это предполагалось ранее [Mollon J. D., Bowmaker J. К.,
1991]. В макуле синие колбочки отсутствуют.
Распределение Ьиполяров и ганглиозных клеток в фовеальной об-
ласти соответствует плотности колбочек. В фовеальной зоне 2° одна
колбочка соединена с 2 ганглиозными клетками. Однако на перифе-
рии распределение этих клеток значительно изменяется, так как мно-
жество палочек и некоторые колбочки конвергируют к меньшему чис-
лу биполяров и к еще меньшему числу больших ганглиозных клеток,
от которых аксоны несут информацию в зрительную кору. На пери-
ферии сетчатки соотношение фоторецепторов и ганглиозных нейро-
цитов составляет 1000:1.
13
Колбочки в фовеоле образуют так называемый ’’карликовый”
(миниатюрный) (midget) путь по принципу ’’один к одному”, когда
колбочка соединена с on- и off-биполяром и on- и off-ганглиозной
клеткой, проецирующейся в клетках парвоцеллюлярного слоя наруж-
ного коленчатого тела. Если на уровне сетчатки этот путь имеет ми-
нимальную конвергенцию, то область коры, перерабатывающая ин-
формацию от макулярной зоны, занимает 60%. Такое увеличение
представительства макулярной зоны, обеспечивающее высокую раз-
решающую способность зрительной системы, получило название
’’фактор магнификации”.
Анатомическое строение сетчатки у различных животных, нали-
чие в ее структуре палочковой и/или колбочковой системы, топогра-
фия и плотность фоторецепторов в разных участках сетчатки от цен-
тра к периферии определяют форму и параметры электроретино-
граммы (ЭРГ).
1.2. НЕЙРОФИЗИОЛОГИЯ СЕТЧАТКИ
Восприятие мира начинается с поглощения света светочувствитель-
ными молекулами зрительного пигмента (родопсина), содержаще-
гося в мембране дистальной части фоторецепторов. Затем следует
цепь молекулярных реакций его последовательного превращения, в
результате которого пигмент обесцвечивается. После этого начина-
ется каскад реакций, отвечающих за процессы фототрансдукции, за-
канчивающейся гиперполяризацией клеточной мембраны наружно-
го сегмента фоторецептора.
Спектр поглощения родопсина состоит из трех основных полос: а
(500 нм) ир (350 нм), за которые ответственна хромофорная группа—
11-цис-ретиналь, и у (278 нм), обусловленной в основном аромати-
ческими аминокислотами: опсинотирозином и триптофаном. Извест-
но, что зрительный пигмент представляет собой хромогликопротеид,
молекула которого содержит одну хромофорную группу, две олиго-
сахаридные цепочки и водонерастворимый мембранный белок опсин.
В механизме зрения молекула зрительного пигмента выполняет
две основные физиологические функции: во-первых, поглощая свет
в некоторой характерной области длин волн, определяет спектраль-
ный диапазон фоторецепторной клетки; во-вторых, запускает фото-
рецепторный процесс. В основе первой функции лежит спектр погло-
щения молекулы зрительного пигмента, который зависит от приро-
ды хромофорной группы и ее ковалентных и нековалентных
взаимодействий с белковой частью молекулы. Вторая функция ос-
нована на способности молекулы зрительного пигмента при погло-
щении света менять свою конформацию: сначала хромофорной груп-
пы, а затем белка; молекула зрительного пигмента на одной из ста-
дий фотопревращения приобретает способность взаимодействовать
с другими белками, участвующими в механизме фоторецепции. Бел-
14
ковая часть обесцвеченной молекулы зрительного пигмента активи-
рует большое количество молекул фермента трансдуцина, который
инактивирует молекулы циклического гуанозинмонофосфата
(цГМФ), участвующего в ионном обмене В мембране фоторецепто-
ров, что в свою очередь приводит к фотопревращению зрительного
пигмента (фотолиз). Ретиналь на последних стадиях отщепляется от
опсина и переносится в пигментный эпителий, где он хранится в виде
эфиров пальмитиновой или стеариновой кислот, а затем возвраща-
ется в наружный сегмент фоторецепторов в виде 11-цис-ретиналя. В
ходе фотолиза происходит фотохимическая реакция — пикосекунд-
ная изомеризация хромофорной группы из 11-цис-формы ретиналя в
транс-форму. Все последующие процессы в молекуле темновые и за-
висят от температуры реакции [Островский М.А., 1992; Shichida Y.,
1986]. Процесс фотолиза родопсина происходит следующим образом.
БАТОРОДОПСИН (543 нм)
30 нс 1
Поглощение
кванта света
(1 - Юфс)
ГЮМИРОДОПСИН (497 нм)
МЕТАРОДОПСИН I (478 нм)
> 10 мс| |
минуты
МЕТАРОДОПСИН II (380 нм)
^^минугы
МЕТАРОДОПСИН III (465 нм)
(па^ародопснн 460 нм)
Полностью транс-РЕТИНАЛЬ+ОПСИН
РОДОПСИН (498 нм)
15
Перестройка в молекуле зрительного пигмента осуществляется
при переходе метародопсина I в метародопсин II, когда происходят
конформационные изменения, ответственные за ее электрогенную ак-
тивность и ряд биохимических процессов.
В ходе распада метародопсина II на ретиналь и опсин или его
превращения в метародопсин III белок возвращается в исходное со-
стояние. Таким образом, на ранних стадиях фотолиза в клетке воз-
никает быстрый потенциал, обусловленный электрогенной активно-
стью самой молекулы родопсина (ранний рецепторный потенциал).
На стадии образования метародопсина II запускается каскад фермен-
тативных реакций, который приводит к ускорению гидролиза цГМФ
и как следствие к блокированию светозависимых Na+- и Са2+-кана-
лов и генерации позднего рецепторного потенциала [Остров-
ский М.А., Говардовский В.И., 1992].
Продукты фотолиза родопсина связаны с генерацией компонен-
тов раннего рецепторного потенциала (РРП) и активацией каскада
ферментативных реакций, участвующих в усилении и внутриклеточ-
ной передаче фоторецепторного сигнала, который превращается в
электромагнитную энергию и в ответ нейронов. Вызванная светом
гиперполяризация мембран фоторецепторов распространяется к
синаптическим окончаниям, связывающим их с другими клеточными
элементами [Бызов А. Л., 1992 ].
Результаты электрофизиологических и психофизических исследо-
ваний, проведенных в последнее время, позволили предположить, что
свойства колбочек, чувствительных к длинно- и средневолновой час-
ти спектра, различны, и временные параметры их ответов зависят от
условий адаптации, которая оказывает слабое воздействие на кине-
тику трансдукции в колбочках человека [Whitmore A.V., Bow-
maker G.K., 1995].
Трансформация и интеграция сигналов происходят как в сетчат-
ке, так и на других уровнях зрительной системы с помощью рецеп-
тивных полей нейронов, которые, имея типичную организацию
центр — периферия, обладают способностью осуществлять латераль-
ное торможение [Глезер В.Д., 1992; Шевелев И.А., 1992]. Техника мар-
кетинга отдельных нейронов по Гольджи, внутриклеточное введение
пероксидазы хрена, электронная микроскопия позволили идентифи-
цировать все типы нейронов в сетчатке и их синаптические связи
[KolbH., 1994,1995].
В сетчатке выделяют дистальный и проксимальный отделы. В ди-
стальный входят фоторецепторы, горизонтальные клетки и биполя-
ры. К проксимальному отделу относится внутренний синаптический
слой, состоящий из аксонов биполярных, амакриновых и ганглиоз-
ных клеток, которые имеют многочисленные связи во внутреннем
плексиформном слое с аксонами ганглиозных клеток, образующих
зрительный нерв. Все нейроны этого слоя образуют сложные синап-
тические переключения во внутреннем синаптическом слое, количе-
16
ство подслоев в котором доходит до 10. Каждый из основных типов
нейронов имеет множество подтипов, образующих палочковый и
колбочковый пути.
Палочковый путь содержит палочковые фоторецепторы, бипо-
лярные и ганглиозные клетки, а также несколько видов амакрино-
вых клеток, являющихся интернейронами. Фоторецепторы переда-
ют зрительную информацию к биполярным клеткам, которые явля-
ются нейронами второго порядка. При этом палочки контактируют
только с биполярными клетками одной категории, называемыми па-
лочковыми биполярными клетками, которые деполяризуются под
действием света. Между палочками и деполяризующимися биполя-
рами имеется инвертирующий синапс, который и осуществляет пере-
дачу сигнала от гиперполяризующихся под действием света палочек
с негативным мембранным потенциалом к палочковым деполяризу-
ющимся биполярам с позитивным мембранным потенциалом. Эти
деполяризующиеся биполяры в литературе названы оп-центр-клет-
ками, или клетками, имеющими рецептивные поля с оп-центром.
Такие палочковые пути называют on-путями. Палочковая система в
высшей степени конвергентна и дивергентна, что определяет обра-
ботку и усиление сигнала для передачи его в мозг.
Колбочковый путь отличается от палочкового тем, что уже в на-
ружном плексиформном слое колбочки имеют более обширные свя-
зи и синапсы связывают их с колбочковыми биполярами различных
типов. Одни из них деполяризуются подобно палочковым деполяри-
зующимся биполярам и формируют колбочковый on-путь с инверти-
рующими синапсами, другие гиперполяризуются, образуя off-путь, в
котором колбочки контактируют с биполярами посредством предох-
раняющих синапсов. Колбочки фовеальной области, каждая из ко-
торых связана с двумя биполярами с on- и off-центром и двумя ганг-
лиозными клетками с on- и off-центром, образуют соответствующие
on- и off-каналы.
Отличие колбочкового пути от палочкового состоит также в том,
что колбочковые биполяры имеют прямые синапсы с дендритами
ганглиозных» клеток без промежуточных амакриновых клеток. Та-
ким образом, колбочковый путь состоит из двух параллельных кана-
лов, идущих от колбочек к биполярным и ганглиозным нейроцитам.
Один канал связан с сигналами, возникающими на стимул, который
ярче фона (on-center channel), другой — на стимул, более темный, чем
фон (off-center channel). Эти каналы обеспечивают восприятие кон-
траста зрительных образов.
Отличительной особенностью горизонтальных клеток разных
типов является их электрическая связь друг с другом через синцити-
альные сети. Различают два основных типа горизонтальных клеток:
L-клетки (ахроматические), реагирующие гиперполяризацией на из-
лучение любой части спектра, и цветовые клетки, знак реакции кото-
рых зависит от длины волны света. Горизонтальные клерки имеют
17
связь с фоторецепторами через тормозящие синапсы обратной связи
в триадах колбочковых терминалей. Эта обратная связь рассматри-
вается как основной механизм, посредством которого организованы
деполяризационные ответы горизонтальных клеток цветового типа,
а также оппонентная периферия рецептивных полей биполяров [Бы-
зов А.Л., 1992].
В сетчатке человека различают два типа биполяров: гиперполя-
ризационные (с off-центром) и деполяризационные (с оп-центром),
названные по знаку электрической реакции, возникающей в ответ на
освещение центра рецептивного поля пятном диаметром 150—
200 мкм, покрывающим область ветвления дендритов. Освещение
периферии рецептивного поля кольцом на фоне освещенного пятна
вызывает ответ противоположного знака, что называют простран-
ственной оппонентностью. Кроме пространственной, у биполяров
известна и цветовая оппонентность. Так, существуют биполяры, от-
вечающие деполяризацией на предъявление красного пятна и гипер-
поляризацией — синего. Биполяры связаны большей частью окон-
чаний ветвящихся дендритов с синаптическими терминалями фото-
рецепторов. Оппонентная реакция периферии рецептивного поля
биполяра передается через горизонтальные клетки, а пространствен-
ная оппонентная реакция — посредством обратной связи от горизон-
тальных клеток к фоторецепторам.
Амакриновые клетки, имеющие отростки длиной до 1 мм, обла-
дают импульсным механизмом и участвуют в передаче сигналов на
большое расстояние внутри сетчатки в горизонтальном направлении,
что объясняет процессы латерального торможения. Различают два
типа амакриновых клеток: фазные, реагирующие короткими депо-
ляризационными пиками с импульсами на включение и выключение
света, и тонические двух типов, отвечающие на воздействие светово-
го раздражителя стойкой деполяризацией (on-тип) или гиперполяри-
зацией (off-тип), дендритные разветвления тонических амакриновых
клеток связаны с реципрокными синапсами, в основном с палочко-
выми биполярами.
’’Ганглиозные клетки сетчатки делятся на Х-, Y- и W-типы, тела
которых отстоят друг от друга на определенные расстояния. Клетки
каждого типа объединены в строгие мозаичные структуры, называе-
мые растрами. Отдельные ячейки в таких растрах по форме близки к
квадратам или шестиугольникам. Растр, образованный телами X-
нейронов, состоит из мелких ячеек, среднее расстояние между тела-
ми которых 150 мкм, тогда как расстояние между клетками растра,
образованного Y-нейронами, составляет 250 мкм. В центральной
области сетчатки больше нейронов Х-типа, а на периферии — Y-типа.
Диаметр центральной части рецептивного поля в угловых градусах
для Y-нейронов 0,5—2,5, для Х-нейронов 0,1—1,3. Однако размер яче-
ек в растрах каждого типа клеток меняется по сетчатке в зависимос-
ти от распределения плотности ганглиозных клеток. Ретинотопичес-
18
кая упорядоченность в афферентных параллельных связях сетчатки
со зрительными структурами промежуточного и среднего мозга со-
провождается тонкой пространственной упорядоченностью, обуслов-
ленной растровым расположением в сетчатке отдельных типов ганг-
лиозных клеток, раздельными растрами — матрицами для on- и off-
клеток каждого типа, и существующими закономерностями
изменения размера и степени перекрытия рецептивного поля (РП)
нейронов различных типов в зависимости от их эксцентриситета по
сетчатке? [Подвигин Н.Ф., 1992]. Х-клетки, кроме тонической импуль-
сации, наблюдающейся в течение всего периода освещения, харак-
теризуются меньшим размером центра поля и большим простран-
ственным разрешением, меньшей скоростью проведения возбужде-
ния по аксонам, линейной суммацией сигналов внутри рецептивного
поля. Они цветооппонентны и передают импульсы в мелкоклеточ-
ные слои наружных коленчатых тел (НКТ). Y-клетки, отвечающие в
основном на изменение интенсивности света, имеют центры рецеп-
тивных полей значительно больших размеров, обладают меньшим
пространственным разрешением, большей скоростью проведения по
аксонам и нелинейным взаимодействием сигналов внутри поля. Y-
клетки цветонезависимы и посылают аксоны в крупноклеточные слои
НКТ (обладают более высокой пространственной чувствительнос-
тью). Клетки W-типа с импульсной активностью характеризуются до-
полнительными специфическими свойствами, например чувствитель-
ностью к направлению движения стимула. Они могут быть фазными
и тоническими, с периферией, оппонентной центру поля. Антагонис-
тическая центру реакция осуществляется посредством горизонталь-
ных клеток.
В классических работах выделяют четыре основных типа гангли-
озных клеток [Wiesel T.N., Hubei D., 1966]. Клетки 1-го типа имеют
круглые рецептивные поля, состоящие из центральной и перифери-
ческой зон с разной спектральной чувствительностью и тормозными
связями в направлении от периферии к центру. Эти поля называют
концентрическими без двойной оппонентности.
Клетки 2-го типа имеют круглые рецептивные поля, в которых
отсутствует периферическая зона. Эти поля избирательно реагиру-
ют на излучение с различной длиной волны, обладают оппонентны-
ми связями внутри своей единственной зоны и дают ответы с проти-
воположным знаком на стимулы различных цветов. Среди нейронов
встречаются не чувствительные к цвету, но обладающие свойствами
пространственной организации полей, сходными со свойствами дру-
гих нейронов. Их называют черно-белыми нейронами или on- и off-
нейронами [Бертулис А.В., Глезер В.Д., 1990].
Клетки 3-го типа — нейроны, не чувствительные к длине волны
излучения, имеющие концентрическую организацию рецептивных
полей с тормозными связями от периферической к центральной зоне.
Нейроны 4-го типа, близкие по строению к нейронам 1-го типа, от-
19
личаются от них более широкой спектральной чувствительностью.
Их рецептивные поля состоят из центральной и периферической зон,
каждая из которых избирательно реагирует на цветовой стимул с
разной длиной волны.
Среди клеток 1-го типа выделяют два класса: красно-зеленые (R-
G) и желто-синие (Y-В). Красно-зеленых клеток в 8 раз больше, чем
сине-желтых. Таким образом, в сетчатке существуют три параллель-
ных канала, связанных с цветовосприятием. Один из них передает
различие сигнала между средне- и длинноволновыми колбочками,
второй — разницу сигнала коротковолновых колбочек из суммы сиг-
налов средне- и длинноволновых колбочек, а третий — суммарный
ответ колбочек всех трех классов. В сетчатке человека обработка сиг-
налов колбочек имеет оппонентный характер. Возникновение ней-
ронов, отвечающих реакциями с противоположными знаками, явля-
ется определяющим фактором функционирования механизмов про-
странственного и цветового зрения. Таким образом, в сетчатке
человека от слоя рецепторов к слою ганглиозных клеток тянется сеть
нейронных связей, образующих множество пострецепторных путей,
или каналов, предназначенных для передачи и обработки сенсорной
информации, которые выполняют разные функции и работают как
параллельные системы.
РП нейрона — это структура, состоящая из выходного нейрона,
промежуточных клеток и совокупности рецепторов сетчатки, в пре-
делах которой изменение светового потока в пространстве или вре-
мени вызывает изменение реакции выходного нейрона. Рецептивные
поля имеют центральную и периферическую зоны, стимуляция кото-
рых вызывает противоположные по знаку реакции, например оп-ре-
акцию — в центре и off-реакцию — на периферии. Между центром и
периферией существуют взаимотормозные отношения так, что осве-
щение обеих зон одновременно приводит к уменьшению как цент-
ральной, так и периферической реакций. Оппонентная центру пери-
ферия ганглиозной клетки организована по механизму обратной связи
от горизонтальных клеток к фоторецепторам.
РП нейронов увеличиваются с эксцентриситетом и имеют систе-
му латерального и возвратного торможения, действующего через
вставочные тормозные нейроны. Латеральное торможение в рецеп-
тивном поле определяет его пространственные характеристики, а
собственное возвратное торможение — временные свойства сигналов,
передаваемых по системе.
Пространственно-временные свойства взаимодействия процессов
возбуждения и торможения в РП нейронов лежат в основе ряда меха-
низмов, осуществляющих преобразование зрительных сигналов на
пути их передачи от сетчатки в кору. К ним относится пространствен-
ная суммация входных сигналов, осуществляющаяся в зонах возбуж-
дения РП. Благодаря конвергенции афферентных связей на один ней-
рон его выходной сигнал увеличивается при увеличении в опреде-
20
ленных пределах площади светового стимула. Такое свойство РП
присуще как нейронам сетчатки, так и нейронам афферентируемых
ею структур мозга, и оно лежит в основе высокой световой чувстви-
тельности зрительной системы. РП подкорковых структур участву-
ют в дифференциации перепадов яркости, усиливают сигналы, несу-
щие в себе сведения о высоких пространственных частотах изобра-
жений, о границах перепада яркости в изображении, служат для
увеличения пространственного разрешения системы.
Пространственно-суммационные свойства РП нейронов опреде-
ляют разрешающую способность зрения. Процессы возбуждения и
торможения развиваются и затухают в РП в течение какого-то вре-
мени после предъявления стимула. Это время составляет 100—200 мс,
т. е. время, сопоставимое с реальным временем зрительного воспри-
ятия. Эти временные изменения пространственных характеристик РП
имеют функциональное значение.
1.3. НЕЙРОМЕДИАТОРЫ
Большинство известных нейротрансмиттеров, нейромодуляторов,
нейропептидов, обнаруженных в сетчатках разных животных, встре-
чается, за небольшим исключением, и в сетчатке человека.
Ауторадиография, иммунология, молекулярная биология, нейро-
химия позволяют маркировать клетки, содержащие любой трансмит-
тер.
В экспериментальных исследованиях доказана роль допамина в
функционировании рецептивных полей, в антагонистических взаи-
модействиях его с мелатонином при дегенеративных изменениях в
сетчатке, а также в возникновении электрофизиологических феноме-
нов, которые их сопровождают. При использовании селективных
рецепторных антагонистов выявлена функциональная роль эндоген-
ных трансмиттеров, таких как глутамат, который является главным
в передаче возбуждения в цепи ретинальных нейронов (фоторецеп-
торов, биполяров, ганглиозных клеток), аспартат, специфичный для
палочек, и ацетилхолин, известный как трансмиттер холинергичес-
ких амакриновых клеток. Установлено, что при деполяризации
фоторецепторы передают глутамат биполярам и горизонтальным
клеткам и от биполярных клеток к ганглиозным и амакриновым. По-
казана роль М-метил-Э-аспартатных (NMDA) рецепторов во взаи-
модействии с ганглиозными клетками, которые в физиологических
условиях остаются интактными, а в случае увеличения содержания
глутамата активируются. Различают два основных типа рецепторов
эндогенного действия: NMDA и HeNMDA. Определена функциональ-
ная роль ГАМК (гаммааминомасляная кислота)-и глицинергичес-
ких путей, которые являются зрительными ингибиторами соответ-
ственно оп-и off-ганглиозных клеток. В клинике получено подтвер-
21
ждение связи рецепторов бензодиазепина с ГАМК-рецепторами, а эк-
спериментально показана активность допаминергических путей в сет-
чатке на различных уровнях. Основной, глутаматовый, путь идет от
фоторецепторов к ганглиозным клеткам через биполяры, а тормоз-
ной путь от ГАМК- и глицинергических амакриновых клеток к ган-
глиозным. Фоторецепторы, деполяризуясь, выделяют в темноте глу-
тамат, который, освобождаясь от них, переносится к горизонталь-
ным клеткам и биполярам и через конвенциональные синапсы к
neNMDA-рецепторам.Воздействие фоторецепторного глутамата на
оп-биполярные клетки осуществляется при посредничестве амино-4-
фосфобутиратчувствительных рецепторов, у которых глутамат зак-
рывает ионные каналы, обусловливая деполяризацию оп-биполяр-
ных клеток на свету. Off-биполяры, деполяризующиеся в темноте, и
оп-биполяры, деполяризующиеся при раздражении светом, выделя-
ют глутамат к HeNMDA-рецепторам off- и оп-ганглиозных клеток
соответственно, вызывая зрительные потенциалы действия. NMDA-
рецепторы оп-ганглиозных клеток активируются избыточно аккуму-
лированным глутаматом. Тормозные пути возникают от ГАМК- и
глицинергических амакриновых клеток, однако выделение ими транс-
миттеров “запускается” через биполярные клетки. Off-биполярные
клетки выделяют глутамат к деполяризованным ГАМК-ергическим
амакриновым клеткам с последующей активацией оп-ганглиозных
клеток. Определенную роль в процессах торможения играют бензо-
диазепиновые рецепторы, которые связаны с ГАМК-рецепторами
через ганглиозные клетки.
Допамин, нейроактивная субстанция сетчатки, содержится в амак-
риновых клетках внутреннего плексиформного слоя и отличается от
возбуждающих и тормозных трансмиттеров, составляя другой, до-
паминовый, путь. Допаминергическая активность проявляется в сет-
чатке на разных уровнях. Допаминергические клетки получают сиг-
нал от оп-биполяров, управляемых колбочками, посылая импульсы
к фоторецепторам, горизонтальным клеткам и пигментному эпите-
лию. Результаты физиологических и биохимических исследований
показали, что допамин и мелатонин, синтезируемый в фоторецепто-
рах, играют реципрокную роль в ускорении процессов их обновле-
ния, а также в адаптивных процессах в темноте и на свету, в наруж-
ных слоях сетчатки. Допаминергические амакриновые клетки явля-
ются пресинапсами к ГАМК- и глицинергическим амакриновым
клеткам. Допаминергическая активность проявляется на различных
уровнях в сетчатке. Несмотря на то что допаминергические клетки
имеются только во внутреннем плексиформном слое, Dj-и D2-рецеп-
торы проходят через всю сетчатку от пигментного эпителия к ганг-
лиозным клеткам. Такое распределение допаминовых рецепторов в
сетчатке подтверждено результатами клинических наблюдейий: до-
паминовые рецепторы могут активироваться под влиянием всех до-
паминергических лекарств, которые проходят кровяной сетчаточный
22
барьер. При проведении электрофореза с использованием допамина
выявлены подавление функции всех ганглиозных клеток и интакт-
ность их рецепторных антагонистов. Вследствие этого допаминерги-
ческие антагонисты (бромокриптин, апоморфин) могут активировать
рецепторы допамина, проходящие через сетчатку, а психотропные
препараты — производные фенотиазинового рада тормозят эндоген-
ную активность допамина.
Из множества пептидов, обнаруженных в сетчатке разных живот-
ных, в сетчатке человека выявлен только нейропептид Y, соматоста-
тин и субстанция Р, которые наряду с регулярными нейротрансмит-
терами и нейрорегуляторами локализуются в различных амакрино-
вых и ганглиозных клетках. Получены морфологические доказательства
того, что два класса трансмиттеров — возбуждающие и тормозящие,
названные ацетилхолином и ГАМК, содержатся в амакриновых клет-
ках одного типа. Каковы их функциональные различия в нейрональ-
ной сети сетчатки еще предстоит понять.
Таким образом, нейроактивные вещества, обнаруженные в сет-
чатке (ацетилхолин, глутамат, ГАМК, глицин и допамин), являются
трансмиттерами, которые взаимодействуют друг с другом, поэтому
функция сетчатки зависит от их тонкого нейрохимического баланса.
Некоторые лекарственные препараты, попадающие экзогенным пу-
тем в сетчатку, могут нарушать этот баланс, вследствие чего изменя-
ется функция сетчатки.
Хорошо известно, что мелатонин и допамин регулируют обнов-
ление наружных сегментов фоторецепторов. В то же время наруше-
ние фагоцитарной функции пигментного эпителия является причи-
ной дегенерации сетчатки. Возникновение дисбаланса между мела-
тонином и допамином может быть одним из факторов, приводящих
к развитию дистрофического процесса в сетчатке, пигментному ре-
тиниту, ретинопатиям лекарственного происхождения. Эти факто-
ры предопределяют новые подходы к терапевтическому лечению
некоторых видов дистрофий. Так, после длительной аппликации
агониста допамина В2-бромокриптина крысам с дистрофией сет-
чатки отмечалось значительное увеличение амплитуды ЭРГ, при
нарастании интенсивности стимулирующего света, по сравнению с
контрольной группой. В то же время, у нормальных гетерозигот
после лечения бромокрипином отмечено значительное снижение ам-
плитуды ЭРГ по сравнению с таковой в контрольной группе. Эти
данные согласуются с представлением о том, что В2-допаминрецеп-
торный механизм способствует длительной адаптации к свету, a D2-
агонист обладает защитным свойством против длительного свето-
вого повреждения. В многочисленных работах показана роль допа-
миновых механизмов в изменении функции сетчатки при болезни
Паркинсона, ретинопатии лекарственного происхождения (мелле-
риловые, фенотиазиновые), побочных действиях лекарственных
препаратов. Тиоридазин и фенотиазин, являющиеся агонистами
23
допамина, широко используются в психиатрической практике. Эти
препараты связывают мелатонин в пигментном эпителии и могут в
течение длительного времени оставаться в клетках. На фоне приме-
нения этих препаратов возникает пигментная ретинопатия, по кли-
ническому течению сходная с наследственной формой. Хотя меха-
низм развития этих лекарственных ретинопатий до конца неясен,
очевидно токсическое воздействие антагонистов допамина на син-
тез мелатонина, которое приводит к нарушению отторжения дис-
ков и фагоцитоза. Не исключено, что точная циклическая модуля-
ция мелатонина и допамина с соответствующими системами связи
в определенных слоях сетчатки может оказывать лечебное
действие [Hankins M.W., 1993; Ikeda Н., 1993]. На крысах KCS с
пигментным ретинитом была подтверждена связь допамина с фун-
кциональными нарушениями, сопровождавшимися высоким содер-
жанием мелатонина, который тормозил освобождение ретинально-
го допамина.
1.4. КЛЕТКИ МЮЛЛЕРА И ИХ ВЗАИМООТНОШЕНИЯ
С НЕЙРОНАМИ
Глиальные клетки до настоящего времени рассматривались как
вспомогательные элементы нервной ткани в центральной нервной
системе и сетчатке. Однако открытия в области нейробиологии, сде-
ланные в последние годы, показали, что глиальные элементы сетчат-
ки обладают различными функциональными свойствами.
Известно, что метаболизм в нейроглие изменяется под влиянием
внешней и внутренней среды и, подобно метаболизму в нейронах,
характеризуется функциональной лабильностью. Наряду с опреде-
ленным сходством существуют и особенности в биохимической орга-
низации нейроглии и нейронов.
Глиальная популяция сетчатки представлена преимущественно
клетками Мюллера, которые, являясь высокоспециализированными
глиальными элементами, выполняют опорно-изолирующую функ-
цию, осуществляют активный транспорт метаболитов на разных уров-
нях сетчатки и участвуют в светоиндуцированном генерировании
волны b ЭРГ. Мюллеровские клетки обладают способностью к по-
ляризации высокой степени. Они образуют все типы контактов, из-
вестных у глиальных клеток: с жидкой средой, базальной мембра-
ной, нейронами и другими глиальными элементами сетчатки. Струк-
турно-функциональная организация отростков радиальных клеток
Мюллера направлена на обеспечение транспорта веществ, поступа-
ющих между наружными сегментами фоторецепторов через гемато-
ретинальный барьер из собственно сосудистой оболочки глазного
яблока (choroidea) [Певзнер Л.З., 1979].
Базальная концевая ножка клеток Мюллера контактирует с ме-
зенхимальными производными и со стороны стекловидного тела
24
покрыта внутренней пограничной мембраной. Плазмолемма нейро-
глии на границе с базальной мембраной обладает высокой проница-
емостью для ионов калия и содержит особые прямоугольные части-
цы, обеспечивающие стабильность мембраны при натяжении на по-
верхности стекловидного тела. Контакты клеток Мюллера в ядерных
и плексиформных слоях обладают свойствами, необходимыми для
нейроглиальных взаимодействий.
Наряду с клетками Мюллера в слое нервных волокон сетчатки
млекопитающих имеются астроциты. Олигодендроциты, участвую-
щие в миелинизации аксонов ганглиозных клеток, описаны лишь у
некоторых млекопитающих. Микроглиоциты обнаруживают глав-
ным образом в сетчатке плодов и в раннем постэмбриональном пе-
риоде, реже — у половозрелых особей. Большинство авторов указы-
вают на расположение микроглии до внутреннего плексиформного
слоя сетчатки. Предполагают, что нарушения структуры и функции
глиальных клеток являются одним из звеньев патогенеза некоторых
заболеваний сетчатки.
Ретинальную нейроглию наряду с клетками пигментного эпителия
относят к основным типам клеток, участвующих в образовании эпи-
ретинальных мембран [Hogg Р., 1990]. Глиальным астроцитам отво-
дят немаловажную роль в процессе неоваскуляризации у пациентов с
пролиферативной диабетической ретинопатией (ДР) и отслойкой сет-
чатки с пролиферативной витреоретинопатией. При ДР установлена
чувствительность цитоплазмы клеток Мюллера, которая трансформи-
руется и приобретает вид луковой шелухи, к длительным гиперглике-
мическим состояниям. При наследственных дегенерациях сетчатки у
животных, схожих с пигментным ретинитом человека, пролифератив-
ная ретинопатия сопровождается процессом неоваскуляризации, по-
добным таковому при ДР. В экспериментах на мышах прослежена ак-
тивация синтеза глиального фибриллярного кислого белка (ГФКБ) в
клетках Мюллера при наследственной дегенерации фоторецепторов в
сетчатке, сопровождающей наследственную ретинальную патологию.
Пролиферация и гипертрофия клеток Мюллера наблюдаются уже на
3—4-е сутки после отслойки сетчатки [Fisher S.K., 1990]. При этом в
нейроглие, так же как и при дегенерации фоторецепторов, возрастает
уровень ГФКБ и виментина. Установлено, что после хирургического
лечения при прилегании сетчатки клетки Мюллера ингибируют реге-
нерацию наружных сегментов фоторецепторов и в наружном плекси-
формном слое развивается глиоз.
Таким образом, повреждение именно глиальных клеток Мюллера
при отслойке сетчатки препятствует последующему восстановлению
ретинальных связей, нормальной структурно-функциональной орга-
низации сетчатки.
Реакция клеток Мюллера на воздействия различных стрессовых
факторов проявляется изменением содержания”белков стресса”, од-
ним из которых считают указанный выше глиальный фибриллярный
25
кислый белок. Подобная реакция нейроглии лежит в основе наруше-
ний, происходящих в сетчатке при травме глаза.
Не исключена связь клеток Мюллера с аутоиммунными механиз-
мами, играющими определенную роль в развитии ретинальной па-
тологии [Roberge 'F.G., 1990].
Известно, что L-глутаминовая аминокислота выполняет ряд функ-
ций в метаболизме нервной ткани. Одна из них состоит в том, что
глутамат является возбудительным нейротрансмиттером. Его ос-
вобождение от синаптических окончаний фоторецепторов происхо-
дит в темноте и прекращается при световой стимуляции. Избыток
глутамата и длительное нахождение его в межклеточном пространст-
ве оказывают ретинотоксическое действие и приводят к блокаде сина-
птической передачи между фоторецепторами и нейронами 2-го поряд-
ка. В экспериментах на сетчатке позвоночных и беспозвоночных
доказано строгое взаимодополняющее пространственное разделение
(компартментализация) в обмене веществ, локализации и потребле-
нии глутамата и глутамина в нейроглии и нейронах [Duce I. R., 1988].
Блокада синаптической передачи между фоторецепторами и ней-
ронами 2-го порядка глутаматом и его аналогами в значительной
степени зависит от pH межклеточной среды. Сетчатка — метаболи-
чески активная и хорошо васкуляризированная ткань, продуцирую-
щая углекислый газ и молочную кислоту, поэтому фоторецепторы
сетчатки нуждаются в механизмах защиты от внутриклеточного аци-
доза. Клетки Мюллера, содержащие карбоангидразу, потребляют
СО2, образующийся в процессе метаболизма, в свою очередь карбо-
ангидраза катализирует реакцию СО2 с водой, в результате которой
образуются Н+ и НСО’ и таким образом глиальные клетки Мюллера
участвуют в регуляции кислотности среды и модуляции палочковой
активности. Аналогичную роль в поддержании ионного гомеостаза
играет нейроглия и в ткани мозга.
Из изложенного выше очевидна роль глиальных клеток Мюлле-
ра в модуляции нейрональной активности в сетчатке и процессах пе-
реработки информации в зрительной системе, а также в развитии ре-
тинальной патологии. В связи с этим определение степени участия
клеток Мюллера в суммарной биоэлектрической активности сетчат-
ки расширяет возможности дифференциальной диагностики нейро-
нальных и нейроглиальных нарушений.
1.5. АНАТОМО-ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ЗРИТЕЛЬНОГО ПУТИ
В настоящем разделе мы лишь отметим те основные структуры зри-
тельного пути, которые принимают участие в обработке сенсорной
информации, и их функция может быть оценена с помощью психо-
физических и электрофизиологических методов исследования.
Анатомия зрительных путей подробно описана в многочислен-
26
Рис. 1.2. Схематическое
изображение зрительных
путей от сетчатки до зри-
тельных центров.
ных монографиях и руководствах [Зальцман М., 1913; Кравков С. В.,
1950; Трон Е. Ж., 1955; Бинг Р., Брюкнер Р.,1959; Меркулов И. И.,
1971; Брошевский Т. И., Бочкарева А. А., 1983; Скоромец А. А., 1989;
Сапин М. Р., 1993; Polyak S. L., 1957].
Третий нейрон зрительного пути начинается с аксонов ганглиоз-
ных клеток сетчатки, которые по морфологическому строению сход-
ны с аксонами нейронов коры головного мозга, так как их аксоны
не имеют шванновской оболочки и по строению близки к нервным
волокнам белого вещества мозга (рис. 1.2).
Аксоны ганглиозных нейроцитов сетчатки образуют зрительный
нерв (n.opticus), который относится к черепным нервам (II пара). Зри-
тельным нервом называется отрезок третьего нейрона зрительного
пути, который начинается от уровня решетчатой пластинки и закан-
чивается в хиазме (chiasma opticum). Длина зрительного нерва 35—
55 мм, толщина (вместе с оболочками) 4 мм. Общее число волокон
от 400 000 до 800 000. Зрительный нерв покрыт твердой, паутинной и
мягкой оболочкой, подобно головному мозгу. В полости черепа зри-
тельные нервы обоих глаз соединяются в области турецкого седла и
образуют частичный перекрест (chiasma opticum) перед воронкой над
гипофизом и пещеристой пазухой, на основании головного мозга
кпереди от серого бугра. Волокна зрительного нерва от височных
(латеральных) отделов сетчатки обоих глаз в области хиазмы не пе-
рекрещиваются и идут снаружи до зрительного тракта, а от носовых
(медиальных) перекрещиваются и переходят в противоположный
27
зрительный тракт. Волокна от нижненосовых квадрантов сетчатой
оболочки переходят на другую сторону ближе к нижнепереднему
краю хиазмы, а от верхненосовых квадрантов — к ее задневерхнему
краю. Волокна от центральной, фовеальной, области сетчатки (так
называемый папиллр-макулярный пучок) в хиазме занимают средин-
ное положение, его волокна перекрещиваются недалеко от верхней
поверхности хиазмы в заднем ее отделе, под дном третьего желудоч-
ка (вопрос о частичном перекресте этих волокон окончательно не ре-
шен).
Дальнейший участок зрительного пути от хиазмы до латераль-
ного коленчатого тела называется зрительным трактом, в состав ко-
торого входят волокна, идущие от сетчаток обоих глаз. Зрительный
тракт, постепенно дивергируя, направляется кверху и кзади, огибает
с обеих сторон серый бугор, проходя по нижней поверхности ножек
мозга, подходит к подкорковым центрам зрения, латеральному (на-
ружному) коленчатому телу (НКТ) и верхним холмикам крыши сред-
него мозга. В НКТ заканчивается третий нейрон зрительного пути.
Из серого слоя верхнего холмика импульсы поступают в ядро гла-
зодвигательного нерва и добавочное ядро (ядро Якубовича), откуда
осуществляется иннервация глазодвигательных мышц, а также мыш-
цы, суживающей зрачок, и ресничной мышцы.
Как в зрительном нерве, так и в тракте существует множествен-
ное проекционное представительство сетчаток обоих глаз. Аксоны,
образующие каждое из этих представительств, направляются в опре-
деленные слои подкорковых структур, где находит отражение рети-
нотопическая мозаичная организация ганглиозного слоя сетчатки.
Третий нейрон зрительного пути осуществляет проведение зри-
тельного возбуждения от сетчатки к латеральному коленчатому телу,
которое находится на нижнебоковой поверхности таламуса, сбоку
от подушки. От НКТ берет начало четвертый нейрон зрительного
пути, проходящий к подушке таламуса. Аксоны четвертого нейрона,
направляющиеся в первичные зрительные центры в затылочной доле
коры, проходя через подчичевицеобразную часть внутренней капсу-
лы, образуют зрительную лучистость (пучок Грациоле).
Система эфферентных связей основных структур промежуточно-
го мозга дает существенную информацию для анализа процессов пе-
реработки зрительных сигналов и функциональной роли отдельных
подкорково-корковых путей их передачи.
“Подкорковый уровень зрительной системы состоит из несколь-
ких параллельных ретинопроекционных путей передачи сигналов от
сетчатки в мозг, каждый из которых обладает определенными функ-
циональными свойствами, что позволяет относить их к так называе-
мым первой, второй и третьей зрительным системам. Каждый из этих
путей имеет многоуровневую организацию последовательно соеди-
ненных нейронов, относящихся либо к отдельным образованиям, либо
к отдельным слоям одного образования мозга” [Подвигин Н.Ф., 1992].
28
Среди структур среднего мозга, имеющих прямое отношение к зри-
тельной и глазодвигательной системам, принимающих аксоны ганг-
лиозных клеток, выделяют претектальную область, ядра крыши сред-
него мозга (tectum), верхние холмики, colliculi superiores (передние буг-
ры четверохолмия), относящиеся ко ’’второй зрительной системе”.
К первичному комплексу зрительной коры в каждом полушарии
относят затылочные цитоархитектонические поля 17—19. При этом
поле 17 называют стриарным, а 18—19 — пара- или экстрастриар-
ной корой. Корковый отдел зрительного пути располагается в обла-
сти верхнего и нижнего краев шпорной борозды (sulcus calcarinus) на
медиальной поверхности затылочной доли (цитоархитектоническое
поле 17). Участок затылочной доли, лежащий между теменно-заты-
лочной и шпорной бороздами, имеющий форму треугольника, назы-
вается клином (cuneus); шпорная борозда ограничивает сверху языч-
ную извилину (gyrus lingualis).
В пределах каждого ретинопроекционного пути существует про-
екционно-топическая упорядоченность связей. Благодаря этому поле
зрения цмеет достаточно точное ретинопроекционное отображение
в нейронном пространстве каждой структуры промежуточного и сред-
него мозга.
Структура ретинотопической организации строго представлена
в первичных зрительных центрах, и каждое из этих полей содержит
почти полное представительство контралатерального полуполя зре-
ния: волокна макулярной области, имея преимущественное предста-
вительство, заканчиваются в окципитальной доле (поле 17), перифе-
рическая область — в более растральной порции sulcus calcarine. Пе-
рекрещенные в хиазме волокна от назальной части сетчатки
достигают поля 17 по Бродману в контралатеральной части окципи-
тальной доли, в то время как неперекрещенные волокна от темпо-
ральной части сетчатки каждого глаза заканчиваются на ипсилате-
ральной стороне. Проводники от верхней половины сетчаток закан-
чиваются в cuneus, а от нижней — в gyrus lingualis.
Пирамидальные клетки от поля 17 проецируются на высшие зри-
тельные области, поля 18 и 19 (по Бродману), которые окружают пер-
вичные зрительные центры. Афферентная информация в зрительную
кору идет четырьмя параллельными путями: 1) ретиногеникулокор-
ковая проекция, 2) ретинотекто(или претекто)-таламокорковая про-
екция, 3) ипсилатеральные корково-корковые проекции, 4) транскал-
лозальные межполушарные связи.
Основной ретиногеникулокорковый путь проходит через дорсаль-
ное ядро НКТ в поле 17—19. Поле 19 снабжается афферентами из
слоев НКТ, медиального интраламинарного ядра и подушки зритель-
ного бугра. Медиальное интраламинарное ядро НКТ снабжает аф-
ферентами поле 18 и латеральную супрасильвиеву зону (Клера — Би-
шопа). Претектум через латеральную часть подушки снабжает вхо-
дами поле 19—20 и зону Клера — Бишопа.
29
В ретиногеникулокорковом пути представлены три параллельных
отдельных функциональных подсистемы, различающихся по свой-
ствам рецептивных полей, типам ответов на свет и скорости проведе-
ния сигналов: Х-, Y- и W- каналы. В отличие от сетчатки и НКТ клет-
ки зрительной коры имеют на свету не круглые, а вытянутые рецеп-
тивные поля (РП) разной ориентации. Соответственно они лучше
всего отвечают не на круглые световые пятна, а на вспыхивающие в
РП или движущиеся через него световые полоски, края или решетки
определенной ориентации. Среди нейронов коры выделяют простые
и сложные. Простые нейроны делятся на on и off (с антагонистичес-
ким взаимодействием и пространственной суммацией) по ответам на
вспышечный стимул. Сложные рецептивные поля характеризуются
тем, что зоны on- и off-ответов в этих полях совпадают или в значи-
тельной степени перекрываются, что позволяет получить смешанный
(on-off) ответ. Максимальный ответ нейрона может быть вызван сти-
мулом оптимальной ориентации не в центре РП, а в некоторой обла-
сти вокруг этого центра. Размеры РП определить трудно, однако их
качественная оценка показала, что корковые нейроны различаются
между собой. Меньший размер у простых корковых РП от 0,3 до 6°,
больший — от 0,5 до 8°; у сложных — от 2 до 30°. Форма корковых
рецептивных полей при световой адаптации вытянута, близка к пря-
моугольной или эллиптической [Шевелев И.А., 1992]. По функ-
ции интенсивности можно оценить порог реакции, характеризующий
световую чувствительность. Ответ корковых зрительных нейронов
(всех трех типов по сложности РП) зависит от размеров стимула.
Большинство простых клеток оптимально реагирует на полосы ши-
риной 0,1—1—2° (для движущихся полос 0,2—0,8°). Сложные нейро-
ны в отличие от простых хорошо отвечают только на узкие полосы,
ширина которых меньше их возбудительной зоны.
В первичной зрительной коре в обработке информации участву-
ют параллельные каналы и системы клеток, которые образованы
афферентами из НКТ, что определяет последовательность передачи
и обработки информации. При этом происходит параллельный от-
бор признаков: пространственная локализация объекта, формы, ско-
рости движения, цвета. Многие нейроны участвуют в кодировании
не одного, а ряда признаков, а многие признаки и их параметры ко-
дируются не одним нейроном, а некоторым их набором.
Разные каналы в коре пересекаются. Ведущая роль в механизме
формирования специфических детекторных свойств РП нейронов
зрительной коры принадлежит процессам торможения. В условиях
световой адаптации РП невелики по размеру, имеют четкую ориен-
тационную и, как правило, дирекциональную специфичность. При
понижении уровня адаптации до темнового РП значительно увели-
чиваются в размере, а их детекторные свойства — ориентационная и
дирекциональная селективность — резко ухудшаются. Центробежные
волокна сетчатки идут от нейронов разных отделов мозга: ретйку-
30
лярной формации, среднего мозга, гипоталамуса, верхнего двухол-
мия, зрительной коры. Центробежные волокна оканчиваются в сет-
чатке, образуя аксосоматическую связь с телами амакриновых клеток;
не исключается связь с горизонтальными клетками и биполярами.
Таким образом, по современным данным переработка зритель-
ной информации на уровне коры мозга осуществляется не только в
поле 17 по Бродману, но также в 18 и 19, где обнаружены зоны с пол-
ным представительством поля зрения. Кроме того, нейроны, реаги-
рующие на зрительную стимуляцию, обнаружены вдоль латеральной
супрасильвиевой борозды, в височной, лобной и теменной коре. Не
менее важную роль в обработке информации играют экстрастриар-
ные зрительные зоны. Физиологические исследования, применение
метода аксонного транспорта красителей, а также топографическое
картирование ЗВКП позволили выделить множественное ретиното-
пическое представительство стриарных и экстрастриарных центров,
связанных с переработкой зрительной информации, но лежащих за
пределами классических ретиногеникулостриарных путей, в связи с
чем и появился термин экстрагеникулостриарное зрение. Корковые
и подкорковые окончания эфферентных волокон идут от стриарной
и престриарной зоны коры в область верхней височной извилины,
верхний зрительный бугор, подушку четверохолмия, нижневисочную
и заднетеменную и лобную области коры. Стриарные и внестриар-
ные зоны находятся в сложном взаимодействии, а множественные аф-
ферентные и эфферентные связи определяют процесс переработки зри-
тельных сигналов, идущих как в первичные зрительные центры, так
и по “вторичному зрительному пути” [Пигарев И.Н., 1992; Подви-
гни Н.Ф.,1992; Пасик Т., Пасик П.,1993; Pirtosek Z., 1993;]. После де-
струкции ретикулостриарной системы в связи с функционированием
указанных зон могут сохраняться разнообразные зрительные реак-
ции: зрачковая, мигательная реакции на свет, следящее движение глаз
и оптокинетический нистагм, дискриминация объектов, различаю-
щихся общим световым потоком и яркостью, формой, цветом, а так-
же способностью локализовать предметы в пространстве. Измене-
ние биоэлектрической активности под влиянием тех или иных пато-
логических нарушений на разных уровнях зрительной системы
является поводом для топографического картирования зрительных
вызванных кортикальных потенциалов (ЗВКП) и локализации источ-
ников биоэлектрической активности.
ГЛАВА 2
ОСТРОТА ЗРЕНИЯ, КОНТРАСТНАЯ
ЧУВСТВИТЕЛЬНОСТЬ И УСТОЙЧИВОСТЬ
К СЛЕПЯЩИМ 3 АСВЕТ AM
2.1. ОПТИКО-АНАТОМО-ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ
ФУНКЦИИ
2.1.1. Острота зрения
Острота зрения (visus) — составная часть более широкого понятия
“разрешающая способность глаза”, поэтому оценивать зрительные
функции только по остроте зрения, как это нередко практикуется,
неправомерно. Однако острота зрения является одним из основных
критериев оценки способности к зрительному анализу форм, разме-
ров, структуры и ориентации объектов в пространстве, а визометрия
относится к числу обязательных методик функциональных исследо-
ваний органа зрения.
Первой ступенью в формировании благоприятных условий для
такого анализа служит оптическая система глаза. Она регулирует
распределение световой энергии в сфере изображений на сетчатке со-
ответственно таковому в сфере объектов. Особенность изображения
заключается в том, что даже в случае идеальной фокусировки точеч-
ного источника световая энергия, концентрируясь в центре (по ходу
главного луча), слегка рассеивается по окружности (рис.2.1). При
расфокусировке это рассеивание усиливается. Следует заметить, что
линейное расстояние, равное на сетчатке эмметропического глаза
1 мм, соответствует угловому отклонению луча около 3,5° [West-
heimer G., 1987]. На распределение световой энергии по сетчатке ока-
зывают влияние следующие оптические факторы.
Дифракция. При очень узком зрачке (меньше 2 мм) благодаря вол-
новой природе света его распределение в изображении точечного
объекта на сетчатке даже в полностью сфокусированной системе ока-
жется иным, чем на рис 2.1. Кривая по форме будет напоминать фи-
гуру колокола, окаймленного дополнительной волной светорассея-
ния.
Аберрации. Смещение фокусов лучей, проходящих через перифе-
рические отделы роговицы и хрусталика, из-за перепадов в прелом-
ляющей силе этих отделов (относительно центральной области) на-
зывается сферической аберрацией. При этом на распределении све-
товой энергии в сетчатке неблагоприятно сказывается не только
многофокальность, но и усиление эффекта Стайлса — Крауфорда (см.
главу 8). Геометрические аберрации (сферическая, астигматизм, дис-
32
сировки и при умеренной
ширине зрачка [Westhei- Ретинальное расстояние
met G., 1987]. в дуговых мин
торсия, кома) особенно ощутимы при широком зрачке (более 5 мм),
поскольку в этом случае увеличивается доля лучей, поступающих
через периферию роговицы и хрусталика. Хроматическая аберрация,
обусловленная различиями в силе преломления и расположения фо-
кусов лучей разной длины волны, в меньшей степени зависит от ши-
рины зрачка.
Рассеивание света. Микроструктура оптических сред глаза тако-
ва, что часть света рассеивается в них. С возрастом выраженность
этого феномена возрастает и он может послужить причиной слепи-
мости от ярких засветов глаза.
Абсорбция. Часть световой энергии поглощается уже при прохож-
дении лучей через оптические среды: чем короче длина волны излу-
чения, тем меньшая доля его достигает сетчатки.
Фокусировка. Механизм аккомодации в определенной мере спо-
собствует уменьшению кругов светорассеяния вследствие фокусиров-
ки изображения на сетчатке. Однако в случае отсутствия в поле зре-
ния остро очерченных мишеней этот фактор может и не сработать
[Westheimer G., 1987].
Второй ступенью, способствующей зрительному восприятию
мельчайшей структуры окружающего пространства, является гекса-
гональное строение ретинальных рецептивных полей [Шелепин Ю.Е.,
1990; Campbell R.W.,1970]. В зоне фовеолы, где колбочки тесно при-
лежат одна к другой, образуется неразбиваемая матрица из 7 элемен-
тов (один — в центре и шесть — по окружности). Угловой размер
такой минимальной распознающей ячейки не выходит за пределы Г,
поэтому если изображение простого объекта занимает большее про-
странство (размещается как минимум на двух гексагональных эле-
33
ментах дискретизации), то можно различить его детали. S. Polyak
(1941) полагал, что для человеческого глаза предельной возможнос-
тью раздельного видения объектов является расположение их один
от другого на расстоянии менее 0,5' по дуге.
В зависимости от эксцентриситета фоторецепторы связаны с со-
ответствующим количеством биполярных клеток с дальнейшей кон-
вергенцией контактов на ганглиозные клетки сетчатки. Таким обра-
зом, образуется множество рецептивных полей, частично перекры-
вающих друг друга. Диаметр каждого такого поля у млекопитающих
составляет в среднем около 1 мм [Kufller S. W., 1950]. К периферии
глазного дна размеры этих полей увеличиваются; в направлении фо-
веолы — уменьшаются, достигая 0,01 мм.
Согласно концепции H.Hartline (1940), так начинается сложный
путь передачи в мозг информации об основных качествах видимого
объекта, в частности его форме. На этой, третьей, ступени зритель-
ного опознания важную роль играет система фильтров различной
пространственной частоты, ориентации, формы. Они функциониру-
ют на уровне ганглиозных клеток сетчатки, наружных коленчатых
тел и в зрительной коре [Глезер В.Д., 1975; Campbell R.W., 1970,1972].
Пространственная дифференциация находится в тесной зависи-
мости от световой. Нейрофизиологические механизмы, включающие
способность к пространственной суммации и латеральному тормо-
жению (см. главу 3), позволяют проводить сравнительную оценку
количества света, падающего на какой-либо участок сетчатки, при
равномерной средней освещенности ее. Этим определяется констан-
тность восприятия белого как белого, а черного как черного, незави-
симо от освещенности. Эти же механизмы позволяют видеть раздель-
но два близко лежащих малых стимула, если между ними имеется до-
статочно глубокая темновая “впадина”. На остроту зрения, кроме
функции светоощущения, оказывают влияние уровень адаптации и
длительность экспозиции объекта.
Качество разрешения зависит и от сложности объекта. В экспе-
риментах, проведенных Ю. Е. Шелепиным и F. W. Campbell, установ-
лено, что для узнавания лица человека требуется участие не менее 5
описанных выше элементов дискретизации рецептивных полей, что
соответствует разрешающему углу зрения в 5'. Лимитирующим фак-
тором, по мнению авторов, является необходимость учитывать осо-
бенности строения глаз человека, в том числе расстояние между зрач-
ками. При наблюдении с расстояния 30—35 м оно составляет в изоб-
ражении на сетчатке 6'. Еще сложнее оценить выражение лица: для
этого необходимо как минимум 8—9 гексагональных элементов дис-
кретизации. Матрицы с таким радиусом расценены авторами как оп-
тимальные для распознавания сложных объектов. Они позволяют при
минимальной нейронной организации видеть максимально удален-
ный стимул.
Казалось бы, большой диаметр ретинальных рецепторов должен
34
создавать ограничения для целостного восприятия протяженных
объектов. Однако мозаичное расположение рецепторов в сетчатке и
наличие нервных механизмов их взаимодействия не только предот-
вращают восприятие фактической прерывистости изображения, но
и создают предпосылки к так называемой гиперостроте зрения.
Мерой остроты зрения служит тот минимальный зрительный угол
(в секундах, минутах, градусах), которым в зависимости от задачи
исследования удается оценить еще различаемые глазом простран-
ственные величины. Чем дальше от глаза расположена эта порого-
вая для различения величина, тем меньше угол, под которым она
видна. На практике для характеристики чувствительности соответ-
ствующих рецепторов чаще используют систему относительных по-
казателей, обратных пороговым, выраженных в дуговых углах. Кро-
ме того, учитывая возрастные нормы, результаты исследования ост-
роты зрения иногда оценивают в процентах сохранности этой
функции.
Ступени форменного зрения Нередко стимулом для фиксации взо-
ра на каком-либо объекте оказывается его внезапное появление в поле
зрения или его продолжающееся движение. Таким образом, сигнал с
периферии сетчатки, которая с помощью палочковой рецепции спо-
собна ответить на вопрос: “Где?”, заставляет повернуть глаз в нуж-
ном направлении, и при включении колбочкового аппарата дать от-
вет на более сложный вопрос: “Что?”.
В этом едином процессе можно выделить три ступени зрительно-
го анализа структуры, ориентации и формы предметов: 1) способ-
ность заметить присутствие объекта (minimum visible); 2) способность
разглядеть структуру объекта в деталях (minimum separable или
minimum resolvable) и 3) способность опознать, идентифицировать
зрительный образ в соответствии с ранее известными представлени-
ями об объектах внешнего мира (minimum cognoscible). На третьей
ступени также анализируется относительное взаиморасположение
видимых образов (spatial minimum discriminable) и оценивается их
форма по контуру (minimum deformable).
Minimum visible — способность заметить стимул минимального
размера, нарушивший непрерывность обозримого гомогенного про-
странства, характеризует абсолютный порог форменного зрения.
Давно известно, что линейные предметы легче заметить, чем точеч-
ные. Так, черный волос толщиной 0,12 мм можно увидеть на белом
фоне с расстояния до 12 м, т.е. при зрительном угле 2" [Burchardt D.,
1869], но если на листе белой бумаги изобразить точку сходного диа-
метра (0,1 мм), то она будет заметна только с 0,6 м, при этом порого-
вый зрительный угол увеличится до 16". Если рядом с первой точ-
кой поместить еще несколько таких же точек с интервалом 0,1 мм, то
с прежнего расстояния уже не удастся различить промежутки между
ними. Для того чтобы увидеть раздельно каждую из этих точек, нуж-
но поднести лист бумаги еще ближе к глазу (до 0,16 м), при этом по-
35
MINIMUM VISIBLE
Рис. 2.2. Схема, поясня-
ющая преимущества яр-
костных характеристик
объекта (1") перед про-
странственными в проис-
хождении minimum visible
[Westheimer G., 1987].
роговый зрительный угол увеличивается до Г, но это уже следую-
щая ступень форменного зрения — собственно острота зрения
(minimum separable).
Парадоксальная способность замечать протяженный объект, тол-
щина которого меньше разрешающей способности колбочковых ре-
цепторов [Westheimer G., 1979], объясняется следующим образом
(рис.2.2). Тончайшая линия шириной 1" по дуге, спроецированная
оптической системой глаза на равномерно освещенной сетчатке, мо-
жет стать видимой, поскольку благодаря расфокусировке ее изобра-
жение достигает значительной ширины. Однако еще важнее контра-
стность. Линия, оставаясь инвариантной по форме, будет заметна
только в том случае, если яркостные различия (Д1) между ее изобра-
жением и адаптационным фоновым освещением сетчатки (I) достиг-
нут пороговой для восприятия контраста величины (AI/I). Таким об-
разом, minimum visibile представляет собой скорее яркостный, чем
пространственный порог, поэтому используется при исследовании
контрастной чувствительности и поля зрения.
Minimum separable — способность разглядеть структуру объекта
или собственно острота зрения. При оценке простой дифференциаль-
ной остроты зрения за единицу измерения принимают минимальный
зрительный угол, характеризующий не размеры всего видимого
объекта, а размер самых малых промежутков, которые еще обеспе-
чивают раздельное видение с данного расстояния близко располо-
женных одна к другой мелких деталей объекта или соседних точек.
Для наблюдателя без патологии зрения в условиях наилучшей
фокусировки минимальный угол разрешения находится в пределах
36
от 30" до Г. Полагают, что усилению этой функции препятствуют,
с одной стороны, ограниченные возможности оптики, с другой — ней-
рональная организация сетчатки в макулярной области глаза чело-
века.
Первыми простейшими оптотипами для определения minimum
separable были полосчатые знаки, состоявшие из чередующихся чер-
ных и белых линий. Эти “решетки” позволили F. W. Campbell и
D. Green (1965) оценить относительную роль оптических и ретиналь-
ных факторов в обеспечении разрешающей способности глаза. Дру-
гим оптотипом были пары квадратов разных размеров с увеличи-
вавшимися промежутками между ними. Однако отсутствие выбора
(каждый раз предъявляли два квадрата) привело к необходимости
изменения этого теста.
Предложенный Снелленом еще в 1862 г. и усовершенствованный
Пфлюгером трехзубчатый оптотип (по типу буквы Е) находит при-
менение и в настоящее время, особенно в детской практике.
Попытки избежать влияния общей конфигурации знака на раз-
личение его деталей привели к созданию так называемой миры Голь-
дмана (рис.2.3). Однако наиболее удачным и признанным во всем
мире оказался оптотип, предложенный Landoldt. Этот оптотип в виде
разорванного на одном из участков кольца имеется и в отечествен-
ных конструкциях таблицы Сивцева.
Minimum cognoscible — способность опознать зрительный образ,
оценив, в частности, его пространственную локализацию. Включаю-
щаяся при этом сверхвысокая острота зрения (hyperacuity) характе-
ризует minimum deformable. Зрительные образы, как простые, так и
сложные, опознаются благодаря не только непосредственному вос-
приятию, но и зрительной памяти человека, а также его умственному
развитию. При исследовании этой интегральной функции обязатель-
но используют психофизические методы.
Набор наиболее простых оптотипов для оценки интегральной ост-
роты зрения у взрослых был предложен J. Casanovas (1958). В набор
входили 4 одинаковые по площади фигуры: звезда, круг, крест и квад-
рат. В тестах, используемых при исследовании зрения у детей, применя-
ют силуэты хорошо знакомых им предметов и домашних животных.
Буквенные и цифровые оптотипы относятся к числу достаточ-
но сложных и не являются оптимальными по точности тестами, хотя
они и получили широкое распространение во всем мире, так как
удобны для контакта с пациентом и дают возможность объективи-
зировать правильность ответов. На результаты исследования ока-
зывает влияние также количество одновременно предъявляемых
тест-объектов.
Способность опознать объект в целом через оценку формы, раз-
меров, ориентации (minimum cognoscible) в отличие от способности
просто заметить присутствие объекта (minimum visible), предпола-
гает видение достаточно четко очерченных контуров. Это “чувство
37
Em epi
a
6
a
Рис. 2.3. Тесты для оценки простой дифференциальной остроты зрения
(minimum separable).
а — «крючки» по Снеллену и Пфлюгеру; б — миры Гольдмана; в — кольцо Ландоль-
та в модификации Вербицкого; г — оригинальное кольцо Ландольта; д — кольца в
модификации Поляка (для целей экспертизы); е — точечно-штриховые оптотипы Вол-
кова для исследования зрения астигматического глаза [Волков В. В. и др., 1976].
линии”, умение отличать правильную окружность от неправильной,
прямой угол от тупого, слегка изогнутую линию от прямой высоко
ценятся в чертежном деле, у дизайнеров, художников, а способность
быстро определить по контурам марку самолета, тип корабля исклю-
чительно важна в военном деле. Существуют тесты, позволяющие
оценить способность к такой “гиперостроте зрения”. Для определе-
ния minimum deformable чаще всего применяют метод проверки но-
ниусной, или верньерной, остроты зрения. Адекватный тест имеется
в таблицах, используемых в США.
От испытуемого требуется указать, на каком отрезке креста име-
ется ступенеобразное смещение прямой линии. У людей с нормаль-
38
Таблица 2.1. Классификация разновидностей понятия остроты зрения в
соответствии с критериальной оценкой (по G. Westheimer, 1974, с некоторыми
модификациями В. В. Волкова)
Оцениваемый показатель Характеристика разновидностей остроты зрения
Критерий функции Минимум ви- димого Минимум разрешае- мого (обычная ост- рота зрения) Минимум прост- ранственно опозна- ваемого, в том числе «гиперсотрота зре- ния»
Решаемая задача Определение присутствия или отутст- вия мишени Определение наи- меньших промежут- ков между элемен- тами стимула, т. е. выявление деталей в зрительной мишени, отличающих ее от других мишеней Определение отно- сительного взаимо- расположения ви- димых деталей, по- зволяющих опознать объект
Формирова- ние типич- ного вопроса при «сило- вом» выборе Имеется ли в поле зрения линейный объ- ект? Как он расположен? Имеется ли в коль- це разрыв? Если да, то где он располо- жен? Верхняя линия рас- положена справа или слева от нижней
Физиологиче- ский базис Локальный яр- костный диф- ференциаль- ный порог (Al) относитель- но фона Наличие яркостных различий между не- сколькими смежны- ми локальными сти- мулами Расположение ка- ких-либо локальных признаков относи- тельно двух или бо- лее надпороговых зрительных образов
Методики измерения Объекты раз- ной величи- ны Объекты или рас- стояние между ними, промежутки между деталями объектов или детали Вариации располо- жения различных об- разов относитель- но друг друга
Величина нор- мальногопорога Около 1" по дуге Около 30” по дуге Около 3" по дуге
Степень влия- ния размыто- сти изобра- жения Умеренная Высокая Низкая
39
ным зрением порог такой пространственной оценки образа уклады-
вается в интервале от 2" до 10", что существенно ниже порога обыч-
ной остроты зрения. Так как для просто остроты зрения имеются
ограничения как оптические, так и анатомо-физиологические, оче-
видно, что процессы пространственной локализации образа обеспе-
чиваются более сложными нервными механизмами.
Таким образом, на различных ступенях форменного зрения реша-
ются различные задачи, что определяет неоднозначность применяе-
мых методов измерения остроты зрения и оценки получаемых резуль-
татов (табл.2.1).
2.1.2. Контрастная чувствительность
Способность улавливать минимальные различия в освещенности двух
соседних областей, а также дифференцировать их по яркости, опре-
деляется контрастной чувствительностью.
Пространственная контрастная чувствительность (ПКЧ) зритель-
ного анализатора является функцией, которая определяет минималь-
ный контраст, необходимый для обнаружения изображений различ-
ных размеров. Она отражает зависимость порогового контраста от
пространственной частоты стимула.
ПКЧ определяется как величина, обратная минимальному кон-
трасту решетки, при котором последняя различима при данной про-
странственной частоте. ПКЧ имеет другую природу, нежели ост-
рота зрения, хотя в известной мере и связана с последней. Если
при снижении остроты зрения вообще теряется возможность опре-
делить ПКЧ на высоких частотах, то сохранение полной остроты
зрения отнюдь не исключает снижения ПКЧ на тех же высоких
частотах.
Острота зрения отражает минимальную величину различимых
глазом символов, имеющих максимальный контраст с фоном. Недо-
статком показателей остроты зрения является их одномерность.
Включение оценки контрастной чувствительности позволяет решать
двухмерные проблемы, а с учетом вариабельности внешней освещен-
ности — и трехмерные.
Подсчитано [Prager Th.C., 1990], что если исследовать остроту зре-
ния в темном помещении, то контраст черных знаков, предъявляе-
мых на белом экране, оценивается в 94%, если же в комнате вклю-
чить освещение, контраст знаков в тестовой таблице снижается
до 31%.
Поскольку порог контрастной чувствительности является фун-
кцией размеров объекта, графически ее можно выразить в виде
кривой контрастно-частотной чувствительности (рис. 2.4). Метод
исследования ПКЧ назван визоконтрастометрией [Волков В.В.,
1985].
Для оценки этой функции используют черно-белые решетки с
40
Пространственная чувствительность
Рис. 2.4. Частотно-контрастная характеристика зрительной системы чело-
века [Wolfe J. М., 1990].Схематически показано, как функция контрастной
чувствительности складывается на основе действия четырех гипотетических
каналов, отличающихся по пространственной частоте пропускаемых частот.
плавным синусоидальным профилем изменения яркости. Толщина
полос, определяющая их пространственную частоту, выражаемую
количеством черно-белых циклов на градус, варьирует. Решетки раз-
личаются также по контрасту, который плавно меняется, например
сверху вниз от 0 до 1 (100%).
Синусоидальный профиль решетки признан предпочтительным,
несмотря на сложность ее изготовления, поскольку, во-первых, со-
гласно теореме Фурье, в серию синусоидальных волн может быть
преобразован любой волнообразный, в том числе прямоугольный,
комплекс, во-вторых, всякая дефокусировка, уменьшая контраст
таких волн, не нарушает их форму, что никак нельзя сказать о
буквенных оптотипах, используемых для проверки остроты зре-
ния.
Имеет значение и величина рецептивных полей ганглиозных кле-
ток сетчатки. В том случае, если размеры их адекватны волновому
профилю тестирующей решетки, то сила ответа увеличивается. Не-
соответствие профиля рецептивных полей волновому профилю те-
стирующей решетки при ее более высоких или низких простран-
ственных частотах приводит к ослаблению ответа [Dowling J.E. et
at, 1984].
В условиях обычного дневного освещения контрастная чувстви-
тельность в норме особенно высока при опознании стимулов сред-
ней пространственной частоты (порядка 4—5 цикл/град). В случае
увеличения яркости объекта сдвиг максимальной чувствительности
происходит в направлении высоких, а с увеличением размеров объек-
та — низких пространственных частот.
41
По мере смещения проекции изображения решеток от центра к
периферии глазного дна контрастная чувствительность на высоких
и средних пространственных частотах, подобно остроте зрения, рез-
ко снижается. Для решеток 20 цикл/град отмечена линейная зависи-
мость этого падения от степени эксцентриситета. Длительное наблю-
дение какой-либо одной из пространственных частот, как и засвет,
снижает чувствительность соответствующего фильтрующего канала;
на фоне этой “адаптирующей” частоты может происходить повыше-
ние чувствительности к другим частотам.
Обнаружены также явления как подпороговой суммации эффек-
та двух близких по частоте решеток [Graham N. et al., 1971], так и
подавления восприятия (“маскировка”) одних частот другими
[Olzak L., Thomas J.P., 1981; Wilson H.R. et al., 1983].
Рецептивные поля ганглиозных клеток наружных коленчатых тел,
таламуса и зрительных нейронов коры обеспечивают функциониро-
вание нескольких каналов, предположительно настроенных на 6—8
пространственных частот и обладающих избирательной чувствитель-
ностью к той или иной ориентации решеток. При воздействии мель-
кающим стимулом количество функционирущих каналов уменьша-
ется в два раза (до 3—4) [Manfield R.J., 1974, 1978; Watson A. et al.,
1981].
Нейрофизиологические основы контрастной чувствительности*
В данном разделе рассмотрена функция пространственной контраст-
ной чувствительности, которая отражает восприятие стимулов, по-
роговых по пространственной частоте, временным параметрам и кон-
трастности. В основе нейрофизиологических процессов такого вос-
приятия лежит структура сетчатки с ее различными по размерам
рецептивными полями в центре и на периферии, со структурой само-
го рецептивного поля, с конкурирующими зонами возбуждения и тор-
можения, со структурой связей отдельно взятой ганглиозной клетки,
с процессами пространственной суммации, присущими рецептивным
полям.
На основании психофизических экспериментов с использовани-
ем синусоидальных и прямоугольных паттернов R.W. Campbell еще
в 1968 г. было высказано предположение, что зрительная система
состоит из множества параллельных каналов-фильтров, каждый из
которых чувствителен к определенным пространственным частотам,
т.е. имеет свою полосу пропускания.
Сложная структура коленчатых тел и более высоких отделов мозга
предопределяет более сложные нейрофизиологические механизмы
пространственной контрастной чувствительности к различным про-
странственным частотам. Каждый нейрон центральных отделов зри-
тельного анализатора реагирует на оптимальную для него простран-
ственную частоту. Так, нейроны Х-типа, располагающиеся в зритель-
ной коре и связанные в основном с макулярной областью сетчатки,
относятся к каналам средней и высокой пространственной частоты;
42
Y-нейроны имеют большие размеры, им не свойственно комбиниро-
вать возбуждающие и тормозящие сигналы с рецептивных полей по
линейному закону. В отличие от Х-нейронов, они лучше реагируют
на стимулы низкой пространственной частоты. Рецептивные поля
многих клеток зрительной коры имеют продолговатую форму и, сле-
довательно, вытянуты в том или ином направлении. R.J.W. Manfield
(1978) полагает, что на восприятие горизонтально ориентированных
решеток “настроены” клетки горизонтальной, а для вертикальных
решеток — вертикальной ориентации; предполагается наличие кле-
ток и косой ориентации.
Таким образом, для нормального зрительного восприятия окру-
жающего мира необходимы не только высокая острота зрения, но и
полноценные пространственно-частотные каналы контрастной чув-
ствительности, которые обеспечивают фильтрацию высоких частот,
информирующих о мелких деталях объекта, низких, без которых не-
возможно восприятие целостного образа даже при различимости мел-
ких деталей, и средних, особенно чувствительных к контрастам и со-
здающих предпосылки для качественного высокочастотного анали-
за контуров предметов. В связи с этим проверка контрастной
чувствительности по низкочастотным (0,72 цикл/град) буквенным те-
стам РеШ — Robson или по высокочастотным (10 цикл/град) тестам
D. Regan, A. Holladay и I. Bailey-Lovie не обеспечивает той полноты
информации, какую предоставляет визоконтрастометрия во всем
диапазоне пространственных частот. Знаки в первой таблице Pelli —
Robson слишком велики, а во второй,— наоборот, малы, чтобы с их
помощью можно было оценить функцию той области, в которой на-
ходится физиологически и клинически наиболее важный пик кон-
трастной чувствительности.
2Л.З. Устойчивость к слепящим засветам
Некоторые пациенты при нормальной, казалось бы, остроте зрения,
установленной по традиционным таблицам, т.е. при высоком кон-
трасте тестовых знаков и в условиях сниженной освещенности поме-
щения, как только оказываются на ярком дневном свету, жалуются
на ’’слепимость”. Здоровым людям знакомо это неприятное ощуще-
ние, возникающее иногда при взгляде на рефлектирующую морскую
гладь или сверкающий белизной снежный покров. В этих случаях
из-за отсутствия на глазном дне контрастирующих различий между
яркостью изображения объекта наблюдения и яркостью фона труд-
но увидеть предметы, хорошо различимые в иной ситуации (напри-
мер, высоко летящий самолет в голубом безоблачном небе, встреч-
ную машину, ослепляющую фарами, и т.п.). Установлено, что более
2000 лет назад эскимосы Аляски и Сибири применяли защитные очки
из слоновой кости с горизонтальными щелочками для защиты от
ослепления.
43
t,c
Рис. 2.5. Зависимость времени (t) восстановления остроты зрения (цифры
на кривых) от яркости фона адаптации (L) и энергетической экспозиции зас-
вета (Н) [Преображенский П. В. и др., 1986].
Ослепление при засветах (glare-effekt) в руководстве для инжене-
ров-осветителей, изданном в США в 1984 г., определяется как ощу-
щение, вызываемое появившимся в поле зрения светом, более силь-
ным, чем тот, к которому глаз адаптирован, и выражающееся в
зрительном дискомфорте, снижении видимости или потере работо-
способности.
В исследованиях, проведенных L.L. Holladay (1926), G. Fry,
М. Alpem (1953), Е. Wolfe (1960), было уставленно, что первопричи-
на зрительных нарушений лежит в усилении “паразитного” свето-
рассеяния в средах глаза (особенно на участках, на которых наруше-
на прозрачность роговицы или хрусталика). Вследствие этого в при-
сутствии слепящего источника света уменьшается контрастность
ретинального изображения в сравнении с реальным, в результате чего
снижается острота зрения. Результаты экспериментов, выполненных
D. Miller, свидетельствуют, что катаракта уже в начальных прояв-
лениях (вакуоли, водяные щели и т.п.), отек эпителия и помутнение
стромы роговицы не столько задерживают, сколько рассеивают свет.
Слепящим, однако, может быть не только боковой (“точечный”) ис-
точник света, например фары встречной машины, но и прямой диф-
фузный засвет по всему полю зрения, также приводящий к сниже-
нию контрастности изображения на сетчатке и уменьшению четкос-
44
ти видения деталей. С учетом этих двух вариантов слепящего дей-
ствия и конструируют приборы для определения устойчивости к за-
светам.
Иную природу имеет ’’временное ослепление”, связанное с обесц-
вечиванием зрительных пигментов под действием сверхъярких источ-
ников света, в частности светового излучения при ядерном взрыве, и
снижение зрения при нарушении темновой адаптации [Преображен-
ский П.В. и др., 1986]. На рис.2.5 представлена двухфакторная зави-
симость времени (t) восстановления остроты зрения от яркости фона
адаптации (L) и энергетической экспозиции засвета (Н). Верхняя плос-
кость фигуры соответствует остроте зрения, которая устанавливает-
ся на фоне адаптации (заштрихована зона, в которой острота зрения
не достигает 0,05).
На рисунке отражены не воздействия ярких источников света в
процессе зрительного восприятия, а их последствия, хотя обычно и
кратковременные, которые приводят к снижению центрального зре-
ния в связи с возникновением интенсивного последовательного об-
раза, что можно определить как ’’фотостресс” для зрительной систе-
мы (см. главу 3) [Кравков С.В., 1950; Луизов А.В., 1983; Schober Н.,
1964; Sliney D., 1994, и др.].
2.2. ОСНОВНЫЕ МЕТОДЫ ФУНКЦИОНАЛЬНЫХ
ИССЛЕДОВАНИЙ
2*2.1* Методы оценки остроты зрения
Скрининговые (ориентировочные) тесты. Маленькому ребенку пока-
зывают две карты: одну — со структурированным изображением,
вторую — с гомогенным серым полем. Предпочтительное внимание
к первой свидетельствует о наличии форменного зрения [Hywarinen L.,
1988].
Испытуемому, в том числе в целях экспертизы, предъявляют дви-
жущийся экран с изображением клеток шахматной доски или черно-
белых решеток. По минимальным размерам деталей, еще вызываю-
щих оптокинетический нистагм, ориентировочно (но методологичес-
ки объективно) судят об остроте зрения.
Пациенту с помутнением оптических сред через две линейные
диафрагмы, установленные перед зрачком, в глаз направляют мало-
интёнсивное излучение гелий-неонового лазера. Образующееся на
глазном дне изображение интерференционной решетки можно варь-
ировать, изменяя расстояние как до источника света, так и между
диафрагмами. По минимальной еще различаемой ширине полос су-
дят о потенциально возможной после устранения помутнений остро-
те зрения.
45
Еще более простыми и надежными методами оценки форменно-
го зрения (после устранения помутнений сред) являются тест При-
мроза и аутоофтальмоскопия по Пуркинье. Первый выполняют с по-
мощью палочки Меддокса, формирующей на глазном дне из света,
направленного через нее в зрачок, прерывистую светящуюся полосу,
направление которой и должен определить обследуемый. При исполь-
зовании второго метода цель больного — уловить непростую для вос-
приятия картинку теней от мельчайших извилистых сосудов, подхо-
дящих к макуле со всех сторон в виде веточек, сеточки, крошек, шаг-
рени, при определенных условиях просвечивания глазного яблока
(через верхнее или нижнее веко прикрытого глаза). Источник света
(например, склеральную лампу) необходимо слегка смещать из сто-
роны в сторону.
При остроте зрения, не позволяющей видеть самые крупные зна-
ки обычных таблиц, прибегают к показу одиночных знаков в виде
полосчатых черных тестов Тибоде— Поляка (на "белом фоне) или
пальцев врача на темном фоне. Учитывают максимальное расстоя-
ние, с которого пациенту удается определить количество показывае-
мых пальцев. Если это расстояние равно 0,5 м, то остроту зрения обо-
значают как 0,01, если это 1м — 0,02,1,5 м — 0,03 и т.д.
В том случае, если обследуемый не может правильно сосчитать
показываемые пальцы, находящиеся у самого лица, выясняют, уда-
ется ли ему определить направление перемещаемого перед глазом
предмета (видение движения), например руки врача. При положи-
тельном результате остроту зрения обозначают как 0,001. Если при
мутных средах такой пациент с расстояния 60 см от глаза сможет уви-
деть два световых пятна от фонариков, находящихся на расстоянии
12 см один от другого, можно предположить, что у него сохранена
функция макулы [Kanski J., 1989].
Наконец, если невозможно решить и эту задачу, то выясняют,
отличает ли пациент свет от тьмы. “Зайчик” от зеркального офталь-
москопа периодически направляют в глаз больного и следят за его
ответами. При ответах невпопад зрение расценивают равным нулю
(амавроз — абсолютная слепота).
Уточняющие тесты. Устройства для традиционных клинических
исследований. Поскольку обычно необходимо иметь представление
об остроте зрения при достаточной освещенности, по мнению
С.В. Кравкова (1934,1945), предпочтительно применение знаков чер-
ного цвета на белом фоне.
Для проверки остроты зрения вдаль широко используют настен-
ные таблицы (по типу таблиц Снеллена, Сивцева и др.). Обычно они
равномерно освещаются спереди (около 700 лк), реже — транспаран-
тно (на просвет). В этих таблицах применяют буквенные или цифро-
вые стимулы; они удобны при обследовании пациента, однако не-
достатком этих стимулов являются их различия по узнаваемости зна-
ков.
46
Наилучшим из известных тестов, используемых для исследова-
ния minimum separable уже более 100 лет, остается тест в виде колец
Ландольта. Единая форма теста почти исключает влияние на показа-
тели обследуемого minimum visible и minimum cognoscible. Толщина
черной полосы, из которой образуется кольцо, выбрана с таким рас-
четом, чтобы она, как и разрыв в кольце (индекс), составляла 1' по
дуге с расстояния, указанного на боковом поле таблицы. При этом
внешний диаметр кольца (базис) с того же расстояния должен состав-
лять 5', а внутренний — 3' (дуговые минуты).
В нашей стране исследование остроты зрения вдаль обычно про-
водят с расстояния 5 м, за рубежом — чаще с 6 м (20 футов). Для того
чтобы при рассмотрении с расстояния 5 м кольцо удовлетворяло при-
нятым условиям, т.е. все оно составляло бы 5', а индексируемый раз-
рыв — 1', его линейные размеры в десятом ряду таблицы должны со-
ставлять 6,25 и 1,25 мм соответственно. Если обследуемый различает
место разрыва в данном кольце именно с 5 м, а в более мелких коль-
цах без приближения к таблице это ему не удается, пороговым углом
разрешения для него и является угол в 1'.
В отечественной офтальмологии это обозначается как vi-
sus = 1,0, за рубежом обычно в числителе указывают и расстоя-
ние, с которого проводилось исследование, а именно 6/6 (или в
футах 20/20). Такую остроту зрения принимают за нижнюю грани-
цу нормы.
Благодаря включению в таблицу рядов с более крупными и мел-
кими кольцами создаются условия для определения как более высо-
кой, так и более низкой остроты зрения без изменения расстояния.
Установив тот ряд, в котором обследуемый распознает разрывы в
наименьших по диаметру кольцах, считывают указанную на полях
таблицы остроту зрения. Этот показатель представляет собой соот-
ношение двух расстояний: того, с которого ведут проверку, к тому,
на котором разрывы в знаках данного ряда видны под углом в 1'. По
принятой в нашей стране системе данный показатель преобразуется
в десятичную дробь, а за рубежом сохраняются значения обеих дис-
танций: в числителе — реально используемой, в знаменателе — рас-
четной, т.е. той, с какой данный тест различим наблюдателем с нор-
мальной остротой зрения.
Проекторы испытательных знаков более удобны в работе офталь-
мологов, так как снабжены дистанционным управлением, позволя-
ют при необходимости демонстрировать знаки отдельными строч-
ками, вращать тест-объект и плавно изменять его размеры. Вместе с
тем их слабым местом является значительное влияние на результаты
исследования оптико-световых характеристик прибора и колебаний
внешней освещенности.
Высокая острота зрения вдаль не означает наличие высокой ост-
роты зрения на близкие расстояния. Наоборот, при высокой остроте
’ зрения сильно близорукого глаза вблизи не удается достичь высокой
47
остроты зрения с коррекцией вдаль. Причина этого, очевидно, в не-
совершенстве применяемой коррекции: обычные очки при высокой
диоптрийности и неизбежном отдалении от роговицы минусового
стекла из-за конструкции оправы резко уменьшают размеры изобра-
жения на глазном дне. Следовательно, специальная проверка остро-
ты зрения для определенных расстояний вблизи так же нужна, как и
проверка остроты зрения вдаль.
Для проверки остроты зрения вблизи чаще всего применяют на-
бор пробных текстов для чтения, в которых различные угловые раз-
меры букв рассчитаны на определенные расстояния от глаза. В на-
шей стране распространены шрифты, оценку по которым проводят с
расстояния 30 см от глаз. Безошибочно читаемый текст характеризу-
ет остроту зрения вблизи.
Известны портативный транспарантный аппарат для проверки
зрения вблизи (ПОЗБ-1), сконструированный в Государственном
оптическом институте им.Вавилова, специальный буклет с набором
тестов для исследований с различных расстояний вблизи (40 см, 1 м),
изданный в 1982 г. в ГДР М. Заксенвегером. Последний представля-
ет большую ценность для исследования остроты зрения и ее коррек-
ции применительно к строго определенному расстоянию от глаза,
неодинаковому в разных профессиях. Ряд тестов имеет вид нот, гео-
графических карт, чертежей и пр.
Оценка результатов исследования. Как в любых
других психофизических испытаниях, при проверке остроты зрения
предъявление объектов, начиная от надпороговых, ведет к заниже-
нию результатов (так называемая ошибка привычки), а начиная от
подпороговых — к завышению результатов (так называемая ошиб-
ка антиципации). Однако эти вариации не столь значимы при исполь-
зовании стандартного объекта (кольца Ландольта) с возможным
выбором 4, а лучше 8 позиций разрыва в кольце.
Для клинических целей за порог достаточно принять 75% пра-
вильных ответов.
С помощью множественных психофизических замеров можно
определить не только средние значения минимального разрешаемо-
го угла (МРУ), но и их стандартную ошибку (АМРУ). По закону Ве-
бера ДМРУ является постоянной частью МРУ:
МРУсредн.
Исходя из этого, для клинической оценки остроты зрения при-
годна и логарифмическая шкала (табл.2.2), если размеры знаков от
ряда к ряду меняются в геометрической прогрессии. В таких log MAR-
таблицах (log. min. acuity resolution), распространенных сейчас в
США, в каждом из 13 рядов имеется только по 5 равноузнаваемых
букв, предложенных L. Sloan (1951), а также J. Bailey-Lovie (1976).
48
Таблица 2.2. Способы спецификации зрения. Эквиваленты остроты зре-
ния в различных обозначениях (по G. Westheimer)
Минималь- ный разре- шаемый угол, дуговые минуты Острота зрения по таблице Снеллена Зрительная эффективно- сть по Shell—Sterling, % Десятичная дробь по Снеллену и Сивцеву Острота зрения в логарифми- ческих единицах по отношению 20/20
футы метры
0,5 20/10 6/3 109 2,0 0,3
0,75 20/15 6/4,5 104 1,33 0,1
1,0 20/20 6/6 100 1,0 0
1,25 20/25 6/7,5 96 0,8 “0,1
1,5 20/30 6/9 91 0,67 -0,18
2,0 20/40 6/12 84 0,5 -0,3
2,5 20/50 6/15 76 0,4 “0,4
3,0 20/60 6/18 70 0,33 -0,5
4,0 20/80 6/24 58 0,25 -0,6
5,0 20/100 6/30 49 0,2 “0,7
6,0 20/120 6/36 41 0,17 -0,78
7,5 20/150 6/45 31 0,133 -0,88
10,0 20/200 6/60 20 0,1 “1,0
20,0 20/400 6/120 3 0,05 -1,3
В наборе имеются три таблицы раздельно для правого и левого глаз
и для подбора очков [Ferris F. et al., 1982].
Н. Snell и J. Sterling (1925) предложили интересный подход к ана-
лизу остроты зрения, который основан на представлении о процен-
тах утраты функций в результате болезни или травмы. Авторы уста-
новили, что любое увеличение МРУ на 1' можно рассматривать как
снижение эффективности зрения до 84% от установленной ранее.
Исходя из этих представлений, при снижении остроты зрения с 1,0 до
0,5, функция зрения ухудшается не в 2 раза, а только на 16%, а при
снижении с 1,0 до 0,1 — не в 10 раз, а на 80%.
Показатели остроты зрения зависят от многих факторов, что при-
вело к созданию большого количества модификаций устройств и
методик, используемых для ее оценки.
Устройства для определения и оценки остроты зрения при реше-
нии специальных задач. Для дифференциальной диагностики нару-
шений остроты зрения, возникающих при макулярной патологии или
в зрительно-нервных проводящих путях, эффективен тест в виде сет-
ки Амслера. Каждый ее квадрат виден с расстояния 30 см под углом
49
Рис. 2.6. Зависимость ос-
троты зрения от дефоку-
сировки типичного глаза.
По оси ординат — острота
зрения, определенная по таб-
лицам Снеллена, представле-
на логарифмической шка-
лой; по оси абцисс — дефо-
кусировка в диоптриях [Lau-
ranceL., 1926].
в 1 °. Патогномоничная для ретинальных изменений метаморфопсия
выявляется без особого труда: определенные звенья сетки представ-
ляются пациенту извилистыми или изогнутыми (метаморфомы), часть
из них может выпадать (скотомы).
Аметропия как причина нарушения фокусировки изображений
на глазном дне отрицательно влияет на остроту зрения. Зависи-
мость остроты зрения от степени дефокусировки представлена на
рис.2.6. График, однако, не отражает особенности остроты зрения
при астигматизме, который является основной причиной ее мери-
диональных вариаций. Последние с высокой точностью можно
оценить с помощью астигмовизометров [Волков В.В., 1976], кото-
рые позволяют определить разрешающую способность астигматич-
ного глаза в каждом из двух главных сечений. Высокой точности
измерений дают возможность добиться точечно-штриховые опто-
типы, “уравновешенные” по принципу оптотипов Вейнберга. При-
мер регистрации результатов исследования: “0,8—0,3 (90°)”, в скоб-
ках указан меридиан, к которому относится минимальная острота
зрения.
Крайние варианты изменений величины зрачка, такие как мид-
риаз (более 6 мм) или миоз (менее 2 мм), отрицательно сказывают-
ся на остроте зрения даже эмметропического глаза. Если же остро-
та зрения снижается и при умеренном расширении зрачка (в пре-
делах 4—5 мм), то это свидетельствует о наличии аметропии.
Применение искусственного зрачка (диафрагмы диаметром 2—3
мм из набора стекол) окажет корригирующее действие и явится
свидетельством необходимости подбирать очки [Кириллов Ю. А.,
1974].
50
В довольно широком диапазоне фотопического освещения (от
уровня в яркий солнечный день и до такового в полнолунную ночь)
острота зрения мало меняется, но, как можно видеть из таблицы
Г. С. Ландсберга (1952), при освещенности, сниженной до 0,1 лк и
более, “угол разрешения” начинает быстро увеличиваться. Правда,
по мере адаптации к сумеркам острота зрения вновь более или менее
быстро восстанавливается. Для исследования сумеречной остроты
зрения созданы специальные приборы, в частности никтометр Ком-
берга, мезоптометр и др. Адаптация остроты зрения к темноте после
предварительной трехминутной дезадаптации к довольно яркому
(порядка 7000 апостильб) свету внутри прибора в норме наступает
в течение около 1,5 мин.
Для оценки способности различать мелькающий объект иссле-
дуется так называемая экспозиционная острота зрения (ЭОЗ). С
уменьшением времени предъявления тестирующего объекта порог
различения возрастает. Эта зависимость особенно выражена во
временном интервале 15—100 мс [Колбанов В.В., 1968]. Однако и
в этом случае возможна компенсация за счет усиления яркости
объекта. Для практических измерений ЭОЗ создан специальный
прибор на основе диапроектора “Этюд” с фотообъективом [Со-
мов Е. Е., Левин Б. М., 1989].
Способность различать детали движущегося объекта называют
динамической остротой зрения. Прибор, сконструированный М.Г.Ко-
зырьковой (1968) для измерения этой функции, напоминает большой
периметр, по дуге которого предъявляется объект (кольцо Ландоль-
та), движущийся с заданной скоростью. Преследование взором в те-
чение 0,5 с мишени, двигающейся с умеренной скоростью (несколько
градусов в секунду), может обеспечить остроту зрения, близкую к
нормальной. Строгая стабильность ретинального изображения, как
известно, не является обязательным условием для оптимального раз-
решения, хотя во время быстрых саккад и при очень быстрых пере-
мещениях изображений по сетчатке зрительные функции ослабе-
вают.
Способность длительно сохранять четкость видения объекта, по-
добранного на пределе разрешающей способности глаза, называ-
ют устойчивостью ясного видения [Ferree С., Rand G.,1913]. На ки-
мографе регистрируют периоды, когда на протяжении 3-минутно-
го непрерывного наблюдения возможность различать разрыв в
кольце Ландольдта исчезает и появляется вновь. В итоге сопостав-
ляют суммарную продолжительность периодов видения и невиде-
ния, а также частоты перерывов и дают оценку профпригодности
испытуемого.
В психофизических исследованиях продуктивности зрительной
работы во времени используют корректурные таблицы Weston, со-
стоящие из большого набора колец Ландольдта с разной ориентаци-
ей разрыва. Обследуемый должен в пределах отведенного времени
51
выбрать кольца определенной ориентации. Продуктивность зрения
(V) оценивают по формуле
у _ Nxn
Nxt ’
где п — количество правильных решений; N — общее количество ко-
лец с заданной ориентацией разрыва; t — время, затраченное обсле-
дуемым.
Тест может быть полезным при оценке зрительного утомления и
эффективности медикаментозной стимуляции зрительной работы
[Вайнштейн Б.И., 1972 ].
В интересах экспертной практики (для выявления симуляции и
диссимуляции) и просто для более детальной оценки изменений ост-
роты зрения предложены таблицы с уменьшенным “шагом” между
рядами [Холина А.А., 1924; Розенблюм Ю.З., 1961; Кириллов Ю.А.,
1972]. Особый интерес представляют контрольные таблицы Б.Л.По-
ляка (1948). В отличие от обычных таблиц в них, во-первых, предна-
меренно нарушена правильность взаимоотношений между рядами,
во-вторых, изменены взаимоотношения размеров базиса и индекса,
принятые для колец Ландольта. Для уточнения остроты зрения у сла-
бовидящих служит набор таблиц Н.В.Серпокрыл (1965). В практике
ВТЭК за норму времени на демонстрацию одного знака принимают
4—5 с, а при обычной клинической работе — 2—3 с.
Т.С.Егоровой (1987) разработаны таблицы подбора средств кор-
рекции для близи лицам с пониженным зрением (от 0,01 до 0,8), с
помощью которых исследуется острота зрения по изолированным
буквенным знакам с расстояний 20,25,33 и 40 см, при этом угловой
размер знака изменяется в геометрической прогрессии с шагом 1,25.
Таблицы позволяют также определять минимальный размер читае-
мого обследуемым типографского шрифта (порог чтения) и скорость
чтения непрерывного текста. В обоих случаях используется литера-
турный полужирный шрифт, размер которого обозначается в “пунк-
тах” и системе “М” единиц.
2.2.2. Методы исследования контрастной чувствительности
Поскольку рецептивные поля зрительного анализатора подобны
системе локальных фильтров разных пространственных частот и ори-
ентаций, лучшими тестовыми стимулами для их возбуждения слу-
жат разноконтрастные черно-белые или хроматические синусоидаль-
ные решетки, т. е. с размытыми контурами, разных направлений и
ширины.
Первой попыткой создания методики для клинической практики
были таблицы, разработанные G.B.Arden (1983). Таблицы (карты)
различались шириной полос, а контраст в них постепенно изменялся
52
Рис. 2.7. Кривая, отражающая контрастную чувствительность во всем диа-
пазоне видимых пространственных частот (ахроматических) при централь-
ной фиксации и фотопической освещенности, полученная с использованием
«Атласа по визоконтрастопериметрии» [Волков В. В. и др., 1987].
вдоль полос: от 100% по нижнему и практически до 0 по верхнему
контуру таблицы. Соответственно модуляционную чувствительность
по логарифмической шкале трактовали, наоборот, как минимальную
(1 отн. ед.) внизу и как максимальную (более 100 отн. ед.) наверху.
Контраст решетки, или глубину модуляции (К), рассчитывают по
формуле [Michelson С., 1891]:
Lmax-Lmin _ 110-90 _ 20
Lmax+Lmin 110 + 90 200
где L — интенсивность (яркость) стимулов.
Модуляционная контрастная чувствительность как величина, обрат-
ная уровню контраста (в данном примере 0,1), составит 10 отн. ед.
Кривая контрастной чувствительности во всем диапазоне види-
мых пространственных частот при центральной фиксации и фотопи-
ческой освещенности представлена на рис.2.7. Стрелкой показан ма-
ленький участок кривой, информацию о которой получают с помо-
щью обычного исследования остроты зрения по Снеллену, т.е. в
условиях высокого контраста.
Основоположником метода исследования ПКЧ в нашей стране
является Ю. Е. Шелепин, по инициативе которого В. Б. Макуловым,
В. Н. Пауком, Ю. И. Левковичем и В.В. Волковым в 1985 г. были
синтезированы синусоидальные изображения тестовых решеток, на
их основе издан “Атлас и пособие по визоконтрастопериметрии” [Вол-
ков В.В. и др. 1987]. Атлас включает отдельные карты, которые раз-
личаются по пространственной частоте решеток и по сочетанию цве-
тов (черно-белые, красно-белые, зелено-белые и сине-белые). В каж-
дой карте при постоянной частоте контраст плавно изменяется, осла-
бевая по направлению снизу вверх. Частота решеток варьирует от 0,37
до 18 цикл/град. Кривая ПКЧ, представляемая на регистрационном
бланке, строится по оси абсцисс в логарифмических единицах.
Для упрощения анализа результатов исследования контрастной
чувствительности Ю.Е. Шелепин предложил ’’выпрямить” кривую
нормы, обозначив для каждой пространственной частоты процент
сохранности зрительных функций в норме, что при патологических
процессах будет изменяться соответственно локализации и степени
его выраженности.
Исследование проводится с расстояния 1,5 м, так как при эммет-
ропической рефракции глаз фокусируется именно на это расстояние
в условиях относительного покоя аккомодации у большинства взрос-
лых людей [Волков В.В., Колесникова Л. Н., 1972]. При аметропии в
пробную оправу вставляют корригирующие линзы, подбираемые с
учетом расстояния до таблиц (на 0,5—0,75 дптр слабее для миопов и
сильнее для гиперметропов в сравнении с их оптимальной коррекци-
ей для дали). Исследование выполняют монокулярно. Карты, поме-
щаемые в аппарат Рота, равномерно освещают и предъявляют в про-
извольном порядке. Тыльной стороной неиспользуемой для тести-
рования карты прикрывают тестируемую. Постепенно, сдвигая
первую вдоль полос второй, в направлении от невидимого (нулево-
го) контраста, определяют момент обнаружения полосчатости. По
боковой шкале определяют пороговый показатель контрастной чув-
ствительности в процентах сохранности относительно нормативов
для данной частоты. При использовании описанного метода для по-
лучения точных результатов на тестирование каждой карты необхо-
димо затрачивать не менее 2 с.
Возможен вариант совмещения в единственной карте решеток,
различающихся как по частоте, так и по контрасту, который реали-
зован в буклете, применяемом для самопроверки зрения [Волков В. В.,
Лебехов П. И., 1993]. Обследуемый должен на прозрачной бумаге,
наложенной на тест-карту, прочертить линию по границе, где теря-
ется структура решетки; полученный результат сопоставляется с ре-
зультатами контрольной схемы.
Предложен также вариант размещения решетчатых тестов, раз-
личающихся по контрастности и частоте, на поле единой настенной
таблицы по типу таблицы Снеллена [Ginsburg А.Р., 1984]. Однако
для этого пришлось уменьшить общие размеры каждой решетки, что
привело к невозможности использования широкополосных тестов,
так как разместить на малой площади требуемые 6 полос было не-
возможно. Методика значительно упростилась, так как не требова-
лось экранировать тесты и искать границу видимого с невидимым,
54
каждый тест постоянен не только по частоте, но и по контрастности.
Правильность ответов обследуемого подтверждалась указанием на ори-
ентацию решетки (вертикально, горизонтально, с наклоном верхних
концов от вертикали вправо и влево). В сходных картах, выпускаемых
фирмой “Vistech” (Dayton), каждый знак размещается таким образом,
что виден с расстояния 3 м под углом всего Г, и при центральной фикса-
ции периферические области сетчатки не стимулируются.
Разработана сходная методика исследования контрастной чув-
ствительности с использованием проектора знаков для проверки зре-
ния вдаль, в котором предъявляется серия слайдов, упакованных в
диск [Волков В.В. и др., 1995].
Первая программа для компьютерной визоконтрастометрии [Ше-
лепин Ю.Е., 1985] в нашей стране реализована для прибора “Эрго-
тест” с видео и регистрирующим устройствами. Прибор работает в
пространственно-частотном диапазоне от 0,4 до 18 цикл/градус, в
пределах контрастов от 0,03 до 0,9% (при разрешающей способности
дисплея 320х 1024 элемента).
Тест для оценки временной контрастной чувствительности
[Lachenmayr, Drance S., 1992] используют при исследовании функции
макулярной зоны. Нормальная кривая этой функции, как и простран-
ственно-частотной функции по Де Ланге, имеет вид параболы. Если
по оси абсцисс обозначить частоту мельканий (цикл/с), а по оси ор-
динат — изменения контраста вспышки (в величине модуляций), то
наибольшая чувствительность и минимальный порог определяют на
средних частотах (7—8 мельканий в 1 с).
Следующим этапом развития метода исследования пространствен-
ной контрастной чувствительности было создание оригинальной ком-
пьютерной программы “ZEBRA” [Шамшинова А. М. и др., 1996,
1997], в которой вертикальные и горизонтальные синусоидальные ре-
шетки выводятся на монитор IBM-совместимого компьютера (VGA
и выше) без использования дополнительных устройств. Программа
предусматривает “накопление кривой” КПЧ для нормального зре-
ния (эталон) в конкретных условиях исследования, благодаря чему
не требуется фотометрирования помещения и монитора. В отличие
от “Атласа по визоконтрастопериметрии” как ахроматические, так и
хроматические решетки различной пространственной частоты (0,5—
22 цикл/градус) предъявляются на черном фоне экрана. Контраст
плавно увеличивается вдоль направления решетки от минимального
0,2% до максимального — 100%, обеспечиваемого данным монито-
ром в принятых фотопических условиях адаптации (средняя освещен-
ность комнаты 10 — 12 лк, яркость экрана 40 — 60 кд/м2). При рас-
стоянии до экрана 2 м и размере изображения 18 см поле стимуля-
ции составляет 5,4°. Для каждой пространственной частоты обследу-
емый устанавливает шторку (которая прикрывает область высокого
контраста), плавно перемещаемую вдоль решетки, в положение по-
рогового различения полос. По результату исследования строится гра-
55
Контрастная чувствительность, усл. ед.
Рис. 2.8. Кривые контрастной чувствительности на цветовые и ахромати-
ческую синусоидальные решетки на черном фоне в норме [Белозеров А. Е.
и др., 1996].
1 — синяя; 2 — зеленая; 3 — красная; 4 — белая.
фик зависимости контрастной чувствительности, равной единице (по-
роговый контраст), от пространственной частоты решетки, а также
график ее сохранности (в дБ), показывающий, во сколько раз кон-
трастная чувствительность испытуемого на различных частотах от-
личается от эталона. Использование этой программы позволяет по-
лучить данные о цветовой контрастной чувствительности наряду с
ахроматической, выявить доли участия цветовых фоторецепторов в
ПКЧ, а также уточнить особенности кривых ПКЧ для той или иной
локализации патологического процесса. В норме наблюдаются ва-
риации ПКЧ (рис.2.8), причем кривые, регистрируемые при исполь-
зовании ахроматического и хроматического паттернов, сходны во
всем диапазоне пространственных частот, за исключением сдвига
максимума кривой ПКЧ на черно-синий паттерн в сторону низких и сни-
жения чувствительности в области высоких пространственных частот.
К настоящему времени методика визоконтрастометрии достигла
такого совершенства, что определенно превосходит обычную визо-
метрию по выявляемое™ различных зрительных нарушений.
56
На основании анатомии и нейрофизиологии сетчатки, наличия
параллельных on- и off-каналов колбочковой системы, участвую-
щих в обработке и передаче информации о контрасте, в последнее
время предложен новый метод исследования топографии контраст-
ной чувствительности в каждой заданной точке поля зрения, кото-
рый позволяет дать функциональную характеристику световых (on)
и темновых (off) каналов колбочковой системы сетчатки (on- и off-
активности колбочковой системы), в норме и при различной пато-
логии [Шамшинова А.М. и др., 1996,1997]. Исследование осуществ-
ляют следующим образом: на экране монитора по схеме предъявля-
ют ахроматические стимулы размером 2 и 3 мм в зависимости от
экцентриситета на сером фоне (8 кд/м2) в любой необходимой для
исследования точке поля зрения (в пределах 40°). Яркость стимулов
ступенчато нарастает от более темных, чем фон, до более
светлых, с шагом 0,5 кд/м2. Различие в яркости стимула и фона
(яркостный контраст) оценивают по величине времени сенсомотор-
ной реакции (СМР), изменение которой регистрируют в процессе
уравнивания яркости стимула и фона. В норме при предъявлении
стимулов, близких к фону по яркости, имеется зона, где время СМР
значительно больше, чем при видимом различии стимула и фона
(при уравненных по яркости стимуле и фоне время реакции беско-
нечно). По мере увеличения различия между стимулом и фоном в
сторону как более темных, так и более светлых стимулов время
СМР симметрично и равномерно уменьшается и выходит на плато.
Данные в каждой исследуемой точке поля зрения представляются
в виде гистограммы, отражающей зависимость времени СМР от
яркости стимула (возможно усреднение данных в любых участках
поля зрения). Для центральной зоны в 2—5° используют стимул в
2 мм. По мере удаления от центра к периферии величина стимула
должна быть увеличена до 3 мм и более, что связано с увеличением
размеров рецептивных полей сетчатки от центра к периферии. При
стимуляции центральных и парацентральных зон сетчатки стиму-
лами малой величины, светлее или темнее, чем фон, возникает ре-
акция в рецептивных полях ганглиозных клеток с on- и off-центра-
ми, сигнализирующими об освещенности на определенных участ-
ках сетчатки, в сравнении с их ближайшим окружением (фон), что
и является основанием для исследования топографии контрастной
чувствительности в каждой точке поля зрения.
2.2.3. Методы оценки устойчивости к слепящим засветам
Тесты на ослепление основаны на измерении зрительных функций
в присутствии слепящего источника света. Ослабление зрения при
этом является функцией позиции и интенсивности источника ослеп-
ления и рассеивающих свойств собственных оптических структур
глаза.
57
Для исследования ослепления (glare) предложено несколько уст-
ройств:
Прибор Аветисова — Вергасовой(\99\) создан на основе фотопро-
ектора ПО-1, в центре круглого экрана которого экспонируется изо-
бражение колец Ландольта (с угловым размером, соответствующим
остроте зрения 0,1) переменного контраста. Источник слепящего света
от галогеновой лампы окружает тестирующий экран. Оценивается
пороговая яркость слепящего засвета, которая препятствует разли-
чению колец Ландольта на каждом уровне контраста между тесто-
вым объектом и фоном.
Тестер яркостной остроты зрения (Brightness Acuity Tester —
ВАТ). В приборе предусмотрены три степени яркости засвета, произ-
водимого в тот момент, когда пациент ведет наблюдение за табли-
цей для проверки остроты зрения вдаль через отверстия диаметром
12 мм в гемисфере диаметром 60 мм с диффузно-белой внутренней
поверхностью. Самый высокий уровень засвета (400 фут-Лб) эквива-
лентен бликам от песка на морском пляже, средний уровень (100 фут-
Лб) подобен яркости листвы в солнечный день и слабый (12 фут-Лб)
по яркости напоминает флюоресцирующий свет.
Тестер слепимости Миллера — Надлера, В качестве теста в при-
боре использованы слайды, на которых изображены черные кольца
Ландольта, соответствующие остроте зрения 20/400 (0,05). По мере
смены слайдов в проекторе фон, на котором предъявляют кольцо и
положение разрыва в кольце могут изменяться. Тест окружен слепя-
щим экраном, яркость которого (420 фут Лб) эквивалентна яркости
свежего снега в солнечный день. Слепимость оценивают в процентах
по таблице, прилагаемой фирмой к прибору.
Vistech VCT-8000. В качестве теста используют синусоидальную
решетку, находящуюся в компактной наблюдательной системе тес-
тера. Ослепление создают путем усиления яркости ореола вокруг те-
ста либо включения центрального осветителя, имитирующего свет
фары приближающегося автомобиля.
Eye Con 5. Сменные оптотипы (кольца, буквы, решетки) предъяв-
ляют на мониторе с расстояния 3,6 м. Они окружены слепящим по-
лем, яркость которого (130 кд/м2) эквивалентна яркости облачного
дня.
При сравнительной оценке прогностических возможностей ука-
занных устройств в отношении “слепимости” пациентов с началь-
ной катарактой в различных условиях пребывания вне дома установ-
лена значительная положительная корреляционная связь между про-
гнозируемой и реальной “внедомашней” остротой зрения [Prager Т.С.
et al., 1989]. При освещении средней яркости исследование с помо-
щью ВАТ оказалось наиболее точным. Если использовать очень силь-
ные засветы, нередко имеет место гипердиагностика, в то время как
при засветах средней интенсивности гипердиагностика редка (30%) и
практически отсутствует (2%) гиподиагностика [Ferris F., 1982]. Ус-
58
тановлено, что слепимость оказывает более выраженное влияние на
контрастную чувствительность, чем на восприятие высококонтраст-
ных букв, т.е. на остроту зрения [Legge G. et al., 1987].
Большинство исследователей полагают, что оценка ослепления с
помощью приборов у части людей с высокой “кабинетной” остро-
той зрения может быть полезна при решении вопроса о лечебной так-
тике при начальной катаракте и выборе способа коррекции близору-
кости (кератотомия, очки, контактные линзы).
Принципиально иную цель преследуют, когда изучают возмож-
ность восстановления остроты зрения, сниженной вследствие дез-
адаптации рецепторов сетчатки при ее сверхъярких засветах. Устой-
чивость центрального зрения к такого рода ослеплениям (к фото-
стрессу) может быть проверена на никтометре Комберга или
макулореадаптометре Балашевича — Шостака [Волков В.В. и др.,
1976].
В литературе имеется немало описаний и более простых вариан-
тов “фотостресстеста”, когда для засвета используются обычный
электроофтальмоскоп или световой карандаш [Товкач В.И., 1977].
Сначала проверяют наилучшее корригируемое зрение для дали,
затем с расстояния 3 см в течение 10 с (или с 8 см в течение 30 с)
воздействуют указанным источником света через зрачок на сетчат-
ку. О реакции судят по времени восстановления остроты зрения.
Исследования проводят поочередно на каждом глазу. У пациента с
макулярной патологией время восстановления остроты зрения пре-
вышает 50 с (иногда достигает нескольких минут), при патологии
зрительного нерва оно обычно не выходит за пределы нормальных
значений. Фотостресстест способствует ранней диагностике пато-
логии макулярной области сетчатки, не обнаруживаемой офталь-
москопически (отек при кистовидной дегенерации, серозном хорои-
ретините и др.).
Остроту зрения, контрастную чувствительность и слепимость, по-
добно многим другим функциям организма, необходимо оценивать
с учетом возрастных вариантов нормы (рис.2.9).
Рассматривая перспективные направления усовершенствования
визоконтрастометрических исследований, следует заметить, что на-
ряду с попытками создать идеальные тесты только для визометрии, в
частности оптотипы, лишенные низкочастотной гармонической со-
ставляющей [Волков В.В. и др., 1987; Коскин С. А., 1994], или только
для контрастометрии — буквенные оптотипы разной контрастнос-
ти, но одинаковой пространственной частоты [РеШ D., Robson J. et
al., 1988], сохраняется тенденция к разработке приемов интеграль-
ной оценки всех факторов, обеспечивающих максимум достоверной
зрительной информации.
J.Thaung с соавт. (1995) на основе теоретического анализа дан-
ных других авторов и результатов собственных модельных экспери-
ментов подтвердили, что устойчивость к светорассеянию в глазу, или
59
а б
Рис. 2.9. Возрастные нормативы остроты зрения.
а—для новорожденных [Dobson V., Teller D., 1978]; б — для взрослых [Weymouth,
1960]. По оси ординат — острота зрения по Снеллену, по оси абсцисс — возраст в ме-
сяцах (а) и годах (б).
LSF (light scattering factor), зависит от многих факторов. L. Paulsson,
J. Sjostrand (1980) выразили эту зависимость формулой:
LSF = L/E х (М/М—1),
где L — усредненная яркость мишени в пространстве; Е — осве-
щенность, создаваемая слепящим источником света; Mj и М2 — мо-
дуляции пороговых контрастов в присутствии и в отсутствиислепя-
щего источника света соответственно.
В приведенную формулу требуется, однако, вводить коррективы.
Во-первых, большую роль, особенно при начальных нарушениях
прозрачности хрусталика, играет геометрия стимула. Если картй с
решетками занимают на глазном дне 5—6°, то буквы всего 5—6*. При
этом распределение яркости на сетчатке, а следовательно, и соотно-
шение реальных контрастов стимула и ретинальных контрастов бу-
дут различными, поэтому при использовании тестовых решеток ав-
торы рекомендуют рассчитывать контраст на глазном дне по Ми-
хельсону, а в случае применения буквенных тестов — по Веберу (см.
главу 3).
Во-вторых, согласно данным М. Abrahamsson и J. Sjostrand (1986),
необходимо учитывать возможность влияния темновой адаптации
при использовании стимулов с малой яркостью (порядка 2 кд/м2), т. е.
в сфере мезопического зрения.
J. Thaung и соавт. (1995) рекомендуют применять в клинической
практике стимулы достаточно высокой яркости (более 10 кд/м2), про-
водить исследование на низких пространственных (2 цикл/град) и
временных (1 Гц) частотах с выбором оценки контраста по Михель-
сону или Веберу в зависимости от размеров стимула. В соответствии
с законом Вебера при низкой пространственной и временной частоте
60
контрастные пороги становятся независимыми от освещенности сет-
чатки. При умеренно высокой пространственной частоте по закону
Де Врие — Розе контрастные пороги обратно пропорциональны квад-
ратному корню от величины ретинальной освещенности [Kelly D.H.,
1972].
2.3. КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ НАРУШЕНИЙ ОСТРОТЫ
ЗРЕНИЯ, КОНТРАСТНОЙ ЧУВСТВИТЕЛЬНОСТИ
И УСТОЙЧИВОСТИ К ОСЛЕПЛЕНИЮ С УЧЕТОМ
ОСОБЕННОСТЕЙ ИХ ЭТИОЛОГИИ И ПАТОГЕНЕЗА
2.3.1. Основные варианты и причины нарушений остроты зрения в
клинической практике
• Снижение центрального зрения только вдаль, корригируемое оп-
тическими средствами, является следствием распространенных раз-
ного рода аметропий.
• Снижение центрального зрения только вблизи, устраняемое опти-
ческими средствами,— следствие нарушения функций аккомода-
ции, в том числе возрастной пресбиопии.
• Снижение центрального зрения вдаль при одновременном улуч-
шении его вблизи у людей пожилого возраста связано с миопиза-
цией в связи с набуханием хрусталика.
• Снижение центрального зрения, вплоть до его потери, не устраня-
емое оптическими средствами и независимое от расстояния: при
наличии на хуже видящем глазу гиперметропии, астигматизма, ко-
соглазия,— обычно свидетельствует об амблиопии.
• Снижение центрального зрения при офтальмоскопически выявля-
емых патологических процессах в макулярной области, и при их
отсутствии, но в случае предъявления больным жалоб на метамор-
фопсию или микропсию со снижением цветоощущения по типу три-
тадефицита, указывает на очаговое поражение в макулярной об-
ласти глазного дна.
• При наличии выявляемых офтальмоскопических признаков вос-
паления диска зрительного нерва и при их отсутствии, но при
предъявлении больным жалоб на центральную скотому (без мета-
морфопсии), сочетающуюся с нарушением цветоощущения, осо-
бенно по типу прото- и дейтердефицита, и со снижением контраст-
ной чувствительности на высоких пространственных частотах при
одностороннем процессе наиболее вероятен неврит или ретробуль-
барный неврит зрительного нерва, при двустороннем — оптико-
хиазмальном арахноидите или проявления осложненного застой-
ного диска.
61
• В отсутствие каких-либо нарушений в структурах глаза, но при
очень неустойчивой рефракции (обычно в сторону ее усиления) сни-
жение остроты зрения может быть признаком спазма аккомода-
ции или других невротических состояний (истерия).
• Стойкое снижение не только центрального, но и периферическо-
го зрения (без метаморфопсий или скотом) с ослаблением реф-
лекса с глазного дна, снижением ПКЧ сначала на низкие, а по-
зднее и на высокие пространственные частоты, наличием трита-
дефицита II степени, повышенной слепимостью от ярких засветов,
нередко является следствием выраженных нарушений прозрачно-
сти преломляющих сред глаза (роговицы, хрусталика, стекловид-
ного тела).
• Снижение центрального зрения, возникающее лишь в определен-
ных условиях при определенной освещенности и сохранении нор-
мальных его значений в обычной обстановке.
• Следствие самых начальных нарушений прозрачности хрустали-
ка (предкатаракта) или ограниченных помутнений роговицы
(в частности, после перенесенной кератотомии или фоторефрак-
ционной кератэктомии). По мнению Th.C. Prager, когда катарак-
та еще мало влияет на обычную остроту зрения, но пациента
беспокоит повышенная слепимость на ярком свету, есть показа-
ния для ускорения хирургического вмешательства по удалению
катаракты.
• При нормальной остроте зрения снижение контрастной чувстви-
тельности на средние пространственные частоты, наличие нару-
шений в парацентральном поле зрения, тритадефицит нередко яв-
ляются следствием достаточно развитой глаукомы.
2.3.2. Варианты и возможные причины изменения
пространственной контрастной чувствительности
при заболеваниях зрительной системы
При патологических изменениях зрительного анализатора всегда
отмечается снижение ПКЧ в той или иной пространственной час-
тоте в зависимости от локализации патологического процесса в
сетчатке, зрительном нерве, тракте или в зрительных центрах. Сни-
жение ПКЧ в диапазоне высоких, средних и низких пространствен-
ных частот происходит при поражении соответствующих им кана-
лов контрастной чувствительности от сетчатки до центральных от-
делов зрительного анализатора, а также при нарушениях оптики
глаза.
Для нормального функционирования контрастной чувствитель-
ности на высоких частотах первые ограничения возникают в случае
снижения центрального зрения. При школьной близорукости обыч-
ные корригирующие линзы не только повышают остроту зрения, но
и выравнивают до нормы показатели ПКЧ. У миопов, пользующих-
62
ся для коррекции контактными линзами и у перенесших кератото-
мию в связи с нарушением прозрачности роговицы высокочастот-
ные показатели ПКЧ нередко снижены, особенно в условиях пони-
женной освещенности [Николаенко В.П., 1994]. Снижение ПКЧ имен-
но на высоких частотах неизбежно во многих других клинических
ситуациях, когда сниженное центральное зрение не корригируется в
связи с нарушениями в зрительно-нервных путях (амблиопия, мио-
пическая болезнь, связанная с возрастом или наследственная дистро-
фия макулярной области, альбинизм и др.) и с изменением прозрач-
ности оптического аппарата глаза (выраженные помутнения хруста-
лика, помутнение его задней капсулы при афакии или артифакии,
рубцовые стадии кератоконуса, отек эпителия роговицы) [Шеле-
пин Ю. Е., 1985; Сомов Е.Е., 1992; Рубцова И.В., 1995; Hess R., 1977;
Zadnik К., Mannis М., 1987]. При указанных патологических процес-
сах механизмы нарушения зрительных функций различны, но сопро-
вождаются одним функциональным симптомом: нарушением про-
странственной контрастной чувствительности по высокочастотному
каналу на том или ином уровне зрительной системы.
На кривой ПКЧ могут наблюдаться ’’провалы” на отдельных
участках кривой [Эскина Э.Н., 1996]. Так, при альбинизме наряду с
отсутствием чувствительности на высокие пространственные часто-
ты и снижением чувствительности на средние частоты отмечен по-
чти вертикальный спад кривой в диапазоне 1,0—2,8 цикл/градус при
тестировании красно- и зелено-черным паттерном. При статической
периметрии в центральной и парацентральной областях поля зре-
ния при этом обнаруживаются скотомы.
Нарушения ПКЧ при амблиопии, обнаруженные и на втором гла-
зу с высокой остротой зрения, не исключают центральный генез ука-
занных нарушений [Шелепин Ю.Е. и др., 1985].
Пространственная контрастная чувствительность на низкие час-
тоты, пороговая чувствительность к которым и в норме невелика,
снижается уже при легком нарушении прозрачности преломляющих
сред глаза. Именно на них нарушается пространственная контраст-
ная чувствительность, несмотря на сохранность остроты зрения, при
самых начальных проявлениях кератоконуса [Carney J., 1982], отло-
жениях на контактной линзе [McClure D. et al., 1977], предкатаракте
[Рубцова И.В., 1995]. Это может быть объяснено тем, что при дефо-
кусировке или равномерном снижении освещенности всей сетчатки
из-за помутнения сред снижается четкость изображения низкочастот-
ного паттерна, который и без того не является адекватным раздра-
жителем для макулярной области. Во многих случаях нормализовать
контрастную чувствительность к низким частотам при начальных по-
мутнениях хрусталика удается при пользовании желто-оранжевыми
фильтрами, поглощающими коротковолновые излучения. При сни-
жении пространственной контрастной чувствительности на низкие
частоты, связанном с патологическими процессами, локализованны-
63
Контрастная OD
чувствительность, усл. ед.
OS
Контрастная
чувствительность, усл. ед.
Рис. 2.10. Кривые контрастной чувствительности у больной Б., 37 лет. Ди-
агноз — оптико-хиазмальный арахноидит. Острота зрения: 0,3 на оба глаза
не корригируется.
Обозначения такие же, как на рис. 2.8.
64
ми в центральных отделах зрительного анализатора, например, при
опухолях гипофиза с битемпоральной гемианопсией, такая коррек-
ция не может привести к желаемому результату.
Снижение ПКЧ в среднем диапазоне частот встречается при мно-
гих процессах, локализованных в макулярной области сетчатки и
зрительном нерве различного происхождения. Например, снижение
ПКЧ в области средних частот отмечено при начальной нестабили-
зированной глаукоме, при которой установлена тесная корреляция
этого показателя с уровнем светочувствительности в зоне Бьеррума,
выявленной при статистической периметрии, и периферическими
границами поля зрения [Симакова И.Л., 1996; Arden G., 1990]. При
нормальной остроте зрения снижение контрастной чувствительнос-
ти на средние пространственные частоты вместе с нарушением в поле
зрения и тритадефицитом часто являются следствием достаточно раз-
витой глаукомы.
ПКЧ может служить критерием улучшения проведения возбуж-
дения по зрительным путям. Так, нормализация офтальмотонуса
медикаментозным или хирургическим способом, как правило, сопро-
вождается улучшением ПКЧ, хотя сразу после гипотензивного вме-
шательства визоконтрастограмма на короткое время может быть
ниже исходного уровня.
Снижение ПКЧ во всем диапазоне частот отмечается при мно-
гих заболеваниях зрительного нерва и вышестоящих отделах зри-
тельного анализатора уже в начальных стадиях заболевания
(рис.2.10). Так, у больных с оптическим невритом (ОН) в стадии
обострения на фоне рассеянного склероза, при ОН другой этиоло-
гии, а также при оптико-хиазмальном арахноидите, при ретрохиаз-
мальных поражениях в зависимости от формы заболевания обнару-
живаются те или иные видеограммы в ответ как на предъявление
ахроматического, так и хроматического синусоидального паттерна.
Важное диагностическое значение могут иметь ’’завалы” в кривой,
отмечаемые в коротком диапазоне разных пространственных час-
тот, что, возможно, обусловлено избирательным поражением тех
или иных ’’частотных каналов”.
Изменения ПКЧ при многих патологических состояниях могут
быть первым признаком заболевания зрительной системы. Однако,
поскольку очень сходные нарушения могут возникать при самой раз-
нообразной патологии как зрительно-нервного, так и оптического
аппарата, многие исследователи [Волков В.В., 1996; Arden G., 1990;
Prager Th.,1990; Wolfe J., 1990] рекомендуют расценивать данные ви-
зоконтрастометрии как скрининговые, с последующим уточнением
происхождения снижения ПКЧ другими методами. Для детей до-
школьного возраста исследование пространственной контрастной
чувствительности затруднено [Сомов Е.Е. и др., 1992]. Однако совре-
менные компьютерные игровые программы для исследования ПКЧ
позволяют решить и эту задачу [Белозеров А.Е. и др. 1997].
65
2.3.3. Клинические примеры нарушений устойчивости зрения к
слепящим засветим
Слепящие эффекты от сильных источников света могут возникать в
глазу с нормально функционирующей зрительно-нервной системой.
Причины их возникновения обычно связаны с чрезмерным рассеива-
нием света в преломляющих средах глаза из-за локальных помутне-
ний. Причем нежные мелкоочаговые помутнения в роговице прино-
сят пациенту гораздо больше неприятностей, чем грубые и обшир-
ные бельма. Поэтому рубцы, возникающие после кератотомии,
помутнения роговицы в связи с кератоконусом являются потенци-
альным источником слепимости от засветов [Аветисов С.Э. и др., 1991;
Николаенко В.П., 1995]. Особенный дискомфорт возникает при оте-
ке эпителия роговицы, в то время как стромальный ее отек перено-
сится значительно легче [Muller D. et al., 1990]. Довольно частым
неблагоприятным фактором повышенной слепимости является на-
чальное помутнение хрусталика еще на стадии предкатаракты [Руб-
цова И.В., 1995]. После, казалось бы, успешного оперативного лече-
ния по поводу катаракты слепимость может возобновиться либо в
связи с вторичным помутнением остающейся в глазу задней капсулы
хрусталика, либо от некоторых деталей интраокулярной линзы: мест
крепления опорных элементов, отверстий для фиксации линзы, края
плохо центрированной линзы [Nadler D., 1990]. Оптические средства
коррекции зрения, в частности плохо адаптированные или загряз-
ненные контактные линзы, также нередко являются причиной повы-
шенной чувствительности глаза к яркому свету [Николаенко В.П.,
1995].
Слепящиц эффект возникает при определенном расстоянии меж-
ду зрительным стимулом и ярким источником света, он зависит от
характеристики источника, фона и окружающей освещенности, ус-
ловий предадаптации и многих других факторов. Пациенты обычно
довольно тяжело переносят слепящие засветы, не всегда определяя
суть возникающих неудобств.
ГЛАВА 3
СВЕТООЩУЩЕНИЕ И ЗРИТЕЛЬНАЯ АДАПТАЦИЯ
3.1. (Х1ЮВНЬШ ФОТОМЕТРИЧЕСКИЕ ПОНЯТИЯ И1ЦЦШ
Фотометрия основана главным образом на тех же принципах, что и
радиометрия. Мощность лучевого потока измеряется в ваттах (Вт),
световой поток — в люменах: 1 лм — световой поток от источника
света силой в 1 канделу (кд) в пределах одного стерадиана, т.е. пря-
мого угла определенной направленности, составляющего четвертую
часть шара. Однако если лучевая энергия поддается физическим из-
мерениям и может быть оценена в ваттах в секунду (Вт • с), т.е. в джо-
улях (Дж), то, выражая “световую энергию” (Q) в люменах в секунду
(лм • с), невозможно обойтись без корректив, основанных на психо-
физической оценке яркости излучений с различной длиной волн ви-
димого диапазона.
При фотометрии спектральную чувствительность глаза опреде-
ляют с помощью достаточно простых опытов (рис.3.1). На фоне (С),
равномерно освещенном, т.е. в условиях световой адаптации, или
равномерно затемненном, т.е. при темновой адаптации, обследуемо-
му предлагается подравнять по яркости два разноокрашенных
полуполя (А и В). Одно из них является эталоном, а мощность излуче-
ния другого (в Вт) изменяют до тех пор, пока их яркостные характе-
ристики полностью не совпадут. Этот процесс называют гетерохром-
ным яркостным сравнением. Количество ватт на единицу площади,
которое потребуется для выравнивания субъективного ощущения
яркости, и определит относительную роль длины волны (рис. 3.2).
В пределах видимого диапазона электромагнитного излучения,
как следует из рис. 3.2, а, мощность лучевого потока (в ваттах) по мере
удлинения волны неуклонно возрастает. Вместе с тем, судя по кривой б,
относительная яркость увеличивается только до уровня 555 нм (в гла-
зу, адаптированном к свету) или 510 нм (в глазу, адаптированном к
темноте), а дальше резко снижается. При комплексной оценке свето-
Рис. 3.1. Схематическое изобра-
жение устройства для измерения
яркости двух сравниваемых по-
луполей А и В на фоне С.
67
Рис. 3.2. Переход от единиц, используемых
в спектрорадиометрии, к единицам фотомет-
рии для оценки яркости [Hart W. М., 1987].
а — мощность лучевого потока в стандартном ос-
ветителе (в Вт); б — относительная яркостная эф-
фективность излучений различных длин волн в
субъективном восприятии; в — световой поток (в
лм), рассчитанный на основе произведения дан-
ных, представленных для а и б, соответственно для
каждой длины волны. По оси абсцисс — длина
волны, нм.
вой энергии (с переводом ее величины из
ватт в люмены) кривая светового пото-
ка (при сложении а и б) примет вид В.
При такой интегральной оценке всех
составляющих световая энергия (Q), по
W.Hart (1987), может быть выражена
формулой:
750
Q = 68sJva Ua Дл,
400
где Ux — спектральное распределение
лучевой энергии; Vx — коэффициент
функции фотопической яркости (с мак-
симумом 1,0 при длине волны 555 нм на
свету); 685 — константа пересчета вели-
чин в люменах на значение, выражен-
ное в ваттах (при длине волны 555 нм).
Таким образом, световой поток (F)
является фотометрическим эквивален-
том лучевого потока (Р); его измеряют
в люменах, оценивают по способности
проводимой световой энергии (лм • с)
вызывать зрительные ощущения.
Интенсивность света (I), которая
является фотометрическим эквивалентом интенсивности излучения
(J), оценивают по световому потоку в рамках прямого угла или сте-
радиана (стр) определенной направленности и выражают в канделах
(кд), рассчитываемых как количество люменов на стерадиан (лм/стр).
Освещенность (Е) — фотометрический эквивалент плотности из-
лучения (Н). Это световой поток, приходящийся на единицу площа-
ди, который измеряют в люксах (лк), рассчитываемых как количе-
ство люменов на квадратный метр (лм/м2).
Яркость (L) — наиболее распространенный фотометрический
критерий, понятие яркости эквивалентно понятию мощности излу-
6Я
чения. Это интенсивность света (I), излучаемого или отражаемого
источником, приходящаяся на единицу площади его поверхности.
Обычно яркость выражают в канделах на 1 м2 (кд/м2), но применяют
и другие единицы, любую из которых можно свести в конечном сче-
те к количеству кандел на единицу площади измеряемого источника
света.
Основные единицы, используемые в фотометрии.
Единицы интенсивности света (I):
1 кд = 1 лм/стр (1 стерадиан — единица прямого угла: со = Ч4 п сферы).
Единицы освещенности (Е):
1 люкс (лк) — метр-кандела= 1 лм/м2.
Единицы яркости (L):
1 нит (нт) = 1 кд/м2.
1 стильб = 1 кд/см2.
1 апостильб (асб) = 1 кд/л м2 (область круга радиусом 1 м).
1 ламберт (лб) = 1 кд/л см2 (область круга радиусом 1 см).
Единицы освещенности сетчатки:
1 троланд — освещенность сетчатки глаза, смотрящего на поверхность,
яркость которой равна 1 кд/м2 через зрачок, искусственно суженный до пло-
щади 1 мм2.
Для упрощения перерасчета одних единиц яркости в другие можно ис-
пользовать следующие данные:
1 нит = 0,3142 миллиламберт.
1 ламберт =10 000 апостильб.
1 ламберт = 0,318 стильб.
1 ламберт = 3183 нит.
Обычно применяемый в статической периметрии апостильб в 3 раза мень-
ше нита, в 10 000 раз меньше ламберта и в 30 000 раз меньше стильба. Виды
и единицы излучения, фотометрические единицы и их взаимотношения пред-
ставлены также в табл. 3.1.
3.2.0 МЕХАНИЗМАХ ЗРИТЕЛЬНОГО ВОСПРИЯТИЯ
И ФИЗИОЛОГИЧЕСКИХ ВОЗМОЖНОСТЯХ ЗРЕНИЯ
Процесс преобразования лучистой энергии, поглощенной рецепто-
рами сетчатки, в качественные зрительные образы обеспечивается
разнообразными физиологическими механизмами, о работе которых
позволяют судить многочисленные специальные психофизические на-
блюдения [Hart W., 1987 ]. Так, детально изучена адаптивная способ-
ность органа зрения к работе при очень низких и очень высоких уров-
нях освещенности. Зрительная система адекватно реагирует как на
быстрые, так и на медленные перепады лучистой энергии, причем
69
Таблица 3.1. Виды излучения и соотношения фотометрических
Единицы излучения
Вид Единицы
Энергия излучения (лучистая энергия) Поток излучения, лучистый поток (мощность излучения) Сила излучения (энергетическая сила света) Плотность излучения энергетическая освещенность, поверхностная плотность потока излучения Лучистость, яркость излучения, энергетическая яркость Количество облучения Джоули (Дж) Ватты (Вт)=Джоули/сек (Дж/с) Ватты/стерадиан (Вт/ср) 1 ср=1 ед. пространственного угла Ватты/метр2 (Вт/м2) Ватты/стерадиан /метр2 (Вт/ср/м2) Дж/м2
Примечание. Адаптировано no:J. С. Armington. The Electroretinogram. New York:
ронных приборов.— M.; Советское радио, 1971.
для нее характерна практически мгновенная реакция на быстро ме-
няющуюся обстановку. Своеобразная “редакторская” способность
позволяет зрительной системе избегать избыточной информации.
Хотя большая часть из того, что мы видим, представляет собой
отобранные порции так или иначе варьирующих изображений,
субъективно зрительный образ представляется цельным и плавно
меняющимся в пространстве и во времени. Видению стабильной
сцены способствует то, что формируемые на сетчатке изображения
одновременно суммируются и дифференцируются, оттеняются и
сглаживаются. При этом конденсируется наиболее полезная и от-
брасывается ненужная информация.
Функционально наиболее важными представляются различные
изменения (вариации образов), происходящие в пространстве и во
времени. В частности, большое значение имеет контраст световых
пятен, возникающих на сетчатке, изменения их локализации и вели-
чины. Зрительная система отвечает подчеркиванием границ и опре-
делением временных различий в их локализации. Она действует по-
добно дифференциатору, разделяя несопоставимые изображения де-
70
единиц
Фотометрические единицы
Вид Единицы
Световая энергия Световой поток Сила света (мощность свечи) Освещенность (характеристика освещаемой по- верхности) Яркость Освещенность сетчатки Количество освещения (световая экспозиция) Люмен-секунды (лм • с) Люмены (лм) (1 кандела излучает 4 л лм) Люмены/стерадианы (1 лм/ср=1 канделе=1 кд) 1 люкс (1 люкс=1 лк=1 лм/м2) Люмен/стерадиан/метр2 (1 лм/1 ср/1 м2= 1 кд/м2=1 нит) Троланд (1 кд/м2, видимый через 1 мм зрачка)=1 Т Лк* с
Academic Press, Inc, 1974; Ю. Г, Якушенков. Основы теории и расчета оптико-элект*
талей, и, подобно интегратору, группируя сходные ощущения. Су-
ществуют механизмы усиления светочувствительности путем уплот-
нения энергии в пространстве и во времени, а также механизма сопо-
ставления и экстраполяции деталей для заполнения зрительным со-
держанием пустоты, возникающей на месте скотом. В результате этого
зрительная система обеспечивает интерпретацию постоянно меняю-
щихся изображений внешнего мира на сетчатке, используя и про-
странственный, и временной базисы в непрерывных поисках инвари-
антных образов и упорядоченных отношений.
3.2.1. Физиологические регуляторы светового потока,
поступающего в глаз
Светочувствительность зрительного анализатора столь же вариабель-
на, сколь разнообразны пространственные и временные характерис-
тики световых раздражителей окружающего нас мира. Необходи-
мость адекватно воспринимать энергию как очень слабых, так и очень
сильных источников света, не подвергаясь необратимым структур-
71
ным повреждениям, обеспечивается способностью к перестройке ре-
жима работы рецепторов. На ярком свету световая чувствительность
глаза снижается, но вместе с тем обостряется реакция на простран-
ственную и временную дифференцировку объектов. В темноте весь
процесс происходит наоборот. Этот комплекс изменений как свето-
чувствительности, так и разрешающей способности глаза в за-
висимости от внешней (фоновой) освещенности и называют зритель-
ной адаптацией.
Диапазон освещенности, в пределах которого осуществляется
зрительная адаптация, огромен; в количественном выражении он
измеряется от миллиарда до нескольких единиц. Для удобства сопо-
ставления данных обычно манипулируют не этими цифрами, а их
десятичными логарифмами. В логарифмических единицах (лог.ед.)
пределы рассматриваемых границ делятся всего на десять уровней
(от 0 до 9), и тогда нулевому уровню будет соответствовать 1g 1, пер-
вому — 1g 10, второму — 1g 100 и т.д. до 9-го уровня, который соот-
ветствует 1g 1 000 000 000.
Известны три физиологических регулятора светового потока,
поглощаемого рецепторами сетчатки, т.е. ее светочувствительнос-
ти. Эти регуляторы различаются в скорости и силе. Первый: вслед-
ствие изменений размеров зрачка количество входящего в глаз света
регулируется в пределах лишь немногим более одной лог.ед. и с весь-
ма умеренной скоростью (примерно за 1 с). Второй значительно бы-
стрее (в пределах нескольких миллисекунд) и эффективнее влияет на
светочувствительную активность нервных элементов сетчатки, из-
меняя ее в тысячу раз, т.е. в пределах 3 лог.ед. Третий, хотя и весьма
медленно (на протяжении десятков минут), но в 100 000 000 раз, т.е.
в пределах 8 лог.ед., увеличивает светочувствительность при изме-
нении концентрации светочувствительных пигментов в рецепторах
и пигментном эпителии сетчатки.
3.2.2. Морфологические основы и субъективные проявления фото-
и скотопического зрения
Каждому известно, насколько беспомощным поначалу чувствуешь
себя, попадая из ярко освещенного помещения в темное. Только спу-
стя 8—10 мин начинаешь различать плохо освещенные предметы, а
для того чтобы достаточно свободно ориентироваться, требуется еще
по крайней мере 20 мин, пока зрительная чувствительность в темно-
те достигает максимума. Обратный процесс адаптации к высокому
уровню освещенности протекает заметно быстрее. Так, по выходе из
кинотеатра зрительный дискомфорт исчезает уже спустя 3—5 мин. В
первом случае — в процессе темновой адаптации проявляется ското-
пическое зрение, а во втором, при световой адаптации,— фотопичес-
кое. Анатомы, установившие, что в сетчатке ночных животных пре-
обладают палочки, а в сетчатке дневных — колбочки, заложили мор-
фологический базис для теории двойственного механизма зрения у
человека. Границы преобладания того или иного зрения определя-
ются уровнем яркости фона адаптации (УЯФА). Фотопическое зре-
ние реализуется при яркости фона больше 3 кд/м2; скотопическое —
меньше 10’3 кд/м2. При УЯФА в интервале от 10’3 кд/м2 до 3 кд/м2,
когда активны в равной мере и палочки, и колбочки, функционирует
мезопическое зрение.
Скотопически (темно) адаптированная сетчатка максимально
чувствительна к световой энергии самого низкого уровня, но при
этом резко снижается ее пространственная разрешающая способность
и исчезает цветоощущение. Фотопически (светло) адаптированная
сетчатка, будучи низкочувствительной, чтобы различать слабые ис-
точники света, вместе с тем обладает высокой пространственной и
временной разрешающей способностью, а также цветоощущением.
По указанным причинам даже в безоблачный день блекнет луна и
гаснут звезды, а ночью без подсвечивания мы теряем способность
читать текст, набранный даже самым крупным шрифтом.
По мере увеличения освещенности сетчатки зрение, определяе-
мое в основном палочковым аппаратом, сменяется колбочковым зре-
нием, причем максимум чувствительности сдвигается в направлении
от коротковолновой к длинноволновой части спектра. Этот фено-
мен, описанный J.E.Purkinje еще в XIX в., хорошо иллюстрируется
бытовыми наблюдениями. Кто не замечал, что в букете полевых цве-
тов в солнечный день выделяются желтые и красные маки, а в сумер-
ках — синие васильки (сдвиг максимума чувствительности от 555 до
510 нм). Изменения яркостной чувствительности глаза в отношении
не столько хроматического, сколько ахроматического света явились
предметом многих психофизических исследований. В них изучались
как пространственные, так и временные аспекты проблемы, причем
именно субъективные ощущения испытуемых с учетом их высокой
чувствительности и специфичности оказывались надежным инстру-
ментом познания.
3.23. Пространственные аспекты световой чувствительности
Абсолютная световая чувствительность и пространственная сумма-
ция. Светочувствительность различных зон нормально функциони-
рующей сетчатки далеко не одинакова. Максимальная плотность
палочек соответствует кольцевой зоне, Примерно в 7° от макулы. Не
случайно именно эта экстрафовеальная область наиболее чувстви-
тельна к очень слабым источникам света, когда лишенная палочек
фовеа находится в состоянии относительной депрессии. Так, тускло
мерцающая звезда в безлунном ночном небе лучше видна, если смот-
реть не прямо на нее, а слегка сдвинув взор. Абсолютная световая
чувствительность лежит в основе всех других функций зрения. Она
подвержена колебаниям в зависимости от очень многих внутренних
73
и внешних факторов, детально изучавшихся школой С.В.Кравкова.
Среди прочих факторов (питание, дыхание, кровоснабжение) особое
место занимают условия освещенности и адаптации. Световой фон
(внешняя освещенность) приводит в действие механизмы, перестраи-
вающие (адаптирующие) абсолютную световую чувствительность в
соответствии с этими условиями.
Абсолютная световая чувствительность определяется минималь-
ным количеством квантов света, достаточным для того, чтобы выз-
вать световое ощущение. Еще в 1875 г. A.Ricco обнаружил, что по-
рог светоощущения находится в обратной зависимости от размеров
светового пятна:
I/S = const,
где: 1п — пороговая интенсивность света; S — площадь светового сти-
мула.
Это отношение получило название “закон полной, или простран-
ственной, суммации”. Суть пространственной суммации состоит в
том, что пороговый эффект вызывается строго определенным чис-
лом квантов независимо от того, как плотно они распределены по
площади. Этому явлению на определенном этапе развития клини-
ческих методов исследования абсолютной световой чувствительно-
сти (адаптометрия, периметрия) уделяли особое внимание. Однако в
последние годы, в связи с тем, что закон Рикко имеет немало ограни-
чений, чаще прибегают к исследованию не абсолютной, а разностной
(различительной) чувствительности, определению дифференциаль-
ных порогов светоощущения в разных областях сетчатки.
Дифференциальная световая чувствительность (психофизическая
оценка прироста яркостной чувствительности). Для каждой области
сетчатки характерен тот или иной порог яркостно-контрастного вос-
приятия. Величиной, обратной порогу, как известно, является чув-
ствительность, в данном случае яркостная чувствительность. О по-
казателях ощутимого увеличения яркостной чувствительности мож-
но судить по результатам несложного эксперимента. В этом и многих
последующих психофизических экспериментах выделяется понятие
фонового светового поля и тестирующего светового объекта.
Интенсивность света в области тестирующего пятна складывает-
ся из суммарной энергии двух световых потоков, один из которых (I)
постоянно освещает фоновое поле, а второй в виде вспышки посту-
пает дополнительно (Д1) только на центральное тестирующее пятно.
Если в сумме I + AI добавка (Д1) слишком мала, то метка не будет
видна. При достаточно высокой силе вспышки можно добиться ви-
дения метки при каждом ее предъявлении. За пороговый уровень
принимают такое увеличение яркости, которое улавливается с веро-
ятностью 0,5, т.е. при каждом втором предъявлении пятна.
Отношение Вебера — Фехнера. Зрительный рецептор, как и лю-
бой другой сенсорный рецептор, обладает следующим свойством: чем
74
Рис. 3.3. Соотношение между яркостными порогами в разных областях сет-
чатки и адаптирующей фоновой освещенностью ее в 1g троландов у одного
и того же обследуемого [Baker Н., 1968].
1 — AI в 2° от фовеа, 2 — Д1 в 7° от фовеа. По оси ординат — Д1 (яркостный порог) в
log троландах; по оси абсцисс — 1g I адаптирующего поля.
слабее адаптационный фон, тем острее ощущается усиление стиму-
ла. При этом в определенных диапазонах энергии сохраняется по-
стоянное соотношение ощущаемого прироста стимула (Д1) к адапта-
ционному фону (I). Иными словами, Д1 представляет собой постоян-
ную долю от I.
Д1/1= const.
Подтверждающие это правило результаты психофизических на-
блюдений представлены на рис.3.3. Для палочковой области (в 7°
от фовеолы) отношение Вебера — Фехнера правомерно в более ши-
роком диапазоне интенсивности адаптационного светового фона
(в троландах), чем для колбочковой (в 2° фовеа). Как видно на
рисунке, “палочковая” кривая в темноадаптированном глазу начи-
нает подъем уже с отметки -2 (1g I), т.е. на подпороговом уровне.
Когда же адаптационный (фоновый) уровень интенсивности света
на сетчатке достигает отметки +1 (1g I), начинается подъем и “кол-
бочковой” кривой зрения. Прямолинейность последней в диапазо-
не по крайней мере 3 лог.ед. позволяет считать отношение Вебе-
ра— Фехнера справедливым в данных условиях и для функции
колбочек.
Левые (горизонтальные) части кривых свидетельствуют о том,
что в условиях полной темновой адаптации в сетчатке сумма I + Д1
поддерживается на постоянном уровне. При увеличении I показатель
Д1 даже несколько снижается (отмечается тенденция кривой опускать-
ся ниже горизонтального уровня). Таким образом, в условиях как
крайне низкой, так и чрезмерно высокой фоновой освещенности сет-
75'
Рис. 3.4. Полосы Маха. На границе двух различных по контрасту полей как
бы подчеркивается чернота одних и светлота других.
чатки возникают ограничения для применения правила Вебера —
Фехнера. По крайне правой части кривых можно прогнозировать
их дальнейший ход не столько вверх, сколько по горизонтали далеко
вправо, так что на них будут признаки утраты способности разли-
чать тестовые пятна от фона. При столь высоких уровнях световой
энергии возможна ее трансформация в тепловую, что вызовет повреж-
дение сетчатки.
Проверка дифференциальной световой чувствительности различ-
ных отделов сетчатки в условиях умеренной яркости фонового осве-
щения получила в последние годы широкое распространение в кли-
нической практике как метод статической периметрии.
Латеральное торможение. Еще одним адаптационным феноме-
ном, способствующим распознаванию зрительных образов, являет-
ся усиление контраста на границах различающихся по яркости зон
изображения (рис.3.4). Как полагают, полосы Маха являются след-
ствием нейрональных взаимодействий. В ответ на локальные вариа-
ции засветов соседствующих зон сетчатки происходит усиление про-
цессов возбуждения нервной активности в одних и процессов тор-
можения в других участках. В результате этого на границе темных и
светлых полей усиливаются чернота темного и белизна светлого, что
и получило название “латеральное торможение”.
Этот феномен можно наблюдать, если один из двух совершенно
одинаковых серых квадратов поместить в светлую рамку, а другой —
в темную. При этом квадрат, окруженный черной рамкой, будет ка-
заться более светлым: фоновая рамка по контрасту усиливает яркость
внутреннего квадрата.
Феномен Трокслера. К числу пространственно-временных меха-
низмов зрительной адаптации можно отнести феномен, выявленный
J.Troxler. Он подметил, что если на периферии поля зрения возника-
ет световое пятно, более интенсивное, чем световой фон, то в случае
неподвижности пятна уже через несколько секунд оно становится
невидимым. Чем слабее и меньше световое пятно, тем быстрее оно
исчезает, особенно при расположении на крайней периферии. Стоит
только пятну сдвинуться при изменении точки фиксации взора, как
оно вновь становится видимым. Мгновенное восстановление свето-
чувствительности предполагает нервный механизм феномена, а роль
выцветания пигментов фоторецепторов практически исключается
[Clarke F.J. et al., 1962]. С учетом находок D.Hubel и Т.Wiesel (1960),
касающихся градиента рецептивных полей сетчатки, которые в на-
76
правлении от центра к периферии закономерно увеличиваются в раз-
мерах, можно предположить наличие обратной зависимости вероят-
ного проявления феномена Трокслера от размеров этих полей. При
этом, как и в случае с физиологическим слепым пятном, ощущение
дефекта в пространстве при феномене Трокслера не отмечено.
Сходным по механизму представляется феномен "видения тени
ретинальных сосудов”, впервые описанный J.E.Purkinje. Феномены
Трокслера и Пуркинье позволяют утверждать, что любой образ, ко-
торый в условиях пристального взгляда неподвижен на сетчатке, по-
степенно теряет четкость и исчезает. Существует, однако, компенса-
торный механизм, поддерживающий в процессе зрительной работы
постоянные и незаметные микродвижения глазных яблок, что и обес-
печивает немедленное восстановление блекнущего образа. Эти дви-
жения состоят из медленных дрейфов и быстрых скачков с угловой
амплитудой не более нескольких долей градуса.
3.2.4. Временные аспекты светочувствительности
В зрительном процессе исключительно велика роль временного фак-
тора. Фотохимические изменения, с которых начинается преобразо-
вание световой энергии в зрительные ощущения, весьма скоротечны
[Говардовский В.И.Л Островский М.А., 1983; Alpem М. et al., 1971].
Начало фотохимической реакции пигмента под влиянием поглощен-
ного света занимает всего несколько микросекунд, как и ранние ре-
цепторные потенциалы, возникающие после вспышки света в процес-
се фотохимических превращений пигмента [Brown К.Т., 1965]. Про-
цесс фотолиза родопсина в полный транс-ретиналь и опсин,
фототрансдукция и адаптация растягиваются на минуты (см. главу 1).
Однако уже самое начальное воздействие световой энергии запус-
кает цепь быстрых электрических реакций, происходящих на фоне
достаточно длительных молекулярных преобразований зрительного
пигмента [Островский М.А., 1993; Wald G. et al., 1953]. Об исключи-
тельно важной роли временного фактора в зрительном процессе сви-
детельствуют результаты психофизических и нейрофизиологических
исследований процессов, лежащих в основе поведенческих реакций.
Временная суммация. Нервный импульс от фоторецепторов мож-
но получить при воздействии одиночного кванта света [Hal-
let Р., 1969]. Однако для того, чтобы вызвать ощущение света в тем-
ноадаптированном глазу, требуется абсорбция в фоторецепторе по
крайней мере нескольких квантов [Hecht S. et al., 1933]. При этом для
достижения пространственной и временной суммации область и дли-
тельность воздействия необходимо строго ограничить. Максималь-
ный срок, в пределах которого еще возможна временная суммация,
называют критической длительностью зрения, обычно он достигает
0,1 с. На этот показатель влияют очень многие факторы: яркость и
размеры светового пятна, проецируемого на темноадаптированную
77
Рис. 3.5. Реципрокная зависи-
мость факторов освещенность—
длительность (Igl • t) и ее наруше-
ние за пределами критической
длительности. Тс находится на
местах изломов графика.
По оси ординат — Igl в троландах, по
оси абсисс — время (t), мс [Roufs J.,
1966].
сетчатку, его спектральный
состав и локализация в поле
зрения, форма пятна и гради-
ент перепада освещенности на
его границах, единообразие
структуры [Hart W.M., 1987].
Важна также скорость вклю-
чения и выключения света, но
главное требование экспери-
мента — изоляция источника
света для определения визу-
ального ответа.
Критическая длительность
зрения (Тс). При анализе это-
го показателя важнейшую
роль играет освещенность те-
стового пятна на сетчатке.
С увеличением освещенности,
например, в 2 раза для достижения критической длительности зре-
ния достаточно также вдвое меньше времени, и, наоборот, при сни-
жении освещенности потребуется увеличение длительности зрения в
такое же количество раз. Важно заметить, что указанная реципрок-
ная зависимость сохраняется лишь в пределах критической длитель-
ности зрения.
Отношение B/t = const, где В — яркость объекта, at — длитель-
ность свечения, известно как закон Блоха. Он означает, что в рамках
Тс один и тот же световой эффект может быть достигнут при строго
определенной степени световой энергии, слагающейся из различных
сочетаний яркости тестового объекта и длительности его воздействия.
При этом пороговую энергию световых вспышек определяют при
обязательном условии, что последние короче, чем Тс [Stewart В., 1972].
На критическую длительность влияет ряд факторов. Так, чем слабее
освещенность сетчатки, тем больше Тс (рис.3.5).
На рисунке в местах изломов графика отражена как обратная за-
висимость освещенности и длительности, так и ее нарушения за пре-
делами критической длительности Тс. Критическая длительность
7Я
возрастает также по мере уменьшения тестируемой области [Roufs
J.A., 1967 ] и усложнения различения предъявляемых тест-объектов
[KahnemanD., 1964].
Физический феномен временной суммации световой энергии ока-
залось возможным объяснить не только на основе квантовой тео-
рии, но также с помощью химических реакций, возникающих в фо-
торецепторах под воздействием света. При этом Bunsen и Roscoe
подтвердили закон Блоха о наличии реципрокной зависимости меж-
ду интенсивностью светового раздражителя и длительностью его
воздействия.
Все это, однако, справедливо только для рассмотренного выше
весьма короткого периода критической длительности зрения. Спус-
тя лишь десятые доли секунды отношения существенно изменяются,
пороговый источник света определяется уже не по его энергетичес-
кой характеристике (произведению яркости на время воздействия),
а только по его яркости. Именно такой принцип лежит в основе кли-
нической периметрии. Время предъявления объекта практически пе-
рестает влиять на его распознавание. Различаемость таких объектов
становится функцией их яркости, т.е. контраста, относительно уров-
ня яркости фона, к которому происходит адаптация.
По отношению к структурированным объектам, имеющим отно-
шение к остроте зрения, также справедливы закономерности крити-
ческой длительности. Однако при решении более сложных задач (рас-
познавание тонких деталей) критическая длительность достигает 0,4 с
(вместо предельной 0,1 с при простых световых стимулах).
Для спектрально различных световых стимулов характерны оп-
ределенные показатели критической длительности (табл.3.2).
Таблица 3.2. Критическая длительность стимула в зависимости от дли-
ны волны и яркости [Regan D., Tyler С. W., 1971]
Освещенность сетчатки, троланды Длина волны, нм Критическая длительность, мс
для АХ для AL
од 527 96
10 600 110 64
580 160 62
527 250 68
480 250 56
110 527 94 40
79
Рис. 3.6. Воспринимаемая яркость вспышек (по оси ординат) различных по
силе источников света (указана над каждой кривой в лк) в зависимости от
длительности вспышки (по оси абсцисс, с) [Broca, Sulzer, 1902].
Как видно из данных, представленных в табл. 3.2, зрительная си-
стема быстрее реагирует на изменения яркости (AL), чем на измене-
ние длины волны (ЛХ). В оценке цветности критическая длительность
возрастает обратно пропорционально длине волны. Так, в фотопи-
ческих условиях освещенности сетчатки (10 троландов) критическая
длительность зрения в длинноволновой части спектра составляет
около 110 мс, что близко к показателям для белого светового стиму-
ла в темноадаптированном глазу (90—96 с); для стимула же из корот-
коволновой части спектра этот период в 2 раза длиннее (250 мс). Это
свойство зрительной системы лежит в основе экспериментальной
методики гетерохроматической мелькающей фотометрии.
Эффект Broca — Sulzer. Явление временной суммации означает
необходимость определенного времени от начала вспышки до мо-
мента ее видения и еще некоторого времени до ощущения максиму-
80
ма яркости. При одной и той же интенсивности света чем короче
вспышка, тем она кажется более яркой (рис.3.6). Оптимальной для
субъективного восприятия максимальной яркости является вспыш-
ка длительностью 50—100 мс. Более длинная вспышка той же силы
мало отличается по яркости от плато, которым характеризуется по-
стоянное свечение. Для того чтобы уловить яркостный эффект от уд-
линяющихся вспышек, потребуется увеличить интенсивность света.
Поскольку эффект Вгоса — Sulzer проявляется в широком диапазо-
не спектрального состава света в различных условиях световой и тем-
новой адаптации для всех областей сетчатки, имеются основания
объяснить его универсальностью зрительно-нервных механизмов
[ Katz M.S., 1964].
Критическая частота слияния мельканий и закон Тальбота, Мель-
кающий свет по мере увеличения частоты мельканий воспринимает-
ся как непрерывный. Этот переходный рубеж называют критичес-
кой частотой слияния мельканий. Talbot показал, что в единицу вре-
мени средняя интенсивность мелькающего света при субъективном
слиянии мельканий и объективно устойчивого света той же длины
волны и яркости одинакова (так называемое плато Тальбота).
Феномен усиления яркости по Brucke, Установлено, что в момент
включения света его воспринимаемая яркость на мгновение возрас-
тает. Это имеет прямое отношение к описанному выше феномену -
Вгоса — Sulzer. Видимую яркость мелькающего света можно опре-
делить путем сравнения ее с яркостью постоянно светящегося стан-
дарта. При этом нужно учесть, что, по Talbot, для мелькающего све-
та с 50% рабочими циклами сила каждой вспышки должна быть вдвое
больше, чем у непрерывно светящегося стандарта, используемого для
сравнения, при стремлении сделать их одинаковыми по видимой яр-
кости. С изменением частоты мельканий изменяется и видимая яр-
кость, которая оказывается максимальной в довольно узком диапа-
зоне частот. Это относится к мельканиям с частотой от 5 до 20 Гц в
зависимости от силы источника света.
Влияние на КЧСМ интенсивности стимула и закон Ферри — Пор-
тера. КЧСМ зависит от яркости вспышки и линейно растет по мере
повышения освещенности в макулярной области сетчатки. Закон
Ферри — Портера справедлив для освещенности в диапазоне 4 лог.
ед. [Hecht S., 1933].
Влияние на КЧСМ спектрального состава света. Для различаю-
щихся по спектральному составу, но одинаковых по яркости источ-
ников света в фотопических условиях закон Ферри — Портера также
/оказался справедливым. При низких уровнях освещенности сетчат-
ки для коротковолнового (“синего”) конца спектра КЧСМ оказа-
лась наибольшей, а для длинноволнового (“красного”) — наимень-
шей.
Влияние на КЧСМ локализации светового пятна на сетчатке.
Согласно закону Ферри — Портера при направлении светового пят-
81
на на область фовеа КЧСМ будет расти с увеличением освещенное*
ти. При смещении светого пятна от центра к периферии соотноше-
ние колбочек и палочек в освещенной зоне изменится и КЧСМ в за-
висимости от экцентриецтета будет иметь иной характер, а кривая
этой зависимости станет двугорбой, поскольку при низкой освещен-
ности сетчатки стимулируются преимущественно палочки, а при вы-
сокой — колбочки.
Влияние на КЧСМразмеров стимулируемой области — закон Гра-
ната — Харпера. Установлено, что КЧСМ линейно возрастает с уве-
личением размеров стимулируемой области сетчатки. Для централь-
ной области это положение справедливо в очень широких пределах
(до 50°). При значительном увеличении размеров мишени повышает-
ся уровень КЧСМ и на периферии, причем он может превзойти тако-
вой для малых стимулов в области фовеолы или вблизи нее. Именно
этим объясняется тот факт, что мелькания на телевизионном экране
сильнее в том случае, если он оказывается в периферической части
поля зрения.
Влияние на КЧСМ яркости фона и адаптации. В условиях, когда
мелькающее тестирующее поле окружено полной темнотой, на по-
казатели КЧСМ могут оказать влияние случайные факторы, в част-
ности ничтожные засветы зон сетчатки, отдаленных от тестируемых
областей. При этом в ореоле от уже стабильного тестового поля мо-
гут возникнуть мелькания. Во избежание этих погрешностей КЧСМ
рекомендуют исследовать на фоне экрана, освещенного таким обра-
зом, чтобы контраст между так называемой яркостью Тальбота в
тестируемой области и фоновой яркостью был минимальным. По-
скольку при световой адаптации уровень колбочковой чувствитель-
ности возрастает, КЧСМ находится в прямой зависимости от адап-
тации к свету.
Таким образом, при оценке функции КЧСМ следует учитывать
возможность влияния на нее очень многих факторов: освещенности
и размеров тестирующего поля, его места на проекции сетчатки, спек-
трального состава света, длительности и глубины модуляции стиму-
лов, их количества при многократном предъявлении.
3,3. КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ НАРУШЕНИЙ
СВЕТООЩУЩЕНИЯ И ЗРИТЕЛЬНОЙ АДАПТАЦИИ
Существует два основных типа нарушений абсолютной световой чувг
ствительностц и зрительной адаптации: гипофункция колбочкового
аппарата сетчатки, или “дневная слепота^ и гипофункция палочко-
вого аппарата сетчатки, или “ночная слепота” — гемералопия
(рис.3.7).
1. Гипофункция колбочкового аппарата, или “дневная слепота”,
характерна для колбочковой дистрофин или дисфункции. Симпто-
82
Рис. 3.7. Типичная двухфазная нормальная кривая темновой адаптации (с
колбочковым и палочковым компонентами) и варианты ее нарушений, ха-
рактеризующиеся исчезновением одной из фаз и изменением уровня всей
кривой.
1 — норма; 2 — болезнь Огуши; 3 — стационарная ночная слепота; 4 — палочковый
монохроматизм.
мами колбочковой дисфункции являются некорригируемое сниже-
ние остроты зрения, снижение фоточувствительности или наруше-
ние адаптации от темноты к свету, т. е. световой адаптации, неразли-
чение оттенков цвета или нарушение цветоощущения в различных
вариациях, улучшение зрения в сумерках и ночью. Характерными сим-
птомами являются нистагм и светобоязнь, ослепление и изменения в
колбочковой макулярной ЭРГ, более высокая, чем в норме, скорость
восстановления световой чувствительности в темноте. Колбочково-
палочковый перегиб на кривой темновой адаптации либо вообще
отсутствует, либо возникает раньше, чем в норме.
Среди наследственных форм колбочковой дисфункции или дист-
рофии выделяют врожденные формы, к которым относят палочко-
вый монохроматизм (ахроматопсия) — аутосомно - рецессивная фор-
ма, голубой колбочковый монохроматизм — форма, наследуемая как
рецессивная сцепленная с полом, и формы с поздним проявлением и
различными формами наследования, а также сцепленная с полом ауто-
сомно-рецессивная колбочковая дистрофия красных колбочек и с
лентовидным блеском. У большинства больных изменения в маку-
лярной области обусловлены атрофическими или дегенеративными
изменениями, некоторые из которых начинаются по типу дистрофии
“бычий глаз“ (Bull’s eye). Характерным признаком является врож-
денный нистагм. Дифференциальную диагностику проводят с кол-
бочково-палочковым пигментным ретинитом или его “инвертиро-
ванной формой” при колбочковом поражении.
83
Офтальмоскопические изменения при колбочковой дегенерации
определяются потерей нервных волокон, атрофией темпоральной
части диска зрительного нерва, гранулярными изменениями в маку-
ле с последующей ее кольцевидной атрофией.
Изменения свето- и цветоощущения наблюдаются и при приоб-
ретенных патологических процессах в макулярной области, обуслов-
ленных токсическими макулопатиями, вызванными длительным при-
менением хлорохина (гидроксихлорохин, делагил), нейролептиками
фенотиазинового ряда.
2. Гипофункция палочкового аппарата, или “ночная слепота” (ге-
мералопия). В настоящее время выделяют два типа гемералопии: про-
грессирующую форму, обусловленную мутацией родопсина, и врож-
денную стационарную. К прогрессирующим формам относят пиг-
ментный ретинит, колбочково-палочковую дистрофию, синдромы,
сопровождаемые пигментным ретинитом (Usher’s, Moon — Bardet —
Biedl, Bassen Komzweig, Leber’s и др.), fundus punctata albescens. Ста-
ционарные формы гемералопии классифицируются следующим об-
разом:
I. Стационарная ночная слепота с нормальным глазным дном
(аутосомно-рецессивная, аутосомно-доминантная и сцепленная с
полом рецессивная):
А — отсутствующая скотопическая ЭРГ ( тип Riggs; тип I);
В — негативная ЭРГ ( Schubert — Bomschein; тип II): 1 — полный;
2 — неполный (соответственно степени сохранности палоч-
ковой функции).
Сцепленная с полом форма стационарной ночной слепоты (тип
II) сочетается с миопией тяжелой или средней степени.
II. Стационарная ночная слепота с патологическим глазным
дном:
А — болезнь Огуши (Oguchi), феномен Мизуо (Mizuo);
Б — fundus albipunctatus;
В — flick retina of Kandory.
В основе этой классификации лежат изменения в ЭРГ, которая
отражает функцию колбочкового и палочкового аппаратов сетчат-
ки. Имеется два типа изменений в ЭРГ:1-й — когда редуцируется фо-
топический ответ, не нарастающий в процессе темновой адаптации
(тип I, тип Нугаре); 2-й — когда отсутствует в-волна темноадапти-
рованной, общей ЭРГ (тип II, тип Шуберта — Борншейна). Патоло-
гические формы ЭРГ отражают различные виды нарушения кривой
темновой адаптации, которая может быть с изменениями колбочко-
вой функции и без них, отсутствием или снижением темновой адап-
тации, пролонгированным временем адаптации с медленным паде-
нием порога до нормального уровня, наличием палочковой адапта-
ции при завышенном конечном пороге (1,0—1,5 лог.ед.).
Врожденная стационарная ночная слепота с патологическими
изменениями на глазном дне, болезнь Огуши, характеризуется свое-
84
образной серо-белой дисколорацией сетчатки в заднем полюсе и эк-
ваториальной зоне, макулярная область при этом темная в контрас-
те с окружающим фоном. Вариацией этой формы является извест-
ный феномен Мизуо (Mizuo), который выражается в том, что после
длительной темновой адаптации необычная окраска глазного дна ис-
чезает и дно выглядит нормальным. После пребывания на свету она
медленно возвращается к своему оригинальному металлическому
цвету. Темновая адаптация пролонгирована с медленным падением
порога до нормального уровня в течение 2—24 ч [Oguchi С., 1907;
Mizuo G., 1913; Nakamura В., 1920].
Большую группу составляют и разнообразные виды ненаслед-
ственной гемералопии, обусловленные общими нарушениями обме-
на веществ (при дефиците витамина А в связи с неполноценным пи-
танием, при хроническом алкоголизме, заболеваниях желудочно-ки-
шечного тракта, гипоксемии и начальном сидерозе).
Прогрессирующие формы гемералопии (ночная слепота) имеют
особенности изменения темновой адаптации в зависимости от вклю-
чения в патологический процесс колбочковой и/или палочковой сис-
темы: темновая адаптация может отсутствовать как для колбочко-
вой, так и для палочковой систем, при снижении темновой адапта-
ции возможен нормальный конец ее порога (fundus albipunctatus) или
замедление темновой адаптации (flick retina Kandor). В основе этих
процессов лежит ненормальная кинетика зрительных пигментов, один
из вариантов которого — значительное запаздывание их ресинтеза.
Однако получены данные [Carr R.E., Ripps Н., 1992] о том, что кине-
тика и регенерация родопсина, его фоточувствительность при стаци-
онарной ночной слепоте нормальны, а нарушение темновой адапта-
ции связано с нарушением нейрональной передачи информации на
уровне биполярных клеток.
Нарушение скотопического зрения нередко проявляется сужени-
ем периферических границ поля зрения и отсутствием спектрально-
го сдвига яркости (по Пуркинье) в процессе темновой адаптации.
Одним из ранних проявлений многих приобретенных заболева-
ний глазного дна может быть нарушение зрения в условиях сниже-
ния освещенности. При этом светоощущение нередко нарушается по
смешанному колбочково-палочковому типу. Так, при отслойке сет-
чатки любого генеза, при которой прерывается связь между фоторе-
цепторами и пигментным эпителием, нарушаются процессы фотохи-
мического превращения зрительных пигментов, фототрансдукция и
электрогенез. Хотя светоощущение при этом длительное время со-
храняется, дифференциальная чувствительность на отслоившихся уча-
стках утрачивается, а способность к адаптации ослабевает как в об-
ласти отслоившейся сетчатки, так и на неотслоившихся участках.
Согласно гипотезе Ферстера — Вильбрандта, при любой патоло-
гии зрительно-нервного пути, сопровождающейся нарушениями в
поле зрения, вероятность снижения темновой адаптации в функцио-
85
пирующей его части тем выше, чем дистальнее локализованы основ-
ные нарушения. Так, адаптация нарушается при миопической болез-
ни, глаукоме и даже при трактусовых гемианопсиях, а при амблио-
пии центрального характера, корковой гемианопсии адаптационных
нарушений обычно не обнаруживают.
Нарушения светоощущения могут быть не связаны с патологией
зрительно-нервного пути. В частности, порог светочувствительнос-
тей возрастает при ограничении поступления света внутрь глаза в слу-
чаях резкого миоза или помутнения оптических сред. Особую форму
нарушения ретинальной адаптации представляет эритропсия. При
афакии, когда сетчатка подвержена воздействию яркого света без
фильтрации хрусталиком коротковолновых лучей, пигмент “синих”
и “зеленых” колбочек выцветает, чувствительность колбочек к крас-
ному цвету увеличивается и красночувствительные колбочки отве-
чают суперреакцией. Эритропсия может сохраняться в течение не-
скольких часов после засвета высокой интенсивности.
3.4. КЛИНИЧЕСКИЕ МЕТОДЫ ТЕСТИРОВАНИЯ
СВЕТОЧУВСТВИТЕЛЬНОСТИ
3.4.1. Темновая адаптометрия
Поскольку светочувствительность весьма изменчива, снижаясь на
свету и повышаясь в темноте, она достигает максимума только при
полной темновой адаптации. Этот абсолютный световой порог уда-
ется определить лишь после 40—60-минутного непрерывного пребы-
вания в полной темноте. Периодические замеры нарастающей в со-
храняющейся темноте светочувствительности (с интервалом, напри-
мер, 5—10 мин) позволяют с помощью как довольно старых
конструкций (адаптометр Нагеля), так и более современных адапто-
метров (типа отечественных АДМ или зарубежных никтометров)
построить кривую темновой адаптации. Угловые размеры тестового
светового пятна и длительность его предъявлений поддерживаются
постоянными. Для того чтобы на кривой темновой адаптации полу-
чить не только палочковый, но и начальный колбочковый компо-
нент, необходимо создать высокий уровень световой предадаптации
(порядка 40 000 кд/м2) (рис. 3.8).
Оценка результатов. На нормальной кривой темновой
адаптации в первые же минуты обнаруживается прирост светочув-
ствительности за счет включения функции колбочек (“колбочковое
колено”). Затем на протяжении 15 мин резко возрастает светочув-
ствительность палочек. В дальнейшем, вплоть до 40-й минуты, све-
точувствительность увеличивается очень медленно и достигает в
конечном итоге стабильного уровня, позволяющего воспринимать
яркость порядка 1Q-6 кд/м2.
86
Рис. 3.8. Нормативы кривых темновой адаптации [Волков В. В. и др., 1989].
а — варианты кривых при различных уровнях предадаптации, кд/м2:1 — 40 000; 2 —
4000; 3 — 2000; 4 — 400; 5 — 30; б — компоненты (ветви) кривой при высоком уров-
не предадаптации (40 000 кд/м2); в — полоса нормы для кривой темновой адаптации.
Световые пороги у вполне здоровых людей широко варьируют,
поэтому при оценке результатов целесообразно пользоваться поло-
сой нормы (см.рис. 3.8). С возрастом светочувствительность изменя-
ется: она максимально высока у 20-летних и после этого возраста
начинает снижаться, достигая к старости минимальных значений.
Введение в клиническую практику ориентировочного быстрого
3-минутного исследования световой чувствительности оказалось по-
лезным особенно в летной экспертизе. Способность различать при
этом форму испытательного объекта (круг, контуры самолета, квад-
рат) не позже чем через 45—50 с после погружения в темноту прибо-
ра (АДМ или камер Кравкова — Вишневского, Семикопного) рас-
ценивали как признак нормальной темновой адаптации. Однако по
87
мере дальнейшей разработки методик оценки центрального сумереч-
ного зрения от сомнительной по точности скрининговой 3-минутной
адаптометрии отказались, так как для ее проведения необходима
предварительная адаптация к освещенности в 4—5 лк в течение
20 мин.
3.4.2. Световая адаптометрия
С помощью очень ярких засветов подавляют функцию фоторецепто-
ров (фотостресс-тест) и по времени последующего восстановления
остроты зрения судят о сохранности колбочковой функции. Адапта-
ция к свету — процесс гораздо более быстрый, чем адаптация к тем-
ноте.
3.4.3. Исследование критической частоты слияния мельканий
В разное время было предложено множество вариантов устройств
для данного исследования в виде вращающихся дисков и электрон-
ных аппаратов, но в клиническую практику ни одно из них так и не
вошло.
Техника исследования. Обследуемому предлагают
смотреть на мелькающий источник света и сообщить, когда по мере
увеличения частоты мельканий свет для него станет постоянным.
И.И.Меркуловым был изготовлен КЧСМ-периметр, с помощью ко-
торого можно оценивать методом слияния мельканий любую точку
в поле зрения. Дозировать можно как силу вспышек и, следователь-
но, амплитуду мельканий, так и их частоту. Для исследования КЧСМ
в наиболее ранимых при глаукоме точках зоны Бьерума В.А.Заха-
ров и И.В. Волкова (1983) создали специальный прибор — глауко-
анализатор.
Оценка результатов. Существование зрительной инерции
в виде последовательного образа позволяет глазу воспринимать пе-
риодически затухающий источник света как непрерывно светящий-
ся, если частота мельканий возрастает до определенного уровня. Наи-
меньшая частота, необходимая для этого, называется критической
частотой слияния мельканий.
КЧСМ не зависит от остроты зрения, рефракции, размера зрач-
ка, но для центральной зоны сетчатки она выше, чем для периферии.
В литературе представлены различные нормативы для этой функ-
ции. Так, большинство авторов установили, что для фовеальной
области КЧСМ составляет 40—55 Гц, для парафовеальной она воз-
растает до 55—60 Гц, а на крайней периферии снижается до 35—40 Гц.
С целью определения КЧСМ для так называемых проблесковых ис-
точников света с яркостью (Ц) не выше 103 кд/м2 В.В. Волков и со-
авт. (1989), принимая во внимание исследования С.В.Кравкова (1950),
рекомендуют использовать формулу
88
v„=121gL,+35,
где: Lo — яркость мелькающего источника, кд/м2. Формула спра-
ведлива при условии, что длительность проблеска занимает поло-
вину периода мелькания. vKp уменьшается с уменьшением глубины
модуляции, т.е. отношения разницы максимальной и минимальной
яркости к средней яркости. Е.Е. Сомов и В.А. Маревский (1982) пред-
ложили для оценки зрительного утомления использовать модифи-
цированную авторами методику определения КЧСМ. Сначала оп-
ределяют минимальное время (в миллисекундах) восприятия про-
блескового светового стимула, а затем, увеличивая частоту его
предъявления, находят показатель КЧСМ, который и рассматрива-
ют как критерий оценки зрительного утомления.
А.В. Луизов (1983) предлагает учитывать феномен инерции зре-
ния при расчете информационной пропускной способности зритель-
ной системы.
По мнению А.И. Богословского (1962), методика исследования
КЧСМ заслуживает большего внимания клиницистов, чем это было
до сих пор, и ее нужно использовать не только при электрофизиоло-
гических исследованиях. КЧСМ может нарушаться при поражении
любого отдела зрительно-нервной системы глаза.
3.4.4. Периметрия
Эта методика является одной из наиболее информативных в офталь-
мологии. Она позволяет судить не только о характере распределения
светочувствительности в каждом глазу по площади рецептора (сет-
чатки), но и об уровне поражений зрительно-нервного пути по сово-
купности нарушений в обоих глазах.
Пространство, одномоментно воспринимаемое глазом при непод-
вижном взоре, называется полем зрения. На протяжении многих лет
ученые А. Грефе, А. Дюбуа-Пульсен и др. придавали большое значе-
ние состоянию поля зрения и возникающим при некоторых заболе-
ваниях сужению его границ и “провалам”(скотомам) в видимом про-
странстве.
Второе “рождение” периметрии произошло, когда, кроме опре-
деления границ контуров видимого и невидимого, с ее помощью
появилась возможность определять распределение световой чувстви-
тельности в каждой точке поля зрения. Важным этапом на пути к
этому явилось представление об объемной модели поля зрения
(рис.3.9) как об “острове видения” (Г. Трекуэр), каждый уровень
которого над “морем невидения” можно было бы оценить количе-
ственно, а границы участков, одинаковых по уровню светочувстви-
тельности и соединенных воображаемой линией, обозначить как
изоптеры.
89
-40 — 0.1
90-
Рис. 3.9. Объемная модель нормальной и патологической дифференциаль-
ной световой чувствительности как функция локализации в поле зрения
[Bebie Н., Fankhauser F., 1983]. В точке р поля зрения стимул В (порядка
20 асб) не воспринимается в отличие от стимула А (100 асб). Нормальная
(N) дифференциальная световая чувствительность в этой точке находится
на уровне 3 асб.
1 — нормальное зрительное поле; 2 — патологическое зрительное поле.
Однако наиболее совершенная тактика периметрии состоит в том,
чтобы сначала получить представление о рельефе светочувствитель-
ности зрительной системы в видимом пространстве, а затем выде-
лить границы интересующих исследователя участков поля зрения.
Техника исследования. Различают кинетическую и
статическую периметрию (рис. 3.10). В обоих случаях взор исследуе-
мого глаза должен быть фиксирован на метке в центре равномерно
освещенной поверхности либо дуги, либо полусферы радиусом 30 см.
При кинетической периметрии тестируемый объект плавно или
ступенчато смещают по поверхности периметра либо от периферии
к центру, либо наоборот. Одновременным вращением дуги перимет-
ра можно создавать и более сложную траекторию движения тест-
объекта. Чаще границу видения определяют в момент появления
объекта в поле зрения, иногда используют критерий его исчезнове-
ния. При любом из этих вариантов кинетической периметрии в тех
случаях, когда исследование проводят с помощью надпорогового
объекта, технически невозможно уловить некоторые скотомы.
В течение многих лет этот вид периметрии был повсеместно един-
ственным; в настоящее время в недостаточно оснащенных учрежде-
90
Рис. 3.10. Принципы
проведения статической
и кинетической пери-
метрии.
1 — при статической пери-
метрии варьирует интен-
сивность стимула (в одной
и той же позиции объекта);
2 — при кинетической из-
меняется местоположение
объекта.
ниях по-прежнему ограничиваются определением границ поля зре-
ния.
Предпринимались попытки повысить информативность кинети-
ческой периметрии. Заслуживают упоминания два следующих направ-
ления.
1. Оценка способности к пространственной суммации (равноэнер-
гетическая периметрия). В основу этого метода положены наблюде-
ния, свидетельствовавшие о том, что при использовании тест-объек-
тов с различными сочетаниями яркости и размеров, если, однако, они
энергетически эквивалентны, т.е. произведения яркости на площадь
представляют какую-либо константу для сравниваемых объектов, то
результаты периметрии такими объектами у здоровых людей прак-
тически совпадают (разброс показателей границы нормы Не превы-
шает 5—7°). При использовании равноэнергетической кинетической
периметрии было установлено [Немцеев Г.И., Сабаева Г.Ф.; 1985;
Organd A., Etienne R., 1961], что уже в самых начальных стадиях гла-
укомы нарушается пространственная суммация. У части пациентов
это проявляется рассогласованием между изоптерами на 10° й более
в каком-либо секторе или по всему полю зрения. Метод, однако, не
получил широкого распространения в клинической практике преж-
де всего из-за длительности каждого исследования.
2. Хронопериметрия — способ одновременного измерения границ
поля зрения и времени сенсомоторной реакции на пересечение их тест-
объектом [Немцеев Г.И.,1988]. Тест-объект автоматически перемеща-
91
ют вдоль любого меридиана поля зрения от его центра к периферии
до исчезновения из поля зрения, затем объект перемещается в обрат-
ном направлении от периферии к центру до его появления в поле зре-
ния. Момент исчезновения и появления объекта отмечается обследу-
емым нажатием на кнопку электромагнитного отметчика. Для каж-
дого глаза получается два поля зрения: измеренное при движении
объекта от центра к периферии (границы поля зрения будут шире
действительных) и от периферии к центру (границы поля зрения бу-
дут уже действительных). Это расхождение в полях зрения соответ-
ствует удвоенному времени сенсомоторной реакции, степень измене-
ния которой и может быть критерием нарушения проводимости зри-
тельно-нервных волокон, наблюдаемое, например, при передней
ишемической нейропатии, при рассеянном склерозе, сопровождаю-
щемся обширным демиелинизирующим процессом [Немцеев Г.И.,
1968—1988; Сабаева Г.Ф., 1981].
При статической периметрии, которую в последние годы исполь-
зуют более широко, чем кинетическую, тестирующий объект не пе-
ремещают и не меняют в размерах, а предъявляют в заданных по той
или иной программе точках поля зрения с переменной яркостью.
Создателями метода являются Н. Goldmann, W. Leydhecker,
Е. AulhornnH. Harms, F. Fankhauser. В стремлении к компромиссу
между максимальным количеством исследуемых точек и минималь-
ными затратами времени возникла идея создания автоматизирован-
ных периметров с компьютерными программами. В итоге появились
скрининговые и пороговые стратегии. Стало возможным в зависи-
мости от предполагаемой патологии проводить исследование в оп-
ределенных участках поля зрения с повторным контролем, сохранять
результаты в памяти прибора и по специальной программе произво-
дить статистический анализ.
Одним из важнейших условий, обеспечивающих точность пери-
метрического исследования, является хорошая фиксация взора. В
современных периметрах (“Rodenstock”, “Humphrey”, “Octopus” и
др.), кроме непосредственного наблюдения за положением глаза па-
циента на телевизионном экране и механической подстройки фикса-
ции, включена система периодической подачи сигналов в зону сле-
пого пятна для учета частоты ошибочно положительных ответов
пациента, свидетельствующих о нарушении фиксации взора (метод
Хейл — Кракау). На характер фиксации оказывают влияние два ос-
новных фактора: длительность стимула и случайность места его
предъявления. С учетом времени нормальной сенсомоторной реак-
ции и временной суммации длительность стимула не должна быть ни
слишком короткой, ни слишком длинной. По данным Е. Aulhorn и
Н. Harris (1972), стимул, который длится 3 с, распознается хуже, чем
стимул продолжительностью 1 с. В то же время распознаваемость
очень коротких стимулов снижается по мере их укорочения.
Распознавание стимулов длительностью более 0,1 с для перифе-
92
рии и более 0,4 с для центральных отделов сетчатки не зависит от
времени предъявления, а определяется только их яркостью. Однако с
учетом скрытого времени произвольных движений глаз (около 0,25 с)
при более длительных стимулах пациент успевает перевести взор от
центра к объекту на периферии и фиксировать его макулярной зо-
ной. В связи с этим в периметре “Humphrey” длительность каждого
стимула уменьшена до 0,2 с.
В большинстве программ периметра “Octopus” предусмотрена
экспозиция стимула 0,1 с, и только при специальных исследованиях
длительность стимула увеличивают до 0,2—0,5 с (программы иссле-
дования зрения у слабовидящих, водителей и blepharoptosis test).
При ручной периметрии легко предсказать направление, в кото-
ром можно ожидать появления тест-объекта, в автоматических же
периметрах это исключено благодаря их случайному (беспорядочно-
му) предъявлению. Данное обстоятельство важно и для исключения
возможных влияний местной адаптации, если предъявлять тест по-
вторно в одном и том же месте.
В большинстве современных периметров с учетом периметричес-
ких стандартов (1979) яркость фона составляет 31,5 асб (10 кд/м2).
Использование такого умеренно яркого фона, близкого к мезопичес-
ким условиям зрения, во-первых, уравнивает вклад палочек и колбо-
чек в светочувствительность, во-вторых, исключает необходимость
предварительной световой или темновой адаптации зрительной сис-
темы пациента. Вместе с тем на этот достаточно яркий фон в пери-
метре не влияют световые помехи, которые могут случайно возник-
нуть при перемещении персонала в помещении, где проводят иссле-
дование.
Целесообразность исследования порога световой чувствительно-
сти в каждой точке поля зрения при любом уровне яркости фона оче-
видна, однако длительность исследования лимитирует использова-
ние такого типа программ. Поэтому следует помнить, что хотя для
широкого ряда адаптирующих освещенностей отношение ДI/I пред-
ставляет собой константу, но в условиях скотопического зрения это
правило нарушается и в профиле нормального поля зрения возника-
ют особенности (рис. 3.11).
Как видно на рис.3.11, в условиях полной темновой адаптации
(U = 0 асб) или очень низкой фоновой освещенности в центральной
зоне сетчатки обнаруживается некоторая локальная депрессия све-
товой чувствительности. По мере повышения адаптирующей ярко-
сти в этой зоне намечается уже небольшой ее пик, а снижение чув-
ствительности от центра к периферии происходит более плавно. На
практике используют постоянство сдвигов яркостной чувствитель-
ности в широком диапазоне освещенностей адаптирующего фона. Это
позволяет в условиях принятой для клинической периметрии осве-
щенности получить информацию о функциональном состоянии по-
давляющего большинства областей сетчатки.
93
40* 20* O’ 20’ 40’ 60’ 80’
Рис. 3.11. Результаты пороговой статической периметрии у одного и того
же субъекта при различных уровнях адаптации (Aulhom Е., 1964].
I — назальная часть поля зрения, II — темпоральная часть поля зрения; 1 —U=0
асб, 2 —U=0,001 асб, 3 —U=0,01 асб, 4—U=0,lac6, 5 —U=lac6, 6 —U=10ac6,
7 —U=100 асб, 8 — U=1000 асб; U — яркость фона.
Периметры, например “Octopus-101”, в которых используется
менее яркий фон (4 асб), т.е. создаются условия мезопического зре-
ния, позволяют более точно оценивать периферические участки поля
зрения. Однако, как свидетельствует опыт проведения статической
периметрии, в клинической практике значительно чаще требуется
точный анализ изменений световой чувствительности, происходящих
В центральной зоне поля зрения (в пределах 30° от точки фиксации).
Как отмечено выше, в центральной зоне сетчатки расположено 66%
рецептивных полей всех ганглиозных клеток, связанных с большей
частью (83%) зрительной коры.
Размеры и интенсивность стимул а. При тради-
ционно проводимой статической периметрии размер стимулов не
изменяется. Выбранный в качестве универсального стимул имеет ди-
аметр 0,43°, что эквивалентно объекту с размером III по Гольдману
(его площадь 4 мм2). Он достаточно велик, чтобы на его распознава-
ние не влияли нарушения рефракции [Sloan L., 1961] и вместе с тем
достаточно мал, что позволяет выявлять даже небольшие скотомы
[Flammer J. et al., 1981]. В проекционных периметрах (“Octopus”,
“Humphrey”) остается возможность изменения размера стимула, что
используется для исследования поля зрения при низком зрении (сти-
мул, эквивалентный объекту с размером V по Гольдману).
Яркость распознаваемого стимула стала единственным количе-
ственным критерием оценки световой чувствительности в исследуе-
мой точке. При возрастании пороговой яркости (от 0,08 до 10 000 асб,
т.е. в пределах 5,1 лог.ед.), оцениваемая в децибелах (дБ) световая
чувствительность снижается с 51 до 0 дБ. На рис. 3.12 представлены
уровни пороговой яркости, которые соответствуют определенным
стандартам по Гольдману. Таким образом, все результаты статичес-
94
Gottnannlll
Рис. 3.12. Шкаля ин-
тенсивностей стимулов,
принятых в современ-
ных отечественных пе-
риметрах, дана в срав-
нении с аналогичной
шкалой (1а — 4е) рас-
пространенного зару-
бежного периметра «Gold-
mann III» [ed. Haley M.
Ь: The Fipld Apaljser
Primer «Humphrey»
Instr. Inc., 1993].
кой периметрии выражаются н единицах светочувствительности (дБ)
на основе единственного критерия — пороговой яркости тест-объекта
(асб). Между указанными показателями существует обратная зави-
симость, причем сдвиг на 1 дБ соответствует изменению яркости на
0,1 лог.ед.
Стратегия исследования. Выбор прибора и программ, адекват-
ных целям исследования, позволит сэкономить время врача и изба-
вить пациента от излишнего утомления.
Для решения скрининговых задач вполне пригодны зарубежные
периметры типа “Ocuplot”, “Peritest”, отечественные анализаторы
поля зрения по Астраленко и простейшие устройства в виде набора
из 20 карт, предъявляемых на фоне негатоскопа или как тест-диск
[Волков В.В. и др., 1985]. Последний вариант можно использовать
для самопроверки в домашних условиях.
Скрининговые программы имеются и в сложных автоматизиро-
ванных периметрах (‘«Humphrey”, “Octopus” др.). При этом исследо-
вание проводят с использованием надпороговых объектов, которые
на 0,6 лог.ед. ярче, чем предполагаемый в норме порог для каждой
точки. В приборе учтена необходимость предъявления на периферии
более ярких стимулов, чем в центре. Если предварительно замерить
индивидуальный порог в 4 парацентральных точках, то в приборе
автоматически перед дальнейшим проведением скрининга по всему
полю зрения вносится поправка на индивидуальную норму (с учетом
возраста, размеров зрачка, прозрачности сред, общего состояния).
Хотя при скрининговом исследовании световая чувствительность сни-
жена на 6 дБ, достоверность отрицательных ответов, свидетельству-
95
5А 10*
15’
20^
3£
40’
50^
Матричное распределение
г
ющих о наличии скотомы, возрастает, и, несмотря на то, что чув-
ствительность теста несколько уменьшается, его специфичность со-
храняется. В тех точках, где надпороговый стимул не воспринимает-
ся, для исключения абсолютной скотомы определяют чувствитель-
ность к самому яркому из имеющихся в приборе стимулов (10 000 асб).
Кроме того, вокруг каждой невидимой точки может быть исследова-
но дополнительно то или иное количество окружающих ее точек.
При проверке порогов световой чувствительности стратегию по-
иска чаще строят по принципу “вилки” или “лестницы”. Первона-
чальный стимул должен быть чуть ярче порогового, затем делают
шаг в сторону его ослабления, в пересчете на децибелы уменьшают
яркость стимула на 4 ’’ступеньки лестницы”, затем при необходимо-
сти еще на 4 дБ, пока он не станет недоступным для восприятия. Пос-
ле этого яркость стимула увеличивают по 2 ’’ступеньки” (до момента
видения). В тех случаях, когда уже первый стимул не виден, с шагом в
4 ’’ступеньки” увеличивают его яркость, а добившись видения, опуска-
ются на 2 ’’ступеньки”. В любом варианте последний видимый стимул
окажется в “вилке” и может быть зарегистрирован как пороговый.
При повторных исследованиях за основу принимают результаты
предыдущей проверки и исходным пунктом тестирования выбирают
стимул, который заведомо на 2 дБ ярче. Затем, как и прежде, спуска-
ются по 4 и поднимаются по 2 ’’ступеньки”.
При еще более ускоренном тестировании обследованных ранее
пациентов иногда ограничиваются исследованием порогов светочув-
ствительности только на те стимулы, которые на 2 дБ ярче установ-
ленного порогового значения для каждой данной точки, и в случае
их распознавания исследование на этом завершают. В периметре
“Octopus” порог светочувствительности определяется с большей дис-
кретностью (пороговая стратегия 4—2—1 дБ). Для уменьшения про-
должительности исследования и нагрузки на пациента применяют ди-
намическую тестовую стратегию. Размер шагов при определении по-
рога светочувствительности автоматически адаптируется к уровню
чувствительности и может изменяться от 2 дб (в области нормаль-
ных значений чувствительности) до 10 дБ (в областях со сниженной
чувствительностью). Окончательное значение пороговой чувстви-
<-------------------------------------------------------------------------
Рис. 3.13. Некоторые варианты расположения в парацентральной части поля
зрения точек, в которых исследуют пороговую светочувствительность в слу-
чаях возникновения подозрения на глаукому.
а — в 12 точках зоны Бьеррума по Волковой; б — в 6 точках глаукотестера, предло-
женного В. В. Волковым и соавт. (1985); в — в 20 точках, расположенных в радиусе до
18° от центра с интервалами в 3°, что создает не только высокую чувствительность,
но и придает пробе специфичность в диагностике глаукомы; г — при исследовании в
поисках назальной “ступеньки” по Армали. Все схемы предназначены для исследова-
ния правого глаза (метка слепого пятна справа от точки фиксации).
97
тельности вычисляют как среднее между интенсивностями двух пос-
ледних стимулов после изменения направления шага. Преимущество
динамической стратегии состоит в уменьшении продолжительности
тестирования на 40—50%.
Как при скрининговой, так и особенно при пороговой стратегии,
минимальное количество точек в схеме исследования должно помочь
быстрому решению клинической задачи. Расстояние между исследу-
емыми точками в центральной или другой зоне специального инте-
реса составляют 4—6°, на периферии (за пределами 30°) — 8—12°.
При тестировании макулярной зоны расстояния между точками мо-
гут быть уменьшены до 1—2°. Например, в периметре “Octopus” про-
грамма М2 для подробного исследования макулярной области по-
зволяет предъявлять в пределах 4° от точки фиксации стимулы, уда-
ленные на 0,7° друг от друга. Для выявления ранних глаукоматозных
изменений созданы паттерны точек, охватывающих зону Бьеррума:
12-точечный тест Волковой для ПРП-60 или полушарового перимет-
ра (фирма “Karl Zeiss, Jena”); 6-точечный тест Волкова и соавт. в гла-
укотестере, тест на так называемую назальную ступеньку по Арма-
ли, 20-точечный тест Хенсона (рис.3.13). В нейроофтальмологии пе-
риметрию проводят при расположении точек строго симметрично
по сторонам от вертикального меридиана (рис. 3.14). Кроме того,
имеются наборы точек для исследования отдельных зон, например
височного полулуния. Если периметрию, в том числе автоматичес-
кую, может с успехом выполнить оптометрист, то выбор программы
для исследования и оценку результатов должен осуществлять только
врач.
Оценкарезультатов. При скрининговой методике оценка
результатов исследования может быть либо положительной (стимул
виден), либо отрицательной (стимул не виден); при многократных
замерах целесообразно использовать принцип 50% вероятности. Эле-
мент количественной оценки результатов в периметрии появляется
при трехступенчатом скрининге: надпороговый стимул виден — нор-
ма; надпороговый стимул не виден, но при увеличении яркости до
максимума (10 000 асб) виден — относительная скотома; стимул не
виден и при максимально используемой яркости — абсолютная ско-
тома.
Пороговая методика предполагает многоступенную количествен-
ную оценку либо в децибелах (при статической периметрии), либо в
градусах отстояния границ от центра (при кинетической периметрии).
Нормативы для количественной кинетической периметрии, прово-
димой разноэнергетическими объектами, были выработаны С.Б. По-
ляк (1968). При равноэнергетической периметрии критерием служит
совпадение или несовпадение границ при повторных исследованиях.
Нормативы для статической периметрии в разных возрастных
группах выработаны R. Brenton и Ch. Phelps (1986). Установлено, что
с возрастом светочувствительность снижается по всему полю зрения
98
а б
Рис. 3.14. Некоторые варианты расположения тестирующих точек при ней-
роофтальмологических исследованиях.
а — паттерн-тест для оценки порогов по обе стороны вертикального меридиана до
50° (с целью выявления гемианопсических нарушений); б — паттерн-тест для оценки
порогов в височном полулунии правого глаза.
равномерно, поэтому рельеф острова зрения (крутизна ската) в нор-
ме сохраняется обычным. Индивидуальные различия увеличивают-
ся по мере усиления эксцентриситета замеров, в частности в перифе-
рической зоне 30—60°, краткосрочные флюктуации были статисти-
чески достоверно более выраженными (2,6 дБ), чем в центральной
зоне (до 30°). Повторное исследование проводили не ранее чем пос-
ле 2 мин отдыха.
На результат периметрии могут повлиять следующие факторы:
слишком глубокое расположение глазного яблока в глазнице из-за
особенностей лицевого черепа, в частности высокой переносицы;
приспущенность верхнего века, в том числе при слишком присталь-
ной фиксации взора; попадание стимула в область крупного сосуда
вблизи диска зрительного нерва или направленное под углом (ано-
мальное) прохождение зрительного нерва через склеру; некачествен-
ная оптическая коррекция зрения, слишком высокая степень амет-
ропии, даже при условии хорошей коррекции, помехи от очковой
оправы. Любой из этих факторов может стать причиной артефакт-
ных дефектов в поле зрения.
Для регистрации результатов периметрии разработано
99
множество схем, некоторые из которых получили международное
признание. Очень распространены схемы в виде пары округлых фи-
гур с 10 концентрическими окружностями, две первые на расстоянии
5—10° от центра и далее через каждые 10°. Окружность пересечена
12 меридианами с угловыми интервалами в 15°. За нулевой меридиан
принят правый конец горизонтального меридиана, отсчет остальных
ведется против часовой стрелки (по системе ТАВО). Внешние грани-
цы каждого из кругов, размещенных на одном уровне, соответству-
ют височным границам поля зрения. В височных половинах поля зре-
ния в 15° от центра на горизонтальном меридиане обозначаются фи-
зиологические скотомы соответственно проекции диска зрительного
нерва. В норме периферические границы поля зрения для белого цве-
та составляют к виску чуть более 90°, к носу и кверху около 60°, кни-
зу до 70°. Иногда на схемах отмечают границы усредненного нор-
мального поля зрения и, очертив границы по результатам кинети-
ческой периметрии, заштриховывают те участки поля зрения, которые
у пациента оказываются “выпавшими”, т.е. с отсутствующей или сни-
женной световой чувствительностью.
Результаты скрининговой статической периметрии регистриру-
ют следующим образом: те участки поля зрения, чувствительность
которых не нарушена, отмечают, например, незаштрихованными
кружками, а на участках сниженной светочувствительности простав-
ляют условные знаки (заштрихованные квадраты или другие фигу-
ры). В результате “вырисовываются” локализация, форма и размер
скотом. Для получения точной информации о расположении ското-
мы предпочтительнее использовать объекты низкой интенсивности,
а для определения глубины поражения необходимы объекты с увели-
чивающейся яркостью. Для регистрации результатов пороговой ста-
тической периметрии применяют либо штриховую, либо цифровую
маркировку (рис. 3.15). Ключ к схеме представляют в нижней части
рисунка в виде таблицы, где указана цена каждой штриховки (в апо-
стильбах и децибелах): чем темнее штриховка, тем ниже светочув-
ствительность. Результаты того же исследования автоматически вы-
даются также в цифровом выражении в децибелах (дБ) на двух дру-
гих схемах, располагаемых на бланке ниже. По ним можно
определить как глубину дефицита, так и пороговые показатели све-
товой чувствительности в каждой исследованной точке. Детали пас-
портной части, состояние зрачка и параметры тестов также представ-
ляются на бланке заключения в децибелах.
При исследовании на глаукотестере [Волков В.В. и др., 1981] ре-
гистрацию порогов световой чувствительности в избранных точках
парацентральной зоны Бьеррума проводят на специально разрабо-
танном приборе, который позволяет выполнять статическую пери-
метрию в условиях дозированной вакуум-компрессии глазного яб-
лока. Этот вариант статической периметрии используют в клиничес-
кой практике для определения устойчивости ДЗН к сдавливанию и
100
Рис. 3.15. Пример изменений в поле зрения при проведении центральной
пороговой периметрии на периметре Хамфри по программе 30-2 в пределах
30° при 2-м варианте расположения точек. Результаты представлены штри-
ховкой разной значимости по прилагаемой к программе шкале.
тем самым для прогностической оценки возможности его дальней-
ших глаукоматозных повреждений.
Современные периметры типа “Humphrey” позволяют автома-
тически проводить статистическую оценку результатов, определяя
среднее отклонение порога световой чувствительности суммарно по
всему полю зрения данного пациента в сравнении с возрастной нор-
мой, разброс ответов (стандартные отклонения) по определенному
паттерну, которые для большинства людей не превышают 4,81 дБ, и
кратковременные флюктуации с учетом индекса постоянства отве-
тов, который у большинства не выходит за пределы 2,32 дБ. Извле-
кая из памяти прибора результаты предшествующих измерений, мож-
но провести линейно-регрессивный анализ реальных отклонений
порогов световой чувствительности от средней девиации; фиксируя
эти отличия (с учетом возрастного снижения показателей), прибор
автоматически отмечает случаи плохой фиксации взора пациента и
низкую надежность статистического анализа в этих случаях.
Основные варианты нарушения поля зрения и интерпретация его
причин. При обнаружении дефектов в поле зрения одного глаза сле-
дует принять за правило обязательно исследовать поле зрения вто-
рого глаза. Из беседы с пациентом нужно выяснить, ощущает ли он
сам недостатки зрения и в чем они выражаются. Одни пациенты не
только видят скотому в собственном поле зрения, но и могут ее обри-
совать, другим она представляется как провал в поле зрения. Ското-
101
мы могут сопровождаться чувством так называемой метаморфопсии
(искажение видимого), микро- или макропсии, фосфена (ощущение
вспышки света). Иногда, подобно физиологическим скотомам (сле-
пое пятно Мариотта в проекции диска зрительного нерва, ангиоско-
томы), приобретенные скотомы не ощущаются вовсе. Вариант клас-
сификации встречающихся дефектов поля зрения и причин их воз-
никновения представлен ниже.
Скотомы разделяют по особенностям локализации, а дефекты в
границах поля зрения — по их моно- или бинокулярности.
I. СКОТОМЫ (особенности локализации в поле зрения; вероятные причи-
ны с учетом некоторых субъективных проявлений).
Центральная зона (с перекрытием точки фиксации — центральная
скотома)
Монокулярная:
• положительная скотома, нередко с метаморфопсией (макулярный отек,
дистрофия Фукса, кисты, вплоть до разрыва сетчатки в макуле, геморра-
гии, экссудат, опухоль, лучевой ожог, сосудистые мембраны и пр);
• положительная скотома с микропсией (центральная серозная хороирети-
нопатия);
• отрицательная скотома (аксиальный неврит, травма или ишемия зритель-
ного нерва).
В редких случаях все перечисленные типы скотом наблюдаются и при би-
нокулярном обнаружении, но с разрывом во времени.
Бинокулярная:
• отрицательная скотома либо сразу на обоих глазах, либо с небольшим вре-
менным интервалом между поражением одного и второго глаза (оптико-
хиазмальный арахноидит).
Зона слепого пятна в проекции диска зрительного нерва (цекаль-
ное пятно Мариотта)
Бинокулярная: физиологическая скотома, незамечаемая в обычной жизни
(около 5° в поперечнике при диаметре диска зрительного нерва около 1,5 мм).
Монокулярная: расширение слепого пятна более 5° в поперечнике, субъек-
тивно обычно незамечаемое (застойный сосок, друзы диска зрительного не-
рва, в меньшей мере глаукома).
Центральная зона и зона слепого п я т н а (центро-цекаль-
ная скотома)
Монокулярная: ремиттирующая скотома (врожденная “ямка” диска зри-
тельного нерва с серозной отслойкой сетчатки).
Бинокулярная: токсическая, леберовская и другие формы оптической ней-
ропатии.
Парацентральная зона (по окружности в пределах 5—15° от
точки фиксации)
Монокулярная: при глаукоме (скотома Бьеррума — первоначально в виде
отдельных островков, расположенных выше либо ниже точки фиксации, в
последующем они сливаются в арочную конструкцию, с одной стороны свя-
занную со слепым пятном, а с другой оканчивающуюся в носовой половине
поля зрения строго по горизонтальному меридиану, образуя два угла (бли-
102
жайший к центру угол скотомы называют выступом Ренне). Уже при моно-
кулярном процессе (глаукома) возможен зрительный дискомфорт, снижение
контрастной чувствительности и темновой адаптации.
Парацентральные боковые зоны(гомонимноправосторон-
ние, гомонимно левосторонние)
Бинокулярная: создает затруднение при чтении: теряется конец слова, строч-
ки или их начало,— при высокой конгруэнтности (сходных скотомах на обо-
их глазах) — корковая патология, при невысокой — трактусовая.
Парацентральные горизонтальные зоны (верхние или
нижние)
Монокулярные: скотомы определяются при наличии чувства “срезания”
верхней или нижней части рассматриваемого объекта (ишемическая опти-
ческая нейропатия).
Срединная зона (между центром и периферией в виде замкнутого
или разомкнутого кольца, кольцевидная скотома, в поздних стадиях за-
болевания кольцо сжимается к центру до 3—5 °)
Монокулярная: при далеко зашедшей глаукоме и др.;
Бинокулярная: при тапеторетинальной дистрофии, медикаментозной ди-
строфии сетчатки и др., обычно сопровождается снижением темновой адап-
тации.
Островковые скотомы (в различных участках периферии поля
зрения)
Монокулярные, реже бинокулярные: часто остаются незамеченными из-за
бинокулярного перекрытия значительной части поля зрения (по горизонта-
ли до 120° ). Встречаются при патологических хороиретинальных очагах,
сопоставимых по диаметру с диском зрительного нерва (кровоизлияния, опу-
холи, воспалительные фокусы и пр).
II. СУЖЕНИЕ ГРАНИЦ ПОЛЯ ЗРЕНИЯ.
Монокулярное:
• сужена часть нижнего поля зрения без четкой квадрантной или гемиано-
пической локализации, ощущение пелены снизу и медиально, ослабеваю-
щей после постельного режима (свежая отслойка сетчатки с разрывом в
верхненаружной или верхней части глазного дна);
• сужена часть верхнего поля зрения; с ощущением нависающей пелены,
усиливающемся при физической активности; с течением времени возника-
ет стойкий горизонтальный уровень дефекта с выключением всей верхней
половины (при свежих отрывах или разрывах сетчатки в нижних отделах);
• постоянное выпадение верхней половины поля зрения, к которому обыч-
но привыкают, оно переносится легче, чем выпадение нижней половины
(при старых отслойках сетчатки);
• клиновидное сужение в верхневнутреннем или нижневнутреннем квадран-
тах, без четкого обозначения границ квадранта по вертикали или гори-
зонтали; может долго не замечаться при бинокулярном зрении из-за нало-
жения внутренних половин полей зрения обоих глаз (развитая или далеко
зашедшая открытоугольная глаукома, в том числе при нормальном оф-
тальмотонусе);
• конусовидное сужение поля зрения с дугообразным (серповидным) пат-
терном, который вершиной конуса связан со слепым пятном, а расширяю-
103
щимся основанием уходит к периферии (скотома Йенсена), возникающая
при юкстапапиллярных патологических очагах , которые чаще всего яв-
ляются очагами хронического продуктивного воспаления хороидеи. Так
как они располагаются по краю диска зрительного нерва, поражаются
также нервные волокна, проходящие по соседству в сетчатке и обеспечи-
вающие функцию определенного сектора в поле зрения;
• выпадение на одном глазу всей верхней или нижней половины поля зре-
ния, или альтитудинальные скотомы (ишемическая оптическая нейропа-
тия).
Бинокулярное:
• выпадение нижних половин с четкой горизонтальной границей (травмы, в
особенности огнестрельные ранения черепа, с поражением обеих затылоч-
ных долей коры больших полушарий головного мозга в области клина);
• выпадение гомонимно правых или гомонимно левых половин поля зре-
ния с четкой границей по вертикальному меридиану — недостаток, кото-
рый может быть компенсирован поворотом головы в сторону нарушения
поля зрения (при решении вопроса об уровне поражения в этих случаях
учитывают данные об отсутствии или наличии гемианопической реакции
зрачка: при сохранении реакции зрачка на очень слабый свет поражен цен-
тральный нейрон одной из гемисфер зрительной коры, отсутствие реак-
ции свидетельствует о поражении зрительного тракта, противоположно-
го гемианопическому дефекту);
• выпадение на обоих глазах и правых, и левых половин поля зрения с со-
хранением лишь островка в центре поля зрения 8—10° (патология преклон-
ного возраста как результат обширной ишемии обеих половин затылоч-
ной коры атеросклеротического генеза; островок зрения сохраняется из-
за двойного кровоснабжения проекции макулярной зоны ветвями
внутренней сонной и позвоночной артериями;
• выпадение гомонимных (правых или левых) верхних либо нижних квад-
рантов полей зрения с четкой границей не только по вертикальному, но и
по горизонтальному меридиану; чаще встречается верхнеквадрантная го-
монимная гемианопсия (признак поражения пучка Грациоле при опухол®
или абсцессе в соответствующей височной доле; зрачковые реакции не на-
рушаются);
• перекрестное (гетерономное) выпадение либо половин, либо квадрантот
поля зрения (патогномонично для хиазмальной патологии):
при биназальной гемианопсии, которая часто сочетается с концентричес
ким сужением поля зрения и центральными скотомами, характерна выра
женная флюктуация изменений (оптико-хиазмальный арахноидит);
при битемпоральной гемианопсии: при выраженной несимметричности де
фектов, появляющихся в первую очередь в нижненаружных квадранта:
(супраселлярные менингиомы бугорка турецкого седла, опухоли III желу
дочка и аневризмы в этой области); при превалировании верхненаружны:
дефектов (аденома гипофиза, аневризмы внутренней сонной артерии и е<
ветвей);
• периферический дефект поля зрения, моно- и бинокулярный, не уклады
вающийся в описание представленных выше вариантов, в отсутствие из
менений глазного дна, объясняющих выпадение поля зрения (может быт:
следствием давления на зрительный нерв в орбите, костном канале ши
полости черепа опухоли, гематомы, обломков кости; так может начинать
ся пре- или постхиазмальный процесс либо проявляться периневрит зри
104
тельного нерва). В основе изменений в поле зрения могут лежать корко-
вые нарушения. Так, при истерии нередко определяется значительное кон-
центрическое сужение границ поля зрения (до 10—30°), что не согласует-
ся с сохранением способности хорошо ориентироваться в окружающей об-
становке. Центральных скотом при истерии не бывает.
Функциональное поле зрения (ФПЗ) в отличие от поля зрения,
исследованного в стандартных условиях, характеризуется тем, что
оно определяется бинокулярно при наличии нестандартных фонов
без фиксации головы, но непременном сохранении единой точки фик-
сации взора. По данным G. Verriest с соавт. (1982), описанное вне-
осевое (периферическое) зрение в обычной жизни, в частности при
решении поисковых задач, играет исключительно важную роль. Эк-
страфовеальная информация в условиях сложного меняющегося фона
управляет саккадическими движениями глаз и способствует оптими-
зации профессиональной деятельности.
Согласно результатам скрининговых исследований, проведенных
у 10 000 водителей автотранспорта, 3—3,5% обследованных в возра-
сте до 60 лет не знали о наличии у них нарушения поля зрения, при-
чем более половины никогда не испытывали затруднений в связи со
зрением [Keltner J., Johnson Ch., 1982]. Примечательно, что если мо-
нокулярные нарушения совсем не влияли на частоту аварий, то при
бинокулярных она была в 2 раза выше в тех же возрастных группах.
Г Л АВ А 4
ЦВЕТООЩУЩЕНИЕ
4.1. АНАТОМО-ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЦВЕТОВОГО
ЗРЕНИЯ
4.1.1. Трихроматичность цветового зрения
Представление о существовании в сетчатке нормального человечес-
кого глаза трех цветовых рецепторов, которые различаются пре-
имущественной чувствительностью к одному из трех основных цве-
тов и вместе с тем способны воспроизвести все многоцветие окру-
жающего мира, связывают с именами М.В. Ломоносова, Т. Юнга и
Г. Гельмгольца [Кравков С.В., 1951].
Экспериментальные подтверждения справедливости гипотезы о
существовании трех классов колбочек получены лишь в середине
текущего столетия благодаря возможностям в то время новых ме-
тодов отраженной денситометрии и микроспектрофотометрии.
В настоящее время установлено наличие трех типов колбочек,
содержащих различные пигменты с пиком спектральной чувстви-
тельности в зоне 570—590,535—555 и 440—450 нм соответственно
[Rushton W., 1963]. Чувствительность каждого из зрительных
пигментов резко снижается на длинноволновой стороне пика, но
уменьшение чувствительности к данной волне компенсируется вклю-
чением соседнего фоторецептора. Достаточно близкие данные спек-
тральной чувствительности были получены при микроспектрофо-
тометрии: 549—570 нм; 522—539 нм; 414—424 нм для каждого клас-
са рецепторов [Dartnall Н. et al., 1983]. Усредненные данные о кри-
вых спектральной чувствительности трех типов колбочек чело-
веческого глаза представлены на рис. 4.1.
Трехрецепторная теория цветового зрения Гельмгольца подтвер-
ждалась тем, что цветность любой длины волны можно составить,
смешивая в определенных пропорциях три монохроматических цве-
та: 400—450, 510—520, 630—700 нм. Так, в уравнении:
а3 Xj + а4 Х4 = aj Aq + Х^,
где а — интенсивность света, X— длина волны, цвет а3 Х3 может
быть составлен из суммы цветов aj \ + а2 Х2 за вычетом а4 Х4.
Правильность представлений о смеси цветов определяется нор-
мальным состоянием как соответствующих колбочек в сетчатке, так
и нейронов зрительных центров. Одни и те же цветовые сочетания
в фовеа и других отделах сетчатки воспринимаются неодинаково,
что объясняется топографией фоторецепторов в сетчатке. Однако
трихроматичность сохраняется и в случае включения палочковых
106
Рис. 4.1. Спектральная чувствительность фотопигментов трех типов кол-
бочек (В — голубой, G — зеленый, R — красный) нормальной сетчатки че-
ловеческого глаза.
а — по оси ординат — световая абсорбция, по оси абсцисс — длина волны, нм [Mollon
J.D., 1982]; б — то же по Е.Н. Юстовой (1993); по оси ординат — спектральная чув-
ствительность в относительных единицах, по оси абсцисс — длина волны, нм.
рецепторов, так как последние замыкаются на те же расположен-
ные выше нейроны сетчатки, что и колбочки. Таким образом, для
проведения информации о цвете имеется только три канала, каж-
дый из которых формируется совокупностью восприятий колбоч-
ковых рецепторов, подразделяющихся у человека на три типа, в
зависимости от особенностей колбочкового пигмента.
4.1.2. Цветовой треугольник и координаты цветов
Различные варианты объемных моделей цветового тела дают пред-
ставление о цветовых смесях и об относительном расположении
неотличимых один от другого цветов.
Особой простотой, но и определенной условностью отличается
модель МКО (колориметрическая система CIE или МКО, разрабо-
танная Международной комиссией по освещению в 1931 г.), в кото-
рой все координаты из объемной модели переведены на плоскость
XYZ (рис.4.2).
В качестве эталонных длин волн, размещенных на вершинах
треугольника XYZ, приняты соответственно 450, 520,650 нм. Пози-
цию искомой длины волны, образующейся в результате составле-
ния смеси цветов, и энергетический вклад формирующих ее цветов
можно определить путем построения цветового треугольника на
основе “правила центра тяжести — W” (см. рис.4.2).
W представляет собой центр тяжести весов (интенсивностей): а,
от X, а2 от Y, а3 от V, а4 от Z до W. Искомым оказался цвет с
107
Рис. 4.2. Диаграмма
цветностей в системе
CIE (Comisson Interna-
tionale de L'Eclairage)
или MKO (1931) no Wa-
shington (1963). Объяс-
нение в тексте.
длиной волны 500 нм. Цветовой треугольник не дает описание
цвета полосы смешиваемых частот (если не определены факторы
фона, адаптации и пр.), он позволяет лишь определить, какими
еще смесями можно воспроизвести один и тот же цвет (точнее,
цветность).
Те длины волн, энергия которых при смешении в определенных
пропорциях дает белый цвет, называют дополнительными. На
рис.4.2 их можно определить, проведя прямую линию через точку Е
(либо А или С при других условиях освещения). Точки пересечения
этой линии с кривой цветности по одну и другую сторону от Е
показывают дополнительные длины волн. Так, например, для 455 нм
это будет 575 нм, а для 650 нм — 497 нм и т.д.
Для создания надежных тестов для оценки нарушений цвето-
ощущения необходимо рассматривать взаимоотношения восприя-
тия цветовых смесей колориметра (КЗС) относительно линий не-
различения их цветослепыми не в плоскостной, а в объемно-вектор-
ной системе (рис. 4.3), т.е. в физиологической системе координат
цветового пространства (RGB) [Юстова Е.Н. и др., 1986 — 1993].
По мнению авторов, лишь в этом случае определяется цвет, а не
цветность. Последняя не содержит полной информации о цвете, так
как определяется только двумя независимыми координатами. Цвет-
ность, являющуюся результатом вычисления, измерить непосред-
ственно при колориметрии невозможно.
Предварительные колориметрические исследования, выполнен-
ные Н.Д. Нюбергом, Е.Н. Юстовой и др. еще в начале 50-х годов на
дихроматах, позволили установить в системе цветовых координат
колориметра (КЗС) взаиморасположение тех линий, по которым
108
Рис. 4.3. Объемное век-
торное совмещение цве-
тов колориметра КЗС и
цветов в физиологичес-
кой системе координат
RGB [Юстова Е.Н. и др.,
1993].
теряется способность к восприятию определенного цвета вследствие
нарушения функции соответствующего приемника (рис.4.4).
Кривые относительной спектральной чувствительности трех
цветоприемников глаза нормального трихромата представлены на
рис. 4. 1. Именно относительно этой системы координат (RGB)
Е. Н. Юстова и ориентировала обнаруживаемые ею нарушения цве-
тового зрения, постоянно испытывая при этом затруднения в трак-
товке цветоаномальности.
Для разграничения понятия цветослабость и цветоаномальность
В.В.Волков (1995) предложил рассматривать снижение уровня той
или иной кривой как проявление слабости приемника (вплоть до
потери чувствительности), а сдвиг пика кривой влево или вправо —
как признак аномальности (без непременной связи с потерей силы
приемника).
4.1.3. Основные параметры цвета
Цветовой тон определяется длиной волны излучения. Так, при раз-
ложении белого цвета призмой на спектр составляющих его цветов
в качестве основных выделяют фиолетовый (430 нм), синий (460 нм),
зеленый (520 нм), желтый (575 нм), оранжевый (600 нм), красный
(650 нм) и пурпурный (> 650 нм). Монохроматический цвет с про-
межуточной длиной волны вызывает ощущение смешанного цвето-
вого тона, например желто-зеленого (при X > 520 нм и < 575 нм).
109
с
'Э—и—и *-*-----
'м—к—но х-х----
M-Х X X—о—
-X—м—О---м—х-х
Л « . О-«< . , 3
20 40 60 80 100
Рис. 4.4. Координаты неразличимых дихроматами цветов в экспериментах
Е.Н. Юстовой и соавт. (1993).
а — при протанопии; б — при дейтеранопии, о — местоположение заданного цвета,
х — соединенные прямой линией координаты других цветовых смесей, не отличаю-
щихся от заданного цвета, К, 3, С — количественные характеристики соответствен-
но красного, зеленого и синего цветов. у
Насыщенность — качество, под которым понимают интенсив-
ность цветового тона, слабую или сильную окрашенность. Цвето-
вой тон определенной спектральной волны теряет насыщенность
при разбавлении его белым цветом или другими составляющими.
Светлоту характеризуют как безразмерную величину, исполь-
зуемую для количественной оценки различий между световыми ощу-
щениями от двух смежных одноцветных поверхностей [Советский
энциклопедический словарь. М.,1980]. Применительно к сравнива-
емым цветам очевидно, что желтый, например, всегда светлее ко-
ричневого. Однако сложность состоит в том, что в условиях фото-
пического зрения на светлоту стимула большое влияние оказывает
ПО
яркость фона. С усилением последнего цветовой стимул становится
темнее и, наоборот, с понижением общей освещенности (в сумер-
ках), например, сине-зеленые цвета становятся светлее, а оранжево-
красные — темнее (феномен Пуркинье). Однако это происходит уже
на рубеже потери ощущения цветности.
Для спецификации светлоты использовали различные модели
[Соколов Е.Н., Измайлов Ч.А., 1984]. Установлено, что в опреде-
ленных пределах (при не слишком больших различиях по светлоте)
можно оперировать логарифмами яркости (согласно закону Вебе-
ра — Фехнера).
Световая энергия, вызывающая ощущение цвета, обладает двумя
основными взаимозависимыми свойствами: интенсивностью и дли-
ной волны. Интенсивность может быть определена по числу фото-
нов, попадающих на определенную область в единицу времени. Элек-
тромагнитные волны различной длины вызывают ощущение цвета
от голубого до красного. Глазом может быть виден только свет с
длиной волны 400—700 нм, абсорбируемый фоторецепторами. Ма-
лейшие квантитативные изменения в физических параметрах цвета
приводят к изменениям в его восприятии (см. главу 1).
Смешение трех монохроматических цветов в синей, зеленой и
красной части спектра вызывает цветоощущение любых произволь-
ных спектральных композиций. Монохроматический свет из разных
областей спектра (коротковолновой, средневолновой и длинновол-
новой) абсорбируется тремя типами колбочковых пигментов (резо-
наторы Юнга), так что каждая клетка генерирует ответ, пропорцио-
нальный числу абсорбированных фотонов. Относительная величина
этих ответов соответствует определенной длине волны, что и оцени-
вается в центральных отделах зрительного анализатора.
Распространенное представление о том, что палочки не участву-
ют в цветовом зрении, справедливо лишь относительно фовеальной
области. Т. Wiesel и D. Hubei (1966) установили, что цветокодирую-
щие нейроны в ЦНС получают импульсы также и от палочек. Уча-
стие палочек в цветовом зрении реализуется при мезопических усло-
виях видения [DawN., HartW., 1987]. По мнению McCann (1969),
цветовой контраст воспринимается главным образом палочками и
только в известной мере красно-колбочковыми рецепторами.
4.1.4. Об одновременном и последовательном цветовом контрасте
Явление одновременного контраста имеет прямое отношение к од-
ному из ’’наиболее достопримечательных и важных факторов в
области физиологической оптики”,— по выражению Э. Геринга,—
константности цветовосприятия. Константность цветовосприятия
определяется способностью зрительной системы правильно узна-
вать окраску предметов в разнообразных условиях освещения [Мак-
симов В.В., 1984]. Цветовое ощущение может определяться только
111
излучением, попадающим на сетчатку, но одно и то же излучение в
разных условиях может вызывать разные цветовые ощущения. В
зрительной системе существуют специальные компенсаторные ме-
ханизмы, благодаря которым наши цветовые ощущения об окраске
предметов остаются неизменными после изменения освещения (по-
правка на освещение), что представляется как обратное преобразо-
вание (трансформация) цветов. В свою очередь трансформация цвета
рассматривается как синоним константности цветовосприятия.
Серое пятно в красной рамке кажется зеленоватым, а в зеленой,
наоборот, — красноватым, т.е. принимает дополнительный цвет.
Об отсутствии строгой корреляции между ощущаемым цветом и
длиной волны излучения свидетельствует следующее наблюдение
[ Daw N., Hart W., 1987 ]. Как известно, излучения с X 585 нм в воз-
духе создают ощущение желтого цвета, однако в окружении X 650 нм
они кажутся зелеными, а при X 540 нм — красными. Если тот же
экран желтого цвета с X 585 нм окружить рамкой того же цвета, но
на 0,7 лог. ед. более яркой, то экран станет серым, а при яркости
рамки на 2 лог. ед. больше — даже черным.
Суть последовательного контраста состоит в сохранении перед
глазом ощущения пятна в течение некоторого времени после при-
стального разглядывания яркого источника света. Это пятно при-
нимает окраску, дополнительную к той, которая вызвала феномен.
В нормальных условиях значение этого феномена невелико из-за
непрерывных движений глаз и частой смены рассматриваемых сцен.
Явление последовательного цветового контраста связывают с
функцией цветооппонентных клеток, которые располагаются уже в
ганглиозном слое сетчатки и наружных коленчатых телах. Клетки
такого типа имеют одну полярность на один цвет и противополож-
ную полярность на другой,, дополнительный, цвет. Существуют
клетки, которые на включение света дают ощущение красного цве-
та, а на выключение — зеленого, причем на белый цвет они вообще
не отвечают.
В центральных отделах зрительного анализатора находятся двой
ные оппонентные клетки, которые по разному реагируют при воз
действии на центральные либо периферические отделы рецептивно
го поля сетчатки. Двойные оппонентные клетки отвечают за функ
цию одновременного цветового контраста.
Современные представления о нейронной организации зритель
но-нервных путей допускают существование в стриарной коре гс
ловного мозга несколько иного, чем в сетчатке, механизмов цвете
восприятия. Как установили M.Livingston и D.Hubel (1981), так не
зываемые пузырьковые клетки образуют оппонентные поля тре
пар цветов: красного — зеленого, синего — желтого, черного -
белого. Таким образом, трехкомпонентная теория Гельмгольца с<
гласуется с оппонентной теорией Геринга.
В исследованиях, проведенных C.Michael (1979) на макаках р
112
Рис. 4.5. Цветовая модель Ман-
селла в виде цилиндрического тела
в трехмерном сенсорном простран-
стве. Образующие цилиндра харак-
теризуют цветовой тон (Н), насы-
щенность (С), светлоту (V).
Н
10РВ
зусах, установлено, что в стриарной коре существуют клетки, спе-
цифичные не только для определения цвета, но и для ориентации.
Эти нейроны могут отвечать на воздействие определенным цветом
при определенной ориентации границ цветовой полосы раздражи-
теля, т. е. анализ цвета осуществляется вместе с восприятием фор-
мы [Livingston М. et al., 1984]. Е. Н. Соколов и Ч. А. Измайлов
(1984) предложили модель цветового пространства в виде "четырех-
мерной сферы, лежащей в четырехмерном эвклидовом простран-
стве". Авторы считают, что три угла, которые образуют полярные
координаты такой сферы, совпадают с субъективными характерис-
тиками цветов: цветовой тон (Н), насыщенность (С) и светлота (V)
(рис.4.5) и коррелируют с координатами цветовой трехмерной мо-
дели, тела Манселла, построенной в 1943 г. для описания отражен-
ного от поверхности света. В модели Харвича и Джемсона цвет
был представлен в виде точек, расположенных в пространстве трех
декартовых координат X (красно-зеленая ось), Y (желто-синяя), Z
(бело-черная). Из модели с помощью расчетов была выведена функ-
ция смешения цветов, порогового цветоразличения и насыщеннос-
ти спектральных цветов.
4.2. ОСНОВНЫЕ ПРИЧИНЫ И ПРИЗНАКИ НАРУШЕНИЙ
ЦВЕТОВОГО ЗРЕНИЯ
В 1986 г. J.Nathans изолировал и идентифицировал ген для трех
колбочковых пигментов, предположив их структурное сходство с
геном родопсина. Локализация одного гена в 7-й хромосоме и двух
других, расположенных очень близко в Х-хромосоме, определила
ИЗ
представление о том, что красно-зеленый дефект цветового зрения
сцеплен с полом. Ген же в 7-й хромосоме обусловливает код для
голубого пигмента. Ген в 6-й хромосоме был идентифицирован у
мужчин, тестирование которых на цветовосприятие показало недо-
статочность в их сетчатке пигмента красных или зеленых колбочек.
Анализ локуса ’’красного и зеленого гена” на Х-хромосоме,
проведенный J.Nathans, позволил предположить механизм возник-
новения различных наследственных дефектов цветового зрения,
включая красно-зеленую цветовую слепоту, впервые описанную
J.Dalton. Врожденные нарушения цветового зрения генетически
обусловлены и рецессивно связаны с полом. Они встречаются у 8%
мужчин и 0,4% женщин. Почти все эти нарушения характеризуются
отсутствием или повреждением одного из трех пигментов фоторе-
цепторов и, таким ообразом, являются причиной дихромазии. По-
вреждение одновременно всех пигментов встречается очень редко и
обусловливает развитие монохроматизма (видение, как в черно-
белом кино). Дихроматы имеют своеобразное цветовое зрение и
нередко узнают о своем недостатке случайно (при специальных
медицинских осмотрах или в некоторых сложных жизненных ситу-
ациях)^ отличие от врожденных приобретенные дефекты цветово-
го зрения, по крайней мере в начале заболевания, проявляются
только на одном глазу. Нарушения цветового зрения при них ста-
новятся более выраженными со временем и могут быть связаны с
нарушением прозрачности оптических сред, но чаще относятся к
патологии макулярной области сетчатки. По мере прогрессирова-
ния заболевания к ним присоединяются снижение остроты зрения,
нарушения в поле зрения, изменяются контрастная чувствительность,
критическая частота слияния мельканий, показатели локальной ЭРГ.
Патологические процессы могут локализоваться и на более
высоком уровне — в зрительном нерве, зрительной коре. Факторы,
вызывающие их, разнообразные: токсические воздействия, сосуди-
стые нарушения, воспалительные, дистрофические, демиелинизиру-
ющие процессы и др.
Несмотря на существующие классификации врожденного и при-
обретенного нарушения цветового зрения, четкого разграничения
клинических проявлений, врожденной и приобретенной цветопато-
логии не существует. Например, при дихромазии встречаются нару-
шения восприятия не одного, а двух цветов; явления цветоаномаль-
ности нельзя рассматривать как предпосылку к возникновению
цветослепоты или ступень ее развития; нет способа избавиться от
врожденных дихромазий, но и приобретенная дихромазия может
оказаться необратимой.
Комитет по международным стандартам унифицировал и пред-
ложил к использованию созданные в разные годы тесты для иссле-
дования цветоощущения [Birch J.G., 1985]. Несмотря на различную
природу врожденных и приобретенных нарушений, было отмечено,
114
что они имеют много общего: характеризуются ошибками в сопо-
ставлении разных цветов, правильном их названии, значительным
уменьшением количества различаемых оттенков и, наконец, смеще-
нием на кривой спектральной чувствительности относительно бо-
лее ярких участков. Так, при врожденных нарушениях первого типа
наряду с потерей способности к различению красно-зеленых оттен-
ков происходит сдвиг относительной яркости к коротковолновой
части спектра; при втором типе наряду с аналогичной потерей спо-
собности к различению красно-зеленых оттенков сдвига яркостной
чувствительности не наблюдается; при третьем типе теряется спо-
собность различать сине-зеленые оттенки, а относительная яркость
сдвигается, как и при первом типе, в коротковолновую часть спек-
тра. Нарушения третьего типа сходны с врожденным тританопи-
ческим дефектом, который встречается только у одного из 13 000
обследованных.
Некоторые приобретенные дефекты проходят от развития фазы
трихроматических через фазу ди- и монохроматических нарушений.
Чаще определяются нарушения третьего типа, при этом расширяется
зона сниженной чувствительности в сине-желтой части спектра.
4.3. ФУНКЦИОНАЛЬНЫЕ ТЕСТЫ НА ЦВЕТОВОЕ ЗРЕНИЕ
Если для выявления врожденных дефектов цветоощущения, особен-
но при массовых профессиональных отборах, с целью экономии
времени допустимо производить проверку двух глаз одновременно
(врожденные нарушения почти всегда двусторонние), то при подо-
зрении на приобретенные изменения цветоощущения тестирование
следует проводить только монокулярно. Нарушения цветового зре-
ния могут быть одним из ранних симптомов приобретенной пато-
логии, поэтому они важны в дифференциальной диагностике.
При скрининговых проверках цветового зрения необходимо
следить за правильным освещением. Комитет по международным
стандартам (1985) рекомендует при проверке цветоощущения у
живущих в северном полушарии земного шара использовать осве-
щенность из окна, ориентированного на север (350—500 лк). Одна-
ко спектральная композиция естественного дневного света непо-
стоянна, что отличает его от искусственного освещения, поэтому
предпочтительно освещение синевато-белым светом "С" (порядка
330 лк), а использование вольфрамовых ламп с желтым светом
должно быть исключено. Во многих светильниках, используемых в
клинических условиях, необходимый свет достигается установкой
на лампы с вольфрамовой нитью в 100 Вт полусферических синих
фильтров (Macbeth easel daylight lamp). Псевдоизохроматические
таблицы стандартизированы для такого света. Можно применять
также высококачественные флюоресцентные лампы. При этом тре-
115
525нм
Рис. 4.6. Хроматические
координаты (в пределах
цветового треугольника)
тех цветов, которые ис-
пользованы: в 15-отте-
ночном тесте Фарнсвор-
та (а) и в 100-оттеночном
тесте Фарнсворта — Ман-
делла (б).
Линии, пересекающие овал
в точке ”с” и касающиеся
его попарно, для цветосле-
пых являются неразличимы-
ми изохромами [Birch J.,
1985].
буются балансировка спектрального распределения и индекс пере-
вода более 90. При скрининговых проверках допускается использо-
вание освещения "С" в диапазоне от 100 до 2000 лк. Источник под-
света важно разместить так, чтобы он не вызывал ни слепящих
рефлексов, ни теней.
При исследовании цветового зрения в вопросах и ответах обыч-
но не используют название цветов. Пробы строятся на определении
116
способности видеть какую-либо фигуру (ее детали), изображенную
одним цветом на другом цветовом фоне, или ранжировать цвета
либо подбирать их под пару.
В настоящее время существует более десятка тестов для диагно-
стики дефектов цветового зрения. Их используют с целью выявле-
ния как врожденных, так и приобретенных нарушений. Различие
состоит лишь в том, что в качестве скрининговых тестов для обна-
ружения врожденных красно-зеленых нарушений наряду с таблица-
ми Ишихары Комитет по международным стандартам рекомендует
использовать спектральный аномалоскоп. В наборе скрининговых
крининговых тестов для диагностики приобретенных цветонаруше-
ний аномалоскоп не упоминается, а, кроме таблиц Ишихары, реко-
мендованы таблицы Американской оптической компании (Hardy—
Rand — Rittler), которые позволяют диагностировать нарушения
сине-желтого приемника, таблицы Японского медицинского кол-
леджа, 15-фишечная панель Farnsworth, в том числе в варианте
объектов ослабленной насыщенности по Лантони (Lanthony).
Т а б л и ц ы. В клинической практике наиболее распростране-
ны псевдоизохроматические таблицы, впервые предложенные Штил-
лингом в 1876 г. Чаще других в настоящее время используются
таблицы Ишихары, Фельхагена, Ишикавы, Рабкина, Флетчера —
Гамблина и др. Таблицы рекомендуется предъявлять на расстоянии
вытянутой руки (0,66 м). При этом таблицы должны освещаться
светом определенного спектрального состава. Исследователь сам
переворачивает страницы, контролируя таким образом время
предъявления символов (около 4 с) и просит обследуемого назвать
цифру или фигуру сразу после переворачивания страницы. Чтение
всех знаков, в том числе и скрытых (псевдоизохроматических), ре-
гистрируется.
Панельные тесты ранжировки цветов. Наи-
большее распространение и признание в диагностике приобретен-
ных нарушений цветового зрения получили панельные тесты, со-
зданные на основе стандартного "атласа цветов” Munsell. За рубе-
жом широко используют 15-, 85- и 100-оттеночные тесты
Фарнсворта (Farnsworth) различных цветов (1940). Цветовые от-
тенки, избранные для тестирования, расположены в цветовом тре-
угольнике (МКО) по контуру овалов вокруг нейтральной точки
(рис. 4.6). В первом случае (рис. 4.б' а) изохроматические линии не-
распознавания пересекают овал в трех направлениях, во втором
(рис. 4.6, б) они по касательным охватывают этот овал. Таким об-
разом, сделана попытка представить замкнутый круг существую-
щих цветовых оттенков как можно более полно. В некоторых вари-
антах теста для усложнения задачи уменьшают насыщенность цве-
тов и для определения нейтральной зоны вводят шкалу серых фишек
(28- и 40-оттеночные тесты Лантони как вариант 15-оттеночного
теста Фарнсворта).
117
В задачу обследуемого входит расставить фишки, начиная с за-
данной (в 15-оттеночном тесте — голубая фишка), чтобы цвет от
фишки к фишке плавно изменялся (по кругу).
В 100(85)-оттеночном тесте фишки хранятся в 4 емкостях, и за-
дача решается в 4 приема: в каждой подгруппе расставляют по
21 фишке, соответствующей цветностям одного из квадрантов (N} —
от розового через оранжевый к желтому; N2 — от желтого к зелено-
голубому; N3 —от зелено-голубого к сине-пурпурному; N4 — от си-
него через красно-пурпурный к розовому), и в конечном счете фор-
мируется замкнутый цветовой круг. Рекомендуется ограничивать
время расстановки фишек из каждой коробки 2 мин. На скорость
выполнения теста и его результаты могут влиять как тренирован-
ность обследуемого, так и его утомление.
В 15-оттеночном дихотомном тесте позиции перепутанных мес-
тами фишек мгновенно замечаются, так как соединяющие их пря-
мые линии не очерчивают, а пересекают тестовый круг (рис.4.7).
Эти линии направлены под прямым углом к линии неразличения
соответствующего цвета в цветовом треугольнике (например, точ-
ки 1 и 14 при протадефиците, точки 4 и 13 при дейтердефиците, 8
и 15 при тритадефиците). Панель Д-15, особенно в усложненном
варианте (с менее насыщенными цветами), широко применяется при
профессиональном отборе. В клинической практике 100(85)-отте-
ночный тест Фарнсворта позволяет более точно определять харак-
тер нарушения цветовосприятия по определенному типу при врож-
денных и приобретенных состояниях. При обработке результатов
размещения фишек каждый объект характеризуется суммой разно-
стей его номера с номерами двух, оказавшихся по соседству. Если
последовательность установлена правильно, сумма разностей номе-
ров составляет 2 (нулевая отметка). При ошибочной установке ре-
зультат всегда будет превышать цифру 2; чем выше окажется иско-
мый показатель, тем тяжелее дефект цветоразличения в направле-
нии соответствующих изохром. Это позволяет судить о типе
нарушения, суммарная же погрешность, с учетом всех меридианов,
свидетельствует о степени цветонарушения. Например, при выра-
женном дефекте восприятия синего цвета на схеме отчетливо видна
полярность нарушений в двух диаметрально противоположных
направлениях от центра цветового круга (рис.4.8).
В последние годы J.D.Mollon значительно упростил тест. В пред-
ложенном им наборе имеются красные, зеленые и синие фишки, отли-
чающиеся не только по цвету, но и по его насыщенности. Перемешан-
ные в беспорядке фишки обследуемый должен разобрать по цветам и
ранжировать по насыщенности. В качестве эталона ему предлагают
установленный в требуемом порядке набор из серых фишек.
Аномалоскопы — приборы, основанные на принципе до-
стижения субъективно воспринимаемого равенства цветов путем до-
зированного составления цветовых смесей. Классическим прибором
118
a
Рис. 4.7. Примеры диагностического использования 15-оттеночного теста
Фарнсворта при слабых и сильных степенях нарушения цветоприемников
[Birch J, 1985].
а — красного; б — зеленого; в — синего. Pilot — голубая фишка.
119
Рис. 4.8. Пример диагностического использования 100-оттеночного теста
Фарнсворта. Отмечается четкое нарушение цветовосприятия синего цвета
[Birch J., 1985].
этого типа, предназначенным для исследования врожденных нару-
шений восприятия красно-зеленых цветов, остается аномалоскоп
Нагеля. По способности уравнять полуполе монохроматического
желтого цвета с полуполем, составленным из смеси чисто красного
и зеленого, дозируемой перемещением щели, судят о наличии или
отсутствии нормальной трихромазии. Последней свойственны стро-
го определенные пропорции смесей (уравнение Релея), хотя некото-
рые различия между показателями парных глаз вполне допустимы.
Аномалоскоп позволяет диагностировать как крайние степени дих-
ромазии (протанопию и дейтеранопию), когда обследуемый прирав-
нивает к желтому чисто красный или чисто зеленый цвета, меняя
лишь яркость желтого полуполя, так и более умеренно выраженные
120
нарушения, заключающиеся в необычно широкой зоне, в пределах
которой смешение красного с зеленым воспринимаются как желтый
цвет (названо это было протаномалией и дейтераномалией).
По тому же принципу, что и аномалоскоп Нагеля, в более по-
зднее время были построены аномалоскопы Мореланда (Moreland),
Найтца (Neitz), Рабкина* аномалометр Безансона (Besancon) и спек-
тротест Рихтера (Richter). Наиболее совершенным из приборов
данного типа является аномалоскоп Гейдельберга фирмы "Oculus".
Попытки создать условия для диагностики тританопии путем
включения тестов на сине-зеленые смеси [вторая модель аномало-
скопа Нагеля, аномалоскоп Пикфорд — Николсон (Pickford —
Nicolson)] пока нельзя признать удачными.
В 1957 г. Г.Н.Раутианом создан аномалоскоп, который в тече-
ние многих лет серийно выпускали в нашей стране (модель АН-59).
Прибор впервые предоставил возможность не только оценки ра-
венства Релея, но и пороговой чувствительности каждого из трех
цветоприемников зрительной системы (RGB). Понятия цветоано-
мальности и цветослабости начали приобретать самостоятельное
значение.
Аномалоскоп Раутиана позволяет определить цветосилу (цвето-
слабость) исследуемого приемника в условных единицах, обознача-
емых как количество порогов цветоразличения (чем больше требует-
ся этих дифференциальных порогов, тем ниже чувствительность).
Пороговые таблицы Юстовой, Алексеевой,
Волкова, Рослякова и Сергеева. Развивая идеи
Г. Н. Раутиана, Е. Н. Юстова (1953.), а затем К. А. Алексеева на базе
вышеописанной физиологической системы цветовых координат RGB
(см. рис.4.3) разработали методику априорного подбора цветов, не-
различаемых дихроматами (см. рис. 4.4). Именно данная система
позволила воспроизводить желаемые стимулы непосредственно в
колориметре. Благодаря этому были разработаны новые оригиналь-
ные таблицы, в основу которых был положен тот же пороговый
принцип оценки цветослабости и дихромазии, что и в аномалоско-
пе Раутиана. Отличие заключалось лишь в том, что пороговые
различия между сравниваемыми цветами в аномалоскопе улавлива-
ются плавно, а в таблицах — дискретно.
Степень сложности различения пар цветов, отобранных для тес-
тирования, измеряемая количеством порогов для сильного нормаль-
ного трихромата, устанавливали в экспериментах на колориметри-
ческой вертушке Максвелла. В набор включено 12 таблиц: по 4 для
испытания красного и зеленого приемников, 3 — для синего и 1 —
контрольная, служащая для исключения симуляции. Таким образом,
предусмотрена трехступенчатая оценка цветослабости каждого при-
емника, а для красного и зеленого — и тест на цветослепоту.
Апробация таблиц на тысячах обследованных показала высо-
кую повторяемость результатов исследования и тем самым четкость
121
г
е
Рис. 4.9. Основные вари-
анты патологических
хроматограмм (по клас-
сификации Гункеля—
Когана).
а — ахроматический "язык”
в направлении зеленого цве-
та; б — красного; в — крас-
но-зеленый ахроматический
’’пояс"; г — ахроматический
"язык" в направлении жел-
того; д — синего; е — сине-
желтый ахроматический
"пояс". Нередко встречается
небольшое расширение цен-
тральной ахроматической
зоны во все стороны.
количественной и качественной оценки порогов цветоразличения,
невозможность заучивания тестов. Кроме того, благодаря простоте
эксплуатации таблиц они доступны практическому врачу.
Более простым вариантом таблиц Юстовой является планшет
[Рубцова И. В., 1994], в котором удалось уменьшить всю их систему
за счет модификаций тестируемого объекта. Планшет позволяет дать
количественную оценку различительной цветосилы, т. е. чувствитель-
ности каждого из трех цветоприемников сетчатки.
Приборы для определения порога воспри-
нимаемой насыщенности того или иного
цвета, протяженности "нейтральной зон ы".
Минимально различимую насыщенность цвета определяют путем
либо добавления к нему белого, как в анализаторе Ловибонда, либо
перемещения от белой "нейтральной" зоны в направлении насыще-
ния исследуемого цветового тона, как в хромографе, созданном
R. Gunkel и D. Cogan (1978), и хромотесте, разработанном
А. В. Луизовым и соавт.(1990). Первые признаки насыщения цвет-
ности основного экрана обследуемому помогает уловить сравнение
его с контрольным, яркость которого синхронно меняется, но он
остается серовато-белым. В хромотесте Луизова на круговой диа-
122
грамме регистрируются по радиусам пороги цветовой чувствитель-
ности (обычно не более чем по 12 цветам), начиная от белого, рас-
положенного в центре круга.
При оценке результатов по классификации Гункеля — Когана
выделяют 6 вариантов диагноза цветослепоты (рис. 4.9). Важным
достоинством методики является возможность графического выра-
жения размеров и направленности цветонейтральной зоны в систе-
ме цветоприемников каждого индивидуума.
При оценке цветоощущения в клинической практике пороговые
таблицы Е. Н. Юстовой и соавт. заняли ведущее место, особенно в
дифференциальной диагностике приобретенных заболеваний зритель-
ных путей, в диагностике начальных нарушений прозрачности хру-
сталика (предкатаракта), при которых одним из наиболее частых
симптомов, выявляемых таблицами, оказался тритадефицит второй
степени [Рубцова И. В., 1995; Волков В. В., 1996]. Таблицы могут
быть использованы и при мутных средах, если сохраняется формен-
ное зрение не ниже 0,03—0,04. При более низком зрении ориентиро-
вочное представление о сохранности цветоощущения можно полу-
чить, направляя в глаз пациента свет от офтальмоскопа или щелевой
лампы через цветные светофильтры. Однако в дифференциальной
диагностике заболеваний зрительно-нервного аппарата предлагает-
ся использовать описанные в данном разделе специальные методики.
Перспективы в улучшении диагностики офтальмологической и
нейроофтальмологической патологии открывает новый метод, раз-
работанный А.М.Шамшиновой и соавт. (1985 — 1997), — цветовая
статическая кампиметрия. Теоретической предпосылкой к его со-
зданию явились наблюдения о том, что время сенсомоторной реак-
ции на зрительный стимул находится в обратной зависимости от
яркостных различий между стимулом и фоном. С помощью дисп-
лея IBM-совместимого компьютера была реализована возможность
определения порога цветовой и световой чувствительности, време-
ни сенсомоторной реакции, приобретенного нарушения цветоощу-
щения в центральной области сетчатки (40°) в любой точке поля
зрения. Программой исследования предусматривается возможность
изменения не только длины волны и яркости стимула и фона, но и
величины стимула в зависимости от топографии рецептивных по-
лей в сетчатке, уравнивание по яркости стимула и фона (рис.4.10).
Время сенсомоторной реакции оказывалось максимальным в про-
цессе уравнивания, когда яркости фона и стимула были одинаковы,
т. е. создавались условия, при которых время реакции становилось
зависимым только от разности в цвете фона и стимула (цветоразли-
чения). Метод цветовой кампиметрии позволяет проводить "топо-
графическое картирование" световой и цветовой чувствительности
зрительного анализатора при начальной диагностике заболеваний
различного генеза.
На базе компьютерной установки (“Sigma Electronics System
123
1.4
1.2
1
0.8
0.6
0.4
1.4
1.2
1
0.8
0.6
0.4
0.01 0.1 1
в
0.01
a
Рис. 4.10. Результаты исследования цветоощущения при уравнивании по
яркости зеленого стимула на пурпурном фоне с использованием дисплея
компьютера [Ендриховский С.Н. и др., 1996].
На рисунке видно значительное увеличение времени сенсомоторной реакции при
нарушении цветоощущения, а—норма; б — больной с дейтеранопией; в — больные
с начальной глаукомой (vis = 1,0); г — больные с альбинизмом (vis = 0,3). По оси
ординат — время сенсомоторной реакции, мс; по оси абсцисс — единицы яркости
стимула по отношению к фону, кд/м2.
5688” с подчиненным дисплеем “Barco CD351”) D.Regan и соавт.
(1994) реализовали новый вариант быстрой диагностики наруше-
ний цветового зрения путем сочетания принципов Штиллинга
(Stilling) и Чибрета (Chibret). На черном фоне с расстояния 2,4 м
демонстрируются составленные из цветовых пятен (как таблицы
Штиллинга) кольца Ландольта с разрывом в 1°. Цветовые парамет-
ры фона и знака изменяют в системе X, Y- или Up V^-координат
(CIE, 1931 или CIE, 1960). С помощью скрининговой методики
(4 мин) определяют пороги по линиям неразличения дихроматов, а
по полной методике (20 мин) строят эллипсы границ неразличения
[McAdam, 1942]. Направленность оси и вытянутость найденного
эллипса характеризуют вид и степень цветослепоты, или цветоано-
мальности, точнее, цветослабости (рис. 4.11). При врожденных по-
роках результаты наблюдений совпадали с показателями как ано-
малоскопа Нагеля, так и 100-оттеночного теста Фарнсворта — Ман-
селла (Farnsworth — Munsell). Примечательно, что распознавание
цветовых сигналов легче удавалось обследуемым при наличии ’’яр-
костных шумов”, возникавших в приборе [Mollon J.D., 1982]. Одна-
124
Рис. 4.11. Эллипсы цвето-
различения у пациентов
с приобретенной трита-
нопией [Regan D. et al.,
1994].
По оси ординат и абсцисс—
координаты цветности.
ко эта модификация имеет определенные недостатки, которые осо-
бенно четко проявляются при обследовании пациентов с демиели-
низирующей патологией зрительного нерва. Лучше ограничиться
пространственной модуляцией шумов, избегая временной. К числу
“слабых мест“ методики относится также ее недостаточная чувстви-
тельность при низких степенях цветослабост#.
4.4.0 КЛАССИФИКАЦИИ НАРУШЕНИЙ ЦВЕТОВОГО ЗРЕНИЯ
Функциональные дефекты колбочковой системы обусловлены на-
следственными факторами и приобретенными патологическими
процессами на различных уровнях зрительной системы.
Врожденные нарушения цветовосприятия выражаются в неспо-
собности отличать световые излучения, различимые человеком с
125
нормальным цветовым зрением. Эти расстройства связаны с гене-
тическими дефектами и наследуются. Хотя у женщин нарушения
цветового зрения наблюдаются значительно реже, они являются
носителями патологического гена и его передатчиками. Врожден-
ные нарушения цветового зрения связаны чаще всего с нарушением
функции одного из фоторецепторов. Различают три вида врожден-
ных расстройств цветового зрения: дефект восприятия красного
цвета (протан-дефект), зеленого (дейтер-дефект) и синего (тритан-
дефект). По степени нарушения каждый из видов подразделяется на
аномальную трихромазию, протанопию, дейтеранопию и монохро-
мазию [Verriest G., 1982; МаггёМ., МаггёЕ., 1987]. Врожденные
нарушения цветового зрения встречаются у 7—8% населения. Ча-
стота цветоаномалий у мужчин в 10—15 раз выше, чем у женщин.
Под приобретенными нарушениями цветового зрения понима-
ют все изменения цветовосприятия, обусловленные патологически-
ми процессами в сетчатке, зрительном нерве, вышележащих отде-
лах зрительного анализатора. Они могут возникать при соматиче-
ских заболеваниях организма и его интоксикации. К приобретенным
относятся также нарушения цветового зрения, которые возникают
как следствие генетически обусловленных и приобретенных заболе-
ваний сетчатки. Приобретенные нарушения цветового зрения все-
гда вторичны, поэтому их определяют случайно. В зависимости от
чувствительности метода исследования эти нарушения могут быть
диагностированы уже при начальном снижении остроты зрения, а
также при ранних изменениях на глазном дне. Если в начале забо-
левания снижение хроматической чувствительности может касаться
либо красного, либо зеленого, либо синего цвета, то при развитии
патологического процесса снижается чувствительность ко всем трем
основным цветам, чаще к зеленому, затем красному и синему.
Е. Б. Рабкин подразделял, например, приобретенные нарушения
цветового зрения на формы, схожие с врожденными цветовыми
расстройствами, формы, не схожие с врожденными цветовыми рас-
стройствами, и смешанные формы.
Первой достаточно стройной клинической классификацией на-
рушений цветового зрения была классификация Криса — Нагеля
(Kries — Nagel). J.V.Kries (1907,1911) предложил обозначать непол-
ную цветослепоту, т.е. слепоту на отдельные цвета, терминами прот-,
дейтер- и тританопия. Термин же аномальность, уже тогда использу-
емый среди специалистов, J.V.Kries применил к ослаблению цвето-
вого чувства. Так, в классификации Криса— Нагеля появились гра-
фы прота-, дейтер- и тританомалии. В прота- и дейтераномалии
Е. Б. Рабкиным (1971) введены подгруппы (АВС): А—резкое сниже-
ние, В — среднее, С — легкое. Основываясь на колориметрических
измерениях в стремлении к этиопатогенетическому обоснованию ме-
ханизмов как врожденных, так и приобретенных нарушений цвето-
ощущения, J. V. Kries (1911) выделил три системы:
126
{.Абсорбция (объясняет цветоаномалию нарушением про-
хождения “цвета” через абсорбционную систему, например пигмент);
2. Альтерация (дейтераномалия и протаномалия, врожден-
ные формы аномальной трихромазии, при которых происходит из-
менение восприятия света в какой-то части зрительной системы,
что интерпретируется как нарушение в системе альтерации);
3. Редукция (дейтер- и протанопия, обе формы врожденной
дихромазии обозначены как редукция нормальной трихроматиче-
ской системы).
При этом система абсорбции реализуется через патологический
пререцепторный фильтр при нормальных ретинальных механизмах.
В спектральной зоне действие фильтра “снижает цветоразличение”
и световую чувствительность.
Для абсорбирующих сред характерно изменение интенсивности,
а не цвета гомогенного спектра света, проходящего через патологи-
ческие среды, эта абсорбционная система характерна для приобре-
тенных цветонарушений, при катаракте, помутнениях стекловидно-
го тела, после инъекции флюоресцеина.
В последующие годы G.Verriest (1958—1983) отнес к альтераци-
онной системе такие формы приобретенной цветоаномалии, при
которых колбочковый пигмент отличается от такового у нормаль-
ных трихроматов. Редукционную систему G.Verriest использовал для
объяснения возникновения таких приобретенных форм нарушений
цветоощущения, при которых встречались нормальные цветовые
уравнения, показатели аномалоскопа соответствовали дейтерано-
пии, а все красно-зеленое принималось одинаковым.
Одна из ранних классификаций [Kdllner Н., 1912] была основана
на результатах колориметрических исследований нарушений цвето-
вого зрения, которые позволили выделить две основные группы: с
изменениями в сине-желтой и красно-зеленой части спектра (с эво-
люцией заболевания), а также прогрессирующую красно-зеленую
слепоту с возможным включением сине-желтых нарушений. Прави-
ло Кельнера гласило: ’’сине-желтые нарушения преимущественно
характерны для болезней сетчатки, красно-зеленые нарушения —
для заболеваний зрительного нерва”. Правда, из этого правила он
делал и исключения (при люэтическом неврите нарушалось воспри-
ятие и сине-желтых, а при макулодистрофиях нередко страдало
восприятие и красно-зеленых цветов). Последующие клинические
наблюдения несколько изменили это представление.
Уточняя справедливость "правила Кельнера", W. Jaeger и
Р. Grutzner (1961) показали, что красно-зеленые нарушения возни-
кают при заболеваниях зрительного нерва, протекающих без нару-
шения кривой относительной спектральной чувствительности, а при
макулодистрофиях отмечается снижение световой чувствительнос-
ти в области красного приемника, т. е. сокращается спектр со сто-
роны его длинноволновой части.
127
Таблица 4.1. Классификация приобретенных нарушений цветового
Тип наруше- ния Классификация нарушений восприятия
смеси цветов цветового тона
по оси дисхроматопсии по степени тяжести
Тип I Красно-зе- леные Трихромазия Монохрома- зия Красно-зеленые Легкие Тяжелые
Тип II Красно-зе- леные Трихромазия Монохрома- зия Красно-зеленые с сопутствующими сине-желтыми Легкие Тяжелые (иногда)
Тип III Сине-жел- тые Трихромазия Дихромазия (редко моно- хромазия) Сине-желтые Легкие Умеренно тяжелые
Для объяснения причин более частых нарушений цветовосприя-
тия в красно-зеленой части спектра, в частности аномальной три-
хромазии, J. D. Mollon (1995) предложил гипотезу о наличии в чело-
веческом глазу двух генетически обусловленных подсистем цветово-
го зрения: древней (сине-желтой) и появившейся позже, относительно
молодой (красно-зеленой). Нормальное функционирование последней
и обеспечивает полноценное цветовое зрение в современном мире.
Последующие классификации цветонарушений, основанные на
кривой спектральной чувствительности, определяемой по 100-отте-
ночному тесту Фарнсворта — Манселла, предложены J. Francois,
G. Verriest, W. Jager, P. Grutzner.
В настоящее время в мировой клинической практике признана
классификация приобретенных нарушений цветового зрения, раз-
работанная G. Verriest, в которой цветонарушения подразделены
на три типа в зависимости от механизмов их возникновения: абсор-
бции, альтерации и редукции (табл. 4.1).
1. Приобретенные прогрессирующие нарушения восприятия
красно-зеленого от трихромазии до монохромазии по оси дисхро-
матопсии, от легких до тяжелых, которые сопровождаются ухудше-
нием зрения. Фотопическая кривая относительной спектральной чув-
ствительности смещается к коротковолновой части спектра, пока
не достигает скотопического уровня, как при врожденной ахрома-
топсии. На аномалоскопе выявляются изменения различной степе-
ни выраженности от протаномалиидопротанопии и ахроматопсии.
Нарушения этого типа характерны для патологии макулярной об-
128
зрения по A. Pinckers, J. Pokorny, V. С. Smith и G. Verriest (1979)
Механизм по Kries Острота зрения Исход
Альтерация и скотопизация Снижена от умерен- ной до резкой степени Ахроматопсия (скотопическая)
Редукция То же Восстановление или прогрессирующее ухудшение
Редукция или абсорбция или альтерация Снижена от легкой до значительной степени Различный
ласти сетчатки и свидетельствуют о нарушениях в колбочковой
системе, в основе которых лежат механизмы альтерации и скотопи-
зации, исход этих нарушений — ахроматопсия (скотопическая).
2. Приобретенные красно-зеленые нарушения, характеризующи-
еся прогрессирующим ухудшением различения цветового тона от
трихромазии до монохромазии по оси дихроматопсии, сопровож-
даемые сине-желтыми нарушениями, от легких до тяжелых форм.
Относительная кривая спектральной чувствительности нормальная.
В ранней стадии нарушения более насыщенные тона кажутся блек-
лыми; на аномалоскопе в равенстве Релея расширен диапазон зеле-
ного. В случае прогрессирования нарушений с изменением остроты
зрения все красно-зеленые уравнения при нормальной установке
яркости принимаются. При тяжелом течении заболевания с эксцен-
тричной фиксацией цветовое зрение приобретает форму ахрома-
топсии и может проявляться скотопизацией. Механизм нарушения
цветового зрения — редукционный, исход нарушения — восстанов-
ление цветового зрения или прогредиентное ухудшение. Наруше-
ния этого типа встречаются при заболеваниях зрительного нерва.
3. Приобретенные сине-желтые нарушения цветового зрения по
сине-желтой оси дисхроматопсии, от легкой до среднетяжелой фор-
мы, изменения от трихромазии к дихромазии (редко монохрома-
зия). В ранних стадиях заболевания больные путают цвета пурпур-
ный, фиолетовый, синий и сине-зеленый, при его прогрессировании
наблюдается дихроматическое цветовое зрение с нейтральной зо-
ной в области около 550 нм. Кривая относительной спектральной
129
чувствительности может быть нормальной или измененной. Пока-
затели аномалоскопа в ряде случаев свидетельствуют о псевдопро-
таномалии. Механизм нарушения цветового зрения — редукция,
абсорбция или альтерация, исход нарушения различный. Наруше-
ния этого типа характерны для заболеваний хороидеи и пигментно-
го эпителия сетчатки, заболеваний сетчатки и зрительного нерва,
встречаются также при бурой катаракте.
Эта классификация была расширена М. Маггё, Е. Маггё (1987),
которые сделали попытку дать более четкую количественную оценку
нарушениям по трем первичным механизмам (синему, зеленому и крас-
ному), они ввели дополнительную информацию о показателях абсо-
лютной и относительной спектральной чувствительностии об особен-
ностях зрительной фиксации (фовеальной, эксцентричной и др.).
Однако, несмотря на полноту и многосторонность классифика-
ции G.Verriest и ее модификаций, вопрос о тождественности поня-
тий цветоаномальности и цветослабости, поставленный еще
J. V. Kries, решен не был. Еще С. В. Кравков (1950—1951), Leo
Hurvich (1981) предлагали разделить эти состояния. Попытки ко-
личественно оценить пороги различения любого цвета по насыщен-
ности от белого предпринимались R.Gunkel, D.Cogan (1978). Наи-
более близко к решению этой проблемы подошел Г.Н. Раутиан
(1956), который создал аномалоскоп, позволивший впервые иссле-
довать светочувствительность каждого из трех приемников раздель-
но. В предложенной Г. Н. Раутианом системе классификации форм
цветового зрения уже отсутствует смешение (отождествление) поня-
тий цветоаномальности и цветослепоты. Применив для исследова-
ния цветослабости у одних и тех же лиц пороговые таблицы Юсто-
вой, а для исследования цветоаномальности — аномалоскоп АН-59
(Раутиана), В.В. Волков и В.П. Сергеев (1987) показали, что цвето-
аномалия и цветослабость не являются ’’жестко сцепленными” меж-
ду собой качествами. Так, при обнаружении одновременно и дейте-
раномальности, и цветослабости зеленого приемника еще можно
было объяснить влияние одного фактора на другой. Но нередко
встречалось и раздельное нарушение этих функций, отсюда возни-
кала необходимость обособленного их толкования. Если вновь
обратиться к кривым чувствительности трех цветоприемников сет-
чатки (см. рис. 4.1), нетрудно представить возможность изменения
любой из кривых как по высоте (сдвиг по ординате), так и по по-
ложению спектрального максимума (сдвиг по абсциссе). В первом
случае это свидетельствовало бы об изменении цветосилы приемни-
ка, а во втором — об аномальности цветовосприятия. Заслуживает
внимания такой на первый взгляд парадоксальный факт: абсолют-
ное неприятие нормального уравнения Релея при легких степенях
аномальности и спокойное принятие его экстрем-аномалами. Са-
мое простое объяснение этому факту состоит в том, что состояния,
относимые по результатам аномалоскопии к крайним степеням
130
аномальности, по сути не являются таковыми, а представляют со-
бой выраженную цветослабость, граничащую с цветослепотой, по-
тому и реагируют они на аномалоскопическую задачу подобно
дихроматам. В. В. Волковым и соавт. (1987—1995) было установле-
но, что цветоаномальность существует независимо от цветослабос-
ти, хотя и является частым спутником последней, что дашло отра-
жение в предложенной автором новой классификации цветового
зрения, являющейся дальнейшим развитием представлений, впер-
вые высказанных Г. Н. Раутианом (табл 4.2).
Таблица 4.2. Классификация уровней и форм цветового зрения [Волков
В. В., 1995]
Уровни цветового зрения по количеству действующих цветоприемников и их силе Формы цветового зрения по расположению максимумов цветочувствительности в спектре
Расположение максимумов
Количе- ство при- емников Цветосила Нормальное Аномальное
Цветосильные
Три Полная Трихромазия нор- мальная, сильная, по всем трем прием- никам Трихромазия сильная, но ано- мальная в виде прото-, дейтер- или тританома- лии
Цветослабые
Три Редуци- рованная I степени II степени III степени Трихромазия нор- мальная (по спек- тру), но редуциро- ванная (по силе) в виде прото-, дейтер- или тритадефицита или их комбинации Различные со- четания цвето- слабости с цве- тоаномалиями
Цветослепые
Два Утрачена В одном из приемников, т. е. частич- ная цвето- слепота Дихромазия в виде прото-, дейтер- или тританопии Различные со- четания цвето- слепоты и цвето- аномальности
Один Утрачена В двух при- емниках, т. е. полная цве- тослепота Монохромазия
Примечание. При нарушениях функции в проводящих путях и зрительных цен-
трах возможны одновременные нарушения красно-зеленого или сине-желтого меха-
низмов цветовосприятия, а также расширение ахроматической зоны на все цвета.
131
Ведущее место в классификации отведено цветосиле приемнийд.
Именно высокой цветочувствительностью, а не отсутствием ано-
мальности определяется, в частности, профессиональная пригод-
ность к вождению транспортных средств, возможность быть летчи-
ком, машинистом, моряком, пожарным, работником органов пра-
вопорядка, электриком, садовником, дизайнером, кондитером.
Только при отборе на работу таких довольно редко встречающихся
специалистов, как колориметристы, художники в текстильной, ла-
кокрасочной или полиграфической промышленности, которым по
роду деятельности необходимо улавливать самые слабые оттенки
цвета, целесообразно исключить малейшую цветоаномалию. В по-
следнем случае для исследования понадобится аномалоскоп, во всех
остальных достаточную информацию можно получить с помощью
таблиц Е. Н. Юстовой и соавт.
ГЛАВА5
ЭЛЕКТРОРЕТИНОГРАФИЯ
5.1. ОБЩАЯ ЭЛЕКТРОРЕТИНОГРАММА
С 1945 г. электроретинография заняла особое место среди функци-
ональных методов исследования в клинике глазных болезней
[Karpe G., 1945 ]. Наряду с общеизвестными физиологическими и
психофизическими методами, с помощью которых получают дан-
ные о функции зрительного анализатора на всем протяжении зри-
тельного пути от сетчатки до центральных отделов, электрорети-
нографию применяют для количественной оценки функционально-
го состояния нейронов сетчатки, более точного определения
локализации патологического процесса.
Электроретинограмма представляет собой графическое отобра-
жение изменений биоэлектрической активности клеточных элемен-
тов сетчатки в ответ на световое раздражение.
Механизмы генерации ЭРГ к настоящему времени хорошо изве-
стны благодаря экспериментальным исследованиям на изолирован-
ных сетчатках различных животных, изучению ЭРГ в различных
условиях стимуляции [Granit R.,1947; YonemuraD., 1977]. В этих
исследованиях изучено происхождение компонентов ЭРГ и их связь
с клеточными элементами сетчатки, образующими нейрональную
сеть.
Общая ЭРГ отражает электрическую активность большинства
клеточных элементов сетчатки и зависимость от количества здоро-
вых функционирующих клеток. Спайковая активность нейронов
различного класса не может быть видна на ЭРГ в связи с домини-
рованием в ответе, превышающем 200 мкв низкочастотных компо-
нентов, однако ганглиозные клетки участвуют в генерации паттерн-
ЭРГ, регистрируемой на реверсивный паттерн-стимул (см.ниже).
Различные типы ЭРГ отражают все многообразие структуры сет-
чатки, а их компоненты, связанные с клеточными элементами сет-
чатки, и являются диагностическим инструментом в клинике.
5.1.1. Современные представления о происхождении ЭРГ
Каждый компонент ЭРГ генерируется различными структурами
сетчатки (рис.5.1). Результатом взаимодействия электрической ак-
тивности нескольких процессов, описанных G. Granit (1947) как
компоненты Рр Рп, Рш (рис.5.2), являются а-, Ь-, с-волны ЭРГ.
ЭРГ глаза человека содержит негативную a-волну, отражающую
функцию фоторецепторов как начальную часть позднего рецептор-
ного потенциала. На нисходящей части a-волны можно видеть две
133
мкВ
Рис. 5.1. Схематическое изображение общей ЭРГ.
а, и а2 — амплитуда a-волны; Ь, и Ь2 — амплитуда b-волны; Ь' — амплитуда Ь'-вол-
ны; D — длительность b-волны; La — латентный период a-волны; Lb — латентный
период b-волны; ТсЬ, — время кульминации Ьр ТсЬ2 — время кульминации Ь2; ТсЬ' —
время кульминации Ь*. По оси ординат — амплитуда волн ЭРГ, мкВ, по оси абс-
цисс — длительность волн ЭРГ, мс.
волночки очень маленькой латентности — ранние рецепторные по-
тенциалы (РРП), отражающие цикл биохимических превращений
родопсина. Волна а имеет двойное происхождение соответственно
двум видам фоторецепторов: более ранняя а^волна связана с ак-
тивностью фотопической системы сетчатки, а2-волна — со скотопи-
ческой системой. Волна а переходит в позитивную b-волну, отража-
ющую электрическую активность биполяров и клеток Мюллера с
возможным вкладом горизонтальных и амакриновых клеток.
Волна Ь, или on-эффект, отражает биоэлектрическую активность
в зависимости от условий адаптации, функции фотопической и ско-
топической системы сетчатки, которые представлены в позитивном
компоненте волнами bj и Ь2. Большинство исследователей, связы-
вая происхождение b-волны с активностью биполяров и клеток Мюл-
лера, не исключают вклад ганглиозных клеток сетчатки. На восхо-
дящей части b-волны отмечается 5—7 волночек, называемых
осцилляторными потенциалами (ОП), которые отражают взаимо-
действие клеточных элементов во внутренних слоях сетчатки, в том
числе амакриновых клеток.
При прекращении действия стимула (выключение света) регис-
трируется d-волна (off-эффект). Эта волна, последняя фаза ЭРГ,
является результатом взаимодействия a-волны и компонента посто-
янного тока b-волны. Эта волна — зеркальное отражение а-волны —
134
Рис.5.2. Схематическое изображение электроретинограммы в условиях тем-
новой и световой адаптации.
а — типичный ответ от палочковой сетчатки в условиях темновой адаптации; б —
то же при световой адаптации; в — типичный ответ колбочковой сетчатки [Granit R.,
1933]. По оси ординат — амплитуда ответа, мкВ, по оси абсцисс — время ответа, с.
имеет фотопическую и скотопическую фазы. Она лучше регистри-
руется в случае преобладания в сетчатке колбочковых элементов.
Таким образом, считается, что главным источником a-волны в
ЭРГ позвоночных являются фоторецепторы, как колбочки, так и
палочки. Однако ранние исследования [TomitaT., 1963; Mura-
kami М., Kaneko А., 1966; Knapp A. G., Schiller Р. Н., 1984;
Evers Н. О., Gouras Р., 1986; Bush R. A., Sieving Р. А., 1994] показа-
ли, что часть a-волны как от темно-, так и от светоадаптированной
сетчатки может происходить после синапсов фоторецепторов, т.е. в
ее генерации принимают участие различные слои сетчатки, в част-
ности гиперполяризующиеся биполяры.
Генерация ЭРГ начинается с выхода К+ из деполяризующихся
ретинальных нейронов в межклеточное пространство в ответ на
включение света. Мембрана клеток Мюллера, проницаемая для К+,
локально деполяризуется [Karwoski C.J., Proenza L.M., 1987] и явля-
135
ется основным источником генерации b-волны ЭРГ. Биоэлектри-
ческая активность распространяется от наружного ядерного слоя
до поверхности сетчатки и коррелирует с количеством нейронов в
дистальной и проксимальной части сетчатки. Данные, полученные
в последние годы, свидетельствуют о том, что возможен прямой
вклад биполярных клеток в развитие b-волны ЭРГ [Xu X., Karwoski
C.J., 1993]. В сетчатке теплокровных в генерации b-волны опреде-
ленное значение имеет активность амакриновых клеток.
В фотопических условиях стимуляции на обезьянах получены
доказательства того, что активность гиперполяризующихся нейро-
нов второго порядка (биполярные и горизонтальные клетки) влияют
на амплитуду и форму фотопической b-волны [Sieving Р. A. et al., 1994].
Выключение стимула приводит к генерации d-волны (off-ответ),
которая начинается с негативного отклонения в преимущественно
палочковых сетчатках и позитивного — в колбочковых. В смешан-
ной палочково-колбочковой сетчатке человека в зависимости от усло-
вий адаптации и стимуляции d-волна может быть представлена двумя
компонентами: dj и d2 (колбочковый и палочковый). При слабых сти-
мулах off-ответ ЭРГ имеет негативную полярность и становится пози-
тивным при увеличении интенсивности стимулирующего света, а при
высокой интенсивности света фотопический off-ответ может иметь
вид позитивного плато.
При высоком уровне световой адаптации позитивный off-ответ
не регистрируется, несмотря на значительную интенсивность стимула
[Kawasaki К., 1984]. Колбочковая и палочковая природа позитивно-
го и негативного off-ответа подтверждена в экспериментах при опре-
делении их спектральной чувствительности, а также при обследова-
нии лиц с палочковым монохроматизмом и стационарной ночной
слепотой. Высказано предположение, что у человека низкопорого-
вый негативный off-ответ должен представлять собой окончание Рп
или компонента постоянного тока (d.c.) [Brown К.Т., 1968], а началь-
ная восходящая фаза высокопорогового позитивного off-ответа —
окончание Рш или позднего рецепторного потенциала. В сетчатке
амфибий d-волна может отражать деполяризацию off-биполяров, а
также активность амакриновых и горизонтальных клеток, отвечаю-
щих на выключение света [Stockton R., Slaughter М., 1989].
Различные свойства колбочек и палочек сетчатки предусматри-
вают использование в электроретинографии коротких стимулов
(несколько микро- и пикосекунд) для регистрации темноадаптиро-
ванной ЭРГ, в которой генерация биопотенциала определяется па-
лочковым on-путем, и стимулов большой длительности для выделе-
ния нейронов светоадаптированной фотопической ЭРГ, которая
содержит ответ колбочковых фоторецепторов и нейронов колбоч-
ковых on- и off- путей.
В исследованиях, проведенных в последние годы, установлено,
что в проксимальной сетчатке позвоночных Ь- и d-волны ЭРГ на
136
большую длительность стимулирующего света при световой адап-
тации отражают активность колбочковых on- и off-путей
[Sieving R.A., 1993; 1994].
Следующее медленное позитивное отклонение с быстрым (45 с)
и медленным (12 мин) пиками осцилляций названо с-волной, кото-
рая может быть выделена лишь при использовании стимулов, не-
прерывно предъявляемых, высокой интенсивности и большой дли-
тельности в темноадаптированном глазу. Это транспигментный по-
тенциал эпителия, медленный позитивный потенциал внеклеточного
тока, образующийся в связи с изменением концентрации калия,
который выделен при введении микроэлектрода в субретинальное
пространство. Регистрация этого медленного потенциала осуществ-
ляется непрямым способом с помощью ЭОГ. Однако выделение
этого постоянного потенциала возможно также в ЭРГ, хотя это
связано с определенными трудностями, обусловленными дрейфом
базовой линии. В настоящее время существует мнение, что позитив-
ный компонент с-волны, генерируемой в слое пигментного эпите-
лия, представляет собой разницу в гиперполяризации между апи-
кальной и базальной мембранами, возникающей в процессе свето-
вой стимуляции, а негативный компонент, значительно меньшей
величины, регистрируется от клеток Мюллера (медленный Рш). Так
как с-волна ЭРГ сохраняется при отсутствии пигментного эпите-
лия, ее происхождение связывают с активностью фоторецепторных
клеток, субстанциями, отвечающими за световой пик (ЭОГ), транс-
миттерами (мелатонин, допамин) фоторецепторов. Однако с-волна
ЭРГ не может быть зарегистрирована без нормальных физических
и биохимических связей между пигментным эпителием и наружны-
ми сегментами фоторецепторов, обновления дисков, фотохимиче-
ских превращений зрительных пигментов и нормального питания
сетчатки. Отделение пигментного эпителия от наружного сегмента
фоторецепторов, отслойка сетчатки, приводит к функциональ-
ной несостоятельности сетчатки, сопровождающейся нерегистри-
руемой ЭРГ.
5.1.2. Факторы, влияющие на ЭРГ
В клинической электроретинографии выделение функций фотопи-
ческой и скотопической систем основано на различии физиологи-
ческих свойств колбочек и палочек сетчатки, поэтому используют
соответствующие условия, в которых доминирует каждая из этих
систем. Колбочки чувствительны к более ярким стимулам света,
палочки — к слабым световым воздействиям. В-волна ЭРГ значи-
тельно изменяется в зависимости от условий адаптации (рис.5.3).
В первые минуты темновой адаптации большей чувствительностью
обладает колбочковая система (фотопическая ЭРГ); к 7-й минуте
световая чувствительность палочковой и колбочковой систем урав-
137
Рис. 5.3. Изменения а- и b-компонентов ЭРГ в течение темновой адаптации
[Auerbach Е., Burian Н.М., 1955].
По оси ординат — 1g I (логарифм интенсивности), по оси абсцисс — время темновой
адаптации, мин.
ниваются и начинает доминировать функция палочковой системы;
после 15-й минуты темновой адаптации в электроретинограмме пре-
обладает биоэлектрическая активность палочковой системы (ско-
топическая ЭРГ). Эта трансформация ЭРГ аналогична процессам
изменения световой чувствительности, наблюдаемым в ходе темно-
вой адаптации, и используется для выделения функции палочковой
системы сетчатки в общей “динамической” ЭРГ, регистрируемой в
процессе темновой адаптации в течение 30 мин с использованием
стимулов низкой интенсивности (белого или голубого). Увеличение
амплитуды b-волны в процессе темновой адаптации является объек-
тивной, более точной характеристикой адаптационных свойств сет-
чатки по сравнению с психофизическими исследованиями темновой
адаптации. Любые факторы, влияющие на регенерацию зрительно-
го пигмента, состояние палочковой системы, медиаторную функ-
цию и нейронную организацию, приводят к изменениям “динами-
ческой” ЭРГ. У пациентов, у которых подозревают ночную слепо-
ту, регистрируется резко измененная ЭРГ. При дефиците витаминов
А, Е и цинка в организме у пациентов с разными формами анемии,
при хроническом алкоголизме, циррозе печени также имеются со-
ответствующие предпосылки к изменениям ЭРГ.
В-волна является постсинаптической волной ЭРГ, отражающей
нейрональную активность как в наружных, так и во внутренних
плексиформных слоях, поэтому она реагирует на некоторые ток-
сичные субстанции и лекарственные препараты, которые блокиру-
138
ют синаптическую передачу или нейрональную модуляцию, что
приводит к снижению соотношения Ь- и а-волн.
Из практики клинической электроретинографии известно, что
поражения макулярной области с локализацией патологического
процесса в пределах до 6—15° не влияют на величину общей ЭРГ,
отражающей функцию всей сетчатки. В то же время на ответ кол-
бочковой (фотопической) ЭРГ, зарегистрированной на предъявле-
ние единичных световых стимулов или частых мельканий (свыше
20—50 Гц), влияют патологические процессы, как локализующиеся
в центральных отделах сетчатки, так и захватывающие периферию,
где имеются колбочки. В связи с этим наличие или отсутствие рит-
мического ответа на световые мелькания с высокой частотой и
амплитуда ритмической ЭРГ позволяет оценить сохранившуюся
функцию колбочковой системы. При этом нужно учитывать значи-
тельный вклад колбочек макулярной области в ритмическую ЭРГ.
Использование световых стимулов возрастающей интенсивности и
световой адаптации приводит к доминированию в ЭРГ функции
колбочковой системы. Зависимость ЭРГ от интенсивности стиму-
лирующего света является тестом в диагностике патологических
процессов в сетчатке.
Для выделения функции колбочковой системы сетчатки необхо-
димы световая преадаптация в течение 5—10 мин и более яркие сти-
мулы длинноволновой части спектра (красные). При этом можно
использовать фоновое освещение до 20 лк с целью подавления фун-
кции палочкового аппарата сетчатки (достаточно 7—10 фут Л б).
С увеличением интенсивности фонового освещения чувствительность
фоторецепторов снижается.
Другой метод выделения активности палочковой и колбочко-
вой систем сетчатки состоит в регистрации ритмической ЭРГ при
воздействии разной частоты мелькающего стимулирующего света.
Это связано с тем, что палочковая система способна воспроизво-
дить ритм мельканий не более 20 Гц, а колбочковая система отве-
чает на значительно большую частоту стимуляции (до 100 Гц).
ЭРГ, зарегистрированная при предъявлении стимула, мелькаю-
щего постоянно с разной частотой, названа мелькающей (ритми-
ческой) ЭРГ. Эту форму ЭРГ чаще всего используют для характе-
ристики функции колбочковой системы сетчатки. Частота света
постоянной интенсивности, при которой b-волна сливается с фоно-
вым шумом на базовой линии, называется фузионной частотой
мельканий. В диагностике начальных поражений колбочковой си-
стемы, при колбочковой дисфункции играет роль как уменьшение
амплитуды мельканий на 30 Гц, так и изменение частоты, которая
способна вызвать ритмический ответ. При дистрофических измене-
ниях в макулярной области мелькающая ЭРГ при предъявлении
стимула в 30 Гц не регистрируется. При этом рекомендуется ис-
пользовать красный стимул высокой частоты, к которому наиболее
чувствительна колбочковая система.
139
Таким образом, на величину и форму ЭРГ влияют различные
факторы: яркость стимула, величина засвета, длительность вспыш-
ки, длина волны стимулирующего света (цвет стимула), условия
адаптации к темноте или свету, частота стимуляции. Величина ЭРГ
зависит от количества стимулируемых фоторецепторов и углового
размера стимула. В случае возникновения патологических процес-
сов в сетчатке при оценке ЭРГ важно учитывать, что амплитуда
а- и b-волн зависит от количества сохранившихся фоторецепторов.
В зависимости от условий исследования ЭРГ может быть заре-
гистрирована от всей сетчатки — общая ЭРГ (ганцфельд-ЭРГ) и от
области на сетчатке различной величины — локальная ЭРГ. Локаль-
ная ЭРГ, зарегистрированная от макулярной области, называется
макулярной или фокальной ЭРГ.
Величина b-волны ЭРГ зависит от интенсивности стимулирую-
щего света. При слабых стимулах регистрируется только Ь-волна
очень низкой амплитуды. Однако это не означает, что волна а во-
обще отсутствует. ЭРГ — сумма как негативных, так и позитивных
потенциалов, поэтому негативная а-волна частично маскируется от-
носительно большей позитивной b-волной. При изучении зависи-
мости ЭРГ от интенсивности стимулирующего света учитывают два
основных критерия — порог логарифма интенсивности, при кото-
ром возможна регистрация ЭРГ (этот порог имеет определенные
границы для пациентов без патологии сетчатки и зрительного не-
рва), и максимум b-волны, который может быть использован в ком-
бинации с порогом интенсивности. Даже при нормальном пороге
незначительное увеличение нейронального тока на каждую едини-
цу интенсивности приводит к значительному увеличению амплиту-
ды ЭРГ, поэтому связь величины b волны ЭРГ и интенсивности
стимулирующего света является очень важным критерием в клини-
ческой электроретинографии, поскольку уровень зрительного раз-
дражения увеличивается пропорционально логарифму интенсивно-
сти или контраста стимула. Именно поэтому в ведущих лаборато-
риях мира для оценки функции сетчатки используется не единичный
мелькающий стимул одной интенсивности, а измеряется функция
ЭРГ от интенсивности. Очевидно, что электроретинография — очень
чувствительный метод оценки функционального состояния сетчат-
ки, позволяющий определить самые незначительные биохимичес-
кие нарушения, которые могут предшествовать начальным клини-
ческим проявлениям.
Нарушение нормального физиологического состояния сетчат-
ки, изменение одного из многочисленных факторов, участвующих
в возникновении ЭРГ, предопределяют изменение временных и/или
амплитудных параметров ЭРГ. К этим факторам относятся состо-
яние хороидальной и ретинальной циркуляции, пигментного эпите-
лия, колбочек и палочек, биполярных, горизонтальных и амакрино-
вых клеток, колбочкового пигмента и родопсина, метаболизм, ион-
140
ный состав вне- и внутриклеточной среды, цепь биохимических
реакций между пигментным эпителием и фоторецепторами, состо-
яние наружного и внутреннего слоев с их синаптической, химиче-
ской передачей зрительного возбуждения и нейромодуляционной
функцией, нормальное взаимодействие между нейронами и глией,
состояние аксонального транспорта ионов. Хотя по виду ЭРГ не
всегда возможно определить, какой из факторов послужил причи-
ной ее изменения, выбор условий регистрации ЭРГ, знание ее
классификации определяют возможность целесообразного исполь-
зования различных электроретинографических тестов для опреде-
ления локализации патологического процесса.
5.1.3. Классификация ЭРГ
Общая ЭРГ при патологических процессах, локализующихся в раз-
личных отделах сетчатки, изменяется в зависимости от патогенеза
заболевания, локализации тяжести и распространенности патоло-
гического процесса, а также от длительности его течения [Шамши-
новаА.М., 1978—1995; CarrR.E., Siegel I.M., 1982]. Если в норме
амплитуда b-волны общей ЭРГ 250—400 мкВ, то при патологиче-
ских состояниях сетчатки возможны изменение отдельных компо-
нентов ЭРГ и полное ее исчезновение. Классификация ЭРГ разра-
ботана еще в 1945 г. G. Кагре и дополнена H.E.Henkes (1953). В ос-
нове этой классификации лежат амплитудные характеристики
основных а- и b-волн ЭРГ. Различают следующие виды ЭРГ: нор-
мальную, супернормальную, субнормальную (плюс- и минус-нега-
тивную), угасшую, или нерегистрируемую (отсутствующую) (рис.5.4).
Супернормальная ЭРГ характеризуется увеличением а- и Ь-волн,
что, возможно, является следствием раздражения фоторецепторов
сетчатки, которое может отмечаться при первых признаках гипок-
сии, медикаментозных интоксикациях, симпатической офтальмии
и пр. Супернормальная биоэлектрическая реакция при травмати-
ческом перерыве зрительного нерва и его атрофии обусловлена на-
рушением проведения возбуждения по ретиноталамическим цент-
робежным тормозящим волокнам. Однако в ряде случаев трудно
объяснить природу супернормальной ЭРГ.
Субнормальная ЭРГ— это наиболее частый вид патологической
ЭРГ, которая характеризуется снижением а- и b-волн. Ее регистриру-
ют при дистрофических заболеваниях сетчатки и хороидеи, отслойке
сетчатки, увеитах с вовлечением в процесс 1—2 нейронов сетчатки,
хронической сосудистой недостаточности с нарушением микроцир-
куляции, при некоторых формах ретиношизиса (х-хромосомный,
сцепленный с полом, тип Вагнера), fundus albipunctatus и т.д.
Негативную ЭРГ характеризует увеличение или сохранность
амплитуды a-волны и небольшое (до изолинии плюс-негативная)
или значительное снижение (ниже изолинии минус-негативная) ам-
плитуды b-волны. Ее хорошо видно при регистрации в условиях
141
200 мкВ
Рис. 5.4. Основные виды
ЭРГ в клинике глазных
болезней.
а — нормальная; б — супер-
нормальная; в — субнор-
мальная плюс-негативная;
г — минус-негативная; д —
нерегистрируемая.
темновой адаптации (скотопическая ЭРГ), хотя ее можно видеть
при многих патологических состояниях в фотопических условиях
(фотопическая ЭРГ). Негативную ЭРГ можно наблюдать при пато-
логических процессах, когда предполагается дисфункция дисталь-
нее фоторецепторного слоя. Степень негативизации может быть
различной — от очень маленькой b-волны до нерегистрируемой. Так,
в некоторых случаях врожденного ретиношизиса и некоторых фор-
мах врожденной стационарной ночной слепоты b-волна отсутству-
ет и ЭРГ состоит только из Рш-компонента, в то время как тем-
ноадаптированная вспышечная ЭРГ имеет большую a-волну и сла-
бовыраженную b-волну. Такого типа минус-негативная ЭРГ
встречается при острых нарушениях кровообращения в сетчатке,
выраженных хороиретинальных поражениях, лекарственных инток-
сикациях (хинином, винкристином), прогрессирующей дегенератив-
ной миопии и врожденной стационарной ночной слепоте (рецессив-
ная форма и сцепленная с полом), болезни Огуши, Х-хромосомном
ювенильном ретиношизисе, металлозах сетчатки, ранних стадиях
доминантной формы тапеторетинальной абиотрофии (пигментный
ретинит), болезни Рефзума (Refsum), Аланда (глазной альбинизм
Forsius — Eriksson), ретинопатии Гольдмана — Фавра (Goldmann —
Favre), врожденном амаврозе Лебера и других ретинальных пато-
логиях. Однако механизмы, приводящие к прогрессивному угнете-
нию b-волны ЭРГ, при указанных заболеваниях различны.
142
Угасшая, или отсутствующая (нерегистрируемая), ЭРГ являет-
ся электрофизиологическим симптомом тяжелых необратимых из-
менений в сетчатке при развитом металлозе, воспалительных про-
цессах в оболочках глаза, тотальной отслойке сетчатки, окклюзии
центральной артерии сетчатки, а также патогномоничным призна-
ком тапеторетинальной абиотрофии сетчатки (пигментного рети-
нита). Отсутствие ЭРГ отмечено при старых тотальных отслойках
сетчатки и грубых необратимых изменениях ее нейронов, которые
могут наблюдаться при воспалительных и травматических пораже-
ниях глазного яблока. ЭРГ этого типа регистрируется в терминаль-
ной стадии диабетической ретинопатии, когда грубый пролифера-
тивный процесс распространяется на дистальные отделы сетчатки,
и ретиношизисе типа Фавра — Гольдмана. Термин “отсутствующая”
ЭРГ не во всех случаях отражает истинное состояние сетчатки и
тем более не несет никакой информации при помутнении оптиче-
ских сред, поэтому в последнее время предлагается использовать
термин нерегистрируемая, неопределяемая, нераспознанная.
J.R.Brunett (1982) классифицировал ЭРГ по формам заболева-
ния сетчатки, используя при этом термины “дегенерация”, “дистро-
фия”, ’’атрофия”. Автор выделил 7 форм ЭРГ.
1. Паттерн врожденной дисфункции, когда отсутствует один из
компонентов ЭРГ (палочковый или колбочковый), соответствую-
щий пораженной системе при сохранности другой. ЭРГ этого вида
регистрируется при первичных, врожденных непрогрессирующих
заболеваниях сетчатки, ахроматопсии, эссенциальной гемералопии,
fundus albipunctatus, болезни Огуши.
2. Дегенеративный паттерн, когда амплитуда b-волны снижена
при увеличенном времени ее пика, когда поражается как палочко-
вая, так и колбочковая система. Такая ЭРГ встречается при пиг-
ментном ретините (палочково-колбочковая дегенерация, пигмент-
ная и беспигментная формы), retinitis punctata albescens, fundus
flavimaculatus, ишемической хороидальной дистрофии, хороирети-
нальном склерозе.
3. Паттерн, характерный для дистрофии и вторичной атрофии с
редуцированным ответом. Он встречается при дистрофиях типа ат-
рофии Гирате (Gyrate), хороидермии, ишемической дистрофии, хо-
роидальном склерозе, первичных процессах в хороидее с вторичным
вовлечением как колбочковой, так и палочковой системы сетчатки.
Среди вторичных атрофических ретинопатий выделяют пигментную
лекарственную ретинопатию, токсическую ретинопатию, металлоз,
инфаркты сетчатки, вторичные системные заболевания.
4. Гипер-паттерн (повышенный ответ ЭРГ), характерный для
гиперметаболических состояний при гипертиреоидизме, расшире-
нии сосудов, альбинизме.
5. Паттерн острой ишемии характеризуется снижением ампли-
туды b-волны и увеличением времени до ее пика, снижением ампли-
туды и уменьшением количества осцилляторных потенциалов.
143
Вспышка 10 мкс
50 мкВ
Вспышка 150 мс
Рис. 5.5. ЭРГ на вспышку различной длительности.
а, б — скотопический палочковый ответ, записанный после темновой адаптации;
виг — колбочковый ответ на яркую вспышку на фоне световой адаптации [Sie-
ving?. А., 1993—1995].
6. Паттерн a-волны, или негативная ЭРГ, характерная для окк-
люзии центральной артерии сетчатой оболочки.
7. Отсутствующая ЭРГ — конечная стадия многих заболеваний
сетчатки.
Классификация, предложенная A.E.Kriil (1970), позволяет более
полно оценить функцию сетчатки и предполагает учет амплитуды а- и
b-волн в зависимости от интенсивности стимулирующего света, нарас-
тание амплитуд а- и b-волн или изменение времени до пика в ходе тем-
новой адаптации, зависимость интервалов до пика а- и b-волн от интен-
сивности стимула, отношение амплитуды b к частоте мельканий.
В последнее время интерес офтальмологов вызывает новый элек-
троретинографический метод исследования фотопической ЭРГ
(рис.5.5) на стимул большой длительности (до 300 мс), позволив-
ший выявлять патологию в on/off-путях колбочковой системы при
колбочковых дистрофиях [Sieving Р.А., 1993].
Выше было отмечено, что гипер- и деполяризация нейронов
отражается на форме и амплитуде суммарной ЭРГ. Как известно,
функция гиперполяризующихся биполярных клеток (ГБК), являю-
щихся частью off-путей сетчатки, отражена в негативных компо-
нентах (d-волна) фотопической ЭРГ приматов, в то время как депо-
ляризующиеся биполярные клетки (ДБК) являются зрительными
on-путями и представлены в b-волне ЭРГ. Функции ГБК и ДБК
144
могут быть разделены в ЭРГ с помощью аналогов трансмиттера
глутамата, блокирующих либо on-, либо off-пути. При использова-
нии этой модели, названной “push-pull”, выделяющей функцию
гипер- и деполяризующихся биполяров в ЭРГ, в экспериментах на
обезьянах зарегистрировали ЭРГ, имеющие форму “гиперполяри-
зационного” и ’’деполяризационного” паттерна, соответствующие
различным заболеваниям сетчатки.
Вариации форм ЭРГ при различных патологических процессах в
палочковой и колбочковой системах сетчатки позволили Р. A. Sie-
ving (1993) предложить электрофизиологическую классификацию
заболеваний сетчатки, в которой выделено 5 типов ЭРГ (рис. 5.6): с
общим снижением амплитуды; тип врожденной стационарной ночной
слепоты; гиперполяризационный паттерн; деполяризационный пат-
терн; патологический off-ответ. На рисунке представлены наряду с
нормальной ЭРГ на длительную вспышку, субнормальная—со сни-
жением амплитуды всех волн a, b, d, i, у больного с пигментным
ретинитом (аутосомно-доминантная форма наследования) в семье с
мутацией гена родопсина, установленной на молекулярном уровне;
тип CSNB у больного с врожденной стационарной ночной слепотой
типа Шуберта — Борншайна (Shubert — Bomshein); гиперполяриза-
ционный паттерн, который сопровождается относительным подав-
лением b-волны у пациента с колбочковой дисфункцией и макулопа-
тией типа ’’бычий глаз”; деполяризационный паттерн, характеризу-
ющийся подъемом плато выше изолинии (этот тип ЭРГ характерен
для палочково-колбочковой дисфункции); патологический off-ответ,
характерный для различных видов дистрофий.
ЭРГ со снижением амплитуды всех ее волн характерна для рети-
нальных дистрофий, в частности колбочково-палочковой. Тип врож-
денной стационарной ночной слепоты, когда угнетена в основном
b-волна при сохранности d-волны ЭРГ, встречается при дисфунк-
ции палочковой системы типа Шуберта — Борншайна (Shubert —
Bomshein), в основе развития которой, как предполагают, лежит
блокада on-путей APB (2-amino-4-phosphonobutyric acid).
Гиперполяризационный паттерн, так же как тип врожденной
стационарной ночной слепоты, характеризуется нарушением соот-
ношения b/а- и b/d-волн, снижением Ь- и d-волн, что является при-
чиной патологического соотношения b/а < 1,0. У пациентов этой
группы в отличие от врожденной стационарной ночной слепоты
отмечается прогрессирующее снижение зрения.
Деполяризационный паттерн ЭРГ относится к новой категории
патологических ЭРГ, для которых характерен общий подъем
плато кривой к позитивному потенциалу над базовой линией. В эк-
сперименте на обезьянах деполяризационный паттерн наблюдался
при блокаде PDA (± cis-2,3 piperidine dicarboxylic acid) или KYN
(kynurenic acid) синаптической передачи к гиперполяризационным
биполярам колбочковых off-путей. В клинике этот вид ЭРГ-паттер-
на встречается при палочково-колбочковой дистрофии.
145
a
Рис. 5.6. Виды ЭРГ на сти-
мул большой длительности
(150 мс) [стимул интенсивно-
стью 185 кд/м2, фоновое осве-
щение (ганцфельд) 43 кд/м2].
а — нормальная ЭРГ; б — суб-
нормальная ЭРГ; в — тип CSNB;
г — гиперполяризационный пат-
терн; д— деполяризационный
паттерн; е — патологический off-
ответ [Sieving Р. А., 1993].
Патологический off-ответ сопровождается снижением и ”нега-
тивизацией” off-компонента и встречается при колбочково-палоч-
ковой дистрофии (например, по типу “бычий глаз”).
Эти категории электроретинографических ответов особенно
146
важны при оценке механизмов снижения зрительных функций, свя-
занных с изменениями в колбочковых on- и off-путях проксималь-
ной сетчатки.
Основным в этих исследованиях было то, что фотопическая ЭРГ,
регистрируемая на стимул большой длительности, у пациентов с
морфологически различными дистрофиями сетчатки может быть
подразделена на несколько различных физиологических категорий.
Так, в одних случаях потеря зрения связана с патологией в on- или
off-путях при сохранности самих фоторецепторов, в других — на-
оборот, с деструкцией последних. Эти новые концепции могут быть
основой выделения подтипов ретинальных дистрофий на основе функ-
ционально-физиологических принципов, а ЭРГ на световой стимул
большой длительности — единственным инструментом дифферен-
циации процессов в палочковых и колбочковых ДБК синапсах.
5.1.4. Метод регистрации ЭРГ (рекомендации ISCEV)
Большое разнообразие технических и методических приемов реги-
страции ЭРГ привело к необходимости стандартизировать условия
ее регистрации, для того чтобы получить возможность сравнивать
результаты исследований, проводимых в различных лабораториях
мира. Большой клинический опыт позволил выбрать условия реги-
страции, наиболее соответствующие современным представлениям
о происхождении ЭРГ и удобные для использования в клинике.
Ниже приведены основные принципы и стандарты регистрации
ЭРГ, рекомендуемые Международным обществом клинических элек-
трофизиологов зрения для исследования зрительных функций у
пациентов с их различными нарушениями. Предлагаются стандар-
ты для пяти основных воспроизведений ЭРГ (рис.5.7):
1) максимальный ответ в темноадаптированном глазу;
2) палочковый ответ в темноадаптированном глазу;
3) колбочковый ответ;
4) осцилляторные биопотенциалы;
5) фликер-ответ, возникает при быстро повторяющемся стимуле
(мелькающая ЭРГ) (рис.5.8).
Для регистрации общей ЭРГ необходимо использовать ганц-
фельд-стимуляцию и равномерное освещение всего поля сетчатки.
Отводящие (активные) электроды могут представлять собой кон-
тактную линзу с возможно большим открытием оптической зоны
роговицы. Поверхность роговицы должна быть защищена не раз-
дражающим и не вызывающим аллергию электропроводимым ра-
створом, например 0,5% метилцеллюлозой. Кожные электроды и
Другие типы корнеальных электродов (крючки, волоски и пр.) ис-
пользовать нежелательно, так как в этом случае амплитуды ответов
снижаются и их невозможно будет сравнить.
Референтный электрод может быть вмонтирован в контактную
147
Рис.5.7. Основные виды ЭРГ в соответствии со стандартами ISCEV.
а — максимальный комбинированный ответ в темноадаптированном глазу; б — па-
лочковый ответ после темновой адаптации; в — колбочковый ответ на фоновом
освещении, подавляющем палочковую активность, после 10 мин предварительной
световой адаптации; г — осцилляторные потенциалы (в темноадаптированном гла-
зу). Стрелками обозначено время начала стимуляции.
линзу со стороны конъюнктивы (биполярный электрод) или исполь-
зован как накожный, который помещается на мочку уха, верхний
край орбиты. Заземление осуществляется отдельным накожным
электродом, как правило, на мочке уха. При этом накожные элек-
троды, используемые в качестве референтного электрода или элек-
трода для заземления, должны иметь сопротивление 10 кОм или
меньше, измеренное при 30—200 Гц. Перед наложением электрода
кожу очищают и на нее наносят электропроводную пасту.
t Базовая линия в отсутствие световой стимуляции должна быть
стабильной, что может быть достигнуто при изготовлении корнеаль-
ных и референтных электродов из неполяризующихся материалов.
После каждого исследования очень важно обработать электро-
ды веществами, предотвращающими передачу инфекционных аген-
148
.цлллмм-
Рис. 5.8. Нормальная ЭРГ на белый стимул, мелькающий с разной час-
тотой.
а — 10 Гц; б — 20 Гц; в — 30 Гц.
тов, так как возможен контакт со слезой и капельками крови, по-
являющимися при очистке кожи.
Длительность стимула должна быть значительно меньше, чем
время интеграции сигнала фоторецепторов, т. е. меньше 5 мс. Та-
кие короткие по продолжительности вспышки света могут быть
получены с помощью газоразрядных трубок, тробоскопов и др.
Длина волны стимулирующего света должна быть в пределах
видимого спектра (белый свет). Цветные стимулы также могут быть
использованы для выделения палочкового и колбочкового ответов,
однако исследования такого типа не определены стандартом. Ин-
тенсивность стимулирующего света на поверхности ганцфельд-шара
1,5—3,0 кд/м2 (3,43 кд/м2 = 1 фут Лб). Стимул такой величины но-
сит название “стандартного”.
Фоновое освещение используют для подавления функций па-
лочковой системы и выделения функции колбочковой. Освещенность
поля должна составлять 17—34 кд/м2 (5—10 фут Лб). Перевод еди-
ниц освещенности представлен в табл. 5.1. Цвет фона белый, однако
можно использовать и цветной фон (например, желтый). В использу-
емых системах стимуляции рекомендуется регулировать интенсив-
ность стимула и фонового освещения так, чтобы интенсивность сти-
мула могла изменяться в диапазоне по крайней мере 3 лог. ед. либо
плавно, либо ступенчато, но не более чем на 0,25 лог. ед. Фоновое
освещение должно быть калибровано соответственно уровню интен-
сивности стимулирующего света, используемого для “колбочкового
149
Таблица 5.1. Перевод единиц освещенности
Единицы освещенности Нит Стильб Апостильб
1 кандела на 1 м2 1 кд/м2 1 10-4 3,14
1 нит (нит) 1 кд/м2 1 104 3,14
1 стильб (сб) 1 кд/см2 104 1 3,14 IO"4
1 апостильб (асб) 1 кд/л см2 0,3183 3,183 • 10 s 1
1 ламберт (Лб) 1 кд/л см2 3,183 103 0,3183 ю-4
1 миллиламберт (мЛб) 1 кд/л см2 3,183 3,183 IO"4 10
1 фут-ламберт (фут-Лб) 1 кд/л фуг 3,426 3,426-10'4 10,764
1 кандела на 1 фут2 1 кд/фут2 10,764 1,0764 IO-3 3,382 10
ответа на единичный стимул”. Желательно, чтобы температура не
повышалась при изменении яркости фона. Для оценки фонового
освещения можно использовать .интеграционный фотометр, который
помещают около глаза.
Электронная регистрирующая система должна состоять из уси-
лителя и предусилителя с фильтрами 0,3 — 300 Гц. Сопротивление
предусилителя должно быть по крайней мере 1 МОм. Пациент дол-
жен быть электрически изолирован от усилителя.
Рекомендуется максимально расширять зрачок и отмечать его
величину, если он максимально не расширен.
Продолжительность темновой адаптации должна быть не менее
20 мин для создания стабильных физиологических условий и полу-
чения максимального скотопического ответа.
Необходимо избегать выполнения флюоресцентной ангиогра-
фии и фотографирования глазного дна перед проведением электро-
ретинографических исследований. Однако в случае необходимости
можно использовать темновую адаптацию в течение часа с после-
дующей регистрацией ЭРГ. Предпочтительно первоначально реги-
стрировать палочковый ответ в темновой адаптации, перед регис-
трацией смешанного колбочкового ответа на более яркие стимулы,
с минимальной световой адаптацией и минимальным временем
нахождения электрода на роговице.
Фиксационная точка должна быть, особенно при использова-
нии структурированных стимулов, однако некоторые пациенты ее
не видят, в связи с чем обследуемому предлагают смотреть прямо
перед собой и глазами не двигать. В строгой фиксации взора нет
необходимости при использовании ганцфельд-стимулятора.
Максимальный ответ регистрируют при использовании стандарт-
ных стимулов в темноадаптированном глазу с интервалом между ними
по крайней мере 5—10 с. Этот ответ состоит из комбинации ответов
палочковой и колбочковой систем. Полосы пропускания: нижняя 75—
100 Гц на нижнем пределе и 300 Гц и выше на верхнем пределе.
150
Ламберт Миллиламберт Фут-ламберт Кандела на 1 м2
3,14 Ю4 3,14 IO4 3,14 ю-4 1 ю-3 1,0764 КГ3 3,382-Ю"3 0,314 0,314 3,14 • 103 101 103 1 1,0764 3,382 0,2919 0,2919 2,919 • 103 9,29 • IO'2 9,29 • IO3 0,929 1 3,14 9,29 • IO’2 9,29 • IO"2 229 2,957 ♦ IO 2 295,7 • IO"2 0,2957 0,3183 1
Палочковый ответ — это первый сигнал после темновой адапта-
ции, так как палочки очень чувствительны к световой адаптации.
Слабый белый стимул интенсивностью 2,5 лог. ед. подают на поверх-
ность ганцфельд-шара. Минимальный интервал между стимулами
2 с. Возможно использование и синего стимула в этих же условиях.
Осцилляторные биопотенциалы регистрируют от темно-адапти-
рованного глаза при использовании того же самого стандартного
белого стимула, однако применяемые фильтры должны быть от 75
до 100 Гц на нижнем конце и 300 Гц или больше в верхнем диапазоне
частот. Осцилляторные потенциалы вариабельны и изменяются пос-
ле первой стимуляции, поэтому первый стимул, подаваемый с интер-
валом 15 с, не следует учитывать, а усреднение полезного сигнала
должно начинаться после второго или следующего стимула.
Простой колбочковый ответ на единичный стимул с интервалом
0,5 с регистрируется при светлом фоне, подавляющем палочковую
активность, с освещенностью 17—34 кд/м2 (5—10 фут Лб), измерен-
ной на поверхности ганцфельд-фона. Рекомендуется высокая осве-
щенность фона при высших пределах яркости стимула и слабое ос-
вещение фона при ее низшем пределе. Стандартное оборудование
должно иметь идентичное соотношение яркости стандартного сти-
мула и фона, подавляющего палочковую активность, эквивалент-
ное Зкд/м2. Для выделения колбочкового ответа рекомендуется
преадаптация в течение 10 мин перед регистрацией ЭРГ, так как в
этот период колбочковый ответ может увеличиться.
Мелькающий (ритмический) ответ регистрируется с помощью
стандартного стимулятора при том же фоновом освещении, подав-
ляющем палочковую активность, после регистрации колбочкового
ответа на единичный стимул. Запись мелькающей ЭРГ при фоно-
вом освещении уменьшает дискомфорт и приводит к стандартиза-
ции условий фотопической адаптации. Частота стимула должна быть
строго 30 стимулов в 1 с, а первые несколько записей пропускают
для стандартизации записи.
151
Рис. 5.9. Нормальные
значения общей и ло-
кальной ЭРГ.
а — нормальная общая
ЭРГ на белый стимул
(0,3 Дж); б — нормальная
локальная ЭРГ на зеле-
ный стимул (18°); в —
нормальная локальная
ЭРГ на красный стимул.
Анализ колбочковой, палочковой и максимальной ЭРГ реко-
мендуется проводить по амплитуде и времени до пика b-волны от
начала стимула; для колбочковой ЭРГ этот анализ проводится как
на единичные вспышки, так и на мелькающий стимул в 30 Гц. Со-
ответственно существующим стандартам a-волну оценивают от
базовой линии до пика, амплитуду b-волны измеряют от пика
a-волны до пика b-волны, а время b-волны (implicit time) измеряет-
ся от времени вспышки до пика этой волны. Значение для клиники
определения времени до пика b-волны ЭРГ очевидно, так как было
установлено, что ранним симптомом до начала снижения амплиту-
ды b-волны, патологических процессов в фоторецепторах сетчатки
является увеличение implicit time. Измерение этого параметра стало
возможным только в последнее время, когда для анализа ЭРГ нача-
ли использовать компьютеры.
Для сравнения пресинаптической активности сетчатки, представ-
152
Таблица 5.2. Нормальное значение клинической ЭРГ на вспышку в
зависимости от возраста при монополизмом отведении (LKC, 1996)
Возраст, годы 20 30 40 50 60 70
Форма ЭРГ мкВ мс мкВ мс мкВ мс мкВ мс мкВ мс мкВ мс
Скотопическая Ь-вол- на ЭРГ 24 дБ Скотопическая Ь-вол- на ЭРГ 0 дБ 185 419 52 13 403 52 141 387 52 119 371 52 97 355 52 75 339 52
Осцилляторный по- тенциал Фотопическая Ь-вол- на ЭРГ (вспышка ПО 102 32 89 92 32 75 82 32 61 72 32 50 62 32 50 52 32
ОдБ) Фотопическая мель- кающая ЭРГ (30 Гц) 70 30 63 30 56 30,5 49 30,5 42 31 35 31
Примечание. Амплитуда ЭРГ в микровольтах; время до пика b — в
миллисекундах.
ленной a-волной с постсинаптической активностью, представлен-
ной b-волной, наряду с оценкой амплитуды и временных парамет-
ров ЭРГ используют соотношение амплитуды b/a-волн. В норме
b-волна в два раза превышает a-волну при использовании насыща-
ющей интенсивности стимулирующего света (максимальная интен-
сивность стимулирующего света, при которой ЭРГ достигает пре-
дельной величины). При заболеваниях сетчатки сосудистого генеза,
сохранности хороидальной циркуляции или воздействии лекарствен-
ных препаратов на сетчатку отношение b/a-волны редуцировано.
Измерение и описание осцилляторных потенциалов не стандар-
тизировано. Их появление зависит от условий стимуляции, адапта-
ции и характеристик фильтров усилителя. Однако большинство
авторов описывают четыре основных пика, которые можно исполь-
зовать в повседневной практике для клинических целей. Усредне-
ние может быть применено для выявления очень слабых ответов.
Рекомендуется, чтобы все лаборатории разрабатывали нормати-
вы ддя каждого используемого ими оборудования и каждого вида
ЭРГ (рис. 5.9). На рисунке представлены данные лаборатории клини-
ческой физиологии зрения им. С.В.Кравкова МНИИ ГБ им. Гельм-
гольца при регистрации общей и локальной ЭРГ с помощью специ-
ализированной системы для электрофизиологических исследований
Neuropto (фирмы “Medelec”, Англия) и электроретинографа (Науч-
но-медицинской фирмы МБН, Россия). Параметры ЭРГ могут изме-
няться как в зависимости от типов электродов, так и в зависимости
от возраста, поэтому расчет стандартного отклонения нормативов
необходимо проводить корректно (табл. 5.2). Все данные о локаль-
153
ных и других нестандартных ответах должны включать пределы нор-
мальных показателей. Для того чтобы можно было сравнивать пока-
затели, полученные в разных лабораториях мира, предлагается в док-
ладах и публикациях представлять ЭРГ с калибровочным сигналом
для амплитудных и временных параметров, указывать интенсивность
стимула и адаптивного фона (в кд/м2), а также условия адаптации.
Показатели приводят в абсолютных величинах.
5.1.5. Роль ЭРГ в диагностике заболеваний сетчатки и зрительного
нерва
Локализация патологических процессов в зрительной системе, в диа-
гностике и дифференциальной диагностике которых используются
электрофизиологические методы исследования, разнообразна. При
этом выбор метода определяется происхождением биоэлектрической
активности в пигментном эпителии, тех или иных слоях и нейронах
сетчатки, центральном отделе зрительного анализатора.
Локализация заболеваний, при которых проводят электрофи-
зиологическое тестирование, может быть следующей: пигментный
эпителий сетчатки (ЭОГ, de ЭРГ, с-волна), наружный сегмент фото-
рецепторов (ранний, рецепторный потенциал, денситометрия), ре-
цепторный слой (а-волна в общей ЭРГ), колбочковая система (фо-
топическая ЭРГ, тесты на цветовое зрение, мелькающая ЭРГ), па-
лочковая система (палочковая изолированная ЭРГ на слабый
голубой или белый стимул ниже колбочкового порога, тесты на
темновую адаптацию), средние слои сетчатки, клетки Мюллера
(b-волна общей ЭРГ), амакриновые и биполярные клетки (осцилля-
торные потенциалы, Р50-компонент паттерн-ЭРГ, порог негатив-
ного ответа), слой ганглиозных клеток сетчатки (паттерн ЭРГ), ма-
кулярная область (локальная, макулярная ЭРГ), зрительный тракт
(зрительные вызванные корковые потенциалы).
Снижение b-волны ЭРГ — один из основных характерных элект-
роретинографических симптомов при заболеваниях сетчатки различ-
ного генеза. Поскольку b-волна отражает активность нейронов второ-
го порядка, вполне естественно ожидать изменение формы ЭРГ при
локализации патологического процесса во внутреннем ядерном слое
сетчатки. При сохранности фоторецепторного слоя, когда амплитуда
a-волны хорошо выражена, наблюдается типичное изменение формы
биоэлектрического ответа сетчатки, известное как негативизация ЭРГ.
Однако не всегда избирательное угнетение b-волны отражает
органические нарушения в сетчатке. Это связано с тем, что фоторе-
цепторы имеют синаптические окончания не на одной, а на несколь-
ких биполярных клетках (за исключением центральных колбочек).
Эта структурная конвергенция сигнала является одной из причин
пространственной суммации в сетчатке. Вследствие этого наруше-
ние процессов пространственной суммации в сетчатке, даже при
отсутствии в ней выраженных органических изменений, неизбежно
154
отражается на амплитудных параметрах ЭРГ, что приводит к изме-
нению соотношения амплитуд а- и b-волн в общей ЭРГ.
В качестве примера рассмотрим наиболее демонстративные
нозологические формы заболеваний сетчатки, при которых резуль-
таты электроретинографических исследований имеют важное зна-
чение в установлении диагноза.
При палочковом монохроматизме скотопическая, палочковая ЭРГ
нормальной амплитуды, в то время как фотопическая, или колбоч-
ковая ЭРГ, а также макулярная ЭРГ отсутствуют.
Формы врожденной колбочковой дисфункции и ее психофизиче-
ские симптомы описаны ранее (см. главу 3). Диагноз колбочковой
дисфункции легко подтверждается при изоляции колбочковой и па-
лочковой систем в ЭРГ. Фотопическая ЭРГ, регистрируемая в усло-
виях световой адаптации, использование частоты мельканий свето-
вого стимула (20—30 Гц) позволяют оценить функциональное со-
стояние колбочковой системы. Фотопическая ЭРГ может быть
нормальной при изолированном поражении макулярной области,
и, наоборот, патологическая фотопическая ЭРГ свидетельствует о
большем распространении патологического процесса, чем это вид-
но офтальмоскопически, т. е. имеет место панретинальная колбоч-
ковая дистрофия или дегенерация.
При поздних проявлениях наследственной колбочковой дистро-
фии всегда наблюдаются изменения в ЭРГ, выраженные в той или
иной степени (от значительных изменений фотопической ЭРГ до
нерегистрируемой), тогда как палочковая ЭРГ нормальна или суб-
нормальна. У пациентов с колбочковой дисфункцией и макулопати-
ей типа “бычий глаз” при достаточно хорошей амплитуде а-волны
ЭРГ может быть значительно снижена b-волна ЭРГ [Yokoyama, 1974].
Макулопатия этого типа встречается при различных видах за-
болеваний сетчатки, таких как хлорохиновая ретинопатия, колбоч-
ковая дистрофия, болезнь Штаргардта, периферическая форма ТРА,
нейрональный кероидный липофусциноз и др. Получаемая с помо-
щью ЭРГ информация позволяет в разных случаях косвенно судить
о характере развивающегося патологического процесса в сетчатке.
М. Kato и I. Watanabe (1991) установили, что при макулопатии
типа “бычий глаз” степень негативизации ЭРГ коррелирует с уров-
нем циклической ГМФ (цГМФ) в плазме. Авторы предположили,
что обнаруженное ими селективное угнетение b-волны ЭРГ на оди-
ночную вспышку яркого света следует рассматривать как результат
аномального метаболизма ц-ГМФ в фоторецепторах. Поскольку
известно, что ц-ГМФ играет важную роль не только в зрительной
трансдукции, но и в синаптической передаче [Barbehen Е.К. et al.,
1988], считается, что у данных больных негативная ЭРГ отражает
синаптическую дисфункцию.
В то же время существует предположение [Gouras Р. et al., 1993],
что при колбочковой дистрофии, характеризуемой супернормаль-
155
1
а б 8
Рис. 5.10. Общая ЭРГ.
а — максимальный колбочково-палочковый ответ на белый стимул; б — палочко-
вая ЭРГ на голубой стимул в темноадаптированном глазу; в — колбочковая ЭРГ на
фоне световой адаптации. 1 — стационарная ночная слепота, 2 — норма, 3 — па-
лочковый монохроматизм.
ними значениями b-волны, в наружных сегментах фоторецепторов
отмечается аномально высокий уровень ц-ГМФ.
Аналогичное нарушение передачи информации от фоторецеп-
торов к палочковым деполяризующимся биполярам было отмечено
при кожной злокачественной меланоме, заболевании, не относящем-
ся к ретинальным дистрофиям.
Эти данные свидетельствуют о большом разнообразии механизмов,
ответственных за потерю зрения у пациентов с дистрофическими забо-
леваниями сетчатки, а также о важной роли электроретинографии в
выявлении звеньев патогенеза разнообразной ретинальной патологии.
Стационарная ночная слепота (гипофункция палочкового аппа-
рата, или гемералопия). Современная классификация гемералопии
представлена в главе 2. Ранние психофизические данные показали,
что имеются две формы врожденной стационарной ночной слепо-
ты: одна форма с нормальной колбочковой функцией и другая с
некоторым ее снижением, что подтверждено в последнее время элек-
трофизиологическими исследованиями. Поэтому классификация
врожденных форм стационарной ночной слепоты основана на раз-
личиях в ЭРГ. Для одного типа характерна редуцированная ампли-
туда фотопического ответа, который не увеличивается в темноте
(I тип Nougaret), в другом типе нарушения функций выражаются в
отсутствии темновой адаптации, b-волны в общей ЭРГ(рис. 5.10), в
связи с чем а-волна выражена больше, чем в норме, при этом фото-
пический ответ снижен (II тип Шуберта — Борншейна). В I типе при
нормальной кинетике зрительного пигмента изменения в ЭРГ ука-
зывали на нарушения в трансмиссии во внутреннем сегменте фото-
рецепторов. При II типе с нормальным зрительным пигментом и
нормальной a-волной ЭРГ отмечено нарушение нейрональной пе-
156
OD OS
a
6
Рис. 5.11. Нерегистрируемая ЭРГ у больного с рецессивно наследуемой
формой пигментного ретинита.
а — на белый стимул; б — красный стимул.
редачи в области биполярных клеток. Электрофизиологические и
психофизические данные предполагают наличие первичного дефек-
та в интерплексиформных клетках внутреннего ядерного слоя, что
подтверждено, с одной стороны, отсутствием структурных наруше-
ний в фоторецепторах, а с другой — тем, что нормальный палочко-
вый сигнал не достигает биполярных клеток. Работы последних лет
показали, что при этом виде заболевания нарушается синаптиче-
ская передача на инвертирующем глутаматном синапсе между па-
лочками и деполяризующимися биполярными нейронами [Carr R.E.,
1992]. При болезни Огуши (врожденная стационарная ночная сле-
пота с изменением глазного дна) b-волна ЭРГ отсутствует
[HiroseT.,1956], хотя в некоторых работах была показана сохран-
ность скотопической функции [Nagata М., 1963]. При использова-
нии красного фона низкой интенсивности найден нормальный кол-
бочковый ответ, но сниженный по амплитуде и пролонгированный
палочковый ответ. Р. Guras (1970) зарегистрировал нормальный
скотопический ответ в ЭРГ при этом заболевании на первый сти-
мул и исчезновение этого скотопического ответа при последующей
стимуляции. В связи с найденной нормальной регенерацией родоп-
сина и нормальной его фоточувствительностью было высказано
предположение, что это заболевание связано с нарушениями в адап-
тивных сетях [CarrR.E., Ripps Н., 1966], а при fundus albipunctata
изменение ЭРГ и ЭОГ связывается с замедлением регенерации па-
лочкового и колбочкового зрительного пигментов. Врожденная
стационарная ночная слепота типа Шуберта — Борншейна (Schu-
bert — Bomschein) подразделяется на два подтипа: полный (с мио-
пией) и неполный (по Miyake) соответственно степени сохранности
палочковой и колбочковой функции в ЭРГ (либо отсутствие ЭРГ,
либо частичная ее сохранность; рис 5.10).
Тапеторетинальная дистрофия (пигментныйретинит). Класси-
ческим патогномоничным признаком этого тяжелого наследствен-
ного заболевания, многообразная клиническая картина которого
обусловлена мутациями гена родопсина, является отсутствие ЭРГ
уже в начальной стадии (рис. 5. 11) [Franceschetti A. et al., 1963;
157
BersonE.L., 1981; Francois J., 1984; Jacobson S.G., 1996]. В самых
ранних стадиях дистрофии с доминантной формой наследования
ЭРГ может быть субнормальной, что дает возможность разделить
колбочковые и палочковые компоненты ЭРГ, изучить эволюцию и
локализацию заболевания. При этом отмечено увеличение времени
кульминации b-волны, что служит дифференциальным признаком
между стационарной ночной слепотой и секторальной формой пиг-
ментного резинита, при которых пиковое время в пределах нор-
мы. Электрофизиологические изменения при пигментном ретините
предшествуют изменениям на глазном дне и другим функциональ-
ным находкам. Делаются попытки, по данным ЭРГ, разделить типы
наследования при пигментном ретините [Berson E.L., 1981—1991;
Apfelstedt-Sylla Е., 1996; Cideciyan A.V., et al., 1996]. Обнаружено,
что при доминантной форме с полной пенетрантностью гена Ь-вол-
на колбочковой ЭРГ имеет нормальную или несколько сниженную
амплитуду с нормальным временем кульминации, в то время как
палочковая ЭРГ значительно редуцирована и время ее кульмина-
ции увеличено. При доминантном типе с редуцированной пенет-
рантностью палочковая и колбочковая ЭРГ редуцированы с про-
лонгированным временем кульминации. Митахондриальная форма
пигментного ретинита по функциональным симптомам сходна с до-
минантной формой. Уже в начальной стадии пигментный ретинит
с аутосомно-рецессивным типом наследования и сцепленным с по-
лом характеризуются очень слабым колбочковым ответом и значи-
тельным увеличением латентного периода b-волны. Если палочко-
вый ответ определяется, время кульминации также значительно уве-
личено. Весьма важным является обследование родственников
больных пигментным ретинитом по восходящей и нисходящей ли-
ниям. Снижение скотопической b-волны ЭРГ, пролонгированное
время ее кульминации, отсутствие или редуцирование осциллятор-
ных потенциалов характерны не только для больных с клинически
установленными формами пигментного ретинита, но и для сибсов,
гетерозигот, носителей патологического гена пигментного ретини-
та (рис.5.12). Изменения ЭРГ, выявленные в раннем детском возра-
сте (до 6 лет), могут помочь как в уточнении диагноза и генетиче-
ской формы заболевания, так и в выявлении носителей патологи-
ческого гена и генетических консультациях. ЭРГ отсутствует при
беспигментной и мелкопигментной унилатеральной форме пигмент-
ного ретинита. [Francois J., 1984; Marques М. et al., 1988; Lorenz В.,
Andrassi М.,1996]. При атипичной односторонней форме заболева-
ния следует повторить обследование через определенный промежу-
ток времени, чтобы установить подлинную унилатеральность. Элек-
троретинографические ответы зависят от распространенности па-
тологического процесса. Так, ЭРГ имеет субнормальный характер
и всегда присутствует при секторальных формах пигментного рети-
нита (квадрантных, половинных, центральных и парацентральных).
158
200 мкВ
30 мс
Рис. 5.12. ЭРГ у больного пигментным ретинитом и его здоровых ближай-
ших родственников.
а—нормальная общая ЭРГ у здорового отца; б — субнормальная ЭРГ у клиниче-
ски здорового сына (носителя патологического гена); в — отсутствующая ЭРГ у ма-
тери, больной пигментным ретинитом.
ЭРГ отсутствует при пигментном ретините, который входит в сим-
птомокомплекс (также включающий мутацию гена родопсина) не-
которых наследственно-атаксических поражениях мозжечка, туго-
ухости, синдроме Ушера (Usher), синдроме Муна — Барде — Бидля
(Moon — Bardet — Biedl), мукополисахаридозе Хурбера — Хунте-
ра — Шеи (Hurber — Hunter — Sheie), некоторых миотонических ди-
строфиях. При Retinitis punctata albescens (тяжелая форма) ЭРГ зна-
чительно снижена, в то время как при retinitis albipunctatus с геме-
ралопией ЭРГ частично сохранена.
Поскольку колбочковый зрительный сигнал в сетчатке прохо-
дит через on- и off-биополярные клетки, Р.А. Sieving (1993) исполь-
зовал для записи фотопической ЭРГ у пациентов с пигментным
ретинитом более длительные, чем другие авторы, стимулы (150—
300 мс), что позволило даже с помощью суммарной ЭРГ выявить
нарушения в on- и off-путях передачи зрительной информации в
сетчатке. Результаты выполненных им исследований показали, что
одной из причин ослабления зрения у этих пациентов является па-
тологическое прохождение сигнала в проксимальной сетчатке, а фун-
кциональный дефект локализован постсинаптически по отношению
к колбочковым фоторецепторам.
Генетические находки последнего времени, свидетельствующие
о мутации родопсина при пигментном ретините аутосомно-доми-
нантного типа наследования [DryjaT.P. et al., 1990], позволяют
предположить, что при многих дистрофиях сетчатки первичный
дефект находится на фоторецепторном уровне. Однако неясно, по-
чему в некоторых случаях при палочково-колбочковой дистрофии
159
(традиционно называемой за рубежом “пигментным ретинитом”)
снижение b-волны ЭРГ не соответствует достаточно хорошо сохра-
нившемуся полю зрения и при этом лишь незначительно повышает-
ся порог темновой адаптации.
С помощью рефлексоденситофотометрии in vivo было показа-
но, что у пациентов с пигментным ретинитом, у которых значи-
тельно снижено сумеречное зрение, содержание зрительного пиг-
мента родопсина в фоторецепторах достаточно велико [Ripps Н. et
al., 1978]. На основании этого сделали вывод о том, что снижение
сумеречного зрения обусловлено нарушениями в клеточных элемен-
тах, расположенных проксимальнее фоторецепторов [Arden G.B.,
1982; Sieving Р.А., 1993].
К редким формам пигментного ретинита с характерными изме-
нениями ЭРГ относятся синдром Бассена — Корнцвейга (Bassen —
Komzweig), проявляющийся наследственной аР-гипопротеинемией,
и болезнь Рефзума (Refsum), характеризующаяся нарушением мета-
болизма, выражающимся в аккумуляции экзогенной фитановой
(phytanic) кислоты. Выявление этих форм заболевания уже в ран-
нем детском возрасте (до 6 лет) с помощью ЭРГ позволит своевре-
менно начать раннее корректирующее метаболизм лечение.
При смешанной форме колбочково-палочковой дегенерации запись
ЭРГ в условиях, позволяющих выделить палочковые и колбочко-
вые компоненты, может показать преимущественное поражение тех
или других компонентов. Если фотопическая b-волна больше реду-
цирована, чем скотопическая b-волна, можно предполагать колбоч-
ково-палочковую дисфункцию, если же изменения скотопической
ЭРГ выражены в большей степени, то речь идет о палочково-кол-
бочковой дисфункции.
Близким заболеванием к пигментному ретиниту является атро-
фия Гирате (атрофия хороидеи и сетчатки), имеющая аутосомно-
рецессивный тип наследования, в основе которого лежит дефект
энзима орнитинаминотрансферазы (гиперорнитинемия) [Takki К.,
1974]. На глазном дне обширные круговые очаги полной атрофии
хороидеи и сетчатки. В ЭРГ обнаруживаются значительные измене-
ния уже в раннем детском возрасте от субнормальной до нерегис-
трируемой. Время до пика фотопической ЭРГ пролонгировано. ЭОГ
также изменяется от субнормальной до значительно субнормаль-
ной.
Хороидеремия — наследственное заболевание (тип наследствен-
ности, сцепленный с полом), характеризуется прогрессивной диф-
фузной дегенерацией пигментного эпителия и хороикапилляров и
отличается от пигментного ретинита и атрофии гирате не только
клинической картиной, которая может быть сходной, но также
типом наследования. ЭРГ, если регистрируется, то имеет вид, ха-
рактерный для палочково-колбочковой дегенерации.
Врожденный амавроз Небера, по данным некоторых авторов,
160
50 мкВ
Рис.5.13. ЭРГ при Х-хро-
мосомном ювенильном
ретиношизисе (больной
Ш., 36 лет., vis: OD=0,2;
OS=0,3).
а — резко субнормальная
общая ЭРГ (OD); б — суб-
нормальная ЛЭРГ (OD).
30 МС
является причиной слепоты у детей в 10—20 % случаев. С рождения
нерегистрируемая ЭРГ является патогномоничным диагностическим
признаком этого тяжелого заболевания.
Ретиношизис. При идиопатической форме Х-хромосомного юве-
нильного ретиношизиса ЭРГ помогает в создании нозологической
классификации заболевания. Так, патогномоничным признаком X-
хромосомного ювенильного ретиношизиса является минус-негатив-
ный тип ЭРГ и значительно сниженная локальная ЭРГ (рис.5.13);
при гиалоидной форме ретинита типа Вагнера (доминантный тип
наследования) ЭРГ имеет субнормальный характер; при гиалоид-
ной тапеторетинальной дегенерации типа Фавра — Гольдмана (ре-
цессивная форма наследования) и макрофибриллярной гиалоидной
дегенерации Фавра (аутосомно-доминантный тип наследования)
ЭРГ не регистрируется.
Сенильный дегенеративный ретиношизис не имеет типичного
изменения ЭРГ.
Отслойка сетчатки. При этой патологии ЭРГ всегда снижена,
и по степени этого снижения можно судить о давности процесса,
его распространенности, прогнозе течения заболевания (рис.5.14).
Если площадь отслойки больше 35%, то ЭРГ изменяется значитель-
но или не регистрируется. Особый интерес представляет исследова-
ние “здоровых” глаз у больных с односторонней отслойкой сетчат-
161
в
Рис. 5.14. ЭРГ при отслойке сетчатки (больной А., 50 лет, отслойка сетчат-
ки на левом глазу, vis: OS = 0,03; правый глаз здоров, vis OD = 1,0).
а — ОП отсутствуют; б — общая ЭРГ субнормальна; в — общая ЭРГ не регистриру-
ется.
ки, в которых отмечаются витреоретинальные изменения дистро-
фического характера на периферии глазного дна. ЭРГ на этих гла-
зах может иметь субнормальный характер, что подтверждает нали-
чие патологического процесса на периферии. Локальная ЭРГ на
таких глазах в пределах нормы, что коррелирует с высокой остро-
той зрения. Мало изменяется ЭРГ при локализованной отслойке
сетчатки. Функциональные результаты после хирургического лече-
ния лучше в тех случаях, когда значения ЭРГ в предоперационном
периоде были удовлетворительными. Даже после с успехом прове-
денного оперативного вмешательства ЭРГ восстанавливается очень
медленно.
Острые циркуляторные нарушения в сетчатке. При острой не-
проходимости центральной артерии сетчатки вместе с появлением
функциональных и клинических симптомов происходят изменения
в ЭРГ. При этом b-волна общей ЭРГ практически не регистрирует-
ся, так как она генерируется во внутренних слоях сетчатки, крово-
снабжение которых нарушено.
При острой закупорке центральной вены сетчатки изменения в
палочковой ЭРГ предшествуют изменениям в колбочковой ЭРГ:
а-волна ЭРГ сохраняется, так как она генерируется фоторецептора-
ми, питающимися от хороидеи. При закупорке ветви центральной
вены сетчатки ЭРГ носит негативный характер, однако изменения
в ЭРГ менее выражены, так как степень изменения b-волны зависит
от ишемии внутренних слоев сетчатки (рис. 5.15, 5. 16). В последнее
время установлено [Matsui Y., 1994], что именно отношение ампли-
туд b/a-волн ЭРГ является индикатором степени ишемизации сет-
чатки и прогноза восстановления зрительных функций. При острых
162
OD
Рис. 5.15. ЭРГ при тромбозе ветви центральной вены сетчатки (больной
Р., 61 года, тромбоз центральной вены сетчатки на правом глазу, vis=0,2;
левый глаз — здоров, vis=l,0).
OD: а — ОП отсутствуют; б - общая ЭРГ резко субнормальна; OS: в — ОП отсут-
ствуют; г — общая ЭРГ в пределах нормы.
а б
Рис.5 Л 6. Фотопический ответ на вспышку большой длительности у паци-
ента с односторонним тромбозом центральной вены сетчатки (выраженная
редукция Ь- и d-волн по сравнению с нормой) [Sieving Р.А., 1993].
а — здоровый глаз; б — тромбоз центральной вены сетчатки.
163
тромбозах сосудов сетчатки этому соотношению придается прогно-
стическое значение, так как по отношению b/a-волн ЭРГ можно
судить о патогенетически различных стадиях острой сосудистой
катастрофы (при геморрагической форме тромбоза b/a-волн боль-
ше 1, при венозном стазе меньше 1). При незначительных изменени-
ях b-волны прогноз лучше, при негативной ЭРГ b-волна ниже изо-
динии, прогноз плохой и является фактором риска для образования
неоваскулярной мембраны, хотя нормальное соотношение Ь/а-волн
не исключает возможность возникновения пролиферативных изме-
нений.
Хронические циркуляторные нарушения в сетчатке, значительные
изменения в хориокапиллярах приводят к снижению ЭРГ, что отра-
жает степень метаболических нарушений.
Диабетическая ретинопатия. Большое значение имеет ЭРГ в
оценке тяжести диабетической ретинопатии, при которой патоло-
гический процесс сосудистой системы, сопровождающийся ишеми-
ей, приводит к метаболическим изменениям в сетчатке.
В клинических и электрофизиологических исследованиях уста-
новлено, что в доклинической стадии диабетической ретинопатии,
когда лишь при ангиографии можно отметить микроаневризмы со-
судов сетчатки, уже имеются изменения осцилляторных потенциа-
лов. ЭРГ, как общая, так и локальная, может иметь супернормаль-
ный характер, отражая начальные метаболические нарушения в
сетчатке. По мере развития клинических признаков заболевания
(облитерация сосудов, возникновение геморрагий, ишемических
инфарктов, появление хлопковидных пятен и неоваскуляризации,
нарушение венозного оттока, наличие липопротеинового экссуда-
та) во II и III стадиях заболевания диабетической ретинопатии от-
мечаются изменения фотопической и скотопической ЭРГ, выражен-
ность которых зависит от патологических изменений на глазном
дне. В IV стадии диабетической ретинопатии ЭРГ может не регис-
трироваться вследствие функциональной несостоятельности наруж-
ных и внутренних слоев сетчатки.
Травмы органа зрения. Исследование общей ЭРГ необходимо
проводить во всех случаях повреждений глаза, особенно когда глаз-
ное дно не видно из-за наличия изменений в роговице, хрусталике,
стекловидном теле для оценки функционального состояния сетчат-
ки и прогнозирования возможных исходов травм и оперативных
вмешательств. ЭРГ необходимо исследовать для оценки функцио-
нального состояния сетчатки при субатрофии глазного яблока трав-
матического и воспалительного генеза, для решения вопроса о воз-
можности хирургического вмешательства.
Гемофтальм. Значительные изменения ЭРГ при использовании
красного стимула свидетельствуют о тяжелой патологии сетчатки,
сопровождающей гемофтальм.
В случае наличия инородного тела в глазу именно при анализе
164
ЭРГ обнаруживают или подтверждают первые проявления сидероза
или халькоза. Признаки металлической интоксикации определяют
достаточно быстро в зависимости от природы инородного тела.
При сидерозе первые нарушения биопотенциала сетчатки можно
отметить уже через месяц, при халькозе — спустя 2—3 мес. При
этом общая ЭРГ сначала супернормальна, что является признаком
раздражения нейронов сетчатки, затем b-волна ЭРГ прогрессивно
снижается, а а-волна остается увеличенной и ЭРГ носит сначала
плюс-, а затем минус-негативный характер. При развитии процесса
а-волна также уменьшается и ЭРГ становится резко субнормальной
или отсутствует. При негативном характере ЭРГ прогноз сомните-
лен, а нерегистрируемая ЭРГ является свидетельством необрати-
мых изменений в сетчатке. Исследование ЭРГ при глазном травма-
тизме — наиболее точный способ контроля за эффективностью про-
водимого консервативного и хирургического лечения.
При заболеваниях зрительного нерва ЭРГ, как правило, нормаль-
на, что является дифференциально-диагностическим признаком за-
болеваний сетчатки и зрительного нерва. Однако отмечаются слу-
чаи ретроградного вовлечения ретинальных нейронов в патологи-
ческий процесс в сетчатке. Если причиной заболевания зрительного
нерва являются биохимические нарушения в организме, ЭРГ может
исчезать или значительно редуцироваться.
ЭРГ может быть использована при некоторых заболеваниях для
определения степени вовлечения сетчатки в патологический про-
цесс. К ним относятся системные заболевания с нарушением пита-
ния, в частности, обусловленные дефицитом цинка, анемия, хрони-
ческий алкоголизм и цирроз печени, диабет, наследственные и врож-
денные заболевания детей, кожные болезни, болезнь Рефзума,
синдром Бассена — Корнцвейга, нейромускулярные нарушения (ми-
тохондриальная миопатия, атаксия Фридрейха, синдром Этона —
Ламберта (Eaton — Lambert), миастения gravis, неврологические на-
рушения — хорея Хантингтона (Huntington, синдром Лауренса —
Муна — Бидля (Laurence — Moon — Biedl), болезнь Паркинсона,
синдромы, включающие слабоумие.
Симпатическая офтальмия. Изменения ЭРГ информативны для
диагностики поражений сетчатки при симпатической офтальмии.
Изменения b-волны ЭРГ, ее супернормальный, а затем субнормаль-
ный характер предшествуют появлению клинических признаков
заболевания на глазном дне. Изменения ЭРГ нарастают по мере
развития и генерализации воспалительного процесса. При этом
фотопическая, колбочковая, локальная и ритмическая ЭРГ не изме-
няются до конечных стадий заболевания.
Сетчатка может реагировать также на токсичные вещества и
лекарственные препараты [Шамшинова А. М., 1971; Shamshi-
nova А. М. et al., 1990]. ЭРГ является индикатором интоксикаций
сетчатки, а также побочных действий лекарственных препаратов.
165
ЭРГ изменяется на фоне таких токсичных веществ, как алкоголь,
табак, карбон моноксид (пренатальная диагностика), карбон
бисульфид (производство вискозы), ртуть, метил, глютамат и др.
К лекарственным препаратам, вызывающим побочные действия, от-
носятся хинин и его производные, винкристин, этамбутол, феноти-
азин, ингибиторы моноаминоксидазы, трициклические антидепрес-
санты, бензодиазепин. ЭРГ изменяется до появления клинических
признаков дистрофических процессов. Особое внимание привлека-
ют препараты, длительно применяемые в клинике, такие, как анти-
малярийные (хинин), индометацин, нейролептики фенотиазиново-
го ряда, цепарин, которые вызывают необратимые патологические
изменения в сетчатке и зрительном нерве. ЭРГ изменяется от супер-
нормальной до субнормальной на стадии функциональных обрати-
мых нарушений и полностью исчезает при эволюции дистрофиче-
ских процессов на глазном дне. В связи с этим исследование ЭРГ в
терапевтической, психиатрической и других клиниках целесообраз-
но периодически проводить при многолетнем курсовом лечении, по
крайней мере 3 раза в год. Хроническая табачная и алкогольная
интоксикация вызывает значительные изменения как палочковой,
так и колбочковой ЭРГ. Курс лечения, включающий назначение
витамина А, приводит к полному восстановлению биопотенциалов
сетчатки.
* Существует ряд критериев, обусловливающих необходимость
проведения электрофизиологических исследований в клинике глаз-
ных болезней:
1. Необходимость оценки функционального состояния сетчатки
в тех случаях, когда определить зрительные функции обычным
методом невозможно, а глазное дно не офтальмоскопируется, при
помутнении сред глаза, гемофтальме. Проведение электроретино-
графических исследований особенно ценно для решения вопроса о
целесообразности хирургического лечения заболевания.
2. Диагностика заболеваний сетчатки, так как в ряде случаев
изменения ЭРГ являются патогномоничными симптомами заболева-
ния. (
3. Оценка глубины, распространенности, степени поражения
сетчатки и его локализации.
4. Изучение звеньев патогенеза заболеваний сетчатки и зритель-
ного нерва.
5. Дифференциальная диагностика заболеваний сетчатки и зри-
тельного нерва различного генеза.
6. Диагностика начальных функциональных изменений сетчат-
ки, предшествующих клиническим проявлениям заболевания (меди-
каментозная интоксикация, диабетическая ретинопатия, сосудистые
нарушения и пр.).
7. Необходимость определения прогноза течения патологиче-
ского процесса, контроль за его эволюцией.
166
5.2. ЛОКАЛЬНАЯ ЭРГ
5.2.1. Происхождение локальной ЭРГ
Если метод общей ЭРГ в настоящее время достаточно унифициро-
ван, то единой методики для регистрации локальной ЭРГ пока не
существует [Говардовский В.И. и др., 1985; Шамшинова А.М.,1990;
Sandberg М.А., 1977; Miyake Y., 1988,1990; Biersdorf W.R.,1990].
Регистрация биопотенциала от макулярной области очень важна
для оценки функционального состояния колбочковой системы и
диагностики процессов, локализующихся в центральной области сет-
чатки. Выделение локального макулярного биопотенциала связано
с физиологическими свойствами палочковой и колбочковой систем
и условиями регистрации. Очевидно, что величина общей ЭРГ за-
висит от количества стимулируемых фоторецепторов. Однако па-
тологический процесс, локализующийся в макулярной области, не
влияет на величину общей ЭРГ. Это объясняется тем, что колбочки
макулярной ЭРГ составляют лишь 10% от общего количества кол-
бочек сетчатки и при макулярной дегенерации (дистрофия нейро-
нов первого, второго порядка), часто в процесс вовлекаются не все
фоторецепторы макулярной области. Кроме того, макулярная деге-
нерация нередко ограничена фовеальным рефлексом в 5° зритель-
ного угла. Эта область содержит 100 000 колбочек, которые состав-
ляют 1,5% всех колбочек сетчатки, поэтому трудно предположить,
что изменения в этой области влияют на общую ЭРГ, которая от-
ражает функцию как колбочковой, так и палочковой системы
всей сетчатки [Шамшинова А.М., 1989, 1990].
При выборе условий регистрации биоэлектрического потенциа-
ла важно выделить функцию колбочковой системы в макулярной
области. Колбочки чувствительны к длинноволновой части спект-
ра, стимулам высокой интенсивности, световой адаптации, способ-
ны реагировать на частоту мельканий свыше 20—30 Гц, в то время
как палочки чувствительны к коротковолновой части спектра, сти-
мулам низкой интенсивности, их чувствительность повышается при
темновой адаптации, достигая максимума к 45—60 мин, они не спо-
собны отвечать на ритмические стимулы с частотой свыше 20 Гц.
5.2.2. Методы регистрации локальной (макулярной) ЭРГ
Для регистрации локального (макулярного) биопотенциала необ-
ходимо решить три основные задачи: 1) выделить маленький по
величине биопотенциал из фонового шума; 2) нивелировать эффект
светорассеяния, всегда возникающий при стимуляции любой обла-
сти сетчатки; 3) контролировать фиксацию взора.
Метод усреднения позволяет выделить небольшой по величине
биопотенциал (меньше 5 мкВ) из шума. Величина ЭРГ зависит от
167
плотности и числа функционирующих колбочек. Нормальный ло-
кальный биопотенциал свидетельствует о нормальной функции
макулярной области, а снижение макулярной локальной ЭРГ при
использовании методов исследования без контроля фиксации взора
не является достоверным, так как при смещении взора стимул по-
падает в зону доминирования палочковой системы сетчатки.
Если учесть, что фотопическая ЭРГ, т.е. электроретинограмма,
зарегистрированная на ганцфельд-стимул в фотопических условиях
освещения, отражает функцию общего количества функционирую-
щих и стимулируемых колбочек, а нормальная острота зрения сви-
детельствует о высокой плотности функционирующих колбочек в
макулярной области, то очевидно, что определенная связь этих
функций существует, хотя биопотенциалы колбочек и острота зре-
ния имеют различное происхождение.
Известны устройства для локальной световой стимуляции сет-
чатки [Sandberg М. А., 1977; Brunette J. R., 1982; Miyake У., 1988,
Biersdorf W. R., 1990; 1990]. Некоторые из них позволяют исследо-
вателю наблюдать глазное дно испытуемого и подавать световые
вспышки, когда “нужный” участок сетчатки ’’смотрит” на источ-
ник стимулирующего света. Однако вследствие неконтролируемых
движений глаз обследуемого воспроизводимая стимуляция одного
и того же участка сетчатки затруднена. Кроме того, для исследова-
ния глазного дна необходим яркий источник света, в связи с чем
его проведение невозможно в условиях темновой и умеренной све-
товой адаптации, а при многократной стимуляции возникает опас-
ность повреждающего действия света на сетчатку. Устройство для
автоматического контроля за направлением взора, позволяющее
подавать световые стимулы в момент, когда испытуемый надежно
фиксирует взгляд на небольшом вспомогательном источнике, не
может быть использовано при наличии у пациента патологических
изменений в макулярной области. Между тем именно в этих случа-
ях электроретинографические методы исследования в диагностике
заболеваний сетчатки являются особенно важными.
Некоторые методы выделения функций колбочковой системы
предусматривают использование фонового освещения, которое при
определенной яркости подавляет палочковую активность, но при
этом снижается и без того небольшой по величине биопотенциал
макулярной области. Для выделения колбочковой и палочковой
фокальной ЭРГ применяют модификацию ТВ-фундускамеры
[Miyake У., Horiguchi М., 1992], в которой контроль за местом сти-
муляции на глазном дне осуществляется в инфракрасном свете.
Сочетание фонового освещения (2,2 лог. скотопических троландов)
для подавления палочковой активности и стимула 5°, 10°, 15° в
3,2 лог. скотопических троландов позволило выделить фокальную
макулярную колбочковую ЭРГ (ФМК ЭРГ) после 526 усреднений.
Регистрации фокальной макулярной палочковой ЭРГ (ФМП ЭРГ)
168
проводили после 30 мин темновой адаптации на слабом красном
фоне белым стимулом в 2,2 лог. ед. с такой же величиной стимула,
как для регистрации колбочковой фокальной ЭРГ, после 128 усред-
нений. Различная постоянная времени 0,03 и 0,003 была использо-
вана для регистрации фокальной ЭРГ и осцилляторных потенциа-
лов (ОП) соответственно.
На основании результатов исследований, проведенных у боль-
ных с пигментным ретинитом, стационарной ночной слепотой,
диабетической макулопатией, наследственной макулярной дистро-
фией типа Штаргардта, макулярная дистрофия разделена по значе-
ниям фокальной макулярной колбочковой и палочковой ЭРГ на
колбочко-палочковый и палочко-колбочковый синдромы. Оба ме-
тода выявляют патологию, не обнаруживаемую с помощью общей
ЭРГ. С помощью компонентного анализа фокальной ЭРГ можно
установить локализацию дисфункции в определенных слоях сетчат-
ки [Horiguchi М., Miyake У., 1992].
Большой опыт клинических электрофизиологов, накопленный в
течение последних 20 лет, позволил сформулировать основные ус-
ловия для регистрации локального макулярного биопотенциала
сетчатки. Наличие рассеянного света в глазу и физиологические
свойства самой сетчатки ограничивают возможность регистрации
локальной ЭРГ. Известно, что при регистрации ЭРГ с помощью
роговичных электродов макулярная ЭРГ прямо пропорциональна
освещенной площади сетчатки. ’’Каждый участок сетчатки с актив-
ными нейронами можно рассматривать как генератор внеклеточно-
го тока, нагруженный на относительно низкое сопротивление окру-
жающих тканей. Падение напряжения на этом сопротивлении, пред-
ставляющее собой ЭРГ, пропорционально силе тока, т. е. площади
возбужденной части сетчатки” [Говардовский В.И. и др., 1985]. Та-
ким образом, световой стимул с угловыми размерами 15°, возбуж-
дающий 1,5% площади всей биоэлектрически активной сетчатки
(полное поле зрения 160—180°), должен вызывать ЭРГ амплитудой
около 1,5% от максимальной, получаемой при засвете всего поля
зрения, т. е. 3—5 мкВ. При стимуле 1,5° амплитуда локальной ЭРГ
должна быть меньше в 100 раз (меньше 0,05 мкВ). Размер тестиру-
емого поля должен быть настолько мал, насколько это возможно
для получения надежного ответа, поэтому многие исследователи
выбирают для фовеального теста размер 2—5°.
По данным ряда авторов, ФМК на меньший стимул лучше кор-
релирует с остротой зрения при условии хорошей фиксации взора.
В то же время расчеты показывают, что надежно регистрируемая
ЭРГ с амплитудой в несколько микровольт (мкВ) может быть полу-
чена при любых размерах стимула, если яркость его достаточно
высока. Однако для стимула в 5° истинно локальный ответ состав-
ляет не более 15%, а для стимула 1° — менее 1% от регистрируемо-
го. Остальные 85—99% генерируются всей сетчаткой. Удовлетвори-
169
тельная степень локальности (70% локального ответа) может быть
получена только для стимулов умеренной интенсивности, величи-
ной не менее 10—15°. При попытке повысить амплитуду ответа
путем дальнейшего увеличения яркости вспышки локальность бу-
дет потеряна. При правильном выборе яркости стимула минималь-
ный размер области на сетчатке, от которой можно отвести локаль-
ную ЭРГ, определяется возможностями регистрации малых отве-
тов. ЭРГ с амплитудой 5 мкВ, условно принятая нами в качестве
пороговой, уже находится на уровне шума, который даже в хоро-
ших условиях регистрации составляет примерно ту же величину.
В связи с этим для получения удовлетворительного отношения сиг-
нала к шуму приходится накапливать ответы на 25—500 вспышек,
т. е. использовать метод усреднения. Положение резко ухудшается
при дальнейшем уменьшении угловых размеров стимула. Посколь-
ку амплитуда локального ответа обратно пропорциональна квад-
рату углового размера стимула, отношение сигнал — шум растет
как корень квадратный из числа накоплений; необходимое число
накоплений обратно пропорционально четвертой степени углового
размера стимула, поэтому стимул в 5° требует 2—4 тыс. накопле-
ний, что находится за пределами допустимой длительности иссле-
дования.
При регистрации локальной ЭРГ у человека необходимо учиты-
вать неравномерное распределение колбочек в сетчатке, поэтому
ЭРГ, регистрируемая в условиях, подавляющих палочковую актив-
ность с учетом соответствующих описанных выше физиологичес-
ких свойств колбочковой и палочковой систем, будет генерироваться
главным образом центральной, богатой колбочками, областью сет-
чатки. Вклад периферических отделов сетчатки, где доминируют
палочковые компоненты, относительно мал, в связи с этим относи-
тельная локальность макулярной ЭРГ автоматически обеспечива-
ется неоднородностью свойств сетчатки.
Итак, использование красного, длиноволнового стимула в 5 раз
Таблица 5.3. Средние значения общей и локальной ЭРГ, полученные с
помощью специализированной электрофизиологической системы «Nueropta»
фирмы «Medelec», Англия [Шамшинова А. М., 1989]
Значение ЭРГ а-Волна Ь-Волна
амплитуда, мкВ время, мс амплитуда, мкВ время, мс
Общая ЭРГ для ин- тенсивности сти- мула Q3 Дж 96±1,3 20±0,4 310±6,9 43±0,7
Локальная ЭРГ 18°: красный стимул зеленый стимул 4,9±0,4 7,8±1,6 28±0,5 32,5±0,9 14,4±ЦЗ 6Qtl4,5 53±0,5 78,7±3,4
170
Рис. 5.17. Электрод-присоска для регистрации общей и локальной ЭРГ
[Шамшинова А.М., 1989).
ярче полунасыщающей яркости длительностью Юме позволяет
выделить в макулярной области ответ преимущественно колбочко-
вой системы. Применение зеленого стимула в тех же условиях, урав-
ненного с красным стимулом по яркости, выделяет функцию кол-
бочковой и палочковой систем в стимулируемой области. Чувстви-
тельность палочковой системы к зеленому стимулу определяется
спектром поглощения родопсина.
Средние значения макулярной ЭРГ на красный и зеленый сти-
мулы в норме представлены в табл. 5.3.
Имеется прямая корреляционная связь ЛЭРГ с остротой зрения
(коэффициент корреляции 1,0, коэффициент регрессии 0,5).
Очевидно, что центральные и периферические колбочки различ-
ны по своим функциональным свойствам. Колбочки макулярной
области имеют более пролонгированный ответ, чем перифериче-
ские. Способ локальной ЭРГ, предложенный в лаборатории клини-
ческойфизиологии зрения С.В.Кравкова, обеспечивает возможность
электроретинографического исследования макулярной области для
диагностики локальных поражений, сопровождающихся нарушени-
ем фиксации взора, при использовании источника локальной свето-
вой стимуляции (светодиод), установленном непосредственно на
электроде-присоске с вмонтированной оптической системой, кото-
рая вместе с оптической системой глаза создает неподвижное изоб-
ражение источника света на центральной области сетчатки (рис.5.17).
Так как при всех движениях глаза присоска передвигается вместе с
ним, изображение источника всегда попадает на одно и то же место
171
A- A* A* A*- W- л-
Рис. 5.18. Мультифокальная запись ЭРГ [Sutter Е.Е., 1985]. Топография
биоэлектрической активности сетчатки в норме (vis= 1,0) и трехмерное
представление плотности ответа стимулируемой области.
сетчатки. Этим обеспечивается многократная воспроизводимость
стимуляции, заданной в макулярной области без необходимости
контроля фиксации взора. Использование светодиода с двумя цве-
тами 660 и 550 нм позволяет изучать функцию колбочковой и па-
лочковой систем в макулярной области (зона раздражения 18°).
Исследование колбочковой функции макулярной области дополня-
ет традиционный метод регистрации общей ЭРГ.
Предложенная к использованию конструкция электрода-присос-
ки, ее форма, величина, комфортность, атравматичность, быстрота
и надежность фиксации на глазном яблоке позволяют использовать
электрод одного диаметра и применять его для регистрации ЭРГ у
взрослых и детей, нежность крепления — при свежих проникающих
ранениях для характеристики функций сетчатой оболочки, при
клинической необходимости и выборе тактики вмешательства.
С помощью электрода-присоски возможна регистрация как ган-
цфельд, так и макулярной локальной ЭРГ при нистагме, когда не-
произвольные движения глаза мешают фиксации взора. Вмонтиро-
172
1 мкВ
200 мс
Рис. 5.19. Мультифокальная запись ЭРГ. Топография биоэлектрической
активности сетчатки в центральном поле зрения ( 23°) у пациента с дист-
рофией макулярной области, связанной с возрастом (vis = 0,2); трехмерное
представление плотности ответа стимулируемой области.
ванная оптическая система и соответствующий расчет фокусных
расстояний позволяют подавать стимул на макулярную область
размером 10—20°, различной длины волны 660 и 560 нм, длитель-
ности (10—200 мс) от светодиода с минимальным оптическим иска-
жением. Плотность контакта с лимбом роговицы обусловливает
минимальное сопротивление до 0—8 кОм с минимальной потерей
информации. В светодиодном стимуляторе заложены широкие воз-
можности регулировки интенсивности, частоты, длительности ло-
кального стимула для макулярной области.
Последним достижением клинической электроретинографии
является мультифокальное топографическое исследование биоэлек-
трической активности зрительных путей, идентификация специфи-
ческих источников ЭРГ и ЗВКП [Sutter Е.Е., 1992]. Метод позволя-
ет проводить топографический анализ зрительного поля и изучать
нелинейные ответы, связанные с адаптивными механизмами зри-
тельных путей, и изолировать биоэлектрические ответы от наруж-
ных и внутренних слоев сетчатки.
173
Для этих целей используют мультифокальную ЭРГ-систему
(’’VERIS” фирма ’’Тотеу”, Япония), которая позволяет проводить
стимуляцию сетчатки специальными, сдвоенными стимулами гек-
согональной формы (61 элемент), предъявляемыми псевдослучай-
ным образом виртуально бесконечное число раз с частотой 75 Гц от
CRT-монитора. Для регистрации ЭРГ используются электроды типа
крючков, волосков, электроды Буриана — Алена (Burian — Allen) с
прозрачной передней поверхностью, не искажающие изображение
стимула. Перекрывая область сетчатки в 23°, паттерн-элемент сти-
мула увеличивается от центра к периферии соответственно разме-
рам рецептивных полей сетчатки. Локализация ответа соответству-
ет каждому гексагональному элементу, и топография ЭРГ пред-
ставляется на дисплее компьютера. При использовании системы
анализа multi-input, метода кросскорреляции, оценивается плотность
распределения биоэлектрической активности сетчатки в каждой
стимулируемой области при регистрации ЭРГ и ЗВКП (рис. 5.18),
которая представляется в виде цветных карт и трехмерных графи-
ков. Пик амплитуды в норме доминирует в центре диаграммы и
отсутствует при дистрофических изменениях в макулярной области
(рис.5.19). По мнению авторов, пространственное распределение
патологических биоэлектрических ответов на диаграмме соответ-
ствует области дистрофических очагов, найденных офтальмоскопи-
чески или периметрически.
5.2.3. Локальная ЭРГ в диагностике заболеваний
макулярной области сетчатки
В зависимости от патогенетических механизмов заболевания и ло-
кализации патологического процесса в различных структурах сет-
чатки общая и локальная ЭРГ будут изменяться по-разному. При
этом необходимо учитывать как амплитуду а- и b-волн, так и вре-
менные параметры: время до пика a-волны, до пика b-волны и
кинетику биопотенциала, время от пика a-волны до пика Ь-волны.
При пигментном ретините отсутствие общей ЭРГ при рецессив-
ной форме и резкое снижение общей ЭРГ(микро-ЭРГ) при доми-
нантной форме наследования являются патогномоничными призна-
ками заболевания. Однако острота зрения у этой категории боль-
ных, несмотря на концентрическое сужение полей зрения, может
оставаться достаточно высокой. Развитие изменений в макулярной
области при пигментном ретините коррелирует с остротой зрения и
может сопровождаться нормальной, субнормальной, резко субнор-
мальной и отсутствующей локальной ЭРГ (рис. 5.20), которая име-
ет определенную связь с регистрируемой микро-ЭРГ, отражающей
при пигментом ретините функцию сохранившейся колбочковой си-
стемы. Достоверной разницы между ЛЭРГ на красный и зеленый
стимул при концентрическом до 10—20° сужении поля зрения при
174
Рис. 5.20. Общая и ло-
кальная, макулярная, ЭРГ
при пигментном ретини-
те (больной X., 35 лет,
OU: vis OD = 0,6; поле
зрения концентрически
сужено до 30°).
а — субнормальная общая
ЭРГ; б — субнормальная
ЛЭРГ на зеленый стимул;
в — субнормальная локаль-
ная ЭРГ на красный стимул.
пигментном ретините нет, что говорит об участии в ЭРГ колбочко-
вой системы сетчатки центральной области. Можно представить
пигментный ретинит как модель колбочковой функции при сохран-
ности остроты зрения, в которой равенство биоэлектрических от-
ветов на красный и зеленый стимул в одинаковых условиях регис-
трации подтверждает патогенетическое звено преимущественного
поражения палочкового аппарата сетчатки. Данные ЛЭРГ, которая
в той или иной степени всегда регистрируется при пигментном ре-
тините, могут служить прогностическим тестом эволюции патоло-
гического процесса.
Наследственные юношеские макулярные дистрофии сетчатки
часто представляют трудности для дифференциальной диагно-
стики.
Наиболее часто встречается и лучше других изучена макулярная
дегенерация — тип Штаргардта, наследуемая по рецессивному типу.
175
Рис. 5.21. Общая, ло-
кальная, макулярная,
ЭРГ при дисциформной
макулярной дистрофии
типа Кунта — Юниуса
(больной П., 70 лет, vis:
OD == 0,8; OS = 0,1, в поле
зрения центральные ско-
томы разной степени вы-
раженности).
а — ЛЭРГ в норме; б —
ЛЭРГ субнормальна; в —
ЛЭРГ практически отсут-
ствует.
Благодаря электроретинографическим исследованиям можно выде-
лить чисто центральную форму поражения глаз, при которой ган-
цфельд ЭРГ имеет нормальную амплитуду, а снижаются в первую
очередь локальный, макулярный, биопотенциал и фотопическая
ЭРГ. По мере прогрессирования процесса ЛЭРГ закономерно сни-
жается до полного исчезновения. При смешанной дистрофии кол-
бочек наблюдают изменения как локальной, так и скотопической
ЭРГ. Болезнь Штаргардта может сочетаться с fundus flavimaculatus,
а также с тапеторетинальной дистрофией, проявляющейся в класси-
ческой и атипичной форме. В этом случае скотопическая, фотопи-
ческая и локальная ЭРГ имеют патологический характер.
Уже в начальных стадиях Х-хромосомного ювенильного ретино-
шизиса в отличие от дистрофии типа Штаргардта общая ЭРГ все-
гда значительно снижена, резко субнормального характера, изме-
нения в ЛЭРГ также более выражены.
При вителиформной дистрофии типа Беста общая и локальная
176
Рис.5.22. ЭРГ при нистаг-
ме (больной Р., 10 лет, с
врожденным нистагмом,
альбинизмом, гипоплазией
макулярной области, vis:
OD = 0,4; OS = 0,5).
а — общая ЭРГ нормальна
(OD так же, как на OS); б —
ЛЭРГ супернормальна; в —
ЛЭРГ нормальна.
ЭРГ в пределах нормы, что подтверждает интактность нейронов
1 — 2-го порядка макулярной области на первых этапах заболева-
ния.
ЛЭРГ при хороиретинальной дистрофии, связанной с возрастом,
может служить прогностическим критерием эволюции заболевания,
так как в начальной стадии процесса при друзах макулярной обла-
сти локальный потенциал не изменен (рис.5.21). По мере развития
заболевания, появления отслойки пигментного эпителия в макуле,
фиброзных и экссудативных изменений в сетчатке вместе со сниже-
нием остроты зрения нарушается электрогёнез в макулярной обла-
сти, что свидетельствует о вовлечении в патологический процесс
фоторецепторов, а также слоя биполяров. При связанной с возра-
стом хороиретинальной дистрофии чаще отмечают нормальные по-
казатели общей ЭРГ, однако возможны ее изменения, являющиеся
177
результатом более генерализованных изменений в сетчатке, чем те,
которые выявляют офтальмоскопически. Значительная вариабель-
ность локальной ЭРГ (от нормальных значений до отсутствую-
щей) — результат различного электрогенеза в разных стадиях хо-
роиретинальной дистрофии, связанной с возрастом.
При вторичных изменениях в макулярной области ЛЭРГ играет
важную дифференциально-диагностическую роль. Так, сопоставле-
ние данных ЛЭРГ со световой чувствительностью глаза в поле зре-
ния, статической периметрией позволяет уточнить уровень пораже-
ния: нормальные показатели ЛЭРГ и наличие центральной ското-
мы в поле зрения — признак патологического процесса в зрительном
нерве или вторичности дистрофических изменений в макуле; суб-
нормальный характер ЛЭРГ при наличии центральной скотомы в
поле зрения — свидетельство локализации патологического процесса
в первом-втором нейронах макулярной области. Супернормальные
значения ЛЭРГ при наличии центральной скотомы возможны из-за
изменения тормозного влияния подкорковых структур при пораже-
нии центробежных волокон зрительного нерва.
Еще в 1990 г. утверждали, что запись ЛЭРГ при нистагме не-
возможна из-за движения глазных яблок, отсутствия фиксации взо-
ра. Представленный метод регистрации ЛЭРГ позволяет регистри-
ровать макулярный биопотенциал у больных с нистагмом (рис.5.22).
Наличие или отсутствие изменений в сетчатке и зрительном нерве,
выявленные с помощью электрофизиологических методов исследо-
вания, позволяет дифференцировать нистагм центрального или
периферического происхождения, а выраженная асимметрия общей
или ЛЭРГ свидетельствует о различном функциональном состоя-
нии сетчатки, ее макулярной области, а также зрительного нерва в
исследуемых глазах.
5.3. ПАТТЕРН-ЭРГ
5.3.1. Происхождение паттерн-ЭРГ
Другая форма электроретинограммы — паттерн-ЭРГ, ретинальный
биопотенциал, который является ответом, вызываемым паттерн-
стимулом (ПЭРГ) в виде полосчатого или шахматного поля при
временном его изменении и постоянной общей освещенности. ПЭРГ
чаще всего регистрируется при реверсивных изменениях ’’шахмат-
ных” паттернов и имеет очень маленький сигнал (0,5—8 мкВ), зави-
сящий от характера стимула, поэтому регистрация ПЭРГ техниче-
ски более сложна, чем обычной ЭРГ [Berninger Т., Arden В., 1988—
1991].
Предложенная L.A.Riggs в 60-х годах ПЭРГ нашла примене-
ние в диагностике заболеваний, при которых поражен третий
178
нейрон сетчатки. ПЭРГ имеет несколько иное происхождение по
сравнению с ЛЭРГ, поскольку всегда снижается при заболеваниях
зрительного нерва, ретробульбарном неврите, травматическом раз-
рыве зрительного нерва, его атрофии, токсической амблиопии,
в то время как общая ЭРГ на вспышку света остается нормаль-
ной.
Происхождение ПЭРГ дискутируется в литературе. Результаты
многочисленных экспериментальных, морфологических и электрон-
но-микроскопических исследований свидетельствуют, что ПЭРГ
генерируется не только ганглиозными клетками, часть ПЭРГ, в
основном позитивный компонент, вызывается другими структура-
ми сетчатки. Дискриминация пространственного контраста пред-
ставлена в большей степени в ПЭРГ, чем в общей ЭРГ на вспы-
шечный стимул. Хотя первый этап пространственной дискримина-
ции контраста происходит уже на уровне биполярных клеток, она
более четко представлена на уровне on- и off-ганглиозных клеток
сетчатки, выделяемых по их способности реагировать на стимулы,
более или менее яркие, чем фон. Эта деполяризация и гиперполя-
ризация биполяров, on- и off-ганглиозных клеток проявляется
именно при чередовании предъявления черных и белых клеток
паттерна в виде шаматного поля или решетки. Аналогичная реак-
ция наблюдается в наружных коленчатых телах и зрительной коре.
Нейроны зрительной коры, которые более чувствительны к стиму-
лам определенной ориентации (горизонтальным или вертикальным),
дают максимально выраженный ответ на паттерн в виде шахмат-
ного поля. Высказано также предположение, что ПЭРГ включает
в себя on- и off-ответ фоторецепторов при их локальной стимуля-
ции [Ikeda Н., 1987].
Параметры стимулов и ПЭРГ. Клинически доказано, что ПЭРГ
отражает активность внутренних слоев сетчатки. Количественная
зависимость между ПЭРГ и активностью внутренних слоев сетчат-
ки устанавливается соотношением размеров элементов паттерна и
амплитудой ответа. Ганглиозные клетки, как известно, реагируют
на соответствующие размеры паттерна, так как имеют концентри-
ческие рецептивные поля, центр и периферия которых находятся в
антагонистических отношениях. Если центр рецептивного поля пе-
рекрыт темной ячейкой паттерна, то в этом случае наиболее веро-
ятно, что периферия рецептивного поля освещена. При этом клет-
ки, имеющие on-центр, в процессе реверсии паттерна, после макси-
мального торможения, будут возбуждаться. При увеличении или
уменьшении величины паттерна заметного изменения возбуждения
не происходит, так как центр и периферия стимулируются частично
светлыми или темными областями. Таким образом, при реверсив-
ном паттерне эффективность стимуляции меньше оптимальной.
Данное положение меняется с изменением размеров рецептивных
полей от центра к периферии. При этом нужно принимать во вни-
179
мание, что биполярные клетки организованы по принципу центр —
периферия, как и большинство ганглиозных клеток, а последние
воспринимают контраст между центром и периферией в отличие от
фоторецепторов, реагирующих просто на освещение. Контраст пред-
ставляется, как C=(Lmax— Lmin)/(Lmax+Lmiii), и в большинстве гангли-
озных клеток сетчатки черно-белая граница вызывает удлиненную
реакцию (см.главу 2). Это изменение (реверс) паттерна, который
является причиной временных изменений контраста, и является более
мощным стимулом по сравнению с вспышкой света. Таким обра-
зом, ПЭРГ отличается от общей ЭРГ тем, что отражает реакцию на
контраст, а не на освещение.
Экспериментально была выявлена связь между амплитудой
ПЭРГ и размером паттерна. Структуры, продуцирующие ПЭРГ,
зависят от пространственной настройки (spatial tuning) и величины
решетки или шахматного элемента, что определяется понятием
’’пространственная частота”, которая измеряется в циклах на гра-
дус зрительного угла. Установлено, что реверсивный паттерн, со-
провождаемый изменениями освещенности, вызывает малые сигна-
лы, которые содержат все обычные компоненты ЭРГ, но другого
происхождения [Spekreijse Н. et al., 1973; Sokol S. et al., 1983]. Пер-
вые количественные измерения ПЭРГ с использованием тонкой
техники показали, что амплитуда ПЭРГ не зависит от контраста
в отличие от зрительных вызванных кортикальных потенциалов.
Последующие работы [Riemslag, 1983, 1985; Arden G. В., Vae-
gan P. P. T., 1983; Baker H.D., 1984] подтвердили некоторые резуль-
таты H. Spekriejse с соавт. и показали, что если сетчатка стиму-
лируется стандартными световыми решетками с появлением и ис-
чезновением паттерна, при которых меняется освещенность,
результирующей ответа не может быть только ПЭРГ, а оптималь-
ный размер ячеек, который вызывает ответ большей амплитуды,
можно установить лишь для рецептивных полей фовеа.
Зависимость ПЭРГ от пространственной частоты была показа-
на для негативного компонента ПЭРГ [Odom J. et al., 1983, 1986;
Berninger T.A., 1988; Korth M., 1989], что и определило противоре-
чивость данных литературы [OdomJ., 1982; Baker Н. D., 1983;
Sokol S., 1983]. Большинству исследователей связь позитивного ком-
понента ПЭРГ с пространственной частотой установить не уда-
лось.
Взаимосвязь между амплитудой ПЭРГ, размером паттерна и
контрастом трудно объяснима на основе только того факта, что
локальные области сетчатки отвечают независимо друг от друга.
Не исключено, что латеральные взаимодействия модифицируют
ответ наружных слоев сетчатки при изменении стимулирующего
света [Baker H.D., 1984; Korth М., 1983; Veagan Р.Р.Т., 1987].
N. Drasdo и D. A. Thompson (1992) рассчитали изменение осве-
щения в сетчатке в связи с каждым воздействием паттерна и опре-
180
делили соотношение амплитуды и пространственной частоты для
маленького диска, включающего фовеа, и серии концентрических
колец. При этом они предположили, что большие паттерн-элемен-
ты, каждый из которых покрывает область исследования, могут вы-
зывать ЭРГ за счет изменений освещенности — “ответ ретинальной
освещенности”. При маленьком размере паттерна контрастирую-
щий край на сетчатке является причиной дополнительного ответа,
называемого “паттерн-специфическим ответом”. Однако, посколь-
ку на границе при переходе светлого квадрата в темный и наоборот
изменение ретинального освещения не так велико для маленьких
паттернов, как для больших, “ответ ретинальной освещенности”
уменьшается. Так же редуцируется “паттерн-специфический ответ”
со снижением контраста. Результирующая амплитуда “паттерн-спе-
цифического ответа” может быть скорректирована для сниженного
контраста.
Таким образом, амплитуда ПЭРГ зависит от пространственной
частоты паттерна, стимулируемой области сетчатки, а так же от
позитивности или негативности компонентов ПЭРГ. Максималь-
ная амплитуда “паттерн-специфического ответа” достигается как с
увеличением высоких пространственных частот, так и с перемеще-
нием исследуемой области в сторону фовеа. При этом “ответ рети-
нальной освещенности” может быть больше при увеличении усред-
ненной освещенности.
Отмечена взаимосвязь между относительным объемом слоев
сетчатки и амплитудой различных компонентов ПЭРГ: в перифери-
ческой сетчатке больше “ответ ретинальной освещенности”, проис-
ходящий от наружных слоев сетчатки, в то время как в экстрафо-
веальной области, в которой увеличен плексиформный слой и слой
ганглиозных клеток, больше “паттерн-специфический ответ”
[Steinberg R. Н., 1970; Sieving Р. А., 1984, 1987].
5.3.2. Методологические основы регистрации паттерн-ЭРГ
(рекомендации ISCEV)
Методики регистрации ПЭРГ, приводимые различными авторами,
очень разнообразны как по технике, так и по способам регистра-
ции. Стандарты для ПЭРГ, как и для общей ЭРГ, основываются
на современных знаниях и опыте множества экспериментальных
офтальмологических и электрофизиологических лабораторий ми-
ра. Общие принципы для исследования детей и взрослых одина-
ковы.
Обозначение волн ПЭРГ. Форма ПЭРГ зависит от
пространственной частоты стимула. Выделяют два вида ПЭРГ в
зависимости от частоты реверсии паттерна (рис. 5.23). При частоте
стимула 1—3 Гц регистрируется “преходящая, скоротечная”
(transient) ПЭРГ. При более высоких пространственных частотах,
181
б
Рис. 5.23. ПЭРГ в нор-
ме.
а — ПЭРГ быстрая, фа-
зическая (transient), Np Рр
N2— ее компоненты; б —
ПЭ^Г устойчивого состо-
яния (steady-state).
около 6—8 Гц, последовательно записываются волны одна за дру-
гой — ПЭРГ устойчивого состояния (steady-state), при которой фор-
ма волны синусоидальная, так что для определения амплитуды и
фазы относительно стимула используют анализ Фурье. ПЭРГ имеет
первый негативный компонент, приблизительно на 35 мс (N35), сле-
дующий более поздний и длительный позитивный компонент на
45—60 мс (Р50), за этой позитивной частью ПЭРГ следует большой
негативный компонент на 90—100 мс (N95). Для transient ПЭРГ ам-
плитуду измеряют от пика N35 до пика Р50, амплитуду пика N95 от
пика Р50 до основания N95. Измерение латентного периода проводят
от начала стимула до пика компонента. В случае мышечной актив-
ности и при других артефактах, изменяющих форму ПЭРГ, невоз-
можно выделение ее компонентов.
Электроды. Для регистрации ПЭРГ используют тонкие во-
локна или крючки, закладываемые за нижнее веко, не искажающие
сигнал, которые можно применять без анестезии. В связи с этим не
рекомендуется использовать контактные линзы и корнеальные элек-
троды. Крючковидные электроды необходимо устанавливать точ-
но по центру зрачка, чтобы свести к минимуму их смещение при
моргании. Референтный электрод помещают на ипсилатеральной
182
стороне наружной спайки. Установка электродов на мастоидаль-
ный отросток, ухо или лоб может привести к искажению сигнала
ПЭРГ. Если производят монокулярную регистрацию ПЭРГ, то
корнеальный электрод на закрытом глазу можно использовать как
референтный. Накожный электрод после предварительной очистки
кожи помещают на лоб и заземляют. Сопротивление между рефе-
рентным и земляным электродами не должно превышать 5 кОм после
установки их на пациента.
Параметры стимула. Стандарты предлагают общие по-
ложения по использованию параметров стимулов для регистрации
ПЭРГ. Однако лаборатории могут выбирать те или иные условия
или параметры стимулов, отличающиеся от описанных в соответ-
ствии с техническими возможностями и задачами исследования. Ре-
комендуется использовать черно-белые реверсирующие квадраты с
общим размером поля стимуляции 10—16° и размером каждого квад-
рата около 40'. Контраст между черными и белыми квадратами
должен быть максимальным (но не менее 80%).
Освещенность. ПЭРГ трудно воспроизвести при низкой
освещенности, поэтому рекомендуется уровень фотопической осве-
щенности белых ячеек 80 кд/м2. В процессе реверсии квадратов общая
освещенность экрана не меняется. Фоновое освещение должно быть
слабым или обычным комнатным и не влиять на освещенность
Экрана, при этом в поле зрения обследуемого не должно быть ярких
источников света.
Transient ПЭРГ. Выделение Р50- и М95-компонентов может быть
полезным в оценке динамики заболеваний зрительного нерва и
макулярной области. Steady-state ПЭРГ используют при глаукоме
для оценки функционального состояния ганглиозного слоя сетчат-
ки. Однако необходимо помнить, что данный тип записи применя-
ют для измерения амплитуды и фазы 2-й гармоники с помощью
анализа Фурье, поэтому не рекомендуется производить эту запись
без соответствующих инструментов для подобного анализа. Часто-
та реверсий, рекомендуемая для регистрации transient ПЭРГ 2—
6 реверсий в 1 с (1—3 Гц), для устойчивой steady-state ПЭРГ —16 ре-
версий в 1 с (8 Гц).
Электронная система записи Необходимо ис-
пользовать предусилитель и усилитель с полосой пропускания от 1
до 100 Гц, с входным импедансом усилителя не менее 10 МОм. Си-
стема должна быть изолирована от пациента. Использование сете-
вых фильтров не рекомендуется. В связи с тем, что ПЭРГ имеет
маленькую амплитуду, необходимо усреднение сигналов. Для
transient ПЭРГ анализируемый период должен составлять не менее
150 мс.
Для проведения анализа Фурье steady-state ПЭРГ период анали-
за должен быть больше стимульного интервала. Удаление артефак-
тов (не более 100 мкВ) осуществляется программным методом, чаще
183
всего автоматически. Дисплейные системы должны иметь адекват-
ное разрешение для точного представления характеристик сигна-
лов маленькой амплитуды.
Клинический протокол. ПЭРГ регистрируют без рас-
ширения зрачков для сохранения аккомодации. Точка фиксации рас-
полагается в центре экрана. При плохой фиксации пациенту дают
установку смотреть в центр экрана или через него. Зрение корриги-
руют.
Запись ПЭРГ можно проводить как монокулярно, так и биноку-
лярно. Бинокулярная запись ПЭРГ как более стабильная рекомен-
дуется в качестве “базовой ПЭРГ” во всех случаях асимметричного
снижения зрения. Монокулярную стимуляцию применяют при од-
новременной записи ПЭРГ и ЗВКП. Усреднение (не менее 150 отве-
тов) ПЭРГ необходимо производить до получения стабильной фор-
мы волны. Заключение по результатам исследования включает ам-
плитуду пиков Р50 и N95, латентный период Р50 и нормативы
лаборатории. При регистрации steady-state ПЭРГ записывают амп-
литуду и фазу второй гармоники.
Факторы, влияющие на ПЭРГ. Величина зрачка
существенно не влияет на позитивный компонент паттерн-ЭРГ, од-
нако при миозе негативный компонент N95 редуцируется и увеличи-
вается время латентности компонентов Р50 и N95. Этот факт важно
учитывать в связи с необходимостью сохранения аккомодации при
исследовании больных глаукомой. ПЭРГ очень чувствительна к де-
фокусировке паттерна: уже при 0,5 дптр. отмечено снижение ПЭРГ
на 50%.
Анализ ПЭРГ. В зависимости от вида предъявляемого
паттерн-стимула измеряют амплитуду одного или двух пиков. Если
регистрируется transient ПЭРГ, то первый позитивный пик возни-
кает на 50 мс и его измеряют от базовой линии или раннего нега-
тивного отклонения (иногда на 35 мс). Следующая негативная вол-
на N95 может быть измерена от базовой линии или пика Р50. Если
увеличивать частоту стимулов или имеется синусоидальная вре-
менная модуляция, то ближайшая синусоида steady-state ПЭРГ
может быть измерена от пика до пика как амплитуда второй гар-
моники ответа.
Латентность ПЭРГ постоянна и редко изменяется при заболева-
ниях. Однако мнения ученых по этому вопросу различны, если одни
не находили изменений латентности пика Р50 при заболеваниях зри-
тельного нерва, отслойке сетчатки, ретинопатии воспалительной
природы, тромбозе ветви центральной вены сетчатки, то другие
отмечали значительное увеличение времени латентности этого пика
у пациентов с глазной гипертензией и глаукомой.
Амплитуда ПЭРГ значительно уменьшается у пожилых пациен-
тов, что объясняется уменьшением числа ганглиозных клеток в сет-
чатке (связанная с возрастом редукция ПЭРГ).
184
Цвет и ПЭРГ. В последние годы сформировалась концепция
существования параллельных зрительных путей на основании зна-
ний о сложном, слоистом строении наружных коленчатых тел: пар-
во- и магноцеллюлярные пути. Магноцеллюлярные пути отходят
от а-ганглиозных клеток, которые слабо чувствительны к цвету, но
очень чувствительны к движению, мельканиям и ахроматическому
пространственному контрасту. Парвоцеллюлярная система идет от
b-ганглиозных клеток, которые кодируют цвет и имеют высокую
пространственную разрешающую способность, слабую чувствитель-
ность к движению и ахроматическому пространственному контра-
сту. В связи с этим идеальным стимулом для магноцеллюлярной
системы являются черно-белые паттерны или полосы, представляе-
мые в реверсивной ’’моде”, а для парвоцеллюлярной системы — цве-
товые контрастные полосы, в которых все цвета одной яркости.
Th.Beminger и B.Adler(1992) зарегистрировали близкий к нега-
тивному ответ около 2 мкВ на красно-зеленые полосы с яркостью
40 кд/м2, уравненные по яркости (при 1000 усреднениях), в то время
как на сине-желтый паттерн ответа совсем не было, что объясня-
лось небольшим количеством ретинальных ганглиозных клеток (5%)
и синих колбочек в сетчатке, реагирующих на сине-желтый стимул.
Регистрируемая ПЭРГ такого типа является отражением цветового
контраста [Korth М. et al., 1983]. Определенные различия в проис-
хождении ахроматической и хроматической ПЭРГ могут быть ос-
нованием для использования их в диагностике заболеваний зри-
тельного анализатора.
5.3.3. Паттерн-ЭРГ в диагностике заболеваний сетчатки
и зрительного нерва
ПЭРГ изменяется при дисфункции макулярной области, которая, как
правило, не отражается на ганцфельд-ЭРГ. Поэтому регистрацию
ПЭРГ, ЛЭРГ и ганцфельд-ЭРГ целесообразно использовать в диф-
ференциальной диагностике.
В клинических наблюдениях было отмечено, что компоненты
ПЭРГ Р50 и N95 изменяются по-разному при заболеваниях различно-
го происхождения [Arden G.B., 1984; Holder G.E., 1989, 1991]. При
макулярных заболеваниях снижается компонент Р50, а при пораже-
нии зрительного нерва — компонент N95
Отсутствие ПЭРГ было отмечено при ювенильной дисциформ-
ной макулярной дегенерации, при болезни Тея — Сакса (Тау —
Sach). Ценность исследования ПЭРГ при макулярных заболеваниях
связана еще и с тем, что в ПЭРГ находят отражение изменения
контрастной чувствительности, изменяющейся при различной па-
тологии сетчатки и зрительного нерва. Однако изменения контраст-
ной чувствительности в большей степени проявляются в изменении
паттерн-ЗВКП. Паттерн-электроретинография удачно дополняет
185
другие методы исследования макулярной области, так как ПЭРГ
изменяется в ряде случаев, когда имеет место нормальная общая и
макулярная ЭРГ, что указывает на связь ПЭРГ со слоем ганглиоз-
ных клеток сетчатки.
Дисфункция зрительного нерва. В генерации ПЭРГ участвуют
ганглиозные клетки сетчатки, поэтому ее изменение можно ожи-
дать при патологии зрительного нерва. Действительно, снижение
ПЭРГ или ее отсутствие, в основном N95, отмечено у пациентов с
атрофией зрительного нерва, при сохранности макулярной и общей
ЭРГ [Berninger Т.А., 1988; Vaegan Р.Р.Т., 1990], доминантной наслед-
ственной атрофии зрительного нерва, имеющей большое количе-
ство фенотипических вариаций.
При оптическом неврите в острой стадии процесса на фоне
рассеянного склероза прежде всего увеличивается время латентно-
сти ЗВКП, что обусловливается выраженностью и распространен-
ностью демиелинизации зрительных волокон. Однако у этой кате-
гории больных через несколько недель после острой стадии процес-
са обнаруживались изменения в сетчатке [Arden G.B., 1983], что
подтверждалось данными ПЭРГ в течение года после обострения
процесса. Компонент N95 ПЭРГ был снижен значительнее, чем Р50.
Несмотря на то что ПЭРГ всегда изменяется при оптическом не-
врите, ЗВКП на паттерн-стимуляцию остается более чувствитель-
ным клиническим тестом при этом виде патологии.
Глаукома. Известно,что при хронической глаукоме повышение
внутриглазного давления, сопровождающееся ишемическим процес-
сом, вызывает поражение зрительного нерва с последующей его
атрофией. Изменения в полях зрения возникают у больных в случае
потери 40—50% волокон зрительного нерва. При начальном глау-
коматозном процессе, когда наблюдаются изменения пространствен-
ной контрастной чувствительности, на фоне нормальной общей ЭРГ
отмечено снижение ПЭРГ, что коррелировало с изменениями в поле
зрения. При наличии циркулярной скотомы ПЭРГ является наибо-
лее чувствительным тестом в диагностике глаукоматозных наруше-
ний.
Амблиопия. Снижение остроты зрения при амблиопии связано,
как известно, с изменениями в корковых центрах, что было под-
тверждено в экспериментальных исследованиях^а животных
[Ikeda Н. et al., 1993]. В целом ряде работ показано, что ПЭРГ
редуцирована при амблиопии и в большей степени изменен компо-
нент N95. У взрослых амблиопов снижение остроты зрения, контра-
стной чувствительности и редуцированная ПЭРГ составляют опре-
деленный комплекс симптомов, по которым можно дифференциро-
вать тип амблиопии, а также судить об эффективности лечения.
Снижение ПЭРГ при амблиопии хотя и подтверждает ее кортикаль-
ное происхождение, но может быть связано с нарушением рети-
нальной функции на уровне ганглиозных клеток.
186
5.4. РИТМИЧЕСКАЯ ЭРГ
5.4.1. Происхождение ритмической ЭРГ
Следующая важная модификация ЭРГ — это мелькающая (ритми-
ческая) ЭРГ, т.е. ЭРГ, зарегистрированная на стимул постоянной
интенсивности, предъявляемый с различной частотой.
С момента внедрения электроретинографии в глазную клинику
ритмическую электроретинографию (РЭРГ) на высокие частоты
длинноволнового (красного) света использовали для оценки функ-
ционального состояния колбочковой системы. При этом, чем ниже
частота мелькающего света, вызывающая ретинальный ответ, тем
большая площадь макулярной области была вовлечена в патологи-
ческий процесс.
Частота мельканий света постоянной интенсивности, при кото-
рой b-волна сливается с шумом базовой линии, называется фузион-
ной частотой мельканий. В диагностике начальных поражений кол-
бочковой системы, при колбочковой дисфункции, дистрофических
изменениях в макулярной области играют роль как способность
сетчатки к усвоению ритма световых мельканий с частотой 30 Гц,
так и снижение амплитуды ритмического ответа. При дистрофиче-
ских изменениях в макулярной области ЭРГ на световые мелькания
с частотой 30 Гц не регистрируется.
Выделение фотопической и скотопической активности в ритми-
ческой ЭРГ (РЭРГ) основано на различной способности нейронов
сетчатки воспроизводить световой ритм. Максимально возможная
частота мельканий, воспроизводимая колбочками,— 50—100 пери-
одов в 1 с, а палочками — 10—20 периодов в 1 с.
Критическая частота слияния мельканий в ЭРГ зависит от мно-
гих факторов, в том числе от соотношения уровня интенсивности
адаптирующего фона и стимулирующего света. Для патологии па-
лочковой системы характерны отсутствие усвоения светового рит-
ма на низкие частоты и усвоение его на высокие в фотопических
условиях регистрации. При врожденных дефектах цветового зре-
ния, а также при макулярных заболеваниях сетчатки с вовлечением
в патологический процесс колбочковой системы отмечается низкая
способность к воспроизведению ритма на мелькания высокой час-
тоты.
В исследованиях, проведенных в последние годы, установлено,
что РЭРГ на высокие частоты стимуляции при большой яркости
стимулирующего света имеет главным образом фоторецепторное
происхождение [BaronW. S., BoytonR. М., 1975; AbrahamF. А.,
1984], в то время как при умеренных интенсивностях стимулирую-
щего света (10—20 кд/м2) в генерации высокочастотной РЭРГ воз-
можно участие внутреннего ядерного слоя сетчатки [Зуева М. В.,
Цапенко И. В., 1993]. Однако при снижении яркости вспышек до
187
АдкВ
100 ‘‘
-100 L
о
Рис. 5.24. Ритмическая
ЭРГ (РЭРГ) в норме
(частота — 8 Гц, R3,
».. * R4, ее компоненты) [Зу-
510 <1 Ж ева М. В., 1996].
10—20 кд/м2 на высоких частотах стимуляции изменение природы
генерации ритмических ответов сетчатки приводит к тому, что вклад
активности нейронов макулярной зоны в суммарную РЭРГ на 40 Гц
становится доминирующим, вследствие чего ее можно с большей
степенью достоверности использовать в диагностике заболеваний
макулярной области сетчатки.
РЭРГ на 5—15 Гц в отличие от высокочастотных ритмических от-
ветов полифазна. Она не является синусоидальным сигналом, а имеет
сложную форму, т. е. состоит из ряда хорошо идентифицируемых
волн или субкомпонентов (рис. 5.24). При раздражении сетчатки
мелькающими стимулами (10—20 кд/м2) частотой ниже 20 Гц био-
электрический ответ состоит из комплекса волн, выраженность ко-
торых зависит от условий адаптации и стимуляции. Выделяют пять
субкомпонентов РЭРГ, обозначаемых Rp R2, R3, R4 и R5 [Зу-
ева M. В., Цапенко И. В., 1992, 1993].
При ахроматических мельканиях умеренной яркости в низкоча-
стотной РЭРГ наиболее хорошо выражены и стабильно регистри-
руются два позитивных отклонения, которые обозначают R3 и R4.
Регистрация РЭРГ на хроматические (красный, оранжевый, сине-
зеленый) стимулы позволяет селективно выделять субкомпоненты
низкочастотной РЭРГ, имеющие колбочковое (Rj и R2) и палочко-
188
вое (R3 и R^) происхождение. Волна Я5 носит смешанный (палочко-
колбочковый) характер. Предполагается, что волна Я3 отражает
преимущественно нейрональную оп-активность, а волна Я4 зависит
от функционирования off-нейронов и ее проявление в ритмическом
ответе связано с ослаблением в сетчатке процессов латерального
торможения.
5.4.2. Методологические основы регистрации
ритмической ЭРГ
Оборудование, необходимое для реализации метода, включает в себя
стандартную диагностическую электрофизиологическую аппарату-
ру, применяемую в офтальмологии и предусматривающую усиле-
ние и усреднение биопотенциалов, возможность синхронизации и
анализа регистрируемого ответа, вариацию частоты, цветности и
яркости стимулирующего света и фонового освещения.
Для регистрации РЭРГ используют различные корнеальные элек-
троды, с помощью которых можно получить максимальный био-
электрический ответ. Источником может служить ганцфельд-сти-
мулятор или локальный стимулятор, который включается в специ-
ализированную электрофизиологическую систему.
Для регистрации РЭРГ рекомендуется использовать низкие и
высокие частоты с энергией вспышек 10—20 кд/м2. Применение яр-
ких (насыщающих) стимулов света возможно при селективном ис-
следовании функционального состояния колбочковых рецепторов.
Амплитуду РЭРГ, так же как волн, входящих в состав низкоча-
стотной РЭРГ, рассчитывают по наиболее типичной (или средней)
волне от ее отрицательного пика до положительного.
Помимо амплитудных параметров, в РЭРГ учитываются времен-
ные параметры, измеряемые от момента стимуляции до пика волны,
а также от пика до пика ритмических волн. Не менее важной является
оценка степени усвоения сетчаткой частоты светового ритма.
Для регистрации хроматической РЭРГ с помощью стандартных
широкополосных светофильтров: сине-зеленого (сзс7), оранжевого
(ос14), а также сине-зеленого (сзс3) и красного (ксп), попарно урав-
ненных по яркости с помощью нейтральных светофильтров, исполь-
зуют мезопические условия стимуляции (световая преадаптация в
течение 10 мин при фоновом освещении 5—10 лк). При этом рас-
считывается коэффициент пропускания (плотности) цветных свето-
фильтров и их комбинаций с нейтральными светофильтрами.
5.4.3. Роль ритмической ЭРГ в диагностике заболеваний сетчатки
Ритмическую ЭРГ наряду с общей и локальной ЭРГ используют в
клинике глазных болезней для установления локализации патоло-
гического процесса в различных слоях сетчатки и его распростра-
189
ценности. Хотя РЭРГ не имеет самостоятельного значения в клини-
ческой электрофизиологии, ее исследование дополняет представле-
ние о биоэлектрической активности, генерируемой различными
нейронами сетчатки в норме и при патологии.
Сравнение локальной макулярной ЭРГ и РЭРГ на высокие ча-
стоты стимуляции показало, что изменения этих биоэлектрических
обетов сетчатки не всегда совпадают при разных патологиях. При
дистрофии типа Штаргардта, центральной связанной с возрастом
хороиретинальной дистрофии (ЦХРД) ЛЭРГ является более чув-
ствительным тестом, хотя РЭРГ также снижается по мере прогрес-
сирования процесса, но в меньшей степени. РЭРГ снижается (в 2—
3 раза) раньше М-ЭРГ при экссудативных процессах в макулярной
области [Зуева М. В. и др., 1991; Цапенко И. В. и др., 1992], напри-
мер при синдроме Ирвина — Гасса и диабетической экссудативной
макулопатии. Макулярная ЭРГ при этих формах поражения маку-
лярной области меняется по-разному — от супернормальной до суб-
нормальной. В связи с этим патогномоничным симптомом отечно-
го компонента в макулярной области, сопровождающего ту или
иную патологию, является субнормальная РЭРГ на 40 Гц при нор-
мальных или супернормальных значениях макулярной ЭРГ. Чув-
ствительность этого симптома достаточно высока, так как патоло-
гическая РЭРГ наблюдается в 80% верифицированных случаев ма-
кулярного отека.
При одиночных световых стимулах в генерации b-волны ЭРГ
принимают участие как нейроны II порядка, так и глиальные клет-
ки Мюллера [Stockton R., Slaughter М., 1987]. Известно, однако, что
глиальные мюллеровские клетки неспособны воспринимать свето-
вой ритм свыше 2—4 Гц [Бызов А. Л., 1979; Miller R. F., Dowling J. Е.,
1970], поэтому РЭРГ на мелькающие стимулы высокой частоты
является сигналом, отражающим суммарную результирующую ак-
тивность только нейрональных элементов сетчатки, не опосредо-
ванную нейроглией.
Различие в генерации ЭРГ и РЭРГ на частоту свыше 5—15 Гц
определяет возможность получения нового диагностического тес-
та в клинической электроретинографии, позволяющего выявлять
долю участия глиальных элементов сетчатки в патологическом про-
цессе.
Сидероз сетчатки, окклюзия ветвей центральной артерии сет-
чатки характеризуются значительно более выраженным снижением
амплитуды b-волны ганцфельд-ЭРГ по сравнению с ритмическими
ответами сетчатки (рис.5.25), свидетельствуя о большей выражен-
ности изменений в клетках Мюллера по сравнению с нейронами
второго порядка.
Выраженное доминирование в РЭРГ субкомпонента R4 и изме-
нение соотношения волн R3 и R4 характерны для ранней (функци-
ональной) стадии диабетической ретинопатии (рис. 5.26) и травм
190
25 мкВ |__
20 мс
15 мкВ |__
10 мс
Рис. 5.25. Сопоставление общей ЭРГ (а) и РЭРГ (б).
1 — при пигментном ретините (vis=0,7, поле зрения концентрически сужено до 20°);
2 — тромбозе ветви центральной вены сетчатки (vis = 0,3); 3 — диабетической рети-
нопатии в ранней стадии (vis - 1,0) [Зуева М. В., 1996].
Рис. 5.26. РЭРГ у боль-
ного с диабетической
ретинопатией. Измене-
ние формы и увеличе-
ние компонента R4 [Зу-
ева М. В., 1996].
191
органа зрения как в травмированном, так и в парном глазу. Изби-
рательное угнетение скотопических субкомпонентов РЭРГ и сохран-
ность фотопических отмечены при прогрессировании сидероза сет-
чатки.
Раннее селективное угнетение волны R3 РЭРГ наблюдается при
дистрофии типа Штаргардта и пигментном ретините [Зуева М.
В., Цапенко И. В., 1992].
5.5. ЭЛЕКТРОРЕТИНОГРАФИЯ ПРИ КАТАРАКТЕ
В последнее десятилетие электрофизиологические методы исследо-
вания нашли широкое применение в диагностике заболеваний сет-
чатки и зрительного нерва не только при прозрачных средах, но и
при их помутнениях. В связи с совершенствованием микрохирурги-
ческой техники, внедрением в практику новых операций на рогови-
це, радужке, хрусталике, стекловидном теле и сетчатке повысились
требования к точности прогнозирования зрительных функций, в
связи с чем особое значение приобрели и функциональные методы
исследования.
В настоящее время при катаракте наиболее информативными
методами исследования, позволяющими оценить функцию сетчат-
ки, ее центральных и периферических отделов, функциональное
состояние зрительного нерва, зрительных путей и центральных
отделов зрительного анализатора, являются электрофизиологиче-
ские, которые включают электроретинографию (ЭРГ), электрооку-
лографию (ЭОГ), запись зрительных вызванных корковых потен-
циалов (ЗВКП). Кроме того, при катаракте для оценки функции
сетчатки и уточнения локализации процесса в различных структу-
рах внутренних отделов глаза применяют ультразвук, исследова-
ние энтоптических феноменов, определение критической частоты
слияния мельканий (КЧСМ), ретинальной остроты зрения,
порога электрической чувствительности и лабильность зрительно-
го нерва.
Регистрация общей ЭРГ с использованием общепринятых ин-
тенсивностей стимулирующего света при плотных помутнениях в
хрусталике и гемофтальме приводит к снижению яркости стимули-
рующего света и соответственно к снижению амплитудных пара-
метров, удлинению латентности и увеличению времени до пиков а-
и b-волны ЭРГ. В связи с этим регистрацию общего и макулярно-
го биопотенциалов сетчатки при катаракте осуществляют в пер-
вые минуты темновой адаптации с использованием стимулов нара-
стающей интенсивности до насыщающего уровня, при котором
общая ЭРГ перестает увеличиваться в размере, и красного стимула
с угловым размером 10—20°, используемого для макулярной ЭРГ,
но удвоенной яркости [Шамшинова А.М., 1991]. Выбор такой ин-
192
тенсивности определяется тем, что при катаракте отмечается раз-
личное светопоглощение в зависимости от характера, степени вы-
раженности и локализации помутнения в хрусталике, от типа ката-
ракты. При ступенчатом увеличении интенсивности стимулирую-
щего света у больных со зрелой катарактой показатели общей ЭРГ
достоверно отличаются от нормальных значений при прозрачных
средах глаза при использовании низких и средних (до 0,3 Дж)
интенсивностей и совпадают с нормативами общей ЭРГ в
случае применения насыщающих интенсивностей стимулирующего
света.
При патологических процессах в периферических отделах сет-
чатки достоверность различия отмечена на всех интенсивностях
стимулирующего света, включая насыщающую. Для оценки функ-
ционального состояния макулярной области при катаракте исполь-
зование стимулов насыщающей интенсивности красного цвета
оправдано тем, что нормативы локальной ЭРГ, полученные в тех
же условиях регистрации при прозрачных средах глаза, совпадают
с биопотенциалом, регистрируемым при зрелой катаракте. Полу-
ченные данные свидетельствуют об одинаковом количестве рецеп-
торов, принимающих участие в ответе, несмотря на бесспорно име-
ющий место эффект светорассеяния.
Многочисленные клинические исследования, проведенные у
больных с катарактой различного происхождения, подтвердили это
предположение.
При регистрация общей ЭРГ используют усреднение 8—10 ре-
акций, а для макулярной ЭРГ 50—60, что при ярких стимулах ис-
ключает необходимость предварительной световой адаптации, так
как ее включает сама система усреднения. Электроретинографи-
ческие данные при катаракте различного происхождения оценива-
ют путем сопоставления биоэлектрических ответов, получаемых
при катарактах с данными нормальных показателей здоровых ис-
пытуемых и с показателем парного глаза. При этом экстраполиру-
ют известные электрофизиологические симптомы заболеваний сет-
чатки при прозрачных средах глаза на оценку функций сетчатки
при катаракте, когда видеть глазное дно не представляется воз-
можным. Проведение электроокулографии лимитировано вслед-
ствие значительного снижения остроты зрения и отсутствия фикса-
ции взора. Использование ЗВКП ограничено тем, что послед-
ние характеризуют зрительный путь в целом от макулярной
области до зрительной коры. При некоторых видах катаракты
нередко отмечается увеличение биопотенциала сетчатки при нор-
мальном состоянии зрительного анализатора,что объясняется ря-
дом авторов ганцфельд-эффектом, обусловленным мутным хруста-
ликом. При катаракте у взрослых и детей целесообразно использо-
вать комплексное исследование общей и макулярной ЭРГ, а также
ЗВКП.
193
Таким образом, нарушение нормального физиологического со-
стояния сетчатки, изменение одного из многочисленных факторов,
участвующих в возникновении ЭРГ, предопределяет изменение ее
временных и/или амплитудных параметров. К этим факторам отно-
сятся состояние хороидальной и ретинальной циркуляции, пигмент-
ного эпителия, колбочек и палочек, биполярных горизонтальных и
амакриновых клеток, колбочкового пигмента и родопсина, метабо-
лизм, ионный состав и биохимическое звено между пигментным
эпителием и фоторецепторами, состояние наружного и внутреннего
синаптических слоев с их синаптической передачей, состоянием
нейротрансмиттеров и нейромодуляционной функцией, нормальное
-взаимодействие между нейронами и нейроглией. С помощью элек-
троретинографии возможно разделение колбочковой и палочковой
функции и, следовательно, диагностика патологических процессов,
локализованных в колбочковой и палочковой системах. Поскольку
заболевания сетчатки наследственного, врожденного, сосудистого
и другого генеза имеют характерную электрофизиологическую симп-
томатику, с помощью электроретинографии возможна дифферен-
циальная диагностика заболеваний сетчатки различного происхож-
дения и прогнозирование их течения. Электроретинографию исполь-
зуют в предоперационном периоде при помутнении оптических сред,
например, при катаракте и гемофтальме, для характеристики функ-
ций сетчатки как центральных, так и периферических отделов, а
также для прогнозирования послеоперационного восстановления
зрительных функций.
Международное общество клинических электрофизиологов зре-
ния рекомендует проводить электрофизиологические исследования
при следующих заболеваниях: наследственных заболеваниях сет-
чатки, глазной гипертензии и глаукоме, сидерозе, отслойке сетчат-
ки (особенно при помутнении оптических сред), рассеянном скле-
розе, некоторых воспалительных заболеваниях глаз, ишемии сет-
чатки при окклюзии центральной артерии и вены сетчатки, а также
при диабетической ретинопатии, хороидальной меланоме и невусе,
симпатической офтальмии, эндофтальмите. Исследования ЭРГ ре-
комендовано проводить при профилактических осмотрах в золах
экологических катастроф, интоксикации промышленными ядами и
лекарственными препаратами, неожиданной потере центрального
зрения и поля зрения, затруднениях при вождении автомобиля, по-
мутнении оптических сред, определении носителей патологического
гена, оценке функции сетчатки и зрительного нерва при травме.
При заболеваниях сетчатки изменения ЭРГ могут возникать рань-
ше клинических признаков заболевания, до офтальмоскопически
видимых изменений. В противоположность электроокулографии,
которая характеризует пресинаптическую функцию сетчатки, элек-
троретинограмма отражает и постсинаптическую функцию.
При заболеваниях, при которых в патологический процесс во-
194
влекаются только ганглиозные клетки сетчатки (зрительный нерв),
наблюдается нормальная ЭРГ. Таким образом, с помощью элект-
роретинографии возможна дифференциальная диагностика
заболеваний, локализующихся в первом и втором нейронах сет-
чатки, третьем нейроне (зрительном нерве) и центральных
отделах зрительного анализатора.
Для того чтобы понять механизмы нарушений зрительных функ-
ций и звеньев патогенеза заболеваний сетчатки, необходимо сопо-
ставлять ЭРГ с результатами других современных психофизических
исследований зрительного анализатора, генетических исследований,
офтальмоскопии глазного дна, фундусрефлектометрии, флюорес-
центной ангиографии, ЭОГ и ЗВКП.
ГЛАВА6
ЭЛЕКТРООКУЛОГРАФИЯ
6.1. ПОСТОЯННЫЙ ПОТЕНЦИАЛ ГЛАЗА
Существование постоянного потенциала глаза открыл Emil Du Bois-
Reymond в 1849 г. В дальнейшем было установлено, что движения
глаз сопровождаются биоэлектрической активностью, которая мо-
жет быть зарегистрирована с помощью накожных электродов
[Meyer J.J., 1929]. Если сначала связывали регистрируемые потен-
циалы с мышечными биотоками, то позднее было показано [Mowrer,
Д936] существование постоянного потенциала глаза около 6 мВ меж-
ду роговицей и задней поверхностью глаза.
Колебания электроокулограммы (ЭОГ) у человека при измене-
нии световых условий первым сделал W.R.Miles (1940), который
применял электроокулографию для регистрации глазных движений.
В 1954 г. L.A.Riggs зарегистрированы изменения этого потенциала
при пигментной дегенерации сетчатки. Начало применения ЭОГ в
качестве объективного электрофизиологического теста при различ-
ных заболеваниях сетчатки относится к 1956 г., когда были опубли-
кованы работы В.Я.Эскина и J.Francois, в которых авторы описали
изменения ЭОГ при различных заболеваниях сетчатки. J.Francois и
соавт. (1956) также показали, что постоянный потенциал глаза уве-
личивается при переходе из темноты к свету и снижается при тем-
новой адаптации.
Электроокулограмма — электрофизиологический метод исследо-
вания, используемый для регистрации постоянного потенциала (ПП)
глаза, который можно зарегистрировать при длительной стимуля-
ции сетчатки, в отличие от ЭРГ, регистрируемой на короткую
вспышку света, при использовании различных условий адаптации
[MarmorM.F., 1982, 1989, 1991].
G.B.Arden в серии работ показал, что наиболее важную инфор-
мацию дают световые и темновые колебания постоянного потенци-
ала, что позволило автору сделать следующие выводы.
1. От каждого глаза обследуемого могут быть получены незави-
сящие друг от друга записи. ЭОГ, т.е. светотемновые колебания
потенциала каждого глаза, отражают процессы регулирования, за-
мыкающиеся в пределах данного глаза.
2. Уменьшение освещенности сетчатки, в том числе переход к
полной темноте, вызывает уменьшение ПП до самого низкого уров-
ня (уровень “темновой впадины”), который не зависит ни от пред-
шествующих световых условий, ни от того, в какой фазе находился
процесс светотемновых колебаний ПП к моменту снижения осве-
щенности сетчатки.
3. Засвет глаза, адаптированного к темноте, вызывает значи-
196
тельный подъем ПП, так называемый световой пик, амплитуда
которого в определенных пределах пропорциональна логарифму
освещенности сетчатки.
Эти данные, полученные при физиологических исследованиях и
анализе патологических состояний, позволили G.B.Arden предло-
жить ряд методических приемов и критериев, благодаря которым
ЭОГ стал надежным параклиническим методом диагностики.
Необходимыми условиями для реализации нормальных свето-
темновых колебаний ПП являются нормальное функционирование
фоторецепторов и пигментного эпителия, контакт между этими
слоями, а также адекватное хориоидальное кровоснабжение.
6.2. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ И ПРОИСХОЖДЕНИЕ
ЭОГ
Исследования, выполненные A.Kris (1958) и H.E.Kolder (1974), по-
зволили выявить многие важные изменения постоянного потенциа-
ла в условиях световой и темновой адаптации глаза и дать им
физиологическое объяснение. Установлено, что резкий'переход от
одного стабильного уровня освещенности к другому вызывает мед-
ленные осцилляции с периодом около 25 мин. Если освещение из-
меняется только один раз, то амплитуда осцилляций прогрессивно
уменьшается, приближаясь к среднему (базовому) значению после
длительного интервала затухающих колебаний (около 90 мин). Если
же фазы освещения и затемнения следуют друг за другом каждые
12 мин, то наблюдается феномен резонанса.
Исследованы начальные быстрые отклонения ЭОГ, возникаю-
щие в момент смены освещенности. Эти отклонения выявляются не
всегда, невелики по амплитуде, имеют период около 2 мин и на-
правлены в противоположную сторону по отношению к медленно-
му колебанию. Высказано мнение, что положительная начальная
часть быстрых колебаний соответствует с-волне ЭРГ, а следующая
за ней отрицательная волна — Рш-компоненту ЭРГ, описанному
R.Granit (1957).
Таким образом, в условиях темновой адаптации ПП глаза умень-
шается, достигает минимума и затем медленно повышается. Если
включить свет, то сначала возникает электроретинографический
ответ, после чего потенциал уменьшается в течение 2 мин, имея
форму осцилляций, которые называют быстрыми.
Быстрые (transient) осцилляции отмечаются на 60—75-й, затем
потенциал начинает увеличиваться [Heilig Р., 1977], достигая через
4—7 мин максимальной величины (“световой ответ” или медлен-
ные осцилляции), после чего начинает уменьшаться, несмотря на
наличие света [Kris А., 1958]. Если свет выключить, то корнеорети-
нальный потенциал снова уменьшается до темнового минимума.
197
Небольшой позитивный пик можно отметить сразу после выключе-
ния света. Он соответствует негативному transient-потенциалу в
начале светового подъема. При использовании слабого стимула
постоянный потенциал может быстро превратиться в 2—3 осцилля-
ции, при более ярком стимуле осцилляции могут продолжаться в
течение длительного периода.
В норме величина ПП варьирует от одного до нескольких мил-
ливольт в зависимости от распределения освещения на сетчатке,
потому что свет вызывает деполяризацию базальной мембраны ре-
тинального пигментного эпителия, переходящую в изменения транс-
эпителиального биопотенциала [Marmor M.F., 1982].
Обнаружение быстрых колебаний, особенно начального поло-
жительного участка при обычной методике регистрации ЭОГ, зат-
руднено, однако в отсутствие светового подъема, например в слу-
чае закупорки центральной артерии сетчатки, выявление их облег-
чается. Снижение ПП обнаружено под влиянием внутривенного
введения гипертонического раствора 20% маннитола [Kawasaki К.,
1986], ацетазоламида и диамокса [Yonemura D., 1977]. Эти данные
свидетельствуют о гиперосмолярной зависимости ПП, подтверж-
дая его происхождение в пигментном эпителии. Регистрация посто-
янного потенциала представляет уникальную возможность иссле-
дования изолированной функции пигментного эпителия у пациен-
тов с патологией в заднем полюсе глаза, хотя не исключено участие
в нем нескольких потенциалов.
Ответ при воздействии ацетазоламида отражает первичную
функцию пигментного эпителия сетчатки заднего поля глазного дна,
поэтому этот ответ может быть патологическим у некоторых паци-
ентов с изменениями глазного дна сетчатки, связанными с процес-
сом в пигментном эпителии. При этом индуцированный ацетазол-
амидом и гиперосмолярный ответы свидетельствуют о патологи-
ческих изменениях пигментного эпителия и позволяют отметить его
отличие от нормы. Изменение ПП авторы связывают со снижением
концентрации натрия и калия в субретинальном пространстве вслед-
ствие метаболических нарушений натрий-калиевого насоса во внут-
реннем сегменте фоторецепторов [Linsenmeier R.A., Steinberg R.H.,
1984].
Таким образом, в ЭОГ отмечается два основных компонента:
светонечувствительный, зависящий от интеграции функции пигмент-
ного эпителия, роговицы, хрусталика и ресничного тела, независя-
щий от функции фоторецепторов, определяющий ’’темновое паде-
ние”; и светочувствительный — медленный ’’световой подъем” в
ЭОГ, обусловленный деполяризацией базальной мембраны пигмент-
ного эпителия сетчатки и опосредованно связанный с изменениями
концентрации калия и натрия в субретинальном пространстве. Де-
поляризация базальной мембраны является результатом увеличе-
ния электрического потенциала, проходящего через клетки пигмен-
198
тного эпителия (трансретинальный потенциал) и участия фоторе-
цепторов. Необходимым условием возникновения этого потенциа-
ла является контакт между фоторецепторами и пигментным эпите-
лием, что объясняет его отсутствие при отслойке сетчатки. Сниже-
ние потенциала при окклюзии центральной артерии сетчатки
предполагает участие в его возникновении внутренних слоев сет-
чатки [Thaler А., 1977, 1989]. В эксперименте показана связь сниже-
ния светового подъема с повышением концентрации внеклеточного
кальция.
Гиперполяризация базальной мембраны пигментного эпителия
сетчатки в ЭОГ определяется по слабому, быстрому и скоротечно-
му отклонению трансэпителиального потенциала после включения
света, который отличается от светового подъема и медленных ос-
цилляций. Эта быстрая осцилляция имеет отношение к изменению
концентрации калия в сетчатке [Griff Е.К.,1983]. Амплитуда свето-
чувствительного компонента ЭОГ увеличивается с повышением
уровня освещенности сетчатки (от 1 до 1000 фут Лб), зависящей от
многих факторов, в том числе от яркости экрана. Более высокий
уровень освещения приводит к большей вариабельности коэффици-
ента Ардена. Световой подъем ЭОГ не связан только с колбочко-
вой или палочковой системой, поскольку нормальный подъем заре-
гистрирован как у больных с врожденной ахромазией, так и у па-
циентов со стационарной ночной слепотой. Каковы доказательства
этого? ЭОГ заметно снижается в темноте, под влиянием субстан-
ций, избирательно воздействующих на пигментный эпителий; све-
товой подъем исчезает при окклюзии центральной артерии сетчат-
ки как у людей, так и в эксперименте у животных.
В работах, выполненных в последние годы, отмечено увеличе-
ние светового подъема ЭОГ под воздействием синего и красного
света, в то же время у цветослепых при воздействии красного света
световой подъем был ниже нормы, а при синем свете — в пределах
нормы. Эти факты свидетельствуют об участии в генерации ЭОГ
колбочек и палочек сетчатки. Не исключено, что световой подъем
в световой фазе исследования отражает химические процессы све-
товой адаптации, а медленный подъем корнеоретинального потен-
циала в течение нескольких минут является признаком регенерации
фотопигмента.
6.3. МЕТОДЫ РЕГИСТРАЦИИ ЭОГ
6.3.1. Стандартная методика регистрации ЭОГ
Постоянный потенциал сетчатки распространяется по глазному
яблоку и в ткани, окружающие глаз. Он может быть зарегистриро-
ван с поверхности кожных покровов по краю глазницы при движе-
199
Рис. 6.1. Один цикл ЭОГ.
hp h2 и h3 — амплитуды ЭОГ, мм.
Рис. 6.2. ЭОГ и импульсная ЭОГ.
Вверху — ЭОГ: а — тестовый сигнал 500 мкВ; б — результат его усреднения; в —
один цикл ЭОГ; г — результат усреднения ЭОГ; д — светотемновые изменения:
Аб — базовая амплитуда, Ат — темновая, Ас — световая; Тт и Тс — соответствующие
им временные параметры. Внизу — импульсная ЭОГ, записанная синхронно с ЭОГ.
нии глаз в горизонтальном направлении при неподвижной голове
(рис. 6.1). Такая методика отведения потенциала положена в основу
клинической электроокулографии (рис. 6.2).
Величина отводимого от кожных покровов постоянного потен-
циала зависит от угла поворота глаз, особенно в пределах угла
±30° от центрального положения взора. Из этого факта следуют
два основополагающих для методики электроокулографии вывода.
1. При постоянном (стандартном) угле поворота глаз (обычно
±20° от центрального положения взора) можно сравнивать величи-
ну постоянного потенциала разных людей (здоровых, больных) и
таким образом оценивать интенсивность обменных процессов в
наружных слоях сетчатки обследуемых.
2. При сопоставлении изменения величины постоянного потен-
циала при движении глаз (произвольном или непроизвольном) с
величиной потенциала для стандартного угла поворота глаза об-
следуемого можно количественно охарактеризовать функции гла-
зодвигательного аппарата.
200
Обе эти методики применяют в клинической практике: первую—
для диагностики нарушений функции сетчатки, вторую — с целью
выявления нарушений функции глазодвигательного аппарата.
Базовый потенциал э лектр о оку ло гр а По-
стоянный потенциал сетчатки, измеренный у пациента, длительно
(более 1 ч) находящегося в условиях неизменной освещенности, ус-
танавливается на определенном уровне, который назван базовым.
Базовый потенциал, зарегистрированный в начале исследова-
ния при умеренном освещении, является одним из диагностических
показателей. Его величина, соотнесенная с одним градусом угла
поворота глаза (в микровольтах на градус — мкВ/градус), обозна-
чается Аб — амплитуда базового потенциала.
Светотемновой тест. При резком изменении свето-
вых условий (от умеренного освещения к темноте, от темноты к
яркому свету) происходят характерные изменения величины посто-
янного потенциала сетчатки. Это позволяет выделить ряд ампли-
тудных и временных параметров, которые используют в диагнос-
тике, так как они закономерно изменяются у здоровых и больных
людей (см. рис. 6.2).
Потенциал темнового с п а д а. Резкий переход от уме-
ренной освещенности к темноте приводит к возникновению серии
затухающих колебаний постоянного потенциала (’’спад” кривой),
достигающего минимума на 10—12-й минуте (а при патологии —
позже), а затем сменяющегося увеличением (подъемом кривой).
Величина потенциала, измеренная в самой низкой точке спада, на-
звана амплитудой темнового спада, обзначается Ат и измеряется
так же, как и базовый потенциал, в микровольтах на 1 градус.
Диагностическое значение имеет и время (в минутах) достижения
темнового спада от момента погружения исследуемого в темноту
(обозначается Т).
Понятие о потенциале светового подъема.
Резкий переход от темноты к яркому освещению экрана, находяще-
гося перед обследуемым, также приводит к возникновению серии
затухающих колебаний постоянного потенциала, которые в отли-
чие от темновых начинаются с подъема, достигают максимума на
8 — 10-й минуте, а затем сменяются спадом. Величина потенциала,
измеренная в самой высокой точке подъема, названа амплитудой
светового подъема (световым пиком), обозначается Ас и измеряется
в микровольтах на 1 градус. В число диагностически важных пока-
зателей входит и время (в минутах) достижения светового пика от
момента включения яркого света (обозначается Т^.
Перечисленные параметры ЭОГ как в приведенном здесь виде,
так и в форме отношений (коэффициент Ардена, коэффициенты
Глиема и др.) используют в клинической диагностике.
Комбинированные и безразмерные парамет-
ры оценки клинической ЭОГ. Уже в первых работах по
201
клинической электроокулографии был отмечен значительный раз-
брос величины постоянного потенциала сетчатки у разных пациен-
тов. Хотя была выявлена достоверная зависимость величины по-
тенциала от функциональных и морфологических сдвигов в зри-
тельном аппарате, они носили статистический характер, т.е.
проявлялись только при вариационно-статистической обработке и
сопоставлении средних данных, полученных при обследовании групп
здоровых и больных людей.
G.B.Arden и соавт. (1962) первыми предложили заменить пока-
затель величины потенциала безразмерным показателем — отно-
шением потенциала светового пика к потенциалу темнового спа-
да —так называемый коэффициент Ардена (КА):
КА = А/А • 100%.
с т
Для оценки патологических состояний сетчатки хороидеи КА
подразделяется на субнормальный (от 135 до 180%); анормальный
(от 130 до 110%); погасший (от 110 до 100%); извращенный (ниже
100%). Коэффициент Ардена, превышающий 185%, считается нор-
мальным, однако следует обратить внимание на состояние глаза с
меньшим КА.
H.Gliem (1971) предложены коэффициенты, учитывающие базо-
вый потенциал, раздельно для световой и темновой фазы исследо-
вания:
КГ, = А/А • 100%; КГ, = А/А • 100%.
1 СО 7 2 ТО
О преимущественном поражении фотопического или скотопи-
ческого компонента ЭОГ можно судить по этим двум относитель-
ным показателям — первому и второму коэффициентам Глиема.
Коэффициент Глиема представляет диагностически ценное до-
полнение к коэффициенту Ардена, так как позволяет раздельно
оценить фотопический и скотопический компоненты ЭОГ и таким
образом установить механизм нарушения светотемновых колеба-
ний ЭОГ. Однако для коэффициента Глиема была получена боль-
шая дисперсия, чем для коэффициента Ардена [Krogh Е., 1977].
Оснащение электро окулографического ка-
бинета. Проведение электроокулографического исследования по
стандартной методике предполагает соответственно подготовлен-
ное помещение и электронное и светотехническое оборудование.
Электроокулографический кабинет включает специально обо-
рудованную камеру для пациента и рабочее место оператора. Ми-
нимальный комплект аппаратуры, необходимый для проведения ис-
следования, состоит из 5 слабополяризующихся электродов, усили-
теля, регистратора, источников умеренного (50 кд/м2) и яркого
(800 кд/м2) света, белого экрана с установленными на нем тремя
точками фиксации взора (одна — в центре и две — под углом ±20°),
а также устройство, позволяющее управлять точками фиксации, ис-
202
точниками освещения, усилителем и регистратором в соответствии
с программой исследования.
Процесс регистрации и обработки данных может быть значи-
тельно облегчен, если в распоряжении исследователя имеется спе-
циализированная автоматизированная ЭОГ — система или персо-
нальная ЭВМ, которую в последние годы все чаще используют в
клинической диагностике.
Стандартизация условий исследования. Для
того чтобы обеспечить доверие врача к получаемой информации и
дать возможность сравнить результаты исследования с данными дру-
гих авторов, исследование должно быть метрологически коррект-
ным, т.е. выполненно в стандартных условиях.
Для повышения точности исследования ЭОГ в световой фазе
необходимо учитывать не только освещенность экрана, но и коли-
чество света, попадающего на сетчатку через зрачок (стандартиза-
ция световых режимов). В офтальмологической практике нередко
происходит отключение естественной зрачковой реакции на свет
как в результате применения фармакологических средств (миотики,
мидриатики), так и вследствие патологического процесса (спайки,
нарушение иннервации) или оперативного вмешательства (колобо-
ма радужки). Сохранение диагностической ценности ЭОГ в этих
случаях должно основываться на учете реального состояния опти-
ческих сред глаза, особенно зрачка. В табл. 6.1 приведены значения
поправочного зрачкового коэффициента (Кз), на который умножа-
ют амплитуду светового подъема, установленную у пациента с не-
реагирующим зрачком:
А = К . А ,
С з сз7
Таблица 6.1. Определение зрачкового коэффициента для некоторых зна-
чений диаметра зрачка и уровней яркости экрана Рекин В. Я., 1984]
Диаметр зрачка, мм Яркость экрана, Кд/м2
400 600 800 1000
1,0 1,49 1,39 1,33 1,28
1,5 1,28 1,21 1,16 1,12
2,0 1,16 1,11 1,06 1,04
2,5 1,09 1,04 1,00 0,97
3,0 1,03 0,99 0,95 0,93
3,5 0,99 0,95 0,92 0,90
4,0 0,96 0,92 0,89 0,87
5,0 0,90 0,87 0,84 0,82
6,0 0,87 0,83 0,81 0,79
х 7,0 0,84 0,81 0,78 0,77
8,0 0,81 0,78 0,76 0,75
203
где А — приведенное значение амплитуды светового пика; Асз —
амплитуда светового подъема ЭОГ глаза с нереагирующим зрач-
ком.
В табл. 6.1 величины поправочных коэффициентов даны для
четырех уровней яркости экрана — от 400 до 1000 кд/м2. Оптималь-
ной для ЭОГ-исследования считается яркость экрана в диапазоне
от 600 до 800 кд/м2, поскольку она соответствует регулирующим
возможностям зрачковой реакции [Эскин В.Я., 1984].
Продолжительность световых фаз электроокулограммы. Для
корректного измерения временных и амплитудных характеристик
первых медленных волн темновой и световой фаз электроокуло-
граммы у больных людей длительность исследования в темновой
фазе должна быть не менее 16 мин, а в световой — не менее 12 мин.
В специальных тестах продолжительность темновой и световой фаз
исследования может быть увеличена.
Дискретность измерения. Частые отсчеты, требующие непрерыв-
ного движения глазных яблок, утомительны для пациента, редкие
содержат опасность утери информации об истинном значении ам-
плитудных и временных параметров ЭОГ. В диагностической прак-
тике рекомендуют использовать интервалы длительностью 1 мин
для молодых лиц и 2 мин для пожилых лиц, у которых отмечается
повышенная утомляемость.
Оптимальная частота переключения точек фиксации. Этот эле-
мент методики клинической электроокулографии определяется сум-
марной продолжительностью латентного периода движения глаз-
ного яблока и длительностью самой саккады. Рекомендуется уста-
новить время переключения точек фиксации 1 с, а частоту
переключения 0,5 Гц.
Влияние на ЭОГ аметропии и оптической коррекции. Проведение
ЭОГ-исследований у слабовидящих лиц имеет методические осо-
бенности: если острота зрения вдаль без коррекции не ниже 0,1, то
исследование можно проводить без очков. При аметропии (близо-
рукость или дальнозоркость высокой степени) во время исследова-
ния можно применять очки.
Для получения истинных значений амплитуды потенциала (А)
его измеренную у пациента в очках величину (Аок) нужно умножить
на коэффициент оптической коррекции (Кок) в соответствии с фор-
мулой:
а = к011.ам,.
Значения Кок для аметропии различной степени приведены в табл.
6.2. При использовании контактных линз и наличии ИОЛ не требу-
ется введение поправки.
Патологические изменения пигментного эпителия проявляются
в снижении КА и КГр в то же время КГ2 повышается и при “плос-
кой” ЭОГ становится равным 100%.
204
Таблица 6.2. Поправочный коэффициент для амплитуды ЭОГ при опти-
ческой коррекции очками [Эскин В. Я., 1984]
Миопия Гиперметропия
оптическая сила линзы коэффициент коррекции (Кок) оптическая сила линзы коэффициент коррекции (Кок)
-20 1,64 +1 0,97
-18 1,52 +2 0,93
-16 1,41 +3 0,89
-14 1,32 +4 0,85
-12 1,23 +5 0,82
-10 1,18 +6 0,78
-8 1,11 +7 0,74
-6 1,08 +8 0,70
-4 1,05 +9 0,68
-2 1,03 +10 0,67
Р.А.Толмачевым (1977) предложен так называемый темновой
коэффициент КТ, характеризующий состояние скотопических про-
цессов в сетчатке:
КТ = А /А • 100%,
тм т 7
где Атм — максимальная амплитуда ЭОГ в темноте (амплитуда тем-
нового подъема, следующего за темновым спадом при длительном
пребывании в темноте).
Приведенные в табл. 6.3 данные получены методом стандарт-
ной ЭОГ при исследовании 54 здоровых лиц. Между тем рядом
авторов делаются попытки усовершенствовать метод ЭОГ как с це-
лью сделать его более независимым от поведения пациента во вре-
мя исследования, так и с целью получения дополнительной диагно-
стической информации.
Таблица 6.3. Амплитудные, временные и относительные показатели ЭОГ
здоровых глаз [Эскин В. Я. 1984]
Статистиче- ские показатели Показатели ЭОГ
А с А т Т Т т с К а кг, кгг
мкВ/град мин %
k М 19,0 12,5 32,3 10,2 7,6 266 175 67
8 6,36 4,16 9,31 1,86 1,43 49,9 34,4 11,3
0,87 0,57 1,27 0,25 0,20 6,79 4,55 1,54
Примечание. М — среднее арифметическое значение, 5 — среднее квадратиче-
ское отклонение, |1 — ошибка среднего арифметического.
205
6.3.2. Импульсная электроокулография
Для проведения исследований у детей, слабовидящих, а также лиц
с установочным поведением в МОНИКИ им. М.Ф.Владимирского
разработана объективная методика — импульсная электроокуло-
графия [Эскин В.Я., 1984]. Особенностью данной методики являет-
ся возможность получения полноценной ЭОГ при недозированных
и даже непроизвольных движениях глазных яблок. В основе мето-
дики импульсной электроокулографии лежит дифференцирование
электроокулосигнала при саккадических движениях глаз. Отличие
в методе измерения величины потенциала при обычной электро-
окулографии от такового при импульсной электроокулЪграфии со-
стоит в том, что при первой измеряют размах потенциала в стати-
ческих положениях взора при визуальной фиксации точек, а при
второй информацию о величине потенциала получают в переход-
ных состояниях при смене точек фиксации. Величина максималь-
ной угловой скорости движения глазных яблок человека при смене
точек фиксации Wmax составляет 400—600% и для углов поворота
глаза более 20° почти не зависит от их величины.
Более объективной, независимой от воли и желания обследуе-
мого импульсная электроокулография становится при использова-
нии оптокинетической стимуляции движения глазных яблок. Фи-
зиологической основой в этом случае является идентичность скоро-
стных характеристик быстрого компонента оптокинетического
нистагма (ОКН) и саккад произвольных движений. Оптимальные
характеристики оптокинетического поля, эффективные при опто-
кинетической стимуляции как в условиях яркого света (600—800 кд/
м2), так и в скотопических, а точнее, в мезопических условиях ис-
следования темновой фазы ЭОГ (0,1 кд/м2), имеют такие значения:
видимая пациентом угловая ширина белых и черных полос рав-
на 4,5°, угловая скорость их перемещения относительно глаза паци-
ента — 30%.
Применение оптокинетической стимуляции движения глаз по-
зволяет определить изменение постоянного потенциала глаз у па-
циентов, для которых стандартная методика электроокулографии
неприменима из-за возрастных особенностей (дети) или установоч-
ного поведения (аггравация, симуляция).
6.3.3. Другие методические подходы в электроокулографии
Диагностические данные, которые можно получить при обработке
ЭОГ, зарегистрированной по стандартной методике (засвет всего
поля зрения белым светом после пребывания в темноте), отражают
состояние наружных слоев всей сетчатки. Так же как в электроре-
тинографии, очаговые поражения сетчатки и ее пигментного эпите-
лия не отражаются на ЭОГ.
206
Рядом исследователей предприняты попытки модифицировать
методику электроокулографии при изменении цвета фонового осве-
щения, применяя локальный засвет центральных отделов сетчатки
и фармакологические пробы, сопоставляя характеристики быстрых
и медленных компонентов ЭОГ. Так, при электроокулографичес-
ком исследовании центральных отделов сетчатки в световой фазе
использовали слабый диффузный сине-зеленый фоновый засвет все-
го поля зрения и засвет макулярной области локальным белым
светом большой яркости на красном фоне и регистрировали так
называемую макулярную или центральную ЭОГ для разных углов
движения глаз [Миронова Э.М. и др., 1976; Hache J.C. et al., 1987].
R.W.Massof (1986) наряду с ганцфельд-ЭОГ исследовал так называ-
емую фокальную ЭОГ, осуществляя засвет центральной области
сетчатки (в 25°) белым светом при яркости экрана 1388 кд/м2. Сле-
дует отметить, что различие методик и разнообразие заболеваний,
при которых их применяют, пока не позволяют сделать вывод о
метрологической корректности и клинико-диагностической ценно-
сти макулярной ЭОГ.
Быстрые колебания в ЭОГ видны в начальной фазе снижения
потенциала, следующего за световой или темновой стимуляцией.
Однако при стандартной записи они неразличимы и сливаются со
световым подъемом. В течение 90 мин темновой адаптации ЭОГ
осциллирует медленными колебаниями, которые достигают устой-
чивого (steady-state) состояния. Быстрые осцилляции с периодом
150 с (2,5 мин), которые накладываются на медленные осцилляции,
можно выделить, если ЭОГ регистрировать при нарастающей (с
определенным шагом) интенсивностью света, и привести к резонан-
су, варьируя частоту (каждые 70—75 с) включения и выключения
стимулирующего света интенсивностью 68 кд/м2. Оптимальная про-
должительность повторяющихся стимулов составляет 1—1,25 мин.
Амплитуда быстрых осцилляций зависит от интенсивности стиму-
лирующего света [Weleber R.G., 1989; Kolder Н.Е., 1991].
Происхождение быстрых потенциалов связано с ионными изме-
нениями, возникающими в пигментном эпителии при воздействии
света. Установлено, что быстрые и медленные колебания потенци-
ала не зависят друг от друга, генерируются разными структурными
элементами сетчатки и по-разному проявляются при различной
глазной патологии. У здоровых лиц хорошо выражены и быстрые,
и медленные колебания ЭОГ, быстрые осцилляции более резистен-
тны к гипоксии, чем медленные потенциалы; при пигментной деге-
нерации сетчатки, передающейся по аутосомно-рецессивному типу,
вителиформной макулярной дистрофии Беста значительно сниже-
ны медленные колебания при нормальных и субнормальных быст-
рых осцилляциях.
207
6.3.4. Методологические основы регистрации ЭОГ (рекомендации
ISCEV)
Во многих лабораториях мира используют различную технику элек-
троокулографии, поэтому для того чтобы можно было сравнивать
результаты исследований, ISCEV предложены стандарты [Маг-
mor M.F., 1993] для двух альтернативных методов регистрации ЭОГ:
1) отношение светового пика к темновому падению (коэффициент
Ардена); 2) отношение светового пика к темноадаптированной ба-
зовой линии (коэффициент Глиема). Не исключена возможность
использования в лабораториях оригинальных методов регистрации
и анализа ЭОГ, измерения дополнительных параметров.
Световой стимуляцией должна быть равномерно охвачена вся
площадь сетчатки, так как ЭОГ включает множество ретинальных
ответов. Система фиксации должна индуцировать движения глаз на
постоянный угол зрения приблизительно в 30° по горизонтальному
меридиану. (При отнесении потенциала к 1° угла поворота величи-
на общего угла теряет принципиальное значение в пределах линей-
ной зависимости до ±30°.) Наиболее часто используют красные
светодиоды, достаточно яркие, чтобы быть видимыми в темновой
и световой фазе исследования. Предполагается, что направление дви-
жения глаза от одной точки фиксации до другой должно меняться
каждые 1—2,5 с, что эквивалентно полному циклу 2—5 с (минимум
10 раз). Накожные электроды должны быть изготовлены из неполя-
ризующегося материала, такого, как серебро, хлорированное сереб-
ро, золото. Межэлектродное сопротивление должно быть меньше
10 кОм в диапазоне частот от 30 до 200 Гц. Электроды (два для
каждого глаза) накладывают на кожу со стандартной электропро-
водной пастой очень близко к назальной и темпоральной спайкам
глаза. При этом нужно иметь в виду, что при близком расположе-
нии электродов к углу глаза (менее 5 мм) возможно подтекание
слезы, что затруднит исследование и может вызвать артефакты.
Заземленный электрод помещают на фронтальную часть головы (на
лоб).
Источники света. Для освещения можно использовать
одну или несколько ламп, продуцирующих видимый белый свет,
которые должны быть расположены так, чтобы равномерно осве-
щалось все поле зрение пациента. В поле зрения должны быть ус-
транены тени и более светлые пятна (hot spots). Освещенность дол-
жна быть калибрована в канделах на 1 м2 стандартным фотомет-
ром, имеющим в основе кривую фотопической освещенности. При
обследовании пациентов с расширенными и нерасширенными зрач-
ками предусматривается использование регулирующих фильтров.
Электронное регистрирующее оборудование.
Основой регистрации ЭОГ является то, что глаз представляет со-
бой диполь с положительным полюсом на роговице по отношению
208
к заднему негативному полюсу глаза, саккадические движения глаз,
результатом которых являются потенциалы вокруг глазницы, про-
порциональные величине стандартного биопотенциала каждого
глаза. Это изменение вольтажа может быть измерено от кожных
электродов, расположенных в области назальной и темпоральной
спаек дек.
Система усиления. Непосредственное усиление тока воз-
никает при изменении взора пациента в горизонтальном направле-
нии, когда регистрируются квадратоподобные волны. Для практи-
ческих целей предлагают использовать стандартные системы, в ко-
торых решена проблема минимизации дрейфа и стабильности,
низкие частоты 0,1 Гц или ниже, а высокие не ниже 20 Гц (предпоч-
тительно выше 50—60 Гц для минимизации интерференции). Важно
вывести оригинальную запись на дисплей во время регистрации
ЭОГ, чтобы видеть артефакты, неожиданные саккады, нехарактер-
ные наложения и пр. — все, что может послужить причиной нало-
жения электродов заново, повторной инструкции пациента или иной
интерпретации результатов. Усилитель должен быть электрически
изолирован от пациента.
Клинический протокол. ЭОГ можно регистрировать
как при расширенном, так и при нерасширенном зрачке. С одной
стороны, расширение зрачка позволяет более равномерно освещать
сетчатку, с другой — эта процедура неудобна при исследовании
определенной группы пациентов. Освещение сетчатки играет очень
важную роль в исследовании ЭОГ, в возникновении светового от-
вета в постоянном потенциале глаза в зависимости от состояния
зрачка, поэтому его необходимо специально корректировать. Пе-
ред темновой фазой преадаптация пациента должна проходить при
обычном комнатном освещении, а перед световой — при 35—70 лк
в течение 15 мин. После пребывания на солнце, офтальмоскопии,
ангиографии исследование ЭОГ следует проводить не ранее чем
через 60 мин.
Тем новая фаза. Эта часть исследования делится на две фазы.
1. Определение отношения светового пика к темновому паде-
нию (коэффициент Ардена), когда запись проводят в течение 15 мин
в темноте, а минимальную амплитуду зарегистрированных в этот
период волн обозначают как темновое падение, чаще всего отмеча-
ющееся между 11-й и 12-й минутами.
2. Отношение светового пика к темноадаптированной базовой
линии. Темновая адаптация в течение 40 мин приводит к стабили-
зации базовой линии. Запись необязательно осуществлять на про-
тяжении всего этого периода, однако необходимо начать ее в пос-
ледние 5 мин перед световой фазой. В этот период темновой адап-
тации электроды можно установить при насыщенном красном
освещении. Возможно использование этого периода для клиничес-
кой адаптометрии.
209
Световой ответ. После включения светового стимула
запись проводят до того момента, когда определяется световой пик
и амплитудный сигнал начинает снижаться (точка, на которой мож-
но остановить исследование). Если световой пик неясен, то исследо-
вание продолжают еще 20 мин, в течение которых необходимо убе-
диться в том, что световой пик действительно не отмечается. Вы-
бор уровня освещенности исследования зависит от того, расширен
зрачок или нет. Интенсивность света должна быть фиксированной
величиной для расширенного зрачка 50—100 кд/м2, дЛя нерасши-
ренного — 400—600 кд/м2. Эти величины рекомендуется соотносить
с размером расширенного и нерасширенного зрачка. При этом для
стандартизации ЭОГ лучше использовать уровень ретинальной
освещенности в троландах (кд/м2 на 1 мм2 площади зрачка в едини-
цу времени), что позволяет гарантировать одинаковый уровень
ретинальной освещенности независимо от размера зрачка, оптималь-
ный уровень которой ISCEV считает от 1000 до 3000 троландов
(3—3,5 лог. троландов). Однако на практике трудно измерять диа-
метр зрачка в процессе исследования, когда пациент смотрит в рас-
сеивающую сферу, калиброванную неточно. Поэтому рекомендуют
разделять фиксированную интенсивность света для расширенного
и нерасширенного зрачка, что является лучшим компромиссом меж-
ду регулируемым уровнем освещенности и реальными условиями
клинического тестирования.
Измерение осцилляций ЭОГ осуществляют по наиболее стабиль-
ному уровню амплитуды. Если имеются выраженные отклонения
сигнала, то рекомендуется измерять начальное или заднее колено
первого стабильного сигнала.
Отношение светового подъема к темново-
му падению. Берут наименьший потенциал при темновой адап-
тации и наибольший — при световой адаптации и рассчитывают
коэффициент Ардена.
Отношение светового пика к базовой линии в фазе темновой
адаптации ниже, чем коэффициент Ардена. Латентный период све-
тового пика определяется от момента включения света до пика све-
тового ответа. Очень важно измерять как стандартный потенциал
(в мкВ), так и амплитуду темнового падения или темноадаптиро-
ванной базовой линии, если используется коэффициент Ардена, по-
скольку уменьшение этих компонентов может иметь самостоятель-
ное клиническое значение, а также использоваться при вычислении
других коэффициентов. Эти амплитуды базовой линии рассчитыва-
ются в микровольтах на 1°(мкВ/градус), если зрительный угол не-
стандартный (стандартный зрительный угол 30°).
Каждая лаборатория должна иметь свои нормативы для ЭОГ,
как для оборудования, так и для популяций людей.
Рекомендуется представлять заключение по ЭОГ, содержащее
коэффициент Ардена, отношение светового пика к темновой базо-
210
вой линии, латентный период и амплитуду светового пика, темно-
вого падения или темновой базовой линии (в микровольтах на 1°
зрительного цикла). Необходимо указать соответствие оборудова-
ния международным стандартам. В заключение должны также
содержаться данные о величине зрачка, пространственных отноше-
ниях, интенсивности световых стимулов, условий преадаптации и
темновой адаптации, продолжительности стимуляции и характери-
стика фильтров регистрирующего оборудования.
Быстрые осцилляции. Это сигнал, отражающий гипер-
поляризацию базальной ретинальной мембраны пигментного эпи-
телия между ЗО-й и 40-й секундой освещения. Исследователи уже
начали включать быстрые осцилляции как в нормативы ЭОГ, так
и изучать их изменения при патологии.
Для выделения быстрых осцилляций используют то же самое
оборудование, что и для записи ЭОГ. Главное отличие быстрых
осцилляций состоит в том, что их регистрация осуществляется
во время цикла ’’свет — темнота”. Рекомендуется проводить сме-
ну светового и темнового периодов каждые 60—80 с для получе-
ния минимум шести полных циклов ’’свет — темнота”. Поскольку
каждый цикл короткий, продолжением его может стать исследова-
ние ЭОГ.
Быстрые осцилляции могут регистрироваться как независимый
тест или как начальный компонент ЭОГ (между преадаптацией и
темновой фазой). Опыт большинства исследователей ЭОГ показал,
что коэффициент Ардена незначительно изменяется при регистра-
ции быстрых осцилляций в начале темновой фазы. Для определе-
ния отношения светового пика к базовой линии важно не нарушить
период темновой адаптации. Представление быстрых осцилляций
должно включать отношение усредненного быстрого пика к темно-
вому падению и среднюю их латентность или фазу смены пика, а
также абсолютную величину постоянного потенциала при его паде-
нии в мкВ на величину зрительного угла.
J.Babel (1977) провел сопоставление результатов электроокуло-
графии в возрастных группах до 50 и после 50 лет и установил, что
существенные различия в этих группах отмечаются в базовом по-
стоянном потенциале и световом подъеме. Темновое падение и от-
ношение светового подъема к темновому падению менее вариабель-
ны. Базовая величина постоянного потенциала, темновое падение и
световой подъем несколько ниже у людей старше 50 лет, а соотно-
шение светового подъема и темнового падения выше у обследован-
ных моложе 50 лет. Межиндивидуальная дисперсия по всем показа-
телям ЭОГ в группе до 50 лет невелика, световой подъем и темно-
вое падение постоянно возникали на 7—9-й и 9—10-й минуте
соответственно. Между показателями правого и левого глаза зна-
чительных различий не отмечено. Нормальный коэффициент Арде-
на 180—250%.
211
6.4. РОЛЬ ЭЛЕКТРООКУЛОГРАФИИ В ДИАГНОСТИКЕ
ЗАБОЛЕВАНИЙ ПИГМЕНТНОГО ЭПИТЕЛИЯ СЕТЧАТКИ
И ХОРОИДЕИ
Электроокулография имеет большое значение в исследовании функ-
ции и первичных дефектов пигментного эпителия при дифференци-
альной диагностике заболеваний сетчатки и зрительного нерва раз-
личной локализации и генеза.
ЭОГ изменяется при различных заболеваниях сетчатки воспали-
тельной и токсической природы, при циркуляторных нарушениях
и т.д. с вовлечением в патологический процесс фоторецепторов и
хороидеи.
Несмотря на то что ЭОГ и ЭРГ имеют разное происхождение,
оба потенциала изменяются при патологии одинаковой локализа-
ции и природы. Это, возможно, обусловлено тем, что патологиче-
ские процессы не ограничиваются одним слоем, а распространяют-
ся более широко. Однако при одних заболеваниях изменяется толь-
ко ЭРГ, при других — только ЭОГ, что является важным моментом
диагностики. Примером служит вителиформная макулярная дист-
рофия Беста, при которой ЭРГ нормальная, а ЭОГ патологическая.
Одновременная регистрация ЭРГ и ЭОГ может облегчить диффе-
ренциальную диагностику наследственных функциональных нару-
шений сетчатки, при которых ЭРГ субнормальная, а ЭОГ нормаль-
ная, и пигментном ретините, характеризующемся изменением обо-
их потенциалов.
При центральных дистрофиях сетчатки патологическая ЭОГ
свидетельствует о вовлечении в патологический процесс пигмент-
ного эпителия сетчатки. Это общее положение установлено в ре-
зультате сопоставления ЭРГ и ЭОГ при центральных дистрофиях
сетчатки различного генеза, очень важно в оценке распространен-
ности и локализации патологического процесса.
При врожденных стационарных функциональных дефектах све-
тоощущения, таких как аутосомно-рецессивная форма с миопией и
сцепленная с полом стационарная ночная слепота типа Шуберта —
Борншейна (негативный тип ЭРГ: при нормальной скотопической
a-волне редуцированная b-волна), ЭОГ, как правило, не изменяет-
ся, и только при аутосомно-доминантной форме типа Риггс (ЭРГ:
скотопическая b-волна редуцирована, но превышает a-волну, ско-
топическая а-волна нормальна либо снижена) отмечен редуциро-
ванный световой подъем в ЭОГ. При болезни Огуши одни исследо-
ватели не обнаружили изменений ЭОГ, другие отмечали снижение
светового подъема [Carr R.E., 1990]. При белоточечной дистрофии
сетчатки (fundus albipunctatus) как при стационарной, так и про-
грессивной форме световой подъем отсутствует при стандартной
записи с периодом темновой адаптации продолжительностью 15 мин,
однако нормальный световой подъем и коэффициент Ардена уста-
212
Рис. 6.3. Электроокулограм-
ма при пигментном рети-
ните (больная 40 лет, vis
OU = 0,6, поле зрения кон-
центрически сужено до 10°,
КА: OD 101%, OS 112% при
норме больше 185%; КГ :
OD 100%, OS 107%; КГ,:
OD 99%, OS 96%).
новлены в случае, когда период темновой адаптации предшество-
вал световой экспозиции.
Таким образом, характерные изменения ЭРГ и ЭОГ при нор-
мальной картине глазного дна и снижении зрения в темноте явля-
ются определенным симптомокомплексом нарушения светоощуще-
ния, дифференциально-диагностическим признаком гемералопии.
При нарушении цветоощущения, врожденной ахромазии и при
прогрессивной колбочковой дистрофии изменений ЭОГ не отмеча-
ется.
Дистрофические изменения в сетчатке; периферические хорой-
ретинальные изменения. При пигментном ретините (тапеторетиналь-
ная абиотрофия с преимущественным поражением палочкового
аппарата сетчатки) исследование ЭОГ имеет высокую диагности-
ческую ценность. Патологические признаки в ЭОГ при пигментном
ретините возникают рано, часто до появления выраженных офталь-
москопически обнаруживаемых изменений и функциональных на-
рушений (рис. 6.3). Патологическая ЭОГ, снижение базового потен-
циала (в среднем до 9 мкВ/градус), уменьшение коэффициента Ар-
дена (в среднем до 113%) и отсутствие общей ЭРГ являются
патогномоничными признаками пигментного ретинита и свидетель-
ствуют об органических изменениях фоторецепторов и пигментно-
го эпителия сетчатки. ЭОГ при этом виде патологии во всех фазах
исследования патологическая и характеризуется межвидовыми раз-
личиями. Отношение свет/темнота резко снижено в 90% случаев: не
отмечено светового подъема в фотопической фазе, базовый потен-
циал изменен в 83%, темновое падение отсутствует в 63% случаев.
При нормальных показателях базового потенциала отмечается из-
вращение светотемновой реакции (коэффициент Ардена < 100%), а
амплитуда постоянного потенциала в фазе темновой адаптации
выше, чем в фазе световой адаптации.
При палочково-колбочковой дистрофии, при которой отмече-
ны значительные изменения ЭРГ, ценность ЭОГ не так велика.
Корреляции ЭОГ с остротой зрения найдено не было.
213
Так или иначе, изменения ЭОГ при пигментном ретините пред-
шествуют клиническим проявлениям и свидетельствуют о наличии
патологического процесса в пигментном эпителиц уже в начальных
стадиях заболевания. При одностороннем процессе изменения ЭОГ
были типичными для пигментного ретинита, хотя вопрос возмож-
ности одностороннего поражения остается открытым.
При секторальном пигментном ретините ЭОГ патологическая,
отношение световой подъем/темновое падение как и базовый по-
тенциал значительно снижены; в фазе световой адаптации световой
подъем отсутствует, хотя в фазе темновой адаптации отмечалось
темновое падение. Отсутствие ЭРГ и значительные изменения ЭОГ
показывают важность участия в генерации ЭОГ нейроэпителия сет-
чатки наряду с пигментным эпителием.
При пигментном ретините заднего поля (инверсная форма), со-
провождающемся центральной или кольцевой парацентральной
скотомой, когда типичная пигментация отмечается в заднем полю-
се глаза, регистрируется патологическая ЭОГ: базовый потенциал и
коэффициент Ардена значительно снижены, темновое падение в фазе
темновой адаптации отсутствует в 30%.
При леберовском амаврозе до появления клинических призна-
ков заболевания у одних детей установлена патологическая ЭОГ, у
других — нормальная.
Дистрофия пигментного эпителия; макулярные дистрофии. При
дистрофии макулярной области типа Штаргардта и при ее сочета-
нии с желтопятнистым глазным дном (fundus flavimaculatus), кото-
рое встречается в более поздних стадиях заболевания, у 67—74%
пациентов отмечена патологическая ЭОГ (снижен коэффициент
Ардена до 120—166%). У этих же больных отмечалась патологичес-
кая ЛЭРГ и ЭРГ на красный свет уже в начальных стадиях забо-
левания. В то же время при офтальмоскопически видимых измене-
ниях, которые ограничивались макулярной областью, общая ЭРГ и
ЭОГ были в пределах нормы.
Изменения ЭОГ предшествуют функциональным и видимым
офтальмоскопически нарушениям и являются первым симптомом
патологии пигментного эпителия. A.Pinckers (1979) отметил пато-
логическую ЭОГ у большинства наблюдаемых больных с fundus
flavimaculatus, однако он не обнаружил четкой корреляционной
зависимости между изменениями ЭОГ и остротой зрения, наличием
скотом в поле зрения, темновой адаптацией [Babel J., 1977; Krill А.Е.,
1977].
При вителиформной дистрофии типа Беста в течение
многих лет сохраняются нормальные зрительные функции, несмотря
на значительные изменения в макулярной области, видимые офталь-
москопически. Тем не менее уже в ранних стадиях отмечается сни-
жение коэффициента Ардена( около 100%), что является патогно-
моничным электрофизиологическим признаком заболевания. Ба-
214
зовые значения потенциала, как правило, нормальные, но могут
быть и значительно сниженными. ЭОГ нередко извращена, т.е. на-
блюдается снижение амплитуды электроокулосигнала в интервале
яркого света до значений, меныпих, чем в темноте. У пациентов с
односторонней локализацией патологического процесса ЭОГ изме-
нена на обоих глазах.
Патологическую ЭОГ регистрируют у гетерозигот с нормаль-
ным глазным дном. Этот признак очень важен в тех случаях, когда
нет типичных проявлений макулярной дегенерации [Deutmann A.F.,
1971; Francois J., 1972]. Изменения ЭОГ связываются авторами с
генерализованными метаболическими нарушениями пигментного
эпителия (дисфункция энзимов). Нормальное отношение светового
подъема и темнового падения позволяют отличить вителиформную
макулярную дистрофию Беста от других изменений в макулярной
области, таких как дистрофия псевдо-Бест, макуло'дистрофия, свя-
занная с возрастом, и др.
Для бабочковидной пигментной дистрофии, при которой в па-
тологический процесс вовлекается пигментный эпителий, характер-
на патологическая, иногда плоская ЭОГ (снижено отношение све-
товой подъем/темновое падение), однако возможны и нормальные
ее значения при хорошо сохранившихся функциях центрального
зрения и нормальных показателях общей ЭРГ.
У больных с пигментной ретикулярной дистрофией заднего
полюса Съегрена изменений в ЭОГ не отмечено, хотя гистологичес-
ки показано, что процесс локализуется в пигментном эпителии;
этот факт пока не имеет объяснения.
При центральной серозной хороиретинопатии коэффициент
Ардена обычно не ниже 185%, однако по сравнению с его значени-
ем для здорового глаза этого же пациента разница составляет 20—
30%. В части случаев обнаруживают патологическую ЭОГ как ре-
зультат генерализованного процесса. После клинического выздо-
ровления асимметрия показателей ЭОГ не исчезает.
Макулярная дистрофия, связанная с возрастом. При доминант-
ных наследственных друзах мембраны Бруха некоторыми авторами
отмечено снижение соотношения светового подъема к темновому
падению (ниже 143%), другими — нормальные значения коэффици-
ента Ардена. При перераспределении пигмента на глазном дне,
наличии друз в мембране Бруха ЭОГ была патологической (коэф-
фициент Ардена ниже 143%) [Babel J., 1977; Gubta L.Y., 1994]. Воз-
можны различные показатели ЭОГ на двух глазах при сходной
офтальмоскопической картине заболевания. В любом случае элек-
трофизиологические изменения были более выраженными и рас-
пространенными, чем выявленные офтальмоскопически, а в сочета-
нии с перераспределением и миграцией пигмента они свидетель-
ствуют о распространенном поражении пигментного эпителия при
этом виде патологии.
215
OD -
1111111ПППпгт1тпП11ппнвор11П)П
Рис. 6.4. ЭОГ с изменениями в глазу вследствие увеита (больной М., 6 лет,
OD — последствия увеита, КА: OD 142%, OS — здоров, 218%).
Дисциформная дегенерация типа Кунта — Юниуса характери-
зуется значительными изменениями базового потенциала, коэффи-
циента Ардена, отсутствием светового подъема.
Дистрофия и дисфункция фоторецепторов. Палочково-колбоч-
ковая дистрофия характеризуется патологической ЭОГ и отсутстви-
ем ЭРГ в ранних стадиях заболевания.
Супернормальный или нормальный характер ЭОГ отмечен при
альбинизме, дискутируется вопрос о происхождении этих измене-
ний, которые могут быть объяснены либо большим количеством
света, проникающего в глаз в отсутствие пигментного экрана, либо
увеличением клеточного метаболизма под действием интенсивной
световой стимуляции.
Хороидальная дистрофия (хороидеремия, атрофия гирате, диф-
фузная атрофия хороидеи) сопровождается характерной патологи-
ческой ЭОГ (КА<100%) уже в ранних стадиях заболевания, что
свидетельствует о том, что патологический процесс более обширен,
чем это видно офтальмоскопически. При диффузной абиотрофии
хороидеи ЭОГ, как правило, значительно изменена (КА<100%).
При этом наблюдается и субнормальная ЭРГ.
При воспалительных заболеваниях ЭОГ изменяется по-разному.
При этом отмечаются отсутствие как светового подъема (рис.6.4),
так и темнового падения, снижение базового потенциала. Патоло-
гический коэффициент Ардена отмечен при отечной форме заболе-
вания, в то время как при рубцовой форме этот коэффициент был
нормальным. Локальные патологические процессы не изменяют
ЭОГ. При генерализованных воспалительных заболеваниях сосуди-
стого тракта изменения в ЭОГ определяются распространенностью
и клиническим течением болезни. При остром переднем увеите (ири-
доциклите) показатели ЭОГ, как правило, нормальны. При хрони-
ческих передних увеитах обнаруживалось снижение базового по-
тенциала, а также снижение светотемновых колебаний ЭОГ. При
216
задних увеитах (хороидитах) изменения ЭОГ зависят от локализа-
ции и распространенности патологического процесса. При хорои-
дитах электроокулография — более чувствительный метод в оценке
функции сетчатки, чем электроретинография, которая при этом виде
патологии долго остается нормальной.
Сосудистые заболевания сетчатки, В литературе отмечено быс-
трое изменение ЭОГ при окклюзии центральной вены и артерии
сетчатки или их ветвей. Электроокулографические данные при цир-
куляторных расстройствах в системе центральной артерии и цент-
ральной вены сетчатки неодинаковы: острое прекращение кровото-
ка в центральной артерии сетчатки существенно не влияет на вели-
чину базового потенциала, однако сравнение ЭОГ здорового и
больного глаза при односторонней окклюзии центральной артерии
сетчатки позволяет выявить асимметрию базового значения потен-
циала и коэффициента Ардена. При тромбозе отдельных ветвей
центральной вены сетчатки изменение показателей ЭОГ менее вы-
ражено. При закупорке центральной вены сетчатки ЭОГ обычно
патологическая.
Выраженный артериосклероз сосудов сетчатки вызывает сниже-
ние показателей ЭОГ до субнормальных значений.
При отслойке сетчатки снижение базового потенциала ЭОГ и
коэффициента Ардена зависит от площади отслойки и ее давности.
Отслойка, занимающая более Ч3 площади сетчатки, приводит к за-
туханию светотемновых колебаний в ЭОГ. Восстановление нормаль-
ных характеристик ЭОГ после оперативного вмешательства по по-
воду отслойки сетчатки, даже если операция была успешной, на-
блюдается редко.
При близорукости слабой степени ЭОГ нормальна, а при высо-
кой степени, сопровождающейся дегенеративными изменениями
сетчатки и хоройдеи, выявляется уменьшение светового подъема и
коэффициента Ардена, а также ухудшение временных показателей—
времени темнового спада.
Ценные диагностические данные получены у больных с трав-
мой глаз. При легких контузиях сетчатки, когда офтальмоскопичес-
ки выявляются лишь региональный отек и небольшие кровоизлия-
ния, снижается световой подъем ЭОГ. При тяжелых контузиях ЭОГ
уплощается и может принять извращенный характер. Поэтому при
проникающих ранениях глазного яблока ценность исследования
ЭОГ в оценке функции сетчатки и хороидеи при непрозрачности
глазных сред очевидна. Особенно эффективным оказалось приме-
нение исследования ЭОГ при химических ожогах глаз, при которых
химический фактор повреждает сосудистый тракт, наружные и внут-
ренние слои сетчатки [Винц И.М.и др., 1975].
Патологическая ЭОГ зарегистрирована при сидерозе, однако не
указывается, на каком этапе эволюции заболевания изменения ЭОГ
могут быть обнаружены. Некоторыми авторами изменения ЭОГ
отмечаются уже в начальных стадиях заболевания.
217
Функциональное состояние зрительно-нервного аппарата при
некоторых общих заболеваниях. Сахарный диабет и сопровождаю-
щие его патологические изменения сетчатки проявляются в сниже-
нии коэффициента Ардена не только при диабетической ретинопа-
тии в разной стадии, но и при нормальном глазном дне [Байтеря-
кова Л.С. и др., 1975].
При острой, хронической почечной недостаточности, сопровож-
дающейся нарушениями электролитного состава крови и уремиче-
ской интоксикацией, выявляется снижение коэффициента Ардена
до субнормальных значений [Архангельская Г. И. и др., 1975].
Большое количество работ посвящено прогностическому значе-
нию ЭОГ и ее роли в начальной диагностике, так как изменения
ЭОГ являются одним из ранних признаков побочного действия
лекарственных препаратов. Методом ЭОГ успешно выявляется ре-
тинотоксическое действие противотуберкулезных (этамбутола и
изониазида), синтетических антималярийных противовоспалитель-
ных и аналгезирующих (индометацин, метиндол) и противоревма-
тических, (хлорохин, делагил, резохин и плаквенил) препаратов, ди-
гиталиса, нейролептиков фенотиазинового ряда [Варновицкий А.М.
и др., 1975]. При побочных действиях лекарственных препаратов от-
мечается снижение базового потенциала, коэффициента Ардена и
темнового падения в фазе темновой адаптации. Изменения ЭОГ
опережают возникновение патологических изменений в пигментном
эпителии на периферии сетчатки, изменения ЭРГ и других функци-
ональных тестов. Начальные изменения ЭОГ являются первым сиг-
налом для отмены или замены препарата. Однако нормальная ЭОГ
регистрируется у 37% пациентов с ретинальной интоксикацией и
макулопатией типа ’’бычий глаз”, возникшей в результате лечения
хлороквином и гидрохлороквином; патологическая ЭОГ у 20%
больных системными заболеваниями, такими как ревматоидный
артрит, независимо от использования антималярийных лекарствен-
ных средств. Препараты накапливаются в пигментном эпителии,
подавляют протеиновый синтез, нарушают ионный обмен и изме-
няют структуру клеточных мембран, что приводит к снижению
коэффициента Ардена. Поэтому лечение этими препаратами долж-
но проводиться под контролем ЭОГ. Снижение коэффициента Ар-
дена на 10% от исходного (до лечения) требует снижения дозы пре-
парата, а более 20% — немедленной его отмены.
При злокачественной меланоме хороидеи в ЭОГ отмечается реду-
цированный световой пик и сниженный коэффициент Ардена, что
очевидно в связи с локализацией патологического процесса в пиг-
ментном эпителии сетчатки. Изменения в ЭОГ могут служить диф-
ференциальным симптомом между невусом и меланомой [Ponte F.,
Lauricella N., 1977; Jones R.M., 1981].
Как видно из приведенных данных, методом электроокулогра-
фии можно оценить функциональное состояние сетчатки, однако
218
диагностическая информативность электрофизиологического иссле-
дования значительно возрастает, если одновременно проводится и
электроретинография (табл. 6.4).
Таблица 6.4. Дифференциально-диагностическая таблица электрофизио-
логических симптомов заболеваний сетчатки и хороидеи
Патология ЭОГ Общая ЭРГ ЛЭРГ
Наследственные стационарные дефекты
Стационарная ночная слепота 4П 1 ±н
Болезнь Огуши 1 i 44 ± 4
Ахроматопсия Наследственные дистрофии 4Н 4Н 4
сетчатки
Пигментный ретинит п -4 ±4Н
Хороидеремия п 4 ± 4
Ретиношизис Фавре — Гольдмана п — - ±
Х-хромосомный ретиношизис п 4 4
Приобретенный ретиношизис н 4 Н Н
Наследственные макулярные заболевания сетчатки
Дистрофия типа Штаргардта 1 н 4 Н -4
Дистрофия типа Беста п Н ±Н
Дистрофия, связанная с возрастом Друзы мембраны Бруха н Н Н
Центральная хороиретинальная дистрофия 1 н 4Н -4Н
Дисциформная дистрофия типа Кунта — Юниуса 4 Н -4
Сосудистые заболевания сетчатки Тромбоз центральной вены сет- г п ± - 4 ± 4
чатки
Эмболия центральной артерии сет- 1 п ± - 4 -4
чатки
Диабетическая ретинопатия 1 п - 4 Т Н -4ТН
Воспалительные заболевания сетчатки и хороидеи Острый передний увеит н Н Н
Хронический передний увеит 1 4 4 Н
Генерализованный увеит п 4 Н 4Н
Нейроувеит н -НТ Н
219
Продолжение табл. 6.4.
Патология ЭОГ Общая ЭРГ ЛЭРГ
Хороиретинит диссеминированный 4 ±н н
Хороиретинит центральный 4Н н хн
Центральная серозная хороире- 4Н н ±н
тинопатия
Симпатическая офтальмия П 4Т X н
Другие заболевания
Отслойка сетчатки П -4 -X
Врожденная близорукость Н4 ±Н ±н
Дегенеративная миопия П 4 ± 4 X ±
Глаукома 4 Н Т н
Лекарственная ретинопатия 4 П -4 - X ±
Паранеопластическая меланома П 4- +
Атрофия зрительного нерва Н ±НТ ±н т
Амавроз Лебера 4 4- X-
Примечание. Н — норма, ± амплитуда вариабельна, Т — амплитуда супернор-
мальна, i — амплитуда выраженно субнормальна, — отсутствующая ЭРГ.
Таким образом, данные литературы [GliemH., 1971; MoriT.,
1991] и наш клинический опыт позволили обобщить результаты
исследования ЭОГ при вовлечении в патологический процесс пиг-
ментного эпителия сетчатки фоторецепторов и хороидеи. Очевид-
но, что процент патологической ЭОГ высок при рассматриваемой
патологии; в патологической ЭОГ базовый потенциал, отношение
светового подъема к темновому падению обычно снижены, хотя
при этом возможно изменение и одного из параметров ЭОГ. В еди-
ничных случаях отмечено повышение коэффициента Ардена, что
объяснялось изменением метаболизма в слое пигментного эпите-
лия, предшествующим деструктивным изменениям (например, при
макулярной дегенерации, связанной с возрастом). Выраженные из-
менения в ЭОГ появляются в случае распространенности патологи-
ческого процесса в пигментном эпителии, небольшие изменения в
макулярной области не влияют на параметры ЭОГ (даже в случае
значительного снижения зрения). Ангиографические исследования
при небольших изменениях сетчатки, видимых офтальмоскопичес-
ки, подтверждали данные патологической ЭОГ о распространенно-
сти изменений в пигментном эпителии на периферии сетчатки.
Однако нет корреляции между патологической ЭОГ и степенью дис-
семинации патологического процесса, выявляемого ангиографичес-
ки, что, например, наблюдается при вителиформной дистрофии типа
Беста и при друзах макулярной области, при которых характер
220
изменения пигментного эпителия и мембраны Бруха разный. Опи-
санная J.D.Zanen(1968) плоская ЭОГ как патогномоничный при-
знак вителиформной дистрофии типа Беста, отмеченная на самых
ранних ее стадиях и у носителей патологического гена, присуща
лишь одной из ее форм. ЭОГ играет ведущую роль в диагностике
отека макулярной области при воспалительных заболеваниях и
симптоме Ирвина — Гасса [Миронова Э.М., 1987]. Есть предполо-
жение [Deutman A.F., 1971; Krill А.Е., 1977], что патологическая ЭОГ
возникает лишь в случае отслойки пигментного эпителия. Это мне-
ние нуждается в морфологическом подтверждении.
Современная электроокулография позволяет получить объектив-
ные данные о ранних, доклинических признаках нарушения функ-
циональной активности сетчатки и пигментного эпителия, опреде-
лить тяжесть патологического процесса, что способствует вынесе-
нию научно обоснованного суждения о клиническом и трудовом
прогнозе.
Основные амплитудные, временные и относительные показате-
ли электроокулограммы изменяются по-разному при различно ло-
кализованных и отличающихся по генезу патологических состояни-
ях. Результаты электроокулографических исследований или их со-
поставление с данными других электрофизиологических и
клинических методов исследования позволяют решить следующие
диагностические задачи:
— подтвердить диагноз заболеваний, для которых изменения
ЭОГ являются патогномоничными;
— провести дифференциальную диагностику заболеваний с не-
ясной или сходной клинической картиной;
— дать оценку функционального состояния сетчатки при не-
прозрачных глазных средах (например, при катаракте) для прогно-
зирования зрительных функций после хирургического лечения;
— осуществлять динамический контроль за течением заболева-
ния и эффективностью проводимого лечения;
— как объективный метод исследования ЭОГ может быть ши-
роко использована в экспертизе трудоспособности, для выявления
профессиональной пригодности, в диагностике профпатологии, а
также для оценки функционального состояния органа зрения при
профотборе и при решении вопроса о воинской пригодности в слож-
ных для диагностики случаях.
ГЛ АВА7
ЗРИТЕЛЬНЫЕ ВЫЗВАННЫЕ КОРКОВЫЕ
ПОТЕНЦИАЛЫ
7.1. ПРОИСХОЖДЕНИЕ ЗРИТЕЛЬНЫХ ВЫЗВАННЫХ
КОРКОВЫХ ПОТЕНЦИАЛОВ
M.L.Ciganek в 1960 г. предложил использовать зрительные вызван-
ные корковые потенциалы (ЗВКП) на вспышку света в клинической
практике, a A.M.Halliday в 1976 г. применил контрастную стимуля-
цию на реверсивный паттерн. В дальнейшем эти методы начали
широко применять в клинике для диагностики заболеваний зри-
тельных путей, при патологии зрительного нерва, его отеке, воспа-
лении, атрофии, компрессионных повреждениях травматического и
опухолевого генеза, при локализации патологического процесса в
хиазме, зрительном тракте, коре головного мозга, рефракционных
изменениях, амблиопии, заболеваниях сетчатки.
Для интерпретации ЗВКП важна топографическая анатомия
зрительных путей (см.гл.1).
Механизмы и источники генерации ЗВКП.
В отличие от электроэнцефалограммы, отражающей активность
коры головного мозга, зрительные вызванные корковые потенциа-
лы (ЗВКП) представляют собой суммарный ответ больших популя-
ций нейронов коры на приходящий к ним синхронный поток
импульсов, возникающий под воздействием афферентного раздра-
жителя. Разность потенциалов, возникающая между вне- и внутри-
клеточной средой, регистририруемая на мембране нейрона, обус-
ловлена разностью концентрации ионов Na+, К+ и СГ в экстракле-
точной среде и в протоплазме нейрона. Этот потенциал называют
потенциалом покоя. При воздействии света сопротивление на мем-
бране изменяется, она деполяризуется или гиперполяризуется и тогда
возникает потенциал действия. Нейрон отвечает короткими импуль-
сами, с помощью которого аксоны нейронов передают информа-
цию на большие расстояния. Импульсы в миелиновых волокнах
перепрыгивают скачками от одного перехвата Ранвье к другому,
что значительно увеличивает скорость проведения возбуждения. Си-
наптическая передача, так же как и в сетчатке, в центральных от-
делах зрительной системы осуществляется с помощью нейротранс-
миттеров, под воздействием которых изменяется проницаемость
постсинаптической мембраны и в зависимости от типа нейротран-
смиттера возникает ее возбуждение или торможение. Различают выз-
ванные биопотенциалы ближнего и дальнего поля. Если ЗВКП от-
ражают реакцию генераторов, находящихся в коре, а электроды
расположены в непосредственной близости от них на скальпе, реги-
222
N
Рис. 7.1. Схематическое изображение ЗВКП (римскими цифрами обозначе-
ны волны первичного ответа).
стрируется биоэлектрическая активность ближнего поля, например
компонент Р100 ЗВКП. Слуховые потенциалы от ядра ствола мозга
являются примером биопотенциалов дальнего поля, когда электрод
располагается на значительном расстоянии от генераторов сигна-
лов.
ЗВКП отражают электрическую активность макулярной облас-
ти, что связано с ее большим представительством в шпорной бороз-
де, чем периферических отделов сетчатки — ’’кортикальный фак-
тор магнификации”, который может быть выражен линейно в мил-
лиметрах коркового пространства, соответствующего 1° зрительного
угла. Поэтому величина ЗВКП будет уменьшаться с увеличением
скотом в поле зрения. В связи с этим в методах исследования ЗВКП
предусмотрена возможность изменения величины шахматных квад-
ратов для стимуляции центральной и парацентральной областей
сетчатки. Например, для стимуляции фовеа оптимальной является
величина квадрата 10 — 15', а парафовеальная область стимулиру-
ется квадратом в 50' [Bodis-Wollner I. et al., 1986].
Величина ЗВКП значительно меньше (до 40 мкВ), чем волны
энцефалограммы (до 100 мкВ), поэтому для их выделения необхо-
дима техника усреднения. ЗВКП регистрируются в виде последова-
тельных колебаний, или компонентов (рис.7.1), различающихся
полярностью (позитивный — Р, негативный — N) и пиковой латен-
тностью — время от момента включения стимула до достижения
максимума того или иного колебания.
Несмотря на широкое использование ЗВКП в клинике [Leh-
mann D., 1984; Harding G.F.A., 1991], вопрос о механизмах их воз-
никновения и источниках генерации дискутируется в литерату-
ре. Для определения места происхождения электрической активно-
сти используют технику моделирующего диполя, с помощью
которой установлено, что первые три компонента могут генериро-
ваться одной и той же областью коры. Анализ распределения от-
дельных компонентов ЗВКП по скальпу, изменений их полярности
и амплитуды показал, что локализация источников ЗВКП на кон-
векситальной или медиальной поверхности затылочной доли, ради-
альная или тангенциальная ориентация этих источников зависят от
положения стимула в зрительном поле, его размера, пространствен-
юй частоты, эксцентриситета, а также от способа предъявления
223
стимула [Sokol S., 1980, 1981; CelesiaG., 1982; OnofryM., 1990;
Srebo R., 1990; Ossenblo K.P. et al., 1994].
Методом главных компонент [Ossenblo K.P. et al., 1994] показа-
но, что ЗВКП на вспышку, мелькания, выключение паттерна, его
реверсию и движение генерируется преимущественно в стриарной
коре. При этом подчеркивается близость источников и сходство
формы ответов на паттерн-реверсию и движение. По данным
Л.Р.Зенкова (1991), сверхранние компоненты ЗВКП относятся к до-
корковым источникам и связаны с активностью зрительного нерва
и тракта, подкорковых ядер (латеральное коленчатое тело) и тала-
мокортикальных путей. Представление о стриарном источнике
(поле 17) раннего компонента ЗВКП с латентным периодом 93 мс и
престриарном (поля 18 и 19) — более позднего компонента с латен-
тным периодом 130 мс было подтверждено при изучении топогра-
фии компонентов ЗВКП на включение паттерна, предъявляемого в
разных частях поля зрения, и использовании в анализе электричес-
ких полей скальпа уравнения Лапласа.
Ранние компоненты ЗВКП на стимуляцию паттерном верхнего
поля генерируются во вторичных зрительных зонах (поля 18 и 19), а
более поздние компоненты при стимуляции паттерном нижнего поля
имеют более распространенный источник в поле 17 и 18 по Бродма-
ну. Происхождение более поздних компонентов (Р200) и следующих
остается неясным, однако предполагается лобно-центральная лока-
лизация источников генерации, являющихся преимущественно не-
специфическими ответами таламуса и стволовых структур мозга. Эти
компоненты, чувствительные к состоянию сознания и уровню комы,
имеют меньшее значение в оценке зрительных нарушений.
Генерация ЗВКП связана также с общими механизмами синхро-
низации, определяющими спонтанную активность мозга, регистри-
руемую на ЭЭГ. Считается, что неспецифические образования ре-
тикулярной формации оказывают общее модулирующее воздействие
на ранние и поздние компоненты ЗВКП. Источником ЗВКП, реги-
стрируемых вне затылочной коры, являются независимые таламо-
кортикальные проекции, отличающиеся по своим характеристикам
и физиологическим механизмам [Зенков Л.Р., 1991].
Некоторые исследователи исходили из представления о том, что
отдельные компоненты ЗВКП являются отражением активности не
единичного диполя, а нескольких зрительных зон. Еще в 60-х годах
было установлено, что при отведении биопотенциалов от нейронов
стриарной коры (первичных зрительных центров) кошки многие из
нейронов не реагировали на диффузный засвет, который, как пред-
полагается, не ^является оптимальным для нейронов коры
[Kuffler S.W., Hubei D., 1960]. В связи с этим признано существова-
ние множества различных нейронов со специфическими функциями
обработки сигнала. Очевидно, что клетки коры с различной ориен-
тацией оказывают тормозящее влияние друг на друга.
224
О 100 200 мс О 100 200 мс о 100 200 мс
1 > L I . ,.L > i > .i > I, - I I I ,1,— 1 1 1
Рис. 7.2. ЗВКП на паттерн-реверсивный стимул при стимуляции у нор-
мальных субъектов.
а — всего поля зрения; б — назальной части поля зрения; в — темпоральной части
поля зрения [Brecelj J., 1993]. N, Р — анализируемые компоненты ЗВКП. 1—5 —
схема локализации электродов в затылочной области и соответствующая им кри-
вая ЗВКП.
Одновременная послойная регистрация нейронной активности
различных отделов мозга, проведенная у обезьяны при вспышеч-
ной стимуляции и на реверсивный шахматный паттерн, анализ плот-
ности источников тока в стриарной коре, сопоставление компонен-
тов ЗВКП обезьяны и человека позволили предположить, что на-
чальный компонент имеет субкортикальный источник, a N70
возникает в IV слое со звездчатыми клетками, однако не исключа-
ется вклад в генерацию этого компонента активности таламокорти-
кальных аксонов. Р100, возможно, является суммарным отражением
постсинаптической активности в сети пирамидных нейронов III слоя
и тормозных процессов в звездчатых нейронах в IV слое. Несмотря
на внешнее сходство компонентов ЗВКП на вспышку и реверсию
паттерна, ЗВКП на паттерн-реверсивный стимул вызывает более
выраженную синаптическую активность нейронов II и III слоев,
которая отражается в Р100. Более поздние компоненты N145 и Р200
происходят из внестриарного источника. Однако высказывается
мнение о том, что ЗВКП, регистрируемые от скальпа в проекции
затылочной коры, отражают активность, имеющую относительную
связь с прилежащими структурами.
При стимуляции всей сетчатки паттерн-стимулом в 16° над пра-
вой и левой гемисферами регистрируется потенциал, состоящий из
компонента N70, Р100 и N145. При стимуляции половины поля зрения
(рис.7.2) над ипсилатеральной гемисферой регистрируются волны
N7o, P1oo, N145, а над контралатеральной — волны Р75, N105, Р1зг Не-
смотря на стимуляцию перекрещенных и неперекрещенных воло-
225
Рис. 7.3. Фазический, прехо-
дящий (transient) паттерн
ЗВКП.
а — при стимуляции централь-
ного поля сетчатки 14° (пат-
терн-элементы размером 12' с
частртой 2 с); б — при экрани-
ровании центрального поля
зрения в 3° (снижение амплиту-
ды и увеличение латентности);
в — при экранировании цент-
рального цоля зрения в 9°.
кон, активный генератор виден в каждой гемисфере [Blumhardt L.,
Halliday А.М., 1979,1993]. Предполагается, что регистрация компо-
нентов N70, Р100 и N143 с ипсилатеральной поверхности скальпа пр
отношению к стимулируемой половине поля зрения, представляю-
щей противоположную сторону активированной зрительной норы,
объясняется так называемой парадоксальной литерализацией, явля-
ющейся результатом заднемедиальной ориентации нейронов-гене-
раторов-
Подтверждением того, что ЗВКП отражают зрительную актив-
ность в основном центрального поля зрения, которое имеет боль-
шее представительство в коре головного мозга, чем периферичес-
кие области (рис. 7.3), является окклюзия центральной части поля
зрения как модель наличия центральной скотомы в поле зрения.
При этом у здоровых субъектов после окклюзии центральной зоны
сетчатки редуцируются ИДИ исчезают макулярные компоненты N70,
Р)00, N143, при этом в ипсидатеральной гемисфере волна Р100 приоб-
ретает W-подобный тип, а над контралатеральной гемисферой
регистрируются парамакулярные компоненты Nl03, Р133. В этой мо-
дели не исключается и сложение некоторых компонентов, происхо-
дящих от проекции макулярной области (N^, Pf00, N143), с компонен-
тами (Р73, N103, Р|35), генерируемыми в проекции неэкрацированной
парамакулярной области.
ЗВКП на паттерн-стимул генерируются в проекционной зоне
макулярной области, чувствительной к изменениям яркости, гра-
ниц контуров, контраста и пространственной частоты элементов
шахматного поля, а также в экстрастриарных областях. Макуляр-
ный пучок зрительного нерва и тракта и соответствующая ему груп-
па таламокортикальных волокон относятся к быстропроводящим
миелинизированным аксонам, поэтому ЗВКП может быть индика-
226
тором поражения зрительных нервов и показателем их демиелини-
зации.
Аналогичная (скотоматозная) форма ЗВКП наблюдается при
стимуляция всего поля и половины поля зрения у больных, в анам-
незе которых имелся оптический неврит. В ЗВКП отсутствует маку-
лярный Р.ю-компонент, регистрируется W-подобная волна, обуслов-
ленная объединением Р100- с Р135-компонентом, в то время как пара-
макулярные N103- и Р135-компоненты могут быть увеличены при
отведении потенциала от контралатеральной гемисферы.
С помощью анализа потенциальных полей и трехмерной лока-
лизации источников этих полей можно получить объяснение неко-
торым встречаемым в ЗВКП изменениям. Например, парадоксаль-
ное увеличение амплитуды компонента Р|Юна электродах, находя-
щихся ипсилатерально или ПО средней линии по отношению к
стимулируемому полуполю зрения, парадоксальная латералцзация
источников этого компонента возникают из-за того, что ответ дей-
ствительно может генерироваться на контралатеральной стороне
соответственно анатомическим представлениям, но источники ори-
ентированы в ипсилатеральном направлении.
Согласно детекторной гипотезе, по которой нейроны зритель-
ной коры избирательно реагируют на различные элементы контура
изображения: линии, полосы разной ширины и ориентации, кон-
трастные границы между темным и светлым полями, углы И др-,
оптимальными стимулами для исследования зрительной системы
являются структурированные стимулы с прямоугольным распреде-
лением освещенности [Jeffreys D., 1977; Spefcreije Н., 1986]. В связи с
ЭТИМ в клинико-физиологических исследованиях в качестве стиму-
лов обычно используют диффузные вспышки света и пространствен-
но-структурированные стимулы в виде шахматных паттернов и
решеток с прямоугольным профилем освещенности. Решетки с си-
нусоидальным распределением освещенности, которые редко при-
меняют в клинических исследованиях, используют при изучении
центральных механизмов переработки зрительной информации- При
этом авторы основываются на представлении о существовании в
зрительной системе независимых пространственно-частотцых кана-
лов и возможности с помощью синусоидальных стимулов, каждый
из которых содержит одну пространственную частоту, изолирован-
но активировать эти каналы [Глезер В.Д., 1985; Хьюбел Д-, 1990].
Сопоставление топографии электрических потенциалов и маг-
нитных полей при изолированной стимуляции половины поля зре-
ния шахматными паттернами показало, что при комбинации этих
методов информация, полученная с помощью одного из них, до-
полняет данные, установленные При использовании другого. При
картировании электрических полей лучше выражены радиально ори-
ентированные источники, при картировании магнитных полей —
тангенциальные [Bodis Wollner I., 1986,1990; Branam M.S. et al., 1994].
227
7.2. ВИДЫ ЗРИТЕЛЬНЫХ ВЫЗВАННЫХ КОРКОВЫХ
ПОТЕНЦИАЛОВ
Состав компонентов и последовательность ЗВКП устойчивы. Вре-
менные характеристики и амплитуда значительно варьируют в за-
висимости от условий исследования, локализации электродов, спо-
соба предъявления и характера стимула.
Виды ЗВКП зависят от характера стимула. ЗВКП на вспышку
света называется вспышечным ЗВКП. Он может регистрироваться
при разных интенсивностях и длинах волн стимулирующего света.
ЗВКП на паттерн-стимул называют паттерн-ЗВКП. При регистра-
ции этой формы ЗВКП стимулы предъявляют либо в режиме вклю-
чения — выключения, когда средняя освещенность паттерна и сме-
няющего его гомогенного поля постоянна, либо в режиме реверсии,
когда в постоянно присутствующем на экране монитора изображе-
нии шахматного поля белые квадраты сменяются черными, а чер-
ные — белыми.
Установлено, что оптимальный размер квадрата для паттерн-
стимуляции макулярной области должен составлять 10—15', а для
парафовеальной области — 50' [Bodis-Wollner I., Kubova K.Z., 1992].
При частоте реверсии от 1 до 4 раз в 1 с регистрируется фазический,
transient-ЗВКП (рис.7.4), в котором идентифицируются последова-
тельные компоненты N70, Р100 и N150; в случае увеличения частоты
реверсии — более 4 раз в 1 с (6—14 реверсий в 1 с) из фоновой ЭЭГ
можно выделить суммарный ритмический ответ в виде синусоидаль-
ной кривой, так называемый ЗВКП устойчивого состояния steady-
state (см. рис.7.4), отличающийся от фазического ЗВКП отсутстви-
ем последовательных компонентов [Kremmer S. et al., 1992]. Таким
образом, различия между этими видами ЗВКП состоят в том, что
transient-ЗВКП вызываются редкими стимулами, предъявляемыми с
интервалом, достаточным для восстановления постоянной актив-
ности мозга, а steady-state-ЗВКП — быстро повторяющимися сти-
мулами, при этом электрическая активность мозга не успевает вер-
нуться в исходное состояние. Адекватным методом анализа фази-
ческого ЗВКП является анализ Фурье, с помощью которого
выделяют гармонические составляющие, различающиеся функцио-
нальными свойствами и распределением по коре [Regan В., 1989].
Параметры ЗВКП. ЗВКП на диффузную вспышку света
большой интенсивности состоит из ранней позитивной волны не-
большой амплитуды — Р40, за которой следуют негативный N70,
доминирующий в комплексе волн Р100, и поздние волны N130 и Р170
[Иваницкий А.М., 1976]. Временные характеристики компонентов
фазического transient-ЗВКП на включение шахматного паттерна,
по данным разных авторов, значительно варьируют: — от 36 до
60 мс, Р100 — от 65 до 110 мс, N150 — от 90 до 160 мс, Р200 — от 160
до 200 мс [Зислина Н.Н., 1987; Бетелева Т.Г., 1990] в зависимости от
228
Рис.7.4. Паттерн-реверсивный
ЗВКП на паттерн с величиной
элемента в 12’ (изменение от
transient-ЗВКП до stady-state-
ЗВКП) при различной частоте
предъявления стимула.
а — 2 Гц; б — 4 Гц; в — 6 Гц; г —
8 Гц; д—10 Гц; е—12 Гц; ж —
14 Гц. N и Р — компоненты ЗВКП.
размеров стимульного поля и составляющих его шахматных клеток
[Фильчикова Л.И., 1989; Шпак А.А., 1993; Jonson, 1987].
В фазическом ЗВКП на низкочастотную реверсию шахматного
паттерна наиболее постоянно регистрируется доминирующий в
комплексе волн позитивный компонент Р100, время латентного пе-
риода которого находится в диапазоне от 95 до 120 мс; у предше-
ствующего ему компонента N70 время латентности составляет от 60
до 80 мс, а компонента N150— от 150 до 200 мс. Менее постоянно
регистрируется поздний позитивный компонент Р200 [Celesia G., 1985].
7.3. МЕТОДЫ РЕГИСТРАЦИИ ЗРИТЕЛЬНЫХ ВЫЗВАННЫХ
КОРКОВЫХ ПОТЕНЦИАЛОВ
Для регистрации ЗВКП применяют стандартные специализирован-
ные системы на основе современных компьютеров, которые позво-
ляют выделять потенциалы малой величины из шума, использовать
усреднение, нивелировать различные артефакты.
При регистрации ЗВКП активный электрод (чашечки из золота,
229
F—фронтальное; О —окципитальное; Р — париетальное; Т — темпоральное; Fz,
Pz, Oz— центральное расположение электродов во фронтальной, париетальной и
затылочной областях соответственно; а, b — активные электроды; с — индиффе-
рентный электрод.
серебра и др.) помещают на 1,5—2 см выше затылочного бугра по
средней линии над областью проекции зрительной зоны (поле 17),
часть которой, выходящая на конвекситальную поверхность полу-
шария, располагается у затылочного полюса. Для лучшего контак-
та используют абразивную пасту. В случае необходимости зареги-
стрировать ответы вторичных зрительных зон (поля 18 и 19) элек-
троды смещают на 2 см вправо и влево от средней линии.
Индифферентный электрод помещают на мочке уха или сосцевид-
ном отростке, заземляющий электрод — на фронтальной части
скальпа (рис.7.5). Поскольку амплитуда ЗВКП составляет всего не-
сколько микровольт, выделение его из фоновой электроэнцефало-
граммы (ЭЭГ) осуществляют с помощью метода многократной сум-
мации ЗВКП, синхронизированных со стимулом (метод усреднения).
Количество накоплений может варьировать от 25 до 100 и больше,
что определяется условиями регистрации, характером стимулов,
задачами исследования. Размер стимулируемого поля около 15°.
Использую! две величины квадратов реверсивного шахматного
Ноля — 12 — 20 мин (14 мм) и 48—60 мин (35 мм) с расстояния 1 м.
Предполагается, что использование квадратов малой и большой
Величины позволяет исследовать фовеальную и парафовеальную
области сетчатки. Зрачок не расширяют, нормативы латентных
периодов должны быть получены в возрастных группах.
Нормативы латентности Р100, определенные в стандартных усло-
виях регистрации, представлены в табл. 7.1. Амплитуда ЗВКП очень
вариабельна, поэтому, не представляя клинического интереса, ред-
ко используется в анализе (в таблице не приведена).
Анализ ЗВКП начинают с идентификации компонентов N75,
наиболее Вариабельного и наиболее устойчивого Р100. При анализе
ЗВКП учитываются амплитуда в микровольтах от изолинии или от
230
Таблица 7.1. Нормальные значения ЗВКП [LKC, 1996]
ПАТТЕРН-ЗВКП
Размер стимулирующего поля, мм Размер шахматного поля, дуговые мин Латентность Р100, мс
4 х4 6 119
8 х8 12 ПО
16 х 16 25 109
32 х 32 50 118
64 х 64 100 123
128 х 128 199 141
ЗВКП НА ВСПЫШКУ, ЛАТЕНТНОСТЬ Р100
Волна Нижняя граница Верхняя граница
36 60
Р* 50 90
N„ 70 120
Р.м 95 155
115 200
Примечание. Амплитуда ЗВКП вариабельна, поэтому редко используется в
клинике.
предыдущего пика противоположной полярности (“от пика до
пика”) и время до пиков волн в миллисекундах. Учитывается раз-
ность величины латентности при стимуляции правого и левого гла-
за — ’’межокулярная разность латентностей” и амплитудная асим-
метрия — “межокулярные амплитудные соотношения”. Межполу-
шарная асимметрия оценивается по отношению максимальной
амплитуды Р1в0 к ее минимальному значению (Р^ к Pmin); в норме
это соотношение меньше 2,5. Латентность считается патологичес-
кой, если она отличается от нормы на 2,5 стандартных отклонения,
абсолютные значения латентности используются при интерокуляр-
ном сравнении. Важна и оценка формы ЗВКП, при нормальной
амплитуде и латентности изменение формы компонентов ЗВКП
может указывать на необходимость дополнительного обследования
Или динамического наблюдения. W-подобная форма пика Р100 со-
здает определенные трудности для интерпретации ЗВКП. При этом
могут рассматриваться два основных варианта: наличие централь-
ной скотомы в поле зрения или частичной атрофии зрительного
йерва. В клинике чаще всего измеряют первый позитивный компо-
нент Рр который в норме составляет около 100 мс, латентность его
колеблется в пределах 2 — 5%, в то время как амплитуда варьирует
в 25%. Как правило, в связи со значительной вариабельностью ЗВКП
231
на вспышку, его амплитуду не рекомендуется учитывать в повсед-
невной клинической практике. Однако, хотя в клинике большее
значение имеют временные параметры при сопоставлении их с воз-
растными нормативами, в ряде случаев амплитуду также можно
принимать во внимание при ее межокулярном сопоставлении, а
также если потенциал следует за увеличением стимула, так как это
предполагает сохранность по крайней мере восприятия света.
На латентность ЗВКП влияет ряд факторов, в том числе диа-
метр зрачка, возраст обследуемого, изменение рефракции. Умень-
шение диаметра зрачка ведет к снижению освещенности сетчатки,
что в свою очередь определяет удлинение латентного периода. Он
уменьшается до 20 мс, если диаметр зрачка увеличивается от 1 до
9 мм, и достаточно постоянен при диаметре зрачка 4 мм. При пат-
терн-стимуляции большими и маленькими квадратами латентность
ЗВКП экспоненциально уменьшалась у обследуемых в возрасте от
2 мес до 40 лет, а затем медленно увеличивалась до 80 лет. Среднее
время латентности для маленьких квадратов больше, чем для боль-
ших, что объясняется различием их освещенности и контраста
[Spekreijse Н., 1981]. При эмметропической рефракции отмечена
более низкая латентность ЗВКП, чем при миопической и гипермет-
ропической.
При проведении исследования у маленьких детей необходимо
учитывать быстроту изменения амплитудных и временных показа-
телей ЗВКП в первые 6 мес жизни, поэтому у них предпочтительнее
регистрировать transiente-, а не steady-state-ЗВКП. Исследование
следует проводить в темной комнате для лучшей фиксации груд-
ным ребенком паттерн-стимула. Усреднение ЗВКП нужно контро-
лировать в связи с изменениями фиксации при движении головы
ребенка, а артефакты, связанные с этими движениями, должны уда-
ляться специальными алгоритмами. У малоконтактных детей ис-
следование можно проводить с использованием седативных средств
(например, хлоралгидрата). Оптимальный размер квадратов в шах-
матном поле при исследовании ЗВКП варьирует в течение первых
6 мес жизни ребенка: в возрасте 1 мес — 120*—240', 2 мес — 60' —
120', 3—5 мес — 30—60', 6 мес и старше — 15'. Если при неисполь-
зовании шахматного квадрата, размер которого соответствует груд-
ному возрасту, ответа не получено, то величину шахматного квад-
рата увеличивают в 2 раза.
При бинокулярном тестировании особенно важно учитывать
возраст пациента и выбирать больший или меньший квадрат шах-
матного поля в зависимости от величины регистрируемых ЗВКП.
Использование квадратов шахматного паттерна различной ве-
личины способствует повышению чувствительности метода в опре-
делении функциональных нарушений зрения. Маленькие квадраты
(10 — 30') используют для выявления увеличивающихся дефектов в
макулярной области, большие квадраты более адекватны для ис-
232
Рис. 7.6. Различия ЗВКП на паттерн-стимул по амплитуде и латентному
периоду при использовании паттерн-элементов различной величины у ре-
бенка 7 лет с амблиопией [Brecelj J., 1993].
а — 49’; б — 24’; в— 12*. Наиболее значительные отличия в ответе на паттерн-эле-
менты в 12'.
следования периферии. Вопрос о физиологических механизмах чув-
ствительности ЗВКП к паттернам различной величины, их связи с
организацией рецептивных полей ганглиозных клеток сетчатки и
на кортикальном уровне дискутируется в литературе. Известно, что
размеры рецептивных полей ганглиозных клеток фовеа меньше в
сравнении с парафовеальными; аналогичные различия в рецептив-
ных полях наблюдаются в первичной зрительной коре. Предпола-
гается, что изменение размера паттерна позволяет активизировать
различные классы нейронов. От величины паттерна зависят ампли-
туда и время латентности компонентов ЗВКП, поэтому при различ-
ных заболеваниях рекомендуется использовать паттерны разных
размеров для выявления патологического состояния в зрительной
системе. Примером могут служить паттерн-ЗВКП при амблиопии
(рис.7.6), выраженные изменения которых отмечены при использо-
вании паттернов малой величины.
Часто в одной серии исследований используют квадраты раз-
ной величины [Brecelj J., 1993], например 7,5', 15', 30', 60', 120'.
У детей исследование начинают с применения шахматного квадра-
та размером 60'. Если сигнал получен, то размер квадрата умень-
шают (30'), в случае отсутствия сигнала величину квадрата увели-
чивают (120'). Если ЗВКП не регистрируется ни при одной ве-
личине квадрата, используют различную частоту светового
раздражителя— 1, 2, 6, 10 и 20 вспышек света в 1 с. Перед моноку-
лярным тестированием пациентов старше 6 мес нужно проводить
бинокулярное тестирование, начиная с применения квадрата разме-
ром 15'. В случае необходимости величину квадрата шахматного
233
поля увеличивают до регистрации видимого сигнала. После полу-
чения бинокулярного сигнала монокулярное исследование начина-
ют с лучше видящего глаза. При этом особое значение приобретает
различие амплитуды ЗВКП, зарегистрированных на обоих глазах,
на одну и ту же величину шахматного квадрата.
Таким образом, в паттерн-ЗВКП определенную ценность имеют
три параметра: происхождение волн, время латентности компонен-
тов И их амплитуда. Время латентности ЗВКП — олень чувстви-
тельный тест в определении субклинических поражений зритель-
ных путей у взрослых, а также при наблюдении за развитием зри-
тельной системы и выявлении ее патологических процессов у детей.
Амплитуда паттерн-ЗВКП также может быть важна в анализе при
Проведении исследований у детей, так как имеется определенная
корреляция ее величины с остротой зрения, особенно для квадра-
тов малой величины. Абсолютные же амплитуды в отличие от ла-
тентности учесть трудно в связи с их различиями у разных индиви-
дуумов. Так или иначе, ЗВКП дают квантитативную информацию
о функции зрительной системы, которую нельзя Получить с помо-
щью других клинических методов исследования.
Рекомендации ISCEV (1996). Так же как в клиничес-
кой электроретинографии, для унифицирования метода и методоло-
гии регистрации ЗВКП и обеспечения возможности сравнения ре-
зультатов исследования, полученных в разных офтальмологических
диагностических лабораториях и клиниках, Международное обще-
ство клинических электрофизиолйгов предлагает рекомендации по
регистрации и анализу ЗВКП в клинике глазных болезней. Для реги-
страции ЗВКП необходимо использовать стимулы трех типов: вспыш-
ку, реверсивный стимул и on/set- и off/set-паттерн. Вспышечный сти-
мул с угловыми размерами 20° и максимальной длительностью 5 мс
должен давать диффузную вспышку 3 кд/м2. Как альтернативная мо-
жет быть использована вспышка для стандартной ЭРГ.
Паттерн-реверсивный стимул состоит из черно-белых квадра-
тов или полос, которые постоянно меняются местами, так что осве-
щенность экрана не изменяется. Величина стимула определяется зри-
тельным углом для каждого квадрата (1° и 15’) или пространствен-
ной частотой для полос (1 и 4 цикл/градус).
Паттерн-on/set И паттерн-off/set для регистрации вызванных
биопотенциалов представляют собой полосы на фоне с равномер-
ной освещенностью, которая Должна оставаться постоянной при
появлении И исчезновении паттерна. Паттерн-ofl/set- и паттерн-off/
set-стимулы имеют те же параметры, что и паттерн-реверсивные
стимулы, за исключением дополнительной равномерной освещен-
ности экрана, предъявляемого в течение 400 мс между паттерн-
стИМуляцией, длительностью 200 мс.
Клинический протокол. Регистрацию ЗВКП проводят без изме-
нения величины зрачка мидриатиками или миотиками при опти-
234
мальной рефракционной коррекции для сохранения единого рас-
стояния до стимула. Необычная величина зрачка должна отмечать-
ся в протоколе. Исследование ЗВКП при паттерн-стимуляции реко-
мендуется проводить монокулярно, когда пациент находится в рас-
слабленном состоянии для минимизации мышечных и других
артефактов. Изучение ЗВКП на вспышку позволяет получить ори-
ентировочную информацию о проведении возбуждения, сохранно-
сти светоощущения и какого-либо форменного зрения у пациентов,
у которых трудно выполнить исследование, при помутнении опти-
ческих сред, изменяющем четкость изображения на сетчатке.
В случае использования для стимуляции паттерн-on/set и пат-
терн-off/set отмечаются значительные межиндивидуальные разли-
чия! Однако on/set-стимул более эффективен при определении ост-
роты зрения и объективной рефракции.
При определении прехиазмальных нарушений наиболее инфор-
мативна монокулярная стимуляция реверсивным паттерном
(transient-паттерн) при локализации активных электродов на О,
O4h°3.
Хиазмальные и постхиазмальные нарушения определяются при
регистрации ЗВКП с обеих гемисфер мозга, так что активные элек-
троды должны располагаться на Oz, О4 и О3 с общим референтным
электродом Fz. Для этих целей может быть использован как ревер-
сиёный паттерн, так й on/set- и off/set-паттерн. Стимуляция полови-
ны поля зрения в 15° должна быть использована при исследовании
одной гемисферы.
Диагностические центры должны иметь свои нормативы для
временных и амплитудных параметров каждого компонента ЗВКП
и для межокулярных различий этих компонентов. Для выработки
нормативов рекомендовано использовать описательную статисти-
ку, которая не принимает во внимание нормальное распределение,
а основывается на расчете средних значений и процента распреде-
ления (вариабельности ряда).
В ЗВКП, зарегистрированных в стандартных условиях, рекомен-
довано учитывать две основные волны и их полярность (к90 и Р12о).
Амплитуду transient-ЗВКП измеряют от пика до пика, а латен-
тность до пика (implicit time) — время от начала стимуляции до
пика волны.
Протокол должен содержать нормальные значения измеряемых
параметров, границы нормы с отметкой о проведении исследова-
йия в стандартных условиях, а также описание трех стандартных
ответов.
ЗВКП на вспышечный стимул, регистрируемые на стандартном
оборудовании, содержат комплекс негативных и позитивных волн,
регистрируемых в диапазоне от 30 до 300 мс. При вспышечных ЗВКП
наиболее часто принимают во внимание следующие компоненты:
Негативный — N2 (на 90 мс) и позитивный — Р2 (на 120 мс). Латен-
235
тность вспышечных ЗВКП зависит от возраста пациента. Ранний
позитивный компонент около 50 мс регистрируется наиболее часто
у пожилых пациентов.
В ЗВКП, регистрируемых на паттерн-реверсивный стимул, ана-
лизируют негативные N75-, N135- и позитивный Р1оо- компоненты
(обозначения пиков негативный и позитивный соответствуют зна-
чениям латентности), а при стимуляции паттерн-on/set и паттерн-
off/set— компоненты Cj (позитивные около 75 мс), С2 (негативный
около 125 мс), С3 (позитивный около 150 мс).
7.4. КАРТИРОВАНИЕ ЗРИТЕЛЬНЫХ ВЫЗВАННЫХ
КОРКОВЫХ ПОТЕНЦИАЛОВ
Новым этапом в развитии клинической электрофизиологии зрения
является использование топографического картирования ЗВКП для
оценки уровня нарушения передачи и обработки информации в
зрительной системе, а также в диагностике патологических процес-
сов различного происхождения и локализации.
Топографическое картирование биоэлектрической активности
мозга появилось почти 20 лет назад, однако в клинике этот метод
не использовали, что было связано с определенными техническими
трудностями. В последнее десятилетие после появления быстродей-
ствующих персональных компьютеров количественные методы ана-
лиза биоэлектрической активности мозга (ЭЭГ и ЗВКП) начали
широко применять в клинике [Иваницкий А.И., 1976; Зенков Л.Р.,
1991; Ньюер М.Р., 1992; Шпак А.А., 1993). Если ранее считали, что
ЗВКП происходят из стриарной коры, и пытались выявить их ис-
точники и локализацию в различных структурах мозга, не имея
представления о сложной топографии зрительного анализатора, то
к настоящему времени очевиден сложный путь, который проходит
зрительный сигнал от сетчатки к центральным отделам зрительной
системы в процессе его обработки (см. гл.1).
С помощью топографического картирования стало возможным
проследить весь путь последовательного возбуждения анатомиче-
ских структур, принимающих участие в обработке информации во
временном диапазоне, так как топографическое картирование пре-
дусматривает регистрацию ЗВКП на уровне не только затылочной,
но также теменной, височной и лобной областей. Топографические
карты, кодируемые цветом (вверх — негативная, вниз — позитив-
ная полярность), отражают амплитуду ЗВКП и область распрост-
ранения биоэлектрической активности в разные промежутки време-
ни от момента начала стимуляции либо вспышкой света, либо ре-
версивным паттерном. Особенно заметны различия паттерн- и
вспышечных ЗВКП при анализе топографии обоих ответов. Более
широкая топография ЗВКП на вспышечный стимул и максималь-
236
ная биоэлектрическая активность проявляются в центральных об-
ластях. На шахматный паттерн максимум ответа приходится на за-
тылочно-теменную область, особенно для ранних компонентов. При
обоих видах стимуляции по-разному проявляется соотношение спе-
цифических и неспецифических ответов. Для ЗВКП на реверсивный
шахматный паттерн характерна более четкая выраженность специ-
фической афферентации. Более широкая топография с большей
выраженностью неспецифических ответов представлена на вспышеч-
ный паттерн.
Топографическое картирование позволяет проводить простран-
ственный анализ и прямое сравнение ЗВКП, регистрируемых в раз-
ных отведениях. Сопоставление с нормативами является критерием
в оценке топографического картирования ЗВКП при патологиче-
ских процессах в зрительной системе.
Методология исследования. Для картирования
ЗВКП используют программу для компьютера IBM PC [Пирожен-
ко А.В., 1995], которая позволяет выделять из шума и анализиро-
вать ЗВКП, зарегистрированные при монополярном отведении от
10 электродов и больше, располагаемых по международной си-
стеме “10—20” [Jasper Н., 1957] на поверхности скальпа в проек-
ции затылочной, височной, теменной и лобной долей на правом и
левом полушарии коры головного мозга; координата электродов
по оси X — от левого уха к правому, по оси Y — от лба к затылку
[Зенков Л.Р., Ронкин М.А. 1991]. Оценивают амплитудные и вре-
менные параметры компонентов ЗВКП, анализируют частотный
спектр по картам распределения мощности частот в определенном
спектральном диапазоне, характерном для ЗВКП (по аналогии с
ЭЭГ: 1—3, 4—7, 8—13, 14—24), в зависимости от топографии био-
электрической активности. Возможны анализ межполушарной асим-
метрии биоэлектрической активности мозга, измерение когерент-
ности, фазовых различий для каждой частоты и диапазона частот
в различных отведениях по скальпу. При стимуляции правого и
левого глаза наличие Отчетливой асимметрии на одной стороне
дает основание предположить хиазмальную или постхиазмальную
локализацию патологического процесса, амплитудная асимметрия,
выявляемая при стимуляции одного глаза, указывает на прехиаз-
мальный уровень поражения. Наличие инверсии Р100-компонента в
лобных отведениях является критерием правильного его опреде-
ления. Эти данные представляют с помощью топографических карт,
на которых цветом закодированы анализируемые параметры
ЗВКП.
Регистрацию и обработку сигнала осуществляют с помощью
специализированной системы, например “МБН-нейрокартограф”
(фирма “МБН”, Россия) для картирования ЗВКП, включающего уси-
литель, стимулятор, аналогово-цифровой преобразователь и ком-
пьютер. Система предусматривает цифровую фильтрацию записи,
237
удаление артефактов, выбор участков записи произвольной дли-
тельности для последующей обработки, получение цифровых дан-
ных по каждому из каналов для любого момента времени относи-
тельно нулевой линии, помеченного курсором.
Картирование амплитудных значений ЗВКП проводят по
топографическим картам, представляющим собой схематическое
изображение поверхности головы, на котором разными цветами обо-
значены различные значения потенциала, определяемые по изобра-
женной на схеме цветовой шкале (каждому цвету карты соответ-
ствует определенная величина биопотенциала).
Локализация источников биоэлектричес-
кой активности мозга. Новым шагом в развитии топо-
графического картирования ЗВКП является локализация источни-
ков биоэлектрической активности мозга. Очаги патологической ак-
тивности можно установить путем трехмерной локализации
источников биоэлектрической активности мозга методом решения
обратной задачи по потенциалам, регистрируемым с поверхности
головы. Для этцх целей служит компьютерная программа
“Brainlock” (фирма ’’МБН”), предназначенная для определения и
отслеживания в объеме мозга человека центров электрического воз-
буждения, представленных в виде одного или двух эквивалентных
дипольных источников, в качестве входной информации использу-
ют одномоментные значения ЗВКП (не менее 8 отведений для опре-
деления однодипольной модели и це менее 16 отведений для двух-
дицольной). Система отведений может быть произвольной.
Поиск минимума суммы квадратов отклонений осуществляют с
помощью комбинированных алгоритмов минимизации с ограниче-
ниями. При этом область изменения пространственных координат
дипольных источников ограничивается поверхностью эллипсоида,
аппроксимирующего поверхность мозга. Для пересчета потенциа-
лов ЗВКП с поверхности головы на поверхность коры полушарий
большого мозга используют неоднородно проводящую восьмислой-
ную модель головы- кроме того, для построения потенциальных
карт применяют алгоритмы сферического анализа, аналитическо-
го продолжения потенциала и линейной интерполяции [Копте-
лов Ю.М-, 1988; Коптелов Ю.М., Гнездицкий В-В-, 1989; Щамши-
нова А.М., Коптелов Ю.М., 1998].
Программа “Brainlock” может быть использована как в клини-
ке для определения локализации патологических очагов возбужде-
ния В глубине мозга воспалительного и опухолевого генеза или
эпиактивности, при патологических состояниях зрительной систе-
мы, так и в научных исследованиях, поскольку позволяет просле-
дить пути биоэлектрической активности в структурах мозга при
различных воздействиях световых и звуковых стимулов, регистра-
ции вызванных потенциалов различной модальности, исследовании
динамики развития волновых комплексов и т. п.
238
МБН
сек
У= 6,78 мкВ
Т= 8.226 сек
8.228 от f
8.128 от f
7Х stiwuls
Ч. ЗЛОХ*. 1
Эпоха N* 1
Рзм 1.824 сек
Чувс 18 мкВ
Част 588.8 Гц |
Зап 1.828 сек
Файл SERC J& |
Внврд SERCJ) J
Рис. 7.7. Топографическое картирование ЗВКП на вспышку в норме (OD).
Хорошо выражен первичный ответ, курсор на PJ00. Доминирующий очаг
биоэлектрической активности в затылочной доле (Ор О2). В этом же вре-
менном диапазоне отмечается очаг активности в теменно-височных и фрон-
тальных долях.
Предварительны? результаты картирования ЗВКП в клинике
гдаэных болезней показали, что у здоровых обследуемых и боль-
ных с различными патологическими состояниями зрительного ана-
лизатора имеется несколько источников биоэлектрической актив-
ности зрительных центров наряду с классическим центром а заты-
лочной области. С определенной закономерностью выявлена
доминирующая активность в лобной, теменной и височной облас-
тях, возникающая в различных временных диапазонах (рис-7.7).
Сопоставление результатов топографического картирования
ЗВКП в норме и при некоторых патологических процессах: ретро-
бульбарном неврите, оптохиазмальном арахноидите, атрофии зри-
тельного нерва, диэнцефальном синдроме, амблиопии, альбинизме
и т.д., показало, что при патологии зрительного анализатора изме-
нения топографии ЗВКП были различными — от уменьшения апп-
роксимированных зон И снижения амплитуды ЗВКП ДР уменьше-
ния колйчесгва доминирующих очагов электрической активности
ца поверхности скальпа, которые и прослеживались в процессе
Развития ЗВКП. ЗВКП в норме и при патологии имеют различия в
частотном спектре. На уровне доминирующих ЗВКП В различных
Областях коры в норме были выделены частоты от 10 до 12 Гц. При
239
патологических состояниях зрительного анализатора низкие и вы-
сокие частоты (0,5—6 и 14—24 Гц) проявлялись в областях, где в
норме они отсутствовали. Степень указанных изменений коррели-
ровала с глубиной поражения тех или иных структур (нейронов) в
зрительном анализаторе, а также с распространенностью патологи-
ческого процесса, изменением остроты и поля зрения. При значи-
тельном снижении остроты зрения отсутствовали характерные для
ЗВКП доминирующие источники биоэлектрической активности и
ос-ритм регистрировался над всей поверхностью коры. При диэнце-
фальном синдроме (острота зрения 1,0), наоборот, отмечался об-
ширный сливной очаг от затылочной доли к лобной в правом и
левом полушарии, что могло быть результатом непрямой реакции
на световое раздражение других не зрительных нейронов.
Картирование ЗВКП позволяет определить динамику и топо-
графию распространения потенциала в разных временных диапазо-
нах на различных участках скальпа, которые, возможна, соответ-
ствуют проекции источников биопотенциала. При картировании
ЗВКП у больных с амблиопией (рис.7.8) не обнаружено характер-
ных для нормы дифференцированных очагов активности в правой
и левой гемисфере затылочной доли при стимуляции амблиопично-
го глаза вспышкой света. В случае паттерн-реверсивной стимуля-
ции (величина элемента шахматного поля 56*) отмечается значи-
тельное снижение амплитуды и увеличение латентности в затылоч-
ной и теменной областях. Уровень источников биоэлектрической
активности мозга при амблиопии показал, что при стимуляции
здоровых глаз у большинства больных очаг возбуждения локализу-
ется в течение всего времени ответа на ипсилатеральной стороне
неперекрещенных зрительных путей (рис.7.9), в проекции затылоч-
ной и заднетеменной областей. При стимуляции амблиопичного
глаза очаги возбуждения располагались на контралатеральной сто-
роне мозга в крайнем нижнем положении схемы, что не исключает
вклад активности подкорковых структур. Значительная дисперсия
очагов возбуждения в ипси- и контралатеральном полушариях, по-
видимому, обусловлена включением большого количества неспеци-
фических нейронов.
Важный дифференциальный электрофизиологический признак
отмечен при сопоставлении топографических карт, полученных у
больных с оптохиазмальным арахноидитом и острым ретробуль-
барным невритом (оптическим невритом), возникающим на фоне
рассеянного склероза. Если в первом случае доминировали очаги
биоэлектрической активности в лобной доле при почти полном их
отсутствии в затылочной, то во втором отмечалась активность в
ипси- и/или контралатеральной гемисфере (в зависимости от лока-
лизации процесса в зрительном нерве) затылочной области при
значительном снижении амплитуды, удлинении латентности и по-
чти полное отсутствие очагов возбуждения в лобной доле в случае
240
ИБН
U~ 5.63 мкВ
8.205 сек
8.205 от ▼
0.105 от |
62 stiMuls
Ч. эпох * 1
Эпоха № 1
Рэм 0.488 сек
Чувс 58 мкВ
Част 1288. Гц |
Зап 8.401 сек
Файл HARCJM34
Внвод HftRC J>04
ft.,1985г. Амблиопия Vis OS 1.8 OS 8,84
1
1
5.88
мкВ
0.24 мкВ
Т= 0,284 сек
0.204 от Т
0.104 от f
62 stiwuls
Ч. эпох:
Эпоха К*
Рэм 0.480 сек
Чувс 50 мкВ
Част 1280. Гц |
Зал 0.401 сек
Файл HARC_S04
Вывод HARC-S04
Рис. 7.8. Топографическое картирование ЗВКП на вспышку (больной X.,
10 лет, с дисбинокулярной амблиопией, vis: OD=1,0; OS=0,04).
а — здоровый глаз (OD): Р100 нормальной амплитуды и латентности выражен на
контралатеральной гемисфере в отведении Ор на ипсилатеральной гемисфере он
практически отсутствует; б — амблиопичный глаз (OS): то же, однако доминирую-
щий очаг нормальной амплитуды и латентности выражен на ипсилатеральной ге-
мисфере, а на контралатеральной стороне амплитуда ЗВКП значительно снижена.
МБН
ээг
241
Рис.7.9 Источники ЗВКП у больной с рефракционной амблиопией (на схе-
ме обозначены точками) при стимуляции амблиопичного глаза.
значительного снижения остроты зрения. При картировании ЗВКП
у альбиносов выявлены межполушарная амплитудная и временная
асимметрия, раздвоение и расширение пика Р100, локализация ис-
точников в одном полушарии (определяемая по двухдипольной мо-
дели) в проекции лобной и затылочной коры с уменьшением зоны
биоэлектрической активности в проекции теменных и височных
долей. Во всех случаях преобладала частота 14—24 Гц.
Очевидно, что картирование ЗВКП и определение локализации
источников биоэлектрической активности в мозговых структурах
как в норме, так и при патологических состояниях зрительной си-
стемы расширило диагностические возможности общепринятой
методики регистрации зрительных вызванных биопотенциалов на
вспышечный стимул и реверсивный паттерн. Появилась возмож-
ность количественно интерпретировать ЗВКП, определять те уров-
ни зрительной системы, в которых располагаются их источники, а
также наблюдать за динамикой их развития.
Результаты топографического картирования ЗВКП подтверж-
дают, что обработка и передача информации происходят в разных
отделах мозга. По мере передачи информации расширяются зоны
242
биоэлектрической активности, появляются или исчезают новые ис-
точники возбуждения.
Таким образом, появилась возможность изучения в динамике
доминирующих очагов ЗВКП в различных временных диапазонах.
7.5. ЗРИТЕЛЬНЫЕ ВЫЗВАННЫЕ КОРКОВЫЕ ПОТЕНЦИАЛЫ
И ОСТРОТА ЗРЕНИЯ
Данные о генезе различных типов ЗВКП позволяют более точно
оценить возможности метода при его использовании в клинической
офтальмологии и для определения состояния зрительных функций.
Так, многократно отмечавшееся в литературе расхождение в оцен-
ке остроты зрения по ЗВКП и поведенческим тестам [Dobson V.,
1978] не может быть полностью объяснено различиями используе-
мых стимулов и оценочных критериев. Главная причина, по-види-
мому, в том, что структурную основу, ответственную за форменное
зрение, кроме первичной зрительной коры (поле 17), составляют
вторичные (поля 18,19) и ассоциативные зоны, тогда как в качестве
критерия для определения остроты зрения используют амплитуду
компонента Р100 ЗВКП на реверсию шахматного поля, генерируемо-
го стриарной корой.
Vilson (1988), сопоставив теоретическую остроту зрения, рассчи-
танную на основе меняющейся с возрастом ребенка плотности фо-
веальных колбочек, с остротой зрения, установленной методами
регистрации ЗВКП и предпочтительной фиксации у младенцев и
детей 1-го года жизни, пришел к выводу, что ЗВКП отражают сте-
пень зрелости фовеальной области сетчатки. Значительно более
низкая острота зрения, полученная методом предпочтительной
фиксации, обусловлена незрелостью высших зрительных центров.
Можно предположить, что одной из причин расхождений в оценке
остроты зрения по ЗВКП и поведенческим критериям является то
обстоятельство, что в генерации ЗВКП на реверсию паттерна, по-
мимо нейронов, реагирующих на контраст и световое мелькание,
важную роль играют детекторы движения. Более низкие пороги об-
наружения световых мельканий и движения могут быть причиной
переоценки остроты зрения по ЗВКП [Taylor V. et al., 1991].
В аспекте обсуждаемого вопроса о соотношении между паттерн-
ЗВКП и зрительными функциями определенный интерес представ-
ляют результаты исследования ЗВКП при корковой слепоте
[Frank Y., 1992]. Несмотря на значительный разброс в полученных
результатах, отмечается возможность появления нормальных ЗВКП
на вспышку и шахматные паттерны у части тотально слепых детей
и взрослых, что было показано при сопоставлении ЗВКП с данны-
ми компьютерной томографии, выполненной при обширном била-
теральном разрушении стриарной коры (поле 17) и частичном по-
243
ражении полей 18 и 19. Результаты анализа позволяют сделать вывод
о том, что для осуществления зрительной функции необходима
сохранность критической величины поля 17. Наличие ЗВКП отра-
жает приход в кору афферентного потока, однако при значитель-
ной деструкции стриарной коры субкортикальные структуры, уча-
ствующие в генерации ЗВКП, не способны обеспечить реализацию
зрительной функции. Авторы указали на прогностическую ценность
ЗВКП: у большинства детей, у которых наблюдалась корковая сле-
пота в острой стадии заболевания, когда регистрировались нор-
мальные ЗВКП на вспышку, зрительные функции полностью вос-
станавливались независимо от этиологии заболевания; измененные
ЗВКП, наоборот, предшествовали неблагоприятным исходам.
Приведенные данные свидетельствуют о том, что ЗВКП на ре-
версию паттерна не являются надежным критерием оценки остроты
зрения у отдельных индивидуумов, но могут быть использованы
для определения степени зрелости и характеристики структурно-
функционального состояния ретинокортикального пути. Хотя ост-
роту зрения нельзя определить по ЗВКП, их можно использовать
для клинических целей для получения важной информации о каж-
дом глазе.
7.6. РОЛЬ ЗРИТЕЛЬНЫХ ВЫЗВАННЫХ КОРКОВЫХ
ПОТЕНЦИАЛОВ В ДИАГНОСТИКЕ ЗАБОЛЕВАНИЙ
ЗРИТЕЛЬНЫХ ПУТЕЙ
ЗВКП широко используются в офтальмологии, нейроофтальмоло-
гии, неврологии, нейрофизиологии для оценки функционального
состояния и степени сохранности зрительных путей, для локализа-
ции уровня патологического процесса, оценки развития зрительной
системы.
ЗВКП дополняют данные ЭРГ, а также являются единственным
источником информации в случаях, когда ЭРГ зарегистрировать
по тем или иным причинам невозможно.
В клинико-физиологических исследованиях наиболее широко
используют метод регистрации фазического ЗВКП на реверсию
шахматных паттернов. Основное преимущество ЗВКП этого типа
по сравнению с другими состоит в большей стабильности компо-
нентного состава и временных характеристик в норме, лучшей
воспроизводимости, чем ответы на вспышечный паттерн, мень-
шей межиндивидуальной вариабельности и в большей чувстви-
тельности к патологическим изменениям в зрительной системе.
Многие исследователи наблюдали сдвиги амплитудно-временных
характеристик ЗВКП при изменении четкости изображения при
катаракте, снижении контрастной чувствительности, заболевани-
ях зрительного нерва и сетчатки [Celesia G., 1985, 1990;
244
Chiappa K.N., 1990; Brecelj J., 1992; Kavender et al., 1992; Halliday A.
M., 1993; Cohen S.Y., 1994].
В клинике глазных болезней паттерн-ЗВКП рекомендовано ис-
пользовать:
1) при оценке функционального состояния зрительного нерва и
макулярной области сетчатки;
2) для ориентировочной оценки остроты зрения и степени раз-
вития зрительной системы у детей;
3) для объективных периметрических исследований, например
при гемианопсии, травмах головного мозга;
4) при амблиопии;
5) при корковой и психогенной слепоте;
6) во всех случаях, когда поведение и реакция больного не по-
зволяют проводить другие психофизические исследования для оцен-
ки функции зрительной системы.
В отличие от ЗВКП на реверсию паттерна ЗВКП на вспышку
значительно более вариабелен, но при этом менее чувствителен к
нарушениям в зрительной системе, в связи с чем его можно исполь-
зовать лишь в случаях более грубых поражений, когда применение
паттерн-ЗВКП оказывается невозможным, например при отсутствии
фиксации взора, значительном снижении остроты зрения, при пол-
ном помутнении оптических сред, нистагме и т.д. [Лысков Е.Б, 1985;
Sokol S., 1977; Halliday А.М., 1979, 1982, 1993; Kriss А., 1982; Bass,
1985; Blumhardt L.D., 1986; Bodis-Wollner I., 1986; Holopigian K.,
1990; Desmedt J.E., 1990; Smith, 1990; Holder G.E., 1992; Denislic M.,
1993; Lowitzsch K., 1993, и др.].
В настоящее время регистрацию ЗВКП на вспышку применяют:
1) в оценке состояния зрительного нерва при травматическом
его повреждении, при черепно-мозговых травмах;
2) при глаукоме, атрофиях зрительного нерва наследственной и
другой этиологии;
3) токсических и ишемических оптических нейропатиях;
4) при рассеянном склерозе и оптохиазмальном арахноидите;
5) мониторинге при хирургии аденомы гипофиза или других
операциях на селлярной области;
6) при локализации процесса в хиазме и зрительном тракте, в
оценке способности к видению при различных зрительных агнози-
ях, повреждениях зрительной коры и при церебральной патологии,
ее компрессионных повреждениях, в том числе в связи с новообра-
зованиями;
7) в оценке состояния зрительных функций при нарушении со-
знания;
8) при помутнении оптических сред;
9) при альбинизме.
Критериями клинически значимых отклонений при оценке ЗВКП
являются отсутствие ответа или значительное снижение амплиту-
245
ды, удлинение латентности всех пиков, значительные различия в
амплитуде и латентности при стимуляции правого и левого глаза
со снижением амплитуды и удлинении латентности при стимуляции
худшего глаза. В литературе встречаются указания на то, что пара-
доксальное увеличение амплитуды и изменение формы ответа ЗВКП
с проявлением его в виде пик-волнового комплекса является свиде-
тельством дисфункции мозга и снижения порога пароксизмальной
готовности. Другие изменения формы ЗВКП, латентности, числа
компонент следует интерпретировать с осторожностью. У новорож-
денных или неконтактных больных нормальные ЗВКП не указыва-
ют еще на наличие сознания и зрительного восприятия. Они могут
лишь показывать на сохранность неспецифической восходящей аф-
ферентации [Гнездицкий В.В., 1997].
Хиазмальный уровень поражений. При ЧАЗН
вследствие оптохиазмального арахноидита наблюдалось снижение
амплитуды и выпадение компонентов ЗВКП, увеличение латентно-
сти. Изменения ЗВКП нарастали с увеличением тяжести пораже-
ния, при этом повреждение прехиазмального участка зрительного
нерва сопровождалось манифестацией клинических проявлений за-
болевания, а поражение супрахиазмальных отделов зрительного
пути — межполушарной асимметрией ЗВКП.
Распределение ЗВКП на поверхности скальпа связано с перекре-
щиванием зрительных путей в хиазме. Среди хиазмальных пораже-
ний, при которых используют ЗВКП в диагностике, выделяют опу-
холь, аневризмы, воспаления, демиелинизирующие процессы, трав-
му. С учетом анатомических особенностей зрительного пути в хиазме
целесообразно проводить исследование ЗВКП при многоканальном
их отведении. Для хиазмальных поражений характерна перекрещен-
ная (crossed) асимметрия ЗВКП, которая выражается в патологи-
ческом ответе, например, над левой гемисферой при стимуляции
всего поля левого глаза и в более значительных изменениях над
правой гемисферой при стимуляции правого глаза. При стимуля-
ции темпоральной половины поля зрения имеет место отсутствие
или патологический ответ на обоих гемисферах, а при стимуляции
назальной половины поля зрения — нормальный ответ. Неожидан-
ностью является большая выраженность изменений ЗВКП на ипси-
латеральной стороне по отношению к дефекту в поле зрения, что
названо “парадоксальной” латерализацией. В то же время при сти-
муляции всего поля паттерн-ЗВКП были всегда изменены на кон-
тралатеральной стороне по отношению к дефектам в поле зрения.
При этом патологические паттерн-ЗВКП могут быть зарегистриро-
ваны и при нормальном поле зрения. Дисфункция перекрещенных
волокон, обнаруженная при стимуляции темпоральной половины
поля, может сочетаться с изменениями в темпоральной части поля
зрения, а дисфункция неперекрещенных волокон может выявляться
при стимуляции назальной половины. Таким образом, при хиаз-
246
Рис. 7.10. Реверсивная асимметрия ЗВКП на вспышку света у больного с
альбинизмом [Brecelj J., 1993].
Различия между двумя глазами в распределении ЗВКП при отведении от
затылочной области (1—3 — схема локализации электродов в затылочной
области и соответствующие им кривые ЗВКП):
а — при стимуляции левого глаза; б — при стимуляции правого глаза; виг — ре-
версивная асимметрия (отмечена стрелками) между правым и левым глазом видна
на результирующих кривых, где различия потенциалов представлены при вычи-
тании левого окципитального затылочного ответа из правого.
мальных поражениях стимуляция половины поля зрения повышает
чувствительность метода в выявлении различий между дисфункци-
ей в волокнах, идущих от темпоральной и назальной частей поля
зрения.
Анатомические исследования, проведенные у альбиносов, пока-
зали, что большинство волокон одного глаза перекрещивается в
хиазме и проецируется на контралатеральной гемисфере. Этот оши-
бочный перекрест темпоральных ретинальных волокон отражается
на форме ЗВКП, регистрируемых при стимуляции вспышкой
(рис.7.10). Асимметрия видна около 80 мс и выражается в негативи-
зации компонентов на контралатеральной стороне и позитивности
ответа на ипсилатеральной стороне головы по отношению к стиму-
лируемому глазу. Таким образом, стимуляция левого глаза приво-
дит к выраженной негативизации компонентов, зарегистрирован-
ных при отведении от поверхности головы над правой окципиталь-
ной областью, и наличию маленького позитивного компонента той
же латентности в проекции левой окципитальной коры. При стиму-
ляции правого глаза эти нарушения носят обратный характер. Та-
кая асимметрия ЗВКП позволяет предположить, что они происхо-
дят от концов общего диполя генератора, который расположен на
медиальной стороне гемисферы, контралатерально к стимулируе-
247
мому глазу. Использование паттерн-реверсивного стимула ограни-
чивается наличием нистагма у этих пациентов. При on/set-стимуля-
ции, так же как и при вспышечной, выявлена реверсивная асиммет-
рия компонентов Ц и С2, характерная для альбиносов.
Реверсивная асимметрия у альбиносов отличается от перекре-
щенной асимметрии ЗВКП при хиазмальных и ретрохиазмальных
поражениях зрительных путей, сопровождающихся поражением
перекрещивающихся волокон в хиазме. Если у альбиносов компо-
нент Р100 виден при стимуляции правого глаза над левой гемисфе-
рой, а при стимуляции левого глаза — над правой гемисферой, то
при хиазмальных поражениях Р100 регистрируется при стимуляции
правого глаза в правой гемисфере, а при стимуляции левого гла-
за — в левой гемисфере. Таким образом, кортикальная репрезента-
ция ЗВКП у альбиносов отражает первичную активность перекре-
щенных ретинальных волокон, а при хиазмальных поражениях —
дисфункцию перекрещенных волокон [Brecelj J., 1993].
Выявленная при альбинизме характерная межполушарная асим-
метрия раннего компонента ЗВКП на вспышку в сочетании с супер-
нормальной ЭРГ позволяет дифференцировать альбинизм от врож-
денной колбочковой дисфункции [Russel-Eggit, 1990; Apkarian Р., 1991].
Ретрохиазмальный уровень поражения. При
ретрохиазмальных дисфункциях зрительного тракта и окципиталь-
ной коры чувствительность паттерн-ЗВКП ниже, чем периметрия,
так как не установлено специфических изменений ЗВКП при пора-
жениях на разных уровнях зрительной системы. Однако типичным
проявлением односторонней ретрохиазмальной дисфункции явля-
ется неперекрещенная (uncrossed) асимметрия, выражающаяся в па-
тологических ЗВКП, сходных для каждого глаза. Гомонимные де-
фекты в поле зрения являются причиной снижения биоэлектриче-
ской активности нейронов центральных отделов зрительного
анализатора и приводят при стимуляции всего поля сетчатки к ти-
пичной неперекрещенной (uncrossed) асимметрии ЗВКП, при этом
нормальный Р100-компонент регистрируется на стороне гомонимно-
го дефекта,что является проявлением парадоксальной латерализа-
ции. Если гомонимные дефекты включают макулу, то при стимуля-
ции половины поля зрения ЗВКП изменены и имеют форму, харак-
терную для скотом в центральном поле зрения. ЗВКП не отличаются
от нормы, если макула не включена в гомонимный дефект поля
зрения, что не исключает наличие органических изменений, приво-
дящих к зрительным нарушениям. Выделение макулярных и пара-
макулярных компонентов при различных нарушениях в поле зре-
ния возможно при использовании стимуляции правой и левой по-
ловин поля зрения.
При двустороннем поражении затылочной доли с двусторонней
гомонимной гемианопсией, корковой слепотой описаны различные
изменения ЗВКП. При интактности первичных зрительных центров
248
OS
a — засвет всего поля зрения (1—5 схема локализации электродов в затылочной
доле и соответствующие им кривые ЗВКП): волны W-подобной формы (OS) наибо-
лее выражены при отведении 3, интактный OD имеет нормальный Р100; б, в —
патологически измененные кривые ЗВКП, соответствующие скотомам, обнаружива-
ются на OS. Р135 -компонент (отмечен открытым кружком) хорошо представлен, Р100-
компонент отсутствует или прерывается в связи с взаимодействием компонентов Р100
ЗВКП могут быть нормальными. Они могут быть нормальными и
при полной корковой слепоте. Предполагается при этом участие
экстрагеникулокалькариновых путей. Несмотря на существование
более чувствительных тестов, таких как компьютерная томография,
магнитный резонанс, исследование ЗВКП для оценки функциональ-
ного состояния центральных зрительных путей рекомендуется как
объективный мониторинг с контролем за развитием заболевания.
Заболевания зрительного нерва. Наиболее ха-
рактерным признаком заболевания зрительного нерва (острый не-
врит, ретробульбарный неврит на фоне рассеянного склероза, ише-
мическая оптическая нейропатия и др.) является увеличение перио-
да латентности основного позитивного компонента (Р100) ЗВКП
[Фильчикова Л.И. и др., 1991; Harding G.F.A., 1991]. При неврите
зрительного нерва наряду с увеличением периода латентности отме-
чается изменение компонентного состава и снижение амплитуды
249
ЗВКП, W-образная форма Р1оо (рис.7.11). При стимуляции полови-
ны поля зрения выявляется отсутствие макулярного компонента Р105
или его соединение с компонентом Р135, в то же время компоненты
Р105 и Р145 парамакулярной области регистрируются на контралате-
ральной гемисфере. Эти данные сопровождаются изменениями цен-
трального поля зрения, наличием относительных или абсолютных
скотом в поле зрения. Таким образом, W-образная форма компо-
нента Р1оо отражает снижение функции аксиального пучка зритель-
ного нерва, а наиболее устойчивым и ранним признаком, возника-
ющим еще до клинических проявлений заболевания, является уве-
личение латентности компонента Р100. При прогрессировании
болезни период латентности увеличивается, превышая нормальные
показатели на 30 — 35%, с одновременным снижением амплитуды
и изменением конфигурации ответа. При восстановлении зритель-
ных функций амплитуда и конфигурация ЗВКП постепенно норма-
лизуются, тогда как временные характеристики продолжают оста-
ваться увеличенными иногда в течение 2—3 лет после нормализа-
ции клинических показателей [Sanders Е., 1986].
Причиной увеличения латентности при оптическом неврите яв-
ляется демиелинизация нервных волокон, приводящая к снижению
скорости проведения возбуждения в пораженных волокнах зритель-
ного нерва на корковом и сетчаточном уровнях. Изменение ампли-
туды ЗВКП отражает полную блокаду проведения возбуждения в
пораженных волокнах. Поскольку при этом заболевании поража-
ются главным образом волокна, связанные с ганглиозными клетка-
ми фовеальной области сетчатки, наиболее адекватными для обна-
ружения патологии являются соответствующие величине фовеаль-
ной области стимулы размером 4 угловых градуса и клетки
шахматного поля величиной 30 угловых минут. При оптическом
неврите, возникающем на фоне рассеянного склероза, ЗВКП на
паттерн-стимул изменяются до появления клинических признаков
заболевания, что свидетельствует о субклиническом вовлечении в
патологический процесс зрительных путей.
Диагностическая ценность ЗВКП увеличивается при подозрении
на рассеянный склероз, когда в процессе исследования изменяются
параметры реверсивного паттерна, частота реверсии, величина поля
стимуляции и шахматных квадратов, яркость стимула и ориента-
ция решеток.
Описаны также характерные для поражения зрительного нерва
при рассеянном склерозе, сосудистой нейропатии и др. нарушения
конфигурации ЗВКП, которые выявляются в форме расщепленного
компонента Р100, образующего трехфазный комплекс волн — Р2, N2,
Р3 вместо нормального Np Р2, N2. В этих случаях период латентно-
сти Р2 меньше, а период латентности Р3 больше нормальных значе-
ний периода латентности Р100. По мнению исследователей, аномаль-
ная форма ЗВКП может быть обусловлена подавлением макуляр-
250
ного компонента Р100 и доминированием парамакулярного компо-
нента Р|35, что приводит к видимому увеличению периода латентно-
сти позитивной волны. Истинное увеличение периода латентности
макулярного компонента Р100 обычно не превышает 55 мс; при боль-
шем его увеличении можно предполагать наличие позитивной вол-
ны парамакулярного генеза. Таким образом, увеличение периода
латентности компонента Р100 ЗВКП является характерным, но не
патогномоничным признаком демиелинизации зрительных путей.
А. М. Halliday (1982) подчеркивает, что изменения ЗВКП не являются
специфичными для какого-либо поражения зрительной системы.
При частичной атрофии зрительного нерва наблюдается сниже-
ние амплитуды и увеличение периода латентности компонента Р|00
ЗВКП; эти изменения могут возникать независимо друг от друга
[Sanders Е., 1986].
Роль исследования ЗВКП в оценке функционального состояния
зрительных путей, идущих от макулярной области, можно просле-
дить на примере ишемической оптической нейропатии, в последнее
время подразделяемой на две группы: с гигантоклеточным артери-
итом и без него. Причиной является острое нарушение кровоснаб-
жения передней порции зрительного нерва, обусловленное наруше-
нием циркуляции в задних цилиарных артериях, глазничной арте-
рии. ЗВКП на паттерн и вспышечный стимул, как правило, в обоих
группах имеют сниженную амплитуду и небольшие изменения ла-
тентности Р|00-компонента, на клинически здоровых глазах патоло-
гических изменений ЗВКП не отмечено. Сниженная амплитуда
ЗВКП, замедленный или трехфазный ответ на вспышечный стимул
были отмечены при темпоральном артериите [Holder G.E., 1992].
Очевидно, что паттерн-ЗВКП более чувствительны к сосудистым
нарушениям в системе зрительного нерва у пациентов без артерии-
та по сравнению со вспышечным ЗВКП. Типичным для ишемичес-
кой оптической нейропатии была незначительная разница в латен-
тности пика Р100 между больным и здоровым глазом (3 мс), в то
время как при демиелинизирующем процессе в зрительном нерве
эта разница была значительной (21 мс). Отмечается важность реги-
страции ЗВКП от двух гемисфер, а не от одного центрального элек-
трода, накладываемого в области затылочного бугра над inion
[Сох Т.А. at al., 1982; Weinstein J.M., 1982; Moschos M„ 1984].
Заболевания сетчатки. При различных формах ма-
кулодистрофий и макулопатий, центральной серозной ретинопатии
также наблюдаются увеличение латентности и снижение амплиту-
ды компонентов ЗВКП, однако увеличение пиковой латентности
компонентов ЗВКП не коррелирует со снижением амплитуды. При
поражении центральных областей сетчатки увеличение латентности
ЗВКП может наблюдаться на паттерны малых угловых размеров,
при поражении периферических отделов — больших размеров [Bodis-
Wollner, 1987; Kato М„ 1991].
251
Значение критической частоты воспроизведения ритма световых
мельканий, используемых в регистрации ЗВКП устойчивого состо-
яния, показано в диагностике поражений аксиального пучка зри-
тельного нерва и макулярной области сетчатки у детей с помутне-
ниями оптических сред глаза. Снижение амплитуды ЗВКП на рит-
мически предъявляемые световые вспышки наблюдали при глаукоме.
Редукция амплитуды была наиболее выраженной при использова-
нии средних и высоких временных частот (от 10 до 50 Гц). Авторы
полагают, что эти данные свидетельствуют о селективном пораже-
нии оптических волокон, проводящих высокие временные частоты
при повышении внутриглазного давления.
Дифференциальный диагноз заболеваний сетчатки и зрительного
нерва с помощью электрофизиологических методов может быть уточ-
нен при сопоставлении данных ЭРГ и ЗВКП. Значительное (> 130 мс)
увеличение периода латентности Р100 и снижение амплитуды ЗВКП
при сохранных общей и локальной ЭРГ указывает на поражение зри-
тельного нерва. Заболевания макулярной области, как правило, со-
провождаются снижением локальной ЭРГ и изменением амплитуд-
но-временных параметров ЗВКП, при этом общая ЭРГ может оста-
ваться сохранной. Рекомендуется также одновременная регистрация
ЭРГ и ЗВКП на реверсию паттерна, при которой возможно опреде-
ление ретинокортикального времени, т.е. времени между активацией
структур сетчатки и приходом сигнала в зрительную кору. Увеличе-
ние латентности b-волны в паттерн-ЭРГ и Р100-компонента ЗВКП при
сохранном ретинокортикальном времени характерно для поражений
макулярной области. Увеличение ретинокортикального времени при
сохранной паттерн-ЭРГ и увеличение временных параметров ЗВКП
указывают на демиелинизацию зрительного нерва. При грубых по-
ражениях аксонов с ретроградной дегенерацией волокон зрительно-
го нерва может наблюдаться резкое снижение амплитуды или отсут-
ствие паттерн-ЭРГ в сочетании с выраженными изменениями ампли-
тудно-временных параметров ЗВКП. Однако четкость приведенной
схемы может нарушаться из-за двойственной природы паттерн-ЭРГ—
ее связи с активностью как преганглионарных элементов, так и ган-
глиозных клеток сетчатки.
Одна из важных проблем дифференциальной диагностики, в
которой метод регистрации ЗВКП может иметь большое значение,—
разграничение функциональных и органических поражений в зри-
тельной системе.
Многочисленные исследования показали, что наиболее харак-
терной чертой ЗВКП при амблиопии является снижение амплитуды
компонентов преимущественно при предъявлении шахматных пат-
тернов с клетками малых размеров (<30 угл.мин); при использова-
нии паттернов с клетками больших размеров амплитуда компонен-
тов ЗВКП может не отличаться от нормы или даже превосходить
ее. При этом, несмотря на значительную вариабельность, в среднем
252
снижение амплитуды ЗВКП при амблиопии пропорционально
снижению остроты зрения. Отмечается также небольшое увеличе-
ние периода латентности Р100-компонента, которое, по данным раз-
ных авторов, в пределах от 5 до 15 мс [Зислина Н.Н., 1987; Толсто-
ва В.А., 1989; Фильчикова Л.И., 1991]. Анализ распределения зна-
чений периода латентности Р100-компонента ЗВКП на реверсию
шахматных полей с клетками размером 28 угл.мин показал, что при
рефракционной и обскурационной амблиопии это значение не пре-
вышает 130 мс.
При ЧАЗН в 70% случаев период латентности Р100 превышает
130 мс, что позволяет с определенной степенью вероятности исполь-
зовать временные характеристики ЗВКП наряду с ЗВКП устойчи-
вого состояния на вспышки света с частотой 33 Гц для дифференци-
альной диагностики амблиопии и частичной атрофии зрительного
нерва. Нозологические формы заболеваний сетчатки и зрительного
нерва, при которых рекомендуется использовать метод исследова-
ния ЗВКП, представлены в табл. 7.2
Таблица 7.2. Изменение ЗВКП при патологии зрительного нерва и зри-
тельных путей
Патология зрительного нерва и зрительного пути Амплитуда Латентность
Ишемическая оптическая нейропатия 11 т
Оптический неврит (рассеянный скле- нт ттт
роз)
Токсическая амблиопия ни нттт
Доминантная атрофия зрительного нерва 1 нт
Леберовская атрофия зрительного нерва 11 т
Гипоплазия зрительного нерва 11 т
Глаукома Н1 НТ1
Друзы зрительного нерва 1 т
Отек зрительного нерва Н1 нт
Опухоль передних зрительных путей 1 т
Амблиопия Н1 нт
Макулодистрофия 'L'L'L тт
Таким образом, приведенные данные свидетельствуют о том,
что, хотя ни одна из аномалий ЗВКП не обладает нозологической
специфичностью, метод исследования ЗВКП может быть использо-
ван для раннего выявления демиелинизирующих поражений зри-
тельного нерва. Включение же этого метода в комплекс электрофи-
зиологических (ЭОГ, ЭРГ) и психофизиологических (компьютерная
периметрия, измерение световой и контрастной чувствительности и
др.) методов увеличивает возможности выявления и уточнения уров-
ня и степени поражения ретинокортикального пути при различных
заболеваниях органа зрения.
253
ГЛАВА8
ЭНТОПТИЧЕСКИЕ ФЕНОМЕНЫ В ДИАГНОСТИКЕ
ФУНКЦИОНАЛЬНЫХ НАРУШЕНИЙ
Под энтоптическими явлениями (образами) понимают необычные
зрительные ощущения, обусловленные не только некоторыми пато-
логическими, но и нормальными структурами глаза, которые либо
мешают обычному распределению лучей света, либо являются не-
адекватным (не световым) раздражителем фоторецепторов.
Целесообразно рассматривать энтоптические явления примени-
тельно прежде всего к анатомии глаза, локализации того морфоло-
гического субстрата, который при определенных условиях лежит в
основе того или иного феномена [Hart W.H., 1987].
8.1. РОЛЬ ОСОБЕННОСТЕЙ СТРУКТУРЫ СЛЕЗНОЙ
ПЛЕНКИ, РОГОВИЦЫ И ХРУСТАЛИКА
Если, прищурившись, рассматривать через точечную диафрагму
(Д = 0,1 мм), установленную в переднем фокусе глаза (на расстоя-
нии примерно 17 мм от роговицы), ярко освещенный экран (белую
стену, небо и т. п.), то можно заметить горизонтально расположен-
ную полосу с яркими медленно плавающими по ней (при движении
век) пятнами с темным ореолом. A. Esser (1926) объяснил это явле-
ние изменениями, происходящими в слезной пленке при движениях
век, особенно при наличии на роговице комочков слизи.
При широко раскрытой глазной щели в тех же, что описаны
выше, условиях обнаруживается мозаичная картина из серии пере-
крещивающихся горизонтальных и вертикальных плавно извиваю-
щихся световых полосок. Полагают, что морфологическим субстра-
том феномена являются складки эпителиального покрова рогови-
цы под слезной пленкой. Образованию складок и бороздок между
ними способствует тонус круговой мышцы век. Увеличение выра-
женности феномена можно вызвать, несильно надавив пальцем на
глазное яблоко через веко. М. Dangle и соавт. (1984) установили,
что сходный эффект вызывают жесткие контактные линзы, упло-
щающие роговицу.
Поскольку аналогичная мозаика может возникать и в присут-
ствии витреальных треков нормального стекловидного тела, для
дифференциальной диагностики закапывают флюоресцеин, который
подчеркивает роговичную природу мозаики.
В условиях высокой освещенности при рассматривании окружа-
ющих предметов люди, у которых в роговице или хрусталике име-
254
ются множественные полупрозрачные помутнения, отмечают сни-
жение контрастности изображения и слепимость, если не прибег-
нуть к диафрагмированию. В основе этого явления лежат “парази-
тическое” рассеивание (“разбрызгивание”) света на полупрозрач-
ных участках и в результате снижение контрастности изображения
на глазном дне.
В коллимированном свете пациент с предкатарактой может
видеть спицевидные помутнения в хрусталике; облегчение возмож-
но при использовании желто-оранжевых светофильтров, отсекаю-
щих коротковолновое излучение.
При нарушении прозрачности роговицы грубое полностью мут-
ное пятно (бельмо) субъективно легче переносится, чем группа едва
заметных полупрозрачных облачковидных помутнений. При их
парацентральном расположении альтернативой кератопластике, как
это ни парадоксально, может явиться татуаж (превращение полу-
прозрачного участка в непрозрачный).
Видение ореолов (нимбов, halo, радужных кругов). Причины об-
разования ореолов разнообразны, и некоторые детали феномена мо-
гут оказаться полезными в дифференциальной диагностике. Орео-
лы, независимо от причины, их вызывающей, легче всего анализиру-
ются при рассматривании белого точечного источника света.
Существует физиологическое halo, обусловленное лучистым
расположением нормальных волокон и швами в хрусталике. Види-
мая благодаря этому лучистость вокруг светящейся точки послужи-
ла поводом для названия небесных светил звездами.
Округлый край зрачка действует наподобие дифракционной
решетки и создает ореол. Он едва заметен из-за небольших разме-
ров (не более 2—3°), но в нем можно уловить хроматичность: ко-
ротковолновые (синие) окружности расположены ближе к центру,
чем длинноволновые (красные).
Если на поверхности слезной пленки скапливается много слизи
и детрита, то хроматическая дисперсия белого света через слизь
усиливается. При этом радужные круги вокруг белого источника
света становятся отчетливо видимыми, однако они тотчас исчезают
при протирании глаз. Чаще всего указанные явления отмечаются
при пробуждении ото сна у больных, страдающих конъюнктиви-
том.
Стойкие радужные круги, не исчезающие при протирании глаз,
или возникают время от времени, или видны постоянно при взгляде
на белый источник света. Периодическое появление стойких радуж-
ных кругов свидетельствует о транзиторном повышении офтальмо-
тонуса и связанном с этим накоплением в роговице, кпереди от
боуменовой мембраны, между клетками эпителия, отечной жидко-
сти. Последняя через растянутый вследствие офтальмогипертензии
слой эндотелия роговицы проникает в ее строму из передней каме-
ры. По мере нормализации офтальмотонуса лишняя влага уходит
255
из роговицы, она становиться прозрачной, а радужные круги исче-
зают.
Стойкие радужные круги, возникающие всякий раз при взгляде
на белый точечный источник света, являются проявлением хрома-
тической дисперсии последнего при диффузном помутнении рого-
вицы или хрусталика и при эндотелиально-эпителиальной дистро-
фии роговицы.
8.2. РОЛЬ ОСОБЕННОСТЕЙ СТРУКТУРЫ СТЕКЛОВИДНОГО
ТЕЛА
"Летающие мушки"— видение на фоне ярко освещенного гомоген-
ного экрана (например, безоблачного неба) темных, иногда слегка
рефлектирующих тончайших цепочек, похожих на ожерелье и со-
стоящих из отдельных фрагментов. В норме они чаще одиночные,
но могут быть и множественными. Характерно их продолжающее-
ся перемещение и после остановки движения глаза.
В основе феномена лежит огрубление отдельных структур стек-
ловидного тела. Чем ближе к поверхности сетчатки находятся эти
структуры (например, задняя гиалоидная мембрана при задней от-
слойке стекловидного тела), тем заметнее для пациента тени от них.
Для задней отслойки стекловидного тела характерно видение по-
мутнения в форме кольца чуть кнаружи от точки фиксации. Часто
эти постепенно появляющиеся в пожилом возрасте (из-за сморщи-
вания стекловидного тела) плавающие помутнения безобидны и об-
ращающиеся к врачу пациенты в лечении не нуждаются, однако не
всем удается избавиться от возникающего при этом зрительного
дискомфорта и тревоги.
'Золотой или серебряный дождь"(астероидный хиалоз). Он прак-
тически не беспокоит пациентов, хотя в светлое время суток у части
из них может немного снижаться острота зрения. Вместе с тем
офтальмоскопия часто затруднена из-за большого количества мель-
чайших золотистых или серебристых включений, плавающих в стек-
ловидном теле и отбрасывающих на сетчатку из-за ее отдаленности
очень слабые тени. Указанные включения нередко представляют
собой кристаллы кальциевых солей.
Плавающие помутнения. При миопической болезни грубые, иног-
да хлопьевидные плавающие помутнения могут существовать дли-
тельно как результат хронической деструкции стекловидного тела.
Особую тревогу они должны вызывать при внезапном возникнове-
нии, поскольку в этом случае они являются следствием либо выпо-
та экссудата в стекловидное тело при увеитах, либо кровоизлияния
в стекловидную полость per diapedesis и при разрыве сосудистой
стенки (при этом необходимо исключить и разрыв сетчатки с пред-
посылками к ее отслойке).
256
8.3. РОЛЬ ОСОБЕННОСТЕЙ ВИТРЕОРЕТИНАЛЬНЫХ
ОТНОШЕНИЙ
Светящиеся полоски Moore (Муре). В 1935 г. R.F. Мооге впервые
описал феномен мелькающих световых полос, возникающих при
движении глазных яблок как на свету, так и в темноте. Полоски
чаще ориентированы вертикально и ощущаются на крайней пери-
ферии поля зрения со стороны виска. Автор объяснил их происхож-
дение витреоретинальными тракциями, предшествующими задней
отслойке стекловидного тела в здоровых глазах пожилых людей.
Отсутствие назальной локализации светящихся полосок связывали
с “оптической недеятельностыо” крайней височной периферии сет-
чатки при проекции на нее части пространства, расположенного со
стороны носа. В 1954 г. Berens указал на необходимость более вни-
мательного отношения к жалобам на полоски Мооге, наличие ко-
торых свидетельствует о витреоретинальной дегенерации.
Мелькающий поворотный фосфен. Сходный с предыдущим по
происхождению и клиническим проявлениям феномен описан в
1957 г. В. Nebel. В темноадаптированном глазу при пробуждении
ото сна (еще до включения света) быстрое движение глазных яблок
из стороны в сторону может сопровождаться видением вспышек
ярко-желтой лучистости, напоминающих сноп соломы. Комель снопа
ориентирован либо к точке фиксации, либо к слепому пятну. Вспыш-
ки могут ощущаться как раздельно, так и одновременно в обоих
глазах, синхронно с движением глазных яблок. Первая вспышка,
самая яркая, длится доли секунды, последующие быстро угасают.
Феномен объясняется ранним старением организма, сопровож-
дающимся синерезисом стекловидного тела, которое перестает за-
щищать сетчатку от гидродинамических противоударов в момент
саккадических движений глазных яблок.
8.4. ФАКТОР НАЛИЧИЯ НЕПРОЗРАЧНЫХ КРОВЕНОСНЫХ
СОСУДОВ В НОРМАЛЬНО ФУНКЦИОНИРУЮЩЕЙ
ПРОЗРАЧНОЙ СЕТЧАТКЕ
Древовидные фигуры Пуркинье. Если необычным образом осветить
сетчатку рассеянным светом (например, через верхнее веко, приста-
вив к нему кончик светящегося трансиллюминатора или направив
в глаз широкий пучок света от щелевой лампы), то на фоне светя-
щегося поля большинство людей увидят темные извилистые, иног-
да ветвящиеся полоски, сравниваемые с веточкой дерева, паутиной,
растрескавшейся землей. При фиксации взора картинка быстро
исчезает, а при постоянном смещении источника света, наоборот,
сохраняется.
257
Фигуры Пуркинье представляют собой не что иное, как зри-
тельное восприятие тени, которую отбрасывают собственные сосу-
ды сетчатки на фоторецепторы. Благодаря близости тенеобразую-
щих объектов к экрану (рецепторам) они достаточно четко видны.
Иногда, кроме полосок (сосудов), удается видеть “шагреневость”
структуры сетчатки.
Возможность вызвать феномен в глазу с непрозрачными среда-
ми, в частности при гемофтальме, катаракте и другой.патологии,
обусловливает благоприятный прогноз относительно достижения
достаточно высокого форменного зрения после операции. Однако
не каждый пациент может описать быстро гаснущую картинку “ауто-
офтальмоскопии” сосудов сетчатки и особенно “макулошагрени”
{Пивоваров Н.Н., 1973; Сосновский В.В., 1990].
Бегущие светлые пятнышки. Если рассматривать яркий неструкту-
рированный экран, освещаемый сине-голубым светом (например, небо
в безоблачный день), то вблизи зоны фиксации взора можно заметить
передвижение одного за другим ярких светящихся пятнышек. В обра-
зуемой ими цепочке отчетливо видны перерывы. С. Marshall (1935)
объяснил происходящее видением передвижения эритроцитов в ка-
пиллярах парафовеальной области сетчатки, а наличие промежутков —
прохождением невидимых лейкоцитов, что позволяет судить о цирку-
ляции крови в концевых отделах региональных капилляров парафове-
альной зоны. Иногда удается наблюдать пульсацию, синхронную с
сердечным ритмом. В центральной зоне, точке фиксации, феномен в
норме не проявляется, так как фовеальная зона диаметром 0,4—0,5 мм
(1°30*) не содержит капилляров.
8.5. ФАКТОР НАЛИЧИЯ ДУГООБРАЗНОСТИ
В НОРМАЛЬНОЙ ТОПОГРАФИИ НЕРВНЫХ ВОЛОКОН
СЕТЧАТКИ
Синие дуги сетчатки возникают в условиях хотя бы частичной
адаптации к свету при рассматривании прямоугольных предметов с
четкими гранями, светящихся любым, но нагляднее длинноволно-
вым светом (например, тлеющие угли из костра). Когда наиболее
вытянутая грань оказывается параллельной ходу каких-либо не-
рвных волокон в сетчатке, то в поле зрения появляются дуги синего
света (Пуркинье), идущие от изображения самого стимула в на-
правлении слепого пятна. J. Moreland (1968) предположил, что эти
дуги не что иное, как восприятие рисунка части собственных зри-
тельно-нервных волокон сетчатки. Направление стимула со сторо-
ны носа к височной части сетчатки дает ощущение наличия двух
дуг, которые наподобие рогов окаймляют изображение предмета
сверху и снизу. По мере приближения изображения к макулярной
зоне дуги все более сходятся.
258
Феномен используют в нейроофтальмофизиологических иссле-
дованиях, так как он позволяет установить распределение нервных
волокон в сетчатке. С. Ingling и В. Drum (1977) в эксперименте ус-
тановили, что, если воспроизводить феномен в глазу, в котором
часть дуги попадает на темноадаптированную зону сетчатки, эта
часть дуги вместо синей будет казаться серой. Авторы полагают,
что биоэлектрические влияния с непосредственно возбужденных не-
рвных волокон сетчатки передаются на подлежащие палочковые и
колбочковые элементы. В некоторых сине-желтых цветооппонент-
ных нейронах светлоадаптированной сетчатки обычно тонус “оп-
активности” центра рецептивных полей к желтому цвету повышен.
Под влиянием указанных биоэлектрических влияний наступает ла-
теральное торможение восприятия желтых сигналов, и тогда в свет-
лоадаптированном глазу все другие сигналы воспринимаются как
цветооппонентные к желтому, т.е. как синие.
В темноадаптированных глазах, когда тонус к восприятию жел-
того стимула в центрах рецептивных полей ослаблен, “молчащее
окружение” способно только на скотопический ответ и “синие дуги”
не возникают.
8.6. ФАКТОР НАЛИЧИЯ В НОРМЕ СТРОГО ОПРЕДЕЛЕННОЙ
(ОТНОСИТЕЛЬНО ЦЕНТРА МАКУЛЫ) ОРИЕНТАЦИИ
МОЛЕКУЛ КСАНТОФИЛЬНОГО ПИГМЕНТА В НАРУЖНОМ
ПЛЕКСИФОРМНОМ СЛОЕ СЕТЧАТКИ
Щетки (кисти) Гайдингера. Известно, что в фовеальной области сет-
чатки происходит плоскостная поляризация света [De Vries N. et al.,
1953; Stanworth A., Nayler E., 1955]. С помощью макулотеста мож-
но увидеть распределение света в форме мальтийского креста (ще-
ток или кистей). Установлено, что у приматов это явление обуслов-
лено характером ориентации молекул ксантофила в слое Генле
центральной области сетчатки [Snodderly D.M. et al., 1984]. Жел-
тый пигмент блокирует преимущественно синий цвет, что и приво-
дит к плоскополяризованному распределению света. Исчезновение
щеток Гайдингера свидетельствует о патологическом процессе в
фовеальной зоне (например, о макулярном отеке) даже в тех случа-
ях, когда слой фоторецепторов не поврежден, острота зрения оста-
ется высокой и офтальмоскопически патологические процессы не
обнаруживают.
Физиологическая центральная скотома на синий цвет. В присут-
ствии желтого (ксантофильного) пигмента в macula lutea эта об-
ласть в обычных условиях дезадаптирована к синему цвету.
Пятно Максвелла описано автором как темное пятно в точке
фиксации (2—3°) в виде горизонтального овала, окруженного яр-
259
ким синим ореолом. Субъективно эта скотома не ощущается, ее
определяют только в случае использования чистого монохромати-
ческого синего цвета при центральной периметрии..
8.7. РОЛЬ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ
ФОТОРЕЦЕПТОРОВ СЕТЧАТКИ В ПРОИСХОЖДЕНИИ
ЭНТОПТИЧЕСКИХ ЯВЛЕНИЙ
Ощущение светимости собственной сетчатки. Давно замечено, что
при пребывании в полной темноте не возникает зрительного ощу-
щения таксой черноты, как, например, при рассматривании на свету
абсолютно^ерных матовых поверхностей, отражение от которых
близко к нулю. В темноадаптированных глазах постоянно присут-
ствует ощущение серого. Этот “темный свет” рассматривают как
эквивалент фона темноадаптированной сетчатки, свидетельствую-
щий о существовании спонтанной нейрональной активности, веро-
ятно, центрального происхождения [Hart W., 1987].
Фосфены. Этим термином объединяют ощущения свечения,
вспышки в глазу в ответ на воздействие неадекватным для фоторе-
цепторов раздражителем (механическим, термическим, электричес-
ким и др). Светящееся округлое пятно локализуется в поле зрения
на участке, противоположном зоне контакта. Фосфен давления был
открыт в 1819 г. Пуркинье, детально изучен Гельмгольцем (1867) и
назван механофосфеном [Пивоваров Н.Н., 1973]. Феномен механо-
фосфена вызывается после капельной анестезии легким давлением
кончиком стеклянной палочки непосредственно на склеру или че-
рез веко в 13—14 мм от лимба [Волков В.В., Шиляев В.Г., 1976].
При продолжительном давлении на глазное яблоко центр светяще-
гося овала представляется бесцветным, а вокруг него формируются
дуги синего цвета, сливающиеся в сплошной синий пояс [Fried-
man В., 1942]. Феномен более отчетливо заметен при воздействии
на области с максимальной плотностью палочек.
Особым образом феномен проявляется в полностью темноадап-
тированном глазу, например после ночного сна в условиях
сохраняющейся темноты при кратковременном негрубом механи-
ческом воздействии на глазное яблоко. Возникающий при этом
механофосфен в поле, противоположном области давления, прояв-
ляется в виде яркого синего круга, который мгновенно появляется
и столь же быстро исчезает, несмотря на продолжающееся давле-
ние.
Локальные транссклеральные засветы сетчатки с определением
фотопорога [Пивоваров Н.Н., 1973; Сосновский В.В., 1990]. При
контакте (после капельной анестезии) наконечника трансиллюми-
натора с поверхностью глазного яблока в различных квадрантах и
на разном удалении от лимба можно ориентировочно судить о сте-
260
пени функциональной сохранности, наличии или отсутствии отслой-
ки сетчатки в глазу с мутными средами. При этом критерием явля-
ется пороговое ощущение светового пятна в секторе поля зрения,
противоположном засвету. Если транссклеральный фотопорог на-
ходится в пределах от 6 до 24 кд/м2, то поле зрения в исследуемой
зоне/не изменено [Сосновский В.В., 1990]. При повышении порога
до 25 кд/м2, вплоть до отсутствия световосприятия, можно ожидать
сужения поля зрения в исследуемом квадранте до 40° от точки фик-
сации и более.
Полосчатый тест Примрозе (Primrose). Если ’’палочку Мед-
докса” расположить в переднем фокусе глаза и через нее зеркалом
офтальмоскопа направить пучок света на зрачок, то даже при мут-
ных средах в макулярной области формируется изображение светя-
щейся прерывистой линии, расположенной перпендикулярно к оси
призматических элементов. Исследователь может видеть эту линию
на поверхности глаза. Правильное определение пациентом положе-
ния светящейся полоски свидетельствует о хорошем прогнозе вос-
становления форменного зрения после хирургического восстанов-
ления прозрачности сред глаза [Волков В.В., Шиляев В.Г., 1976]. При
положительной полосчатой пробе прогнозируемая острота зрения
может составить не менее 0,03 [Сосновский В.В., 1990].
Эффект Стайлса — Крауфорда (Stiles — Crawford). Этот фено-
мен свидетельствует о различном распределении относительной чув-
ствительности ретинальных фоторецепторов в зависимости от их
топографии и места прохождения света через зрачок. Один и тот же
пучок света кажется неодинаково ярким в зависимости от того, как
он проникает в глаз: через центр зрачка или его периферические
отделы. Параллельные лучи света, попадающие на центральную зону
глазного дна под прямым углом к поверхности сетчатки, вызывают
ощущение большей яркости, чем лучи, которые входят в глаз по
краю широкого зрачка и, преломившись, достигают фоторецепто-
ров под острым углом. Таким образом, чувствительность сетчатки
значительно снижается при вхождении в глаз светового луча не через
центр зрачка, а с периферии.
По характеру фовеального пятна от наблюдаемого источника
света можно установить, что в норме чувствительность глаза рав-
номерно снижается от центра к периферии.
Как проявление эффекта Stiles — Crawford возможна асиммет-
рия яркостной чувствительности в фовеальной зоне у больных с
миопическими стафиломами на глазном дне и некорригированны-
ми аномалиями рефракции. Даже при небольшом угле наклона па-
дающего в глаз луча света по отношению к оси фоторецептора све-
точувствительность последнего заметно снижается.
G. Westheimer (1968) изучал феномен на себе при рассматрива-
нии яркого источника света (звезды на фоне темного неба) с пред-
намеренно расфокусированным изображением. По распределению
261
яркости ощущаемого в пятне светорассеяния автор мог судить о
появляющейся асимметрии светочувствительности в макулярной
области. В условиях искусственной аметропии ее максимум мог
сдвигаться либо к височной, либо к носовой половине макулы (с
учетом смещения направления основного пучка преломленных лу-
чей). Следовательно, светочувствительность определяется не толь-
ко плотностью размещения фоторецепторов, но и их ориентацией
относительно направления падающих световых лучей.
Таким образом, энтоптические феномены не только способству-
ют лучшему пониманию физиологических процессов в зрительной
системе, но также могут быть полезны клиницистам в оценке функ-
ционального состояния сетчатки, объяснении природы нарушения
зрения и прогнозировании перспектив его восстановления при со-
ответствующем лечении.
ЧАСТЬП
ГЛАВА 1
ВЕКИ КАК ЗАЩИТНЫЙ АППАРАТ ГЛАЗА
1.1. СТРОЕНИЕ И ФУНКЦИЯ ВЕК В НОРМЕ
Веки выполняют защитную функцию благодаря наличию ресниц и
бровей, выделению сального секрета мейбомиевыми (железы хряща век)
и другими железами, согласованной работе двигательного аппарата.
Ресницы и б р о в и. Ресницы образуют двухрядный нежный
заслон для глазной щели из 100—150 коротких волосков на верх-
нем веке и вдвое меньшего количества на нижнем. Через каждые
3—5 мес они заменяются новыми. Брови усиливают контур верхне-
го края глазницы. Как и ресницы, они состоят из коротких волос-
ков. В какой-то мере брови защищают глаз от неблагоприятных
воздействий с поверхности лба.
Мейбомиевы железы. Располагаясь в толще хрящевой
пластинки параллельными рядами, эти железы (около 30 в каждом
веке) открываются позади корней ресниц, в межреберном простран-
стве (рис. 1.1). Выделяемый ими сальный секрет участвует в форми-
ровании поверхностного слоя пленки, препятствующего как испа-
рению слезы, так и скатыванию ее через край века на кожу лица.
Слезная пленка предохраняет глаз от высыхания.
Двигательный аппарат век является решающим фактором защит-
ной функции век. Он представлен мышцей, поднимающей верхнее
веко, круговой мышцей глаза, частью которой является мышца Ри-
олана, и гладкомышечными волокнами верхнего и нижнего век (мыш-
ца Мюллера). Веки могут открываться и смыкаться как произволь-
но, так и рефлекторно, и мигание осуществляется с определенной
периодичностью с помощью указанных механизмов и спонтанно.
Поднимание верхнего века и раскрывание глазной щели. Основной
мышцей является мышца, поднимающая верхнее веко, вспомога-
тельную роль играет лежащая под ней мышца Мюллера, а при взгля-
де вверх — лобная и верхняя прямая мышцы. Ослабление функции
последней может повлиять на действие мышцы, поднимающей вер-
хнее веко [Moses R.A., 1987]. Эта мышца, как и верхняя прямая
мышца, иннервируется III парой черепных нервов, лобная мыш-
ца — VII парой, а мышца Мюллера имеет симпатическую иннерва-
цию. В норме диапазон раскрытия глазной щели от 8 до 12 мм.
263
1
Рис. 1.1. Сечение через верхнее веко и передний отдел глазного яблока.
1 — мышца, поднимающая верхнее веко; 2 — мышца Мюллера; 3 — орбитальная
мышца; 4 — периферическая аркада; 5 — нисходящая ветвь; 6 — мейбомиева железа
в тарзальной части; 7 — восходящая ветвь; 8 — краевая аркада; 9 — мышца Риола-
на; 10 — малый круг кровообращения; И — конъюнктива; 12 — большой круг кро-
вообращения; 13 — передняя цилиарная артерия; 14 — верхняя прямая мышца [Moses
R., 1987].
Опускание верхнего века и закрывание глазной щели. Глазная щель
закрывается благодаря сокращению круговой мышцы глаза, иннер-
вируемой VII парой черепных нервов. В ней выделяют две основ-
ные порции: пальпебральную, лежащую претарзально, и орбиталь-
ную, расположенную пресептально. Первая функционирует при
мигании, вторая поддерживает постоянный тонус и используется
при произвольном смыкании век. Орбитальная порция, сокращаясь
в 2 раза медленнее, действует сильнее [ Hoyt W. et al., 1965 ]. Край
века прижимается к глазному яблоку волокнами, известными как
мышца Риолана. Глубокая часть пальпебральной порции круговой
мышцы играет важную роль в механизме слезоотведения.
264
Рис. 1.2. Оценка силы мышцы, под*
нимающей верхнее веко [Kanski J.,
1989]. Объяснение в тексте.
Мигательные движения век. Электромиографически установле-
но, что при произвольных мигательных движениях мышца, подни-
мающая верхнее веко, и круговая мышца глаза действуют рецип-
рокно: активность одной сопровождается пассивностью другой
(рис. 1.1). Если верхнее веко медленно опускается, то не только сни-
жается активность поднимающей его мышцы, но остается пассив-
ным и антагонист (круговая мышца). Минимальный интервал меж-
ду повторными произвольными миганиями 0,3 с.
Рефлекторное мигание возникает по разным причинам: при воз-
действии яркого света, внезапном появлении объекта перед глаза-
ми, сильном шуме, прикосновении к ресницам и т.д. Дозированные
яркие засветы, оценку тактильного корнеального рефлекса исполь-
зуют для диагностики уровня поражения в нейроофтальмологии
[HoytW. et al., 1965].
Спонтанные мигания возникают без очевидных стимулов в, пери-
од бодрствования, сохраняются у слепых. У большинства взрослых
частота мигания достигает 15 раз в 1 мин, при длительности каждого
мигания 0,3—0,4 с. Эти мигания редки у маленьких детей. Зритель-
ное ощущение не прерывается при мигании, как и при быстрых сак-
кадических движениях глазного яблока. Между тем искусственное
прерывание видимости объекта даже на более короткое время обыч-
но вызывает дискомфорт. Этот факт заслуживает внимания при изу-
чении пилотирования высокоскоростных летательных аппаратов.
Согласованность действий различных мышц век и глазного ябло-
ка. Результаты электромиографии свидетельствуют о том, что к
моменту начала сокращения круговой мышцы глаза ее антагонист
265
(мышца, поднимающая верхнее веко) приходит в состояние полной
релаксации (см. рис. 1.1). Это обстоятельство, а также способность
претарзальных волокон круговой мышцы сокращаться с очень
высокой частотой (порядка 390 в 1 мин) создают условия для высо-
кой стартовой скорости непроизвольного смыкания век, остающе-
гося незамеченным. Мышцы, поднимающие верхние веки обоих глаз,
работают синхронно, поскольку, согласно закону Геринга, парные
мышцы одновременно получают нервные импульсы равной силы.
Не случайно, для того чтобы облегчить осмотр больного глаза,
пациента просят открыть оба глаза.
Наконец, наблюдаются ассоциированные движения век и глаз-
ного яблока. Так1 при переводе взора круто вверх в норме движе-
ния век и глазного яблока сочетаются.
Полагают, что за координацию движения век отвечает особый
’’моторный центр”, расположенный в лобной доле коры больших
полушарий головного мозга. Каков путь от этого центра к стволу
мозга до конца неизвестно.
1.2. КЛИНИЧЕСКИЕ ВАРИАНТЫ НАРУШЕНИЯ
ФУНКЦИИ ВЕК
• Снижение защищенности глаз от случайных травм, проявляюще-
еся в частом попадании в конъюнктивальный мешок инородных
тел вследствие облысения краев век или ослабления тактильного
мигательного рефлекса, проявляющегося также в более высокой
вероятности других травм глаза в связи с потерей остальных ми-
гательных рефлексов (на яркий свет, внезапно обнаруженные ря-
дом с глазом опасные объекты).
• Слезотечение, связанное с гипофункцией мейбомиевых желез (не-
качественность слезной пленки), из-за атонии век, обусловлен-
ной слабостью круговой мышцы глаза, особенно ее пальпебраль-
ной порции и мышцы Риолана.
• Невозможность полноценного зрения вследствие нарушения функ-
ции открывания глазной щели (парез мышцы, поднимающей верх-
нее веко, или выраженный блефароспазм).
• Подсыхание роговицы с возможным стойким ее помутнением из-
за нарушения функции закрывания глазной щели (гипофункция
круговой мышцы или гиперфункция мышцы, поднимающей верх-
нее веко).
• Рассогласование нормальных и появление патологических ассо-
циированных движений век и глазного яблока, век и челюстно-
лицевой мускулатуры (нарушение связей в центральной нервной
системе).
266
1.3. ПРИЕМЫ ФУНКЦИОНАЛЬНОЙ ДИАГНОСТИКИ
И ТРАКТОВКА РЕЗУЛЬТАТОВ
1.3.1. Тестирование мигательного рефлекса
На основе тактильного мигательного рефлекса разработаны тесты
оценки чувствительности роговицы. В упрощенном варианте тест
состоит в следующем. Пациента просят слегка запрокинуть голову,
открыть пошире глаза и смотреть прямо перед собой. Врач, не
придерживая век, сбоку подносит ватку, скрученную на конус, и
осторожно касается ее кончиком роговицы. Наличие или отсутствие
ответного мигания, хотя и ориентировочно, свидетельствует о чув-
ствительности роговицы. Элементы неожиданности и грубого трав-
мирования роговицы должны быть исключены. Усовершенствова-
ние теста шло по пути более строгого выбора зоны касания и силы
прилагаемого воздействия.
Техника количественной пробы. Согласно мето-
дике Фрея — Самойлова, для контакта с роговицей используют
волоски, установленные под прямым углом, калиброванные по ди-
аметру торцовой части с силой давящего воздействия 0,3,1,0 и 10 г/
мм2. В другом варианте алгезиметра волоски заменены проволоч-
кой диаметром 0,2 мм, силу воздействия которой дозируют массой
упорной муфты (2, 10 и 50 мг) [Меркулов И.И., 1964].
В алгезиметре Добромыслова — Ромашевского, который при-
знан наиболее совершенным, использована капроновая нить диа-
метром 0,25 мм, которую помещают на роговицу подобно тономет-
ру Маклакова с переменной муфтой, осуществляющей воздействие
весом 50, 100, 200 и 400 мг. При этом исключен сложный момент
улавливания прогиба нити. С учетом ощущаемого пациентом по-
рогового давления на единицу площади по таблице легко рассчи-
тать показатель относительной чувствительности роговицы (в про-
центах от нормы), суммарный для всех исследуемых точек, количе-
ство которых у разных авторов варьировало от 13 до 33 при
различных схемах их распределения по роговице.
Оценка результатов. О степени снижения чувствитель-
ности (анестезии) в исследуемой точке на роговице судят не только
по отсутствию субъективного чувства касания, но и объективно —
по отсутствию мигательного рефлекса.
При определении причин снижения чувствительности роговицы
обычно обращают внимание на возможное поражение тактильных
рецепторов, которые поражаются в ходе и исходе различных забо-
леваний роговицы. Однако отсутствие мигания в ответ на прикос-
новение к роговице может быть следствием патологии как по ходу
афферентных (V пара черепных нервов), так и эфферентных
(VII пара) путей рефлекторной дуги. О наличии у человека связи
тактильного мигательного рефлекса с корой свидетельствует воз-
267
никновение боли и блефароспазма одновременно с ощущением при-
косновения. Мигательный рефлекс отсутствует при локализации по-
ражения в роландовой борозде коры; потеря тактильной чувстви-
тельности роговицы на стороне опухоли мостомозжечкового угла —
наиболее ранний симптом заболевания.
Мигательные рефлексы на яркий засвет и внезапную угрозу для
глаза берут начало в сетчатке и по афферентным путям зрительно-
го нерва замыкаются: первый — на подкорковом уровне (в ядре VII
пары черепных нервов), второй — в коре больших полушарий го-
ловного мозга.
Отсутствие мигательного рефлекса на яркие засветы может быть
признаком поражения мезенцефальной области. Повышенная чув-
ствительность к засветам, сопровождающаяся чувством боли в гла-
зу, иногда в голове, слезотечением, блефароспазмом (в отсутствие
симптомов раздражения глазного яблока), наоборот, позволяет
заподозрить синдром Бурденко — Крамера (раздражение мозговых
оболочек с вовлечением тенториальных веточек тройничного не-
рва).
Отсутствие рефлекса на внезапную угрозу для глаза при сохра-
нении тактильного и светового мигательных рефлексов свойствен-
но корковой природе поражения.
1.3.2. Функциональная оценка края век
Защитная функция края век определяется наличием и качеством
ресниц, характером секреции мейбомиевых желез и положением
краевых структур века относительно глазного яблока.
Простого осмотра достаточно, чтобы обнаружить обломанность
ресниц, их небольшое количество, а иногда и неправильный рост,
в частности по направлению к глазному яблоку.
Для оценки функции мейбомиевых желез пациента просят при-
крыть глаза. Расположив I и II пальцы на каждом из век, края их
форсированно сближают, добиваясь легкого выворота. При этом
веки соприкасаются уже не своими краями, а слизистыми оболоч-
ками, покрывающими хрящи. Смещая верхнее и нижнее веко в
разные стороны и продолжая сдавливать хрящевые пластинки, на-
блюдают за протоками мейбомиевых желез. Отсутствие секрета или
его водянистость расценивают как проявление гипосекреции липи-
дов, необходимых для формирования защитной слезной пленки,
обильный желтовато-белый вязкий секрет — признак хронического
мейбомиевого блефарита.
Положение края века относительно сферичной поверхности глаз-
ного яблока в отсутствие рубцовых изменений определяется состо-
янием тонуса пальпебральной порции круговой мышцы глаза и
особенно мышцы Риолана. При их избыточном тонусе на фоне
сглаживания заднего ребра края века возникает entropion (заворот
268
нижнего века), что легче всего определить по положению ресниц,
которые, подвернувшись, касаются роговицы. Чаще из-за недостат-
ка тонуса этой мышцы край века отходит от глазного яблока, слез-
ные точки теряют контакт со слезным озером, глаз начинает сле-
зиться.
Состояние тонуса претарзальной порции круговой мышцы гла-
за легко проверить, если оттянуть пальцем нижнее веко, а затем
отпустить, наблюдая за положением его края. У пожилых людей,
особенно при слабости мышцы, веко очень медленно возвращается
в исходную позицию, а иногда не приходит в полное соприкосно-
вение с глазным яблоком. По наблюдениям М.Ю.Султанова (1987),
еще одним тонким признаком атонии мышцы Риолана является
неспособность слезных точек и канальцев к сплющиванию (и при
сомкнутых, и при открытых веках они остаются зияющими).
1.3.3. Тестирование функции открывания глазной щели
Роль мышцы, поднимающей верхнее веко, Ослабление или утрата
функции мышцы, поднимающей верхнее веко, проявляется в птозе
(опущении) верхнего века разной степени выраженности.
Техника исследования силы мышцы, под-
нимающей верхнее веко. Для того чтобы исключить вли-
яние лобной мышцы, на бровь пациента помещают I палец и плот-
но прижимают им подлежащие ткани к кости. Пациента просят
посмотреть вниз, насколько это возможно. Нулевое деление верти-
кально расположенной линейки совмещают с ресничным краем
верхнего века в средней его части (см. рис. 1.2). Затем просят паци-
ента с усилием посмотреть вверх и по линейке, остающейся непод-
вижной, устанавливают величину экскурса края верхнего века.
Оценка результатов определения силы мышцы, поднимающей
верхнее веко: при амплитуде 15 мм — нормальная сила; 8—14 мм —
достаточно высокая сила; 5—7 мм — небольшое уменьшение, 4 мм
и менее — выраженная слабость.
Техника определения степени выраженно-
сти птоза. При одностороннем птозе достаточно сопоставить
по ширине (высоте) глазные щели парных глаз — разница в милли-
метрах и составит искомый показатель. При двустороннем птозе
исходят из представлений о средней норме. Нулевую отметку ли-
нейки помещают на край нижнего века (рис. 1.3), который практи-
чески у всех в меридиане 6 ч совпадает с нижним лимбом роговицы.
Далее принимают во внимание, что вертикальный диаметр рогови-
цы в среднем равен 11 мм, а при первичной позиции глазного яб-
лока (взгляд прямо перед собой) верхняя часть ее в норме на 2 мм
прикрыта верхним веком. Следовательно, в представленном случае
на долю птоза приходится также 2 мм.
Оценка результатов исследования птоза [KanskiJ., 1989]: до
269
Рис. 1.3. Оценка степени птоза [Kanski J., 1989].
2 мм — птоз легкий, веко не прикрывает зрачок; 3 мм — птоз уме-
ренный, зрачок наполовину прикрыт; 4 мм и более — птоз выра-
женный, зрение нарушено, так как прикрыт весь зрачок.
Особенности клинической диагностики в зависимости от проис-
хождения птоза. При врожденном птозе следует исключить нали-
чие синкинетических связей мышцы, поднимающей верхнее веко, с
другими группами мышц. Очень редко птоз является всего лишь
составной частью генетически обусловленного синдрома, включа-
ющего прогрессирующую наружную офтальмоплегию, пигментную
ретинопатию и сердечную блокаду (синдром Kearns — Sayre). Врож-
денный птоз может входить также в симптомокомплекс миопати-
ческой дистрофии, при которой в процесс вовлекаются лицевые
мышцы, создавая вид печальной экспрессии, наблюдаются потеря
волосистого покрова головы и пресенильная катаракта.
При приобретенном птозе важно исключить системную патоло-
гию мышц, в частности myasthenia gravis. Последняя относится к
числу редких аутоиммунных заболеваний, чаще развивается у жен-
щин в возрасте 20—40 лет. Характеризуется повышенной утомляе-
мостью всей скелетной мускулатуры, однако в 60% случаев первым
признаком является легкий, обычно двусторонний, хотя и необяза-
тельно симметричный птоз, усиливающийся к исходу дня, позже
нередко присоединяется диплопия.
Для выявления слабости мышцы, поднимающей верхнее веко,
проще всего попросить пациента несколько раз закрыть и открыть
глаза, однако существует и специальный тест на ’’подергивания” по
Когану [Kanski J., 1989].
270
Техника выполнения проб и оценка ре-
зультатов. Пациента просят посмотреть вниз и затем быстро
перевести взор в обычную (взгляд прямо вперед) позицию. При по-
ложительной пробе верхнее веко начинает подергиваться, после чего
медленно опускается в привычную позицию.
Применяют также специальный тензилонный тест (тензилон —
ингибитор холинэстеразы), который выполняют следующим обра-
зом.
1. Определяют исходную степень выраженности птоза и/или силу
мышцы, поднимающей верхнее веко.
2. Внутривенно вводят 0,3 мг атропина (для смягчения действия
тензилона).
3. В туберкулиновый шприц набирают 1 мл тензилона (10 мг).
4. В локтевую вену вводят 0,2 мл (2 мг) тензилона и, не извлекая
иглу из вены, наблюдают за состоянием пациента.
5. Через 60 с, если не появилось обильное слюноотделение, пот,
слезотечение, медленно вводят остальные 0,8 мл тензилона.
При положительном результате пробы уже через 30—60 с птоз
исчезает и эффект сохраняется до 5 мин. Для документации резуль-
татов пробы может быть использована электромиография.
Роль мышцы Мюллера. Волокна этой мышцы очень тонкие и
маломощные, поэтому недостаточность ее функции может быть
причиной лишь птоза легких степеней, придающего пациенту сон-
ный вид (на обоих глазах при трахоме, на одном глазу при синдро-
ме Горнера).
При синдроме Горнера парез гладкой мышцы верхнего века
обычно сочетается с парезом аналогичной мышцы нижнего века.
Вследствие этого не только слегка опускается верхнее веко, но и
приподнимается край нижнего века, освободившийся от тяги соб-
ственных гладкомышечных структур. В конечном счете из-за суже-
ния глазной щели создается впечатление энофтальма.
Роль круговой мышцы глаза. Открыванию глазной щели не столь
уж редко может препятствовать временный спазм круговой мышцы
глаза, который возникает при острых воспалительных заболевани-
ях и травмах переднего отдела глазного яблока. Этот блефаро-
спазм усиливается при ярких засветах и осложняется иногда заво-
ротом век.
Значительно реже встречается эссенциальный блефароспазм
обычно трудно распознаваемого неврологического происхождения.
Приступы непроизвольного сокращения круговой мышцы глаза у
больных с выраженным склерозом сосудов головного мозга неред-
ко двусторонние, и тогда больные на время становятся практически
слепыми. Между тем при осмотрах вне приступа каких-либо откло-
нений от нормы в состоянии функции круговой мышцы глаза най-
ти не удается. При одностороннем затруднении открывания глаз-
ной щели прежде всего нужно исключить раздражение лицевого
271
нерва вследствие сдавления его артериальной петлей в зоне выхода
корешка. Эту патологию необходимо дифференцировать от миоки-
мии, которой свойственно постоянное трепетание мышц лица (по
причине рассеянного склероза, глиомы и других заболеваний го-
ловного мозга), а также от апраксии, т.е. нарушения целенаправ-
ленного действия, на почве экстрапирамидной дисфункции (при
сохранении нормальной силы всех мышц). Какой бы ни была эти-
ология блефароспазма, в патологический процесс всегда вовлека-
ются все порции круговой мышцы, включая мышцы брови
[Hoyt W.F., Loeffler J.D., 1965].
1.3.4. Тестирование функции закрывания глазной щели
Роль круговой мышцы глаза. Во время сна веки не просто смыкаются
в результате расслабления мышц, поддерживающих глазную щель
открытой. Наряду с торможением функции мышцы, поднимающей
верхнее веко, в состояние определенного тонуса приходит круго-
вая мышца глаза. При пробуждении все указанные процессы идут
с ’’обратным знаком”.
Смыкание век становится невозможным при ослаблении или
утрате функции круговой мышцы глаза, например, вследствие
паралича лицевого нерва (паралич Белла). Образующийся лаго-
фтальм (“заячий глаз”) нередко осложняется подсыханием и по-
мутнением роговицы, отвисанием нижнего века (ectropion) и сле-
зотечением.
Роль мышцы Мюллера. Попытка нормально сомкнуть веки, что-
бы полностью закрыть глазную щель, иногда затруднена при повы-
шении тонуса гладкомышечных волокон мышцы Мюллера [Вол-
ков В.В., Шиляев В.Г., 1985]. Так, у больных тиреотоксикозом пре-
обладание тонуса симпатической иннервации нередко проявляется
в ретракции верхних век обоих глаз. При этом глазная щель оказы-
вается максимально широкой, в ее пределах заметна белая полоска
склеры над роговицей (симптом Дальримпля). Чрезмерный тонус
мышцы Мюллера лежит в основе и другого симптома гипертирео-
за— симптома Грефе, проявляющегося в нарушении ассоцииро-
ванного движения глазного яблока и верхнего века (при взгляде
вниз веко отстает от яблока).
Роль мышцы, поднимающей верхнее веко. Очень редко, когда
тиреотоксикоз исключен, ретракция век может быть связана с по-
вышением тонуса не симпатической, а парасимпатической иннерва-
ции в связи с патологией в области задней комиссуры мозга. Гипер-
тонус мышцы, поднимающей верхнее веко, приводит к подворачи-
ванию верхнего века — симптом Кольера(СоШег). Однако
патологический очаг в зоне ствола мозга чаще вызывает не возбуж-
дение, а парез мышцы, поднимающей верхнее веко, и птоз.
272
1.3.5. Тестирование ассоциированных движений
феномен Маркуса — Гунна, У некоторых пациентов с врожденным
птозом движения нижней челюсти (открывание-закрывание рта,
смещение в стороны) сопровождаются синхронными движениями
опущенного века. Так, при открывании рта птоз исчезает, а при
закрывании — появляется вновь. Полагают, что причина указан-
ных синкинезий в наличии патологических связей между центрами
этих функций в коре мозга.
Феномен псевдо-Грефе, Наблюдается в процессе регенерации
после паралича III пары черепных нервов. При взгляде прямо, а
также при отведении глазного яблока на стороне бывшего пораже-
ния сохраняется легкий птоз. Однако стоит перевести взгляд кнут-
ри (состояние приведения глазного яблока), как не только исчезает
птоз, но и возникает ретракция верхнего века. Полагают, что при
регенерации часть волокон III пары черепных нервов изменяет на-
правление роста в сторону мышцы, поднимающей верхнее веко.
В итоге стимул к аддукции и создает эффект поднимания верхнего
века.
Физиологический симптом Белла. Нормальное смыкание век, как
во время сна, у 90% здоровых людей сопровождается “закатывани-
ем” глазного яблока кверху. Примечательно, что даже при наруше-
нии произвольного взгляда вверх (например, вследствие супранук-
леарного паралича) симптом Белла может сохраняться [Moses R.A.,
1987].
ГЛАВА2
СЛЕЗОПРОДУЦИРУЮЩИЙ
И СЛЕЗООТВОДЯЩИЙ АППАРАТ
2.1. СТРОЕНИЕ И ФУНКЦИИ СЛЕЗНОГО АППАРАТА
В НОРМЕ
В конъюнктивальном мешке постоянно присутствует около 6—7 мкл
слезной жидкости. При раскрывании глазной щели она не излива-
ется из мешка, а в виде пленки распространяется по поверхности
глазного яблока [Сомов Е.Е., Бржеский В.В., 1994].
2.1.1. Слезная пленка
В слезной пленке выделяют 3 слоя (рис.2.1), различающихся по
составу и функции [Holly F.J. и др., 1977; Milder В., 1987].
Первый, поверхностный, слой—липидный: замедляет испарение
влаги из подлежащего водянистого слоя и уменьшает теплоотдачу;
увеличивая поверхностное натяжение, способствует стабильности
пленки, что особенно важно в вертикальном положении тела, так
как пленка препятствует перекатыванию слезы через край века;
служит смазкой для век при скольжении их по поверхности глазно-
го яблока.
Второй, средний, самый широкий, слой — водянистый: благода-
ря непрерывному обновлению способствует доставке к роговично-
му эпителию кислорода из атмосферы, ферментов, аминокислот и
других продуктов метаболизма из желез и кровеносных сосудов;
служит защитной средой, препятствующей инфицированию, благо-
даря наличию в нем лактоферрина, лизоцима, иммуноглобулинов и
других биохимических ингредиентов; обеспечивает гладкость опти-
ческой поверхности роговицы, устраняя все неровности на ней и
как бы формируя контактную линзу; смывает шлаковые метаболи-
ты, детрит и инородные тела.
Третий, глубокий, очень тонкий, слой —муциновый: преобразу-
ет поверхность роговицы из гидрофобной в гидрофильную.
В отсутствие муцина роговица теряет способность смачиваться
слезой, так как ее поверхность становится гидрофобной. Попав на
такую поверхность, частицы влаги как бы отталкиваются от нее,
собираясь в капли.
2.1.2. Слезопродукция
Для формирования липидного слоя пленки необходима нормаль-
ная функция мейбомиевых желез, а также железок Цейсса и Молля,
274
Рис. 2.1. Структура и состав слез-
ной пленки [Holly F.J., Lemp М.А.,
1977].
1 — поверхностный липидный слой'
(0,1 мкм), состоящий из воска и холес-
териновых эфиров, а также поляризо-
ванных липидов; 2 — водянистый
слой (7 мкм), содержащий в, раство-
ренном виде неорганические соли, глю-
козу, мочевину, поверхностно-актив-
ные биополимеры, белки и гликопро-
теины; 3 — микровиллы (ворсинки);
4 — слизистый слой (0,02—0,05 мкм);
5 — гидратированный слой мукопроте-
инов, обогащенных в саломуцине.
открывающихся на краю века. Липиды составлены из молекул хо-
лестерина, которые обеспечивают связь липидного слоя с подлежа-
щим водянистым слоем пленки, и молекул воска, создающих на ее
поверхности условия гидрофобности.
Основная масса слезной жидкости, которая формирует водяни-
стый слой пленки, секретируется слезными железами, главной (Кра-
узе) и добавочными (Вольфринга). При этом именно добавочные
железы функционируют постоянно, а главная железа обеспечивает
преимущественно рефлекторное слезотечение (при действии разно-
образных раздражителей на слизистую оболочку глаз или носа, а
также во время плача).
В образовании муцинового слоя участвуют эпителиальные и
бокаловидные клетки роговицы, а также железки Генле и Манца,
имеющиеся в конъюнктиве. Синтезируемый и выделяемый рогович-
ным эпителием муцин состоит из молекул мукопротеинов, которые
275
Рис. 2.2. Функция муци-
нового слоя слезной плен-
ки, эпителиальные клет-
ки (схема).
а — недостаточная; б — нор-
мальная.
создают на поверхности эпителиальных ворсинок (microvilli) базаль-
ную прослойку. Она водонерастворима и, следовательно, гидрофоб-
на. При контакте такого слоя с водянистым слоем слезной пленки
последняя распадается на отдельные капли. Только тогда, когда в
пространство между ворсинками эпителия проникает еще одна со-
ставляющая муцинового слоя, секрет бокаловидных клеток, и об-
разуется дополнительная прослойка толщиной всего 0,02 — 0,05 мкм,
поверхность эпителия становится гидрофильной, а слезная пленка
приобретает структурное единство (рис. 2.2).
Основные условия нормальной смены слезной пленки на повер-
хности роговици следующие [Kanski J., 1985]:* достаточная продук-
ция всех трех основных компонентов слезной пленки; нормальный
мигательный рефлекс (при его нарушении, например, в случае ла-
гофтальма, страдает механизм перемещения муцина бокаловидных
клеток из нижнего свода конъюнктивы на поверхность эпителия
роговицы); хорошее соответствие по кривизне контактирующих
поверхностей (внутренней на веках и наружной на глазном яблоке).
2.1.3. Слезоотведение
Примерно 25% слезы, поступившей в. конъюнктивальный мешок,
испаряется с поверхности глаза, остальная оттекает через слезоот-
водящие пути в полость носа.
В конъюнктивальной полости слеза смещается в направлении
внутреннего угла глазной щели. Этому способствуют сила тяжес-
ти, благодаря которой слезный ручей формируется именно вдоль
края нижнего века; капиллярное притяжение слезных точек, актив-
но всасывающих слезу в вертикальную ампулу слезных канальцев
(преимущественно нижних); мигательные движения век, обеспечи-
вающие дальнейшее продвижение слезы.
При сокращении пресептальных волокон круговой мышцы гла-
276
за веки по типу застежки-молнии плавно смыкаются в направлении
от виска к носу и смещают в соответствующем направлении слез-
ную жидкость, находящуюся в конъюнктивальном мешке; часть этой
жидкости проталкивается в слезные канальцы.
На завершающем этапе смыкания век слезные точки и верти-
кальные отделы слезных канальцев сплющиваются, горизонталь-
ные их отделы укорачиваются, а наружная стенка слезного мешка
оттягивается от внутренней, благодаря чему порция слезы из слез-
ных канальцев перемещается в слезный мешок.
При раскрывании глазной щели после мигательного движения
и релаксации упомянутых выше мышц стенки слезного мешка спа-
даются, а содержащаяся в нем слеза выдавливается через слезно-
носовой канал в полость носа.
В процессе размыкания век слезные точки и ампулы слезных
канальцев также раскрываются и заполняются новыми порциями
слезы.
Стоит моргнуть — и цикл слезоотведения повторится.
Скоростная фотография мигательных движений век, выполнен-
ная M.G. Doane (1981), показала, что до завершения смыкания век
верхняя й нижняя слезные точки успевают встретиться и блокиро-
вать друг друга. В таком положении они остаются не только при
сомкнутых веках, но и до почти полного раскрытия глазной щели.
Наконец, в последний момент они вдруг расходятся (при уже пол-
ностью открытых глазах) и в канальцы, а также в мешок ex vacuo
устремляется слеза. Согласно этой теории, слезному мешку отво-
дится пассивная роль в слезоотведении.
Тот факт, что частым морганием и усилением носового дыха-
ния можно компенсировать избыток слезопродукции, не вызывает
сомнений. Однако это не исключает наличия и других механизмов
слезоотведения. Известно, что с уменьшением количества мигатель-
ных движений в 10 раз (от 30 в 1 мин во время беседы до 3 в 1 мин
при чтении) слезотечение не появляется. Возможно, это связано со
снижением слезопродукции.
2.2. КЛИНИЧЕСКИЕ ВАРИАНТЫ НАРУШЕНИЙ ФУНКЦИИ
УВЛАЖНЕНИЯ ГЛАЗА
• Чувство сухости, засоренности, а периодически и жжения в глазу,
возникающее в связи с гипофункцией слезных желез, проявляю-
щееся в виде сухого кератоконъюнктивита, синдрома Шегрена,
болезни Микулича либо по причине неполноценности слезной
пленки из-за нарушения механизмов образования муцина в конъ-
юнктиве или роговице, регулируемых симпатической иннерваци-
ей, а также из-за дефицита липидов, выделяемых железами, от-
крывающимися на краю века.
277
• Отсутствие слез во время плача при блокировании иннервирую-
щих слезную железу парасимпатических волокон лицевого нерва
и ganglion sphenopalatinum.
• Рефлекторное слезотечение при гиперфункции основной слезной
железы как периферического генеза в случае раздражения окон-
чаний тройничного нерва в слизистой оболочке носа, глотки,
конъюнктивальной полости, так и центрального психогенного
происхождения.
• Постоянное слезотечение, нередко с наличием серозного или даже
гнойного отделяемого из слезных точек обычно вследствие пол-
ной облитерации слезоотводящих путей в нижнем их отделе (дак-
риостенозы, дакриоциститы).
• Слезотечение без отделяемого, возникающее периодически только
на ветру или в холодное время года, чаще обусловлено нарушени-
ем функции верхнего отдела слезоотводящих путей (дислокация,
атония, сужение или закупорка слезных точек или канальцев).
2.3. ПРИЕМЫ ФУНКЦИОНАЛЬНОЙ ДИАГНОСТИКИ
И ТРАКТОВКА РЕЗУЛЬТАТОВ
2.3.1. “Сухой” глаз, тестирование слезопродукции и качества
слезной пленки
Слезопродукция даже в норме отличается высокой вариабельно-
стью. Однако ее стойкое снижение удается надежно объективизиро-
вать с помощью тестов Ширмера.
Тест Ширмера I. Техника выполнения
пробы. Тестовые полоски, например из набора фирмы “Alcon”,
имеют ширину 5 мм и длину 35 мм (для пробы пригодна обычная
фильтровальная бумага такого размера). На расстоянии 5 мм от
одного из концов полоску перегибают, создавая угол порядка 45°.
Короткий конец полоски закладывают за нижнее веко на границе
наружной и средней третей его края. Пациента просят прикрыть
глаза, разрешая при необходимости моргать. Через 5 мин полоску
извлекают и, начиная от изгиба, измеряют длину увлажненной ча-
сти длинного конца полоски.
Оценка результатов. Большинство исследователей
считают слезопродукцию нормальной, если влага продвинулась до
10 — 25 мм. При увлажнении менее 5 мм появляются основания
подозревать гипопродукцию слезы, наблюдающуюся при послеожо-
говых и других рубцовых изменениях в конъюнктиве, однако сле-
дует иметь в виду, что увлажнению может мешать и просто скопив-
шаяся в сводах конъюнктивы слизь.
Результаты проверки на парных глазах могут не совпадать, но
клинически значимой является лишь разница, превышающая 27%.
278
0-15 16-30 31-45 46-60 61-75 76-90 лет
Рис. 2.3. Слезопродукция в зависимости от возраста [Бржеский В.В., 1990].
1 — базисная; 2 — рефлекторная.
Суточные колебания слезопродукции несущественны, но снижение
ее с возрастом, по данным Е.Е. Сомова и В.В. Бржеского, следует
принимать в расчет (рис. 2.3).
Поскольку в описанном варианте пробу (тест Ширмера I) вы-
полняют без предварительной анестезии, ее результаты характери-
зуют тотальную секрецию слезы. Для того чтобы разграничить долю
“рефлекторной” (на контакт глаза с бумажкой) секреции и порцию
выделившейся за то же самое время “базисной” слезы, L. Jones (1966)
модифицировал пробу, назвав ее тестом Ширмера II.
Тест Ширмера II. Варианты техники выполне-
ния п р о б ы. L. Jones предложил модифицировать описанный выше
классический тест Ширмера, введя элементы предварительной ка-
пельной анестезии и отсасывания остатков влаги из сводов ватным
тампоном. В. Milder (1987) дополнил тест, применив специальный
раздражитель рефлекторной секреции слезы (вдыхание паров раствора
аммиака или введение в носовые входы сухого ватного “банничка”).
Оценка результатов. В варианте, предложенном
L. Jones, для суждения о рефлекторной секреции из показателей теста
Ширмера! вычитают показатели теста ШирмераII. По данным
В.В. Бржеского (1990), в норме эта разница составляет в среднем
11,1 ±2,8 мм, снижаясь с возрастом (см. рис. 2.3). В варианте Миль-
дера (при более сильном раздражителе) данные о рефлекторной
секреции слезы получают непосредственно при проведении пробы,
причем она вместо 5 мин длится только 2 мин.
Следует отметить, что в клинической практике тесты на гипер-
продукцию слезы, регулируемую парасимпатической иннервацией,
используют мало, так как собственно рефлекторная секреция ее на-
рушается исключительно редко.
279
Тест на стабильность прек орнеа льной
слезной пленки [Norn М.,1969]. Техника выполне-
ния пробы. В конъюнктивальный мешок закапывают 1%
раствор флюоресцеина, исключая, естественно, касания роговицы.
Пациенту предлагают несколько раз моргнуть, а затем просят воз-
держаться от мигания и засекают время секундомером. В синем свете
щелевой лампы (с кобальтовым фильтром) широким пучком скани-
руют роговицу.
Оценка результатов. На фоне равномерной желтовато-
зеленой окраски поверхности роговицы на том или ином ее участке
вдруг возникают черные пятна или полоски. Они соответствуют
участкам подсыхания эпителия и разрыва над ним окрашенной
слезной пленки. По секундомеру определяют время, прошедшее от
последнего мигания до появления черных пятен. Пробу желательно
провести трижды. Время разрыва слезной пленки (ВРСП) в норме
довольно вариабельно. По данным М. Norn, ВРСП у здоровых
людей оно колеблется от 3 до 132 с, по сведениям В.В. Бржеского
(1990), — от 7 до 52 с. Если у молодых людей стабильность пленки
сохраняется в среднем на уровне 20 с, то в пожилом возрасте ВРСП
уменьшается до 16 — 2 с. При ВРСП до 10 с и меньше пленку рас-
ценивают как неполноценную. В данном случае обязательно следу-
ет исключить сухой кератоконъюнктивит и другую патологию ро-
говицы, а также нарушения механизмов формирования слезной
пленки (при недостаточной выработке муцина или липидов ВРСП
патологически уменьшается).
Тест прижизненного прокрашивания рого-
вицы [KanskiJ., 1985]. После закапывания в конъюнктивальный
мешок 1% раствора бенгальского розового оценивают участки про-
крашивания, которые представляют собой не дефекты покровного
эпителия, обнаруживаемые с помощью флюоресцеина, а скопления
погибших клеток и слизи. Краситель проникает внутрь этих струк-
тур. Для сухого кератоконъюнктивита характерно прокрашивание
межпальпебральных участков конъюнктивы глазного яблока (в пре-
делах открытой глазной щели) в виде двух треугольников.
Недостатком теста является возможность выраженной индиви-
дуальной непереносимости в виде длительной (до 1 сут) ирритации.
Предварительное использование дикаина нежелательно, так как
может привести к ошибочно положительному результату (токси-
ческое действие дикаина на клетки).
Синдром “сухого глаза” относится к числу патологических со-
стояний, довольно распространенных, особенно среди женщин в
климактерическом периоде, и вместе с тем в их диагностике зачас-
тую не уделяют должного внимания описанным тестам. Выполне-
ние лишь количественных тестов на слезопродукцию может ока-
заться недостаточным для установления правильного диагноза, так
как наличие сухих пятен на роговице стимулирует рефлекторное
280
слезотечение, вследствие чего результат такой пробы будет смазы-
ваться. Только качественная оценка слезной пленки в ряде случаев
помогает установить правильный диагноз.
2.3.2. “Переувлажненный” глаз, тестирование слезоотведения
Иногда глаз может быть влажным от переполняющей конъюнкти-
вальный мешок слезы вследствие гиперпродукции последней, но
значительно чаще — в связи с затрудненным ее оттоком по слезо-
отводящим путям [Волков В.В., 1975; Султанов М.Ю., 1983]. Эти
состояния можно сразу дифференцировать с помощью цветовой слез-
но-носовой пробы с флюоресцеином.
Цветовая слезно-носовая проба. Техника выпол-
нения пробы. При необходимости освободить носовые ходы от за-
бивающего их содержимого пациента прежде всего просят высмор-
каться. В оба глаза закапывают по 1 — 2 капле 1% раствора флю-
оресцеина-натрия, просят пациента посидеть, наклонив голову
вперед, и засекают время.
Оценка результатов. Спустя 3, а затем 5 мин либо
вводят в носовые ходы влажные “баннички”, либо просят пациента
высморкаться во влажную марлевую салфетку (из каждой ноздри
отдельно!). Зеленовато-желтое окрашивание свидетельствует о нор-
мальной функции слезоотведения и гиперпродуктивном характере
слезотечения.
Согласно результатам многочисленных исследований, проведен-
ных М.Ю. Султановым, в норме краситель проходит в нос за 1
5 мин.
При отрицательной цветовой слезно-носовой пробе приступают
к зондированию и промыванию.
Зондирование. После капельной анестезии, расширив слез-
ную точку коническим зондом, заменяют его на тупоконечный
цилиндрический зонд Боумена. Оттянув нижнее веко, зонд прово-
дят сверху вниз в вертикальную, а затем, развернув зонд на 90°, и
в горизонтальную часть нижнего слезного канальца. Возможно, зонд
не встретит по пути никакого сопротивления и сразу упрется в слез-
ную косточку (через медиальную стенку слезного мешка) либо сна-
чала натолкнется на “мягкий стопор”, и тогда, только преодолев
это пружинящее “мягкое сопротивление”, можно довести зонд до
“твердого сопротивления” кости.
Оценка результатов. “Мягкое пружинящее сопротив-
ление” зонду свидетельствует о наличии стриктуры (частичной или
полной) в системе слезных канальцев. Для уточнения ее уровня и
степени выраженности целесообразно провести промывание слезо-
отводящих путей.
Промывани е.После капельной анестезии по тому же пути
вместо зонда вводят прямолинейную тупоконечную канюлю, наса-
281
женную на шприц с промывной жидкостью (изотонический раствор
натрия хлорида, новокаин, левомицетин). Пациента просят накло-
нить голову вперед, под подбородком помещают почковидный
тазик, а врач, слегка придавливая указательным пальцем область
слезного мешка, продвигает канюлю до участка сопротивления, а
затем, ослабив давление на ткани, начинает вводить промывную
жидкость.
Оценка результатов. Если жидкость свободно
проходит в нос, значит, стриктуры нет. Вытекание жидкости через
нос каплями свидетельствует о сужении, причем в том случае, если
зонд не дошел до кости, — в системе канальцев, если же дошел, —
в слезно-носовом канале.
В некоторых случаях жидкость совсем не проходит в нос и воз-
вращается через противоположную или ту же самую слезную точ-
ку. Возвращение жидкости через противоположную точку свиде-
тельствует о сохранившемся сообщении между канальцами и более
низком расположении стриктуры, через ту же слезную точку — о
высоком расположении стриктуры (в том же самом канальце, через
который проводят промывание). Для уточнения уровня поражения
целесообразно выполнить рентгенографию слезоотводящих путей с
введением контрастного вещества.
Насосная (канальцевая) пр о б а, В случае полного
отсутствия пассивной проходимости слезоотводящих путей для пре-
доперационной оценки перспектив возможного вмешательства вы-
ясняют наличие насосной функции слезных канальцев и мешка.
Техника выполнения пробы. В конъюнктивальный
мешок закапывают 2% раствор колларгола. Пациента просят слег-
ка запрокинуть голову и поморгать. Выжидают 1 — 2 мин, избыток
колларгола осторожно, не надавливая на веки, отсасывают ваткой.
Затем, слегка оттянув нижнее (или верхнее) веко, наблюдают за со-
ответствующей слезной точкой, однрвременно надавливая на слез-
ный мешок.
Оценка результатов. Если при надавливании на
слезный мешок из слезной точки появляется капля колларгола, то
проба положительная— насосная функция существует. В случае
отсутствия выделений из слезной точки при давлении на мешок
проба отрицательная. Очень важно до постановки пробы освобо-
дить мешок от возможного содержимого (путем промывания или
хотя бы выдавливания).
Тесты Д ж о не с а. За рубежом принята несколько иная си-
стема исследования слезоотводящих путей [Kanski J., 1985].
Первичный тест Джонеса. Техника пробы практически очень
близка к таковой при проведении цветовой слезно-носовой пробы.
В глаз также закапывают флюоресцеин, но для оценки результатов
не прибегают к сморканию, а вводят в нижний носовой ход ват-
ный “банничек”, увлажненный 4% раствором кокаина (важно иметь
282
в виду, что от входа в нос до устья слезно-носового канала около
Зсм).
Оценка результатов. Обнаружение окраски на конце
банничка не позднее чем через 5 мин свидетельствует о положи-
тельной пробе и отсутствии необходимости других проверок.
В отсутствие окраски и, следовательно, при отрицательной про-
бе выполняют вторичный тест Джонеса.
Вторичный тест Джонеса. Техника выполнения пробы анало-
гична по существу описанной выше, но промывание слезоотводя-
щих путей осуществляется с несколько иной интерпретацией ре-
зультатов, поскольку пробе предшествует инстилляция флюоресце-
ина.
Оценка результатов. Если из носа появляется
окрашенный флюоресцеином раствор, то, значит, имеется активная
насосная функция верхнего отдела слезоотводящей системы и толь-
ко расположенное ниже сужение путей не позволило краске выйти
в нос при проведении первичного теста. Результат теста указывает
на благоприятный прогноз возможной дакриоцисториностомии.
Если же из носа вытекает неокрашенный раствор, значит, при вы-
полнении первичного теста засасывания окрашенной слезы не про-
изошло. В этом случае следует искать нарушения в верхнем отделе
дренажной системы слезоотведения (атоническая эверсия или дис-
локация слезных точек, стриктуры канальцев, в частности их устья
при входе в мешок).
ГЛАВА 3
ЦИРКУЛЯЦИЯ КРОВИ И ТКАНЕВЫХ
ЖИДКОСТЕЙ В ГЛАЗУ
3.1. ПРИКЛАДНАЯ АНАТОМИЯ СОСУДИСТОЙ
СИСТЕМЫ ГЛАЗА
В кровоснабжении глазного яблока участвуют две раздельные сосу-
дистые системы: ретинальная и увеальная. Если первая образует в
основном два слоя (в заднем полюсе до 3—4 слоев) капиллярных
сетей, располагающихся в 2/3 внутренних слоев сетчатки, то вторая,
находясь вне сетчатки, обеспечивает питание бессосудистой V3 на-
ружных слоев сетчатки единственным слоем хороикапилляров.
Между последними и фоторецепторами сетчатки имеются тончай-
шая прослойка, называемая мембраной Бруха, и слой клеток пиг-
ментного эпителия. Поскольку в направлении центральной зоны,
именуемой желтым пятном (fovea), внутренние слои сетчатки ис-
тончаются, в норме на участке диаметром 0,5 мм капилляров вооб-
ще нет (рис. 3.1).
В ретинальной системе артериолы и венулы расходятся от диска
зрительного нерва (ДЗН) в слое нервных волокон сетчатки, подоб-
но ветвям дерева, стволом которого служат центральные вена и
артерия. Последние, в свою очередь, являются ветвями более круп-
ных орбитальных сосудов (глазничная артерия, верхняя и нижняя
орбитальные вены). Важно, что глазничная артерия (a. ophthalmica)
берет начало непосредственно от внутренней сонной артерии, а
орбитальные вены связаны с кавернозным синусом и венами лица.
Увеальная сосудистая система представлена в заднем отделе
мощным слоем многочисленных хороикапилляров, а в переднем
отделе — тремя уровнями сосудистых колец: эписклеральным, ин-
трамускулярным (в ресничной мышце) и большим кругом кровооб-
ращения (в корне радужки). Артериальные и венозные коллекторы
в увеальной системе регионарно разобщены.
Сосуды, несущие артериальную кровь, проходят сквозь склеру
вблизи заднего полюса (задние ресничные артерии), но имеются и
такие, которые входят внутрь глаза впереди мест прикрепления
наружных прямых мышц (передние ресничные артерии). Среди зад-
них ресничных артерий выделяют короткие (обычно 3—4 стволика,
каждый из которых внутри глаза образует 10—20 веточек, обрыви-
сто переходящих в слой густой капиллярной сети) и длинные, меди-
альную и латеральную, которые направляются в супрахороидаль-
ном пространстве кпереди и вместе с веточками передних реснич-
ных артерий участвуют в формировании трех упомянутых выше
сосудистых колец.
284
Рис. 3.1. Флюоресцентная ангиограмма нормального глазного дна. Темное
пятно в центре соответствует зоне фовеа, которая в норме лишена капил-
лярной сети.
Сосуды, отводящие кровь из увеальной системы, обычно представ-
лены четырьмя вортикозными венами, проходящими через склеру в
косых меридианах. В оттоке крови из переднего отдела глаза, как и в
оттоке камерной влаги, участвуют эписклеральные вены, в том числе
описанные Ашером водяные вены. В зависимости от позы человека
кровь далее поступает либо в орбитальные, либо в лицевые вены.
Довольно сложной является система кровоснабжения ДЗН
[HayrehS.S., 1995]. Его внутриглазная часть снабжается кровью
ветвями задних коротких ресничных артерий, однако без участия
хороикапилляров, сеть которых обрывается в перипапиллярной зоне;
ретинальные капилляры питают только поверхностные слои ДЗН.
В ретроокулярную часть зрительного нерва входят веточки, отде-
лившиеся от центральной артерии сетчатки (еще до ее проникнове-
ния в полость глаза), и от сосудов, питающих мягкую оболочку
зрительного нерва. В слоях склеры непосредственно вокруг ДЗН из
ветвей задних коротких ресничных артерий формируется полный
или частичный артериальный круг. Небольшая область сетчатки,
расположенная вблизи фовеа, имеет изолированное кровоснабже-
ние, осуществляемое за счет тонкой самостоятельной цилиорети-
нальной артерии (у каждого третьего). Роль этого сосуда трудно
Переоценить, когда нарушается кровообращение в центральной
артерии сетчатки.
285
Ретинальные, как и крупные хороидальные артерии, являются
по структуре концевыми, поэтому при обтурации какой-либо из
указанных артерий, несмотря на обилие в сетчатке и хороидее ка-
пиллярных связей, определенный сектор внутренних или наружных
слоев сетчатки полностью лишается питания и гибнет, не имея ана-
стомозов. При обтурации ретинальной артерии необратимые изме-
нения в сетчатке наступают уже к исходу 1-го часа. При обтурации
увеальной артерии сетчатки патологический процесс проходит не
столь быстро, и возникает дегенерация по типу булыжной мосто-
вой или многофокусной плакоидной эпителиопатии.
3.2. СТРУКТУРЫ И ФУНКЦИИ ГЕМАТООФТАЛЬМИЧЕСКИХ
БАРЬЕРОВ
Внутренний и внешний барьеры. Капилляры сетчат-
ки и хороидеи существенно различаются между собой по строению
и функциональным возможностям стенки [Kanski J., 1989]. Капил-
ляры радужки функционально близки к капиллярам сетчатки, а ка-
пилляры отростков ресничного тела — к капиллярам хороидеи.
Внутренним тематическим барьером, для сетчатки являются ее
капилляры с плотной внутренней выстилкой из эндотелиальных
клеток, которые хорошо пропускают только жирорастворимые
структуры (кислород, СО2); водорастворимые структуры проходят
через стенку ретинальных капилляров с большим затруднением. Их
транспорт осуществляют микропиноцитарные пузырьки эндотелия
(путем заполнения эндотелиальных пор водой). Сетчатка, таким
образом, подобно мозгу, защищена от проникновения из кровяно-
го русла через стенку собственных сосудов различных токсинов,
нейромедиаторов и других активных веществ.
Внешним барьером капилляры хороидеи быть не могут, так как
их стенка очень тонка, в ней много дырчатых мембран, подобных
имеющимся в капиллярах почек и различных желез. В них происхо-
дит беспрепятственный обмен интра- и экстраваскулярных ком-
понентов, в том числе крупных молекул белка. Таким путем посту-
пают к пигментному эпителию провитамин А и глюкоза. Вторым
внешним гематическим барьером для сетчатки является слой кле-
ток пигментного эпителия. В области ДЗН этот барьер ослаблен,
но непигментированный эпителий ресничного тела, а также эндоте-
лий капилляров радужки выполняют барьерную функцию в полной
мере.
Для изучения функции гематоофтальмических барьеров служат
устройства для флюорофотометрии сред глаза и флюоресцентной
ангиографии глазного дна.
Методы исследования барьерных функций. Флюорометрия (флю-
орофотометрия). Молекулы флюоросцеина и ряда других веществ
286
обладают свойством преобразовывать световую энергию таким
образом, что, возбуждаясь от засвета относительно коротковолно-
выми излучениями (например, синим цветом с X 490 нм), они не
отражают, а преобразуют световую энергию в более длинноволно-
вую (например, в желто-зеленый цвет с X 530 нм).
В приборах это свойство реализуется таким образом, что свет,
направляемый в глаз через синий светофильтр, изучается после
возвращения из глаза через желто-зеленый светофильтр.
Для оценки функции гематоофтальмического барьера существу-
ют приборы, позволяющие точно определить время выхода флюо-
ресцеина, введенного внутривенно, из сосудов в полость глаза (в
стекловидную полость, во влагу передней камеры) [Hochheimer В.,
1979; Miyake J., 1983; Sanders D., 1994]. Барьерная функция наруша-
ется при гипертонической болезни, диабете, многих воспалитель-
ных и дегенеративных заболеваниях глаза, после хирургических
операций со вскрытием глазного яблока.
Флюоресцентная ангиография. Для оценки рети-
нальной и увеальной циркуляции, а также состояния тематических
барьеров в глазу исключительно ценным методом является флюо-
ресцентная ангиография (ФАГ).
Физиологические основы метода. Большая
часть (80%) флюоресцеина-натрия, введенного в кровяное русло,
связывается альбумином и другими белками сыворотки крови; ос-
тальные 20% остаются несвязанными (“свободными”).
В норме гематоретинальные барьеры, как внутренний (эндоте-
лий капилляров сетчатки), так и внешний (пигментный эпителий
сетчатки), не пропускают не только связанные, но и свободные
молекулы флюоресцеина. Аналогичным свойством обладают стен-
ки крупных ретинальных и хороидальных сосудов.
Иные свойства у хороикапилляров. Через их очень тонкие стен-
ки свободные молекулы флюоресцеина беспрепятственно выходят в
экстраваскулярное протранство, а далее через вполне нормальную
мембрану Бруха они поступают к ретинальному пигментному эпи-
телию, но он из-за плотно прилежащих одна к другой клеток и
наличия в норме “запирательных” связок оказывается непроницае-
мым для флюоресцеина.
Техника выполнения пробы. Пациенты, как пра-
вило, хорошо переносят внутривенное введение флюоресцеина. Лег-
кие побочные эффекты проявляются в виде последовательных эрит-
ро- или ксантопсических образов, кратковременного поташнива-
ния, изменения цвета кожи и мочи. Случаи анафилактического шока,
сопровождающегося бронхоспазмами и обмороками, исключитель-
но редки. Для выявления индивидуальной непереносимости препа-
рата из анамнеза выявляют склонность к аллергическим реакциям
и внутрикожно вводят 0,1—0,2 мл подготовленного для внутривен-
ного вливания флюоресцеина. В кабинете для ФАГ необходимо
287
иметь запас медикаментозных средств для борьбы с анафилакти-
ческим шоком.
Для получения качественной ангиограммы зрачок должен быть
максимально расширен, а преломляющие среды глаза — прозрач-
ны. Глазное дно пациента фокусируют фотофундускамерой. В лок-
тевую вену вводят ’’толчком” 5 мл 10% раствора очищенного флю-
оресцеина (при недостаточной прозрачности сред глаза 3 мл 25%
раствора). В исходном состоянии глазное дно фотографируют с
зеленым фильтром, т.е. в бескрасном свете. Компактно введенная
доза позволит получить снимки высокого качества. По данным
A. Wessing (1969), время “рука — сетчатка” в норме варьирует от 5
до 24 с, поэтому фотографирование глазного дна начинают на
5-й секунде и продолжают эту процедуру ежесекундно до 25-й се-
кунды. Время от времени, быстро перемещая камеру на другой глаз,
для сравнения делают контрольные фотоснимки. Глазное дно ис-
следуемого глаза при необходимости фотографируют дополнитель-
но в поздние сроки (спустя 10 и 20 мин).
Флюоресцентную ангиоиридографию выполняют с использова-
нием сходной техники.
Трактовка результатов ФАГ. Через глазную ар-
терию флюоресцеин поступает в различные ее ветви, в частности в
задние короткие ресничные артерии и центральную артерию сет-
чатки, одновременно. Однако, поскольку путь в систему хороидаль-
ной циркуляции несколько короче, она заполняется на 1 с раньше,
чем система ретинальной циркуляции. Как уже отмечалось, свобод-
ный флюоресцеин легко преодолевает стенку хороикапилляров и,
оказавшись вне их, “смазывает” детали хороидальной циркуляции.
Осмотр затрудняет также меланин пигментного эпителия.
Для оценки хороидальной циркуляции при ФАГ целесообразно
применять красители, стимулирующие флюоресценцию в более длин-
новолновом диапазоне, например индоцианин зеленый с пиком
флюоресценции на 720 нм [Hochheimer B.F., 1979]. Этот же краси-
тель используют для усиления эффекта лечебного лазерного воз-
действия ИК-излучателями на хороидальные сосуды [ReichelE.,
PuliafitoC., 1994].
Ретинальная циркуляция в отличие от хороидальной прослежи-
вается значительно отчетливей.
В нормальной флюоресцентной ангиограмме глазного дна вы-
деляют четыре фазы:
1) хороидальную (для сетчатки — преартериальная): флюорес-
цеин заполняет только сосуды хороидеи;
2) артериальную: флюоресцеин поступает в артерии сетчатки;
3) артериовенозную: еще присутствуя в артериях, флюоресцеин
заполняет капиллярную сеть сетчатки и обозначает пристеночный
ток в венах;
4) венозную: флюоресцеин находится в основном в венозной сети
288
сетчатки (эту фазу подразделяют иногда на раннюю, срединную и
позднюю венозные фазы).
При оценке ангиограммы необходимо учитывать длительность
перечисленных фаз, характер распределения флюоресцеина (по ин-
тенсивности флюоресценции) в каждой из анатомически дифферен-
цируемых областей глазного дна, обращая внимание на участки как
гипер-, так и гипофлюоресценции, специально наблюдая за измене-
ниями, происходящими на этих участках с течением времени.
Свойственная макулярной зоне гипофлюоресценция и ее темная
окраска обусловлены не только отсутствием в норме капилляров,
но и более сильной пигментацией клеток пигментного эпителия,
расположенных в фовеальной и парафовеальной областях. Мела-
нин и ксантофильные пигменты сетчатки экранируют в этой зоне и
хороидальную флюоресценцию.
Поскольку вышедший из сосудов глаза флюоресцеин спустя 3—
5 мин вновь появляется в них благодаря рециркуляции крови, отме-
ченные выше фазы его прохождения повторяются, но с каждым
разом интенсивность свечения уменьшается. Длительное послесве-
чение ДЗН относится к числу часто встречающихся в норме состо-
яний.
3.3. ГЛАЗНОЙ КРОВОТОК В НОРМЕ. МЕХАНИЗМЫ
РЕГУЛИРОВАНИЯ КРОВОТОКА В ГЛАЗУ И КЛИНИЧЕСКИЕ
МЕТОДЫ ЕГО ИССЛЕДОВАНИЯ
Степень кровотока и оксигенация сетчатки. Среднее время цирку-
ляции крови в сетчатке у здорового человека по данным ФАГ со-
ставляет 4,7 ± 1,1 с [Hyckam J.B. et al., 1965]. Об объеме крови, про-
текающей в единицу времени через определенную массу различных
тканей, легко судить по результатам экспериментов на обезьянах
[Alm А., 1973; Folkov В., 1973], в которых установлен ретинальный
и хороидальный кровоток, в 10 раз превышающий кровоток в моз-
ге (из расчета на 100 г соответствующей ткани). Поскольку подоб-
ных различий в метаболической активности сравниваемых тканей
не существует, предполагают, что из большой массы крови, прохо-
дящей через хороидею, утилизируется сравнительно небольшая доля
питательных веществ (в частности, 5—8% кислорода). Из ретиналь-
ных сосудов кислород усваивается гораздо интенсивнее: разница в
его концентрации на входе и выходе достигает 38%; как свидетель-
ствуют результаты экспериментов на анестезированных животных,
снабжение сетчатки кислородом на 60—80% зависит от кровотока
не в сетчатке, а в хороидее.
Важность обильного кровотока в хороидее исследователи ус-
матривают не только в необходимости поддержания высокого на-
пряжения кислорода, чем обеспечивается его диффузия в бессосуди-
289
стую наружную треть сетчатки, но и в создании своеобразного за-
щитного механизма [Alm A., Bill А., 1987]. Кровоток защищает сет-
чатку от перегрева или переохлаждения: при локальном преобразо-
вании, например, лазерной энергии в механическую или тепловую
температура может достигать десятков тысяч градусов; световая
энергия операционного микроскопа, фокусированного на область
макулы, при длительных воздействиях также может представлять
опасность.
Глаз в отличие от многих других органов, в которых степень
раскрытия капиллярной сети определяется изменением калибра пре-
капиллярных артериол с гладкомышечной стенкой, таким свойством
не обладает. Ни в сетчатке, ни в хороидее прекапиллярных сфинкте-
ров не обнаружено, ток крови в норме здесь не прерывается.
В сетчатке и в хороидее, как и в любой другой ткани, кровоток
определяется следующими факторами: давлением, создающим этот
кровоток; сопротивлением сосудистых стенок; вязкостью крови.
A.Alm и A. Bill предложили выражать глазной кровоток (blood
flow - BF) в случае постоянной вязкости крови, формулой:
BF = (РА - Pv)/R,
где (РА - Pv) — разница между артериальным давлением (РА) на
входе в глаз и венозным давлением (Pv) на выходе из него, т.е.
перфузионное давление; R — резистентность сосудистой стенки.
Этот показатель в значительной степени зависит не только от
уровня ригидности самой стенки, но и разности давлений внутри и
вне сосуда (трансмуральное давление); для внутриглазных сосудов
давление вне сосуда и есть истинное внутриглазное давление.
3.3.1. Артериальное давление и методы исследования активной
(моторной) части глазного кровотока
Офтальмодинамометрия. Цель этого исследования —
определение уровня давления в центральной артерии сетчатки
(ЦАС). Метод основан на офтальмоскопической регистрации появ-
ления пульсаций и последующей остановки кровотока в ЦАС по
мере дозированного сдавливания глазного яблока.
Техника выполнения пробы. При медикаментозно
расширенных зрачках обоих глаз после капельной анестезии один
из исследователей прикладывает к склере пружинящий плунжер
офтальмодинамометра Байара и начинает сдавливать глазное яб-
локо. В то же время другой исследователь путем непрямой офталь-
москопии устанавливает непрерывное наблюдение за сосудами на
диске зрительного нерва. Регистрируют уровень склеральной ком-
прессии, при которой начинается отчетливая пульсация ЦАС, и
второй уровень, при котором она прекращается. По таблицам
Мажито — Байара производят необходимые перерасчеты.
290
Оценка результатов. Исследовать необходимо оба гла-
за; если разница показателей, установленных на парных глазах, не
превышает 20%, то она не может свидетельствовать о патологии с
какой-либо стороны. Хотя пульсация ЦАС и контролируется, ре-
зультаты исследования отражают уровень артериального давления
в глазной артерии и состояние циркуляции крови в системе внут-
ренней сонной артерии. Таким путем диагностируют некоторые сте-
нозирующие и аневризматические нарушения циркуляции крови в
данной системе.
Для диагностики по результатам офтальмодинамометрии цереб-
ральных форм гипертонической болезни иногда используют рети-
нобрахиальный индекс по Байару. За норму принимают такое соот-
ношение, при котором давление в глазничной артерии примерно в
2 раза ниже, чем в плечевой (для диастолического давления этот
показатель составляет 0,48, а для систолического — 0,59). Более
высокий индекс многие годы расценивали как патологический.
Коррективы к нему внесли A.Alm и A.Bell, оперировавшие показа-
телем среднего давления, понимая под этим сумму из диастоличес-
кого давления +Ч3 пульсового давления. Так, при кровяном давле-
нии в плечевой артерии 140/80 мм рт.ст. среднее давление в ней
составит 100 мм рт.ст. [80 + (140 - 80)/3]. В норме при вертикальном
положении туловища среднее давление в артериях, входящих в глаз,
составляет 65—70 мм рт.ст. и за норматив ретинобрахиального
индекса принимаются цифры 0,6—0,7.
Разность между показателем среднего давления в глазной арте-
рии и истинным внутриглазным давлением называют градиентом
давления [Lobstein A., Herr F., 1966]. В норме при замерах офталь-
модинамометром Мюллера с использованием калибровочных таб-
лиц Бедованья градиент давления составляет около 55 мм рт.ст.
Более низкий градиент является плохим прогностическим призна-
ком в отношении сохранности зрительных функций при глаукоме
[Бунин А.Я. и др., 1974].
Снижение этого градиента может происходить не только вслед-
ствие повышения офтальмотонуса, но и в связи с низким уровнем
артериального давления [Волков В.В. и др., 1970, 1974]. Общая ар-
териальная гипотензия и связанное с ней снижение показателей
внутричерепного давления могут стать причинами прогрессирова-
ния глаукомы на фоне невысоких показателей офтальмотонуса.
Офтальмодинамографи я. Для графической регистра-
ции давления в глазной артерии используют специальные приборы
(М. Уемуры, Хагера и др.). Пульсация сосудов глазницы вызывает
синхронные колебания глазного яблока в переднезаднем направле-
нии. Если приложить к роговице капсулу с тонкостенной мембра-
ной, то эти колебания можно записать (рис. 3.2). По мере усиления
компрессии кривая глазоорбитального пульса снижается. Это, в ча-
стности, наблюдается при стенозирующих поражениях в системе
291
1
2
Рис. 3.2. Офтальмодинамограмма.
1 — запись компрессионной нагрузки; 2 — запись глазного пульса.
внутренней сонной артерии. При аневризме же в пещеристом сину-
се амплитуда пульса, наоборот, возрастает.
К недостаткам метода относится невозможность избавиться от
помех, связанных с мигательными движениями век, со сдавлением
отдельных ветвей наружной сонной артерии краями капсулы. При
очень высоком артериальном давлении проводить исследование
опасно.
Ю.С. Астахов (1990) с помощью усовершенствованного вакуум-
ного офтальмодинамографа конструкции ВНИИМП установил, что
давление в глазной артерии в норме составляет 73,5 ± 10 мм рт.ст.,
а ретинобрахиальный индекс — 0,59.
Ультразвуковая или лазерная допплерография. Метод позволяет
судить о скорости кровотока в исследуемом сосуде на основании
изменения отраженных ультразвуковых колебаний или длины вол-
ны. X. Допплер (1842) сформулировал принцип, который позволяет
оценивать направление и скорость движения любого объекта по
изменениям отраженного от него эхосигнала: если объект переме-
щается с определенной скоростью, то время, через которое эхосиг-
нал возвращается к источнику излучения, отлично от времени воз-
врата эхосигнала от неподвижного объекта. ’’Эффект Допплера”
сделал возможным не только расчет линейной систолической, диа-
столической, средней скорости и определение направления крово-
тока в сосудах, но и оценки состояния стенки исследуемого сосуда
[Борисова С. А., 1997; Johnson J.M. et al., 1982; Ficher G.G., 1985].
Исследование кровотока в глазной артерии можно проводить с
помощью транскраниальной допплерографии при использовании
импульсного режима, лоцирования сосудов. Возможность лоциро-
вания в непрерывном режиме глазной артерии, задних цилиарных
сосудов и сосудов хороидеи, что часто проводится в клинической
практике, весьма сомнительна, поскольку в этом случае спектро-
грамма представляет собой суммарную производную от спектров
всех сосудов, встретившихся на пути ультразвукового луча. Для
локации сосудов мозга, тонких орбитальных сосудов, глазной и
заднецилиарной артерии можно применить транскраниальный доп-
плерограф (флуометр ТК-264 В с диаметром датчика 20 мм), исполь-
зовав 10% его максимальной мощности, или компьютерные доп-
292
плеровские системы при использовании глубины локации 30 —
50 мм [Hayreh S.S.,1962] и объема локации 7 — 13 мм, что позволя-
ет проводить прямое исследование гемодинамики [Никитин Ю.М.,
1995; Шахнович и др., 1996; Galassi F. et al., 1994; Belden C.J. et al.,
1995; Karczewicz D. et al., 1995; Michelson G.et al., 1995; Valli A et al.,
1995]. В последние 5 лет появились сообщения об использовании
лазерной допплерографической техники, которая позволяет не толь-
ко вычислять скорость кровотока во внутриглазных сосудах, диа-
метр которых меньше 1 мм, например центральной артерии сетчат-
ки, но и оценить капиллярное кровообращение участков диска зри-
тельного нерва при расчете скорости передвижения эритроцитов в
капиллярах [Dennis K.J., 1995]. О применении дуплексного метода в
диагностике глазных заболеваний имеются единичные сообщения
[GuthoffR.G. et al., 1991]. При использовании В-сканирующего
ультразвукового зонда (в 5 и 7,5 Мгц) авторами были отмечены
определенные трудности в оценке кровотока в сосудах заднего от-
резка орбиты. Новый подход к оценке кровотока в сосудах орбиты,
который состоит в измерении и сопоставлении параметров крово-
тока по надблоковой и глазной артерии с использованием компрес-
сионных проб, осуществлен при исследовании кровотока по внут-
риглазным сосудам у больных открытоугольной глаукомой [Бори-
сова С.А. и др., 1997].
3.3.2. Венозное давление и методы исследования “стопорной” части
глазного кровотока
В норме при офтальмоскопическом исследовании всегда можно
заметить пульсацию крупных ветвей центральной вены сетчатки
(ЦВС), что свидетельствует о близости показателей давления внут-
ри и вне этих сосудов. Приняв давление в окружающих внутриглаз-
ные вены структурах (внутриглазное давление), равным 15 мм рт.ст.,
с определенной долей ошибки можно считать, что и венозное дав-
ление (Pv) равно 15 мм рт.ст. Исходя из этого, в норме перфузион-
ное давление в глазу составит
РА- Pv = 65 - 15 = 50 мм рт.ст.,
где РА — среднее давление в ЦАС; Pv — давление в ЦВС.
Перепады давления по ходу внутриглазных венозных стволов
ничтожны, поэтому сами пульсовые волны мало влияют на дина-
мику объема протекающей крови и связанные с пульсом колебания
офтальмотонуса. Так, офтальмотонус колеблется синхронно с пуль-
сом в пределах от 1 до 2 мм рт.ст. (по А.П.Нестерову — 1,2 мм рт.ст.),
а объем крови — в пределах от 1 до 4 мкл (по Г.А.Киселеву —
1,55 мм3).
Офтальмосфигмография. Этот метод предназначен
для измерения и регистрации пульсовых колебаний офтальмотону-
293
Рис. 3.3. Пульсовые колебания офтальмотонуса и волны Геринга — Трау-
бе на тонограмме.
са (рис. 3.3) в ходе топографии, в частности, электронным тоногра-
фом. Амплитуду волн первого порядка (Геринга — Траубе) оцени-
вают с помощью шкалы, прилагаемой к тонометру Шиотца. При
использовании ручного сфигмоманометра начальное давление на
роговицу составляло всего 2 г, по мере его повышения до 18 г реги-
стрировали пульсовую волну [Bynke, 1964], амплитуда которой по-
степенно снижалась. Пьезоэлектрические кристаллы и плунжерная
система служили датчиком. С помощью таблиц (нормограмм) Фри-
денвальда на основании пульсовых колебаний офтальмотонуса
можно составить представление о пульсовых изменениях объема
глазного яблока.
На состояние венозного оттока из глаза и давление во внутри-
глазных венах выраженное влияние может оказать давление в экст-
рабульбарных венах (эписклеральных, вортикозных, глазничных).
Между внутри- и внеглазными частями венозного русла в здоровых
глазах существует значительный (порядка 5 — 10 мм рт.ст.) пере-
пад давления. При таких условиях внутриглазной вене, примыкаю-
щей к выходному отверстию в склере, с одной стороны, угрожает
коллапс, а с другой — переполнение кровью. Развитию коллапса
противостоит ауторегуляторный механизм, обеспечивающий (за счет
изменений тонуса соседних мышечных структур) поддержание дав-
ления во внеглазных венах выше критического [Alm A., Bill А., 1973].
Последствия переполнения внутриглазной вены кровью регулиру-
ются с большим трудом. При сдавлении экстраокулярных вен по-
вышенное давление сразу распространится на интраокулярные вены,
а они лишены компенсаторных механизмов. В норме внутриглаз-
ные вены из-за тонкости стенки функционируют почти при нуле-
вом трансмуральном давлении, и когда произойдет даже незначи-
тельное повышение давления в просвете такого венозного сосуда,
трансмуральное давление в нем возрастет в несколько раз. След-
ствием этого будет не только расширение вен сетчатки, которые
становятся извитыми, но и, возможно, появление микроаневризм и
кровоизлияний.
Методы исследования давления в эписклеральных венах. Специ-
альный веноманометр укрепляют на щелевой лампе. После капель-
ной анестезии датчик с латексной мембраной на торце касается
глазного яблока. Эписклеральную вену (эти вены расположены
глубже, темнее и прямолинейнее конъюнктивальных) сдавливают
латексной мембраной (предпочтительнее в 3 — 4 мм от лимба вра-
294
щая диск прибора) под контролем биомикроскопии, добиваясь
прекращения кровотока в вене. Оценку результатов проводят после
троекратного замера. В норме давление в эписклеральных венах
составляет в среднем 7,6 мм рт.ст. [Zeimer R., 1983].
3.3.3. Ауторегуляция кровотока в глазу и методы его исследования
В глазу, как и в мозге, существуют механизмы ауторегуляции кро-
вотока.
Нервный механизм. При нарушении симпатической иннервации
ослабляется кровоток во всех отделах увеального тракта, что мож-
но рассматривать как защитную реакцию (при стрессовых ситуаци-
ях, рабочей перегрузке), предохраняющую гематоофтальмический
барьер от ’’поломки”. При этом роль парасимпатической иннерва-
ции незначительна.
Воздействие симпатической системы проявляется в изменении
просвета увеальных и ретинальных сосудов, что коррелирует с уров-
нем артериального давления, хотя его небольшое понижение со-
провождается не снижением, а увеличением увеального кровотока
[Астахов Ю.С., 1972].
В норме регулирующее влияние офтальмотонуса на кровоток,
очевидно, невелико. Однако следует учитывать, что в эксперименте
снижения перфузионного давления можно достичь путем не только
снижения артериального давления, но и повышения офтальмотонуса.
Метаболический (химический) механизм. По данным A.Alm и
A.Bill (1987), уменьшение хороидального кровотока вследствие
умеренной офтальмогипертензии не отражается на снабжении внут-
ренних слоев сетчатки питательными веществами. Необходимое
напряжение кислорода в тканях поддерживается на постоянном
уровне благодаря метаболической регуляции кровотока. Дефицит
кислорода в тканях глаза (гипоксия) и избыток углекислоты (ги-
перкапния) компенсируются увеличением в них кровотока вслед-
ствие расширения сосудов. Повышение парциального давления
кислорода приводит к сужению внутриглазных сосудов и уменьше-
нию кровотока.
Офтальмоплетизмография. По результатам опре-
деления динамики объема переднего сегмента глазного яблока в
ходе сердечного цикла можно судить об изменениях объема всего
глазного яблока.
Техника исследования. Больной находится в
положении лежа, датчик пульса устанавливают на анестезирован-
ной роговице, как при проведении топографии. С помощью оф-
тальмоплетизмографа ОПГ-Т можно производить запись глазного
пульса (рис.3.4). В условиях постепенно наращиваемой компресси-
онной нагрузки на перилимбальную область глазного яблока амп-
литуда пульсовых волн увеличивается.
295
a
15 20 30 40 50 60 70 80
—---S----------Л- —*--*-------*—'—
6
Рис. 3.4. Офтальмоплетизмограмма [Нестеров А.П. и др., 1974].
а — элементы волны глазного пульса: А — амплитуда, а — время анакротической
фазы, Р — время катакротической фазы; б — нормальная кривая. При повышении
компрессионной нагрузки (цифры сверху) амплитуда пульсовых волн возрастает.
Оценка результатов. В норме увеличение пульсового
объема при каждой систоле составляет около 2 мм3, что в пересчете
на объем всего глазного яблока должно быть примерно в 10 раз
больше, т.е. 20 мм3, а за период времени в 1 мин составит 1100—
1200 мм3 (минутный артериальный объем крови). С возрастом этот
показатель снижается. Увеличение пульсового объема наряду со
снижением офтальмотонуса расценивают как благоприятный при-
знак в отношении сохранности зрительных функций при глаукоме.
По свидетельству авторов метода [Бунин А.Я. и др., 1974], с учетом
динамики изменений возможности его ограничены* получаемые
данные носят лишь относительный характер.
Реоофтальмография. В отличие от плетизмографии
реография позволяет оценивать не только объемные пульсовые из-
менения кровотока, но и его скорость. В основе метода лежит ре-
гистрация изменений импеданса (общего электрического сопротив-
ления ткани при указанных выше изменениях кровотока) [Несте-
ров А.П. и др., 1974]. С помощью РОГ оценивается кровенаполнение
главным образом 'в зоне задних длинных цилиарных артерий, так
как располагают электроды перилимбально в области цилиарного
тела. При этом нельзя исключить вклад в РОГ хороидального кро-
вообращения в связи с большим количеством анастомозов между
задними длинными и короткими цилиарными артериями.
Техника исследования. Регистрацию проводят с
помощью усилителя с реографической приставкой и специального
электрода для реоофтальмографических исследований конструкции
И.Чибирене (1971), представляющего собой пластмассовую прозрач-
ную чашечку, изогнутую по кривизне глазного яблока, к внутрен-
296
2
3
Рис. 3.5. Нормальная реограмма.
1 — угол наклона реограммы, определяющий форму анакротической части кривой;
2 — пик, соответствующий максимальному кровенаполнению; 3 — дикротический
зубец на дикротической части кривой; 4 — максимальная амплитуда реограммы.
ней стороне которой прикреплены 2 серебряных бобовидных элек-
трода, снаружи — резиновая присоска, позволяющая с помощью
вакуума фиксировать датчик на поверхности глазного яблока.
Оценка результатов. В реоофтальмограмме (РОГ)
оценивают форму реографической волны, ее амплитуду, время вос-
ходящей (а) и нисходящей (р) частей РОГ (рис. 3.5). При анализе
кривых началом цикла сердечной деятельности считают начальную
точку зубца Q на ЭКГ, через которую проводят ортогональную
систему координат, в цикле кривой РОГ измеряют различные вре-
менные интервалы, а также принятые в кардиологии показатели
центральной гемодинамики, что позволяет анализировать глазной
кровоток в зависимости от фазовой деятельности левого желудочка
сердца [Чибирене И.О., 1984]. В процессе математической обработ-
ки РОГ принимают во внимание калибровочный сигнал, величина
которого находится в обратной зависимости от показателя относи-
тельного пульсового объема.
При анализе кривых реоофтальмограммы используется рео-
графический коэффициент по Jantch (1958), который характеризует
максимальный объем крови в сосудистом тракте в процентах.
А-Еом-1000
REMM
где RQ — реографический коэффициент, который характеризует
максимальный объем крови в сосудистом тракте; А — амплитуда
реографической волны и дополнительная амплитуда а, мм; R —
297
сопротивления между электродами (балансировочное сопротивле-
ние), Ом; ЕОм — калибровочный импульс в Ом, Емм — калибровоч-
ный импульс в мм.
Для оценки растяжимости сосудистой стенки принято использо-
вать отношение амплитуды РОГ к времени анакротической части
цикла РОГ A/а (коэффициент Hadjiev — Tzenov), где А — ампли-
туда волны, а — время анакротической части кривой цикла РОГ.
С помощью реографического метода можно оценить не только
степень кровенаполнения, но также и степень влияния вазогенных
медикаментозных средств на кровообращение в увеальном тракте
глаза [Кацнельсон Л.А., 1976].
Гамма-резонансная офтальмовелосимметрия. Метод разработан
Ю. С. Астаховым (1990). Помещаемая на роговицу таблетка массой
20 мг с радиоактивным источником практически не оказывает дав-
ления на глазное яблоко, вместе с тем ее микроперемещения, обус-
ловленные глазоорбитальной пульсацией, точно регистрируются
специальным детектором.
Установлено, что у здоровых людей амплитуда глазоорбиталь-
ного пульса составляет в среднем 22,0 ± 0,6 мкм„а скорость пульсо-
вой волны — 0,294 ± 0,01 мм/с.
Этот неинвазивный метод позволяет с высокой точностью выяв-
лять нарушения проходимости брахиоцефальных артерий, т.е. су-
дить, в частности, о кровоснабжении глазницы и глазного яблока.
3.4. ЭТИОЛОГИЯ, ПАТОГЕНЕЗ И КЛИНИЧЕСКИЕ
ВАРИАНТЫ НАРУШЕНИЯ ГЛАЗНОЙ ГЕМОЦИРКУЛЯЦИИ
3.4.1. Нарушения артериального кровотока
В основе нарушений артериального кровотока могут лежать возра-
стные (атеросклеротические, артериогипертензионные), инфекцион-
ные (эндокардиты), ангиоспастические заболевания, а также трав-
мы глазного яблока, глазницы, трубчатых костей (жировая эмбо-
лия). Нередко причиной этих нарушений является патология
краниобазилярных артерий (стенозы глазной артерии, височный
периартериит и др.).
Основные механизмы острого нарушения кровотока в сосудах
сетчатки состоят в следующем.
1. Эмболизация, которой способствует короткий путь для попа-
дания из сердца в ветвь внутренней сонной артерии, глазную арте-
рию, элементов атероматозных бляшек. Желтые (холестероловые)
или белые (содержащие кальций) элементы, перекрывающие крово-
ток в соответствующем сосуде сетчатки, либо цепочка из белова-
тых элементов (скопления фибрина и тромбоцитов), слегка переме-
щающихся в сосудистом русле, резко нарушают зрение (с ощущени-
298
ем занавески, закрывающей верхнюю или нижнюю половину поля
зрения (amaurosis fugax).
2. Тромбообразование как следствие атероматозного артериита—
одного из проявлений системных коллагенозов (красной волчанки,
склеродермии, узелкового, в том числе височного, периартериита).
При этом острота зрения снижается постепенно.
3. Спазмирование артериол, в стенке которых имеются мышеч-
ные волокна, возникает при ангионеврозах у молодых людей (чаще
у женщин) и характеризуется возможностью восстановления зри-
тельных функций и более стойкими их изменениями при гиперокси-
генации у недоношенных детей.
4. Длительное механическое'сдавление ЦАС в случаях очень рез-
кой артефициальной офтальмогипертензии (при операциях по по-
воду отслойки сетчатки с циркляжем, проводимых под наркозом).
5. Вирусное поражение концевых ветвей ретинальных артерий,
которому нередко предшествует иридоциклит, с развитием синд-
рома острого некроза сетчатки (множественные ее разрывы и от-
слойка).
Аналогичные этиологические факторы могут приводить к нару-
шениям кровообращения и в хороидальных сосудах. Несмотря на
обилие анастомозирующих капилляров, компенсация недостатка
кровотока через венулы невозможна из-за резких различий в уров-
не давления, поскольку крупные хороидальные артерии функцио-
нируют как концевые. В связи с прекращение^ кровотока в рети-
нальных артериях возникает острый ишемический отек внутренних
слоев сетчатки с последующей восходящей атрофией нервных воло-
кон диска зрительного нерва, а обтурация хороидальной артерии
приводит к медленно развивающейся хороиретинальной дистрофии
в зоне ее разветвления.
Попытки повысить напряжение кислорода в сетчатке и стекло-
видном теле путем усиления кровотока с помощью таких сосудо-
расширяющих средств, как изопротеренол, гистамин, никотиновая
кислота, ксантинола никотинат, в опытах на кошках были малоэф-
фективны [Alm А., 1972], в то же время активнее оказался папаве-
рин, что связано с его высокой жирорастворимостью, позволявшей
ему легко преодолевать тематический барьер.
Нарушение кровотока может возникнуть не только вследствие
обтурации сосуда, но и в связи с разрывом его стенки, возникнове-
нием обширных кровоизлияний внутрь глаза (так называемое экс-
пульсивное кровотечение).
3.4.2. Нарушения венозного кровотока
Факторами риска нарушения венозного кровотока являются пожи-
лой возраст (старше 70 лет), артериальная гипер- или гипотензия,
частые и значительные подъемы офтальмотонуса, многие бо-
299
лезни крови, перифлебиты и другие васкулиты (саркоидоз, болезнь
Бехчета).
Венозная окклюзия в сетчатке нередко становится причиной
формирования порочного круга, в котором затруднение и останов-
ка кровотока приводят к гипоксии сетчатки, вызывающей отек и
геморрагии на глазном дне, что становится причиной повышения
экстравазального тканевого давления, еще более затрудняющего
кровоток, и т. д.
В зависимости от степени окклюзии ветвей и основного ствола
ЦВС острота зрения изменяется по-разному. В начальной стадии
заболевания больных беспокоит лишь дискомфорт, они предъявля-
ют жалобы на “мазки грязи” в поле зрения, при этом продолжи-
тельность острой фазы, в ходе которой зрение постепенно снижает-
ся, увеличивается до 6 мес. Причиной нарушения остроты зрения
считают макулярный отек, который может протекать как в ишеми-
ческой, так и в неишемической форме при нарушении и сохранно-
сти функции капилляров соответственно. Если в первом случае про-
гноз в отношении восстановления зрения плохой, то во втором —
достаточно благоприятный. В хронической фазе (после 6 мес) при
офтальмоскопии отмечаются коллатерали и шунты; появляется
рубеоз радужки, возможна офтальмогипертензия.
Причиной нарушения циркуляции в ЦВС может служить дисба-
ланс: ВГД = 2 ВЧД. В диске зрительного нерва в случае стойкого
снижения ВГД (при посттравматической офтальмогипотонии) или
повышения ВЧД (при церебральной гипертензии) расширяются
крупные ветви ЦВС и замедляется кровоток, что сопровождается
отеком ткани ДЗН (застойный сосок). При тромбозе ЦВС различ-
ного происхождения по ходу крупных венозных ветвей появляются
интраретинальные геморрагии (картина “раздавленного помидора”).
У лиц молодого возраста на фоне циркуляторных нарушений иног-
да появляются признаки воспаления (папиллофлебит).
Симптомом нарушения кровотока в эпибульбарных венах явля-
ется их расширение. Для эндокринной офтальмопатии характерно
расширение вен в меридианах прикрепления к склере пораженных
прямых мышц. Патогномоничным признаком внутриглазных опу-
холей, исходящих из ресничного тела, является расширение эпи-
бульбарных вен в точном соответствии с сектором локализации
новообразования. Расширение эпибульбарных вен по всему пери-
метру переднего отдела глаза встречается при терминальной глау-
коме (“голова медузы”), но может быть также признаком наруше-
ния кровообращения в венах глазницы. При сопутствующем экзо-
фтальме, жалобах на дующий шум в половине головы, синхронный
с пульсом (врач может уловить этот шум фонендоскопом через
прикрытые веки), возникают основания для подозрения на наличие
каротидно-кавернозного соустья, представляющего угрозу для жизни
больного.
300
3.4.3. Нарушения капиллярного кровотока
Неблагоприятная роль большей части и артериальных, и венозных
нарушений гемоциркуляции в глазу обусловлена патологией капил-
лярной сети. Однако при некоторых заболеваниях, в частности при
, диабете, уже в самых ранних стадиях процесс начинается с микро-
ангиопатии, охватывающей не только капилляры, но также прека-
пиллярные артериолы и посткапиллярные венулы.
Нарушение постоянства содержания кислорода в сетчатке ока-
зывает неблагоприятное влияние на функцию внутреннего гемато-
офтальмического барьера, проявляющееся в закупорке или пропо-
тевании капилляров.
1. Закупорка прекапилляров (вследствие утолщения базисных мем-
бран, пролиферации эндотелия, усиления склеивания тромбоцитов,
деструкции эритроцитов, теряющих способность переносить кисло-
род) приводит к ретинальной ишемии (кардинальный симптом),
затем к ретинальной гипоксии, освобождению ’’вазоформатной суб-
станции”, что, в свою очередь, обусловливает артериовенозное шун-
тирование, неоваскуляризацию сетчатки и рубеоз радужки, кото-
рые приводят к внутриретинальному фиброзу и отслойке сетчатки.
2. Пропотевание (“протечка”) капилляров (в связи с уменьше-
нием количества перицитов в стенке сосудов) приводит к геморра-
гиям и плазморрагиям. Результатом геморрагии является фиброз, а
плазморрагии обусловливают ретинальноый отек: диффузный вслед-
ствие просачивания плазмы и локальный в результате образования
микроаневризм. Первый заканчивается “мягким экссудатом” и ки-
стовидными дегенерациями, а второй — “твердым экссудатом” (ли-
посодержащие макрофаги и холестерол).
Второй характерной, хотя и значительно более редкой, чем
диабет, клинической формой нарушений капиллярного кровотока
сетчатки является ретинопатия недоношенных детей, звенья патоге-
неза которой таковы: наблюдается избыток кислорода в крови и
сужение ретинальных артерий с облитерацией их русла, которая
последовательно приводит к гипоксии и отеку сетчатки, вазопроли-
ферации, рубцеванию и отслойке сетчатки.
3.5. ТЕСТИРОВАНИЕ НАРУШЕНИЙ ВНУТРИГЛАЗНОЙ
ГЕМОДИНАМИКИ В КЛИНИЧЕСКОЙ ПРАКТИКЕ
(ПО ДАННЫМ ФЛЮОРЕСЦЕНТНОЙ АНГИОГРАФИИ
И ДРУГИХ МЕТОДИК)
3.5.1. Оценка кровотока при симптомах гипофлюоресценции
Возможные причины патологической гипофлюоресценции при ФАГ
гдазного дна:
1. Флюоресцеин не поступает либо поступает с задержкой в ре-
301
тинальное или хороидальное русло вследствие закупорки артерии,
медленнее выводится из-за обтурации вены. По локализации зоны
незаполнения можно судить о месте нахождения эмбола или тромба.
2. Аномальное кровоснабжение какой-либо части глазного дна
(при хороидеремии, миопической болезни).
3. Экранирование хороидальной и ретинальной флюоресценции:
а) увеличение плотности пигментации либо в нейросенсорных
слоях сетчатки (ксантофил), либо в пигментном эпителии (мела-
нин);
б) отложение патологических субстратов (липофусцина при
дистрофии типа Беста, твердых экссудатов при диабете и пр.);
в) кровоизлияния в различные слои сетчатки, пре- и пострети-
нальные.
Прогноз заболевания в значительной мере определяется запол-
няемостью капиллярной сети. Если после рассасывания маскирую-
щих кровоизлияний, отека, экссудатов становится очевидным, что
в сетчатке или хороидее сформировалась бессосудистая зона, то
прогноз в отношении восстановления утраченной зрительной фун-
кции при такой ишемической патологии (в том числе макулопа-
тии) плохой (неэкссудативные формы “географической” и сухой
атрофии хороикапилляров, экссудативные формы других макуло-
патий).
Используя офтальмодинамометрию при установлении причины
артериальных нарушений, целесообразно определить перфузионное
давление (РА — Pv), рассчитать градиент давления (РА — ВГД). При
стенозах внутренней сонной артерии и ее ветви — глазной артерии
РА значительно снижено. Градиент ниже 55 мм рт.ст. при повышен-
ном офтальмотонусе является плохим прогностическим признаком.
В то же время имеются клинические наблюдения, свидетельству-
ющие о том, что у больных диабетом в глазу, в артериальной сис-
теме которого ослаблен кровоток, ангиоретинопатия может быть
менее выраженной, чем на парном глазу. На основании этого сде-
лан вывод, что уменьшение трансмурального давления в венах и
капиллярах как следствие ослабления кровотока защищает сосуди-
стую стенку от характерных для диабета повреждений [Gay A.J.,
Rosenbaum R.L., 1965]. Вместе с тем наиболее тяжелые формы от-
слойки сетчатки возникают на фоне ухудшения кровотока, выра-
женного снижения реографического коэффициента и пульсового
объема крови [Малашенкова Е.Н. и др., 1973, 1975].
Для того чтобы определить минутный объем крови, можно про-
вести офтальмоплетизмографию или офтальмореографию, но луч-
ше — допплерографию. Две последние методики позволяют также
получить представление о скорости кровотока, с их помощью можно
контролировать степень спазмирования сосудов и влияние фармако-
логических препаратов, ингаляций карбогена (7% СО2 + 21% О2) на
калибр сосудов сетчатки [Яковлев А.А., 1970; Alm A., Bill А., 1987].
302
3.5.2. Оценка тематических барьеров при симптомах
гиперфлюоресценции
Возможные причины патологической гиперфлюоресценции при ФАГ
глазного дна:
1. Образование атрофических “окон” в пигментном эпителии
(рис.3.6), позволяющих отчетливо видеть нормальную флюоресцен-
цию хороидеи (например, из-за токсической макулопатии).
2. Просачивание красителя под пигментный эпителий в местах
нарушения мембраны Бруха (рис.3.7).
3. Просачивание красителя под фоторецепторы не только сквозь
мембрану Бруха, но и через пигментный эпителий, т.е. при наруше-
нии наружного тематического барьера (центральная серозная рети-
нопатия).
4. Просачивание красителя под сетчатку из проросших под
нее новообразованных сосудов хороидеи (неоваскулярные мемб-
раны).
5. Просачивание красителя сквозь стенку измененных сосудов
сетчатки, т.е. при нарушении внутреннего тематического барьера
(при диабете, кистовидном макулярном отеке, застойных дисках)
(рис. 3.8).
6. Задержка выхода красителя из некоторых патологических
образований (длительное прокрашивание друз, опухолей хороидеи)
(рис. 3.9).
Таким образом, в основе образования очагов гиперфлюорес-
ценции лежит либо задержка флюоресцеина в тканях (прилипание
молекул флюоресцеина к друзам, захват их клетками злокачествен-
ных опухолей), либо значительно чаще — просачивание его через
нарушенные тематические барьеры.
Современные методики количественной флюорофотометрии
позволяют диагностировать нарушения функции внутреннего тема-
тического барьера в случаях диффузного просачивания красителя в
среды глаза, например при диабете, еще до появления очаговых
изменений, видимых при флюоресцентной ангиографии. Иногда
участок диффузного просачивания определяют уже при офтальмо-
скопии по скоплению микрокист в макулярной области в форме
тычинок и пестика цветка. Однако особенно отчетливо кистовид-
ный отек макулярной области идентифицируется при ФАГ. Весьма
тонкий в норме слой Генле (наружный плексиформный слой) утол-
щается в несколько раз вследствие повышения проницаемости ре-
тинальных капилляров парамакулярной области (при синдроме
Ирвина — Гасса, диабете и других заболеваниях).
Нарушение внутреннего гематоретинального барьера позволя-
ет не только свободным, но и связанным молекулам флюоресцеи-
на выходить в экстраваскулярное пространство парафовеолярной
зоны.
303
Рис. 3.6. Усиление флюоресценции на участке атрофии пигментного эпите-
лия при хлороквиновой макулопатии. Объяснение в тексте.
Отдельные фокусы гиперфлюоресценции, появившиеся в арте-
риовенозной фазе, в поздние сроки сливаются, а затем долго ярко
светятся, распадаясь на мелкие капельки (задержка красителя в мик-
рокистах).
ФАГ позволяет обнаруживать также зоны нарушения внешнего
гематоретинального барьера, в частности при различных экссуда-
тивных формах макулодистрофии, и оценивать их не только по
площади, но и по глубине, что исключительно важно для правиль-
ного выбора метода лазерной терапии.
При изолированной отслойке только пигментного эпителия сет-
чатки участок гиперфлюоресценции намечается уже в хороидаль-
ной фазе в виде слабо светящегося округлого очажка. В артериове-
нозной фазе интенсивность свечения очажка увеличивается, при этом
четкость его границ сохраняется на протяжении всего цикла про-
хождения флюоресцеина через сосуды глаза. Это обусловлено со-
храняющейся прочной связью пигментного эпителия с мембраной
Бруха вокруг зоны отслойки (эта связь прочнее, чем связь пигмен-
тного и сенсорного нейроэпителия).
304
Рис. 3.7. Скопление флуоресцеина под отслоенным пигментным эпите-
лием.
При центральной серозной хороиретинопатии, когда из-за “по-
ломки” внешнего гематоретинального барьера свободные молеку-
лы флюоресцеина проникают не только под пигментный эпителий,
но и сквозь него — под нейроэпителий, встречаются два варианта
ФАГ-проявлений [Kanski J., 1989]. Первый вариант — по типу ды-
мовой трубы (рис. 3.10). Уже в хороидальной фазе на ограничен-
ном участке (в зоне отслойки пигментного эпителия) замечают то-
чечное свечение. В поздней венозной фазе краситель просачивается
в субретинальное пространство и поднимается в нем, подобно дыму
из трубы, прямо вверх. По достижении верхней границы отслоен-
ной зоны краситель “обтекает” ее с обеих сторон, формируя в этой
фазе картину “гриба”, и иногда постепенно занимает всю обычно
округлую полость расслоения.Второй вариант — по типу черниль-
ной кляксы: характерное для венозной фазы нарастание флюорес-
ценции распространяется равномерно во всех направлениях от пер-
воначальной точки.
Разнообразные патологические состояния макулярной области
могут стать причиной развития субретинальных неоваскулярных
305
Рис. 3.8. Кистовидный макулярный отек, вызванный нарушением внутрен-
него гематоофтальмического барьера.
мембран (рис. 3.11). Они берут начало из хороикапилляров сосуди-
стой оболочки и прорастают через мембрану Бруха сквозь щели в
пигментном эпителии под нейроэпителий. Это прежде всего экссу-
дативные макулодистрофии, в том числе диабетические, друзы
(очажковое скопление гиалиновой субстанции между базальной
мембраной пигментного эпителия, с одной стороны, и коллагено-
вым слоем мембраны Бруха — с другой), пятно Фукса при миопиче-
ской болезни, ангиоидные полосы при болезни Гренблата — Странд-
306
Атрофия и д епигментация РПЭ
Друзы
Рис. 3.9. Гистопатологическая схема формирования друз и флюоресцент-
ная ангиограмма глазного дна с множеством друз.
берга, опухоли хороидеи, участки травматических разрывов, чрез-
мерной коагуляции, поствоспалительные очаги.
Причинно-следственной связи между отслойкой пигментного
эпителия и развитием хороидальной неоваскулярной мембраны не
установлено. В ранних стадиях при ФАГ можно не обнаружить эти
мембраны, хотя они уже видны офтальмоскопически при локализа-
ции под пигментным эпителием в виде серовато-зеленоватых слег-
ка выступающих над поверхностью сетчатки очагов, а при прора-
стании в субретинальное пространство они воспринимаются блед-
но- или желтовато-розовыми. Поскольку капилляры хороидеи в
отличие от капилляров сетчатки не выполняют барьерную функ-
цию, субретинальные мембраны легко пропускают свободные мо-
лекулы флюоресцеина. При ФАГ по контуру очага неоваскулярной
307
Рис. 3.10. Серия флюоресцентных ангиограмм при центральной серозной
хороидопатии по типу дымовой трубы.
мембраны уже в хороидальной фазе возникает гиперфлюоресцен-
ция, напоминающая кружевной воротничок. На высоте флюорес-
ценции нормальных сосудов сетчатки (через 20 с) довольно ярко
светится и “воротничок”, состоящий из сливающихся пятен. Свече-
ние его еще более усиливается через 1—2 мин, когда флюоресцен-
ция нормальных структур существенно гаснет.
Позднее свечение объясняется, во-первых, тем, что фиброзная
ткань самой мембраны задерживает краситель, а во-вторых, тем
что нередко мембрана окружена жидкостью, отслаивающей пиг-
ментный эпителий, где также задерживается краситель.
Субретинальная мембрана не только непосредственно ухудшает
зрение, но и может быть причиной следующих тяжелых осложне-
ний:
308
Рис. 3.11. Серия флюорес-
центных ангиограмм при
хороидальной неоваскуляр-
ной мембране.
1) геморрагической отслойки пигментного эпителия при разры-
ве собственного сосуда мембраны;
2) геморрагической отслойки нейроэпителия (в первое время под
отслоившейся сетчаткой виден очаг сине-черного цвета, который
спустя 1,5—2 нед окружает ореол ярко-красного цвета);
3) прорыва крови в стекловидное тело (при сниженной актив-
ности свертывающей системы крови, например на фоне лечения ан-
тикоагулянтами);
4) формирования рубца дисковидной формы с необратимой ут-
ратой зрительных функций;
5) экссудативной отслойки сетчатки, выходящей за пределы ма-
кулярной области.
Для профилактики указанных осложнений с целью сохранения
макулярной функции прибегают к лазерной коагуляции сосудистой
мембраны, предварительно вводя внутривенно индоцианин зеленый
[Reichel Е., 1994]. Некоторые офтальмологи считают лазерную коа-
гуляцию противопоказанной, если какой-либо край мембраны на-
309
ходится на расстоянии менее 200 мкм от центра фовеолы (диаметр
фовеолы 350 мкм), а также при наличии непосредственно под зо-
ной фовеа геморрагической отслойки пигментного эпителия. По
этой причине лишь 1 из 10 пациентов с рассматриваемой патологи-
ей отбирают для лазерного лечения.
Задержка флюоресценции по краям “обстрелянной” мембраны
не свидетельствует о рецидиве, а является признаком новообразо-
ванной соединительной ткани.
При ишемической макулопатии, когда капиллярное кровообра-
щение в макулярной зоне отсутствует (в норме, как уже отмечалось,
аваскулярная зона занимает участок диаметром не более 0,5 мм), не
приходится рассчитывать на эффект от лазерной терапии [Kanski J.,
1989].
глABA 4
аппарат, регулирующий гидродинамику
И ОПТИЧЕСКУЮ УСТАНОВКУ ГЛАЗА
4.1. АНАТОМО-ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
ИРИДОХРУСТАЛИКОВОЙ ДИАФРАГМЫ, РЕСНИЧНОГО
ТЕЛА, КАМЕРНОГО УГЛА И РЕШЕТЧАТОЙ МЕМБРАНЫ
ДИСКА ЗРИТЕЛЬНОГО НЕРВА
Рассматриваемые структуры, кроме решетчатой мембраны диска
зрительного нерва (ДЗН), в совокупности составляют передний отдел
глаза. Их объединяет также участие в регулировании таких важных
функций, как продукция и отток внутриглазной жидкости, управле-
ние оптическим аппаратом глаза. По зрачковым реакциям можно
судить также о сохранности и об уровне поражения зрительно-не-
рвного пути. Расположение решетчатой мембраны ДЗН способствует
гидродинамическому балансу между тканевыми структурами, рас-
положенными перед и за мембраной.
4.1.1. Продукция и отток камерной влаги, офтальмотонус
Внутриглазная гидродинамика необходима не только для регули-
рования обменных процессов во внутриглазных структурах, в час-
тности в оптических средах, но и для поддержания в глазу тонуса
в определенных пределах. Последний обеспечивает сохранность
формы глазного яблока, в частности предотвращает образование
складок роговицы и склеры, создает условия для нормального кро-
вообращения во внутриглазных сосудах, поддерживает на оптималь-
ном уровне перфузионное и трансмуральное давление.
Офтальмотонус, или внутриглазное давление (ВГД), — это дав-
ление глазного содержимого, составляющее у большинства здоро-
вых людей в среднем около 15 мм рт.ст. Индивидуальные вариации
этого так называемого истинного ВГД (Ро) в норме достаточно ве-
лики (от 8 до 21 мм рт. ст.). В клинической практике отечественных
офтальмологов чаще используют показатели тонометрического
давления (Pt).
С учетом веса тонометра Pt всегда выше, чем Р0(рис. 4.1). Хотя
в медицинской документации принято фиксировать строго опреде-
ленные величины ВГД, эти цифры всегда выходят за рамки точно-
сти метода. Даже колебания офтальмотонуса в зависимости от фазы
артериального пульса достигают ±1 мм рт. ст. (рис. 4.2). Кроме того,
имеют значение фаза дыхания и степень напряжения глазных мышц,
особенно мышц века, и многие другие факторы, в том числе долго-
срочные (время суток, сезонность). Основными факторами, опреде-
311
Рис. 4.1. Истинное (Ро) и тонометрическое (Pt) внутриглазное давление
(приведенные цифровые значения свидетельствуют об офтальмогипертен-
зии).
ляющими уровень ВГД, являются: продукция камерной влаги (F),
сопротивление оттоку ее из глаза (R).
Продукция и перемещение влаги. Известны два механизма появ-
ления влаги в задней камере глаза.
Секреторный механизм реализуется благодаря насосно-метаболи-
ческой функции эпителия ресничных отростков ресничного тела, ко-
торый избирательно “нагнетает” в глаз аскорбаты, действуя незави-
симо от разности давления в капиллярах крови и задней камере глаза.
Фильтрационный механизм, особенно ультрафильтрационный
(в норме препятствующий выходу крупных молекул белков плаз-
мы из капилляров), зависит от разности гидростатического и онко-
тического давления в плазме крови и камерной влаге.
Перемещение влаги. Из задней камеры большая часть влаги через
зрачок, омывая хрусталик, поступает в переднюю камеру; неболь-
шая порция просачивается в стекловидное тело и направляется к
заднему полюсу глаза; часть влаги подвергается реабсорбции на месте
ее выделения ресничным телом. По мере продвижения влаги внутрь
глаза ее состав изменяется под влиянием метаболических процессов.
Само существование движения влаги свидетельствует о наличии
разницы давления в задней и передней камерах. Хотя за счет этого
возможен легкий физиологический ’’бомбаж” корня радужки, заме-
рить указанную разницу из-за ее малой величины пока не удалось.
Вместе с тем известно, что благодаря эластичности и упругости
хороидеи давление в супрахороидальном пространстве на 1 —
2 мм рт. ст. ниже, чем в полости глаза.
Отток влаги из глаза. Изучены два основных пути
оттока.
1. Путь через сеть корнеосклеральных трабекул в углу передней
камеры. Жидкость сквозь межтрабекулярные микропоры и щели
312
24
22
12 3 4
16 1 1 1 LUJ И ,1.,1.1.1,1.1 И 1 1 1 1 i.l 1J1 1 1..1 1 1 1 1 11 XI 1 i II 1 1
0 1 2 3 Время (с) 4 5
Пульс ВГД min, ВГД max, АП, па ПКГ,
mmHg mmHg mmHg p.l/min
1 18,0 20,9 2,86 5,46 576
2 18,0 20,9 2,97 5,66 612
3 18,0 20,6 2,63 5,04 561
4 18,0 21.2 3,17 5,99 5,54 651
средн. 18.0 20,9 2,91 600
Рис. 4.2. Типичная запись пульсовых колебаний офтальмотонуса и рассчи-
танные компьютером результаты измерений [Langham М., 1989].
ВГД — внутриглазное давление, АП — амплитуда глазного пульса, ПО — пульсо-
вой объем, ПКТ — пульсирующий глазной кровоток.
шириной 1,5 мкм проникает между клетками эндотелия, выстилаю-
щими стенки венозного синуса склеры, именуемого иногда круго-
вым венозным синусом (хотя в норме он не содержит крови), и
попадает в его просвет (диаметр 0,28 мм). С наружной стороны
канала имеется от 17 до 35 тонких (диаметром от 5 до 160 мкм)
выпускников. Эти коллекторные каналы, анастомозируя между со-
бой, образуют интрасклеральное сплетение. Из него влага идет по
так называемым водяным венам Ашера (каждая шириной 0,01—
0,1 мм) в эписклеральные вены. Здесь происходит смешение влаги с
кровью. Давление в эписклеральных венах обычно сохраняется по-
стоянным (в среднем 8—12 мм рт. ст.). Описанным путем в веноз-
ную систему глазницы оттекает не менее 80% камерной влаги. Ве-
личина оттока по этому пути зависит от разности давления внутри
глаза и в эписклеральных венах. Однако, по данным R. Brubaker
(1975), прямой зависимости между повышением ВГД и усилением
оттока (С) не существует, поскольку при офтальмогипертензии из-
за уменьшения глубины передней камеры и сужения угла сопротив-
ление оттоку (R) может возрастать.
313
Ресничная мышца в процессе аккомодации оказывает опреде-
ленное влияние на степень фильтрации жидкости через трабекулу
[Allen L., Burian Н., 1965]. Та часть наружных волокон ресничной
мышцы, которая вплетается в склеральную шпору и трабекулы, при
своем сокращении оттягивает и распрямляет трабекулярную сеть,
тем самым не только предотвращается ее коллапс, но и улучшается
проходимость [Moses R., 1987]. При резком сокращении циркуляр-
ной порции ресничной мышцы отток может даже быть затруднен.
В то же время и возрастное ослабление активности мышцы также
оказывает отрицательное влияние на функцию трабекулярной ди-
афрагмы, нарушается ее метаболизм, забиваются поры, уменьшает-
ся эластичность [Нестеров А.П., 1995].
2.Увеосклеральный путь. По данным A. Bill (1966), около 20%
камерной влаги из угла передней камеры просачивается через ту
часть ресничного тела, которая выступает в камеру между скле-
ральной шпорой и корнем радужки. Независимо от уровня ВГД
жидкость просачивается по межклеточным пространствам реснич-
ного тела и далее— хороидеи. На своем пути к заднему отделу
глаза она частично резорбируется сосудами указанных структур и
из полости глаза просачивается непосредственно через ткань скле-
ры в глазницу. Получены данные, согласно которым атропин уси-
ливает, а пилокарпин ослабляет этот механизм оттока. В генезе пос-
леоперационных цилиохороидальных отслоек следует учитывать
роль увеосклеральных взаимоотношений [Волков В. В., 1973; Van
Alfhen G. W. H., 1961; Moses R., 1965].
Имеются некоторые основания считать, что существует третий
путь оттока жидкости из глаза через структуры диска зрительного
нерва в его межоболочечные пространства. М.Я. Фрадкин с соавт.
(1961), а в последние годы Е. Г. Рапис установили, что частицы
китайской туши, введенной в полость стекловидного тела глаза
кролика, спустя несколько часов обнаруживаются в межоболочеч-
ных пространствах зрительного нерва. На существование этого пути
оттока указывают и случаи развития посттромботической глауко-
мы, как известно, плохо поддающейся традиционному лечению.
При сопоставлении возможностей различных путей оттока жид-
кости из глаза очевидно, что решающий вклад в этот процесс вно-
сит транстрабекулярный путь [Goldmann Н., 1951; Grant М., 1958],
где локализуется и основной механизм регуляции сопротивления
оттоку, позволяющий поддерживать офтальмотонус на необходи-
мом уровне. Подсчитано, что сопротивление движению влаги в глазу
в 100 000 раз превосходит *то сопротивление, которое испытывает
кровоток по сосудам, хотя вязкость крови почти в 5 раз выше вяз-
кости камерной влаги [Нестеров А.П., 1967].
Принципы расчетов показателей гидродинамики глаза. Первона-
чально, по предложению Н. Goldmann (1951), за сопротивление от-
току (R) принимали давление (в миллиметрах ртутного столба),
314
которое требуется, чтобы, преодолевая сопротивление, вытеснить
из глаза за 1 мин 1 мм3 влаги. Результат оценивали в мм рт. ст./мин
. мм3. Однако в широкую клиническую практику вошел обратный
показатель (I/R), обозначенный как коэффициент легкости оттока
(С) с размерностью мм3/мин на 1 мм рт. ст.
На основе закона Пуазейля, касающегося истечения жидкостей
через цилиндрическую трубку, R. Moses, М. Grant, А. П. Нестеров
и др. для расчетов объема камерной влаги (F), оттекающей за 1 мин
через трабекулярную систему глаза, использовали формулу:
F=aP.C или AP = F/C,
где ДР — разница между истинным ВГД (Ро) и давлением в эпи-
склеральных венах (Pv), т.е. Ро — Pv; С — коэффициент легкости от-
тока.
Согласно результатам электротонографии, коэффициент легко-
сти оттока (С) в норме варьирует от 0,15 до 0,55 мм3/мин на 1 мм рт.
ст. [Нестеров А.П. и др., 1967]. При построении математической
модели гидродинамики глаза С принимают равным 0,3 мм3/мин на
1 мм рт. ст., a Pv— 9 мм рт. CT.[Moses R., 1987].
Для случаев устойчивого офтальмотонуса (Ро) и с учетом высо-
кой стабильности уровня Pv показатель F часто рассматривают не
только как минутный объем (мм3/мин) оттекающей из глаза влаги,
но и как минутный объем продуцируемой внутриглазной жидкости.
Как повышение, так и понижение уровня ВГД относительно нор-
мы свидетельствует о дисбалансе между объемами продуцируемой и
оттекающей влаги. В клинической практике значительно чаще при-
чиной указанных сдвигов является нарушение оттока или затрудне-
ния в циркуляции жидкости в полости глаза (так называемые блоки,
из которых наиболее распространенным является зрачковый).
Отмечаемые у здоровых людей суточные колебания офтальмо-
тонуса обычно не превышают 5 мм рт. ст. Значительно чаще утром
(сразу после пробуждения, т.е. в 6 — 7 ч) давление оказывается
выше, чем вечером. Можно было бы предположить, что во время
ночного сна ВГД постепенно повышается, однако это не согласует-
ся с данными Р. Henkind и соавт. (1973), которые установили наи-
более низкие показатели именно в ночное время. Исследователя
должна насторожить заметная стойкая асимметрия показателей оф-
тальмотонуса парных глаз (более 3 мм рт. ст.).
4.1.2. Взаимоотношения офтальмотонуса с артериальным
и внутричерепным давлением, а также с тканевым давлением
в зрительном нерве
Принято считать, что внутриглазное давление сохраняет выражен-
ную автономность и практически не зависит от давления в сосудах
глаза и тканях зрительного нерва. Изложенные в данном разделе
315
Таблица 4.1. Влияние среднего артериального давления (АД) на базовое
внутриглазное давление кролика
Планируемое АД Фактическое АД ВГД
100 98 ± 0,9 21 ± 1,7
80 80 ± 0,7 16 ± 1,4
60 62 ± 0,8 14 ± 0,9
40 41 ± 1,5 11 ± 0,6
20 20 ± 1,3 10 ± 0,7
Посмертно 9 ± Q6
результаты достаточно новых и пока мало известных эксперимен-
тов на животных свидетельствуют в пользу наличия связей между
ними.
Офтальмотонус и артериальное давление. Интегральным вы-
ражением существующих представлений об офтальмотонусе могут
служить тонообъемные взаимоотношения в глазу [Moses R. S., 1987].
Коэффициент ригидности, лежащий, по Фриденвальду (Frieden-
wald, 1937), в основе этих взаимоотношений, до недавнего времени
считали относительно постоянным и независящим от общей и мест-
ной гемодинамики. Основанием для этого послужили результаты
многочисленных работ, в которых не находили существенной
разницы между коэффициентами ригидности живого и свеже-
энуклеированного глаза.
Ошибочность такого подхода показана в экспериментах (прове-
денных.на 23 животных), в которых было установлено, что, во-пер-
вых, артериальное давление влияет на офтальмотонообъемные со-
отношения, а значит, и на офтальмотонус (табл. 4.1) [Kiel J.W., 1994,
1995].
Данные, представленные в табл. 4.1, свидетельствуют, что уро-
вень офтальмотонуса экспоненциально варьирует в зависимости от
среднего артериального давления.
Во-вторых, когда по мере наполнения глаза жидкостью повы-
шается и офтальмотонус, видно, что чем выше АД, тем круче под-
нимается начальная часть соответствующей кривой. Как только ВГД
приближается к АД, эта зависимость исчезает.
При сопоставлении офтальмотонообъемных соотношений при
разных уровнях АД показана зависимость коэффициента ригидно-
сти глаза от АД. Коэффициент ригидности рассчитывается из ли-
нейной корреляции офтальмотонообъемных соотношений со сред-
ним АД.
В-третьих, после того, как офтальмотонус приблизится к уров-
ню АД, дальнейшее наполнение глаза жидкостью не сразу приво-
дит к повышению ВГД (рис.4.3). Видимое на рисунке плато объяс-
316
йяется компенсаторным вытеснением части крови из сосудов хоро-
идеи. Снижение перфузионного давления (АД — ВГД) приводит сна-
чала к ослаблению кровотока, а затем и к уменьшению концентра-
ции движущихся клеток крови.
Можно предположить, что неблагоприятное течение глаукома-
тозного процесса у пациентов с низким артериальным давлением
оказывается следствием низкого перфузионного давления, когда при
умеренных показателях офтальмотонуса существенно затрудняется
кровообращение в глазу (эффект Старлинга). Таким образом, гид-
родинамические процессы в глазу, гомеостаз и офтальмотонус на-
ходятся в тесной связи с гемодинамикой.
Офтальмотонус, внутричерепное давление (ВЧД) и тканевое
давление (ТД) в зрительном нерве. Взаимоотношения между внутри-
глазным и внутричерепным давлением давно привлекают внимание
исследователей [Ноишевский К. И., 1912, и др.]. В эксперименте после
костной орбитотомии впервые удалось измерить давление жидко-
сти в межоболочечных пространствах зрительного нерва (у кроли-
ка), которое составило 110—140 мм вод. ст., что эквивалентно 8,1—
0,5 мм рт. ст. [Волков В. В., Коровенков Р. И., 1973]. Сходные дан-
ные были получены и в опытах на кошках [Hedges Т., ZarenH.,
1973]. Тканевое давление в зрительном нерве и межоболочечных
пространствах оказалось близким к внутричерепному давлению
(последнее в зависимости от положения тела колеблется от 6 —
10 мм рт. ст. до нуля) и ориентировочно на 10 мм ниже истинного
внутриглазного давления, составляющего в норме 12—20 мм рт. ст.
(в среднем 16 мм рт. ст., по F. Hollows и др., 1966).
Спустя 20 лет рассмотренные исследования получили дальней-
шее развитие [Morgan W. et al., 1995]. Для изучения влияния давле-
ния цереброспинальной жидкости на градиент тканевого давления
в решетчатой мембране ДЗН применено трансвитреальное введе-
ние микропипетки прямо в ДЗН глаза собаки (решетчатая мембра-
на собаки напоминает таковую у человека). Основной перепад от
ВГД к тканевому давлению в зрительном нерве происходит, как
оказалось, в слое решетчатой мембраны толщиной всего 400 мкм.
Причем на глубине первых 200 мкм на уровне тканевого давления
ДЗН еще сказывалось искусственное изменение офтальмотонуса. На
большей глубине уровень тканевого давления в зрительном нерве,
составлявший в среднем 8,6±3,5 мм рт. ст., уже не зависел от внут-
риглазного давления, однако зависел от окружающего давления
жидкостей в межоболочечных пространствах зрительного нерва,
которое практически соответствовало давленииэ в боковых желу-
дочках мозга. Таким образом, чем более высокими были индивиду-
альные показатели давления цереброспинальной жидкости, тем
выше были и показатели ретроламинарного тканевого давления в
зрительном нерве.
Полученные данные свидетельствуют о том, что ретроламинар-
317
Рис. 4.3. Типичный ход кривых, отражающих хороидальный кровоток и на
основе лазерной допплерографии концентрацию движущихся по сосудам
клеток крови (КДКК), а следовательно, и объем крови во время и после
прекращении вливания в глаз изотонического раствора натрия хлорида
[Kiel J.W., 1995].
318
ное тканевое давление в ДЗН определяется главным образом уров-
нем давления цереброспинальной жидкости. Следовательно, внут-
ричерепное давление наряду с внутриглазным формирует транс-
мембранный градиент тканевого давления в зрительном нерве. Ре-
зультаты исследований, проведенных W. Morgan и соавт.,
свидетельствуют о постоянстве гидростатического давления по ходу
зрительного нерва при плавном его снижении в направлении от
глаз к мозгу.
Вероятно, аксоны зрительного нерва по выходе из глаза на
уровне решетчатой мембраны ДЗН даже в норме испытывают на
себе резкий перепад давления. Согласно закону Ла Пласа, этот
градиент создает механические усилия в направлении, поперечном
тому, в каком действует градиент, т.е. усилия приходятся на плос-
кость решетчатой мембраны. Не исключено, что именно рассмат-
риваемый градиент давления в норме оказывается движущей си-
лой, во-первых, для медленного аксоплазматического тока, суще-
ствующего в аксонах всякого нормального зрительного нерва, а
во-вторых, для перемещения жидкостей в его межоболочечных про-
странствах от глаз к мозгу.
Известно, что повышение ВЧД затрудняет или останавливает
ток жидкости во внутри— и межоболочечном пространстве; клини-
чески это проявляется симптомами застойного диска зрительного
нерва. Если из-за отека его ткани прерывается не только медлен-
ный, но и быстрый аксоплазматический ток, то начнется ухудше-
ние зрительных функций, которое может принять необратимый
характер. Примечательно, что причиной застойного диска может
быть не только повышение ВЧД, но и выраженное стойкое пониже-
ние ВГД (например, вследствие рецессии угла передней камеры после
контузии или оперативного вмешательства).
Меньше единства в толковании офтальмологами изменений в
ДЗН, связанных с противоположной ситуацией, когда либо абсо-
лютно, либо относительно более высоким оказывается ВГД по срав-
нению с ВЧД, т.е. с офтальмогипертензией или глаукомой. Многие
авторы объясняют глаукоматозную атрофию ДЗН сосудистыми
нарушениями, оставляя без внимания очевидную роль механичес-
кого гипертензионного фактора. Их не смущает тот факт, что ни
понять, ни смоделировать патогномоничную для глаукомы атро-
фию ДЗН с прогибанием решетчатой мембраны на основе ишеми-
ческих факторов невозможно. Почему достаточно часто при незна-
чительном повышении офтальмотонуса и при “нормальных” его
<------------------------------------------------------------
Плато кривых офтальмотонуса при разных уровнях АД в глазничной артерии: а —
при 80 мм. рт. ст.; б — при 60 мм рт ст.; в — при 40 мм рт. ст. Как только офталь-
мотонус начинает превышать уровень АД в глазничной артерии — кровоток осла-
бевает и объем циркулирующей крови уменьшается. 1 — среднее АД, мм рт. ст., 2 —
хороидальный кровоток, мм3 в 1 мин, 3 — КДКК, мкл, 4 — ВГД, мм рт. ст.
319
значениях глаукоматозный процесс прогрессирует, остается неяс-
ным.
Для объяснения представленных выше противоречивых, на пер-
вый взгляд, клинических ситуаций, а именно возможности развития
застойного диска зрительного нерва при нормальном ВЧД и глау-
комы при нормальном ВГД, предлагается [ Волков В. В., 1974,1976,
1995 ] оценивать не столько их абсолютные, сколько относительные
значения, т.е. принимать во внимание наличие градиента давления
в области решетчатой мембраны ДЗН (норма до 10 мм рт. ст). Ва-
риант так называемой глаукомы низкого (’’нормального”) давле-
ния особенно характерен для людей преклонного возраста с ’’юно-
шеским” артериальным давлением. При такой выраженной артери-
альной гипотонии вероятной представляется и внутричерепная
гипотония. Отсутствие щадящих способов исследования ВЧД не по-
зволяет проводить широкую клиническую проверку справедливос-
ти указанного предположения. D. Liu с соавт. (1995) с помощью
микроманометра измеряли давление жидкостей в стволе зрительно-
го нерва на операционном столе у людей непосредственно перед
тем, как произвести энуклеацию по поводу какого-либо заболева-
ния глаза, когда это было необходимо. Мы сопоставили средние
показатели, полученные этими исследователями у больных глауко-
мой и больных других категорий, и обнаружили, что при глаукоме
они были на 25% ниже (соответственно 6 и 8,5 мм рт.ст.). Возникает
важный вопрос: может быть, не только при глаукоме низкого дав-
ления, а и вообще при неблагоприятно текущей глаукоме привыч-
ный в норме градиент порядка 10 мм рт. ст. нарушается вследствие
как повышения первого, так и снижения второго?
Накопилось уже немало фактов, подтверждающих неблагоприят-
ное влияние механического воздействия на зрительно-нервные волок-
на ДЗН именно в зоне решетчатой мембраны при ее прогибании.
1. В некоторых клинических наблюдениях отмечено, что при
остром приступе глаукомы по мере снижения ВГД экскавация ДЗН
исчезает и появляется вновь при рецидивах приступа [Neumann Е.,
Hyams S., 1973].
2. В экспериментах на энуклеированных глазах [ Нестеров А.П.,
Егоров Е.А., 1977 ] и в наблюдениях за добровольцами [ Волков В.В.
и др., 1981 ] отмечалось прогибание ДЗН при искусственной оф-
тальмогипертензии.
3. Деформация отверстий решетчатой мембраны при ее проги-
бании сопровождается уменьшением площади сечения в местах
прохождения зрительных нервных волокон [Волков В. В., 1976;
Quigley Н., Addicks Е., 1982].
4. При искусственной острой офтальмогипертензии в зритель-
но-нервных волокнах у обезьян прекращается быстрый аксоплаз-
матический ток как раз на уровне деформируемых отверстий в ре-
шетчатой мембране ДЗН [ Minckler D. и др., 1978 ].
320
5. Отрицательное воздействие нагрузочной вакуум-периметри-
ческой пробы используемой для диагностики глаукомы у людей
[Волков В.В. и др., 1981 ].
Если у взрослых наименее прочным участком капсулы глаза,
неспособным противостоять стойкой абсолютной или относитель-
ной офтальмогипертензии, оказывается решетчатая мембрана ДЗН,
то у маленьких детей прежде всего растягиваются податливые ро-
говица и склера, что является причиной мегалокорнеса, а возможно,
и миопической болезни [Кондратенко Ю. Н., Шаповалова В. М.,
1995, и др.]
4.1.3. Зрачковые реакции
Зрачок способен реагировать не только на свет, но и на установку
оптики глаза на близкое расстояние в связи со стимулом, что реа-
лизуется с помощью аккомодации и конвергенции.
Световой рефлекс — это сложная четырехнейронная дуга:
1-й нейрон: от фоторецепторов сетчатки до претектальных ядер
в среднем мозге;
2-й нейрон: от каждого претектального ядра к обоим ядрам
Якубовича — Эдингера — Вестфаля (межнунциальные нейроны
обеспечивают содружественную реакцию зрачков);
3-й нейрон: от указанных выше ядер идет в толще III пары
черепных нервов до ресничного узла в глазнице. Важно знать, что
эти волокна при выходе III пары из среднего мозга располагаются
поверхностно, поэтому могут сдавливаться аневризмой внутренней
сонной артерии, однако, проходя через боковую стенку пещеристо-
го синуса, они располагаются центральнее, в связи с чем даже при
полной наружной офтальмоплегии обычно не страдают; в глазнице
зрачковые вегетативные волокна от нижней ветви глазодвигатель-
ного нерва отходят, образуя глазодвигательный (парасимпатичес-
кий) корешок, волокна которого направляются к ресничному узлу.
4-й нейрон: от ресничного узла (который, хотя и содержит ряд
других волокон, лишь для парасимпатических является синапсом)
пупиллярные волокна вместе с короткими ресничными нервами
достигают сфинктера зрачка.
Рефлекс на близкие расстояния (“на аккомодацию и конверген-
цию”). Он состоит не только в сужении зрачка, но и в стимуляции
аккомодации вблизь и конвергенции зрительных осей. Когда свето-
вой рефлекс присутствует, нет надобности проверять рефлекс на близь
(он будет в наличии). Световая короткодистанционная диссоциация
зрачкового рефлекса относится лишь к тем редким случаям, когда
реакция на свет нарушается (исчезает), а рефлекс на близь, для кото-
рого наличие зрения не является необходимым, сохраняется.
Конечные пути рассмотренных рефлексов одинаковы (в III паре
черепных нервов). Различия заключаются в двух видах супранукле-
321
арных влияний на центры в среднем мозге: претектальные ядра,
обеспечивающие рефлекс на близь, лежат глубже (вентральнее), а
ведающие рефлексами на свет ядра Якубовича — Эдингера — Вес-
тфаля — дорсальнее. Вследствие этого последние раньше поража-
ются (как и рефлекс на свет) при росте пинеаломы и других процес-
сах в зоне дорсального водопровода задней комиссуры.
Время реакции зрачка на свет (0,26—0,3 с) заметно меньше, чем
время его реакции на близкий стимул, по-видимому, в связи с более
короткой рефлекторной дугой в первом случае.
4.1.4. Структурно-функциональные элементы аккомодационного
аппарата
Аккомодацией глаза называют его способность к рефлекторной пе-
рефокусировке своей оптической системы применительно к меняю-
щимся расстояниям до объекта зрительной фиксации. Стимулом к
аккомодации служит потеря четкости видения в зоне интереса из-за
неадекватной сходимости лучей и возникновения хроматической
аберрации.
Положение фокусной зоны на глазном дне может варьировать
вследствие следующих факторов: кривизны роговицы при прищу-
ривании век; длины оси глазного яблока при конвергенции — ди-
вергенции; позиции хрусталика относительно роговицы при накло-
нах головы; вариаций ’’кривизны” поверхностей хрусталика и оп-
тической плотности его вещества вдоль зрительной оси при
напряжении — расслаблении ресничных мышц; размеров зрачка,
причем в норме оптическая установка глаза на близкие расстояния
синкинетически сопровождается сужением зрачка.
Наиболее значимым из перечисленных и вместе с тем довольно
сложным является хрусталиковый механизм аккомодации. Прежде
чем попытаться его рассмотреть, необходимо остановиться на осо-
бенностях строения и функции участвующих в этом акте тканевых
структур.
Ресничная мышца. Она представляет собой кольцо гладкомы-
шечных волокон на внутренней поверхности склеры в ее переднем
отделе. Часть волокон вплетается в склеральную шпору, другие
берут начало от трабекулярной сети. Самый наружный слой распо-
лагается вдоль склеры меридионально (продольно) в виде идущих
кзади групп волокон и вплетается в хороидею. Волокна следующе-
го слоя направлены как бы радиально к оси глаза. Наконец, внут-
ренний слой не имеет связи со склерой, а формирует кольцо
сфинктера ресничной мышцы. По Е. F. Fincham (1937), волокна
мышцы в целом образуют изящную сетку. J. Rohen (1953) предста-
вил ее состоящей из волокон, которые во всех трех измерениях
ромбовидно перекрещиваются между собой. Углы перекреста, ме-
няющиеся в процессе аккомодации, в каждый данный момент нео-
322
динаковы в разных отделах мышцы, что и побуждало исследовате-
лей делить ее на порции.
Ресничная мышца обладает двойной иннервацией. Подобно
всякой гладкой мышце, она высокопластична, поэтому степень ее
напряжения не определяется длиной волокна. Известно, что глад-
кие мышцы многих полостных органов (мочевого пузыря, кишеч-
ника) работают не только при сокращении, но и при растяжении
мышечного волокна. В случае преобладания парасимпатических
влияний волокна укорачиваются, при перевесе симпатических вли-
яний — удлиняются. Даже в средней позиции, в состоянии равнове-
сия всей системы, гладкомышечные структуры, как известно, со-
храняют определенный тонус. Правда, энергозатраты при этом ми-
нимальны.
Хороидея. Предположения Т. Henderson (1926) относительно того,
что хороидея может быть не только пассивной, но и активной струк-
турой, воздействующей при аккомодации на ресничную мышцу, не
лишены оснований. Она, хотя и очень тонкая, в определенных пре-
делах, подобно резине, эластичная [ Moses R., 1965]. Ее связи со
склерой довольно нежны.
Смещаемость комплекса ресничное тело — хороидея в процессе
аккомодации установлена экспериментально [Hensen V., Volkers С.,
1973] и подтверждена клинически при трансиллюминации и опреде-
лении границы механофосфена. По данным R. Moses (1987), у мо-
лодых людей ширина диафаноскопической тени от ресничного тела
на склере на каждую 1 дптр аккомодации уменьшается на 0,05 мм,
т. е. при 10 дптр сокращение достигает 0,5 мм.
Зонулярная пластинка. В противоположном от хороидеи направ-
лении ресничное тело испытывает натяжение тончайшей мембраны
(зонулы), состоящей из множества бесклеточных волокон, одна
группа которых веером идет к передней, а другая — к задней пре-
экваториальной поверхности хрусталика. Волокна исходят из крипт
ресничного тела (между ворсинками), и только часть из них на-
правляется к хрусталику. Кроме них, имеются волокна, которые
формируют сетку на поверхности ресничного тела, ориентирован-
ную меридионально в направлении хороидеи и плоской части рес-
ничного тела. Эти волокна участвуют в формировании стекловид-
ного тела. Зонула состоит не из коллагена, а представляет собой
эластичную микрофибриллярную ткань. Последняя не только пере-
дает усилия ресничной мышцы на капсулу хрусталика, поддержива-
ющие его позицию, в частности предотвращающие хрусталиковый
зрачковый блок, но и вместе с капсулой образует барьер для камер-
ной влаги. Убедительно доказано неблагоприятное воздействие
последней на макулярную область сетчатки после интракапсуляр-
ной экстракции катаракты [Worst J.C., 1978].
Капсула и вещество хрусталика. Форма хрусталика определяет-
ся эластичностью его капсулы и содержимого. Старческий хруста-
323
лик со склерозированным ядром почти не способен к изменению
формы. В молодом возрасте, когда аккомодирующий хрусталик
способен округляться, из-за особой тонкости его капсулы в области
переднего полюса именно здесь он приобретает наиболее выражен-
ную кривизну (по Е. Fincham, 1937, ее радиус уменьшается с 10 до
6 мм). При этом, сокращаясь в размере по диаметру, хрусталик утол-
щается в направлении оптической оси (почти исключительно вслед-
ствие смещения его передней поверхности кпереди). Кроме тдго, в
процессе аккомодации по оптической оси возрастает показатель
преломления собственно эластичного вещества хрусталика в связи
с его анизотропностью [WealeR., 1962; Fischer R., 1982].
При значительном округлении хрусталика, в частности под воз-
действием аккомодативно-конвергентных усилий, хрусталик из-за
расслабления зонулы может “провисать”, что встречается у пресби-
опов при чтении с наклоном головы [Hess С., 1910]. В результате
этого блокируется зрачок, нарушается циркуляция камерной влаги и
повышается офтальмотонус у людей с предрасположенной к этому
анатомо-функциональной структурой рассматриваемой области.
Особый интерес представляет наблюдение W. O’Neill и J. Doyle
(1968), установивших, что в “неаккомодирующем” глазу содержи-
мое хрусталика находится под несколько более высоким давлени-
ем, чем окружающее его внутриглазное давление. Это обстоятель-
ство следует принимать во внимание при рассмотрении механизма
аккомодации.
Скорость аккомодативных реакций. При переводе взгляда с да-
леко расположенных предметов на близлежащие затрачивается
0,64 с, а в обратном направлении этот процесс происходит быстрее
(0,56 с). Уже этот факт заставляет усомниться в том, что при пере-
ключении зрения вдаль аккомодация выполняет пассивную роль. В
ответ на предъявление стимула аккомодация реагирует в среднем
через 0,36 с, т. е. медленнее, чем зрачок на свет (0,26—0,3 с)
[Lovenstein О., 1958; Campbell F., Westheimer G., 1959].
4.1.5. Аккомодация и оптическая установка глаза
Преломляющую способность оптической системы глаза принято вы-
ражать не в абсолютных значениях физической рефракции, а через
отношение главного фокуса системы к сетчатке. В прошлом веке
Ф. Дон дере для упрощения расчетов и рассуждений предложил
выделить три вида клинической рефракции в зависимости от то-
го, где собираются в главный фокус поступающие в глаз парал-
лельные лучи: 1) эмметропию (главный фокус на сетчатке), 2) ги-
перметропию (главный фокус за сетчаткой), 3) миопию (главный
фокус перед сетчаткой). Уже эта теоретическая предпосылка от-
водила эмметропии особую роль оптимальной нормы. Позже воз-
никло даже представление о целенаправленной эмметропизации глаза
324
в процессе формирования детского организма. Между тем такие
“соразмерные” (эмметропические) глаза имеются менее чем у тре-
тьей части всей популяции взрослого населения.
Нефизиологичность утвердившегося подхода к определению
того, как реагирует глаз на параллельные лучи, проявилась, напри-
мер, при оценке возможностей так называемой относительной
аккомодации. Вместо проведения ее прямых замеров в каждом
конкретном случае глаз, оцененный как миопический или гипер-
метрический, сначала искусственно уподобляют эмметропическому
с помощью корригирующих линз и только после этого определяют
возможности его аккомодации. При таком подходе в угоду физи-
ческой чистоте наблюдений (в исходной позиции на сетчатке сфо-
кусированы параллельные лучи) не учитывают тот факт, что еще
до начала основных физиологических испытаний на аккомодацию
глаза больного с миопией искусственно с помощью линз возложена
дополнительная нагрузка, а с глаза пациента с гиперметропией также
посредством линз она частично снята.
Существуют две принципиальные возможности определения
клинической рефракции глаза в условиях относительного покоя
аккомодации:
1) оценка отношения глаза к линзам, корригирующим остроту
зрения вдаль, т.е. при наличии стимула к аккомодации, по крайней
мере к аккомодации вдаль, в диоптрийном исчислении;
2) определение расстояния до той зоны в пространстве, на кото-
рую фокусирована оптическая система глаза в отсутствие стимула
к аккомодации (использование инфракрасных приборов, предъяв-
ление тусклых, лишенных каких-либо деталей стимулов в условиях
низкой освещенности).
Несовпадение результатов исследований с использованием раз-
ных методик принято объяснять несовершенством методик второй
группы. Между тем скорее методики первой группы не в полной
мере отвечают задачам исследования; при их применении не прямо,
а косвенно определяется оптическая установка. На точность резуль-
татов исследования отрицательное влияние оказывает сила корри-
гирующих линз (особенно при высокой диоптрийности). Наконец,
наличие в ходе проверки адекватного для включения аккомодации
стимула (таблицы) не позволяет считать, что исследование ведется
в условиях покоя аккомодации, хотя бы и относительного.
В то же время достоинства методик второй группы позволили
установить и объяснить ряд, казалось бы, парадоксальных явлений,
в частности так называемую аккомодацию в прибор.
1. В условиях общего физического переутомления у так называ-
емых эмметропов возникает миопическая установка глаз [Шапова-
лов С. Л., 1972].
2. При низкой освещенности “эмметропический” на свету глаз
преобразуется в миопический (приобретает “темновой фокус”).
325
Таблица 4.2. Кобальтометрическая оценка оптической установки в гла-
зах с различной клинической рефракцией
Клиническая рефракция (в ее обычном понимании) Количество исследован- ных глаз Оптическая установка при использовании кобальтового светофильтра
Н Em миопия, дптр ответы в > широком диапазоне
до 0,5 0,75— 1,5 2,0— 3,0 3,0
Н 83 11 8 26 25 6 2 5
Em 159 — 3 31 90 24 1 10
М 78 — — 11 11 32 15 9
Всего... 320 11 11 68 128 62 18 14
3. В условиях безориентирного пространства, так называемого
“пустого поля”, у летчиков определяется миопизация.
В этом перечне указаны как раз те ситуации, в которых миопи-
зацию естественнее было бы объяснять не действием чрезмерных
стимулов к аккомодации, как это иногда делают, а ослаблением ее
активности из-за отсутствия стимулов. Именно в подобных услови-
ях относительного покоя аккомодации оптический аппарат глаза
принимает такую оптическую установку (“позу”), которая соответ-
ствует его устройству.
При использовании кобальтового светофильтра, подавляюще-
го стимул к аккомодации, в условиях низкой освещенности и на-
блюдения за слабым неструктурированным источником света при
приближении и удалении от него до исчезновения либо красного,
либо синего ореола найденная позиция характеризовала оптичес-
кую установку в описанных условиях (табл. 4.2) [Волков В. В., Ко-
лесникова Л. Н., 1972]. У большинства пациентов, у которых в обыч-
ных условиях определялась эмметропия, в отсутствие стимулов к
аккомодации глаз фокусировался к зоне, удаленной на 0,6—1,3 м
(0,75—1,5 дптр), у большинства других, по обычным понятиям
близоруких, этот показатель составлял 2,0—3,0 дптр, а у дально-
зорких — он варьировал от 0 до 1,5 дптр (изредка более). У неболь-
шой части гиперметропов оптическая установка имела отрицатель-
ное значение.
На основании результатов этих исследований представляется
более рациональным подразделять глаза не на эмметропические и
аметропические, а на имеющие среднюю (0,75—1,5 дптр), сильную
(> 1,5 дптр) и слабую (<0,75 дптр) клиническую рефракцию в зави-
симости от оптической установки в условиях относительного покоя
аккомодации. Расположение темнового фокуса в большинстве глаз
с эмметропической, гиперметропической и слабой миопической реф-
ракцией на расстоянии около 1 м биологически значимо. Благода-
326
ря этому глаз надежно защищен от наиболее часто возникающей
потенциальной угрозы травмирования близко расположенными
объектами, которые могут внезапно возникать в сумерках или в
безориентирном пространстве. Острота фокусировки обычно не
выходит за пределы 0,5 дптр, реже ширина зоны соразмерности
(сохранения фокуса на сетчатке без напряжения аккомодации) рас-
тягивается до 1,0—2,0 дптр.
Во время сна, когда активное взаимодействие организма с ок-
ружающей средой ослабевает и наступает “царство вагуса”,— эм-
метропический глаз, согласно результатам экспериментов на обезь-
янах, принимает миопическую установку в 2,5—3,0 дптр [West-
heimer G., Blair S., 1973]. Аналогичным образом действует наркоз.
Во время бодрствования, но при медикаментозном подавлении па-
расимпатической иннервации атропиноподобными препаратами,
наоборот, доминирует симпатическая иннервация, и оптическая
установка глаза, как известно, сдвигается в пространство дальнего
видения, приближаясь к ’’дальнейшей точке ясного видения”
(punctum remotum).
Таким образом, очевидно, что позиция “функционального по-
коя аккомодации” в интервале между первой и второй промежу-
точной точкой (punctum intermediunij и punctum intermedium2) мо-
жет сдвигаться либо к глазу в случае преобладания статического
тонуса парасимпатической иннервации (сон, наркоз, миотики),
либо от глаза, если доминируют влияния симпатической нервной
системы (циклоплегия, симпатомиметики). На основании того
факта, что в условиях функционального покоя аккомодации глаз
большинства людей, особенно молодых, фокусируется не в беско-
нечность, а на какое-либо определенное конечное расстояние,
следует сделать ряд важных для клинической практики заключе-
ний.
1. Результаты рефрактометрии, особенно основанной на исполь-
зовании в приборах инфракрасного (ИК) излучения, заслуживают
самой высокой оценки. Получаемые при этом исследовании более
’’сильные” показатели клинической рефракции, чем при проверке
по таблицам для дали (субъективным способом), отнюдь не явля-
ются артефактом, а отражают действительную оптическую установ-
ку глаза в условиях относительного покоя аккомодации.
2. Отклонения от оптической установки, занимаемой глазом в
условиях относительного покоя аккомодации, возможны не только
в сторону усиления его рефракции за счет напряжения аккомода-
ции вблизь, но и в сторону ослабления — за счет напряжения акко-
модации вдаль. Найдя зону покоя, несложно оценить и рабочие
зоны аккомодации [Волков В. В., Парпаров А. Б., 1976]. В норме
для близи зона работы в диоптрийном исчислении всегда больше,
чем для дали. Если зона работы для дали совсем исчезает (паралич
аккомодации вдаль), а для близи суживается, то это свидетельству-
327
ет о тоническом напряжении аккомодации вблизь, которое клини-
чески проявляется так называемой ложной миопией. Если, наобо-
рот, зона работы для близи пропадает (например, при медикамен-
тозном параличе аккомодации вблизь), а для дали сохраняется, но
суживается, то это признак тонического напряжения аккомодации
вдаль, которое клинически иногда проявляется так называемой
скрытой гиперметропией. При расслаблении аккомодации и вблизь,
и вдаль, когда рабочие зоны исчезают полностью, оптическая уста-
новка глаза соответствует его клинической рефракции.
z 3. При подборе корригирующих очков для постоянного ноше-
ния следует ориентироваться не на показатели рефракции в даль-
нейшей точке ясного видения, как это довольно широко принято, а
на оптическую установку глаза в покое аккомодации [Волков В. В.,
Парпаров А. Б., 1976].
4. Проблемы медикаментозной циклоплегии также следует рас-
сматривать с учетом преимущественного влияния того или иного
препарата на аккомодацию вблизь либо вдаль.
4.1.6. О механизмах аккомодации
Новые факты, не укладывающиеся в рамки классической теории
Гельмгольца, согласно которой аккомодация рассматривается как
односторонне активный процесс, служат поводом к корректировке
привычных представлений. Одна из версий возможного механизма
активной аккомодации не только вблизь, но и вдаль основана на
представлении о динамическом равновесии нескольких противобор-
ствующих сил. Эластичность вещества и капсулы молодого хруста-
лика, да еще более высокое тканевое давление в нем (в сравнении
с давлением окружающих жидкостей), с одной стороны, и эласти-
ческие силы также молодой хороидеи — с другой, противостоят
друг другу. Первые направлены на усиление, вторые — на ослабле-
ние рефракции. Третьей силой, как и в модели Гулыптранда, выс-
тупает ресничная мышца. Обладая двойным антагонизмом (к пер-
вой и второй силам), она уравновешивает всю систему на требуе-
мом для зрения уровне.
В условиях относительного покоя аккомодации вся уравнове-
шенная система находится в состоянии минимального тонического
напряжения. Как только с сетчатки поступит импульс на подстрой-
ку аккомодации для близи или дали, прежде всего нарушается
мышечное равновесие вследствие укорочения одних и удлинения
других гладкомышечных волокон ресничной мышцы. Благодаря
этому в ту или иную сторону сдвигается основание цинновой связ-
ки. Мышца, помогая одной из противоборствующих эластичных
структур, действует против другой и в итоге удерживает их на од-
ном уровне тонического напряжения. Так, для аккомодации вблизь
328
под влиянием парасимпатической иннервации сокращаются мери-
диональные и кольцевые волокна ресничной мышцы. Благодаря
этому ресничное тело, а вместе с ним и основание цинновой связки
смещаются кпереди и кнутри; ослабевает натяжение капсулы хрус-
талика, что позволяет ему округлиться, снижается внутрихрустали-
ковое давление, натягивается хороидея, которая, подобно растяну-
той пружине, приходит в состояние повышенного тонуса.
Для реализации аккомодации вдаль сокращаются преимуще-
ственно радиальные волокна ресничной мышцы, возможно, при
участии симпатической иннервации; благодаря этому ресничное
тело, а вместе с ним и основание цинновой связки смещаются не-
сколько кзади и эксцентрично от оптической оси; снимается напря-
жение с натянутых до того эластических волокон хороидеи, но
приходит в состояние тонического напряжения капсула уплощен-
ного хрусталика, давление внутри которого возрастает.
Тот факт, что аккомодация вдаль и вблизь далеко не равнознач-
ны по силе, не может служить доводом против принципиальной
возможности существования описанного механизма их действия. В
организме многие мышцы-антагонисты также существенно разли-
чаются между собой по силе (сгибатели и разгибатели конечностей,
мышцы, ведающие конвергенцией и дивергенцией и др.), но вместе
с тем они обеспечивают некое равновесное состояние.
Не отступая от теории Гельмгольца об односторонней активно-
сти ресничной мышцы, В.Ф.Ананин (1992) выдвинул концепцию
механизма аккомодации, отводя важную роль в процессе аккомо-
дации наряду с ресничной мышцей изменениям переднезадних раз-
меров глазного яблока. Основанием для этого явилась частичная
сохранность способности к аккомодации в глазах, лишенных хрус-
талика. Между тем нельзя не учитывать возможность действия на
рефракцию при афакии и других факторов, особенно в случае на-
личия в глазу искусственного хрусталика (ИОЛ). Усилению реф-
ракции при работе на близком расстоянии (вблизи) могут способ-
ствовать увеличение кривизны роговицы при неполном смыкании
век (прищуривании), легкое смещение ИОЛ кпереди при наклоне
головы и, наконец, рефлекторное сужение зрачка при взгляде на
предметы, расположенные вблизи.
Прямые измерения переднезадних размеров глазного яблока при
аккомодации и конвергенции на небольших расстояниях от глаза
были выполнены с помощью компьютерной томографии и А-скан-
ультразвуковой эхобиометрии [Волков В. В., 1988]. При этом уста-
новлена принципиальная возможность удлинения глазного яблока
при релаксации отводящих и сокращении приводящих прямых
мышц. Возможно, этот механизм играет определенную роль в фор-
мировании глазного яблока в школьные годы, но у взрослых с
потерей склерой эластичности его значение невелико.
329
4.1.7. Попытки биомеханического моделирования одновременного
действия внутренних мышц глаза на оптическую установку
и дренажную систему
Участие сократительно-релаксационных элементов радужки и рес-
ничного тела в акте аккомодации очевидно; о роли тех же элемен-
тов в регуляции оттока камерной влаги, особенно на уровне трабе-
кулярного аппарата, известно мало [Нестеров А. П., 1995; Allen L.,
Burian Н., 1965; Moses R., 1987]. Между тем, как полагают
О. В. Светлова и Б. А. Смольников (1993), именно нарушения неко-
торых структур аккомодационного аппарата являются основной
причиной офтальмогипертензии, развивающейся после контузион-
ного отрыва части цинновых связок при подвывихе хрусталика. Смо-
делированы варианты взаимодействия иридоцилиарных мышечных
элементов с эластичными структурами трабекулярного аппарата в
зависимости от биологических особенностей строения и локализа-
ции венозного синуса склеры (шлеммова канала) [Котляр К. Е. и
др., 1995]. Установлено, что натяжение трабекул, возможное при
сокращении сфинктера зрачка и круговой порции ресничной мыш-
цы, может повлиять на площадь сечения венозного синуса склеры.
При одном и том же векторе тяги в зависимости от позиции более
длинной оси овального сечения канала последний может либо сплю-
щиваться, а по А. П. Нестерову — даже полностью закрываться,
либо раскрываться. То же происходит с отверстиями в трабекуле.
Благодаря этому, как свидетельствует математическое моделирова-
ние, по мере нарастания мышечной тяги в первом случае сопротив-
ление оттоку увеличивается, а во втором — уменьшается (рис. 4.4).
Наличие статистически достоверной прямой корреляции между
некоторыми параметрами венозного синуса склеры (площадью его
поперечного сечения, длиной внутренней стенки и общим перимет-
ром) и коэффициентом легкости оттока убедительно показано в
экспериментах по перфузии энуклеированных человеческих глаз
R. Allingham с соавт. (1996).
Смоделированы биологические механизмы взаимодействия
аккомодационной и дренажной регуляторных систем глаза. Их, как
известно, объединяет один и тот же активный исполнительный
механизм: работа мышечных элементов радужки и ресничного тела.
Аккомодационная и дренажная системы с точки зрения теории
управления могут быть рассмотрены как биологические многокон-
турные системы связанного автоматического регулирования [Кот-
ляр К. Е. и др., 1996]. В частности, суточные колебания офтальмо-
тонуса, для которого в норме характерны более высокие показате-
ли по утрам, вероятно, связаны с особенностями деятельности
аккомодации. Аккомодационный аппарат бодрствующего организ-
ма, пребывая в течение дня в тонусе, способствует более активной
функции дренажной системы глаза, что проявляется в самых низ-
330
Рис. 4.4. Результаты математического моделирования изменений коэффи-
циента легкости оттока (С) по мере увеличения тяги трабекулярного аппа-
рата ресничной мышцей в зависимости от особенностей расположения внут-
ренней стенки венозного синуса склеры (шлеммова канала).
а — при совпадении вектора тяги и плоскости расположения стенки (угол 0 = 0); б —
при ориентации вектора тяги под прямым углом к плоскости расположения внут-
ренней стенки канала (угол 0 = 90°). С — мм3/мин • мм рт. ст.; Р — процент от мак-
симальной результирующей мышечного усилия.
ких показателях офтальмотонуса к исходу дня. Вместе с тем при
’’поломке” аппарата аккомодации (травматическое смещение хрус-
талика, афакия, хронические воспалительные или дистрофические
процессы в радужке и ресничном теле) более характерны вечерние
подъемы офтальмотонуса.
Еще В. П. Филатов считал полезной для больных глаукомой
зрительную нагрузку, связанную с напряжением аккомодации. Од-
нако у некоторых пациентов чтение и напряжение аккомодации
вблизь закономерно приводит к повышению офтальмотонуса. Пос-
леднее не исключает наличия механизмов сочетанного действия
аккомодационной и дренажной систем глаза, не всегда, однако, при
совместном функционировании работающих с максимальной
пользой для организма.
331
4.2. ОСНОВНЫЕ ПРИЧИНЫ, КЛИНИЧЕСКИЕ ПРИЗНАКИ
И МЕТОДИКИ ИССЛЕДОВАНИЯ НАРУШЕНИЙ ГЛАЗНОЙ
ГИДРОДИНАМИКИ, ФУНКЦИИ ЗРАЧКА,
АККОМОДАЦИИ И ОПТИЧЕСКОЙ УСТАНОВКИ ГЛАЗА
4.2.1. Нарушения глазной гидродинамики и их последствия
Функциональные проявления и причины нарушений гидродинами-
ки глаза представлены в табл. 4.3.
Неглаукоматозная офтальмогипертензия полиэтилогична. В ее
основе лежат как гиперпродукция камерной влаги (при эндокрин-
ной, диэнцефальной и другой общей патологии), так и затруднения
при его перемещении либо внутри глаза (’’блоки”), либо за его
пределы. Для изучения механизмов гипертензии целесообразно про-
водить офтальмотонографию. Считать офтальмогипертензию глау-
коматозной нет оснований до тех пор, пока подъемы ВГД не при-
ведут к повреждению ДЗН и связанным с этим специфичным для
глаукомы нарушениям зрения. Офтальмогипертензия в восьми из
десяти случаев является симптомом различных местных или общих
заболеваний, а отнюдь не глаукомы. Встречающиеся при высоких
подъемах офтальмотонуса (при глаукомоциклических кризах) крат-
ковременные нарушения зрения и затуманивание обусловленны
преходящим отеком эпителия роговицы, который встречается при
многих других заболеваниях.
Таблица 4.3. Участие отдельных структурных и функциональных про-
явлений нарушенной гидродинамики глаза и головного мозга в симптомо-
комплексе различных патологических состояний
Диагноз (симптомокомплекс) Структуры или функции
ВГД ДЗН поле зрения ВЧД (АД) оболоч- ки глаза
Офтальмогипертензия неглаукоматозная + — — — —
Преглаукома + - + - + — —
Глаукома + + + ? —
Глаукома низкого ("нормального") давления — + + + —
Глаукома юношеская (детская) + ± ± — +
Офтальмогипотония + ± ? — ±
Примечание. + нарушены, - не нарушены, ± нарушений может не быть,
- <-> + нарушения обратимы, ? — состояние функций недостаточно изучено.
332
Преглаукома характеризуется не только офтальмогипертензией,
но и снижением устойчивости опорных структур ДЗН к сдавлива-
нию. Это периодически проявляется в его обратимых изменениях в
виде временно расширяющейся экскавации, что сопровождается
появлением скотом в парацентральной части поля зрения. Указан-
ные симптомы, стойко сохраняющиеся при глаукоме, у больных
преглаукомой носят временный характер. Они могут быть спрово-
цированы специальной нагрузочной пробой или неблагоприятной
жизненной ситуацией. Однако в стадии преглаукомы все выявлен-
ные нарушения полностью обратимы.
Глаукома в типичных случаях может проявляться в форме зак-
рыто- или открытоугольной (выделяют и смешанные формы). Не-
зависимо от особенностей угла передней камеры и обусловленного
ими клинического течения для установления достоверного диагно-
за необходима триада симптомов: офтальмогипертензия, прогрес-
сивно расширяющаяся экскавация ДЗН (особенно достоверна асим-
метрия экскавации, которая более широкая, чем на парном глазу)
и специфические нарушения в поле зрения (парацентральные ско-
томы, назальная ’’ступенька” и др.).
Для закрытоугольной глаукомы характерны высокие подъемы
офтальмотонуса (иногда с сильными болями вокруг глаза, т.е. в
соответствующей половине головы).
При открытоугольной глаукоме, особенно в начальных стади-
ях, офтальмотонус мало отличается от верхней границы нормы;
патологические подъемы чаще регистрируют в ранние утренние
часы.
В запущенных стадиях как закрыто-, так и открытоугольной
глаукомы наблюдаются выраженные дистрофические изменения не
только в ДЗН, но и во многих других внутриглазных структурах и
его оболочках (дистрофия радужки и роговицы, помутнение хрус-
талика и пр.).
Глаукома низкого (“нормального”) давления. Характеризуется
классическими для глаукомы изменениями ДЗН и поля зрения,
однако повышения офгальмотонуса по сравнению с принятыми для
популяции нормативами выявить не удается. Эту редкую форму
глаукомы чаще выявляют у лиц преклонного возраста, как прави-
ло, с относительно низким (с учетом возраста) артериальным дав-
лением. Артериальная гипотензия у них предположительно сочета-
ется с низким внутричерепным давлением. На основании этого
можно предположить, что даже нормальное для других ВГД у этих
больных оказывается относительно высоким (в норме, как отмеча-
лось, градиент ВГД=2 ВЧД). Именно такой механизм при глаукоме
низкого давления может лежать в основе сниженной толерантности
ДЗН к, казалось бы, нормальному офтальмотонусу.
Юношеская (детская) глаукома чаще развивается вследствие
органической блокады угла передней камеры зародышевой мезо-
333
дермальной тканью, не претерпевшей обратного развития. Посколь-
ку в этот период, когда все ткани не только продолжают расти, но
и сохраняют податливость к механическим воздействиям, стойко
повышенный офтальмотонус растягивает и роговицу (до керато-
глобуса), и склеру, приводя к миопизации или даже буфтальму, ука-
занные изменения до определенного момента могут предотвращать
или ’’смягчать” типичные для глаукомы нарушения в ДЗН. Однако
и при этой форме глаукомы наряду с дистрофией всех оболочек
глаза в конечном счете происходит нарушение зрения вследствие
патологической экскавации ДЗН и атрофии его волокон.
Офтальмогипотензия может быть преходящей или стойкой.
Стойкая гипотензия возникает вследствие либо недостаточной про-
дукции камерной влаги (например, после травматической деструк-
ции ресничного тела), либо чрезмерного дренирования влаги (через
супрацилиарную щель при посттравматической рецессии угла пере-
дней камеры, через нерасчетливо выполненный хирургом слишком
большой фистулизирующий ход). Чаще всего нестойкой бывает
гипотензия, обусловленная воспалением ресничного тела и времен-
ным подавлением продукции камерной влаги. При этом, однако,
периоды гипотензии могут сменяться периодами гипертензии вслед-
ствие закупорки дренажной системы фибрином, лейкоцитами и
другими элементами воспаления. При длительной офтальмогипо-
тензии развивается застойный сосок ДЗН, белковые элементы из
кровяного русла проникают в камерную влагу, возможна субатро-
фия глазного яблока.
4.2.2. Методики и устройства для исследования офтальмотонуса
и компонентов глазной гидродинамики
Офтальмотонометрия. Существуют тонометры двух видов: аппла-
национные и импрессионные (вдавливающие). К настоящему вре-
мени принцип аппланации роговицы является ведущим в констру-
ировании офтальмотонометров. Принцип вдавливания сохранил
свое значение для разработчиков глазных тонографов. Простота
конструкции и хорошая повторяемость получаемых результатов
обусловили большую популярность среди офтальмологов нашей
страны аппланационного тонометра Маклакова, созданного уже
более 100 лет назад. Тонометр подобного типа в несколько видоиз-
мененном виде [Posner и Inglima,1962] используют и в США.
Однако методика тонометрии по Маклакову нуждается в уточне-
нии, так как глаз не является идеальным тонкостенным сферическим
телом. В связи с этим правило Имберта — Фика, согласно которому
о давлении (Р) внутри сферического тела можно судить по силе (F),
необходимой для того, чтобы ’’уплостить” сферу в расчете на едини-
цу площади (А), откуда P=F/A, неприменимо к глазному яблоку из-
за упругих свойств роговйцы. При проведении тонометрии с исполь-
334
зованием рассматриваемой методики тонометрическое давление (PJ
оказывается существенно выше истинного внутриглазного давления,
причем разница может достигать 30% (см. рис. 4.1).
В стремлении создать более точный прибор Goldmann и Schmidt
(1957) решили не только стандартизировать, но и значительно умень-
шить зону сплющивания роговицы. Влияние упругих сил роговицы
при этом уменьшилось настолько, что Pt оказывалось выше Ро все-
го на 3%, причем оценивали не диаметр кружка сплющивания, а то
усилие, которое необходимо приложить, чтобы сплющить рогови-
цу до определенной степени, до пятна диаметром 3,06 мм. По рас-
четам Goldmann, именно при таких размерах пятна сплющивания
взаимно нейтрализуются два фактора, неблагоприятно влияющих
на точность замеров. Имеются в виду, с одной стороны, сопротив-
ление роговичных структур их деформации, а с другой — то допол-
нительное давление на роговицу, которое оказывает грузик на не-
посредственно своей площадкой, а через окружающий ее мениск
слезной жидкости, неизбежно возникающий в ходе исследования.
Для улучшения визуального контроля используют щелевую лам-
пу, а также в конъюнктивальный мешок закапывают раствор флюо-
ресцеина-натрия. При этом на общем синеватом фоне становятся
отчетливо видны мениски слезной жидкости, окрашенные в желто-
зеленый цвет. Поскольку площадка тонометра составлена из двух
призм, окрашенное кольцо слезного мениска представляется наблю-
дателю разделенным на два полукольца (верхнее и нижнее). Как
только в процессе компрессии концы полуколец соприкоснутся,
роговица сплющивается до пятна диаметром 3,06 мм. Умножив
показатели шкалы тонометра на 10, определяют истинное внутри-
глазное давление (Ро)
Современными тонометрами Гольдмана оснащены многие ще-
левые лампы. Близкий по конструкции прибор был создан в нашей
стране А.Я.Буниным с соавт. (1964). Известно немало других моди-
фикаций, основанных на том же принципе.
Ручной тонометр П е р к и н с а не нужно крепить
к щелевой лампе, он адаптируется к маленькому источнику света,
например к электроофтальмоскопу, и позволяет исследовать оф-
тальмотонус у лежачих больных, в том числе у находящихся в со-
стоянии наркотического сна.
Воздухоструйные тонометры основаны на прин-
ципе Гольдмана, но роговица при их использовании аппланируется
не призмой, а струей сжатого воздуха. При проведении исследова-
ния по этой методике уменьшается точность измерения, но не тре-
буется применения анестетиков и полностью устраняется опасность
заноса инфекции от одного пациента другому.
Тонометр Маккея — Марта (Mackay — Marg) —
портативный электронный прибор, в котором объединены как ап-
планационный, так и импрессионный принципы тонометрии. Ак-
335
тивной частью тонометра, устанавливаемой на роговицу, служит
тонкий (1,5 мм) плунжер. Подобно импрессионному тонометру
Шиотца, прибор может быть полезен при асферичной рубцово-из-
мененной роговице.
По этому же принципу построен микроэлектронный тонокаран-
даш, снабженный иглоподобным трансдюсером, который при со-
прикосновении с роговицей преобразует механический сигнал в
электрический. Результаты нескольких касаний записываются в
цифровом или графическом виде. Тонокарандаш может быть ис-
пользован в ранние сроки после кератопластики или кератотомии.
Наиболее совершенным из всех существующих тонометров яв-
ляется компьютеризированный прибор мембранного типа конст-
рукции Langham, позволяющий проводить, кроме тонометричес-
ких, тонографические исследования с учетом различных парамет-
ров глазного пульса.
Для скрининговых исследований создавались менее точные, но
более простые в обращении устройства типа индикатора ВГД Бе-
ренса — Толмана, глаукотеста Брунна — Йенсена (фирмы ’’Heine”),
прибора Цаймера и др. Последний в США рекомендуют использо-
вать для самоконтроля внутриглазного давления в домашних усло-
виях. Прибор состоит из двух основных элементов: оптоэлектрон-
ного зонда и электронной схемы. Лампа накаливания излучает свет,
который после линзы идет параллельным пучком. Этот коллимиро-
ванный свет попадает на эластичную и прозрачную мембрану, ук-
репленную на торце поршня с помощью кольцевой подставки. Часть
света отражается от мембраны и на обратном пути фокусируется
линзой после отражения от стеклянной пластинки фотодетектора.
Давление воздуха внутри цилиндра, в котором перемещается пор-
шень, контролируется нажатием на эластичную педаль и измеряет-
ся манометром.
Для правильной ориентации исследуемого глаза относительно
прибора необходимо увидеть мишень и центрировать на ней изоб-
ражение диафрагмы. При низком зрении это не каждому удается.
Нажатием на педаль пациент повышает давление воздуха в прибо-
ре и приводит в движение оптоэлектронный зонд до контакта его
торцовой мембраны с роговицей, предварительно анестезирован-
ной. По мере аппланации роговицы свет, отраженный от ее повер-
хности, усиливается. Это происходит до тех пор, пока роговица не
начнет вдавливаться. Электрическая схема точно улавливает мо-
мент максимального уровня света, т. е. аппланации роговицы, до ее
прогибания и регистрирует потребовавшееся для этого усилие (в
миллиметрах ртутного столба). По окончании измерения срабаты-
вает звуковой сигнал, и пациент прекращает давление на педаль, а
зонд сразу же отводят от глаза назад. При неправильной юстировке
прибор не срабатывает. Для контроля ориентировки пациентом
мишени на ней имеется одна светлая точка. Пациент сообщает о
336
положении этой точки. В ходе тонометрии возможна коррекция
аметропии в пределах от +14,0 до -6,0 дптр. Стандартные откло-
нения показателей домашнего тонометра от результатов обычной
аппланационной тонометрии достигают 2,5 мм рт.ст. Пациенты даже
преклонного возраста, разводя веки пальцами, могли без больших
усилий приспосабливаться к самостоятельному проведению тоно-
метрии. В настоящее время изучается возможность уменьшения
диаметра рабочей части тонометра.
Топография. Для клинической топографии используют
либо импрессионные, либо аппланационные устройства. В нашей
стране применяли импрессионные электротонографы Сахарова, Не-
стерова и др., длительное время использовали тонометр Шиотца,
удерживаемый на роговице в течение определенного времени, при-
меняли также перилимбальные присоски.
А. И. Дашевским (1961), С. Ф. Кальфа и М. Б. Вургафтом (1962),
А. П. Нестеровым (1963) были разработаны оригинальные методи-
ки для аппланационной топографии. В.С. Красновидов (1970) наря-
ду с достоинствами отметил недостатки этих методик. Наибольшее
распространение в нашей стране в свое время получила упрощен-
ная методика, предложенная А. П. Нестеровым.
В начале измеряют офтальмотонус с помощью тонометров
Маклакова массой 5 и 15 г, затем на 4 мин на роговицу помещают
грузик массой 15,0 г. По окончании компрессии измеряют давление
в глазу, используя тот же грузик. После сопоставления размеров
кружков сплющивания до и после компрессии по таблицам находят
коэффициент легкости оттока (С). Минутный объем камерной вла-
ги (F) определяют расчетным путем, принимая уровень давления в
эписклеральных венах, равным 10 мм рт.ст.
Хотя современная топография и позволяет установить некото-
рые патофизиологические механизмы течения глаукомы, ее клини-
ческое значение в оценке статуса конкретного больного невелико,
получаемые показатели довольно вариабельны. Не случайно даже
А.П. Нестеров, потративший много усилий для детальной разра-
ботки и пропаганды метода, в настоящее время относит эту мето-
дику во “второй эшелон” диагностических приемов, используемых
при глаукоме. Привлекательной представляется американская тех-
нология офтальмотонометрии и топографии, предложенная
М. Langham (1995). Его пневматический компьютеризированный
тонометр позволяет благодаря контакту податливой мембраны,
поддавливаемой изнутри потоком газа, а снаружи контактирую-
щей с анестезированной роговицей, вести непрерывную запись
офтальмотонуса на фоне пульсовых колебаний крови. При этом
также регистрируют амплитуду глазного пульса, измеряют пульсо-
вой и минутный объем крови (см. рис. 4.2).
В приборе Лангхама предусмотрена также возможность прове-
дения топографии с использованием лимбальных присосок; можно,
337
кроме того, измерять давление в глазной артерии, выявляя, в час-
тности, относительную недостаточность глазного кровообращения,
например, вследствие стеноза внутренней сонной артерии. Однако,
как отметил J. Kiel (1995), в приборе Лангхама заложен показатель
ригидности оболочек глаза по Фриденвальду, вычисленный без учета
влияния на результаты исследования индивидуального среднего
уровня артериального давления.
Прогностическая нагрузочная вакуум-пе-
риметрическая проба. В. В. Волков и соавт. (1974) пер-
выми предложили использовать дозированную компрессию глазно-
го яблока сжатым воздухом через очки гермошлема пилота с одно-
временным контролем за состоянием зрительных функций в
центральном поле зрения.
По мере усовершенствования метода были созданы специальные
устройства, одним из вариантов является глаукотестер Волкова —
Сухининой — Тер-Андриасова, компрессия в котором создается раз-
режением воздуха в подочковом пространстве. Искусственное
повышение ВГД в обоих случаях (как при компрессии нагнетанием
воздуха, так и при вакуумировании) создается за счет деформации
глазного яблока и уменьшения его объема. Вариант с использова-
нием вакуума оказался предпочтительным, так как меньше запоте-
вали стекла и желаемый нагрузочный эффект создавался при мень-
ших изменениях давления в подочковом пространстве (до 45—
50 мм рт. ст.). При использовании вакуума, правда, требуется
временная фиксация верхнего века полоской липкого пластыря для
предотвращения сужения глазной щели. В очковой оправе предус-
мотрены гнезда для установления заслонки перед одним из глаз и
корригирующего стекла перед исследуемым глазом. В конструкции
вакуум-компрессора предусмотрена защита от передозировки на-
грузочного эффекта.
4.2.3. Нарушения зрачковых реакций, их диагностика и последствия
Варианты патологической зрачковой симптоматики в зависимости
от уровня поражения рефлекторной дуги можно проследить на
следующей схеме:
Уровень поражения
Сетчатка или зритель-
ный нерв
Нейрон, связующий пре-
тектальное ядро с ядром
Якубовича — Эдинге-
ра — Вестфаля
При грубом
нарушении
При умеренном
нарушении
При нейросифилисе
Название симптома
Амавротический зра-
чок
Симптом Маркуса —
Гунна
Симптом Аргайла Ро-
бертсона
338
По ходу III пары череп-
ных нервов
При патологической
регенерации ветвей III
пары черепно-мозго-
вых нервов
Постганглионарно
Псевдосимптом
Аргайла Робертсона
П реганглионарно
По ходу одного из трех
нейронов симпатиче-
ской иннервации глаза
П реганглионарно
(ниже верхнего шейно-
го ганглия)
П остганглионарно
Тонический зрачок
Эди — Холмса
Симптом нарушения
среднемозговых струк-
тур
Центральный симптом
Горнера (паредрин
расширяет зрачок)
Постганглионарный
симптом Горнера (па-
редрин не влияет на
зрачок)
Основные признаки и причины наруше-
ний. При амаврозе, когда восприятие света отсутствует из-за по-
вреждения сетчатки или зрительного нерва, оба зрачка одинаковы
по размеру, при засвете слепого глаза ни один из зрачков не реаги-
рует, при засвете здорового (парного) оба зрачка реагируют, сохра-
нена также реакция на ближний стимул.
При симптоме Маркуса — Гунна, когда сетчатка или зрительный
нерв повреждены лишь частично (например, при неврите), а острота
зрения может быть ненарушенной, ответная реакция обоих зрачков
при засвете больного глаза замедлена. Для того чтобы легче уловить
разницу, глаза поочередно засвечивают зеркальным офтальмоско-
пом, пытаясь уловить наличие парадоксальной реакции: при быст-
ром переносе света со здорового глаза на больной его зрачок не только
не суживается, а расширяется. Полагают, что это результат более
сильной, чем прямая, содружественной реакции (зрачок здорового
глаза при перенесении света на больной глаз начинает расширяться).
Симптом Аргайла Робертсона патогномоничен для нейросифи-
лиса. Для него характерна диссоциация реакций на свет и ближний
стимул: реакция на свет отсутствует, а на “близь” — живая. Обыч-
но реагируют оба зрачка, хотя не исключена асимметрия реакций,
зрачки узкие и плохо расширяются под влиянием мидриатиков.
Диссоциация связана с нарушением нейрональной связи между
претектальным ядром и ядром Якубовича — Эдингера — Вестфаля
(при утрате зрения диссоциацию реакций уловить не удается).
При тонической реакции зрачка (Эди — Холмса) нарушения свя-
зывают с патологией постганглионарной части III пары черепных
нервов. Это наиболее частая причина анизокории у женщин в воз-
расте 30—40 лет, перенесших вирусную инфекцию (как правило,
ослабевают также сухожильные рефлексы).
Более широкий, т.е. поврежденный, зрачок слабее реагирует на
339
свет, а на “близь” его реакция очень замедлена. Так же замедлена
обратная реакция расслабления сфинктера (он, а также ресничная
мышца как бы находятся в состоянии повышенного тонуса). Для
подтверждения диагноза проводят инстилляцию 0,1% раствора пи-
локарпина в конъюнктивальный мешок обоих глаз, при этом зрачок
здорового глаза почти не реагирует, а больного — резко сужается.
Данный симптом не является признаком тяжелого течения про-
цесса, хотя эффективного лечения нет, но нарушения аккомодации
со временем могут уменьшаться.
При среднемозговой (тектальной) природе нарушений зрачковых
реакций причиной их возникновения считают компрессию III желу-
дочка (например, пинеаломой). Наряду с расширением зрачка и ос-
лаблением его реакции на свет характерна длительная сохранность
реакции на “близь”, так как обеспечивающие ее волокна лежат более
вентрально, чем те, от которых зависит реакция на свет. Этот симптом
необходимо дифференцировать от симптома Аргайла Робертсона.
Нейросифилис не является единственной причиной диссоциаций
реакций зрачка на свет и близко расположенный раздражитель (дис-
социации “свет — близь”). Последние встречаются при юношеском
диабете, миотонической дистрофии, неполноценной регенерации во-
локон III пары черепных нервов, синдроме Парино, при котором,
кроме зрачковых нарушений, наблюдаются ограничения взора вверх
и конвергентно-ретракционный нистагм.
При повреждении III пары черепных нервов, например, вследствие
сдавления мозговой аневризмой нарушается эфферентный путь зрач-
кового рефлекса и, естественно, на поврежденной стороне исчезают
реакции на свет и ”близь”. В процессе восстановления функции
поврежденного нерва регенерация идет аберрантным путем и тогда
в веточку зрачковых волокон вплетаются волокна приводящей, т.е.
внутренней, прямой мышцы глаза. При этом может наблюдаться
ложная диссоциация “свет — близь” (зрачок псевдо-Аргайла Роберт-
сона). В связи с повреждением веточки III пары черепных нервов,
иннервирующей сфинктер зрачка, реакция на свет невозможна (или
очень ослаблена), но реакция на ’’близь” наблюдается. Она связана
с напряжением внутренней прямой мышцы, в частности, при кон-
вергенции (синкинетические движения зрачка этой мышцы явились
следствием патологической регенерации). Дополнительными при-
знаками внутричерепной аневризмы являются симптом псевдо-Гре-
фе., выражающийся в ретракции верхнего века в ответ на аддукцию
глаза, и сегментарные подергивания сфинктера зрачка при движе-
ниях глаз [Chamecki J.S., Thompson Н., 1978].
Особняком стоит синдром Горнера (глазосимпатический пара-
лич), поскольку при нем, кроме зрачковых нарушений, проявляю-
щихся лишь в виде миоза, более заметного при сниженной освещен-
ности (все зрачковые реакции сохранены), наблюдаются умерен-
ный птоз верхнего века (в связи с парезом мышцы Мюллера), а
340
также приподнятость нижнего века (вследствие пареза гладкой
мускулатуры, прислоняющей в норме “хрящевую” пластинку ниж-
него века к глазу). По указанным причинам сужается глазная щель,
в связи с чем ошибочно диагностируют энофтальм, при котором
обнаруживают усиление аккомодативной способности глаза к близ-
ким расстояниям.
Необходимо дифференцировать пре— и постганглионарные (от-
носительно шейного ганглия) поражения. Первые (бронхиальная
карцинома, аневризма грудной аорты и пр.) менее благоприятны,
вторые, при которых нарушается также и потоотделение, чаще
имеют сосудистый генез, являются причиной головной боли, но
протекают более благоприятно.
Синдром Горнера подтверждается при инстилляции в конъюн-
ктивальный мешок 4 % раствора кокаина, когда зрачок больного
глаза не реагирует на капли, а здоровдго глаза — расширяется.
Нормальные зрачковые реакции являются благоприятным про-
гностическим признаком улучшения зрительных функций в различ-
ных условиях внешней освещенности, а стойкий мидриаз на разных
расстояниях — причиной повышенной слепимости при ярких засве-
тах. Благодаря мидриазу уменьшается глубина фокусной зоны и,
следовательно, поля, в пределах которого одновременно можно
четко видеть предметы, удаленные на разные расстояния. Стойкий
миоз затрудняет ориентацию в условиях сниженной освещенности,
а при крайних степенях сужения зрачка вследствие дифракции ста-
новится причиной снижения остроты зрения.
Общие методические рекомендации по ис-
следованию зрачков. Исследование проводят в слабоос-
вещенном помещении при взгляде пациента вдаль (например, на
таблицу Сивцева), при этом подсвечивают его лицо так, чтобы оба
глаза были равномерно освещеньг^косыми лучами. Диаметр зрачка
измеряют непосредственно миллиметровой линейкой или пристав-
ленным со стороны виска на исследуемой стороне пупиллометром,
на котором рядом с линейкой представлены кружки черного цвета
диаметром от 1,5 до 8 мм с интервалом 0,5 мм. Поскольку у каждо-
го пятого обследуемого в норме бывает легкая анизокория, в поис-
ках патологии следует изменять освещение. Так, у больных с син-
дромом Горнера разница проявляется гораздо отчетливее при сни-
женном освещении.
Нужно учитывать, что одностороннее снижение зрения само по
себе не влияет на размеры зрачка, но при патологии именно зри-
тельно-нервных путей зрачковая реакция может нарушаться, на-
пример, по типу симптома Гунна.
Для проверки светового рефлекса зрачка можно воспользовать-
ся зеркалом офтальмоскопа или осветителем щелевой лампы. В
случае возникновения подозрения на одностороннюю слабость зрач-
ковой реакции при прямом освещении проверяют содружествен-
341
ную реакцию зрачка другого глаза. При одинаковой выраженности
прямой и содружественной реакций афферентную дугу рефлекса
признают нормальной. Для того чтобы выявить гемианопсические
нарушения в реакции зрачка, удобнее всего воспользоваться точеч-
ным источником света от щелевой лампы, переводя его поочередно
то в правую, то в левую позиции и наблюдая при этом через бино-
куляр за выраженностью реакции зрачка.
Для оценки “ближнего” рефлекса зрачка пациента просят сна-
чала посмотреть вдаль, а затем перевести Ьзор на кончик собствен-
ного пальца, приставленного к носу. Подбородок желательно дер-
жать слегка приподнятым, так как многие люди легче конвергиру-
ют при опущенном взоре. Иногда приходится придерживать верхнее
веко, чтобы легче было проследить за реакцией зрачка. Степень
сужения можно оценивать по трех- или четырехбалльной системе.
Установки для регистрации движений зрачка предлагались еще
в прошлом веке (Л.Г. Беллярминов и др.); в нашей стране извест-
но устройство Самойлова — Шахновича, но в широкой клиниче-
ской практике обходятся без записывающих устройств.
4.2.4. Нарушения аккомодации, их связь с “неудачными”
вариантами оптической установки глаза. Основные признаки
и причины нарушений
Варианты нарушений аккомодации и отправные пункты для их ко-
личественной оценки представлены на следующей схеме:
Варианты нарушений
Дефицит аккомода-
ции для зрения на
близких расстояниях
Дефицит аккомода-
ции для зрения на
далеких расстояниях
Нарушение тонуса Чрезмерная
покоя аккомодации возбудимость
Инертность
Количественная пространственная
х арактеристика
Укорочение зоны действия аккомо-
дации в диапазоне от установки
оптики глаза в промежуточную по-
зицию покоя {Punctum intermedium I)
к установке на предельно близкое
{Punctum proximum) расстояние
Укорочение зоны действия аккомо-
дации в диапазоне от установки
оптики глаза в промежуточную по-
зицию покоя {Punctum intermedium
II) к установке на предельно дале-
кое {Punctum remotum) расстояние
Сужение (< 0,5 дптр.) интервала
между ближайшей {Punctum interme-
dium I) и ближней {Punctum interme-
dium II) точками оптической установ-
ки в зоне покоя аккомодации
Расширение (> 2,0 дптр) интервала
между теми же точками в зоне покоя
аккомодации
342
Дефицит аккомодации вблизи. Он в основном проявляется зат-
руднениями при зрительной работе на близких расстояниях, в час-
тности при чтении, работе с экранами индикаторов и чертежами, с
сохраняющейся на прежнем уровне остротой зрения вдаль. Возмож-
ные причины:
• возрастное склерозирование хрусталика;
• индивидуальный вариант слишком отдаленной или даже отрица-
тельной оптической установки глаза, когда P.intermedium гра-
фически находится за глазом, характеризуя степень сходимости,
которой должны обладать лучи, чтобы, преломившись в глазу,
сфокусироваться на сетчатке (в привычном понимании — гипер-
метропия);
• общая астенизация организма (при инфекционных болезнях, фи-
зическом переутомлении), проявляющаяся, в частности, аккомо-
дативной астенопией;
• слабость конвергенции;
• паралич ресничной мышцы, обусловленный действием токсинов
(ботулизм) или фармакологических препаратов (холинолитики);
• чрезмерная стимуляция аккомодации вдаль Р-адренергическими
препаратами (неосинэфрин, фенилэфрин и др.).
Дефицит аккомодации вдаль. Основным признаком является
вначале преходящее, а затем стойкое ухудшение зрения вдаль при
сохранении его на близких расстояниях. Возможные причины:
• привычное, обусловленное зрительной работой напряжение ак-
комодации вблизь, ведущее в период формирования рефракции к
перемещению всей оптической установки глаза в зону ближнего
видения (школьная и другая профессиональная миопизация мо-
лодых людей);
• слабость всех наружных мышц глаза при общей стойкой астени-
зации организма и неполноценности склеры, проявляющаяся про-
грессирующим удлинением переднезадней оси глазного яблока и
другими симптомами миопической болезни;
• спазм конвергенции, в частности так называемый эксцесс кон-
вергенции при высоком отношении АК/А;
• спазм ресничной мышцы с более выраженным Спазмом тех ее
порций, которые “ведают” аккомодацией вблизь, медикаментоз-
ной (холиномиметики, антихолинэстеразные препараты), невро-
тической (истерия) и другой природы;
• одно из проявлений синдрома Горнера, а также действия Р-адре-
ноблокаторов (пропранолол и др.).
Нарушения тонуса покоя аккомодации и сдвиги оптической уста-
новки (Р. intermedium I и II), Следствием таких нарушений могут
быть затруднения при перестройке аккомодации с далеких стиму-
лов на близкие и наоборот.
• Слишком широкая зона покоя аккомодации, являющаяся свиде-
тельством инертности рефлекса, что проявляется медленным рас-
343
сеиванием тумана при переводе взгляда с одних расстояний на
другие.
• Практически неулавливаемая (при определении зоны покоя) спо-
собность к физиологической релаксации всех порций ресничной
мышцы даже в условиях отсутствия стимулов. По-видимому, при
таком варианте работающей без устали аккомодации даже ско-
рее, чем при инертном варианте, возможны истинные спазмы ак-
комодации от переутомления (с болевым синдромом и потерей
способности к аккомодации как вблизь, так и вдаль).
С.Л.Шаповалов (1976) установил, что при длительной напря-
женной зрительной работе аккомодационная способность сначала
возрастает, а по мере утомления снижается.
• Перестройка привычного тонуса покоя аккомодации со стойким
изменением оптической установки (равно рефракции) глаза. Ра-
нее уже была отмечена возможность стойкого изменения рефрак-
ции глаза при дефиците аккомодации вдаль. Иногда ложная ми-
опия по мере ослабления аккомодации вдаль становится истин-
ной, а скрытая гиперметропия по мере ослабления аккомодации
вблизь — явной. В исследованиях, проведенных С.Л.Шаповало-
вым (1976), отмечена возможность временных сдвигов зоны по-
коя аккомодации по мере зрительного утомления. Когда эти сдви-
ги становятся привычными, реальным оказывается изменение оп-
тической установки, которую приобретает глаз в условиях нового
уровня равновесия аккомодации вблизь и вдаль.
4.3. МЕТОДЫ И УСТРОЙСТВА ДЛЯ ФУНКЦИОНАЛЬНЫХ
ИССЛЕДОВАНИЙ АККОМОДАЦИИ И ОПТИЧЕСКОЙ
УСТАНОВКИ ГЛАЗА
О те ст-объе ктах для исследования аккомодации.
Не всякий видимый глазом объект может возбудить аккомодацию.
По данным F. Campbell (1954), световой минимум, способный выз-
вать рефлекс аккомодации, должен быть несколько сильнее, чем по-
роговый раздражитель, воспринимаемый макулярной областью сет-
чатки. Кроме того, контуры объекта должны быть достаточно четко
очерченными, чтобы был стимул для фокусировки. Без конкретной
структуры поля зрения стимул к аккомодации легко затухает.
Тест-объекты, применяемые для возбуждения аккомодации,
могут быть простыми и сложными, черными и цветными, на белом
фоне и светящимися на темном фоне. Казалось бы, лучше всего
использовать простейший тест-объект. Например, рассматривать ряд
линий, постепенно приближая до появления первых признаков на-
рушения четкости видения, как предлагал Грефе, или цепочку све-
тящихся точек, как рекомендовал Ландольт. Однако у этих тест-
объектов имеются недостатки. Дело не только в том, что контроль
344
Рис. 4.5. Некоторые тест-объекты, применяемые для исследования аккомо-
дации.
а — по Дуане; б — по Шенку; в — по Парпарову.
врача за субъективными ощущениями обследуемого при использо-
вании таких объектов практически невозможен. Важнее то, что сами
пациенты затрудняются выбрать момент для ответа: один сообща-
ет о нечеткости видения объекта при появлении ее ничтожных при-
знаков, а другой лишь тогда, когда фактически совсем перестает
видеть объект.
По существу все изложенное относится и к современному зна-
ку, применяемому для исследования аккомодации, который был
предложен Duane (рис.4.5). Правда, имеющиеся в этом знаке жир-
ные черные полосы, расположенные по сторонам от тонкой тесто-
вой линии, облегчают ориентировку. Благодаря им обследуемый не
тратит усилия на поиски белого поля, к которому должно быть
приковано его внимание. Это важно на пределе видимости тесто-
вой линии, чтобы не выпустить ее из-под наблюдения.
Если для исследования аккомодации использовать такой эле-
ментарно простой объект, как одиночная линия или даже точка, то
для анализа возникающих при этом зрительных ощущений лучше
всего обратиться к феномену Шейнера.
Простотой в эксплуатации, доступностью для понимания зада-
чи обследуемым и достаточной чувствительностью при исследова-
нии аккомодации у лиц с нормальным чувством цвета привлекают
двухцветные тест-объекты. При приближении такого объекта к глазу
обследуемый замечает уменьшение четкости видения сначала тех
линий и точек, которые изображены на красном фоне, а при про-
должающемся приближении — на зеленом; при удалении объекта
все наблюдается в обратной последовательности. Этот феномен обус-
ловлен тем, что красные лучи преломляются несколько слабее, чем
зеленые. После того как определены пороги для красного и зелено-
го цветов, взяв среднее между ними, просто определить показатель
для промежуточного желтого цвета.
Тест-объекты сложной конфигурации. Чем сложнее объект, тем
активнее работает аккомодация, однако сложные объекты с трудом
поддаются стандартизации.
345
Наиболее доступными и удобными, хотя и не самыми точными,
являются тест-объекты в виде букв, точнее, в виде литературных
текстов. Обследуемый легко усваивает, что от него требуется чте-
ние вслух до тех пор, пока различается текст. Однако смышленый
человек продолжит чтение и тогда, когда, не видя отдельных букв
и слов, он воспроизводит их, угадывая. Следовательно, буквенные
тесты приемлемы лишь для ориентировочного определения возмож-
ностей аккомодации.
В случае необходимости проведения более тонких йСследований
аккомодации используют тест-объект в виде кольца Ландольта. Его
применяют в ряде аккомодометров, в частности в глазных эргогра-
фах. Психологически пробы с кольцом создают лучшие возможно-
сти для более четких ответов обследуемого. Он должен уловить не
столько исчезновение разрыва, сколько появление в кольце пере-
мычки (“превращение” буквы “С” в букву “О”), а появление стиму-
лов в поле зрения улавливается острее, чем исчезновение. Кроме
того, если сделать кольцо вращающимся, то создаются условия для
проведения врачом контрольных исследований.
Поскольку различение букв и колец основано на принципе
minimum separable, в ходе исследования аккомодации важно обес-
печить соответствие угловых размеров знаков остроте зрения (про-
веряемого глаза) и расстояниям, с которых их предъявляют. По
мере изменения расстояния величину объекта следует менять.
Особого рассмотрения заслуживают тест-объекты, пригодные
для исследования аккомодации при астигматизме. В ориентировоч-
ных пробах можно ограничиться тест-объектом Шенка. Он с доста-
точной надежностью позволяет определить возможности аккомо-
дации в каждом из двух главных сечений астигматической системы.
По мере приближения или удаления тест-объекта линия тестового
(тонкого) кольца прерывается сначала в каких-либо двух противо-
лежащих секторах, затем — в двух остальных.
Для более тонких исследований целесообразно использовать
тесты, ориентация которых легко меняется в строгом соответствии
с положением главных сечений астигматизма. При этом оправды-
вают себя двухразрывные кольца Ландольта, однако еще перспек-
тивнее тест, предложенный А.Б. Парпаровым. Тест состоит из двух
параллельных линий с меняющимся по мере удаления или прибли-
жения к глазу расстоянием между ними. Важно, что, вращая тест,
его легко ориентировать в любом меридиане.
Таким образом, существует немало разнообразных тест-объек-
тов для исследования аккомодации. Выбор наиболее подходящего
из них определяется в первую очередь задачей исследования: для
ориентировочных проб нужны одни, для уточняющих — другие.
В ориентировочных пробах, не добиваясь высокой точности,
проверяют лишь отдельные стороны процесса аккомодации: сте-
пень ее максимального напряжения для близи и дали; резервные
346
возможности уже работающей аккомодации при зрении вблизь или
вдаль; долю участия хрусталика в акте аккомодации; правильность
подбора корригирующих очков с учетом свобода действия аккомо-
дации; ответные реакции аккомодационного рефлекса на пристав-
ление к глазу пробных очковых стекол.
При уточняющих пробах важно установить прежде всего исход-
ный пункт аккомодации, ее зону покоя, после чего становится
возможной более правильная оценка всех ее рабочих зон. Цель
уточняющих проб — оценка аккомодационного аппарата в целом,
с учетом его работоспособности и сложных связей с другими систе-
мами, в частности с конвергенцией.
Проксиметрия и ремотометрия. Названные методики в сово-
купности позволяют измерить объем абсолютной аккомодации. Этот
показатель, выражаемый в диоптриях, характеризует диапазон воз-
можных изменений оптической установки глаза за счет действия
аккомодации от ближайшей (Рр) до дальнейшей (Рг) точек ясного
видения. Иными словами, объем абсолютной аккомодации есть
разница между оптическими установками глаза при максимальном
напряжении аккомодация вблизь и вдаль. Э.С. Аветисов (1973) на-
звал эту функцию динамической рефракцией глаза, полагая, что
статической является рефракция, определяемая при медикаментоз-
ном (атропиновом) параличе ресничной мышцы, т.е. при установке
глаза к дальнейшей точке ясного видения. Такая позиция совпадает
с классическими представлениями, но при этом не учитываются
новые данные о существовании активной аккомодации при зрении
не только вблизь, но и вдаль. Обычно полагают, что плюсовые
линзы замещают аккомодацию, а минусовые стимулируют (нагру-
жают) ее действие. Однако из признания двусторонней активности
аккомодации следует, что линзы одного и того же знака в зависи-
мости от зрительной задачи могут оказывать на аккомодационный
аппарат прямо противоположное действие. Так, плюсовые линзы
при зрении вдаль уже не замещают, а нагружают (стимулируют)
аккомодацию вдаль. Именно по этой причине в упражнениях по
Шерду за счет подобной стимуляции аккомодации вдаль иногда
удается достичь повышения остроты зрения близорукого глаза пу-
тем приставления плюсовых линз возрастающей силы.
Таким образом, нулевой точкой для отсчета действия аккомо-
дации, т.е. статической рефракцией, нужно считать оптическую
установку глаза не при медикаментозном параличе ресничной мыш-
цы, а при ее физиологическом расслаблении в отсутствие стимулов
для включения как на близкие, так и на далекие расстояния. Исходя
из этого, при определении ближайшей и дальнейшей точек ясного
видения на практике проще использовать тесты с реальным измере-
нием расстояний от глаза до объекта, а не прибегать к косвенной
оценке этих расстояний по реакции глаза на линзы из набора сте-
кол. Как бы ни измерять положения указанных точек, оценивать
347
объем абсолютной аккомодации нужно от оптической установки
глаза (в условиях покоя аккомодации) в случаях как приближения,
так и удаления от нее.
В ходе исследования можно смещать объект как из зоны его
неразличения, так и в обратном направлении. При этом важно со-
блюдать условия единообразия исследования. В идеале угловые
размеры деталей объекта, которые необходимо распознать, долж-
ны были бы сохраняться, несмотря на изменения расстояния, но,
как правило, этим пренебрегают, так как размеры объекта не меня-
ются и, следовательно, условие постоянства его угловых значений
не соблюдается.
Для простейшей проксиметрии используют миллиметровую ли-
нейку, конец которой с нулевым делением упирают в наружный
край глазницы со стороны исследуемого глаза. Подсветку распо-
лагают над головой пациента, направляя свет на экран, пристав-
ленный к линейке перед глазом. Это могут быть пробные шрифты
для чтения вблизи (обычно соответственно остроте зрения 0,7—
0,8 с расстояния 35 см), лучше — кольцо Ландольта. В последнем
случае обследуемого просят уловить момент, когда становится раз-
личимым разрыв в кольце, если экран смещают от глаза, или ког-
да разрыв уже не виден (“С” превращается в “О”) при смещении
экрана к глазу. Рассчитывают средние результаты повторных про-
верок, переводят линейные меры в диоптрии и получают значения
ближайшей точки ясного видения. В разное время предлагались
многочисленные варианты специальных проксиметров [Волков В. В.
и др., 1976].
Несколько меньше внимания было уделено в литературе прибо-
рам для ремотометрии. Необходимым элементом для этих устройств
является редуцирующая линза, которая искусственно приближает
дальнейшую точку ясного видения к глазу. С этой целью обычно
используют линзу +3,0 дптр. При очень слабой рефракции, в частно-
сти при афакии, требуется более сильная линза, а при собственной
оптической установке глаза в пределах 40—50 см редукция вообще
не нужна. При ремотометрии, которую проводят как и во время
проксиметрии, с помощью миллиметровой линейки определяют гра-
ницу возможности различения деталей стандартного объекта (либо
на появление, либо на исчезновение). Из полученной величины, вы-
раженной в диоптриях, вычитают силу редуцирующей линзы.
По разнице между показателями, полученными при проксимет-
рии и ремотометрии, судят об объеме абсолютной аккомодации,
нормативы для которой меняются с возрастом. Максимальный (10—
15 дптр) объем аккомодации наблюдается во втором десятилетии,
затем он постепенно уменьшается с возрастом и после 50 лет оказы-
вается в рамках 1 дптр (рис. 4.6), главным образом за счет сохране-
ния аккомодации вдаль.
Для определения положения ближайшей и дальнейшей точек
348
Рис. 4.6. Возрастные нормативы аккомодации.
а — по Радзиховскому: 1 — усредненные, 2 и 3 — крайние варианты максимального
напряжения для близи; б — по Лау — Мютце: 1 — для аккомодации вдаль, 2 — для
оптической установки глаза в покое аккомодации, 3 — для аккомодации вблизь.
ясного видения очень удобен прибор АКА-01. В приборе имеется
редуцирующая линза силой 10 дптр, что позволяет определять не
только ближайшую, но и дальнейшую точку ясного видения. Тесто-
вое кольцо Ландольта вращением рукоятки плавно перемещают
вдоль оптической оси прибора. Шкала прибора градуирована в
диоптриях от +6,0 до -5,0 дптр.
Инфракрасный сканирующий аккомодометр [Анании В.Ф. и др.,
1970—1971 ]. Прибор регистрирует с точностью 0,93 дптр измене-
ния аккомодации в процессе перемещения тест-объекта из дальней-
349
шей точки ясного видения в ближайшую. В приборе записывается
не сам процесс фокусировки глазом предъявляемого ему стимула (в
виде трех параллельных коротких линий), постепенно удаляемого
от глаза или приближаемого к нему в диапазоне от 100 до 700 мм,
а степень расфокусировки глаза относительно другого неподвиж-
ного и невидимого (из-за его ИК-излучения) стимула, изображение
которого до начала аккомодации было сфокусировано точно на
сетчатке. По мере потери резкости указанного изображения (в свя-
зи с расширением круга светорассеяния) прибор регистрирует уве-
личение времени сканирования, которое необходимо для регистра-
ции увеличивающегося по площади пятна. Таким путем можно
количественно оценить аккомодацию. С помощью этого метода
осуществляли исследование аккомодации вблизь (исследование ак-
комодации вдаль авторы не проводили).
Портативный аккомодометр [Мяло Н.М., 1985]. Прибор прост в
устройстве и обращении. Основу его составляет раскручивающаяся
линейка длиной до 100 см. На одном конце линейки расположены
упор и редуцирующая линза, которую можно откидывать на 90°, на
другом — подвижный стандартный тест-объект в виде кольца Лан-
дольта (размеры соответствуют остроте зрения 0,7—0,8 с расстояния
33 см). Прибор позволяет определять положение в пространстве трех
точек: дальнейшей (Рг), ближайшей (Рр) и точки, находящейся в про-
межутке между ними, ближе ко второй,— срединной точки (Pinterm),
соответствующей оптической установке глаза при минимальном на-
пряжении аккомодации.
Клиническая рефрактометрия, проводимая для определения опти-
ческой установки глаза в условиях относительного покоя аккомода-
ции. Исходя из представленных выше данных, мы рассмотрим толь-
ко те методики, которые позволяют определять оптическую уста-
новку глаза непосредственно, не прибегая к корригирующим линзам,
в условиях использования тест-объектов, лишь минимально стиму-
лирующих аккомодацию.
Обычные приемы, используемые для исключения влияний акко-
модации, не выдерживают критики. Исследование в условиях
предъявления тест-объектов на далеких расстояниях, если и исклю-
чает стимул для аккомодации вблизь, то в полной мере является
таковым для аккомодации вдаль. Циклоплегические средства пара-
лизуют аккомодацию (главным образом вблизь), но их действие
никак не может быть адекватным ее физиологическому расслабле-
нию, при котором определенный минимальный тонус сохраняется
во всех обеспечивающих аккомодацию структурах.
Следующие условия могут способствовать исследованию акко-
модации без побуждений к ее активности: предъявляемый стимул
должен быть лишен каких-либо деталей, в идеале не должен иметь
формы и четких граней; яркость его должна быть на пределе разли-
чения; в окружающем стимул пространстве не должно быть допол-
350
нительных ориентиров (“пустое поле”). В идеале обследуемого нуж-
но было бы лишить возможности даже косвенно судить об удален-
ности стимула, в том числе по степени расходимости идущих от
него лучей. Соблюсти эти условия в полной мере весьма сложно, а
тем более создать соответствующий прибор. Однако имеется нема-
ло устройств, основанных на разных принципах, но используемых
с одной целью — определить оптическую установку глаза в усло-
виях относительного покоя аккомодации.
Растровый диоптриметр Лау — Мутце. Это, по-видимому, один
из первых приборов, с помощью которого еще в 1952 г. были про-
изведены исследования интерференционной карты, позволившие
получить представление об оптической установке глаза без включе-
ния аккомодации. Основными элементами прибора являются две
стеклянные пластины с изображенными на каждой из них фотогра-
фическим путем решетками в виде множества вертикальных полос
черного цвета. Прозрачные пространства между полосами имеют
строго постоянные размеры и составляют константу прибора. За
матовым стеклом и диафрагмой размещен источник света в виде
лампы. Если пластины расположены параллельно, то лучи, прохо-
дящие в промежутках между полосами сначала на первой, а затем
на второй пластине, попадают в глаз наблюдателя также парал-
лельным пучком. Повернув входную решетку, которая расположе-
на ближе к источнику света, вокруг вертикальной оси, уменьшают
константу этой решетки. Пучок света, достигающий глаза наблю-
дателя, становится теперь расходящимся. С помощью дозирован-
ных поворотов другой, т.е. выходной, решетки, делают пучок света
сходящимся. Таким образом, поворотами решеток на определен-
ный угол формируется соответствующая степень ди— или конвер-
генции лучей.
Для аккомодирующего глаза изображения решеток будут на-
кладываться одно на другое и представляться все время движущи-
мися и потому нечеткими. Однако как только в глаз начнут посту-
пать лучи под углом, не требующим напряжения аккомодации,
изображение решеток сразу становится отчетливым и кажущееся их
движение прекращается. Возникающая картина является следстви-
ем интерференции лучей.
В приборе имеется призма Аббе, которая позволяет поворачи-
вать изображение решеток как бы вокруг зрительной оси и тем
самым проводить описанные выше исследования при астигматизме
по разным меридианам. Авторы установили, что с возрастом при
так называемой эмметропии отдаляется не только ближайшая точ-
ка ясного видения, но и зона оптической установки глаза (см.
рис. 4.6). Так, в возрасте 20 лет эта зона расположена на расстоянии
в среднем 25 см, в возрасте 40 лет — 50 см, а в 50 лет — на удалении
1 м. При субъективной проверке рефракции с помощью тестов для
дали определялась эмметропия. Следовательно, обычно применяе-
351
мая субъективная рефрактометрия не позволяла выявить те значи-
тельные перемены, какие с возрастом происходят в оптической ус-
тановке глаза, исследуемой в условиях физиологического покоя ак-
комодации.
Лазерскиаскопия по Аветисову [Аветисов Э. С. и др., 1972]. По
данным Ю. 3. Розенблюма и Е. И. Шапиро (1976), разработанный в
НИИ глазных болезней им. Гельмгольца лазерный скиаскоп (на базе
гелий-неонового лазера-75) позволяет на основе интерференции ко-
герентных лучей судить о клинической рефракции глаза. Жаль, ко-
нечно, что исследования традиционно проводили с расстояния 5 м,
т.е. сохранялся стимул для аккомодации вдаль. По-видимому, по этой
причине показатели миопии оказались более низкими, а гиперметро-
пии более высокими, чем при обычной скиаскопии. На основе ре-
зультатов лазерной рефрактометрии удалось получить более точное
представление об оптической установке глаза, чем с помощью скиа-
скопии, проведенной в условиях циклоплегии.
Кобальтовый рефрактометр. Идея использовать кобальтовое
(Со) стекло, пропускающее только длинные (красные, X 750 нм) и
короткие (синие, X 480 нм) лучи, для определения зоны покоя акко-
модации принадлежит В.В.Волкову и соавт. (1972). Расчет строился
на том, что практически неструктурированный слабо светящийся
стимул в условиях скотопического зрения не способен активно сти-
мулировать аккомодацию. Вместе с тем особенности преломления
исходящих из него только синих и красных лучей, отмеченные еще
Н. von Helmholtz, позволяли довольно четко поймать в “вилку” их
фокусировку на сетчатке глаза. Если точечный стимул казался ро-
зовым, а ореол был синим, то фокус располагался перед сетчаткой,
если же соотношения были обратными, то фокус “уходил” за сет-
чатку. Только в отсутствие ореолов и при равномерной сине-розовой
окраске светящегося стимула, наблюдаемого через кобальтовое
стекло, можно было считать, что его изображение фокусируется
именно на сетчатке.
По мнению В.В. Волкова и соавт. (1972,1973), стимул к включе-
нию аккомодационного рефлекса в описанной обстановке был
минимальным, и потому у подавляющего большинства испытуе-
мых удавалось определить, какую оптическую установку принима-
ет глаз в условиях физиологического покоя аккомодации.
Первая стационарная кобальторефрактометрическая установка
была создана в Военно-медицинской академии им. Кирова
В. В. Волковым с соавт. в 1975 г.
Собственно, имея кобальтовое стекло, можно проводить иссле-
дование и без специальной установки, хотя и с меньшими удобства-
ми. В слегка затемненном помещении обследуемый, держа кобаль-
товое стекло перед глазом, должен медленно приближаться к
точечному источнику света или удаляться от него. В обстановке
сниженной освещенности и практически в безориентирном простран-
352
стве, т.е. в условиях, исключающих наличие стимулов к аккомода-
ции, светящаяся точка будет видна обследуемому без красного или
синего ореола именно на том удалении (и только на нем), на кото-
ром глаз установлен при бездействующей аккомодации. При этом,
хотя аккомодация и не работает, фиксируемая точка оказывается
сфокусированной на сетчатке. Задача исследователя состоит в том,
чтобы отыскать данную установку глаза. С целью проведения ак-
комодометрии непосредственно в производственных условиях
Е.Е.Сомовым и Н.М.Мяло был предложен портативный кобальто-
рефрактометр. Прилагаемая к прибору номограмма позволяет бы-
стро рассчитывать (в диоптриях) как положение оптической уста-
новки глаза в состоянии покоя аккомодации, так и ширину этой
установки. В норме ширина этой зоны варьирует от 0,5 до 2,0 дптр.
Дуохромный рефрактометр [Сомов Е.Е., Баранов Ю.А., 1983].
Прибор создан на основе раздельного раздражения сетчатки крас-
ным и фиолетовым пучками света щелевидной формы, что позво-
лило использовать прибор для определения не только моно-, но и
бинокулярной оптической установки глаза.
Устройства, основанные на феномене Шейнера. Суть явления
заключается в оценке монокулярной диплопии, возникающей при
рассматривании нефокусированного на сетчатке объекта через два
точечных отверстия, расположенных настолько близко, что оба
умещаются в проекции зрачка. В условиях, когда фиксируемый
объект оказывается в фокусе оптической системы глаза, диплопия
исчезает. Для использования феномена при определении оптичес-
кой установки глаза Н. Пикулева (1973) предложила применить
фильтры (красный перед одним и зеленый перед другим глазом)
либо призму, которую установливают основанием кверху или кни-
зу перед одним из отверстий. С целью использования феномена для
определения зоны покоя аккомодации С.Л. Шаповалов (1976) со-
здал специальное устройство. На феномене Шейнера основано це-
лое поколение высокочувствительных приборов, каковыми явля-
ются рефрактометры Хартингера.
Современные автоматические рефрактометры (диоптроны, оф-
тальметроны, авторефракторы). Приборы этого класса основаны
на сканировании глазного дна невидимым инфракрасным источни-
ком света и регистрации той оптической установки, при которой в
исследуемом глазу исчезает монокулярная диплопия. Они освобож-
дают практикующего офтальмолога или оптометриста от весьма
трудоемких процедур. Важно, что с помощью данных приборов
определяют показатели, соответствующие именно клинической реф-
ракции глаза в зоне относительного покоя аккомодации. При этом
никаких поправок “на аккомодацию в прибор” не требуется, хотя
все показатели, получаемые при объективной рефрактометрии,
подлежат проверке на субъективную переносимость применитель-
но к конкретным задачам оптической коррекции зрения. Это осо-
353
бенно касается астигматизма и бинокулярной переносимости пред-
полагаемой коррекции анизометропии.
Субъективные тесты на астигматизм. Среди множества тес-
тов, позволяющих выявить наличие астигматизма по субъектив-
ным ощущениям, обследуемого, следует выделить лучистую фигуру
Снеллена, вращающийся крест Паскаля, стрелу Раубичека, фигу-
ру-циферблат Кривенковой. Неодинаковая четкость видения по-
разному ориентированных деталей перечисленных фигур объясня-
ется различиями в близости к сетчатке одной из двух фокальных
линий коноида Штурма, описывающего преобразования точечно-
го источника света в астигматической системе. В. И. Доброволь-
ский, И. П. Кричагин, К. А. Адигезалова-Полчаева в разное время
выполнили важные исследования, подтверждавшие существование
способности хрусталика к частичной аккомодации, компенсирую-
щей роговичный астигматизм у молодых людей.
Аккомодометрия с графической регистрацией и расчетом пара-
метров рабочих зон и зоны относительного покоя абсолютной акко-
модации. Возможность получить наглядное представление о работе
и покое аккомодации появилась благодаря детальной разработке
методики глазной эргографии и эргометрии.
Эргометр Волкова — Парпарова—Лопаева (1975). В отличие от
всех ранее предлагавшихся вариантов прибора (Н. В. Зимкин,
Э. С. Аветисов и др.) в эргометре, созданном В.В. Волковым и со-
авт., впервые был применен объект, в котором сохраняется посто-
янство угловых размеров, несмотря на изменения расстояния до
глаза, имеется возможность менять ориентацию объекта с учетом
главных сечений астигматизма, впервые реализована возможность
графической регистрации аккомодации не только вблизь, но и вдаль
(благодаря удлинению регистрирующего барабана и введению ре-
дуцирующих линз).
Эргограмма близорукого глаза с нанесенной на график зоной
покоя аккомодации, которая была получена при кобальторефрак-
тометрии, представлена на рис. 4.7. Две эргограммы, выполненные
в условиях полной медикаментозной циклоплегии и при афакии,
изображены на рис. 4.8. На рис. 4.9 можно видеть, как меняется
эргограмма при выраженном утомлении аккомодации для близи.
О ценности эргометрии при подборе рациональных корригирую-
щих очков можно судить из расчетов [Волков В. В., Парпаров А. Б.,
1976] показателя утомления для близи (ПУ):
ПУ = 2[(a5+b^ - (a,+b,)]/(a1+b,)(a5+b5)-T,
где а — ближняя, b — ближайшая точки ясного видения, цифровой
индекс указывает на время от начала эргометрии, Т — продолжитель-
ность зрительной нагрузки.
ПУ, превышающий 0,2 дптр в 1 мин, свидетельствует о выра-
женном зрительном утомлении при выполнении заданной работы.
354
20 r-
1 2 3 4 Ъмин
Рис. 4.7. Компоненты эргограммы нормально функционирующей аккомо-
дации.
Внизу — эргограмма для близи (а,, а5 — ближние, bp Ь5 — ближайшие точки ясного
видения соответственно в начале и на 5-й минуте исследования); вверху — эрго-
грамма для дали, при необходимости записываемая с редуцирующим стеклом (с,,
с5—дальние, d,, d5 — дальнейшие точки ясного видения).
19
18
17
16
15
14
в
11
И
NNHnuavvi и rnfrirnrrim
fiJii'irivaiiiini.iniiin uiMiii нншиштмиш ti
ytr a b i г 1ЛА mil 141 111 i n i i
'пгт нищими! ммп'иш
яай'1'.ии IU Ilk'll ill 11 nil ШЙГЛ1Н
цДвцииииилниимямиимииии
0 1 мин 2 мин 3 мин 4 t, мин
Рис. 4.8. Эргограмма в случае полной потери способности аккомодации к
работе в зоне как ближнего, так и дальнего видения.
а — в условиях глубокой медикаментозной циклоплегии всех порций ресничной мыш-
цы; б — при афакии.
355
Рис. 4.9. Эргограмма, свидетельствующая о выраженной слабости аккомо-
дации для близи (ее утомление развивается в ходе кратковременной на-
грузки).
Амплитуда аккомодации, роль различных факторов, влияющих на
аккомодацию, некоторые рекомендации к назначению корригиру-
ющих очков с учетом оптической установки глаза.
Оценивать амплитуду аккомодации следует раздельно для зре-
ния вблизи и вдаль, ведя отсчеты в обе стороны от зоны, на кото-
рую фокусирован глаз в состоянии физиологического покоя акко-
модации. Эта зона должна была бы стать исходной и для установки
тестов, используемых с целью проверки индивидуальной абсолют-
ной (без коррекции) остроты зрения. Всякое смещение тестов из
этой зоны ближе к глазу потребует нагрузки на аккомодацию вблизь.
Ее затраченную часть можно уподобить размещению перед глазом
положительной линзы соответствующей силы, а еще не израсходо-
ванную часть оценивать силой отрицательной линзы, которую спо-
собна преодолеть аккомодация без снижения остроты зрения при-
менительно к конкретному расстоянию. Обратные соотношения
складываются при отдалении тестов от нулевой точки отсчета. При
этом затраченная часть аккомодации вдаль подобна размещению
отрицательной линзы, а резервная часть выявляется нагрузкой по-
ложительными линзами, подобно тому, как это происходит в тре-
нировочных упражнениях по Шерду.
Амплитуда аккомодации, как уже отмечалось, является прежде
всего функцией возраста. Если отбросить влияние глубины поля,
определяемой размерами зрачка, и некоторую устойчивость акко-
модационного рефлекса к размытости изображений, то реальная
амплитуда аккомодации должна быть уже, чем на рис. 4.6. Она
существенно расширяется с увеличением фоновой освещенности. Рас-
356
пространенное суждение о том, что затуманиванием зрения при
подборе очков можно снять стимул к аккомодации, ставится под
сомнение. Вообще, как и многие другие функции, аккомодация
обладает способностью к флюктуациям амплитуды. Это следует
рассматривать как вариации тонического напряжения, свойствен-
ные любой двигательной системе организма.
Влияние аккомодации на конвергенцию рассматривается в сле-
дующей главе, здесь же уместно отметить, что существуют и обрат-
ные влияния. Конвергентная аккомодация, по данным Е. Finchan и
I. Walton (1957), характерна для молодых людей. Из наблюдений
указанных исследователей видно, что влияние конвергенции на
аккомодацию ослабевает с возрастом, а также при использовании
циклоплегических средств.
Принято считать, что на угол конвергенции к объектам на
расстоянии 1 м приходится 1,0 дптр аккомодации, но это соотноше-
ние не столь уж абсолютно справедливо, а самое главное, в этих
расчетах относительной аккомодации много условностей.
Выше отмечалось, что нельзя признать методически правомер-
ным проведение всех отсчетов от нулевой рефракции (с главным
фокусом в бесконечности) при параллельном расположении зри-
тельных осей обоих глаз. Это удобно математически, но физиоло-
гически неприемлемо, так как противоречит изложенным в данном
разделе фактам. Если в состоянии покоя аккомодации оптика глаза
фокусирует изображение предметов, например, на удалении 1 или
2 м, а при проверке и расчетах относительной аккомодации этот
фокус сначала с помощью корригирующих линз ’’перенести” в бес-
конечность и лишь после этого начинать дозированные нагрузки
на аккомодацию и конвергенцию, то при этом изначально для бли-
зорукого глаза искусственно создаются менее выгодные условия,
чем для дальнозоркого.
При подборе корригирующих очков для постоянного ношения
нужно стремиться переместить оптическую установку неаккомоди-
рующего глаза с помощью линз в зону, удаленную от него на 1,5—
2,5 м. Исходя из такой установки в подобранных очках создаются
благоприятные условия для активности аккомодации и вблизь, и
вдаль. В случае недостаточной активности аккомодации в каком-
либо из направлений приходится подбирать специальные очки толь-
ко для близи или только для дали, но и в этом случае, чем сильнее
стекло, выбранное из равноулучшающих зрение, тем меньше оста-
ется свободы для подключения аккомодации и меньше глубина поля
четкого видения в очках.
В распоряжении офтальмологов имеется достаточный выбор
тестовых таблиц для проверки остроты зрения с самых разных
расстояний. Наиболее распространены таблицы Головина — Сив-
цева для проверки с 5 м и их же шрифты для исследования с рассто-
яния 33 см. Кроме того, существуют оптотипы Серпокрыл, рассчи-
357
Таблица 4.4. Влияние основных современных фармакологических препаратов на офтальмотонус, зрачок и аккомодацию
Группы препаратов Характер действия на
офтальмотонус зрачок ресничную мышцу (оптическую установку)
Холинамилдопики: пилокарпин, ацетилхолин и др. Антихолинэстеразные веще- ства: физостигмин, эхотиофат и др. Снижают ВГД вследствие спа- стического сокращения зрачка и ресничной мышцы Суживают на несколько часов. Суживают на несколько дней Усиливают рефракцию (приближают фокус опти- ческой установки к глазу), аккомодация вблизь (+) Вызывают контрактуру ресничной мышцы (спазм аккомодации)
Xa/iuHOi/iumuKu: атропин и другие циклоплеги- ки в том числе быстрого действия: тропикамид (мидриажил), циклопентолат (цикложил) При анатомической предраспо- ложенности могут повысить ВГД вследствие блокады угла передней камеры Расширяют на несколько дней Расширяют на несколько часов Ослабляют рефракцию (отдаляют фокус оптиче- ской установки от глаза) Аккомодация вблизь (-)
Симпатомиметики: Адреномиметики: Снижают ВГД, угнетая продук- цию внутриглазной жидкости и улучшая ее отток Расширяют особенно ак- тивно при симпатической денервации Слегка ослабляют ре- фракцию (отдаляют фо- кус оптической уста- новки)
адреналин (действует на а- и fl- ap енорецепторы), эпинефрин, глаукон, кокаин (способствует накоплению адреналина) дипивалил эпинефрина, дипи- вефрин обладают высокой ли- пофильностью) (а2) Адреноагонисты: клофелин, изоглаукон, апра- клонидин, бримонидин Снижают ВГД вследствие умень- шения секреции внутриглазной жидкости на фоне вазоконст- рикции цилиарных отростков, улучшения оттока водянистой влаги Не влияют Аккомодация вдаль (+) fl-Адренергический ме- ханизм активирует неко- торые порции ресничной мышцы
Симпагпслитики fl-Адреноблокаторы: тимолол, пропранолол (несе- лективно блокируют fl^ и р2- рецепторы); ос-Адреноблокаторы: тимоксамин, гуанетидин (исме- лин, актафен вызывают симпа- тическую денервацию, как при синдроме Горнера Снижают ВГД, угнетая продук- цию камерной влаги Не влияют Суживают зрачок, рас- ширенный эпинефрином Приближают фокус опти- ческой установки к глазу Аккомодация вдаль (-)
Примечание: значок «+» или «-» указывает на тенденцию к усилению или ослаблению аккомодации.
тайные для дистанции 3 м, оптотипы Захсенвегера (1982) — для 1 м
и его же шрифты — для 40 см. Достаточно широкие возможности
открываются при использовании современных проекторов знаков.
Подбирая очки для постоянного ношения, можно воспользо-
ваться распространенной таблицей Сивцева, разместив пациента
не в 5 м, а в 2,5 м от нее. При этом потребуется единственная по-
правка: уменьшение в 2 раза получаемых показателей остроты зре-
ния. В частности, при способности читать знаки последней строки
остроту зрения нужно считать равной не 2 ед., а 1 ед.; при чтении,
например, 5-й строки сверху — не 0,5, а 0,25 и т.п. От применения
циклоплегиков при подборе корригирующих очков, как правило,
воздерживаются. Они нужны лишь в крайне редких случаях при
патологически спастическом состоянии ресничной мышцы с клини-
чески проявляющимся болевым синдромом. Что же касается обыч-
ного физиологического тонуса ресничной мышцы, то никаких спе-
циальных приемов для его преодоления не требуется. Скрытая ги-
перметропия и ложная миопия (порядка 0,5—1,0 дптр) по сути
являются результатом недооценки существования аккомодации
вдаль, проявляющейся после медикаментозного паралича аккомо-
дации вблизь. Не случайно очки, подобранные в условиях цикло-
плегии, касающейся только аккомодации вблизь, больные плохо
переносят. Такие очки либо создают чрезмерную нагрузку на акко-
модацию вдаль и сужают амплитуду аккомодации вблизь (при из-
бытке преломляющей силы корригирующего стекла, т.е. в случае
выбора слишком сильной линзы convex), либо сужают амплитуду
аккомодации вдаль и перегружают аккомодацию вблизь при недо-
статке преломляющей силы корригирующего стекла, т.е. в случае
выбора слишком сильной линзы concave.
Аналогичным образом подбираются очки к любому расстоянию,
которое определяется целью коррекции. Лишь в тех случаях, когда
предполагается специально нагрузить для тренировочных целей или
разгрузить с лечебной целью какую-либо из “чаш весов” аккомода-
ции, при выборе оптической установки отступают от принципа рав-
новесия тонических напряжений аккомодации вблизь и вдаль.
4.4. СВОДНЫЕ МАТЕРИАЛЫ ОБ ОТВЕТНОЙ РЕАКЦИИ
МЕХАНИЗМОВ РЕГУЛЯЦИИ ОФТАЛЬМОТОНУСА, ЗРАЧКА
И РЕСНИЧНОЙ МЫШЦЫ НА ФАРМАКОЛОГИЧЕСКИЕ
ВОЗДЕЙСТВИЯ
Многие лекарственные препараты, проявляющие активность в окон-
чаниях парасимпатической или симпатической иннервации, оказы-
вают комплексное влияние на зрачок, аккомодацию и офтальмото-
нус. Имеющиеся в литературе сведения об этом действии представ-
лены в табл. 4. 4.
360
ГЛАВА5
ГЛАЗОДВИГАТЕЛЬНЫЙ АППАРАТ
И БИНОКУЛЯРНОЕ ЗРЕНИЕ
5.1. УСТРОЙСТВО И РАБОТА ГЛАЗОДВИГАТЕЛЬНОГО
АППАРАТА В НОРМЕ
Движения глазного яблока осуществляются шестью экстраокуляр-
ними мышцами, из которых четыре прямые и две косые. Верхняя
косая и наружная прямая мышцы имеют особую иннервацию: пер-
вая — за счет блоковидного нерва, т.е. IV пары черепных нервов,
вторая — отводящего нерва, т.е. VI пары черепных нервов. Все ос-
тальные мышцы иннервируются ветвями глазодвигательного не-
рва, т. е. III пары черепных нервов.
5.1.1. Глазницы и биомеханика глазодвигательных мышц
Глазницы напоминают пару конусов (рис.5.1, а) с углами при вер-
шинах порядка 45°, обращенными кзади и к срединной плоскости
черепа. Как видно на рисунке, чтобы глаз принял позицию для
взгляда прямо вперед (в бесконечность), его переднезадняя ось долж-
на отклониться от оси глазницы кнутри на 22,5—23°. Эту позицию
глазного яблока называют базисной, и максимально возможные
действия любой из мышц глаза, именно исходя из этой позиции,—
также базисными. Оси, мысленно проведенные через центр враще-
ния глазного яблока (рис. 5.1, б), принято обозначать так: X —
горизонтальная, Y — переднезадняя и Z — вертикальная.
Для наружной и внутренней прямых мышц, ориентированных
относительно глазного яблока горизонтально, единственным базис-
ным действием является вращение глазных яблок вокруг вертикаль-
ной оси Z вправо и влево, т.е. по горизонтали.
Прямые мышцы, расположенные выше и ниже глазного яблока,
т.е. верхняя и нижняя, ориентированы вдоль оси глазницы, поэтому
в базисной позиции глазного яблока эти мышцы образуют с его
переднезадней осью угол порядка 23° (рис. 5.2, а).
Для верхней прямой мышцы, прикрепляющейся впереди эква-
тора глаза, базисным действием является поднимание взора вра-
щением глазного яблока вокруг оси X, проходящей через центр
вращения глаза и его экватор (рис. 5.1, б). Оптимальные условия
для оценки действия верхней прямой мышцы исключительно как
поднимателя возникают при отклонении глазного яблока от ба-
зисной позиции кнаружи на 23° (рис.5.2, б): при этом оси глазного
яблока и глазницы совпадут. Если же из этой позиции повернуть
глазное яблоко на 90° кнутри, т.е. на 23° до базисной позиции и
361
Рис. 5.1. Некоторые геометрические параметры зрительной системы,
а — углы, формируемые стенками глазницы; б — оси и плоскости глазного яблока.
еще на 67° (рис. 5.2, в), то верхняя прямая мышца окажется спо-
собной действовать только как вращатель глазного яблока (будет
смещать верхний конец вертикального меридиана кнутри и книзу).
Верхняя косая мышца, прикрепляющаяся к глазному яблоку
позади экватора, в базисной позиции образует с оптической осью
глаза угол порядка 50° (рис. 5.3,а). При этом она действует глав-
ным образом как вращатель, поворачивая глазное яблоко вокруг
оси Y, проходящей сагиттально спереди назад через зрачок, и сме-
щая верхний лимб кнутри. Дополнительно глазное яблоко немного
опускается и отмечается его отведение. Если же оптическую ось
Рис. 5.2. Варианты действия
верхней прямой мышцы в за-
висимости от исходной пози-
ции глазного яблока.
а — в базисной позиции (взгляд
прямо кпереди); б — при совпаде-
нии оптической и глазной осей;
в — в позиции аддукции (приве-
дения).
362
a
Рис. 5.3. Варианты действия
верхней косой мышцы в зави-
симости от исходной позиции
глазного яблока.
а — в базисной позиции (взгляд
прямо кпереди); б — в позиции
аддукции (приведения); в — в по-
зиции абдукции (отведения).
глазного яблока развернуть на 50° к носу, т.е. привести в состояние
приведения, и тем самым совместить эту ось с послеблоковой ори-
ентацией брюшка верхней косой мышцы (рис. 5.3,6), то она начнет
действовать исключительно как опускатель. При повороте глазно-
го яблока из данной позиции на 90° кнаружи (на 50° до базисной
позиции и еще на 40°) (рис.5.3, в), когда верхняя косая мышца ока-
жется расположенной под прямым углом к оптической оси глазно-
го яблока, действие данной мышцы сведется к разворотам верхнего
лимба кнутри.
На основании приведенных схем и описаний, касающихся фун-
кции как прямой, так и косой верхних мышц глаза, по аналогии
можно без особого труда составить представление о биомеханике
прямой и косой нижних мышц.
5.1.2. Виды произвольных движений здоровых глаз
Выделяют три вида произвольных движений глаз: Аукционные, вер-
зионные (торзионные) и вергентные.
Аукционные движения подразделяют на приводящие (аддукция),
отводящие (абдукция), поднимающие, опускающие, вращающие
(судят по смещению верхнего конца вертикального меридиана ро-
говицы кнутри или кнаружи). В зависимости от роли той или иной
мышцы в Аукционном движении исследуемого глаза рассматрива-
ют агониста (основного двигателя в заданном направлении), синер-
гистов (вспомогательных двигателей в том же направлении) и анта-
гониста (действующего в противоположном направлении). Как ус-
363
ВКос'
НижПР*
Рис. 5.4. Основные позиции взора (кроме двух: прямо вверх и прямо вниз)
с указанием для каждого глаза соответствующих взору мышц-антагонис-
тов (по первым буквам их названий).
тановил Шеррингтон, при напряжении мышцы-агониста мышца,
находящаяся в реципрокной зависимости от первой, автоматически
расслабляется.
Синхронные и симметричные смещения взора обоих глаз в од-
ном и том же заданном направлении называют верзионными. Со-
гласно правилу Геринга, к попарно работающим мышцам обоих
глаз в норме направляется одинаковый по силе нервный импульс.
Выделяют девять кардинальных позиций взора. На рис.5.4, кроме
первичной, представлены еще шесть позиций (две вторичные пози-
ции — прямо вверх и прямо вниз — исключены). Стрелками пока-
зано, благодаря каким попарно работающим мышцам осуществля-
ются эти верзионные движения.
Extorsion Intorsion
Intorsion
Extorsion
Рис. 5.5. Первичная и третичная позиции взора. В представленном вариан-
те эксторзия и инторзия не являются истинными [Listing J.B., 1854].
364
При рассмотрении четырех третичных позиций взора (рис. 5.5)
создается впечатление вращения (торзии) глазных яблок вокруг оси
Y. Действительно, на рис. 5.5 верхний конец вертикального мери-
диана представляется наклоненным либо к носу (intorsion), либо к
виску (extorsion). Однако, по мнению R.M. Burde и S.E. Feldon (1987),
следует согласиться с J.B. Listing, который еще в 1854 г. показал,
что это является результатом смещения глаза только относительно
координат X и Z (см.рис. 5.1,6), т.е. вращения глаза вокруг некой
косой оси, а потому описанный эффект является псевдоторзион-
ным. Истинная торзия вокруг оси Y возможна лишь в небольших
пределах при непроизвольной циклодукции.
Вергентными называют такие бинокулярные движения глазных
яблок, при которых они синхронно и симметрично смещаются по
горизонтали в противоположных направлениях. Движения навстречу
друг другу называют конвергенцией, в обратных направлениях —
дивергенцией. Конвергенция и дивергенция могут быть произволь-
ными, но чаще они происходят рефлекторно, т.е. непроизвольно.
5.1.3. Виды непроизвольных движений здоровых глаз
Группу, состоящую из некоторых непроизвольных движений глаз,
иногда называют спонтанным физиологическим нистагмом. В эту
группу входят тремор, дрейф и скачки (саккады). Эти движения
неизбежно присутствуют даже при стремлении обследуемого сохра-
нить взором непрерывную фиксацию какого-либо неподвижного
точечного стимула. Именно благодаря им преодолеваются те тор-
мозные процессы, которые невольно возникают в длительно стиму-
лируемом участке сетчатки. О некоторых количественных характе-
ристиках непроизвольных микродвижений глаз можно судить по
данным, приводимым A.Levy-Shoen (1969), которые основаны на
результатах исследований различных авторов (табл. 5.1).
Таблица 5.1. Параметры физиологического нистагма
Вид непроизвольных периодических движений здорового глаза Основные характеристики
пространственные дуговые минуты временные
по частоте, Гц по интервалам
Тремор 0,2—2 30—200 —
Скачки (саккады) 1—30 — 0,03—5
Дрейфы 1—10 2—5 —
Перечисленные виды микродвижений здоровых глаз представ-
ляют интерес для психологов и физиологов, в частности в связи с
365
решением проблем эргономики зрительного утомления [ Ярбус А.Л.,
1965; Волков В.В., 1989; Сомов Е.Е., 1992 ].
К непроизвольным движениям глаз, обеспечивающим биноку-
лярное зрение, относятся вергентные (не только произвольные, но
и непроизвольные) и фузионные движения глаз, а также циклодук-
ция. В вергентном рефлексе выделяют четыре компонента:
— тонический — результат врожденной способности сразу пос-
ле пробуждения от сна благодаря нервной импульсации придавать
мышцам глаза определенный тонус;
— проксимальный — результат психогенного осознания близо-
сти или удаленности объекта фиксации;
— фузионный — проявление оптомоторной реакции в ответ на
диспаратность ретинальных изображений; в итоге без изменений в
рефракции глаза изображения оказываются на корреспондирующих
точках сетчатки (при битемпоральной диспаратности возбуждается
конвергенция, при биназальной — дивергенция);
— аккомодативный — результат нормального влияния аккомо-
дации на конвергенцию (проявление синкинетической реакции на
близко расположенные стимулы). В норме на 1,0 диоптр, аккомода-
ции приходится прирост конвергенции в 4 призменных диоптрии.
Отношение аккомодативной конвергенции к аккомодации обозна-
чают АК/А. Результат выражают в призменных диоптриях на еди-
ницу аккомодации, измеренной в диоптриях.
В этой группе отдельно стоит циклодукция, которая проявляет-
ся в том, что при наклоне головы к одному или другому плечу оба
глазных яблока испытывают истинный торзионный эффект: ком-
пенсаторно вращаются наподобие рулевого колеса. При этом каж-
дый глаз верхним концом вертикального меридиана разворачива-
ется в сторону, противоположную стороне наклона головы. На-
клон к носу называют инциклодукциёй, к виску — эксциклодукцией.
По данным R.S. Jampel (1970), в этом случае вокруг оси Y враща-
ется не столько все глазное яблоко, сколько его передний отдел
(вокруг точки на наружном лимбе). Таким способом поддерживает-
ся сенсорное постоянство расположения вертикали швов сетчатки
относительно горизонта. При наклоне головы на 30° глаз непроиз-
вольно компенсаторно отклоняется на 7—8° в противоположную
сторону [Кириллов Ю.А., 1976].
Если сопоставить амплитуду непроизвольных движений глаз при
фиксации неподвижного стимула, то, как установил Э.С. Аветисов
(1977), при наличии бификсации глазные яблоки заметно устойчи-
вее. Величина поля бификсации по горизонтали составляет 5—
10 мин, а по вертикали не выходит за пределы 4 мин. Удержание
объекта фиксации в пределах узкого фузионного поля является
результатом непроизвольной статической работы глазодвигатель-
ных мышц, координируемой сенсорным аппаратом бинокулярного
зрения.
366
5.2. БИНОКУЛЯРНОЕ ЗРЕНИЕ
Под нормальным бинокулярным зрением понимают такую совмест-
ную работу сенсорных и моторных систем правого и левого глаза,
которая обеспечивает одновременную ориентацию обеих зритель-
ных осей на объект фиксации, формирует пару сходных монокуляр-
ных изображений этого объекта на сетчатке парных глаз, способ-
ствует слиянию их в единый зрительный образ, правильному опре-
делению местоположения стимула, в том числе его удаленности от
глаз в видимом пространстве [Аветисов Э.С., 1977; Bishop Р.С. et
al., 1987].
При бинокулярной фиксации точечного объекта значительная
часть окружающего его пространства воспринимается одновремен-
но одним и другим глазом, образуя таким образам общее (биноку-
лярное) поле зрения. Однако и в зоне бинокулярного перекрытия
есть немало деталей “сцены”, которые видны только монокулярно.
Таким образом, в естественных условиях неизбежно параллельное
согласованное функционирование бинокулярных и монокулярных
механизмов. Возможность совместного одиночного или раздвоен-
ного видения изображений, возникающих в парных глазах, опреде-
ляется рядом условий.
5.2.1. Совместное одиночное видение
Одиночное видение при бинокулярном восприятии формируется уже
в первые годы жизни. В нормальной работе механизмов, обеспечи-
вающих слияние двух различных изображений, которые формиру-
ются в парных глазах, и ощущение трехмерности (объемности)
пространства, важную роль играют три фактора.
Качество изображений в каждом глазу. Четкость изображения
обеспечивается не только состоянием оптического аппарата глаза,
но также особенностями строения и функцией макулярной области
сетчатки. Эта область шире, чем macula lutea. Периферической гра-
ницей макулярной области, занимающей центральную зону сетчат-
ки диаметром 5,5 мм (9,2°), является зона, где плотность ганглиоз-
ных клеток значительно ниже. Центром макулярной области явля-
ется фовеола диаметром 0,35 мм (0,6°). В этой области сетчатка очень
тонкая и представлена лишь слоем фоторецепторов. Фокусировка
стимула точно в этой области обеспечивает наиболее высокую ос-
троту зрения. В пределах фовеолы находится и зона Панума, име-
ющая прямое отношение к фузионным механизмам бинокулярного
зрения. В окружающей фовеолу области фовеа диаметром 1,5 мм
(5°) сетчатка в 2 раза тоньше, чем в остальной ее части. В этой зоне
почти нет биполярных клеток, а слой ганглиозных клеток с аксона-
ми отсутствует вовсе. С участием именно этой области осуществля-
ется тонкий стереопсис.
367
Бинокулярные рецептивные
поля
'Ч | Диплопия
Бинокулярное
_/\ одиночное зрение
А
Диплопия
Бинокулярные ответы
а б
Рис. 5.6. Схема основного нервного механизма, лежащего в основе биноку-
лярного одиночного видения и стереопсиса [Bishop P.O., 1987].
а — накладывающиеся одно на другое монокулярные рецептивные поля парных глаз
соотнесены с фронтопараллельными плоскостями, мысленно проходящими в види-
мом пространстве через обе линии взора вблизи места их пересечения; б — гисто-
граммы усиления или подавления бинокулярных ответов на стимулы, которые ло-
кализуются в соответствующих фронтопараллельных плоскостях.
Пара- и перифовеальный отделы макулярной области участву-
ют в формировании стереоскопического зрения. Однако возможно-
сти разрешающей способности сетчатки в этих областях по мере
эксцентриситета быстро и значительно ухудшаются.
Способность зрительных центров мозга к слиянию двух изобра-
жений от парных глаз. Бинокулярное взаимодействие двух рецеп-
тивных полей парных глаз как в тех случаях, когда эти поля накла-
дываются одно на другое, так и в тех, когда они все больше расхо-
дятся, представлено на рис.5.6. Область, возбужденная стимулом,
имеет дид вертикальной полоски, а зоны торможения вокруг за-
штрихованы. В каждой из пяти фронтопараллельных плоскостей, вы-
деленных вдоль линии взора, представлены соответствующие пары
рецептивных полей. Там, где поля отражают пересечение зрительных
осей на объекте фиксации (полоска), поля точно совпадают. Кпере-
ди и кзади зона возбуждения в одном из полей накладывается на
зону торможения в другом. На правой части рисунка видна депрес-
сия бинокулярного ответа. При значительном удалении от поля
слияния рецептивные поля правого и левого глаза все больше
разделяются и благодаря этому стимулируются уже независимо одно
от другого.
На схеме весьма упрощенно представлены сложные процессы,
происходящие в мозге, но она хорошо объясняет одиночность би-
нокулярного видения вследствие усиления ответа при совпадении
полей и гашения сигналов при незначительном несоответствии.
368
Рис. 5.7. Круг гороптера (Вайса—
Мюллера). Точка фиксации (Р) и дру-
гая точка в пространстве (Q) лежат
на общей для них окружности, прове-
денной также через узловые точки
обоих глаз. Благодаря равенству уг-
лов 01 = 02 изображения точек Р и Q
на глазном дне совпадают с коррес-
пондирующими точками сетчаток
обоих глаз (qj и q2).
Когда же рецептивные поля становятся полностью независимыми,
возникают условия для возникновения диплопии.
Роль фузионных и вергентных движений глаз в поддержании нор-
мальной корреспонденции сетчаток. В первой половине прошлого
века сформировалось представление о круге гороптера. Согласно
концепции Вайса и Мюллера, при бинокулярном зрении каждым
глазом можно видеть одновременно и четко лишь те точечные сти-
мулы, которые располагаются в пространстве по окружности, охва-
тывающей точку фиксации и узловые точки обоих глаз. Стимулы
гороптера (например, стимул Q) (рис.5. 7) будут изображаться на
парных сетчатках на одинаковом расстоянии и в одном направле-
нии от фовеолярных областей. Точки qt и q2 на двух сетчатках
оказываются геометрически корреспондирующими. Их одновремен-
ная стимуляция приведет к формированию в сознании представле-
ний о том, что одиночный стимул Q расположен правее от точки
фиксации Р. Из данной схемы следует, что все точечные стимулы,
расположенные как за пределами круга гороптера, так и внутри
него, будут изображаться не в корреспондирующих, а диспаратных
точках сетчатки, что не позволит им сливаться.
Дальнейшие исследования показали, однако, что свойство сет-
чаток к корреспонденции в процессе жизни поддается тренировкам
и благодаря этому гороптер фактически не является точечным, а
приобретает определенную глубину. На рис.5.8, кроме эмпиричес-
ки рассчитанного точечного гороптера и круга Вайса — Мюллера,
представлена заштрихованная зона одиночного видения. Если то-
чечный объект фиксировать на расстоянии 40 см перед глазом, то
все точки гороптера будут видны как одиночные, а точки, распо-
ложенные в 5—10 мм кпереди или кзади от гороптера, сольются в
единый образ. Чем дальше расположен объект фиксации, т.е. чем
больше радиус круга гороптера, тем большую глубину имеет и эта
зона одиночного видения, получившая название зоны Панума.
369
Рис. 5.8. Относительное расположение зон в бинокулярно видимом про-
странстве.
1 — эмпирического гороптера; 2 — фузионной области Панума (зоны одиночного
видения); 3 — круга Вайса — Мюллера и зон, обозначенных слева: дальней и ближ-
ней диплопии, частично прекрывающей их зоны стереопсиса.
Фузионные движения глаз определяются наличием такой зоны.
Исследование зоны Панума с помощью стереомишеней (рис. 5.9,а)
и при перемещении вдаль или вблизь одной из вертикальных пало-
чек (рис.5.9,б) позволяет получить объемное представление о про-
ксимальных и дистальных пределах фузионной зоны Панума.
В опыте, представленном на рис.5.9,б, взором ведущего (левого) гла-
за фиксируется расположенная слева палочка. При смещении вто-
рой (правой) палочки кпереди или кзади от местоположения ее в
круге гороптера (с сохранением угла эксцентриситета между палоч-
ками) в определенный момент появится диплопия. На стереомише-
ни, изображенной на рис.5.9,а, это обозначено пунктирными лини-
ями. Угловое расстояние между линиями (± 8S) отражает фузион-
ную зону Панума, т.е. протяженность по горизонтали зоны
одиночного бинокулярного зрения при данном эксцентриситете.
Когда диспаратность точек выходит за пределы зоны Панума, по-
является физиологическая диплопия, которую в обычной жизни не
замечают. Однако именно ее наличие позволяет правильно опреде-
лить взаиморасположение различных предметов в видимом про-
странстве.
K.Ogle (1962) установил, что при умеренной диспаратности
объекта, изображаемого на сетчатках парных глаз, т.е. при диспа-
ратности, возникающей сразу за пределами зоны Панума (когда
фузионные движения уже не способны привести к слитному воспри-
ятию), появляется картина стереопсиса, в которой одни детали вид-
ны более четко, чем другие, и изображение приобретает черты
объемности. По мере усиления диспаратности стереопсис становит-
370
Левая Правая
l S±8SJ угол
Мишени эксцентРисит^
Точка
фиксации
Проксимально
Дистально
Sx
а
Левый глаз
б
Правый глаз
Рис. 5.9. Приемы определения зоны Панума (схема) [Bishop P.O., 1987].
Область
' бинокулярного
одиночного видения
Размеры угла (зона Панума)
фузионной зоны Панума
а — стереомишени парных глаз; б — объемное изображение опыта, в котором ле-
вый вертикальный объект (точка фиксации) неподвижен, правый изображен в двух
переменных позициях (проксимальной и дистальной), за пределами которых по-
явится двоение. На левой стереомишени: S — угол эксцентриситета; на правой ми-
шени — ± 6S — угол между вертикальными пунктирными линиями соответствует
горизонтальной ширине зоны Панума.
ся все более грубым (рис.5.10). Тонкий стереопсис называют безус-
ловным, а грубый, результаты которого не поддаются количествен-
ной оценке,— качественным (сохранено ощущение того, что ближе,
а что дальше, но насколько ближе, определить невозможно). Для
фузионных движений основным и достаточным стимулом является
очень небольшая диспаратность изображений и совсем не обяза-
тельно возникновение диплопии и других проявлений стереоскопи-
ческого эффекта. Для преодоления более значительной диспаратно-
сти по горизонтали необходимы вергентные движения глаз. По мере
удаления стимула от линии фиксации к периферии стереопсис в
отличие от фузии позволяет заметно активнее преодолевать диспа-
ратность.
Следует подчеркнуть, что ощущение стереопсиса вызывается
только горизонтальной диспаратностью ретинальных изображений.
Вертикальная диспаратность не оказывает влияния на эффект сте-
реопсиса. Фузия же осуществляется независимо от направления
диспаратности. Порог стереоскопичности в значительной степени
определяется уровнем адаптации зрительной системы к свету. I.Le
Grand установил, что на кривой стереоскопической чувствительно-
сти имеется излом, аналогичный тому, который определяется при
переходе кривой скотопического зрения в кривую фотопического
[Bishop Р.В., 1987]. Это свидетельствует об определенной общности
механизмов этих явлений.
371
400-j
-6° а Пределы для дали
о Пределы для близи
Безусловный
стереопсис
Отклонение к периферии, градусы
Рис. 5.10. Пределы диспаратности для фузии, стереопсиса (безусловного и
только качественного) в зависимости от ретинального эксцентриситета
[OgleK.N., 1962].
По данным R.N.Berry (1948), стереоострота и верньерная ост-
рота зрения, исследованные в одних и тех же условиях, нередко
хорошо совпадают, хотя у некоторых субъектов при наличии высо-
кой остроты зрения бинокулярное зрение может быть плохим. Ос-
трота стереоскопичности представляет собой величину, обратную
глубинному порогу. Последний же определяется тем минимальным
сагиттальным расстоянием между двумя объектами, которое еще
удается уловить как различие в их относительной удаленности.
Остроту стереоскопичности измеряют угловыми порогами биноку-
лярной диспаратности (в дуговых секундах). H.Howard (1919) уста-
новил, что в норме стереопорог составляет 2 дуговые секунды. В не-
которых условиях, когда в качестве объектов используют точки
(вместо вертикальных линий), порог возрастает до 40 дуговых се-
кунд. Нормативы варьируют с возрастом [Simons К., 1981]. На ре-
зультаты исследования оказывает влияние также наличие анизомет-
ропии свыше 0,5%. Иногда хорошо переносится разница до 3%, но
при 5% качество стереопсиса всегда ухудшается. E.Levinson и
Е.Blake (1979) установили, что если в качестве теста использовать
синусоидальную решетку, то при различиях в восприятии простран-
ственной частоты в парных глазах (например, при односторонней
афакии) до 20% способность к фузии нарушается. При определении
372
диспаратности на основе цветовых различий может потребоваться
удлинение контуров стереограмм по сравнению с обычными [Lu С.,
Fender F., 1972].
При перемещении объекта по глубине возможности стереоост-
роты особенно снижаются. Примечательно, что по мере возраста-
ния эксцентриситета до 5° порог стереоостроты сначала увеличива-
ется медленно, а затем круто поднимается.
Существует понятие абсолютной диспаратности. Поскольку по
кругу гороптера порог самый низкий, абсолютную диспаратность
расценивают как нулевую. Она возрастает по мере приближения
объекта к точке фиксации и удаления от нее. В случае приближения
объекта определяют порог конвергентной абсолютной диспаратно-
сти, в случае отдаления — порог дивергентной абсолютной диспа-
ратности. Теоретически предельное расстояние, на котором еще
возможно различать предметы по глубине, составляет 500 м. Раз-
личия в показателях абсолютной диспаратности двух объектов на-
зывают относительной диспаратностью между ними. При абсолют-
ной дивергентной диспаратности величиной в 2 дуговых градуса не
только может сохраняться стереопсис, но благодаря присутствию
фузии еще возможно различение относительной диспаратности в <£'
[Fender A. et al., 1967]. При этом поражает не способность к тонко-
му различению глубины, а возможность фузии при дивергенции
изображений от центра фовеолы на целый дуговой градус. Таким
образом, бинокулярное одиночное видение возможно только при
бинокулярной стабилизации ретинальных изображений. Следова-
тельно, фузия и стереопсис существуют не потому, а несмотря на
то, что глазные яблоки подвижны.
5.2.2. Совместное раздвоенное видение
Очевидными причинами нарушения слияния парных изображений
в одиночное является рассогласование установки зрительных осей
на единый объект, скрытое или явное косоглазие. Последнее прояв-
ляется в одном из двух функциональных вариантов: либо ухудша-
ется качество фокусировки и фиксируемый объект воспринимается
как в тумане, хотя и слитно двумя глазами (рис.5. 11), либо четко,
но только одним глазом, фокусирующим объект. В первом случае
из-за наслоения изображений разных объектов, информация о ко-
торых исходит от макулярных областей парных глаз, возникает
“неурядица”. Во втором случае другой глаз вынужденно отклоня-
ется от точки фиксации и в итоге возникает двоение.
При сходящемся косоглазии двоение носит одноименный ха-
рактер, при расходящемся — разноименный. Иными словами, если
при косоглазии зрительные оси сходятся, изображения не перекре-
щиваются, если расходятся — перекрещиваются.
Механизмы компенсации раздвоенного видения. Подавление (суп-
373
a
Рис. 5.11. Варианты совместного видения двумя глазами при косоглазии.
а — нормальная бификсация с одиночным видением в отсутствие косоглазия; б —
’’неурядица”: на изображение объекта (кружок), фиксируемого лучше видящим гла-
зом, в зрительных центрах накладывается изображение другого объекта (треуголь-
ник), попавшего на макулу косящего глаза, оба объекта проецируются в зону пере-
креста зрительных осей и видны нечетко, поскольку там нет ни одного, ни другого
объекта; в — диплопия: в косящем (правом) глазу сформировалась ложная макула,
которая, будучи диспаратной по отношению к макуле другого глаза, посылает в
центр ложную информацию о проекции объекта в пространстве.
рессия) зрительного образа косящего глаза. Т.П.Кащенко (1978)
считает этот механизм бинокулярной сенсорной адаптации наибо-
лее важным, особенно у детей, поскольку он встречается у 70% всех
пациентов с косоглазием. Суть его в том, что изображение, переда-
ваемое в центр от косящего глаза, активно отвергается (тормозит-
ся) зрительной корой головного мозга. Примечательно, что меха-
низм функционирует только тогда, когда оба глаза открыты
(рис.5.12). Стоит прикрыть лучший (фиксирующий) глаз, как зри-
тельная ось косящего глаза переводится на объект фиксации; толь-
ко что косивший глаз как бы освобождается от супрессии.
374
5.12. Механизмы компенсации диплопии.
а, б — супрессия; в — амблиопия; г — аномальная корреспонденция сетчаток.
В происхождении супрессии, по-видимому, участвуют тормоз-
ные процессы, возникающие как в зрительных центрах, так и в
пределах самой сетчатки косящего глаза. Подавление изображения
может приобрести стойкий характер, и тогда зрение становится
монокулярным, а в косящем глазу развивается амблиопия
(см.рис.5.12). В этом случае зрение косящего амблиопичного глаза
приобретает следующие особенности: острота зрения уже не повы-
шается при фиксации предметов этим глазом; ущербность остроты
зрения особенно выражена при одновременном показе группы зна-
ков (феномен кучности); на остроту зрения почти не влияют нейт-
ральные фильтры, как и при исследовании здорового глаза. Он
таким образом отличается от глаз, в которых острота зрения сни-
жена в связи с органическими изменениями в зрительно-нервном
аппарате. В последнем случае нейтральные светофильтры законо-
мерно ухудшают остроту зрения.
375
Кроме амблиопии, развивающейся в связи с косоглазием, встре-
чаются еще два вида амблиопии: обскурационная (при нарушении
прозрачности сред и при птозе) и анизометропическая (при низком
качестве изображения в глазу с большей аметропией).
Аномальная корреспонденция сетчаток. Этот вид бинокулярной
сенсорной адаптации при косоглазии встречается нечасто (в 5—
7% случаев). У маленьких детей при стойком косоглазии может сфор-
мироваться подобный неполноценный вариант асимметричного
бинокулярного зрения с ложной макулой [Кащенко Т.П., 1978]. Ре-
тинальные элементы косящего глаза устанавливают корреспондиру-
ющие отношения с фовеа и другими областями сетчатки некосящего
глаза, развивается аномальная корреспонденция сетчаток гармони-
ческого типа. Она особенно характерна для микрострабизма.
Спровоцированная диплопия. Ее выявляют у 20—25% лиц с косо-
глазием. Э.С. Аветисов и Т.П. Кащенко (1977, 1993) полагают, что
наличие скрытой диплопии или специальное ее вызывание в есте-
ственных условиях создают дополнительный шанс для “воспита-
ния” способности к ее преодолению “путем саморегулирующегося
механизма бификсации”. Эта система лечения получила название
диплоптики [Аветисов Э.С., 1977].
Давно отмечено, что при слабости какой-либо из экстраокуляр-
ных мышц дети с потенциально хорошим бинокулярным зрением и
взрослые, не способные к супрессии, могут подсознательно и созна-
тельно подавлять диплопию, придавая определенное положение
голове. Общим правилом является то, что голова должна привыч-
но разворачиваться в сторону действия ослабленной мышцы, так
как при этом глаз автоматически поворачивается в противополож-
ную сторону и нагрузка на слабую мышцу снимается.
Поскольку горизонтальная девиация заключается в развороте
головы, например, влево (если поражена соответствующая мыш-
ца), глаза автоматически поворачиваются вправо. При вертикаль-
ной девиации (при слабости поднимателей) пациент будет подни-
мать подбородок и смотреть свысока, а при слабости опускате-
лей — опускать его и смотреть исподлобья. Торзионная девиация
заключается в наклоне головы к тому плечу, в сторону которого
должна была бы поворачивать верхний лимб ослабленная косая
мышца.
При параличе одйого из инторзеров, т.е. мышц, вращающих
верхний лимб кнутри, глаз примет положение эксторзии. Для того
чтобы компенсировать ее и вернуть глаз в положение инторзии,
голову следует наклонить к тому плечу, в сторону которого дол-
жна была бы действовать ослабленная мышца. Так, слабость
правой верхней косой мышцы компенсируется наклоном головы
влево.
376
5.3. КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ НАРУШЕНИЙ
ФУНКЦИИ ГЛАЗОДВИГАТЕЛЬНОГО АППАРАТА
И БИНОКУЛЯРНОГО ЗРЕНИЯ С УЧЕТОМ
РАЗЛИЧИЙ ЭТИОЛОГИИ И ПАТОГЕНЕЗА
Сходящееся косоглазие (детская эзотропия):
1) младенческое косоглазие. Обычно обнаруживают в первые
6 мес жизни. Угол очень большой, фиксация взора при боковых
предъявлениях перекрестная, в прямой позиции — альтернирующая;
наличие горизонтального нистагма — предвестник развития верти-
кальной девиации;
2) аккомодативное косоглазие характеризуется повышенным
аккомодационным рефлексом, но если глаз не отклоняется за пре-
делы фузионной дивергентной амплитуды, то косоглазие остается
скрытым; варианты:
• рефракционный (гиперметропия > 4 диоптр.),
• нерефракционный (АК/А превышает 10 призменных диоп-
трий), косоглазие усиливается при зрении вблизи и обна-
руживается только при разглядывании мелких предметов
на близких расстояниях,
• смешанный (высокое АК/А + гиперметропия). Если соче-
тается высокая гиперметропия с низким АК/А, косоглазие
не развивается;
3) неаккомодативное косоглазие обусловлено “поломкой” ди-
вергентно-фузионного механизма; возможные причины: стрессовые
ситуации, общие заболевания; сенсорная депривация (при помутне-
нении сред, атрофии зрительного нерва и других заболеваниях);
слабость дивергенции без паралича отводящего нерва или при его
наличии; “спазм” рефлекса на близкие расстояния (при истерии,
иногда с выраженным миозом и спазмом аккомодации).
Расходящееся косоглазие (детская экзотропия):
1) перемежающееся косоглазие (по величине угла), в отличие от
сходящегося, характеризуется более широкими пределами фузион-
ной дивергенции, при которой скрытое косоглазие (фория) еще не
переходит в явное (тропия); обнаруживают в возрасте около 2 лет,
усиливается при ярком свете, утомлении, в состоянии задумчивос-
ти, иногда отмечается тенденция к стойкому усилению девиации;
ретинальная корреспонденция при выравнивании зрительных осей
становится нормальной, а с появлением явного косоглазия возни-
кает темпоральная гемисупрессия сетчаток; амблиопия развивается
нечасто.
2) постоянное косоглазие:
• врожденное, нередко сочетающееся с неврологическими на-
рушениями. Часто обнаруживают уже при рождении, так
как угол достаточно большой. Фиксация взора гомоним-
377
ная (правым глазом справа, левым слева); амблиопия неха-
рактерна;
• декомпенсированно-перемежающееся, V-синдром. При
взгляде вверх и прямо — экзотропия, при взгляде вниз —
эзотропия;
• сенсорно-депривационное, развивающееся у детей старше
5 лет в связи с прерыванием бинокулярного рефлекса из-за
помутнения преломляющих сред;
• последовательное, возникающее как следствие гиперкоррек-
ции эзотропии или при глубокой амблиопии (глаз, перво-
начально косящий кнутри, постепенно отходит кнаружи).
Выраженная гетерофория может быть причиной быстрого утом-
ления органа зрения, тошноты, диплопии и головной боли [Шапо-
валов С.Л. и др., 1985]. При отборе кандидатов на летные профес-
сии следует осуществлять проверку этой функции и включить ее в
экспертизу [Сомов Е.Е., 1992].
Паралитическое косоглазие. Это приобретенное косоглазие,
развивается после травм, хирургических вмешательств, как ослож-
нение нейроинфекций и других заболеваний. Характеризуется огра-
ничением подвижности глазного яблока в сторону действия пара-
лизованной мышцы, скрытой или явной диплопией, усиливающей-
ся в том же направлении. При измерении угла косоглазия вторичное
отклонение (здорового глаза) превышает первичное (косящего гла-
за). При наличии тортиколлиса (кривошеи) необходимо обнаружить
ослабленную глазную мышцу; о давности процесса судят, сравни-
вая старые фотографии больного с имеющимся статусом.
Редкие дефекты подвижности глаз. Следствием врожденных
аномалий являются ретракционный синдром при болезни Дауна,
синдром врожденного фиброза поднимателей глазного яблока и век
или внутренних прямых мышц обоих глаз (фиксированный страбизм).
5.4. МЕТОДИКИ ФУНКЦИОНАЛЬНЫХ ИССЛЕДОВАНИЙ
ГЛАЗОДВИГАТЕЛЬНОГО АППАРАТА И БИНОКУЛЯРНОГО
ЗРЕНИЯ
5.4.1. Измерение угла косоглазия
Тест Гиршберга весьма ориентировочен, но пригоден при обследо-
вании лиц, плохо фиксирующих взор. Свет (от зеркала офтальмо-
скопа, лампочки, свечи) направляют прямо (спереди) в глаза и оце-
нивают место расположения его рефлекса на роговице. Угол косо-
глазия принимают равным 15° при расположении рефлекса по краю
зрачка средней ширины (4 мм), 45° при его локализации на лимбе.
С помощью этого теста легко исключается ложное косоглазие, на-
поминающее сходящееся при широкой переносице или расходяще-
еся при укорочении глазной щели со стороны виска.
378
+5‘
Ортофория
(угол Каппа=+5°) ;+5<
Рис. 5.13. Варианты распо-
ложения роговичных реф-
лексов при зрачке диамет-
ром 4 мм и их использова-
ние для оценки угла косо-
глазия по Гиршбергу.
глаз
В норме рефлекс часто слегка смещен от центра роговицы к
носу (рис.5.13). Рефлекс обозначает положение зрительной оси, а
оптическая ось при положительном угле Каппа (0,25°) проходит
темпоральнее. При сходящемся косоглазии рефлекс смещается кна-
ружи, при расходящемся — кнутри. При высокой миопии, когда ма-
кула сдвигается ближе к диску, зрительная ось расположена под
отрицательным углом Каппа по отношению к оптической оси и
возникает ощущение ложного сходящегося косоглазия. Угол Кап-
па — важный фактор и в оценке эффективности операции, поэтому
большинство авторов считают косметический результат операции
удовлетворительным при отклонении глаза по горизонтали в пре-
делах ±5°, а по вертикали ±3°. R.Hugonnier и S.Clayette-Hugonnier
(1969) полагают, что при сходящемся косоглазии (эзотропии) пре-
делы допустимого по косметическим критериям угла Каппа могут
быть расширены до +14° (см. рис.5.13). Нужно иметь в виду, что
смещение роговичного рефлекса на 1 мм соответствует углу в 6—
7°. В качестве ориентировочной шкалы используют размеры диа-
метра зрачка (на рис. 5.13 он равен 4 мм).
Методика определения угла косоглазия на дуговом периметре по
результатам оценки роговичных рефлексов с использованием двух
свечей достаточно точна, но, по-видимому, уже мало кем применяется.
Тесты с перекрытием одного глаза предназначены для выявле-
379
ния явного и скрытого косоглазия по установочному движению, а
также для измерения его угла страбометром или миллиметровой
линейкой по степени смещения (сдвиг на 1 мм принимается соот-
ветствующим повороту на 5—6°).
В том случае, если при перекрытии ведущего (фиксирующего)
глаза второй (неприкрытый) глаз совершает установочное движе-
ние, то это — признак явного косоглазия (при перемещении к носу —
расходящегося, к виску — сходящегося, по вертикали — гипо- или
гипертропии). После устранения перекрытия ведущего глаза кося-
щий глаз чаще сразу же возвращается в исходное положение. Если
же он продолжает сохранять приданную позицию (прямо кпереди,
то его прикрывают и, убедившись в том, что после этого восстано-
вились первоначальные отношения, расценивают косоглазие как
альтернирующее.
Для выявления гетерофории, т.е. скрытого косоглазия, следят за
поведением уже не того глаза, который оставляют открытым, а того,
который на короткое время (на 2 с) перекрывают. Освобожденный
от экрана, он может оставаться неподвижным: скрытого косогла-
зия нет (оргофория), либо при наличии гетерофории совершит не-
большое движение. Если движение направлено к носу, то значит,
глаз при кратковременном его выключении из бинокулярного зре-
ния отклонился к виску (экзофория), а затем возвратился в прямое
положение. При смещении глаза после открывания к виску — эзо-
фория, книзу — гиперфория, кверху — гипофория. Нужно заметить,
что обнаруживаемая при тестировании гетерофория не исключает,
а подтверждает наличие бинокулярного зрения.
Для скрининговой оценки бинокулярности зрения у маленьких
детей используют простой прием. К одному из глаз пациента при-
ставляют сильную призму (20 призматических диоптрий) основани-
ем к виску и, если, наблюдая через нее, улавливают компенсирующее
смещение глаза кнутри, делают вывод о наличии у ребенка стимула
к одиночному видению предметов, к бинокулярности зрения.
5.4.2. Измерение мышечного баланса, в том числе
конвергентно-аккомодационных отношений
Проба Грефе. Это простейшая проба из числа предложенных в раз-
ное время для указанной цели, и достаточно точная. Рисуют верти-
кальную линию, пометив ее середину точкой или крестиком, и про-
сят обследуемого посмотреть на точку (крестик). Затем, поместив
перед одним из глаз призму, например, силой 6 призменных диоп-
трий, основанием книзу (или кверху), создают второе изображение
рисунка, сдвинутое по вертикали. При наличии дисбаланса мышц,
работающих в горизонтальном направлении, линия удваивается и
одна из образовавшихся линий оказывается правее или левее дру-
гой. Величину этого смещения можно определить с помощью призм
380
последовательно возрастающей силы, если их устанавливать осно-
ванием к виску или носу (в зависимости от вида гетерофории) до
момента слияния двух изображений в одно.
Аналогичные измерения проводят с целью оценки степени вы-
явленной фории в вертикальном и при необходимости в косых на-
правлениях.
Тесты Меддокса (с разделением изображений парных глаз для
выявления скрытого косоглазия). В устройстве для близи (33 см)
благодаря разделению полей зрения испытуемый одним глазом
может видеть только две (горизонтальную и вертикальную) кресто-
видно пересекающиеся шкалы цифр, а другим — вертикально и го-
ризонтально ориентированные стрелки. В результате бинокулярно-
го наложения одного изображения на другое и невозможности сли-
яния столь разных картин в единый образ обнаружится скрытое
косоглазие. Степень и вид его с учетом возможной циклофории
показывают стрелки, накладывающиеся на шкалы.
С целью определения наличия гетерофории вдаль перед одним
из глаз обследуемого (на рис.5.14. перед правым глазом) помеща-
ют взятую из набора пробных очковых линз “палочку” Меддокса
и предлагают обоими глазами смотреть на световое пятно, находя-
щееся на расстоянии 3—5 м. В случае горизонтальной ориентации
осей призм “палочки” пятно для правого глаза представится вытя-
нутым в вертикальную светящуюся линию. При наличии скрытой
девиации тот глаз, который прикрыт “палочкой”, отклонится в со-
ответствующую сторону и видимые одновременно пятно левым
глазом и линия правым не смогут слиться воедино. При экзофории
для глаза, отклонившегося за “палочкой” кнаружи, и линия ока-
жется сместившейся кнаружи, при эзофории восприятие линий бу-
дет обратным (см.рис.5.14). Величина отстояния линии от пятна
соответствует степени скрытого косоглазия. Для измерения угла
косоглазия применяют призмы, помещая их основанием в направ-
лении, противоположном девиации. По силе призмы, при которой
удастся достигнуть совпадения пятна и линии, определяют степень
гетерофории.
Тесты с очковыми сферическими линзами (для оценки влияния
привычного тонуса аккомодации на положение зрительных осей
при явном косоглазии). При явном косоглазии в оценке мышечного
баланса особое место занимают взаимоотношения между аккомо-
дацией и конвергенцией. Это прежде всего касается сходящегося
косоглазия (эзотропии), в основе которого лежит необходимость в
чрезмерно высокой активности аккомодации вблизь, что и опреде-
ляет четкость видения.
В ряде случаев имеет значение не просто наличие высокой ги-
перметропии, а отсутствие оптимальных взаимоотношений акко-
модации и конвергенции. Нормативы прироста конвергенции на
единицу усиления аккомодации вблизь приведены выше. Следует
381
Рис. 5.14. Тест с “палочкой” Меддокса.
отметить, что если выявляются высокие значения показателя АК/А,
то это следует расценить как предпосылку к так называемому экс-
цессу конвергенции. Иными словами, при потребности напрягать
аккомодацию для работы на близких расстояниях, как это свой-
ственно лицам с гиперметропией, возникают предпосылки к эзо-
тропии, т.е. чрезмерному схождению зрительных осей (среди про-
чих механизмов конвергенции у таких людей велика роль аккомо-
дативной конвергенции). В этих случаях снятие нагрузки на
аккомодацию вблизь с помощью положительных линз приводит к
ослаблению импульсов на конвергенцию и устранению эзофории.
Последняя при зрении вдаль исчезает и без очков.
382
Таким образом, причиной сходящегося косоглазия может по-
служить избыточное напряжение аккомодации, обусловленное ак-
комодативной эзофорией высокой гиперметропией и высоким от-
ношением АК/А, эзотропией смешанного типа. Низкие значения
АК/А могут приводить при рассматривании предметов, находящихся
на близком расстоянии, к экзотропии и расхождению зрительных
осей.
5.4.3. Определение слабофункционирующей (парализованной)
мышцы и оценка ее силы
Оценка амплитуды движений. Наиболее простым способом оценки
функции наружных мышц глаза является измерение амплитуды дви-
жений глазного яблока. В направлении действия парализованной
мышцы движение глаз более или менее ограничено. Правда, для
уточнения диагноза необходимо исключить влияние контрактуры
антагониста. С этой целью после местной анестезии выполняют
пробу с насильственным (при пинцетном захвате) поворотом глаз-
ного яблока в требуемом направлении.
Роль степени вторичного отклонения. При параличе какой-либо
наружной мышцы глаза для суждения о силе направляемых к ней (и
к другим мышцам) импульсов прибегают к оценке взаиморасполо-
жения фиксирующего и косящего глаз.
Предположим, имеется парез правостороннего отводящего не-
рва (n. abducens). В этом случае при спокойном взгляде прямо, когда
нервная импульсация распространяется равномерно на все мышцы,
на точку фиксации устанавливается здоровый (левый) глаз, а кося-
щий (правый) из-за слабости наружной прямой мышцы (и перевеса
тонуса внутренней) может слегка отклониться кнутри. Это так на-
зываемое первичное отклонение, иногда улавливаемое с трудом.
Стоит, однако, предложить пациенту усилием воли фиксировать
взор больным (правым) глазом (здоровый глаз прикрывают), как
положение резко меняется. Для того чтобы установить больной
глаз прямо, потребуется очень сильный нервный импульс к ослаб-
ленной наружной прямой мышце этого глаза. При этом, согласно
правилу Геринга, неадекватно высокий импульс направится и к
синергисту в верзионном движении другого глаза, т.е. к внутренней
прямой мышце левого (здорового) глаза. В итоге вторичное откло-
нение здорового глаза при фиксации больным оказывается заметно
более выраженным, чем первичное.
Роль характера диплопии. Исследование диплопии по Гаабу в
отсутствие коордиметра играет важную роль в определении слабо-
функционирующей мышцы. Для этого требуются красное стекло и
свеча (фонарик, лампа). Исследование проводят при открытых
глазах. Перед одним из них помещают красное стекло. Источник
света располагается прямо по средней линии на уровне глаз.
383
Зел
Правый глаз фиксирующий Левый глаз фиксирующий
б в
Рис. 5.15. Принцип теста Гесса с красным и зеленым светофильтрами. Объяс-
нение в тексте.
При наличии диплопии, не всегда осознаваемой пациентом до
пробы, в ходе исследования он замечает удвоение источника света
(один красного, а другой белого цвета). Взаиморасположение двой-
ных изображений в пространстве изменяется в зависимости от на-
правления взора. Задача обследуемого состоит в том, чтобы пра-
вильно оценить происходящие перемены (расхождение изображе-
ний, их сближение, слияние, какое из них правее, левее, какое выше
или ниже, не появляется ли в определенном положении взора на-
клон) и сообщить врачу. Это касается каждой из 9 основных пози-
ций взора. Создавать нужные позиции проще не путем перемеще-
ния источника света, а посредством вращения головы пациента в
направлениях, противоположных тем, которые нужны для созда-
ния нагрузки на определенную мышцу.
Оценка силы мышц по данным коордиметрии (тесты Гесса, Лан-
кастера). Пациенту надевают очки с красным и зеленым фильтра-
384
ми, в руки дают зеленую световую указку. У врача световая указка
красного цвета. Свет от нее врач направляет на экран и просит
пациента своим зеленым “зайчиком” накрыть красное пятно
(рис.5.15). При нормальной функции всех наружных мышц обоих
глаз это без труда удается, хотя каждый глаз видит только одну
метку такого цвета, который совпадает с цветом соответстствую-
щего фильтра.
В том случае, если имеется парез, например, наружной прямой
мышцы правого глаза, то, согласно правилу Геринга, чрезмерный по
силе импульс направляется не только к указанной слабой мышце, но
и к верзионно работающей с ней внутренней прямой мышце другого
(левого) глаза. В результате этого пациент уже не сможет попасть
зеленой меткой на красную метку врача и обнаружится смещение
проекционной фиксации левого глаза в правую сторону. Если поме-
нять стекла местами и пациент будет пытаться фиксировать указку
врача своим здоровым (левым) глазом, то это ему опять не удастся.
При этом будет отмечено недостаточное смещение проекционной
фиксации больного глаза. Результаты обоих этапов исследования
наносят на бланк с коордиметрической сеткой. В представленном
выше примере на сетке для правого глаза обозначится дефицит фун-
кции паретичной мышцы в главном для ее действия направлении
(при взгляде вправо). На сетке для левого глаза, наоборот, проявится
избыток функции при действии в том же направлении.
При повторных исследованиях по изменениям размеров и фор-
мы коордиметрических фигур судят о динамике патологического
процесса.
5.4.4. Выявление сенсорной аномалии, в том числе аномальной
корреспонденции сетчатки
Четырехточечный цветовой тест типа Уорса (Worth). При оче-
видной простоте устройства тест обладает хорошими диагности-
ческими возможностями. Если перед одним глазом (например, пра-
вым) поместить красный фильтр, а перед другим (левым) — зеле-
ный и предложить пациенту смотреть на экран устройства с че-
тырьмя светящимися окнами, одно из которых красное, два зеленых
и одно белое, то в зависимости от восприятий обследуемого воз-
можны следующие диагнозы:
• при правильном указании цветности всех окрашенных окон в
отсутствие явного косоглазия — бинокулярное зрение с нор-
мальной фузией (белое окно кажется окрашенным в цвет стекла,
расположенного перед ведущим глазом);
• при правильном указании цветности всех окрашенных окон, но
при наличии явного косоглазия — асимметричное бинокулярное
зрение с аномальной корреспонденцией сетчаток (белое окно при-
обретает цвет стекла, расположенного перед некосящим глазом);
385
• если видны только два окна и оба кажутся красными,— моноку-
лярное зрение с супрессией изображения левого глаза;
• если видны только три окна и. все кажутся зелеными,— моноку-
лярное зрение с супрессией изображения правого глаза;
• если видны то три зеленых, то два красных окна,— монокуляр-
ное зрение с альтернирующей супрессией;
• при одновременном видении не четырех, а пяти окон (три зеленых
и два красных) — одновременное зрение с диплопией.
Имеется вариант устройства, в котором тест Уорса (Worth)
реализуется путем поляризации света. Четыре светящихся объекта
рассматривают через лорнет со стеклами, поляризованными в двух
взаимно перпендикулярных направлениях. Одним глазом при этом
видны верхний поляризованный и нижний неполяризованный объек-
ты, а вторым — оба средние, горизонтально спаренные поляризо-
ванные в противоположном первому направлении.
При «наличии бинокулярного зрения обследуемый видит все
четыре светящихся объекта, при монокулярном зрении число их
уменьшается до двух или трех (по ведущему глазу), а при одновре-
менном зрении увеличивается до пяти, так как неполяризованный
объект удваивается.
Тестирование последовательных образов по Чермаку — Бильшов-
скому (Tschermak— Bielschowsky). Перед одним глазом включают
вертикальную, а перед другим — горизонтальную вспышку. Если
последовательный образ выступает в виде креста — корреспонден-
ция сетчаток нормальная, при расхождении перекреста — аномаль-
ная.
Тест Баголини (Bagolini). Данный тест реализуется еще проще,
чём предыдущий. Две линзы Баголини, которые представляют со-
бой стекло, покрытое тончайшими параллельными полосками, раз-
мещают в пробной оправе таким образом, чтобы полоски на стекле
находились под углом 45 и 135°. Лучи от точечного источника све-
та, проходящие через указанные линзы, преобразуются в две пере-
крещивающиеся линии.
Тестирование характера зрения на приборе Маревского — Сомо-
ва. В этом приборе предусмотрено полное разделение полей зрения
правого и левого глаза. В торцовой части тубуса имеется табло с
цифровыми индикаторами, дискретно включаемыми и выключае-
мыми, возможны изменения комбинации предъявляемых цифр (от
О до 9). Индикаторы могут включаться как раздельно, так и син-
хронно. Методика выгодно отличается от других возможностью
контроля правильности ответов обследуемого.
Исследования на синоптофоре. Прибор позволяет измерить угол
косоглазия, определить характер зрения двумя глазами, установить
наличие одновременного зрения, выявить аномальную корреспон-
денцию сетчаток, супрессию и амблиопию, оценить состояние фу-
зионной способности.
386
Если предъявлять разным глазам две сходные картинки, в каж-
дой из которых нет какой-либо детали, при наличии фузии удается
слить их воедино. Оценивая фузию, важно, в частности, опреде-
лить, в каком диапазоне вергенции удается сохранять ее действие.
В норме фузионная вергенция, измеренная на синоптофоре или
просто с помощью призм, достигает при конвергенции 15 призма-
тических диоптрий для дали и 25 призматических диоптрий для
близи. При дивергенции показатели ниже. Тренировки фузионной
конвергенции могут быть полезными при экзофории, а фузионной
дивергенции — при эзофории. При ослаблении этого механизма, в
частности при утомлении, существует угроза перехода скрытого
косоглазия (фории) в явное (тропия).
Для количественной оценки прочности фузии Э.С.Аветисов и
Т.П.Кащенко (1971) рекомендовали использовать анаглифный фу-
зиометр. Прибор основан на создании искусственной дозирован-
ной анизоэйконии.* Методика применения с той же целью поляри-
зационного фузиоэйдометра усовершенствована И.И.Дитрихом
(1989).
Т.П.Кащенко (1978) критически оценивала результаты гапло-
скопических исследований на синоптофоре, полагая, что считать
нормальной корреспонденцию сетчаток, если этот результат полу-
чен у больного с косоглазием при бифовеальном слиянии и вместе
с тем в отсутствие бинокулярного зрения, неправомерно. “Прибор-
ным артефактом”, по ее мнению, является выделение понятия “не-
гармонического” типа аномальной корреспонденции сетчаток, так
как сравниваемые при этом показатели объективных и субъектив-
ных углов косоглазия фактически получены в разных условиях.
5.4.5. Оценка глубинного зрения и стереопсиса
Высшим проявлением слитного бинокулярного восприятия являет-
ся ощущение объемности предметов — стереопсис.
Для количественной оценки реального глубинного зрения клас-
сическим остается прибор Говарда — Долмана. В этом приборе име-
ются три вертикальные стойки, одна из которых подвижна. Обсле-
дуемый должен уловить момент, когда смещаемая стойка сравня-
ется по удаленности с остальными двумя (неподвижными). За порог
принимают то максимальное различие, еще сохраняющееся по глу-
бине, которое обследуемый уже не способен ощутить. При проф-
отборе предлагали использовать ряд приборов для исследования
глубинного зрения, например, у операторов-микроскопистов, лет-
ного состава [Колосова С.А., Колосов В.В., 1977; Сомов Е.Е., Ба-
ранов В.А., 1982]. Нормативы по остроте глубинного зрения у
людей зрелого возраста для близи составляют всего 0,075 мм, а
для дали — 8,5 мм (соответственно 10,8" и 4,4").
Для исследования стереопсиса, кроме синоптофора, в клиничес-
387
кой практике применяют немало других гаплоскопических уст-
ройств, находивших применение [Sachsenweger R., 1963]. К таким
приборам относится Titmus stereotest, с помощью которого можно
измерить пороги от 3000 до 14" при средней норме применительно
к данному прибору 24" [Romano Р.Е., Romano J.A., 1973]. При
микрострабизме порог повышается до 70". В некоторых приборах
используют поляроидные вектографические карты или проекцион-
ные слайды [Parks М.М., 1971]. Одним из многих вариантов таких
стереотестов (фирма Titmus) является тест “летающая муха”. Тест
выпускают в виде буклета с поляроидными вектограммами, созда-
ющими трехмерные картинки. Их изготавливают сложным путем
фотографирования одной и той же области с двух позиций. Фото-
графии представляют в разных плоскостях поляризации, поэтому
они видны только через соответствующие фильтры, размещаемые
один перед правым, другой перед левым глазом. При слиянии изоб-
ражений возникает стереоэффект. Стереоскопические картинки вид-
ны обследуемому без каких-либо условностей, нарушающих про-
странственные отношения в нормальном зрительном поле (линзы,
призмы, перегородки не требуются).
Для проверки стереоостроты зрения у детей в возрасте 3—5 лет
пригодным оказался неупорядоченно-точечный Е-стереотест
[Simons К.,1981]. Применение этого прибора позволило установить,
что частота косоглазия у детей с учетом микрострабизма может
достигать 3—5%.
Для скрининговых исследований стереопсиса можно использо-
вать обычный проектор испытательных знаков, снабженный специ-
альным тестом и измерительной линейкой (фирма “Carl Zeiss”). Тест
состоит из двух вертикальных штрихов и округлого светящегося
пятна над ними. Если смотреть на этот тест через лорнет с поляри-
зованными стеклами (при разной направленности поляризации), то
при наличии стереопсиса пятно и штрихи воспринимаются как рас-
положенные на разной глубине (каждый из штрихов виден моноку-
лярно, пятно — обоими глазами)
5.5. О ТЕНДЕНЦИЯХ В РАЗВИТИИ ПРЕДСТАВЛЕНИЙ
О МЕХАНИЗМАХ БИНОКУЛЯРНОГО ЗРЕНИЯ И ВЫБОРЕ
АДЕКВАТНЫХ МЕТОДОВ ЕГО ИССЛЕДОВАНИЯ
Выделяют две основные тенденции в совершенствовании диагнос-
тики бинокулярных функций [Аветисов Э. С., Кащенко Т. П., 1993]:
приоритет методов, которые, в отличие от гаплоскопических, по-
зволяют проводить исследование в условиях, близких к естествен-
ным; совершенствование методов количественной оценки резуль-
татов.
Основной смысл нового направления, названного авторами
388
диплоптикой, состоит в стремлении разными способами в естествен-
ной обстановке спровоцировать у пациента диплопию, а затем с
помощью лечебных тренировок ее преодолевать. К средствам, спо-
собным вызвать диплопию, относят призмы, положительные и от-
рицательные, сферические линзы, приставляемые поочередно, све-
тофильтры возрастающей плотности и меняющейся окраски
[Тарасцова М.М., 1985; Белькова А.Г., 1991]. Использование нейт-
ральных и цветных светофильтров позволило выработать приемы
количественной оценки устойчивости бинокулярного зрения: чем
слабее устойчивость, тем выраженнее бинокулярная амблиопия (“ам-
блиобинопия” по Аветисову).
Количественно оценивая феномен “бинокулярного соперниче-
ства”(с красным фильтром перед одним глазом и зеленым перед
другим) по частоте смены циклов преобладания то одного, то дру-
гого глаза, предлагается судить о степени утомления зрительного
анализатора, а у школьников — о тенденции к миопизации [Тете-
рина Т.П. и др., 1987]. Для оценки тренировки бинокулярного зре-
ния в естественных условиях используют пространственные зритель-
ные эффекты, основанные на стимуляции, физиологического
двоения (за пределами зоны Панума) [Могилев Л. Н., 1978; Соловь-
ева В.В., 1984]. Метод, названный бинариметрией, включает не толь-
ко оценку, но и тренировку компонентов бинокулярного зрения в
реальных условиях, когда расстояние межде парами тестов изменя-
ется как во фронтальной, так и сагиттальной плоскостях.
Многокомпонентность моторных и сенсорных факторов обес-
печивает нормальное функционирование такой сложной системы,
какой является бинокулярное зрение. Исходя из этого, чтобы упро-
стить практическому врачу подход к решению диагностических
задач в данной области, И.И.Дитрих (1991), апробировавший все
существующие методы, рекомендовал в качестве самого первого
(скринингового) этапа производить оценку характера зрения двумя
глазами, используя, в частности, разделитель полей зрения по Баго-
лини. Если в результате проверки выявляется монокулярное или
одновременное зрение, то по профессиональным показаниям неза-
висимо от результатов исследования на первом этапе — приступа-
ют ко второму (уточняющему) этапу. Бинокулометрия, по И.И. Дит-
риху, должна как минимум включать: повторную проверку харак-
тера зрения по Уорсу (Worth) (на четырехоконном цветотесте или
поляризаторе света), оценку мышечного баланса (в призменных ди-
оптриях), проверку устойчивости бинокулярного зрения (в процен-
тах), измерение порога глубинного зрения (в угловых секундах или
линейных мерах).
Совершенно очевидно, что разные тесты позволяют оценивать
различные стороны бинокулярного зрения. Даже тесты одной на-
правленности очень различаются по эффективности. Так, Bagolini
(1980) установил, что если с помощью предложенного им теста
389
Левый глаз
Правый глаз
Рис. 5.16. Гипотетическая схема бинокулярного зрения человека [Рожко-
ва Г. И., Плосконос Г. А., 1988].
MCL (левая), МСГ (правая) монокулярные подсистемы, ЧБС — чисто бинокулярная
подсистема, МБС — монобинокулярная подсистема, ПМС — постмонокулярная под-
система.
признаки бинокулярного зрения выявляли у 90% обследованных,
то при использовании теста Уорса (Worth) в той же группе пациен-
тов их удавалось обнаружить только у 40%, а на синоптофоре —
лишь у 15%. При оценке одной и той же функции, а именно воспри-
ятия глубины на основе диспаратности, одна часть пациентов ус-
пешнее решает задачи по фузионному слиянию контурных стерео-
грамм, а другая — случайно-точечных [Frisby J., 1975]. Нуждается в
объяснении более успешное решение задач по мере удлинения вре-
мени показа тестов; обнаружение неравномерности распределения
аномальной стереоостроты по полю зрения [Richards W., 1970]. Для
390
объяснения этих и других фактов, не укладывающихся в существу-
ющие представления, предпринимаются попытки создания теории
множественности механизмов бинокулярного синтеза. Одно из пред-
ложений (рис.5.16) принадлежит Г.И.Рожковой и Г.А.Плосконос
(1988). Авторы полагают, что в зрительной системе человека име-
ется пять подсистем, из которых две монокулярные и три биноку-
лярные. Однако из числа бинокулярных только одна подсистема
чисто бинокулярная (ЧБС): она функционирует независимо от мо-
нокулярных подсистем; еще одна подсистема монобинокулярная
(МБС) и, наконец, предполагается существование постмонокуляр-
ной (ПМС) подсистемы. Именно последняя формирует трехмерные
образы на основе синтеза тех мелких деталей и признаков, которые
выделяются обеими монокулярными подсистемами зрительного
анализа (MCL и МСг).
ЧБС очень чувствительна к относительной диспаратности, если
сигналы поступают из определенной области пространства и обяза-
тельно через оба глаза. Эта подсистема обеспечивает максимально
высокую остроту бинокулярного стереоскопического зрения.
МБС срабатывает при поступлении сигналов как через оба гла-
за, так и через один. Она уменьшает площади слепых зон в поле
зрения.
ПМС в отличие от ЧБС оперирует обобщенными абстрактны-
ми характеристиками объектов, которые не поддаются в деталях
слиянию, поскольку в их изображениях, передаваемых от правого и
левого глаз, могут быть различия по цвету и по знаку контраста.
Таким образом, ПМС обеспечивает бинокулярный синтез “конф-
ликтующих изображений” как бы с высоты третьего измерения,
формируя стереопрозрачность, ощущение бликов [Рожкова Г.И.,
1992]. Очень важной и нерешенной задачей остается согласование
теоретических представлений о бинокулярном зрении с информа-
цией, которую на практике позволяют получить многочисленные
методики исследования разных сторон проявления этой сложней-
шей функции.
СПИСОК ОСНОВНОЙ ЛИТЕРАТУРЫ
Аветисов Э.С. Содружественное косоглазие.— М.:Медицина, 1977.— 312 с.
Аветисов Э.С, Диплоптика — принципиально новая система лечения ко-
соглазия // Вести, офтальмол.— 1977.— № 6.— С. 15.
Аветисов Э.С. Близорукость.— М.: Медицина, 1986.—239 с.
Аветисов Э.С., Кащенко Т.П. Бинокулярное зрение: клинические методы
исследования и восстановления // Клиническая физиология зрения:
Сборник трудов МНИИГБ им.Гельмгольца.— М.: Руссомед, 1993.—
С. 199 —209.
Ананин В.Ф. Аккомодация и близорукость.— М.: Изд. Рос. университета
дружбы народов, 1992.— 135 с.
Арефьева Ю.А., Еричев В.П., Шамшинова А.М., Лелекова А.П. Функцио-
нальная топография on- off-каналов колбочковой системы сетчатки в
диагностике начальной глаукомы // Вести. Офтальмол.— 1997.— Т. 113,
№ 1.—С. 28—30.
Астахов Ю.С. Глазоорбитальный пульс и клиническое значение его иссле-
дования: Автореф. дисс. ... д-ра мед. наук. — Л., 1990.
Баголини Б. Некоторые аспекты анормального бинокулярного зрения при
эзотропиях с малым углом // Нарушения бинокулярного зрения и ме-
тоды его восстановления.— М.: Медицина, 1980.— С. 20—29.
Балашевич Л.И., Шостак В.И. Динамика восстановления остроты зрения
после воздействия на глаз кратковременных световых раздражителей
// Вестн.офтальмол.— 1970.— № 6.— С. 63 — 65.
Белькова А.Г. Использование светофильтров в исследовании и восстанов-
лении бинокулярных функций: Автореф. дисс. ... канд. мед. наук.—
М., 1991.—22 с.
Бетелева Т.Г. Структурно-функциональная организация развивающегося
мозга.— Л.: Наука, 1990.— 65 с.
Богословский А.И. Основные закономерности зрительных ощущений //
Многотомное руководство по глазным болезням.— М.: Медгиз, 1962.—
Т. 1, кн. 1.—С. 400—411.
Богословский А.И. Различительная световая чувствительность // Многотом-
ное руководство по глазным болезням.— М.: Медгиз, 1962.— Т. 1,
кн. 1.—С. 421—425.
Богословский А.И. Клиническая электрофизиология зрительной системы в
практике офтальмологии//Вести, офтальмол.—1982. — № 6.— С. 56—63.
Борисова С.А., Никитин Ю.М., Еричев В.П. Ультразвуковое допплерогра-
фическое исследование кровотока в орбитальных сосудах у больных
первичной глаукомой // Ультразвуковая диагностика.— 1997.— Вып. 2,
№2.—С. 8.
392
Борисова С,А. Ультразвуковая допплерография в офтальмологии//Вести,
офтальмол.— 1997.—Т. 113, № 6.—С. 43—45.
Бржеский В.В, Слезная жидкость в диагностике некоторых повреждений и
заболеваний глаз: Автореф. дисс. ... канд. мед. наук. — Л., 1990.—
230 с.
Бунин А.Я. Гемодинамика глаза и методы ее исследования.— М.: Медици-
на, 1971.
Бызов А.Л. Нейрофизиология сетчатки // Физиология зрения / Под ред.
А.Л.Бызова. — М.: Наука, 1992.— С.115 — 162.
Водовозов А.М. Толерантное и интолерантное внутриглазное давление.—
Волгоград, 1993.— 240 с.
Волков В.В., Султанов М.Ю. Наружная дакрио-цисториностомия.— Л.:
Медицина, 1975.— 104 с.
Волков В.В, Существенный элемент глаукоматозного процесса, неучитыва-
емый в клинической практике // Офтальмол. журн.— 1976.— № 7.—
С. 500—505.
Волков В.В., Горбань А.Н., Джалиашвили О.А. Клиническая визо- и рефрак-
тометрия.— Л.: Медицина, 1976.— С. 5—65, 161—202.
Волков В.В., Шиляев В.Г. Комбинированные поражения глаз.— Л., 1976.—
С. 54 — 62.
Волков В.В., Парпаров А.Б. Аномалии рефракции и аккомодации как при-
чина ограничения зрительной работоспособности и эргономические
основы очковой коррекции зрения. Офтальмоэргономика.— М.:
МЗРСФСР, МНИИ ГБ им. Гельмгольца, 1976.— С. 20—42.
Волков В.В. Прицельная периметрия — важнейший метод диагностики
глаукомы // Офтальмол. журн.— 1979.— № 6.— С. 331—334.
Волков В.В., Сухинина Л.Б., Тер-Андриасов Э.Л. О применении вакуума в
компрессионно-периметрической пробе при диагностике глаукомы //
Вести, офтальмол.— 1981.— № 2.— С. 22—25.
Волков В.В., Колесникова Л.Н., Шелепин Ю.Е. Частотно-контрастная ха-
рактеристика и острота зрения в офтальмологической практике И Оф-
тальмол. журн.— 1983.— № 3.— С. 148—151.
Волков В. В., Колесникова Л.Н., Шелепин Ю.Е. Методика клинической визо-
контрастометрии // Вести, офтальмол.— 1983.— № 3.— С. 59—61.
Волков В.В., Сергеев В.П., Бугрова А.Ф. Простая методика множественной
центральной статической периметрии и первые результаты ее приме-
нения // Вести, офтальмол.— 1983.— № 4.— С. 48—50.
Волков В.В., Сухинина Л.Б., Устинова Е.И. Глаукома, преглаукома, оф-
тальмогипертензия.— Л.: Медицина, 1985.— С. 59—203.
Волков В.В., Шиляев В.Г. Методы исследования и дифференциальная диаг-
ностика при экзофтальмах.— Л.: Воен.-мед. акад., 1985.— С. 16—17,
58—60.
Волков В.В. Острота зрения и рефракция как критерий трудовой пригодно-
сти // Офтальмоэргономика операторской деятельности: Материалы
симпозиума.— Л., 1986.— С. 39—40.
Волков В.В., Шелепин Ю.Е., Колесникова Л.И. Атлас и пособие по визокон-
трастопериметрии.— Л.: ЦВМУ, 1987.
393
Волков В.В., КолесниковаЛ.И., Макулов В.Б. и др. Новые буквенные тесты
для измерения остроты зрения И Офтальмол. журн.— 1987.— № 5.—
С. 294—296.
Волков В.В., Сергеев В.П. О взаимоотношении цветослабости и цветоанома-
лии по данным исследований пороговыми таблицами Юстовой и ано-
малоскопом Раутиана // Офтальмол. журн.— 1987.— № 1.— С. 24—27.
Волков В.В. Визометрия и визоконтрастометрия: состояние, перспективы //
Вести, офтальмол.— 1988.— Т. 104, № 4.— С. 3—7.
Волков В.В., Луизов А.В., Овчинников Б.В. и др. Эргономика зрительной
деятельности человека.— Л.: Машиностроение, 1989.— С. 21—22,
31—34.
Волков В.В. Мышечно-адаптивный механизм миопизации глаза у школь-
ников // Близорукость.—М., 1990.— С. 20—24.
Волков В.В., Луизов А.В., Федорова Н.С. и др. Новый прибор для выявления
аномалий цветового зрения// Оптико-электрон. промышленность.—
1990.— №12.—С. 44—47.
Волков В.В., Бржеский В.В, Гладких А.Ф. Диагностика и лечение синдрома
“сухих” глаз ожоговой этиологии И Офтальмол. журн.—1990.— № 6.—
С. 328—330.
Волков В.В., Лебехов П.П. Тесты и упражнения для проверки и тренировки
зрения.— Л.: Гиппократ, 1993.— 6 с.
Волков В.В. Психофизиология зрительных процессов и методы его изуче-
ния: Сб. научных трудов.— М.: Руссомед, 1993.— С. 158—179.
Волков В.В., Шамшинова А.М., Розенблюм Ю.З. и др. Физиологические
основы цветового зрения и его нарушения: Сб. научных трудов.— М.:
Руссомед, 1993.— С. 224—260.
Волков В.В., Юстова Е.Н. О диагностике и классификации вариантов цве-
тового зрения в свете новых методических подходов//Вестник оф-
тал.—1995.— № 2.— С. 19—24.
Волкова М.В. Нормальные показатели статической периметрии в зоне
Бьеррума И Вести, офтальмол.— 1980.— № 1.— С. 55—58.
Гиппенрейтер Ю.Б. Движения человеческого глаза.— М.: Изд-во Моск, ун-
та, 1978.—256 с.
Глезер В.Д. Зрение и мышление.— Л.: Наука, 1985.—246 с.
Глезер В.Д. Модели нейронов зрительной коры // Физиология зрения / Под
ред. АЛ.Бызова.— М.: Наука, 1985.— С. 315—345.
Говардовский В.И., Шамшинова А.М., Голубцов КВ. К методике регистра-
ции локальной электроретинограммы // Современные аспекты клини-
ческой физиологии зрения: Респ. сборник научных работ.— М., 1985.—
С. 33—41.
Голубев С.Ю. Визоконтрастометрические и электрофизиологические пока-
затели при острых отравлениях //Дисс. ... канд. мед. наук.— СПб.,
1995.—149 с.
Дитрих И.И. Сравнительная оценка различных компонентов бинокуляр-
ного зрения и выбор наиболее информативных способов его исследо-
вания: Автореф. дисс. ... канд. мед. наук.— Л., 1991.—20 с.
394
Ендриховский С.Н., Шамшинова А.М., Соколов Е.Н. и др. Время сенсомо-
торной реакции человека в современных психофизических исследова-
ниях // Сенсорные системы.— М., 1996.— Т. 10, № 2.— С. 13—29.
Зенков Л.Р., Ронкин М.А. Функциональная диагностика нервных болез-
ней.— М.: Медицина, 1991.— 2-е изд.—640 с.
Зислина Н.Н. Нейрофизиологические механизмы нарушения зрительного
восприятия у детей и подростков.— М.: Педагогика, 1987.— 168 с.
Зуева М.В., Цапенко И. В. Электрофизиологическая характеристика гли-
альнонейрональных взаимоотношений при ретинальной патологии //
Сенсорные системы.— 1992.— № 3.— С. 58—63.
Иваницкий А.М. Мозговые механизмы оценки сигналов.— М.: Медицина,
1976.—263 с.
Карлова ИЗ., Шамшинова А.М., Белозеров А.Е., Лоскутов И.А. Простран-
ственная контрастная чувствительность в дифференциальной диагно-
стике оптических невритов // Вести, офтальмологии.— 1996.— Т. 112,
№ 3.—С. 21—24.
Катаргина Л.А., Шамшинова А.М., Орловская Л. С. Функциональное состо-
яние сетчатки у детей с увеитами и постувеальными осложнениями
// Патологии глазного дна и зрительного нерва: Респ. сборник науч-
ных трудов.— М.: МНИИГБ им.Гельмгольца, 1991.— С. 58—68.
Кацнельсон Л.А. Реоофтальмография. Клинико-экспериментальные иссле-
дования: Дисс. ... д-ра мед. наук.— М., 1968.— 426 с.
Кащенко Т.П. Бинокулярная зрительная система при содружественном ко-
соглазии: Автореф. дисс. ... д-ра мед. наук.— М., 1978.— 31 с.
Кириллов Ю.А. Циклодукция и зрачок в исследовании астигматизма глаза:
Автореф. дисс. ... канд. мед. наук. — Л., 1976.— 23 с.
Колосова С.А. Бинокулярное глубинное зрение человека в норме и при
воздействии некоторых экстремальных факторов: Автореф. дисс. ...
канд. мед. наук. — М., 1981.— 18 с.
Коптелов Ю.М. Исследование и численное решение некоторых обратных
задач электроэнцефалографии: Автореф. дисс.... канд. физ.-мат. наук.—
М., 1988.— 14 с.
Коптелов Ю.М., Гнездицкий В.В. Анализ “скальповых потенциальных по-
лей” и трехмерная локализация источников эпилептической активно-
сти мозга человека” // Журн. нервопатол. и психиатр.— 1989.— Т.89,
№ 6.— С. 16—20.
Коскин С.А. Диагностические возможности и пути усовершенствования
визоконтрастометрии: Дисс.... канд. мед. наук.— СПб., 1994.— 242 с.
Котляр К.Е., Кошиц И.Н., Смольников Б.А. Биомеханическая модель регу-
ляции внутриглазного давления // Инновационные наукоемкие техно-
логии для России.— СПб.: Изд-во СПб. гос. техн, ун-та, 1995.— Ч. 5.—
80 с.
Котляр К.Е., Светлова О.В., Смольников Б.А. О возможном механизме
функциональной блокады венозного синуса в случае анатомической
предрасположенности к открытоугольной глаукоме И Глаукома.— М.:
МНИИГБ им.Гельмгольца, 1996.— С. 7—16.
395
Кравков С.В. Глаз и его работа.— М.: Медицина, 1945.— С.85—168.
Кравков С.В. Цветовое зрение.— М.: Академия наук СССР, 1951.— 175 с.
Луизов А.В. Цвет и свет.— Л.: Энергоатомиздат, 1989.— 256 с.
Луизов А.В. Глаз и свет.— Л.: Энергия, 1983.— 140 с.
Лысков Е.Б. Динамика вызванной биоэлектрической активности мозга при
поражениях зрительных путей: Автореф. дисс. ... канд. мед. наук. —
Л., 1985.— 21 с.
Максимов В.В. Трансформация цвета при изменении освещения.— М.:
Наука, 1984.— 160 с.
Меркулов И. И. Введение в клиническую офтальмологию.— Харьков: Харь-
ковский гос. ун-т, 1964.— С. 14—93.
Миронова Э.М., Егорова Э.В., Куман Н.Г. и др. Электрофизиологическая
диагностика и патогенез макулярного отека //Офтальмол. журн.—
1987.—№ 1.—С. 21—24.
Могилев Л. Н., Рабичев И. Э. и др. Восстановление бинокулярного зрения в
естественных условиях // Охрана зрения детей и подростков.— М.,
1984.—С. 7—18.
Мяло Н.М. Кобальторефрактометрия и визоконтрастометрия в интересах
профотбора и в клинической практике: Автореф. дисс. ... канд. мед.
наук. — Л., 1985.— 20 с.
Нейроофтальмология / Под.ред. С. Лессела, Дж.Т.У. Ван Далена.— М.:
Медицина, 1983.— 464 с.
Немцеев Г.И. Современные способы исследования поля зрения И Вести,
офтальмол.— 1977.— № 1.— С.82—85.
Немцеев Г. И., Сабаева Г.Ф. Клиническое применение ППУ: Методические
рекомендации.— М., 1988.— 18 с.
Немцеев Г.И., Шамшинова А.М., Сабаева Г.Ф. Дифференциальная диагно-
стика ранних стадий макулодистрофий и патологий зрительного не-
рва с помощью современных электроретинографических и периметри-
ческих тестов // Вести, офтальмол.— 1988.— № 5.— С. 50—52.
Немцеев Г. И. Актуальные вопросы современной клинической периметрии //
Клиническая физиология зрения: Сборник научных трудов МНИИГБ
им. Гельмгольца.— М.: Руссомед, 1993.— С. 277— 295.
Немцеев Г.И. Автоматическая компьютерная периметрия и хроноперимет-
рия в диагностике заболеваний зрительного пути: Метод, рекоменд.—
М„ 1996.— 18 с.
Нестеров А.П. Гидродинамика глаза.— М.: Медицина, 1967.— 142 с.
Нестеров А.П., Бунин А.Я., Кацнельсон Л.А. Внутриглазное давление. Фи-
зиология и патология.— М.: Наука, 1974.— 381. с.
Нестеров А.П. Первичная глаукома.— М.: Медицина, 1982.— 287 с.
Никитин Ю.М. Ультразвуковая допплерография в диагностике поражений
магистральных артерий головы и основания мозга. Институт Невро-
логии РАМН.— АО Спектромед.— 1995.— 45 с.
Нюер М.Р. Количественный анализ и топографическое картирование ЭЭГ:
методики, проблемы, клиническое применение // Успехи физиол.
наук.— 1992.— Т.23, № 1.— С.20—39.
396
Островский М.А., Говардовский В.И. Механизмы фоторецепции позвоноч-
ных // Физиология зрения / Под ред. А.Л. Бызова.— М.: Наука, 1992.—
С. 5—59.
Павлов Н,Н„ Волков В.В., Шелепин Ю.Е. и др. Принципы и перспективы
построения систем измерения возможностей бинокулярного и стерео-
скопического зрения человека. Автоматизированные системы реаль-
ного времени для эргономических исследований.— Тарту, 1988.—
С. 118—120.
Певзнер Л.3. Функции нейроглии.— Тбилиси: Мецниерба, 1979.— С. 165—251.
Пивоваров Н.П. Аутоофтальмоскопия и ее диагностические возможности //
Веста, офтальмол.— 1974.— № 5*— С. 64—64.
Пивоваров Н.Н. Энтоптические феномены в исследовании функции сетчат-
ки и зрительного нерва // Методические рекомендации М3 СССР (Все-
союзный НИИ глазных болезней).— М., 1974.— 14 с.
Пивоваров Н.Н. Диагностическое значение зрительных сенсорных феноме-
нов в патологии оптического и нервного аппарата глаз: Дисс. ... fl-
pa мед. наук.— М., 1982.— 352 с.
Пигарев И.Н. Экстрастриарные зрительные зоны коры мозга // Физиоло-
гия зрения / Под ред. А.Л.Бызова.— М.: Наука, 1992.— С. 345—401.
Преображенский П.В., Шостак В.И, Балашевич Л.И. Световые поврежде-
ния глаз.— Л.: Медицина, 1986.— 198 с.
Раутиан Г.Н. Новый аномалоскоп// Биофизика.—1957.— Т. 2., вып.16.—
С. 10.
Рожкова Г.И. Бинокулярное зрение // Физиология зрения.— М.: Наука,
1992.— С. 586—664.
Розенблюм Ю.З. Основные тенденции в исследовании остроты зрения //
Новые методы функциональной диагностики в офтальмологии.— М.:
МНИИ ГБ им. Гельмгольца, 1973.— С. 161—184.
Розенблюм Ю.З. Рефракция, аккомодация, зрение // Клиническая физиоло-
гия зрения: Сборник научных трудов МНИИГБ им. Гельмгольца.—
М.: Руссомед, 1993.— С. 180—198.
Розенблюм Ю.З. Оптометрия.— СПб.: Гиппократ, 1996.— 247 с.
Рубцова И. В. Ранняя диагностика и лечение нарушений прозрачности хру-
сталика: Дисс. ... канд. мед. наук.— СПб., 1994.— 223 с.
Сапин Б.А. Анатомия человека.— М.: Медицина, 1993.— Т. 2.— С. 68—
498.
Симакова И.Л. Видеограмма и диск зрительного нерва при разных стадиях
глаукомы и в оценке эффективности ее оперативного лечения // Дисс.
... канд.мед. наук.— СПб., 1997.— 198 с.
Смольянинова И.Л. Врожденный нистагм и возможное улучшение зритель-
ных функций при нем // Клиническая физиология зрения: Сборник
научных трудов МНИИГБ им. Гельмгольца.— М.: Руссомед, 1993.—
С. 210—223.
Соколов Е.Н., Измайлов ИА. Цветовое зрение.— М.: Московский универ-
ситет, 1984.— 175 с.
Соловьева В.В. Метод бинариметрии в диплоптическом лечении содруже-
397
ственного косоглазия: Автореф. дисс.... канд. мед. наук. — М., 1988.—
23 с.
Сомов Е.Е. Методы офтальмоэргономики.— Л.: Наука, 1989.— 157 с.
Сомов Е.Е. Офтальмоэргономика операторской деятельности летного со-
става.— СПб.: Политехника, 1992.— 176 с.
Сомов Е.Е. Введение в клиническую офтальмологию.— СПб.: Петербург,
пед. ин-т, 1993.— С. 133—176.
Сомов Е.Е., Бржеский В.В. Слеза. Физиология, методы исследования, кли-
ника.— СПб.: Наука, 1994.— С. 13—21.
Сосновский В.В. Методика энтоптических исследований, возможные при-
чины нарушений и нормативные критерии результатов И Труды Воен-
но-мед. академии.— Л., 1989.— Т. 226.— С. 57—66.
Сосновский В.В. Энтоптические и электрофизиологические исследования в
оценке сохранности зрительных функций при тяжелых повреждениях глаз
и их последствиях: Автореф. дисс.... канд. мед. наук.— Л., 1990.— 26 с.
Султанов М.Ю., Алиева З.А. Физиология слезоотведения.— Баку: Азер-
нешр, 1983.— 60 с.
Султанов М.Ю., Алиева З.А. Щадящая хирургия слезоотводящих путей.—
Баку: Азер. гос. издат., 1987.— С. 11—14.
Тарасцова М.М. Диплоптическое лечение содружественного косоглазия у
детей раннего и дошкольного возраста: Автореф. дисс. ... д-ра. мед.
наук.— М., 1985.— 26 с.
Тетерина Т.П., Волков В.В., Кочеткова Л.П. Биоритмы бинокулярного
зрения // Физиология человека.— Л., 1987.— Т.13, № 5.— С. 779—782.
Товкач В.И. Фотостресс обычным электроофтальмоскопом в диагностике
заболеваний глаз И Воен.-мед. журн.— 1977.— № 8.— С. 40—44.
Толмачев Р.А., Эскин В.Я. Адапто-электроокулография у лиц с высокой
миопией // Веста, офтальмол.— 1977.— Вып. 2.— С. 53—56.
Фильчикова Л.И., Новикова Л.А., Гусева М.Р. и др. Зрительные вызванные
потенциалы на реверсию шахматных полей у детей с оптическим не-
вритом И Веста, офтальмол.— 1991.— Т. 107, № 3.— С. 65—69.
ХьюбелД. Глаз, мозг, зрение: Пер. с англ.— М., Мир, 1990.— 297 с.
Шамшинова А.М., Щербатова О.И. Новые достижения в электрофизиоло-
гии глаза и их роль в диагностике заболеваний сетчатой оболочки //
Актуальные вопросы патологии сетчатки.— М., 1978.— С. 64—89.
Шамшинова А.М. Локальная электроретинограмма: проблема регистрации
и роль в клинике глазных болезней. Обзор литературы // МРЖ.—
1985.— Разд. 8, № 5.— С. 11—18.
Шамшинова А.М. Электрофизиологические методы исследования в клини-
ке глазных болезней: Обзор литературы // МРЖ.— 1986.— Разд. 8,
№56.—С. 1—11.
Шамшинова А.М. Локальная ЭРГ и ее роль в клинике глазных болезней //
Реабилитация; Сборник научных трудов МНИИГБ им. Гельмголь-
ца.— М., 1988.— С. 137—148.
Шамшинова А.М. Локальная электроретинограмма в клинике глазных
болезней: Дисс. ... д-ра мед. наук. — М., 1989.— 333 с.
398
Шамшинова А.М. Локальная электроретинограмма в клинике глазных
болезней: Метод, рекомендации.— М., 1991.— 24 с.
Шамшинова А.М., Смольянинова И.Л., Мац К.А. и др. Роль биопотенциалов
сетчатки в изучении механизмов возникновения нистагма И Вести, оф-
тальмол.— 1991. —№ 6.— С. 46—51.
Шамшинова А.М., Говардовский В.И., Голубцов К.В. К методике регистра-
ции локальной ЭРГ//Вестн. офтальмол.— 1989.— № 6.— С. 47—49.
Шамшинова А.М. Электрофизиологические методы исследования в диаг-
ностике заболеваний органа зрения у детей и оценка их реабилита-
ции//Функциональная реабилитация в офтальмологии^ Сборник науч-
ных работ МНИИ ГБ им. Гельмгольца.— М., 1990.— С. 122—127.
Шамшинова А.М., Яковлев А.А. Оценка функций сетчатки при катаракте с
помощью электроретинографии И Вести, офтальмол.— 1991.— № 1.—
С. 36—42.
Шамшинова А.М., Нестерюк Л.И., Ендриховский С.Н. Возможности компь-
ютерной обработки результатов исследования зрительной системы //
Вести, офтальмол.— 1992. —№ 2.— С. 29—35.
Шамшинова А.М., Ендриховский С.Н., Еричев В.П. и др. Цветовая кампи-
метрия — новый метод исследования функций зрительного анализа-
тора И Глаукома: Сборник научных трудов МНИИ ГБ им. Гельмголь-
ца.— М., 1994.— С. 26—36.
Шамшинова А.М. Электроретинография в клинике глазных болезней //
Сборник работ научной конф., посвящ. 100-летию со дня рождения
С.В.Кравкова и 90-летию со дня рождения А.И.Богословского.— М.,
1993.— С. 57—82.
Шамшинова А.М., Нестерюк Л.И., Ендриховский С.Н. и др. Цветовая кам-
пиметрия в диагностике заболеваний зрительного нерва: Методичес-
кие рекомендации.— М., 1994.— 10 с.
Шамшинова А.М., Нестерюк Л.И., Ендриховский С.Н. и др. Цветовая кампи-
метрия в диагностике заболеваний сетчатки и зрительного нерва //
Вести, офтальмол.— 1995.— № 2.— С. 24—28.
Шамшинова А.М., Белозеров А.Е., Шапиро В.М. и др. Новый метод ис-
следования контрастной чувствительности в клинике глазных болезней//
Вести, офтальмол.— 1997.— Т. 113, № 1.— С. 22—25.
Шаповалов С.Л. Клинико-физиологические особенности абсолютной акко-
модации глаз человека и методы ее исследования: Дисс. ... д-ра мед.
наук.—М., 1977.— 279 с.
Шаповалов С.Л. Методы исследования динамической рефракции глаза в
норме и при патологии.— М., 1981.— 34 с.
Шаповалов С.Л., Шелепин Ю.Е., Милявская Т.Н. и др. Пособие по диагно-
стике, экспертизе, восстановлению нарушений зрительной работоспо-
собности у лиц летного и диспетчерского состава гражданской авиа-
ции.— М.: Воздушный транспорт, 1990.— 104 с.
Шахнович А.Р., Шахнович В.А. Диагностика нарушений мозгового крово-
обращения. Транскраниальная допплерография.— М., 1996.—446 с.
Шевелев И.А. Нейроны зрительной коры. Адаптивность и динамика рецеп-
тивных полей.— М.: Наука, 1984.— 232 с.
399
Шевелев И.А. Зрительная кора // Физиология зрения / Под ред. А.Л.Бызо-
ва.— М.: Наука, 1992;— С. 243—315.
Щекутьев Г.А., Коптелов Ю.М. Метод дипольной локализации в анализе
акустического стволового вызванного потенциала // Журн. высш,
нервн. деят.— 1992.— Вып. 1, т. 42.— С. 176—185.
Шелепин Ю.Е., Колесникова Л.Н., Левкович Ю.И. Визоконтрастометрия.—
Л.: Наука, 1985.— 102 с.
Шелепин Ю.Е., Волков В.В., Колесникова Л.Н. и др. Измерение функцио-
нальных возможностей зрительной системы человека // Вести. АН
СССР.— 1987.— № 9.— С. 63—72.
Шелепин Ю.Е., Глезер В.Д., Бондаренко В.М. и др. Пространственное зре-
ние // Физиология зрения / Под ред. А.Л.Бызова.— М.: Наука, 1992.—
С. 528—586.
Шелепин Ю.Е., Павлов Н.Н., Амельянова С.Г. и др. Поляризованный свет,
контрастная чувствительность и структурная организация зрительной
системы человека//Сенсорные системы.— 1989.— № 4.— С. 390—
395.
Шпак А. А. Исследования зрительных вызванных потенциалов в офтальмо-
логии и офтальмохирургии.— М., 1993.— 191 с.
Эскин В.Я. Электрографическая регистрация корнеоретинальных потенци-
алов // Сборник трудов КТМИ.— Фрунзе, 1956.— Т.8.— С. 365—370.
Эскин В.Я. Исследование электрического поля глаза в клинической оф-
тальмологии: Дисс. ... д-ра мед. наук. — М., 1984.— 462 с.
Эскин В.Я., Щербатова О.И., Толмачев Р.А. Клиническая электроокуло-
графия // Клиническая физиология зрения: Сборник трудов МНИИГБ
им.Гельмгольца — М.: Руссомед, 1993.— С. 130—145.
Юстова Е.Н., Алексеева КА., Волков В.В. и др. Пороговые таблицы для
исследования цветового зрения: Метод, рекомендации.— М., 1993.—
47 с.
Ярбус А.Л. Роль движений глаз в процессе зрения.— М.: Наука, 1965.—
166 с.
Abraham F.A., Alpem М., Kirk D.B. Electroretinograms evoked by sinusoidal
excitation of human cones // J. Physiol. Lon.— 1985.— Vol. 363.— P.
135—150.
Adashi-Usami E., Lehmann D. Monocular and binocular evoked average potential
field topography: upper and lower hemiretinal stimuli // Exp. Brain Res.—
1983.— Vol. 50.— P. 341—346.
Alm A., Bill F. Ocular circulation. In Adler’s Physiology of the Eye/Ed by Moses
and Hart.— Mosby Co., 1987.— P. 183—203.
Allingham R.R., de Kater A. W., Ethier C.R. Schlemm’s canal and primary open
angle glaucoma: correlation between Schlemm’s canal dimensions and
outflow facility // Exp. Eye Res.— 1996.— Vol. 62, № 1.— P. 101—1Ю.
Alpem V. The kinetics of cone visual pigments in man // Vis. Res.— 1971.—
Vol. 11.—P. 539.
400
Apkarian P., Spekreijse H. The use of the electroretinogram and visual evoked
potentials in ophthalmogenetics // Visual evoked potentials.— Amsterdam:
Elsevier, 1990.— P. 169—223.
Apkarian P. Methodology of testing for albinism with visual evoked cortical
potentials // Principles and practice of clinical electrophysiology of vision.—
London: Mosby Year Book, 1991.— P. 425—434.
Arden G.B., Barrada A., Kelsey J.H. New clinical test of retinal function based
upon the standing potential of the eye // Brit. J. Ophthal.— 1962.—
Vol. 46.— P. 449—467.
Arden G. B„ Gunduz K, Perry S. Color vision testing with a computer graphics
system//Clin. Vision Sci.— 1988.— № 2.— P. 303.
Arden G.B., Carter R.M., Hogg C. et al. A gold foil electrode extending the
horizons for clinical electroretinography // Invest. Ophthalmol. Vis. Sci.—
1979.— Vol. 18.— P. 421-426.
Arden G.B., Vaegan P.P. T. Electroretinigrams evoked in man by local uniform
of pattern stimulation // J. Physiol.— 1983.— Vol. 341.— P. 85—104.
Arden G.B. Recent development in clinical contrast sensivity testing // Advances
in diagnostic vizual optics.— Berlin: Springer — Verlag, 1983.
Armaly M.F. Selective perimetry for glaucomatous defects in ocular hypertension //
Arch. Ophthalmol.— 1970.— Vol. 9, № 6.— P. 425—429.
Armington J.C. The Electroretinogram.— New York: Academic Press, 1974.—
262 p.
Attwell D., Barbour B., Brew H. et al. Modulation of glutamate uptake in
salamander Muller cells by potassium and arachidonic acid // Proceeding
of the International Society for Eye Research.— Helsinki, 1990.— Vol.
6.— P. 243—249.
Aulhorn E. Uber die Beziehung Zwischen Lichtsinn und Sehsharfe//Gr. Arch.
Ophthahn. —1964.— Bd.167.— № 4.— P. 71—74.
Aulhorn E. Einfache perimetrische Methoden fur die Erfassung der glaucomatosen
Fruhaus falle // Glaucoma.— .Wurzburg, 1974.— S. 117—130.
Babel J., Stangos N., Korol S. et al. Ocular Electrophysiology.— Stuttgart.:
Georg Thieme Publishers, 1977.— 125 p.
Baker H.D. Night vision and visual sensitivity // In Whitcomb M.A. and Benson
W. (Ed.). The measurment of visual function.— Washington D.C.: Armed
Forces NRC Committee on vision, National Academy Science, National
research Conncil, 1968.— 240 p.
Barbehen E.K., Gagnon C., Noelker D.M. et al. Inherited rod-cone dysplasia:
abnormal distribution of cyclic GMP in visual cells of affected Irish setters //
Exp. Eye Res.— 1988.— Vol. 46.— P. 149—159.
Barch W.S., Boyton R.M. Responses of primate cones to sinusoidally flickering
homohromatic stimuli // J. Physiol.— 1975.— Vol. 246.— P. 311—331.
Barret G, Blumhardt L., Halliday E. et al. A paradox in the lateralisation of the
visual evoked responce.— London: Nature, 1976.— Vol. 261.— P. 253—255.
Bartl J., Van Lith G.H.M., Van Marie G. W. Cortical potentials evoked by a TV
pattern reversal stimulus with varying check sizes and stimulus field // Brit.
J. Ophthalmol.— 1978.— Vol. 62.— P. 216—219.
401
Baron W.S., Boynton R.M. Response of primat cones to sinusoid alle flickering
monochromatic stimuli // J. Physiol.— 1975.— Vol. 246.— P. 311—337.
Bass S., Sherman J., Bodis-Wollner I. et al. Visual evoked potentials in macular
diseases // Inv. Ophthalmol. Vis. Sci.— 1985.— Vol. 26, № 8.— P. 1071.
Bearse M.A., Sutter J., Sutter E.E. et al. Imaging retinal damage with the multi-input
electroretinogram // Vis. Sc. and its appl.— 1994.— Vol. 2.— P. 358—361.
Bebie H., Fankhauser E, Jenn A. et al. Perimeter Digest // Interzeag Ag., Schlieren,
Switzerland, 1983.— P. 49.
Berhman J., Nissim S, Arden G.B. A clinical method for obtaining pattern
visual evoked responses // Advances in experimental medicine and biology.—
New York: Plenum Press, 1972.— Vol. 24.— P.199—206.
Berninger T.A., Arden G.B. The pattern electroretinogram // Eye.— 1988.—
Vol. 2.— P. 257—283.
Berson E.L. Hereditary retinal diseases. Classification with the full-field
electroretinogram // Doc. Ophthalmol. Proc. Ser.— 1977.— Vol. 13.—
P. 149.
Berson E.L. Retinitis pigmentosa and allied diseases: applications of
electroretinographic testing // Int. Ophthalmol.— 1981.— Vol. 4.—
P. 7—22.
Berson E.L. Retinitis Pigmentosa:Some Observations on the Course //Principles
and Practice of Clinical Electrophyssiology of Vision.— Mosby Year Book:
St. Louis.— 1991.— P. 501—509.
Biersdorf W.R. The clinical utility of the foveal electroretinogram: A review //
Doc. Ophthalmol.— 1990.— Vol. 73.— P. 313 —325.
Bill A. The uveal venous pressure // Arch. Ophthalin.— 1963.— Vol. 69 —
P. 780.
Birch D.G. Clinical electrophysiology // Ophthalmol. Clin. North. Amer.—
1989.— Vol. 2.— P. 469-497.
Bishop P.O. Binocular vision. In Adler’s Physiology of the Eye. Ed. Moses and
Hart. — Mosby Co.— St. Louis.—1987.— P. 619—689.
Bjork A. Electromyographic Studies on coordination of antagonistic muscle in
cases of abducens and facial pulsy//Brit. J. Ophthalm.— 1954.— Vol.38.—
P. 605.
Blumhardt L.D., Halliday A.M. Hemisphere contributions to the composition of
the pattern evoked potential waveform // Exp. Brain Res.— 1979.— Vol.
36.— P. 53—69.
Blumhardt L.D. The abnormal pattern visual eviked response in neurology //
A textbook of clinical neurophysiology.— Chichester: John Wiley & Sons.,
1987.— P. 307—342.
Blumhardt L„ Barett G., Halliday A. et al. The effect of the field size on the
pattern-reversal visual evoked responce // Clin. Vis. Sci.— 1989.— Vol.
4.— P. 27—40.
Bodis-Wollner I., Brannan J.R., Chillardi M.F. et al. The importance of physiology
visual evoked potentials // Visual evoked potentials.— Amsterdam — New
York —Oxford: Elsevier, 1990.—P. 1—24.
Bodis-Wollner I„ Ghialardi M.F., Mylin L.H. The importance of stimulus selection
402
in VEP practice: the clinical relevance of visual physiology // Frontiers of
Clinical Neuroscience: Evoked Potentials.— New York: AR Liss, 1986.—
P. 15—27.
Bowmaker J.K., Dartnall H.J. Visual pigments of rods and cones in a human
retina // J. Physiol.— 1980.— Vol. 298.— P. 501—511.
Bradnam M.S., Evans A.L., Montgomery D.N.I. et al. A personal computer-
based visual evoked potential stimulus and recording system // Doc.
Ophthalmol.— 1994.— Vol. 86, № 1.— P. 81—95.
Brecelj J., Strucl M., Skrbek M. Visual evoked potentials in compressive lesions of
the optic chiasm // Neuroophthalmology.— 1992.— Vol. 12.— P. 207—214.
Brecelj J., Strucl M. Some anatomical and physiological aspects of clinical visual
electrophysiology // Zdrav. Vestn.— 1993.— Suppl. 1.— P. 47—57.
Brigel M„ Kaufman D.I., Bobak P. et al. The pattern visual evoked potential: A
multicenter study using standardized techniques // Doc. Ophthalmol.—
1994.— Vol. 86, № 1.— P. 65—81.
Brindley G.S., Westheimer G. The spatial properties of the human electro-
retinogram // J. Physiol.— 1965.— Vol. 179.— P. 518—537.
Broca A., Sulzer D. La sensation luminense en fonction du temps // J. Physiol.
Path. Gen — 1902.— Vol.4.— P. 632.
Brown К. T. The electroretinogram: its components and their origins // Vis. Res.—
1968.— Vol. 8.— P. 633—677.
Brunette J.R. Clinical electroretinography // Appl. Canad. J. Ophthalmol.—
1982.— Vol. 17, № 6.— P. 239—244.
Bush R.A., Sieving P.A. A proximal retinal component in the primat fotopic
ERG a-wave // Inv. Ophthalmol. Vis. Sci.— 1994.— Vol. 735, № 2.— P.
635—645.
Bynke H. The visual field // Neuroophthalmology.— Amsterdam — Oxford—
Princenton, 1980.— P. 352—362.
Campbell F. W., Green D. C. Optical and retinal factors affecting visual resolution//
J. Physiol. Lond.— 1965.— Vol. 181.—P. 576.
Campbell F. W., Kulikowski J. J. An electrophysiological measure of the
psychophysical contrast threshold//!. Physiol.— 1997.— V. 217, № 1.—
P. 54—55.
Carr R.E., Siegel I.M. Electrodiagnostic testing of the visual system: a clinical
guide.— Philadelphia: Davis, 1990.— 252 p.
Cavender, Hobson S.A., Hobson R.R. et al. Comparison of preoperative 10-Hz
visual evoked potentials to contrast sensitivity and visual acuity after cataract
extraction // Doc. Ophthalmol.— 1992.— Vol. 81, № 2.— P. 181—189.
Celesia G., Kaufman D. Pattern ERG and visual evoked potential in
maculopathies and optic nerve diseases // Inv. Ophthalmol. Vis. Sci.—
1985.— Vol. 26.— P. 726.
Celesia G, Ghilardi M.I., Brannan J.R. et al. Clinical application of pattern
visual evoked potentials // Visual evoked potentials.— Amsterdam: Elsevier,
1990.— P. 121—146.
Chiappa KN. Evoked Potentials in Clinical Medicine.— New York: Raven Press,
1990.— P. 111—172.
403
Ciganek L. The EEG response (evoked potentials) to the light stimulus in man //
Electroenceph. Clin. Neurophysiol.— 1961.— Vol. 13.— P. 165—168.
Cohen S. Y., Le Gagasson J.-E, Guez J.-E. et al. Focal visual evoked potentials
generated by scanning laser ophthalmoscope in patients with age-related
macular degeneration treated by perifoveal photocoagulation // Doc.
Ophthalmol.— 1994.— Vol. 86, № 1.— P. 55—65.
Cracco R.Q., Bodis-Wollner I. Frontiers of clinical neuroscience: evoked
potential.— New York: A.R. Liss., 1986.
Dawson W. W., Trick G.L., Maida T.M. Evaluation of the DTL corneal electrode //
Doc. Ophthalmol. Proc. Ser.— 1982.— Vol. 31.— P. 81—88.
Denislic M. Multimodal evoked potentials and central motor latencies in
evaluation of patients with optic neuritis // Zdrav. Vestn.— 1993.— Vol.
62, Suppl. 1.— P. 73—79.
Dennis K.J., Dixon R.D., Winsberg F. et al. Variability in measurement of central
retinal artery velocity using color doppler imaging//!. Ultrasound-Med.—
1995.—Vol. 14, № 6.— P. 463—466.
Desmedt J.R. Visual evoked potentials.— Amsterdam.: Elsevier, 1990.— Vol.
61.— P. 121—146.
Deutman A.F. The hereditary dystrophies of the posterior pole of the eye.—
Springfild, III.: Charles C. Thomas, 1971.— 457 p.
Van Dijk B. W., Spekreijse H. Localisation of electric and magnetic sources of
brain activity // Visual evoked potentials.— Amsterdam—New York —
Oxford: Elsevier, 1990.— P. 57—74.
Doane M.G. Blinking and the mechanics of the lacrimal drainage system//
Ophthalmology.— 1981.— Vol. 88.— P.844.
Dobson V., Teller D. Visual acuity in human infants: a rewier and comparison
of behavioral and electrophysiological studies // Vis. Res.— 1978.— Vol.
18.— P. 1469—1475.
Dowling J.E. Organization of vertebrate retinas // Inv. Ophthal.— 1970.— Vol.
9.— P. 655—680.
Dowling J.E. Electroretinogram and glial responses in retina.— Cambridge:
Belknapp Press, Mass., 1987.— P. 164—186.
Dome P.A., Espiard J.F. L’electro-oculogramme. Principe et technique; son interet
dans 1’etude des maculopathies // Arch. Ophthal.— 1971.— Vol. 31.— P.
217—224.
Drasdo N., Thompson D.A., Thompson S.N. An optical stimulator for studying
the topography of electrical and magnetic visual evoked responses // Doc.
Ophthalmol.— 1992.— Vol. 81, № 2.— P. 219—227.
Dryja T.P., VcGee T.L., Reichel E. et al. A point mutation of the rhodopsin gene
in one form of retinitis pigmentosa // Nature.—1990.— Vol. 343.— P. 364—
366.
Duce I.R. Glutamate // Comparative Invertebrate Neurochemistry.— London—
Sydney: Croom Helm, 1988.— P. 43—89.
Evers H. O., Gouras P. Three cone mechanism in the primate electroretinogram:
two with one without off-center bipolar responses // Vis. Res.— 1986.—
Vol. 26.— P. 245—254.
404
Fagin R.R., Griffin G.R. Stereoaquity tests: comparison of mathematical
equivalents // Amer. J. Optom. Physiol. Opt.— 1982.— Vol. 59.— P. 427.
Falsini B., Porciatti V., Fadda A. et al. The first and second harmonics of the
macular flicker electroretinogram: differential effects of retinal diseases //
Doc. Ophthalmol.— 1995.— Vol. 90.— P. 157—167.
Fender D., Julesz B. Extension of Pannum’s fusional area in binocular stabilized
vision// J. Opt. Soc. Amer.— 1967.— Vol.57.— P. 819.
Ferris III F.L., Kassoff A. et al. New visual acuity charts for clinical research //
Amer. J. Ophthalm.— 1982.— Vol. 94, № 1.— P. 91—96.
Ferris III F.L., Sperduto R.D. Standardized Illumination for visual acuity testing
in clinical research // Amer. J. Ophthalm.— 1982.— Vol. 94, № 1.— P. 97—
98.
Fisher S.K., Anderson D.H^ Erickson P.A. The response of Muller cells in
experimental detachment and reattachment // Proc. Int. Soc. Eye Res.—
1990.— Vol. 6.— P. 76.
Fishman G.A., Cunha-Vaz J.G., Salzano T. Vitreous fluorophotometry in patients
with retinitis pigmentosa//Arch. Ophthalmol.— 1981.— Vol. 99.— P.1202.
Fishman G.A. Basic principles of clinical electrophysiology // Retina.— 1985.—
Vol. 5.— P. 123—126.
Fishman G.A., Sokol S. Electrophysiologic testing in disorders of the retina,
optic nerve and visual pathway // Amer. Acad. Ophthalmol.— 1990.— Vol.
2.— P. 164.
Franceschetti A., Francois J., Babel J. Les Heredo-Degenerescences Chorio-
Retiniennes // Masson Cie.— 1963.— Vol. 2.— P. 1205—1250.
Francois J., Verriest G., De Rouck A. Electro-oculography as a functional test
in pathological conditions of the fundus. First results // Brit. J.
Ophthalmol.— 1956.— Vol. 40.— P. 108—112, 305—312.
Francois J., De Rouck A., Fernandez-Sasso D. Electroretinography and electro-
oculography in diseases of the posterior pole of the eye // Adv. Ophthal.—
1969.— Vol. 21.— P. 132—163.
Francois J. Importance of electrophysiology in ophthalmogenetics //
Ophthalmol.— 1984.— Vol. 188.— P. 14—27.
Frank Y., Kurtzberg D., Kreuzer I. et al. // Dev. Med. Child. Neurol.— 1992.—
Vol. 34, № 4 — P. 305.
Fulton A.B., Hartmann E.E., Hansen R.M. Electrophysiologic testing techniques
for children // Doc. Ophthalmol.— 1989.— Vol. 71.— P. 341—354.
Galloway N.R. Ophthalmic Electrodiagnosis.— London: Lloyd-Luke, 1981.—
180 p.
Gallemore R.P., Griff E.R., Steinberg R.H. Evidance in support of a
photoreceptoral origin for the “light-peake” substance // Invest. Ophthalmol.
Vis. Sci.— 1988.— Vol. 29.— P. 566—571.
Gliem H. Das Elektrookulogramm.— Leipzig: Thieme-Verlag, 1971.— 106 s.
Godde-Jolly D., Larmande A. Les Nystagmus.— Paris, 1973.
Goede P., Kolb H. Identification of the Synaptic Pedicles Belonging to the
differen Spectral Types of Photoreceptor in the Turtle Retina // Vis. Res.—
1994.— Vol. 34, № 21.— P. 2801—2811.
405
Gouras P. Relationships of electro-oculogram to the electroretinogram // The
Clinical value of electroretinography / ISCERG Symp.— Ghent, 1968.—
P. 66—73.
Gouras P. Electroretinography: same basic principles // Invest. Ophtalmol.—
1970.— Vol. 9, № 8.— P. 557—569.
Granit R. The components of the retinal action potential and their relation to
the discharge in the optic nerve // J. Physiol.— 1933.— Vol. 77.— P. 207—
240.
Granit R, Sensory mechanisn of the retina.— London: Oxford Univ. Press,
1947.— P.265.
Grant W.M. Tonographic method for measuring the facility and rate of aqueous
flow in human eyes // Arch. Ophthalm.— 1950.— Vol. 44.— P. 204.
Griff E.R., Linsenmeier R.A., Steinberg R.H. The cellular origin of the fast
oscillation // Doc. Ophthalmol. Proc. Ser.— 1983.— Vol. 37.— P. 13—20.
Gubta L. К, Marmor M.F. Sequental recording of photic and nonphotic electro-
oculogram responses in patients with extensive extramacular drusen // Doc.
Ophthalmol.— 1994.-- Vol. 88, № 1.— P. 49—57.
Gunkel R.D., Cogan D.G. Colorimetry by a new Principle // Arch.Ophthalm.—
1978.— Vol. 96.— № 2.— P. 331—334.
Haley W.J. The field analyzer primer // Humphrey instr.— California, 1993.—
P. 167.
Halliday A. Evoked potentials in clinical testing.— Edinburgh, London: Churchill
Livingstone, 1993.— 357 p.
Hankins M. W., Ikeda H. Dopamine pathways in the outer mammalian retina
and abnormalities expressed in hereditary retinal distrophy // Zdrav. Vestn.—
1993.— Vol. 62, Suppl. 1.— P 33—38.
Harding G.F.A. Origin of visual evoked cortical potentials components //
J.Heckenlively, G.B.Arden. Principles and practice of clinical electrophy-
siology of vision.— Mosby Year Book, 1991.— P. 132—144.
Hart W.M. Visual adaptation // Adler’s Physiology of the Eye / Clinical
application.— Saint Louis: Mosby Co., 1987.— P. 389—414.
Hart W.M. The temporal responsiveness of vision. // Adler’s Physiology of the
Eye / Clinical application.— Saint Louis: Mosby Co, 1987.— P. 429—457.
Hart W.M. Entopic imagery // Aider’s physiology of the Eye. Clinical
application.— St. Louis, 1987.— P.373—388.
Hawlina M., Ikeda H. The role of melatonin and dopamine in retinal renewal
and degeneration // Zdrav. Vestn.— 1993.— Suppl. 1.— P. 23—31.
Hay reh S.S. The optic Nerve Head circulation in health and diseases // Exp. Eye
Res.— 1995.— Vol. 61, № 3.— P. 259—272.
Heckenlively J., Arden G.B. Principles and practice of clinical electrophysiology
of vision.— Mosby Year Book, 1991.— 493 p.
Henkes HE., Lith F, Van Legein C. Electrodiagnosis of the visual system of the
young child with defective vision // Trans. Ophthal. Soc.—1973.— Vol. 93.—
P. 17—20.
Hinchliffe H.N. Clinical evaluation of stereopsis// Brit. Orthopt. J.— 1978.—
Vol. 35.— P. 46..
406
Hobley A. J., Harding G.F.A. The topography of the Pl component of the flash
visual evoked response // Doc. Ophthalmol.— 1989.— Vol. 73.— P. 119—
125.
Hofmann H„ Niemeyer G. Calcium blocks selectively the EOG light peak // Doc.
Ophthalmol.— 1985.— Vol. 60.— P. 361—368.
Hogg P., Grirson L, Willis J. et al. Signalling between cells ivolved in epiretinal
membranes: an in vitro study // Proc. Inter. Soc. Eye Res.— 1990.—
Vol. 6.— P. 35—46.
Hochheimer B.F. A dye for experimental choroidal angiography// Exp. eye res.—
1979.— Vol.29.— p. 141.
Holder G.E, Pattern electroretinography in patients with delayed pattern visual
evoked potentials due to distal anterior visual pathway disfunction //
J. Neurol. Neurosurg. Psychiat.— 1989.— Vol. 52.— P. 1364—1368.
Holder G.E. Chiasmal and retrochiasmal lesions // Principles and practice of clinical
electrophysiology of vision.— Mosby Year Book, 1991.— P. 557—564.
Holly F.J., Lemp M.A. Tear physiology and dry eyes// Surv. Ophthalm.—
1977.— Vol. 22.— P. 69.
Holopigian K., Snow J., Seiple W. et al. Variability of the pattern
electroretinogramm // Doc. Ophthalmol.— 1978.— Vol. 70.— P. 103—115.
Horiguchi M., Miyake Y. Electroretinographic analysis of macular cone and rod
systems in macular diseases // Chibret Int. J. Ophthalmol.— 1992.— Vol.
9, № 2.—P. 25.
Houchin K.W., Purple R.L., Wirtschafter J.D. X-linked congential stationary
nightblindness and depolarizing bipolar system dysfunction // Inv.
Ophthalmol. Vis. Sc.— 1991.— Vol. 32.— P. 1229.
Howard H.J. A test for the judment of Distance // Amer J. Ophthalm.— 1919.—
Vol. 2.— P. 656.
Hoyt W.F., Daroff R.B. Supranuclear disorders of ocular contor system in man:
clinical, anatomical and physiological correlations//The control of eye
movements/Eds. P. Bach-y-Rita, С. C. Collins.— New-York, 1971.— P.175.
Ikeda H. Electrophysiology of the retina and visual pathway // Med.
Ophthalmol.— London: Chapman & Hall, 1976.— P. 38—55.
Ikeda H. Clinical electroretinography // Evoked potentials in clinical testing.—
London: Churchill Livingstone, 1993.— P. 115—139.
Ikeda H. Electrodiagnosis of the primary afferent visual system. Past, present,
future // Zdrav. Vestn.— 1993.— Vol. 62, Suppl. 1.— P. 57—65.
Ikeda H. Mammalian retinal neurotransmitters — as seen throgh the eyes of a
neurophysiologist // Zdrav. Vestn.— 1993.— Vol. 62, Suppl. 1.— P. 7—16.
Ingling C.R., Drum D.A. Why the blue arcs of the retina are blue / Vis. res.—
1977.— Vol. 17.— P. 498.
International standardisation committee. Standard for clinical electroretinography//
Arch.Ophthalmol.— 1989.—Vol. 107.—P. 816—819.
Jacobson J.H. Clinical Electroretinography.— Springfield, Illinois USA: Charles
C Thomas, 1961.—P. 170.
Jacobson S.G., Parel J.-M., Knighton R.W. Hereditary retinal degeneration:
Advances in methodes of testing visual function // Neuro-Ophtalmology
Now.— New-York: Field-Rich Associates, 1986.— P. 98—108.
407
Jacobi P.C., Miliczek K.-D., Zrenner E. Experiences with the international
standard for clinical electroretinography: Normative values for clinical
practice, interindividual and intraindividual variations and possible
extensions // Doc. Ophthalmol.— 1993.— Vol. 85, № 2.— P. 95-—115.
Jayle G.E., Boyer R.L., Saracco J.B. L’electro-oculographie (en tant qu’examen
foncsionel sensoriel) // L’electroretinographie.— Paris: Masson, 1965.—
1259 p.
Jones L. T. The lacrimal secretory System and its treatment // Amer. J. Opht-
halm.— 1966.— Vol. 62, № 1. — P. 47—60.
Jones R .M., Klein R., De Venecia G. et al. Abnormal electro-oculograms from
eyes with a malignant melanoma of the choroid // Inv. Ophthalmol. Vis.
Sci.— 1981.— Vol. 20.— P. 276—279.
Joseph D., Miller S. Alpha-adrenergic receptors mediate basal membrane voltage
and resistahse changes in bovine and retinal pigment epithelium (RPE) //
Inv. Ophthalmol. Vis. Sci.— 1988.— Suppl. 29.— P. 20.
Kahneman D. Temporal summation in an acuity task at different energy levels—
a study of the determination of summation // Vis. Res.— 1964.— Vol. 4.—
P. 557.
Kanski I. Clinical Ophthalmology.— London: Butterworth, 1989.
Karpe G. The Basis of clinical electroretinography // Acta Ophthalmol.— 1945.—
Vol. 24.— P. 1—118.
Karwoski C.J., Proenza L.M. A sources and sinks of light-evoked [K+]o in the
vertebrate retina // Can. J. Phisiol. Pharmacol.— 1987.— Vol. 65.—
P. 1009—1017.
Kato M„ Nakagami T., Watanabe I. Visual evoked potential in macular hole //
Doc. Ophthalmol.— 1991.— Vol. 76, № 4.— P.341—351.
Kato M., Watanabe I. Bull’s eye makulopathy, negative electroretinogram and
low plasma cyclic guanosine monophosphate level // Doc. Ophthalmol.—
1990.— Vol. 75, № 1.— P. 23—32.
Kawasaki K, Madachi-Yamamoto S., Yonemura D. Hyperosmolarity response
of ocular standing potential as a clinical test for retinal pigment epithelium
activity: rhegmatogenous retinal detachment // Doc. Ophthalmol.— 1984.—
Vol. 57.— P. 175—180.
Kawasaki K, Tsuchida K, Jacobson J.H. Positive and negative Deflection in the
off-response of the ERG in man // Amer. J. Ophthalmol.— 1984.— Vol.
72.— P. 367—375.
Kawasaki K, Mukoh S., Yonemura D. et al. Acetazolamide-induced changes of
the membrane potential of the retinal pigment epithelial cell // Doc.
Ophthalmol.— 1986.— Vol. 63,— P. 375—381.
Kiel J. W. The effect of arterial pressure on the ocular pressure // Exp. Eye
Res.— 1995.— Vol. 60, № 3.— P.267—278.
Knapp A.G., Schiller P.H. The contribution of on-bipilar cells to the
electroretinogram of rabbits and monkeys // Vis. Res.— 1984.— Vol. 24.—
P. 1841— 1846.
Kolb H. The architecture of functional neural circuits in the cat retina // Basic
and Clinical Perspectives in Vision Research / Ed. J. Robbins et al.— New
York: Plenum Press, 1995.— P. 3—51.
408
Kolb H., Fernandes E., Ammermuller J., Cuenca N. Substance P: A
neurotranmitter of amacrine and ganglion cells in the vertebrate retina//
J. Histol. Histopathol.— 1995.— Vol. 10.— P. 947—968.
Kolder H.E. Electrooculography // 4th Congress of the European Society of
Opthalmology.— Budapest, 1974.— Part I.— Vol. 169.— P. 127—140.
Korth M. Pattern evoked responses and luminance evoked responses in the human
electroretinogramm // J. Physiol.— 1983.— Vol. 337.— P. 451—469.
Kremmer S., Stodtmeister R., Tolksdorf A. et al. Averaged steady-state visual
evoked cortical potentials at raised intraocular pressure // Doc.
Ophthalmol.— 1992.—Vol. 81, № 2.— P. 189 —197.
Krill A.E. The electroretinogram and electro-oculogram: clinical applications //
Inv. Ophthalmol.—» 1970.— Vol. 9.— P. 600—617.
Laurance L. Visual optics and sight testing// London. Ed3, 1926.
Levinson E., Blake R. Stereopsis by harmonic analysis // Vis. Res.— 1979.—
Vol. 19.— P. 73.
Levy-Schoen A. L’etude des mouvments oculares.— Paris, 1969.
Krill A.E. Hereditary Retinal and Choroidal Diseases // New York: Harper &
Row, 1977.— Vol. 2.— P. 739—824.
Kriss A., Jeffrey B., Taylor D. The electroretinogram in infants and young
children // J. Clin. Neurophysiol.— 1992.— Vol. 9.— P. 373—393.
Kriss A., Russell-Eggitt I. Electrophysiological assessment of visual pathway
function in infants // Eye.— 1992.— Vol. 6.— P. 145—153.
Kubova M„ Kuba Z., Kuba M. Clinical application of motion-onset visual evoked
potentials // Doc. Ophthalmol.— 1992.— Vol. 81, № 2.— P. 209— 219.
Kuffler S. W. The single cell approach in the visual system and the study of
receptive fields // Inv. Ophthalmol.— 1973.— Vol. 12.— P. 794—813.
Langham M.E., Farrell R.A., O’Brien V. et al. Blood flow in the human eye //
Acta Ophthalmol.— 1989.— Vol. 67, suppl. 191.— P. 9—13.
Lehmann D., Skrandies W. Spatial analysis of evoked potentials in man—a
review // Progress in Neurobiology.— 1984.— Vol. 23.— P. 227—250.
Leydhecker W. Perimetry update. // Amer. Ophthalm.— 1983.— Vol. 15, № 6.—
P. 511—541.
Van Lith G. Quantitative evaluation of the electroretinogram // Ophthal-
mologica.— 1981.— Vol. 182.— P. 218—223.
Lee В. B. Parallel pathways in primate retina/ZJohn Dalton’s Colour Vision
Legacy: Selected proceedings of the international conference.— UK: Taylor
& Francis, 1997.—P. 65—72.
Lobstein A., Herr F, Gethard G. Factors effecting the susceptibility of the
glaucomatous eye to raised intraocular pressure// Mod. Probl. Ophthalm.,
Basel.— 1968.— Vol. 6.— p. 73—93.
Lowitzsch K. Visual evoked potentials in neurology: clinical applications in pre —
and post-chiasmal lesions // Zdrav. Vestn.— 1993.— Vol. 62, Suppl. 1.—
P. Gl—TL
Macaluso C., Barrata G, Lamedica A. et al. Visual evoked cortical potentials and
psychophysical determinition of color contrast thresholds along different
chromatic axes // Doc. Ophthalmol.— 1995.— Vol. 90, № 2 .— P. 201—209.
409
Marmor M.F. Clinical electrophysiologic tests for evaluating the retinal pigment
epithelium // Retinal pigment epithelium / Proceedings of the International
Meetings of S. Margherita Ligure.— Amsterdam.: Kugler & Ghedini
Publications, 1989.— P. 9—15.
Marmor M.F., Lurie M. Light-induced electrical responses of the pigment
epithelium: Physiological propities and clinical significance of the c-wave,
standing potential changes (EOG) and melanin response // The retinal
pigment epithelium.— Cambridge.: Harvard University Press, 1979.—
P. 226—244.
Marmor M.F., Hock P.A. A practical method for c-wave recording in man //
Doc. Ophthalmol. Proc. Ser.— 1982.— Vol. 31.— P. 67—72.
Marmor M.F. Clinical electrophysiology of the retinal pigment epithelium //
Doc. Ophyhalmol.— 1991.— Vol. 76, № 4.— P. 301—315.
Marmor M.F., Zrenner E. Standard for clinical electro-oculography // Doc.
Ophthalmol.— 1993.— Vol. 85, № 2.— P. 115—125.
Marmor M.F., Zrenner E. Standard for clinical electroretinography // Doc.
Ophthalmol.— 1995.— Vol. 89, № 3.— P. 199—210.
Marre M., Marre E. Erworbene Storungen des Farbensehens.— VEB Georg
Thieme Leipzig.—1986.—190 p.
Marshall J. The Ageing Retina: Physiology or Pathology // Eye.— 1987.—
Vol. 1.— P. 282—295.
Massof R. W., Finkelstein D. Subclassification of retinitis pigmentosa from two-
color scotopic static perimetry // Doc. Ophthalmol. Proc. Ser.— 1984.—
Vol. 26.— P. 219—225.
Matsui K, Katsumi O., Sakave H. et al. Electroretinogram b/a wave ratio
improvement in central retinal vien obstruction // Brit. J. Ophthalmol.—
1994.— Vol. 78, № 3.— P. 191—198.
Marquez M.F., Shamshinova A.M. Value of electroretinogram (ERG) in
pigmentary retinosis // Revista Cubana Ophthalmol.— Cuba.— 1988.—
№ 1.— P. 33—41.
Michaelson G, Groh M.J., Groh M.E. et al. Advanced primary open-angle
glaucoma is associated with decreased ophthalmic artery blood-flow velocity//
Ger. J. Ophthal.—1995.— Vol. 4, № 1.— P. 21—24.
Miller R.F., Dowling J.E. Intracellular responses of the Muller (glial) cells of
mudpuppy retina: their relation to b-wave of the electrretinogramm //
J. Neurophysiol.— 1970.— Vol. 33.— P.323—341.
Miyake Y. Vitreous fluorophotometry in congential stationary night blindness //
Arch, ophthalmol.— 1983.—Vol. 101.—P.112.
Miyake Y. Studies of local macular ERG // Acta Soc. Ophthalmol. Jpn.— 1988.—
Vol. 92.— P. 1419—1449.
Miyake Y. Macular oscillatory potentials in humans // Doc. Ophthalmol.—
1990.— Vol. 75.— P. 111—124.
Miller N.R. Vascular lesions and circulatory Disorders of the Nervous System
(vasculitis).— In: Walsh and Hoyt’s., Clinical Neuro-Ophthalmology, 4th/
Ed. Williams and Wilkins.— Vol. 4.-1991.— P. 2575—2761.
Morgan W.H., Dao-Yi Yu, Cooper R.L.et al. The influence of cerebrospinal fluid
410
pressure on the lamina cribrosa tissue.Pressure Gradient// Invest. Ophthal.
and Vis. Res—1995 — Vol. 36, № 6.— P. 1163—1173.
Mori T., Marmor M.F., Miyoshi K. et al Combined photic and nonphotic electro-
oculographic responses in the clinical evaluation of the retinal pigment
epithelium // Doc. Ophthalmol.— 1991.— Vol. 76, № 4.— P. 315—323.
Motion J.D., Bowmaker J.K. The spatial arrangement of cones in the primate
fovea // Nature.— 1992.— Vol. 360.— P. 677—679.
Motion J.D. Seeing colour // Colour: art and science / Ed. T. Lamb and
J.Bourrian.— Cambridge University Press, 1995.— 230 p.
Moreland J.D. Threshold measurement of the blue arcs phenomenon // Vis.
res — 1968.— Vol. 8.— P. 1093.
Moses R.A. Intraocular Pressue. Adler’s Physiology of the eye. (Clinical
application).— Saint Louis: The C.V. Mosby Company, 1970.— P. 223—
245.
Moses R.A. Detachment of ciliary body — anatomical and physical considerations //
Invest. Ophthalm — 1965.— Vol. 4.— №5.— p. 930—941.
Miitze K. Die Akkomodation des menschlichen Auges. Berlin.— 1956.
Nadler M.P., Miller D., Nadler D.J. Glare and Contrast Sensitivity for
clinicians.— New York: Springer-Verlag, 1990.— 150 p.
Nilsson S.E., Andersson B.E., Corneal D.C. Recordings of slow ocular potential
changes such as the ERG c-wave and the light peak in clinical work // Doc.
Ophthalmol.— 1988.—Vol. 68.— P. 313—325.
Van Norren D. The technical limitations of clinical electroretinography // Doc.
Ophtalmol.— 1982.— Vol. 31.— P. 3—12.
Odom J., Dawson W., Romano P. et al. Human pattern evoked retinal response
(PERR): spatial tuning and development // Doc. Ophthalmol. Proc. Ser.—
1983.—Vol. 37.— P. 265—271.
Onofry M. Generators of pattern visual evoked potentials in normals and in
patients with retrochiasmatic lesions // Visual evoked potentials.—
Amsterdam: Elseiver, 1990.— P. 87—113.
Opremcak E.M. Uveitis a clinical manual for ocular inflammation.— New York.:
Springer-Verlag, 1995.— 278 p.
Orden V.K.F., Sturr J.F. Age related amplitude changes of the pattern reversal
evoked potential depand on stimulus spatial frequency and contrast // Clin.
Vis. Sci.— 1990.— Vol. 6, № 1.— P. 31—39.
Osborn N.N., Chader G.J. Progress in retinal research.— Oxford: Pergamon
press, 1985.— Vol 4.— 250 p.
Ossenblok P. The sourses of the pattern VEP in man // Academish proefschrift.—
Netherlands: University of Amsterdam, 1992.— P. 1—141.
Ossenblok P., Reits T., Spekreijse H. Check size dependency of the sources of
the hemifield-onset evoked potentials // Doc. Ophtalmol.— 1994.— Vol. 88,
№ 1.— P. 77—89.
Osterberg G. Topography of the layer ofrods and cones in the human retina //
Acta. Ophthalmol.— 1935.— Suppl. 6:1—103.— P. 62—92.
Pallas S., Schmidt W., Dodt E. Scotopic versus photopic pattern onset-offset
electroretinograms // Doc. Ophthalmol.—1992.—Vol. 81, № 2.— P. 239—253.
411
Park M.M. Sensory adaptation in strabismus // Sympos. on strabismus / Transact,
of N-Orleans Acad. Ophthalmol.— St.Louis.: C.N.Mosby, 1971.— P. 121.
Pirtosek Z., Barret G, Lees A J. Seeing not just watching: poststriate cognitive
processing of visual signals // Zdrav. Vestn.— 1993.— Vol. 62, Suppl. 1.—
P. 39—47.
Potts A.M. Electrophysiological measurements // The Assessment of Visual
Function.— St Louis: CV Mosby Co, 1972.— P. 187—206.
Regan D., Tyler C. W. Temporal summation and its limit for wavelength changes:
an analog of Bloch’s law for color vision // J. Opht. Soc. Amer.— 1971.—
Vol. 61.— P. 1414.
Regan D. Human brain electrophysiology: evoked potentials and evoked magnetic
fields in science and medicine.— New York: Elsevier, 1989.
Reichel E., Pulliafito C.A., Ducker J.S. et al. Indocianin green dye-enchanced
diod laser photocoagulation poorly defined subfoveal choroidal
neovascularisation // Ophthal. Surg.— 1994.— Vol. 25.— P.195—201.
Rimmer S, Katz B. The pattern electroretinogramm: technical aspects and clinical
significance // J. Clin. Neurophysiol.— 1989.— Vol. 6.— P. 85—99.
Ripps H., Witkovsky P. Neuron-glia interaction in the brain and retina // Progress in
Retinal Research.— Oxford: Pergamon Press, 1985.— Vol. 5.— P. 181—219.
Roberge F.G. T-cell activation by eye tissue specific cells // Proc. Int. Society Eye
Res. —1990.— Vol. 6.— 73 p.
Rodeick R. W. The Vertebrate retina: priciples of structure and function.— San
Francisko: W.H. Freeman & Co., 1973.
Roufs I.A.I. On the relation between the threshold of short flashes, the flicker
fusion frequency and the visual latency // Inst, for Perception research.
Eindhoven.— The Netherlands Annual Progress Report.— 1966.— № 1.—
P. 69— 77.
Rovamo J., Virsu V. An estimation and application of the human cortical
magnification factor // Exp. Brain Res.— 1979.— Vol. 37.— P. 495—510.
Sanders E., Volkers A., Van Der Poel I. et al. Estimation of visual function after
optic neuritis: a comparison of clinical tests // Brit. J. Ophthalmol.— 1986.—
Vol. 70, № 12.— P. 918—924.
Sanders D.R. Quantitative assessment of post-surgial breakdown of the blood-
aqueous barrier // Arch. Ophthalmol.— 1983.— Vol. 101.— P. 103.
Salzman J., Seipie W., Carr R. et al. Electrophysiologic assessment of aphakic cystoid
macular edema // Brit. J. Ophthalmol.— 1986.— Vol. 70.— P. 819—824.
Sandberg M.A., Ariel M. A hand-held, two-channel stimulator-ophthalmoscope //
Arch. Ophthalmol.— 1977.— Vol. 95.— P. 1881—1882.
Schiller P.H. On and off channels of the visual system // Vision and the Brain.—
Raven Press, 1990.— P. 35—41.
Schippers V, Silny J., Rau G. et al. Results with multichannel topographical
bipolar recordings of the visual evoked potential in normals and amlyopes //
Doc. Ophthalmol.— 1990.— Vol. 76, № 1.— P. 91—101.
Shamshinova A.M., Ivanina T.A., Yakovlev A.A. et al. Electroretinography in the
diagnosis of tallium intoxication // J. Hyg., Epidemiol., Microbiol.,
Immunol.— 1990.— Vol. 34, № 2.— P. 113—121.
412
Shamshinova А.М. Local ERG for clinical examination of eye diseases // Doc.
Ophthalmol.— 1990.— Vol. 76.— P. 1—11.
Shamshinova A.M., Katargina L.A., Orlovskaya L.S. Electroretinographyc
findings in children with uveitis. Recent advances in uveitis.— Amsterdam—
New York, 1992.— P. 397-400.
Shamshinova A. M., Endrichovsky S. N., Nesteruk L. I., Yacovlev A. A. New
Method for Investigation of Colour Sensitivity//John Dalton’s Colour Vision
Legacy: Selected proceedings of the international conference.— UK: Taylor
& Francis, 1997.—P. 317—324.
Sieving P.A. Photopic on- and off-pathway abnormalities in retinal dystrophies //
Trans. Amer. Ophthalmol. Soc.— 1993.— Vol. 91.— P. 701—773.
Sieving P.A., Murayama K., Naarendorp F. Push-pull model of the primat
photopic electroretinogram: A role for hyperpolarizing neurons in sharping
the b-wave // Vis. Neurosc.— 1994. —Vol. 2.— P. 515—532.
? Silver D.M., Farrell R.A., Langham M.E. et al. Estimation of pulsative ocular
blood flow from intraocular pressure // Acta Opthalmol.— 1989.— Vol.
67.— P.25 —29.
Simons K. Stereaquity norms in young children // Arch. Ophthalmol.— 1981.—
Vol. 99.— P. 439.
Simons K. A comparison of the Frisby, Random-Dot E., TNO and Randot
circles stereotests in screening and office use // Arch. Ophthalmol.— 1981.—
Vol. 99.— P. 446.
Skarf B. Clinical use of visual evoked potentials // Ophthalmol. Clin. North
Amer.— 1989.— Vol. 2.— P. 499—518.
Skrandes W., Lehmann D. Spatial principle components of multichannel maps
evoked by lateral visual half-field stimuli // Electroenceph. Clin.
Neurophysiol.— 1982.— Vol. 54.— P. 662—667.
Smith E.L. Ill, Harwerth R.S., Crawford M.L.J. et al. Contribution of the retinal
Ochannels to Scotopic and photopic spectral sensitivity // Vis. Neurosc.—
1991.— Vol. 7.— P. 75—86.
Smith R.G., Brimlow G.M., Hardman L.S.J. et al. Evoked responses in patients
with macular holes // Doc. Ophthalmol.— 1990. — Vol. 75.— P. 135—144.
Snodderly D.M., Auran J.D., Delor F.C. The macular pigment. Spatial distribution
in primat retinas // Inv. Ophthalmol. Vis. Sci.— 1984.— Vol. 25.— P. 674.
Sokol S., Nadler D. Simultaneous electroretinograms and visual evoked potentials
from adult amblyopes in response to pattern stimulus // Inv. Ophthalmol.
Vis. Sc.— 1979.— Vol. 18.— P. 848—855.
Sokol S. Electrophysiology and psychophysics; their use in ophthalmic diagnosis //
Int. Ophthal. Clin.— Boston: Little Brown & Co, 1980.—Vol. 31.—P. 34—36.
Sokol S. Abnormal evoked potential latencies in ambliopia // Brit. J.
Ophthalmol.— 1983.— Vol. 67.— P. 310—314.
Sorsby A. Modem ophthalmology.— London: Butterworth, 1972.
Spekreijse H., Apkarian P. The use of a system analysis approach to
electrodiagnostic (ERG and VEP) asesstment // Vis. Res.— 1986.— Vol.
26, № 1.— P. 195—219.
Spekreijse H., Van der Tweel L.H., Zuidema T. Contrast evoked responses in
man // Vis. Res.— 1973.— Vol. 13.— P. 1577—1601.
413
Srebro R. Realistic modeling of VEP topography // Vis. Res. 1990.— Vol. 30,
№ 7.— P. 1001—1009. ,
Steinberg R.H., Linsenmeier R.A., Griff E.R. Retinal pigment epithelial cell
contributions to the electroretinogram and electrooculogram // Progress in
retinal research.— Oxford: Pergamon Press, 1985.— Vol. 4.— P. 33—66.
Stockton R., Slaughter M. В-wave of the electroretinogram. A reflection of ON-
bipolar cell activity // J. Gen. Physiol.— 1989.— Vol. 93.— P. 101—122.
Strucl M. Electrophysiological basis of diagnostics at different levels of the
visual system // Neurologia.— 1990.— Vol. 39, Suppl. 1.— P. 203—210.
Sutter E.E., Tran D. Imaging of visual function using ERG and VEP responses //
J. Noninvasive assessment of the visual system.— Nevada: Incline Village,
1990.— Vol. 3.— P. 54—57.
Sutter E.E., Tran D. The field topography of ERG components in man-1. The
photopic luminance. Response // Vis. Res.— 1992.— Vol. 32, № 3.—
P. 433—446.
Thaler A., Lessel M.R. Ischemic retinopathy: reduced light-peak and dark trough
amplitudes in electrooculography, ERG, VER, and Psychophysics // Doc.
Ophthalmol. Proc. Ser.— 1977.— Vol. 13.— P. 119—121.
Thaler A.R.G., Lessel M.R., Heiling P. et al. The fast oscillation of the electro-
oculogram // Ophthalmol. Res.— 1982.— Vol. 14.— P. 210—214.
Thaung J., Beckman C., Abrahamsson M. et aL The “light scattering factor”
(importance of stimulus geometry contrast definition and adaptation) //
Inv. Ophthalmol. Vis. Sci.— 1995.— Vol. 36, № 11.— P.2313—2317.
Thimmins N., Marmor M.F. Studies on the stability of the clinical electro-
oculogram // Doc. Ophthalmol.— 1992.— Vol. 81, № 2.— P. 163—173.
Tomita T. Electrical activity in the vertebrate retina // J. Optical Soc.— 1963.—
Vol. 53.— P. 49—57.
Thompson H.S. Light — near dissotioation of the pupil // Ophthalmologica.—
1984.— Vol. 189.— P. 21.
Trick G.L. Visual disfunction in normotensive glaucoma // Doc. Ophthalmol.—
1993.— Vol. 85, № 2— P. 125—135.
Usui S, Nagasaka E. Spatial distribution of local flash electroretinogram by multi-
input stimulation // Doc. Ophthalmol.—1994.— Vol. 88.— P. 57—63.
Vaegan P.P. T., Satter EE. Fundamental differences between the nonlinearities
of pattern and focal electroretinogramms // Doc. Ophthalmol.— 1990.—
Vol. 76, № 1.— P. 13—27.
Vaegan P.P. T Amine transmitters and their associated second messenger systems //
Compar. Invert. Neurochemistry.— London, Sydney: Croom Helm, 1988.—
P. 124—174.
Verriest G. The occupational visual field. — Sacramento, USA, 1982.
Weleber R.G^ Eisner A. Retinal function and physiological studies // Retinal
Dystrophies and Degeneration.— New York: Raven Press, 1988.— P. 21—69.
Westheimer G. Entoptic visualization of Styles-Crawford effect // Arch.
Ophthalmol.— 1968.— Vol. 79.— P. 584.
Westheimer G., Hauskey. Temporal and spatial interference with vernier acuity //
Vis. Res.— 1975.— Vol. 15.— P. 1137—1146,
414
Westheimer G. The scaling of visual acuity measurement // Arch. Ophthalmol.—
1979.— Vol. 97.— P. 327—329.
Westheimer (7. Visual acuity // Adler’s physiology of the Eye.— St. Louis: Moslos
Courp.— 1987.— P. 415—428.
Whitmore A. V, Bowmaker J.K. Short communication. Differeces in the Temporal
Properties of Human Longwave- and Middlwave sensitive Cones // Europ.
J. Neurosci — 1995.— Vol. 7.— P. 1420—1423.
Williamson Т.Н., Baxter G.M., Pyott A. et al.A. comparison of colour Doppler
imaging of orbital vessels and other methods of blood low assessment//
Graefes Arch. Clin. Exp. Ophthal.— 1995.—Vol. 233.— № 2.— P. 80—84.
Wolfe J.M. In Introduction to contrast Sensitivity Testing.— In: Nader M.P. et
al. (Ed) Glare and Contrast Sensitivity for Clinicians.— New York — Berlin:
Springer-Verlag, 1990.— P. 5—23.
Xu X, Karwoski C.J. Current sourses and sinks associated with the ERG
b-wave // Inv. Ophthalmol. Vis. Sci.— 1993.— Vol. 34.— P. 1272—1275.
Yokoyama M., Vi K, Youshida T. The spectral response in the retina with
progressive cone dystrophy // Jap. J. Clin. Ophthalmol.— 1974.— Vol. 28.—
P. 805—813.
Volkov V. V., Kolesnikova L.N. Use of cobalt glass in Accomodation studies. In
Functional Examination in Ophthalmology/Ed. J. Francois.— Basel: S.
Karger, 1974.—Pt. IL— P. 303—306.
Yonemura D., Kawasaki K. Electrophysiological study on activities of neuronal
and nonneuronal retinal elements in man with reference to its clinical
application // Jap. J. Ophthalmol.— 1977.— Vol. 22.— P. 195—213.
Yoshii M., Yanashima K, Kawara T. et al. Luminance-unbalanced pattern onset-
ofset electroretinogram and visual evoked cortical potencial // Doc.
Ophthalmol.— 1991.— Vol. 76, № 4.— P. 359—367.